/











































































































































































































































































































































































































































































































































































Автор: Стерки П.
Теги: физиология животных общая физиология обмен энергией иммунология физиология
Год: 1984




Текст
BASIC PHYSIOLOGY
Edited by P. D. Sturkie
Springer — Verlag
New York Heidelberg Berlin
1981
основы
ФИЗИОЛОГИИ
Под редакцией П. СТЕРКИ
Перевод с английского
канд. биол. наук н. Ю. АЛЕКСЕЕНКО
и Н. Н. АЛИПОВА
под редакцией
чл.-корр. АМН СССР г. И. КОСИЦКОГО
Москва «Мир» 1984
ББК 28.073
075
УДК 591.1
Основы физиологии: Пер. с англ. — М.: Мир; 1984. —
О 75 556 с, ил.
В коллективной монографии, написанной известными американскими
физиологами, представлены все разделы современной физиологии позвоночных.
Последовательно рассмотрены: строение органов и тканей, задачи общей и клеточной
физиологии, функция и организация нервной системы и органов чувств, высшие
функции головного мозга, работа мышц, кровеносных и лимфатических сосудов,
сердца, органов дыхания, почек, эндокринной, репродуктивной и выделительной
систем, процессы обмена веществ, питание и система крови.
Предназначена для студентов биологических и медицинских специальностей,
для физиологов и врачей.
2001040000— ББК 28.073
О 134—84, ч. 1
041(01)—84 57.04
Редакция литературы по биологии
© 1981 by Springer-Verlag New York Inc.
© Перевод на русский язык, «Мир», 1984
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА
Книга «Основы физиологии» написана группой
преподавателей Ратгерсовского университета (штат Нью-Джерси, США)
и задумана как учебное пособие по физиологии человека и
животных для студентов — биологов, зоологов, учащихся
сельскохозяйственных вузов и школ медицинских сестер.
Это пособие выгодно отличается от других учебников и
руководств тем, что в нем в значительной мере реализовано
стремление авторов излагать физиологические закономерности в
тесной связи с соответствующими данными по макро- и
микроанатомии органов и систем, а также анализировать
физиологические механизмы на основе современных биохимических данных.
Таким образом, в книге предпринята попытка синтезировать
сведения о структуре и функции, а сам анализ функций
довести до молекулярного уровня. Такой характер изложения
соответствует стремлениям многих педагогов излагать курс
физиологии в виде комплексных лекций; в некоторых вузах такие
лекции читаются несколькими специалистами — анатомом,
гистологом, физиологом, биохимиком, а иногда и патологом.
Подобная тактика призвана дать студенту всестороннее
представление о структуре и функции изучаемого органа или
системы. В этом бесспорно проявляется прогрессивная тенденция
интегрировать данные смежных наук.
Однако на примере настоящей книги можно видеть и
теневую сторону такого педагогического приема. Ввиду огромного
фактического материала отдельные разделы приходится
излагать весьма сжато, конспективно.
Далеко не все разделы равноценны. Данные по анатомии и
гистологии органов и тканей изложены отрывочно, иногда
поверхностно. При этом авторы нередко придерживаются своей,
оригинальной, классификации, не совпадающей с
общепринятой. Не везде анатомические данные увязаны с собственно
физиологическими.
Немногим больше повезло биохимическим разделам книги.
Здесь приводятся современные сведения по биохимии и
молекулярной биологии, на основе которых предпринимается попытка
изложить фактический материал о функциях органов и систем.
Однако, если учесть сравнительно небольшой объем книги, а
также то, что в пей нередки экскурсы в область патологии и
клиники, то станет ясно, что для изложения на современном
уровне многих физиологических закономерностей порой не
остается места.
6
Предисловие редактора перевода
Из сказанного следует, что данная книга не может служить
учебником физиологии, скажем, для студентов-медиков, но
может быть полезным дополнительным пособием, в частности
конспектом для повторения некоторых основ курса. Этому
бесспорно помогут и вопросы, приведенные в конце каждой главы.
Главы снабжены списками рекомендуемой литературы с
тем, чтобы студент мог работать самостоятельно. Однако, к
сожалению, в этих списках нет работ советских авторов и
вообще книг на русском языке. Мы приводим ниже небольшой
список, чтобы хоть в какой-то мере восполнить этот пробел.
Хотелось бы подчеркнуть, что данная книга полезна, кроме
того, и для инженеров, психологов и тех, кто самостоятельна
изучает основы физиологии. Этим читателям сведения по
анатомии, гистологии и особенно биохимии, а также экскурсы в
клинику не просто полезны, а настоятельно необходимы.
Все сказанное позволяет надеяться, что книга, в которой
сжато и на современном уровне излагаются основы курса
физиологии, найдет у нас широкого читателя.
Сеченов И. М. Рефлексы головного мозга. — М.: изд-во АМН СССР, 1952.
Павлов И. П. Полное собрание сочинений, т. 1—6. — М.: изд-во АН СССР,
1951—1952.
Анохин П. К. Биология и нейрофизиология условного рефлекса. — М.:
Медицина, 1968.
Аршавский И. А. Очерки по возрастной физиологии кровообращения. — М.:
Медицина, 1967.
Асратян Э. А. Очерки по физиологии условных рефлексов. — М.: Наука,
1971.
Бабский Е. £., Зубков А. А., Косицкий Г. И., Ходоров Б. И. Физиология
человека.— М.: Медицина, 1972.
Бериташвили И. С. Общая физиология мышечной и нервной системы, т. I—II. —
М., Л.; изд-во АН СССР, 1947—1948.
Бехтерева Н. П. Нейрофизиологические аспекты психической деятельности
человека.— Л.: Медицина, 1974.
Быков К. М. Кора головного мозга и внутренние органы. — М., Л.: Медгиз,
1947.
Костюк П. Г. Физиология центральной нервной системы. — М.: Высшая
школа, 1977.
Орбели Л. А. Лекции по физиологии нервной системы. — М., Л.: изд-во АН
СССР, 1938.
Парин В. В., Меерсон Ф. 3. Очерки клинической физиологии
кровообращения.— М.: Медгиз, 1965.
Симонов П. В. Теория отражения и психофизиология эмоций. — М.: Наука,
1970.
Уголев А. М. Мембранное пищеварение. — Л.: Наука, 1972.
Ухтомский А. А. Собрание сочинений, т. I—V. Изд-во ЛГУ, 1950—1954.
Черниговский В. Н. Интерорецепторы. — Л.: Наука, 1968.
Шаповалов А. И. Клеточные механизмы синаптической передачи. — М.:
Медицина, 1966.
Штерн Л. С. Непосредственная питательная среда органов и тканей. — М.:
изд-во АН СССР, 1960.
ПРЕДИСЛОВИЕ
Настоящая книга представляет собой введение в
физиологию позвоночных, в котором содержатся необходимые
морфологические понятия, соотнесенные с функцией. Особое внимание
уделено физиологии человека. В одной из глав кратко
излагаются основы морфологии на макро- и микроуровне. В других
главах обсуждаются вопросы общей и клеточной физиологии,
физиологии нервно-мышечной системы, жидких сред организма,
кровообращения и сердца, дыхания, пищеварения и
всасывания, обмена веществ и энергии, терморегуляции, питания, почек
и эндокринной системы (в этот последний раздел входит
учение о функциях гипофиза, репродуктивной системы, щитовидной,
паращитовидной и поджелудочной желез и надпочечников).
В книге нашли отражение все основные концепции, которые
достаточно хорошо обоснованы. Изложение рассчитано на
студентов, получивших предварительные сведения по биологии,
физике, химии и математике, а также на специалистов, желающих
освежить свои знания в области основ физиологии.
Книга может быть использована в качестве пособия
учащимися биологических, ветеринарных, медицинских,
сельскохозяйственных учебных заведений, в которых преподается курс
физиологии животных и человека.
Книга «Основы физиологии» написана коллективом из семи
специалистов, имеющих опыт преподавания этой дисциплины.
Пол Д. Стёрки
СПИСОК АВТОРОВ
Casey Т. М. (гл. 18, 19) Department of Physiology, Rutgers
University, New Brunswick, New Jersey 08903, U. S. A.
Frankel H. M. (гл. 17) Department of Physiology, Rutgers
University, New Brunswick, New Jersey 08903, U. S. A.
Griminger P. (гл. 23) Department of Nutrition, Rutgers
University, New Brunswick, New Jersey 08903, U. S. A.
Hazelwood R. L. (гл. 22) Department of Biology, University of
Houston, Central Campus, Houston, Texas 77004, U. S. A.
Page С. Н. (гл. 3—10) Department of Physiology, Rutgers
University, New Brunswick, New Jersey 08903, U. S. A.
Sturkie P. D. (гл. 1, 2, 11—16, 20, 21, 25—30) Department of
Physiology, Rutgers University, New Brunswick, New
Jersey 08903, U. S. A.
Zambraski E. J. (гл. 24) Department of Physiology, Rutgers
University, New Brunswick, New Jersey 08903, U. S. A.
Глава 1
ОБЩИЙ ПЛАН СТРОЕНИЯ ТКАНЕЙ, ОРГАНОВ
И ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Живой организм состоит из систем, несущих определенные
функции и образованных органами, тканями и клетками.
Системы эти следующие: костная, мышечная, нервная,
сердечнососудистая, пищеварительная, дыхательная, мочеполовая и
эндокринная.
Клетке — структурно-функциональной единице всех органов
и тканей — посвящена гл. 2.
Макро- и микроскопическое строение органов и систем
является предметом изучения морфологии. Анатомия исследует
форму и строение органов и систем на макроуровне.
Микроскопическая анатомия, к которой относятся гистология и
цитология, изучает соответственно структуру тканей и клеток.
Настоящая глава представляет собой короткий экскурс в
гистологию, а также в нормальную и топографическую анато-
Таблица 1.1. Термины, определяющие положение частей тела у человека
и четвероногого животного
Человек Четвероногое
Верхний, или краниальный Передний, или краниальный
Передний, или вентральный Нижний, или вентральный
Задний, или дорсальный Верхний, или дорсальный
Медиальный — находящийся ближе к срединной плоскости (например, нос
расположен медиально относительно глаз)
Латеральный — находящийся дальше от срединной плоскости (например,
глаза расположены латерально относительно носа)
Проксимальный — расположенный ближе (например, проксимальным
называют тот сустав пальца ноги, который ближе к плюсне)
Дистальный — расположенный дальше (например, палец ноги расположен
дистально по отношению к плюсне)
Периферический — удаленный от центра (спинномозговые нервы
расположены периферически по отношению к головному мозгу)
Висцеральный — относящийся к внутренностям
10
Глава 1
Краниальный
Верхний
v Фронтальная плоскость
Дорсальный
Задний
Вентральный
Передний
Каудальный
Задний
Дорсальный
Верхний
Краниальный
Передний
Вентральный
Нижний
I
Каудальный
Нижний
Рис. 1.1. Термины, принятые для обозначения расположения частей тела
человека и четвероногого животного.
мию. Более подробные анатомические сведения приурочены к
обсуждению физиологии различных систем в отдельных главах.
ТЕРМИНОЛОГИЯ
Анатомические термины, определяющие положение частей
тела, приведены в табл. 1.1 и на рис. 1.1.
На рис. 1.2 показаны плоскости тела — сагиттальная,
поперечная (горизонтальная); фронтальная.
Общий план строения
11
ТКАНИ
Отдельные органы и системы организма образованы одной
тканью или группами тканей, обладающих характерным
строением. К ним относятся: 1) эпителий, 2) кожа, 3) оболочки,
4) мышцы, 5) нервы, 6) соединительная ткань, 7) эластическая
ткань, 8) хрящевая ткань, 9) костная ткань. В процессе
эмбриогенеза эти ткани развиваются из трех зародышевых
листков— эктодермы, энтодермы и мезодермы. Происхождение
некоторых органов и тканей из этих листков указано на рис. 1.3.
ЭПИТЕЛИИ
Эпителий состоит из тесно связанных друг с другом клеток,
образующих покровы на наружных поверхностях тела и
выстилки внутренних поверхностей органов и полостей. Кровеносных
Рис. 1.2. Плоскости тела.
12
Глава 1
и их расположению
сосудов в эпителии нет. По типу клеток
различают следующие виды эпителия.
Однослойный плоский. Представляет собой один слой плоских
клеток; выстилает альвеолы легких, хрусталик и часть
внутреннего уха.
Многослойный плоский. Состоит из плоских клеток,
расположенных в несколько слоев; образует эпидермис (рис. L4,Л). Со
Покровный эпителий тела
Хрусталик глаза,
слуховой пузырек,
внутреннее ухо,
глазной пузырек
Волосы, кожа, зубы, эмаль,,
эпителий рта и анального
отверстия, потовые и молочные
железы
( Энтодерма J
ч
с
Эпителий
Мезодерма
ч
Опорные ткани органов
Нервная система
Нервная трубка,
спинной мозг,
ганглии,
головной мозг
Пищеварительной трубки,печени,
поджелудочной железы, мочевого
пузыря, глотки, трахеи, бронхов,
легких, щитовидной, паращито-
видных и вилочковой желез
Кости, хрящи, соединительная и
эластическая ткань, эндотелий
кровеносных сосудов
Рис. 1.3. Производные трех зародышевых листов.
временем плоские ороговевающие клетки слущиваются, а на
смену им приходят новые, образующиеся в глубоком
(ростковом) слое.
Цилиндрический. Образован высокими цилиндрическими
клетками с ядром у основания, многочисленными митохондриями и
развитым аппаратом Гольджи в цитоплазме (рис. 1.4, В). Такой
эпителий характерен для слизистых оболочек желудка и
кишечника; его клетки бывают с гладкой поверхностью или
имеют выросты — реснички, а также микроворсинки — крохотные
выросты, увеличивающие поверхность всасывания.
Кубический. Этот эпителий сходен с цилиндрическим, однако
клетки его имеют кубическую форму; представлен в печени.
Эндотелий. Представляет собой один слой клеток,
выстилающий внутреннюю поверхность кровеносных и лимфатических
сосудов.
Общий план строения
13
ЖЕЛЕЗЫ И ОБОЛОЧКИ
В отличие от желез, не обладающих протоками (см. гл. 25),
экзокринные железы выделяют свой секрет через выводной
проток. Протоки бывают самой различной формы и выстланы
цилиндрическим, кубическим или плоским эпителием. При
классификации желез учитывается их тип и форма (рис. 1.5).
Эпителий и примыкающие к нему ткани образуют оболочки,
покрывающие различные органы (см. гл. 2). Эти оболочки
выстилают полости и органы и могут вырабатывать жидкий
серозный или густой слизистый секрет.
Кровеносные сосуды выстланы оболочкой, состоящей из
эндотелия и соединительной ткани. К оболочкам относятся также
перикард, брюшина и плевра.
С костно-мышечным аппаратом тесно связаны синовиальные
оболочки. Они образованы плотным наружным фиброзным
слоем и внутренним слоем рыхлой соединительной ткани,
содержащей эластические и коллагенрвые волокна. Эти оболочки
вырабатывают синовиальную жидкость (тканевая жидкость и
муцин), которая играет роль смазки при движении суставов
(например, коленного и локтевого).
Слизистые оболочки выстилают пищеварительную трубку
(желудочно-кишечный тракт), трахею, бронхи и многие другие
органы; эти оболочки вырабатывают слизь. Слизистая
оболочка состоит из трех слоев: 1) эпителия, 2) собственной
пластинки и 3) мышечной пластинки (см. рис. 20.2). Слизистые
оболочки служат опорой и защитой для кровеносных и
лимфатических сосудов; они создают обширную всасывающую
поверхность (например, в кишечнике).
СИСТЕМА СОЕДИНИТЕЛЬНОЙ ТКАНИ
Ткани этой системы обеспечивают опору для различных
структур и связывают их между собой. Соединительная ткань
богато васкуляризирована, содержит большое количество
межклеточного вещества или жидкости, тогда как клеточных
элементов в ней мало. Строение волокон и межклеточного
вещества, в значительной степени определяющих тип соединительной
ткани, широко варьирует.
КЛАССИФИКАЦИЯ
Ниже приведена классификация видов соединительной
ткани (СТ).
у рыхлая неоформленная
Волокнистая коллагеновая
^ плотная ^
^ эластическая
Слой плоских клеток
Промежуточн
слой
Переходной
слой
Слой
цилиндрических
клеток
Рыхлая
соединительная
Блестящий
слой
Роговой слой
Зернистый
слой
5
CL
Сосочки дерм
Нерв
Жировые клетки
Ростковый s
слой о
Осязательное
тельце
СО
5
Потовая железа
Микроворсинки
В
14
Общий план строения
15
,ретикулярная
С особыми '„,„„„„,,„
свойствами ~жиР°вая
\
пигментная
СТ
Собственно СТ
/
гиалиновая
Хрящевая < коллагеново-волокнистая
эластическая
Костная
/(компактная
\1 губчатая
Межклеточное вещество собственно соединительной ткани —
пластичное, хрящевой — твердое, но гибкое, костной — твердое
из-за наличия в его структуре солей кальция.
Широко распространенная рыхлая неоформленная СТ
состоит из клеток и волокон, взвешенных в студнеобразном
основном веществе; благодаря такой структуре эта ткань мягкая,
полупрозрачная и легко смещаемая (рис. 1.6,Л). Волокна (кол-
лагеновые и эластические) образуют строму многих органов и
оболочку кровеносных сосудов и нервов. Клеточные элементы
Трубчатая Альвеолярная Альвеолярно - трубчатая Сложная железа
Рис. 1.5. Типы желез и разветвления их секреторной части и протоков.
соединительной ткани представлены в основном фибробластами
и гистиоцитами; последние обладают способностью к
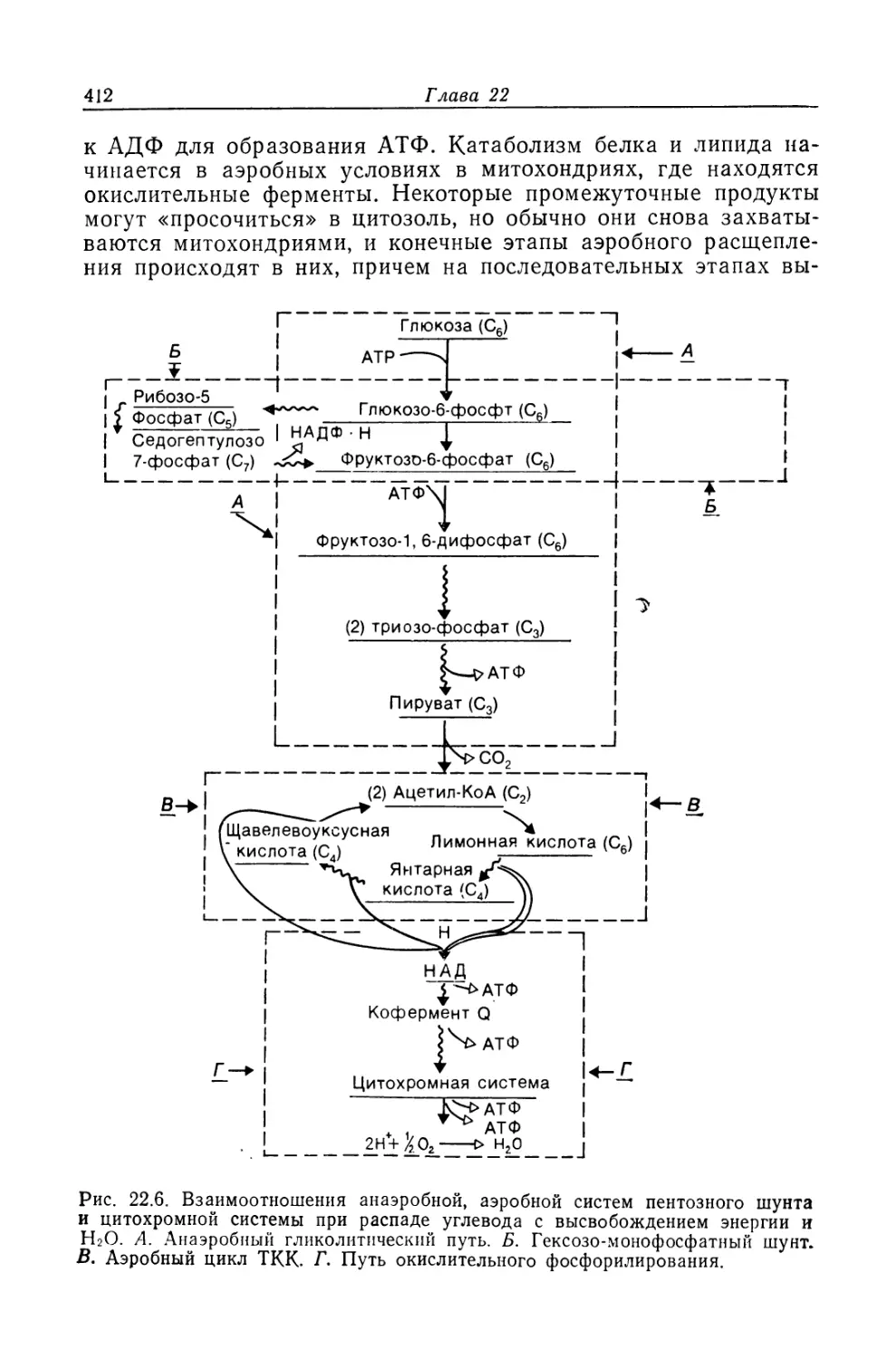
фагоцитозу, т. е. к поглощению различных веществ и их
перевариванию.
Ретикулярная СТ состоит из сети, образованной клетками и
ретикулярными и коллагеновыми волокнами (рис. 1.6,5). Она
Рис. 1.4. Клеточное строение некоторых тканей. А. Плоский эпителий. Б. Тот
же эпителий в коже; видны три слоя эпидермиса и дерма с нервными
волокнами, осязательными тельцами и потовыми железами. В. Простой,
мерцательный и цилиндрический эпителий (последний встречается в кишечнике) и
расположенная под ним соединительная ткань.
Краевой синус
Коллагеновые.
волокна
Внутренняя
оболочка
Средняя
оболочка
у Адвентициальная
оболочка
в
Общий план строения
17
составляет строму лимфатических узлов, селезенки и костного
мозга.
Эластическая СТ— это ткань, в составе которой
преобладают эластические волокна. Эта растяжимая ткань встречается
в легких, некоторых хрящах, кровеносных сосудах (рис. 1.6,5),
бронхах и эластических связках.
В фиброзной СТ преобладают коллагеновые волокна. Эти
крупные беловатые волокна плотны, но не растяжимы. Из
такой ткани состоят: 1) связки, подкрепляющие места соединения
костей, и 2) сухожилия, служащие для прикрепления мышц к
костям (сухожилия имеются также в некоторых оболочках,
обеспечивающих защиту таких органов, как сердце и почки).
Жировая ткань содержит крупные овальные жировые
клетки, в цитоплазме которых имеются капельки жира. На
обезжиренных препаратах (рис. 1.7,Л) эти клетки выглядят как
пустые колечки или овалы. Жировая ткань распространена
повсеместно в составе подкожной жировой клетчатки.
ХРЯЩЕВАЯ ТКАНЬ
Твердая, но гибкая хрящевая ткань состоит из клеток,
погруженных в межклеточное вещество, состав которого бывает
различным, и волокон (рис. 1.7). Гиалиновый хрящ
представляет собой голубовато-белую однородную массу, содержащую
клетки и коллагеновые волокна. Он покрывает суставные
поверхности костей и образует хрящи ребер (вентральную часть
ребер). Гиалиновый хрящ участвует также в формировании
носа, гортани, трахеи и бронхов.
Эластический хрящ представляет собой соединительную
ткань с преобладанием эластических волокон (рис. 1.7). Из
такого хряща состоят гортань, надгортанник и внутреннее ухо;
эластический хрящ способствует сохранению формы этих
органов и в то же время придает им гибкость.
Волокнистый хрящ образован клетками, вкрапленными в
межклеточное вещество, характерное для коллагеновой ткани.
Этот хрящ образует прочное и гибкое соединение между
некоторыми костями в тех участках, где необходимы жесткость и
прочность.
Рис. 1.6. Типы соединительной ткани (СТ). Л. Плотная СТ; /—эластические
волокна, 2 — коллагеновые волокна, 3 — фибробласты, 4 — плазматические
клетки, 5 — гистиоциты, 6 — тучные клетки, 7 — лимфоциты. Б. Ретикулярная
СТ; видны ретикулярные и коллагеновые волокна и клетки. В. Эластическая
СТ во внутренней оболочке артерии (темные линии) на поперечном разрезе
2-1705
18
Глава 1
КОСТНАЯ ТКАНЬ
Костная ткань — это разновидность соединительной ткани с
межклеточным веществом, пропитанным слоями кальция и
фосфора. Неорганический компонент составляет около 2/з массы
кости. Органический компонент образуют костные клетки (ос-
теоциты, остеобласты, остеокласты), остеоидыое межклеточное
Гиалиновый
Эластический
Волокнистый
Губчатая ткань
с красным
костным мозгом
Ядро
Надкостница^
Костномозговая
полость
Суставной хрящ
Эпифиз
Компактная кость
Рис. 1.7. Жировая, хрящевая и костная ткани. Л. Жировая ткань с
вакуолями (препарат обезжирен) и волокнами. Б. Разные виды хряща. Видны
хрящевые клетки в межклеточном веществе, содержащем эластические и колла-
геновые волокна. В, Поперечное сечение компактной кости; видны клетки
(остеоциты) в межклеточном веществе. Г. Продольное сечение кости; видно
компактное и губчатое вещество.
вещество, кровеносные сосуды и нервы (см. гл. 28). Костная
ткань подразделяется на компактную и губчатую (рис. 1.7).
На ранних стадиях эмбриогенеза образуется хрящевой
скелет; в дальнейшем начинается его оссификация (замена
хрящевой ткани костной), которая продолжается и после рождения.
Общий план строения
19
В длинных трубчатых костях (рис. 1.7) центры окостенения
находятся в диафизе, построенном из компактного вещества, и
эпифизах. Между диафизом и эпифизами остаются тонкие
прослойки, хрящевой ткани. В норме костное вещество постоянно
разрушается остеокластами и восстанавливается остеобластами
(см. гл. 28). '
МЫШЕЧНАЯ ТКАНЬ
В зависимости от физиологических свойств и структуры,
определяемой при световой микроскопии, различают три типа
мышечной ткани: скелетную, сердечную и гладкую.
Сокращение скелетных мышц осуществляется произвольно
через посредство соматических нервов в отличие от сердечной
и гладких мышц, управляемых вегетативной нервной системой.
Как следует из названия, скелетные мышцы прикрепляются к
костям скелета (рис. 1.13 и 1.14); сердечная мышца образует
основную массу ткани сердца, а гладкие мышцы — мышечные
слои внутренних органов (пищеварительного тракта,
кровеносных сосудов и т. д.).
Скелетные мышцы состоят из пучков, образуемых
множеством волокон (рис. 1.8,Л). Пучки отделены друг от друга
соединительнотканными прослойками, по которым проходят сосуды
и нервы. Отдельные волокна заключены в трубчатую оболочку
(сарколемму) и отличаются характерной поперечной исчерчен-
ностью (рис. 1.8,5). Длина волокон варьирует от 1 до 40 мм,
а диаметр — от 0,01 до 0,15 мм; они состоят из плотно
упакованных, ориентированных по длинной оси волокна миофибрилл.
Более подробно структура волокон представлена в гл. 11.
Поперечная исчерченность образована чередующимися светлыми
(изотропными) I-дисками и темными (анизотропными) А-дис-
ками. Оба диска образуют саркомер. Каждый диск пересекает
тонкая темная полоска — Z-линия.
Сердечная мышца, так же как и скелетная, имеет
поперечную исчерченность, однако в отличие от скелетной мышцы здесь
в области Z-линии имеются участки, где волокна смыкаются,
образуя вставочные диски (рис. 1.8,5). Благодаря этим дискам
волокна прочно сцеплены, вследствие чего сокращение одного
из них передается другому. Хотя волокна сердечной мышцы
ветвятся и переплетаются, каждое из них заключено в
отдельную мембрану. Сердечная мышца получает питание через
венечные артерии. Более подробно структура сердечной мышцы
описана в гл. 11 и 15.
Узкие и длинные (но более короткие, чем скелетные) глад-
комышечные волокна не имеют поперечной исчерченности
(рис. 1.8,Г). В каждом таком волокне всего одно ядро в отли-
2*
Сарколемма
Поперечная
исчерченность
Ядро
Вставочные
диски
Ядра
Гладкомышечныв
клетки
Ядро
Кровеносные
капилляры
20
Общий план строения
21
чие от многоядерных волокон скелетных ^ышц. Гладкомышеч-
ные клетки веретенообразны; длина их составляет около
0,015—0,5 мм, диаметр — 0,002—0,02 мм. В стенках внутренних
органов и кровеносных сосудов гладкие мышцы обычно
образуют два слоя — толстый наружный циркулярный и тонкий
внутренний продольный. Питающие мышцу кровеносные сосуды
проходят между мышечными пучками параллельно гладкомы-
шечным волокнам (рис. 1.8,Г).
НЕРВНАЯ ТКАНЬ
Нервная ткань образована нервными клетками (нейронами),
которые по своей структуре и функции отличаются от других
клеток. Нейроны обладают особыми физиологическими
свойствами— раздражимостью, возбудимостью и проводимостью;,
иными словами, они способны принимать сигналы и передавать
ответные импульсы. Более подробно анатомия и гистология
нервной системы обсуждается в соответствующих главах.
Топография периферических нервов, их выход из спинного мозга и
отношение к сплетениям и позвоночным ганглиям
представлены схематически на рис. 1.9.
АНАТОМИЯ
Опорными структурами организма служат костная и
мышечная системы, ограничивающие полости, в которых
расположены внутренние органы (рис. 1.10 и 1.11).
КОСТНАЯ СИСТЕМА
Скелет взрослого человека состоит примерно из 200 костей,,
в том числе
Позвоночник 26
Мозговой череп 8
Лицевой череп 14
Подъязычная кость, ребра, грудина 26
Верхние конечности 64
Нижние конечности 62
Всего 200
Рис. 1.8. Типы мышечной ткани. Л. На поперечном сечении мышцы видны три'
мышечных пучка (1, 2, 3), состоящие из волокон, и соединительнотканные
прослойки между волокнами и пучками. Б. Продольное сечение скелетной-
мышцы; видны отдельные волокна, сарколемма и поперечная исчерченность
(темные и светлые полосы). В. Сердечная мышца; видны поперечная
исчерченность и области соединения волокон (вставочные диски). Г. Гладкая
мышца; видны отдельные клетки без поперечной исчерченности.
Полушария
головного мозга
Спинномозговой
нерв
Спинномозговой
узел
Задняя ветвь
Передняя ветвь
Плечевое
сплетение
Подмышечный
нерв
Срединный
нерв
Локтевой
нерв
Первый поясничный
позвонок
Поясничное
сплетение
Крестцовое
сплетение
Копчиковое
сплетение
.Срамное
сплетение
Седалищный нерв
Рис. 1.9. Спинномозговые нервы и сплетения.
22
Полость
глазницы
Носовая_
полость
Полость рта
. Полость
черепа
&
Спинномозговая
-полость
Грудная
полость"
Брюшная,
полость
Диафрагма;
Полость таза
Рис. 1.10. Полости тела.
24
Глава 1
Рис. 1.11. Основные внутренние органы, вид спереди.
На рис. 1.12 и 1.13 показано расположение и приведены
названия важнейших костей.
МЫШЕЧНАЯ СИСТЕМА
Скелетные мышцы прикрепляются к костям и другим
структурам либо непосредственно, либо при помощи фиброзных
сухожилий (апоневрозов). Размеры и форма мышц бывают
самыми разными в зависимости от связанных с ними костей.
Мышцы ног длинные, а мышцы туловища плоские и широкие;
икроножная мышца очень толстая, а волокна портняжной
мышцы достигают в длину почти 60 см.
Архитектура мышечных волокон также бывает различной в
зависимости от выполняемой мышцей функции (см. гл. 11).
Лобная кость
Глазница.
Ключица
Грудина
Мечевидный,
отросток
Реберный хрящ
Свободное^
ребро
Межпозвоночный
диск
Безымянная
кость
Подвздошная
кость
Лонное сочленение
Таранная кость
Плюсна
Фаланги
Рис 1.12. Скелет человека, вид спереди.
Череп
Верхняя челюсть
Нижняя челюсть
Яремная вырезка
Ребра
Плечевая кость
Позвоночник
Локтевая кость
Лучевая кость
Большой вертел
Запястье
Пясть
Фаланги)
Малый вертел
Крестец
Бедренная кость
Коленная чашечка
Большеберцовая кость
Малоберцовая кость
25
Теменная кость
Затылочная кость
Шейные позвонки (7)
Ключица
Акромион
Плечевая кость
Лучевая кость
Локтевая кость
Шиловидный
отросток "~
Бедренная кость
Малоберцовая кость
Болыиеберцовая кость
Рис. 1.13. Скелет человека, вид сзади.
26
Грудные
позвонки (12)
Поясничные
позвонки (5)
Медиальный надмыщелок
плечевой кости
Латеральный надмыщелок
плечевой кости
Подвздошная кость
Крестец
Лонное сочленение
Седалищная кость
Латеральный надмыщелок
бедренной кости
Медиальный мыщелок
бедренной кости
Латеральная лодыжка
Пяточная кость
Фаланги
Лобная
Круговая глаза
Круговая рта
Грудино - щитовидная
Дельтовидная
Большая грудная
Широчайшая спины
Передняя зубчатая
Плечелучевая
Влагалище прямой
мышцы живота
Длинный лучевой
разгибатель
запястья
Подвздошно-
поясничная
Камбаловидная
Жевательная
но-ключично-сосцевидная
Грудино-подъязычная
Трапециевидная
Трехглавая плеча
Двухглавая плеча
Плечевая
Круглый пронатор
Наружная косая
Прямая живота
евой сгибатель
запястья
линная ладонная
Натягивающая
Портняжная
Четырехглавая
бедра
широкую фасцию
бедра
Икроножная
Длинная малоберцовая
Длинный оазгибатель пальцев
Передняя эолыиеберцовая
Короткая малоберцовая
Рис. 1.14. Мышцы человека, вид спереди.
27
Грудино-ключично
сосцевидная
Ременная
Большая
ромбовидная
Широчайшая
спины
Средняя
ягодичная
Длинный лучевой
разгибатель
запястья
Локтевая
Короткий лучевой
разгибатель
запястья -"""^1
Длинная отводящая,
большого пальца
Короткий разгибатель'
большого пальца
Подошвенная
Икроножная
Трапециевидная
Подостная
Малая круглая
Большая круглая
Трехглавая плеча
Поясничный
апоневроз
Плечелучевая
Наружная косая
Локтевой сгибатель
запястья
7/ Локтевой разгибатель
:j_ запястья
/ Разгибатель пальцев
Большая ягодичная
Подвздошно-больше-
берцовый тракт
Большая приводящая
Полусухожильная
Двуглавая бедра
Полуперепончатая
Ахиллово сухожилие
Камбаловидная
Короткая малоберцовая
Рис. 1.15. Мышцы человека, вид сзади.
2а
Затылочная-
Внутренняя сонная _
Наружная сонная
Позвоночная"
Правая общая сонная-
Правая подключичная-
Плечеголовная-
Дуга аорты-
Правая венечная-
Аорта-
Чревный ствол-
Верхняя брыжеечная-
Брюшная аорта-
Общая подвздошная,
Внутренняя
подвздошная
Наружная
подвздошная
Глубокая бедренная
Подколенная.
Задняя большеберцовая
Дорсальная плюсневая
Тыльная стопы
Левая общая сонная
Левая подключичная
-Легочная
"С
-Подмышечная
"Левая венечная
-Плечевая
- Селезеночная
-Почечная
Нижняя
' брыжеечная
-Лучевая
-Локтевая
^Ладонные
дуги
-Пальцевые
-Бедренная
Передняя большеберцовая
Малоберцовая
Дугообразная
Рис. 1.16. Главные артерии человека.
29
Наружная яремная
Внутренняя яремная
Правая
плечеголовная
Верхняя полая
Нижняя полая
Печеночная
Почечная
Общая
подвздошна)
Срединная
крестцовая
Наружная \
подвздошная
Внутренняя
подвздошная
Поверхностная височная
~j Лицевая
Левая подключичная
Подмышечная
Плечевая
Диафрагмальная
Надпочечная
Яичковая
Поясничные
Лучевая
Бедренная -х—,
\
Большая подкожная
Малая подкожная
Подколенная
Задняя большеберцовая
Передняя большеберцовая
Тыльные пальцевые
ве^ы стопы
Рис. 1.17. Главные вены человека.
30
Общий план строения
31
Название и расположение главных наружных мышц тела
указаны на рис. 1.14 и 1.15.
СИСТЕМА КРОВООБРАЩЕНИЯ
К этой системе относятся сердце, кровеносные и
лимфатические сосуды и их содержимое. Подробно об анатомии и
гистологии этих структур сказано в главах, посвященных
физиологии сердца и кровообращения.
Топографическая анатомия главных артерий и вен
приведена на рис. 1.16 и 1.17 и в гл. 15, а анатомия лимфатических
сосудов и лимфатических узлов обсуждается в гл. 14.
ЛИТЕРАТУРА
Ebe T.t Kobayashi S. (1973). Fine Structure of Human Cells and Tissues, John
Wiley and Sons, New York.
Miller M. A., Drakontides А. В., Leavell L. C. (1979). Kimber-Gray-Stackpoles-
Anatomy and Physiology (17th ed.), MacMillan, New York.
Pick T. P., Howden R. (1977). Gray's Anatomy (revised from 15th English ed.),
Bounty Books, New York.
Weiss L. (1977). The cell. In: Weiss L., Greep R. (eds.), Histology (4th ed.),
McGraw Hill, New York.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Назовите системы организма.
2. Назовите главные виды тканей организма.
3. Дайте определение анатомии и гистологии.
4. Назовите по крайней мере два органа или ткани,
производные эктодермы, энтодермы, мезодермы.
5. Дайте определение и охарактеризуйте: 1) плоский эпителий,
2) разветвленную трубчатую железу с протоками
(изобразите в виде схемы), 3) экзокринные и эндокринные железы.
6. Что такое рыхлая неоформленная соединительная ткань и
где она встречается?
7. Что такое эластическая соединительная ткань и где она
встречается?
8. В чем разница между гиалиновым и эластическим хрящом?
9. Из чего состоит кость?
10. В чем разница между скелетной, сердечной и гладкой
мышцей?
11. Назовите главные мышцы рук и ног и укажите их
расположение.
12. Каково расположение костей пясти и запястья?
Глава 2
ОБЩАЯ И КЛЕТОЧНАЯ ФИЗИОЛОГИЯ
Хотя Роберт Гук впервые описал клетку еще в 1665 г.,
клеточная теория была провозглашена Шлейденом и Шванном
лишь в 1839 г. Согласно этой теории, элементарной структурно-
функциональной единицей всех живых организмов является
клетка. В своих исследованиях Шлейден и Шванн использовали
световой микроскоп. Изобретение электронного микроскопа
позволило изучить строение клеток более детально. До появления
электронной микроскопии многие субклеточные частицы (орга-
неллы) оставались неизвестными: различить их было
невозможно. К настоящему времени эти частицы выделены, изолированы
и идентифицированы с помощью различных методик, в том
числе ультрацентрифугирования, при котором органеллы
разделяются в соответствии с их размером и плотностью. Подробный
физико-химический анализ некоторых органелл показал, что
они выполняют важные метаболические и биохимические
функции.
Органы и ткани состоят из скопления клеток, размеры,
форма и число которых различны в зависимости от органа и
выполняемой им функции. Существует несколько типов клеток:
1) эпителиальные, 2) мышечные, 3) нервные, 4) клетки крови,
5) костные и 6) клетки соединительной ткани. Эти типы клеток
и образуемые ими ткани описаны в гл. 1.
Содружественная деятельность клеток, тканей и органов и,
следовательно, организма в целом определяется обменом и
транспортом — внутри- и внеклеточными процессами.
Регуляция этих процессов частично зависит от структуры и функции
клеточной оболочки, или мембраны.
СТРОЕНИЕ КЛЕТКИ
Главными компонентами клетки являются ядро и
цитоплазма (рис. 2.1). Размеры клеток варьируют в широких пределах.
ЯДРО
Ядро, управляющее всей жизнедеятельностью клетки,
обычно располагается в центре. Микрохирургическое удаление ядра
Сэнуклеация) приводит к дезорганизации функций цитоплазмы.
Общая и клеточная физиология
33
Ядерная мембрана
Рис. 2.1. Схема электронно-микроскопической картины типичной клетки.
(Anthony С. P., Kolthoff N. J., Anatomy and Physiology, 1975.)
Ядро ограничено мембраной и содержит одно или несколько
ядрышек — небольших сферических телец, а также
многочисленные гранулы хроматина; из хроматина построены
хромосомы— носители вещества наследственности (генов). Хромосома
состоит из молекулы дезоксирибонуклеиновой кислоты (ДНК)
и структурного белка; каждый ген соответствует отдельному
участку молекулы ДНК.
В процессе митоза — нормального клеточного деления —
клетка и находящиеся в ней хромосомы делятся на две равные
половины, каждая из которых получает полный (диплоидный)
3-1705
34
Глава 2
набор хромосом. Первичные половые клетки претерпевают
редукционное деление (мейоз), в результате которого образуются
сперматозоиды и яйцеклетки, содержащие половинный
(гаплоидный) набор хромосом. Слияние гамет (сперматозоида и
яйцеклетки) приводит к образованию оплодотворенной
яйцеклетки, или зиготы, которая в дальнейшем превращается в
зародыш и плод, содержащие диплоидный набор хромосом (см.
гл. 26). Лишенные мембран ядрышки состоят преимущественно
из рибонуклеиновой кислоты (РНК) и белка. Больше всего
ядрышек в растущих клетках; вероятно, в них синтезируется
РНК, обнаруживаемая в рибосомах. Более подробно синтез
РНК и ДНК обсуждается в гл. 22.
Ядро окружено двухслойной мембраной с пространством
между слоями. Она проницаема даже для таких крупных
молекул, как молекулы РНК, которые свободно проходят из ядра
в цитоплазму.
ЦИТОПЛАЗМА
К цитоплазме относятся все компоненты клетки, кроме ядра
и плазматической мембраны. Важнейшие из органелл клетки —
это 1) эндоплазматическии ретикулум, 2) рибосомы, 3)
митохондрии, 4) аппарат Гольджи, 5) центросома и 6) лизосомы.
ЭНДОПЛАЗМАТИЧЕСКИИ РЕТИКУЛУМ
Эндоплазматическии ретикулум (ЭР), имеющийся в
цитоплазме почти всех клеток, представляет собой совокупность
образованных мембранами полостей, размер и форма которых
значительно варьируют в зависимости от физиологического
состояния. В некоторых клетках, например в эритроцитах,
эндоплазматическии ретикулум полностью отсутствует. Одно время
считали, что эндоплазматическии ретикулум имеется лишь в
эндоплазме, однако теперь установлено, что он располагается и
в периферических отделах клетки (эктоплазме) и тесно связан
с плазматической мембраной.
Эндоплазматическии ретикулум представляет собой систему
взаимосвязанных каналов и полостей, пронизывающих всю
цитоплазму. Существуют два типа эндоплазматического ретику-
лума — шероховатый (гранулярный) и гладкий (агрануляр-
ный). Ограничивающая его двойная мембрана содержит белки,
фосфолипиды и ферменты.
РИБОСОМЫ
Рибосомы — это электроноплотные частицы размером 10—
25 нм (1-10-6 мм), содержащие РНК и белок. В большинстве
.своем они связаны с гранулярным эндоплазматическим ретику-
Общая и клеточная физиология
35
лумом, но могут и свободно располагаться в цитоплазме.
Рибосомы, связанные с эндоплазматическим ретикулумом,
синтезируют белок «на экспорт»; свободные рибосомы синтезируют
белок, используемый на месте синтеза.
МИТОХОНДРИИ
Диаметр митохондрий варьирует от 0,5 до 1,0 мкм, а длина
составляет до 0,7 мкм. Их конфигурация и структура различны
в зависимости от функционального состояния.
В некоторых клетках (в частности, делящихся) митохондрии
!^огут быть подвижными, а в ряде тканей, особенно
обладающих секреторной активностью, они образуют отчетливые
скопления. У митохондрий существует наружная и внутренняя
мембраны; внутренняя мембрана выпячивается, образуя
складки (кристы), в каждой из которых имеется множество
элементарных частиц. Эти частицы представляют собой скопления
ферментов, и главным образом аденозинтрифосфата (АТФ) —
основного источника энергии для большинства биохимических
реакций, протекающих в организме.
Поскольку в результате ферментативных реакций в
митохондриях высвобождается большое количество энергии, эти
органеллы называют «силовыми станциями» клетки. Чем
интенсивнее активность клетки, тем больше в ней митохондрий; так,
в активной клетке печени число митохондрий достигает 2500, а
в сравнительно неактивной клетке — их всего 25.
АППАРАТ ГОЛЬДЖИ
Аппарат Гольджи (АГ) образован системой канальцев и
мешочков, или цистерн. Хотя форма канальцев бывает
различной, как правило, аппарат Гольджи представляет собой стопку
уплощенных мешочков, связанных с мелкими пузырьками.
В некоторых клетках (например, в мышечных) аппарат Гольд*
жи представлен слабо, в других (секреторные клетки, нервы)
он развит хорошо. Гранулы, образуемые в аппарате Гольджи,
содержат гормоны и ферменты преимущественно белковой
природы.
Последними данными установлено, что в аппарате Гольджи
происходит синтез углеводов и их связывание с белками, в
результате чего образуются гликопротеиды.
ЛИЗОСОМЫ
Лизосомы — это окруженные мембраной частицы размером
от 0,25 до 0,8 мкм; на электронных микрофотографиях они
выглядят плотными, зернистыми (рис. 2.1). Лизосомы содержат
3*
36
Глава 2
многочисленные ферменты, участвующие в расщеплении белков,
нуклеиновых кислот, полисахаридов, жиров и даже клеток и
бактерий. Известны заболевания, в основе которых лежит
снижение активности лизосом или их разрушение. В образовании
лизосом участвуют либо эндоплазматический ретикулум, либо
пузырьки аппарата Гольджи.
ЦЕНТРОСОМА
Как показывает само название, центросома расположена
вблизи центра клетки у ядра (рис. 2.1). Она содержит две
центриоли, которые в световом микроскопе выглядят как точки,
а на электронной микрофотографии — как маленькие цилиндры.
Эти органеллы принимают участие в образовании веретена в
процессе митоза.
КЛЕТОЧНАЯ МЕМБРАНА
Толщина клеточной мембраны, часто называемой также
плазматической, составляет около 75 А (7,5-10~6 мм); она
имеет складки, вдающиеся внутрь клетки. В состав клеточной
мембраны входят липиды и белкит относительное расположение
которых окончательно не выяснено. Существуют, однако,
некоторые гипотезы, основанные на косвенных данных. В течение
многих лет наиболее общепринятой была модель Даниэлли —
Дэвсона, предполагающая наличие двух слоев радиально
расположенных липидных молекул, полярные группировки которых
направлены наружу, а неполярные углеводные цепи — внутрь,
по направлению друг к другу. К каждому из слоев,
образованному полярными группировками липидов, примыкает слой
глобулярных белков.
Согласно этой модели, белки расположены в виде
протяженных слоев. Однако последние данные, полученные путем
спектрографического анализа, свидетельствуют в пользу
глобулярного строения белков, свернутых в виде спирали. Эти
данные позволили предложить жидкомозаичную модель мембраны
(рис. 2.2). Согласно этой модели, в мембране имеется двойной
слой фосфолипидных молекул, соединенных с цепями жирных
кислот (на рисунке — тонкие линии). Белки (ломаные
линии) на рисунке частично свернуты в виде спирали, причем их
заряженные группировки (Н ) обращены в сторону
внутренней и наружной поверхностей мембраны. В трехмерном
изображении, белки выглядят как крупные, беспорядочно
расположенные глобулы; последние данные свидетельствуют о том, что
одни глобулы обращены внутрь, а другие — наружу
относительно двойных фосфолипидных слоев.
Общая и клеточная физиология
37
Некоторые клетки снабжены микроворсинками —
пальцевидными выростами наружной мембраны, увеличивающими
всасывающую поверхность клетки.
В тканях клетки связаны между собой посредством
межклеточных соединений. К таким соединениям относятся плотные
контакты, где мембраны двух клеток примыкают друг к другу
и сливаются. При другом типе соединения мембраны разделены
относительно широкой щелью — 15—35 нм. Есть и третий тип
соединения, когда между мембранами клеток имеется щель
шириной 2 нм (щелевой контакт), через которую возможно про-
Рис. 2.2. Модель клеточной мембраны как жидкой мозаики из липидов и
глобулярных белков. А. Поперечный срез. Б. Трехмерное изображение
(Singer S. J., Nicolson G. L., Science, 175, 720, 1972).
хождение ионов и небольших молекул из одной клетки в
другую, минуя межклеточное пространство.
На основании данных о скорости перехода растворенных в
воде солей в клетку предположили существование в мембране
маленьких отверстий, или пор, диаметром около 7 нм. Однако
электронная микроскопия таких образований не обнаружила.
Главная функция плазматической мембраны заключается в
том, что сквозь нее движутся различные вещества из клетки и
в клетку. Это движение осуществляется при помощи целого
ряда механизмов, обсуждаемых в следующем разделе настоящей
главы и в гл. 12. Для одних веществ мембрана служит
барьером, для других — входными воротами. Проницаемость
мембраны высоко избирательна.— она препятствует прохождению
(диффузии) одних веществ, свободно пропуская другие.
Мембрана осуществляет также активный транспорт ряда веществ,
т. е. выполняет функцию насоса.
Проницаемость мембраны зависит от ее структуры и
размеров проходящих через нее молекул. Она практически
непроницаема для крупных белковых молекул и органических
анионов (А-), составляющих большинство внеклеточных анионов,
ограниченно проницаема для Na+ и свободно проходима для
К+ и С1+. Мембрана проницаема также для жирорастворимых
веществ.
38
Глава 2
ДВИЖЕНИЕ ВЕЩЕСТВ ЧЕРЕЗ КЛЕТОЧНЫЕ
МЕМБРАНЫ
Различные вещества могут перемещаться через мембрану в
том или ином направлении путем: 1) фильтрации, 2) пассивной
диффузии и осмоса, 3) облегченной, или опосредованной
переносчиком, диффузии, 4) диализа, 5) активного транспорта и
6) пиноцитоза.
ФИЛЬТРАЦИЯ
Процесс фильтрации зависит от разности давлений (АР)
снаружи и внутри кровеносного сосуда и его проницаемости
для жидкой части плазмы и низкомолекулярных веществ. Мак-
ромолекулярные соединения не проходят через поры мембраны.
В гл. 12 обсуждается роль фильтрации в образовании тканевой
жидкости.
ДИФФУЗИЯ
Пассивной диффузией называется перемещение молекул из
области высокой концентрации в область низкой концентрации»
Это передвижение носит вероятностный характер. Скорость
диффузии определяется: 1) концентрационным (химическим)
градиентом раствора (концентрацией); 2) электрическим
градиентом, т. е. электрическими зарядами ионов; 3)
проницаемостью мембраны, т. е. соотношением между размерами пор
мембраны и молекул вещества; 4) размерами молекул
вещества: скорость диффузии обратно пропорциональна размеру
молекул. Так, диффузия воды гораздо выше, чем других веществ»
РАВНОВЕСИЕ ДОННАНА
Обмен жидкостью и некоторыми способными к диффузии
ионами (К+, С1+, Na+) между кровью и тканями и между
внеклеточным и внутриклеточным пространствами
осуществляется путем диффузии, при которой ионы свободно проходят через
мембрану в соответствии с их концентрационными градиентами
(от большей концентрации к меньшей). Крупные белки плазмы
не диффундируют через мембрану, и их электрические
(отрицательные) заряды препятствуют движению и распределению»
способных к диффузии ионов. Наблюдающееся в результате
Общая и клеточная физиология
39
этого разделение ионов можно точно вычислить и предсказать
следующим образом:
А Мембрана Б
к+
С1-
Белок-
К+
С1-
Мембрана частично проницаема для К+ и С1~, но не для
белка, и его отрицательный заряд (—) препятствует диффузии
катионов ( + ). В результате частички с большей осмотической
активностью скапливаются по одну сторону мембраны (А);при
этом [К+]а+ [С1-]А+;[Белок-]А> [К+]б +[С1-]б. Однако
диффундирующие ионы (К+ и С1~) распределятся по обе стороны
мембраны и будут удовлетворять равенству
[К+]А _ [СГ]Б
[К+]Б ~~ [С1-]А "
Это и есть так называемое равновесие Доннана.
Белки — важнейшие компоненты плазмы — редко
диффундируют через капилляры в тканевую жидкость, поэтому эффект
Доннана играет в организме важную роль. Количественное
распределение ионов по обе стороны мембран и их роль в
генерации электрической активности клеток (потенциала
действия) и изменения мембранного потенциала обсуждаются
в гл. 4 и 5.
ВЫЧИСЛЕНИЕ СКОРОСТИ ДИФФУЗИИ
Уравнение Фика. Скорость диффузии растворенного вещест-
ва (-37-) равна количеству этого вещества, перемещающегося
из области высокой концентрации (Si) в область низкой
концентрации (52) за единицу времени
ds _ _ п А (Si — S2)
dt u* dX, или Т '
где D — коэффициент диффузии, постоянный для данного
растворенного вещества и обратно пропорциональный размеру
молекул (см. гл. 1), S\—52 — разность концентраций
растворенного вещества (диффузионный поток), А — площадь
поперечного сечения, dX — диффузионное расстояние, или толщина
{Т) мембраны.
Таким образом, скорость диффузии прямо
пропорциональна D, А и разности концентраций (Si — S2) и обратно пропор-
fv7l"?\:.:,J
s -
88
.'•'.'Д4-:-:
—>
76
NaCI
H20
•; 18-. ;-
s
4
<--
82
:•.': 18 ':'•;'•
s
•*
82
NaCI
Рис. 2.З. Диффузия. А. 12%-ный и 24%-ный растворы разделены мембраной;
для NaCI и воды существуют концентрационные градиенты; диффузия
осуществляется из области высокой концентрации в область низкой
концентрации (по концентрационному градиенту); преимущественные направления
диффузии NaCI (—*) и воды ( >-) взаимно противоположны. Б. Диффузия
по градиенту концентрации прекращается, и между растворами
устанавливается равновесие: концентрации NaCI и воды по обе стороны мембраны
выравниваются и диффузия идет в обоих направлениях (двойные стрелки).
Цитоплазма клетки
Клеточная мембрана
Просвет кишечника
Рис. 2.4. Схема всасывания глюкозы в кишечнике — пример опосредованного
переносчиком (активного) транспорта. Na+ облегчает всасывание глюкозы.
Энергия, необходимая для работы натриевого насоса, поступает от АТФ
(Crane R. К., Fed. Ргос, 24, 1000, 1965).
40
Общая и клеточная физиология 4£
циональна толщине мембраны, или диффузионному
расстоянию. На рис. 2.3 приведен пример диффузии вещества из
раствора большей концентрации в раствор меньшей концентрации.
Мембрана в данном случае проницаема как для растворенного
вещества (NaCl), так и для растворителя (Н20).
ОБЛЕГЧЕННАЯ ДИФФУЗИЯ
Об облегченной диффузии говорят в том случае, когда
какое-либо вещество способно свободно диффундировать, но этот
процесс облегчается другим веществом, или молекулой,
переносящим первое. Такая диффузия идет по градиенту
концентрации. В качестве примера можно привести диффузию, или
транспорт, глюкозы из крови, где ее концентрация высока, в клетки
тканей, где концентрация глюкозы низка. Эта диффузия
облегчается (опосредуется) молекулой-переносчиком. Транспорт же
глюкозы против концентрационного градиента (всасывание из
:шшечника)—это активный процесс, требующий затраты
энергии (рис. 2.4).
ДИАЛИЗ
Диализ — это диффузия вещества через полупроницаемую
мембрану, пропускающую мелкие и задерживающую крупные
молекулы (рис. 2.5).
ОСМОС
Осмос — это диффузия
растворителя (воды) через
полупроницаемую мембрану, т. е.
мембрану, не пропускающую
некоторые растворенные
вещества. Так, стенка
кровеносных капилляров относительно
непроницаема для белков,
молекулы которых слишком
крупны и не могут выходить
из сосудов. В результате
белки плазмы создают
осмотическое давление (см. гл. 12),
противодействующее
гидростатическому, или
фильтрационному, давлению и движению
воды и некоторых веществ
через стенку сосуда. Осмоти-
p-:o-o-ofo'
о;о'ор'0-
Рис. 2.5. Диализ. В банку с водой
помещен мешочек с раствором
кристаллоидов (глюкоза) и коллоидов
(белок). Глюкоза диффундирует
через полупроницаемую мембрану в
воду, тогда как молекулы белка не
проходят через нее и остаются в
мешочке.
42
Глава 2 .
II
\{::§#
щщ
-■»
Рис. 2.6. Осмос. /. Два различных раствора глюкозы (6%- и 12%-ный)
разделены мембраной, проницаемой для воды, но не пропускающей глюкозу.
Следовательно, через мембрану может проходить только вода из того раствора,
где содержание ее больше, т. е. из 6%-ного раствора глюкозы (указано
стрелкой). Концентрация глюкозы по обе стороны мембраны может сравняться
только в результате разбавления водой более концентрированного раствора.
Это приводит к уменьшению (—) объема воды в менее концентрированном
растворе и увеличению (+) в более концентрированном. //. Одновременно
давление на той стороне, где объем увеличивается, повышается, а на той, где
объем уменьшается,— понижается (подразумевается, что отсек, в который
переходит вода, способен растягиваться); диффузия воды через мембрану идет
в обе стороны (двойные стрелки).
ческое давление удерживает жидкость в сосудистом русле и
оказывает влияние на всасывание тканевой жидкости. Пример
осмоса приведен на рис. 2.6.
ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ
Осмотическое давление зависит от числа частиц
растворенного вещества в растворе. Формулируя закон осмотического
давления, Вант-Гофф использовал аналогию сильно
разбавленного раствора со смесью идеальных газов (закон Авогадро).
В соответствии с этим законом одна грамм-молекула любого
вещества в 1 л растворителя (одномолярный раствор)
содержит одинаковое число молекул и при 0°С может создавать
давление 22,4 атм (17 024 мм рт. ст.), а при 37°С —19 300 мм
Общая и клеточная физиология
43
рт. ст. Таким образом,
осмотическое давление
зависит от числа частиц в
1 л раствора, что
определяется молярной
концентрацией раствора и
числом ионов в молекуле.
Осмотическое
давление — это то давление,
которое оказывал бырас-
-твор, отделенный от
воды мембраной с
избирательной проницаемостью.
Давление,
препятствующее при этих условиях
переходу растворителя
через мембрану (рис. 2.7),
называется эффективным
осмотическим давлением.
Объем отсека А,
содержащего раствор
сахарозы (рис. 2.7), постоянен;
•отсек замкнут и соединен
с манометром. Вода
будет переходить из
отсека Б в отсек А до тех пор, пока давление в манометре не
сравняется с осмотическим давлением раствора сахарозы.
Когда вода переходит (диффундирует) в раствор вещества,
«е проникающего через мембрану, давление и объем в отсеке,
содержащем более концентрированный раствор, повышается.
Рис. 2.7. Осмотическое давление. Резервуар
с водой разделен на два отсека
полупроницаемой мембраной, пропускающей воду, но
не сахарозу. В отсек Л, заполненный водой
и закрытый, помещен кубик сахара. Отсек
£, содержащий воду, открыт. Вода
перемещается из отсека Б в отсек А до тех пор,
пока давление водяного столба в открытой
манометрической трубке не сравняется с
осмотическим давлением раствора сахара
(Dowben R. W. в кн.: Goldstein L.,
Comparative Physiology, 1977).
Расчет осмотического давления при 37 °С
Осмотическое давление неэлектролита в мм рт. ст. равно:
Молярность раствора= 19 300
10%-ный раствор глюкозы = 100г/л
Молекулярный вес глюкозы =180
100
— =0,555; 0,555-19 300=10 711 (мм рт. ст.)
180
Для раствора электролита:
4%-ный раствор NaCl=40 г/л
Молекулярный вес NaCl = 58
Число ионов в одной молекуле = 2
40
58
= 0,69; 0,69-2-19 300=26 634 (мм рт. ст.)
44
Глава 2
Изоосмотические, гипоосмотические и гиперосмотические растворы
Осмотическое давление изоосмотических растворов
одинаково; осмоса между ними не происходит, и они считаются
изотоническими.
Осмотическое давление гипоосмотических, гипотонических
растворов ниже осмотического давления некоего эталонного
раствора; давление же гиперосмотических, гипертонических,
растворов выше давления такого раствора. Более подробно
осмотическое и фильтрационное давление крови обсуждается
в гл. 12 и 14.
АКТИВНЫЙ ТРАНСПОРТ
При активном транспорте вещество переносится против
концентрационного или электрохимического градиента.
Необходимая для этого энергия образуется в ходе метаболических
реакций, обычно при гидролизе АТФ i (аденозинтрифосфата). Этот
гидролиз катализируется специфическим ферментом —
АТФазой; полагают, что АТФаза осуществляет активный
транспорт Na+, т. е. может считаться натриевым насосом. После
деполяризации клетки Na+ выкачивается из нее через мембрану
против высокой внеклеточной концентрации этого иона.
Гидролиз 1 моля АТФ сопровождается выкачиванием из клетки
около 2—3 молей :Na+; при этом удалению каждых трех ионов
Na+ сопутствует накачивание в клетку двух ионов К+ («натрий-
калиевый насос»). Так как при переносе в клетку одного
иона К+ из нее удаляется более одного иона Na+, в конечном
итоге имеет место выход положительных зарядов из клетки, что
приводит к гиперполяризации. Точные механизмы действия
натриевого насоса в последнее время привлекли внимание
многих исследователей (см. также гл. 3 и 4).
Облегчающее влияние работы натриевого насоса на
транспорт глюкозы, опосредованный переносчиком, схематически
показано на рис. 2.4.
пиноцитоз, экзоцитоз, фагоцитоз
Эти разновидности активного транспорта связаны с
деятельностью самой клетки.
Фагоцитоз — способность лейкоцитов «заглатывать»
бактерии (см. гл. 13). При пиноцитозе вокруг того или иного тельца,,
находящегося вне клетки, в результате углубления мембраны
1 В биохимической литературе в настоящее время принято для этого
соединения латинское сокращение АТР. — Прим. ред.
Общая и клеточная физиология
45
образуется «кармашек»; затем он отделяется от остальной
мембраны и передвигается внутрь клетки в виде замкнутого
пузырька.
ООО00'
Экзоцитоз — процесс, обратный пиноцитозу. При экзо-
цитозе та или иная внутриклеточная гранула перемещается
по направлению к плазматической мембране;- мембрана этой
гранулы сливается с плазматической; в конечном счете гранула
выталкивается из клетки.
О О с?
МЕМБРАННЫЕ ПОТЕНЦИАЛЫ
В клетках имеется электрохимический градиент,
обусловленный разницей концентрации внеклеточных и
внутриклеточных ионов. Это так называемый мембранный потенциал. У
разных клеток и тканей, а также в разных физиологических
условиях он различен. Когда клетка находится в покое, ее мембрана
поляризована: внутренняя поверхность мембраны заряжена
отрицательно по отношению к наружной, несущей
положительный заряд.
Мембранный потенциал измеряется в милливольтах (мВ).
Величина потенциала покоя колеблется от —10 до —100 мВ;
для клеток миокарда характерен потенциал от —80 до —90 мВ.
При деполяризации клетки (например, при раздражении
мышцы или нерва) ионы перемещаются через мембрану и возникает
электрический импульс; это приводит к распространению
возбуждения и потенциала действия. Мембранный потенциал
изменяется и становится менее отрицательным. Подробнее о
потенциале действия сказано в гл. 4.
КИСЛОТНО-ЩЕЛОЧНОЕ РАВНОВЕСИЕ
Термин «кислотно-щелочное равновесие» относится к под-
Держанию постоянства (гомеостаза) концентрации водородных
ионов [Н+] в жидкостях организма. Кислотность обычно
выражается в виде концентрации водородных ионов, или рН.
46
Глава 2
рН является мерой концентрации свободных водородных
ионов [Н+]. Кислотность, определяемая титрованием,
обусловлена свободными и связанными водородными ионами; она
выражается в миллиэквивалентах/л (мэкв/л), или в граммах
способного к обмену Н+ на 1 л. Один эквивалент кислоты
нейтрализует один эквивалент основания.
Степень диссоциации кислоты отражает константа
диссоциации, или константа равновесия (Ка):
Ка прямо пропорциональна степени ионизации и силе кислоты;
рКа обратно пропорционален силе кислоты и величине Ка.
Ионизация и диссоциация сильной кислоты типа 0,1 н. НО
гораздо сильнее, чем слабой.
Сильная кислота (серная) H2S04 1,2-Ю-2 1,9
Слабая кислота (угольная) Н2С03 7,9-Ю-7 6,1
pH=-lg[H+]=lg^|
Чем выше кислотность, тем меньше значение рН. Если при
25 °С рН = 7, то концентрации ионов Н+ и ОН~ равны и такой
раствор нейтрален.
Если рН=1, то [Н+] = 10-1, или —
L J 10
Если рН=7, то [Н+] = 10-7, или
10 000 000
Если рН больше 7, то раствор щелочной.
БУФЕРНЫЕ СИСТЕМЫ
Большинство процессов в организме протекает при реакции
среды, близкой к нейтральной; поддержание этой реакции
обеспечивается целой системой буферов, т. е. веществ,
препятствующих значительным сдвигам рН при добавлении в среду
сильных кислот или оснований (щелочей).
Буферы представляют собой смесь слабой кислоты и
основания или соли. Основные буферы крови и тканей следующие:
, г Л „ , ^ К-Hb К-НЬ02 Соль
1. Гемоглобиновыи буфер и
НЬ НЬ02 Кислота
НЬ (восстановленный)
НЬ02 (окисленный)
Общая и клеточная физиология
47
л _ Протеинат Na
2. Белки плазмы крови
Белки (слабые кислоты)
Na2HP04 (основной фосфат)
3. Фосфатный буфер
NaH2P04 (кислый фосфат)
_ , NaHC03 КНСОз Соль
4. Бикарбонатныи буфер или •
Н2С03 Н2С03 Кислота
В буферных системах происходит замена сильной кислоты
слабой, при диссоциации которой образуется меньше ионов Н+,
и, следовательно, рН раствора снижается в меньшей степени
(см. гл. 17). В наибольших количествах в организме образуется
угольная кислота, однако кислота эта слабее, чем молочная,
которая буферируется, или нейтрализуется, бикарбонатом
(НС03~) и замещается угольной кислотой (Н2С03).
Проницаемость, всасывание, транспорт и выделение
различных веществ в организме зависят от степени ионизации и
диссоциации, которая в свою очередь определяется значением
рН и температурой окружающей среды (см. гл. 24).
УРАВНЕНИЕ ГЕНДЕРСОНА — ГАССЕЛЬБАХА
рКа кислоты соответствует тому значению рН, при котором
концентрации ее ионизированных и неионизированных форм
равны. рКа различных веществ и буферных растворов известны,
и для удобства составлены их таблицы. Уравнение Гендерсо-
на — Гассельбаха выглядит следующим образом:
pH = Ptfa+lg- соль
кислота
При помощи этого уравнения можно определить: 1)
соотношение соли и кислоты в буферной системе, если известны рН
и р/(а, 2) рН, если известны рКа и соотношение соли и кислоты.
Содержание НС03~ и Н2С03 в крови в норме составляет около
26 и 1,3 мэкв/л соответственно.
Следовательно, диссоциация наступает при рН среды,
равном:
u is i i^ HCOo" 26 мэкв
pH=6,l + lg-o- = 6,l+lg 20 = 6,1 + 1,3 = 7,4.
Роль дыхания и почек в регуляции кислотно-щелочного
равновесия подробно обсуждается в гл. 17 и 24.
48
Глава 2
ЛИТЕРАТУРА
Davson H. (1970). A textbook of general physiology (4th ed.), Williams and Wil-
kins, Baltimore.
Dowben R. (1977). Membrane physiology. In: Goldstein L. (ed.), Introduction to
Comparative Physiology, Holt, Rinehart and Winston, New York.
Oanong W. F. (1977). Review of Medical Physiology (8th ed.), Lange Medical,
Los Altos, Cal.
Giese Л. С. (1973). Cell Physiology (4th ed.), Saunders. Philadelphia.
Singer S. J., Nicolson J. L. (1972). The fluid mosaic model of the structure of
cell membranes, Science, 175, 720.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Нарисуйте схему строения клетки и укажите основные ор-
ганеллы в ядре и цитоплазме.
2. Что такое рибосомы и какова их функция?
3. Что такое митохондрии и какова их функция?
4. Что такое лизосомы и какова их функция?
5. Что такое плазматическая мембрана и почему она так
важна для жизнедеятельности клетки?
6. Назовите механизмы или пути, при помощи которых
различные вещества переносятся через клеточную мембрану.
7. В чем разница между пассивной диффузией и активным
транспортом?
8. Что такое равновесие Доннана?
9. Что означают термины: «изоосмотический», «гипоосмотиче-
ский», «гиперосмотический»?
10. Чему равно осмотическое давление 10%-ного раствора
глюкозы? Сделайте расчет.
11. Откуда поступает энергия для активного транспорта?
12. Дайте определение диализа.
13. Дайте определение рН.
14. Что такое буферные системы и в чем их главная роль?
15. Из чего состоят бикарбонатные буферы (приведите пары
соединений).
16. Что такое р/( кислоты?
Глава 3
ОРГАНИЗАЦИЯ НЕРВНОЙ СИСТЕМЫ
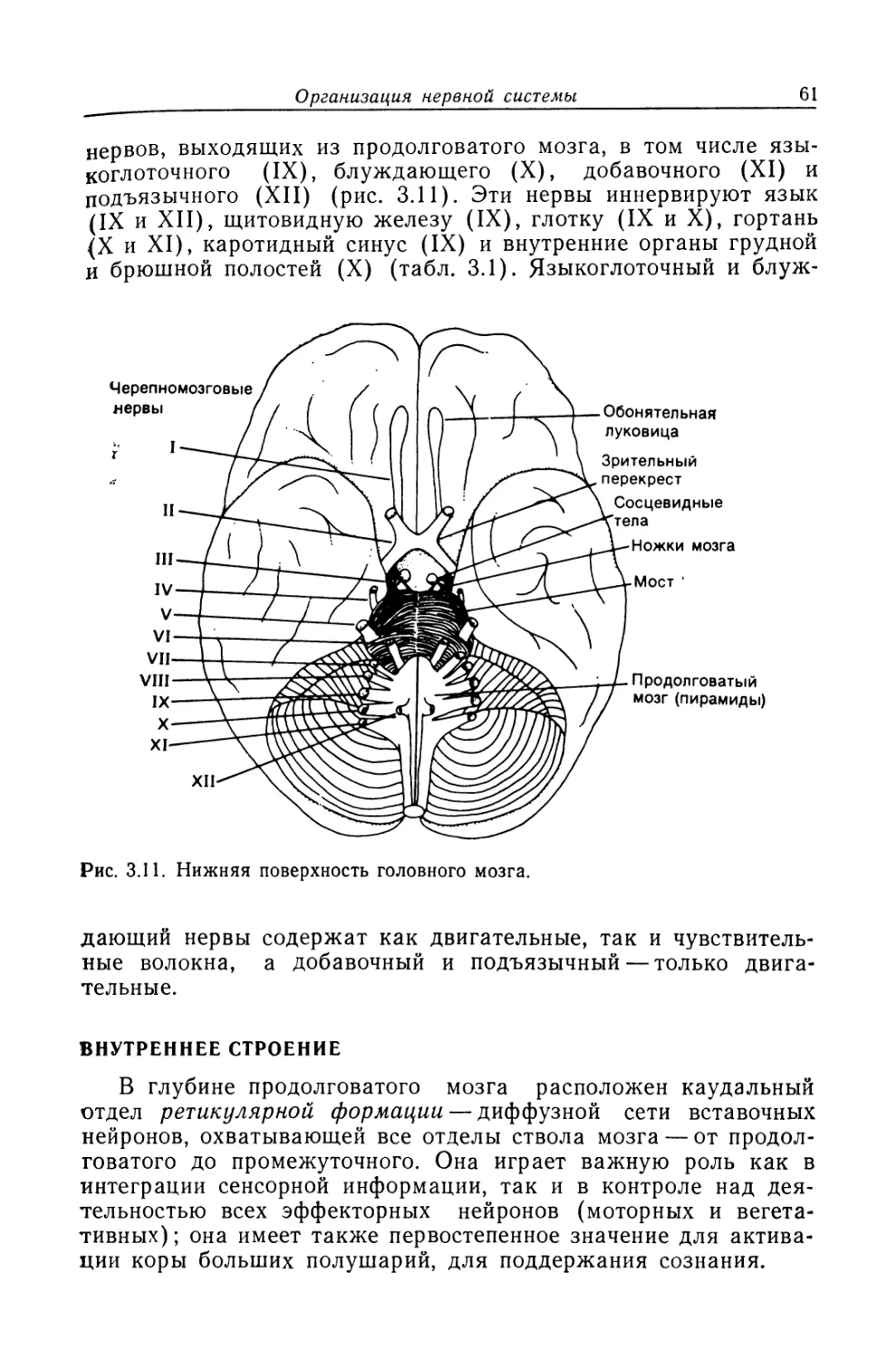
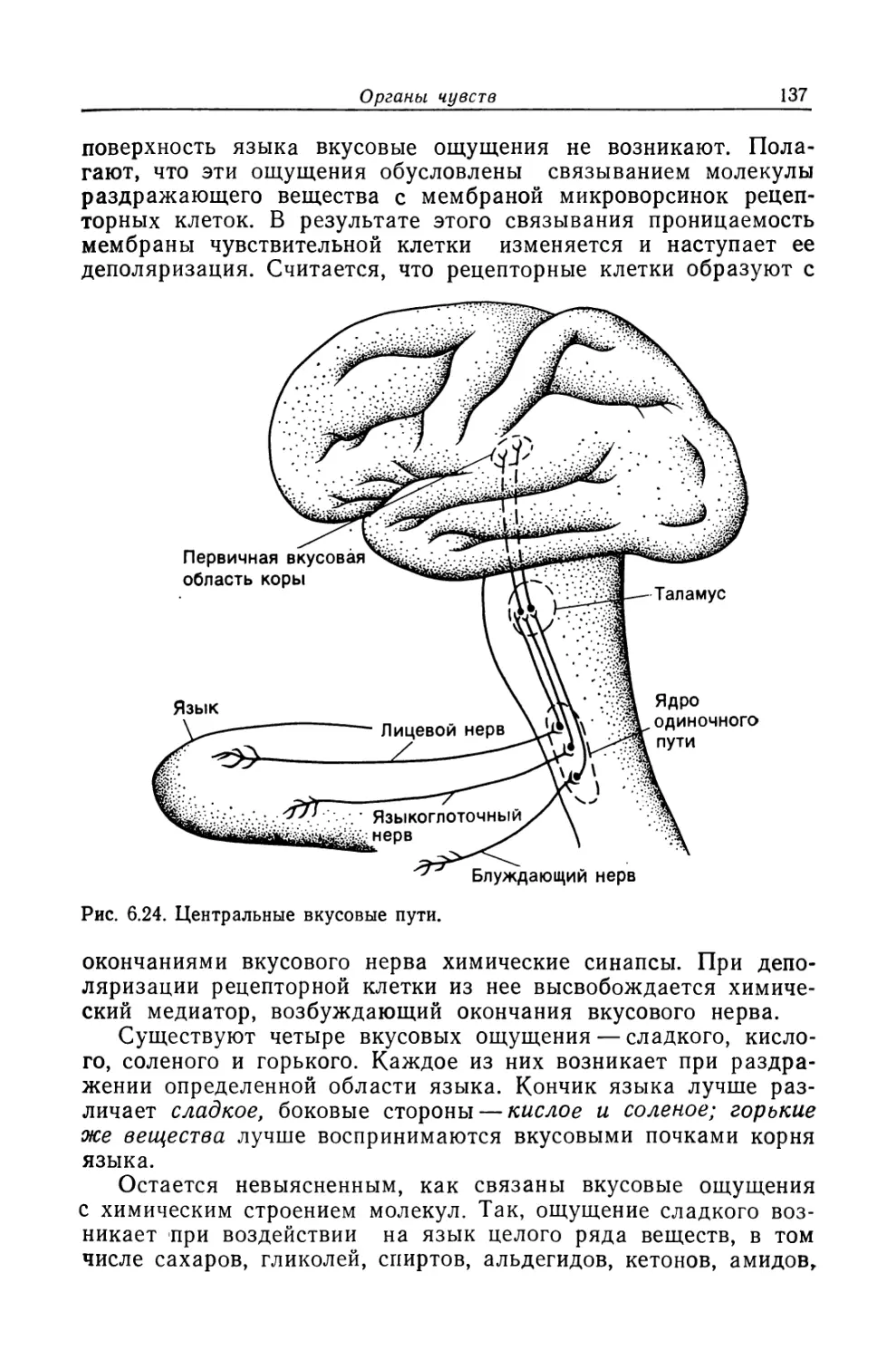
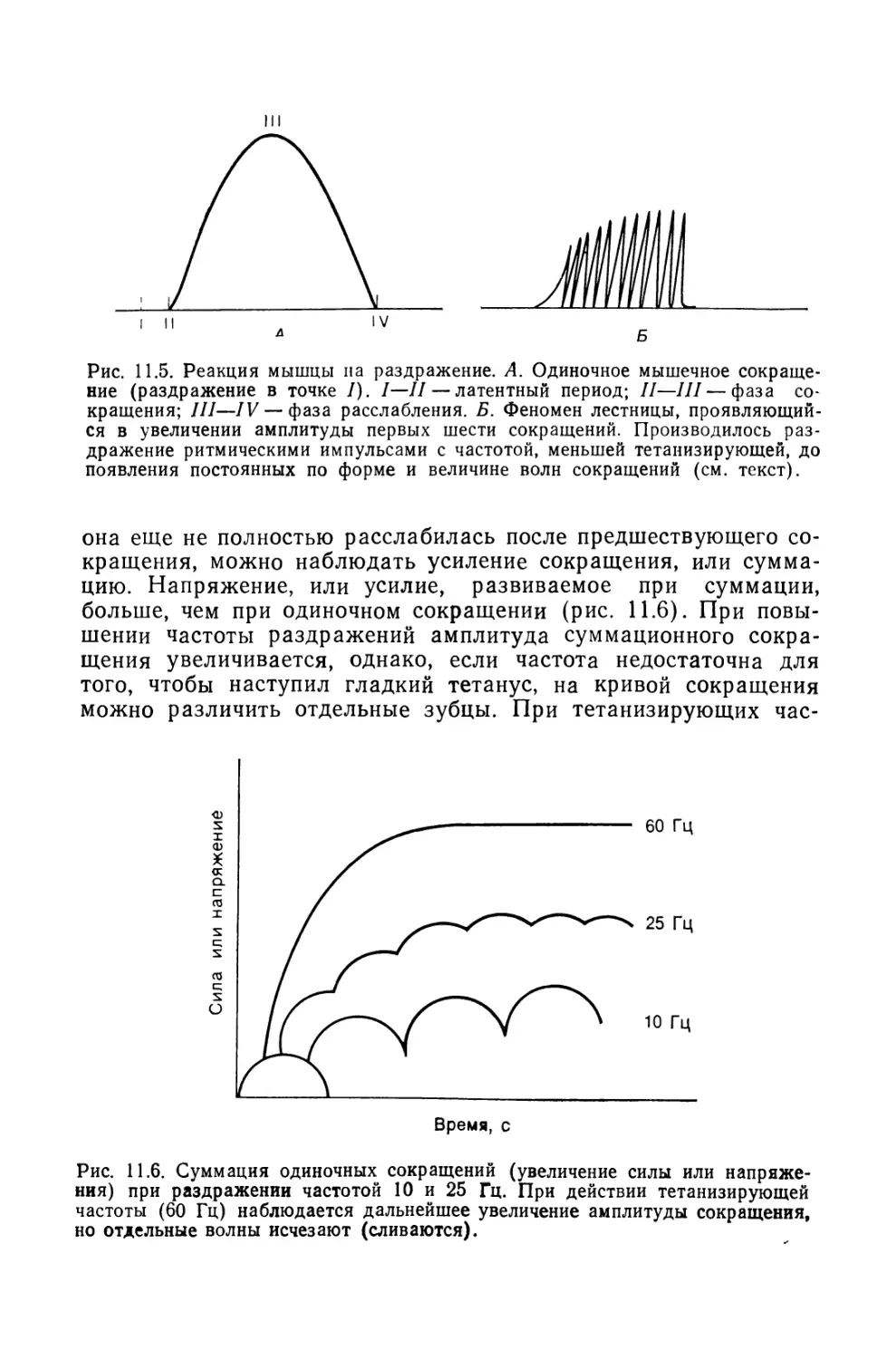
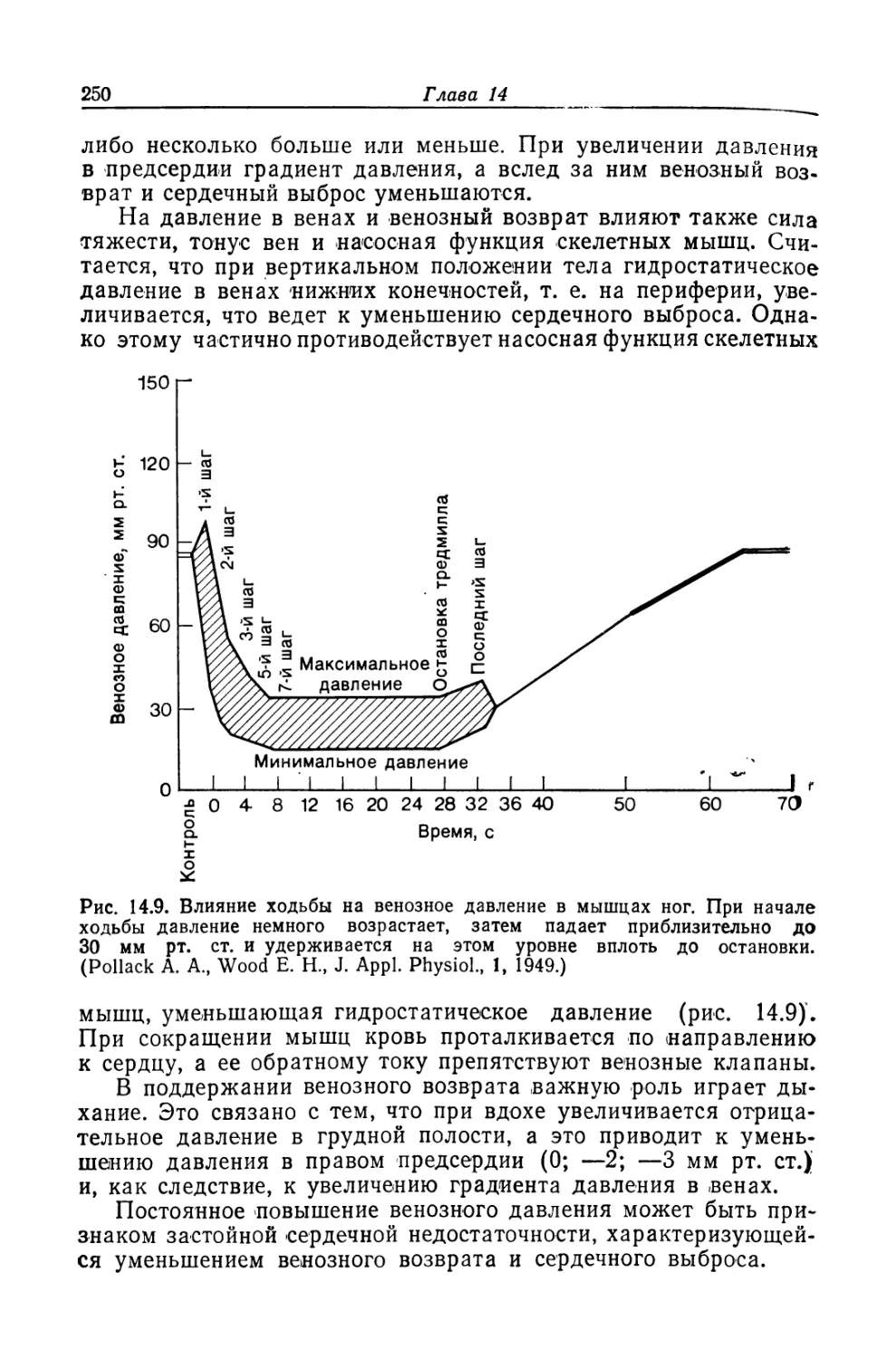
Из всех известных человеку структур его собственная
нервная система — самая сложная. Она содержит 50 миллиардов
нервных клеток, объединенных в невероятно сложную сеть.
В мозгу находятся чувствительные центры, анализирующие
изменения, которые происходят как во внешней, так и во
внутренней среде. Он управляет всеми функциями организма,
включая мышечные сокращения и секреторную активность желез.
НЕЙРОН
Нейрон, или нервная клетка, — это функциональная единица
нервной системы, строение и функции которой приспособлены
к передаче и интеграции информации. В каждом нейроне
различают четыре различные области: тело, дендриты, аксон и ак-
сонные окончания (терминали) (рис. 3.1). Все эти области
выполняют строго определенные функции.
Центр процессов синтеза в нервной клетке — ее тело, или
сома, — содержит ядро, рибосомы, эндоплазматический ретику-
лум и другие органеллы. Здесь синтезируются медиаторы,
клеточные белки и другие важнейшие компоненты. Сома имеет
первостепенное значение для существования и целостности
нейрона. При ее разрушении дегенерирует вся клетка, включая
аксон с его терминалями и дендриты. Главная функция аксона
состоит в проведении нервных импульсов к другим клеткам —
нервным, мышечным или секреторным. Большинство аксонов
представляет собой длинные нитевидные отростки, исходящие
из сомы. До переключения на воспринимающие отростки других
нейронов они проходят путь от нескольких миллиметров до
нескольких метров. Ряд аксонов соединяет центральную нервную
систему (головной и спинной мозг) с периферической. Аксоны
чувствительных (сенсорных) нейронов передают информацию
от расположенных на периферии рецепторов к центральной
нервной системе (ЦНС). Аксоны двигательных (моторных)
нейронов проводят нервные импульсы от ЦНС к мышцам
туловища и конечностей. Другие аксоны соединяют ЦНС с рецеп-
4-1705
50
Глава 3
торами, мышечными и секреторными клетками внутренних
органов.
Специфической функцией аксона является проведение
нервных импульсов. Эти импульсы возникают в результате
небольших изменений проницаемости мембраны аксона, приводящих
к возникновению электрического потенциала; последний,
подобно волне, пробегает по всей длине аксона — от сомы до
окончаний.
Ближе к окончанию аксон ветвится и образует тонкую
кисточку из конечных ветвей (аксонных терминалей, или оконча-
Рис. 3.1. Нейрон.
ний). На конце каждая терминаль образует
специализированный контакт, или синапс, с постсинаптической клеткой
(нервной, мышечной или железистой). Подавляющее большинство
синапсов ЦНС образовано окончаниями аксонов одних
нейронов на дендритах других.
Специальная функция синапса заключается в передаче
информации от клетки к клетке. Когда к окончанию аксона
приходит нервный импульс, в этом окончании образуется
небольшое количество особого химического вещества—нейромедиато-
ра. Высвобождаясь из окончания, медиатор связывается с
мембраной дендрита постсинаптического нейрона и изменяет ее
проницаемость, что приводит к сдвигу ее электрического
потенциала. Возникающий в результате этого синаптический
потенциал может быть возбуждающим или тормозным. В первом
случае он увеличивает вероятность генерации нервного
импульса в постсинаптическом нейроне; тормозный же постсинаптиче-
ский потенциал, напротив, этому препятствует.
Дендриты образуются в результате древовидного
разветвления отростков нервной клетки, отходящих от ее тела; их
специальная функция заключается в восприятии синаптнческих
Организация нервной системы
51
влияний. На дендритах типичной нервной клетки оканчиваются
терминали аксонов сотен или тысяч нейронов. Эти терминали
покрывают всю поверхность дендрита. В активном состоянии
каждая терминаль высвобождает медиатор, вызывающий
местное изменение проницаемости мембраны дендрита. В результате
этого меняется ее электрический потенциал. Это изменение
потенциала (синаптический потенциал) передается от дендритов
к начальному сегменту аксона. Если синаптический потенциал
возбуждающий, то частота генерации нервных импульсов
возрастает, если же он тормозный — уменьшается.
КЛЕТКИ ГЛИИ
Хотя именно нервные клетки являются функциональными
единицами, обрабатывающими информацию, на их долю
приходится лишь 10% общего числа клеток в нервной системе.
Периваскуляр' ый \ \-—-капилляр
отросток
Аксоны
Астроцит Олигодендроиит
Миелиновая оболочка
Д,
L- Аксон
Микроглиальная клетка Шванновская клетка
Рис. 3.2. Четыре основных вида клеток глии.
Большинство же здесь составляют глиальные клетки,
заполняющие все пространство между нейронами. Существуют четыре
основных разновидности глиальных клеток (рис. 3.2): астроциты,
олигодендроциты и микроглия, находящиеся в головном и
спинном мозгу, и шванновские клетки, расположенные в перифери-
4*
52
Глава 3
ческих нервах. Многие клетки глии — олигодендроциты в ЦНС
и шванновские клетки периферических нервов — тесно связаны
с длинными нервными путями, образованными пучками аксонов.
Многие крупные аксоны как бы заключены в футляр из
мембранных выростов глиальных клеток, образующих миели-
новую оболочку (рис. 3.1). Последняя изолирует мембрану
аксона, что играет очень важную роль, так как способствует
повышению скорости проведения нервного импульса. Другие
глиальные клетки — астроциты — расположены между
кровеносными сосудами и телами нейронов. Некоторые их отростки
контактируют со стенкой капилляров. Эти периваскулярные
отростки служат компонентом гематоэнцефалического барьера.
Многие нейробиологи считают, что клетки глии регулируют
транспорт питательных веществ от капилляров к нейронам.
Предполагают, что между клетками глии и связанными с ними
нейронами осуществляется обмен белками, нуклеиновыми
кислотами и другими важными веществами. Ряд данных
свидетельствует о том, что активность нейронов способна влиять на
мембранный потенциал глиальных клеток путем увеличения
концентрации К+ во внеклеточном пространстве.
Клетки микроглии — это клетки-мусорщики, или фагоциты
мозга. Они входят в состав ретикулоэндотелиальной системы
(см. гл. 13). Клетки микроглии редки в неповрежденном мозгу,
в области же повреждений ткани мозга они всегда
представлены в изобилии.
СХЕМА СТРОЕНИЯ НЕРВНОЙ СИСТЕМЫ
Все нейроны можно разделить на 3 класса: чувствительные
(сенсорные), вставочные и эффекторные (рис. 3.3).
Чувствительные и эффекторные нейроны связывают структуры,
расположенные на периферии (рецепторы, мышцы и железы), с ЦНС
(головным и спинным мозгом). Чувствительные нейроны
представляют собой афферентные пути, по которым импульсы
передаются от рецепторов в ЦНС, а эфферентные нейроны проводят
импульсы от ЦНС к эффекторам (мышцам и железам). К эф-
фекторным нейронам относятся двигательные (моторные)
нейроны, иннервирующие скелетные мышцы, и нейроны
вегетативной нервной системы, осуществляющие центральную регуляцию
мышц и желез внутренних органов.
Отростки вставочных нейронов не выходят за пределы ЦНС.
Почти все нейроны ЦНС, за исключением сенсорных и эффек-
торных, являются вставочными. В центральной нервной системе
вставочные нейроны образуют цепи, осуществляющие анализ
входной сенсорной информации, хранение опыта в виде памяти
и формирование соответствующих нервных команд.
Организация нервной системы
53
Варианты структуры вставочных нейронов почти
бесконечны (рис. 3.3). Амакриновые клетки сетчатки или клетки-зерна
обонятельной луковицы представляют собой мелкие нервные
клетки без аксонов, тогда как другие нейроны, например
крупные пирамидные клетки двигательной области коры, обладают
аксонами длиной в 1 м и более. Отростки некоторых нейронов
Чувствительные
Вставочные
Двигательные-
У ^
ОбН АмН
■ж*тт
J^<
Рис. 3.3. Три класса нейронов. Обратите внимание на большое разнообразие
строения вставочных нейронов. В качестве сенсорных нейронов изображены:
нейрон, отросток которого идет в составе слуховых волокон преддверно-улит-
кового нерва (VIII пара), нейрон, реагирующий на стимуляцию кожи (КН).
Вставочные нейроны представлены амакриновой (АмН) и биполярной (БН)
клетками сетчатки, нейроном обонятельной луковицы (ОбН), нейроном
голубоватого места (ГМН), пирамидной клеткой коры головного мозга (ПН) и
звездчатым нейроном (ЗН) мозжечка. В качестве двигательного нейрона
изображен мотонейрон спинного мозга.
(биполярных клеток сетчатки и звездчатых клеток коры)
разветвляются в очень ограниченной области нервной системы,,
других же (клетки голубоватого места в стволе
мозга)—распространяются по большей части массы мозга.
Нервная система организована таким образом, что
вставочные нейроны, выполняющие одинаковые функции (т. е. с
одинаковыми входами и выходами), сгруппированы в виде так
называемых ядер (рис. 3.4). В мозгу имеются сотни различных,
ядер, каждое из которых содержит тысячи нейронов,
участвующих в интеграции тесно связанных между собой функций.
В дальнейшем будут обсуждаться, в частности, ядра базальных
ганглиев, мозжечка, гипоталамуса и таламуса.
Наиболее сложна нервная организация коры. Все ее
отделы— неокортекс, кора мозжечка и гиппокампа — состоят из
нескольких слоев нейронов и их отростков (рис. 3.4). В больший-
54
Глава 3
Вход
Выход
Наружный
плексиформный слой
Средний
плексиформный слой
Внутренний
плексиформный слой
->
Выход
Рис. 3.4. Сложные формы организации нейронов. Л. Ядро — скопление
нейронов с одинаковыми входами и выходами: все нейроны, входящие в состав
какого-либо ядра, выполняют одну общую функцию. Б. Кора — многослойная
структура, состоящая из чередующих ядерных и волоконных слоев. Нейроны,
расположенные в одной и той же области коры, получают одинаковую
информацию.
стве областей кора образована чередующимися ядерными
(содержащими тела клеток) и плексиформными (содержащими
дендриты и синаптические окончания) слоями. Разные слои
коры состоят из разных нейронов. Центростремительные пути
к коре обычно оканчиваются в одном или двух плексиформных
слоях; аксоны же, передающие сигналы из коры к другим
отделам ЦНС, как правило, отходят от основания одного из
ядерных слоев. Подробнее кора большого мозга и мозжечка
описана в гл. 6 и 8.
Организация нервной системы
55
АНАТОМИЧЕСКАЯ ОРГАНИЗАЦИЯ НЕРВНОЙ СИСТЕМЫ
Нервная система подразделяется на две основные части:
центральную и периферическую. Центральная нервная система
состоит из нейронов, их отростков и глии, расположенных в
головном и спинном мозгу. Периферическая нервная система,
напротив, образована нейронами, отростками и глией,
находящимися за пределами ЦНС. К ней относятся все нервные
отростки, идущие в составе периферических нервов (черепномоз-
- Боковой желудочек
-Третий желудочек
Сильвиев водопровод
Четвертый желудочек.
Рис. 3.5. Развитие нервной системы. А. На 3-й неделе внутриутробного
развития. Б. На 7-й неделе.
говых, спинномозговых и вегетативных), а также
расположенные на периферии скопления нервных клеток — чувствительные
и вегетативные ганглии.
Анатомически ЦНС состоит из четырех отделов,
образующихся у человека на третьей неделе внутриутробного развития
(рис. 3.5). К ним относятся передний мозг (прозенцефалон),
средний мозг (мезенцефалон), задний мозг (ромбенцефалон) и
спинной мозг. На седьмой неделе эмбриогенеза происходит
дальнейшее разделение переднего и заднего мозга, в результате
чего образуются пять отделов, имеющихся и в зрелом мозгу:
конечный мозг (теленцефалон), промежуточный мозг (диенце-
фалон), средний мозг (мезенцефалон), задний мозг (метенце-
фалон) и продолговатый мозг (миеленцефалон). Конечный мозг
и промежуточный мозг являются производными переднего
мозга, а задний мозг и продолговатый — ромбовидного (заднего).
Внутри каждого из этих отделов находится пространство,
заполненное жидкостью, или желудочек (рис. 3.5); всего таких
Передний мозг
Средний мозг
Задний -
мозг
Спинной мозг
Конечный мозг
Промежуточный
мозг
Средний мозг
Задний мозг
Продолговатый
мозг
Спинной мозг
56
Глава 3
1-й шейный (С1)
1-й грудной (Т1)
т 12
1-й поясничный (L1).
1-й крестцовый (S1)
1-й копчиковый
.^Кора головного
мозга
Мозжечок
Продолговатый мозг
Шейные позвонки
Спинной мозг
Грудные
позвонки
Поясничные
позвонки
Крестец
Концевая нить
Копчик
Рис. 3.6. Спинной мозг и спинномозговые нервы (31 пара).
желудочков четыре. Боковые (правый и левый) желудочки
находятся внутри долей конечного мозга, третий желудочек
локализуется в промежуточном мозге, а четвертый расположен
между задним и продолговатым мозгом. Третий и четвертый
желудочки сообщаются между собой посредством сильвиева
водопровода, проходящего в виде канала в среднем мозгу.
Организация нервной системы
57
СПИННОЙ МОЗГ И СПИННОМОЗГОВЫЕ НЕРВЫ
Спинной мозг лежит в позвоночном канале. У человека он
располагается между продолговатым мозгом и вторым
поясничным позвонком, и длина его составляет примерно 45 см
(рис. 3.6). Краниальный отдел спинного мозга переходит в
продолговатый мозг; от каудального отдела отходит концевая
нить, прикрепляющаяся к первому сегменту копчика. От
спинного мозга отходит 31 пара спинномозговых нервов (от каждого
сегмента по одной паре), в том числе 8 шейных, 12 грудных,
5 поясничных, 5 крестцовых и одна копчиковая (рис. 3.6).
Задний корешок /^': '' '■'•'"'.'••'.: • :V—■
1;V Задний рог
•*•".;. (Промежуточная
^£*! 7 "область
■./ Передний рог
Передний корешок ^-—^ W Восходящие пути
Нисходящие пути
Рис. 3.7. Внутреннее строение спинного мозга (поперечное сечение).
Каждый спинномозговой нерв образуется в результате слияния
двух спинномозговых корешков — заднего (сенсорного) и
переднего (моторного) (рис. 3.7). Нервы выходят через
межпозвоночные отверстия соответствующих позвонков и направляются
к периферическим структурам — рецепторам и мышцам.
31 пара спинномозговых нервов иннервирует все
рецептивные поля кожи, за исключением лица и передней поверхности
волосистой части головы (рис. 3.8). В составе каждого нерва
имеются сенсорные волокна, идущие от рецептора
соответствующего сегмента. Распределение зон спинномозговых нервов
по сегментам было установлено путем определения размеров и
границ участков кожи (дерматомов), иннервируемых каждым
нервом. Дерматомы расположены на поверхности тела по
сегментарному принципу. К шейным дерматомам относятся задняя
поверхность головы, шея, плечи и передняя поверхность
предплечий. Грудные сенсорные нейроны иннервируют оставшуюся
поверхность предплечья, грудь и большую часть живота.
Сенсорные волокна поясничных, крестцовых и копчиковых
сегментов подходят к остальной части живота и ногам.
Спинномозговой
нерв
Рис. 3.8. Схема дерматомов. Иннервация поверхности тела 31 парой
спинномозговых нервов (С — шейные, Т — грудные, L — поясничные, S —
крестцовые) .
Организация нервной системы
59
ВНУТРЕННЕЕ СТРОЕНИЕ
Спинной мозг состоит из серого и белого вещества (рис. 3.7).
Серое вещество, расположенное в центре, участвует в
интеграции сенсорных и двигательных функций спинного мозга. В нем
находятся тела, дендриты и аксонные окончания
спинномозговых нейронов. Белое вещество образовано совокупностью мие-
линизированных волокон (трактами), служащих для
проведения возбуждения между различными сегментами спинного-
мозга и между спинным и головным мозгом.
Три основных отдела спинного мозга — задний рог,
промежуточное вещество и передний рог (рис. 3.7)—соответствуют
трем его основным функциям, а именно: 1) восприятию
чувствительных импульсов (задние рога); 2) формированию
эфферентной импульсации к внутренним органам (промежуточное
вещество грудных и поясничных сегментов); 3) посылке
двигательных импульсов к скелетным мышцам (передние рога).
Спинномозговые пути (тракты), составляющие белое веще*
ство, можно разделить на восходящие (проводящие импульсы к
головному мозгу) и нисходящие (проводящие импульсы от
головного мозга) (рис. 3.7). Восходящие пути образованы
аксонами сенсорных и вставочных нейронов спинного мозга. Они
являются проводниками разных видов чувствительности —
тактильной, температурной и болевой — к различным отделам
головного мозга, включая продолговатый мозг, зрительные
бугры и мозжечок. Аксоны вставочных нейронов коры больших
полушарий и двигательных ядер ствола мозга образуют
нисходящие спинномозговые пути. Благодаря этим путям центры
головного мозга осуществляют регуляцию спинальных сенсомо-
торных функций.
ПРОДОЛГОВАТЫЙ МОЗГ
Продолговатый мозг расположен между спинным мозгом,
мостом и мозжечком (рис. 3.9). Каудальный отдел
продолговатого мозга, переходящий в спинной, напоминает последний
по строению. Краниальный же отдел продолговатого мозга
представляет собой пирамидальную структуру, дорсальная
поверхность которой ограничена четвертым желудочком
(рис. 3.10). Большинство путей, соединяющих спинной мозг с
высшими отделами головного, проходит через продолговатый
мозг без переключений. Волокна кортикоспинального пути,
идущие от моторных областей коры головного мозга к
спинному, образуют на вентральной поверхности продолговатого
мозга два продольных валика — пирамиды (рис. 3.11). К
другим внешним ориентирам относятся корешки четырех черепных
Центральная борозда Теменная доля
Лобная доля-
Латеральная борозда
Височная доля
Мост
Продолговатый мозг
Рис. 3.9. Головной мозг, вид сбоку.
Затылочно-теменная
борозда
Мозжечок
Затылочная
доля
Гипоталамус [
Зрительный
лерекрест
Гипофиз
Центральная борозда
Таламус
Затылочно-теменная
борозда
Шишковидное
тело
Четверохолмие
Сосцевидное тело
Средний мозг
Сильвиев водопровод
Четвертый желудочек
Листки мозжечка
1Рис. 3.10. Главные образования медиальной поверхности головного мозга на
ерединно-сагиттальном разрезе.
«60
Организация нервной системы
61
нервов, выходящих из продолговатого мозга, в том числе язы-
коглоточного (IX), блуждающего (X), добавочного (XI) и
подъязычного (XII) (рис. 3.11). Эти нервы иннервируют язык
(IX и XII), щитовидную железу (IX), глотку (IX и X), гортань
(X и XI), каротидный синус (IX) и внутренние органы грудной
и брюшной полостей (X) (табл. 3.1). Языкоглоточный и блуж-
Черелномозговые
нервы
Обонятельная
луковица
Зрительный
перекрест
Сосцевидные
тела
Ножки мозга
Мост
Продолговатый
мозг (пирамиды)
Рис. 3.11. Нижняя поверхность головного мозга.
дающий нервы содержат как двигательные, так и
чувствительные волокна, а добавочный и подъязычный — только
двигательные.
ВНУТРЕННЕЕ СТРОЕНИЕ
В глубине продолговатого мозга расположен каудальный
отдел ретикулярной формации — диффузной сети вставочных
нейронов, охватывающей все отделы ствола мозга — от
продолговатого до промежуточного. Она играет важную роль как в
интеграции сенсорной информации, так и в контроле над
деятельностью всех эффекторных нейронов (моторных и
вегетативных) ; она имеет также первостепенное значение для
активации коры больших полушарий, для поддержания сознания.
62
Глава 3
Таблица 3.1. Функции черепномозговых нервов
Нерв
Иннервируемый орган
Функция
Обонятельный (I)
Зрительный (II)
Глазодвигательный (III)
Блоковый (IV)
Тройничный (V)
Отводящий (VI)
Лицевой (VII)
Улитковый (VIII)
Языкоглоточный
(IX)
Блуждающий (X)
Добавочный (XI)
Подъязычный
(XII)
Обонятельный эпителий
Сетчатка
Мышцы глаза
Мышцы глаза
Голова и челюсть
Мышцы глаза
Мышцы лица, слюнные
и слезные железы,
вкусовые сосочки языка
Слуховые и
вестибулярные рецепторы
Глотка, слюнные железы,
вкусовые сосочки
языка, каротидный синус
Глотка, гортань, органы
грудной и верхней
половины брюшной
полости
Мышцы шеи, плечевого
пояса, гортани и
глотки
Мышцы языка
Сенсорная: обоняние
Сенсорная: зрение
Моторная: движения глаз,
сужение зрачка и изменение
формы хрусталика
(аккомодация)
Моторная: движения глаз
Моторная: жевание и глотание
Сенсорная: соматическая
чувствительность (тактильная и
температурная) передней
части головы и лица
Моторная: движения глаз
Моторная: мимика,
слюноотделение и слезообразование
Сенсорная: вкус
Сенсорная: слух и чувства
равновесия
Моторная: глотание и
слюноотделение
Сенсорная: вкус, кровяное
давление и содержание газов в
крови
Моторная: глотание, движения
гортани, парасимпатическая
иннервация внутренних
органов
Сенсорная: висцеральная
чувствительность
Моторная: движения головы*
плеч, гортани и глотки
Моторная: движение языка
Вокруг диффузной, расположенной в центре продолговатого
мозга ретикулярной формации в нем имеются также
многочисленные отдельные скопления нейронов или ядра. К ним
относятся: ядра четырех черепномозговых нервов (языкоглоточногоу
блуждающего, добавочного и подъязычного); нежное и
клиновидное ядра в задних столбах спинного мозга (по этим путям
информация передается к соматосенсорной зоне коры);
улитковые ядра слуховой системы; ядра нижних олив, играющие
важную роль в поступлении информации в мозжечок; наконец,,
так называемые «жизненные центры», регулирующие
деятельность сердечно-сосудистой и дыхательной систем.
Организация нервной системы
63
ЗАДНИЙ МОЗГ
Задний мозг состоит из двух отделов — моста и мозжечка
(рис. 3.9). Мост представляет собой часть ствола мозга,
расположенную между продолговатым и средним мозгом.
Мозжечок образуется в результате интенсивного разрастания
крыши передней части заднего мозга и покрывает дорсальную
поверхность моста и продолговатого мозга (рис. 3.10). Мост
соединен с мозжечком посредством передних ножек
мозжечка— широких пучков нервных волокон, обусловливающих
характерную выпуклость на вентральной поверхности моста
(рис. 3.11).
МОСТ
Задняя часть моста является непосредственным
продолжением продолговатого мозга. Многие структуры продолговатого
мозга, в частности восходящие и нисходящие пути и
ретикулярная формация, проходят через мост не прерываясь. В
области соединения моста и продолговатого мозга выходит вестибу-
локохлеарный нерв (VIII). (рис. 3.11). Из моста выходят
также тройничный (V), отводящий (VI) и лицевой (VII)
нервы. Они иннервируют лицо, рот и кожу волосистой части головы
(V и VII), язык (VII), слуховые и вестибулярные рецепторы
внутреннего уха (VIII) и боковые прямые мышцы глаза (VI)
(см. табл. 3.1). Вестибулокохлеарный нерв является чисто
чувствительным, тройничный и лицевой — смешанными, а
отводящий— чисто двигательным.
В ткани моста разбросаны некоторые важные
чувствительные и двигательные ядра. Кроме ядер тройничного, отводящего,
лицевого и вестибулокохлеарного нервов здесь расположены
также ядра моста, дающие начало средним ножкам мозжечка,
соединяющим мост и мозжечок. К другим важнейшим ядрам
моста относятся: слуховое ядро верхней оливы и вегетативные
центры, регулирующие деятельность сердечно-сосудистой и
дыхательной систем.
МОЗЖЕЧОК
Мозжечок представляет собой главную структуру заднего
мозга. Он расположен над дорсальной поверхностью моста и
продолговатого мозга непосредственно за большими
полушариями конечного мозга (рис. 3.9). Мозжечок связан со стволом
мозга посредством трех пар ножек (нижних, средних и
верхних) ; нижние ножки соединяют его с продолговатым и спинным
64
Глава 3
мозгом, средние — с мостом, а верхние — со средним мозгом и
таламусом.
Мозжечок состоит из двух отделов — поверхностно
расположенной коры и глубинных ядер. На поверхности мозжечка
имеется множество складок, или листков, образованных сложными
изгибами его коры (рис. 3.10).
Выходные импульсы от коры мозжечка передаются на его
ядра, расположенные в глубине мозжечковых ножек. От этих
ядер отходят волокна, часть которых оканчивается в
двигательных ядрах ствола мозга, а часть идет через таламус в
двигательные зоны коры больших полушарий.
СРЕДНИЙ МОЗГ
Средний мозг соединяет передний мозг с задним. К
характерным внешним ориентирам среднего мозга относятся:
четверохолмие с его округлыми верхними и нижними бугорками
(рис. 3.10), корешки глазодвигательного (III) и блокового (VI)
нервов (рис. 3.11) и ножки мозга — два крупных пучка нервных
волокон, в которых проходят прямые нисходящие пути от коры
больших полушарий (рис. 3.11).
Средний мозг образует переднюю часть ствола мозга,
поэтому многие образования задних отделов ствола (моста и
продолговатого мозга), в том числе восходящие и нисходящие пути
и ретикулярная формация, проходят через него не прерываясь.
В среднем мозгу расположен ряд важных чувствительных и
двигательных центров. Четверохолмие, находящееся на его
дорсальной поверхности, имеет большое значение как интегратив-
ный центр зрительной (верхние бугорки) и слуховой (нижние
бугорки) систем. К важным двигательным центрам относятся
также черная субстанция (входящая в систему базальных ядер)
и красное ядро. Большую роль играют также ядра
глазодвигательного и блокового нервов, управляющих мышцами глаз.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг расположен у переднего конца ствола
мозга и с трех сторон (передней, дорсальной и боковой)
окружен нервными пучками внутренней капсулы и долями коры
больших полушарий (рис. 3.10 и 3.12). На вентральной
поверхности мозга переднюю и заднюю границу промежуточного
мозга обозначают два четких ориентира (рис. 3.11): передняя
граница проходит в области хиазмы (зрительный перекрест), а
задняя — по заднему краю хорошо заметных мамиллярных тел.
