

























































































































































































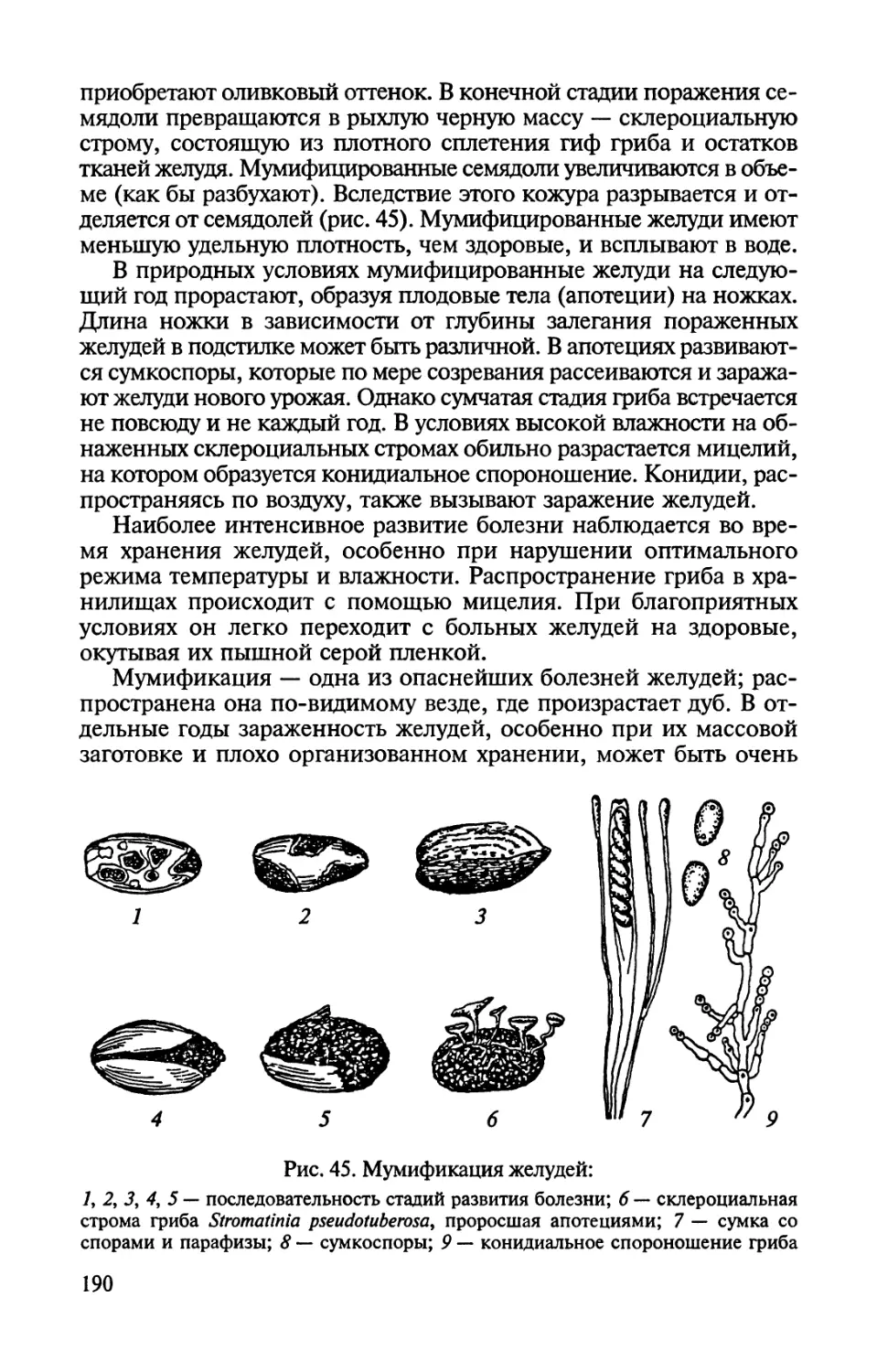











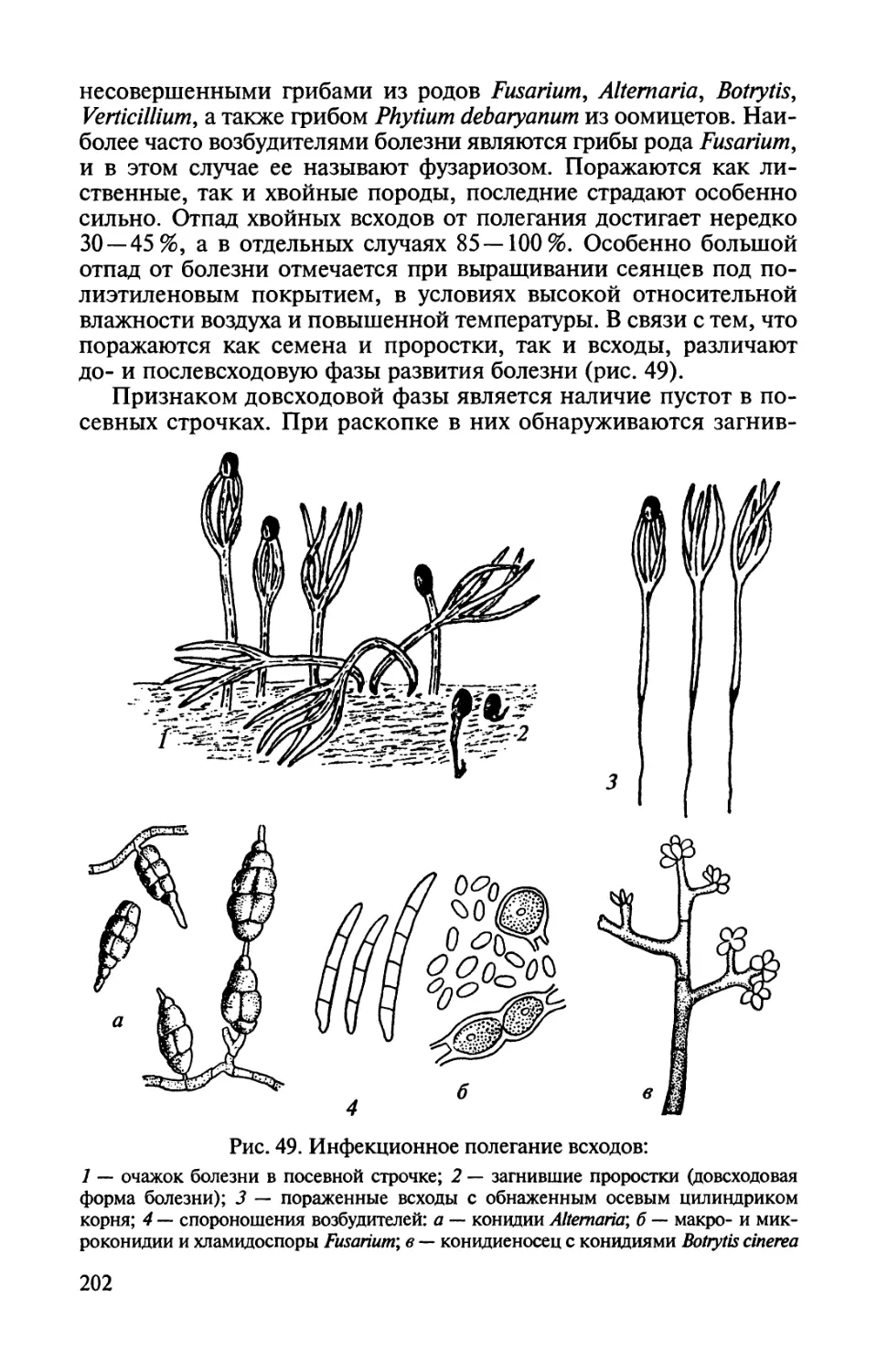





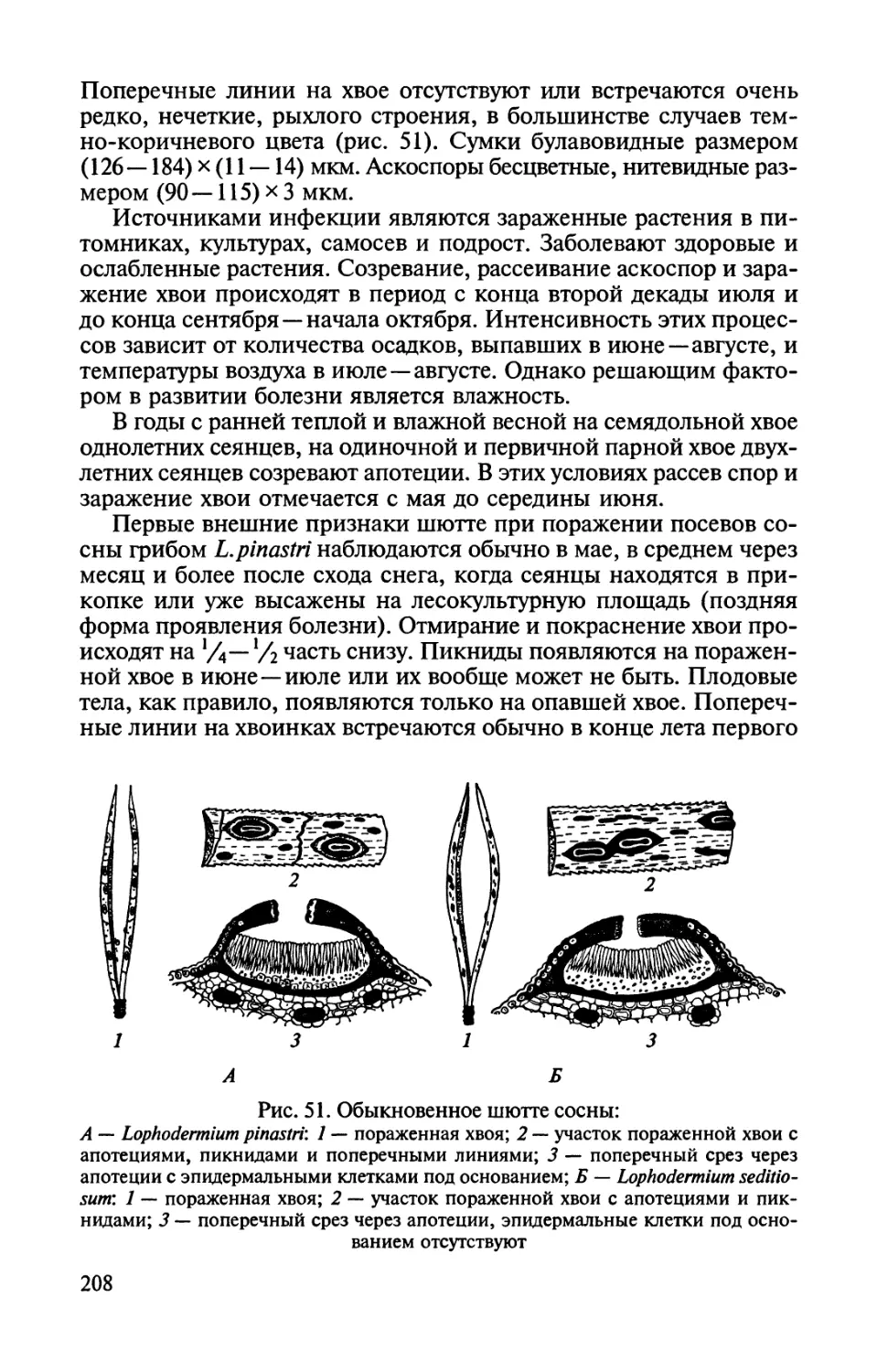


























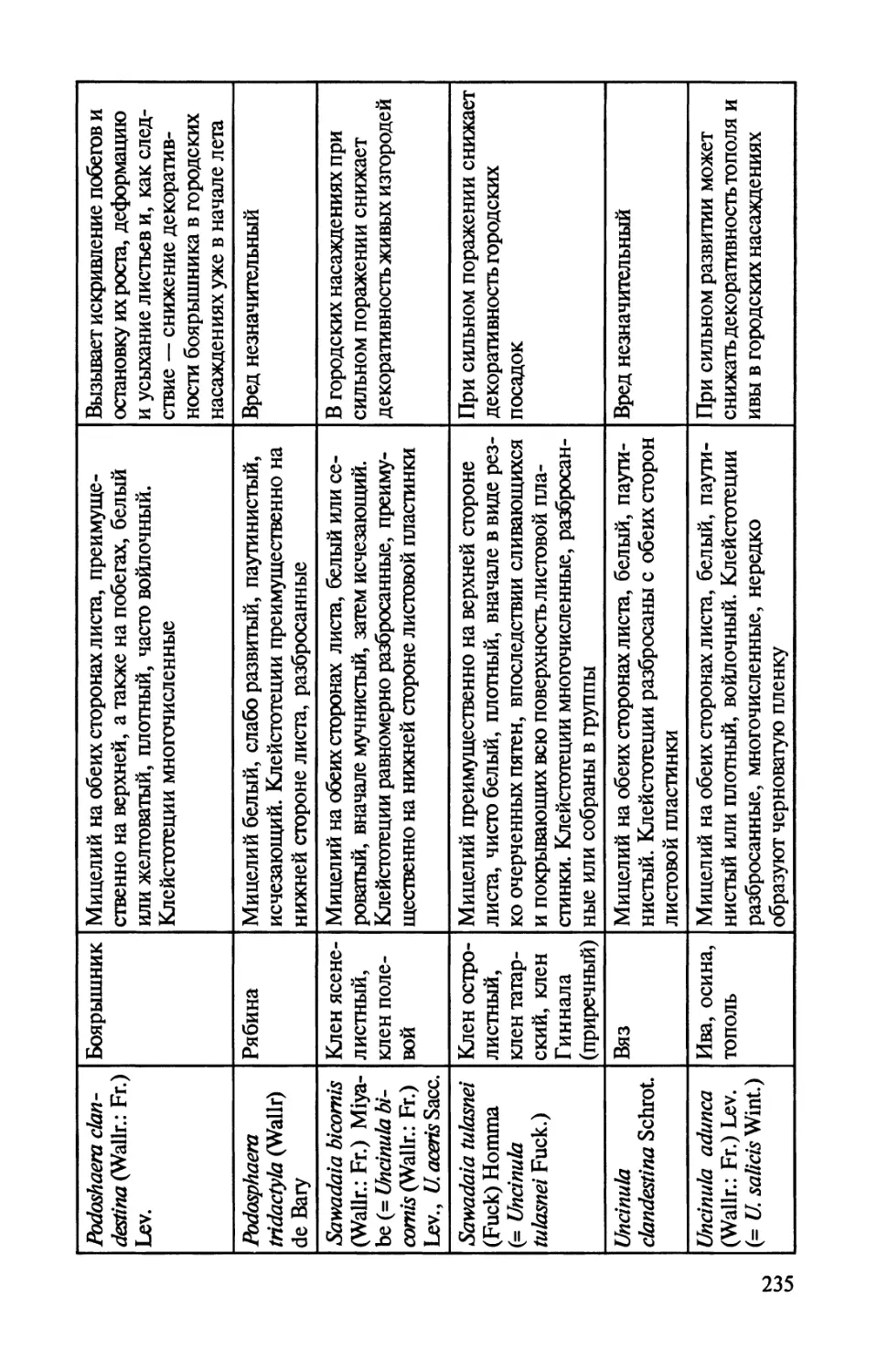































































































































































































































































Автор: Соколова Э.С. Семенкова И.Г.
Теги: лесное хозяйство лесоводство болезни растений и борьба с ними биология экология ландшафт систематика издательский центр академия учебник для студентов
ISBN: 5-7695-1259-8
Год: 2003




• :
U
^ь
ВЫСШЕЕ ПРОФЕССИОНАЛЬНОЕ ОБРАЗОВАНИЕ
И.Г.СЕМЕНКОВА, Э.С.СОКОЛОВА
ФИТОПАТОЛОГИЯ
Допущено
Министерством образования Российской Федерации в качестве
учебника для студентов высших учебных заведений, обучающихся
по направлению подготовки дипломированных специалистов
«Лесное хозяйство и ландшафтное строительство»
Москва
ACADEMA
2003
УДК 630.44(075.8)
bliK 44.7я73
сзо
Рецензенты:
д-р биол. наук, проф. А.М.Жуков (Всероссийский научно-исследог ггсльский
институт лесоводства и механизации лесного хозяйства);
д-р биол. наук, проф. Ю. Л.Демаков; д-р с.-х. наук, проф. И. А. Алексеев
и канд. с.-х. наук, доц. Н.И.Гаврицкова
(кафедра управления природопользованием и лесозащиты Марийского
государственного техническою университета)
Семенкова И. Г.
СЗО Фитопатология: Учебник дли студ. вузов /
И.Г.Семенкова, Э.С.Соколова. — М.: Издательский центр «Академия»,
2003. — 480 с, [16] с цв. ил.: ил.
ISBN 5-7695-1259-8
В учебнике изложены основы биологии, экологии и систематики
грибов и грибоподобных организмов, бактерий, вирусов, фитонематод и
других возбудителей болезней растений, симптомы и типы этих болезней.
Рассмотрены основные методы и средства борьбы с болезнями растений,
диагностические признаки, особенности развития и условия
распространения наиболее опасных болезней деревьев, кустарников и цветочных
культур, изложены меры борьбы с ними. Приводится описание наиболее
вредоносных видов складских и домовых грибов и методов Защиты древесины.
Для студентов высших учебных заведений, обучающихся по
направлению «Лесное хозяйство и ландшафтное строительство».
УДК 630.44(075.8)
ББК 44.7я73
© Семенкова И. Г., Соколова Э.С., 2003
ISBN 5-7695-1259-8 © Издательский центр «Академия», 2003
ПРЕДИСЛОВИЕ
Лес — это ценнейшее достояние народа. Все леса подлежат
охране от пожаров, незаконных порубок, нарушений установленного
порядка лесопользования и других действий, причиняющих вред
лесу, а также защите от вредных насекомых и болезней.
Болезни наносят значительный ущерб лесным насаждениям,
препятствуют выращиванию посадочного материала, вызывают
гибель семян древесных пород и кустарников. Поэтому в научных
учреждениях нашей страны и за рубежом ведется интенсивная
работа по изучению основных болезней древесных пород,
разрабатываются меры борьбы с ними. Ежегодно возрастает поток
научной информации, издаются пособия по лесной фитопатологии и
защите леса. За 10 лет, прошедших после выхода в свет учебника
лесной фитопатологии, значительно расширился объем знаний о
систематике, биологии и экологии грибов и других возбудителей
болезней растений, эпифитотиологии, иммунитете растений.
Выявлены новые для нашей страны болезни лесных пород.
Разработаны методы и средства защиты леса от болезней. Все это
обусловило необходимость издания предлагаемого учебника.
Учебник написан на основе курса лекций, которые авторы
читают в течение многих лет на лесном факультете и факультете
ландшафтной архитектуры Московского государственного
университета леса. В нем рассматриваются все основные вопросы,
предусмотренные примерной программой дисциплины «Защита
растений», рекомендованной Министерством образования
Российской Федерации для подготовки специалистов по направлению
«Лесное хозяйство и ландшафтное строительство».
Введение и главы 1 — 5, 9, 12—14 написаны И. Г. Семенковой,
главы 6 — 8, 10 и 11 — Э.С.Соколовой. Указатель латинских
названий составлен И. Г. Семенковой, указатели русских названий
болезней растений и фитопатогенных организмов, наименований
химических и биологических средств защиты растений и
древесины, предметный — Э.С.Соколовой.
ВВЕДЕНИЕ
Фитопатология — наука о болезнях растений, их причинах и
мерах борьбы с ними. В ее названии соединены корни трех греческих
слов: phyton — растение, pathos — болезнь, logos — слово, учение.
Различают общую, сельскохозяйственную и лесную фитопатологию.
Общая фитопатология рассматривает причины болезней
растений, закономерности взаимоотношений между возбудителями
болезней, пораженными растениями и окружающей средой,
факторы устойчивости растений к болезням и другие общие
теоретические вопросы. Сельскохозяйственная фитопатология
занимается изучением болезней сельскохозяйственных культур и
разработкой мер борьбы с ними.
Лесная фитопатология изучает болезни древесных растений и
процессы биологического разрушения древесины на складах, в
сооружениях и постройках, разрабатывает меры борьбы с ними.
В центре внимания фитопатолога находятся чаще всего не
отдельные растения, а большие группы растений или их сообщества:
питомник, сад, лесное насаждение в целом. В таком сложном
биогеоценозе, каким является лес, массовое заболевание одного или
нескольких видов древесных пород обычно влечет за собой
нарушение сбалансированных связей между популяциями и внутри
популяций организмов, населяющих лес, т. е. нарушение всей
экосистемы. Лесная фитопатология в этих случаях имеет дело с болезнями
не только отдельных видов древесных растений, но и леса в целом.
Большинство наиболее распространенных и вредоносных
болезней растений, в том числе лесных пород, вызывается грибами,
бактериями, вирусами и другими микроорганизмами.
Биологические свойства, особенности развития и распространения
возбудителей во многом определяют сущность и характер развития самих
болезней растений. Поэтому совершенно естественно, что
фитопатология возникла и развивалась на базе соответствующих наук:
микологии (науки о грибах), бактериологии (науки о бактериях),
вирусологии (науки о вирусах), микробиологии (науки о микроорганизмах
вообще). Несмотря на то что фитопатология уже давно оформилась
в самостоятельную отрасль биологии, она и сейчас опирается на
эти науки, используя их методы и новейшие достижения.
Существует тесная взаимосвязь между фитопатологией и
другими биологическими и естественными науками. Так, при
исследовании изменений анатомического строения, обмена веществ и
физиологических функций больного растения фитопатология со-
4
прикасается с ботаникой, физиологией растений и биохимией, а
при изучении механизмов восприимчивости и устойчивости
растений к болезням — с генетикой, цитологией и молекулярной
биологией. В то же время фитопатологические наблюдения и
исследования являются важным звеном в работах по селекции
новых сортов и форм культурных растений, в том числе цветочных и
древесных пород.
Распространение и развитие болезней в лесах и цветочных
хозяйствах, причиняемый ими ущерб в значительной степени
зависят от почвенных и других условий, организации и технического
уровня ведения лесного хозяйства, соблюдения правил высокой
агротехники. Поэтому фитопатология непосредственно связана с
многими другими дисциплинами, особенно с такими, как
экология, почвоведение, лесные культуры, лесоводство, лесная
таксация, цветоводство, экономика и организация производства.
Современная фитопатология немыслима без использования
эффективных химических средств борьбы с болезнями растений; отсюда
очевиден ее контакт с химией.
Неразрывна связь фитопатологии с лесной энтомологией —
наукой о вредных и полезных для леса насекомых. Поражение и
ослабление насаждений грибными болезнями, например
корневыми гнилями, часто приводят к увеличению численности и
вспышкам массового размножения вредных насекомых, особенно
стволовых вредителей. В свою очередь, повреждение насаждений
хвое- и листогрызущими насекомыми может понизить их
устойчивость к различным заболеваниям. Кроме того, некоторые
насекомые являются переносчиками возбудителей болезней растений,
способствуют их размножению и сохранению в природе.
Велико значение фитопатологии и защиты растений в
решении экологических проблем, в деле охраны окружающей среды.
Процесс возникновения, становления и развития
фитопатологии как науки связан с именами многих отечественных и
зарубежных ученых. Основоположником лесной фитопатологии
считается немецкий ботаник, лесовод и фитопатолог Роберт Гартиг
(1839-1901).
В 1874 г. вышел в свет учебник Р.Гартига «Болезни древесных
пород», который был переведен на все европейские языки. В 1894 г.
он был издан и в России. В этой и других работах ученый дал
подробное описание болезней лесных пород и биологии их
возбудителей, сопроводив текст превосходными иллюстрациями.
После Гартига в Германии и других странах Европы и Америки в
области лесной фитопатологии работали многие известные ученые.
Классические исследования грибов — возбудителей
заболеваний растений были выполнены выдающимся русским ученым,
основателем отечественной микологии и фитопатологии
академиком М.С.Ворониным (1838 — 1903).
5
В 1897 г. в Петербурге вышел «Краткий очерк микологии с
указанием грибов, наиболее вредных в сельском хозяйстве и
лесоводстве» профессора И. П. Бородина. Эту книгу можно считать
первым в России учебником микологии, ибо в ней самое большое
место отведено описанию морфологии и биологии грибов,
паразитирующих на древесных растениях и сельскохозяйственных
культурах.
Неоценимый вклад в развитие отечественной и мировой
микологии и фитопатологии внес А.А.Ячевский (1863—1932) — один
из талантливейших учеников М. С. Воронина. Он написал около
500 научных работ, им были созданы такие фундаментальные
труды, как «Паразитные и сапрофитные грибы русских лесных
пород» (1897), двухтомный «Определитель грибов» (1913, 1917),
«Основы микологии» (1933), «Бактериозы растений» (1935) и многие
другие. Ячевский был также неутомимым организатором фитопа-
тологических исследований, инициатором создания в нашей стране
службы защиты растений. В 1902 г. он организовал и возглавил
Центральную фитопатологическую станцию, где издавал
«Ежегодник сведений о болезнях и повреждениях культурных и
дикорастущих полезных растений». Эти выпуски имели важное
значение в распространении и популяризации фитопатологических
знаний, организации фитопатологических наблюдений на местах.
В 1907 г. Ячевский организовал в Петербурге Бюро по микологии
и фитопатологии; впоследствии оно было преобразовано в
Лабораторию микологии им. А. А.Ячевского, вошедшую в состав
Всесоюзного института защиты растений (ВИЗР).
В 1920 г. в Петроградском лесном институте
(Санкт-Петербургская государственная лесотехническая академия) была основана
первая в нашей стране кафедра лесной фитопатологии, которую
в течение 26 лет возглавлял ученик А. А.Ячевского, крупнейший
ученый и педагог С.И.Ванин (1890—1951).
С. И. Ванин вошел в историю науки как создатель
отечественной школы лесных фитопатологов и автор первого в стране
учебника лесной фитопатологии для вузов (первое издание вышло в
свет в 1931 г., а последнее, пятое, — в 1951 г.). Ученым были
опубликованы многочисленные работы по различным вопросам
лесной фитопатологии. Среди них наиболее известны такие
книги, как «Болезни сеянцев и семян лесных пород» (1931),
«Домовые грибы» (1931), «Методы исследований грибных болезней леса
и повреждений древесины» (1934) и др. Плодотворная научно-
педагогическая деятельность Ванина явилась стимулом к
развертыванию фитопатологических исследований в других лесных вузах и
научных учреждениях, оказала огромное влияние на весь ход
последующего развития отечественной лесной фитопатологии, имела
большое значение в подготовке высококвалифицированных
специалистов лесного хозяйства и воспитании научных кадров лес-
6
ных фитопатологов. Многие ученики и последователи С. И. Ванина
(А. Т. Вакин, Д. В. Соколов, И. Я. Шемякин и др.) также внесли
значительный вклад в развитие лесной фитопатологии и лесозащиты.
В ряду ученых, наиболее ярко вписавших свои имена в
историю лесной фитопатологии, стоит имя А. С. Бондарцева (1877 —
1969), одного из крупнейших в мире специалистов по
систематике и биологии дереворазрушающих грибов, автора капитального
труда «Трутовые грибы европейской части СССР и Кавказа» (1953),
книги «Пособие для определения домовых грибов» (1950) и др.
Работы В.Ф.Купревича (1897—1969), среди которых
выделяется монография «Физиология больного растения в связи с
общими вопросами паразитизма» (1947), положили начало физио-
лого-биохимическому направлению в отечественной
фитопатологии. Исследования по патологической физиологии больного
растения успешно развиваются в последние десятилетия как в нашей
стране, так и за рубежом,
На современном этапе развития лесной фитопатологии
большое внимание уделяется изучению факторов иммунитета
растений, причин возникновения эпифитотий и их прогнозированию,
разработке систем мероприятий по борьбе с наиболее опасными
болезнями и комплексами заболеваний в определенных эколого-
производственных объектах (питомниках, теплицах, молодых
культурах, взрослых насаждениях, городских посадках и т.п.),
внедрению новых способов повышения устойчивости растений к
инфекционным болезням и другим неблагоприятным факторам
окружающей среды.
По ряду важнейших вопросов лесной фитопатологии, в
частности по проблемам корневой губки, сосудистых болезней,
проводятся комплексные исследования, в которых вместе с фитопато-
логами участвуют представители смежных наук — физиологи,
биохимики, лесоводы и другие специалисты.
Глава 1
ОБЩИЕ СВЕДЕНИЯ О БОЛЕЗНЯХ
РАСТЕНИЙ
1.1. ПОНЯТИЕ О БОЛЕЗНЯХ РАСТЕНИЙ
И ИХ ПРИЧИНАХ
Болезнь растения — это сложный патологический процесс,
который возникает под влиянием внешних факторов, протекает
во взаимодействии с окружающей средой и проявляется в
нарушениях физиологических функций и анатомо-морфологических
изменениях всего растения или отдельных его органов. Болезнь
ведет к отмиранию пораженных тканей, ослаблению, снижению
продуктивности или гибели всего растения.
На протяжении всей жизни растение тесно связано с
окружающей средой и постоянно взаимодействует с ней. Его рост,
развитие, продуктивность зависят от температуры и влажности,
химического состава почвы и воздуха, разнообразных живых
организмов и множества других факторов. Когда условия окружающей
среды соответствуют требованиям растения и во
взаимоотношениях растения с населяющими среду живыми организмами
наблюдается равновесие, его обмен веществ протекает нормально,
оно растет и развивается, не обнаруживая функциональных и
морфологических нарушений. Если же какой-либо фактор среды
резко отклоняется от оптимума или на растение нападают другие
организмы, то происходят нарушения обмена веществ и
физиологических функций. Это, как правило, ведет к изменениям
анатомического строения и внешнего вида растения, т.е. возникает
патологический процесс, или болезнь.
Различают инфекционные и неинфекционные болезни
растений. Неинфекционные болезни возникают без участия фи-
топатогенных организмов, под влиянием неблагоприятных для
нормального развития растений факторов окружающей среды.
Таковы, например, болезни, вызываемые недостатком или избытком
воды, нарушениями режима питания, недостатком кислорода,
воздействием слишком высокой или низкой температуры,
недостатком света, наличием в воздухе или почве токсичных для растений
веществ и т.д.
8
Важнейшая особенность неинфекционных болезней — их
неспособность передаваться от больного растения к здоровому.
Инфекционные болезни растений вызываются
микроорганизмами (преимущественно грибами, бактериями, вирусами),
паразитическими нематодами или цветковыми
растениями-паразитами. Организм, вызывающий болезнь, называют ее
возбудителем, или патогеном. Организмы, способные вызывать
болезни растений, называются фитопатогенными. Растение, на
котором поселяется и за счет которого питается патоген,
называют растением-хозяином.
Инфекционные болезни растений весьма многочисленны, и
многие из них причиняют большой ущерб, так как могут
передаваться от больных растений здоровым. Они разнообразны по
характеру вызываемых ими патологических процессов и внешним
признакам.
Несмотря на существенные различия между инфекционными
и неинфекционными болезнями, их нельзя рассматривать как
явления обособленные. В природе между ними наблюдается
определенная взаимосвязь: часто инфекционные заболевания
возникают на фоне предварительного поражения и ослабления растений
неинфекционными болезнями. Во многих случаях
неинфекционный патологический процесс обусловливает саму возможность
проникновения патогена в растение, облегчает его заражение,
способствует более интенсивному развитию инфекционного
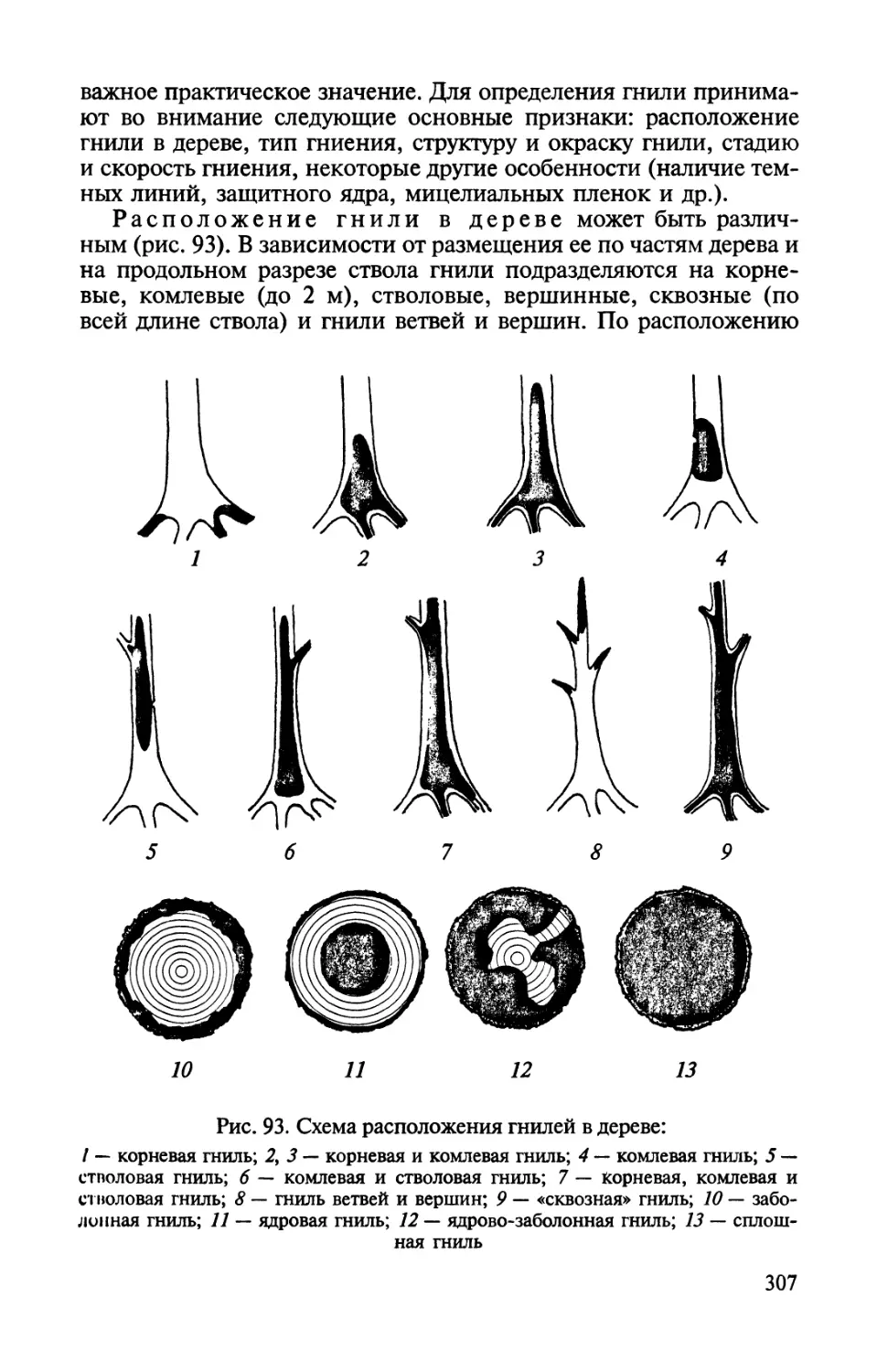
патологического процесса.ГТак, морозобоины и ожоги коры могут
явиться первой ступенью в развитии инфекционных некрозно-
раковых болезней стволов. Ослабление физиологических
процессов и отмирание мелких корней вследствие недостатка кислорода
при уплотнении почвы способствуют поражению хвойных пород
корневой губкой. Инфекционным полеганием и черной ножкой
чаще всего поражаются всходы растений, ослабленные вследствие
плохого ухода, неблагоприятных почвенных или погодных
условий. Нарушения обмена веществ в результате одностороннего
удобрения повышают восприимчивость растений к мучнистой росе и
другим болезням, способствуя их развитию. Такие
взаимосвязанные болезни, одна из которых предопределяет или стимулирует
развитие другой, получили название сопряженных.
1.2. СИМПТОМЫ БОЛЕЗНЕЙ РАСТЕНИЙ
Патологические реакции клеток и тканей растения при
различных заболеваниях весьма разнообразны. От их характера во
многом зависят и внешние проявления болезни. Видимые
признаки болезней, доступные невооруженному глазу, называют
симптомами.
9
Различают симптомы типичные (регулярно появляющиеся при
данном заболевании) и нетипичные; главные (наиболее
характерные для данной болезни) и сопутствующие; специфические
(свойственные лишь данной болезни) и общие (наблюдаемые при
различных заболеваниях); первичные (появляющиеся первыми, сразу
после окончания инкубационного периода) и вторичные
(возникающие при дальнейшем развитии болезни). Постоянство симптомов,
появляющихся у растений при определенном заболевании,
обусловливает типичное течение болезней и делает возможной их
диагностику. Наиболее простыми с точки зрения диагностики
являются случаи, когда разные возбудители вызывают различные
симптомы. Однако часто разные возбудители (как и разные абиотические
факторы) вызывают сходные и даже одинаковые проявления
болезни. Такое совпадение признаков при различных заболеваниях
получило название конвергенции симптомов. Например, усыхание
кроны деревьев наблюдается при засухе, гнилевых болезнях
корней, сосудистых и некрозно-раковых болезнях. Иногда, напротив,
один и тот же патоген вызывает у различных видов растений или
на различных органах одного и того же растения-хозяина
появление разных симптомов. Поэтому специфическими для той или иной
конкретной болезни часто являются не отдельные симптомы, а их
совокупность (симптомокомплекс, или синдром) и для
диагностики болезней важны не отдельные признаки, а их определенное
сочетание, последовательность появления, связь между ними.
Наряду с болезнями, сопровождающимися появлением
симптомов, встречаются болезни, протекающие без видимых внешних
признаков (например, некоторые вирусные заболевания). Такие
болезни называют бессимптомными. Бессимптомность может быть
обусловлена выносливостью (толерантностью) пораженного растения по
отношению к возбудителю, слабой патогенностью самого возбудителя
и другими причинами. Болезни, которые вначале протекают
бессимптомно, а проявляются через какое-то время, получили название л а-
тентных. Иногда под влиянием определенных внешних факторов
уже появившиеся симптомы временно исчезают (например,
мозаичная расцветка листьев, характерная для некоторых вирусных
болезней, в жаркую погоду пропадает, а при похолодании возникает вновь).
Это явление называют маскировкой симптомов. В других случаях
определенные условия окружающей среды могут задержать появление
симптомов болезни или повлиять на степень их выраженности.
1.3. ТИПЫ БОЛЕЗНЕЙ РАСТЕНИЙ
Типом болезни называют группу заболеваний,
характеризующихся комплексом сходных симптомов и объединяемых общим
названием. Тип болезни обусловливается характером взаимодей-
10
ствия патогена и хозяина и совокупностью происходящих в
растении патологических изменений, а они, в свою очередь, зависят от
биологических свойств патогена, особенностей поражаемых
органов и тканей, иногда от возраста и состояния растений. Наиболее
часто встречаются следующие типы болезней растений.
Гниль. Болезни этого типа вызываются грибами или бактериями.
Для них характерны резкие нарушения структуры, распад тканей
пораженных органов растений. Гнилью могут поражаться
различные части растений: плоды, семена, луковицы, корни, стволы и др.
В зависимости от строения и химического состава растительной
ткани, характера воздействия на нее ферментов возбудителя, от
степени разрушения отдельных компонентов пораженной ткани
и других особенностей патологического процесса возникают
мягкие или твердые, мокрые или сухие гнили. Более подробно
признаки и особенности развития разных типов гнилей изложены в
соответствующих главах учебника.
Рак. Этим термином обозначают болезни, характеризующиеся
развитием тканевых новообразований: наростов (опухолей),
образующихся вследствие усиленного деления или разрастания клеток
или незарастающих, окруженных наплывами язв, смолоточащих
ран и т. п. Раковые новообразования могут возникать на стволах,
корнях и других органах растений. Большинство раковых болезней
древесных растений вызывается грибами или бактериями, однако
первопричиной развития рака могут быть механические
повреждения (при раневом раке), резкая смена температур (при морозо-
бойном раке) и другие факторы.
Вилт. Широко распространенный тип сосудистых болезней,
связанных с поражением проводящей системы растений и
проявляющихся в увядании (усыхании) всего растения или его части,
например отдельных ветвей кроны. Инфекционные болезни этого
типа можно отличить от неинфекционного усыхания (в
результате засухи, механических повреждений и других причин) по
характерному потемнению сосудов на поперечном срезе увядшего
стебля, засохшей ветви, ствола усохшего дерева.
Некроз коры. Тип грибных болезней, которые характеризуются
поражением коры и камбия с изменением их цвета, отмиранием
тканей и формированием в них специфических грибных
образований (стром, спороношений и др.).
Ржавчина. Под этим названием известны многочисленные
болезни, вызываемые ржавчинными грибами. В типичных случаях
ржавчина характеризуется образованием пустул, т.е. скоплений
оранжево-желтых, ржаво-бурых или темно-бурых спор,
выступающих наружу через разрывы покровных тканей: эпидермиса и
кутикулы листьев, коры стволов и ветвей.
Шютте. Название этого типа болезней хвойных пород произошло
от немецкого глагола schutten, что значит осыпаться. Болезни типа
11
шютте вызываются различными видами грибов и проявляются в
изменении цвета, отмирании и опадении хвои. Характерным
признаком инфекционных болезней этого типа (в отличие от
неинфекционного засыхания и опадения хвои) служит появление на
пораженной хвое спороношений возбудителя.
Мучнистая роса. Болезни этого типа вызываются грибами
порядка мучнисторосяных. Их своеобразие заключается в том, что
мицелий патогена развивается на поверхности растений, образуя
тонкий паутинистый или более плотный ватообразный налет
белого цвета. При массовом образовании спор гриба налет
становится как бы присыпанным мукой (отсюда название этих болезней и
грибов-возбудителей).
Пятнистость. Это один из самых распространенных в природе
типов болезней растений. Пятнистости вызываются грибами,
бактериями, вирусами, а также неблагоприятными абиотическими
факторами. Обычно пятнистостями поражаются листья, но
иногда они встречаются на плодах, молодых побегах. Болезни этого
типа проявляются в отмирании (некрозе) тканей отдельных
участков пораженного органа, что сопровождается изменением их
окраски, структуры, а при грибных пятнистостях — появлением
на них спороношений патогена. Пятнистости очень разнообразны
по цвету, величине, форме, структуре пятен. Они бывают белые,
черные, бурые, мелкие или крупные, округлые, угловатые или в
виде штрихов, окаймленные, зональные, плоские или выпуклые,
точечные, дырчатые и т.д.
Мумификация. Так называются грибные болезни, при которых
мицелий паразита, обильно разрастаясь в тканях пораженного
органа, превращает его в плотное черное образование — «мумию»,
или склероциальную строму, состоящую из сплетения гиф гриба
и остатков растительной ткани. Чаще всего встречается
мумификация плодов и семян, но мумифицироваться могут также
отдельные участки листьев и других органов растений.
Парша. Поражение покровных тканей листьев, плодов и
побегов, сопровождающееся пятнистостью, растрескиванием и стру-
пьевидным шелушением соответствующих участков. Парша
вызывается некоторыми видами грибов. Встречается на плодовых
деревьях, осине, тополе, иве.
Антракноз. Грибное заболевание, проявляющееся в
образовании углубленных язв или сухой гнили при поражении плодов,
семян и других мясистых органов растений, а также в форме пят-
нистостей при поражении листьев.
Ведьмина метла. Представляет собой множество тесно
скученных тонких побегов, образовавшихся на небольшом отрезке ветви
в результате интенсификации ростовых процессов под влиянием
патогена. Ведьмины метлы чаще всего возникают при поражении
растений грибами и вирусами.
12
Деформация. Нарушения формы тех или иных органов
растений могут возникать под влиянием различных факторов, но чаще
всего вызываются патогенами. Типы деформаций весьма
разнообразны. Так, деформации листьев проявляются в виде курчавости,
пузыревидных вздутий, нитевидное™, морщинистости. Это связано
с неравномерным ростом отдельных частей листовой пластинки.
Мешковидное или листовидное разрастание пораженных тканей
происходит при деформациях плодов и семян (образуются так
называемые кармашки, или дутые плоды). При деформациях
цветков наблюдаются превращение генеративных частей в
вегетативные (махровость), непомерное разрастание или, наоборот,
недоразвитие цветка. Встречаются также деформации побегов, стеблей
в виде искривлений, утолщений, деформации корней и стволов в
виде опухолей.
Ожог. Болезни типа ожога могут быть вызваны грибами или
бактериями. Они проявляются на различных органах древесных
растений: молодых побегах, цветках, коре стволов и ветвей, иногда
на почках и молодых листьях. Цветки и побеги внезапно отмирают
и чернеют. Листья также чернеют, но не опадают, а свертываются
и остаются на ветвях. Кора покрывается пузырями и
растрескивается, поэтому дерево выглядит, как опаленное огнем. При
бактериальных ожогах из пораженных листьев и трещин коры иногда
вытекает мутная жидкость, содержащая массы бактерий, и
застывает на поверхности ветвей в виде капель и подтеков.
Плесень. На поверхности пораженных органов растений
образуются паутинистые или порошащие налеты, состоящие из
мицелия и спор грибов различного цвета (зеленая, розовая, серая и
другие плесени).
Чернь. Так называют черные, напоминающие сажу налеты на
зеленых частях растений, образуемые мицелием и спороношенй-
ями сапротрофных сажистых грибов, которые питаются не за счет
тканей растения-хозяина, а за счет посторонних поверхностных
наносов (различных выделений насекомых или самого растения,
пыли и др.).
Мозаика. Неравномерная окраска листьев, чередование темно-
зеленых участков листа с более светлыми, создающее мозаичность
расцветки, характерны для многих вирусных болезней растений,
но могут быть связаны и с недостатком отдельных элементов
питания.
Хлороз. Хлорозом называют побледнение или пожелтение
листьев в результате снижения содержания в них хлорофилла.
Причиной хлороза чаще всего являются нарушения минерального
питания растений; иногда хлороз возникает при вирусных болезнях
растений.
Наряду с ярко выраженными типами болезней встречаются
заболевания (например, полегание всходов), характеризующиеся
13
сложным комплексом признаков, присущих разным типам
болезней. Другие примеры таких заболеваний рассмотрены в
соответствующих главах учебника.
Кроме классификации болезней древесных растений по типам
заболеваний, их классифицируют также по возрастным группам
(болезни всходов, сеянцев, молодняков, приспевающих, спелых
и перестойных насаждений) и по поражаемым органам (болезни
плодов, семян, листьев, хвои, корней, стволов и ветвей).
Болезни цветочных культур чаще всего классифицируют по группам
(болезни многолетников, однолетников, оранжерейных культур)
и родам поражаемых растений (например, болезни пиона, астры,
хризантемы и т.п.).
У растений различают местные и общие болезни. При местных
заболеваниях поражение ограничивается лишь отдельными
участками или органами растения. Типичными примерами местных
болезней могут служить пятнистости, гнили, раковые и многие
другие болезни растений. При общих заболеваниях растение
бывает поражено полностью, хотя заражение могло произойти в
каком-то одном месте. Таковы сосудистые болезни, большинство
вирусных болезней растений.
В зависимости от скорости развития патологического процесса и
характера его внешних проявлений, различают две формы
болезни: острую (т. е. быстро протекающую, с резко выраженными
симптомами) и хроническую (медленно развивающуюся, с неясно
выраженными, иногда малозаметными признаками). В острой или
хронической форме могут протекать, например, сосудистые болезни
древесных пород. Некоторые болезни всегда развиваются как
острые (например, шютте, некрозы ветвей и стволов), другие — как
хронические (например, гнилевые и раковые болезни стволов).
1.4. УЩЕРБ, ПРИЧИНЯЕМЫЙ БОЛЕЗНЯМИ
РАСТЕНИЙ И ДЕРЕВОРАЗРУШАЮЩИМИ
ГРИБАМИ
Экономическое значение болезней культурных растений и
лесных пород исключительно велико. История знает примеры, когда
массовое распространение болезней важных
сельскохозяйственных культур приводило к переселению народов, гибели от голода
миллионов людей. Известны случаи, когда в результате
распространения болезней на огромных территориях полностью
прекращалось возделывание поражаемых растений. Так произошло,
например, с кофейным деревом в Юго-Восточной Азии, где в
конце прошлого столетия из-за опустошительной эпифитотии
ржавчины были ликвидированы плантации этой ценнейшей куль-
14
туры. Эпифитотическое распространение ржавчины в Бразилии
также в свое время поставило под угрозу выращивание там
кофейного дерева.
В современных условиях, несмотря на известные достижения в
выведении устойчивых сортов, создание и применение
различных средств борьбы с болезнями растений, многие заболевания
еще приносят ущерб лесному и сельскому хозяйству. В лесных
питомниках нередко наблюдается массовая гибель сеянцев
древесных пород от полегания и болезней типа шютте. Усыхание и
распад хвойных насаждений на больших площадях вызывает
корневая губка. Существенный вред насаждениям сосны причиняет
смоляной рак, а ильмовым — голландская болезнь. Насаждения липы
почти повсеместно усыхают от тиростромоза. Огромные убытки
связаны с болезнями цветочных культур.
Ущерб, причиняемый болезнями растений, складывается из
прямых и косвенных потерь. Прямые потери непосредственно
определяются снижением количества и качества получаемой
продукции, например уменьшением выхода стандартного посадочного
материала или деловой древесины, ухудшением качества семян
или цветочной продукции и т. п. Прямые потери могут быть с той
или иной степенью точности оценены и в денежном выражении
по разнице в стоимости продукции, получаемой от здорового и
больного растения, с гектара здорового и больного леса и т.п.
Так, например, в Западной Европе ежегодные потери деловой
древесины в результате поражения насаждений корневой губкой
оцениваются в 15 — 20 млн долл. Косвенные потери связаны с
последствиями болезни, которые могут выражаться в снижении
зимостойкости растений и их устойчивости к другим болезням, в
затратах на пересев в случае гибели всходов от полегания, на
дополнение молодых культур при усыхании их от шютте, ржавчины
или мучнистой росы, лесовосстановительные работы в очагах
корневых гнилей, некрозно-раковых, сосудистых заболеваний и т.д.
Появляется необходимость ежегодных затрат и на проведение
профилактических мероприятий по защите растений, на
химические, биологические и другие меры борьбы с болезнями леса.
Огромный ущерб причиняют и сапротрофные дереворазруша-
ющие грибы, которые поражают лесоматериалы, хранящиеся на
складах, вызывают гниение деревянных конструкций в
различных сооружениях и зданиях. По подсчетам специалистов, не
менее 20 % от общего количества заготавливаемой в стране деловой
древесины расходуется для восполнения ее потерь от
преждевременного разрушения грибами. Известно также, что складские и
домовые грибы ежегодно уничтожают миллионы кубометров
древесины, а общие затраты на периодически проводимый противо-
грибной ремонт зданий приближаются к затратам на их
строительство.
Глава 2
ГРИБЫ И ГРИБОПОДОБНЫЕ
ОРГАНИЗМЫ - ВОЗБУДИТЕЛИ БОЛЕЗНЕЙ
РАСТЕНИЙ
Эта обширная группа бесхлорофилльных организмов
объединяет более 100 тыс. видов. Богатство видового состава, своеобразие
происхождения, морфологии и биологии грибов послужили
основанием для выделения их в самостоятельное царство живой
природы (Mycota). Некоторые грибоподобные организмы
отнесены к царствам Protozoa и Chromista. Однако в дальнейшем как
собственно грибы, так и грибоподобные организмы мы будем для
краткости изложения условно называть грибами. Грибы очень
разнообразны по форме, строению, физиологическим и
экологическим особенностям, их роли в природе и жизни человека. Наряду
с широко известными видами съедобных и ядовитых шляпочных
грибов, в том числе грибов-микоризообразователей, существует
огромное число видов микроскопических грибов. Многие из них
являются возбудителями болезней древесных растений, а также
сельскохозяйственных и цветочных культур. Известны грибные
болезни животных и человека. Грибы вызывают также разрушение
древесины в лесу, на складах, в зданиях и сооружениях, порчу
продуктов и промышленных изделий. Некоторые виды грибов
находят применение в медицине и различных отраслях
промышленности.
2.1. ВЕГЕТАТИВНОЕ ТЕЛО ГРИБОВ
И ЕГО ВИДОИЗМЕНЕНИЯ
Вегетативное тело большинства грибов состоит из тонких
простых или разветвленных нитей — гиф. Гифы растут вершинами
и, сплетаясь между собой, образуют мицелий (грибницу). Одна
из специфических особенностей грибов — способность к
неограниченному росту мицелия. Гифы могут быть одноклеточными (без
перегородок) или многоклеточными (с поперечными перегород-
16
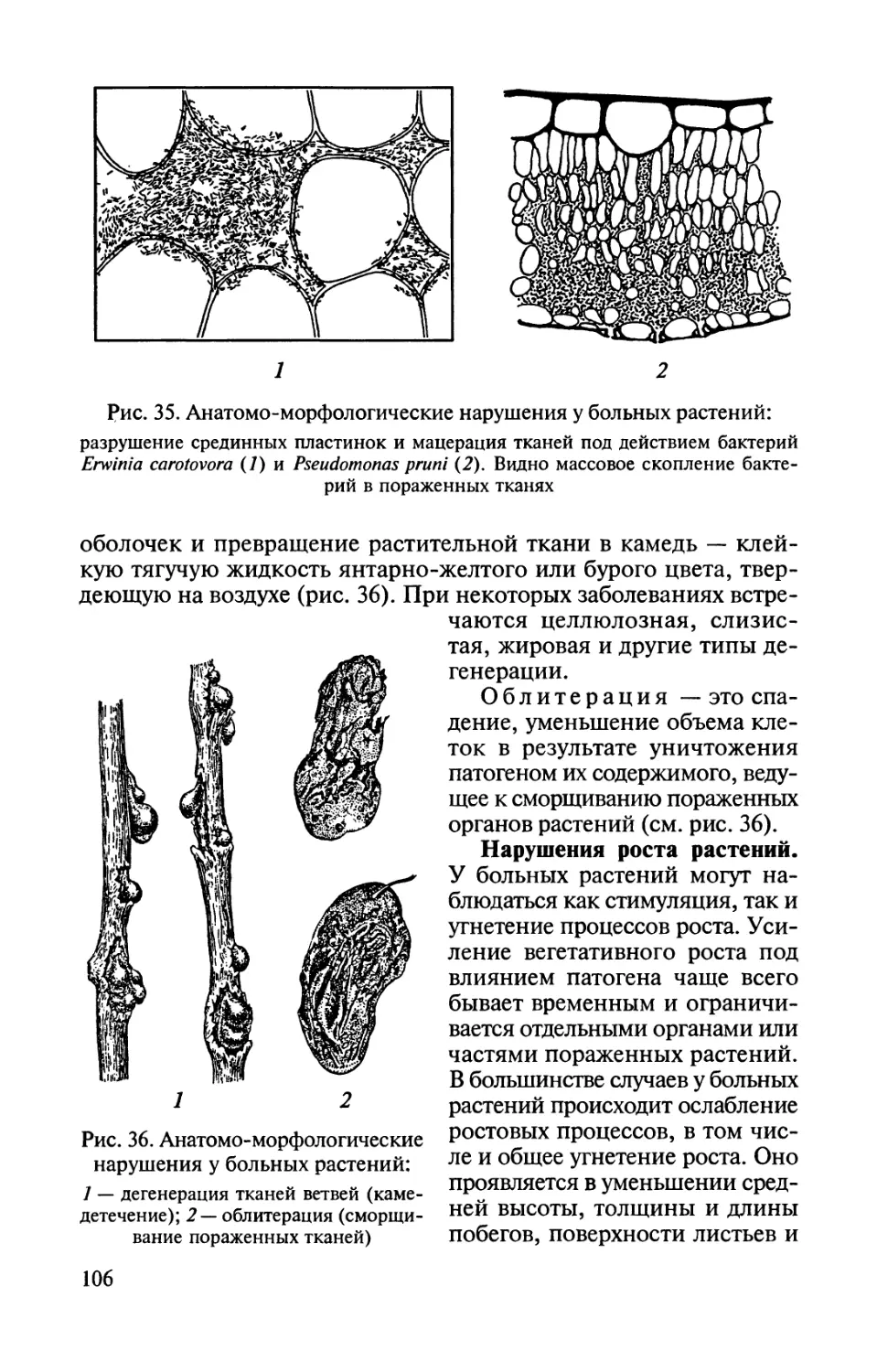
Рис. 1. Типы вегетативного тела грибов и грибоподобных организмов:
/ — плазмодии в клетках растения; 2 — одноклеточный мицелий; 3 —
многоклеточный мицелий
ками — септами). Многоклеточный мицелий называют септи-
рованным; мицелий без перегородок — несептированным
(рис. 1).
У небольшого числа простейших грибоподобных организмов
вегетативное тело представляет собой голый комочек
цитоплазмы (плазмодий). Грибы, имеющие плазмодий или
одноклеточный мицелий, считаются низшими, а грибы с многоклеточным
мицелием — высшими. В поперечных перегородках септирован-
ных гиф имеются отверстия — поры, через которые происходит
движение цитоплазмы по направлению к растущим концам гиф.
Гифы грибов чаще всего бесцветны, но иногда имеют зеленый,
серый, розовый оттенок или бурую окраску различной
интенсивности.
Грибы, мицелий которых наделен свойствами обоих полов,
называют гомоталличными, а грибы, у которых каждый мицелий
обладает свойствами только одного пола, — гетероталличными.
У многих грибов в связи с особенностями их биологии или
условиями развития на мицелии образуются специальные
органы, выполняющие определенные функции (рис. 2). Например, для
паразитных грибов характерны гаустории — различной
формы боковые отростки гиф, внедряющиеся внутрь клеток
пораженного растения. С помощью гаусториев гриб извлекает из тканей
растения-хозяина растворы питательных веществ. У некоторых
грибов формируются пучки тонких разветвленных гиф,
напоминающих корни, — ризоиды, посредством которых гриб
прикрепляется к субстрату и осуществляет питание. Нередко наблюдается
образование анастомозов — соединительных «мостиков» между
соседними гифами. Через анастомозы происходит обмен
содержимым, в том числе и ядерным. На диплоидном мицелии некоторых
базидиальных грибов образуются пряжки — особые канальцы,
соединяющие соседние клетки одной и *ой же рифм и эбеспечи-
17
Рис. 2. Специализированные органы мицелия:
1 — ризоиды; 2 — столон; 3 — гаустории (а — гифа гриба, б — гаусторий, в —
зараженная клетка растения); 4 — анастомозы; 5 — пряжки
вающие переход ядер из клетки в клетку (см. рис. 2). Известны и
другие видоизменения гиф.
Мицелий, развивающийся на поверхности пораженного
растения или другого питательного субстрата, называют
поверхностным, или воздушным (экзофитным). Такой мицелий обычно
имеет вид нежного пушистого или паутинистого налета, тонких
пленочек или ватообразных скоплений. Мицелий,
распространяющийся внутри субстрата, называют субстратным (эндофитным).
Субстратный мицелий фитопатогенных грибов в свою очередь бывает
межклеточным, но может распространяться также внутри клеток
пораженной ткани. Плазмодии фитопатогенных видов обычно
развиваются только внутри живых клеток растения.
Распространение мицелия по поверхности или внутри субстрата
чаще всего ограничено определенными небольшими участками.
Такой мицелий называют местным, или локальным. Если же
мицелий полностью пронизывает субстрат или все органы
пораженного растения, то его называют диффузным.
В процессе эволюционного развития у грибов выработалась
способность видоизменять мицелий соответственно изменениям
условий среды или при переходе в состояние покоя (рис. 3).
Например, в трещинах древесины, пораженной грибами-трутовиками,
или под корой часто развиваются довольно толстые и плотные
мицелиальные пленки, напоминающие замшу. Ткань та-
18
Рис. 3. Видоизменения мицелия:
/ — простые мицелиальные шнуры; 2 — сложные мицелиальные шнуры
(ризоморфы); 3 — ризоморфы под корой ствола; 4 — мицелиальная пленка в гнилой
древесине; 5— склероции произвольной формы; б— шаровидные склероции; 7—
склероции в виде рожков; 8, 9 — микроскопическое строение склероциев (на
срезах видна утолщенная наружная ткань — кора склероция)
ких пленок образована плотным равномерным переплетением гиф
гриба.
Другое видоизменение мицелия — тяжи, или шнуры,
сложенные из параллельно идущих, частично сросшихся, обычно
однородных по строению и функциям гиф (см. рис. 3).
Мицелиальные тяжи представляют собой разветвленные образования
различной толщины (от нескольких микрометров до нескольких
миллиметров). Они характерны для некоторых базидиальных грибов,
например домовых.
Ризоморфы — более мощные темные ветвящиеся шнуры,
длина которых может достигать нескольких метров при толщине в
несколько миллиметров. Гифы, образующие ризоморфы,
различны по строению и выполняемым функциям. Наружная часть ризо-
морф сложена из темных толстостенных гиф, а внутренняя — из
бесцветных тонкостенных гиф и гиф типа сосудов: неветвящих-
ся, без перегородок, с широкими просветами и кольцевыми
утолщениями на оболочках. Типичным примером могут служить ризо-
19
морфы опенка. Ризоктонии в отличие от ризоморф имеют
более рыхлую структуру и меньшие размеры, они встречаются у
некоторых сумчатых грибов. Тяжи и ризоморфы грибов играют
роль проводящих органов. По ним поступают вода и питательные
вещества к развивающимся плодовым телам. Кроме того, тяжи и
ризоморфы способствуют распространению гриба.
Своеобразным видоизменением мицелия являются с к л е р о -
ции (см. рис. 3). Это плотные твердые тела различной формы и
величины. Они образуются в результате тесного переплетения гиф,
Рис. 4. Видоизменения мицелия:
А — мягкие яркоокрашенные мицелиальные стромы: / — головчатые
фиолетовые стромы, спорыньи, выросшие на склероции; 2 — внешний вид головчатой
стромы с устьицами плодовых тел гриба; 3 — продольный разрез через
головчатую строму с погруженными в нее плодовыми телами; 4 — красные шаровидные
стромы; 5 — схематический разрез через такую строму; Б —- твердые черные
мицелиальные стромы: 1 — плоская трещиноватая строма с точечными
устьицами плодовых тел; 2 — разрез через эту строму; 3 — разрез через подушковидную
строму с плодовыми телами; 4 — многолетние шаровидные стромы на стволе
березы; 5 — схематический разрез через такую строму (видны «годичные слои» с
плодовыми телами); В — склероциальные стромы (мумии): 1 — строма гриба
Stromatinia pseudotuberosac плодовыми телами; 2— строма гриба Moniliniafructigena;
3 — стромы с развивающимися в них плодовыми телами на листе клена,
пораженном черной пятнистостью; 4 — разрез через такую строму
20
богатых запасными питательными веществами, и предназначены
для сохранения гриба при неблагоприятных условиях и для его
распространения. Внутренняя часть склероция обычно
формируется из бесцветных гиф. Наружная часть (кора) состоит из
толстостенных темноокрашенных элементов, поэтому склероции
снаружи чаще всего выглядят черными. По окончании периода покоя
склероции прорастают, образуя грибницу или органы спороно-
шения. Склероции известны у многих грибов, главным образом
сумчатых, некоторых базидиальных и несовершенных. Они
образуются, например, на сеянцах, погибших от выпревания,
цветочных растениях, пораженных белой гнилью.
Внутри пораженных тканей растений иногда образуются мик-
росклероции — очень мелкие склероции, сложенные из
однородных бурых клеток.
Кроме настоящих склероциев, которые формируются только
из гиф гриба и легко отделяются от ткани пораженного растения,
встречаются склероциальные стромы, или мумии, в
формировании которых участвуют не только гифы гриба, но и
пораженная ткань растения. Примерами подобных образований могут
служить мумифицированные желуди и яблоки (рис. 4).
Многие грибы (особенно сумчатые) образуют мицелиаль-
ные стромы — мясистые сплетения гиф, на поверхности или
внутри которых закладываются плодовые тела или другие
органы спороношения. Стромы могут быть однолетние или
многолетние, мягкие или деревянистые, различной формы и окраски (см.
рис. 4).
Ткани грибов. Формирование тканей у грибов в отличие от
растений происходит не путем деления клеток в разных
направлениях, а в результате переплетения и срастания «готовых» гиф
мицелия. Таким образом, грибная ткань является ложной; она
называется плектенхимой. Как и у высших растений, у
грибов различают несколько типов тканей: образовательную,
механическую, покровную, проводящую.
2.2. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ КЛЕТКИ
У большинства грибов клетка по своему строению и
выполняемым ею функциям в целом аналогична клетке растений. Она
состоит из твердой оболочки и внутреннего содержимого,
представляющего собой цитоплазменную систему, окруженную ци-
топлазматической мембраной и содержащую митохондрии,
рибосомы, ядро (или ядра), вакуоли и различные включения. Однако
грибная клетка имеет ряд специфических особенностей,
отличающих ее от растительной клетки и послуживших в числе других
аргументов основанием для выделения грибов в самостоятельное
21
царство живой природы. Клеточная оболочка у грибов выполняет
роль защитного барьера и, кроме того, непосредственно
участвует в процессах питания гриба и обмена веществ между клеткой и
внешней средой. Оболочка клетки может быть однослойной или
многослойной, разнообразной по химическому составу. Строение,
состав и свойства клеточной оболочки зависят от вида гриба и
функций клетки. Они могут изменяться с возрастом, при
переходе из одной фазы развития в другую, под влиянием условий
питания и других факторов.
Основу оболочки составляют полисахариды (например
целлюлоза), простые сахара, белки, липиды и фосфаты. Кроме того, в ее
состав входят лигниноподобные вещества, производные
нуклеиновых кислот, аминокислоты, различные соли, смолы, а также
хитин, свойственный покровным тканям насекомых, хитозан, (3-глю-
кан. Эти и другие компоненты содержатся в оболочках клеток
грибов в самых разнообразных сочетаниях, образуя сложные
комплексы, характерные для определенных систематических групп грибов.
Оболочки молодых клеток обычно тонкие, бесцветные,
однородные по структуре. По мере старения оболочка может утолщаться,
ослизняться, становиться более темной благодаря отложению
пигментов. Наружные слои оболочки клеток (особенно спор) многих
грибов кутинизированы, пропитаны воском и жиром, что делает
их несмачиваемыми. У трутовых грибов, особенно часто в плодовых
телах, наблюдаются лигнификация и опробковение оболочек гиф.
По строению ядерного аппарата грибы относятся к эукариотам.
Ядро в клетках грибов четко обособлено, снабжено оболочкой и
содержит ядрышко. У грибов разных систематических групп число
ядер в клетке неодинаково. Хорошо развитый несептированный
мицелий низших грибов содержит много ядер. У большинства
сумчатых грибов (за исключением мучнисторосяных) и базидиоми-
цетов клетки одно- или двуядерны, в зависимости от фазы
развития. Ядра обычно мелкие, в среднем 2—3 мкм (в сумках и базиди-
ях — более крупные), круглой, овальной или веретеновидной
формы, однако форма их не постоянна.
Своеобразная особенность грибов — отсутствие в цитоплазме
их клеток растительного крахмала. В то же время важнейшая роль
принадлежит гликогену, который обычно содержится в тканях
животных. Гликоген является основным запасным веществом
грибной клетки и равномерно распределяется по всей цитоплазме в
виде мелких гранул. Клетки грибов содержат также большое
количество метахроматина (волютина). Он относится к полифосфатам
и играет важную роль в процессах обмена. Из других включений в
клетках многих грибов содержатся жировые вещества; особенно
богаты ими споры, плодовые тела, склероции, старые части
мицелия. Жиры находятся в цитоплазме в мелкораспыленном
состоянии или образуют более крупные капли (липосомы).
22
В состав клеток мицелия, репродуктивных органов,
покоящихся структур грибов могут входить и многие другие вещества:
пигменты, органические кислоты и их соли, витамины, терпены
(ароматические эфирные масла), токсины, смолы и др. Некоторые из
них играют роль запасных питательных веществ клетки,
участвуют в физиологических процессах, выполняют защитную
функцию, другие являются вредными для клетки продуктами ее
метаболизма.
2.3. РАЗМНОЖЕНИЕ ГРИБОВ
Известны три типа размножения грибов: вегетативное,
бесполое и половое.
Вегетативное размножение осуществляется непосредственно
частями вегетативного тела или спорами, которые образуются
клетками вегетативных гиф (рис. 5).
Рис. 5. Вегетативное размножение грибов:
/, 2, 3 — различные формы почкования мицелия с образованием бластоспор; 4, 5 —
формы образования оидий; 6 — образование гемм; 7, 8, 9 — различные
формы образования хламидоспор; 10 — хламидоспоры
23
Простейшая форма вегетативного размножения —
размножение частицами гиф (а также обрывками шнуров, ризоморф, скле-
роциями); будучи отделены от материнского мицелия и попав в
благоприятную среду, они могут дать начало новому
самостоятельному мицелию. Этот способ размножения весьма
распространен среди сапротрофных грибов.
Функцию вегетативного размножения у некоторых грибов,
например дрожжевых, выполняет почкующийся мицелий. Его
клетки на определенном этапе развития округляются, обособляются,
и на их поверхности появляются небольшие выросты, которые
постепенно увеличиваются в размерах, затем отделяются от
материнской клетки и в свою очередь начинают почковаться. Такие
клетки называют бластоспорами.
Особая форма вегетативного размножения — образование оидий
и хламидоспор. Оидии, или артроспоры, образуются в
результате дробления гиф, начиная с их концов, на короткие
обособленные членики, которые в дальнейшем дают начало новому
мицелию. Они снабжены тонкой оболочкой и недолговечны.
Разновидностью оидий являются геммы, отличающиеся от них
более плотной темной оболочкой и способностью дольше
сохраняться (например, в зимний период). Оидии и геммы известны у
многих грибов разных классов.
Хламидоспоры возникают путем уплотнения и
обособления содержимого отдельных клеток мицелия, которое при этом
покрывается толстой темноокрашенной оболочкой.
Освободившиеся из клеток материнских гиф хламидоспоры способны долго
сохраняться при неблагоприятных условиях. Прорастая, они
образуют органы спороношения или мицелий. Хламидоспоры широко
распространены в природе. Они встречаются у многих видов бази-
диомицетов, оомицетов, несовершенных грибов.
Артроспоры и хламидоспоры представляют собой как бы
переходные формы от вегетативного к бесполому размножению.
Бесполое размножение осуществляется спорами, которые
образуются без оплодотворения на особых органах, отличающихся по
форме и строению от вегетативных гиф мицелия.
Бесполые спороношения (анаморфы) обычно образуются на
хорошо развитом мицелии, имеющем достаточный для
формирования спор запас питательных веществ. Спороносными органами
при бесполом размножении грибов служат специализированные
ответвления или участки гиф, внутри которых (эндогенно) или
снаружи (экзогенно) созревают споры (рис. 6).
Самый примитивный орган бесполого размножения низших
грибов — зооспорангий, представляющий собой
расширенное окончание гифы. Его содержимое распадается на подвижные
споры с одним или двумя жгутиками — зооспоры. Зооспоры
могут передвигаться и сохранять жизнеспособность только в воде.
24
Рис. 6. Основные формы бесполого размножения грибов:
/ — зооспорангий (а — двужгутиковая зооспора, б — одножгутиковые
зооспоры); 2 — спорангий со спорангиоспорами; 3 — конидиеносцы с конидиями
Поэтому они свойственны, главным образом, водным и
некоторым почвенным грибам. У наземных видов, сохранивших связь с
водной средой, зооспоры образуются во время дождя, обильной
росы или густого тумана.
Более совершенной формой бесполого размножения низших
грибов являются спорангии — шаровидные вместилища на
концах ответвлений грибницы. Ветвь, несущая спорангий,
называется спорангиеносцем. Внутри спорангия формируются
неподвижные одноклеточные споры — спорангиоспоры. Они покрыты
твердой оболочкой, образуются у мукоровых грибов.
Наиболее распространенная форма бесполого размножения,
свойственная почти всем высшим и некоторым низшим грибам, —
конидиальное спороношение. Конидиями называют споры,
образующиеся на конечных ответвлениях специальных органов —
конидиеносцев. Конидиеносцы и конидии очень
разнообразны по форме, размерам, строению и окраске, а также по
характеру их развития и размещения. Своеобразными формами кониди-
ального спороношения грибов являются коремии, конидиальные
ложа (ацервулы) и пикниды.
Коремии — это пучки плотно соединенных друг с другом
конидиеносцев, на концах которых образуются конидии.
Скопления конидий на коремиях могут быть порошащими или
слизистыми. Спороношения коремиального типа часто встречаются у
грибов класса гифомицетов, у сумчатых грибов.
Конидиальные ложа и пикниды (так называемые конидио-
м ы) характерны для грибов класса целомицетов.
25
Ложа представляют собой подушковидные, более или менее
плотные мицелиальные образования, формирующиеся на
поверхности пораженного субстрата или погруженные в него, но при
созревании выступающие наружу. Конидиеносцы располагаются
на поверхности ложа сплошным слоем. Такая форма конидиаль-
ного спороношения характерна для грибов порядка Acervulales.
Пикниды — это шаровидные (иногда немного
приплюснутые) или грушевидные вместилища с плотной, обычно темной
оболочкой. Они бывают одиночными (иногда их называют
однокамерными) или образуются группами в общей строме
(многокамерные пикниды). Конидии в пикнидах, или пикноспоры,
образуются на коротких конидиеносцах, покрывающих внутреннюю
стенку пикниды. Созревая, они выделяются из пикниды через
специальное отверстие (устьице) сплошной слизистой массой,
застывающей на воздухе в виде капель или скрученных нитей,
нередко имеющих яркую окраску. Как правило, пикниды
частично или полностью погружены в питающий субстрат и наружу
выступают лишь их верхние части или устьица. Спороношение типа
пикнид — основной признак грибов порядка пикнидальных.
У многих фитопатогенных грибов в течение вегетационного
периода наблюдается несколько генераций бесполого
спороношения. Этим обеспечиваются массовое распространение
инфекционного начала и повторное заражение растений.
Половое размножение. При этом способе размножения
образованию спор предшествует половой процесс. Формы полового
процесса у грибов весьма разнообразны (рис. 7). Их можно разделить
на три группы.
Гаметогамия — это слияние специализированных половых
клеток (гамет), образующихся в особых половых структурах (гаме-
тангиях). Простейшей формой гаметогамииявляется изогамия,
т.е. слияние разнополых, но одинаковых по строению и размерам
зооспор. В результате образуется циста. У некоторых наиболее
примитивных грибов (например, слизевиков) продуктом полового
процесса типа изогамии является диплоидный плазмодий.
Более сложная форма гаметогамии — о о г а м и я, т. е. слияние
различных по строению и размерам половых клеток: женской (оого-
ния) и мужской (антеридия). В результате образуется ооспора.
Гаметангиогамия — это слияние двух
специализированных половых структур (гаметангиев), отличающихся по строению
от вегетативных гиф, на которых они образуются. Примерами га-
метангиогамии могут служить зигогамия с образованием
зигоспоры и половой процесс у сумчатых грибов.
В последнем случае сливается только цитоплазма, а ядра
группируются попарно, образуя так называемые дикарионы,
располагающиеся в аскогенных гифах. В результате полового процесса
аскомицетов образуются сумки (аски). Это различной формы меш-
26
Рис. 7. Основные формы полового размножения грибов:
/ — изогамия с образованием цист; 2 — оогамия с образованием ооспор в тканях
растения; 3 — зигогамия (процесс формирования зигоспоры); 4 — типы сумок и
сумкоспор; 5 — типы базидий и базидиоспор
ковидные клетки, внутри которых (эндогенно) развиваются сум-
коспоры (аскоспоры). Обычно сумка содержит восемь сумкоспор.
Соматогамия — это слияние обычных клеток
вегетативного мицелия. Типичная форма соматогамии характерна для бази-
диальных грибов. В процессе соматогамии также образуются дика-
рионы, а содержащий их мицелий называют дикариотическим. По-
27
ловой процесс завершается формированием базидий. Базидия
представляет собой орган булавовидной или цилиндрической
формы, на котором экзогенно образуются базидиоспоры (обычно их
четыре).
И сумки, и базидий чаще всего образуются не открыто на ди-
кариотическом мицелии, а в развивающихся на нем плодовых телах
разнообразного строения (см. рис. 14, 15, 19).
Переход к половому размножению с образованием половых
спороношений (телеоморф) у грибов часто бывает связан с
окончанием вегетативного роста, наступлением неблагоприятных условий,
перезимовкой. У фитопатогенных видов половое спороношение,
как правило, наблюдается один раз в течение года, а споры,
образующиеся половым путем, осуществляют первичное заражение
растений весной или в начале лета. Однако у многих трутовиков,
имеющих многолетние плодовые тела, половые споры
(базидиоспоры) могут формироваться и заражать субстрат на протяжении
длительного периода времени, иногда в течение многих месяцев.
Таким образом, грибам присуще свойство плеоморфизма, т.е.
способность формировать в цикле развития1 разные типы
спороношений.
Обычный (простой) цикл развития включает два спороноше-
ния: бесполое и половое. Однако есть немало видов, имеющих
наряду с половым несколько различных бесполых спороношений.
Такой цикл развития называют сложным. Примером грибов со
сложным циклом развития могут служить ржавчинные грибы.
Многие митоспоровые (несовершенные) грибы имеют только
бесполые спороношения (анаморфы). В то же время у некоторых
сумчатых и базидиальных грибов известно лишь половое
спороношение. В ряде групп грибов имеются виды, цикл развития которых до
сих пор неясен.
Споры грибов, предназначенные для распространения и
быстрого прорастания в течение вегетационного периода, называются
пропагативными. Они имеют тонкие, обычно бесцветные
оболочки и не могут долго сохранять жизнеспособность. Таковы
зооспоры, конидии и другие анаморфы.
Споры, служащие для длительного сохранения при
неблагоприятных условиях, называются покоящимися. Они бывают
снабжены толстой и, как правило, пигментированной оболочкой
(например, ооспоры, зигоспоры, телиоспоры ржавчинных грибов).
Спорообразующая способность большинства грибов очень
велика. Например, у мукоровых грибов один спорангий может со-
1 Циклом развития у грибов называют последовательное прохождение
различных стадий и спороношений, завершающееся образованием исходных спор.
У большинства грибов в цикле развития происходит чередование анаморфных
и телеоморфных стадий.
28
держать несколько десятков тысяч спорангиоспор. В каждой
пустуле ржавчинных грибов созревает несколько сотен спор. Число сум-
коспор в плодовом теле некоторых сумчатых грибов исчисляется
миллионами, а плодовые тела многих труговых грибов выделяют
миллиарды базидиоспор в течение всего периода споруляции.
Количество продуцируемых грибами спор многократно
увеличивается соответственно числу генераций бесполого спороношения.
2.4. РАСПРОСТРАНЕНИЕ СПОР ГРИБОВ
Отделение спор от спорообразующих органов грибов может
происходить пассивно или активно. Так, у большинства мукоро-
вых грибов оболочки созревших спорангиев естественно
разрушаются, и спорангиоспоры, пассивно освобождаясь, попадают в
окружающую среду. Пассивно, вместе со слизистым содержимым
пикниды, выделяются наружу и пикноспоры грибов. Отчленение
зрелых конидий от конидиеносцев на воздушном мицелии также
носит чаще всего пассивный характер. Активное освобождение спор
наблюдается у различных групп грибов. Оно особенно типично
для аскомицетов: при созревании сумок их содержимое благодаря
особому механизму с силой выбрасывается наружу, тем самым
облегчается дальнейшее распространение сумкоспор.
Освободившиеся споры частично оседают в непосредственной
близости от спорообразующих органов гриба, частично
распространяются на то или иное расстояние от них. Это распространение в
свою очередь может быть пассивным или активным. В
подавляющем большинстве случаев оно происходит пассивно, при
помощи различных агентов, таких, как воздушные течения (особенно
ветер), вода, животные (в том числе насекомые), человек.
Важнейшим фактором, определяющим пути и способы
распространения грибов в природе, является физическое состояние
споровых масс в момент их созревания. В одних случаях
созревшие споры сухие и легко распыляются (их называют ксероспоры).
В других случаях споры выделяются из споровместилищ вместе со
слизистым веществом (это так называемые миксоспоры).
Соответственно этому различают и два типа грибов: сухоспоро-
вые и слизистоспоровые. Первые переносятся
преимущественно по воздуху, вторые — с помощью воды и насекомых.
Анемохория. Распространение спор по воздуху встречается в
природе наиболее часто. Анемохорным способом
распространяются ржавчинные, мучнисторосяные, плесневые, трутовые и
многие другие грибы. Их споры образуют порошащие скопления,
выбрасываются из споровместилищ в виде пылевидных масс
(«облачков») или свободно выпадают из плодовых тел. Благодаря
ничтожно малым величине и массе споры грибов подхватываются
29
ветром, восходящими теплыми потоками воздуха и другими
воздушными течениями. В то же время чрезвычайно малая скорость
падения спор у грибов-анемохоров позволяет им долго
удерживаться в воздухе, подниматься в верхние слои атмосферы и
переноситься на огромные расстояния от мест их образования,
сохраняя при этом способность к прорастанию. Так, жизнеспособные
споры различных видов грибов, в том числе фитопатогенных,
неоднократно находили в сотнях миль от берега над акваторией
Океана, горными массивами, в Арктике.
Особенно много спор обнаруживается над лесами и
плантациями сельскохозяйственных и цветочных культур, при этом их
количество в воздухе и видовой состав подвержены значительным
колебаниям в зависимости от времени года, направления и силы
господствующих ветров.
Хотя с увеличением расстояния от места спорообразования
количество спор в воздухе резко снижается (основная масса спор
оседает в радиусе 250 — 300 м), однако потенциальная угроза
распространения на большие расстояния и накопления
инфекционного начала постоянно существует, и этот фактор имеет важное
значение для прогноза и профилактики эпифитотий. Есть
основания считать, что возбудители ряда опасных грибных болезней
растений попали из Америки в Европу и, наоборот, именно анемо-
хорным путем.
Зоохория. Роль животных в распространении спор грибов
достаточно велика. При этом чаще всего наблюдается эпихория,
т.е. перенос гриба на поверхности тела животного, реже
встречается эндохория — перенос внутри тела, но иногда эпихорию
и эндохорию трудно разграничить. Многие грызуны (белки, мыши-
полевки и др.), запасая на зиму плодовые тела шляпочных
грибов, способствуют рассеиванию базидиоспор. Моллюски,
питаясь плодовыми телами шляпочных грибов, также разносят их споры.
В распространении трюфелей участвуют кабаны, выкапывающие
плодовые тела этих грибов. Землеройные позвоночные животные,
домашние и дикие копытные считаются возможными
разносчиками спор дереворазрушающих грибов, в том числе корневой
губки. При скармливании скоту растений, пораженных грибными
болезнями, споры возбудителей, пройдя через пищеварительный
тракт животных, часто сохраняют жизнеспособность. Так, с
навозом могут распространяться возбудители килы крестоцветных, рака
картофеля и других опасных заболеваний, а также споры копро-
фильных грибов. Известны случаи переноса плодовых тел и спор
грибов птицами (например, распространение дятлами спор
возбудителя эндотиевого рака каштана). Грибную инфекцию могут
переносить дождевые черви, фитонематоды, клещи.
Энтомохория. Важное значение в природе имеет
распространение инфекционного начала грибов насекомыми. В большинстве
30
случаев насекомые механически переносят споры грибов на
поверхности тела. Иногда между насекомым-переносчиком и
грибом наблюдается более тесная биологическая взаимосвязь. Так,
ильмовые заболонники являются переносчиками возбудителя
голландской болезни. Дубовый заболонник, желтопятнистый усач,
непарный шелкопряд, златогузка переносят возбудителей
сосудистого микоза дуба. Многие виды стволовых вредителей
распространяют возбудителей синевы древесины. Различные виды мух и
жуков, личинки которых (а иногда и имаго) питаются
плодовыми телами шляпочных и трутовых грибов, также способствуют их
распространению. Насекомые (в частности, мухи) играют
важную роль в распространении, половом размножении и
гибридизации ржавчинных грибов, способствуя тем самым появлению
новых физиологических рас этих паразитов.
В большинстве случаев насекомых-переносчиков привлекают
сахаристые или пахучие выделения плодовых тел грибов, их
яркая окраска. У некоторых видов грибов на спорах имеются
своеобразные приспособления (крючки, выступы и т.д.),
облегчающие перенос их насекомыми. Обычно насекомые распространяют
инфекционное начало грибов на ограниченной территории
(например, в пределах зараженного насаждения), но в периоды
миграций могут переносить его и на значительные расстояния.
Гидрохория. Вода также может служить распространению грибов.
Гидрохорным способом распространяются в основном грибы,
споры которых образуют слизистые скопления или выделяются из споро-
вместилищ в виде склеенных масс (например, грибы-целомицеты).
Важная роль при гидрохории принадлежит дождям. Во время дождя
слизь, склеивающая споры, набухает, разжижается, споры
разъединяются и смываются с пораженных частей растений, попадая на их
здоровые части и соседние растения. Особое значение дожди и росы
имеют ддя грибов, спороношение которых возможно лишь при
наличии капельно-жидкой влаги (например, ложномучнисторосяных).
С каплями и брызгами дождя конидии и зооспоры этих грибов
рассеиваются, попадают на растения и, тут же прорастая, заражают их.
Смывая с ветвей и стволов деревьев осевшие на них споры
фитопатогенных грибов, занося их в трещины и раны на стволе и
корнях, создавая при этом благоприятные условия увлажнения,
дожди способствуют распространению в насаждении также гни-
левых и некрозно-раковых болезней.
Таким образом, гидрохория обеспечивает главным образом
местное распространение инфекции — в пределах растения, поля
или насаждения. Однако бурные ливневые потоки, паводки,
оросительные воды, реки могут переносить плодовые тела и споры
самых различных грибов и на большие расстояния.
Антропохория. Распространению грибов, в том числе
фитопатогенных, нередко способствует хозяйственная деятельность че-
31
ловека. Перенос мицелия, спор, склероциев паразитных грибов
может происходить при транспортировке семян, посадочного
материала, сельскохозяйственной продукции, декоративных
растений, древесины, причем не только внутри страны, но и в
мировом масштабе. Установлено, что за последнее столетие из
Америки в Европу были завезены десятки возбудителей болезней
культурных растений и древесных пород, в том числе таких опасных,
как мучнистая роса дуба, крыжовника, винограда, фитофтороз
картофеля. В свою очередь в Америку были завезены парша
яблони, эндотиевый рак каштана, пузырчатая ржавчина веймутовой
сосны и многие другие болезни растений.
Инфекционное начало грибов распространяется человеком
также в процессе выполнения различных лесохозяйственных работ,
при уходе за растениями в питомниках и теплицах — на руках,
одежде, с частицами почвы и растений, приставшими к обуви,
орудиям и инструментам. Особенно часто это происходит при
нарушении необходимых санитарно-гигиенических правил.
Аутохория. Это активное, самостоятельное распространение спор.
Оно свойственно грибам, в цикле развития которых образуются
зооспоры: слизевикам, многим оомицетам. Благодаря наличию
жгутиков зооспоры могут свободно передвигаться в воде, при этом
часто наблюдаются явления фототаксиса (движение в
направлении источника света) и хемотаксиса (привлечение зооспор
выделениями восприимчивого хозяина).
2.5. ПИТАНИЕ, ПАРАЗИТИЗМ И СПЕЦИАЛИЗАЦИЯ
ГРИБОВ
Грибы и грибоподобные организмы характеризуются
гетеротрофным типом питания. Они не способны, в отличие от зеленых
растений, самостоятельно синтезировать органические вещества
(углеводы, белки, жиры и др.), а извлекают их в готовом виде из
питательного субстрата, на котором поселяются. Органические и
минеральные соединения поступают в организм гриба
непосредственно через оболочки гиф (осмотрофное питание). Поэтому
грибы могут поглощать их только в виде водных растворов. Высокая
поглощающая способность хорошо развитого мицелия
обеспечивается очень большой суммарной поверхностью гиф.
Питание грибов осуществляется с помощью мощного
ферментного аппарата, включающего экзоферменты, выделяемые грибом
в питательный субстрат, и эндоферменты, действующие внутри
грибной клетки. При участии экзоферментов грибы превращают
сложные органические соединения растительных тканей в более
простые водорастворимые соединения, а при помощи эндофер-
ментов протекает как бы обратный процесс — синтез высокомо-
32
лекулярных соединений, необходимых для нормального обмена
веществ, роста и размножения гриба.
Грибам свойственны ферменты всех основных групп:
гидролитические, окислительно-восстановительные, бродильные. Среди
гидролитических ферментов грибов наибольшим разнообразием
отличаются карбогидразы, т.е. ферменты, осуществляющие
гидролиз углеводов — основных источников питания фитопатоген-
ных грибов. Из них широко распространены целлюлазы и геми-
целлюлазы, гидролизующие клетчатку и гемицеллюлозу —
соединения, которые являются важнейшей составной частью
клеточных оболочек высших растений; пектиназы, разрушающие
пектиновые вещества растительных тканей; амилазы, гидролизующие
крахмал, и др. Гидролиз белковых соединений растительных и
животных тканей обеспечивается комплексом протеолитических
ферментов — протеиназ; расщепление жиров осуществляют
липазы. Окислительные ферменты грибов представлены оксидазами
и дегидразами, а восстановительные — редуктазами. Бродильные
ферменты (зимазы) свойственны некоторым сумчатым
(например, дрожжевым), несовершенным и мукоровым грибам,
благодаря чему они находят применение в пищевой промышленности.
У отдельных групп и видов грибов имеется набор
специализированных ферментов, участвующих в гидролизе специфических
компонентов питательного субстрата и промежуточных
продуктов их расщепления. Кроме того, они являются катализаторами
процессов энергетического обмена и биосинтеза сложных
соединений.
Наличие ферментов определенной группы и их состав тесно
связаны с образом жизни и степенью специализации гриба. Так,
неспециализированные грибы-сапротрофы, поселяющиеся на
отмерших частях растений и животных, органических остатках и
других мертвых субстратах, обладают наиболее богатым и
разнообразным набором ферментов, что позволяет им использовать
самые различные источники питания. Чем больше выражена
специализация гриба в отношении питающего субстрата, тем
обычно беднее и специфичнее ассортимент вырабатываемых им
ферментов. Активность ферментов у грибов может изменяться под
влиянием температуры, влажности, величины рН и других
условий окружающей среды.
Для нормальной жизнедеятельности грибы нуждаются в
многочисленных элементах питания, среди которых первостепенное
значение имеют углерод, азот, некоторые зольные элементы,
биологически активные вещества, микроэлементы.
Образ жизни грибов обусловливается их биологическими
свойствами, которые вырабатывались и изменялись в процессе
эволюции. Среди грибов различают сапротрофы, которые заселяют
мертвые органические субстраты, и паразиты, развивающих-
33
ся на живых органах растений или животных. Грибы-паразиты в
свою очередь различаются по характеру воздействия на ткань
растения-хозяина.
Некротрофы сначала убивают клетки растения, грубо
воздействуя на них своими выделениями (ферментами и
токсинами), а затем питаются их содержимым. Зараженные некротрофа-
ми участки тканей часто представляют собой островки мертвых
клеток в живом организме хозяина (например, при пятнистостях
листьев). Иногда некротрофы полностью убивают растение, а
потом уже развиваются в нем. К этому типу грибов относятся и так
называемые токсигенные паразиты; они выделяют
сильнодействующие токсины, которые, распространяясь по проводящей
системе растения-хозяина, быстро вызывают его гибель. Таковы,
например, грибы рода Fusarium, вызывающие увядание сеянцев
древесных пород и цветочных растений.
Биотрофы обычно не выделяют токсинов и осторожно
воздействуют ферментами на зараженную ими ткань, извлекая
питательные вещества непосредственно из живых клеток растения,
которые при этом могут сравнительно долго оставаться
жизнедеятельными.
Сапротрофный образ жизни считается первичной, наиболее
древней формой существования грибов. Переход к паразитизму
происходил у отдельных групп грибов в ходе длительного
эволюционного развития и сопровождался выработкой у них особых
приспособительных качеств (наличие специальных ферментов и
токсинов, способность внедряться в живую клетку, жить в
условиях недостатка кислорода, избытка углекислоты и т.д.).
В ходе эволюции возникли и обособились группы грибов,
различающиеся по степени паразитической активности, форме
существования и, соответственно, располагающиеся на разных
ступеньках «эволюционной лестницы» (рис. 8).
Облигатные сапротрофы — это грибы, способные
поселяться и развиваться только на отмерших тканях растений или
животных и других органических остатках. Они хорошо растут на
различных искусственных питательных средах. К ним относится
большинство известных видов грибов. Особенно большую группу
составляют сапротрофные почвенные грибы, играющие весьма
важную роль в почвообразовательных процессах. Разлагая
растительные остатки, они способствуют накоплению гумуса и
повышению плодородия почв. Некоторые из них являются
продуцентами антибиотиков. Не представляя опасности для живых растений,
грибы-сапротрофы, однако, причиняют большой ущерб,
разрушая древесину на складах и в открытых сооружениях (складские
грибы), в отапливаемых зданиях (домовые грибы). Многие виды
сапротрофных грибов вызывают порчу плодов и семян,
продуктов питания, бумаги и других изделий.
34
Грибы, у которых единственная
или преобладающая форма
существования сапротрофная
1 ступень
Облигатные
(обязательные)
сапротрофы
2 ступень
Факультативные
(условные)
паразиты
Грибы, у которых единственная
или преобладающая форма
существования паразитическая
3 ступень
Факультативные
(условные)
сапротрофы
4 ступень
Облигатные
(обязательные)
паразиты
Рис. 8. Условная схема ступенчатой эволюции паразитизма у грибов
Факультативные паразиты — это грибы, для которых
обычным в природе является сапротрофный образ жизни. Однако
при определенных условиях (поэтому их называют также
условными паразитами) они способны переходить к паразитированию
на живых, чаще всего ослабленных растениях. Все
факультативные паразиты характеризуются некротрофным способом питания,
который у них ярко выражен. Эти грибы также легко
культивируются на искусственных питательных средах. К факультативным
паразитам относятся, например, возбудители полегания всходов
древесных растений, многие трутовые и плесневые грибы. В
отсутствие подходящих живых растений они нормально
развиваются и размножаются на мертвой древесине или органических
остатках в почве.
Факультативные сапротрофы обладают более
высокой степенью паразитической активности. Среди них имеются как
некротрофы, так и биотрофы. Некоторые могут переходить от не-
кротрофного к биотрофному способу питания, и наоборот.
Обычно они ведут паразитический образ жизни, развиваясь в течение
вегетации на живых растениях и образуя на них бесполые споро-
ношения. Однако завершают свой цикл развития они нередко как
сапротрофы. Половое спороношение этих грибов чаще всего
формируется на отмерших частях растения (опавших листьях и хвое,
засохших ветвях и т.п.). Факультативные сапротрофы могут быть
выращены и на определенных искусственных субстратах. К этой
группе относятся очень многие фитопатогенные грибы из числа
сумчатых и несовершенных, например возбудители пятнистостей
листьев, парши, антракнозов, шютте и др.
35
Среди факультативных паразитов и сапротрофов выделяют
группу раневых паразитов. Эти грибы, поражающие живые
органы растений, способны проникнуть в них только через раны или
отмершие участки покровных тканей. После предварительного
этапа сапротрофного развития на поврежденных тканях раневые
паразиты обычно переходят на соседние живые ткани. Таковы,
например, возбудители ступенчатого рака стволов древесных
пород.
Облигатные паразиты составляют группу грибов,
характеризующихся наивысшей степенью паразитической
активности. Они полностью утратили способность к сапротрофному
образу жизни и могут питаться только за счет живых клеток растения-
хозяина, т. е. всегда являются биотрофами. Облигатные паразиты
обычно поражают вполне жизнеспособные, без каких-либо
признаков ослабления, экземпляры растений. Их не удается (за
исключением отдельных немногих случаев) культивировать на
искусственных питательных средах. Свойствами облигатных
паразитов обладают мучнисторосяные, плазмодиофоровые, тафриновые,
большинство ржавчинных грибов.
Выделяюттакжегруппуэкологически облигатных
паразитов, которые в природе всегда существуют только как
облигатные паразиты. Однако в лабораторных условиях они могут
быть выделены в чистую культуру на специально приготовленную
питательную среду. Таковы головневые грибы, некоторые виды
аскомицетов и ржавчинных грибов.
В зависимости от того, где развивается вегетативное тело
паразитических грибов, среди них различают эндо- и эктопаразитов.
У первых мицелий распространяется внутри тканей пораженных
органов растений. Он может разрастаться как в межклетниках, так
и в самих клетках, или только в межклетниках (в этом случае от
него отходят внутрь клеток отростки — гаустории). Если
вегетативное тело гриба — плазмодий, то его развитие происходит только
внутри клеток. Мицелий эндопаразитов бывает местный или
диффузный, однолетний или многолетний. Эндопаразитизм наблюдается
у подавляющего большинства фитопатогенных грибов. У
эктопаразитов мицелий развивается на поверхности растений, посылая в
клетки эпидермиса гаустории, с помощью которых гриб питается
и прочно удерживается на субстрате. Эктопаразитизм характерен
для мучнисторосяных грибов.
В отличие от облигатных сапротрофов, большинству которых
свойственны неспециализированное питание и способность
заселять самые разнообразные органические субстраты,
паразитические грибы характеризуются специализацией, т.е.
приспособленностью к поражению лишь известного круга
растений-хозяев или определенных органов и тканей, либо
растений и тканей только данного возраста. Различают следующие
36
типы специализации: филогенетическую, онтогенетическую,
органотропную и гистотропную. Степень (широта)
специализации любого типа у разных возбудителей неодинакова и
специфична.
Филогенетическая специализация означает
приспособленность патогена к паразитированию на одном или нескольких
(иногда многих), но всегда определенных питающих растениях.
Она является результатом длительной сопряженной эволюции
(филогенеза) паразитов и их растений-хозяев.
Широкоспециализированных, малоразборчивых в отношении питательного субстрата
патогенов называют полифагами, узкоспециализированных
паразитов, отличающихся строгой избирательностью в отношении
растений-хозяев, — монофагами.
Существуют патогенные грибы, способные поражать многие
виды растений из разных ботанических семейств. Таковы,
например, Sclerotinia graminearum и Typhula graminearum, вызывающие
выпревание различных злаков, некоторых других травянистых
растений, а также сеянцев хвойных и лиственных пород. В этом
случае мы имеем дело с очень широкой филогенетической
специализацией.
Менее широкая специализация у паразитов, поражающих
растения разных родов в пределах одного семейства. Примером
может служить Plasmodiophora brassicae, возбудитель килы
крестоцветных, паразитирующий на многих растениях семейства.
Узкая специализация наблюдается у многих фитопатогенных
грибов, относящихся к различным систематическим группам.
Она может быть родовой, когда патоген поражает все или
отдельные виды одного рода растений-хозяев. Так, виды рода Lopho-
dermium, возбудители обыкновенного шютте сосны, поражают
хвою у разных видов рода Pinus. Rhytisma acerinum, возбудитель
черной пятнистости листьев клена, паразитирует на разных видах
рода Acer.
Среди облигатных паразитов (мучнисторосяных, ржавчинных
грибов) имеются виды (их иногда называют сборными),
представленные совокупностью форм, различающихся по своей
специализации, но морфологически неотличимых друг от друга.
Такие формы внутри одного и того же вида гриба получили
название специализированных форм (латинское название forma specialis,
сокращенно/ sp, или просто/). Например, вид мучнисторосяного
гриба Phyllactinia suffulta, встречающийся на лещине, ясене,
березе, буке и других породах, включает ряд специализированных
форм, каждая из которых приурочена к одному из названных
родов растений-хозяев: Ph. suffultaf. coryli, Ph. suffulta f fraxini, Ph.
suffultaf. betulae, Ph. suffultaf. carpini и т.д.
Видовая специализация наблюдается в тех случаях, когда
паразит способен развиваться на растениях только одного вида. Такой
37
специализацией обладает, например, Peridermium pint —
возбудитель смоляного рака сосны обыкновенной.
Существует еще более узкая — сортовая специализация,
свойственная физиологическим расам паразитных грибов.
Физиологические расы способны поражать лишь определенные сорта
какого-либо вида растений. Они известны у многих грибов из
разных систематических групп. Физиологические расы
обозначаются цифрами.
Онтогенетическая специализация патогенов означает их
приуроченность к определенному возрастному этапу
индивидуального развития (онтогенеза) растений или к определенному воз-
растно-физиологическому состоянию поражаемых органов.
Например, возбудители полегания сеянцев поражают всходы лишь в
первые недели или месяцы жизни. Melampsora pinitorqua,
возбудитель соснового вертуна, поражает сосну лишь в возрасте до 10 —
12 лет. Microsphaera alphitoides, возбудитель мучнистой росы дуба,
как и многие мучнисторосяные, поражает только молодые,
растущие листья и побеги. Phacidium infestans, возбудитель снежного шют-
те сосны и ели, а также Lophodermium seditiosum — один из
возбудителей обыкновенного шютте сосны поражают в основном
молодую хвою, тогда как Lophodermium pinastri, другой возбудитель
обыкновенного шютте сосны, наоборот, специализирован к
физиологически старой хвое и поражает ее на нисходящем отрезке
кривой онтогенеза.
Рассмотренные типы специализации (филогенетическая и
онтогенетическая) имеют большое практическое значение, поскольку
непосредственно связаны с устойчивостью растений к
инфекционным болезням. Они могут быть использованы в селекционной
работе, при подборе пород и форм древесных растений, при
обосновании конкретных лесохозяйственных и химических мер
борьбы в системах мероприятий по защите растений и повышению
устойчивости насаждений.
Органотропная специализация — это приспособленность
патогенов к определенным органам, а гистотропная —к
определенным тканям питающих растений. Например, некоторые виды
возбудителей парши (представители рода Venturid) обладают
сравнительно широкой органотропной специализацией, так как
поражают листья, побеги, а иногда еще и плоды. В
противоположность им, например, Septoria populi, возбудитель белой
пятнистости листьев тополя, поражает только листья, а в самих листьях —
только паренхимную ткань. Возбудители некоторых гнилевых и
некрозно-раковых болезней древесных пород могут иметь
одинаковую органотропную специализацию, поражая стволы деревьев,
но при этом их гистотропная специализация будет совершенно
разной: первые развиваются в древесине ствола, а вторые
поражают кору и камбий.
38
2.6. ТРЕБОВАНИЯ ГРИБОВ К УСЛОВИЯМ
ОКРУЖАЮЩЕЙ СРЕДЫ
Нормальный рост, развитие и жизнедеятельность грибов, их
паразитическая активность в значительной мере зависят от условий
окружающей среды и прежде всего от влажности, температуры,
кислотности, наличия кислорода, освещенности. Влияние каждого
из этих факторов может меняться в зависимости от изменения
других условий. Важнейшее значение при этом имеют определенное
сочетание отдельных факторов и продолжительность их воздействия.
Влажность. Наличие воды в той или иной форме — одно из
главных условий жизни грибов. Отношение к этому фактору
определяется принадлежностью гриба к определенной экологической
группе, а у фитопатогенных грибов связано также с
особенностями их паразитизма. От влажности питательного субстрата во
многом зависит возможность его заселения и использования
определенными видами грибов. Влажность окружающего воздуха часто
обусловливает интенсивность спорообразования и
распространения инфекционного начала грибов.
Водные грибы требуют полного погружения в воду как во
время вегетативного роста, так и в период размножения, поскольку
оно осуществляется зооспорами. Многие представителя
почвенной микофлоры, в цикле развития которых образуются
зооспоры, лучше всего развиваются при насыщенной влажности почвы.
Однако для грибов, требовательных к кислороду, избыточное
увлажнение почвы неблагоприятно, так как при этом резко ухудшается
ее аэрация.
Для большинства дереворазрушающих грибов оптимальна
умеренная влажность древесины (в пределах 30 — 80 %), но у
отдельных видов потребность в увлажнении субстрата выходит за эти
пределы. Почти все наземные грибы требуют повышенной
влажности субстрата во время роста мицелия, т.е. в период активного
накопления вегетативной массы.
Высокая относительная влажность воздуха (а для грибов,
образующих зооспоры, — капельно-жидкая влага) чаще всего
необходима при образовании органов бесполого спороношения. При
формировании же органов полового размножения потребность в
воде иногда снижается. Особенно резко она уменьшается при
переходе гриба в покоящуюся стадию, например при образовании
склероциев. Высокая насыщенная влажность воздуха или наличие
капельно-жидкой влаги необходимы большинству грибов при
освобождении спор из споровместилищ, их рассеивании и
прорастании. Способность развивать мицелий и обильно спороносить
при низкой относительной влажности воздуха присуща немногим
грибам (например, мучнисторосяным).
39
Фитопатогенные грибы, особенно облигатные паразиты,
обычно находят необходимую для них воду в живых тканях растения-
хозяина. Поэтому их развитие, прежде всего в фазе вегетативного
роста, в меньшей степени, чем у грибов-сапротрофов, зависит от
колебаний влажности окружающего воздуха.
Температура. Как и влажность, температура окружающей
среды оказывает очень сильное влияние на рост, размножение и
физиологическую активность грибов. Температурные требования
разных видов и групп грибов неодинаковы. Наряду с грибами,
температурный оптимум которых лежит около 25... 3 О °С (а иногда
и выше), известно немало видов, для которых оптимальны
температуры от 0 до 10 °С. Есть виды, активное развитие которых
происходит при температурах от 1 до 3 °С (под снегом), например
возбудители снежного шютте сосны и выпревания сеянцев. Для
большинства грибов оптимальны температуры в пределах от 18 до 25 °С.
При оптимальной температуре процессы обмена веществ,
роста и спорообразования обычно протекают наиболее интенсивно.
Однако иногда температурный оптимум для вегетативного роста
не совпадает с оптимумом для спороношения или
физиологической активности того же самого гриба. Далеко не всегда и
максимальная патогенность паразитного гриба проявляется при
оптимальной для него температуре: решающую роль здесь часто играет
возрастное или физиологическое состояние поражаемого
растения в сочетании с другими факторами среды. Большое значение
имеет температура среды для прорастания спор грибов. Иногда от
температуры зависят не только сама возможность прорастания
спор, но и скорость прорастания и его характер.
Если для наилучшего развития вида необходима оптимальная
температура, то для выживания и сохранения его в природе
нередко более важны крайние (минимальная и максимальная)
температуры, которые называют критическими. При минимальной
температуре начинаются процессы жизнедеятельности гриба, при
максимальной они резко ослабевают или почти
приостанавливаются. Покоящиеся споры, склероции, плодовые тела некоторых
грибов способны в течение определенного срока выдерживать
экстремальные температуры, при которых полностью
прекращаются все жизненные процессы. Так, многолетние плодовые тела
трутовых грибов сохраняют жизнеспособность во время сильных
(до -40 °С) морозов.
Фунгицидное действие высоких температур используется для
дезинфекции зараженных грибами материалов, изделий и
субстратов, для обеззараживания семян и посадочного материала. На
использовании высокой температуры в сочетании с химической
обработкой основан термохимический метод стерилизации.
Кислород. Все грибы относятся к числу аэробных организмов,
однако потребность в кислороде у разных видов различна. Боль-
40
шинство грибов требует достаточно высокого содержания
кислорода в среде обитания. В то же время многие грибы могут развиваться
при слабом доступе кислорода. Таковы водные и некоторые
плесневые грибы, живущие в насыщенных водой субстратах, а также
облигатные паразиты, развивающиеся в живых, активно
дышащих тканях растения-хозяина. Известны также виды (например,
некоторые дрожжевые грибы), способные в течение
определенного периода времени выдерживать полное отсутствие кислорода.
Кислотность. Реакция среды имеет важное значение для роста
и жизнедеятельности грибов. Она является одним из главных
факторов, обусловливающих возможность поражения грибами
определенных видов или органов растений, заселения ими различных
природных субстратов, пищевых продуктов, промышленных
материалов.
Большая часть грибов предпочитает слабокислую реакцию
среды (рН 4,0 — 6,0), но есть виды, для которых благоприятны более
кислые, нейтральные или даже щелочные субстраты. Щелочная
среда необходима, например, грибам, паразитирующим на
животных (в частности, энтомопатогенным видам) или
разлагающим остатки животного происхождения.
Оптимум и критические значения рН для вегетативного роста,
процессов метаболизма, спорообразования, прорастания спор чаще
всего несколько различаются. Требования грибов к реакции среды
могут изменяться в зависимости от состава питательных веществ в
субстрате, температуры, аэрации. Некоторые виды грибов сами
способны подкислять или подщелачивать среду, выделяя
соответствующие ферменты или продукты метаболизма.
Свет. Большинство грибов наилучшим образом развивается на
рассеянном свету. Мицелий грибов обычно малочувствителен к
свету, но для нормального развития органов спороношения свет,
как правило, необходим. Многие грибы в темноте не спороносят
или слабо спороносят, другие (например, некоторые трутовики)
образуют уродливые, иногда стерильные плодовые тела. Есть и
такие виды (настоящий домовый гриб, шампиньон, трюфели),
которые вообще не нуждаются в освещении.
Грибам свойственны реакции фототаксиса и фототропизма,
причем они бывают положительными или отрицательными. Так,
плазмодии и зооспоры могут активно передвигаться к свету или в
противоположную сторону. Явление фототропизма чаще всего
наблюдается у репродуктивных органов грибов и проявляется в
росте или изгибании конидиеносцев, спорангиеносцев,
плодовых тел по направлению к источнику света, или наоборот.
Чередование освещения и затемнения нередко стимулирует процессы
роста и спорообразования грибов.
Прямой солнечный свет обычно задерживает рост мицелия, а
при длительном воздействии вызывает его отмирание. Не выдер-
41
живают прямых солнечных лучей и пропагативные споры многих
грибов, особенно неокрашенные. Исключение составляют мучни-
сторосяные грибы. Не случайно склероции, покоящиеся споры и
споровместилища грибов, выносливые к прямому освещению,
имеют, как правило, толстые, интенсивно пигментированные
оболочки.
Наличие в оболочках спор, плодовых тел и других органов
грибов оливково-черных пигментов (меланинов) оказывает также
защитное действие против ультрафиолетовых, инфракрасных,
космических и других излучений. Ионизирующее облучение в
зависимости от дозы, экспозиции, физиологического состояния
гриба, температуры, влажности и других факторов может
стимулировать рост и спороношение, вызывать изменение
генетических свойств (появление мутаций) или гибель гриба.
2.7. СИСТЕМАТИКА ГРИБОВ
Современная систематика грибов и грибоподобных
организмов означает распределение их по царствам живого мира,
отделам, классам, порядкам, семействам, родам и видам.
За последние 10—20 лет традиционные представления о
царстве грибов, взгляды на их происхождение, эволюцию и
филогенетические связи подверглись основательному пересмотру. В связи
с накоплением обширного материала по физиологии и биохимии
грибов и грибоподобных организмов, строению и составу их
клеточной стенки, генетическим особенностям, ультраструктуре
клетки произошли существенные изменения в объеме и содержании
царства грибов и таксонов (прежде всего — отделов и классов).
Так, из прежнего царства грибов были исключены организмы,
имеющие подвижные стадии: все миксомицеты, наряду с
амебами, вошли в царство Protozoa, а оомицеты и некоторые другие
псевдогрибы — в царство Chromista.
Таким образом, в последних системах грибы и грибоподобные
организмы разделены на три самостоятельных эволюционных ствола
и распределены по трем царствам живого мира: Protozoa (= Proto-
ctista), Chromista и Mycota. Однако единой общепризнанной
системы грибов пока еще нет.
Современный этап в формировании взглядов на филогенез и
систематику грибов характеризуется интенсивным изучением их
методами молекулярной биологии. Это позволило получить новые
данные, используемые при построении филогенетической
системы грибов.
В настоящее время на основании анализа результатов
многочисленных исследований наиболее значимыми
таксономическими признаками считаются строение подвижных стадий (зооспор,
42
Таблица 1
Распределение грибов и грибоподобных организмов по царствам, основным отделам и классам
Царство Protozoa
(Простейшие)
Отдел
Мухо-
mycota
(Слизевики)
Класс
Мухо-
тусе-
tes
Отдел
Plasmodio-
phoromycota
(Плазмо-
диофоро-
вые)
Класс
Plasmodio-
phoromyce-
tes
Размножение:
половое — изогамия;
бесполое —
зооспорами
Вегетативное тело —
плазмодий
Царство
Chromista
(Псевдогрибы)
Отдел
Oomycota
(Оомицеты)
Класс
Oomycetes
Спороношения:
половое —
ооспоры,
бесполое —
зооспоры,
конидии
Царство Mycota, Fungi (Настоящие грибы)
Отдел
Zygomycota
(Зигомицеты)
Класс
Zygomycetes
Спороношения:
половое —
зигоспоры,
бесполое — спо-
рангиоспоры,
конидии
Вегетативное тело —
одноклеточный мицелий
Низшие грибы
Отдел Dicaryomycota
Подотдел
Ascomycotina
(Сумчатые)
Класс
Archae-
asco-
mycetes
Класс
Euas-
comy-
cetes
Класс
Locu-
loasco-
mycetes
Спороношения:
половое — сумкоспо-
ры, бесполое —
конидии
Подотдел
Basidiomycotina
(Базидиальные)
Класс
Telio-
myce-
tes
Класс
Usto-
myce-
tes
Класс
Basi-
diomy-
cetes
Спороношения:
половое — базидио-
споры, бесполое —
споры типа
конидий
Mitosporis fungi,
Fungi imperfecti
(Несовершенные)
Класс
Ago-
nomy-
cetes
Класс
Hyp-
homy-
cetes
Класс
Сое-
lomy-
cetes
Спороношения:
половое —
отсутствует, бесполое —
конидии или оно
отсутствует
Вегетативное тело — многоклеточный мицелий
Высшие грибы
гамет), состав и строение клеточной стенки, физиолого-биохи-
мические свойства, ультраструктурные особенности клетки. В
характеристике таксонов важное значение имеют также
особенности вегетативного тела и размножения грибов.
В учебнике приведена сокращенная система грибов и грибопо-
добных организмов. В ней рассмотрены основные
таксономические группы (табл. 1) и те из более мелких таксонов, в состав
которых входят фитопатогенные виды — возбудители болезней
лесных древесных растений и цветочных культур и некоторые
другие грибы, имеющие практическое значение.
2.7.1. Царство Protozoa, или Protoctista
К этому царству отнесены простейшие грибоподобные
организмы, которые включены в два основных отдела: Myxomycota и
Plasmodiophoromycota. Вегетативное тело — плазмодий, бесполое
размножение осуществляется зооспорами, половой процесс —
изогамия, в результате которой образуется диплоидный
плазмодий.
Отдел Myxomycota
Сюда относятся типичные миксомицеты (собственно
слизевики), имеющие сапротрофный плазмодий. Клеточные стенки
спороносных органов и покоящихся спор содержат целлюлозу.
Основной класс этого отдела — Myxomycetes. Плазмодий миксоми-
цетов не имеет ни оболочки, ни постоянной формы и поэтому
способен к более или менее активному амебообразному
передвижению во влажной среде.
Большая часть слизевиков — лесные сапротрофы, плазмодии
которых обитают в почве, на лесной подстилке, всевозможных древесных
остатках (валежнике, пнях и т. п.). Слизевики образуют особые,
разнообразные по форме и строению органы спороношения — эта-
лии, при созревании которых формируются покоящиеся споры.
Наиболее часто в наших лесах встречаются миксомицеты,
относящиеся к родам Fuligo, Spumaria, Lycogala, Stemonitis, Physarum
и др.
Отдел Plasmodiophoromycota
К этому отделу относится только один класс — Plasmodiophoro-
mycetes. Плазмодиофоровые — облигатные паразиты со сложным
циклом развития, имеющие внутриклеточный плазмодий.
Развиваясь в клетках растения-хозяина, плазмодии вызывают
увеличение их размеров (явление гипертрофии). К концу вегетации плаз-
44
модий распадается на множество спор, покрытых плотной
оболочкой. Попадая в почву при сгнивании пораженных тканей
растений, они выполняют роль покоящихся спор, которые могут
сохраняться в течение нескольких лет. С наступлением
благоприятных условий они прорастают амебообразными зооспорами (микс-
амебами), последние вновь заражают растения.
Характерным признаком болезней, вызываемых паразитами
этого класса, является разрастание пораженных тканей,
образование на подземных частях растений (корнях, клубнях) наростов
различной формы и величины. Наибольшее хозяйственное
значение среди плазмодиофоровых имеет Plasmodiophora brassicae —
возбудитель килы крестоцветных растений, в том числе
цветочных культур.
2.7.2. Царство Chromista (Псевдогрибы)
Царство включает три отдела, два из которых представлены
сапротрофами или паразитами водных растений и простейших.
Практическое значение имеет отдел Oomycota, в состав которого
входит лишь один класс — Oomycetes.
Класс Oomycetes — Оомицеты
Вегетативное тело — в основном одноклеточный диплоидный
мицелий. В составе клеточных стенок преобладает глюкан-целлюло-
за; изредка содержится небольшое количество хитина. Основные
запасные вещества — Р-глюкан и миколаминоран. Бесполое спо-
роношение — двужгутиковые зооспоры с гетероморфными
жгутиками (гладким и перистым) или конидии. Половой процесс —
оогамия, в результате которой образуется телеоморфа — ооспора.
Класс включает девять порядков, важнейшими из которых
являются порядки Peronosporales, Pythiales и Saprolegniales.
Порядок Peronosporales — Пероноспоровые. В его
состав входят фитопатогенные виды, относящиеся к семействам
Phytophtoraceae, Peronosporaceae и Albuginaceae. Все они имеют
хорошо развитый несептированный мицелий. Покоящиеся споры
(ооспоры) сохраняются в почве или на растительных остатках, а
при наступлении благоприятных условий прорастают, образуя
органы бесполого размножения, чаще всего зооспорангии.
К семейству фитофторовых относятся фитопатогенные виды
рода Phytophthora: Ph. cactorum — возбудитель фитофторозной
гнили сеянцев лиственных пород и декоративных растений и Ph.
infestans — возбудитель фитофтороза картофеля и томата.
Виды семейства пероноспоровых (ложномучнисторосяные) —
облигатные паразиты, вызывающие болезни растений под
названием ложные мучнистые росы. Они проявляются в образовании
45
Рис. 9. Ложная мучнистая роса:
/ — на листе винограда; 2 — на листе подсолнечника
на верхней стороне листьев хлоротичных или бурых
некротических пятен, а на нижней — беловатого мучнистого налета,
состоящего из зооспорангиев или конидиеносцев и конидий
возбудителя. Ложной мучнистой росой поражаются многие
сельскохозяйственные и цветочные культуры (рис. 9).
Виды семейства альбугиновых — также облигатные паразиты
травянистых растений, в том числе некоторых возделываемых
культур (например, левкоя, хрена), вызывающие болезни под
названием «белая ржавчина».
Порядок Pythiales — Питиевые. Представители порядка —
в основном почвенные сапротрофы. Из фитопатогенных видов
наиболее известным является Pythium debaryanum —
факультативный паразит, один из возбудителей полегания всходов древесных
пород и декоративных кустарников, а также черной ножки
цветочной рассады.
Порядок Saprolegniales — Сапролегниевые. В основном
это сапротрофы, обитающие на остатках водных растений и
животных, преимущественно в загрязненных водоемах. Среди сапролег-
ниевых есть и паразиты, вызывающие опасные заболевания водных
животных, особенно рыб, и причиняющие ущерб рыбоводству.
2.7.3. Царство Mycota, Fungi (Настоящие грибы)
Царство Mycota содержит огромное количество видов самых
разнообразных грибов, в том числе фитопатогенных, —
возбудителей многих опасных болезней древесных растений, цветочных и
сельскохозяйственных культур.
46
К этому царству относятся два неравных по объему и значению
отдела: Zygomycota и Dicaryomycota, а также группа без
определенного таксономического статуса — Mitosporis fungi
(Несовершенные грибы).
Отдел Zygomycota
Вегетативное тело — преимущественно несептированный
мицелий. Септы встречаются лишь у некоторых
специализированных групп (например, энтомофторовых). В составе клеточных
стенок — хитин в комплексе с хитозаном. Запасное вещество —
гликоген. Половой процесс — зигогамия, в результате которой
образуется телеоморфа (зигоспора). Важнейший класс отдела —
Zygomycetes.
Класс Zygomycetes — Зигомицеты
Класс делится на несколько порядков, различающихся по
морфологии анаморф (типу бесполого спороношения), циклам
развития и экологическим особенностям. Важнейшие порядки — му-
коровые и энтомофторовые.
Порядок Mucorales — мукоровые. У большинства
мукоровых грибов имеется разветвленный паутинистый мицелий,
обьино стелющийся по субстрату. Он состоит из довольно толстых
гиф, на которых образуются органы бесполого размножения —
спорангиеносцы со спорангиями. Спорангии мукоровых имеют вид
темных, хорошо различимых простым глазом головок, благодаря
чему эти грибы получили название головчатых плесеней.
Освобождающиеся из спорангиев спорангиоспоры, прорастая, дают
начало новому мицелию. Для некоторых видов характерно
наличие ризоидов — корнеподобных разветвлений, которыми гриб
укрепляется на субстрате, и воздушных гиф — столонов (рис. 10).
Особенностью мукоровых является гетероталлизм, т.е. разнопо-
лость мицелиев. При копуляции гаметангиев разного пола
происходит половой процесс — зигогамия (см. рис. 7).
Мукоровые грибы весьма широко распространены в природе.
Большинство этих грибов — почвенные сапротрофы, в изобилии
встречающиеся в пахотном слое, особенно в почвах, богатых
органическими веществами (число их спор в 1 г почвы нередко
достигает нескольких десятков тысяч). Они принимают активное
участие в процессах разложения органических веществ и накопления
гумуса. В условиях повышенной влажности мукоровые могут быть
причиной порчи кормов, продуктов и материалов.
Многие представители мукоровых грибов вызывают плесне-
вение семян и плодов при хранении. Таковы, например, виды
рода Mucor, Rhizopus nigricans, Thamnidium elegans и др. (см. рис. 10).
47
Рис. 10. Грибы порядка мукоровых:
А — разветвленный одноклеточный мицелий и спорангии гриба рода Мисог; Б —
строение вегетативного тела и органов бесполого спороношения гриба Rhizopus
nigricans: 1 — спорангиеносцы; 2 — спорангии; 3 — ризоиды; 4 — столоны; 5 —
спорангиоспоры; В — спороношение гриба Thamnidium elegans
Некоторые виды вызывают микозы теплокровных, в том числе
человека.
Порядок Entomophthorales — энтомофторовые.
К этому порядку относятся паразиты насекомых. Во внутренних
органах насекомого развивается мицелий гриба; на нем
образуются органы бесполого размножения — конидиеносцы, выходящие
наружу через дыхательные отверстия и тонкие участки
хитинового покрова. На конидиеносцах формируются конидии; при
созревании они отбрасываются и попадают на здоровых насекомых.
Прорастая, конидия дает ростковую гифу, которая проникает в тело
насекомого, где вновь развиваются мицелий и органы
спороношения гриба (рис. 11).
Практический интерес представляют энтомофторовые,
паразитирующие на насекомых, вредных для леса и сельскохозяйствен-
Рис. 11. Насекомые, пораженные грибами порядка энтомофторовых (7, 2,3)
и формы конидиального спороношения возбудителей (4, 5)
48
ных культур. Так, Entomophaga aulicae вызывает на больших
площадях сильные эпизоотии у сосновой совки, златогузки. Другие виды
этого рода поражают пядениц, листоверток, прямокрылых, мух,
пилильщиков. Виды рода Zoophthora паразитируют на различных
насекомых, например, вызывают эпизоотии у тлей; виды рода
Tarichium поражают многих вредных чешуекрылых и
представителей других отрядов. Изредка встречаются факультативные
паразиты теплокровных и сапротрофы.
Энтомофторовые грибы могут являться причиной массовых
заболеваний и снижения численности насекомых. Это позволяет
использовать их в рамках биологического метода борьбы с
вредными насекомыми в лесном и сельском хозяйстве.
Представители порядка Zoopagales— в основном
паразиты животных, имеющие простой ловчий аппарат (ветви гиф с
каплями слизи на их кончиках).
Виды порядка Dimargaritales— паразиты грибов.
Некоторые виды порядка Kickxellales — также
паразиты на грибах.
Представители порядка Endogonales — в основном
симбиотрофы, образующие эндотрофную микоризу с корнями
растений.
Отдел Dicaryomycota
Самый большой отдел не только царства грибов, но и всех
грибоподобных организмов, насчитывающий огромное
количество видов. Главный признак отдела — наличие септированного
дикариотического (двуядерного) мицелия. Отдел включает два
подотдела: Ascomycotina и Basidiomycotina.
Подотдел Ascomycotina (Сумчатые грибы)
Подотдел включает три основных класса, 46 порядков и
264 семейства грибов, что составляет около 75 % всех известных
видов. Сумчатые грибы весьма разнообразны по морфологии,
образу жизни, значению в природе и практической
деятельности человека. Однако всех их объединяет важнейший
диагностический признак — наличие сумок с эндогенными сумкоспорами
(аскоспорами), которые образуются в результате полового
процесса.
Второй важный диагностический признак — наличие
двуслойных клеточных стенок, в состав которых входят хитин и
Р-глюкан.
Подотдел Ascomycotina состоит из четырех классов,
различающихся в основном по способу формирования и размещения
сумок: Archaeascomycetes, Hemiascomycetes, Euascomycetes и Loculo-
ascomycetes.
49
Класс Archaeascomycetes
Относительно новый класс, выделенный на основании
результатов биохимических исследований. В него вошли некоторые
грибы, прежде относившиеся к Hemiascomycetes, в частности грибы
порядка тафриновых.
Порядок Taphrinales — тафриновые. Мицелий
тафриновых — в основном септированный. Сумки образуются
непосредственно на мицелии, причем сумкоспоры способны
почковаться. Бесполое спороношение отсутствует. Порядок включает
около 100 видов, относящихся к роду Taphrina. Все тафриновые —
облигатные паразиты с узкой специализацией. Они поселяются в
молодых растущих органах и тканях многих древесных лиственных
пород. Развиваясь в них, эти грибы усиливают ростовые процессы
или нарушают фенологический цикл растений, вызывая
деформации пораженных органов. На поверхности последних
образуется розоватый, желтоватый или золотистый налет, состоящий из
сумок гриба, выступающих наружу из-под кутикулы и чаще всего
расположенных слоем (рис. 12). Тафриновые поражают листья,
завязи и побеги (рис. 13).
Поражение листьев проявляется в неравномерном росте
отдельных участков листа, в изменении его формы и окраски,
образовании вздутий. Болезни такого типа получили название курчавости
или пузырчатки листьев (например, пузырчатка листьев тополя
вызывается грибом Т. аигеа).
Поражение завязей наблюдается у многих плодовых и
лиственных пород. Вследствие разрастания стенок завязей последние
превращаются в вытянутые мешковидные образования, совсем не
похожие на нормальные плоды. Примерами болезней этого типа могут
служить кармашки сливы и черемухи (возбудитель T.pruni),
деформация плодиков ольхи (Т. aini =
incanae) и др.
При поражении ветвей
заболевание проявляется в
многократном ветвлении побегов, так
как под действием гриба резко
сокращается период покоя, и
спящие почки преждевременно
пробуждаются. В результате
патологического процесса на
зараженных участках ветвей
образуются ведьмины метлы,
состоящие из тонких, тесно скученных
побегов с хлоротичными
недоразвитыми листьями. Ведьмины
метлы, вызываемые грибами
Рис. 12. Открытое размещение сумок
у грибов порядка тафриновых.
Сумки (а) образуются на мицелии под
кутикулой и выступают на
поверхность пораженного органа растения
50
Рис. 13. Типы болезней, вызываемых грибами порядка тафриновых:
1 — ведьмины метлы ольхи; 2 — деформация листьев клена татарского; 3 — пузырчатка листьев тополя: а — пораженный лист, б —
— сумки гриба; 4 — деформация плодов ольхи; 5 — деформация плодов осины; 6 — кармашки сливы; 7 — кармашки черемухи
Т. betulina и Т. turgida, образуются, например, на березе. Ведьми-
ны метлы на клене вызывает T.acerina, на ольхе — T.epiphylla.
Аналогичные заболевания встречаются и на других лиственных.
Класс Hemiascomycetes
Сумки образуются открыто на почкующемся мицелии.
Главный порядок класса — Saccharomycetales, включающий
семейство Saccharomycetaceae — дрожжевые грибы. К этому семейству
относится важнейший в практическом отношении вид — Saccharo-
myces cerevisae, известный под названием «пекарские дрожжи». Он
содержит множество штаммов, очень разнообразных по физиоло-
го-биохимическим свойствам и широко используемых в пищевой
промышленности.
Класс Euascomycetes — Плодосумчатые
К плодосумчатым относится около 90 % всех сумчатых грибов,
что подтверждается данными молекулярно-филогенетических
исследований. Вегетативное тело — хорошо развитый септирован-
ный мицелий, образующий гифальную систему. В результате
полового процесса формируются настоящие плодовые тела (аско-
мы), содержащие сумки с сумкоспорами.
Различают следующие типы плодовых тел (рис. 14).
Клейстотеций — плодовое тело шаровидной формы,
совершенно закрытое. Сумки располагаются внутри плодового тела
в беспорядке или пучком. Созревшие сумкоспоры пассивно
освобождаются в результате постепенного разрушения оболочек
сумок и плодового тела или активно выбрасываются при набухании
сумок через разрыв оболочки плодового тела. Клейстотеций
всегда образуются на мицелии.
Перитеций — плодовое тело шаровидной, грушевидной или
колбообразной формы с узким выводным отверстием на вершине.
Сумки расположены в плодовом теле чаще всего пучком,
прикрепленным к его основанию, но иногда в беспорядке.
Освобождение сумкоспор в большинстве случаев активное; иногда
сумкоспоры пассивно выделяются через отверстие плодового тела вместе
со слизью. Перитеции образуются на мицелии или погружены в
стромы. Перитеции и стромы могут различаться по строению и
окраске.
Апотеций — плодовое тело чаще всего воронковидной,
чашеобразной или блюдцевидной формы, в зрелом возрасте —
открытое, на ножке или без ножки. Сумки располагаются внутри или на
поверхности плодового тела плотным слоем — гимением.
Освобождение сумкоспор — активное. Апотеций образуются на
мицелии, стромах или склероциях.
52
10 11
Рис. 14. Типы плодовых тел у плодосумчатых грибов:
7, 2, 3 — клейстотеции; 4, 5, б, 7, 8 — перитеции; Р, /0 — гистеротеции; 77, 72, 7J, 74 — настоящие апотеции
В циклах развития плодосумчатых грибов, кроме полового споро-
ношения в виде плодовых тел с сумками, обычно имеется и
бесполое (конидиальное) спороношение. У фитопатогенных видов
именно конидиальная стадия, как правило, является
паразитической.
В зависимости от типа, внутреннего строения, места
формирования плодовых тел, расположения в них сумок, характера и
способа освобождения сумкоспор, строения сумок и других
признаков класс делится на большое число (до 17) порядков. Порядки,
сходные по важнейшим признакам (типу плодового тела,
расположению в нем сумок и др.), для удобства объединены в три
группы порядков: плектомицеты, пиреномицеты и дискомицеты.
Плектомицеты
Плодовые тела плектомицетов — в основном клейстотеции,
реже перитеции. Сумки в плодовых телах располагаются
беспорядочно, освобождение сумкоспор — пассивное. Из нескольких
порядков, относящихся к плектомицетам, наибольшее значение в
фитопатологическом плане имеет порядок Eurotiales.
Порядок Eurotiales — Эвроциевые. Порядок
включает многочисленные и широко распространенные виды родов
Penicillium и Aspergillus, характеризующихся образованием клей-
стотециев. Однако в связи с тем, что в циклах развития этих
грибов ведущая роль принадлежит анаморфам (конидиальным
стадиям и спороношениям), а не телеоморфам (плодовым телам
сумчатого спороношения), их чаще всего относят не к аскомицетам,
а к митотическим (несовершенным) грибам. В настоящем
учебнике они также рассматриваются как представители группы митоти-
ческих грибов (см. рис. 22).
Пиреномицеты
Плодовые тела пиреномицетов — клейстотеции или
перитеции, сумки в плодовом теле располагаются беспорядочно или
пучком. При созревании сумкоспор происходит их пассивное
освобождение или активное выбрасывание из плодового тела.
Конидиальная стадия фитопатогенных пиреномицетов является
паразитической: она развивается летом на живых тканях растений,
служит для массового размножения и распространения гриба в
течение вегетационного периода. Сумчатая же стадия чаще
развивается на отмирающих или мертвых тканях (на опавших
листьях и хвое, отмершей коре и т.п.) и служит для перезимовки.
Эту же роль выполняют склероции, характерные для некоторых
видов.
Среди пиреномицетов немало грибов, вызывающих болезни
древесных растений, кустарников, цветочных культур. Типы
заболеваний разнообразны. Это мучнистые росы, некрозно-раковые и
54
другие болезни. Практическое значение для лесного хозяйства
имеют представители семи порядков.
Порядок Erysiphales — Мучнисторосяные.
Плодовые тела — клейстотеции, образующиеся на мицелии.
Мучнисторосяные грибы однотипны по своей биологии. Все они —
узкоспециализированные биотрофы, облигатные паразиты растений.
Характерная особенность большинства мучнисторосяных грибов —
поверхностный, экзофитный мицелий. Гриб прикрепляется к
растению и питается с помощью гаусториев, внедряющихся в
полость клеток эпидермиса. Летом на мицелии образуется бесполое
спороношение в виде коротких конидиеносцев, на вершине
которых развиваются овальные бесцветные конидии, соединенные
в цепочки.
Благодаря обилию конидий налет гриба приобретает
порошащий, мучнистый вид. Конидии служат для широкого
распространения гриба: они вызывают массовые повторные заражения
растений, давая несколько генераций в течение лета. Во второй
половине вегетационного периода на том же мицелии развивается
сумчатое спороношение — клейстотеции, имеющие вид черных
точек. Плодовые тела зимуют на почве, опавших листьях и
пораженных побегах. Весной или в начале лета созревшие клейстотеции
растрескиваются, сумкоспоры высвобождаются из сумок и
осуществляют первичное заражение молодых листьев и побегов.