Промежуточный мозг подразделяется на отделы: 1) тала-
мус, 2) гипоталамус, 3) эпиталамус. Первые два отдела
являются важнейшими нервными центрами. Эпиталамус содержит
Организация нервной системы
65
два образования: шишковидное тело — секреторный орган
(рис. ЗЛО) и поводок — обонятельный центр, связанный с лим-
бической системой.
ТАЛАМУС
Таламус (зрительный бугор) расположен в центромедиаль-
ной области переднего мозга (рис. 3.10). В его состав входит
множество ядер, каждое из которых передает импульсацию в
определенный участок коры больших полушарий. Нейроны
этих ядер являются последним этапом на пути импульсов,
направляющихся к коре. Все поступающие к ней сигналы, за
исключением обонятельных, должны пройти через интегративные
и релейные ядра таламуса. Большая часть волокон зрительного
нерва оканчивается в латеральном коленчатом теле зрительного
бугра. Ядра таламуса можно подразделить либо в зависимости
от их общего назначения (сенсорные, моторные и
ассоциативные), либо в соответствии с теми областями коры, с которыми
они связаны. Важнейшие сенсорные ядра — это вентробазаль-
ный комплекс, обрабатывающий информацию, поступающую в
соматосенсорную и вкусовую области коры, медиальное
коленчатое ядро, связанное со слуховой областью коры, и
латеральное коленчатое ядро, передающее информацию в зрительную
кору. В переднем вентральном и вентролатеральном ядрах
переключаются импульсы, идущие от базальных ганглиев и
мозжечка к двигательным зонам коры. К другим важным ядрам
относятся передние, передающие сигналы от лимбической
системы в кору, и межпластинчатые, служащие промежуточными
этапами для проведения в кору недифференцированной болевой
информации.
ГИПОТАЛАМУС
Гипоталамус расположен в вентральной части
промежуточного мозга (рис. 3.10) непосредственно над гипофизом,
деятельностью которого он управляет.
Гипоталамус содержит восемь мелких ядер, управляющих
большинством функций внутренних органов. Кроме секреторной
активности гипофиза эти ядра регулируют температуру тела,
водный баланс и чувство голода и насыщения. Гипоталамус,
играет также первостепенную роль в эмоциональном и половом
поведении.
КОНЕЧНЫЙ МОЗГ
Конечный мозг состоит из двух анатомически обособленных
частей — коры головного мозга и базальных ганглиев. Кора —
самый большой отдел мозга — разделена на два полушария,
5—1705
66
Глава 3
соединенных очень крупными пучками нервных волокон; эти
пучки образуют мозолистое тело (рис. 3.12). Каждое полушарие
расположено над передней дорсальной поверхностью ствола
мозга (рис. 3.9). Базальные ганглии образуют комплекс из
взаимосвязанных ядер, локализующихся в глубине каждого
полушария (рис. 3.12).
Рис. 3.12. Внутреннее строение переднего мозга на поперечном разрезе. На
врезке изображен уровень разреза.
КОРА ГОЛОВНОГО МОЗГА
Хотя кора головного мозга образует множество складок
(рис. 3.9), расположение важнейших извилин и борозд
отличается постоянством. В связи с этим главные извилины и
борозды используются в качестве ориентира при разграничении
областей коры. Наружная поверхность коры разделена тремя
бороздами на четыре доли: лобную, теменную, затылочную и
височную (рис. 3.9 и 3.10). Борозды эти следующие: 1)
центральная борозда, разделяющая лобную и теменную доли;
2) затылочно-теменная борозда, образующая границу между
теменной, затылочной и височной долями; 3) латеральная
борозда, отделяющая височную борозду от лобной и теменной.
К каждой из этих долей подходят волокна от одного или
нескольких таламических ядер. К сенсорным зонам — сомато-
сенсорной и вкусовой в теменной доле, зрительной в
затылочной доле и слуховой в височной •— поступают импульсы от вен-
тробазального комплекса, латерального и медиального ядер
соответственно. Моторные зоны коры связаны с передним вен-
Организация нервной системы
67
тральным и вентролатеральным ядрами таламуса.
Ассоциативные зоны получают преимущественно информацию от нейронов
других зон коры.
По обонятельному нерву (I пара) импульсы направляются
непосредственно в кору головного мозга. Его волокна
заканчиваются в обонятельных луковицах, расположенных на вентро-
медиальной поверхности коры (рис. 3.11). Вход обонятельных
сигналов в кору тесно связан с лимбической извилиной,
расположенной в виде пояса в глубине больших полушарий
(рис. 3.10). Лимбическая кора и ряд соединенных с ней
подкорковых и гипоталамических структур образуют лимбическую
систему, содержащую высшие центры регуляции деятельности
внутренних органов.
Кора больших полушарий связана с нижележащими
отделами мозга (базальными ядрами, таламусом, средним мозгом
и мостом) крупными пучками волокон, образующими
внутреннюю капсулу (рис. 3.12). Эти широкие пласты белого вещества
содержат миллионы нервных волокон, часть из которых
(аксоны нейронов таламуса) служат для передачи нервных сигналов
к коре, а часть (аксоны корковых нейронов)—от коры к
нижележащим нервным центрам.
БАЗАЛЬНЫЕ ГАНГЛИИ
Базальные ганглии — это комплекс подкорковых ядер —
бледного шара, скорлупы, хвостатого ядра и миндалины, —
расположенный в глубине больших полушарий и окруженный
волокнами внутренней капсулы (рис. 3.12). Первые три из этих
ядер связаны с некоторыми стволовыми двигательными ядрами
и вместе с ними образуют систему базальных ядер — важный
центр координации движений. Миндалины же, морфологически
входящие в состав базальных ганглиев, функционально
относятся к вегетативным центрам лимбической системы.
ЛИТЕРАТУРА
Bodian D. (1967). Neurons, circuits and neuroglia. In: Quarton G. C, Melne-
chuk Т., Schmitt F. O. (eds.), The neurosciences, Rockefeller University
Press, New York.
Eyzaguirre C, Fidone S. J. (1975). Physiology of the nervous system. 2nd edn.,
Yearbook Medical, Chicago.
Heitner L. (1971). Pathways in the brain. Sci. Amer., 225, 48.
Kuffler S. W., Nicolls J. G. (1976). From neuron to brain: a cellular approach
to the function of the nervous system. Sinauer, Sunderland, Mass. [Имеется
перевод: Куффлер С, Николе Дж. От нейрона к мозгу. — М.: Мир, 1979.]
Noback С, Demarst R. /. (1975). The human nervous system, 2nd edn., McGraw-
Hill, New York.
5*
68
Глава 3
Рарау S. L., Chan-Palay V. (1977). Morphology of neurons and neuroglia, In:
Kandel E. R. (ed.), Handbook of physiology: Cellular biology of neurons,
Amer. Physiol. Soc. Bethesda, Maryland.
Truex R. C, Carpenter M. B. (1975). Human neuroanatomy, Williams and Wil-
kins, Baltimore.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Каковы четыре функциональных отдела нейрона? Опишите
вкратце их функции.
2. Что такое глиальные клетки? Чем они отличаются от
нейронов?
3. Опишите коротко две высшие формы нейронной
организации — ядра и кору.
4. Приведите схему поперечного разреза спинного мозга и
обозначьте его основные структуры.
5. Составьте таблицу анатомических и функциональных
характеристик черепномозговых нервов. В таблицу должны войти
данные об основных функциях этих нервов, области выхода
их из головного мозга и данные о важнейших из иннерви-
руемых ими органах.
6. Приведите схему поперечного разреза головного мозга и
укажите на ней область расположения продолговатого мозга,
моста, мозжечка, среднего мозга, гипоталамуса, таламуса,
мозолистого тела и коры.
7. Составьте схему анатомических взаимоотношений между
корой головного мозга, мозолистым телом, внутренней
капсулой, таламусом и базальными ядрами.
Глава 4
ФИЗИОЛОГИЯ НЕЙРОНОВ
Главная функция нервной системы состоит в быстрой и
точной передаче информации. Сигнал от рецепторов к сенсорным
центрам, от этих центров — к моторным центрам и от них к
эффекторным органам — мышцам и железам—должен
передаваться быстро и точно. Выживание организма зависит от его
способности улавливать опасные изменения окружающей среды
и быстро на них реагировать.
Нервные клетки специализированы для передачи
информации как морфологически, так и функционально (см. гл. 3).
У большинства нейронов главным отростком является аксон,
служащий для быстрой передачи возбуждения от одного
пункта нервной системы к другому. Именно в проведении нервных
импульсов (потенциалов действия) и заключается специальная
функция аксона.
Межнейронная передача нервных импульсов происходит в
области специализированных соединений — синапсов,
образованных аксонами одних нейронов на дендритах и телах других.
В синапсе происходит как передача информации от нейрона к
нейрону, так и ее .интеграция. Импульсы передаются через
синапс посредством выделения из пресинаптического окончания
аксона специфического химического передатчика (медиатора).
Взаимодействуя с постсинаптическими участками дендритов
и сомы, медиатор вызывает кратковременные изменения их
мембранных потенциалов (синаптические потенциалы).
Как проведение нервных импульсов, так и генерация синап-
тических потенциалов зависят от способности мембран нервных
клеток быстро изменять свой электрический потенциал. Хотя
мембранные потенциалы существуют у многих клеток, лишь на
мембранах нервных, чувствительных и мышечных клеток могут
генерироваться потенциалы действия и синаптические
потенциалы. Эти клетки называют «возбудимыми» из-за их
способности временно изменять мембранный потенциал; все остальные
клетки организма относятся к невозбудимым. Именно эта
особенность определяет уникальную способность нейрона
передавать и обрабатывать информацию.
70
Глава 4
МЕМБРАННЫЙ ПОТЕНЦИАЛ
Трансмембранное распределение ионов. Концентрации
основных одновалентных ионов — хлора (О-), калия (К+) и натрия
(Na+) —внутри клетки существенно отличаются от их
содержания в омывающей клетки внеклеточной жидкости. Как видно
из рис. 4.1, главным внутриклеточным катионом (положительно
\ \ J \ I \ /1 Дендриты
Na+: 142
к+ : 5
or : 11°
А- ; 5
Насос
\[ Аксон
Рис. 4.1. Распределение ионов по обе стороны мембраны нейрона. Приведены
типичные значения вне- и внутриклеточного содержания Na+, К+, С1~ и
органических анионов (А-). Низкий внутриклеточный уровень Na+
поддерживается благодаря работе Na+/K+-Hacoca, «выкачивающего» из клетки Na+ в
обмен на внеклеточный К+. Низкая концентрация С1~ в нейроне обусловлена
отталкивающим действием отрицательно заряженных органических молекул,
включенных в мембрану. Концентрации приведены в мэкв/л.
заряженным ионом) является К+; внутриклеточные же анионы
(отрицательно заряженные ионы) представлены
преимущественно остатками аминокислот и других органических молекул.
Основной внеклеточный катион — это Na+, а анион — С1~.
Такое распределение ионов создается в результате двух
факторов: 1) наличия отрицательно заряженных органических
молекул внутри клетки и 2) существования в клеточной
мембране систем активного транспорта, «перекачивающих» Na+ из
клетки, а К+ — в клетку (см. гл. 2). Если такие небольшие
ионы, как С1~, К+ и Na+, клеточная мембрана легко пропускает,
то органические анионы, например аминокислот и органических
кислот цитоплазмы, слишком крупны и не могут пройти через
мембрану. В связи с этим в клетке накапливается значитель*
Сома
Физиология нейронов
71
ный избыток отрицательных зарядов (органических анионов).
Эти заряды препятствуют проникновению в клетку
отрицательных ионов, например С1~, но притягивают в нее положительно
заряженные катионы (К+ и Na+); однако большая часть
поступающего в клетку Na+ немедленно удаляется натрий-калиевым
насосом. Быстрое удаление Na+ приводит к тому, что в клетке
накапливается только К+, который притягивается
отрицательными зарядами органических анионов и накачивается Na+/K+-
насосом.
ИЗМЕРЕНИЕ КЛЕТОЧНЫХ ПОТЕНЦИАЛОВ
Между наружной и внутренней поверхностью всех клеток
существует разность потенциалов. Потенциал покоя варьирует
от —40 до —95 мВ в зависимости от особенности той или иной
клетки. Потенциал покоя нервных клеток обычно равен от —60
до —80 мВ.
Мембранный потенциал быстро определяют, измеряя
разность потенциалов между двумя одинаковыми электродами,
один из которых введен в клетку, другой помещен в омываю-
+ + + + + + + + + ++ + + + + + + + +// + 4- + + + + +
-70 мВ
+ + ++ + + + + +
+ + + + + + +++ + + + + + +
0 мВ
Рис. 4.2. Внутриклеточное измерение мембранного потенциала аксона.
Величина усиленного потенциала регистрируется при помощи осциллоскопа.
72
Глава 4
Вертикальные пластины
Горизонтальные пластины
Экран
Регистрируемый
потенциал
стимулятор
d
YL
1
к
Рис. 4.3. Электронно-лучевой осциллоскоп. В месте встречи пучка электронов
с флуоресцирующим экраном образуется световое пятно. Движение пучка на
экране регулируется потенциалами, создаваемыми на вертикальных и
горизонтальных пластинах. Блоком развертки генерируется потенциал,
управляющий движением луча с постоянной скоростью вдоль горизонтальной оси.
Скорость развертки служит основой отсчета времени на кривой изменения
вольтажа во времени, регистрируемой на экране. Вертикальное отклонение луча
управляется потенциалами, приложенными к горизонтальным пластинам. Так
как именно к этим пластинам через усилитель подаются сигналы отводящих
электродов, вертикальное отклонение луча указывает на величину
потенциала. Кривая на экране отражает амплитудно-временные характеристики
потенциала действия, возникающего под влиянием раздражающих импульсов.
Запись осуществляется через внутриаксонные микроэлектроды.
щую ее жидкость (рис. 4.2). Электроды соединены с
усилителем, увеличивающим амплитуду регистрируемого потенциала;
эта амплитуда определяется при помощи измерителя
напряжения типа осциллоскопа (рис. 4.3).
Для измерения мембранного потенциала обычно используют
микроэлектроды. Благодаря малому диаметру кончика
микроэлектрод может быть введен через мембраны любых клеток,
кроме самых маленьких, без повреждения этих клеток.
Физиология нейронов
73
ИОННЫЕ ОСНОВЫ МЕМБРАННОГО ПОТЕНЦИАЛА
Потенциал покоя возникает прежде всего в связи с
асимметричным распределением К+ по обе стороны мембраны. Так
как концентрация его в клетке примерно в 30 раз выше, чем
эо внеклеточной среде, существует трансмембранный
концентрационный градиент, способствующий диффузии К+ из клетки
(рис. 4.1). Выход каждого положительного иона К+ из клетки
приводит к тому, что в ней остается несбалансированный
отрицательный заряд (органические анионы). Эти заряды и
обусловливают отрицательный потенциал внутри клетки (см.
также гл. 1).
УРАВНЕНИЕ НЕРНСТА
Когда химический концентрационный градиент,
способствующий выведению К+ из клетки, уравновешивается
отрицательным внутриклеточным потенциалом, вовлекающим К+ в клетку,
потоки калия становятся сбалансированными. В этом состоянии
реальный калиевый ток через мембрану равен нулю.
Диффузионный потенциал в состоянии равновесия называется
равновесным потенциалом (Е^.+). Его можно вычислить по
уравнению Нернста (при 25 °С):
£K+ = _591g£T.
14 out
Уравнение Нернста можно также применить для
определений разновесных потенциалов С1~ и Na+. Типичные значения
этих потенциалов для нервных клеток равны:
Ек+ = —86 мВ; Еа-= —78 мВ; £Na+ = +66 мВ.
Потенциал покоя большинства клеток почти равен Ек+. Ее-
ли бы мембрана была проницаема лишь для К+, т. е. не
пропускала бы С1~ и Na+, то потенциал покоя в точности
соответствовал бы £к+. Однако, хотя проницаемость для К+ намного
выше, чем для С1~ и Na+, последние все же могут проходить
через мембрану и участвуют в создании мембранного
потенциала. Относительный вклад каждого иона в генерацию
потенциала покоя пропорционален проницаемости мембраны для
этого иона; чем выше проницаемость для него, тем его
вклад больше. В связи с тем что проницаемость
мембран нервных клеток для К+ примерно в 7 раз выше, чем
для С1~ и в 25 — чем для Na+, их потенциал покоя
определяется главным образом значением Ек+.
74
Глава 4
ПОТЕНЦИАЛ ДЕЙСТВИЯ
Функция аксона заключается в проведении нервных
импульсов. Потенциал действия возникает в результате
кратковременной реверсии мембранного потенциала, волнообразно
распространяющейся вдоль аксолеммы. Обычно потенциал
действия зарождается в начальном, ближайшем к телу клетки
I I У
I \
I I
' \
У,-
дкт
Рис. 4.4. Потенциал действия.
А. График развития потенциала действия. Б. Изменения проницаемости
мембраны в процессе развития потенциала действия.
Пороговая деполяризация мембраны приводит к увеличению натриевой
проницаемости (gNa+), обусловливающему восходящую фазу потенциала
действия. Нисходящая фаза связана с уменьшением натриевой и
увеличением калиевой (g K+) проницаемости. Потенциал покоя равен —70 мВ, а
пороговый потенциал — 50 мВ. Проницаемость измеряется в ммо/см2 (единица
проводимости мо — величина, обратная единице сопротивления — Ому).
сегменте аксона, и пробегает по аксону к его окончаниям. На
препарате изолированного аксона потенциал действия можно
вызвать, приложив к его мембране короткий толчок
электрического тока (рис. 4.3 и 4.4). Амплитуда потенциала действия
(100—125 мВ) и его длительность (1—2 мс) мала, и поэтому,
для того чтобы получить изображение, его необходимо усилить
и вывести на экран осциллоскопа.
Потенциал действия состоит из двух фаз (рис. 4.4). Фаза
деполяризации (/) соответствует быстрому изменению
мембранного потенциала (деполяризации мембраны) примерно на
110 мВ. Мембранный потенциал изменяется от уровня покоя
(около —70 мВ) до значения, близкого к ENa+ (примерно
40 мВ). Во время фазы реполяризации (2) мембранный
потенциал вновь достигает уровня покоя (мембрана реполяризует-
ся) после чего наступает гиперполяризация до значения при-
мерно на 10 мВ меньшего (более отрицательного), чем
потенциал покоя, т. е. примерно —80 мВ.
Фаза деполяризации потенциала действия обусловлена
временным повышением проницаемости мембраны аксона для нат-
Физиология нейронов
75
рия (рис. 4.4). В этот момент открываются специфические
натриевые каналы, и Na+ лавинообразно устремляется в
клетку. Этот приток положительных ионов приводит к
деполяризации мембраны.
Фаза реполяризации потенциала действия связана с
закрытием натриевых и открытием калиевых каналов (рис. 4.4).
Вход натрия в аксон снижается из-за падения натриевой
проницаемости; повышение же калиевой проницаемости приводит
к увеличению выхода К+. Так как по мере выхода К4
удаляются положительные заряды, мембрана реполяризуется.
Гиперполяризация мембраны до уровня большего (более
отрицательного), чем потенциал покоя, обусловлена очень высокой
калиевой проницаемостью в фазу реполяризации. Закрытие калиевых
каналов приводит к восстановлению исходного уровня
мембранного потенциала; значения проницаемости для К+ и Na+
при этом также возвращаются к прежним.
ВОЗНИКНОВЕНИЕ НЕРВНОГО ИМПУЛЬСА
Все возбудимые клетки характеризуются неким пороговым
значением мембранного потенциала, при котором возникает
потенциал действия. Когда в результате деполяризации
потенциал мембраны достигает этого значения (у нервных клеток
он равен примерно —50 мВ), генерируется нервный импульс.
Пороговый уровень соответствует потенциалу, при котором
входящий поток натрия равен выходящему потоку калия; это
приводит к быстрой деполяризации мембраны нейрона до
величины £Na+ (около 40 мВ), т. е. к возникновению потенциала
действия.
Потенциал действия возникает только в том случае, когда
под действием раздражителя мембрана деполяризуется до
порогового уровня. Генерация потенциала действия подчиняется
закону «все или ничего». Это означает, что лишь тогда, когда
сила раздражения достаточна для пороговой деполяризации
мембраны нейрона, может возникнуть нервный импульс. Хотя
на практике обычно в качестве раздражителя используют
короткие толчки деполяризующего тока, любой стимул,
способный снизить потенциал клетки до порогового (например,
механическая деформация мембраны или кратковременное
воздействие лучом лазера), может вызвать потенциал действия.
РЕФРАКТЕРНОСТЬ НЕРВА
В момент генерации потенциала действия мембрана аксона
пребывает в состоянии рефрактерности, т. е. пониженной
возбудимости. Длительность периода рефрактерности можно опре-
76
Глава 4
делить, раздражая аксон парными толчками тока. Если
интервал между парными стимулами достаточно велик (больше
10 мс), каждый из них приводит к возникновению потенциала
действия. Если интервал между стимулами сократить, сделав
его менее 10 мс, то под действием второго из них нервный
импульс не возникнет. Этот период сниженной возбудимости
называется фазой относительной рефрактерности, так как
увеличение силы второго стимула может привести к генерации нерв-
+40
ш
5
пГ
<0
.X
ZT
X
ф
}-
о
п.
•70
L
I
□и
\
и и и и и и i
Пороговый
потенциал
и:
з
Фаза Фаза относительной
абсолютной рефрактерности
рефрактерности
О
ПП
Первый
стимул
ш
Второй стимул
10
Рис. 4.5. Фазы абсолютной и относительной рефрактерности. Для определения
длительности этих фаз аксон раздражают парными толчками электрического
тока. Величина первого из них достаточна для того, чтобы возник потенциал
действия. Эффект второго стимула зависит от промежутка времени между
стимулами. Если второй стимул наносится в фазу абсолютной
рефрактерности, то он не может вызвать потенциал действия. В фазе же относительной
рефрактерности, хотя порог возбуждения и повышен, потенциал действия
может возникнуть, если увеличить силу стимула.
Физиология нейронов
17
ного импульса (рис. 4.5). Во время фразы относительной
рефрактерности порог возникновения потенциала действия
повышен; следовательно, для того чтобы мембранный
потенциал достиг порога, необходимо приложить ток большей силы.
Фаза относительной рефрактерности соответствует большей
части нисходящего колена потенциала действия и периоду
следовой гиперполяризации; в это время проницаемость для К1"
очень высока, а проходимость натриевых каналов только
восстанавливается.
Если же интервал между парными стимулами сделать еще
короче (меньше 3 мс), то второй стимул будет попадать в фазу
абсолютной рефрактерности, когда аксон полностью
невозбудим (рис. 4.5). Она соответствует фазе деполяризации
потенциала действия, когда проницаемость для Na+ высока.
Рефрактерпость играет важную роль в деятельности аксона,
так как из-за нее минимальный интервал между двумя
последовательными потенциалами действия ограничен. В результате
максимальная частота импульсов, которую способно провести
большинство нервных волокон, составляет 200 в 1 с.
ВОССТАНОВЛЕНИЕ ИОННЫХ ГРАДИЕНТОВ
Хотя в ходе одиночного потенциала действия в аксон
входит лишь около 1 • 10-12 экв (1 пкмоль) 'Na+ (и примерно такое
же количество К+ выходит из него), при генерации большего
количества импульсов эти ионные потоки могут привести к
значительному изменению содержания внутриклеточного Na+ и
внеклеточного К+. Первостепенную роль в предупреждении
длительных сдвигов градиентов в концентрации Na+ и К+ по
обе стороны мембраны играет Na/K-насос, который быстро
выкачивает все вошедшие в аксон ионы натрия и обменивает их
на внеклеточный калий; однако в генерации потенциала
действия он непосредственно не участвует. Крупные аксоны,
«отравленные» уабаином (сердечным гликозидом, подавляющим
деятельность Na/K-насоса), способны провести тысячи нервных
импульсов до того, как концентрации внутриклеточного Na+ и
внеклеточного К+ достигнут уровня, при котором проведение
становится невозможным. Напротив, специфический яд тетро-
дотоксин, блокирующий открытие натриевых каналов,
подавляет генерацию потенциалов действия, не влияя на функцию
Na+/K+-Hacoca и трансмембранные градиенты Na+ и К+.
ПРОВЕДЕНИЕ НЕРВНОГО ИМПУЛЬСА
Проведение нервного импульса по волокну можно сравнить
с распространением пламени вдоль фитиля. Оба этих процесса
подчиняются закону «все или ничего» и обладают конкретны-
78
Глава 4
ми порогами: для того чтобы открылись натриевые каналы
аксона, необходимо деполяризовать его до порогового
потенциала, и точно так же фитиль может загореться, лишь будучи
нагретым до критической температуры. Пламя
распространяется по фитилю благодаря тому, что тепло передается от горя-
+ 40 г
-70
+ + + Ч- +
+ + +^Гк- - | + ^ + + + + + + + +
)
+ 40
-70
1^ф-_*?\ +
+ + + + + + Е> + А 1+Л+ + + +
Рис. 4.6. Распространение потенциала действия. На каждой схеме показаны
потенциалы и ионные токи в последовательные промежутки времени по мере
распространения нервного импульса по аксону. Следует обратить внимание
на то, что вход Na+ сопровождается «местными токами», деполяризующими
прилежащие участки мембраны до порогового уровня. Выход же К+
приводит к генерации токов, гиперполяризующих мембрану.
щего участка к соседнему негорящему. Когда этого тепла
достаточно для того, чтобы нагреть негорящии участок до
критической температуры, он воспламеняется.
Точно так же распространяется нервный импульс по аксону.
Этому распространению способствуют небольшие «местные
токи», создаваемые входящим током Na+ во время фазы
деполяризации потенциала действия (рис. 4.6). В результате входа
Na+ возникает ток положительных зарядов, текущих вдоль
мембраны непосредственно перед фронтом потенциала действия.
Этот ток приводит к открытию натриевых каналов, тем самым
деполяризуя мембрану от уровня потенциала покоя до поро-
Физиология нейронов
79
гового потенциала, при котором генерируется потенциал
действия, и способствует распространению нервного импульса
вдоль аксона.
САЛЬТАТОРНОЕ ПРОВЕДЕНИЕ
Большинство аксонов миелинизировано (см. гл. 2).
Благодаря миелиновой оболочке потенциал действия проводится по
волокну быстрее: в миелинизированных волокнах скорость
прохождения импульса достигает 30—100 м/с, тогда как в безмя-
котных волокнах млекопитающих она не превышает 3 м/с.
Такая большая разница связана с тем, что в безмякотных
волокнах для распространения нервного импульса необходимо,
чтобы под действием местных токов последовательно
деполяризовался каждый участок волокна; в миелиновых же волокнах
деполяризующий ток не протекает через участки, покрытые
изолирующей миелиновой оболочкой (участки между
перехватами). Потенциал действия здесь распространяется
скачкообразно (сальтаторно). Он как бы «прыгает» лишь по небольшим
обнаженным участкам аксона (перехватам), разделяющим со-
Направление распространения импульса
>
Рис. 4.7. Сальтаторное проведение. В каждом перехвате поочередно
возникают следующие процессы: 1) пороговая деполяризация; 2) вход Na+; 3)
выход К+; 4) реполяризация. Вход Na+ сопровождается деполяризующим
током, проходящим через участок между перехватами к следующему перехвату.
При деполяризации перехвата до порогового уровня проницаемость его
мембраны для Na+ возрастает, что приводит к входу Na+.
седние миелииизированные участки (рис. 4.7). Поскольку
импульс возникает только в небольших областях аксона
(перехватах), проведение его по миелинизированному аксону
осуществляется с высокой скоростью.
Сальтаторное проведение обусловлено тем, что в области
мембраны перехвата течет ток положительных зарядов,
распространяющихся к следующему перехвату, деполяризуя его
мембрану до порогового уровня и способствуя открытию натриевых
каналов. В результате потенциал действия перемещается вдоль
аксона на один перехват.
80
Глава 4
СМЕШАННЫЕ НЕРВЫ
Любой периферический нерв состоит из сотен или тысяч
аксонов. В смешанном нерве все эти аксоны, за исключением
наиболее тонких, миелинизированы: диаметр их колеблется от
0,1 мкм у самых тонких безмякотных волокон, до 20 мкм у
наиболее толстых. Скорость проведения потенциала действия
по аксону пропорциональна его диаметру. В волокнах
смешанного нерва она варьирует от 120 м/с (толстые миелинизирован-
ные волокна) до 0,5 м/с (самые тонкие безмякотные волокна).
Рис. 4.8. Запись составного потенциала действия. Периферический нерв
раздражают короткими толчками тока. Возникающий в результате этого
составной потенциал действия записывается при помощи пары проволочных
электродов, на которые положен нерв. При прохождении нервного импульса над
каждым электродом на экране осциллоскопа возникает отклонение.
Двухфазная форма волны связана с тем, что электроды подсоединены к разным
вертикальным пластинам.
Если раздражать смешанный нерв коротким толчком тока,
по нему будет распространяться составной потенциал действия
(рис. 4.8), имеющий форму двухфазной волны. Такая волна
регистрируется при использовании двух отводящих электродов.
По мере распространения вдоль нерва потенциал действия
поочередно улавливается каждым электродом. При приближе-
Физиология нейронов
81
нии импульса под первым электродом возникает отрицательный
потенциал по отношению ко второму; когда же импульс
проходит от первого электрода ко второму, более отрицательный
потенциал регистрируется уже под вторым. В результате
записывается двухфазное отклонение.
Л.
V
Раздражающие
электроды
r€h
Отводящие*
электроды
^^
Порог
Субмаксимальный
стимул
Максимальный стимул
Рис. 4.9. Связь между величиной составного потенциала действия и силой
раздражителя. Периферический нерв стимулируется импульсами тока
возрастающей силы. Максимальная сила стимула — это такая сила раздражителя,
при которой возбуждаются все аксоны, входящие в состав нерва.
Составной потенциал действия — это суммарный потенциал,
образующийся в результате сложения потенциалов действия
всех аксонов, входящих в состав нерва (рис. 4.9). Амплитуда
его зависит от силы раздражителя. Наиболее низким порогом
возбудимости обладают самые толстые волокна, поэтому
именно они возбуждаются при нанесении минимальных стимулов,
способных вызвать реакцию нерва. При увеличении силы
стимула растет и амплитуда потенциала действия, так как при
6-1705
*2
Глава 4
этом последовательно возбуждаются волокна все меньшего
диаметра. Когда сила раздражителя с?ановится достаточной
для того, чтобы возбудились все волокна нерва, говорят о
максимальном значении стимула.
СИНАПТИЧЕСКАЯ ПЕРЕДАЧА
Синапсом называется функциональное соединение между
пресинаптическим окончанием аксона и мембраной
постсинаптической клетки. Синапсы — это те участки, где нервные
импульсы могут влиять на деятельность постсинаптической клетки,
тормозя или возбуждая ее. Существуют две разновидности
синапсов: в пресинаптическом окончании химических синапсов (1)
выделяется медиатор, генерирующий синаптический потенциал
в постсинаптической клетке; в электрическом синапсе (2) от
пресинаптического нейрона к постсинаптическому идет
электрический ток.
Несмотря на то что общие принципы строения синапсов
одинаковы, синапсы, образованные разными нейронами,
различаются: 1) структурой аксонных окончаний, 2) природой
медиатора и постсинаптической клетки и 3) характером постси-
наптического потенциала. В простейшем синапсе постсинапти-
ческая клетка иннервируется лишь одним аксоном. В качестве
примера можно привести нервно-мышечный синапс, или
соединение, в котором каждое мышечное волокно иннервируется
одним мотонейроном. В более сложных синапсах на постсинап-
тическом нейроне оканчиваются тысячи аксонов. У таких
нейронов (например, у клеток Пуркине мозжечка и пирамидных
нейронов коры головного мозга), как правило, имеются
чрезвычайно сложные древовидные разветвления дендритов.
НЕРВНО-МЫШЕЧНОЕ СОЕДИНЕНИЕ
Каждое волокно скелетной мышцы иннервируется одним
мотонейроном. Его окончания образуют синаптические
контакты со специализированной областью мышечной мембраны —
двигательной концевой пластинкой (рис. 4.10). Мембрана
пресинаптического окончания отделена от постсинаптической
концевой пластинки промежутком в 30 нм — синаптической щелью.
ПРЕСИНАПТИЧЕСКОЕ ВЫДЕЛЕНИЕ И ПОСТСИНАПТИЧЕСКОЕ
ДЕЙСТВИЕ МЕДИАТОРА
В пресинаптическом окончании медиатор ацетилхолин
хранится в мелких сферических пузырьках диаметром 50 нм
(рис. 4.10). Эти синаптические пузырьки скапливаются вдоль
Физиология нейронов
83
внутренней поверхности пресинаптической мембраны. В
окончании содержатся также многочисленные митохондрии,
служащие поставщиками аденозинтрифосфата, необходимого для
синтеза медиатора и других важнейших компонентов. Когда
под действием нервного импульса наступает деполяризация
окончания аксона, синаптические пузырьки сливаются с
пресинаптической мембраной и содержимое их выбрасывается в
ПД нервного волокна
Синаптическая Рецепторы АХ
щель v j
Двигательная концевая пластинка
Рис. 4.10. Нервно-мышечная передача. К моторному окончанию приходит
потенциал действия (ПД); синаптические пузырьки высвобождают ацетилхолин
в синаптическую щель; ацетилхолин (АХ) связывается с рецепторами,
встроенными в постсинаптическую мембрану (двигательную концевую пластинку);
потенциал двигательной концевой пластинки снижается от —85 мВ до
—10 мВ (возникает ПКП); под действием тока, идущего от
деполяризованной двигательной концевой пластинки к граничащей с ней мембране
мышечного волокна, в последней возникает потенциал действия.
синаптическую щель. Роль пускового механизма в выделении
медиатора играет повышение концентрации кальция (Са2+)
внутри окончания, наступающее в свою очередь в результате
деполяризации последнего.
Ацетилхолин диффундирует через синаптическую щель и
связывается с рецепторными белками, встроенными в мембрану
постсинаптической двигательной концевой пластинки (рис. 4.10),
что приводит к повышению проницаемости для натрия и калия.
В результате этого возникает входящий поток Na+ и
выходящий— К+, под действием которых концевая пластинка
деполяризуется примерно до —10 мВ (т. е. до среднего значения
между £к+ и ENa+ (см. выше). Эта деполяризация концевой
пластинки называется возбуждающим постсинаптическим
потенциалом (ВПСП) или потенциалом концевой пластинки (ПКП).
6*
84
Глава 4
ВОЗНИКНОВЕНИЕ ПОТЕНЦИАЛА ДЕЙСТВИЯ
В МЫШЕЧНОМ ВОЛОКНЕ
Нервно-мышечное соединение работает как повторитель:
в ответ на каждый импульс, исходящий от мотонейроиа, в
мышце всегда возникает потенциал действия. В отличие от ВПСП
в межнейронном соединении одиночный ПКП всегда вызывает
генерацию потенциала действия в мышечном волокне.
Потенциал действия мышечного волокна возникает под влиянием
деполяризующего тока, направленного от деполяризованной
концевой пластинки к соседним участкам мембраны мышечного
волокна (рис. 4.10). Величина тока, порождаемого ПКП, более
чем достаточна для того, чтобы мышечная мембрана
деполяризовалась до порогового уровня, при котором генерируется
потенциал действия. Последний распространяется по мембране
волокна и запускает мышечное сокращение (см. гл. 11).
ВОССТАНОВИТЕЛЬНЫЕ ПРОЦЕССЫ В НЕРВНО-МЫШЕЧНОМ
СОЕДИНЕНИИ
На наружной поверхности двигательной концевой
пластинки расположены два белковых комплекса, связывающиеся с
медиатором ацетилхолином: рецепторный белок, при помощи
которого регулируется ионная проницаемость мембраны, и
фермент ацетилхолинэстераза, инактивирующий ацетилхолин путем
расщепления его на составные части — ацетат и холин. Холин
переносится в пресинаптическое окончание, где используется
для ресинтеза ацетилхолина. Активность ацетилхолиэнстеразы
очень высока. Время полураспада молекулы ацетилхолина
составляет всего несколько тысячных долей секунды. За 20 мс
расщепляется весь выделившийся в синаптическую щель
ацетилхолин. По мере уменьшения количества ацетилхолина,
способного связываться с рецепторным белком, падает и ионная
проницаемость мембраны концевой пластинки, и, когда эта
проницаемость возвращается к исходному уровню, ПКП
заканчивается.
Восстановление пузырьков в аксонном окончании происходит
путем отщепления участков мембраны от ее краев. В этих
пузырьках накапливается ацетилхолин, который при помощи
фермента холинацетилазы синтезируется в окончании из
предшественников — ацетилкофермента А, образующегося в
митохондриях, и холина.
ФАРМАКОЛОГИЧЕСКИЕ ИНГИБИТОРЫ
НЕРВНО-МЫШЕЧНОЙ ПЕРЕДАЧИ
Многие фармакологические агенты блокируют
синаптическую передачу в области нервно-мышечного соединения. Наи-
Физиология нейронов
85-
более широко известный яд — кураре, впервые
использовавшийся южноамериканскими индейцами, блокирует
нервно-мышечную передачу, связываясь с рецепторными белками постсинап-
тической мембраны и тем самым препятствуя взаимодействию
с ними ацетилхолина. Кураре в зависимости от концентрации
уменьшает или полностью подавляет ПКП.
Некоторые фосфорорганические соединения — физостигмин
и неостигмин — влияют на нервно-мышечную передачу, инак-
тивируя ацетилхолинэстеразу. В результате ПКП удлиняется,
что приводит к генерации в мышце множественных
потенциалов действия. В ответ на каждый импульс мотонейрона в ней
возникает целый залп потенциалов действия, и вследствие этого
наступает сильное спастическое сокращение мышцы.
Одним из наиболее сильных среди известных ядов является
ботулин-токсин, выделяемый бактерией Clostridium botulinum.
Этот яд угнетает нервно-мышечную передачу, блокируя выход
ацетилхолина из пресинаптических терминалей. Он вызывает
смертельное пищевое отравление.
МЕЖНЕЙРОННЫЙ СИНАПС
На одном нейроне могут образовывать синаптические кон-
такты аксонные окончания сотен или тысяч других нервных:
клеток (рис. 4.11). Большинство этих окончаний неоднократно
ветвится. Синаптические контакты почти всегда образуются на
дендритах нейрона, и лишь около 15% всех аксонных
терминалей оканчивается на теле клетки. В конце каждого аксона
имеется маленькое расширение, или синаптическая бляшка
(рис. 4.12). Эта пресинаптическая терминаль содержит
синаптические пузырьки, заполненные медиатором; она отделена от
мембраны постсинаптического нейрона синаптической щелью
шириной 30 нм.
Последовательность событий, приводящих к возникновениям
постсинаптического потенциала в межнейронном синапсе, почти
такая же, как и при генерации ПКП в нервно-мышечном
соединении (см. выше). Однако межнейронные синапсы
отличаются от нервно-мышечных двумя основными особенностями:.
1) многие межнейронные синапсы являются тормозными, т. е.
из их синаптических бляшек высвобождается медиатор,
вызывающий генерацию тормозного постсинаптического потенциала
(ТПСП) в мембране постсинаптического нейрона; 2) ВГ1СП,
возникающий при деполяризации одиночной синаптической
бляшки, слишком мал (1—2 мВ) для того, чтобы вызвать по-
Синаптические
бляшки
Аксон
Рис. 4.11. Синаптические входы нейрона. Синаптические бляшки окончаний
пресинаптических аксонов образуют соединения на дендритах и теле (соме)-
постсинаптического нейрона.
ПД нервного волокна
Митохондрия
Синаптическая
бляшка
Постсинаптическая--
мембрана
Синаптические
пузырьки
Синаптическая.
щель
Постсинаптические рецепторы
Рис. 4.12. Межнейронный синапс. Когда под действием нервного импульса
пресинаптическое окончание (бляшка) деполяризуется, синаптические
пузырьки высвобождают медиатор в синаптическую щель. В результате связывания
медиатора с постсинаптическими рецепторами возникает либо местная
деполяризация (ВПСП), либо местная гиперполяризация (ТПСП) постсинаптиче-
ской мембраны.
86
Физиология нейронов
8Т
тенциал действия, и импульсы в постсинаптическом нейроне-
возникают при суммации нескольких, накладывающихся друг
на друга ВПСП.
ПОСТСИНАПТИЧЕСКИЕ РЕАКЦИИ
Медиатор в зависимости от его природы и специфики пост-
синаптической клетки может либо деполяризовать, либо гипер-
поляризовать ее мембрану (рис. 4.13). Если под действием
медиатора растет проницаемость мембраны для Na+, K+ и С1~^
то возникает ее деполяризация — ВПСП. Если же увеличивает-
Возбуждающие
А синапсы ^
Микроэлектрод
синапс
о
1=
-70
Аксон
Временная Пространственная Подавление
суммация суммация ВПСП
ВПСП
г
/N
А
г т
^
А А
А + Б
А
А •- В
Рис. 4.13. Взаимодействие ВПСП и ТПСП в межнейронном синапсе. Под
окончаниями А и Б возникают ВПСП, а под окончанием В — ТПСП;
потенциалы регистрируются при помощи микроэлектрода. Суммарный ВПСП может
возникнуть в результате либо временной (при повторных разрядах одного и
того же волокна), либо пространственной (при одновременном возбуждении
двух или нескольких волокон) суммации. Тормозное действие ТПСП
проявляется в уменьшении величины ВПСП.
-88
Глава 4
ся проницаемость лишь для К+ и С1~, то мембрана
гиперполяризуется— генерируется ТПСП.
Потенциал действия впервые возникает в области аксонного
холмика нейрона (рис. 4.14)—начального сегмента аксона в
месте его отхождения от тела клетки. Аксонный холмик — это
самый возбудимый участок нейрона с наиболее низким поро-
Рис. 4.14. Генерация потенциала действия в аксонном холмике. Под
действием ВПСП в дендритах и соме возникают деполяризующие токи, направленные
к холмику. Если наступающая в результате этого деполяризация холмика
^превышает пороговый уровень, генерируется потенциал действия.
гом. Для того чтобы в постсинаптическом нейроне возник
нервный импульс, необходимо деполяризовать мембрану аксонного
холмика на величину от —10 до —25 мВ.
ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ НЕЙРОНА
ВПСП, генерируемый в одном возбуждаемом синапсе,
приводит лишь к незначительному колебанию мембранного
потенциала в аксонном холмике (1 мВ или меньше); это связано с
тем, что ВПСП возникает в отдалении от аксонного холмика,
а амплитуда его мала. Потенциал действия может появиться
лишь в случае повторных разрядов в одной синаптической
бляшке (временная суммация) либо при одновременном
возбуждении нескольких окончаний (пространственная суммация).
Отдельные ВПСП, генерируемые при повторных или
одновременных разрядах, складываются и образуют суммарный ВПСП
(рис. 4.13). Если местные деполяризующие токи, возникающие
под влиянием суммарного ВПСП, достаточно велики для того,
чтобы мембрана аксонного холмика деполяризовалась до
порогового уровня, возникает потенциал действия.
ТПСП тормозит генерацию потенциала действия, уменьшая
.величину суммарного ВПСП (рис. 4.13). Возникновение нерв-
Физиология нейронов
89>
ного импульса зависит от того, достаточна ли амплитуда
суммарного синаптического потенциала, образующегося в
результате сложения всех ВПСП и ТПСП, для деполяризации
мембраны аксонного холмика до порогового уровня.
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
В межнейронных синапсах возможны два типа торможения:
1) постсинаптическое торможение, связанное с генерацией ги-
перполяризующего ТПСП, и 2) пресинаптическое торможение,
при котором уменьшается выброс возбуждающего медиатора.
Пресинаптическое торможение наблюдается в тех случаях,
когда окончания аксона образуют синапс на другой синапти-
t Т t
_, А Б А
Рис. 4.15. Пресинаптическое торможение. При возбуждении окончания Б
уменьшается количество медиатора, высвобождающегося из окончания А.
Судя по микроэлектродной записи от окончания А, окончание Б оказывает
на него тормозное действие, деполяризуя его и тем самым уменьшая
амплитуду возникающего в нем потенциала действия.
ческой бляшке (рис. 4.15). Когда по тормозному пресинапти-
ческому окончанию пробегает нервный импульс, из этого
окончания выделяется медиатор, деполяризующий «постсинаптиче-
скую» бляшку. Это приводит к тому, что количество медиатора,
которое может выделяться этой бляшкой, уменьшается, так
как снижается амплитуда генерируемого в ней потенциала
90
Глава 4
действия; амплитуда же этого потенциала действия
уменьшается потому, что постсинаптическая бляшка уже частично
деполяризована пресинаптическим тормозным окончанием.
ВОЗБУЖДАЮЩИЕ И ТОРМОЗНЫЕ МЕДИАТОРЫ
Предполагается, что роль медиаторов играют некоторые
низкомолекулярные органические вещества. В окончаниях
мотонейронов спинного мозга выделяется ацетилхолин. Он служит
также возбуждающим медиатором в вегетативной нервной
системе (ВНС) и, возможно, в некоторых областях мозга. Нор-
адреналин играет роль возбуждающего медиатора как в ВНС,
так и в ряде отделов мозга. Дофамин является возбуждающим
медиатором для базальных ганглиев. К другим возможным
возбуждающим медиаторам относятся аспартат, глутамат и
серотонин. Есть данные о том, что некоторые белки —
субстанция Р, рилизинг-факторы гипоталамуса и энкефалины — могут
влиять на деятельность нейронов, возможно, выступая в роли
медиаторов.
В ЦНС есть по меньшей мере два тормозных медиатора:
гамма-аминомасляная кислота и глицин. Ацетилхолин,
выделяющийся парасимпатическими нейронами, тормозит активность
сердечной и ряда гладких мышц. Норадреналин,
высвобождающийся в окончаниях симпатических нейронов, также оказывает
тормозное влияние на некоторые гладкие мышцы (эти эффекты
парасимпатических и симпатических нервов обсуждаются
в гл. 11).
ЭЛЕКТРИЧЕСКИЕ СИНАПСЫ
В некоторых областях мозга млекопитающих обнаружены
межнейронные синаптические соединения, в которых передача
осуществляется нехимическим путем. К ним относятся щелевые
контакты (нексусы). В этих соединениях промежуток между
пре- и постсинаптическим нейронами равен 2 нм; через него
проходят тончайшие (диаметром 1 нм) цитоплазматические
каналы. Эти каналы служат проводниками тока, идущего от
пресинаптического нейрона к постсинаптическому. В таких
соединениях от нейрона к нейрону распространяются
деполяризующие и гиперполяризующие токи. Щелевой контакт
характеризуется двусторонним проведением (в химическом синапсе
проведение одностороннее — от пресинаптического окончания к
постсинаптическому нейрону).
Физиология нейронов
91
ЛИТЕРАТУРА
Aidley D. J. (1971). The physiology of excitable cells, Cambridge University
Press, New York.
Axelrod J. (1974). Neurotransmitters, Sci. Amer., 230, 59.
Baker P. F. (1966). The nerve axon, Sci. Amer., 214, 74.
Eccles J. C. (1965). The synapse, Sci. Amer., 212, 56.
Eccles J. C. (1977). The understanding of the brain, McGraw-Hill, New York.
Hodgkin A. L. (1964). The ionic basis of nerve conduction, Science, 145, 1148.
Katz B. (1968). Nerve, Muscle and synapse, McGraw-Hill, New York. [Имеется
перевод первого издания: Катц Б. Нерв, мышца и синапс. — М.: Мир,
1969.]
Katz В. (1971). Quantal mechanisms of neural transmitter release, Science, 172,
123.
Kmjevic K. (1974). Chemical nature of synaptic transmission in vertebrates,
Physiol. Rev., 54, 418.
Lester H. A. (1977). The response to acetylcholine, Sci. Amer., 236, 106.
Mountcastle V. B. (1974). Medical physiology, Mosby. St. Louis.
Shepherd G. M. (1974). The synaptic organization of the brain: An introduction,.
Oxford University Press, New York.
Shepherd G. M. (1978). Microcircuits in the nervous system, Sci. Amer., 238, 92.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Что такое возбудимая клетка? Приведите несколько при-
меров.
2. Составьте таблицу распределения основных катионов и
анионов по обе стороны мембраны; объясните, почему они
так распределяются.
3. Раскройте ионные основы мембранного потенциала покоя.
Почему он не соответствует равновесному потенциалу для
калия, вычисляемого из уравнения Нернста?
4. Нарисуйте график потенциала действия. Укажите его
длительность и амплитуду (вольтаж).
5. Опишите последовательность изменений ионной
проницаемости нервной мембраны в процессе развития потенциала
действия, т. е. в фазе деполяризации, реполяризации и
гиперполяризации.
6. Что такое закон «все или ничего»? Объясните, почему
возникновение потенциала действия подчиняется этому закону.
7. В чем разница между фазами абсолютной и относительной
рефрактерности? Как их измеряют?
8. В чем состоит значение Na+/K+-Hacoca для поддержания
возбудимости нерва? Как изменится проницаемость при
подавлении активности этого насоса?
9. Расскажите, как нервный импульс распространяется вдоль
аксона. Что такое «местные токи»?
10. Что такое сальтаторное проведение?
11. Нарисуйте схему двигательной концевой пластинки.
^2
Глава 4
12. Опишите последовательность событий, происходящих при
передаче возбуждения через двигательную концевую
пластинку, начиная с прихода нервного импульса к окончанию
мотонейрона.
13. В чем основные морфологические и физиологические
различия между нервно-мышечным и межнейронным
синапсами?
14. Дайте определение пространственной и временной сумма-
ции.
15. Сравните и укажите разницу между пресинаптическим и
постсинпатическим торможением. Опишите морфологический
субстрат, особенности выделения медиатора и влияние на
возбудимость нейрона для каждого из этих видов
торможения.
16. Расскажите, как нейрон выполняет свою интегративную
функцию. В чем состоит роль аксонного холмика в этом
процессе передачи информации?
17. Что такое щелевой контакт (нексус)? Как через него
передается возбуждение с нейрона на нейрон?
Глава 5
РЕЦЕПЦИЯ.
СОМАТИЧЕСКАЯ ЧУВСТВИТЕЛЬНОСТЬ
Центральная нервная система (ЦНС) получает
информацию о состоянии окружающей среды от рецепторов. Каждый
рецептор воспринимает определенный раздражитель —
химический, электромагнитный (световые волны), механический или
температурный. Рецепторы — это датчики, преобразующие
энергию раздражителя в электрохимический потенциал.
Информация о раздражителе кодируется в виде импульсов в
чувствительных (сенсорных) нервах. Эта информация поступает в
сенсорные структуры нервной системы, где подвергается
декодированию и анализу.
РЕЦЕПТОРЫ
Морфологически и физиологически каждый рецептор
приспособлен для восприятия раздражителя строго определенной
модальности. Это так называемые «адекватные» раздражители,
т. е. раздражители, к которым рецептор наиболее чувствителен.
В основу одной из общепринятых классификаций
рецепторов положена модальность адекватных раздражителей. По
этому признаку все рецепторы обычно делят на пять групп:
1) фоторецепторы, воспринимающие свет; 2) механорецепторы,
воспринимающие механическое перемещение (прикосновение,
давление, звуковые волны); 3) терморецепторы, чувствительные
к температуре (холоду и теплу); 4) хеморецепторы,
воспринимающие химические вещества внешней и внутренней среды
(к ним относятся рецепторы, чувствительные к напряжению
дыхательных газов и уровню глюкозы в крови, а также вкусовые
и обонятельные); 5) ноцицептивные рецепторы, реагирующие на
повреждения ткани, сопровождающиеся болью.
Рецепторы можно также подразделить в зависимости от
того, где находится воспринимаемый ими раздражитель. В
соответствии с такой классификацией рецепторы делятся на четыре
группы: 1) дистантные экстероцепторы, реагирующие на
отдаленные раздражители (зрительные, обонятельные и слуховые);
2) контактные экстероцепторы, воспринимающие раздражение
поверхности тела (рецепторы прикосновения, давления, темпе-
94
Глава 5
ратурные и вкусовые); 3) йнтероцепторы, воспринимающие
раздражители от внутренних органов и уровень химических
веществ в крови, и 4) проприоцепторы, сигнализирующие о
положении тела в пространстве (о расположении суставов, длине
мышц).
ПРЕОБРАЗОВАНИЕ ЭНЕРГИИ РАЗДРАЖИТЕЛЯ
Первичная реакция любого рецептора состоит в генерации
рецепторного потенциала, возникающего в результате
взаимодействия между раздражителем и мембраной рецептора
'Мембрана рецептора Начальный сегмент
Рис. 5.1. Возникновение рецепторного потенциала в чувствительном нервном
окончании. При взаимодействии раздражителя с мембраной рецептора ее
ионная проницаемость возрастает. Это вызывает вхождение Na+ в окончание
рецептора, его деполяризацию и генерацию рецепторного потенциала. Под
действием последнего возникают местные токи, деполяризующие начальный
сегмент чувствительного нерва. Когда эта деполяризация достигает
порогового уровня, возникает потенциал действия.
(рис. 5.1). В зависимости от характера адекватного
раздражителя это взаимодействие может сопровождаться: 1)
механической деформацией мембраны (механорецепторы); 2)
возбуждением связанного с мембраной фотопигмента под действием
световых волн (фоторецепторы); 3) изменением проницаемости
мембраны под влиянием температуры (терморецепторы);
4) связыванием химических веществ мембраной рецептора (хе-
морецепторы); 5) связыванием пептидов, высвобождающихся
при повреждении ткани, с мембраной рецептора (ноцицептив-
ные рецепторы). Любой из этих процессов приводит к
повышению общей ионной проницаемости мембраны,
сопровождающейся вхождением Na+ в чувствительное окончание. В результате
этого входящего тока окончание деполяризуется и возникает
рецепторный потенциал; в фоторецепторах глаза вместо
деполяризации наступает гиперполяризация (см. гл. 6).
Рецепция. Соматическая чувствительность
95
ВОЗНИКНОВЕНИЕ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Нервные импульсы возникают в начальном сегменте
чувствительного нерва в результате возбуждающего действия рецеп-
торного потенциала (рис. 5.1). Последовательность процессов,
приводящих к генерации потенциала действия в чувствительном
нерве, зависит от анатомических взаимоотношений между этим
нервом и рецептором, в котором возникает рецепторный
потенциал. Этот рецептор может представлять собой либо окончание
чувствительного нерва, выполняющее функцию преобразования
Энергия раздражителя
Энергия
Энергия
Преобразование
\
Рецепторный
(генераторный)
потенциал
Потенциал
действия
Рецепторная
клетка >
Синапс
Сенсорный
нейрон
< И
Преобразование
Рецепторный
потенциал
Синаптическая
передача
Ф
Генераторный
потенциал
I
Потенциал
действия
Рис. 5.2. Схема возникновения возбуждения в чувствительном нерве,
иллюстрирующая преобразование рецептором энергии раздражителя в нервную им-
пульсацию. А. Рецептор представляет собой окончание чувствительного
нерва. Б. Рецептор — отдельная клетка, иннервируемая чувствительным
окончанием.
сенсорной информации (рис. 5.2,Л), либо отдельную клетку,
образующую с чувствительным окончанием химический синапс
(рис. 5.2,Б). В первом случае рецепторный потенциал,
возникающий в специализированных сенсорных окончаниях
афферентных нервов, играет роль генераторного потенциала
(рис. 5.2,Л). Во втором же случае под действием рецепторного
потенциала из «пресинаптической» чувствительной клетки
высвобождается химический медиатор, вызывающий возникнове-
96
Глава 5
ние деполяризующего генераторного потенциала в «ггостсинап-
тическом» сенсорном окончании (рис. 5.2,5).
Как в первом, так и во втором случае деполяризующие
токи, возникающие под действием генераторного потенциала,
приводят к возникновению нервных импульсов в
чувствительных нервах. Эти токи направляются к начальному сегменту
аксона, где порог возбуждения наиболее низок (рис. 5.1). По-
+ 30
ю
а*
х
Q)
I-
О
С
О h
-30
II
Порог
V 1
-70 £_Ё 1_^
Время, мс
Рис. 5.3. Возникновение нервного импульса под действием рецепторного
(генераторного) потенциала. А. Сила раздражителя достаточно велика, и
амплитуда рецепторного (генераторного) потенциала превышает пороговый
уровень для генерации нервного импульса в начальном сегменте. Б. Сила
раздражителя меньше; возникающий рецепторный (генераторный) потенциал
слишком мал, чтобы деполяризовать мембрану до порогового значения.
следовательность событий, приводящих к возникновению
потенциала действия в чувствительных нервах, такая же, как и при
возбуждении несенсорных нейронов и мышц: потенциал
действия возникает в том случае, если генераторный потенциал
достаточно велик для того, чтобы деполяризовать начальный
сегмент до порогового значения (рис. 5.3; подробнее см. гл. 2).
КОДИРОВАНИЕ СЕНСОРНОЙ ИНФОРМАЦИИ.
АДАПТАЦИЯ
Главный принцип кодирования сенсорной информации
состоит в том, что характер ощущений, возникающих при
возбуждении чувствительных нервов, зависит от того, в какой области
ЦНС эти нервы оканчиваются.
Рецепция. Соматическая чувствительность
97
Интенсивность раздражения кодируется амплитудой рецеп-
торного потенциала. Величина этого потенциала
пропорциональна логарифму силы раздражителя. Так как в свою очередь
частота разрядов в чувствительных нервах пропорциональна
величине рецепторного потенциала, частота сенсорной импуль-
сации тоже пропорциональна логарифму силы раздражителя.
200
150 И-
2
S
Рецептор растяжения мышцы
100
Рис. 5.4. Адаптация рецепторов. При постоянном раздражении рецептора
уровень импульсации снижается. Рецепторы давления и прикосновения
относятся к фазным — они быстро адаптируются к постоянному раздражению.
Рецепторы же растяжения мышц тонические: при постоянном растяжении мышцы
они адаптируются медленно (лишь через несколько часов).
Недавно было показано, что логарифмическая зависимость
между силой раздражения и сенсорным разрядом
приблизительна. Более точно эта зависимость описывается степенными
уравнениями типа R = KIA, где R — величина сенсорного
разряда, / — сила раздражения, К и А — константы.
Если на любой рецептор в течение продолжительного
времени действовать постоянным раздражителем, то реакция
постепенно уменьшится (рис. 5.4). Это явление называется
адаптацией. По мере адаптации снижаются оба параметра
возбуждения — частота импульсации и величина рецепторного
потенциала. Считается, что рецепторный потенциал
уменьшается в результате постепенного падения ионной проницаемости
7—1705
98
Глава 5
мембраны рецептора. Само собой разумеется, что адаптивные
изменения уровня сенсорной импульсации являются прямым
следствием «адаптации» рецепторного потенциала: по мере
уменьшения этого потенциала частота разряда в
чувствительных нервах падает. '
Хотя адаптация свойственна всем рецепторам, скорость ее
у разных рецепторов различна (рис. 5.4). Некоторые из них
(например, тактильные рецепторы) независимо от длительности
раздражения отвечают на него всего несколькими импульсами.
В других же (рецепторы растяжения мышц и хеморецепторы
каротидных телец) в ответ на постепенно действующие
раздражители возникает импульсация, постепенно уменьшающаяся
лишь через несколько часов. В зависимости от скорости
адаптации рецепторы могут быть разделены на быстро
адаптирующиеся — фазные — и медленно адаптирующиеся — тонические.
СОМАТОСЕНСОРНАЯ СИСТЕМА
Виды чувствительности, сигнализирующие о состоянии тела,
называются соматестезией. К соматосенсорным рецепторам
относятся кожные рецепторы, реагирующие на прикосновение,
давление, температуру и боль, а также проприоцепторы,
воспринимающие движения в суставах и мышцах.
Соматосенсорным рецепторам можно противопоставить другую обширную
группу рецепторов — специальные сенсорные рецепторы, или
органы чувств, включающие зрительные, слуховые,
обонятельные, вкусовые и вестибулярные рецепторы (см. гл. 4). Все эти
рецепторы расположены в области головы и иннервируются
черепномозговыми нервами; соматосенсорные же рецепторы
находятся во всех частях тела — в конечностях, туловище и
голове. Подавляющее большинство соматосенсорных рецепторов
локализуется в туловище и конечностях и иннервируется
спинномозговыми нервами. Соматосенсорные рецепторы головы
иннервируются черепномозговыми нервами.
ЧУВСТВИТЕЛЬНЫЕ ВОЛОКНА СПИННОМОЗГОВЫХ
НЕРВОВ
В зависимости от размеров и скорости проведения все
чувствительные волокна спинномозговых нервов можно разделить
на четыре группы (табл. 5.1). Различные типы соматосенсорных
рецепторов иннервируются волокнами разного размера. Самые
крупные волокна (группа I) отходят от проприоцепторовмышц,
аннулоспиральных окончаний мышечных веретен и
сухожильных органов Гольджи. Волокна группы II идут от кожных
рецепторов прикосновения и давления, а также от гроздевидных
Рецепция. Соматическая чувствительность
99
Таблица 5.1. Чувствительные нервные волокна млекопитающего
Группа
I
II
III
IV
Рецепторы, характер чувствительности
Аннулоспиральные окончания
мышечных веретен, сухожильные органы
Гольджи
Гроздевидные окончания мышечных
веретен; тактильная
Болевая, температурная, тактильная
Болевая, температурная
Диаметр
волокон, мкм
12—20
4—12
1—4
0,5—1
Скорость
проведения,
м/с
75—120
24-72
6-24
<6
окончаний мышечных веретен. Тактильные, а также болевые и
температурные рецепторы иннервируются волокнами группы III.
Самые тонкие волокна группы IV несут импульсы от болевых
и температурных рецепторов.
Существуют два вида болевых ощущений — острая колющая
боль, возникающая при порезах или уколах, и жгучая боль,
наблюдаемая при ожогах. При повреждающих воздействиях,
сопровождающихся возбуждением болевых рецепторов
группы III, появляется острая колющая, строго локализованная
боль. Раздражение же болевых рецепторов группы IV приводит
к возникновению разлитой жгучей боли.
ЧУВСТВИТЕЛЬНЫЕ ОКОНЧАНИЯ И РЕЦЕПТИВНЫЕ
ПОЛЯ
Для многих соматосенсорных рецепторов характерны
специализированные окончания (рис. 5.5). Некоторые из наиболее
крупных окончаний, отвечающих за тактильную и болевую
чувствительность, окружены соединительнотканной сумкой; так
построены тельца Мейсснера, колбы Краузе и тельца Панины,
Механорецепторные окончания либо образуют утолщения
(диски Меркеля, тельца Руффини), либо оплетают основание
волосяного фолликула. Мышечные веретена и сухожильные
органы Гольджи воспринимают изменения длины и напряжения
в мышце (подробнее см. гл. 7). Наиболее тонкие соматосенсор-
ные волокна образуют свободные окончания, расположенные в
разных областях тела. К ним относятся все холодовые,
тепловые и болевые рецепторы, а также механорецепторы, иннерви-
рующие те области, в которых нет более сложно построенных
окончаний. Так, в роговице, чувствительной к механическим
воздействиям, находятся только свободные окончания.
Рецептивным полем чувствительного нейрона называется
поверхность тела, раздражение которой приводит к
возбуждению этого нейрона. Размеры рецептивного поля примерно соот-
7*
100
Глава 5
ветствуют величине области распространения волокон
соответствующего нейрона. Чувствительные волокна группы II
отходят от инкапсулированных рецепторов и тактильных окончаний
волосяных луковиц и возбуждаются лишь при раздражении
непосредственно прилегающих к ним тканей. Их рецептивные
( Ж '
Рис. 5.5. Виды чувствительных нервных окончаний. К инкапсулированным
окончаниям относятся тельца Мейсснера (Л), колбы Краузе (Б) и тельца
Пачини (В). Расширенные окончания, воспринимающие механические
раздражения, представлены дисками Меркеля (Г) и тельцами Руффини (Д).
Окончания чувствительных нервов могут оплетать основания волосяных
фолликулов (Е) или быть свободными (Ж).
поля строго ограничены. Свободные же окончания
чувствительных нейронов (группы III и IV) охватывают обширные области;
эти нейроны обладают большими диффузными рецептивными
полями.
ЦЕНТРАЛЬНЫЕ СОМАТОСЕНСОРНЫЕ ПУТИ
Волокна от соматосенсорных рецепторов туловища и
конечностей идут к спинному мозгу в составе спинномозговых нервов
и вступают в него через задние корешки. В спинном мозгу они
Рецепция. Соматическая чувствительность
101
либо заканчиваются в сером веществе на нейронах, дающих
начало соответствующим трактам (волокна групп I, III, IV),
либо образуют восходящие пути в белом веществе (волокна
Группы II).
Существуют четыре основных спинальных афферентных
пути (рис. 5.6): 1) задние столбы, 2) спинно-мозжечковый, 3)
латеральный спинно-таламический и 4) передний спинно-талами-
Рис. 5.6. Главные чувствительные пути спинного мозга (объяснение см. в
тексте).
ческий. Задние столбы образованы аксонами чувствительных
нейронов группы II. Нейроны группы I переключаются на
спинномозговых вставочных нейронах, аксоны которых образуют
спинно-мозжечковый путь. Латеральный спинно-таламический
путь состоит из аксонов вставочных нейронов спинного мозга,
на которых оканчиваются отростки нейронов группы IV.
Передний спинно-таламический путь образован аксонами вставочных
нейронов, возбуждаемых чувствительными волокнами группы III.
Соматосенсорные волокна головы и лица идут к головному
мозгу в составе тройничного нерва и оканчиваются в его
чувствительном ядре.
СИСТЕМА ЗАДНИХ СТОЛБОВ
Задние столбы спинного мозга образованы аксонами
чувствительных нейронов группы II, восходящими к продолговатому
мозгу в составе спинного (рис. 5.7). В продолговатом мозгу
эти аксоны переключаются на вставочных нейронах ядер
задних столбов (клиновидного и нежного ядра). Отростки нейро-
102 Глава 5
Рис. 5s.7(. Задние столбы (объяснение см. в тексте). Эта цепь, образованная
тремя нейронами, передает информацию от рецепторов давления и
прикосновения к соматосенсорной коре.
нов этих ядер переходят на противоположную сторону
продолговатого мозга и восходят в составе медиальной петли к задне-
вентральному ядру таламуса. Здесь отростки нейронов
продолговатого мозга переключаются на нейроны таламуса,
аксоны которых идут к соматосенсорным областям коры. Таким
образом, задние столбы представляют собой трехнейронную
цепь, проводящую импульсы от рецепторов группы II к
соматосенсорной коре; в эту цепь входят: 1) чувствительные нейроны
группы II; 2) нейроны продолговатого мозга и 3) нейроны
Рецепция. Соматическая чувствительность
103
таламусэ. Эти пути служат для быстрой передачи тактильных
сигналов к коре.
Чувствительные нейроны задних столбов обладают
небольшими, четко ограниченными рецептивными полями. Многие из
них возбуждаются при слабом прикосновении или
надавливании на кожу, на которые реагируют инкапсулированные
окончания. Нейроны задних столбов передают информацию о
точной локализации раздражителя на поверхности тела.
Существует соответствие между положением нейрона в
задних столбах и проекцией его рецептивного поля на поверхности
тела. Нейроны, воспринимающие раздражения соседних
участков кожи, расположены в задних столбах рядом друг с другом.
Место каждого волокна в заднем столбе определяется
локализацией его окончания. Волокна нейронов первого порядка,
идущие от рецепторов ног и туловища, занимают в задних столбах
медиальное положение, волокна же от рук и шеи —
латеральное. Чем краниальнее локализовано окончание волокна, тем
латеральнее это волокно расположено в задних столбах.
СПИННО-ТАЛАМИЧЕСКИЕ ПУТИ
Система спинно-таламических путей состоит из двух
основных трактов — переднего и латерального. Передний спинно-та-
ламический тракт образован аксонами спинномозговых
вставочных нейронов задних рогов; на этих нейронах переключаются
сенсорные нейроны группы III, воспринимающие
прикосновение, давление, температуру и острую боль (рис. 5.8). Волокна
нейронов группы IV, реагирующие на температурные
раздражители или воздействия, сопровождающиеся жгучей болью,
заканчиваются на спинальных вставочных нейронах, входящих
в состав латерального спинно-таламического тракта (рис. 5.9).
Перед тем как войти в передний и латеральный спинно-та-
ламический пути, восходящие по спинному мозгу и стволу
мозга к таламусу, отростки обеих групп вставочных нейронов
перекрещиваются. При прохождении через продолговатый мозг,
мост и средний мозг аксоны спинно-таламического пути отдают
многочисленные коллатерали. По этим коллатералям
возбуждение направляется в срединные структуры ствола мозга, и в
частности в ретикулярную формацию, которая играет важную
роль в поддержании состояния активации (см. гл. 7).
Волокна нейронов второго порядка, идущие в составе
разных спинно-таламических путей, оканчиваются в разных
отделах таламуса. Аксоны переднего тракта переключаются в зад-
нелатеральном вентральном ядре, а отростки нейронов второго
104
Глава 5
Соматосенсорная кора
Вентробазальный
комплекс таламуса
Средний мозг
Медиальная петля
Продолговатый мозг
Передний спинно-таламический
путь
Рис. 5.8. Передний спинно-таламический тракт. По этому пути в соматосен-
сорную кору поступает информация от рецепторов давления и прикосновения,
а также от болевых и температурных.
порядка, образующие латеральный тракт, оканчиваются в
заднем и межпластиночном ядрах таламуса. Аксоны нейронов
третьего порядка идут от таламуса к коре головного мозга.
Отростки нейронов, воспринимающих тактильные,
температурные и ноцицептивные (острую боль) раздражители,
заканчиваются в соматосенсорной коре; окончания же нейронов,
отвечающих за жгучую боль, широко ветвятся во всех областях коры.
Рецепторы нейронов групп III, IV образованы ветвящимися
окончаниями чувствительных нервов. Рецептивные поля этих
Рецепция. Соматическая чувствительность
105
Диффузные
проекции на кору
Межпластиночные
ядра таламуса
•Средний мозг
Продолговатый мозг
Латеральный
спинно-таламический путь.
Рис. 5.9. Латеральный спинно-таламический тракт. По этому пути информация
от температурных и болевых рецепторов поступает к обширным областям
коры головного мозга (подробнее см. в тексте).
нейронов диффузны и обширны. В связи с этим нейроны спин-
но-таламических путей не способны к тонкой пространственной
дифференцировке локализации раздражителя. Эти нейроны
возбуждаются при тактильных, температурных или болевых
воздействиях на обширные области тела. Напротив, волокна
задних столбов воспринимают только тактильное раздражение
небольших локализованных областей. По спинно-таламической
системе чувствительные сигналы передаются в кору медленно,
тогда как волокна задних столбов обеспечивают быстрое
проведение в кору информации о локализации раздражения.
106
Глава 5
ВЕНТРОБАЗАЛЬНЫИ КОМПЛЕКС ТАЛАМУСА
В заднелатеральный вентральный отдел таламуса импульса-
ция поступает от двух групп нейронов второго порядка — от
системы задних столбов и от образующих передние спинно-та-
ламические тракты. Области их проекции в таламусе
перекрываются и организованы соматотопически: проекция нейрона в
таламусе определяется расположением его рецептивного поля
на поверхности тела. Волокна нейронов второго порядка,
рецептивные поля которых граничат друг с другом (эти волокна
возбуждаются при раздражении смежных участков кожи),
переключаются на соседних нейронах таламуса.
Соматосенсорные волокна от области лица идут в составе
тройничного нерва и переключаются на нейроны второго
порядка в его ядре, расположенном в среднем мозгу. Волокна
нейронов второго порядка заканчиваются в заднемедиальном
вентральном ядре таламуса. Это ядро вместе с заднелатеральным
вентральным ядром образует вент роб азальный комплекс
таламуса.
Соматотопическая организация, характерная для заднела-
терального вентрального ядра таламуса, распространяется и на
его заднемедиальное вентральное ядро. Чем медиальнее
расположен нейрон в вентробазальном комплексе, тем краниальнее
его рецептивное поле.
СОМАТОСЕНСОРНАЯ КОРА
Волокна нейронов вентробазального комплекса таламуса
оканчиваются в соматосенсорнои коре (рис. 5.7 и 5.8).
Первичная соматосенсорная область коры расположена в
постцентральной извилине непосредственно позади центральной
борозды (рис. 5.10). Сзади и сбоку от этой области находится
вторичная зона соматосенсорнои интеграции — вторичная соматосен-
сорная область.
Первичная соматосенсорная область организована
соматотопически. Проекция каждого таламического нейрона в этой
зоне коры определяется его положением в таламусе. Волокна,
отвечающие на раздражение стопы, заканчиваются в дорсоме-
диальной части постцентральной извилины, в то время как
рецепторы головы проецируются в области соматосенсорнои коры,
занимающие самое вентролатеральное положение (рис. 5.10).
Расположение нейрона в соматосенсорнои коре четко
соответствует локализации его рецептивного поля на поверхности тела.
Поля всех нейронов постцентральной извилины находятся на
контрлатеральной стороне тела. Нейроны же вторичных сома-
тосенсорных областей получают сигналы от обеих сторон —
как ипсилатеральной, так и контрлатеральной.
Рецепция. Соматическая чувствительность
107
Рис. 5.10. Соматосенсорная кора.
СТРОЕНИЕ КОРЫ
Кора состоит из шести слоев нейронов. Импульсы от вентро-
базального комплекса таламуса приходят к нейронам слоя IV,
отростки которых идут в двух направлениях: к поверхности,
в слои I и II, и в глубину, к слоям V и VI. От соматосенсорной
области сигналы посылаются по аксонам нейронов слоев V, VI,
соединяющим эту область с другими зонами коры, в частности
ассоциативной, соматосенсорной и двигательной.
Отростки нейронов коры ориентированы главным образом
вертикально. Поэтому взаимодействие между нейронами
осуществляется в основном в пределах вертикальных столбиков
(колонок), расположенных перпендикулярно к поверхности
коры и захватывающих все шесть ее слоев. Такая организация
коры в виде колонок обеспечивает обработку соматосенсорной
информации. Каждая колонка отвечает за информацию о сома-
тосенсорном раздражителе определенной модальности,
действующем на определенную область. Так, многие колонки
определяют качество стимула (остроту, шероховатость и интенсивность)
и его локализацию: для каждой точки поверхности тела
существует своя колонка. Другие колонки реагируют на движения
или положение суставов.
Первичная соматосенсорная область отвечает как за
определение локализации раздражения на поверхности тела, так
и за различение локализации и силы двух одновременно
наносимых раздражителей. В этой области происходит оценка каче-
108
Глава 5
ства стимула (остроты, шероховатости или температуры) и
интеграция информации о расположении и движениях тела в
пространстве (происходящая на уровне сознания).
Деятельность вторичной соматосенсорной области
недостаточно изучена. Эта область получает сигналы от обеих половин
тела и участвует в интеграции соматосенсорной информации от
всего организма.
ЛИТЕРАТУРА
Catton W. Т. (1970). Mechanoreceptor function, Physiol. Rev., 50, 297.
Eyzaguirre С, Fidone S. J. (1975). Physiology of the nervous system, 2nd edn.
Year Book Medical, Chicago.
Lim R. K. S. (1970). Pain, Ann. Rev. Physiol., 32, 269.
Lowenstein W. R. (I960). Biological transducers, Sci. Amer., 203, 98.
Lynn B. (1975). Somatosensory receptors and their CNS connections, Ann. Rev.
Physiol., 37, 105.
Mountcastle V. B. (1974). Medical physiology, 13th edn., Mosby, St. Louis.
Sensory Receptors (1965). Cold Spring Harbor symposium on quantitative
biology. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Wall P. D., Dubner R. (1977). Somatosensory pathways, Ann. Rev., Physiol., 34,
315.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Какие стимулы называются «адекватными» для данного
рецептора?
2. Что такое 1) дистантные экстероцепторы; 2) контактные
экстероцепторы; 3) интероцепторы; 4) проприоцепторы?
3. Опишите последовательность событий, приводящих к
возникновению разряда в чувствительном нерве под действием
раздражителя.
4. Составьте таблицу различных групп чувствительных волокон
спинномозговых нервов. Включите в нее данные об
относительных размерах этих волокон, связанных с ними
спинномозговых путях и о том, какие виды чувствительности они
проводят.
5. Что такое рецептивные поля?
6. Назовите два спинномозговых пути, проводящих соматосен-
сорную информацию к вентробазальному комплексу таламу-
са. Коротко расскажите, в чем состоят различия в нейро-
анатомической организации этих путей и в передаваемой
ими информации.
7. Опишите нервные пути, передающие соматосенсорную
информацию от области лица к коре.
8. Для соматосенсорной коры характерны как соматотопиче-
ская организация, так и строение в виде колонок. Опишите
коротко сущность этих двух типов организации.
Глава 6
ОРГАНЫ ЧУВСТВ
Органами чувств называются сенсорные системы,
расположенные в области головы. К ним относятся зрительная,
слуховая, вестибулярная, вкусовая и обонятельная системы.
ЗРИТЕЛЬНАЯ СИСТЕМА
Функция глаза состоит в получении зрительной информации
от окружающей среды и передаче ее в сенсорные области
головного мозга. Зрительный образ проецируется на рецепторы
сетчатки глаза благодаря сложной оптической системе.
Сетчатка образована густой сетью рецепторов и связанных с ними
нейронов, извлекающих информацию о таких параметрах
зрительного раздражителя, как, в частности, интенсивность, цвет,
размер, кривизна и скорость перемещения. Эта информация
передается по зрительному нерву к зрительным областям
мозга, где происходит ее переработка.
СТРОЕНИЕ ГЛАЗА
Глаз представляет собой сферический орган, покрытый
плотной фиброзной соединительнотканной оболочкой —
склерой (рис. 6.1). Спереди склера переходит в прозрачную
роговицу. Внутренняя поверхность склеры выстлана двумя тонкими
оболочками — сосудистой и сетчаткой. Сосудистая оболочка,
содержащая многочисленные сосуды, питающие глаз,
расположена между склерой и сетчаткой. Сетчатка — это слой,
образованный нервными элементами; здесь расположены
фоторецепторы и вставочные нейроны. Аксоны ганглиозных клеток
сетчатки образуют зрительный нерв.
Хрусталик делит глаз на два отсека с жидким содержимым:
передняя камера заполнена водянистой влагой; позади
хрусталика находится студенистая масса — стекловидное тело. Перед
тем как попасть на сетчатку, свет должен пройти через
прозрачную роговицу, водянистую влагу, зрачок, хрусталик и
стекловидное тело.
по
Глава 6
ЗРАЧКОВЫЙ РЕФЛЕКС
Поступающий в глаз световой поток регулируется
радужкой, изменяющей размер зрачка. В радужке имеются две
группы мышечных волокон — суживающие и расширяющие зрачок
(рис. 6.2). Мышца, суживающая зрачок (сфинктер), состоит
из окружающих отверстие циркулярных волокон, тогда как
Рис. 6.1. Строение глаза; передняя камера глаза заполнена водянистой
влагой; позади хрусталика находится стекловидное тело. Перед тем как попасть
на фоторецепторы сетчатки, свет последовательно проходит через роговицу,
водянистую влагу, хрусталик и стекловидное тело.
мышца, расширяющая зрачок (дилятатор), образована
радиальными волокнами, отходящими от зрачка подобно спицам
колеса. Сфинктер иннервируется парасимпатическими нервами,
а дилятатор — симпатическими (см. гл. 10).
На ярком свету радужка сокращается, и входящий в глаз
световой поток уменьшается. При увеличении освещения
нейроны претектального ядра посылают импульсы по
парасимпатическим волокнам, иннервирующим сфинктер зрачка. В
результате сфинктер сокращается, зрачок суживается и световой
поток, идущий к сетчатке, уменьшается. Если уровень
освещения снижается, то нейроны претектального ядра тормозят
Органы чувств
111
активность парасимпатических волокон, что приводит к
расслаблению сфинктера и расширению зрачка. Возбуждение
симпатических нервов при эмоциональной или физической
нагрузке сопровождается активным расширением зрачка в
результате сокращения волокон дилятатора (см. гл. 10).
ФОРМИРОВАНИЕ ИЗОБРАЖЕНИЯ
Оптическая система глаза сходна с фотоаппаратом.
Роговица и хрусталик фокусируют лучи на сетчатке, содержащей
слой фоторецепторов; последние запечатлевают зрительный
образ в виде изменений электрических потенциалов.
Возбуждение фоторецепторов световыми лучами аналогично
экспозиции пленки в фотоаппарате.
Прохождение световых лучей через искривленную
поверхность, разграничивающую две среды с различной оптической
Зрачок
Циннова
связка
Хрусталик Зрачок
Сфинктер
Ресничная
мышца
Дилятатор
зрачка
А
Хрусталик
Рис. 6.2. Механизмы зрачкового рефлекса и аккомодации. А. Строение
радужки и хрусталика. Б. Зрачковый рефлекс; видны две группы мышечных
волокон в радужке; при сокращении сфинктера зрачок сужается, а при
сокращении дилятатора — расширяется. В. Аккомодация; хрусталик подвешен
на цинновых связках; при сокращении ресничной мышцы натяжение этих
связок уменьшается и эластичная капсула хрусталика становится почти
сферической.
плотностью, сопровождается преломлением лучей, или
рефракцией. Если лучи от отдаленного источника проходят через
двояковыпуклую линзу, то в результате преломления они
сходятся в некой точке сзади этой линзы — фокусе (рис. 6.3).
Преломление зависит от угла падения световых лучей на
поверхность линзы: чем больше угол падения, тем сильнее
преломление луча. Лучи, падающие на края линзы, значительно
преломляются по направлению к центральной оси; лучи же,
112
Глава 6
проходящие через центр линзы перпендикулярно к ней, не
преломляются вовсе.
Общая преломляющая сила глаза составляет примерно
66,7 диоптрии. Диоптрия — это величина, обратная фокусному
расстоянию (Z)=l:/); фокусным расстоянием (/) называется
расстояние от линзы до ее фокуса в метрах (рис. 6.3).
При прохождении лучей через глаз они преломляются на
четырех поверхностях раздела: 1) между воздухом и
роговицей, 2) между роговицей
и водянистой влагой,
3) между водянистой
влагой и хрусталиком и
4) между хрусталиком и
стекловидным телом.
Наибольшей
преломляющей силой (около 45
диоптрий) обладает
поверхность раздела между
воздухом и роговицей. Это
связано с большой
кривизной роговицы и
значительной разницей между
оптической плотностью
воздуха (1,0) и
роговицы (1,38).
Преломляющая сила хрусталика
равна приблизительно 20 диоптриям, что обусловлено относительно
небольшой разницей между его оптической плотностью (1,39)
и оптической плотностью водянистой влаги (1,33).
АККОМОДАЦИЯ
Если, не напрягая глаз, рассматривать предмет, удаленный
не более чем на 6 м, то изображение его будет размытым. Это
связано с тем, что преломляющая сила глаза оказывается
недостаточной, чтобы сфокусировать изображение на сетчатке
(рис. 6.4). Для того чтобы изображения близлежащих
предметов могли фокусироваться на сетчатке, существует
аккомодационный рефлекс, под влиянием которого преломляющая сила
глаза может увеличиваться на 14 диоптрий.
При аккомодации преломляющая сила глаза возрастает в
результате увеличения кривизны хрусталика. Эта кривизна
изменяется вследствие сокращения кольцевой ресничной
мышцы, окружающей хрусталик (рис. 6.2). Хрусталик как бы
подвешен в центре этого кольца на радиальных волокнах цинновой
связки.
Фокус
Фокусное расстояние
Рис. 6.3. Построение изображения в
двояковыпуклой линзе. Лучи от удаленного
источника сходятся позади линзы в фокусе.
Расстояние от линзы до фокуса называется
фокусным расстоянием.
Органы чувств
113
Рис. 6.4. Аккомодация. Если, не напрягая глаз, смотреть на близко
расположенный предмет, то его изображение будет проецироваться позади сетчатки
(а). В результате аккомодации преломляющая сила глаза увеличивается и
четкое изображение строится в области сетчатки (б).
Ресничная мышца сокращается под действием
парасимпатических волокон, идущих в составе глазодвигательного нерва.
Когда взгляд переводится с отдаленного предмета на
близлежащий, эти волокна возбуждаются и ресничная мышца
сокращается. При этом уменьшается диаметр образованного ею
мышечного кольца. Так как волокна цинновой связки
вплетаются в это кольцо, при уменьшении его диаметра они
расслабляются. Снижение натяжения связок позволяет эластическому
хрусталику принять более сферическую форму; при этом
преломляющая сила глаза увеличивается.
Возбуждение парасимпатических волокон при
рассматривании близлежащего предмета приводит также к сужению
зрачка в результате сокращения его сфинктера.
Периферические лучи не попадают на сетчатку, и глубина резкости
увеличивается. На сетчатке формируется более четкое изображение.
С оптической точки зрения сужение зрачка при аккомодации
аналогично уменьшению отверстия диафрагмы фотоаппарата с
целью увеличения глубины резкости.
ОСТРОТА ЗРЕНИЯ
Острота зрения отражает способность оптической системы
глаза строить четкое изображение на сетчатке. Она
измеряется путем определения наименьшего расстояния между двумя
точками, при котором их изображения не сливаются. Это
расстояние должно быть достаточным для того, чтобы лучи от
обеих точек попали на разные рецепторы сетчатки. В норме
острота зрения равна углу в 1'; если расстояние между двумя
точками таково, что они просматриваются под углом в одну
минуту, их можно различить. Если же этот угол меньше, чем Г
8-1705
114
Глава 6
(или если изображение на сетчатке размыто из-за плохой
фокусировки), то изображения точек сливаются в одно.
Остроту зрения можно проверить с помощью таблиц Снел-
лена. На этих таблицах нанесено несколько строчек букв
разного размера. Строчки составлены таким образом, что детали,
отличающие одну букву от другой (например, С от О),
просматриваются с расстояния в 6 м (20 футов) под углами в 1',
2', 3', 4' и т. п. Определение остроты зрения сводится к оценке
у испытуемого способности различать последовательность таких
букв с одинаковыми размерами деталей. При остроте зрения
20/20, т. е. при нормальном зрении, человек может различать
буквы, детали которых просматриваются под углом в V. Если
острота зрения равна 20/40 (детали просматриваются лишь
под углом 2') или 20/60 (3'), то она считается сниженной;
таким людям необходимы очки.
АНОМАЛИИ РЕФРАКЦИИ
У многих людей изображение на сетчатке всегда получается
нечетким. Это бывает связано либо с необычной формой
глазного яблока (рис. 6.5), либо с неправильной кривизной
роговицы или хрусталика.
Дальнозоркость, или гипер метр опия, возникает в
результате укорочения глазного яблока. Расстояние между
хрусталиком и сетчаткой слишком мало, и фокус оказывается позади
Дальнозоркость
Близорукость
Рис. 6.5. Аномалии рефракции. А. При дальнозоркости (гиперметропии) глаз
укорочен и изображение располагается за сетчаткой. Для коррекции этого
дефекта перед глазом помещается двояковыпуклая линза. Б. При
близорукости (миопии) глаз чрезмерно удлинен и изображение располагается перед
сетчаткой. Для коррекции перед глазом помещается двояковогнутая линза.
Органы чувств
115
Рис. 6.6. Строение сетчатки. В наружном слое расположены пигментные
клетки (ПК). Рецепторы — палочки (П) и колбочки (К)—состоят из четырех
отделов: наружного сегмента, внутреннего сегмента, ядерной зоны и синап-
тической зоны. Наружный слой нейронов образован биполярными (Б),
горизонтальными (Г) и амакриновыми (А) клетками; внутренний слой образован
ганглиозными клетками (ГК), из отростков которых состоит зрительный нерв
(ЗН).
сетчатки. Этот дефект исправляется ношением очков с
выпуклыми линзами, увеличивающими преломляющую силу глаза.
Люди с удлиненными глазными яблоками страдают
близорукостью (миопией). В этом случае четкое изображение
формируется впереди сетчатки. Для исправления этого нарушения
применяют вогнутые линзы, уменьшающие преломляющую силу
глаза. Неправильная кривизна роговицы лежит в основе астиг-
8*
116
Глава 6
матизма. Изображение на сетчатке искажено: одни его части
находятся в фокусе, а другие — нет. В этом случае используют
линзы, корригирующие неправильную кривизну роговицы.
ФОТОРЕЦЕПЦИЯ
Сетчатка состоит из четырех слоев клеток (рис. 6.6):
пигментного слоя, слоя фоторецепторов и двух слоев нейронов
сетчатки. Наружный (ближайший к склере) слой образован
пигментными клетками. Слой фоторецепторов расположен между
пигментными и нервными клетками. Во внутреннем
(прилегающем к стекловидному телу) слое находятся ганглиозные
нервные клетки, аксоны которых образуют зрительный нерв.
Вследствие такого «обратного» расположения слоев
сетчатки позвоночных животных свет у них, прежде чем попасть на
фоторецепторы, должен пройти оба слоя нервных клеток. По
мере прохождения через эти слои многие лучи рассеиваются на
нейронах. Из-за этого рассеяния качество изображения на
сетчатке страдает.
Верхняя прямая
Рис. 6.7. Наружные мышцы глаза.
Лишь в небольшом участке сетчатки — центральной ямке —
формируется четкое изображение. Это связано с тем, что здесь
слои нейронов смещены к периферии и фоторецепторы открыты
для восприятия световых лучей. Свет непосредственно падает
на рецепторы центральной ямки, не рассеиваясь на нейронах.
В этом участке содержится большое количество маленьких
фоторецепторов в виде колбочек, что повышает остроту зрения.
Так как четкое изображение может формироваться только
в центральной ямке, важно, чтобы лучи от любого рассматри-
Органы чувств
117
Таблица 6.1. Наружные мышцы глаза
Мышца
Верхняя прямая
Нижняя прямая
Внутренняя прямая
Наружная прямая
Верхняя косая
Нижняя косая
Нерв
Глазодвигательный
>
Отводящий
Блоковый
Глазодвигательный
Движение глазного
яблока
Вверх и внутрь
Вниз и внутрь
Кнутри
Кнаружи
Вниз и кнаружи
Вверх и кнаружи
ваемого предмета попадали именно в эту область. В том,
чтобы обеспечить это, состоит важнейшая функция трех пар
наружных мышц глаза (рис. 6.7 и табл. 6.1).
ПАЛОЧКИ И КОЛБОЧКИ
В сетчатке имеется два вида фоторецепторов — палочки и
колбочки. Палочки, чувствительность которых выше при
слабом освещении, ответственны за «сумеречное зрение»; колбочки
же воспринимают различные цвета и отвечают за «дневное
зрение». В сетчатке более 100 млн. палочек и около 5 млн.
колбочек. Последние сосредоточены преимущественно в центре
сетчатки, в частности в центральной ямке. На периферии
сетчатки большую часть рецепторов составляют палочки.
Как палочки, так и колбочки представляют собой
продолговатые клетки, которые морфологически можно разделить на
четыре зоны: 1) наружный сегмент (членик), содержащий
фотопигмент; 2) внутренний сегмент, в котором расположены
митохондрии и другие органеллы; 3) ядерная зона и 4) синап-
тическая зона (рис. 6.6). У палочек наружный сегмент тонкий
и длинный, у колбочек — короткий и расширенный (как у
колбы). Мембрана наружного сегмента образует складки в виде
пластин (дисков), содержащих фотопигмент.
Палочки и колбочки обладают уникальной среди всех
рецепторов электрической активностью: их рецепторные
потенциалы— гиперполяризующие, и потенциалы действия под их
влиянием не возникают. Поглощение света молекулами
фотопигмента, расположенными на мембранных пластинах,
приводит к снижению проницаемости для натрия мембраны
наружного сегмента, что сопровождается гиперполяризацией
фоторецептора. Гиперполяризующие рецепторные потенциалы
наблюдаются только в палочках и колбочках; во всех других
рецепторах в ответ на раздражение генерируются
деполяризующие рецепторные потенциалы (см. гл. 5).
118
Глава 6
ФОТОХИМИЧЕСКИЕ СВОЙСТВА
Во всех видах фотопигментов содержится ретиналь
(ретинен) и опсин. Ретиналь представляет собой альдегид
витамина А, а опсин — белок. В палочках и колбочках содержатся
разные формы опсина. Фотопигмент палочек родопсин состоит
из ретиналя и скотопсина. Этот пигмент наиболее чувствителен
к синему и зеленому свету с максимумом поглощения при
505 нм.
Фотопигмент колбочек иодопсин образован ретиналем и
опсином колбочек. Существует три формы опсинов колбочек
(фотопсинов) соответственно трем типам колбочек. Эти три
формы опсинов обладают максимумами поглощения при 430 нм
(«синие» колбочки), 535 нм («зеленые» колбочки) и 575 нм
.Фиолетовый Синий Зеленый Желтки Оранжевый Красный
Рис. 6.8. Три типа пигмента в колбочках. Следует отметить, что их кривые
поглощения охватывают следующие области спектра: у «синих» колбочек —
фиолетовую и синюю; у «зеленых» колбочек — синюю, зеленую и желтую;
у «красных» колбочек — зеленую, желтую, оранжевую и красную.
(«красные» колбочки) (рис. 6.8). Ии энно наличие этих трех
типов колбочек (воспринимающих синий, зеленый и красный
цвета) обусловливает цветовое зрение. Так как синий, зеленый
и красный —это основные цвета, при возбуждении трех типов
колбочек в различной комбинации могут возникать все
разнообразные цветовые ощущения.
В родопсине содержится ретиналь в виде П-цис-изомера.
При попадании света на молекулу родопсина 11-цис-рътиналь
Органы чувств
119
превращается в полностью-граяс-ретиналь (рис. 6.9). В
результате этой фотоизомеризации ретиналя в полностью-траяс-фор-
му из родопсина образуется крайне нестойкий прелюмиродоп-
син, быстро распадающийся через ряд промежуточных стадий
до транс-решналя и скотопсина. В ходе его распада
образуются два нестойких промежуточных продукта: люмиродопсин и
метародопсин. Этот распад не зависит от световых лучей и
может идти как на свету, так и в темноте.
Родопсин
Свет
_Ж_
Прелюми родопсин
Люмиродопсин
Метародопсин
11-Цис-ретиналь +
Опсин
Изомераза
Пол ноетью-транс-
ретиналь - Опсин
Ж
Полностью-
транс-витамин А
Рис. 6.9. Синтез и распад родопсина.
Для ресинтеза родопсина полностью-граяс-ретиналь должен
превратиться в 11-^ис-изомер. Эта изомеризация
катализируется ферментом ретиненизомеразои. В результате соединения
И-^ис-ретиналя с опсином образуется родопсин. Еще одним
источником 11-цис-ретиналя служит витамин А. 11-цис-ретиналь
образуется из витамина А в результате окисления последнего
до полностью-ту?бшс-ретиналя, который в дальнейшем
превращается в И-цис-изомер.
ПЕРЕРАБОТКА ИНФОРМАЦИИ НЕЙРОНАМИ СЕТЧАТКИ
Посредством сети нейронов сетчатки фоторецепторы
соединяются с ганглиозными клетками (рис. 6.6). Аксоны ганглиоз-
ных клеток образуют зрительный нерв, по которому от сетчатки
к головному мозгу передается зрительная информация.
Существуют три типа нейронов сетчатки — биполярные,
горизонтальные и амакриновые клетки. Биполярные клетки
непосредственно связывают фоторецепторы с ганглиозными
клетками, т. е. осуществляют передачу информации через сетчатку в
120
Глава 6
вертикальном направлении. Напротив, горизонтальные и ама-
криновые клетки, соединяя рецепторы с биполярными клетками,
передают информацию по горизонтали.
Ганглиозные клетки обладают концентрическими
рецептивными полями (рис. 6.10): каждой ганглиозной клетке
соответствует маленький круглый участок сетчатки, при освещении
которого клетка подвергается либо возбуждению, либо
торможению. В рецептивном поле различаются две противоположные
по функции области — центр и периферия. Примерно половина
всех ганглиозных клеток возбуждается при попадании света в
•^нмишм пни пиши •
1 II mil iiiiiiHiiiiiiitt
t 4
ON OFF
Рис. 6.10. Возбуждение ганглиозных клеток при раздражении центра и
торможение— при раздражении периферии рецептивного поля. А. Раздражение
центра рецептивного поля световым пятном размером 0,5 мм сопровождается
возбуждением ганглиозной клетки. Б. Раздражение периферии рецептивного
поля световым кольцом размером 3 мм приводит к торможению ганглиозной
клетки.
центр рецептивного поля; напротив, при освещении периферии
световым кольцом эти клетки подвергаются торможению.
Другие же ганглиозные нейроны возбуждаются при освещении
периферии и тормозятся при освещении центра рецептивного
поля.
Такая антагонистическая организация рецептивных полей
ганглиозных клеток обусловлена наличием двух путей
проведения возбуждения в сетчатке (см. рис. 6.6). Реакции на
возбуждение центра связаны с вертикальным путем (1), по
которому информация передается от фоторецепторов к биполярным
клеткам и от них — к ганглиозным. Реакции же на
раздражение периферии рецептивного поля обусловлены горизонтальным
путем проведения возбуждения (2). Сигналы от
фоторецепторов периферических зон передаются к ганглиозным клеткам
через сеть, образованную отростками амакриновых, биполярных
и горизонтальных клеток.
Разница в остроте фовеального (центрального) и бокового
зрения соответствует различию в размерах рецептивных полей
периферических и центральных ганглиозных клеток. В
периферических областях сетчатки к каждой такой клетке
поступают сигналы примерно от 600 палочек; поэтому острота зре-
о
Органы чувств
121
ния в этих областях низка. В фовеальной же зоне центры
рецептивных полей многих ганглиозных клеток образованы всего
одной колбочкой. Такое соотношение, при котором на одну
ганглиозную клетку приходится лишь одна колбочка,
обусловливает высокую остроту зрения в центральной ямке.
ЦЕНТРАЛЬНЫЕ ЗРИТЕЛЬНЫЕ ПУТИ
По зрительному нерву информация от глаза передается к
зрительным областям головного мозга. Оба зрительных нерва
соединяются в области основания мозга, образуя зрительный
Зрительная кора
Рис. 6.11. Путь от сетчатки к зрительной коре (вид снизу). В сетчатке
выделяют назальную (Н) и темпоральную (Т) половины. В результате
перекреста волокон в области хиазмы импульсы от правых половин обоих глаз
поступают в правые отделы коры, а от левых половин — в левые. Волокна
зрительных нервов оканчиваются также в претектальном ядре и верхнем
двухолмии (здесь не показано).
перекрест (хиазму) (рис. 6.11). Здесь половина всех волокон
зрительных нервов, а именно волокна от назальных половин
сетчатки, перекрещивается и переходит на противоположную
сторону; волокна же от темпоральных половин не
перекрещиваются и остаются на ипсилатеральной стороне. В результате
этого перекреста в правом таламусе оканчиваются волокна от
правых половин каждого глаза, а в левом — от левых.
122
Глава б
Зрительные волокна оканчиваются преимущественно в
латеральных коленчатых телах таламуса (рис. 6.11). Волокна же,
не прерывающиеся в таламусе, идут кзади, оканчиваясь вверх-
нем двухолмии и претектальных ядрах среднего мозга. В
верхнее двухолмие поступают также волокна от зрительной коры.
Нейроны верхнего двухолмия и претектальных ядер
управляют деятельностью трех пар черепномозговых нервов, иннерви-
рующих глазные мышцы (глазодвигательного, блокового и
отводящего), и тем самым регулируют движение глазного
яблока. Претектальные ядра отвечают также за аккомодационные
рефлексы.
ЗРИТЕЛЬНАЯ КОРА
Латеральное коленчатое тело играет главным образом роль
станции переключения при передаче информации к зрительной
коре, расположенной в задней половине затылочной доли
(рис. 6.12). Зрительная кора организована ретинотопически:
расположение какого-либо нейрона в этой коре соответствует
локализации его рецептивного поля в сетчатке. К центральным
областям зрительной коры поступают сигналы от центральной
Рис. 6.12. Расположение первичен, вторичной и третичной зрительных зон.
А. Латеральная поверхность полушария. Б. Медиальная поверхность
полушария.
ямки — области наибольшей остроты зрения. Световое
раздражение дорсальных зон сетчатки приводит к возбуждению
нейронов дорсальных областей зрительной коры; импульсы от
вентральных участков сетчатки поступают к вентральным зонам
зрительной коры. Локализация фоторецепторов в сетчатке
(поточечно) соответствует расположению нейронов зрительной
области.
Органы чувств
123
АКТИВНОСТЬ ЗРИТЕЛЬНОЙ КОРЫ
Нейроны зрительной коры отвечают за извлечение
информации о форме зрительного образа. Большая часть этих нейронов
возбуждается при воздействии на отдельный маленький
участок сетчатки (рецептивное поле) световыми раздражителями в
виде линии или полоски. Каждый нейрон реагирует лишь на
очень ограниченное число зрительных раздражителей. В
зависимости от того, насколько сложным должно быть изображение
на сетчатке, чтобы вызвать возбуждение зрительного нейрона,
эти нейроны были разделены на три типа.
Простые клетки реагируют на темные полоски на светлом
фоне или светлые полоски на темном фоне. Каждый такой
нейрон возбуждается лишь при определенном положении
полоски в поле зрения. Согласно последним данным, каждому
возможному расположению и ориентации полоски в поле
зрения соответствует по меньшей мере один простой нейрон.
Сложные клетки также лучше воспринимают определенным
образом ориентированные полоски (вертикальные или
горизонтальные). Однако расположение полоски имеет значительно
меньшее значение для возбуждения сложных клеток, чем
простых. В связи с этим сложные клетки способны реагировать на
полоски, проецирующиеся в самые различные области
сетчатки. Сверхсложные клетки сходны со сложными, однако в
отличие от них эти клетки реагируют не только на ориентацию
и расположение полоски в рецептивном поле, но и на ее длину.
Если полоска слишком коротка или длинна, то сверхсложная
клетка не подвергается возбуждению.
Все нейроны зрительной коры сгруппированы в колонки,
каждая из которых играет роль интегративной единицы (в гл. 5
обсуждается роль колонок в соматосенсорной коре). Каждая
колонка состоит из нейронов, воспринимающих полоски
определенной ориентации, проецирующиеся на небольшие участки
сетчатки. Каждой возможной ориентации полоски соответствует
отдельная колонка, и для любого рецептивного поля на
сетчатке существует группа колонок.
В зрительной коре обычно различают первичную, вторичную
и третичную области (рис. 6.12). Вторичная и третичная
области представляют собой ассоциативные зоны зрительной
коры. Сигналы к ним поступают от латерального коленчатого
тела и от первичной зрительной области. В ассоциативных
зонах, отвечающих за более сложное абстрактное зрительное
восприятие, содержатся почти все сверхсложные нейроны.
124
Глава 6
СЛУХОВАЯ СИСТЕМА
Орган слуха состоит из трех отделов — наружного,
среднего и внутреннего уха (рис. 6.13). Наружное и среднее ухо —
это вспомогательные сенсорные структуры, обеспечивающие
Рис. 6.13. Строение уха. Ушная раковина и слуховой проход направляют
звуковые волны к барабанной перепонке. Внутреннее ухо состоит из
вестибулярного аппарата и улитки. К вестибулярному аппарату относятся
мешочек, маточка и три полукружных канала, ориентированных соответственно
трем плоскостям тела. Евстахиева труба соединяет полость среднего уха
с глоткой.
проведение звука к слуховым рецепторам в улитке (внутреннее
ухо). Во внутреннем ухе содержатся два типа рецепторов —
слуховые (в улитке) и вестибулярные (в структурах
вестибулярного аппарата).
ЗВУК
Ощущение звука возникает, когда волны сжатия, вызванные
колебаниями молекул воздуха в продольном направлении,
попадают на слуховые органы. Волны из чередующихся участков
сжатия (высокой плотности) и разрежения (низкой плотности)
молекул воздуха распространяются от источника звука
(например, камертона или струны) наподобие ряби на поверхности
воды. Звук характеризуется двумя основными параметрами —
силой и высотой.
Органы чувств
125
Высота звука определяется его частотой, или числом волн
за одну секунду. Частота измеряется в герцах (Гц). 1 Гц
соответствует одному полному колебанию в секунду. Чем больше
частота звука, тем этот звук выше. Человеческое ухо
различает звуки в пределах от 20 до 20 000 Гц. Наибльшая
чувствительность уха приходится на диапазон 1000—4000 Гц.
Сила звука пропорциональна амплитуде колебаний звуковой
волны и измеряется в логарифмических единицах — децибелах.
Один децибел равен 10 lg I/ISy где Is — пороговая сила звука.
За стандартную пороговую силу принимается 0,0002 дин/см2 —
величина, весьма близкая к пределу слышимости у человека.
НАРУЖНОЕ И СРЕДНЕЕ УХО
Ушная раковина служит как бы рупором, направляющим
звук в слуховой канал (рис. 6.13). Для того чтобы попасть на
барабанную перепонку, отделяющую наружное ухо от среднего,
звуковые волны должны пройти через этот канал. Колебания
барабанной перепонки передаются через заполненную воздухом
полость среднего уха по цепи из трех маленьких слуховых
косточек: молоточка, наковальни и стремечка. Молоточек
соединяется с барабанной перепонкой, а стремечко — с перепонкой
овального окна улитки внутреннего уха. Таким образом,
колебания барабанной перепонки передаются через среднее ухо на
овальное окно по цепи из молоточка, наковальни и стремечка.
Среднее ухо играет роль согласующего устройства,
обеспечивающего передачу звука от среды низкой плотности (воздух)
к более плотной (жидкость внутреннего уха). Энергия,
требующаяся для сообщения какой-либо перепонке колебательных
движений, зависит от плотности окружающей эту перепонку
среды. Колебания в жидкости внутреннего уха требуют в
130 раз больших затрат энергии, чем в воздухе.
При передаче звуковых волн от барабанной перепонки к
овальному окну по цепи слуховых косточек звуковое давление
увеличивается в 30 раз. Это связано прежде всего с большой
разницей в площади барабанной перепонки (0,55 см2) и
овального окна (0,032 см2). Звук от большой барабанной перепонки
передается по слуховым косточкам к маленькому овальному
окну. В результате звуковое давление на единицу площади
овального окна по сравнению с барабанной перепонкой
возрастает.
Колебания звуковых косточек уменьшаются (гасятся) при
сокращении двух мышц среднего уха: мышцы, напрягающей
барабанную перепонку, и мышцы стремечка. Эти мышцы
присоединяются соответственно к молоточку и стремечку. Их
сокращение приводит к увеличению ригидности в цепи слуховых
126
Глава 6
косточек и к уменьшению способности этих косточек проводить
звуковые колебания к улитке. Громкий звук вызывает
рефлекторное сокращение мышц среднего уха. Благодаря этому
рефлексу слуховые рецепторы улитки предохраняются от
повреждающего воздействия громких звуков.
ВНУТРЕННЕЕ УХО
Улитка образована тремя спиральными каналами,
заполненными жидкостью, — вестибулярная лестница (лестница
преддверия), средняя лестница и барабанная лестница
(рис. 6.14). Вестибулярная и барабанная лестницы соединяются
Круглое окно Барабанная лестница Основная мембрана
Рис. 6.14. Улитка в развернутом виде.
в области дистального конца улитки посредством отверстия —
геликотремы, а средняя лестница расположена между ними
(рис. 6.15). Средняя лестница отделена от вестибулярной
тонкой рейснеровой мембраной, а от барабанной — основной (ба-
зилярной) мембраной.
Улитка заполнена двумя видами жидкости: в барабанной и
вестибулярной лестницах содержится перилимфа, а в средней —
эндолимфа. Состав этих жидкостей различен: в перилимфе
много натрия (150 мэкв), но мало калия (5 мэкв), в эндолимфе
же мало натрия (16 мэкв), но много калия (114 мэкв). Из-за
этих различий в ионном составе между эндолимфой средней
лестницы и перилимфой барабанной и вестибулярной лестниц
возникает эндокохлеарный потенциал величиной около +80мВ.
Поскольку потенциал покоя волосковых клеток равен
примерно— 80 мВ, между эндолимфой и рецепторными клетками
создается разность потенциала в 160 мВ (от —80 мВ до +80 мВ).
Эта разность потенциалов имеет большое значение для
'поддержания возбудимости волосковых клеток.
В области проксимального конца вестибулярной лестницы
расположено овальное окно (рис. 6.14). При низкочастотных
колебаниях перепонки овального окна в перилимфе вестибу-
Органы чувств
127
лярной лестницы возникают волны давления. Колебания
жидкости, порожденные этими волнами, передаются вдоль
вестибулярной лестницы и затем через геликотрему в барабанную
лестницу, на проксимальном конце которой находится круглое
окно. В результате распространения волн давления в барабан-
Рис. 6.15. Поперечное сечение витка улитки. Над волосковыми клетками,
расположенными на основной мембране, нависает покровная мембрана.
ную лестницу колебания перилимфы передаются на круглое
окно. При движениях круглого окна, играющего роль
демпфирующего устройства, энергия волн давления поглощается.
КОРТИЕВ ОРГАН
Слуховыми рецепторами являются волосковые клетки
(рис. 6.16). Эти клетки связаны с основной мембраной; в
улитке человека их около 20 тысяч. Апикальные поверхности клеток
снабжены ресничками типа волосков, располагающимися в
эндолимфе средней лестницы. Над волосками нависает особый
лоскут основной мембраны — покровная (текториальная)
мембрана. С базальной поверхностью каждой волосковой клетки
образуют синапсы окончания кохлеарного нерва. С этими
волокнами соединяются волокна вестибулярного нерва, образуя
вестибулокохлеарный нерв (VIII пара черепномозговых нер-
128
Глава 6
bob). Слуховой нерв образован волокнами кохлеарного нерва.
Волосковые клетки, окончания кохлеарного нерва, покровная
и основная мембраны образуют кортиев орган.
ВОЗБУЖДЕНИЕ РЕЦЕПТОРОВ
При распространении звуковых волн в улитке покровная
мембрана смещается, и ее колебания приводят к возбуждению
волосковых клеток. Обычно покровная мембрана слегка сопри-
Рис. 6.16. Основная мембрана. Снабженные ресничками поверхности
волосковых клеток обращены к эндолимфе средней лестницы. Над этими
поверхностями нависает покровная мембрана. Волосковые клетки иннервируются
волокнами кохлеарного нерва.
касается с ресничками этих клеток, и при ее перемещениях
реснички сгибаются. Это сопровождается изменением ионной
проницаемости и деполяризацией. Возникающий при этом рецеп-
торный потенциал возбуждает окончания кохлеарного нерва.
Механизм генерации потенциалов действия в кохлеарном нерве
точно не известен. На этот счет существуют две гипотезы:
1) волосковые клетки образуют с окончаниями кохлеарного
нерва химические синапсы; при деполяризации из клеток
высвобождается химический медиатор, возбуждающий постсинапти-
ческие окончания кохлеарного нерва; 2) гипотеза электрической
передачи: окончания кохлеарного нерва непосредственно
возбуждаются высокоамплитудными рецепторными потенциалами,
возникающими при раздражении волосковых клеток.
Органы чувств
129
РАЗЛИЧЕНИЕ ВЫСОТЫ ЗВУКА
Колебания основной мембраны зависят от высоты (частоты)
звука (рис. 6.17). Эластичность этой мембраны постепенно
возрастает по мере удаления от овального окна. У
проксимального конца улитки (в области овального окна) основная
мембрана уже (0,04 мм) и
жестче, а ближе к гелико-
треме — шире и более
эластична. Поэтому
колебательные свойства
основной мембраны
постепенно изменяются по
длине улитки:
проксимальные ее участки более
восприимчивы к звукам
высокой частоты, а дисталь-
ные реагируют лишь на
низкие звуки.
Согласно
пространственной теории различения
высоты звука, основная
мембрана действует как
анализатор частоты
звуковых колебаний. От
высоты звука зависит,
какой участок основной
мембраны будет
отвечать на этот звук
колебаниями наибольшей
амплитуды (рис. 6.17). Чем
звук ниже, тем больше и
расстояние от овального
окна до участка с
максимальной амплитудой
колебаний. Вследствие этого та частота, к которой наиболее
чувствительна какая-либо волосковая клетка, определяется ее
расположением. Клетки, реагирующие преимущественно на высокие
тона, локализуются на узкой, туго натянутой основной мембране
близ овального окна; рецепторы же, воспринимающие низкие
звуки, расположены на более широких и менее туго натянутых
дистальных участках основной мембраны.
Информация о высоте низких звуков кодируется также
параметрами разрядов в волокнах кохлеарного нерва; согласно
«залповой теории», частота нервных импульсов соответствует
частоте звуковых колебаний. Частота потенциалов действия б
9—1705
\
20 000
Рис. 6.17. Звуковые частоты,
воспринимаемые различными участками улитки.
Колебательные характеристики основной
мембраны постепенно изменяются вдоль ее
длины.
130
Глава 6
волокнах кохлеарного нерва, реагирующих на звук ниже 2000 Гц,
близка к частоте этих звуков: так, в волокне, возбуждающемся
при действии тона в 200 Гц, возникает 200 импульсов в 1 с.
ЦЕНТРАЛЬНЫЕ СЛУХОВЫЕ ПУТИ
Волокна кохлеарного нерва идут в составе вестибулокохле-
арного нерва к продолговатому мозгу и заканчиваются в его
кохлеарном ядре (рис. 6.18). От этого ядра импульсы переда-
Рис. 6.18. Центральные слуховые пути. Волокна кохлеарного (улиткового)
нерва оканчиваются в ипсилатеральных ядрах продолговатого мозга —
верхней оливе, дорсальном кохлеарном ядре и вентральном кохлеарном ядре.
Большая часть волокон от этих ядер идет к контрлатеральной верхней оливе.
Отростки нейронов всех этих ядер направляются к нижнему двухолмию;
волокна от двухолмия оканчиваются в медиальном коленчатом теле таламуса;
отсюда сигналы поступают в слуховую кору.
Органы чувств
131
ются в слуховую кору по цепи вставочных нейронов слуховой
системы, расположенных в продолговатом мозгу (кохлеарные
ядра и ядра верхних олив), в среднем мозгу (нижнее двухол^-
мие) и таламусе (медиальное коленчатое тело). «Конечный
пункт назначения» слуховых каналов — это дорсолатеральный
край височной доли, где расположена первичная слуховая
область (рис. 6.19). Эту область в виде полосы окружает
ассоциативная слуховая зона.
Рис. 6.19. Слуховая кора занимает дорсолатёральные отделы височной доли
и подразделяется на первичную и вторичную области.
Слуховая кора отвечает за распознавание сложных звуков.
Здесь соотносятся их частота и сила. В ассоциативной
слуховой области интерпретируется смысл услышанного. Нейроны
нижележащих отделов — средней части оливы, нижнего
двухолмия и медиального коленчатого тела — осуществляют
извлечение и переработку информации о высоте и локализации звука.
ВЕСТИБУЛЯРНАЯ СИСТЕМА
Лабиринт внутреннего уха, содержащий слуховые
рецепторы и рецепторы равновесия, расположен в пределах височной
кости и образован системой наполненных жидкостью
перепончатых каналов (рис. 6.20). В состав лабиринта входят
вестибулярный аппарат и улитка (cochlea). Вестибулярный аппарат
подразделяется на два функциональных отдела: 1) маточку
(utriculus) и мешочек (sacculus), воспринимающие положение
в пространстве и линейное ускорение, и 2) три полукружных
канала, реагирующие на угловое ускорение. В улитке
расположены слуховые рецепторы. Вестибулярный и слуховой отделы
9*
132
Глава 6
лабиринта сообщаются посредством канала, соединяющего
среднюю лестницу с мешочком. Благодаря этому каналу
средняя лестница и вестибулярный аппарат заполнены общей эн-
долимфой.
Вестибулярными и слуховыми рецепторами являются
волосковые клетки. Эти клетки расположены на стенке лабиринта;
Рис. 6.20. Лабиринт. Вестибулярные рецепторы расположены в ампулах
полукружных каналов и в пятнах мешочка и маточки.
их покрытые ресничками поверхности погружены в эндолимфу.
По всей видимости, ионный состав эндолимфы обеспечивает
поддержание возбудимости волосковых клеток.
МАТОЧКА И МЕШОЧЕК
Часть лабиринта, содержащая полукружные каналы и
улитку, расширяется и образует две полости — маточку и мешочек
(рис. 6.20). На внутренней поверхности каждой из них
расположено так называемое пятно (macula) —скопление рецепторов
(рис. 6.21). В каждом пятне содержится несколько тысяч
волосковых клеток, реснички которых погружены в плотную
студенистую массу. В этой массе расположены мелкие кристаллы
углекислого кальция (отолиты, или отоконии). Волосковые.
клетки воспринимают отклонение ресничек под действием веса
покрывающей их студенистой массы.
Волосковые клетки реагируют на перемещение в
определенном направлении. Это обусловлено наличием двух видов рее-*.
Органы чувств
133
ничек (рис. 6.22). На каждой клетке расположена одна
крупная ресничка (киноцилия), окруженная многочисленными:
мелкими (стереоцилиями). В зависимости от того, в каком
направлении действует сгибающая реснички сила, в волосковой
клетке возникает возбуждение или торможение. Если стерео-
цилии сгибаются по направлению к киноцилии, возникает воа-
буждение; если же они отклоняются от киноцилии — торможе-
Студенистая
масса
Z: J* • • Z'.Z.Z]Z-Z.Z Z •*.—"Отолиты /.'• .';'.; • '• "\
~~'~. т-••• ••••••• •■?':' /.; ;.'.:.".-".V"-V/:;-.:. •"•*.•/ "."• л
••'••'.'.'.• • ' -Студенистая г-.'..".'. '^ ••'.'»№"•'•'.• "v •'.'•'
••••'•"-•'•.■.•'.'" ..'.'..• '• масса ■'•;. \^-'.%^M^'f-:.:--'.^>^
1. . .'.%• JPC»! \^Рк: -i/s^--'
•',н • ,'i "••'•■ л."•■■'.'•'■' V'• • •'«(•I'Afo) f \•»■•'■ '•'•'•
•A'' &,-■ ■ ■ UW ли '* Vv^-A\\.&')V [0)Л\У/' ■ 1
■III ■ "'11- '■■ -• Jit ■ '•■ ш • "'■' •В—-—"Реснички Х-%£!У§/л^^
JLijJL-'' JlL_^^M^i^JiL-l \^у\ \ 1 //ш^у
1 \ о 1 \ ° I \ о
f о \ fa J
\ / V /\7\\1///V^m
АО/ U—Волосковые Л^у \\у/// v$^v
0 ) | 0 ] клетки /O^V \tf/ \^<
.V
^Реснички
^.Волосковые
клетки
— Опорные
v клетки
Волокна
вестибулярного
Волокна
вестибулярного '
нерва
Рис. 6.21. Вестибулярные рецепторы. А. Пятна маточки и мешочка.
Волосковые клетки покрыты студенистой массой, в которую погружены их реснички.
В наружных слоях этой массы расположены отолиты. Б. Рецепторы
полукружных каналов; реснички апикальных поверхностей волосковых клеток
погружены в студенистую массу — купулу.
ние. В каждом пятне насчитывается несколько тысяч
волосковых клеток, ориентированных во всех возможных направлениях.
Благодаря этому общая картина возбуждений и торможений в
области пятна отражает направление действующей силы.
Маточка и мешочек воспринимают положение головы в
пространстве и линейное ускорение. При изменении положения
головы студенистая масса, содержащая отолиты, смещается под
действием собственного веса; при этом возбуждается новая
группа волосковых клеток. Эти отделы вестибулярного
аппарата крайне чувствительны: они реагируют на изменение
положения даже на 0,5°.
Маточка и мешочек воспринимают также линейное
ускорение, вызванное внезапным изменением скорости движения
вперед или назад. При внезапном смещении головы возникает
ускорение, под действием которого изменяется сила давления
134
Глава 6
отолитов на волосковые клетки. Информация о линейном
ускорении кодируется характером (паттерн) импульсации
волосковых клеток, возникающей в ответ на это ускорение.
ПОЛУКРУЖНЫЕ КАНАЛЫ
От маточки под прямыми углами друг к другу отходят три
полукружных канала (рис. 6.20). Расположение их таково, что
каждый из них реагирует на угловое ускорение головы, т. е.
ее внезапный поворот в одной из трех плоскостей —
фронтальной, сагиттальной и горизонтальной.
В каждом канале имеется расширенный участок с
чувствительными волосковыми клетками — ампула (рис. 6.21).
Реснички этих клеток покрыты студенистым колпачком (cupula) —
Смещение
Киноцилия ' I
/ I К киноцилии I От киноцилии
Стереоцилия / I ^ I ^
Импульсация в вестибулярном нерве
I I И I IIIIIIIIIIIII Illlllllj 1111
В покое | Возбуждение i Торможение
Рис. 6.22. Схема, иллюстрирующая избирательную чувствительность
волосковых клеток к движениям в определенном направлении.
купулой. Купула выступает в просвет канала и легко
смещается при движениях эндолимфы, заполняющей канал. Смещение
купулы приводит к возбуждению погруженных в эндолимфу
волосковых клеток. На каждой такой клетке расположена одна
крупная киноцилия и множество мелких — стереоцилий.
Ориентация всех волосковых клеток в пределах каждой ампулы
одинакова, и поэтому при смещении купулы в одном
направлении возникает возбуждение клеток, а в противоположном —
торможение.
Органы чувств
135
Волосковые клетки реагируют на угловое ускорение,
возникающее при внезапных поворотах головы. При движении
полукружного канала перемещение заполняющей его эндолимфы в
силу инерции запаздывает. Во время вращения головы, а
значит, полукружных каналов и купулы, эндолимфа стремится
сохранить прежнее положение в пространстве, и вследствие
этого она давит на купулу, смещая ее. В результате
этого смещения стереоцилии сгибаются по направлению к ки-
ноцилии и волосковые клетки подвергаются возбуждению. При
внезапном прекращении вращения головы полукружные каналы
испытывают отрицательное угловое ускорение. В течение
короткого времени после остановки головы, полукружных каналов и
связанных с ними купул эндолимфа продолжает двигаться и
вновь смещает купулу. Так как при этом стереоцилии
отклоняются от киноцилий, волосковые клетки в ответ на
отрицательное угловое ускорение подвергаются торможению.
От полукружных каналов поступает информация о
плоскости, в которой происходит угловое ускорение (положительное
или отрицательное) и его величине. Информация о плоскости
вращения кодируется в виде сигналов от какого-либо
определенного канала (верхнего, заднего или горизонтального), так
как каждый канал реагирует на угловое ускорение только в
одной из плоскостей. Степень смещения купулы и,
следовательно, частота импульсации в вестибулярном нерве, иннервирую-
щем волосковые клетки, зависит от величины ускорения.
ЦЕНТРАЛЬНЫЕ ВЕСТИБУЛЯРНЫЕ ПУТИ
Волосковые клетки вестибулярного аппарата иннервируют-
ся волокнами вестибулярного нерва. Эти волокна идут в
составе вестибулокохлеарного нерва к продолговатому мозгу, где
и заканчиваются в вестибулярных ядрах (см. рис. 6.21).
Отростки нейронов этих ядер идут к мозжечку, ретикулярной
формации и спинному мозгу — двигательным центрам,
управляющим положением тела при движениях благодаря
информации от вестибулярного аппарата, проприоцепторов шеи и
органов зрения.
Поступление вестибулярных сигналов к зрительным центрам
имеет первостепенное значение для важного
глазодвигательного рефлекса — нистагма. Благодаря нистагму взор при
движениях головы фиксируется на неподвижном предмете. Во время
вращения головы глаза медленно поворачиваются в обратную
сторону, и поэтому взор фиксирован на определенной точке.
Если угол вращения головы больше, чем тот, на который могут
повернуться глаза, то они быстро перемещаются в направлении
вращения и взор фиксируется на новой точке. Это быстрое
136
Глава 6
движение и есть нистагм. При повороте головы глаза
попеременно совершают медленные движения в направлении поворота
и быстрые в противоположном направлении.
ВКУСОВАЯ СИСТЕМА
Вкусовые ощущения возникают в результате химического
раздражения различными веществами вкусовых почек в
слизистой оболочке полости рта. На языке, нёбе и стенках глотки
расположено около 10 000 вкусовых почек.
ВКУСОВЫЕ ОЩУЩЕНИЯ
Поверхность языка покрыта множеством мелких выростов,
или сосочков, на апикальных концах которых расположена
большая часть вкусовых почек (то 100 почек на сосочке).
Каждая почка образована примерно 40 продолговатыми клетками,
окружающими в виде долек апельсина вкусовую пору
Эпителий языка Вкусовая пора
клетка клетка
Рис. 6.23. Типичная вкусовая почка.
(рис. 6.23). Среди этих клеток различают два вида — опорные
и рецепторные. На апикальной поверхности рецепторных клеток
расположено несколько микроворсинок в виде волосков,
выступающих во вкусовую пору; к базальным поверхностям
рецепторных клеток подходят окончания вкусового нерва.
Для того чтобы то или иное вещество могло вызвать
возбуждение вкусовой почки, оно должно раствориться в жидкой
среде полости рта. При наложении сухих кристаллов на сухую
Органы чувств
137
поверхность языка вкусовые ощущения не возникают.
Полагают, что эти ощущения обусловлены связыванием молекулы
раздражающего вещества с мембраной микроворсинок рецеп-
торных клеток. В результате этого связывания проницаемость
мембраны чувствительной клетки изменяется и наступает ее
деполяризация. Считается, что рецепторные клетки образуют с
Рис. 6.24. Центральные вкусовые пути.
окончаниями вкусового нерва химические синапсы. При
деполяризации рецепторной клетки из нее высвобождается
химический медиатор, возбуждающий окончания вкусового нерва.
Существуют четыре вкусовых ощущения — сладкого,
кислого, соленого и горького. Каждое из них возникает при
раздражении определенной области языка. Кончик языка лучше
различает сладкое, боковые стороны — кислое и соленое; горькие
же вещества лучше воспринимаются вкусовыми почками корня
языка.
Остается невыясненным, как связаны вкусовые ощущения
с химическим строением молекул. Так, ощущение сладкого
возникает при воздействии на язык целого ряда веществ, в том
числе Сахаров, гликолей, спиртов, альдегидов, кетонов, амидов.
138
Глава 6
эфиров, аминокислот, сульфокислот, галогеноводородных
кислот, солей свинца и бериллия. Структура сахарина, который в
600 раз слаще сахарозы (сахара), существенно отличается от
строения Сахаров. Другие вкусовые ощущения более
соответствуют химическому строению веществ. Соленый вкус всегда
возникает при действии ионизированных неорганических солей,
кислый — при нанесении кислот. Чем ниже рН раствора, тем
он кислее. Горькие вещества — это либо алкалоиды, либо
органические соединения, имеющие молекулу с длинной цепью.
Неприятный горький вкус может служить предупреждением об
опасности — токсины многих ядовитых растений являются
алкалоидами.
ЦЕНТРАЛЬНЫЕ ВКУСОВЫЕ ПУТИ
Вкусовые волокна идут от языка к ядру солитарного пучка
продолговатого мозга в составе трех черепномозговых нервов —
лицевого, языкоглоточного и блуждающего (рис. 6.24). Лицевой
нерв содержит волокна от передних двух третей, а языкогло-
точный — от задней трети языка. Волокна от вкусовых почек
глотки идут в составе блуждающего нерва. Вкусовые волокна
нейронов второго порядка восходят от ядра солитарного пучка
продолговатого мозга к отделам таламуса, примыкающим к
вентробазальному комплексу. Волокна нейронов третьего
порядка направляются от таламуса к постцентральной извилине
коры. Они заканчиваются во вкусовой области, непосредственно
примыкающей к зоне соматосенсорной коры, возбуждающейся
при раздражении соматосенсорных рецепторов языка.
ОБОНЯТЕЛЬНАЯ СИСТЕМА
Обонянием называют способность ощущать запахи.
Обонятельные рецепторы расположены в обонятельном эпителии,
выстилающем верхнюю поверхность полости носа. В этом эпителии
содержится около 100 млн. рецепторов, расположенных среди
опорных клеток (рис. 6.25). На апикальной поверхности
каждого рецептора имеется несколько ворсинок, или чувствительных
волосков; эти волоски погружены в слой слизи, покрывающий
обонятельный эпителий. Обонятельные рецепторы представляют
собой чувствительные нейроны, аксоны которых образуют
обонятельный нерв.
Для того чтобы возникло ощущение запаха, вещество
должно быть летучим (благодаря чему оно попадает с вдыхаемым
воздухом в полость носа) и растворимым в воде (что
позволяет ему проникать через слой слизи, покрывающий рецепторы).
Считается, что возбуждение обонятельных рецепторов про-
Органы чувств
139
исходит при взаимодействии молекулы пахучего вещества с
особыми воспринимающими участками, расположенными на
покрытой ресничками поверхности рецептора. Хотя механизм
этого взаимодействия неясен, известно, что рецепторная клетка
деполяризуется и возникает генераторный потенциал.
Последний и служит тем деполяризующим стимулом, под влиянием
Слизь _
Рис. 6.25. Обонятельный рецептор.
которого генерируются потенциалы действия в обонятельном
нерве.
По имеющимся данным, существует целый ряд первичных
обонятельных ощущений — примерно от 50 до 100. Многие
люди не воспринимают определенные запахи; это так
называемая аносмыя. Описано более 70 видов аносмий, что заставляет
думать о не меньшем числе различных типов обонятельных
рецепторов.
Волокна обонятельного нерва заканчиваются в обонятельной
луковице (рис. 6.26). Волокна, идущие от луковицы, делятся
на латеральный и медиальный пути, направляющиеся к
различным, плохо изученным областям переднего мозга. К этим
областям относятся гиппокамп и лобная и височная доли коры.
Обонятельная информация поступает также в таламус,
гипоталамус и ретикулярную формацию. Большинство центральных
структур, получающих обонятельную информацию, отвечают за
интеграцию соматической и вегетативной деятельности. Эти
140
Глава 6
Медиальная обонятельная область
Рис. 6.26. Центральные обонятельные пути.
структуры, образующие лимбическую систему, участвуют в
регуляции пищевого, полового и сложного эмоционального
поведения, например страха и удовольствия (см. гл. 10).
ЛИТЕРАТУРА
Brindley G. S. (1970). Central pathways of vision, Ann. Rev. Physiol., 32, 259.
Daw N. W. (1973). Neurophysiology of color vision, Physiol. Rev., 53, 571.
Eldredge D. //., Miller J. D. (1971). Physiology of hearing, Ann. Rev. Physiol.,
33, 281.
hyzagulrre C, Fidone S. J. (1975). Physiology of the nervous system, 2nd edn.,
Year Book Medical, Chicago.
Goldberg J. M., Fernandez C. (1975). Vestibular mechanisms, Ann. Rev. Physiol.,
37, 129.
Hodgson E. S. (1961). Taste receptors, Sci. Amer., 204, 135.
Hubel D. H. (1963). The visual cortex of the brain, Sci. Amer., 209, 54.
Mcllwain J. T. (1972). Central vision: Visual cortex and superior colliculus, Ann.
Rev. Physiol., 34, 291.
Moulion D. G., Beidler L. M. (1967). Structure and function in the peripheral
olfactory system., Physiol. Rev., 47, 1.
Mounlcastle V. B. (1974). Medical physiology, 13th edn., Mosby, St. Louis.
Oakleij В., Benjamin R. M. (1966). Neural mechanisms of taste, Physiol. Rev.,
46, 173.
von Bekesy G. (1957). The ear, Sci. Amer., 197, 66.
Wald G. (*1968). Molecular basis of visual excitation, Science, 163, 230.
Werblin F. S. (1973). The control of sensitivity in the retina, Sci. Amer., 228, 69.
Witkovsky P. (1971). Peripheral mechanisms of vision, Ann. Rev. Physiol., 33,
257.
Органы чувств
141
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Нарисуйте схему поперечного сечения глаза и обозначьте
его структуры.
2. Как регулируется размер зрачка? Почему эта регуляция
так важна для нормального зрения?
3. Расскажите, как изменяется преломляющая сила глаза при
аккомодации. Почему аккомодация важна для четкого
различения предметов?
4. Что такое центральная ямка и какое отношение она имеет
к остроте зрения?
5. Проведите в кратких чертах сравнение двух типов
фоторецепторов. Укажите на сходство и различия в их строении,
функции и фотохимических свойствах.
6. Опишите последовательность этапов синтеза и распада
родопсина.
7. Как кодируется зрительная информация в зрительной
коре? Расскажите об особенностях ответов простых, сложных
и сверхсложных нейронов.
8. Каковы две важнейшие характеристики звука? Чем они
измеряются?
9. Нарисуйте схему соединения слуховых косточек среднего
уха. Расскажите об их роли в передаче звука к
внутреннему уху.
10. Нарисуйте схему поперечного сечения витка улитки.
11. Расскажите кратко, как кортиев орган преобразует
слуховые раздражения. Опишите все этапы преобразования
звуковых волн в импульсы улиткового нерва.
12. Расскажите о механизме кодирования высоты звука во
внутреннем ухе.
13. Нарисуйте схему вестибулярного аппарата.
14. Расскажите о морфологических и физиологических
особенностях маточки и мешочка, позволяющих этим
образованиям воспринимать положение в пространстве и линейное
ускорение»
15. Нарисуйте схему строения ампул полукружных каналов.
Как они реагируют на угловое ускорение?
16. Какие особенности вестибулярных волосковых клеток
обусловливают их чувствительность к перемещениям в
определенном направлении?
17. Нарисуйте схему строения вкусовой почки.
18. Назовите четыре вкусовых ощущения. Соотнесите их с
химическим строением веществ.
19. Расскажите о нервных путях, передающих информацию от
вкусовых почек к коре головного мозга.
20. Нарисуйте схему строения обонятельного рецептора. Чем
он отличается от вкусовой почки?
Глава 7
СПИННОМОЗГОВЫЕ РЕФЛЕКСЫ
Рефлексом называется, ответная реакция на раздражение
рецептора, опосредованная нервной системой и
характеризующаяся постоянством. Рефлексы — это простейшие реакции
нервной системы, возникающие в результате последовательного
возбуждения чувствительных, нервных и двигательных
структур. Эти структуры образуют рефлекторную дугу (рис.
7.1, Л)—нервный путь, соединяющий чувствительные
рецепторы с мышцами, участвующими в рефлекторной реакции.
В простейших рефлекторных дугах чувствительные нейроны
образуют синапсы непосредственно на двигательных (рис. 7.1).
Такие дуги называются моносинаптическими, так как в их
составе всего один синапс. Большинство же дуг является поли-
синаптическими, т. е. содержат чувствительные, вставочные и
двигательные нейроны (рис. 7.1,5). В полисинаптической
рефлекторной дуге отростки чувствительных нейронов не
оканчиваются непосредственно на двигательных: возбуждение от них
проводится через один или несколько рядов вставочных
нейронов и лишь затем поступает к двигательным. В центральной
части полисинаптических дуг имеются два синапса или более.
ВИДЫ РЕФЛЕКСОВ
Рефлексы осуществляются на многих уровнях нервной
системы. Рефлексы спинного мозга играют важную роль в
регуляции движений туловища и конечностей. К ним относятся
рефлексы, контролирующие длину мышц (рефлексы
растяжения), отвечающие за уход от вредных воздействий (сгибатель-
ные рефлексы) и движения (перекрестные разгибательные
рефлексы). Другие рефлексы — например отвечающие за
поддержание вертикального положения и регуляцию зрения —
замыкаются на уровне ствола мозга. Многие висцеральные
рефлексы, в том числе участвующие в половом возбуждении,
опорожнении мочевого пузыря и регуляции артериального давления,
опосредованы рефлекторными дугами автономной нервной
системы.
Спинномозговые рефлексы
/ 143
СПИННОЙ МОЗГ
Изучение рефлексов спинного_ мозга—это первая ступень
к пониманию того^аким образом нервная система регулирует"
д^и£ател±>дую _активно.ст,ь. Возможности спинного мбзТаТГ'конт-
роле над двигательными функциями можно выявить, наблюдая
за реакциями животного с перерезанным спинным мозгом.
После исчезновения явлений спинального шока такое спинальное
животное способно к поразительно сложным двигательным
реакциям. Оно может стоять, если конечности его контактируют
Чувствительный
Рецептор нейрон^ Двигательный Мышца
нейрон
А
> } О
> L О О
Рис. 7.1. Рефлекторная дуга. А. Моносинаптическая дуга: окончания
чувствительных нейронов образуют синапсы на двигательных. Б. Полисинаптическая
дуга: между чувствительным и двигательным нейронами имеется один или
несколько вставочных.
с твердой опорой, отдергивать конечность от ноцицептивного
раздражителя, почесывать какую-либо область поверхности
тела или конечности, если ее «пощекотать», а также производить
попеременное движение конечностями, как при ходьбе.
ОРГАНИЗАЦИЯ СПИННОГО МОЗГА
Способность спинного мозга осуществлять двигательные
реакции обусловлена взаимосвязями нейронов в его сером
веществе. Это вещество состоит из окончаний чувствительных
нейронов, а также из вставочных и двигательных нейронов
(рис. 7.2). Двигательные нейроны расположены в передних
рогах, а вставочные — в задних рогах и в промежуточном
веществе. Отростки чувствительных нейронов входят в спинной
мозг через задние корешки и оканчиваются на вставочных
нейронах задних рогов и промежуточного вещества. Аксоны
некоторых чувствительных нейронов входят в состав восходящих
или нисходящих спинномозговых путей и достигают других
уровней нервной системы (см. гл. 5). Лишь один вид
чувствительных волокон, принадлежащих к группе IA и отходящих от
рецепторов мышечных веретен, оканчивается непосредственно
па двигательных нейронах. На вставочных нейронах спинного
мозга образуют синапсы чувствительные волокна, другие вста-
144
Глава 7
вочные спинномозговые нейроны и восходящие и нисходящие
волокна спинномозговых путей. На двигательные нейроны
переключаются вставочные нейроны спинного мозга, восходящие
и нисходящие спинномозговые волокна и чувствительные
нейроны группы IA.
Было подсчитано, что в одном сегменте спинного мозга
собаки (L5) содержится 2000 чувствительных, 6000 двигательных
и 360 000 вставочных нейронов. Однако на самом деле слож-
Вставочные нейроны
спинного мозга
Рис. 7.2. Спинной мозг (поперечный разрез). Изображены контакты между
чувствительными, вставочными и двигательными нейронами в сером
веществе спинного мозга.
ность организации спинного мозга определяется не
количеством нейронов в одном сегменте, а числом синапсов. Число это
поистине астрономическое, так как на «типичном» вставочном
нейроне расположено около 650 синапсов, а на «типичном»
двигательном — около 5500.
МОНОСИНАПТИЧЕСКИЕ РЕФЛЕКСЫ РАСТЯЖЕНИЯ
Моносинаптические рефлексы растяжения — это простейшие
рефлексы спинного мозга. Их можно вызвать путем быстрого
растяжения мышцы, например ударяя по ее сухожилию.
Растяжение мышцы приводит к возбуждению заложенных в ней
чувствительных к растяжению рецепторов — мышечных
веретен. Сенсорные волокна, иннервирующие эти рецепторы, входят
в спинной мозг в составе задних корешков и заканчиваются в
передних рогах. Здесь они образуют синаптические контакты
на двигательных нейронах, иннервирующих растягиваемую
мышцу. Так как в центральной части (в ЦНС) дуги рефлекса
Спинномозговые рефлексы
14S
растяжения имеется лишь один синапс (между чувствительным
и двигательным нейронами), рефлекс этот относится к моноси-
наптическим.
Центральное время рефлекса растяжения можно измерить,,
наложив на передний корешок отводящие электроды для
регистрации разряда мотонейронов, вызванного электрическим раз-
1 1 1 l I I I 1 1 i I I I l I
мс
Рис. 7.3. Анализ дуги рефлекса растяжения. Раздражение чувствительных
волокон заднего корешка (ЗК) вызывает ранний и поздний ответы,
регистрируемые от переднего корешка (ПК). Ранний ответ является моносинаптическим,
а поздний — полисинаптическим. Раздражение волокон группы IA, идущих от
аннулоспиральных окончаний, сопровождается моносинаптическим ответом, а
чувствительных волокон групп II, III, IV — полисинаптическим.
дражением чувствительных нейронов в заднем корешке
(рис. 7.3). При записи активности переднего корешка
обнаруживаются два компонента — ранний с кратковременной
задержкой (около 1,5 мс) и поздний. Если подобрать величину
раздражения так, чтобы возбуждались только волокна группы
IA (сделать это несложно, так как это самые крупные из всех
чувствительных волокон), то при отведении активности
переднего корешка можно зарегистрировать только ранний ответ.
Задержка раннего ответа (1,5 мс) складывается из
центрального времени рефлекса (0,5 мс) и времени, необходимого для
проведения потенциала действия по чувствительным волокнам
к мотонейрону (около 1,0 мс). Центральное время рефлекса в
данном случае мало, и за это время возбуждение может пере-
10—1705
146
Глава 7
даться лишь через один синапс. Следовательно, часть
чувствительных волокон группы IA оканчивается непосредственно на
тех мотонейронах, которые они возбуждают. Этот вывод можно
проверить, введя микроэлектрод в двигательный нейрон перед-
Раздражающие
электроды
Рис. 7.4. Микроэлектродный анализ дуги рефлекса растяжения. А. Слабое
раздражение волокон IA вызывает генерацию ВПСП в мотонейроне. Б. При
усилении раздражения величина ВПСП превышает пороговое значение; в
мотонейроне возникает потенциал действия.
него рога спинного мозга (рис. 7.4). Слабое раздражение
волокон IA приводит к возникновению в нем возбуждающих пост-
сииаптическпх потенциалов (ВПСП). При усилении
раздражения, когда увеличивается число возбужденных волокон IA, эти
ВПСП суммируются и становятся достаточными для того,
чтобы в мотоненропе возник потенциал действия.
Спинномозговые рефлексы
147
ЧУВСТВИТЕЛЬНЫЕ РЕЦЕПТОРЫ И МЫШЕЧНЫЕ
ВЕРЕТЕНА
Мышечные веретена — это специализированные
поперечнополосатые мышечные волокна, иннервируемые
чувствительными окончаниями, воспринимающими растяжение (рис. 7.5).
Речь идет о мелких тонких интрафузальных волокнах,
отличающихся от экстрафузальных волокон, составляющих основную
мышечную массу. Сокращения интрафузальных волокон слишком
слабы, чтобы повлиять на деятельность всей мышцы: развивае-
Рис. 7.5. Мышечное веретено и его иннервация. Обратите внимание на
параллельное расположение интрафузальных и более крупных экстрафузальных
волокон.
мые ею усилия связаны с сокращением крупных экстрафузальных
волокон. Эти два типа волокон иннервируются различными
видами мотонейронов. К экстрафузальным волокнам подходят
отростки крупных альфа-мотонейронов, а эфферентная
иннервация интрафузальных волокон осуществляется мелкими гамма-
мотонейронами.
В каждом интрафузальном волокне имеется центральный
несокращающийся чувствительный участок, в котором
концентрируются ядра волокна (рис. 7.5). Этот участок иннер-
вируется двумя разновидностями чувствительных нервов: ан-
нулоспиральными окончаниями волокон группы IA (первичные
рецепторные окончания) и гроздевидными терминалями
волокон группы II (вторичные рецепторные окончания). Аннулоспи-
ральные окончания волокон IA окружают в виде спирали
ядерную сумку, а волокна группы II, иннервирующие мышечное
веретено, образуют в нем множество гроздевидных
разветвляющихся окончаний.
ВОЗБУЖДЕНИЕ МЫШЕЧНЫХ ВЕРЕТЕН
Как первичные, так и вторичные рецепторные окончания
возбуждаются при растяжении центральной чувствительной
зоны интрафузального волокна. Это растяжение может про-
10*
Л 48
Глава 7
Сила стимула
.! А
II
Рис. 7.6. Реакция афферентов
мышечного веретена на растяжение.
Волокна группы IA реагируют как
на скорость растяжения веретена
(динамические параметры), так и
на изменение его длины
(статические параметры); волокна же
группы II реагируют лишь на
изменение длины веретена
(статические параметры).
изойти либо под действием
внешних сил (например, при покола-
чивании по сухожилию), либо
при сокращении самого интрафу-
зального волокна.
Интрафузальное волокно
сокращается под влиянием
импульсов, исходящих от
гамма-мотонейронов, которые иннервируют
периферические сократительные
участки волокна. Длина волокна
в целом постоянна, так как оно
расположено параллельно более
крупным экстрафузальным
волокнам и может укоротиться
лишь при сокращении последних.
Поэтому при сокращении
сократительных участков интрафузаль-
ного волокна его центральная
чувствительная область
растягивается.
Возбуждение первичных и
вторичных рецепторных
окончаний приводит к усилению импульсной активности в
чувствительных волокнах групп IA и II (рис. 7.6). Волокна группы IA
воспринимают как скорость растяжения мышечного веретена (ди-
^ «амические параметры), так и само это растяжение (статиче-
'-£ <жие параметры); волокна же группы II чувствительны лишь к
изменению длины волокна (статический параметр).
РЕГУЛЯЦИЯ ДЛИНЫ МЫШЦЫ
Мышечные ведетена и ос^ще^^^ рефлексы
растяжения лежат в основе механизма регуляции длины мышц.
Главная функция рефлекса растяжения состоит в
противодействии изменениям длины мышцы, возникающим под
влиянием приложенных к ней внешних сил. Эти изменения
наступают, когда мышца растягивается: 1) при поднятии груза; 2) при
ударе по сухожилию; 3) при сокращении мышцы-антагониста
(например, при сокращении сгибателя какого-либо сустава
разгибатель этого же сустава растягивается). Вследствие того
что интра- и экстрафузальные волокна расположены
параллельно друг другу, при растяжении мышцы удлиняются и те
и другие. Импульсация, возникающая при этом в первичных и
вторичных рецепторных окончаниях, передается в спинной
:Мозг, где вызывает рефлекторный разряд альфа-мотонейронов,
Спинномозговые рефлексы
149
иннервирующих экстрафузальные волокна растянутой мышцы.
Сокращение последних приводит к укорочению мышцы,
противодействуя тем самым удлинению ее под влиянием внешней
силы.
РЕГУЛЯТОРНЫЕ ВЛИЯНИЯ ГАММА-МОТОНЕИРОНОВ
Чувствительность , MJ?LU.If-4F£!rP,.-.5^J?£I^^^JL.par-Tff^K^4Hlff. °JIPe"
деляется ритидц^тктл >гяММ^--М^Т0Н£Йрон.0^-.Ш5ЛРрВИруЮФ,¥У вгп
интрафузальные^^волокна. При усилении этой активности
растет сила сокращения интрафузальных волокон и, как
следствие, напряжение в этих волокнах и чувствительность веретена
к растяжению. Увеличение напряжения в интрафузальных
волокнах приводит к повышению уровня чувствительной импуль-
сации, что сопровождается рефлекторным разрядом
альфа-мотонейронов; под действием этого разряда экстрафузальные
волокна сокращаются и мышца укорачивается. Напротив, при
снижении активности гамма-мотонейронов уменьшается
напряжение в интрафузальных волокнах и разряд мышечных
веретен, что приводит к снижению активности альфа-мотонейронов,
расслаблению экстрафузальных волокон и удлинению мышцы.
СОВМЕСТНАЯ АКТИВАЦИЯ МОТОНЕЙРОНОВ
Считается, что нервная система регулирует мышечные
сокращения, изменяя активность как альфа-, так и
гамма-мотонейронов. Обычно эти нейроны возбуждаются одновременно:
повышение активности альфа-мотонейронов сопровождается
усилением активности гаммй-мотонейронов. Благодаря такой
совместной активации (коактивации) сокращение мышцы не
влияет на чувствительность мышечных веретен к растяжению.
Если этого нет (при увеличении разряда только
альфа-мотонейронов), укорочение экстрафузальных волокон приводит к
снижению напряжения в интрафузальных волокнах и
чувствительности мышечного веретена к растяжению.
ТОРМОЗНЫЙ СУХОЖИЛЬНЫЙ РЕФЛЕКС См ;Jr
Сильное растяжение сухожилия мышцы, например при ее
моиЩо^Тбкращении"* или действии внешней силы, приводит к
расслаблению этой мышцы. При таком растяжении
возбуждаются сухожильные органы J дльджи (рис. 7.7) —окончания
чувствительных волокон группы 1Б, локализующиеся в сухожилиях
и воспринимающие растяжение. Эти органы расположены
последовательно с экстрафузальными волокнами и активизиру-
ются при увеличении напряжения в^м^ище.
150
Глава 7
Волокна, идущие от органов Гольджи, входят в спинной
мозг и оканчиваются на вставочных нейронах; последние
образуют тормозные синапсы на альфа-мотонейронах, иннервиру-
ющих растянутую мышцу (рис. 7.7). При разряде этих
тормозных вставочных нейронов в мотонейронах возникают тормозные
постсинаптические потенциалы (ТПСП), подавляющие
активность мотонейрона, препятствуя возникновению потенциала
действия в аксонном холмике (см. гл. 4).
Рис. 7.7. Дуга сухожильного рефлекса.
Сухожильные рефлексы играют двоякую роль. Способствуя
быстрому расслаблению мышцы, они защищают ее от
повреждения при сильных сокращениях. Они участвуют также в
регуляции напряжения в мышце точно так же, как мышечные
веретена регулируют ее длину. Когда напряжение достигает
значения, при котором возбуждаются сухожильные органы
Гольджи, альфа-мотонейроны, иннервирующие сокращающуюся
мышцу, тормозятся, что приводит к ее расслаблению и падению
напряжения в ней.
ПОЛИСИНАПТИЧЕСКИЙ СГИБАТЕЛЬНЫЙ РЕФЛЕКС
Сгибательный рефлекс, или рефлекс отдергивания,
относится к защитным рефлексам. Если на поверхность конечности
нанести повреждающее или болевое раздражение (щипок,
прикосновение горячим или холодным предметом), то эта конеч-
Спинномозговые рефлексы 151
ность быстро согнется — отдернется от раздражителя,
предотвращая дальнейшее повреждение ее тканей. Сгибание
обусловлено сокращением флексоров и расслаблением экстензоров
соответствующего сустава.
Сила двигательной реакции зависит от интенсивности
раздражителя: при увеличении интенсивности растет как число
возбужденных рецепторов, так и уровень их возбуждения.
Слабую двигательную реакцию можно вызвать путем
прикосновения к коже или легкого надавливания на нее. Интенсивные но-
цицептивные раздражители могут приводить к широко ирра-
диирующим реакциям: сильному сгибанию раздражаемой (ип-
силатеральной) конечности и разгибанию противоположной
(контрлатеральной) (этот рефлекс часто называют
перекрестным разгибательным); возбуждение может распространяться
вверх или вниз по спинному мозгу к другой паре конечностей,
и в этом случае наступает разгибание ипсилатеральной и
сгибание контрлатеральной.
Возбуждение чувствительных к прикосновению и давлению
волокон группы II может вызвать лишь слабую сгибательную
реакцию. Сильное сгибание возникает при раздражении
наиболее мелких сенсорных волокон (группы III и IV), реагирую-
и разгибателям и разгибателям
Рис. 7.8. Дуги сгибательного и перекрестного разгибательного рефлекса.
щих на ноцицептивные стимулы (см. гл. 5). Эти волокна
оканчиваются на сети вставочных спинальных нейронов,
образующих синапсы на соответствующих мотонейронах (рис. 7.8).
Непосредственно сииаптического переключения чувствительных
нейронов на двигательные в этом случае нет. Электрическое
раздражение волокон группы II, III п IV в области заднего
корешка вызывает поздний полисинаптическип ответ, регистри-
152
Глава 7
руемый в отведениях от переднего корешка (см. рис. 7.3); эти
волокна не участвуют в генерации раннего моносинаптического
ответа, возникающего при возбуждении волокон IA.
РЕЦИПРОКНАЯ ИННЕРВАЦИЯ
Сеть вставочных нейронов, возбуждающаяся при
раздражении чувствительных ноцицептивных волокон, организована по
принципу реципрокности, так что нейроны ипсилатериального
сгибателя возбуждаются, а ипсилатерального разгибателя —
тормозятся (рис. 7.8). При достаточной интенсивности
раздражителя возбуждаются интернейроны, отростки которых
пересекают спинной мозг и заканчиваются на мотонейронах
контрлатерального переднего рога, активируя разгибатели и тормозя
сгибатели.
Реципрокное возбуждение и торможение мотонейронов
мышц-антагонистов — важнейшие принципы организации
нейронных сетей спинного мозга. Как правило, при возбуждении
двигательных нейронов какой-либо мышцы мотонейроны, ин-
нервирующие мышцу-антагониста, тормозятся. Такая реципрок-
ная иннервация предотвращает одновременное сокращение
противоположных групп мышц, которое, препятствовало бы
любому движению в суставе или конечности.
ДРУГИЕ ПОЛИСИНАПТИЧЕСКИЕ РЕФЛЕКСЫ
Раздражение кожных рецепторов давления на стопе
приводит к одновременному сокращению разгибателей и сгибателей
и разгибанию конечности. Это разгибательный рефлекс,
имеющий важное значение для регуляции ригидности конечности,
что необходимо для поддержания веса тела, находящегося в
вертикальном положении.
Слабое тактильное раздражение кожи вызывает чесатель-
ный рефлекс. В этом рефлексе два компонента: 1) нахождение
раздраженного участка и 2) ритмичные чесательные движения.
Тактильное раздражение стопы у зафиксированного в
станке спинального животного может вызвать ритмичные
попеременные шагательные движения всех четырех конечностей. Эти
движения возникают вследствие поочередного сокращения и
расслабления разгибателей и сгибателей. Такое поочередное
возбуждение группы мотонейронов и торможение их
антагонистов отражает реципрокную иннервацию мотонейронов сетью
вставочных нейронов. Ритмичные шагательные движения
обусловлены колебаниями активности спинномозговых вставочных
нейронов, так как эти движения не прекращаются при
перекрытии всех чувствительных каналов.
Спинномозговые рефлексы
153
СПИННОМОЗГОВОЙ МОТОНЕЙРОН КАК ОБЩИЙ
КОНЕЧНЫЙ ПУТЬ
Спинномозговой мотонейрон служит эфферентным путем
всех рефлекторных дуг. Для того чтобы мышцы сократились и
возникла поведенческая реакция, все нервные импульсы от
спинного и головного мозга должны пройти через эти нейроны.
Импульсная активность мотонейрона в каждый момент
времени определяется результирующей величиной действующих на
него возбуждающих и тормозных синаптических влияний. На
мотонейроне переключаются чувствительные волокна группы
IA, возбуждающие и тормозные вставочные нейроны спинного
мозга и внутрицентральные нисходящие пути от двигательных
областей головного мозга. Активность мотонейрона
формируется в результате возбуждающих и тормозных влияний,
поступающих по всем этим каналам. Редко бывает так, чтобы один
из них доминировал. Например, одновременно с возбуждением
мотонейрона задней конечности волокнами мышечных веретен
может наблюдаться его торможение вследствие раздражения
кожи этой конечности и под действием спускающихся из
двигательной коры вызванных болевым раздражением передней
конечности импульсов от вставочных нейронов.
ЛИТЕРАТУРА
Burke R. Е., Rudomin Р. (1977). Spinal neurons and synapses. In: Kandel E. R.
(ed.), Handbook of physiology: cellular biology of neurons, American
Physiological Society, Bethesda, Maryland.
Eyzaguirre C, Fidone S. J. (1975). Physiology of the nervous system, 2nd ed.,
Year Book Medical, Chicago, Chaps, 13—17.
Hunt C. C, Perl E. R. (1960). Spinal reflex mechanisms concerned with skeletal
muscle, Physiol. Rev., 40, 538.
Merton P. A. (1972). How we control the contraction of our muscles, Sci. Amer.,
226, 30.
Mountcastle V. B. (1974). Medical physiology, 13th edn. Mosby, St. Louis,
Chaps. 22—24.
Pearson K. (1976). The control of walking, Sci. Amer., 235, 72.
Stein R. B. (1974). Peripheral control of movement, Physiol. Rev., 54, 215.
Wilson V. J. (1966). Inhibiton in the central nervous system, Sci. Amer., 214,
102.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Что такое рефлекторная дуга? Нарисуйте схему
рефлекторных дуг сгибательного и перекрестного разгибательного
рефлекса, а также рефлекса растяжения.
154
Глава 7
2. В чем состоят морфологические и функциональные различия
между экстра- и интрафузальными мышечными волокнами?
3. Почему одновременная активация альфа- и
гамма-мотонейронов имеет важное значение? Что произойдет при
раздельном возбуждении альфа- и гамма-мотонейронов?
4. Коротко расскажите, как можно доказать, что рефлекс
растяжения является моносинаптическим.
5. Что такое реципрокная иннервация и в чем ее значение?
6. Почему альфа-мотонейрон называют общим конечным
путем?
Глава 8
ЦЕНТРАЛЬНАЯ РЕГУЛЯЦИЯ ДВИЖЕНИЙ
Движения, которые может выполнять человек и животное,
практически бесконечно разнообразны, и каждое из них
обусловлено специфическим комплексом разрядов мотонейронов.
Лишь наиболее п_£ость1е дви^е_ни[^^например, отдергивание ко-
нечности или почесывание) осуществляются изолированным
спинн'ым"' 1УЮз7омГГ"Все разнообразие двигательных актов, на ко-
т^фьГ^ Способны"" мотонейроны и вставочные нейроны спинного
мозга, сводится к рефлекторным реакциям (см. гл. 5). Все
остальные сложные движения — от координированной ходьбы до
исполнения бетховенских сонат — требуют участия
центральных областей головного мозга. Эти области регулируют
активность мотонейронов спинного мозга через нисходящие
спинномозговые пути. К высшим центрам регуляции движений
относится кора головного мозга, осуществляющая контроль как над
пирамидной, так и над экстрапирамидной системами, базаль-
ные ганглии и мозжечок. Хотя многие функции этих центров
еще не выяснены, считается, что каждый из них вносит свой
особый вклад в осуществление движений. Комплекс
двигательная кора — пирамидная система отвечает за тонкие
произвольные движения. Грубые непроизвольные движения
осуществляются блоком двигательная кора — экстрапирамидная система.
Базальные ганглии и мозжечок участвуют в координации
движений. С базальными ганглиями связана координация
медленных (червеобразных) движений, а с мозжечком — быстрых
(баллистических).
ДВИГАТЕЛЬНЫЕ ФУНКЦИИ КОРЫ
К двигательным областям коры головного мозга относятся
первичная и вторичная моторная и премоторная кора
(рис. 8.1). Первичная двигательная моторная область
локализована в прецентральной извилине непосредственно кпереди от
центральной борозды. Премоторная и вторичная двигательная
области расположены в виде широкой полосы перед
прецентральной извилиной. Премоторная область занимает всю
латеральную поверхность этой полосы, а вторичная моторная соот-
156
Глава 8
ветствует ее медиальной поверхности, скрытой в глубине меж-
полушарной борозды.
Локальное электрическое раздражение какого-либо
участка этих областей воспроизводимо сопровождается дискретным
движением. Каждый участок коры характеризуется тем, какое
движение возникает при его электрическом раздражении. Пер-
Первичная
Рис. 8.1. Двигательные области коры головного мозга. Большая часть
вторичной двигательной области расположена на медиальной поверхности коры,
которая на данном рисунке не видна.
вичная двигательная область отвечает за сокращения
отдельных мышц. Раздражение вторичной двигательной области
сопровождается менее дискретными и локализованными
двигательными реакциями; к ним относятся сложные движения
головы, шеи, туловища и конечностей. Премоторная кора
контролирует локомоторные акты, в том числе движения рта и языка
при словообразовании, координированные движения глаз и
головы и тонкие движения рук и пальцев.
ПЕРВИЧНАЯ ДВИГАТЕЛЬНАЯ ОБЛАСТЬ
Эта область образована совокупностью вертикальных
колонок нейронов. Каждая из них отвечает за возбуждение или
торможение одной группы мотонейронов, иннервирующих отдель-
Центральная регуляция движений
157
ную мышцу. Колонки, возбуждающие и тормозящие
какую-либо определенную группу мотонейронов, находятся по
соседству.
Многие нейроны двигательной коры отвечают на
раздражение соматосенсорных рецепторов. Главными источниками
чувствительной информации двигательной коры служат связи с
соматосенсорной корой и ретикулярной формацией. Каждая
колонка двигательной области получает импульсацию от про-
приоцепторов мышц и суставных рецепторов, возникающую при
осуществлении регулируемого этой колонкой движения. Так,
если какая-либо колонка отвечает за сокращение
мышцы-сгибателя определенного сустава, то к ней поступают импульсы от
рецепторов этого сустава и проприоцепторов относящихся к
нему мышц. Колонки, регулирующие движения кистей рук,
получают также импульсацию от расположенных в них рецепторов^
давления и осязания.
СОМАТОТОПИЧЕСКАЯ ОРГАНИЗАЦИЯ
Колонки двигательной коры характеризуются соматотопи-
ческим расположением (рис. 8.1); подобным же образом
построена и соматосенсорная кора (см. гл. 5). Местонахождение
колонки в двигательной коре непосредственно связано с
локализацией иннервируемой ею мышцы. Колонки, регулирующие,
деятельность соседних мышц, располагаются по соседству.
Чем краниальнее расположена мышца, тем более вентролате-
ральное положение занимают соответствующие колонки.
Раздражение колонок дорсомедиального края двигательной коры
сказывается на движениях мышц ноги. Стимулируя колонки
вентролатерального края, можно вызвать движение рта к.
языка. \
Размер корковой области, отвечающей за регуляцию какой-
либо группы мышц, определяется степенью участия этих мышц
в тонких движениях. Относительно обширные участки
двигательной коры регулируют деятельность пальцев рук, губ и
языка — структур, осуществляющих крайне тонкие движения..
Мышцам же спины и ног, участвующим в поддержании позы^
соответствует лишь небольшая область двигательной коры.
ПИРАМИДНАЯ СИСТЕМА
Пирамидный, или кортикоспинальный, путь служит прямым;
каналом регуляции двигательной активности спинного мозга со
стороны коры (рис. 8.2). Эти пути идут от двигательной коры
Двигательная кора
Средний мозг_
Пирамидный путь__
Продолговатый -^
Кюзг
4L
0^ \
\ t
ft 1
3>
^3
1 Боковой
/^кортико-
спинальный
путь
1 Передний
кортико-
спинальный
путь
Рис. 8.2. Пирамидный, или кортикоспи-
нальный, путь.
через ствол мозга к
продолговатому мозгу, где
большая часть составляющих их
волокон перекрещивается и
переходит на
противоположную сторону. После этого
пирамидный путь
разделяется на латеральный и
передний пучки, которые
входят в спинной мозг,
спускаясь по нему в составе
латерального и переднего корти-
коспинальных путей (рис.
8.3). Большинство волокон
этих путей оканчивается на
вставочных нейронах
спинного мозга, но около 20%
(наиболее крупные волокна)
образуют синапсы
непосредственно на мотонейронах.
Все волокна
пирамидного пути начинаются либо в
первичной двигательной
(60%), либо в соматосен-
сорной (40%) коре.
Последние служат каналом для
корковой регуляции
чувствительных функций
спинного мозга: они тормозят
афферентную сенсорную
активность в задних рогах.
ФУНКЦИИ ПИРАМИДНОЙ СИСТЕМЫ
Главная функция пирамидной системы состоит в
осуществлении тонких движений — например при игре на фортепиано,
продевании нитки в иголку, беге с препятствиями или при
разговоре. Считается, что таким движениям предшествует
возникновение возбуждения в соседних областях премоторной и
вторичной двигательной коры. После того как сформируется
«идея» движения, в двигательной коре образуется сложный
комплекс возбуждений, необходимый для осуществления
тонкого движения. Для образования такого комплекса в нервных
сетях двигательной коры соматосенсорная информация
(тактильная, температурная и болевая) от ретикулярной формации
Центральная регуляция движений
159
и соматосенсорной коры соотносится с импульсацией от базаль-
ных ганглиев и мозжечка.
Следует отметить, что для многих основных двигательных
актов, таких, как стояние, ходьба, бег, прыгание и потребление
пищи, участие пирамидной системы необязательно. Эти акты
сохраняются у обезьян и после перерезки пирамидных путей.
Передний кортикоспинальный путь
Рис. 8.3. Поперечное сечение спинного мозга. Изображены главные
нисходящие спинномозговые пути пирамидной (латеральный и передний кортикоспи-
нальные пути) и экстрапирамидной (руброспинальный, ретикулоспинальный
и вестибулоспинальный пути) систем.
Пирамидная система играет важную роль в поддержании
мышечного тонуса, так как при перерезке пирамидных путей
мышечный тонус существенно снижается. Нисходящие
пирамидные влияния на спинной мозг являются преимущественно
возбуждающими. Устранение этих влияний приводит к
угнетению активности спинного мозга.
ЭКСТРАПИРАМИДНАЯ СИСТЕМА
К экстрапирамидной системе относятся все двигательные
ядра и пути, по которым двигательные команды коры
проводятся к спинному мозгу, за исключением пирамидных. В
отличие от пирамидной системы экстрапирамидная система не
организована в виде одиночного дискретного двигательного пути:
160
Глава 8
Двигательная кора
Средний мозг
Продолговатый
мозг
Спинной мозг
Красное ядро
Ретикулярная
формация
Руброспинальный
путь
Ретикуло-.
спинальный путь
Рис. 8.4. Экстрапирамидная система. Руброспинальный и ретикулоспинальныи
пути, входящие в состав мультинейронного экстрапирамидного пути, идущего
от коры больших полушарий к спинному мозгу.
она скорее представляет собой сложную сеть двигательных
ядер и соединяющих их между собой трактов, по которым
сигналы из коры больших полушарий передаются на
мотонейроны спинного мозга (рис. 8.4). В состав ее входят пучки,
соединяющие кору с двигательными ядрами ствола мозга,
важнейшими из которых являются красное и вестибулярное ядра и
ретикулярная формация (рис. 8.4 и 8.5).
От этих промежуточных ядер начинается ряд нисходящих
спинномозговых путей, оканчивающихся на вставочных и дви-
Центральная регуляция движений
161
гательных нейронах спинного мозга, — руброспинальный, ре-
тикулоспинальный и вестибулоспинальный (рис. 8.3).
Некоторые нейроанатомы относят к экстрапирамидной системе и ба-
зальные ганглии, однако последние данные говорят о том, что
эти ганглии, как и мозжечок, участвуют в кординации движе-
Ядра мозжечка
Вестибуляр
ядро
ядро \фЛм'££*У~*;^ ,
нижней - - f у :^.\\
оливы ~ ^ — " ""^^
. N Ретикулярная
Продолговатый. \ формация
МОЗГ \_
Мост
Красное ядро
Средний мозг
Рис. 8.5. Сагиттальный разрез ствола мозга и мозжечка. Показано строение
мозжечка и расположение главных двигательных центров ствола мозга.
ний, осуществляемых и пирамидной, и экстрапирамидной
системами.
Волокна экстрапирамидных путей идут почти от всех
областей коры головного мозга. Большинство из них представляет
собой аксоны нейронов, расположенных либо в двигательных
областях коры (первичной, вторичной и премоторной), либо в
соматосенсорной коре, однако многие волокна берут начало в
других чувствительных зонах коры — слуховой, зрительной и
вкусовой, или в высших ассоциативных зонах лобной, теменной
и височных долей.
ДВИГАТЕЛЬНЫЕ ЯДРА СТВОЛА МОЗГА
Промежуточные двигательные ядра участвуют в регуляции
позы и в поддержании вертикального положения тела. Они
расположены "в глубине ствола мозга от продолговатого мозга до
11—1705
162
Глава 8
центральной части зрительных бугров (рис. 8.5). Часть
нейронов, относящихся к этим образованиям, сгруппирована в виде
специализированных ядер, однако большинство этих нейронов
входит в состав ретикулярной формации — диффузной сети
вставочных нейронов, расположенной в глубине ствола мозга.
На этих ядрах переключаются экстрапирамидные волокна
нейронов коры, базальных ганглиев и мозжечка (рис. 8.6). В ре-
/
1—7
Мозжечок
Двигательная
кора
^
S
V
Красное
ядро
s
ьны
Вестибулярное
ядро
ве^5;
Нижняя
олива
лиаг
уброст
LL.
N
\
^
.о
<н
N
/
Ретикулярная
формация
/ N
v
t/.
Базальные
ганглии
Ретикуло-
спинальный путь
Спинной мозг
Рис. 8.6. Главные связи между базальными ганглиями, мозжечком,
двигательными ядрами ствола мозга и двигательной корой.
тикулярной формации и связанных с ней ядрах сигналы,
поступающие по этим волокнам от высших двигательных центров,
интегрируются с соматосенсорной информацией, передаваемой
по спинно-таламическим путям и с импульсами от
вестибулярной системы. В результате формируются двигательные акты,
необходимые для поддержания вертикального положения.
Для сохранения вертикального положения тела силе
тяжести должно противодействовать сокращение разгибателей.
Передние две трети двигательных срединных структур ствола
мозга служат источником мощной облегчающей импульсации
к мотонейронам разгибателей. На этот разгибательный тонус в
норме оказывают тормозное влияние сигналы, идущие от
высших двигательных центров коры и базальных ганглиев.
Величина этого тонуса хорошо видна у животного после перерезки
ствола мозга, отделяющей кору и базальные ганглии от
двигательных ядер ствола. У такого децеребрированного животного
Центральная регуляция движений
163
резко повышен тонус разгибателей; в типичных случаях
наблюдается ригидность разгибателей, обусловленная их сильным
возбуждением.
ФУНКЦИИ ЭКСТРАПИРАМИДНОЙ СИСТЕМЫ
Экстрапирамидная система участвует в регуляции позы и
осуществлении таких локомоторных актов, как ходьба, стояние,
прыжки, бег и плавание. У интактного животного эти
двигательные акты запускаются в результате возбуждения нейронов
коры, посылающих отростки к двигательным ядрам ствола
мозга. Для осуществления соответствующего двигательного
акта информация о его временных параметрах, поступающая от
мозжечка и базальных ганглиев, интегрируется в
промежуточных ядрах с чувствительными сигналами о состоянии
организма (от ретикулярной формации).
От коры головного мозга к двигательным ядрам ствола по
экстрапирамидным путям направляются преимущественно
тормозные влияния. Это нисходящее торможение играет важную
роль в регуляции активности двигательных экстрапирамидных
ядер ствола, так как от этих ядер к двигательным центрам
спинного мозга поступает мощная возбуждающая импульсация.
В отсутствие тормозных влияний коры — например при
перерезке ствола мозга — движения превращаются в ряд
судорожных мышечных сокращений.
МОЗЖЕЧОК
Мозжечок, образованный в результате разрастания
дорсальной поверхности ствола мозга, соединяется с последним
посредством трех крупных нервных пучков — ножек мозжечка
(рис. 8.5). Мозжечок состоит из двух частей: 1) коры,
образующей складки и покрывающей большую часть дорсальной
поверхности ствола мозга, и 2) мелких мозжечковых ядер,
расположенных в глубине ножек мозжечка.
Мозжечок, участвующий в координации движений и
распределении их во времени, играет важную роль как
сравнивающее устройство. Когда в двигательной коре принимается
решение о каком-либо движении, то в мозжечок направляется
информация о природе и ожидаемых результатах этого
движения. В мозжечке эта информация хранится и сличается с
чувствительной импульсацией от проприоцепторов и других
рецепторов, возбуждающихся при совершении движения. Если
сигналы, поступающие в мозжечок в ходе двигательного акта,
свидетельствуют о том, что последний выполняется неправильно,
то от мозжечка в ствол мозга и в корковые двигательные
!!•
164
Глава 8
центры посылаются импульсы, благодаря которым
осуществляется необходимая коррекция.
Мозжечок имеет особенно большое значение для построения
и осуществления баллистических движений. Скорость
выполнения таких движений слишком велика, чтобы во время
двигательного акта в него вносились какие-либо исправления; к ним
относится быстрая игра на фортепиано, бросание мяча в цель,
прыжок через препятствие и речь. Коррекция по ходу
движения в таких случаях невозможна, так как время, необходимое
для 1) передачи сенсорной информации к мозжечку, 2)
анализа этой информации и 3) построения корректирующего
движения, гораздо больше, чем длительность самого двигательного
акта. Следовательно, баллистические движения должны быть
запрограммированы заранее. Мозжечок имеет первостепенное
значение для такого программирования, поскольку в нем
хранится чувствительная и двигательная информация,
позволяющая пирамидной и экстрапирамидной системам избрать тот
комплекс двигательных импульсов, под действием которого
будет успешно выполнено необходимое баллистическое движение.
ВХОДНЫЕ КАНАЛЫ МОЗЖЕЧКА
Большинство волокон, несущих импульсацию в мозжечок,
заканчиваются как в его ядрах, так и в коре (рис. 8.7). К ним
относятся важные проприоцептивные афферентные каналы от
спинного мозга, вестибулярной системы и ядра нижней оливы.
Чувствительные пути идут также от верхнего (зрительное) и
нижнего (слуховое) двухолмия. К мозжечку приходит также
важнейшая информация от двигательной и соматосенсорной
областей коры больших полушарий.
Все эти импульсы поступают в мозжечок по двум типам
волокон: лазящим и моховидным (рис. 8.7). Лазящие волокна,
на активность которых влияют различные проприоцептивные
стимулы, начинаются от ядер нижних олив. О том, как
происходит их активация, в настоящее время известно мало, хотя
полагают, что они активизируются под влиянием импульсов от
двигательных центров коры. По моховидным волокнам
передается основная часть поступающей в мозжечок чувствительной
информации (проприоцептивная, тактильная, вестибулярная,
слуховая и зрительная), а также импульсы от коры больших
полушарий. По этим волокнам в мозжечок приходят сигналы
от всех чувствительных и двигательных систем организма.
ВЫХОДНЫЕ КАНАЛЫ МОЗЖЕЧКА
Выходные сигналы мозжечка передаются через четыре его
ядра — зубчатое, шаровидное, пробковидное и ядро шатра.
Центральная регуляция движений
165
Кора
мозжечка
Ядра
мозжечка
Рис. 8.7. Нервная цепь мозжечка. К коре и ядрам мозжечка поступают
возбуждающие импульсы как по лазящим, так и моховидным волокнам. Из
мозжечка же сигналы идут лишь от клеток Пуркине (П), угнетающих
активность нейронов в ядрах мозжечка (Я). К собственным нейронам коры
мозжечка относятся возбуждающие клетки-зерна (3) и тормозные корзинчатые
нейроны (К), нейроны Гольджи (Г) и звездчатые нейроны (Зв). Стрелками
указано направление движения нервных импульсов. Имеются как
возбуждающие (+), так и тормозные (—) синапсы.
Импульсы от этих ядер направляются как к двигательным
центрам ствола мозга (красному ядру, ретикулярной формации
и вестибулярным ядрам), так и — через вентролатеральный
отдел таламуса — к двигательным и соматосенсорным областям
коры больших полушарий (рис. 8.6).
Вся информация от коры мозжечка передается к его ядрам
через клетки Пуркине — крупные тормозные вставочные
нейроны, образующие средний слой коры мозжечка. Таким образом,
вся активность коры мозжечка в конечном счете преобразуется
в тормозные влияния на нейроны его ядер.
166 Глава 8
АКТИВНОСТЬ МОЗЖЕЧКА
Кора мозжечка обладает стереотипной структурой. В
состав ее входит пять типов нейронов: клетки Пуркине, клетки-
зерна, корзинчатые клетки, клетки Гольджи и звездчатые
клетки (рис. 8.7). Клетки Пуркине — главные нейроны,
выполняющие интегративную функцию, — возбуждаются под влиянием
как лазящих, так и моховидных волокон. К каждому такому
нейрону подходит лишь одно лазящее волокно, образующее
на нем крупные возбуждающие синапсы. Одного импульса,
поступающего по лазящему волокну, достаточно для того, чтобы
вызвать разряд нейрона Пуркине. Напротив, моховидные
волокна оказывают на нейроны Пуркине слабое и непрямое
возбуждающее действие. Эти волокна не оканчиваются
непосредственно на клетках Пуркине, но возбуждают вставочные
нейроны (клетки-зерна), образующие на дендритах нейрона
Пуркине небольшие возбуждающие синапсы. Тозмозные
корзинчатые нейроны, нейроны Гольджи и звездчатые нейроны
оказывают модулирующее влияние на возбуждение клеток Пуркине
клетками-зернами.
Полагают, что кора мозжечка функционирует как блок
памяти, т. е. в ней в доступной для пирамидной и
экстрапирамидной систем форме хранится информация о состоянии про-
приоцепторов и общем состоянии чувствительных и
двигательных систем организма, необходимая для координации
движений. Единицей памяти служит нейрон Пуркине, в котором в
виде небольших ВПСП запасается информация об активности
моховидного волокна. Функцию считывающего устройства
выполняет лазящее волокно. Как уже указывалось, одиночного
импульса в лазящем волокне достаточно для того, чтобы
вызвать разряд иннервируемого им нейрона Пуркине. В то же
время величина этого разряда, т. е. количество и частота
нервных импульсов, зависит от возбудимости нейрона Пуркине в
данный момент времени. Если иннервирующие этот нейрон
моховидные волокна возбуждены, то на одиночный импульс,
пришедший по лазящему волокну, он отвечает мощным залпом.
Если же активность моховидных волокон минимальна, то
возбуждение лазящего волокна вызывает лишь одиночный
потенциал действия в нейроне Пуркине.
В коре мозжечка человека содержится 15 млн. клеток
Пуркине и к каждой из них подходит группа моховидных волокон,
в чем-то отличающаяся от остальных таких групп. Поэтому в
каждом нейроне Пуркине хранится обособленная часть
существенной для координации движений информации о состоянии
чувствительных и двигательных систем организма — например
положении того или иного сустава, степени сокращения мышц,
Центральная регуляция движений
167
расположении образа в поле зрения, и о только что
сформировавшемся в двигательной коре комплексе возбуждений. Эта
информация передается к двигательным областям коры
больших полушарий и к двигательным ядрам ствола мозга
посредством тормозно-модулирующего влияния на активность ядер
мозжечка.
ПОВРЕЖДЕНИЯ МОЗЖЕЧКА
Повреждения мозжечка сопровождаются рядом
характерных двигательных устройств, в том числе интенционным
тремором, дисметрией и атаксией. Изучение этих расстройств
служит важнейшим источником познания функций мозжечка.
ИНТЕНЦИОННЫЙ ТРЕМОР
Одна из функций мозжечка в координации мышечной
деятельности состоит в прекращении или затормаживании
движения. Для успешного совершения какого-либо движения
необходимо участие двух групп мышц, одна из которых
продвигает конечность к той точке в пространстве, которую необходимо
достичь, а вторая прекращает движение по достижении этой
точки. При повреждениях мозжечка плавная остановка
движений нарушается или становится невозможной, и объем
движений становится либо чрезмерным, либо недостаточным.
Пытаясь исправить чересчур размашистое движение, человек или
животное с поврежденным мозжечком обычно делает это в
большей степени, чем необходимо, и объем движения
становится недостаточным. В результате возникает интенционный
тремор. При остановке движения конечность совершает ряд
затухающих колебательных движений в ту или иную сторону и
лишь затем занимает постоянное положение. -
АТАКСИЯ
Следующая важнейшая функция мозжечка состоит в
координации движений, требующих последовательного сокращения
многих мышц. Атаксия — это невозможность координирование
выполнить последовательность движений. У человека или-
животного с поврежденным мозжечком нарушено распределение
движения во времени, поэтому движения, требующие точной
последовательности сокращений различных мышц, становятся
невозможными. Человеку с атаксией требуются огромные усилия
для совершения многих обычных двигательных актов типа
бега, письма и речи.
168
Глава 8
ДИСМЕТРИЯ
Для успешного выполнения быстрых (баллистических)
движений нервная система должна иметь возможность
«предугадывать» как ход, так и длительность движения, вызванного
неким комплексом двигательных возбуждений. Такой
способностью обладает именно мозжечок. Его поражение
сопровождается дисметрией — неспособностью предугадать последствия
движения. Желая достичь цели или перешагнуть через
препятствие, человек с поврежденным мозжечком совершает
движения в избыточном объеме. Дисметрия, как и другие описанные
выше мозжечковые расстройства, существенно влияет на
способность человека к координированным движениям. Особенно
сильно страдают при поражениях мозжечка баллистические
движения.
БАЗАЛЬНЫЕ ГАНГЛИИ
К базальным ганглиям относятся: хвостатое ядро, скорлупа
и бледный шар — образования, располагающиеся под корой
больших полушарий в глубине конечного мозга (рис. 8.8).
Деятельность их тесно связана с некоторыми двигательными
ядрами ствола мозга — гипоталамусом, черной субстанцией и
красным ядром. Все эти структуры, объединенные в сложную
нейронную сеть, образуют систему базальных ганглиев.
Рис. 8.8. Система базальных ганглиев.
Центральная регуляция движений
169
Основная часть импульсации поступает к базальным
ганглиям от двигательных областей коры головного мозга и
мозжечка. Сигналы от базальных ганглиев направляются к
двигательной коре, мозжечку и ретикулярной формации (рис. 8.6).
Следует отметить, что благодаря этим связям образуются две
важные нейронные петли, одна из которых связывает базаль-
ные ганглии с двигательной корой, а другая — с мозжечком.
ЗНАЧЕНИЕ БАЗАЛЬНЫХ ГАНГЛИЕВ В РЕГУЛЯЦИИ
ДВИЖЕНИЯ
Двигательные функции базальных ганглиев в настоящее
время не ясны. Считается, что они обеспечивают выполнение
сложных стереотипных движений при ходьбе, осуществлении
акта еды и поддержании позы. Согласно распространенной
гипотезе, деятельность базальных ганглиев связана с
выполнением медленных, или червеобразных, движений. Такие
движения осуществляются достаточно медленно, и в них могут
вноситься изменения через отрицательную афферентную обратную
связь от рецепторов, возбуждающихся в ходе самого движения.
Примером таких двигательных актов может быть медленная
ходьба, перешагивание через препятствие, вдевание нитки в
иголку. Когда по каналу афферентной обратной связи
поступает информация о том, что какое-то движение не достигает
цели, или, напротив, совершается в чрезмерном объеме, от
базальных ганглиев к двигательным областям коры больших
полушарий и ствола мозга поступают корректирующие сигналы.
Поскольку червеобразные движения совершаются относительно
медленно, пирамидная и экстрапирамидная системы успевают
по ходу их выполнения внести исправления в непрерывный
поток двигательных возбуждений.
ПОРАЖЕНИЯ БАЗАЛЬНЫХ ГАНГЛИЕВ
Как ни странно, основная информация о роли базальных
ганглиев в регуляции движений получена при изучении
двигательных расстройств, возникающих при поражении этих
ганглиев. К таким расстройствам относятся атетоз, хорея, баллизм
и болезнь Паркинсона.
Атетоз, наблюдающийся при поражениях хвостатого ядра и
скорлупы, характеризуется медленными извивающимися
непроизвольными движениями конечностей. При этом нарушении
возникают попеременные порывистые сокращения сгибателей и
разгибателей. Как и другие нарушения функции базальных
ядер, атетоз сопровождается невозможностью удержать
конечность в постоянном положении.
170
Глава 8
При хорее, сопровождающей поражения хвостатого ядра,
наблюдаются постоянные непроизвольные порывистые
движения головы и конечностей.
Для поражений субталамической области характерны
сильные неконтролируемые движения конечностей: если такие
движения возникают постоянно, то наблюдается картина бал-
лизма.
Главным симптомом болезни Паркинсона является
постоянный низкочастотный тремор, сочетающийся с мышечной
ригидностью. Из-за этой ригидности больной с трудом может начать
движение. Постоянный непроизвольный тремор препятствует
успешному выполнению медленных движений. Болезнь
Паркинсона возникает при поражениях черной субстанции.
Современные данные свидетельствуют о том, что в норме черная
субстанция оказывает тормозное влияние на хвостатое ядро,
скорлупу и бледный шар. При ее разрушении эти тормозные
влияния устраняются. В результате усиливаются возбуждающие
влияния базальных ганглиев на кору головного мозга и
ретикулярную формацию и возникают характерные симптомы
болезни Паркинсона — тремор и ригидность.
ЛИТЕРАТУРА
Asanuma H. (1975). Recent developments in the study of the columnar
organization of neurons within the cerebral coretx. Physiol. Rev., 55, 143.
Brooks V. В., Stoney S. D. (1971). Motor mechanisms: the role of the pyramidal
system in motor control, Ann. Rev. Physiol., 33, 337.
Eccles J. C. (1977). The understanding of the brain, McGraw-Hill, New York,
Chap. 4.
Evarts E. V. (1973). Brain mechanisms in movements, Sci. Amer., 230, 96.
Eyzaguirre С Fidone S. J. (1975). Physiology of the nervous system, 2nd edn.,
Year Book Medical, Chicago. Chaps. 18—21.
Kornhuber H. H. (1974У?СегеЬга1 cortex, cerebellum and basal ganglia: An
introduction to their motor function. In: Schmitt F. O. and Worden F. G. (eds.),
The neurosciences III, MIT Press, Cambridge.
Llinas R. R. (1975). The cortex of the cerebellum,* Sci. Amer., 232, 56.
Mountcastle V. B. (1974). Medical physiology, 13th edn., Mosby st. Louis,
Chaps. 26, 28 and 29.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Коротко опишите строение колонок первичной двигательной
области коры.
2. В чем состоят основные различия в анатомической
организации пирамидной и экстрапирамидной систем?
3. Сравните функциональное значение пирамидной и
экстрапирамидной систем в регуляции движений.
4. Что такое баллистические движения и чем они отличаются
от медленных движений?
Центральная регуляция движений
171
5. Нарисуйте схему нервной сети мозжечка и обозначьте ее
основные нервные элементы.
6. Используя схему, нарисованную при ответе на вопрос 5,
расскажите, как мозжечок действует в качестве
сравнивающего устройства при осуществлении движений. Укажите
механизмы, которые предположительно участвуют в хранении
и считывании информации.
7. Сравните двигательные нарушения, возникающие при
поражении мозжечка и базальных ганглиев. Свидетельствует ли
характер этих нарушений в пользу гипотезы, согласно
которой базальные ганглии участвуют в осуществлении
медленных, а мозжечок — баллистических движений?
Глава 9
ВЫСШИЕ ФУНКЦИИ КОРЫ ГОЛОВНОГО МОЗГА
Высшие функции нервной системы — способность к
мыслительной деятельности, осознанию сигналов из окружающей
среды, к абстрактному мышлению и запоминанию — в
значительной мере связаны с деятельностью коры больших
полушарий. Возбуждение нейронных сетей коры обусловливает осо-~
знание личностью своих взаимоотношений с окружением.
Именно кора служит структурной основой сознания и интеллекта.
ОРГАНИЗАЦИЯ КОРЫ
В коре головного мозга содержится около 14 миллиардов
нейронов, большая часть которых (примерно 90%)
сгруппирована в шесть слоев и образует неокортекс — высший интегра-
тивный отдел соматической нервной системы. Неокортекс
отвечает за переработку и интерпретацию чувствительной
информации (слуховой, вкусовой, соматосенсорной и зрительной), а
также за управление сложными мышечными движениями. Здесь
расположены центры, участвующие в процессах абстрактного
мышления, речи и хранения памяти. Большая часть процессов
в неокортексе является нейрофизиологической основой сознания.
Второй большой отдел коры головного мозга — это палео-
кортекс. В отличие от неокортекса эта часть коры обладает
более простой трехслойной структурой. Процессы, протекающие
в палеокортексе, не всегда отражаются в сознании. К палео-
кортексу относятся отделы коры, связанные с лимбической
системой (см. гл. 10). Здесь расположены высшие вегетативные
центры.
Неокортекс состоит из четырех долей — лобной, теменной,
затылочной и височной (рис. 9.1). Каждая из них
подразделяется на проекционные и ассоциативные области. К
проекционным областям коры сигналы поступают преимущественно от
специфических сенсорных ядер таламуса — латерального
коленчатого тела (зрительное), медиального коленчатого тела
(слуховое) и вентробазального комплекса (соматосенсорное и
вкусовое), а также от его специфических двигательных
(передних) ядер. Основным источником импульсов, поступающих
Высшие функции коры головного мозга
173
к ассоциативным областям коры, служат другие корковые —
проекционные и ассоциативные — зоны. Как проекционные, так
и ассоциативные области коры получают также сигналы от
неспецифических ядер таламуса. Эти неспецифические сигналы
определяют состояние активации коры.
НЕОКОРТЕКС
Проекционные облает небкортекса обладают
стереотипными функциями. Электрическое раздражение какой-либо точки
чувствительных зон коры при Нейрохирургических операциях
сопровождается специфическими ощущениями, например
1) чувством прикосновения или температурного воздействия на
кожу при раздражении соматосенсорной коры, 2) вкусовыми
ощущениями (вкусовая кора), 3) видением светящихся полос
или пятен (зрительная кора), 4) звучанием простых тонов и
Затылочная доля *=С
Рис. 9.1. Проекционные (отмечены точками) и ассоциативные (светлые)
зоны коры головного мозга. К проекционным зонам относятся двигательная
область (лобная доля), соматосенсорная область (теменная доля), зрительная
область (затылочная доля) и слуховая область (височная доля).
шумов (слуховая кора). На уровне специфических
чувствительных областей коры определяются общие параметры
раздражения (например, цвет, ориентация и положение образа,
высота и длительность звука). Двигательная кора управляет
деятельностью мышц. Функциональная организация этих областей
коры головного мозга рассматривается в гл. 5
(соматосенсорная область), гл. 6 (слуховая, вкусовая и зрительная области)
и гл. 8 (двигательная область).
174
Глава 9
АССОЦИАТИВНЫЕ ОБЛАСТИ КОРЫ
Существуют три ассоциативные области коры: лобная,
височная и затылочно-теменная. Информацию об их функции
можно получить двумя основными путями. Первый из них —
электрическое раздражение, возможное при некоторых
нейрохирургических операциях. Наблюдая внимательно за
реакциями на это раздражение, можно получить ценные сведения,
позволяющие глубже понять сложные функции коры. Вторым
источником информации служит изучение нарушений поведения
у больных с повреждениями ассоциативных зон в результате
инсульта или травмы.
Считается, что затылочно-теменная область участвует в
интерпретации поступающих соматосенсорных, вкусовых и
зрительных сигналов. Здесь соотносятся друг с другом физические
параметры раздражителя (например, характер поверхности,
вес, размер, форма и цвет) и формируется его интегральный
образ. Больной с поражением затылочно-теменной области не
может узнать предмет, исходя из соматосенсорной и
зрительной информации о нем. Так, он может осознавать, что некий
объект оранжевый, круглый и имеет кисло-сладкий вкус, но не
способен понять, что это апельсин.
Поражения области стыка теменной, затылочной и височных
долей сопровождаются словесной слепотой (алексией). Такие
больные узнают буквы, составляющие какое-либо слово, но не
способны объяснить его значение. Алексия может также
проявляться в понимании отдельных слов, но не фразы в целом.
У больных с поражениями задних областей височной доли
наблюдается словесная глухота. Они легко понимают значение
написанной фразы, но, если ту же фразу произнести вслух,
они не в состоянии объяснить ее значение. Полагают, что
обширные области височной доли участвуют в долговременной
памяти. Раздражение этих долей, в частности их задних
отделов, сопровождается возникновением сложных картин из
прошлого. Обычно эти картины бывают связаны с каким-либо
моментом из жизни: игрой в детстве, посещением концерта,
беседой с конкретным человеком. Воспоминания эти бывают очень
яркими и обычно включают комплекс соматосенсорных,
слуховых, зрительных и вкусовых ощущений, относящихся к
всплывающему в памяти событию. Так, воспоминание о концерте
может сопровождаться звучанием исполнявшейся музыки,
зрительным образом сцены с оркестром и ощущением
температуры в зале.
В лобных долях расположены главные центры речи и
письма. При поражениях задне-боковых отделов ассоциативных
областей лобной коры теряются все речевые навыки — больной
' Высшие функции коры головного мозга
175
не может выразить свои мысли ни в устной, ни в письменной
форме.
Самый передний отдел лобной доли (префронтальная кора),
участвует в формировании личностных качеств, творческих
процессов и влечений. При поражении этой области наступают
глубокие изменения личности, интересов и способности к
концентрации внимания. Такие люди утрачивают социальные
«тормоза», интерес к работе и собственной внешности и становятся
некоммуникабельными.
МЕЖПОЛУШАРНАЯ АСИММЕТРИЯ
Многие высшие функции ассоциативных областей коры
выполняются каким-либо одним полушарием, так что в
отношении различных сторон высшей нервной деятельности между
полушариями существует асимметрия. Ведущее полушарие
(обычно это бывает левое) отвечает за интерпретацию и
формирование устной и письменной речи. Речевые центры
расположены в лобной и теменной долях этого полушария. Другое
Сосцевидное тело
Рис. 9.2. Кора головного мозга на поперечном разрезе. Оба полушария
соединены пучками белого вещества, образующими мозолистое тело.
полушарие, так сказать «подчиненное», участвует в
пространственных построениях и определении временных
взаимоотношений, а также содержит центры музыкального и
художественного творчества.
Важнейшая функция мозолистого тела (рис. 9.2) — крупных
пучков белого вещества, соединяющих полушария, — состоит в
координации деятельности полушарий. В некоторых случаях
для того чтобы предупредить распространение тяжелых эпи-
176
Глава 9
лептических припадков, производят хирургическое рассечение
мозолистого тела. На первый взгляд поведение таких больных
с «расщепленным мозгом» кажется нормальным. Однако
тщательное наблюдение показывает, что оба полушария
функционируют независимо друг от друга: речь и письмо
осуществляется ведущим, а способность к узнаванию предметов исходя из
их зрительных и соматосенсорных характеристик —
подчиненным полушарием.
АКТИВАЦИЯ КОРЫ ГОЛОВНОГО МОЗГА. СОЗНАНИЕ
Для сознательного (бодрствующего) состояния характерно
восприятие субъектом физических и психических факторов
окружающего мира. Сознание, обусловленное активностью
миллионов нейронов коры, зависит от степени их возбуждения.
Это возбуждение создается постоянным притоком импульсов от
ретикулярной формации ствола мозга, поступающих в кору по
диффузной нервной сети — активирующей ретикулярной
системе. Уровень активации в значительной мере определяется
степенью возбуждения этой системы.
ЭЛЕКТРОЭНЦЕФАЛОГРАММА
Уровень активации коры обычно определяют путем записи
электроэнцефалограммы (ЭЭГ). ЭЭГ отражает колебания
потенциалов мозга, зарегистрированные через черепную коробку
посредством электродов, на-
Альфа vtoMWfyw^^AAAMtowvvwl ложенных на кожу головы.
ЭЭГ можно записать также
непосредственно с коры го-
Бета w»»*^^ ??ртиес?иТоп^
в опытах над животными.
Существует связь между
уровнем активации коры и
Тэта >wnv^W\AAAAAvH^A^W частотой колебаний на ЭЭГ.
Обычно типы
электроэнцефалограмм, полученные при
^, переходе от бодрствования
s\ /V /Ч /V /\ к бессознательному состоя-
Делыа / \г ^^ ^v \/ нию или сну, разделяют на
альфа-, бета-, тэта- и
дельта-ритмы (рис. 9.3).
Альфа-ритм характерен
1 с t для нормального бодрству-
Рис. 9.3. Четыре основных ЭЭГ-ритма. ющего человека или живот-
Высшие функции коры головного мозга
177
ного в состоянии покоя. Особенно хорошо он выявляется, если
глаза испытуемого закрыты. Частота колебаний альфа-ритма
варьирует в пределах от 8 до 14 Гц. Если на человека,
находящегося в состоянии покоя, внезапно подействовать каким-либо
достаточно интенсивным раздражителем — например включить
свет в темной комнате, неожиданно издать тот или иной звук
или нанести кожное раздражение, то на ЭЭГ будет
зарегистрирован переход альфа-ритма в более высокочастотный (15—50
Гц) бета-ритм (рис. 9.4). Этот ритм, связанный с сознательной
концентрацией внимания на каком-либо внешнем объекте,
характерен для активного бодрствующего состояния. Бета-ритм
наблюдается также при умственном напряжении во время
решения задачи или формулирования мысли.
г
Звуковой стимул —
Рис. 9.4. Активация коры. Предъявление стимула испытуемому,
находящемуся в спокойном, расслабленном состоянии, сопровождается десинхронизаци-
ей ЭЭГ — переходом альфа-ритма в бета-ритм.
У взрослого переход от состояния бодрствования к сну
соответствует переходу альфа-ритма ЭЭГ в дельта-ритм,
характерный для глубокого сна. Низкочастотный ритм ЭЭГ
наблюдается у бодрствующих субъектов с незрелой нервной системой.
Так, у новорожденных преобладает дельта-ритм (частота менее
4 Гц). Для нормальных детей характерен тэта-ритм (4—8 Гц).
НЕЙРОФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЭЭГ
Главные нервные клетки коры — это пирамидные и
звездчатые нейроны (рис. 9.5). Хотя тела пирамидных нейронов
заложены в глубине коры (слой IV), их дендриты располагаются в
наиболее поверхностных ее слоях — I и II. Тела и отростки
звездчатых нейронов находятся в слое IV. Волокна нейронов
таламуса заканчиваются в двух слоях коры. От нейронов
специфических ядер таламуса эти волокна идут к дендритам
звездчатых нейронов слоя IV, аксоны которых подходят к ба-
зальным дендритам пирамидных нейронов. Отростки же
нейронов неспецифических ядер таламуса оканчиваются в слоях I и
II на апикальных дендритах пирамидных нейронов.
ЭЭГ отражает уровень синаптической активности внешних
слоев коры головного мозга. Ритм ЭЭГ определяется
величиной синаптических потенциалов — ВПСП и ТПСП —этих слоев.
12—1705
178
Глава 9
Большая часть синапсов во внешних слоях коры образована
окончаниями восходящих волокон от неспецифических ядер
таламуса на апикальных дендритах пирамидных нейронов.
Величина ритмических колебаний потенциалов,
создаваемых в результате активности нейронов поверхностных слоев
коры, будет достаточна для того, чтобы отразиться на ЭЭГ\
лишь в том случае, если эта активность синхронизирована.
Tain
IV
V
VI
Нейроны
коры
Апикальные | Афферентные
дендриты | волокна таламуса
3L-L —
Пирамидный
нейрон
Звездчатый
нейрон
Базальные Специфи- Неспеци -
дендриты ческие фические
Рис. 9.5. Клеточное строение коры головного мозга. Афферентные волокна от
специфических ядер таламуса оканчиваются на звездчатых нейронах слоя IV.
У пирамидных нейронов имеются два вида дендритов: расположенные в слое
V базальные дендриты, образующие синапсы со звездчатыми нейронами, и
расположенные в слое I апикальные, получающие волокна от
неспецифических структур таламуса. Пирамидные нейроны — это эфферентные нейроны
коры, передающие по аксонам ее команды.
кая синхронизация возникает в результате двух факторов,
1) собственной ритмической активности нервных сетей
поверхностных слоев коры и 2) ритмического характера нервных
разрядов неспецифических ядер таламуса, посылающих сигналы к
этим слоям.
Пирамидные нейроны — это главный путь передачи команд
от одних отделов коры к другим, а также к нижележащим
центрам, и поэтому деятельность этих нейронов имеет
первостепенное значение для бодрствующего состояния. По всей
видимости, для поддержания возбудимости пирамидных
нейронов необходим постоянный приток к ним импульсов от
неспецифических ядер таламуса. Если в результате пересечения
путей от указанных ядер этот поток прекращается, то
возбудимость пирамидных нейронов снижается: животное после такой
операции постоянно пребывает в состоянии ступора.
Высшие функции кори головного мозга
179
АКТИВАЦИЯ КОРЫ: РОЛЬ АКТИВИРУЮЩЕЙ
РЕТИКУЛЯРНОЙ СИСТЕМЫ
Если человеку, находящемуся в состоянии покоя,
предъявить сильный раздражитель, то он моментально
настораживается и концентрирует внимание на этом раздражителе. Этому
переходу умственной деятельности от состояния покоя к
активности соответствует переход от альфа-ритма ЭЭГ к бета-
ритму и другим более частым колебаниям (см. рис. 9.4). Этот
переход, или де синхронизация ЭЭГ, возникает в результате
поступления в кору сенсорных возбуждений от неспецифических
ядер таламуса. ^
~Неспецифические
ядра таламуса
Продолговатый \ У-VN \
мозг х v •'—\
A V;:.\'
/ \ V
Ретикулярная \
, формация
Рис. 9.6. Активирующая ретикулярная система (АРС)—нервная сеть, по
которой сенсорное возбуждение передается от ретикулярной формации ствола
мозга к неспецифическим ядрам таламуса. Волокна от этих ядер регулируют
уровень активности коры.
Неспецифические ядра образуют диффузную нервную сеть
в медиальных отделах таламуса. Они составляют самый
передний отдел активирующей ретикулярной системы (АРС),
также расположенной в виде диффузной нервной сети между
ретикулярной формацией и таламусом и ответственной за
регуляцию возбудимости коры (рис. 9.6). АРС активируется
самыми различными сенсорными сигналами, поступающими в
12*
180
Глава 9
ретикулярную формацию — слуховыми, обонятельными, сома-
тосенсорными, вестибулярными и зрительными. Активирующая
ретикулярная система служит каналом, по которому эти
сигналы передаются через неспецифические ядра таламуса к
поверхностным слоям коры.
Возбуждение АРС необходимо для поддержания
бодрствующего состояния. Разрушение этой системы у подопытных
животных неминуемо приводит к сноподобному коматозному
состоянию. Если же у спящего животного произвести
электрическое раздражение АРС, то оно немедленно проснется.
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ СЕНСОРНЫЕ СИСТЕМЫ
Следует отметить существенные различия между функцией
АРС — не специфической сенсорной системы, служащей
диффузным путем к поверхностным слоям всех областей коры, и
функциями специфических сенсорных систем, образующих
строго локализованные проекции в средних слоях соответствующих
сенсорных областей коры. В отличие от поражений
неспецифической системы, сопровождающихся стойкой потерей сознания,
нарушения в области какого-либо специфического сенсорного
пути не влияют на уровень активации коры. При таких
нарушениях утрачивается способность к оценке той или иной
специфической сенсорной информации. Электрическое
раздражение этих путей также не влияет на уровень активации коры, но
приводит к возбуждению в ее сенсорных зонах (например,
слуховой или зрительной), что сопровождается специфическими
ощущениями типа шумов или световых вспышек.
СОН
Сон — это состояние, характеризующееся угнетением
сознания и восприятия окружающей действительности. Считается,
что сон играет роль восстановительного процесса. Во сне
снижены многие функции организма, в том числе мышечный тонус,
частота сокращений сердца, интенсивность обмена веществ,
дыхание, симпатический тонус и температура тела.
Сон необходим для нормальной умственной деятельности.
Длительное лишение сна приводит к психическим
расстройствам общего характера. У таких людей снижается
работоспособность и умение решать задачи. Эти общие явления часто
сопровождаются повышенной раздражительностью, приводящей
иногда к аномальному поведению.
Высшие функции коры головного мозга
181
СМЕНА СНА И БОДРСТВОВАНИЯ:
ЭЛЕКТРОЭНЦЕФАЛОГРАФИЧЕСКИЕ ХАРАКТЕРИСТИКИ
При нормальной смене сна и бодрствования у взрослого на
каждые 16 часов бодрствования приходится около 8 часов сна.
Эти состояния сопровождаются характерными изменениями
ЭЭГ.
При приближении сна бета-ритм ЭЭГ, типичный для
активного бодрствования, сменяется альфа-ритмом (рис. 9.7).
Альфа-ритм наблюдается у людей в расслабленном состоянии с
закрытыми глазами, когда сознание сохранено, но целостные
реакции на раздражители угнетены.
Покой
4WWV4Y^
ДpeмoтнoevлV^*^\vлr•^^^^v^
состояние
Легкий
сои
глубокий ^rJ\J\/\^^^J\^
Парадоксальный
сон
1 с
Рис. 9.7. ЭЭГ, характерные для различных фаз цикла сон — бодрствование.
На наступление сна и полное подавление сознания
указывает дальнейшее замедление ритма ЭЭГ — альфа-ритм переходит
в тэта- или даже низкоамплитудный дельта-ритм (рис. 9.7). На
волны доминирующего ритма (бета или дельта)
накладываются короткие вспышки, или веретена, альфа-ритма,
характерные для ЭЭГ при поверхностном сне.
В дальнейшем сон углубляется, что сопровождается
появлением высокоамплитудного дельта-ритма ЭЭГ (рис. 9.7). Этот
медленный ритм занимает около 80% всего времени сна. Так
182
Глава 9
называемый медленноволновый сон характеризуется
снижением всех функций организма, отсутствием сновидений и быстрых
движений глаз (БДГ).
Хотя сознание у человека в состоянии медленноволнового
сна угнетено, он может быстро проснуться под действием особо
важных для него раздражителей — например звука шагов,
скрипа открываемой двери или детского плача. Малозначимые
же стимулы такой же или даже большей интенсивности не
оказывают подобного действия: так, люди, живущие близ
железной дороги, спокойно спят под шум проходящих поездов.
Для того чтобы отличить медленноволновый сон от
второй важнейшей формы сна — сна с быстрыми движениями глаз
(БДГ), его иногда называют сном без БДГ. Обычно ритм ЭЭГ
5—6 раз за ночь резко переходит от высокоамплитудного
дельта-ритма, характерного для медленноволнового сна, к
низкоамплитудным бета-подобным волнам, наблюдающимся при сне
с БДГ, или парадоксальном сне (рис. 9.7). Эти эпизоды
высокочастотного ритма на ЭЭГ длятся от 5 до 20 мин и
сопровождаются появлением сновидений и быстрых маятникообразных
движений глаз.
Высокочастотные ритмы ЭЭГ, наблюдающиеся при сне с
БДГ, названы парадоксальными, так как точно такие же
ритмы характерны для человека в активном бодрствующем
состоянии. В то же время в фазе сна с БДГ и сознание, и
восприятие раздражителей резко угнетены. Разбудить человека в этот
момент гораздо труднее, чем во время медленноволнового сна.
Для того чтобы человек проснулся в фазу сна с БДГ,
раздражитель— звук шагов или плач ребенка — должен быть
гораздо сильнее, чем в фазу медленноволнового сна. Предполагают,
что характерные для сна с БДГ «активные»
электроэнцефалографические ритмы (а также сами быстрые движения глаз)
отражают психические процессы, связанные со сновидениями.
ИНДУКЦИЯ СНА
Механизмы, приводящие к возникновению сна, не
выяснены. В настоящее время существуют две основные теории
индукции сна: теория утомления нервных клеток и теория
возбуждения центра сна. Не исключено, что справедлива и та и другая:
сложные нервные процессы, которые мы называем сном, могут
возникать как в результате утомления центров, связанных с
бодрствованием, так и под действием возбуждения центров
сна.
Высшие функции коры головного мозга
183
УТОМЛЕНИЕ НЕРВНЫХ КЛЕТОК
Наступление сна характеризуется переходом
высокочастотного бета-ритма ЭЭГ в низкочастотный дельта-ритм. Этот
переход отражает снижение активности АРС, а крупные дельта-
волны, наблюдающиеся при глубоком сне, возникают в
результате ее торможения. Сходные волны наблюдаются у
подопытных животных после перерезки волокон, соединяющих АРС с
корой. Как уже указывалось, раздражение АРС у нормального
спящего животного приводит к его мгновенному пробуждению.
Считают, что угнетение активности АРС и сопутствующий
этому угнетению медленноволновый сон связаны с утомлением
нейронов АРС. Как в спинном мозгу, так и в других отделах
ЦНС интенсивная деятельность нервных клеток приводит к их
утомлению, т. е. угнетению синаптической передачи и
снижению возбудимости нейронов. Так как ретикулярная формация
образована диффузными мультинейронными сетями с большим
количеством синапсов, вполне возможно, что ее постоянная
интенсивная деятельность может привести к утомлению
образующих ее нейронов и, как следствие, к снижению ее активности.
ЦЕНТРЫ СНА
В мозгу было обнаружено несколько центров, раздражение
которых вызывает сон. Эти центры расположены в базальных
отделах переднего мозга, таламусе и каудальных отделах
ретикулярной формации. При раздражении таламического центра
сна импульсами частотой 8—10 Гц бодрствующее животное
впадает в глубокий медленноволновый сон. Повреждение
центров сна в базальных областях переднего мозга и
каудальных отделах ретикулярной формации сопровождается
бессонницей.
Индукция медленноволнового и парадоксального сна
осуществляется двумя различными нейрохимическими системами
ствола мозга (рис. 9.8). Считается, что медленноволновый сон
обусловлен системой вырабатывающих серотонин нейронов,
находящихся в ядрах области срединного шва. Хирургическое
разрушение этих ядер приводит к постоянной бессоннице,
исчезающей при введении подопытному животному серотонина.
За парадоксальный сон ответственна другая
нейрохимическая система, образованная норадренэргическими нейронами
моста (рис. 9.8). После хирургического разрушения
голубоватого участка (locus ceruleus), расположенного в дорсальной
части моста, фаза парадоксального сна выпадает. Считается, что
эта фаза обусловлена периодическим возбуждением норадре-
нэргических нейронов.
184
Глава 9
ПАМЯТЬ
Одна из важнейших функций нервной системы заключается
в ее способности накапливать и хранить прошлый опыт.
Памятью называются мысли или элементы прошлого опыта,
отложенные в нервной системе в форме, доступной для извлече-
Рис. 9.8. Гипногенные нейрохимические структуры ствола мозга. Норадрен-
эргические нейроны голубоватого места отвечают за парадоксальный сон.
Предполагают, что серотонинэргические нейроны ядер области срединного
шва вызывают медленноволновый сон.
ния. Существуют по крайней мере три вида памяти —
кратковременная, долговременная и промежуточная. Физиологические
основы памяти в настоящее время не известны. Несмотря на
интенсивную нейрофизиологическую разработку проблемы
памяти, у нас еще слишком мало достоверной информации о
механизмах запечатления следов памяти и о тех отделах нервной
системы, где эти следы хранятся. Поэтому дальнейшее
обсуждение памяти и ее механизмов носит предварительный и
гипотетический характер.
В кратковременной памяти информация хранится менее
одной секунды. С ней связана способность кратковременного
запечатления сенсорного опыта. За тот небольшой срок, в
течение которого «действует» кратковременная память, корковые
и подкорковые центры успевают обработать сенсорную
информацию и оценить ее значимость для организма. Если эта инфор-
Высшие функции коры головного мозга
185
мация важна, то она запечатлевается в промежуточной памяти:
в противном случае она быстро забывается.
Промежуточной памятью называют способность к хранению
подробной информации в течение нескольких минут или часов.
Такая память включается, например, при разговоре,
формулировании мыслей и последовательностей слов или чисел. Эта
форма памяти запечатлевает также важные события,
происходившие в последние несколько часов.
В долговременной памяти информация может храниться всю
жизнь. В ней могут откладываться удивительно подробные
обстоятельства — например расположение комнат и обстановка
в доме, в котором прошло детство, или слова песен,
услышанных много лет тому назад. Важные события запечатлеваются
в долговременной памяти особенно живо. (В память автора на
всю жизнь врезались подробности обстановки, время суток,
погода, окружающие люди — словом все обстоятельства того
момента, когда он узнал об убийстве Джона Кеннеди.)
МЕХАНИЗМЫ ЗАПОМИНАНИЯ
В настоящее время существуют две теории формирования
долговременной памяти — гипотеза изменения межнейронных
связей и гипотеза образования специфических макромолекул
(нуклеиновых кислот и белков).
Эти теории не являются взаимоисключающими: возможно,
что для изменений в области синапсов необходим синтез
специфических макромолекул, и, наоборот, этот синтез может
вызывать перестройку синаптических структур.
Повторные возбуждения в нейронных цепях способны
привести к изменениям входящих в их состав синапсов. Эти
изменения могут заключаться в увеличении (уменьшении) размеров
синапсов или образовании (разрушении) новых (старых)
контактов. Большое количество экспериментальных данных
свидетельствует о том, что длительное проведение возбуждения в
синапсе приводит к возрастанию постсинаптических
потенциалов. Эта потенциация обусловлена увеличенным выходом
медиатора из пресинаптических терминалей.
Многочисленные экспериментальные данные подтверждают,
что угнетение синтеза РНК или белков блокирует запечатле-
цие информации в форме долговременной памяти. При
обучении синтез белков и РНК усиливается. Некоторые
нейрофизиологи выдвигают гипотезу о хранении памяти в виде
специфической последовательности аминокислот, образующей
особую белковую энграмму (или специфической
последовательности нуклеотидов, образующей рибонуклеиновую энграмму).
Однако большинство исследователей считают, что связь между
186 Глава 9
хранением следов памяти и синтезом белков и РНК
объясняется важностью этого синтеза для роста и формирования
синапсов.
ЛОКАЛИЗАЦИЯ СЛЕДОВ ПАМЯТИ
В запечатлении и извлечении следов памяти участвуют
тысячи нейронов коры головного мозга, лимбической системы, та-
ламуса и других нервных центров. Эти следы, как известно,
распределены диффузно, но в хранении и извлечении памяти
особую роль играют два отдела коры — гиппокамп и височная
доля неокортекса.
Для образования долговременной памяти требуется
несколько часов. Процесс запечатления следов памяти легко может
быть нарушен — например электрошоком, охлаждением мозга
или наркозом. Особенно четко эти нарушения выявляются,
если воздействовать на гиппокамп. Больные с двусторонними
поражениями гиппокампа не способны к длительному
запоминанию, хотя опыт, запечатленный до поражения гиппокампа, не
страдает.
Деятельность нейронов височной доли тесно связана с
хранением и извлечением следов долговременной памяти. При
электрическом раздражении височных долей возникают живые
картины из прошлого — например занятия в школе или
просмотренный кинофильм. Эти картины очень ярки: так, если у
женщины возникают воспоминания о родах, то она как бы
снова переживает все обстоятельства этого события — разговор
врача с акушеркой, обстановку в родильной комнате,
возбуждение и боль.
Несмотря на то что височной доле приписывают функцию
извлечения следов памяти, ее не считают хранилищем этих
следов. Большинство нейрофизиологов полагают, что память
обусловлена деятельностью большого количества нейронов,
локализованных диффузно в коре головного мозга, а также в
таких подкорковых образованиях, как лимбическая система,
таламус и гипоталамус.
ЛИТЕРАТУРА
Agranoff В. W. (1967). Memory and protein synthesis, Sci. Amer., 216, 115.
Atkinson R. C, Shiffrin R. M. (1971). The control of short-term memory, Sci.
Amer., 225, 82.
Buser P. (1976). Higher functions of the nervous system, Ann. Rev. Physiol.,
38, 217.
Eccles J. C. (1977). The understanding of the brain, McGraw-Hill, New York,
Chapt. 6.
Eyzaguirre C, Fidone S. J. (1975). Physiology of the nervous system, 2nd edn.,
Year Book Medical, Chicago, Chapt. 24.
Высшие функции коры головного мозга
187
Gazzaniga M. S. (1967). The split brain in man, Sci. Amer., 217, 24.
Harris A. J. (1974). Inductive functions of the nervous system, Ann. Rev.
Physiol., 36, 251.
Kandel E. R. (1977). Neuronal plasticity and the modification of behavior. In:
Handbook of physiology: cellular biology of neurons, American
Physiological Society, Bethesda, Maryland p. 1137.
Luria A. R. (1970). The functional organization of the brain, Sci. Amer., 222, 66.
Mountcastle V. B. (1974). Medical physiology. Bosby, St. Louis, Chaps. 8 and 19.
Penfield W. (1958). The excitable cortex in conscious man, CC Thomas,
Springfield, Illinois.
Sperry R. W. (1974). Lateral specialization in the surgically separated
hemispheres. In: Schmitt F. O. and Worden I. G. (eds.), The neurosciences, MIT Press,
Cambridge, p. 1.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. В чем состоят функциональные различия между
проекционными и ассоциативными областями коры?
2. Какова функция мозолистого тела?
3. Нарисуйте схему соединений между главными нейронами
коры головного мозга. Сравните влияния специфических и
неспецифических структур таламуса на функцию этих
нейронов.
4. Опишите изменения ЭЭГ во время перехода ко сну.
5. Что такое сон с БДГ (парадоксальный)? В чем его отличие
от сна без БДГ («ортодоксального»)?
6. Каковы нейрохимические системы, обусловливающие
парадоксальный и «ортодоксальный» сон?
7. Перечислите три вида памяти.
8. Расскажите о современных теориях образования и хранения
следов памяти.
Глава 10
НЕРВНАЯ РЕГУЛЯЦИЯ ФУНКЦИЙ
ВНУТРЕННИХ ОРГАНОВ
ЦЕНТРЫ РЕГУЛЯЦИИ ФУНКЦИИ
Функции внутренних органов регулируются нервными
центрами продолговатого мозга, гипоталамуса и лимбической
системы, импульсы из которых доходят до внутренних органов
через волокна и узлы вегетативной (автономной) нервной системы
(АНС). Все эти структуры участвуют в поддержании
постоянства внутренней среды организма и влияют на работу всех его
систем — сердечно-сосудистой, эндокринной, пищеварительной,
выделительной, половой, дыхательной; они влияют также на
обмен веществ.
Вегетативная нервная система регулирует как сокращения
висцеральных мышц (сердечной и гладких), так и деятельность
желез. Эта система является непроизвольной, и ей часто
противопоставляют «произвольную» соматическую нервную систему,
управляющую сокращениями скелетных мышц. Регуляторные
влияния соматической нервной системы подчинены сознанию.
К ней относятся главные чувствительные и двигательные пути
и их центры, кора головного мозга, базальные ганглии и
мозжечок.
Различные отделы вегетативной нервной системы — лимби-
ческая система, гипоталамус, продолговатый мозг и АНС —
отвечают за регуляцию деятельности внутренних органов на
разных уровнях. Лимбическая система, образованная
совокупностью нескольких структур переднего мозга, участвует в
осуществлении сложных поведенческих реакций — пищевого,
родительского, полового и территориального поведения. Главным
образованием, поддерживающим постоянство внутренней
среды организма, служит гипоталамус, в котором расположены
центры регуляции температуры тела и водного баланса, а
также пищевого, полового и эмоционального поведения.
Гипоталамус играет также важную роль в регуляции эндокринных
функций, так как именно он управляет деятельностью передней и
задней долей гипофиза. В продолговатом мозгу находятся
регуляторные центры сердечно-сосудистой и дыхательной систем.
В АНС, главной задачей которой является проведение
импульсов от центральной нервной системы (ЦНС) к внутренним
органам, расположены эфферентные вегетативные нейроны.
Глаз
Слезная железа
Слюнные железы
Глотка, трахея
Бронхи; легкие
Сердце
Желудок
Тонкий кишечник
Печень, желчный;
пузырь
Поджелудочная
железа
Мозговой слой
надпочечника
Толстый кишечник
Прямая кишка
Почки
Мочевой пузырь
Внутренние
половые органы
Наружные
половые органы
К потовым железам
и кровеносным
сосудам кожи
Рис. 10.1. Организация автономной нервной системы (АНС).
189
190
Глава 10
АНС подразделяется морфологически и функционально на два
отдела — парасимпатический и симпатический (рис. 10.1).
Парасимпатический отдел имеет краниальную и сакральную
части. Нейроны его расположены в вегетативных ядрах III, VII,
IX и X черепно-мозговых нервов и в крестцовых сегментах
спинного мозга. Тела нейронов симпатического отдела
локализуются в грудных и первых трех поясничных сегментах.
Большинство внутренних органов имеет как парасимпатическую, так и
симпатическую иннервацию. Между этими отделами обычно
существует антагонизм: под действием парасимпатических
нервов железы или мышцы внутренних органов подвергаются
возбуждению, а под действием симпатических — торможению.
АВТОНОМНАЯ НЕРВНАЯ СИСТЕМА
Сигналы от ЦНС к внутренним органам передаются по
эфферентному пути, состоящему из двух нейронов — преганглио-
нарного и постганглионарного (рис. 10.2). Эти два нейрона
образуют двигательную часть вегетативной рефлекторной дуги.
Тела преганглионарных нейронов расположены либо в боковых
рогах спинного мозга, либо в вегетативных ядрах
черепно-мозговых нервов. По преганглионарным нейронам импульсы
передаются от ЦНС к постганглионарным нервным клеткам,
расположенным за пределами центральной нервной системы в
вегетативных ганглиях. Постганглионарные нейроны — последнее
звено вегетативной рефлекторной дуги. Их аксоны
оканчиваются на соответствующих эффекторных внутренних органах.
СИМПАТИЧЕСКИЙ ОТДЕЛ
Нейроны симпатической нервной системы расположены в
16 сегментах спинного мозга — начиная от первого грудного до
четвертого поясничного включительно (рис. 10.1). К ним
относятся как преганглионарные нейроны боковых рогов спинного
мозга, так и постганглионарные нейроны симпатических
ганглиев. Большая часть этих ганглиев соединена в цепочку,
находящуюся вблизи спинного мозга. Постганглионарными
нейронами этой цепочки осуществляется симпатическая иннервация
головы (глаз, слюнных желез и сосудов), органов грудной
полости (бронхов и сердца) и кожи (сосудов и потовых желез).
Большинство органов брюшной полости (желудок, тонкий и
толстый кишечник, печень, мочевой пузырь и половые органы)
иннервируются постганглионарными нейронами,
расположенными в одном из трех коллатеральных симпатических ганглиев —
чревном либо верхнем или нижнем брыжеечном. Все эти
ганглии находятся в брюшной полости.
Ядра блуждающего нерва
Парасимпатическая дуга
Блуждающий
Чувствительное
волокно
Продолговатый
мозг
Преганглионарнь
нейрон
Постганглионарный'
нейрон
Чувствительное волокно Симпатическая дуга
инно мозговой
нерв /
К потовым железам
и кровеносным
сосудам кожи
Постганглионарные
нейроны / иг
Преганглионарные ' — /
нейроны
Грудной отдел
спинного мозга
Чувствительное волокно
Симпатический
ганглий
Парасимпатическая дуга
Преганглионарный
нейрон
Крестцовый, отдел
спинного мозга
Тазовый нерв
Постганглионарный
нейрон
Рис. 10.2. Вегетативные рефлекторные дуги. Постганглионарные нейроны
расположены либо в вегетативных ганглиях (симпатические и часть
парасимпатических), либо во внутренних органах (большинство парасимпатических
нейронов).
191
192
Глава 10
ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ
Парасимпатический отдел вегетативной нервной системы
состоит из двух частей — краниальной и сакральной (см.
рис. 10.1). Преганглионарные парасимпатические нейроны
расположены как в вегетативных ядрах III, VII, IX и X черепно-
мозговых нервов, так и в боковых рогах крестцового отдела
спинного мозга. Постганглионарные же нейроны
локализуются вблизи иннервируемого органа, а чаще — в нем самом;
отростки их очень коротки.
Главным путем передачи парасимпатических влияний
служат волокна блуждающего нерва, который иннервирует
большинство органов грудной и брюшной полости, в том числе
сердце, трахею, бронхи, тонкий кишечник и печень.
Парасимпатическая иннервация структур головы (слюнных и слезных
желез, кровеносных сосудов и пр.) осуществляется волокнами
других черепно-мозговых нервов — III, VII и IX.
Парасимпатические волокна сакральной части иннервируют прямую кишку,
почки, мочевой пузырь и половые органы.
МЕДИАТОРЫ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Два главных медиатора вегетативных нейронов — это
ацетилхолин и норадреналин. Нейроны, выделяющие ацетилхолин,
называются холинэргическими, а норадреналин — адренэргиче-
скими. Все преганглионарные нейроны (как симпатические, так
и парасимпатические), парасимпатические постганглионарные
нейроны, а также симпатические постганглионарные нейроны,
иннервирующие кровеносные сосуды кожи, скелетных мышц
и потовых желез, являются холинэргическими.
Большинство же постганглионарных симпатических нейронов ад-
ренэргические, так как их окончания высвобождают
норадреналин.
Норадреналин и его производное адреналин выделяются не
только большей частью постанглионарных симпатических
нейронов, но также секреторными клетками мозгового слоя
надпочечников. Эти клетки, иннервируемые преганглионарными
симпатическими волокнами, родственны постганглионарным
симпатическим нейронам и сходны с ними по эмбриогенезу.
Действие медиаторов на эффекторные органы определяется
свойствами постсинаптических мембран клеток этих органов.