Важнейшие виды мучнисторосяных грибов: Microsphaera alphi-
toides — на листьях и побегах дуба; Uncinula aceris — на листьях
клена; Phyllactinia suffulta — на ясене, березе, лещине, буке,
фисташке, грабе, шелковице и других породах; Sphaerotheca pannosa — на
розах; Uncinula salicis — на иве, тополе, осине; Microsphaera lonice-
гае — на жимолости.
Порядок Hypocreales — Гипокреевые. Плодовые
тела — перитеции — яркоокрашенные или светлые, мягкие или
мясистые. Перитеции иногда образуются непосредственно на
субстрате, но чаще располагаются на строме (погружены в нее
своими основаниями или целиком). Стромы такой же консистенции и
окраски, как и перитеции. Среди представителей порядка немало
сапротрофов, развивающихся в почве, на древесных и других
растительных остатках. Есть и фитопатогенные грибы, относящиеся к
семействам Нуросгеасеае и Clavicipitaceae. Из семейства Нуросгеасеае
для лесного хозяйства имеют значение виды рода Nectria: N. cinnaba-
rina — возбудитель некроза ветвей и стволов различных
лиственных деревьев и кустарников; Kgalligena — возбудитель ступенчатого
рака лиственных пород. На плодовых телах трутовиков
паразитирует гриб Hypocrea pulvinata. К семейству Clavicipitaceae относятся
виды рода Claviceps, вызывающие спорынью злаков (в том числе
газонных трав), и гриб Cordyceps militarise паразитирующий на
гусеницах и куколках чешуекрылых.
55
Порядок Diatrypales — Диатриповые. Плодовые
тела — перитеции, погруженные в строму. Стромы однолетние,
черные, плоские, твердые, иногда широко распростертые, с
многочисленными мелкими трещинками, как бы обугленные.
Устьица перитециев на поверхности стромы имеют вид мельчайших
бугорков, хорошо видимых в лупу. Среди фитопатогенных видов
наиболее часто встречается Diatrype stigma с конидиальной стадией
Naemospora croceola — возбудитель диатрипового (немоспорово-
го) некроза ветвей и стволов дуба.
Порядок Xylariales ( = Sphaeriales) — Ксиляри-
евые. Представители порядка образуют перитеции,
располагающиеся обособленно или группами на мицелии (в этом случае они
обычно погружены в субстрат) или в строме. Перитеции и стромы
темного цвета, чаще черные, твердые, углистой или деревянистой
структуры, различной формы (распростертые, шаровидные, подуш-
ковидные, разветвленные наподобие оленьих рогов и др.). В
составе порядка преобладают сапротрофы, но имеются и фитопатоген-
ные виды, например Hypoxylon pruinatum — возбудитель черного
рака осины и тополя, Nummularia bulliardii — возбудитель некроза
ветвей и стволов дуба; сюда же относится Daldinia concentrica —
гриб, вызывающий белую гниль стволов березы и ясеня и
образующий на усыхающих стволах крупные (до 6 см в диаметре)
шаровидные многолетние стромы черного цвета с погруженными в
них многочисленными перитециями.
Порядок Diaporthales — Диапортовые. Грибыэтого
порядка очень сходны с представителями предьщущего порядка
по строению, окраске и расположению перитециев. Стромы —
распростертые или шаровидные, черные, отличаются от Xylariales
характером развития перитециев и строением сумок. Конидиаль-
ные стадии фитопатогенных видов диапортовых обычно
развиваются на живых частях растений, вызывая болезни типа антракно-
зов, пятнистостей, некрозов коры. Сумчатое спороношение
закладывается на отмерших тканях в конце вегетации. Важнейшие
представители: Endothia parasitica — возбудитель эндотиевого рака
каштана съедобного, Valsa sordida (конидиальная стадия — Cytospora
chrysosperma) — возбудитель бурого цитоспороза тополя.
Порядок Ophiostomatales — Офиостомовые.
Порядок характеризуется образованием перитециев с длинным
«хоботком», через устьице которого созревшие сумкоспоры выходят
наружу вместе с сахаристыми слизистыми выделениями,
привлекающими насекомых. К этому порядку относятся виды рода
Ophiostoma, вызывающие сосудистые болезни лиственных
древесных пород: О. ulmi — возбудитель голландской болезни ильмовых,
О. roboris — возбудитель сосудистого микоза дуба.
Порядок Microascales — Микроасковые.
Плодовые тела микроасковых того же типа, что и у офиостомовых. Пред-
56
ставители порядка — дереворазрушающие сапротрофные виды рода
Ceratocystis (С.coeruleum, C.piceae и др.), вызывающие синеву за-
болонной древесины хвойных пород.
Дискомицеты
Плодовые тела дискомицетов — апотеции, различающиеся по
степени развития, строению и форме. Кроме сумчатой стадии, в
цикл развития некоторых дискомицетов входят конидиальная
стадия или склероции, однако у многих видов они отсутствуют.
Среди дискомицетов имеются как сапротрофы, так и паразиты
растений, характеризующиеся различной степенью паразитизма. Типы
болезней растений, вызываемых дискомицетами, также очень
разнообразны. Большинство фитопатогенных дискомицетов,
вызывающих болезни древесных растений, относится к порядку Leotiales.
Порядок Leotiales (=Helotiales) — Леоциевые.
Важнейшие семейства этого порядка — Phacidiaceae и Scleroti-
niaceae.
К семейству Phacidiaceae относятся представители
дискомицетов, имеющие черты сходства с пиреномицетами. Их плодовые
тела по форме и строению напоминают перитеции (они обычно
плотные, черные или в черной строме). Вначале они закрыты
сплетением гиф, а при созревании открываются щелью или лопасте-
видным разрывом, однако сумки в плодовом теле располагаются
уже слоем (гимением), как и у других дискомицетов. Такие
нетипичные апотеции называют гистеротециями. У некоторых
видов имеются конидиальные стадии, развивающиеся на живых
частях растений.
В состав семейства входит большое число фитопатогенных
видов, вызывающих весьма распространенные и опасные болезни
древесных пород: Lophodermium seditiosum и L.pinastri —
возбудители обыкновенного шютте сосны, Phacidium infestans —
возбудитель снежного шютте сосны и ели, Rhytisma acerinum и Rh.salici-
пит — возбудители черных пятнистостей листьев клена и ивы,
Clithris quercina — возбудитель клитрисового некроза ветвей и
стволов дуба и др.
Семейство Sclerotiniaceae характеризуется образованием
типичных, хорошо развитых апотециев в виде мягких
светло-коричневых чашечек, воронок или дисков, чаще всего на ножке.
Апотеции обычно образуются при прорастании склероциев или склеро-
циальных стром гриба после периода покоя. Сюда относятся виды
родов Sclerotinia и Stromatinia, вызывающие ряд болезней
растений: Scl.betulae и Str.pseudotuberosa — возбудители мумификации
семян березы и дуба, ScLgraminearum — возбудитель выпревания
сеянцев, Dasyscypha willkommii — возбудитель ступенчатого рака
стволов лиственницы, Sclsclerotiorum — возбудитель белой гнили
цветочных культур и др.
57
Порядок Pezizales — Пезизовые. Порядок содержит
многочисленные виды почвенных сапротрофных грибов,
образующих апотеции, разнообразные по форме, строению, окраске и
размерам, в том числе такие широко известные съедобные
грибы, как сморчки (виды Morchella и Verpa). Из фитопатогенных
грибов к данному порядку относится Rhizina undulata (ризина
волнистая) — возбудитель корневой гнили хвойных деревьев.
Порядок Tuberales — Трюфелевые. Характеризуется
довольно крупными подземными клубневидными плодовыми
телами. Гимений образуется на лабиринтообразных складках внутри
плодового тела. Трюфелевые грибы являются микоризообразова-
телями, съедобны. Наиболее известный род — Tuber.
Класс Loculoascomycetes — Полостносумчатые
У полостносумчатых сумки формируются в особых полостях
(локулах) стром, которые получили название аскостром, или
псевдотециев (рис. 15). В зависимости от особенностей
развития и строения аскостром, количества и расположения локул,
количества и размещения сумок в локуле подкласс делится на
порядки.
Фитопатогенные представители полостносумчатых относятся к
трем основным порядкам. Важнейшие виды порядка доти-
д е е в ы х (Dothideales) — Dothidella betulina и D. ulmi, вызывающие
черные пятнистости листьев березы и вяза. К порядку плео-
споровых (Pleosporales) относятся виды рода Venturia,
вызывающие болезни типа парши у древесных растений (яблони, груши,
осины, тополя). Представителем порядка гистериевых
(Hysteriales) является Hysterographiumfraxini — возбудитель
некроза ветвей и стволов ясеня.
Подотдел Basidiomycotina (Базидиальные грибы)
К базидиальным относится около одной трети всех описанных
видов грибов. Основной диагностический признак базидиальных
грибов — наличие базидии с экзогенными базидиоспорами,
образующимися в результате полового процесса — соматогамии. Типичная бази-
дия — несептированная с четырьмя одноклеточными гаплоидными
базидиоспорами. У некоторых видов базидия двух- или четырехкле-
точная (имеет продольные или поперечные перегородки), с
числом базидиоспор меньше или больше четырех. В цикле развития
преобладает септированный дикариотический мицелий с пряжками.
Среди базидиальных грибов встречаются паразиты растений,
симбиотрофы (микоризообразующие грибы) и сапротрофы.
В современной системе грибов подотдел включает 3 класса
(Teliomycetes, Ustomycetes, Basidiomycetes), 41 порядок, 165 семейств.
58
Рис. 15. Типы аскостром у полостносумчатых грибов.
Видны многообразие и особенности строения аскостром, количество в аскост-
ромах и различное количество сумок в локулах
Класс Teliomycetes — Телиомицеты
Мицелий телиомицетов — без пряжек. Базидии четырехклеточ-
ные, образуются при прорастании покоящихся спор — телиоспор.
Важнейший порядок — Uredinales.
Порядок Uredinales — Ржавчинные. Порядокпред-
ставлен исключительно облигатными паразитами — биотрофами —
59
возбудителями широко распространенных и вредоносных
болезней растений. Ржавчинные грибы обладают узкой специализацией.
Они поражают очень многие виды растений и вызывают болезни
под названием «ржавчина». Основной признак ржавчины —
ржаво-бурые, оранжево-желтые или темно-бурые скопления спор,
которые в большинстве случаев выступают наружу через разрывы
покровных тканей растений.
Одна из важнейших биологических особенностей ржавчинных
грибов — сложный цикл развития, включающий несколько стадий
и спороношений, последовательно сменяющих друг друга (рис. 16).
Другой важной особенностью этих грибов является разнохозяй-
ность, т. е. свойство развиваться в одной из стадий на одном, а в
Рис. 16. Спороношения ржавчинных грибов:
1 — спермогоний и спермации; 2 — эций и эциоспоры; 3 — урединиоспоры; 4, 5,
6— несросшиеся телиоспоры на ножках; 7, 8, 9, 10— сросшиеся телиоспоры без
ножек; 11 — базидии с базидиоспорами
60
других стадиях — на другом виде растений. Существуют и однохо-
зяйные ржавчинные грибы, у которых все стадии протекают на
растениях одного вида.
Эциостадия ржавчинных грибов чаще всего развивается
весной или в начале лета, в результате первичного заражения
растений базидиоспорами. В этой стадии гриб образует два спороноше-
ния: спермогонии и эции. Спермогонии — это маленькие
незаметные вместилища, в которых формируются очень мелкие
споры — спермации; они не вызывают заражения растений, но
играют важную роль в половом процессе. Эции — более крупные
вместилища, имеющие форму бокальчиков, корзиночек,
продолговатых разрывов или пузыревидных вздутий, заполненных
порошащей золотисто-желтой или оранжевой массой эциоспор. По мере
созревания эциоспоры высыпаются из эциев и служат (у разнохо-
зяйных видов) для заражения другого хозяина.
Урединиостадия ржавчинных грибов развивается летом и
слагается обычно из нескольких генераций урединиоспор. Они
имеют вид порошащих светло-желтых или оранжевых скоплений,
образующихся в урединиях, представляющих собой округлые или
продолговатые разрывы эпидермиса и кутикулы (пустулы). Уреди-
ниоспоры вновь заражают растения данного вида, обеспечивая
тем самым быстрое распространение паразита и массовое
развитие болезни.
Телиостадия характеризуется наличием двух спороношений: те-
лиоспор и базидиоспор. Телиоспоры обычно образуются к концу
вегетационного периода и представляют собой покоящиеся
споры ржавчинных грибов. Они снабжены толстыми оболочками,
имеют темную окраску и чаще всего предназначены для
сохранения в зимний период. Телиоспоры выходят на поверхность
растений в виде порошащих темно-бурых или черных скоплений через
разрывы покровных тканей (открытые телиопустулы) либо
размещаются под эпидермисом (закрытые телиопустулы). У
некоторых видов телиоспоры располагаются на поверхности растений в
виде студенистых скоплений или тонких волосовидных столбиков.
После перезимовки телиоспоры прорастают, образуя базидии с
базидиоспорами; базидиоспоры вновь осуществляют переход
гриба на другой вид растения-хозяина и вызывают первые весенние
заражения. У некоторых видов прорастание телиоспор,
образование базидиоспор и заражение ими растений происходят в конце
вегетационного периода.
Таким образом, в цикле развития ржавчинных грибов
возможны три стадии и пять спороношений. Такой цикл называют
полным. Цикл развития, в котором отсутствуют те или иные стадии и
спороношения, называют неполным.
Особенности строения телиоспор положены в основу деления
порядка ржавчинных грибов на два семейства: Pucciniaceae и Ме-
61
Рис. 17. Виды ржавчины,
вызываемые грибами рода Gymnosporangium:
1 — эции гриба G.mali-tremelloides на
листе яблони; 2— эции гриба G.sabinae
на листе груши; 3 — эции гриба G.juni-
perinum на листе рябины; 4 — телиоспо-
ры Gymnosporangium на стволе
можжевельника
lampsoraceae. Для грибов
первого семейства характерны тели-
оспоры на ножках, не
соединенные между собой. У грибов
второго семейства телиоспоры без
ножек, сросшиеся друг с
другом.
К семейству пукциниевых
(Pucciniaceae) относятся виды
рода Puccinia, паразитирующие
в эциостадии на кустарниках, а
в урединио- и телиостадии
вызывающие ржавчину злаков.
Виды рода Gymnosporangium,
имеющие неполный цикл, в
эциостадии развиваются на плодовых, а
в телиостадии — на можжевель-
никах (рис. 17). Однохозяйный
вид Phragmidium distiflorum
вызывает ржавчину роз, Uromyces
caryophyllinus — ржавчину
гвоздики и других цветочных
растений.
К семейству мелампсоровых
(Melampsoraceae) относятся
многие разнохозяйные виды с
полным циклом развития,
вызывающие болезни древесных
пород. Например, Melampsora pinitorqua в эциостадии вызывает
ржавчину всходов и побегов сосны, а в урединио- и телиостадии —
ржавчину листьев осины (см. рис. 17). Thekopsorapadi является
возбудителем ржавчины шишек ели и листьев черемухи. Есть в
составе семейства и однохозяйные виды с неполным циклом развития:
Peridermium pini, возбудитель смоляного рака сосны, имеет только
эциостадию.
Класс Ustomycetes — Устомицеты
К устомицетам относятся семь порядков, два из которых
имеют практическое значение: Exobasidiales и Ustilaginales.
Порядок Exobasidiales — Экзобазидиальные. Ба-
зидии одноклеточные, образуются непосредственно на мицелии
рыхлым слоем.
Экзобазидиальные грибы — облигатные паразиты
вечнозеленых растений. Все они относятся к роду Exobasidium и вызывают
болезни под названием «экзобазидиозы», проявляющиеся в харак-
62
терных деформациях пораженных
органов, на поверхности которых
образуется налет, состоящий из
базидий и базидиоспор. В наших
лесах повсеместно встречается эк-
зобазидиоз брусники
(возбудитель — Exobasidium vaccina).
Заболевание проявляется в
деформации стеблей, листьев, цветков,
изменении окраски пораженных
частей (они становятся розовато-
красными) и появлении на них
беловатого налета базидиально-
го спороношения гриба (рис. 18).
Известны также экзобазидиозы
чайного дерева, лавра
благородного, азалии.
К порядку Ustilaginales
(головневые) относятся возбудители
болезней зерновых злаков под
названием «головня». Они
проявляются в разрушении пораженных
частей растений с образованием
черных пылящих масс, состоящих
из огромного количества хламидоспор гриба. При прорастании хла-
мидоспор образуются базидий с базидиоспорами, которые вновь
заражают растения. Известны также головня виолы, головня лука
и другие виды.
Класс Basidiomycetes — Базидиомицеты
Базидий образуются в более или менее крупных плодовых
телах — базидиомах. В зависимости от типа базидий и особенностей
строения плодовых тел класс делится на два подкласса: Heterobasi-
diomycetidae и Homobasidiomycetidae.
Подкласс Heterobasidiomycetidae —
Гетеробазидиальные
Базидий двух- или четырехклеточные образуются в
студенистых, желатинообразных, слизистых или хрящевидных плодовых
телах различной формы и окраски. Из-за своеобразной
структуры плодовых тел многие из этих грибов получили название «дро-
жалковых». Большинство гетеробазидиальных — лесные сапрот-
рофы (ксилотрофы) на валежнике, пнях и других древесных
остатках. Основные порядки: Auriculariales, Tremellales, Dacrymy-
cetales.
Рис. 18. Экзобазидиоз брусники:
1 — деформация листьев; 2 —
деформация стебля; 3 — деформация
цветка; 4 — открытое размещение
мицелия и слоя базидий (гимения) на
поверхности пораженного органа
растения
63
Подкласс Homobasidiomycetidae —
Хомобазидиальные
Подкласс включает большое число порядков, которые по типу
плодовых тел объединены в две группы порядков: гименомицеты
и гастеромицеты.
Гименомицеты
Гименомицеты — самая большая группа порядков в классе ба-
зидиомицетов. Характерная особенность гименомицетов —
наличие крупных плодовых тел (базидиом), развивающихся на
многолетней грибнице. Базидии в плодовых телах образуются плотным
слоем — гимением. Плодовые тела различаются по величине,
форме, строению и окраске. Гимений располагается непосредственно
на поверхности плодового тела или в особой его части,
называемой гименофором. Гименофор также может иметь различное
строение (рис. 19).
В зависимости от макроморфологических особенностей
базидиом и гименофора (формы, строения и др.) и
микроморфологических признаков (типа гифальной системы плодовых тел,
строения гимения) гименомицеты делят на афиллофороидные и ага-
рикоидные, а те, в свою очередь — на порядки и семейства.
Афиллофороидные гименомицеты: плодовые тела
имеют различную форму и структуру и обычно не загнивают при
старении. Гименофор разнообразного строения, плотно
соединенный с тканью плодового тела.
К афиллофороидным гименомицетам относятся порядки
Thelephorales, Clavariales, Hericiales, Poriales, Hymenochaetales,
Ganodermatales и другие, в состав которых входит большое число
семейств.
Многие афиллофороидные гименомицеты составляют
обширную группу трутовых грибов.
Среди трутовиков встречаются как сапротрофные грибы —
разрушители мертвой древесины (в том числе складские и домовые
грибы), так и фитопатогенные виды с различной степенью
паразитической активности и специализации, вызывающие гнилевые
болезни леса.
Плодовые тела трутовых грибов — одно- или многолетние,
разнообразные по форме (копытовидные, распростертые, желвако-
образные, в виде шляпок и т.д.). По структуре они могут быть
деревянистыми, пробковидными, мясистыми, кожистыми.
Разнообразны и типы гименофора. У большинства трутовиков
гименофор трубчатый, у других — лабиринтообразный,
пластинчатый, ирпексовидный и т.д. (см. рис. 19).
К трутовикам относится множество видов из родов Fomitopsis,
Phellinus, Inonotus, Daedalea, Gloeophyllum и многих других. Глав-
64
10 И 12
Рис. 19. Формы плодовых тел (/—9) и типы гименофора (10—13) у ги-
меномицетов:
/ — копытообразное; 2 — черепицеобразная группа сидячих (боковых) шляпок;
3 — одиночная боковая шляпка; 4 — желвакообразное; 5 — воронковидное; 6 —
распростертое; 7— коралловидное; 8— шляпка с ножкой; 9 — булавовидные; 10 —
трубчатый; // — лабиринтообразный; 12 — пластинчатый; 13 — игольчатый
нейшие виды трутовых грибов и вызываемые ими гнили живых
деревьев и мертвой древесины рассматриваются в
соответствующих главах.
Агарикоидные гименомицеты: их плодовые тела
имеют форму шляпок с центральной, эксцентрической или боковой
ножкой, иногда без нее, при старении они загнивают. Гименофор
пластинчатый или трубчатый, легко отделяющийся от ткани
плодового тела.
К агарикоидным гименомицетам относятся порядки Agaricales,
Boletales, Polyporales, Russulales и другие, включающие большое
число семейств.
Среди агарикоидных гименомицетов большая часть видов —
почвенные грибы — микоризообразователи, в том числе съедобные
(белый, подосиновик, масленок, рыжик, сыроежка и др.) и ядови-
65
тые (мухомор, бледная поганка и др.). Есть ксилотрофы,
разрушающие мертвую древесину в лесу, на складах и в различных
сооружениях. Фитопатогенных видов немного, важнейший из них — опенок
осенний (Armillaria me Ilea), вызывающий белую корневую и
комлевую гниль многих древесных растений.
Гастеромицеты
Гастеромицеты характеризуются особым типом развития и
строения базидиом. К ним относится около тысячи видов грибов,
большинство которых — почвенные сапротрофы, в том числе и
микоризообразующие. Некоторые виды поселяются на мертвой
древесине. Среди гастеромицетов (рис. 20) широко
распространены представители порядка Lycoperdales: Дождевики (род Lycoper-
dori) и Порховки (род Bovista) с крупными плодовыми телами
грушевидной или шаровидной формы, а также звездовики (род
Geastrum) с плодовыми телами, по форме напоминающими
морские звезды. Из грибов порядка Nidulariales (нутревики) часто
встречаются бокальчики (род Cyathus), гнездовки (род Nidularia)
и круцибулум гладкий {Crucibulum laeve). В наших лесах есть также
виды порядков Phallales и Sclerodermatales. При созревании
плодового тела гастеромицетов споры выходят из отверстия или
разрыва оболочки на его вершине в виде темной пылящей или
слизистой массы.
Рис. 20. Плодовые тела гастеромицетов:
1 — дождевики {а — род Lycoperdon, б — род Bovista); 2 — нутревики (в — род
Cyathus; г — род Crucibulum); 3 — звездовики (род Geastrum)
66
Митоспоровые, или несовершенные, грибы —
Mitosporis fiingi (= Fungi imperfecta Deuteromycetes,
Asexual fungi, Conidial fungi)
Группа без определенного таксономического статуса. К ней
отнесены высшие грибы, имеющие только бесполые (конидиаль-
ные) стадии. Сюда входят виды, утратившие способность к
образованию сумок или базидий, а также грибы, у которых половое
спороношение (сумчатое, базидиальное) не известно. У многих
фитопатогенных видов несовершенных грибов найдены
соответствующие телеоморфы. Однако некоторые из этих видов числятся
в классе несовершенных, поскольку анаморфная стадия у них
играет ведущую роль, а телеоморфная встречается редко или не
имеет значения в развитии гриба и вызываемого им заболевания.
К митоспоровым отнесены также грибы, не образующие никаких
спороношений и существующие только в форме вегетативного
мицелия и склероциев.
Группа включает около 25 тыс. видов, разнообразных по
строению, образу жизни и типам вызываемых ими болезней растений.
По типу конидиального спороношения (рис. 21)
митоспоровые грибы делятся на два класса: Hyphomycetes и Coelomycetes.
Третий класс — Agonomycetes — включает виды, в цикле
развития которых имеются лишь мицелий и склероции.
Рис. 21. Типы конидиального спороношения митоспоровых грибов:
/ — одиночный конидиеносец и конидии; 2 — коремия; 3 — конидиальное ложе
и конидии; 4 — однокамерная пикнида и выделяющаяся из нее масса склеенных
конидий; 5 — многокамерная пикнида и конидии
67
Класс Hyphomycetes — Гифомицеты
Это самый большой класс несовершенных грибов. Конидие-
носцы с конидиями развиваются прямо на мицелии и выходят на
поверхность субстрата одиночно или пучками — коремиями,
поэтому спороношение чаще всего имеет вид порошащего налета.
Гифомицеты отличаются исключительным разнообразием
формы, строения и окраски конидиеносцев и конидий. Эти
особенности лежат в основе деления класса на порядки, семейства, роды
и виды.
В состав класса входит множество фитопатогенных грибов,
вызывающих болезни древесных растений, которые приносят
значительный вред лесному хозяйству. Болезни, вызываемые гифо-
мицетами, проявляются в усыхании хвои, увядании растений, а
также в виде гнилей и плесеней. Один из важнейших видов —
Meria lands, возбудитель шютте лиственницы. Большое
практическое значение имеют грибы рода Fusarium — возбудители
полегания всходов, увядания растений, плесневения и загнивания
семян и луковиц. Аналогичные заболевания сеянцев и семян
вызывают грибы родов Alternaria, Botrytis и др. Двоякое значение имеют
грибы рода Penicillium: многие из них являются возбудителями
плесневения плодов и семян при хранении; некоторые виды
находят широкое применение в медицине и промышленности. К
этому классу относятся и виды, используемые для биологической
борьбы с вредными насекомыми (род Beauveria), фитопатоген-
ными грибами (Trichoderma lignorum, Trichothecium roseum), фито-
гельминтами (хищные грибы) (рис. 22).
Рис. 22. Грибы-гифомицеты, используемые для производства
биопрепаратов:
1 — мицелий гриба Beauveria bassiana, пронизывающий тело погибшего
долгоносика; 2 — конидиальное спороношение гриба Trichothecium roseum; 3 — кониди-
альное спороношение гриба Trichoderma lignorum
68
Класс Coelomycetes — Целомицеты
У грибов этого класса конидии образуются в особых спорообра-
зующих органах — конидиомах. По типу конидиом класс делится
на два порядка: Acervulales (= Melanconiales) и Pycnidiales
(= Sphaeropsidales).
Порядок Acervulales — сравнительно небольшая,
однородная группа грибов. Они сходны по строению и развитию и
вызывают у растений однотипные заболевания под названием ант-
ракнозы. Антракнозы проявляются в изъязвлении плодов и семян,
растрескивании стеблей и в виде пятнистостей листьев. Конидии
образуются на коротких конидиеносцах в конидиальных ложах,
выступающих на поверхность субстрата в виде плоских или
выпуклых подушечек (см. рис. 21). К этому порядку относятся,
например, Marssonina populi — возбудитель бурой пятнистости
листьев (марссониноза) тополя, Marssonina juglandis — возбудитель марс-
сониноза ореха грецкого, Gloeosporium quercinum — возбудитель
антракноза желудей и бурой пятнистости листьев дуба.
Порядок Pycnidiales— конидии формируются в пикни-
дах, которые могут быть расположены на поверхности субстрата,
погружены в субстрат или в общую строму. К этому порядку
относятся как сапротрофные грибы, так и фитопатогенные виды из
числа факультативных сапротрофов и факультативных паразитов.
Типы болезней, вызываемых пикнидиальными грибами,
довольно разнообразны: пятнистости листьев, гнили плодов и
семян, некрозные и раковые заболевания древесных пород.
Характерный общий признак всех этих болезней — появление на
пораженных частях растений многочисленных пикнид возбудителя в
виде бугорков или черных точек (см. рис. 21).
Порядок Пикнидиальные включает большое число родов,
различающихся строением пикнид, строением, формой и окраской
конидий. Среди представителей порядка могут быть названы
такие виды, как Septoria populi — возбудитель белой пятнистости
листьев тополя, виды рода Cytospora — возбудители некрозных
заболеваний ветвей и стволов тополя и других древесных пород,
виды родов Phoma, Phyllosticta, Phomopsis — возбудители
пятнистостей листьев цветочных культур, болезней плодов и семян
древесных растений.
Класс Agonomycetes (Mycelia sterilia) — Агономицеты
Грибы этого класса вызывают болезни растений,
проявляющиеся в виде гнилей, пятнистостей, увядания. На пораженных
частях растений образуются склероции возбудителя, что является
типичным признаком заболеваний, вызываемых мицелиальными
грибами — агономицетами.
69
Класс включает два рода: Sclerotium (характеризуется белым
мицелием и мелкими округлыми склероциями) и Rhizoctonia
(мицелий темноокрашенный, скяероции неправильной формы). Виды
рода Sclerotium паразитируют на многих сельскохозяйственных и
цветочных растениях. Виды рода Rhizoctonia (например, R. solani)
могут вызывать полегание сеянцев, черную ножку и корневую
гниль цветочных культур, черную паршу картофеля (рис. 23).
Глава 3
БАКТЕРИИ, ВИРУСЫ, НЕМАТОДЫ
И ЦВЕТКОВЫЕ ПАРАЗИТЫ -
ВОЗБУДИТЕЛИ БОЛЕЗНЕЙ РАСТЕНИЙ
Среди фитопатогенных организмов важная роль принадлежит
бактериям, вирусам, микоплазмам, фитонематодам и
паразитическим цветковым растениям. Однако болезни древесных пород,
вызываемые патогенами этих групп, встречаются реже и имеют
меньшее хозяйственное значение, чем грибные болезни.
3.1. ФИТОПАТОГЕННЫЕ БАКТЕРИИ
3.1.1. Общие сведения о бактериях
Бактерии относятся к царству дробянок (Mychota), надцарству
прокариот (Procariota). В отличие от эукариот, к которым
относятся растения и грибы, в клетках прокариот нет настоящего
обособленного ядра, клеточная стенка не содержит ни целлюлозы, ни
хитина, а ее каркас составляет мукопептид муреин. Ядерное
вещество (нуклеоид) распределено в цитоплазме клетки диффузно
или в виде мелких зерен.
Бактерии представляют собой одноклеточные организмы, не
имеющие хлорофилла и живущие, как и все гетеротрофы, за счет
готовых органических веществ растений и животных. Среди
бактерий имеются сапротрофы и паразиты, однако случаи облигатного
паразитизма отсутствуют: все бактерии способны в тех или иных
условиях развиваться вне живого организма.
Размеры фитопатогенных бактерий незначительны: от 0,5 до
4,5 мкм в длину и от 0,3 до 0,6 мкм в поперечнике.
Бактериальная клетка окружена жесткой, довольно толстой
многослойной оболочкой, придающей клетке определенную
постоянную форму. По форме клеток различают бактерии
шаровидные и палочковидные; последние могут иметь вид коротких или
длинных, прямых или в различной степени изогнутых палочек
71
(рис. 24). Болезни растений вызываются преимущественно
бактериями, имеющими форму коротких прямых палочек. Чаще они
одиночные, но иногда соединены попарно или в цепочки*
У некоторых фитопатогенных бактерий имеется ослизненная
оболочка (капсула), защищающая бактериальную клетку от
воздействия прямых солнечных лучей и высыхания. При болезнях
растений, вызываемых такими бактериями, на пораженных органах
обычно образуются скопления слизи (экссудата).
Бактерии бывают неподвижные и подвижные, т.е. активно
передвигающиеся при помощи жгутиков. Количество и
расположение жгутиков могут быть различными. Известны бактерии с
одним, двумя и большим количеством жгутиков. У одних бактерий
жгутики находятся на концах клетки, тогда их называют
полярными; у других они расположены по всей поверхности клетки — в
этом случае их называют перитрихальными. Большинство
фитопатогенных бактерий — подвижные формы.
Размножение бактерий происходит путем деления или
перешнуровывания клеток. При делении посредине клетки возникает
поперечная перегородка, которая разделяет материнскую клетку
на две дочерние. При перешнуровывании посредине материнской
клетки появляется перетяжка, которая постепенно утончается и в
I/, ?3л
2
1_
Рис. 24. Основные формы бактерий:
1— кокки; 2 — собственно бактерии; 3— бациллы; 4— вибрионы; 5— спирохеты;
6 — подвижные формы со жгутиками
72
конце концов разделяет клетку на две дочерние. Иногда
наблюдаются дробление материнской клетки на несколько дочерних и
размножение почкованием. У некоторых бактерий установлен половой
процесс типа конъюгации. При оптимальных условиях бактерии
размножаются очень быстро: примерно через каждые 20 — 30 мин
их число удваивается. Быстро размножаясь на одном месте,
бактерии могут образовать довольно крупные скопления (колонии),
имеющие у разных видов различную форму и окраску. На
искусственной питательной среде фитопатогенные бактерии обычно
развивают мелкие, блестящие, слизистые колонии округлой формы,
чаще всего с ровными краями, белого, сероватого или
желтоватого цвета.
Некоторые виды бактерий образуют споры. При этом
содержимое клетки теряет воду и концентрируется в одном месте. Затем
этот участок уплотняется и покрывается собственной, более
плотной оболочкой, превращаясь в спору. Споры бактерий очень
устойчивы к различным внешним воздействиям и могут сохранять
жизнеспособность при неблагоприятных условиях, например, в течение
длительного времени выдерживать нагревание до 100 °С и выше, а
также сильное охлаждение, высушивание, воздействие
химических веществ и т. д. Попав в благоприятную среду, спора
прорастает, образуя новую вегетативную клетку, которая в дальнейшем
размножается путем деления. Большинство фитопатогенных
бактерий спор не образует.
Питание бактерий осуществляется осмотическим путем:
питательные вещества поступают внутрь бактериальной клетки
непосредственно через ее оболочку. Поселившись в растении,
фитопатогенные бактерии выделяют ферменты, с помощью которых они
разрушают ткани пораженных органов и используют их в пищу.
Набор ферментов у фитопатогенных бактерий достаточно широк;
в него входят ферменты, гидролизующие углеводы, белки и другие
компоненты растительной ткани. Под действием хлорофиллазы
происходит расщепление хлорофилла, вследствие чего на
пораженных участках листьев и других зеленых частей растений обычно
появляются характерные для бактериозов светлые маслянистые
пятна. Побурение и почернение пораженных бактериями тканей
связаны с воздействием окислительного фермента тирозиназы.
Заражение растений фитопатогенными бактериями
осуществляется только через естественные отверстия (устьица, чечевички,
гидатоды, рыльца, нектарники) или повреждения (ранки,
срезы, морозобойные трещины и т.п.). Внедряться в растение через
неповрежденную кутикулу или кору бактерии не могут.
Фитопатогенные бактерии различаются по степени
специализации: наряду с узкоспециализированными монофагами и
бактериями, поражающими растения нескольких видов или родов,
имеются типичные полифаги. Таким почти «всеядным» является,
73
например, возбудитель корневого рака — Agrobacterium twnefaciens,
который поражает растения более чем 60 родов из различных
семейств.
Одним из важных свойств бактерий является их большая
изменчивость. Новые формы и биотипы, возникающие в результате
мутаций, рекомбинаций генов и другими путями, нередко
отличаются от исходных форм более высокой или, наоборот,
меньшей патогенностью в отношении растений-хозяев.
Условия окружающей среды оказывают большое влияние на
развитие и распространение фитопатогенных бактерий. Так, наиболее
активное развитие большинства бактерий наблюдается при
температуре от 20 до 35 °С, избыточной влажности и нейтральной или
слабощелочной реакции среды. По отношению к кислороду все фитопа-
тогенные бактерии являются типичными аэробами. Свет
неблагоприятно действует на большинство бактерий; прямые же
солнечные лучи их быстро убивают. Поэтому солнечный свет является
мощным фактором, ограничивающим развитие бактерий в природе.
Все фитопатогенные бактерии относятся к классу Eubacteria,
порядку Eubacteriales, включающему несколько семейств.
Основными признаками семейств являются форма бактериальных
клеток, способность к активному передвижению, количество и
расположение жгутиков, наличие или отсутствие спор.
Принадлежность бактерий к тому или иному роду определяется, кроме того,
формой и цветом колоний, биохимическими свойствами,
например способностью к образованию определенных пигментов,
отношением к прижизненному окрашиванию определенными
красителями («окраска по Граму») и некоторыми другими
признаками. При определении ввдов фитопатогенных бактерий прежде всего
учитываются особенности ферментной системы, специализации,
патогенность и другие паразитические свойства. Большинство
фитопатогенных бактерий относится к родам Pseudomonas, Xanthomonas,
Erwinia, Agrobacterium и др.
3.1.2. Типы бактериальных болезней растений
Характер внешнего проявления бактериальных болезней
растений во многом зависит от того, какие органы и ткани
поражены бактериями и какие патологические процессы в них
происходят. Можно выделить следующие основные группы и типы
бактериальных болезней растений (рис. 25):
I. Болезни, связанные с отмиранием паренхимных тканей —
паренхиматозные бактериозы. Они обычно носят
местный характер. Среди паренхиматозных бактериозов встречаются
пятнистости, ожоги, гнили.
Проникая в межклеточные пространства листьев,
фитопатогенные бактерии вызывают быстрое отмирание окружающих кле-
74
Рис. 25. Типы бактериальных болезней растений:
7, 2 — угловатые пятнистости листьев; 3 — ожог побегов; 4 — сосудистый
бактериоз; 5 — кольцевая гниль; б — корневой рак
ток. Это проявляется в образовании пятен чаще всего угловатой
формы. Примерами бактериальных пятнистостей могут служить:
пятнистость листьев и плодов ореха грецкого (возбудитель —
Xanthomonas juglandis), пятнистость листьев смородины
(возбудитель — Xanthomonas heterocea), дырчатая пятнистость косточковых
(Xanthomonas pruni) и др.
Среди болезней типа ожога практическое значение имеют:
бактериальный ожог груши (возбудитель — Pseudomonas piri), ожог
сирени (возбудитель — Pseudomonas syringae), ожог шелковицы
(возбудитель — Pseudomonas mori). К важнейшим объектам внешнего
карантина относится ожог плодовых деревьев (возбудитель — Erwinia
amylovora).
Бактериальные гнили возникают при поражении луковиц,
клубней, плодов и семян. Под действием пектолитических ферментов
бактерий происходит разрушение срединных пластинок.
Пораженные ткани размягчаются или превращаются в слизистую, дурно
пахнущую массу (например, при поражении желудей бактериями
рода Erwinia).
II. Болезни, связанные с разрастанием тканей —
гиперпластические бактериозы. При заболеваниях этой группы
бактерии вызывают ускоренное беспорядочное деление клеток, иногда
сопровождающееся увеличением их размера, что приводит к
образованию раковых опухолей. Опухолевидный рак встречается у
многих древесных растений.
75
Важнейшие виды: поперечный рак стволов дуба (возбудитель —
Pseudomonas quercus), опухолевидный рак стволов ясеня
(Pseudomonas fraxini), бактериальный рак тополя (Pseudomonas remifaciens),
бугорчатый рак сосны (Pseudomonas pini), корневой рак плодовых
деревьев и лесных древесных пород (Agrobacterium tumefaciens),
туберкулез маслины и ясеня (Pseudomonas savastanoi).
III. Болезни, связанные с поражением сосудов — сосудистые
бактериозы. Обычно характеризуются общим поражением
растений и проявляются в их увядании (усыхании). Размножаясь в
сосудах ксилемы, заполняя и закупоривая их густой слизистой
массой, бактерии нарушают подачу воды от корней к надземным
частям. Кроме того, бактерии выделяют токсины, отравляющие
ткани растения. Все эти нарушения приводят к быстрому
отмиранию пораженных частей, а затем и всего растения. Сосудистыми
бактериозами поражаются преимущественно
сельскохозяйственные и цветочные культуры. На древесных растениях они
встречаются реже. В качестве примера можно привести бактериальное
увядание ивы — весьма опасное карантинное заболевание,
вызываемое бактерией Erwinia salicis.
Среди бактерий имеются виды, обладающие способностью
растворять мицелий и спороношения грибов. Они получили
название миколитических бактерий. Их используют в лесном
и сельском хозяйстве для борьбы с грибными болезнями
растений (мучнистой росой, полеганием сеянцев и др.).
3.1.3. Источники инфекции и пути распространения
фитопатогенных бактерий
Одним из основных источников бактериальной инфекции
являются остатки пораженных растений. Фитопатогенные бактерии
способны сохраняться в растительных тканях до тех пор, пока не
произойдет их полное разложение под влиянием сапротрофных
микроорганизмов. Возбудители многих бактериозов могут
сохраняться на поверхности и внутри семян, саженцев, черенков и
другого посадочного материала древесных и кустарниковых
растений. При использовании зараженного посевного и
посадочного материала бактерии переходят на развивающиеся молодые
растения. Источником инфекции нередко служат зимующие части
пораженных растений: луковицы, клубнелуковицы, корневища.
В распространении фитопатогенных бактерий важную роль
играют дождевые брызги и потоки, ирригационные воды. Велика
роль антропогенного фактора: бактерии распространяются при
транспортировке пораженных растений и семян, при уходе за
растениями, а также с различными орудиями, на одежде, обуви и
т.д. Переносчиками фитопатогенных бактерий могут быть различ-
76
ные насекомые, нематоды, грызуны. Некоторые виды бактерий
сохраняются в почве.
Меры борьбы с фитопатогенными бактериями в основном
сводятся к уничтожению остатков пораженных растений или
созданию условий для их быстрейшего разложения. Необходимы также
использование здорового посадочного материала, дезинфекция
семян и почвы.
3.2. ФИТОПАТОГЕННЫЕ ВИРУСЫ
3.2.1. Общие сведения о вирусах
Вирусами называются субмикроскопические возбудители
инфекционных болезней, характеризующиеся отсутствием
клеточного строения, относительной простотой химического состава и
способностью жить и размножаться только в живых клетках
организма хозяина.
Существование вирусов было открыто в 1892 г. русским
ученым-физиологом Д.И.Ивановским, который по праву
считается основоположником вирусологии — науки, изучающей
вирусы и вызываемые ими болезни. В настоящее время известно около
600 фитопатогенных вирусов, хотя точное их число указать трудно.
Вопрос о происхождении и природе вирусов до сих пор не
решен окончательно, хотя такие свойства вирусов, как инфекци-
онность, способность к размножению, паразитизму и
сохранению наследственных признаков, свидетельствуют об их живой
природе. В то же время многие вирусы способны вступать в
реакции и кристаллизоваться подобно минеральным соединениям.
Химический состав и строение вирусов достаточно хорошо
изучены. Частица фитопатогенного вируса (вирион) состоит из
одиночной или двойной нити нуклеиновой кислоты (почти всегда
РНК), окруженной множеством симметрично расположенных
молекул белка (рис. 26). Размеры вирусных частиц чрезвычайно
малы (в большинстве случаев — менее 200 нм). Они проходят через
бактериальные фильтры и недоступны для наблюдения даже при
самых сильных увеличениях обычных оптических микроскопов. Лишь
с появлением электронных микроскопов, увеличивающих в
десятки тысяч раз, стало возможным изучение и измерение вирусных
частиц. Форма частиц фитопатогенных вирусов чаще всего
шаровидная или палочковидная, реже изогнуто-нитевидная.
В фазе покоя вирусные частицы образуют в зараженных клетках
скопления в виде аморфных тел (х-тел) или кристаллов.
Вирусные кристаллы могут иметь форму восьмигранников, тонких игл,
шестиугольных пластинок. Некоторые вирусы образуют веретено-
видные и петлеобразные кристаллы (см. рис. 26).
77
Рис. 26. Строение вирусов:
1 — модель структуры вирусной частицы (видна нить нуклеиновой кислоты,
окруженная молекулами белка); 2 — вирусные частицы шаровидной формы; 3 —
палочковидные вирусные частицы; 4, 5— различные формы кристаллов вирусов
в клетках пораженных растений
Размножение вирусов коренным образом отличается от
размножения грибов и бактерий. Попав в клетку растения, вирусная
частица распадается на белок и нуклеиновую кислоту. Под
действием вирусной нуклеиновой кислоты обмен веществ
зараженной клетки перестраивается таким образом, что в ней начинается
синтез новых молекул вирусного белка и вирусной нуклеиновой
кислоты, идущих на построение новых вирионов.
Перемещение вирусов из клетки в клетку происходит по
плазменным тяжам — плазмодесмам, а в тканях типа меристемы — в
процессе деления клеток. Передвижение вирусов из одних
органов растения в другие осуществляется с током питательных
веществ по сосудам, преимущественно по флоэме, т. е. от верхних
78
частей растения к нижним. Перемещение вирусов по растению в
направлении снизу вверх наблюдается реже и происходит с
меньшей скоростью.
Вирусы могут по-разному реагировать на воздействие
факторов внешней среды. По этому признаку вирусы делят на стойкие и
нестойкие. Стойкие вирусы выдерживают подкисление,
высушивание, нагревание до 80...90°С, воздействие света и различных
химических веществ. Такие вирусы могут быть выделены из
растений в чистом (кристаллическом) виде с сохранением их
жизнеспособности. Нестойкие же вирусы под воздействием
неблагоприятных факторов быстро разрушаются (инактивируются).
Например, большинство фитопатогенных вирусов инактивируется при
значениях рН выше 8,0, т.е. в щелочной среде. Гибель некоторых
вирусов происходит при температурах 37... 50 °С. На этом
основано термическое оздоровление зараженного вирусами посадочного
материала (саженцев, черенков и т.д.).
3.2.2. Типы вирусных болезней растений
По характеру поражения и внешним признакам вирусные
болезни растений можно отнести к двум типам: мозаики и желтухи
(рис. 27).
Мозаики проявляются главным образом в изменении
окраски различных органов растений. На листьях, стеблях, лепестках
цветков возникают бледно-зеленые, желтые, иногда бурые
(некротические) участки в виде пятен, колец, полос различной
формы и величины. Чередуясь с нормально окрашенными участками,
они создают пеструю, мозаичную расцветку. Эти симптомы
появляются в результате повреждения хлоропластов, снижения их фо-
Рис. 27. Типы вирусных болезней растений:
1 — мозаика (побеление жилок) и деформация листьев; 2 — кольцевая мозаика
листьев; 3 — пестролепестность цветка; 4 — пролиферация цветка; 5 — ведьмины
метлы; 6 — курчавость листьев
79
тосинтетической активности, непосредственного разрушения
вирусом хлорофилла, а также вследствие изменения активности
некоторых ферментов и нарушения метаболизма клеток в связи с
синтезом вирусных частиц. Иногда мозаичная окраска
сопровождается деформацией листовых пластинок: они становятся
курчавыми, морщинистыми или нитевидными.
Большое распространение и хозяйственное значение имеют
многие мозаичные болезни цветочных культур, например
вирусная мозаика гладиолуса, пестролепестность тюльпана и гвоздики.
Эти болезни приводят к деформации надземных частей,
измельчанию цветков и луковиц и другим проявлениям вырождения.
В южных районах часто встречаются мозаики листьев клена ясене-
листного, вяза, ясеня, жимолости и других древесных пород и
кустарников.
Желтухи характеризуются более сильным и глубоким
воздействием вируса на организм хозяина и, как следствие этого,
более тяжелыми и разнообразными нарушениями обмена веществ,
роста и формы пораженных растений. При вирусных болезнях
желтушного типа часто наблюдаются деформации отдельных органов
или всего растения, связанные с угнетением или, наоборот, со
стимуляцией ростовых процессов. В первом случае могут
наблюдаться общее недоразвитие, карликовость растений, различные
дегенеративные изменения поражаемых частей; во втором —
разрастание тканей (опухоли и галлы), усиленное
побегообразование (ведьмины метлы), пролиферация тканей, превращение
генеративных органов в вегетативные (например, превращение
частей цветка в листовидные образования). Для вирусных желтух,
возбудители которых вызывают некроз флоэмы, характерны
утолщение и скручивание листьев в результате нарушения
нормального оттока ассимилятов из листьев в другие части растения. Лишь
сравнительно немногие болезни этой группы проявляются в
резком пожелтении листьев (общий хлороз). К болезням желтушного
типа относятся ведьмины метлы робинии лжеакации, розеточная
болезнь яблони и фисташкового дерева, ряд весьма
распространенных и вредоносных заболеваний сельскохозяйственных и
цветочных культур.
3.2.3. Сохранение и распространение вирусов
в природе
Большинство фитопатогенных вирусов способно сохраняться
только в живых клетках хозяина. У вегетативно размножаемых
растений вирус может из года в год сохраняться в луковицах,
клубнях, корневищах и передаваться с ними. Распространение
вирусов, поражающих древесные и кустарниковые растения, час-
80
то происходит с черенками и
отводками, взятыми от больных
маточников. Вирусы, вызывающие
поражение плодовых деревьев,
передаются главным образом
прививкой при использовании зараженных
подвоев или привоев. Подобным
образом вирусы могут
распространяться и при естественном
срастании корней растений.
Основной способ
распространения фитопатогенных вирусов в
природе — передача их насекомыми и
клещами. Переносчиками вирусов
являются преимущественно насеко- рИс. 28. Тля — переносчик мо-
мые с колюще-сосущим ротовым ап- заичных вирусов. Виден ход сти-
паратом: тли, цикады, клопы, трип- лета насекомого в тканях рас-
сы и некоторые другие. Питаясь на тения
больном растении, насекомое
всасывает вирус вместе с клеточным содержимым. Попадая затем на
здоровое растение, прокалывая его покровные ткани своим
стилетом, вводя в растение слюну, насекомое вносит в него и вирус
(рис. 28).
Различают два типа передачи фитопатогенных вирусов
насекомыми. В одних случаях насекомые приобретают вирофорность (т. е.
способность передавать вирус) сразу после питания на больном
растении, но сохраняют ее недолго. Такой способ передачи часто
называют механическим, а вирусы, передающиеся этим путем —
неперсистентными. Неперсистентные вирусы вызывают, как
правило, мозаичные болезни растений; они распространяются
чаще всего тлями, причем в организме переносчика вирус не
перемещается и не размножается. При механической передаче обычно
наблюдается взаимно широкая специализация вирусов и их
переносчиков: один и тот же вид тлей может переносить большое
число (иногда несколько десятков) различных вирусов, а один и тот
же вирус нередко передается разными видами тлей.
В других случаях насекомые становятся вирофорными не сразу
после питания на зараженном растении, а лишь по истечении
скрытого (инкубационного) периода, во время которого вирус
перемещается (циркулирует) и размножается в теле насекомого.
В этом случае насекомое, воспринявшее вирус, долго, иногда в
течение всей жизни, сохраняет вирофорность. Этот способ
передачи называют биологическим, а вирусы, передающиеся таким
образом, — персистентными, или циркулятивными.
Большинство персистентных вирусов вызывает болезни типа желтух и
переносится цикадами, реже трипсами, тлями, белокрылками и
81
другими насекомыми, а также клещами. Переносчики персистент-
ных вирусов, как правило, характеризуются более узкой
специализацией: некоторые виды цикад способны передавать только один
определенный вирус. Соответствующую высокую степень
избирательности в отношении переносчиков обнаруживают и вирусы этой
группы.
Имеются сведения о распространении некоторых вирусов и
грызущими насекомыми — жуками, прямокрылыми, уховертками.
При этом наблюдается малоспециализированная механическая
передача вируса. Переносчиками фитопатогенных вирусов в
природе могут быть также нематоды. Установлена передача вирусов
грибами, в частности фитопатогенными видами хитридиевых, и
паразитическими цветковыми растениями, например повиликой.
С семенами больных растений передаются сравнительно
немногие вирусы. Зараженные вирусом семена часто не отличаются
внешне от здоровых, и это создает особую угрозу распространения с
семенами карантинных вирусных заболеваний.
Стойкие фитопатогенные вирусы могут передаваться
контактным способом непосредственно от растения к растению при их
соприкосновении через мельчайшие ранки, например через
обломанные волоски, при трении друг о друга листьев или стеблей.
Возможна контактная передача вируса через загрязненные соком
больных растений сельскохозяйственные машины, инструмент,
руки и одежду работающих. Некоторые наиболее стойкие вирусы
способны передаваться через почву, где они адсорбируются на
почвенных коллоидах или сохраняются некоторое время в
зараженных растительных остатках.
В системе профилактических мероприятий против вирусных
болезней растений первостепенное значение имеют отбор
здорового семенного и посадочного материала, борьба с насекомыми-
переносчиками, соблюдение санитарно-гигиенических правил,
культивирование вирусоустойчивых видов и сортов растений.
3.3. МИКОПЛАЗМЫ
Микоплазмы как возбудители болезней растений впервые
описаны в 1967 г. японскими вирусологами Исии, Дои, Асуяма и др.
При электронно-микроскопическом исследовании тканей
шелковицы, пораженной карликовостью, были выявлены частицы, во
многом сходные с бактериями, но в то же время отличающиеся
от них некоторыми чертами строения и другими особенностями.
Микоплазмы представляют собой мельчайшие тельца (лишь
немного крупнее вирусов) неопределенной формы (часто
округло-продолговатые и гантелевидные), окруженные трехслойной
мембраной, но не имеющие клеточной стенки (рис. 29). Мико-
82
Рис. 29. Микоплазмы:
1 — формы микоплазменных частиц; 2 — возбудитель микоплазменной болезни
в тканях пораженного растения
плазменные частицы содержат два типа нуклеиновых кислот (РНК
и ДНК) и способны размножаться делением. Среди микоплазм
имеются как паразиты, так и сапротрофы. Их можно
культивировать на специальных искусственных средах.
Фитопатогенные микоплазмы передаются цикадами по типу
персистентных вирусов, т. е. биологическим способом, хотя
особенности их передачи могут варьировать, и они еще недостаточно
хорошо изучены. Микоплазмы вызывают патологические
изменения в проводящей системе (чаще всего во флоэме) пораженных
растений. Симптомы микоплазменных болезней очень сходны с
симптомами вирусных желтух. Исследованиями последних лет
доказано, что многие болезни желтушного типа, прежде
считавшиеся вирусными, в действительности вызываются микоплазмами.
Таковы желтуха астр, махровость (реверсия) смородины,
карликовость шелковицы, ведьмины метлы некоторых растений.
3.4. ФИТОПАТОГЕННЫЕ НЕМАТОДЫ
3.4.1. Общие сведения о фитонематодах
Нематоды относятся к типу круглых червей (Nemathelminthes),
которые широко распространены в природе и представлены как
свободно живущими, так и паразитическими видами.
Фитонематодами (фитогельминтами) называются
микроскопические черви класса Nematoda — возбудители нематодных бо-
83
лезней растений (фитогельминтозов). Подавляющее большинство
фитонематод относится к отрядам Tylenchida и Dorylaimida.
Зараженность фитонематодами некоторых почв очень велика и может
достигать нескольких тысяч особей в 100 м3 верхнего слоя почвы и
нескольких сотен особей в 1 г корней.
Тело у фитонематод — преимущественно нитевидное или ве-
ретеновидное, реже лимоновидное или шарообразное, длиной
0,5—3,5 мм, толщиной 0,01 — 0,6 мм, покрытое оболочкой
(кутикулой), чаще гладкой, но иногда снабженной щетинками или
чешуйками. В ротовой полости фитонематод расположен колюще-
сосущий иглообразный орган — стилет, которым нематода
прокалывает ткани растений и вводит в них ферменты и токсины.
В средней части пищевода имеется мускулистое расширение —
бульбус, выполняющий функцию насоса при поглощении
нематодой жидкого содержимого растительных тканей (рис. 30).
Фитонематоды размножаются яйцами, откладывая их в
субстрат в свободном виде (стеблевая нематода) или выделяя в жела-
тинозном мешке (галловая нематода); у нематод рода Heterodera
яйца остаются в теле самки, которое после ее смерти
превращается в твердую цисту. Отрождающиеся из яиц личинки заражают
Рис. 30. Фитонематоды и вызываемые ими болезни растений:
1 — внутреннее строение тела нематоды (а — стилет, б — бульбус, в — железы, г —
кишка); 2 — внешний вид нематоды; 3 — самка нематоды, набитая яйцами
(циста); 4 — корневая система, пораженная галловой нематодой; 5 — цисты нематоды
на корне растения; 6 — растение, пораженное хризантемной нематодой
84
растения, внедряясь преимущественно в мелкие корни.
Некоторые виды продолжают развиваться в корнях до взрослой стадии,
другие переходят из корней в надземные части. Немногие виды
заражают проростки и затем распространяются в тканях
развивающихся растений. Известны виды, которые передвигаются из почвы
вверх по растению в пленке воды и поражают надземные органы:
стебли, листья, цветки, семена.
Плодовитость фитонематод очень велика. Например, самка
галловой нематоды откладывает за сезон до 3 тыс. яиц. В течение
вегетационного периода многие нематоды дают несколько генераций,
поэтому потомство одной самки за год может составить
несколько миллиардов особей.
Среди факторов окружающей среды, влияющих на энергию
размножения, скорость развития, распространение и динамику
численности фитонематод, главную роль играют влажность и
температура почвы, так как нематоды активны только в пленке воды
и очень чувствительны к охлаждению или перегреву. Важное
значение имеют также кислотность почвы, ее механический состав,
сезонные физиологические изменения у растений-хозяев и
другие факторы. Например, максимальная численность фитонематод,
вызывающих болезни сеянцев хвойных пород, наблюдается в
питомниках, расположенных на легких песчаных и супесчаных
почвах, при температуре почвы 18... 20 °С и влажности 18... 28 %,
особенно в годы с теплым дождливым летом. Фитогельминтозы
цветочных культур чаще встречаются и наиболее вредоносны в
условиях защищенного грунта.
При неблагоприятных условиях фитонематоды могут впадать в
состояние анабиоза, но чаще сохраняются на фазе яйца.
Большинство их зимует в почве, некоторые виды — в корнях, клубнях,
луковицах, галлах. Они распространяются с зараженным
растительным материалом, почвой, поливными и дождевыми водами.
Степень паразитической специализации различных видов
фитонематод неодинакова. Большинство их имеет ограниченный круг
растений-хозяев, однако известны и полифаги: например,
корневая галловая нематода Meloidogyne marioni поражает свыше 1700 видов
растений, а нематода Xiphinema americanum — более 70 видов
деревьев и кустарников, в том числе 12 видов сосны.
3.4.2. Симптомы и типы нематодных болезней
растений
Нематодные болезни часто встречаются в питомниках,
теплицах, лесных культурах и естественных насаждениях различного
возраста. Известны фитогельминтозы многих лиственных и хвойных
пород, плодовых деревьев и декоративных кустарников. Широко рас-
85
пространены нематодные болезни цветочных и
сельскохозяйственных культур. Однако их диагностика затруднена тем, что симптомы
фитогельминтозов во многом сходны с симптомами других
болезней. Например, при нематодных заболеваниях всходов часто
наблюдаются увядание, полегание и другие общие симптомы, характерные
для некоторых грибных и неинфекционных болезней.
Неспецифичны и многие признаки поражения нематодами взрослых деревьев.
Для древесных растений нематодные болезни наиболее опасны
в первые годы их жизни. В зараженных нематодами питомниках
часто наблюдаются запоздалые всходы, резко увеличивается
отпад (отмечались случаи 50%-й и даже 100%-й гибели посевов),
ухудшается качество посадочного материала: у пораженных
растений, вследствие нарушения физиологических функций,
наблюдаются общее ослабление, ухудшение приживаемости, снижение
морозостойкости и устойчивости к другим патогенам.
Признаки нематодных болезней, проявляющиеся на
надземных частях молодых древесных растений, — отставание в росте,
деформация стволиков и побегов, увядание верхушек, изменение
цвета хвои, некротические пятна на листьях и др. Иногда
наблюдаются короткоиглость, образование галлов.
Патологические изменения корней более разнообразны. Самые
обычные из них — общее недоразвитие и деформация корневой
системы. При поражении сеянцев хвойных пород нематодами,
относящимися к родам Tylenchorhynchus и Trichodorus, корни
становятся укороченными, утолщаются, искривляются, иногда
отмирают, наблюдается комковатость корневой системы,
сопровождающаяся хлоротичностью и увяданием надземных частей. Нематода
Paralongidorus maximus вызывает у саженцев сосны деформацию
корневых окончаний с одновременным угнетением роста растений. При
этом хвоя осенью приобретает фиолетово-красный оттенок, а на
следующий год желтеет, отмирает, и саженцы погибают.
Корневые галловые нематоды рода Meloidogyne образуют на
корнях многих лиственных пород типичные сферические галлы, а у
хвойных вызывают утолщение корней (см. рис. 30). Галлообразование на
корнях древесных растений вызывают также нематоды родов Xiphine-
та и Longidorus. Из-за оттока питательных веществ к формирующимся
галлам растения сильно отстают в росте и нередко засыхают.
Нематоды рода Heterodera вызывают массовое образование
вторичных боковых корешков, благодаря чему корневая система
приобретает «бородатый» вид.
Под влиянием токсичных выделений эктопаразитических фито-
нематод (например, родов Xiphinema, Tylenchorhynchus, Paratylenchus,
Dolichodorus, Rotylenchus) происходит образование на корнях бурых
некротических пятен, обесцвечивание тканей, усыхание
кончиков сосущих корней, некроз и шелушение коры, отмирание
молодых корешков. Эндопаразитические нематоды рода Pratylenchus,
86
проникая внутрь корней, образуют глубокие некротические
ранки и полости; пораженные корни отмирают.
У взрослых деревьев нематодные болезни характеризуются
разнообразными признаками: снижением прироста, частичной сухокрон-
ностью, увяданием молодых побегов, ослаблением цветения и
плодоношения, преждевременным опадением плодов, недоразвитием
и утолщением корней. При резком ухудшении почвенных,
метеорологических или других условий на фоне высокой численности
нематод может произойти быстрое усыхание пораженных деревьев.
Одной из наиболее опасных болезней считается бурсафеленхоз
сосны, вызываемый нематодой Bursaphelenchus xylophilus Stein, et
Buhr. Заболевание распространено в США, Канаде, Китае и
особенно в Японии, где оно приобрело характер эпифитотии. В
нашей стране бурсафеленхоз не обнаружен и является объектом
внешнего карантина.
Поражаются деревья разного возраста. Первый признак
болезни — обесцвечивание ветвей с последующим их ярким красно-
коричневым окрашиванием. Пораженные ветви отмирают, а
затем усыхают и сами деревья. Эта нематода может размножаться и
накапливаться в сосновой щепе. Ее переносчиками являются жуки-
усачи рода Monochamus.
Фитонематоды вызывают тяжелые заболевания многих
цветочных культур (см. гл. 14); это приносит очень большой вред
промышленному цветоводству.
Повреждая корневые волоски, кору корней, многие
фитонематоды способствуют проникновению в них вторичной грибной или
бактериальной инфекции, загниванию корней. Вызывая
биохимические изменения в тканях пораженных растений,
фитонематоды часто снижают их устойчивость к грибным и бактериальным
болезням или усиливают развитие последних. В свою очередь, грибы
и другие патогены также могут вызывать определенные сдвиги в
физиолого-биохимических реакциях клеток хозяина, создавая
более благоприятные условия для нематод. Такие взаимосвязи
приводят к возникновению комплексных заболеваний. Например,
причиной загнивания корней у сеянцев хвойных пород может быть
одновременное заражение их нематодами и грибами рода Fusarium;
поражение сеянцев сосны обыкновенным шютте часто сочетается
с более высокой зараженностью их паразитическими нематодами.
Некоторые фитонематоды отряда Dorylaimida способны быть
переносчиками фитопатогенных вирусов.
Для защиты растений от нематод применяют комплекс
агротехнических, лесохозяйственных, физико-механических, химических
и биологических мер борьбы. Он включает: выращивание
устойчивых к фитонематодам видов и сортов растений; использование
здорового посадочного материала или его пропаривание; специальные
севообороты, сбалансированное удобрение, использование минераль-
87
ных субстратов; уничтожение больных растений и сорняков;
термическое или химическое обеззараживание почвы, применение
системных нематицидов; выращивание ловчих культур, внесение в
почву биопрепаратов хищных грибов, а также паразитирующих на
нематодах грибов и бактерий; карантинные и другие мероприятия.
3.5. ПАРАЗИТИЧЕСКИЕ ЦВЕТКОВЫЕ РАСТЕНИЯ
3.5.1. Общие сведения о цветковых
растениях-паразитах
Большинство высших (цветковых) растений обладает
способностью к самостоятельному (автотрофному) питанию. Эти
растения, как правило, имеют хорошо развитую корневую систему,
дающую им возможность добывать из почвы воду и минеральные
вещества, и надземный ассимилирующий аппарат, с помощью
которого они синтезируют сложные органические соединения,
идущие на построение всех тканей и органов растений. Однако в
составе некоторых семейств есть растения, частично или
полностью утратившие способность к автотрофному питанию и
перешедшие к паразитическому существованию за счет дфугих растений.
Переход к паразитизму осуществлялся у них путем
приспособления к жизни на корнях или на надземных органах других растений.
Соответственно этому среди паразитических цветковых растений
сформировались группы корневых и стеблевых (стволовых)
паразитов. У тех и у других наблюдаются значительное видоизменение,
недоразвитие или полная деградация корневой системы. Те и
другие берут у растения-хозяина воду и минеральные вещества. Вместе
с тем в обеих группах есть виды, резко различающиеся по
способности ассимилировать углекислоту и создавать органические
вещества своего тела. Одни в полной мере сохранили эту способность:
они имеют зеленые листья и стебли, что дает основание называть
их полупаразитами, или частичными паразитами. Другие
полностью утратили это свойство высших растений (а вместе с ним —
хлорофилл и зеленую окраску). Они извлекают из растения-хозяина
не только воду и элементы минерального питания, но и
органические вещества. Такие растения являются абсолютными паразитами.
Иногда растение-паразит почти целиком находится внутри
растения-хозяина (например, между древесиной и корой корня или
стебля), причем его вегетативные органы настолько
видоизменены, что напоминают тонкие нити мицелия. Они пронизывают ткани
растения-хозяина, а наружу выходят лишь цветки и плоды.
Некоторые виды наряду с присосками (гаусториями) имеют настоящие
корни, способные извлекать воду и питательные вещества из
почвы; другие на начальных этапах своего развития ведут паразитиче-
88
ский образ жизни, но в дальнейшем переходят к
самостоятельному питанию. Однако, несмотря на столь существенные различия
между отдельными группами и видами паразитических цветковых
растений в степени и форме паразитизма, все они являются об-
лигатными паразитами, поскольку не в состоянии развиваться (на
протяжении всей жизни или на определенных стадиях) без
растения-хозяина. Степень специализации цветковых паразитов также
различна: наряду с узкоспециализированными видами
встречаются виды с широкой, но четко выраженной специализацией.
Экономическое значение паразитических цветковых растений
в целом по сравнению с другими группами фитопатогенных
организмов (например, грибами или вирусами) не столь велико,
однако некоторые виды, особенно повилики, заразихи, омела,
причиняют значительный вред.
Большинство паразитических цветковых относится к семействам
норичниковых (Scrophulariaceae), ремнецветниковых (Lorantha-
сеае), повиликовых (Cuscutaceae), заразиховых (Orobanchaceae),
орхидных (Orchidaceae).
3.5.2. Корневые полупаразиты
Типичным представителем корневых полупаразитов является иван-
даг-марья (Melampyrum nemorosum Z,.), широко известное растение
из семейства норичниковых, часто встречающееся на опушках леса
и лесных полянах. Корни иван-да-марьи снабжены особыми
присосками, которые присасываются к корням других растений, в
основном деревьев и кустарников, и таким путем извлекают из
растения-хозяина растворы минеральных питательных веществ. Другие
представители семейства норичниковых (погремок — Rhinanthus major
Ehrb., очанка — Euphrasia officinalis L., мытник — Pedicularis pa-
lustris L.) являются полупаразитами луговых трав. Они
отрицательно влияют на густоту и высоту травостоя, снижают качество сена.
3.5.3. Стволовые полупаразиты
Среди стволовых полупаразитов наибольшее хозяйственное
значение имеет омела белая (Viscum album L.), растение из
семейства ремнецветниковых. Омела широко распространена в
южных районах нашей страны и на Дальнем Востоке. Она поражает
многие лиственные и хвойные породы. На пихте паразитирует
близкий вид — пихтовая омела (Viscum abietis Beck.Stank).
Омела — двудомное растение, имеющее форму куста, с ярко-
зелеными кожистыми листьями и зеленым дихотомически
ветвящимся стеблем (рис. 31). Плоды омелы — шаровидные сидячие
ягоды белого цвета. Созревающие в них зимой семена покрыты
клейким веществом — висцином. Они распространяются птица-
89
Рис. 31. Паразитические цветковые растения (стволовые полупаразиты):
1 — кустики омелы в кроне дерева; 2 — омела белая на стволе; 3 — начальная
стадия заражения ствола омелой; 4 — можжевелоядник
ми, особенно дроздами, и, попадая на ветви и стволы деревьев,
прилипают к ним, а весной прорастают. Кончик проростка при
соприкосновении с корой образует плоскую присоску. От нее берет
начало первичное сосальце (гаусторий), которое внедряется в кору,
а затем по мере роста проникает в древесину. На следующий год
первичное сосальце образует корнеподобные боковые ветви —
ризоиды, они продвигаются от основания первичного сосальца в
стороны по коре. Из них перпендикулярно отходят вторичные
сосальца, постепенно погружающиеся в глубь древесины.
Вред, причиняемый омелой, очень велик. Он заключается
прежде всего в том, что паразит отнимает у растения-хозяина большое
количество воды, а также азота, фосфора, калия и других
зольных элементов. Вследствие этого деревья, в сильной степени
пораженные омелой, плохо растут, слабо плодоносят, частично
или полностью усыхают. Кроме того, пораженные омелой стволы
обесцениваются с технической точки зрения: из них нельзя
получить высококачественные деловые сортименты.
К семейству Loranthaceae относятся также можжевеловая
омела и ремнецветник. Можжевеловая омела, или
можжевелоядник (Arceuthobium oxycedri M. В.), часто встречается в
Крыму, на Кавказе и в Средней Азии, поражает различные виды
можжевельника. Растения можжевелоядника имеют вид
небольших кустиков с разветвленными побегами и мелкими
чешуйчатыми листьями. Обильное ветвление делает эти кустики похожими
на ведьмины метлы. Плод — мелкая продолговатая голубовато-
зеленая ягода, содержащая одно семя. При созревании плодов се-
90
мена «выстреливаются» из них на расстояние до 10 м и благодаря
висцину прочно приклеиваются к чему-либо, в том числе и к
коре ветвей и стволов можжевельника, а затем прорастают, давая
начало новым особям паразита. Распространению семян
способствуют птицы, поедающие мякоть плодов. На одном растении
можжевельника иногда развивается более 100 кустиков паразита,
продолжительность жизни которых достигает нескольких
десятилетий. В местах поражения ветвей и стволов можжевельника
возникают патологические изменения тканей, нарушается
сокодвижение, появляются утолщения и другие деформации. У больных
деревьев снижается прирост, отмирают отдельные ветви, а при
сильной степени поражения деревья полностью усыхают.
У стволовых полупаразитов в распространении и прорастании
семян, благодаря участию птиц и наличию в ягодах висцина,
существует много общего. Внедрение же паразита в ствол растения-
хозяина, дальнейшее развитие в нем и механизм питания у
разных видов имеют свои особенности, причем многие моменты этих
процессов еще не вполне ясны.
3.5.4. Корневые паразиты
К абсолютным корневым паразитам древесных растений
относится петров крест (Lathraea squamaria L.) из семейства
норичниковых. Свое название это растение получило благодаря
наличию у него мясистых подземных стеблей с характерными
крестовидными разветвлениями (рис. 32). У паразита есть многолетнее
Рис. 32. Паразитические цветковые растения (корневые и стеблевые
паразиты):
1 — петров крест; 2 — заразиха; 3 — повилика; 4 — соцветие повилики и
присоски на ее стебле
91
корневище, от которого отходят особые корни,
присасывающиеся к корням деревьев. На поверхность земли ранней весной
выходит невысокий желтоватый стебель с чешуевидными розоватыми
листьями и малиново-красными цветками. Петров крест поражает
березу, осину, ольху, лещину, черемуху, бук, ель и другие
древесные растения.
Очень большой вред сельскохозяйственным культурам
причиняют заразихи — виды рода Orobanche (см. рис. 32).
3.5.5. Стеблевые паразиты
Из паразитов этой группы наиболее широко распространены и
хорошо известны повилики — виды рода Cuscuta (семейство
повиликовые). Это бесхлорофилльные растения, не имеющие ни
корней, ни настоящих листьев. Тонкие, иногда ветвящиеся
стебли повилики обвиваются вокруг стеблей растения-хозяина,
прочно прикрепляются к ним с помощью особых присосков (гаусто-
риев) и извлекают из сосудов пораженного растения воду,
минеральные и органические вещества (см. рис. 32).
Повилика — наиболее опасный и трудно искоренимый
паразит среди цветковых растений. Она очень плодовита (каждое
растение дает несколько тысяч мелких, как пыль, семян). У
большинства видов повилик резерватором семян служит почва, но у
некоторых видов они сохраняются только с семенами растения-
хозяина. Семена повилики не теряют всхожести, проходя через
пищеварительный тракт животных. Распространяется повилика с
семенами растений, с навозом, с оросительными водами и другими
путями. При этом распространяться могут не только ее семена, но
и кусочки стеблей. Различные виды повилик поражают ценные
технические и плодово-ягодные культуры, многолетние травы.
На деревьях и кустарниках чаще всего паразитируют два вида:
повилика европейская (С. europaeaL.) и повилика од-
ностолбиковая (С. monogyna Vahl.), принадлежащие к числу
карантинных объектов. Они поражают иву, робинию лжеакацию,
тополь, ольху, клен, бузину, сирень, смородину и другие
деревья и кустарники, а также многие культурные растения.
Вред, наносимый повиликами, исключительно велик. В лесном
хозяйстве он особенно ощутим при распространении повилик в
пойменных лесах, а также в полезащитных насаждениях,
питомниках, молодых культурах и парках. Кроме того, повилики могут
служить переносчиками фитопатогенных вирусов.
Глава 4
ПАТОГЕНЕЗ И ДИНАМИКА
ИНФЕКЦИОННЫХ БОЛЕЗНЕЙ РАСТЕНИЙ
Патогенезом, или патологическим процессом, называют
процесс развития инфекционной болезни, протекающий во
взаимодействии растения-хозяина, патогена и окружающей среды.
Патологический инфекционный процесс может возникнуть
лишь в том случае, если произойдет нападение патогена на
восприимчивое к нему растение. Для этого необходимо, чтобы
патоген был способен проникать в растение и распространяться в нем;
чтобы он обладал способностью убивать зараженную ткань или
питаться содержимым живых клеток, преодолевая их защитные
механизмы и изменяя обмен веществ хозяина соответственно своим
потребностям. Иными словами патоген должен обладать
определенными свойствами, обеспечивающими заражение растения и
дальнейшее развитие болезни.
4.1. СВОЙСТВА ПАТОГЕНОВ
Важнейшими свойствами возбудителей болезней являются па-
тогенность (вирулентность) и агрессивность.
Патогенностью называется болезнетворная способность
патогена, его свойство вызывать у растения-хозяина
патологические реакции, т. е. болезнь. Патогенность проявляется в
повреждениях клеток и тканей, нарушениях обмена веществ и
физиологических функций растения.] Она обусловливается воздействием на
растение ферментов, токсинов, регуляторов роста и других
биологически активных веществ. Эти вещества служат основными
средствами нападения патогенов на растения. Именно они
определяют механизмы патогенности, характерные для определенных групп
и видов возбудителей болезней. С их помощью патоген выводит из
строя защитные механизмы растения-хозяина и использует его
ткани в качестве питательного субстрата.
93
Важнейший фактор патогенности грибов и бактерий
—ферменты. Они играют основную роль в патогенезе инфекционных
болезней растений. Ферменты разрушают покровную ткань,
оболочки и содержимое клеток растения, делая возможным
внедрение патогена в растение, его распространение во внутренних
тканях хозяина и питание за его счет. Особенности ферментного
аппарата, качественный состав ферментов и характер их воздействия
на растительную ткань зависят от типа питания, уровня
паразитической активности и специализации возбудителей.
Ферменты облигатных паразитов-биотрофов обычно
оказывают умеренное воздействие на живые клетки растения, не вызывая
(особенно на начальных этапах патогенеза) сильного
повреждения их структур, заметных нарушений обмена веществ и
физиологических функций хозяина. Мягкое воздействие некоторых био-
трофов на ткани и обмен веществ растения-хозяина связаны с
отсутствием у них оксидаз, амилаз и протеолитических
ферментов или низкой их активностью. Облигатные паразиты иногда даже
стимулируют у пораженных растений обменные процессы, что
выражается во временном усилении фотосинтеза, дыхания и
других физиологических функций.
Ферменты некротрофных паразитов, как правило, грубо
воздействуют на ткани растения-хозяина, вызывая более полное
разрушение клеточных структур и резкие нарушения
физиологических функций.
Второй фактор патогенности грибов и бактерий, присущий
главным образом некротрофам (в основном факультативным
паразитам и факультативным сапротрофам) — токсины, т. е.
вредные для растений продукты жизнедеятельности возбудителей
болезней. Они вызывают у растений специфические повреждения
клеток, ведущие к быстрому отмиранию (некрозу) тканей, что
делает их доступными для заселения патогеном.
Патогенность ряда грибов обусловлена воздействием таких
токсинов, как ликомаразмин, патулин, диапортин, альтернариевая
и фузариевая кислоты и др. Нарушения водного режима, ведущие
к усыханию деревьев при сосудистых микозах, связаны главным
образом с действием токсинов, вызывающих отмирание тканей,
окружающих сосуды, и закупорку сосудов образующимися при
этом камедеобразными веществами, слизью, тиллами. Токсины
увядания (например, гликопептиды), выделяемые некоторыми
фитопатогенными бактериями, вызывают повреждение клеточных
мембран, что приводит к большой потере воды клетками
больного растения.
Некоторые биотрофы также образуют токсины, но в отличие
от некротрофов они повреждают ткань растения-хозяина не во
время ее заселения, а после завершения в ней своего
вегетативного роста и размножения.
94
Третий фактор патогенности грибов и бактерий — ростовые
вещества. Вещества, регулирующие рост тканей (индолилук-
сусная и гибберелловая кислоты, гиббереллины и др.), играют
важную роль в механизмах патогенности многих биотрофных
паразитов. Обычно они вырабатываются патогеном или самим
растением-хозяином под влиянием патогена на ранних этапах
инфекционного процесса, в период вегетативного роста и
распространения возбудителя болезни в живых тканях растения.
Дисбаланс ростовых веществ вызывает временную
стимуляцию процессов роста (это проявляется в патологическом
разрастании зараженных тканей) или, наоборот, угнетает эти
процессы, что приводит к недоразвитию пораженных органов или всего
растения.
Патогенность вирусов связана с воздействием вирусной
нуклеиновой кислоты на зараженные клетки растения-хозяина. Она
проявляется прежде всего в повреждении ферментного аппарата
растения и как следствие этого — в разнообразных нарушениях
обмена веществ и физиологических функций, процессов роста и
развития, разрушении хлорофилла, а также в использовании
азота, фосфора и других элементов содержимого клеток хозяина для
построения новых вирусных частиц.
Патогенность цветковых растений-паразитов обусловливается
главным образом выносом из тканей растения-хозяина больших
количеств воды, органических и минеральных веществ, что ведет
к истощению, угнетению роста или гибели пораженных растений.
Патогенность некоторых видов цветковых паразитов (например,
повилики, омелы) усиливается выделением ими токсинов.
Вирулентность — понятие, тесно связанное с
филогенетической специализацией возбудителей болезней. Это патогенность
определенного таксона (вида, специализированной формы, расы)
паразита по отношению к определенному таксону (семейству, роду,
виду, сорту) растений. Например, патогенность гриба Ophiostoma
ulmi — возбудителя голландской болезни, проявляется только в
отношении ильмовых; следовательно, этот гриб вирулентен
только для древесных пород этого семейства.
Если вид паразита физиологически неоднороден и состоит из
специализированных форм, то он неоднороден и по
вирулентности. Примером может служить мучнисторосяной гриб Phyllactinia
suffulta: каждая из специализированных форм этого вида вирулентна
только для растений определенного рода (ясеня, березы,
лещины, граба и т.д.). Точно так же определенные физиологические
расы патогенов (в пределах специализированных форм)
вирулентны лишь для определенных видов, сортов или форм растений.
Агрессивностью называют способность фитопатогенных
организмов вызывать массовые заболевания растений (эпифи-
тотии).
95
Агрессивный патоген должен обладать определенным
комплексом свойств: высокой энергией размножения (плодовитостью),
способностью быстро распространяться на далекие расстояния и
накапливаться в природе, способностью заражать хозяина
минимальным количеством инфекционного начала, малой
продолжительностью процесса заражения и инкубационного периода,
высокой экологической пластичностью (приспособляемостью к
различным условиям внешней среды).
Высокой агрессивностью характеризуются многие ржавчинные,
мучнисторосяные, пероноспоровые грибы. Они способны не только
заразить растение малым количеством спор (нередко одной
спорой), но и быстро осуществить заражение, имеют короткий
инкубационный период, образуют в течение лета несколько
генераций бесполых спороношений, которые легко распространяются
ветром и служат для массовых повторных заражений. Таким
образом увеличивается запас инфекции и создается предпосылка для
возникновения эпифитотии.
Возьмем для сравнения два гриба: Microsphaera alphitoides и
Rosellinia quercina. Оба гриба патогенны и вирулентны для дуба.
Однако первый, возбудитель мучнистой росы, обладает всеми
качествами высокоагрессивного паразита, поэтому развитие
мучнистой росы часто носит характер эпифитотии. Второй,
возбудитель корневой гнили сеянцев, лишен этих качеств, имеет
незначительное распространение и массового заболевания растений
обычно не вызывает.
Патогенность и вирулентность — постоянные, не
подверженные колебаниям свойства возбудителей болезней. Уровень же
агрессивности одного и того же патогена может изменяться в
зависимости от условий развития и других причин.
4.2. ИНФЕКЦИОННЫЙ ПРОЦЕСС
Развитие патологического процесса при инфекционных
болезнях растений обычно проходит несколько этапов,
последовательно сменяющих друг друга. Основные этапы патогенеза:
заражение, инкубационный период, собственно болезнь и
выздоровление (или отмирание) пораженных частей либо всего растения.
Каждый из этапов характеризуется определенными
взаимоотношениями между патогеном и растением-хозяином, причем
развитие этих взаимоотношений в значительной степени зависит от
условий окружающей среды.
Заражение. Этот этап инфекционного процесса, в свою
очередь, включает как бы несколько фаз: попадание возбудителя на
поверхность растения, прорастание инфекционного начала,
проникновение возбудителя в растение.
96
Попадание возбудителя на поверхность растения
обеспечивается способностью патогенов распространяться в природе
самостоятельно или с помощью различных агентов (воздушных
течений, воды, животных или человека). В большинстве случаев этот
процесс протекает неспецифично: одним и тем же способом
(например, с помощью воздушных течений) могут
распространяться как различные паразитические, так и непатогенные сапро-
трофные микроорганизмы. На поверхность растения могут
попадать любые сапротрофы и возбудители не свойственных данному
растению болезней. Однако в некоторых случаях наблюдается
специфичность распространения возбудителя (например, при
передаче персистентных вирусов специализированными насекомыми-
переносчиками, распространении возбудителей сосудистых
болезней определенными видами стволовых вредителей).
Прорастание инфекционного начала наблюдается у грибов и
паразитических цветковых растений: у первых прорастают споры,
у вторых — семена. Прорастание спор фитопатогенных грибов может
происходить не только на поражаемых органах восприимчивых
растений, но и на других растениях и даже на мертвых субстратах,
если имеются подходящие внешние условия. В ряде случаев,
однако, прорастание спор патогенных грибов и семян паразитических
растений стимулируется специфическими выделениями
восприимчивого растения-хозяина. Иногда прорастание инфекционного
начала у специализированных паразитов вообще возможно
только при наличии выделений восприимчивых растений.
Для прорастания спор и семян фитопатогенных организмов
необходим комплекс условий внешней среды: определенные
значения температуры, влажности, кислотности, аэрации и т.д.
Важное значение среди факторов окружающей среды имеет также
присутствие других микроорганизмов, которые могут
стимулировать или, наоборот, подавлять прорастание патогенов. Факторы
внешней среды обусловливают не только возможность
прорастания инфекционного начала, но влияют и на скорость этого
процесса, что имеет иногда решающее значение для заражения
растений. При этом разные группы и виды возбудителей болезней
предъявляют к факторам среды различные, подчас очень строгие
требования.
Проникновение возбудителя в растение может происходить
различными путями. Способ внедрения патогена в растение
зависит от его биологических особенностей и во многих случаях
является специфичным для той или иной группы фитопатогенных
организмов (рис. 33).
Грибы могут проникать в растение по-разному. Например,
ростковые трубки (инфекционные гифы) базидиоспор ржавчинных
грибов, конидий мучнисторосяных, некоторых гифомицетов и
других паразитных грибов обладают способностью внедряться в
97
Рис. 33. Процесс заражения растений фитопатогенными организмами:
7 — заражение хвои через устьице возбудителем шютте и развитие мицелия в
зараженных тканях; 2 — процесс заражения листа грибом Botrytis cinerea через
неповрежденные покровные ткани (а — спора гриба в капле воды, б — прорастание
споры и проникновение инфекционной гифы через кутикулу, в — проникновение
инфекционной гифы сквозь эпидермис и внедрение патогена в ткань листа); 3 —
заражение листа через устьице бактерией Pseudomonas pruni и начало размножения
бактериальных клеток; 4 — внедрение сосальца растения-паразита петров крест в
корень дерева; 5 — внедрение гаусториев повилики в сосуды стебля
растение через неповрежденные наружные покровы. При таком
прямом проникновении в растение гриб оказывает на кутикулу и
наружные стенки клеток эпидермиса как механическое давление,
так и химическое воздействие, размягчая и растворяя их
соответствующими ферментами. Ростковые гифы сумкоспор многих ас-
комицетов, урединиоспор ржавчинных грибов, зооспоры низших
грибов проникают внутрь растений через устьица, чечевички, ги-
датоды, рыльца пестиков и другие естественные отверстия, а
зооспоры — также через органы растений, не покрытые кутикулой,
например корневые волоски. Многие грибы, относящиеся к
факультативным паразитам и факультативным сапротрофам (так
называемые раневые паразиты), способны заражать растения
через какие-либо ранки, повреждения коры и кутикулы или
отмершие части. Таковы, например; возбудители ступенчатого рака,
некрозных и гнилевых болезней древесных пород.
Бактерии, вирусы и микоплазмы внедряются внутрь растений без
прорастания, непосредственно после попадания на их поверхность.
98
Бактерии проникают во внутренние части растений только через
естественные ходы или различные повреждения. Вирусы и микоплаз-
мы чаще всего вносятся в ткани растений насекомыми во время их
питания, а также через мелкие ранки, возникающие при трении
соприкасающихся частей растений или при уходе за растениями.
Проростки семян паразитических цветковых растений
углубляются в ткани стеблей, стволов или корней растения-хозяина
путем прямого внедрения через неповрежденную кутикулу или
кору. Фитонематоды проникают в растения преимущественно
через корневые волоски.
Внедрение патогена в растение еще не означает, однако, что
заражение осуществилось. Через устьица и другими путями внутрь
растения могут проникнуть и случайные микроорганизмы, не
способные к дальнейшему развитию в данном растении. Только в том
случае, когда внедрившийся патоген начинает питаться и расти
за счет тканей хозяина, т. е. когда между партнерами
устанавливаются стабильные паразитические взаимоотношения, заражение
можно считать осуществившимся.
Инкубационный период. Инкубационным (или скрытым)
периодом называется отрезок времени от момента заражения до
появления первых симптомов болезни. Продолжительность
инкубационного периода зависит прежде всего от паразитических свойств
возбудителей болезней, поэтому при разных заболеваниях она
различна. Например, при некоторых вирусных болезнях
инкубационный период может составлять несколько часов, при
поражении растений мучнистой росой — несколько дней, при болезнях
типа шютте — несколько недель или месяцев, а при поражении
сосен возбудителями пузырчатой ржавчины (смоляного рака) —
более полутора лет.
В пределах, характерных для определенных групп или видов
заболеваний, продолжительность инкубационного периода может
колебаться в зависимости от степени устойчивости или выносливости
растения-хозяина (т. е. от типа реакций растения на воздействие
патогена), а также от многих внешних факторов, в первую очередь от
температуры и влажности. Если возбудитель способен проникать в
растение разными путями, то период инкубации может изменяться
и в зависимости от способа заражения. При оптимальных для
развития паразита условиях инкубационный период сокращается, при
неблагоприятных — соответственно увеличивается. В течение
инкубационного периода патоген активно воздействует на обмен
веществ растения-хозяина, полностью переходит к питанию за счет
его тканей, распространяется, а иногда (например, при
бактериозах и вирусных болезнях) и размножается в нем, заселяя
подходящие органы и ткани. В этот период в растении постепенно, пока
еще скрыто, накапливаются те патологические изменения,
которые к концу инкубационного периода становятся явными.
99
Собственно болезнь. Появление видимых внешних признаков
болезни означает начало третьего, основного этапа
инфекционного процесса. В этот период взаимодействие растения-хозяина и
патогена достигает наибольшей остроты и проявляется в
характерных для данной болезни физиологических, анатомических и
морфологических нарушениях. Продолжительность этого этапа
может быть различной и зависит от многих факторов. Чаще всего
он завершается отмиранием пораженных частей или всего
растения. Выздоровление больных растений наблюдается реже.
Выздоровление растений. Процесс восстановления и
нормализации нарушенных болезнью физиологических функций может
происходить естественным путем или при помощи человека.
Естественное выздоровление наблюдается в тех случаях, когда
изменение внешних условий (например, резкое колебание
температуры окружающей среды) подавляет жизнедеятельность патогена в
тканях растений. Выздоровление растений иногда бывает связано
и с накоплением в пораженных тканях токсичных для патогена
веществ, под влиянием которых возможны угнетение роста и
развития, дегенерация или гибель возбудителя болезни.
Изменением условий минерального питания пораженных
растений (путем внесения
удобрений, микроэлементов),
введением антибиотиков и других
веществ, воздействием на
пораженные растения различных
физических факторов человек
может резко изменить обмен
веществ растения-хозяина, создать
тем самым неблагоприятные
условия для развития патогена и
вызвать у него регрессивные
изменения (например,
прекращение роста, дегенерацию и лизис
мицелия паразитных грибов),
что ведет к прекращению
патологического процесса и
выздоровлению растений (рис. 34).
Выздоровление деревьев,
пораженных некрозно-раковыми
болезнями, опенком, может быть
достигнуто применением
системы соответствующих лечебных
мероприятий (зачисткой,
дезинфекцией и пломбированием
ран, обрезкой больных ветвей и
корней, аэрацией корней и т.п.).
Рис. 34. Разрушение мицелия фито-
патогенного гриба в тканях
пораженного растения под влиянием
микроэлементов:
1 — нормальный мицелий; 2, 3 —
распад и лизис гиф
100
4.3. ПАТОЛОГИЧЕСКИЕ ИЗМЕНЕНИЯ БОЛЬНОГО
РАСТЕНИЯ
В процессе заболевания паразиты механически повреждают
покровные ткани растения-хозяина, клеточные оболочки и
мембраны, воздействуют на них своими ферментами, токсинами и
другими биологически активными выделениями, отнимают воду
и питательные вещества, закупоривают сосуды растений. В
результате у растений нарушаются нормальные связи между
отдельными структурами внутри клетки и между самими клетками,
различными тканями и органами, нарушается синтез ферментов,
контролирующих и координирующих биохимические реакции
клеток. Это приводит к дальнейшим нарушениям обмена веществ
и физиологических функций больного растения. Как следствие
физиолого-биохимических нарушений возникают отклонения от
нормы в росте и развитии растений, а также разнообразные ана-
томо-морфологические изменения, создающие внешнюю
картину (симптомокомплекс) болезни.
4.3.1. Физиолого-биохимические нарушения
Нарушения биохимических реакций, обмена веществ и
физиологических процессов, наблюдаемые у больных растений,
возникают уже на самых первых этапах патогенеза и оказывают
решающее влияние на дальнейшее развитие болезни и состояние
растения-хозяина, хотя внешние симптомы болезни могут долго
не проявляться. К наиболее важным и характерным физиолого-
биохимическим нарушениям относятся нарушения ферментного
аппарата и функций клеточных мембран, нарушения
фотосинтеза, дыхания, углеводного и белкового обмена, водного режима
растений и другие патологические изменения.
Нарушения ферментного аппарата. Нарушения синтеза и
деятельности ферментов наблюдаются при любых болезнях растений.
Они приводят к весьма тяжелым последствиям, поскольку
именно ферменты управляют всем ходом биохимических процессов
клетки. От них зависят синтез структурных элементов, запасных и
биологически активных веществ, координация энергетических
процессов, скорость обмена веществ, интенсивность
физиологических функций растения. Особенно резко деятельность
ферментов нарушается при инфекционных заболеваниях. При поражении
фитопатогенными грибами, вирусами, бактериями может быть
нарушена деятельность различных групп ферментов.
Наиболее изучены изменения активности
окислительно-восстановительных ферментов больных растений. Эти изменения
специфичны: они зависят от видовой принадлежности патогена, вида
101
растения-хозяина и других факторов. В одних случаях активность
ферментов заметно повышается, в других — понижается.
Например, в листьях дуба, пораженных мучнистой росой, активность
каталазы снижается до 50 %, тогда как активность пероксидазы,
наоборот, возрастает.
Поскольку ферментные системы растения тесно
взаимосвязаны, нарушения в деятельности ферментов одной группы влекут
за собой изменения не только непосредственно регулируемых ими
процессов, но и многих других. С нарушениями в ферментном
аппарате в той или иной степени связаны почти все физиолого-
биохимические нарушения, наблюдаемые у больных растений.
Нарушения функций клеточных мембран. Повреждение плазма-
леммы и других клеточных мембран в результате механического
или химического воздействия патогена приводит к нарушению
нормального взаимодействия между отдельными компонентами
клетки, вследствие чего в ней возникают патологические реакции
с образованием токсичных веществ. Повреждение
полупроницаемых мембран ведет также к нарушению нормальных
взаимоотношений клетки с окружающей средой: в клетку начинают
проникать вредные для нее вещества и в то же время усиливается отток
из клетки необходимых для ее жизнедеятельности соединений и
запасных питательных веществ; значительно повышается и
водоотдача. Все эти нарушения влекут за собой патологические
изменения концентрации клеточного сока, осмотического давления,
сосущей силы, тургора клеток. При большинстве инфекционных
болезней эти показатели у пораженных растений по сравнению со
здоровыми снижаются.
Понижение осмотического давления клеточного сока у
больных растений связано также с усиленным потреблением
паразитом питательных веществ. Однако при некоторых болезнях в связи
с нарушением оттока продуктов фотосинтеза из листьев и
одновременным нарушением притока воды концентрация клеточного
сока и осмотическое давление могут повышаться. Нарушения
осмотического давления у пораженных растений имеют очень
важное значение, поскольку от этого показателя в значительной мере
зависят такие свойства растений, как засухоустойчивость, соле-
выносливость, морозостойкость.
Нарушения фотосинтеза. В большинстве случаев у пораженных
растений наблюдается значительное снижение интенсивности
фотосинтеза, вплоть до прекращения процесса ассимиляции. Это
связано прежде всего с разрушением токсинами возбудителя
хлорофилла и ферментов, управляющих фотосинтезом. При
поражении растений облигатными паразитами (чаще всего
ржавчинными грибами) и некоторыми факультативными сапротрофами
(например, грибом Rhytisma acerinum) на начальных этапах
инфекционного процесса нередко отмечается усиление фотосинтеза. У боль-
102
ных растений иногда повышается и содержание хлорофилла:
зараженные участки листьев (места формирования пустул или
пятен) бывают в этих случаях окружены кольцом ткани более яркой
зеленой окраски. Однако со временем стимуляция фотосинтеза
сменяется резким снижением или полным торможением этого
процесса.
Нарушения дыхания. Нарушения дыхания происходят в
результате воздействия патогена на деятельность дыхательных
ферментов. Начальные этапы инфекционного процесса, как правило,
характеризуются повышением активности дыхания пораженных
органов растений. По мере развития болезни интенсивность
дыхания обычно снижается. Усиление дыхания приводит соответственно
к усиленному расходованию растением запасных питательных
веществ. Если повышение активности дыхания, при котором
увеличивается количество освобождающейся энергии, сопровождается
нарушениями в системе переноса энергии, может возникнуть
явление гипертермии — повышение температуры пораженных
органов растений.
Нарушения углеводного и белкового обмена. При
инфекционных болезнях растений наблюдаются как количественные, так и
качественные изменения процессов обмена веществ, в том числе
основных органических веществ клетки — белков и углеводов.
Количественные изменения в большинстве случаев сводятся к
снижению общего содержания углеводов и белковых соединений
пораженных тканей в результате ослабления процессов
ассимиляции и синтеза, усиленного расходования растением
органических веществ на дыхание и непосредственного потребления их
патогеном.
Качественные изменения выражаются в нарушении
нормального соотношения отдельных компонентов углеводного и
азотсодержащего комплексов. Так, при поражении растений ржавчиной
содержание общего и белкового азота, как правило, снижается,
но при этом резко увеличивается содержание небелкового азота,
представленного в основном вредными для растения продуктами
жизнедеятельности патогена. Изменяется также состав белков. При
вирусных болезнях наряду с общим снижением содержания
растительных белков происходит накопление вирусного белка.
При многих болезнях растений значительно изменяются
состав и процентное соотношение моносахаридов и полисахаридов.
Нарушения обмена веществ проявляются также в ненормальном
распределении углеводов и белков по растению. Особенно ярко
это проявляется при заболеваниях, сопровождающихся некрозом
флоэмы. Вследствие нарушения нисходящего тока ассимилятов
листья бывают перегружены крахмалом, в то же время
содержание углеводов в других частях растений, как правило, резко
снижается.
103
Нарушения водного режима. Все болезни растений
сопровождаются нарушениями водного режима, причем по мере развития
патологического процесса и увеличения степени поражения
растений эти нарушения усиливаются и проявляются в более острой
форме. У пораженных растений, как правило, изменяется водо-
удерживающая способность, нарушается транспирация. В
большинстве случаев (при поражении ржавчиной, мучнистой росой,
паршой и многими другими болезнями) больные растения
характеризуются повышенной транспирацией, но при некоторых
заболеваниях (например, при сосудистых болезнях, поражении
корневой губкой) интенсивность транспирации уменьшается.
Нарушения водного режима при инфекционных болезнях
растений обусловливаются различными причинами. Так, усиление
водоотдачи при поражении листьев паршой или ржавчиной
бывает связано с разрывом покровных тканей, спороношениями
возбудителя и нарушением динамики движения устьиц, при других
болезнях — с повышением водопроницаемости цитоплазматиче-
ских мембран под действием токсинов патогена.
Нарушение подачи воды к надземным частям растений может
быть следствием отмирания корней (например, при корневых гни-
лях древесных пород, полегании сеянцев), результатом
патологических изменений проводящей системы (при сосудистых болезнях
растений), отравления растений токсинами патогена (при фуза-
риозном увядании растений, при поражении опенком). Во всех
случаях нарушения водного баланса приводят к снижению или
полной потере тургора клеток, увяданию листьев, постепенному
или быстрому усыханию растений.
Среди биохимических нарушений в клетках пораженных
растений следует назвать также нарушения синтеза нуклеиновых
кислот и других жизненно важных соединений, нарушения баланса
ростовых веществ, витаминов и гормонов, изменения
кислотности клеточного сока, накопление вредных метаболитов или не
свойственных растению пигментов (явление метаплазии) и
другие патологические изменения.
4.3.2. Анатомо-морфологические нарушения
Анатомо-морфологические изменения растений, возникающие
на основе физиолого-биохимических нарушений, определяют
характер симптомов и тип болезни. Они проявляются в нарушениях
строения и структуры тканей, роста и формы пораженных
органов или растения в целом.
Нарушения строения и структуры тканей. Анатомические
изменения происходят в клетках и тканях пораженных растений при
самых различных заболеваниях. Наиболее распространенные сре-
104
ди них — гипертрофия, гиперплазия, гипоплазия, некроз,
мацерация, дегенерация, облитерация.
Гипертрофия — это увеличение размера и изменение
формы клеток под влиянием патогена (например, при поражении
ветвей и стволов можжевельника ржавчинными грибами рода
Gymnosporangium, при экзобазидиозах растений).
Гиперплазия — увеличение количества клеток вследствие
их усиленного беспорядочного деления под воздействием
патогена (например, при бактериальном раке корней и стволов
древесных растений, ржавчинном раке пихты).
Гипертрофия часто сочетается с гиперплазией, например, при
деформациях плодов, листьев и ветвей, вызываемых грибами
порядка тафриновых, при поражении корней крестоцветных килой.
Как гипертрофия, так и гиперплазия ведут к патологическому
разрастанию тканей, образованию на пораженных органах
растений вздутий, опухолей и других деформаций.
Гипоплазия — это уменьшение размера или количества
клеток, иногда качественное изменение содержимого клеток,
ведущее к уменьшению объема, патологическому недоразвитию
какой-либо ткани, отдельных органов или всего растения.
Гипоплазия может проявляться в образовании перетяжек на стеблях,
мелколистное™ растений, редукции листовых пластинок, атрофии
пораженных органов, карликовости растений.
Некроз — отмирание отдельных клеток или участков ткани
под действием токсинов патогена или различных абиотических
факторов (высоких или низких температур, химических веществ,
механических повреждений и т.п.). Некроз паренхимных тканей
листьев характерен для пятнистостей, некроз коры и камбия — для
некрозно-раковых болезней стволов и ветвей, некроз флоэмы —
для вирусных заболеваний. Некротические процессы в
пораженных тканях сопровождаются структурными и биохимическими
изменениями цитоплазмы клеток.
Мацерация — это разъединение клеток ткани вследствие
разрушения межклеточного вещества, что ведет к ее размягчению
и распаду (рис. 35). Мацерация, наблюдаемая при поражении
плодов, семян, луковиц, корней и других органов растений
различными видами гнилей, происходит в результате разрушения
грибами или бактериями срединных пластинок растительной ткани.
Дегенерация — перерождение тканей, связанное с
нарушениями обмена веществ и глубокими структурными
изменениями, которые могут быть вызваны различными биотическими и
абиотическими факторами. В результате разрушения протопластов
и клеточных оболочек отдельные клетки или целые участки
растительной ткани превращаются в скопления веществ, не
свойственных нормальным тканям. Например, при камедетечении в
стволах древесных растений происходит разжижение клеточных
105
Рис. 35. Анатомо-морфологические нарушения у больных растений:
разрушение срединных пластинок и мацерация тканей под действием бактерий
Erwinla carotovora (7) и Pseudomonas pruni (2). Видно массовое скопление
бактерий в пораженных тканях
оболочек и превращение растительной ткани в камедь —
клейкую тягучую жидкость янтарно-желтого или бурого цвета,
твердеющую на воздухе (рис. 36). При некоторых заболеваниях
встречаются целлюлозная,
слизистая, жировая и другие типы
дегенерации.
Облитерация — это
спадение, уменьшение объема
клеток в результате уничтожения
патогеном их содержимого,
ведущее к сморщиванию пораженных
органов растений (см. рис. 36).
Нарушения роста растений.
У больных растений могут
наблюдаться как стимуляция, так и
угнетение процессов роста.
Усиление вегетативного роста под
влиянием патогена чаще всего
бывает временным и
ограничивается отдельными органами или
частями пораженных растений.
В большинстве случаев у больных
1 2 растений происходит ослабление
Рис. 36. Анатомо-морфологические Ростовых процессов, в том чис-
нарушения у больных растений: ле и оби*ее угнетение роста. Оно
1 - дегенерация тканей ветвей (каме- проявляется В уменьшении сред-
детечение); 2 - облитерация (сморщи- неи ВЫСОТЫ, ТОЛЩИНЫ И ДЛИНЫ
вание пораженных тканей) Побегов, ПОВерХНОСТИ ЛИСТЬев И
106
хвои, значительном снижении продуктивности больных растений
по сравнению со здоровыми. При некоторых болезнях сильно
выраженное угнетение процессов роста приводит к общему
недоразвитию, карликовости растений. Это явление иногда наблюдается
при диффузном поражении растений ложной мучнистой росой,
при вирусных болезнях.
Нарушения формы растений. Изменения формы (деформации)
отдельных органов, а иногда всего растения, весьма
разнообразны и встречаются при различных болезнях. Таковы опухоли,
наросты, галлы, образующиеся на корнях или надземных частях
растений при некоторых бактериозах, фитогельминтозах и грибных
болезнях, а также курчавость листьев и ведьмины метлы,
вызываемые тафриновыми грибами и вирусами (рис. 37).
При вирусных заболеваниях часто наблюдаются изменения
формы цветков и листьев, утолщение и скручивание листовых
пластинок, различные новообразования. Характерными
особенностями определенных болезней древесных пород являются
ступенчато разрастающиеся наплывы и язвы (ступенчатый рак ство-
Рис. 37. Нарушения формы пораженных органов растений:
7 — развитие ступенчатой язвы на стволе; 2 — курчавость листьев; 3 —
скручивание листьев; 4 — ненормальное ветвление; 5 — образование опухолей; 6 —
искривление побегов
107
лов), искривление побегов (сосновый вертун) и другие
деформации, возникающие в результате нарушения физиологических
процессов и патологических изменений строения тканей.
4.4. ПОНЯТИЕ ОБ ЭПИФИТОТИЯХ
Инфекционные болезни растений существенно различаются
между собой не только по особенностям патогенеза и внешнему
проявлению, но и по характеру их развития в природе. Некоторые
болезни распространены повсеместно, но встречаются редко, и
уровень их встречаемости почти не меняется. Другие болезни
встречаются чаще, но также мало подвержены колебаниям; массовое
развитие этих болезней на больших площадях и сильное
поражение растений, представляющее угрозу посевам или насаждениям,
не наблюдается. К таким болезням относятся многие виды
стволовых гнилей древесных пород, выпревание сеянцев и ряд других
заболеваний.
Вместе с тем существуют болезни, распространение и
развитие которых в определенной местности или в пределах всего
ареала непостоянно, подвержено резким колебаниям. Таковы многие
виды ржавчины и мучнистой росы, некоторые виды сосудистых
болезней и корневых гнилей древесных пород, ряд вирусных и
других болезней.
Массовое развитие инфекционной болезни растений на
определенной территории в течение определенного периода времени
называется эпифитотией. Изучениемэпифитотийзанимается
специальный раздел фитопатологии — эпифитотиология. Это
учение о развитии популяций патогена внутри популяций хозяина и
о болезнях растений, возникающих в результате их
взаимодействия под влиянием окружающей среды или вмешательства
человека.
4.4Л. Роль патогена, растения-хозяина
и окружающей среды в развитии эпифитотий
Возникновение, развитие и затухание эпифитотий, как и
динамика болезней вообще, подчиняются определенным
закономерностям и зависят от взаимодействия трех компонентов,
участвующих в эпифитотическом процессе: популяции возбудителя
болезни, популяции растения-хозяина и окружающей среды. Если
это взаимодействие оказывается благоприятным для развития
болезни, она прогрессирует и возникает эпифитотия. Если же в
ходе эпифитотий создаются условия, препятствующие ее
дальнейшему развитию, происходит ее постепенное затухание, и
вспышка болезни прекращается. Каждый из названных компо-
108
нентов играет определенную роль и имеет одинаково важное
значение.
Роль патогена. Роль возбудителя болезни исключительно
велика. Для возникновения эпифитотии необходимо, чтобы
возбудитель обладал высокой агрессивностью и вирулентностью по
отношению к произрастающему в данной местности растению-
хозяину и чтобы запас инфекции был достаточно велик.
Решающей предпосылкой эпифитотии может оказаться появление
нового для данной местности высокоагрессивного патогена,
обладающего большой плодовитостью и способностью накапливаться в
природе.
Чем выше скорость размножения патогена, чем легче и
быстрее он распространяется, чем дольше способен сохраняться, не
теряя жизнеспособности, тем больше угроза возникновения
эпифитотии. И, наоборот, сокращение запаса инфекции,
уменьшение энергии размножения и скорости распространения
возбудителя, снижение его агрессивности — важнейшие предпосылки
затухания эпифитотии.
Роль растения-хозяина. Она также весьма существенна.
Массовое развитие болезни происходит только в том случае, если на
определенной площади сосредоточено множество восприимчивых
растений. Значение этого фактора возрастает, когда цикл
развития возбудителя протекает на двух разных питающих растениях,
что наблюдается у разнохозяйных ржавчинных грибов. В таком
случае для возникновения эпифитотии необходимо наличие в
достаточном количестве обоих растений-хозяев. Поэтому удаление
промежуточного хозяина может явиться решающим условием
прекращения дальнейшего развития ржавчины.
Очень важную роль в накоплении инфекции и развитии
эпифитотии играют сорные растения, на которых могут
размножаться и сохраняться возбудители болезней культурных растений и
лесных пород. Особенно велика роль растения-хозяина при
интродукции: если ввезенный вид растения оказывается
восприимчивым к местным возбудителям болезней, то со временем
развитие этих болезней на новом хозяине может приобрести характер
эпифитотии. То же самое происходит, если патоген, попадая в
новые районы, находит там новых восприимчивых хозяев.
Растение-хозяин может играть важную роль и в затухании
эпифитотии. Если эпифитотия носит сезонный характер, то ее
затуханию могут способствовать возрастные изменения растений или
их тканей, благодаря чему растения приобретают устойчивость,
как это наблюдается, например, при полегании сеянцев или
мучнистой росе дуба.
Фактором затухания эпифитотии может явиться и общее
noli ышение устойчивости насаждений под влиянием изменения
внешних условий или в результате естественного отбора, поскольку
109
в популяции выживают наиболее устойчивые или выносливые
индивидуумы. Наконец, эпифитотия может прекратиться при
введении в культуру устойчивых пород или форм растений.
Роль окружающей среды. Эта роль часто оказывается
решающей. Особенно большое значение имеют климатические условия
местности и погодные условия данного года, иногда нескольких
последних лет. Определяющим моментом при этом является, как
правило, не один какой-либо фактор (например, оптимальная
температура или влажность), а оптимальное сочетание многих
факторов, благоприятствующих массовому размножению
патогена, его накоплению и распространению, заражению растений.
Факторы внешней среды могут способствовать возникновению
эпифитотий и в том случае, если они оказывают неблагоприятное
влияние на растение-хозяина, вызывая его ослабление и снижая
тем самым его устойчивость к болезни. И, наоборот, любые
условия, которые препятствуют ускоренному размножению и
сохранению патогена в природе, его быстрому распространению и
заражению растений, равно как и все факторы, повышающие
жизнеспособность и устойчивость растения-хозяина, могут являться
факторами затухания эпифитотий.
Таким образом, эпифитотия представляет собой сложный
комплекс взаимосвязанных элементов, непрерывно изменяющихся
под влиянием многих факторов: генетических, экологических,
экономических и т.д. Эти элементы образуют во времени и
пространстве как бы непрерывную цепь, причем отдельные ее звенья и
характер связей между ними определяются спецификой
взаимоотношений в системе патоген—хозяин и особенностями
окружающей среды.
4.4.2. Динамика эпифитотий
Эпифитотия — это динамический процесс, в развитии
которого выделяют ряд последовательно сменяющих друг друга стадий:
1) подготовительную стадию, или предэпифитотию; 2) вспышку
болезни, или собственно эпифитотию; 3) стадию депрессии, или
затухания эпифитотий.
Во время первой стадии в природе происходят те изменения в
популяциях патогена и растения-хозяина, которые затем
приводят к вспышке заболевания: появление новых, более агрессивных
рас возбудителя или более активное, в связи с оптимальными
погодными условиями, размножение и накопление уже
имеющегося в данной местности патогена, сосредоточение на больших
площадях восприимчивых растений (например, при создании
чистых лесных культур) или снижение по тем или иным причинам
устойчивости насаждений; возникновение благоприятных
условий для заражения растений (например, в связи с хозяйственной
ПО
деятельностью человека, повышением рекреационных нагрузок
или воздействием абиотических факторов и т.д.).
Продолжительность этой стадии может быть различной, но чаще всего она
длится несколько лет.
Вторая стадия (вспышка) характеризуется одновременным
поражением большого количества растений, сильной степенью
поражения и гибелью значительной части больных растений,
высоким уровнем причиняемого болезнью ущерба. Момент
кульминации вспышки может быть приурочен к определенному возрасту
растений или к периоду с наиболее благоприятными для
развития болезни погодными условиями.
Во время третьей стадии (депрессии) наблюдается
постепенное снижение количества больных растений и степени их
поражения. Обычно уменьшается и зона распространения эпифитотии.
Продолжительность эпифитотии, как и отдельных ее стадий,
зависит от многих факторов и может сильно колебаться.
Эпифитотии некоторых болезней развиваются в течение одного сезона,
другие могут продолжаться много лет.
Процессы, происходящие во взаимодействующих популяциях
патогенов и растений-хозяев, описываются и объясняются
математическими моделями. Математическое моделирование
эпифитотии рассматривается в работах Ван дер Планка и других
авторов.
В качестве простейшего примера можно привести модель
изменения популяции патогена во времени для болезней,
распространяющихся анемохорным способом: D = А(сР)т/\ где D — скорость
нарастания болезни за генерацию (цикл); А — первичный запас
инфекции или начальная пораженность; с — число спор,
образующихся в очаге поражения; Р — вероятность заражения (0 — ни
одна из спор не вызывает заражения, 1 — все споры вызывают
заражение); Т— время; / — инкубационный период.
Модели изменения популяций во времени используются для
анализа параметров популяций, оценки развития и
прогнозирования эпифитотии.
4.4.3. Типы эпифитотии
В зависимости от особенностей развития и масштабов
распространения в природе различают следующие основные типы
эпифитотии.
Местные эпифитотии, или энфитотии. Характеризуются
ежегодным (в течение нескольких лет) сильным развитием болезни на
ограниченной территории, иногда в виде отдельных очагов.
Возбудители местных эпифитотии, как правило, постоянно
присутствуют в данной местности. Они способны долго сохраняться в
почве, на растительных остатках, семенах, сорняках и т.п. Ин-
111
фекционное начало таких патогенов обычно медленно
накапливается в природе и сравнительно медленно распространяется.
Однако, если запас инфекции достигает высокого уровня, то при
наличии восприимчивых растений и благоприятных внешних условиях
нередко возникают эпифитотии. Примером местных эпифитотий
могут служить энфитотии полегания всходов, ежегодно
наблюдаемые в питомниках многих районов страны.
Прогрессирующие эпифитотии. Эпифитотии этого типа
начинаются как местные, но со временем охватывают более обширные
территории. Они обычно вызываются наиболее агрессивными
патогенами, которые имеют высокую энергию размножения,
образуют в течение лета несколько генераций бесполого спороноше-
ния и способны быстро распространяться по воздуху или с
помощью насекомых (например, эпифитотии ржавчины, мучнистой
росы, некоторых сосудистых и вирусных болезней).
Причиной возникновения прогрессирующих эпифитотий
может оказаться переброска из одних районов в другие зараженного
посадочного материала или попадание патогена в новые для него
районы, где имеются значительные площади восприимчивых
растений-хозяев. Примером такой эпифитотии может служить эпи-
фитотия пузырчатой ржавчины веймутовой сосны, возникшая и
быстро охватившая огромные площади, занятые этой сосной в
США, после того как возбудитель болезни был завезен в Америку
из Европы.
Прогрессирующие эпифитотии часто развиваются в течение
многих лет. Так, сильные прогрессирующие эпифитотии
голландской болезни ильмовых, распространившиеся на больших
территориях в лесостепной, степной и полупустынной зонах нашей
страны, были отмечены в 1935—1940 и 1955—1959 гг. В молодых
культурах сосны, создаваемых на обширных площадях
концентрированных вырубок в северных и северо-западных районах России,
наблюдаются прогрессирующие эпифитотии снежного шютте и
ржавчины побегов сосны.
Повсеместные эпифитотии, или панфитотии, характеризуются
массовым развитием болезни на территории целой страны,
иногда нескольких стран или континентов. Панфитотии — явление
довольно редкое, но они могут принимать размеры
национального бедствия, как это случилось во время панфитотии фитофторо-
за картофеля в середине XIX в. В начале XX в. характер панфитотии
носило массовое распространение мучнистой росы дуба и
мучнистой росы крыжовника, завезенных из Америки в Европу.
Повсеместное распространение корневой губки во многих странах
Европы и Северной Америки в течение последних десятилетий также
достигло уровня панфитотии.
Кроме того, различают медленно развивающиеся, или тар-
див н ы е, и быстро развивающиеся, или эксплозивные, эпи-
112
фитотии. Первые чаще всего наблюдаются при поражении
многолетних растений (например, древесных) заболеваниями типа
голландской болезни ильмовых или корневой губки на хвойных. Они
характеризуются плавным ходом нарастания вспышки и
постепенным ее затуханием. Вторые вызываются в основном
патогенами с высокой скоростью размножения и характеризуются резким
нарастанием вспышки и быстрым ее затуханием. Ход эпифитотии
этого типа часто подчинен сезонным изменениям и в
значительной степени определяется факторами внешней среды. Примерами
могут служить эпифитотии парши яблони, полегания сеянцев,
мучнистой росы, ржавчины, шютте и др.
Знание особенностей различных типов эпифитотии позволяет
предвидеть их возникновение, ход дальнейшего развития и
использовать эти данные для составления более точных прогнозов и
планирования лесозащитных мероприятий.
Глава 5
ИММУНИТЕТ РАСТЕНИЙ
К ИНФЕКЦИОННЫМ БОЛЕЗНЯМ
5.1. ОСНОВНЫЕ ПОНЯТИЯ И КАТЕГОРИИ
ИММУНИТЕТА
Иммунитетом называется невосприимчивость (устойчивость)
организма к инфекционной болезни при контакте с ее
возбудителем и наличии условий, способствующих заражению. Например,
хвойные породы никогда не поражаются мучнистой росой, а
лиственные — болезнями типа шютте. Ель абсолютно
невосприимчива к ржавчине побегов, а сосна — к ржавчине шишек. Такой
абсолютный иммунитет обусловлен биологическим
несоответствием этих растений свойствам и требованиям
возбудителей данных болезней. Чаще всего он объясняется неспособностью
патогена проникнуть в растение и развиваться в нем даже при
самых благоприятных внешних условиях.
Наряду с абсолютной невосприимчивостью к одним болезням
у растений может наблюдаться относительная
устойчивость (или, соответственно, относительная восприимчивость)
к другим заболеваниям. Она зависит от индивидуальных свойств
растения, его анатомо-морфологических или физиолого-биохи-
мических особенностей, уменьшающих возможность заражения или
ограничивающих распространение патогена в тканях
растения-хозяина. Степень устойчивости растений к болезням может быть
различной: от весьма высокой (близкой к полной
невосприимчивости) до очень низкой.
У растений различают неспецифический и специфический
иммунитет. Неспецифическим (или видовым)
иммунитетом называется устойчивость определенного вида растений к тем
возбудителям, которые вообще неспособны поражать этот вид.
Неспецифический иммунитет обеспечивает недоступность растений
для основной массы сапротрофной и патогенной микрофлоры,
населяющей среду обитания этих растений. Специфическим
(или сортовым) иммунитетом называется устойчивость
отдельных сортов или форм какого-либо вида растений к возбудителям,
способным поражать этот вид.
114
Различают также иммунитет врожденный (естественный) и
приобретенный (искусственный). Врожденным иммунитетом
называется наследственная невосприимчивость к болезни,
сформировавшаяся в результате длительной совместной эволюции
(филогенеза) растения-хозяина и патогена или направленной
селекции. Приобретенным иммунитетом называют устойчивость
к болезни, приобретаемую растением в процессе его
индивидуального развития (онтогенеза) под влиянием определенных
внешних факторов или в результате перенесения данной болезни.
Приобретенный иммунитет не передается по наследству.
Устойчивость растений (обычно какого-либо сорта) лишь к
определенным физиологическим расам патогена называют
вертикальной, ату или иную степень устойчивости ко всем расам
данного патогена — горизонтальной. Устойчивость какого-
либо вида или сорта растений одновременно к нескольким
болезням называют групповой, или комплексной, устойчивостью.
Врожденный иммунитет растений бывает пассивным и
активным. Пассивный иммунитет, или аксения,
—этоустойчивость к болезни, которая обеспечивается свойствами,
проявляющимися у растений независимо от угрозы заражения. Таким образом,
свойства, обусловливающие пассивный иммунитет, не являются
защитными реакциями растения на нападение патогена.
Активный иммунитет — это устойчивость к болезни,
которая обеспечивается свойствами растений, проявляющимися у них
только в случае нападения патогена, т.е. в виде защитных реакций
растения-хозяина на внедрение возбудителя.
5.2. ПАССИВНЫЙ ВРОЖДЕННЫЙ ИММУНИТЕТ
Пассивный иммунитет может быть связан с особенностями
формы и анатомического строения растений или с их
функционально-физиологическими и биохимическими особенностями.
Свойства растений, обусловливающие пассивный иммунитет, —
это, как правило, проявления горизонтальной устойчивости. Они
весьма многочисленны, разнообразны и могут быть объединены в
две основные группы: анатомо-морфологические и физиолого-
биохимические.
5.2.1. Анатомо-морфологические факторы пассивного
иммунитета
Защитные приспособления растений, выражающиеся в
особенностях их формы или строения, широко распространены в
природе и играют важную роль в естественном иммунитете растений.
Эти особенности растений в основном препятствуют заражению,
115
не давая возможности возбудителю болезни прорасти и
проникнуть внутрь растения. Иногда они повышают устойчивость
растений, препятствуя распространению паразита в тканях хозяина,
если заражение уже осуществилось. Иммунитет, основанный на
анатомических или морфологических особенностях растений,
называют также структурным.
Анатомо-морфологическими факторами пассивного
иммунитета могут служить раскидистая форма кроны, малое количество
и особое строение устьиц, закрытый тип цветка, наличие на
поверхности поражаемых органов густого опушения или воскового
налета, толстая кутикула или пробковый слой, мощное развитие
склеренхимной ткани и другие особенности строения.
При рыхлой, раскидистой форме куста или кроны в нее
беспрепятственно проникают прямые солнечные лучи, она лучше
проветривается, в ней почти не задерживается влага, вследствие
чего создается неблагоприятный микроклимат для сохранения
инфекционного начала возбудителей болезней, прорастания спор
фитопатогенных грибов. Поэтому при прочих равных условиях
деревья и кустарники с такой кроной меньше поражаются
болезнями, чем деревья с густой, компактной кроной.
Количество и размеры устьиц и чечевичек, форма устьичной
щели могут быть факторами устойчивости растений против
патогенов, проникающих в растение через эти естественные ходы. Чем
меньшее число устьиц и чечевичек приходится на единицу
поверхности поражаемых органов, чем меньше устьичные щели, тем
меньше у патогена шансов на заражение, тем выше устойчивость
растения.
Строение цветка и характер цветения могут определять
устойчивость или восприимчивость растений к заболеваниям,
возбудители которых внедряются через рыльца, нектарники и другие
части цветков. Виды и сорта растений, характеризующиеся
закрытым типом цветения, обычно поражаются такими болезнями
меньше, чем виды или сорта с открытым цветением.
Неблагоприятные условия для прорастания спор грибов
создаются при наличии на хвое, листьях, плодах воскового налета
или обильного опушения, так как это делает их несмачиваемыми.
Например, сизые («голубые») формы некоторых хвойных пород,
хвоя которых покрыта восковым налетом, как правило, более
устойчивы к шютте и ржавчине по сравнению с обычными
формами.
Устойчивость некоторых видов и сортов растений к заражению
грибами, которые внедряются непосредственно через кутикулу,
часто обусловливается большей, чем у восприимчивых видов,
толщиной кутикулярного слоя. Так, у сильно поражаемого
ржавчиной вида барбариса Berberis dictyophylla общая толщина кутикулы и
наружной стенки эпидермиса составляет 0,82 мкм, а у невоспри-
116
имчивого Berberis thunbergii — 1,57 мкм, т. е. она почти вдвое
больше. С этим фактором связана и возрастная устойчивость листьев и
побегов дуба к мучнистой росе. Молодые растущие листья,
имеющие тонкую, нежную кутикулу, сильно поражаются мучнистой
росой. По мере увеличения толщины и прочности кутикулярного
слоя повышается и устойчивость листьев. Листья же,
закончившие рост, практически невосприимчивы к болезни. Такую же
защитную роль играет одревеснение побегов. Кутикула может
служить для патогенов не только механическим, но и химическим
барьером, так как содержащиеся в ней воск и кутан обладают
фунгицидными свойствами.
Важным фактором устойчивости могут быть анатомические
особенности и физико-механические свойства внутренних тканей
растений: более плотная паренхимная ткань, мощное развитие
склеренхимы и расположение ее в периферической части стебля
или вокруг сосудисто-проводящих пучков препятствуют
распространению и нормальному развитию патогена внутри растения.
Так, устойчивые (или выносливые) к корневой губке
экземпляры сосны в очагах болезни обычно характеризуются большей
толщиной годичных слоев и стенок трахеид, более высоким
процентом поздней древесины и другими особенностями. Исследования
показали, что утолщение клеточных стенок в древесине
устойчивых сосен обусловлено повышенным содержанием в них гемицел-
люлозы и лигнина.
5.2.2. Физиолого-биохимические факторы
пассивного иммунитета
К этой группе факторов относятся специфические
особенности обмена веществ растений, высокое содержание или
определенный качественный состав углеводов, белков и продуктов их
распада, наличие в клетках растений веществ, выполняющих
защитную роль, физико-химические особенности тканей,
некоторые функциональные особенности растений.
Устойчивость растения к болезни может быть связана с
отсутствием в его тканях необходимых для возбудителя элементов
питания или физиологически активных веществ, несоответствием
обмена веществ растения-хозяина обмену веществ патогена,
угнетением патогена токсичными продуктами метаболизма растения,
другими неблагоприятными для патогена факторами.
Одним из важнейших факторов устойчивости к
инфекционным болезням является неблагоприятный для патогенов характер
углеводного и белкового обмена растений. Устойчивость,
связанная с этим фактором, в значительной мере зависит от типа
питания патогена, степени его паразитической активности и специализа-
117
ции. Так, растения, устойчивые к некротрофам (факультативным
паразитам и факультативным сапротрофам), обычно
характеризуются более высоким общим содержанием углеводов по
сравнению с восприимчивыми. Это установлено, например, для вязов,
устойчивых к голландской болезни. Качественный состав углеводов
в этом случае не имеет большого значения, поскольку некротро-
фы обладают богатым ассортиментом гидролитических
ферментов, в том числе карбогидраз. На устойчивость же растений к
узкоспециализированным биотрофам, имеющим ограниченный набор
ферментов, в большей степени влияет качественный состав
углеводов в тканях растения-хозяина. В то же время к облигатным
паразитам особенно восприимчивы наиболее жизнеспособные и
хорошо развитые растения, характеризующиеся активным
течением процессов фотосинтеза и, следовательно, высоким общим
содержанием углеводов в их тканях.
Фактором устойчивости растений к факультативным
паразитам может служить высокое содержание в тканях белков и
промежуточных продуктов белкового обмена. Устойчивость же к
облигатным паразитам определяется в основном качественными
особенностями белкового комплекса растений, несходством строения
белков растения-хозяина и патогена, отсутствием в тканях
растения белковых соединений, доступных для питания патогена.
Наличие в тканях растений определенных аминокислот,
токсичных для паразита (или, наоборот, отсутствие аминокислот,
необходимых для его жизнедеятельности), высокое содержание
токсичных продуктов распада белков — аммиака и мочевины —
также могут обеспечивать устойчивость растений к
инфекционным болезням.
Среди химических соединений, обусловливающих устойчивость
растений, большое значение имеют фенолы, алкалоиды,
эфирные масла, пигменты, смолы, терпены, дубильные и другие
вещества. Некоторые из них сами по себе токсичны для паразитов,
и наличие их в растении служит как бы «химическим барьером»,
препятствующим заражению.
Защитная роль других веществ более сложна и связана с их
активным участием в биохимических реакциях клетки, ведущих
к образованию токсичных соединений. В тех случаях, когда
подобные реакции происходят под влиянием возбудителя, их
следует рассматривать уже как защитные реакции активного
иммунитета.
Одним из факторов устойчивости сосны к корневой губке
является содержание в древесине веществ фенольной природы (в
частности, резорцина), оказывающих на гриб сильное ингибиру-
ющее и токсическое действие. Доказана также связь устойчивости
луба у ели к распространению корневой губки с содержанием в
нем фенольных соединений.
118
Антибиотические вещества, свойственные определенным
видам высших растений и всегда содержащиеся в их тканях,
называют фитонцидами. Фитонциды обусловливают
неспецифический иммунитет растений к сапротрофным микроорганизмам,
сдерживая процесс их приспособления к обмену веществ
растений-хозяев. В некоторых случаях фитонциды могут служить одним
из факторов устойчивости растений к факультативным
паразитам, однако их роль в комплексе защитных средств растений
невелика. В иммунитете растений к облигатным паразитам и
факультативным сапротрофам фитонциды не имеют значения, хотя и
могут в какой-то мере препятствовать прорастанию спор
патогена, его внедрению в растение.
Определенную роль в пассивном иммунитете растений к фито-
патогенным организмам играют такие физико-химические
показатели растительных клеток, как проницаемость цитоплазмы,
осмотическое давление и кислотность клеточного сока.
Устойчивость к внедрению патогенов может быть обусловлена
некоторыми функциональными особенностями растений,
например суточным ритмом движений устьиц, характером прорастания
семян, способностью к интенсивному образованию каллюса,
быстрому заживлению ран и др.
53- АКТИВНЫЙ ВРОЖДЕННЫЙ ИММУНИТЕТ
Проявления активного иммунитета носят характер
специфических защитных реакций растения, возникающих в ответ на
заражение. Они могут быть направлены на подавление самого
возбудителя болезни, разрушение и обезвреживание его
выделений или на восстановление нарушенных болезнью
физиологических функций и обмена веществ пораженного растения.
Различают антиинфекционные и антитоксические защитные реакции
растений (однако между ними не всегда можно провести четкую
грань).
Антиинфекционные защитные реакции направлены
непосредственно против патогена. Они препятствуют распространению
паразита в тканях растения, подавляют его развитие, приводят к его
локализации и гибели. Примерами таких реакций могут служить
реакция сверхчувствительности, явление фагоцитоза, синтез фи-
тоалексинов.
Реакция сверхчувствительности, возникающая в
ответ на внедрение патогена, наиболее ярко проявляется при
заражении растений облигатными паразитами, которые
характеризуются биотрофным типом питания, например ржавчинными и
мучнисторосяными грибами, вирусами, микоплазмами. Суть этой
реакции заключается в быстром отмирании клеток устойчивого
119
растения в местах внедрения возбудителя. Оказавшись в зоне
мертвой ткани, паразит-биотроф локализуется в ней, лишается
питания и погибает.
Внешне реакция сверхчувствительности выражается в
появлении на листьях небольших хлоротичных, а затем некротических
пятен (так называемых защитных некрозов). У высокоустойчивых
растений образуются мелкие, точечные некрозы, которые
практически не влияют на работу ассимиляционного аппарата.
Реакция сверхчувствительности — наиболее характерное проявление
вертикальной устойчивости.
Фагоцитозом называют внутриклеточное переваривание
патогенных микроорганизмов. В наиболее характерной форме он
проявляется при развитии эндотрофной микоризы в корнях растений.
Когда гифы гриба-микоризообразователя проникают в клетки
первичной коры, в цитоплазме этих клеток возникают ответные
защитные реакции, направленные на ослабление и уничтожение
мицелия паразита или на предотвращение дальнейшего
распространения гиф в живых тканях корней. В результате этих реакций
гифы микоризного гриба сплетаются в клубки (везикулы),
которые затем постепенно перевариваются клеткой, или образуют
древовидные разветвления — ар бус кул ы, не способные
активно расти и переходить в соседние клетки корня. Благодаря
фагоцитозу развитие и распространение гриба в корнях растения-
хозяина поддерживается на уровне симбиоза, обеспечивающего
определенную выгоду для обоих партнеров.
Фитоалексины — особые липидоподобные защитные
вещества, которые, как и фитонциды, обладают антибиотическим
действием. Они задерживают развитие возбудителей болезни или
подавляют синтез патогенами ферментов и токсинов.
Однако в отличие от фитонцидов, фитоалексины в здоровых
тканях отсутствуют и образуются только в случае заражения
растений микроорганизмами. В очень малых количествах
(следы) они обнаруживаются также при механическом повреждении
тканей.
Фитоалексины вырабатываются зараженным или
поврежденным растением в результате взаимодействия метаболитов
патогена и метаболитов растения-хозяина, причем образование фи-
тоалексинов может быть вызвано не только паразитирующими
на данном растении видами возбудителей, но и не патогенными
для него микроорганизмами. Растениям определенного рода или
вида свойственны определенные фитоалексины. Характерно, что
фитоалексины продуцируются как устойчивыми, так и
восприимчивыми к патогену формами и сортами растений. Степень
устойчивости (или восприимчивости) определяется в этом случае
количеством фитоалексинов и скоростью их накопления в
тканях зараженного растения. Устойчивые растения могут вы-
120
рабатывать в 2 — 3 раза больше фитоалексинов, чем
восприимчивые.
Антитоксические защитные реакции растений направлены
главным образом на обезвреживание ферментов, токсинов и других
вредных продуктов жизнедеятельности патогенов. К реакциям этого
типа можно отнести перестройку и активизацию ферментных
систем растения-хозяина, образование механических барьеров и др.
Важнейшим фактором активного иммунитета является
перестройка ферментных систем растения, прежде всего
окислительной системы. При заражении устойчивых растений деятельность
окислительной системы у них резко активизируется и
приобретает характер защитной реакции, направленной на уменьшение
причиняемого патогеном вреда.
Окислительные ферменты устойчивого растения инактивиру-
ют гидролитические ферменты возбудителя или подавляют
синтез этих ферментов. Кроме того, они нейтрализуют токсины
возбудителя, разрушая их или окисляя до безвредных для растения
соединений. Следует отметить, что у растений устойчивых (в
отличие от восприимчивых) преобладают окислительные
ферменты, не чувствительные к токсинам патогена и даже
активизирующиеся под их действием.
Защитная роль окислительных ферментов растения-хозяина
заключается и в том, что они активно участвуют в процессах
синтеза веществ, необходимых для восстановления разрушенных
паразитом клеточных структур и тканей и нормализации
физиологических процессов.
Активное сопротивление растения может проявляться в
утолщении оболочек клеток, окружающих зону заражения, образовании
вокруг зараженного участка пробкового слоя или слоя вторичной
перидермы. Такие механические барьеры задерживают
распространение патогена и его токсинов, ведут к его локализации или
вытеснению из растения вместе с участком пораженной ткани
(как это наблюдается при дырчатой пятнистости листьев). Еще
один пример антитоксической защитной реакции этого типа —
образование в корнях деревьев, зараженных опенком, вторичной
перидермы, препятствующей распространению токсинов гриба в
вышележащие части дерева. Процессы образования защитных
тканевых барьеров также тесно связаны с активизацией ферментных
систем растения-хозяина.