И ацетилхолин, и норадреналин могут в зависимости от органа
оказывать на него как возбуждающий, так и тормозный
эффект. Если какой-либо орган иннервируется и симпатическими,
и парасимпатическими нервами, то влияния ацетилхолина и
норадреналина обычно бывают противоположными.
Нервная регуляция функций внутренних органов 193
Общее возбуждение симпатической нервной системы
помогает организму справиться со стрессовой ситуацией.
Симпатическая система ускоряет ритм сердца, повышает артериальное
давление, кровоток в мышцах и уровень глюкозы в крови; на
деятельность же пищеварительной системы она оказывает
угнетающее влияние. Все эти эффекты помогают организму
справиться со стрессом. Парасимпатическая система
отвечает за восстановление ресурсов; именно на это направлены
основные эффекты парасимпатических нервов — снижение
частоты сердечных сокращений и артериального давления и
стимуляция функции пищеварительной системы.
На отдельные органы парасимпатические и симпатические
нервы оказывают сложные и разнообразные влияния (табл.
10.1). В следующем разделе будут кратко рассмотрены
влияния этих нервов на главные системы организма; более
подробно эти влияния описаны в главах, посвященных отдельным
системам.
ГЛАЗ
Парасимпатические волокна иннервируют сфинктер зрачка,
цилиарную мышцу хрусталика и слезные железы.
Возбуждение этих волокон сопровождается: 1) сужением зрачка и
вследствие этого уменьшением падающего на сетчатку
светового потока (зрачковый рефлекс на свет), 2) фокусировкой
хрусталика на близко расположенный предмет
(аккомодационный рефлекс) и 3) секрецией слезной жидкости. Мышца,
расширяющая зрачок, иннервируется симпатическими волокнами,
и их возбуждение при стрессе приводит к расширению зрачка.
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Влияние парасимпатических нервов на желудочно-кишечный
тракт сводится к усилению пищеварения. При возбуждении
этих нервов моторика пищеварительных органов и секреторная
активность входящих в их состав железистых клеток
усиливается, а сфинктеры расслабляются. Симпатическая же система
оказывает на пищеварительный тракт угнетающее щдействие:
уменьшает моторную и секреторную активность и усиливает
сокращения сфинктеров. Под влиянием симпатических нервов
уменьшается также кровоснабжение органов пищеварительной
системы.
СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Симпатические нервы увеличивают частоту и силу
сокращений сердца, в результате чего увеличивается минутный объем.
При активации симпатической системы наступает перераспре-
13—1705
Таблица 10.1. Влияние вегетативных нервов на внутренние органы3
Орган
Действие симпатической
системы
Действие
парасимпатической системы
Глаз
Зрачок
Цилиарная мышца
Слезные железы
Слюнные железы
Легкие
Бронхи
Бронхиальные
железы
Сердце
Мышца
Кровеносные сосуды
Коронарные
Кожные
Скелетных мышц
Брюшной полости
Желудок
Моторика и тонус
Сфинктеры
Секреция
Кишечник
Моторика и тонус
Сфинктеры
Секреция
Поджелудочная железа
Печень
Желчные пути
Мочевой пузырь
Мышца,
выталкивающая мочу
Мочепузырный
треугольник и
сфинктер
Моторика и тонус
мочеточников
Половые органы
Мозговой слой
надпочечников
Основной обмен
Расширение
Расслабление для фикса
ции отдаленных
предметов
Выделение вязкого
секрета
Расширение
Торможение
Ускорение
Увеличение силы
сокращения
Расширение
Сужение
Торможение
Сокращение
Торможение
Торможение
Сокращение
Торможение
Высвобождение глюкозы
Расслабление
Расслабление
Сокращение
Усиление
Эякуляция
Секреция адреналина
Увеличение
Сужение
Сокращение для
фиксации близко
расположенных предметов
Возбуждение секреции
Выделение водянистого
секрета
Сужение
Возбуждение секреции
Замедление
Уменьшение силы
сокращений
Усиление
Расслабление
Возбуждение
Усиление
Расслабление
Возбуждение
Возбуждение секреции
Сокращение
Сокращение
Расслабление
Эрекция
а) Прочерком обозначено отсутствие влияния.
194
Нервная регуляция функций внутренних органов 195
деление кровотока от внутренних органов и поверхности тела
к скелетным мышцам. Под влиянием симпатических нервов
кровеносные сосуды органов брюшной полости и кожи
суживаются, а скелетных мышц—расширяются. Парасимпатические
же нервы вызывают снижение частоты и силы сокращений
сердца. На кровоток в сосудах органов брюшной полости,
кожи и скелетных мышц эти нервы оказывают минимальное
влияние.
ЧУВСТВИТЕЛЬНЫЕ ВЕГЕТАТИВНЫЕ ПУТИ
Большинство рецепторов внутренних органов реагирует на
ноцицептивные стимулы или изменения давления. Волокна от
них идут к ЦНС в составе симпатических и парасимпатических
нервов и заканчиваются либо в задних рогах спинного мозга,
либо в чувствительных вегетативных ядрах черепно-мозговых
нервов.
Хотя на уровне ЦНС область нанесения кожного
раздражения может быть определена очень точно, локализация
воздействия, нанесенного на внутренние органы, зачастую
невозможна. Болезненное состояние этих органов может вызвать
болевые ощущения в самых отдаленных областях тела. Так, при
нарушении коронарного кровообращения возникают очень
резкие боли в области шеи, плеча и руки; раздражение болевых
рецепторов червеобразного отростка, расположенного в
брюшной полости, сопровождается сильными болями в нижних
отделах грудной клетки; при ноцицептивных раздражениях
желудка наблюдается «жжение в сердце», воспринимаемое как
боли в груди.
Болевые ощущения в какой-либо области при
ноцицептивных воздействиях на другую область называются
отраженными болями; возникновение их объясняется особым ходом
большинства чувствительных вегетативных волокон в спинном
мозгу.
Большая часть вегетативных афферентных волокон идет от
внутренних органов к спинному мозгу в составе симпатических
нервов. Затем они входят в симпатический ствол, поднимаются
по нему на несколько сегментов и лишь после этого вступают
в спинной мозг. В результате такие волокна входят в спинной
мозг на несколько сегментов выше, чем расположены
соответствующие рецепторы внутренних органов. Из-за сегментарного
строения спинного мозга раздражение этих рецепторов
проецируется в область иннервации того сегмента, в который
вступают упомянутые выше волокна, а так как этот сегмент
расположен выше, чем соответствующий внутренний орган,
раздражение этого органа также может проецироваться на несколько
сегментов выше его истинного положения.
13*
196
Глава 10
ЦЕНТРЫ РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ
Главные нервные центры, регулирующие деятельность
сердечно-сосудистой и дыхательной систем, расположены в про-
долговатом мозгу (рис. 10.3). Через эти жизненно важные
центры осуществляется большинство сердечно-сосудистых и
дыхательных рефлексов. В них поступают сигналы от бароре-
Рис. 10.3. Главные вегетативные центры ствола мозга.
цепторов и хеморецепторов систем кровообращения и дыхания.
Некоторые клетки продолговатого мозга реагируют
непосредственно на уровень С02 и Н+ в крови. Сердечно-сосудистые и
дыхательные рефлексы, замыкающиеся на уровне
соответствующих центров продолговатого мозга, имеют первостепенное
значение для выживания организма; они описаны в главах,
посвященных регуляции кровообращения и дыхания.
Кроме перечисленных выше центров, в продолговатом
мозгу расположены также центры кашлевого, чихательного,
глотательного и рвотного рефлексов, для осуществления которых
необходима координированная деятельность глотки, верхних
дыхательных путей и верхних отделов пищеварительного
тракта.
ГИПОТАЛАМУС
Гипоталамус — это главный центр регуляции вегетативных
функций. Он непосредственно управляет как деятельностью
АНС, так и секреторной активностью передней и задней долей
гипофиза. В нем расположены центры регуляции температуры
Нервная регуляция функций внутренних органов
\97
тела, потребления пищи, водного баланса, полового и
эмоционального поведения. Благодаря связям с жизненными
центрами продолговатого мозга гипоталамус влияет также на
сердечно-сосудистые и дыхательные рефлексы.
Анатомически гипоталамус состоит из скопления ядер,
расположенных в промежуточном мозгу вентральнее таламуса
(рис. 10.3). Его можно разделить на два отдела (рис. 10.4).
Паравентрикулярное
ядро
Переднее ядро
ХЛ // )Г~\ Г
Заднее
ядро
/J-JI lLy<( / \
Преоптическое
ядро
Сосцевидное
тело
Супраоптическое
ядро
нтро^О^'Иальное
ядро
Гипофиз
Рис. 10.4. Главные ядра гипоталамуса.
В переднем отделе находятся передние, паравентрикулярное,
преоптическое и супраоптическое ядра, в заднем — заднее и
вентромедиальное, а также сосцевидные тела.
Основная часть импульсации поступает в гипоталамус от
лимбической системы таламуса и ретикулярной формации. От
гипоталамуса сигналы направляются к лимбической системе
и —через ретикулярную формацию —к спинному мозгу.
Гипоталамус не только посылает нервные сигналы, но также
вырабатывает гормоны задней доли гипофиза и рилизинг-факторы,
управляющие деятельностью передней доли гипофиза.
ТЕМПЕРАТУРА ТЕЛА
В гипоталамусе расположены центры, управляющие
теплообразованием, теплоотдачей и сохранением тепла (см. гл. 19).
При электрическом раздражении переднего отдела гипоталаму-
198
Глава 10
са возрастает теплоотдача, а при возбуждении заднего —
теплообразование.
Все эти воздействия, направленные на регуляцию
температуры, возникают при непосредственном раздражении
термочувствительных нейронов гипоталамуса. Некоторые из них
возбуждаются при повышении, а другие — при понижении
температуры крови. Раздражение рецепторов, чувствительных к теплу,
приводит к возбуждению центра теплоотдачи в переднем
гипоталамусе; стимуляция же холодовых рецепторов
сопровождается активацией влияний заднего гипоталамуса, усиливающих
теплообразование и способствующих сохранению тепла. В эти
центры поступают также сигналы от кожных терморецепторов.
Подробнее терморегуляция обсуждается в гл. 19.
ПОТРЕБЛЕНИЕ ПИЩИ
В гипоталамусе расположены два пищевых центра — центр
голода и центр насыщения. При электрическом раздражении
латеральных отделов гипоталамуса у подопытного животного
возникает пищевое поведение. При разрушении центра голода
животное отказывается от еды и, если его не кормить насильно,
погибает от истощения. В вентромедиальном ядре расположен
центр насыщения, являющийся антагонистическим по
отношению к центру голода: его разрушение сопровождается гипер-
фагией (постоянным потреблением пищи) (см. также гл. 21).
В центры голода и насыщения поступает импульсация от
нескольких различных типов рецепторов. К ним относятся глю-
корецепторы, рецепторы растяжения желудка (механорецепто-
ры) и терморецепторы. Глюкорецепторы — это гипоталамиче-
ские нейроны, чувствительные к изменениям уровня глюкозы в
крови. Механорецепторы стенок желудка воспринимают
изменения объема желудочного содержимого. Импульсация,
поступающая к центрам насыщения и голода от терморецепторов,
также имеет большое значение. У голодавшего животного
температура и интенсивность обмена низки, причем температура
особенно • снижается непосредственно перед потреблением
пищи. После кормления эти два показателя повышаются.
ВОДНЫЙ БАЛАНС
Раздражение дорсального гипоталамуса у собаки или козы
вызывает жажду. После разрушения этого центра питье
полностью прекращается и животное погибает от обезвоживания.
Считается, что основная информация поступает в центр питья
от осморецепторов гипоталамуса — нейронов, возбуждающихся
при изменении осмотического давления крови.
Нервная регуляция функций внутренних органов 199
ЭМОЦИОНАЛЬНОЕ И ПОЛОВОЕ ПОВЕДЕНИЕ
При раздражении вентромедиальных ядер возникает бурный
приступ ярости, часто сопровождающийся немотивированным
нападением на какой-нибудь расположенный поблизости
предмет. Напротив, животные с разрушенными вентромедиальными
ядрами становятся совершенно ручными.
Электрическое раздражение некоторых областей
гипоталамуса влияет на половое поведение. После введения в
гипоталамус тестостерона у кастрированных животных половое
поведение нормализуется. При поражениях переднего гипоталамуса
половая активность угнетается как у самцов, так и у самок.
Введение эстрогенов в гипоталамус животных с удаленными
яичниками приводит к характерным для периода течки
поведенческим реакциям, в частности к активному поиску и
завлеканию партнера.
ПЕРЕДНЯЯ И ЗАДНЯЯ ДОЛИ ГИПОФИЗА
Гипоталамус — главный нервный центр, управляющий
деятельностью гипофиза и регулирующий секрецию гормонов
обеими его долями.
Передняя доля гипофиза сообщается с гипоталамусом
посредством сосудистой сети, образующей гипоталамо-гипофизар-
ную портальную систему (рис. 10.5). По этим сосудам рили-
зинг-факторы гипоталамуса поступают в гипофиз. Благодаря
влияниям на секрецию гормонов передней доли гипофиза
гипоталамус может регулировать высвобождение половых гормонов
(фолликулостимулирующего, лютеинизирующего и пролакти-
на), а также тиреотропного, соматотропного и адренокортико-
тропного (более подробно рилизинг-факторы и гормоны
передней доли гипофиза обсуждаются в гл. 25). В отличие от
передней доли гипофиза задняя доля иннервируется нейронами су-
праоптического и паравентрикулярного ядер гипоталамуса
(рис. 10.5). По отросткам этих нейронов в заднюю долю
гипофиза из гипоталамуса поступают гормоны окситоцин и вазо-
прессин. Подробнее функции задней доли гипофиза
обсуждаются в гл. 25.
ЛИМБИЧЕСКАЯ СИСТЕМА
Лимбическая система, содержащая высшие центры
интеграции деятельности внутренних органов, образована несколькими
взаимосвязанными корковыми и подкорковыми отделами
конечного мозга. Между этой системой и гипоталамусом
имеются тесные анатомические и функциональные связи; некоторые
авторы даже относят гипоталамус к лимбической системе.
200
Глава 10
Паравентрикулярное/^"у
ядро /
Супраоптическое
ядро
/
Гипоталамо-
гипофизарная
портальная
система
Передняя
доля
гипофиза.
Вена
Ядра
гипоталамуса
Задняя -n
доля
гипофиза
Рис. 10.5. Пути, соединяющие гипоталамус и гипофиз. Рилизинг-факторы
переносятся от гипоталамуса к передней доле гипофиза по гипоталамо-гипофи-
зарной портальной системе. Волокна нейросекреторных нейронов паравентри-
кулярного и супраоптического ядер гипоталамуса оканчиваются в задней
доле гипофиза.
Лимбическая система является высшим центром регуляции
деятельности автономной нервной системы и гипофиза. В ней
осуществляется интеграция трех видов информации: 1) о
деятельности внутренних органов; 2) обонятельная и 3) о
деятельности чувствительных и двигательных ассоциативных зон
коры больших полушарий. Лимбическая система отвечает за
мотивацию и выработку сложных поведенческих актов,
успешное выполнение которых требует координации вегетативных и
соматических рефлексов. В настоящее время конкретных
сведений о роли корковых и подкорковых образований лимбиче-
ской системы мало. Это связано с тем, что их нейроанатомиче-
ская организация и регулируемые ими поведенческие акты
очень сложны.
Нервная регуляция функций внутренних органов 201
АНАТОМИЯ
Лимбическая кора расположена в виде кольца в области4
нижних отделов коры головного мозга, окружающих верхнюю
часть ствола мозга (рис. 10.6). В состав ее входят
обонятельные области коры и гиппокамп. Лимбическая кора тесно
связана с грушевидной корой и фронтоорбитальными
ассоциативными областями коры.
Лимбическая кора образована палеокортексом —
«примитивной» корой, содержащей лишь три слоя нейронов (чувствитель-
Грушевидная кора Миндалина
Рис. 10.6. Лимбическая система.
ные, двигательные и ассоциативные зоны коры больших
полушарий относятся к неокортексу, содержащему шесть слоев
нейронов).
К основным связанным с лимбической системой
подкорковым образованиям относятся миндалина и перегородка
(конечный мозг), а также переднее ядро таламуса и гипоталамуса
(промежуточный мозг) (рис. 10.6). Лимбическая система
соединена также с обонятельной луковицей.
Прямых связей между лимбической системой и
остальными областями коры мало (возможно, поэтому так трудно
сдерживать свои эмоции). Импульсы от чувствительных,
двигательных и ассоциативных отделов коры поступают в лимбиче-
скую систему через передние ядра таламуса. Единственное
исключение — это связи между лимбической системой и корой
лобных долей. К важнейшим сигналам, поступающим в лимби-
ческую систему, относятся также обонятельные (от обонятель-
202
Глава 10
ных луковиц) и висцеросенсорные (от гипоталамуса и
вегетативных ядер ствола мозга).
От лимбической системы импульсы направляются прежде
всего к вегетативным центрам гипоталамуса и через него к
гипофизу, а также к автономной нервной системе. Благодаря
своим связям с базальными ганглиями, передними отделами
таламуса и ретикулярной формацией лимбическая система
может влиять на деятельность скелетных мышц.
ПОВЕДЕНЧЕСКИЕ АКТЫ
При нанесении электрических раздражений на различные
области лимбической системы возникают самые различные
сложные поведенческие акты, связанные с пищевым и
половым поведением, нападением и бегством; наблюдаются также
сопровождающие эти акты эмоции удовольствия, ярости,
отвращения и страха.
Активность лимбической системы тесно связана с
гипоталамусом. Есть данные о том, что различные отделы лимбической
системы регулируют секрецию рилизинг-факторов
гипоталамусом. В настоящее время мало известно о функциях,
выполняемых лимбической системой или приписываемых ей.
Поведенческие акты, протекающие при участии лимбической
системы, связаны с сохранением особи или вида. Данные,
полученные при раздражении или разрушении различных
областей лимбической системы, свидетельствуют о том, что
миндалина и грушевидная кора отвечают прежде всего за
сохранение особи, тогда как перегородка и гиппокамп участвуют в
выработке поведения, направленного на сохранение вида.
При раздражении миндалины возникают поведенческие
акты, связанные с потреблением пищи — жевание, глотание и сли-
зывание. Стимуляция других отделов миндалины приводит к
яростному нападению на животных или экспериментатора. При
поражениях миндалины животное становится ручным,
прожорливым и гиперсексуальным. При раздражении перегородки
агрессивное животное превращается в ручное; при поражениях
же этого отдела животное становится яростным и агрессивным.
Поражения гиппокампа сопровождаются приступами ярости.
При раздражении многих областей перегородки возникает
половое и родительское поведение.
ЛИТЕРАТУРА
Axelsson J. (1971). Catecholamine functions, Ann. Rev. Physiol., 33, 1.
Barchas J. D., Akil H., Elliott G., et al. (1978). Behavioral neurochemistry: neu-
roregulators and behavioral states, Science, 200, 964.
Нервная регуляция функций внутренних органов 203
Blackwell R. E.f Guillemin R. (1973). Hypothalamic control of adenohypophysi-
cal secretions, Ann. Rev. Physiol., 35, 357.
DiCara L. V. (1970). Learning in the autonomic nervous system, Sci. Amer.,
222, 30.
Eyzaguirre C, Fidone S. J. (1975). Physiology of the nervous system, 2nd edn.,
Year Book Medical Chicago, Chaps, 22 and 23.
Guillemin R., Burgus R. (1972). The hormones of the hypothalamus, Sci. Amer.,
227, 24.
Schally A. V., Arimura A., Kastin A. J. (1973). Hypothalamic regulatory
hormones, Science, 179, 341.
Snyder S. H. (1977). Opiate receptors and internal opiates, Sci. Amer., 236, 44..
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Что такое вегетативный нейрон? По каким признакам
определяют его принадлежность к парасимпатической или
симпатической системе?
2. Опишите влияние парасимпатической системы на желудочно-
кишечный тракт. Как действует на него симпатическая
система?
3. Каковы анатомические основы отраженных болей?
4. Какие регуляторные центры расположены в продолговатом
мозгу? В гипоталамусе?
5. Каким образом гипоталамус регулирует секреторную
деятельность гипофиза?
6. Расскажите о роли лимбической системы в регуляции
деятельности внутренних органов.
Глава 11
МЫШЦЫ
Существуют три типа мышц — скелетные, гладкие и
сердечная. Их основные свойства, локализация и анатомическое
строение обсуждались в гл. 1.
СКЕЛЕТНЫЕ МЫШЦЫ
Скелетные мышцы состоят из отдельных многоядерных
волокон, обладающих поперечной исчерченностью (см. рис. 1.8
и ИЛ). Эта исчерченность образована чередованием темных
(анизотропных) А-дисков и светлых (изотропных) 1-дисков
(рис. 11.1 и 11.2). Через середину I-диска проходит Z-линия;
две соседние Z-линии ограничивают саркомер, структурно-
функциональную сократительную единицу. При электронной
микроскопии изолированных волокон видно, что в составе А-ди-
ска имеется более светлый участок (так называемая Н-зона),
а в центре этот диск пересекает темная полоска — М-линия.
САРКОПЛАЗМАТИЧЕСКИИ РЕТИКУЛУМ И Т-ТРУБОЧКИ
Волокно скелетной мышцы имеет сарколемму и состоит из
миофибрилл. Саркоплазматический ретикулум и сеть
поперечных Т-трубочек образуют вокруг миофибрилл как бы решетку,
пронизанную отдельными фибриллами (рис. 11.2). Т-трубочки
расположены перпендикулярно фибриллам, а
саркоплазматический ретикулум — параллельно. Участки соприкосновения
Т-трубочек и саркоплазматнческого ретикулума состоят из
небольшой трубочки в центре и двух цистерн ретикулума по
бокам — так называемые триады. В скелетной мышце триады
прилегают к каждому участку, где перекрываются актиновые
и миозиновые нити (А- и I-диски). На каждый саркомер
приходятся две триады.
Главная функция Т-системы состоит в быстрой передаче
потенциала действия от клеточной мембраны к миофибриллам.
Саркоплазматический ретикулум обеспечивает также
внутриклеточный ток кальция.
Мышцы
205
Мышца
Рис. 11.1. А. Скелетная мышца; мышечные волокна прикреплены к
сухожилиям. Б. Отдельное волокно, состоящее из миофибрилл. В. Отдельная мио-
фибрилла; видно чередование светлых актиновых I-дисков и темных миози-
новых А-дисков. Внутри А-диска видна более светлая Н-зона, в центре
которой имеется темная М-линия. Саркомер ограничен двумя соседними Z-ли-
ниями. Г. Поперечные мостики между толстыми миозиновыми и тонкими ак-
тиновыми нитями.
СОКРАТИТЕЛЬНЫЕ БЕЛКИ
Чередующиеся I- и А-диски состоят соответственно из
молекул актина и миозина, образующих протофибриллы (рис.
11.1, В и 11.2). При электронной микроскопии на поперечном
разрезе А- и I-дисков видно миозиновое волокно (точка
большего размера), окруженное шестью актиновыми (точки
меньшего размера) (рис. 11.3, Д). В мышечном волокне содержатся
также тропомиозин и тропонин; последний состоит из трех
субъединиц — I, Т и С. По современным представлениям тро-
206 Глава 11
Рис. 11.2. Объемное изображение скелетной мышцы. Волокно, состоящее из
фибрилл, окружено саркоплазматическим ретикулумом и Т-трубочками,
открывающимися в области сарколеммы (с любезного разрешения G. Hoyle).
помиозиновые и тропониновые нити, прикрепляющиеся к
гораздо более крупным актиновым нитям, регулируют
взаимодействие толстых и тонких нитей в процессе мышечного
сокращения. Тропонин обладает высоким сродством к кальцию,
и реакция между этим белком и кальцием может быть
пусковым механизмом для мышечного сокращения.
МЕХАНИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
Механическому сокращению мышцы предшествует ее
электрическое возбуждение, вызываемое разрядом двигательных
нейронов в области нервно-мышечного соединения
(двигательной концевой пластинки), т. е. в месте контакта нерва и
мышцы (см. гл. 4 и 7). Здесь высвобождается медиатор аце-
тилхолин, который взаимодействует с постсинаптической
мембраной и вызывает электрическое возбуждение мышцы —
потенциал действия. Под влиянием потенциала действия
высвобождается кальций, запускающий механическое сокращение
(см. также гл. 4) в следующей последовательности;
Мышцы
2)7
Актин
Миозин
Г\ Поперечный
1 [Г мостик I
2 0
• •
• • •
• •
д
Рис. 11.3. Саркоплазматический ретикулум и Т-трубочки. Миозиновые
(толстые) и актиновые (тонкие) нити в состоянии покоя (Л), сокращения (Б) и
растяжения (В). Укорочение мышцы при ее сокращении связано со
скольжением актиновых нитей (Г). Прикрепление поперечных миозиновых
мостиков к нитям актина. Благодаря этим мостикам, большинство которых
участвует в процессе сокращения, актиновые нити скользят по направлению к
центру саркомера, что приводит к укорочению мышцы (2 и 3). Д. Поперечный
разрез через А- и I-диски (электронная микроскопия); видна толстая миози-
новая нить, окруженная шестью тонкими актиновыми.
1. Кальций выходит из цистерн, или пузырьков, саркоплаз-
матического ретикулума и перемещается к толстым и тонким
нитям.
2. Кальций связывается с тропонином.
3. Между миозиновыми и актиновыми нитями образуются
поперечные мостики, и актиновые нити скользят вдоль
миозиновых, что приводит к сокращению мышцы (рис. 11.2 и 11.3).
Основным источником энергии, необходимой для
мышечного сокращения, служит аденозинтрифосфат (АТФ). Энергия
высвобождается в результате реакции: АТФ—^Энергия+
<+АДФ + Фосфат.
Потенциал покоя мембраны мышечного волокна равен
примерно 85 мВ (см. гл. 4), а длительность потенциала действия —
208
Глава 11
около 1—5 мс, т. е. больше, чем у нерва. Скорость проведения
возбуждения в скелетных мышцах составляет 3—5 м/с
(значительно медленнее, чем в миелинизированных нервах).
ТЕОРИЯ «СКОЛЬЗЯЩИХ НИТЕЙ»
В сократительной единице мышц — саркомере — длина
миофибрилл изменяется в результате скольжения актиновых
нитей вдоль миозиновых, но сами нити при этом не
укорачиваются. В соответствии с этим в расслабленной, а тем более
растянутой мышце актиновые нити располагаются дальше от
центра саркомера, и длина саркомера больше (рис. 11.3, Л и
В). В сокращенной же мышце актиновые нити скользят по
направлению к центру саркомера, примыкают друг к другу (рис.
11.3, Б) и могут даже взаимно перекрываться в зависимости от
степени сокращения и укорочения мышцы. Изменение
ориентации миозиновых мостиков и их роль в движении молекул
актина показаны на рис. 11.3, Г.
При изотоническом (происходящем .при постоянной силе)
сокращении мышца укорачивается. При изометрическом
сокращении мышца не может преодолеть приложенную к ней
нагрузку и сократиться, однако она развивает усилие,
напряжение в ней растет и затрачивается энергия. При этом
сохраняется постоянная длина (изометричность) мышечных волокон.
При совершении работы по перемещению груза мышца
обычно сокращается сначала изометрически, затем
изотонически. Максимальный коэффициент полезного действия при
изотонических сокращениях равен приблизительно 25%. Мышцы-
сгибатели типа двуглавой мышцы плеча сокращаются
изотонически, т. е. укорачиваются, а четырехглавая мышца бедра при
стоянии напрягается и сокращается в изометрическом режиме.
ДЛИНА, СИЛА И СКОРОСТЬ СОКРАЩЕНИЯ МЫШЦ
Длина, сила и скорость сокращения — наиболее важные
механические свойства мышц. Существует некая оптимальная
длина мышцы, при которой сокращение максимально. Это
можно показать в опыте по изучению изометрических сокращений
изолированной мышцы, фиксируемых при различных
значениях исходной длины (рис. 11.4). Если исходная длина мышцы
мала, то и усилие, развиваемое ею при сокращении, невелико;
при растяжении ее до определенного уровня (точка 2 на рис.
11.4, Л) это усилие достигает максимального значения. Если
же мышца перерастянута (точка 3), сила ее сокращения вновь
падает. Для скелетных мышц это взаимоотношение между
длиной и силой не имеет большого значения, однако в сердечной
Мышцы
209
мышце оно играет важную роль; именно это взаимоотношение
лежит в основе закона Франка—Старлинга (см. гл. 15).
Увеличение нагрузки на мышцу снижает скорость ее сокращения
(см. рис. 11.4, £ и гл. 15).
РЕАКЦИЯ МЫШЦ НА РАЗДРАЖЕНИЕ
На одиночный стимул мышца отвечает одиночным
сокращением (рис. 11.5,Л). Раздражение, наносимое на мышцу,
характеризуется следующими параметрами: 1) интенсивностью
(В или мВ), 2) длительностью (с или мс), 3) частотой (имп/с).
Длительность единичного мышечного сокращения составляет
примерно 0,1 с. Если на мышцу наносить повторные
раздражения постоянной силы и длительности с частотой, несколько
ниже той, при которой возникает тетанус, то можно наблюдать
феномен лестницы (зубчатый тетанус) (рис. 11.5,5). Это
явление заключается в том, что амплитуда сокращений в
ответ на первые несколько раздражений увеличивается, а затем
Длина
А
Рис. 11.4. Зависимость между силой, напряжением и длиной мышцы. А. При
увеличении длины мышцы до точки 2 напряжение и сила ее сокращения
возрастают, а при дальнейшем растяжении мышцы — падают (3). Б. Зависимость
скорости сокращения мышцы от нагрузки; чем больше нагрузка, тем меньше
скорость сокращения.
устанавливается на постоянном уровне. Полагают, что
феномен лестницы обусловлен увеличением количества кальция,
способного связываться с тропонином.
Электрический ответ мышцы на раздражение (потенциал
действия) характеризуется периодом рефрактерности, когда
мышца не отвечает на раздражение, в механическом же
сокращении скелетной мышцы такого периода нет. Поэтому если на
мышцу наносить повторное раздражение в тот момент, когда
Н—1705
Ill
IV Б
Рис. 11.5. Реакция мышцы на раздражение. А. Одиночное мышечное
сокращение (раздражение в точке /). /—// — латентный период; //—/// — фаза
сокращения; ///—IV — фаза расслабления. Б. Феномен лестницы,
проявляющийся в увеличении амплитуды первых шести сокращений. Производилось
раздражение ритмическими импульсами с частотой, меньшей тетанизирующей, до
появления постоянных по форме и величине волн сокращений (см. текст).
она еще не полностью расслабилась после предшествующего
сокращения, можно наблюдать усиление сокращения, или сумма-
цию. Напряжение, или усилие, развиваемое при суммации,
больше, чем при одиночном сокращении (рис. 11.6). При
повышении частоты раздражений амплитуда суммационного
сокращения увеличивается, однако, если частота недостаточна для
того, чтобы наступил гладкий тетанус, на кривой сокращения
можно различить отдельные зубцы. При тетанизирующих час-
60 ГЦ
Время, с
Рис. 11.6. Суммация одиночных сокращений (увеличение силы или
напряжения) при раздражении частотой 10 и 25 Гц. При действии тетанизирующей
частоты (60 Гц) наблюдается дальнейшее увеличение амплитуды сокращения,
но отдельные волны исчезают (сливаются).
Мышцы
211
тотах (60 Гц) отдельные волны исчезают, и сокращения
сливаются. Суммация наблюдается также при увеличении числа
одновременно сокращающихся моторных единиц.
МЫШЕЧНЫЙ ТОНУС И ФИЗИЧЕСКАЯ НАГРУЗКА
Под тонусом мышцы понимают состояние ее частичного
сокращения, когда мышца напрягается, но не производит
движения. При потере тонуса мышцы становятся вялыми. При
интенсивной физической нагрузке мышцы увеличиваются
(гипертрофируются), что связано с увеличением размеров мышечных
волокон. При неинтенсивной физической нагрузке гипертрофии
мышц не наступает. Сила и размер мышц значительно
увеличиваются в результате коротких статических упражнений.
Скелетные мышцы подразделяются в зависимости от
выполняемой ими функции, т. е. сгибания, разгибания, вращения в
суставе, отведения и приведения. Агонистами называются
мышцы, производящие движения в одном направлении, например
сгибатели руки или ноги; их антагонисты осуществляют
движение в обратном направлении (разгибатели руки или ноги). При
повышении тонуса сгибателя падает тонус соответствующего
разгибателя. Синергистами называются мышцы,
способствующие деятельности агонистов.
БЫСТРЫЕ И МЕДЛЕННЫЕ МЫШЦЫ
Скорость сокращения мышц различна в зависимости от их
функции. Так, икроножная мышца сокращается быстрее, чем
камбаловидная, отвечающая за осуществление медленных
реакций, а глазные мышцы — еще быстрее. Как правило, в
быстрых мышечных волокнах более развит саркоплазматический
ретикулум, что способствует быстрому выбросу кальция, и они
менее богато васкуляризированы. Их называют «белыми»
мышечными волокнами. Медленные мышцы построены из более
мелких волокон. Такие мышцы часто называют «красными» из-
за их красноватой окраски, связанной с высоким содержанием
миоглобина.
Группа мышечных волокон, иннервируемых одним нервным
окончанием, называется моторной единицей. В моторных
единицах небольших быстрых мышц, как правило, меньше
волокон, чем в более крупных медленных мышцах: в некоторых
быстрых мышцах на одну моторную единицу приходится всего
три волокна, тогда как в крупных медленных мышцах это
число достигает 1000.
14*
212 Глава 11
УТОМЛЕНИЕ МЫШЦ И НАРУШЕНИЯ ИХ ФУНКЦИИ
Длительное напряжение мышцы приводит к ее утомлению
и неспособности совершать дальнейшую работу. Поступление
нервных импульсов к мышце может быть нормальным, однако
механическое сокращение мышцы в ответ на эту импульсацию
угнетено вследствие истощения основного источника энергии —
АТФ.
Нервный паралич (например, при полиомиелите) приводит
к утрате способности к сокращению и атрофии мышц. Многие
заболевания центральной нервной системы сопровождаются
тяжелыми нарушениями мышечной деятельности — судорожным
сокращением, тремором и тетанусом, хотя сами мышцы при
этом не изменены (см. гл. 8).
Мышечная дистрофия, заболевание неизвестной этиологии,
характеризуется нарастающей слабостью и утратой способности
к сокращению. При заболевании, известном как myasthenia
gravis, отмечается нарушение нервно-мышечной передачи; при
этом заболевании количество ацетилхолина, выделяющегося в
области двигательной концевой пластинки и действующего на
рецепторы, мало, и передача возбуждения либо нарушена,
либо не осуществляется вовсе.
СЕРДЕЧНАЯ И ГЛАДКИЕ МЫШЦЫ
Сердечная мышца, как и скелетная, имеет поперечную ис-
черченность (см. рис. 1.8); эти два вида мышечной ткани во
многом сходны, однако в миокарде в области Z-линий имеются
участки слияния (переплетения) волокон. В этих участках
образуются вставочные диски. Благодаря этой особенности
сердечная мышца представляет собой сеть волокон, что
способствует более быстрому проведению возбуждения от волокна к
волокну. Деятельность сердечной мышцы осуществляется
непроизвольно. Т-система кардиомиоцитов локализована в области
Z-линий (а не на месте слияния А- и I-дисков, как в скелетной
мышце). В ответ на раздражение сердечная мышца в
соответствии с законом «все или ничего» либо сокращается с
максимальной силой, либо не сокращается вовсе. В отличие от
скелетной мышцы для миокарда характерен период рефрактерно-
сти, во время которого он не реагирует на раздражение. Более
подробно структура и функция сердечной мышцы описаны в
гл. 15.
Строение гладких мышц обсуждалось в гл. 1. Длинные и
тонкие волокна этих мышц лишены поперечной исчерченности,
они короче волокон скелетных мышц и содержат по одному
ядру. Гладкие мышцы присутствуют во внутренних органах и
кровеносных сосудах.
Мышцы
213
Гладкие мышцы разделяются на две основные группы: муль-
тиунитарные и унитарные. Мультиунитарные волокна
функционируют независимо друг от друга, и каждое такое волокно
может иннервироваться отдельным нервным окончанием. Такие
волокна обнаружены в ресничной мышце глаза, мигательной
перепонке и мышечных слоях некоторых крупных сосудов.
К этому типу относятся и мышцы, поднимающие волосы.
Унитарные же волокна настолько тесно переплетены друг с другом,
что их мембраны могут плотно примыкать или сливаться. При
раздражении одного унитарного волокна импульс (потенциал
действия) быстро распространяется на соседние волокна, так
как электрическое сопротивление между ними мало. Такие
мышцы имеются в большинстве органов, в том числе в
пищеварительном тракте, матке и мочеточниках.
ЛИТЕРАТУРА
Gauthier G. F. (1977). In: Weiss L. and Greep R. 0. (eds.), Histology, 4th edn.,
McGraw-Hill, New York, Chap. 7.
Guyton A. C. (1977). Basic human physiology, 2nd edn., Saunders,
Philadelphia, Chaps. 9 and 10.
Hoyle G. (1970). How is muscle turned on and off, Sci. Amer., 222, 84.
Julian F. J., Mos P. L., Sillins M. R. (1978). Mechanism for vertebrate striated
muscle contraction (a review), Circ. Res., 42, 1.
Meiss R. A. (1976). In: Selkurt E. E. (ed.), Phisiology, 4th edn., Little, Brown,
Boston, Chap. 3.
Murray M. M., Weber A. (1974). The cooperative action of muscle and proteins,
Sci. Amer., 230, 58.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Назовите и охарактеризуйте основные типы мышц в
соответствии с особенностями их строения.
2. Подробно раскройте теорию скользящих нитей,
объясняющую мышечное сокращение.
3. Назовите основные сократительные белки и расскажите об
их роли в мышечном сокращении.
4. Дайте определение: а) саркомера, б) саркоплазматического
ретикулума, в) Т-трубочек, г) А-диска, д) I-диска, е) Z-ли-
нии, ж) мышцы-сгнбателя, з) мышцы-разгибателя.
5. Какие мышцы являются произвольными, какие
непроизвольными? Что означают эти термины?
6. В чем сходство и различие между скелетной и сердечной
мышцей?
7. Где и в каких органах имеются скелетные и гладкие мышцы?
8. Дайте определение изотонических и изометрических
сокращений.
9. Чем запускается потенциал действия в скелетных мышцах?
Глава 12
ЖИДКИЕ СРЕДЫ ОРГАНИЗМА.
КРОВООБРАЩЕНИЕ В КАПИЛЛЯРАХ.
ЛИМФООБРАЩЕНИЕ
Жидкие среды организма находятся в различных органах
в виде внутриклеточной,внеклеточной и межклеточной (интер-
стициальной) жидкости. Таким образом, вся жидкость в
организме распределена между внутриклеточным и внеклеточным
пространствами (компартментами), состав которых представлен
в табл. 12.1. Объем внутриклеточной воды составляет в
среднем 70% всей жидкости в организме, варьируя в зависимости
от возраста и количества тканей, не содержащих жира (табл.
12.2). Содержание воды в этих тканях выше, чем в жировой;
ее больше также у новорожденных и растущих детей и
животных, чем у взрослых.
У молодых особей доля внеклеточной жидкости выше. У
человека по сравнению с другими видами объем и соотношение
жидкости во внеклеточных пространствах относительно
постоянны. Способность поддерживать постоянство внутренней
среды организма называется гомеостазом\ это понятие, хотя и
достаточно общее, весьма плодотворно для учения о функциях
организма.
ИЗМЕРЕНИЕ ОБЪЕМА ЖИДКОСТНЫХ ПРОСТРАНСТВ
ОРГАНИЗМА
Это измерение основано на принципе разведения или
распределения определенного объема (V) введенного индикатора:
Количество введенного Количество выделившегося
,г индикатора индикатора
Концентрация индикатора в исследуемой жидкости
Исследуемой жидкостью может быть жидкость любого из
пространств организма — общая вода (внутриклеточная и
внеклеточная), внеклеточная жидкость, плазма крови (методы
измерения плазмы крови подробно описаны в гл. 13). Количество
внутриклеточной жидкости вычисляется путем вычитания
объема внеклеточной жидкости из общего объема воды в
организме. Выбор индикатора зависит от того, объем какого именно
Жидкие среды организма
215
Таблица 12.1. Внутриклеточные и внеклеточные жидкостные пространства*1
Пространство
Объем, л
Общее количество жидкости в организме
Внеклеточное6*
Плазма
Внутриклеточное
а' Приведены цифры для взрослого человека весом 70 кг.
40
15
3
25
б)
В состав внеклеточного пространства входит также лимфа, количество которой
-составляет около 2 л.
Таблица 12.2. Внеклеточная и внутриклеточная жидкости в организме
человека
Пространство
Внеклеточное
Вода плазмы
Межклеточная вода
Внутриклеточное
Всего
Доля общего веса
тела, %
13.1 } 17'5
40,6
58,1
Доля общего
количества воды, %
22,1 } 30'°
70,0
100,0
пространства измеряется. Так, для измерения общего объема
жидкости используются вещества типа антипирина, которые,
будучи введенными в кровоток, проникают как во
внеклеточное, так и во внутриклеточное пространства. Измерив
концентрацию антипирина в крови после его распределения,
вычисляют количество жидкости в организме, необходимое для
разведения его до этой концентрации. Этот метод применим при
условии, что индикатор не выводится с мочой; в противном
случае необходимо вносить соответствующую поправку.
Для определения внеклеточной жидкости необходимо
использовать индикатор, распределяющийся только во
внеклеточном пространстве — тиоцианат натрия, маннитол или
инулин.
СОСТАВ ЖИДКИХ СРЕД ОРГАНИЗМА
В табл. 12.3 перечислены основные компоненты
внутриклеточной, внеклеточной и плазматической жидкости. Тканевая,
или интерстициальная, жидкость по своему составу сходна с
плазмой крови, однако последняя гораздо богаче белками.
216
Глава 12
Таблица 12.3. Главные компоненты жидких сред организма, мэкв/л
Компонент
Na+
К+
Са2+
С1-
ро4з-
НС03-
Mg2 +
Белки
Плазма крови
152
5
5
113
2
27
13
16
Внеклеточная
жидкость
143
4
5
117
2
27
3
2
Внутриклеточная
жидкость
12
157
—
15
113
10
26
74
Внутриклеточная жидкость отличается от внеклеточной в
основном содержанием Na+ и К+: во внеклеточной жидкости
концентрация Na+ высока, а К+— низка, а во внутриклеточной —
наоборот. Ионы Na+ и К+ играют важную роль в процессах
деполяризации и реполяризации клеток.
Лимфа сходна по составу с внеклеточной жидкостью,
отличаясь от нее более высоким (около 2 г%) содержанием
белков.
ОБРАЗОВАНИЕ ТКАНЕВОЙ ЖИДКОСТИ
Тканевая жидкость образуется благодаря переходу
(фильтрации) жидкой части крови из капилляров в ткани. На уровне
венозного конца капилляра происходит обратное всасывание
жидкости.
Рис. 12.1. Схема капилляров мышцы, иллюстрирующая строение микроцирку-
ляторной единицы; ABA — артерио-венозный анастомоз. (Zweifach В. W.
Report from the Josiah Macy jr. Foundation Conference, 1950.)
Жидкие среды организма
217
Анатомия капилляров. На рис. 12.1 представлена схема
капилляра с прилежащими к нему артериолами и венулами.
Подобный комплекс называется микроциркуляторной единицей
(см. также гл. 1 и 14). Истинный капилляр представляет собой
тонкостенную выстланную эндотелием трубку, относительно
проницаемую для воды и низкомолекулярных солей (рис. 12.2),
но не пропускающую белки и некоторые крупные молекулы.
Рис. 12.2. Две эндотелиальные клетки стенки капилляра. Через щели
осуществляется ток жидкости из просвета капилляра и обратно.
Это связано с тем, что такие вещества не могут пройти через
мелкие поры в капиллярной стенке (вероятно, расположенные
у места соединения эндотелиальных клеток). Однако
некоторые капилляры (например, в печени) проницаемы для белков
(см. также гл. 2). Это заставило предположить наличие пор,
достаточно крупных для прохождения таких макромолекул;
однако при электронной микроскопии подобные поры или щели
обнаружены не были (рис. 12.2). В то же время были найдены
многочисленные пузырьки, накапливающие меченые белки в
большом количестве, что свидетельствует о возможности
переноса белков посредством пиноцитоза и экзоцитоза (см. гл. 2).
ДВИЖЕНИЕ ЖИДКОСТЕЙ ЧЕРЕЗ КАПИЛЛЯРЫ
Если прекапиллярные сфинктеры открыты, то кровь через
окончания артериол и метартериолы поступает
непосредственно в истинные капилляры (рис. 12.1). Если же эти сфинктеры
закрыты, то кровь может течь через главный канал в венулу,
218
Глава 12
минуя истинные капилляры. Кроме того, из артериолы кровь
может поступать непосредственно в венулу через артерио-ве-
нозный анастомоз — шунт (ABA). Переход жидкости в ткани
осуществляется путем транскапиллярного обмена в истинном
капилляре. Обратное же всасывание жидкости происходит как
в венозном конце капилляра, так и в венуле.
ФИЛЬТРАЦИЯ И ГИПОТЕЗА СТАРЛИНГА
Переход жидкости в ткани зависит от разности между
гидростатическим давлением в капилляре (ГДК), под действием
которого жидкость как бы проталкивается через стенку
капилляра (фильтрация), и осмотическим давлением белков плазмы,
называемым также онкотическим давлением (ОДК),
удерживающим жидкость в просвете капилляра. Этим силам (ГДК
и ОДК), действующим внутри капилляра, противодействуют
силы гидростатического (ГДТ) и онкотического (ОДТ) давления
в тканях, однако они относительно невелики и влияние их
незначительно. Следовательно, фильтрационное давление (ФД),
способствующее переходу жидкости в ткани (образованию
тканевой жидкости), равно:
ФД= (ГДК+ОДТ)-(ГДТ+ОДК).
Проиллюстрируем гипотезу Старлинга примером:
Гидростатическое давление в капилляре (ГДК) =35 мм рт. ст.
Гидростатическое давление в тканях (ГДТ) = 1 мм рт. ст.
Онкотическое давление плазмы (в капилляре) (ОДК) =
= 24 мм рт. ст.
Онкотическое давление в тканях (ОДТ) =2 мм рт. ст.
При движении жидкости из капилляров в ткани (направление
действующих сил указано стрелками):
Тканевая
жидкость
'Кровеносный сосуд / Л
(артериола) I I ( | (35 мм
v v v V V гя\
Эффективное фильтрационное давление составляет: 34 мм
рт. ст. — 22 мм рт. ст.= 12 мм рт. ст. В этом случае жидкость
движется из капилляров в ткани. При движении жидкости из
Жидкие среды организма
219
ткани в кровеносные сосуды (всасывание):
Тканевая ГДТ °АТ
жидкость I (1 мм) (2 мм)
Сила всасывания = 14 мм рт. ст. — 22 мм рт. ст. = —8 мм
рт. ст.; жидкость переходит из ткани в венулы и вены. Таким
образом, главную роль в процессе всасывания играет онкотиче-
ское давление белков плазмы, которое обычно превышает ГДК
и поэтому способствует перемещению жидкости в кровеносное
русло.
ПРОНИЦАЕМОСТЬ
Проницаемость для какого-либо вещества зависит от его
природы, а также от эффективного фильтрационного давления,
размеров пор и наличия пузырьков в эндотелии капилляров.
Чем меньше молекула вещества, тем выше проницаемость для
Таблица 12.4. Коэффициенты проницаемости для различных веществ
Вещество
Поваренная соль
Глюкоза
Инулин
Альбумин
Молекулярный вес
58
180
5 500
67 000
Диффузия, смЗ/с
2,0
9-10-1
2,4-10-1
8,5-102
Проницаемость,
см/с на 100 г ткани
15
6
з.ю-1
1-Ю"3
него. В табл. 12.4 приведены коэффициенты проницаемости
для различных веществ в соответствии с их молекулярным
весом.
ФИЛЬТРАЦИЯ И ОБРАЗОВАНИЕ ТКАНЕВОЙ ЖИДКОСТИ
На фильтрацию и образование тканевой жидкости влияют
любые факторы, изменяющие давление в капиллярах (Рк).
К ним относятся:
Артериальное давление Ра;
Венозное давление Рв;
220
Глава 12
Сопротивление прекапилляров (артериальный конец) Ra;
Сопротивление посткапилляров (венозный конец) RB.
Давление в капилляре
р (Кв/Ка)Ра + Рв
1 + (Ra/Ra) e
Из этой формулы следует, что:
1. Рк возрастает при повышении Ра или Рв.
2. При повышении Рв Рк возрастает в 5—10 раз больше, чем
при таком же повышении Ра.
3. Рк возрастает при повышении RB.
4. Рк уменьшается при повышении Ra.
5. Рк в большинстве капилляров равно 32—35 мм рт. ст. на
артериальном и 15 мм рт. ст. — на венозном конце.
6. Повышение у взрослого человека центрального венозного
давления на 10 см водного столба приводит к выходу из
плазмы 250 мл жидкости за 10 мин.
ВОДНЫЙ ОБМЕН
В норме скорости фильтрации и всасывания таковы, что
чрезмерного накопления жидкости в тканях (отеков) нет.
Отеки возникают в случае, если скорость перехода жидкости в
ткани выше, чем скорость всасывания. Такая ситуация может
создаться, когда: 1) онкотическое давление в капиллярах
падает вследствие снижения уровня белков в крови, что приводит к
уменьшению интенсивности всасывания; 2) венозное давление
(ГДК) начинает превышать онкотическое давление плазмы
(ОДК), что также сопровождается снижением всасывания,
уменьшением оттока лимфы и отеками.
Увеличению фильтрации и объема тканевой жидкости
способствуют такие факторы, как 1) расширение прекапилляров
(уменьшение Ra); 2) снижение уровня белков в крови
(уменьшение ОДК).
К снижению же фильтрации приводят: 1) обезвоживание;
2) сужение прекапилляров (увеличение Ra). Ткани таких
органов, как, например, легкие, лишены воды, и образование
межклеточной жидкости в них незначительно, так как давление в
их капиллярах меньше, чем осмотическое давление плазмы.
ПОСТУПЛЕНИЕ И ПОТЕРЯ ЖИДКОСТИ
В связи с тем что поступление и потеря жидкости
непосредственно влияют на общее количество ее в организме,
соотношение между этими двумя процессами направлено на
поддержание равновесия между различными жидкостными пространств
Жидкие среды организма 221
вами. Вода выводится главным образом почками и путем
испарения легкими и кожей. При избыточном поступлении воды
выделение ее почками возрастает, при жажде и
обезвоживании — падает. В среднем взрослый человек теряет в сутки около
1 л воды в виде паров через легкие и с поверхности кожи и
около 1,5 л с мочой через почки; эти показатели зависят от
температуры окружающей среды и других факторов.
На потребление воды влияют жажда, обезвоживание,
необходимость восполнить объем циркулирующей плазмы и —
в конечном счете — объем других водных пространств
организма. Потеря воды в количестве, приблизительно равном 0,8%
веса тела, сопровождается чувством жажды и увеличенным
потреблением жидкости. В среднем за сутки в организм
поступает около 1,2 л воды при питье и примерно столько же при
потреблении и метаболизме пищи.
ДИФФУЗИЯ
В капиллярах фильтруется лишь 2% плазмы, и в норме-
столько же подвергается реабсорбции. Это относительно
небольшое количество имеет ключевое значение для поддержав
ния постоянного соотношения между жидкостными
пространствами организма, т. е. одного из параметров гомеостаза.
Гораздо большие (почти в 5000 раз) количества жидкости
переходят из крови в ткани и обратно путем диффузии —
равновесного процесса, при котором обмен жидкостью между кровью
и Тканями зависит от концентрации в ней солей. Более
подробно механизм диффузии разбирается в гл. 2.
осмоляльность жидкости во внутриклеточном
И ВНЕКЛЕТОЧНОМ ПРОСТРАНСТВАХ
Сразу же после потребления поваренной соли возрастает
осмоляльность жидкости внеклеточного пространства. При этом
вода начинает поступать из внутриклеточного пространства,
где осмоляльность ниже, во внеклеточное, что способствует
уменьшению концентрации солей в последнем. Объем воды во
внеклеточном пространстве при этом растет, во
внутриклеточном — уменьшается.
Напротив, при потреблении гипотонических растворов
осмоляльность (концентрация поваренной соли) внеклеточного-
пространства быстро падает, и вода перемещается из него во-
внутриклеточное пространство с более высокой осмоляль-
ностью, способствуя ее уменьшению. В результате объем водьь
во внутриклеточном пространстве растет. В конечном счете кон-
222
Глава 12
центрации хлорида натрия во внутриклеточном и внеклеточном
пространствах уравниваются и устанавливается равновесие
(см. рис. 2.3).
ОБРАЗОВАНИЕ ЛИМФЫ
Лимфа представляет собой особую ткань, или тканевую
жидкость, собираемую специальными сосудами (рис. 12.3); эти
сосуды впадают в правый и левый лимфатические протоки, ко-
Фис. 12.3. Схема лимфооттока и лимфатических сосудов.
Жидкие среды организма
223
торые в свою очередь открываются в крупные центральные
вены. Лимфа отличается от тканевой жидкости главным
образом более высоким содержанием белков (2 г%). За сутки
образуется 2—4 л лимфы.
Лимфатическая система образована преимущественно
околовенозными, т. е. идущими вдоль вен (а также вдоль
артерий) лимфатическими сосудами. Эти сосуды бывают
глубокими и поверхностными и по структуре напоминают вены,
отличаясь от них меньшей толщиной стенки и большей
проницаемостью. Как и вены, они имеют клапаны, препятствующие
обратному току лимфы.
Лимфатические сосуды пронизывают почти все органы, за
исключением ногтей, кожи, волос, роговицы и некоторых
других. В печени и тонком кишечнике они особенно
многочисленны, высоко проницаемы, и уровень белка в содержащейся в
них лимфе довольно высок (5—6 г%). Лимфатические сосуды
способствуют оттоку жидкости от тканей (рис. 12.3).
Лимфа от верхней половины туловища оттекает в грудной
проток, открывающийся в венозное русло в области слияния
левой внутренней яремной и подключичной вен. В грудной
проток перед местом впадения его в венозную сеть открываются
также сосуды, собирающие лимфу от левой половины головы
и груди.
Лимфа от правой половины головы, шеи и правой руки
оттекает в правый лимфатический проток, впадающий в правую-
подключичную вену. В среднем ток лимфы у человека
составляет 1,4 мл/кг веса в 1 ч, или около 2 л за сутки. Каждые 24 ч
в лимфу переходит от lU ДО половины всех белков плазмы
крови.
Току лимфы способствуют как сокращения мышц, так и
сокращения непосредственно самих лимфатических сосудов. На*
него влияют также изменения давления в тканях и
капиллярах.
Закупорка лимфатических сосудов в результате
воспалительных изменений вызывает заболевание, называемое
слоновостью (элефантиаз), характеризующееся прекращением
оттока лимфы и непомерным увеличением и утолщением тканей
конечности.
Так же как и в венах, в лимфатических сосудах
существует градиент давления, направленный от периферии к
центральным сосудам (грудному и правому лимфатическим протокам).
Лимфатические узлы представляют собой мелкие овальные*
образования размером от едва различимой крупинки до
величины миндального ореха; они группируются вокруг
лимфатических сосудов (рис. 12.3). В организме эти узлы
распространены повсеместно; особенно много их в области нижних и верх-
224
Глава 12
них конечностей. Они выполняют функцию фильтра и защиты
от инфекции. В них содержатся лимфоциты, участвующие в
борьбе с инородными телами и инфекционными агентами. При
патологии лимфатические узлы, например подмышечные и
паховые, могут увеличиваться и становиться болезненными при
пальпации.
ЛИТЕРАТУРА
Alt man P. L., Dittmer D. S. (1961). Blood and other body fluids, Fed. Amer.
Soc. Exp. Bio., Washington.
Alt man H. L., Dittmer D. S. (1974). Biology data book, 2nd edn., vol. Ill, Fed.
Amer. Soc. Exp. Biol., Washington.
Burke S. R. (1972). The composition and function of body fluids. Mosby, St.
Louis.
Guyton A. C. (1977). Basic human physiology, 2nd edn., Saunders, Philadelphia.
Selkurt E. E. (1975). Basic physiology for the health sciences. Little, Brown,
Boston.
Williams W. /., Beutler E., Erslev A. /., et al. (1972). Hematology, McGraw-Hill,
New York.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Что такое жидкостные пространства организма?
Перечислите их и укажите их объемное соотношение.
2. Как можно измерить объем крови, используя принцип
разведения?
3. Как образуется тканевая жидкость? Покажите на примере,
как в соответствии с гипотезой Старлинга происходит а)
выход жидкости из кровеносного русла в ткани и б)
всасывание жидкости из тканей.
4. Какие факторы определяют проницаемость кровеносных
сосудов?
5. Как влияет на образование тканевой жидкости а) снижение
содержания белков в крови; б) уменьшение
гидростатического давления крови; в) повышение венозного давления?
8. Каковы главные лимфатические сосуды, собирающие и
отводящие лимфу?
Глава 13
кровь
Цельная кровь состоит из жидкой части (плазмы),
содержащей соли и ряд других компонентов, и форменных элементов,
или кровяных телец. К последним относятся красные кровяные
тельца (эритроциты) и белые кровяные тельца (лейкоциты). На
долю форменных элементов приходится около 45% общего
объема крови, а остальные 55% составляет плазма. К основным
компонентам плазмы относятся Na+, K+, P, Са2+, глюкоза и
белки — альбумины, глобулины и фибриноген, составляющие
6—7% объема плазмы. Фибриноген принимает участие в
свертывании крови. Альбумины и глобулины представляют собой
высокомолекулярные соединения, практически не проходящие
через полупроницаемую стенку капилляров, благодаря чему они
способствуют поддержанию онкотического давления в
кровеносном русле (см. гл. 2 и 12). Это препятствует избыточному
выходу жидкости в межклеточное пространство и является
одним из механизмов поддержания водного баланса между
кровью и тканями (см. гл. 12).
Поскольку содержание альбуминов в крови почти вдвое
больше, чем глобулинов (А/Г), они играют важную роль в
поддержании онкотического давления крови, а также в связывании
переносимых плазмой веществ — лекарственных препаратов,
витаминов, гормонов и пигментов. Глобулины же представляют
собой антитела и другие защитные факторы организма. Белки
плазмы выполняют также функцию буферов, корректируя
сдвиги кислотно-щелочного равновесия (рН) крови.
Важными компонентами крови и многих тканей являются
электролиты — Na, К, Са и Р, играющие существенную роль в
возникновении биоэлектрических явлений.
Кроме особых функций, выполняемых эритроцитами,
лейкоцитами и тромбоцитами, речь о которых пойдет ниже, кровь
играет еще роль в процессах: 1) всасывания и переноса
питательных веществ от пищеварительного тракта к тканям, 2)
переноса газов от легких к тканям и обратно, 3) удаления
метаболитов, 4) транспорта гормонов, 5) регуляции водного баланса
тканей, рН и температуры тела, 6) образования антител и
других веществ, участвующих в борьбе с инфекцией.
15—1705
226
Глава 13
Плазма
Лейкоциты
т
I
Эритроциты
-0\
-Ъ
-3
-Ъ\
__|-6
-8
Кровь
Объем эритроцитов,
отделяющихся путем центрифугирования крови
при 1500 g в течение 30 мин (или при
более высокой скорости в течение
меньшего времени), называется гема-
токритом. Для определения гемато-
крита в кровь, взятую из артерии
или вены, добавляют антикоагулянт,
предупреждающий ее свертывание
(например, оксалат или цитрат
натрия либо калия). Кровь помещают в
пробирку и центрифугируют до тех
пор, пока эритроциты не осядут на
дно, а плазма останется над осадком
Цифры на трубке при этом указывав
ют на объемное соотношение плазмы
и форменных элементов (рис. 13.1).
Так как в составе эритроцитарной
массы еще остается 4% объема
плазмы (кажущийся гематокрит),
истинный гематокрит вычисляют, умножая
полученный на 0,96. Факторы,
влияющие на форму и количество
эритроцитов, изменяют гематокрит.
При центрифугировании непосредственно над слоем
эритроцитов образуется тонкий желтоватый слой, содержащий
лейкоциты.
-9п
10
Рис. 13.1. Плазма и цельная
кровь с эритроцитами и
лейкоцитами.
ФИЗИЧЕСКИЕ СВОЙСТВА КРОВИ
Кровь представляет собой вязкую жидкость с большим,
чем у воды, удельным весом. Обмен жидкостью между тканью
и кровью зависит от ее осмотического давления.
ОБЪЕМ КРОВИ
Из всех жидкостных пространств организма кровь после
лимфы является самым небольшим, однако она выполняет
важнейшие функции по регуляции состава других пространств,
кровяного давления, венозного возврата и сердечного выброса.
Количество плазмы крови у взрослого человека составляет
в среднем 4,3—5% веса тела, т. е. около 3 л; на долю
эритроцитов приходится примерно 2 л. Около 60—80% общего объема
крови находится в венах, остальная часть — в полостях сердца,
артериях и капиллярах.
Объем крови, как и других водных пространств, достаточно
постоянен. Кровопотеря сопровождается резким снижением
Кровь
227
объема крови, однако это снижение компенсируется выходом
жидкости из тканевого пространства в кровь. При кровопотере
включаются и другие механизмы, направленные на сохранение
жидкости, например уменьшение испарения и выделения мочи.
Потеря жидкой крови может быть восполнена обильным питьем,
однако нормальная численность эритроцитов восстанавливается
лишь через несколько дней.
ИЗМЕРЕНИЕ ОБЪЕМА КРОВИ
Для измерения объема крови (плазмы) в кровоток вводят
известное количество красителя (типа синьки Эванса — Т 1824)
и определяют его концентрацию после того, как он полностью
смешается с циркулирующей кровью, но до того как он из нее
исчезнет. При этом
г^<. Количество введенного красителя
Объем плазмы = -т7 —: •.
Концентрация красителя в крови
У человека краситель перемешивается с кровью в среднем за
10 мин.
~^ „ ^ Объем плазмы* 100
Общий объем крови = Г/ .
^ r % iwia3Mbij
Если объем плазмы равен 3000 мл, а гематокрит — 40%, то
Общий объем крови = ^ = 5000 мл = 5 л.
Вязкость крови примерно в 5 раз больше вязкости воды; на
нее оказывает сильное влияние содержание эритроцитов и
белков плазмы. При повышении вязкости увеличивается как
сопротивление кровотоку, так и работа сердца. Удельный вес цельной
•крови зависит в основном от числа эритроцитов и у человека
колеблется от 1,0520 до 1,0610; он является одним из факторов,
определяющих скорость оседания эритроцитов в цельной крови,
помещенной в специальную пробирку. Скорость оседания
эритроцитов (СОЭ) измеряют в мм/ч; она варьирует от 2 до 10. При
•некоторых заболеваниях (ревматизм, туберкулез, артриты,
токсемии) СОЭ повышается.
ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ
К форменным элементам крови относятся эритроциты,
лейкоциты и кровяные пластинки (тромбоциты) (рис. 13.2). В 1 мм3
Цельной крови содержится примерно 5 млн. эритроцитов,
9000—10 000 лейкоцитов и 300 000 тромбоцитов (табл. 13.1).
Изображенные на рис. 13.2 форменные элементы образуются в
костном мозгу, где они проходят несколько стадий клеточного
Деления (рис. 13.3). Большинство исследователей признают, что
15*
Глава 13
Эритроциты
Тромбоциты
Лейкоциты
Гранулоциты
Нейтрофил
Базофил
Эо*инофил
Моноцит
Рис. 13.2. Форменные элементы крови.
Лимфоцит
форменные элементы происходят от одной
недифференцированной мультипотентной стволовой клетки (монофилетическая
теория). Эта клетка дает начало четырем типам коммитировацных
стволовых клеток, от которых в свою очередь происходят бласт-
ные клетки — предшественники зрелых форменных элементов.
Из бластных клеток образуются юные формы клеток крови
(«циты»), превращающиеся в конечном счете в зрелые клетки.
Зрелые эритроциты не имеют ядра, однако их
предшественники — нормобласты, а также эритроциты более
низкоорганизованных животных (птиц и рептилий) являются ядерными
клетками. Нарушение развития эритроцитов может привести к
состоянию, называемому эристобластозом плода, при котором
у плода и новорожденного преобладают эритробласты, не
развивающиеся в зрелые эритроциты.
Кровь
229
Таблица 13.1. Нормальные показатели
(в 1 мм3)
Форменные
элементы
Общее число
лейкоцитов
Эозинофилы
Базофилы
Нейтрофилы
Моноциты
Лимфоциты
Эритроциты
Тромбоциты
В среднем
9000
275
25
5000
400
2000
4,7.10е
300 000
форменных элементов крови
Границы нормы
5000—11000
100—400
3000—7000
100—600
1000—3000
4—6-10е
200 000—400 000
Процент общего
числа в среднем
3,0
0,27
55,50
4,44
22,22
Образование красных кровяных телец в костном мозгу
называется эритропоэзом. Эритропоэз регулируется гуморальными
факторами; дефицит кислорода (гипоксия) вызывает повышение
образования особого вещества, стимулирующего эритропоэз —
эритропоэтина (ЭРП). ЭРП представляет собой гликопротеин,
вырабатываемый почками и циркулирующий в крови; действуя
непосредственно на костный мозг, он стимулирует образование
клеток эритроидного ряда (ретикулоцитов) и ускоряет их
превращение в зрелые эритроциты.
Эритропоэз усиливается под действием андрогенов и
гормонов коры надпочечников и уменьшается под влиянием
эстрогенов. Образование эритроцитов зависит от достаточного поступ-
Монобласт
кск-
Миелобласт
Некоммитированная
стволовая клетка
Коммитированная КСК
стволовая клетка (КСК)
КСК
Пром'иелоцит
Промоноцит Миелоциты
Лимфобласт Проэритробласт МегакариобласГ
I I
Пролимфоцит Пронормобласт
Базофил Нейтрофил Эозинофил
Нормобласт
Большой лимфоцит I
| Ретикулоцит
Малый лимфоцит . I
Эритроцит
Моноцит
Рис. 13.3. Развитие клеток крови в костном мозгу
Мегакариоцит
Тромбоцит
230
Глава 13
ления железа и белков, необходимых для синтеза гемоглобина,
а также других веществ, например витаминов группы В и
особенно Bi2. Недостаток этих веществ и ряд других нарушений
может приводить к анемиям различного типа.
ЭРИТРОЦИТЫ
Средняя продолжительность жизни эритроцитов человека
составляет 120 дней. Старые погибающие формы эритроцитов
становятся хрупкими, мембрана разрывается, и клетка
распадается; освобождающееся при этом железо вновь используется в
костном мозгу. Входящий в состав молекулы гемоглобина гем
превращается в желчный пигмент билирубин, который
впоследствии выводится печенью.
ПРОЧНОСТЬ ЭРИТРОЦИТОВ
Гемолизом называется разрыв эритроцитов с выходом
содержащегося в них гемоглобина в плазму. Гемолиз наступает при
замораживании, оттаивании и изменении осмотического
давления крови. Гемолиз не происходит в растворе, осмотическое
давление которого равно осмотическому давлению крови; такие
растворы называются изотоническими. Растворы с более низким
осмотическим давлением называются гипотоническими, а с более
высоким — гипертоническими. В гипотонических растворах
эритроциты поглощают воду и разрываются; в гипертонических они
теряют воду и сморщиваются.
Показателем прочности эритроцитов может быть их
осмотическая резистентность в гипотонических растворах с известной
концентрацией NaCl (концентрация изотонического раствора
NaCl равна примерно 0,9%) (рис. 13.4).
Эритроциты представляют собой двояковогнутые диски
диаметром около 8,5 мкм (см. рис. 13.2). Такая форма и площадь
поверхности способствуют максимальной диффузии и
транспорту 02 и С02. У взрослого мужчины число эритроцитов в
кубическом миллиметре крови колеблется от 4 до 6 млн., составляя в
среднем 5 млн., у женщин — в среднем около 4,5 млн.
(табл. 13.1).
На численность эритроцитов влияет ряд факторов, например
поступление железа, витаминов группы В, пищевого белка,
а также гипоксия и мышечная работа.
АНЕМИИ
В зависимости от числа и размеров эритроцитов и
содержания в них гемоглобина анемии можно классифицировать
следующим образом.
Кровь
231
Нормоцитарные и нормохромные
Размеры эритроцитов и содержание в них гемоглобина не
изменены, однако число эритроцитов уменьшено;
наблюдаются после острой кровопотери в результате того, что плазма
восполняется гораздо быстрее, чем число эритроцитов.
Микроцитарные и гипохромные (со сниженным содержанием
гемоглобина)
Кривая осмотической
1Qq резистентности эритроцитов
0,55
0,50 0,45 0,40 0,35
Концентрация NaCI, %
0,30
Рис. 13.4. Хрупкость эритроцитов. По мере того как концентрация NaCI
падает и раствор становится гипотоническим, все больше эритроцитов
разрывается (гемолиз).
Размеры и число эритроцитов и содержание в них
гемоглобина снижено. Такие анемии называются железодефицит-
ными.
3. Макроцитарные и гиперхромные
Число эритроцитов уменьшено, но их размеры и
содержание гемоглобина повышены — типичная картина пернициоз-
ной анемии, связанной с дефицитом внешнего фактора
(витамин Bi2) или внутреннего фактора, вырабатываемого
слизистой оболочкой желудка.
4. Апластическая анемия
Связана с нарушением костномозгового кроветворения;
может появиться в результате чрезмерного рентгеновского
облучения или под действием неизвестных причин.
5. Серповидноклеточная анемия
232
Глава 13
Характеризуются неправильной формой эритроцитов и
нарушением транспорта кислорода; эта форма анемии является
наследственной.
Из-за пониженного количества гемоглобина при анемиях
ухудшается снабжение тканей кислородом. Для больных
анемией характерна слабость, быстрая утомляемость. При полици-
темии, наблюдающейся в результате обезвоживания или
гиперфункции костного мозга, число эритроцитов повышено. Оно
значительно увеличивается также при гипоксии или пребывании в
высотных районах, однако в норме при адаптации к высоте
число эритроцитов уменьшается и может стать таким же, как и
на уровне моря.
ТРОМБОЦИТЫ И СВЕРТЫВАНИЕ КРОВИ
Тромбоциты (кровяные пластинки) млекопитающих
представляют собой безъядерные цитоплазматические образования
диаметром 2—4 мкм (рис. 13.1), развивающиеся из мегакарио-
цитов; у птиц и рептилий эти клетки имеют ядро. Содержание
тромбоцитов в крови человека варьирует от 200 тыс. до 400 тыс.
Внутренние факторы Внешние Факт°РЫ
Тканевой промбопластин,
высвобождающийся при
повреждении или разрыва
сосуда
Протромбин
Плотный
->сгусток
фибрина
Рис. 13.5. Этапы процесса свертывания крови.
в 1 мм3 (табл. 13.1). Тромбоциты играют роль в остановке
кровотечения (свертывании крови). Скапливаясь в области разрыва
сосуда (агрегация тромбоцитов), они вместе с тромбопластина-
ми способствуют образованию кровяного сгустка.
При свертывании крови образуется плотный сгусток из нитей
фибрина (тромб). Фибрин представляет собой нерастворимый
белок, образующийся из фибриногена в результате его
активации ферментом тромбином; последний в свою очередь образует-
Тромбоциты
| Рыхлый
Фибриноген > Тромбин >сгусток >Х1Н
фибрина
Кровь
233
ся из протромбина под действием ряда ускоряющих факторов.
Подробно фазы и факторы свертывания крови представлены на
рис. 13.5 и в табл. 13.2.
Таблица 13.2. Факторы свертывания крови
I Фибриноген
II Протромбин
III Тромбопластин
IV Са2+
V Лабильный фактор, проакцелерин, АК-глобулин
VI Акцелерин
VII Стабильный фактор, проконвертин, сывороточный ускоритель
превращения протромбина
VIII Антигемофильный фактор А, антигемофильный глобулин (АГГ),
тромбоцитарный кофактор I, тромбопластиноген А
IX Антигемофильный фактор Б, плазменный компонент тромбопла-
стина (ПКТ), тромбоцитарный кофактор II, фактор Кристмаса
(ФК)
[ X Фактор Стюарта — Прауэра
XI Антигемофильный фактор В, предшественник плазменного тром-
бопластина (ППТ)
XII Фактор Хагемана
XIII Фибрин-стабилизирующий фактор
Прозрачная сыворотка крови, лишенная, как и плазма,
форменных элементов, не содержит фибриногена, израсходованного
на образование сгустка.
ВРЕМЯ СВЕРТЫВАНИЯ
Под временем свертывания понимают промежуток времени,
в течение которого кровь, помещенная в пробирку при
температуре 37 °С, образует сгусток; в норме оно равно 3—8 мин.
Лучшим показателем свертываемости служит протромбиновое
время, т. е. время, за которое происходит свертывание крови при
добавлении к ней известного количества кальция и тромбоплас-
тина (предварительно из крови удаляется Са2+). В норме
протромбиновое время составляет 12—17 с. У лиц с недостатком
некоторых факторов (V, VII, X) этот показатель увеличен.
Свертыванию крови препятствуют антикоагулянты.
Некоторые вещества (оксалаты и цитраты) в пробирке предупреждают
образование сгустка, удаляя (осаждая) кальций из крови.
Образующийся в организме гепарин в норме препятствует
свертыванию, подавляя активность тромбина, протромбина и тромбо-
пластина. Для нормального свертывания необходим витамин К.
Дикумарин — вещество, впервые найденное в доннике,
препятствует утилизации витамина К и тем самым нарушает
образование протромбина. Гепарин и дикумарин не дают при введении
никаких побочных эффектов, тогда как введение в организм
оксалатов и цитратов, осаждающих кальций, может приводить
К серьезным осложнениям.
234
Глава 13
ГРУППЫ КРОВИ
При переливании крови от человека к человеку возможны
тяжелые реакции, иногда со смертельным исходом. Это
происходит потому, что эритроциты человека содержат антигены, или
агглютиногены, вступающие при переливании в реакцию с
антителами (агглютининами). В результате наступает склеивание
эритроцитов. Тяжесть реакции зависит от групповой
принадлежности крови донора и реципиента. В системе АВО существуют
четыре группы крови: О, А, В и АВ.
Таблица 13.3. Определение группы крови
Сыворотка3'
А и В
А
В
А и В
Агглютинация
есть
есть
есть
нет
Группа крови
АВ
В
А
О
Исследуемая кровь добавляется к сыворотке указанной группы.
Люди с группой АВ являются универсальными
реципиентами, так как в крови у них нет агглютининов, и им можно
переливать любую кровь, не опасаясь реакции агглютинации
(табл. 13.3 и 13.4). Люди с группой крови О — это
универсальные доноры: в их крови нет агглютиногенов, и переливание ее
другим лицам не сопровождается реакциями групповой
несовместимости.
Приведем пример групповой (АВО) несовместимости.
Эритроциты группы В в плазме группы В — агглютинации нет.
Эритроциты группы А в плазме группы В — агглютинация
(склеивание).
Таблица 13.4. Группы крови и реакции при переливании
Группа
крови
А
В
АВ
О
Переливание
( Эритроциты к сыворотке В и О
1 Сыворотка к эритроцитам В и АВ
f Эритроциты к сыворотке А и О
\ Сыворотка к эритроцитам А и АВ
Г Эритроциты к сыворотке А, В, О
1 Сыворотка к эритроцитам А, В, АВ, О
( Эритроциты к сыворотке А, В, АВ, О
\ Сыворотка к эритроцитам А, В, АВ
Агглютинация
Есть
Есть
Есть
Есть
Есть
Нет
Нет
Есть
Кровь
235
Группу крови конкретного человека определяют путем
смешивания его эритроцитов со стандартными группоспецифиче-
скими сыворотками (табл. 13.4).
РЕЗУС-ФАКТОР
В крови большинства людей найдена еще одна группа агглю-
тиногенов (Rh); носители таких агглютиногенов называются
резус-положительными. Если перелить резус-положительную кровь
резус-отрицательному человеку, то видимой реакции не
произойдет, однако в его крови появятся резус-антитела, что приведет
к осложнениям при повторном переливании Rh+-KpOBH.
Гены, определяющие Rh+, являются доминантными по
отношению к определяющим Rh-; следовательно, потомство от
резус-положительного и резус-отрицательного родителей будет
Rh+. В том случае, когда отец Rh+, а мать —Rh~, кровь плода
будет Rh+. При проникновении агглютиногенов плода через
плаценту в кровь матери в организме матери начнется образование
антител (агглютининов) к Rh+ эритроцитам; если эти антитела
поступят через плаценту обратно в кровь плода, произойдет
агглютинация и наступит эритробластоз. Гибель плода в этих
случаях наступает обычно к моменту рождения. Первый
ребенок у Rh--MaTepH может быть здоровым, тогда как при
последующих беременностях возможна внутриутробная смерть плода.
Проявление резус-несовместимости можно свести к минимуму
путем назначения беременным Rh''-женщинам анти-резус гам-
маглобулина, нейтрализующего Rtr-антигены плода.
М- И N-ФАКТОРЫ
В зависимости от присутствия этих агглютиногенов
различают три группы крови: М, N и MN, однако сывороточные
антитела к ним не вырабатываются, и поэтому для переливания
крови они значения не имеют.
ГЕМОГЛОБИН
Гемоглобин —это кислородпереносящий пигмент
эритроцитов. В легких он связывает кислород (насыщается), а в
тканях—отдает его (см. гл. 17). Гемоглобин представляет собой
белок с мол. весом около 60 тысяч. Его молекула состоит из
четырех субъединиц, каждая из которых содержит гем
—железосодержащее производное порфирина. Это производное связано
с более сложной полипептидной структурой, часто называемой
глобиновой частью молекулы. В этой части имеются две а- и
Две р-полипептидные цепи (рис. 13.6), каждая из которых со-
236
Глава 13
держит более 140 аминокислотных остатков. В нормальной
крови человека присутствуют преимущественно два типа
гемоглобина: тип А у взрослых и тип F у плода. Найдены и другие
(аномальные) разновидности гемоглобина.
При присоединении 02 к железу (Fe2+) гемоглобина (НЬ)
образуется оксигемоглобин, полностью насыщенный
кислородом. При отщеплении кислорода гемоглобин восстанавливается.
Образование и диссоциация оксигемоглобина будут подробно
Рис. 13.6. Молекула гемоглобина (НЬ). А. Четыре субъединицы (а и Р),
образующие молекулу гемоглобина. Каждая субъединица состоит из
полипептидной цепи и гема (Н). Б. Отдельная субъединица гемоглобина. Гем встроен
в полипептидную цепь, представляющую собой линейную последовательность
аминокислот (светлые кружки). В начале цепи — аминогруппа (NH3), в
конце— карбоксильная группа (СООН). (Perutz M. F. Scientific American, 239,
№ 6, 92, 1978.)
рассмотрены в гл. 17. Окись углерода присоединяется к
гемоглобину активнее, чем кислород, образуя карбоксигемоглобин; при
этом гемоглобин утрачивает способность связываться с
кислородом и переносить его, что может оказаться смертельным.
Содержание гемоглобина в крови здорового человека колеблется
от 15 до 16 г%; у мужчин оно несколько выше, чем у женщин,
1 г НЬ способен связать 1,3 мл 02.
ЛЕЙКОЦИТЫ
Лейкоциты развиваются из одной родоначальной стволовой
клетки костного мозга. От этой клетки происходят
соответствующие клетки-предшественники (см. рис. 13.3), из которых в
конечном счете развиваются одноядерные моноциты и
полиморфноядерные базофилы, нейтрофилы и эозинофилы. У
животных, не относящихся к млекопитающим, например у птиц,
вместо нейтрофилов имеются гетерофилы. Полиморфноядерные
Кровь
237
клетки обладают цитоплазматическими гранулами характерной
формы, которые отчетливо окрашиваются. Крупные гранулы
базофилов окрашиваются основными красителями в пурпурные
ШЙ1 фиолетовые тона, а гранулы эозинофилов воспринимают
кислые красители (эозин) желто-розового цвета. Реакция нейтро-
филов по отношению к красителям нейтральна.
Моноциты обладают крупным ядром и относительно
небольшим количеством цитоплазмы. Лимфоциты — тоже одноядерные
клетки; некоторое их число обнаруживается в костном мозгу,
однако основная их часть образуется в лимфатических узлах,
селезенке и вилочковой (зобной) железе (тимусе) из
первичных стволовых клеток костного мозга.
Число лейкоцитов в крови изменчиво и в норме колеблется
от 5000 до 11 000 в 1 мм3; при ряде заболеваний, а также при
интенсивной физической нагрузке число лейкоцитов резко
возрастает (лейкоцитоз). Содержание различных клеток в крови,
а также процентный состав лейкоцитов приведены в табл. 13.1.
Основная функция лейкоцитов — это борьба с чужеродными
агентами, в том числе токсинами и бактериями, и выработка
антител. Лейкоциты фагоцитируют (поглощают) чужеродные
тела и уничтожают их. Они способны передвигаться к участку
повреждения и внедрения инфекции. Самыми многочисленными
из всех лейкоцитов являются нейтрофилы, и поэтому роль их в
фагоцитозе наиболее значительна. Содержание эозинофилов и
базофилов мало, и функция их до конца не ясна.
Лимфоциты, на долю которых приходится более 20% общего
числа лейкоцитов, играют важнейшую роль в защите организма
от инфекций. Им не присуща способность к фагоцитозу, однако
они играют важную роль в иммунных механизмах, в частности
синтезируют антитела. Моноциты способны к фагоцитозу.
При патологических состояниях число лейкоцитов может
увеличиваться в 5—20 раз; это лейкоцитоз. Злокачественный
процесс, проявляющийся резко выраженной пролиферацией грану-
лоцитов, называется лейкемией (лейкозом). Снижение общего
числа лейкоцитов — лейкопения — может быть вызвано
угнетением костного мозга под действием рентгеновских лучей или
токсичных веществ.
ЛИТЕРАТУРА
Altman P. L., Dittmer D. S. (1961). Blood and other body fluids, Fed. Amer.
Soc. Exp. Biol. Washington.
Altman P. L., Dittmer D. S. (1974). Biology data book, 2nd edn., vol. III. Fed.
Amer. Soc. Exp. Biol., Washington.
Guy ton A. C. (1977). Basic human physiology, 2nd edn., Saunders, Philadelphia.
Perutz M. F. (1978). Hemoglobin structure and respiratory transport, Scientific
American, 239, No. 6, p. 92.
238
Глава 13
Selkurt E. E. (1975). Basic physiology for the health sciences, Little Brown,
Boston.
Williams W. J., Beutler E., Erslev A. J., Rundles R. W. (1972). Hematology,
McGraw-Hill, New York.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Каковы основные компоненты цельной крови? Что такое
плазма? Что такое сыворотка?
2. Сколько эритроцитов в крови мужчин и женщин?
3. Каково содержание гемоглобина в крови и его основная
функция?
4. Что такое гематокрит?
5. Что такое эритропоэз и эритропоэтин?
6. Что представляет собой: а) микроцитарная гипохромная
анемия, б) макроцитарная анемия, в) серповидноклеточная
анемия, г) пернициозная анемия?
7. Что такое а) полицитемия, б) тромбоциты?
8. Каковы конечная и главная фазы свертывания крови?
9. Назовите основные группы крови.
10. Кто считается универсальным донором, а кто —
универсальным реципиентом?
11. Группы А и О встречаются чаще, чем другие группы
(правильно это или нет?).
12. Что произойдет при групповой несовместимости эритроцитов
и сыворотки?
13. Дайте определение агглютиногенов и агглютининов.
14. Дайте определение резус-фактора. Чем опасно для
женщины быть Rh~?
15. Перечислите и дайте определение разных видов лейкоцитов.
16. Что такое а) лейкоцитоз, б) лейкопения?
\
Глава 14 \
КРОВООБ^ЩЕНИЕ
\
ОБЩИЕ ПРЕДСТАВЛЕНИЯ
К системе кровообращения относятся: сердце, выполняющее
функцию насоса, и периферические кровеносные сосуды —
артерии, вены и капилляры. Выбрасываемая сердцем кровь
разносится к тканям через артерии, артериолы (мелкие артерии) и
капилляры, и затем возвращается к сердцу по венулам (мелким
венам) и крупным венам. На рис. 14.1 приведена схема
кровообращения в важнейших органах и системах (подробнее см. гл. 1).
Насыщенная кислородом артериальная кровь выбрасывается из
левого желудочка и через аорту направляется к органам;
венозная кровь возвращается к правому предсердию, затем поступает
в правый желудочек и далее через легочные артерии к легким,
где вновь насыщается кислородом. После этого кровь по
легочным венам возвращается в левое предсердие. Давление крови
в легочных артериях и венах меньше, чем артериальное
давление в большом кругу.
В артериальной системе давление высокое, а в венозной —
низкое; особенности строения артерий и вен соответствуют этим
условиям.
АНАТОМИЧЕСКИЕ СВЕДЕНИЯ
Стенка артерии состоит из нескольких слоев. Внутренний,
ближайший к просвету, слой называется эндотелием; к нему
прилегает эластическая оболочка (рис. 14.2), толщина которой
различна в зависимости от типа сосуда. Далее следует слой
гладких мышц, обусловливающий способность сосудов к
расширению и сужению. Существуют два типа гладкомышечных
волокон— циркулярные и продольные. Сокращение циркулярных
волокон сопровождается сужением коротких ограниченных
отрезков сосуда. Под мышечными слоями проходят сосуды (vasa
vasorum) и нервы. Раздражение симпатических нервов приводит
к сокращению гладких мышц и сужению сосудов. В наружной
оболочке имеются коллагеновые волокна; эти волокна способны
растягиваться, но не обладают упругостью эластических
волокон.
Голова и шея>
Вены верхней
половины тела
Руки и грудь
Верхняя
полая вена'
Легкое
WY
Легочные артерии
Артерии верхней
-половины тела
Левое
предсердие
Легочные
вены
Легкое
Правый
желудочек -
Печеночные^
вены
Нижняя
полая
вена
Правое
^предсердие4
Печень
Пищеварительный-
тракт
Почки.
\\^Ш
-£"
.Воротная вена
-Левый
желудочек
.Печеночная
артерия
-Аорта
Вены нижней
половины тела"
^cs
Артерии нижней
половины тела
Рис 14.1. Схема кровообращения (артериальная кровь показана светлым,
венозная— темным),
240
Кровообращение
241
Диаметр крстеносных сосудов и тканевой состав их стенок
различны в зависимости от типа сосуда (рис. 14.2). Как
правило, в стенках артерий больше эластической ткани и меньше кол-
нагеновых волокон,\чем в стенках вен; вены же, напротив, более
богаты коллагеновыми волокнами, нежели эластическими.
Капилляры имеют эндотелиальный слой, но их стенки
лишены мышечной и соединительной ткани. Они относительно пассив,
ны, и их поведение определяется преимущественно процессами.
Аорта Артерии Сфинктеры Венулы Вены
Артериолы Капилляры
О П
Полые вены
Диаметр 25 мм
Толщина
стенки 2 мм
Эндотелий
Эластическая
оболочка
Мышечная
оболочка
Фиброзная
оболочка
<§> ®
4мм
1 мм
О
ЗОмкм 35мкм
20мкм ЗОмкм
Р
ушшт
в®»
8мкм 20мкм 5мм
1мкм 2мкм i0,5mm
30 мм
Рис. 14.2. Строение кровеносных сосудов различных отделов сосудистой
системы. Вся сосудистая сеть выстлана слоем эндотелиальных клеток;
количество же мышечной и соединительной ткани в стенках различных сосудов
варьирует. (Rushmer R. F. Cardiovascular Dynamics, 1970.)
происходящими в примыкающих артериолах и венулах (см.
гл. 12).
Лимфатические сосуды, выполняющие особую функцию, по
строению сходны с венами, отличаясь от них меньшей толщиной
и большей проницаемостью (см. также гл. 12).
ФИЗИЧЕСКИЕ ЗАКОНЫ ДВИЖЕНИЯ КРОВИ
(ГЕМОДИНАМИКА)
Кровь, которую выбрасывает сердце, движется по сосудам
разного калибра, растяжимости и сопротивления. Так как в
состав крови входят форменные элементы, она вязкая, и ее
гидродинамическое сопротивление больше, чем у воды. Объем крови,
поступающей к какому-либо органу за определенное время
(объем/время), равен отношению разности давлений (АР) к
16-1705
242
Глава 14
гидродинамическому сопротивлению: /
Р
Q (объемная скорость кровотока) =/g-;
р
отсюда P = Qg, a R=-q-.
Эти отношения иллюстрирует рис. 14.3. Разница (градиент)
давления между точками Pi и Рг зависит от расстояния между
ними, или длины отрезка трубки (1). Радиус трубки (г) по всей
ее длине одинаков, поэтому сопротивление кровотоку
постоянно. Для жестких трубок существует линейная зависимость меж-
Рис. 14.3. Зависимость между давлением, сопротивлением и объемной
скоростью тока жидкости в трубке. Объемная скорость прямо пропорциональна
градиенту давления (АР) и четвертой степени радиуса трубки (г4) и
обратно пропорциональна ее длине (1) и соответственно гидравлическому
сопротивлению (см. текст).
ду расходом жидкости и давлением, для сосудов же,
обладающих эластичностью, эта зависимость не линейна (рис. 14.4).
Объемная скорость (Q) прямо пропорциональна четвертой
степени радиуса сосуда (г4); так, при увеличении радиуса на
16% объемная скорость тока жидкости возрастает «а 100%.
Поэтому незначительные изменения ширины просвета кровеносных
сосудов сильно отражаются на кровотоке.
Сопротивление току жидкости (R) зависит от ее вязкости.
Вязкость воды, относящейся к ньютоновским жидкостям,
постоянна. Вязкость крови зависит от числа эритроцитов,
содержания белка в плазме и прочих факторов. Чем больше вязкость,
тем меньше величина кровотока.
Уравнение Пуазейля описывает все факторы, определяющие
гидродинамическое сопротивление:
P-.L- 8гИ
14 ~" О ^4~»
ЯГ4
где г\— вязкость (в Пуазах), 8 — коэффициент
пропорциональности.
л.
Кровообращение
243
Взаимоотношение между давлением, объемной скоростью и
сопротивлениемчможно обобщить следующим образом.
1. Объемная скорость прямо пропорциональна высоте
гидростатического напора и радиусу сосуда (г4).
2. Объемная скорость обратно пропорциональна длине сосуда
(1) и вязкости жидкости (г\).
3. Гидродинамическое сопротивление прямо пропорционально
длине сосуда и вязкости жидкости.
4. Гидродинамическое
сопротивление обратно про- , .а
порционально радиусу
сосуда (г4).
ЕДИНИЦЫ
ПЕРИФЕРИЧЕСКОГО
СОПРОТИВЛЕНИЯ
Гидродинамическое
сопротивление измеряют в
динах на с/см5;
периферическое сопротивление
сосудистой сети человека,
выраженное в этих единицах, равно
1700.
Для сопоставления
изменений сосудистого
сопротивления предложены более
удобные относительные
величины — единицы
периферического сопротивления
(ЕПС). Периферическое
сопротивление в этих
единицах вычисляется следующим
образом:
Давление■
Рис. 14.4. В жесткой трубке (Л) между
давлением и объемной скоростью тока
жидкости существует прямая
зависимость; в эластичном и растяжимом
кровеносном сосуде (Б) равноценному по
сравнению с жесткой трубкой
увеличению давления соответствует меньший
прирост объемной скорости (зависимость
не линейная). Сокращение гладких
мышц сосуда в результате их
раздражения (В) приводит к росту давления и
менее выраженному увеличению
объемной скорости.
ЕПС =
Кровяное давление (мм рт. ст.)
Минутный объем (л/мин или мл/мин)
Чем больше величина, выраженная в ЕПС, тем больше
сопротивление кровотоку; возрастание этой величины может
свидетельствовать (хотя и не всегда) о повышении сосудистого тонуса.
ЛИНЕЙНАЯ СКОРОСТЬ КРОВОТОКА
Ток крови так же, как и поток воды в струе, может быть
либо ламинарным, либо турбулентным. Можно представить, что
текущая жидкость состоит из тонких скользящих друг
относительно друга слоев (laminae). На каждый из этих слоев действу-
16*
244 Глава 14
ет напряжение, или усилие сдвига, замедляющее скорость их
передвижения.
Распределение слоев в кровеносном сосуде с ламинарным
течением определяется линейной скоростью кровотока, зависящей
в свою очередь от таких факторов, как размер сосуда и
гидродинамическое сопротивление (рис. 14.5). При известных
значениях гидростатического напора и вязкости линейная скорость
обратно пропорциональна радиусу или площади поперечного се-
А
Рис. 14.5. А. Распределение скоростей в струе с ламинарным потоком
жидкости. Скорость возрастает от нуля в пристеночном слое до максимального
значения в центре трубки. Б. Турбулентный поток, характеризующийся
завихрениями и воронками.
чения (S) сосуда. Таким образом, линейная скорость кровотока
..__ Q (объемная скорость кровотока)
S (площадь поперечного сечения сосуда)
выше в сосудах малого диаметра.
Средняя линейная скорость тока крови в аорте человека
(диаметр — 2 см, площадь сечения — 3 см2, объемная скорость
кровотока — 84 мл/с) вычисляется следующим образом:
V = |-=84MfCM(rS/C)=28cM/c.
В более мелких артериях линейная скорость значительно выше,
в венах большего диаметра ниже.
При возрастании линейной скорости до некоторой величины
в струе образуются завихрения (как в быстром потоке воды),
сопровождающиеся шумом — течение превращается из
ламинарного в турбулентное (рис. 14.5,Б). Эта величина определяется
числом Рейнольдса (Re):
где V —линейная скорость тока жидкости, D — диаметр сосуда,
р — плотность жидкости, ц — вязкость жидкости.
В местах разветвления сосудов завихрения образуются легче,
поэтому вероятность сосудистых поражений (артериосклероза)
в этих участках выше.
Кровообращение
245
ЗАКОН ЛАПЛАСА
Хорошо известно, что мелкие артерии и даже капилляры
разрываются гораздо реже, чем более крупные сосуды, что на
первый взгляд кажется парадоксальным; если исходить из закона
Лапласа, то это можно объяснить их малым диаметром. Закон
гласит, что давление (Р) в полом сосуде равно отношению
напряжения в его стенке (Т) к радиусу сосуда (г) (рис. 14.6).
Таким образом для кровеносных
•сосудов справедлива
зависимость:
р--Ь
Т = Рг.
Рис. 14.6. Зависимость между
давлением внутри сосуда (Р) и
напряжением в его стенке (Т), т. е.
силой, предохраняющей его от
разрыва (закон Лапласа).
.Это означает, что:
1. Повышение давления (Р)
приводит к росту напряжения
(Т).
2. Поскольку давление (Р)
обратно пропорционально
радиусу, более мелкие сосуды
могут выдерживать большее
давление.
3. Напряжение (Т) прямо
пропорционально радиусу (г)
(Т=Р-г): чем больше радиус,
тем больше напряжение, и
наоборот.
В соответствии с законом Лапласа мелкие сосуды, а также
сердца небольших размеров способны выдержать большее
давление, чем более крупные образования, и вероятность их
разрыва меньше.
В законе Лапласа речь идет о пассивном напряжении, т. е.
напряжении, зависящем от структурных особенностей самого
сосуда, таких, как количество эластических и коллагеновых
волокон. Активное напряжение связано с сокращением гладких
мышц сосуда, приводящим к его сужению и уменьшению
кровотока в нем. Если нервы, оканчивающиеся на этих мышцах,
раздражать с возрастающей частотой, давление в сосудах будет
увеличиваться, а кровоток падать (см. рис. 14.4,В).
Трансмуральное давление равно разнице между давлением,
действующим на сосуд извне, а именно со стороны окружающих
тканей и тканевой жидкости, и изнутри (кровяным давлением).
Так, при сокращении мышцы кровоток в ее сосудах может
временно прекратиться в связи с тем, что действующая извне
сдавливающая сосуд сила будет больше давления внутри сосуда.
246
Глава 14
СЕРДЕЧНЫЙ ВЫБРОС И РАСПРЕДЕЛЕНИЕ КРОВИ
Около 80—85% общего объема циркулирующей крови
находится в большом 'кругу кровообращения, остальная часть —
в малом (легочном). Распределение крови показано на рис. 14.7;
более половины всей крови, циркулирующей в сосудах большого
круга кровообращения, находится в венах.
Мелкие артерии,
5%
Артериолы,
2%
Капилляры,
5%
Рис. 14.7. Количество крови в различных отделах сосудистой системы в
процентах от общего объема.
Сердечный выброс — это общее количество крови,
выбрасываемой сердцем в единицу времени; обычно оценивают выброс
за 1 мин (минутный объем). Объем крови, выбрасываемой за
одно сокращение, называется ударным объемом; минутный
объем равен ударному объему, помноженному на частоту
сокращений сердца.
Существует несколько методов измерения сердечного
выброса (СВ), в том числе прямой и непрямой метод Фика, методы
разведения красителей или радиоактивных веществ и тепловые
методы (термодилюция) (см. гл. 2). Сердечный выброс можно
также измерить непосредственно при помощи электромагнитных
датчиков, накладываемых под прямым углом на артерию; эти
датчики определяют линейную скорость кровотока, зная которую
можно вычислить объемную скорость.
При использовании прямого метода Фика измеряется объем
потребленного (поглощенного в легких) кислорода, и
полученная величина делится на разницу между объемным
содержанием кислорода (в процентах) в артериальной (А02) и смешанной
Кровообращение 247
венозной (V02) крови:
PR __ Поглощенный 02 (мл/мин)
^Ь- A02-V02 •
При использовании непрямого метода Фика (метода
разведения индикатора) определенное количество красителя или
какого-либо другого индикатора вводят в вену; по вене краситель
поступает в правое сердце, где и перемешивается. Через
канюлю, вставленную в артерию, сразу после введения красителя
начинают собирать кровь. Далее, кровь с разведенным в ней
/ \ R
12 3 4 5 6
0 6 12 18 24 30
Время, с
Рис. 14.8. График сердечного выброса, полученный методом разведения
индикатора, R — волна рециркуляции (полулогарифмический масштаб).
красителем можно либо пропускать через денситометр (прибор
с фотоэлементом), измеряющий концентрацию индикатора и
вычерчивающий кривую его содержания, либо собирать порции
крови через определенные промежутки времени и определять
концентрацию красителя в них. В любом случае необходимо
построить кривую разведения (рис. 14.8). Прежде чем весь
введенный краситель шройдет мимо места забора проб, первые
порции его рециркулируют, что сопровождается подъемом
кривой разведения (волна R, рис. 14.8). Эту часть кривой не
учитывают, а нисходящее колено экстраполируют до пересечения
с нулевой линией (на рисунке изображено прерывистой
линией).
Сердечный выброс (СВ) определяют либо путем прямого
планиметрического измерения площади, ограниченной^ кривой,
либо вычисляя среднюю концентрацию красителя (С) за ис-
с;
"lT
2
к
с;
Q)
Ь
X
°
<з
Q-
X
к
S
J
(Л
CL
V-
I
ф
3
I
о
248
Глава 14
следуемый промежуток времени:
рр> А (количество введенного красителя, мг)
С*Время исследования (на площади, ограниченной кривой)'
pR Количество введенного красителя (12 мг)
С • Время (30 с)
После определения концентраций красителя в пробах (Сь С2,.
С3,... Сп) полученные значения наносят на ось ординат и по ним
строят график (рис. 14.8). Однако кривую разведения можно
получить и непосредственно, при помощи регистрирующего
прибора (денситометра). Средняя концентрация равна:
с=с1+с2+с3+с4+с5+св=
= 0+4,2+8,5+7,5+3,8+0 = -^-= 4 мг/л;
отсюда
пт. 12 мг введенного красителя „ ,on c ,
СВ = 4 мг/л-30 с = 3 Л/3° °' ИЛИ 6 Л/МИН-
ФАКТОРЫ, ВЛИЯЮЩИЕ НА СЕРДЕЧНЫЙ ВЫБРОС
В среднем у большинства взрослых С В составляет 5 л/мин,
варьируя в зависимости от массы и конституции. Более точным
показателем является сердечный индекс, равный сердечному
выбросу, отнесенному к площади поверхности тела (в м2). У
человека средней массы площадь поверхности тела составляет
приблизительно 1,7 м2, а сердечный индекс соответственно равен
3 л/мин/м2.
Как и следовало ожидать, физическая нагрузка влияет на СВ
и частоту сокращений сердца (табл. 14.1). Факторы,
увеличивающие частоту сокращений сердца (физическая нагрузка,
эмоциональное возбуждение и т. п.), как правило, увеличивают и
СВ. У тренированных людей (например, у спортсменов) при
физической нагрузке частота сокращений сердца возрастает не
в такой сильной степени, как у нетренированных, при таком же
Таблица 14.1. Влияние физической нагрузки на сердечный выброс
и частоту сокращений сердца у человека
Состояние
Покой
Умеренная нагрузка
Интенсивная нагрузка
Частота
сокращений сердца в 1 мин
60
100
138
Сердечный выброс,
л/мин
5,5
10,9
15,0
Кровообращение
249
приросте сердечного выброса. Это означает, что при физической
нагрузке у этих лиц увеличивается ударный объем.
За достаточно длительный отрезок времени выброс правого
^сердца, из которого кровь поступает в легочные сосуды и затем
возвращается к левому сердцу (предсердию), должен быть
таким же, как и выброс левого сердца. Это действительно имеет
место, если учитывать выброс за несколько сокращений, однако
при отдельных сокращениях ударные объемы правого и левого
сердца могут быть различными.
От момента рождения и приблизительно до десятилетнего
возраста сердечный индекс быстро возрастает, а затем к
старости постепенно уменьшается (табл. 14.2).
Таблица 14.2. Изменение сердечного
индекса с возрастом
Возраст, годы
10
20
40
60
80
Сердечный индекс,
л/мин/м2
4,3
3,6
3,0
2,7
2,5
На сердечный выброс влияют заболевания и нарушения, прл
которых уменьшается приток крови к сердцу по венам
(венозный возврат). При значительном уменьшении объема крови
(например, при кровотечениях) венозный возврат и вследствие
этого сердечный выброс падают. При расширении и ослаблении
сердца (например, при застойной сердечной недостаточности)
сердечный индекс также уменьшается из-за снижения
сократимости миокарда.
ВЕНОЗНЫЙ ВОЗВРАТ
На сердечный выброс влияют сократимость миокарда и
венозный возврат. Сократимость и факторы, от которых она
зависит, подробно обсуждаются в гл. 15. Выброс нормально
функционирующего сердца определяется объемом притекающей к
нему по венам крови (венозным возвратом). Потеря крови
(кровотечение) ведет к снижению венозного возврата и сердечного
выброса. На венозный возврат влияет также градиент давления
и сопротивление току крови в венах. Градиент давления равен
разности между давлением на периферическом конце венозной
сети и более низким давлением на центральном ее конце (в
правом предсердии). Давление в предсердии может быть равно нулю,
250
Глава 14
либо несколько больше или меньше. При увеличении давления
в предсердии градиент давления, а вслед за ним венозный воз-
врат и сердечный выброс уменьшаются.
На давление в венах и венозный возврат влияют также сила
тяжести, тонус вен и насосная функция скелетных мышц.
Считается, что при вертикальном положении тела гидростатическое
давление в венах 'нижних конечностей, т. е. на периферии,
увеличивается, что ведет к уменьшению сердечного выброса.
Однако этому частично противодействует насосная функция скелетных
£ 0 4- 8 12 16 20 24 28 32 36 40 50 60 10
8. Время, с
Рис. 14.9. Влияние ходьбы на венозное давление в мышцах ног. При начале
ходьбы давление немного возрастает, затем падает приблизительно до
30 мм рт. ст. и удерживается на этом уровне вплоть до остановки.
(Pollack A. A., Wood E. H., J. Appl. Physiol., 1, 1949.)
мышц, уменьшающая гидростатическое давление (рис. 14.9)'.
При сокращении мышц кровь проталкивается по (направлению
к сердцу, а ее обратному току препятствуют венозные клапаны.
В поддержании венозного возврата важную роль играет
дыхание. Это связано с тем, что при вдохе увеличивается
отрицательное давление в грудной полости, а это приводит к
уменьшению давления в правом предсердии (0; —2; —3 мм рт. ст.)
и, как следствие, к увеличению градиента давления в ,венах.
Постоянное повышение венозного давления может быть
признаком застойной сердечной недостаточности,
характеризующейся уменьшением венозного возврата и сердечного выброса.
Кровообращение
251
РАСПРЕДЕЛЕНИЕ КРОВОТОКА
Распределение кровотока по органам и тканям может быть
оценено либо путем прямого измерения кровотока, либо
непрямым методом с использованием разведения красителя или
других индикаторов.
В табл. 14.3 представлено распределение общего объема
крови в различных органах человека, за исключением легких, через
которые проходит вся выбрасываемая сердцем кровь. Кровоток
в пересчете на 100 г ткани выше всего в почках, затем следуют
печень, сердце и головной мозг. В процентах же от общего
объема крови лучше всего кровоснабжаются органы
пищеварительного тракта (22%), затем мышцы (18%), головной мозг (14%),
сердце (5%) и кожные покровы (4%). Кровь переносит
кислород к тканям, и количество кислорода, извлеченного 'из крови
при прохождении ее через тот или иной орган, равно разнице
между содержанием кислорода в артериальной (А) и венозной
(В) крови в сосудах этого органа (артерио-венозной разнице по
Ог). Эта разница (см. табл. 14.3) наиболее высока для ткани
сердца (9 мл на 100 г/мин). На втором месте стоят головной
мозг и печень.
Физическая нагрузка, усиливающая деятельность сердца и
кровоснабжение органов, стимулирует экстракцию кислорода.
Уменьшение гидравлического сопротивления сосудов также
способствует усиленному кровоснабжению органов.
Некоторые вещества, попавшие в сосуды головного мозга, не
проходят через стенки этих сосудов (гематоэнцефаличеокий
барьер). Лишь вода, кислород и углекислый газ легко
проникают через капилляры; другие же вещества либо проходят через
них медленно, либо не проходят вовсе.
КРОВЯНОЕ ДАВЛЕНИЕ
Текущая жидкость оказывает на стенку сосуда давление,
измеряемое обычно в миллиметрах ртутного столба (торр) или
(реже) в дин/см2. Давление, равное ПО мм рт. ст., означает,
что, если бы сосуд был соединен с ртутным манометром,
давление жидкости на конце сосуда сместило бы непрерывный
столбик ртути на высоту ПО мм. При использовании водного
манометра перемещение столбика было бы примерно в 13 раз
большие. Давление в 1 мм рт. ст.= 1330 дин/см2.
В мелких тонкостенных сосудах давлению внутри сосуда
частично противодействует давление снаружи; эта разница
между внутренним и наружным давлением называется трансмураль-
ным давлением. В артериях с высоким давлением крови этот
фактор играет незначительную роль.
Таблица 14.3. Распределение общего объема сердечного выброса по различным органам и тканям
Орган
Мозг
Сердце
Печень
Желудочно-кишечный тракт
Почки
Мышцы
Кожа
Прочие органы (кости,
костный мозг, жировая и
соединительная ткань)
Всего
Вес
органа,
кг
1,4
0,3
1,5
2,5
0,3
35
2
27
70
Кровоток в покое (при максимально
расширенных сосудах)
мл/мин
750(1500)
250 (1200)
1300 (5000)
1000 (4000)
1200(1800)
1000 (20 000)
200 (3000)
800 (4000)
6500 (6,5 л)
мл/мин
на 100 г
55
80
85
40
400
3
10
3
% общего
сердечного
выброса
14
5
23
22
18
4
14
100
Потребление кислорода в покое
А —В
разница по 02,
мл на 100 мл
крови
6
10
6
1,3
5
2,5
5
мл/мин
45
25
75
15
50
5
35
250
мл/мин
на 100 г
3
8
2
5
0,15
0,2
0,15
i
% общего
потребления
О*
18
10
30
6
20
2
14
100
В таблице приведены округленные цифры, лишь приближенно характеризующие соответствующие параметры у «среднего» человека в
покое. В скобках даны приблизительные значения крозотока в различных органах при максимально расширенных сосудах (по Folkow B.f
Neil P., Circulation, 1971),
Кровообращение
253
Давление и кровоток в легких меняются в зависимости от
положения тела (см. гл. 17).
Существует градиент давления, направленный от артерий к
артериолам и капиллярам и от периферических вен к
центральным (рис. 14.10). Таким образом, кровяное давление уменьша-
Артериальное
Артерии Вены
Рис. 14.10. Среднее давление в различных областях сосудистого русла в
состоянии покоя (/), при расширении (//) и сужении (///) сосудов. В крупных
венах, расположенных около сердца (полые вены), давление при вдохе
может быть несколько ниже атмосферного. (Sampson Wright's Applied Physiol-
ogy, Keele С A., Neil E., 1971.)
ется в следующем направлении: аорта-)-артериолы-^капилля-
ры-^венулы-)-крупные вены-*полые вены. Именно благодаря
этому градиенту кровь течет от сердца к артериолам, затем к
капиллярам, венулам, венам и обратно к сердцу (см. рис. 14.1
и 14.10). На рис. 14.10 показано также влияние на градиент
кровяного давления расширения и сужения сосудов.
СИСТОЛИЧЕСКОЕ И ДИАСТОЛИЧЕСКОЕ ДАВЛЕНИЕ
Максимальное давление, достигаемое в момент выброса
крови из сердца в аорту, называется систолическим (СД). В
диастолу же, когда -после изгнания крови из сердца аортальные кла-
254
Глава 14
паны захлопываются, давление падает до величины,
соответствующей так называемому диастолическому давлению (ДД).
Разница между систолическим и диастолическим давлением
называется пульсовым давлением (рис. 14.11). Среднее давление
(Ср.Д) можно определить, измерив площадь, ограниченную
кривой давления, и разделив ее на длину этой кривой
/р ГП Площадь под кривой
л Р-м; — длина кривой *
Колебания кровяного давления обусловлены пульсирующим
характером кровотока и высокой эластичностью и
растяжимостью кровеносных сосудов. У человека форма и амплитуда
120
со 80
Пульсовое
давление
Систолическое
давление
Среднее давление
Диастолическое
давление
Рис. 14.11. График давления в аорте с указанием систолического, диастоличе-
ского, пульсового и среднего артериального давления.
пульсовой волны меняется от аорты к периферии: амплитуда
волны в периферических сосудах выше (рис. 14.12) (у птиц
этого не наблюдается). Амплитуда же кривой объемной
скорости кровотока уменьшается от аорты к периферии. Большинство
исследователей считают, что эти изменения связаны с
отражением волн, хотя, по мнению ряда авторов, они обусловлены не
отражением волн, а геометрией сужения и эластичностью
сосудов.
Скорость распространения пульсовой волны зависит от
размера и упругости сосуда. В аорте она составляет 3—5 м/с,
в средних артериях (подключичной и бедренной) —7—9 м/с, а в
мелких артериях конечностей — 15—40 м/с.
ИЗМЕРЕНИЕ КРОВЯНОГО ДАВЛЕНИЯ
При измерении кровяного давления прямым методом в
исследуемый сосуд вводится игла или катетер, соединенный с
концом соответствующего U-образного манометра; в этом случае
о давлении судят по перемещению столбика ртути. Можно
также измерять давление при помощи гибкой мембраны,
соединенной с тензодатчиком. Однако чаще используют «епрямой ме-
Кровообращение
255
тод измерения давления при помощи сфигмоманометра
(рис. 14.13). При использовании непрямого метода на плечо
накладывается резиновая манжета соответствующего размера,
раздуваемая при помощи груши (рис. 14.13). Когда давление в
манжете начинает превышать давление в плечевой артерии,
последняя спадается и кровоток в ней прекращается. Давление,
оказываемое манжетой на руку, регистрируется манометром»
100
Q.
2
2
80
60
\
Аорта
Восходящая Грудная Брюшная Бедренная Подкожная
артерия артерия
Рис. 14.12. Давление (Р) и объемная скорость кровотока (Q) в центральных
артериях и артериях нижних конечностей. Следует отметить, что по
направлению к периферии амплитуда волн давления растет, а объемная скорость
падает.