Резюмируя сказанное о врожденном иммунитете растений,
необходимо подчеркнуть, что устойчивость растений к
инфекционным болезням в большинстве случаев обусловливается не
отдельными особенностями или свойствами организма, а
комплексом сложнейших защитных механизмов, тесно взаимосвязанных
между собой и действующих на определенных этапах
патологического процесса.
121
5.4. ГЕНЕТИЧЕСКИЕ ОСНОВЫ ИММУНИТЕТА
РАСТЕНИЙ
Наследуемые защитные свойства растений, являющиеся
факторами врожденного иммунитета, контролируются генами
устойчивости, а свойства патогенов, обусловливающие их способность
вызывать болезни, — генами патогенности (вирулентности).
Наследование устойчивости растений, как и других свойств,
контролируемых генами, подчиняется законам Менделя.
Одной из теорий, объясняющих генетические механизмы
иммунитета, является теория «ген на ген», разработанная
американским фитопатологом X. Флором. Согласно этой теории каждому
гену растения-хозяина, определяющему его устойчивость,
соответствует ген паразита, определяющий его вирулентность. Такие
взаимосвязанные гены хозяина и паразита называют
комплементарными. Совершенно очевидно, что гены устойчивости имеют
большую селекционную ценность.
Механизм эволюции и саморегуляции взаимоотношений в
системе паразит—хозяин заключается в постоянном появлении (при
мутациях и естественном отборе) новых генов устойчивости в
популяциях растения-хозяина и новых генов вирулентности в
популяциях паразита. Результатом такой изменчивости паразита и
хозяина является генетическая неоднородность популяций как
возбудителя болезни, так и поражаемых растений. Столь сложное
генетическое взаимодействие в системе паразит—хозяин,
обеспечивающее выживание обоих партнеров и их равновесие в
природе, могло сложиться лишь в результате их длительной
совместной эволюции.
Теория сопряженной эволюции паразита и хозяина на их
совместной родине была выдвинута и блестяще аргументирована
основоположником учения об иммунитете растений к
инфекционным болезням Н. И. Вавиловым. Сущность этой теории состоит
в том, что географические центры формирования видов растений
в большинстве случаев являются одновременно и центрами
формирования рас их паразитов. Исходя из того, что именно в этих
генетических центрах происходит интенсивный процесс
естественного отбора устойчивых форм растений, Н.И.Вавилов обосновал
положение о том, что устойчивые виды и формы растений,
которые могут служить исходным материалом для селекции на
устойчивость, следует искать на первичной родине хозяина и паразита.
Искусственный отбор устойчивых форм растений является
одним из основных методов селекционной работы. Наилучшие
результаты дает отбор на инфекционном фоне, т.е. при наличии
контакта популяции растений с возбудителями болезней. Так,
отдельные устойчивые экземпляры хвойных пород нередко
встречаются в очагах корневой губки, а устойчивые экземпляры осины —
122
в очагах осинового трутовика. Семена, собранные с таких
деревьев, могут служить исходным материалом для дальнейшего отбора
на устойчивость к гнилевым болезням.
Второй не менее важный метод селекции на устойчивость —
гибридизация. Этот метод дает возможность получать новые
формы растений, сочетающие высокие хозяйственные качества,
унаследованные от одного родителя, с устойчивостью к болезням,
унаследованной от другого. Таким путем, например, были
получены гибридные формы ильмовых пород, устойчивые к
голландской болезни, и формы тополей, устойчивые к стволовой гнили
и некрозам стволов и ветвей.
5.5. ВЫНОСЛИВОСТЬ РАСТЕНИЙ
Под выносливостью (толерантностью) понимают свойство
растений, не обладающих устойчивостью к болезни и не способных
противостоять заражению, оказывать сопротивление
дальнейшему развитию инфекционного процесса, сохраняя при этом
жизнеспособность и удовлетворительную продуктивность.
Наиболее часто выносливость проявляется в том, что
симптомы болезни (т. е. видимая реакция растения на воздействие
патогена) бывают плохо выражены. Таким образом, выносливые
растения, как правило, отличаются слабой поражаемостью. Высшая
степень выносливости характеризуется отсутствием внешних
признаков болезни у зараженного растения.
Понятие выносливость употребляют и в тех случаях, когда
растения характеризуются сильной поражаемостью, однако
удовлетворительно растут и развиваются, как это иногда наблюдается
при поражении пятнистостями, некоторыми видами ржавчины,
гнилевых и других болезней. Такая выносливость может объясняться
несинхронностью развития растения-хозяина и патогена,
способностью растения развивать новые побеги, листья или корни
взамен пораженных (компенсаторный рост), вегетативной мощью
растения и т. п.
Например, поражение растений многими видами пятнистостей
листьев происходит во второй половине лета, когда растения в
основном уже закончили рост, сформировали плоды и семена.
Растения, выносливые к болезням корней, обычно обладают
способностью к образованию придаточных корней. В очагах корневой
губки часто встречаются деревья, пораженные грибом, но при
этом отличающиеся нормальным приростом и отсутствием явных
признаков ослабления; их выносливость обусловлена мощным
развитием корневой системы.
В природных популяциях растений выносливость является
одним из факторов естественного отбора: растения, сильно стра-
123
дающие от болезни, не выдерживают конкуренции с более
выносливыми экземплярами, вследствие чего повышается
выносливость популяций в целом. Однако выносливые растения могут
стать накопителями патогенов и источниками инфекции для других
растений, особенно в том случае, если патоген обладает
широкой специализацией. Кроме того, в больших популяциях
патогенов возрастает вероятность появления новых рас возбудителей
болезней.
5.6. ПРИОБРЕТЕННЫЙ ИММУНИТЕТ И ПУТИ
ПОВЫШЕНИЯ УСТОЙЧИВОСТИ РАСТЕНИЙ
К БОЛЕЗНЯМ
У растений различают инфекционный и неинфекционный
приобретенный иммунитет. Инфекционный приобретенный
иммунитет может появиться в результате перенесенной болезни, если
она закончилась выздоровлением растения. Проявления такого
иммунитета у растений встречаются крайне редко, и его
практическое значение невелико.
Гораздо большее значение имеет неинфекционный
приобретенный иммунитет, возникающий под влиянием каких-либо
внешних факторов, под действием определенных химических
веществ. Растения могут также приобрести устойчивость к
инфекционным болезням в результате обработки их специальными
биологическими препаратами (вакцинами).
Приобретение растениями искусственного иммунитета,
повышение их устойчивости к болезням с помощью различных
приемов называют иммунизацией.
5.6.1. Химическая иммунизация растений
Методы химической иммунизации получили наиболее
широкое распространение и признание. В качестве химических имму-
низаторов используют основные удобрения (азотные, калийные,
фосфорные), микроэлементы, антиметаболиты.
Применение удобрений. Использование азота, калия и фосфора
основано на том, что они оказывают существенное влияние на
анатомическое строение, обмен веществ и физиологические
функции растений. Изменяя их в направлении, неблагоприятном для
фитопатогенных организмов, основные удобрения могут
повышать устойчивость или выносливость растений к болезням.
Особенно важное значение для повышения устойчивости
растений имеют калийно-фосфорные удобрения. Калий и фосфор
активизируют деятельность ферментов, снижают скорость гидро-
124
литических процессов, увеличивают вязкость цитоплазмы, тургор
клеток, механическую прочность тканей. В результате повышается
общая сопротивляемость растений воздействию неблагоприятных
факторов среды, их устойчивость или выносливость к
инфекционным болезням. Например, внесение в питомниках повышенных
доз калия и фосфора уменьшает поражаемость сеянцев
полеганием, выпреванием, мучнистой росой, ржавчиной.
Азотные удобрения, в сочетании с калийно-фосфорными,
усиливают рост, повышают продуктивность, устойчивость и
выносливость растений. Однако относительный избыток азота или
внесение его в поздние сроки могут понизить устойчивость
растений к болезням. Так, например, одностороннее азотное
удобрение заметно усиливает поражение сеянцев и молодых культур дуба
мучнистой росой. Это объясняется тем, что под влиянием азота
удлиняется период образования и роста новых побегов и листьев.
Они дольше остаются восприимчивыми к возбудителю болезни,
так как он поражает именно молодые, растущие органы растений.
Избыток азота в почве снижает также устойчивость сосны к
корневой губке.
Применение микроэлементов. Медь, цинк, железо, марганец и
другие микроэлементы играют очень важную роль в
биохимических реакциях клеток, синтезе структурных элементов
растительных тканей и других процессах. Сдвигая их в неблагоприятном для
патогена направлении, микроэлементы могут повысить
болезнеустойчивость растений.
Микроэлементы способствуют утолщению кутикулы и
клеточных стенок, повышению прочности тканей, т.е.
формированию механических защитных барьеров, препятствующих
заражению растений и распространению в них возбудителей
болезней.
Многие микроэлементы входят в состав окислительных и
других ферментов, непосредственно участвующих в защитных
реакциях растений. Кроме того, микроэлементы могут инактивировать
ферменты и токсины патогенов, вызывать у них регрессивные
изменения: угнетение роста, лизис и дегенерацию клеток.
Например, цинк, кобальт, медь, бор, молибден угнетают рост мицелия
корневой губки, других фитопатогенных грибов.
Велика роль микроэлементов и в профилактике
неинфекционных болезней, связанных с недостатком этих веществ в
почве.
Существуют различные способы применения основных
удобрений и микроэлементов: внесение в почву, предпосевная
обработка семян и посадочного материала, внекорневая подкормка
вегетирующих растений. Наилучший результат может быть
достигнут при сбалансированном сочетании микроэлементов,
минеральных и органических удобрений. Эффективность иммунизации за-
125
висит также от применяемых концентраций и доз макро- и
микроэлементов, способов и сроков их применения (с учетом
биологических особенностей патогенов и обрабатываемых растений),
почвенных и других условий.
Применение антиметаболитов. Антиметаболитами называют
органические вещества, очень близкие по химическому составу,
структуре и свойствам к соединениям, участвующим в нормальном
обмене веществ растений.
Антиметаболиты настолько сходны с нормальными
метаболитами, что могут замещать их в обменных процессах. Будучи
безвредными для самого растения, антиметаболиты вызывают в его
тканях биохимические изменения, неблагоприятные для
патогена: увеличение количества белков, дубильных и других защитных
веществ, усиление активности ферментов и т.д. В результате
растение приобретает устойчивость к инфекционной болезни.
Например, гидрохинон и паранитрофенол повышают устойчивость
ильмовых к голландской болезни. К антиметаболитам относят также
аргинин, аденозин, пиритиамин. Предполагают, что свойствами
антиметаболитов обладают и некоторые фунгициды, например
коллоидная сера и др.
Антиметаболиты применяют разными способами:
замачиванием семян в их растворах, внесением в почву, путем инъекций и
др. Иммунизирующее действие антиметаболитов может
проявляться не только в год обработки растений, но и в последующие 1 —
3 года.
5.6.2. Биологическая иммунизация (вакцинация)
растений
В качестве биологических иммунизаторов (вакцин)
используют ослабленные или убитые культуры фитопатогенных
микроорганизмов, вытяжки из них, продукты их жизнедеятельности
(токсины), антиинфекционные сыворотки. Механизм
иммунизирующего действия вакцин изучен еще недостаточно. Под влиянием
вакцины в тканях растений изменяется обмен веществ,
образуются соединения, подавляющие развитие патогена или
нейтрализующие его токсины, возникают защитные реакции типа
фагоцитоза, подобные тем, какие наблюдаются при врожденном
активном иммунитете растений.
Действие вакцин специфично: растения, обработанные
вакциной, приготовленной из определенного патогена,
приобретают устойчивость только к этому патогену. Однако известны
случаи и перекрестной защиты, когда введение в ткани растения
одного патогена повышает его устойчивость и к другим
возбудителям.
126
Основные способы применения вакцин: обработка семян,
опрыскивание вегетирующих растений, введение внутрь растений
путем инъекций.
Устойчивость и выносливость насаждений можно повысить
направленным изменением условий их роста, применением
комплекса эффективных лесохозяйственных мероприятий. Важное
значение имеют использование высококачественных семян и
здорового посадочного материала, правильный уход за
насаждениями на всех фазах развития, профилактика повреждений,
регулирование рекреационных нагрузок, строгое соблюдение
санитарных правил и т. п.
Глава 6
МЕТОДЫ БОРЬБЫ С БОЛЕЗНЯМИ ЛЕСА
6.1. ОБЩИЕ СВЕДЕНИЯ
Все мероприятия по защите леса могут быть
классифицированы по разным признакам. Так, ранее было принято делить их на
две группы — предупредительные (или профилактические) и
истребительные; иногда дополнительно выделяли и
организационные. Первые объединяли методы, направленные на создание
условий, препятствующих размножению и распространению вредных
организмов, ко вторым относились методы, направленные на их
непосредственное уничтожение. В настоящее время методы
защиты леса чаще всего подразделяют на следующие группы: 1 —
методы надзора и прогноза, к ним обычно относят и методы
лесопатологического обследования; 2 — карантинные
мероприятия; 3 — лесохозяйственные методы защиты леса; 4 —
биологические методы; 5 — генетические методы борьбы с насекомыми;
6 — химические методы; 7 — использование аттрактантов; 8 —
физико-механические методы; 9 — интегрированный метод
защиты леса. Многие из них имеют многоцелевой характер и
выполняют одновременно несколько функций профилактического и
истребительного плана.
Эффективность защиты леса возможна лишь при
использовании против вредных организмов не какого-либо одного, а
обязательно комплекса методов — системы мероприятий,
предусматривающей одновременное создание условий, неблагоприятных для
развития очагов вредителей и болезней, в сочетании с методами
их непосредственного уничтожения или подавления.
Под системой лесозащитных мероприятий
понимают сочетание методов, приемов и средств, используемых для
защиты от вредителей и болезней лесов определенных природных
территориально-производственных комплексов, эколого-производ-
ственных лесных объектов и объектов озеленения.
Под природным
территориально-производственным комплексом подразумеваются леса или зеленые
128
насаждения на определенной территории, где руководство всеми
хозяйственными мероприятиями осуществляется администрацией
предприятия, объединения, кооператива, арендодержателями или
органами власти республики, края, района, населенного пункта.
Эколого-производственные объекты — этоотдель-
ные участки леса или лесной площади, отличающиеся по
экологической обстановке, целевому назначению и обитающему там
комплексу популяций живых организмов. Это семенные хозяйства,
плантации, склады-хранилища семян и плодов древесных пород,
питомники, лесные культуры, лесные площади, подлежащие закульти-
вированию, лесные насаждения, вырубки, склады древесины и
сооружения из древесины, объекты озеленения в городах и
населенных пунктах (лесопарки, парки, уличные и внутридворовые
посадки, бульвары и скверы), дендрологические и ботанические сады,
полезащитные полосы и прочие объекты, защита которых требует
специфических методов, средств и технологических приемов.
Под очагами вредных организмов понимают
участки леса (лесной площади) или объекта озеленения,
характеризующиеся повышенной концентрацией патогенных организмов,
наносящих ощутимый экологический и экономический ущерб. Обычно
образование очага сочетается с массовым поражением леса. Часто
площадь очагов может достигать многих сотен и даже тысяч
гектаров, а иногда очаги распространяются на леса целых регионов.
Системы лесозащитных мероприятий разрабатывают отдельно
для каждой физико-географической зоны. В их пределах эти
системы создают для отдельных лесорастительных и лесоэкономичес-
ких районов.
Система лесозащитных мероприятий включает:
организованную службу надзора за появлением и массовым распространением
вредителей и болезней; мероприятия по повышению
биологической устойчивости насаждений; активные меры борьбы с
вредителями и болезнями, включающие все способы использования
средств защиты растений; экономическую и экологическую оценку
результатов мероприятий до и после их применения.
На основании данных службы надзора составляют прогноз
численности вредителей и болезней, позволяющий планировать
активные меры борьбы, сроки их проведения и осуществлять
определенные лесохозяйственные мероприятия.
6.2. НАДЗОР ЗА ПОЯВЛЕНИЕМ БОЛЕЗНЕЙ
И ВРЕДИТЕЛЕЙ
Надзор включает постоянные и периодические наблюдения и
учеты появления, распространения и развития очагов вредителей
и болезней, состояния леса в целях своевременного планирова-
129
ния и осуществления лесозащитных мероприятий. В лесах России
осуществляют общий и специальный надзор.
Общий надзор заключается в выявлении случаев массового усы-
хания и повреждения (поражения) леса, массового
распространения вредителей или болезней. Надзор осуществляется всеми
лесными специалистами и работниками лесных предприятий в
процессе их повседневной деятельности. О наблюдаемых явлениях
немедленно оповещаются службы защиты леса. Основную работу по
общему надзору выполняет лесная охрана под непосредственным
руководством лесничих. В случае обнаружения признаков
неблагополучного состояния леса или его повреждения заполняют
листок сигнализации или передают по каналам связи (рации,
телефону) фиксируемое в специальном журнале сообщение. Сигнал
проверяет лесничий, затем заполняет специальный акт проверки
сигнализации и передает его специалистам службы защиты леса.
После этого акт поступает в вышестоящую организацию (лесхоз,
леспромхоз, объединение) для принятия соответствующих мер.
Специальным (лесопатологическим) надзором называют
систему дистанционных и наземных наблюдений, анализов и учетов.
Надзор устанавливают за опасными видами вредителей и
болезней или их комплексами и за состоянием леса с целью
получения данных для прогноза и планирования лесозащитных
мероприятий.
Специальный надзор может быть рекогносцировочным и
детальным в зависимости от методов и точности наблюдений и учетов.
Рекогносцировочный лесопатологический надзор — это
визуальный дистанционный и наземный надзор за состоянием, поражен-
ностью (поврежденностью) леса, развитием болезней и
численностью вредителей. Он позволяет выявить повреждение и усыха-
ние леса на определенных маршрутах, проложенных с учетом
прошлогодних повреждений, потенциальные, возникающие и
действующие очаги опасных вредителей и болезней леса. Этот вид
надзора проводят на специально отобранных маршрутах и
участках насаждений, где были замечены, предполагаются или
действуют очаги. Их наносят на схематическую карту и отмечают в
специальной ведомости. Осмотр насаждений осуществляют в
периоды наибольшего проявления признаков повреждения леса. Цель
рекогносцировочного надзора — выявить степень поражения,
повреждения, усыхания леса в поднадзорных участках, установить
ориентировочный уровень численности насекомых или степень
развития болезни, границы и площади очагов.
Рекогносцировочный надзор — наиболее гибкая и удобная
форма контроля за появлением и распространением вредителей и
болезней, так как он не требует больших затрат времени и сил и
позволяет быстро выявить очаги вредителей и болезней, заметить
тенденции изменения уровня численности и масштаба распрост-
130
ранения вредных организмов. Рекогносцировочный надзор
выполняют среднее и высшее звенья лесной охраны, инженер по
охране и защите леса при участии или руководстве специалистов
станций защиты леса.
Детальный лесопатологический надзор — это надзор с
применением методов детального анализа состояния леса, учета
плотности структуры и жизнеспособности популяции вредителей и
установлением характера распространения и степени развития
болезней леса на основании наземного выборочного обследования.
Так же как и рекогносцировочный, детальный
лесопатологический надзор устанавливают за определенными вредителями и
болезнями или их комплексами. Для этого выбирают постоянные
участки или маршруты, на которых проводят соответствующие
наблюдения и учеты. Детальный надзор включает в себя, кроме
наземных обследований, применение специальных технических и
биологических средств — светоловушек, токсических и ловчих
поясов, клеевых колец, феромонных ловушек и приманок с ат-
трактантами.
Специальный надзор дополняют данными лесопатологическо-
го обследования.
6.3. МЕТОДЫ ДИАГНОСТИКИ БОЛЕЗНЕЙ
ДРЕВЕСНЫХ РАСТЕНИЙ
Диагностика — это распознавание причин болезней с
применением различных методов. Постановка диагноза включает
определение типа и характера болезни (инфекционный или
неинфекционный), возбудителя или причины; оценку вредоносности,
определение давности поражения, выявление условий,
способствовавших развитию болезни.
Правильная постановка диагноза позволит своевременно
определить и предупредить угрозу массового поражения растений
болезнями, осуществить профилактические и истребительные
защитные мероприятия, не допустить потерь и неоправданных затрат.
В настоящее время для диагностики болезней леса применяют
следующие методы: макроскопический (патографический),
микроскопический, микологический, химический и физический.
Часто их используют в комплексе.
Макроскопический (патографический) метод дает
возможность диагностировать болезнь невооруженным глазом или при
помощи лупы, бинокля в полевых условиях. Этот метод широко
применяется в производственных условиях и до настоящего
времени остается ведущим. Для правильного распознавания болезней
при помощи этого метода необходимо проанализировать большое
количество экземпляров или органов растений. Такая необходи-
131
мость вызвана тем, что совокупность признаков болезней может
встречаться не на всех растениях или симптомы болезней могут
быть не всегда ярко выражены.
Другим условием правильной диагностики является
тщательный осмотр больного растения от вершины до корней. У взрослых
деревьев крону и ствол осматривают с помощью бинокля, а
корневую систему исследуют, обнажая 2—3 корня. Для того, чтобы
сделать правильный вывод, необходимо тщательно
проанализировать условия местопроизрастания исследуемых объектов и
установить причины, способствующие заболеванию. Далеко не всегда
факт наличия болезни говорит о том, что она является причиной
ослабления или усыхания растений. Например, в Очагах
массового усыхания дубрав на стволах и ветвях ослабленных и усыхающих
деревьев часто встречаются некрозы (клитрисовый, вальзелловый
и др.), которые не являются первопричинами усыхания. Они
развиваются на предварительно ослабленных деревьях и лишь
ускоряют процесс усыхания.
При осмотре растения обращают внимание на наличие
плодовых тел, спороношений, различных видоизменений мицелия (ри-
зоморфы, пленки, склероции и т.д.), типов новообразований
(раны, опухоли, трещины и т.д.), гнилей. Легче всего определить
возбудителя болезни по хорошо развитым, характерным
плодовым телам и спороношениям. Например, окаймленный трутовик
распознают по плодовым телам, имеющим характерную красную
или желто-красно-коричневую кайму; клитрисовый некроз дуба
определяют по струпьевидно изогнутым, сероватым апотециям;
достоверный диагностический признак смоляного рака — эци-
альные спороношения в виде крупных оранжевых вместилищ.
Если плодоношения отсутствуют, то в некоторых случаях
можно определить возбудителя по характерным грибным
образованиям. Так, поражение опенком распознают по темным шнуровид-
ным ризоморфам или белым веерообразным пленкам грибницы,
а возбудителя эндотиевого рака каштана — по веерообразным
пленкам желтого или оранжевого цвета, которые являются
диагностическим признаком болезни. Особенности структуры, цвет и
другие признаки пораженной трутовиками древесины дают
возможность установить вид возбудителя гнили.
Сложнее поставить правильный диагноз, когда есть только
признаки патологии, т.е. раны, опухоли, пятна, увядание и т.д. По
ним можно установить тип болезни, а для определения
возбудителя необходимо применять другие методы. Макроскопический
метод дает возможность распознавать возбудителей наиболее
распространенных и известных болезней на тех стадиях их развития,
когда появляются ясно выраженные внешние признаки. Если по
макроскопическим признакам диагноз поставить нельзя,
пользуются микроскопическим методом.
132
Микроскопический метод заключается в
исследовании под микроскопом спороношений возбудителей или
пораженных тканей растений. Его применяют для определения вида
возбудителя или установления наличия патогена в тканях растения.
Только микроскопирование дает возможность обнаружить
характерные признаки, присущие патогену (у грибов форму и цвет спор,
форму сумок, наличие парафиз, характер размещения сумок в
плодовых телах и др.). При микроскопическом анализе делают
временные или постоянные препараты. В зависимости от характера
объектов препараты готовят разными способами.
Если мицелий или спороношения возбудителей (налеты,
пленки, пустулы, подушечки) располагаются поверхностно, их
соскабливают препаровальной иглой или скальпелем на чистое
предметное стекло в каплю воды. Концами препаровальных игл
материал расправляют и накрывают чистым покровным стеклом,
помещая его на край капли под острым углом. Лишнюю воду с
предметного стекла убирают фильтровальной бумагой.
Для установления наличия мицелия в тканях хвои и листьев,
определения типов плодовых тел и т.д. лезвием производят очень
тонкие продольные или поперечные срезы, чтобы на препарате
были хорошо видны признаки возбудителя. С этой целью
применяют новые или малоиспользованные лезвия.
Срезы хвои и листьев с наиболее характерным поражением
делают в сердцевине бузины, кукурузы, моркови или пенопласте
(рис. 38). Из нескольких срезов выбирают наиболее тонкие для
просмотра под микроскопом. Плодовые тела рассекают
продольно или поперечно. По срезам можно установить тип плодовых тел
(апотеции, перитеций, пикниды). Сначала проводят срез через
середину плодового тела, отсекая одну половину. По
вертикальной поверхности оставшейся половины делают несколько срезов,
выбирая затем наиболее удачные для просмотра. Если хвоя или
кора сухие, то их предварительно размачивают или кипятят в воде.
Препараты просветляют или
окрашивают. Просветляют их в том случае, если
просмотру подлежат толстые
непрозрачные срезы. Для этого фильтровальной
бумагой с препарата оттягивают воду и
на объект наносят две капли молочной
кислоты или раствора едкой щелочи
(NaOH, КОН). Затем препарат
подогревают на спиртовке до кипения,
накрывают покровным стеклом и только после
этого просматривают под микроскопом.
В качестве просветлителей кроме выше- рИс. 38. Этапы приготовле-
указанных можно применять хлоралгид- ния тонких срезов хвои и
рат, лактофенол, бергамотовое масло. листьев
133
Окрашивание применяют для обнаружения в тканях растения
бесцветных образований возбудителей (мицелий и др.).
Приготовленные срезы вначале рассматривают в капле воды и выбирают
наиболее удачные. Затем воду оттягивают фильтровальной
бумагой и заменяют ее красителем, в котором выдерживают объект в
течение 5 мин. После этого краситель удаляют фильтровальной
бумагой, а препарат промывают водой. На объект пипеткой
переносят капли воды и удаляют их до тех пор, пока вода не станет
бесцветной.
Для окрашивания препаратов можно использовать 1 %-й
водный или молочно-кислый раствор хлопчатобумажного синего
красителя, 1 %-й молочно-кислый раствор анилинового синего, 1 %-й
водный раствор водного синего. Перед окрашиванием срезы
просветляют указанными выше способами. Просветление и
окрашивание лучше получать путем фиксации препаратов. Наиболее
простой способ фиксации — нагревание срезов в капле кипящей воды
или молочной кислоты на предметном стекле.
Исследование тканей с дифференцированным окрашиванием
применяют при диагностике обыкновенного шютте в ранний
период его развития, когда еще нет характерных внешних
признаков инфекционного полегания всходов.
При необходимости проводят измерения элементов гриба с
помощью окуляр-микрометра или микрометра окулярного
винтового MOB-1-15. Определение размеров спор необходимо
проводить в первую очередь при разделении видов грибов в пределах
рода. При этом используют окуляр-микрометр, вложенный в
окуляр и имеющий 100 делений.
Цена деления окуляр-микрометра устанавливается следующим
образом. Окуляр-микрометр вводят в окуляр, затем на столик
микроскопа вместо препарата помещают объектив-микрометр и,
выбрав требуемое увеличение, устанавливают на резкость.
Объектив-микрометр представляет собой шкалу со 100 делениями,
расположенными в 10 мкм (0,01 мм) друг от друга. С его помощью
определяют цену деления окуляр-микрометра. Сначала
совмещают два любых деления обеих шкал, затем ищут отрезок, на котором
опять совпадают два деления обеих шкал, и рассчитывают
соотношения. Например, 17 делений объектив-микрометра совпадают с
40 делениями окуляр-микрометра. Следовательно, 40 делений
окуляр-микрометра соответствуют 170 мкм (17-10 мкм). Расстояние
между двумя делениями на окуляр-микрометре составляет 4,25 мкм
(170:40). Этот расчет повторяют еще два раза, совмещая любые
другие деления. Из трех значений, которые в данном примере
лежат между 4,25 и 4,30 мкм, рассчитывают среднее, которое и
используют в качестве окончательной цены деления. Этот
показатель действителен только для данного микроскопа при
применении объектива с увеличением, используемым для калибровки.
134
Для измерения спор сначала готовят водный препарат
(суспензию спор). Затем его помещают на предметный столик таким
образом, чтобы линейка окуляр-микрометра проходила над спорой
по продольной или поперечной ее оси (длина или ширина),
подсчитывают деления окуляр-микрометра от одного конца споры до
другого и умножают их число на цену деления. Изогнутые споры,
например конидии Fusarium, измеряют от верхушки до верхушки
(рис. 39). Чтобы получить среднее значение, а также определить
размах вариабельности от самой маленькой до самой большой
споры, находят размеры не менее чем 100 спор.
Микологический метод заключается в выделении
гриба из пораженных частей растений, в его изоляции и
выращивании на искусственной или естественной среде. Довольно просто
можно выделить грибы из пораженных частей растений,
пользуясь методом влажной камеры. Он основан на способности
грибницы, находящейся внутри тканей растения, во влажных
условиях прорастать наружу и образовывать спороношения. Метод не
требует стерильных условий и позволяет в довольно короткий срок
получать спороношения возбудителей. Он широко применяется
для распознавания инфекционного полегания, болезней хвои,
семян и др.
Чаще применяют влажные камеры следующего устройства. На
дно чашек Петри или Коха помещают кружок фильтровальной
бумаги, равный по диаметру дну чашки. Готовые влажные камеры
стерилизуют в сушильном шкафу. Чашки можно стерилизовать
кипятком или спиртом, а кружки фильтровальной бумаги —
пламенем горелки или спички, быстро проводя над ним 2—3 раза.
Перед загрузкой камеры фильтровальную бумагу смачивают
дистиллированной или охлажденной кипяченой водой.
Объекты исследования (всходы, хвою, стволики, корни) перед
закладкой в камеру промывают под струей воды и затем
стерилизуют различными способами (погружением в 0,5 %-й раствор мар-
135
ганцовокислого калия, обработкой спиртом, проведением через
пламя). После этого их раскладывают на фильтровальную бумагу в
чашки так, чтобы они не соприкасались друг с другом (рис. 40).
Загруженные камеры оставляют в условиях комнатной
температуры или помещают в термостат с температурой 20...25 °С. Камеры
периодически просматривают и по мере необходимости бумагу в
них увлажняют стерильной водой из пипетки. В зависимости от
особенностей объекта мицелий, спороношения или
плодоношения образуются через несколько дней или недель, после чего
проводят микроскопический анализ.
Более сложное выделение патогена из тканей растений
осуществляют методом чистых культур, основанным на использовании
искусственных или естественных питательных сред. Для
получения чистых культур грибов и поддержания их жизнеспособности в
целях дальнейшего изучения применяют различные питательные
среды (твердые и жидкие, естественные и искусственные). Выбор
того или иного питательного субстрата зависит от потребностей
патогена и целей исследований.
Для получения спороношения большинства видов грибов
(факультативных сапротрофов и факультативных паразитов)
используют суслоагаровую среду, содержащую 7 %-е неохмеленное
пивное сусло и агар-агар, а также агаровую среду, компонентами
которой являются вода и агар-агар. Специфические среды для
отдельных видов грибов отбирают экспериментальным путем.
Рецепты приготовления наиболее широко применяемых
питательных сред приводятся в специальных пособиях по технике и
методам фитопатологических исследований. При выборе
питательных сред для грибов необходимо учитывать, что они лучше растут
в среде, богатой углеводами, и предпочитают слабокислую
реакцию.
Для выделения патогенов, поражающих кору и древесину,
объекты дезинфицируют в 96 %-м этиловом спирте в течение 2 мин или
путем фламирования, т.е. стерилизации над пламенем спиртовки.
Затем стерильным скальпелем срезают поверхностные слои с
образца, а из внутренних слоев вырезают небольшие кусочки
размером не более 5x5 мм и стерильным пинцетом переносят их на
питательную среду в чашки Петри или пробирки.
При выделении патогена из хвои ее поверхностную
дезинфекцию осуществляют путем погружения в одно из следующих
веществ: в 70 %-й или 96 %-й спирт на 2—3 мин, в 0,5 %-й раствор
марганцовокислого калия на 30—40 мин. После этого хвою
трижды промывают в стерильной воде, разрезают на кусочки
размером 5—7 мм и переносят в питательную среду.
При определении поверхностной инфекции семян их
раскладывают рядами в чашки Петри на агаровую среду без
предварительной стерилизации. Для определения внутреннего заражения
136
семена дезинфицируют в 0,5 %-м растворе марганцовокислого
калия или 96 %-м спирте, а затем раскладывают на питательную
среду. Крупные семена разрезают стерильным скальпелем
пополам.
Для выявления патогенов, обитающих в почве, берут образцы
из характерных почвенных разностей на глубине, где
расположена основная масса корней. Собранную почву помещают в
бумажные пакеты и в течение нескольких дней высушивают до
воздушно-сухого состояния. Затем ее измельчают и высевают на
агаровую питательную среду в чашки Петри. По мере развития
колоний грибов их переносят в пробирки на косо разлитую агаровую
среду.
Важнейшим требованием при использовании метода чистых
культур является соблюдение стерильности посуды,
инструментов, одежды, помещения. Нарушение этого требования может
привести к загрязнению питательной среды посторонней
инфекцией и ошибочным выводам при постановке диагноза. Работы с
чистыми культурами грибов осуществляют в специальных
помещениях-боксах. Конструкция и размеры боксов могут быть
различны в зависимости от общей площади лабораторного
помещения. Бокс должен быть герметичен, изготовлен из легко
моющихся материалов (стекла, окрашенных масляной краской фанеры
или металла). Для работы при естественном освещении одна
стенка бокса должна быть застеклена не менее чем на половину.
Однако это не исключает применения электрического освещения.
Химический метод основан на использовании цветных
индикаторов, изменяющих окраску водных вытяжек из хвои или
листьев, пораженных разными грибами. Подробно химический
метод диагностики изложен в книге И.И.Журавлева (1962).
Физический метод основан на различии физических
свойств органов и тканей у здоровых и больных растений.
Качества семян определяют по их плотности. Больные семена,
имеющие меньшую плотность, всплывают при погружении их в
различные жидкости (пресная или соленая вода). Для установления
причин поражения пользуются одним из описанных выше
способов (выделением в чистую культуру, влажной камерой).
Для выявления скрытой гнили древесины применяют
звуковую пробу; при выстукивании здоровых и фаутных стволов
получают разный звук. Этот метод используют как при определении
фаутности древостоев, так и при обследовании зданий на
зараженность домовыми грибами. Однако способ выстукивания не дает
точных результатов, так как характер звука зависит не только от
наличия гнилей в древесине, но и от других факторов.
В последнее время все более широкое распространение при
диагностике болезней древесных пород получает
люминесцентный анализ. Он основан на разном свечении в ультрафиолетовых
137
лучах пораженных тканей растений. Как источник света можно
использовать лабораторные и медицинские кварцевые лампы. Этим
методом устанавливают жизнеспособность семян. Например,
жизнеспособные семена хвойных пород на разрезе при облучении
светятся фиолетовым цветом, клена — голубым, а
нежизнеспособные семена этих пород не светятся. По-разному светятся на
срезах черенки тополей здоровых и пораженных цитоспорозом.
Люминесцентный анализ позволяет установить причину
специфической окраски древесины, определить возбудителей сходных
по внешним признакам гнилей (гнили ствола ели, вызванные
еловой и корневой губкой). Е. И. Ладейщикова применила
люминесцентный анализ для ранней диагностики корневой губки.
6.4. ПРОГНОЗ РАЗВИТИЯ БОЛЕЗНЕЙ
Прогнозирование болезней — это предвидение их появления,
массового распространения и размеров вредоносности. Прогноз
болезней основан на изучении связи развития и распространения
болезней с метеорологическими, экологическими и другими
факторами. Прогноз имеет огромное значение, поскольку позволяет
заранее планировать необходимые виды и объем лесозащитных
мероприятий и дает возможность предотвратить или снизить
возможный ущерб от болезней.
В зависимости от целей различают краткосрочный,
долгосрочный и многолетний прогноз.
Краткосрочный прогноз — это предвидение массового
развития болезней на относительно короткий срок (вегетационный
период, месяц, декада). Этот прогноз основан на определении
сроков массовой споруляции патогена, его инкубационного
периода, динамики болезни. По результатам анализа и обобщения
этих данных строятся математические модели, на основе которых
устанавливают оптимальные сроки проведения лесозащитных
мероприятий, в том числе химических обработок. Для составления
краткосрочного прогноза необходимо проводить наблюдения за
фенологией растений-хозяев, состоянием патогена и запасом
инфекции, погодными условиями. Кроме того, определяют сроки
появления первых признаков болезни и ее динамику.
Долгосрочный прогноз — это предвидение развития
болезней на продолжительный срок (вегетационный период
следующего года). Для составления долгосрочного прогноза анализируют
данные о запасе инфекции и состоянии растения-хозяина в
текущем году и метеорологические факторы предшествующего года. Из
погодных факторов используют температуру воздуха и почвы,
относительную влажность воздуха, сумму осадков, число дней с
осадками, количество солнечных дней, высоту снежного покрова и др.
138
Многолетний прогноз — это предсказание массового
развития болезни на длительный срок (год, несколько лет).
Многолетний прогноз болезней, способных принимать характер эпифито-
тий (некрозы, раки, сосудистые), основан на использовании
многолетних данных о поражении и состоянии деревьев в конкретной
зоне или конкретных экологических условиях. При этом
учитываются средние показатели поражения и количество лет с
благоприятными для развития болезни условиями и их повторяемость,
ежегодный отпад растений. По результатам анализа многолетних
данных строится математическая модель. Многолетний прогноз дает
возможность составлять перспективные планы различных
лесозащитных мероприятий.
В лесном хозяйстве прогноз болезней недостаточно изучен, он
разработан пока для отдельных видов болезней в питомниках, в
том числе для обыкновенного, снежного шютте и ржавчины
побегов сосны.
6.5. КАРАНТИН РАСТЕНИЙ
Карантин растений — это система государственных
мероприятий, направленных на охрану территории Российской Федерации
от проникновения из других государств карантинных и других
опасных болезней растений, вредителей и сорняков, способных
причинить значительный вред народному хозяйству страны. К
карантинным объектам относятся возбудители болезней, виды
вредителей и сорняков, которые не встречаются в пределах
государства или имеют ограниченное распространение на территории
страны, но могут попасть извне и нанести значительный ущерб.
В настоящее время в связи с интенсивной хозяйственной
деятельностью человека и значительным расширением
международных торговых связей в нашу страну ввозится большое количество
семенного и посадочного материала, плодов, овощей, ценной
древесины, вместе с которыми могут проникать возбудители
болезней, не имевшие до этого распространения в РФ. Часто это
приводит к возникновению эпифитотий. В связи с этим
организации карантина уделяется большое внимание. В нашей стране
охрану территории от проникновения из других государств
карантинных объектов, а также проведение карантинных мероприятий
осуществляют органы государственной службы по карантину
растений Министерства сельского хозяйства Российской Федерации.
Руководство всеми работами по карантину осуществляет
Государственная инспекция по карантину растений Российской
Федерации (Росгоскарантин). В государственную службу карантина
входят государственные органы карантина растений в
автономных республиках, краях, областях (государственные, областные,
городские, межрайонные, районные инспекции и карантинные
139
пункты; пограничные пункты карантина).
Научно-исследовательская и методическая работа по карантину растений проводится
Всероссийским научно-исследовательским институтом карантина
растений (ВНИИКР). Лесной карантин относится к службе
карантина растений, действующей в системе сельского хозяйства. В
настоящее время отдел лесного карантина входит в состав Росгоска-
рантина и обеспечивает карантинный контроль над экспортной
древесиной и охрану территории Российской Федерации от
заноса и распространения опасных для страны видов болезней и
вредителей леса.
Одна из основных задач карантинной службы — ликвидация
возникших очагов болезней, являющихся объектами карантина.
В зависимости от поставленных задач различают внешний и
внутренний карантин.
Задача внешнего карантина заключается в
предупреждении ввоза в нашу страну отсутствующих карантинных объектов
и предотвращении вывоза с экспортируемым материалом
карантинных объектов, обусловленных договорами стран-импортеров.
Ввоз из других государств продукции растительного
происхождения разрешается только при наличии специальных сертификатов,
характеризующих карантинное состояние ввозимых в страну
объектов. Ценные семена и посадочный материал, ввозимые в нашу
страну, направляются для оздоровления и выявления скрытой
зараженности в карантинные питомники и оранжереи.
Мероприятия по внешнему карантину распространяются на
семена, живые растения, коллекции возбудителей болезней
растений, гербарии растений, неокоренную древесину,
поделочную древесину и изделия из нее, образцы почв. Болезни лесных
пород, плодовых и цветочных культур, относящиеся в настоящее
время к числу объектов внешнего карантина, и их возбудители
представлены в табл. 2.
Задача внутреннего карантина заключается в
предупреждении распространения болезней внутри страны. С этой
целью систематически проводятся обследования
сельскохозяйственных угодий и лесных насаждений, мест хранения и переработки
продукции растительного происхождения. К объектам
внутреннего карантина относятся болезни, имеющие очень
ограниченное распространение, но при проникновении в другие районы
они могут вызвать эпифитотии. Объектами внутреннего
карантина являются: голландская болезнь ильмовых пород (Ophiostoma
ulmi), сосудистый микоз дуба (грибы родов Ceratocystis и Fusarium),
крифонектриевый рак каштана съедобного {Cryphonectria
parasitica), побеговый рак хвойных пород (Scleroderris lagerbergii),
бактериоз сеянцев сосны (Pseudomonas fluorescens), повилика
европейская (Cuscuta europaea) и одностолбовиковая (C.monogyna),
паразитирующие на лиственных породах и кустарниках. При уста-
140
Таблица 2
Объекты внешнего карантина
Группа фитопатогенных организмов.
Вид возбудителя
Вызываемая
болезнь
Древесная порода,
цветочная культура
Грибы
Ceratocystisfagacearum (Т. W. Bretz)
J.Hunt
Cronartium sp. p. (неевропейские
виды)
\Chrysomyxa arctostaphyliDietel
\Endocronartium sp. p.
(неевропейские виды)
llnonotusweirii (Murill.) Kotlalaet
Pouzar (= Phellinus weirii (Murill.)
R. L. Gibberston
Mycosphaerella populorum G. E.
Tompson
\Phyllosticta solitaria Ellis, exEverh
\Puccinia horiana Henn
Trechispora brinkmannii (Bres.) D. P.
Rogers et H. Iacke (= Sistotrema
brinkmannii (Bress.) I. Eriksson
Сосудистое
увядание
Рак
Ржавчина хвои
Рак
Гниль
Пятнистость
листьев
Пятнистость
листьев
Белая ржавчина
Гниль
Дуб
Сосна
Ель
Сосна
Хвойные
Тополь
Боярышник,
груша, яблоня
Хризантема
Сосна, пихта,
дуб, клен, ива,
тополь
Бактерии
\Erwinia amylovora(Burill.)
Corn. S.A.B.
\Pseudomonas caryophylli Starr, et
Burkholder
\Xanthomonascampestris Dows
Xanthomonas hyacinthi (Wakker.)
Dowson
Бактериальный
ожог
Бактериальное
увядание
Сосудистый
бактериоз
Желтая гниль
Груша
Гвоздика
Левкой
Гиацинт
Нематоды
Bursaphelenchus xylophilus Stein, et
Buhr
Бурсафеленхоз
(усыхание)
Сосна
141
новлении зараженности принимаются меры по локализации
выявленных очагов.
Карантинные мероприятия распространяются на семена и
живые растения, образцы почв, живые культуры грибов и бактерий,
коллекции возбудителей болезней, неокоренную древесину.
6.6. ЛЕСОХОЗЯЙСТВЕННЫЕ МЕТОДЫ
Лесохозяйственные методы включают мероприятия,
направленные на повышение биологической устойчивости насаждений
и их оздоровление путем своевременного применения
существующих правил ухода за лесом и использования современных
достижений лесозащиты. Например, использование при
лесоразведении здорового посевного и посадочного материала. Нарушение
этого правила может привести к возникновению очагов болезней.
Саженцы, имеющие механические повреждения, пораженные
болезнями или вредителями, нередко отстают в росте и теряют
устойчивость к воздействию абиотических неблагоприятных
факторов. Механические повреждения на стволиках являются
воротами инфекции некрозно-раковых болезней. Саженцы, пораженные
болезнями, при высадке их на лесокультурную площадь часто
становятся центрами возникновения очагов. Ярким примером этому
служит использование для посадки на лесокультурную площадь
сеянцев сосны, пораженных снежным шютте. При высадке
сеянцев, пораженных болезнью на 25 %, через 2 — 3 года в культурах
их сохраняется менее 50 % от общего числа высаженных.
Выращивание здорового посадочного материала и ограничение
распространения болезней в питомниках и культурах возможно
только при правильной агротехнике. Своевременное проведение
посева, рыхления, прополки сорняков и прочих мер по уходу за
посевами создает оптимальные условия для роста и развития
всходов и сеянцев, косвенно повышая их устойчивость к болезням.
Большое значение имеет правильное внесение удобрений с
учетом почвенных условий. В хорошо удобренной почве создаются
неблагоприятные условия для развития паразитной микрофлоры.
Сеянцы, обеспеченные всеми необходимыми элементами
питания в оптимальных соотношениях, обладают высокой
устойчивостью к болезням. При выращивании в питомнике дуба, клена и
других лиственных пород, восприимчивых к мучнистой росе,
необходимо регулировать внесение азотистых и фосфорных
удобрений. Избыток азота вызывает бурный рост зеленой массы,
задерживает созревание листьев, увеличивая период восприимчивости
их к болезни. Фосфорные удобрения способствуют созреванию
листьев и одревеснению побегов, повышая их устойчивость к
мучнистой росе. При избытке азотистых удобрений и недостатке фос-
142
форных замедляется одревеснение стволиков и всходов хвойных
пород и повышается их восприимчивость к полеганию. При
посевах очень важно соблюдать установленные нормы высева,
учитывая, что загущенные посевы более сильно страдают от полегания,
шютте и других болезней.
Одним из мероприятий, направленных на предупреждение и
ограничение распространения болезней, является применение
рациональных севооборотов в питомниках с подбором
соответствующих предшественников. Это мероприятие препятствует
накоплению почвенных патогенов и снижает запас почвенной
инфекции, в частности возбудителей полегания сеянцев.
Монокультура в питомниках, особенно хвойных пород, приводит к
накоплению в почве большого количества инфекции и
систематическому отпаду всходов от полегания.
Во избежание заражения болезнями посевов текущего года их
следует пространственно изолировать от прошлогодних посевов
той же породы.
Большое значение имеет правильный подбор пород с учетом
почвенно-климатических условий, поражаемости их болезнями,
повреждаемости вредителями, аллелопатических отношений между
ними. Известно, например, что сосна хорошо растет с березой и
рядом кустарников, в том числе со скумпией, с акацией желтой
и другими, а сочетание дуба, ясеня, бука является
неблагоприятным. Некоторые древесные породы поражают одни и те же
возбудители болезней и поэтому породы заражаются друг от друга. Так,
береза и лиственница поражаются ржавчинным грибом Melampso-
ridiwn betulaeArth., осина и сосна — ржавчинным грибом Melampsora
pinitorqua Rostr., что следует учитывать при создании культур и
размещении питомников.
Одним из мероприятий, ограничивающих распространение
болезней, является подбор устойчивых пород и форм, отбор
семян с устойчивых форм, гибридизация. В настоящее время
довольно хорошо изучена устойчивость древесных пород к
главнейшим болезням. Некоторые виды лиственниц, произрастающих на
Дальнем Востоке, устойчивы к шютте; вяз перистоветвистый
более устойчив к голландской болезни, чем другие ильмовые
породы; белые тополя обладают более высокой устойчивостью к доти-
хициевому некрозу, чем бальзамические и черные, ясень зеленый
меньше поражается эндоксилиновым и цитофомовым раком, чем
обыкновенный, и т.д.
Сбор и заготовку семян проводят с деревьев, которые
оказались непораженными в очагах болезней. Такие деревья при рубках
оставляют как семенники.
Создание смешанных разновозрастных насаждений —
важнейшее условие, обеспечивающее их биологическую устойчивость и
ограничивающее распространение болезней. В чистых одновозраст-
143
ных насаждениях создаются условия для доминирования
отдельных видов возбудителей болезней, что приводит к
возникновению эпифитотий. Создание чистых культур сосны вызвало их
гибель на огромных площадях от корневой губки; в чистых
ильмовых насаждениях возникают очаги голландской болезни. Введение
в состав хвойных насаждений лиственных пород ограничивает
возможность передачи инфекции через корни. Кроме того,
некоторые лиственные породы (черемуха, аморфа, робиния
лжеакация) выделяют активные вещества, подавляющие рост корневой
губки, а береза улучшает плодородие почвы. Введение в состав
ильмовых насаждений других лиственных пород препятствует
распространению спор возбудителя голландской болезни и
ограничивает передачу инфекции через корни.
Важную роль в оздоровлении насаждений играет правильный
и своевременный уход за молодыми культурами и лесом,
предусматривающий своевременное удаление больных и ослабленных
деревьев.
Большое значение для ограничения распространения болезней
имеет правильный выбор системы рубок и их своевременное
проведение. У большинства возбудителей главнейших болезней леса
споруляция происходит в весенне-летний период. В связи с этим
рубки в очагах болезней проводят в осенне-зимний период.
Важную роль в создании биологически устойчивых насаждений
играют рубки ухода. При всех видах этих рубок в первую очередь
выбирают деревья, пораженные болезнями, заселенные стволовыми
вредителями, сухостойные, суховершинные. Рубки ухода
способствуют созданию смешанных насаждений, обладающих высокой
биологической устойчивостью по сравнению с чистыми.
Прореживание при рубках ухода в молодняках улучшает условия роста,
повышает устойчивость к снеголомам и снеговалам,
ограничивает распространение болезней и стволовых вредителей. Однако
сильное изреживание приводит к распространению в насаждениях
болезней (смоляной рак сосны), возникновению очагов
стволовых вредителей с повышенной плотностью популяции, усиливает
подверженность ветровалу и бурелому.
Важную роль в ликвидации очагов болезней и улучшении
санитарного состояния насаждений играют санитарные рубки, при
которых удаляют деревья, пораженные опасными болезнями
(корневой губкой, смоляным раком, голландской болезнью и т.д.),
заселенные стволовыми вредителями, имеющие механические
повреждения и ослабленные по разным причинам. Способ и объем
санитарных рубок определяют по результатам лесопатологических
обследований с учетом хозяйственной целесообразности.
При слабой пораженности древостоя болезнями проводят
выборочные санитарные рубки. Если пораженность насаждения
превышает 40 % или выборка пораженных деревьев может привести к
144
расстройству насаждений, назначают сплошные санитарные
рубки. В лесах III группы сплошные санитарные рубки включают в
план рубок главного пользования. Несвоевременное проведение
санитарных рубок в очагах болезней после полного усыхания
деревьев приводит к массовому распространению инфекции и
возникновению новых очагов.
Основным требованием профилактики является тщательное
выполнение элементарных санитарных правил в лесах. Участки
ветровала, бурелома, усохшие от болезней, должны
разрабатываться в кратчайший срок во избежание потери технических
качеств древесины и распространения возбудителей болезней и
стволовых вредителей. Эти площади должны тщательно очищаться от
порубочных остатков и захламленности. Поскольку порубочные
остатки, кора и щепа могут служить источниками инфекции, их
необходимо сжигать. Заготовленную древесину окоряют и вывозят
из леса в зависимости от сроков заготовки, а также споруляции
возбудителей болезней, в очагах которых проводится рубка.
Повышение биологической устойчивости насаждений и их
оздоровление достигается также путем реконструкции насаждений с
учетом типа лесорастительных условий. Реконструкция
осуществляется изменением состава насаждений и введением почвоулуч-
шателей, какими являются некоторые травянистые растения и
кустарники. При реконструкции сосновых насаждений,
пораженных корневой губкой, в их состав вводят лиственные породы.
Лучшим компонентом является береза, которая не только
препятствует распространению болезни через корни, но и улучшает
плодородие почвы, изменяет состав микрофлоры в ризосферно-при-
корневой зоне. В ризосферно-прикорневой зоне березы
преобладают микроорганизмы, проявляющие сильную степень
антагонистической активности по отношению к корневой губке. В
сосновых культурах, созданных на бедных песчаных почвах с
недостатком азота, проводят посев многолетнего люпина, который
улучшает аэрацию почвы, повышает запасы гумуса и азота,
активизирует круговорот органических веществ. Кроме того, он
способствует более глубокому проникновению корней в почву, ускоряет
плодоношение деревьев.
6.7. БИОЛОГИЧЕСКИЙ МЕТОД
Биологический метод борьбы с болезнями основан на
использовании антагонистических связей между различными
микроорганизмами и аллелопатических отношений растений. В настоящее
время первостепенное внимание уделяется проблемам
окружающей среды. В этой связи велико значение биологического метода.
Его применение снижает загрязнение биосферы пестицидами и
145
уменьшает их вредное последействие. В борьбе с болезнями
древесных пород биологический метод пока применяется весьма
ограниченно, но испытания различных биологических средств в
последние годы проводятся в больших масштабах.
Одним из направлений, в которых ведется разработка
биологического метода, является использование антагонистов,
подавляющих развитие возбудителей болезней или вызывающих их
гибель. Установлено, что обитающие в пахотном слое почвы грибы
рода Trichoderma являются антагонистами некоторых возбудителей
болезней древесных пород, в том числе корневой губки и
возбудителей полегания.
Большой интерес представляют дереворазрушающие грибы-
сапротрофы, находящиеся в активных антагонистических связях
с корневой губкой. Эти грибы называют пневыми конкурентами.
Среди них наиболее эффективны: Peniophora gigantea, Fomitopsis
pinicola, которые способны быстро проникать в глубь древесины и
защищать пни не только от проникновения внешней споровой
инфекции, но и препятствовать развитию мицелия в пораженной
древесине корней. Обработка пней осуществляется водными
суспензиями плодовых тел или чистых культур грибов.
Кроме грибов-антагонистов известно много видов бактерий,
проявляющих высокую антагонистическую активность по
отношению к фитопатогенным грибам, в том числе к корневой губке,
возбудителям полегания и мучнистой росы.
Высокой антагонистической активностью к возбудителям
полегания обладают антибиотики, продуцируемые некоторыми де-
реворазрушающими грибами: окаймленным, настоящим, ложным
дубовым, осиновым трутовиками и др.
Большой интерес представляет использование в борьбе с
болезнями растений сверхпаразитов (гиперпаразитов) —
микроорганизмов, паразитирующих на других фитопатогенных
микроорганизмах. В настоящее время известен целый ряд грибов-сверхпаразитов,
паразитирующих на фитопатогенных грибах. Например, Cicinnobolus
cesati развивает мицелий внутри гиф мучнисторосяных грибов. Он
был использован для борьбы с мучнистой росой яблони и дал
положительные результаты. Tuberculina persicina подавляет
развитие ржавчинных грибов, паразитирующих на груше, барбарисе,
крушине; Т. maxima паразитирует на Cronartium ribicola —
возбудителе ржавчины веймутовой сосны.
Особое внимание при разработке биологического метода
уделяется использованию аллелопатических отношений растений.
Сущность этих взаимоотношений заключается в том, что одни
растения оказывают влияние на другие через выделения активных
веществ, поступающих в окружающую среду при жизни растений
и после их отмирания.'Корни древесных пород выделяют большое
количество разнообразных по химическому составу веществ, ока-
146
зывая различное влияние на компоненты биогеоценоза. Корневые
выделения могут стимулировать или ингибировать рост
некоторых грибов и бактерий* Например, фитонциды, выделяемые
некоторыми деревьями (черемухой), кустарниками (желтой
акацией, смородиной черной) и травянистыми растениями (лютиком
едким, геранью луговой, пижмой), оказывают токсическое
действие на корневую губку.
Аллелопатические отношения растений представляют большой
практический интерес и требуют глубокого дальнейшего
изучения — это одно из перспективных направлений в разработке
биологических мер борьбы с корневой губкой и другими болезнями.
6.8. БИОФИЗИЧЕСКИЙ И МЕХАНИЧЕСКИЕ
МЕТОДЫ
Эти методы включают мероприятия, направленные на
предупреждение распространения болезней и уничтожение источников
инфекции. Данные методы, особенно в сочетании с другими,
эффективны, но применение их экономически оправданно
только на небольших площадях. К ним относятся: удаление больных
растений, обрезка больных частей растений, изоляция растений,
уничтожение промежуточных хозяев грибов, сбор и уничтожение
плодовых тел грибов, сбор и уничтожение опавшей пораженной
листвы и хвои, просушка и аэрация корневой системы, обжиг и
окорка пней, термическая обработка почвы и древесины.
Наличие в насаждении или в питомнике больных экземпляров
может привести к образованию очагов болезни. Так, чтобы
предупредить возникновение очагов снежного шютте, весной перед
выкопкой сеянцев или во время выкопки удаляют из посевов все
пораженные экземпляры. Это ограничивает дальнейшее
распространение болезни в питомниках и предупреждает ее появление в
культурах. В городских посадках, парках и скверах полностью
удаляют деревья ильмовых пород, пораженные голландской
болезнью. В связи с тем, что она может передаваться через корни,
больные пни выкорчевывают с корнями. Это мероприятие имеет
большое значение в условиях закрытого грунта, где несвоевременное
удаление пораженных растений приводит к быстрому
распространению болезни.
В городских, садовых, парковых и лесопарковых насаждениях
для предупреждения распространения болезней часто прибегают
к удалению пораженных органов: ветвей, корней, частей ствола.
При поражении некрозами обрезают ветви с плодоношениями
возбудителя или с другими признаками болезни. Лучше это делать
весной или осенью до начала сокодвижения и споруляции
возбудителей. В случае образования на стволах ран их удаляют, вырезая
147
древесину до здорового слоя. У деревьев, пораженных опенком,
обрезают больные корни, что предотвращает дальнейшее
распространение в них гнили и передачу болезни соседним здоровым
деревьям.
Для ограничения распространения болезней, передающихся
через корни, больные растения изолируют от здоровых. Так, для
изоляции пораженных корневой губкой или опенком молодых
культур или отдельных деревьев их окапывают канавами,
локализуя очаги или отдельные источники болезни.
В борьбе с болезнями, вызываемыми разнохозяйными
ржавчинными грибами, уничтожают промежуточных хозяев, если они
являются малоценными растениями. Например, чтобы
ограничить распространение ржавчины побегов сосны, уничтожают
поросль осины, а для предупреждения ржавчинного рака веймуто-
вой сосны — разные виды смородины, являющиеся
промежуточными хозяевами. Однако эта мера неприемлема в борьбе со
ржавчиной лиственницы и березы, так как они относятся к ценным
породам.
Чтобы ограничить распространение гнилевых болезней в
парках, садах и лесах I группы, целесообразно проводить сбор и
уничтожение плодовых тел дереворазрушающих грибов. Сроки сбора
зависят от видового состава грибов. Основную массу грибов
собирают в летний период. Обнаруженные плодовые тела срубают
топориком, собирают и сжигают.
В питомниках, молодых культурах, парках и садах проводят сбор
и сжигание опавшей листвы или хвои, на которых может
сохраняться зимующая стадия возбудителей болезней. На опавших
листьях перезимовывают возбудители различных пятнистостей,
мучнистой росы, парши, ржавчины тополя и побегов сосны.
Опавшая хвоя является источником инфекции таких заболеваний, как
шютте сосны, ели, лиственницы.
В садово-парковых условиях для предупреждения
распространения опенка производят просушку и аэрацию корневой системы
больных деревьев. Корни освобождают от земли и держат
открытыми в течение 3—4 мес. При этом грибница подвергается
действию высоких температур, иссушению и погибает.
В борьбе с опенком, корневой губкой часто обжигают пни. При
этом образуется щелочная среда, неблагоприятная для развития
опенка и корневой губки, но пригодная для поселения
дереворазрушающих грибов — пневых конкурентов корневой губки и
опенка. На обожженных пнях развиваются пениофора гигантская,
столбовой гриб и др.
Окорка пней является действенным мероприятием в борьбе с
болезнями, распространяющимися через поросль, и проводится
для того, чтобы предупредить ее образование. Это мероприятие
целесообразно в условиях садово-парковых хозяйств как профи-
148
лактика распространения вилта клена, черного некроза тополя,
сосудистого микоза дуба и др.
В борьбе со многими грибами используют высокие
температуры, губительные для мицелия, например с почвообитающими
грибами в питомниках и в теплицах. Термическая обработка
обеззараживает древесину, убивает мицелий и покоящиеся споры
возбудителей. Прогревание почвы можно осуществлять различными
способами. Термическую обработку древесины производят
горячим паром или воздухом. Температура внутренних слоев
древесины при этом должна быть не ниже 70 °С. Высокая температура не
только убивает грибницу, но и создает условия, неблагоприятные
для поселения других дереворазрушающих грибов.
6.9. ХИМИЧЕСКИЙ МЕТОД
6.9.1. Общие сведения
Химический метод защиты леса основан на использовании
органических и неорганических веществ, токсичных для вредных
организмов. Химические вещества наносят непосредственно на
вредные организмы, на поверхность различных органов растений
или вносят в среду обитания (почву, древесину, воздушную
среду). Химические средства защиты растений отличаются большой
универсальностью, их можно применять против большинства
вредителей и болезней леса на разных эколого-производственных
лесных объектах, в том числе складах, теплицах.
Ассортимент химических средств защиты растений в нашей
стране постоянно пополняется более эффективными и менее
опасными в экологическом отношении препаратами. Ведутся
активные поиски их оптимальных форм, удобных для хранения,
применения и менее опасных для работающих. Разрабатываются и более
эффективные способы применения пестицидов.
Одно из важных преимуществ этого метода перед другими —
возможность механизации. Использование новой совершенной
аппаратуры позволяет значительно повысить производительность
лесозащитных работ, снизить затраты и время на их проведение.
Кроме того, применение химических веществ, действующих
непосредственно на вредителей и возбудителей болезней, дает
возможность в короткий срок ликвидировать их очаги на больших
площадях.
Основной недостаток химического метода — отрицательное
влияние пестицидов на полезную фауну леса, микрофлору
почвы, их токсичность для человека и теплокровных животных. Обычно
при химических обработках лесных насаждений гибнут не только
вредные организмы, но и полезные — опылители растений, эн-
149
томофаги, микроорганизмы в почве и др. После многократных
химических обработок вредные организмы приобретают
устойчивость к химическим веществам, что снижает эффективность их
применения. Кроме того, результат использования химического
метода зависит от погодных условий. Сильные осадки смывают и
растворяют химические вещества, а ветер препятствует их
оседанию на растения во время обработки насаждений. Химические
вещества оказываются опасными для человека не только при
работе с ними, но и в связи с постепенным накапливанием в
организме, куда они попадают с зараженной пищей и из воздуха.
В районах интенсивной химической обработки растений резко
сократилась численность их опылителей (пчел, шмелей и др.).
Отрицательные последствия использования химических веществ
заставляют ограничивать их применение. Однако без химической
обработки невозможно ликвидировать очаги многих вредителей и
болезней. Поэтому изыскиваются наименее опасные химические
вещества избирательного действия, быстро разрушающиеся в
окружающей среде, совершенствуются методы и способы применения
пестицидов в целях уменьшения потерь активных ингредиентов и
сокращения норм их расхода. Одной из важнейших задач является
также совершенствование методов химического анализа
микроколичеств пестицидов в пищевых продуктах, фураже, почве, воде
и атмосферном воздухе.
6.9.2. Классификация химических средств защиты
растений
Химические вещества, используемые для защиты растений,
называются пестицидами (от лат. pestis — зараза,
разрушение, caedo — убиваю). Пестициды классифицируются по
химическому составу, объектам применения, а также по характеру
действия и способам проникновения в организм.
По химическому составу выделяют три основные группы:
неорганические соединения (соединения меди, серы, фтора,
бария и т.д.);
органические соединения (хлорорганические, фосфороргани-
ческие соединения, синтетические пиретроиды, производные
карбаминовой, тио- и дитиокарбаминовой кислот, нитрофенолы
и т.д.);
препараты растительного, бактериального и грибного
происхождения (пиретрины, антибиотики).
Объектами применения пестицидов являются вредители,
возбудители болезней растений, нежелательная растительность.
Для защиты растений от вредителей, относящихся к разным
классам животных, используют следующие основные группы Цести-
150
цидов: инсектициды (от лат. insectum — насекомое и caedo —
убиваю) — средства для борьбы с насекомыми; акарициды
(от греч. acari — клещ и лат. caedo — убиваю) — средства для
борьбы с клещами; моллюскоциды (от греч. mollusca —
моллюски и лат. caedo — убиваю) — для борьбы с моллюсками; з о о ци-
д ы (от греч. zoon — животное) — средства для борьбы с
вредными животными, в частности — с грызунами.
Гербициды (от лат. herba — трава и caedo — убиваю)
применяют для уничтожения травянистой растительности; ар бори-
циды (от лат. arbor — дерево и caedo — убиваю) — для
уничтожения древесно-кустарниковой растительности; а л ь г и ц и д ы (от
лат. algae — водоросль и caedo — убиваю) — для уничтожения
водорослей.
Пестициды, применяемые для защиты растений от
инфекционных болезней, подразделяются на следующие группы:
фунгициды (от лат. fungus — гриб и caedo — убиваю) —
используют для борьбы с грибными болезнями; бактерициды
(от греч. bacterion — палочка) — для борьбы с бактериальными
болезнями; вирусоциды (от лат. virus — яд) — для борьбы с
вирусными болезнями; нематоциды (от лат. nematodes —
круглые черви, фитогельминты) — для борьбы с нематодными
болезнями (фитогельминтозами); антисептики (отгреч. anti —
против, septikos — гнойный) — для защиты древесины от гнилей.
Классификация по объектам применения в известной степени
условна, так как многие пестициды обладают универсальностью
действия, т.е. способностью поражать разные группы вредных
организмов.
По характеру действия все пестициды подразделяют на две
большие группы: контактного и системного действия. К
контактным относят химические вещества, вызывающие гибель или
подавление вредных организмов при контакте с ними. Системные
пестициды способны проникать в растения, перемещаться в их
тканях и вызывать гибель возбудителя болезни, вредителя.
Пестициды должны отвечать требованиям ГОСТа, т. е.
соответствовать стандартам. Они предусматривают точное название
пестицида с указанием состава действующего химического вещества,
наполнителей, тонины помола для дустов и смачивающихся
порошков, влажности, способов отбора проб для анализа, а также
вида упаковки, условий и сроков хранения, технических условий
на изготовление пестицида.
Пестициды обычно изготовляют из дешевого недефицитного и
неогнеопасного сырья. Их применение должно быть экономически
выгодным. Пестициды перевозят в легкой и простой таре и
используют с малой нормой расхода на единицу обрабатываемой площади.
Госхимкомиссией при Министерстве сельского хозяйства
Российской Федерации совместно с Минздравом РФ и Госкомэколо-
151
гией РФ ежегодно утверждается «Государственный каталог
пестицидов, разрешенных к применению на территории Российской
Федерации», содержащий перечень пестицидов, используемых в
сельском, лесном, коммунальном хозяйствах, включая
фермерские и приусадебные. Он устанавливает основные регламенты
использования и контроля пестицидов. Препараты, не указанные в
Каталоге, запрещены к ввозу, продаже и применению на
территории Российской Федерации. Данный перечень пестицидов
постоянно пополняется более эффективными и менее опасными в
экологическом отношении препаратами с указанием форм их
применения.
Глава 7
ХИМИЧЕСКИЕ СРЕДСТВА БОРЬБЫ
С БОЛЕЗНЯМИ ЛЕСА
7.1. ОСНОВЫ ТОКСИКОЛОГИИ
Токсичность пестицидов. Способность химического вещества в
определенном количестве вызывать отравление организма
называется токсичностью. Мерой токсичности является доза —
количество вещества (в граммах или миллиграммах), вызывающее
отравление. Доза выражается в единицах массы пестицида по
отношению к единице массы, объема, площади. Различают
летальную, сублетальную и пороговую дозы пестицидов.
Летальная, или смертельная, доза — это наименьшее
количество ядовитого вещества, вызывающее гибель организма.
Сублетальная доза — это количество яда, вызывающее
серьезные нарушения функций организма, но не приводящее к смерти.
Пороговая доза — это количество яда, вызывающее
значительные изменения в организме.
Эффект действия пестицидов для исследуемых организмов
(спор, насекомых и т.д.) определяют по их гибели или по
наиболее характерным признакам отравления (изменению массы,
роста, снижению репродуцирующей способности и т.д.) и
выражают в процентах. Показатели токсичности обозначают буквенными
символами: СД (смертельная доза), ЛД (летальная доза), СК
(смертельная концентрация) и ЕД (эффективная доза) с указанием
эффекта. Если эффект действия пестицида учитывают по количеству
погибших объектов, то используют показатели СД, ЛД, СК. В
практике о токсичности пестицидов судят по усредненной
характеристике, чаще всего по дозам, вызывающим 50 %-й эффект (СД5о,
СК50). Токсичность пестицидов определяют опытным путем.
Показатели токсичности используют при определении норм
расхода пестицидов и при их гигиенической характеристике. На
токсичность пестицидов влияют многие факторы. Прежде всего
токсичность препарата определяется наличием в нем отдельных
химических элементов или групп (меди, ртути, серы и др.). Токсичность
пестицидов зависит от количества в них ядовитого (действующего)
153
вещества. Чем больше в препарате содержится действующего
вещества, тем выше его токсичность. На токсичность пестицидов
оказывает влияние строение молекул, изменение которого может
повысить или ослабить токсичность. Одним из факторов, влияющих на
токсичность, является продолжительность контакта действующего
вещества с организмом. Токсичность пестицидов зависит также от
их физико-химических свойств: размера и формы твердых частиц,
растворимости, стойкости, летучести, гигроскопичности.
На токсичность пестицидов влияют и условия внешней среды:
температура и влажность. С повышением температуры токсичность
химических веществ в большинстве случаев возрастает. Увеличение
влажности приводит, как правило, к понижению токсичности.
Токсичность пестицидов в значительной мере зависит от
биологических особенностей вредных организмов. Один и тот же фунгицид
может оказывать высокое токсическое действие на одни виды
возбудителей болезней и быть совершенно нетоксичным для других.
Например, фунгициды, содержащие в составе медь, токсичны для
многих видов ржавчинных грибов и нетоксичны для мучнисторосяных.
Вредные организмы в разных стадиях развития проявляют
неодинаковую чувствительность к пестицидам. Так, одни и те же
фунгициды являются токсичными для конидиальной стадии
грибов и малотоксичны или совсем нетоксичны для покоящихся
стадий (хламидоспор, склероциев, телиоспор).
Все факторы, влияющие на токсичность пестицидов,
используют при установлении их ЛД, норм расхода и концентраций.
Концентрация и норма расхода. В практике защиты растений
применяют и часто путают понятие дозы, или дозировки, с
нормой расхода и концентрацией. Рабочие составы, применяемые для
защиты растений, редко представляют собой чистое действующее
вещество. Чаще всего они содержат в своем составе определенное
количество действующего вещества и различные
вспомогательные вещества (наполнители, смачиватели, прилипатели и т.д.).
Содержащееся в рабочем составе количество действующего
вещества должно обеспечивать гибель вредного организма и не
оказывать отрицательного действия на защищаемое растение.
Концентрация — это количество действующего вещества
или препарата в рабочем составе, вызывающее гибель вредного
организма. Она выражается в процентах, массовых или объемных
единицах действующего вещества (концентрация по
действующему веществу) или препарата (концентрация по препарату),
содержащихся в определенном количестве рабочего состава.
Норма расхода— это количество действующего вещества,
препарата или рабочего состава, необходимое для обработки
единицы площади, объема или отдельного объекта. Нормы расхода
пестицидов указываются в ежегодно составляемом Каталоге, о
котором говорилось выше. Завышение рекомендованных норм рас-
154
хода препаратов может оказать отрицательное действие на
растения, а также привести к накоплению пестицидов в окружающей
среде и получаемой продукции.
7.2. ДЕЙСТВИЕ ПЕСТИЦИДОВ
НА ЖИВЫЕ ОРГАНИЗМЫ И ОКРУЖАЮЩУЮ
СРЕДУ
Действие пестицидов на биоценозы, в том числе и на лесные
биогеоценозы, сложно и многообразно. Систематическое
применение пестицидов вызывает частичное уничтожение насекомых-
опылителей, муравьев, водных беспозвоночных, птиц,
почвенной флоры и фауны, способствует появлению у вредных
организмов (фитопатогенных грибов, насекомых) устойчивости к
действию пестицидов.
Попадая в почву, пестициды изменяют состав ее микрофлоры.
Пестициды в основном быстро разлагаются в почве и вследствие
этого их влияние на почвенную флору и фауну незначительно.
Установлено, что почвенные фунгициды и фумиганты
отрицательно действуют на большинство представителей почвенной
микрофлоры. Но, подавляя одни виды, они могут стимулировать
развитие других. Например, карбатион подавляет развитие всех
почвенных микроорганизмов, кроме актиномицетов. Хлорпикрин
снижает количество грибов в почве, которое впоследствии
восстанавливается, но при этом изменяется их видовой состав. После
фумигации почвы преобладающим видом в ней часто становится
гриб Trichoderma viride — это сильный антагонист возбудителей
полегания и корневой губки.
Действие пестицидов на почвенные микроорганизмы
(положительное и отрицательное) изменяет плодородие почвы, что
сказывается на последующих звеньях лесного геоценоза.
Действие пестицидов на растения. Оно может быть
положительным или отрицательным в зависимости от свойств пестицида,
применяемой нормы расхода, видовых и возрастных
особенностей защищаемых растений, условий внешней среды.
Отрицательное действие на растения проявляется при
неправильном применении пестицидов. Повышение норм расхода
пестицидов приводит к серьезным нарушениям обмена веществ, что
выражается в угнетении роста и развития, повреждении и даже гибели
растений. Повреждающее (фитонцидное) действие пестицидов на
растения проявляется в образовании ожогов. Поврежденные листья
и хвоя приобретают бурый или коричневый цвет и отмирают.
Применение пестицидов в небольших дозах оказывает
стимулирующее действие на растения: повышает всхожесть и энергию
прорастания семян, усиливает рост и развитие растений.
155
Пестициды должны быть токсичными для вредных организмов
и безвредными для растений. Для определения пригодности
препарата к применению пользуются хемотерапевтическим
индексом, который характеризует сравнительную токсичность
пестицидов для вредных организмов и защищаемых растений.
Хемотерапевтический индекс — это отношение
минимальной дозы действующего вещества, убивающей вредный
организм (С), к максимальной дозе, безвредной для
защищаемого растения (/). Препарат пригоден в том случае, если отношение
С:/<1.
Действие пестицидов на человека и теплокровных животных.
Химические вещества, применяемые для защиты растений, в
большинстве случаев токсичны для человека и теплокровных
животных. Они могут проникать в их организм через дыхательные пути,
кожу, желудочно-кишечный тракт, накапливаться в отдельных
органах тела (печени, почках, легких, сердце). В местах
накопления пестициды подвергаются существенным изменениям
(метаболизму). Продукты метаболизма, как правило, менее токсичны,
чем исходные вещества, но могут быть и более токсичными. От
скорости метаболизма зависит концентрация пестицидов в
организме и степень их влияния на него. Пестициды вызывают острые
или хронические отравления в тех случаях, когда организм не
способен обеспечить своевременный вывод или разрушение
пестицида.
Острое отравление происходит при поступлении в организм
сразу больших доз действующего вещества. Оно сопровождается
резким нарушением функций и нередко кончается смертью.
Хроническое отравление наблюдается при систематическом
поступлении малых доз действующего вещества, способных
накапливаться в организме. Оно не приводит организм к гибели, но
вызывает его ослабление и нарушение функций. Хроническому
отравлению способствует загрязнение внешней среды пестицидами,
наличие остаточных количеств пестицидов в продуктах питания.
В действии пестицидов на человека различают пороговую,
токсическую и смертельную дозы. Токсичность пестицидов для
человека устанавливается в опытах на крысах. Доза, вызывающая
гибель 50 % животных, называется средней смертельной и
обозначается как СД5о или ЛД5о- По этому показателю все пестициды
делятся на четыре группы (определенное количество
миллиграммов на 1 кг массы):
I — сильнодействующие — ЛД5о меньше 50;
II — высокотоксичные — ЛД5о от 50 до 200;
III — среднетоксичные — ЛД5о от 200 до 1000;
IV — малотоксичные — ЛД5о превышает 1000.
Препараты, относящиеся к группе сильнодействующих,
используются очень редко и с большими ограничениями.
156
7.3. ПРЕПАРАТИВНЫЕ ФОРМЫ ПЕСТИЦИДОВ
Для борьбы с вредителями и болезнями леса применяют
следующие формы препаратов: дусты, смачивающиеся порошки,
гранулированные препараты, растворы в воде и органических
растворителях, концентраты эмульсий, аэрозоли, фумиганты.
Дусты — это порошки очень тонкого размола,
представляющие собой смесь действующего вещества (д. в.) и наполнителя.
В качестве наполнителя используют вещества, которые не
изменяют химических свойств пестицида: тальк, пирофилит, мел,
каолин, трепел, силикагель и различные глины. Предпочтение
отдают пирофилиту и тальку, так как они имеют слоистую структуру
и поэтому лучше прилипают к растениям. Для уменьшения
непроизводительной распыляемое™ и потерь в связи со сносом
мелких и мельчайших частиц к дустам добавляют 3 — 5 %
минерального масла. Оптимальный диаметр частиц дуста при наземных
обработках 15 — 25 мкм, а при авиаопыливании 25 — 50 мкм более
грубые дусты (с диаметром частиц 70 мкм и более) плохо
удерживаются на листьях растений.
Смачивающиеся порошки — порошковидные
пестициды, содержащие действующее вещество, наполнители и
поверхностно-активные вещества (ингредиенты). При разбавлении водой
они дают устойчивые суспензии. Их применение имеет
значительные преимущества: уменьшаются непроизводительные потери, так
как суспензии лучше прилипают к растениям и дольше
удерживаются на них. Смачивающиеся порошки высокодисперсны и
содержат 80 % частиц диаметром 30 мкм. В состав смачивающихся
порошков наряду с действующим веществом и наполнителем
входят поверхностно-активные вещества и прилипатели. Обычно
смачивающиеся порошки содержат 30 — 80% д.в., 15—60%
наполнителя, 2 —4 % поверхностно-активного вещества и прилипателя.
Смачивающиеся порошки часто используют для приготовления
водных суспензий, представляющих устойчивые взвеси твердых
частиц в воде. Для этого порошок сначала смешивают с небольшим
количеством воды до получения сметанообразной массы, которую
затем при постоянном помешивании соединяют с оставшимся
количеством воды. Более удобной формой порошковидных
пестицидов являются растворимые порошки, образующие в воде и
органических соединениях однородные стабильные составы — растворы.
Гранулированные препараты— это пестициды
зернистой формы, состоящие из действующего вещества и
наполнителя. Средний диаметр гранул составляет 0,25 — 5 мм.
Гранулированные препараты готовят путем пропитки пестицидом
гранул из минералов перлита, вермикулита или гранулированием
порошковидных препаратов. Такие препараты вносят в почву в
качестве защиты от обитающих в ней вредителей, для интоксика-
157
ции растений через корневую систему. Применение
гранулированных препаратов уменьшает опасность загрязнения
окружающей среды.
Растворы пестицидов в воде и органических
растворителях применяют редко, так как они имеют большое
поверхностное натяжение, вследствие чего плохо смачивают
обрабатываемую поверхность, а также неудобны при хранении и
транспортировке. Наиболее совершенны, эффективны и удобны в
применении и хранении масляные растворы.
Суспензии — это взвеси твердых частиц в воде. Для
приготовления суспензий используют смачивающиеся порошки. Чем
дольше твердые частицы находятся во взвешенном состоянии, чем
медленнее они оседают, тем равномернее распределяются
пестициды по обрабатываемой поверхности. Для повышения
стабильности суспензий в них добавляют вспомогательные вещества —
бонификаторы. Они способствуют лучшему покрытию и
удержанию раствора на растениях.
Концентраты эмульсий — жидкие или пастообразные
пестициды, содержащие действующее вещество, растворитель,
эмульгатор и смачиватель. При разбавлении водой образуют
устойчивые, долго не расслаивающиеся эмульсии, дисперсную фазу
которых составляют капельки масла с растворенным в нем
пестицидом, а дисперсную среду — вода. Концентраты эмульсий
готовят с применением гомогенизаторов.
Различают два типа концентратов эмульсий. Концентраты
первого типа получают диспергированием в воде раствора пестицида
в не смешивающемся с водой растворителе. Концентраты второго
типа состоят из пестицида, растворителя и эмульгатора. Для
получения концентрата эмульсии пестициды растворяют и
смешивают с эмульгаторами при нагревании до 40...80°С.
Применяют также обратные эмульсии, в которых дисперсной
фазой является пестицид, растворенный в воде, а дисперсной
средой — масло. При этом мелкие капельки водного раствора
пестицида как бы вмонтированы в крупные капли масла. Обратные
эмульсии используют для малообъемного опрыскивания, они
лучше прилипают к растениям и не испаряются.
Фумиганты представляют собой газо- или парообразные
пестициды, применяемые для обеззараживания почвы, складских
помещений, хранилищ.
7.4. СПОСОБЫ ПРИМЕНЕНИЯ ПЕСТИЦИДОВ
В борьбе с вредителями и болезнями леса, а также при защите
срубленной древесины пестициды применяют путем
опрыскивания растений, использования аэрозолей, путем фумигации по-
158
чвы, помещений, семян и посадочного материала, интоксикации
растений, протравливания семян, черенков и почвы,
изготовления отравленных приманок и токсичных поясов, антисептирова-
ния древесины.
Опрыскивание — нанесение пестицида в виде раствора,
суспензии или эмульсии на поверхность растений или тела
вредителя. К достоинствам этого способа относят сравнительно
небольшой расход пестицидов и равномерное покрытие ими
обрабатываемых поверхностей, хорошую прилипаемость, к недостаткам —
сложность приготовления рабочих составов, большой расход воды,
порчу аппаратуры в результате коррозии.
Эффективность опрыскивания зависит от величины,
количества и распределения капель жидкости на обрабатываемую
поверхность. При авиационных обработках различают крупнокапельное
(диаметр капель более 300 мкм), среднекапельное (151 — 300 мкм),
мелкокапельное (51 — 151 мкм) опрыскивание. В первых двух
случаях расходуется большое количество рабочей жидкости, что в
условиях лесного хозяйства экономически нецелесообразно.
Наиболее приемлем способ мелкокапельного опрыскивания с
расходом рабочей жидкости до 50 л/га. В зависимости от нормы расхода
при мелкокапельном опрыскивании различают малообъемное и
ультрамалообъемное опрыскивание.
Малообъемным называют опрыскивание со сравнительно
небольшой нормой расхода рабочей жидкости 5 — 50 л/га. Для этого
используют водные эмульсии и суспензии, мелкие капли которых
в наибольшей степени подвержены сносу и испарению. Поэтому
опрыскивание с расходом менее 20—25 л жидкости на 1 га
водными препаратами не применяется.
Ультрамалообъемное опрыскивание (УМО) производится не
разбавленными водой концентратами эмульсий с нормами расхода
0,5—10 л/га. По сравнению с малообъемным это опрыскивание
увеличивает производительность самолета более чем в четыре раза
и значительно удешевляет стоимость работ, не требует
предварительной подготовки рабочих растворов, уменьшает контакт
рабочих с химическими веществами. Концентраты для УМО должны
обладать текучестью при комнатной температуре, плотностью не
менее единицы, низкой токсичностью для теплокровных
животных, высокой биологической активностью и не ожигать растения.
При наземных обработках различают многолитражное,
малообъемное и ультрамалообъемное опрыскивание с расходом
рабочей жидкости соответственно 600—2000, 200 — 500 и 0,5—10 л/га.
При многолитражном опрыскивании размер капель составляет
120 — 300 мкм, а при малообъемном и ульрамалообъемном — не
более 100 мкм.
Аэрозоли — это дисперсные системы, содержащие частицы
во взвешенном состоянии. Аэрозоли делятся на туманы и дымы.
159
Туманы содержат капли жидкости, дымы — твердые частицы.
Аэрозоли при обработке зараженных насаждений оказывают
кратковременное воздействие на насекомых при непосредственном
контакте и остаточное последействие при накоплении инсектицида в
биомассе. В первом случае достигается высокая эффективность в
борьбе с насекомыми во взрослой фазе (летающие бабочки,
пилильщики). Аэрозоли хорошо проникают в кроны деревьев, щели
и трещины коры. При работе с аэрозолями значительно
уменьшается расход рабочей жидкости, увеличивается производительность
работы, улучшаются равномерность покрытия поверхностей
жидкостью и прилипаемость. Однако применение аэрозолей связано
с некоторыми осложнениями: обычно на туман воздействуют
воздушные потоки и разносят его. Определенные затруднения в
лесхозах вызывает также подготовка масляных растворов,
сопряженная с нагреванием масла для растворения в нем инсектицидов.
Фумигация — процесс введения пестицидов в паро- и
газообразном состоянии в воздушную среду, окружающую
вредителей и возбудителей заболеваний растений. Производится
фумигация почвы, складов и семенохранилищ, крон деревьев.
Своеобразным методом фумигации является введение отравляющих веществ
(парадихлорбензола и др.) в ходы скрытностволовых вредителей.
Интоксикация растений— внедрение в растение
безвредных для него химических веществ, растение становится
ядовитым для вредных насекомых и возбудителей болезней. Такими
веществами являются инсектициды внутрирастительного действия,
обладающие способностью проникать в растение и быстро
распространяться по его сосудистой системе. Этим методом можно
пользоваться для борьбы с сосущими и скрытноживущими
насекомыми.
Протравливание семян— это способ обработки семян,
он осуществляется в специальных аппаратах или вручную
порошковидными препаратами. При небольшом объеме семян их
перемешивают с пестицидом в плотно завязанных полиэтиленовых
мешках. Протравливание можно проводить непосредственно
перед посевом или заблаговременно. С целью повышения
эффективности сухого протравливания применяют клеящие вещества:
сульфитно-спиртовую барду, обезжиренное молоко, мучной клейстер.
Антисептирование — обработка поверхности древесины
химическими веществами, защищающими ее от разрушения
грибами, насекомыми, морскими древоточцами. Существуют разные
способы антисептирования древесины. При капитальном ремонте
зданий, пораженных домовыми грибами, применяют глубокую
диффузионную пропитку и поверхностную обработку древесины.
Антисептирование столбов и строительных деталей осуществляют
заводским (в автоклаве под давлением, методом горяче-холодных
ванн) и полевым (длительное вымачивание, суперобмазка, бан-
160
даж) способами. Выбор способов и средств антисептирования
зависит от свойств антисептика, назначения сооружений,
местонахождения защищаемых конструкций, влажности древесины,
наличия соответствующего оборудования и других условий.
7.5. ФУНГИЦИДЫ
К фунгицидам относятся органические и неорганические
вещества, обладающие фунгицидным и бактерицидным действием.
Распространение грибной и бактериальной инфекции
происходит главным образом по воздуху с помощью ветра, дождя,
посредством насекомых и человека в процессе ухода за растениями.
Большинство паразитов проникает в них через естественные
отверстия в растительных тканях (устьица, водные поры,
нектарники, чечевички в коре), и через повреждения механического
характера.
Есть и такие паразиты, которые внедряются в растение
непосредственно через эпидермис. Таковы, например, возбудители
настоящей мучнистой росы. Попав на растение, споры этих грибов
прорастают и внедряются в ткань, обеспечивая питание и
удерживаясь на пораженной поверхности. Гриб в этом случае
развивается на поверхности растений (экзопаразит). В большинстве же
случаев инфекция, попав в растение, развивается либо в
межклетниках, либо в клетке (эндопаразит). Развитие паразитов
внутри растения затрудняет их уничтожение, поэтому защитные
мероприятия чаще направлены на предупреждение заражения, чем
на уничтожение возбудителей.
Из всех групп пестицидов фунгициды наименее токсичны для
человека и теплокровных животных. Многие фунгициды не
оказывают отрицательного действия на защищаемые растения, а,
наоборот, стимулируют их рост и развитие. По характеру действия
на патоген фунгициды делят на защитные (профилактические) и
лечебные (искореняющие).
Защитные фунгициды воздействуют на возбудителя до
того, как произойдет заражение, и предотвращают развитие
болезни, но не способны уничтожить возбудителей, уже
внедрившихся в растительные ткани. Применяют такие препараты в
периоды, предшествующие массовому распространению инфекции.
Лечебные фунгициды (искореняющие) влияют на
вегетативные, репродуктивные органы возбудителей заболевания, а
также на их зимующие стадии, угнетая или вызывая гибель
патогена после того, как произошло заражение растения.
Эффективность лечебных фунгицидов зависит от времени, прошедшего с
момента внедрения возбудителя в ткани растений до начала
обработки их фунгицидами.
161
Таблица 3
Перечень фунгицидов, разрешенных для применения в лесных
и городских зеленых насаждениях в 2000—2001 гг.
Название препарата,
препаративная форма
Фундазол, СП1
Кемикар, СП
Медный купорос,
РП
Бордоская смесь, П
Действующее
вещество
Беномил
Карбоксин
Меди
сульфат
Меди
сульфат +
кальция гидро-
ксил
Норма
расхода
препарата
0,5-0,8
кг/га
6 кг/т
6 кг/т
~-
10—20 кг/га
30-60 кг/га
Объект защиты
Сосна
Хвойные породы
Тоже
Хвойные и
лиственные породы
Лиственные
породы в городских
насаждениях
Лиственные и
хвойные породы
в питомниках и
молодняках
Объект борьбы
Снежное шют-
те
Полегание
Тоже
Раны, дупла,
срезы
Парша, мони-
лиоз,
пятнистости
Ржавчина,
парша тополя и
ивы, шютте
лиственницы
Способ и время
обработки
Опрыскивание
осенью
Протравливание
семян
Тоже
Дезинфекция ран,
дупел, срезов 3—5 %-м
раствором
Опрыскивание
в период вегетации
Ранневесеннее
опрыскивание до или в
период распускания
почек
Кратность
обработок
1
1
1
1-2
1
Бордоская смесь, П
Картоцид, СП
Оксихлорид меди,
СП
Абига-Пик, ВС
1 Кумулус ДФ, ВДГ
Сера коллоидная,
|пс
|Топсин-М, СП
Меди
сульфат +
кальция гидро-
ксил
Меди трикап-
ролактам +
+ дихлорид
моногидрат
Меди
хлорокись
Тоже
Сера
Сера
Тиофанат-
метил
6 —12 кг/га
6 —15 кг/га
6 кг/т
3,6—4,8 кг/га
2—8 кг/га
2,4—9,6 кг/га
7—10 кг/га
12—15 кг/га
6 кг/т
Лиственные
породы в городских
насаждениях
Лиственные и
хвойные породы
в питомниках и
молодняках
Хвойные породы
Тоже
Лиственные и
хвойные породы
в питомниках и
молодняках
Тополь
Тоже
Лиственные
породы
Лиственные
породы
Хвойные породы
Ржавчина,
пятнистости,
парша
Ржавчина хвои
и листьев,
пятнистости
листьев
Полегание
Тоже
Ржавчина
листьев и хвои
Цитоспороз
Тоже
Мучнистая роса
Мучнистая
роса
Полегание
Опрыскивание
в период вегетации
Опрыскивание
в период вегетации
Протравливание семян
Опрыскивание
посевов при появлении
отдельных очагов
болезни
Опрыскивание
в период вегетации
Тоже
Тоже
Опрыскивание
в период вегетации
Опрыскивание
в период вегетации
Протравливание
семян
1-2 1
1-2
—
1-2
1
1
1 J
2
1
1
Окончание табл. 3
Название препарата,
препаративная форма
Топсин-М, СП
|ТМТД, СП
Байлетон, СП
Привент, СП
Действующее
вещество
Тиофанат-
метил
Тирам
Триадимефон
Тоже
Норма
расхода
препарата
2—4 кг/га
6 кг/т
1,5 кг/га
1,5 кг/га
2,4 кг/га
0,15-0,4
кг/га
Объект защиты
Сосна в
питомниках и
культурах до
5-летнего
возраста
Хвойные породы
Сосна в
питомниках
Тоже
Лиственница в
питомниках
Дуб
Лиственные
породы в городских
насаждениях
Объект борьбы
Снежное
шютге
Обыкновенное
шютге
Полегание
Снежное
шютге
Обыкновенное
шютге
Тоже
Шютге
Мучнистая роса
Мучнистая
роса, парша,
ржавчина
Способ и время
обработки
Опрыскивание
в осенний период
Опрыскивание
в летний период
Протравливание семян
Опрыскивание
осенью
Опрыскивание
в летний период
Тоже
Опрыскивание
в летний период
Тоже
Тоже
Кратность
обработок
1-2
1-2
1
1-2
1-2
1
1
1
2
1 Условные обозначения препаративных форм фунгицидов: ВС — водная суспензия; ВДГ — вододиспергируемые гранулы;
ПС — паста; П — порошок; РП — растворимый порошок; СП — смачивающийся порошок.
По характеру распределения фунгицидов в растении их
подразделяют на фунгициды контактного действия и системные (внут-
рирастительные).
Контактные фунгициды не проникают в растения. Они
могут ограниченно передвигаться с одной поверхности листа на
другую и действовать на возбудителя болезни при
непосредственном контакте. К этой группе относится большинство применяемых
в настоящее время фунгицидов. Продолжительность действия
контактных фунгицидов определяется временем нахождения их на
поверхности растений и зависит от метеорологических условий.
Системные фунгициды — соединения или продукты
их распада, которые быстро поглощаются растением,
переносятся из корней в листья, из старых листьев в молодые и т.д.; в
концентрациях, не причиняющих вреда растению,
предупреждают заражение всего растения или уничтожают уже внедрившихся
в него возбудителей заболеваний. Продолжительность действия
системных фунгицидов в незначительной степени зависит от
метеорологических условий и составляет от 20 до 70 дней.
Современные фунгициды и особенности их применения для
борьбы с разными болезнями древесных пород в лесных и
городских зеленых насаждениях представлены в табл. 3, составленной
на основе государственного каталога.
7.6, ПРАВИЛА ТЕХНИКИ БЕЗОПАСНОСТИ
ПРИ РАБОТЕ С ПЕСТИЦИДАМИ
Все работы с пестицидами, их отпуск, прием,
транспортировка должны проводиться в строгом соответствии с
существующими инструкциями по технике безопасности. Лица, работающие с
пестицидами, должны быть проинструктированы и снабжены
защитными средствами.
При работах с порошкообразными препаратами и
фунгицидами, обладающими высокой летучестью, следует надевать
спецодежду, защитные очки, респираторы и резиновые перчатки.
Использование при этом медицинских резиновых перчаток запрещается.
После окончания работ спецодежду необходимо вытряхнуть на
открытом воздухе. При сильном загрязнении ее замачивают в 1 %-м
растворе марганцовокислого калия или в 2 %-м растворе соды и
стирают в горячей воде. Резиновую обувь и перчатки тщательно
обливают водой, обрабатывают хлорной известью и снова моют
теплой водой. Лицевые части респираторов следует промыть
теплой водой, обработать 0,5 %-м раствором марганцовокислого
калия и еще раз промыть водой.
Во время работы с пестицидами нельзя курить, пить и
принимать пищу. Перед курением, едой и питьем необходимо вымыть с
165
мылом руки, а после работы лицо. Участки, использовавшиеся
для приготовления рабочих составов, после окончания работ
должны быть перекопаны. Тару из-под пестицидов, непригодную для
дальнейшего использования, сжигают, а золу закапывают. После
окончания работ вся аппаратура должна быть промыта и сдана на
склад.
Перевозка пестицидов производится в исправной таре, после
перевозки транспорт должен быть тщательно очищен.
Категорически запрещается перевозить пестициды вместе с пищевыми
продуктами и кормами. В местах, где проводилась обработка
пестицидами, в течение 25 дней запрещается выпас скота, косьба, сбор
грибов и ягод. Продолжительность рабочего дня при работе с
пестицидами не должна превышать 6 ч. Занятость лиц, работающих
с сильнодействующими пестицидами, не должна превышать 4 ч.
Все работы по защите растений производятся под руководством
специалистов или лиц, прошедших специальную подготовку.
Глава 8
НЕИНФЕКЦИОННЫЕ БОЛЕЗНИ
ДРЕВЕСНЫХ ПОРОД
Неинфекционные болезни — это поражения древесных
растений, вызываемые абиотическими факторами. К ним относятся
неблагоприятные для растений почвенно-климатические условия,
воздействие промышленных выбросов, хозяйственная деятельность
человека, рекреационный пресс, механические повреждения.
Нередко древесные растения подвергаются воздействию комплекса
факторов, и в этом случае физиологические изменения
ускоряются, проявляются внешние признаки болезни, процессы
ослабления и гибель растений протекают быстрее.
8Л. ОТРИЦАТЕЛЬНОЕ ВЛИЯНИЕ ПОЧВЕННЫХ
УСЛОВИЙ
Почва — одна из составных частей лесной экосистемы,
поэтому различные изменения в почве приводят к нарушению ее устой-
чивости. Отрицательное влияние на лесные биогеоценозы
оказывают избыточное увлажнение и дефицит влаги в почве,
недостаток или избыток питательных веществ, уплотнение почвы.
Избыток влаги. Причинами избыточного увлажнения почвы
могут быть: обильные продолжительные дожди, быстрое таяние
снега весной, заболачивание почвы, подъем уровня грунтовых
иод. В этих случаях в почве образуется недостаток кислорода,
происходит нарушение нормальных окислительных процессов и
образование различных, вредных для растений продуктов, какими
ииляются органические кислоты, углекислота, соли закиси желе-
аа. В результате деятельность корневой системы нарушается, сни-
м ается поглощение корнями питательных веществ, деревья
слабеют или погибают.
Избыточное увлажнение почвы может быть приурочено к
местам естественного заболачивания и поднятия уровня грунтовых
167
вод вследствие выпадения осадков в количестве, превышающем
норму на протяжении нескольких лет. Оно наблюдается на
участках со строящимися и эксплуатируемыми дорогами,
преграждающими сток воды, и в местах с загрязненными осушительными
каналами, которые не обеспечивают сток воды.
Древесные породы устойчивы к весенним и осенним
затоплениям, но большинство их не выносит длительного летнего
затопления. Наиболее устойчивы к затоплению сосна, береза, дуб,
менее устойчивы ель, вяз.
Переувлажнение почвы приводит к ослаблению и усыханию
насаждений, а в питомниках — к снижению грунтовой всхожести
и загниванию корневой системы всходов. В ослабленных
насаждениях нередко возникают очаги стволовых вредителей.
Дефицит влаги в почве. Он может возникнуть из-за регулярно
повторяющихся засух, а также падения уровня грунтовых вод,
вызываемого строительством гидросооружений. Недостаток воды
в почве приводит к нарушениям жизнедеятельности деревьев,
которые проявляются в увядании кроны, засыхании листьев, усы-
хании вершин, и к гибели растений.
Последняя волна усыхания дубрав, начавшаяся в 1964 г.,
охватила почти весь ареал дуба. Такой же периодический характер носит
и усыхание ельников, его грандиозный размах отмечался после
засух 1936, 1938, 1939 гг. (А.И.Воронцов). Более позднее усыхание
еловых насаждений наблюдалось в период 1963 — 1971 гг. в
Калининградской области и отмечалось не повсеместно, а только в
отдельных районах, где количество осадков было ниже нормы. Усыхание
ели сопровождалось массовым размножением короеда-типографа.
Увядание и засыхание листьев от недостатка воды в почве
наблюдается у многих лиственных пород. Массовое увядание и
засыхание листьев в городских посадках, защитных лесных полосах
отмечали во время летней засухи 1972 г., охватившей несколько
районов страны. У многих древесных пород преждевременный
листопад привел к ослаблению деревьев, а в отдельных случаях и
к их гибели.
Для питомников и молодых культур большую опасность
представляют весенние засухи. При этом снижается грунтовая
всхожесть, всходы появляются недружно и теряют устойчивость к
болезням, в частности к полеганию. Так, в питомниках Бузулукско-
го бора после весенней засухи 1975 г. пораженность всходов сосны
фузариозом достигала 50—70 %. Недостаток влаги в почве при
высокой температуре воздуха может вызывать массовое усыхание
насаждений.
Недостаток и избыток питательных веществ в почве. Для
нормального роста и развития растений требуется достаточное
количество питательных веществ и их определенное соотношение.
Недостаток какого-либо одного элемента питания не может быть
168
восполнен избытком другого. При недостатке питательных веществ
у растений нарушаются жизненные функции, что влечет за собой
различные патологические изменения. Для нормального роста
растению необходимы основные элементы питания: азот, калий,
фосфор, кальций, железо, сера и микроэлементы: бор, медь,
марганец, молибден, цинк и др. Избыток питательных веществ
также может привести к патологическим изменениям.
Азот — один из основных элементов питания растений. Он
входит в состав аминокислот, из которых строятся белки. Для
обеспечения нормального роста и развития азот необходим растениям
в большом количестве. С его недостатком связано угнетение роста
растений. При продолжительном азотном голодании побеги у
древесных пород становятся тонкими, короткими, твердыми, а
листья мелкими с желтовато-зеленой окраской. У некоторых пород в
листьях накапливается красноватый пигмент. Иногда наблюдается
преждевременное их опадение. Недостаток азота наиболее сильно
проявляется на переувлажненных почвах, особенно после
ливневых дождей, когда легкорастворимые соединения азота
вымываются в более глубокие слои почвы, недоступные корням.
Недостаток в связанном азоте восполняют почвенные бактерии,
осуществляющие процессы нитрификации. В холодную засушливую
погоду эти процессы замедляются, в таких условиях наблюдается
азотное голодание.
Избыток азота вызывает усиленное нарастание зеленой массы
с мягкими, рыхлыми тканями, долго не вызревающими и менее
устойчивыми к болезням и заморозкам. Например, при
избыточном количестве азота в почве сеянцы дуба, клена, березы и
других древесных пород сильно поражаются мучнистой росой. Нео-
древесневшие побеги дуба часто побиваются ранними
заморозками и отмирают. У цветочных культур избыток азота замедляет фазу
цветения, уменьшает количество цветков, снижая их сортность.
Калий — необходимый элемент питания растений. Он
находится в почве в виде ортоклаза, который растениями не может
усваиваться. В доступную для растений форму калий переводят
силикатные бактерии. Недостаток калия вызывает угнетение роста
древесных пород, при этом наблюдается слабое развитие
побегов, они часто искривляются. Края листьев и участки между
жилками желтеют, затем буреют и отмирают. У некоторых пород
листья становятся морщинистыми. Устойчивость растений к холоду
понижается. У цветочных культур недостаток калия вызывает
хлороз листьев, т. е. желто-зеленую окраску, побурение и отмирание
ткани около листового побега. При избытке калия наблюдается
задержка роста цветочных культур, побурение листьев,
ухудшается картина цветения.
Фосфор — является составной частью нуклепротеидов и фос-
фолипидов и служит главнейшим источником энергии у растений.
169
Недостаток его вызывает угнетение роста растений, листья
приобретают темно-зеленую или бронзовую окраску. Хвоя сеянцев
сосны приобретает фиолетовый цвет, который обнаруживается
осенью. При длительном фосфорном голодании формируются
тонкие побеги, листья становятся мелкими. У цветочных культур при
фосфорном голодании наблюдается укорачивание стеблей, побу-
рение и отмирание листьев.
Кальций входит в состав клеточных оболочек растений и
служит связующим веществом клеток. Кроме того, он
нейтрализует некоторые органические кислоты, вырабатываемые в
процессе жизнедеятельности, которые могут быть вредными для
растений. Недостаток кальция отрицательно влияет на развитие
корневой системы древесных пород, наблюдается отмирание
верхушек побегов, отмирают также ткани по краям или вдоль средней
жилки листьев.
Железо — важный элемент, обеспечивающий нормальную
жизнедеятельность растений. Оно способствует образованию
хлорофилла и в больших количествах содержится в листьях. В кислых
почвах железо находится в доступной для растений форме. При
повышении значения рН оно переходит в труднорастворимые
соединения, малодоступные для растений. Недостаток железа
препятствует образованию хлорофилла и приводит к возникновению
хлороза листьев и хвои, который выражается в бледной, иногда
белой окраске хвои и листьев. Следствием хлороза является
нарушение ассимиляции, что в свою очередь приводит к гибели
растения. У сеянцев, пораженных хлорозом, отмечается угнетение
роста корневой системы, хвои, листьев, приостанавливается рост,
и в итоге сеянцы отмирают.
Магний входит в состав хлорофилла и некоторых
органических соединений, образующихся в растении. При недостатке
магния у древесных пород на листьях появляются желто-бурые пятна.
Вскоре все листья опадают, за исключением розетки
темно-зеленых листьев на верхушках побегов. Раннее опадение листвы
приводит к ослаблению растений, особенно молодых. У цветочных
культур магниевое голодание вызывает пожелтение нижних листьев,
угнетает образование боковых корешков, что отрицательно
сказывается на поступлении питательных веществ и воды в растение.
Медь— потребность в ней испытывают растения,
произрастающие на почвах, имеющих высокую концентрацию
органических веществ и щелочную реакцию. Недостаток меди вызывает
снижение содержания белкового азота, органического фосфора,
падение интенсивности фотосинтеза. При медном голодании
древесные растения отстают в росте и не плодоносят. Избыток меди
токсичен для растений.
Бор потребляется растениями в небольших количествах.
Поскольку он находится в равновесии с кальцием, то его недостаток
170
связан с избытком кальция в почве. При обратном соотношении
образуется избыток бора. Недостаток его вызывает отмирание у
растений верхушечных почек, нарушает цветение и
плодоношение. При избытке бор накапливается в листьях, они желтеют и
преждевременно опадают, что приводит к ослаблению растения.
Цинк является составной частью карбоангидразы —
фермента, активизирующего дыхание растений и способствующего
синтезу гетероауксина. Недостаток цинка вызывает пожелтение
листьев, розеточность и мелколистность, образование мелких
уродливых плодов.
Вредное действие на растения оказывает избыток солей в
почве. Соли повышают осмотическое давление почвенного
раствора, всасывающая сила корней уменьшается, происходит
нарушение водоснабжения растений. Кроме того, некоторые соли в
высоких концентрациях токсичны для растений. Наиболее вредны
для древесных пород: сода (Na2C03), сернокислый (Na2S04) и
хлористый натрий (NaCl). Избыток этих солей вызывает
пожелтение и засыхание листьев и хвои сеянцев.
8,2. ОТРИЦАТЕЛЬНОЕ ВЛИЯНИЕ
МЕТЕОРОЛОГИЧЕСКИХ УСЛОВИЙ
Влияние ветра. Под влиянием ветра в лесах возникают
ветровалы и буреломы, причиняющие вред лесному хозяйству,
происходит обеднение почвы, нарушается нормальный рост и развитие
посадок.
Ветровал — это отдельные деревья и древостой,
поваленные с корнями, что происходит при слабой связи корней с
почвой. Прежде всего страдают деревья со слаборазвитой и
поверхностной корневой системой. Из хвойных пород наиболее устойчивы
к ветровалу сосна, кедр и лиственница, из лиственных — дуб, граб,
ильм, ясень. Сильнее других пород ветровалу подвергается ель.
Степень ветровальности деревьев в значительной мере зависит
от почвенных условий. Чаще всего ветровал происходит в
насаждениях на мелких, рыхлых переувлажненных почвах, где сила
сцепления корней с почвой недостаточна для противостояния силе
ветра. Большие площади ветровалов отмечаются в горах с
насаждениями на мелких почвах с каменистой подстилающей породой,
у деревьев развивается поверхностная корневая система, не
способная противостоять силе ветра, особенно на ветроударных
склонах. Степень устойчивости к ветру одних и тех же пород может
меняться в зависимости от условий произрастания.
Ветроустойчивая сосна в горных условиях и на сухих песчаных, мелких и
заболоченных почвах в значительной степени подвергается ветровалу.
Неустойчивая ель хорошо противостоит ветру на богатых, хоро-
171
шо развитых почвах. Более устойчивы к ветровалу смешанные
насаждения. Ветровалу в значительной степени способствует
поражение деревьев корневыми гнилями, вызываемыми корневой
губкой {Heterobasidion annosum), опенком {Armillaria mellea) и др.
Бурелом — это деревья, сломанные ветром. Он
характеризуется разломом ствола ниже кроны, что происходит в случае,
когда корни хорошо укреплены в почве, а сила ветра превосходит
сопротивление ствола по излому. Устойчивость к бурелому зависит
от породы деревьев, обладающих древесиной различной
прочности. Наименее устойчивы к буреломам хвойные породы. Степень
устойчивости к буреломам тесно связана с возрастом
насаждений: больше страдают спелые и перестойные насаждения,
которые сильнее поражаются гнилями и раковыми болезнями.
По нашим данным, в Мурманской области 100% буреломных
деревьев ели поражены гнилями, вызванными еловым комлевым
трутовиком {Onnia triqueter), еловой губкой {Phellinus chrysoloma),
окаймленным трутовиком {Fomitopsis pinicola). В Бузулукском бору
75 % бурелома осины было поражено черным раком (Hypoxylon pruina-
tum). В Ростовской области 95 % бурелома в чистых насаждениях
ясеня оказалось пораженным эндоксилиновым раком (Endoxylina
stellulata). В Тебердинском заповеднике среди бурелома пихты
преобладали деревья, пораженные ржавчинным раком (Melampsorella
caryophyllacearum). Несвоевременное удаление поваленных
деревьев приводит к массовому размножению стволовых вредителей,
которые впоследствии могут заселять соседние здоровые деревья.
Кроме ветровала и бурелома, в насаждениях, подвергающихся
постоянному действию ветра (морские побережья, ветроударные
склоны и т.д.), у деревьев наблюдаются анатомические
изменения древесины, проявляющиеся в свилеватости и крени стволов,
которые затрудняют ее механическую обработку. В городских
насаждениях ветровал и бурелом представляет большую опасность
для населения, транспорта, жилых зданий и других помещений.
Массовому ветровалу и бурелому в годы с ураганными ветрами в
городских насаждениях в большинстве случаев способствуют
гнили, вызываемые ложным {Phellinus igniarius), настоящим {Fomes
fomenatarius), серно-желтым {Laetiporus sulphureus), чешуйчатым
{Polyporus squamosus), плоским {Ganoderma applanatum)
трутовиками, феллинусом точечным {Phellinus punctatus).
Ветер оказывает отрицательное влияние и на почву. На
опушках, горных хребтах, морских побережьях вследствие постоянных
ветров почва осушается, сдуваются ее верхний тонкий слой и
почвенная подстилка. Это вызывает снижение прироста у деревьев,
препятствует естественному возобновлению, приводит к
расстройству молодых насаждений. Таким образом, вред, причиняемый
ветром, заключается в снижении продуктивности насаждений,
выхода и качества деловой древесины.
172
Влияние осадков. Осадки в виде снега, ожеледи, града и дождя
могут причинять существенный вред лесным насаждениям. Снег,
задерживаясь на кронах в большом количестве, вызывает
снеголомы и снеговалы.
Снеголом — это деревья, у которых стволы или вершины
сломаны под тяжестью снега. При снеговале деревья бывают
повалены с корнями под тяжестью снега. Скоплению снега
способствует резкая смена температур. При теплой, безветренной
погоде снег падает крупными, пушистыми, влажными хлопьями
и легко примерзает к стволам и ветвям во время резкого
понижения температуры. Большой вред причиняют снеголомы и снеговалы
молодым насаждениям. От снеголома особенно сильно страдает
сосна, ель, осина. При снеговале чаще ломаются деревья с кривыми
стволами, слабо развитые, пораженные болезнями и
поврежденные вредителями. Снеголому сосновых культур и подроста может
способствовать смоляной (Cronartium flaccidum) и биаторелловый
(Biatorella difformis) рак.
В горных условиях массовые снеголомы и снеговалы часто
вызываются снежными лавинами. Участки снеголомов и снеговалов
нередко становятся очагами стволовых вредителей, ускоряющих
усыхание деревьев. Более высокой устойчивостью к навалу снега
обладают неоднородные по возрасту и составу насаждения. Резкая
смена погоды зимой вызывает образование на стволах и ветвях
ледяного слоя, называемого ожеледью. Под ее действием
обламываются ветви и молодые стволики. Повреждения ожеледью
большей частью наблюдаются в степных лесах. Но в отдельные годы
массовое повреждение ожеледью отмечается в городских
насаждениях, где она наносит большой вред декоративным посадкам.
Наиболее сильно страдают акация белая, ясень, берест, вяз, дуб.
Существенный вред может причинять деревьям град. При этом
чаще страдают их нежные части: листья, хвоя, цветки, плоды,
побеги и тонкая кора. В результате преждевременной потери
листьев или хвои у молодых деревьев отмечается потеря прироста,
отмирают молодые побеги. Сильное повреждение градом может
вызвать массовое усыхание деревьев. Наибольшую опасность
градобой представляет для лесных и декоративных питомников и
городских посадок, особенно молодых. Поврежденные градом
деревья заселяются стволовыми вредителями, которые ускоряют
процесс усыхания. Кроме того, через повреждения градом (градобои-
ны) часто проникают возбудители болезней. Значительный вред
древесным растениям могут причинять дожди ливневого
характера, вызывающие опадение цветков, завязи, вымывание семян и
плохо укоренившихся всходов и сеянцев в лесных питомниках.
Влияние температуры. Все высшие растения развиваются при
определенных температурах, которые неодинаковы для
различных видов растений, а также для их органов. Для главнейших дре-
173
весных пород установлены оптимальные, минимальные и
максимальные температуры. Отклонение температур за пределы
минимума или максимума, а также резкая смена температур
отрицательно влияют на растения, вызывая патологические изменения,
которые нередко приводят к гибели растений.
Низкие температуры вызывают обмерзание кроны и
подмерзание корней, образование морозобойных трещин, выжимание
сеянцев в питомниках. Температура зимой до -30 °С и ниже (до
-40...-45°С), сохраняющаяся в течение продолжительного
времени, вызывает обмерзание побегов, а при малоснежных зимах —
и корней деревьев. Наиболее сильно от морозов страдают
молодые растения. Деревья старшего возраста чаще полностью
оправляются на второй год после обмерзания, реже погибают.
Суровые зимы 1939/40, 1941/42 и 1978/79 гг. явились одной из
причин массового усыхания дубрав в центральных районах,
Среднем и Нижнем Поволжье. В эти же периоды отмечалась массовая
гибель садов на европейской части России. Уцелевшая часть
деревьев к началу вегетации была сильно ослаблена. Впоследствии
такие деревья заселяются стволовыми вредителями или поражаются
грибными болезнями, вызывающими их усыхание.
Значительный вред древесным растениям, особенно молодым,
могут причинять поздние весенние и ранние осенние заморозки.
Особенно опасны поздние заморозки в годы с необычно теплыми
веснами. Сильнее страдают от заморозков растения в пониженных
участках, где застаивается холодный воздух. Поздние весенние
заморозки повреждают молодые листья, побеги, бутоны, вызывая
их отмирание. Пораженные части растений чернеют, засыхают и
обламываются. Как правило, более устойчивы к весенним
заморозкам поздно распускающиеся
формы.
Часто после повреждения
дуба поздними весенними
заморозками происходит
поражение вторичных побегов
мучнистой росой, которые не
успевают одревеснеть и
побиваются ранними заморозками.
Засыхание побегов у молодых
дубков вызывает потерю
прироста и ослабление.
При резком понижении
температуры зимой на стволах
Рис. 41. Повреждение стволов низкими деревьев появляются мороз-
температурами: ные трещины (рис. 41). По дли-
1 - морозобойная трещина; 2 - морозо- н^ ОНИ могут достигать 1 м И
бойныйрак более, по глубине распро-
174
страняются до сердцевины. Они образуются вследствие того, что
наружные слои древесины при внезапном понижении
температуры сжимаются, а внутренние остаются менее сжатыми. Морозо-
бойные трещины образуются как на хвойных, так и на
лиственных породах, но чаще на последних. Наиболее сильно
поражаются твердолиственные породы: клен, дуб, бук, ясень, вяз. Но
довольно часто морозные трещины встречаются и на мягких
лиственных породах: липе, каштане конском, тополе.
Однородные древостой поражаются неодинаково в разных
типах лесорастительных условий. Наиболее сильно страдают
древостой в пойменных условиях, здесь увеличивается не только
количество пораженных деревьев, но и количество морозобойных
трещин на дереве. Высокополнотные насаждения меньше страдают
от морозобоя, чем низкополнотные. Поражение морозными
трещинами увеличивается на опушках леса и открытых участках в
городских насаждениях в связи с повышением перепадов
температур. С возрастом устойчивость деревьев к морозобою снижается.
На образование морозобойных трещин влияют особенности
анатомического строения и физиологического состояния
древесины, которые определяются типом лесорастительных условий.
Наибольшее значение в снижении прочности древесины на
растяжение в тангентальном направлении, что происходит при морозо-
бое, имеет высота многорядных сердцевинных лучей, изменяющаяся
в зависимости от типа лесорастительных условий. С увеличением
количества таких лучей в древесине и ж высоты прочность
древесины на растяжение уменьшается. Так, у дуба в пойменных
условиях высота сердцевинных лучей значительно больше, чем в
плакорных условиях. Этим и объясняется сильное поражение
морозобоинами деревьев в пойменных условиях.
В случае систематического повреждения морозом на месте
морозобоиной трещины образуется раковая рана, которая может
разрастаться из года в год (см. рис. 41). Деревья с морозобойными
трещинами и ранами часто бывают ослаблены, суховершинят,
реже — усыхают. Повреждение морозобойными трещинами снижает
выход деловой древесины, способствует проникновению
возбудителей гнилей, снижает устойчивость деревьев к бурелому. Пора-
женность морозобоинами дуба в спелых древостоях центральной
лесостепи превышает 30%, а в хвойно-широколиственных лесах
Среднего Поволжья достигает 50 % и выше.
Опасность появления морозных трещин и ран возрастает в ур-
боэкосистемах, где состояние пораженных морозобоем деревьев
усугубляется действием многих антропогенных факторов. По
данным обследования, в городских насаждениях г. Москвы и
Подмосковья поражение морозобоем разных древесных пород
составляет: у каштана конского — 10—50 %, у вяза гладкого — 5—20 %,
у клена остролистного — 10—20%, у липы — 2—10 %.
175
Низкие температуры вызывают выжимание сеянцев и
саженцев, которое чаще всего отмечается на переувлажненных
почвах. При понижении температуры пары воды конденсируются
в верхнем слое почвы, а из нижних слоев почвы поднимаются
новые пары воды. Если температура воздуха понижается постепенно
до точки замерзания и ниже, то в верхнем слое почвы образуются
кристаллы льда, которые вытягивают воду из нижележащих
слоев почвы. При этом лед нарастает снизу, примерзает к корневой
шейке и поднимает растение за собой. Корни, находящиеся в
промерзшей почве, обрываются. При оттаивании почва оседает, а
растения, оказавшиеся на ее поверхности, засыхают.
Выжимание может вызываться и резким понижением
температуры, когда теплая дождливая погода сменяется температурой ниже
О °С. При этом верхний слой почвы быстро промерзает, а на
границе с замерзшей почвой образуются кристаллы льда, которые
начинают оттягивать воду из нижележащих слоев. Увеличивающийся
слой льда приподнимает верхний замерзший слой вместе с
растением. При последующем потеплении оттаявший верхний слой
земли оседает, а растения оказываются на поверхности и погибают.
Так, в Мурманской области поврежденность саженцев сосны и
ели достигала 66—74 %, лиственницы — 45 %. Значительная
разница в поражении разных пород объясняется особенностями
строения корневой системы. У лиственницы корневая система
проникает глубоко в почву и сильно ветвится, что обеспечивает прочное
сцепление корней с почвой и большую устойчивость к
выжиманию по сравнению с сеянцами сосны и ели. В питомниках
Среднего Поволжья от выжимания в отдельные годы погибает до 20 %
сеянцев ели.
Высокие температуры, превосходящие максимум, также
вызывают патологические изменения у растений. Длительные
засухи с температурой выше 30...35°С, сопровождающиеся
дефицитом влаги в почве, приводят к массовому усыханию
насаждений. Смена дождей солнечной погодой с высокой температурой
воздуха вызывает ожог листьев, причиной которого являются
задерживающиеся между жилками листьев капли дождя,
концентрирующие солнечные лучи. Ткани листа в этих участках
перегреваются, а затем отмирает и весь лист.
Действие сильных солнечных лучей у деревьев с тонкой
гладкой корой вызывает ожог коры. Пораженная кора
приподнимается, в ней появляются продольные и поперечные трещины. Позже
отмершие участки коры опадают, обнажая древесину, которая
подсыхает и растрескивается. Через трещины в ткани дерева
проникают фитопатогенные грибы, в том числе возбудители гнилей и
некрозно-раковых болезней. Деревья с ожогом коры
ослабляются, у них отмечается падение прироста, нередко они заселяются
стволовыми вредителями.
176
В питомниках у всходов хвойных пород под действием высокой
температуры происходит ожог или опал корневой шейки. При опале
у корневой шейки образуется хорошо заметная перетяжка,
вследствие чего всходы падают. В питомниках Среднего Поволжья гибель
всходов от опала корневой шейки при температуре на поверхности
почвы до 42 °С составляла на посевах лиственницы 25 %, а на
посевах ели 20 %. При температуре почвы 44 °С гибель всходов сосны
достигала 48 %'. Чаще всего опал корневой шейки встречается в
южных районах, где температура поверхности почвы повышается
до 70 °С и выше. Наибольший опал всходов отмечается на темно-
окрашенных почвах, которые нагреваются особенно сильно.
Резкие колебания температур вызывают у деревьев
образование отлупных трещин, ожог коры, усыхание сеянцев.
Отлупные трещины (отлупы) образуются у деревьев при
резком повышении температуры после сильных морозов. В данном
случае наружные слои древесины нагреваются и расширяются, а
внутренние остаются сжатыми, поэтому между годичными
слоями развивается кольцевая трещина. Отлупы образуются у всех
древесных пород в нижней части ствола и распространяются по его
длине на несколько метров. Отлупные трещины нарушают
целостность древесины, снижают выход высокосортных сортиментов.
Резкие колебания температур в зимний период вызывают ожог
коры древесных пород. Продолжительные оттепели в конце зимы
или ранней весной преждевременно выводят из периода покоя
отдельные органы или все дерево. Следующие за оттепелями
резкие понижения температуры вызывают отмирание коры, луба или
камбия, чаще всего с южной стороны ствола. Пораженная кора
Рис. 42. Повреждение молодых побегов ели поздними весенними
заморозками
177
опадает, обнажая древесину. Наиболее сильно от зимних ожогов
страдают плодовые деревья.
В ранневесенний период после малоснежных зим резкие
колебания температуры в дневное и ночное время вызывают
пожелтение и опадение хвои: температура воздуха днем повышается до
15...18°С, а температура почвы остается низкой, не выше 4°С.
При этом у растений на солнце происходит активное испарение,
а корни функционируют еще очень слабо и не способны
обеспечить растения необходимым количеством влаги. В результате хвоя
становится красно-бурой и опадает. Чаще всего это явление
наблюдается в питомниках и на открытых местах (рис. 42).
S3. ОТРИЦАТЕЛЬНОЕ ВЛИЯНИЕ
АНТРОПОГЕННЫХ ФАКТОРОВ
К антропогенным факторам неблагоприятного воздействия на
лесные и урбоэкосистемы относятся: загрязнение атмосферы,
поверхностных и грунтовых вод, почвы промышленными и
автотранспортными выбросами; хозяйственные мероприятия,
проводимые в лесных и городских насаждениях; возрастающие с
каждым годом масштабы рекреации насаждений.
Загрязнение окружающей среды. Промышленные эмиссии и
выхлопные газы автотранспорта приводят к значительным
стрессам, которые испытывают лесные и урбоэкосистемы. Однако из
всех компонентов экосистем от загрязнения атмосферы и почвы
наиболее сильно страдает растительность. Лесные и городские
насаждения, находящиеся в зоне атмосферного загрязнения,
ослабляются и усыхают.
Различные древесные породы обладают неодинаковой
газоустойчивостью и газочувствительностью.
Газоустойчивость — это способность растения
противостоять вредному действию газов, сохраняя свою жизнеспособность.
Газочувствительность — это скорость и степень
появления у растений патологической реакции на токсическое
действие газов. Например, лиственница более газочувствительна, чем
сосна и ель, и тем не менее она обладает большей
газоустойчивостью, чем указанные породы. Различают три вида газоустойчивости
растений: физиологическую, морфологическую и биологическую.
Физиологическая устойчивость определяется низкой окисляе-
мостью клеточного содержимого. Двуокись серы и другие кислые
газы, проникая в клетки, связывают активное железо, без
которого невозможен фотосинтез. Так как солнечная энергия
продолжает поступать в листья, то хлорофилл, обладающий
флуоресцирующей способностью, проявляет фотодинамическое действие,
которое выражается в фотоокислении. Окисленные вещества раз-
178
рушаются, что приводит к отмиранию клеток. Поэтому чем
меньше окисляемость протоплазмы, тем выше газоустойчивость
растений. В связи с этим хвойные породы, имеющие большую
окисляемость, менее устойчивы к действию газов. Лиственные породы, у
которых общая окисляемость меньше, обладают более высокой
газоустойчивостью.
Морфолого-анатомическая газоустойчивость обусловливается
особенностями строения листьев, которые препятствуют
поступлению газов в растение. Биологическая газоустойчивость связана
со способностью растений быстро восстанавливать пораженные
газами органы.
Растение может обладать одновременно различными видами
газоустойчивости. При этом какой-либо один вид
газоустойчивости может доминировать и определять степень газоустойчивости
данного вида растения. Газоустойчивость древесных пород зависит
от химического состава соединений, которые есть в
промышленных отходах, от условий внешней среды и характера задымления.
Основными примесями, содержащимися в выбросах
промышленных предприятий и автотранспорта, являются соединения серы,
фтора, хлора, азота, магния и др. Ниже приводятся
диагностические признаки поражения под действием опасных соединений.
Диоксид серы (S02) — этот бесцветный газ
выбрасывается в атмосферу коксохимическими заводами, горнорудными и
целлюлозно-бумажными предприятиями. Характерные признаки
поражения появляются на хвое (листьях) только при сильном
действии выбросов, когда разрушаются клеточные структуры и
пигмент. При действии высоких концентраций газа четкие признаки
поражения проявляются через несколько дней. Хвоя приобретает
рыже-бурую окраску. У сосны и пихты изменения окраски хвои
часто начинаются с концов. Причем у сосны оно протекает
постепенно, тогда как у ели вначале идет медленно, а затем очень
бурно. На листьях под влиянием сернистого газа появляются пятна
красно-бурого цвета различного размера. Высокие концентрации
сернистого газа вызывают образование пятен, часто
охватывающее более половины площади листовой пластинки (рис. 43).
Поражение диоксидом серы приводит к преждевременному
опадению листьев и хвои, а высокие концентрации газа вызывают
искривление и отмирание молодых побегов. Лиственные породы
более устойчивы к действию S02, чем хвойные. Предельно
допустимая концентрация его составляет (мг/м3): для лиственницы
0,25, для сосны 0,40, для ели 0,70.
Ослабление деревьев сопровождается нарушением обмена
веществ, падением активности окислительных ферментов,
ослаблением фотосинтеза и разрушением хлорофилла. В то же время
происходят деформация и разрушение клеток и тканей коры, луба,
камбия, хвои и листьев.
179
Рис. 43. Повреждение древесных пород диоксидом серы:
1 — хвои пихты; 2 — листьев клена; 3 — листьев дуба
Фтор и его соединения — в твердом или газообразном
состоянии они выбрасываются в атмосферу заводами по
производству алюминия, кирпича, керамических изделий, фосфатных
удобрений; выделяются при выплавке стали, попадают в
атмосферу из дымовых труб и фабричных установок.
Поражение растений происходит через листья (хвою) и корни.
При концентрации, равной 0,01 мг/м3, поражения образуют по
периферии листа узкие некротические полосы светло-желтого цвета.
У хвойных пород происходит побеление, а затем потемнение
концов хвои, которое распространяется к основанию игл. Действие
фтора в высоких концентрациях выражается в прекращении
фотосинтеза, нарушении роста и развития, отмирании завязей,
загнивании плодов. Наиболее восприимчивы к соединениям фтора
хвойные породы, среди них менее устойчива сосна.
Хлор и хлористый водород применяют в
производстве пластмасс и инсектицидов. Эмиссии соляной кислоты
встречаются на заводах по изготовлению калийных солей. Пары хлора и
хлористого водорода быстро оседают на землю и поэтому
повреждают растительность только вблизи источника эмиссии. Их
содержание в воздухе в концентрации менее 1 мг/м3 вызывает сильное
поражение листьев. Вначале листья приобретают темный цвет с
хорошо заметным серебристым оттенком, затем на них
появляются обесцвеченные участки разных размеров. По мере отмирания
тканей они крошатся, образуя отверстия. При длительном
действии низких концентраций хлора краснеют края листьев.
Нитрозные газы. Это смесь окисей азота, которая
выбрасывается в атмосферу заводами по производству азотной, серной
кислот и нитратных удобрений, а также — с отработанными
газами автотранспорта. Окиси азота вызывают сильное поражение ли-
180
стьев (хвои) в концентрации более 2 мг/м3. На вершинах и по
краям листьев образуются буровато-черные участки. У хвойных
пород происходит покраснение кончиков хвои.
Выхлопные газы автотранспорта. В их состав входят
фумиганты окиси углерода, нитрозные газы, ненасыщенный
водород, полициклические ароматические углеводороды, сажа и
свинцовые соединения. Выхлопные газы вызывают образование
некрозов на листьях, преждевременное усыхание и опадение
листвы, ослабление и усыхание деревьев.
Пылевидные эмиссии выбрасываются топками,
металлургическими и цементными заводами.
Цементная пыль представляет собой смесь минералов,
содержащих калий, кальций, алюминий. Такая смесь выбрасывается
печами цементных заводов, пылевидные эмиссии осаждаются
вблизи источников загрязнения. Оседающая на листьях и хвое пыль
снижает ассимиляцию и эффективность солнечного излучения,
способствует повышению температуры. При попадании на почву
пыль изменяет ее кислотность и содержание в ней
микроэлементов. Все это приводит к усыханию хвои и листьев, нарушению
роста корневой системы и как следствие — к ослаблению и
гибели деревьев.
Магнезитовая пыль образуется при обжиге магнезитовой руды.
Основным компонентом магнезитовой пыли является оксид
магния. Она оказывает отрицательное действие как на лиственные,
так и на хвойные породы, однако последние страдают сильнее.
Магнезитовая пыль повреждает только молодую хвою и листву.
Поэтому в начале вегетации деревья менее устойчивы к действию
магнезитовой пыли. В связи с этим хвоя, уцелевшая от
токсического действия магнезитовой пыли в первые годы жизни, в
дальнейшем не погибает. В результате действия магнезитовой пыли хвоя
на побегах текущего года принимает светло- или
желтовато-зеленую окраску. Двухлетняя хвоя краснеет, буреет и частично
погибает. У лиственных пород между жилками листа образуются
светло-зеленые или желтовато-зеленые пятна.
В результате постоянного действия магнезитовой пыли
хвойные породы усыхают, особенно сосна и ель. Их гибель наступает
при сильном изреживании кроны, которое происходит вследствие
поражения молодой хвои магнезитовой пылью и под влиянием ее
естественного старения. Отмирание пораженной сосны часто
связано со стволовыми вредителями, поселяющимися на
ослабленных деревьях.
Степень повреждения растений промышленными эмиссиями
зависит от их концентрации в окружающей среде и длительности
действия. Длительное воздействие выбросов в концентрациях ниже
предельной нормы вызывает хронические поражения насаждений,
проявляющиеся в постепенном изменении физиологических и
181
биохимических функций. Аварийные выбросы,
характеризующиеся высокими концентрациями и кратковременным действием,
приводят к острым поражениям, которые проявляются в
массовом образовании некрозов на листьях и хвое и сравнительно
быстром отмирании деревьев.
Усыхание насаждений в зоне промышленных выбросов
зависит от следующих факторов: возраста, состава и полноты
насаждений, близости источника выбросов, направления ветра,
рельефа, погодных условий, концентрации токсичных веществ.
В зоне промышленных выбросов процесс усыхания более
интенсивен в насаждениях старшего возраста, в изреженных и высо-
кополнотных древостоях. Это объясняется тем, что в изреженных
насаждениях газы беспрепятственно проникают вглубь, а при
слишком плотном смыкании крон они застаиваются под пологом
длительное время, не изменяя концентрации. Значительно
меньше усыхают среднеполнотные насаждения, в которых воздух
максимально перемешивается и концентрация токсичных веществ
уменьшается. Смешанные насаждения в меньшей степени
страдают от дымовых газов, чем чистые хвойные, так как лиственные
породы поглощают часть газов и уменьшают их вредное влияние
на хвойные породы.
Сохранность и повышение устойчивости лесных и зеленых
городских насаждений в зонах воздействия промышленных
выбросов достигается комплексом технических и лесохозяиственных
мероприятий. Для того чтобы уменьшить влияние промышленных
выбросов, необходимо прежде всего постоянно совершенствовать
пыле- и газоочистительные установки, а также технологии
промышленных процессов. Сокращение выбросов автотранспорта
может быть достигнуто совершенствованием двигателей
внутреннего сгорания, переводом их на другие малотоксичные виды
топлива. Большое значение имеет организация мониторинга за
состоянием лесных и зеленых насаждений в зонах промышленных
выбросов. Из числа лесохозяиственных мероприятий, направленных
на снижение степени воздействия промышленных выбросов,
можно назвать следующие: создание смешанных насаждений с
опушками из стойких пород, расположение насаждений с учетом
рельефа и направления господствующих ветров, определяющих
распространения выбросов. Кроме того, при озеленении
промышленных центров необходимо учитывать различную степень
газоустойчивости древесных пород. Г.М.Илькуном (1978) дана
следующая оценка газоустойчивости деревьев и кустарников:
очень устойчивые — белая акация, боярышник, ива
белая, роза, сирень, тополь бальзамический и канадский, ясень
зеленый;
устойчивые — ель колючая, можжевельники казацкий,
сибирский и обыкновенный, вяз, дуб, карагана древовидная, раз-
182
ные виды клена, липа крупнолистная и войлочная, рябина
обыкновенная, сирень, тополь белый, черный, крупнолистный,
яблоня, ясень американский, обыкновенный и пушистый;
относительно устойчивые — можжевельник
виргинский, береза пушистая, граб, конский каштан, клен
остролистный, липа мелколистная, орех, тополь китайский, лавролистный;
малоустойчивые — ель восточная, сибирская, пихта
белая, сибирская, барбарис обыкновенный, береза бородавчатая;
неустойчивые — лиственница, сосна обыкновенная,
Банкса, веймутова.
Хозяйственная деятельность человека наряду с положительным
влиянием (уход за лесом, создание насаждений, борьба с
болезнями и вредителями) в ряде случаев отрицательно сказывается на
насаждениях, вызывая их ослабление, отмирание, способствуя
развитию очагов вредителей и болезней. Негативными сторонами
хозяйственной деятельности, способствующими развитию
болезней, являются следующие мероприятия: порослевое
возобновление, монокультура, неудачные типы и конструкции посадок,
выпас скота, травматизм растений, применение пестицидов и
других химических веществ.