Исследующий накладывает стетоскоп ниже манжеты и
выслушивает тоны, создаваемые текущей кровью. При спадении
артерии тоны не прослушиваются.
Затем давление в манжете медленно снижают, и когда
давление снаружи сосуда становится меньше, чем внутри его, ток
крови в отрезке артерии, расположенном ниже манжеты,
возобновляется. При этом возникают так называемые тоны Коротко-
ва, выслушиваемые при помощи стетоскопа. Показания
манометра в этот момент соответствуют систолическому давлению.
По мере того как давление в манжете продолжает падать,
кровоток в артерии растет и таны меняются; непосредственно перед
исчезновением тонов или их резким ослаблением в артерии вое-
256 Глава 14
Рис. 14.13. Непрямое измерение кровяного давления при помощи сфигмомано-
метра. Изображены манометр, манжета на плече и стетоскоп, наложенный
ниже манжеты для выслушивания тонов, создаваемых током крови.
станавливается нормальный кровоток. Давление в манжете в
этот момент равно диастолическому.
Точность этого метода зависит как от исследователя, так и
в особенности от соотношения размеров манжеты и плеча
исследуемого. Если манжета чересчур узка, то показания будут
завышенными, если же она широка — заниженными. Без поправки
на размер манжеты у тучных людей с толстыми руками
показатели артериального давления могут оказаться завышенными.
При измерении давления у взрослых используется манжета
шириной в среднем 12 см.
В табл. 14.4 даны приблизительные поправки на длину
окружности руки при использовании стандартной /манжеты.
Кровообращение
257
Таблица 14.4. Поправки на окружность плеча при использовании манжеты
стандартной ширины для определения артериального давления
Окружность плеча,
см
15-18
27—30
35—38
42—45
Поправка к показателю систолического давления
Занижено; добавить 15 мм рт. ст.
Поправка не требуется
Завышено; вычесть 10 мм рт. ст.
Завышено; вычесть 20 мм рт. ст.
НОРМАЛЬНЫЕ ПОКАЗАТЕЛИ АРТЕРИАЛЬНОГО
ДАВЛЕНИЯ
На рис. 14.14 приведены значения артериального давления у
здоровых людей в возрасте от 15 до 60 лет и старше. С
возрастом у мужчин как систолическое, так и диастолическое давление
растет равномерно, у женщин же зависимость давления от воз-
160 г-
-о Женщины
150
140
L 130
12Q
2 11°
со
а:
§100
а. 90
<
80
70
60
ф- —« Мужчины
LiT^ Систолическое
Диастолическое
J L
15 20 25 30 35 40 45 50 55 60 65
Возраст, годы
Рис. 14.14. Половые и возрастные различия в систолическом и диастоличе-
ском давлении. (Morris J. N. Modern Concepts of Cardiovascular Disease. 30,
635, 1961.)
17-1705 ^
258
Глава 14
раста сложнее: от 20 до 40 лет давление у них увеличивается
незначительно, и величина его меньше, чем у мужчин; после
40 лет с наступлением менопаузы показатели давления быстро
возрастают и становятся выше, чем у мужчин.
У тучных людей (даже после соответствующих поправок)
артериальное давление выше, чем у людей с нормальным весом.
Если при измерении давления испытуемый лежит, то в
левом сердце, артериях головы и ног давление практически
одинаково (95—100 мм рт. ст.), что связано с отсутствием
гидростатического давления, создаваемого силой тяжести; в венах же
головы и ног давление больше, чем в правом сердце (5 и 2 мм
рт. ст. соответственно). Этот градиент давления способствует
току венозной крови от ног и головы к сердцу. Показатели
давления в положении стоя (но не при ходьбе) приведены в
табл. 14.5.
Таблица 14.5. Артериальное и венозное давление при вертикальном
положении, мм рт. ст.
Область
Голова
Сердце
Нижние конечности
Артериальное давление
51
100
188
Венозное давление-
—40*>
2
90
а^ Следует отметить, что давление в венах головы, равное —40 мм рт. ст., может
Привести к спадению этих сосудов, так как давление в них в этом случае будет
отрицательным (меньше, чем давление снаружи сосуда).
При вертикальном положении тела току венозной крови к
сердцу препятствует гидростатическое давление; это приводит к
уменьшению сердечного выброса; вот ^почему длительное
стояние иногда вызывает обморок. —• * ~г>. . ,г"-л:,Т *"
Ходьба или даже 'незначительные движения ног приводят в
действие «мышечный насос» нижних конечностей, уменьшающий
влияние гидростатического давления и разницу давления между
венами ног и сердцем. В результате венозное давление
снижается (см. рис. 14.9).
При физической нагрузке повышается систолическое и диа-
столическое давление, сердечный выброс и частота сокращений
сердца. При ходьбе в умеренном темпе артериальное давление
растет (приведены значения в мм рт. ст.) :
Давление До ходьбы После ходьбы
Систолическое 117 132
Диастолическое 74 67
Кровообращение
259
При курении систолическое давление может возрасти на 10—
20 мм рт. ст. Артериальное давление повышается также при
кашле и во время полового акта; в покое и во время сна оно
существенно снижается.
На артериальное давление оказывает влияние дыхание: при
вдохе давление снижается, при выдохе — повышается; это
связано главным образом с изменениями сердечного выброса. В
момент вдоха увеличивается выброс из правого сердца в легочные
сосуды, но количество крови, возвращающейся из них в левое
сердце, уменьшается; это приводит <к уменьшению выброса из
левого желудочка. При выдохе же повышается артериальное
давление и увеличивается сердечный выброс.
Артериальное давление зависит также от поведения. Оно
повышается в состоянии тревоги и эмоционального стресса; на
него влияют стрессорные факторы современной жизни.
Определенную роль играют также генетические факторы.
Есть семьи с явным наследственным предрасположением к
гипертонии. Такое предрасположение отмечено также у некоторых
видов животных.
Норадреналин (НА), высвобождающийся из окончаний
симпатических волокон и выделяемый надпочечниками, повышает
артериальное давление. Известна опухоль надпочечников (фео-
хромоцитома), при которой усилено образование норадренали-
на, что приводит к гипертонии. Повышение давления
наблюдается и при повышенном образовании гормонов коры
надпочечников. При заболеваниях почек, когда в крови возрастает
уровень ангиотензина II, также происходит сужение сосудов >и
повышается артериальное давление.
РЕГУЛЯЦИЯ АРТЕРИАЛЬНОГО ДАВЛЕНИЯ
Выше отмечалось, что на уровень артериального давления
влияют главным образом три фактора: а) частота сокращений
сердца (ЧСС), б) изменение периферического сопротивления
сосудистого русла и в) изменение ударного объема, или
сердечного выброса (рис. 14.15).
При увеличении частоты сокращений сердца от 60 до 120 в
1 мин (рис. 14.15,2,А) незначительно повышается
систолическое и в большей степени диастолическое давление; пульсовое
давление уменьшается. Снижение частоты сокращений сердца
До 40 в 1 мин (рис. 14.15, 3) сопровождается снижением и
систолического и диастолического давления (по сравнению с
рис. 14.15, 1 и 2), однако пульсовое давление (по сравнению
с рис. 14.15, 1) несколько возрастает. При низкой частоте
сокращений сердца увеличиваются промежуток времени между
сердечными сокращениями и количество крови, оттекающей за
17*
260
Глава 14
В
л
1 1
А
ЧСС 60
с
А
+
+
120 40
Изменение ЧСС
+
I — I Ускореь
+
I
ние
"^
Изменения периферического сопротивления
лри постоянной ЧСС (60 в 1 мин)
п г
—I +
Ускорение
— Замедление
Д
Изменение ударного об'ема
при постоянной ЧСС (60 в 1 мин)
1
Рис. 14.15. Влияние изменения частоты сокращений сердца (Л),
периферического сопротивления (Б) и ударного объема (В) на показатели
артериального давления. Квадратики второй вертикальной колонки отражают давление
в покое, первой и третьей — его изменения. Повышение или снижение
систолического (С), диастолического (Д) и пульсового (П) давления указано
значками «+» и «—» наверху, внизу и в центре квадратика соответственно (см.
также текст).
этот промежуток в периферические сосуды. При высокой частоте
сокращений сердца пульсовое давление меньше, так как
вследствие уменьшения интервала между сокращениями меньше и
количество оттекающей крови.
Увеличение периферического сопротивления (Б) при
постоянной частоте сокращений сердца (60 в 1 мин) сопровождается
значительным повышением систолического и диастолического
давления; время оттока крови и пульсовое давление
уменьшаются. Снижение периферического сопротивления (рис. 14.15, 3,
Кровообращение
261
Б) приводит к существенному снижению систолического и ди-
астолического и увеличению пульсового давления.
При увеличении ударного объема (УО) (рис. 14.15, 2, В\
наблюдается значительное повышение систолического давления
и небольшое повышение диастолического; в результате
пульсовое давление возрастает. Уменьшение УО (рис. 14.15, 5, В)
Приводит к снижению систолического и диастолического
давления; при этом систолическое давление снижается в большей
степени, что сопровождается значительным уменьшением
пульсового давления.
НЕРВНАЯ РЕГУЛЯЦИЯ
Частота сокращений сердца, периферическое сопротивление,
ударный объем — все эти факторы, определяющие артериальное
давление, регулируются нервной системой. Периферическое
сопротивление изменяется в результате либо сокращения гладких
мышц сосудов (при этом сопротивление возрастает), либо их
расслабления (при этом оно уменьшается) (рис. 14.16).
Тела нейронов симпатических нервов расположены в боковых
рогах спинного мозга на уровне грудного и поясничного
отделов; их преганглионарные волокна подходят к паравертебраль-
ным или висцеральным ганглиям, где образуют синапсы. Пост-
ганглионарные волокна оканчиваются на гладких мышцах
кровеносных сосудов (рис. 14.16). Большинство этих волокон при
раздражении либо прямым путем, либо через сосудодвигатель-
ный центр продолговатого мозга вызывают сужение сосудов
путем высвобождения из своих окончаний норадреналина (адрен-
эргические волокна). Существуют, однако, и симпатические ва-
зодилятаторы, иннервирующие кровеносные сосуды скелетных
мышц и кожи. Сосудосуживающие я сосудорасширяющие
волокна, как правило, перемешаны, и (раздражение симпатических
нервов обычно сопровождается сужением сосудов ib связи с
преобладанием вазоконстрикторов; однако при помощи ряда
фармакологических агентов можно выявить и наличие
сосудорасширяющих волокон. Эти волокна, называемые холинэргическими,
при раздражении выделяют ацетилхолин, который вызывает
расширение сосудов. Они шшервируют также некоторые вены,
К парасимпатическим нервам относятся нервы головного, или
Краниального, отдела парасимпатической нервной системы
(в том числе многие черепно-мозговые нервы), посылающие
волокна к сосудам головы и шеи, и нервы сакрального отдела,
иннервирующие сосуды половых органов, мочевого пузыря и
толстого кишечника. Их длинные преганглионарные волокна
отходят от нейронов, расположенных в краниальном и сакральном
отделах. Волокна сакрального отдела выходят из спинного мозга
Афференты
языкоглоточного
нерва (IX)
Внутренняя
Наружная
сонная
артерия
Кора
головного мозга
Гипоталамус
Средний мозг
Дыхательный
центр
Продолговатый
мозг
Прессорный
Депрессорный
Отделы
Эфференты
блуждающего
нерва
СА-узел
(—Желудочек
Симпатические нервы
Кровеносные
сосуды
Парасимпатические
Ч^ / нервы
Рис. 14.16. Прессорецепторы дуги аорты и каротидного синуса,
воспринимающие артериальное давление, и афферентные волокна черепно-мозговых нервов
(IX и X), передающие информацию от них в депрессорный отдел сосудодви-
гательного центра (расположен медиально в виде треугольника). Кроме
этого изображены пути от коры и гипоталамуса, по которым также могут
передаваться команды в депрессорный и прессорный отделы. А. Хеморецепторы
каротидных и аортальных телец воспринимают изменение рСОг (повышение)
и р02 (снижение) и через нервы (IX и X) передают эту информацию в
прессорный отдел. Б. Импульсация от депрессорного и прессорного отделов идет—
через спинной мозг и соответствующие симпатические ганглии и нервы и
блуждающий нерв — к сердцу и — через нервы поясничного (П) и
крестцового (К) отделов — к кровеносным сосудам. В части А изображены
афферентные волокна обоих языкоглоточных нервов и одного блуждающего.
262
Кровообращение 263
на уровне соответствующих позвонков и заканчиваются в
ганглиях, расположенных внутри иннервируемых органов
(рис. 14.16). Из окончаний коротких постганглионарных
волокон высвобождается ацетилхолин, вызывающий расширение
сосудов. Нервы краниального отдела выходят из головного мозга
и направляются к соответствующим органам (см. также гл. 3
и Ю).
СОСУДОДВИГАТЕЛЬНЫЕ ЦЕНТРЫ
Главный сосудодвигательный центр расположен в
продолговатом мозгу. Он состоит из двух основных отделов, называемых
депрессорным и прессорным; локализация их изображена на
рис. 14.16. Раздражение более обширного латерального отдела
приводит к сужению сосудов; эфферентные импульсы при этом
идут вниз по спинному мозгу к сосудодвигательным нервам.
При раздражении же депрессорного отдела, расположенного
медиально в виде треугольника, артериальное давление падает
в результате торможения активности вазоконстрикторов или
выделения ацетилхолина симпатическими вазодилятаторами.
И сосудосуживающие и (Сосудорасширяющие нервы влияют
на артериальное давление путем изменения сосудистого
сопротивления, однако давление зависит еще от частоты сокращений
сердца и ударного объема. Эти параметры также регулируются
как симпатическими -нервами, увеличивающими частоту
сокращений сердца (кардиоакцелераторами); так и парасимпатиче*
скими (блуждающими) нервами, уменьшающими ее (кардиоин-
гибиторами). Симпатические нервы, оканчивающиеся в
желудочках, увеличивают силу сокращений и ударный объем.
СОСУДОДВИГАТЕЛЬНЫЕ НЕРВЫ И ЧАСТОТА
СОКРАЩЕНИЙ СЕРДЦА
Для осуществления рефлекса необходимо, чтобы сигнал был
воспринят рецептором и послан центром к эффекторному
органу (см. гл. 7). При этом рефлекс может затрагивать рецептор
и центр, расположенный только на уровне спинного мозга (спи-
нальный рефлекс), или же замыкаться также на уровне
головного мозга и сосудодвигательного центра; такие рефлексы
влияют на артериальное давление и частоту сокращений сердца.
К рецепторам, участвующим в регуляции гемодинамики,
относятся: 1) прессорецепторы (барорецепторы), расположенные в
каротидном синусе и дуге аорты, и 2) хеморецепторы,
локализующиеся в области разветвления каротидного синуса и дуги
аорты (аортальные тельца) (рис. 14.16).
Реакции депрессорного и прессорного отделов на
раздражения показаны на рис. 14.17 и 14.18 соответственно.
Стимул I
гПовышенное давление
в области барорецептороЕ.
По афферентным
волокнам
IX и X нервов
2
Передается в
Депрессорный центр
продолговатого мозга
Снижению
кровяного давления
Вследствие
Снижения тонуса
симпатических
вазоконстри кторов
Повышения
тонуса
вазодилята-
торов
Повышения тонуса
блуждающего нерва
(торможение
сердечной
деятельности)
Рис. 14.17. Раздражители, приводящие к возбуждению депрессорного центра
(см. текст).
Стимул
Снижение рО.
(Повышение
рС02) .
|Дыхательный|
центр
Кора больших полушарий «4-
Гипоталамус ^-
Повышению
кровяного давления
Под
влиянием
I
Действия
симпатических
вазоконстрикторов
на сосуды
Прессорный центр
Приводит к
1
Увеличению частоты
сердечных
сокращений
I
Действия
(симпатических нервов
на СА-узол
(ускорение ,
деятельности сердца)}
Стимул
Эмоции
I Возбуждение
Повышению
сократимости'
1
Действия
симпатических
нервов .
на желудочки
Рис. 14.18. Действие некоторых раздражителей на прессорный центр (см.
текст).
264
Кровообращение
265
АЛЬФА- И БЕТА-РЕЦЕПТОРЫ
Влияния симпатических нервов на частоту и силу
сокращений сердца и артериальное давление опосредованы в конечном
счете рецепторами, расположенными в соответствующих
органах и тканях.
Норадреналин (НА) и адреналин (Адр) повышают
артериальное давление и вызывают сужение большей части
кровеносных сосудов путем возбуждения альфа-рецепторов,
расположенных в сосудистой стенке. Действие альфа-стимулятора
норадреналина всегда сопровождается сужением сосудов. Адреналин же
может возбуждать бета-рецепторы некоторых сосудов (в
частности, скелетных мышц), вызывая их расширение (см. также
гл. 15).
В сердечной мышце также имеются бета-рецепторы,
возбуждение которых под действием адреналина, норадреналина и
изопротеренола приводит к увеличению частоты и силы
сокращений сердца.
Многие фармакологические препараты блокируют действие
агентов, которые в норме стимулируют альфа- и бета-рецепторы.
МЕСТНАЯ РЕГУЛЯЦИЯ КРОВООБРАЩЕНИЯ
В местной регуляции периферического кровообращения
играют роль такие факторы, как соотношение давления внутри и
снаружи сосуда и реакции гладких мышц артериол и веяул на
сосудосуживающие и сосудорасширяющие агенты. О некоторых
из этих агентов уже упоминалось в связи с обсуждением нейро-
медиаторов, альфа-, бета- и холинорецепторов.
На тонус периферических сосудов влияют также другие
факторы, в том числе вырабатываемые организмом гормоны и
биологически активные вещества. Приведем ниже некоторые из
них1:
Серотонин (С, Р) Глюкагон (Р)
Гистамин (Р) Холецистокинин (Р)
Ацетилхолин (Р) Секретин (Р)
Ангиотензин (С) Простагландины (С, Р)
Кинины (Р) Гипоксемия (Р)
Окситоцин (С, Р) Гиперкапния (Р)
Вазопрессин (С) Н+ (Р)
Аденозин (С, Р) К+ (Р)
В принципе как в физических системах, так и в некоторых
кровеносных сосудах при повышении давления растет и
объемная скорость тока жидкости, однако в ряде органов и тканей
С — сужение сосудов, Р — расширение.
266
Глава 14
организма изменение давления не сопровождается
дальнейшими изменениями кровотока. Это явление, называемое
саморегуляцией, ярко выражено в почках. К механизмам его относятся
1) деятельность самих гладких мышц (миогенная активность) и
2) влияние перечисленных выше метаболитов и биологически
активных веществ (метаболический эффект) (подробнее
см. гл. 24).
ЛИТЕРАТУРА
Alt man P. L., Dittmer D. S. (1974). Biology data book, 2nd edn., vol. Ill, Fed.
Amer. Soc. Exper. Biol.
Berne R. M., Levy M. N. (1977). Cardiovascular physiology, 3rd edn., Mosby.
St. Louis.
Folkow В., Neil E. (1971). Circulation, Oxford University Press, London.
Guy ton A. C. (1977). Basic human physiology, Saunders, Philadelphia.
Guyton A. C, Young D. B. (1978). Cardiovascular physiology III: International
Review of Physiology, vol. 18, University Park Press, Baltimore.
Little R. C. (1977). Physiology of the heart and circulation, Yearbook Medical,
Chicago.
Selkurt E. E. (1975). Basic physiology for the health sciences, Little, Brown,
Boston.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Как соотносятся между собой давление, объемная скорость
кровотока и сопротивление? Напишите формулу.
2. Какие факторы определяют линейную скорость кровотока?
3. Что такое закон Лапласа, отражающий соотношение между
напряжением в стенке сосуда, давлением в нем и его
радиусом? Напишите формулу.
4. В каких артериях и каких их участках наиболее 'высока
вероятность возникновения турбулентного потока крови?
5. Перечислите факторы, влияющие на сердечный выброс.
6. Как влияет на сердечный выброс физическая нагрузка?
7. Назовите два органа, получающие наибольшее количество
крови.
8. Что такое сфигмоманометр и как с ним обращаться? ]
9. Дайте определение а) систолического давления, б) диасто-
лического давления, в) пульсового давления, г) среднего
давления.
10. Как изменяется кривая артериального давления от аорты к
артерии ноги?
11. Каково систолическое и диастолическое давление у
мужчины 20 и 60 лет?
12. Как влияет вес тела на артериальное давление?
13. Что такое прессорецепторы, и каким образом они
способствуют повышению и понижению артериального давления?
14. Что такое рефлекторная регуляция артериального давления?
Глава 15
СЕРДЦЕ КАК НАСОС
АНАТОМИЧЕСКИЕ СВЕДЕНИЯ
Сердце представляет собой полый мышечный орган
размером приблизительно с кулак взрослого человека; вес его
составляет от 220 до 300 г у мужчин и от 180 до 220 г у женщин.
Сердце находится в грудной полости между легкими; большая
часть его расположена непосредственно слева от срединной
линии тела (рис. 15.1). Сердце состоит из четырех камер — двух
предсердий и двух желудочков. Стенки предсердий тонкие,
желудочков же — значительно толще, особенно левого, давление
в котором велико.
Более подробно анатомическое строение сердца представлено
на рис. 15.2 и 15.3, где изображены области впадения и выхода
главных вен и артерий, а также коронарных сосудов.
Коронарные артерии снабжают кровью сердечную мышцу (миокард);
венозный отток от миокарда осуществляется через широкий ко*
ронарный синус, открывающийся в правое предсердие.
Закупорка коронарных артерий или прекращение тока крови по ним
(нарушение коронарного кровообращения) приводит к уменьше-
Aojtfa
Верхняя полая вена
Легочный клапан
Правое
предсердие
Правый
атриовентрикулярный
клапан
Правый желудочек
Левое предсердие
Аортальный
клапан
Левый
атриовентрикулярный
клапан
Левый желудо^К
Рис. 15.1. Расположение сердца и его клапанов в грудной полости.
Левая общая сонная артерия
Плечеголовной ствол —
Аорта
Верхняя полая вена
Легочный ствол
Ушко правого
предсердия
Правая венечная
артерия
Задняя нисходящая
ветвь левой
венечной артерии
Левая подключичная
артерия
Левое предсердие
Левая венечная
артерия
Огибающая ветвь
левой венечной
артерии
Передняя
нисходящая ветвь
левой венечной
артерии,
проходящая в
межжелудочковой
борозде
Левая общая сонная артерия
Плечеголовной ствол
Аорта
Верхняя полая вена
Легочный ствол
Правое предсердие
(оттянуто)
Коронарный синус
<впадает в правое
лредсердие)
Передняя вена
сердца
Малая вена
сердца
Левая подключичная
артерия
Левое предсердие
(оттянуто кверху)
Большая вена
сердца
Средняя вена сердца
Рис. 15.2. Внешний вид сердца с изображением главных артерий (Л) и вен
(Б).
Плечеголовной ствол
Верхняя полая вена
Левая общая сонная артерия
Левая подключичная
артерия
Аорта
©етви правой
легочной артерии
Правые <^"
легочные вены^^--^<
Легочный
лолулунный клапан
Правое предсердие
Трехстворчатый (правый
атриовентрикулярный)
клапан'
Сухожильные струны
Нижняя полая вена*
> Ветви левой
л
легочной артерии?
Левые легочные
вены
Левое предсердие
Двустворчатый (левый
атриовентр икулярный)
клапан
Аортальный
полулунный клапан
Левый желудочек
Межжелудочковая
перегородка
Сосочковая мышца
Верхушка сердца
Нисходящая аорта
Рис. 15.3. Внутреннее строение сердца; видны клапаны и места впадения в
выхода основных артерий и вен.
Левое предсердие
Ветви левой
легочной вены
Правое
предсердие
Трехстворчаты^
«лапан "^
Правый желудочек
Межжелудочковая
перегородка
Париетальный
листок
перикарда
Полость перикарда
Митральный
клапан
Висцеральный листок
перикарда (эпикард)
Миокард
Эндокард
Левый желудочек
Верхушка
Рис. 15.4. Поперечный разрез сердца с изображением перикарда и слоев
сердца — наружного (эпикарда), среднего (миокарда) и внутреннего
(эндокарда).
269
270
Глава 15
нию кровоснабжения сердечной мышцы (инфаркт миокарда) к
нарушению функции сердца.
Сердце одето околосердечной сумкой — перикардом
(рис. 15.4); внутренний листок перикарда (эпикард) тесно
прилежит к сердечной мышце — миокарду, внутренний слой сердца
называется эндокардом. Строение сердечной мышцы подробно
описано в гл. 11.
Среди клапанов сердца различают правый атриовентрику-
лярный трехстворчатый (трикуспидальный) клапан и левый
атриовентрикулярный двустворчатый (бикуспидальный)
клапан, называемый митральным. Эти клапаны отделяют
предсердия от желудочков. Аортальный клапан называется
полулунным (рис. 15.1 и 15.3); подобный клапан имеется и в легочном
стволе. Полулунные клапаны открываются под действием
высокого давления лишь в одном направлении; при понижении
давления они вновь закрываются. В открытом и закрытом виде
трехстворчатый клапан выглядит примерно так
СЕРДЕЧНЫЙ ЦИКЛ
Фазы сердечного цикла следующие (рис. 15.5): 1) систола
предсердий; 2) фаза изометрического напряжения желудочков;
3) фаза быстрого изгнания; 4) фаза медленного изгнания;
5) фаза изометрического расслабления желудочков; 6) фаза
быстрого наполнения и 7) фаза медленного наполнения.
Как указывалось ранее (гл. 14), венозная кровь поступает
в правое предсердие, затем — через правое атриовентрикулярное
отверстие — в правый желудочек и далее через легочный ствол
оттекает к легким. Здесь кровь насыщается кислородом и по
легочным венам возвращается к левому предсердию, откуда
через левое атриовентрикулярное отверстие она поступает в левый*
желудочек. Наконец, через аорту кровь выбрасывается в
артерии большого круга кровообращения.
Сердечный цикл начинается с систолы предсердий (1).
В этой фазе кровь проталкивается через атриовентрикулярные
отверстия. Затем следует фаза изометрического напряжения
желудочков (2), происходящая при закрытых атриовентрикуляр-
ных и аортальных клапанах. Наполненный кровью желудочек
сокращается без изменения объема (см. кривую объема
желудочка), однако внутрижелудочковое давление быстро возрастает
ТО Q.
5 ®
о о
Оф
s Q.
О с
. Ф
Its* -и II si
'Ills ss || И
sums 5ssq.iiq i
5?
2
2
ф
о
'5 о
л -о
\
120
\
100
80
60
40
20
0
38
32
26
20
'
-
-'
-
-
У
<1 1
2^
,зч
/ Открытие
j аортального
V V Закрытие
1 л митрального
а
f
]
J
0
с
0
1
с
I 1
i 1
1
li
1
1
1
\
t
С
Сист
<елу£
1 1
^ 4 Ф 5Ф "6 sL- 7 s
<. ^fs ^ }^ у>
Захлопывание
^*^\ аортального
\| клапана 1 „
Yv-—Ц Давление
\ П^\ в а°рте
\ к
1 ^-^
\ 1 I ^^-
\ Давление в левом
\ j желудочке
^.
\г"
От
МИТ
Давление
в левом
пре/
Т
ола
,очков
_L J
сер/
2
j
ЦИИ
3
J
крытие
рального
лапана
J I
- А
А
А
-1
А
А
^
=>
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8
Время, с
Рис. 15.5. Кривые внутрижелудочкового давления, объема желудочка,
венозного пульса, фонокардиограмма и электрокардиограмма за один сердечный
Цикл.
271
272 Глава 15
до величины, при которой открываются аортальное клапаны
(конец второй и начало третьей фазы); вслед за этим
наступает фаза быстрого изгнания, и давление достигает максимума,
после чего оно начинает падать (фаза медленного изгнания).
Во время фазы быстрого изгнания (3) объем желудочков
быстро уменьшается, однако желудочки никогда не
опорожняются полностью. В ходе фазы медленного изгнания (4) внутриже-
лудочковое давление падает, а объем желудочков уменьшается
до минимальной величины; в этот момент аортальный клапан и
клапан легочного ствола захлопываются, что препятствует как
обратному забросу крови в желудочки, так и значительному
падению давления в аорте и легочной артерии. Захлопывание
полулунных клапанов знаменует начало фазы изометрического
(без изменения объема) расслабления желудочков (5) и
приводит к быстрому падению внутрижелудочкового давления до
определенного низкого значения, при котором открываются
правый и левый атриовентрикулярные клапаны, и кровь начинает
поступать из предсердий в желудочки (фаза быстрого
наполнения) (6). За этим следует фаза медленного наполнения (7).
Во время шестой и седьмой фаз объем желудочков возрастает,,
и, когда он достигает исходного уровня, начинается сокращение
предсердий — новый цикл.
Описанные изменения внутрижелудочкового давления и
объема тесно коррелируют с тонами сердца, венным пульсом и
электрокардиограммой.
Как правило, существуют четыре тона сердца, из которых
лишь два удается выслушать при помощи стетоскопа; однако-
эти тоны можно усилить и зарегистрировать в виде зубцов. Так
получают фонокардиограмму (рис. 15.5). Первый тон
соответствует началу систолы желудочков и закрытию атриовентрикуляр-
ных клапанов (вторая фаза). Этот тон, связанный с
многократными колебаниями крови у стенки желудочка и вибрацией
последней, наиболее звучный. Начало его примерно соответствует
пику зубца R на ЭКГ. Второй тон возникает при закрытии
полулунных клапанов, приводящем к колебаниям крови вследствие
«отдачи». Этот тон характеризуется меньшей длительностью и
большей высотой по сравнению с первым; начало его
соответствует концу зубца Т ЭКГ.
Третий тон возникает в ранней диастоле вследствие
быстрого притока крови к желудочкам; он характеризуется слабой
интенсивностью и низкой частотой. У некоторых людей
прослушивается четвертый тон; он вызван колебаниями крови во
время систолы предсердий.
Комплекс QRS на ЭКГ несколько предшествует
механическому сокращению сердца. Начало зубца Т совпадает с началом
фазы медленного изгнания, а конец примерно соответствует мо-
Сердце как насос
273*
менту захлопывания аортального клапана. Зубцы ЭКГ,
отражающие электрическое возбуждение сердца, знаменуют начала
его механического сокращения.
СИЛА СОКРАЩЕНИЙ
Когда сердечная мышца сокращается, отмечается тенденция
к укорочению (изотоническое сокращение). Это связано с
наличием в миокарде сократительных элементов (СЭ), которые
укорачиваются, и эластического компонента (ЭК), который
растягивается. Если к мышце не приложена нагрузка (нагрузка
равна 0), мышца сокращается с максимальной скоростью
о 1U h-
СО
X
Нагрузка, г
Рис. 15.6. График зависимости скорости укорочения от нагрузки на мышцуч
Vmax соответствует изотоническому сокращению, Р0 — максимальному напря*
зкению при изометрическом сокращении.
(рис. 15.6). При возрастании нагрузки от 0 до бесконечности
скорость сокращения падает и становится равной 0 при такой
нагрузке, когда мышца уже не способна укоротиться (на
рисунке: Ро). В этом случае говорят об изометрическом сокращении
мышцы. При таком сокращении в мышце развивается
напряжение. Когда сердце сокращается при замкнутых клапанах, то со-
18—1705
574 Глава 15 /
Г ~
кращение происходит именно в изометрическом режиме в
условиях постоянного объема; кровь из него не /выбрасывается
(фаза изометрического сокращения). /
НАСОСНАЯ ФУНКЦИЯ СЕРДЦА
Насосная функция сердца зависит от силы сокращения
сердечной мышцы, достигающей максимума при выбросе крови из
левого желудочка в аорту. Количество выброшенной крови, или
ударный объем, зависит от объема крови, находящейся в
желудочке в момент его сокращения, и от силы сокращения.
ЗАКОН ФРАНКА —СТАРЛИНГА
Этот закон был впервые открыт немецким ученым Отто
Франком и позднее английским исследователем Эрнестом Стар-
лингом. Закон гласит, что сила сокращения мышцы сердца тем
больше, чем сильнее растянуты ее волокна, т. е. количество
крови, выбрасываемой при сокращении, тем больше, чем больше ее
накапливается в сердце во время диастолы. Увеличение объема
крови в сердце сопровождается его растяжением и ростом
внутрисердечного давления и объема в диастоле (конечно-диас-
толического давления и объема).
Благодаря этому механизму выброс сердца может
(изменяться в зависимости от объема притекающей к сердцу крови, т. е.
венозного притока. Механизм Франка — Старлинга служит для
согласования выбросов правого и левого сердца, ударный объем
которых может изменяться от сокращения к сокращению,
однако общий объем в единицу времени должен быть одинаковым.
Так, если ударный объем левого сердца во время какого-либо
сокращения будет повышенным из-за значительного конечно-
диастолического давления или объема, при следующем
сокращении ударный объем уменьшится и будет таким же, как и
выброс правого сердца.
Таким образом, если составить график зависимости ударного
объема (выброса) от конечно-диастолического давления в левом
предсердии или длины мышечных волокон (рис. 15.7), то из него
будет видно, что по мере увеличения этих параметров ударный
объем возрастает (каждая кривая представляет собой кривую
Франка — Старлинга). Это так .называемая гетерометрическая
(связанная с изменением длины волокон) саморегуляция. Так
«сердце управляет своим выбросом.
Сердце как насос
275
X
ф
3"
(О
Q.
О
<0
сс
Q.
О)
О
о
о
(О
о.
\
60
50
40
30
20
10
_L
J_
J_
J_
0 5 10 15 20 25
Среднее давление в левом предсердии,
см вод. ст.
Рис. 15.7. Кривые, иллюстрирующие закон Франка — Старлинга (гетеромет-
рическую регуляцию) и гомеометрическую регуляцию (увеличение ударного
объема при постоянном давлении в предсердии или длине мышечных
волокон). На каждой из кривых, отражающих зависимость Франка — Старлинга,
видно, что по мере увеличения давления в предсердии и длины мышечных
волокон растет ударный объем, или сердечный выброс. Производилось
раздражение сердечных нервов с нарастающей частотой (от 0 до 4 Гц); видно, что
в результате произошел сдвиг полученных кривых (так называемых кривых
Сарнова) влево (вверх). Это означает, что при одних и тех же значениях
внутрипредсердного давления ударный объем (и соответственно
сократимость) возрастает при увеличении частоты стимуляции от 0 до 4 Гц (пример
гомеометрической регуляции).
ГОМЕОМЕТРИЧЕСКАЯ САМОРЕГУЛЯЦИЯ СИЛЫ
СОКРАЩЕНИЯ
Термин «сократимость» отражает способность сердечной
мышцы сокращаться и совершать работу при определенном
растяжении или длине ее волокон. В этих условиях действует гомео*
метрическая (при постоянной длине волокон) саморегуляция
сердца, отраженная на рис. 15.7 семейством кривых Сарнова;
при одних и тех же значениях внутрипредсердного давления
или длины волокон эти кривые сдвигаются влево, что
указывает на большую сократимость. На сократимость влияют такие
факторы, как раздражение симпатических волокон или действие
норадреналина, повышение концентрации кальция или
воздействие другими агентами типа наперстянки, применяемыми в
клинике для усиления деятельности ослабленного сердца. Сила со*
18*
276
Глава 15
кращений возрастает также при увеличении нагрузки «а сердце
вследствие повышения давления в аорте, или при увеличении
частоты сокращений.
РАБОТА СЕРДЦА
Работа сердца (А) состоит в выбросе крови и
проталкивании ее под давлением в различные части тела. Для того чтобы
переместить некий объем крови (V) на определенное
расстояние, необходимо приложить силу, или создать давление (Р).
Работа равна произведению среднего давления на объем, т. е.
A = P-V, где Р выражено в граммах, а V в см3.
В связи с тем что давление в левом желудочке в 5 раз
больше, чем в правом, а минутный объем (V) обоих желудочков
одинаков, работа левого желудочка в 5 раз превышает работу
правого. Для вычисления работы, выполняемой сердцем в
единицу времени, необходимо умножить работу, совершаемую за
один цикл, на частоту сокращений сердца.
Работа левого желудочка сердца человека, перекачивающего
при среднем давлении 100 мм рт. ст. (135 Г/см2) 5 л (5000 см3)
крови в 1 мин, составляет: 5000-135 = 675 000 Г-см = 6,75 кГ-м
(за 1 мин).
КОЭФФИЦИЕНТ ПОЛЕЗНОГО ДЕЙСТВИЯ СЕРДЦА
Коэффициент полезного действия (к.п.д.) сердца, равный
отношению совершенной работы к затраченной энергии,
составляет всего 14—25%, что говорит о значительных потерях энергии.
Большая часть ее преобразуется в тепловую энергию, т. е. не
затрачивается на совершение работы. Источником энергии для
сердца служит кислород; сердце весом 300 г может потреблять
около 27 мл кислорода в минуту, однако большая часть этого
количества расходуется на образование тепла. При физической
нагрузке и тренировке к.п.д. сердца может увеличиться.
При повышении кровяного давления нагрузка на сердце
увеличивается, а к.п.д. уменьшается. Поэтому для облегчения
работы сердца желательно, чтобы кровяное давление было
сравнительно низким, а сердечный выброс — большим.
РИТМ СЕРДЦА
Ритм сердца человека, находящегося в состоянии
физического и эмоционального покоя, т. е. частота сокращений сердца
колеблется, но в среднем составляет 70 ударов в 1 мин. Во время
сна, а также у тренированных спортсменов ритм сердца равен
приблизительно 50—60 ударам в 1 мин.
Сердце как насос
277
При физической нагрузке частота сокращений сердца и
кровяное давление увеличиваются пропорционально интенсивности
нагрузки. От 50—70 ударов в 1 мин в покое ритм сердца может
возрастать до 100 при слабой, 120 умеренной и 138 сильной
физической нагрузке. Значительно повышается ритм сердца и во
время полового акта.
ЭКСТРАКАРДИАЛЬНАЯ РЕГУЛЯЦИЯ
Ритм сердца контролируется как интракардиальными, так и
экстракардиальным1и механизмами. К последним относятся
симпатические (ускоряющие) и парасимпатические (тормозные)
сердечные нервы (рис. 14.16 и 15.8). Тела преганглионарных
симпатических нейронов находятся в боковых рогах спинного
Рис. 15.8. Схема симпатической иннервации сердца; видно, что постганглио-
нарные волокна берут начало в шейных и грудных ганглиях. Эти волокна
образуют сплетение, затем идут к сердцу, иннервируя синоатриальный и ат-
риовентрикулярный узел и желудочки (см. в тексте). Парасимпатические
(блуждающие) нервы также иннервируют синоатриальный и атриовентрику-
дярный узел.
278
Глава 15
мозга; отростки их выходят по направлению к паравертебраль-
ным ганглиям на уровне верхнего, среднего и нижнего шейных
и 1—5 грудных сегментов. Отходящие от этих ганглиев пост-
ганглионарные волокна идут к сердечному сплетению. У
человека верхний, средний и нижний сердечные нервы образованы
отростками постганглионарных нейронов, тела которых
находятся в указанных шейных ганглиях, хотя в состав этих нервов
могут входить и отростки нейронов из грудных ганглиев.
Постганглионарные волокна правых сердечных нервов иннер-
вируют преимущественно синоатриальный узел, а левых — ат-
риовентрикулярный узел; к желудочкам идут волокна с обеих
сторон. Волокна, иннервирующие синоатриальный и атриовент-
рикулярный узлы, участвуют в регуляции частоты сокращений
сердца, волокна же, идущие к желудочкам, влияют на силу
сокращений. В окончаниях сердечных симпатических нервов
высвобождается медиатор норадреналин, стимулирующий бета-
рецепторы сердца (см. гл. 14); эти рецепторы возбуждаются
также бета-адреномиметиками типа изопротеренола и
блокируются бета-блокаторами (например, пропранололом). Адреналин
также стимулирует бета-рецепторы. Возбуждение этих
рецепторов приводит к повышению частоты и силы сокращений
сердца.
В рефлекторной регуляции ритма сердца и кровяного
давления (см. гл. 14) участвуют высшие нервные центры головного
мозга (кора больших полушарий, гипоталамус и продолговатый
мозг); в ответ на афферентную импульсацию эти центры
посылают сигналы по волокнам, идущим в составе спинного мозга к
соответствующим симпатическим ганглиям и нервам (см.
рис. 14.16).
ПАРАСИМПАТИЧЕСКИЕ НЕРВЫ
Парасимпатическая иннервация сердца осуществляется
через блуждающие нервы. Тела первых нейронов этих нервов
заложены в дорсальном двигательном ядре продолговатого мозга;
их отростки идут на шее рядом с общей сонной артерией.
Длинные преганглионарные волокна направляются к внутрисердеч:
ным ганглиям, откуда исходят отростки постганглионарных
нейронов (рис. 15.8). Правый блуждающий нерв влияет
преимущественно на синоатриальный узел, вызывая замедление
(торможение) деятельности сердца. Левый блуждающий нерв
действует главным образом на проведение в атриовентрикулярном узле,
хотя часть волокон его иннервирует синусный узел. Холинэрги-
ческие (высвобождающие ацетилхолин) волокна блуждающих
нервов оказывают на сердце отрицательный хронотропный
эффект, что доказывается, в частности, тем, что пересечение блуж-
Сердце как насос
279
дающих нервов или блокада их атропином приводит к
ускорению ритма сердца вследствие устранения тормозного влияния
блуждающего нерва.
ТОНУС СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ НЕРВОВ
Так как симпатические нервы ускоряют ритм сердца, а
блуждающие замедляют, выраженность хронотропного влияния этих
нервов является показателем их тонуса. При пересечении
блуждающих нервов или их блокаде атропином наблюдается
значительное повышение частоты сокращений сердца, при
перерезке же симпатических нервов ритм сердца замедляется, но
незначительно. Это свидетельствует о преобладании
отрицательного хронотропного эффекта блуждающих нервов над
положительным хронотропным эффектом симпатических. При блокаде
или перерезке и симпатических, и блуждающих нервов у
взрослого человека ритм сердца возрастает примерно до 105
сокращений в 1 мин, что значительно выше нормального ритма
покоя. Эта частота называется собственным ритмом сердца.
РЕФЛЕКТОРНАЯ РЕГУЛЯЦИЯ РИТМА СЕРДЦА
Этот вопрос обсуждался в гл. 14, где указывалось, что
колебания кровяного давления улавливаются прессорецепторами,
что приводит к рефлекторным изменениям ритма сердца через
сердечно-сосудистый центр. Так, при повышении кровяного
давления импульсация от прессорецепторов (вызывает снижение
давления и замедление ритма сердца, при понижении же
давления частота сокращений сердца повышается. Было показано
также, что эмоции и состояние 'возбуждения, связанное с
высшей нервной деятельностью, также приводят к активации
сердечно-сосудистого центра (см. рис. 14.16 и 14.18).
ИНТРАКАРДИАЛЬНАЯ РЕГУЛЯЦИЯ
Под интракардиальной регуляцией подразумевается
способность сердца управлять -собственной деятельностью независимо
от внешних (нервных и гуморальных) влияний. В опытах на
полностью денервированном или выделенном из организма
сердце было показано, что такое сердце способно регулировать
свой ритм в соответствии с нагрузкой. Борзые с денервирован-
ными сердцами могли бежать столь же быстро, как и
нормальные собаки, и при этом частота сокращений сердца у них
увеличивалась, хотя (И не так быстро, как у собак с -ненарушенной
иннервацией сердца. Другим примером интракардиальной
регуляции может быть закон Франка — Старлинга, в соответствии
280
Глава 15
с которым при растяжении мышцы сердца увеличивается сер*
дечный выброс. На собственный ритм сердца влияет также
температура тела: при повышении температуры и энергетического
обмена этот ритм возрастает, при снижении температуры —
уменьшается.
ЛИТЕРАТУРА
Altman P. L., Dittmer D. 5. (1974). Biology data book, 2nd edn. Vol. Ill, Fed.
Amer. Soc. Exper. Biol.
Berne R. M., Levy M. N. (1977). Cardiovascular physiology, Mosby, St. Louis.
Guyton A. C. (1977). Basic human physiology, Saunders, Philadelphia.
Guyton Л. C, Young D. B. (1978). Cardiovascular physiology III: International
Review of Physiology, vol. 18, University Park Press, Baltimore.
Jensen D. (1971). Intrinsic cardiac rate regulation. Appleton-Century Crofts,
New York.
Little R. С (1977). Physiology of the heart and circulation, Year Book Medical,
Chicago.
Selkurt E. E. (1975). Basic physiology for the health sciences, Little, Brown,
Boston.
Sturkie P. D. (1976). Avian physiology. Chap 5, Springer Verlag, New York.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. К какому типу принадлежат правый и левый атриовентри-
кулярные клапаны? Опишите их строение.
2. Какая фаза сердечного цикла протекает при закрытых
клапанах?
3. В какой фазе сердечного цикла открыты атриовентрикуляр-
ные клапаны?
4. Что такое изометрическое и изотоническое сокращения?
5. Что такое механизм Франка — Старлинга?
6. Что такое гетерометрическая регуляция?
7. Что такое гомеометрическая регуляция?
8. Каков к.п.д. сердца (в процентах)?
9. Как связано потребление сердцем кислорода с его к.п.д.?
10. Назовите несколько факторов, влияющих на ритм сердца.
11. Что понимают под экстракардиальной и интракардиальной
регуляцией функции сердца?
12. Какие нервы участвуют в регуляции ритма сердца, каковы '
механизмы их регуляторных влияний?
13. Приведите пример рефлекторной регуляции ритма сердца.
Глава 16
ЭЛЕКТРОКАРДИОГРАФИЯ
ПРОВОДЯЩАЯ СИСТЕМА СЕРДЦА
В сердце человека и животных существует
специализированная, анатомически обособленная проводящая система (рис. 16.1),
Она состоит из синоатриального (СА) и атриовентрикулярного
(АВ) узлов, пучка Гиса с его левой и правой ножками и
волокон Пуркине. Эта система образована специализированными
мышечными клетками, обладающими свойством автоматизма и
высокой (по сравнению с неспециализированными мышечными
клетками предсердий и желудочков) скоростью проведения
возбуждения. Импульс зарождается в пейсмекерных клетках
СА-узла (пейсмекер первого порядка), который в норме
определяет ритм сердца. Затем возбуждение распространяется по
поверхности предсердий, вызывая их деполяризацию, после чего
переходит к атриовентрикулярному узлу (пейсмекер второго
порядка) и возбуждает (деполяризует) его. От атриовентрикуляр-
ного узла импульс следует по пучку Гиса и распространяется
вправо и влево, возбуждая мышцу желудочков.
Специализированная ткань проводящей системы составляет лишь
незначительную часть общей массы миокарда предсердий и
желудочков.
ДЕПОЛЯРИЗАЦИЯ И РЕПОЛЯРИЗАЦИЯ
Распространение электрического импульса (потенциала
действия) по проводящей системе и мышце предсердий и
желудочков сопровождается деполяризацией ih реполяризацией.
Регистрируемые в результате этого волны, или зубцы, называются
волнами деполяризации (QRS) и реполяризации (Т) желудочков.
В покое, т. е. в невозбужденном состоянии, мышца'
поляризована: наружная поверхность ее несет положительный, а
внутренняя— отрицательный заряд (рис. 16.2, 1). На рис. 16.2, 2
изображена мышца в процессе деполяризации (уничтожение
положительных и отрицательных зарядов), а на рис. 16.2, 3 —
полностью деполяризованная мышца. На записи в этот момент
регистрируется законченная волна деполяризации. Затем
следует волна реполяризации, начинающейся с того же участка,
что и деполяризация (рис. 16.2, 4). На рис. 16.2, 1 изображена
полностью реполяризованная мышца; этот момент соответствует
282
Глава 16
завершению отрицательного отклонения на записи и возврату
мышцы к состоянию покоя. Деполяризация и реполяризация
сердечной мышцы предшествуют ее механическому сокращению
(рис. 15.5).
ЭЛЕКТРОКАРДИОГРАММА
Электрокардиограмма (ЭКГ) —это запись электрической
активности (деполяризации и реполяризации) сердца,
зарегистрированная при помощи прибора — электрокардиографа, элек-
Синусный узел
Правая ножка
Рис. 16.1. Специализированная проводящая система сердца (Lipman В. S.,
Massie E., 1959.)
троды которого помещены не непосредственно на сердце, а на
различные участки тела (отведения).
. ЭКГ может быть записана непосредственно чернильным пис-
чиком или подогреваемым лером на теплочувствительной
бумаге. Полоса пропускания частот для приборов с механическим
писчиком ограничена (около 80—100 Гц) вследствие его
инерционности, однако для регистрации ЭКГ человека этого
достаточно. Можно также зарегистрировать ЭКГ при помощи
высокочастотного (до 500 Гц) прибора с фотографической или
электронной записью; при этом более точно воспроизводятся форма
и крутизна зубцов ЭКГ у животных с большой частотой
сокращений сердца (свыше 300 ударов в 1 мин) —у крыс, птиц и
других мелких животных.
ОТВЕДЕНИЯ
Запись, получаемая при отведении электрической активности
непосредственно от сердца, называется электрограммой (ЭГ).
При наложении электродов на различном расстоянии от сердца
Электрокардиография
283
1
Рис. 16.2. Деполяризация и реполяризация миокарда (желудочек). Мышца
в состоянии покоя (1)\ положительные и отрицательные заряды
сбалансированы и не передвигаются. Затем импульс начинает распространяться по
миокарду (2 — сердечная мышца деполяризована частично, 3— деполяризована
вся мышца сердца, при этом регистрируется полный комплекс QRS). Затем
следует реполяризация сердечной мышцы (4 — начало реполяризации, 1 —
реполяризация завершена). Обратите внимание на форму зубцов ЭКГ на
?азличных стадиях деполяризации и реполяризации (см. текст). На рис. 16.2
/) Т-зубец отрицателен; таким он был бы в том случае, если бы скорости
деполяризации и реполяризации были равны, однако у большинства
здоровых людей эти скорости различны и зубец Т направлен вверх. (Katz L. N.
Electrocardiography, 1947.)
получают непрямые отведения, в том числе отведения от
конечностей и грудные, обозначаемые V. Эти отведения могут быть
как униполярными, та.к -и биполярными. При биполярных
отведениях электрическая активность улавливается обоими
электродами, а при униполярных используется один активный и один
неактивный (нулевой, индифферентный) электрод. Разность по-
284
Рис. 16.3. Треугольник Эйнтхове-
на и положение электродов в
стандартных отведениях (I, II и
III).
Глава 16
тенциалов, регистрируемая при
униполярном отведении,
примерно в два раза меньше, чем при
биполярном. Гольдбергер
предложил усилить стандартные
униполярные отведения таким
образом, чтобы величина
регистрируемой разности потенциалов
составляла около 70% амплитуды
биполярного отведения. Такие
отведения называются усиленными
униполярными отведениями от
конечностей (aVR — от правой
руки, aVL — от левой руки,
aVF — от левой ноги).
СТАНДАРТНЫЕ ОТВЕДЕНИЯ
ОТ КОНЕЧНОСТЕЙ
К этим биполярным
отведениям относятся: первое (I)
отведение (правая рука, ПР — левая
рука, ЛР); второе (II) отведение
(ПР — левая нога, ЛН) и третье
(III) отведение (ЛР — ЛН). С
известным допущением можно
считать, что эти три отведения
образуют равносторонний
треугольник, около центра которого
расположено сердце (рис. 16.3).
ГРУДНЫЕ, ИЛИ ПРЕКАРДИАЛЬНЫЕ,
ОТВЕДЕНИЯ
При снятии этих
униполярных отведений активный
электрод накладывают на различные
точки грудной клетки (рис. 16.4)1„
обозначаемые цифрами (Vb V3,
V3, V4, V5, V6). Эти отведения
отражают электрические процессы
в более или менее
локализованных участках и помогают
выявлять ряд сердечных заболеваний.
Рис. 16.4. Прекардиальные, или грудные,
униполярные отведения, обозначаемые V.
Изображены отведения Vi, V2, V3, V4,
V5 и V6. Второй электрод —
индифферентный.
Электрокардиография
285
СКОРОСТЬ ПРОТЯЖКИ БУМАГИ И УСИЛЕНИЕ
При снятии ЭКГ человека используется стандартная скорость-
протяжки бумаги — 25 мм/с. Промежуток между двумя
горизонтальными линиями на бумаге, равный 1 мм, служит для отсчета
временных интервалов и соответствует 0,04 с. Вертикальные ли-
1
1
1
1
1
1
f
/
/
С(
э
К
\
р
г -
р
D
эгмент
h
1
1
J_
р
- п
-м
1
1
1
1
1
1
1
1
i
1-
l
\
i
1
ч
N
Q
eQR
RI
<ч
| Сегм
1 1 >
1 \^~
\Г
\f
i'<
ь
s [
Г
т
ент
i
1
1
1
i
с
. ..1
/
ч
S-"
г
-0,2
г
с-
т
—>
/
с
с
| <
i
\
•J
Л \
1
J
ч
Э J
и—
/ 1
\
Рис. 16.5. Типичная ЭКГ человека во втором отведении (фронтальная
плоскость). Обозначены зубцы Р, Q, R, S и Т и сегменты Р—R, S—Т и Q—Т#
Промежутки между линиями на бумаге составляют 1 мм (здесь увеличено)!
нии, также нанесенные через 1 мм, являются масштабом ампли^
туды, -или вольтажа (рис. 16.5). За стандартный вольтаж при
снятии ЭКГ человека принят 1 мВ. Если приложить напряжение
•в 1 мВ ко входу прибора, на бумаге регистрируется
вертикальное отклонение, равное 10 мм. Отводящие электроды
накладываются или привязываются на соответствующие участки с
использованием специальной пасты, обеспечивающей надежный
электрический контакт. На приборе имеются обозначения для
различных отведений.
286
Глава 16
ЗУБЦЫ И ИНТЕРВАЛЫ ЭКГ
На рис. 16.5 изображена типичная нормальная ЭКГ человека
в одном из стандартных отведений; конфигурация, длительность
и амплитуда зубцов приведены в табл. 16.1. Зубец Р
соответствует деполяризации предсердия, комплекс QRS—иачалу
деполяризации желудочков, зубец Т—реполяризации желудочков.
Зубец U обычно отсутствует.
Таблица 16.1. Зубцы нормальной ЭКГ человека
Обозначение
Р
0
R
S
QRS
Т
Характеристика
Отражает деполяризацию предсердий;
обычно положительный '
Отражает начало деполяризации
желудочков, отрицательный, может
отсутствовать
Главный зубец деполяризации
желудочков, положительный
(направлен вверх)
Отражает окончание деполяризации,
отрицательный, может
отсутствовать
Совокупность зубцов, отражающих
деполяризацию желудочков
Отражает реполяризацию, у человека
положительный (направлен вверх)
Диапазон
длительности, с
0,08—0,12
см. QRS
0,08—0,10
колеблется
в среднем
0,28
Диапазон
амплитуды
в I, II и
III
отведении, мм
0,55—1,25
0,36—0,61
5,5—11,5
1,5-1,7
1,2—3,0
Таблица 16.2. Интервалы ЭКГ
Обозначение
интервала
P-R
Q —Т
S —Т
| Характеристика интервала
От начала Р до начала R (или Q, если таковой
имеется)
От начала R (или Q, если таковой имеется) до
конца S (или R, если S отсутствует)
От начала Q до конца Т; соответствует
деполяризации и реполяризации желудочков
От конца S до начала Т
Длительность, с
0,18—0,20
0,08
0,38—0,31
При анализе ЭКГ большое значение имеют временные
интервалы между некоторыми зубцами (табл. 16.2). Отклонение
длительности этих интервалов за пределы нормы может
свидетельствовать о нарушениях функции сердца.
Электрокардиография
28Т
ЭЛЕКТРИЧЕСКАЯ ОСЬ СЕРДЦА
В 1908 г. Эйнтховен предложил регистрировать
электрическую активность сердца при помощи трех удаленных от него
электродов, накладываемых на конечности таким образом, что
получаемые в результате три отведения образуют стороны
примерно равностороннего треугольника, в центре которого распо-
Рис. 16.6. Отношение электрической оси сердца к отведениям. Методика оп~
ределения оси приведена на рис. 16.6, Г. Комплекс QRS (представленный в<
основном зубцом R) в отведении 1 направлен вверх и величина его состав*-
ляет два деления; в третьем отведении он равен трем делениям и во втором—*•
пяти. Таким образом, 11 = 1+111 (5=2+3), что справедливо также для
каждого изображенного здесь треугольника (закон Эйнтховена). Подробнее см.
текст. (Greenspan К., 1977, с изменениями.)
ложено сердце (рис. 16.3). Согласно закону Кирхгофа,
алгебраическая сумма всех электродвижущих сил (э.д.с.) в
замкнутой цепи равна нулю. В треугольнике, изображенном на
рис. 16.6, Л, э.д.с. в первом отведении направлена от ПР
(отрицательный полюс) к ЛР (положительный полюс) (указано
стрелками). В третьем отведении э.д.с. направлена от ЛР
(отрицательный полюс) к ЛН (положительный полюс), а во
вторам— от ПР (отрицательный полюс) к ЛН (положительный
полюс). Таким образом, алгебраическая сумма э.д.с. первого и
третьего отведений, направленных к ЛН, равна э.д.с. второго
отведения, также направленной к ЛН, но с другой стороны
(т. е. 1 + 1П = П или 1 + 111—11 = 0. Отсюда 1 = 11—III и 111 =
= 11—1). О величине э.д.с. можно судить по пропорциональной
288
Глава 16
ей высоте зубца в соответствующем отведении (рис. 16.6, А—Г).
На рис. 16.6, В зубцы направлены вверх (положительны) во
всех трех отведениях, так как возбуждение впервые возникает
^ближе к отрицательным электродам: ПР в первом и втором
отведениях и ЛР в третьем.
Электрическая ось сердца — это вектор, который отражает
среднюю величину и направление э.д.с, действующей во время
электрической систолы сердца. Электрическая ось указывает, в
каком направлении действует максимальная э.д.с. в течение
наибольшего времени. Методика определения электрической оси
показана на рис. 16.6, Г. Для этого используются любые два
отведения (обычно I и III). В первом отведении на рис. 16.6, Г
величина комплекса QRS (представленного преимущественно
зубцом R) соответствует двум делениям, в третьем—трем. Эти
значения откладываются от нулевых точек (на рисунке из этих
точек исходят сплошные линии) на ось соответствующего —
первого и третьего — отведения.
Из концов отрезков, отложенных на осях отведений и равных
2 и 3 единицам соответственно в отведениях I и III,
восстанавливаются перпендикуляры (прерывистые линии). Рисуется
стрелка, идущая от точки пересечения сплошных (нулевых)
линий до точки пересечения прерывистых линий, исходящих из
концов отрезков на осях отведений I >и III. Эта стрелка (вектор)
идет под углом между 0 >и 90°, или примерно 65°. Следует
отметить, что амплитуда зубца во втором отведении равна пяти
делениям, что соответствует правилу 11 = 1 +III (пять делений во
втором отведении равны сумме двух делений в первом и трех —
в третьем).
Если электрическая ось сердца параллельна или почти
параллельна линии того или иного отведения, например первого
отведения на рис. 16.6, Б, амплитуда зубцов в этом отведении
будет наибольшей. Если же ось сердца направлена
перпендикулярно отведению (отведение I на рис. 16.6, Л), то в этом
отведении зубец не запишется. Это связано с тем, что электрическая
активность возникает приблизительно у центра отведения и
распространяется в дальнейшем в обе стороны, так что
возникающие импульсы взаимно гасятся и разность потенциалов
равна 0.
Амплитуда зубцов отведения III на рис. 16.6, В мала, в связи
с тем что электрическая ось сердца почти перпендикулярна
этому отведению.
ОТКЛОНЕНИЯ ЭЛЕКТРИЧЕСКОЙ ОСИ СЕРДЦА
В норме направление электрической оси сердца колеблется
от 0 до 90° (рис. 16.6, Г). Если направление оси находится в
интервале от 0 до —90 °С, говорят об отклонении оси влево
Электрокардиография
289
{рис. 16.6, Г). Такое направление часто встречается у
низкорослых, тучных людей и свидетельствует не столько о патологии,
сколько о смещении анатомической оси сердца влево.
Если же направление оси колеблется в промежутке от +90
до +180° (рис. 16.6, Г и 16.7), речь вдет об отклонении оси
вправо, что уже более подозрительно в смысле патологии серд-
Нормальное
положение
оси в = 60° .
'А
А
■1
in _Д_
Отклонение оси
влево в = 0°
■ А
«А
„, -у"
Отклонение оси
вправо в = 120°
■ Т
■ А
|"1
Рис. 16.7. Изменение конфигурации желудочковых комплексов в отведениях
I, II и III под влиянием отклонения электрической оси влево и вправо.
Обратите внимание на инвертированный (отрицательный) зубец в отведении III
при отклонении оси влево и в отведении I — при отклонении оси вправо.
ца. На рис. 16.7 показано, как отражается отклонение
электрической оси вправо и влево на амплитуде желудочковых
комплексов в отведениях I, II и III.
ПАТОЛОГИЧЕСКИЕ ИЗМЕНЕНИЯ ЭКГ
Существуют два основных типа патологических изменений
ЭКГ: к первому относятся нарушения ритма и возникновения
возбуждения, ко второму — нарушения проведения возбуждения
и искажения формы и конфигурации зубцов.
Аритмии, или нарушения ритма сердца, характеризуются
нерегулярным поступлением импульсов из СА-узла, что и
приводит к аритмии. Ритм, или частота сокращений сердца, может
быть низким (брадикардия), неправильным или очень высоким
(предсердная тахикардия, рис. 16.8). Предсердные
преждевременные сокращения (экстрасистолы) характеризуются
укороченным Р—Р интервалом, после которого следует длинный
Р—Р интервал (рис. 16.8, А). При этом может наблюдаться
значительное замедление ритма сердца, характеризующееся
длительными паузами и увеличенным, но нерегулярным интерва-
19—1705
290
Глава 16
лом Р—Р. Это нарушение возникновения (импульса в СА-узле.
При желудочковых экстрасистолах, когда возбуждение
возникает в эктопическом очаге, локализованном в стенке желудочка,
преждевременное сокращение характеризуется искаженным
комплексом QRS (рис. 16.8, В). Желудочковая тахикардия
проявляется быстрыми регулярными разрядами эктопического
очага, расположенного в желудочке (рис. 16.8, Д).
Предсердная тахикардия (230 в 1 миьЛ
я vYvV
Желудочковая тахикардия
Мерцание предсердий
Трепетание предсердии
Желудочковая экстрасистола
Рис. 16.8. Аритмии сердца (подробнее см. текст). (Greenspan K-, Т966.)
Фибрилляция предсердий или желудочков характеризуется
нерегулярными аритмичными сокращениями, неэффективными в
гемодинамическом отношении. Особую опасность представляет
фибрилляция желудочков, которая приводит к летальному
исходу. Фибрилляция предсердий не столь опасна и проявляется
нерегулярными аритмическими сокращениями, при которых
частота сокращения предсердий в 2—5 раз выше, чем желудочков
(рис. 16.8, Е). При этом на каждый зубец R приходится 2,1 или
3 нерегулярных зубца Р (называемых в этом случае зубцами F).
При трепетании предсердий наблюдаются более регулярные и
менее частые предсердные комплексы, частота которых все же
в 2—3 раза превышает частоту сокращения желудочков
(рис. 16.8, Ж).
Мерцание предсердий может вызываться множественными
эктопическими очагами в их стенке, тогда как разряды одиноч-
Электрокардиография
291
ного эктопического очага, очевидно, сопровождаются
трепетанием предсердий. Нормальный предсердный ритм может быть
восстановлен соответствующей медикаментозной терапией или
толчком электрического тока (дефибрилляция предсердий).
НАРУШЕНИЯ ПРОВЕДЕНИЯ
Коронарная болезнь, и в частности свежий инфаркт
миокарда, возникает вследствие нарушения кровоснабжения миокарда,
что влечет за собой изменение последнего. На рис. 16.9
приведены изменения комплекса QRS при свежем инфаркте миокарда
и через некоторое время после выздоровления. В острой стадии
наблюдаются выраженные изменения зубцов Q и Т и сегмента
Отведения
! II III aVR aVL aVF
Норма
Острый
период
-JU J\f—L^ -y^r _J\—a^
Выздоровление
Рис. 16.9. Эволюция ЭКГ в отведениях I, II, III, aVL и aVF при нарушении
коронарного кровообращения (инфаркт миокарда). При свежем инфаркте в
ряде отведений наблюдается патологический зубец Q, отрицательный зубец Т
и смещение кверху сегмента S—Т. Через несколько недель ЭКГ почти
восстанавливается до нормы — стадия выздоровления. (Lippman В. S., Mas-
sie E. 1959, с изменениями.)
S—Т. Следует отметить, в частности, подъем сегмента S—Т и
инвертированный зубец Т в некоторых отведениях. Прежде
всего наступает ишемия миокарда (нарушение его кровоснабжения,
болевой приступ), повреждение ткани с последующим
образованием омертвевшего участка. Эти нарушения кровообращения в
сердечной мышце сопровождаются изменениями проведения.
Нарушения коронарного кровотока приводят также к аритмиям.
19*
292
Глава 16
БЛОКАДЫ СЕРДЦА
Блокаду сердца при нарушениях в области СА-узла (СА-бло-
када) не следует смешивать с замедлением ритма, или
остановкой сердца. Такая блокада характеризуется длительными
паузами и увеличением интервала Р—Р, кратным обычному интервалу
Р—Р; при этом наблюдается выпадение предсердных зубцов (Р)
и желудочковых комплексов. Возникновение возбуждения в
СА-узле не нарушено, однако это возбуждение не выходит за
пределы синусного узла, а блокируется в нем.
Блокада
1 -й степени
Блокада 2-й степени
Полная блокада
Рис. 16.10. Атриовентрикулярная блокада (см. текст). (Greenspan K-, 1966.)
Атриовентрикулярная блокада может быть разной степени —
первой, второй и третьей (рис. 16.10). При блокаде первой
степени наблюдается постепенное увеличение интервала Р—R с
возможным последующим выпадением одного желудочкового
сокращения.
При блокаде второй степени наблюдается нормальное
чередование зубцов Р с периодическим выпадением желудочкового
комплекса. Частота выпадения желудочковых комплексов может
быть самой различной: от каждого второго до любой другой
периодичности.
При полной поперечной блокаде зубцы Р следуют с более
высокой частотой, чем желудочковые комплексы. Это
свидетельствует о том, что желудочки и предсердия сокращаются
независимо друг от доуга, и о том, что проведение импульсов к
желудочкам блокировано в области А—В-узла и имеют место лишь
эктопические желудочковые сокращения. Больным с полной
атриовентрикулярной блокадой часто вживляют искусственные
электрические водители ритма (пейсмекеры), задающие
желудочкам ритм, соответствующий ритму предсердий.
Электрокардиография
293
ЛИТЕРАТУРА
Altman P. S., Dittmer D. S. (1974). Biology data book, 2nd edn, vol. Ill, Fed.
Amer. Soc. Exper. Biol.
Berne R. M., Levy M. V. (1977). Cardiovascular physiology, 3rd edn., Mosby,
St. Louis.
Burch G., Windsor T. (1972). A primer of electrocardiography, 6th edn., Lea and
Febiger, Philadelphia.
Greenspan K. (1977). Physiology, 2nd edn. (1966) and 4th edn. (1977) edited
by E. E. Selkurt, Little, Brown and Co., Boston.
Lipman B. S., Massie E. Clinical scalar electrocardiography, 4th edn. (1959) and
Lipman, Massie and Kleiger (1972) 6th edn. Year Book Medical Publishers,
Inc., Chicago.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Нарисуйте схему проводящей системы сердца человека.
2. Что такое пейсмекеры первого и второго порядка, почему их
так называют?
3. Нарисуйте схему треугольника Эйнтховена <и
взаимоотношений между электрическими процессами в сердце и зубцами
стандартных (I, II и III) отведений.
4. Что такое закон Эйнтховена, и каково его значение?
5. При снятии стандартных отведений у больного обнаружено,
что в первом отведении комплекс QRS почти не
регистрируется, а во втором и третьем отведениях величина его
одинакова. Что это означает? Каково направление электрической оси
сердца?
6. Из трех стандартных отведений наибольшую величину
зубец R имеет во втором. Каково направление электрической оси
сердца?
7. При снятии ЭКГ человека обычно используют бумагу с
нанесенными через 1 мм линиями. Чему соответствуют эти линии
в вертикальном и горизонтальном направлениях?
8. Каково направление электрической оси, если зубец R равен
двум делениям в первом и трем — в третьем отведении?
9. Изобразите схематично атриовентрикулярную блокаду второй
степени.
Глава 17
ДЫХАНИЕ
Химическое восстановление молекулярного кислорода с
образованием воды служит для млекопитающих основным
источником энергии. Эта реакция происходит главным образом в
митохондриях. Без нее жизнь не может продолжаться дольше
нескольких секунд. Восстановлению кислорода сопутствует
образование С02 преимущественно в результате окислительного фосфо-
рилирования глюкозы, которое тоже протекает главным образом
в митохондриях.
Кислород С02 не происходит непосредственно из
молекулярного кислорода. Использование 02 и образование С02 связаны
между собой промежуточными метаболическими реакциями;
теоретически каждая из них длится короткое время.
Обмен 02 и С02 между организмом и средой называется
дыханием. У высших животных процесс дыхания осуществляется
благодаря ряду последовательных процессов. 1. Обмен газов
между средой и легкими, что обычно обозначают как «легочную
вентиляцию». 2. Обмен газов между альвеолами легких и кровью
(легочное дыхание). 3. Обмен газов между кровью и тканями.
Наконец, газы переходят внутри ткани к местам потребления
(для 02) и от мест образования (для С02) (клеточное дкхание).
Выпадение любого из этих четырех процессов приводит к
нарушениям дыхания и создает опасность для жизни животного.
Движение газов в дыхательной системе и между средой и
тканями происходит в результате разницы давлений.
Пониженное давление 02 в ткани заставляет газ двигаться к ней. Для
С02 градиент давления направлен в обратную сторону, и С02
переходит в окружающую среду (табл. 17.1). Поэтому изучение
физиологии дыхания, по существу, представляет собой изучение
этих градиентов и того, как они поддерживаются. (Обратите
внимание в табл. 17.1 на то, что давление водяных паров в
организме выше, чем в окружающей среде, и, таким образом, при
дыхании организм теряет воду.)
АНАТОМИЯ
Дыхательная система млекопитающих состоит из тканей и
органов, обеспечивающих легочную вентиляцию и легочное
дыхание (воздухоносные пути, легкие и элементы костно-мышеч-
Дыхание 2%
Таблица 17.1. Среднее давление (торры) и содержание газов (%)
в окружающей среде и в организме человека при барометрическом давлении
760 торр
Газ
Кислород
Двуокись
углерода
Водяные пары
Окружающая среда
О)
к
а
О)
га
с*
159
0,3
6
Я
Я
га
о.
О)
«=С
о
о
20,94
0,04
0,78
Альвеолы
0)
к
а
О)
га
100
40
47а)
О)
Я
а
га
о,
О)
еС
О
13,6
5,26
6,18
Артериальная кровь
большого
круга
4>
К
а
CJ
и
ч~
95
40
47
О)
Я
а
га
*
о,
о
еС
О
и
20б)
1,2В>
Ткани
0)
к
а
о
д
<40
^46
Артериальная кровь
легочного
круга
Я
а
О)
ч
CQ
га
40
46
47
«и
га
с*
о
и
15^)
1,4В)
а; При 37 °С.
б| В мл 02 на 100 мл крови.
в) В мМ.
ной системы). К воздухоносным путям относятся: нос, полость
носа, носоглотка, гортань, трахея, бронхи и бронхиолы. Легкие
состоят из бронхиол и альвеолярных мешочков, а также из
артерий, капилляров и вен легочного круга кровообращения. К
элементам ко сто-мышечной системы, связанным с дыханием,
относятся ребра, межреберные мышцы, диафрагма и
вспомогательные дыхательные мышцы.
ВОЗДУХОНОСНЫЕ ПУТИ
Нос и полость носа служат проводящими каналами для
воздуха, в которых он нагревается, увлажняется и фильтруется
(рис. 17.1, А). В полости носа заключены также обонятельные
рецепторы. Наружная часть «оса образована треугольным
костно-хрящевым остовом, который покрыт кожей; два овальных
отверстия на нижней поверхности — ноздри — открываются
каждое в клиновидную полость носа. Эти полости разделены
перегородкой. Три легких губчатых завитка (раковины) выдаются из
боковых стенок ноздрей, частично разделяя полости на четыре
незамкнутых прохода (носовые ходы). Полость носа выстлана
богато васкуляризованной слизистой оболочкой. Сальные
железы и многочисленные жесткие волоски, а также снабженные
ресничками эпителиальные и бокаловидные клетки служат для
очистки вдыхаемого воздуха от твердых частиц. В верхней части
296
Глава 17
полости лежат обонятельные клетки (обонятельные рецепторы).
Носовые ходы открываются в носоглотку.
Гортань лежит между трахеей и корнем языка. Полость
гортани разделена двумя складками слизистой оболочки, не
полностью сходящимися по средней линии. Пространство между
этими складками — голосовая щель — защищено пластинкой
волокнистого хряща — надгортанником (рис. 17.1, А). По краям
Лобная пазуха
Турецкое седло
Клиновидная пазуха
Глоточная
миндалина
Отверстие —■
евстахиевой труб!
Носоглотка
Мягкое небо
Ротовая часть
глотки
Гортанная част
глотки
Надгортанник
Голосовая склад
Гортань
Перстневидны
хрящ
Пищевод
Трахея
Преддверие
Твердое небо
Язык
Подбородочно-
язычная мышца
Подбородочно-
подъязычная
мышца
Подъязычная кость
Щитовидный хрящ
Перстневидный
хрящ
Перешеек
щитовидной железы
Рис. 17.1. Дыхательные пути. А. Анатомия воздухоносных путей; продольный
разрез через полость носа, рот и гортань. Б. Анатомия легких, гортани,
трахеи, бронхов и бронхиол; части лежащей над ними легочной ткани удалены,
чтобы показать разветвление.
Дыхание
297
Гортань
Щитовидная железа,
Верхушка легкого
Правый бронх
Трахея
Вентральные ветви
аерхнедолевого
бронха
Среднедолевой
бронх
Вентральные
ветви нижне
долевого бронха
Левый бронх
Внутренняя, или
медиастинальная
поверхность легкого
Вентральные ветви
верхнедолевого
бронха
Вентральные
ветви
нижнедолевого
бронха
Дорсальные
бронхиальные ветви
Рис. 17.1. (Продолжение).
голосовой щели в слизистой оболочке лежат фиброзные
эластичные связки, которые называются нижними, или истинными,
голосовыми складками (связками). Над ними находятся ложные
голосовые складки, которые защищают истинные голосовые
складки и сохраняют их влажными; они помогают также
задерживать дыхание, а при глотании препятствуют попаданию пищи
в гортань. Специализированные мышцы натягивают и
расслабляют истинные и ложные голосовые складки. Эти мышцы
играют важную роль при фонации, а также препятствуют попаданию
каких-либо частиц в дыхательные пути.
Трахея начинается у нижнего конца гортани (рис. 17.1, Б)
и спускается ;в грудную полость, где делится на правый и левый
бронхи; стенка ее образована соединительной тканью и хрящом.
У большинства млекопитающих хрящи образуют неполные
кольца. Части, примыкающие к пищеводу, замещены фиброзной
связкой. Правый бронх обычно короче и шире левого. Войдя
в легкие, главные бронхи постепенно делятся на все более
мелкие трубки (бронхиолы), самые мелкие из которых — конечные
298
Глава 17
бронхиолы — являются последним элементом воздухоносных
путей. От гортани до конечных бронхиол трубки выстланы
мерцательным эпителием.
ЛЕГКИЕ
В целом легкие имеют вид губчатых, пористых конусовидных
образований, лежащих в обеих половинах грудной полости.
Наименьший структурный элемент легкого—долька (рис. 17.2)
состоит из конечной бронхиолы, ведущей в легочную бронхиолу
и альвеолярный мешок. Стенки легочной бронхиолы и
альвеолярного мешка образуют углубления — альвеолы. Стенки
альвеол состоят из одного слоя эпителиальных клеток типа I и
окружены легочными капиллярами. Внутренняя поверхность
альвеолы покрыта поверхностно-активным веществом сурфак-
тантом, состоящим из фосфолипопротеинов или липо-полисаха-
ридов. Как полагают, сурфактант является продуктом секреции
Рис. 17.2. Долька легкого; видны бронхиола, альвеолярные структуры и их
отношение к сосудам малого и большого круга кровообращения.
Дыхание
299
гранулярных пневмоцитов (клетки типа II). Отдельная
альвеола, тесно соприкасающаяся с соседними структурами, имеет
форму неправильного многогранника и приблизительные
размеры до 250 мкм. Принято считать, что общая поверхность
альвеол, через которую осуществляется газообмен, экспоненциально
зависит от веса тела. С возрастом отмечается уменьшение
площади поверхности альвеол.
ПЛЕВРА
Каждое легкое окружено мешком, образованным серозной
оболочкой — плеврой (рис. 17.3). Наружный (париетальный)
листок плевры примыкает к внутренней поверхности грудной
стенки и диафрагме, внутренний (висцеральный) покрывает
легкое. Щель между листками называется плевральной полостью.
При движении грудной клетки внутренний листок обычно легко
Рис. 17.3. Изменения внутрилегочного и внутриплеврального давления на
протяжении цикла вдох — выдох. Показаны также изменения скорости тока
воздуха и объема легких.
скользит по наружному. Давление в плевральной полости всегда
меньше атмосферного (отрицательное). В условиях покоя внут-
риплевральное давление у человека в среднем на 4,5 торр ниже
атмосферного (—4,5 торр). Межплевральное пространство
между легкими называется средостением; в нем находятся трахея,
. зобная железа (тимус) и сердце с большими сосудами,
лимфатические узлы и пищевод.
300
Глава 17
КРОВЕНОСНЫЕ СОСУДЫ ЛЕГКИХ
Легочная артерия несет кровь от правого желудочка сердца.
Она делится на правую и левую ветви, которые направляются
к легким. Эти артерии ветвятся, следуя за бронхами, снабжают
крупные структуры легкого и образуют капилляры, оплетающие
стенки альвеол (рис. 17.2).
Воздух в альвеоле отделен от крови в капилляре 1) стенкой
альвеолы, 2) стенкой капилляра и в некоторых случаях 3)
промежуточным слоем между ними. Из капилляров кровь
поступает в мелкие вены, которые в конце концов соединяются и
образуют легочные вены, доставляющие кровь в левое предсердие.
Бронхиальные артерии большого круга тоже приносят кровь
к легким, а именно снабжают бронхи и бронхиолы,
лимфатические узлы, стенки кровеносных сосудов и плевру. Большая часть
этой крови оттекает в бронхиальные вены, а оттуда — в
непарную (спра/ва) и в полунепарную (слева). Очень небольшое
количество артериальной бронхиальной крови поступает в
легочные вены.
ГРУДНАЯ ПОЛОСТЬ
Грудная полость ограничена сзади первыми 10 грудными
позвонками (рис. 1.8—1.11); последние два грудных позвонка
функционально относятся к брюшной полости и не принимают
активного участия в дыхании (см. также гл. 1). Переднюю
стенку грудной клетки образует грудина — плоская узкая кость,
расположенная по средней линии груди. Самая нижняя часть
грудины называется мечевидным отростком (рис. 1.10). Боковая
стенка грудной клетки образована ребрами и реберными
хрящами. Ребра представляют собой костные дуги, соединенные сзади
суставами с телами и отростками грудных позвонков. Ребра
лежат парами по обе стороны позвоночника. Каждое следующее
из первых семи ребер длиннее предыдущего,, а из следующих
трех (VIII—X) —короче. Пары XI и XII спереди не
фиксированы (подвижные ребра). Ребра I, X, XI и XII сочленены с одним
позвонком (рис. 1.11), остальные с телами двух смежных
позвонков. Каждое ребро наклонено вниз, от уровня своего
сочленения с позвонком и прикреплено к грудине ниже. Пространства
между ребрами называются межреберными.
ДЫХАТЕЛЬНЫЕ МЫШЦЫ
Дыхательные мышцы — это те мышцы, сокращения которых
изменяют объем грудной клетки. Мышцы, направляющиеся от
головы, шеи, рук и некоторых верхних грудных и нижних шей-
\
Дыхание
301
ных позвонков, а также наружные межреберные мышцы,
соединяющие ребро с ребром, приподнимают ребра и увеличивают
объем грудной клетки. Диафрагма — мышечно-сухожильная
пластина, прикрепленная к позвонкам, ребрам и грудине, —
отделяет грудную полость от брюшной (см. рис. 1.8 и 1.9). Это
главная мышца, участвующая в нормальном вдохе. При
усиленном вдохе сокращаются дополнительные группы мышц. При
усиленном выдохе действуют мышцы, прикрепленные между
ребрами (внутренние межреберные мышцы), к ребрам и
нижним грудным и верхним поясничным позвонкам, а также мышцы
брюшной полости; они опускают ребра и прижимают брюшные
органы к расслабившейся диафрагме, уменьшая таким образом
емкость грудной клетки.
ЛЕГОЧНАЯ ВЕНТИЛЯЦИЯ
Пока внутриплевральное давление остается ниже
атмосферного, размеры легких точно следуют за размерами грудной
полости. Движения легких совершаются в результате сокращения
дыхательных мышц в сочетании с движением частей грудной
стенки и диафрагмы.
ДЫХАТЕЛЬНЫЕ ДВИЖЕНИЯ
Расслабление всех связанных с дыханием мышц придает
грудной клетке положение пассивного выдоха. Соответствующая
мышечная активность может перевести это положение во вдох
или же усилить выдох.
Вдох создается расширением грудной полости и всегда
является активным процессом. Благодаря своему сочленению с
позвонками ребра движутся вверх и наружу, увеличивая
расстояние от позвоночника до грудины, а также боковые размеры
грудной полости (реберный или грудной тип дыхания).
Сокращение диафрагмы меняет ее форму из куполообразной в более
плоскую, что увеличивает размеры грудной полости в
продольном направлении (диафрагмальный или брюшной тип дыхания).
Обычно главную роль во вдохе играет диафрагмальное дыхание.
Поскольку люди — существа двуногие, при каждом движении
ребер и грудины меняется центр тяжести тела и возникает
необходимость приспособить к этому позные мышцы.
При спокойном дыхании у человека обычно достаточно
эластических свойств и веса переместившихся тканей, чтобы
вернуть их в положение, предшествующее вдоху. Таким образом,
выдох в покое происходит пассивно вследствие постепенного
снижения активности мышц, создающих условие для вдоха.
Активный выдох может возникнуть вследствие сокращения внут-
302
Глава 17
ренних межреберных мышц в дополнение к другим мышечным
группам, которые опускают ребра, уменьшают поперечные
размеры грудной полости и расстояние между грудиной и
позвоночником. Активный выдох может также произойти вследствие
сокращения брюшных мышц, которое прижимает внутренности к
расслабленной диафрагме и уменьшает продольный размер
грудной полости.
Расширение легкого снижает (на время) общее внутрилегоч-
ное (альвеолярное) давление. Оно равно атмосферному, когда
воздух не движется, а голосовая щель открыта. Оно ниже
атмосферного, пока легкие не наполнятся при вдохе, и выше
атмосферного при выдохе. Внутриплевральное давление тоже
меняется на протяжении дыхательного движения; но оно всегда ниже
атмосферного (т. е. всегда отрицательное) (рис. 17.3).
ИЗМЕНЕНИЯ ОБЪЕМА ЛЕГКИХ
Объем легкого меняется при вдохе не всюду одинаково. Для
этого имеются три главные причины. Во-первых, грудная полость
увеличивается неравномерно во всех направлениях. Во-вторых,
не все части легкого одинаково растяжимы. В-третьих,
предполагается существование гравитационного эффекта, который
способствует смещению легкого книзу (рис. 17.4).
Объем воздуха, вдыхаемый при обычном (неусиленном)
вдохе и выдыхаемый при обычном (неусиленном) выдохе,
называется дыхательным воздухом. Объем максимального выдоха после
т
о
О
100
5 10 15 20 25 30
Расстояние от верхушки легкого, см
Верхушка
Основание
Рис 17 4. Объем части легкого, способный к расширению, т. е. жизненная
емкость части (ЖЕЧ). Верхушка легкого показана в левой части графика
ФОЕ — функциональная остаточная емкость.
\
Дыхание
303
предшествовавшего максимального вдоха называется жизненной
емкостью. Она не равна всему объему воздуха в легком
(общему объему легкого), поскольку легкие полностью не спадаются.
Объем воздуха, который остается в неспавшихся легких,
называется остаточным воздухом. Имеется дополнительный объем,
который можно вдохнуть при максимальном усилии после нор-
Общая
легочная
емкость
1 6 6
$ 4,2
Жизненна'я
емкость
6 4,8
? 3,3
Остаточный
объем
'<? 1,2
$ 1,1
Резервный
объем вдоха
S 3,3
? 1,9
Дыхательный объем
6 и ? 0,5
Резервный
объем
выдоха
<Л,0?0,7
Емкость
вдоха
Функциональная
остаточная
емкость
Рис. 17.5. Распределение объема и емкости легких у взрослых. Средние
величины (в литрах) у мужчин (d) и женщин (9).
мального вдоха. А тот воздух, который выдыхается
максимальным усилием после нормального выдоха, это резервный объем
выдоха. Функциональная остаточная емкость состоит из
резервного объема выдоха и остаточного объема. Это тот находящийся
в легких воздух, в котором разбавляется нормальный
дыхательный воздух (рис. 17.5). Вследствие этого состав газа в
легких после одного дыхательного движения обычно резко не
меняется.
Минутный объем V — это воздух, вдыхаемый за одну минуту.
Его можно вычислить, умножив средний дыхательный (tidal)
объем (Vt) на число дыханий в минуту (f), или V = fVt. Часть
Vt, например воздух в трахее и бронхах до конечных бронхиол
и в неперфузируемых альвеолах, не участвует в газообмене, так
304
Глава 17
I
как не приходит в соприкосновение с активным легочным
кровотоком— это так называемое «мертвое» (dead) пространство
(Vd). Часть Vt, которая участвует в газообмене с легочной
кровью, называется альвеолярным объемом (Va). С
физиологической точки зрения альвеолярная вентиляция (Va) —наиболее
существенная часть наружного дыхания VA = f(Vt—Vd), так как
она является тем объемом вдыхаемого за минуту воздуха,
который обменивается газами с кровью легочных капилляров.
МЕХАНИКА ДЫХАТЕЛЬНЫХ ДВИЖЕНИЙ
Перемещение воздуха в легкие и из них требует совершения
работы. Для того чтобы воздух вошел в легкие, должны быть
преодолены силы трех типов, а именно: 1) эластическое
сопротивление, 2) сопротивление воздушного потока в трахео-брон-
хиальном дереве и 3) сопротивление неэластичных тканей,
например ребер.
РАСТЯЖИМОСТЬ
Работа, необходимая для преодоления эластического
сопротивления легких и грудной стенки, как полагают, не зависит от
времени. Максимум работы .производится тогда, когда
дыхательный объем также максимален. Эту форму сопротивления
можно вычислить, определив давление, необходимое для
изменения объема легких и грудной клетки. Эта величина
называется растяжимостью (С),
где AV — изменение объема, а АР — изменение давления (в см
Н20).
Общую растяжимость легкого и грудной стенки можно
определить, составив график, выражающий внутрилегочное давление,
необходимое для поддержания в легком известных объемов газа.
Экспериментально это производится путем наполнения легких
известными объемами, расслабления всех дыхательных мышц и
измерения давления во рту (при закрытых ноздрях).
Растяжимость легкого равна величине внутриплеврального давления и
может быть определена таким же образом (рис. 17.6).
Установлено, что от 3Д до 7/s общего эластического
сопротивления создается поверхностным натяжением пленки жидкости,
выстилающей внутреннюю поверхность альвеолы, а остальная
часть эластическими свойствами ткани. Чем выше поверхностное
натяжение, тем больше нужно энергии для преодоления его
сопротивления. Поверхностное натяжение снижается за счет сур-
Дыхание 30S
100
80
\-
о
2 60
О)
о
| 40
ф
X
со
0
-20
- 20 0 +20
Легочное давление, мм рт. ст.
Рис. 17.6. Общее давление Р0, создаваемое суммой эластических свойств
грудной клетки (Ргр) и легких Рл на разных уровнях расширения груди (% ЖЕ).
Наклон кривых соответствует растяжимости. Обратите внимание на то, что
конечный дыхательный объем в покое (Vn) приходится на точку, где
отрицательное Ргр равно положительному Рл. При изменении эластических
свойств легкого или грудной клетки Vn должно сместиться. Любой другой,
объем, кроме Vn, требует напряжения мышц для создания нужной силы
(Ро).
фактанта. Как полагают, сурфактант стабилизирует легочные
альвеолы, так что они не спадаются при выдохе.
Показано, что сопротивление воздушному потоку создается
главным образом в бронхах среднего размера (рис. 17.7). На
основании уравнения Пуазейля (см. гл. 14) следовало бы
ожидать, что местом наибольшего сопротивления будут самые
мелкие бронхиолы, но на самом деле это не так. Воздушные пути
с диаметром меньше 2 мм создают менее 20% измеренного
сопротивления воздушному потоку. Обилие мелких воздушных
путей создает большое суммарное поперечное сечение для
воздушного потока. Для очень малого объема легких описано явление
«закрытия воздухоносного пути», т. е. обратимого спадения
мелких бронхиол. В таких условиях некоторое количество энергии
затрачивается при вдохе на открывание спавшихся бронхиол.
Сопротивление воздушному потоку зависит от времени; оно наи-
20-1705
ее
О
о
О
0,08
0 06Н
0,04
0,02h-
• 1
•
•
•
•
•
Сегментарные
бронхи #
"i
1
1 1 1
Ч
Терминальные
бронхиолы
\ 1 1
1 # • Р ••
0 5
Трахея
10
15 20-
Терминальная
бронхиола
Рис. 17.7. Сопротивление в разных частях воздухоносных путей. Обратите
внимание на то, что сопротивление выше всего в крупных бронхах и самое
низкое в мелких бронхиолах. (Pedley et al., Resp. Physiol., 1970, 9, 387.)
P —>
Рис. 17.8. Схема зависимости между давлением и объемом при одном
дыхательном объеме (сплошная линия, направленная в сторону возрастающих
значений, обозначает вдох; сплошная линия, направленная в сторону снижения
объема, обозначает выдох). Суммированные площади А и Б соответствуют
общей работе эластических компонентов, совершаемой при вдохе. Площадь В
соответствует всей работе неэластических компонентов, производимой при
вдохе, т. е. работе ткани и воздушной струи. Площадь Б соответствует
работе, совершаемой неэластическими компонентами при выдохе.
306
t
о
со о
Ф Q
S|.
® ф
-«?
СО О
о. с
Общая
Частота дыханий в 1 мин
>
(tfE=K)
Рис. 17.9. Гипотетическая кривая работы эластических и неэластических
компонентов и суммарной работы, производимой при разной частоте дыхания у
человека при постоянном минутном объеме. Обратите внимание на
оптимальную частоту, когда общая сумма работы эластических (/) и неэластических
(//) компонентов дает минимум общей потребной работы. Работа выражена
в количестве потребляемого Ог (Vo2).
2 200 п
£ 100U-
Эмфизема
о
•>
20 40 60 80
Вентиляция VE, л/мин
100
Рис. 17.10. Потребление кислорода дыхательными мышцами при разной
частоте дыхания в норме и при пониженной растяжимости легких (эмфизема).
20*
307
308
Глава 17
большее лри частом дыхании и достигает максимума, даже если
объем вдоха не максимален.
Работа по перемещению грудной стенки и легкого против
сопротивления неэластичных тканей тоже зависит от времени.
У взрослых молодых людей она составляет около 20% общего
расхода энергии при дыхании.
Общую работу, затрачиваемую на перемещение воздуха в
легкое и из него, включая движение грудной стенки, можно вы-
v
числить по графику давление — объем (рис. 17.8): W=JVdp.
о
Эта работа складывается из работы против эластических сил
(площади Л и Б на рис. 17.8) и против неэластических
(площадь В на рис. 17.8). Для данного минутного объема существует
интенсивность работы, при которой сумма эластического и
зависимого от времени неэластического компонентов минимальна
(рис. 17.9). При нормальном дыхании для перемещения воздуха
в легкие и из них требуется менее 5% общего потребления 02
(рис. 17.10).
ЛЕГОЧНОЕ КРОВООБРАЩЕНИЕ
В легочный круг кровь поступает из правого желудочка по
легочным артериям, проходит по капиллярной сети, лежащей
между альвеолами, и возвращается в левое предсердие по
легочным венам. Помимо его роли в газообмене легочный
кровоток служит также резервуаром крови для левой половины
сердца; он осуществляет питание альвеолярной ткани, удаляет из
альвеол избыточную жидкость и служит фильтром венозной
крови большого круга. Он может также активировать и инакти-
вировать ряд фармакологических препаратов.
Роль легочного кровообращения в газообмене составляет
основной предмет настоящего изложения. По сравнению с
большим кругом кровообращения малый круг представляет собой
систему с низким давлением. Давление в легочных артериях
равно примерно 25/10 мм рт. ст. при среднем давлении около
15 мм. Поскольку объем крови в правой и левой половинах
сердца должен быть одинаковым, сопротивление в малом круге
(R= КрОВгОТок ) составляет приблизительно от 1/4 до 1/5
сопротивления в системе большого круга (см. также гл. 14).
Емкость артерий и вен легочного круга может быть
увеличена ценою очень малых изменений кровяного давления (высокая
растяжимость). Из-за их высокой растяжимости на емкость
сосудов сильно влияют гравитационные силы (т. е. высота
столба крови). Из-за гравитационного эффекта у человека при
нормальной вентиляции нижняя часть легких получает
сравнительно больше крови, чем верхняя (рис. 17.11).
Дыхание
309
Рис. 17.11. Распределение кровотока в легком определяется расстоянием от
основания. В зоне I (верхушка) внутрилегочное давление (Ра) превышает
давление артериальной крови (Ра) в течение некоторой части дыхательного
цикла, и кровоток при этом ограничен. В зоне II Ра больше Ра, а Ра^Рв
(давление в легочной вене). Кровоток регулируется разностью Ра—Ра
(кровоток слегка усилен по сравнению с зоной I). В зоне III Ра меньше Ра и Рв.
Кровоток в этой зоне определяется градиентом Ра—Рв (значительно
усиленный кровоток). В основании легкого, в зоне IV, кровоток снова ограничен, и
одной из возможных причин этого считается интерстициальное давление Ри.
Капилляры легких образуют на поверхности альвеол густую
сеть, так что по одной альвеоле может проходить несколько
капилляров. В условиях покоя кровь в капиллярах находится в
контакте с альвеолярным газом примерно в течение 0,75 с, а при
легкой физической работе в течение 0,34 с. Давление в
капиллярах низкое (в пределах 7—9 мм рт. ст.). Согласно гипотезе Стар-
линга о движении жидкости между капилляром и тканями
(см. также гл. 12), существует перепад давления, который
перемещает жидкость из альвеол обратно в кровяное русло.
НЕРВНЫЕ И ГУМОРАЛЬНЫЕ ВЛИЯНИЯ
Легочные сосуды обильно снабжены сосудосуживающими
симпатическими нервными волокнами. Как полагают,
возбуждение этих волокон участвует в мобилизации крови из легочного
резервуара. Симпатические и парасимпатические сосудорасши-
310
Глава 17
ряющие нервные волокна встречаются лишь в ограниченном
количестве. Их физиологическая роль еще не совсем ясна.
Адреналин и ангиотензин II вызывают сужение легочных артериол;
серотонин (5 ОТ) и гистамин — сужение легочных вен. В
отличие от реакции сосудов большого круга локальное повышение
парциального давления С02 или понижение парциального
давления 02, или же одновременно и то и другое, вызывает сужение
сосудов. Таким образом ограничивается приток крови к плохо
вентилируемым альвеолам и достигается преимущественная
перфузия хорошо вентилируемых альвеол.
ЛЕГОЧНОЕ ДЫХАНИЕ
Газ является таким состоянием вещества, при котором оно
равномерно распределяется по ограниченному объему. В
газовой фазе взаимодействие молекул между собой незначительно.
Когда они сталкиваются со стенками замкнутого пространства,
их движение создает определенную силу; эта сила, приложенная
к единице площади, называется давлением газа и выражается в
миллиметрах ртутного столба, или торрах; давление газа
пропорционально числу молекул и их средней скорости. При
комнатной температуре давление какого-либо вида молекул,
например 02 или N2, не зависит от присутствия молекул другого газа
(см. также гл. 2). Общее измеряемое давление газа равно сумме
давлений отдельных видов молекул (так называемых
парциальных давлений) или Pb = Pn2+Po2 + Ph2o + Pz, где Рв —
барометрическое давление. Долю (F) данного газа (х) в сухой газовой
смеси можно вычислить по следующему уравнению:
F- *
х Рв — Рн20(паР)
И наоборот, парциальное давление данного газа (х) можно
вычислить из его доли: Px = Fx(Pb—Рн2о). Сухой атмосферный
20 94
воздух содержит 20,94% 02-Po2=-txq--760 торр (на уровне
моря) = 159,1 торр (см. табл. 17.1).
Газообмен в легких между альвеолами и кровью происходит
путем диффузии. Диффузия возникает в силу постоянного
движения молекул газа и обеспечивает перенос молекул из
области более высокой их концентрации в область, где их
концентрация ниже (закон Фика; см. гл. 2).
ГАЗОВЫЕ ЗАКОНЫ
На величину диффузии газов между альвеолами и кровью
влияют некоторые чисто физические факторы. 1. Плотность
газов. Здесь действует закон Грэма. Он гласит, что в газовой фазе
Дыхание
311
при прочих равных условиях относительная скорость диффузии
двух газов обратно пропорциональна квадратному корню из их
плотности. 2. Растворимость газов в жидкой среде. Здесь
действует закон Генри: согласно этому закону, масса газа,
растворенного в данном объеме жидкости при постоянной
температуре, пропорциональна растворимости газа в этой жидкости и
парциальному давлению газа, находящегося в равновесии с
жидкостью. 3. Температура. С повышением температуры растет
средняя скорость движения молекул (повышается давление) и
падает растворимость газа в жидкости при данной температуре.
4. Градиент давления. К газам в дыхательной системе приложим
закон Фика.
КОЭФФИЦИЕНТЫ ДИФФУЗИИ
Исходя из растворимости и величины молекул, коэффициент
диффузии для СОг приблизительно в 2,7 раза больше, чем для
Ог. Поскольку эта величина постоянная и температура в легких
обычно тоже остается постоянной, то только парциальные
давления этих газов определяют направление газообмена между
легкими и альвеолами (см. табл. 17.1). При рассмотрении
физиологических аспектов газообмена в легких следует учитывать
1) легочное кровообращение в альвеолах, 2) доступную для
диффузии поверхность, 3) характеристики альвеолярной и
капиллярной тканей и 4) расстояние, на которое происходит
диффузия.
Определить диффузионную способность легких,
обозначаемую как коэффициент переноса (Tlx, или Dlx некоторых
исследователей), можно, измерив количество газа (х), переносимое
каждую минуту на каждый торр разницы парциального давле«
ния в альвеолах (Рах) и капиллярах (Рсар), или: Тх =
=^5 ^5—'» Tlx варьирует в зависимости от изучаемого газа и
г\Ах — *сар
его места в легком. Tlx кислорода во всем легком человека в
состоянии покоя колеблется от 19 до 31 мл/мин на 1 торр. При
легкой физической работе оно возрастает до 43 мл/мин
СООТНОШЕНИЕ МЕЖДУ ВЕНТИЛЯЦИЕЙ И ПЕРФУЗИЕЙ
Эффективность легочного дыхания варьирует в разных
частях легкого. Эта вариабельность в значительной мере
объясняется представлением о соотношении между вентиляцией и
перфузией (Va/Q). Указанное соотношение определяется числом
вентилируемых альвеол, которые соприкасаются с хорошо
перфорируемыми капиллярами. При спокойном дыхании (эупное)
312 Глава 17
у человека верхние отделы легкого расправляются полнее, чем
нижние отделы (см. рис. 17.4), но при вертикальном положении
нижние отделы перфузируются кровью лучше, чем верхние
(рис. 17.11). По мере увеличения дыхательного объема нижние
части легкого используются все больше и все лучше
перфузируются. Соотношение V/Q в нижней части легкого стремится к
единице (рис. 17.12).
ТРАНСПОРТ ДЫХАТЕЛЬНЫХ ГАЗОВ
Около 0,3% Ог, содержащегося в артериальной крови
большого круга при нормальном Ро2, растворено в плазме. Все
остальное количество находится в непрочном химическом
соединении с гемоглобином (НЬ) эритроцитов. Гемоглобин
представляет собой белок (глобин) с присоединенной к нему
железосодержащей группой (тем). Fe2+ каждой молекулы гемоглобина
соединяется непрочно и обратимо с одной молекулой 02.
Полностью насыщенный кислородом гемоглобин содержит 1,39 мл
4
з
v/6
2
1
5 Ю 15 20
Верхушка см Основание
Рис. 17.12. Региональная вентиляция (V) и кровоток (Q), выраженные в
процентах общей для легких величины (на основании данных рис. 17.4 и 17.11),
и их соотношение (V/Q) на разных расстояниях от верхушки легкого у
стоящего человека. Обратите внимание на изменение V/Q, вызванное понижением
кровотока в зоне IV.
150
100
50
\ Vй'
i i
_
6
ч
Дыхание
313
02 на 1 г НЬ (в некоторых источниках указывается 1,34 мл).
Если Fe2+ окислен до Fe3+, то такое соединение утрачивает
способность переносить 02.
Полностью насыщенный кислородом гемоглобин (НЬ02)
обладает более сильными кислотными свойствами, чем
восстановленный гемоглобин (НЬ). В результате в растворе, имеющем
20 40 60 80 100 120
Рис. 17.13. Кривые диссоциации оксигемоглобина, выражающие
отношение давления кислорода (Ро2) к проценту гемоглобина, связывающему
кислород. А. Влияние изменения давления Рсо2 при 38 °С. Б. Влияние
изменения рН при 38°С. В. Влияние температуры при постоянном Рсо2 (40 торр).
рН 7,25, освобождение 1 мМ 02 из НЬ02 делает возможным
усвоение 0,7 мМ Н+ без изменения рН; таким образом, выделение
02 оказывает буферное действие.
Соотношение между числом свободных молекул 02 и числом
молекул, связанных с гемоглобином (НЬ02), описывается
кривой диссоциации 02 (рис. 17.13). НЬ02 может быть представлен
в одной из двух форм: или как доля соединенного с кислородом
гемоглобина (% НЬ02), или как объем 02 на 100 мл крови во
взятой пробе (объемные проценты). В обоих случаях форма
кривой диссоциации кислорода остается одной и той же.
314
Глава 17
ВЛИЯНИЕ рН, УГЛЕКИСЛОТЫ И ТЕМПЕРАТУРЫ
НА ТРАНСПОРТ КИСЛОРОДА
Легкость, с которой 02 присоединяется к гемоглобину и
образует оксигемоглобин (НЬ + 02ч^НЬ02), зависит от Рсо2,
температуры и концентрации в крови Н+ (или рН), а также
органических фосфатов, в особенности 2,3-дифосфоглицерата (ДФГ)
(рис. 17.13). Повышение [Н+] (понижение рН), Рсо2,
концентрации ДФГ или температуры вызывает смещение равновесия
реакции диссоциации в сторону дезоксигемоглобина, что
соответствует правой части кривой, где представлены более низкие
значения [Н+], Рсо2, ДФГ или температуры; этот эффект
называется «смещением вправо». Смещение влево соответствует
более высокому содержанию НЬ02 при данном Ро2. В иной форму*
лировке для достижения того же уровня НЬ02 требуется более
низкое Ро2.
Поскольку артериальная кровь обладает более НИЗКИМ Jrco«
и более высоким рН, содержание НЬ02 в ней будет выше, чем
в венозной крови с таким же Ро2 (табл. 17.1 и рис. 17.13).
В тканях повышение Рсо2 и понижение рН приводит к тому, что
даже при неизменном Ро2 некоторое количество 02, связанное
гемоглобином в артериальной крови, будет освобождаться и
использоваться тканью. Таким образом, повышение содержания Н+*
или Рсо2, или того и другого, способствует выходу 02 в ткани
(эффект Бора).
Коэффициентом использования кислорода называется
разность концентраций 02 в артериальной [02]а и смешанной
венозной [02]в крови, деленная на [02]д:
[02]а - [02]в
[02]А *
Обычно эта средняя величина составляет около 0,25 и
определяется в пробе смешанной венозной крови большого круга.
Коэффициенты использования в ограниченных участках ткани
могут быть выше или ниже этой величины. Общий коэффициент
использования снижается вследствие шунтирования.
ТРАНСПОРТ УГЛЕКИСЛОТЫ
Более 90% углекислоты, переносимой кровью, находится в
химически связанном состоянии, остальная часть (<5%)
растворена в плазме. Двумя химическими формами транспорта С02
является ион бикарбоната (60—70%) и аминогруппы белков
крови, например глобина в составе гемоглобина (10—30%).
Дыхание
315
Бикарбонат образуется в крови в результате реакции С02
с водой, т. е. перехода ее в угольную кислоту, которая
диссоциирует на водород и ион бикарбоната:
Н20 + С02 +=+ Н2С03 =<=* H+ + HCO3-.
Угольная кислота образуется медленно. В присутствии
фермента карбоангидразы этот процесс протекает в несколько
тысяч раз быстрее. Поскольку карбоангидраза содержится в
эритроцитах, а не в плазме, образование и диссоциация Н20 и СОг
совершаются главным образом в клетках. На тканевом уровне
при добавлении С02 к крови концентрация бикарбоната в
эритроцитах повышается. Бикарбонат свободно диффундирует через
мембрану эритроцита в окружающую плазму в силу разницы
концентраций этого аниона. Благодаря избирательной
проницаемости мембраны эритроцита диффузия бикарбоната создает
трансмембранную разность потенциалов (катионов внутри
больше, чем анионов). В соответствии с равновесием Гиббса — Дон-
нана ион хлора диффундирует внутрь клетки (см. гл. 2). В
легких после выхода из крови С02 и поступления 02 происходит
противоположный процесс, т. е. бикарбонат диффундирует
внутрь, а хлор наружу. Это перемещение иона хлора,
сохраняющее электрохимическое равновесие, называется «сдвигом хлора».
Образование карбаминовых соединений (Белок—NH2 +
+ С02^Белок—NHCOO-+H+) происходит быстро и,
по-видимому, не нуждается в присутствии ферментов. Восстановленная
форма гемоглобина может связать больше С02, чем окисленная,
что облегчает транспорт как 02, так и С02.
РОЛЬ ДЫХАНИЯ В ПОДДЕРЖАНИИ
КИСЛОТНО-ЩЕЛОЧНОГО РАВНОВЕСИЯ
Поскольку, соединяясь с водой, С02 переходит в Н2С03,
транспорт С02 оказывает сильное влияние на
кислотно-щелочное равновесие циркулирующей крови, а также всего
организма. Отношение между раствором С02 в крови и рН крови
выражается уравнением Гендерсона — Гассельбаха (см. гл. 2):
pH = p/fa+bg 'Соль11,
г i'^a 1 &» [Кислота] '
где рКа — это отрицательный логарифм константы диссоциации
для всей реакции.
н2о + со2 -<—>- н2со3 <—>• н+ + нсо3-.
При 37 °С рКа составляет приблизительно 6,1; [НС03"~] и
{Н2СОз] выражены в мМ/л. Поскольку концентрация Н2С03
пропорциональна концентрации С02 в растворе, мы можем рассчи-
316
Глава 17
тать [Н2С03], определив давление С02 в растворе (Рсо2). При
37°С на каждый торр РСо2 образуется 0,03 мМ/л угольной
кислоты, и уравнение Гендерсона — Гассельбаха можно записать
следующим образом:
Пример такого отношения: если рН артериальной крови
равно 7,4, а Расо2 — 40 торр, то вычисленная [НС03"~] составит
24 мМ/л. Если Рсо2 снижается до 20 торр, то соответственное
изменение рН вычисляется следующим образом:
Р D,1^0g (0,03-40)— (0,03-20) ~~
= 6,l + logff =
= 6,l + log39 =
= 6,1+1,6=
= 7,7.
В реальных условиях содержащиеся в крови буферы
изменяют отношение между Рсо2 и рН. Конкретные реакции лучше
всего можно определить по имеющимся графикам, которые
описывают отношение между Рсо2, НС03~ и рН для разных жидкостей
тела.
Следует помнить, что НС03~ крови, кроме Н+, связана еще
и с другими катионами. Изменение Н+ не обязательно меняет
концентрации других катионов. Существует равновесие между
поглощением и выведением отдельных катионов. Выведение
катионов регулируется главным образом почками (см. гл. 24).
Изменение рН, наступающее при повышении или понижении
содержания С02 в крови (алкалоз для повышенного и ацидоз для
пониженного рН), обозначается как «дыхательное». Если же
изменяется концентрация бикарбоната, то происходящее при этом
изменение рН называют «метаболическим» (рис. 17.14).
Возможна компенсация дыхательного ацидоза метаболическим
алкалозом.
НАСЫЩЕНИЕ ТКАНЕЙ КИСЛОРОДОМ
Транспорт Ог из крови в те участки ткани, где он
используется, происходит путем простой диффузии (см. гл. 2). Поскольку
кислород используется главным образом в митоховдриях, рас-
1 Из концентрации бикарбоната следует вычесть число мМ/л,
поставляемое угольной кислотой.
Дыхание
31>
бОторр
40торр
20 торр
О
О
I
Рис. 17.14. График зависимости между [НС03_] и рН для водного раствор»
при Рсо2, показанном изобарами. Обозначенный в центре шестиугольник
соответствует диапазону величин, найденных для здоровых людей на уровне
моря. Смещение этих величин за пределы нормального диапазона
представляет собой изменение кислотно-щелочного равновесия у данного индивидуума.
рН меньше 7,4 и [НС03~] больше 25 мМ/л соответствуют состоянию
первичного дыхательного ацидоза; рН больше 7,4 и [НС03~] меньше 25 мМ/л —
состоянию первичного дыхательного алкалоза. Первичный метаболический
ацидоз представлен частью графика ниже изобары Расо„—40 торр и рН
меньше 7,4. Первичный метаболический алкалоз представлен частью графика
выше изобары РаСо2 — 40 торр и рН больше 7,4. Другие части графика
(штриховка) показывают роль дыхания и обмена в нарушении равновесия.
Основная причина нарушенного кислотно-щелочного равновесия может быть
компенсирована его нарушением с обратным знаком из другого источника,
например при компенсации дыхательного алкалоза метаболическим ацидозом.
(Woodbury /. W., 1974. In: Ruch Т., Patton H., eds., Physiology and Biophysics,
v. 11, pp. 480—524.)
стояния, на которые происходит диффузия в тканях,
представляются большими по сравнению с обменом в легких. В
мышечной ткани присутствие миоглобина, как полагают, облегчает
диффузию 02. Для вычисления тканевого Ро2 созданы
теоретические модели, которые предусматривают факторы, влияющие.
318
Глава 17
на поступление и потребление Ог, а именно расстояние между
капиллярами, кровоток в капиллярах и тканевой метаболизм.
Самое низкое Ро2 установлено в венозном конце и на полпути
между капиллярами, если принять, что кровоток в капиллярах
одинаковый и что они параллельны.
РЕГУЛЯЦИЯ ДЫХАНИЯ
ЦЕНТРАЛЬНАЯ НЕРВНАЯ РЕГУЛЯЦИЯ
Уровни СО2, [H+] и Ог в артериальной крови, как правило,
регулируются в узких пределах через легочную вентиляцию.
В центральной нервной системе (ЦНС) имеются специальные
области, которые участвуют в создании каждого
вентиляционного усилия дыхательных мышц, а также регулируют общую
деятельность дыхательной системы. Участие ЦНС складывается из
двух функционально раздельных элементов: 1) автоматическое
дыхание, связанное главным образом со структурами ствола
мозга, и 2) произвольное дыхание, связанное со структурами
высших уровней мозга, главным образом с корой больших
полушарий.