Систематическое возобновление насаждений порослью
приводит к их вырождению. Порослевые насаждения второго и
последующих поколений, как правило, ослаблены и менее
производительны, чем насаждения семенного происхождения, что
объясняется резким нарушением развития корневой системы и кроны.
Ослабленность порослевых насаждений является предпосылкой для
поражения их болезнями. В порослевых насаждениях особенно
часто возникают очаги сосудистых болезней, передающихся через
зараженные пни: сосудистый микоз дуба, голландская болезнь
ильмовых пород, вертициллезное усыхание клена. Приспевающие
и спелые насаждения порослевого происхождения сильнее, чем
семенные, поражаются гнилями. По данным А. И. Воронцова
(1978), пораженность Останкинской дубравы ложным дубовым
трутовиком (Phellinus robustus) в среднем составляла 25,3%, а на
отдельных участках достигала 43 %. В Тульских засеках в
средневозрастных дубравах порослевого происхождения плодовые тела
ложного дубового трутовика имели 15 — 20% деревьев, а 35—40%
модельных деревьев были с гнилью, вызванной этим трутовиком.
Создание монокультур является причиной развития опасных
болезней, принимающих характер эпифитотии. Это объясняется
тем, что однопородные насаждения характеризуются бедным
видовым составом патогенов, но некоторые из них, способные
поражать породу, образующую насаждение, оказываются в
оптимальных условиях. Создание чистых сосновых культур, особенно на
нелесных почвах, привело к развитию эпифитотии корневой губки
(Heterobasidion annosum) в разных регионах России. В очагах корне-
183
вой губки развиваются очаги стволовых вредителей. Чистые
сосновые культуры сильнее страдают от снежного шютте (Phacidium infe-
stans), побегового рака (Scleroderris lagerbergii). В чистых культурах
и посадках тополя в благоприятные для возбудителей годы
наблюдается массовое развитие ржавчины (Melampsora allii—populina),
бурой пятнистости (Marssonina populi), парши (Pollaccia radiosa).
Неудачно подобранные типы и конструкции посадок,
особенно из пород, не соответствующих условиям местопроизрастания,
приводят к их ослаблению и массовому распространению
болезней. Важное значение при этом имеет густота посадок, оптимум
которой определяется в каждом конкретном местообитании.
Известно, что загущенные культуры и посадки ели в сильной
степени поражаются шютте (Lophodermium macrosporum). В
перегущенных посадках школьных отделений древесных питомников и в
культурах тополя чаще возникают очаги мокрого язвенно-сосудистого
рака и цитоспороза.
Отрицательное воздействие на насаждения оказывает
перевыпас, от которого наиболее страдают насаждения на слаборазвитых
почвах с редким травяным покровом. Протаптываемые скотом
тропы сливаются, на этих участках исчезают травяной покров и
лесная подстилка. На оголенных, уплотненных участках почвы
корни деревьев в сильной степени повреждаются копытами
животных. Через повреждения в корни проникают возбудители
гнилей, в частности корневая губка. Ослабленные деревья заселяются
стволовыми вредителями, ускоряющими процесс отмирания.
Лесохозяйственные мероприятия травмируют растения, что если
и не приводит к их гибели, то ослабляет их и способствует
заражению болезнями. При культивации посевов в питомниках
нередко наблюдается повреждение корневой системы у сеянцев в
крайних рядах. Поврежденные сеянцы засыхают или ослабляются,
отстают в росте и становятся непригодными к посадке. Посадочный
материал в питомниках, культуры и молодые посадки в городских
насаждениях травмируются при уходе за ними. Через нанесенные
повреждения в ткани деревьев легко проникают возбудители
болезней. В 1991 — 1992 гг. при обследовании Московского совхоза
декоративного садоводства было обнаружено значительное
поражение (43 и 57 %) нектриозом клена остролистного,
предназначенного для реализации. Анализ пораженных деревьев показал,
что все они имели механические повреждения в нижней части
стволов, полученные при культивации. В культурах и городских
насаждениях деревья часто травмируются во время посадки и пос-
лепосадочного ухода, при небрежной обрезке сучьев, когда
вместе с обрезанными ветвями сдирается кора, а иногда повреждается
древесина стволов. В насаждениях массовые механические
повреждения стволов связаны с проведением рубок, трелевками,
прокладкой визиров и др. В таких насаждениях отмечается высокий
184
уровень пораженности гнилевыми болезнями, возбудители
которых проникают через повреждения стволов.
Интенсивное развитие химической промышленности
приводит к тому, что постоянно в биосферу поступает большое
количество различных химических веществ, в том числе и пестицидов.
Пестициды обладают различной физиологической активностью и
в зависимости от физико-химических свойств, доз, способов и
сроков применения могут оказывать фитотоксическое или
стимулирующее действие на защищаемые растения.
Способность пестицидов оказывать токсическое (отравляющее)
воздействие на растение называют фитотоксичностью. Она
проявляется при неправильном применении пестицидов, когда
завышают допустимые концентрации или нарушают технологию
обработки растений. При этом пестициды могут вызвать
повреждение защищаемых древесных пород, а также соседних с ними
деревьев и подлеска.
При быстром распространении по растению пестициды
вызывают общее отравляющее действие на весь организм. Если
пестицид распространяется медленно и локализуется в местах
проникновения в растения, он оказывает местное действие. Оно чаще
всего проявляется в виде ожогов листьев с образованием бурых и
коричневых пятен.
В городских насаждениях негативное влияние на древесные
растения оказывают хлористые соли, в том числе поваренная соль,
применяемые для борьбы с наледями и снегом на
автомагистралях, на улицах и во дворах. Длительное применение противолед-
ных препаратов приводит к постепенному засолению почв.
Отрицательное действие хлоридов на древесные растения
проявляется в более позднем и неравномерном распускании листьев, су-
хокронности и снижении интенсивности роста, образовании на
листьях некрозов, преждевременном их усыхании и опадении.
Особенно страдают от комплексного воздействия засоления,
промышленных и автотранспортных выбросов деревья на
автомагистралях крупных городов и насаждения вблизи промышленных
предприятий.
Рекреационная нагрузка. Увеличивающиеся с каждым годом
масштабы городского строительства и рост городского населения
приводят к возникновению больших рекреационных нагрузок в
зеленых зонах. Особенно высокий рекреационный пресс
создается в наиболее посещаемых населением пригородных лесах и
лесопарках. Избыточное рекреационное воздействие на пригородные
лесные насаждения сопровождается уплотнением почвы,
нарушением естественного живого покрова, уничтожением и
повреждением подроста и подлеска, изреживанием древостоя,
обнажением корневых лап, нанесением многочисленных механических
повреждений стволов и корней.
185
Все изменения природных комплексов в зонах рекреации
приводят к увеличению уровня пораженности древостоев болезнями.
В результате изреживания сосняков под действием
рекреационного пресса возрастает их пораженность смоляным раком. В ельниках
пригородных лесов механические повреждения корней
способствуют заражению ели самыми опасными корневыми гнилями,
вызываемыми корневой губкой и опенком осенним. С повышением
рекреационной нагрузки и количества механических повреждений на
стволах в ельниках заметно увеличивается распространение
раневого рака. По мере возрастания рекреационной дигрессии
повышается уровень пораженности деревьев стволовыми гнилями.
Для ограничения вреда, причиняемого лесным насаждениям
рекреацией, необходимо проводить комплекс лесохозяйственных
мероприятий, включающий организацию мониторинга состояния
пригородных лесов и зеленых городских насаждений, проведение
лесоустройства, регулирование допустимой нормы посетителей в
разных экологических объектах, рациональное устройство дорож-
но-тропиночной сети, эстетически улучшать открытые участки для
отдыха, периодически или длительно ограждать наиболее ценные
участки насаждений, устанавливать маршруты для лыжных
прогулок и пр.
Глава 9
БОЛЕЗНИ ПЛОДОВ И СЕМЯН
И МЕРЫ БОРЬБЫ С НИМИ
Получение необходимого количества здоровых сеянцев, рост
лесных культур, формирование высокопродуктивных устойчивых
насаждений в значительной мере зависят от качества посевного
материала. Качество семян играет важную, подчас решающую роль
в естественном возобновлении леса, в развитии благонадежного
подроста. Одним из главных факторов, снижающих качество
семян и влияющих на дальнейшее развитие сеянцев, является
поражение плодов и семян микроорганизмами, чаще всего грибами.
Грибы — возбудители болезней семян существенно
различаются между собой по уровню паразитической активности и
специализации. Они весьма многочисленны, а вызываемые ими болезни
разнообразны по характеру развития и симптомам.
Грибные болезни плодов и семян причиняют лесному
хозяйству немалый ущерб: они вызывают гибель семян или снижают их
посевные кондиции, приводят к заболеваниям всходов и
посадочного материала в питомниках, ухудшают развитие самосева
под пологом леса. Иногда наблюдается гибель всего урожая семян
на дереве или под пологом насаждения, потеря всхожести
больших партий семян во время хранения и транспортировки.
Заражение фитопатогенными грибами может происходить еще
на дереве, на различных этапах развития и созревания семян. В
других случаях семена заражаются грибами во время сбора или
хранения.
Многие болезни плодов и семян, особенно болезни, связанные
с весенним заражением завязей паразитными грибами, развиваются
в летний период, вызывая характерные изменения формы, цвета,
размеров или структуры семян, и поэтому легко обнаруживаются
во время их заготовки. В этом случае больные семена могут быть тем
или иным способом отделены от здоровых и уничтожены. Таковы
мумификация семян березы, болезни, вызываемые голосумчатыми
и ржавчинными грибами, пятнистости плодов и семян.
187
При заболеваниях, связанных с более поздним заражением
семян (особенно после их созревания и опадения), внешние
признаки поражения в момент заготовки еще не проявляются в
полной мере. Поэтому зараженные семена могут попасть в хранилища
и явиться источником инфекции для здоровых семян. К болезням
такого типа относятся различные гнили, мумификация желудей и
заболевания, вызываемые плесневыми грибами.
Болезни плодов и семян характеризуются внутренней
инфекцией, при которой мицелий гриба проникает внутрь семени и
развивается в тканях оболочки, зародыша или семядолей,
используя их как питательный субстрат. Встречается и поверхностное
загрязнение семян спорами фитопатогенных грибов или так
называемая внешняя инфекция, при которой сами семена обычно не
поражаются, но при их высеве могут возникнуть заболевания
проростков, всходов и посадочного материала. Бактериальные
болезни плодов и семян встречаются сравнительно редко и не имеют
большого практического значения. По характеру заражения семян,
срокам развития и особенностям проявления можно выделить
следующие основные группы и типы болезней плодов и семян.
9.1. БОЛЕЗНИ, РАЗВИВАЮЩИЕСЯ В ТЕЧЕНИЕ
ВЕГЕТАЦИОННОГО ПЕРИОДА
Большинство болезней, поражающих семена во время
вегетации, вызывается узкоспециализированными грибами из числа
облигатных паразитов и факультативных сапротрофов. Это
ржавчина шишек, деформации плодов и семян, болезни типа
мумификации. Исключение составляют лишь некоторые виды пятни-
стостей, вызываемые факультативными паразитами или
факультативными сапротрофами с более широкой специализацией.
Болезни этой группы разнообразны по внешнему проявлению. Как
правило, они характеризуются глубокими специфическими
изменениями пораженных тканей.
9.1.1. Мумификация семян
Болезни этого типа вызываются сумчатыми грибами порядка
Leotiales. Они характеризуются интенсивным развитием в тканях
зараженных семян мицелия гриба и превращением семян в
черную губчатую массу (склероциальную строму) или образованием
на пораженных семенах склероциев возбудителя.
Мумифицированные семена теряют всхожесть. Мумификацией поражаются
семена березы, ольхи, дуба и других пород.
Мумификация семян березы. Заболевание вызывается грибом
Sclerotinia betulae Woron. Весной, во время цветения березы, сум-
188
коспоры заражают завязи.
Мицелий паразита развивается в
тканях зараженных семян и
образует на границе между семянкой и
крылаткой хорошо заметные
склероции в виде черного
подковообразного ободка. Склероции
являются покоящейся формой
паразита и служат для его
перезимовки. Пораженные семянки
отличаются от здоровых темной
окраской. При сильном развитии
болезни в сережках могут быть
поражены все семена.
Мумифицированные семена опадают.
Весной следующего года
склероции на этих семенах
прорастают, образуя плодовые тела — апо-
теции в виде неглубокой чашечки
или воронки диаметром 1—4 мм,
сидящей на тонкой длинной
ножке (рис. 44). Основание ножки
имеет бурый цвет и покрыто
темно-бурыми волосками. На
внутренней стороне апотециев
формируется слой сумок (гимений) грязновато-белого или охряно-
желтого цвета. Сумкоспоры в сумках созревают ко времени
цветения березы, разносятся ветром и, попадая на женские сережки,
заражают завязи. Мумифицированные семена легче здоровых и
поэтому легко отделяются при сортировке.
Мумификация желудей дуба. Вызывается грибом Stromatinia
pseudbtuberosa Rehm. В лесу желуди заражаются сумкоспорами из
апотециев, образующихся на мумифицированных желудях. Сроки
заражения желудей сумкоспорами обычно растянуты: частично
желуди могут заражаться еще на деревьях, но в основном это
происходит после опадения их на землю. При непосредственном
контакте опавших здоровых желудей с прошлогодними
мумифицированными происходит также заражение их мицелием,
проникающим внутрь желудя через трещины в кожуре и сосудистые пучки
в местах прикрепления плюски. Эта форма заражения,
по-видимому, является главной.
В местах заражения семядолей сначала появляются небольшие
рыжевато-коричневые пятна с резко очерченными краями.
Постепенно увеличиваясь в размерах, пятна сливаются, охватывая
всю семядолю. На поверхности семядолей образуется сероватая
пленочка грибницы. Внутренние ткани семядолей буреют, затем
Рис. 44. Мумификация семян
березы:
/ — пораженная крылатка со склеро-
циями возбудителя; 2 — прорастание
склероциев апотециями; 3 — сумка с
сумкоспорами; 4 — парафиза
189
приобретают оливковый оттенок. В конечной стадии поражения
семядоли превращаются в рыхлую черную массу — склероциальную
строму, состоящую из плотного сплетения гиф гриба и остатков
тканей желудя. Мумифицированные семядоли увеличиваются в
объеме (как бы разбухают). Вследствие этого кожура разрывается и
отделяется от семядолей (рис. 45). Мумифицированные желуди имеют
меньшую удельную плотность, чем здоровые, и всплывают в воде.
В природных условиях мумифицированные желуди на
следующий год прорастают, образуя плодовые тела (апотеции) на ножках.
Длина ножки в зависимости от глубины залегания пораженных
желудей в подстилке может быть различной. В апотециях
развиваются сумкоспоры, которые по мере созревания рассеиваются и
заражают желуди нового урожая. Однако сумчатая стадия гриба встречается
не повсюду и не каждый год. В условиях высокой влажности на
обнаженных склероциальных стромах обильно разрастается мицелий,
на котором образуется конидиальное спороношение. Конидии,
распространяясь по воздуху, также вызывают заражение желудей.
Наиболее интенсивное развитие болезни наблюдается во
время хранения желудей, особенно при нарушении оптимального
режима температуры и влажности. Распространение гриба в
хранилищах происходит с помощью мицелия. При благоприятных
условиях он легко переходит с больных желудей на здоровые,
окутывая их пышной серой пленкой.
Мумификация — одна из опаснейших болезней желудей;
распространена она по-видимому везде, где произрастает дуб. В
отдельные годы зараженность желудей, особенно при их массовой
заготовке и плохо организованном хранении, может быть очень
Рис. 45. Мумификация желудей:
1, 2, 3,4, 5 — последовательность стадий развития болезни; б — склероциальная
строма гриба Stromatinia pseudotuberosa> проросшая апотециями; 7 — сумка со
спорами и парафизы; 8 — сумкоспоры; 9 — конидиальное спороношение гриба
190
высокой (20—30% и более). S.pseudotuberosa вызывает также
мумификацию плодов каштана.
Мумификация, вызываемая грибами Stromatinia aucupariaeWoron.
и Stromatinia crataegi Magn., встречается соответственно на рябине
и боярышнике.
Меры борьбы разработаны лишь против мумификации
желудей. Для профилактики болезни необходима правильная
организация сбора, транспортировки и хранения желудей. Сбор их
следует проводить в сжатые сроки. При сборе и перевозке желудей
нельзя допускать их механического повреждения. Перед закладкой
на зимнее хранение желуди рекомендуется обсушить, не
допуская потери ими более 5 — 8 % влаги, и произвести сухое
протравливание эффективными фунгицидами. Следует избегать перегрева
и подмерзания желудей. Зимнее их хранение осуществляют теми
способами, которые наиболее целесообразны в конкретных
условиях хозяйства и исключают возможность развития грибной
инфекции и снижения посевных качеств желудей. Во время
хранения рекомендуется поддерживать температуру от 0 до 4 °С; более
низкая температура неблагоприятна для желудей, а при более
высокой возможно развитие болезни.
9.1.2. Ржавчина шишек
Ржавчиной поражаются шишки ели. Их легко отличить от
здоровых по широко раскрытым чешуям, на которых располагаются
эции ржавчинных грибов в виде оранжевых или коричневых
шаровидных вместилищ (рис. 46). Поражение ржавчиной резко
уменьшает выход семян. Кроме того, семена, развивающиеся в таких
шишках, обычно щуплые, мелкие, легковесные. Они обладают
низкой всхожестью. При сильном развитии болезни пораженные
шишки иногда совсем не дают всхожих семян. Поэтому на семена
не следует собирать шишки, даже в незначительной степени
пораженные ржавчиной. Это заболевание причиняет вред лесному
хозяйству, препятствуя естественному возобновлению леса и
получению полноценных семян для искусственного разведения ели.
Ржавчину шишек ели вызывают два вида разнохозяйных грибов с
полным циклом развития.
Thekopsora padi Kleb. образует на внешней стороне кроющих
чешуи малозаметные спермогонии, имеющие вид плоских
корочек, а на внутренней стороне — многочисленные тесноскученные
шаровидные эции диаметром 1 — 2 мм темно-коричневого цвета.
Высыпающиеся из них эциоспоры заражают листья черемухи, на
которых развиваются сначала урединиоспоры, а затем телиоспо-
ры гриба. Поражение листьев черемухи проявляется в виде
характерных угловатых фиолетовых или буровато-малиновых пятен. После
перезимовки на опавших листьях телиоспоры прорастают и обра-
191
Рис. 46. Ржавчина шишек ели:
1 — пораженная шишка; 2 — чешуйка с эциями гриба Thekopsora padi\ 3 — разрез
через эций с эциоспорами; 4 — эциоспоры; 5 — урединио- и телиостадия гриба на
листе черемухи; 6 — урединиоспоры; 7 —- телиоспоры в клетках эпидермиса
зуют базидии с базидиоспорами; последние, распространяясь по
воздуху, вновь заражают шишки ели.
Chrysomyxa pirolae Rostr. также образует на кроющих чешуях ели
спермогонии и эции. Но в отличие от T.padi у C.pirolae эции
располагаются обычно по два на чешуйке; они более крупные (3—4 мм
в диаметре), в виде округлых оранжевых субэпидермальных
подушечек, наполненных эциоспорами. При созревании эции быстро
разрушаются, и освобождающиеся эциоспоры покрывают
поверхность пораженных шишек оранжевым порошком. Эциоспоры
заражают листья грушанок, на котбрых развиваются урединио- и телио-
стадии гриба. Меры борьбы с ржавчиной шишек ели пока не
разработаны. Существуют рекомендации об уничтожении на лесосемен-
ных участках ели и в прилегающих к ним насаждениях
пораженных шишек и черемухи — промежуточного хозяина гриба T.padi
9.1.3. Деформация плодов
Деформация плодов наблюдается при заболеваниях,
вызываемых сумчатыми грибами рода Taphrina. У пораженных плодов
резко изменяются форма, цвет, анатомическое строение. Семена в
деформированных плодах обычно совсем не образуются, иногда
остаются недоразвитыми и невсхожими. Наиболее часто встреча-
192
ется деформация плодов косточковых древесных пород и плоди-
ков ольхи. Иногда болезнь поражает осину и некоторые виды
тополя (см. рис. 13).
Деформация плодов (кармашки) косточковых пород.
Кармашки, или дутые плоды, образуются в результате поражения
косточковых пород (черемухи, сливы, алычи и др.) грибом Taphrinapruni
Fuck. Мицелий паразита многолетний, он зимует в ветвях и
почках деревьев, а во время цветения проникает в завязи. Гриб
вызывает сильное разрастание стенок завязи, подавляя в то же время
развитие косточки. В результате из зараженных завязей
формируются уродливые мешковидные бурые тела, внутри полые (отсюда
название болезни). Снаружи они покрываются как бы восковым
сероватым налетом, состоящим из сумок возбудителя. Сумкоспо-
ры дозревают во второй половине лета и, попадая на чешуйки
почек и ветви, заражают их. Поскольку мицелий из года в год
сохраняется в ветвях, на них ежегодно вместо нормальных плодов
образуются кармашки. Заболевание распространено повсеместно.
Развитию его способствует влажная погода весной и летом.
Деформация плодов ольхи. Болезнь вызывается грибом Т. alni-
incanae (Ktihn.) Magn. Она проявляется в деформации семян и
разрастании чешуек, превращающихся в уродливые листовидные
образования, на которых развиваются сумки. Мицелий
возбудителя зимует в почках, откуда после цветения ольхи переходит в
женские сережки.
Меры борьбы с деформацией плодов должны быть
направлены на уничтожение зимующей инфекции и
предупреждение новых заражений. Это достигается обрезкой ветвей, на
которых наблюдалась деформация плодов, сбором и уничтожением
пораженных плодов, осенним или ранневесенним
профилактическим опрыскиванием деревьев фунгицидами. Чаще всего
применяют 3 %-й раствор железного купороса.
9-2. БОЛЕЗНИ, РАЗВИВАЮЩИЕСЯ
ПРИ ХРАНЕНИИ СЕМЯН
Болезни этой группы вызываются главным образом
факультативными паразитами из числа сумчатых и несовершенных грибов.
В большинстве случаев активное развитие возбудителей
происходит во время транспортировки и хранения семян, хотя их
заражение могло осуществиться еще в лесу, в период созревания или
сбора. Болезни плодов и семян при хранении обычно
проявляются в форме гнили или плесени. Заражению семян способствуют их
повреждение и ослабление. Массовое развитие этих заболеваний,
как правило, является следствием нарушения правил сбора и
перевозки семян или неправильного режима их хранения.
193
9.2.1. Гнили плодов и семян
Гнили характеризуются нарушением структуры, размягчением
и распадом тканей пораженных плодов и семян, которые при этом
полностью теряют всхожесть. Гнили чаще поражают мясистые
семена, богатые водой и запасными питательными веществами
(желуди, плоды каштана, ореха грецкого и др.). Особенно
благоприятным субстратом для развития грибов являются желуди. Гнили
желудей приносят большой ущерб лесному хозяйству, при
заготовке и хранении крупных партий желудей создаются
предпосылки для массового развития болезней этого типа.
Сухая гниль (антракноз) желудей вызывается несовершенным
грибом порядка Acervulales Gloeosporium quercinum West.
Заболевание характеризуется появлением на пораженных семядолях
серовато-бурых или темно-бурых, иногда почти черных, резко
очерченных пятен неправильной формы. Они постепенно
углубляются, превращаясь в язвы, и увеличиваются в размерах. Полностью
пораженные семядоли имеют неровную, изъязвленную, как бы
обугленную поверхность, и ссыхаются. Во влажной среде на
пораженных местах образуются тонкие грязновато-белые или желтоватые
пленки грибницы, в которых формируются конидиомы гриба. Они
имеют вид мелких бугорков или подушечек желтовато-бурого цвета,
расположенных концентрическими кругами. Конидии выступают
наружу в виде беловатых слизистых капелек.
Антракноз желудей распространен повсеместно и при хранении
желудей нередко является преобладающим заболеванием. GL quercinum
поражает также сеянцы дуба, вызывая бурую пятнистость листьев.
Белая гниль желудей. Возбудитель болезни — пикнидиальный
гриб Phomopsis quercella (Sacc.) Died. На пораженных семядолях
образуются сероватые пятна, которые со временем темнеют,
сливаются между собой и охватывают всю поверхность семядолей. В условиях
повышенной влажности на них развивается пышная белая пленка
мицелия (отсюда и название болезни). В толще грибницы
закладываются многочисленные черные довольно крупные (до 1,5 мм в
диаметре) пикниды возбудителя (рис. 47). По мере развития пик-
ниды надавливают на кожуру, вследствие чего поверхность
желудя становится бугристой, а затем звездообразно разрывается. Из
созревших пикнид выходят склеенные массы конидий в виде
оранжевых капель или извитых нитей.
Первичное заражение желудей происходит в лесу, но
интенсивное развитие болезни наблюдается уже в хранилищах в том
случае, если влажность и температура поднимаются выше нормы.
Заболевание распространено во многих областях России. Сумчатая
стадия гриба развивается на коре и древесине ветвей дуба.
Цитоспороз желудей вызывается грибами рода Cytospora (чаще
всего С. intermedia Sacc). При цитоспорозе на семядолях появля-
194
Рис. 47. Гнили желудей:
1 — черная; 2, 3 — белая; 4 — цитоспороз; 5 — многокамерная пикнида гриба
Cytospora intermedia; 6 — пикнида с выходящей из нее ленточкой склеенных
конидий; 7 — конидии
ются отчетливые темно-коричневые пятна, покрытые тонкой
белой мицелиальной пленкой. Со временем пленка приобретает
желтоватый или оливково-бурый оттенок, разрастается и покрывает
всю поверхность семядолей. На пораженных семядолях
развиваются крупные, до 3—4 мм в диаметре и 2 — 3 мм в высоту, олив-
ково-черные стромы с пикнидами гриба, выступающие на
поверхность желудей через продольные трещины и разрывы в кожуре
(см. рис. 47). При созревании конидий они выходят наружу
склеенными массами. Мицелий может легко переходить на соседние
здоровые желуди, поэтому в хранилищах обычно наблюдается
распространение болезни в виде очагов. Цитоспороз — одна из
наиболее опасных болезней желудей, часто встречающаяся в
различных районах нашей страны.
195
Черная гниль желудей вызывается грибами Ophiostoma roboris Geoig.
et Teod. и O.valachicum G. et Teod., которые поражают все части
желудя: черешок, плюску, кожуру и семядоли. Заражению
желудей в лесу способствует повреждение их насекомыми. Заболевание
проявляется в образовании черных пятен, локализующихся
главным образом у основания желудя, и сопровождается быстрым
размягчением семядолей и почернением кожуры желудей. На
пораженных желудях формируются конидиальные спороношения,
иногда коремиального типа, а после отмирания желудя — и плодовые
тела возбудителя. Перитеции грушевидные, черные, с длинным
хоботком. Поражение желудей этой гнилью отмечалось главным
образом в Краснодарском крае, где нередко заболевание
приводило к массовой гибели желудей во время зимнего хранения.
Возбудители болезни представляют особую опасность в связи с
тем, что они поражают также проростки желудей и сеянцы дуба,
а во взрослых насаждениях вызывают сосудистый микоз ветвей и
стволов дуба.
Бактериальные гнили плодов и семян древесных пород не
имеют широкого распространения и большого хозяйственного
значения. Чаще всего поражаются желуди. Возбудителем бактериоза
желудей является Erwinia quercina Held, et Schr. Пораженные
семядоли приобретают темно-бурую окраску и размягчаются. Распад
тканей сопровождается образованием дурно пахнущей бурой
слизи, состоящей из скоплений бактериальных клеток и их
выделений. Развитию болезни способствуют механические повреждения,
подмерзание и перегрев желудей.
9.2.2. Плесневение плодов и семян
Плесневение плодов и семян всегда служит показателем плохой
организации их сбора или хранения. Грибы, вызывающие
плесневение семян, относятся к числу сапротрофов или факультативных
паразитов и способны развиваться лишь на мертвых или сильно
ослабленных тканях. Их ослабление или отмирание могут вызвать
механические повреждения, подмерзание, чрезмерное увлажнение
или, наоборот, пересыхание семян, повреждение насекомыми,
повышенная температура, недостаточная аэрация и другие факторы.
Споры плесневых грибов обычно попадают на семена во время
их заготовки и перевозки, иногда еще при созревании семян на
деревьях. Для прорастания же спор, развития мицелия и спороноше-
ний, внедрения гриба в ткань семени необходимы условия,
которые чаще всего возникают при неправильном хранении семян.
Развиваясь вначале на покровных тканях семян, плесневые
грибы не оказывают существенного влияния на их посевные качества.
Однако в дальнейшем гифы грибов могут разрушить оболочки
семян, проникнуть в их внутренние ткани и вызвать гибель зародыша.
196
Рис. 48. Спороношения грибов — возбудителей плесневения семян:
/ — Penicillium glaucum; 2 — Aspergillus glaucus (возбудители зеленой плесени); 3 —
Fusarium moniliforme; 4 — Trichothecium roseum (возбудители розовой плесени); 5 —
Rhizopus nigricans (возбудитель головчатой плесени); б — Cladosporium herbarum; 7—
Alternaria tenuis (возбудители черной плесени); 8 — Botrytis anthophila; 9 — Botrytis
cinerea (возбудители серой плесени)
Под влиянием этих грибов возможно снижение энергии
прорастания и силы роста семян. Семена, пораженные плесенью, нередко
слеживаются, загнивают и становятся непригодными для высева.
Наиболее распространенные плесневые заболевания плодов и
семян вызываются грибами-гифомицетами (например зеленая,
розовая, черная, серая плесень) и грибами порядка мукоровых
(головчатая плесень) (рис. 48).
Зеленая плесень вызывается многими видами грибов рода
Penicillium, а также грибом Aspergillus glaucus Link.
На пораженных семенах образуются яркие, рыжевато-бурые или
красноватые, резко очерченные пятна, постепенно сливающиеся.
Мицелий гриба сравнительно быстро проникает во внутренние
ткани семян, которые становятся рыхлыми и приобретают
коричневую окраску. На поверхности пораженных семян появляются
зеленые, зеленовато-сизые, голубоватые порошащие налеты. Они
состоят из гиф, конидиеносцев и конидий возбудителя. Кониди-
альное спороношение часто имеет форму коремий.
Вследствие обильного образования пылящих масс конидий
распространение инфекции в местах хранения семян происходит
очень быстро. Пораженные семена полностью теряют всхожесть.
197
Зеленой плесенью особенно часто поражаются сочные семена
(например, желуди), подвергшиеся чрезмерному подсушиванию в
период заготовки. Заболевание широко распространено, и степень
поражения семян иногда бывает очень высокой.
Розовая плесень чаще всего вызывается грибом Trichothecium
roseum Link. Развитию гриба способствует высокая влажность. На
зараженных семенах образуются матовые, темно-коричневые,
иногда почти черные, резко очерченные, слегка углубленные пятна.
На их поверхности возникает розовый с оранжевым оттенком
мучнистый налет конидиального спороношения гриба.
Заболевание встречается на семенах различных видов деревьев и
кустарников (клена, ясеня, дуба, сосны, ели и др.), которые могут
заражаться еще на дереве.
Поражение семян, проявляющееся в виде розовой (с
оттенками от нежно-розового до малинового цвета) плесени,
вызывается также грибами рода Fusarium. Поражение мясистых семян
сопровождается загниванием внутренних тканей, приобретающих при
этом красноватую окраску, и отмиранием зародыша. Грибы рода
Fusarium при высеве зараженных ими семян могут вызвать
опаснейшее заболевание — загнивание проростков и полегание всходов.
Черная плесень. Поражение характеризуется появлением на
семенах сажистых темно-бурых или оливково-черных налетов и дер-
новинок, состоящих в основном из конидиеносцев и конидий
гриба. Виды рода Alternaria (особенно часто A. tenuis Nees.)
встречаются повсеместно на семенах многих древесных пород и
кустарников. Эти грибы обладают сравнительно высокой патогенностью.
Они ухудшают посевные качества семян и, кроме того, поражают
прорастающие семена и всходы, вызывая загнивание проростков
и полегание сеянцев.
Из грибов рода Cladosporium наиболее распространен С. herbarum
Link., поражающий семена лиственных и хвойных деревьев. Такие
семена имеют низкую всхожесть. Некоторые виды этого рода
развиваются на листьях и хвое древесных растений. Черная плесень
семян часто вызывается грибом Aspergillus niger Link.
Серая плесень. Возбудителем чаще всего является гриб Botrytis
cinerea Pers., образующий на семенах густой пушистый
темно-серый налет, как бы присыпанный светло-серой мукой. Налет состоит
из гиф и конидиеносцев, на которых в огромном количестве
образуются конидии. При длительном развитии плесени во время
хранения пораженные семена загнивают, и на них появляются
плотные черные склероции гриба. Заболевание наиболее интенсивно
протекает в условиях повышенной влажности. В. cinerea поражает
семена многих лиственных и хвойных пород и также входит в
комплекс возбудителей полегания сеянцев.
Головчатая плесень чаще всего вызывается грибами Rhizopus
nigricans Ehr., Thamnidium elegans Link, и видами рода Мисог. При
198
высокой влажности воздуха и субстрата эти грибы образуют
обильный паутинистый мицелий, на котором формируются
многочисленные спорангии в виде черных головок. Головчатая плесень
обычно развивается на мертвых семенах. Однако при неправильном
режиме хранения мукоровые грибы поражают и живые семена.
Обычно они почти не снижают всхожести семян, но при длительном
развитии на семенах могут заметно задерживать их прорастание,
что в дальнейшем отражается на росте сеянцев.
Меры борьбы с гнилями и плесневением плодов и семян:
мероприятия по профилактике заражения семян грибами
(правильная организация сбора и перевозки семян, дезинфекция тары
и хранилищ, химическое протравливание семян перед зимним
хранением), создание оптимального режима хранения семян.
9.3. СИСТЕМА МЕРОПРИЯТИЙ
ПО ЗАЩИТЕ ПЛОДОВ И СЕМЯН
ОТ БОЛЕЗНЕЙ
Система мероприятий по защите плодов и семян включает
проведение надзора, выполнение правил лесной профилактики в
лесосеменных хозяйствах, правильную организацию сбора и
хранения плодов и семян, фитопатологическую экспертизу семян,
химические меры борьбы.
Надзор за появлением и распространением болезней плодов и
семян проводится с целью выявления их видового состава,
степени пораженности семян, динамики их развития в насаждениях и
причиняемого вреда. Для проведения надзора в лесосеменных
хозяйствах отводят таксационные выделы сроком на 10 лет. Надзор
осуществляют путем периодического сбора и анализа плодов
(желудей, шишек и т.д.). Сбор плодов проводят не менее двух раз в
год в соответствии со сроками развития плодов и семян, а также
возбудителей главнейших заболеваний.
В каждом отведенном для надзора выделе с 3 — 5 плодоносящих
деревьев методом случайной выборки снимают не менее 300
плодов и определяют их зараженность болезнями. На основании этих
данных делают заключение о необходимости тех или иных
мероприятий. Анализ данных надзора за несколько лет позволяет
судить о динамике развития болезней и угрозе поражения ими
плодов и семян в последующие годы.
Для получения здоровых семян с высокими генетическими и
посевными качествами необходимо производить их заготовку на
специальных лесосеменных плантациях или семенных участках. Под
лесосеменные участки отводят совершенно здоровые, лучшие по
всем показателям древостой. При закладке новых участков следует
отбирать экотипы и формы древесных пород, наиболее устойчи-
199
вые к инфекционным болезням и воздействию абиотических
факторов. На лесосемейных плантациях и в окружающих насаждениях
необходимо строго выполнять санитарные правила, осуществлять
надзор за распространением вредителей и болезней, проводить
борьбу с теми из них, которые могут снизить урожай и качество
семян.
На территории лесосеменных участков и в прилегающих к ним
насаждениях следует уничтожать деревья и кустарники,
являющиеся промежуточными хозяевами возбудителей болезней семян.
Сбор семян на лесосеменных участках следует проводить
только с абсолютно здоровых, лучших по росту и развитию деревьев.
Важное значение имеет выбор оптимальных сроков сбора и
способов заготовки семян с учетом биологических особенностей
древесных пород и кустарников, а также местных условий. Для
предупреждения заражения плодов и семян фитопатогенными грибами
необходимо во время сбора, транспортировки и переработки
плодов и шишек оберегать их от загрязнения, механических
повреждений, самонагревания или подмерзания, чрезмерного увлажнения
и пересушивания.
Способ и режим хранения семян определенных пород имеют
очень большое значение. При этом особое внимание следует
уделять поддержанию оптимальных температуры и влажности
воздуха, аэрации семенохранилищ. Так, во время хранения семян
допускаются колебания температуры лишь в пределах от 0 до 5 °С, а
влажности — от 65 до 70 %. Хранящиеся семена рекомендуется
периодически перемешивать, а семенохранилища — проветривать.
Для контроля за состоянием и качеством семян в процессе
хранения периодически берутся пробы.
Химическая защита семян включает следующие мероприятия:
дезинфекцию 3 %-м раствором формалина орудий труда и тары
для семян как перед сбором, так и по окончании сбора каждой
партии семян; дезинфекцию семенохранилищ перед закладкой
семян на хранение путем фумигации — сжигания серы из расчета
30—40 г на 1 м3 помещения; протравливание семян химическими
препаратами. Для предотвращения развития грибных болезней во
время зимнего хранения рекомендуется сухое протравливание
семян, особенно желудей, перед закладкой их в хранилища.
Наиболее эффективно применение ТМТД, фундазола, топсина-М и
некоторых других фунгицидов.
Семена всех древесных растений, предназначенные для
посева, перед посевом должны пройти специальную проверку (фито-
патологическую экспертизу) на лесосеменной станции. По
результатам этой проверки лесосеменная станция делает заключение о
наличии грибной или бактериальной инфекции и степени
зараженности семян, дает рекомендации по предпосевной обработке
семян соответствующими препаратами.
Глава 10
БОЛЕЗНИ ВСХОДОВ, СЕЯНЦЕВ,
МОЛОДНЯКОВ И МЕРЫ БОРЬБЫ С НИМИ
Питомники и культуры представляют собой эколого-производ-
ственный объект, включающий определенные возрастные
группы растений: всходы, сеянцы, молодняки. В этих возрастных
периодах растения наиболее сильно поражаются болезнями. Одни из
них являются специфичными для растений определенного
возрастного периода. Например, полеганием поражаются только
всходы, а выпревание встречается только на однолетних сеянцах.
Другие болезни поражают как растения в питомниках, так и
взрослые насаждения, однако представляют опасность только для
молодых растений. К ним относятся шютте, ржавчина побегов
сосны (сосновый вертун), мучнистая роса и др.
Болезни, развивающиеся в питомниках, имеют разное
хозяйственное значение. Например, полегание сеянцев распространено
повсеместно и ежегодно вызывает отпад сеянцев в среднем на
20 — 30 %. Большой ущерб в разных географических зонах
причиняет снежное шютте. Такие болезни, как выпревание сеянцев,
шютте лиственницы, развиваются лишь в отдельные годы,
наиболее благоприятные для развития возбудителей, но вред
растениям в эти годы они могут принести большой.
В настоящее время создают крупные базисные питомники, в
широком масштабе проводят работы по лесовосстановлению,
защитному лесоразведению и озеленению городов, что требует
большого количества высококачественного посадочного материала.
Однако серьезным препятствием для выращивания стандартного
посадочного материала являются болезни.
ЮЛ. ПОЛЕГАНИЕ ВСХОДОВ И СЕЯНЦЕВ
Болезнь поражает семена, проростки и всходы в возрасте до
3—4 нед. В связи с этим ее полное название — загнивание семян и
проростков, полегание и увядание всходов. Полегание вызывается
201
несовершенными грибами из родов Fusarium, Alternaria, Botrytis,
Verticillium, а также грибом Phytium debaryanum из оомицетов.
Наиболее часто возбудителями болезни являются грибы рода Fusarium,
и в этом случае ее называют фузариозом. Поражаются как
лиственные, так и хвойные породы, последние страдают особенно
сильно. Отпад хвойных всходов от полегания достигает нередко
30—45%, а в отдельных случаях 85—100%. Особенно большой
отпад от болезни отмечается при выращивании сеянцев под
полиэтиленовым покрытием, в условиях высокой относительной
влажности воздуха и повышенной температуры. В связи с тем, что
поражаются как семена и проростки, так и всходы, различают
до- и послевсходовую фазы развития болезни (рис. 49).
Признаком довсходовой фазы является наличие пустот в
посевных строчках. При раскопке в них обнаруживаются загнив-
Рис. 49. Инфекционное полегание всходов:
1 — очажок болезни в посевной строчке; 2 — загнившие проростки (довсходовая
форма болезни); 3 — пораженные всходы с обнаженным осевым цилиндриком
корня; 4 — спороношения возбудителей: а — конидии Alternaria; б — макро- и
микроконидии и хламидоспоры Fusarium; в — конидиеносец с конидиями Botrytis cinerea
202
шие семена и проростки. Для послевсходовой фазы характерно
очаговое расположение больных всходов. При благоприятных
условиях очаги быстро сливаются и могут охватить всю посевную
площадь.
У пораженных всходов около корневой шейки образуется
перетяжка темно-бурого цвета, вследствие чего всходы падают на
землю и усыхают. Нередко во влажную погоду в местах перетяжек
образуется мицелий возбудителей. Часто больные всходы долго не
сбрасывают семенные колпачки. У пораженных всходов загнивают
и отмирают боковые корешки, поэтому больные растения легко
вынимаются из земли вместе с обнаженным осевым
цилиндриком корня (см. рис. 49, 3). При поражении всходов старше 4-не-
дельного возраста, с начинающими одревесневать стволиками,
перетяжка у корневой шейки не образуется, и в этом случае
растения усыхают стоя. Один из типов проявления болезни —
увядание верхушек всходов, происходящее вследствие закупорки
сосудов гифами возбудителей. Пораженные всходы теряют тургор и
падают, но корневая система при этом не загнивает.
Источниками инфекции являются зараженные семена и почва.
В почве возбудители полегания, в частности фузариумы, долгое
время могут существовать сапротрофно на растительных остатках.
Кроме того, у фузариумов образуются хламидоспоры, которые
длительное время сохраняются в почве и обладают высокой
устойчивостью к действию неблагоприятных факторов внешней
среды и пестицидов.
Фузариумы наряду с всходами древесных пород поражают
многие сельскохозяйственные культуры, в том числе бахчевые,
кукурузу, картофель. В связи с этим почвы, вышедшие из-под
указанных культур, могут служить мощным источником инфекции и
представляют большую угрозу для посевов древесных пород.
Болезнь распространяется посредством конидий, заносимых в
почву с семенами, и мицелием. Скорость распространения
грибницы в почве составляет 2 —5 см в сут, поэтому при
благоприятных условиях очаги полегания могут за 2 мес охватить более 60 %
площади посевов.
Развитие болезни связано с почвенно-климатическими
условиями. Наиболее активно распространение болезни происходит
на тяжелых по механическому составу кислых, влажных почвах,
на которых образуется корка, препятствующая появлению
всходов. Благоприятные условия для развития возбудителей полегания
создаются и на сухих песчаных почвах. Большое влияние на
развитие и распространение болезни оказывают погодные условия.
Отпад от болезни увеличивается при сочетании низких температур с
большим количеством осадков и высоких температур с
дефицитом влаги в первые 3—4 нед после появления всходов. Кроме того,
развитию болезни в значительной степени способствуют наруше-
203
ния правил агротехники, приводящие к ослаблению всходов и
снижающие их устойчивость.
Полегание может быть вызвано и абиотическими факторами, в
том числе избыточным увлажнением почвы, чрезмерным отене-
нием посевов, перегревом верхних слоев почвы и т.д., но в этом
случае распространения очагов, как при инфекционном
полегании, не происходит.
Для правильного планирования мероприятий, направленных на
предотвращение и ограничение полегания, важно установить
характер заболевания — инфекционный или неинфекционный. С этой
целью используют различные методы диагностики болезни:
макроскопический, микроскопический, микологический.
Для инфекционного полегания характерна очаговость,
наличие в пустотах посевных строчек загнивших семян и проростков,
наличие у больных всходов перетяжки и обнаженного осевого
цилиндрика корешка. В сомнительных случаях дополнительно
проводят анализ больных всходов методом влажной камеры. При
помещении в нее всходов грибница возбудителя, находящаяся
внутри тканей, прорастает наружу через 3 — 6 дней. По цвету
мицелия, а также по форме и цвету конидий можно определить род
возбудителя (см. рис. 49, 4).
Грибы из рода Fusarium образуют на стебельках у корневой
шейки всходов пушистый белый или розовато-белый мицелий,
на котором формируются конидии двух типов. Макроконидии
серповидные или веретенообразные, бесцветные, с 3—5
перегородками размером (28 — 60) х (3 — 6,5) мкм. Они являются
характерным симптомом фузариоза. Микроконидии овальные, бесцветные,
одноклетные размером (5—9)х(2 —5) мкм.
Грибы из рода Alternaria характеризуются образованием у
корневой шейки всходов бархатистых оливковых, темно-бурых или
почти черных дерновинок мицелия. Конидии обратно-булавовидные,
оливково-бурые, с несколькими поперечными и 1 — 3
продольными перегородками, размером (30—60) х (14—18) мкм. Они могут
быть одиночными или собраны в легко распадающиеся цепочки.
У гриба Bottytis cinerea на всходах развивается мицелий в виде
серого налета или дернинок. Конидии округлые или яйцевидные,
бесцветные, одноклетные размером (9—12) х (7 — 9) мкм,
собраны гроздьями на разветвленных вершинах конидиеносцев.
Грибы рода Verticillium проявляются в виде белого, пушистого
мицелия, на котором формируются конидиеносцы с конидиями.
Конидии удлиненно-овальные или удлиненно-яйцевидной формы,
бесцветные или буроватые, одноклетные размером (5 — 12) х 3 мкм.
Если полегание носит неинфекционный характер, налет
грибницы на всходах во влажных камерах будет отсутствовать.
По способу, предложенному И.И.Журавлевым, берут кусочек
стебелька в области корневой шейки, размачивают его в капле
204
воды и раскатывают до образования тонкой полоски ткани. На эту
полоску переносят кагуш 3 — 5 %-го раствора марганцовокислого
калия. Через 2 — 3 мин раствор удаляют пипеткой или
фильтровальной бумагой, препарат промывают водой и рассматривают
под микроскопом. При инфекционном характере болезни в
клетках будут видны окрашенные гифы гриба.
Болезнь распространена повсеместно в европейской части
России, на Урале, в Сибири, на Дальнем Востоке.
10.2. ГНИЛЬ СЕЯНЦЕВ
Гниль надземных частей сеянцев вызывается грибом оомице-
том Phytophthora cactorum (Lebert.et Kohn.) Schrot.). Гриб поражает
сеянцы лиственных и хвойных пород, особенно часто — всходы и
сеянцы бука. Первичное заражение осуществляется весной
посредством ооспор, сохраняющихся в почве. У пораженных всходов на
стеблях, семядолях или листочках (хвое) появляются темные
пятна. Во влажную погоду пятна быстро разрастаются и охватывают
все загнивающее растение. В сухую погоду растения буреют и
выглядят, как обожженные. В местах поражения в межклеточных
пространствах образуются скопления мицелия, а внутрь клетки
проникают гаустории. На пораженных органах вскоре образуются ко-
нидиеносцы с конидиями в виде серовато-белого налета. Попав
во влажную среду, конидии дают мицелиальный росток или их
содержимое распадается на зооспоры. Конидии и зооспоры
образуются в течение весны и лета и осуществляют массовые
повторные заражения. Заражение здоровых растений может происходить
и посредством грибницы через почву. Осенью внутри зараженных
тканей образуются ооспоры, которые с отмершими частями
сеянцев попадают в почву, где способны сохраняться в течение
нескольких лет. При благоприятных условиях ооспоры прорастают и
заражают сеянцы и всходы.
Болезнь не имеет широкого распространения. Наиболее часто
она встречается в условиях Черноморского побережья Кавказа.
Гниль корней сеянцев вызывается сумчатым грибом Rosellinia
quercina Hart. Он поражает чаще всего сеянцы дуба, значительно реже
другие лиственные и хвойные породы. Заражение растений
осуществляется мицелием, ризоктониями, аскоспорами. У больных
сеянцев поражаются и загнивают корни, поэтому они засыхают
начиная с вершины (рис. 50). На пораженных корнях образуется
грибница с тонкими ветвящимися светлыми ризоктониями. В местах
соединения боковых корешков с главным корнем развиваются
черные склероции. После перезимовки склероции прорастают, особенно
активно во влажную погоду, и образуют серовато-беловатый
мицелий, который распространяется в почве или по ее поверхности и
205
Рис. 50. Гниль корней сеянцев дуба (дубовый корнедушитель):
1 — пораженный сеянец; 2 — конидиальное спороношение гриба; 3 —
пораженная часть корня со склероциями и ризоктониями возбудителя; 4 — перитеции
гриба; 5 — зрелая сумка со спорами; 6 — аскоспоры
заражает здоровые сеянцы. У корневой шейки усохших растений
образуются шаровидные перитеции диаметром до 1 мм, в которых
формируются сумки со спорами. Сумки цилиндрические на
длинных ножках. Аскоспоры эллипсоидальные, одноклетные, темно-
бурые, с двумя каплями масла. Распространению аскоспор
способствуют насекомые, повреждающие корневую систему сеянцев.
Источниками инфекции являются почва и пораженные сеянцы.
Чаще болезнь развивается на отдельных растениях или
проявляется небольшими очажками. В более редких случаях наблюдается
массовое поражение и усыхание посевов дуба в питомниках и
самосева в насаждениях.
10.3. БОЛЕЗНИ ТИПА ШЮТТЕ
Обыкновенное шютте сосны. Возбудители болезни — сумчатые
грибы Lophodermium seditiosum Mint., Stal. et Millar и Lophodermium
pinastri Chev. По данным Н. М. Ведерникова, шютте сеянцев,
саженцев, подроста, культур и молодняков сосны до пятилетнего
206
возраста вызывается преимущественно (96—100% случаев)
грибом L.seditiosum и лишь изредка на сеянцах встречается L.pinastri
Подрост, культуры и молодняки сосны в 6 — 14-летнем возрасте
поражаются одновременно двумя грибами: L. seditiosum в основном
до восьмилетнего возраста и L.pinastri главным образом с восьми
лет. Начиная с 15-летнего возраста сосна заражается только
грибом L.pinastri.
Наибольший ущерб шютте причиняет посевам сосны в
питомниках, в годы эпифитотий поражение достигает 60—-100 %
двухлетних сеянцев. При условиях, благоприятных для развития
болезни, пораженные сеянцы гибнут в питомниках или после
высадки их в культуры. Больные растения, оставленные в
питомниках, частично восстанавливаются, хотя в последующие годы, как
правило, опять заболевают. У больных сосенок в культурах
прирост снижается.
При благоприятных условиях для развития шютте,
вызываемого L. seditiosum, первые его признаки появляются уже в конце
октября — начале ноября, чаще всего в виде пятен желтого цвета по
всей длине хвои или с верхних ее концов, в нижней части охвое-
ния (ранняя форма проявления болезни). Иногда в это время на
части хвоинок можно обнаружить пикниды гриба.
В большинстве случаев первые внешние признаки болезни в
питомниках (обычная форма проявления болезни) наблюдаются
через 3—9 дней после стаивания снега (при солнечной погоде
раньше, а при холодной и пасмурной — позднее). Хвоя сеянцев
отмирает и краснеет. Для шютте типично равномерное
распространение болезни по площади, а на сеянцах — снизу вверх.
Нередко растения поражаются почти на 100 %. У однолетних сосенок
хвоя не опадает, может опадать частично в сезон проявления
болезни или остается на них еще в течение года. С больных
трехлетних сеянцев хвоя опадает большей частью в течение первого лета.
В период с середины апреля до конца второй декады мая на
пораженной хвое появляются пикниды гриба. Апотеции образуются в
конце июня — начале июля. Пикниды и апотеции являются
наиболее характерными признаками болезни.
Пикниды L. seditiosum на одиночной хвое овально-удлиненные,
на парной хвое, как правило, округло-линейные, реже
эллиптические, длиной 300 — 500 мкм, часто сливающиеся (в цепочку) друг
с другом. Конидии одноклеточные, бесцветные, цилиндрические
длиной 6—8 мкм. Апотеции расположены под эпидермисом, в
большинстве случаев серого цвета, нередко соединяющиеся на концах
друг с другом, эллиптические или удлиненно-эллиптические,
заостренные. Плодовые тела размером (901 —1281) х (435 — 554) мкм,
при созревании раскрывающиеся продольной щелью,
окаймленной серо-голубыми или грязно-зелеными губами. Под
основанием апотециев на поперечном разрезе эпидермальных клеток нет.
207
Поперечные линии на хвое отсутствуют или встречаются очень
редко, нечеткие, рыхлого строения, в большинстве случаев
темно-коричневого цвета (рис. 51). Сумки булавовидные размером
(126—184) х (11 —14) мкм. Аскоспоры бесцветные, нитевидные
размером (90— 115) х 3 мкм.
Источниками инфекции являются зараженные растения в
питомниках, культурах, самосев и подрост. Заболевают здоровые и
ослабленные растения. Созревание, рассеивание аскоспор и
заражение хвои происходят в период с конца второй декады июля и
до конца сентября—начала октября. Интенсивность этих
процессов зависит от количества осадков, выпавших в июне—августе, и
температуры воздуха в июле —августе. Однако решающим
фактором в развитии болезни является влажность.
В годы с ранней теплой и влажной весной на семядольной хвое
однолетних сеянцев, на одиночной и первичной парной хвое
двухлетних сеянцев созревают апотеции. В этих условиях рассев спор и
заражение хвои отмечается с мая до середины июня.
Первые внешние признаки шютте при поражении посевов
сосны грибом L.pinastri наблюдаются обычно в мае, в среднем через
месяц и более после схода снега, когда сеянцы находятся в
прикопке или уже высажены на лесокультурную площадь (поздняя
форма проявления болезни). Отмирание и покраснение хвои
происходят на У4— !/2 часть снизу. Пикниды появляются на
пораженной хвое в июне—июле или их вообще может не быть. Плодовые
тела, как правило, появляются только на опавшей хвое.
Поперечные линии на хвоинках встречаются обычно в конце лета первого
Рис. 51. Обыкновенное шютте сосны:
А — Lophodermium pinastri: 1 — пораженная хвоя; 2 — участок пораженной хвои с
апотециями, пикнидами и поперечными линиями; 3 — поперечный срез через
апотеции с эпидермальными клетками под основанием; Б — Lophodermium seditio-
sum: 1 — пораженная хвоя; 2 — участок пораженной хвои с апотециями и
пикнидами; 3 — поперечный срез через апотеции, эпидермальные клетки под
основанием отсутствуют
208
или в начале второго года после заражения, незадолго до
появления апотециев. На хвое сеянцев, за редким исключением,
поперечных линий и апотециев гриба не бывает.
Пикниды гриба L.pinastri темно-серые или черные, округлые
длиной 300—400 мкм. Пикниды при созревании раскрываются
продольной щелью на боковой стороне, конидии одноклеточные,
бесцветные, цилиндрические размером (4,5x6,25) мкм. Апоте-
ции своим основанием погружены под эпидермис, они черного
цвета, овальные или эллиптические размером (825—1099) х (432 —
563) мкм. Плодовые тела при созревании раскрываются
продольной щелью, окаймленной красноватыми, реже
темно-коричневыми губами. Под основанием апотециев на поперечном разрезе
находится, как правило, более пяти эпидермальных клеток.
Поперечные линии на хвое, особенно на парной, многочисленные,
четкие, в большинстве случаев черного цвета (см. рис. 51, А, 2).
Сумки булавовидные размером (90—148) х(Ю—12) мкм. Аско-
споры нитевидные, бесцветные размером (80—105) х (2 — 3) мкм.
Источниками инфекции являются культуры, подрост и
насаждения старше 15-летнего возраста. На пораженной хвое, как
правило, на следующий год после ее опадания образуются апотеции.
Наиболее активный разлет аскоспор и заражение растений
наблюдается в июле — августе, но возможен и весной, в мае.
Поражаются ослабленные сеянцы. Причинами ослабления бывают
неблагоприятные условия роста растений, поражение их другими
болезнями, повреждения пестицидами и механические.
Болезнь широко распространена в ареале сосны.
Снежное шютте сосны. Возбудитель болезни — сумчатый гриб
Phacidium infestans Karst. Его называют «снежным» грибом из-за
способности развиваться под снежным покровом, а болезнь —
снежным шютте. Большую опасность болезнь представляет для сеянцев
и самосева, а также для культур и подроста, которые зимой почти
полностью находятся под снегом. Отпад в питомниках составляет
10—20 %, а в отдельных случаях достигает 80 % и более. В такой же
степени поражаются самосев и подрост сосны. В питомниках
устойчивее однолетние сеянцы, потому что они не так тесно
соприкасаются, как двухлетние, кроме того, хвоя однолетних сеянцев
покрыта восковым налетом, с низкой смачиваемостью. Кроме сосны,
болезнь может поражать кедр сибирский, ель, можжевельник.
Первые признаки болезни появляются в январе, когда в толще
снежного покрова устанавливается температура от 0 до -5 °С. На
хвое бледно-зеленого цвета образуется нежный, паутинистый
мицелий. В феврале пораженная хвоя приобретает мраморную окраску с
характерным чередованием пятен зеленого, желтого и коричневого
цветов. В марте с повышением температуры в толще снежного
покрова до 0,5 °С начинается активное развитие наружного мицелия,
который распространяется на здоровую хвою и заражает ее. По мере
209
таяния снега мицелий уплотняется в беловато-серые пленки,
являющиеся важным диагностическим признаком болезни. Однако
пленки очень быстро разрушаются, и от них остаются лишь грязно-
серые обрывки на сеянцах и почве. В первые дни после схода снега
пораженная хвоя отмирает и приобретает яркую красноватую или
рыжую окраску. В этот период куртины пораженных сеянцев четко
выделяются на зеленом фоне здоровых растений. Вскоре после
схода снега пораженная хвоя светлеет, и на ней появляются зачатки
апотециев в виде мелких, темных, слегка расплывчатых пятнышек
(рис. 52). Летом пораженная хвоя становится сероватого цвета, а
апотеции имеют вид темных бугорков. К началу осени хвоя
приобретает характерную пепельную окраску, становится ломкой,
хрупкой. Апотеции раскрываются, разрывая эпидермис хвои
звездообразными лопастями, и при этом обнажается округлый гимениаль-
ный слой серовато-розового цвета. Сумки булавовидной формы,
содержащие восемь спор эллипсоидальной, реже яйцевидной
формы размером (72—140) х (12—24,8) мкм, с хорошо заметной
оболочкой размером (11,5 — 27,6) х (5,2—9,4) мкм (см. рис. 52, 2—5).
Созревание и распространение аскоспор начинается не сразу
после раскрытия апотециев, а спустя некоторое время. Эти процес-
Рис. 52. Снежное шютте (фацидиоз) сосны:
1 — пораженная хвоя с зачатками апотециев; 2 — хвоя со зрелыми апотециями; 3 —
участок пораженной хвои с полностью раскрывшимися во влажных условиях
апотециями; 4 — срез через апотеции; 5 — сумки со спорами; 6 — аскоспоры
210
сы наиболее активно происходят при большом количестве
выпадающих осадков и температуре воздуха выше средней многолетней.
Особым спорообразовательным толчком являются осенние
заморозки, чередующиеся с дождливой погодой. Начало рассеивания
спор отмечается обычно в конце сентября—начале октября,
массовый разлет происходит в середине октября. Наиболее
благоприятные условия для разлета спор и заражения хвои создаются в
октябре — начале ноября, когда чередуются дни с дождями,
выпадением и таянием снега, моросями, рассев спор заканчивается
до установления снежного покрова. Если же количество
выпадающих осадков и температура не превышают многолетних данных,
рассев спор может продолжаться вплоть до установления
снежного покрова. В этих условиях споры в апотециях могут встречаться в
январе и даже весной, однако они не способны к прорастанию, а
пораженная хвоя не является источником инфекции.
Попадая на хвою, аскоспоры прорастают в ткани хвои. После
установления снежного покрова начинается развитие гриба
внутри тканей. Главнейшая роль при этом принадлежит
температурному режиму снежного покрова. Снежный покров обладает большой
теплоизолирующей способностью, которая изменяется в
зависимости от его глубины и плотности. Благоприятные условия для
развития возбудителя создаются в тех случаях, когда высокий
снежный покров ложится на непромерзшую почву, она остается талой
в течение всей зимы. При этом в толще снежного покрова
устанавливается температура в пределах О °С, а иногда и выше.
По данным И. С. Коссинской, развитие гриба не происходит
совсем, если даже глубокий снежный покров ложится на
промерзшую землю. В этом случае в толще снежного покрова
сохраняются низкие температуры, препятствующие развитию болезни. Если
в зимний период снежный покров не промерзает, то повышение
температуры весной приводит к значительной активизации роста
наружного мицелия и заражению здоровой хвои посредством
этого мицелия. После схода снега наступает сапротрофная фаза
развития гриба, длящаяся весь вегетационный период. В этой фазе
плодовые тела гриба формируются и созревают при любых условиях.
Паразитическая фаза гриба начинается с раскрытия апотециев
и продолжается до схода снега; освобождение аскоспор,
заражение и развитие гриба происходит под снежным покровом.
Выше указывалось, что все эти процессы находятся в тесной
связи с метеорологическими условиями. В зависимости от них было
выделено три типа развития паразитической фазы гриба:
европейский, сибирский и промежуточный.
Европейский тип — осенью происходит распространение спор,
а зимой — развитие болезни. Болезнь развивается в районах,
характеризующихся влажной осенью и сравнительно мягкой зимой
с высоким снежным покровом.
211
Сибирский тип — освобождение спор, прорастание их и
заражение здоровой хвои происходят одновременно в период
весеннего таяния снега в образующихся в толще снежного покрова
полостях или снежных парничках. Болезнь развивается в условиях резко
континентального климата с сухой осенью, суровой зимой,
теплой, дружной весной.
Промежуточный тип может приближаться к одному из
вышеназванных в зависимости от изменения метеорологических условий.
Знание типов развития паразитической фазы возбудителя имеет
большое практическое значение, так как от этого зависят
способы, средства борьбы и сроки проведения мероприятий по защите
от болезни.
Болезнь распространена в европейской части России, на
Урале, в Сибири.
Снежное шютте ели. Возбудитель — гриб Phacidium infestans Karst.
Болезнь поражает 1 — 3-летние посевы ели обыкновенной. Отпад в
отдельные годы, по данным Н.М.Ведерникова, достигает 60%.
Первые признаки болезни проявляются под снегом в феврале —
марте. В этот период хвоя сеянцев становится бледно-зеленой, на
ней образуются желтоватые и бурые пятна, появляется наружный
мицелий. Во время таяния снега происходят активное развитие на
хвое мицелия и заражение им здоровой хвои. Сразу же после схода
снега на больных сеянцах хорошо заметны беловатые пленки
мицелия; хвоя в этот период оливково-зеле-
ная, с коричневыми пятнами. В течение
нескольких дней пораженная хвоя отмирает
и становится красновато-коричневой;
зачатки апотециев приобретают вид черных
пятнышек. В дальнейшем апотеции
увеличиваются в размере, но сумки со спорами
в них не формируются. Пораженная хвоя
осыпается, но в связи с указанной
особенностью плодовых тел не может служить
источником инфекции. Первичное
заражение хвои ели происходит спорами,
которые образуются на пораженной хвое,
произрастающей поблизости сосны, и только
при высокой инфекционной нагрузке.
Болезнь отмечена в питомниках
Среднего Поволжья.
Серое шютте сосны. Возбудитель — сум-
Рис. 53. Серое шютте чатый гриб Lophodermella sulcigena (Rostr.)
сосны Hohn. (= Hypodermella sulcigena (Rostr.) Tub.)
l-пораженнаяхвоя; 2- c конидиальной стадией - Hendersonia
сумка со спорами; з - ас- acwola Tub. Поражаются культуры сосны в
коспоры возрасте от 3 до 10—12 лет и подрост.
212
Заражается хвоя на побегах прошлых лет. Верхняя часть хвои
желтеет, резко отделяясь от здоровой части бурой полоской
шириной до 2 — 3 мм. Затем на пожелтевшей части развиваются пик-
ниды в виде мелких черных точек. Позже отмершая часть хвои
становится серой и на ней хорошо заметны черные удлиненные
апотеции (рис. 53). Больная хвоя долго не опадает. Развитию
болезни способствует высокая влажность. Наиболее сильное поражение
сосны отмечается на открытых, хорошо освещенных местах.
Болезнь не причиняет существенного вреда, но ее высокий уровень
может вызвать ослабление сосны в возрасте до пяти лет.
В ареале сосны серое шютте распространено повсеместно.
Шютте лиственницы. Возбудитель болезни — несовершенный
гриб Meria laricis Vuil. Он поражает лиственницу в возрасте до 30
лет, но болезнь особенно опасна для питомников, где отпад от
нее в среднем составляет 20—30 %, достигая в отдельных случаях
90 % и более.
Рис. 54. Шютте лиственницы:
/ — пораженная хвоя; 2 — больные сеянцы с полностью опавшей хвоей; 3 — вид
окрашенных спороношений возбудителя под лупой на пораженной хвое; 4 —
пучок конидиеносцев, выходящий из устьица хвои; 5 — конидиеносец с конидиями
213
Первые признаки болезни появляются через 10—14 дней
после охвоения сеянцев. Вначале на кончиках хвои возникают
отдельные бурые пятнышки, которые быстро разрастаются,
охватывая всю хвою. Пораженная хвоя имеет красно-бурую окраску и
слегка закручивается, на ней образуются пучки конидиеносцев с
конидиями, выходящие из устьиц, расположенных с нижней
стороны хвои. Спороношения имеют вид мелких, блестящих песчинок,
которые очень трудно различить невооруженным глазом. Для того
чтобы точно поставить диагноз, пораженную хвою погружают на
15—20 мин в 2 %-й раствор марганцовокислого калия. Пучки
конидиеносцев окрашиваются и становятся хорошо заметными в виде
параллельных рядов черных точек (рис. 54). Конидии
цилиндрические, с закругленными концами и перетяжкой посредине,
бесцветные, с двумя каплями масла, размером (6—10) х (2 —3) мкм.
Гриб зимует в виде мицелия в опавшей пораженной хвое,
которая является источником инфекции. Посевы текущего года
значительно меньше страдают от болезни. На посевах же второго года
жизни по мере накопления инфекции болезнь часто принимает
массовый характер. Сначала поражается и опадает хвоя нижнего
яруса, затем среднего и верхнего. С момента попадания конидий на
хвою и до образования нового спороношения проходит 10—14 дней.
При сильном развитии болезни сеянцы уже к середине
вегетационного периода теряют большую часть ассимиляционного
аппарата. В результате они сильно слабеют, отстают в росте и
становятся непригодными к посадке. Кроме того, у больных сеянцев в
зимний период загнивает корневая система, они погибают. Выход
стандартного посадочного материала в результате заболевания
снижается на 30—40 %, а иногда и больше.
Разные виды лиственниц неодинаково поражаются болезнью.
Наиболее сильно страдают сибирская и даурская лиственницы, несколько
меньше — европейская и Сукачева. Развитие болезни зависит от
погодных условий в летний период. Температура не оказывает
существенного влияния на развитие возбудителя. Мицелий гриба растет
при температуре от 5 до 30 °С, оптимальная температура 18... 20 °С.
Интенсивность разлета спор возбудителя находится в прямой
зависимости от количества выпадающих осадков, поэтому засушливая
погода является серьезным препятствием для развития болезни.
Наиболее благоприятные условия для нее складываются в первой
половине вегетационного периода, когда происходит быстрое
нарастание новой хвои и выпадает наибольшее количество осадков.
Шютте лиственницы встречается в северо-западных и
центральных областях европейской части России, на Урале, в Сибири и на
Дальнем Востоке.
Шютте ели. Возбудители — сумчатые грибы Lophodermium
macrosporum (Hart.) Rehm. (= Linda macrospora (R.Hart.) Dark.) и
Lophodermium piceae (Fuck.) Hohn.
214
Рис. 55. Шютте ели:
/ — побег с пораженной хвоей; 2 — хвоя
с апотециями возбудителя; 3 — сумка со
спорами
L. macrosporum поражает
сеянцы в питомниках,
культуры, самосев, подрост и мо-
лодняки ели естественного
происхождения. Заражение
растений осуществляется
весной сумкоспорами. Первые
признаки болезни
обнаруживаются в начале лета
следующего года на прошлогодних
побегах, хвоя желтеет, затем
становится бурой. К концу
лета с нижней стороны хвои
образуются темно-бурые,
почти черные апотеции,
вытянутые до половины длины
хвои и больше. Отмершая хвоя
остается висеть на побегах.
Весной в апотециях
созревают сумки со спорами. В этот период пораженная хвоя опадает. Сумки
возбудителя булавовидные размером 100 х (15 — 20) мкм, аско-
споры нитевидные размером 75 х 155 мкм (рис. 55). Однако часто
апотеции на хвое не образуются. В таком случае для постановки
диагноза пользуются методом влажной камеры.
Развитию болезни способствует избыточное увлажнение и
факторы, вызывающие ослабление ели. Наиболее сильно поражаются
загущенные посевы, подрост ели под пологом леса и защитные
лесные полосы. При благоприятных для возбудителя условиях
болезнь приводит к уменьшению выхода стандартного посадочного
материала, ослаблению и отставанию в росте ели в культурах,
снижению благонадежности естественного возобновления и реже — к
гибели растений. Источниками инфекции являются еловые
насаждения или вырубки с неубранными порубочными остатками.
Болезнь встречается повсеместно в ареале ели, но особенно
широко распространена в северных районах России.
L.piceae поражает ель чаще в естественных насаждениях, реже —
в культурах. Вначале на хвое появляются красно-бурые пятна, и
такая пестрая хвоя может долго оставаться на побегах.
Впоследствии она вся становится бурой и быстро опадает. На отмершей
хвое образуются черные овальные апотеции длиной 1 — 1,5 мм,
отделенные друг от друга поперечными темно-бурыми или
черными тонкими линиями. В апотециях созревают сумки с
сумкоспорами. Споры нитевидные размером (70—80) х 1 мкм. Гриб L.piceae
поселяется на хвое ослабленных деревьев, встречается
значительно реже, чем L. macrosporum, и не причиняет заметного вреда.
Болезнь отмечена в европейской части России, на Урале.
215
Шютте можжевельника. Возбудитель — сумчатый гриб Lophoder-
mium juniperinum de Not. Признаки болезни проявляются в начале
лета на прошлогодней хвое. Пораженная хвоя приобретает
желтоватую или красно-бурую окраску, с верхней стороны хвои в начале
июля образуются плодовые тела — апотеции. Апотеции округлые
или эллипсовидные, выпуклые черные, 0,5 —1,5 мм длиной,
раскрываются узкой щелью, обнажая светлый гимениальный слой сумок
со спорами. Сумки булавовидные, размером (70—90) х (9—12) мкм;
аскоспоры нитевидные, одноклеточные, бесцветные, размером
(65-75) х (1-2) мкм.
Гриб поражает можжевельник на фоне ослабления, вызванного
разными причинами. Болезнь встречается на можжевельнике
разного возраста в разных природных условиях, но особенно высокий
уровень поражения отмечается в более влажных районах в изре-
женных, низкополнотных насаждениях. По данным наших
исследований, уровень болезни в заповедниках Нижне-Свирском и «Ки-
вач» достиг соответственно 94 и 98 — 100% со средней степенью
пораженности кроны, количество усыхающих от болезни
деревьев колебалось с 3,6 до 14 %.
Болезнь была отмечена в средней полосе и северо-западных
районах европейской части России.
Бурое шютте хвойных пород. Возбудитель — сумчатый гриб
Herpotrichiajuniperi (Duby) Petr. (=#. nigra Hart.). Поражаются
разные хвойные породы, в том
числе сосна, кедр, ель,
пихта, можжевельник. Болезнь
встречается в питомниках,
культурах и на подросте.
Признаки заболевания
обнаруживаются весной, сразу
после схода снега. В этот
период хвоя опутана густым
черно-бурым мицелием. Она
буреет и отмирает, но склеенная
мицелием долго не опадает
(рис. 56). К осени на мицелии
образуются плодовые тела —
перитеции. Они шаровидной
или грушевидной формы
диаметром 0,2—0,3 мм, у
основания покрыты длинными,
Рис. 56. Бурое шютте: бурыми, волосистыми
придатками. Сумки мешковидные
1 — пораженная ветка сосны с плотным жгттттт„тлттттттл ^„™,л^л,, /то
темно-бурым мицелием на хвое; 2-пери- Удлиненные размером (72-
теций возбудителя; 3 — сумка со спорами; M)X(1U 12) МКМ. АСКОСПО-
4 — аскоспора ры веретеновидные, бесцвет-
216
ные, одно-, четырехклеточные размером (15 — 30) х (6—12) мкм,
с капельками масла.
Болезнь развивается под снегом в период весеннего
снеготаяния при температуре 0,5 °С. Наибольший вред она причиняет в
местах, где скапливается и долго не тает снег. Самый высокий
уровень болезни отмечается в горах, на крутых, теневых склонах,
где высокий снежный покров и длительный период его таяния
создают оптимальные условия для развития бурого шютте. Гриб
поражает подрост разной высоты. Степень поражения кроны
уменьшается по мере увеличения высоты подроста. Иногда болезнь
захватывает нижние ветви деревьев основного полога и второго
яруса, находившихся под снегом. Поражение бурым шютте хвойных
пород приводит к их ослаблению и гибели, снижает выход
посадочного материала, препятствует естественному возобновлению.
Болезнь встречается в северных районах европейской части
России, в горных районах Северного Кавказа, на Урале, в Сибири.
Побурение хвои ели. Возбудитель — несовершенный гриб
Rhizosphaera kalkhqffii Bub. Заражение хвои происходит весной.
Конидии распространяются посредством дождя. К концу лета или
осенью на хвое побегов текущего года появляются отдельные
желтые пятна. Постепенно они увеличиваются в размерах, становятся
бурыми или красно-бурыми, и в конце зимы — начале весны вся
хвоя буреет. Весной с нижней стороны хвоинок образуются
черные округлые пикниды, располагающиеся продольными рядами.
Конидии овальные бесцветные размером (7—10) х (3 — 5) мкм,
выходят из пикнид в виде мелких белых капель. В этот период
пораженная хвоя опадает.
Распространение конидий и заражение хвои может
осуществляться только в условиях повышенной влажности. Конидии
возбудителя прорастают очень медленно и поэтому заражение хвои
возможно только при длительном ее увлажнении. Болезнь
развивается в питомниках, культурах и особенно в сильно загущенных
плантациях новогодних елей. Часто болезнь протекает на фоне
предварительного ослабления ели, вызванного разными
факторами. По данным М.Э.Хансо, широкое распространение гриба
отмечалось в районах, подвергающихся воздействию
промышленных выбросов, основным компонентом которых является двуокись
серы. Систематически повторяющиеся поражения ели грибом
Kkalkhoffii приводят к ослаблению, реже — к усыханию ее в
питомниках и молодых культурах.
Болезнь распространена в северо-западных районах
европейской части России.
Побурение хвои пихты. Возбудитель — несовершенный гриб
Rhizosphaera pini (Corda) Maubl. Поражаются культуры и подрост
разных видов пихты под пологом леса. Заражение хвои
осуществляется конидиями, которые созревают и распространяются в конце
217
лета, реже — весной. На пораженной хвое вначале появляются
желтые пятна, а позже вся хвоя буреет. Летом на нижней стороне
хвои, вдоль средней жилки, образуются пикниды гриба,
имеющие вид цепочек из мелких, черных точек. В пикнидах
развиваются одноклеточные бесцветные яйцевидные, цилиндрично-оваль-
ные конидии размером (16 —23) х 7,5 мкм. Отмершая хвоя
остается висеть на побегах до весны.
Болезнь встречается в разных типах условий
местопроизрастания на подросте разной высоты. По нашим данным, поражен-
ность пихтового подроста грибом Я pint в разных заповедниках
составляла: 15 % в Тебердинском (Северный Кавказ), 12,9—34,9 %
в Байкальском (Восточная Сибирь), 82 —87% в «Басегах» (Урал).
На высокую степень пораженности подроста пихты в Верхнем
Приобье (Западная Сибирь) указывает и А. М. Жуков.
Систематически повторяющееся поражение хвои приводит к ослаблению
пихтового подроста, а при сильной степени пораженности кроны —
к его гибели.
Болезнь широко распространена на Кавказе, Урале, в Сибири.
10.4. ВЫПРЕВАНИЕ И ДРУГИЕ БОЛЕЗНИ
СЕЯНЦЕВ ХВОЙНЫХ ПОРОД
Выпревание сеянцев. Возбудителями болезни являются грибы
Sclerotinia graminearum Elen. и Typhula graminearum Tul. Первый
относится к сумчатым, а второй — к афиллофороидным гименомице-
там. S. graminearum поражает дикорастущие и культурные злаки и
однолетние сеянцы древесных пород, чаще всего сосны. Первые
признаки болезни обнаруживаются сразу после схода снега. В этот
период на пораженных сеянцах хорошо заметны серовато-белые
пленки мицелия, которые быстро разрушаются и исчезают.
Больная хвоя отмирает и приобретает красно-бурую окраску. Вскоре
после схода снега вблизи почки или на стволике, а иногда внутри
стволика образуются склероции. Они имеют вид образований
неправильной формы размером 1 — 6 мм в диаметре, вначале белого,
позже черного цвета. К началу июня склероции опадают, а осенью,
в конце сентября, они прорастают в апотеции. Апотеции
возбудителя крупные диаметром до 7 мм, на ножке длиной от 2 до 10 мм
(рис. 57, 4). Аскоспоры, развивающиеся в апотециях, разлетаются
и заражают здоровые сеянцы.
Развитие болезни происходит в зимний период под снежным
покровом. Для заражения сеянцев и развития болезни
необходимы определенные условия температуры и влажности. Склероции
прорастают при повышенной влажности и температуре воздуха
около 12 °С. Созревание, разлет спор и заражение сеянцев
наиболее активно происходит при большом количестве осадков и уме-
218
Рис. 57. Выпревание сеянцев:
/ — сеянец сосны, пораженный Typhula graminearum; 2 — сеянцы сосны,
пораженные Sclerotinia graminearum', 3 — склероции гриба T.graminearum, проросшие
булавовидными плодовыми телами; 4 — склероции S. graminearum, проросшие
апотециями
ренной температуре. Развитию болезни в зимний период
способствуют теплая зима с высоким снежным покровом и затяжная
весна с медленным таянием снега.
Болезнь не имеет массового распространения, но в годы,
наиболее благоприятные для развития возбудителя, может причинить
значительный вред. Пораженные сеянцы хотя и редко гибнут, но
становятся многовершинными, отстают в росте и делаются
непригодными для посадки.
T.graminearum поражает однолетние, реже — двухлетние
сеянцы сосны, а также различную сорную травянистую растительность,
в том числе незабудку, звездчатку, мятлик и др. Признаки болезни
обнаруживаются после схода снега. Пораженные сеянцы в этот
период покрыты сероватым паутинистым мицелием, который быстро
разрушается, хвоя красноватого цвета опущена книзу. На хвое, в
пазухах хвоинок и на почках образуются склероции округлой
формы, черные, до 2 мм в диаметре. К началу лета они опадают на
землю, а осенью прорастают в булавовидные плодовые тела, на
которых формируются базидии с базидиоспорами (см. рис. 57, 7,3).
219
Последние и заражают сеянцы. Этот гриб
встречается значительно реже, чем S.grami-
пеагит. В связи с этим данные об условиях
его развития в литературе практически
отсутствуют.
Выпревание встречается в питомниках
центральной полосы европейской части
России, в Сибири, на Дальнем Востоке.
Склерофомоз сосны. Возбудитель
болезни — несовершенный гриб Sclerophoma pithy a
v.Hohn. Поражаются сеянцы 2 —3-летнего
возраста и культуры до 12-летнего
возраста. На подросте сосны болезнь не
зарегистрирована.
Признаки болезни обнаруживаются с
мая по октябрь. По характеру проявления
симптомов выделено несколько типов
поражения. В одном случае побеги имеют S-
образное искривление, изменения
окраски не наблюдается; иногда на таких
побегах можно обнаружить темно-бурые
некротические участки вытянутой формы
длиной до 1 см. Хвоя не имеет признаков
заболевания. В другом — пораженные побеги
деформируются и приобретают яркую ржаво-рыжую окраску,
становятся как бы стекловидными; на хвоинках образуются бурые
широкие перетяжки, выше которых кончики буреют. Позднее на
пораженных побегах появляются некротические участки
вытянутой формы длиной до 1,5 см, вначале темно-бурого, позднее
сероватого цвета, на его фоне хорошо заметны черные, овальные или
круглые пикниды, выступающие продольными рядами из трещин
коры (рис. 58). Хвоя на таких побегах поражена вся или частично.
Кончики хвоинок бурого цвета. В дальнейшем побеги или их
вершины усыхают, чернеют; поверхность побегов и больная хвоя
сплошь покрываются пикнидами.
Многие пораженные побеги не искривляются, но их
поверхность имеет яркую золотисто-ржавую окраску, на фоне которой
заметны небольшие некротические участки бурого цвета. В
верхней части хвоинок образуется бурая перетяжка, выше которой
кончик хвои буреет. Пикниды гриба выступающие, округлые, до
460 мкм в диаметре, с черными толстыми оболочками. Конидии
размером (6 —7) х (2 —2,5) мкм, яйцевидные, яйцевидно-верете-
новидные, неравнобокие, каплевидные.
Источником инфекции являются пораженные отмершие
побеги, осенью они опадают или остаются на мутовках до следующего
года. Болезнь поражает сосну в разных лесорастительных условиях.
Рис. 58. Склерофомоз
сосны:
1 — пораженный побег с
некротическим участком,
покрытым пикнидами
гриба; 2 — конидии
возбудителя
220
Уровень склерофомоза снижается по мере увеличения возраста
культур. Это связано с тем, что для развития гриба необходимы
высокая температура воздуха, хорошее освещение и дефицит
влаги. Такие условия создаются в питомниках и культурах сосны до
6 —7-летнего возраста. Кроме того, свободное движение воздуха в
питомниках и несомкнувшихся культурах способствует активному
распространению спор возбудителя болезни.
Массовое развитие болезни в питомниках вызывает усыхание
сеянцев и снижение выхода стандартного посадочного материала.
Пораженность культур сосны до 5 —7-летнего возраста приводит
к искривлению побегов, образованию многовершинности в
случае усыхания главного побега, а при повторяющихся эпифитоти-
ях — к заметному ослаблению деревьев.
Болезнь зарегистрирована в Ленинградской и Оренбургской
областях.
Бактериоз сосны. Возбудитель болезни — бактерия Pseudomonas
fluorescens Migula. Болезнь поражает только однолетние сеянцы
сосны обыкновенной. Распространенность бактериоза в питомниках
колеблется от 0,1 до 38 %.
Первые признаки болезни проявляются через 10—15 дней
после схода снега. Поражается верхняя часть стволиков и основания
хвои, часто — верхушечная почка. Они приобретают
угольно-черную с синеватым оттенком окраску. Больные растения или их
небольшие куртины располагаются на посевах диффузно. Болезнь
приводит к гибели сеянцев, реже — к ослаблению. У выживших
сеянцев наблюдается деформация стволиков или многовершин-
ность, вследствие чего они становятся непригодными для
посадки. Инфекция сохраняется в почве на растительных остатках.
Весной происходит заражение семян и проростков.
Распространенность бактериоза сеянцев сосны обыкновенной
зависит от температуры воздуха в период с июля по октябрь и
количества осадков в августе в год, предшествующий его
появлению. Погодные условия этого периода оказывают влияние на
растение-хозяина, определяют его устойчивость. Развитие
инфекционного процесса в растении-хозяине происходит в ранневесен-
ний период, течение болезни зависит от температуры воздуха в
первые 10 — 15 дней после схода снега, причем оптимальной
является температура 6 °С и выше.
По данным Н.М.Ведерникова, болезнь впервые была
обнаружена в 1975 г. на территории Марийской Республики. В
последующие годы она распространилась в Татарии и Ульяновской
области.
Серая плесень. Возбудитель болезни — гриб Bottytis cinerea Pers.
Она широко распространена в лесных питомниках на ели,
лиственнице, сосне, реже — в культурах, созданных посевом.
Особенно сильно страдают посевы под полиэтиленовым покрытием.
221
Кроме того, гриб часто поражает многие сельскохозяйственные
культуры, в том числе картофель, люпин и др.
На поверхности больных сеянцев появляется налет
темно-серого цвета, состоящий из мицелия и конидиеносцев с
конидиями. Осенью на грибнице образуются склероции, весной они
прорастают, образуя мицелий и конидиеносцы с конидиями.
Конидии заражают сеянцы. Конидии одноклеточные, яйцевидные,
заостренные у основания, бесцветные, в массе дымчатые размером
(9—12) х (6,5 — 10) мкм. Сильнее поражаются ослабленные и
загущенные посевы. Развитию болезни в открытом грунте
способствуют высокий снежный покров зимой и теплая влажная погода
в период вегетации. Наиболее широкое распространение серая
плесень имеет в закрытом грунте, где создаются благоприятные
условия для развития гриба: высокие температура и влажность
воздуха, большая густота посева, слабая циркуляция воздуха.
Источниками инфекции являются пораженные сеянцы, сорняки
и многие сельскохозяйственные культуры (картофель, люпин и др.).
По данным Н.М. Ведерникова, при сильном развитии болезни
гибель сеянцев достигает 20—30 %, а в отдельных случаях — 90 %.
Болезнь распространена в ареале поражаемых пород.
Удушье сеянцев. Чаще всего болезнь
вызывается афиллофороидным гимено-
мицетом Thelephora terrestris Ehr. Это
типичный почвенный сапротроф,
развивающийся преимущественно на песчаных
и супесчаных почвах. Плодовые тела
гриба имеют вид тонких кожистых раковин
темно-бурого цвета. Вырастая рядом с
сеянцами, гриб обволакивает их,
механически препятствуя дыханию и росту
ассимиляционного аппарата (рис. 59).
От гриба сильно страдают сеянцы
сосны, ели, березы и других пород.
Побеговый рак («зонтичная болезнь»,
склеродерриоз, круменулез). Болезнь
вызывается сумчатым грибом Scleroderris
lagerbergii Gremm (= Gremmeniella abietina
(Lagerb.) Morelet; Crumenula abietina
Lagerb.) с конидиальной стадией Brun-
chorstia pinea (Karst.) Hohn (= B. destruens
Eriks). Поражаются многие хвойные
породы: разные виды сосны, ель, пихта,
лиственница, многие хвойные экзоты, но
чаще встречается на сосне обыкновенной.
Гриб и вызываемая им болезнь широ-
Рис. 59. Удушье сеянцев ко распространены во многих странах
222
Европы, в Северной Америке, в Японии, где причиняют
значительный вред в лесных питомниках и молодняках сосны. В России
болезнь впервые была отмечена в конце 30-х гг. XX столетия в
лесных питомниках Татарии. По данным В. И. Крутова и М. Э. Хансо,
в 1965 — 1967 гг. побеговый рак повсеместно встречался в
сосновых культурах разного возраста в Мурманской области, а в 1981 г.
с ним связана гибель культур сосны на большой площади в
Карелии. В этот же период болезнь была зарегистрирована в
питомниках Ленинградской области. Несколько позже (1987), массовое
распространение сосны в питомниках Среднего Поволжья было
отмечено Н. М. Ведерниковым.
Поражается сосна в питомниках, молодняках искусственного
и естественного происхождения и в насаждениях разного возраста.
Первые признаки заболевания сосны в питомниках наблюдаются
через несколько дней после схода снега. Гриб проникает в хвою,
верхушечную почку, в камбий молодых побегов, ветвей и
стволиков. Вначале хвоя имеет зеленую окраску и только у основания, в
месте прикрепления к стволику, краснеет. У пораженных 2—3-
летних сеянцев парная хвоя висит косо, «зонтиком» и при
малейшем прикосновении опадает (рис. 60, 7). Поэтому в питомниках
эту болезнь часто называют зонтичной. Сосна поражается
одиночно или куртинами. После схода снега хвоя в куртинах больных
сеянцев кажется как бы примятой. Со временем она буреет,
может долгое время оставаться на сеянцах. Одновременно отмирают
не только хвоя, верхушечная почка, но и камбий стволика. При
попытке выдернуть сеянец кора легко снимается чулком. У
сосенок старше пяти лет в очагах болезни хвоя взъерошенная или
флагообразно повисшая, верхушечные побеги укорочены или
утолщены, с изогнутой или утолщенной хвоей (см. рис. 60, 2).
Растения с диаметром стволика меньше 1 см, пораженные побеговым
раком, чаще всего погибают. Для растений с большим диаметром
характерно появление некротических пятен опоясывающего
некроза ствола и ветвей, отмирание верхушечных побегов. Изредка
отмечается ступенчатый некроз коры с последующим
образованием раковых язв (см. рис. 60, <?).
В основании хвоинок и на коре усыхающих или уже усохших
сеянцев в течение лета появляются зрелые пикниды. Они имеют
вид черных шероховатых бородавочек размером до 2 мм. Конидии
бесцветные, серпообразные, чаще всего 3—4-клеточные, иногда
4—6-клеточные размером (16—32) х (2—3,5) мкм (см. рис. 60,3,5, б).
Сумчатая стадия возбудителя встречается очень редко. Апотеции
черно-бурого цвета диаметром 1 — 1,8 мм, с булавовидными
сумками размером (95 — 110) х (7,5—10) мкм. Аскоспоры
эллипсоидальные, 3—4-клеточные, бесцветные размером (15—24) х (4—4,8) мкм
(см. рис. 60, 4, 7). При отсутствии плодоношений диагностическим
признаком болезни может служить зеленовато-желтая до изум-
223
рудно-зеленой окраска древесины на продольном срезе по
границе между мертвой и живой тканью в местах некроза.
В молодняках 15—20-летнего возраста у пораженных сосен
наблюдается отмирание концов верхушечных побегов, котброе
сопровождается их характерным утончением и опадением хвои. На таких
Рис. 60. Побеговый рак сосны:
/ — пораженный двухлетний сеянец с зонтикообразно опущенной хвоей; 2 —
пораженные ветви в культурах; 3 — скопления пикнид в отмершей коре побегов;
4 — апотеции в трещинах коры отмершего побега; 5 — пораженная хвоя с пик-
нидами; 6 — конидии возбудителя; 7 — сумка со спорами; 8 — раковая рана на
стволике в культурах
224
побегах нередко образуются пикниды возбудителя. Аналогичная
картина поражения в средневозрастных, спелых и перестойных
насаждениях. Кроны больных деревьев приобретают пеструю
окраску из-за множества отмерших побегов. Нижние ветви погибают
полностью. В годы эпифитотий отмирание побегов происходит не
только в нижней, но и в средней, и верхней частях кроны.
Заражение здоровых растений осуществляется конидиями и
аскоспорами, которые разносятся ветром и, возможно,
насекомыми. Рассев конидий происходит в течение всего лета, аскоспор —
во второй половине лета — начале осени.
Развитию побегового рака способствуют различные факторы,
вызывающие ослабление растений и тем самым снижающие их
устойчивость к болезни. К ним относятся неблагоприятные
климатические и почвенные условия: сильные морозы, холодные и
дождливые вегетационные периоды, задерживающие одревеснение
побегов, нарушение баланса питательных веществ. Наиболее
активно болезнь развивается в низинах, где часты туманы и заморозки.
Устойчивость к болезни снижается в посевах и посадках с высокой
густотой. Наиболее восприимчивы к побеговому раку те климати-
пы сосны, которым не соответствуют условия произрастания.
Поражаются сеянцы в питомниках, молодняки естественного
и особенно искусственного происхождения до 25-летнего
возраста, реже — средневозрастные, спелые и перестойные насаждения.
При благоприятных условиях, когда болезнь принимает характер
эпифитотий, пораженность сеянцев в питомниках достигает 85 —
100 % с усыханием до 50 —60 %. При заболевании молодняков и
насаждений более старшего возраста деревья ослабляются,
значительно уменьшается прирост, возрастает вероятность заселения
стволовыми вредителями.
Болезнь зарегистрирована в европейской части России. По
данным А. М.Жукова (1978), в Сибири схожее с этим заболевание на
пихте вызывает гриб Ascocalyx abietis Naum.
10.5. БОЛЕЗНИ, ВЫЗЫВАЕМЫЕ РЖАВЧИННЫМИ
ГРИБАМИ
Ржавчинные грибы поражают многие древесные породы и
кустарники, вызывая преждевременное засыхание и опадение
листьев и хвои. Больные растения сильно ослабляются, отстают в
росте, а иногда гибнут.
Ржавчина побегов сосны (сосновый вертун) вызывается разно-
хозяйным ржавчинным грибом Melampsora pinitorqua (A. Br.) Rostr.
Эции образуются весной и в начале лета на всходах, сеянцах и
молодых побегах сосны. Они имеют вид желтых вздутий,
прикрытых эпидермисом. Созревшие эциоспоры разрывают эпидермис и
225
выступают в виде оранжевой массы, разносятся воздушными
потоками и заражают листья осины или белого тополя, на которых
развивается урединиостадия. Урединии представляют собой
оранжевые или ярко-желтые порошащие подушечки, которые при
сильном развитии болезни покрывают весь лист.
В конце лета на верхней стороне пораженных листьев
образуется телиостадия. Телиопустулы имеют вид темно-коричневых или
черных плотно прилегающих коростинок, сильно набухающих во
влажную погоду. В этой стадии гриб зимует на опавших листьях
осины и тополей. Весной телиоспоры прорастают, образуя бази-
дии с базидиоспорами в виде золотистого налета, хорошо
заметного на серых прошлогодних листьях. Созревшие базидиоспоры
заражают хвою, стволики и молодые побеги сосны, где вновь
развиваются эции.
Поражаются всходы, сеянцы в питомниках, культуры и мо-
лодняки естественного происхождения (рис. 61). У больных
всходов сосны эции образуются как на стволике, так и на хвоинках.
Они имеют вид вздутий желтого цвета, располагающихся
цепочкой. Поражение только хвои не приводит к гибели всходов.
Возникновение же эциев на стволике вызывает усыхание и опадение
хвои, а нередко и гибель всходов, достигающую 20—40 %.
У двухлетних сеянцев сосны поражаются только побеги
текущего года. Скопления эциев имеют вид желтых, выпуклых
образований длиной 1 см. В месте возникновения эциев побег
искривляется (см. рис. 61,2). После созревания и разлета эциоспор на
побегах остаются бурые, часто засмоленные ранки. У больных сеянцев
верхушки чаще всего отмирают, развивается многовершинность,
но гибель сеянцев отмечается в редких случаях. В благоприятные
для развития болезни годы характерно ослабление и сильное
искривление сеянцев, которые становятся непригодными к посадке.
У сосен от 2 до 10-летнего возраста поражаются как боковые
побеги, так и верхушечный (см. рис. 61, 7). В случае усыхания
побегов под ними развиваются дополнительные почки, из которых
образуются новые побеги. Один из этих побегов становится
главным, заменяя усохший верхушечный побег, остальные отстают в
росте и чаще отмирают. Поражение сосны в этот возрастной
период снижает прирост и способствует образованию внутренних
пороков древесины. Сосны старше 10-летнего возраста
поражаются значительно реже, только при наиболее благоприятных для
развития болезни условиях. При этом чаще поражаются боковые
побеги.
Жизнедеятельность гриба зависит от погодных условий. Эциоста-
дия наиболее активно развивается при температуре 18 ...21 °С. В
зависимости от климатических условий эциоспороношение
появляется в разное время, в период с конца мая до третьей декады
июня. В конце августа — начале сентября образуется телиостадия,
226
Рис. 61. Ржавчина побегов сосны (сосновый вертун):
/ — пораженные побеги в культурах; 2 — пораженный всход с эциями на стволике;
3—лист осины с урединиоспороношением; 4— телиоспоры, проросшие базидиями
с базидиоспорами; 5 — искривление стволика у пораженного двухлетнего
сеянца
зимующая на опавших листьях осины и тополей. Для прорастания
телиоспор необходима высокая относительная влажность воздуха
(80—97%,) и температура в пределах 18...20°С. Однако
решающим фактором является влажность. В связи с этим наиболее
сильное поражение болезнью наблюдается в годы с теплыми, влажными
веснами.
На развитие болезни значительное влияние оказывает близость
осиновых молодняков и доля участия осины в составе полога
лиственных пород. Примесь других лиственных пород приводит к
снижению распространения болезни. Пораженность культур и
молодняков сосны зависит от типа лесорастительных условий.
Сильнее поражается сосна на кипрейных вырубках, чем на вей-
227
никовых, что объясняется отсутствием высокого и густого
травяного покрова, препятствующего распространению спор и
защищающего сосну от заражения. Особенно подвержены болезни
культуры, созданные в сосняках-черничниках, кисличниках, где не
только повышена влажность, но и низкий травяной покров не
препятствуют разлету базидиоспор и попаданию их на сосну.
Болезнь широко распространена в центральных и северных
районах европейской части России.
Ржавчина хвои сосны. Возбудителями болезни являются раз-
нохозяйные ржавчинные грибы из рода Coleosporium. Эциостадия
этих грибов развивается на хвое сосны, а урединио- и телиоста-
дии — на различных травянистых растениях. Промежуточным
хозяином C.senecionis Fr. является крестовник, С. tussilaginis Lev., —
мать-и-мачеха, С. sonchiarvensis Lev. — осот, С. campanulae Pers. —
колокольчик, С. melampyri (Reb.) Tul. — марьянник (рис. 62, А).
Поражается хвоя в разных частях кроны. В начале лета на хвое
образуются эции гриба, имеющие вид желто-оранжевых
пузырьков длиной 1—3 мм, шириной 1,5 — 2 мм. При благоприятных для
развития возбудителя условиях эции почти сплошь покрывают
поверхность хвои. В эциях летом созревают эциоспоры, которые
при рассеивании заражают соответствующие каждому виду гриба
растения. После рассеивания эциоспор и разрушения оболочек
эциев на хвое остаются буроватые пятна, а хвоя приобретает
пеструю окраску.
У травянистых растений вначале образуются урединиостадии,
а ближе к осени — телиостадии, покрывающие почти всю
поверхность листа. Весной на листьях травянистых растений —
промежуточных хозяев возбудителей развиваются базидии с базидиос-
порами, которые заражают хвою сосны.
По данным А.М.Жукова (1978), гриб C.senecionis в условиях
Западной Сибири способен существовать только в эциостадии.
В этом случае мицелий зимует в хвое сосны, и на следующий год
на ней вновь образуются эции. Ржавчина поражает посевы в
питомниках, культуры до 15-летнего возраста, самосев и молодняки
сосны естественного происхождения. Чаще отмечается невысокий
уровень болезни, но в отдельные годы она может вызывать
массовое поражение сосны. Эпифитотия ржавчины отмечалась Б.Н. Ни-
колиным в 5 — 10-летних культурах в Ленинградской области. На
сильное развитие ржавчины в культурах и на подросте сосны в
Западной Сибири указывает А. М.Жуков. Ржавчина даже в период
эпифитотии не вызывает гибели сосны, но приводит к ее
ослаблению и снижает устойчивость к другим болезням.
Болезнь распространена в европейской части России, на
Урале, в Сибири.
Ржавчина хвои ели. Возбудители болезни — ржавчинные грибы
Chrysomyxa ledi DB и Ch. abietis (Wallr.) Unger.
228
Рис. 62. Ржавчина хвои:
А — ржавчина сосны: 1 — пораженная хвоя; 2 — зрелые эции на хвое; 3 —
урединиоспороношение на листе мать-и-мачехи; Б — ржавчина ели: 4 — хвоя
с эциями; В — золотистая ржавчина ели: 5 — пораженная хвоя с телиоспоро-
ношением гриба; 6 — срез через телиопустулу
Гриб С. ledi разнохозяйный с полным циклом развития. Эциоста-
дия развивается на хвое ели в виде многочисленных мелких
цилиндрических пузырьков, заполненных желтой массой эциоспор
(рис. 62, Б). Урединио- и телиостадия образуются на нижней
стороне листьев багульника. Весной телиоспоры прорастают в базидии с
базидиоспорами, которые вновь заражают хвою ели. Поражаются
229
взрослые деревья, подрост, реже — ель в культурах и питомниках.
Болезнь встречается в разных типах лесорастительных условий, где
в состав напочвенного покрова входит багульник. С увеличением
доли участия последнего в живом покрове повышается уровень
болезни. Особенно сильно поражается ель в условиях хорошего
освещения. При оптимальных для развития возбудителя условиях болезнь
может принимать характер эпифитотии. Массовое поражение
ржавчиной приводит к опадению хвои, что особенно сильно
отражается на подросте, вызывая его ослабление, а нередко и усыхание.
Болезнь широко распространена в европейской части России,
в Сибири и на Дальнем Востоке.
Гриб С. abietis вызывает золотистую ржавчину хвои.
Возбудитель однохозяйный, в цикле развития которого образуются те-
лио- и базидиостадии. Молодая хвоя поражается весной, сразу после
распускания почек. Заражение осуществляется базидиоспорами,
образующимися на прошлогодней больной хвое. Летом на
пораженной хвое появляются сначала мелкие, желтоватые точечные
пятна, которые затем увеличиваются и, сливаясь, могут
охватывать всю поверхность хвои. Следующей весной на месте пятен
образуется телиостадия гриба, имеющая вид ярко-оранжевых или
желто-бурых, вначале восковидных, вытянутых вдоль подушечек
длиной до 1 см (рис. 62, В), При прорастании телиоспор и
образовании базидий с базидиоспорами подушечки становятся
бархатистыми. После распространения базидиоспор хвоя опадает.
Болезнь поражает ель в возрасте 10—20 лет. Особенно сильно
страдают густые молодняки под пологом леса. В годы наиболее
активного развития болезни происходит массовое опадение хвои.
Систематически повторяющиеся поражения приводят к
ослаблению деревьев, реже — к гибели.
Золотистая ржавчина широко распространена на севере
европейской части России и в Западной Сибири.
Ржавчина побегов и хвои ели. Возбудитель — разнохозяйный
ржавчинный гриб Chrysomyxa woroniniTtaxa. Эциостадия гриба
развивается на ели, урединио- и телиостадии — на видах багульника.
Поражаются молодые побеги, из почек которых на следующий
год появляются укороченные побеги с укороченной хвоей. На
молодой хвое образуются эции возбудителя, вытянутые до
половины длины хвои и больше, шириной до 0,5 мм. Больные побеги с
укороченной, покрытой эциями хвоей, имеют вид характерных
оранжевых или ярко-желтых кисточек и шишечек, резко
выделяющихся на фоне зеленой кроны. Созревающие эциоспоры
заражают багульник, на котором гриб зимует в виде телиоспор.
Болезнь поражает подрост ели разной высоты, значительно реже
встречается на деревьях второго яруса. Наиболее активно она
развивается в годы с теплой, влажной весной и сухим летом,
особенно в условиях хорошего освещения. При сильном развитии ржав-
230
чины, вызываемой C.woronini, происходит массовое отмирание
побегов, образование многовершинности, снижение прироста по
высоте.
Болезнь зарегистрирована на северо-западе европейской части
России, в Сибири, на Камчатке и Сахалине.
Ржавчина листьев тополя. Болезнь вызывается разнохозяйными
ржавчинными грибами из рода Melampsora: M. larici-populina ЮеЬ.,
М. alli-populina Kleb., MAarici-tremulae Kleb., M.pinitorqua (A. Br.)
Rostr.
Урединио- и телиостадии у грибов М. alli-populina и М. larici-
populina развиваются на листьях черных и бальзамических
тополей, эциостадия — соответственно на луке и лиственнице. У
видов Mlarici-tremulae и M.pinitorqua урединио- и телиостадии
протекают на листьях белых тополей, эциостадия — соответственно
на лиственнице и сосне.
Первые урединии на листьях черных и бальзамических тополей
появляются в конце июня — начале июля, а на белом тополе — в
середине или конце мая. Урединиоспоры в течение лета
осуществляют массовое заражение молодых листьев. В конце лета на верхней
стороне пораженных листьев образуется телиостадия, имеющая
вид темно-коричневых, почти черных коростинок, часто сплошь
покрывающих поверхность листовой пластинки. Телиоспоры
зимуют на опавших листьях, а весной прорастают в базидии с бази-
диоспорами, которые заражают промежуточных хозяев.
Поражается тополь в маточных и школьных отделениях питомников, в
культурах и городских насаждениях всех типов, но особенно
опасна болезнь для растений тополя в возрасте до 4—5 лет. Развитию
ржавчины способствует высокая влажность и наличие
промежуточных растений-хозяев.
При сильном развитии ржавчины листья преждевременно
засыхают и опадают, а кроны деревьев становятся ажурными. В
питомниках, молодых культурах и городских посадках у больных
растений задерживается одревеснение молодых побегов, они нередко
повреждаются ранними заморозками. Ослабленные деревья, как
правило, поражаются цитоспорозом, который значительно ускоряет
процесс ослабления и нередко приводит к гибели тополя.
Болезнь широко распространена в ареале тополя и
промежуточных хозяев возбудителей.
Ржавчина лиственницы и березы. Возбудитель болезни — разнохо-
зяйный ржавчинный гриб Melampsoridium betulinum ЮеЬ. (= М betulae
Arth.). Эциостадия развивается на хвое лиственницы и имеет вид
желтых пузырьков, расположенных вдоль средней жилки хвои.
Урединио- и телиостадии образуются на листьях березы. Уредини-
оспороношение появляется в середине лета и имеет вид ярко-
оранжевых пылящих подушечек, расположенных с нижней
стороны листа. При сильном развитии болезни листья сплошь покрыва-
231
ются урединиями. В конце лета на листьях появляются телиоспо-
роношения в виде коричневых подушечек. Телиоспоры зимуют на
опавших листьях, а весной прорастают в базидии с базидиоспо-
рами. Последние заражают хвою лиственницы.
В некоторых случаях гриб может зимовать на опавших листьях в
урединиостадии, при этом урединиоспоры весной вновь
заражают березу. Развитием гриба по неполному циклу объясняется по-
раженность березы в тех местах, где поблизости отсутствует
лиственница.
В годы, благоприятные для развития возбудителя, ржавчина
может причинять большой вред посевам лиственницы и березы.
Пораженные сеянцы преждевременно теряют ассимиляционный
аппарат, ослабляются, а иногда и усыхают.
Болезнь широко распространена в ареале березы.
10.6. МУЧНИСТАЯ РОСА
Болезнь вызывается сумчатыми грибами, относящимися к
семейству Мучнисторосяные. Она поражает многие лиственные
породы и характеризуется образованием на листьях белого
мучнистого налета, состоящего из мицелия и конидиеносцев с
конидиями. Мучнисторосяные грибы заражают только молодые листья и
неодревесневшие побеги. Мучнистая роса встречается как в
районах с умеренно влажным теплым летом, так и в засушливых
условиях. Наиболее широко распространена мучнистая роса дуба.
Мучнистая роса дуба. Возбудитель болезни — сумчатый гриб Mic-
rosphaera alphitoides Griffon et Maubl. (конидиальная стадия Oidium dubium
Jacz.). Болезнь распространена по всему ареалу дуба. Поражаются
разные его виды, но особенно сильно черешчатый и зимний (рис. 63).
В начале лета на больных листьях образуется плохо заметный
беловатый паутинистый налет мицелия. С началом развития кони-
диальной стадии он становится порошистым, мучнистым.
Созревающие конидии в течение всего лета заражают новые листья. Конидии
овальные, бесцветные, одноклетные, расположены цепочками,
размером (20 — 55) х (13 —27) мкм. К тому времени, когда
конидиальная стадия заканчивает свое развитие, налет становится
войлочным. Он может появляться как на верхней, так и нижней
стороне листьев. В конце лета на мицелии образуется сумчатая стадия
гриба — клейстотеции. Они имеют вид вначале коричневых,
позже черных, мелких, многочисленных точек, расположенных вдоль
жилок листа. У клейстотециев есть придатки, дихотомически
разветвленные на концах. В них формируются булавовидные сумки
размером (43 — 85) х (26—550) мкм. Аскоспоры эллипсоидальные
бесцветные одноклетные размером (18 —32) х (9—18) мкм. Клейстотеции
зимуют на опавших листьях. В следующем году, когда среднесуточ-
232
Рис. 63. Мучнистая роса дуба:
/ — пораженный лист с белым налетом мицелия и клейстотециями; 2 —
пораженный молодой побег; 3 — конидиальное спороношение возбудителя; 4 —
зрелый клейстотеций с выходящими сумками; 5 — сумки со спорами
ная температура воздуха достигает 16 °С, в сумках созревают аскоспо-
ры, которые осуществляют первичное заражение молодых листьев.
Заражение мучнистой росой вызывает нарушение
физиологических функций листьев (фотосинтез, дыхание, транспирацию),
степень которого находится в прямой зависимости от степени
поражения листьев. В результате листья засыхают и опадают.
Преждевременное опадение листьев приводит к тому, что побеги не
успевают одревеснеть и отмирают при перезимовке. Пораженные
мучнистой росой побеги отмирают уже после ранних осенних
заморозков. Вследствие засыхания верхушечных почек или
верхушечного побега сеянцы и молодые дубы в культурах становятся
многовершинными, приобретают кустообразную форму.
Жизнедеятельность гриба связана с определенными
погодными условиями. Его конидиальная стадия успешно развивается как
при сухой, так и при влажной погоде, поэтому болезнь встречает-
233
Таблица 4
Характеристика возбудителей мучнистой росы листьев древесных пород
Вид возбудителя
\ Micmsphaera
betulae Magn.
Micmsphaera coryli
(Jacz.) Golow
Micmsphaera
divaricata (Wallr)
|Lev.
Micmsphaera van-
bruntiana Gerard.
\Phyllactinia gut-
\tata (Wallr.iFr.)
Lev. (=P.suffulta
|Sacc.)
Поражаемая
древесная
порода, вид
кустарника
Береза
Лещина
Крушина
ломкая
Бузина
Береза, бук,
лещина,
рябина, ясень
Характер расположения мицелия и клейстотециев
Мицелий в виде слабо развитого налета на обеих
сторонах листа. Клейстотеции немногочисленные,
разбросанные или собраны в небольшие группы
Мицелий хорошо развитый, белый,
паутинисто-мучнистый, на обеих сторонах листа, но преобладает
на верхней в виде резко очерченных пятен.
Клейстотеции разбросанные, чаще на верхней стороне
листа
Мицелий на обеих сторонах листьев, белый,
паутинистый, быстро исчезающий. Клейстотеции
разбросанные или собраны в рыхло-юйлочные группы
Мицелий на обеих сторонах листа, белый,
паутинистый, особенно хорошо заметный на верхней
стороне листовой пластинки. Клейстотеции
разбросанные или скученные, особенно многочисленны
на нижней стороне листа
Мицелий на нижней стороне листа, белый, слабо
развитый, паутинистый, часто исчезающий.
Клейстотеции на нижней стороне листа, разбросанные,
часто многочисленные
Причиняемый вред
Незначительный
Незначительный
Незначительный
Незначительный
Вызывает преждевременное
пожелтение листьев во второй
половине лета
Podoshaera
clandestine (Wallr.: Fr.)
Lev.
Pbdosphaera
tridactyla (Wallr)
|de Вагу
Sawadaia bicornis
(Wallr.: Fr.) Miya-
be (= Uncinula bi-
\comis(Wallr.: Fr.)
Lev., U.acerisSacc.
Sawadaia tulasnei
(Fuck) Homma
(= Uncinula
tulasnei Fuck.)
Uncinula
clandestina Schrot.
Uncinula adunca
(Wallr.: Fr.) Lev.
(= U salicisWmt)
Боярышник
Рябина
Клен ясене-
листный,
клен
полевой
Клен
остролистный,
клен
татарский, клен
Гиннала
(приречный)
Вяз
Ива, осина,
тополь
Мицелий на обеих сторонах листа,
преимущественно на верхней, а также на побегах, белый
или желтоватый, плотный, часто войлочный.
Клейстотеции многочисленные
Мицелий белый, слабо развитый, паутинистый,
исчезающий. Клейстотеции преимущественно на
нижней стороне листа, разбросанные
Мицелий на обеих сторонах листа, белый или
сероватый, вначале мучнистый, затем исчезающий.
Клейстотеции равномерно разбросанные,
преимущественно на нижней стороне листовой пластинки
Мицелий преимущественно на верхней стороне
листа, чисто белый, плотный, вначале в виде
резко очерченных пятен, впоследствии сливающихся
и покрывающих всю поверхность листовой
пластинки. Клейстотеции многочисленные,
разбросанные или собраны в группы
Мицелий на обеих сторонах листа, белый,
паутинистый. Клейстотеции разбросаны с обеих сторон
листовой пластинки
Мицелий на обеих сторонах листа, белый,
паутинистый или плотный, войлочный. Клейстотеции
разбросанные, многочисленные, нередко
образуют черноватую пленку
Вызывает искривление побегов и
остановку их роста, деформацию
и усыхание листьев и, как
следствие — снижение
декоративности боярышника в городских
насаждениях уже в начале лета
Вред незначительный
В городских насаждениях при
сильном поражении снижает
декоративность живых изгородей
При сильном поражении снижает
декоративность городских
посадок
Вред незначительный
При сильном развитии может
снижать декоративность тополя и
ивы в городских насаждениях
ся в засушливых районах и в местах с влажным климатом.
Образование и распространение конидий происходит при температуре в
пределах от 18 до 25 °С; оптимальной для прорастания конидий
является температура 20... 22 °С. Прорастанию конидий
способствует хорошее освещение. Более активное формирование клейстоте-
циев отмечается в сухую погоду. Для их созревания необходима
влажная теплая погода в мае —начале июня. Из всего сказанного
следует, что наиболее широко мучнистая роса распространена в
южных областях и центральной полосе страны. Особенно сильно
поражается дуб мучнистой росой на участках с понижениями. Здесь
дуб очень подвержен действию весенних заморозков, поэтому у
него чаще образуются вторичные побеги, восприимчивые к
болезни. Мучнистая роса может поражать не только сеянцы и
культуры, но и древостой. Это происходит в случае нарушения
нормального развития деревьев, когда образуются поздние побеги с
молодой листвой, восприимчивой к болезни. Такая картина
наблюдается после объедания дуба листогрызущими вредителями. При
этом первичная листва и листва третьего прироста поражаются
слабее, чем листья второго прироста, что объясняется меньшей
суммой эффективных температур и количеством осадков в июне
и августе.
Кроме дуба мучнистая роса поражает многие древесные
породы и виды кустарников в лесных и городских зеленых
насаждениях. Краткие сведения о других возбудителях мучнистой росы
приведены в табл. 4.
Мучнистая роса широко распространена во многих регионах
России, в том числе в европейской части, в Сибири, на Дальнем
Востоке.
10.7. ПЯТНИСТОСТИ И ДРУГИЕ БОЛЕЗНИ
ЛИСТЬЕВ
Пятнистости выражаются в образовании на листьях пятен
различной формы, окраски и размера в зависимости от вида возбудителя.
Пятнистости могут быть вызваны вирусами, бактериями и грибами.
На древесных породах и кустарниках наиболее широко
распространены пятнистости грибного происхождения (рис. 64). Пятнистости
поражают растения в питомниках, культурах, естественных и
городских зеленых насаждениях. Систематически повторяющееся сильное
поражение пятнистостями ослабляет растения, снижает
устойчивость к другим болезням и ухудшает их декоративные качества.
Из других типов болезней листьев часто встречаются парша,
деформация, чернь.
Черная пятнистость клена. Возбудитель — сумчатый гриб Rhytisma
acerinum (Pers.) Fr. Заражение листьев осуществляется в начале
236
Рис. 64. Пятнистости листьев:
/ — бурая пятнистость тополя (Marssonina populi); 2 — черная пятнистость клена
(Rhytisma acerinum); 3 — черная пятнистость ивы (Rhytisma salicinum); 4 — темно-
бурая пятнистость липы (Cercospora microsora); 5 — бурая пятнистость березы
(Ceuthospora betulae); 6— белая пятнистость тополя (Septoriapopuli)
лета. Позже, в июле — августе на листьях появляются
многочисленные светло-желтые пятна, на которых возникают мелкие,
черные, постепенно сливающиеся точечные бугорки. К концу лета
слившиеся отдельные бугорки образуют строму, имеющую вид
черных, выпуклых, округлых пятен с блестящей поверхностью
диаметром 10—15 мм, с хорошо заметной желто-зеленой каймой
(см. рис. 64, 2). Осенью в строме формируются плодовые тела
возбудителя — апотеции, которые зимуют на опавших листьях. На
следующий год весной или в начале лета в апотециях созревают
сумки со спорами, заражающими листья.
Чаще болезнь встречается в естественных насаждениях, парках
и других типах посадок, удаленных от источников атмосферного
загрязнения. Этим объясняется отсутствие черной пятнистости в
крупных промышленных центрах с хорошо развитой сетью
транспортных магистралей.
Гриб не причиняет существенного вреда, но при
систематически повторяющемся поражении клена в школьных отделениях
237
питомников может вызвать снижение выхода стандартного
посадочного материала, а в городских насаждениях — снижение
декоративности клена.
Болезнь широко распространена в европейской части России,
на Урале.
Заболевание, схожее с описанным, вызывает гриб Kpunctatum
Rehm. На листьях клена образуются строматические черные
мелкие пятна. Болезнь причиняет очень незначительный вред.
Черная пятнистость ивы. Возбудителем болезни является
сумчатый гриб Rhytisma salicinum Rehm., образующий на листьях
черные выпуклые блестящие пятна, до 1,5 см в диаметре. Болезнь
часто встречается на ивовых плантациях, где при благоприятных
для развития гриба условиях вызывает преждевременное
засыхание и опадение листьев, а нередко и гибель растений. Болезнь
распространена в ареале ивы (см. рис. 64, 3).
Черная пятнистость вяза вызывается сумчатым грибом Dothidella
ulmi (Duv.) Wint. На листьях вяза образуются серовато-черные
округлые выпуклые пятна диаметром до 2 — 3 мм, представляющие
собой аскостромы, в локулах которых развиваются сумки со
спорами. Сумки цилиндрические, на короткой ножке, размером (60 —
70) х (8—9) мкм. Аскоспоры удлиненно-овальные бесцветные двух-
клеточные, неравноклетные размером (10—12,5) х4,5 мкм.
При сильном развитии болезни многочисленные пятна сплошь
покрывают поверхность листьев, что приводит к их
преждевременному засыханию и опадению. В результате этого значительно
снижаются декоративные качества вяза, который широко
применяется для озеленения городов.
Болезнь распространена в европейской части России.
Черная пятнистость березы. Возбудитель болезни — полостно-
сумчатый гриб Dothidella betulina (Fr.) Sacc. На верхней стороне
листьев образуются многочисленные мелкие черные выпуклые
пятна округлой или угловатой формы. При сильном развитии
болезни пятна сплошь покрывают листовую пластинку, что
приводит к преждевременному опадению листвы. На опавших листьях
внутри пятен, представляющих собой аскострому, образуются
локулы. Весной в них созревают сумки со спорами, заражающими
листья. Сумки цилиндрические размером (38—44) х (12—12,5) мкм.
Аскоспоры эллиптические, с одной перегородкой, зеленоватые
размером 14 х 15 мкм.
Массовая летняя дефолиация снижает декоративность березы
в городских насаждениях.
Болезнь распространена в европейской части России, на
Урале, в Сибири, на Дальнем Востоке.
Белая пятнистость тополя (септориоз). Возбудитель —
несовершенный гриб Septora populi Desm. Во второй половине лета на
листьях образуются двусторонние белые или сероватые пятна, округ-
238
лой или неправильной формы, с темной, почти черной, тонкой
каймой. На верхней стороне пятен развиваются пикниды,
имеющие вид мелких черных точек, хорошо заметных
невооруженным глазом на белом фоне пятна (см. рис. 64, 6). В пикнидах
созревают конидии, осуществляющие заражение листьев. Конидии
цилиндрические, прямые или согнутые, двухклеточные размером
(38-45) х (2,5-3,5) мкм.
Поражаются разные виды и гибриды тополей в питомниках,
культурах, городских насаждениях. В годы, благоприятные для
развития возбудителя, болезнь вызывает массовое преждевременное
опадение листьев; уже к концу июля деревья теряют до 30—50 %
листьев. В питомниках интенсивное развитие болезни в течение
нескольких лет ослабляет растения в маточных плантациях и
школах, снижая выход и качество посадочного материала. В городских
посадках вследствие массового поражения болезнью
декоративные качества деревьев значительно снижаются.
Болезнь распространена повсеместно в ареале тополя.
Бурая пятнистость тополя (марссониоз). Возбудитель болезни —
несовершенный гриб Marssonina populi Kleb. Гриб поражает разные
виды и гибриды тополей. Источником инфекции являются
опавшие пораженные листья. Весной на них развиваются споры,
заражающие молодые листья. Первые пятна на листьях появляются в
конце мая — начале июня. Пятна бурые, серовато-бурые
округлые, расплывчатые, иногда неясно выраженные (см. рис. 64, 7). На
пятнах с обеих сторон листа, но преимущественно с верхней
образуются конидиальные ложа возбудителя, имеющие вид мелких,
желтоватых или белесых, округлых или плоских образований.
Конидии яйцевидные, удлиненно-булавовидные, грушевидные,
бесцветные, прямые или согнутые, вначале одноклетные, затем с
одной перегородкой у основания, размером (14—29) х (5 — 10) мкм.
Инкубационный период возбудителя составляет всего 3 — 5 дней,
поэтому количество пораженных деревьев и степень пораженно-
сти кроны увеличивается очень быстро. В начале заражаются листья
нижних ветвей, находящихся ближе к первоначальному
источнику инфекции, затем от пораженных листьев инфекция
распространяется на ветви средней и верхней частей кроны. Болезнь
наиболее интенсивно развивается в условиях высокой влажности
воздуха и умеренной температуры (13... 18 °С).
Поражается тополь в питомниках, культурах, в естественных и
городских насаждениях. Массовое, систематически
повторяющееся поражение саженцев в питомниках вызывает их ослабление и
снижение выхода стандартного посадочного материала. Высокий
уровень болезни в городских насаждениях приводит к
преждевременному листопаду, к почти полной потере декоративности
тополя и защитных функций. Это особенно отрицательно
сказывается на крупных промышленных центрах, где зеленые насажде-
239
ния прежде всего выполняют роль очистителей атмосферного
воздуха от различных примесей.
Болезнь распространена в европейской части России, на
Урале, в Сибири.
Схожее заболевание на черных тополях вызывает гриб М. brunnea
(Ellis ex Everh. Magn.).
Бурая пятнистость березы вызывается грибами Ceuthospora betulae
(Fuck.) v. Arx. (= Gloeosporium betulinum West.) и Marssonina betulae
(Lib.) Magn. При поражении С. betulae на листьях образуются
пятна оливкового или темно-коричневого цвета, округлые, с
расплывчатыми краями, до 10 мм в диаметре. На нижней стороне
пятен образуются ложа гриба. Конидии бесцветные
цилиндрические размером (4—10) х (1,5 — 2) мкм (см. рис. 64, 5).
Гриб М. betulae вызывает появление бурых, округлых или
неправильной формы пятен, часто с более темной каймой. На верхней
стороне пятен образуются ложа в виде мелких, плоских,
темно-коричневых подушечек. При созревании конидии выступают серовато-
белыми мелкими каплями. Конидии продолговато-овальные или
удлиненно-яйцевидные, прямые или согнутые, вначале одноклет-
ные, позже двуклетные бесцветные размером (15 — 22) х (6,5 —10)
мкм. Поражается береза в питомниках, культурах, естественных и
городских насаждениях. При сильном поражении листья
преждевременно усыхают и опадают, снижается выход стандартного
посадочного материала в питомниках и декоративность березы в
городских насаждениях.
Бурая пятнистость встречается в ареале березы.
Бурая пятнистость дуба (глеоспориоз). Возбудитель болезни —
несовершенный гриб (Discula umbrinella (Berk, ex Broome) Sutton
(= Gloeosporium quercinum West.). На листьях в начале июля
появляются пятна диаметром 2—4 мм неправильной формы или
округлые, вначале желтовато-зеленые, затем бурые. На нижней
стороне пятен образуются конидиальные ложа возбудителя в виде
желтовато-коричневых подушечек. Конидии двух типов:
макроконидии бесцветные, одноклетные, овальные, нередко
булавовидные, суженные к основанию, размером (8—20) х (3,5 — 8) мкм;
микроконидии овальные, палочковидные, клиновидные
размером (4—8) х (1,5 — 2) мкм.
При развитии сумчатой стадии гриба (Gnomonia quercina Kleb.)
пятна увеличиваются в размере и принимают бурую окраску.
Многочисленные пятна сливаются, покрывая всю поверхность листа. На
опавших пораженных листьях весной образуются перитеции.
Созревшие в перитециях аскоспоры осуществляют первичное заражение
листьев дуба. Аскоспоры бесцветные, удлиненно-эллипсоидные или
веретеновидные, двухклетные размером (13—15) х (3,5—4) мкм.
Поражается дуб в питомниках, культурах, естественных и
городских насаждениях. При благоприятных условиях для развития
240
возбудителя бурая пятнистость вызывает преждевременное
опадение листьев, что приводит к ослаблению растений в питомниках,
снижает декоративность дуба в городских насаждениях.
Болезнь широко распространена в европейской части России.
Серая пятнистость осины (глеоспориоз) вызывается
несовершенным грибом Gloeosporium tremulae Pass. Во второй половине
лета на листьях образуются большие пятна диаметром 5 — 15 мм,
неправильной формы, серого или серо-желтого цвета с темной
каймой. На поверхности пятен развивается конидиальное споро-
ношение гриба (ложа) в виде многочисленных темно-бурых или
черных шероховатых плоских подушечек. Сливаясь, пятна часто
охватывают всю поверхность листа. В этом случае деревья
полностью теряют декоративность, листья преждевременно опадают.
Встречается в европейской части России, на Урале.
Кремовая пятнистость липы (глеоспориоз). Возбудитель —
несовершенный гриб Gloeosporium tiliae Oudem (=Discula umbrinella
(Berk, ex Broome) Sutton). В июле на листьях появляются крупные,
до 4—8 мм в диаметре, неправильно-округлые пятна кремового
или охряного цвета с узким темным ободком. На пятнах
развиваются конидиальные спороношения в виде темно-бурых мелких
подушечек. Конидии двух типов: макроконидии бесцветные од-
ноклетные, овальные, яйцевидные размером (10—18) х (4—6) мкм;
микроконидии палочковидные размером (4—8,5) х (1 —1,5) мкм.
Кроме листьев при сильном развитии болезни поражаются
черешки и околоцветники. Во второй половине лета пятна нередко
покрывают всю поверхность листовой пластинки, что приводит
к сильной деформации листьев, вследствие этого крона
становится ажурной.
Болезнь может принимать характер длительной эпифитотии,
поражаются не только листья, но и черешки и соцветия. В таком
случае наблюдается массовое преждевременное опадение
листьев, цветение отсутствует, в более редких случаях происходит
отмирание деревьев в очагах болезни. Кроме того пораженные глео-
спориозом липняки перестают выполнять роль базы пчеловодства.
Эпифитотии глеоспориоза отмечались в липняках Жигулевского
заповедника и в лесном массиве «Липовый остров»,
расположенном на юго-востоке Западной Сибири. В городских насаждениях у
сильно пораженных деревьев теряется декоративность и
снижаются защитные функции.
Болезнь распространена в европейской части России, на
Урале, в Сибири.
Темно-бурая пятнистость липы (церкоспороз) вызывается
несовершенным грибом Cercospora microsora Sacc. Поражается липа в
питомниках, естественных и городских насаждениях разного возраста.
На листьях в начале июля образуются темно-бурые пятна со
светлым центром, округлые или неправильной формы, до 2 — 3 мм в
241
диаметре (см. рис. 64, 4). На пятнах развиваются пучки конидие-
носцев с конидиями, которые осуществляют заражение листьев
в летний период. Конидии палочковидные, веретенообразные,
булавовидные, слегка согнутые, с 3 — 8 поперечными
перегородками, бледно-бурые размером (20—100) х (3—4,5) мкм. На
опавших пораженных листьях образуется сумчатая стадия гриба
(Mycosphaerella microsora Syd.). Созревающие в перитециях аскоспо-
ры осуществляют первичное заражение листьев. При сильном
развитии болезни многочисленные пятна сплошь покрывают
листовую пластинку.
Поражается липа в посевных и школьных отделениях
питомников, в естественных и городских насаждениях. Наибольший вред
церкоспороз причиняет питомникам, вызывая ослабление
всходов и сеянцев. В городских насаждениях при сильном развитии
болезни снижается декоративность липы. Естественным
насаждениям церкоспороз заметного вреда не причиняет.
Болезнь распространена в европейской части России, на
Урале, в Сибири.
Парша тополя и осины. Парша листьев тополя вызывается
несовершенным грибом Pollaccia radiosa (Lib.) Bald, et Cif. (=Fusi-
cladiwn radiosum (Lib.) Lind.). Сумчатая стадия — Venturia tremulae
Aderh. — образуется далеко не всегда. Поражаются разные виды
белого тополя.
На листьях в начале лета образуются сначала фиолетово-бурые
округлые или неправильной формы пятна разных размеров. Позднее
на пятнах появляется
бархатистый оливкового цвета налет,
представляющий конидиаль-
ное спороношение
возбудителя. Конидии
овально-эллипсоидальной формы, с одной,
реже двумя поперечными
перегородками, неравноклет-
ные, желто-зеленого или
светло-оливкового цвета,
размером (17 —26) х (7—11) мкм
(рис. 65, /, 3). Конидии
образуются в течение всего
вегетационного периода. С момента
заражения листьев до
образования на них конидиального
спороношения проходит 8 —
Рис. 65. Парша тополя: *2 Дней- Конидии распростра-
у ™«о^Лг,«^ ™^<т. о K^msiM ^ няются ветром, дождевыми
1 — пораженные листья; 2 — больной по- * ' ^ ^
бег, сломанный в месте образования не- каплями, а также через зара-
кроза; 3 — конидии возбудителя женные листья.
242
Развитие болезни зависит от погодных условий, главным
образом от влажности. Образованию конидий и заражению
способствуют затяжные дожди или часто выпадающие росы, при этом
листья продолжительное время находятся в увлажненном
состоянии. Температурные условия для возбудителя парши большого
значения не имеют, он может развиваться в пределах от 13 до 35 °С.
Наиболее благоприятные условия для возбудителя создаются в
первой половине вегетации, когда выпадает наибольшее количество
осадков и образуется масса молодых листьев, наиболее
восприимчивых к болезни. При сильном поражении листья
преждевременно засыхают и опадают, уже к середине августа опавшие
листья могут составлять 30—50 %.
Кроме листьев, болезнь поражает молодые побеги, они
чернеют, засыхают и загибаются в виде крючков, а иногда ломаются (см.
рис. 65, 2). Побеги заражаются мицелием, проникающим в них
через черешки пораженных листьев. Гриб зимует в виде мицелия
на опавших листьях или пораженных побегах. Весной на мицелии
образуются конидиеносцы с конидиями, которые осуществляют
первичное заражение. В условиях развития сумчатого спороноше-
ния на опавших листьях первичное заражение осуществляется ас-
коспорами, образующимися в перитециях.
Поражается тополь в маточных и школьных отделениях
питомников, в культурах и городских насаждениях. Систематически
повторяющиеся поражения паршой приводят к ослаблению и усы-
ханию тополя в питомниках, вследствие чего снижается выход
стандартного посадочного материала. В городских насаждениях
массовое поражение листьев и побегов паршой снижает
декоративность тополя и его защитные функции.
Болезнь зарегистрирована в европейской части России.
Паршу осины вызывает гриб P. elegans Serv. (сумчатая стадия —
Venturia populina Vuill.). Признаки болезни сходны с таковыми на
тополе белом, но от возбудителя парши на тополе белом гриб
P. elegans отличается микроскопическими признаками. Конидии
P. elegans светло-оливковые, удлиненно-овальные, веретеновид-
ные, преимущественно 3-клетные, реже — 2-й 4-клетные
размером (26,5 —31,2) х (6,5 — 7) мкм. Гриб поражает поросль осины и
крайне редко листья на деревьях.
Парша осины распространена повсеместно в ареале растения-
хозяина.
Парша ивы. Возбудитель — несовершенный гриб Pollaccia saliclperda
(Allesch. ex Tub.) Arx. (=Fusicladium saliciperdwn Lind.). Поражает
разные виды ивы (белую, козью, серую, ломкую и др.) в
питомниках, на плантациях, в городских насаждениях. Заражение
листьев осуществляется весной, в мае, конидиями, которые
образуются на прошлогодних пораженных листьях. Пораженные
молодые листья и побеги чернеют и усыхают (рис. 66). При этом побе-
243
ги загибаются в виде крючка, а усохшие листья повисают.
Почерневшие побеги с усохшими, повисшими, почерневшими
листьями резко вьщеляются на фоне зеленой кроны. На больных побегах
и листьях образуется темно-оливковый, почти черный, плохо
заметный бархатистый налет конидиального спороношения.
Конидии желтоватые, овальные, цилиндрические, грушевидные, с 1 —
2 поперечными перегородками и заметными перетяжками,
размером (12—40) х (6—10) мкм. Конидии заражают молодые листья
и побеги в течение всего вегетационного периода. Зимует гриб в
виде мицелия в пораженных побегах и на опавших листьях.
Наиболее интенсивно болезнь развивается в годы с большим
количеством осадков в весенне-летний период. Массовое
поражение паршой приводит к снижению выхода посадочного
материала в питомниках, качественного материала на ивовых
плантациях, декоративности ивы в городских насаждениях.
Болезнь широко распространена в европейской части России.
Деформация листьев. Возбудители — голосумчатые грибы из рода
Taphrina. Заражение листьев осуществляется сумкоспорами весной.
Мицелий проникает в ткань эпидермиса и, развиваясь внутри
клеток, деформирует листья. На поверхности пораженных
участков листьев образуется налет, состоящий из слоя сумок.
Созревающие сумкоспоры заражают молодые побеги и листья.
Поражаются многие лиственные породы в питомниках,
естественных и городских
насаждениях. Наиболее часто
деформация листьев
встречается на березе, вязе, клене,
ольхе, тополе. Деформацию
листьев березы вызывает гриб
Г. betulina Rostr. На листьях
образуются округлые вздутия,
на внутренней стороне
которых развивается беловатый
или желтоватый слой сумок
с сумкоспорами. Вред от
болезни несущественный. Реже
на листьях березы
встречается гриб Т. carnea Johans.
Деформация листьев вяза
вызывается грибом Г. ulmi
Johans. На листьях
образуются слегка вздутые, округлые
или неправильной формы
Рис. 66. Парша ивы: серые пятна. С нижней сто-
1 — ветки с пораженными листьями и по- РОНЫ ЛИСТЬев на пятнах
бегами; 2 — конидии возбудителя развивается сероватый ИЛИ
244
коричневатый слой сумок со
спорами. В отдельные годы сильно
поражаются листья на поросли, они
преждевременно засыхают и опадают. Вред
от болезни незначительный.