Области ЦНС, особенно ствола мозга, связанные с
дыханием, называют «дыхательными центрами». Термин этот
неправильный, так как речь идет не о «центре», а об анатомических
областях, где находится определенное число нейронов,
связанных с дыханием. В стволе мозга имеется несколько таких
областей. На реакцию каждого центра влияют афферентные
волокна, идущие из разных частей тела, а также локальный уровень
метаболитов в крови и в спинномозговой жидкости (СМЖ).
В спинном мозгу существуют какие-то внутренние связи,
которые влияют на деятельность дыхательных мышц. Но
автономное дыхание невозможно, если спинной мозг отделен от
головного.
Самая каудальная часть головного мозга, управляющая
ритмом дыхания, это продолговатый мозг (medulla). Будучи
связаны со спинным мозгом, медуллярные центры способны
поддерживать дыхательные движения без участия вышележащих
отделов ЦНС. В продолговатом мозгу заложены два
скопления (или группы) дыхательных нейронов: 1) первичные
нейроны вдоха, называемые дорсальной дыхательной группой (ДДГ),
2) нейроны вдоха и выдоха, названные вентральной
дыхательной группой (ВДГ). Относительно происхождения дыхательной
ритмики многое далеко не ясно. Ритм, возможно, создается
автоматической активностью определенных медуллярных
нейронов, например ДДГ, или сетью нейронов, соединенных между
собой таким образом, что их активность на выходе оказывается
ритмической. Такие сети были обнаружены. Предполагается,
Дыхание
31fr
Кора
Мост
Пневмотаксичес-
кий центр
\i /
Центр апное
.(-)
!♦ t
(+,-)!
Другой проприоцеп
тивный афферент
Периферический]
I хеморецептор
Щ
Барорецептор
и
Продолговатый мозг
/ Хемочувстви -\
[тельные клетки 1
Медуллярно-
спинальная
координация
Легочные
рецепторы
растяжения
(-,+)
i i i
Дыхательные мотонейронык-
Проприоцепторы
Диафрагма
Вспомогательные
мышцы |
Межреберные
мышцы
\ i /
I Грудная I
1 клетка I
Рис. 17.15. Основные функциональные компоненты системы регуляции
дыхания. Показаны некоторые проявления возбудительных (+) и тормозных (—)
влияний. На основе имеющейся информации высказывается предположение а
взаимоотношениях между дорсальной и вентральной дыхательными
группами (ДДГ и ВД Г).
что ДДГ либо служит источником, либо расположена близ
источника генерации ритма. Таким образом, ДДГ приводит в
действие ВДГ, а также множество спинномозговых дыхательных
мотонейронов.
Выше медуллярных дыхательных областей, в каудальных
Двух третях моста, лежит область, называемая центром апноэ.
.320
Глава 17
Перерезка головного мозга выше этого центра в сочетании с
двусторонней ваготомией вызывает почти непрерывный вдох.
Таким образом, центр апноэ является областью, откуда
непрерывно вызывается вдох; он тормозится 1) из области в самой
задней части моста, называемой пневмотаксическим центром,
или 2) афферентными волокнами блуждающего нерва, или же и
тем и другими (рис. 17.15). Как полагают, пневмотаксический
центр обеспечивает тонкую настройку автоматического
дыхательного ритма, либо регулируя время вдоха, либо модулируя
реакцию ЦНС на множество афферентных воздействий,
влияющих на дыхание, например на РСо2 крови и на степень
растяжения легких и т. д.
Высший отдел ЦНС, кора больших полушарий, оказывает
влияние на глубину и частоту дыхания. При стимуляции особых
областей коры дыхание или усиливается, или ослабляется. Эти
области находятся под произвольным контролем и проявляют
себя, когда мы едим или говорим.
СЕНСОРНЫЕ РЕЦЕПТОРЫ
В дыхательных путях множество сенсорных рецепторов;
рецепторы имеются также в кровеносных сосудах, соприкасаются
с СМЖ, с тканями ЦНС, суставов и мышц конечностей; они
влияют на ритм дыхания.
Рецепторы в носовых ходах иннервируются обонятельным
(I) и тройничным (V) черепно-мозговыми нервами; они
чувствительны к разным химическим веществам, а также к
механическим раздражителям. Реакция на их стимуляцию варьирует
от апноэ до чихания. Глоточная зона иннервируется веточкой
языкоглоточного нерва (IX). Стимуляция этой области
вызывает резкие вдохи. В гортани и трахее находятся рецепторы
разного типа, отвечающие на химические и механические
раздражения. Иннервируются они преимущественно веточками
блуждающего нерва (X). Их стимуляция оказывает разное действие,
в том числе вызывает кашель, апноэ и медленное глубокое
дыхание. Наблюдается также спазм бронхов.
МЕХАНОРЕЦЕПТОРЫ ЛЕГКИХ
В легких заложены рецепторы трех типов, иннервируемые
блуждающим нервом: 1) медленно адаптирующиеся рецепторы
растяжения легких, 2) рецепторы, чувствительные к
раздражающим влияниям, и 3) рецепторы растяжения типа «J».
Расправление легкого возбуждает медленно адаптирующиеся
рецепторы растяжения, действие которых на ЦНС состоит в том, что
они тормозят разряд эфферентного нерва вдоха и вызывают
Дыхание
321
пассивный выдох. Эта самоограничивающаяся система, впервые
ставшая известной в 1860-х годах, названа рефлексом
Геринга— Брейера. У людей этот рефлекс обычно не вступает в
действие, пока объем легкого не превысит в 1,5—2,0 раза
дыхательный объем в покое. При нормальных условиях нейроны
легочных рецепторов растяжения в блуждающем нерве изменяют
частоту дыхания, как воздействуя на продолговатый мозг, так и
тормозя центр апноэ.
Рецепторы, чувствительные к раздражающим влияниям,
представляют собой быстро адаптирующиеся рецепторы,
воспринимающие химические и механические воздействия,
например аммиака и твердых частиц. Рецепторы этого типа рефлек-
торно вызывают усиленный вдох и спазм бронхов.
Рецепторы типа «J» в норме не реагируют на движение
легких как таковое. Предполагается, что они заключены в стенке
легочных капилляров. Они возбуждаются при чрезмерном
расширении легких. В настоящее время их функция точно не
установлена.
Имеется ряд других механорецепторов, возбуждение
которых влияет на характер дыхания. Среди них рецепторы
давления, расположенные в артериальной и венозной системах
большого круга кровообращения. При их возбуждении возникают
реакции, варьирующие от временного апноэ до значительного
учащения дыхания. Движение суставов и растяжение мышц
конечностей повышают как частоту дыхания, так и
дыхательный объем. Боль тоже действует ,на дыхание.
ХЕМОРЕЦЕПТОРЫ
В результате тканевого метаболизма в артериальной крови
происходят следующие изменения — понижение Ро2, повышение
Рсо2 и повышение [Н+]; все это ведет к стимуляции дыхания.
Наиболее важные дыхательные рецепторы, чувствительные к
химическому составу окружающей их жидкости, так называемые
хеморецепторы, лежат в дуге аорты (аортальные тельца), в
области бифуркации общей сонной артерии на внутреннюю и
наружную сонные артерии (жаротидные тельца) (см. гл. 14) и в
ЦНС близ поверхности четвертого желудочка или в вентролате-
ральных областях продолговатого мозга. Аортальные тельца ин-
нервируются веточками блуждающего нерва, а каротидные —
веточками языкоглоточного. Аортальные и каротидные тельца
считаются периферическими хеморецепторами, а находящиеся в
мозгу — центральными.
Частота импульсации в афферентном нерве, идущем от
периферических хеморецепторов, обратно пропорциональна арте-
21—1705
322
Глава 17
риальному Ро2 в пределах от 30 до 500 торр (рис. 17.16, А).
Легочная вентиляция в норме существенно яе усиливается, пока
артериальное Ро2 не снизится от нормального уровня в 90—
100 торр до 60 торр или меньше (рис. 17.16, Б). Степень
влияния Ро2 на дыхание зависит от других параметров — С02 и рН
'(рис. 17.16, Б). Центральные хеморецепторы не чувствительны
к понижению Ро2. Влияние низкого Роа на дыхание состоит в
60 80 100
Ра02, торр
60
40
20
С
т \
_.
,
с\
Ч Расо,
^Ча (49)
*«»*• 144)
(36)
|| II
100
Ра0^, торр
140
Рис. 17.16. Изменение Расо2 в ответ на изменяющееся Рао2. А Средняя
частота разрядов нейрона из каротидного тельца кошки. Б. Вентиляционная
реакция у человека (изобары Расо2—в торрах).
повышении частоты дыхания или дыхательного объема, или того
и другого.
На периферические и центральные хеморецепторы влияет
СОг. Около 80—85% всех наблюдаемых реакций вызываются
возбуждением центральных хеморецепторов под действием СОг.
Подъем напряжения С02 в артериальной крови (Расо2) до
30 торр повышает частоту импульсации в нервном волокне,
идущем от рецептора. На эту реакцию влияет уровень Ро2
(рис. 17.16,Б). Повышение РаСо2 всегда усиливает
дыхательный объем; его действие на частоту дыхания варьирует и,
вероятно, связано с эффектом Геринга — Брейера. Увеличение [Н+]
в артериальной крови (снижение рНА) при постоянном
Расо2 стимулирует дыхание, а увеличение рН снижает
вентиляцию. Концентрация [Н+] в СМЖ также влияет на дыхание,
Поскольку изменение Расо2 меняет рН (см. рис. 17.14),
действие Расо2 на характер дыхания отчасти связано с действием
Дыхание
323
[Н+] на активность рецепторов. Движение Н+ через гематоэн-
цефалический барьер и в СМЖ происходит гораздо медленнее,
чем диффузия СОг.
гипоксия
Гипоксия возникает при недостаточном снабжении тканей
кислородом. Краткое изложение разных причин гипоксии может
служить и сокращенным обзором всех дыхательных процессов.
Ниже в каждом пункте указаны нарушения одного или более
процессов, которые перечислены в начале главы.
Систематизация их позволяет рассматривать все эти явления одновременно.
I. Недостаточный транспорт Ог кровью (аноксем'ическая
гипоксия) (содержание Ог в артериальной крови большого круга
понижено).
А. Сниженное Ро2:
1) недостаток Ог во вдыхаемом воздухе;
2) снижение легочной вентиляции;
3) снижение газообмена между альвеолами и кровью;
4) смешивание крови большого и малого круга,
Б. Нормальное Ро0:
1) снижение содержания гемоглобина (анемия);
2) нарушение способности гемоглобина присоединять
02.
II. Недостаточный транспорт крови (гипокинетическая
гипоксия).
A. Недостаточное кровоснабжение:
1) во всей сердечно-сосудистой системе (сердечная
недостаточность) ;
2) местное (закупорка отдельных артерий).
Б. Нарушение оттока крови:
1) закупорка определенных вен.
B. Недостаточное снабжение кровью при возросшей
потребности.
III. Неспособность ткани использовать поступающий 02 (ги-
стотоксическая гипоксия).
ЛИТЕРАТУРА
Bouhuys А. (1977). The physiology of breathing, Grune and Stratton, New York.
Slonim N. В., Hamilton L. H. (1976). Respiration physiology, 3rd edn., Mosby,
St. Louis.
West J. B. (1974)). Respiratory physiology — the essentials, Williams and Wil-
kins, Baltimore.
21*
324
Глава 17
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Дайте определение терминам: а) дыхательный объем; б)
резервный объем вдоха; в) функциональная остаточная емкость;
г) минутный объем; д) альвеолярная вентиляция; е)
растяжимость; ж) кривая диссоциации кислорода; з) рефлекс
Геринга— Брейера и) эффект Бора; к) центр апноэ.
2. Какие мышцы участвуют во вдохе?
3. Какие три основных компонента следует иметь в виду при
рассмотрении работы, производимой при дыхании?
4. Какое физиологическое значение имеет соотношение между
вентиляцией и перфузией (V/Q) в легких?
5. Назовите три формы транспорта С02 в крови.
6. Каким образом поглощение кровью 02 в легких облегчает
выход С02 из крови?
7. Какие рецепторы возбуждаются при понижении
парциального давления кислорода?
8. На каком уровне головного мозга формируется основной ритм
дыхания?
Глава 18
ЭНЕРГЕТИЧЕСКИЙ ОБМЕН
Живые организмы отличаются от неодушевленных предметов
сложностью, разнообразием и упорядоченностью своих
компонентов. Всем системам присуща тенденция к уменьшению
упорядоченности, т. е. к увеличению энтропии. Чтобы противостоять
этой естественной тенденции, организмам приходится
непрерывно затрачивать энергию. Согласно первому закону
термодинамики, энергия не исчезает и не возникает вновь. Организмы
должны получать энергию в доступной для них форме из
окружающей среды и возвращать в среду соответствующее количество
энергии в форме, менее пригодной для дальнейшего
использования.
Около столетия тому назад французский физиолог Клод Бер-
нар установил, что животное и среда образуют единую систему,
поскольку между ними происходит непрерывный обмен
веществами и энергией. Организмы находятся в стационарном состоя-
нии (steady state) —переход материи и энергии из среды в
организм точно соответствует переходу материи и энергии из
организма в среду. Нормальная жизнедеятельность поддерживается
регуляцией внутренних компонентов, требующей затраты
энергии. Использование химической энергии в организме называют
энергетическим обменом: именно он служит показателем общего
состояния и физиологической активности организма.
ОПРЕДЕЛЕНИЯ И ЕДИНИЦЫ ИЗМЕРЕНИЯ
Традиционная единица измерения энергии, применяемая, как
правило, в биологии, — это калория (кал). Она определяется как
количество энергии, необходимой для повышения температуры
1 г воды на 1 °С. Однако это количество энергии так мало, что
при изучении энергетических процессов в организме человека
Удобнее пользоваться более крупными единицами. Одна
килокалория (ккал) —это 1000 кал.
Хотя калории и килокалории используются в биологии, не
вызывая особой путаницы, иногда эти единицы оказываются
неудобными, в особенности при междисциплинарных
исследованиях. В работах, касающихся потока энергии, физики, химики и
326
Глава 18
инженеры пользуются разными наборами произвольно
определяемых единиц измерения, что частично затрудняет
взаимодействие физических и биологических наук. Сравнительно недавно
во всем мире принята единая система единиц измерения, легко
переводимых в единицы других систем, применяемых во всех
научных дисциплинах. Международная система единиц (СИ)
определяет семь основных физических величин: длина, масса,
время, электрический ток, сила света, термодинамическая
температура и количество вещества. Из этих единиц прямо
выводятся все остальные единицы измерения. Так, например, сила =
= масса-ускорение, или масса-длина за единицу времени
(скорость) : время. Работа (энергия) =сила-длина; мощность
(поток энергии) = работа в единицу времени.
Единицами СИ, использующимися при измерении
энергетических величин являются: джоуль (1 Дж = 4,187 кал) в качестве
единицы измерения энергии и ватт (1 Вт=1 Дж/с) для измерения
мощности. В табл. 18.1 приведены основные единицы
измерения энергетических величин и коэффициенты перевода единиц,
традиционно применяемых физиологами.
ПРЕВРАЩЕНИЕ И ИСПОЛЬЗОВАНИЕ ЭНЕРГИИ
Организмы получают энергию из окружающей среды в виде
потенциальной энергии, заключенной в химических связях
молекул жиров, углеводов и белков. Сложные органические
молекулы постепенно окисляются, /выделяя энергию,
высвобождающуюся при разрыве химических связей. Молекулы эти распадаются
до трехуглеродных соединений, которые включаются в цикл
Кребса (так называемый цикл лимонной кислоты), окисляясь
дальше до СОг и НгО. Высвободившиеся в этих окислительных
реакциях протоны и электроны вступают в цепь переноса
электронов, в которой молекулярный кислород служит конечным
акцептором электронов с образованием воды. Все процессы,
генерирующие энергию и требующие участия молекулярного
кислорода, образуют систему аэробного обмена. Генерация энергии
без участия кислорода, как при гликолизе, при котором
происходит расщепление глюкозы до молочной кислоты,
называется анаэробным обменом.
Во всех цепях энергия высвобождается при разрыве
химических связей. Накапливается энергия в «высокоэнергетических»
фосфатных связях аденозинтрифосфата (АТФ) и в меньшей
степени— других фосфатных соединений. Биологическое окисление
по существу представляет собой «сгорание» вещества при
низкой температуре. Поскольку теплота не может быть
использована организмом в качестве источника энергии, некоторая часть
энергии, высвобождающаяся при сгорании, сохраняется путем
Таблица 18.1. Коэффициенты перевода для некоторых общепринятых энергетических единиц
Работа, энергия, теплота
ккал Дж
см3 02«)
л 02')
кал
ккал
Дж
л О/
1,0000 (—3)
4,1868 (0)
2,0833 (—4)
1,0000 (+3)
4,1868 (+3)
2,0833 (—1)
2,3885 (—1)
2,3885 (—4)
5,0073 (—5)
4,8000 (0)
4,8000 (—3)
2,0097 (+1)
1,0000 (—3)
4,8000 (+3)
4,8000 (0)
2,0097 (+4)
Вт
Мощность, потребление энергии
кВт ккал-мин-1
ккал-сут-1
Вт
кВт
ккал-мин-1
ккал-ч-1
ккал-сут-1
1,0000 (—3)
1,4331 (—2)
8,5985 (—1)
2,0636 (+1)
1,000 (+3)
1,4331 (+1)
8,5985 (+2)
2,0636 (+4)
6,9780 (+1)
6,9780 (—2)
6,0000 (+1)
1,4400 (+3)
1,1630 (0)
1,1630 (—3)
1,6667 (-2)
2,4000 (+1)
4,8458 (-2)
4,8458 (—5)
6,9444 (—4)
4,16671 (—2)
Вт-кг-1
Интенсивность обмена
ккал-г-'ч-1 ккал-
•сут-1
ккал • кг-1 -ч-1
ккал-кг-1-сут-1
Вт-кг-1
ккал- (г-ч)-1
ккал-сут-1
ккал•кг-ч-1
ккал-(кг сут)-1
—
8,5985 (—4)
2,0636 (—2)
8,5985 (—1)
2,0636 (+1)
1,1630 (+3)
—
2,4000 (+1)
1,000 (+3)
2,4000 (+4)
4,8458 (+1)
4,1667 (—2)
—
4,1667 (+1)
1,0000 (+3)
1,1630 (0)
1,000 (—3)
2,4000 (—2)
—
2,4000 (+1)
4,8458(—2)
4,1667 (—5)
1,0000 (—3)
4,1667 (- 2)
—
') Коэффициенты перевода для потребляемого 02 варьируют в зависимости от ДК; значение ДК принято здесь рапным 0,79.
328
Глава 18
образования АТФ; АТФ служит также средством переноса
энергии, поскольку он диффундирует в те места, где требуется
энергия. Образование и распад АТФ сопряжены с процессами,
требующими затраты энергии. Когда возникает необходимость в
энергии, путем гидролиза разрывается связь концевой
фосфатной группы, и
высвобождается заключенная в ней
химическая энергия. В этой
форме она может быть
использована клетками.
Перенос энергии —
процесс, не сопровождающийся
производством работы, и
при всех таких реакциях
высвобождается некоторое
количество тепла. В таких
случаях говорят, что
энергия «деградировала» в
тепло, поскольку в такой
форме она не может быть
превращена организмом в
полезную энергию. Около
половины всей химической
энергии обмена теряется в
виде тепла в процессе
образования молекул АТФ.
Мышечное сокращение —
процесс еще менее
эффективный. Около 80% энергии,
используемой при мышечном
сокращении, теряется в
виде тепла из-за малой
эффективности ее
превращения, и только 20%
превращаются в механическую
работу (движение мышцы). Если только человек не совершает
работу над окружающей средой, практически вся генерируемая
энергия теряется в форме тепла. Например, когда человек
плывет, он сообщает некоторое количество энергии воде, чтобы
продвинуться в ней. Это создает волну, которая в конечном счете
потеряет всю сообщенную ей энергию в виде тепла, и
поверхность воды снова станет спокойной. Затраченная таким образом
при плавании энергия представляет собой совершенную работу
плюс тепло, потерянное телом пловца. Если обратиться к
человеку, лежащему в постели, то, пока он не совершает никакой
внешней работы, практически вся работа, совершающаяся в его
Рис. 18.1. Калориметрическая бомба для
измерения энергетического эквивалента
вещества, Тепло, высвобождающееся при
взрывном сгорании пробы внутри
бомбы, выделяется в водяную баню, где
оно измеряется чувствительным
термометром.
Энергетический обмен
329
теле, рассеивается в форме тепла. Следовательно, величина
теплопродукции является точным выражением величины
обмена в организме человека, находящегося в состоянии покоя (это
так называемый «основной обмен»).
ЭНЕРГЕТИЧЕСКИЙ ЭКВИВАЛЕНТ ПИЩИ
Количество энергии, выделяемой при сгорании какого-либо
соединения, не зависит от числа промежуточных этапов его
распада. 1 моль глюкозы дает 2867 кДж (686 ккал) независимо от
того, сгорел ли он сразу в пробирке или окислился в организме
в ходе катаболических процессов. Поэтому не составляет труда
определить запас энергии <на единицу массы в различного рода
пищевых продуктах.
Запас энергии в пище измеряется в калориметрической
бомбе (рис. 18.1)—замкнутой камере, погруженной в водяную
баню. Точно взвешенную пробу помещают в эту камеру,
наполненную чистым Ог, под давлением 20 атм. Пробы поджигают
платиновой проволокой так, что они быстро сгорают.
Высвобождаемое при этом тепло точно измеряется чувствительными
калиброванными термометрами по изменению температуры
определенного объема воды, окружающей камеру; зная
теплоемкость воды, ее объем и изменение температуры, можно
определить количество выделившейся энергии.
Углеводы дают в среднем 17,16 кДж/г (4,1 ккал/г)1
(табл. 18.2). Самой высокой энергетической ценностью
обладают жиры: 1 г жира дает вдвое больше энергии, чем 1 г углевода.
Запасание в форме жира является самым экономичным
способом длительного хранения энергии в организме, так как единица
запасенной энергии оказывается в меньшем объеме вещества.
Белки окисляются в организме не полностью. Аминогруппы
отщепляются от молекулы белка и выводятся с мочой в форме
мочевины. Поэтому при сжигании белка в калориметрической бом-
Таблица 18.2. Энергетический эквивалент, коэффициент ДК,
объем потребляемого 02 и выделяемого С02 для разных пищевых веществ
Калориметрическая
бомба
Углеводы
Белки
Жиры
X
X
4,1
5,4
9,3
с*
17,2
22,6
38,9
Окисление
в организме
человека
^
X
tt
4,1
4,1
9,3
17,2
17,2
38,9
о2
ч
ч
5,05
4,46
4,74
ч
с*
21,1
18,7
19,8
со2
2
5,05
5,57
6,67
*
п
х
21,1
23,3
27,9
ДК
1,00
0,80
0,71
о2
~ч
0,81
0,94
1,96
со2
"ч"
0,81
0,75
1,39
330
Глава 18
бе выделяется больше энергии, чем при его окислении в
организме. Разница как раз и приходится на ту энергию, которая
выделяется при сжигании мочевины. Так, при сжигании белка
в калориметрической бомбе выделяется 22,61 кДж/г (5,4 ккал/г),
а при окислении »в организме человека—17,17 кДж/г
(4,1 ккал/г) (см. табл. 18.2).
ОПРЕДЕЛЕНИЕ ИНТЕНСИВНОСТИ МЕТАБОЛИЗМА
(ОБМЕНА)
Поскольку энергия, затрачиваемая человеком в покое,
быстро переходит в тепло, общая теплопродукция эквивалентна
затраченной энергии. Поэтому, измерив теплопродукцию, можно
определить интенсивность обмена непосредственно и
теоретически более правильно, чем любыми другими косвенными
методами.
ПРЯМАЯ КАЛОРИМЕТРИЯ
Такие прямые измерения энергетического обмена провели
еще в 1788 г. Лавуазье и Лаплас. Они помещали животное в
камеру, окруженную водяной рубашкой со льдом и
изолирующим слоем из льда и воды при температуре 0°С. Поскольку
тепло не могло проникать через ледяной слой извне, любое
прибавление его в ледяной рубашке происходило за счет
теплопродукции в организме животного. Зная, какое количество теплоты
требуется для таяния данного количества льда, исследователи
производили прямое измерение теплопродукции по количеству
воды, образовавшейся в ледяной рубашке.
В современных системах прямой калориметрии ледяная
рубашка заменена циркулирующей жидкостью (рис. 18.2). Для
вычисления теплопродукции используются данные о
теплоемкости жидкости, общем ее объеме, протекающем через
изолированную камеру за единицу времени, и разности температур
поступающей в камеру и оттекающей жидкости. Прямая
калориметрия применяется редко, не считая особых случаев, потому
что при ней трудно добиться высокой точности измерений при
отсутствии сложной аппаратуры, косвенные же методы точны,
дешевы и легко осуществимы.
КОСВЕННАЯ КАЛОРИМЕТРИЯ
Энергетический обмен можно оценивать по количеству
потребляемого кислорода или выделяемой углекислоты в
контролируемых условиях. В замкнутых системах (рис. 18.3)
испытуемый вдыхает определенное количество воздуха из воздухонепро-
Энергетический обмен
331
ницаемой системы, и при этом измеряется уменьшение объема
или парциального давления кислорода (газообмен). В
открытой системе испытуемый находится в непрерывно
вентилируемой камере, и потребление кислорода определяется по разности
концентраций газа на входе и по скорости тока воздуха через
камеру. В обеих системах следует удалить водяные пары, до-
Рис. 18.2. Калориметр для исследований, проводимых на человеке.
Суммарная выделяющаяся энергия состоит из: 1) возникающего тепла
(измеряемого по повышению температуры воды, протекающей в змеевике, который идет
через камеру), 2) скрытой теплоты парообразования (измеряемой по
количеству водяных паров, извлекаемых из окружающего воздуха первым
поглотителем НгО), и 3) работы, направленной на объекты вне камеры; во
избежание накопления в камере С02 следует обеспечить ее поглощение. При
этом процессе образуется вода, поэтому требуется второй ее поглотитель.
Потребление 02 измеряется по тому количеству, которое приходится добавлять,
чтобы содержание его в камере оставалось постоянным.
бавляющиеся к выдыхаемому воздуху, так как они изменяют
показатели давления других газов (см. гл. 17 и 19).
Количество энергии на единицу потребляемого 02 или
выделяемого С02 зависит от типа окисляющихся в организме
веществ. Углеводы дают около 21 кДж на 1 л 02 (5 ккал/л),
белки—18,7 кДж (4,5 ккал), а жиры—19,8 кДж (4,74 ккал).
Поэтому для перевода измеренных количеств поглощенного
кислорода в энергетический эквивалент должен быть известен тип
«топлива». Такую информацию дает одновременное измерение
потребления кислорода и выделения СО2 в условиях
стационарного состояния. Отношение выделяемого С02 к потребляемому
С>2, дыхательный коэффициент (ДК) характерен для данного
«топлива» (табл. 18.2). Обычно в условиях покоя человек ис-
332
Глава 18
Мундштук
/ Клапаны
г1"""Т1
Поглотитель
СО,
Ц=
ш.
jj
Колокол
Водяной
затвор
Писчик
Кимограф
Рис. 18.3. Спирометр, приспособленный для измерения количества
поглощаемого О2. В рот испытуемого вставлен мундштук; внешний газообмен
исключается применением носового зажима. Во избежание сильного увеличения
эффективного мертвого пространства для вдыхаемого и выдыхаемого газов
применяются раздельные трубки с клапанами, исключающие смешивание;
поглотитель СОг исключает накопление выдыхаемого С02 в замкнутой системе.
Объем 02, остающегося в спирометре, регистрируется писчиком на бумаге,
укрепленной на кимографе.
пользует разное «топливо», и ДК в этих условиях равен 0,83.
Считается, что при таком ДК энергетический эквивалент
кислорода составляет 20,22 кДж/л 02 (4,83 ккал/л). Пользуясь этим,
можно определить интенсивность энергетического обмена
(табл. 18.3).
ДРУГИЕ КОСВЕННЫЕ МЕТОДЫ
Для определения интенсивности обмена могут быть
использованы некоторые физиологические параметры, коррелирующие
с потреблением кислорода. Частота дыханий, вентиляционный
объем (частота дыханийХдыхательный объем), частота
сокращений сердца и минутный объем (частота сокращений сердцаХ
Хударный объем) — все они связаны с потреблением кислорода
и поэтому в известной мере отражают затраты энергии. Однако,
как правило, эти показатели недостаточно точны, за
исключением тех случаев, когда человек находится в состоянии покоя.
Энергетический обмен
333
Таблица 18.3. Энергетический эквивалент 1 л 02 при разных дыхательных
коэффициентах
Дыхательный коэффициент
0,707
0,75
0,80
0,85
0,90
0,95
1,00
Энергетический эквивалент
килокалории
4,686
4,739
4,801
4,862
4,924
4,985
5,047
килоджоули
19,62
19,84
20,10
20,36
20,62
20,87
21,13
Для измерения интенсивности обмена предложено
использовать изотопы, например цезия и цинка. В общем, чем
интенсивнее метаболизм, тем быстрее выводится метка. Точные
сведения об интенсивности обмена на протяжении длительного
времени можно получить, применив воду с двойной меткой (по
водороду и кислороду).
Методики соотнесения радиотелеметрических
физиологических параметров или скорости выведения изотопов с
энергетическим обменом привлекательны тем, что они позволяют получать
информацию на протяжении длительных периодов, не требуя
ограничения движений испытуемого. В последнее время эти
методики применяются все более широко. Однако присущие им
серьезные недостатки, в том числе использование слишком
упрощающих допущений и недостаточная точность, в какой-то
мере ограничивают их применение.
ОСНОВНОЙ ОБМЕН
Интенсивность энергетического обмена значительно
варьирует и зависит от многих факторов. Относительно постоянный
уровень энергетических затрат в условиях физического и
эмоционального покоя называют основным обменом (ОО).
Измерение ОО производят в строго контролируемых условиях.
Человек должен: 1) находиться при нейтральной
температуре среды, исключающей активность механизмов
терморегуляции; 2) быть в спокойном удобном положении, но не спать, так
как сон понижает энергетический обмен приблизительно на
10%; 3) быть в состоянии душевного покоя, так как
эмоциональный стресс может усилить метаболизм; 4) не есть на
протяжении не менее 12 ч, причем всасывание пищи должно быть
закончено, чтобы снизить теплопродукцию, связанную с
перевариванием пищи.
334 Глава 18
Основной обмен человека определенного роста и веса
можно предсказать с точностью до 5 или 10%. Более значительные
отклонения от предсказанных величин часто свидетельствуют о
нарушениях обмена, обычно связанных с нарушением
гормонального равновесия.
РАЗМЕРЫ ТЕЛА
Основной обмен у крупных людей очевидным образом ниже,
чем у маленьких. Энергетический обмен может быть выражен
величиной общего обмена (ккал/ч, или по системе СИ в
ваттах) или в виде затраты энергии на единицу массы тела
(ккал/кг/ч = Вт/кг), или величины энергии, расходуемой на
единицу поверхности тела (ккал/м2/ч = Вт/м2).
Кал/день
Кал/кг/день
10000
| 1000
-о
о
>х
о
X
ах
§ ЮО
О
10
0,01 0,1 1,0 10 100 1000 500О
Вес тела, кг
Рис. 18.4. Отношение основного обмена к размерам тела у млекопитающих.
(Brody, 1945.)
У млекопитающих отношение величины основного обмена к
массе тела приблизительно одинаково (рис. 18.4). В
логарифмическом масштабе энергетический обмен млекопитающих, в том
числе человека, описывается уравнением
lgE-0,751gM+7,84,
где Е — энергетический обмен, а М — масса тела.
Физиологи-клиницисты часто пишут о величине метаболизма
на единицу поверхности тела. Такой способ восходит к класси-
Лошадь
1 Молочный скот, бык^
Мясной скот, корова ^
Кал = 70,5 кг0-734 Мясной скот, бык-J
^\^ Человек у?/
Слон_^-^и^
чЯА( Мясной
У/\ скот, бык
Молочный скот,
у корова
\ \^ /J(s\ "~~ ^Свинья
Гусь 2&х\ °BL*a
Кошка^ \ уЯ5*^йг* *
L \л\ Ж/'^-"^ Собака
р^^^^^^ Кролик-—__A>£?£s=~Линии 20%-ного отклонения
К, ^*^^-^^1ка^^^=:=^ Курица
Морская свинка^-^4<2^ 1
Голубь ч Лб<2-( ^""^^*^,^
Канарейка ^^^^^ Крыса ' ^^V*r
\ /4^~~ Горлица ^**^
~~\ A^/^i- Воробей
Щяг7^> Мышь
/УуК | | | |
С^5^вв
1
Энергетический обмен
335
ческой работе Рубнера, который в 1868 г. в своем исследовании
на собаках установил, что интенсивность обмена определяется
размерами поверхности тела и необходимостью поддерживать
постоянную температуру; он сделал вывод, что теплопродукция
также должна меняться в зависимости от веса тела. Однако
данные, полученные па многих видах млекопитающих, не
подтвердили этого предположения. Поэтому теплоотдача и
площадь поверхности тела, по-видимому, не являются основными
факторами, определяющими метаболизм у животных разной
величины. Тем не менее обычай относить энергетический обмен к
единице поверхности прочно укоренился в физиологической
литературе. Случилось так, что у человека отношение основного
обмена к поверхности тела оказалось величиной сравнительно
постоянной. Таблицы для определения поверхности тела у
человека на основании роста и веса облегчают выражение
данных об интенсивности метаболизма на единицу поверхности
(рис. 18.5).
01—N,v Г»»"1 LLI^J 1 1 I 1 1 1 1 1 1 1 1 1 1
20 30 40 50 60 70 80 90 100 110
Вес, кг
Рис. 18.5. График для определения поверхности тела человека (в м2) по
отношению к весу (в кг) и рост (в м) в соответствии с формулой для площади
(m2)=W°'42SXH°'725X0,202. (DuBois, DuBois, 1916. Arch. Intern. Med.).
Мышечная работа существенно изменяет интенсивность
обмена. У тренированных спортсменов при кратковременных
интенсивных упражнениях были зарегистрированы величины
метаболизма, в 20 раз превосходящие показатель основного
обмена; отмечены в 10 раз большие величины при продолжительной
работе (табл. 18.4). Потребление кислорода при физической
нагрузке не отражает общего расхода энергии, так как часть
ее тратится на гликолиз (анаэробный) и не требует затраты
кислорода.
336
Глава 18
Таблица 18.4. Влияние разного рода активности на интенсивность обмена1)
Покой
Сон 35
Бодрствование лежачее положение 4а
ьодрствование сидячее положение 5а
Незначительная активность
Письмо, канцелярская работа 60
Стояние 85
Умеренная активность
Умывание, одевание 100
Ходьба (3 км/ч) 140
Домашняя работа 140
Выраженная активность
Езда на велосипеде 250
Плавание 350
Пилка дров 350
Ходьба на лыжах 500
Бег 600
Дрожь до 250
■) Brown А. С. (1973). Energy metabolism. In: Т. С. Ruch and H. D. Patton,
eds., Physiology and Biophysics (v. 3), Philadelphia, Saunders.
На рис. 18.6 показано потребление кислорода при
субмаксимальной физической работе. На нее требуется определенное
количество энергии, но потребление 02 не сразу удовлетворяет
эту потребность. Со временем устанавливается стационарное
состояние, при котором потребность в 02 удовлетворяется его
притоком. Разность между потребностью в 02 и его
потреблением составляет энергию, получаемую в результате
анаэробного распада, и называется кислородным долгом. Теоретически
расход энергии после окончания мышечной работы должен
вернуться к уровню покоя, однако потребление 02 остается
высоким и только медленно возвращается к этому уровню.
Принято говорить, что в это время происходит оплата кислородного
долга (рис. 18.6).
Кислородный долг состоит из двух компонентов. Лактацид-
ный долг представляет собой добавочную энергию,
затрачиваемую на превращение главного побочного продукта анаэробного
метаболизма — молочной кислоты (lactic acid) в пировиноград-
ную, которая может быть использована в цикле Кребса для
аэробного метаболизма. В больших дозах молочная кислота
вредна для организма и отчасти лежит в основе мышечного
утомления непосредственно после бега. Нелактацидный долг
представляет собой дополнительную энергию, необходимую для
рефосфорилирования таких высокоэнергетических резервных
соединений, как креатинфосфат, и для восстановления запасов
02 в истощившемся мышечном миоглобине.
Энергетический обмен
337Г
Стационарное
состояние
при нагрузке
Время, мин
Рис. 18.6. Потребление 02 при умеренной мышечной работе. Величина
кислородного долга вычисляется путем измерения разности между общим
потреблением Ог и уровнем покоя по окончании работы. В идеальном случае
площади обоих секторов равны.
Анаэробный обмен имеет первостепенное значение при
кратковременной максимальной физической нагрузке. Забег на
100 м приблизительно за 10 с осуществляется на 85% за счет
анаэробной энергии. По мере увеличения продолжительности
физической нагрузки отношение аэробной энергии к анаэробной
возрастает. Если максимальная активность длится две минуты».
Рис. 18.7. Потребность в 02 (прерывистые линии) и потребление Ог
(сплошные линии) у человека при трех интенсивностях мышечной работы: I — бег
в течение 30 с; II — бег в течение 2 мин до истощения, с максимальным
потреблением Ог и III — менее интенсивная работа, при которой потребление 02
было равно потребности в нем. Площадь треугольника, выражающего
потребность в Ог, равна сумме площадей для Ог, потребленного до и после
мышечной работы.
22-1705
338 Глава 18
100
80
60
20
0 10 20 30 40 50 60
Мин
Рис. 18.8. Относительный вклад аэробного (/) и анаэробного (//) процессов
при максимальном усилии. При максимальной активности на протяжении
около 2 мин как на аэробную, так и на анаэробную работу приходится
приблизительно по половине затраченной энергии.
аэробная и анаэробная энергия уравниваются, и каждая из них
составляет 50% генерируемой энергии. Максимальное усилие
длительностью 60 мин требует 90% аэробной энергии. При его
дальнейшем продлении общая интенсивность обмена
(аэробного и анаэробного) снижается (рис. 18.7 и 18.8).
ДРУГИЕ ФАКТОРЫ И ИНТЕНСИВНОСТЬ ОБМЕНА
Человек сохраняет постоянную температуру внутренних
областей тела независимо от температуры окружающей среды.
Для этого должна осуществляться регуляция теплопродукции
или теплоотдачи. В покое интенсивность теплопродукции (и,
следовательно, интенсивность метаболизма) прямо зависит от
разности между температурой внутренних областей тела и
окружающей температурой. Дополнительный расход энергии
(теплопродукция) уравновешивает большую теплопотерю при
низкой температуре окружающей среды (подробнее см. гл. 19).
Само поступление пищи усиливает энергетический обмен.
Этот процесс, называемый специфическим динамическим
действием пищи (СДД), не нашел определенного физиологического
объяснения. Величина СДД зависит от характера съеденной
пищи. Белковая пища повышает интенсивность обмена на 25—
•30%, а углеводы и жиры—всего на 10% или меньше.
Энергетический обмен
33£
Таблица 18.5. Нормальные стандарты интенсивности основного обмена,
ккал/мг/ч1)' 2>
Мужчины
Возраст Основной обмен
6
7
8х/2
9
9V2
10
10V2
11
12
13-15
16,
161/!
17
I7V2 I
18,
18^2
19
191/*
20—21
22—23
24—27
28—29
30—34
35—39
40—44
45—49
50—54
55-59
60—64
65—69
53,0
52,5
51,8
51,2
50,5
49,4
48,5
47 7
47,2
46,7
46,3
45,7
45,3
44,8
44,0
43,3
42,7 |
42,3 1
42,0
41,4
40,8
40,2
39,8
39,3
38,7
38,0
37,4
36,7
36,1
35,5
34,8
1 Женщины
1 Возраст
6,
6V2
1 7
7V2
8
8V2
9—10
11
II1/!
12
12х/2
1.3
i3Va
14
I4V2
15
15V2
16Vl
IW
18—19
20—24
25—44
45—49
50—54
55—59
60—64
65—69
J Основной .обмен
50,6
50,2
49,1
47,8
47,0
1 46,5
45,9
45,3
44,8
44,3
43,6
42,9
42,1
41,5
40,7
40,1
39,4
38,9
38,3
37,8
37,4
36,7
36,2
35,7
34,9
34,0
33,2
32,6
32,3
') Boothby W. M., Berkson J., Dunn H. L. (1936). Am. J. Physiol., 116, 468.
2) Нормальный диапазон обычно принимается равным ±10%; более выраженное от*
Клонение считается патологическим.
У людей основной обмен неуклонно меняется с возрастом;
для каждой возрастной группы выведены клинические
стандарты (табл. 18.5). Отклонение от такого стандарта на ±10%
считается нормой. У детей оно больше, в период полового
созревания уменьшается и меньше всего в старости.
Во время сна интенсивность метаболизма приблизительно на
10% ниже ОО. Вероятно, разница между бодрствованием в
состоянии покоя и сном объясняется тем, что во втором случае
мышцы более расслаблены.
Лихорадочное состояние с повышением средней температуры
тела связано с повышением интенсивности энергетического об-
22*
.340
Глава 18
мена. Хорошо известно, что с повышением температуры
химические реакции ускоряются. Поэтому более высокая средняя
температура создает тенденцию к ускорению химических
реакций, связанных с энергетическим обменом. Кроме того, часто
возникает усиленная мышечная активность, аналогичная
дрожи. Сужение периферических сосудов и пониженная
способность к потоотделению уменьшают потерю тепла и приводят
к накоплению большего количества тепла в организме, что
>еще больше повышает температуру тела и теплопродукцию.
ЛИТЕРАТУРА
Bartholomew G. А. (1977). Energy metabolism. In: Gordon M. S. (ed.), Animal
physiology: principles and adaptations, 3rd edn., MacMillan, New York.
.Benedict F. G. (1938). Vital energetics. Carnegie Institution of Washington
Publications, No. 503, Washington.
Brody S. (1945). Bioenergetics and growth, Reinhold, New York.
Hemmingsen A. M. (1960). Energy metabolism as related to body size and
respiratory surfaces and its evolution. Rep Steno Mem Hosp. Nord Insulin Lab.,
9, 7—110.
.Kleiber M. (1961). The fire of life, Wiley, New York.
Lefebvre E. A. (1964). The use of D2018 for measuring energy metabolism in
Columbia livia at rest in flight, Auk, 81, 403—416.
Lusk G. (1928). The elements of the science of nutrition, 4th edn., Saunders,
Philadelphia.
Mechtly E. A. (1973). The international system of units: physical constants and
conversion factors. NASA-7012, National Aeronautics and Space
Administration, Washington.
Miller А. Т., Jr. (1954). Energy metabolism and metabolic reference standards,
Math. Med. Res., 6, 76—84.
Passmore R., Durnin J. (1955). Human energy expenditure, Physiol. Rev., 35,
801—840.
Stahl W. R. (1967). Scaling of respiratory variables in mammals, J. Appl.
Physiol., 22, 453—460.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. При каких условиях количество потребляемого Ог может
служить мерой интенсивности энергетического обмена, и какая
требуется дополнительная информация?
2. Почему потребление Ог точнее отражает интенсивность
энергетического обмена, чем выделение ССЬ?
3. Почему при одинаковом потреблении 02 теплопродукция
будет различной, если используются разные виды «топлива»?
4. Дайте характеристику аэробной и анаэробной генерации
энергии для бегуна на длинную дистанцию а) сразу после
старта, б) на середине дистанции, в) у финиша.
5. Сколько имеется разных видов энергии? Какие формы энергии
являются для организма «полезными»?
Глава 19
ТЕРМОРЕГУЛЯЦИЯ
Температура среды, непосредственно окружающей животное,
оказывает большое влияние на его физиологическую
активность. На Земле температура колеблется от —50° во время
арктической зимы до +60°С летом в некоторых пустынях. Однако
температурный диапазон, в котором способны функционировать
живые клетки, составляет всего около 50°.
Живые клетки замерзают при нескольких градусах ниже
0°С. Само по себе замерзание не всегда является роковым для
живых тканей; ученые получали ткани, которые при
замораживании сохраняли на некоторое время свою жизнеспособность.
Однако обычно при замерзании тканей образующиеся
кристаллы льда разрушают тонкие клеточные структуры. При
температурах выше 45 °С происходит денатурация белков. Поскольку
белки ответственны фактически за все регуляторные функции
животных, их структурная и функциональная сохранность
жизненно необходима для нормального функционирования
организма. Температура сильно влияет на метаболизм живой ткани,
так как скорость биохимических реакций зависит от
температуры окружающей среды и обычно возрастает в два-три раза
«а каждые 10° повышения температуры.
В животном мире существуют несколько основных способов
реагирования на внешнюю температуру. У пойкилотермных
животных, к которым относятся большинство беспозвоночных и
низших позвоночных, температура тела зависит от окружающей
температуры (рис. 19.1). Альтернативой к такой зависимости
служит терморегуляция. В этом случае температура внутренних
областей тела поддерживается на постоянном уровне
независимо от температуры окружающей среды. Терморегуляция
присуща птицам и млекопитающим, которых называют гомойотерм-
ными (или гомеотермными). Терморегуляция позволяет
животным функционировать в температурных условиях, которые
обеспечивают оптимальную активность ферментов. Гетеротермия —
это особое состояние, в котором гомеотермные животные на
время выключают терморегуляцию и позволяют температуре
своего тела снижаться до пределов, отличных приблизительно
на 1 °С от окружающей температуры.
342
Глава 19
Многие наземные животные — рептилии и некоторые
насекомые— сохраняют в дневное время сравнительно
постоянную температуру тела в известных пределах температур
воздуха, используя источники тепла и утечку тепла в окружающей
среде. Поэтому в течение дня такие животные скорее подходят
под определение гомеотермных, чем пойкилотермных (рис. 19.1).
Животных можно также классифицировать по тем источникам
о
о
to"
с;
Q)
\-
<0
О.
>ч
I-
(0
О.
Q)
С
Q)
О 10 20 30 40
Температура воздуха. °С
Рис. 19.1. Соотношение температуры тела у животных и температуры
воздуха. У пойкилотермных животных первая довольно близка второй. Гомойотерм-
ные сохраняют температуру тела на сравнительно постоянном уровне в
широком диапазоне температур воздуха.
тепла, которые они используют для поддержания температуры
тела. Эктотермные, например рептилии, используют для этого
наружное тепло; эндотермные, и в частности человек,
используют тепло метаболического происхождения.
ТЕМПЕРАТУРА ТЕЛА И ЕЕ ИЗМЕРЕНИЕ
Температура тела эндотермных организмов зависит как от
внешней среды, так и от состояния активности. В покое
температура тела человека составляет около 37 °С, хотя она
предсказуемо меняется в течение суток с небольшим снижением во
время сна. При мышечной работе температура тела часто
повышается на несколько градусов вследствие усиленной
теплопродукции.
Температура не одинакова во всем теле, и распределение
внутренней температуры является сложной функцией
теплопродукции в разных тканях, переноса тепла в результате циркуля-
Терморегуляция
343
ции крови и локальных температурных градиентов. Если не
считать условий напряженной мышечной работы, большая
часть метаболической теплопродукции происходит во
внутренних органах (в сердце и внутренностях), а также в мозгу.
Тепло, образованное в глубоких областях тела, или внутреннее
{core) тепло, должно перейти к поверхности тела. Температура
на периферии определяется теплом, перенесенным из глубоких
областей тела, и температурой воздуха. Следовательно, можно
считать, что тело обладает внутренней, относительно
постоянной температурой и изолирующим слоем, оболочкой (shell),
весьма изменчивой в зависимости от энергетического баланса
организма. В холодной среде приток крови к периферии
сокращается, и это вызывает понижение периферической температу-
Рис. 19.2. Температура различных областей тела человека при температуре
воздуха 20°С (Л) и 35 °С (Б). Показаны изотермы (линии, соединяющие
точки с одинаковой температурой). При 20°С между внутренней областью тела,
(заштриховано) и поверхностью существуют резкие перепады температуры,
причем внутренняя область ограничена головой и туловищем. При 35 °С
внутренняя область распространяется далеко на конечности.
344
Глава 19
ры. При высокой температуре окружающей среды теплоотдача
затрудняется, и внутренняя температура может
распространяться почти на все тело (рис. 19.2).
Температура ткани отражает содержание в ней тепла.
Поэтому определение температуры тела по одному измерению
ошибочно, но все же оно удобнее, чем попытки судить по
совокупности измерений температуры во многих областях тела. Этот
Теплообразование
Теплоотдача
Потоотделение
Одышка
Изменение
температурь!
Более легкая
одежда
Усиление
незаметного
испарения •
Усиленное
излучение
Усиленное
j движение
' воздуха
Гипотермия
Рис. 19.3. Равновесие между факторами, усиливающими приток и отдачу
тепла, создающее постоянную температуру тела. Если равновесия нет,
температура тела изменяется.
последний подход дает больше информации об энергетическом
балансе организма, но технически такие измерения трудно
осуществимы. Кроме того, интерпретация их тоже трудна
вследствие динамического характера изменений температуры в
изолирующем слое.
Температуру тела обычно измеряют в заднем проходе
(ректально) или во рту (орально). Ректальная температура —
самый надежный и самый удобный способ измерения, и обычно
принято считать, что она отражает среднюю внутреннюю
температуру. Однако этот способ измерения признан не везде. Часто
приводится температура во рту, под мышкой, в сердце или
печени. Измеренная ректально внутренняя температура меняется
слишком медленно и не дает точной информации при
исследовании терморегуляции. В таких исследованиях желательно
знать температуру мозга, которую у человека и животных хо-
Терморегуляция
345
рошо отражает температура барабанной перепонки.
Температура крови, оттекающей от левого желудочка, тоже используется
в качестве динамического показателя энергетического баланса
организма.
ТЕПЛОВОЙ БАЛАНС
Чтобы температура тела оставалась постоянной, необходимо,
чтобы отдача тепла была равна его притоку (в результате
метаболической теплопродукции или из внешней среды)
(рис. 19.3). Если приток выше отдачи, тепло накапливается,
вызывая повышение температуры (гипертермию). Такой баланс
удобно описывается уравнением
М ± ER ± Ес ± EG—EE ± S = О,
где М — метаболическая теплопродукция, Er — излучение, Ее —
теплопроводность, EG — конвекция, ЕЕ — испарение, S —
накопление тепла; « + » означает приток, а «—»— теплоотдачу,
Тепло может быть получено или отдано путем излучения,
теплопроводности и конвекции в зависимости от условий внешней
среды. Тепло всегда образуется в качестве побочного продукта
химических реакций, протекающих в организме (см. гл. 22), и
поэтому метаболизм всегда имеет положительный знак; испарение
всегда имеет отрицательный знак. Противоположная реакция,
конденсация, обычно мало влияет на тепловой баланс у
человека.
ИЗЛУЧЕНИЕ
Все предметы с температурой выше абсолютного нуля
{—273 °С) отдают энергию путем излучения. Излучение
происходит в форме электромагнитных волн. Электромагнитная
радиация свободно проходит через вакуум; атмосферный воздух
также можно считать прозрачным для электромагнитных волн.
ИЗЛУЧАЮЩАЯ СПОСОБНОСТЬ
Температура поверхности предметов и тип или качество их
поверхности имеют важное значение для движения энергии в
форме излучения. Излучающая способность поверхности связана
с ее свойствами как излучателя. Поверхность, поглощающая и
совсем не отражающая лучистой энергии, обладает
максимальной излучающей способностью, равной 1. Когда поверхность
отражает всю лучистую энергию, ее излучающая способность
равна 0. Очевидными примерами такой поверхности служат
зеркало или хорошо отполированный металл. Обычно предметы
346
Глава 19
почти полностью поглощают волны одной длины и в то же
время сильно отражают волны другой длины. Так, например,
человеческая кожа, как белая, так и пигментированная, поглощает
почти все инфракрасное излучение. В то же время черная
кожа поглощает значительно больше видимого излучения.
В большинстве случаев окружающие предметы, излучение от
которых достигает организма или на которые попадает
излучение от него, имеют температуру, отличающуюся примерно на
20 °С от температуры поверхности тела. Поток энергии в
ваттах описывается уравнением
ER=KR(TS-Ta),
где Kr — это коэффициент передачи тепла излучением (Вт/°К),
a (Ts—Та) выражает разность температур между кожей и
внешней средой. Член Ts—Та часто определяют как движущую силу,
поскольку для существования потока лучистой энергии
необходима разница температур.
ТЕПЛОПРОВОДНОСТЬ
Переход тепла от одного предмета к другому при
соприкосновении их поверхностей называется теплопроводностью. Тепло
перемещается по тепловому градиенту от более теплого к более
холодному предмету. Этот процесс идет без перехода материи
от одной поверхности к другой; энергия переносится через слои
молекул без всякого перемещения самих молекул. Ощущение
прикосновения к «теплому» или «холодному» предмету
связано с направлением движения тепла путем его проведения. Если
температура предмета ниже температуры пальца,
прикасающегося к нему, то тепло переходит от пальца к предмету, а если
предмет теплее пальца, тепло движется в противоположную
сторону.
Переход тепла путем теплопроводности (W) является
линейной функцией разности температур двух соприкасающихся
предметов и описывается уравнением
Ес = Кв(Т1-Т^.
Величина К выражает количество тепла, переходящего путем
теплопроводности между двумя предметами (W/K). Однако
нередко представляющим интерес параметром является не
количество тепла, проходящего через вещество, а удержание тепла
этим веществом. Хорошие изолирующие оболочки плохо
проводят тепло, и наоборот.
Одним из лучших проводников тепла служат такие
металлы, как серебро и медь, а к самым худшим проводникам
относятся разные газы. Биологические ткани служат примерно та-
Терморегуляция
347
кими же изоляторами, как вода; жир в этом отношении вдвое
эффективнее мышечной или костной ткани. У тюленей,
например, толстый слой жира служит для изоляции внутренних
областей тела от кожи, которая соприкасается с ледяной водой.
В результате кожа сохраняет температуру лишь на несколько
градусов выше температуры воды. Не будь жирового слоя,
температура кожи была бы близка внутренней температуре тела
(37 °С), и потеря тепла была бы в 10 раз больше.
У млекопитающих теплопроводность не является основным
способом теплообмена, потому что площадь соприкосновения их
поверхности с твердыми предметами минимальна. При низких
температурах среды такие эктотермные, как пресмыкающиеся и
насекомые, часто прижимаются поверхностью тела к теплым
предметам, чтобы путем теплопроводности повысить
температуру тела.
КОНВЕКЦИЯ
Перенос тепла от предмета, окруженного жидкой или
газообразной средой, происходит посредством конвекции.
Конвекционный теплообмен в отличие от теплопроводности связан с
обменом не только энергии, но и молекул. Это происходит потому,
что вокруг всех предметов существует «нерушимый»
пограничный слой влаги (воздуха или жидкости). Толщина этого слоя
зависит от окружающих условий. Поток тепла идет от более
теплых к более холодным участкам. Если температура воздуха
выше температуры тела, тепло будет передаваться телу.
Когда тело окружено неподвижным воздухом, от кожи
отходит теплый воздух, который, переходя в окружающий воздух,
переносит как молекулы, так и энергию. Такой процесс
называется свободной конвекцией. Если окружающий воздух
движется, толщина пограничного слоя зависит от скорости
движения воздуха. Пограничный слой, равный при неподвижном
воздухе нескольким миллиметрам, при ветре может уменьшиться
до нескольких микронов. Такого типа теплообмен в
значительной степени зависит от движения воздуха и называется
принудительной конвекцией.
Количество переносимого конвекцией тепла описывается
уравнением
Ec = hc(Ts-Ta),
где Ее — количество тепла, передаваемого путем конвекции
(W), Ts и Та — температура кожи и воздуха, a hc —
коэффициент передачи тепла путем конвекции. Коэффициент hc
зависит от величины предмета и скорости ветра. Для гладких ци-
348
Глава 19
Крыса
линдров hc описывается формулой
hc = KV1/3D2/\
где V —скорость движения воздуха, a D — диаметр цилиндра
(рис. 19.4). Влияние скорости ветра на коэффициент конвекции
выражается «показателем охлаждения ветром». Из-за того что
ветер уменьшает пограничный слой воздуха около кожи, отдача
тепла при ветре больше,
12 г- ^пыло чем в спокойном воздухе.
Поэтому при сильном
ветре воздух «ощущается
более холодным», чем он
есть в действительности.
Метеорологические
станции часто сообщают
наряду с истинной
температурой воздуха также
кажущуюся, вызываемую
фактором охлаждения
ветром.
Для регуляции
внутренней температуры тела
большое значение имеет
конвекционная передача
тепла в нем. Большая
часть мышц,
обеспечивающих теплопродукцию,
находятся во внутренних
областях тела. Как было указано в предыдущем разделе, ткань
тела плохо проводит тепло, а кровь выносит большое
количество тепла, образованного внутри тела, на периферию, т. е. к
рукам и ногам, где относительно большие их поверхности и более
низкие температуры облегчают отдачу этого тепла во внешнюю
среду.
ИСПАРЕНИЕ
При испарении воды с какой-либо поверхности поверхность
охлаждается вследствие затраты энергии на переход жидкости
в пар, или газообразное состояние. На каждый грамм
испарившейся воды уходит около 0,59 Вт-ч. Почти в любых условиях
окружающей среды вода непрерывно испаряется с поверхности
тела и составляет важный механизм теплоотдачи. Основные
места испарения — это кожа и дыхательные пути. Объем потери
воды зависит от внешних условий, особенно от температуры и
влажности.
0,25 0,50 0,75
Скорость движения воздуха, м/с
Рис. 19.4. Зависимость коэффициента
конвекции (hc) двух цилиндров от скорости
ветра. D — диаметр.
Терморегуляция
349-
Если воздух насыщен водяными парами (100%
относительной влажности), испарения с поверхности кожи не происходит.
Вдыхаемый воздух нагревается в легких до внутренней
температуры тела и насыщается водяными парами. Даже если
наружный воздух насыщен ими, при выдохе теряется некоторое
количество воды, потому что при повышении температуры
воздуха при выдохе данный объем воздуха удерживает больший
объем воды.
Испарение приобретает чрезвычайно большое значение при
высокой температуре воздуха, поскольку при температуре
воздуха, равной температуре тела (или точнее — температуре
кожи), ни один из обычных механизмов теплоотдачи — излучение,
теплопроводность, конвекция — не функционирует.
У человека главной для терморегуляции формой испарения-
служит пот. У других животных — многих птиц (не имеющих
потовых желез), некоторых млекопитающих и отдельных
рептилий, потеря воды путем испарения усиливается при высокой
температуре благодаря учащенному дыханию — так называемой"
«тепловой одышке» (panting). Тепловая одышка увеличивает
ток воздуха над влажными респираторными поверхностями и
тем самым усиливает испарение.
КОЭФФИЦИЕНТ ПЕРЕДАЧИ ТЕПЛА
Теплообмен путем излучения, теплопроводности и конвекции*
приблизительно линейно зависит от разности температур телаг
и внешней среды. Коэффициенты излучения, теплопроводности
и конвекции могут быть объединены в общий коэффициент
передачи тепла (теплопроводности), который выражает величину
теплообмена на каждый радиус разности температур тела и
воздуха в соответствии с формулой
dH/dt = C(Tb-Ta),
где dH/dt означает потерю тепла (ватты), С — коэффициент-
передачи тепла (Вт/К), а Ть и Та — соответственно внутреннюю
температуру тела и температуру воздуха. Это уравнение
представляет собой упрощение и в такой форме полезно только при-
тщательно контролируемых внешних условиях.
ИНТЕНСИВНОСТЬ МЕТАБОЛИЗМА И ТЕМПЕРАТУРА
ВОЗДУХА
Метаболическая реакция млекопитающих на температуру^
воздуха показана на рис. 19.5. Минимальный уровень
метаболизма приходится на зону температур воздуха, называемую по-
350
Глава 19
разному: зоной физической регуляции, термонейтральной зоной,
или зоной комфорта. В этой температурной зоне
теплопродукция не меняется, и температура тела может регулироваться
незначительными изменениями теплопроводности животного путем
изменения позы, оттоком крови от внутренней части тела к
периферии или усилением потоотделения. При самой низкой
температуре термонейтральной зоны, при наименьшей критической
^6
X
sec 5
«о
Q)
о -о 4
«О Л
^ о 3
о со °
О п
с Я о
О) О. £
TH3
с дополнительной
изоляцией
твк
с дополнительной изоляцией ^^
^х
_L
10
20
30
40
•Рис. 19.5. Соотношение между энергетическим обменом у млекопитающего и
температурой воздуха (сплошная линия). Прерывистая линия — то же
животное с более сильной изоляцией (у животных при смене на более густой мех,
у человека при надевании пальто). Коэффициент минимального переноса
тепла равен наклону кривой, выражающей отношение между обменом и
температурой.
ТНЗ — термонейтральная зона; ТНК — нижняя граница — температуры
комфорта; ТЁК — верхняя граница температуры комфорта.
температуре, животное сводит к минимуму все пути
теплообмена. Если температура внешней среды падает ниже
наименьшей критической, то постоянная температура тела уже не
может быть сохранена путем сокращения передачи тепла, и
поэтому во избежание снижения температуры тела должна
повыситься теплопродукция. Зона температур ниже наименьшей
критической температуры называется зоной химической
регуляции, или зоной метаболической регуляции. Человек может
менять это соотношение искусственно, меняя вес своей одежды.
Надев зимнее пальто, он повышает теплоизоляцию и тем самым
снижает теплоотдачу (рис. 19.5) Такие же сезонные изменения
наблюдаются у животных, у которых зимой после линьки
вырастает более густой и длинный мех (улучшающий
теплоизоляцию).
Терморегуляция
351
СИСТЕМА ТЕРМОРЕГУЛЯЦИИ
Система терморегуляции состоит из ряда элементов со
взаимосвязанными функциями. Термическая информация приходит
от периферических или глубинных температурных рецепторов.
Импульсы от этих рецепторов поступают по афферентным
нервам к центру терморегуляции в гипоталамусе, после чего он
Афферентные нервы Эфферентные нервы Венозный отток Обратная связь*
Терморецепторы | цнс | Эффекторы | Тепловой смеситель |
Обратная связь с терморецепторами через кровь
Рис. 19.6. Система терморегуляции у млекопитающего. Тонкие линии — нерв>-
ные пути, толстые — кровеносные сосуды. (Bligh J., 1973.)
активирует различные механизмы, которые обеспечивают либа
теплопродукцию, либо теплоотдачу. Механизм обратной связи
с участием нервной системы и кровоток изменяют
чувствительность температурных рецепторов, и таким образом создается
замкнутая петля (рис. 19.6).
По своей работе система терморегуляции аналогична
системе автоматизированного контроля с отрицательной обратной
связью. Температура тела поддерживается на определенном
Уровне, и величина реакции эффекторов пропорциональна
отклонению истинной температуры от этой точки. Так, если
внутренняя температура падает на 2°С, то повышение
теплопродукции посредством дрожи будет гораздо больше, чем при падении
внутренней температуры на 0,5 °С. В следующем разделе будут
рассмотрены разные элементы системы терморегуляции.
352
Глава 19
ТЕМПЕРАТУРНЫЕ РЕЦЕПТОРЫ
Температурные рецепторы распределены по всей
поверхности тела и в глубоких областях. Некоторые сенсорные клетки
обладают необычайно высокой температурной
чувствительностью, но отдельного морфологического типа сенсорных клеток,
связанного специально с температурной чувствительностью, не
•существует.
Кожные терморецепторы бывают двух типов — холодовые
и тепловые. Оба типа особенно чувствительны к степени
изменения температуры. Холодовые рецепторы резко снижают
частоту импульсации в ответ на повышение температуры и
повышают частоту, когда температура падает. В тепловых
рецепторах происходят обратные изменения. Если температура
меняется быстро, реакция рецепторов будет гораздо большей, чем в
том случае, когда такое же изменение температуры происходит
медленно.
Температурные рецепторы расположены в разных областях
тела. Они встречаются во внутренних органах (ауэрбахово
сплетение), гипоталамусе, ретикулярной формации, в преоптической
области ствола мозга и спинном мозгу. Они найдены в
дыхательных путях, продолговатом мозгу, в двигательной коре, и
весьма вероятно, что после дальнейших исследований такие
рецепторы будут обнаружены и в других местах.
ДЕНТР ТЕРМОРЕГУЛЯЦИИ
Теплопродукция и теплоотдача организма меняются под
действием многих нервных центров. Поэтому должен
существовать не только центр, интегрирующий различные сенсорные
входы, связанные с тепловым балансом организма, но и центр
регуляции разных двигательных реакций. Такой контроль
составляет одну из функций гипоталамуса, который иногда называют
термостатом организма (см. гл. 10 и 24). После хирургического
отделения гипоталамуса от нижележащих отделов головного
мозга и от спинного мозга способность к регуляции
температуры тела утрачивается. Контроль за разными эффекторами
осуществляют отдельные участки гипоталамуса.
ТЕПЛООТДАЧА
Двигательные центры переднего гипоталамуса контролируют
регуляцию теплоотдачи, главным образом предотвращая
перегревание. После разрушения этого центра физиологическая
активность в холодной среде продолжается, но в теплой среде
отсутствует контроль температуры тела, и температура повы-
,шается.
Терморегуляция
353
ТЕПЛОПРОДУКЦИЯ
Другой центр, лежащий в заднем гипоталамусе,
контролирует величину теплопродукции и тем самым предотвращает
излишнее охлаждение. Разрушение заднего гипоталамуса у
животного, находящегося в холодной среде, нарушает
способность к усилению энергетического обмена, и температура тела
падает. Центры переднего гипоталамуса, регулирующие
расширение сосудов, потоотделение и тепловую одышку,
чувствительны к температуре протекающей через них крови. Кроме того,
имеются данные, позволяющие думать, что для функции
гипоталамуса имеет большое значение приток Na+ и Са2+,
по-видимому определяющий «заданный» уровень температуры.
Изменение движения Na+ и Са2+ приводит к изменению уровня
температуры тела. Механизм этого воздействия и отношение
температуры тела к уровню Na+—Са2+ в крови остаются пока
неясными. Полученные данные показывают, что незначительное
понижение температуры гипоталамуса вызывает у мелких
млекопитающих усиление теплопродукции посредством дрожи и
сохранение тепла путем сужения сосудов.
РЕГУЛЯЦИЯ ТЕПЛООТДАЧИ ПРИ УЧАСТИИ
ЭФФЕКТОРОВ
Конвекция тепла из внутренних областей тела к
конечностям в результате изменения объема кровотока является
важным средством регуляции теплоотдачи (вазомоторный
контроль). Конечности выдерживают гораздо больший диапазон
температур, чем внутренние области тела и образуют
прекрасные температурные «отдушины», т. е. места, которые могут
обеспечить потерю больших или меньших количеств тепла в
зависимости от притока тепла из внутренних областей тела через
кровоток.
Вазомоторный тонус регулируется адренэргическими
симпатическими нервными волокнами, которые меняют приток крови
к конечностям и температуру крови, поступающей в кожу (см.
гл. 14).
В ответ на сильное охлаждение общая вазомоторная
реакция снижает приток крови на периферию посредством
эффективного сужения сосудов. У человека по мере прохождения
крови по крупным артериям рук и ног температура ее значительно
падает. Прохладная венозная кровь, возвращаясь внутрь тела
по сосудам, расположенным близ артерий, захватывает боль-
Шую долю тепла, отдаваемого артериальной кровью (рис. 19.7).
Такая система называется противоточным теплообменником.
Она способствует возвращению большого количества тепла к
23-1705
354
Глава 19
внутренним областям тела при завершении кровотоком круга
через конечности. Суммарным эффектом такой системы
является снижение теплоотдачи.
При температуре воздуха, близкой к нулю, такая система
невыгодна, так как в результате интенсивного теплообмена
между артериальной и венозной кровью температура пальцев
на руках и ногах может упасть ниже точки замерзания (отмо-
Артерии
Рис. 19.7. Кровоток к периферическим тканям, допускающий расширение
сосудов кожи (клапан открыт), которое облегчает отдачу тепла, и сужение
сосудов кожи (клапан закрыт), которое снижает отдачу тепла. Заметьте, что
кровь возвращается к внутренним областям тела по другим сосудам.
рожение). У животных, адаптированных к арктическим
условиям, происходит избирательное расширение сосудов для
поддержания температуры кожных покровов выше точки замерзания.
При сильной жаре резко увеличивается приток крови к коже
и избыток тепла рассеивается от конечностей. Кровь
возвращается к внутренним областям тела по венам, лежащим под
самой поверхностью кожи. Изменяя свой путь, венозная кровь
минует противоточный теплообменник, благодаря чему
снижается количество тепла, которое захватывается из нисходящей
артериальной крови. Близость вен к кожной поверхности сильно
увеличивает охлаждение венозной крови, возвращающейся к
внутренним областям тела.
ТЕПЛООТДАЧА ПУТЕМ ИСПАРЕНИЯ
Высокая температура воздуха создает для системы
терморегуляции млекопитающих серьезные трудности. В большинстве
регионов планеты температура тела млекопитающих значитель-
Терморегуляция
355
но выше, чем окружающей среды, и основная задача состоит в
уменьшении теплопотерь. При повышении температуры воздуха
разница между окружающей температурой и температурой
тела уменьшается, и этим снижается эффективность теплообмена
путем излучения, теплопроводности и конвекции. Если
температура окружающего воздуха выше температуры тела, оно может
стать приемником тепла, поступающего из внешней среды.
Запасы тепла в теле возрастут, пока не будет достигнута для него
высшая точка температурного равновесия. В этих условиях
испарение воды служит главным способом отдачи тепла во
внешнюю среду.
ТЕПЛОВАЯ ОДЫШКА
Самой распространенной формой отдачи тепла путем
испарения у млекопитающих и птиц обычно служит тепловая
одышка— учащенное дыхание. При этом объем воздуха,
проходящего над влажными поверхностями дыхательных путей, резко
возрастает. Например, у собак, у которых частота дыхания в
норме составляет 30—40 в 1 мин, в жару она достигает 300—400.
У некоторых животных такое дыхание имеет частоту,
соответствующую резонансным частотам их дыхательной системы. Это
самая эффективная частота, при которой дыхательная система
в состоянии функционировать и снижать затраты энергии. Она
связана с эластическими свойствами системы, которая
поглощает некоторое количество энергии при вдохе и затем отдает
ее при выдохе.
Тепловая одышка эффективна только на некоторое время,
потому что, если она длится долго, нарушается
кислотно-щелочной баланс организма, поскольку усиленная вентиляция может
привести к избыточному выведению С02, а это приводит к
повышению рН крови (см. гл. 17). Хотя потери С02 могут быть
снижены за счет перехода на поверхностное дыхание
(уменьшение дыхательного объема), в результате чего большая часть
воздушного потока оказалась бы ограниченной мертвым
пространством, тем не менее у большинства млекопитающих этот
механизм эффективно не используется, так что тепловое
дыхание на протяжении длительного времени вызывает алкалоз.
ПОТООТДЕЛЕНИЕ
У человека и других млекопитающих, например у лошади и
коровы, теплоотдача происходит главным образом не путем
учащенного дыхания, а за счет потоотделения. Потоотделение
особенно эффективно у животных со скудным шерстным покровом.
При этом для испарения используется вся поверхность тела.
23*
'56 Глава 19
Такой способ теплоотдачи не сказывается на нормальном
дыхании и не нарушает кислотно-щелочного баланса; испарение
путем потоотделения не связано с большим расходом энергии.
Однако длительное усиленное потоотделение ведет к
обезвоживанию и потере солей, что нарушает баланс электролитов,
восстановление которого требует потребления соли.
В коже человека насчитывается более двух миллионов
потовых желез, функция которых регулируется симпатической
нервной системой. Они чувствительны и к местной температуре
кожи, и к температуре в области мозговых центров (рис. 19.8).
Потоотделение становится максимальным при повышении
внутренней и периферической температуры.
т 1 1 1 1 1 1 1
Температура кожи-~^^^
39°, 38°,370,360,350,340,33°С/
/ 33°С32оСЗГС30оС29°С
' ддл Температура кожи
—I 1 1 I I I I I
36,4 36,8 37,2 37,6
Температура барабанной перепонки, °С
Рис. 19.8. Взаимодействие между температурой барабанной перепонки и
температурой кожи по отношению к потоотделению у человека. При
температуре кожи от 33 до 39 °С потоотделение начинается при строго определенной
температуре барабанной перепонки. При более низких температурах кожи для
начала потоотделения требуются более высокие температуры барабанной
перепонки.
чии
300
I
о
с;
О
СТ
\-
О
о
\-
о
с
200
100
Терморегуляция
357
РЕГУЛЯЦИЯ ТЕПЛОПРОДУКЦИИ ЭФФЕКТОРАМИ
При снижении температуры воздуха для компенсации
увеличившейся теплопотери должна усилиться теплопродукция.
Повышенная активность и мышечные сокращения
сопровождаются теплопродукцией. Эта активность находится под
произвольным контролем. Вторым механизмом усиления теплопродукции
в организме является дрожь.
Дрожь — это автономная реакция на холод, хотя она может
быть преодолена произвольным мышечным контролем. При
дрожи одновременно сокращаются ритмически с большой частотой
и сгибатели и разгибатели. И частота, и сила сокращения могут
варьировать. Дрожь и мышечная активность не аддитивны:
тепло генерируется дрожью, только если мышцы не вовлечены в
другую деятельность. Если, например, человек спокойно сидит
на прохладном воздухе, он может внезапно начать дрожать без
всякого сознательного усилия.
Такие произвольные действия, как ходьба, связаны с
мышечным сокращением, которое преодолевает дрожь. И дрожь, и
ходьба сопровождаются образованием тепла. Но во время
мышечной активности вследствие повышенного конвекционного
теплообмена усиливаются и потери тепла. Небольшое движение
мышц при дрожи повышает эффективность теплопродукции,
понижая конвекционную теплоотдачу.
Центр дрожи, находящийся в заднем гипоталамусе,
по-видимому, влияет на частоту и силу мышечных сокращений при
дрожи. В свою очередь к этому центру поступают импульсы от
центра терморегуляции в переднем гипоталамусе и от рецепторов
мышц. Центр дрожи имеет выходы ко всем уровням спинного
мозга. Однако вызвать дрожь удавалось охлаждением и после
перерезки спинного мозга, хотя сигналы от центра дрожи не
могли достичь спинномозговых нервов. Стало быть,
ритмические сигналы, вызывающие в мышцах дрожь, возникают на
уровне спинного мозга (рис. 19.9). Центр дрожи модулирует
ритмическую выходную активность осциллятора. И к спинальному
осциллятору, и к гипоталамическому центру приходят сигналы
от терморецепторов.
Регуляция дрожи, по-видимому, зависит от активности
терморецепторов— как периферических, так и глубинных. У
некоторых видов животных дрожь возникает при локальном
охлаждении гипоталамуса, при неизменной температуре кожи.
Однако во многих случаях у животного, помещенного в холодную
среду, дрожь начинается слишком быстро, чтобы ее можно
было отнести за счет снижения внутренней температуры тела, чт>
свидетельствует о важном значении периферических
терморецепторов. Такую реакцию следует признать адаптивной, по-
358
Глава 19
Холодовой стимул Тепловой стимул
Пути от внецентральных
терморецепторов
Тепла
Рецепторы
•
Холода
Центр терморегуляции
(преоптическая область)
Другие влияния
на мышечный тремор
Центр дрожи
(задний гипоталамус)
Путь в спинном мозгу
Мышца
Спинальный осциллятор
Рис. 19.9. Взаимодействие между центром дрожи в заднем гипоталамусе и
спинномозговым осциллятором, управляющим дрожью. (Bligh J., 1973.).
скольку она предвосхищает изменение внутренней температуры
тела.
В определении начала и интенсивности дрожи при холоде
играют роль как периферические, так и центральные
терморецепторы. Так, если температура барабанной перепонки человека
(показатель температуры мозга) равна 36,6 °С, а температура
кожи — 28 °С, то теплопродукция составляет около 200 Вт
(рис. 19.10). Но если температура барабанной перепонки
останется 36,6 °С, а температура кожи упадет до 22 °С, то
теплопродукция возрастет приблизительно на 50%. Внутренняя тем-
Терморегуляция
359
пература тела, при которой начинается дрожь, и степень
увеличения теплопродукции при дрожи снижаются при повышении
температуры кожи.
НЕДРОЖАТЕЛЬНЫЙ ТЕРМОГЕНЕЗ
Давно известно, что увеличение теплопродукции не
обязательно сопровождается дрожью. Повсеместное повышение
теплопродукции называют недрожательным термогенезом (НДТ).
Чаще всего (но не исключительно) он происходит у некоторых
животных, подвергающихся действию холода. Этот термогенез
аддитивен по отношению к теплопродукции посредством дрожи
Кожа
36,6 36,8 37,0 37,2 37,4 37,6
Температура барабанной перепонки, °С
Рис. 19.10. Взаимодействие между температурой кожи и температурой
барабанной перепонки по отношению к началу и интенсивности дрожи у
человеку При повышении температуры кожи от 20 до 31 °С требуется большее
падение температуры барабанной перепонки, чтобы началась дрожь (см. текст).
(Benzinger Т. Н., 1969.)
360
Глава 19
или другой активности скелетных мышц: нормально он
начинается до дрожи и снижает температуру, при которой наступает
дрожь. У большинства видов около половины увеличения
теплопродукции в результате НДТ происходит в скелетных
мышцах.
КОРИЧНЕВЫЙ ЖИР
Коричневый жир — ткань, состоящая из многих долек и
расположенная под лопаткой, чаще встречается у животных,
которым свойственна зимняя спячка. В литературе неоднократно
сообщалось о калоригенной роли коричневого жира при выходе
из спячки. Коричневый жир хорошо развит также у многих
новорожденных млекопитающих, в том числе у детей. В течение
первых нескольких дней после рождения недрожательный тер-
могенез (путем теплопродукции в клетках коричневого жира)
составляет главную реакцию на холод. Позднее такой реакцией
становится дрожь, и это коррелирует с превращением
коричневого жира в белый. Белый жир несет главным образом функцию
сохранения тепла, а не его образования.
У некоторых видов, например у крысы, коричневый жир
сохраняется в течение всей жизни и служит источником
недрожательного термогенеза, особенно при воздействии холода. Долю
участия коричневого жира в общей теплопродукции за счет
недрожательного термогенеза оценить трудно. Коричневый жир,
который участвует в термогенезе у крысы, составляет всего 1%
массы тела, тогда как на скелетные мышцы приходится 45%.
Хирургическое удаление всех следов коричневого жира у крыс
приводит к снижению недрожательного термогенеза, но не к
его исчезновению. В общем, в реакции крупных млекопитающих
на холод более важное значение имеет дрожь, а в реакции
мелких млекопитающих, по-видимому, важнее недрожательный тер-
могенез.
ТЕРМОРЕГУЛЯЦИЯ И МЫШЕЧНАЯ РАБОТА
Во время мышечной работы внутренняя температура тела
повышается: повышение ее пропорционально интенсивности
производимой работы и не зависит от окружающей температуры,
во всяком случае у крупных млекопитающих. У мелких
млекопитающих температура тела колеблется в зависимости от
интенсивности работы и температуры окружающей среды, которая
определяет температурный градиент, лежащий в основе тепло-
потери. Связь Ть с температурой воздуха у мелких
млекопитающих (но не у крупных), вероятно, отражает более высокую
теплоотдачу на единицу поверхности у таких животных вслед-
Терморегуляция
361
ствие общего отношения поверхности к объему и менее
эффективной теплоизоляции.
Повышение внутренней температуры тела у человека при
различной интенсивности работы позволяет думать, что
«заданный» уровень температуры тела при мышечной работе
меняется. Однако проще объяснить это тем, что при более
интенсивной продукции тепла внутри тела (в условиях мТлшечной
работы) в нем накапливается больше тепла и устанавливается
новое равновесие между теплопродукцией и теплоотдачей. При
постоянной температуре кожи интенсивность потоотделения у
человека тесно связана с внутренней температурой тела как в
покое, так и при мышечной работе (рис. 19.11).
Многочисленные исследования убедительно свидетельствуют
в пользу того, что внутренняя температура тела является
важным фактором, определяющим функцию эффекторов при термо-
ф
X
Ф
с;
Ф
»-
о
о
I-
о
и.
350
300
250
200
150
100
50
-
-
as °
V °
1
ш
г
}
I- I ~~Т ! I I I J
36,4 36,6 36,8 37,0 37,2 37,4 37,6
Температура барабанной перепонки, °С
ис. 19.11. Величина потоотделения у человека в покое (О) и при мышечной
работе (Л). Температура кожи была между 33 и 38°С, температура внутри
тела (т-ра барабанной перепонки) колебалась от 36,7 до 37,3 °С. Подобные
отношения между состоянием покоя и мышечной работой позволяют
дуть, что во время работы уровень, вокруг которого колеблется регуляция
не изменился. '
362
Глаао, 19
регуляции. Внутренняя температура тела повышается в
линейной зависимости от интенсивности работы (и, следовательно, от
интенсивности обмена). Средняя температура кожи является
функцией температуры воздуха и снижается при испарении.
Она не связана непосредственно ни с внутренней температурой,
ни с интенсивностью обмена. Если при данной внутренней
температуре тела повышается средняя температура кожи,
усиливается также и потоотделение. Как показано на рис. 19.8,
интенсивность потоотделения при любой внутренней температуре
может значительно меняться в зависимости от уровня средней
температуры кожи.
ФИЗИОЛОГИЧЕСКАЯ АДАПТАЦИЯ И ЭКСТРЕМАЛЬНАЯ
ТЕМПЕРАТУРА
Обычные сезонные климатические колебания вызывают у
животных физиологические изменения, называемые
акклиматизацией. Аналогичное явление можно наблюдать и в лаборатории,
если содержать животных длительное время при заданной
температуре и неизменных остальных параметрах среды. Чтобы
отличить адаптивные (приспособительные) изменения,
наблюдаемые в лаборатории, от природной акклиматизации,
физиологические изменения при экспериментальных условиях
называют акклимацией.
ХОЛОД
Эндотермные животные по-разному реагируют на
климатические изменения, особенно на холод. Гетеротермные птицы и
млекопитающие снижают свои метаболические процессы и
допускают снижение температуры тела до точки, отличающейся от
окружающей температуры на один градус или около того. Эта
адаптация уменьшает затраты энергии, необходимые для
выживания. Однако гетеротермные животные не совсем «отданы на
милость» окружающей среды. Если температура воздуха падает
ниже точки замерзания, эти животные могут удерживать
температуру тела на уровне выше этой точки, и, следовательно,
происходит некоторая регуляция. Сильное увеличение
теплоизоляции зимой в результате сезонной линьки у некоторых
животных снижает утечку тепла, и таким образом теплопродукция
удерживается на одинаковых уровнях зимой и летом.
У многих животных теплоизоляция как таковая существенно
не меняется и приходится использовать усиленную
метаболическую теплопродукцию.
Поскольку у человека не так уж много коричневого жира,
усиление теплопродукции должно происходить посредством
Терморегуляция
363
дрожи или мышечной активности. Но эти средства годятся
только на короткое время. Дальнейшая реакция на холод
состоит в генерализованном сужении сосудов, которое помогает
сохранять внутреннюю температуру, снижая утечку тепла в
периферические области тела. Как было указано выше, такая
реакция приносит лишь ограниченную пользу, особенно при очень
низких температурах, так как возникает опасность
отморожений. Главная реакция человека на холод поведенческая, а не
физиологическая. Ощущение человека, что «ему холодно»,
заставляет его утеплять жилище, прибегать к теплой одежде
и т. п. Попытки выявить специфические физиологические
изменения, связанные у человека с акклиматизацией, оказались
несостоятельными.
ЖАРА
Жаркая среда потенциально является большей нагрузкой
для организма, чем холод. Когда температура среды выше
температуры тела, тепло переходит из окружающей среды в
организм. Тепло вырабатывается в процессе метаболизма и, кроме
того, поступает в организм путем излучения, теплопроводности
и конвекции — всеми теми путями, которыми происходит
теплоотдача в холодной среде. В таких случаях единственным
способом отведения тепла служит испарение.
Крупные млекопитающие, приспособленные к жаркому
сухому климату, например верблюд, могут переносить большие
колебания внутренней температуры тела, и этот процесс
называется факультативной гипертермией. При притоке тепла в
организм верблюда испарение не усиливается и тепло
накапливается, вызывая повышение температуры тела. Эта стратегия
способствует выживанию благодаря большим размерам верблюда
и относительно низкому отношению поверхности его тела к
объему. Температура тела повышается так медленно, что к концу
дня она не достигает летального уровня. Ночью, когда воздух
прохладнее, температура тела снова падает до нормального
уровня. Отказываясь от точной регуляции температуры тела,
верблюд снижает потребность в охлаждении путем испарения,
что имеет несомненное адаптивное значение в пустыне с ее
скудными запасами свободной воды.
Хотя человек не может жить в жарком сухом климате без
доступа к воде, он все же обладает определенными
физиологическими приспособлениями к существованию в условиях
избытка внешнего тепла. В начале действия жары ректальная
температура, потребление кислорода, частота сокращений сердца и
минутный объем быстро растут. Потоотделение невелико и
недостаточно для сохранения постоянной температуры тела
364
Глава 19
(рис. 19.12). Через 12 дней пребывания на жаре наступают
очевидные признаки приспособления. Потоотделение удваивается,
а частота сокращений сердца и потребление кислорода падают
примерно на 40%. Усиление испарения и понижение обмена
уменьшают повышение внутренней температуры тела. С более
эффективной регуляцией связано сильное расширение сосудов,
что повышает теплоотдачу на периферии и усиливает
потоотделение. Такая реакция обнаруживается приблизительно через
5 дней и быстро прекращается, если условия изменяются. Ох-
Ректальная температура °F
//V/V/:
Частота сокращений сердца в 1 мин
V4
Потоотделение, мл/ч
123451
Часы
Дни 1-й 2-й
102
101
140
130
10-й 11-й 12-й
Рис. 19.12. Ректальная температура, частота сокращений сердца,
интенсивность потоотделения и потребление кислорода у молодых людей, работающих
в течение 5 ч на жаре (Та = 36°С), в первый день работы и после 12 дней.
лаждение путем испарения решает вопрос лишь ненадолго, так
как чрезмерное потоотделение может нарушить водный баланс
в организме и привести к недостаточности кровообращения из-за
уменьшения объема плазмы.
Тепловой удар — это состояние перегрева, при котором
температура тканей на периферии и в глубоких областях тела
повышается, что приводит к потере сознания; пот не выделяется,
охлаждение путем испарения отсутствует. Кожа часто краснеет,
что указывает на расширение периферических сосудов, но, если
жара продолжается, повышение температуры оказывает
повреждающее действие на ткани головного мозга и всей нервной
системы и в конечном счете приводит к смерти.
Терморегуляция 365
ЛИХОРАДОЧНОЕ СОСТОЯНИЕ
Лихорадка — это физиологическое состояние,
характеризующееся повышением внутренней температуры тела, обычно
вызванным присутствием в крови особых химических веществ —
пирогенов. Пирогены бывают микробного происхождения—
эндотоксины бактерий. Лихорадочное состояние, вызываемое
экспериментальным введением эндотоксина, обычно возникает у
человека не ранее, чем через 30 мин после введения. Существуют
о
40
39
38
37
Дрожь
Сужение
г- 1
Исходная Г
температура 1
1. _ .! 1 1
сосудов
i
/\ Истинная
' ^ цен"
1
Потоотделение
Расширение сосу/
1 \
i \
1\
1 \
1 \
тральная температура\
1 \
1 \
1 \
1 1
1 V
L ^
1 1 1 1
Г
Время, ч
т
Начало
лихорадки
Прекращение
лихорадки
Рис. 19.13. Течение типичного приступа лихорадки. Температура тела отстает
от быстрых изменений того уровня, вокруг которого идет регуляция. Во
время лихорадочного состояния температура регулируется на более высоком
уровне.
эндогенные пирогены, которые образуются в тканях организма;
такие пирогены выделены из лейкоцитов. После введения
эндогенных пирогенов температура повышается почти немедленно.
Под влиянием пирогенных веществ повышается тот
«заданный» уровень, на котором поддерживается постоянная
температура тела. В начале лихорадочного состояния организм
реагирует как бы на холод. Теплоотдача понижается вследствие
сужения сосудов в периферических тканях, а теплопродукция
усиливается. Человек зябнет, и организм реагирует на это
Дрожью, ознобом. Внутренняя температура тела повышается до
Некоторого нового уровня, на котором она регулируется, пока
не прекратится действие пирогена (рис. 19.13). После его
удаления «заданный» уровень понижается, и организм реагирует как
366
Глава' 19
°F
|114-
110-
106-
102-
98-
94-
90-
86-
.
82-
78-
74-
°C
-44
-42
-40
-38
-36
-34
-32
-30
-
-28
-26
-24
Верхний предел
выживания ~)
~) _. Нарушение
I Тепловой удар UepMO-
\ Повреждения регуляции
J мозга
"1 Лихорадочные")
[состояния и
J тяжелая мышеч I
}ная работа
Обычные
пределы нормы
1 Сердечная
j аритмия
Терморегуляции
эффективна
при лихорадке,
в норме и при
мышечной
работе
Нарушение
термо -
регуляции
Потеря
тёрмо -
регуляции
бы на тепловую нагрузку. В
этой фазе обычно сосуды
кожи расширяются, начинается
потоотделение, и температура
постепенно возвращается к
нормальному уровню.
ЛИХОРАДОЧНОЕ СОСТОЯНИЕ
И ИНФЕКЦИЯ
Долгое время считали, что
лихорадка возникает потому,
что повышение температуры
тела чем-то полезно для
борьбы с инфекцией. Однако лишь
отдельные клинические факты
свидетельствуют в пользу
такой точки зрения. Повышенная
температура явно вредна
организму. Все метаболические и
физиологические процессы
протекают при ней на опасно
высоких уровнях, и температура
тела уже приближается к той,
при которой начинается
денатурация белков (рис. 19.14).
Лихорадочное состояние, в
общем, считается вредным для организма, и лечение большинства
болезней обычно предусматривает снижение температуры тела
обычно применением жаропонижающих средств.
ЛИТЕРАТУРА
Blight J. (1973). Temperature regulation in mammals and other vertebrates,
American Elsevier, New York.
Benzinger Т. Н. (1969). Heat regulation: homeostasis of central temperature in
man, Physiol. Rev., 49, 671.
Dubois E. F. (1948). Fever and the regulation of body temperature, С. С.
Thomas, Springfield, 111.
Hammel H. T. (1968). Regulation of internal body temperature, Ann. Rev.
Physiol., 30, 641.
Hardy J. D. (1961). Physiology of temperature regulation, Physiol. Rev., 41,
521.
Heller H. C, Crawshaw L. I., Hammel H. T. (1978). The thermostat of vertebrate
animals, Sci. Amer. August, 102.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Какое значение для теплового баланса человека имеет
снижение потребления кислорода во время акклимации к жаре?
2. Опишите преимущества и недостатки теплоотдачи путем
испарения при потоотделении и при тепловой одышке.
Рис. 19.14. Температура тела
человека: нормальные и крайние точки.
\
Терморегуляция 367
3. Поскольку для поддержаниялостоянной внутренней
температуры тела теплопродукция дол^кна быть равна теплоотдаче
и поскольку теплоотдача пропорциональна разности между
температурой тела и температурой воздуха, каким образом
в пределах термонейтральной зоньг сохраняется постоянная
теплопродукция?
4. Какова физическая основа ощущения холода от ветра?
5. Приняв, что интенсивность обмена у марафонского бегуна
определяется его активностью во время бега, опишите
некоторые механизмы, регулирующие температуру тела.
6. Объясните главное различие тепловой нагрузки при высокой
температуре окружающего воздуха и мышечной работе.
7. Каковы основные источники притока и отдачи тепла у
человека а) в полдень в пустыне и б) в условиях арктической
ночи?
8. Какое значение имеет недрожательный термогенез?
9. Почему терморегуляторная система использует и
периферические, и центральные терморецепторы? Изложите ваши
соображения о взаимоотношениях между периферическими и
центральными терморецепторами при а) низкой температуре
окружающей среды, б) высокой температуре окружающей
среды, в) мышечной работе.
Глава 20 /
МОТОРИКА ЖЕЛУДОЧНО-КИШЕЧНОГО
ТРАКТА
Функции переваривания и всасывания пищи в желудочно-
кишечном тракте (ЖКТ) включают в себя пережевывание
пищи и смешивание ее со слюной (жевание и слюноотделение),
глотание пищи и ее передвижение по пищеводу в желудок, где
начинается пищеварение, и в тонкий кишечник (место
дальнейшего переваривания и всасывания). Из тонкого кишечника
пищевая масса переходит в толстый кишечник, откуда
непереваренные вещества выбрасываются наружу. Строение и функция
этих отделов должны быть такими, чтобы пища проталкивалась
и передвигалась (моторика).
ЭЛЕМЕНТЫ АНАТОМИИ
Расположение и общая анатомия основных частей
пищеварительного тракта показаны на рис. 20.1. Более детально
анатомия будет описана при рассмотрении различных отделов
ЖКТ. Зубы, язык, слюнные железы и рот являются теми
самыми верхними органами пищеварительного тракта, где пища
пережевывается, смешивается со слюной и проглатывается.
Пищевод начинается в конце глотки и идет вниз до
желудка на протяжении 250—300 мм. Детальная структура
(гистология) стенки или выстилки пищевода в основном такая же, как
и остальных частей пищеварительного тракта, за исключением
небольших различий в типе и строении выстилки просвета
(внутренней стороны трубки) и желез в слизистой и подслизи-
стой. Строение стенки пищевода показано на рис. 20.2 (см.
также гл. 1). По направлению от наружной стенки к просвету
лежат соединительная ткань и мышцы двух типов (кольцевые и
продольные), подслизистая, слизистая оболочки и
эпителиальная выстилка внутренней поверхности просвета. Железы лежат
в слизистой и подслизистой оболочках.
Между кольцевыми и продольными мышцами заложены
специализированные нервные клетки, объединенные в сеть, или
сплетение — ауэрбахово, или миоэнтеральное, сплетение. Кнуг-
Моторика же луЬщно-кишечного тракта
Околоушная слюнная железа
Печень
Желчный пузырь
Двенадцатиперстная
кишка
) Правый угол
ободочной кишки
Восходящая
ободочная кишка
Слепая кишка
Аппендикс
Подвздошная кишка
Изгиб сигмовидной кишки
Слюнные железы:
Подъязычная
Подчелюстная
Пищевод
Кардия
Желудок
Поджелудочная
железа .
Левый угол
ободочной кишки
Поперечная
ободочная кишка
Нисходящая
ободочная кишка
Тощая кишка
Сигмовидная кишка
Прямая кишка
Рис. 20.1. Органы желудочно-кишечного тракта.
ри от этих клеток лежат нервные клетки, образующие мейсне-
Рово, или подслизистое, сплетение. Эти внутристеночные (ин-
трамуральные) нервные сплетения находятся на всем
протяжении пищеварительного канала. Сплетения содержат нервные
клетки с отростками, которые идут от рецепторов,
расположенных в стенке кишки или слизистой, и вызывают автоматические
сокращения и движения тракта, независимые от экстрамураль-
ньгх (наружных) нервов.
24—1705
370
Глава/20
' Брыжейка
Серозная оболочка ^
Мезотелий серозной оболочки
Соединительная ткань
Наружный мышечный слой
Кольцевой мышечный слой
Продольный мышечный слой
Подслизистая
Слизистая
Эпителий
Собственная пластинка слизистой
Мышечная пластинка слизистой
Круговая складка
Мейснерово сплетение
Ауэрбахово сплетение
Выводной проток
крупной железы
Подслизистая
железа
Слизистая железа
Рис. 20.2. Поперечный разрез стенки пищеварительного канала.
НЕРВНО-МЫШЕЧНЫЙ АППАРАТ
Кроме интрамуральных нервных сплетений, к большинству
гладких мышц ЖКТ подходят наружные нервы, участвующие
в возбуждении мышц и их последующей моторике. На рис.
20.3, А, Б и В показаны типы мышц и нервные волокна.
В табл. 20.1 приведены данные об иннервации разных
областей или органов (см. также гл. 10). В общем, симпатические
двигательные нервы оказывают тормозное, или расслабляющее,
действие на гладкие мышцы желудка и кишечника, а
парасимпатические нервы оказывают возбуждающее действие и
вызывают большинство моторных эффектов. Преганглионарные
парасимпатические волокна образуют синапсы с ганглиозными
клетками в миоэнтеральном и подслизистом сплетениях (рис.
20.3, Б)у а волокна от этих нервных ганглиев (постганглионар-
ные) иннервируют и возбуждают гладкие мышцы (продольные
и кольцевые волокна), выделяя ацетилхолин (см. гл. 11).
После разрушения или денервации наружных нервов гладкие
мышцы желудка и кишечника еще способны к ритмическим со-
Моторика желудоЧ^о-кишечного тракта 37J_
Таблица 20.1. Иннервация желудочно-кишечного тракта симпатическими (С)
и парасимпатическими (ПС) двигательными (эфферентными) нервами
Иннервируемый орган
}Келудок (мышца)
Сфинктер привратника
Тонкий кишечник (мышца)
Илеоцекальный сфинктер
Мышца слепой кишки
Толстый кишечник
Восходящая ободочная кишка
Нисходящая ободочная кишка
Анальный сфинктер
Внутренний
Наружный
Нервы
С (Чревный)
ПС (Блуждающий)
С (Чревный)
ПС (Блуждающий)
С (Чревный)
ПС (Блуждающий)
С (Чревный)
ПС (Блуждающий)
С (Чревный)
ПС (Блуждающий)
С (Чревный)
ПС (Блуждающий)
С (Подчревный)
ПС (Тазовый)
С (Чревный)
ПС (Тазовый)
Половой нерв
(соматический)
Эффект1)
+
+
—
—
+
—
+
—
+
—
+
—
+
+
—
+
') — торможение; + возбуждение.
кращениям, хотя и более слабым. Такие сокращения
вызываются действием интрамуральной нервной системы (ауэрбахово
и мейснерово сплетения). Даже после блокады интрамуральных
сплетений кокаином гладкая мышца все еще может совершать
некоторые автоматические, но не перистальтические движения;
перистальтика требует интактной интрамуральной нервной
системы. На рис. 20.3 показана афферентная иннервация
пищеварительного тракта. Стимулы, идущие к слизистой, в которой
лежат рецепторы (механо- и хеморецепторы), передаются по
афферентным волокнам блуждающего и чревного нервов в
центральную нервную систему (ЦНС), а ответ передается по
указанной выше эфферентной системе. Те же самые механо- и
хеморецепторы могут включаться в простой рефлекс, в котором
эфферентная, или моторная, реакция осуществляется через
спинной мозг.
Главные кровеносные сосуды ЖКТ показаны на рис. 20.4.
К каждому органу желудочно-кишечного тракта кровь
поступает по своей артерии и оттекает по своей вене (артерия —
афферентный сосуд, а вена — эфферентный). По-иному обстоит
Дело с печенью, у которой имеется одна артерия, а вены двух
типов: воротная, несущая кровь и усвоенные пищевые
вещества от кишечника к печени (афферентная), и печеночная вена,
24*
Рис. 20.3. Наружные и внутренние нервные волокна, идущие к
желудочно-кишечному тракту. Л. Симпатические нервы — эфферентные. Б.
Парасимпатические нервы — двигательные, холинэргические. В. Афферентные нервные
волокна, получающие импульсы от желудочно-кишечного тракта и передающие их
в центральную нервную систему. (Печатается с разрешения Davenport H. W.:
Physiology of the digestive tract, 4th ed., 1977.)
372
'^ Чревный ствол
Желчный пузырь
Печень
Печеночная
артерия
Гастродуоденальная
артерия
Правая желудоч
ная артерия
Верхняя
брыжеечная
артерия
Внутренняя
подвздошная
артерия
Средняя и нижняя
прямокишечные
артерии
Печеночная вена
Желчный пузырь
Печень-
Воротная вена
Верхняя
брыжеечная
вена
Правая
ободочная
вена
Внутренняя.'
подвздошная
вена
Прямокишечные
вены
Брюшная
аорта Левая желудочная
артерия
Селезеночная
артерия
Селезенка
Желудочно-
сальникозая
артерия
Нижняя брыжеечная
артерия
Левая
ободочная
артерия
Сигмовидная
артерия
Верхняя
прямокишечная
артерия
Наружная
подвздошная
артерия
Нижняя полая вена
Левая желудочная вена
Селезеночная вена
Панкреатическая
вена
Желудочно-
сальниковая*
вена
Нижняя
брыжеечная
вена
Сигмовидная
вена
Наружная
подвздошная
вена
Рис. 20.4. Главные кровеносные сосуды ЖКТ.
373
374
Глава 20
Аорта
Нижняя
полая вена
Чревная а.
Селезеночная а.
Верхняя
брыжеечная а.-
Нижняя
брыжеечная а.
ф
т
ф
С
Суммарный
приток:
1500 мл/мин
при 90 мм рт. ст.
Суммарный
отток:
1500 мл/мин
при 3 мм рт. ст.
Рис. 20.5. Кровоток через ЖКТ у человека. (Selkurt E. E., Physiology, 4th ed.,
1976.)
которая несет кровь от печени (эфферентная) в нижнюю полую
вену.
Количество крови, протекающей через ЖКТ у человека
среднего роста, составляет около 1500 мл/мин. Доля общего
кровотока, приходящаяся на каждый кровеносный сосуд, показана на
рис. 20.5. Печень получает 30% всей крови по печеночной
артерии и 70% по воротной вене.
ЕДА И ПИТЬЕ
Когда пища попадает в рот и пережевывается, она
стимулирует вкусовые сосочки и обонятельные рецепторы, от
которых главным образом зависит удовлетворение от еды. Вид пи-
Моторика желудочно-кишечного тракта
375
щи и ее прикосновение к слизистой рта вызывают
рефлекторное выделение слюны; слюна смешивается с пищей, размягчает
и смачивает ее, и в результате жевания образуется комок,
который проглатывается.
Вкус и вид пищи стимулируют рецепторы во рту, откуда
импульсы передаются в мозг, в латеральный гипоталамус, где
лежит пищевой центр, или центр голода (см. гл. 10). Стимуляция
этой области заставляет животных жадно поглощать пищу.
В вентромедиальной части гипоталамуса лежит другая область,
которая оказывает действие, обратное тому, какое производит
центр голода. Это центр насыщения, который сообщает
человеку, что он сыт и больше есть не хочет. Наполненный желудок
или растяжение любой части ЖКТ стимулирует центр
насыщения и угнетает центр голода.
Питье воды частично зависит от качества съеденной пищи;
оно также регулируется центром жажды в гипоталамусе (см.
гл. 10). Другие факторы, влияющие на прием жидкости и
выделение ее из организма, рассмотрены в гл. 12 и 24.
ЖЕВАНИЕ И ГЛОТАНИЕ
Жевание происходит при участии жевательных мышц, губ,
щек и языка. Их работа координируется нервными
импульсами V, VII, IX, X и XII черепно-мозговых нервов. Хотя
жевание— это произвольный акт, но частично оно регулируется
также и рефлекторно, поскольку происходит и в том случае, когда
пища находится во рту у животного с удаленной корой больших
полушарий. Дробящая сила зубов гораздо больше (454—545 кг),
чем нужно для большинства сортов пищи.
Проглоченная пища проходит из полости рта через глотку и
пищевод в желудок. Механика этого процесса такова.
1. Пищевой комок (bolus) направляется к глотке. Пища или
вода скатывается по спинке языка, а кончик прижимает ее
вверх к твердому небу; за этим следует сокращение мышц,
которое проталкивает комок в глотку.
2. Комок перемещается в пищевод.
Пищевод разделен на три функциональные части: 1)
верхний сфинктер пищевода (фарингоэзофагальный), 2) тело и
3) нижний сфинктер пищевода (гастроэзофагальный). Для всех
трех частей характерна своя сократительная активность в
покое и при глотании.
СОКРАТИТЕЛЬНЫЕ ДВИЖЕНИЯ ПИЩЕВОДА
В состоянии покоя верхний сфинктер закрыт и пищевод
расслаблен. Замкнутый сфинктер не позволяет большим объемам
воздуха проникать в желудок при дыхании. При глотании верх-
376
Глава 20
ний сфинктер расслабляется, открывается и пропускает пищу.
Затем он сокращается, после чего следует перистальтическая
сократительная волна, пробегающая по пищеводу к желудку
(рис. 20.6). Непосредственно перед тем как сократительная
волна достигает нижнего сфинктера, он расслабляется
(открывается), пропускает пищу и снова закрывается. Если комок
водянистый, то он перемещается до желудка силой тяжести. Но ес-
Перистальтика Сегментация
(проталкивание) (перемешивание)
Рис. 20.6. Сократимость желудочно-кишечного тракта. А. Перистальтические
волны. Б. Сегментные сокращения тонкого кишечника. В. Мультигаустраль-
ные сокращения толстой кишки. Волны начинаются в точке 1 и
распространяются до точек 2 и 3. За сокращением следует расслабление.
ли он твердый или полутвердый, то проталкивается в желудок
перистальтическими волнами со скоростью от 2 до 4 см/с.
Давление, создаваемое перистальтическим сокращением,
колеблется от 30 до 140 мм рт. ст. Для того чтобы глоток полностью
прошел в желудок, часто нужно больше одной
перистальтической волны.
Начало глотания — это произвольный акт, в котором
главную роль играет кора больших полушарий; оно также может
быть вызвано рефлекторно скоплением во рту жидкости или
слюны. По имеющимся данным в продолговатом мозгу
находится глотательный центр, который посылает моторные импульсы
к мышцам, осуществляющим акт глотания. В регуляции
перистальтических волн пищевода важную роль играет
блуждающий нерв, поскольку его перерезка прекращает перистальтику.
Моторика желудочно-кишечного тракта 377
МОТОРИКА ЖЕЛУДКА
Желудок представляет собой хорошо оформленный орган,
состоящий из дна, кардиальной части (близ входа пищевода),
тела и привратника, который соединяется с двенадцатиперстной
кишкой (рис. 20.7). Объем человеческого желудка составляет
от 50 мл, когда в нем нет пищи, до 750 мл или более, когда
он наполнен или растянут.
Эпителий желудка отличается наличием специализированных
желез. У млекопитающих обкладочные клетки выделяют в
желудочный сок НС1, а главные клетки секретируют гранулы пеп-
синогена, но у птиц они продуцируют и кислоту, и пепсиноген
(рис. 20.8).
Верхний отдел желудка включает дно и небольшую часть
тела, а нижний отдел состоит из тела и антральной области;
эти отделы обладают разной сократимостью. Верхний отдел
желудка малоактивен, а нижний (дистальная часть тела и ант-
ральная область) отличается высокой активностью.
Антральная область
Рис. 20.7. Желудок человека.
378
Глава 20
Поверхностный Желудочные
эпителий ямки
Обкладочные
Собственная
пластинка
слизистой
Мышечный
слой
подслизистой I>J^
Поперечный
разрез железы
Рис. 20.8. Специализированные клетки слизистой желудка,
продуцируют пепсиноген, обкладочные — НС1.
Главные клетки
Сокращения начинаются и обычно усиливаются в средней
области желудка по мере продвижения к месту перехода в
двенадцатиперстную кишку. Эти волны, преимущественно
перистальтические, распространяются с частотой 3 в 1 мин. С
волнами сокращения связаны волны давления разной амплитуды
и длительности. Волны типа I и II — это медленные
ритмические волны давления разной амплитуды. Длительность их от 2
до 20 с, и они возникают с частотой 2—4 в 1 мин. Вероятно, это
давление генерируется перистальтическими сокращениями.
Тип III состоит из сложных волн давления длительностью около
минуты.
Проглоченная твердая пища скапливается преимущественно
в верхнем отделе желудка, почти не перемешивается и лежит
слоями. Проглоченная жидкость стремится стечь над этими
слоями в нижний отдел желудка, где жидкая и твердая пища
перемешивается.
Моторика желудочно-кишечного тракта
379
ОПОРОЖНЕНИЕ ЖЕЛУДКА
Скорость продвижения проглоченной массы из желудка в
кишечник зависит главным образом от ее физико-химического
состава в желудке и двенадцатиперстной кишке. Углеводы
выходят из желудка быстрее всего, белки — медленнее, а жиры
остаются в желудке дольше всего.
Консистенция содержимого желудка тоже влияет на время
эвакуации. Крупные куски мяса остаются в желудке дольше,
чем мелкие. Гипотонические растворы дольше задерживаются
в желудке, чем изотонические, а растворы с рН 5,3 или ниже
задерживают опорожнение.
Эвакуация содержимого желудка зависит от взаимодействия
желудка с двенадцатиперстной кишкой, но точный механизм
этого акта неизвестен. Впрочем, называются несколько
возможностей, а именно: 1) активность пилорического сфинктера,
2) желудочно-кишечные гормоны и 3) координированные циклы
активности входа и проксимальной части двенадцатиперстной
кишки. За сокращением входа идут последовательные
сокращения привратника (пилоруса) и двенадцатиперстной кишки.
На прохождение пищи влияет разница давления в пилориче-
ской части и в двенадцатиперстной кишке. Если давление ниже
со стороны желудка, то пища не проходит, даже если пилориче-
ский сфинктер неактивен или отсутствует в результате
хирургического вмешательства.
Желудочно-кишечные гормоны — гастрин, секретин и холеци-
стокинин — тормозят эвакуацию, но как именно, еще не ясно.
Жир в кишечнике имеет тенденцию тормозить опорожнение
желудка, возможно, через секретин.
НЕРВНАЯ РЕГУЛЯЦИЯ МОТОРИКИ
Как было указано выше, главным внешним нервом,
влияющим на моторику желудка и вызывающим перистальтические
волны, является блуждающий нерв. Раздражающие стимулы,
гипертонические растворы и растяжение двенадцатиперстной
кишки тормозят моторику желудка через посредство кишечно-
желудочного рефлекса, рецепторы которого находятся в
двенадцатиперстной кишке; они посылают импульсы по афферентным
волокнам блуждающего нерва; торможение моторики желудка
вызывается эфферентными волокнами симпатического нерва.
В этот рефлекс могут вовлекаться также и интрамуральные
нервные сплетения.
Эвакуация желудка задерживается, когда подвздошная
кишка наполнена (илеогастральный рефлекс) и когда анус
стимулируется механически (аногастральный рефлекс). Такие состоя-
380
Глава 20
ния, как тревога и возбуждение, усиливают моторику желудка;
страх и гнев тормозят ее.
Рвота представляет собой опорожнение желудка в обратном,
или ненормальном, направлении. Основное усилие создается
сокращением главным образом брюшных, а не желудочных
мышц. Кураре — препарат, парализующий мышцы брюшной
стенки, — прекращает рвоту. Рвота представляет собой сложный
рефлекторный акт и координируется центром, расположенным
в продолговатом мозгу. Афферентные импульсы возникают от
стимуляции глотки и желудка и от стимулов, возникающих в
вестибулярном аппарате при морской болезни.
МОТОРИКА КИШЕЧНИКА
Тонкий кишечник человека состоит из трех частей. Первая
часть, двенадцатиперстная кишка, или дуоденум, лежит кау-
дальнее желудка и служит его продолжением; она имеет длину
около 20 см и образует петлю (рис. 20.1). Вторая часть, тощая
кишка, имеет длину около 3,5 м, а третья — подвздошная
кишка— 4,5 м. Между тощей и подвздошной кишкой нет резкой
границы, хотя имеются структурные различия в эпителиальной
выстилке ворсинок (рис. 20.9), пальцевидных выступов, или
Цилиндрический эпителий
Центральный
лимфатический
сосуд
Ворсинка
^Капиллярная
^сеть
Кишечная железа
(либеркюнова крипта)
Подслизистая
Лимфатический :^^г==- —:—^^=-=^=^=^^
канал ,1ГТ1 И Ш д Круговая
мышца
Продольная
мышца
Серозная
оболочка
Рис. 20.9. Строение ворсинки тонкого кишечника.
Моторика желудочно-кишечного тракта
381
складок, которые чрезвычайно сильно увеличивают поверхность
слизистой, т. е. площадь всасывания (примерно в 30 раз). На
каждой складке, или ворсинке, лежат более мелкие складки
(микроворсинки), которые еще больше увеличивают
поверхность. Диаметр тонкого кишечника составляет около 3—4 см.
Подвздошная кишка кончается у илеоцекального сфинктера
при входе в ободочную кишку.