Деформацию листьев клена
татарского вызывает гриб T.polyspora
(Svrcek) Johans. Пятна неправильной
формы, сначала коричневые, позже
черные, вздутые, морщинистые, со
временем прорываются. Гимениаль-
ный слой на пятнах образуется с
верхней стороны листьев. При сильном
развитии болезни листья
деформируются, чернеют, засыхают и
преждевременно опадают. Болезнь может вы- Рис. 67. Деформация (пузыр-
зывать усыхание сеянцев в питомни- чатка) листьев тополя
ках. В городских насаждениях сильное
поражение клена приводит к значительной потере
декоративности деревьев.
Деформация листьев ольхи вызывается грибом Т. tosquinetii
Magn. На листьях появляются коричневые, морщинистые пятна.
При образовании серого гимениального слоя листья становятся
липкими.
Деформация (пузырчатка) листьев тополя вызывается грибом
Т. aurea (Pers.) Fr. На листьях появляются крупные, округлые
вздутия вначале золотисто-желтого, позже бурого цвета (рис. 67). На
внутренней стороне вздутий образуется желтый слой сумок со
спорами. Болезнь поражает молодые листья тополей в школах,
городских посадках, но заметного вреда не причиняет.
Чернь листьев вызывается несовершенными грибами из родов
Fumago, Dematium и др. На листьях образуются легко стирающиеся
сажистые налеты темно-бурого или черного цвета.
Возбудители черни — сапротрофные грибы, которые питаются
сахаристыми выделениями, образующимися в результате
нарушения обмена веществ или повреждения тлями, щитовками, а
также поверхностными наносами (пыль и др.).
Чернь поражает листья очень многих древесных пород и
кустарников, нарушает процессы ассимиляции, затрудняет доступ
воздуха и света к листьям. Развитию черни способствует
засушливая жаркая погода.
Основной вред причиняется городским насаждениям,
благоприятные условия вызывают массовое поражение деревьев и
кустарников и значительную потерю их декоративности.
Болезнь распространена повсеместно, особенно в городских
насаждениях.
245
10.8. СИСТЕМА МЕРОПРИЯТИЙ ПО ЗАЩИТЕ
ПИТОМНИКОВ, КУЛЬТУР И МОЛОДНЯКОВ
ОТ БОЛЕЗНЕЙ
10.8.1. Общие сведения
В питомниках на сравнительно маленькой площади
концентрируется большое количество ценного посадочного материала, от
сохранности и состояния которого зависит успешность лесокуль-
турных работ. Огромные площади лесных культур, многие из
которых расположены в малолесных районах страны, также требуют
заботы со стороны лесоводов и должны быть сохранены. В отличие
от хорошо возобновляющихся естественных лесов, не
требующих больших вложений в период своего роста и развития, при
создании питомников и выращивании культур вкладываются
большие трудовые и финансовые средства. Тем важнее не допустить
напрасных затрат и предотвратить ущерб от вредных организмов.
В ряде случаев возникает необходимость и в защите естественных
молодняков и подроста на вырубках и под пологом леса,
которые, как и молодые растения в питомниках и культурах, могут
страдать от повреждений насекомых, клещей, позвоночных
животных (грызунов и копытных) и болезней.
К числу наиболее распространенных болезней молодых
растений относятся шютте, ржавчина хвои и побегов, пятнистости,
мучнистая роса, ржавчина листьев, некрозы и раки побегов.
10.8.2. Надзор за появлением и распространением
болезней
В питомниках организуется рекогносцировочный надзор,
который ведется путем систематического наблюдения за
состоянием растений, появлением очагов болезней, их распространением
и степенью пораженности растений. Рекогносцировочный надзор
дополняется детальным лесопатологическим обследованием,
которое проводят три раза в год: весной, после схода снега, в
первой половине лета и осенью (в сентябре) путем учета на пробных
площадях. В питомниках весной, после схода снега проводят
обследование с целью выявления очагов снежного, обыкновенного
и бурого шютте, выпревания и побегового рака сосны. В начале
лета осуществляется надзор за полеганием всходов, шютте
лиственницы, ржавчиной побегов и склерофомозом сосны. При
осеннем обследовании оценивают состояние посадочного материала и
степень пораженности его болезнями типа ржавчина,
пятнистости, мучнистая роса и др.
246
При детальном обследовании питомников проводят учет на
пробных площадках размером 1 м посевной строчки с перечетом
всех растений. Выделяют растения без признаков поражения,
пораженные и погибшие.
У сеянцев оценку степени пораженности хвои производят по
четырехбалльной шкале: 1 — поражено до 25 % хвои; 2 —
поражено 25 — 50% хвои; 3 — поражено 50 — 75% хвои; 4 — поражено
более 75 % хвои. Пораженность сеянцев лиственных пород
оценивают по трехбалльной шкале: 1 — поражено до 25 % листьев; 2 —
поражено 25 —50 % листьев; 3 — поражено более 50 % листьев.
Наряду с оценкой степени пораженности проводят также
оценку сеянцев по состоянию, подразделяя их на следующие
категории: 1 — без признаков ослабления; 2 — ослабленные; 3 —
погибшие.
Для определения необходимого числа пробных площадок на
выделенном для обследования участке предварительно
закладывают шесть пробных площадок, на которых проводят перечет
сеянцев или всходов, подразделяя их на здоровые и больные, и
устанавливают средний процент поражения болезнью. Далее
проводят расчет по формуле:
т2 '
где п — необходимое число пробных площадок; / —
нормированное отклонение при соответствующих уровнях вероятности (обьино
для полевых исследований он принимается за 0,68, при этом
значении нормированное отклонение равно 1); р — средний процент
поражения растений, выраженный в долях единицы; т —
допустимая ошибка в процентах или долях единицы.
Вначале подсчитывают примерный процент пораженности по
5—6 пробным площадкам, расположенным в разных местах
исследуемого участка. Затем полученное значение пораженности
подставляют в формулу и из полученного числа вычитают
количество уже заложенных площадок. При неравномерном
распределении пораженных растений по площади глазомерно выделяют
участки (слои) с разной степенью пораженности, находят
необходимое количество пробных площадок для каждого слоя, а затем
вычисляют средневзвешенное значение (% пораженности) для
участка в целом.
Размещать пробные площадки можно равномерно по участку,
по двум его диагоналям или пользуясь методом случайной
выборки с использованием таблицы случайных чисел. В последнем
варианте две условно выравненные стороны обследуемого участка
разбивают на 100 частей каждую и используют как координатные
оси. Выбранные пары случайных чисел отмеряют по двум сторо-
247
нам участка и по этим координатам выбирают места
расположения пробных площадок. Методика анализа выборочных единиц
зависит от вида болезни и формы ее проявления.
По результатам детального обследования определяют
распространенность болезни, т. е. количество пораженных растений в
однородном сообществе. Ее выражают в процентах и вычисляют по
формуле Р = (аЮ0)/А, где Р — распространенность болезни; А —
общее количество учтенных растений; а — количество
пораженных растений.
Для характеристики усредненной степени пораженности
участка развитие болезни определяют по формуле:
ПК
где R — развитие болезни, %; Х(Л^) ~~ сумма произведения
числа больных растений а на соответствующий балл пораженности Ь;
п — общее количество учтенных растений; к — высший балл учета
принятой шкалы.
В культурах и молодняках надзор ведется за основными
болезнями (снежное шютте сосны и ели, ржавчина побегов сосны,
побеговый рак, шютте ели и лиственницы). Кроме того, проводят
текущие оперативные обследования, а в случае необходимости
назначают специальные лесопатологические обследования.
Детальное обследование проводят путем закладки
безразмерных пробных площадей, чтобы в перечет вошло не менее 100
деревьев. Степень пораженности крон оценивают по приведенным
выше шкалам. При оценке состояния культур деревья делят на
следующие категории: 1 — без признаков ослабления; 2 —
ослабленные; 3 — усыхающие; 4 — усохшие в текущем году; 5 —
усохшие в прошлом году.
Для определения состояния подроста в пределах обследуемого
выдела насаждений выбирают типичный для них участок
насаждения, на котором располагают учетные площадки одного из
выбранных размеров (м): (1 х 1), (1 х 2), (3 х 3) или 2x5. Размер
площадки зависит от высоты и возраста подроста и характера
размещения его на пробной площади. Чем старше и выше подрост и
чем неравномернее его размещение, тем больше должен быть
размер площади. Размещать площадки на пробе наиболее
целесообразно в случайном порядке, используя метод случайных чисел.
Учет естественного возобновления ведется по группам высот,
возрасту и категориям состояния. Для подроста выделяют
следующие группы: всходы — растения до 1 года или появившиеся в год
учета; самосев — в возрасте от 2 до 5 лет высотой до 0,5 см;
подрост — молодые деревца старше 5 лет высотой от 0,5 м до
половины высоты основного полога (0,5—1 м, 1 —1,5 м, выше 1,5 м).
248
Для выяснения степени влияния болезней на рост деревьев
проводят детальный анализ 10 пораженных (из разных категорий
состояния) и 10 здоровых деревьев с измерением прироста по
высоте за последние 3 — 5 лет. При наличии культур разного
возраста, состава, густоты и т.д. учет ведут методом послойной выборки
с последующим сравнением данных о состоянии и степени
пораженное™ в каждом участке (слое). По материалам надзора назначают
необходимые меры борьбы, сроки их проведения и объем работ.
Немедленно после обследования питомников или молодняков
проводят лабораторный анализ образцов хвои, листьев, коры и
древесины с целью определения тех болезней, диагноз которых не
удалось поставить в полевых условиях. При этом пользуются
одним или комплексом методов диагностики, описанных в гл. 6.
Для того чтобы ограничить распространение и ликвидировать
очаги болезней, для планирования необходимых объемов защитных
мероприятий и достижения их высокой эффективности большое
значение имеет прогнозирование наиболее опасных болезней леса.
В лесном хозяйстве прогноз болезней недостаточно изучен и
разработан пока для отдельных видов болезней в питомниках.
Эффективность химической борьбы с обыкновенным и
снежным шютте в значительной степени зависит от правильного
выбора сроков первой обработки, связанной с массовой споруляци-
ей возбудителей. Сроки в зависимости от погодных условий могут
изменяться в пределах не только региона, но и небольшого
района. В связи с этим установление сроков массовой споруляции и
соответственно даты первой обработки имеют большое значение.
Н.М.Ведерников разработал прогноз дат первого
опрыскивания сосны против обыкновенного и снежного шютте для
Среднего Поволжья. Для прогноза даты первого летнего опрыскивания
посевов сосны против обыкновенного шютте предлагается
номограмма с тремя шкалами (рис. 68). На левой шкале откладывают
суммы осадков за период с 1 мая по 16 июля текущего года. На
правой — суммы среднесуточных температур воздуха выше 15 °С
за период от схода снега до 16 июля. При этом включают только ту
часть значения температуры, которая превышает 15 °С. На
средней шкале отмечают фактические даты начала летней споруляции
возбудителя болезни. Для прогноза сведения об осадках и
температуре берут на ближайшей от питомника метеостанции. На левой
шкале откладывают суммы осадков, на правой — суммы
температур и соединяют эти точки прямой линией. На пересечении этой
линии со средней шкалой находят дату первой обработки.
Для установления даты опрыскивания сосны против снежного
шютте используется такая же номограмма, но с другими
значениями (рис. 69). На левой шкале откладывают сумму осадков за
период с 1 июля по 30 сентября. На правой — сумму
положительных среднесуточных температур за период с 1 июня по 30 сентяб-
249
Ключ:
мм-срок-°С 1|
мм °С
18
19
20
21
22 |
23 зЗ
24
26
28
30
5
10 |
13 3
Рис. 68. Номограмма для
определения начала летнего рассева
аскоспор возбудителя
обыкновенного шютте сосны в
питомниках
6*
90 g^
:110S-
130 Sg
150 S 2
170 | |
190 £g
210 | g
230 |2
250 &3
Ключ:
мм-срок-°С j
мм °C
Срок
&
Цзт|
»
25
23
21
19
17
15
14
12
11
10
«i
40
2
b 2210
2190
2170
2150
2130
2110 a£
2090 S3
2070 g <£
2050 I в
2030 g£
2010 й^
1990
1970
1950
1930
1910
1890
1870
1850
1830
1810
1790
1770
U
»U8
Рис. 69. Номограмма для определения
даты первого опрыскивания сосны
против снежного шютте в
питомниках
ря. На средней шкале отмечают фактические даты начала спору-
ляции возбудителя. Соединив эти точки прямой линией, на
пересечении ее со средней шкалой находят нужную дату.
Для долгосрочного прогноза (на весну будущего года)
обыкновенного и снежного шютте Н. М. Ведерниковым составлены
прогностические уравнения регрессии, которые отражают связь
степени развития болезней с погодными условиями.
Для определения ущерба от обыкновенного шютте
необходимо прогнозировать не только распространенность болезни
(процент поражения растений), но и интенсивность ее развития
(степень пораженности растений).
Прогноз распространенности болезни осуществляется с
использованием уравнения У, = 78,50 - 4,19^, + 0,22ДГ2 ± 18,3.
Для прогноза интенсивности болезни применяют уравнение
Y2 = 119,50 - 5,57Х, + 0,18*2 + 21,7, где Хх — средняя из
среднемесячных температур воздуха за июль и август; Х2 — сумма
осадков за июнь—август. Значения Х\ и Х2 берутся за год,
предшествующий прогнозируемому.
При постановке прогноза снежного шютте предсказывают
только распространенность болезни, так как интенсивность болезни в
питомниках равна 100 %. Поскольку развитие снежного шютте зави-
250
сит от количества осадков в летне-осенний период и высоты
снежного покрова, то прогноз устанавливают в два этапа.
Предварительное прогнозирование осуществлялось в ноябре с использованием
формулы Y3 = 1,37^ - 32,43 + 20,0. Окончательный прогноз ставят
в марте, применяя формулу Y4 = 0,54^ + 0,99^ - 52,70 ± 17,9, где
Х\ — число дней с осадками за июнь — октябрь; Х2 —
максимальная высота снежного покрова в данном питомнике.
Предложенные методы прогноза могут быть использованы и для других
регионов.
Метеобиологический метод долгосрочного прогноза
ржавчины побегов сосны (соснового вертуна), разработанный В.И.Кру-
товым для Южной Карелии, основан на зависимости проявления
болезни (распространенность и интенсивность) от различных
погодных факторов предшествующего года. Определяющими в
развитии болезни являются сумма температур воздуха выше 10 °С за
июнь, июль, сентябрь и количество осадков в августе за дни с
температурой воздуха выше 5 °С (табл. 5).
Таблица 5
Прогноз развития ржавчины побегов сосны
Сумма / воздуха выше 10°С
за июнь, июль, сентябрь
900 и более
Более 1100
Менее 900
Сумма осадков за август
при / выше 5 °С
Около 70
Около 50
Более 70
Проявление болезни
Выше нормы
Норма и ниже нормы
Примечание. В числителе — показатели погодных факторов для определения
распространенности болезни, в знаменателе — для развития болезни.
Прогноз ржавчины побегов сосны может осуществляться
разными способами. В первом случае получают приблизительное
представление о проявлении болезни по степени отклонения от
нормы. Норма распространенности болезни колеблется от 30 до 50 %
и средний балл поражения 0,8—1,5; выше нормы соответственно
значения более 50 % и выше 1,5 баллов, ниже нормы — до 30 % и
0,8 балла. Используя табл. 5 и данные о погодных факторах
предшествующего года, можно представить картину проявления
болезни.
Для более точного прогнозирования болезни используют
математические модели. Распространенность и развитие болезни определяют
соответственно по формулам: Yx = 90,11 + 0,21^ - 10,95*2 + 10,26
и Y2 = 0,597 + 0,06*, - 0,197*2 + 0,33, где Хх — сумма температур
воздуха 10 °С за июнь, июль, сентябрь, а Х2 — сумма осадков за
август при температуре воздуха 5 °С. Значения показателей Хх и Х2
берутся за предшествующий год.
251
10.8.3. Лесохозяйственные мероприятия
Своевременное и технически грамотное выполнение лесохо-
зяйственных мероприятий не только предотвращает появление
очагов болезней в питомниках и культурах, но и является
залогом высокой эффективности химических и биологических мер
борьбы.
Для закладки новых, особенно базисных, питомников следует
выбирать по возможности ровные участки с легкими
супесчаными почвами. Питомники нельзя закладывать на площадях,
вышедших из-под картофеля, бахчевых культур и кукурузы, так как
возможно их поражение возбудителями инфекционного полегания.
Питомники рекомендуется закладывать на расстоянии 250 м от
стены леса, чтобы предотвратить занесение инфекции из
насаждений. Для снижения влажности почвы необходимо проводить
дренирование низких сырых мест. Кислые почвы надо известковать,
так как кислая среда активизирует жизнедеятельность
возбудителей полегания. С целью создания неблагоприятных условий для
развития в почве паразитических микроорганизмов и сорняков
следует применять севообороты с черным паром. Применяемые в
питомниках севообороты должны предусматривать высев одной и
той же породы на одном месте не ранее чем через два года.
Большую роль в снижении потерь от болезней в питомниках играют
удобрения. Они улучшают рост и развитие растений, делая их
более устойчивыми к болезням. Кроме того, внесение удобрений
способствует созданию неблагоприятных условий для развития
паразитной микофлоры.
Семена перед высевом следует обязательно проверять в
лабораториях Центрлессема или на зональных лесосеменных станциях
для выяснения степени зараженности их болезнями, определения
энергии прорастания и всхожести. Высев семян проводят в
оптимальные сроки с соблюдением нормы высева и глубины заделки
семян для каждой породы и конкретных условий. Для
предотвращения поражения хвойных посевов болезнями типа шютте их не
следует укрывать ветвями той же породы или мхом из этих
насаждений, так как хвоя может быть источником инфекции.
В питомниках, чтобы предотвратить развитие грибных
болезней, необходимо систематически проводить тщательную
прополку сорняков, которые не только ослабляют сеянцы, но и могут
служить источником инфекции, полегания и выпревания. Для
предупреждения распространения болезней листьев (пятнистостей,
мучнистой росы, ржавчины) в посевных, школьных отделениях
и культурах необходимо осенью сгребать и сжигать опавшую
листву, которая является источником инфекции. С этой же целью
следует ликвидировать имеющиеся в питомниках переросшие
школы.
252
Состояние культур и степень пораженности их грибными
болезнями в значительной мере зависят от методов подготовки
почвы, качества посадочного материала, техники посадки, ухода за
ними, типов смешения. При глубокой зяблевой вспашке споры и
склероции фитопатогенных грибов (возбудителей полегания, вы-
превания) попадают в глубокие слои почвы и теряют способность
к прорастанию и заражению растений. Посадочный материал
перед высадкой необходимо тщательно отсортировать, удаляя
пораженные болезнями, плохо развитые, многовершинные сеянцы. При
посадке следует избегать загиба или механических повреждений
корневой системы, так как в дальнейшем такие саженцы
поражаются болезнями, повреждаются насекомыми и часто гибнут. При
создании культур необходимо подбирать древесные и
кустарниковые породы, устойчивые к болезням, с учетом их влияния друг
на друга применительно к конкретным условиям. При создании
культур необходимо учитывать, что в смешанных насаждениях
создаются неблагоприятные условия для распространения
болезней. Большое значение в повышении устойчивости культур к
болезням имеет своевременный уход за посадками,
обеспечивающий оптимальные условия для их роста и развития. Однако для
предупреждения появления и ликвидации имеющихся очагов
болезней одних только лесохозяйственных мероприятий недостаточно.
Они должны дополняться химическими и биологическими
мерами борьбы.
10.8.4. Химические меры защиты
Они осуществляются путем протравливания семян,
опрыскивания посевов и посадок. В питомниках применяется многолит-
ражное опрыскивание.
Протравливание семян проводят в том случае, если имеется
заключение лесосеменной станции об их зараженности фитопа-
тогенными грибами или при заведомо плохом качестве семян. Этим
способом уничтожается инфекция, находящаяся на поверхности
семян, а также создается защитная зона вокруг проростков.
Семена протравливают сухим способом, эффективность которого
повышается при его сочетании с намачиванием семян в растворах
микроэлементов. Такая обработка повышает энергию
прорастания и всхожесть, устойчивость к полеганию и способствует
появлению дружных всходов.
Борьба с полеганием включает протравливание семян и
опрыскивание посевов. Протравливание производится путем опудривания
семян непосредственно перед посевом или заблаговременно. В
последние годы для этой цели применяют фундазол, кемикар, кар-
тоцид, топсин-М, ТМТД из расчета 6 кг на 1 т семян. При
появлении первых признаков болезни посевы опрыскивают картоцидом
253
из расчета 3,6—4,8 кг препарата на 1 га. В случае дальнейшего
распространения очагов болезни обработку повторяют.
Для защиты посевов и культур сосны до трехлетнего возраста от
обыкновенного шютте рекомендуется проводить 1—2
опрыскивания, начиная с конца второй — начала третьей декады июля,
водными суспензиями топсина-М (2—4 кг/га), байлетона (1,5 кг/га),
привента (1,5 кг/га). Интервал между обработками — 2 — 3 нед.
Норма расхода рабочей жидкости 400 — 800 л на 1 га.
Борьба со снежным шютте осуществляется путем 1 — 2-кратного
опрыскивания посевов и культур сосны до пятилетнего возраста в
середине октября водными суспензиями фундазола (0,5—0,8 кг/га),
топсина-М (2—4 кг/га байлетона (1,5 кг/га), привента (1,5 кг/га).
Нормы расхода рабочих жидкостей 400—800 л на 1 га на
питомниках и до 1000—1200 л/га — в культурах. Интервал между
обработками — 3 нед.
Для защиты лиственницы от шютте рекомендуется проводить
профилактическое и искореняющее опрыскивания посевов и
культур до трехлетнего возраста. Искореняющую обработку следует
проводить в период распускания почек на двухлетних посевах и в
культурах, где имеется опавшая прошлогодняя хвоя, которая
является источником первичной инфекции. Для этой цели
используется бордоская смесь из расчета 30—60 кг препарата на 1 га.
Норма расхода рабочей жидкости 400—600 л на 1 га.
В период вегетации, через 10—14 дней после распускания хвои,
проводят опрыскивание водными суспензиями байлетона и
привента с нормой расхода 2,4 кг препарата на 1 га. При
необходимости через 2 нед обработку повторяют. Нормы расхода рабочих
жидкостей составляют 400 — 500 л/га для однолетних посевов,
800 л/га — для двухлетних, 1000—1200 л/га — в культурах.
Для борьбы со ржавчиной побегов сосны (сосновый вертун) в
первой половине мая проводят опрыскивание водными
суспензиями оксихлорида меди (2,4—8 кг/га), бордоской смеси (6 — 8 кг/га),
Абига-Пик (6 — 9,8 кг/га) с нормой расхода рабочих жидкостей
600 —800 л на 1 га.
Защита сосны от побегового рака целесообразна только в
питомниках, где в середине мая проводят опрыскивание
посевов водными суспензиями фундазола (2,4—3,2 кг/га) и
байлетона (1,8 — 2,4 кг/га) с нормой расхода рабочих жидкостей 600 л
на 1 га.
Химическая защита сосны, ели и лиственницы от ржавчины
хвои целесообразна только в том случае, если болезнь
наблюдается из года в год и причиняет ощутимый вред. Проводится
опрыскивание посевов в начале лета водными суспензиями
бордоской смеси (6—8 кг/га), оксихлорида меди (2,4—8 кг/га), Абига-
Пик (6 — 8 кг/га) с нормой расхода рабочей жидкости 600 — 800 л
на 1 га.
254
При появлении первых признаков мучнистой росы дуба и других
лиственных пород проводят опрыскивание водными суспензиями
байлетона (2,4 кг/га), коллоидной серы (12—15 кг/га), привента
(2,4 кг/га), кумулуса ДФ (7—10 кг/га) с нормой расхода рабочей
жидкости 600 — 800 л на 1 га.
С целью предупреждения пятнистостей листьев рано весной, до
распускания почек, проводят искореняющее опрыскивание по
опавшей листве, которая является источником первичной инфекции. При
этом используют водные суспензии бордоской смбси (6—16 кг/га)
с нормой расхода рабочей жидкости 600—800 л на 1 га.
При появлении пятен на листьях проводят опрыскивание
водными суспензиями бордоской смеси в концентрации 0,5 % или
фундазола — 0,1—0,15 %.
Защита лиственных пород от ржавчины осуществляется путем
опрыскивания в период вегетации водными суспензиями
следующих препаратов: Абига-Пик в концентрации 0,4—1%, бордоская
смесь — 0,5 — 1 %, оксихлорид меди — 0,4— 1 %, привент — 0,01 %,
байлетон — 0,01 %, арцерид — 0,5 %.
Для защиты тополя и ивы от парши проводят опрыскивание
через 10—14 дней после распускания листьев водными
суспензиями бордоской смеси (10—12 кг/га) и привента (0,1 — 0,4 кг/га)
при норме расхода рабочих жидкостей 1000—1200 л на 1 га.
Глава 11
СОСУДИСТЫЕ И НЕКРОЗНО-РАКОВЫЕ
БОЛЕЗНИ ДРЕВЕСНЫХ ПОРОД
И МЕРЫ БОРЬБЫ С НИМИ
Болезни этой группы поражают стволы и ветви различных
древесных пород и кустарников в школах, культурах и насаждениях
различного возраста. Они вызываются грибами разных классов и
бактериями. Возбудители болезней стволов и ветвей отличаются
патогенностью, образом жизни, способом проникновения и
характером распространения в тканях дерева. В связи с этим
вызываемые ими болезни причиняют неодинаковый вред. Одни (вальзел-
ловый некроз дуба, серо-желтый некроз тополя) поражают
только усыхающие или уже отмершие стволы и ветви, ускоряя
процесс разложения древесины, другие, например голландская
болезнь ильмовых пород, смоляной рак сосны, эндоксилиновый рак
ясеня, вызывают ослабление, суховершинность, а нередко и
гибель насаждений на больших площадях.
Болезни стволов и ветвей в зависимости от характера
поражения и внешних признаков условно делят на три группы:
сосудистые, некрозные и раковые.
11.1. СОСУДИСТЫЕ БОЛЕЗНИ
Сосудистые болезни характеризуются поражением проводящей
системы дерева. Пораженные сосуды имеют вид темных сплошных
или прерывистых колец на поперечных срезах и темных штрихов —
на продольных. Вследствие поражения сосудов вся крона или
отдельные ее ветви усыхают, при этом листья желтеют или буреют,
иногда окраска не изменяется.
Для сосудистых болезней характерна очаговость поражения. При
благоприятных условиях очаги быстро разрастаются, и болезнь
принимает характер эпифитотий. Сосудистые болезни могут
протекать в острой или хронической форме. В первом случае усыхание
деревьев происходит в течение одного вегетационного периода,
256
месяца или нескольких дней. При хронической форме болезнь
длится в течение 8—10 лет.
Голландская болезнь ильмовых пород (графиоз). Возбудитель —
сумчатый гриб Ophiostoma ulmi (Buisman) Nannf. (= Ceratocystis ulmi
(Buism.) Moreau) с конидиальной стадией Graphium ulmi Schwarz.
Поражаются все европейские и азиатские виды ильмовых пород.
Впервые болезнь была зарегистрирована во Франции в 1918 г.,
позже ее отмечали в Голландии, Бельгии и других странах. В конце
20-х гг. XX в. возбудитель болезни и европейские виды
переносчиков болезни — ильмовые заболонники с вязовыми бревнами были
завезены в США.
На территории бывшего СССР голландская болезнь впервые
была обнаружена Э. Э. Гешеле в 1935 г. в Одессе; эпифитотии
голландской болезни были зарегистрированы в периоды с 1936 по
1941 гг. и с 1955 по 1959 гг. В последующие годы болезнь охватила
весь ареал ильмовых пород, вызывая их усыхание в лесах,
полезащитных лесных полосах, городских насаждениях. Быстрое
распространение болезни поставило под угрозу существование
ильмовых пород. Во многих регионах они практически исчезли из
основного полога насаждений.
Изучением развития эпифитотии голландской болезни в
нашей стране в разное время занимались Е.С.Квашнина (1941),
Ю. В. Синадский (1960, 1977), Н. П. Озолин и Е. А. Крюкова (1958 —
1991), Р.А.Крангауз (1964-1965), В.А.Зудилин (1968, 1984).
Многолетние исследования специфики развития эпифитотии
голландской болезни в условиях большого города проведены Е. П. Кузь-
мичевым (1985 —1987).
Возбудитель голландской болезни был выделен и описан в 1922 г.
в Голландии. Первые капитальные работы по изучению болезни
принадлежат также голландским исследователям. Эти
обстоятельства и послужили основанием для названия болезни. По
конидиальной стадии болезнь часто называют графиозом. В 1928 г. Х.Бу-
исман установила, что у гриба есть сумчатая стадия.
Сумчатая стадия возбудителя встречается редко, преобладает
бесполое спороношение разных типов: Cephalosporium, Rhinotrichum,
Hyalodendron, Graphium. Однако первые три типа спороношений очень
недолговечны и легко разрушаются при механическом воздействии.
Более долговечно спороношение типа Graphium, представляющее
собой коремии. Коремии имеют вид гладких, коричневых столбиков
высотой от 10 до 100 мкм, на вершине которых образуются округлые
белесые головки из склеенных слизью конидий. Конидии
яйцевидные или грушевидные размером (3—4) х (2—3) мкм (рис. 70, 6).
Коремии образуются весной под отставшей корой и на заболони
усохших от болезни деревьев, особенно обильно — в ходах заболонников.
Заражение деревьев осуществляется спорами возбудителя или при
контакте корневых систем больных и здоровых деревьев. Главная роль
257
Рис. 70. Голландская болезнь ильмовых пород:
/ — поперечный срез через пораженную ветку; 2 — характерное скручивание
(«флажок») засохших листьев на пораженном побеге; 3 — проникновение
инфекции из пораженных побегов поросли в корни; 4 — дополнительное питание
заболонника в развилке веточек; 5 — ранка в месте дополнительного питания
жука, через нее инфекция проникает в ткани дерева; 6 — конидиальное споро-
ношение возбудителя (коремии), образующееся в ходах заболонников
в заражении принадлежит спороношению типа Graphium. Споры гриба
переносятся от дерева к дереву ветром, дождем, насекомыми. В
переносе инфекции первое место принадлежит заболонникам, из которых
наиболее активны: разрушитель (Scolytus scolytus F.), струйчатый
(Scolytus multistriatus Marsham), реже — пигмей (Scolytus pygmaeus F.).
Заражение происходит только при попадании спор на свежие поражения.
258
Заболонники осуществляют заражение в период
дополнительного питания молодых жуков в кронах деревьев. При повреждении
развилок тонких веточек жуки вносят в сосуды инфекцию с
поверхностных покровов (см. рис. 70, 4, 5). После попадания в
проводящие ткани споры распространяются пассивно. При этом
движение спор вверх происходит значительно быстрее, чем вниз.
Образующиеся гифы сразу после заражения распространяются от
сосуда к сосуду. Скорость распространения гриба и степень пора-
женности деревьев находятся в прямой зависимости от размера
сосудов. У деревьев с ббльшим диаметром сосудов споры в
большем количестве и легче проникают в глубь растения.
В зависимости от скорости развития листовых симптомов
(скручивание и изменение окраски) и отмирания деревьев различают
острую и хроническую формы болезни. При острой форме деревья
усыхают быстро, за вегетационный сезон или месяц. Усохшие
листья в кроне часто не изменяют окраску. На поперечных срезах
стволов и ветвей усохших деревьев пораженные сосуды заметны в
виде сплошного темно-бурого кольца (см. рис. 70, /).
Чаще встречается хроническая форма болезни, при которой
деревья болеют и усыхают в течение нескольких лет. У больных
деревьев листва распускается позже, чем у здоровых, листовые
пластинки уменьшаются, поэтому крона имеет ажурный вид.
Листва на таких деревьях опадает раньше, отмирание пораженных
ветвей происходит после зимы. Усыхание начинается с верхней
части кроны и распространяется вниз. Вначале усыхают
отдельные побеги, принимающие форму крючка. Усохшие листья
приобретают красно-бурую окраску, свертываются вдоль средней
жилки и повисают в виде флажков. Пораженные сосуды на
поперечных срезах стволов и ветвей заметны в виде отдельных темно-
бурых точек или прерывистого кольца (см. рис. 70, 7, 2). При
хронической форме болезни пораженные участки ксилемы всегда
обнаруживаются в более старых годичных слоях. Это связано с
тем, что у деревьев радиальный прирост образуется быстрее по
сравнению с ростом мицелия гриба в радиальном направлении.
По данным Е. П. Кузьмичева, экологическая неоднородность
среды большого города определяет разнообразие внешних
симптомов болезни и их встречаемость в городских насаждениях у
деревьев вяза гладкого в возрасте 15 — 40 лет. Наряду с типичным
скручиванием листьев вдоль осевой жилки Кузьмичевым были
отмечены широко распространенные в городских насаждениях вяза
общие симптомы: уменьшение размера листьев, изменение
окраски, преждевременный опад.
Болезнь поражает деревья всех возрастов, но чаще всего очаги
отмечаются в насаждениях от 10 до 40 лет. Быстрота
распространения голландской болезни, образование очагов и их рост
зависят от численности заболонников. Радиус разлета заболонников
259
не превышает 400 м, а чаще он достигает лишь десятков метров.
Поэтому и разрастание очагов голландской болезни идет от
источника инфекции во все стороны, постепенно охватывая новые
деревья и новые участки насаждений. Кроме того, заражение
здоровых деревьев может осуществляться споровой инфекцией,
которая находится в воздухе в очагах голландской болезни. Инфекция
может передаваться от пораженных материнских деревьев пневой
поросли к корневым отпрыскам, большая часть которых усыхает
в ближайшие 2 — 3 г. Массовое распространение спор гриба и
заражение деревьев происходит в годы с теплым влажным летом.
Наиболее активно болезнь развивается в условиях
повышенной почвенной влажности. Этим объясняется тот факт, что
быстрее и интенсивнее усыхают ильмовые в поймах рек. Наиболее
сильно поражаются чистые ильмовые насаждения. Примесь других
пород снижает их заболеваемость. В засушливые годы больные
деревья усыхают интенсивнее.
В условиях города вяз поражается голландской болезнью в
разных типах насаждений, но наиболее высокий уровень пораженно-
сти отмечается в куртинных посадках. Расширение площади
очагов болезни в городских насаждениях происходит за счет уличных
посадок, насаждений, парков, скверов, дворов. Источниками
инфекции являются усохшие и усыхающие деревья, кора, щепа,
порубочные остатки, срубленная древесина. Поражаются все
европейские и азиатские виды ильмовых пород. К наиболее
восприимчивым относятся вяз гладкий (Ulmus laevis) и берест (U.campestris).
Наиболее устойчивым считается вяз мелколистный (U.pumila).
Голландская болезнь широко распространена в центральных,
южных и юго-восточных районах европейской части России.
Сосудистый микоз дуба вызывается сумчатыми грибами из рода
Ceratocystis (C.roboris (Georg. et Y.Teod.) Potl., C.kubanicum (Scz.-
Par.) Potl., C.valachicum C.Georgescu (конидиальные стадии —
Graphium roboris Schw., Verticillium sp., Cephalosporium sp., Hyalodendron
sp., Rhinotrichum sp,,) и несовершенными грибами из рода Fusarium
(F.sporotrichiella BUai var. poae (PK.) Wr., FJavanicum Koord.).
Последние два вида были выделены и описаны в 1983 г. Е. П. Кузьмиче-
вым. Указанные виды могут поражать дуб, действуя в комплексе
или каждый отдельно.
Впервые сосудистый микоз дуба был обнаружен в 1926 г. в
Югославии. Впоследствии болезнь распространилась в другие
европейские страны и в США. На территории нашей страны болезнь и ее
возбудители были описаны А.Л.Щербиным-Парфененко в 1953 г.
В последующие годы изучением сосудистого микоза дуба
занимались Ю.Н.Иванченко (1957), В.И.Потлайчук (1957), И.И.Минке-
вич(1963,1972), Е. А. Крюкова и Т. С. Плотникова (1976,1977,1979),
Э. С. Гусейнов (1979,1986), Е. П. Кузьмичев (1983,1984), Н. Н. Селоч-
ник (1987, 1989).
260
В усыхании дубрав на территории России, которое в XX
столетии приняло широкие масштабы, сосудистый микоз дуба играет
важную роль, что было доказано всеми указанными выше
исследователями. Их работы дают основание считать сосудистый микоз
самым опасным заболеванием дуба. Поражаются все его виды.
Заражение деревьев осуществляется спорами возбудителя,
которые распространяются посредством ветра, дождя, насекомых.
Среди последних главную роль играют стволовые вредители, в
том числе дубовый заболонник {Scolytus intricatus Ratz.), желто-
пятнистый усач (Mesosa myops Dalm.), дубовая узкотелая златка
(Agrilus angustulus HI.).
Инфекция проникает в ткани дерева через свежие поранения,
наносимые стволовыми вредителями и долго незарастающие
мертвые сучья. Решающим фактором для заражения и дальнейшего
распространения инфекции является предварительное ослабление
деревьев. В случае распространения инфекции из ствола в корни
заражение может происходить при срастании корней больных и здоровых
деревьев. От больных пней и корней заражается пневая и корневая
поросль, поэтому пни от сильно зараженных деревьев непригодны
для возобновления дуба. Болезнь может передаваться и с
желудями, заражение которых происходит на деревьях и в период
хранения. Развиваясь в сосудах, возбудители вызывают отмирание
околососудистой паренхимы и интенсивное образование тиллов.
Болезнь носит ярко вьфаженный очаговый характер и может протекать в
острой и хронической форме, последняя встречается чаще.
При острой форме увядание начинается с концов ветвей и
быстро распространяется по всей кроне. Листья в этом случае
закручиваются краями вверх, приобретают желтоватую или бронзовую
окраску и через 3 — 6 нед после заражения опадают. Если
заражение происходит в конце лета, листья буреют и опадают не
скручиваясь, а часть их сохраняется на ветвях зимой. Под корой
усохших деревьев образуются скопления мицелия, выступающие из
трещин коры.
При хронической форме болезни постепенное отмирание
кроны сопровождается ее деформацией. Подобный тип усыхания
наблюдается в дубравах, находящихся под длительным
воздействием неблагоприятных экологических факторов. Усыхание
начинается с отдельных ветвей, листья уменьшаются в размерах,
желтеют, буреют и опадают. Часто листья на больных ветвях не
распускаются или опадают, не достигнув нормальных размеров.
Вследствие измельчения листьев пораженные деревья отличаются
хорошо заметной ажурностью кроны. Нередко у больных деревьев
возникает суховершинность. Сильно ослабленные и усыхающие
деревья образуют водяные побеги.
Внешние признаки сосудистого микоза могут проявляться и
при поражении другими болезнями или под воздействием раз-
261
личных неблагоприятных почвенно-климатических и
антропогенных факторов. Поэтому достоверная диагностика болезни возможна
только по внутренним признакам, главным для сосудистого
микоза. На поперечном срезе пораженных ветвей признаки
обнаруживаются по наличию бурых или темно-оливковых пятен и
точек, а в отдельных случаях по изменению окраски всей заболони.
На тангентальных срезах внутренние симптомы имеют вид темно-
бурых прерывистых тяжей длиной от 0,01 м до 2 м.
Сосудистый микоз поражает дуб в естественных насаждениях и
культурах в разных лесорастительных условиях, охватывая
пойменные и нагорные дубравы. Но в каждой конкретной зоне
наиболее высокий уровень поражения отмечается в пойменных
дубравах. Болезнь распространена в насаждениях дуба разного
возраста, однако при хронической форме течения пораженность
увеличивается по мере их старения. Степень пораженности
сосудистым микозом не зависит от состава, полноты и бонитета
насаждений. Сильнее страдают порослевые насаждения дуба,
характеризующиеся пониженной устойчивостью. Е.П.Кузьмичевым
установлена тесная связь между степенью усыхания насаждений дуба
и уровнем пораженности их сосудистым микозом. Интенсивное
усыхание дубрав может происходить и без участия этой болезни.
Но по мере продвижения на юг влияние пораженности
сосудистым микозом на размеры усыхания дуба возрастает.
В очагах сосудистого микоза нередко встречается опенок
осенний и некрозы, в том числе клитрисовый (колпомовый), вилле-
миниевый, черный немоспоровый и другие, которые могут
ускорять процессы ослабления и усыхания дуба.
Сосудистый микоз дуба распространен в южных и
юго-восточных районах европейской части России.
Вертициллезное усыхание (вилт) клена вызывается
несовершенным грибом Verticillium dahliae Kleb. Он поражает
некоторые виды клена, дуб, ильмовые и другие породы. Особенно сильно
поражается клен остролистный. Усыхание разных видов клена
под названием «вилт» известно в США с 1915 г., в Европе — с
1926 г. В России массовое усыхание сеянцев клена в
питомниках было впервые обнаружено С.И.Ваниным в 1952 г. в
Московской области. Позже массовое усыхание клена остролистного от
вертициллеза отмечалось в насаждениях юго-востока
европейской части России и в центральной зоне смешанных лесов.
Данные об особенностях развития и распространения болезни в
России принадлежат Р.А.Крангауз, изучавшей вилт клена с 1956 по
1960 гг.
Внешние признаки заболевания проявляются в усыхании
кроны, ее отдельных ветвей или всего дерева. При усыхании кроны
иногда появляются водяные побеги, а при усыхании дерева —
поросль, которые затем также усыхают (рис. 71). В некоторых слу-
262
Рис. 71. Вертициллезное усыхание (вилт) клена:
/ — усыхающая поросль, зараженная от пня больного дерева; 2 — конидиенос-
цы с конидиями; 3 — хламидоспоры возбудителя; 4 — потемнение древесины на
поперечных срезах пораженных стволов
чаях крона дерева приобретает ажурный вид в связи с
уменьшением количества и размеров листьев.
Гриб вызывает полное или частичное отмирание корней, оно
распространяется от корневой шейки и сопровождается
разрушением древесины. Поэтому усохшие деревья часто легко
вырываются из почвы. Древесина пораженного дерева приобретает
светло-зеленоватый, зеленовато-черный или оливковый цвет. При этом
сначала появляются полосы или штрихи древесины с
измененной окраской, переходящие затем в сплошное окрашивание.
Наружные слои пораженной заболони до полной гибели дерева
обычно имеют слабое потемнение. При несвоевременной выборке
усохших деревьев кора вдоль ствола отстает и опадает.
263
Наблюдается различное течение болезни. В одних случаях усы-
хание начинается с отдельных ветвей и постепенно охватывает
всю крону. В других случаях дерево без внешних признаков
заболевания внезапно усыхает среди лета или не распускается весной.
В-третьих, довольно редко болезнь принимает как бы
хронический характер, иногда заканчивающийся даже выздоровлением. Это
бывает в том случае, когда заражены глубокие слои древесины и
скорость нарастания годичных слоев ее превышает скорость
распространения гриба по радиусу.
Усыхание деревьев при острой форме заболевания наступает
через 1—4 г. после заражения. Поросль и водяные побеги чаще
всего усыхают в первый и второй годы жизни.
Первичное заражение происходит главным образом через корни
в области корневой шейки. Некоторая часть деревьев заражается через
ствол и ветви. Распространение болезни вверх и вниз по дереву идет
как по наружной, так и по внутренней, в том числе ядровой
древесине, распространение вверх происходит гораздо быстрее. В местах
внедрения возбудителя развивается грибница и микросклероции,
представляющие собой кучки хламидоспор. Возбудитель не является
обитателем почвы и сохраняется только на растительных остатках в
виде микросклероциев. Гриб легко обнаруживается во влажных
камерах: на 4—6 день он образует характерные мутовчатые спороно-
шения. Развитию болезни способствует теплая влажная погода.
Болезнь встречается в питомниках, культурах и насаждениях
европейской части России.
11.2. НЕКРОЗНЫЕ БОЛЕЗНИ
При некрозных болезнях поражаются кора, луб, камбий и
наружные слои древесины. Некрозы протекают сравнительно
быстро, вызывая гибель деревьев за несколько лет, а иногда — за
несколько недель. Возбудителями некрозов чаще всего бывают
грибы, одни поражают уже ослабленные деревья, другие поселяются
на вполне жизнеспособных деревьях.
В зависимости от диаметров стволов и ветвей, а также от
скорости распространения грибницы возбудителей отмирание
тканей дерева происходит по окружности или отдельными участками.
В соответствии с этим различают круговые и локальные некрозы.
Круговые некрозы проявляются в отмирании тканей по всей
окружности стволов и ветвей. Локальные некрозы выражаются в
образовании отмерших участков овальной формы или вытянутых в виде
полос, часто отличающихся от здоровой коры окраской. Нередко
локальные некротические участки отграничиваются от здоровых
валиком каллюса. В отмершей коре развиваются спороношения
грибов, имеющие вид различно окрашенных бугорков, пустул,
264
мелких подушечек и т.д. Многие некрозные болезни
сопровождаются гнилью, которая развивается в древесине уже усохших
стволов и ветвей. Часто на некротических участках образуются
раковые раны, особенно на стволах и толстых деревьях.
Некрозы поражают очень многие древесные породы и
кустарники, причиняя вред лесным питомникам, культурам и
насаждениям. Степень вредоносности некрозов неодинакова в разных
частях ареала определенной древесной породы и в различных типах
лесорастительных условий.
Ценангиевый некроз сосны (ценангиоз). Возбудитель —
сумчатый гриб Cenangium ferruginosum Fr. (= С. abietis (Pers.) Rehm.) с
конидиальной стадией Dothichizaferruginosa Sacc. Гриб часто
встречается как сапрофит на нижних, отмерших ветвях или на
порубочных остатках, но может поражать вполне жизнеспособные
растения. Это объясняется тем, что его популяция представлена
штаммами с различной степенью патогенности, среди которых
есть штаммы, проявляющие высокую патогенность и
агрессивность по отношению к сосне (Н. Н.Арапова, 1986). Болезнь
встречается в насаждениях, культурах, реже — в питомниках.
Заражение сосны осуществляется аскоспорами, реже
конидиями. Массовое созревание и распространение аскоспор гриба и
заражение сосны происходит в конце августа — сентябре.
Мицелий возбудителя развивается в коре и лубе. Первые признаки
болезни обнаруживаются весной. Хвоя на пораженных ветвях и
побегах сначала желтеет, затем краснеет и постепенно опадает, а
крона изреживается. На оставшейся висеть хвое и усохших побегах
образуются пикниды, имеющие вид мелких черных бугорков,
выступающих из разрывов эпидермиса хвои и трещин коры (рис.
72). В пикнидах созревают бесцветные одноклетные конидии
яйцевидной, веретеновидной, эллипсоидальной формы размером
(6,3 — 9,5) х (3—4) мкм. На следующий год на отмерших побегах и
ветвях образуются группы тесно скученных апотециев, имеющих
вид многочисленных темно-бурых, почти черных шероховатых,
бугорков диаметром до 1 — 3 мм, выступающих из трещин коры.
Во влажном состоянии апотеции раскрываются и приобретают
вид чашечек диаметром 1—3 мм с бурой шероховатой
поверхностью и зеленовато-желтым гимениальным слоем, состоящим из
сумок. Аскоспоры одноклетные бесцветные или слегка буроватые
эллипсовидные, яйцевидные размером (10—13) х (5 — 8) мкм.
Иногда на пораженных побегах наблюдается усиленное
выделение смолы, что связано с проникновением мицелия в древесину.
Ценангиевый некроз сосны, как правило, развивается на фоне
предварительного ослабления растений под воздействием
различных неблагоприятных факторов: засухи, сильные морозы,
поражение болезнями (снежное шютте, корневые гнили),
повреждение насекомыми и копытными животными, нарушение правил
265
Рис. 72. Ценангиевый некроз сосны:
1 — пораженный побег с опавшей хвоей; 2 — пораженный стволик с отмершей
корой и многочисленными группами апотециев; 3 — раскрытые апотеции в
трещинах коры; 4 — сумка со спорами
ухода за лесом, введение интродуцированных видов сосны,
промышленные выбросы.
При благоприятных для возбудителя условиях болезнь может
принимать характер эпифитотий, вызьгоая массовое усыхание культур и
подроста сосны. По данным Н.Н.Араповой (1992), в период
эпифитотий ценангиоза в Северном Казахстане отмечалось массовое
поражение самосева сосны под пологом леса и сеянцев в питомниках.
Болезнь распространена в европейской части России, в
Сибири, на Алтае.
Нектриевый некроз (нектриоз) лиственных пород. В цикле
развития у возбудителя образуются конидиальная {Tubercularia vulgaris
Tode) и сумчатая (Nectria cinnabarina (Tode) Seav.) стадии.
Последняя развивается крайне редко. По конидиальной стадии болезнь
называют туберкуляриозом. Поражаются многие лиственные
древесные породы и кустарники: береза, боярышник, бук, дуб, вяз,
каштан конский, клен, липа, рябина, тополь, ясень, жимолость,
карагана, кизильник и др.
Гриб часто встречается как сапротроф на отмерших стволах и
ветвях, но может поражать живые деревья и кустарники. Это
объясняется различной степенью вирулентности штаммов,
составляющих популяцию гриба в природе.
Поражается кора, сосудистая система и древесина. В случае
проникновения гриба в сосуды у больных деревьев вначале происхо-
266
дит увядание листьев, а затем отмирают ветви. При поражении
коры и древесины на стволах и толстых ветвях сначала
появляются локальные некрозы, постепенно преобразующиеся в
неступенчатые раны. Характерным симптомом болезни при всех типах ее
проявления являются стромы гриба, развивающиеся в большом
количестве на пораженных стволах и ветвях (рис. 73, 7, 2). Кони-
диальные стромы имеют вид гладких киноварно-красных или
розовых выпуклых подушечек диаметром 0,5—2 мм, высотой до 1,5 мм,
выступающих из трещин коры. Конидиальные стромы
образуются с ранней весны. С поверхности подушечек в массовом
количестве отчленяются цилиндрические, бесцветные конидии
размером (5 —8) х (1,5— 3) мкм (см. рис. 73, 3). В умеренной зоне
образование конидий происходит весь год, но их прорастание и
заражение растений возможно только в период вегетации при
температуре от 10 до 27 °С.
Сумчатая стадия гриба развивается осенью на тех же конидиаль-
ных стромах. Перитеции закладываются у основания стром. С обра-
Рис. 73. Нектриевый некроз клена:
1 — пораженные стволик и побег, покрытые стромами возбудителя; 2 —
стромы, выступающие из трещин коры; 3 — конидиеносцы с конидиями; 4 — сумки
со спорами
267
зованием перитециев стромы приобретают темно-красную или
коричневую окраску, а поверхность их становится неровной, как бы
зернистой. В перитециях формируются сумки со спорами. Сумки
цилиндрически-булавовидные размером (60—90) х (9 — 12) мкм.
Сумкоспоры бесцветные эллипсоидально-цилиндрические двух-
клеточные размером (12 — 20) х (4—7) мкм (рис. 73, 4).
Распространение гриба чаще всего происходит посредством
конидий, которые очень эффективно разносятся каплями дождя. Разносу
инфекции могут способствовать и насекомые. Споры проникают в
ткани растений через различные поранения коры, обломы сучьев,
морозобоины, ожоговые трещины. Развитию болезни способствуют
природные и антропогенные факторы, ослабляющие растения:
погодные условия, промышленные выбросы, нарушение водного и
воздушного режима почвы.
Развиваясь на фоне предварительного ослабления, течение
болезни ускоряется, что приводит к частичному или полному усы-
ханию кроны. В редких случаях заболевание может принимать
характер эпифитотии. Наибольший вред нектриоз причиняет
растениям в питомниках и молодым городским посадкам.
Ареал болезни охватывает европейскую часть России, Сибирь,
Дальний Восток.
Колпомовый (клитрисовый) некроз дуба. Возбудитель —
сумчатый гриб Colpoma quercinwn (Pers.) Wallr. (= Clithris quercina (Pers.)
Rehm.) поражает растения в питомниках, культуры и
насаждения разного возраста.
На пораженных стволах и ветвях кора вначале становится
красноватой, а после отмирания приобретает желтовато-белесую
окраску, при этом некротические участки четко отграничиваются
от здоровой коры. Некрозы могут быть ленточными или
круговыми. На отмирающих участках вначале образуются пикниды гриба в
виде мелких серовато-белых бугорков, затем в пораженной
грибом коре появляются многочисленные апотеции длиной до 5 мм.
Они имеют вид серовато-белых или темно-серых изогнутых, стру-
пьевидных образований, расположенных поперек стволиков или
ветвей. Во влажную погоду апотеции раскрываются довольно
широкой продольной щелью, обнажая сероватый студенистый гиме-
ниальный слой сумок со спорами. Сумки булавовидные, на
ножке, размером (130—150) х (9—10) мкм. Аскоспоры нитевидные,
бесцветные, с каплями масла, вначале одноклеточные, позже с
одной перегородкой, размером 90 х 1,5 мкм (рис. 74).
Заражение осуществляется аскоспорами через различные
повреждения коры. Кроме отмирания коры гриб вызывает белую за-
болонную гниль.
В насаждениях лесной зоны болезнь поражает нижние,
отмирающие ветви, способствуя самоочищению стволов. В степной и
лесостепной зонах в неблагоприятных, засушливых условиях роста
268
болезнь вызывает ослабление
и усыхание культур до
20-летнего возраста. Пораженность
молодых культур в степной
зоне достигает 10—40 %.
Источником инфекции
являются пораженные деревья и
неубранные порубочные
остатки. Болезнь широко
распространена в ареале дуба.
Черный немоспоровый (диа-
триповый) некроз дуба.
Возбудитель болезни —
несовершенный гриб Naemospora croceola
Sacc. (сумчатая стадия — Dia-
trype stigma (Hoffm.) Wint.).
В начальной стадии
развития болезни кора в местах
поражения приобретает
красноватую окраску, при этом
пораженные участки трудно
отличить от здоровых. Позднее в
отмирающей коре образуется
конидиальная стадия
возбудителя — ложа. Ложа имеют вид
выпуклых мелких пятен
желто-бурого или красно-бурого
цвета, окаймленных тонкой черной линией диаметром 1 — 3 мм.
В центре пятен хорошо заметны красные капельки или жгутики
застывающей массы конидий. Конидии одноклеточные бесцветные
цилиндрические или слегка изогнутые размером (5 — 7) х (0,8 — 1) мкм.
Через 2 — 3 года в толще отмершей коры развивается строма,
часто окольцовывающая стволики и ветви. Строма плоская,
толщиной до 2 мм, темно-бурого цвета, покрытая многочисленными
поперечными и продольными трещинами. На поверхности стро-
мы в лупу заметны многочисленные мелкие точечные бугорки —
выступающие устьица перитециев. После опадания наружной корки
строма обнажается, придавая пораженной отмершей коре обо-
женный вид. Перитеции обнаруживаются на поздней стадии
развития болезни. Сумки булавовидно-цилиндрические, на удлиненной
ножке, размером (30 — 60) х (4—3) мкм. Аскоспоры
одноклеточные, светло-коричневые цилиндрические, прямые или согнутые, с
двумя каплями масла, размером (4—12) х (0,6—1,5) мкм (рис. 75).
Из отмершей коры гриб проникает в заболонь, вызывая белую
гниль. В лесной зоне болезнь встречается на тонких ветвях в
нижней части кроны, а также на сильно ослабленных деревьях. В за-
Рис. 74. Колпомовый (клитрисовый)
некроз дуба:
/ — пораженная ветка с апотециями и
пикнидами возбудителя; 2 — зрелые рас-,
крытые апотеции; 3 — поперечный срез
через апотеции; 4 — сумки со спорами
269
Рис. 75. Черный немоспоровый (диатриповый) некроз дуба:
1 — часть пораженного ствола с обнаженной темно-бурой, покрытой трещинами
стромой возбудителя; 2 — сумка со спорами; 3 — срез через строму с перитециями
сушливых условиях степной и лесостепной зон болезнь поражает
культуры до 25-летнего возраста и может вызвать их усыхание. По-
раженность культур дуба в условиях степей достигает 40—50 %, а
иногда и более 70 %.
Болезнь встречается в европейской части России, особенно
широко распространена в лесостепной зоне.
Нуммуляриевый некроз дуба. Возбудитель — сумчатый гриб Num-
mullaria bulliardiiTul. Поражаются стволы и толстые ветви деревьев.
Видимые признаки болезни проявляются через несколько лет
после заражения. В толще пораженной коры, часто по всей длине
ствола, образуются толстые (до 3 мм), овальные, подушковидные
стромы, достигающие 1 —6 см ширины и 2—40 см длины. Вначале
они имеют табачный цвет и мажущую консистенцию, позже
становятся черными, углистыми (рис. 76). В толще стром развиваются
Рис. 76. Нуммуляриевый некроз дуба:
1 — часть пораженной ветви со стромами возбудителя; 2 — срез через строму с
перитециями; 3 — сумка со спорами; 4 — аскоспоры
270
многочисленные перитеции, устьица которых образуют на
поверхности стромы многочисленные точечные бугорки. Сумки в перитециях
цилиндрические, на короткой ножке, размером (100—120) х 10 мкм.
Аскоспоры эллипсовидные одноклеточные темно-коричневые
размером (12—14) х (6—Ю) мкм. Часто стромы сливаются,
окольцовывая стволы на протяженности до 1 м и более. В это же время
гриб, распространяясь в заболонной части стволов и ветвей,
вызывает белую гниль с черными извилистыми линиями.
Болезнь развивается в культурах и насаждениях II—III классов
возраста на фоне предварительного ослабления. В насаждениях
процесс усыхания под воздействием различных факторов
ускоряется. Поражение нуммуляриевым некрозом молодых культур в
неблагоприятных условиях роста может вызвать их гибель.
Заболевание широко распространено на юго-востоке
европейской части России.
Виллеминиевый некроз дуба вызывается грибом Vuilleminia
comedens Maize., относящимся к афиллофороидным гименомице-
там. Кроме дуба в более редких случаях болезнь поражает бук,
березу, лещину, граб.
Поражаются чаще ветви. Под эпидермисом коры, обычно с
нижней стороны ветвей, образуются плодовые тела гриба, которые
после разрыва и опадения верхней корки обнажаются. Они имеют
вид плотно приросших к субстрату восковидных пленок
беловатого, желтоватого или светло-бурого цвета толщиной 1 — 1,5 мм. В
гимениальном слое развиваются базидии с базидиоспорами.
Образование и распространение спор зависит от погодных условий. В
период сухой погоды споруляция прекращается и возобновляется
после выпадения осадков. В связи с этим у гриба V. comedens в
течение года может наблюдаться несколько периодов массового
образования и рассеивания спор и заражения деревьев. Инфекция
проникает в ткани дерева через различные поранения коры, в
том числе через повреждения насекомыми. Мицелий возбудителя
развивается в заболонной части ветвей, вызывая отмирание коры
и белую заболонную гниль.
В лесной зоне гриб поражает нижние отмирающие ветви. В
засушливых условиях степной зоны развивается в ослабленных
насаждениях, часто в очагах сосудистого микоза, способствуя более
быстрому усыханию. При неблагоприятных условиях роста болезнь
может вызывать массовое усыхание культур дуба в возрасте до 25 лет.
Болезнь встречается в европейской части России, на Южном
Урале.
Цитоспороз лиственных пород. Это общее название некрозных
болезней, вызываемых грибами из рода Cytospora. Поражаются
многие лиственные породы и кустарники.
Бурый цитоспоровый некроз тополя. Возбудитель болезни —
несовершенный гриб Cytospora chrysosperma (Pers.) Fr., (сумчатая ста-
271
дия — Valsa sordida Nitschke). Гриб поражает разные виды и
гибриды тополя в маточных плантациях и школах питомников, в
культурах, естественных и городских насаждениях.
Заражение тополя осуществляется конидиями, которые
распространяются посредством дождя или насекомых. Возбудитель попадает
в ткани дерева через различные повреждения, трещины у
основания ветвей; гриб выделяет токсины, вызывающие отмирание коры.
В мертвой коре развивается темно-серая или бурая строма, в
которой образуются пикниды. На пораженных ветвях и стволах они имеют
вид многочисленных конических бугорков, которые вначале
прикрыты перидермой, а затем выступают устьицами из разрывов коры.
Созревающие конидии выходят из устьиц и застывают на воздухе в
виде золотисто-красноватых капель или тонких спиралек. Конидии
бесцветные, в массе серовато-беловатые, одноклеточные
цилиндрические, с закругленными концами, часто изогнутые, размером
(3—5) х 1 мкм (рис. 77). Массовое спороношение возбудителя
происходит в мае — начале июня и в конце августа—сентябре.
Очаги цитоспороза возникают, как правило, на фоне
предварительного ослабления тополя, в связи с неблагоприятными по-
чвенно-климатическими условиями (засуха, длительное
затопление, подмерзание, нарушение баланса питательных веществ в
почве), нарушение правил ухода за питомниками и посадками,
загрязнением воздуха и почвы.
Рис. 77. Бурый цитоспоровый некроз тополя:
/ — пораженный стволик с выходящими в виде спиралек конидиями
возбудителя; 2— поперечный срез через пикниду; 3— конидии; 4 — споры, выходящие из
пикниды, застывают в виде тяжей
272
Источниками инфекции являются черенки и пораженные
деревья. С черенками, заготовленными в пораженных маточных
плантациях, инфекция проникает в школьные отделения питомников.
Вместе с зараженным посадочным материалом болезнь
распространяется в культурах и городских посадках.
При длительном течении болезни происходит постепенное, в
течение нескольких лет усыхание кроны. Усыхают и
появляющиеся водяные побеги. При быстром течении болезни деревья могут
усохнуть еще до распускания почек или через 1 — 2 нед после
появления листьев.
Болезнь распространена в ареале тополя.
Черный цитоспоровый некроз тополя. Возбудителем болезни
является несовершенный гриб Cytosporafoetida VI. et Rr. Болезнь
поражает различные виды и гибриды тополя, в том числе белый,
серебристый, канадский, осокорь, тополь Болле и др.
Гриб заражает кору и луб. В толще пораженной коры
развивается черная строма, в которой образуются пикниды возбудителя. На
участках стволов и ветвей с тонкой гладкой корой пикниды
заметны в виде черных округлых, выпуклых пятен диаметром до 2 мм.
Весной, в конце апреля—начале мая, из пикнид выходит
слизистая масса спор, застывающая на воздухе в виде кроваво-красных
капель или жгутиков. Конидии бесцветные одноклеточные
цилиндрические с закругленными концами, слегка согнутые, размером
(5—7) х (1,5—2) мкм (рис. 78). При поражении толстой коры
пикниды не видны, и гриб обнаруживается по кроваво-красным
жгутикам спор, выступающим из трещин коры. Строма гриба в свежем
состоянии издает неприятный
запах испорченной селедки,
который служит одним из
характерных симптомов болезни.
Источниками инфекции
являются больные деревья и кора
порубочных остатков.
Заражение тополя осуществляется
конидиями, которые
распространяются посредством дождя,
насекомых, реже — ветра.
Инфекция проникает в ткани дерева
через сухие сучья, различные
поранения коры, повреждения
насекомыми.
Возникновению очагов
болезни способствуют различные
факторы, влияющие на
ослабление деревьев: длительные
затопления, засухи, сильные моро-
Рис. 78. Черный цитоспоровый некроз
тополя:
/ — пораженный ствол с тяжами спор;
2 — поперечный срез через пикниду с
выходящими конидиями; 3 — конидии
возбудителя
273
зы, ожог коры, нарушение баланса питательных веществ,
загрязнение воздуха и почвы, повреждение насекомыми.
Болезнь вызывает массовое усыхание культур, насаждений II—
III классов возраста и поросли в очагах заражения. У молодых
тополей поражаются преимущественно стволики под кроной и в
нижней части кроны, что приводит к их быстрому усыханию в
течение 1 — 2 лет. В насаждениях более старшего возраста чаще всего
стволы поражаются в средней и нижней, реже — в подкроновой
части. В этом случае процесс усыхания длится в течение
нескольких лет, пораженные деревья становятся частично сухокронными
и постепенно усыхают.
Черный цитоспоровый некроз распространен в юго-восточных
районах европейской части России.
Цитоспороз на других породах мало изучен, хотя встречается
часто и приносит существенный вред, приводит к ослаблению и
усыханию деревьев, особенно в городских насаждениях.
Цитоспороз липы вызывает гриб Cytospora leucosperma Fr. (= С. carphosperma
Fr.), рябины — C.rubescens Fr., ясеня — С. pruinosa (Fr.) Defago (=
C.pulchella Sacc), яблони — C.schulzeri Sacc. et Syd. (=C.capitata
Schulz.).
Дотихициевый (дискоспориевый) некроз тополя. Возбудитель —
несовершенный гриб Sutton (Dothichiza populea Sacc.) (= Discosporium
populeum (Sacc). Поражаются разные виды и гибриды черных и
бальзамических тополей в питомниках* культурах, городских
насаждениях. Гриб заражает кору, луб, камбий конидиями, которые
распространяются дождем, насекомыми, в меньшей степени —
ветром. Возбудитель проникает в ткани дерева через различные
повреждения, а также через естественные ворота инфекции:
чечевички, основания почек, места соединения побегов, трещины
у основания ветвей и сучьев.
Мицелий развивается в коре стволов и ветвей, образуя
локальные или круговые некрозы, реже — раковые раны. Вначале на
коре стволов и ветвей появляются вдавленные некротические
участки овальной формы, до нескольких сантиметров в диаметре. Они
появляются по всей длине стволов и побегов, но чаще всего в
местах прикрепления ветвей к стволам, побегов — к ветвям. На
живых стволах и ветвях пораженные участки выделяются более
темным цветом, но по мере отмирания кора приобретает
желтоватый цвет. Вокруг некротических участков образуются валики
каллюса толщиной в несколько миллиметров. Постепенно
отдельные некротические участки сливаются, окольцовывая ствол или
ветвь. При поражении толстых стволов грибница
распространяется в тканях дерева в течение 2—3 лет, вследствие чего на стволах
развиваются раковые раны. Пораженные деревья имеют ажурную
крону с мелкими листьями, на стволах образуются
многочисленные водяные побеги.
274
Оа
о
Рис. 79. Дотихициевый (дискоспо-
риевый) некроз тополя:
1 — пораженный стволик с пикнида-
ми возбудителя; 2 — срез через зрелую
пикниду; 3 — конидии
На отмирающих и отмерших
участках коры весной
образуются пикниды гриба в виде
бугорков до 2 мм в диаметре.
Пикниды располагаются чаще всего
продольными рядами, реже —
беспорядочно. Зрелые споры в
пикнидах сохраняются большую
часть года, однако массовое
созревание конидий происходит в
первой половине вегетации.
Выходящие из пикниды споры
имеют вид черновато-белых или
светло-оливковых жгутиков
длиной до 2 —4 мм. При
сильном поражении во время
массовой споруляции стволы
становятся белесо-серыми от
массы тяжей выходящих спор.
Конидии одноклеточные
бесцветные, яйцевидные, суженные на одном конце, реже —
эллипсовидные, шаровидные размером (9 —13) х (7—9) мкм (рис. 79).
Заражение деревьев может происходить с апреля по сентябрь.
Инкубационный период длится 6 — 7 мес. Развитию болезни
способствуют разные факторы, вызывающие ослабление тополя (см.
цитоспороз). Болезнь нередко приводит к массовому ослаблению
и усыханию тополя в питомниках, культурах, городских
насаждениях. Особенно сильно страдают загущенные посадки в школьных
отделениях питомников при неблагоприятных условиях роста.
В культуры и городские насаждения инфекция проникает с
зараженным посадочным материалом.
Дотихициевый некроз распространен в центральных районах
европейской части России.
Гистерографиевый некроз ясеня. Возбудитель болезни — поло-
стносумчатый гриб Hysterographium fraxini (Pers.) De Not.
Поражается ясень зеленый и ясень обыкновенный. Гриб развивается в
коре и лубе толстых ветвей и стволов молодых деревьев. На
зараженных стволах и толстых ветвях образуются некротические
участки, вытянутые на 0,5 м и более. При заболевании тонких ветвей и
побегов развиваются круговые некрозы. Вначале кора в местах
поражения краснеет, позже некротические участки становятся
желтовато-белесыми и покрываются мелкими продольными и
поперечными трещинами. На светлых отмерших участках коры хорошо
заметны плодовые тела возбудителя — гистеротеции, имеющие
вид мелких черных кофейных зерен до 3 мм длиной. При созревании
они раскрываются продольной щелью, обнажая гимениальный слой,
275
состоящий из сумок со спорами. Сумки цилиндрические, на
короткой ножке, размером (250—400) х (24—40) мкм. Аскоспоры олив-
ково-бурые овальные яйцевидные, имеющие 6—9 поперечных и
2 — 3 продольных перегородки размером (30—48) х (10—20) мкм.
Гистерографиевый некроз заражает преимущественно порослевые
насаждения I —II классов возраста в неблагоприятных условиях
роста и нередко вызывает их усыхание. Особенно сильно
поражаются изреженные насаждения.
Болезнь зарегистрирована на юго-востоке европейской части
России.
11.3. РАКОВЫЕ БОЛЕЗНИ
Раковые болезни представляют собой поражения коры, луба,
камбия, древесины. Патологический процесс может развиваться в
течение нескольких десятков лет. Раковые болезни чаще
вызываются грибами, бактериями, реже — абиотическими факторами.
Внешне болезни этой группы проявляются в образовании на
стволах и ветвях язв, ступенчатых ран или опухолей. В зависимости от
характера поражения и внешних признаков различают следующие
типы раковых болезней: язвенный рак, ступенчатый рак,
смоляной рак, опухолевидный рак. Раковые болезни поражают хвойные
и лиственные породы, вызывая их ослабление и усыхание.
Смоляной рак (серянка) сосны. Возбудителями болезни
являются ржавчинные грибы Cronartium flaccidum Wint. и Peridermiwn pint
(Willd) Lev. et Kleb. Оба гриба вызывают одинаковое по внешним
признакам заболевание, но отличаются циклом развития. C.flaccidum
разнохозяйный, с полным циклом развития. Эциостадия
развивается на сосне, урединио- и телиостадии — на различных
травянистых растениях: ластовне обыкновенном (Cynanchum vincetoxicum),
мытнике болотном (Pedicularis palustris), недотроге (Impatiens sp.)9
вербене (Verbena sp.) и др. P. pini имеет только эциальную стадию,
развивающуюся на сосне.
Время появления эциев зависит от погодных условий. Их
развитию и созреванию способствует теплая погода. Массовое развитие
эциев и разлет эциоспор происходят в июне. Поражаются спелые
насаждения и сосновый подрост. В разных районах страны пора-
женность подроста смоляным раком колеблется от 5 до 25 %, а во
взрослых насаждениях достигает 40—60 %. Заражение
осуществляется через молодые охвоенные побеги. Мицелий возбудителя
проникает в клетки древесины и смоляные ходы, разрушает их,
вследствие чего смола пропитывает близлежащие слои древесины и
вытекает наружу. Развиваясь в клетках камбия, грибница прекращает
рост древесины через 2 — 3 года после заражения. В это же время на
пораженных местах развиваются эции возбудителей. Они выступают
276
Рис. 80. Смоляной рак (серянка) сосны:
/ — пораженный стволик подроста с эциями возбудителя; 2 — поперечный срез
через пораженный, деформированный участок ствола; 3 — телиоспороношение
на листе ластовня обыкновенного — промежуточного хозяина гриба С. flaccidum;
4 — эциоспоры; 5— урединиоспоры; 6— разрез через пораженный лист ластовня:
а — урединиопустулы, б — столбик телиоспор
из трещин коры в виде оранжевых пузырьков высотой до 3 — 5 мм,
заполненных массой эциоспор и сплошь покрывают пораженный
участок ветви или ствола (рис. 80, 7). После рассеивания эциоспор
в местах поражения остаются очень тонкие, хрупкие белесые
оболочки эциев, которые быстро разрушаются и опадают, а на их
месте остаются вдавленные желтовато-белые пятна различной
формы, сохраняющиеся 1 — 2 года, хорошо заметные на гладкой
коре.
Пораженные молодые побеги после 1 —2-кратного
плодоношения часто погибают вместе с мицелием возбудителя, который
277
не успевает при этом перейти в ствол. При проникновении
мицелия в ствол он распространяется в нем из года в год вдоль и по
окружности, вызывая отмирание камбия и образование раны. Кора
в местах поражения шелушится и опадает, вытекающая смола
застывает в виде серовато-желтых желваков или подтеков.
Вследствие усиленного притока питательных веществ в непораженную
часть ствола ширина годичных колец значительно увеличивается,
что приводит к деформации ствола, выражающейся в резкой
эксцентричности (см. рис. 80, 2).
Средняя скорость распространения грибницы составляет 11 см
в год по длине ствола и 2,1 см в год по окружности. Болезнь может
длиться от 2—3 до 100 лет в зависимости от скорости
распространения мицелия, возраста и жизнеспособности дерева. При
поражении подроста в возрасте от 3 до 20 лет болезнь продолжается
несколько лет. Во взрослых насаждениях она протекает в течение
нескольких десятков лет.
Состояние дерева зависит от расположения и количества ран
на стволе (рис. 81). При их возникновении в вершинной части
наблюдается суховершинность. Если усохшая вершина меньше
половины длины кроны, то такие деревья могут жить в течение
длительного времени. В противном случае пораженные деревья
заметно ослабляются. Возникновение ран в нижней части кроны и под
ней приводит к частичному или полному ее усыханию.
Рис. 81. Расположение ран смоляного рака на стволах сосны:
1 — под кроной; 2 — в нижней части кроны; 3 — в верхней части кроны
278
Смоляной рак вызывает нарушение фотосинтеза. Раковые раны,
охватывающие ствол более чем на 2/з его окружности,
препятствуют поступлению воды и питательных веществ, что приводит
к резкому снижению текущего прироста у больных деревьев.
Наиболее сильно поражаются изреженные насаждения. Особенно
страдают деревья по опушкам, около дорог и просек. Это объясняется
тем, что возбудители смоляного рака относятся к свето- и
теплолюбивым видам. Развитие болезни в условиях интенсивного
освещения и сильного нагревания стволов происходит в 2,5 раза
быстрее, чем в затененных местах.
Пораженные смоляным раком ослабленные деревья
заселяются стволовыми вредителями, видовой состав которых изменяется
в зависимости от полноты насаждений, типов леса и от других
факторов. Наиболее распространенными видами стволовых
вредителей в очагах смоляного рака являются: вершинный короед
(Ips acuminatus Gyll.), малый сосновый лубоед (Tomicus minor Hart.),
четырехзубый гравер {Pityogenes quadridens Hart.), восточный
гравер {Pityogenes irkutensis Egg.), большой сосновый лубоед {Tomicus
piniperda L.) и др. Стволовые вредители в значительной степени
ускоряют процесс отмирания деревьев в очагах болезни.
Смоляной рак широко распространен в ареале сосны
обыкновенной в естественных и городских насаждениях.
Ржавчинный рак (пузырчатая ржавчина) сосны. Возбудитель
болезни — ржавчинный гриб Cronartium ribicola Dietr. Поражаются
кедровая сосна сибирская, веймутова сосна, реже — другие виды
сосны и разные виды смородины. Эциостадия возбудителя
развивается на видах сосны, урединио- и телиостадии — на
смородине.
Заражение сосны происходит в конце лета или осенью
посредством базидиоспор, образующихся на листьях смородины. Через
2 — 3 г. весной в местах поражения образуется эциостадия. Эции
имеют вид желто-оранжевых пузырьков длиной до 10 мм,
высотой 1—2 мм, заполненных массой оранжевых эциоспор. Эциоспо-
ры заражают листья разных видов смородины; в начале лета на
них появляется урединиостадия. Урединии образуются на нижней
стороне листьев и имеют вид оранжевых подушечек диаметром
1 — 3 мм (рис. 82). Урединиоспоры осуществляют 2—3-кратное
заражение смородины в течение лета. В конце лета на листьях
смородины образуется телиостадия. Телиоспоры после короткого
периода покоя прорастают в базидии с базидиоспорами, которые
разлетаются и заражают сосну.
Вначале заражаются молодые побеги, откуда мицелий
проникает в древесину ветвей и стволов. На пораженных ветвях и
стволах возникают утолщения, которые постепенно разрастаются и
превращаются в многолетние, ступенчатые, смолоточащие раны.
Ветви выше мест поражения усыхают. Многократное поражение
279
сосны приводит к
постепенному усыханию кроны,
ослаблению и гибели деревьев.
Наиболее восприимчива к
болезни сосна Веймутова.
Существенный вред этому виду
сосны болезнь причиняет в
питомниках, культурах и в
городских посадках, поблизости от
которых есть смородина.
Кедровая сосна сибирская,
хотя и более устойчива к
пузырчатой ржавчине, но в молодом
возрасте может поражаться
достаточно сильно. В
Байкальском заповеднике ослабление и
гибель от пузырчатой
ржавчины кедровой сосны сибирской
отмечались только у подроста
высотой до 1 м в пойменных
участках кедрово-пихтовых
насаждений, где в состав
подлеска входит красная смородина.
В России болезнь
распространена в ареале кедровой сосны сибирской, а также в регионах,
где в городских насаждениях культивируют Веймутову сосну.
Ржавчинный рак пихты. Возбудитель болезни — ржавчинный
гриб Melampsorella caryophyllacearum G.Schrot. (=M.cerastii Wint.).
Эциостадия развивается на пихте, урединио- и телиостадии — на
растениях семейства гвоздичных: звездчатке (Stellaria), ясколке
{Cerastium), мягковолоснике (Malachium).
Весной базидиоспоры, развивающиеся на листьях указанных
растений, заражают пихту. Вначале поражаются молодые ветви и
побеги, на которых образуются муфтообразные утолщения.
Следующей весной из почек зараженных побегов вырастает
вертикально ведьмина метла с укороченной желто-зеленой хвоей (рис.
83, 7). С середины лета на хвое ведьминых метел образуются эции,
а к осени эта хвоя опадает. В последующие годы на них образуются
новые побеги, на которых развиваются эции. Ведьмины метлы могут
жить в течение 20 лет.
Из пораженных ветвей грибница проникает в ствол, где
вызывает отмирание камбия, в результате чего на стволе образуется
утолщение, покрытое корой с продольными трещинами. С
течением времени кора растрескивается, опадает, обнажая открытую
ступенчатую рану (см. рис. 83, 2). Наросты появляются по всей
длине ствола, нередко по несколько штук. Раны разрастаются мед-
Рис. 82. Ржавчинный рак (пузырчатая
ржавчина) сосны:
1 — пораженный стволик подроста
кедровой сосны с эциями возбудителя; 2 —
лист смородины — промежуточного
хозяина гриба, покрытый урединиопустулами
280
Рис. 83. Ржавчинный рак пихты:
1 — ведьмина метла с опавшей пораженной хвоей; 2 — пораженный ствол с
опухолью, превращающейся в открытую рану
ленно. Скорость их роста составляет в среднем 0,6 см в год по
окружности и от 0,7 до 1,3 см по высоте ствола.
Пораженные раком деревья могут не иметь признаков
ослабления в течение нескольких десятков лет. Состояние пораженных
деревьев зависит от расположения ран на стволе. Образование ран
в кроне вызывает отмирание вершины или большей части кроны.
Наиболее опасны раны, развивающиеся в подкроновой части ствола.
В этом случае при распространении раны более чем на 1/2
окружности ствола деревья усыхают. Трещины в коре утолщений и раны
являются местами проникновения грибов, вызывающих гниль
древесины: чешуйчатки жирной, трутовика Гартига и др. Такие
деревья в сильной степени подвергаются бурелому. Образование
на больных стволах ран и дальнейшее развитие гнили приводят к
значительному снижению выхода деловой древесины.
Поражение ржавчинным раком способствует росту численности
стволовых вредителей в насаждении. Большую роль в отмирании
пихтарников в очагах болезни играет пихтовая смолевка (Pissodes piceae
111.) Она поселяется на живых участках ствола вокруг раковых ран,
увеличивая отмирающую часть ствола и ускоряя гибель деревьев.
Болезнь поражает пихту белую, кавказскую, сибирскую в
насаждениях разного возраста. Наиболее высокий процент пораженно-
сти отмечается во влажных условиях и в тех участках, где в
травяном покрове преобладают промежуточные хозяева возбудителя, в
чистых пихтарниках. Ржавчинный рак пихты является одной из
причин накопления сухостоя и расстройства пихтовых насаждений.
281
Болезнь широко распространена в ареале пихты.
Ступенчатый рак лиственницы. Возбудитель — сумчатый гриб
Lachnellula willkommii (R. Hartig.) Dennis {-Dasyscypha willkommii
Hartig.). Гриб поражает разные виды лиственницы, в более редких
случаях — кедровую сосну сибирскую и пихту.
Заражение осуществляется сумкоспорами, которые
проникают в ткани дерева через сухие ветви или обломы сучьев, где гриб
развивается как сапротроф. Позже мицелий возбудителя попадает
в ствол и живые ветви, поражая кору и камбий. При заражении
лиственниц до 15-летнего возраста чаще поражаются ветви, на
которых вначале образуются небольшие вздутия с обильно
выделяющейся смолой. Позже на месте вздутий развиваются ранки, а
вокруг них возникают апотеции. У деревьев старше 15 лет в
первые годы развития болезни на пораженных ветвях и стволах
образуются овальные засмоленные вмятины, которые, разрастаясь из
года в год, превращаются в открытые ступенчатые раны.
Вследствие усиленного притока питательных веществ в здоровую часть,
окружающую рану, стволы и ветви в местах повреждения
односторонне утолщаются. Поверхность пораженных мест
покрывается вытекающей смолой, которая, окисляясь на воздухе, чернеет.
Рис. 84. Ступенчатый рак лиственницы:
1 — рана с апотециями возбудителя на стволе; 2 — некроз на ветке в месте
обломанного сучка; 3 — срез через пораженную деформированную часть ствола;
4 — срез через апотеции возбудителя; 5 — сумка со спорами
282
На одном стволе нередко образуется несколько ран, вследствие
чего весь ствол становится сильно деформированным (рис. 84).
На коре в местах поражения появляются апотеции
возбудителя, имеющие вид чашечек на ножке, диаметром 2—4 мм.
Снаружи апотеции покрыты белыми густыми волосками, внутренняя
поверхность покрыта оранжевым гимениальным слоем сумок со
спорами. Сумки цилиндрично-булавовидные размером (90 —
180) х (8 —12) мкм. Аскоспоры бесцветные овальные
одноклеточные, реже двухклеточные размером (12—28) х (6—9) мкм.
Созреванию и рассеиванию спор способствует влажная, теплая погода.
Развитие мицелия в тканях дерева происходит в зимний период,
особенно активно во время оттепелей.
Поражается лиственница разного возраста, но наиболее
сильно — саженцы в питомниках и молодняки естественного и
искусственного происхождения до 30 лет. Значительно реже рак
встречается на деревьях более старшего возраста. Болезнь приводит к
снижению выхода посадочного материала в питомниках,
ослаблению и гибели молодых культур. Наиболее восприимчивы к
ступенчатому раку лиственница европейская и сибирская, более
устойчива лиственница японская.
Болезнь зарегистрирована на
северо-западе европейской части России, на Урале, в
Сибири.
Биаторелловый рак сосны. Возбудитель
болезни — сумчатый гриб Biatorella difformis (Fries.)
Rehm. (конидиальная стадия — Biatoridina pinastri
Golow. et Schzedr.). В России болезнь впервые
обнаружена и описана в 1955 г. в Карелии
В. И. Щедровой. Значительно позже она была
зарегистрирована в разных регионах страны:
на Кольском п-ове, на Северном Кавказе, на
Урале, в Сибири, в Центральном районе
европейской части России. Во всех указанных
регионах биаторелловый рак отмечен только на
сосне обыкновенной. Поражается сосна в
возрасте от 10 до 80 лет, но наиболее опасна
болезнь для подроста и культур.
Гриб поражает кору, камбий, древесину. В
начале на стволах и ветвях образуются
засмоленные, вдавленные язвы. По мере развития они
превращаются в глубокие, ступенчатые раны
широкоовальной, почти ромбовидной формы риС 85. Биаторелло-
с сильно засмоленными краями (рис. 85). На вый рак сосны: от-
одном стволике у молодых деревьев образует- крытая ступенчатая
ся от 3 до 10, иногда до 20 ран. Они разраста- рана на стволике
ются очень медленно, некоторые со временем подроста
283
зарастают, поэтому на взрослых деревьях количество ран не
превышает 2—4. Раны возникают по всей длине ствола, но большее
их количество сосредоточено в средней и нижней частях стволов,
иногда у корневой шейки. Преобладающее количество ран
приурочено к северной стороне стволов.
На поверхности ран образуются пикниды, имеющие вид
мелких черных бугорков и черные восковидные апотеции,
расположенные одиночно или группами. Возбудитель проникает в ткани
дерева через трещины в коре, а также через повреждения,
наносимые насекомыми, в том числе побеговьюном-смолевщиком
(Petrova resinella L.), сосновым долгоносиком (Hylobius abietis L.),
смолевками рода Pissodes.
Болезнь приурочена к неблагоприятным лесорастительным
условиям. По данным некоторых исследователей, наиболее высокий
уровень поражения биаторелловым раком отмечается на подросте
сосны под пологом леса. В Мурманской области он составлял 86 %,
а в Байкальском заповеднике — 75 %. По данным И.Г.Семенко-
вой (1987), пораженность биаторелловым раком подроста сосны в
Кабардино-Балкарском заповеднике была одинаково высока как
под пологом леса, так и на открытых местоположениях и
колебалась от 35 до 60 %. При сильном развитии болезнь приводит к
ослаблению и усыханию подроста и культур сосны.
Болезнь широко распространена в европейской части России,
на Урале, в Сибири.
Язвенный рак сосны. Возбудитель — гриб Lachnellula pini (Brunch.)
Dennis. Поражается сосна обыкновенная, кедровая сибирская,
кедровый стланик. Первые признаки болезни обнаруживаются по
слабому засмолению отдельных участков на стволах и ветвях. Позже в
местах поражения образуются небольшие вмятины с
трещиноватой, засмоленной поверхностью. Впоследствии трещины
расширяются, и образуются открытые, засмоленные ступенчатые раны.
Характерным симптомом болезни являются многочисленные
апотеции, выступающие из разрывов эпидермиса коры. Апотеции
возбудителя чашевидные диаметром 1 — 5 мм на короткой ножке,
снаружи покрыты густыми, длинными коричневыми волосками
с ярко-оранжевым гимениальным слоем. Сумки цилиндрические
или булавовидные размером (100—122) х (7,7—20) мкм. Споры
бесцветные, эллипсовидные или яйцевидные, молодые
одноклеточные, зрелые двухклеточные размером (10 —19) х (5—6) мкм. В
сухом состоянии апотеции закрываются и приобретают вид
мелких бурых треугольников, плохо заметных на коре.
Распространение аскоспор происходит во влажную погоду,
когда апотеции открыты. Разнос инфекции осуществляется
посредством дождя и насекомых. Инфекция проникает в ткани
древесины через естественные отверстия в коре и механические
повреждения, в том числе — поранения от насекомых. На одном
284
стволике или ветви часто образуется по нескольку ран,
сливающихся по мере развития, вследствие чего возникает характерная для
раковых болезней деформация пораженных стволиков и ветвей.
Болезнь развивается в разных лесорастительных условиях, но
ее массовое распространение приурочено к открытым
местоположениям. По нашим данным пораженность подроста кедровой
сосны в Байкальском заповеднике колебалась от 11 до 56 % под
пологом леса и достигала 34 —85 % на открытых участках.
Сильное поражение язвенным раком приводит к ослаблению и
усыханию указанных видов сосны. По данным В. И. Крутова,
гибель сосны обыкновенной от болезни в Мурманской области не
превышает 10 %. В то же время данные Т. В. Галасьевой,
полученные при обследовании Байкальского заповедника, свидетельствуют
о более значительном размере усыхания от болезни разных видов
сосны: на подросте кедра в среднем — 38 %, а на кедровом
стланике усыхание достигает 80 %. Помимо непосредственного,
язвенный рак причиняет косвенный вред, способствуя заселению
ослабленных растений стволовыми вредителями, ускоряющими и
часто довершающими процесс усыхания.
Болезнь зарегистрирована в Мурманской области, на Урале, в
Сибири, на Камчатке и Сахалине.
Бугорчатый рак сосны. Возбудитель — бактерия Pseudomonas pini
Vuil. Болезнь поражает разные виды сосны, но чаще встречается
на сосне обыкновенной. Характерным симптомом болезни
является образование на стволах вздутий. Вначале они небольшие,
диаметром до 10 см. Разрастаясь из года в год, опухоли достигают
больших размеров, до 1 м в диаметре, а в центре наростов
образуется пустота. С течением времени опухоли покрываются
глубокими трещинами и засмоляются.
На одном стволе образуется от 1 до 30 опухолей, которые при
разрастании могут окольцевать ствол на большом протяжении. Они
возникают по всей длине ствола, но преимущественно в средней
части. Ветви в зоне расположения опухолей усыхают. Кроны
деревьев с большим количеством раковых наплывов становятся
ажурными и часто приобретают флагообразную форму.
Болезнь редко приводит к усыханию деревьев, но при
большом количестве опухолей на стволе вызывает ослабление сосны.
Поражается сосна с 10 — 20 лет, но наиболее высокий уровень
болезни отмечается в сосняках старше 60-летнего возраста.
Болезнь распространена в европейской части России, на
Урале, в Сибири.
Ступенчатый рак лиственных пород (обыкновенный рак, не-
ктриевый рак). Возбудитель — сумчатый гриб Nectria galligena Bres.
(конидиальная стадия — Cylindrocarpon willkommii (Lind.) Wr.).
Болезнь поражает бук, дуб, клен, липу, яблоню, ясень и другие
породы.
285
Рис. 86. Ступенчатый рак лиственных пород:
1 — открытая ступенчатая рана на стволе; 2 — конидии возбудителя; 3 — сумка
со спорами; 4 — аскоспоры; 5 — раковая рана в развилке ветвей
Гриб поражает кору, луб, камбий и заболонь. Заражение
деревьев осуществляется аскоспорами и конидиями. Споры
распространяются посредством ветра, дождя и насекомых. Инфекция
проникает в ткани дерева через различные повреждения коры, в
местах обрезки сучьев, ожогов коры. Мицелий развивается в тканях
дерева в течение многих лет, вызывая ежегодное отмирание
камбия и образование все новых наплывов каллюса, вследствие чего
на стволах возникают многолетние, ступенчатые раны. Кора в
местах поражения отмирает и опадает, обнажая древесину раны
(рис. 86, 7, 5). Раны образуются с разных сторон ствола, нередко
по нескольку штук, чаще в местах облома сучьев или соединения
ветвей со стволом. При развитии ран на ветвях наплывы каллюса
почти полностью закрывают их, оставляя лишь небольшую щель,
поэтому поражение ветвей обнаруживается по наличию на них
опухолевидных образований. Ежегодно по краю раны возникают
вначале конидиальные, а позже — сумчатые стромы. Конидиальные
стромы имеют вид желтоватых подушечек, на которых
развиваются конидиеносцы с конидиями. Конидии бесцветные
цилиндрические, прямые или слегка изогнутые, с 2 —5 перегородками,
размером (10—65) х (4—7) мкм. Осенью стромы становятся кир-
пично-красными, и в них формируются перитеции. Сумки
булавовидные размером (90—110) х( 12 —15) мкм. Сумкоспоры бес-
286
цветные овальные двухклеточные размером (14—22) х (6 — 9) мкм
(см. рис. 86, 2, J, 4).
Температура не играет существенной роли в развитии болезни.
Созревание и распространение спор возбудителя может
происходить при температуре от 2 до 30 °С. Но главным фактором,
определяющим сроки наиболее интенсивного рассеивания конидий и
аскоспор, является влажность. Этим объясняется максимум спо-
руляции возбудителя в наиболее влажные весенние и осенние
месяцы.
Поражаются естественные насаждения I — IV классов возраста
и разные типы городских насаждений. Ступенчатый рак приводит
к ослаблению деревьев, а при сильном поражении может вызвать
их усыхание. Кроме того, через раны проникают возбудители
гнилей, которые ухудшают состояние деревьев и еще больше
снижают их устойчивость к ветру. Схожее по признакам заболевание на
буке и клене вызывает сумчатый гриб N. ditissima Tul.
Болезнь распространена в европейской части России, на
Урале, в Сибири, на Дальнем Востоке.
Эндоксилиновый рак ясеня. Возбудитель — сумчатый гриб
Endoxylina stellulata Rom. Поражается ясень обыкновенный. Гриб
поражает кору, луб, камбий и древесину. Заражение происходит через
отмершие сучья, преимущественно в нижней части ствола.
Многолетний мицелий распространяется в тканях дерева из года в год,
в результате чего образуются ступенчатые раны (рис. 87, 7). Они
возникают по 1 — 2, реже больше, часто с разных сторон ствола.
Рис. 87. Эндоксилиновый рак ясеня:
/ — открытая ступенчатая рана с темной древесиной и поперечными
трещинами; 2 — срез через строму с перитециями; 3 — сумка со спорами; 4 — аскоспоры
287
Разрастание ран может происходить в течение 7—10 лет и
достигать в длину 0,7 м, и охватывать 2/3 окружности ствола. Под
отмершей корой образуется темная, черная строма. Позже пораженная
кора и частично строма опадают, обнажая темную с поперечными
трещинами древесину. Поверхность раны покрыта сплошным слоем
мелких черных точечных бугорков, представляющих собой
устьица погруженных в древесину перитециев (см. рис. 87, 2). Перитеции
развиваются на второй год после отмирания заболони. Сумки
булавовидные, с длинными ножками, размером (135—150) х (10—12)
мкм. Аскоспоры цилиндрические, с закругленными концами,
согнутые двухклеточные оливково-бурые размером (14—20) х (4—6)
мкм (см. рис. 87, 3, 4). Болезнь сопровождается образованием ядро-
во-заболонной гнили. При этом гнилая древесина приобретает
желтоватый мраморный цвет. Гниль может распространяться выше и
ниже раковой раны. Вследствие образования ран
преимущественно в комлевой части, с разных сторон ствола, болезнь приводит к
сравнительно быстрому усыханию деревьев. В зависимости от
количества и расположения ран на стволе гибель деревьев может
наступить через 5—12 лет. Пораженные эндоксилиновым раком
насаждения ясеня, особенно чистые, нередко страдают от
бурелома, что объясняется снижением устойчивости к ветру у
больных деревьев, имеющих многолетние раны.
Поражаются ослабленные деревья в приспевающих и спелых,
особенно в изреженных насаждениях И —III бонитетов на сухих
склонах, участках с солонцовыми пятнами. Наиболее сильно
поражаются порослевые насаждения ясеня, где количество больных
деревьев может достигать 40—60 %.
Болезнь распространена в южных регионах России.
Черный рак осины и тополя. Возбудителем болезни является
сумчатый гриб Hypoxylon pruinatum (Kl.) Cooke (= H. mammatum
(Wahlenberg) H.Miller). Гриб поражает кору, камбий и древесину
стволов и ветвей. Первые признаки проявляются в образовании на
коре неправильной формы вдавленных участков буроватого
цвета, нерезко отграниченных от здоровой коры. Позже в местах
поражения возникают мокнущие вздутия, кора покрывается
мелкими трещинами, из которых при надавливании вытекает
белесоватая жидкость. Постепенно пораженные участки превращаются в
раковые раны. В коре и заболони развивается черная мажущая
строма толщиной в несколько миллиметров. На поверхности стромы
вначале образуется конидиальное спороношение, имеющее вид
темно-серых, почти черных зубовидных выростов высотой до 1 мм.
Они располагаются в виде частой щетки по краям раны и
приподнимают кору, вызывая ее растрескивание и опадение. Конидии
гриба одноклеточные бесцветные размером (5,5—8) х (1,5—4) мкм.
На третий год после заражения и опадения зубовидных выростов
конидиального спороношения на той же строме образуется сумча-
288
тая стадия гриба. Группы перитециев имеют вид серовато-черных
образований многоугольной формы диаметром до 1 см,
расположенных вдоль раны. В перитециях поздней осенью и ранней весной
созревают удлиненно-эллиптические темно-коричневые
одноклеточные аскоспоры размером (20—33) х (9 —12) мкм (рис. 88, 2—5).
Заражение осуществляется конидиями и аскоспорами,
проникающими в ткани дерева через старые поранения коры, сучья.
Споры распространяются посредством ветра, дождя и насекомых.
Переносчиками инфекции могут быть большой осиновый усач
{Saperda carcharias L.), зеленая узкотелая златка (Argilus viridis L.),
осиновый древоточец (Cossus terebra F.), большая тополевая
стеклянница (Aegeria apiformis CL).
Мицелий распространяется в тканях дерева очень быстро,
поэтому наплывы вокруг пораженной части не успевают образоваться,
в результате раны не имеют ясно выраженной ступенчатости (см.
рис. 88, 7). Скорость распространения мицелия вдоль ствола
значительно больше, чем по окружности, вследствие чего раны
достигают длины 1,5 — 2 м. Они образуются в средней, реже в нижней
части ствола. Болезнь сопровождается развитием в стволе белой
смешанной гнили. Гибель дерева происходит постепенно, за
несколько лет. Разные виды и формы рода Populus обладают неоди-
Рис. 88. Черный рак осины и тополя:
/ — вытянутая рана с черной стромой гриба; 2 — участок пораженной коры с
группами перитециев; 3— конидии; 4— конидиенисцы с конидиями; 5— сумка
со спорами; 6 — аскоспоры
289
наковой устойчивостью к болезни, что зависит от влажности,
физических свойств коры и химического состава клеточного сока.
Наиболее сильно поражаются черным раком осина и тополь
белый. Значительно реже болезнь встречается на черных и
бальзамических тополях. Поражаются деревья I —IV классов возраста в
естественных насаждениях, культурах и в городских посадках.
Для естественных насаждений осины черный рак является
одним из самых опасных заболеваний, он вызывает усыхание
осинников, снижает выход деловой древесины, способствует
массовому бурелому. Черный рак осины встречается в разных типах
условий местопроизрастания. Наиболее высокий уровень болезни
отмечается на открытых, хорошо освещенных местах. В городских и
парковых насаждениях больные деревья теряют декоративность и
защитные функции.
Болезнь широко распространена в ареале осины.
Крифонектриевый (эндотиевый) рак каштана съедобного.
Возбудитель — сумчатый гриб Cryphonectria parasitica (Murril.) Barr. (=Endothia
parasitica (Murr.) P. And. et H. And.). Впервые эпифитотии
крифонектриевого рака были зарегистрированы в Японии и Китае. Оттуда
в 1900 г. возбудитель вместе с древесиной был завезен в Северную
Америку. За сравнительно короткий период гриб вызвал эпифи-
тотию в насаждениях каштана зубчатого (Castanea dentata),
который в ряде штатов являлся основной лесообразующей породой.
Из Северной Америки возбудитель проник в Западную Европу.
Здесь болезнь распространялась медленнее, чем в США,
поскольку произрастающий в Европе каштан посевной (C.sativa)
оказался более устойчивым, а использование его только как садового
дерева исключало концентрацию этой породы на больших
площадях. Тем не менее, с 1930 г. в отдельных странах Европы
началась эпифитотия крифонектриевого рака, а к 1950 г. она охватила
все страны, где произрастает каштан посевной. В России болезнь
впервые была зарегистрирована в 80-х гг. XIX столетия на каштане
съедобном, а массовое его поражение было отмечено в 1915 —
1920 гг. в районе г. Сочи.
Кроме каштана съедобного болезнь поражает разные виды
дуба, бук, граб. Гриб развивается в коре и камбии стволов,
ветвей, корней. На стволах и ветвях с тонкой гладкой корой
пораженные участки вначале слегка темнеют, а затем приобретают
красновато-бурую окраску, резко выделяясь на оливково-зеленом
фоне здоровой коры. В толще пораженной коры образуются кони-
диальные стромы, имеющие вид оранжевых бугорков,
выступающих из трещин коры. Во влажную погоду из пикнид,
развивающихся в строме, выходит масса конидий, застывающих в виде
желтых, впоследствии красновато-бурых усиков. Конидии
бесцветные одноклеточные овально-цилиндрические, прямые или
слегка согнутые размером 3,6 х 1,3 мкм. Позже, когда кора отмирает,
290
строма увеличивается в несколько раз, становится плоской, и в
ней образуются перитеции. Цвет стромы становится оранжевым
или красно-бурым. На фоне такой окраски хорошо заметны
черные точки устьиц перитециев. Из перитециев выходят зрелые
сумки с сумкоспорами. Сумки гриба удлиненно-эллипсоидальные
размером (20—38) х (6 — 8) мкм; сумкоспоры бесцветные
эллипсоидальные двухклеточные с перетяжкой у перегородки
размером (5 — 7) х (3—4) мкм (рис. 89, 2—6).
На стволах с толстой корой признаки болезни
обнаруживаются только по подушечкам стром, выступающим из трещин в коре.
Пораженная кора вначале как бы присыхает к заболони, но затем
отстает, и на ее внутренней поверхности видны веерообразные
пленки мицелия от белого до оранжевого цвета. Позже на стволах
и ветвях образуются раны различных типов: закрытые, открытые,
в виде опухолевидных вздутий с трещинами. Раны достигают
нескольких метров в длину (см. рис. 89, 7). Одновременно с ранами
на стволах появляется масса водяных побегов. Больные деревья
отличаются ажурностью кроны, наличием усохших ветвей. К
концу лета листья засыхают и буреют или засыхают, сохраняя
зеленую окраску. Бурая листва резко выделяется на фоне зеленой
кроны, и такие деревья хорошо видны на значительном расстоянии.
Рис. 89. Крифонектриевый (эндотиевый) рак каштана съедобного:
/ — раковая рана на стволе; 2 — срез через строму с перитециями; 3 — сумки со
спорами; 4 — аскоспоры; 5 — срез через строму с пикнидой; 6 — конидии
291
Засохшая листва опадает только на следующий год. В древесине
стволов и корней пораженных деревьев происходит закупорка
проводящих сосудов тиллами. Такие сосуды хорошо заметны на
поперечных срезах в виде темно-бурых точек. Заражение деревьев
осуществляется конидиями и реже — сумкоспорами. Конидии
распространяются посредством дождя, насекомыми, птицами, сум-
коспоры переносятся по воздуху. Инфекция проникает в ткани
дерева через различные поранения коры, ожоговые и морозобой-
ные трещины.
После гибели деревьев гриб в течение продолжительного
времени продолжает развиваться как сапротроф. Источниками
инфекции являются больные деревья, пни, кора, порубочные остатки
и лесоматериалы, заготовленные из зараженных деревьев.
Каштан разного возраста поражается независимо от условий
местопроизрастания. При проникновении инфекции в корни
заболевает и поросль. Наиболее высокий уровень поражения
отмечается у опушек, в сильно разреженных древостоях, чистых
насаждениях каштана и в смешанных с большой долей его участия.
Развитию болезни способствует мягкий, влажный климат. Болезнь
приводит к ослаблению и усыханию насаждений каштана
съедобного. В зависимости от количества ран, их расположения, возраста
деревьев, условий окружающей среды гибель деревьев
происходит в возрасте от года—двух до 10 лет.
В России болезнь распространена на Черноморском побережье.
Инфекционное усыхание липы и вяза (стигминиоз, тиростромоз,
стеганоспориоз). Возбудителем болезни является несовершенный
гриб Stigmina compacta (Sacc.) M.B. Ellis (= Thyrostroma compactum Sacc,
Steganosporium compactum Sacc). Гриб поражает липу и вяз.
Первые сведения об инфекционном усыхании липы на
территории бывшего СССР появились в Литве в 1974 г. В 1976 г.
заболевание было обнаружено в городских насаждениях Московской
области. А в период с 1985 по 1992 гг. болезнь зарегистрировали в
естественных насаждениях липы в Самарской и Тульской
областях и в зеленых городских насаждениях Москвы и Московской
области. В 1999 г. единичные случаи инфекционного усыхания липы
были отмечены в насаждениях Санкт-Петербурга.
Заражение липы осуществляется конидиями, которые
распространяются ветром. Инфицирование ветвей может происходить как
в период покоя дерева, так и во время вегетации, во второй
половине лета. Возбудитель проникает в ткани дерева в основном через
почки, реже — через другие естественные ворота: чечевички,
места соединения побегов. После заражения активное развитие
патогена в тканях дерева происходит в фазе покоя, т.е. с начала
прекращения вегетации и до распускания листьев. Это
объясняется тем, что оптимальные условия для интенсивного
вегетативного роста гриба создаются при низких температурах, минимум ко-
292
торых лежит в пределах -2 °С. От заражения до появления
характерных симптомов болезни проходит больше года, чаще два.
Первые признаки поражения обнаруживаются на тонких
веточках — приростах последнего года. При позднелетнем заражении
следующей весной почки на этих ветвях не распускаются. В конце
лета или следующей весной на них образуется спороношение
гриба (рис. 90, 2). Из тонких пораженных ветвей инфекция проникает
в более крупные элементы кроны — приросты предыдущего года,
имеющие также тонкую гладкую кору. Заражение новых ветвей
может осуществляться и путем проникновения возбудителя из
более толстых ветвей в тонкие.
На ветвях и стволах с тонкой гладкой корой появляются
некротические, слегка вдавленные участки, выделяющиеся на фоне
здоровой коры более темной окраской. Нередко пораженный
участок отграничивается от здоровой части хорошо заметной темной
каймой. Впоследствии на ее месте образуется валик каллюса, а
еще позже — трещина. В некоторых случаях в этот период
пораженная кора светлеет, приобретая желтоватую или сероватую
окраску. Некротический участок коры покрывается многочисленными
темно-бурыми, почти черными бархатистыми подушечками ко-
нидиальных стром, выступающих из разрывов эпидермиса коры.
На поверхности стром образуется спороносящий слой. Конидии
Рис. 90. Инфекционное усыхание липы:
/ —- пораженная ветка с пучком укороченных побегов; 2 — часть пораженного
побега со спороношением возбудителя; 3 — конидии; 4 — открытая рана на
ветке
293
продолговатые, булавовидные, с 3—6 поперечными и одной
продольной перегородками, с перетяжками у перегородок, бурые или
оливковые размером (18,5 — 24,5) х (50—58) мкм (см. рис. 90, 3).
На стволах и ветвях с более толстой корой сначала образуются
некротические участки, не так резко выделяющиеся, как на
тонкой коре, а затем закрытые раны. Кора, покрывающая раны, по
мере их развития натягивается по центру раны в виде ремешка,
поверхность которого покрыта спороношениями гриба.
Впоследствии кора опадает, обнажая древесину раны. Открытые раны
продолговатые, часто веретенообразной формы, неступенчатые. В
большинстве случаев на ветвях и стволах образуется несколько ран,
чаще в местах соединения ветвей со стволом или побегов с
ветвями (см. рис. 90, 4).
При отмирании приростов последних лет, образующих
основную массу листвы, дерево компенсирует утрату фотосинтезирую-
щей поверхности образованием побегов и листьев из спящих
почек. У пеньков обломившихся пораженных ветвей, на скелетных
ветвях и стволе образуются пучки водяных побегов с крупными
листьями. Появившаяся «вторичная» крона тоже постепенно
отмирает, что сопровождается отрастанием новых пучков водяных
побегов (см. рис. 90, 7). За короткий период времени дерево
приобретает характерный, растрепанный внешний вид. Постепенно
усыхают более крупные ветви. Через несколько лет кроны и
стволы старых деревьев деформируются, в городских насаждениях они
полностью утрачивают декоративность. Молодые деревья за это
время усыхают.
Поражается липа всех возрастов в разных типах леса и разных
уровнях вертикальной структуры древостоев. Встречаемость
болезни в разных экологических условиях достигает 92—100 %. Уровень
встречаемости болезни повышается с увеличением доли участия
липы в составе насаждения и его полноты. В естественных
насаждениях стигминиоз приводит к постепенному ослаблению липы и
является главной причиной гибели подроста.
В насаждениях Москвы встречаемость стигминиоза составляет
74—90 %. В условиях города болезнь поражает липу разного
возраста в насаждениях, созданных разными способами на главных
улицах и магистралях, в скверах, парках, на бульварах и во дворах.
Вместе с тем было установлено, что загрязнение воздуха в
городских условиях снижает встречаемость болезни и интенсивность
спорообразования (Кузьмичев). Поражаются разные виды липы.
Массовое поражение стигминиозом вяза впервые было
обнаружено в 1958 г. в полезащитных лесных полосах Воронежской
области. Позже массовое усыхание вяза мелколистного (Ulmus
pumila) отмечалось в полезащитных полосах Ростовской области в
1971 г. В 1994 г. болезнь была зарегистрирована в естественных
насаждениях НПП «Самарская лука». С 1992 г. поражение вяза мел-
294
колистного отмечалось в городских посадках Москвы и
Подмосковья. Симптомы болезни на вязе сходны с таковыми на липе.
Поражаются разные виды ильмовых пород, но наиболее
восприимчив к болезни вяз мелколистный (Ulmus pumila). Эпифитотия
болезни в полезащитных лесных полосах приводит к тому, что
они перестают выполнять важную природоохранную роль, что
влечет за собой снижение урожайности ценных
сельскохозяйственных культур. Массовое усыхание вяза в городских насаждениях
снижает декоративность посадок, которые при этом утрачивают
частично или полностью и свои защитные функции.
Болезнь распространена в европейской части России.
Мокрый язвенно-сосудистый рак тополя (бурое слизетечение,
раково-язвенное заболевание). Возбудители болезни — бактерии
Pseudomonas cerasi Griffin и P.syringae Van Hall, f.populi Sabet et
Dowson. Это одна из наиболее распространенных и вредоносных
болезней тополя в странах Европы. Изучение его было начато в
30-х гг. XX столетия. Дальнейшее распространение болезни и
увеличение масштабов введения тополя в лесные культуры
обусловили неослабевающий интерес исследователей многих
европейских стран к заболеванию. Мнения ученых о причинах массового
усыхания тополя разделились. Одни считали, что причиной
заболевания являются вредные насекомые, другие называли
различные абиотические факторы (резкая смена дневных и ночных
температур в весенний период, засухи, подтопления, длительные
затопления, высокая плотность посадок и т.д.). В то же время
многие исследователи утверждали, что усыхание тополя вызвано
бактериями. Бактериальная природа болезни была подтверждена
экспериментально. Однако вопрос о виде возбудителя до сих пор не
решен. Из пораженных тканей тополя исследователями были
выделены разные бактерии, в числе которых доминировал вид
Pseudomonas rimaefaciens Коп. Однако Р. И. Гвоздяк и Л. М.
Яковлева установили, что возбудителями бактериального рака тополя
являются два вида P. cerasi, P.syringae f.populi, из которых главная
роль принадлежит первому.
Бактерии поражают кору, луб, камбий и заболонь. Признаки
болезни появляются в конце апреля — начале мая. На стволах и
ветвях с тонкой гладкой корой образуются округлые или
овальные вздутия до 1 — 2 см в диаметре. При надавливании из них
вытекает прозрачная жидкость, которая под воздействием
бактерий приобретает бурый цвет. На стволах с трещиноватой корой
таких вздутий не образуется, и первые признаки болезни
обнаруживаются по наличию мокнущих пятен и подтеков на коре.
Несколько позже на месте вздутий появляется продольная трещина.
Пораженные участки постепенно разрастаются и за несколько
месяцев достигают 3 — 3,5 см в диаметре, приобретая вид
типичной раны. По краям ран образуются наплывы древесины толщи-
295
ной до 2—3 мм. По всей длине ствола за год может возникнуть до
10—25 ран. Разрастаясь, они сливаются в одну большую рану
длиной до 1 м, нередко полностью окольцовывающую ствол. Чаще
всего раны образуются на наиболее освещенных сторонах стволов.
На следующий год на пораженных стволах возникают новые раны,
которые к концу вегетационного периода тоже сливаются. В таком
случае пораженные деревья сильно ослабляются и за 3—4 г. усыхают.
Весной и осенью на них видны бурые потеки, часто
появляющиеся в местах прикрепления сучьев. Больные стволы сильно
деформируются из-за утолщений, образующихся с разных сторон ствола.
Инфекция распространяется дождевой водой, насекомыми,
человеком при обрезке сучьев.
Болезнь встречается в школьных отделениях питомников, в
культурах и городских насаждениях. Пораженность тополя
мокрым раком в разных экологических объектах колеблется от 5 до
90 % и выше. Развитию болезни способствуют различные
факторы, вызывающие ослабление тополя (засуха, затопление,
морозы, промышленные выбросы, высокая рекреационная нагрузка,
нарушение технологии посадки и выращивания тополя).
Разные виды и гибриды тополя обладают неодинаковой
устойчивостью к болезни. Сильнее поражаются бальзамические и черные
тополя и их гибриды. Наиболее устойчивы к мокрому раку белые
тополя. Мокрый язвенно-сосудистый рак вызывает ослабление и
усыхание тополя. В городских насаждениях сильное поражение
приводит к потере декоративности этой породы и ослаблению ее
защитных функций. Кроме того, деревья тополя
с большим количеством ран на стволах теряют
устойчивость к ветру и подвергаются
бурелому, что в городских условиях особенно опасно.
Широкое распространение болезни
отмечено в центральных районах европейской части
России и на Алтае.
Сходное заболевание тополя и осины
вызывает бактерия Pseudomonas rimaefaciens Kon.
Поперечный рак дуба. Возбудитель —
бактерия Pseudomonas quercina Schem. Поражаются
разные виды дуба, но наиболее восприимчив
к болезни дуб черешчатый.
Болезнь проявляется в образовании
опухолей, расположенных поперек стволов и ветвей.
Вначале они имеют вид небольших, гладких
наплывов с одной стороны ствола (ветви) или
муфт, охватывающих весь ствол (ветвь) (рис. 91).
По мере роста деревьев опухоли увеличивают-
Рис. 91. Поперечный ся, на них появляются глубокие поперечные
рак дуба трещины, а позже образуются раны с неровны-
296
ми, отогнутыми краями. На одном дереве может возникать от 1 —
2 до 10 опухолей. Наибольшее их количество отмечается на
молодых дубках. По данным Р.И.Гвоздяка и Л.М.Яковлевой, новые
наросты образуются у молодняка дуба, чаще у 2—5-летних
растений. У деревьев старше 20 лет новые опухоли не появляются.
Переносчиком болезни считается пестрая дубовая тля {Lachnus roboris
L.), однако экспериментально это не доказано. В местах
образования опухолей стволы сильно деформируются, а выше и ниже
прирост древесины падает. Болезнь поражает дуб разного возраста в
различных лесорастительных условиях, но наиболее активное ее
развитие отмечается на бедных, сухих почвах. В чистых дубовых
насаждениях уровень болезни выше, чем в смешанных. Особенно
сильно страдают насаждения порослевого происхождения.
Именно в связи с систематическим порослевым возобновлением дуба
значительно возросла в последние десятилетия его пораженность
поперечным раком, достигающая в отдельных случаях 60—80 %.
В дубравах семенного происхождения зараженность редко
превышает 5 %. Молодые культуры при сильном поражении
ослабляются, отстают в росте, а иногда усыхают. Широкое распространение
болезни в насаждениях старших возрастных категорий снижает
выход и качество деловой древесины. Кроме того, через трещины в
опухолях проникают споры дереворазрушающих грибов {Phellinus
robustus, Inonotus dryophilus и др.), гниль от которых еще больше
снижает выход деловой древесины и устойчивость деревьев к
бурелому. Поперечный рак встречается в ареале дуба повсеместно,
особенно в лесостепной и степной зонах России.
Бактериальный рак ясеня вызывается бактерией Pseudomonas
syringae van Hall. (= Д/Уго/ш" Wuill). Поражается ясень
обыкновенный. На стволах и ветвях вначале появляются овальные или
округло-овальные гладкие опухоли. Позже в центре опухоли образуется
узкая, неглубокая, прямая или изогнутая трещина. По мере
развития болезни трещины расширяются, удлиняются, появляются
новые и постепенно образуются характерные раковые раны. Кора
в местах поражения растрескивается, шелушится, отмирает и
опадает, обнажая древесину раны. Раны возникают в разных частях
ствола. По данным Р.И.Гвоздяка и Л.М.Яковлевой, их
количество на одном дереве достигает несколько десятков и сотен. При
сильном поражении многочисленные раны сливаются по длине
ствола, что приводит к образованию сухобочин. По данным
Ф. Е. Маленева, это заболевание может поражать и соцветия
ясеня, каждый цветок превращается в слегка приплюснутую опухоль
с неровной поверхностью размером 0,5 — 0,9 см.
Заражение дерева происходит через различные повреждения
коры, надломы побегов, листья и почки. Переносчиками
инфекции могут быть малый ясеневый лубоед {Leperisinus fraxini Panzer)
и ясеневая моль {Praus curtisellus Don.).
297
Болезнь поражает ясень всех возрастов, но особенно опасна
для молодняков. Она встречается в разных лесорастительных
условиях. Наиболее интенсивные поражения ясеня наблюдаются в
неблагоприятных условиях роста, на бедных, плохо аэрируемых
почвах, в загущенных куртинах, в насаждениях с большой долей
его участия.
При сильном развитии болезни части стволов и ветвей выше ран
усыхают, реже усыхает все дерево. Поражение бактериальным
раком приводит к снижению сортности и выхода деловой древесины.
Кроме того, через раны в опухолях легко проникают споры
возбудителей гнилей {Phellinus igniarius, P.punctatus, Oxyporus
populinus, Polypoms squamosus, Ganoderma applanatum), которые в
значительной степени снижают устойчивость ясеня к бурелому.
Болезнь распространена в лесостепной и степной зонах
европейской части России.
11.4. СИСТЕМА МЕРОПРИЯТИЙ ПО ЗАЩИТЕ
ДРЕВЕСНЫХ ПОРОД ОТ СОСУДИСТЫХ
И НЕКРОЗНО-РАКОВЫХ БОЛЕЗНЕЙ
Борьба с сосудистыми и некрозно-раковыми болезнями
включает надзор за их появлением и распространением, лесохозяй-
ственные и химические мероприятия.
Надзор организуют за главнейшими болезнями, причиняющими
существенный вред лесному хозяйству. Рекогносцировочный
надзор проводят путем обследования насаждений по ходовым
линиям, в качестве которых используют визиры, просеки, лесные
дороги и тропы. Ходовые линии прокладывают с таким расчетом,
чтобы были сделаны заходы во все выделы с преобладанием хвойных
и твердолиственных пород (дуба, ясеня, ильмовых, клена и др.)
независимо от площади выдела. Выделы с преобладанием мягко-
лиственных пород обследуют только в случае, если их площадь
составляет не менее 5 га. Обязательному осмотру подлежат также
все неблагополучные по состоянию участки леса. На все выделы,
намеченные к обследованию, делают выборку из таксационного
описания. Ходовые линии и выявленные очаги болезней или
неблагополучные в санитарном отношении участки наносят на план
насаждений.
Рекогносцировочное обследование включает глазомерную
оценку санитарного и лесопатологического состояния участков леса.
При оценке санитарного состояния учитывают сухостой,
захламленность, ветровал, бурелом, снеголом, невывезенную
древесину с примерным указанием занимаемой ими площади (га) и их
массы. Запас сухостоя дается в м3/га или в процентах от общего
числа деревьев. Указывают время образования сухостоя (свежий,
298
старый) и его состояние (пораженный болезнями, заселенный
или не заселенный стволовыми вредителями).
Лесопатологическое состояние насаждений оценивают по трем
классам биологической устойчивости:
I класс (устойчивые) — размер усыхания до 5%, величина
текущего отпада (усыхающие деревья и свежий сухостой)
составляет не более 2 %. Поражение болезнями отсутствует или
единичное, состояние лесной среды не нарушено;
II класс (с нарушенной устойчивостью) — размер усыхания
6—40 %, величина текущего отпада может в два раза и более
превышать размер естественного для данного возраста и типа лесорасти-
тельных условий, болезни могут иметь массовое распространение,
лесная среда часто нарушена, полнота неравномерная;
III класс (утратившие устойчивость) — размер усыхания более
40 %, отпад намного превышает естественный, болезни могут иметь
массовое распространение, лесная среда нарушена, полнота
неравномерная или насаждения полностью расстроены.
При рекогносцировочном обследовании отмечают характер
распределения сухостоя и пораженных болезнями деревьев. При этом
выделяют следующие категории распределения сухостоя:
единичный — пораженные болезнями и сухостойные деревья встречаются
единично; групповой — небольшими группами (до 10 деревьев);
куртинный — наблюдается поражение или усыхание деревьев
куртинами разной величины, до 0,25 га; сплошной — усыхание
деревьев или пораженность их болезнями наблюдается сплошь на
участках площадью более 0,25 га.
По возможности устанавливают первопричины усыхания или
ослабления деревьев: поражение болезнями, повреждение
вредителями, нарушение санитарных правил, пастьба скота, действие
метеорологических факторов и т.д.
Общая оценка пораженности насаждений болезнями дается
по проценту пораженных деревьев: слабая — до 10 %, средняя —
10 —25 %, сильная — свыше 25 %.
Детальный надзор проводят на постоянных или временных
пробных площадях. Наиболее удобными являются безразмерные
пробные площади вытянутой формы. При слабой степени
зараженности древостоя в перечет включают не менее 200 деревьев,
при средней — 100 деревьев, при сильной — от 50 до 20 деревьев.
На пробных площадях определяют состояние насаждений,
особенности поражения болезнями, их распространение и
стадию развития.
Определение состояния насаждений проводят путем перечета
деревьев по породам, категориям состояния, ступеням толщины
с выделением заселенных и отработанных стволовыми
вредителями. В табл. 6 приведена характеристика основных категорий
состояния деревьев хвойных и лиственных пород.
299
Таблица 6
Шкала категорий состояния деревьев
Категория деревьев
Основные признаки
Хвойные породы
1 — без признаков
ослабления
2 — ослабленные
3 — сильно
ослабленные
4 — усыхающие
5 — сухостой
текущего года
6 — сухостой
прошлых лет
Хвоя зеленая блестящая, крона густая, прирост
текущего года нормальный для данной породы,
возраста, условий местопроизрастания и сезона
Хвоя часто светлее обычного, крона
слабоажурная, прирост уменьшен не более чем наполовину
по сравнению с нормальным
Хвоя светло-зеленая или сероватая матовая,
крона ажурная, прирост уменьшен более чем
наполовину по сравнению с нормальным
Хвоя серая, желтоватая или желто-зеленая, крона
заметно изрежена, прирост текущего года еще
заметен или отсутствует
Хвоя серая, желтая или бурая, крона часто
изрежена, мелкие веточки сохраняются, кора
сохранена или осыпалась лишь частично
Хвоя осыпалась или сохранилась лишь частично,
мелкие веточки, как правило, обломились,
большая часть ветвей и коры осыпалась
Лиственные породы
1 — без признаков
ослабления
2 — ослабленные
(в кроне до 25 %
сухих ветвей)
3 — сильно
ослабленные (сухих
ветвей 50-75 %)
4 — усыхающие су-
хокронные (в
кроне более 75 % сухих
ветвей)
Листва зеленая, блестящая, крона густая,
прирост текущего года нормальный для данной
породы, возраста, условий местопроизрастания и
сезона
Листва зеленая, крона слабоажурная, прирост
может быть ослаблен по сравнению с
нормальным
Листва мельче или светлее обычной,
преждевременно опадает, крона изрежена
Листва мельче, светлее или желтее обычной,
преждевременно опадает или увядает, крона сильно
изрежена
300
Окончание табл. 6
Категория деревьев
5 — сухостой
текущего года
6 — сухостой
прошлых лет (старый)
Основные признаки
Листва усохла, увяла или преждевременно
опала, мелкие веточки и кора сохранились
Листва и часть ветвей опали, кора разрушена или
опала на большей части ствола
Ветровал, бурелом и снеголом учитываются отдельно с
указанием времени их образования.
Учеты на пробных площадях дополняют данными анализа
модельных деревьев. Из каждой категории состояния берут по три
модельных дерева. При анализе деревьев, пораженных
сосудистыми болезнями, исследуют ветви, ствол и корни. Для обнаружения
признаков болезни делают поперечные срезы через ствол и
корни, отмечают наличие потемневших сосудов в годичных кольцах
и характер потемнения (в виде сплошного или прерывистого
кольца, в виде отдельных темных точек). При анализе модельных
деревьев, пораженных раковыми болезнями, отмечают характер
новообразований (ступенчатая рана или без ясно выраженных
ступеней, язва, опухоль), расположение их на стволе (под кроной, в
нижней, средней и верхней частях кроны), степень охвата ствола
по окружности (на У3, У2, 3/4, кольцевая), экспозицию на стволе
(южная, северная сторона и т.д.).
Сроки проведения надзора за болезнями определяются их
биоэкологическими особенностями. Надзор за появлением и
распространением сосудистых болезней лучше проводить с середины июня
по август, когда четко проявляются внешние симптомы болезней
этой группы (характерное усыхание отдельных побегов и ветвей в
кроне или целиком кроны) и прослеживается динамика усыха-
ния кроны деревьев.
Надзор за большинством видов некрозно-раковых болезней
осуществляется в летний период, когда хорошо выражены их
основные симптомы, в том числе характерные для возбудителей
грибных болезней спороношения. К этим объектам надзора относятся:
нектриевый некроз лиственных пород, некрозы дуба, ясеня;
ступенчатый рак лиственных пород, эндоксилиновый рак ясеня,
черный рак тополя, смоляной рак и пузырчатая ржавчина сосны,
ржавчинный рак пихты, ступенчатый рак лиственницы и др.
За отдельными видами некрозно-раковых болезней (цитоспо-
роз и дискоспориевый (дотихициевый) некроз тополя,
инфекционное усыхание (стигминиоз) липы и вяза, мокрый
язвенно-сосудистый рак тополя) надзор рекомендуется проводить сразу после
распускания листьев, когда резко выделяются деревья с нераспу-
301
стившейся полностью кроной или с отдельными побегами с
нераспустившимися листьями, особенно хорошо заметны спороно-
шения возбудителей, появляются бурые потеки экссудата на
стволах и ветвях.
На основании данных надзора с учетом санитарного состояния
насаждений планируют мероприятия по борьбе с болезнями и
оптимальные сроки их проведения.
Лесохозяйственные меры борьбы включают мероприятия,
направленные на предупреждение очагов болезней, ограничение их
распространения и причиняемого ими вреда. Все проводимые
лесохозяйственные мероприятия должны обеспечить биологическую
устойчивость насаждений.
В существующих насаждениях необходимо своевременно
проводить санитарные рубки, при которых выбирают сильно
ослабленные, усыхающие и свежезаселенные деревья. После этого
следует тщательно собрать и сжечь все порубочные остатки, которые
могут быть источниками инфекции. Заготовленная древесина
должна быть своевременно вывезена из леса. Рубки рекомендуется
проводить в осенне-зимний период, когда нет споруляции
возбудителей. В противном случае делают антисептирование больных
пней. При проведении рубок необходимо поддерживать
оптимальную для конкретных условий и насаждений полноту; очень важно
строго соблюдать возраст рубок, так как перестойные деревья
часто становятся источниками инфекции. В зависимости от характера
очага и степени пораженности древостоя назначают выборочные
или сплошные санитарные рубки. Если пораженность невелика
(до 10 %) и больные деревья располагаются группами или единично
и их удаление не повлечет за собой расстройства насаждений,
назначают выборочные санитарные рубки. В том случае, когда
пораженность древостоя превышает 25 % или больные деревья
расположены рассеянно, проводят сплошную санитарную рубку.
Одновременно с санитарными мероприятиями следует
проводить реконструкцию насаждений, вводя в их состав более
устойчивые к тем или другим болезням породы или изменяя схемы
смешений. При создании насаждений следует использовать
породы, приспособленные к местным условиям. С целью
предупреждения болезней и ограничения их дальнейшего распространения
необходимо создавать смешанные насаждения. Схемы смешения,
а также размещения посадочных мест должны выбираться в
зависимости от конкретных типов лесорастительных условий. В
дальнейшем необходимо поддерживать оптимальную густоту посадок
путем своевременных прореживаний. Во избежание
распространения инфекции обрезку сучьев рекомендуется проводить ранней
весной, до начала споруляции возбудителей. После
прореживаний и обрезки сучьев все порубочные остатки необходимо
удалить и сжечь как источники инфекции.
302
В связи с тем, что стволовые вредители могут быть в одних
случаях переносчиками инфекции (голландская болезнь
ильмовых, сосудистый микоз дуба), в других — способствовать
проникновению инфекции, в третьих — ускорять процесс усыхания в
очагах болезней, с ними следует вести систематическую борьбу
путем выборки свежезаселенных и выкладки ловчих деревьев.
Система защиты городских насаждений от
болезней включает мониторинг состояния древостоев и загрязнения
среды, надзор за появлением и распространением фитофагов и
болезней, мероприятия по повышению устойчивости растений и
насаждений, интегрированные методы истребительных и
профилактических мероприятий. Неотъемлемые элементы такой
системы — банк экологических данных и стратегия принятия решений
о целесообразности и методах борьбы с вредными организмами с
помощью современных безопасных для человека средств. Большое
значение имеют организационные мероприятия по
совершенствованию службы озеленения и методы повышения устойчивости
насаждений.
В городских и парковых насаждениях, где проведение
санитарных рубок ограничено, большое внимание уделяется лечению ран,
обрезке пораженных ветвей, локализации корневых систем при
появлении голландской болезни.
Химический метод. Химическая защита древесных пород от
сосудистых и некрозно-раковых болезней применяется очень
ограниченно, что связано прежде всего с трудностями уничтожения
их возбудителей, развивающихся внутри тканей стволов и ветвей.
Поэтому большое значение имеет дезинфекция срезов ветвей,
механических и морозобойных ран, через которые проникают
возбудители многих некрозно-раковых болезней (нектриевый и ци-
тоспоровый некрозы, нектриевый ступенчатый, черный гипо-
ксилоновый, мокрый язвенно-сосудистый раки и др.).
Дезинфекция мест проникновения инфекции проводится сразу же после их
зачистки путем опрыскивания 3 —5 %-м раствором медного
купороса.
Профилактические опрыскивания проводятся только против
цитоспороза тополя в маточных и школьных отделениях
питомников, в молодых культурах и городских посадках при наличии
болезни на участке или вблизи него. Обработка проводится в
конце августа — начале сентября с использованием 0,4—1 %-х
водных суспензий оксихлорида меди или Абига-Пик.
Глава 12
ГНИЛЕВЫЕ БОЛЕЗНИ ДРЕВЕСНЫХ ПОРОД
И МЕРЫ БОРЬБЫ С НИМИ
Гнили корней и стволов растущих деревьев составляют одну из
самых больших и важных групп болезней леса. При поражении
деревьев гнилевыми болезнями у них может произойти резкое
нарушение физиологических процессов, ведущее к снижению
прироста, общему ослаблению и усыханию деревьев. В насаждениях,
пораженных этими болезнями, часто наблюдаются ветровал и
бурелом, что в конечном счете приводит к распаду насаждений, утрате
лесом его ценнейших свойств и функций. Вред, причиняемый гни-
левой болезнью дереву как живому организму и насаждению как
биогеоценозу, можно рассматривать как биологический. Но гнили
причиняют еще и технический вред. Он заключается в разрушении
и обесценивании основного продукта леса — древесины,
снижении выхода и качества деловых сортиментов. Кроме того,
распространение гнилевых болезней в древостоях, не достигших возраста
естественной спелости, приводит к колоссальным потерям
(недобору) древесины из-за вынужденных преждевременных рубок.
Гнилевые болезни древесных растений вызываются дереворазру-
шающими грибами, подавляющее большинство которых относится
к классу базидиомицетов, группе порядков гименомицеты. Грибы-
возбудители гнилей различаются по биологическим особенностям,
степени паразитизма, специализации, характеру воздействия на
древесину и на дерево в целом. Среди них отсутствуют облигатные
паразиты, мало облигатных сапротрофов; большая часть этих грибов
относится к числу факультативных паразитов и факультативных
сапротрофов, обладающих способностью, в зависимости от условий
развития, питаться живой или мертвой тканью, переходить с живых,
растущих деревьев на мертвые древесные остатки, и наоборот.
Среди грибов, поражающих растущие деревья, в свою очередь
имеются виды, которые питаются живой тканью заболонной
древесины, виды, заселяющие только мертвую (ядровую) древесину
центральной части ствола, и виды, способные развиваться как в
304
живой, так и в мертвой древесине. Наряду с широко
специализированными представителями дереворазрушающих грибов, которые
поражают многие хвойные и лиственные породы, существуют виды
с более узкой специализацией, вплоть до типичных монофагов.
Заражение деревьев возбудителями стволовых гнилей в
большинстве случаев происходит через различные повреждения коры,
вызванные абиотическими факторами (морозобоины и др.),
животными (копытными, грызунами, насекомыми) или
хозяйственной деятельностью человека (механические повреждения, ожоги
и т. д.). Заражение возбудителями корневых гнилей
осуществляется через повреждения корней, отмершие мелкие корешки и при
непосредственном контакте (или срастании) здоровых и
пораженных корней. Заражению деревьев гнилевыми болезнями и
интенсивному развитию их в насаждении способствуют любые
факторы, ведущие к общему ослаблению древостоя, нарушению
сложившихся экологических связей, снижению биологической
устойчивости насаждения (засухи, неправильное ведение хозяйства,
повышенные рекреационные нагрузки и т.д.).
12.1. СУЩНОСТЬ ПРОЦЕССА ГНИЕНИЯ
ДРЕВЕСИНЫ
Гниением древесины называется ее биологическое разложение.
Суть этого процесса заключается в разрушении ферментами грибов
оболочек древесных клеток. В зависимости от того, какими
ферментами гриб воздействует на
клеточные стенки, какие их
компоненты, в какой степени и
последовательности он
разрушает, в древесине происходят
те или иные нарушения
анатомической структуры,
изменения ее химического состава и
физических свойств.
Главным критерием,
характеризующим особенности
гниения древесины и
происходящих в ней изменений,
является тип гниения. От него, в свою
очередь, зависят структура и
окраска гнили. Различают
деструктивный и коррозионный
типы гниения (рис. 92).
При деструктивном
типе гниения гриб воздейству-
о а
Рис. 92. Схема разложения древесины
дереворазрушающими грибами:
/ — при деструктивном типе гниения
(развитие бурой трещиноватой гнили); 2 —
при коррозионном типе гниения
(развитие пестрой ямчатой гнили); 3— при
коррозионном типе гниения (развитие белой
слоисто-волокнистой гнили)
305
ет на всю древесную массу, не оставляя не затронутых
разложением частей древесины. В этом случае разлагается целлюлоза
клеточных оболочек, а лигнин остается нетронутым. По мере
разрушения целлюлозы и освобождения лигнина пораженная
древесина темнеет, ее объем уменьшается, она становится хрупкой,
растрескивается, распадается на отдельные куски, а в конечной
стадии гниения легко растирается в порошок. Поэтому
деструктивные гнили характеризуются трещиноватой, призматической,
кубической или порошкообразной структурой и бурой (различных
оттенков) окраской — бурая гниль.
При коррозионном типе гниения разлагаются как
целлюлоза, так и лигнин. Однако при поражении разными видами
грибов этот процесс протекает неодинаково. В одних случаях гриб
одновременно разлагает целлюлозу и лигнин, полностью разрушая
клеточные оболочки, а затем и целые группы клеток. В пораженной
древесине появляются отверстия, ямки, пустоты, заполненные
остатками белой неразложившейся целлюлозы; так возникает
пестрая гниль. При коррозионном гниении, в отличие от
деструктивного, разложению подвергается не вся пораженная
древесина: отдельные группы разрушенных клеток чередуются с
совершенно нетронутыми участками древесины. Поэтому гниль
расщепляется на волокна, крошится, но долго сохраняет вязкость, и ее
объем не уменьшается.
В других случаях сначала полностью разлагается лигнин, а
затем постепенно разрушается целлюлоза. Однако при этом
разлагается не вся целлюлоза: часть ее остается в пустотах древесины в
виде белых скоплений (выцветов). Пораженная древесина
равномерно или полосами светлеет, приобретает белую,
светло-желтую рши «мраморную» окраску (белая гниль). Коррозионные
гнили на разных стадиях разрушения древесины характеризуются
ямчатой, ямчато-волокнистой, волокнистой и
слоисто-волокнистой структурой.
В любом случае биологическое разложение древесины
возможно лишь при определенных условиях, допускающих развитие де-
реворазрушающих грибов. Например, содержание свободной воды
в древесине должно составлять не менее 18—20 %, а
минимальный объем воздуха в зависимости от экологических требований
гриба — от 5 до 20 %.
12.2. КЛАССИФИКАЦИЯ И ПРИЗНАКИ ГНИЛЕЙ
Пораженная древесина, утрачивая свои нормальные
биологические свойства и технические качества, приобретает новые
признаки, характерные для отдельных групп и видов гнилевых
болезней. Диагностические признаки и классификация гнилей имеют
306
важное практическое значение. Для определения гнили
принимают во внимание следующие основные признаки: расположение
гнили в дереве, тип гниения, структуру и окраску гнили, стадию
и скорость гниения, некоторые другие особенности (наличие
темных линий, защитного ядра, мицелиальных пленок и др.).
Расположение гнили в дереве может быть
различным (рис. 93). В зависимости от размещения ее по частям дерева и
на продольном разрезе ствола гнили подразделяются на
корневые, комлевые (до 2 м), стволовые, вершинные, сквозные (по
всей длине ствола) и гнили ветвей и вершин. По расположению
10 И 12 13
Рис. 93. Схема расположения гнилей в дереве:
/ — корневая гниль; 2, 3 — корневая и комлевая гниль; 4 — комлевая гниль; 5 —
стволовая гниль; 6 — комлевая и стволовая гниль; 7 — корневая, комлевая и
стволовая гниль; 8 — гниль ветвей и вершин; 9 — «сквозная» гниль; 10 — забо-
лонная гниль; 11 — ядровая гниль; 12 — ядрово-заболонная гниль; 13 —
сплошная гниль
307
гнили на поперечном срезе корня, ствола или ветви различают
ядровые, заболонные и ядрово-заболонные гнили.
Гнили, различающиеся по расположению в дереве или в
стволе, по-разному влияют на жизненные функции и состояние
дерева, а также на выход деловой древесины; следовательно, они
характеризуются различной степенью причиняемого ими
биологического и технического вреда. Так, наибольший биологический
вред приносят корневые гнили и заболонные гнили стволов,
наибольший технический вред — ядровые и ядрово-заболонные
гнили стволов.
Тип гниения (см. рис. 92) отражает особенности процесса
разрушения древесины, связанные с биологическими свойствами
гриба и характером его воздействия на клеточные оболочки
пораженной ткани (рис. 94).
Окраска гнили зависит от стадии ее развития и типа
гниения. При деструктивном типе гниения обычно возникает бурая,
красновато-бурая или серовато-бурая окраска, при коррозионном —
пестрая или белая (светло-желтая, полосатая, мраморная).
Структура гнили свидетельствует об изменениях
анатомического строения и физических свойств древесины в
зависимости от типа гниения. Деструктивные гнили характеризуются
призматической, кубической или порошкообразной структурой;
коррозионные — ямчатой, волокнистой, ямчато-волокнистой и
слоисто-волокнистой структурой. По структуре и окраске гнили в
конечной стадии разрушения древесины можно определить тип
гниения. Зная тип гниения, нетрудно предвидеть, какую окраску и
структуру будет иметь гниль в конечной стадии.
Стадия гниения является показателем степени
разрушения древесины в процессе гниения. Каждая стадия
характеризуется определенными изменениями окраски и структуры поражен-
Рис. 94. Основные диагностические признаки гнилей:
2 _ бурая призматическая деструктивная гниль; 2 — пестрая ямчатая
коррозионная гниль; 3 — белая слоисто-волокнистая гниль коррозионного типа
308
ной древесины. Различают I (начальную), II и III (конечную)
стадии развития гнили. Образование дупла (IV стадия) — признак
прекращения процесса гниения древесины и начала ее
механического распада естественным путем или при участии насекомых,
птиц, других животных или человека. Определение стадии
развития гнили имеет важное практическое значение, особенно в тех
случаях, когда речь идет о возможностях технического
использования пораженной древесины.
Скорость гниения характеризует продолжительность
отдельных стадий процесса гниения и позволяет определить время
наступления конечной стадии. Различают медленное, быстрое и
очень быстрое гниение древесины. Большое практическое
значение, особенно при оценке влияния гнили на выход деловых
сортиментов, имеет и скорость распространения
гнили в различных частях дерева, в бревнах или деревянных
конструкциях зданий и сооружений в единицу времени (сутки, месяц,
год). Так, скорость распространения гнили, вызываемой
корневой губкой, в стволе ели достигает в среднем 48 см в год.
Быстрота гниения и скорость распространения гнили зависят
от биологических особенностей гриба — возбудителя гнили и
условий его развития, от свойств живого дерева, физического
состояния и технических качеств древесины.
Независимо от быстроты гниения древесины распространение
гнили в пределах дерева может быть как медленным, так и
быстрым. Например, гниль от еловой губки распространяется по
стволу ели очень быстро, а гниль дуба, вызываемая дуболюбивым
трутовиком, — медленно, хотя в обоих случаях наблюдается быстрое
гниение древесины.
12.3. КОРНЕВЫЕ ГНИЛИ
Корневые гнили древесных пород принадлежат к числу
наиболее распространенных и вредоносных болезней леса. Возбудители
корневых гнилей заражают деревья спорами (главным образом
через поврежденные или отмершие корни) и мицелием — при
соприкосновении или срастании здоровых и пораженных корней.
Вследствие распространения инфекции по корням от дерева к
дереву развитие корневых гнилей в насаждениях обычно носит
куртинный характер и проявляется в групповом ослаблении и
отмирании деревьев. Иногда возникают крупные очаги,
охватывающие большие площади леса.
Поражение и разрушение корней очень сильно влияет на
состояние дерева, так как нарушается поступление в его надземные
части воды и питательных веществ. Поэтому корневые гнили
приводят к быстрому ослаблению и усыханию деревьев, ветровалу,
309
заселению деревьев стволовыми вредителями, изреживанию
древостоя, а при сильной степени поражения насаждений — к их
полному распаду.
Некоторые виды гнилей из корней переходят в ствол и,
поражая комлевую, а иногда и большую часть ствола, приводят к
значительным потерям деловой древесины.
Среди болезней этой группы наибольшую опасность
представляют гнили, вызываемые корневой губкой и опенком осенним.
Менее распространены гнили корней, вызываемые трутовиком
Швейница, ризиной волнистой. Из ствола в основания корней
могут распространяться гнили, вызываемые еловой губкой,
северным, чешуйчатым и некоторыми другими трутовиками.
Корневая губка (Heterobasidion annosum (Fr.) Bref., (= Fomitopsis
annosa Karst.) Гриб относится к классу базидиомицетов, группе
афиллофороидных гименомицетов. Вызывает пеструю ямчато-во-
локнистую корневую и стволовую гниль. Корневая губка — один
из самых распространенных в мире грибов. Заболевание охватило
огромные площади хвойных насаждений земного шара и приобрело
характер глобальной эпифитотии (панфитотии). Во многих
странах гниль, вызываемая корневой губкой, считается самым
вредоносным заболеванием леса.
Корневая губка может заражать многие хвойные породы и
некоторые мягколиственные (например, березу), однако
лиственные породы поражаются редко. Гриб представляет большую
опасность лишь для хвойных насаждений, прежде всего для сосны,
ели, пихты и в меньшей степени для лиственницы.
Описан ряд морфологических форм или разновидностей
корневой губки, которые различаются по географическому
распространению, уровню патогенности и специализации к различным
древесным породам.
Первичное заражение деревьев осуществляется базидиоспора-
ми и конидиями гриба. Базидиоспоры образуются в плодовых
телах, а конидии — на мицелии в тех местах, где гниль выходит на
поверхность зараженных пней или корней. Корневая губка
способна сохраняться и развиваться не только в древесине живых
деревьев, но и на мертвых корнях, пнях, древесных остатках, в
подстилке, где часто формируются ее плодовые тела. Споры гриба
переносятся потоками воздуха, водой, различными животными.
Попадая на поверхность корней, особенно при наличии
механических повреждений, они заражают их. Затем в корнях
распространяется мицелий гриба и развивается гниль. При попадании
спор на свежие срезы пней (например, после рубок ухода) они
прорастают на них, и мицелий сначала распространяется в
древесине пней, а затем переходит в корни. Дальнейшее
распространение инфекции и вторичное заражение корней живых деревьев
осуществляются грибницей при непосредственном контакте здоро-
310
вых корней с пораженными. Этим объясняется групповое, или
куртинное, поражение древостоя. Заражение деревьев может
осуществляться и через отмершие мелкие корешки или мертвые
окончания глубинных корней.
Характер развития болезни и ее признаки у разных древесных
пород заметно различаются. Так, при поражении сосны гниль
развивается только в корнях. Поэтому для ее обнаружения
необходимо обследовать корневую систему. На начальной стадии развития
гнили происходит обильное выделение живицы из
разрушающихся смоляных ходов. Древесина корней пропитывается смолой,
приобретает красновато-оранжевый, иногда чуть лиловатый
оттенок, становится как бы стекловидной и издает специфический
запах скипидара. Смола скапливается под корой пораженных
корней, затем вытекает наружу и склеивает окружающие частицы
почвы, образуя на корнях твердые желваки. По мере развития гнили
просмоленность постепенно исчезает, древесина принимает
более светлую, равномерно желтую окраску, иногда с едва заметными
белыми вкраплениями целлюлозы. На последней стадии гниения
в древесине образуются многочисленные мелкие пустоты; гниль
распадается на отдельные волокна, становится мочалистой,
трухлявой.
По мере отмирания корней нарушается водный баланс дерева,
снижается интенсивность транспирации, фотосинтеза, других
физиологических функций, происходит общее ослабление дерева,
отчетливо проявляющееся в изменении состояния кроны.
Первые признаки ослабления сосны — уменьшение прироста
по высоте, наличие укороченных побегов, на которых образуется
укороченная хвоя. Значительная часть двух- и трехлетней хвои
опадает, крона постепенно изреживается, становится как бы
ажурной. Оставшаяся на побегах хвоя собрана в виде кисточек, она
бледная, тусклая. Такие деревья резко выделяются среди здоровых.
В дальнейшем хвоя постепенно желтеет, а затем и полностью
засыхает.
В сосновых насаждениях действующие очаги корневой губки
можно узнать по наличию ослабленных и усыхающих деревьев,
свежего и старого сухостоя, а также характерных наклонившихся
деревьев и ветровала. Групповое усыхание деревьев и их
повышенная ветровальность, последующие санитарные рубки
приводят к образованию «окон» и прогалин. Куртины усохших деревь-
си и «окна» в сосняках имеют более или менее четкие очертания.
С каждым годом они расширяются, по их краям появляются все
новые усыхающие деревья, отдельные прогалины сливаются, и в
конце концов насаждение превращается в редину (рис. 95).
При поражении ели и пихты мицелий гриба сначала
распространяется в корнях, затем переходит в ствол, вызывая ядровую
гниль, окаймленную лиловато-серым кольцом. Она поднимается
311
Рис. 95. Куртина усыхания и
ветровал сосны в очаге
корневой губки
по стволу в среднем на высоту 3 —
4 м, иногда до 8—10 м и более. На
первой стадии развития гнили
древесина приобретает
серовато-фиолетовую окраску; затем она становится
красновато-коричневой, а на
последней стадии гниения — типично
пестрой: в ней появляются отчетливые,
довольно крупные белые выцветы
целлюлозы и весьма характерные
черные штрихи. Гниль имеет ямчато-
волокнистую структуру, в сухом
состоянии легко крошится. Наличие в
стволе ядровой гнили с типичными
для корневой губки признаками
можно установить с помощью
возрастного бурава. Со временем в нижней
части ствола образуется дупло.
Деревья ели и пихты, пораженные
корневой губкой, даже при значительном развитии гнили в корнях и
стволах могут долго не усыхать, хотя признаки ослабления
бывают хорошо выражены: снижение прироста по высоте, изрежен-
ная крона, тусклая с буроватым оттенком хвоя,
деформированные побеги. В связи с тем, что заболевание у ели часто носит
скрытый характер, а отпад происходит в основном за счет ветровала, в
ельниках не образуются столь ясно выраженные и равномерно
увеличивающиеся по радиусу куртины усыхания и «окна», как в
сосновых насаждениях.
Самый верный признак поражения дерева корневой губкой —
наличие на корнях плодовых тел гриба. Обычно они образуются
в затененных местах, на нижней поверхности гнилых корней
ветровальных деревьев, иногда у корневой шейки усохших
деревьев, на полуразрушенных пнях. Плодовые тела корневой губки
имеют различную форму и величину, они многолетние, тонкие,
распростертые, обращенные гименофором кнаружи (рис. 96). Края
плодовых тел немного отстают от корня. Их поверхность
коричневая, с более светлым краем и концентрическими бороздками.
Гименофор вначале белый, позднее желтоватый, с
шелковистым блеском. Поры мелкие, округлые или угловатые, иногда
скошенные.
Корневая губка встречается почти во всех типах лесораститель-
ных условий, за исключением заболоченных местообитаний. Очень
редко поражаются сосняки сфагновые и лишайниковые. Наиболее
сильное развитие болезни и наибольший вред от нее
наблюдаются при поражении высокобонитетных насаждений в свежих типах
леса. Поражаются насаждения разного возраста, причем первые
312
Рис. 96. Корневая губка:
1 — распространение ядровой гнили из корней в ствол ели; 2 — плодовое тело
гриба на пораженном корне; 3 — конидиальное спороношение гриба
признаки болезни могут обнаруживаться уже в 15—20-летних древо-
стоях. Самосев хвойных пород, появляющийся в очагах корневой
губки, также заражается грибом и погибает. В наибольшей степени
страдают чистые хвойные насаждения, особенно культуры,
созданные на бывших пашнях, пустырях или площадях, вышедших
после рубки древостоев, пораженных корневой губкой. В
естественных насаждениях сосны корневая губка встречается реже. Ель и
пихта сильно поражаются не только в культурах, но и в
естественных лесах. Смешанные хвойно-лиственные насаждения
более устойчивы к болезни. Чрезмерная густота насаждений при
наличии в почве тесно переплетающихся и сросшихся корней
способствует распространению гриба и быстрому развитию очагов.
В пригородных лесах и лесопарках развитию очагов корневой
губки благоприятствуют повышенные рекреационные нагрузки,
следствием которых являются повреждения корней, уплотнение
почвы и ухудшение ее аэрации. Это в свою очередь влечет за собой
отмирание корней и создает условия для перехода гриба от сап-
ротрофного питания на древесных остатках к паразитизму на
живых деревьях.
Ущерб, причиняемый корневой губкой, очень велик.
Заболевание приводит к массовому усыханию деревьев и распаду
насаждений. Поражение ели и пихты приносит, кроме того, большой
технический вред, так как у этих пород гниль из корней поднимается
в ствол; в результате резко уменьшается выход деловых
сортиментов из наиболее ценной части ствола. Потери деловой древесины
могут составлять у ели около 50 %, а у пихты свыше 75 %.
Ослабление и усыхание пораженных деревьев, как правило, влечет за
собой усиленное размножение насекомых-ксилофагов. Поэтому
313
очаги корневой губки обычно превращаются и в очаги стволовых
вредителей, которые ускоряют процесс усыхания насаждений.
Меры борьбы: система мероприятий, направленных на
ограничение массового развития болезни и формирование
устойчивых насаждений с помощью оптимального режима лесовыра-
щивания. Эта система включает обследование насаждений с
целью выявления и учета очагов болезни, лесоводственный уход,
лесовосстановление и санитарно-оздоровительные мероприятия,
назначаемые с учетом прогноза развития болезни, а также
контроль качества лесохозяйственных мероприятий.
Выявление и учет очагов корневой губки проводят при
лесоустройстве и лесопатологических обследованиях. В процессе
рекогносцировочного обследования дают приблизительную оценку
состояния и степени поражения насаждений, определяют
площадь очагов корневой губки. За очаг болезни принимают весь
таксационный выдел, в котором наблюдается куртинное усыхание
или вывал пораженных деревьев, т. е. отпад является
патологическим и превышает естественную норму.
В зависимости от давности развития очагов, их структуры и
внешних признаков различают следующие категории очагов:
возникающие, действующие и затухающие.
Возникающие очаги представляют собой небольшие (до
10 деревьев) группы сильно ослабленных и усыхающих деревьев,
свежего сухостоя или ветровала, чаще в насаждениях I—II
классов возраста. В очагах, как правило, еще нет ни прогалин («окон»),
ни пней от санитарных рубок, поскольку их еще не проводили.
Действующие очаги характеризуются наличием хорошо
выраженных куртин усыхания и прогалин с пнями различного
состояния от санитарных рубок различной давности. В окружающем
«окна» древостое (который, как правило, уже поражен корневой
губкой) присутствуют деревья всех категорий состояния: от
ослабленных в разной степени до свежего и старого сухостоя и
ветровала. В окнах начинается смена хвойных пород лиственными,
обычно березой или осиной.
Затухающие очаги характеризуются отсутствием
усыхающих деревьев, свежего сухостоя, свежего ветровала, что
свидетельствует об окончании активной фазы развития очага. Вокруг окон
может оставаться еще не вырубленный старый сухостой. В окнах
преобладают полуразрушенные или трухлявые пни от давних
рубок, имеется хорошо развитый подрост лиственных пород.
Степень поражения сосновых насаждений считается слабой,
если куртины поражения или прогалины суммарно составляют до
5 % площади выдела в насаждениях до 20 лет, до 10 %—в
насаждениях от 21 года до 50 лет и до 15 % — в насаждениях старше 50 лет.
Степень поражения считается средней, если куртины поражения
и прогалины суммарно составляют, соответственно по возраст-
314
ным группам, до 15 %, до 25 % и до 33 % площади выдела.
Степень поражения сосняков считается сильной, если куртины
поражения и прогалины суммарно составляют соответственно 16% и
более, 26 % и более, 34 % и более площади выдела.
Степень поражения еловых и пихтовых насаждений считается
слабой, если деревья, зараженные корневой губкой, составляют
до 20 %; средней, если таких деревьев 21—40 %, и сильной, если
их более 40%.
При детальном обследовании, во время которого проводят
закладку пробных площадей со сплошным перечетом деревьев,
уточняют данные о состоянии и степени пораженности насаждений.
По результатам рекогносцировочного и детального обследований
составляют карту очагов корневой губки, разрабатывают
конкретный план санитарно-оздоровительных мероприятий,
определяют их очередность и объем.
В пораженных и предрасположенных к болезни насаждениях в
зависимости от их происхождения, возраста, состояния и уровня
экологической устойчивости назначают рубки ухода или
санитарные рубки. Их проводят в соответствии с «Санитарными
правилами в лесах Российской Федерации», «Основными положениями
по защите сосны, ели и пихты от корневой губки» и
действующими инструкциями.
Рубки ухода в молодняках следует ориентировать на
формирование древостоев с густотой, оптимальной для каждой возрастной
группы и местных условий лесопроизрастания. Возраст, в котором
следует начинать рубки ухода, и их интенсивность зависят от
состава и состояния молодняков, густоты и схемы посадки. При
рубках ухода в чистых культурах хвойных пород необходимо сохранять
естественную примесь лиственных пород. К возрасту 20—25 лет
полноту насаждений рекомендуется довести до 0,7—0,8 и
поддерживать ее при последующих рубках.
При санитарных рубках объем вырубаемой массы определяют
по сумме запасов усохших, усыхающих и сильно ослабленных
деревьев на пробных площадях.
Выборочные санитарные рубки назначают в более взрослых
насаждениях со слабой степенью поражения. В этом случае удалению
подлежат сухостой, усыхающие, сильно ослабленные и
ветровальные деревья. Интенсивность и периодичность таких рубок зависят
от целевого назначения насаждений, их полноты, возраста,
общего состояния и других факторов. В возникающих и действующих
очагах болезни рекомендуются более интенсивные рубки, чем в
затухающих. При средней степени поражения насаждений с
наличием явно выраженных небольших куртин усыхания
рекомендуются рубки изолирующих полос или так называемые группово-
выборочные санрубки. При этом вырубают все деревья в пределах
«окна», а также в 4—6-метровой полосе вокруг него (в зоне скры-
315
того поражения). При наличии в насаждении больших участков с
различной степенью поражения проводят частично сплошные или
выборочно-сплошные рубки: наиболее пораженную часть выдела
вырубают полностью, а на участках со слабой степенью
поражения проводят выборочную санитарную рубку.
Сплошные санитарные рубки назначают в насаждениях с
сильной степенью поражения корневой губкой. На вырубках рекомендуется
выкорчевать пни, «вычесать» из почвы корни, пни и корни сжечь.
Все виды рубок следует проводить поздней осенью и зимой —
в период зимнего покоя деревьев. При проведении рубок в другие
сроки одновременно с рубкой или в течение 4—5 дней после нее
рекомендуется проводить химическую обработку (антисептирова-
ние) пней и корневых лап или удалять их. Срубленную древесину
следует сразу же вывозить из леса. Оставленную древесину
необходимо окорять или обрабатывать инсектицидами против
стволовых вредителей.
Для химической обработки пней рекомендуются
водорастворимые антисептики: 20%-й раствор карбамида (мочевины), 10%-й
раствор нитрафена, 10 %-й раствор сульфата аммония, 5 %-й
раствор хлористого цинка, 4 %-й раствор марганцовокислого калия,
4 %-й раствор буры и др. Обработку проводят с помощью
ранцевых опрыскивателей с таким расчетом, чтобы вся поверхность
пней и корневых лап была тщательно покрыта антисептиком.
С целью локализации возникающих очагов усыхания
рекомендуют обработку почвы 1 %-м раствором фундазола, которую
проводят одновременно с санитарной рубкой. Для этого по
периферии куртин усыхания в зоне шириной до 1 м разрыхляют почву и
вносят в нее препарат при норме расхода 1—2 л/м2. Рекомендуют
также применение биопрепаратов, например микоризина.
Лесовосстановление на вырубках после сплошных и частично
сплошных санитарных рубок, а также облесение площадей,
находившихся под сельскохозяйственным пользованием, проводят
путем создания чистых лиственных или смешанных культур с
учетом типа леса, характера вырубки, инфекционного фона,
наличия подроста и других местных условий. Во всех случаях хвойные
породы не должны занимать в составе более 30 %, а число
посадочных мест не должно превышать 5000 на 1 га. Схемы смешения
и размещения пород подбирают в соответствии с условиями
местопроизрастания.
При создании культур необходимо использовать
высококачественный посадочный материал с хорошо развитой корневой
системой и микоризой. На нелесных площадях и бедных песчаных
почвах для улучшения роста и повышения устойчивости культур
необходимо вносить органо-минеральные удобрения.
Рекомендуется также посев многолетнего люпина. В условиях пригородных
лесов проводят мероприятия по регулированию рекреационных
316
нагрузок. В насаждениях с преобладанием хвойных пород
запрещается выпас скота.
Опенок осенний (Armillaria mellea (Vahl. ex Fr.) Karst.). Вызывает
белую заболонную гниль корней и стволов хвойных и лиственных
деревьев. Гриб относится к классу базидиомицетов, группе агари-
коидных гименомицетов. Он почти повсеместно встречается во всех
частях света и является типичным полифагом: поражает более
230 видов растений, в том числе многие древесные породы,
кустарники, сельскохозяйственные, цветочные культуры. Он
способен как к паразитическому образу жизни, так и к длительному
сапротрофному развитию на мертвой древесине.
В составе популяций A. mellea выделяют формы,
различающиеся по эколого-морфологическим, культуральным и другим
признакам.
В последнее время A. mellea нередко рассматривают не как один
вид, а как комплекс видов, различающихся по морфологическим
признакам, экологическим особенностям и географическим
ареалам. Семь из них выявлены в Европе и не менее трех в нашей
стране.
Важнейшие диагностические признаки поражения деревьев
опенком — наличие на корнях и стволах сильно ветвящихся
темно-бурых мицелиальных шнуров (ризоморф) и пленок (рис. 97).
На поверхности корней гриб образует корнеподобные, круглые в
Рис. 97. Опенок осенний:
/ — мицелиальная пленка гриба под корой пораженного пня; 2 — мицелиальные
пленки под корой ствола; 3 — группа плодовых тел у корневой шейки дерева; 4 —
образование плодовых тел на ризоморфах; 5— фрагмент плодового тела (а —
пластинчатый гименофор, б — кольцо на ножке под шляпкой); 6 — белая заболонная
гниль ствола с характерными темными линиями; 7 — участок гимения с базидия-
ми и базидиоспорами гриба; 8 — базидиоспоры; 9 — прорастание базидиоспор
317
сечении ризоморфы, которые могут, распространяясь в лесной
подстилке и почве, переходить на корни соседних здоровых
деревьев и заражать их через отмершие мелкие корешки,
повреждения коры, чечевички. Под корой пораженных корней и стволов
развиваются плоские ризоморфы, нередко многометровой длины.
Именно на таких ризоморфах формируются хорошо знакомые всем
плодовые тела гриба.
Плодовые тела опенка осеннего образуются преимущественно
в августе — октябре большими группами, чаще всего на пнях
(отсюда название гриба), валежнике, сухостое, реже на корнях и
основаниях стволов пораженных живых деревьев. Шляпка
диаметром до 15 см, мясистая, вначале выпуклая, затем плоская, с
подвернутым краем, часто с бугорком в центре,
желтовато-коричневая или серовато-бурая, с более темными (или того же цвета)
многочисленными чешуйками. Внутренняя ткань белая, рыхлая,
с приятным запахом, сладковато-вяжущая. Пластинки гименофо-
ра слегка нисходящие, белые, со временем темнеющие. Ножка
центральная, цилиндрическая, длиной до 10—15 см, толщиной
до 1 — 1,5 см (у основания иногда немного вздутая),
мелкочешуйчатая, беловатая или светло-бурая, книзу более темная, с белым
толстым пушисто-шелковистым кольцом под шляпкой.
Базидиоспоры, созревающие в плодовых телах,
распространяются ветром, дождевой водой, животными и, попадая на пни и
корни деревьев, прорастают и заражают их.
От мест заражения мицелий гриба разрастается под корой
корней и ствола, нередко поднимаясь на высоту до 2 — 3 м (иногда
еще выше). Под действием токсинов патогена живые ткани луба,
камбия и заболони отмирают, после чего мицелий гриба
внедряется в них и вызывает в периферических слоях заболонной
древесины мягкую волокнистую белую или светло-желтую гниль с
характерными извилистыми тонкими черными линиями. Из
зараженных участков токсины опенка по сосудам могут
распространяться в другие части дерева, ускоряя его ослабление и гибель.
Между корой и древесиной пораженных корней и стволов
развиваются белые веерообразные пленки, которые со временем
уплотняются, становятся кожистыми, желтеют и, частично
расщепляясь, дают начало плоским ризоморфам.
Наибольший вред опенок причиняет насаждениям хвойных
пород, дуба, ясеня, ильмовых, осины, различных видов тополя,
шелковицы, плодовых деревьев, вызывая корневую и комлевую
белую заболонную гниль. В чистых хвойных насаждениях и
дубравах распространение опенка часто приобретает характер эпифи-
тотий.
Опенок поражает насаждения различного возраста.
Распространение гриба от дерева к дереву по корням обусловливает куртинный
характер заболевания. У молодых деревьев болезнь часто протека-
318
ет в острой форме, ведущей к их быстрому (за 1 — 2 года) усыха-
нию. При поражении взрослых деревьев болезнь развивается
медленнее (6—10 лет), вызывая их постепенное ослабление.
Усыхающие деревья часто заселяются стволовыми вредителями. Для
пораженных деревьев характерны изреженность кроны, мелкие
листья, короткая бледно-зеленая или буроватая хвоя, резкое падение
прироста по высоте, растрескивание коры в нижней части ствола.
При поражении хвойных пород смола пропитывает кору; между
корневыми лапами, у оснований стволов и на корнях образуются
скопления смолы.
Интенсивному развитию очагов опенка осеннего
способствуют загущенность древостоев, переплетение и срастание корневых
систем, ослабление деревьев абиотическими и другими
факторами, а также теплая влажная погода, благоприятная для массового
формирования плодовых тел, рассеивания базидиоспор и
заражения ими свежих пней, на которых вновь образуются мицелий,
пленки и, наконец, ризоморфы, обеспечивающие дальнейшее
распространение гриба.
Меры борьбы: комплекслесохозяйственныхмероприятий,
химических и биологических мер борьбы, направленных на
повышение устойчивости насаждений, устранение источников
инфекции, профилактику заражений, локализацию очагов болезни и
оздоровление насаждений.
Для уменьшения угрозы поражения опенком необходимо
создавать смешанные насаждения из более устойчивых к болезни
древесных пород. При подборе пород следует учитывать
почвенные и климатические условия местности. Перед посадкой
предусматривается известкование кислых почв, внесение основных
удобрений и микроэлементов, способствующих лучшему росту и
повышению устойчивости молодых насаждений.
Создавая культуры на вырубках, весьма желательно с целью
снижения запаса инфекции предварительно выкорчевать пни
вместе с корнями или обработать их фунгицидами (10 %-м раствором
КМп04, фундазолом или топсином-М). Рекомендуется также
окорять пни и корневые лапы или обжигать их.
В молодых насаждениях следует регулярно проводить рубки
ухода, удаляя больные, усохшие и угнетенные деревья, регулируя
состав и полноту древостоя. Локализацию возникающих очагов
болезни в молодых культурах проводят путем удаления
пораженных деревьев с корнями.
Во взрослых насаждениях, пораженных опенком, необходимо
проводить санитарные рубки: выборочные — при слабой степени
поражения (до 20%); группово-выборочные — при средней
степени поражения (до 40%); сплошные — при сильной степени
поражения (свыше 40 %). Эффективность санитарных рубок будет
более высокой, если пни срубленных деревьев выкорчевать с кор-
319
нями, обработать фунгицидами или, в крайнем случае, окорить
либо обжечь.
Химическую защиту пней можно заменить обработкой их
биопрепаратами или споровыми суспензиями грибов-конкурентов
(пениофоры гигантской, окаймленного трутовика, вешенки
обыкновенной и др.).
В лесопарках, парках, городских посадках, мемориальных и
других особо ценных насаждениях целесообразны меры по
индивидуальному лечению больных деревьев (подсушивание корневой
системы, обрезка или кольцевание зараженных корней), а также
создание «химических барьеров» (путем внесения в почву
фунгицидов) для локализации очагов опенка или окапывание куртин
усыхания после санитарной рубки канавами глубиной не менее
0,5 м и шириной 0,4—0,5 м.
Трутовик Швейница (Phaeolus schweinitzii (Fr.) Pat.) вызывает
бурую призматическую ядровую корневую и комлевую гниль
хвойных пород. Чаще всего он поражает ель, сосну, лиственницу, реже
пихту и кедр, но иногда встречается и на лиственных породах:
дубе, лещине и некоторых других.
Плодовые тела образуются у оснований стволов и на корневых
лапах старых деревьев (рис. 98). Они однолетние, имеют вид
плоских или воронковидных, желтых или коричневых толстых
шляпок до 40 см в диаметре, без ножки или с короткой
клубневидной ножкой. Поверхность шляпок бархатисто-волосистая с
неясными зонами. Ткань мягкопробковидная или губчатая,
ржаво-коричневая. Трубочки гименофора ржаво-бурые с зеленоватым
оттенком, короткие, с крупными угловатыми, иногда
расщепленными порами.
Рис. 98. Возбудители корневых гнилей:
1 — плодовое тело трутовика Швейница; 2 — ризина волнистая (а — плодовое
тело, б — разрез через плодовое тело с ризоидами, в — сумка с сумкоспорами и
парафизы, г — сумкоспора)
320
Меинфекиионный некроз
листьев березы в зоне
сильного техногенного
нозлействия
Морозобойная трешина
in стволе дуба
Краевой некроз неинфекционного
характера на листьях деревьев,
произрастающих в зоне воздействия
автомагистралей
Молодые побеги ели, поврежденные
поздними весенними заморозками
Массовое поражение всходов сосны полеганием
в питомнике
Снежное шютте сосны Серое шютте сосны
У Шютте сеянцев лиственницы
Бурое шютге на подросте пихты
\
V
Ржавчина хвои пихты
Ржавчина
хвои ели
Усыхание хвои ели
колючей от шютте
Ржавчина побегов
и хвои ели
Склерофоллоз побегов сосны
Мучнистая роса
Темно-бурая пятнистость липы Парша тополя белого
Серая пятнистость осины
• • /
Бурая пятнистость (глеоспориоз) березы
Дерево, пораженное
голландской болезнью
Характерное скручивание листьев на
усохших побегах вяза, пораженного
голландской болезнью
Действующий очаг
голландской болезни
в вязовых насаждениях
Прерывистые темно-бурые кольиа
на поперечном срезе ветвей вяза,
пораженного хронической формой
голландской болезни
4*'
t
4~
'Л
Очаг
сосудистого
микоза в
пойменной
дубраве
Конидиальное спороношение
возбудителя черного немо-
спорового некроза на ветвях
дуба
Распростертое плодовое тело
возбудителя виллеллиниевого
некроза на ствоЛИке дуба
Цитоспороз на стволе
молодого тополя
Дотихиииевый (дискоспорие
вый) некроз на стволике
молодого тополя
Инфекиионное усыхание липы (стигллиниоз, тиростроллоз)
\-
Обший вил кроны липы,
пораженной болезнью
Неступенчатые раны на
стволе липы, пораженной
болезнью
Закрытые ранки на тонких
ветвях липы
Открытая рана на ветке
липы
\
^
t*
Черный гипоксилоновый рак осины
Рана в полкроновой части
ствола
Пузырчатая ржавчина
сосны вейллутовой
Группы плодовых тел
возбудителя в месте поражения
Эиии пузырчатой ржавчины
на стволике кедра
Ржавчинный рак пихты
I
П
* /
4* '*
.V<
Вельмина метла на подросте Раковая опухоль на стволе
пихты
Муфтообразное утолшение
на тонкой ветке пихты
Ветровал и бурелом
в Пихтарнике
в комплексном очаге
ржавчинного рака и гнилей
■J -.
и •: «.-"■
\л
\
Суховершинность сосны,
вызванная смоляным раком
(серянкой)
Плодовые тела возбудителя
рака побегов хвойных
пород на подросте кедра
Рак побегов хвойных пород
на кедровом стланике
Раны биатореллового рака
на стволике сосны
Корневая губка
Действующий очаг болезни Действующий очаг
в культурах сосны болезни в ельниках
Плодовые тела корневой губки
ч
болезни жзн
Гнилевые болезни корней
Плодовые тела опенка
осеннего на стволе
сухостойной березы
Мииелиальные пленки опенка
под корой сухостоя дуба
Плоские ризоллорфы опенка
под корой усыхаюшего дуба
Плодовое тело трутовика
Швейнииа у основания
ствола сосны
»
■ I
Плодовые тела елового
комлевого трутовика
на стволе ели
Плодовое тело трутовика
Гартига на стволе пихты
N
\
Плодовые тела серно-
желтого трутовика на
стволе живого дуба
Плодовые тела плоского
трутовика
Плодовые тела ложного трутовика
на стволе березы
Гниль от ложного трутовика на поперечном
срезе ствола березы
Плодовые тела настоящего
трутовика на березе
Чага на стволе березы
t>
*
ч
Плодовое тело дуболюбивого
трутовика на стволе дуба
Л
^ -
4| *
I
1*
" I
Плодовые тела шелестника
обыкновенного на стволе
сильно ослабленного
дерева каштана конского
* V
i f
Плодовые тела березовой
губки на сухостое березы
Плодовые тела столбового
(заборного) гриба на старом
ветровале сосны
Плодовые тела чешуйчатки жирной
на стволе ели
На цветной вклейке представлены фотографии Е.Г.Мозолевской, А.М.Жукова,
А.С.Рожкова, Т.В.Галасьевой, Т.В.Шарапа, ВХТузова, Н.М.Ведерникова, Г.Б.Колганихиной.
Заражение растущих деревьев базидиоспорами и грибницей
происходит через корни. Из корней гриб переходит в ствол и вызывает
ядровую гниль, которая может подниматься на высоту до 2 — 3 м.
Пораженная древесина сначала немного темнеет и приобретает
красноватый оттенок, затем становится бурой, и в ней появляются
трещины, располагающиеся по радиусу и годичным слоям. В
трещинах образуются тонкие белые мицелиальные пленки. Гниль имеет
скипидарный запах. Деревья при сильной степени поражения
корней усыхают или вываливаются ветром, иногда значительны и
потери древесины.
Трутовик Швейница — типичный обитатель старых
насаждений, обычно поражающий древостой старше 60 лет. В некоторых
лесных массивах он встречается довольно часто. Этот гриб широко
распространен в лесах европейской части страны и в Сибири, но
встречаемость его сравнительно невелика и крупных очагов он не
образует.
Ризина волнистая (Rhizina undulata Fr.) (=Rh. inflata (Schaeff.)
Rehm). Гриб-дискомицет, вызывающий корневую гниль молодых
культур сосны, но иногда он поражает также ель, пихту,
лиственницу. Как сапротроф этот гриб живет в подстилке на песчаных
почвах, но нередко переходит к паразитическому существованию
на корнях живых деревьев.
Чаще всего ризина поражает сосновые культуры, созданные в
борах и суборях после сплошных рубок в сосняках. Гриб часто
встречается на гарях и в местах, где во время лесозаготовок сжигали
ветки и другие порубочные остатки. Мицелий гриба развивается
на корнях 2 —5-летних растений, затем вокруг них на
поверхности почвы образуются плодовые тела — однолетние апотеции.
Диаметр плодовых тел 2 — 9 см, толщина 2—4 мм. Они имеют
мясистую сухую структуру и выпуклую неровную (бугристую)
поверхность каштаново-бурого цвета, вначале с более светлым краем.
Гимений располагается на поверхности апотециев. Сумки
удлиненно-булавовидные, с утолщенной красновато-бурой на
вершине оболочкой (как и у парафиз), что обусловливает темный цвет
плодового тела. Снизу плодовые тела беловато-желтые, с
отходящими от них тонкими ризоидами, которые внедряются в почву, а
затем и в корни (см. рис. 98).
У пораженных сосенок сначала желтеет хвоя, а затем они
полностью усыхают.
Плоский трутовик {Ganoderma applanatum (Pers. ex Wallr.) Pat.)
вызывает белую ядрово-заболонную корневую и комлевую гниль
лиственных деревьев.
Плодовые тела этого гриба — многолетние, в виде крупных (до
40 см в диаметре) плоских, довольно толстых шляпок без ножки,
иногда в виде низкого копыта, чаще одиночные, реже
соединенные черепицеобразными группами. Поверхность плодовых тел
321
матовая, шоколадно-коричневая, иногда серовато-бурая, обычно
неровная, бугристая или с концентрическими бороздками. На
разрезе плодового тела хорошо видна тонкая твердая темно-бурая
корка. Ткань пробковидная или войлочная коричневая. Гимено-
фор сначала белый, затем сероватый или буроватый, с очень
мелкими порами.
Плоский трутовик обычно встречается на пнях, сухостое, но
иногда поражает и живые деревья различных лиственных пород,
чаще всего липу, клен, иву, осину, тополь, вяз. Изредка
встречается на хвойных породах. Деревья заражаются через всевозможные
раны и повреждения поверхностных корней и основания ствола.
Мицелий распространяется в корнях и комлевой части дерева.
Гниль от плоского трутовика сначала ядровая, светло-желтого
цвета, с продолговатыми углублениями, заполненными белой
грибницей. Со временем гниль переходит и в заболонную часть
ствола. В конечной стадии гниль становится белой, волокнисто-
трухлявой, нередко образуется дупло. Пораженные деревья
подвержены ветровалу и бурелому. Плоский трутовик распространен
повсеместно, особенно в лесопарках, парках, городских
насаждениях.
12.4. СТВОЛОВЫЕ ГНИЛИ
Стволовые гнили составляют обширную группу гнилевых
болезней хвойных и лиственных пород. Это в основном адровые и
ядрово-заболонные гнили, возбудители которых (трутовые
грибы) заражают деревья базидиоспорами через обломанные ветви,
морозобойные трещины, ошмыги коры, затески и другие
повреждения стволов и ветвей. Ядровые гнили развиваются в стволах
живых деревьев и обычно мало влияют на их состояние, однако при
сильном развитии могут вызвать ослабление и даже усыхание
деревьев. Ядрово-заболонные гнили чаще поражают усыхающие, сильно
ослабленные деревья, развиваются на сухостое, валежнике.
Поражение стволовыми гнилями приводит к бурелому, снеголому,
захламленности, изреживанию древостоев (рис. 99). Это ухудшает
условия роста оставшихся деревьев, способствует размножению
стволовых вредителей. Все стволовые гнили причиняют
значительный технический вред, снижая выход деловых сортиментов. При
большой протяженности гнили в стволе потери деловой
древесины могут достигать 100 %. Во многих случаях пораженная
древесина не годится даже на дрова, так как ее теплотворная способность
резко снижается.
Иногда стволовые гнили развиваются на фоне некрозно-рако-
вых заболеваний, являясь в этом случае типичным примером
сопряженных болезней.
322
Рис. 99. Бурелом и захламленность в лиственном насаждении, пораженном
стволовыми гнилями
Стволовые гнили весьма разнообразны по специализации
возбудителей, особенностям развития и диагностическим признакам.
Они известны у всех лиственных и хвойных пород.
12.4.1. Стволовые гнили хвойных пород
Сосновая губка (Phellinuspini (Thore ex Fr.) Pil.). Гриб вызывает
пеструю ядровую гниль стволов у живых деревьев: сосны, кедра,
лиственницы. Плодовые тела многолетние, копытовидные или жел-
вакообразные, реже плоские. Поверхность темно-бурая или
серовато-черная шероховатая, грубощетинистая или голая, с
концентрическими бороздками и многочисленными радиальными
трещинами, часто покрытая лишайниками. Ткань деревянистая, ржаво-
бурая. Гименофор желтовато-коричневый, с неравновеликими
округлыми, угловатыми и лабиринтообразными порами. Плодовые тела
появляются на стволах уже при значительном развитии гнили.
Заражение стволов базидиоспорами обычно происходит через
обломанные ветви и раны на стволах. Пораженная древесина
приобретает красновато-бурую окраску. В дальнейшем развивается
ядровая ямчато-волокнистая гниль с многочисленными белыми
пятнами целлюлозы, которые позднее превращаются в пустоты
(«ситовина»). По годичным кольцам часто образуются трещины.
Гриб наиболее интенсивно разрушает ядровую часть ствола на
высоте от 2 до 10 м. Сосновая губка заражает древостой не моложе
40 лет. Пораженность старых насаждений иногда достигает 60 %.
Развитие гнили в стволе почти не влияет на состояние дерева,
однако приводит к большим потерям деловой древесины.
323
Еловая губка (Phellinus pini Pil. var. abietis Karst.) (=Phellinus
chrysoloma Karst.) вызывает пеструю ядровую гниль стволов и
ветвей у деревьев не моложе 40 лет. Плодовые тела многолетние,
пробково-деревянистые, распростертые или полураспростертые
(рис. 100). Они образуются на ветвях и, сливаясь, обволакивают их
с нижней стороны. Иногда плодовые тела имеют вид
полуотогнутых шляпок, расположенных на стволе черепицеобразными
группами. Поверхность их неровная, с радиальными трещинками,
часто складчатая, с волнистым краем, темно-бурая или серовато-
черная. Ткань ржаво-бурая, очень тонкая. Гименофор
серовато-бурый, с неравновеликими угловатыми или извилисто-лабиринто-
образными (дедалеевидными) порами.
По характеру заражения деревьев и особенностям паразитизма
этот гриб сходен с сосновой губкой. В насаждениях ели
обыкновенной пораженность редко превышает 16%, значительно сильнее
поражаются древостой ели аянской и ели Шренка (до 70 — 80 %).
Гниль развивается очень быстро. Она типично коррозионная,
ноздреватая («ситовина»), ямчато-волокнистая, окруженная
темно-бурым кольцом, с ярко выраженными пустотами и белыми
выцветами целлюлозы. Чаще она распространяется в средней и
нижней частях ствола, но иногда поднимается довольно высоко,
захватывая до 50 % его объема. Поэтому пораженные деревья
почти не дают деловой древесины и часто целиком идут на дрова.
Лиственничная губка {Fomitopsis officinalis (Will.) Bond, et Sing.)
вызывает красно-бурую ядровую гниль лиственницы. Плодовые тела
крупные, многолетние, копытообразные,
продолговато-цилиндрические или желвакообразные (рис. 101). Поверхность плодовых
тел без корки, серовато-белая или желтоватая, покрытая
трещинами. Ткань белая, мягкая, с возрастом твердеющая, в сухом
состоянии легко крошится, на вкус горькая, используется в
фармацевтической промышленности. Гименофор желтовато-белый, поры
мелкие округлые. Пораженная древесина вначале приобретает
светло-бурую, а затем красновато-бурую окраску. В конечной стадии
Рис. 100. Распростертые и
полураспростертые плодовые
тела еловой губки на стволе Рис. 101. Плодовое тело
и ветви ели лиственничной губки
324
гниения появляются широкие трещины, в которых образуются
кремово-белые толстые пленки, похожие на замшу, или ватооб-
разные скопления грибницы. Гниль распадается на крупные
кубики и призмочки. Она развивается в центральной части ствола, но
часто захватывает и заболонь, вызывая суховершинность деревьев.
Лиственничная губка встречается главным образом в
ослабленных, спелых и перестойных насаждениях Сибири, Урала,
Дальнего Востока.
Комлевой еловый трутовик (Onnia triqueter Bres.) является
возбудителем пестрой крупноямчатой комлевой гнили ели. Плодовые
тела гриба — однолетние, в виде шляпок без ножки (иногда с
зачатком ножки), расположенные поодиночке или в виде чере-
пицеобразных групп. Шляпки тонкие, плоские,
желтовато-коричневые, с острым краем, в молодом возрасте — грубоволосистые.
Ткань темно-коричневая. Гименофор трубчатый. Трубочки
короткие, с маленькими сероватыми или коричневыми порами.
Плодовые тела чаще всего располагаются на высоте до 70 см, а на
сильно ослабленных деревьях — до 1 —1,5 м от земли.
Комлевой еловый трутовик распространен во всех таежных
ельниках. В европейской части страны он встречается и в зоне
смешанных лесов, почти до южной границы произрастания ели. Он
поражает стволы живых деревьев ели, изредка — сосны и
лиственницы, вызывая в их комлевой части ядровую или одностороннюю
ядрово-заболонную гниль.
Вначале зараженная древесина становится желтоватой, затем в
ней появляются светло-бурые овальные пятна, внутри которых со
временем образуются довольно крупные пустоты, заполненные
белым веществом, с коричневым окаймлением. Оно особенно
хорошо заметно на продольных разрезах гнили. На последних стадиях
разрушения древесина растрескивается по годичным слоям. В
трещинах развиваются темно-коричневые тонкие разветвленные
шнуры грибницы.
В местах образования плодовых тел гниль выходит к
периферии, поражает заболонь и вызывает отмирание камбия. Обычно
гниль распространяется вниз, иногда поражая также и корни. Вверх
по стволу гниль может подниматься на высоту до 2—4 м от
корневой шейки. На пораженных деревьях часто наблюдаются
сильные смолотечения.
Комлевым трутовиком могут поражаться деревья от III до VIII,
но чаще до IV класса возраста. Первичное заражение стволов
осуществляется базидиоспорами через различные механические
повреждения.
Вред, наносимый грибом, весьма значителен: резко
уменьшается выход наиболее ценных сортиментов, так как нижние части
пораженных стволов пригодны только на дрова. Потери деловой
древесины могут составлять от 17 до 50 %.
325
Рис. 102. Плодовые тела чешуйчатки
жирной
Чешуйчатка жирная (Pholiota
adiposa Fr.) вызывает бурую ям-
чато-волокнистую ядровую гниль
хвойных и лиственных пород.
Наибольший вред чешуйчатка
причиняет древостоям пихты, но
поражает также ель,
лиственницу, реже березу, тополь, ольху,
липу, бук. Встречается на живых
и отмерших деревьях.
Гниль, вызываемая чешуйчат-
кой, — стволовая, а у
лиственницы иногда и корневая.
Древесина в начальной стадии гниения
становится желтоватой, затем приобретает бурую окраску, и в ней
появляются небольшие углубления или узкие канальцы,
заполненные рыжевато-бурым мицелием. В конечной стадии разрушения
древесины иногда образуется дупло.
Плодовые тела имеют форму шляпок с центральной или
боковой ножкой; часто образуются группами. Шляпки округлые, обычно
выпуклые, с бугорком в центре, мясистые; поверхность клейкая,
золотисто-желтая или коричневато-желтая, с
немногочисленными бурыми чешуйками, которые со временем исчезают. Ткань
желтовато-белая. Гименофор пластинчатый. Пластинки частые,
широкие, буровато-желтого цвета. Ножка плотная, беловатая, внизу
более темная и утолщенная, с перепончатыми кольцами и
чешуйками (рис. 102).
Чешуйчатка жирная широко распространена в Сибири, на
Урале, встречается на Кавказе.
Окаймленный трутовик (Fomitopsis pinicola (Sw. et Fr.) Karst.)
вызывает светло-бурую ядрово-заболонную призматическую гниль
хвойных, реже — лиственных пород. Плодовые тела многолетние,
копытообразные, подушковидные, плоские или
полураспростертые. Поверхность молодых плодовых тел глянцевая, охряно-жел-
тая или красная, старых — матовая, от буровато-серой до почти
черной, неровная, с кремово-белой, оранжевой,
вишнево-красной или красновато-бурой каймой по краю (отсюда название
трутовика). Гименофор желтовато-белый или буроватый, трубчатый,
с довольно крупными округлыми порами. Ткань желтоватая или
светло-коричневая, пробково-деревянистая.
Окаймленный трутовик встречается в основном на валежнике,
пнях, сухостое, буреломных деревьях, срубленной древесине в
лесу и на складах, сухобочинах живых деревьев.
Пораженная древесина становится розоватой, затем желтовато-
или красновато-бурой, с белесыми пятнами и полосами. В
конечной стадии гниль приобретает более темную окраску, и в ней обра-
326
зуются многочисленные трещины,
заполненные беловатыми пленками мицелия.
Трутовик Гартига {Phellinus hartigii (All.
et Schnab.) Bond.) вызывает белую
ядровую гниль пихты. Плодовые тела гриба
многолетние, твердые, копытообразные, с
утолщенно-закругленным краем,
широкими, иногда неясно выраженными
концентрическими зонами и единичными
радиальными трещинами или без них.
Поверхность плодовых тел желтовато-бурая,
часто серая или черноватая, с более
светлым краем. Гименофор трубчатый,
ржаво-коричневый, с мелкими округлыми Рис. 103. Плодовое тело
порами. Ткань плодового тела ржаво-бу- трутовика Гартига
рого цвета, деревянистая (рис. 103).
Заражение происходит чаще всего через механические
повреждения ствола, трещиноватые опухоли ржавчинного рака, места
облома ветвей; поэтому гниль нередко имеет раневой характер.
Протяженность гнили в стволе составляет в среднем 6—8 м, гниль
обычно ядровая, иногда ядрово-заболонная.
В начальной стадии развития гнили древесина темнеет, затем
постепенно становится более светлой. В конечной стадии гниль
светло-желтого цвета с белесоватыми пятнами и черными
извилистыми линиями на границе между пораженной и здоровой
древесиной. Развитие гнили способствует бурелому.
Трутовик Гартига поражает различные виды пихты: сибирскую,
белокорую, кавказскую, европейскую. Зараженность пихтовых дре-
востоев местами весьма значительна. Гриб распространен в
Сибири, на Дальнем Востоке, Кавказе.
12.4.2. Стволовые гнили лиственных пород
Ложный дубовый трутовик {Phellinus robustus (Karst.) Bourd. et
Galz.) вызывает белую полосатую гниль живых деревьев дуба.
Плодовые тела многолетние, бесформенно желвакообразные, подуш-
ковидные или копытообразные, с широким закругленным
желтовато-охряным или рыжеватым краем и гименофором того же
цвета с очень мелкими округлыми порами. Поверхность серовато-
бурая или черноватая, бугристая, с трещинами. Ткань очень
твердая, деревянистая, желтовато-бурая. Плодовые тела ложного
дубового трутовика внешне очень похожи на плодовые тела
трутовика Гартига (см. рис. 103), хотя филогенетическая специализация
этих грибов совершенно разная.
Заражение стволов происходит через морозобоины, раны от
затесок и другие повреждения. Пораженная древесина буреет, за-
327
тем в ней появляются светлые полосы. В конечной стадии гниль
желтовато-белая, с тонкими извилистыми темными линиями. В
пораженной древесине часто образуются скопления рыжеватого
мицелия.
Гниль в основном ядровая, но местами заходит в заболонь,
камбий и луб, вызывая образование сухобочин и ран. Поражение
ложным дубовым трутовиком приводит к суховершинности,
бурелому, в старых древостоях — к усыханию деревьев. Развиваясь в
нижней части ствола и поднимаясь до 2 — 3 м, гниль приводит к
значительным потерям деловой древесины. Ложный дубовый
трутовик широко распространен и является одним из наиболее
вредоносных трутовиков, поражающих дуб.
Дуболюбивый (дубовый) трутовик (Inonotus dryophilus (Pers.)
Murr.) вызывает пеструю ядровую гниль живых стволов дуба.
Плодовые тела однолетние, толстые, копытообразные или желвако-
образные, сначала мягкие, затем твердеющие. Поверхность их
ржаво-коричневая, шероховатая или ямчатая, щетинистая или
голая, с тонкой кожицей. Гименофор сложен из длинных
коричневых трубочек с округлыми или угловатыми неравновеликими
порами, на которых выделяются капли желтоватой жидкости. Ткань
ржаво-бурая грубоволокнистая, в месте прикрепления к стволу с
желтовато-белыми прожилками, песчанистой структуры.
Плодовые тела, как правило, быстро разрушаются насекомыми.
Заражение происходит в местах облома живых ветвей,
имеющих ядро, откуда гриб переходит в ядровую часть ствола.
Древесина сначала буреет, потом в ней появляются белые пятна и полосы
целлюлозы, постепенно сливающиеся. Со временем образуются
ямки и пустоты. В конечной стадии гниль становится рыхлой,
пористой, волокнистой. Вдоль ствола и по диаметру гниль
распространяется быстро, а ее протяженность в стволе может достигать
6—14 м. Это приводит к большим потерям деловой древесины.
Дубовая губка (Daedalea quercina (L.) Fr.). Плодовые тела
многолетние, в виде сидячих полукруглых копытообразных, иногда
плоских шляпок с утолщенным основанием и тонким краем, реже
полураспростертые, часто расположенные черепицеобразными
группами (рис. 104). Поверхность голая, неровная, бугорчатая или
морщинистая, с неясными зонами, серовато-коричневого или
бежевого цвета. Гименофор лабиринтообразный
того же цвета. Ткань пробковидная.
Дубовая губка обычно встречается на
сухостое, буреломе, валежнике, пнях,
заготовленной древесине. Изредка поражает старые,
поврежденные, сильно ослабленные деревья,
вызывая ядровую (и частично заболонную) гниль
Рис. 104. Плодовое комлевой части ствола. Из разрушающихся
тело дубовой губки пней мицелий гриба иногда переходит в ос-
328
нования стволов порослевого дуба, поднимаясь на высоту 1 — 3 м.
Гниль деструктивная, темно-бурая, с сероватым оттенком. В
конечной стадии образуются трещины, гниль разделяется на
пластинки. В трещинах развиваются желтоватые или серовато-белые
пленки мицелия.
Серно-желтый трутовик (Laetiporus sulphureus (Bull.) Bond, et
Sing.) вызывает красно-бурую ядровую гниль стволов дуба и
других лиственных пород, а также некоторых хвойных, например
лиственницы. Плодовые тела однолетние, собранные черепицеоб-
разными группами, имеют вид плоских, округлых или
веерообразных шляпок, с волнистым или лопастным краем. Поверхность
голая, складчатая, розовато-оранжевая; ткань желтоватая, почти
белая (рис. 105). Гименофор серно-желтого цвета, с короткими
трубочками. Молодые плодовые тела мягкие, мясистые,
съедобные. С возрастом они обесцвечиваются, становятся хрупкими.
Серно-желтый трутовик поражает деревья разного возраста, но
чаще старые, ослабленные. Заражение происходит через морозобой-
ные трещины и обломанные сучья. Гниль на начальных стадиях
розоватая, с белыми штрихами и точками; позднее она
приобретает красновато-бурую окраску, растрескивается по годичным
слоям и распадается на крупные призмочки. В трещинах образуются
желтовато-белые, плотные, замшевидные пленки мицелия. Обычно
поражается нижняя часть ствола, но иногда протяженность гнили
в стволе может достигать 10 —15 м. Сильное ее развитие приводит
к ослаблению и постепенному усыханию деревьев, образованию
дупел, бурелому, большим потерям деловой древесины, так как
наиболее ценные отрезки ствола идут на дрова или в отходы.
Чешуйчатый трутовик (Polyporus squamosus Huds. ex Fr.)
вызывает белую раневую ядровую гниль лиственных пород (ильмовых,
клена, липы, дуба, тополя, березы и других).
Плодовые тела гриба однолетние, в виде шляпок с немного
загнутыми краями и чаще всего с боковой ножкой. Поверхность
Рис. 105. Плодовые тела возбудителей стволовых гнилей лиственных пород:
/ — серно-желтого трутовика; 2 — чешуйчатого трутовика (а — поверхность
шляпки, б — гименофор)
329
шляпки немного вдавленная, желтоватая, с крупными
коричневыми чешуйками. Ножка мясистая коричневая, с черным
основанием. Ткань плодового тела в свежем состоянии белая, мягкая,
при высыхании желтеет и крошится. Гименофор трубчатый с очень
большими, в виде ячеек, угловатыми, часто расщепленными
порами (см. рис. 105).
Заражение ствола происходит через раны в коре,
морозобоины и другие повреждения. Гниль, вызываемая чешуйчатым
трутовиком, ядровая, но распространяется и на заболонь. Развивается
чаще всего в нижней части ствола, иногда заходит в корни. В
конечной стадии гниль становится белой, часто с темными
линиями, и в ней появляются многочисленные трещинки, идущие в
разных направлениях; в них часто образуются скопления белого
мицелия.
Чешуйчатый трутовик — типичный раневой паразит, наиболее
часто встречается в поврежденных старых древостоях, лесопарках
и других насаждениях с высокой рекреационной нагрузкой.
Ложный трутовик (Phellinus igniarius (L. ex Fr.) Quel.) —
возбудитель белой ядровой гнили многих лиственных пород. Это
сборный вид, который включает ряд специализированных форм,
поражающих разные породы (березу, ольху, иву, клен, тополь и др.).
Плодовые тела многолетние, чаще копытообразные, иногда жел-
вакообразные или подушковидные, реже распростертые (рис. 106).
Их поверхность темно-серая или буровато-черная, трещиноватая,
с концентрическими бороздками. Ткань очень твердая,
деревянистая, рыжевато-коричневая. Гименофор трубчатый, ржаво-бурый,
с очень мелкими, едва заметными округлыми порами.
Наибольшее распространение имеют формы ложного трутовика,
поражающие осину и березу.
Заражение стволов базидиоспорами происходит через
механические повреждения, морозобоины, места облома ветвей.
Пораженная древесина вначале буроватая, потом в ней появляются
светло-желтые пятна и полосы, которые со временем сливаются.
В последней стадии гниль желтовато-белая, с черными
извилистыми замкнутыми линиями и скоплениями рыжеватого
мицелия. Она отделяется от здоровой части
темно-бурым кольцом. Гниль распространяется
главным образом в средней части ствола, но
иногда тянется по всему стволу и толстым
ветвям, вызывая потери деловой древесины
до 100%.
Ложный трутовик повсеместно
встречается в насаждениях всех классов возраста; по-
Рис. 106. Плодовые раженность спелых древостоев иногда дости-
телаложноготруто- гает 80—100% и может носить скрытый ха-
вика рактер (без образования плодовых тел).
330
Осиновый трутовик (Phellinus tremulae (Bond.) Bond, et Bor.)
поражает только осину, вызывая белую ядровую гниль живых стволов.
Плодовые тела многолетние, относительно небольшие, полукопы-
товидные или желвакообразные, полураспростертые или
распростертые. Поверхность серовато-черная, с мелкими трещинками,
выступающая из-под наплыва коры. Ткань очень твердая,
деревянистая, ржаво-коричневая. Гименофор скошенный или
округло-выпуклый, рыжевато-бурый, с очень мелкими округлыми порами.
Заражение стволов осуществляется базидиоспорами через
обломанные ветви и повреждения коры, а также мицелием через
корневые отпрыски и пневую поросль. Древесина центральной
части ствола вначале приобретает серовато-буроватую или бледно-
розовую окраску, затем становится желтовато-белой. Развитая гниль
имеет волокнистую структуру, окружена темно-бурой линией и
буровато-оливковым широким кольцом.
Осиновый трутовик — один из самых вредоносных трутовых
грибов. Пораженность средневозрастных, приспевающих и спелых
насаждений может достигать 70—90 %. Гниль развивается главным
образом в средней части ствола, но иногда распространяется по всей его
длине, что влечет за собой огромные потери деловой древесины.
Чага. Чагой называют образующиеся на живых стволах березы
крупные бесформенные наросты черного цвета, очень твердые,
деревянистые, изрезанные многочисленными глубокими
трещинами. Внутренняя ткань наростов неоднородна по цвету и
структуре: в основном твердая, ржаво-коричневого цвета, но
местами более мягкая и светлая, с тонкими желтоватыми
прожилками. Наросты чаще всего образуются в местах облома ветвей,
механических и других повреждений ствола (рис. 107).
Плодовые тела возбудителя чаги — гриба Inonotus obliquus (Pers.)
Pil. — появляются под корой около
наростов. Они распростертые, тонкие, бурого
цвета, с трубчатым гименофором,
довольно быстро разрушающиеся; поэтому стадия
плодовых тел часто проходит незамеченной.
Чага поражает главным образом березу,
но встречается и на других лиственных
породах: ольхе, клене, буке, ясене, рябине. В
стволах пораженных деревьев развивается
ядровая белая гниль, сходная по своим
признакам с гнилью от ложного трутовика. Чага
широко распространена и часто встречается
в насаждениях поражаемых пород.
«Березовая» чага обладает лечебными свойствами и
ее заготавливают как лекарственное сырье.
Кленовый трутовик (Oxyporuspopulinus (Fr.) Рис. 107. Нарост чаги
Donk.) вызывает желтовато-белую ядровую на стволе березы
331
гниль стволов живых деревьев. Плодовые тела многолетние, собраны
черепицеобразными группами на общем основании, реже —
одиночные. Они имеют вид небольших шероховатых шляпок с белой,
желтовато-серой или черноватой поверхностью, обычно
зарастающей зеленым мхом. Иногда плодовые тела полураспростертые.
Ткань плодового тела белая или светло-желтая, пробковидная или
деревянистая. Трубочки гименофора короткие с очень мелкими
округлыми желтовато-белыми порами.
Кленовый трутовик особенно часто встречается на клене, но
может поражать также вяз, ясень, липу, тополь и другие
лиственные. Пораженность клена в насаждениях, особенно городских
парках и лесопарках, может быть высокой. Заражение происходит через
морозобойные трещины и механические повреждения, главным
образом в комлевой части ствола.
На первых стадиях развития гнили древесина становится
зеленовато-бурой, затем несколько светлеет, приобретая желтоватый
оттенок. В конечной стадии гниль распадается вдоль сердцевинных
лучей. В пораженных стволах часто образуются дупла.
Настоящий трутовик (Fomes fomentarius (L.) Gill.) поражает
лиственные породы (березу, бук, ольху, осину, реже дуб, ясень,
липу и др.), вызывая белую мраморную ядрово-заболонную гниль
стволов. Плодовые тела многолетние,
копытообразные, обычно одиночные (рис. 108).
Поверхность голая, гладкая, с
концентрическими бороздками, пепельно-серая
(иногда желтоватая) до черно-серой,
покрытая твердой коркой. Гименофор
сероватый, с возрастом буреющий, с
округлыми порами, заполненными белым
мицелием. Ткань желто-коричневая,
плотно-войлочная или
клочковато-волокнистая.
Настоящий трутовик заражает
отмирающие, старые, поврежденные и
ослабленные, реже здоровые деревья. Заражение
происходит через обломанные ветви,
раны, сухобочины. Мицелий быстро
распространяется из заболони во внутренние
части ствола, интенсивно разрушая
древесину, которая в начальной стадии гниения
буреет, потом становитея светло-желтой
Рис. 108. Плодовые тела и> наконец, почти белой, с буровато-чер-
возбудителей гнилей ли- ными штрихами и тонкими извилистыми
ственных пород: линиями, похожими на графики. В конеч-
/ — настоящего трутовика; н°й стадии гниль расслаивается на тонкие
2 — березовой губки ' пластинки, расщепляется на волокна и
332
превращается в очень легкую мягкую труху. Настоящий трутовик
наиболее опасен в буковых и березовых лесах, так как может стать
причиной массового бурелома. Поражая заготовленную
древесину, приносит большой технический вред.
Березовая губка (Piptoporus betulinus (Bull, ex Fr.) Karst.) поражает
только березу, вызывая красно-бурую гниль стволов. Плодовые
тела — однолетние боковые шляпки без ножки или с ее зачатком,
округлые или почковидные, слегка выпуклые, с тупым, иногда
подогнутым вниз краем (см. рис. 108). Поверхность ровная, гладкая,
в молодом возрасте белая, позднее коричневая или сероватая,
покрытая тонкой, легко отделяющейся кожицей. Ткань однородная,
белая, мягкопробковидная. Трубочки гименофора белые, с
возрастом темнеющие. Поры мелкие, неравновеликие. Встречается
главным образом в насаждениях с неблагоприятными условиями
роста, на валежнике, сухостое, редко на живых ослабленных березах.
Гниль ядрово-заболонная. Вначале древесина приобретает
желтовато-бурую, затем красновато-коричневую окраску,
растрескивается и распадается на отдельные куски. В трещинах часто
образуются кремово-белые пленки мицелия. В конечной стадии гниль
очень хрупкая и легко растирается в тонкий порошок.
12.4.3. Защита насаждений от стволовых гнилей
Мероприятия по защите насаждений от стволовых гнилей
должны планироваться и проводиться дифференцированно по
группам лесов, а в пределах каждой группы — с учетом целевого
назначения и возраста насаждений.
В лесах I группы необходимо прежде всего строго
выполнять санитарные правила, осуществлять комплекс санитарно-оздо-
ровительных мероприятий. В молодняках следует своевременно
проводить рубки ухода с целью формирования здоровых, биологически
устойчивых насаждений оптимальных полноты и состава. В
средневозрастных и более старых древостоях необходимо регулярно
проводить выборочные санитарные рубки с выборкой сухостоя,
бурелома, усыхающих и ветровальных деревьев, а также сильно
ослабленных деревьев с плодовыми телами трутовых грибов —
возбудителей наиболее опасных гнилевых болезней. Сильно изре-
женные древостой (с полнотой ниже 0,3 — 0,4) целесообразно
назначать в сплошную санитарную рубку с последующим закульти-
вированием площадей. Срубленную древесину следует
своевременно вывозить из насаждений. Чтобы предупредить заселение
древесины стволовыми вредителями, надо осуществлять ее
химическую защиту (она запрещена для древесины, идущей на сплав)
или окорять. Не следует допускать захламленности насаждений;
порубочные остатки, бурелом, валежник надо регулярно
вывозить из леса и уничтожать.
333
Большое значение имеет профилактика заражения деревьев
стволовыми гнилями. Для предотвращения механических
повреждений стволов при рубках необходимо строго соблюдать правила
их проведения. Чтобы уменьшить опасность возникновения моро-
зобойных трещин, рекомендуется оставлять густые опушки. Также
следует проводить биотехнические мероприятия, направленные
на регулирование численности копытных (особенно лосей) и
уменьшение угрозы повреждения ими стволов и молодого
подроста. Не менее важно создание более устойчивых к болезням
смешанных хвойно-лиственных насаждений с правильным подбором
и размещением пород. Схемы смешения должны составляться с
учетом лесорастительного района, типа леса и реальной угрозы
тех или иных болезней.
В парках, мемориальных насаждениях,
городских посадках и особо ценных участках лесопарков наряду с
тщательным проведением общих санитарно-оздоровительных
мероприятий важное значение приобретает индивидуальная защита
деревьев: лечение ран, обрезка усохших или пораженных ветвей,
удаление плодовых тел трутовых грибов, пломбирование дупел и т. п.
Лечение ран и сухобочин лучше всего проводить ранней весной.
Прежде всего необходимо очистить рану до здоровой древесины
ножом или стамеской. Очищенная поверхность должна быть слегка
вогнутой, гладкой. Затем поверхность обрабатывают
антисептиком (5 %-м раствором медного купороса, 20 %-м карболинеумом
или креозотовым маслом) и после просыхания покрывают
водонепроницаемым составом: масляной краской, садовой замазкой
(10 частей нигрола, 6 частей канифоли, 3 части золы, 1 часть
воска) или петролатумной замазкой (петролатума 80 %,
канифоли 10 %, растительных масел 10 %). Последняя особенно
эффективна, так как способствует заживлению и зарастанию ран.
Если на сухобочине наблюдается гниение древесины, лечение
проводят аналогичным способом. Если гнили нет, то для
профилактики надо промазать сухобочину антисептиком и закрасить
масляной краской под цвет коры.
Обрезку усохших или больных ветвей, водяных побегов и
пасынков проводят заподлицо с последующим антисептированием
поверхности среза и покрытием его петролатумной или садовой
замазкой либо масляной краской. Так же обрабатывают места
среза плодовых тел трутовиков.
Пломбирование дупел проводят летом в сухую погоду. Дупло
очищают, срезая ножом или стамеской пораженную древесину, и
удаляют весь мусор. После просушки поверхность дупла
дезинфицируют антисептиком, вновь просушивают и покрывают масляной
краской, каменноугольным (кузбасским) лаком, асфальтобитум-
ным лаком или битумной эмульсией. Затем заполняют дупло
цементирующей смесью: 1 часть цемента и 2 части песка или 1 часть
334
расплавленного битума и 3—4 части опилок. Еще не остывшую
смесь закладывают в дупло, заглаживают ее верхний слой и после
остывания окрашивают под цвет коры. Рекомендуют и другие
лечебные составы. Дупло заполняют до каллюса или камбиального
слоя, иначе цементирующая смесь будет мешать естественному
зарастанию дупла.
Исключительно важное значение в насаждениях лесопарковых
зон, мемориальных насаждениях и парках имеют мероприятия по
регулированию рекреационных нагрузок, профилактике
механических и других повреждений деревьев антропогенного характера.
В лесах II и III групп, где ведут рубки главного
пользования и в раде случаев невозможны санитарно-оздоровительные
мероприятия, важнейшая роль принадлежит периодическим
обследованиям. Их проводят для выявления видового состава гниле-
вых болезней, степени пораженности насаждений и снижения под
влиянием гнилей товарности древостоев в связи с изменениями
их возрастного состава. Эти работы особенно актуальны для тех
районов, где проектируется создание крупных
лесопромышленных комплексов.
Глава 13
гАЗРУШЕНИЕ ДРЕВЕСИНЫ НА СКЛАДАХ,
В ТЕХНИЧЕСКИХ СООРУЖЕНИЯХ
И ЗДАНИЯХ
Живые ткани растений, в том числе живые слои заболони
растущих деревьев, обладают свойствами, обеспечивающими их
устойчивость к сапротрофной микрофлоре. После рубки дерева его
защитные механизмы выходят из строя, в отмирающих клетках
древесины происходят необратимые изменения, и она постепенно
заселяется сапротрофными дереворазрушающими грибами.
Биологическое разложение древесины, которое начинается еще
в лесу и продолжается в процессе ее хранения, транспортировки
и эксплуатации, обычно проходит несколько этапов,
характеризующихся последовательной сменой комплексов грибов, каждый
из которых заселяет древесину лишь на определенном этапе и при
определенных условиях внешней среды (влажности древесины и
воздуха, температуры, аэрации и др.). Таким образом, эти
комплексы представляют собой экологические группы грибов, тесно
связанные с режимом хранения лесоматериалов и эксплуатации
зданий и сооружений.
Первый комплекс грибов, раньше всех заселяющих свежесруб-
ленную древесину в лесу или на складах, составляют
деревоокрашивающие и плесневые грибы. Вслед за ними идут складские
грибы, разрушающие древесину на складах, а также деревянные
части открытых (холодных) сооружений; эти грибы составляют
второй комплекс. Наконец, к третьему комплексу относятся
домовые грибы, разрушающие деревянные конструкции
преимущественно в отапливаемых зданиях. Однако не всегда между этими
экологическими группами можно провести четкую грань. Иногда
срубленную древесину поражают грибы, вызывающие гнили
растущих деревьев (например, корневая губка, окаймленный
трутовик и др.). Некоторые деревоокрашивающие грибы могут вести
себя как складские дереворазрушающие грибы. Складские грибы
при подходящих условиях могут продолжать свое развитие в
закрытых постройках и т.д. Внутри экологических групп отдельные
336
виды также заметно различаются по своим требованиям к
условиям среды.
13.1. ПОРАЖЕНИЕ ДРЕВЕСИНЫ
ДЕРЕВООКРАШИВАЮЩИМИ И ПЛЕСНЕВЫМИ
ГРИБАМИ
Неестественная окраска древесины (в виде цветных полос,
пятен или сплошного окрашивания) — весьма распространенное
явление, связанное с деятельностью деревоокрашивающих и
плесневых грибов.
Деревоокрашивающие грибы поражают в основном свежезаго-
товленную древесину при ее хранении (в лесу или на складах) и
транспортировке, но часто встречаются также на валежнике,
сухостое, а иногда и на растущих деревьях. Эти грибы могут заселять
лесоматериалы в широком диапазоне температур (от 5 до 30 °С) и
влажности древесины (22 — 75 %), однако экологические
требования отдельных видов бывают ограничены более узкими рамками.
Древесина заражается спорами или мицелием при контакте
пораженных и здоровых лесоматериалов. Развитие
деревоокрашивающих грибов может продолжаться до тех пор, пока древесина
сохраняет естественную влажность. После ее подсыхания
жизнедеятельность этих грибов практически прекращается, а при
нагревании древесины до 80 °С они погибают.
Деревоокрашивающие грибы
Среди этих грибов имеются представители различных
систематических групп — сумчатых, базидиальных, несовершенных.
Некоторые из них поражают древесину лишь одной-двух
определенных пород, другие — многих лиственных и хвойных.
Различаются они и по отношению к тем или иным лесоматериалам и
сортиментам.
В биологии этих грибов много общего. Все они — сапротрофы,
питающиеся в основном содержимым отмирающих клеток
заболони. Мицелий гриба разрастается в паренхимных клетках
сердцевинных лучей, богатых питательными веществами, а затем
переходит в трахеиды, распространяясь в глубь древесины. Клеточные
стенки при этом почти не разрушаются, и, следовательно, не
разлагается лигниноцеллюлозный комплекс, составляющий
основную массу древесины.
Окраски древесины, вызываемые деревоокрашивающими
грибами, разнообразны по расположению, рисунку, цвету и
интенсивности. Обычно они проявляются в виде цветных пятен или
полос, реже — в виде сплошного окрашивания. Характер окраски
337
зависит от вида гриба, глубины его проникновения в древесину,
количества и цвета его гиф и спороношений, а также выделяемых
грибом пигментов.
Различают внутренние и наружные окраски древесины.
Внутренние, или ядровые, окраски, как правило, возникают
еще при жизни дерева. Они вызываются грибами, проникающими
внутрь ствола через обломанные сучья или раны, и
обусловливаются накоплением в центральной части ствола грибных
пигментов. После рубки дерева развитие этих грибов обычно
прекращается. Внутренние окраски наблюдаются сравнительно редко и почти
не имеют практического значения.
Наружные, или заболонные, окраски древесины (так
называемая цветная заболонь) иногда обнаруживаются вскоре
после отмирания дерева на корню (в сухостое) или на ветровальных
деревьях, но особенно часто появляются в древесине свежесруб-
ленных стволов при ее медленном просыхании. В большинстве
случаев эти окраски захватывают лишь заболонные слои древесины,
но некоторые из них переходят и в ядровую часть ствола.
Заболонные окраски, распространяющиеся на глубину до 2 мм, называют
поверхностными; окраски, проникающие более глубоко, —
глубокими. К заболонным окраскам относятся синева, кофейная тем-
нина, краснина, желтизна, оранжевая, зеленая и др.
Синева. Так называют глубокую патологическую окраску
голубовато-серых, синеватых, зеленовато-синих, серо-коричневых и
синевато-черных тонов, которая наиболее часто встречается на
заболонной древесине как сухостойных, так и срубленных
деревьев. Синева поражает преимущественно окоренную древесину
многих хвойных, реже лиственных пород. Неокоренная древесина
обычно поражается только на торцах, так как кора препятствует
проникновению грибов — возбудителей синевы. Повреждение коры
стволовыми вредителями (короедами, златками, усачами и др.),
механические повреждения стволов способствуют заражению
заболони деревоокрашивающими грибами.
В круглых лесоматериалах может развиваться боковая, торцовая
и подслойная синева. При боковой синеве, которая
наблюдается в основном у хвойных пород, на боковой поверхности
древесины вначале образуются пятна, позднее сливающиеся. Затем
окраска проникает по сердцевинным лучам в глубь заболони. На
поперечных срезах бревен она имеет форму клиньев, отдельных
сегментов или сплошного кольца, охватывающего наружные слои
древесины (сплошная синева). Торцовая синева,
встречающаяся на любых породах, распространяется от торцов во
внутренние части сортиментов. На поперечных срезах она проявляется в
виде пятен, а на продольных — в виде полос различной ширины.
Подслойная синева окрашивает более глубокие слои
древесины, располагаясь под неокрашенным наружным слоем заболони.
338
При хранении пиломатериалов и шпона может появиться
налетная синева, которая имеет вид серых или
синевато-черных пятен и чаще всего поражает боковые поверхности
сортиментов, но иногда проникает и в глубь древесины. При летней сушке
досок в штабелях, в местах соприкосновения их с прокладками,
встречается так называемая прокладочная синева.
Для большинства возбудителей синевы оптимальны
температура 20...27°С, влажность древесины 35—90% и наличие в ней
воздуха (не менее 15 % от ее объема).
Синеву вызывают около 70 видов деревоокрашивающих
грибов. Лесоматериалы хвойных пород, в основном сосны и ели, чаще
всего поражают сумчатые грибы рода Ceratocystis: C.pini (Miinch)
H.et Syd., Cpiceae (Miinch) H.et Syd., Ccoeruleum (Miinch) H.et
Syd. (рис. 109) и др. Реже встречаются другие представители
сумчатых и несовершенных грибов. Видовой состав грибов, вызываю-
Рис. 109. Синева древесины:
/ — продвижение гиф гриба рода Ceratocystis сквозь стенки трахеид сосны; 2 —
пораженная древесина; 3 — конидиальное спороношение гриба; 4 — коремии
(а) и перитеции (б) возбудителя; 5 — сумка с сумкоспорами; 6 — перитеций
339
щих синеву древесины лиственных пород, также представлен в
основном грибами этих групп.
Кофейная темнина. Глубокая кофейно-коричневая окраска
заболони в большинстве случаев вызывается несовершенным
грибом Discula brunneo-tingens Mejer и часто появляется в бревнах и
непросохших пиломатериалах хвойных пород, преимущественно
сосны, лиственницы и ели. В круглых лесоматериалах она из
заболони клиньями (в основном по сердцевинным лучам)
распространяется в сторону ядра. На боковых поверхностях бревен эта
окраска имеет вид полос или продолговатых пятен. Кофейная
темнина нередко обнаруживается вместе с синевой или красниной.
Коричневые, бурые и коричнево-черные окраски, вызываемые
другими видами сумчатых, базидиальных и несовершенных
грибов, встречаются на древесине различных хвойных и лиственных
пород.
Краснина. Этим термином обозначают красную, розовую,
малиновую окраску разнообразных тонов и оттенков. Краснина
также вызывается многими высшими грибами различных классов и
может встречаться на древесине любых лиственных и хвойных
пород.
Наиболее часто краснину вызывает базидиальный гриб Corticium
laeve Fr., образующий небольшие мягкокожистые плодовые тела
розовато-желтого или светло-коричневого цвета, сливающиеся в
тонкие пленки. Этот гриб поражает в основном окоренную
древесину сосны, ели и других хвойных пород, реже осины, вызывая
красноватую или нежную розовато-оранжевую окраску в виде
полос, глубоко проникающую в заболонь. По мере просыхания
древесины окраска бледнеет.
Интенсивную кровяно-красную окраску древесины хвойных
пород и дуба вызывает базидиомицет Peniophora sanguinea Bres. Он
выделяет красный пигмент, который накапливается в клетках
сердцевинных лучей и трахеидах. На поверхности пораженных
лесоматериалов гриб образует тонкие пленчатые плодовые тела и шнуры
красновато-оранжевого и красного цвета.
В красные, фиолетовые и розовые тона древесину хвойных и
лиственных пород окрашивают некоторые грибы-гифомицеты
родов Penicillium и Fusarium. Розовое, в виде полос, окрашивание
заболони вокруг карр у подсоченных сосен обычно связано с
заселением древесины сумчатым грибом Biatorella resinae Mudd.
Желтизна. Часто встречающаяся патологическая окраска как
грибного, так и негрибного происхождения. Например, глубокую
лимонно-желтую окраску (в виде прерывистых полос и пятен)
заболони хвойных и лиственных пород вызывает гифомицет
Verticillium glaucum Bon., обычно развивающийся при очень
высокой (близкой к 100%) влажности древесины. При обработке
пораженных лесоматериалов 10 %-м раствором едкого натра (NaOH)
340
желтая окраска, вызванная этим грибом, переходит в оранжево-
красную. Глубокая желтая окраска древесины дуба и ореха
грецкого возникает в результате окрашивания клеточных оболочек
золотисто-желтым пигментом, который выделяется несовершенным
грибом Eidamia catenulata Will., и образования желтых спороноше-
ний гриба. Известны и другие окраски этой группы.
Зеленая окраска также встречается довольно часто. Она может
быть вызвана многими видами сумчатых и несовершенных грибов.
Так, например, глубокое окрашивание древесины березы и
других лиственных пород в интенсивный сине-зеленый цвет
вызывают грибы-дискомицеты рода Chlorosplenium; зеленые окраски
разных оттенков возникают при заселении лесоматериалов грибами-
гифомицетами родов Penicillium, Trichoderma и др.
Деревоокрашивающие грибы могут поражать не только
древесину, но также искусственные материалы, вырабатываемые из
древесной массы или целлюлозы (бумагу, картон,
древесностружечные и древесноволокнистые плиты, ткани и др.), вызывая их
побурение, покраснение, пожелтение, появление пятен
коричнево-черного, розового или зеленоватого цвета.
Как правило, деревоокрашивающие грибы слабо воздействуют
на клеточные стенки и срединные пластинки и поэтому не
оказывают заметного влияния на химический состав и
физико-механические свойства древесины. Однако при длительном воздействии
некоторых видов этих грибов (например, вызывающих синеву)
технические свойства древесины ухудшаются: на 10—30 %
снижается ударная вязкость, понижается прочность при продольном
сжатии. Использование древесины, пораженной желтизной, в
клееных конструкциях может привести к разрушению клея. Поражение
деревоокрашивающими грибами (например, при синеве, красни-
не) увеличивает проницаемость древесины, делая ее
непригодной для многих целей. Кроме того, присутствие деревоокрашива-
ющих грибов свидетельствует о неправильном режиме хранения
древесины, при котором она может быть поражена и более
опасными дереворазрушающими грибами. Отдельные виды деревоок-
рашивающих грибов даже стимулируют развитие складских и
домовых грибов, а некоторые (например, Peniophora sanguinea (Fr.)
Bres., Hericium diversidens (Fr.) Pers. и др.) могут вызывать не
только окрашивание, но и гниль древесины. Кстати, изменение
нормального цвета древесины служит одним из первых признаков
гниения, вызываемого трутовыми грибами.
По этим причинам патологические окраски считаются
серьезным пороком древесины, снижающим ее ценность и сортность.
Значительная часть древесины с грибными окрасками (особенно
с синевой) бракуется, сбыт ее затруднен. Наличие окрасок не
допускается в древесине, предназначенной для несущих
конструкций зданий и сооружений, в других сортиментах ответствен-
341
ного и специального назначения (например, для
судостроительной, авиационной, оборонной промышленности), в фанерном
кряже и пиломатериалах, идущих на изготовление мебели,
музыкальных инструментов, бочек. Древесина с грибными окрасками
не допускается на. экспорт, что наносит огромный ущерб
внешней торговле и в целом экономике стран — экспортеров леса.
Иногда использование окрашенной древесины ограничивается из-
за утраты ее нормального цвета.
Неестественные окраски древесины могут возникать и без
участия грибов, являясь следствием внешних химических воздействий,
а также внутренних биохимических процессов, протекающих в
древесине. Например, на поверхности пиломатериалов и фанеры
из древесины пород, богатых дубильными веществами, в
результате реакций между этими веществами и солями железа могут
появиться пятна или полосы синевато-серого или черного цвета
(так называемые чернильные пятна). При обработке таких пятен
растворами роданидов калия или аммония они становятся ярко-
красными. Этого не происходит, если пятна имеют грибное
происхождение. У сплавной древесины хвойных пород поверхностная
лимонно-желтая окраска (сплавная желтизна) может быть
вызвана биохимическими процессами, связанными с недостатком
кислорода. При смачивании 10 %-м раствором NaOH сплавная
желтизна, в отличие от грибной, не изменяет цвета. Древесина и
изделия из нее могут приобретать не свойственную им бурую,
сероватую и другую окраску также под действием прямых солнечных
лучей и прочих абиотических факторов.
Плесневые грибы
Виды этих грибов развиваются в основном на поверхности
субстрата, где они образуют тонкие паутинистые пленки и
порошащие налеты, состоящие из мицелия и спороношений гриба.
Большая часть плесневых грибов относится к классу гифоми-
цетов или к порядку мукоровых. Это главным образом
представители родов Penicillium, Botrytis, Fusarium, Aspergillus^ Alternaria,
Cladosporium, Mucor и др. Образуемые ими налеты плесени
обычно имеют определенную, иногда довольно яркую окраску,
характерную для каждого рода или вида гриба: зеленую,
грязно-голубую, бурую, серую, розово-красную, желтую, черную и т.д.
Развиваясь на лесоматериалах, плесневые грибы выделяют
пигменты, которые проникают в поверхностный слой заболони (на
глубину не более 1 мм), вызывая ее неестественное окрашивание.
Плесневые грибы заселяют свежесрубленную древесину
одновременно с деревоокрашивающими грибами или вслед за ними.
Поражению лесоматериалов плесневыми грибами способствуют
те же условия, при которых развиваются деревоокрашивающие и
342
более опасные дереворазрушающие грибы: высокая влажность
самой древесины, увлажненная поверхность лесоматериалов, сырая
умеренно теплая погода. Появление плесени на хранящихся
сортиментах свидетельствует о неправильном режиме хранения
древесины и об угрозе ее заражения складскими и домовыми
грибами. При этом сами плесневые грибы, как и большинство дерево-
окрашивающих, активного разрушения древесины не вызывают.
Поскольку плесневые и деревоокрашивающие грибы очень
сходны по способности окрашивать древесину, экологическим
требованиям, характеру и уровню причиняемого вреда, четко
разграничить эти группы грибов довольно трудно.
13.2. РАЗРУШЕНИЕ ДРЕВЕСИНЫ НА СКЛАДАХ,
В ОТКРЫТЫХ СООРУЖЕНИЯХ И ХОЛОДНЫХ
ПОСТРОЙКАХ
В процессе хранения древесины на смену деревоокрашиваю-
щим и плесневым грибам приходят дереворазрушающие грибы
второго экологического комплекса — так называемые складские,
или штабельные. Вытеснение ими грибов-первопоселенцев
обусловливается происходящими в древесине изменениями
(увеличением содержания в ней кислорода, уменьшением влажности
и запаса легкодоступных питательных веществ в содержимом
клеток), а также угнетением грибов первого комплекса
антибиотиками, выделяемыми грибами второго комплекса. Увлажненная
поверхность лесоматериалов и наличие на них трещин, высокая
атмосферная влажность окружающего воздуха способствуют
прорастанию спор и проникновению складских грибов в
поверхностные слои древесины. Дальнейшее развитие этих грибов может
происходить в широком диапазоне влажности, но не ниже 30—35 %, и
в условиях достаточной аэрации.
Складские грибы разрушают также холодные постройки
(сараи, ангары и т.п.), деревянные части различных открытых
сооружений: опоры мостов, телеграфные столбы, ограды и пр.
Благоприятные условия для жизнедеятельности складских
грибов создаются при нарушении правил заготовки, перевозки и
хранения древесины, правил строительства и эксплуатации
сооружений, имеющих деревянные конструкции.
Большинство складских грибов относится к гименомицетам.
Среди них различают слабых и сильных разрушителей древесины.
Представители первой группы (субдеструкторы) в начальный
период заселения ими древесины питаются, как и
деревоокрашивающие грибы, в основном содержимым клеток, но в
дальнейшем воздействуют и на клеточные стенки; это приводит к
медленному и неполному разрушению наружных слоев древесины
343
(возникаетзаболонная гниль). Представители второй группы
(деструкторы) потребляют преимущественно вещества клеточных
оболочек, что приводит к более интенсивному гниению
древесины. Они начинают развиваться в наружных слоях
лесоматериалов, но потом проникают и во внутренние их части, вызывая
сплошную ядрово-заболонную гниль, быстрое и полное
разрушение пораженных сортиментов.
Твердую заболонную гниль круглого леса, преимущественно
хвойных пород, чаще всего вызывают грибы-субдеструкторы: пе-
ниофора гигантская, щелевик обыкновенный, стереум кровяно-
красный. При поражении этими грибами древесина разрушается
слабо, и ее технические свойства изменяются незначительно.
Мягкую заболонную гниль, при которой происходит более
активное разрушение древесины и существенно снижается ее
прочность, вызывают валежный еловый, буро-фиолетовый, розовый
и другие трутовики-субдеструкторы.
Ядрово-заболонную гниль лесоматериалов хвойных пород при
хранении, а также поражение деревянных конструкций в
открытых сооружениях вызывают грибы-деструкторы. Чаще всего это
шпальный гриб, столбовые грибы, трутовики групповой и
душистый. Под воздействием этих грибов обычно развивается бурая
трещиноватая гниль, происходит интенсивное и полное
разрушение древесины.
Ядрово-заболонную гниль лесоматериалов лиственных пород
при длительном хранении вызывают очень многие
трутовики-деструкторы: разноцветный, утолщенный, волосистый, серый,
березовый пластинчатый и др. При поражении этими грибами обычно
развивается белая коррозионная гниль.
Пениофора гигантская (Peniophora gigantea (Fr.) Mass.)
встречается на древесине хвойных пород (особенно неокоренной) уже
в первый сезон хранения на лесных биржах и складах, а также в
постройках из сырого леса и подвалах. Гриб вызывает потемнение
и сравнительно медленное разрушение поверхностных слоев
древесины, но иногда проникает и в более глубокие ее слои. Гниль
имеет волокновидную, неясно коррозионную структуру.
Пораженная древесина сначала почти не изменяет свою окраску, затем
слегка буреет, в ней иногда появляются мелкие неясные трещины.
На поверхности пораженных частей образуются беловатые, вато-
образные, иногда довольно мощные пленки грибницы и очень
тонкие веерообразные шнуры.
Плодовые тела имеют вид длинных (до 50 см длиной)
молочно-белых или серовато-кремовых восковидных пленок. В сырую
погоду они разбухают, а при высыхании становятся как бы
пергаментными и легко отделяются от субстрата. Гименофор
совершенно гладкий, матовый, бледный, грязно-белый или
желтовато-сероватый с розовым оттенком (рис. 110).
344
Рис. 110. Плодовые тела складских грибов:
1 — пениофоры гигантской; 2 — щелевика обыкновенного; 3 — шпального гриба;
4 — столбового (заборного) гриба
Щелевик, или щелелистник, обыкновенный {Schizophyllum
commune Fr.) поражает древесину как хвойных, так и лиственных
пород; часто встречается на пнях, валежнике, сухостое, сухобо-
чинах растущих деревьев и различных лесоматериалах при
хранении. Плодовые тела обычно располагаются в щелях (отсюда
название) и имеют вид маленьких, 1 —4 см в диаметре, тонких
округлых сидячих шляпок (см. рис. 110), иногда с зачатком боковой ножки.
Поверхность шляпок светло-серая, войлочная, со слегка
загнутыми вниз краями. Пластинки гименофора расположены
веерообразно. Они сероватые или лиловато-коричневые, иногда
расщепленные, кожистые. Гриб вызывает медленно развивающуюся
поверхностную бурую гниль древесины.
Валежный еловый трутовик {Hirschioporus abietinus (Fr.) Donk)
поражает лесоматериалы, валежную древесину, пни, сухостой
хвойных пород, поселяется на сухобочинах стволов. Гниль
развивается преимущественно в заболони, но у ели и пихты может
проникать и в более глубокие слои древесины. Пораженная
древесина красновато-бурая с небольшими белыми выцветами
целлюлозы и черными штрихами. В дальнейшем появляются
удлиненные ямки, гниль становится мягковолокнистой, трухлявой, легко
крошится. Плодовые тела тонкие, кожистые, в виде небольших
345
сидячих шляпок, полуотогнутые или распростертые, собранные
черепицеобразными группами, часто сплошь покрывающими
лесоматериалы. Поверхность шляпок пепельно-серая, войлочная, ги-
менофор фиолетовый, трубчатый, поры округлые, угловатые или
вытянутые, часто с расщепленными краями.
Буро-фиолетовый трутовик (Hirschioporus fusco-violaceus (Fr.)
Donk) поражает преимущественно обработанную древесину
сосны и лиственницы. Встречается также на стволах и ветвях
сухостойных и ветровальных деревьев этих пород. Вначале гниль
светло-бурая, позднее пестрая, с белыми вытянутыми пятнами
целлюлозы и пустотами. В конечной стадии гниль мягкая, легко
расщепляется на волокна. Плодовые тела в виде небольших боковых
шляпок, полураспростертые или распростертые, часто
расположенные черепицеобразными группами. Поверхность мягковойлоч-
ная, серовато-буроватая. Гименофор фиолетово-бурый, в виде
радиально расходящихся зубчатых пластинок.
Стереум кровяно-красный (Stereum sanguinolentum Fr.)
встречается на круглых лесоматериалах, пнях и стволах ели, реже —
других хвойных пород. Поражает заболонную древесину, вызывая
вначале светло-бурую, затем более темную, коричнево-бурую гниль
с мелкими пустотами, которая часто развивается в местах
механических повреждений ствола. На пораженных материалах
образуются полураспростертые тонкокожистые плодовые тела диаметром
до 3 см. Отогнутая поверхность плодовых тел
шелковисто-волосистая, серая или желтовато-коричневая. Гименофор гладкий,
слабобугорчатый или слегка волнистый, коричневый с более светлым
кремовым краем, краснеющий при нажатии или повреждении.
Розовый трутовик (Fomitopsis rosea Karst.) вызывает бурую
призматическую гниль заготовленной древесины, преимущественно
сосны и ели, реже — лиственных пород. Встречается на сухостое,
валежнике, пнях, нередко разрушает деревянные столбы, опоры
мостов и других сооружений. Плодовые тела в виде небольших
(диаметром до 7 см) толстых шляпок или копытообразные, чаще
одиночные. Поверхность коричнево-бурая или черноватая, ткань
розоватая, деревянисто-пробковая, гименофор розоватый, со
временем буреющий, трубчатый, с мелкими округлыми или
угловатыми порами.
Шпальный гриб (Lentinus lepideus Fr.) вызывает очень сильное и
быстрое гниение древесины, является главным разрушителем
железнодорожных шпал. Шпальный гриб поселяется на древесине
различных хвойных пород, но предпочитает сосновую древесину.
Разрушает пни, валежник, хранящиеся лесоматериалы; встречается в
погребах, сараях, шахтах; разрушает основания столбов, опоры
мостов и т. п. Изредка поселяется и на сухобочинах живых деревьев.
Гниль бурая, трещиноватая, легко распадающаяся на
продолговатые куски, рассыпчатая. В трещинах гнилой древесины часто
346
образуются белые налеты и тонкие пленки мицелия. Со временем
на пленках появляются желтовато-коричневые пятна.
Плодовые тела в виде довольно толстых плотных твердеющих
шляпок до 12 см в диаметре, с плотной центральной или немного
смещенной вбок (эксцентрической) ножкой. Поверхность
шляпки кремово-желтая или охряно-рыжая, покрытая темными
чешуйками. Края шляпки тонкие, извилистые. Ножка чешуйчатая,
желтоватая, у основания деревянистая. Пластинки гименофора низ-
бегающие, желтоватые, зубчато-рассеченные (см. рис. 110).
Столбовой, или заборный, гриб (Gloeophyllum sepiarium (Wulf.)
Karst.) свое название получил в связи с тем, что часто заселяет
столбы, заборы, шпалы, древесину различных открытых
сооружений и холодных построек (мостов, сараев и т.п.). Встречается
также на лесных складах, где поражает лесоматериалы
длительного хранения, и в лесу — на валежнике, пнях, срубленной
древесине. В жилых постройках иногда обнаруживается на чердаках.
Заборный гриб имеет очень широкое распространение. Он
заражает древесину хвойных пород, главным образом сосны, и
вызывает бурую гниль деструктивного типа, характеризующуюся
весьма интенсивным развитием. Пораженная древесина вначале
желтеет, потом приобретает красноватый оттенок, в ней
появляются мелкие трещины. На более поздней стадии гниения
древесина становится светло-коричневой и растрескивается по годичным
слоям. На последних стадиях разрушения гниль становится темно-
бурой, и в ней появляются крупные радиальные трещины. В
трещинах иногда образуются скопления желтовато-коричневой
грибницы. На поверхности пораженных частей появляются тонкий
войлочный налет и тонкие шнуры грязновато-белого цвета,
которые затем принимают желтую, а позднее буроватую окраску.
Пораженная древесина имеет приятный запах.
Плодовые тела обычно развиваются на следующий год после
заражения древесины, располагаясь в трещинах разрушающихся
материалов. Они имеют вид тонких пробковидно-кожистых
шляпок, прикрепленных боком или полураспростертых (см. рис. 110).
Поверхность темно-коричневая, у основания бугристая, иногда
грубоволосистая с концентрическими зонами. Края более
светлые, желтовато-бурые. Гименофор светло-коричневый или
ржаво-бурый, в виде невысоких разветвленных частых пластинок,
расходящихся в радиальном направлении. Ткань плодового тела
1 — 2 мм толщиной, пробковая, рыжевато-коричневая.
Столбовой елово-пихтовый гриб (Gloeophyllum abietinum (Butt.)
Karst.) в отличие от предыдущего вида предпочитает древесину
ели и пихты, заселяя хранящиеся на складах лесоматериалы, а
также пни, валежник, бурелом. Гниль, вызываемая этим грибом,
очень сходна с гнилью от G. sepiarium как по внешним признакам,
так и по интенсивности разрушения древесины.
347
Плодовые тела по форме, структуре и расположению на
субстрате похожи на плодовые тела G. sepiarium, но они немного
мельче, имеют более тусклую окраску и гименофор с более
редкими, почти не ветвящимися и несросшимися пластинками.
Групповой трутовик {Coriolellus serialis (Fr.) Murr.) часто
встречается в лесах на мертвой древесине. На складах поражает
лесоматериалы хвойных пород, вызывая бурую призматическую гниль,
в трещинах которой образуются скопления беловатого мицелия.
Плодовые тела полураспростертые или в виде небольших желва-
кообразных шляпок, сливающихся в черепицеобразные группы с
общим основанием. Поверхность шляпок желтовато-бурая, ткань
пробковидная, белая, гименофор кремово-белый, с округлыми
или слегка угловатыми порами.
Душистый трутовик (Osmoporus odoratus (Fr.) Sing.) разрушает
лесоматериалы, обработанную древесину, пни, валежник ели и
пихты, а также деревянные части открытых сооружений. На
первой стадии гниения древесина становится красновато-бурой,
затем растрескивается, и в трещинах образуются коричневые
шнуры и скопления мицелия со специфическим запахом аниса.
Полностью разрушенная древесина приобретает темно-бурый цвет и
распадается на кубики.
Плодовые тела подушковидные или в виде толстых шляпок
неправильной формы с темно-бурой бугристой грубовойлочной
или голой поверхностью и более светлым утолщенным краем.
Гименофор ржаво-коричневый, с округлыми крупными порами.
Внутренняя ткань мягкопробковая, красновато-коричневая, с
сильным ванильно-анисовым запахом.
Трутовик серый, или опаленный (Bjerkandera adusta Karst.)
относится к числу тех видов, которое наиболее часто встречаются в
лесах (на сухостое, пнях, валежнике, реже на стволах живых
ослабленных деревьев) и первыми заселяют лесоматериалы
лиственных пород на складах. Гриб довольно активно разрушает
древесину, вызывая белую волокнистую гниль.
Плодовые тела — в виде тонких сидячих шляпок,
полуотогнутые или распростертые, расположенные черепицеобразными
группами. Поверхность шляпки грязно-белая, пепельно-серая или
желтовато-буроватая, бархатисто-волосистая, слабощетинистая или
голая; край острый, часто черноватый, снизу
беловато-желтоватый, стерильный. Ткань волокнисто-кожистая, кремово-белая или
бледно-буроватая; на границе с гименофором проходит тонкая
черная линия. Трубочки от пепельно-серых до серовато-черных, с
очень мелкими округлыми или округло-угловатыми порами.
Трутовик разноцветный, или многоцветный (Coriolus versicolor (L.)
Quel.) — один из самых обычных, широко распространенных де-
реворазрушающих грибов. Встречается на складах, где активно
разрушает круглые неокоренные лесоматериалы различных ли-
348
ственных, реже хвойных пород. Нередко разрушает мосты,
столбы, заборы. Пораженная древесина вначале буреет, затем
размягчается, становится рыхлой и приобретает светло-желтую окраску.
В ряде стран этим грибом специально заражают древесину бука,
используемую при производстве карандашей, для ее размягчения.
В лесах разноцветный трутовик встречается на пнях, валежнике,
сухостое, буреломе. Он образует очень тонкие кожистые, округлые
или половинчатые шляпки, иногда полураспростертые плодовые
тела, обычно расположенные в виде черепицеобразных групп или
розеток на боковой поверхности или торцах пораженных
лесоматериалов. Поверхность шляпок бархатистая, блестящая, с
концентрическими зонами желтовато-коричневых, серовато-бурых, олив-
ково-черных тонов, с более светлым бесплодным неподгибающимся
краем. Ткань белая, замшевидная, при разрыве клочковато-вато-
образная. Гименофор трубчатый, белый, кремово-желтый или
буроватый, с мелкими округлыми или округло-угловатыми порами.
Трутовик утолщенный, или зональный (Coriolus zonatus (Fr.)
Quel.) встречается так же часто, как и разноцветный трутовик.
Поражает преимущественно круглые лесоматериалы, реже —
обработанную древесину лиственных пород, но разрушает ее не столь
интенсивно, как предыдущий вид. В условиях леса активно
заселяет сухостой, пни, бурелом, валежную древесину. Иногда
встречается в постройках. Вызывает белую гниль, сходную с гнилью от
разноцветного трутовика.
Плодовые тела по форме и расположению на субстрате очень
похожи на С, versicolor, однако шляпки, тонкие по краям,
заметно утолщены у основания, поверхность матовая, с более
светлыми тусклыми охряно-желтыми, сероватыми или
ржаво-буроватыми зонами. Гименофор беловато-кремовый, серовато-желтоватый
или охряно-буроватый, с округлыми или округло-угловатыми
мелкими (но чуть крупнее, чем у С. versicolor) порами.
Трутовик волосистый (Coriolus hirsutus (Wulf.) Quel.) — один из
наиболее распространенных видов трутовых грибов. Поражает
бревна, другие лесоматериалы, а также сухостой, пни, валежник
лиственных пород. Вызывает белую гниль. Пораженная древесина
расщепляется по годичным слоям на тонкие пластинки, затем
становится трухлявой, легко крошится.
Плодовые тела имеют вид гибких утолщенных, округлых или
почковидных, часто сросшихся боками шляпок, образующих че-
репицеобразные группы, иногда полураспростертые. Поверхность
густоволосистая, буровато-желтая, рыжеватая или
пепельно-серая, с более или менее однотонными, слегка выпуклыми
концентрическими зонами. Ткань упругая, пробково-кожистая, при
разрыве ватообразно-клочковатая, кремово-белая. Гименофор
пепельно-серый, желтовато-серый или буроватый, с округлыми
довольно мелкими порами.
349
Трутовик березовый пластинчатый (Lenzites betulina (L. ex Fr.) Fr.)
повсеместно встречается на складах, где разрушает
лесоматериалы различных лиственных пород (чаще всего березы), а также в
лесах — на отмерших стволах, валежнике, пнях. Гриб вызывает
белую ядрово-заболонную гниль. В конечной стадии гниения
древесина становится легкой и распадается на отдельные волокна.
Плодовые тела имеют вид полукруглых сидячих шляпок,
образующихся поодиночке или группами. Поверхность шляпки
бархатистая или волосисто-войлочная, грязновато-желтая, пепельно-серая
или буроватая, обычно с неясными зонами. Ткань кремово-белая
или желтоватая, пробково-кожистая, при разрыве ватообразно-
волокнистая. Гименофор пластинчатый, с радиально
расположенными, слегка извилистыми, слаборазветвленными и местами
сросшимися пластинками желтоватого или серовато-белого цвета.
13.3. ЗАЩИТА ЛЕСОМАТЕРИАЛОВ
ОТ ПОРАЖЕНИЯ ГРИБАМИ НА СКЛАДАХ
Поражение древесины грибами обычно происходит вследствие
грубого нарушения санитарных правил при заготовке и
транспортировке лесопродукции, а также нерационального и небрежного
хранения ее на лесных складах. Чтобы предупредить разрушение
древесины, необходимо правильно организовать хранение лесоматериалов,
создать условия, неблагоприятные для активной
жизнедеятельности дереворазрушающих грибов и насекомых. При этом очень
важно выбрать наиболее подходящий способ хранения древесины.
Существуют два основных способа хранения лесоматериалов:
сухой и влажный. Несмотря на существенные технологические
различия (характер предварительной подготовки лесоматериалов,
особенности укладки сортиментов, размещение штабелей и т.п.),
оба способа хранения имеют общую экологическую основу и
единую цель — обеспечение такого режима влажности, температуры,
затенения и воздухообмена, при котором складские грибы и на-
секомые-ксилофаги не способны проникать в древесину и
развиваться в ней. Однако в первом случае этой цели достигают
интенсивным просушиванием лесоматериалов, а во втором — их
максимальным увлажнением, охлаждением и затенением.
Выбор того или иного способа хранения древесины в каждом
конкретном случае зависит от породного и сортиментного
состава лесоматериалов, их целевого назначения, продолжительности
хранения, климатических и погодных условий, времени года,
технической оснащенности лесозаготовительных и
деревообрабатывающих предприятий и других факторов.
Необходимая предпосылка правильного хранения
лесоматериалов — рациональное размещение склада, его хорошее санитарное
350
состояние. Склады, предназначенные для сухого хранения
древесины, рекомендуется размещать на сухих, открытых, хорошо
продуваемых ветрами местах, а склады для влажного хранения — в
понижениях, защищенных от ветра, но в любом случае не ближе
чем в 100 м от стены леса. Территорию склада следует регулярно
очищать от щепы, коры, опилок и других древесных остатков. Всю
древесину (хранящиеся сортименты, прокладки, щиты и др.) с
плодовыми телами дереворазрушающих грибов, налетами,
пленками, другими грибными образованиями и признаками гнили
необходимо уничтожать, почву в местах локализации инфекции
дезинфицировать 5 %-м раствором хлорной извести из расчета 5 л/м2
почвы. Дровяную древесину следует хранить отдельно от здоровых
лесоматериалов.
13.3Л. Сухое хранение древесины
Сухой способ хранения древесины основан на неспособности
деревоокрашивающих и подавляющего большинства
дереворазрушающих грибов и насекомых развиваться в древесине с
влажностью ниже 20 — 25%. Поэтому главными задачами сухого
хранения являются снижение влажности лесоматериалов до этого
уровня путем ускоренной сушки и последующее их содержание в
хорошо просушенном состоянии. Этот способ применяется для
хранения пиломатериалов и широко используется (наряду с
влажным способом) для хранения круглого леса, в основном круглых
лесоматериалов хвойных пород.
Сухое хранение пиломатериалов. Доски и другая пилопродук-
ция при небрежном или неправильном хранении на складах в
наибольшей степени подвержены поражению деревоокрашивающи-
ми, плесневыми и дереворазрушающими грибами. Поэтому сушка
пиломатериалов с последующим их антисептированием —
единственный надежный способ сохранения качества древесины и
необходимое условие ее дальнейшего эффективного использования.
Чаще всего применяется атмосферная (на открытом
воздухе) сушка пиломатериалов. В этом случае их укладывают в
штабеля высотой от 4—5 до 7 — 8 м. Штабеля располагают на подшта-
бельных фундаментах (высотой примерно 0,5 м) рядами с таким
расчетом, чтобы расстояние между соседними штабелями внутри
ряда было не менее 1 —1,5 м, а между соседними радами
штабелей — 2 м. Внутри штабеля доски по одной кладут друг на друга,
причем между соседними стопками досок оставляют
пространства (шпации) шириной не менее половины ширины доски.
Через каждые 1 —1,5 м по вертикали располагают поперечные
прокладки — длинные доски или рейки.
Для защиты пиломатериалов от осадков и растрескивания под
действием солнечных лучей, для повышения сохранности древе-
351
сины над штабелями устраивают односкатные крыши из
низкосортных досок или щитов. С боков штабеля прикрывают щитами
или досками. Для сушки и хранения пиломатериалов иногда
устраивают навесы.
Скорость просыхания пиломатериалов при атмосферной
сушке, в зависимости от климатических и погодных условий, времени
штабелевки, конструкции штабелей, породы древесины,
толщины лесоматериалов и других факторов, может колебаться от 1 нед
до 2 — 3 мес.
Самый эффективный и современный способ просушивания
пиломатериалов — так называемая камерная сушка,
позволяющая надежно и быстро защитить пилопродукцию от
заражения грибами. В этом случае свежий пиловочник помещают в
особые сушильные установки (камеры), где он подвергается
воздействию высокой температуры в сочетании с усиленной
циркуляцией специального состава, ускоряющего процесс просыхания
древесины. Продолжительность камерной сушки может составлять
от нескольких часов до нескольких дней в зависимости от
заданного режима, который устанавливается по таблицам с учетом
древесной породы, толщины пиломатериалов и требуемого уровня
их просушивания.
Высушенные пиломатериалы, влажность которых не
превышает 22%, укладывают на хранение в плотные штабеля,
обеспечивая их защиту от атмосферных осадков, так как при повторном
увлажнении древесины вновь возникает угроза ее поражения
грибами.
Если пилопродукция уже заражена грибами и насекомыми,
рекомендуется ее пропарка, т.е. прогрев до 80... 120°С
насыщенным водяным паром в специальных парильных камерах, после
чего древесину подвергают сушке.
Химическая защита пиломатериалов от поражения грибами при
атмосферной сушке проводится путем их антисептирования. Для
этого древесину в течение 2 — 3 дней после распиловки
опрыскивают или погружают в ванны с рабочим составом антисептика.
В качестве антисептиков применяют водные растворы препаратов
ГР-48 и ПБТ (в концентрации 1 — 2%), пентахлорфенола (1 %),
буры (5 %) и др.
Сухое хранение круглых лесоматериалов. Сухим способом
хранят в основном круглые сортименты хвойных пород и некоторые
сортименты лиственных, используемые в дальнейшем без
продольной распиловки: строительные бревна, балансы, рудничную
стойку и др.
Перед укладкой на хранение круглые лесоматериалы
подвергают окорке, благодаря этому они быстрее просыхают, не
заселяются насекомыми-ксилофагами и лучше пропитываются
антисептиками. В этом плане наиболее предпочтительна чистая, или сплош-
352
ная, окорка, при которой кора удаляется полностью. Широко
применяют и так называемую лубяную окорку: удаляют лишь
наружный корковый слой коры, а внутренний (лубяной) оставляют. Этот
способ окорки имеет свои преимущества, так как лубяной слой
предохраняет древесину от растрескивания. Лесоматериалы,
предназначенные для сухого хранения, при заготовке в теплое время
года следует окорять не позже чем через 3 — 5 дней после рубки, а
при заготовке в зимний период — как только установится теплая
погода (со среднесуточной температурой не ниже 5 °С).
Для круглых окоренных лесоматериалов применяется в
основном атмосферная сушка. После окорки их укладывают в рыхлые
сушильные штабеля, которые размещают на прочных бетонных
или деревянных, обработанных антисептиками
подставках-фундаментах высотой не менее 0,5 м. Высота штабелей обычно не
превышает 2 —3 м, а длина — 50 м. Между штабелями оставляют
проходы шириной 1 —1,5 м при хранении древесины хвойных
пород и 0,6—0,7 м для лиственных.
При благоприятных для сушки условиях (например, в южных
районах, в годы с сухим жарким летом) лесоматериалы
укладывают в нормальные штабеля. В этом случае бревна кладут
плотными слоями без промежутков (шпаций) между бревнами, а
слой от слоя отделяют поперечными горизонтальными
прокладками из более тонких сортиментов (подтоварника, жердей). При
неблагоприятных условиях (например, в холодные дождливые
периоды) лесоматериалы укладывают в разреженные штабеля:
между соседними бревнами оставляют шпации шириной 5 — 6 см.
Поперечные прокладки между слоями бревен в таких штабелях
делают более толстыми, обычно используя для этого те же
укладываемые бревна.
Короткие сортименты (балансы, рудничную стойку и др.)
укладывают в штабеля-клетки (отличающиеся от обычных
сушильных штабелей чередованием в смежных слоях направления
укладки бревен) или в невысокие (до 2 м) рыхлые
поленницы. После просушивания их можно хранить в плотных
поленницах.
Над штабелями для защиты их от дождя и снега необходимо
устраивать крыши из горбыля, щитов, низкосортных досок или бревен.
При сухом хранении круглых лесоматериалов, как правило,
наблюдается их растрескивание, что ухудшает качество
древесины. Для предохранения торцов бревен от воздействия прямых
солнечных лучей и сильного растрескивания применяют белую краску
или специальные влагозащитные замазки. Вещества,
используемые в качестве торцовых покрытий, способы нанесения и нормы
расхода при сухом хранении — те же, что и при влажном.
Влагозащитные покрытия наносят на торцы лесоматериалов перед их
укладкой на сушку. Это позволяет снизить интенсивность растрески-
353
вания или даже исключить его. Применение торцовых покрытий
особенно целесообразно для защиты лесоматериалов лиственных
пород, которые наиболее подвержены сильному растрескиванию:
дуба, бука, березы и др. Для лесоматериалов хвойных пород
(кроме лиственницы) торцовые покрытия не применяют.
Для антисептирования круглых лесоматериалов используют те
же химические соединения, что и для обработки пиломатериалов.
Расход рабочего состава при антисептировании древесины
зависит от толщины сортимента, вида антисептика, его
концентрации и колеблется от 0,05 до 0,7 кг/м3 лесоматериалов.
Нижние рады штабелей рекомендуется антисептировать
растворами пентахлорфенолята натрия, железного купороса, препарата
ГР-48.
Сроки атмосферной сушки круглых лесоматериалов зависят от
вида древесной породы, толщины и длины сортиментов, способа
окорки, времени заготовки и укладки на сушку, типа штабелей,
климатических и погодных условий (температуры и
относительной влажности воздуха, силы ветра и т.д.).
13.3.2. Влажное хранение древесины
Этим способом хранят в основном круглые лесоматериалы
лиственных и хвойных пород, предназначенные для дальнейшей
переработки, например пиловочные и фанерные кряжи. Главная
задача влажного хранения — постоянное поддержание в
древесине высокой влажности (на уровне, свойственном живому дереву
или еще более высоком). Такой режим хранения исключает или
затрудняет просыхание лесоматериалов, обеспечивая их
охлаждение и затенение.
Наиболее подходящий вариант влажного хранения древесины
выбирают в зависимости от вида лесопродукции, древесной
породы, размера лесоматериалов и их назначения, предполагаемого
срока хранения и технической оснащенности лесного склада.
Для лучшего сохранения влаги в свежезаготовленных или
сплавных лесоматериалах их хранят неокоренными в плотных
(беспрокладочных), плотнорядовых (с небольшим количеством
прокладок) или пачковых штабелях.
В плотном штабеле бревна укладывают вплотную друг к
другу, без прокладок, и только по краям штабеля слои бревен
разделяют тонкими (до 0,5 их диаметра) горизонтальными прокладками.
В плотнорядовом штабеле бревна кладут так же тесно,
как и в плотном штабеле, но между слоями лесоматериалов
(толщина слоя обычно составляет 3—4 бревна) по всей длине
штабеля помещают тонкие прокладки.
Пачковый штабель отличается от плотнорядового тем,
что в нем делают не только горизонтальные прокладки, но и вер-
354
тикальные стойки; поэтому такой штабель состоит как бы из
отдельных пачек по 9—16 бревен, в зависимости от их толщины.
Штабеля делают длинными (не менее 25 м), высотой не менее
2 м для лиственных и 3 м для хвойных пород. Расстояние между
штабелями должно быть минимальным. Важное значение имеет
также время закладки штабелей. Наиболее предпочтительна
осенняя штабелевка.
Для защиты круглых лесоматериалов от высыхания,
растрескивания и заселения грибами торцовые поверхности колод
покрывают влагозащитными замазками. Их применяют в основном для
бревен лиственных пород. В качестве замазок используют
вещества, которые образуют на поверхности древесины плотную водо-
и воздухонепроницаемую пленку: парафин, петролатум,
синтетические смолы и латексы, препараты карбафен-16 и ПФК-У-12
(светлые замазки), а также нефтяные битумы и приготовленные
на их основе пасты, эмульсии и лаки, пекосмоляные смеси,
солидол, каменноугольный деготь (темные замазки). Влагозащитные
покрытия в горячем или холодном состоянии наносят кистями
или опрыскивателями на чистые, слегка подсушенные свежие
торцы. Применяют также побелку боковых поверхностей
лесоматериалов и затенение штабелей. При весенне-летних заготовках
лесоматериалы надо обрабатывать не позже чем через 3 — 5 дней
после раскряжевки, при зимних — после оттаивания торцов, но
не позже чем за 10—15 дней до распускания листвы у той
породы, древесину которой защищают. В средней полосе европейской
части страны обработку лесоматериалов зимней заготовки надо
заканчивать до начала мая.
Химическую защиту (антисептирование) свежезаготовленных
неокоренных лесоматериалов проводят одновременно с укладкой
их в штабеля или после ее завершения. Для противогрибной
профилактики используют креозот, 3 — 5 %-е растворы пентахлорфенола
или нафтената меди в нефтепродуктах (дизельном топливе и
керосине), водные растворы пентахлорфенолята натрия (1 — 2 %),
антисептиков ГР-48 (1-2%), ПБТ (2-3%), ББК-3 (3%), 5-10%-й
раствор сульфата меди и другие препараты.
Круглые неокоренные лесоматериалы весенне-летней
заготовки следует антисептировать не позже чем через 3 — 5 дней после
их раскряжевки, лесоматериалы зимней заготовки — после
оттаивания поверхности древесины, но не позже чем за 10—15 дней до
появления листвы у той породы, древесину которой защищают.
Примерный расход антисептика для защиты 1 м3 лесоматериалов —
от 0,05 до 0,75 кг, в зависимости от вида антисептика и
концентрации применяемых растворов.
Круглые окоренные лесоматериалы и пилопродукцию также
антисептируют водными растворами пентахлорфенолята натрия,
препаратов ГР-48, ПБТ или ББК-3. Древесину обрабатывают опрыски-
355
ванием или погружением в антисептический раствор в теплое время
года (со среднесуточной температурой не ниже 5°С) и не
позднее чем через 12 ч после ее окорки или распиловки.
К наиболее эффективным методам влажного хранения
неокоренных круглых лесоматериалов относится их искусственное
увлажнение, или дождевание, которое заключается в регулярном, на
протяжении всего теплого времени года, поливе штабелей
древесины водой. Его применяют на деревообрабатывающих
предприятиях и лесоскладах, оснащенных специальными дождевальными
установками.
Для хранения небольших партий древесины можно применять
замораживание: при укладке в штабеля лесоматериалы
пересыпают снегом; затем штабель обильно поливают водой, а когда он
покроется слоем льда, засыпают опилками или покрывают
хвойным лапником.
Круглые неокоренные лесоматериалы хранят также
погружением в воду. Этот способ применяют на лесоскладах заводов,
располагающих естественными или искусственно созданными
водоемами, а также на деревообрабатывающих предприятиях.
Например, на заводах по производству фанеры фанерные кряжи хранят
в специальных бассейнах, что надежно защищает их от грибов и
насекомых, а в дальнейшем облегчает обработку.
13.4. РАЗРУШЕНИЕ ДРЕВЕСИНЫ В ЗДАНИЯХ
И СООРУЖЕНИЯХ С ПОСТОЯННЫМ
ТЕМПЕРАТУРНЫМ РЕЖИМОМ
Грибы, разрушающие деревянные конструкции в жилых
домах, административных, фабрично-заводских и других
отапливаемых зданиях, в промышленных и хозяйственных сооружениях с
относительно постоянным режимом температуры и влажности
(шахтах, семенохранилищах и т.п.), составляют особую
экологическую группу домовых грибов. Домовые грибы широко
распространены как в городах, так и в сельской местности. Вызывая
разрушение деревянных конструкций, они иногда быстро выводят из
строя относительно новые здания, причиняя очень большой ущерб.
Домовые грибы могут заражать древесину мицелием и базидио-
спорами, которые при благоприятных условиях образуются в
огромном количестве. Базидиоспоры легко распространяются по
воздуху и сохраняют способность к прорастанию в течение
нескольких лет. Гифы домовых грибов могут более года сохранять
жизнеспособность в древесине с влажностью 11%. Еще более
стойки и живучи шнуры и плодовые тела этих грибов.
Инфекционное начало домовых грибов (споры, обрывки
мицелия и шнуров) может попасть в здание из соседних зараженных
356
помещений по воздуху или с предметами домашнего обихода,
при использовании на дрова старой зараженной древесины.
Домовые грибы могут быть занесены в новую постройку и с
зараженным строительным материалом. При наличии очага инфекции
домовый гриб проникает из зараженных помещений в соседние,
распространяясь при помощи шнуров через междуэтажные перекрытия,
по поверхности стен и полов, под плинтусами и штукатуркой.
Прорастание спор, рост мицелия домовых грибов и активное
разрушение ими древесины происходят лишь при определенных
условиях окружающей среды, прежде всего при подходящей
температуре. В большинстве случаев для домовых грибов оптимальна
температура в пределах от 20 до 27 °С, минимальная температура
составляет 5... 9 °С, а максимальная 35... 37 °С. Только у
настоящего домового гриба максимум лежит несколько ниже: 26...27°С.
Вторым важным фактором является влажность древесины и
окружающего воздуха. Для большинства видов домовых грибов
благоприятна высокая относительная влажность воздуха — не ниже
80 — 85 %. Что касается влажности древесины, то здесь
наблюдаются более резкие различия между отдельными видами. Так, для
роста настоящего домового гриба достаточна влажность 19—22 %,
для белого и пластинчатого домовых грибов она необходима в
пределах 50—70 %. Развитие же пленчатого домового гриба и
наиболее сильное разрушение этим грибом древесины происходит
при ее влажности от 34 до 46 %.
Большое значение имеют наличие кислорода и углекислоты, а
также кислотность субстрата. Домовые грибы лучше всего
развиваются при значениях рН от 3,0 до 6,0, т. е. при слабокислой
реакции среды.
Домовые грибы чаще и сильнее поражают древесину хвойных
пород. Некоторые из них (например, белый) никогда не
встречаются на древесине лиственных. Другие (например, настоящий и
пленчатый) могут наряду с хвойными поражать и лиственные
породы, однако разрушение древесины во втором случае идет
медленнее. Древесина дуба, робинии лжеакации, каштана
съедобного устойчива к этим грибам.
Занос инфекции и дальнейшее развитие домовых грибов чаще
всего происходят при нарушении правил строительства и
эксплуатации зданий. Наиболее часто встречается такое серьезное
нарушение, как использование при строительстве плохо
просушенной неантисептированной и даже неокоренной древесины.
Развитию домовых грибов особенно благоприятствует сырость
в зданиях вследствие течи в крышах, неисправности
водопровода, отсутствия вентиляции, недостаточного отопления и других
причин. Неправильная эксплуатация зданий, отклонения от
нормального режима температуры и влажности, застойность воздуха
приводят к быстрому разрушению древесины.
357
Поражение зданий домовыми грибами обычно становится
заметным, когда процесс разрушения древесины зайдет уже
сравнительно далеко. Об этом свидетельствуют такие признаки, как
наличие влажных расплывчатых, иногда цветных пятен на стенах
и потолках, увлажнение досок пола, характерный неприятный
запах, осадка стен, прогиб балок, перекос дверных коробок,
зыбкость полов. Местами наблюдаются отлуп и обрушивание
штукатурки и даже появляются плодовые тела и мицелиальные пленки
гриба. Чаще всего они образуются в сырых углах комнат,
санузлах, неотапливаемых помещениях, плохо проветриваемых
подвалах. Сильное поражение может привести к перекосу полов,
обвалу потолков и перекрытий.
Известно около 70 видов домовых грибов, но наиболее
распространены и опасны настоящий, белый, пленчатый и
пластинчатый домовые грибы. При благоприятных условиях они могут в
течение 1 — 2 лет полностью разрушить деревянные конструкции
зданий. Все эти грибы вызывают бурую деструктивную
трещиноватую гниль, легко растирающуюся в порошок.
Настоящий домовый гриб (Serpula lacrimans (Wulf.) Gray.)
образует пышные ватообразные скопления грибницы. Вначале она
белая, затем местами приобретает канареечно-желтую или
розоватую окраску. Под конец мицелий съеживается, превращаясь в
грязно-серые пленки или подушки.
Плодовые тела (рис. 111) широко распростертые, иногда очень
большие (до 0,5 м и более в диаметре), толстые, губчатые, до
1—4 см толщиной, чаще приросшие к субстрату, реже
полуотогнутые или свободные. Край плодового тела утолщенный
войлочный, в молодом состоянии белый, отчетливо выделяющийся. Гиме-
нофор в виде невысоких извилистых переплетающихся складочек,
ячеистый, иногда крупносетчатый или извилисто-зубчатый. Цвет
его охряно-желтый, затем ярко-ржавый до ржаво-коричневого.
Ячейки до 2 — 3 см в диаметре и примерно такой же глубины.
Плодовое тело настоящего домового гриба выделяет за сутки
около 1 млн спор. Во время споруляции
гриба 1 м3 воздуха зараженного подполья
может содержать до 80 тыс. спор, а 1 м3
первого этажа — до 16 тыс. спор. При
обильном образовании базидиоспор они
имеют вид тонкого пылящего ржаво-
бурого порошка, покрывающего
плодовые тела и оседающего внутри
зараженных помещений. Споры разносятся
также на обуви, одежде и т.д. На скоплениях
мицелия и по краям плодовых тел вы-
Рис. 111. Плодовое тело на- деляются капли водянистой жидкости
стоящего домового гриба (Serpula lacrimans от лат. lacrimo — пла-
358
кать, покрываться влагой). В связи с этим древесина в процессе
гниения еще более увлажняется.
Настоящий домовый гриб способен интенсивно разрушать
древесину различных хвойных и мягколиственных пород.
Пораженная древесина буреет и покрывается глубокими продольными и
поперечными трещинами. На последней стадии разрушения она
распадается на отдельные куски и легко растирается в очень
тонкий шелковистый порошок.
Распространение гриба по поверхности стен с этажа на этаж
часто происходит при помощи шнуров, длина которых может
достигать нескольких метров. Шнуры довольно толстые (до 6—7 мм),
иногда плоские беловатые или пепельно-серые деревянистые, в
сухом состоянии — ломкие. С деревянных конструкций мицели-
альные шнуры и пленки нередко переходят на граничащие с ними
кирпичные и каменные кладки (фундаменты, стены), которые
могут стать местом сохранения инфекции.
Настоящий домовый гриб встречается преимущественно в
замкнутых конструкциях жилых, фабрично-заводских и
административных зданий, а также в подвалах, семенохранилищах, плохо
вентилируемых складах и других постройках. Это самый
распространенный и вредоносный вид среди домовых грибов.
Белый домовый гриб (Coriolus vaporarius (Fr.) Bond, et Sing.)
назван так потому, что развивает пышный, хлопьевидный или
ватообразный, всегда белый мицелий. В сырых замкнутых
пространствах образуются особенно мощные скопления рыхлой
грибницы, волокнистых прядей и шнуров (рис. 112). Шнуры толстые (до
4—6 мм), мягкие, неломкие, слабоветвящиеся, тоже белые. В
узких пространствах гриб образует белые пленки
лучисто-веерообразного строения, иногда в виде слоев прессованной ваты.
Плодовые тела появляются очень редко. Они обычно
небольшие, распростертые, приросшие, вначале мягкие, иногда в виде
корочки с узким, неясно выраженным краем. Цвет плодового тела
беловатый, впоследствии желтоватый до рыжевато-желтого. Ги-
Рис. 112. Белый домовый гриб:
/ — слившиеся плодовые тела; 2 — мицелиальные шнуры на разрушенной грибом
древесине
359
менофор трубчатый. Трубочки вначале белые, затем желтоватые
или светло-коричневые. Поры крупные, неравновеликие, в
среднем (1 — 2) х 1 мм, округлые или угловатые, сначала с цельными,
а затем мелкозубчатыми краями.
Белый домовый гриб встречается чаще всего в бесплодной
(мицелиальной) стадии. Его находят в междуэтажных перекрытиях
жилых домов, реже на лесных складах и в надворных постройках.
Разрушение древесины под действием этого гриба происходит
очень быстро, он считается весьма опасным, особенно при
наличии определенной влажности и отсутствии вентиляции.
Зараженная древесина вначале принимает бурую окраску, затем
растрескивается поперечными и продольными трещинами на крупные
призматические участки и на конечной стадии гниения легко
рассыпается в порошок. Белым домовым грибом поражается только
древесина хвойных пород, в основном сосны и ели.
Пленчатый домовый гриб (Coniophora cerebella (Pers.) Schrot)
характеризуется слаборазвитым, иногда пушистым мицелием с
желтоватым оттенком. Старый мицелий спадается, становится
буроватым и потом почти исчезает. На поверхности древесины гриб
развивает очень тонкие, почти нитевидные, стелющиеся по
субстрату, веерообразно ветвящиеся шнуры коричневатого, позднее
темно-бурого, почти черного цвета (рис. 113). Снаружи и в
трещинах древесины также образуются пленки. Они тонкие, вначале
почти белые, затем соломенно-желтого и к старости коричневого
цвета. Плодовые тела довольно мясистые, вначале беловатые,
затем желтовато-бурые.
При благоприятных условиях гриб быстро разрушает
древесину. Она буреет, покрывается многочисленными трещинками и
затем распадается на мелкие призмочки. Такая мелкотрещиноватая
структура гнили — характерный признак поражения пленчатым
домовым грибом.
Пленчатый домовый гриб также относится к числу наиболее
часто встречающихся и опасных разрушителей древесины. Как и
^ г^ г>Ф Рис-113- Домовые грибы:
г^ Ф\С\ ^ "~ мицелиальные шнуры пленчатого домового гриба; 2, 3 —
^ ^ плодовые тела пластинчатого домового (шахтного гриба); 4 —
^ базидиоспоры пластинчатого домового гриба
360
настоящий домовый гриб, он активно разрушает древесину не
только хвойных, но и лиственных пород, за исключением дуба и
робинии лжеакации. В зданиях он поражает деревянные
перегородки и перекрытия. Кроме того, разрушает мосты, шпалы,
изгороди, лесоматериалы на складах. Встречается также на валежнике
и пнях.
Пластинчатый домовый, или шахтный, гриб (Paxillus panuoides
Fr.). Мицелий вначале очень скудный бесцветный паутинистый,
позднее желтеющий, иногда с фиолетовым или лиловатым
оттенком, веерообразно разрастающийся. Старый мицелий
сероватого цвета. Гриб развивает также тонкие, нитевидные,
разветвленные, переплетающиеся шнуры, сначала светлые, затем
зеленовато-желтые, иногда образующие как бы рыхлую пленку.
Плодовые тела в виде шляпок без ножки или с короткой
боковой ножкой (см. рис. 113). Шляпки достигают 2—6 см в диаметре,
тонкие, мясистые, различной формы: часто веерообразные или
раковинчатые, раздвоенные, суженные к основанию, иногда
распростертые. Поверхность шляпки желтовато-охряная или грязно-
желтая, иногда с фиолетовым оттенком или коричневатая,
опушенная или нежно-войлочная, в старости голая и гладкая. Край
шляпки тонкий, острый, подвернутый или прямой, волнисто-
лопастный. Ткань шляпки мягкая, губчатая, без запаха. Ножка
зачаточная или очень короткая, боковая, такого же цвета, как и
шляпка; иногда она отсутствует. Пластинки радиально расходящиеся,
узкие, волнистые мягкие, дихотомически ветвящиеся, в молодом
состоянии беловатые, затем яично-желтые или буроватые.
При благоприятных условиях (влажность древесины 50 — 70 %
и окружающего воздуха не ниже 100 %) пластинчатый домовый
гриб вызывает быстрое и сильное разрушение древесины,
преимущественно сосновой. Пораженная древесина на первых
стадиях развития гнили становится зеленовато-желтой, потом буреет,
приобретает волокнистое строение и, наконец, растрескивается,
главным образом в продольном направлении.
Гриб часто встречается в подпольях, каркасных домах и
перекрытиях с торфяной засыпкой, в погребах и особенно в шахтах. В
связи с этим он и получил название шахтного гриба.