Толстая, или ободочная, кишка сравнительно короткая
(110 см); диаметр ее сечения — 7—10 см. Она начинается у
слепой кишки (рис. 20.1) и тянется до прямой кишки и
заднего прохода — последнего отдела ЖКХ Ободочная кишка
делится на 1) илеоцекальную область; 2) главную часть, состоящую
из восходящей, поперечной, нисходящей ободочной и сигмоид-
ной кишки, переходящей в прямую кишку (rectum) и задний
проход (anus). Ободочная кишка лишена ворсинок.
ДВИЖЕНИЕ ТОНКОГО КИШЕЧНИКА
Пища, частично переваренная в желудке, поступает в
тонкий кишечник, где она полностью переваривается и где
питательные вещества всасываются. Отходы и непереваренная пища
переходят в толстый кишечник. Этим процессам помогают
движения тонкого кишечника — волны, или сокращения, двух
типов, а именно сегментация, иначе обозначаемая как
сокращение типа I, и перистальтика (рис. 20.6).
Сегментация, кольцеобразные сокращения повторяются
через довольно правильные интервалы (около 10 раз в 1 мин) и
служат для перемешивания химуса. Участки сокращения
(рис. 20.6) сменяются участками расслабления, и наоборот.
Перистальтические волны распространяются вдоль
кишечника в направлении от полости рта. Такая волна обычно
движется медленно — со скоростью 1—2 см/с, но в патологических
случаях может двигаться и гораздо быстрее, со скоростью
25 см/с (перистальтический «бросок»). Перистальтические
волны проталкивают перевариваемую пищу; ритмические сегмен-
тационные сокращения перемешивают пищевые массы, движут
их взад и вперед, но не проталкивают.
РЕГУЛЯЦИЯ МОТОРИКИ
Сегментационные и перистальтические движения
продолжаются даже после разрушения внешних нервов (блуждающего
и симпатического), хотя сила сокращений снижается, что
говорит о регуляции этих сокращений интрамуральными нервами;
это справедливо в отношении перистальтики. Устранение
внешней и внутренней нервной активности введением кокаина тоже
прекращает перистальтику, но не сегментацию. Сегментация
инициируется самой гладкой мышцей кишечника, которая мо-
382
Глава 20
жет реагировать на локальные механические и химические
стимулы. Одним из таких химических веществ является серотонин
(5-ОТ), который образуется в кишечнике и стимулирует
моторику. Вероятно, он представляет собой нейромедиатор сенсомотор-
ных нейронов в перистальтическом рефлексе.
Перистальтика состоит сначала из сокращений продольной
мышцы, за которыми следует сокращение циркулярной; эта
последняя в норме возбуждается внутренними нервными
сплетениями, на которые в свою очередь воздействуют внешние нервы.
Растяжение кишки вызывает перистальтический рефлекс.
Итак, сокращения тонкого кишечника регулируются и
контролируются 1) активностью внешних и интрамуральных нервов,
2) активностью самой гладкой мышцы и 3) локальными
химическими и механическими факторами.
ИЛЕОЦЕКАЛЬНЫЙ СФИНКТЕР
Обычно у человека через 3!/г ч после эвакуации желудка
содержимое его достигает конца подвздошной кишки, где
находится клапан, или сфинктер, который, открываясь и
закрываясь, способствует или препятствует прохождению
перевариваемой массы в толстый кишечник.
Сфинктер расслабляется, когда перистальтическая волна
достигает дистальнои части подвздошной кишки, пропуская
перевариваемые массы в толстый кишечник, после чего сфинктер
закрывается.
Одним из факторов, вызывающих закрывание сфинктера,
является мышечно-кишечный рефлекс; механическая стимуляция
или растяжение слизистой слепой кишки вызывает сокращение
клапана даже после блокады экстрамуральных нервов.
Расслабляется сфинктер гастрально-подвздошным рефлексом после
попадания пищи в желудок под влиянием секреторной
активности желез желудка и последующей эвакуации.
МОТОРИКА ТОЛСТОГО КИШЕЧНИКА
Масса, переходящая из подвздошной кишки в ободочную
(ежедневно около 1500 мл), не переварена до конца. В толстом
кишечнике ежедневно всасывается около 1,3 л воды,
содержащей электролиты — количество сравнительно небольшое, но
достаточное для того, чтобы образовались твердые фекальные
массы.
У человека мышцы толстого кишечника большую часть
времени бездействуют, но изредка происходят сильные сокращения,
проталкивающие массу из проксимальной кишки к дистальнои.
После выхода пищи из желудка требуется 6—8 ч, чтобы она
достигла дистальнои кишки, в которой удерживается в течение
различного времени до дефекации.
Моторика желудочно-кишечного тракта
383
Перевариваемые массы проталкиваются по толстому
кишечнику комбинацией трех типов движений, или сокращений, а
именно сегментация, мультигаустральное проталкивание,
перистальтика.
Гаустральное сокращение представляет собой крайний тип
сегментации, при котором слизистая толстой кишки
складывается в мешки, или гаустры (рис. 20.6).
Нервная регуляция активности толстой кишки сходна с
регуляцией остальной части кишечника, с той разницей что
блуждающий нерв иннервирует только часть толстого кишечника
(вероятно, проксимальную треть поперечной ободочной кишки).
Нижняя часть кишки иннервируется парасимпатическим
тазовым нервом (рис. 20.3). В кишке находятся мышечно-кишечное
и подслизистое сплетения (интрамуральные нервы).
ДЕФЕКАЦИЯ
Дефекация — функция произвольная и непроизвольная;
конечный акт контролируется внутренним и наружным
анальными сфинктерами. Внутренний сфинктер гладкомышечный. Он
иннервируется волокнами подчревного симпатического нерва и
находится под непроизвольным контролем, а также под
контролем парасимпатических волокон (произвольным) тазового
нерва. Возбуждение симпатических волокон вызывает
сокращение (замыкание) сфинктера. Активация парасимпатических
волокон тазового нерва приводит к его расслаблению
(открыванию).
Наружный «анальный сфинктер состоит из скелетной мышцы
и иннервируется моторными волокнами полового
(соматического) нерва. В периоды бездействия внутренний и наружный
сфинктеры находятся в состоянии тонической активности и
сокращены или закрыты под действием импульсов соответственно
от симпатического и полового нервов. Наружный сфинктер
может сокращаться и расслабляться произвольно.
Растяжение прямой кишки вызывает рефлекторное
сокращение толстой кишки, расслабление внутреннего анального
сфинктера и позыв к дефекации. Рефлекс осуществляется по
тазовым парасимпатическим волокнам и вызывает расслабление
внутреннего анального сфинктера, его открывание и понижает
частоту импульсов в половом нерве, который вызывает
сокращение наружного анального сфинктера. Боль или страх,
действуя через симпатический нерв, тормозят рефлекс дефекации.
Растяжение желудка при перенасыщении может вызвать
сокращения прямой кишки и позыв к дефекации
(желудочно-кишечный рефлекс). Это верно особенно в отношении детей, у
которых часто после еды происходит дефекация.
384
Глава 20
ЗАПОР И ПОНОС
У многих людей дефекация наступает нерегулярно — один
раз в 2—3 дня или даже реже, без непроизвольных позывов.
При этом может возникнуть легкий дискомфорт и чувство
растяжения, но не бывает аутоинтоксикации, которую можно было
бы приписать всасыванию токсичных веществ.
Главной причиной запоров являются отсутствие привычки к
регулярному стулу и произвольное подавление или торможение
нормальных рефлексов дефекации. Это может ослабить
рефлексы и в конце концов привести толстую кишку к утрате тонуса
и способности к моторике.
Понос случается при нарушении способности тонкого и
толстого кишечника к всасыванию воды. Если такое нарушение
распространяется главным образом на тонкий кишечник, то
большие количества жидкости, поступающие в толстый
кишечник, превосходят его способность к всасыванию. Если в
содержимом подвздошной кишки оказываются какие-либо плохо
всасывающиеся вещества, например сульфат магния, то это
препятствует нормальному всасыванию воды в толстой кишке.
Воспаление тонкого кишечника, вызванное токсинами и бактериями,
нарушает всасывание воды и также вызывает понос.
Наиболее существенные статьи и монографии по
рассмотренным вопросам читатель найдет в списке к гл. 21.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Постройте схему, назовите и разместите на ней главные
части жкт.
2. Что означает кровоснабжение печени по воротной вене?
3. Назовите центры голода и насыщения.
4. Дайте определение и опишите перистальтику. Где она
происходит?
5. Каковы функции главных и обкладочных клеток желудка?
6. Назовите факторы, тормозящие или вызывающие моторику
желудка. Какие гормоны тормозят опорожнение желудка?
7. Назовите отделы кишечника (толстого и тонкого). Каковы
их размеры?
8. Как называется основная всасывающая поверхность в
тонком кишечнике?
9. Какого типа волны, или сокращения, обнаружены в тонком
кишечнике, и какие функции они выполняют?
10. Что такое мышечно-кишечный рефлекс?
11. Что такое гастроилеальный рефлекс?
12. Какие типы движения обнаруживаются в толстом
кишечнике?
13. Что такое дефекационный рефлекс?
Глава 21
СЕКРЕЦИЯ, ПЕРЕВАРИВАНИЕ,
ВСАСЫВАНИЕ
СЛЮННЫЕ ЖЕЛЕЗЫ
Взрослый человек секретирует за сутки в среднем 1—2 л
слюны, которая создает во рту влажную среду, помогает
говорить и смачивает пищу, облегчая ее проглатывание. Если
слюны мало, рот пересыхает. Содержащийся в слюне фермент
птиалин секретируется слюнной железой в виде зимогеновых
гранул (рис. 21.1).
У человека имеются слюнные железы трех типов: 1)
околоушные, 2) подчелюстные и 3) подъязычные. Свойства этих
желез суммированы в табл. 21.1. В слюне содержатся Na+, K+, С1~
и НС03~ в концентрациях, которые обычно выше, чем в плазме
крови; однако эти концентрации варьируют в зависимости от
уровня секреции. рН слюны близко к 7,0. Слюна
вырабатывается в ответ на импульсы, возникающие в слюнных центрах
продолговатого мозга. Эти импульсы инициируются
раздражением рецепторов полости рта и носа и желудочно-кишечного
тракта (ЖКТ). Ответ яа эти импульсы, приходящие в слюнные
центры, посылается к слюнным железам по симпатическим и
парасимпатическим волокнам. Возбуждение парасимпатических
нервов (VII и IX) усиливает серозную (водянистую) секрецию
и расширяет сосуды. Эти холинэргические нервы выделяют аце-
тилхолин, и их действие может быть выключено атропином,
который блокирует секрецию и вызывает сухость во рту.
Симпатические нервы приходят к слюнным железам от
верхних шейных ганглиев. Их возбуждение вызывает выделение
малого количества секрета, содержащего много слизи, но
механизм этого эффекта точно не установлен.
. Вид, запах и вкус пищи вызывают рефлекторную секрецию
слюны. Секреция, возникающая в ответ па нахождение пищи во
рту, является безусловным рефлексом. Вид и запах пищи или
звуки, связанные с пищей, вызывают выделение слюны. Это
условный рефлекс, резко выраженный у собак и более слабый у
человека. На рис. 21.1 показаны типичные клетки слюнных
желез, секретирующие зимогеновые гранулы и жидкость,
поступающую в слюнные протоки и капилляры.
25—1705
386
Глава 21
Капилляр
Лимфатический
сосуд
Зимогеновые^
гранулы
Слюнной
ацинус
Проток
Капилляр с эритроцитами
Рис. 21.1. Серозная слюнная железа с протоками, кровеносными и
лимфатическими сосудами.
ЖЕЛУДОЧНАЯ СЕКРЕЦИЯ
Желудок секретирует НС1, гранулы пепсиногена, воду,
гормоны, муцин и другие вещества. Специализированные
желудочные клетки слизистой у млекопитающих — обкладочные, или
выделяющие кислоту, секретируют НС1 или Н+, а более
мелкие главные клетки секретируют гранулы пепсиногена. У
некоторых видов, не относящихся к млекопитающим (птицы), нет
обкладочных клеток, а только главные, которые выделяют и
кислоту и гранулы пепсиногена. Другие клетки слизистой
выделяют серозный и слизистый секреты (см. рис. 20.7). Секреция
регулируется нейрогуморальными или гормональными
факторами, гастрином, гистамином и другими веществами.
Концентрация кислоты в желудочном соке зависит от
интенсивности секреции и от буферного или разбавляющего действия
пищи и других веществ.
Таблица 21.1. Слюнные железы
Железа
Околоушная
Подчелюстная
Подъязычная
Иннервация
(парасимпатическая)
Языкоглоточный (IX)
Лицевой (VII)
»
Тип секрета
Серозный
Серозный и
слизистый
Слизистый
Общее
количество
секрета, %
25
70
5
Секреция, переваривание, всасывание
387
Поверхность слизистой желудка у человека составляет
около 800 см2 и делится на зоны по распределенным в ней типам
желез. Имеются 1) кардиальные железы, 2) железы,
выделяющие кислоту, и 3) пилорические железы. Секрет кардиальных
желез в основном содержит слизь, некоторые электролиты и
немного (или совсем не содержит) НС1 или пепсиногена.
Выделяющая кислоту область дна и тела желудка составляет около
75% всей поверхности слизистой желудка. Выделяющие
кислоту железы содержат три основных типа секреторных клеток, а
именно 1) выделяющие кислоту, или обкладочные, 2) слизистые
и 3) главные клетки. Пилорические железы содержат клетки,
которые секретируют слизь и щелочной сок.
Слизистая желудка выделяет кислоту, достаточно сильную,
чтобы переварить саму слизистую, однако в норме этого не
происходит. Слизистый барьер желудка препятствует быстрому
проникновению кислоты в клетки. Некоторые вещества и
лекарственные препараты, например аспирин (салициловая кислота),
желчные соли и высокие концентрации спирта, ослабляют этот
барьер. Физиологическая природа сопротивления слизистой
желудка проникновению и вредоносному действию кислоты
неизвестна, но, возможно, она заключена в самих слизистых
клетках; впрочем, их защищенность отчасти может быть связана с
тем, что слизистая покрыта муцином.
СОСТАВ И ИЗМЕРЕНИЕ
Желудочный сок состоит, из кислотного компонента, НС1 и
щелочного, содержащего пепсиноген и некоторые
электролиты— Na4*, K+, C1". Он также содержит муцин, воду, клетки и
некоторые ферменты.
Концентрация [Н+] зависит от интенсивности секреции: при
усилении секреции [Н+] растет, a [Na+] падает. Кислота
желудочного сока выражается в миллиэквивалентах на 1 л или на
заданное время (ч). Максимальная [Н+] составляет около
150 мэкв/л.
Базальную секрецию относят к секреции после голодания и
обычно собирают сок в течение четырех 15-минутных периодов.
Максимальной секрецией считается количество сока,
собранного за первый час после применения стимула, вызывающего
секрецию (пища, гистамин или гастрин), или после стимуляции
блуждающего нерва.
У взрослого человека за сутки выделяется 2—3 л
желудочного сока. Интенсивность базалькой и максимальной секреции
кислоты показана в табл. 21.2. Естественным стимулом
выделения желудочного сока и кислоты служит пища. Секреция
стимулируется также гистамином, пентагастрином, гастрином, ин-
25*
388
Глава 21
Таблица 21.2а). Секреция
кислоты (НС1) в желудке человека, мэкв/ч
Интенсивность базальной и максимальной секреции
Базальная
1,4
Максимальная
После еды После гистамина
30 34,5
До и после еды
Часы
Концентрация, мэкв
0 2,0
0,5 17,0
1,0 27,0
1,5 30,0
2,0 26,5
3,0 20,0
4,0 7,0
а) По Davenport H. W. 1977. Physiology of digestive tract, 4th ed., Yearbook
Medical, Chicago.
сулином и действием блуждающего нерва. Желудочная
секреция начинается при виде, запахе и вкусе пищи (цефалическая
фаза) и продолжается в течение 3—4 ч после приема пищи
(желудочная фаза).
ГИСТАМИН И ГАСТРИН
Долгое время считали, что локально продуцируемый гиста-
мип усиливает секрецию кислоты, стимулируя обкладочные
клетки, и что действие гастрина, выражающееся в усилении
секреции кислоты, объясняется стимулирующим эффектом
гастрина, высвобождаемого действием гистамина. В последнее
время было показано, что гастрин стимулирует обкладочные
клетки и что гистамин не является физиологическим
медиатором. Стимулирующее действие гастрина на желудочную
секрецию (НС1) во много раз сильнее (на молярной основе), чем
действие гистамина, но он лишь слабо стимулирует секрецию
пепсина.
Однако роль гастрина и гистамина в желудочной секреции
точно не выяснена, хотя имеющиеся данные показывают, что
рецепторы для гистамина, стимулирующего секрецию кислоты,
можно заблокировать фармакологическими препаратами. Эти
Секреция, переваривание, всасывание
389
антагонисты блокируют желудочную секрецию независимо от
того, вызвана ли она гастрином, гистамином или же
стимуляцией блуждающего нерва. Продолжаются споры относительно
того, имеются ли для каждого агента, вызывающего секрецию
кислоты, свои отдельные рецепторы.
Гастрин выделяется клетками антрального отдела желудка.
Это пептид из 17 аминокислотных остатков с низким
молекулярным весом. Гастрин не только усиливает желудочную секрецию,
но также стимулирует моторику желудка, пищевода,
кишечника, желчного пузыря и матки.
РЕГУЛЯЦИЯ ЖЕЛУДОЧНОЙ СЕКРЕЦИИ
В регуляции желудочной секреции участвуют гуморальные
и нервные механизмы. Секреция делится на три фазы: цефали-
ческую, желудочную и кишечную.
Цефалическая фаза вызывается факторами, действующими
через головной мозг, — видом, вкусом, запахом и жеванием
пищи. Они вызывают безусловные рефлексы, но здесь могут
участвовать и условные рефлексы, сочетание пищи с
индифферентными стимулами или звуком (павловский рефлекс). В цефали-
ческой фазе секреции выделяется желудочный сок с высоким
содержанием кислоты и пепсина. Эту фазу, опосредованную
исключительно блуждающим нервом (вагусом), можно
подразделить на стимуляцию гастрином, выделяющимся под действием
вагуса, прямую активацию вагусом желез, выделяющих
кислоту, и на сенситизацию гастрином желез к действию вагусных
импульсов. Раздражение блуждающего нерва высвобождает
гастрин в антральном отделе, поскольку оперативное удаление
этого последнего прекращает или сильно снижает секрецию.
Желудочная фаза начинается при поступлении пищи в желудок
или ее соприкосновении со слизистой оболочкой желудка; фаза
длится 3—4 ч и действует даже в изолированном денервирован-
ном малом желудочке независимо от цефалической фазы.
Некоторые виды пищи, например белки, эффективно вызывают
кислую секрецию. Желудочная фаза, вероятно, запускается хо-
линэргическими рефлексами, которые могут быть вызваны
механической (растяжением) или химической стимуляцией
локальных рецепторов 1) стенки антрума (где продуцируется гастрин)
или 2) области слизистых желез (где продуцируется кислота)^
Это показано на рис. 21.2. Считается, что ацетилхолин (АХ)
является конечным стимулятором выделения гастрина, а также
пепсина и кислоты (Н+).
В кишечной фазе стимуляция слизистой кишечника массами,
поступающими из желудка, или пищей, вводимой прямо в
кишку, изолированную хирургическим путем, вызывает желудочную
390
Глава 21
Область желез
привратника
ЦНС
Рис. 21.2. Действие стимуляции нерва и выделения гастрина на секрецию
кислоты. Локальная стимуляция рецепторов 1 путем стимуляции внутреннего
нерва, или вагуса, вызывает выделение ацетилхолина (АХ), который
стимулирует секрецию гастрина клетками антральной области, а тот в свою очередь
вызывает секрецию кислоты обкладочными клетками. Афферентные и
эфферентные пути обозначены стрелками. Стимуляция рецептора 2 локально или
через вагус также вызывает выход АХ и секрецию кислоты обкладочными
клетками. Реакция на цефалические стимулы — зрительные, обонятельные,
вкусовые — тоже может осуществляться по эфферентным вагусным путям.
(Grossman M. I., Physiologist, 1963, 6, 349.)
секрецию. Это свидетельствует о том, что кишка выделяет
непосредственно в кровоток гастрин или какой-то другой гормон,
который, всасываясь, попадает в желудок и вызывает секрецию
желудочного сока. Холецистокинин, тоже выделяемый
кишечником, обладает тем же физиологическим действием, что и
гастрин, и может частично обусловливать кишечную фазу
желудочной секреции.
ТОРМОЖЕНИЕ ЖЕЛУДОЧНОЙ СЕКРЕЦИИ
Различные вещества, приходя в соприкосновение со
слизистой двенадцатиперстной кишки, тормозят желудочную
секрецию. Это жиры, кислоты и продукты переваривания белков;
кислота, соприкасаясь со слизистой антралыюго отдела,
тормозит или замедляет выделение гастрина. Нарушение аппетита
эмоциональными воздействиями подавляет или устраняет
рефлекторную вагусную стимуляцию цефалической фазы секреции.
Антацидные препараты значительно варьируют по своей
способности нейтрализовать кислоты. Так, например, фосфат
алюминия сравнительно мало эффективен по сравнению с препаратом,
Секреция, переваривание, всасывание
391
содержащим гидроокись алюминия, карбонат кальция и
гидроокись магния.
Секреция пепсина стимулируется большинством тех
факторов, которые стимулируют выделение кислоты, а именно пищей,
гастрином, гистамином, секретином и возбуждением
блуждающего нерва. Мотилин — полипептид, содержащий 22
аминокислоты,— стимулирует секрецию пепсина и кислоты и
обнаруживается в слизистой двенадцатиперстной и верхней части тощей
кишки. Это вещество, предположительно являющееся гормоном.,
также вызывает сильные сокращения желудка и, возможно,
участвует в процессе эвакуации его содержимого.
Пептид, оказывающий тормозное действие на желудок,
тормозит желудочную секрецию и стимулирует секрецию
кишечного сока и панкреатического гормона. Он выделяется главным
образом под действием пищи, в частности жиров и углеводов,
и обнаруживается в двенадцатиперстной и тощей кишках.
В 1975 г. он был синтезирован. Секреция внутреннего фактора
слизистой желудка происходит в обкладочных клетках. Этот
фактор необходим для процесса всасывания витамина Bi2
в подвздошной кишке.
ПАНКРЕАТИЧЕСКАЯ И КИШЕЧНАЯ СЕКРЕЦИЯ
Панкреатический секрет выделяется эндокринными и экзо-
кринными железами. Экзокринный секрет содержит главным
образом ферменты, воду и электролиты. Экзокринная часть
поджелудочной железы состоит из сложных альвеолярных желез,
напоминающих слюнные; ее ацинарные клетки тоже выделяют
зимогеновые гранулы, поступающие в протоки, которые выводят
их в просвет двенадцатиперстной кишки (рис. 21.3). Меньше
известно о том, как секретируется жидкий сок, если не считать
того, что предположительно он тоже выделяется ацинарными
клетками.
СОСТАВ И РЕГУЛЯЦИЯ
Панкреатический сок выделяется в разных количествах, но
в среднем оно составляет у человека около 2 л в сутки. Его рН
колеблется от 7,6 до 8,2 (щелочное), и он содержит
бикарбонаты в высокой концентрации. Содержание ферментов в соке
варьирует в зависимости от стимула в пределах от 0,1% ДО
10%. Входящие в него ферменты представляют собой трипсин,
химотрипсин, карбоксипептидазы (протеолитические ферменты),
липазу (расщепляющую жир), амилазу (переваривающую
углеводы) и другие, о которых будет сказано ниже в разделе
о пищеварительных ферментах. Два из них — трипсиноген и хи-
392
Глава 21
мотрипсиноген — в момент выделения неактивны и
активируются позднее.
Секрецию регулируют нервные и гормональные механизмы,
главным образом последние. Раздражение блуждающего
нерва вызывает выделение небольших количеств панкреатического
сока, богатого ферментами. Этот эффект блокируется
атропином. При растяжении желудка, вызывающем возбуждение аф-
Правый
печеночный
Ампула желчного Двенадцатиперстная
протока кишка
Рис. 21.3. Поджелудочная железа, ее протоки, печеночные и желчные
протоки и место их впадения в двенадцатиперстную кишку.
ферентных волокон блуждающего нерва, возникает гастропан-
креатический рефлекс. Симпатические волокна стимулируют
выделение небольшого объема сока, богатого ферментами.
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ
Секретин — гормон, продуцируемый кишечной слизистой, —
вызывает обильное выделение панкреатического сока с низким
содержанием ферментов. Впервые обнаруженный в 1902 г., он
был синтезирован лишь в 1966 г. Секретин содержит 27
аминокислот. Единственным стимулом, эффективным для его
выделения, является действие кислоты на слизистую верхнего
отдела кишечника.
Обнаруженный в кишечной слизистой холецистокинин был
выделен, очищен и синтезирован. Хотя он содержит 33
аминокислоты, вся его активность определяется восемью С-концевы-
Секреция, переваривание, всасывание
393
ми аминокислотами; последние 5 кислот те же, что в гастрине,
и действительно, действие холецистокинина в некоторых
отношениях такое же, как гастрина. Главное его действие состоит
в стимуляции желчного пузыря, но он вызывает также
секрецию ферментов из поджелудочной железы.
Панкреатические полипептиды, впервые обнаруженные в
поджелудочной железе у птиц, были выделены и очищены у
многих млекопитающих, включая человека, и все они
обладают одинаковой структурой. Эти гормоны тормозят сокращение
желчного пузыря, кишечную моторику и выделение
панкреатических ферментов, но усиливает тонус желчного протока и
панкреатическую секрецию воды и НС03~.
Секреция в тонком кишечнике осуществляется главным
образом бруннеровыми и либеркюновыми железами. Бруннеровы
железы двенадцатиперстной кишки выделяют густую щелочную
слизь, которая, возможно, защищает дуоденальную слизистую
от кислоты желудочного сока. Либеркюновы железы секретиру-
ют слизь и изотоническую жидкость. Раздражение
блуждающего нерва стимулирует секреторную активность бруннеровых
желез.
ПЕЧЕНОЧНАЯ И ЖЕЛЧНАЯ СЕКРЕЦИЯ
Печень — самый крупный внутренний орган, выполняющий
множество функций, в том числе J) образование желчи, 2)
метаболизм многих веществ и пищевых ингредиентов,
всасывающихся в кишечнике, 3) синтез и депонирование некоторых
соединений и 4) расщепление и детоксикация лекарственных
средств и других веществ (см. также гл. 22 и 23).
На рис. 20.4, 21.3 и 21.4 показаны рассмотренная ранее
анатомия печеночных кровеносных сосудов и расположение
печеночного и желчного протоков. Печень лежит в брюшной
полости (рис. 20.1), она состоит из правой и левой доли (рис. 21.3 и
21.4), общего желчного протока, пузырного протока и
желчного пузыря. Некоторые виды животных, например крыса, голубь
и лошадь, лишены желчного пузыря. Желчный проток и
желчный пузырь выделяют желчь, которая изливается в
двенадцатиперстную кишку (рис. 21.3).
Доли печени разделены на дольки, функциональные единицы
которых показаны на рис. 21.4. Долька обладает кровеносными
сосудами и специализированными печеночными клетками,
образующими балки. Между этими клетками лежат синусы,
получающие как артериальную кровь, так и кровь воротной вены.
Оттекающая кровь поступает в печеночную вену.
394
Глава 21
СОСТАВ ЖЕЛЧИ
Печеночная желчь содержит около 97% воды; остальные ее
компоненты и их количества в процентах следующие: соли
желчных кислот (0,7), желчные пигменты (0,2), минеральные
соли (0,8), жирные кислоты (0,14), лецитин (0,02) и холесте-
Печеночные балки
Синусоиды
Желчные
канальцы
Центральная вена
Соединительная
ткань
Желчный
проток
Ветвь воротной
вены
Внутри - Ветвь
дольковая печеночной
артериола артерии
Рис. 21.4. Трехмерная схема печеночной дольки.
рин (0,06). Ее рН 7,4. Соли желчных кислот синтезируются в
печени из холестерина.
Пузырная желчь более концентрирована, чем печеночная, и
содержит меньше воды и больше солей; это обстоятельство
позволяет думать, что вода и некоторые минеральные соли
всасываются в желчном пузыре. В желчном пузыре желчь
более кислая (рН 5—6). Желчные кислоты являются важными
компонентами и играют большую роль в пищеварении.
Основные желчные кислоты у человека и их процентное содержание
следующие: холевая кислота (50), хенодезоксихолевая
кислота. (30),, дезоксихолевая кислота (15) и литохолевая
кислота1 (;5).
Секреция, переваривание, всасывание
395
соли желчных кислот
Соли желчных кислот соединяются в кишечнике с частицами
жира и образуют мицеллы, из которых жиры легче
транспортируются и всасываются. Если желчь не попадает в кишечник,
то по меньшей мере 25% жира остается неусвоенным и
появляется в кале. Больше 90% солей желчных кислот всасываются
в подвздошной кишке (активный транспорт), а остальные
попадают в кал. Всосавшись, соли переносятся в печень и снова
выводятся с желчью. Этот цикл движения желчи повторяется
за сут^и 6—8 раз.
Билирубин представляет собой пигмент, образующийся при
расщеплении гемоглобина, и большая часть его связана в
плазме с альбумином. Свободный билирубин возвращается в печень
и вновь используется при синтезе. Цвет желчи определяется
пигментом: она желтая, когда содержит билирубин, и зелено-
черная, если в ней содержится пигмент биливердин.
Желтуха — болезнь, характеризующаяся желтизной кожи и
слизистых оболочек, вызывается накоплением свободного или
связанного билирубина.
ЖЕЛЧНЫЙ ПУЗЫРЬ
У тех животных, у которых есть желчный пузырь, желчь в
нем накапливается и выделяется в кишечник время от
времени; однако хирургическое удаление желчного пузыря вполне
совместимо с жизнью; непрерывно секретируемая желчь
поступает прямо в кишечник.
Желчный пузырь начинает сокращаться вскоре после
приема пищи, и в течение нескольких минут происходит
нерегулярное и неполное опорожнение пузыря. При сокращении пузыря
повышается давление в пузырном протоке, открывается
сфинктер, и желчь выбрасывается в двенадцатиперстную кишку.
Это сокращение находится под нейрогормональным и
гуморальным контролем. Эфферентные нервные пути к
желчному пузырю и сфинктеру идут в составе блуждающего
нерва, и сокращение желчного пузыря составляет часть цефа-
лической фазы пищеварительной секреции. Афферентные
импульсы возникают в двенадцатиперстной кишке и других
органах и передаются по эфферентным волокнам блуждающего
нерва.
Гуморальная регуляция запускается пищевыми веществами,
в том числе жиром, вызывающим выделение гормона из
слизистой кишечника (холецистокинина), который в свою очередь
вызывает сокращение желчного пузыря. Вследствие
избыточного накопления в пузыре различных веществ, главным образом
396
Глава 21
холестерина, образуются желчные камни. Это происходит при
стазе (прекращении или уменьшении тока) желчи или при
закупорке пузырного протока; в пузыре при этом всасывается
вода, а не холестерин.
ПЕРЕВАРИВАНИЕ
Переваривание складывается из ряда физико-химических
процессов, в результате которых поглощаемая пища
расщепляется на более простые компоненты и пищевые вещества,
способные всасываться и проникать в кровоток. Переваривание
начинается во рту, где слюна содержит фермент птиалин,
участвующий в расщеплении крахмала.
Продолжается переваривание в желудке, где образуется
пепсин, который при оптимальном для него рН (2,0) действует на
нативные белки; они расщепляются на протеозы и пептоны,
(и реже, на аминокислоты).
Панкреатический сок содержит ферменты трипсин, химо-
трипсин и карбоксипептидазы (табл. 21.3); ферменты,
расщепляющие жиры (липазы), и ферменты, которые действуют на
углеводы (амилазы). Протеолитические ферменты разрушают
сложные белки до аминокислот, которые всасываются. Липаза
Таблица 21.3. Ферментативное переваривание в желудочно-кишечном тракте
Орган
Полость рта
(слюна)
Желудок
Двенадцатиперстная кишка,
панкреатический сок
Двенадцатиперстная кишка,
панкреатический сок
Двенадцатиперстная кишка,
панкреатический сок
Двенадцатиперстная кишка,
панкреатический сок
Двенадцатиперстная кишка,
панкреатический сок
Слизистая тонкого
кишечника
■ I
Главный
переваривающий фермент
Птиалин (амилаза)
Пепсин, рН 2—3
Трипсин, рН 6—8
Химотрипсин,
рН 6—8
Карбоксипептида-
за, рН 6—8
Липаза, рН 8
Амилаза
Рибонуклеаза, де-
зоксирибонукле-
аза
Субстрат действия
фермента
Крахмал
Белки
Белки,
полипептиды
То же
Конечная СОО-
группа пептидов
Жиры
Крахмал, гидроли-
заты, 1,4-альфа-
связи
Рибонуклеиновая и
дезоксирибону-
клеиновая
кислоты I
Основной продукт
(продукты)
действия фермента
Мальтоза
Протеозы, пептоны
Аминокислоты и
полипептиды
То же
Аминокислоты
Жирные кислоты,
глицерин, моно-
и диглицериды
Мальтоза,
декстрины и глюкоза
Нуклеотиды
Секреция, переваривание, всасывание
397
превращает сложные липиды в более простые формы, а именно
глицерин и жирные кислоты, которые могут всасываться.
Съедобные углеводы — это полисахариды, олигосахариды, диса-
хариды и более простые моносахариды (см. также гл. 23).
Примерами поли-, ди- и моносахаридов являются
соответственно крахмал и гликоген, мальтоза и глюкоза.
Предшественником трипсина служит трипсиноген, который
в кишечнике превращается в активный трипсин под действием
энтерокиназы — фермента кишечного сока. Трипсин
переваривает денатурированные и частично переваренные белки,
поступающие из желудка. При этом конечным продуктом его действия
являются аминокислоты и полипептиды. Продуцируемый
поджелудочной железой ингибитор трипсина препятствует
повреждающему действию трипсина на слизистую кишечника.
Химотрипсин действует на белки и полипептиды (так же,
как трипсин); он образуется в форме химотрипсиногена,
который под действием трипсина или энтерокиназы в кишечнике
переходит в свою конечную форму. Оба протеолитических
фермента расщепляют связи в средней части молекул (эндопеити-
дазы).
Карбоксипептидаза расщепляет аминокислоты пептидов со
свободными карбоксильными (СОО~) группами. Липаза
действует на жиры, превращая их в свободные жирные кислоты,
глицерин и глицериды. Образующаяся в печени желчь при
добавлении к содержимому двенадцатиперстной кишки
активирует липазу. Амилаза расщепляет крахмал и гликоген на
мальтозу, декстрины и глюкозу.
Слизистая кишечника продуцирует ферменты, действие
которых сходно с действием панкреатических ферментов, но
роль их по сравнению с последними невелика. Здесь имеются,
однако, и некоторые специфические амилазы, а именно лактаза
(действует на лактозу), сахараза (действует на сахарозу), дек-
стриназа и дисахаридаза.
ВСАСЫВАНИЕ
Продукты переваривания продвигаются сквозь слизистую
ЖКТ и всасываются в кровоток при помощи нескольких
механизмов, а именно 1) активного транспорта, 2) диффузии и
3) пиноцитоза (см. также гл. 2).
УГЛЕВОДЫ
Сложные полисахариды и некоторые дисахариды в конечном
счете расщепляются в кишечнике на более простые формы, а
именно на легко всасывающиеся моносахариды. Некоторые
дисахариды всасываются, не изменяясь.
398
Глава 21
Глюкоза и другие гексозы (сахара с 6 атомами углерода),
а также пентозы (сахара с 5 атомами углерода) легко
всасываются через слизистую двенадцатиперстной и подвздошной
кишки в капилляры кровеносной системы, которые впадают в
воротную вену печени. Другие углеводы вместе с некоторыми
пептозами могут всасываться путем диффузии, но глюкоза и
галактоза транспортируются через слизистую путем активного
процесса, называемого транспортом с участием переносчика
(см. гл. 2). Переносчиком является Na+; глюкоза переносится
в соединении с Na+.
липиды
Более сложные липиды (см. предыдущий раздел) в
конечном счете расщепляются на жирные кислоты, глицерин и моно-
глицериды и усваиваются с небольшой затратой энергии или
совсем без нее.
Всасывание жирных кислот зависит от величины
соединения (числа атомов углерода). Те соединения, которые содержат
меньше 12 атомов углерода, переносятся через клетки слизистой
непосредственно в кровь воротной вены. Жиры с большим
числом атомов углерода всасываются в лимфатические сосуды и
транспортируются по ним. Большая часть такого жира
переносится в виде триглицеридов; но в кишечнике триглицериды
расщепляются на моноглицериды, всасываются, ресинтезируют-
ся обратно в триглицериды клетками слизистой, а затем
попадают в лимфатические сосуды.
Как полагают, жир усваивается в виде тонкодисперсной
эмульсии и в виде мицелл — скопления молекул, содержащих
жиры и соли желчных кислот. Основное всасывание жира
происходит в двенадцатиперстной кишке, но частично также и в
подвздошной.
БЕЛКИ
Продуктами переваривания белков являются свободные
аминокислоты и некоторые ди- и трипептиды. Белки всасываются
главным образом в виде аминокислот посредством различных
активных процессов с участием переносчика, каким является
Na~!\ Некоторые аминокислоты — L-кислоты — всасываются
быстрее D-кислот; скорость всасывания в двенадцатиперстной и
тощей кишках выше, чем в подвздошной.
ВОДА И ЭЛЕКТРОЛИТЫ
Около 8 л воды всасывается за сутки в тонком кишечнике
и около 1,3 л — в толстом. Эта вода всасывается главным
образом путем осмоса и быстрее всего из гипотонических раство-
Секреция, переваривание, всасывание
399
ров. Осмотически активные частицы продуктов переваривания
всасываются в кишечнике; вода перемещается пассивно
(всасывается) вместе с осмотически активными частицами.
Электролиты всасываются из двенадцатиперстной и тощей кишки
легче, чем из подвздошной. Такие моновалентные ионы, как
Na+, K+, С1~ и НСОз~, всасываются скорее поливалентных
Са2+ и Mg2+. Всасывание Na+, Ca2+ и Fe2+ происходит путем
активного механизма. Для того чтобы произошло всасывание,
железо должно быть двухвалентным (Fe2+); его всасывание
усиливается, когда запасы железа в организме невелики, а
образование эритроцитов повышено.
ЛИТЕРАТУРА
Code С. F. (ed.) (1968). Handbook of physiology: alimentary canal, vol. V,
American Physiological Society, Washington.
Crane R. K. (ed.). (1977). Gastrointestinal Physiology, 1977 International
Review Physiology, vol. 12, Chaps. 2, 3, 5, 6, University Park Press, Baltimore.
Davenport H. W. (1977). Physiology of digestive tract, 4th edn., Year Book
Medical, Chicago.
Jacobson E. D., Shanbour W. L. (eds.) (1974). Gastrointestinal physiology,
vol. 4, Chaps. 1, 4, 5—10, International Review of Physiology, University
Park Press, Baltimore.
Ganong W. F. (1977). Review of medical physiology, 8th edn., Lange Medical,
Los Altos, California.
Knoebil L. K. (1977). In: Selkurt E. E. (ed.), Physiology, 4th edn., Little, Brown,
Boston.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Назовите слюнные железы и их расположение.. Которая из
них секретирует больше всего слюны?
2. Что подразумевается под слизистым барьером желудка?
3. Сколько желудочного сока выделяется у человека за сутки?
4. На какие фазы делится желудочная секреция? Назовите и
опишите их.
5. Назовите три агента, стимулирующие желудочную
секрецию.
6. Опишите местоположение поджелудочной железы и
функцию экзокринных ацинарных клеток.
7. Назовите типы ферментов, вырабатываемых экзокринной
частью поджелудочной железы, и типы субстратов, на
которые они действуют.
8. Какой гормон стимулирует экзокринную функцию
поджелудочной железы?
9. Какой нерв наиболее эффективно действует на экзокринную
функцию поджелудочной железы?
Ю. Каковы главные функции печени?
400
Глава 21
И. Где вырабатывается желчь?
12. Какова функция солей желчных кислот?
13. Что лежит в основе образования желчных камней?
14. Определите следующие вещества и укажите, на что они
действуют: а) карбоксипептидазы, б) трипсин.
15. Какое рН оптимально для пищеварения в тонком
кишечнике?
16. Что такое энтерокиназа, и каково ее воздействие?
17. Как всасываются следующие вещества: а) углеводы, б)
белки и в) жиры?
18. Что такое всасывание при помощи переносчика? Приведите
пример.
Глава 22
ПРОМЕЖУТОЧНЫЙ ОБМЕН
Блоки, из которых строятся три главных энергетических
субстрата организма, могут быть объединены (синтезированы, ана-
болизированы) в более крупные сложные структуры. Точно так
же крупные (сложные) органические соединения могут быть
расщеплены (катаболизированы) на исходные, основные
строительные блоки. Основные единицы, из которых строится
организм, это моносахариды (для построения сложных Сахаров»
или углеводов), аминокислоты (для построения сложных
полипептидов и белков) и глицерин и свободные жирные кислоты
(для построения липидов, или жира). Промежуточный
метаболизм, или обмен, представляет собой итог химических
превращений (обычно катализируемых ферментативными белками),
происходящих одновременно путем анаболических и катаболи-
ческих процессов.
АНАБОЛИЗМ И КАТАБОЛИЗМ
Принято считать, что анаболизм (синтез), но не катаболизм
отличается видовой специфичностью. С одной стороны, все
организмы могут усваивать одни и те же основные компоненты,,
например аминокислоты, и из них строить разные сложные
соединения, например полипептиды или белки. Конечный продукт
будет специфичным для данного организма, для тканевых
процессов в этом организме и специфических реакций,
осуществляемых при помощи ферментов, обеспечивающих химические
взаимодействия.
С другой стороны, каждый организм может поглотить
любой полипептид или белок и расщепить его. Но какого бы типа
ни был этот организм и какие бы катаболические процессы при
этом ни происходили, в результате образуются те же самые
аминокислоты. Они были соединены по-разному, но при их
расщеплении создаются те же строительные блоки. Таким образом,
катаболизм неспецифичен.
Поскольку анаболизм и катаболизм складываются из
химических реакций, происходящих в организме, и поскольку эти
реакции обычно регулируются катализаторами — ферментами
26—1705
402
Глава 22
(среди которых есть ферменты, лимитирующие скорость всего
процесса), — одновременное течение этих процессов может
приводить к динамическому равновесию, росту или частичному
разрушению структур тела. Если в результате анаболических
процессов создается такое же количество ткани (или образуется
такое же количество энергии), какое разрушается (или
выделяется) в результате катаболических процессов, то можно
говорить о динамическом равновесии, при котором общее
количество ткани не увеличивается и не уменьшается (рис. 22.1).
Анаболизм — катаболизм
Основной
i
Потери ткани
(анаб. < катаб.)
Динамическое
равновесие
(анаб. = катаб)
Рост
(анаб. > катаб)
Рис. 22.1. Взаимоотношения между анаболизмом и катаболизмом в условиях
„динамического равновесия, роста и «истощения».
Если же анаболические процессы превалируют над
одновременно происходящими катаболическими, то в итоге происходит
накопление ткани — истинный рост. Наконец, если
разрушительные, или расщепляющие (катаболические), реакции
преобладают над одновременно протекающими синтетическими
процессами, то выделяется энергия, а ткань разрушается. Иначе говоря,
катаболизм преобладает над анаболизмом, и в результате
«содержимое» организма уменьшается: он теряет свою истинную
массу (рис. 22.1). Обычно у взрослых анаболические реакции
уравновешиваются расщеплением или катаболизмом
(равновесие). Дополнительные сведения об этом см. в гл. 2.
ИСТОЧНИКИ ЭНЕРГИИ
Энергетический метаболизм клеток (образование, перенос и
превращение) происходит главным образом в митохондриях.
Цитоплазма животных клеток —«жидкая» часть клетки
—служит растворителем для пищевых веществ и их
предшественников. Кроме того, высокая концентрация белка в цитоплазме
Промежуточный обмен
403'
служит «банком» для многих ферментов, играющих основную
роль в высвобождении энергии, накопленной в структуре
пищевых веществ. Предшественники — это вещества с очень
низким молекулярным весом (обычно меньше 50), которые
воспринимаются из окружающей среды для синтеза определенных,
более крупных (с мол. весом 50—300) промежуточных
соединений; эти последние образуют затем основные строительные
блоки (мол. вес 200—400), а именно аминокислоты, жирные
кислоты, глицерин и моносахариды. Исходными предшественниками
для этих последовательных этапов служат Н20, С02 и другие
простые соединения. Эти же самые простые соединения
являются конечными продуктами полного катаболизма указанных
пищевых веществ.
Еще одним продуктом катаболизма является энергия
(хранящаяся в виде потенциальной энергии в структурных связях
каждого пищевого вещества), выделяемая или в виде тепла
(кинетической энергии, связанной с движением молекул) или в
виде энергии, которая может быть перенесена на имеющиеся
вещества. Эти последние включают энергию в структурные
связи биомолекул. Такая «пойманная» энергия может быть
использована в дальнейшем (потенциальная энергия) в качестве
источника кинетической энергии, как это происходит в энергетике
мышечного сокращения и локомоции (см. гл. 11). Энергия,
рассеиваемая в виде тепла, в значительной мере способствует
поддержанию температуры, оптимальной для ферментативных
процессов, и общей температуры тела (см.-гл. 18).
АКТИВНОСТЬ ФЕРМЕНТОВ
Слово «энзим» (иначе «фермент») происходит от
греческого и буквально значит «дрожжи», или «закваска». Такое
название оправдано, потому что само существование энзимов стало
известным благодаря знаменитым исследованиям Луи Пасте-
ра, касающимся той роли, какую играют дрожжевые клетки
в процессе сбраживания винограда.
Ферменты представляют собой белки, которые действуют
как органические катализаторы. Это впервые показал Освальд
в 1893 г. Ферменты, таким образом, являются регуляторами
скорости химических реакций; обычно они ускоряют реакции
Даже без изменения температуры. Лучше всего они действуют
при специфическом рН, при определенных температурах и
только на специфические субстраты (оптимальные условия).
Подобно небелковым катализаторам, ферменты
объединяются в комплексы или взаимодействуют с субстратом, но не
участвуют в реакции. Напротив, они обычно выходят из
комплекса субстрат — фермент интактными, неповрежденными и спо-
26»
404
Глава 22
собными регулировать скорость другой реакции в том же
субстрате. Таким образом:
•Фермент (Enzyme) -(-Субстрат (Substrate) —
(Е) (S)
> Комплекс Фермент—Субстрат
(ES)
Продукт-)-Фермент.
(F) (Е)
Молекулярная активность фермента выражается числом
молекул субстрата, которые могут реагировать с одной молекулой
фермента за единицу времени, обычно за 1 мин. Ферменты
специфичны относительно реакций, которые они регулируют,
потому что должны физически взаимодействовать с субстратом,
подобно тому как взаимодействуют ключ с замком (см. гл. 25).
Поскольку ферменты не разрушаются при регулируемых ими
Углеводы, )
белки, жиры, /
кислород у4*
-/ Катаболизм
\ Энергия
уЛНА со2
АДФили
креатин
Ч^ Мышечное
АДФ сокращение
Синтез
АТФ
АДФ
АДТ
▲
X
Секреторная
активность
желез
Синтез белка
Ч^ Осмотическая
работа
Рис. 22.2. Роль катаболических процессов в обеспечении энергией для
работы клетки. Рн — неорганический фосфор, АТФ — аденозинтрифосфат, АДФ—
аденозиндифосфат.
Промежуточный обмен
405
реакциях, они могут существовать неделями, месяцами и даже
годами. Отсюда также следует, что, поскольку в
ферментативных реакциях нет побочных продуктов, их выход составляет
всегда 100%. Одновременно активность сотен ферментов (как
анаболических, так и катаболических) создает для организма
многообразие высокоспецифичных конечных продуктов
(рис. 22.2). Таким образом, ферменты осуществляют за секунды
или доли секунды то, что без них заняло бы в лаборатории
биохимика дни или недели.
Эффективность ферментативного действия в организме
может быть изменена молекулами субстрата, ингибиторами и
гормонами. Из-за того что молекула белкового фермента очень
велика по сравнению с молекулами субстрата, ограниченная
зона на катализаторе, где происходит физический контакт и
образуется комплекс ES, представляет собой лишь очень
небольшую часть всей поверхности. Это — активный центр, и
комплексы ES образуются только здесь. Ясно, что при избытке
субстрата все активные центры оказываются занятыми и
активность фермента максимальна. Избыточные молекулы
субстрата не могут взаимодействовать с участками фермента, пока
эти последние не выйдут из ES вследствие образования
продукта реакции (Р). Таким образом, реакция, катализируемая
ферментом, максимальна, когда заняты все активные центры.
Избыток субстрата не может более ускорить эту реакцию.
ИНГИБИТОРЫ
Кинетику ферментативного процесса изменяют также
ингибиторы. Конкурентное ингибирование представляет собой
одну из самых важных форм ингибирования в биологических
системах. Возможно, что конфигурации активного центра на
ферменте соответствует конфигурация двух или более других
молекул. Молекула субстрата конкурирует с молекулой не-суб-
страта за контакт с активным центром, так как и та и другая
обладают конфигурацией, нужной для того, чтобы вставить
«свой ключ в замок фермента». Естественно, что при
повышении концентрации не-субстрата (конкурентного ингибитора)
статистическая вероятность, что субстрат войдет в контакт с
активным центром, ниже. Таким образом, скорость химической
реакции может быть понижена ингибиторами, т. е. теми
веществами, которые конкурируют с субстратом за активные центры
белка-фермента. К другим формам ингибирования относятся
неконкурентные и необратимые формы. Для более подробного
рассмотрения этого вопроса читателю следует обратиться к
списку литературы, приведенному в конце настоящей главы.
406
Глава 22
ЭНЕРГИЯ
Строго говоря, энергия не синтезируется и не может быть-
синтезирована живой клеткой. Согласно первому закону
термодинамики (закону сохранения энергии), энергия не исчезает
и не возникает вновь; в организме она переносится или
превращается. Химическая энергия, накопленная в клетке, — это
потенциальная энергия, подобная энергии растянутой резиновой
ленты. Растягивание ленты потребовало затраты энергии,
которая при укорочении ленты высвобождается в виде тепла или
работы (кинетической энергии), или же и того и другого.
Энергия требуется также для синтеза сложных питательных
веществ, или макромолекул, внутри клетки.
Энергия высвобождается также при каталитическом
разрушении сложного химического вещества, когда оно распадается
на свои промежуточные формы или в конечном счете на
исходные основные компоненты Н20, С02, NH3, №. К счастью, при
ферментативно управляемых реакциях энергия связей
высвобождается постепенно; связи распадаются поэтапно, пока не
высвободится вся энергия. Однако поэтапный выход
молекулярной энергии при расщеплении сложных субстратов позволяет
другим молекулам «захватывать» или запасать энергию порцию
за порцией и тем самым сохранять ее в высокоэнергетических
связях в других химических формах. Поэтому запасы энергии
могут накапливаться в уже существующем соединении без
существенного изменения вещества; энергия может быть также
использована для образования совершенно нового вещества,
обладающего высокой потенциальной энергией.
Анаболизм требует затраты энергии, происходящей чаще
всего от разрыва высокоэнергетических фосфатных связей,—
для биосинтеза сложных молекул из основных строительных
блоков с образованием клеточных компонентов, заряженных
высокой энергией. Катаболизм дробит богатые энергией
сложные молекулы, высвобождая поэтапно ее порции, причем до
42% их могут быть захвачены или отложены в веществах
клетки для будущего превращения (рис. 22.2).
ПОТЕНЦИАЛЬНАЯ И КИНЕТИЧЕСКАЯ ЭНЕРГИЯ
Энергия химической связи является потенциальной. Она
подобна энергии шара на вершине горы. Разрыв связей
сопровождается высвобождением энергии (как при скатывании шара
с горы), когда движение молекул (теплота) превращается в
работу (как при сокращении мышцы) или передается в виде
потенциальной энергии в иное семейство внутриклеточных
химических соединений.
Промежуточный обмен
407
ОБРАЗОВАНИЕ ВОДЫ
В реакции H2 + V2O2—^Н20 на каждый моль образованной
Н2О высвобождается около 68 ккал энергии (см. также гл. 18).
Таким образом, вода содержит на 68 000 калорий меньше, чем
исходные вещества. И когда молекула воды расщепляется на
Аденин
Д-рибоза Остаток фосфорной кислоты
NH,
ОН ОН
I I
• О
с—с—с—с—с—о—р—о—р—о—р—он
I I I I I I I
н н н н н он он он
АТФ (Аденозинтрифосфат)
\
АДФ(Аденозиндифосфат)
О
I
0—Р—О—P—ОН
I I
он -он
--r^VV ,
АМФ(Циклический аденозин-3, 5-монофосфат)
—О
^N^^N н Н
Н
3 41 б!
-С—С—С
Аденин Д-рибоза о-
I I
" о" >
р
ОН Циклический фосфат
Рис. 22.3. Структурные формулы АТФ, АДФ и циклического 3',5'-АМФ.
водород и кислород, энергия поглощается. Для начала
большинства внутриклеточных химических реакций или — чаще —
Для их ускорения нужны небольшие количества энергии. Так,
Для образования определенного количества воды при 20 °С из
€меси водорода и кислорода требуются минуты. Но если
кислород и водород нагреть, то молекулы соединяются быстро с
образованием воды и высвобождением упомянутых 68 ккал/моль.
Энергия, добавленная для облегчения процесса синтеза,
называется «энергией активации». В основном тепло ускоряет
столкновение молекул, что делает их соединение более вероятным.
408
Глава 22
МОЛЕКУЛЫ ВЫСОКОЭНЕРГЕТИЧЕСКОГО ФОСФАТА
Соединение аденозинтрифосфат (АТФ) состоит из аденози-
на, сахара с 5 атомами углерода (пентозы, рибозы) и трех
фосфатных групп, последовательно соединенных с сахаром
(рис. 22.3). При разрыве концевой фосфатной связи (О—Р)
высвобождается значительно больше энергии (7 ккал/моль
АТФ), чем при разрыве обычных низкоэнергетических связей
С—О, Н—О или Н—Н. Высвобожденные 7 ккал/моль сразу же
могут служить для нагревания окружающей клетку среды,
превращения в работу или энергию или перехода на
воспринимающие молекулы для последующего использования. АТФ,
резервуар высокой энергии, предпочтительно используется в таких
постоянно активных тканях, как скелетные мышцы. Переход
энергии от АТФ к скользящим филаментам скелетной мышцы
рассмотрен в гл. 11.
Поскольку АТФ служит депо энергии, перешедшей от других
связей, для него требуется, чтобы в какой-то момент в истории
клетки произошел перенос энергии к воспринимающей
молекуле. Затем эта молекула должна была измениться и образовать
АТФ. Воспринимающей молекулой предпочтительно является
аденозиндифосфат (АДФ), а при некоторых условиях в мышце
это может быть также молекула креатина (К). В обоих
случаях высокая энергия локализована в концевой фосфатной
группе. Отсюда:
АДФ+Фн+Энергия > АТФ+Н20
(Аденозиндифосфат)-[-(Неорганический фосфат)+ 7 ккал/моль >
► (Аденозинтрифосфат) -f- (Вода)
или
К+Фн+Энергия —* КФ+Н20
(Креатин)-[-(Неорганический фосфат)-]-10 ккал/моль ►
► (Креатинфосфат)-[-(Вода)
и затем
КФ+АДФ —* АТФ + К
(Креатинфосфат)-]- (Аденозиндифосфат) >
> (Аденозинтрифосфат) -f- (Креатин)
В мышце концентрация КФ в 5 раз больше концентрации
АТФ, а креатин действует как временное депо энергии, быстро
передавая ее «ожидающей» молекуле АДФ. Этот перенос
облегчается ферментом креатинкиназой, освобождающей креатин,
который вновь используется для переноса энергии. При этом
Промежуточный обмен
409
переносе в качестве депо могут быть использованы и другие
метаболические вещества (в частности, у беспозвоночных).
Энергия постоянно совершает циклы в клетке, а АТФ
обычно служит общим фактором, который поддерживает процессы,
протекающие с затратой энергии. Такими процессами являются
активный транспорт (см. гл. 2), мышечное сокращение и локо-
моция (см. гл. И), секреция гормонов (см. гл. 25) и биосинтез
таких сложных структур, как полипептиды и белки. Хотя общее
количество энергии, заключенной в АТФ, невелико, но
повсеместное наличие этого вещества, а также легкость, с какой
энергия переносится, подчеркивают его важную роль в активности
клетки. Надо также иметь в виду, что катаболизм углеводов,
жиров и белков поставляет энергию для ресинтеза АТФ.
Таким образом, катаболизм приводит к накоплению энергии,
и передаче ее общему носителю энергии — АТФ (рис. 22.2).
АНАЭРОБНЫЙ И АЭРОБНЫЙ КАТАБОЛИЗМ
Большая часть энергии, высвобождаемой при катаболизме
пищевых веществ, образуется в митохондриях клеток
в'присутствии кислорода (аэробные реакции), который действует как
акцептор водорода. Но само высвобождение энергии
происходит и при катаболизме в отсутствие кислорода, например при
начальном распаде глюкозы в цитоплазме клетки. Такие
реакции называются анаэробными («без кислорода»). Следует
подчеркнуть, что путь от предшественника к продукту реакции
(анаболизм) и от продукта к предшественнику (катаболизм) —
не один и тот же.
Анаболизм — это не просто «обращенный» катаболизм.
В большинстве случаев эти пути отличны друг от друга из-за
разной специфичности ферментов. Анаболические процессы
требуют участия АТФ (с образованием АДФ и неорганического
фосфата), тогда как катаболические реакции небольшими
порциями выделяют энергию, которая помогает ресинтезу АТФ из
АДФ и фосфата. Окислительные процессы (аэробные) дают
при катаболизме питательных веществ больше энергии, чем
анаэробные реакции распада.
Аэробный распад пищевых веществ лучше всего можно
описать на примере начальных этапов катаболизма глюкозы,
т. е. превращения СеН^Ое + бОг—^6С02 + 6Н20 + Энергия.
Если бы один моль (180 г) распадался мгновенно, то общая
высвободившаяся энергия составила бы около 673 ккал.
Подобное расщепление буквально превратило бы клетку в печь, и
выделившаяся энергия была бы потеряна в виде тепла. Но
вместо этого анаэробными средствами — путем тщательно
регулируемого и медленного расщепления — энергия высвобожда-
410
Глава 22
Глюкоза
I 2 3 4 5 6
С-С-С-С-С-С
АТФ—> —>ADO + Ph
Гл юкоза-6-фосфат
С-С-С-С-С-С.
А1 Ф—><—> ADO+ Рн
Фруктозо-1,6- дифосфат
С-С-С-С-С-С
р О S р
г 2Рн—D>< K
(2) триозо-фосфаты
i 2 3 или 4,5,6
с-с-с
г
р
2НАДН>Р-0 2НАД-Н2
2ADOH>C—>2АТФ
(2) Фосфоенолпируват"
ется при точно
контролируемых условиях и передается
акцепторам нуклеотидам,
таким, как АДФ, для ре-
синтеза АТФ. Этим
избегается взрывной выход 673
ккал даже несмотря на то,
что распад глюкозы
продолжается в последующих
аэробных реакциях, и в
конце концов выделяются все
673 ккал/моль.
Чаще всего анаэробный
распад происходит в
цитоплазме животных клеток,
потому что ферменты,
регулирующие каждую катабо-
лическую реакцию,
ограничены цитозолем.
Катаболизм молекулы
сахара с 6 атомами
углерода при относительном
отсутствии кислорода дает
молочную кислоту —
промежуточный продукт с 3
атомами углерода (рис. 22.4).
При этом превращении,
которое обычно называется
анаэробным гликолизом,
требуются 2 молекулы АТФ,
чтобы «зарядить» молекулу
глюкозы (глюкозу до
уровня глюкозо-6-фосфата и
фруктозо-6-фосфат до
уровня фруктозо-1,6-дифосфата).
Однако непосредственно
перед образованием пирувата
или лактата или того и
другого 2 молекулы АТФ регенерируют на каждом из двух этапов.
Следовательно, энергетическим выходом анаэробного
гликолиза являются 4 молекулы АТФ за вычетом двух молекул АТФ,
потребовавшихся первоначально, чтобы «зарядить» молекулу
глюкозы и обеспечить «чистый» выход двух молекул АТФ на
одну распавшуюся молекулу глюкозы. Такой выход энергии
невелик и не мог бы обеспечивать клеточную активность у
млекопитающих или птиц сколько-нибудь продолжительное
2АОФ—О/
Пировиноград-
ная кислота
С-С-С=0
Н. II |
3 о о
н
2 АТФ
С-С-С=0
Н3 0 I
н 8
2 АТФ на молекулу гексозы
Рис. 22.4. Анаэробный распад глюкозы.
Извилистые линии означают
многоэтапные процессы. Цифры на атомах
углерода относятся к исходной молекуле
глюкозы; ~Р означает
высокоэнергетический фосфат.
Промежуточный обмен
411
время. Однако у некоторых рыб, амфибий и рептилий он может
обеспечивать потребность в АТФ.
По существу, анаэробный гликолиз — это просто процесс
брожения, требующий совместного последовательного действия
11 ферментов. Все промежуточные продукты фосфорилированы,
а конечным продуктом является лактат.
Аэробный распад пищевых веществ, по-видимому, служит
основным источником восстановленной энергии АТФ, т. е.
энергии, выделенной, захваченной акцептором и затем перешедшей
Пировиноградная кислота
1 2 3
с-с-с=о
Но И I
3 о о
Аминокислоты I > 2Н+ Липиды
(2) Ацетил-КоА
C-C-S-KoA
ЬЦ II
Лимонная кислота (С6)
Этап 1,
Щавелевоуксусная кислота (С4)
Этап 8
3 АТФ.
<кД^нн
Этап 2
| Цикл трикарбоновых
Этап 7 кислот
\
Этап 6
7Н+
ЭтапЗ
Аэ СО, У эт,л4 ЗАТ»
ЗАТФ Этап 5^2^
3 АТФ
Рис. 22.5. Аэробный распад глюкозы до пировиноградной кислоты с
высвобождением энергии на АТФ и С02. Заметьте, что ресинтез АТФ связан с
удалением водорода (см. также рис. 22.6). Конечный выход после одного цикла:
12 молекул АТФ, образованных при конденсации молекулы ацетил-КоА с
одной молекулой щавелевоуксусной кислоты. (Полное расщепление гексозы да-
«т 30 молекул АТФ.)
412
Глава 22
к АДФ для образования АТФ. Катаболизм белка и липида
начинается в аэробных условиях в митохондриях, где находятся
окислительные ферменты. Некоторые промежуточные продукты
могут «просочиться» в цитозоль, но обычно они снова
захватываются митохондриями, и конечные этапы аэробного
расщепления происходят в них, причем на последовательных этапах вы-
Глюкоза (С6)
Б
.1.
ATP
Ч-
j Рибозо-5
| { Фосфат (С5) *+~~
I * Седогептулозо I ^Д*7* JT
-<~^ Фруктозо-6-фосфат (Сб)
Глюкозо-6-фосфт (С6)
7-фосфат (С7)
-4--
Т
АТФ
Фруктозо-1, 6-дифосфат (С6)
(2) триозо-фосфат (С3)
-—\ -t—Л
Б
-J
h
АТФ
| Пируват (С3)
I
Ts>co2
,
в-*
(2) Ацетил-КоА (С2)
Лимонная кислота (Сб)
НАД
^-^АТФ
Кофермент Q
|\^АТФ
Цитохромная система
1ч2>АТФ
tN> АТФ
__2Н^-^0_2_—->_Н^0 j
Рис. 22.6. Взаимоотношения анаэробной, аэробной систем пентозного шунта
и цитохромной системы при распаде углевода с высвобождением энергии и
Н20. А. Анаэробный гликолитический путь. Б. Гексозо-монофосфатный шунт.
В. Аэробный цикл ТКК. Г. Путь окислительного фосфорилирования.
Промежуточный обмен 413
свобождаются порции энергии (рис. 22.5). Таким образом ре-
синтезируется АТФ, который способен переместиться в цито-
золь. Из-за сложности и многообразия белковых структур
количеству АТФ, восстанавливающемуся при белковом катаболизме,,
нельзя придать определенного значения. Однако окисление уме-
|: ренного количества липида до С02 и Н20 в митохондриях дает
§ около 48—58 АТФ на моль субстрата, тогда как аэробный рас-
| пад лактата до С02 и Н20 дает около 30 АТФ. Для полного
| расщепления исходной гексозы весь цикл трикарбоновых кис-
% лот (ТКК) должен повториться 2!/2 раза.
Чистый выход АТФ из глюкозы или гликогена (полимер
глюкозы) в анаэробных условиях составляет соответственно две
и три молекулы АТФ. При аэробном катаболизме начинается
распад белков и липидов и завершается распад углеводов,
начатый анаэробным гликолизом. Несмотря на разные пути
расщепления каждого типа пищевых веществ, эти пути сходятся
на общей промежуточной форме ацетил-КоА, которая ведет к
конечному аэробному распаду каждого из этих веществ в
общей последовательности этапов (рис. 22.5). Эта общая
последовательность называется циклом трикарбоновых кислот, или
циклом Кребса; в этом цикле происходит конечный катаболизм
всех трех типов субстратов с образованием С02, Н20 и энергии
(рис. 22.5 и 22.6). Энергия, высвобождающаяся при распаде
белков и жиров, выделяется только на этом окончательном
уровне промежуточного обмена. Только распад углеводов
высвобождает энергию выше уровня цикла ТКК, представленного
небольшим количеством АТФ, восстановленного в результате
анаэробных реакций.
МЕТАБОЛИЗМ УГЛЕВОДОВ, ЖИРОВ И БЕЛКОВ
Промежуточный обмен складывается одновременно из
анаболизма и катаболизма всех трех пищевых субстратов или их
предшественников; осуществляется он путем координированных,
строго контролируемых химических реакций, регулируемых
ферментами. Катаболизм углеводов начинается анаэробно в
цитоплазме, но на уровне глицерол-3-фосфата или пирувата их
распад завершается в митохондриях (рис. 22.5 и 22.6).
МЕТАБОЛИЧЕСКИЕ ПУТИ
Гликолиз совершается, как уже описано и показано на
рис. 22.6, в блоке, обозначенном А. Посредством как
анаэробных (до уровня пирувата), так и аэробных механизмов (от
пирувата до С02, Н20) непосредственные потребности клетки в
^энергии удовлетворяются метаболизмом сахара. Исходным ис-
414
Глава 22
точником глюкозы обычно служит или нефосфорилированная
глюкоза крови, или глюкоза, фосфорилированная в положении
1-С, из углеводных депо — гликогена печени или мышц. При
относительном отсутствии кислорода образуется молочная
кислота, которая действует как временное депо Н+. Все эти гликоли-
тические реакции совершаются в цитозоле клеток и
регулируются ферментами.
ФОСФОГЛЮКОНАТНЫЙ ПУТЬ
Известный также под названием гексозо- (или пентозо-)
монофосфатного шунта, этот путь действует у большинства
позвоночных во многих тканях (рис. 22.6,Б). Хотя это не главный
путь распада глюкозы, но он служит альтернативным способом
ее окисления и дает С02 и пентозы. Эти сахара с 5 атомами
углерода необходимы для синтеза нуклеиновых кислот. Кроме
того, шунт позволяет пентозам перейти в гексозы для
последующего окислительного распада. Наконец (что очень важно для
тканей, активно синтезирующих жирные кислоты и стероидные
соединения), пентозный шунт производит восстановленную
форму НАДФ, необходимую для описанного выше синтеза. Все
важные основные ферменты, связанные с этим альтернативным
путем катаболизма глюкозы, содержатся в цитоплазме
большинства животных клеток, в печени, молочных железах,
надпочечниках и особенно в гонадах.
Цикл трикарбоновых кислот (рис. 22.5 и 22.6,6) известен
также под названием цикла лимонной кислоты, или цикла
Кребса (по имени нобелевского лауреата Ганса Кребса). Девять
ферментативно регулируемых стадий, составляющих этот
цикл, — это те реакции, которые все вместе уже были нами
рассмотрены как часть аэробного катаболизма глюкозы. Весь этот
процесс с выходом энергии в качестве конечной стадии
происходит в митохондриях. Его окислительная сущность связана с
поэтапным удалением Н+ (рис. 22,6, Г) из промежуточных
продуктов.
За исключением одного пути катаболизма некоторых
аминокислот, минующего промежуточный ацетил-КоА, последний
действует как общий промежуточный этап для всех пищевых
веществ независимо от их происхождения. Таким образом,
конденсация ацетил-КоА с щавелевоуксусной кислотой дает
возможность белкам (дезаминированным аминокислотам),
углеводам (декарбоксилированной пировиноградной кислоте) и
жирам (2-С-фрагментам реакций |3-окисления) войти в этот
последний ряд регулируемых реакций, которые ведут к
образованию С02, Н20 и энергии (рис. 22.7).
Промежуточный обмен
415
Потребляемая I Мясо || Сладости, и Масло сливочное,—)
| (постное) 11 сахар || растительное |
пища:
V V
В желудочно- Белки и Полисахариды
кишечном тракте: полипептиды (крахмал и т.п.)
Компоненты
крови:
Липиды,
жиры
Жирные кислоты/
глицерин
Промежуточные
соединения
в цитоплазме:
Митохондрии:
Митохондрии:
Цитохромный
уровень:
Выделяется
в цитозоль:
<7
Мочевина
t^Окислительное*? ph
фосфори- 1С
лирование У «
Рн3, АДФ
АТФ
V2 О,
Рис. 22.7. Взаимосвязанные процессы при переваривании, всасывании и
метаболизме трех типов пищевых веществ. Обратите внимание на ключевую роль
ацетил-КоА; NH3 служит формой выделения азота (в форме мочевины с
мочой).
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
Роль кислорода в аэробном метаболизме в значительной
степени связана с той ролью, какую он играет в удалении Н+ из
митохондрий. Как показано на рис. 22.6,6, С02 удаляется в две
стадии, а Н30+ по меньшей мере в четыре стадии в процессе
распада в цикле ТКК. Водород, возникающий при разрыве
связей в промежуточных ТКК-соединениях, взаимодействует с
акцептором водорода никотинамидадениндинуклеотидом (НАД),
416
Глава 22
(рис. 22.6,Г). Когда это происходит, не только атом водорода,
но также некоторая часть потенциальной энергии из исходной
молекулы тоже переносится на НАД. Этот исходный перенос
происходит в митохондриях внутри обеих мембран; все
последующие реакции тоже протекают здесь.
Небольшое количество этой потенциальной энергии
выделяется в виде электронов из атомов водорода, переносимых через
ряд «дыхательных белков» — цитохромную систему. Перенос
электронов от НАД-Н на кислород служит непосредственным
источником энергии для окислительного фосфорилирования.
Таким образом энергия переносится с транспортом электронов от
цитохрома к цитохрому, которые все отличаются друг от друга.
Такой транспорт электронов не только оставляет позади себя
свободный НАД и Н+, но и высвобождает поэтапно энергию.
Эта энергия, высвобождаемая небольшими порциями, в
присутствии свободного неорганического фосфата и АДФ,
способствует образованию АТФ (как уже было описано). Ионы Н+
легко связываются в митохондриях с наличным кислородом и
образуют Н20 (рис. 22.6,Г).
Таким образом, окислительное фосфорилирование
обеспечивает большую часть ресинтеза АТФ: при использовании одного
атома кислорода на АДФ переносится 3 молекулы фосфата.
При окислении двух молекул пировиноградной кислоты до С02
и Н20 регенерируют приблизительно 30 молекул АТФ. 6
молекул АТФ образуются в дыхательной цепи окисления НАД-Н,
образованного в результате анаэробного гликолиза, и две
молекулы АТФ — при окислении на уровне субстрата (вне
митохондрий) путем превращения глюкозы в пировиноградную
кислоту. Получающиеся в итоге 38 молекул АТФ оказываются
«запасенными» при расщеплении одного моля глюкозы.
Каждая концевая высокоэнергетическая связь АТФ при
разрыве дает 7—7,5 ккал/моль, а высвободившаяся потенциальная
энергия составляет 266—285 ккал/моль глюкозы. Поскольку не-
расщепленная молекула глюкозы содержит 673 ккал/моль, то
конечный ресинтез 38 молекул АТФ означает, что клетка
(главным образом путем окислительного фосфорилирования в
митохондриях) способна на 39—42% (266—285/673-100) вновь
захватить энергию из молекул глюкозы.
ПРОМЕЖУТОЧНЫЙ ОБМЕН
Промежуточный обмен представляет собой поэтапный
метаболизм всех трех типов пищевых веществ. На рис. 22.7
показаны взаимоотношения этих трех основных типов. Ацетил-КоА
играет важную роль общей стадии всех трех путей
катаболизма, хотя выше этого уровня только анаэробный гликолиз созда-
Промежуточный обмен 417
ет АТФ. Высвобождение и связывание энергии происходит
большей частью в переплетающихся реакциях циклов ТКК и
окислительного фосфорилирования, связанных с транспортом
электронов. Ацетил-КоА является необходимым промежуточным
этапом в синтезе жиров и некоторых белков, но не играет
существенной роли в образовании полисахаридов. Потребность в
притоке энергии (в форме АТФ с высвобождением АДФ и Рн)
возникает на начальных стадиях, когда предшественники
(NH3, C02 и Н20) соединяются и образуют промежуточные
продукты метаболизма; затем эти последние становятся
основными структурными блоками (аминокислоты, сахара, жирные
кислоты и глицерин). В отличие от катаболизма анаболическая
последовательность реакций на всех трех главных
анаболических путях (рис. 22.7) требует энергии между всеми уровнями
суперорганизации над уровнем ацетил-КоА. Важно еще раз
подчеркнуть, что анаболический и катаболический пути — это
не просто прямое и обратное течение процесса по одному и
тому же пути. Большая часть реакций не повторяет в обратном
порядке ту же последовательность (предшественников и
образующихся продуктов), и каждая из них подчиняется
независимой тонкой регуляции. Результатом является
координированное катаболическое и анаболическое взаимодействие,
приводящее к высвобождению энергии и тепла, независимо от природы
предшественника или питательного субстрата. Обычно все
пищевые вещества подвергаются превращению одновременно, что
подчеркивает необходимость выхода энергии частыми
небольшими порциями, находящимися под ферментативным контролем.
НУКЛЕИНОВЫЕ КИСЛОТЫ
Метаболизмом клетки в значительной степени управляет
активность ядра, где синтезируются ферменты, регулирующие
отдельные стадии катаболизма и анаболизма. Особенно важен
синтез белков, связанных со структурными элементами клеток
или продуктами секреторной активности некоторых типов
клеток. Например, клетки печени синтезируют только белки,
присущие структуре этих клеток (мембране), или такие продукты
секреторной активности печеночных клеток, как альбумин,
глобулин и протромбин.
Клетки печени и коры надпочечников различаются,
например в отношении «сил», направляющих как синтез ферментов,
так и качество белковых конечных продуктов. Ядро с
содержащейся в нем нуклеиновой кислотой в значительной степени
направляет синтез белка (и, следовательно, фермента),
происходящий в цитоплазме на рибосомах. Без ядра синтез белка почти
немедленно прекращается и может возобновиться, только если
27—1705
418
Глава 22
Углеводы,
липиды, белки
Метаболизм
Структурный белок Новообра^!0^
зованный °^
белок /Секретируемый
+ f-^ ^" / белок
Рис. 22.8. Управление цитоплазматическими механизмами со стороны ядра.
Обратите внимание на «расщепление» ДНК, при котором происходит
комплементарный синтез мРНК на одной нити ДНК.
в клетку вновь будет имплантировано нормальное во всем
остальном ядро. Очевидно, должен существовать «контакт»
между ядром и рибосомой в форме молекул посредника, которые
переносят информацию от ядра к местам синтеза белка. Это
влияние ядра показано на рис. 22.8.
ДЕЗОКСИРИБОНУКЛЕИНОВАЯ КИСЛОТА
Содержащиеся в ядре макромолекулы ДНК несут в своей
структуре закодированную информацию (рис. 22.9). Именно
эта информация диктует, какие ферменты и структурные белки:
с
с^—I—^ с
Рис. 22.9. Типичная двойная спираль молекулы ДНК с последовательностями
оснований. Обратите внимание на то, что азотистые основания связаны с
остатком дезоксирибозы (сахар-С) в молекуле ДНК, а также на то, что
связи А—Т представляют собой двойные водородные связи, а Ц—Г — тройные..
А — аденин, Ц — цитозин, Г — гуанин, Т — тимин.
Промежуточный обмен
419
будут синтезироваться, какой белковый продукт будет секретом
данной клетки и какой наследственный признак (конфигурация,
окраска, размер) будет заложен в память или передан
потомству.
Дезоксирибонуклеиновая кислота (ДНК)—это молекула,
которая состоит из повторяющихся последовательностей 5-С-са-
хара (пентозы, лишенной одного атома кислорода и поэтому
«дезокси»), одной молекулы фосфорной кислоты и
гетероциклического производного пурина или пиримидина. Эти три
субъединицы— пентоза, фосфат и азотистое основание (пурин или
пиримидин)—называются нуклеотидами и повторяются на
протяжении всей длинной цепи молекулы ДНК. Нуклеотиды
соединены друг с другом связями между сахаром и кислородом
сфосфатной группы; ДНК — это, вероятно, самые крупные
молекулы в животных клетках; хотя они сконцентрированы в
клеточном ядре, они не ограничены этой областью. Они очень
стабильны и относятся к «долгоживущим».
Информация, содержащаяся в ДНК, определяется в
совокупности, во-первых, длиной цепи ДНК, во-вторых, типом
азотистого основания, включенного в каждый нуклеотид, и, в-третьих,
положением нуклеотида в последовательности. Каждый
нуклеотид в ДНК может содержать аденин или гуанин (пурины) либо
цитозин или тимин (пиримидины). Длинная цепь каждой
молекулы ДНК в действительности состоит из двух цепей,
переплетающихся друг с другом через правильные интервалы — это
так называемая «двойная спираль» (рис. 22.9). Силами,
соединяющими обе цепи, являются связи между пуринами и пири-
мидинами, включенными в каждую нуклеотидную субъединицу.
В любой молекуле ДНК концентрация пуринов равна
концентрации пиримидинов; количество тимина равно количеству
аденина, а количество гуанина — количеству цитозина. Пурино-
во-пиримидиновое взаимодействие между двумя цепями ДНК,
как полагают, состоит в связях тимин — аденин и гуанин —
цитозин (Т—А и Г—Ц).
Хотя ДНК была открыта еще в 1870 г., но лишь в 1962 г.
Дж. Уотсон и Ф. Крик получили Нобелевскую премию за
раскрытие трехмерной структуры ее молекулы. Их работа была
подтверждена и продолжена, что позволило понять, каким
образом в структуре молекулы ДНК записывается генетическая
информация и каким образом эта информация переводится на
рибосомы для синтеза белков.
РИБОНУКЛЕИНОВАЯ КИСЛОТА
В животных клетках существует несколько типов
рибонуклеиновой кислоты (РНК), и каждый из них выполняет свою
особую функцию в процессе передачи сообщения от ядерной
27*
20
Глава 22
ДНК к рибосомам, где осуществляется синтез белка.
Приблизительно 15% всей РНК находится в митохондриях, около
25%—в цитоплазме, 10%—в ядре и 50%—в рибосомах. По-
химическому составу РНК сходна с ДНК; она содержит
фосфорную кислоту, рибозу и какое-либо из четырех азотистых
оснований. Но в РНК содержится рибоза (а не дезоксирибоза,
как в ДНК). Кроме того, наряду с аденином, гуанином или ци-
тозином в ее нуклеотидной субъединице вместо тимина
содержится урацил. Молекулы РНК состоят из одиночных цепей и
обычно короче молекулы ДНК, а поэтому обладают меньшим
молекулярным весом (соответственно от 2,8-104 до МО6 и от
Ь106 до 5-Ю9). Кроме того, в животных клетках РНК
преобладает (в 2—6 раз) над ДНК; она, по-видимому, быстрее
расщепляется и ресинтезируется, т. е. является короткоживущей.
ИНФОРМАЦИОННАЯ (МАТРИЧНАЯ) РНК
Эта форма РНК (мРНК) синтезируется главным образом в
клеточном ядре (небольшое ее количество возникает в
митохондриях), когда последовательность нуклеотидов одной цепи
хромосомной ДНК транскрибируется в одиночную цепь
молекулы мРНК. Таким образом, она содержит код той же самой
последовательности оснований, но в форме цепи,
комплементарной цепи ДНК, участвующей в этом процессе (рис. 28.8). Затем
мРНК «проскальзывает» через поры ядра в цитоплазму и
переходит в рибосомы, где она служит матрицей для
формирования соответствующей последовательности аминокислот при
синтезе данного белка. Меньше 20% всей РНК относится к мРНК-
ТРАНСПОРТНАЯ РНК
Этот вид РНК (тРНК) раньше назывался растворимой
РНК; он составляет около 15% всей РНК в животных клетках.
Это мелкие структуры, содержащие меньше 90 оснований, и их
действие чрезвычайно специфично. Иными словами, каждая
тРНК рассматривается как переносчик специфической
аминокислоты; тРНК переносит отдельные аминокислоты на рибосомы в
процессе биосинтеза белка, сочетая их антикодоны с кодонами
мРНК. Поскольку идентифицировано больше 50 тРНК, то
очевидно, что данная аминокислота может соединяться с
несколькими тРНК. Известно однако, что каждая из 20 аминокислот,
встречающихся в белке, имеет в качестве переносчика по
меньшей мере одну тРНК (рис. 22.10).
Промежуточный обмен
421
РИБОСОМНАЯ РНК
Хотя эта форма РНК (рРНК) составляет больше 75%—85%
клеточной РНК, ее функция остается неясной. Известны по
меньшей мере четыре разные формы рРНК, и все они тесно
связаны с рибосомными частицами. Возможно, что они принимают
непосредственное участие в конечных стадиях синтеза белка, но,
«Свободная» тРНК
Растущая (ДЛЯ аланина)
полипептидная
цепь
Активированный
аланин,
связанный с тРНК,
для следующего
добавления
мРНК-матрица
Кодон для аланина Область кодона для валина
Движение рибосомы вдоль цепи мРНК
Рис. 22.10. Синтез белка на рибосоме (см. текст). (А — аденин, Ц — цитозин,
Г — гуанин, Н — неопределенное основание, У — урацил).
как именно, пока еще не известно. По-видимому, они не несут
закодированной информации; может быть, они являются
промежуточным звеном между тРНК и мРНК, причем отдельные
аминокислоты расположены в надлежащей последовательности,
продиктованной «посредником».
СИНТЕЗ БЕЛКА
В биосинтезе белка из аминокислот АТФ прямо поставляет
большую часть энергии, которая нужна для активации каждой
аминокислоты перед включением в растущую пептидную цепь
на рибосоме. При биосинтезе нуклеиновых кислот из нуклеоти-
Дов большую долю энергии доставляет АТФ, а также УТФ,
ГТФ, ЦТФ. Эти последние являются богатыми энергией три-
фосфатными формами соответствующих нуклеотидов РНК. Как
Полагают, в биосинтезе полипептидов (и белков) существует
422
Глава 22
определенная последовательность стадий, включающая
генетический код и его транскрипцию и трансляцию на рибосомные
элементы в цитоплазме.
ГЕНЕТИЧЕСКИЙ КОД
Генетическая информация хранится в спиральных
молекулах ДНК в хромосомах. Каждая аминокислотная
последовательность, составляющая определенный белок, кодируется
конкретной последовательностью нуклеотидов в молекуле ДНК.
Термин «ген», собственно говоря, относится к той части цепи
ДНК, которая является специфичной для одной полной
полипептидной цепи. Ген остается в хромосоме, а закодированное
сообщение «транскрибируется» в форме мРНК, которая
переносит инструкцию на рибосомы для трансляции в процессе
биосинтеза.
Три смежных нуклеотида в ДНК обеспечивают кодирование
каждой из 20 аминокислот, участвующих в синтезе белка.
В 1961 —1965 гг. тремя лабораториями — Ниренберга, Очоа и
Корана — были опубликованы сообщения о «словах»
специфического генетического кода для каждой аминокислоты.
Поскольку нуклеотиды ДНК содержат только четыре разных основания,
для управления всем кодированием имеющихся аминокислот
требуются разные последовательности оснований (в триплетах).
Кодирующие триплеты (нуклеотидных оснований ДНК),
перешедшие на мРНК в качестве комплементарных генетических
инструкций, называются кодонами и действуют как
РНК-матрица, которая определяет точное положение и
последовательность аминокислот при синтезе белка (рис. 22.10).
Для передачи генетической информации от одного
организма к другому из поколения в поколение требуется репликация
ДНК; ДНК является единственным веществом, которое для
своего биосинтеза, т. е. для репликации, не нуждается во
«внешней» матрице. Молекула ДНК «воспроизводится» после
разделения цепей двойной спирали путем разрыва связей
между комплементарными основаниями (пуринами и пиримидина-
ми). Таким путем в двух «половинах» ДНК обнажаются
пурины и пиримидины нуклеотидов, которые теперь могут
соединяться с соответствующими доступными трифосфорилированны-
ми основаниями АТФ, ГТФ, ЦТФ и ТТФ. После сочетания
соответствующих оснований (А — Т, Г — Ц) нуклеотиды
оказываются соединенными, а пирофосфат освобождается. Таким
образом, при репликации обе цепи ДНК действуют как матрицы,
тогда как в синтезе мРНК действует только одна цепь.
Реплицированная ДНК содержит исходную цепь и
сочетавшуюся с ней нуклеотидную цепь, при этом образуются две
Промежуточный обмен
423
идентичные спиральные молекулы ДНК. Репликация ДНК
может занять 8—12 ч. Когда приближается деление клетки, две
удвоенные идентичные цепи ДНК уплотняются или тесно
скручиваются и образуют хромосомы. Копии ДНК переходят в
дочерние клетки; каждая из них — это точная копия
генетического кода в исходной родительской клетке (см. гл. 26).
СИНТЕЗ БЕЛКА НА РИБОСОМАХ
Как только на одной из двух цепей ДНК синтезируется
мРНК (в виде молекулы, в которой последовательность нуклео-
тидов в зеркальном порядке повторяет последовательность в
ДНК), она выходит из ядра в цитоплазму и достигает
рибосом. Здесь мРНК, содержащая кодоны для специфических
аминокислотных последовательностей, прикрепляется к субъединице
рРНК- Кроме этой роли «прикрепления» мало что известно о
функции рРНК; она не содержит закодированных данных.
Одновременно свободные аминокислоты в цитозоле активируются
под действием ферментов, причем активация зависит от АТФ;
в результате возникает эфирная связь с определенными тРНК.
Комплекс из активированной аминокислоты и тРНК
связывается с рибосомой особым образом: пары оснований в тРНК (ан-
тикодоны) сочетаются с соответствующими кодонами мРНК
(рис. 22.10). При этом может добавиться только специфическая
аминокислота, соответствующая коду (продиктованная кодом).
Теперь ферменты катализируют образование пептидной связи,
в то время как очередная аминокислота все еще прикреплена к
соответствующей тРНК.
К растущей пептидной цепи с помощью различных тРНК
одна за другой добавляются аминокислоты, начиная от амин-
ного конца цепи; цепь синтезируется и растет по направлению
к карбоксильному концу (рис. 22.10). Для завершения
построения цепи требуется специальный кодон. Окончательно
синтезированная белковая структура теперь отделяется от рибосомы и
переходит в область Гольджи, где происходит «упаковка» и
включение ее в образованные мембранами пузырьки, или же
она может выйти в цитозоль и затем распределиться по клетке.
Во время этого процесса тРНК не меняется, не разрушается и
может быть использована повторно.
Первичной структурой белка считается специфическая
аминокислотная последовательность; вторичная структура — это
спираль полипептидной цепи, закрученная вокруг продольной
оои, а третичная структура — это трехмерный сложенный в
складки белок; как это можно видеть у глобулярных, или
сферических, белков; конечный белковый продукт, отделившийся от
рибосомы, может быть поэтому спиралью со случайной ориен-
424
Глава 22
тацией, свернутой при помощи дисульфидных связей
{—S—S—). Обычно структурные белки клетки бывают
вытянутыми, длинными — фибриллярными; белки же, которые
обладают физиологической активностью — ферменты или
гормоны, — обычно глобулярные.
Некоторые белки соединены с металлами или
органическими реактивными группами. Многообразие биосинтетического
процесса усугубляется тем, что диапазон исследованных белков
охватывает молекулы с мол. весом от 5000 до 1000 000. Синтез
белка в животных клетках происходит и на «свободных»
рибосомах— не прикрепленных к эндоплазматическому ретикулуму
(так синтезируется, например, гемоглобин), но, как было
указано выше, большей частью синтез происходит на
«фиксированной» рибосоме.
ЛИТЕРАТУРА
Ganong W. F. (1979). Review of medical physiology, 8th edn., Lange Medical,
Los Gatos, California.
Gordon M. S. (1977). Animal physiology, 3rd edn., MacMillan, New York.
Harper H. A. (1977). Review of physiological chemistry, 16th edn., Lange
Medical, Los Gatos, California.
Lehninger A. L. (1975). Biochemistry, 2nd edn., Worth, New York. [Имеется
перевод: Ленинджер А. Биохимия. М.: Мир, 1976.]
Vander A. J., Sherman J. U., Luciano D. S. (1980). Human physiology, 3rd edn.,
McGraw-Hill, New York.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Определите, что такое ген.
2. Чем синтез мРНК отличается от репликации ДНК?
3. Из каких компонентов состоят нуклеиновые кислоты, и
какое специальное значение имеют азотистые основания для
генетического кода?
4. Какую специфическую роль в метаболизме пищевых
веществ играет кислород?
5. Достаточно ли эффективно анаэробный метаболизм
поставляет энергию для поддержания жизненных процессов у
млекопитающих? У птиц? У других позвоночных?
6. В чем основное количественное преимущество углеводного
аэробного обмена над анаэробным (или наоборот)?
7. Какое промежуточное вещество в метаболизме является
центральным для всех форм субстратов?
8. Как ферменты взаимодействуют с соединениями,
участвующими в биологической реакции? При каких условиях?
9. Какое отношение удаление Н+ имеет к повторному
присоединению высокоэнергетического фосфата?
10. Что служит источником энергии для образования связей
между основаниями в структуре ДНК?
Глава 23
ПИТАНИЕ
Питательные вещества делятся на углеводы, жиры, белки,
минеральные вещества, витамины и (последнюю по порядку, но
не по значению) воду. Организму необходимы энергия,
некоторые аминокислоты, витамины и минеральные соли.
Теоретически источниками энергии могут служить углеводы, жиры, белки;
последние в том случае, если их потребление превысило
потребность организма в белке. Однако при такой «свободе выбора»
существуют некоторые ограничения. 1) Совершенно лишенная
жира пища неприятна на вкус, а очень высокое содержание
жира многие люди переносят плохо. 2) Белок в качестве главного
источника энергии имеет тот недостаток, что нуждается в
эффективном механизме для удаления избыточного азота. 3) Что
касается питательности, то небольшие количества углеводов
(около 50 г глюкозы в сутки) и жиров требуются для
получения незаменимых жирных кислот (в особенности линолевой)*
4) Углеводы вообще являются самым дешевым источником
энергии. Исходя из изложенного ясно, что организм человека
черпает энергию из всех трех типов питательных веществ.
Подробнее об образовании энергии сказано в гл. 18 и 22.
Источником энергии может служить также спирт и
некоторые синтетические углеводороды. Но первый считается вредным
для здоровья, а о вторых пока еще мало известно.
ЖИРЫ, УГЛЕВОДЫ И БЕЛКИ
При расщеплении углеводов и белков выделяется
приблизительно 4 ккал/г, жиров — в 2*Д раза больше. Поэтому человек,
получающий 40% калорий за счет жиров, потребляет рацион,
содержащий 23% жира, если не считать некалорийные
питательные вещества (минеральные соли и воду). Энергетические
потребности взрослых людей варьируют от 1800 до 3000 ккал
при умеренной активности. Около двух третей этого количества
покрывают основную потребность организма в энергии. При
тяжелой работе или занятиях спортом потребность в калориях
может заметно возрасти.
н он
н-с-он
I
но-с-н о
н-с-он
1
н-с
I
Н2СОН
А
СН2ОН
СН2ОН
СН2ОН
сн2он
сн2—о—с--(сн2)7—сн=сн—сн2—сн=сн—(сн2)5н
СН—О—С—(СН2)7—СН=
1 /°
СН2—О—С^-(СН2)17 Н
сн2—сн—соон
1 1
OH NH2
Е
=сн-
-(СН2)8 Н
д
О
. СН2 —СН—СООН
н ж
Рис. 23.1. Химическое строение отдельных пищевых веществ. Разные
изображения глюкозы (a-D-глюкопиранозы). Л. По Фишеру; Б. По Ховорту.
В. «Стул», стереохимически наиболее точное изображение. Г. Сахароза (1-а-
D-глюкопиранозид-Р-фруктофуранозид). Д. Триглицерид (линео-олео-стеарин).
Е. Серии, гидроксизамещениая алифатическая аминокислота. Ж. Триптофан,
ароматическая аминокислота.
426
Питание
427
Главным источником углеводов служит крахмал злаков —
пшеницы, кукурузы и риса, а также картофеля. Фрукты и
овощи содержат крахмал, а также сахар, преимущественно диса-
хариды — сахарозу (рис. 23.1) и мальтозу, а также
моносахариды— глюкозу и фруктозу. Молоко является источником ди-
сахарида лактозы. В некоторых экономически более развитых
странах до одной трети углеводов потребляются в виде
сахарозы. В последнее время основной группой углеводов в пищевом
рационе американцев вместо злаков стали сахара (табл. 23.1).
Большую часть потребляемых с пищей жиров составляют
триглицериды — соединения, в которых молекула глицерина
связана с тремя жирными кислотами (рис. 23.1). Кислоты эти
бывают с цепью разной длины и разной степени
ненасыщенности. Фосфолипиды как источник липидных калорий играют
второстепенную роль. Как и углеводы, жиры могут
синтезироваться iB организме, за исключением полиненасыщенной лино-
левой кислоты, которая является предшественницей арахидоно-
вой кислоты и необходима для синтеза простагландинов. Лино-
левая кислота должна давать 1—2% всех калорий —
количество, легко покрываемое средним рационом американцев.
Таблица 23.1. Содержание основных пищевых веществ в продуктах3*,
поставляемых гражданскому населению США
Мясо, птица, рыба
Яйца
Молочные продукты
(кроме масла)
Жиры, растит, масло,
слив, масло
Фрукты
Картофель (включая
батат)
Овощи
Сухие бобы, горох,
орехи
Мучные продукты,
каши
Сахар, сладости
Разное
Энергия
,б)
18,1
2,6
13,5
16,0
3,3
3,0
2,7
2,9
21,4
15,8
0,8
и
20,0
1,8
11,1
18,1
1 3,0
2,9
2,7
3,1
19,2 1
17,3
0,7
Белки
I
35,7
6,8
24,4
0,2
1,2
2,5
3,8
5,2
19,9
в)
0,4
и
42,6
4,8
22,0
0,2
1,1
2,3
3,6
5,4
17,6
в)
0,4
Жиры
I
32,9
4,0
16,6
39,5
0,4
0,1
0,4
3,3
1,6
0
1,4
и
34,1
2,7
12,5
43,3
0,4
0,1
0,4
3,8
1,3
0
1,2
Углеводы
I
0,1
0,1
7,7
а)
7,1
5,5
4,7
2,3
37,6
34,2
0,7
.11
0,1
0,1
6,7
в)
6,6
5,4
5,3
2,1
34,7
38,5
0,5
В процентах от среднего общего потребления продукта. По данным National Food
lew, USDA, 1978.
Период исследования: 1 — 1957—1959 гг. (средние данные); II — 1977 г.
В) Менее 0.05%.
428
Глава 23
Рекомендуемую норму белка могут удовлетворить разные
продукты. Реально существует потребность в небольшом числе
незаменимых аминокислот, которые не синтезируются
организмом или синтезируются в недостаточном количестве (табл. 23.2).
Таблица 23.2. Главные аминокислоты, встречающиеся в белках
Аминокислоты
I. Алифатические
Глицин
Серии
Алании
Треонина)
Цистеинб)
Аспарагиновая кислота
Валина>
Метионина>
Глутаминовая кислота
Лейцина>
Изолейцина)
Аргинин
Лизина)
Гидроксилизин6*
II. Неалифатические
Пролин
Гидроксипролин
Гистидина)
Фенилаланина)
Тирозинб)
Триптофана)
Число
атомов С
2
3
3
4
3
4
5
1 5
5
6
1 6
6
6
6
5
5
6
9 I
9
11
Отличительные признаки
Гидроксильная группа (ОН)
То же
Содержит серу (SH)
Кислая
Боковая цепь (СН3)
Содержит серу (S)
Кислая
Боковая цепь (СН3)
То же
Основная
»
»
Иминокислота (без
аминогруппы)
То же
Основная
Ароматическое ядро
ОН-фенилаланин,
ароматическое ядро
Ароматическое ядро
Незаменимые аминокислоты, не синтезирующиеся в организме; гистидин,
вероятно, не является незаменимым для взрослых.
Организм может синтезировать цистеин только из метионина, гидроксилизин — из
лизина и тирозин — из фенилаланина.
Качество белка в значительной степени определяется долей
этих аминокислот в общем белковом рационе. В продуктах
животного происхождения — мясе, рыбе, яйцах и молоке — состар
аминокислот ближе соответствует потребностям человека.
Впрочем, правильное сочетание источников растительного
белка может дать смесь аминокислот, сходную с той, какую дает
мясо. Немалое число строгих вегетарианцев говорят об
успешном использовании белка из растительных источников.
Предпочтение, отдаваемое бифштексу перед блюдом из чечевицы,
объясняется скорее свойствами жиров, содержащихся в мясе,
нежели входящими в него белками.
Питание\
429
МИНЕРАЛЬНЫЕ СОЛИ И ВИТАМИНЫ
Главным хранилищем минеральных веществ в организме
является скелет (см. гл. 1). В кости содержится большая часть
кальция, фосфора и магния. Но эти и другие элементы играют
также важную роль как кофакторы разных ферментативных
реакций в организме в поддержании постоянства внутренней
среды (осмотического давления и кислотно-щелочного
равновесия) и в нервно-мышечной активности (см. гл. 22). Железо
играет особую роль как компонент гемоглобина, а иод как
неотъемлемая часть гормонов щитовидной железы.
Необходимость в некоторых микроэлементах стала очевидной только
недавно, и, возможно, найдутся еще и другие незаменимые
элементы, хотя и в очень малых количествах.
Смешанный рацион, содержащий достаточное количество
энергии, белка и витаминов, должен также включать нужные
минеральные вещества. Чаще всего в предельно малых или в
недостаточных количествах в пище содержатся железо и
кальций. В некоторых районах США во избежание развития у
населения заболевания щитовидной железы (зоба) в столовую соль
приходится добавлять иод. В табл. 23.3 перечислены
минеральные вещества и микроэлементы, которые считаются
необходимыми, а также указана их установленная или предполагаемая
функция.
Как теперь определено, витамины являются добавочными
факторами, необходимыми для жизни отдельных видов
животных, в организме которых они не синтезируются; они не
поставляют энергии и не становятся постоянной частью организма.
Восемь витаминов группы В полностью отвечают этому
определению. Они действуют в анаболических и катаболических
ферментативных системах, большинство которых подробно
изучено (см. гл. 22). Они растворимы в воде, так же как холин и
аскорбиновая кислота. Холин, собственно говоря, становится
частью организма; так, например, фосфатидилхолины
(называемые также лецитинами) являются структурными
компонентами клеток, и в первую очередь клеток нервной системы.
Включение холина в описок витаминов объясняется главным
образом тем, что он не подходит ни к какой другой категории,
и там, где он не синтезируется организмом, требуется лишь в
сравнительно малых количествах, но больших, чем любые
другие витамины. Хотя аскорбиновая кислота полностью
соответствует только что упомянутому определению витаминов и хотя
хорошо известно, каким образом сказывается ее дефицит,
механизм ее действия точно еще не установлен.
За последние годы стало известно многое о функции
четырех жирорастворимых витаминов—А, Д, Е и К, но чтобы по-
Таблица 23.3. Незаменимые факторы питания человека: минеральные
веществаа)
Название
Кальций (Са)
Фосфор (Р)
Магний (Mg)
Натрий (Na)
Хлор (С1)
Калий (К)
Железо (Fe)
Медь (Си) ,
Марганец (Мп)
Цинк (Zn)
Иод (I)
Молибден (Мо)
Хром (Сг)
Основные функции
Содержится в костях
и зубах, участвует в
реакциях нервной
системы, кофактор
ферментов
Содержится в костях
и зубах, участвует в
промежуточном
обмене
Содержится в костях,
участвует в
реакциях нервной системы,
кофактор ферментов
Поддержание
осмотического равновесия
и объема жидкости
Поддержание уровня
жидкости и
электролитов
Ферментативная
функция в клетке
Содержится в
гемоглобине,
дыхательных ферментах
Кофактор ферментов
(цитохром-с-окси-
даза)д>
Кофактор ферментов
содержится в кости,
участвует в
процессах воспроизведения
Кофактор ферментов
(карбоангидраза)д)
Синтез гормона
щитовидной железы
Кофактор ферментов
(ксантиноксидаза)А)
Регуляция
метаболизОсновные источники
Молочные
продукты, овощи —
зеленые листья
Молочные
продукты, злаки, мясо
Целые зерна, мясо,
молоко
Столовая сольв)
То же
Овощи, мясо,
сухофрукты, орехи
Мясо, печень,
бобы, орехи,
сухофрукты
Optixn, печень,
почки, сухие овощи,
виноград
Орехи, целые зерна
Крабы, мясо,
бобы, яичный
желток
Столовая соль с
добавлением
иода
Почки, некоторые
злаки и овощи
Ограниченные све-
ма (фактор толе- дения |
рантности к глюкозе))
1
Симптомi»i
дефицита б)
Кальциевая
тетания
Цеминерализация
костей
Анорексия,
тошнота,
неврологические симптомы
Слабость, апатия*
подергивание
мышц
_ г)
Слабость,
летаргия, гипорефлек-
сия
Анемия
Анемия, нейтропе-
ния, дефекты
скелета
_ г)
Отставание в
росте, задержка
полового созревания
Зоб
_ г)
_ г)
а' Потребность человека в следующих микроэлементах, возможно, существует, но
точно не установлена: селений (Se), фтор (F), кремний (Si), никель (Ni), ванадий (V)
и олово (Sn). Потребность в сере (S) удовлетворяется метионином и цистином, а
потребность в кобальте (Со) — витамином Bj2.
б' За исключением Са, Fe и I, дефицит в рационе у человека или маловероятен,
или встречается редко.
в' Многие подвергаемые обработке пищевые продукты содержат значительные
количества хлористого натрия.
г)
' У человека не обнаружено каких-либо определенных синдромов дефицита.
д' Примеры активности в качестве кофакторов ферментов.
Питание^ 431
нять, как именно они действуют на молекулярном уровне,
нужны дальнейшие исследования. ч\
Краткий обзор витаминов дан в табл. 2&4.
ДЕФИЦИТ И ИЗБЫТОК \
Нарушения, вызываемые полным отсутствием или
недостатком определенного пищевого вещества, в общем хорошо
установлены; но вредным может оказаться и его избыточное
потребление. Излишнее поглощение энергии приводит к
ожирению. В некоторых частях света, в том числе в США и Западной
Европе, тучность чисто эстетически воспринимается как
отрицательное явление; важнее, однако, то обстоятельство, что, как
показано, ожирение коррелирует с высокой частотой различных
заболеваний. Избыточное потребление насыщенных жиров и
холестерина положительно коррелирует с заболеваниями
сердечно-сосудистой системы.
Токсичность минеральных веществ и микроэлементов в
большой степени зависит от того, насколько организм способен
их выводить. Среди микроэлементов, нужных человеческому
организму, к наиболее токсичным относят медь, кобальт и фтор.
Синдромы гипервитаминоза описаны для витаминов A, D и в
меньшей степени для никотиновой кислоты; симптомы
чрезмерного потребления описаны также и для некоторых других
витаминов. Хотя избыточное поглощение калорий — это
распространенное явление, но маловероятно, чтобы человек,
потребляющий обычную пищу в разумных или даже слегка излишних
количествах, страдал от последствий избытка минеральных
веществ или витаминов. Такой избыток обычно возникает из-за
нарушений обмена, непривычной пищи или приема
концентрированных источников витаминов и минеральных веществ в дозах,
в несколько раз превышающих нормальные. В качестве
примеров можно указать: на токсичность нормальных доз
витамина D, связанную с тем, что в организме человека не происходит
катаболизма этого витамина; на развитие миопатии под
влиянием пива, к которому для получения избытка пены добавляют
вещество, содержащее кобальт; на токсичность больших доз
витамина Л, который назначают по поводу угрей.
В руководствах по питанию обычно описывают довольно
подробно симптомы недостаточности витаминов. Так, например,
в качестве основных симптомов недостаточности тиамина у
крыс указывают потерю аппетита, потерю веса, судороги,
замедление ритма сердца, понижение температуры тела;
симптомами недостаточности пантотеновой кислоты у человека
являются: рвота, слабость, боли в животе, мышечные судороги,
болезненность пяток, утомляемость и бессонница. Совершенно
Таблица 23.4. Незаменимые факторы питания человека: витамины
Название
Основные функции
Основные источники
а)
Симптомы дефицита*'
Ретинол (А)
Кальциферол (D)
Токоферолы (Е)
Витамин К
Незаменимые жир
ные кислоты
Тиамин (ВО
Рибофлавин (В2)
Никотиновая
кислота (ниацин)
Входит в состав зрительного
пурпура, поддерживает целостность
эпителиальных тканей
Участвует в процессе всасывания Са
и Р; образование костей и зубов
Антиоксидант
Участвует в синтезе факторов
свертывания крови
Синтез простагландинов
Энергетический метаболизм — декар-
боксилирование
Перенос водорода и электронов
(ФМН, ФАД)
Перенос водорода и электронов
(НАД, НАДФ)
Яичный желток, рыбий жир, кароти-
ноиды
Рыбий жир, печенка, облученные сте-
риныв)
Растительные масла, зеленые листья
овощей
Зеленые листья овощей,
бактериальный синтез
Ненасыщенные масла с большим
содержанием линолевой кислоты
Целые зерна, потроха
Целые зерна, молоко, яйца, печень
Дрожжи, мясо, печеньд>
кКуриная слепота»,
повреждения роговицы и кожи
Рахит, остеомаляция
У животных: мышечная
дегенерация, бесплодие,
повреждения мозга, отекг)
Замедление свертывания
крови
Дерматит, нарушение
транспорта жировг>
Бери-бери, полиневрит
Хейлоз, глоссит, светобоязнь
Пеллагра
/
g Пиридоксин (Вб) Метаболизм аминокислот
Пантотеновая
кислота
Биотин
Фолиевая кислота
Цианокобаламин
(В12)
Аскорбиновая
кислота
Холин
Перенос ацетильной группы (КоА)
Перенос С02
Перенос одного углерода
Одноуглеродный синтез;
молекулярная перестройка
Гидроксилирование, синтез
коллагена
Транспорт жиров, синтез фосфолипи-
дов
Целые зерна, дрожжи, печень
Широко распространена
Яйца, печень; бактериальный синтез
Зеленые листья овощей, мясо
Продукты животного происхождения,
особенно печень; бактериальный
синтез
Цитрусовые, картофель, перец
Продукты животного происхождения,
а также синтетические
Судороги, повышенная
раздражимость
Нейромоторные и
желудочно-кишечные нарушения
Себорейный дерматит
Анемия, спру
Пернициозная анемия
Цинга
— г)
а' Большинство витаминов, особенно из группы В, содержатся во множестве пищевых продуктов и во всех клетках тела.
б' При недостатке некоторых витаминов возникает множество симптомов: дефицит витаминов иногда имеет множественную природу, и
симптомы, сходные с описанными, могут возникнуть не в связи с питанием.
в' Некоторые каротины, содержащиеся в зеленых и желтых овощах, являются предшественниками витамина Л. Некоторые стерины,
включая 7-дегидрохолестерин, синтезируемый в организме, служат предшественниками витамина D.
г' У человека не описано четко определенного синдрома.
д' Ниацин является одним из конечных продуктов нормального метаболизма триптофана.
434
Глава 23"
ясно, что большая часть этих симптомов может быть
следствием множества других причин, а не только недостаточности
этого витамина. Установить здесь причинную связь с
недостаточностью пантотеновой кислоты у человека — дело далеко не
легкое: пришлось бы применить такие крайние средства, как прием
антивитаминов.
Несмотря на эти факты, описание симштомов
недостаточности может создать впечатление, во всяком случае у широкой
публики, что, какова бы ни была их причина, их можно снять
усиленным потреблением соответствующих витаминов.
Положительная реакция на такое лечение в отсутствие настоящего
дефицита скорее означала бы действие плацебо
(воображаемый эффект), чем действительное устранение гиповитаминоза.
ПИТАНИЕ НАСЕЛЕНИЯ
Первое широкое обследование питания населения в США
было предпринято в 1967 г. в десяти штатах и охватило 70 000
человек из бедных слоев населения. Были обнаружены анемия,
болезни зубов, отставание в росте и случаи недостаточности
витаминов А, С, D и белка. Причиной анемии могут быть
разные виды алиментарной недостаточности, но чаще всего — это
результат недостатка железа. Наблюдаемые симптомы
дефицита витаминов в большинстве случаев ограничивались низким
уровнем данного витамина в крови. Точно так же часто
встречались случаи низкого уровня белка в плазме крови, тогда как
клинические симптомы недостатка белков в пище были
сравнительно редкими. Другие обследования показывают, что кальций
часто потребляется в количествах ниже рекомендуемого уровня.
Однако некоторые исследователи считают этот уровень
завышенным, и, за исключением крайних случаев, трудно связать
низкое потребление кальция с подлинным синдромом его
дефицита. Можно думать, что явления алиментарной
недостаточности, описанные у представителей малоимущих слоев,
встречаются также, хотя и реже, в других группах населения США.
В глобальном масштабе, недоедание распространено очень
широко, и клинические симптомы, связанные с неполноценным
питанием, встречаются часто. Самой серьезной проблемой
считается белково-калориевая недостаточность, которая в наиболее
тяжелой форме называется квашиоркором и маразмом. Другие
часто встречающиеся нарушения — это анемия и слепота —
последняя как результат дефицита витамина А. Описываются
также случаи рахита (недостаток витамина D), цинги
(недостаток витамина С), бери-бери и пеллагры (недостаток тиамина и
ниацина) и симптомы недостатка рибофлавина. Эту проблему
Питание
435
можно обобщить как проблему недостатка пищи, ее выбора и
информации.
Население США и других развитых стран не страдает от
широко распространенной алиментарной недостаточности;
здесь нередки симптомы переедания, которое, как полагают,
ведет к разным хроническим болезням. Хотя случаи употребления
токсичных доз некоторых витаминов встречаются нечасто,
избыточное потребление калорий и соли широко распространено.
Сердечно-сосудистые заболевания, сахарный диабет и
поражение желчного пузыря — таковы некоторые из тех хронических
болезней, которые, как считают, связаны с перееданием.
Излишнее потребление соли, по-видимому, способствует развитию
гипертонии.
ПОТРЕБНОСТЬ В ПИЩЕВЫХ ВЕЩЕСТВАХ
И РЕКОМЕНДУЕМЫЕ СУТОЧНЫЕ НОРМЫ
Отдельные животные в группе, которая получает
достаточно пищи, удовлетворяющей все потребности, растут
неодинаково. Это можно объяснить индивидуальными различиями их
генетически обусловленной способности к росту. Если пищу
видоизменяют, слегка ограничивая один или несколько
ингредиентов, то члены группы тоже растут ло-разному, что говорит о
разной способности использовать данное пищевое вещество и,
следовательно, о разной потребности в нем для оптимальной
активности. Если кормить животных, исходя из среднего
уровня потребностей, то половина животных получит достаточно
питательных веществ, а другая половина получит их меньше, чем
нужно для сохранения оптимального состояния.