13.5. ЗАЩИТА ЗДАНИЙ ОТ РАЗРУШЕНИЯ
ДОМОВЫМИ ГРИБАМИ
В защите строений от домовых грибов важнейшую роль играет
профилактика. Она заключается прежде всего в создании
условий, исключающих возможность попадания инфекции и развития
домовых грибов. Основные моменты противогрибной
профилактики — правильный выбор места под застройку, рациональная
361
конструкция зданий, правильное устройство вентиляционных,
отопительных, гидро-, тепло- и пароизоляционных систем,
строгое соблюдение правил строительства и эксплуатации зданий,
обеспечивающих защиту деревянных конструкций от увлажнения,
систематический надзор за техническим состоянием строений.
Очень важно правильно выбрать место для застройки. Дома
желательно строить в местах с низким уровнем грунтовых вод, на
повышениях, обеспечивающих сток поверхностных вод. Однако
решающее значение имеет конструкция фундамента, устройство
подполья, полов и крыши.
Фундамент лучше делать из кирпича или камня с
использованием высококачественного цемента. Деревянные столбы и стулья
фундамента необходимо предварительно просмолить или
пропитать антисептиками.
Балки, лаги и переводы должны быть проантисептированы.
Прежде чем их укладывать на фундамент, он должен быть хорошо
просушен и покрыт гидроизоляционной прокладкой из двух
слоев рубероида с заливкой петролатумом или асфальтом.
Перед укладкой полов надо тщательно очистить подполье от
щепы и мусора, обработав его формалином, 3 %-м раствором
фторида натрия или 5 %-м раствором медного купороса. В полах
необходимо устанавливать вентиляционные розетки. В подвальных,
полуподвальных, чердачных помещениях, междуэтажных
перекрытиях также должны быть предусмотрены вентиляционные
устройства.
Плохо проветриваемые или постоянно увлажняемые элементы
закрытых конструкций (концы деревянных балок в стенах, стыки
в междуэтажных перекрытиях под санузлами) рекомендуется
просмолить, обмазать нефтебитумом или маслянистым антисептиком,
покрыть рубероидом. Кирпичные кладки, прилегающие к
деревянным частям, следует промазать антисептиком или прокалить
пламенем паяльной лампы.
Конструкция крыши и материал для нее должны
соответствовать местным условиям и надежно защищать перекрытие и
верхнюю часть стен от увлажнения. Балконы и открытые веранды
необходимо делать с небольшим уклоном для стока воды. С целью
отвода талых и дождевых вод устанавливают водосточные трубы,
вокруг дома устраивают бетонированную или асфальтированную
отмостку.
Для строительства следует использовать только новую, хорошо
просушенную (оптимальная влажность 18 — 20%) древесину без
признаков гнили, грибных окрасок, налетов и червоточин,
обработанную соответствующими антисептиками. Последовательность
выполнения строительных работ необходимо строго соблюдать.
Отопительная, водопроводная и канализационная системы
должны соответствовать местным условиям, конструкционным
362
особенностям зданий и обеспечивать постоянный температурно-
влажностный режим, не допускающий активного развития дере-
воразрушающих грибов.
При эксплуатации зданий необходимо постоянно следить за
состоянием кровли, чердачных помещений, канализационных,
водопроводных и водосточных труб, своевременно устранять
неисправности; счищать снег с крыш, отводить от дома талые и
дождевые воды.
Нельзя допускать длительного увлажнения помещений
(особенно кухонь, санузлов), засорения раковин, умывальников,
унитазов и увлажнения полов и стен. При мытье полов не следует обильно
заливать их водой; в жилых помещениях не рекомендуется стирать
и сушить белье, хранить овощи и дрова, загромождать наружные
углы и стены мебелью, затрудняющей циркуляцию воздуха.
В зимнее время дома необходимо хорошо отапливать, а летом
проветривать и просушивать чердаки, подвалы и подполья;
вентиляционные отверстия закрывать и утеплять до наступления
холодов.
Для выявления очагов развития домовых грибов, причин их
возникновения и своевременного проведения защитных
мероприятий необходимо ежегодно (а в новых домах два раза в год)
проводить специальное обследование зданий.
Начинают с общего осмотра объекта, а затем, в случае
обнаружения явных признаков поражения (наличие грибных
образований, гнили, оседание или деформация деревянных конструкций,
специфический запах и др.) или выявления условий,
способствующих развитию домовых грибов (сырость в помещениях,
застойность воздуха, неисправность сантехники, повреждения крыши и
т.д.), — проводят детальное обследование здания, начиная с
подвальных и кончая чердачными помещениями и крышей. При этом
особое внимание обращают на конструкционные дефекты
(например, в устройстве вентиляции, отопления, гидро- и
теплоизоляционных систем, кровли), которые могли быть причиной
развития домовых грибов. В случае необходимости, при подозрении
на скрытую форму поражения, проводят вскрытие конструкций.
Для более точного определения вида домового гриба, а также в
сомнительных случаях из пораженных элементов постройки и тех
мест, где наиболее велика опасность заражения (доски пола в углах,
у наружных стен, под водопроводными трубами и раковинами,
концы балок, лаг, подвальные помещения и др.), берут образцы
древесины для лабораторного анализа.
При выявлении очагов гниения определяют степень
разрушения древесины (в процентах или баллах), а по завершении
обследования составляют акт, содержащий перечень выявленных
очагов поражения с указанием причин развития домовых грибов, а
также конкретные рекомендации по борьбе с ними.
363
При обнаружении очагов развития домовых грибов
необходимо принять срочные меры. Прежде всего следует устранить
источники увлажнения, после чего вскрыть и просушить пораженные
конструкции. Если развитие домовых грибов находится в
начальной стадии и древесина еще не затронута гниением (имеются лишь
поверхностные налеты мицелия) или слабо затронута, нужно
удалить грибные налеты, стесать наружные слои древесины и
обильно промазать антисептиком поверхность пораженных грибом
элементов, захватывая при этом незараженные части не менее чем
на 50—70 см от края зараженного участка. Промазку повторяют
2 — 3 раза с перерывами в 2—3 дня. Рекомендуют и такой способ:
просверливают в древесине этих элементов (после стесывания)
отверстия диаметром 1,5 см и глубиной не менее половины
толщины элемента, располагая отверстия через каждые 10—20 см, и
заливают в них горячий раствор фторида или кремнефторида
натрия. Эту операцию повторяют несколько раз, а затем
обработанные части обмазывают антисептической пастой.
Если обнаружено более глубокое гнездовое поражение и
загнивание древесины в ответственных конструкциях (лаги, балки,
перекрытия, венцы, столбы фундамента и т.п.), а также при
любой степени поражения легко сменяемых конструкций (доски
пола, плинтусы, обшивки, перегородки и т.д.), надо удалить и
сжечь (на костре или в котельной) все загнившие части и
прилегающую к ним здоровую на вид древесину на расстоянии не
менее 50—70 см от пораженного участка. Использовать эту
древесину в качестве топлива и для других хозяйственных нужд
недопустимо.
Следует также тщательно удалить и сжечь все грибные
образования (плодовые тела, налеты, шнуры, пленки) и весь
древесный мусор. Негорючий мусор (куски обвалившейся штукатурки,
засыпку и т.д.) необходимо закопать или вывезти на свалку.
Здоровые части конструкций, прилегающие к пораженным,
обязательно проантисептировать. Каменные или кирпичные
кладки, бетонные и металлические поверхности, примыкавшие к
пораженной древесине, должны быть тщательно очищены,
обеззаражены с помощью паяльной лампы, а затем также обработаны
антисептиками.
Землю или другую засыпку (шлак, керамзит), находившуюся под
пораженными полами, необходимо вынуть (20—30 см) и удалить,
заменив новой. Новую засыпку рекомендуется полить 3 %-м
раствором фторида натрия или 5 %-м раствором медного купороса.
После этого все помещение дезинфицируют парами
формалина (из расчета 10 г формалина на 1 м3) или путем сжигания серы
при плотно закрытых дверях и окнах. Затем помещение
проветривают и тщательно просушивают. Только после этого можно
приступать к замене изъятых деревянных частей новыми.
364
Для ремонта используют новую сухую (не более 18—20 %
влажности) здоровую древесину, предварительно обработанную
фторидом натрия или другим подходящим для этой цели
антисептиком — наиболее целесообразен способ «прогрев — холодная
ванна» или метод автоклавной пропитки.
Ремонт зданий, пораженных домовыми грибами, лучше всего
делать летом, когда легче просушить ремонтируемые помещения
и уберечь стройматериалы от увлажнения. Одновременно с проти-
вогрибным ремонтом обычно проводят ремонт кровли, замену
изношенных элементов сантехники, вносят необходимые
изменения в планировку и техническое оснащение зданий, устраняют
те конструкционные недостатки (например, в устройстве
вентиляционных, отопительных, гидроизоляционных систем), которые
способствовали развитию домовых грибов.
13.6. АНТИСЕПТИРОВАНИЕ ДРЕВЕСИНЫ
Антисептированием называется поверхностная обработка или
глубокая пропитка древесины веществами биоцидного действия
(антисептиками) для защиты ее от разрушения грибами и насекомыми.
Антисептики различаются по химическому составу,
консистенции, стойкости к вымыванию из древесины, токсичности для
микроорганизмов, насекомых и теплокровных, особенностям их
применения и другим показателям. Среди них ^сть соединения
водорастворимые, растворимые в легких органических
растворителях, растворимые в маслах и тяжелых нефтепродуктах, а также
различные пропиточные масла, комбинированные антисептики
и антисептические пасты.
Водорастворимые и растворимые в легких органических
растворителях антисептики применяют для пропитки как сырой, так и
подсушенной древесины различного назначения. Раствор наносят
кистью с помощью гидропульта или ранцевого опрыскивателя.
Маслами и другими антисептиками маслянистой консистенции
обрабатывают только сухую древесину. Антисептики этой группы
обладают высокой стойкостью против вымывания их водой и в то
же время имеют резкий запах, поэтому используются для
пропитки древесины открытых сооружений, контактирующих с грунтом:
оснований столбов, стоек и опор, подвергающихся сильному
увлажнению элементов зданий (стульев фундамента, нижних венцов,
деревянных подкладок под лаги и концы балок и т. п.).
Комбинированные антисептики, как и масла, из-за сильного
неприятного запаха применяют только для обработки древесины
открытых сооружений.
Антисептические пасты, или суперобмазки, рассчитаны на
долговременную защиту древесины и поэтому применяются в ос-
365
новном для глубокой пропитки древесины, используемой для
наиболее ответственных частей деревянных конструкций. В момент
обработки влажность древесины должна быть достаточно высокой
(не менее 40 — 60%).
Антисептические пасты имеют полужидкую (вязкую)
консистенцию и состоят из водорастворимого антисептика (чаще всего
фторида или кремнефторида натрия), клеящей основы,
наполнителя и воды. В качестве клеящей (вяжущей) основы используются
экстракты сульфитных щелоков, битум, жидкое силикатное
стекло, различные клеи, жирная глина и другие вязкие вещества,
благодаря которым паста прочно приклеивается к поверхности
древесины. В зависимости от этого различают экстрактные,
битумные, силикатные, глинистые и другие пасты. Наполнителем
чаще всего служит торфяная мука. Для приготовления пасты
клеящее вещество растворяют в горячей воде, а затем при
непрерывном помешивании добавляют антисептик и торфяную муку.
Экстрактные и силикатные антисептические пасты рекомендуются для
обработки балок в чердачных перекрытиях и санузлах. Битумные
пасты и пасты на каменноугольном лаке, а также маслянистые
антисептики применяют для обработки деревянных частей,
соприкасающихся с грунтом.
По химическому составу действующего вещества выделяют
следующие группы и виды антисептиков: неорганические
соединения (фторид натрия, кремнефторид натрия, кремнефторид
аммония, сульфат меди); фенолы и их производные (2-оксидифе-
нил, оксидифенолят натрия, динитрофенолят натрия, пентахлор-
фенол, пентахлорфениллаурат, пентахлорфенолят натрия);
углеводороды (дифенил, я-дихлорбензол, креозот, каменноугольные
пропиточные масла, карболинеум, сланцевые пропиточные
масла); металлоорганические соединения (этилмеркурфосфат, гекса-
бутилдистанноксан, оксофин); производные карбоновых кислот
(салициланилид, нипагин-М, фталан, каптан);
гетероциклические соединения (купроцин, трилан); комбинированные
антисептики (препараты ГР-48, ПБТ, П-2Т, ББК-3, хромово-мед-
ный препарат ХМ-5, фторохромово-мышьяковый препарат ФХМ,
медно-хромово-цинковый препарат МХХЦ, хромхлорид цинка
ХХЦ, паста ФН-П, уралит, триолит, или препарат ФДХ, и др.);
влагозащитные антисептические покрытия (карбафен-16, ПФК-
У-12идр.).
Существуют разные способы антисептирования древесины.
Выбор того или иного метода зависит от свойств антисептика,
типа и назначения лесоматериалов и строительных деталей,
способа и срока их хранения, влажности древесины, наличия
соответствующего оборудования и других условий.
Поверхностная обработка чаще всего проводится на
лесоскладах при закладке лесоматериалов на непродолжительное
366
хранение, так как при этом способе антисептирования биоцид-
ные вещества проникают только в наружные слои древесины (на
глубину от 1 до 3 мм) и сравнительно легко вымываются.
Поверхностную обработку обычно проводят путем опрыскивания
сортиментов растворами антисептиков с помощью ранцевого
опрыскивателя или гидропульта. При защите небольших партий
лесоматериалов их обмазывают кистью. В зависимости от породы древесины
обработку повторяют от 2 до 4 раз с интервалами 3—4 ч. Для
двукратной обработки требуется 0,6—0,8 л рабочего раствора на 1 м2
поверхности древесины. Небольшие партии малогабаритных
строительных деталей иногда погружают в емкости (ванны, бочки и т. п.)
с горячим (80...90°С) раствором антисептика и выдерживают в
нем в течение нескольких минут. С целью контроля глубины и
качества пропитки в бесцветные растворы антисептиков добавляют
анилиновую краску (4—5 г на 10 л). Для поверхностной обработки
сырой или немного подсушенной древесины применяют
водорастворимые антисептики и соединения, растворимые в легких
органических растворителях, а сухую древесину обрабатывают
маслянистыми антисептиками. Креозот и карболинеум лучше
впитываются в древесину, если их перед употреблением подогреть до
50...60°С.
Глубокая диффузионная пропитка обеспечивает
долговременную противогрибную защиту древесины. Поэтому она
особенно рекомендуется для антисептирования строительных
материалов, телефонных и телеграфных столбов, круглых
лесоматериалов длительного хранения. С этой целью применяют
различные типы антисептиков: водорастворимые, растворимые в легких
органических растворителях и комбинированные, но наиболее
эффективны антисептические пасты для обработки древесины,
имеющей влажность не ниже 40 %. После опрыскивания или обмазки
антисептиком древесину укладывают в плотные штабеля на
мягкую подстилку, укрывают со всех сторон полиэтиленовой
пленкой и оставляют на 3—4 мес, чтобы антисептик проник в
заболонь по всей ее толщине.
Глубокую пропитку древесины можно осуществить путем
длительной выдержки в ванне с антисептическим составом. Сырые
лесоматериалы в зависимости от толщины сортиментов и
температуры раствора выдерживают в течение нескольких суток (в среднем
около недели). Сухие строительные материалы, предназначенные
для замены пораженных домовыми грибами деталей при
капитальном ремонте зданий, выдерживают примерно в течение месяца
(капиллярная пропитка). Для капиллярной пропитки
древесины, предназначенной для использования в сухих (например,
жилых) помещениях, применяют водорастворимые малотоксич-
иые препараты. Для пропитки строительных элементов, идущих
на лаги, нижние венцы, столбы фундамента и другие конструк-
367
ции, эксплуатируемые в условиях постоянного увлажнения,
применяют маслянистые и другие невымываемые и трудновымывае-
мые антисептики, которые перед употреблением рекомендуется
подогреть.
Бандажный метод применяют для защиты деревянных
деталей, контактирующих с грунтом (различных опор, столбов,
стоек). Вкапываемые в землю основания столбов обмазывают
антисептической пастой, а затем обертывают полиэтиленовой
пленкой или покрывают водонепроницаемым составом (например, неф-
тебитумом). В качестве антисептического бандажа можно
использовать два слоя мешковины или другой технической ткани с
прослойкой антисептической пасты. Таким бандажом обертывают
защищаемую деталь, а снаружи ее покрывают гидроизолирующим
материалом. Бандаж должен на 20 — 30 см выступать над
поверхностью земли.
Для противогрибной защиты строительных деталей, столбов,
опор линий электропередачи, других конструкций применяют в
основном пропитку под давлением: через торец, способом
«прогрев — холодная ванна», автоклавную и др.
Пропитка через торец с использованием
водорастворимых соединений рекомендуется для антисептирования свеже-
срубленных лесоматериалов. При этом способе раствор
антисептика под давлением через торец бревна поступает в заболонь и,
вытесняя клеточный сок и воду, постепенно пропитывает ее по
всей толщине.
Способ «прогрев —холодная ванна» — один из
самых эффективных для антисептирования сухой древесины (с
влажностью до 25 %). При выдерживании круглых сортиментов
сначала 3—5 ч в горячем, а затем 2 —3 ч в холодном растворе
антисептик не только пропитывает заболонь, но со стороны торцов
проникает и в ядровую часть бревен.
Наилучшее качество антисептирования обеспечивает глубокая
автоклавная пропитка, осуществляемая в заводских
условиях. Ее применяют для воздушно-сухой древесины наиболее
ответственного назначения (идущей на производство шпал, деталей
мостов, морских свай, мачт и других важных деталей открытых
сооружений). В этом случае древесину попеременно выдерживают
то в вакууме, то под давлением, регулируя его уровень. При этом
применяют различные режимы антисептирования:
вакуум—давление, вакуум—давление —вакуум, давление —вакуум,
давление—давление—вакуум и др. Наиболее известна автоклавная
пропитка древесины маслянистыми антисептиками в режиме
давление—давление —вакуум.
Еще более эффективен применяемый на шпалопропиточных
заводах комбинированный метод пропитки
древесины: сначала водорастворимым антисептиком, проникающим
368
в центральные части чурок, а затем — маслянистым,
пропитывающим наружные слои. Этим достигается сплошная пропитка,
которая надежно защищает древесину от поражения дереворазру-
шающими грибами.
При правильном антисептировании срок службы стандартных
деревянных домов увеличивается в среднем с 15 до 50 лет, шпал —
с 10 до 25, столбов и опор линий электропередачи и связи — с 12
до 50, деревянных мостов и гидросооружений — с 10 до 40,
кузовов грузовых автомобилей и товарных железнодорожных вагонов —■
с 5 до 20 лет.
Антисептирование фанеры, древесностружечных и
древесноволокнистых плит осуществляют путем добавления фунгицида в
состав клея, применяемого для склеивания листов шпона, или
пропитки готовых материалов раствором препарата, содержащего
4 % салициланилида и 0,4 % пентахлорфенолята натрия.
Обычно защиту древесины от биоповреждений совмещают с
защитой от возгорания. Для этого применяют биоогнезащитные
препараты, в которые наряду с антисептиками вводят антипире-
ны: хлорид цинка, буру, карбонат натрия, сульфат аммония и др.
Глава 14
БОЛЕЗНИ ЦВЕТОЧНЫХ КУЛЬТУР
И МЕРЫ БОРЬБЫ С НИМИ
Цветочные культуры занимают очень важное место в
озеленении городов и других населенных пунктов. Они служат
украшением парков, скверов, площадей, мемориальных комплексов и
старинных усадеб. Цветочные растения широко используются в
озеленении приусадебных участков, в интерьерах административных
зданий, учреждений здравоохранения, образования и культуры и
др. Однако многие виды цветочных растений поражаются
различными инфекционными (грибными, бактериальными,
вирусными, нематодными) и неинфекционными болезнями, которые
снижают декоративные качества и продуктивность этих культур,
вызывают их ослабление и гибель, причиняя большой ущерб
цветоводству.
Ниже рассматриваются наиболее опасные и распространенные
болезни важнейших цветочных культур защищенного грунта,
луковичных и клубнелуковичных растений, многолетников и
однолетников открытого грунта.
14.1. БОЛЕЗНИ ЦВЕТОЧНЫХ КУЛЬТУР
ЗАЩИЩЕННОГО ГРУНТА
14 Л Л. Болезни гвоздики садовой ремонтантной
Ржавчина. Болезнь вызывается разнохозяйным грибом с
полным циклом развития Uromyces caryophyllinus (Schr.) Wint. Однако
полный цикл возможен только в условиях открытого грунта, при
наличии промежуточного хозяина — молочая {Euphorbia gerardiana
Jacq.), на котором развивается весенняя стадия гриба.
Пораженные ржавчиной растения молочая имеют укороченные
междоузлия, деформированные листья, на нижней стороне которых
образуются многочисленные, в виде оранжевых порошащих язвочек,
370
эции с эциоспорами. Эциоспоры заражают растения гвоздики, и
на них развивается сначала урединиостадия, а затем и телиоста-
дия гриба (рис. 114).
В урединиостадии на стеблях и нижней стороне листьев
пораженных растений появляются светло-коричневые подушечки —
урединиопустулы, прикрытые эпидермисом и заполненные уре-
диниоспорами. Урединии иногда образуются и на чашечках
цветков. По мере созревания пустул эпидермис разрывается, уреди-
ниоспоры высыпаются наружу и, рассеиваясь, вновь и вновь
заражают растения гвоздики.
В результате многочисленных разрывов эпидермиса
усиливается отдача воды растением, что приводит к постепенному усыха-
нию листьев и угнетению всего растения.
К осени на пораженных органах гвоздики появляются более
темные телиопустулы, содержащие темно-бурые, с толстыми
оболочками, телиоспоры. Последние зимуют на пораженных частях
растений и весной прорастают базидиями с базидиоспорами, ко-
Рис. 114. Ржавчина гвоздики:
/ — пораженный лист гвоздики с урединиями и телиопустулами возбудителя; 2 —
здоровое растение молочая; 3 — пораженное растение молочая с эциями гриба
на деформированных листьях
371
торые заражают молочай, на нем развивается эциостадия гриба,
и цикл развития повторяется. Может зимовать также мицелий гриба
в корнях молочая.
При выращивании гвоздики в теплицах, где нет
промежуточного хозяина (молочая) и не требуется перезимовки телиоспор,
ржавчина развивается по неполному циклу. В течение всего года
гриб образует урединиоспоры, которые обеспечивают
прогрессирующее распространение болезни. При сильном развитии болезни
листья усыхают полностью, растения увядают и гибнут. Телио-
споры в теплицах тоже образуются, но не играют никакой роли в
дальнейшем развитии гриба.
В условиях защищенного грунта ржавчина особенно
интенсивно развивается в осенне-зимний период, когда температура
понижается до 18... 20 °С и при этом влажность воздуха
увеличивается до 80—90 %. Летом, при высокой температуре воздуха в
теплицах (30... 35 °С) болезнь нередко затухает. Развитию ржавчины
способствуют также недостаточное освещение и одностороннее
азотное питание.
Ржавчина — одна из наиболее вредоносных болезней гвоздики
ремонтантной, резко снижающая выход товарной продукции и
декоративные качества растений. Болезнь представляет большую
опасность и для семенников гвоздики. Больные растения слабо
цветут и не завязывают семян. Даже при невысокой степени
поражения количество цветков уменьшается на 25—30%.
Инфекция чаще всего распространяется с черенками, по
воздуху, а также через инвентарь, одежду персонала и другими путями.
Меры борьбы при выращивании гвоздики в
закрытом грунте: незагущенные посадки; регулярное
проветривание теплиц; полив растений под корень; в зимний и ран-
невесенний период — понижение температуры до 10... 12 °С. При
выращивании гвоздики в открытом грунте: уничтожение
пораженных растительных остатков и растений молочая. Проведение
внекорневых подкормок основными удобрениями (N.P.K) в
соотношении 1:2,5:3,5, а также микроэлементами: бором,
марганцем, цинком и др. (0,05 % по препарату) с целью повышения
устойчивости растений к ржавчине. Профилактические
опрыскивания с интервалом 14 дней, а при появлении ржавчины —
через 5—7 дней. Рекомендуется использование байлетона, пенкона-
зола (топаза), оксихлорида меди, препарата Абига-Пик,
биопрепарата «Бактофит» и др.
Фузариоз. Возбудители — различные виды несовершенных
грибов рода Fusariwn, и прежде всего — Fusarium oxysporum Schl. f.
dianthi (Pril. et Del.) Bilai.
Фузариоз поражает гвоздику на всех стадиях ее развития, но
особенно сильно — в период бутонизации. Промышленные
масштабы выращивания гвоздики, широкое распространение фузариоза
372
на больших площадях и массовая гибель пораженных растений
делают это заболевание одним из наиболее вредоносных. Как
правило, болезнь развивается очагами.
Поражаются все органы взрослых растений (корни, стебли,
листья, бутоны), а также укорененные и неукорененные черенки.
Черенки заражаются через почву. Ткани их оснований буреют, затем
загнивают, и на них появляются розовые или белые подушечки
конидиального спороношения возбудителя. Чаще поражаются
черенки, передержанные на стеллажах и при заглубленной (более
1,5 см) посадке. В этом случае отмирание корней и нижней части
стебля происходит в первые 2—4 нед после посадки. Черенки,
взятые с пораженных маточников, обычно уже содержат
инфекцию. Они плохо укореняются и быстро погибают, становясь
причиной заражения соседних черенков.
Главным источником инфекции является пахотный слой
почвы, где возбудители фузариоза годами сохраняются на
пораженных остатках растений в виде мицелия или хламидоспор. Поэтому
заражение взрослых растений также чаще всего происходит через
корневую систему. При таком типе развития болезни корневая
шейка буреет, затем чернеет и подгнивает, нижние части
растений загнивают и размочаливаются, после чего болезнь быстро
распространяется на вышележащие части. Листья из-за потери тур-
гора внезапно увядают, становятся сначала бледно-зелеными,
потом соломенно-желтыми. Отмершие листья обычно не опадают.
Стебли буреют или приобретают красноватый оттенок. На
поперечных срезах отмерших стеблей видны бурые пораженные
сосуды. Бутоны не образуются или плохо раскрываются, их
сердцевина буреет.
Палево-розовые подушечки конидиального спороношения
возбудителя, образующиеся при влажной погоде (в условиях
открытого грунта) или при высокой влажности воздуха в теплицах,
служат источником заражения все новых растений. Конидии легко
переносятся воздушными течениями, брызгами воды,
насекомыми, людьми при уходе за растениями и заражают надземные
части гвоздики. При этом типе инфекции болезнь чаще всего
распространяется от верхних частей растения к нижним, а в дальнейшем
переходит на корни, которые также загнивают, растения валятся
на землю и легко вынимаются из почвы (рис. 115).
На новые участки возбудители фузариоза могут быть занесены
с посадочным материалом, с растительными остатками и с
семенами. Заражению растений способствуют механические
повреждения (особенно корней и корневой шейки), возникающие при
посадке растений, а также повреждения насекомыми.
Интенсивное развитие болезни с ярко выраженными
признаками поражения наблюдается при оптимальных для патогена
условиях: высокой влажности почвы и воздуха, плохой вентиля-
373
ции в теплицах, температуре
воздуха 25...28°С, температуре
почвы 20... 24 °С. По мере
повышения температуры и влажности
почвы и воздуха степень
развития фузариоза возрастает. В
зимний и ранневесенний период,
когда в теплицах температура
воздуха не превышает 15°С, а
почвы 10... 12 °С, болезнь
проявляется слабо.
Ослабление растений и
снижение устойчивости к фузарио-
зу вызывают и такие факторы,
как дефицит в почве фосфора и
калия, высокая кислотность кор-
необитаемой среды, загущенные
посадки, низкая агротехника,
нарушение технологии выращи-
Рис. 115. Фузариозное увядание вания гвоздики (например, сро-
гвоздики ков укоренения черенков и др.).
Фиалофороз. Возбудитель —
гриб Phyalophora cinerescens (Wr.) van Beyma. Типичные признаки
болезни обнаруживаются у растений на плантациях и в теплицах в
возрасте 6—8 мес. Заражение корневых систем происходит через
почву. Мицелий проникает в корни, откуда переходит в
надземные части растений.
Признаки фиалофороза сходны с признаками фузариоза,
однако в отличие от последнего корневая система в целом выглядит
нормально, долго не загнивает, и поэтому растения трудно
выдернуть из почвы. Иногда можно обнаружить потемнение и
отмирание отдельных мелких корешков.
Поражение распространяется снизу вверх по одному или по
нескольким стеблям, при этом растения сравнительно долго
остаются живыми и зелеными. Затем на нижних листьях появляются
соломенно-желтые и серо-зеленые пятна, желтые участки
чередуются с красноватыми. Ближе к верхней части стебля иногда
наблюдается усыхание половины листовой пластинки по жилке. При
высокой температуре пятна становятся вишнево-красными или
имеют ржавый оттенок. Потом листья желтеют, увядают,
повисают. В большинстве случаев отмечается частичное усыхание стеблей.
На них вначале появляются темно-синие зоны, в дальнейшем
стебли приобретают светло-соломенный цвет с сероватым оттенком,
иногда растрескиваются.
При увядании всего растения кора легко отделяется от стеблей.
На продольном и поперечном срезах нижней части стеблей и кор-
374
невой шейки обнаруживается характерная интенсивная
коричнево-бурая окраска проводящих сосудов в виде кольца или
полукольца. Изменение окраски тканей иногда наблюдается до высоты
30 — 50 см.
На листьях боковых побегов старых растений появляется
характерный для этой болезни красно-фиолетовый или лиловато-
красный оттенок. Одновременно увядают бутоны, изменяется и
их окраска.
Оптимальные температуры для развития фиалофороза 14... 18 °С,
влажность воздуха — 75—80 %. Оптимальная температура для спо-
руляции возбудителя 18... 23 °С. Резкие перепады температуры (до
10... 15 °С) стимулируют спороношение. При заражении растений
инкубационный период колеблется от одного до трех месяцев, в
зависимости от температуры, возраста и сорта гвоздики.
Возможна передача возбудителя болезни и с черенками, взятыми от
пораженных растений.
По имеющимся данным, инфекция фиалофороза может
сохраняться в почве на глубине до 1 м, что затрудняет ее
химическое или термическое обеззараживание.
Ризоктониоз. Возбудитель болезни — несовершенный гриб
Rhizoctonia solani Kiihn. Заболевание проявляется в виде корневой
гнили, полегания, черной ножки, поражения корневой шейки.
Ремонтантной гвоздике ризоктониоз наносит значительный вред.
Болезнь особенно опасна в форме корневой гнили: на фоне
здоровых растений хорошо заметны растения с
этиолированными, серо-зелеными, со свинцовым оттенком, увядающими и слегка
морщинистыми листьями. Эти признаки наиболее отчетливо
проявляются в жаркое время дня. После полива или прохладной ночи
тургор восстанавливается.
На пораженной корневой шейке появляется
бледно-коричневое маслянистое пятно. Ткани корневой шейки темнеют,
утончаются и размочаливаются. Растение увядает полностью, тургор не
восстанавливается, листья становятся свинцово-белесыми или
серо-зелеными, стебли поникают. При достаточном количестве
влаги мицелий от больного растения быстро распространяется во
всех направлениях и хорошо заметен как на тканях растения, так
и на поверхности почвы. Он паутинистый,
желтовато-коричневого цвета. Больные растения полегают. При выдергивании их из
почвы пораженные ткани корневой шейки разрываются, хотя
корневая система в целом выглядит здоровой.
Кроме мицелия, в зоне поражения обнаруживаются светло- или
темно-коричневые склероции разной величины (от 0,5 до 1,5 мм),
большей частью округлые или удлиненные.
При укоренении гвоздики на стеллажах ризоктониоз часто
поражает значительный процент черенков. Заболевание носит
массовый характер и развивается очагами в местах, где температу-
375
pa превышает 24...25°C, влажность более 90—95%, густота
посадки — свыше 700—800 шт./м2 и глубина заделки черенков
более 1 см.
На нижней части черенков развивается войлочный или
паутинистый мицелий светло-коричневого цвета. У оснований и в
пазухах листьев формируются склероции гриба. Черенки
становятся вялыми, белесыми, не укореняются и погибают. Болезнь
передается от больных черенков к здоровым. Зараженные
черенки становятся источником инфекции при пересадке их на
постоянное место.
Ризоктониоз является наиболее опасным заболеванием как в
защищенном, так и в открытом грунте в условиях повышенной
температуры и избыточной влажности. Одностороннее азотное
питание, внесение больших доз органики, заглубленная посадка
растений, высокая температура воздуха, частые бесконтрольные
поливы — вот те факторы, которые способствуют развитию ри-
зоктониоза. Наиболее благоприятные условия для развития
болезни создаются в весенне-летний период. Гриб способен
развиваться в диапазоне 6...35 °С (при оптимуме 26...30 °С).
Rh.solani относится к числу высокоагрессивных патогенов.
Инкубационный период длится 11 — 14 дней.
Мицелий гриба сохраняется в почве на глубине до 80 см
(оптимальная глубина — 40—60 см). Находясь в верхних слоях почвы,
он легко распространяется по всему пахотному слою. Резерватора-
ми инфекции служат многие виды сорных растений, на которых
сохраняются мицелий и склероции гриба.
Меры борьбы с болезнями типа увядания.
Самое главное — соблюдение правил высокой агротехники и
технологии выращивания гвоздики. Своевременная диагностика, удаление
и уничтожение больных растений с последующим их сжиганием
на всех этапах выращивания гвоздики; уничтожение растительных
остатков; хорошая отрегулированность режимов температуры и
влажности воздуха, вентиляции, поливов почвы;
сбалансированное минеральное питание; создание высокого карантинного и
фитосанитарного фона. Не рекомендуется заглубленная посадка
укорененных черенков в летний период, когда температура
воздуха достигает 28... 30 °С, а почвы — 25 °С, при влажности 80 % и
выше. Не следует применять полив напуском воды, так как это
способствует распространению инфекции (зооспор, хламидоспор,
конидий и др.) и массовому заражению растений. Отбор черенков
только со здоровых маточников. Перед укоренением черенков
дезинфекция их в течение 15 — 20 мин в суспензиях фундазола (0,2 %),
ТМТД (0,6 %), бактофита и др. Профилактический двукратный
полив с интервалом в один месяц названными фунгицидами (8 —
10 л/м2) с последующими рыхлениями. Недопустимость
загущенных посадок и механических повреждений черенков. Смена культур
376
в открытом грунте. Дезинфекция почвы фунгицидами за 3—4 нед
до посадки растений или пропаривание почвы на глубину 25 — 30 см
при температуре 100... 105 °С и экспозиции 3—4 ч. Изоляция
новых почвосмесей от зараженного грунта полимерными и другими
материалами либо замена зараженной почвы на новую, не
содержащую инфекции. Использование легких и хорошо дренируемых
почвосмесей.
Гетероспориоз. Возбудитель — гриб Heterosporium echinulatum
(Berk.) Cke. Болезнь проявляется в форме пятнистости.
Поражаются все части растений: стебли, листья, чашелистики, бутоны,
цветки и даже семена.
Первым признаком болезни служит появление на листьях
буровато-коричневых пятен с хорошо выраженным темно-красным
ободком. Позже такие же пятна появляются на стеблях и
чашелистиках. Постепенно разрастаясь, пятна сливаются, что сильно
уменьшает ассимилирующую поверхность листьев и часто
приводит к их отмиранию и опадению.
На поверхности пятен образуется бархатистый или пушистый
оливково-зеленоватый налет, состоящий из мицелия и кониди-
ального спороношения возбудителя. Рассеиваясь, конидии вновь
заражают гвоздику.
При сильной степени развития болезни бутоны развиваются
плохо или совсем не раскрываются. Больные растения, успевшие
зацвести, дают зараженные семена, в которых инфекция
сохраняется до следующего года, вызывая новые заражения. Заражение
растений может происходить также через почву, если на одном и
том же участке гвоздика выращивается несколько лет подряд.
В теплицах болезнь бурно развивается на горшечных культурах;
при высадке их в открытый грунт они могут служить источником
распространения болезни.
Развитию болезни способствуют высокая влажность воздуха,
чрезмерно загущенные посадки и обильные поливы. Гриб способен
развиваться до глубокой осени, и его спороношение нередко
наблюдается даже, в ноябре на растениях, погибших от заморозков.
Альтернариоз. Возбудитель — гриб-гифомицет Alternaria dianthi
Stev. et Hall. Болезнь проявляется во второй половине лета.
Поражаются листья и стебли как молодых, так и взрослых растений.
Первоначально заболевание обнаруживается на нижних частях
растений, произрастающих в условиях затенения, повышенной
влажности и пониженной температуры воздуха.
Пораженные листья и стебли буреют, а в дальнейшем
обесцвечиваются. На листьях нижней части растений вначале появляются
бледно-зеленые пятна. Затем вдоль главной жилки образуются
округлые или удлиненные пепельно-серые пятна. На стеблях местами
происходит некроз тканей, что приводит к загниванию стеблей и
общему увяданию растений.
377
На поверхности пораженных частей растений появляется
темно-коричневый бархатистый налет мицелия и конидиального спо-
роношения гриба. Конидии распространяются ветром,
насекомыми, другими способами и, попадая на растения гвоздики, вновь
их заражают.
Черноватый налет появляется также на чашелистиках цветков
и бутонах. Пораженные бутоны часто не раскрываются или
образуют уродливые цветки (рис. 116).
Гриб сохраняется в виде мицелия в зимующих остатках
пораженных растений. Весной он вновь образует конидии, которые
осуществляют первичное заражение здоровых растений. Споры
гриба могут сохраняться и с семенами гвоздики, являясь, таким
образом, одним из источников заражения молодых растений в
следующем году.
Развитию болезни способствуют повышенная влажность
воздуха и загущенная посадка растений. Наиболее интенсивное
заражение гвоздики альтернариозом наблюдается при дождливой
погоде и температуре воздуха от 10 до 27 °С. Заболевание может
встречаться повсеместно.
Септориоз. Возбудитель — пикнидиальный гриб Septoria dianthi
Desm. На листьях и стеблях появляются желтоватые пятна округ-
Рис. 116. Альтернариоз гвоздики Рис. 117. Септориоз гвоздики
378
лой или неправильной формы, окруженные пурпуровой каймой.
На поверхности пятен заметны пикниды гриба в виде
темно-коричневых мелких точек, погруженных в ткань. При сильном развитии
болезни происходит отмирание пораженных листьев (рис. 117).
Из других пятнистостей на гвоздике встречаются аскохитоз
(возбудитель — Ascochyta dianthi Berk.) и филлостиктоз (возбудитель —
Phyllosticta dianthi West.). При аскохитозе на листьях образуются
сероватые пятна с темной каймой и черными точечными пикни-
дами в центре пятен. При филлостиктозе — пятна беловатые, в
ткань которых погружены пикниды, имеющие вид черных точек.
Меры борьбы с пятнистостями: в открытом грунте —
смена культур и уничтожение растительных остатков; в
защищенном грунте — дезинфекция почвы или замена почвенного субстрата.
Заготовка черенков только от здоровых растений; хорошее
проветривание теплиц; опрыскивание растений фунгицидами: фундазо-
лом, байлетоном, охсихлоридом меди, бордоской жидкостью и др.
Мозаика. Возбудитель — Carnation mosaic virus (вирус мозаики
гвоздики). Болезнь поражает молодые листья, на которых
отчетливо проступают длинные бледно-зеленые пятна и штрихи,
идущие параллельно центральной жилке листа. Эти участки со
временем отмирают, листья скручиваются. У сортов с красными и
темно-розовыми цветками появляются симптомы пестролепестности
в виде светлых штрихов и полос. Иногда происходит
растрескивание чашечек. Вирус передается тлями, а также механически с
соком при уходе за растениями.
Кольцевая пятнистость. Возбудитель — Carnation ringspot virus
(вирус кольцевой пятнистости гвоздики). Характерный признак
болезни — появление на молодых листьях хлоротичных
концентрических колец, желтых и серых пятен — линейных или
округлых, светлой крапчатости. Края листьев деформируются.
Пораженные вирусом старые листья мельчают, краснеют, концы листьев
и середина пятен часто некротизируются. На стеблях также
образуются коричневые некротические пятна. Рост и развитие
растений замедляются. Цветки также мельчают, деформируются, их
чашечки растрескиваются, возникает пестролепестность.
Переносчиками являются фитонематоды Xiphinema diversicaudatum и
Longidorus macrosoma.
Пожилковая крапчатость. Возбудитель — Carnation vein mottle
virus (вирус пожилковой крапчатости гвоздики). Жилки на
молодых листьях светлеют, вдоль них образуются хлоротичные пятна и
крапчатость. На цветках, особенно темноокрашенных,
появляется пестролепестность в виде светлых штрихов, наблюдается
деформация (морщинистость) лепестков. Вирус передается тлей
Myzus persicae, иногда механически. Данный вирус один из
наиболее распространенных в цветоводческих комбинатах; встречается
практически на всех сортах.
379
Меры борьбы с вирусными болезнями:
использование только здорового посадочного материала; уничтожение
пораженных черенков и взрослых растений; уничтожение
переносчиков вирусной инфекции; применение методов термотерапии.
Нематодные болезни гвоздики весьма многочисленны. Чаще
других встречаются стеблевая нематода, галловые нематоды,
клеверная цистообразующая нематода и тиленхоринх.
Стеблевая нематода— Ditylenchus dipsaci (Kiihn.) Filip.
Пораженные растения отстают в росте и развитии, плохо цветут.
Стебли утолщаются, укорачиваются, листья деформируются,
цветки остаются недоразвитыми. Часто наблюдается израстание
верхушек стеблей, образуются искривленные, гофрированные или
нитевидные листья. При сильной степени поражения растения
погибают.
Меры борьбы: уничтожение больных растений и сорняков;
севооборот с возделыванием бархатцев; размножение гвоздики
здоровыми черенками; обеззараживание почвы нематицидами. При
температуре воздуха выше 18 °С — предпосадочное вымачивание
кустов в 0,2 %-м растворе гетерофоса (с температурой не ниже
15°С) — экспозиция 30 мин. Последующий полив уже
посаженных гвоздик 0,2 %-м раствором гетерофоса при норме расхода 0,2—
0,5 л на каждый куст; опрыскивание заболевших растений гетеро-
фосом и полив раствора препарата под куст.
Галловые нематоды рода Meloidogyne образуют на
пораженных корнях многочисленные опухоли (галлы). Паразит
угнетает растения — они отстают в развитии и плохо цветут. В зоне
умеренного климата с многоснежной и часто морозной зимой
распространена северная галловая нематода (M.hapla Chit.), а в
более южных районах, с теплыми бесснежными зимами —
арахисовая галловая нематода (М arenaria (N.) Chit.) и яванская галловая
нематода (M.javanica (Trenb.) Chit.).
Меры борьбы: уничтожение корней больных растений, а в
случае поражения всей корневой системы — выбраковка и
уничтожение самих растений; удаление сорняков; чередование культур с
выращиванием хвойных древесных пород; обработка почвы
нематицидами.
Клеверная цистообразующая нематода — Hetero-
dera trifolii Goff.
Во второй половине вегетационного периода на корнях хорошо
видны бурые цисты лимоновидной формы размером около 1 мм;
это отмершие самки нематоды, внутри которых содержатся
жизнеспособные яйца и личинки паразита. Пораженные растения
отстают в развитии, плохо цветут, утрачивают декоративность.
Меры борьбы: смена культур с исключением из
севооборота поражаемых растений семейств гвоздичных, губоцветных,
крестоцветных, бобовых, пасленовых, гречишных, тыквенных,
380
норичниковых. Удаление сорняков из этих семейств;
выращивание гвоздики на прежних участках не ранее, чем через 6—7 лет;
обеззараживание почвы нематицидами.
Тиленхоринх — Tylenchorhynchus dubius (Butsch.) Filip. Эта
нематода паразитирует на корнях, вызывая редукцию
(недоразвитие) корневой системы. Пораженные растения угнетены, отстают
в росте и развитии, слабо цветут, при сильной степени
поражения погибают.
Меры борьбы: уничтожение корней больных растений, а
при сильной зараженности — уничтожение растений целиком,
тщательная прополка; чередование культур с выращиванием
бархатцев; возврат гвоздики на прежние участки через 3—4 г.;
обработка почвы нематицидами.
На гвоздике паразитируют и другие фитонематоды: парати-
ленх {Paratylenchus projectus Jen.), пратиленх {Pratylenchus neglectus
(Rensch) Filip. et Sch.St.), проникающая короткотелая нематода,
или проникающий пратиленх {Ppenetrans (Cobb.) Filip. et Sch.
St.) и др.
14.1.2. Болезни розы
Мучнистая роса. Возбудитель — гриб Sphaerotheca pannosa Lev. van
rosae Woronich. (конидиальная стадия — Oilium leucoconium Desm.).
Одна из наиболее распространенных и вредоносных болезней,
поражающая розы как в защищенном (в том числе горшечные
розы), так и в открытом грунте.
Рис. 118. Мучнистая роса розы:
/ — пораженный лист; 2 — плодовое тело возбудителя: а — придатки, б — клей-
стотеций, в —- сумка с сумкоспорами
381
На поверхности листьев, зеленых побегов и бутонов вначале
появляется паутинистый белый налет мицелия, который затем
становится мучнистым в результате массового образования
конидий и со временем приобретает сероватый оттенок (рис. 118).
Конидии, распространяясь по воздуху, вновь заражают побеги и
листья (особенно молодые).
При выращивании роз в теплицах болезнь развивается
круглый год, причем плодовые тела сумчатой стадии (клейстотеции)
обычно не формируются, и гриб из года в год сохраняется на
растениях в виде мицелия (в пораженных ветвях и спящих
почках) или в конидиальной стадии. В условиях открытого грунта в
отдельных районах гриб образует клейстотеции, зимующие на
почве или опавших листьях. Считается, однако, что сумчатая
стадия не имеет большого значения ни в перезимовке, ни в
весеннем возобновлении цикла развития гриба и первичном
заражении растений.
Оптимальные условия для развития мучнистой росы:
температура 18...20°С, относительная влажность воздуха 60—78%, но
гриб способен развиваться и при влажности 50—60%. Наиболее
интенсивно конидии прорастают при температуре 12... 25 °С и
влажности 60—65 %. В оптимальных условиях прорастание конидий и
заражение растений происходит за 24 ч, через 48 ч появляется
новая генерация.
При сильной степени поражения мучнистой росой листья
сплошь покрываются белым налетом, деформируются
(приобретают морщинистость, утолщаются, скручиваются) или остаются
недоразвитыми, уродливыми и преждевременно опадают.
Молодые побеги искривляются, их рост приостанавливается, и они
постепенно отмирают. Бутоны мельчают, часто не раскрываются,
а распустившиеся цветки и лепестки утрачивают нормальную
форму, приобретая «растрепанный» вид. Все это резко снижает
декоративность и товарную ценность роз.
Мучнистая роса может вызвать настолько сильное ослабление
растений, что они не переносят зимовки и погибают.
Болезнь особенно сильно поражает розы в период жаркой и
влажной погоды (чаще всего это конец июля—начало августа) и
нередко полностью приостанавливает их цветение.
Мучнистая роса роз распространена повсеместно. Появлению и
развитию болезни способствуют загущенные посадки, иссушение
почвы возле корней, недостаток в ней питательных веществ,
сочетание низкой ночной и высокой дневной температуры воздуха.
Меры борьбы: уничтожение сорняков на плантациях роз;
обрезка и уничтожение пораженных ветвей и побегов; осенью —
глубокая обработка почвы; выращивание устойчивых сортов;
поддержание в теплицах правильного (неблагоприятного для
развития мучнистой росы) режима температуры, влажности, аэрации.
382
Обработка по спящим почкам 5%-м раствором медного купороса;
перед посадкой роз фумигация оранжерей серой.
При первых признаках болезни рекомендуется опрыскивание
растений байлетоном, фундазолом (10—15 г СП на 10 л воды) —
пятикратная обработка, вектрой (2 — 3 мл СК на 10 л воды) —
четырехкратная обработка, скором (2 г КЭ на 10 л воды) —
четырехкратная обработка, пенконазолом (топазом), тиовитом,
трифорином (сапролем), коллоидной серой (1 %) — 25 —50 г СК
на 10 л воды, а также бактофитом (опрыскивание 0,7%-м
рабочим расвором 1 — 2 раза в мес) или средством «Multirose».
Эффективен биологический метод борьбы — опрыскивание настоем
навоза.
Ложная мучнистая роса. Возбудитель — оомицет Peronospora sparsa
Berk. Поражаются листья, побеги, цветоножки. Болезнь наиболее
вредоносна в теплицах.
На верхней стороне листьев образуются расплывчатые бледно-
желтые, со временем буреющие и высыхающие пятна,
соответственно которым на нижней стороне листьев появляются
«островки» беловатого, нежного, слабовыраженного мучнистого налета,
состоящего из пучков конидиеносцев с конидиями, выходящих
из устьиц. Пораженные листья часто приобретают
гофрированную форму, преждевременно засыхают и опадают, побеги также
усыхают, на них иногда образуются глубокие трещины.
Инфекция сохраняется в пораженных побегах и опавших листьях.
Развитию ложной мучнистой росы в теплицах способствуют высокая
влажность воздуха, резкие колебания температуры, избыток
азота, а в условиях открытого грунта — теплая погода с моросящими
дождями.
Меры борьбы: дезинфекция оранжерей; уничтожение
опавшей листвы и пораженных побегов; соблюдение постоянного
температурного режима; умеренные азотные удобрения;
систематическое проветривание оранжерей; опрыскивание растений дита-
ном М-45, байлетоном, оксихлоридом меди, бордоской
жидкостью, вектрой, фундазолом, топазом; опыливание смесью серы с
известью (1:1).
Ржавчина. Вызывается однохозяйными видами рода Phragmidium:
Ph.distiflorum (Tode) James, Ph. tuberculatum Mull., Ph.rosae-pimpinelli-
foliae (Rabenh.) Diet. Два первых вида поражают преимущественно
культурные сорта роз, третий вид — как культурные, так и
дикорастущие шиповники, главным образом в умеренном влажном
климате. В связи с тем, что шиповники (особенно Rosa canina)
используются в качестве подвоев, Ph. rosae-pimpinellifoliae
представляет особую опасность. Ржавчина роз встречается повсеместно в
теплицах и открытом грунте.
Все стадии ржавчины развиваются только на розах.
Спермогонии формируются на верхней стороне листьев под кутикулой мел-
383
кими группами, они имеют вид плоских желтых подушечек. Эции
образуются на молодых побегах, нижней стороне листьев,
черешках, цветоножках и цветочных почках в виде ярко-оранжевых
порошащих подушечек. У шиповников эции образуются также на
плодах (рис. 119). На одревесневших частях (штамбах и ветках)
эции развиваются по типу цеомы — в виде продолговатых
вздутий, прикрытых эпидермисом. При созревании эциев эпидермис
разрывается, и эции превращаются в глубокие многолетние язвы,
заполненные эциоспорами. Последние заражают растения в
течение всего вегетационного периода.
На пораженных листьях появляются крупные желтые пятна.
Урединиопустулы образуются на нижней стороне листьев в виде
порошащих желтых подушечек. В отличие от многих других
ржавчинных грибов, ведущая роль в распространении возбудителя
принадлежит эциоспорам. Урединиоспоры в этом плане большого
значения не имеют.
К концу вегетации на различных органах растений образуются
многоклеточные телиоспоры — в открытых пустулах
буро-черного цвета (рис. 120). При сильной степени развития ржавчины
телиоспоры могут сплошь покрывать пораженные органы растений,
которые при этом выглядят, как обугленные. После периода
покоя телиоспоры прорастают базидиями, и базидиоспоры
осуществляют первичное заражение растений.
Пораженные ржавчиной растения плохо развиваются,
выглядят угнетенными, хуже цветут. Побеги деформируются,
искривляются, листья скручиваются, засыхают и в массе осыпаются.
Зимуют телиоспоры на опавших листьях, других пораженных
органах. В пораженных ветвях зимует также мицелий.
Особенно сильно ржавчина поражает многие виды
шиповников (розу собачью, желтую, краснолистную), некоторые сорта
Рис. 119. Ржавчина розы:
1 — эциостадия ржавчины на молодых деформированных плодах шиповника; 2 —
пораженный лист с эциями гриба
384
Рис. 120. Ржавчина розы:
/ — телиостадия гриба на листе розы; 2 — пустула ржавчины с урединиоспора-
ми (а) и телиоспорами (б)
парковых и ремонтантных роз. Более устойчивы чайные, чайно-
гибридные, полиантовые и плетистые розы. Роза морщинистая и
многие ее гибриды иммунны.
В защищенном грунте развитие ржавчины происходит и в
зимний период. Мицелий гриба развивается в покровных тканях
стволиков и ветвей, часто вызывая образование на них глубоких ран,
что препятствует правильному формированию кустов.
Развитию болезни способствуют высокая температура воздуха,
недостаток влаги в почве, дефицит калия.
Меры борьбы: осенью и весной — обрезка и сжигание
пораженных ветвей и побегов. Зачистка язв на штамбах с
последующим антисептированием медным купоросом и замазкой; сбор и
уничтожение опавшей листвы. Осенью или весной (до набухания
почек) перекопка почвы с оборотом пласта; уничтожение ши-
повников на участках, занятых розами, и в радиусе 300 м вокруг
них. Опрыскивание кустов весной по спящим почкам 5 %-м
раствором железного или медного купороса. Опрыскивание в период
вегетации: трехкратное — 1 %-й бордоской жидкостью (первое —
во время распускания листовых почек, второе — в начале
бутонизации, третье — перед цветением); четырехкратное — раствором
арцерида (50 г СП на 10 л воды); двукратное — картацидом (50 г
СП на 10 л воды); двукратное — купроксатом (50 г СК на 10 л
воды). Рекомендуются также охсихлорид меди, топаз, препарат
Лбига-Пик. Каждые три недели желательна обработка нижней
стороны листьев медьсодержащими фунгицидами.
385
Инфекционный ожог (стеблевой рак). Возбудители — грибы рода
Coniothyrium: С. rosarum Sacc. (= С. fuckelii Sacc.) и С. wernsdorffiae
Laub. Первый из них встречается преимущественно в теплицах,
второй — в открытом грунте. Особенно сильно розы поражаются в
открытом грунте, под зимним укрытием, в условиях избыточной
влажности, когда растения ослаблены. Признаки болезни
обнаруживаются сразу же после снятия укрытий. На коре ветвей и
побегов образуются вначале небольшие красноватые пятнышки. Со
временем пятна становятся буровато-черными вдавленными,
окруженными красно-коричневой каймой. Пятна могут появляться
и на листьях. На второй год пятна сереют. Постепенно
разрастаясь, пятна окольцовывают побег, и он отмирает. В отмершей коре
формируются черные пикниды гриба в виде мелких бугорков,
выступающих через разрывы коры своими устьицами. Края
пораженных участков ветвей утолщаются, кора растрескивается,
отваливается кусками или полосами, образуются глубокие раны,
окруженные раневыми валиками.
На однолетних побегах в открытом грунте болезнь проявляется
как ожог — в виде удлиненно-ланцетовидных, слегка вдавленных
пятен темно-коричневого цвета с малиновым ободком, иногда
кольцующих побег; в этом случае вышележащие части отмирают.
В теплицах болезнь часто встречается в случае укоренения
черенков и при выгонке роз на срез. При укоренении черенков
отрастающие молодые побеги буреют и засыхают. Верхние части
черенков также буреют и покрываются мелкими черными пикнида-
ми. У некоторых сортов на стеблях появляются темно-зеленые
мелкие пятна, на которых затем развиваются пикниды.
Возбудитель проникает в ткани растений, как правило, через
ранки, нанесенные насекомыми, или через механические
повреждения, например при рыхлении почвы. Распространение
инфекции, ведущей к заражению растений, происходит также при
обрезке — на секаторах, руках, перчатках и т. п. Мицелий зимует в
зараженных побегах. В открытом грунте особенно сильно
подвержены болезни плетистые сорта роз. В основном поражаются розы,
слишком долго находившиеся под укрытием при плюсовой
температуре (весной или зимой во время оттепелей). На парковых
розах, которые обычно не укрывают на зиму, инфекционный ожог
встречается редко.
Развитию болезни способствуют высокая влажность воздуха,
слабая вентиляция, недостаточное одревеснение побегов, плохое
укрытие на зиму, запоздалое снятие укрытий, различные
повреждения, несбалансированное удобрение (избыток азота),
размножение роз зелеными черенками.
Меры борьбы: своевременное удаление укрытий весной;
обрезка и уничтожение больных побегов; вырезка на стволиках
крупных ран с последующей дезинфекцией срезов и покрытием их са-
386
довой замазкой; подкормка роз комплексным удобрением.
Дезинфекция инструментов (секаторов) после употребления;
опрыскивание роз до распускания почек 5 %-м раствором CuS04, а во
время вегетации — фундазолом, топсином-М, купроксатом,
медьсодержащими фунгицидами; осенью перед укрытием роз
удаление всех листьев и невызревших побегов, обработка 2%-й
бордоской жидкостью, оксихлоридом меди; укрытие роз в сухую погоду.
Черная пятнистость. Возбудитель болезни — гриб-целомицет
Marssonina rosae (Lib.) Diob. с сумчатой стадией Diplocarpon rosae
Wolf.
По имеющимся данным, у возбудителя черной пятнистости
известно не менее 50 физиологических рас, что делает
проблематичным выведение новых устойчивых к этой болезни сортов роз.
Болезнь широко распространена во всех странах, где
выращивают розы, т. е. фактически во всем мире. Черная пятнистость
характеризуется исключительно высокой вредоносностью, так как
поражает очень многие сорта роз из разных групп и наносит
культуре роз большой ущерб. Наиболее восприимчивы к болезни сорта
ремонтантных и чайно-гибридных роз, имеющих бледно-зеленые
листья. Есть данные о более сильной поражаемости красных и
розовых сортов и о повышенной устойчивости белых роз.
Наибольшее распространение болезни отмечается в годы с большим
количеством осадков. Гриб заражает растения в начале вегетации,
но признаки болезни чаще всего становятся заметными во второй
половине вегетационного периода (в южных районах иногда и
раньше), однако конкретные сроки проявления болезни в период
с июля по август могут заметно различаться в зависимости от
экологических условий и сортовых особенностей роз.
На листьях садовых роз образуются преимущественно крупные
черные резко очерченные округлые пятна с желтыми краями, но
нередко наблюдаются и многочисленные мелкие пятна, иногда с
нечеткими бахромчатыми краями. Характер пятен (их форма,
размеры, расположение на листовой пластинке) зависят от
ботанической группы и сортовых особенностей роз (рис. 121). Пятна
могут появляться и на стеблях, которые позже усыхают.
На пятнах формируются конидиальные ложа гриба, имеющие
вид еле заметных округлых или слегка продолговатых
припухлостей (подушечек).
Листья с сильной степенью поражения (наличием
многочисленных, особенно крупных, пятен) быстро желтеют и опадают и
иногда уже в конце июля кусты оголяются, что отрицательно
сказывается на дальнейшем развитии растений. Преждевременная
потеря листьев в открытых экспозициях роз приводит к их
подмерзанию, сильному отставанию в росте и развитии в следующем
году: у роз снижается прирост, обмерзают спящие почки, резко
снижается урожай цветков эфиромасличных сортов. Слабая сте-
387
Рис. 121. Черная пятнистость розы: пораженные листья
пень поражения листьев тоже небезобидна, так как наличие даже
немногочисленных черных пятен делает растения
непривлекательными, снижается их декоративность и товарная ценность
(особенно в тех случаях, когда розы идут на срезку).
На опавших листьях, пораженных пятнистостью, после
перезимовки развивается сумчатая стадия гриба. Плодовые тела (апо-
теции) образуются между палисадной паренхимой и
эпидермисом листа (субэпидермально).
Сумкоспоры осуществляют первичное весеннее заражение
здоровых листьев. Иногда оно может быть вызвано конидиями,
образующимися на перезимовавших листьях. Массовое же конидиаль-
ное спороношение обычно совпадает с длительным периодом
созревания и рассеивания сумкоспор.
Оптимальными условиями для заражения роз в средней полосе
являются дождливая погода и умеренные (12...20°С)
температуры в мае—июне. Если эти условия реализуются в более поздние
сроки (июль—август), то и массовое развитие (эпифитотия)
болезни будет наблюдаться именно в этот период. Появлению и
развитию черной пятнистости способствуют недостаток в почве
калия, загущенная посадка, но особенно часто от болезни страдают
розы в годы с теплым и влажным летом.
Меры борьбы. Бороться с черной пятнистостью довольно
трудно. Прежде всего надо собирать и сжигать опавшие больные
листья. Необходима глубокая перекопка почвы с оборотом пласта.
В начале распускания листьев — дважды, с промежутком в одну
неделю, опрыскивают листья систаном, медно-мыльным
раствором (30 г CuS04 и 200 г мыла на 10 л воды), средством «Multirose»,
388
оксихлоридом меди, бордоской жидкостью, байлетоном). При
появлении первых признаков болезни и по мере необходимости
опрыскивание растений повторяют. Осенью (перед укрытием роз на зиму)
растения обрабатывают 5 %-м раствором медного купороса.
Пурпуровая пятнистость (филлостиктоз) вызывается пикниди-
альным грибом Phyllosticta rosarum Pass, (сумчатая стадия — Sphaceloma
rosarum (Pass.) Jenk.). Поражаются в основном листья. На их
верхней стороне образуются довольно крупные серовато-белые пятна
неправильной формы с четкими очертаниями, окруженные
широкой каймой багряно-пурпурового цвета. На нижней стороне
листьев пятна мелкие буровато-коричневые округлые или
неправильной формы, при сильной степени поражения сливающиеся.
В центре пятен формируются скученные, погруженные в ткань
светло-коричневые пикниды.
Поражаются также цветоножки и молодые зеленые побеги, на
которых появляются мелкие темно-коричневые пятна с белой
серединкой. Больные листья преждевременно опадают, цветение
задерживается, часто образуются недоразвитые цветки. В связи с
этим резко снижается декоративность растений.
Церкоспороз. Возбудитель — гриб Cercospora rosicola Pass. На
поверхности листьев появляются округлые диаметром 1—5 мм
бурые пятна с темным или пурпуровым ободком и нежным
бархатистым налетом оливкового цвета (конидиальное спороноше-
ние гриба).
Септориоз. Возбудитель — гриб Septoria rosae Desm. На листьях
появляются округлые беловатые или буроватые пятна,
окруженные темно-красной каймой, часто сливающиеся. На поверхности
пятен, в их середине, образуются черные точечные пикниды
гриба. Пораженные участки листьев засыхают. У некоторых видов роз
септориоз проявляется и на стеблях. Паразит зимует на остатках
больных растений.
Меры борьбы с пятнистостями - см. «Меры борьбы
с черной пятнистостью роз».
Серая гниль. Возбудитель болезни — гриб-гифомицет Botrytis
cinerea Pers. Поражает многие виды растений многих семейств, в
том числе сорта роз, относящиеся к разным группам. Наиболее
восприимчивы к болезни и сильнее всего поражаются розы из
групп флорибунда и чайно-гибридных. К высоковосприимчивым
относится, например, сорт Карина. Болезнь развивается как в
открытом, так и в защищенном грунте.
В теплицах после обрезки, в период покоя растений, на
нижних частях стеблей появляются бурые мелкие пятна, которые,
увеличиваясь, целиком охватывают стебли и вызывают
отмирание коры и почек.
В период вегетации листья желтеют без явных признаков
грибного поражения и опадают. Бутоны поникают, не раскрываются,
389
1 — пораженные бутоны; 2 — пораженный цветок; 3 — здоровый цветок
и на них развивается серый пушистый налет конидиеносцев,
который вскоре становится порошащим из-за массы образующихся
конидий гриба. На лепестках роз болезнь проявляется в виде
мелких бурых пятен или язвочек. Пораженные лепестки быстро
осыпаются. Декоративность растений резко снижается (рис. 122).
Очень часто в оранжереях серая гниль встречается
одновременно с инфекционным ожогом (стеблевым раком) роз.
Меры борьбы: рыхление и мульчирование почвы;
создание в оранжереях и на открытых участках условий максимальной
сухости для растений, т. е. прекращение полива, усиление
вентиляции и освещения; систематическое удаление и сжигание
пораженных органов растений (листьев, цветков, побегов) и целых
растений; химические меры борьбы.
Из химических препаратов наиболее эффективны ровраль
(2,5 %-й раствор), фундазол (0,2 %-й раствор). Опрыскивание ра-
390
стений указанными препаратами через каждые 3 — 5 дней до
полного исчезновения серого налета. Необходимо чередование
препаратов. При сильном поражении роз возможен их полив под
корень одним из указанных препаратов в 0,2 %-й концентрации (но
в этом случае водой не поливать).
Устойчивость роз к серой гнили повышают марганцевые
микроудобрения, усиливающие в растении окислительные
процессы, неблагоприятные для гриба.
Условия минерального питания оказывают большое влияние
на обмен веществ, процессы роста и развития, продуктивность
растений, их устойчивость к болезням, декоративные качества.
Песталоциоз. Заболевание вызывается грибом-целомицетом
Pestalotia adusta Ell. et Ev. Особенно часто поражаются розы,
привитые на подвои Rosa canina и R. multiflora. У них наблюдается отмира-
ние стебля в местах прививки.
Болезнь очень вредоносна, так как поражает все надземные
части растений. Она проявляется в виде искривления и
сплющивания побегов. Листья приобретают фиолетовый оттенок и опадают.
Деформация стеблей и опадение листьев приводят к
отмиранию бутонов, вследствие чего резко снижается выход товарной
продукции (при срезке цветков). Затрудняется также заготовка
стандартных здоровых черенков. Обычно на пораженных частях
растений отсутствуют четкие симптомы грибного заболевания:
пятнистости, налеты, спороношения и т. п. признаки. Лишь в
условиях высокой влажности воздуха на листьях могут появиться круп-
ные черные конидиальные ложа, на которых образуются
желеобразные скопления конидий гриба (рис. 123).
Рис. 123. Песталоциоз розы:
/ — пораженный лист с конидиомами гриба; 2 — конидии
391
Чернь листьев. Возбудитель — гриб Meliola Penzigi Sacc. На
листьях, черенках и отчасти ветвях образуется черный сажистый
налет в виде тонкой пленки (в основном на верхней стороне
листьев). Гриб не является паразитом: он не поражает ткани растений и
не питается за их счет. Гриб развивается на сахаристой жидкости,
выделяемой тлями и червецами, паразитирующими на нижней
поверхности листьев. Выделяемая ими жидкость падает на
расположенные ниже листья и покрывает их верхнюю поверхность
клейкой сахаристой массой, известной под названием «медвяной росы».
На этой жидкости и развивается гриб, покрывая ее черным
налетом, состоящим из темноокрашенных гиф и спор. Образование
подобного налета на поверхности листьев затрудняет свободный
доступ к ним воздуха и света, нарушает нормальный рост
растений и приостанавливает их развитие. Болезнь встречается в
оранжереях и в открытом грунте. Гриб поражает многие цветочные и
плодовые растения, декоративные кустарники.
Меры борьбы: уничтожение тлей и червецов; удаление с
поверхности листьев сажистого налета мокрой губкой или
сильной струей воды из брандспойта.
Мозаика. Возбудитель — Rosae mosaic virus (вирус мозаики розы).
Внешние признаки болезни различаются в зависимости от сорта
роз. У многоцветных роз на нижних частях стеблей появляется крап-
чатость, образуются мелкие деформированные листья. У чайно-
гибридных роз возникает карликовость растений, цветочные
почки — недоразвитые, бледные. При сильной степени развития
болезни цветки становятся почти белыми. По средней жилке или на
одной половине листа образуются многочисленные хлоротичные
пятна, наблюдается деформация черешков и листовых пластинок,
иногда ткань листа вдоль жилок светлеет. Переносчик вируса не
выявлен.
Увядание. Возбудитель — Rosae wilt virus (вирус увядания розы).
Признаки болезни — пролиферация растений и образование
множества молодых побегов с остроконечными листьями. Рост
побегов приостанавливается, корневая система остается недоразвитой.
У некоторых сортов листья скручиваются и очень рано опадают, а
концы побегов отмирают. Больные растения погибают.
Меры борьбы с вирусными болезнями:
уничтожение пораженных растений, использование для размножения
только здоровых маточников, термическое обеззараживание
посадочного и прививочного материала.
Нематодные болезни роз вызываются галловыми нематодами,
ксифинемой американской, короткотелыми нематодами.
Галловые нематоды рода Meloidogyne паразитируют на
корнях роз, вызывая образование некрозов и галлов. Больные
растения отстают в росте и развитии, при сильной степени
поражения погибают.
392
Меры борьбы: уничтожение корней пораженных растений;
чередование культур с выращиванием хвойных; удаление
сорняков, обеззараживание почвы нематицидами.
Ксифинема американская — XiphinemaamericanumCobb.
Нематода паразитирует на кррнях, вызывая по всей их длине
образование бурых некрозов, а на концах корней — мелких галлов.
Корневая система остается недоразвитой, загнивает. Наблюдается
значительная задержка роста растений. При сильной степени
поражения розы гибнут. X. americanum является переносчиком
многих фитопатогенных вирусов.
Меры борьбы: уничтожение больных растений и
сорняков, обеззараживание почвы нематицидами (фитовермом, окса-
милом, тиабендазолом).
Ранящая короткотелая нематода — Pratylenchusvulrus
All. et Jen. Паразитирует на корнях, вызывая редукцию корневой
системы, хлороз и карликовость растений, усыхание побегов.
Меры борьбы: уничтожение больных растений;
чередование культур с выращиванием бархатцев, обеззараживание почвы
нематицидами.
Проникающая короткотелая нематода —
Pratylenchus penetrans (Cobb.) Filip. et Sch. Stekh. Проникает в корни,
на которых образуются темные некротические пятна. Пораженные
корни загнивают. Больные растения отстают в росте и развитии,
при сильной степени заражения погибают.
Меры борьбы: перед посадкой роз — проверка почвы на
зараженность пратиленхами; предпосадочное обеззараживание
почвы нематицидами; выращивание в междурядьях бархатцев;
уничтожение сорняков и больных растений.
14.1.3. Болезни хризантемы
Черная ножка. Возбудители — оомицет Pythium debaryanum Hesse
и несовершенный гриб Rhizoctonia solani Kuhn. Поражаются
молодые корнесобственные растения и черенки. Область корневой
шейки у рассады и основания черенков при их укоренении чернеют
и загнивают. Верхушки больных растений поникают, растения
увядают, полегают и гибнут в течение 3—4 дней. Возбудители
болезни сохраняются в почве, которая служит источником
инфекции. Наиболее часто черная ножка появляется на участках,
бессменно используемых для выращивания поражаемых
растений, особенно одних и тех же видов. Развитию болезни в
парниках и теплицах способствуют загущенные посевы, сырые
тяжелые почвы, нарушения режима влажности, температуры,
освещения, ослабление растений другими факторами. Заболеванию
подвержены многие цветочные культуры в защищенном и
открытом грунте.
393
Меры борьбы: регулярное проветривание теплиц;
соблюдение оптимального режима температуры и влажности, оптимальной
густоты; использование органических удобрений. Замена или
дезинфекция зараженной почвы (ТМТД, биопрепаратами). Полив
почвы после посева семян и при появлении всходов суспензиями
фундазола, топсина-М, ТМТД, другими фунгицидами с
последующим рыхлением.
Фузариоз. Возбудитель — гриб-гифомицет Fusarium oxysporum
Schl. Растение поражается полностью. В период бутонизации и
цветения листья, начиная с нижних, скручиваются и увядают, часто
не теряя зеленой окраски. Основание стебля чернеет и загнивает,
затем гниль распространяется и в корни. Сосуды пораженных
стеблей заполнены мицелием гриба. Во влажных условиях на корневой
шейке часто образуется розоватый налет спороношения гриба.
См. также «Фузариоз гвоздики».
Вертициллез. Возбудитель — гриб-гифомицет Verticillium dahliae
Kleb. Листья, начиная с нижних, желтеют и усыхают, на них иногда
образуются коричневые пятна. При сильной степени поражения
соцветия блекнут, поникают и увядают, часто с одной стороны
растения. Середина соцветий буреет. Стебель внешне обычно
выглядит здоровым, но на его поперечном срезе обнаруживается
побурение сосудов. В условиях влажной погоды у основания
стеблей появляется беловатый налет. В целом болезнь развивается
менее интенсивно, чем фузариоз.
Меры борьбы с болезнями типа увядания:
исключение в качестве предшественников хризантем других культур,
поражаемых сосудистыми болезнями (астры, гвоздики, левкоя и
др.); замена почвы или ее дезинфекция, если обнаружены очаги
фузариоза или вертициллеза. Немедленное удаление больных
растений; использование черенков, взятых только от здоровых
растений в период цветения; обработка черенков перед их
укоренением суспензиями фундазола или топсина-М, а в период
бутонизации — полив черенков и корнесобственных растений теми же
фунгицидами.
См. также «Меры борьбы с болезнями типа увядания гвоздики».
Серая гниль. Возбудитель — гриб-гифомицет Botrytis cinerea Pers.
Наиболее опасная форма болезни — поражение цветков. Они
буреют, загнивают, покрываются серым (мышиного цвета)
пушистым налетом, который при массовом развитии конидиального
спороношения становится порошащим и приобретает более
светлую, пепельно-серую окраску. Поражаются также листья и
цветоносы. Серая гниль встречается как в открытом, так и в
защищенном грунте, причиняя культуре хризантемы значительный вред.
Появлению и интенсивному развитию болезни способствуют
высокая влажность, низкая температура, недостаточное
освещение, избыточное азотное питание.
394
Меры борьбы: в открытом грунте и в теплицах — незагущен-
ные посадки, умеренное увлажнение; применение
сбалансированного удобрения, микроэлементов; регулярное
проветривание теплиц. Во время вегетации — опрыскивание растений
фунгицидами: байлетоном, фундазолом или БМК, картоцидом, ров-
ралем и др. Немедленное уничтожение пораженных частей
растений.
Мучнистая роса. Болезнь вызывается грибом Oidium chrysanthemi
Rab. На молодых листьях, стеблях, цветоножках и бутонах
появляется белый (иногда грязно-белый) паутинистый налет мицелия,
который затем становится мучнистым благодаря образованию ко-
нидиального спороношения. Сумчатое спороношение (клейстоте-
ции) не образуется. Пораженные части растений засыхают.
Распространению болезни способствуют жаркая погода с росами,
избыток азотных удобрений. Источником инфекции являются
растительные остатки. Болезнь распространена повсеместно,
встречается как в открытом грунте, так и в теплицах. Особенно опасна
в условиях защищенного грунта, где приносит наибольший вред.
Меры борьбы: незагущенные посадки, свободное
размещение растений в теплицах; недопущение резких колебаний
температуры и пересушивания растений, избыточного азотного
удобрения. Внесение фосфорно-калийных удобрений и
микроэлементов. Для профилактики болезни и при первых ее признаках —
многократные опрыскивания растений байлетоном, бромуконазолом,
вектрой, фундазолом, топазом, коллоидной серой; при слабой
степени поражения — удаление больных и засохших листьев, при
сильном развитии болезни — уничтожение пораженных растений.
Ложная мучнистая роса. Возбудитель — оомицет Peronospora
schachtii Fuck. Болезнь чаще встречается в защищенном грунте. На
листьях появляются желтоватые пятна, с нижней стороны
которых образуется беловатый налет, представляющий собой
спороношение гриба в виде выходящих из устьиц конидиеносцев с
конидиями. При интенсивном развитии болезни поражаются также
стебли и цветки. Пораженные органы растений засыхают. Развитию
ложной мучнистой росы способствуют излишняя влажность и
высокая температура воздуха, недостаточная вентиляция теплиц.
Меры борьбы: соблюдение севооборота с возвращением
хризантемы на прежнее место не ранее 4—5 лет; сбор и сжигание
осенью остатков растений; глубокая обработка почвы на участках,
вышедших из-под мака. Удаление из посевов больных растений с
последующим закапыванием их в землю на глубину не менее 30 см.
Опрыскивание растений в период их вегетации растворами
фунгицидов (байлетона, фундазола, топаза). При опрыскивании
необходима тщательная обработка нижней поверхности листьев.
Опрыскивание проводят при первых признаках появления болезни, что в
большинстве случаев совпадает с серединой или окончанием фор-
395
мирования розетки. Последующие
опрыскивания проводят с
промежутком в 10—12 дней.
Ржавчина. Возбудитель — гриб
Puccinia chrysanthemi Roze — одно-
хозяйный гриб с неполным
циклом развития, развивающийся
только в урединиостадии.
Поражаются лишь хризантемы, на
листьях которых с нижней стороны
образуются мелкие оранжево-бурые
урединиопустулы (рис. 124).
Больные листья желтеют и отмирают.
Если ржавчина появляется на
стеблях, растения отмирают
целиком. Инфекция сохраняется на
пораженных растительных остатках.
В оранжереях болезнь может
развиваться из года в год, причиняя
значительный ущерб. Ее развитию
способствуют холодная влажная
погода и одностороннее азотное удобрение.
Меры борьбы: заготовка черенков только от здоровых
маточников; сбалансированное удобрение без избытка азота; неза-
гущенные посадки в открытом грунте и разреженное размещение
растений в теплицах. При обнаружении ржавчины в период
вегетации или угрозе ее появления — опрыскивание растений дита-
ном М-45, байлетоном, байтаном, оксихлоридом меди,
бордоской жидкостью или другими системными и медьсодержащими
фунгицидами. Замена в теплицах верхнего слоя почвы.
Септориоз. Возбудитель — гриб Septoria chrysanthemella Sacc.
Вначале на листьях появляются мелкие черно-бурые, почти черные,
округлые пятна, окруженные широким коричневым ободком.
Увеличиваясь, пятна сливаются и покрывают большую часть
поверхности листьев. На пятнах образуются черные точечные пикниды
гриба. Пораженные листья засыхают и опадают. Заболевание
распространено повсеместно, но наиболее вредоносно при
выращивании хризантем в теплицах и оранжереях, особенно при
высокой влажности и недостаточном освещении. Инфекция
сохраняется из года в год на пораженных опавших листьях.
Бурая пятнистость листьев. Вызывается грибом Cylindrosporiwn
chrysanthemi Stev. Пятна темно-бурые, нерезко очерченные,
диаметром 10 мм и больше. При сршьной степени поражения листья
деформируются, усыхают и повисают на стеблях, не опадая.
Меры борьбы: для профилактики пятнистостей —
свободное размещение растений, умеренное увлажнение и сбалансиро-
Рис. 124. Ржавчина хризантемы:
пораженный лист с пустулами гриба
396
ванное азотное питание.
Регулярное проветривание теплиц. При
появлении первых симптомов
болезни — систематическое
уничтожение пораженных листьев и
многократное опрыскивание
растений оксихлоридом меди,
бордоской жидкостью, байлетоном,
препаратом Абига-Пик и
другими фунгицидами.
Бактериальная пятнистость.
Возбудитель — бактерия Pseudo-
monas syringae van Hall. На
листьях, начиная с краев,
появляются желтоватые маслянистые
просвечивающие пятна, которые
позднее сливаются и становятся
коричнево-черными (рис. 125).
Пораженные листья
скручиваются и отмирают. Заражение
происходит через механические
повреждения и устьица при
высокой влажности воздуха. Оптимальной' для развития бактерии
является температура 25...28°С. Инфекция сохраняется в
пораженных листьях. Болезнь распространена в южных районах
страны, поражает также сирень.
Меры борьбы: для профилактики бактериальной
пятнистости хризантемы — недопущение переувлажнения воздуха и
попадания влаги на надземные части растений, регулярное
проветривание теплиц. При выращивании в открытом грунте — соблюдение
пространственной изоляции от сирени. Опрыскивание растений
оксихлоридом меди, бордоской жидкостью и другими
медьсодержащими фунгицидами с обязательной обработкой нижней
поверхности листьев. Уничтожение появившихся больных растений.
Кольцевая пятнистость. Возбудитель — Chrysanthemum ringspot
virus (вирус кольцевой пятнистости хризантем). Болезнь
проявляется на листьях, которые сильно деформируются и покрываются
крупными хлоротичными пятнами и кольцами. Передача вируса
происходит при прививке или с соком больных растений.
Карликовость. Возбудитель — Chrysanthemum disease virus.
Главный симптом болезни — приостановка роста и, как следствие, —
карликовость растений, сопровождающаяся посветлением темно-
окрашенных цветков. Лепестки больных растений часто короче,
чем у здоровых, иногда свертываются. Пораженные растения
обычно цветут раньше здоровых. У некоторых сортов наблюдаются
деформации листьев: «курчавость», морщинистость, скручивание
Рис. 125. Бактериальная пятнистость
листьев хризантемы
397
Рис. 126. Курчавость хризантемы
(рис. 126). В зависимости от сорта
хризантем и условий их
выращивания внешние проявления
болезни могут сильно варьировать.
Вирус передается повиликой,
при срезке с соком больных
растений, прививкой.
Крапчатость. Возбудитель —
Chrysanthemum vein mottle virus
(вирус крапчатости жилок хризантем).
Первый признак болезни —
малозаметный хлороз жилок. Позднее
появляются желтые пятна между
жилками и крапчатость листьев.
Хлоротичные пятна со временем
превращаются в некрозы,
отмершие участки выпадают, и на
листьях образуются дырки. Вирус
передается тлей Macrosiphoniella
sanborni GUI. и прививкой.
Бессемянность. Возбудитель —
Chrysanthemum aspermy virus
(вирус бессемянности хризантем). На
пораженных листьях образуются
вначале бледные желтовато-зеленые пятна (которые затем часто
маскируются), а также кольца и характерный линейный узор. При
поражении цветков их лепестки скручиваются в виде трубочек;
язычковые цветки становятся как бы курчавыми. Наблюдается
измельчание соцветий, резкое уменьшение количества цветков. Все
эти симптомы могут видоизменяться и варьировать в зависимости
от сортовых особенностей и условий выращивания хризантем.
Переносчики вируса — тли.
Желтуха — микоплазменная болезнь. Характеризуется общим
хлорозом, отставанием в росте, посветлением жилок на листьях.
Меры борьбы против вирусных и микоплазменных
болезней: уничтожение больных растений, сорняков и переносчиков
болезни; соблюдение санитарно-гигиенических правил при
прививке и уходе за растениями; использование метода термотерапии.
Нематодные болезни хризантемы вызываются листовыми
нематодами (хризантемной и земляничной), галловыми
нематодами, проникающей и ранящей короткотелыми нематодами и др.
Среди них наиболее широко распространена и вредоносна хри-
зантемная нематода, которая поражает также астру, дельфиниум,
флокс, бегонию и другие цветочные растения.
Хризантемная нематода— Aphelenchoides ritzemabosi
(Schw.) Stein.
398
В июне на нижних листьях между жилками образуются сначала
желтые, а затем бурые некротические пятна. В дальнейшем
пораженные листья полностью буреют, скручиваются, поникают и
отмирают, но при этом долго не опадают. Постепенно болезнь
переходит на верхние листья и цветки. Пораженные цветки
мелкие уродливые, деформированные. Часто они отмирают уже на
стадии бутонов. Молодые листья остаются недоразвитыми,
деформируются, рост побегов прекращается. При сильной степени
поражения растения полностью погибают (см. рис. 30).
Из опавших листьев и других пораженных остатков растений
нематоды переселяются в почву, где они нередко сохраняют
жизнеспособность до 6 —7 мес, а иногда и дольше. Источниками
инфекции могут служить черенки, взятые от больных растений,
иногда семена и сорняки. Заражение распространяется также с водой.
Нематоды способны перезимовывать в надземных частях
пораженных растений. В состоянии анабиоза они могут находиться до
двух лет.
С поверхности почвы и опавших листьев нематоды проникают
в здоровые растения через корневую систему и стебли, а также
непосредственно в листья через устьица.
Самки откладывают яйца в ткани листьев, и там проходит
весь цикл развития нематоды. При благоприятных условиях
(высокой влажности воздуха и температуре 14... 18 °С) полный цикл
развития — от яйца до половозрелой самки — заканчивается за
13—16 дней, при температуре 8°С размножение прекращается.
В течение года хризантемная нематода способна дать до 10
генераций. Нематод можно обнаружить в корнях, стеблях, цветках и
семенах, но наибольшее их количество обычно сосредоточено в
листьях. Особенно сильно болезнь проявляется в годы с
дождливым летом.
Меры борьбы: чередование культур с возделыванием
злаков; выбраковка и уничтожение больных растений. В осенний
период (сентябрь—октябрь) — пропаривание растений в течение 5 мин
в воде, подогретой до 46 °С (корни надо предварительно обрезать,
их длина не должна превышать 10—12 см). Пропаривание
надземной части маточных растений в течение 5 мин в воде с при 50... 55 °С.
Опрыскивание пораженных растений растворами оксамила, тиа-
бендазола. Обеззараживание почвы нематицидами (гетерофосом и
фитовермом).
Меры борьбы с галловыми нематодами,
проникающей короткотелой и ранящей короткотелой нематодами — см.
нематодные болезни роз.
Хлороз — неинфекционная болезнь. Причины — недостаток в
почве железа или избыток в ней кальция. Рекомендуется
подкислять почву, вносить в нее подкормку в виде солей железа, не
поливать растения жесткой водой.
399
14.2. БОЛЕЗНИ ЛУКОВИЧНЫХ
И КЛУБНЕЛУКОВИЧНЫХ КУЛЬТУР
14.2Л. Болезни тюльпана
Серая гниль. Возбудитель — гриб Botrytis tulipae (Lib.) Lind.
Серая гниль может развиваться в течение всего вегетационного
периода до глубокой осени и поражать все части растений: побеги,
листья, бутоны, цветки и луковицы. Вначале гриб развивается на
полузасохших, отмирающих и отмерших тканях, но затем при
благоприятных условиях способен перейти и на здоровые части
растений.
Основной признак болезни — появление на пораженных
частях растений грибного налета: вначале темного пушистого,
состоящего из тесно расположенных прямостоячих древовидно
разветвленных конидиеносцев оливково-бурого цвета, а затем серого
порошащего, представляющего собой массу одноклеточных конидий,
образующихся на конечных веточках конидиеносцев. Конидии легко
распространяются ветром, брызгами дождя, насекомыми и
другими путями.
При благоприятных условиях конидии, попав на поверхность
листьев или стеблей, прорастают, и образующийся мицелий
проникает в ткани растения главным образом через мельчайшие
ранки, солнечные ожоги и другие повреждения кутикулы.
При поражении листьев на них образуются большие бурые,
вначале сухие, а затем загнивающие пятна, постепенно
охватывающие большую часть поверхности листьев. В этом случае листья
отмирают и во влажных условиях покрываются серым налетом.
Бурые пятна появляются и на цветоносах.
В том случае, если к этому времени сформировались цветки,
на их лепестках образуются мелкие округлые желтоватые или
бесцветные, почти прозрачные пятнышки, со временем буреющие,
и на них также появляется серый налет спороношения
возбудителя. При сильной степени поражения растений бутоны вовсе не
образуются или не раскрываются полностью, и цветки остаются
недоразвитыми. Рост растений приостанавливается.
Верхние части луковиц чаще всего заражаются еще во время
вегетации от пораженных надземных органов. На них появляются
желтовато-бурые, резко очерченные, вдавленные пятна с серым
грибным налетом. Со временем эти части чернеют. Внутренние
ткани луковиц пронизываются коричневым мицелием гриба,
который позднее переходит и в поверхностные ткани. Луковицы
размягчаются, буреют, а впоследствии сморщиваются и загнивают,
покрываясь все тем же серым налетом. На чешуях и на
поверхности пораженных луковиц образуются многочисленные черные скле-
400
роции диаметром до 1 — 1,5 мм, часто
сливающиеся в сплошную черную корочку с
блестящей поверхностью (рис. 127).
Иногда на наружных тканях пораженных
луковиц гниль внешне не проявляется,
однако при надавливании на днища таких
луковиц их ткани проваливаются, так как
внутренние части бывают сильно разрушены.
Луковицы со слабой степенью поражения
могут попасть в хранилища, где, как
правило, полностью сгнивают. Чаще всего это
происходит в сырых холодных хранилищах.
При высадке в грунт луковиц со слабо
выраженными малозаметными признаками
серой гнили всходы часто вообще не
появляются или оказываются нежизнеспособными.
Появлению и интенсивному развитию
болезни способствуют высокая влажность и Рис. 127. Серая гниль
пониженная температура как во время ве- луковицы тюльпана
гетации, так и при хранении, механические
и другие повреждения луковиц и надземных частей тюльпанов,
ослабление растений неблагоприятными почвенными условиями,
несбалансированным удобрением, плохим уходом и другими
факторами. Гриб сохраняется в виде мицелия и склероциев в почве и
луковицах.
Серой гнилью поражаются все луковичные и клубнелукович-
ные культуры, многолетники (пионы и др.), гвоздика, астра,
левкой и другие цветочные растения. Гриб может развиваться на
семенах при их хранении, а также на проростках семян. Внешние
проявления болезни могут несколько различаться в зависимости
от особенностей поражаемых культур и органов растений, их
возраста.
Болезнь распространена повсеместно.
Меры борьбы: высадка луковиц тюльпанов на участках с
рыхлой дренированной плодородной почвой, имеющей
нейтральную или слабокислую реакцию; известкование кислых почв.
Недопущение загущенных посадок, выращивание на хорошо освещенных
солнцем участках; нормирование азотных удобрений, умеренный
полив. Внесение фосфорно-калийных удобрений и микроэлементов.
При появлении признаков болезни введение севооборота с
возвратом на прежние участки тюльпанов и других поражаемых серой
гнилью культур не ранее, чем через пять лет. Обеззараживание
почвы или замена ее другой, не содержащей инфекции. Во время
вегетации растений удаление больных экземпляров вместе с
почвой и их уничтожение; засыпка образовавшихся лунок хлорной
известью или золой. Трехкратное профилактическое опрыскива-
401
ние здоровых растений ОД %-м процимидоном (сумилексом),
0,5 %-м беномилом или 0,1 %-м сапролем.
Рекомендуется возможно ранняя выкопка луковиц и их
протравливание в суспензиях фундазола, топсина-М (0,2—0,5 %), ров-
раля (0,2 %), дитана М-45 (0,2 %), ТМТД (1 —1,5 %), тиабендазо-
ла (0,4 %), либо опудривание их дустами ТМТД или тачигарена с
последующей тщательной просушкой в течение 3—4 нед при
температуре 20...30°С, но не на солнце. Во время уборки, сушки и
транспортировки предохранение луковиц от повреждений,
ушибов и солнечных ожогов. Отбор на хранение только здоровых
луковиц. Перед уборкой и закладкой луковиц на зимнее хранение —
обязательная дезинфекция хранилищ, лотков, ящиков и другого
инвентаря.
Обязательное соблюдение оптимального температурно-влаж-
ностного режима хранения луковиц (10...20 °С и хорошее
проветривание), периодическая проверка их состояния и выбраковка
экземпляров с признаками поражения серой гнилью. За 1 — 3 дня
перед высадкой в грунт проведение повторного протравливания
луковиц и отбор здоровых экземпляров для высадки в грунт и
выгонки.
Склероциальная гниль. Возбудители — грибы Sclerotium tuliparwn
Kleb. или Sclerotinia bulborum (Wakk.) Rehm.
Болезнь поражает луковицы в почве. Гниль начинает развиваться
с верхних частей луковиц, и на них образуется белый ватообраз-
ный мицелий. В толще мицелия на больных луковицах и в
прилегающей к ним почве образуются склероции: у Scletrotium
tuliparwn — светлые мелкие (диаметром до 2 мм), округлые, у Sclerotinia
bulborum — черные, более крупные (до 1,5 см в поперечнике),
неправильной формы. Склероции могут сохраняться в почве до
пяти лет и служить источником инфекции.
Пораженные луковицы загнивают и при сильной степени
поражения нередко полностью сгнивают в почве. В этом случае
всходы вообще не появляются.
При слабой степени поражения больные луковицы иногда
трудно отличить от здоровых, так как они покрыты снаружи
неповрежденными гладкими чешуями. Однако такие луковицы,
высаженные в почву, иногда не прорастают или дают слабые
деформированные всходы, которые, едва образовав первые мелкие
листочки, быстро желтеют, увядают и засыхают. На корнях больных
луковиц в почве образуются склероции, которые, развиваясь в
тканях корешков, разрывают их кожицу, выступая на поверхность
в виде белых или темно-коричневых бугорков. Заражение
здоровых луковиц и проростков происходит от больных луковиц или
через зараженную почву. На новые участки болезнь попадает при
посадке пораженных луковиц. Заболевание развивается очагами
на переувлажненных и кислых почвах.
402
Склероциальная гниль распространена повсеместно и
поражает все луковичные цветочные растения.
Меры борьбы— см. «Серая гниль тюльпана».
Пенициллезная гниль. Возбудители — грибы рода Penicillium: P.
corymbiferum West, и др. Болезнь проявляется преимущественно в
хранилищах. Вначале на луковицах появляются небольшие
оранжевые или желтовато-бурые вдавленные пятна. Со временем они
разрастаются, сливаются и охватывают всю луковицу. На
поверхности зараженных луковиц и между чешуйками образуется
пылящий голубовато-зеленый (бирюзового цвета) налет конидиаль-
ного спороношения гриба. Пораженные ткани загнивают.
Заражению и развитию болезни способствуют механические
повреждения луковиц и плохие условия их хранения.
При высокой влажности почвы в период вегетации
пенициллезная гниль может поражать не только луковицы, но и
надземные части растений.
Меры борьбы: предохранение луковиц от механических
повреждений при выкопке, очистке и транспортировке; тщательная
просушка луковиц перед хранением; протравливание луковиц
фунгицидами (ТМТД, фундазолом, топсином-М); соблюдение
оптимального режима влажности и аэрации в хранилищах.
Опрыскивание растений фунгицидами во время вегетации (см. также
«Болезни гладиолуса»).
Фузариоз. Возбудитель — гриб Fusarium oxysporum Schl. / tulipae.
Болезнь обнаруживается на луковицах чаще всего при их выкопке:
в области донца можно заметить небольшое светло-коричневое
пятно, резко отграниченное от здоровой ткани темно-бурой
каймой. Постепенно пятно разрастается, распространяясь на верхние
части луковиц. Ткань в области пятна загнивает и покрывается
белым или розоватым налетом конидиального спороношения гриба.
Пораженные луковицы обычно полностью сгнивают, выделяя
клейкое тягучее вещество, имеют специфический запах.
В условиях сухого хранения гниющие луковицы превращаются
в труху. В полевых условиях фузариоз обычно проявляется к концу
вегетации в виде пожелтения и увядания надземных частей
вследствие загнивания корней. Однако при высоких температурах
гибель растений может произойти уже в мае.
Инфекция в форме мицелия может долго сохраняться в
луковицах и в почве и распространяется с зараженным посадочным
материалом.
Меры борьбы: ранняя выкопка луковиц и их
протравливание в суспензиях фундазола, топсина-М или ТМТД с
последующей тщательной просушкой. Дезинфекция хранилищ,
ящиков, лотков и другого инвентаря. Хранение луковиц при
температуре 14... 16°С и хорошей вентиляции хранилищ; выбраковка
луковиц с признаками поражения; повторное протравливание
403
луковиц перед посадкой; уничтожение больных растений во
время вегетации.
Пестролепестность. Возбудитель — Tulipa virus 1 (вирус пестро-
лепестности тюльпана). Главный признак болезни — пестрая
окраска лепестков цветка: наличие на них хлоротичной крапчато-
сти, зеленовато-желтых или беловатых штрихов, полос и пятен
(рис. 128). Цветовые вариации пестролепестности зависят от сорта
тюльпанов и степени поражения растений.
Ежегодная высадка луковиц, полученных от растений,
пораженных пестролепестностью, постепенно приводит к
вырождению растений — оно проявляется прежде всего в измельчании
луковиц, из которых вырастают недоразвитые растения с
искривленными стеблями. Цветки год от года мельчают, деформируются
и, наконец, полностью обесцвечиваются. Бутоны часто почти не
раскрываются. Существует мнение, что сильнее всего
поражаются сорта, имеющие белую, желтую, голубую, синюю и лиловую
окраску оснований лепестков. Болезнь может проявиться в
первый год или через несколько лет после заражения. Вирус
передается механически с соком больных растений, при прививке и
тлями.
Некротическая пятнистость. Возбудитель — Tobacco necrosis virus
(вирус некроза табака). На листьях, цветоносах и цветках
образуются некротические штрихи, полосы и пятна. Обычно отмирание
тканей начинается с одной стороны листа, а потом
распространяется по всему растению. Иногда, при заражении в середине
вегетации, поражается только половина растения. Больные
тюльпаны сильно отстают в росте, деформируются, листья засыхают, и
растения погибают. В лучшем случае растения не погибают, но не
цветут или дают недоразвитые цветоносы, деформированные
цветки. На срезах пораженных луковиц видны пятна — вначале
прозрачные, позднее буреющие. Вирус передается инокуляцией сока,
переносится зооспорами Olpidium brassicae. Вирус может сохраняться
в почве в течение нескольких лет.
Рис. 128. Пестролепестность тюльпана
404
Некроз жилок. Возбудитель — Tobacco ringspot virus (вирус
кольцевой пятнистости табака). На жилках листьев образуются
некротические, постепенно сливающиеся участки. Жилки отмирают.
Междужилковые ткани приобретают хлоротичную окраску.
Пораженные растения отстают в росте, деформируются. Вирус
передается механически с соком больных растений, нематодой Xiphinema
sp., клещом Tetranichus sp. и другими переносчиками.
Меры борьбы с вирусными болезнями: выбраковка
и уничтожение больных растений (во время вегетации) и луковиц
с признаками вирусных болезней (при хранении), недопущение
чрезмерно загущенных посадок; уничтожение переносчиков
вирусов, выращивание тюльпанов на одном месте не более 2—3 лет,
смена участка.
Нематодные болезни вызываются стеблевой нематодой Ditylen-
chus dipsaci (Kiihn) Filip. и нематодой Aphelenchoides subtenuis (Cobb)
St. et Buhr.
D. dipsaci поражает все части растений. Больные луковицы —
мягкие на ощупь. На разрезе таких луковиц обнаруживается
кольцевая гниль, развивающаяся вследствие паразитирования
нематод в чешуях. Зараженные растения отстают в росте и развитии,
плохо цветут, а при сильной степени поражения погибают.
Интенсивному развитию этой нематоды благоприятствуют
прохладное дождливое лето и поражение луковиц бактерией Erwinia
carotovora.
Меры борьбы: чередование культур с выращиванием
бархатцев; выбраковка больных растений и луковиц; обеззараживание
почвы оксамилом или другими нематицидами. Через три недели
после выкопки луковиц пропаривание их в воде при температуре
43 °С в течение 2—4 ч (в зависимости от размера луковиц).
A subtenuis поражает как луковицы, так и надземные части
растений. Чешуи луковиц буреют и загнивают. На поперечном срезе
больных луковиц, как и при поражении их стеблевой нематодой,
видна кольцевая гниль. На поверхности больных луковиц, обычно
у донца, образуются бурые некротические пятна.
Меры борьбы: выбраковка больных растений и луковиц;
смена культур с выращиванием древесных пород;
обеззараживание почвы тиабендазолом или другими нематицидами.
Неинфекционные болезни
Посинение луковиц. На луковицах появляются сначала
мелкие водянистые пятна, позднее они увеличиваются и
приобретают синевато-бурую окраску. Причиной болезни является
просушка луковиц на солнце.
Камедетечение луковиц. Причиной камедетечения
считается повышенная влажность почвы в конце вегетации
тюльпанов.
405
Замирание бутонов (слепота). Причины болезни:
ранняя высадка луковиц в теплую погоду (в этом случае развитие
корневой системы отстает от роста надземной части); избыточная
влажность и высокая температура при выгонке; хранение луковиц
при высокой температуре.
Образование наплывов на наружных чешуях
луковиц происходит в результате слишком ранней выкопки
луковиц и воздействия солнечной радиации на невызревшие
луковицы.
Накопление извести в луковицах проявляется в их
побелении и затвердении вследствие хранения луковиц
(особенно невызревших) в условиях повышенной влажности.
Опрокидывание цветков. При выгонке наблюдается
размягчение цветоноса под цветком, в результате чего цветок
повисает и выглядит, как обломанный. Причина — нарушение
правил выгонки.
Во избежание этих явлений для выгонки необходимо
использовать хорошо вызревшие, правильно хранившиеся луковицы,
соблюдать оптимальные сроки посадки растений и выкопки
луковиц, избегать слишком высоких температур и резких ее
перепадов во время вегетации, не допускать полива растений при ярком
солнечном освещении и застаивания воды в почве, соблюдать
правила выгонки тюльпанов.
14.2.2. Болезни гиацинта
Склероциальная гниль. Возбудитель — сумчатый гриб Sclerotinia
bulborum (Wakk.) Rehm. Болезнь поражает луковицы и листья.
Чешуи луковиц обесцвечиваются, между ними развиваются тонкие
белые пленки мицелия, а затем и черные сплюснутые склероции
различной формы и величины. Листья желтеют, увядают и легко
отделяются от луковицы. Растения полностью погибают.
Основными источниками инфекции служат склероции,
сохраняющиеся в почве в течение нескольких лет, и пораженные растительные
остатки. Развитию болезни благоприятствуют высокая влажность
и кислая реакция почвы. Болезнь встречается повсеместно.
Меры профилактики: смена культур с возвратом
гиацинта на прежние участки через 5—6 лет; глубокая перекопка
почвы с оборотом пласта; известкование почвы; уничтожение
пораженных растений.
Пенициллезная гниль. Возбудитель — несовершенный гриб
Penicillium cyclopium West. Поражаются все органы растения. На
луковицах и цветоносах, реже на листьях сначала образуются
небольшие красно-коричневые пятна, со временем сливающиеся.
Луковицы, цветоносы и цветки загнивают, листья желтеют и
отмирают. В условиях повышенной влажности пораженные части рас-
406
тений покрываются порошащим зеленым налетом конидиального
спороношения гриба. Поражению луковиц способствуют
механические повреждения, высокая влажность, плохое проветривание
помещений для хранения луковиц.
Меры борьбы — см. «Пенициллезная гниль тюльпана».
Серая гниль. Возбудитель — несовершенный гриб Botrytis
hyacinthi West, et Beyma. Поражаются луковицы и листья. На
луковицах образуются расплывчатые бурые пятна, быстро
охватывающие всю луковицу. Ткани луковиц размягчаются и загнивают. Во
влажных условиях на пораженных луковицах появляется
обильный серый налет, который затем уплотняется, и в нем
развиваются мелкие черные склероции. На листьях вначале образуются
мелкие желтоватые пятна. Позднее пятна увеличиваются в
размерах и приобретают бурую окраску. При повышенной влажности на
пораженных участках листьев также появляется серый налет
конидиального спороношения возбудителя. Болезнь имеет широкое
распространение.
Меры борьбы — см. «Серая гниль тюльпана».
Фузариоз. Возбудители — грибы рода Fusarium: F. culmorum
(W. G. Sm.) Sacc, F. oxysporum Schl., F. solani (Mart.) App. et Wr.
Поражаются корни, луковицы и надземные части растений.
Луковицы и корни загнивают. Листья желтеют, начиная с концов, и
отмирают. По мере развития болезни растения отстают в росте,
соцветия остаются недоразвитыми. На пораженных луковицах в
области донца появляется беловато-розоватый налет
спороношения возбудителя.
Меры борьбы — см. «Фузариозтюльпана».
Ржавчина. Возбудитель — гриб Uromyces scillarum (Grev.) Wint.
Вначале на листьях появляются мелкие диаметром до 2 мм
бесцветные пятна, со временем желтеющие. В дальнейшем под
эпидермисом на нижней стороне (иногда с обеих сторон)
развиваются пустулы, содержащие светло-коричневые урединиоспоры, при
созревании распыляющиеся. Споры образуются и на чешуях
луковиц. Пораженные листья засыхают и преждевременно опадают.
Растения выглядят ослабленными, плохо цветут. Гриб зимует в
луковицах и на пораженных остатках надземных частей.
Меры борьбы— см. «Ржавчина гвоздики садовой».
Мягкая бактериальная гниль. Возбудитель — бактерия Erwinia
carotovora (Jones.) Wald. Чаще всего болезнь обнаруживается при
выгонке гиацинтов. Растения вначале отстают в росте и не цветут,
затем желтеют и увядают. При поражении луковиц развивается
мокрая дурно пахнущая гниль, которая прогрессирует во время
хранения луковиц при высоких значениях температуры и влажности.
Меры борьбы: соблюдение нормального режима
температуры и влажности. Применение только азотных удобрений.
Уничтожение больных растений во время вегетации, после их удаления —
407
присыпка лунок хлорной известью. Тщательная просушка и
правильное хранение луковиц.
Мозаика. Возбудитель — Grijs mosaic virus (вирус мозаики
гиацинта). Листья больных растений становятся более мелкими и
узкими. На них появляются светло-зеленые пятна, штрихи и полосы.
В дальнейшем эти участки желтеют и отмирают. На цветоносах
также образуются беловатые штрихи и полосы, которые к концу
вегетации превращаются в некрозы. На цветках появляются тонкие
желтовато-белые полосы, пестролепестность. Нижние цветки часто
остаются недоразвитыми. Пораженные растения отстают в росте.
Пестростебелыюсть. Возбудитель — Tobacco rattle virus (вирус
курчавой полосатости табака). На срезах больных луковиц
обнаруживаются некротические участки тканей, на листьях вдоль жилок —
некротические пятна. Переносчики вируса — нематоды рода
Trichodorus.
Меры борьбы с вирусными болезнями:
уничтожение больных растений, сорняков и переносчиков вирусов —
насекомых и нематод.
Нематодные болезни вызываются стеблевой нематодой и
проникающей короткотелой нематодой.
Меры борьбы: смена культур с выращиванием бархатцев.
Дезинфекция почвы нематицидами. Во время вегетации выбраковка
и уничтожение больных растений. Через 3 нед после выкопки про-
паривание луковиц в воде при температуре 43 °С в течение 2—4 ч.
Неинфекционные болезни
Сидячие соцветия. Соцветия остаются недоразвитыми при
слишком ранней посадке луковиц, при посадке невызревших
луковиц, при высокой температуре в период укоренения, при
избыточной влажности.
Гниль верхушки ц в е т о н о с а наблюдается при
выгонке, если растения преждевременно вносят в оранжерею, и при
высоких температурах в период выращивания.
Остановка в росте всходов отмечается при высадке
ослабленных луковиц и при резких колебаниях температуры и влажности.
Для профилактики этих явлений необходимо выкапывать и
высаживать хорошо вызревшие луковицы. Выгонку производить в
условиях постоянной оптимальной температуры и влажности, а посадку
луковиц — в оптимальные сроки, избегая слишком ранней посадки.
14.2.3. Болезни гладиолуса
Фузариоз. Возбудитель — несовершенный гриб Fusarium oxysporum
f. gladioli (Mass.) Sm. et Hans. Болезнь поражает как
клубнелуковицы, так и надземные части растений. На клубнелуковицах
развивается сухая гниль. Она начинается с донца, а в дальнейшем охва-
408
тывает всю клубнелуковицу. Болезнь проявляется в образовании
на клубнелуковицах красновато-бурых сухих пятен, которые при
повышенной влажности покрываются розовато-белым налетом
мицелия и конидиального спороношения гриба. Клубнелуковицы
загнивают как при хранении, так и в грунте.
Сильно пораженные клубнелуковицы сгнивают зимой при
хранении, а пораженные в слабой степени внешне выглядят
здоровыми и могут быть весной высажены в грунт. При благоприятных
для развития болезни условиях такие клубнелуковицы не
прорастают совсем, сгнивая в почве, или из них вырастают слабые
растения, которые преждевременно желтеют и погибают до
образования цветочных стрелок. Стебель полностью сгнивает
(остаются только волокна пораженных сосудистых пучков) и у
оснований листьев переламывается. Больные растения почти не
образуют деток.
Корневая система больных клубнелуковиц развивается слабо,
потом загнивает, и растения легко выдергиваются из почвы. На
срезах клубнелуковиц, пораженных фузариозом, обнаруживается
потемнение сосудов или гниль сердцевины.
Заражение молодых клубнелуковиц и деток происходит от
больной материнской клубнелуковицы или из почвы, где возбудитель
может сохраняться в течение нескольких лет. Проникая в
сосудистую систему молодых клубнелуковиц и деток, гриб вызывает
увядание и засыхание надземных частей, а сами клубнелуковицы и
детки при сильной степени поражения сгнивают.
Развитию фузариоза способствуют многие факторы: удобрение
почвы свежим навозом, загущенная посадка растений, уборка на
хранение невызревших, поврежденных и плохо просушенных
клубнелуковиц, неправильный режим их хранения и др.
Болезнь встречается во всех районах выращивания
гладиолусов, являясь одной из самых распространенных и вредоносных.
Преждевременное пожелтение и усыхание листьев гладиолусов
может быть также вызвано абиотическими факторами: засухой,
вымоканием, плохим уходом и т.д.
Меры борьбы: смена культур, недопущение возврата
гладиолусов на прежний участок в течение 4—5 лет или
дезинфекция почвы; высадка только здоровых клубнелуковиц.
Обеззараживание посадочного материала перед посадкой фунгицидами: ров-
ралем (0,2 %), тиабендазолом, ТМТД, фундазолом, топсином-М
и др. Выбраковка и уничтожение пораженных растений;
оздоровительные поливы фунгицидами; ранняя выкопка клубнелуковиц
и предохранение их от повреждений и ушибов при выкопке,
просушке и транспортировке. Послеуборочное прогревание
клубнелуковиц при температуре 30...35°С и относительной влажности
80% в течение 5 — 7 дней или химическое протравливание
указанными фунгицидами с последующей просушкой. Дезинфекция
409
хранилищ, ящиков, лотков и другого инвентаря. Хранение
клубнелуковиц при температуре 4...5 °С.
Коричневая сердцевинная гниль. Возбудитель —
несовершенный гриб Botrytis gladiolorum Timm. Эта гниль считается наиболее
опасной болезнью клубнелуковиц при хранении. Гриб переходит
на клубнелуковицу из тканей основания стебля и затем
проникает в глубь клубнелуковицы, поражая всю ее сердцевинную часть
(рис. 129).
При надавливании на донце внешне здоровой луковицы
обнаруживается, что ее внутренние ткани уже сгнили. Иногда сгнивает
только часть клубнелуковицы, отделяющаяся от здоровых частей
пробковым слоем. Клубнелуковица, загнивающая сбоку, в
дальнейшем поражается полностью, превращаясь в мягкую
коричневую массу. Во влажных условиях гнилая клубнелуковица
покрывается серым пушистым налетом, в толще которого образуются
черные округлые мелкие (диаметром до 5 мм) склероции.
Даже при малой степени поражения из высаженных в грунт
клубнелуковиц развиваются слабые растения, которые вскоре
погибают. У больных растений в области шейки образуются склероции
и спороношение возбудителя.
В открытом грунте болезнь
чаще проявляется после
цветения (на листьях, стеблях и
шейке растений) или во время
цветения (на цветках). При
заражении листьев на них сначала
появляются мелкие буроватые
пятна; со временем они темнеют,
сливаются, и листья
преждевременно засыхают. Такие же пятна
образуются и на стеблях —
цветоносах. На лепестках цветков
появляются водянистые пятна.
При повышенной влажности
воздуха пораженные части
растений (особенно нижние,
затененные) покрываются серым
пушком. Иногда болезнь
поражает шейку гладиолуса — в этом
случае заболевшее растение
загнивает и полностью отмирает.
Из надземных частей гриб
проникает в клубнелуковицу и таким
путем попадает в хранилище.
Заражение здоровых клубнелуковиц
может происходить и при сопри-
Рис. 129. Коричневая сердцевинная
гниль гладиолуса:
1 — пораженный лист; 2 — поражение
основания стебля и шейки
клубнелуковицы; 3 — сердцевинная гниль на
продольном разрезе клубнелуковицы
410
косновении их с больными во время уборки, просушки перед
зимним хранением, во время транспортировки и хранения, так
как при этом часто образуются различные повреждения и ранки,
в большинстве случаев настолько мелкие, что их трудно
обнаружить невооруженным глазом. Споры гриба, попадая на
поврежденные клубнелуковицы, прорастают, мицелий внедряется во
внутренние ткани клубнелуковиц и вызывает их загнивание.
В поле возбудитель сохраняется на пораженных остатках
растений, а на новые участки заносится чаще всего с посадочным
материалом.
В годы с обильными осадками интенсивность развития болезни
обычно возрастает.
Меры борьбы с коричневой гнилью: тщательный
отбор и просушка клубнелуковиц при температуре выше 20 °С сразу
же после уборки их с поля; предохранение клубнелуковиц от
механических и других повреждений; протравливание клубнелуковиц
перед закладкой на хранение и перед высадкой в поле ровралем
(0,2 %), фундазолом, топсином-М (0,2—0,5 %), ТМТД (1 — 1,5 %),
тиабендазолом (0,4 %) или другими фунгицидами; во время
вегетации — опрыскивание
здоровых растений 0,1 %-м сумилексом,
0,5%-м беномилом или 0,1%-м
сапролем.
См. также «Меры борьбы с
серой гнилью тюльпана».
Твердая гниль. Возбудитель —
пикнидиальный гриб Septoria gladioli
Pass. Болезнь поражает
клубнелуковицы и надземные части
гладиолусов. На клубнелуковицах
образуются мелкие водянистые красновато-
коричневые пятна, которые со
временем увеличиваются, сливаются,
становятся вдавленными, затем
чернеют, приобретают угловатые
контуры и при благоприятных
условиях охватывают всю
клубнелуковицу. Ее ткань твердеет,
сморщивается, мумифицируется (рис. 130).
Обнаружить заболевание в
начальной стадии можно еще осенью,
сразу же после уборки
клубнелуковиц, если снять с них чешую. На
чешуях бывают заметны сероватые
пятна с мелкими черными пикни-
дами. При влажном хранении клуб-
4
Рис. 130. Твердая гниль
гладиолуса:
1, 2> 3 — пораженные
клубнелуковицы (последовательность стадий
развития гнили); 4 — пораженный
лист
411
нелуковиц болезнь развивается очень интенсивно и приводит к
гибели большого количества посадочного материала, причиняя
значительный ущерб культуре гладиолуса.
Гриб поражает также листья гладиолусов во время вегетации.
Они покрываются округлыми пурпурово-коричневыми пятнами
со светло-серой серединой. Вначале пятна бывают мелкими, затем
увеличиваются в диаметре. Со временем в центре пятен
образуются черные точечные пикниды возбудителя. Созревающие в них
конидии заражают надземные части новых растений. Заражение
здоровых клубнелуковиц происходит не только при контакте с
больными в хранилищах, но и в полевых условиях через почву, где
гриб сохраняется на растительных остатках до четырех лет. В почву
гриб попадает с больными клубнелуковицами, высаженными без
отбора и обеззараживания.
Особенно сильно твердая гниль развивается в годы с
прохладным летом и обильными осадками, на бедных, истощенных почвах.
Болезнь распространена повсеместно.
Пенициллезная гниль. Возбудитель — гриб-гифомицет Penicillium
gladioli Mc Cull et Thom. Болезнь проявляется только на
клубнелуковицах во время хранения. В открытом грунте гладиолусы не
поражаются.
Первые признаки болезни сходны с симптомами твердой или
сухой склероциальной гнили. На клубнелуковицах появляются
шероховатые, несколько вдавленные коричневые пятна с
сероватым оттенком. В дальнейшем развиваются мелкие
светло-коричневые шаровидные склероции диаметром около 1 мм. При
повышенной температуре склероции образуются чаще. В условиях
высокой влажности и низкой температуры в хранилищах на
поверхности пятен появляется голубовато-зеленоватый (бирюзового
цвета) порошащий налет конидиального спороношения гриба. При
сильной степени поражения таким налетом покрывается вся
клубнелуковица. Из года в год мицелий сохраняется в пораженных
клубнелуковицах. При нарушении режима температуры и
влажности во время хранения клубнелуковиц пенициллезная гниль
может привести к массовой гибели посадочного материала.
Заражению клубнелуковиц способствуют их небрежная уборка,
плохая очистка и недостаточная осенняя просушка, а также
механические (даже малейшие!) повреждения во время выкопки,
перевозки и хранения. Болезнь распространяется конидиями, а также
мицелием при соприкосновении больных и здоровых клубнелуковиц.
Сухая склероциальная гниль. Возбудитель — аскомицет Sclerotinia
gladioli (Mass.) Dray — гриб, узкоспециализированный к этой
культуре. Диагностические признаки сухой склероциальной гнили
сходны с симптомами твердой гнили. Эти виды гнили различаются
лишь по микроскопическим признакам и по характеру
проявления болезни на растениях в открытом грунте.
412
В период хранения гниль развивается на клубнелуковицах.
Сначала болезнь проявляется на их внутренних чешуйках в виде
коричневых пятен с размягченной тканью. При сильной степени
поражения гниль переходит и на наружные части клубнелуковиц.
На верхних частях, в местах прикрепления листьев, образуются
вдавленные серовато-бурые кольцевые пятна. В конечном счете
клубнелуковицы мумифицируются.
Во время вегетации гриб от пораженных клубнелуковиц
переходит в надземные части растений. Прежде всего болезнь
поражает нижние листья и основание стебля. Вначале листья желтеют, а
затем буреют, размочаливаются и засыхают под воздействием гриба.
Ткань основания стебля размягчается и загнивает, что часто
приводит к его обламыванию и гибели всего растения. На
поверхности пораженных клубнелуковиц, стеблей и листьев образуются
мелкие черные или темно-коричневые склероции гриба.
Из года в год мицелий и склероции вместе с пораженными
растительными остатками сохраняются в почве (до пяти лет), а
также на больных клубнелуковицах в хранилищах, откуда гриб
при их высадке вновь попадает в поле.
Сухая склероциальная гниль распространена повсеместно, но
особенно интенсивно развивается при выращивании гладиолусов
на кислых, влажных, тяжелых почвах, а также при хранении
клубнелуковиц в сырых, плохо проветриваемых помещениях.
Меры борьбы с твердой, пенициллезной и
сухой склероциальной гнилями: правильная агротехника
выращивания гладиолусов; уничтожение в период вегетации
больных растений и растительных остатков, своевременная уборка и
тщательная просушка клубнелуковиц при 20... 25 °С, отбор на хранение
только здоровых экземпляров, соблюдение оптимального режима
хранения, выбраковка больных клубнелуковиц и протравливание их
в суспензиях ТМТД, фундазола, топсина-М и других фунгицидов.
Парша. Возбудитель — бактерия Pseudomonas marginata (Mc Cull.)
Stapp. Болезнь поражает клубнелуковицы и листья. На
клубнелуковицах, чаще вокруг донца, образуются округлые мелкие
(диаметром до 5 мм) углубленные коричневые или бурые язвочки, со
временем сливающиеся в более крупные пятна. Они часто
растрескиваются, и из трещин выступают прозрачные или
желтовато-коричневые выделения бактерий. Глубоко внутрь
клубнелуковиц бактерии не проникают, так как вскоре вокруг пораженных
участков возникает пробковый слой, отделяющий их от здоровых
тканей. Язвочки обнаруживаются только после снятия чешуи. При
сильной степени поражения болезнь проявляется и на чешуях в
виде черных пятен. Иногда пятна коричневые с черными краями.
В условиях повышенной влажности парша на клубнелуковицах
выглядит, как мокрая гниль. Сильно пораженные паршой
клубнелуковицы часто не прорастают совсем. На листьях, главным образом
413
у их оснований, образуются мелкие красновато-коричневые
пятнышки, которые постепенно разрастаются, становятся темными и
похожими на ожоги. В сырую погоду ткань в области пятен загнивает.
Бактерии заносятся в почву с больными клубнелуковицами и
могут сохраняться в ней долгое время. Развитию болезни
способствуют различные повреждения клубнелуковиц.
Основной вред парша причиняет во время вегетации
растений, так как они теряют декоративность. При хранении
клубнелуковиц парша особого вреда не приносит и не является причиной
отпада посадочного материала в хранилищах.
В целях профилактики болезни рекомендуется протравливание
клубнелуковиц перед посадкой.
Мозаика. Возбудитель — Gladiolus mosaic virus (вирус мозаики
гладиолуса). На листьях появляются отчетливые зеленовато-желтые
пятна, полосы и кольца. Цветение растений запаздывает. Цветоносы
вытягиваются, на них образуются мелкие цветки, у которых
наблюдается пестролепестность в виде желтоватых или серых пятен и
штрихов. У бледно окрашенных сортов на лепестках появляются
пурпуровые полоски, лепестки закручиваются. Пораженные растения имеют
угнетенный вид, иногда вообще не дают цветочных стрелок,
клубнелуковицы также год от года мельчают. Болезнь распространяется
тлями, с больными клубнелуковицами, с соком пораженных растений.
Из других вирусных болезней на гладиолусах встречаются
белая разрушающая мозаика, карликовость, растрескивание
листьев; микоплазменная болезнь «травянистость верхушки».
Меры борьбы— обычные против вирусов и микоплазм.
Нематодные болезни гладиолусов вызывают стеблевые
нематоды рода Ditylenchus, галловые рода Meloidogyne, проникающая ко-
роткотелая нематода.
Меры борьбы с нематодами— см. «Нематодные
болезни тюльпана».
Неинфекционное отмирание (внезапное пожелтение и
отмирание листьев, засыхание корней) связано с выращиванием
гладиолусов га гырых тяжелых почвах, поздней глубокой посадкой
клубнелуковиц, избытком азотных удобрений.
14.3. БОЛЕЗНИ МНОГОЛЕТНИКОВ
ОТКРЫТОГО ГРУНТА
14.3.1. Болезни георгины
Черная ножка. Чаще всего возбудителями болезни являются
оомицет Pythium debaryanum Hesse и несовершенный гриб Rhizoctonia
solani Kuhn. Болезнь поражает черенки, сеянцы, рассаду.
Основания черенков (при их укоренении) и стеблей молодых растений
414
вначале немного темнеют, потом становятся черными и
загнивают. В области корневой шейки у больных растений образуется
перетяжка. Часто происходит поражение корешка: он буреет,
загнивает, и сеянец легко выдергивается из почвы. Пораженные
черенки усыхают, молодые сеянцы увядают и погибают.
При повышенной влажности почвы и недостаточном
проветривании болезнь прогрессирует очень быстро, и растения гибнут
в течение нескольких дней. Заболевание носит очаговый
(куртинный) характер.
Меры борьбы: для предупреждения дальнейшего
распространения болезни необходимо немедленное удаление
пораженных растений вместе с комом земли и сжигание. Обеззараживание
участков, где были обнаружены очаги заболевания, путем
двукратного полива почвы с интервалом в 10 — 12 дней фундазолом,
топсином-М или их смесями в равных пропорциях из расчета 5 —
10 л/м2. Необходимы дезинфекция стеллажей, проветривание
теплиц и посадок в открытом грунте, умеренный полив.
Фузариозное увядание и сухая гниль корнеклубней.
Возбудители — грибы рода Fusarium. Поражаются листья, стебли и корне-
клубни георгин. Во время вегетации растений инфекция может
распространяться на большие расстояния конидиями, а во время
хранения корнеклубней — мицелием гриба при
непосредственном контакте здоровых и больных экземпляров. Особенно сильно
поражаются слабые, плохо вызревшие корнеклубни.
В период вегетации корневая шейка растений загнивает, и на
ней при высокой влажности появляются розовато-охряные
подушечки конидиального спороношения возбудителя. Листья и
стебли желтеют, и все растение целиком или отдельные его части
отмирают, корнеклубни становятся вялыми, во время хранения
сморщиваются, их ткань ссыхается и при разломе крошится. На
пораженных участках появляются плоские буроватые пятна.
Вначале они покрываются беловатым, а позднее розоватым налетом,
состоящим из мицелия и конидиального спороношения паразита.
Фузариозная гниль обычно наблюдается в том случае, когда на
хранение убирают слабо развитые, недозрелые, плохо
просушенные или поврежденные корнеклубни.
Сильному развитию гнили способствуют высокая температура
и повышенная влажность воздуха. При отсутствии
профилактических мероприятий и неправильном режиме хранения
корнеклубней они полностью сгнивают и разрушаются с образованием
сухой гнили и появлением розоватого налета. Заболевание
распространено повсеместно.
Меры борьбы: правильная агротехника выращивания
георгин; своевременная уборка на хранение хорошо развитых и
вызревших корнеклубней; выкопка их в сухую солнечную погоду,
раскладка на 2—3 ч на освещенных солнцем местах, затем просушка
415
в затененном месте. При укладке корнеклубней на хранение
рекомендуется пересыпка их песком или торфом; место хранения
должно быть сухим и прохладным с температурой воздуха 4...5°С;
проверка корнеклубней на пораженность перед их посадкой и
высадка только здоровых экземпляров. Больные растения подлежат
немедленному удалению и сжиганию.
Загнивание листьев, стеблей и корнеклубней. Возбудитель
болезни — гриб Bottyosporium pulchrum Corda. Чаще поражаются
растения ослабленные, а также растущие в тени или имеющие
механические повреждения. Вначале на пораженных органах растений
появляются буроватые, расплывчатые, с размягченной тканью
пятна. При затенении и плохом проветривании на пораженных
тканях образуется беловатый пушистый налет, состоящий из
грибницы и органов конидиального спороношения гриба. Заболевание
обычно наблюдается к концу вегетации георгин и значительно
раньше в дождливое холодное лето.
Меры борьбы: создание условий, благоприятствующих
нормальному развитию георгин, — незагущенные посадки, хорошо
разрыхленная почва с содержанием извести, подвязывание
георгин к колышкам. Рядовая посадка георгин с расстоянием между
рядами в 1 — 1,5 м, а между растениями в ряду не более 0,75—1,0 м.
При уходе за георгинами и их обрезке оставление на каждом
кусте не больше 1—2 основных побегов. Своевременная уборка
корнеклубней в сухую погоду.
Вертициллезное увядание и гниль корнеклубней. Возбудители —
несовершенные грибы рода Verticillium: V. dahliae Kleb. и V. lateritium
Berk. Болезнь поражает георгины во время вегетации, а корне-
клубни — также при хранении. Проявляется обычно в жаркие
солнечные дни. Гриб сохраняется в почве и сначала заражает корни и
корнеклубни. От больных подземных частей заболевание быстро
распространяется по стеблям, черешкам листьев, и переходя от
нижних частей к верхним поражает все растение.
Пораженные части сначала желтеют, потом становятся
бурыми, растение увядает и погибает. Корни и корнеклубни больных
георгин также вначале увядают, затем буреют и загнивают. В
тканях луба образуются многочисленные микросклероции.
Вертициллезная гниль отличается от фузариозной тем, что на
поверхности пораженных корнеклубней образуются бархатистые
красновато-бурые или коричневые дерновинки, представляющие
собой конидиальное спороношение гриба.
Развитию вертициллезного увядания способствуют те же
условия, что и развитию фузариоза.
Меры б о р ь б ы те же, что и против фузариоза.
Белая гниль (склеротиниоз). Возбудитель болезни — сумчатый
гриб Sclerotinia sclerotiorum Fuck. Заражение георгин происходит
через повреждения тканей стебля, нанесенные при позднем пасын-
416
ковании или во время обработки растений. Вначале болезнь
проявляется в виде водянистых размягченных пятен на основаниях
стеблей. В дальнейшем пораженные ткани желтеют, потом
буреют, загнивают и разрушаются, покрываясь белым ватообразным
налетом мицелия. Стебель вместе с листьями и соцветиями
увядает и отмирает. Внутри стеблей также образуются скопления
мицелия, а затем и темно-коричневые склероции гриба. Заболевание
может привести к отмиранию только отдельных стеблей, но в
случае поражения центрального стебля погибает все растение.
В хранилищах болезнь продолжает развиваться на
корнеклубнях. Последние покрываются белым пушистым мицелием, на
котором образуются многочисленные плотные черные склероции
различной формы и величины.
В зимний период гриб сохраняется в виде склероциев и
мицелия на зараженных корнеклубнях и остатках надземных частей. При
благоприятных условиях склероции прорастают апотециями, и
сумкоспоры осуществляют первичное заражение георгин.
Склеротиниоз поражает многие цветочные и
сельскохозяйственные растения и обычно проявляется на кислых тяжелых сырых
почвах в форме очагов. В поверхностных слоях почвы склероции
сохраняются 5—7 лет, в более глубоких — погибают значительно
быстрее. Развитию болезни способствуют повышенные
температура и влажность воздуха.
Меры борьбы: соблюдение севооборота, недопущение
возврата георгин на прежние участки в течение 3—5 лет;
известкование почвы; ранняя осенняя глубокая перекопка почвы с оборотом
пласта; удаление и сжигание пораженных растений; содержание
почвы в разрыхленном состоянии, без сорняков; незагущенные
посадки и умеренный полив. Обрезка стеблей на 10—15 см ниже
пораженного участка и сжигание удаленных частей. Полив почвы в
очагах болезни суспензиями фунгицидов (ТМТД, фундазола, топ-
сина-М и др.); присыпка золой, известью. При начальной стадии
развития болезни (водянистые, мокнущие пятна на стеблях или
корнеклубнях) — вырезка пораженных участков и присыпка мест
срезов гашеной известью, золой, смесью толченого древесного угля
с молотой серой. Дезинфекция хранилищ; отбор на хранение
только здоровых корнеклубней; опудривание корнеклубней перед
закладкой на хранение фундазолом, топсином-М, ТМТД. В
хранилищах — обработка их известью-пушонкой с примесью молотой серы.
Серая гниль. Возбудитель — гриб Bottytis cinerea Pers. Чаще и
сильнее болеют ослабленные растения, а также георгины,
растущие в тени или имеющие механические повреждения.
Серая гниль поражает всходы, черенки при их укоренении,
листья, стебли, бутоны и цветки вегетирующих растений, а также
корнеклубни при хранении в условиях высокой влажности и
пониженной температуры. Вначале на зараженных органах растений
417
появляются коричневые пятна, увеличивающиеся в размерах.
Всходы и ростки часто поражаются целиком. Пораженные части
буреют, ткани размягчаются и загнивают, покрываясь обильным
дымчато-серым порошащим налетом конидиального спороношения
гриба. В дальнейшем образуются склероции.
Меры борьбы включают мероприятия, проводимые
против белой гнили. Кроме того, рекомендуется опрыскивание
растений в период вегетации фунгицидами: картоцидом, ровралем и др.
Бурая пятнистость. Возбудитель — пикнидиальный гриб Phyllos-
ticta dahliecola Brun. Поражаются в основном листья, иногда стебли.
На них образуются крупные буро-серые пятна с более темной
серединой, где развиваются черные точечные пикниды гриба.
Листья преждевременно засыхают. Инфекция сохраняется на
растительных остатках. Болезнь встречается повсеместно, но наиболее
интенсивно развивается в периоды с теплой влажной погодой.
Меры борьбы: чередование культур; внесение фосфорно-
калийных удобрений; незагущенные посадки. Во время вегетации —
опрыскивание растений фунгицидами: байлетоном, бордоской
смесью, оксихлоридом меди и др. Осенью — уничтожение
растительных остатков.
Мучнистая роса. Возбудители — сумчатые грибы Sphaerotheca
fuliginea Poll./ dahliae Movs. и Erysiphe cichoracearum D. С. Мучнистая
роса георгин имеет признаки, характерные для всех болезней
этого типа: вначале на молодых листьях появляются небольшие
«островки» белого паутинистого налета мицелия, в дальнейшем —
поражаются бутоны, происходит разрастание и уплотнение ми-
целиального налета, появляется порошащий (мучнистый) налет
конидиального спороношения гриба, а к концу вегетации
образуются плодовые тела паразита. При сильной степени развития
болезни пораженные части растений засыхают. Клейстотеции
зимуют на почве и отмерших остатках пораженных растений. Весной
или в начале лета созревающие в плодовых телах сумкоспоры
осуществляют первичное заражение молодых побегов и листьев.
Распространению болезни в открытом грунте
благоприятствуют жаркая погода с обильными росами, избыток азотных
удобрений. Мучнистая роса георгин встречается часто и повсеместно.
Меры борьбы: удаление пораженных частей и
растительных остатков, свободное размещение растений, применение
сбалансированного комплекса NPK с преобладанием калийно-фос-
форных удобрений и микроэлементов; при выращивании в
теплицах — оптимальный режим температуры и влажности, без
резких колебаний их значений.
Опрыскивание растений (в случае необходимости
многократное) фундазолом, топазом, кумулусом, байлетоном, коллоидной
серой, кальцинированной содой с мылом, трихотецином и
другими препаратами.
418
Бактериальный корневой рак. Возбудитель — бактерия Agrobac-
terium tumefaciens (Sm. et Towns.) Conn. На корнеклубнях, корнях и
в области корневой шейки образуются сначала небольшие
наросты, не отличающиеся по цвету от здоровых частей, но со
временем опухоли увеличиваются и становятся коричневыми. К концу
лета они загнивают, разрушаются и отваливаются. У пораженных
растений нарушается режим питания, они имеют угнетенный вид
и в конце концов погибают. Болезнь особенно опасна для
молодых растений.
Бактерии на остатках пораженных растений и опухолей могут
сохраняться в почве до 3—4 лет. Распространяясь с каплями воды,
они попадают на корни, корнеклубни и корневую шейку здоровых
растений и заражают их. Этому способствует наличие
механических травм и повреждений корнеклубней насекомыми. Болезнь
распространяется также дождевыми червями. Бактерии заражают
новые участки при высадке на них больных растений. Заражение
здоровых корнеклубней может произойти и при совместном
хранении их с больными экземплярами. Развитию болезни способствует
высокая влажность почвы, особенно наличие капельно-жидкой
влаги. Высаженные для черенкования корнеклубни, пораженные
раком, дают слабые черенки, непригодные для выращивания.
Болезнь встречается повсеместно и поражает, кроме георгин,
очень многие цветочные, сельскохозяйственные, плодовые
культуры и лиственные деревья и кустарники.
Меры борьбы: выбраковка посадочного материала;
обрезка больных корешков у саженцев при незначительном их
поражении; удаление наростов с последующей дезинфекцией ран 1 %-м
раствором медного купороса; дезинфекция корневой системы.
Недопущение в течение 3—4 лет посева и посадки на зараженных
участках георгин и других растений, восприимчивых к
бактериальному раку.
При выкопке корнеклубней осенью — тщательная проверка их
на зараженность и немедленное уничтожение больных
экземпляров. Во время вегетации выкопка вместе с землей и уничтожение
сильно пораженных растений. Обеззараживание почвы хлорной
известью, медным купоросом, марганцевокислым калием.
Своевременная уборка созревших корнеклубней, недопущение их
подмерзания или механического повреждения при уборке.
Соблюдение оптимального режима температуры и влажности
при хранении и чистоты хранилищ, их предварительная
дезинфекция. Запрет на использование даже частично пораженных
растений в качестве маточников.
Мозаика. Возбудитель — Dahlia mosic vims (вирус мозаики
георгин). Поражает листья, цветки и корнеклубни. Развитию болезни
способствуют плохие условия выращивания растений. Мозаика
передается с корнеклубнями, черенками, инструментами, кото-
419
Рис. 131. Мозаика георгин
рые использовались при черенковании и срезке георгин, а также
насекомыми-переносчиками.
Болезнь характеризуется появлением на молодых листьях
желто-зеленых, неравномерно расположенных пятен неправильной
формы, посветлением околожилковой ткани, измельчанием,
деформацией (в частности морщинистостью) листовых пластинок
(рис. 131). Наблюдаются также укорочение междоузлий,
деформация цветков (у некоторых сортов встречается пестролепестность),
а также задержка роста с образованием многочисленных боковых
побегов, придающих растению кустистый вид. Цветоносы сильно
укорачиваются, цветки становятся непригодными для срезки.
Переносчиками вируса мозаики являются тли.
Меры борьбы: немедленное удаление с участка и
уничтожение всех пораженных растений; уничтожение
насекомых-переносчиков; дезинфекция инструментария; изоляция наиболее ценных
сортов; использование метода термотерапии.
Нематодные болезни георгин вызываются хризантемной
нематодой, галловыми нематодами, стеблевой нематодой картофеля —
Ditylenchus destructorTh., земляничной нематодой — Aphelenchoides
fragariae (Ritz. Bos.) Christ, и др.
Меры борьбы — см. «Нематодные болезни ириса».
14.3.2. Болезни ириса
Ржавчина. Возбудитель — разнохозяйный гриб Puccinia iridisVfmL
Ржавчина сильно поражает листья, на которых летом образуются
многочисленные ржаво-коричневые продолговатые порошащие
подушечки, выступающие через разрывы эпидермиса, — уреди-
420
нии (урединиопустулы). Созревающие в них урединиоспоры в
течение всего вегетационного периода заражают листья ириса.
Ближе к осени на пораженных листьях появляются черные
бархатистые выпуклые спороношения гриба в виде штрихов — телиопус-
тулы. Образующиеся в них телиоспоры зимуют на отмерших
частях растений, а затем прорастают базидиями с базидиоспорами.
Последние заражают валериану, на листьях которой развивается
эциостадия гриба.
Пораженные ржавчиной листья ириса желтеют и
преждевременно засыхают. Ежегодное сильное развитие ржавчины может
привести к ослаблению, слабому цветению и гибели растений.
Это опасное заболевание распространено главным образом в
южных районах России.
Меры борьбы: сбор и уничтожение крупных растительных
остатков; заделка мелких остатков в почву при ее обработке или
мульчирование участка торфом, перегноем, песком слоем в 2—3
см; возвращение ирисов на прежнее место не ранее чем через 3 г.;
применение фосфорно-калийных подкормок; опрыскивание
растений фунгицидами (коллоидной серой, дитаном М-45).
Пятнистость листьев (гетероспориоз). Возбудитель — гриб
Heterosporium gracile Sacc. При гетероспориозе на листьях ириса
образуются округлые или слегка продолговатые светло-желтые
пятна, окруженные более темной каймой и усеянные оливково-
черными дерновинками, представляющими собой конидиальное
спороношение гриба. Со временем пятна приобретают сероватый
или буроватый оттенок. В случае сильного поражения (при
массовом образовании пятен) листья буреют, начиная с верхушки, и
засыхают. Паразит распространяется конидиями, а зимует
мицелием в пораженных остатках листьев. Развитию болезни
благоприятствует влажная теплая погода.
Наиболее часто заболевание встречается в средней полосе
России.
Меры борьбы: высокая агротехника выращивания ирисов.
Создание условий, способствующих нормальному росту и развитию
растений. Уборка и уничтожение всех пораженных частей во
время вегетации и особенно осенью, перед зимовкой. Уничтожение
сорной растительности. Опрыскивание растений в летний период
фунгицидами: байлетоном, бордоской смесью, оксихлоридом
меди, препаратом Абига-Пик и др.
Серая гниль. Возбудитель — гифомицет Botrytis cinerea Pers. Гриб
поражает цветоносы и концы листьев, которые сначала
обесцвечиваются, затем буреют, загнивают и покрываются дымчато-
серым налетом конидиального спороношения возбудителя. Серая
гниль развивается на растениях в условиях высокой влажности,
при избытке в почве азота, подмерзании корневищ, наличии
механических повреждений. Болезнь распространена повсеместно.
421
Меры борьбы: выращивание ирисов на легких
дренированных почвах с достаточным содержанием фосфора и калия, на
хорошо освещенных солнцем участках; дезинфекция корневищ и
почвы перед посадкой растений; удаление и уничтожение
пораженных частей ирисов и больных растений целиком при сильной
степени поражения. Опрыскивание с добавлением прилипателей
и полив растений фундазолом, БМК, ровралем, топсином-М и
другими фунгицидами.
Мокрая гниль. Возбудители — бактерии Erwinia carotovora (Jones)
Waldee, Erwinia aroidea (Town.) Holl., Pseudomonas iridis Holl.
Весной основания побегов темнеют и засыхают. Если заражаются также
и корневища, то их верхние части и основания стеблей загнивают.
В течение лета пораженные ткани разлагаются, превращаясь в
кашицеобразную дурно пахнущую массу. К концу лета внутренние
ткани корневищ разрушаются полностью, хотя их наружные
оболочки иногда сохраняются. Листья буреют и засыхают.
Пораженные растения погибают. Болезнь развивается в условиях высокой
влажности на тяжелых почвах, при глубокой посадке растений,
при подмерзании корневищ или их повреждении во время
пересадки, при недостатке в почве фосфора и кальция, при
удобрении растений свежим навозом.
Меры борьбы: смена участка для посадки ирисов,
улучшение условий произрастания (особенно почвенных), защита
корневищ от подмерзания в холодные периоды, оголение корневищ
в летний период. Уничтожение больных растений, а при слабой
степени развития болезни — зачистка пораженных участков
корневищ до здоровой ткани с последующей дезинфекцией срезов.
При пересадке растений — удаление пораженных частей и
обработка корневищ суспензией дитана М-45 в течение одного часа
или растворами КМп04, тиабендазола, медного купороса.
Мозаика. Возбудитель — Iris mosaic virus (вирус мозаики
ириса). На листьях образуются светлые пятна и полосы. В период
бутонизации и цветения у основания листьев появляются
светло-зеленые или голубые пятна неопределенной формы. На листьях
возникает мозаика в виде сетки из желтых полос. Пораженные
растения сильно отстают в росте («карликовость»), имеют
укороченные цветоносы и недоразвитые, ломкие, пестролепестные цветки.
Переносчики вируса — тли.
У сортов Iridis germanica L. могут появляться лишь симптомы
мозаичности (светло-желтые пятна различной формы на
листьях), а цветоносы и цветки развиваются нормально.
Крапчатая мозаика. Возбудитель — Iris virus 1 (вирус 1 ириса).
На листьях появляется крапчатость, они желтеют и засыхают.
Цветки деформируются, покрываются пятнами неопределенной
формы. Вирус передается с соком больных растений и переносится
теми же тлями, что и обычная мозаика ириса.
422
Меры борьбы сводятся к выбраковке и уничтожению
больных растений, насекомых-переносчиков.
Нематодные болезни
Земляничная нематода— Aphelenchoides fragariae (Ritz.
Bos.) Christ, (паразит почек, листьев, цветков). Больные растения
отстают в развитии, имеют угнетенный вид.
Меры борьбы: выбраковка больных растений,
уничтожение сорняков, чередование культур с возделыванием злаков,
обработка почвы нематицидами.
Стеблевая нематода картофеля —
DitylenchusdestructorTh. Паразит корневищных и луковичных ирисов. Поражает
корневища и луковицы, на которых образуются некротические
бурые пятна. Впоследствии они загнивают, и растения гибнут.
Меры борьбы: выбраковка больных растений, удаление
сорняков; севооборот с выращиванием бархатцев; прогревание
пораженных корневищ и луковиц в воде с температурой 43 °С в
течение трех часов; затем их охлаждение и обсушивание в хорошо
вентилируемом помещении.
Галловая нематода — Meloidogyne sp. Поражает корни с
образованием на них галлов. Больные растения угнетены, отстают
в развитии, теряют декоративность (рис. 132).
Меры борьбы: уничтожение корней больных растений и
сорняков, чередование культур с выращиванием хвойных,
обеззараживание почвы нематицидами.
Проникающая короткотелая нематода {Pratylenchus
penetrans (Cobb.) Filip. et Sch. St.) — паразит корней. На первых
стадиях болезни на корнях появляются бурые и черные некроти-
Рис. 132. Галловая нематода:
/ — самка; 2 — пораженные корни ириса с галлами
423
ческие пятна. В дальнейшем наблюдается почернение, а затем и
отмирание корневой системы. Пораженные растения отстают в
развитии. Для них характерны хлороз, задержка цветения,
потеря декоративности.
Меры б о р ь б ы: во время вегетации — удаление и
уничтожение пораженных корней, а осенью — уничтожение всех
растительных остатков в почве. Уничтожение сорняков. Чередование
культур с выращиванием бархатцев, гайллардии, которые не
поражаются этой нематодой. Обеззараживание почвы нематицида-
ми: препаратом ДД и др.
14.3.3. Болезни пиона
К числу наиболее распространенных болезней этой цветочной
культуры можно отнести серую гниль, ржавчину, пятнистости
листьев, фитофтороз, стеблевую гниль, а также мучнистую росу,
вертициллезное увядание, нематодные и вирусные болезни.
Серая гниль. Возбудитель — гриб-гифомицет Botrytispaeoniae Oud.
Это наиболее опасная болезнь пионов, так как она поражает все
части растения: корневища, молодые проростки, стебли, листья
и бутоны (рис. 133).
Первичным источником инфекции служат пораженные
корневища и отмершие надземные части растений, на которых зимуют
склероции гриба. В загнивших корневищах может сохраняться и
мицелий возбудителя. Отрастающие весной из пораженных
корневищ молодые побеги внезапно увядают, и на них появляется
порошащий налет серой плесени — конидиальное спороношение
возбудителя. На стеблях возле корневой шейки образуются
коричневые пятна, окольцовывающие стебель и быстро
распространяющиеся на вышележащие части. Побеги и стебли увядают и
отмирают. В дальнейшем пораженные ткани разрушаются,
превращаясь в гниль черного цвета. Конидии гриба по воздуху переносятся
на другие органы растения, которые также поражаются серой
гнилью и становятся вторичным источником инфекции для здоровых
частей и соседних здоровых растений.
На кончиках листьев появляются крупные расплывчатые
коричневые пятна. Во влажную погоду они покрываются серым
грибным налетом. Пораженные листья загнивают, скручиваются и
засыхают. Бутоны поникают, не распускаются, буреют, загнивают
и покрываются серым налетом. На пораженных частях растений
со временем образуются округлые или немного вытянутые
черные склероции размером 1 — 1,5 мм.
В период вегетации пионов болезнь распространяется
конидиями гриба, которые переносятся дождевой водой и муравьями. В
зимний период гриб сохраняется в корневищах и отмерших остатках
растений.
424
Рис. 133. Серая гниль пиона:
/ — часть корневища и основание молодого побега со склероциями возбудителя;
2 — пораженный лист; 3 — цветонос и бутон с признаками гнили и налетом
спороношения возбудителя; 4 — конидиеносец и конидии гриба
Развитию болезни, особенно на подземных органах,
способствуют тяжелые, глинистые, затопляемые почвы, повышенная
кислотность, избыток азотных удобрений, слишком загущенные
посадки, затрудняющие вентиляцию, а также укрытие растений
на зиму опавшими листьями и навозом.
Заболевание распространено повсеместно, особенно часто
встречается в годы с дождливым летом и наиболее сильно
поражает пионы в фазе бутонизации и после цветения.
Меры борьбы: агротехнические мероприятия по
профилактике болезни, предусматривающие хорошее освещение посадок,
дренаж почвы, ограничение азотного и усиление фосфорно-калий-
ного питания. Обязательны осенняя низкая обрезка стеблей и
уничтожение всех растительных остатков. После появления молодых
побегов — профилактическое опрыскивание растений системными
фунгицидами (байлетоном, картоцидом, купроксатом и др.), а также
полив растений под корень фундазолом или 0,6 %-й суспензией
ТМТД из расчета 0,5 л суспензии на одно растение с предвари-
425
тельным удалением больных стеблей. В случае повторного
появления признаков болезни — повторение обработки.
Обрезка больных корней при пересадке растений и
погружение корневой системы в 1 %-й раствор медного купороса или в
суспензию ТМТД. В связи с тем, что распространению болезни
способствуют муравьи, которые питаются сахаристыми
выделениями бутонов и разносят споры гриба, необходимо их
уничтожение. В период вегетации рекомендуется опрыскивание
системными и медьсодержащими фунгицидами (бордоской смесью, ок-
сихлоридом меди и др.).
Гниль корней и черенков. Возбудители — грибы родов Fusarium,
Sclerotinia, Rhizoctonia. Болезнь проявляется в основном при
размножении пионов делением куста. Корни пораженных растений
буреют, размягчаются, загнивают и отмирают (рис. 134). На
поверхности зараженных частей образуется розоватый или белый
налет в виде плесени.
Грибы, вызывающие эту болезнь, хорошо приживаются на
мертвых участках поврежденных корней и оснований стеблей,
откуда переходят на живые ткани, вызывая увядание надземных
частей. Возбудители болезни сохраняются в виде мицелия, скле-
роциев или ризоктоний (тонких шнуровидных сплетений гиф) на
зимующих остатках растений в поверхностных слоях почвы в
течение нескольких лет.
После перезимовки мицелий грибов, вызывающих гниль
корней пионов, возобновляет свое развитие. Кроме того, прорастают
склероции и ризоктоний.
Развитие корневой гнили наблюдается при глубокой посадке
пионов, плохом уходе за ними и при наличии поранений.
Заболевание усиливается при повышенной влажности и кислой реакции
почвы.
Особенно частое и сильное проявление болезни наблюдается
при размножении кустов делением, так как корневой системе
наносятся значительные повреждения и при высадке черенков.
Обнаженная ткань на срезах черенков способствует проникновению
возбудителей внутрь растения. При
недостаточно плотном соприкосновении
посадочного материала с песком срез черенка
зарастает медленно и почвенные грибы,
являющиеся факультативными паразитами,
имеют свободный доступ к обнаженным
тканям растений.
Заболевание распространено в
большинстве цветочных хозяйств и кроме пионов
может поражать астру, гвоздику и многие дру-
Рис. 134. Корневая гниль гие культуры, особенно рассаду, сеянцы и
пиона саженцы этих растений.
426
Меры борьбы: при размножении пионов делением куста
рекомендуется тщательная вырезка загнивших участков;
дезинфекция срезов на корневищах раствором медного купороса;
присыпка толченым углем или ТМТД. Внесение фосфорно-калийных
удобрений и микроэлементов, повышающих устойчивость
растений. При посадке корневищ и черенков заделка их на
оптимальную глубину (2,5 — 3,5 см) для профилактики загнивания тканей.
При этом необходимо, чтобы песок плотно прилегал к
погруженным в субстрат частям черенков и корневищ. Рекомендуется
полив растений смесью фундазола (0,2 %) с ТМТД (0,6 %), дитаном
М-45, опрыскивание пионов биопрепаратом «Бактофит».
Белая стеблевая гниль. Возбудитель — сумчатый гриб Sclerotinia
sclerotiorum Fuck. Болезнь проявляется в быстром увядании
растений. Гриб зимует в форме склероциев, которые образуются внутри
пораженных стеблей. Мицелий пронизывает ткани пораженного
растения и в отдельных местах, особенно в сердцевине стеблей,
образует скопления в виде белых хлопьев, из которых со
временем формируются склероции. Последние сразу прорастают (при
благоприятных условиях) или сохраняются до появления
благоприятных условий. На проросших склероциях образуются
маленькие дискообразные плодовые тела (апотеции) на длинных
ножках. Особенно сильно болезнь развивается в годы с дождливым
летом. Заболевание распространено повсеместно.
Меры борьбы: удаление больных растений вместе с комом
земли и сжигание их.
Вертициллезное увядание. Возбудитель — несовершенный гриб
Verticillium dahliae Kleb. При этом заболевании наблюдается
внезапное увядание листьев и побегов во время цветения растений.
На поперечных срезах внешне здоровых стеблей и черешков
листьев бывает заметно потемнение сосудов. При микроскопическом
анализе в сосудах обнаруживается мицелий.
Возбудитель болезни — почвенный факультативный паразит,
который через корневую систему проникает в сосуды растения и
отравляет его своими токсинами. Гриб зимует в корневищах и
тканях корневой шейки растений. Возбудитель имеет весьма широкую
специализацию и поражает растения разных ботанических семейств.
Вертициллезное увядание широко распространено, но
наиболее часто встречается при выращивании пионов на сырых
тяжелых глинистых почвах, что резко снижает устойчивость растений
к этой болезни.
Меры борьбы: применение агротехники, повышающей
устойчивость растений; удаление пораженных растений с комом
земли и сжигание их; протравливание лунок из-под больных
растений марганцовокислым калием или хлорной известью.
Мучнистая роса. Возбудитель — сумчатый гриб Sphaerotheca
fuliginea Poll./ paeoniae Jacz. Гриб образует на верхней стороне ли-
427
Рис. 135. Мучнистая роса пиона:
/ — пораженный лист; 2 — плодовое тело гриба (клейстотеций); 3 — сумка с
сумкоспорами возбудителя
стьев налет белого паутинистого мицелия, на котором
развиваются цепочки конидий. Созревая, конидиальные цепочки
рассыпаются, и налет становится порошащим (мучнистым). Плодовые тела
(клейстотеций) — разбросанные светло-коричневые с одной
сумкой. Встречается на листьях Paeonia anomala L. и других диких и
культурных видах Paeonia (рис. 135).
Меры борьбы: опрыскивание пораженных растений
фунгицидами сразу же после появления первых признаков мучнистой
росы; двукратное повторение обработки через 8—12 дней после
предыдущего опрыскивания. Эффективны препараты серы (куму-
лус, коллоидная сера и др.), системные (байлетон и др.), брому-
коназол, биопрепарат бактофит.
Пятнистости листьев. Наиболее часто встречаются следующие
виды грибных пятнистостей.
Коричневая пятнистость. Возбудитель — гриб-гифо-
мицет Cladosporium paeoniae Pass. Во второй половине лета на
верхней стороне листьев появляются вначале небольшие
темно-пурпуровые или красновато-коричневые пятна. Постепенно пятна
увеличиваются, становятся темно-коричневыми, и пораженные
участки листьев выглядят, как обожженные. Часто они полностью
покрывают всю листовую пластинку.
Для этой болезни характерна особая структура пятен: они
разрастаются в виде зон со слабо выраженной концентрической
складчатостью; еще одна особенность состоит в том, что пораженные
ткани не засыхают и не загнивают, как это бывает при
поражении серой гнилью.
428
Во влажную погоду на нижней стороне листьев в середине
пятен появляется дымчатый или буровато-оливковый паутинисто-
войлочный налет мицелия и конидиального спороношения гриба.
Болезнь часто переходит на молодые побеги и бутоны. Они также
покрываются дымчатым налетом, бутоны плохо раскрываются,
лепестки цветков осыпаются. Попадая на листья, пораженные
лепестки могут вызвать их заражение.
Септориоз. Один из возбудителей болезни — гриб Septoria
paeoniae West. На листьях и стеблях образуются
желтовато-коричневые пятна, более светлые в центральной части и окруженные
пурпуровой каймой. На пятнах развиваются группы пикнид. Второй
возбудитель — Septoria macrospora Sacc. На листьях появляются крупные
округлые серовато-бурые пятна, усеянные многочисленными
мелкими (едва заметными) черными точками — пикнидами. В обоих
случаях пораженные листья преждевременно засыхают и опадают.
При широком распространении и сильной степени развития
болезни септориозы причиняют значительный вред культуре пиона.
Меры борьбы с пятнистостями: осенняя обрезка и
сжигание надземных частей пораженных растений. В период вегетации
при появлении первых признаков пятнистостей удаление
пораженных частей растений и опрыскивание пионов фунгицидами
(бордоской смесью, оксихлоридом меди, кумулусом, коллоидной
серой, препаратом Абига-Пик).
Кольцевая пятнистость. Возбудитель — Paeonia ringspot vims
(вирус кольцевой пятнистости пиона). Вначале между жилками
листьев появляются светло-зеленые пятна неправильной формы.
Потом пятна желтеют и вокруг них образуются более яркие желтые
кольца или полукольца. Они могут возникать и вокруг зеленых
участков листовой пластинки. Обычно за желтым кольцом следует
светло-зеленое кольцо, а затем опять желтое (рис. 136).
К осени правильность расположения концентрических колец
нарушается, и мозаичный рисунок приобретает вид
извивающихся незамкнутых желтых колец или линий. Возможны и другие
отклонения от типичного рисунка, например образование хлоро-
тичных пятен, среди которых остаются зеленые участки, окру-
Рис. 136. Кольцевая мозаика пиона
429
женные желтыми кольцами. Особенности мозаичного узора
зависят от сорта пионов.
Пораженные растения могут более или менее нормально
развиваться и цвести, хотя иногда несколько отстают в росте от
здоровых. Вирус переносится тлями.
Курчавость листьев. Возбудитель не выявлен. Листья на стеблях
располагаются очень близко друг к другу, их края закручиваются
кверху. Листовая пластинка становится морщинистой. Бутоны не
образуются. Одновременно наблюдается карликовость растений.
Мозаика. Возбудитель не идентифицирован. Как и при
обычных вирусных мозаиках других культур, болезнь проявляется в виде
желтовато-зеленых расплывчатых пятен или точек. Больные
растения имеют угнетенней вид, плохо цветут, теряют декоративность.
Меры борьбы с вирусными болезнями пионов:
уничтожение переносчиков инфекции; правильное и своевременное
проведение всех агротехнических мероприятий; внесение в почву бора
и марганца, повышающих устойчивость растений.
Нематодные болезни вызываются листовыми и галловыми
нематодами, пратиленхами.
14.3.4. Болезни флокса
Мучнистая роса. Возбудитель — сумчатый гриб Etysiphe cichora-
cearum D. C.f.phlogis Jacz. Мучнистая роса — одна из наиболее
распространенных и вредоносных болезней флокса. Поражаются все части
растения: листья, стебли, соцветия (в том числе цветоножки), а
при интенсивном развитии болезни — даже лепестки цветка.
На растениях обычно во второй половине лета сначала
появляются небольшие островки тонкого белого порошащего налета,
состоящего из мицелия и конидиального спороношения гриба.
Затем, в случае сильного поражения растений, налет мучнистой росы
разрастается и постепенно может полностью покрыть всю
листовую пластинку, верхушки зеленых стеблей и зонтики соцветий.
Налет становится плотным, войлочным и приобретает
сероватый оттенок. На его поверхности обычно образуется множество
мелких черных шаровидных плодовых тел сумчатой стадии (клей-
стотециев), содержащих сумки с сумкоспорами. Пораженные
листья скручиваются и опадают. Соцветия часто остаются
недоразвитыми, цветки мельчают, становятся бледными, иногда
деформируются. Пораженные флоксы утрачивают декоративность,
имеют неприглядный вид, не годятся для срезки.
Гриб зимует в форме клейстотециев на опавших листьях,
почве, пораженных стеблях. На стеблях, не срезанных вовремя, при
определенных условиях может сохраняться и мицелий гриба.
Мучнистая роса флокса имеет широкое распространение и
встречается повсеместно.
430
Меры борьбы: сбор и уничтожение опавших пораженных
листьев; обязательная осенняя обрезка и сжигание всех стеблей;
перекопка почвы; осеннее или ранневесеннее опрыскивание
флоксов и почвы вокруг растений раствором медного (1 %) или
железного (1,5%) купороса. Во время вегетации, при появлении
первых симптомов болезни, опрыскивание растений фунгицидами
(байлетоном, дерозалем, коллоидной серой и др.), а также бакто-
фитом. В дальнейшем — при необходимости — повторное
опрыскивание с интервалом в 10 —12 дней.
Вертициллезное увядание. Возбудитель — гриб-гифомицет
Verticillium sp. Болезнь поражает корни и основания стеблей. Их
ткани буреют, размягчаются и загнивают. Это приводит к нарушению
поступления воды и питательных веществ в проводящую систему
надземных частей. На пораженных частях растений образуется
беловатый налет, состоящий из мицелия и конидиального спороноше-
ния гриба. В последующем налет приобретает коричневый оттенок.
Загнивание корней приводит к увяданию стеблей и листьев,
при этом листья отмирают и опадают, часто без пожелтения. Гриб
сохраняется в почве на остатках пораженных растений в виде
мицелия и конидий и заражает корневую систему флоксов. Возбудитель
болезни может распространяться и с посадочным материалом.
Болезнь носит очаговый характер. Особенно часто она
поражает флоксы на влажных участках и кислых почвах.
Меры борьбы: строгое соблюдение севооборота,
возвращение флоксов и других поражаемых вертициллезом цветочных
культур на прежние участки не ранее чем через 3—4 г. Высадка
цветочной рассады со здоровой корневой системой. При посадке
растений и разделении кустов предохранение корней от
повреждений. При наличии в посадках единичных больных растений их
удаление и уничтожение. Дезинфекция почвы в тех местах, где
росли больные растения, 3 %-м раствором железного купороса
или раствором КМп04.
Фомоз. Возбудитель — пикнидиальный гриб Phomaphlogis (Roum.)
Speg. Болезнь листьев и стеблей, имеющая некоторое сходство с
физиологическим засыханием листьев и растрескиванием стеблей. На
пораженных частях растений образуются бурые расплывчатые пятна
с многочисленными черными пикнидами (в отличие от
неинфекционного заболевания). Листья скручиваются и засыхают, ткани
стеблей растрескиваются. Растения отстают в росте, плохо цветут.
Развитию болезни способствуют механические повреждения
оснований стеблей, избыточная влажность, заглубленная посадка.
Белая пятнистость (септориоз). Возбудитель болезни —
пикнидиальный гриб Septoria phlogis Sacc. et Speg. Заболевание проявляется
на листьях в виде очень мелких (2—3 мм), но часто сливающихся
пятен округлой формы. Сначала пятна имеют
пурпурно-фиолетовую окраску, потом становятся белыми с темно-красной каймой
431
Рис. 137. Белая пятнистость (септо-
риоз) флокса
и едва заметными в центре
темными точками — пикнидами
гриба. Первыми заражаются
нижние листья, затем инфекция
распространяется с каплями
воды или ветром на здоровые
верхние листья (рис. 137).
Пораженные листья желтеют,
скручиваются, отмирают и опадают.
Ткани стеблей растрескиваются.
При сильном развитии болезни
растения очень рано
сбрасывают листья, плохо цветут,
утрачивают декоративность. Гриб
зимует на опавших листьях, и
поэтому заражение новых,
молодых листьев начинается
весной при первом появлении
отрастающих побегов.
Септориоз поражает флоксы
в открытом грунте и в парниках
в период укоренения черенков. На ослабленных растениях и
черенках болезнь развивается очень интенсивно.
Пятнистости листьев флокса вызывают также Phyllosticta decussata
Syd., Cercospora omphacodes Ell. et Holw. Но они встречаются реже.
Меры борьбы с пятнистостями: уничтожение
пораженных листьев, опавших в течение вегетации и осенью;
профилактическое, а при появлении болезни — и повторное
опрыскивание фунгицидами: бордоской
смесью, оксихлоридом меди,
препаратом Абига-Пик.
Вирусные и микоплазменные
болезни
Курчавость. Возбудитель —
Phlox necrosis vien virus (вирус
некроза жилок флоксов). Листья
пораженных растений сильно
деформируются, на них образуются
многочисленные желтые и бурые
некротические пятна неправильной
формы. Стебли становятся
ломкими, на них также образуются
участки отмершей ткани.
Наблюдается деформация стеблей. Из-за
резкого уменьшения длины междоуз-
Рис. 138. Курчавость флокса
432
лий растения приобретают кустистый вид («карликовость»).
Цветки или вообще не образуются или слабо развиты. Курчавостью
могут быть поражены как молодые, так и старые листья (рис. 138).
Желтуха. Вызывается микоплазменным организмом. Цветки
утрачивают нормальную окраску, у них возникает белое
окаймление. Иногда лепестки и тычинки становятся зелеными и
превращаются в листовидные образования типа чашелистиков. В
дальнейшем происходит пролиферация (израстание) цветков,
превращение их в вегетативные органы, развитие цветков из завязей.
Растения кустятся и значительно отстают в росте, листья мелкие,
узкие, хлоротичные. Переносчиками желтухи являются цикады.
Нематодные болезни
Стеблевая нематода — Ditylenchus dipsaci (Kiihn.) Filip.
Пораженные растения отстают в росте и развитии, образуют
уродливые листья, поздно и плохо цветут. Стебли сильно утолщаются,
становятся ломкими. Больные растения полностью теряют
декоративность (рис. 139).
Меры борьбы — см. «Стеблевая нематода гвоздики».
Северная галловая нематода —MeloidogynehaplaChit.—
паразит корней, вызывает образование на корнях галлов
различной формы и величины. Больные
растения отстают в развитии, имеют
угнетенный вид и утрачивают
декоративность.
Меры борьбы: уничтожение
корней с галлами, сорняков;
чередование культур с выращиванием
хвойных; обеззараживание почвы
нематицидами.
Неинфекционные болезни
Засыхание и опадение
листьев. Усыхание начинается с
нижних листьев и при длительном
недостатке влаги поднимается до
самой верхушки растения.
Усыхание нижних листьев может
наблюдаться также и при
несвоевременном летнем делении и
пересадке кустов. Неправильное внесение
удобрений также может вызвать это
заболевание. Во время подкормок на
нижние листья часто попадают
капли раствора удобрений. При
испарении воды концентрация раствора Рис. 139. Стеблевая нематода
433
повышается и может вызвать ожог или
усыхание листьев. Это особенно ярко
проявляется при внесении подкормок
в жаркое время дня, поэтому их лучше
проводить в пасмурную погоду или
вечером, а после этого смыть с листьев
попавшие на них удобрения.
Растрескивание стеблей. Во
влажные годы во время сильного
роста растений, при внесении с весны
только азотных удобрений и
недостатке в почве извести, у флоксов в
нижней части стеблей наблюдается
продольное растрескивание тканей (рис.
140). Со временем трещины зарастают
каллюсом и стебли развиваются
нормально, но в местах образования
трещин часто переламываются. Для пре-
Рис. 140. Растрескивание ДУпреэдения растрескивания стеблей
стеблей флокса весной надо вносить под флоксы
полное минеральное удобрение и не
допускать загущенных посадок; не поливать флоксы слишком
холодной водой, особенно днем в жаркую погоду.
14.4. БОЛЕЗНИ ОДНОЛЕТНИКОВ И ДВУЛЕТНИКОВ
ОТКРЫТОГО ГРУНТА
14.4.1. Болезни астры садовой
Фузариозное увядание. Возбудители болезни — грибы-гифоми-
цеты рода Fusarium: R oxysporumf. callistephiWr., F. conglutinansWr.
/ callistephi Beach, и др. Фузариоз считается самым опасным
заболеванием этой культуры. Астра поражается в любом возрасте — от
рассады до взрослых растений, но особенно сильно болезнь
проявляется в фазе бутонизации и цветения.
Возбудители фузариоза сохраняются главным образом в почве
на пораженных растительных остатках, а также на семенах. Поэтому
первичное заражение растений обычно начинается с корневой
системы, откуда гриб постепенно распространяется по сосудам и
другим тканям в надземные органы (рис. 141).
Пораженные корни становятся ломкими и впоследствии
загнивают. Поэтому больные растения в посадках полегают и легко
вынимаются из почвы практически без корней. Загнивают и ткани
стеблей в области корневой шейки, где очень часто во влажных
условиях образуются розоватые подушечки или налеты мицелия
434
и конидиального спороношения
гриба. Конидии с каплями
дождя и потоками воздуха
переносятся на здоровые растения и
заражают их.
Увядание надземных частей
обусловлено закупоркой
проводящей системы. Сосуды стеблей
окрашиваются в коричневый цвет
и заполняются мицелием, что
хорошо видно на поперечных
срезах пораженных стеблей.
У молодых растений листья
быстро бледнеют, затем
поникают и усыхают, у взрослых —
сначала желтеют нижние листья, по- Рис. 141. Фузариозное увядание
том остальные. Они скручивают- сеянцев астры
ся, увядают и повисают. На
стеблях у корневой шейки появляются коричневые некротические
полосы, и в этих местах со временем могут образоваться
продольные трещины. Постепенно болезнь распространяется на
вышележащие части стеблей, боковые веточки, цветки и семена.
Растение желтеет и погибает.
Заболевание сильнее и быстрее поражает растения,
ослабленные какими-либо факторами (например, после пикировки,
пересадки или при наличии поврежденных корней).
Наиболее часто поражение фузариозом наблюдается при
разведении астр на тяжелых глинистых и суглинистых почвах с
повышенной влажностью и кислотностью, а также при бессменном
выращивании этой культуры на одних и тех же участках.
Развитие болезни также в значительной мере зависит от
температуры воздуха: при температурах 20...27°С болезнь проявляется
особенно сильно, при 17...20°С — она развивается медленнее, а
при температурах ниже 12 и выше 35 °С вообще не проявляется.
Нередко наблюдается очаговый характер развития болезни.
Среди множества сортов астр некоторые более устойчивы к
фузариозу (например Виктория, Комета, Исполинская и др.). Сорта
группы Страусово перо, Розовидные, Триумф, Уникум,
некоторые сорта группы Американская красавица и другие, наоборот,
поражаются сильно.
Болезнь распространена повсеместно и часто встречается в
хозяйствах, выращивающих астру, причиняя значительные убытки
цветоводству.
Меры борьбы: соблюдение чередования культур;
недопущение возврата астры на прежние участки в течение 4—5 лет;
внесение в почву извести (30—50 г на 1 м2); сбор семян только со
435
здоровых растений, химическое или термическое
обеззараживание семян; подзимний или ранневесенний посев в грунт.
Использование обеззараженной почвы для выращивания рассады. Перед
высадкой растений в грунт оздоровительный полив почвы фунда-
золом; высадка только здоровой рассады. Внекорневые
подкормки калием и микроэлементами. Уничтожение пораженных
растений и замена почвы в очагах болезни; выращивание устойчивых
сортов. См. также «Фузариоз гвоздики».
Ризоктониоз. Возбудитель — несовершенный гриб Rhizoctonia
solani Kuhn. Ризоктониоз поражает растения любого возраста и
проявляется на всех органах растений: корнях, корневой шейке,
стеблях и листьях. Корневая система загнивает; ткани корневой
шейки и вышележащих частей стебля вначале становятся
водянистыми, затем темнеют и тоже загнивают. Листья желтеют,
увядают и отмирают. Больные растения значительно отстают в
росте.
В отличие от других болезней, вызывающих увядание, при ри-
зоктониозе у основания стеблей пораженных растений
появляется бурый войлочный налет мицелия возбудителя. Местами
мицелий еще более уплотняется, образуя мелкие темные склероции.
Гриб чаще поражает растения при их глубокой посадке и
наличии механических повреждений на корнях и корневой шейке. Это
облегчает проникновение гриба сначала в камбий, а затем в
сосуды и другие ткани растений.
Развитию болезни способствуют кислые, влажные и бедные
органическими веществами почвы. Скорость развития ризоктони-
оза зависит и от погодных условий. При дождливой погоде полное
увядание пораженных растений затягивается на 2 — 3 нед, а в
сухую и жаркую погоду астры гибнут значительно быстрей. Однако
слишком низкая температура почвы и воздуха тоже
благоприятствует развитию болезни, особенно у молодых растений.
Гриб, вызывающий ризоктониоз астры, поражает также
гвоздику, картофель, томаты и многие другие растения, сохраняясь
годами в почве на их остатках. Болезнь широко распространена и
встречается в разных районах.
Меры борьбы: высокая агротехника выращивания астр;
проведение мероприятий, ускоряющих рост и развитие растений
(сбалансированное удобрение, микроэлементы); посадка
растений на легких рыхлых почвах, хорошо пропускающих дождевую и
поливную воду. Недопустимы посадки астр на участках из-под
других поражаемых ризоктониозом культур; обеззараживание почвы
в парниках и теплицах сухим паром или фунгицидами;
чередование культур с возвратом астры на прежние участки не ранее чем
через 3 — 4 г. См. также «Ризоктониоз гвоздики».
Серая гниль. Возбудитель — гриб Bottytis cinerea Pers. Чаще всего
болезнь в открытом грунте обнаруживается во второй половине
436
вегетации, причем поражаются все органы растений. В теплицах
развивается в любое время года.
Вначале наблюдается побурение краев листьев, которое потом
распространяется вдоль жилок. На листовой пластинке
появляются коричневые пятна. Пораженные ткани загнивают, а затем
подсыхают. На стеблях, листьях и цветках появляется серый налет,
состоящий из мицелия и конидиального спороношения гриба.
Инфекция сохраняется в виде мицелия и склероциев в почве —
на остатках пораженных растений, а также в виде спор на рамах,
стеллажах и стенах теплиц.
Серая гниль особенно интенсивно развивается при
повышенных влажности и температуре воздуха. Передается при контакте
больных и здоровых растений или конидиями по воздуху. Это
заболевание поражает очень многие цветочные культуры открытого
и защищенного грунта, а также плодово-ягодные и древесные
растения.
Меры борьбы: высокая агротехника выращивания астр —
глубокая обработка почвы с оборотом пласта; разреженные
посадки растений; рыхление почвы и умеренные поливы;
сбалансированное удобрение (без избытка азота); хорошее освещение и
проветривание, особенно в теплицах и парниках; обрезка
пораженных частей и уничтожение сильно пораженных растений;
опрыскивание растений фундазолом, ровралем, дйтаном М-45,
другими эффективными против серой гнили фунгицидами.
Ржавчина. Возбудитель — разнохозяйный гриб с полным
циклом развития Coleosporium solidaginis Thumen. Поражает разные виды
астр. В первой половине лета на нижней стороне листьев
появляются мелкие желто-оранжевые порошащие подушечки — уреди-
ниопустулы, заполненные урединиоспорами, которые,
рассеиваясь по воздуху, вновь заражают листья астр в течение всего лета.
К осени на этих же листьях под эпидермисом развиваются телио-
споры гриба в виде плоских коричневых коростинок. При
сильной степени поражения листья засыхают и преждевременно
опадают.
Телиоспоры зимуют в отмерших листьях, а весной
прорастают, образуя базидии с базидиоспорами, и последние заражают
всходы и хвою различных видов сосны. В конце весны или в начале
лета на хвоинках образуются беловатые пузыревидные эции.
Высыпающиеся из них эциоспоры гриба разносятся ветром и,
попадая на листья астр, заражают их.
Меры борьбы: участок, на котором выращиваются астры,
должен быть удален от сосновых насаждений или лесных
питомников не менее чем на 250 — 300 м; профилактическое
опрыскивание астр фунгицидами в начале вегетации, а при появлении
первых пустул ржавчины — через каждые 7 — 10 дней топазом
(0,05—0,1 %-м раствором), оксихлоридом меди, бордоской сме-
437
сью (0,5 %-й суспензией), препаратом Абига-Пик (0,5 %-м
раствором).
Пятнистости листьев. Возбудители — несовершенные грибы
Septoria callistephi Gloyer, Alternariaflorigena Nelen.
Септориоз имеет более широкое распространение и
встречается чаще. На листьях во второй половине лета появляются
пятна — вначале светло-серые, позднее с более темным центром и
коричневой каймой, почти округлые, диаметром от 3 до 10 мм.
Пятна быстро разрастаются и часто охватывают значительную часть
листовой пластинки. На пятнах с верхней стороны листа
образуются рассеянные, погруженные в ткань пикниды с сосочковид-
ным устьицем, содержащие конидии гриба. Пораженные листья
засыхают, при сильной степени развития болезни может
погибнуть все растение. Гриб сохраняется в зараженных растительных
остатках.
Альтернариоз характеризуется образованием более
крупных бурых пятен с концентрической зональностью. Во влажную
погоду на них появляются оливково-черные дерновинки кониди-
ального спороношения гриба.
Меры борьбы с пятнистостями: соблюдение
севооборота с возвратом поражаемых растений на прежние участки не
ранее чем через 3—4 года; уничтожение остатков пораженных
растений глубокой зяблевой вспашкой или перекопкой почвы с
оборотом пласта; обрезка пораженных частей растений и их
уничтожение; сбор и уничтожение опавших листьев и других мелких
остатков осенью или весной (закопка в ямы или закладка в
компостные кучи на 3—4 года); сбор семян только со здоровых растений.
Во время вегетации, при появлении первых признаков болезни,
опрыскивание растений фунгицидами: байлетоном (0,01—0,02%),
оксихлоридом меди, фундазолом, бордоской смесью и др.
Черная ножка (гниль корневой шейки рассады). Возбудители —
оомицет Pythium debatyanum Hesse, миксомицет Olpidium brassicae
Woron., несовершенные грибы Rhizoctonia aderholdii (Ruhl.) Kolosh.,
Fusarium sp. Болезнь проявляется в побурении или почернении
тканей, а в дальнейшем — в загнивании основания стебля и
образовании перетяжки в области корневой шейки. Листья и стебли
пораженных растений вначале бледнеют, затем растения
постепенно увядают, а иногда и полегают. Возбудители болезни сапротрофно
живут и размножаются в поверхностных слоях почвы на отмерших
остатках растений, но при соприкосновении мицелия с корнями
или корневой шейкой живых (особенно — ослабленных)
растений переходят на их ткани, становясь факультативными
паразитами. Распространение этих грибов с больных растений на
здоровые осуществляется мицелием через почву, а также спорами,
которые разносятся почвенной водой и насекомыми. Разные виды
возбудителей черной ножки зимуют в виде мицелия и спор или в
438
виде различных видоизменений мицелия (мицелиальных шнуров,
склероциев и др.).
Черной ножкой поражаются всходы и рассада очень многих
видов цветочных, сельскохозяйственных и древесных растений.
Меры борьбы — см. «Болезни виолы».
Гниль корней и основания стебля — см. «Болезни виолы».
Желтуха. Возбудитель — микоплазменный организм. У молодых
растений заболевание начинается с посветления жилок или
хлороза, проявляющихся сначала на одном листе. Постепенно хлороз
охватывает новые листья, а со временем — и все зеленые части
растений. На листовых пластинках возникают некрозы. Молодые
листочки становятся почти белыми и растут вертикально, их края
загибаются кверху.
Листовые пластинки у пораженных растений — мельче, чем у
здоровых, а черешки листьев — длиннее. Иногда наблюдаются
укорачивание жилок, образование на листовых пластинках пузыревидных
вздутий или трещин, неровность краев и другие деформации.
У больных растений, особенно молодых, задерживается рост
главного побега, в тканях стебля появляются некротические
участки. Из пазух листьев развиваются тонкие хлоротичные побеги
второго порядка с удлиненными междоузлиями, нередко
возникает усиленная кустистость растений типа «ведьминых метел».
У растений, пораженных желтухой, образуются цветки с резко
деформированными, сильно укороченными зелеными
лепестками, которые могут превращаться в обычные листочки. Иногда
бутоны не распускаются (рис. 142).
Заболевание передается прививкой и насекомыми: цикадами,
листоблошками.
Кроме астры желтуха поражает тагетес, циннию, календулу и
ряд других цветочных культур.
1 2
Рис. 142. Желтуха астры:
1 — здоровый цветок; 2 — два больных цветка
439
Кольцевая пятнистость. Возбудитель — Aster ringspot virus (вирус
кольцевой пятнистости астр). На листьях образуются хлоротичные
кольца и зигзагообразные линии. Растения при сильной степени
поражения отстают в росте, у листьев появляется морщинистость.
Астры хуже цветут и теряют декоративность. Инфекция
передается насекомыми и с соком больных растений при уходе за ними.
Меры борьбы с вирусными и микоплазменны-
ми болезнями: уничтожение больных растений, сорняков и
насекомых-переносчиков.
Нематодные болезни вызываются хризантемной нематодой,
стеблевой нематодой и северной галловой нематодой.
Два первых вида являются паразитами стеблей, листьев и
соцветий. Вызывают угнетение растений, увядание листьев,
деформацию цветков. Северная галловая нематода — паразит корней,
на которых образуются галлы. Растения отстают в росте, плохо
развиваются и цветут.
Меры борьбы: уничтожение больных растений,
растительных остатков, сорняков; обеззараживание почвы нематицидами.
Для профилактики заражения стеблевыми нематодами — смена
культур с выращиванием злаков, против галловой нематоды —
смена культур с выращиванием хвойных.
14.4.2. Болезни левкоя
Черная ножка. Возбудители болезни — оомицет Pythium
debaryanum Hesse, миксомицет Olpidium brassicae Wor., гриб
Rhizoctonia aderhodii (Ruhl.) Kolosh.
Поражаются всходы и рассада. Область корневой шейки и
основания черенков при их укоренении чернеют, загнивают, и
растения погибают. Обычно болезнь
носит очаговый характер и
развивается в условиях высокой
влажности (рис. 143).
Более подробное описание
болезни и меры борьбы с ней
см. «Черная ножка астры
садовой», «Черная ножка виолы».
Кила. Возбудитель болезни —
миксомицет Plasmodiophora bras-
sicae Wor. Кила — одна из
самых опасных и широко
распространенных болезней,
поражающая растения семейства
крестоцветных в любом возрасте.
Болезнь встречается как в от-
Рис. 143. Черная ножка сеянцев левкоя крытом, так и в защищенном
440
грунте. Возбудитель килы — обли-
гатный внутриклеточный паразит.
Он способен сохраняться в почве
с пораженными остатками
растений в течение 4—6 лет в виде
покоящихся спор. При
благоприятных условиях они прорастают аме-
бообразными зооспорами — мик-
самебами. Миксамебы заражают
корневые волоски или молодые
окончания корней. В клетках
зараженных корней развиваются
плазмодии паразита. Под влиянием
паразита эти клетки быстро
увеличиваются в размерах (явление
гипертрофии), вследствие чего на
пораженных корнях образуются
опухоли. Разрастаясь и сливаясь, Рис. 144. Кила левкоя
опухоли охватывают всю корневую
систему и полностью ее деформируют, превращая в сплошные
твердые наросты (рис. 144).
Пораженные корни не могут снабжать надземные части
растений водой и питательными веществами, что приводит к
угнетению роста и увяданию растений. Больные растения левкоя
образуют недоразвитые деформированные соцветия, а зачастую вообще
не цветут и засыхают.
На определенной стадии развития плазмодии в пораженных
клетках распадаются на покоящиеся споры, опухоли со временем
загнивают, разрушаются, и споры попадают в почву, которая
становится источником инфекции для вновь высаживаемых растений.
Кила чаще всего встречается и особенно сильно поражает
растения на влажных тяжелых кислых почвах, преимущественно в
северных районах страны.
Меры борьбы: чередование культур с исключением из
севооборота всех крестоцветных растений или протравливание
почвы; известкование кислых почв; умеренный полив. В период
вегетации — удаление и уничтожение больных растений с корнями и
комом земли. Засыпка оставшихся лунок известью, толченым
древесным углем или золой, либо их опудривание препаратами ТМТД,
фундазола, сумилекса, картоцида и др.).
Белая гниль, склеротиниоз. Возбудитель — гриб Sclerotinia mattiolae
Beiss. Вначале поражаются основания стеблей и нижние листья.
Они буреют, загнивают, и на них развивается ватообразный
налет мицелия с черными склероциями. Постепенно мицелий
заполняет внутренние части стеблей, растения желтеют, их
верхушки поникают. В конце концов растения погибают.
441
Обычно очаги склеротиниоза появляются во второй половине
вегетационного периода. Развитию болезни способствуют бедные
тяжелые кислые влажные почвы, повышенная влажность воздуха.
Источником инфекции является почва, где склероции
сохраняются в течение 5 — 7 лет. Болезнь распространена повсеместно.
Меры борьбы: смена культур с исключением из севооборота
поражаемых видов растений; известкование кислых почв,
применение фосфорно-калийных удобрений; содержание почвы в рыхлом
состоянии; разреженные посадки и умеренный полив;
уничтожение сорняков. Во время вегетации — удаление пораженных
растений с комом земли. После уборки культуры — глубокая перекопка
почвы с оборотом пласта; дезинфекция почвы дитаном-45, фун-
дазолом (0,2 %), ТМТД (0,5 %) и другими фунгицидами.
Ложная мучнистая роса. Возбудитель — оомицет Peronospora
mattiolae Gaum. На верхней стороне листьев появляются
желтоватые пятна. На нижней стороне, соответственно, образуется слабо-
выраженный серовато-фиолетовый налет спороношения
возбудителя. Пораженные стебли искривляются, бутоны на них не
раскрываются или образуют деформированные уродливые цветки.
Пораженные части растений преждевременно засыхают.
Болезнь интенсивно развивается в периоды дождливой погоды.
Возбудитель сохраняется ооспорами на пораженных растительных
остатках и мицелием в семенах, собранных с больных растений.
Ложная мучнистая роса левкоя распространена повсеместно.
Меры борьбы: чередование культур с возвратом
поражаемых (крестоцветных) растений на прежний участок через 3—4 г.;
разреженные посевы и посадки. Протравливание семян перед
посевом препаратами ТМТД (4 г/кг), фундазола, топсина-М и др.
Во время вегетации — уничтожение пораженных растений и
сорняков. При первых признаках болезни — 2—3-кратное
опрыскивание растений оксихлоридом меди, бордоской жидкостью, бай-
летоном и другими фунгицидами, предпочтительно системного
действия, с обязательной обработкой нижней поверхности
листьев.
Серая гниль. Возбудитель — гриб Botrytis cinerea Pers.
Поражаются всходы, рассада и взрослые растения левкоя. Ткани в области
корневой шейки становятся водянистыми, стебель утончается,
темнеет и загнивает. Растения усыхают, и на пораженных частях во
влажных условиях появляется серый налет мицелия и конидиально-
го спороношения гриба. В толще налета часто образуются мелкие
черные плоские склероции. Могут поражаться также цветоносы,
цветки и стручки, которые загнивают и покрываются таким же
серым пушистым налетом. В этом случае семена левкоя теряют
всхожесть.
Развитию серой гнили способствуют высокая влажность
воздуха, недостаточное освещение и проветривание, избыток азота в
442
почве. Гриб сохраняется главным образом в виде склероциев на
растительных остатках, в почве и (в качестве механической
примеси) в сборах семян.
Меры борьбы: регулярное проветривание парников и
теплиц при выращивании рассады; выбраковка больных растений;
обрезка пораженных частей и их уничтожение; разреженные
посадки; умеренный полив, сбалансированный комплекс
удобрений. Опрыскивание вегетирующих растений байлетоном, фунда-
золом, дитаном М-45, картоцидом и другими фунгицидами.
Сортировка собранных семян с целью удаления склероциев.
Фузариоз. Возбудитель — гриб Fusarium oxysporum Schl.
Поражается все растение в целом (прежде всего стебли и листья, а в
дальнейшем и корни). Листья желтеют, скручиваются и увядают,
начиная с нижних. Основание стебля темнеет, и постепенно
поражение распространяется вверх. На стеблях появляются темные
полосы и продольные трещины. На поперечных срезах стеблей
обнаруживаются побуревшие сосуды. На корнях и корневой
шейке развивается сухая гниль. На корневой шейке образуется
розоватый налет конидиального спороношения. Корни отмирают, и
растение легко выдергивается из земли. Особенно сильно увядание
растений проявляется в фазе бутонизации и цветения.
Меры борьбы с фузариоз ом: выращивание
устойчивых сортов; использование новой или обеззараженной почвы для
посева семян и выращивания рассады; перед посевом семян и
высадкой рассады в грунт оздоровительный полив почвы фунда-
золом; известкование кислых почв; посев семенами, собранными
со здоровых растений и протравленными ТМТД (4 г/кг), фунда-
золом (2 г/кг), беномилом или обеззараженными термическим
способом. Высадка только здоровой рассады; подзимний или ран-
невесенний посев в грунт; внекорневые подкормки калием и
микроэлементами. Во время вегетации — удаление и сжигание
пораженных растений. Обработка оставшихся лунок известью,
толченым древесным углем, золой.
«Белая ржавчина». Возбудитель — оомицет Albugo Candida Lev.
Болезнь встречается на левкое и других растениях семейства
крестоцветных. Гриб поражает все надземные органы растений,
особенно в годы с холодной весной. Пораженные части покрываются
как бы каплями белой масляной краски, затем искривляются или
местами вздуваются под давлением большого количества спор,
образующихся под эпидермисом. Резерватором инфекции обычно
являются дикорастущие растения, от которых болезнь может
передаваться цветочным культурам. Инфекция сохраняется также на
пораженных растительных остатках.
Меры борьбы— см. «Ложная мучнистая роса левкоя».
Сосудистый бактериоз. Возбудитель — бактерия Xanthomonas
campestris Dows.
443
Основные признаки болезни — пожелтение листьев, увядание
отдельных побегов или всего растения. На срезах увядших стеблей
видны потемневшие сосуды. Бактериальная инфекция
сохраняется на пораженных растительных остатках и передается с семенами.
Развитию болезни способствуют повышенные температура и
влажность. Сосудистый бактериоз широко распространен в
европейской части России.
Меры борьбы: чередование культур с возвратом левкоя на
прежний участок через 3—4 г., смена почвы или ее
протравливание перед выращиванием рассады; уничтожение больных
растений с корнями; сбор семян только со здоровых семенников;
протравливание семян термическим способом.
Мозаика. Возбудитель — Stock mosaic virus (вирус мозаики
левкоя). На листьях появляются хлоротичные бледно-зеленые пятна
различной формы и величины. Наблюдаются деформация
(складчатость) листовых пластинок и пестролепестность цветков,
угнетение растений, задержка в росте и развитии. Вирус передается
тлями.
Кольцевой некроз. Вирус, вызывающий кольцевой некроз, еще
плохо изучен. На пораженных листьях наблюдаются хлоротичная
крапчатость, посветление околожилковой ткани, а в дальнейшем —
образование некротических участков и пестролепестность
цветков. Растения сильно отстают в росте, становятся карликовыми.
Меры борьбы: выбраковка и уничтожение больных
растений, сорняков и переносчиков вирусов.
Галловая нематода — Meloidogyne arenaria (Neal.) Chitw.
(арахисовая галловая нематода) — паразитирует на левкоях в
районах с теплым климатом, вызывая на корнях образование галлов.
Пораженные растения имеют угнетенный вид, теряют
декоративность.
Меры борьбы: уничтожение корней больных растений и
сорняков, смена культур с выращиванием хвойных,
обеззараживание почвы нематицидами (нематофагином, авертином N и др.).
14.4.3. Болезни сальвии
Ложная мучнистая роса. Возбудитель — оомицет Peronospora
swinglei Ell. et Kell. В основном поражаются нижние листья, на
которых снизу появляется сероватый налет конидиального спороно-
шения. На верхней стороне, соответственно, образуются
желтоватые, слегка выпуклые, со временем буреющие пятна — некрозы.
Инфекция сохраняется в растительных остатках (ооспоры). Болезнь
распространена повсеместно.
Меры борьбы — см. «Ложная мучнистая роса виолы».
Корневой рак. Возбудитель — бактерия Agrobacterium tumefaciens
S.et Towns. Болезнь проявляется в виде наростов или галлов с бу-
444
горчатой поверхностью, образующихся на корневой шейке и
корнях растения. Вначале наросты мелкие, затем они разрастаются и
образуют большие опухоли. К осени наросты загнивают и
разрушаются.
Больные растения отстают в росте, имеют угнетенный вид,
листья свертываются и засыхают. На влажных тяжелых почвах
болезнь развивается сильнее.
Меры борьбы: тщательный осмотр сеянцев перед
высадкой в грунт и выбраковка больных растений с последующим
сжиганием; глубокая обработка почвы (вспашка на глубину 25 — 27 см);
чередование культур; обеззараживание почвы пропариванием
перед посевом семян; протравливание семян перед посевом в
растворах бактерицидов.
Мучнистая роса. Возбудитель — гриб Erysiphe labiatarun Chev./
salviae Jacz. — специализированная форма, поражающая только
шалфей. На листьях и побегах образуется беловатый налет,
который чаще всего появляется перед цветением или в самом его начале.
В конце вегетации на мицелии образуются точечные плодовые тела
гриба — клейстотеции. При сильном развитии болезни листья и
побеги засыхают. Зимуют мицелий гриба в тканях пораженных
побегов или клейстотеции на остатках растений и на почве.
Меры борьбы: удаление и уничтожение пораженных
остатков растений и глубокая обработка почвы; двукратное
опрыскивание растений системными фунгицидами или препаратами,
содержащими серу (например, 0,2 %-й суспензией коллоидной серы).
Пятнистости: септориоз (возбудитель Septoria salviae Pass.), pa-
муляриоз (возбудитель Ramularia salviae Pass.), церкоспороз
(возбудитель Cercospora salvicola Tharp.) и др.
При септориозе на листьях с
обеих сторон образуются мелкие
бурые пятна. Со временем их середина
светлеет, и на ней появляются
черные точечныепикниды. Рамуляри-
о з характеризуется образованием на
верхней стороне листа четко
очерченных бурых пятен; на нижней стороне
пораженных участков наблюдается
слабо выраженный грибной налет. При
церкоспорозе — пятна
светло-серые, окруженные пурпуровой
каймой, на них образуются черные дер-
новинки конидиального спороноше-
ния гриба. Пятнистости листьев
встречаются повсеместно (рис. 145).
Меры борьбы — см. «Пятни- Рис. 145. Пятнистость (цер-
стости виолы», «Пятнистости астры», коспороз) листьев сальвии
445
Ржавчина. Возбудитель — гриб Puccinia nigrescens Kirch. Летом на
листьях развиваются урединии в виде ржаво-оранжевых
порошащих пустул, а к осени образуются коричневые телиопустулы.
Меры борьбы— см. «Ржавчина виолы».
Фузариозное увядание. Возбудители — грибы-гифомицеты рода
Fusarium. Болезнь проявляется чаще всего в фазе бутонизации и
цветения. Растения желтеют и быстро увядают. На основаниях
стеблей развивается плотный беловато-розоватый налет мицелия и
спороношения гриба.
Меры борьбы — см. «Фузариозастры».
Ризоктониоз (бурая гниль). Возбудитель — несовершенный гриб
Rhizoctonia solani Kuhn. Болезнь поражает корни и прилегающие к
ним надземные части растений. Они постепенно темнеют,
становятся бурыми и загнивают. На пораженных органах образуется
плотный налет бурого мицелия с погруженными в него мелкими
черными склероциями. Обычно болезнь приводит к гибели растений.
Ризоктониоз может развиваться в течение всего периода
вегетации, чаще и сильнее проявляется на участках с тяжелыми
кислыми холодными, чрезмерно влажными почвами, а также при
слишком глубокой посадке растений и наличии на корнях
механических повреждений. Возбудитель может несколько лет
сохраняться в виде склероциев в почве или в виде мицелия на
пораженных растительных остатках. Болезнь распространена
повсеместно и встречается на очень многих цветочных культурах.
Меры борьбы — см. «Ризоктониоз астры».
Пожилковый хлороз. Возбудитель — Salvia virus 1 (вирус саль-
вии). Вирус вызывает побледнение жилок и общий хлороз
надземных частей растений. Распространяется тлей Myzus persicae.
Кольцевая пятнистость. Возбудитель болезни не
идентифицирован. Болезнь проявляется в образовании на листьях хлоротич-
ных колец и пятен. Позднее пораженные участки отмирают.
Меры борьбы с вирусными болезнями:
уничтожение больных растений, сорняков и переносчиков инфекции.
Нематодные болезни представлены листовыми нематодами (хри-
зантемной и земляничной), галловыми нематодами (северной,
арахисовой и яванской), а также ксифинемой американской и
проникающей короткотелой нематодой (проникающим пратиленхом).
Xiphinema americanum Cobb. — паразит корней, вызывающий их
отмирание и сильное угнетение растений, выражающееся в
задержке роста надземных частей. Эта нематода является
переносчиком многих фитопатогенных вирусов (кольцевой пятнистости
гвоздики, мозаики некоторых злаковых трав, желтой
карликовости малины и др.).
Pratylenchus penetrans (Cobb.) Filip. et Sch. St. поражает
корневую систему растений. Вначале на корнях образуются бурые и
черные некротические пятна. В дальнейшем корневая система полно-
446
стью чернеет и отмирает. У пораженных растений развивается
хлороз и наблюдается задержка цветения. Они сильно отстают в
росте, утрачивают декоративные качества.
Меры борьбы с фитонематодами: во время
вегетации — уничтожение пораженных корней или выбраковка
больных растений, а осенью — сжигание всех растительных остатков,
уничтожение сорняков; чередование культур с выращиванием
хвойных (против галловых нематод) или злаков (против листовых
нематод). Обеззараживание почвы нематицидами (тиабендазолом,
нематофагином, оксамилом и др.).
14.4.4. Болезни виолы
Головня. Возбудители — грибы Urocystis violae (Sow.) Fisch. v.
Waldh., Ur. kmetiana Magn. Первый возбудитель поражает
вегетативные органы растений. На листьях, черешках и стеблях
образуются вздутия различной формы и величины, заполненные
черной массой хламидоспор (рис. 146). Последние, распыляясь,
заражают новые растения. Второй возбудитель поражает только
генеративные органы — цветки и завязи. Больные цветки не
раскрываются, завязи вздуваются. Цветки и завязи заполняются
черными клубочками хламидоспор гриба. Во время заготовки семян,
а иногда и раньше, оболочки пораженных цветков и завязей
разрываются, хламидоспоры освобождаются и рассеиваются.
Зимуют споры на поверхности семян или почвы. Весной они
прорастают и осуществляют первичное заражение растений. Головня
виолы встречается во многих областях России.
Меры борьбы: немедленное удаление с плантации и
сжигание больных растений; чередование культур или смена почвы;
на участках с проявлением головни
перекопка почвы с оборотом пласта;
присыпка почвы известью. Перед
посевом протравливание семян фунда-
золом, БМК, топсином-М или
другими эффективными фунгицидами.
Сбор семян только со здоровых
семенников.
Ржавчина. Возбудители — гриб
Puccinia violae (Schum.) DC и другие
виды. На листьях и черешках
появляются ржаво-коричневые урединиопу-
стулы, а затем темно-коричневые те-
лиопустулы. Гриб сохраняется в виде
телиоспор на пораженных остатках
растений. Поражаются многие виды
фиалок. Болезнь обычно проявляется Рис. 146. Головня виолы
447
во второй половине вегетации растений при влажной погоде.
Распространение болезни совпадает с ареалом культуры.
Меры борьбы: сбор и уничтожение растительных
остатков, смена культур с возвратом фиалок на прежний участок через
3—4 г. Усиление фосфорно-калийного питания. Возделывание
устойчивых сортов. Опрыскивание растений во время вегетации
фунгицидами: бордоской смесью (0,5 %), оксихлоридом меди (0,2 %),
топазом (0,05—0,1 %), препаратом Абига-Пик (0,5 %).
Мучнистая роса. Возбудители — сумчатые грибы Erysiphe
cichoracearum DC/ violarum (Died.) Jacz., Oidium violae Sacc.
Поражаются все надземные части растений: стебли, листья обертки
цветочных корзинок. Вначале на них появляется тонкий серовато-
белый паутинистый налет мицелия, который иногда выражен очень
слабо. Позднее налет приобретает мучнистый вид за счет
образования конидиального спороношения гриба. К концу вегетации у
первого вида появляются многочисленные черные точечные
плодовые тела — клейстотеции, которые зимуют на остатках
пораженных растений. Созревающие в них сумкоспоры на следующий
год осуществляют первичное заражение растений.
Мучнистая роса обычно появляется в фазе бутонизации и
цветения. Растения отстают в росте, хуже цветут (при этом
сокращается период цветения), теряют декоративность. Урожай семян резко
снижается.
Развитию болезни способствуют сухость воздуха, теплая
солнечная погода с обильными росами, недостаточный полив,
избыток азотных удобрений.
Меры борьбы: смена культур; сбор и уничтожение
пораженных остатков растений; глубокая перекопка почвы с оборотом
пласта; сбалансированное минеральное питание; использование
более устойчивых сортов; протравливание семян фундазолом (5 г/кг);
опрыскивание растений в период вегетации через 12—14 дней
фундазолом, через 7 — 10 дней — дерозалем, коллоидной серой и
другими серосодержащими и системными фунгицидами,
биопрепаратами — бактофитом (0,7 %) или трихотецином.
Пятнистости вызываются несовершенными грибами Colletotrichum
violae-tricoloris, Septoria violae West., Phyllosticta tricoloris Sacc,
Ascochyta violae Sacc. et Speg., Pleosphaerulina violae P. Magorn.,
видами Ramularia и Cercospora.
На листьях образуются пятна различного цвета, формы и
величины, на которых развиваются столь же разнообразные по
характеру и внешнему виду спороношения возбудителей. При сильной
степени поражения листья преждевременно засыхают. Наиболее
распространенные и вредоносные из перечисленных пятнисто-
стей — антракноз и филлостиктоз.
Антракноз (Colletotrichum violae-tricoloris). На листьях
появляются желтовато-коричневые пятна с ясно выраженной темной
448
каймой. Вначале пятна мелкие округлые, затем они
увеличиваются. При рассматривании пятен в лупу видны темные подушечки —
конидиальные ложа гриба.
Филлостиктоз. На листьях образуются одиночные или
сливающиеся округлые некротические пятна. На их поверхности
развиваются черные точечные пикниды гриба. При наличии
многочисленных пятен листья засыхают целиком. Инфекция
сохраняется на пораженных опавших листьях. Обе пятнистости встречаются
часто и повсеместно.
Меры борьбы с пятнистостями: чередование культур с
возвращением виолы на прежнее место не ранее, чем через 3—4 года;
осенью — сжигание растительных остатков и глубокая перекопка
почвы; протравливание семян ТМТД (2 г/кг); во время вегетации —
опрыскивание через 12—14 дней фундазолом, дерозалем, байле-
тоном, дитаном М-45, хлорокисью меди, коллоидной серой и
другими фунгицидами.
Черная ножка. Возбудители — несовершенный гриб Rhizoctonia
aderholdii (Ruhl.) Kolosh., оомицет Pythium debaryanum Hesse, мик-
сомицет Olpidium brassicae Wor. Поражаются всходы и рассада.
Пораженные участки стеблей темнеют и утончаются. Больные
растения поникают и в течение 3—4 дней погибают. Взрослые
растения не болеют. Развитию болезни в теплицах и парниках
способствуют нарушения режима температуры, влажности и освещения,
загущенность посевов, поздняя пикировка, избыток азотных
удобрений, кислые почвы.
Заболевание обычно носит очаговый характер. Накопление
инфекции происходит при высадке на одном и том же участке в
течение нескольких лет поражаемых культур. Болезнь
распространена повсеместно.
Меры борьбы: соблюдение агротехнических норм —
правильный режим температуры и влажности, сбалансированное
удобрение; незагущенный посев и умеренный полив; систематическое
рыхление почвы и проветривание теплиц; замена или
дезинфекция зараженной почвы. Внесение в почву триходермина, трихоте-
цина, бактофита или других биопрепаратов. После высева семян и
при появлении всходов полив почвы фунгицидами (фундазолом,
топсином-М, ТМТД) с последующим рыхлением.
Серая гниль. Возбудитель — гриб Botrytis cinerea Pers. Описание
болезни и меры борьбы — см. «Серая гниль левкоя».
Гниль корней и основания стебля. Возбудители — оомицет
Phytophthora cryptogea Peth. et Laff., грибы родов Rhizoctonia и Sclerotinia.
У заболевших растений темнеют и загнивают корни, загнивают и
размягчаются нижние части стеблей. При фитофторозе на стеблях
образуется белый паутинистый налет мицелия, при ризоктониозе —
налет бурый и более плотный (войлочный). При поражении склеро-
тиниозом — налет мицелия (белый, ватообразный) развивается
449
не только на основаниях стеблей, вызывая «белую гниль», но
переходит и на вышележащие части растений, при этом внутри
стеблей образуются скопления мицелия и крупные черные склероции
различной формы. Во всех случаях листья желтеют,
скручиваются, часто загнивают. Растения обычно полностью отмирают.
Развитию гнили способствуют кислые, чрезмерно влажные
почвы. Возбудители сохраняются на остатках погибших растений
и непосредственно в почве. Все три возбудителя этой гнили
поражают многие цветочные культуры: астру, петунию, наперстянку,
иберис, алиссум, львиный зев и др.
Меры борьбы: чередование культур; глубокая осенняя или
весенняя перекопка почвы, ее известкование; протравливание
семян 0,2 %-м фундазолом; удаление больных растений с комом
земли и их сжигание; опрыскивание растений 0,2—0,3 %-ми
суспензиями фундазола, топсина-М, ТМТД и другими фунгицидами.
Ложная мучнистая роса. Возбудитель — оомицет Peronospora violae
de By. Поражаются все надземные части растений. На верхней
стороне листьев образуются желтоватые, постепенно буреющие
расплывчатые пятна, на нижней стороне, под пятнами, возникает едва
заметный беловато-сероватый налет конидиального спороношения
возбудителя. Стебли в местах поражения искривляются, пораженные
бутоны не раскрываются или из них получаются
уродливо-деформированные цветки. Пораженные органы преждевременно засыхают.
Массовое развитие ложной мучнистой росы совпадает с
дождливыми периодами. Источниками инфекции являются
растительные остатки и семена пораженных растений.
Болезнь распространена повсеместно.
Меры борьбы: чередование культур с возвратом виолы на
прежнее место через 3—4 г.; разреженные посадки; уничтожение
сорняков и пораженных растений; протравливание семян ТМТД
(4 г/кг), фундазолом, топсином-М; при первых признаках
болезни 2 —3-кратное опрыскивание растений байлетоном, купрокса-
том, бордоской смесью, оксихлоридом меди, топазом, вектрой и
другими эффективными фунгицидами (особое внимание обращать
при этом на обработку нижней поверхности листьев).
Пестролепестность. Возбудитель — Cucumis mosaic virus 1
(вирус огуречной мозаики). Главный симптом — пестролепестность
цветков. Листья становятся хлоротичными, их концы отмирают.
Переносчики — тли.
Кольцевая мозаика. Возбудитель — Tobacco ringspot virus (вирус
кольцевой пятнистости табака). На листьях образуются
многочисленные бурые некротические кольца с темно-зеленой серединой.
Переносчики — тли.
Меры борьбы с вирусными болезнями:
выбраковка и уничтожение больных растений, насекомых-переносчиков и
сорняков.
450
Нематодные болезни вызываются земляничной нематодой,
галловой нематодой, паралонгидорусом большим и проникающим
пратиленхом (см. нематодные болезни других цветочных культур).
14.4.5. Болезни гвоздики турецкой
Ржавчина. Возбудитель — гриб Puccinia arenariae (Schum.) Wint.
На листьях с обеих сторон (чаще с нижней) образуются темно-
коричневые рассеянные, иногда сливающиеся плотные телиопу-
стулы, окруженные кольцом хлоротичной ткани. Телиоспоры
сохраняются на остатках пораженных растений. Развитию болезни
способствует высокая влажность воздуха. Ржавчина поражает
многие культивируемые и дикорастущие растения семейства
гвоздичных, распространена повсеместно.
Меры борьбы — см. «Ржавчинавиолы».
Пятнистости. На гвоздике турецкой встречаются в основном два
вида болезней этого типа: гетероспориоз и альтернариоз
(возбудители — грибы-гифомицеты, соответственно Heterosporium echinulatum
Cke. и Alternaria dianthi Stev. et Hall.). Поражаются все надземные
части растений в течение всего вегетационного периода, сильнее —
во второй половине лета. При гетероспориозе образуются
округлые серые пятна, окруженные темно-красной каймой. На них
развивается конидиальное спороношение гриба в виде оливково-зе-
леного налета. При альтернариозе пятна округлые или
продолговатые, спороношение гриба — в виде черного бархатистого налета.
При сильной степени поражения растений листья засыхают,
стебли переламываются, бутоны не распускаются или образуются
уродливые однобокие цветки. Инфекция сохраняется на
растительных остатках.
Массовому развитию и распространению пятнистостей
способствуют повышенные температура и влажность, плохая
вентиляция, избыток азота. Гетероспориоз и альтернариоз гвоздики
встречаются повсеместно.
Меры борьбы — см. «Пятнистость астры садовой».
Вирусные болезни гвоздики турецкой — мозаика, пожилковая
крапчатость, кольцевая пятнистость. Возбудители,
диагностические признаки и меры борьбы — см. «Вирусные болезни гвоздики
садовой».
Нематодные болезни вызываются стеблевой нематодой Ditylenchus
dipsaci (Kuhn) Filip., галловыми нематодами рода Meloidogyne,
клеверной цистообразующей нематодой Heterodera trifolii Goff. и др.
Диагностические признаки и меры борьбы — см. «Нематодные
болезни гвоздики садовой».
Ломкость стеблей — неинфекционное заболевание. Оно
характеризуется тем, что цветоносные стебли ломаются под тяжестью
соцветий. Профилактика — подкормка растений солями кремния.
451
14.4.6. Болезни мальвы
Ржавчина. Возбудитель — однохозяйный гриб с неполным
циклом развития Puccinia malvacearum Mont. На нижней стороне
листьев образуются телиопустулы в виде коричневых бородавочек,
которым на верхней стороне листа соответствуют мелкие,
несколько вдавленные желтоватые пятна. При сильной степени
поражения листьев они засыхают. Болезнь обычно развивается во второй
половине вегетационного периода при влажной погоде, причем
сильнее страдают растения второго года жизни. Ржавчина
поражает все культивируемые сорта мальвы и алтей. Телиоспоры
сохраняются зимой на остатках больных растений.
Болезнь встречается во всем ареале мальвы.
Меры борьбы: уничтожение крупных растительных
остатков и заделка мелких частей в почву при обработке междурядий;
чередование культур с возвратом мальвы на прежний участок
через 3 г.; внесение фосфорно-калийных удобрений; опрыскивание
растений фунгицидами; возделывание устойчивых сортов.
Пятнистости листьев. Среди пятнистостей наиболее
распространен аскохитоз (возбудитель — пикнидиальный гриб Ascochyta
malvicola Sacc). Пятна округлые, охряного цвета с
темно-коричневой каймой; пикниды образуются на пятнах с верхней стороны
листа и располагаются концентрическими кругами.
Реже встречаются церкоспороз (возбудитель — гриб-гифоми-
цет Cercospora althaeina Sacc.) и антракноз (возбудитель —
несовершенный гриб Colletotrichum malvacearum Sacc).
Меры борьбы: — см. «Пятнистости виолы».
Мозаика. Возбудитель — Malve mosaic virus (вирус мозаики
мальвы). Болезнь характеризуется посветлением жилок и мелкой крап-
чатостью листовых пластинок. Растения отстают в росте и
развитии, хуже цветут. Переносчики вируса — тли.
Меры борьбы: уничтожение насекомых-переносчиков;
выбраковка и уничтожение больных растений и сорняков.
Галловые нематоды рода Meloidogyne. Чаще встречается
М ritzemabosL
Меры борьбы: уничтожение сорняков, выбраковка и
уничтожение пораженных растений; обеззараживание почвы немати-
цидами; смена культур с введением хвойных.
СПИСОК ЛИТЕРАТУРЫ
Бактериальные болезни растений / Под ред. В. П. Израильского. — М.:
Колос, 1979. - 288 с.
Бейлин И.Г. Цветковые полупаразиты и паразиты. — М.: Наука, 1968. —
88 с.
Белоселъская 3. Г., Сильвестров АД Вредители и болезни цветочных и
оранжерейных растений. — М.; Л.: Сельхозгиз, 1953. — 200 с.
Белоселъская 3. Г., Сильвестров АД Зашита цветочных растений от
вредителей и болезней. — Л.; М.: Сельхозгиз, 1960. — 147 с.
Влияние загрязнения воздуха на растительность / С. Бёртитц и др. /
Под ред. Х.-Г. Десслера. — М.: Лесная промышленность, 1981. — 184 с.
Булатов В. А Гвоздика ремонтантная. — Л.: Агропромиздат, 1987. — 95 с.
Вавилов Н. И. Избранные произведения. Работы по иммунитету
растений. - М.: Наука, 1956. - Т. IV.
Вакин А Т., Полубояринов О, И., Соловьев В. А, Альбом пороков
древесины. — М.: Лесная промышленность, 1980. — 125 с.
Ван дер Планк Я. Устойчивость растений к болезням: Пер. с англ. — М.:
Колос, 1972. - 285 с.
Василяускас А. Корневая губка и устойчивость экосистем хвойных
пород. — Вильнюс: Мокслас, 1989. — 98 с.
Ведерников Н. М, Яковлев В. Г. Защита хвойных сеянцев от болезней. —
М.: Лесная промышленность, 1972. — 89 с.
Власов А А, Крангауз Р. А. Негнилевые болезни стволов и ветвей
лиственных пород. — М.: Гослесбумиздат, 1963. — 48 с.
Воронцов А И, Патология леса. — М.: Лесная промышленность, 1978. —
269 с.
Воронцов А. И., Мозолевская E.L, Соколова Э.С Технология защиты
леса. — М.: Экология, 1992. — 304 с.
Вредители и болезни цветочно-декоративных растений. — М.: Наука,
1987. - 591 с.
Гарибова Л. В. Обзор и анализ современных систем грибов. —
Петрозаводск: Карельский научный центр РАН, 1999. — 28 с.
Гарибова Л, В., Сидорова Я. К Грибы. Энциклопедия природы России. —
М.: Наука, 1999. - 350 с.
Гвоздяк Р. И., Яковлева Л.М. Бактериальные болезни лесных
древесных пород. — Киев: Наукова думка, 1979. — 244 с.
Государственный каталог пестицидов и агрохимикатов, разрешенных
к применению на территории Российской Федерации. — М.: Госхимко-
миссия Минсельхозпрода РФ, 2001. — 330 с.
Грот В. А. Лилии и их культура — М.: Изд-во МГУ, 1966. — 90 с.
Губина В, Г Нематоды хвойных пород. — М.: Наука, 1980. — 190 с.
Жизнь растений. — Т. 2. — Грибы. — М.: Просвещение, 1976. — 479 с.
453
Жуков AM. Грибные болезни лесов Верхнего Приобья. —
Новосибирск: Наука, 1978. — 246 с.
Жуков А.М., Гордиенко П. В. Научно-методическое пособие по
диагностике грибных болезней лесных деревьев и кустарников. — М.: Изд-
во Министерства природных ресурсов России, 2001. — 69 с.
Журавлев К В. Диагностика болезней леса. — М.: Сельхозиздат, 1962. —
192 с.
Журавлев К И. Болезни цветочных культур. — Л.: Изд-во ЛГУ, 1973. —
76 с.
Журавлев И. К, Крангауз Р. А., Яковлев В. Г. Болезни лесных деревьев
и кустарников. — М.: Лесная промышленность, 1974. — 160 с.
Защита леса от вредителей и болезней: Справочное издание. — М.:
Агропромиздат, 1988. — 413 с.
Зирка Т. И. Атлас вирусных и микоплазменных болезней
декоративных растений. — Киев: Наукова думка, 1984. — 150 с.
Ипполитова Н.Я., Васильева М. Ю. Пионы. Альбом-справочник. — М.:
Россельхозиздат, 1985. — 222 с.
Карантин растений в СССР / Л.В.Воронкова и др. — М.:
Агропромиздат, 1986. — 205 с.
Коссинская И. С Фацидиоз сосны. — Новосибирск: Наука, 1974. — 91 с.
Краснова Н. С Пионы. — М.: Колос, 1971. — 90 с.
Кругов В. И. Грибные болезни хвойных пород в искусственных ценозах
таежной зоны европейского Севера СССР. — Петрозаводск: Карельский
филиал АН СССР, 1989. - 206 с.
Крутое В. И. Учет и долгосрочный прогноз соснового вертуна. —
Петрозаводск: Карельский филиал АН СССР, 1987. — 20 с.
Крутое В. И., Хансо М.Э. Побеговый рак (склеродерриоз) сосны:
диагностика, профилактика и меры борьбы. — Петрозаводск: Карельский
филиал АН СССР, ЭстНИИЛХОТ, 1989. - 14 с.
Кулибаба Ю. Ф., Сапов С К Болезни ремонтантной гвоздики и меры
борьбы с ними. — М.: Изд-во Министерства сельского хозяйства СССР,
1983. - 5 с.
Ладейщикова Е. И. Устойчивость хвойных пород и перспективы
селекции сосны / Корневая губка. — Харьков, УкрНИИ ЛХА, 1974. — 150 с.
Лесная энциклопедия. — М.: Советская энциклопедия, 1985, 1986. —
590 с. - 605 с.
Матвеева М.А. Защита растений от нематод. — М.: Наука, 1989. — 150 с.
Методы фитопатологии / 3. И. Кирай и др. — М.: Колос, 1974. — 344 с.
Минкевич И. И. Эпифитотиология грибных болезней лесных пород. —
Л.: ЛТА, 1977. - 208 с.
Миско Л. А. Розы. Болезни и защитные мероприятия. — М.: Наука,
1986. - 246 с.
Многолетние цветы открытого грунта: Справочник цветовода / Н. А. Ба-
зилевская и др. — М.: Изд-во Министерства коммунального хозяйства
РСФСР, 1959. - 439 с.
Мозолевская Е.Г., Катаев О. А., Соколова Э. С. Методы лесопатологи-
ческого обследования очагов стволовых вредителей и болезней леса. —
М.: Лесная промышленность, 1984. — 152 с.
454
Негруцкий С. Ф. Корневая губка. — М.: Агропромиздат, 1986. — 200 с.
Номеров Б. А. Садовые розы. — М.: Изд-во МГУ, 1973. — 145 с.
Общая и сельскохозяйственная фитопатология / М.И.Дементьева и
др. — М.: Агропромиздат, 1985. — 495 с.
Однолетние цветочные растения / Составители: В.В.Вакуленко и
Т. М. Алейникова. — М.: Изд-во Министерства коммунального хозяйства
РСФСР, 1961.-259 с.
Основные методы фитопатологических исследований / А. Е.Чумаков
и др. — М.: Колос, 1974. — 191 с.
Попкова К. В. Общая фитопатология. — М.: Агропромиздат, 1989.
Рожков А. А, Козак В. Т. Устойчивость лесов. — М.: Агропромиздат,
1989. -239 с.
Ролл-Хансен Ф., Ролл-Хансен X. Болезни лесных деревьев / Под ред.
В. А. Соловьева. — СПб: ЛТА, 1998. — 120 с.
Сборник руководящих и инструктивных документов по карантину
растений в Российской Федерации. — М.: Министерство сельского
хозяйства и продовольствия Российской Федерации, 1999. — 395 с.
Сборник руководящих документов по лесному карантину. — М.:
Министерство сельского хозяйства и продовольствия Российской
Федерации, 1998. — 103 с.
Семенкова И, Г. Лесная фитопатология. — М.: Изд-во МГУЛ, 2001. —
197 с.
Скипина К. П. Неинфекционные заболевания ремонтантной гвоздики. —
М.: Изд-во Министерства сельского хозяйства СССР, 1984. — 6 с.
Соколов Д.В. Корневая гниль от опенка и меры борьбы с ней. — М.:
Лесная промышленность, 1964. — 181 с.
Тарр С. Основы патологии растений: Пер. с англ. — М.: Мир, 1975. —
588 с.
Уход за декоративными растениями: Справочник «Брунс парланцен». —
М.: Глобус, 2000. - 112 с.
Федоров Н И. Лесная фитопатология. — Минск: Вышэйшая школа,
1989. -316 с.
Федоров Н И. Корневые гнили хвойных пород. — М.: Лесная
промышленность, 1984. — 161 с.
ХессайонД.Г. Все о розах: Исчерпывающее руководство по
выращиванию роз и уходу за ними. — М.: Кладезь-Букс, 2001. — 290 с.
Черемисинов НА., Негруцкий С.Ф., Лешковцева НИ. Грибы и
грибные болезни деревьев и кустарников. — М.: Лесная промышленность,
1970. - 392 с.
Шевченко СВ., Цилюрик А.В. Лесная фитопатология. — Киев: Вища
школа, 1986. — 382 с.
Щербин-Панфененко А.Л. Бактериальные заболевания лесных пород. —
М.: Гослесбумиздат, 1963. — 148 с.
Эпифитотии болезней растений. Математический анализ и
моделирование: Пер. с ангд. / Под ред. К.М.Степанова и Б.И.Гуревича. — М.:
Колос, 1979.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
А
Агрессивность 95
Аксения 115
Антракноз 12
Аски 26
Анаморфы 28
Анастомозы 17
Анемохория 29
Антисептики 151
Антисептирование древесины 160
Антропохория 31
Арбускулы 120
Артроспоры 24
Аутохория 32
Ацервулы 26
Аэрозоли 159
Б
Базидии 28
Базидиомицеты 63
Бактерии миколитические 76
— фитопатогенные 71
Бактериозы гиперпластические 75
— паренхиматозные 74
— сосудистые 76
Бактерициды 151
Бластоспоры 24
Болезни 8
— инфекционные 9
—- местные 14
— неинфекционные 9, 167
— нематодные 380, 408, 420, 430,
446
— общие 14
— острые 14
— хронические 14
Бурелом 172
В
Вакцинация растений 126
Ведьмины метлы 12, 79
Везикулы 120
Ветровал 171
Вещества ростовые 95
Вилт И
Вирулентность 95
Вирусоциды 151
Вирусы неперсистентные 81
— персистентные 81
— фитопатогенные 77
Влажная камера 135, 204
Выздоровление растений 100
Выносливость растений 123
Г
Газоустойчивость растений 178
биологическая 179
морфологическая 178
физиологическая 178
Газочувствительность растений 178
Гаметангиогамия 26
Гаметогамия 26
Гастеромицеты 66
Гаустории 17
Гены патогенности 122
— устойчивости 122
Гидрохория 31
Гименомицеты 64
— агарикоидные 65
— афиллофороидные 64
Гиперплазия 105
Гипертрофия 105
Гипоплазия 105
Гифомицеты 105
Гифы 16
— одноклеточные 16
— многоклеточные 16
Гниль И, 15
Градобоины 173
Грибы 16
— базидиальные 58
— деревоокрашивающие 337
456
Грибы дереворазрушающие 304
— деструкторы 344
— домовые 356
— микоризообразователи 65
— митоспоровые 67
— мучнисторосяные 55
— настоящие 46
— несовершенные 67
— плесневые 342
— плодосумчатые 52
— полостносумчатые 58
— ржавчинные 59
— складские 349
— субдеструкторы 343
—- трутовые 64
— штабельные 343
д
Дегенерация 105
Деформация 13
Дискомицеты 57
Доза 153
— летальная 153
— пороговая 153
— сублетальная 153
Дусты 157
Ж
Желтизна 340
Желтуха 80
3
Заражение 96
Зигогамия 26
Зигомицеты 47
Зооспорангии 24
Зоохория 30
И
Иммунизация биологическая 126
— химическая 124
Иммунитет абсолютный 114
— активный 115
— врожденный 115
— неинфекционный 124
— неспецифический 114
— пассивный 115
— приобретенный 115
— специфический 114
Иммунитет инфекционный 124
— структурный 116
Индекс хемотерапевтический 156
Интоксикация растений 160
К
Камедетечение 106
Карантин растений 139
внешний 140
внутренний 140
Карликовость 107
Конвергенция симптомов 10
Конидиеносцы 25
Конидии 25
Конидиомы 25
Концентрация рабочих составов
154
Коремии 25
Краснина 340
Л
Ложа конидиальные 26
М
Мацерация 105
Метод диагностики болезней
макроскопический 131
микологический 135
микроскопический 133
патографический 131
физический 137
химический 137
Методы защиты леса 128
биологические 145
биофизические 147
лесохозяйственные 142
химические 149
Микоплазмы 82
Микросклероции 21
Мицелий 16
— несептированный 17
— септированный 17
Мозаика 79
Мумификация 12
Мучнистая роса 12
Н
Нагрузка рекреационная 185
Надзор 129
457
Надзор детальный 131
— лесопатологический 130
— общий 130
— рекогносцировочный 130
— специальный 130
Нарушения анатомо-морфологи-
ческие 104
— физиолого-биохимические 101
Некротрофы 34
Нематициды 151
Норма расхода пестицидов 154
О
Облитерация 106
Ожеледь 173
Ожог 13
Оидии 24
Оогамия 26
Оомицеты 45
Опрыскивание 159
— авиационное 159
— крупнокапельное 159
— малообъемное 159
— мелкокапельное 159
— многолитражное 159
— наземное 159
— среднекапельное 159
— ультрамалообъемное 159
Отлупы 177
П
Панфитотии 112
Паразиты 33
— облигатные 36
— раневые 36
— факультативные 35
— экологически облигатные 36
Парша 12
Патоген 9
Патогенность 93
Период инкубационный 99
Пестициды 150
— высокотоксичные 156
— малотоксичные 156
— сильнодействующие 156
— среднетоксичные 156
Пикниды 26
Пиреномицеты 54
Плазмодий 17
Плектенхима 21
Плектомицеты 54
Пленки мицелиальные 18
Плесень 13
Порошки смачивающиеся 157
Препараты гранулированные 157
Прогноз болезней 138
долгосрочный 138, 249, 250,
251
краткосрочный 138
многолетний 139
Псевдогрибы 45
Протравливание семян 160
Процесс инфекционный 96
Пряжки 17
Пятнистость 12
Р
Развитие болезни 248
Размножение грибов 23
бесполое 24
вегетативное 23
половое 26
Рак 11
Растворы 158
Растение-хозяин 9
Реакции антитоксические
защитные 121
Реакция сверхчувствительности 119
Ржавчина 11
Ризоиды 17
Ризоктонии 20
Ризоморфы 19
С
Сапротрофы 33
— облигатные 34
— факультативные 35
Септы 17
Симптомы болезней растений 9
Синдром 10
Системы лесозащитных
мероприятий 128
Склероции 20
Снеговал 173
Снеголом 173
Соматогамия 27
458
Специализация грибов 36
гистотропная 37
онтогенетическая 37
органотропная 37
филогенетическая 37
Спорангий 25
Споры пропагативные 28
Стромы мицелиальные 21
— склероциальные 21
Сумки 26
Суспензии 158
Т
Телеоморфы 28
Телиомицеты 59
Темнина кофейная 340
Типы болезней 15
— эпифитотий 111
Ткань грибов 21
механическая 21
образовательная 21
покровная 21
проводящая 21
Токсичность пестицидов 153
Трещины отлупные 177
Тяжи 19
У
Устойчивость растений 114
вертикальная 115
горизонтальная 115
групповая 115
Устомицеты 62
Ф
Фагоцитоз 120
Ферменты 94
Фитоалексины 120
Фитотоксичность 185
Фумигация 160
Фунгициды 151, 161
— защитные 161
— искореняющие 161
— контактные 165
— лечебные 161
— системные 165
X
Хламидоспоры 24
Хлороз 13
ц
Целомицеты 69
Цитоспороз 271
Ч
Чернь 13
Ш
Шнуры 19
Шютте 11
Э
Эндохория 30
Энтомохория 30
Энфитотии 111
Эпифитотий 108
— местные 111
— повсеместные 112
— прогрессирующие 112
— тардивные 112
— эксплозивные 112
Эпихория 30
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ БОЛЕЗНЕЙ РАСТЕНИЙ
И ФИТОПАТОГЕЕ
А
Альтернариоз гвоздики 377
Антракноз виолы 448
— желудей 194
Б
Бактериоз желудей 196
— сеянцев сосны 211
Бессемянность хризантемы 398
Бурсафеленхоз сосны 141
В
Вертициллез хризантемы 394
Ведьмины метлы березы 52
клена 52
ольхи 52
пихты 280
Вилт клена 262
Выжимание сеянцев 176
Выпревание сеянцев 218
Г
Гетероспориоз гвоздики 377
Глеоспориоз дуба 240
— липы 241
Гнили бактериальные плодов и
семян 196
— желудей 194
Гниль белая георгины 416
левкоя 441
пиона 427
полосатая дуба 327
— ядровая березы 331
— осины 331
— ядровая дуба 327
пихты 327
— желтая гиацинта 141
— корней астры 449
виолы 442
пиона 426
— корнеклубней георгины 416
460
ЫХ ОРГАНИЗМОВ
Гниль коричневая сердцевинная
гладиолуса 410
— красно-бурая призматическая
лиственницы 324
— ядровая дуба 329
— ядрово-заболонная березы 333
— мокрая ириса 422
— мягкая бактериальная гиацинта
407
— пенициллезная гиацинта 406
гладиолуса 412
тюльпана 403
— серая астры 436
— ириса 421
— розы 389
— тюльпана 400
— хризантемы 394
— склероциальная гиацинта 406
гладиолуса 412
— тюльпана 402
— сухая георгины 415
желудей 194
— твердая гладиолуса 411
— черная желудей 196
Голландская болезнь ильмовых
257
Головня виолы 447
Гриб домовый белый 359
настоящий 358
пластинчатый 361
пленчатый 360
— снежный 209
— столбовой 347
елово-пихтовый 347
— шахтный 361
— шпальный 346
Губка березовая 333
— дубовая 328
— еловая 172, 324
— корневая 172, 183, 310
— лиственничная 324
д
Деформация листьев березы 244
вяза 244
Деформация листьев клена 245
ольхи 245
тополя 245
— плодов косточковых 193
ольхи 193
Ж
Желтуха астры 439
— флокса 433
— хризантемы 398
3
Заразиха 92
К
Карликовость хризантемы 397
Курчавость листьев пиона 430
флокса 432
Крапчатость пожилковая гвоздики
379
— хризантемы 398
Круменулез хвойных 222
М
Микоз сосудистый дуба 260
Мозаика гвоздики 379
— георгины 419
— гиацинта 408
— гладиолуса 414
— ириса 422
— кольцевая виолы 450
— крапчатая ириса 422
— левкоя 444
— мальвы 452
— пиона 430
— розы 392
Мумификация желудей 189
— семян березы 188
Мучнистая роса березы 234
боярышника 235
бузины 234
виолы 448
вяза 235
георгины 418
дуба 232
Мучнистая роса ивы 235
клена 235
крушины 234
лещины 234
осины 235
пиона 427
розы 381
рябины 235
сальвии 445
тополя 235
флокса 430
хризантемы 395
ясеня 234
ложная виолы 450
левкоя 442
розы 383
сальвии 444
Н
Некроз бурый цитоспоровый
тополя 271
— виллеминиевый дуба 262, 271
— гистерографиевый ясеня 275
— диатриповый дуба 269
— дискоспориевый тополя 274
— дотихициевый тополя 274
— жилок тюльпана 405
— клитрисовый дуба 262, 268
— колпомовый дуба 268
— кольцевой левкоя 444
— нектриевый 266
— нуммуляриевый дуба 270
— ценангиевый сосны 265
— черный немоспоровый 269
— цитоспоровый тополя 262, 273
Нематода галловая 423
левкоя 444
— земляничная ириса 423
— клеверная цистообразующая 380
— проникающая короткотелая 393
— ранящая короткотелая 393, 423
— северная галловая 433
— стеблевая 380, 433
картофеля 423
— хризантемная 398
Нематоды галловые гвоздики 380
мальвы 452
розы 392
461
о
Ожог бактериальный груши 75, 141
плодовых 75
сирени 75
шелковицы 75
— всходов 177
— инфекционный розы 386
Омела 89
— белая 89
— пихтовая 89
— можжевеловая 90
Опенок осенний 172, 262, 317
П
Парша гладиолуса 413
— ивы 243
— осины 243
— тополя 184, 242
Пениофора гигантская 146, 344
Песталоциоз розы 391
Пестролепестность виолы 450
— тюльпана 404
Пестростебельность гиацинта 408
Петров крест 91
Плесень головчатая 198
— зеленая 197
— розовая 198
— серая 198, 221
— черная 198
Побурение хвои ели 217
пихты 217
Полегание всходов 201
Пятнистость бактериальная
хризантемы 397
— белая тополя 238
флокса 431
бурая березы 240
георгины 418
дуба 240
тополя 184, 239
хризантемы 396
— ириса 421
— кольцевая астры 440
гвоздики 379
пиона 429
сальвии 446
хризантемы 397
— коричневая пиона 428
Пятнистость кремовая липы 241
— некротическая тюльпана 389
— пурпуровая розы 389
— серая осины 241
— темно-бурая липы 241
— черная березы 238
вяза 238
ивы 238
клена 236
розы 387
точечная клена 238
Р
Рак бактериальный ясеня 297
— сосны биаторелловый 173, 283
бугорчатый 285
— корневой сальвии 444
— крифонекриевый каштана
съедобного 290
— мокрый язвенно-сосудистый
тополя 295
— побеговый хвойных 184, 222
— поперечный дуба 296
— ржавчинный пихты 172, 280
— сосны смоляной 173, 276
ржавчинный 279
язвенный 284
ступенчатый лиственницы 282
— черный осины, тополя 172, 288
— эндоксилиновый ясеня 172, 287
Ржавчина листьев астры 437
хризантемы 141, 396
березы 231
осины 231
сальвии 446
тополя 184, 231
— виолы 447
— гвоздики садовой 370
— гвоздики турецкой 451
— гиацинта 407
— мальвы 452
— розы 383
— побегов ели 230
сосны 225
— хвои ели 141, 228, 230
лиственницы 231
пихты 280
сосны 228
462
Ржавчина шишек ели 191, 192
Ризина волнистая 321
Ризоктониоз гвоздики 375
— сальвии 446
С
Септориоз астры 438
— виолы 448
— гвоздики 378
— пиона 429
— розы 389
— сальвии 445
— тополя 238
— флокса 431
Склеротиниоз левкоя 441
Склеродерриоз хвойных 222
Склерофомоз сосны 220
Стеганоспориоз липы 292
— вяза 292
Стереум кровяно-красный 346
Стигминиоз липы 292
Т
Тиростромоз липы, вяза 292
Трутовик березовый пластинчатый
350
— буро-фиолетовый 346
— валежный еловый 345
— волосистый 349
— Гартига 281, 327
— групповой 348
— дубовый 328
— дуболюбивый 328
— душистый 348
— зональный 349
— кленовый 298, 331
— комлевой еловый 172, 325
— ложный 172, 298, 330
дубовый 183, 297, 327
— многоцветный 348
— настоящий 172, 332
— окаймленный 146, 172, 326
— опаленный 348
— осиновый 331
— плоский 172, 298, 321
— разноцветный 348
— розовый 346
— серно-желтый 172, 329
Трутовик серый 348
— утолщенный 349
— чешуйчатый 172, 298, 329
— Швейница 320
У
Увядание вертициллезное
георгины 416
пиона 427
флокса 431
— розы 392
— фузариозное астры 434
георгины 415
Удушье сеянцев 222
Усыхание вертициллезное клена
462
— инфекционное липы 292
вяза 294
Ф
Феллинус точечный 172
Фиалофороз гвоздики 374
Фузариоз гвоздики 372
— гладиолуса 408
— левкоя 443
— сеянцев 202
— тюльпана 403
— хризантемы 394
X
Хлороз пожилковый сальвии 446
— хризантемы 399
ц
Ценангиоз сосны 265
Церкоспороз липы 241
— мальвии 452
— розы 389
— сальвии 445
Цитоспороз желудей 194
Ч
Чага 331
Черная ножка астры 438
виолы 449
георгины 414
левкоя 440
хризантемы 393
463
Чернь листьев 245
розы 392
Чешуйчатка жирная 281, 326
Ш
Шютте ели 184, 214, 215
— лиственницы 213
— можжевельника 216
Шютте сосны обыкновенное 206
бурое 216
серое 212
снежное 184, 209
— снежное ели 212
щ
Щелелистник обыкновенный 345
УКАЗАТЕЛЬ НАИМЕНОВАНИИ ХИМИЧЕСКИХ
И БИОЛОГИЧЕСКИХ СРЕДСТВ ЗАЩИТЫ РАСТЕНИЙ
И ДРЕВЕСИНЫ
Абига-Пик 163, 254, 372, 385, 432,
438, 448
Арцерид 255, 385
Байлетон 164, 254, 372, 383, 418,
425, 428, 431, 438, 450
Бактофит 372, 376, 428, 431, 448
ББК-3 355, 366
Беномил 402, 411, 443
Бромуконазол 428
Вектра 383
Гексабутилдистанноксан 366
ГР-48 355, 366
Дерозаль 431, 448
Дитан М-45 402, 427, 437, 442,
443
Дифенил 366
Калий марганцовокислый 427
Каптан 366
Карбафен-16 355, 366
Карболинеум 334, 366, 367
Картоцид 163, 253, 385, 425, 441,
443
Кемикар 162, 253
Кремнефторид аммония 366
— натрия 366
Креозот 355, 366
Кумулус 163, 255, 418, 428
Купорос железный 385, 431
— медный 162, 303, 334, 364, 385,
387, 426, 431
Купроксат 425
Купроцин 366
Лак каменноугольный 366
Масла каменноугольные
пропиточные 366
— сланцевые пропиточные 366
МХХЦ 366
Нафтенат меди 355
Нипагин-М 366
2-оксидифенил 366
Оксидифенолят натрия 366
Оксихлорид меди 163, 254, 372,
383, 385, 387, 432, 437, 438,
448
Оксофин 366
Паста битумная 366
— глинистая 366
— силикатная 366
— экстрактная 366
ПБТ 355, 366
Пенконазол 383, 385
Пентахлорфениллаурат 366
Пентахлорфенол 355, 366
Пентахлорфенолят натрия 355,
366
Привент 164, 254
Процимидон 402
П-2Т 366
ПФК-У-12 355, 366, 367
Ровраль 395, 402, 409, 411, 437
Салициланилид 366
Сапроль402, 411
Сера 200
— коллоидная 163, 255, 383, 418,
428, 431, 445
Смесь бордоская 162, 254, 383, 387,
432, 437, 448
Сумилекс 402, 411, 441
Сульфат меди 355, 366
Тачигарен 402
Тиабендазол 409, 411
Тиовит 383
ТМТД 164, 200, 253, 376, 394, 402,
403, 409, 411, 413, 417, 425, 426,
427, 441, 442, 443, 449, 450
Топаз 437, 448, 450
Топсин-М 163, 200, 253, 254, 387,
394, 402, 403, 409, 411, 413, 417,
442, 449, 450
Трилан 366
Триолит 366
Трифорин 383
465
Трихотецин 418, 448 Фундазол 162, 200, 253, 254, 376,
Уралит 366 383, 387, 390, 394, 402, 403, 409,
ФДХ 366 2 411, 413, 425, 427, 437, 438, 441,
ФН-П 366 442, 449, 450
Формалин 200, 364 ФХМ 366
Фталан 366 ХМ-5 366
Фторид натрия 364, 366 ХХЦ 366
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
А
Acervulales 26, 69, 194
Agaricales 65
Agonomycetes 43, 67, 69
Agrobacterium 74
— tumefaciens (Sm. et Towns.) Conn.
74, 76, 419, 444
Albuginaceae 45
Albugo Candida Lev. 443
Alternaria 68, 198, 202, 204, 342
— dianthi Stev. et Hall. 377, 451
— florigena Nelen 438
— tenuis Nees. 197, 198
Anisomyces odoratus (Fr.) Sing. 348
Aphelenchoides fragariae (Ritz. Bos.)
Christ. 420, 423
— ritzemabosi (Schw.) Stein. 398
— subtenuis (Cobb) St. et Buhr. 405
Arceuthobium oxycedri M. B. 90
Archaeascomycetes 43, 49, 50
Armillaria mellea (Vahl. ex Fr.) Karst. 317
Ascocalyx abietis Naum. 225
Ascochyta malvicola Sacc. 452
— violae Sacc. et Speg. 448
Ascomycotina 43, 49
Asexual fungi 67
Aspergillus 54, 342
— glaucus Link. 197
— niger Link. 198
Aster ringspot virus 440
Auriculariales 63
В
Basidiomycetes 43, 58, 63
Basidiomycotina 43, 49, 58
Beauveria 68
Biatorella difformis (Fries) Rehm. 173,
283
Biatorella resinae Mudd. 340
Biatoridina pinastri Golow. et Schzedr.
283
Hjcrkandera adusta Karst. 348
Bolctales 65
Rotryosporium pulchrum Corda. 416
Botrytis 68, 98, 202, 342
— cincrea Pers. 197, 204, 221, 389,
394, 417, 436, 442
— hyacinthi West, et Beyma. 407
— gladiolorum Timm. 410
— paeoniae Oud. 424
— tulipae (Lib.) Lind. 400
Bovista 66
Brunchorstia destruens Er. 222
— pinca (Karst.) Нцпп. 222
Bursaphelenchus xylophilus Stein, ct
Buhr. 87, 141
С
Carnation mosaic virus 379
— ringspot virus 379
— wcin mottle virus 379
Cenangium abietis (Pers.) Rehm. 265
— fcrruginosum Fr. 265
Cephalosporium 257, 260
Ccratocystis 57, 140, 339
— cocruleum (Munch.) H. et Syd. 57,
339
— kubanicum (Scz.-Par.) Potl. 260
— piceae (Munch.) H. et Syd. 57, 339
— pini (Munch.) H. et Syd. 339
— roboris (Georg. et I. Teod.) Potl. 260
— ulmi (Buism.) Moreau 257
— valachicum С Georgescu 260
Cercospora 448
— althaeina Sacc. 452
— microsora Sacc. 237, 241
— omphacodes Ell. et Holw. 432
— rosicola Pass. 389
— salicicola Tharp. 445
Ceuthospora betulae (Fuck) v. Arx 237,
240
Chlorosplenium 341
467
Chromista 16, 42, 43, 45
Chrysanthemum aspermy virus 398
— disease virus 397
— ringspot virus 397
— wein mottle virus 398
Chrysomyxa abietis (Wallr.) Unger
228, 230
— arctostaphyli Dietel 141
— ledi D.B. 228, 229
— pirolae Rostr. 192
— woronini Tranz. 230, 231
Cladosporium 198, 342
— herbarum Link. 197, 198
— paeoniae Pass. 428
Clavariales 64
Claviceps 55
Clavicipitaceae 55
Clithris quercina (Pers.) Rehm. 57, 268
Coelomycetes 43, 67, 69
Coleosporium 228
— campanulae Pers. 228
— melampyri (Reb.) Tul. 228
— senecionis Fr. 228
— solidaginis Thumen. 437
— sonchiarvensis Lev. 228
— tussilaginis Lev. 228
Colletotrichum malvacearum Sacc. 452
— violae-tricoloris 448
Colpoma quercinum (Pers.) Wallr. 268
Conidial fungi 67
Coniophora cerebella (Fr.) Karst. 360
Coniothyrium 386
— fuckelii Sacc. 386
— rosarum Sacc. 386
— wernsdorffiae Laub. 386
Cordyceps militaris (Fr.) Link. 55
Coriolellus serialis (Fr.) Murr. 348
— vaporarins (Fr.) Bond, et Sing. 359
Coriolus hirsutus (Wulf.) Quel. 349
— versicolor (L.) Quel. 348, 349
— zonatus (Fr.) Quel. 349
Corticium laeve Fr. 340
Cronartium flaccidum Wint. 173, 276,
277
— ribicola Dietr. 146, 279
— sp. 141
Crucibulum 66
Crucibulum laeve (D. C.) Kambly 66
Cryphonectria parasitica (Murrill.) Barr.
140, 290
Cucumis mosaic virus 450
Cuscuta 92
— europaea L. 92, 140
— monogyna Vahl. 92, 140
Cuscutaceae 89
Cyathus 66
Cylindrocarpon willkommii (Lind.) Wr.
285
Cylindrosporium chrysanthemi Stev. 396
Cytospora69, 271
— capitata Schulz. 274
— carphosperma Fr. 274
— chrysosperma (Pers.) Fr. 56, 271
— foetidaVl.etRr.273
— intermedia Sacc. 194, 195
— leucosperma Fr. 274
— pruinosa (Fr.) Defago 274
— pulchella Sacc. 274
— rubescens Fr. 274
— schulzeri Sacc. et Syd. 274
D
Dacrymycetales 63
Daedalea 64
— quercina (L.) Fr. 328
Dahlia mosaic virus 419
Daldinia concentrica de Not 56
Dasyscypha willkommii Hartig. 57, 282
Dematium 245
Deuteromycetes 67
Diaporthales 56
Diatrypales 56
Diathrype stigma (Hoffm.) Wint. 56,
269
Dicaryomycota 43, 47, 49
Dimaigaritales 49
Diplocarpon rosae Wolf. 387
Discosporium populeum (Sacc.)
Sutton. 274
Discula brunneo-tingens Meyer 340
— umbrinella (Berk, ex Broome) Sutton.
240, 241
Ditylenchus 414
— destructor Th. 420, 423
Ditylenchus dipsaci (Kiihn.) Filip.
380, 405, 433, 451
468
Dolichodorus 86
Dorylaimida 84, 87
Dothichiza ferruginosa Sacc. 265
— populea Sacc. 274
Dothideales 58
Dothidellabetulina (Fr.) Sacc. 58, 238
— ulmi Wint. 58,238
E
Eidamia catenulata Will. 341
Endocronartium sp. 141
Endogonales 49
Endothia parasitica (Murr.) P. And. et
H. And. 56, 290
Endoxylina stellulata Rom. 172, 287
Entomophaga aulicae Batko 49
Entomophthorales 48
Erysiphales 55
Erysiphe cichoracearum D.C. 418
— f. phlogis Jacz. 430
— f. violarum (Died.) Jacz. 448
— labiatarum Chev. f. salviae Jacz. 445
Erwinia 74, 75
— amylovora (Burill.) Corn 75, 141
— aroidea (Town.) Holl. 422
— carotovora (Jones.) Wald. 106, 407,
422
— quercina Held, et Schr. 196
— salicis (Day) Chest. 76
Euascomycetes 43, 49, 52
Eubacteria 74
Eubacteriales 74
Euphrasia officinalis L. 89
Eurotiales 54
Exobasidiales 62
Exobasidium 62
— vaccinii Woron. 63
F
Fomes fomentarius (L.) Gill. 432
Fomitopsis 64
— annosa Karst. 310
— officinalis (Will.) Bond, et Sing. 324
— pinicola (Schwartz, et Fr.) P. Karst.
146, 326
— rosea Karst. 346
Fuligo 44
Fumago 245
Fungi 43, 46
— imperfecti 34, 43, 68, 87
Fusarium 135, 140, 198, 202, 260,
340, 342, 372, 407, 414, 426, 434,
446
— conglutinans Wr. f. callistephi Beach.
434
— culmorum (W. G. Sm.) Sacc. 407
— javanicum Koord. 260
— oxysporum Schl. 394, 407, 443
— f. callistephi Wr. 434
— f. dianthi (Pril. et Del.) Bilai. 372
— f. gladioli (Mass.) Sm. et Hans. 408
— f. tulipae. 403
— solani (Mart.) App. et Wr. 407
— sp. 438
— sporotrichiella Bilai var. poae (Pk.)
Wr. 260
Fusicladium radiosum (Lib.) Lind. 242
— saliciperdum Lind. 243
G
Ganoderma applanatum (Pers.) Pat.
298, 321
Ganodermatales 64
Geastrum 66
Gladiolus mosaic virus 414
Gloeosporium betulinum West. 240
— quercinum West. 69, 194, 240
— tiliae Oudem. 241
— tremulae Pass. 241
Gloeophyllum 64
— abietinum (Butt.) Karst. 347
— sepiarium (Wulf.) Karst. 347
Gnomonia quercina Kleb. 240
Graphium roboris Schw. 260
— ulmi Schwarz. 257
Gremmeniella abietina (Lagerb.)
Morelet. 222
Grijs mosaic virus 408
Gymnosporangium 62, 105
H
Helotiales 57
Hemiascomycetes 49, 50, 52
Hendersonia acicola Tub. 212
Hericiales 64
Hericium diversidens (Fr.) Pers. 341
469
Herpotrichia juniperi (Duby.) Petr. 216
— nigra Hart. 216
Heterobasidiomycetidae 63
Heterobasidion annosum (Fr.) Bref.
183, 310
Heterodera 84, 86
— trifolii Goff. 380, 451
Heterosporium echinulatum (Berk.)
Cke. 377,451
— gracile Sacc. 421
Hirschioporus abietinus (Fr.) Donk
345
Homobasidiomycetidae 63, 64
Hyalodendron 257, 260
Hymenochaetales 64
Hyphomycetes 43, 67, 68
Hypocrea pulvinata Fuck. 55
Hypocreaceae 55
Hypocreales 55
Hypodermella sulcigena (Rostr.) Tub.
212
Hypoxylon mammatum (Wahlenbeig)
Miller 288
— pruinatum (Kl.) Cooke. 56, 172,
288
Hysteriales 58
Hysterographium fraxini (Pers.) de Not
58, 275
I
Inonotus 64
Inonotus dryophilus (Pers.) Murr.
297, 328
— obliquus (Pers.) Pil. 331
— weirii (Mur.) Kotl. et Pouzar 141
Iris mosaic virus 422
— virus 1 422
К
Kickxellales 49
L
Lachnellula pini (Brunch.) Dennis 284
— willkommii (R. Hartig) Dennis 282
Laetiporus sulphureus (Bull, et Fr.)
Bond, et Sing. 329
Lathraea squamaria L. 91
Lentinus lepideus Fr. 346
Lenzites betulina (L. ex Fr.) Fr. 350
Leotiales 57, 188
Lirula macrospora (R. Hart.) Dark. 214
Loculoascomycetes 43, 49, 58
Longidorus 86
Lophodermella sulcigena (Rostr.) Hoh.
212
Lophodermium 37
— juniperinum (Fr.) de Not. 216
— macrosporum (Hart.) Rehm. 184,
214, 215
— piceae (Fuck.) Hohn. 214, 215
— pinastri Chev. 38, 57, 206, 207,
208, 209
— seditiosum 38, 57, 206, 207, 208
Loranthaceae 89, 90
Lycoperdales 66
Lycoperdon 66
M
Malve mosaic virus 452
Marssonina betulae (Lib.) Vagn. 240
— juglandis (Lib.) P. Magn. 69
— populi (Lib.) Sacc. 69, 184, 237,
239
— rosae (Lib.) Diob. 387
Melampsora allii-populina ЮеЬ. 184,
231
— larici-populina ЮеЬ. 231
— larici-tremulae ЮеЬ. 231
— pinitorqua (A. Br.) Rostr. 38, 62,
143, 225, 231
Melampsoraceae 61, 62
Melampsorella caryophyllacearum I.
Schrot. 172, 280
— cerastii Wint. 280
Melampsoridium betulae Arth. 143,
231
— betulinum Kleb. 231
Melampyrum nemorosum L. 89
Melanconiales 69
Meliola penzigi Sacc. 392
Meloidogyne 86, 380, 392, 414, 451
— arenaria (Neal.) Chitw. 380, 444
— javanica (Trenb.) Chit. 380
Meloidogyne hapla Chit 380, 433
— marioni Cornu 85
-sp.423
470
Meria laricis Vuill. 68, 213
Microascales 56
Microsphaera alphitoides Griffon
Maubl. et 38, 55, 96, 232
— betulae Magn. 234
— coryli (Jacz.) Golow. 234
— divaricata (Wallr.) Lev. 234
— lonicerae Wint. 55
— van-bruntiana Gerard. 234
Mitosporis fungi 43, 47, 67
Morchella 58
Mucor 47, 48, 198, 342
Mucorales 47
Mycelia sterilia 69
Mychota71
Mycosphaerella microsora Syd. 242
— populorum G. E. Tompson 141
Mycota 16, 42, 46
Myxomycetes 43, 44
Myxomycota 43, 44
N
Naemospora croceola Sacc. 56, 269
Nectria 55
— cinnabarina (Tode: Fr.) Fr. 55, 266
— ditissimaTul. 287
— galligena Bres. 55, 285
Nemathelminthes 83
Nematoda 83
Nidularia 66
Nidulariales 66
Nummularia bulliardii Tul. 56, 270
О
Oidium chrysanthemi Rab. 395
— leucoconium Desm. 381
— violae Sacc. 448
Olpidium brassicae Woron. 404, 438,
440, 449
Onnia triqueter Bres. 325
Oomycetes 43, 45
Oomycota 43, 45
Ophiostoma 56
— roboris G. et Teod. 56, 196
— ulmi (Buism.) Мог. 56, 95, 140,
257
— valachicum G. et Teod. 196
Ophiostomatales 56
Orchidaceae 89
Orobanchaceae 89
Orobanche 92
Osmoporus odoratus (Fr.) Sing. 348
Oxyporus populinus (Fr.) Donk 331
P
Paeonia ringspot virus 429
Paralongidorus maxumus (Butsch.)
Syd. 86
Paratylenchus 86
— projectus Jen. 381
Paxillus panuoides Fr. 361
Pedicularis palustris L. 89
Penicillium 54, 68, 197, 340, 341,
342, 403
— corymbiferum West. 403
— cyclopium West. 406
— gladioli Mc Cull et Thorn. 412
Peniophora gigantea (Fr.) Mass. 146,
344
— sanguinea (Fr.) Bres. 340, 341
Peridermium pini (Willd.) Lev. et Kleb.
38, 62, 276
Peronospora mattiolae Gaum. 442
— schachtii Fuck. 395
— sparsa Berk. 383
— Swinglei Ell. et Kell. 444
— violae de By. 450
Peronosporaceae 45
Peronosporales 45
Pestalotia adusta Ell. et Ev. 391
Pezizales 58
Phacidiaceae 57
Phacidium infestans Karst. 38, 57,
184, 209, 212
Phaeolus schweinitzii (Fr.) Pat. 320
Phellinus 64
—- chrysoloma (Fr.) Donk. 324
— igniarius (L. et Fr.) Quel. 298, 330
— pini (Thore ex Fr.) Pil. 323
— pini (Thore ex Fr.) Pil. var. abietis
Karst. 324
— punctatus (Fr.) Pil. 298
— robustus (Karst.) Burd. et Galz. 183,
297, 327
— tremulae (Bond.) Bond, et Bor. 331
— weirii (Murill.) Gibb. 141
Phlox necrosis vien virus 432
471
Pholiota adiposa Fr. 326
Phoma 69
— phlogis (Roum.) Speg. 431
Phomopsis 69
— quercella (Sacc.) Died. 194
Phragmidium 383
— distiflorum (Tode) James 62, 383
— rosae-pimpinellifoliae 383
— tuberculatum Mull. 383
Phyalophora cinerescens (Wr.) van
Beyma 374
Phyllactinia guttata (Wallr. et Fr.) Lev.
234
— suffulta Sacc. 37, 55, 95, 234
f. betulae 37
f. carpini 37
f. coryli 37
f. fraxini 37
Phyllosticta 69
— dahliecola Brun. 418
— decussata Syd. 432
— rosarum Pass. 389
— solitaria Ellis, ex Everh. 141
— tricoloris Sacc. 448
Physarum 44
Phytophthora 45
— cactorum (Lebert. et Kohn.) Schrot.
45, 205
— cryptogea Peth. et Laff. 449
— infestans D.B. 45
Phytophthoraceae 45
Piptoporus betulinus (Bull, ex Fr.) Karst.
333
Plasmodiophora brassicae Woron 37,
44, 440
Plasmodiophoromycetes 43, 44
Plasmodiophoromycota 43, 44
Pleosphaerulina violae P. Magorn. 448
Pleosporales 58
Podosphaera clandestina (Wallr. et Fr.)
Lev. 235
— tridactyla (Wallr.) de Bary 235
Pollaccia elegans Serv. 243
— radiosa (Lib.) Bald, et Cif. 184, 242
— saliciperda (Allesch. ex Tub.) Aix. 243
Polyporales 65
Polyporus squamosus Huds. ex Fr. 298,
329
Poriales 64
Pratylenchus 86
— neglectus (Rensch.) Filip. et Sch.
St. 381
— penetrans (Cobb.) Filip. et Sch.
Stekh. 381, 393, 423, 446
— vulrus All. et Jen 393
Procariota71
Protoctista 42, 44
Protozoa 16, 42, 43, 44
Pseudomonas 74
— cerasi Griff. 295
— fluorescens Migula 140, 221
— fraxini Vuill. 76, 297
— iridis Holl. 422
— marginata (Mc Cull.) Stapp. 413
— mori (B. et L.) St. 75
— quercina Schem. 76, 296
— pin Vuill. 76, 285
— remifaciens Kon. 76, 295, 296
— savastanoi 76
— syringae van Hall. 75, 297, 397
— syringae van Hall. f. populi Sabet. et
Dowson 295, 297
Puccinia 62
— arenariae (Schum.) Wint. 451
— chrysanthemi Roze 396
— horiana Henn. 141
— iridis Wint. 420
— malvacearum Mont. 452
— nigrescens Kirch. 446
— violae (Schum.) DC 447
Pucciniaceae 61, 62
Pycnidiales 69
Pythiales 45, 46
Pythium debaryanum Hesse 46, 202,
393, 414, 438, 440, 449
R
Ramularia 448
— salviae Pass. 445
Rhinanthus major Ehrb. 89
Rhinotrichum 257, 260
Rhitisma acerinum (Pers.) Fr. 37, 57,
102, 236, 237
— punctatum Rehm. 238
— salicinum Rehm. 237, 238
Rhizina inflata (Schaeff.) Rehm. 321
472
Rhizina undulata Fr. 58, 321
Rhizoctonia 70, 426, 449
— aderholdii (Ruhl.) Kolosh. 438,
440, 448
— solani Kuhn. 70, 375, 376, 393,
414, 436, 446
Rhizopus nigricans Ehr. 47, 48, 197,
198
Rhizosphaeria kalkhoffii Bub. 217
— pini (Corda) Maubl. 217, 218
Rosae mosaic virus 392
— wilt virus 392
Rosellinia quercina Hart. 96, 205
Rotylenchus 86
Russulales 65
S
Saccharomyces cerevisae 52
Saccharomycetaceae 52
Saccharomycetales 52
Salvia virus 446
Saprolegniales 45, 46
Sawadaia bicomis (Wallr.: Fr.) Miyabe
235
— tulasnei (Fuck.) Homma 235
Schizophyllum commune Fr. 345
Scleroderris lagerbeigii Gremm. 140,
184, 222
Sclerophoma pithya v. Hohnk. 220
Sclerotinia 57, 426, 449
— betulae Woron. 57, 188
— bulborum (Wakk.) Rehm. 402, 406
— gladioli (Mass.) Dray 412
— graminearum Elen. 37, 57, 218,
219, 220
— mattiolae Beiss. 441
— sclerotiorum Fuck. 57, 416, 427
Sclerotiniaceae 57
Sclerotium 70
— tuliparum Kleb. 402
Scrophulariaceae 89
Septoria callistephi Gloyer 438
— chrysanthemella Sacc. 396
— dianthi Desm. 378
— gladioli Pass. 411
— macrospora Sacc. 429
— paeoniae West. 429
— phlogis Sacc. et Speg. 431
Septoria populi Desm. 38, 69, 237, 238
— rosae Desm. 389
— salviae Pass. 445
— violae West. 448
Serpula lacrimans (Wulf.) Gray. 358
Sistotrema brinkmannii (Bres.) I.
Eriksson 141
Sphaceloma rosarum (Pass.) Jenk. 389
Sphaeriales 56
Sphaeropsidales 69
Sphaerotheca fuliginea Poll. f. dahliae
Movs. 418
— fuliginea Poll. f. paeoniae Jacz. 427
— pannosa Lev. 55
— pannosa Lev. var. rosae Woronich. 381
Spumaria 44
Steganosporium compactum Sacc. 292
Stemonitis 44
Stereum sanguinolentum Fr. 346
Stigmina compacta (Sacc.) M. B. Ellis
292
Stock mosaic virus 444
Stromatinia 57
— aucupariae Woron. 191
— crataegi Magn. 191
— pseudotuberosa Rehm. 20, 57, 189,
190, 191
T
Taphrina 50, 192, 244
— acerina Elias 52
— aini-incanae (Kuhn) Magn.) 50,
193
— aurea (Pers.) Fr. 50, 245
— betulina Rostr. 52, 244
— carnea Iohans. 244
— epiphylla Sacc. 52
— polyspora (Svr6ek) Iohans. 245
— pruni Fuck. 50, 193
— tosquinettii Magn. 245
— turgida Giesh. 52
— ulmi Joh. 244
Taphrinales 50
Tarichium 49
Teliomycetes 43, 58, 59
Thamnidium elegans Link. 47, 48, 198
Thekopsora padi (Kze. et Schm.) Kleb.
62, 191, 192
Thelephora terrestris Ehr. 222
Thelephorales 64
Thyrostroma compactum Sacc. 292
Tobacco necrosis virus 404
— rattle virus 408
— ringspot virus 405, 450
Trechispora brinkmannii (Bres.) Rogers
et Jacke 141
Tremellales 63
Trichoderma 146, 341
— lignorum Harz. 68
Trichodorus 86, 408
Trichothecium roseum Link. 68, 197,
198
Tuber 58
Tuberales 58
Tubercularia vulgaris Tode 266
Tulipa virus 404
Tylenchida 84
Tylenchorhynchus 86
— dubius (Biitsch.) Filip. 381
Typhula graminearum Tul. 37,218, 219
U
Uncinula aceris Sacc. 55, 235
— adunca (Wallr. et Fr.) Lev. 235
— bicornis (Wallr. et Fr.) Lev. 235
— clandestina Schrot. 235
— salicis (D. C.) Wint. 55, 235
— tulasnei Fuck. 235
Uredinales 59
Urocystis kmetiana Magn. 447
— violae (Sow.) Fisch. v. Waldh. 447
Uromyces caryophyllinus (Schrank)
Wint. 62, 370
Uromyces scillarum (Grev.) Wint. 407
Ustilaginales 62
Ustomycetes 43, 58, 62
V
Valsa sordida Nits. 56, 272
Venturia 38, 58
— populina Vuill. 243
— tremulae Aderh. 242
Verpa 58
Verticillium 202, 204, 416
— dahliae Kleb. 262, 394, 416, 427
Verticillium glaucum Bon. 340
— lateritium Berk. 416
— sp. 260, 431
Viscum abietis (Beck.) Stank. 89
— album L. 89
Vuilleminia comedens Maire 271
X
Xanthomonas 74
— campestris Dows. 141, 443
— heterocea (Wsor.) Gorl. 75
Xanthomonas juglandis Piers. 75
— pruni (Sw.) Dows. 75
Xiphinema 86
— americanum Cobb. 85, 393
— sp. 405
Xylariales 56
Z
Zoopagales 49
Zoophthora 49
Zygomycetes 43, 47
Zygomycota 43, 47
ОГЛАВЛЕНИЕ
Предисловие 3
Введение 4
Глава 1. Общие сведения о болезнях растений 8
1.1. Понятие о болезнях растений и их причинах 8
1.2. Симптомы болезней растений 9
1.3. Типы болезней растений 10
1.4. Ущерб, причиняемый болезнями растений
и дереворазрушающими грибами 14
Глава 2. Грибы и грибоподобные организмы — возбудители
болезней растений 16
2.1. Вегетативное тело грибов и его видоизменения 16
2.2. Строение и химический состав клетки 21
2.3. Размножение грибов 23
2.4. Распространение спор грибов 29
2.5. Питание, паразитизм и специализация грибов 32
2.6. Требования грибов к условиям окружающей среды 39
2.7. Систематика грибов 42
2.7.1. Царство Protozoa, или Protoctista 44
2.7.2. Царство Chromista (Псевдогрибы) 45
2.7.3. Царство Mycota, Fungi (Настоящие грибы) 46
Глава 3. Бактерии, вирусы, нематоды и цветковые паразиты —
возбудители болезней растений 71
3.1. Фитопатогенные бактерии , 71
3.1.1. Общие сведения о бактериях 71
3.1.2. Типы бактериальных болезней растений 74
3.1.3. Источники инфекции и пути распространения
фитопатогенных бактерий 76
3.2. Фитопатогенные вирусы 77
3.2.1. Общие сведения о вирусах 77
3.2.2. Типы вирусных болезней растений 79
3.2.3. Сохранение и распространение вирусов
в природе 80
3.3. Микоплазмы 82
3.4. Фитопатогенные нематоды 83
3.4.1. Общие сведения о фитонематодах 83
3.4.2. Симптомы и типы нематодных болезней растений 85
3.5. Паразитические цветковые растения 88
3.5.1. Общие сведения о цветковых растениях-паразитах 88
3.5.2. Корневые полупаразиты 89
3.5.3. Стволовые полупаразиты 89
3.5.4. Корневые паразиты 91
3.5.5. Стеблевые паразиты 92
Глава 4. Патогенез и динамика инфекционных болезней растений 93
4.1. Свойства патогенов 93
4.2. Инфекционный процесс 96
4.3. Патологические изменения больного растения 101
4.3.1. Физиолого-биохимические нарушения 101
4.3.2. Анатомо-морфологические нарушения 104
4.4. Понятие об эпифитотиях 108
4.4.1. Роль патогена, растения-хозяина и окружающей
среды в развитии эпифитотий 108
4.4.2. Динамика эпифитотий 110
4.4.3. Типы эпифитотий 111
Глава 5. Иммунитет растений к инфекционным болезням 114
5.1. Основные понятия и категории иммунитета 114
5.2. Пассивный врожденный иммунитет 115
5.2.1. Анатомо-морфологические факторы пассивного
иммунитета 115
5.2.2. Физиолого-биохимические факторы пассивного
иммунитета 117
5.3. Активный врожденный иммунитет 119
5.4. Генетические основы иммунитета растений 122
5.5. Выносливость растений 123
5.6. Приобретенный иммунитет и пути повышения
устойчивости растений к болезням 124
5.6.1. Химическая иммунизация растений 124
5.6.2. Биологическая иммунизация (вакцинация)
растений 126
Глава 6. Методы борьбы с болезнями леса 128
6.1. Общие сведения 128
6.2. Надзор за появлением болезней и вредителей 129
6.3. Методы диагностики болезней древесных растений 131
6.4. Прогноз развития болезней 138
6.5. Карантин растений 139
6.6. Лесохозяйственные методы 142
6.7. Биологический метод 145
6.8. Биофизический и механические методы 147
6.9. Химический метод 149
6.9.1. Общие сведения 149
6.9.2. Классификация химических средств защиты
растений 150
Глава 7. Химические средства борьбы с болезнями леса 153
7.1. Основы токсикологии 153
7.2. Действие пестицидов на живые организмы и окружающую
среду 155
476
7.3. Препаративные формы пестицидов 157
7.4. Способы применения пестицидов 158
7.5. Фунгициды 161
7.6. Правила техники безопасности при работе
с пестицидами 165
Глава 8. Неинфекционные болезни древесных пород 167
8.1. Отрицательное влияние почвенных условий 167
8.2. Отрицательное влияние метеорологических
условий 171
8.3. Отрицательное влияние антропогенных
факторов 178
Глава 9. Болезни плодов и семян и меры борьбы с ними 187
9.1. Болезни, развивающиеся в течение вегетационного
периода 188
9.1.1. Мумификация семян 188
9.1.2. Ржавчина шишек 191
9.1.3. Деформация плодов 192
9.2. Болезни, развивающиеся при хранении семян 193
9.2.1. Гнили плодов и семян 194
9.2.2. Плесневение плодов и семян 196
9.3. Система мероприятий по защите плодов и семян
от болезней 199
Глава 10. Болезни всходов, сеянцев, молодняков и меры борьбы
сними 201
10.1. Полегание всходов и сеянцев 201
10.2. Гниль сеянцев 205
10.3. Болезни типа шютте 206
10.4. Выпревание и другие болезни сеянцев хвойных
пород 218
10.5. Болезни, вызываемые ржавчинными грибами 225
10.6. Мучнистая роса 232
10.7. Пятнистости и другие болезни листьев 236
10.8. Система мероприятий по защите питомников, культур
и молодняков от болезней 246
10.8.1. Общие сведения 246
10.8.2. Надзор за появлением и распространением
болезней 246
10.8.3. Лесохозяйственные мероприятия 252
10.8.4. Химические меры защиты 253
Глава 11. Сосудистые и некрозно-раковые болезни древесных пород
и меры борьбы с ними 256
11.1. Сосудистые болезни 256
11.2. Некрозные болезни 264
11.3. Раковые болезни 276
11.4. Система мероприятий по защите древесных пород
от сосудистых и некрозно-раковых болезней 298
477
Глава 12. Гнилевые болезни древесных пород и меры борьбы
с ними 304
12.1. Сущность процесса гниения древесины 305
12.2. Классификация и признаки гнилей 306
12.3. Корневые гнили 309
12.4. Стволовые гнили 322
12.4.1. Стволовые гнили хвойных пород 323
12.4.2. Стволовые гнили лиственных пород 327
12.4.3. Защита насаждений от стволовых гнилей 333
Глава 13. Разрушение древесины на складах, в технических
сооружениях и зданиях 336
13.1. Поражение древесины деревоокрашивающими
и плесневыми грибами 337
13.2. Разрушение древесины на складах, в открытых
сооружениях и холодных постройках 343
13.3. Защита лесоматериалов от поражения грибами
на складах 350
13.3.1. Сухое хранение древесины 351
13.3.2. Влажное хранение древесины 354
13.4. Разрушение древесины в зданиях и сооружениях
с постоянным температурным режимом 356
13.5. Защита зданий от разрушения домовыми
грибами 361
13.6. Антисептирование древесины 365
Глава 14. Болезни цветочных культур и меры борьбы
с ними 370
14.1. Болезни цветочных культур защищенного грунта 370
14.1.1. Болезни гвоздики садовой ремонтантной 370
14.1.2. Болезни розы 381
14.1.3. Болезни хризантемы 393
14.2. Болезни луковичных и клубнелуковичных культур 400
14.2.1. Болезни тюльпана 400
14.2.2. Болезни гиацинта 406
14.2.3. Болезни гладиолуса 408
14.3. Болезни многолетников открытого грунта 414
14.3.1. Болезни георгины 414
14.3.2. Болезни ириса 420
14.3.3. Болезни пиона 424
14.3.4. Болезни флокса 430
14.4. Болезни однолетников и двулетников
открытого грунта 434
14.4.1. Болезни астры садовой 434
14.4.2. Болезни левкоя 440
14.4.3. Болезни сальвии 444
14.4.4. Болезни виолы 447
14.4.5. Болезни гвоздики турецкой 451
14.4.6. Болезни мальвы 452
478
Список литературы 453
Предметный указатель 456
Указатель русских названий болезней растений
и фитопатогенных организмов 460
Указатель наименований химических и биологических
средств защиты растений и древесины 465
Указатель латинских названий 467
Учебное издание
Семенкова Ирина Григорьевна,
Соколова Элла Сергеевна
Фитопатология
Учебник
Редактор А. Я. Сашина
Технический редактор Е. Ф. Коржуева
Компьютерная верстка: М. Ф. Фомина
Корректоры А. П. Сизова, В.А.Жилкина
Изд. № A-665-I/1. Подписано в печать 16.05.2003. Формат 60x90/16. Бумага тип. № 2.
Печать офсетная. Гарнитура «Тайме». Усл. печ. л. 31,0 (в т.ч. цв. вкл. 1,0).
Тираж 20000 экз. (1-й завод 1-5100 экз.). Заказ № 1598.
Лицензия ИД № 02025 от 13.06.2000. Издательский центр «Академия».
Санитарно-эпидемиологическое заключение № 77.99.02.953.Д.002682.05.01 от 18.05.2001.
117342, Москва, ул. Бутлерова, 17-Б, к. 223. Тел./факс: (095)334-8337, 330-1092.
Отпечатано с готовых диапозитивов
на ОАО «Альянс «Югполиграфиздат» ИПК «Офсет»
400001, г. Волгоград, ул. КИМ, 6.