Рекомендуемые нормы питания (РНП) не означают средних
■потребностей человека. Это те уровни потребления основных
питательных веществ, относительно которых Управление
питания при Национальном исследовательском совете считает на
основании имеющихся научных данных, что они удовлетворяют
известным потребностям «в пище практически всех здоровых
людей. Разумеется, РНП для разных веществ должны быть
значительно выше, чем действительные средние потребности
людей данного возраста, веса и пола. Единственное исключение
составляет рекомендуемая норма энергии, которая установлена
на самом низком значении, соответствующем, как полагают,
здоровому состоянию средних представителей каждой
возрастной группы. В табл. 23.5 приведены РНП некоторых
питательных веществ для молодых взрослых женщин и мужчин (для
Других возрастов см. публикацию Национальной Академии наук,
приведенную в списке литературы в конце настоящей главы).
28*
436
Глава 23
Таблица 23.5. Рекомендуемая суточная норма некоторых питательных
вешества)
Питательное вещество
Энергия
Белок
Витамин А
Витамин D
Витамин Е
Аскорбиновая кислота
Тиамин (Bi)
Рибофлавин (В2)
Пиридоксин (В6)
Ниацин
Фолацин
Кобаламин (Bj2)
Кальций
Фосфор
Магний
Железо
Цинк
Иод
Единицы
ккал
г
мкг Р.Э.В>
мкгг)
мкг Т. Э.д>
мг
мг
мг
мг
мг Н. Э.е>
мкг
мкг
мг
мг
мг
мг
мг
мкг
1
Для мужчин '
2900
56
1000
7,5
10
60
1,5
1,7
2,2
19
400
3
800
800
350
10 ;
15
150
Для женщин6'
2100
44
800
7,5
8
60
1 1
1,3
2,0
14
400
3
800
800
300
18
15
150
Питательные вещества, для которых Управление питания при Национальном
исследовательском совете Национальной Академии наук определило рекомендуемые
суточные нормы (РНГГ); список признанных безвредными и соответствующими потребностям
суточных норм для витаминов и минеральных солей. «Рекомендуемые суточные нормы»
Национальной Академии наук (Recommended Daily Allowances, National Academy of
Sciences, 1980, 9-е изд.).
Возраст мужчин от 19 до 22 лет, вес 70 кг, рост 177 см; возраст женщин от 19
до 22 лет, вес 55 кг, рост 163 см; для других возрастов см. «Рекомендуемые суточные
нормы».
в)
' Р. Э. — ретиноловый эквивалент; 1 Р. Э. = 1 мкг ретинола (спирта витамина А)
или 6 мкг бета-каротина.
г)
' В виде холекальциферола; 10 мкг холекальциферола=400 ед. витамина D.
д{ Альфа-токофероловые эквиваленты; 1 мг d-a-токоферола^ Т. Э.
1 Н. Э. (ниациновый эквивалент) равен 1 мг ниацина или 60 мг пищевого
триптофана.
ОБЩИЕ СООБРАЖЕНИЯ
В 1977 г. Сенатский комитет по питанию опубликовал
рекомендации для населения США. Комитет рекомендует
американцам потреблять меньше жиров, особенно насыщенных,
меньше холестерина, сахара и соли, а также повысить потребление
овощей, фруктов, злаков и ненасыщенных масел.
Теперь американцы получают 42% своих калорий из жиров
и 20% из сахара. Общее потребление калорий растет
(табл. 23.6). Среднее потребление холестерина превышает
500 мг, а соль потребляется за сутки в количестве 6—18 г. Но
природа потребляемых жиров за последние годы изменилась:
Питание
437
Таблица 23.6. Среднее суточное потребление основных питательных веществ
гражданским населением СШАа)
Питательное вещество
Энергия, ккал
Белки, г
Жиры, г
Углеводы, г
1957—1959 гг., в среднем
3140
95(12)6)
143(41)
375(47)
1977 г.
3380
103(12)
159(42)
391(46)
а) По National Food Review, USDA, январь, 1978.
Цифра в скобках — количество питательного вещества в процентах к общему
потреблению калорий.
сливочного масла и свиного сала едят меньше, а использование
менее насыщенных масел, применяемых для салатов и
приготовления пищи, резко возросло. Эта качественная перемена
привела также к значительному сокращению потребления
холестерина за последние 40 лет. Некоторые ученые считают, что
снижение смертности от ишемической болезни сердца,
наметившееся в конце 60-х годов, может быть связано с этими
изменениями в характере потребляемых жиров и что возможно еще
дальнейшее улучшение. Однако другие исследователи критикуют
«пищевые программы» и считают, что они разработаны без
достаточных оснований и не опираются на современную науку.
Критика подчеркивает также отсутствие объективных
доказательств того, что следование этим программам действительно
принесет населению пользу. Хотя такую критику нельзя
полностью игнорировать, все же взгляды, содержащиеся в «пище-
мых программах», представляют собой лучшие из возможных
в настоящее время советов. Как и все рекомендуемые нормы
питания, эти советы будут изменены и улучшены, когда будут
получены новые данные.
ЛИТЕРАТУРА
Baker И., Frank О. (1968). Clinical vitaminology, Wiley, New York.
Coodhart R. S., Shils M. E. (eds.) (1979). Modern nutrition in health and disease,
6th edn., Lea and Febiger, Philadelphia.
Harper A. E. (1978). Dietary goals —a sceptical view, Amer. J. Clin. Nutr., 31.
310.
Hegsted D. M. (1978). Dietary goals — a progressive view, Amer. J. Clin. Nutr.,
31, 1504.
Mitchell H. S., Rynbergen H. J., Anderson L., Dibble AT. V. (1976). Nutrition in
health and disease, 16th edn., Lippincott, Philadelphia.
The Nutrition Foundation, Inc. (1976). Present knowledge in nutrition, The
Nutrition Foundation, Inc., Washington.
Recommended Dietary Allowances (1980). 9th revised, Food and Nutrition Board,
National Research Council, National Academy of Sciences,
Washington.
438
Глава 23
Select Committee on Nutrition and Human Needs (1977). U. S. Senate. Dietary
goals for the United States, 2nd edn., U. S. Government Printing Office,
Washington.
Underwood E. J. (1977). Trace elements in human and animal nutrition, 4th edn.,
Academic, New York.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Энергия может быть получена из углеводов, белков и
жиров. Чем ограничено использование этих питательных
веществ в качестве источников энергии для человека?
2. Какие моно- и дисахариды имеют значение для питания?
3. Какие аминокислоты являются незаменимыми в питании
человека?
4. Какие аминокислоты могут быть синтезированы только из
определенной незаменимой аминокислоты, а не из
аминокислот, которые не являются незаменимыми?
5. Назовите минеральные вещества, необходимые человеку, и
укажите по одной основной функции каждого из них.
6. Какие условия могут вызывать у человека гипервитаминоз?
7. Почему опасно описывать симптомы недостатка
питательных веществ в публикациях, которые читают люди, не
обладающие научными знаниями?
8. Дефицит каких основных питательных веществ существует
в США и других странах?
9. Что такое РНП, и чем они отличаются от действительных
потребностей?
10. Каковы основные рекомендации Сенатского комитета по
питанию?
11. Как изменилось потребление некоторых питательных
веществ в США за последние 20 лет?
12. Если 40% калорий, получаемых данной группой населения,
поступает из жиров, каков процент жиров, потребляемых
этой группой (без учета некалорийных питательных
веществ— минеральных солей и воды)?
Глава 24
ПОЧКА
Чтобы объяснить и выделить роль почек как жизненно
важных органов, нужно указать на несколько обстоятельств.
Прежде всего они получают около 20—25% всей выходящей из
сердца крови — больше крови на единицу веса, чем любой другой
из основных органов тела. Путем образования мочи почки
1) удаляют из плазмы конечные (или побочные) продукты
обмена, такие, как мочевина; 2) контролируют во всем организме
и в плазме уровни разных электролитов — натрия, калия,
хлора, кальция и магния; 3) способствуют регуляции рН
организма (устанавливая уровень бикарбонатов в плазме и выводя
кислую мочу). Они также контролируют количество воды в
плазме и других областях тела и этим поддерживают
постоянство внутренней жидкой среды. Кроме того, почки
продуцируют два вида гормонов, ренин и простагландины, которые
воздействуют на клетки и изменяют физиологические процессы во
всем организме.
ФУНКЦИОНАЛЬНАЯ АНАТОМИЯ
Почки представляют собой пару бобовидных органов,
лежащих в брюшной полости (рис. 24.1,Л). У человека каждая
почка весит около 150 г. В брюшной полости почки окружены и
поддерживаются соединительной тканью, называемой почечной
фасцией, и жировой тканью. На поперечном разрезе
(рис. 24.1, Б) видны их основные анатомические особенности.
Вдоль медиального края почки идет углубление, так
называемые ворота почки. Через них проходят главные кровеносные
сосуды, почечные нервы, лимфатические сосуды и мочеточник.
Наружную часть почки составляет гладкий,
красновато-коричневый корковый слой, или кора. Он по виду заметно отличается от
беловато-серого мозгового вещества, которое тянется от
внутренней стороны коры к воротам почки. Функции этих двух
слоев, мозгового и коркового, различны.
В мозговой области лежат конические структуры,
называемые почечными пирамидами. Верхушки пирамид, или сосочки,
направленные в сторону ворот, окружены малыми чашечками,
440
Глава 24
которые объединяются в большие чашечки, в свою очередь
образующие почечную лоханку. Мочеточник каждой почки
начинается в лоханке, выходит из почки через ворота и идет через
брюшную полость к мочевому пузырю. Колонки
красновато-коричневой корковой ткани, которые лежат между пирамидами и
проникают в мозговую часть, называются бертиниевыми колон-
ками.
Гистологическими исследованиями установлено, что почка
состоит приблизительно из 1,25 млн. единиц, называемых
Рис. 24.1. А. Анатомические соотношения между почками, мочеточниками и
мочевым пузырем в брюшной полости. Б. Поперечный разрез через почку.
нефронами. Совместная деятельность всех нефронов составляет
общую функцию почки. Перейдем теперь к процессам,
протекающим в одном нефроне (рис. 24.2).
НЕФРОН
Каждый нефрон состоит из двух главных частей:
кровеносных сосудов и почечного канальца. Через каналец протекает та
жидкость, которая затем превращается в мочу. Первую
структуру нефрона составляет клубочек — мелкая сеть, или
скопление капилляров, образованных из афферентной артериолы.
Каждый клубочек почти полностью заключен в структуру,
названную боуменовой капсулой. В общем виде ее описывают как
вдавление слепого конца почечного канальца; функция ее
состоит в том, чтобы 1) собирать жидкость и молекулы твердых
веществ, проходящие через клубочковые капилляры, и 2)
направлять жидкость в систему, называемую почечным каналь-
Почка
441
цем. Сочетание клубочка и окружающей его боуменовой
капсулы представляет собой почечное тельце.
Почечный каналец обладает несколькими основными
свойствами. Недалеко от своего начала у почечного тельца он
образует ряд изгибав и летель. Эта его часть называется прокси-
Клубочек
Афферентная
артериола
Боуменова капсула
Корковый нефрон
Проксимальный
извитой каналец
Эфферентная
артериола
Около -
канальцевые
капилляры
Дистальный
извитой <N
каналец \
Моча
Моча
Рис. 24.2. Нефрон. Показаны клубочковые капилляры в боуменовой капсуле
и околоканальцевые капилляры. (С изменениями по Smith H., 1951. The
Kidney.)
мальным извитым канальцем. Далее каналец выпрямляется и
приобретает и-обрааную форму, образуя так называемую
петлю Генле. Та часть, которая идет от извитого отрезка до
середины петли, составляет нисходящее колено, а та, которая идет
от дна петли параллельно нисходящему колену, составляет
восходящее колено. Далее следует новый ряд изгибов и петель,
так называемый дистальный извитой каналец. Наконец, ди-
Афферентная
артериола
Эфферентная
артериола
Эфферентная
артериола
Юкстамедуллярный
клубочек
03
О.
О
3"
CD
Корковый
клубочек
Капсула
Сплетение
капилляров
в коре
Дуговые
артерия
и вена
Прямые
артериолы
Прямые
венулы
Прямые
канальцы
Петля Генле
Собирательная
трубочка
Рис. 24.3. Vasa recta, капилляры, проникающие в мозговое вещество и
окружающие петлю Генле и собирательную трубочку.
442
Почка
443
Клубочек
Боуменова капсула
Фильтрация
Проксимальный
извитой каналец
Реабсорбция <
Околоканальцевые
кровеносные сосуды
Дистальный
извитой
каналец
Секреция
Рис. 24.4. Нефрон; показаны процессы фильтрации, реабсорбции и секреции.
стальные извитые канальцы от нескольких нефронов
объединяются в собирательные трубки, которые сходятся в еще более
крупные собирательные протоки. Почечный каналец
представляет собой непрерывную структуру, идущую от боуменовой
капсулы до собирательных протоков.
Чтобы понять функциональное значение разных частей неф-
рона, надо представить себе его положение и ориентацию
внутри почки. Клубочек, боуменова капсула, проксимальный и
дистальный извитые канальцы и вторичная капиллярная сеть — все
444
Глава 24
они находятся преимущественно в наружных, корковых,
областях почки (рис. 24.2 и 24.3). Петли Генле и собирательные
протоки лежат в мозговом веществе. Небольшая доля всех нефро-
нов расположена глубже в коре. Из-за их положения эти неф-
роны обозначаются как юкстамедуллярные. Разные части
коркового нефрона, включая приносящую и выносящую артериолы,
проксимальный и дистальный канальцы, иннервируются
симпатическими нервными волокнами, которые также проникают в
корковые области по бертиниевым колонкам.
КРОВОСНАБЖЕНИЕ
Каждый нефрон содержит также сосудистый компонент.
После клубочковой капиллярной сети эти мелкие сосуды
соединяются (сливаются) и образуют выносящую артериолу, которая
выходит из почечного тельца (рис. 24.2 и 24.3). В дальнейшем
выносящая артериола разветвляется и образует вторичную
капиллярную сеть, или околоканальцевую капиллярную систему,
оплетающую проксимальные и дистальные извитые канальцы.
Почка представляет собой одно из немногих образований в
организме, которое содержит две последовательно расположенные
капиллярные сети. Околоканальцевая капиллярная система
вновь объединяется и образует вены, которые затем впадают в
почечную вену.
Кровь поступает в мозговое вещество по особым сосудам —
прямым сосудам (рис. 24.3). Это длинные капиллярные петли,
которые ответвляются от корковых кровеносных сосудов,
спускаются в глубокий мозговой слой, а затем возвращаются в
кору, перестраиваются и образуют вены. Хотя и корковые, и
мозговые области содержат кровеносные сосуды, но больше
90% всего почечного кровотока поступает «в кору.
В почке в каждом нефроне происходят три основных
процесса: клубочковая фильтрация, канальцевая реабсорбция и ка-
нальцевая секреция (рис. 24.4).
КЛУБОЧКОВАЯ ФИЛЬТРАЦИЯ
Клубочковая фильтрация представляет собой процесс
перехода жидкости и растворенных в ней веществ из кровеносной
системы, или клубоч:ковых капилляров, в боуменову капсулу,
или почечный каналец (рис. 24.2 и 24.4). Клубочковые
капилляры можно определить как ультрафильтры, через которые
вода и мелкие молекулы плазмы переходят в почечный каналец.
Функционально они ведут себя так, как если бы содержали
цилиндрические поры диаметром 75—100 А, хотя анатомически
эти поры не обнаружены и, возможно, не существуют. Основной
Почка 445
особенностью клубочковой капиллярной мембраны является
наличие эпителиальных клеток, примыкающих к просвету
основной мембраны капилляра, и эндотелиальных клеток,
примыкающих к просвету боуменовой капсулы. Вода, электролиты и
различные мелкие молекулы свободно проникают через клубочко-
1вый капилляр.
Более крупные молекулы, например белки, не проходят
через него свободно. А для очень крупных молекул — сложных
белков или форменных элементов крови — клубочковая
мембрана в норме непроницаема. Главными участками, где
проникновение промежуточных молекул ограничено, являются основная
мембрана и пространства между смежными эндотелиальными
клетками, называемые щелевыми контактами. На способность
к проникновению крупных молекул влияет их электрический
заряд. Проницаемость клубочкового капилляра не всегда
одинакова, ее могут изменять циркулирующие в крови гормоны и
другие вещества.
Скорость клубочковой фильтрации (СКФ) у человека, или
количество фильтрата, поступающего в почечные канальцы в
обеих почках, составляет около 125 мл/мин; ток плазмы через
обе почки составляет, около 650 мл/мин.
Термином фильтрационная фракция обозначается отношение
125
количества фильтрата к току плазмы (-^-=0,20). Это значит,
что из всей плазмы, проходящей через клубочковые
капилляры, только 20% фильтруется через клубочки ih попадает в
почечные канальцы. Остальные, не фильтрующиеся 80% текут
дальше в выносящей артериоле, проходят через вторичную
капиллярную сеть (или околоканальцевые капилляры) и
возвращаются в общее кровяное русло по венозной системе.
МЕТОДИКА КЛИРЕНСА
Измерение количества жидкости, проходящей через клубоч-
ковую мембрану (СКФ), долгие годы считалось трудной
задачей. Решение ее связано с тем, что называется методикой кли-
рееса. Клиренсом обозначают количество вещества, удаляемого
из плазмы за единицу времени.
Клиренс какого-либо вещества (соединения А) можно
выразить следующим простым уравнением или отношением:
1. Концентрация А в плазмеХКоличество плазмы,
фильтрующейся в почечный каналец= Концентрация А в моче X объем
мочи.
Это просто означает, что произведение концентрации А в
плазме и скорость, с какой она фильтруется в почечный
каналец, равны количеству, выходящему в мочу, или: Объем мочиХ
446
Глава 24
X Концентрация А в моче. Очевидным допущением является,
что почка не добавляет А к моче, не удаляет его из мочи. Это
уравнение можно написать иначе:
2. Количество плазмы, фильтрующейся в почечный каналец
Концентрация А в мочеХОбъем мочи
Концентрация А в плазме
Поскольку количество фильтрующейся плазмы равно СКФ, то
уравнение принимает такой окончательный вид:
о /-т^/ь / / \ А в моче (мг/мл) XОбъем мочи „„„. г-т^/ь
3. СКФ (мл/мин) = т—-—-——.—-,—г , или: СКФ =
47 А в плазме (мг/мл)
___UAXV
~~ Ра *
ИНУЛИНОВЫЙ КЛИРЕНС
Вещество, избираемое для измерения клиренса, должно
удовлетворять определенным критериям. Во-первых, оно
должно свободно фильтроваться через клубочковую мембрану.
Во-вторых, оно не должно ни добавляться в почечном канальце,
ни удаляться из него. Наконец, выбранное вещество должно
быть относительно инертным, не оказывать других
физиологических эффектов и быть легко измеримым в плазме и моче.
Всем этим требованиям отвечает инулин — вещество с
небольшой молекулой, которое обычно в плазме не обнаруживается.
Для измерения клубочковой фильтрации инулин следует
вводить внутривенно.
Имеется еще одно вещество, креатинин, которое в норме
содержится в плазме, фильтруется, выводится и которое по
своему клиренсу приближается к клиренсу инулина. Для
определения скорости клубочковой фильтрации можно также
пользоваться клиренсом мочевины, но ценность такого приема
сомнительна, так как содержание мочевины в крови весьма
вариабельно.
ТОК ПЛАЗМЫ В ПОЧКЕ
Принцип клиренса служит для измерения тока плазмы в
почке (ТПП), что является применением принципа Фика (см.
также гл. 2 и 14). Существует следующее отношение:
Ток плазмы в почке (мл/мин) (ТПП) =
Концентрация А в моче (мг/мл) ■ Объем мочи (мл/мин)
Концентрация А (мг/мл) в Концентрация А (мг/мл) в*
почечной артерии почечной вене
В этом случае в качестве А можно выбрать любое вещество
€ тем только ограничением, что сама почка не должна ни при-
Почка
447
бавлять его к моче, ни удалять из нее. Таким идеальным
веществом является парааминогиппурат (ПАГ), потому что он
свободно фильтруется, и у человека больше 90% ПАГ может
быть полностью удалено из почечной плазмы, так что его
содержанием в почечной венозной крови можно пренебречь. Это
позволяет упростить уравнение следующим образом:
ТПП (МЛ/МИН) = ПАГ мочи (мг/мл).Объем мочи (мл/мин)
7 ПАГ в почечной артерии (мг/мл)
Кроме того, ПАГ, который нужно вводить внутривенно в
кровоток, не участвует в метаболизме; поэтому его уровень в
артериальной и венозной плазме один и тот же. Это
обстоятельство позволяет измерять ПАГ в пробах периферической венозной
крови, избегая трудности получения проб артериальной крови.
Величина для цельной крови, содержащей плазму и
эритроциты (ТКП), вычисляется следующим образом:
ТКП = ТПП 71—=-! г.
(1 — Гематокрит)
Поскольку фильтрационная фракция (ФФ)= =^, то
^- _ Инулиновый клиренс
Клиренс ПАГ
С измерениями клиренса инулина и ПАГ связаны
следующие проблемы: 1) эти вещества должны быть введены в
кровоток; 2) через определенные отрезки времени нужно собирать
мочу в течение по меньшей мере 20—30 мин, и 3) получаемые
значения клиренса отражают величины СКФ и ТПП не в
данный момент, а средние за тот отрезок времени, пока собиралась
моча.
Ток плазмы в почке можно определить также
непосредственно при помощи электромагнитного расходомера.
Имплантированный на почечную артерию, он регистрирует скорость
потока, что позволяет определить объем кровотока (см. гл. 14).
Ценность этого прибора в том, что он измеряет ток плазмы в
почке в данный момент и не требует ни введения ПАГ, ни
измерения его в плазме и моче. Поскольку расходомер
имплантируется на кровеносный сосуд, применение его ограничено
исследованиями на животных, и он не используется в клинической
практике.
ФИЗИЧЕСКИЕ И ЭЛЕКТРОХИМИЧЕСКИЕ ВЛИЯНИЯ
Скорость клубочковой фильтрации регулируется
гидростатическими силами, а также физическими и электрохимическими
ограничениями проницаемости клубочковой мембраны. Физиче-
448
Глава 24
ские силы здесь те же, которые определяют фильтрацию или
всасывание в любом капилляре (см. гл. 12). Эти факторы
входят в следующее уравнение:
СКФ = (Коэффициент фильтрации) х
X (Эффективное фильтрационное давление).
Коэффициент фильтрации (Кф) —это термин, описывающий
параметры проницаемости клубочковой мембраны. Эффективное
фильтрационное давление (ЭФД) —это движущая сила,
возникающая из разности между существующим гидростатическим
(Р) и осмотическим (я) давлением плазмы капилляров и
первичной мочи в почечном канальце.
^1\ч^ \*\^/~Г\*клубочк. ^канальц. г "^канальцев ой жидкости ^плазмы,) •
Эффективное фильтрационное давление определяет
движение воды и растворенных веществ через клубочковую мембрану
в почечный каналец. Подставив соответствующие значения для
давлений (мм рт. ст.), можно вычислить эффективное давление.
Знак плюс (, + ) означает положительное давление,
проталкивающее жидкость из кровеносного сосуда в каналец; знак минус
{—) означает отрицательное давление, проталкивающее
жидкость из канальца в, клубочковый капилляр:
СКФ = (Кф)-(+45—10+0—25) = (Кф)-(Ю мм рт. ст. ЭДФ).
Этот расчет показывает, что при любом состоянии
клубочковой проницаемости, или Кф, 10 мм рт. ст. будет
благоприятствовать фильтрации. Важно понимать, что на СКФ влияет
изменение любого из указанных параметров.
Коэффициент фильтрации принято считать относительно
постоянным, но недавние опыты показали, что при некоторых
болезнях или под влиянием определенных гормонов он может
заметно меняться. Существенный способ регулировать СКФ
состоит в изменении гидростатического давления в клубочковом
капилляре. Приносящая и выносящая артериолы клубочков
содержат гладкие мышцы, которые сокращаются при действии
симпатического нерва или под влиянием гормонов. Сужение
приносящих артериол снижает гидростатическое давление клу-
бочковых капилляров и снижает СКФ. Напротив, сужение
выносящей артериолы повышает гидростатическое давление и
повышает СКФ. Этот регулирующий механизм играет
первостепенную роль в регуляции выведения почками жидкости и
электролитов.
Почка
449
АУТОРЕГУЛЯЦИЯ СКФ И ТКП
На основе сказанного выше можно было бы ожидать, что
СКФ изменяется пропорционально величине артериального
давления в большом кругу, или перфузионного давления почки.
Поскольку кровоток также является функцией давления, можно
было бы ожидать, что кровоток в почке (ТКП) увеличивается
или уменьшается при повышении или понижении давления. Но
на самом деле это не так. Существует широкий диапазон пер-
з;о
2,5
1 2 0
ii,5
"с
1,0
0,5
О 40 80 120 160 200 240 280
Почечное перфузионное давление >
Рис. 24.5. Ауторегуляция тока плазмы в почке (I) и скорости клубочковой
фильтрации (II). ТПП и СКФ увеличиваются с повышением почечного
перфузионного давления до определенной точки, а затем при дальнейшем
повышении давления выравниваются (ауторегуляция).
фузионного давления в почке (80—180 мм рт. ст.), в пределах
которого и СКФ и ТКП остаются относительно постоянными
(рис. 24.5). Это свидетельствует об активном приспособлении
сопротивления почки току и степени сужения или расширения
сосудов. Такое явление называется ауторегуляцией.
Факторы, определяющие ауторегуляцию, пока еще не
выяснены. Она происходит и в изолированной почке, которая пер-
фузируется искусственно — с помощью насоса, что
свидетельствует против участия в ауторегуляции нервных и гормональных
факторов. Получены данные, указывающие на роль в
ауторегуляции гормона ренина (образующегося в почке). Однако
большинство авторов считают, что она является следствием
сокращения гладких мышц сосудов в ответ на изменения давления
на их стенки. При повышении перфузионного давления в почке
артериолы сокращаются, увеличивая сопротивление, и таким
29-1705
450
Глава 24
образом ток остается постоянным. Если же давление
снижается, то стимул, вызывающий сокращение, ослабевает, сосуды
расширяются и сопротивление уменьшается.
Хотя почка обладает способностью к ауторегуляции СКФ и
ТКП, многие обстоятельства оказываются сильнее этой
регуляции и резко сказываются на обеих этих величинах. Так,
например, при кровоизлиянии или кровопоте(ре под влиянием
симпатических импульсов происходит сужение сосудов почки, что
приводит к снижению СКФ и ТКП. Они понижаются также при
повышении уровня норадреналина и адреналина в крови. При
напряженной мышечной активности оба этих параметра
уменьшаются на 75—80%. Такое интенсивное сужение почечных
сосудов приводит к перераспределению кровотока в сторону
работающих скелетных мышц.
КАНАЛЬЦЕВАЯ РЕАБСОРБЦИЯ
У человека СКФ составляет в норме 125 мл/мин, или 180 л
в сутки (1440 мин). Поскольку в организме человека образуется
за сутки примерно 1,5 л мочи, т. е. 1,5/180 (около 1%), 99%
жидкости, профильтровавшейся через клубочковые капилляры,
подвергается обратному всасыванию в кровяное русло — реаб-
сорбируются (см. рис. 24.4). Большие количества воды,
электролитов, глюкозы, аминокислот и других жизненно важных
веществ возвращаются в кровь путем реабсорбции.
ОТНОШЕНИЕ: КАНАЛЬЦЕВАЯ ЖИДКОСТЬ/ПЛАЗМА
Для исследования процесса реабсорбции введен полезный
термин, а именно отношение канальцевая жидкость/плазма
(КЖ/П). Отношение КЖ/П для Na+ показано на рис. 24.6.
Концентрация натрия (Na+) в плазме сравнительно
постоянна. Поскольку Na+ свободно фильтруется, концентрация Na+
в КЖ внутри боуменовой капсулы такая же, как в плазме
(КЖ/П=1). Можно также наблюдать, что в точках дистальнее
боуменовой капсулы, в конце проксимального извитого
канальца, [Na+] в КЖ снижена, и отношение КЖ/П для Na+ меньше
единицы. Следовательно, можно сказать, что если отношение
КЖ/П для вещества меньше 1,0, то это (вещество должно было
реабсорбироваться. Если по мере дальнейшего движения по
почечному канальцу реабсорбируется все больше вещества, то
определяемые в специальных точках отношения КЖ/П будут
все время снижаться.
Процесс, посредством которого вещество движется из
общего кровотока в почечный каналец не через клубочек, называется
секрецией. Следовательно, если КЖ/П для данного вещества
Почка
451
больше 1, это значит, что в почечном канальце происходит
секреция этого вещества. КЖ/П можно вычислить для любого
фильтрующегося электролита или вещества с тем, чтобы
количественно определить степень канальцевой реабсорбции или
секреции.
Проксимальный
каналец, длина, %
Дистальный
каналец, длина, %
Моча
Na
гКонцентрация-
в канальцевой
жидкости
Концентрация
в плазме
1,01-
0,51-
0,2
0,1
I I I I I
20 40 60 80 100
'///////Ш/А
I I I I
20 40 60 80
' 1,0
0,5
Na/инулин
-Концентрация-) 0,1
в канальцевой
жидкости
Р,05к
Концентрация
в плазме
0,01k
0,005k
0,001k
0,0005l_
Диапазон
: Диапазон
100
Диапазон
Рис. 24.6. Отношения КЖ/П для Na+ и Na+Дшулин. Л. Показаны участки
нефрона, в которых определялись эти отношения. Если не вносится
поправка на реабсорбцию воды в проксимальном извитом канальце, отношение
КЖ/П для Na+ не меняется. Б. В любой точке проксимального или дисталь-
ного извитого канальца, где внесена поправка на реабсорбцию воды путем
деления КЖ/П для Na на КЖ/П для инулина, отношение постепенно
снижается, что указывает на реабсорбцию Na+. (Netter F. H., 1973. Kidneys, ureters
and urinary bladder, v. 6.)
Применение отношений КЖ/П связано с проблемой
удаления воды. Если почечный каналец содержит фиксированное
количество вещества, например Na4", и реабсорбируется только
вода, [Na+] в КЖ увеличится, и отношение КЖ/П для натрия
станет больше 1,0. Это означало бы (неправильно), что Na+
секретировался. Чтобы внести коррекцию в эффекты удаления
воды из почечного канальца, вычисляется отношение КЖ/П
для инулина. Поскольку инулин не реабсорбируется и не секре-
тируется, на его концентрацию в КЖ влияет только удаление
воды. В табл. 24.1 приведены отношения КЖ/П для инулина в
29*
452
Глава 24
Таблица 24.1. Отношения КЖ/П для инулина (14С-) в разных отделах
почечного канальцаа)
Локализация
Боуменова капсула
Место соединения
средней и дистальной
третей проксимального
канальца
Конец проксимального
канальца (вычислено)
Начало дистального
канальца
Конец дистального
канальца
Мочеточник
Отношение
кж/п
1
з
4
5
20
690
Остаточный
клубочковый
фильтрат, %
100
33
25
20
5
0,14
Реабоорбированная в
отделе профильтрованная
вода, %
75 (в проксимальном
канальце)
5 (в петле)
15 (в дистальном
канальце)
4,86 (в собирательных
трубочках)
а' Gottschalk (I961). Micropuncture studies of tubular function in the mammalian
kidney. Physiol., 4, 35.
разных точках в канальце и количество реабсорбированной
профильтровавшейся воды.
Делением отношения КЖ/П для вещества на отношение
КЖ/П для инулина вносится 'поправка на изменения,
связанные с удалением воды. Такой пример показан для Na+ на
рис. 24.6. Чтобы измерить отношение КЖ/П (для инулина или
любого другого вещества), надо получить пробу жидкости,
содержащейся в определенных участках почечного канальца,
посредством микропункции. Микропункция требует введения очень
маленькой микропипетки с тонким кончиком через стенку
канальца в его просвет у какого-либо животного (крысы) так,
чтобы через пипетку можно было взять пробу канальцевой
жидкости.
МЕХАНИЗМЫ РЕАБСОРБЦИИ
На реабсорбцию оказывают существенное влияние
электрохимические градиенты в почечном канальце. На рис. 24.7
показаны просвет почечного канальца и клетки стенок канальца,
окружающие просвет, околоканальцевая жидкость и
пространство вокруг канальца. При реабсорбции происходит движение
веществ из просвета в околоканальцевое пространство.
Благодаря своей проницаемости и способности к переносу веществ
мембраны создают существующие электрохимический и
концентрационный градиенты.
Почка
453
Электрохимически цитоплазма клеток канальцев заряжена
отрицательно по отношению и к просвету канальца (—52 мВ),
и к околоканальцевои жидкости (—72 мВ). Следовательно,
просвет слегка отрицателен по отношению к околоканальцевои
жидкости (—20 мВ). Мембрана, отделяющая цитоплазму
клеток канальца от просвета, в общем избирательно проницаема
Пространство
вокруг канальца
Пассивный// К +
"7 <*J
145
\ мМ\
v \ \
*
Пространство
вокруг канальца
^-^Клетка эпителТ^"4^
У\ канальца \.
/т1росвет канальца >ч \
Активный 4-У^\активный
Л 145мМ\ Л*
ЗмМ
Nv .г
^420 MB
Пассивный ^7|^аФ4^
/40/
/ мМ / )
-
1—±!
бгмв^х/
72мВ^^
Рис. 24.7. Условия в проксимальном канальце; активные и пассивные этапы
реабсорбции Na+ и К+.
для Na+, а мембрана, окружающая каналец, избирательно
проницаема для К+. Химические концентрационные градиенты
таковы, что концентрация К+ в клетках канальцев высокая, а
концентрации Na+ и С1~ низкие.
ТРАНСПОРТ ЭЛЕКТРОЛИТОВ
На рис. 24.7 показано, каким образом реабсорбируется Na+.
Он пассивно движется вдоль концентрационного и
электрического градиентов из просвета в клетки канальца. Na+ внутри
клетки активно переносится из нее в околоканальцевую
жидкость против концентрационного и электрического градиентов.
Этот второй этап требует затраты энергии. Энергия
поставляется метаболизмом и последующим гидролизом АТФ.
Потребности почки в кислороде, или потребление кислорода, пропорцио-
454
Глава 24
нально количеству активно переносимого Na+. Некоторое
количество реабсорбированного Na+ может «просочиться обратно»
в каналец через межклеточные пространства. Это особенно
заметно в проксимальном извитом канальце, где контакты между
клетками «рыхлы». В дистальных же извитых канальцах
клетки образуют так называемые «плотные контакты», которые не
допускают сильной «утечки».
Стадией, ограничивающей скорость транспорта Na+,
по-видимому, является его пассивное вхождение в клетки канальцев,
а не стадия активного транспорта, или натриевый насос. При
помощи отношений КЖ/П рис. 27.6 показывает, где реабсорби-
руется профильтровавшийся Na+.
РЕАБСОРБЦИЯ КАЛИЯ В ПРОКСИМАЛЬНОМ ИЗВИТОМ КАНАЛЬЦЕ
Хотя переход К+ в клетки канальцев идет по электрическому
градиенту (рис. 24.7), очень большой противодействующий
концентрационный градиент требует, чтобы происходил
активный транспорт; затем К+ пассивно диффундирует из клеток
канальцев против электрического градиента, но при
благоприятном концентрационном градиенте. При высоком общем
содержании К+ в организме возможна его активная секреция в
дистальных извитых канальцах.
Что касается транспорта хлора из просвета канальца в око-
локанальцевую жидкость, то можно было бы предсказать его
направление по электрохимическому концентрационному
градиенту. Но на самом деле С1~ входит в клетки канальцев против
выраженного электрического градиента (—52 мВ). Для этого
существуют два пути. Первый состоит в соединении С1~ с
другим ионом, например с натрием, и при этом образуется
нейтральная молекула. Другой — это движение С1~ через
межклеточные контакты. Большая часть транспорта С1~ происходит
пассивно, хотя неопровержимо показано, что в некоторых
отделах канальцев, например в восходящем колене петли Генле,
происходит активный транспорт С1~.
РЕАБСОРБЦИЯ ГЛЮКОЗЫ
Глюкоза, которая свободно фильтруется в клубочке,
обнаруживается в канальцевой жидкости (рис. 24.8). Как известно,
в норме глюкозы в моче не бывает, что свидетельствует о
полной реабсорбции всей профильтрованной глюкозы. Реабсорбция
глюкозы представляет собой активный процесс, требующий
энергии. По-видимому, для глюкозы, галактозы и фруктозы
существует общий транспортный механизм или
молекула-переносчик. Системе реабсорбции глюкозы присущ так называемый
О 200 400 600 800 1000
Концентрация глюкозы в плазме, мг %
Рис. 24.8. Реабсорбция глюкозы. А. Для глюкозы показаны процессы,
протекающие, когда нефрон работает ниже или выше Ттах- Б. Графически
показаны конечные результаты. Если вещество фильтруется в количестве меньше
Ттах, оно не выделяется. Когда фильтрация превышает Ттах, вещество
появляется в моче. (Netter F. Н. 1973. Kidney, ureters and urinary bladder, v. 6.)
456
Глава 24
транспортный максимум (Ттах). Это значит, что транспортная
система глюкозы имеет конечную пропускную способность.
При нормальных уровнях глюкозы в крови система
функционирует намного ниже своего Ттах реабсорбируя всю
глюкозу, не оставляя ее следов в моче. При повышении уровня
глюкозы в крови система достигает своего Ттах, или точки
насыщения. Если глюкоза плазмы поднимается выше Ттах, то
количество профильтрованной глюкозы превосходит то, какое может
быть реабсорбировано. В результате глюкоза появляется в
моче. Примером этого служит глюкоза в моче, или глюкузурия,
при нелеченном диабете, когда уровень глюкозы в крови
превышает Тщах, который составляет приблизительно 200 мг на
100 мл.
Почки обладают способностью реабсорбировать многие
вещества. Происходит активная реабсорбция Р04, креатина,
сульфата, мочевой кислоты, аскорбиновой кислоты, кальция и
магния. Существуют также транспортные механизмы для реабсорб-
ции аминокислот и фильтрующихся белков с мелкими
молекулами.
КАНАЛЬЦЕВАЯ СЕКРЕЦИЯ
Канальцевая секреция представляет собой 'Процесс,
посредством которого вещества переносятся из околоканальцевой
жидкости в почечный каналец (рис. 24.4). Секреция сходна с
фильтрацией в том отношении, что оба процесса приводят к
проникновению веществ в почечный каналец. Но фильтрация
происходит только в клубочке, а секреция во всех частях нефрона дис-
тальнее клубочка. Секреция может быть 'пассивной или
активной, происходящей с затратой энергии. Большая часть активных
секреторных механизмов имеет ограниченную транспортную
способность, т. е. обладает Ттах.
Существует общий путь секреции органических кислот. К
соединениям, выводимым из кровотока этим механизмом,
относятся феноловый красный, ПАГ, пенициллин и глюкурониды.
Тестом для определения секреторной способности служит
измерение экскреции фенолового красного — соединения, которое
можно вводить в кровь.
Второй секреторный механизм переносит сильные
органические основания. Сюда относятся гуанидин, тиамин, холин, гис-
тамин и тетраэтиламмоний. Пассивная секреция перемещает
вещества в почечный каналец по электрохимическому градиенту.
Этим способом переносятся такие соединения, как слабые
основания и слабые кислоты. Кроме того, по электрохимическому
градиенту в дистальном канальце может пассивно секретиро-
ваться К+.
Почка
457
РЕГУЛЯЦИЯ ЭКСКРЕЦИИ ЭЛЕКТРОЛИТОВ
Действуя как гомеостатический орган, почка поддерживает
постоянство уровня жидкости в плазме и общей концентрации
электролитов в организме. Отсутствие натрия в .пище или
снижение его общего количества в организме заставляет почку
сохранять Na, сводя к минимуму количество, выделяемое с мочой.
Почка весьма эффективно снижает экскрецию Na+, понижая
объем фильтрации. Если вещество не фильтруется или не секре-
тируется, оно не появится в моче. Поскольку существуют
способы изменения СКФ, фильтрующееся количество воды,
электролитов или любого другого вещества может быть резко
изменено
200
Наружный
мозговой
Внутренний
мозговой
слой
100
100
200
400
А
300
300
*сг
400
600
Na+*
Н20
Na+«
800
Мочевина
1000 Н2О<{-1000
НпО«е-'
Мочевина^}-—
1200
300
|*Н20
300
400
600
► К +
н+
800
1200
Рис. 24.9. Сводные данные по обмену воды, ионов и мочевины в почке.
Цифры показывают осмоляльность (в мосмолях) канальцевой мочи и
околоканальцевой жидкости. Обратите внимание на увеличение околоканальцевой
осмоляльности по мере продвижения в глубокие слои мозгового вещества:
кора — 300; мозговое вещество—1200. Сплошные стрелки — активный
транспорт; прерывистые стрелки — пассивный транспорт. Жирные линии вдоль
восходящего колена петли Генле показывают, что этот отрезок сравнительно
непроницаем. (С изменениями по Gottschalk С. W., Mylle M., 1959. Am. J.
Physiol., 196, 927.)
30—1705
458
Глава 24
но. Другая возможность состоит в увеличении реабсорбции Na+
(рис. 24.9). Но количественно именно снижение СКФ
определяет главным образом уменьшение экскреции Na+. Хотя
усиление его реабсорбции действует благоприятно и действительно
происходит, это процесс активный, требующий затраты энергии.
Он менее эффективен, чем простое сокращение экскреции Na+
путем уменьшения его фильтрации.
РЕАБСОРБЦИЯ НАТРИЯ
Реабсорбции Na+ способствуют несколько механизмов. В
основе одного из них лежит действие гормона альдостерона,
выделяемого корой надпочечников. Этот минералокортикоид
усиливает реабсорбцию Na+, главным образом в дистальных
извитых канальцах. Работы последнего времени позволяют думать,
что альдостерон стимулирует синтез пермеазы натрия, которая
способствует пассивному вхождению Na+ в клетки эпителия
канальцев. Альдостерон контролирует лишь малую долю (2—
3%) всей реабсорбции натрия. Поэтому считается, что он
обеспечивает «тонкую настройку» тех процессов, которым Na+
подвергается в почке. Вместе с тем альдостерон имеет большое
значение, если учесть то огромное количество натрия, которое
переносится через почку за несколько дней.
НЕРВЫ ПОЧКИ И РЕАБСОРБЦИЯ НАТРИЯ
Хорошо известно, что стимуляция почечных симпатических
нервов заметно снижает экскрецию <Na+. Это их действие
приписывали исключительно гемодинамическим изменениям, т. е.
понижению ТКП и снижению СКФ. Однако работы последних
лет показывают, что нервы почек приходят в непосредственный
контакт с основной мембраной как проксимального, так и ди-
стального извитых канальцев. Слабая стимуляция этих нервов,
которая не меняет СКФ, усиливает канальцевую реабсорбцию
Na+. Следовательно, существует прямой нервный механизм
регуляции реабсорбции Na+.
РЕАБСОРБЦИЯ ВОДЫ
Из 180 л воды, фильтруемых ежесуточно клубочками, 99%
реабсорбируются в канальцах. Реабсорбция воды — это
пассивный процесс, осуществляемый силами осмотического давления.
На рис. 24.9 показана осмолярность окружающей каналец ин-
терстициальной, или околоканальцевой, жидкости;
осмолярность постепенно увеличивается от 300 мосмоль/л в наружном
Почка
459
корковом слое до 1200 мосмоль/л ©о внутреннем мозговом слое
близ сосочка.
Значения КЖ/П для инулина (табл. 24.1) свидетельствуют
о том, что реабсорбция воды происходит как в проксимальном,
так и в дистальном извитых канальцах. Реабсорбция эта изо-
осмотическая; это значит, что реабсорбирующаяся жидкость и
жидкость, остающаяся в канальце, обладают одинаковой
осмотической концентрацией. Следовательно, ионы и вода
удаляются с одинаковой скоростью. Удаление ионов, например Na+,
происходит активно, а вода следует за ними вследствие
возникающей осмотической силы. Около 80% профильтрованной
воды реабсорбируется посредством этого механизма в
проксимальном извитом канальце, а остальное — из более дистальных
сегментов нефрона.
КОНЦЕНТРАЦИЯ МОЧИ
Осмолярность плазмы составляет около 300 мосмоль/л.
Осмолярность нормальной мочи равна 600—800 мосмоль/л, и
поэтому моча гипертонична по отношению к плазме. Почка
обеспечивает это следующим образом. Фильтрат, который
проходит через клубочковые капилляры, имеет ту же осмолярность,
что и плазма (рис. 24.9). По мере того как жидкость спускается
по петле Генле, наступает равновесие между все возрастающей
осмотической концентрацией среды вокруг канальца и
содержимого канальца. Это происходит потому, что нисходящее колено
проницаемо и для воды, и для электролитов. В верхушке петли
осмолярность канальцевой жидкости составит около 1200
мосмоль/л. Движению жидкости по восходящему колену
сопутствует понижение осмолярности из-за того, что это колено
непроницаемо для воды, и электролиты (т. е. Na+ и С1~)
переносятся из канальца активно (рис. 24.9). В результате жидкость,
дошедшая до дистального канальца, гипотонична по отношению
к плазме, и ее осмолярность составляет около 100 мосмоль/л.
На первый взгляд это невыгодно, поскольку задача заключается
в образовании гипертонической мочи. Но решающим
обстоятельством является то, что канальцевая жидкость должна
спуститься в собирательную трубочку обратно через мозговой
осмотический слой. То, что происходит при этом окончательном
оттоке, и определяет содержание воды или осмолярность
выводимой мочи.
Существуют две возможности. 1. Если собирательная
трубочка проницаема для воды, между окружающей интерстициальной
тканью и содержимым трубочки наступает осмотическое
равновесие. Это приводит к образованию мочи с такой же гипертонич-
ностью, как у жидкости в глубоких участках мозгового вещест-
30*
460
Глава 24
ва (1200 мосмоль/л). 2. Если собирательная трубочка
непроницаема для воды, то осмотического равновесия не наступает.
В этом случае гипотоничная жидкость в дистальном канальце
не подвергается концентрированию, и выделяется гипотоничная
моча.
АНТИДИУРЕТИЧЕСКИЙ ГОРМОН
Нормально собирательная трубочка непроницаема для
воды. Гормон вазопрессин, или антидиуретический гормон (АДГ),
делает ее проницаемой для воды, вследствие чего образуется
концентрированная моча. АДГ выделяется нейрогипофизом (см.
гл. 25). Усиление осмолярности, означающее дефицит воды,
повышает выделение АДГ, что приводит к уменьшению
количества выводимой воды. При некоторых условиях, связанных с
уменьшением объема мочи, например при обезвоживании или
мышечной работе, уровни АДГ бывают повышены. У больных
несахарным диабетом отмечается либо низкий уровень АДГ,
либо почки не реагируют на гормон. Количество выделяемой
мочи может достигать 5—10 л в сутки.
ПРОТИВОТОЧНЫИ УМНОЖИТЕЛЬ
Хотя детальный механизм создания корково-мозгового
осмотического градиента неясен, известно, что это состояние
связано с противоточной умножающей системой (рис. 24.9).
Большая осмолярность в мозговой области объясняется повышением
концентрации Na+ и мочевины. Развитие этой осмолярности
предположительно основано на том, что между восходящим и
нисходящим коленами петли Генле может образоваться
небольшой (200 мосмоль/л) осмотический градиент и что в разных
сегментах почечного канальца проницаемость неодинакова.
Осмолярность начального восходящего сегмента повышена
вследствие того, что по изгибу петли идет более
концентрированная жидкость. Постепенно осмолярность становится гораздо
выше, чем в плазме, где она равна 300 мосмоль/л. Вода и
электролиты в канальце приходят в равновесие с околоканальцевой
жидкостью, образующей осмотическую среду вокруг канальца.
Кокко и Ректор (Kokko, Rector) предложили формальную
модель, использующую принцип противоточного умножения.
Они исходят из того, что мочевина, концентрация которой в
дистальном канальце очень высока, диффундирует из
собирательной трубочки и iB значительной степени определяет
извлечение воды из нисходящего колена. Поскольку большая доля
осмолярности в мозговом интерстиции обусловлена мочевиной,
существует градиент Na+, который благоприятствует пассивно-
Почка
461
Кора
NaCI
Наружный \ w /
мозговой
слой
Внутренний
мозговой
слой
Рис. 24.10. Модель Кокко и Ректора (Kokko, Rector). Жирные стрелки —
активный транспорт хлора, X — нереабсорбируемое растворенное вещество.
В этой модели большая доля осмолярности во внутреннем мозговом слое
создается мочевиной. (С изменениями по Kokko J. P. et al. 1974. Fifth
International Congress of Nephrology.)
му движению Na+ из начального сегмента восходящего колена.
Затем в освободившийся участок диффундирует мочевина,
которая повторяет цикл, выходя в интерстиций мозгового
вещества из собирательной трубочки (рис. 24.10).
ПРОТИВОТОЧНЫИ ОБМЕН
Необходимо, чтобы система концентрации мочи, в которой
участвует гипертоническая область мозгового слоя,
обеспечивала возвращение реабсорбированных из канальца Na+ и воды в
кровоток, но чтобы это не снижало высокой концентрации
молекул веществ, растворенных в мозговом слое. Это
достигается, во-первых, чрезвычайно медленным кровотоком в области
мозгового слоя по vasa recta (рис. 24.3). Во-вторых, здесь
происходит процесс, называемый противоточным обменом, при
котором из мозгового слоя не удаляются большие количества
растворенных веществ. Между двумя коленами петли Генле в
мозговом слое происходит обмен растворенными веществами. В ко-
462
Глава 24
Растворенные
вещества —
300 и О «
Растворенные
вещества
525 Н20 <
Растворенные
вещества
750 Н,0 4
Растворенные
вещества
975
Н20
Растворенные
вещества
I
285
300
525
750
1200
Н20
%
t
315
525
750
'975'
1200
Растворенные
вещества
,2,- 30°
Н20
Растворенные
вещества
Н90
525
Растворенные
вещества
1н2о 75°
Растворенные
вещества
► 975
- Н20
1200
Рис. 24.11. Противоточный обмен. Осмолярность крови, поступающей в
мозговой слой, составляет 285 мосмоль/л, а оттекающей — 315 мосмоль/л. Этот
процесс, благодаря которому оттекающая кровь не содержит много
растворенных веществ, препятствует устранению гипертоничности мозгового интер-
стиция. (Neller F. Н., 1973. Kidneys, ureters and urinary bladder., v. 6.)
нечном счете плазма, выходящая из области мозгового слоя,
лишь ненамного более концентрирована, чем входящая плазма.
Этим предотвращается вымывание растворенных веществ из
мозгового слоя (рис. 24.11).
ПОЧЕЧНЫЕ ГОРМОНЫ
Известны две главные гормональные системы почки.
Первая— это ренин-ангиотензиновая система, основные
компоненты которой описаны в гл. 29. Ренин вырабатывается и выде-
Почка
463
ляется почкой. Он синтезируется в специальной группе эпите-
лиодных клеток, называемых юкстагломерулярными (ЮГ). Эти
клетки выстилают афферентную артериолу и расположены
между ней и специализированным сегментом дистального
канальца, именуемым macula densa (рис. 24.12). Разные стимулы
(в том числе понижение почечного перфузионного давления,
усиление активности симпатической нервной системы,
повышение содержания катехоламинов в крови, уменьшение общего
Афферентная
артериола
Рис. 24.12. Юкстагломерулярный аппарат, состоящий из macula densa и
специфических клеток. (С изменениями по Davis I. 1971. Circ. Res, 28, 301.)
объема внеклеточной жидкости в организме и изменения
электролитного состава жидкости в дистальном канальце), к
которым чувствительна macula densa, усиливают выход ренина.
Ренин действует на глобулин плазмы, синтезируемый в
печени, и образует декапептид ангиотензин I. В присутствии
особого фермента, который обнаружен в легких и почке,
ангиотензин I расщепляется с образованием октапептида ангиотензина II
(АН). Биологически активный АН является мощным
сосудосуживающим агентом (см. гл. 14). Кроме того, он усиливает
выход альдостерона, снижает синтез ренина и может
непосредственно повысить реабсорбцию Na+. Ренин-ангиотензиновая
система участвует в возникновении и поддержании некоторых
типов гипертонии.
Вторым главным эндокринным комплексом в почке является
простагландиновая (ПГ) гормоноподобная система. ПГ
синтезируются из фосфолипидов и арахидоновой кислоты посредством
464
Глава 24
ферментативного комплекса ПГ-синтетазы, или циклооксигена-
зы. Известно целое семейство ПГ-гормонов с многими
различными видами физиологической активности.
Хотя ПГ синтезируются во всем организме, особенно много
их образуется в мозговом слое почки; главные ПГ — это ПГЭ2
и ПГФ2. Кроме воздействий на сердечно-сосудистую систему ПГ
способны изменять эффекты симпатической нервной системы,
усиливать синтез ренина, ослаблять сосудосуживающие
эффекты АН и подавлять активность АДГ. Изменения синтеза про-
стагландинов наблюдаются при многих болезнях. Интересно
отметить, что аспирин, мощный ингибитор ПГ-синтетазы,
ослабляет образование всех ПГ.
ПОЧКИ И КИСЛОТНО-ЩЕЛОЧНОЕ РАВНОВЕСИЕ
Почка участвует в регуляции рН плазмы или поддержании
общего кислотно-щелочного равновесия организма несколькими
способами. Эти способы следующие: 1) регуляция уровня
НСОз" в плазме; 2) регенерация НС03~~ и 3) секреция Н+ в
мочу.
РЕГУЛЯЦИЯ УРОВНЯ НС03- В ПЛАЗМЕ
Как было указано в гл. 2 и 17, роль почек состоит в
регуляции уровня НСОз-. При уровне НСОз" в плазме 28 мМ/л или
меньше отфильтрованный НСОз" целиком реабсорбируется.
Если же уровень его в плазме превышает 28 мМ/л, т. е. выше
почечного порога, НСОз" начинает появляться в моче. Реаб-
Na+<r
Ж
<
Интерстициальная
жидкость
Н20 + С02
Угольная
ангидраза
Н2С03
<-—I Na +
Активный
■нсо3- + н+ тРансп°Рт > н +
Канальцевая
Клетка эпителия канальца ] м°ча
Рис. 24.13. Механизм реабсорбции НС03™ в почке.
Почка
465
сорбция бикарбоната происходит в проксимальных и дисталь-
ных канальцах.
Большая часть профильтрованного НСОз" (приблизительно
90%) реабсорбируется в проксимальном канальце. На рис. 24.13
показан механизм реабсорбции НСОз" в клетках эпителия
канальца. Натрий движется в клетку по электрохимическому
градиенту. Ионы водорода, активно секретируемые в просвет
канальца, реагируют с НСОз" и образуют Н2С03, которая ^ссо-
1рЛФует на С02 и Н20. С02 движется в клетки эпителия
канальца; реакция между С02 и Н20, при которой возникает Н2С03,
катализируется ферментом карбоангидразой. Вновь
образованная Н2СОз диссоциирует на Н+, подвергающийся секреции, и на
активно реабсорбируемый НС03"~.
Кроме реабсорбции НС03~, канальцы также его
регенерируют, возмещая утраты в качестве буфера сильных кислот. Как
НС03~ превращают сильные кислоты в нейтральную соль и
способную к диссоциации угольную кислоту, видно из
следующего уравнения:
Н3Р04 + 2NaHC03
Сильная
кислота
■* NaaHP04 + 2Н20 + 2С02.
Нейтральная
соль
Как показано на рис. 24.14, динатрийфосфат (Na2HP04)
превращается в мононатрийфосфат (NaH2P04), следствием чего
является возвращение Na+ и НС03~ в кровоток.
Канальцевая
моча
Клетка эпителия дистального канальца
Na2HP04
NaH2P04
Метаболит + 02 —> Н20 + С02
Угольная
ангидраза
НР0
->Ыа+НСОз-
Кровь в
капиллярах
канальца
-> С02
NaHC03
Рис. 24.14. Механизмы перехода динатрийфосфата в мононатрийфосфат и
последующей реабсорбции бикарбоната натрия.
466
Глава 24
СЕКРЕЦИЯ ИОНОВ ВОДОРОДА
Ионы водорода могут активно секретироваться в мочу.
Количество секретируемой таким образом кислоты зависит от рН
мочи. Максимальный градиент для секреции Н+ достигается
при рН мочи, равном 4,5. Ионы водорода могут соединяться с
НС03-, образуя С02 и Н20, и с НР042", образуя Н2Р04-. Еще
один способ снизить число свободных Н+ в просвете канальца
состоит в их реакции с NH3, при которой образуется NH4+.
Аммиак секретируется почечными канальцами. Небольшое его
количество может поступать из артериальной крови. Будучи
растворимым в липидах, NH3 способен диффундировать через
мембрану канальца в его просвет. Там он реагирует с секрети-
руемым свободным Н+ и образует NH4+, который нерастворим
в липидах, остается в почечном канальце и выводится с мочой.
ОБРАЗОВАНИЕ МОЧИ
Почки играют чрезвычайно важную роль в поддержании го-
меостаза. Химический состав мочи и ее количество служат
отражением водно-электронного статуса организма. В табл. 24.2
приведены изменения выделения количества и состава мочи,
связанные с дегидратацией.
Таблица 24.2. Количество и состав мочи у человека в норме
и при дегидратацииа>
Норма
Дегидратация6)
Объем
мочи,
(мл/сутки)
1500
300
Удельный
вес
1,022
1,028
Осмоляр-
ность,
мосмоль/л
800
1 100
РН
6,2
5 9
N+
мэкв/л
138
по
мэкв/л
40
120
а> По Zambraski et al. 1975. Med. Sci. Sports. 7, 217.
e' Во время обезвоживания образуется меньше мочи; выделяемая
концентрированная и кислая моча отражает усиленную реабсорбцию Н20 и Na+.
МОЧЕИСПУСКАНИЕ
Выйдя из почечного канальца, моча через собирательный
проток поступает в почечную лоханку, а оттуда по
мочеточникам в мочевой пузырь (рис. 24.1). Продвижение мочи по
мочеточникам облегчается ритмическими перистальтическими
сокращениями, возникающими с частотой 1—5 раз в 1 мин.
Мочевой пузырь представляет собой полый орган с
мышечной стенкой. Двумя главными частями пузыря, составляющими
его тело, являются мышца, выталкивающая мочу, и небольшой
Почка 467
участок, так называемый треугольнику образованный устьями
правого и левого мочеточника и внутренним отверстием
мочеиспускательного канала. Мочевой пузырь инициируется
афферентными и эфферентными нервными волокнами.
Симпатические волокна идут через нижние брыжеечные узлы;
парасимпатические волокна от тазового нерва иннереируют тело пузыря и
внутренний сфинктер. Волокна к наружному сфинктеру отходят
от половых нервов. Мочевой пузырь растяжим. Стенка его
растягивается до максимума, лишь когда его объем достигает у
человека около 400 мл; давление в пузыре при этом резко
возрастает.
Мочеиспускание — выход мочи из пузыря — происходит при
сокращении выталкивающей мышцы (m. detrusor) и брюшных
мышц, а также при расслаблении внутреннего и наружного
сфинктеров. Эти процессы подчинены как автономной, так и
произвольной регуляции. В начальной фазе мочеиспускания
происходит постепенное наполнение пузыря, пока напряжение
его стенки не достигнет уровня, который вызовет позыв к
мочеиспусканию. У человека позыв возникает при объеме пузыря
около 200 мл. Когда афферентные стимулы достигнут
критического уровня, происходит рефлекторное сокращение m. detrusor
и расслабление обоих сфинктеров. Замыкание дуги этого
рефлекса приходится на II—IV крестцовые сегменты спинного
мозга. Влияние высших центров, лежащих в коре, определяют
возможность торможения или облегчения этого рефлекса.
ЛИТЕРАТУРА
Brenner В. М., Rector F. С. (1976). The kidney, Saunders, Philadelphia.
Handbook of Physiology (1973). Renal physiology, American Physiological
Society, Washington.
MTP International Review of Science (1974). Kidney and urinary tract
physiology, vol. 6, University Park Press, Baltimore.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Перечислите основные анатомические признаки нефрона и
связанных с ним специфических структур, а также их
функциональную роль.
2. Опишите главные процессы — фильтрацию, реабсорбцию и
секрецию.
3. Дайте определение клиренса и укажите, как он
используется для количественной оценки скорости клубочковой
фильтрации и тока почечной плазмы.
4. Перечислите факторы, которые регулируют ТПП.
5. Определите отношение КЖ/П и укажите, чем оно полезно
для изучения функции нефрона.
468
Глава 24
6. Опишите концентрацию и электрохимические условия в
проксимальном извитом канальце и стадии реабсорбции
Na+ и К+.
7. Какие различные механизмы или регулирующие факторы
определяют экскрецию электролитов?
8. Опишите процессы, определяющие реабсорбцию 98% воды,
профильтровавшейся в клубочках.
9. Опишите ренин-ангиотензиновую и простагландиновую
гормональные системы.
10. Как благодаря почкам восстанавливается нормальное рН
плазмы при развитии ацидоза?
11. Перечислите все процессы, происходящие при
мочеиспускании.
Глава 25
Эндокринология, функции гипофиза
и гипоталамуса
Эндокринология — это учение о функциях секретов (гормонов)
эндокринных (или беспротоковых) желез. Каждая секреторная
клетка такой железы одной своей поверхностью контактирует с
венозным синусом или капилляром, что способствует быстрому
проникновению гормона в кровь. Гормоны, регулирующие
деятельность отдельных органов или организма в целом, обычно
вырабатываются в небольших количествах и с током крови
переносятся к месту назначения. О существовании какого-либо
гормона думают прежде всего тогда, когда в результате
удаления или заболевания железы возникают нарушения, связанные
с отсутствием этого гормо'на и выпадением его функции.
С целью восстановления нарушенных функций организма в
целом или отдельного органа животным с удаленными
эндокринными железами иногда вводят гормоны, полученные путем
экстрагирования из желез (так называемая заместительная
терапия). Неочищенный экстракт желез может содержать
несколько гормонов; их можно разделить, получить в
очищенном виде, установить их химическое строение и — <в
дальнейшем — синтезировать.
МЕХАНИЗМ ДЕЙСТВИЯ ГОРМОНОВ
Существуют по крайней мере два механизма, посредством
которых гормон, проникший в кровоток и доставленный к месту
назначения, может подействовать на клетки органов и тканей.
Гормоны могут либо 1) усиливать образование циклического
аденозинмонофосфата (цАМФ), влияющего на специфические
функции клеток, 2) активировать гены, вызывая тем самым
синтез внутриклеточных белков, также способствующий
специфическим клеточным функциям (см. гл. 22).
Предполагают, что действие гормонов обусловлено их
связыванием и взаимодействием со специальными участками
клетки— рецепторами. Строение рецепторов до конца не выяснено,
хотя они и были выделены и специфичность их показана
соответствующими иммунохимическими методами.
470
Глава 25
РОЛЬ цАМФ
Считается, что цАМФ играет основную роль в действии
гормонов. Это вещество образуется из АТФ под действием
фермента аденилатциклазы (АЦ), расположенного на клеточной
мембране: АТФ+АЦ—*цАМФ. Механизм сопряжения процессов в
области рецептора с конечной реакцией до конца не выяснен,
однако ряд исследователей полагают, что цАМФ является^
вторым посредником в действии некоторых гормонов; первый
посредник—это сам гормон (рис. 25.1). Реакция образования
цАМФ относится к очень распространенным реакциям
организма; она протекает также в области ряда негормональных рецеп-
Гормон
тУг
Первый
посредник
Рецептор
АТФ
Аденилатциклаза
Клеточная
мембрана
ц-АМФ ■
Второй% посредник
Рис. 25.1. Роль рецепторов клетки и цАМФ в действии гормонов.
1
„^L Г 1. Механ
г | 2. Инакт
изм обратной связи
нактивация
Ответная реакция
органа-мишени
Связывание
с рецептором
Поступление
к тканям
Рис. 25.2. сЖизненный цикл» гормона.
Эндокринология 471
горов. «Жизненный цикл» гормона от момента его синтеза в
клетке до использования или инактивации включает ряд стадий
и.™ процессов (рис. 25.2).
ОПРЕДЕЛЕНИЕ СОДЕРЖАНИЯ ГОРМОНОВ
Гормон может быть извлечен из органа, в котором он
продуцируется; затем «после очистки первоначального экстракта он
может быть введен в организм. Содержание гормона в крови
определяется биологическими пробами, химическими методами
или иммунологическими пробами.
В основе биологических проб лежат реакции органа или
ткани на специфическое действие исследуемого гормона. Так,
тестом на фолликулостимулирующий гормон (ФСГ) может быть
изменение веса яичек неполовозрелых животных в ответ на его
введение. Мерой концентрации гормона служит отношение веса
яичек животных, которым он был введен, к весу яичек
контрольных животных. Биологические пробы сравнительно неточны
и малочувствительны.
Относительно эффективны химические методы, применимые
в том случае, если известен состав гормона, и он может быть
обнаружен химическим путем. Однако эти методы
недостаточно чувствительны в тех случаях, когда требуется определить
низкие концентрации гормонов, а многие из гормонов (белки и
пептиды) практически невозможно обнаружить химическим
путем.
Наилучшим методом для определения пептидов, белков и
стероидных гормонов, когда необходима высокая
чувствительность, является радиоиммунологический. Этот метод основан на
иммунологических свойствах гормонов, или реакции антиген —
антитело. Он состоит в следующем.
1. Сначала гормон, подобный тому, который
вырабатывается данной эндокринной железой, очищается и в течение
определенного промежутка времени вводится животному. После
повторных введений в крови животного появляются антитела,
специфичные в отношении этого гормона.
2. В небольшое количество очищенного гормона вводится
метка — обычно изотоп (чаще всего радиоактивный иод). После
этого известное количество гормона помещается в пробирку,
содержащую небольшой известный объем плазмы с антителами.
3. В пробирку добавляется определенный объем плазмы
подопытного животного, в крови которого измеряют уровень
гормона. Антитела реагируют как с мечеными, так и с
немечеными антигенами, связывая их. Так как количество антител ог-
4/Z
глава 2b
раничено, соотношение связанных и несвязанных меченых
антигенов варьирует в зависимости от содержания немеченых
антигенов в плазме исследуемого животного.
Таким образом, доля связываемых антителами меченых
гормонов тем меньше, чем больше добавляется немеченых
гормонов (антигенов), конкурирующих с мечеными за антитела.
Содержание какого-либо гормона в крови можно определить по
кривой зависимости
отношения связанных меченых
гормонов к несвязанным (с/н) от
содержания немеченого
гормона— антигена (рис. 25.3).
Радиоиммунологические методы
определения разработаны
почти для всех гормонов, в том
числе ТТГ, ФСГ, ЛГ, МТГ,
СТГ, тестостерона, эстрогенов
и простагландинов.
Гормоны , о которых
пойдет речь в данной главе,
подразделяются на следующие
группы.
1. Рилизинг-факторы
гипоталамуса.
2. Тропные гормоны адено-
гипофиза, действующие а) на
гормональную активность
органов-мишеней — щитовидной
железы, половых желез, коры
и мозгового слоя
надпочечников и б) непосредственно на
органы и ткани.
3. Гормоны, секреция которых не управляется аденогипофи-
зом: а) гормоны поджелудочной железы (инсулин и глюкагон);
б) гормоны паращитовидных желез и кальцитонин; 2) прочие
(см. гл. 20).
4. Гормоны нейрогипофиза (окситоцин, вазопрессин).
О 0,2 0,4 0,6 0,8
Немеченый гормон, нг/мл
Рис. 25.3. Радиоиммунологический
метод определения неизвестного
гормона. Представлена кривая
зависимости отношения связанных гормонов,
меченных I131, к несвязанным от
концентрации немеченых гормонов
(антигенов) .
ГОРМОНЫ ГИПОТАЛАМУСА И ГИПОФИЗА
В гипоталамусе вырабатывается ряд гормонов, вызывающих
выделение тропных гормонов передней долей гипофиза (pars
distalis, или аденогипофиз) и задней его долей (pars nervosa,
или нейрогипофиз) (рис. 25.4 и 25.5). Под действием этих так
называемых рилизинг-факторов гипоталамуса происходит
высвобождение из аденогипофиза 1) адренокортикотропного гор-
Г"
кона
Эндокринология
473
лона (АКТГ); 2) фолликулостимулирующего гормона (ФСГ);
3^ лютеинизирующего гормона (ЛГ); 4) тиреотропного гормона
(ТТГ); 5) пролактина; 6) гормона роста, или соматотропного
гормона (СТГ) и 7) меланоцитстимулирующего гормона
(МСГ).
Под влиянием гипоталамуса происходит также выделение
из задней доли гипофиза 1) антидиуретического гормона (АДГ)
Гипоталамус
Ж
\У
V
Аденогипофиз
Ж.
ТТГ
V
ФСГ
у
ЛГ
_ж_
АКТГ
т
V ^
Пролактин
Щитовидная
железа
Гонады
i
Надпочечники Молочные
железы
i
Эстрогены
Андрогены
Прогестерон
Кортикостероиды
ЖЛ_
-Ж
±
СТГ
Рост
МТГ
V \к !£■ \|/ Ф v Ф
Меланин
кожи
Рис. 25.4. Действие рилизинг-факторов — либеринов (—) ингибирующих
факторов статинов ( ) гипоталамуса на тройные гормоны аденогипофиза.
(у людей аргинин-вазопрессина, у птиц — аргинин-вазотоцина)
и 2) окситоцина.
Подробнее высвобождение и действие тройных гормонов
будет обсуждаться в главах, посвященных действию гормонов на
ткани и органы-мишени.
РИЛИЗИНГ-ФАКТОРЫ ГИПОТАЛАМУСА
Выделение каждого из тропных гормонов гипофиза
регулируется по крайней мере одним рилизинг-фактором
гипоталамуса (рис. 25.4). Исключение составляют ФСГ и Л Г: в настоящее
время известен лишь один рилизинг-фактор, вызывающий
высвобождение обоих этих гормонов. Три рилизинг-фактора были
получены от овец и свиней и выделены в чистом виде; их
химическая структура окончательно ©ыяснена (табл. 25.1).
31—1705
474
Глава 25
Преоптические
ядра
Супраоптические
ядра и пути
Срединное возвышение
Портальные сосуды
Аденогипофиз
<pars distalis)
Гипоталамус
-Паравентрикулярные ядра
Преоптический и прочие
•* нервные пути
Нейросекреторные гранулы
К аденогипофизу по
портальным сосудам
Йнфундибулярная ножка
Рис. 25.5. Регуляция активности гипофиза со стороны гипоталамуса. А. Па-
равентрикулярное и супраоптическое ядра и отходящие от них волокна
переносят окситоцин и вазопрессин к задней доле гипофиза. Другие ядра и
волокна гипоталамуса участвуют в транспорте нейросекрета и рилизинг-фак-
торов к аденогипофизу по системе портальных сосудов (вен). Б.
Расположение гипофиза (показано черным).
НЕИРОСЕКРЕЦИЯ
В состав гипоталамо-гипофизарной системы входят: 1)
аденогипофиз; 2) ядра гипоталамуса и срединного возвышения и
направляющиеся от них волокна, импульсы по которым
передаются нейрогипофизу, выделяющему окситоцин и вазопрессин,
а также (непрямым путем) к аденогипофизу, вызывая
высвобождение в нем различных гормонов (рис. 25.4 и 25.5).
Существуют нервные связи между гипоталамусом и нейроги-
пофизом (рис. 25.5), но не между гипоталамусом и аденогипо-
физом. Согласно многочисленным имеющимся данным, нейро-
Эндокринология 475
аблица 25.1. Рилизинг-факторы гипоталамуса
Рилизинг-фактор
ТТГ—РФа> (ТРФ)
ФСГ ч
>РФ
лг /
СТГ-РФ
СТ-ИФб> (сомато-
статин)
АКТГ-РФ
Пролактин-РФ
Пролактин-ИФв)
МСГ-РФ
МСГ-ИФ
Характеристика
Трипептид
Декапептид
Тетрадекапептид
Частично очищен
Трипептид
Пентапептид
Аминокислотная последовательность
(аминокислоты не обведены)
|Пиро| -Глу-Гис-Про- NH2 I
|Пиро 1 -Глу-Гис-Три-Сер-Тир-Гли-
Лей-Арг-Про-Гли-ЫНг!
Структура не известна
|Н|-Ала-Гли-Цис-Лиз-Асн-Фен-Фен-
Три-Лиз-Тре-Фен-Тре-Сер-Цис—
/°н|
Структура не известна
То же
»
Про-Лей-Гли-|ЫН2|
|н|-Цис-Тир-Илей-Гли-Асн-|Ш|
РФ — рилизинг-фактор.
ИФ — ингибирующий фактор.
Доказано, что медиатор гипоталамуса дофамин тормозит выделение пролактина.
секрет гипоталамуса и срединного возвышения (гормоны)
поступает в аденогипофиз по портальным сосудам (венам),
проходящим в ножке воронки (рис. 25.5). При нарушении этого
пути оттока (например, в результате перерезки или остановки
кровотока в области ножки воронки и срединного возвышения)
поступление рилизинг-факторов гипоталамуса в гипофиз
прекращается.
РЕГУЛЯЦИЯ НЕИРОСЕКРЕЦИИ ПО МЕХАНИЗМУ
ОБРАТНОЙ СВЯЗИ
Хорошо известно, что циркулирующие в крови гормоны
могут действовать либо на гипофиз, ингибируя высвобождение
тропных гормонов, либо на гипоталамус, снижая
высвобождение рилизинг-факторов (рис. 25.6). Это так называемая
отрицательная обратная связь, обеспечивающая поддержание
необходимого уровня гормонов. Стероидные гормоны (эстрогены,
андрогены и прогестерон) оказывают ингибирующее действие
преимущественно на уровне гипоталамуса, а гормоны щитовид-
31*
476
Глава 25
Короткая
петля
-=г^
Гипоталамус
=7^6
Л^
Аденогипофиз
_Ж_
Тропные гормоны
Ч/
Гормоны
желез-мишеней
Длинная
петля
Рис. 25.6. Короткая (3) и длинные (/ и 2) петли обратной связи,
обусловливающей торможение синтеза и высвобождения гормонов аденогипофиза и
гипоталамуса гормонами желез-мишеней и тройными гормонами
аденогипофиза. Стероидные гормоны оказывают тормозящее действие преимущественно по
типу петли /, а гормоны щитовидной железы — петли 2.
ной железы — на уровне гипофиза. Некоторые тропные
гормоны гипофиза (АКТГ, СТГ, ФСГ и ЛГ) действуют на уровне
гипоталамуса и, возможно, возвращаются в гипофиз путем
обратной диффузии (короткая петля обратной связи).
Действие гормонов, поступающих в гипоталамус с
кровотоком, реализуется через длинную петлю обратной связи. В
некоторых случаях гормон какой-либо эндокринной железы
стимулирует высвобождение соответствующего тропного гормона
или рилизинг-фактора. Это так называемая положительная
обратная связь.
ГОРМОНЫ АДЕНОГИПОФИЗА
Аденогипофиз расположен в углублении клиновидной
кости— в так называемом «турецком седле». У человека он весит
около 500 мг, а максимальный его диаметр равен примерно
1,4 см (рис. 25.5). При микроскопическом исследовании в
зависимости от реакции на красители в аденогипофизе выявляются
три основных типа клеток: 1) хромофобные, составляющие
около половины всех клеток аденогипофиза; их ядра
окрашиваются синими (основными) красителями; 2) ацидофильные (около
35% всех клеток); их цитоплазматические гранулы
окрашиваются кислыми красителями в розовый и оранжевый тона; 3) базо-
фильные (около 10%); их цитоплазматические гранулы окра-
Эндокринология 477
фиваются в синий цвет. Точное число тех или иных клеток
зависит от возраста и ряда других факторов.
Размеры и форма этих клеток и содержащихся в их
цитоплазме гранул варьируют; возможно, это отражает их
секреторную активность. Существуют убедительные данные о том, что
ацидофильные клетки синтезируют СТГ и пролактин, хромо-
фобные — МСГ, а базофильные клетки различного типа с
гранулами разной формы — большинство других гормонов адено-
гипофиза. Лучший метод определения секреторной активности
клеток — иммуноцитологический.
Все гормоны гипофша были выделены и очищены, а часть
из них — синтезирована. ТТГ, Л Г и ФСГ представляют собой
гликопротеины с мол. весом около 30 000; они состоят из двух
полипептидных цепей и присоединенных к ним углеводов.
Каждый из этих гормонов образован двумя гликопротеиновыми
субъединицами (аи р), представляющими собой
последовательность аминокислот. Альфа-субъединица ТТГ и бета-субъедини-
ца ЛГ почти идентичны и содержат 'по 96 аминокислот.
Альфа-субъединица ЛГ содержит 120 аминокислот, а бета-субъеди-
ница ТТГ 113. Активность субъединиц проявляется только при
их соединении, но не ло отдельности, причем характер
биологического действия определяется бета-субъединицей, например:
а-ЛГ+р-ЛГ=Активный ЛГ
а-ТСТ+Р-ТТГ-Активный ТТГ
а-ТТГ+Р-ЛГ = Активный Л Г
а-ЛГ+Р-ТТГ = Активный ТТГ
Пролактин и СТГ представляют собой белки с высоким
молекулярным весом (около 22 000), состоящие примерно из
190 аминокислот. На рис. 25.7 приведена структурная формула
СТГ; видна последовательность из 188 аминокислот. СТГ
человека сходен по структуре с пролактином овец. Пролактин
обладает значительной соматотропной активностью, а СТГ —
некоторым лактотропным действием.
АКТГ и МСГ— это полипептиды, обладающие
определенным сходством строения. В их состав входит одинаковое ядро
из семи аминокислот, к которому присоединены сходные
боковые аминокислоты. АКТГ образован цепочкой из 39
аминокислот и имеет мол. вес около 4600. Аминокислотная
последовательность в этой цепочке незначительно варьирует в
зависимости от вида животного. Первые 23 аминокислотных остатка в
последовательности у всех видов одинаковы и образуют
центральную и наиболее активную часть молекул; аминокислотные
остатки 24—29 у разных видов различны.
>зо.
Рис. 25.7. Аминокислотная последовательность соматотропного гормона человека.
Эндокринология 479
ДЕЙСТВИЕ ГОРМОНОВ АДЕНОГИПОФИЗА
Здесь будут раскрыты лишь общие механизмы действия
гормонов гипофиза; дальнейшие подробности мы рассмотрим
при обсуждении соответствующих желез-мишеней.
Тиреотропный гормон (ТТГ) стимулирует рост и созревание
клеток щитовидной железы, запускает синтез и высвобождение
гормонов из этой железы.
АКТГ стимулирует рост и созревание клеток коркового слоя
надпочечников, синтез и высвобождение соответствующих
гормонов.
ФСГ стимулирует рост и развитие фолликулов яичников и
в последующем выход из них эстрогенов, а также рост яичек
и сперхматогенез.
Лютеинизирующий гормон (ЛГ) вызывает периодический
выход яйцеклетки из яичника (овуляцию), а также развитие
после этого желтого тела, секретирующего прогестерон; кроме
того, ЛГ способствует росту и развитию интерстициальных
клеток яичка (клеток Лейдига), вырабатывающих андрогены.
Соматотро1Пный гормон (СТГ, гормон роста) способствует
росту и развитию костей и других тканей в раннем возрасте.
Действие его опосредовано низкомолекулярными
полипептидами — соматомединамщ вырабатывающимися в печени в ответ на
действие СТГ. Действие СТГ отличается видовой
специфичностью.
МСГ стимулирует отложение меланина в коже. У
млекопитающих этот гормон находится в промежуточной доле гипофиза,
а у птиц — в передней.
Пролактин поддерживает существование желтого теда-JL
(I крыс или мышей (лютеотропное действие), но не у свиней, ко-
• ров, овец и ч"ёл^ёкТ.~ТНмекопитающих основная функция его
cocfOHT в иници&Цйи и поддержании лактации; у птиц
пролактин вызывает секреторную активность зоба и родительское
поведение. У некоторых видов этот гормон стимулирует рост.
Пролактин-ингибирующий фактор угнетает секрецию про-
лактина у млекопитающих. У птиц этого фактора нет.
СОДЕРЖАНИЕ ГОРМОНОВ ГИПОФИЗА
Содержание тропных гормонов в тканях гипофиза и в
крови отражает динамику их высвобождения и использования. При
высвобождении гормонов уровень их в гипофизе обычно падает,
а в крови — растет. Однако это наблюдается не всегда, так как
к повышенному выходу гормонов может приводить их
усиленный синтез без значительных изменений уровня гормонов в ги-
480
Глава 25
пофизе. При удалении или повреждении гипофиза или
гипоталамуса выход гормонов снижается или полностью подавляется
и содержание их в крови резко падает.
СОМАТОТРОПНЫЙ ГОРМОН
Содержание СТГ в плазме крови взрослого человека
составляет 3—4 нг/мл; период его полураспада равен примерно
30 мин, а рассчитанная суточная секреция—1—4 мг. Недавно
путем усреднения интегрированных значений в суточных
пробах были получены следующие цифры: у юношей в препубер-
татном возрасте — 5,10 нг/мл; у юношей в периоде полового
созревания — 7,37 нг/мл; у молодых взрослых мужчин —
3,47 нг/мл.
Как и следовало ожидать, у растущих организмов уровень
СТГ выше. Есть тенденция к снижению содержания СТГ в
первую половину дня и повышению — вечером. У женщин в
среднем уровень СТГ несколько выше. В гипофизе взрослого
человека содержание СТГ колеблется от 4 до 15 мг. При голодании
и истощении белковых запасов организма (квашиоркор)
содержание СТГ в крови повышается до 40—50 нг/мл. Факторы,
влияющие на уровень СТГ, перечислены в табл. 25.2.
Таблица 25.2. Факторы, влияющие на уровень гормона роста (СТГ)
Повышение уровня СТГ
Отход ко сну
Физическая нагрузка
Умственное напряжение, стресс
Гипогликемия, инсулин
Голодание
Некоторые аминокислоты
Опухоли гипофиза
L-ДОФА
Введение кальция
Снижение уровня СТГ
Парадоксальный сон
Свободные жирные кислоты
Введение глюкозы
Введение СТГ
Соматостатнн
Нарушение секреции СТГ. Карликовость (нанизм)
наблюдается в том случае, если в детстве снижена секреция СТГ и
содержание его в плазме (до 1—3 нг/мл). Нанизм,
обусловленный недостатком СТГ, характеризуется пропорциональным
уменьшением размеров всех органов; если же нанизм связан с
гипофункцией щитовидной железы, то отмечается
непропорциональное телосложение. У гипофизарных карликов наблюдается
снижение уровня других гормонов и задержка полового
развития.
Эндокринология
481
При повышенной секреции СТГ
у детей наблюдается патологически
быстрый рост, приводящий к
гигантизму. Такие люди чрезвычайно
высоки (до 2,4—2,8 м);
усиливается рост всех тканей, но в
особенности длинных костей. Гигантизм
обычно наблюдается при опухолях
гипофиза у детей.
Акромегалией называют
заболевание, обусловленное повышенной
секрецией СТГ (уровень в плазме
от 20 до 100 нг/мл) у взрослых,
когда рост длинных костей уже
завершен. В этом случае СТГ уже не
может вызывать рост костей в
длину, однако может приводить к
их утолщению и росту мягких
тканей. Особенно увеличиваются кисти,
стопы, нос, надбровья и нижняя
челюсть, которая при этом выдается
вперед (рис. 25.8).
Механизм действия СТГ связан
с обменными процессами и
опосредован соматомединами —
полипептидами, вырабатывающимися в
печени под влиянием СТГ. СТГ
стимулирует синтез и снижает распад
белков (1), способствует
мобилизации жиров и использованию их в качестве источника
(2) и снижает потребление углеводов (3).
Подробнее уровни ТТГ, АКТГ, ФСГ, ЛГ и пролактина и ме
ханизмы их действия будут обсуждаться в главах,
посвященных соответствующим железам-мишеням — щитовидной, молоч
ным, половым и надпочечникам.
Рис. 25.8. Акромегалия.
Обратите внимание на выдающуюся
нижнюю челюсть н увеличение
носа.
энергии
ГОРМОНЫ ЗАДНЕЙ ДОЛИ ГИПОФИЗА
Основные гормоны задней доли гипофиза у человека и
большинства млекопитающих — это окситоцин и аргинин-вазо-
прессин (у птиц и некоторых других животных — окситоцин и
аргинин-вазотоцин). У свиней обнаружен лизин-вазопрессин.
Каждый из этих гормонов образован цепью из 9
аминокислот, расположенных в определенной последовательности и
связанных боковыми мостиками (рис. 25.9). Эти гормоны запасены
в задней доле гипофиза, где содержание их значительно: около
432
Глава 25
25 мкг аргинин-вазопрессина и примерно в 1,5 раза меньше
окситоцина. Окситоцин и вазопрессин образуются в супраопти-
ческом и паравентрикулярном ядрах гипоталамуса; отсюда
гранулы нейросекрета поступают по нервным волокнам в нейроги-
пофиз (рис. 25.5), где и хранятся вместе с
белками-переносчиками— нейрофизином I (для окситоцина) и нейрофизином II
(для вазопрессипа). Согласно последним данным, у многих ви-
I 1
Цис-Тир-Иле-Глн -Асы -Цис-Про-Лей-Глн
1234 5 6789
Окситоцин
фен Apr .
8-аргинин- вазопрессин
Apr
8-аргинин-окситоцин (вазотоцин)
Рис. 25.9. Строение гормонов нейрогипофиза у млекопитающих (окситоцин и
вазопрессин) и у птиц и низших позвоночных (вместо вазопрессина —
вазотоцин). Сплошными линиями обозначены аминокислоты, общие у
вазопрессина и вазотоцина с окситоцином.
дов млекопитающих окситоцин образуется в основном в
паравентрикулярном, а вазопрессин — преимущественно в супраоп-
тическом ядре.
АКТИВНОСТЬ ГОРМОНОВ ЗАДНЕЙ ДОЛИ ГИПОФИЗА
Аргинин-вазопрессин представляет собой антидиуретический
гормон (АДГ) млекопитающих, а вазотоцин — птиц. Он
действует на дистальные канальцы и собирательные трубочки почек,
увеличивая реабсорбцию воды и снижая диурез. Он активнее
и играет более важную роль у пустынных животных
(верблюдов, кенгуру, крысы) в местах, где воды значительно меньше и
сохранение ее имеет большое значение.
На секрецию АДГ влияет содержание воды и электролитов
в крови и тканях. При обезвоживании и уменьшении количества
воды в организме секреция АДГ увеличивается. Уровень АДГ
в крови, отражающий интенсивность его синтеза, зависит от
степени гидратации организма (табл. 25.3). При уменьшении
количества воды в организме и объема крови секреция АДГ
значительно возрастает, а при потреблении воды — снижается.
Повышение осмоляльности (содержания электролитов) плазмы
служит мощным раздражителем для выброса АДГ у
млекопитающих (но не у птиц).
На секрецию АДГ влияют и другие факторы. К ним
относится возбуждение областей центральной нервной системы
Эндокринология
483
Таблица 25.3. Зависимость скорости секреции и содержания АДГ
от водного баланса у человека
Состояние водного баланса
Норма
Дегидратация
Значительная кровопотеря
Водная нагрузка (гидратация)
Содержание ЛДГ в
плазме, мкЕД/мл
2
6-8
900
<1
Скорость секреции
ЛДГ, мл/ч
10
20—40
Нет данных
То же
(ЦНС), расположенных выше гипоталамуса при боли, тревоге,
нервном напряжении во время хирургических операций, а
также при еведении ряда лекарственных средств. Потребление
алкоголя угнетает секрецию АДГ и приводит к увеличению
диуреза и обезвоживанию.
При патологическом снижении секреции АДГ возникает
несахарный диабет, характеризующийся повышенным диурезом
и потреблением воды. Для его лечения можно вводить аргинин-
вазопрессин.
окситоцин
Уровень окситоцина в плазме человека составляет от 1 до
5 мкЕД/мл. Период его полураспада невелик (1—4 мин); он
быстро подвергается метаболизму и выводится с мочой.
Наиболее мощным стимулом для выработки окситоцина у
млекопитающих является кормление грудью и растяжение матки и
влагалища. При кормлении грудью возбуждаются рецепторы
молочных желез, импульсы от которых .поступают в головной мозг
и гипоталамус и вызывают выброс содержащегося в последнем
окситоцина в кровь (рис. 25.5). Окситоцин выделяется
порциями в различных количествах.
Окситоцин переносится к молочной железе и вызывает
сокращение миоэпителиальных клеток ее альвеол и протоков; при
этом происходит выделение молока из железы. Окситоцин
влияет лишь на выброс молока, уже находящегося в млечных
протоках, но не на его секрецию, или лактацию (см. гл. 16).
Сосательный рефлекс имеет большое значение в стимуляции
лактации. Существует еще ряд факторов, кроме самого акта
кормления, способствующих лактации. При страхе или
возбуждении отделение молока угнетается.
РОДЫ
Значение окситоцина для сокращений мускулатуры матки
в конце беременности и при родах не ясно. Известно, что во
время родов содержание окситоцина в крови женщин достигает
484
Глава 25
максимального значения — 200 мкЕД/мл. Этот высокий уровень
окситоцина может способствовать усилению сокращений матки
и облегчить роды, особенно после того, как они уже начались.
Очевидно, для родовой деятельности большее значение имеет
растяжение матки. Так, роды возможны у женщин, перенесших
удаление гипофиза и не имеющих гормонов его задней доли.
Действие вазотоцина на матку курицы аналогично действию
окситоцина у млекопитающих: непосредственно перед
откладыванием яйца уровень этого гормона у курицы резко возрастает.
ЛИТЕРАТУРА
Bentley P. J. (1976). Comparative vertebrate endocrinology, Cambridge
University Press, London and New York.
Catt K. J. (1971). An ABC of endocrinology, Little, Brown, Boston.
Ganong W. F. (1977). Review of medical physiology, 8th edn., Lange Medical,
Los Altos, California.
Greep P. 0., Koblinsky M. A. (1977). Frontiers in reproduction and fertility
control, MIT Press, Cambridge.
Greep R. 0. (ed.) (1977). Reproductive physiology, International Review of
Physiology, vol. 13, Chaps. 1 and 8, University Park Press, Baltimore.
Ingbar S. H. (ed.) (1976). The year in endocrinology 1975—1976, Plenum, New
York.
McCann S. M. (ed.) (1977). Endocrine physiology II, International Review of
Physiology, vol. 16, Chapter 1—3, University Park Press, Baltimore.
Sturkie P. D. (1976). Avian physiology, 3rd edn., Chap. 15, Springer Verlag,
New York.
Tepperman J. (1973). Metabolic and endocrine physiology, 3rd edn., Year Book
Medical, Chicago.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Дайте определение эндокринных и экзокринных желез.
2. Расскажите о механизмах действия гормонов.
3. Назовите гормоны передней доли гипофиза (аденогипофиза)
и их основные функции.
4. Какие факторы влияют на секрецию и высвобождение
гормонов передней доли гипофиза?
5. Что такое нейросекреторная система, и как она
функционирует?
6. Какие секреторные функции аденогипофиза выполняются
различными типами его клеток?
7. Что происходит при гиперсекреции СТГ у взрослых?
8. Что наблюдается при недостаточном образовании ФСГ?
9. Какие гормоны вырабатываются в задней доле гипофиза, и
какова их функция?
Глава 26
РАЗМНОЖЕНИЕ
К мужским половым органам (рис. 26.1) относятся яички
(семенники), половой член и добавочные железы и протоки, по
которым секреты предстательной железы, яичек и семенных
пузырьков поступают к половому члену.
Яички представляют собой парные органы. Они состоят из
семенных канальцев, в которых происходит сперматогенез, и
выводных протоков, по которым сперматозоиды поступают в
придаток яичка; здесь они хранятся и затем по семявыносяще-
му протоку направляются к половому члену. Моча,
поступающая из мочевого пузыря, и секреты яичек и добавочных
половых желез выбрасываются из организма через уретру
(мочеиспускательный канал) —общий выводной проток полового члена.
Половой член состоит из чувствительного отдела — головки
и большого количества кавернозной ткани, образующей
губчатые и кавернозные тела. При эрекции эта ткань переполняется
кровью (рис. 26.1,5), в результате чего длина и диаметр
полового члена значительно увеличиваются.
эрекция, эякуляция и эмиссия
Эрекция полового члена наступает в результате расширения
его артериол. При этом кавернозные и губчатые тела (отделы
полового члена, образованные кавернозной тканью)
переполняются кровью, вследствие чего вены сдавливаются и отток крови
затрудняется; это способствует поддержанию эрекции.
Эфферентные (двигательные) нервы, вызывающие расширение
артериол, относятся к парасимпатическим сакральным волокнам
(nervi erigentes).
К эрекции приводят физические и психические
раздражители. К первым относятся прикосновения к половому члену и его
массаж; при этом афферентные импульсы поступают по
половым нервам к центрам спинномозговых рефлексов и к высшим
Центрам головного мозга. Эрекция усиливается при действии
Других стимулов — например при раздражении заднего прохода
и мошонки. К половому возбуждению может также привести
растяжение мочевого пузыря и раздражение предстательной
486
Глава 26
Семенной пузырек
Лобковый симфиз
Уретра
Венчик головки
полового члена
Головка
полового Члена
СемявыносяЩий проток
емявыбрасывающий проток
■Предстательная железа
Мужская маточка
Прямая кишка
Мужская
маточка
Предстательная
железа
Дорсальные вены Дорсальная артерия и неов
___ Кожный покров
Белочная
оболочка
Уретра
Рис. 26.1. Мужские половые органы. А. Срединный разрез области таза.
Б. Продольный разрез полового члена. В. Яичко на разрезе: 1 — белочная
оболочка, 2— перегородка яичка, 3 — семенные канальцы, 4 — сеть яичка, 5—
выносящие канальцы, 6 — придаток яичка, 7 — семявыносящий проток.
Г. Поперечный разрез полового члена.
железы и уретры. Половое поведение и влияние на него со
стороны гипоталамуса рассматриваются в гл. 10.
К психическим раздражителям, вызывающим половое
возбуждение и эрекцию, относятся сексуальные мысли, вид
обнаженного тела и эротические сновидения. У юношей часто
наблюдаются ночные семяизвержения (поллюции).
Эмиссия начинается с сокращений придатка яичка и семя-
выносящего протока, в результате которых сперма
выбрасывается в уретру. Сокращение семенных пузырьков и
предстательной железы и выброс их секретов способствуют
дальнейшему продвижению спермы по уретре. Семенная жидкость
состоит из всех перечисленных секретов, а также выделений куперо-
вых желез. Заключительный этап выброса семенной жидкости
из мочеиспускательного канала полового члена называется
эякуляцией.
Размножение
487
Афферентные импульсы при эмиссии и эякуляции идут
преимущественно от тактильных рецепторов головки полового
члена и по внутренним половым нервам поступают в спинной
мозг. Эмиссия наступает в результате поступления к половому
члену эфферентных импульсов по тюдчревным нервам (Li и L2
сегменты спинного мозга). Эякуляция связана с ритмическими
сокращениями скелетных мышц, окружающих эректильную
ткань полового члена (рис. 26.1). Спинномозговые центры
эякуляции расположены в верхних крестцовых и нижних
поясничных сегментах; эфферентные импульсы от этих центров
поступают по 1—3 крестцовым корешкам и внутренним половым
нервам.
Объем эякулята обычно колеблется от 2 до 6 мл, составляя
в среднем 3,5 мл. Реакция эякулята слабощелочная (рН 7,0—
7,5). В семенной жидкости содержится много простагланди-
нов — гормонов, вырабатываемых семенными пузырьками.
СПЕРМАТОГЕНЕЗ И СПЕРМАТОЗОИДЫ
На рис. 26.2 изображено формирование сперматозоидов в
яичках. Зрелый сперматозоид в своем развитии проходит
следующие стадии: 1) сперматогония; 2) сперматоцит первого
порядка; 3) сперматоцит второго порядка; 4) сперматида
(незрелый сперматозоид) и 5) сперматозоид.
Сперматогонии
Рис. 26.2. Поперечный разрез яичка. Видны семенные канальцы, клетки Лей-
Аига, секретирующие тестостерон, и сперматозоиды, образующиеся из
сперматогонии через стадии сперматоцитов первого и второго порядка, и спер-
матид.
488
Глава 26
Очевидно, процесс созревания сперматозоидов связан с
функцией клеток Сертоли. Для образования зрелого
сперматозоида из сперматогоний требуется около 70 дней.
Форма и размер сперматозоидов у разных видов различна,
однако общий план их строения одинаков (рис. 26.3).
Сперматозоид состоит из головки,
содержащей ядро средней части,
и длинного хвоста, похожего
на кнут. При оплодотворении
головка сперматозоида
проникает в яйцеклетку.
Акросома
Ядро
Клеточная
мембрана
Цснтриоль
М и то хо и л у я ал ь -
ная оболочка
Осезая нить
\Головка
Фибриллы
в оболочке
хвоста
ОПЛОДОТВОРЕНИЕ
Перифери-
Яйцеклетка может быть
оплодотворена в течение 24—
26 ч после выхода ее из
яичника. Большая часть
сперматозоидов сохраняется в
женских половых путях до 24—
48 ч, хотя некоторые из них
жизнеспособны до 72 ч. В
связи с этим период, в течение
которого возможно
оплодотворение, ограничен несколькими
j- ческий отдел днями,
хвоста g эякуляте обычного
объема (3,5 мл) содержится в
среднем 120 млн.
сперматозоидов, однако эта величина
может варьировать от 40 млн. до
400 млн. и более. В
фаллопиевы трубы проникает всего 100
сперматозоидов или немногим
больше, и лишь один из них
оплодотворяет яйцеклетку,
остальные сперматозоиды погибают в женских половых путях.
Так как для оплодотворения достаточно всего одного
сперматозоида, может создаться впечатление, что общее количество этих
клеток в семенной жидкости не имеет значения, однако это не
так. Когда содержание сперматозоидов в семенной жидкости
менее 20 млн./мл, способность к оплодотворению резко
снижена. Напротив, высокое содержание сперматозоидов в семенной
жидкости в известной мере отражает их жизнеспособность и
оплодотворяющую способность.
Рис. 26.3. Схема строения
сперматозоида человека. (Ganong W. F., 1977.
Medical Physiology.)
Размножение
489
КАПАСИТАЦИЯ И УГНЕТЕНИЕ ОПЛОДОТВОРЯЮЩЕЙ СПОСОБНОСТИ
У некоторых видов животных сперматозоиды, покинувшие
придаток яичка, приобретают способность к оплодотворению
яйцеклетки лишь после того, как некоторое время пробудут в
организме самки; это явление называется капаситацией. У
кролика время капаситации составляет около 6 ч, у человека ка-
паситация не обязательна.
Семенная жидкость имеет слабую щелочную реакцию, а
содержимое влагалища — кислую (рН 3—4). Активность
сперматозоида при рН<6,5 снижена. При температуре тела
сперматозоиды жизнеспособны в течение 24—72 ч, а в придатках
яичка срок их жизни значительно больше. Если семенную жидкость
быстро заморозить, то ее можно хранить в течение нескольких
месяцев и использовать для искусственного оплодотворения.
Способность самцов к оплодотворению может быть снижена
или подавлена агентами, угнетающими нормальное созревание
сперматозоидов. Применение их по отношению к человеку
обычно не рекомендуется.
МУЖСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Основным гормоном семенников (андрогеном) является
тестостерон, вырабатываемый интерстициальными клетками
яичек (клетками Лейдига). Основные пути синтеза тестостерона
у человека следующие:
холестерин
I
прегненолон;
t
активируется 17а-гидроксилазой и образует:
I
17-оксипрегненолон
Y
дегидроэпиандростерон
\
андростендиол
{
тестостерон
Существует и другой путь образования тестостерона, имеющий
У человека меньшее значение:
прогестерон
т
17а-оксипрогестерон
I
андростендиол
Т
тестостерон
490
Глава 26
Строение ряда андрогенов приведено на рис. 26.6. Более
половины общего количества тестостерона крови связывается
белками плазмы — альбумином и глобулином. Большая часть
циркулирующего в крови тестостерона превращается в печени в
17-кетостероиды и выводится с мочой.
ДЕЙСТВИЕ ТЕСТОСТЕРОНА
Действием тестостерона обусловлено развитие в
пубертатном периоде вторичных мужских половых
признаков—огрубление голоса, появление волос в подмышечных впадинах и на
лобке, рост бороды и увеличение размеров наружных половых
органов. Под его влиянием изменяется телосложение — плечк
становятся шире, мышцы увеличиваются, наступает изменение
в характере оволосения. У подопытных животных андрогены
достоверно вызывают агрессивное поведение; у мужчин это
влияние выражено в меньшей степени.
Облысение, при наличии к нему наследственного
предрасположения, связано с выработкой андрогенов. У мужчин,
кастрированных в препубертатном периоде, облысения не наступает.
Под действием андрогенов в организме усиливается синтез
и угнетается распад белков (анаболическое действие). Введение
экзогенных андрогенов может привести к усилению полового
влечения (либидо). При крипторхизме яички не опускаются из
брюшной полости и функционируют при температуре тела
более высокой, чем в мошонке. Эта температура препятствует
нормальному протеканию сперматогенеза, и поэтому криптор-
хизм обычно сочетается с бесплодием, хотя тестостерон может
вырабатываться. Введение гонадотропного гормона может
способствовать опусканию яичек.
Состояние, обусловленное нарушением функции клеток Лей-
дига или их отсутствием в результате либо врожденных
аномалий, либо кастрации в препубертатном периоде, называется
евнухоидизмом. Для лиц, страдающих евнухоидизмом,
характерны узкие плечи, плохо развитая мускулатура, женственная
внешность, небольшие размеры половых органов, высокий голос
и редкий или вовсе отсутствующий волосяной покров на лобке.
Введение таким больным андрогенов в течение 3—6 мес может
привести к увеличению и развитию половых органов и росту
волос на лобке (рис. 26.4).
РЕПРОДУКТИВНАЯ ФУНКЦИЯ У ЖЕНЩИН
У женщин детородный период начинается примерно с 14—
16 лет и длится до наступления менопаузы, т. е. до 40—50 лет.
Изменения, связанные с наступлением половой зрелости, назы-
Размножение
491
Рис. 26.4. Больной крипторхизмом с евнухоидным синдромом в возрасте
167г лет. А. До лечения. Б. После лечения андрогеном (6 мес).
ваются пубертатными, а период, предшествующий половой
зрелости, называется препубертатным. У женщин в пубертатном
периоде изменяются размеры и контуры тела, особенно бедер,
а также первичные и вторичные половые 'признаки:
увеличиваются наружные половые органы (рис. 26.5), появляются волосы
на лобке, развиваются молочные железы. Во внутренних
органах— яичниках, матке и влагалище также наступают
изменения, знаменующиеся появлением менструаций. Эти изменения
обусловлены выработкой и действием гонадотропных гормонов
гипофиза (ФСГ и ЛГ) и гормонов яичника (эстрогенов, аидро-
генов и прогестерона) (рис. 26.6).
В препубертатном периоде ФСГ, ЛГ и гормоны яичников
вырабатываются в небольших количествах. После наступления
половой зрелости и во время всего детородного периода
секреция этих гормонов повышается и принимает циклический
характер. В менопаузе же реакция яичников на действие ФСГ и
ЛГ снижается, и, хотя секреция ФСГ и ЛГ по сравнению с до-
климактерическим периодом даже выше, гормоны яичников
(эстрогены и прогестерон) вырабатываются в малых
количествах или не вырабатываются вовсе. Повышенная секреция ФСГ
Матка
Фаллопиева труба
Шейка матки
Отверстие уретры
Девственная
плева
Отверстие
влагалища
Головка клитора
Большая
половая губа
Малая
половая губа
Заднепроходное
отверстие
Лобковый
симфиз
Уретра
Сфинктер
заднего прохода
В
Рис. 26.5. Женские половые органы. А. Верхние отделы женской половой системы; видны яичники, фаллопиевы
трубы, матка и влагалище. Б. Срединный разрез области таза. В. Наружные половые органы. Видны клятор и
вульва; к последней относятся большие и малые половые губы,
17-/5- эстрадиол
Тестостерон
Эстриол
Прогестерон
Рис. 26.6. Структурные формулы некоторых половых гормонов.
с=о
Женщина
Мужчина
(44+ХХ) Ооцит (44+XY) Первичный
Г J первого порядка V J сперматоцит
Первое деление N.—У Первое деление)
созревания (мейоз) / \ созревания
Ооцит S~\ V3fX (мей03) 1^р + у^22+У>1Вторичные
второго порядка (22 + Х ) (А Редукционное Г /V У сперматоциты
^ тельце Второе деление)
созревания
Второе деление
созревания
Зрелый ооцит
Сперматиды и
22+Х (22+Х )(22 + Y J(22+Y J сперматозоиды
Женские зиготы (XX)
Мужские зиготы (ХУ)
Рис. 26.7. Схемы овогенеза и сперматогенеза.
493
494
Глава 26
и ЛГ в гипофизе обусловлена устранением действия
отрицательной обратной связи.
В состав репродуктивного цикла входят такие стадии, как
1) овогенез и оплодотворение (рис. 26.7), 2) овуляция и
менструация (рис. 26.8), 3) беременность.
Денегерирующее
желтое тело
Менструальная
(яичниковая)
фаза
Растущий Зрелый
фолликул фолликул Овуляция
Дегенерирующее
желтое тело
Под влиянием
Эстрогена
Постменструальная
(фолликулярная) фаза
Прогестерона
Пременструальная фаза
(фаза желтого тела)
Дни
4-5
I
14
I
I
28
Рис. 26.8. Рост яйцеклетки, овуляция и изменение эндометрия в ходе
менструального цикла. (Schottelius В. A., Schottelius D. А., 1978. Textbook of
Physiology.)
АНАТОМИЯ ЖЕНСКИХ ПОЛОВЫХ ОРГАНОВ
Половая система взрослой женщины включает внутренние
половые органы (яичники, фаллопиевы трубы, матку и
влагалище) и наружные гениталии — вульву (малые и большие
половые губы, клитор и вход во влагалище, который у девственниц
может быть прикрыт девственной плевой).
ЯИЧНИКИ
У женщин яичники представляют собой парные органы,
расположенные в глубине малого таза с двух сторон то бокам от
матки. Размеры зрелого яичника составляют около 3,5x2 см,
вес —4 г. К моменту рождения в яичниках содержится
примерно 500 тысяч яйцеклеток различного размера, большинство их
сморщиваются, становятся атретическими и не достигают зре-
Размножение
495
лой стадии (овуляции). В течение детородного периода у
женщины обычно овулируют лишь 300—400 яйцеклеток (по одной
каждый месяц в течение 30—35 лет). После овуляции, т. е.
разрыва зрелого фолликула и выхода яйцеклетки, последняя
захватывается близлежащей фаллопиевой трубой, имеющей форму
воронки, и переносится в матку (рис. 26.5). Оплодотворение
яйцеклетки сперматозоидом происходит до того, как она
поступает в матку (в пределах фаллопиевой трубы).
СОЗРЕВАНИЕ ЯЙЦЕКЛЕТКИ
На поперечном сечении зрелого яичника видны яйцеклетки
на различных стадиях развития (рис. 26.9): зачатковый
эпителий, примордиальные фолликулы, растущие фолликулы, преову-
{ Наружная оболочка
Внутренняя оболочка
Зернистый слой
Рис. 26.9. Поперечное сечение яичника. Изображены различные стадии
развития яйцеклетки вплоть до овуляции и образования желтого тела. Обратите
внимание на увеличенные изображения клеток желтого тела на разных
стадиях, а также на срез зрелого фолликула.
ляторный граафов пузырек с яйцеклеткой, яйцеклетка после ее
выхода (овуляции), атретические фолликулы и желтое тело,
образовавшееся из лопнувших фолликулов. Часть желтых тел
активна, другие находятся в состоянии регрессии. В зрелом
граафовом пузырьке (рис. 26.9) видна крупная яйцеклетка,
образованная одной клеткой и расположенная внутри пузырька,
496
Глава 26
и полость, содержащая жидкость. Сам пузырек состоит из
следующих слоев клеток (от наружной поверхности к внутренней):
наружной оболочки (theca externa), внутренней оболочки (theca
interna) и слоя зернистых клеток, достигающего полости
пузырька. Ежемесячно происходит овуляция, когда эти слои
фолликула лопаются и яйцеклетка выбрасывается.
МАТКА
Матка представляет собой мышечный орган треугольной
формы, расположенный между мочевым пузырем и прямой
кишкой (рис. 26.5). Длина ее у взрослых достигает 6—8 см,
толщина стенок — 3 см, а ширина в наиболее узкой части
(перешейка)— 3 см. Матка соединяется с влагалищем посредством
шейки; во время беременности шейка матки закрыта слизистой
пробкой.
Полость матки выстлана слизистой оболочкой —
эндометрием, под которой расположен мышечный слой, или миометрий.
Эндометрий очень богато кровоснабжается и в ходе
менструального цикла претерпевает значительные изменения. Во время
месячных (рис. 26.8) эндометрий вместе с поверхностными
сосудами отторгается: в дальнейшем под действием вырабатываемого
активным желтым телом прогестерона происходит его
регенерация (пролиферация). Периодически желтое тело
дегенерирует, и эндометрий вновь отторгается (возникает менструация).
В том случае, если наступает беременность, желтое тело под
действием хорионического гонадотропина (ХГТ) сохраняется
(персистирует), ХГТ стимулирует выработку в желтом теле
прогестерона; последний совместно с эстрогенами
подготавливает эндометрий к имплантации оплодотворенной яйцеклетки и
развитию зародыша, или эмбриона.
ВЛАГАЛИЩЕ
Влагалище состоит из эпителия, выстилающего его полость,
среднего мышечного и наружного соединительнотканного слоев.
Под действием гормонов яичника происходит пролиферация,
дифференциация и десквамация (отторжение) клеток эпителия
влагалища. В препубертатном и постклимактерическом
периодах, когда синтез эстрогена понижен, эти клетки плоские и ат-
рофичные. Под действием эстрогена наступает их пролиферация
и утолщение эпителия влагалища. Прогестерон способствует
десквамации клеток поверхностного слоя.
ОВОГЕНЕЗ И СПЕРМАТОГЕНЕЗ
Овогенезом и сперматогенезом называются процессы,
благодаря которым из поколения в поколение передается постоянное
число хромосом: 44 аутосомы и 2 половые хромосомы (44 + ХХ
у женщин и 44 + XY у мужчин).
Как показано на рис. 26.7, клетки зачаткового эпителия
яичников делятся и созревают, т. е. происходит овогенез. Эти
клетки представляют собой овогонии, содержащие 44 + ХХ хромосом.
Некоторые из овогонии развиваются в овоцит первого порядка,
который затем претерпевает редукционное деление (мейоз).
В результате мейоза образуются овоциты второго порядка,
несущие половинный набор хромосом (22 аутосомы+1 X
хромосома). Это происходит на стадии зрелого фолликула
непосредственно перед овуляцией. Затем овоциты второго порядка делятся
нередукционным делением (митотически), и в результате
образуется зрелая яйцеклетка, способная к овуляции и
оплодотворению, а также редукционные тельца, которые в дальнейшем
дегенерируют.
В яичках протекает аналогичный процесс — сперматогенез
(см. рис. 26.2). Оперматогонии непрерывно растут, делятся и
образуют сперматоциты первого порядка (44 + XY). Затем
последние претерпевают редукционное деление, в результате
которого образуются сперматоциты второго порядка с гаплоидным
набором хромосом (N). В зависимости от типа половой
хромосомы сперматоциты второго порядка бывают двух видов — 22 + Х
и 22 + Y. В дальнейшем они развиваются сначала в сперматиды,
а затем в зрелые сперматозоиды, или гаметы (рис. 26.7).
Оплодотворение наступает в результате слияния
сперматозоида с яйцеклеткой; это происходит обычно в фаллопиевых
трубах. При соединении женской гаметы (22 + Х) с мужской,
содержащей набор хромосом 22 + Х, образуется женская зигота
(44 + ХХ). Если же с женской гаметой (22 + Х) соединяется
мужская гамета (22+ Y), то образуется мужская зигота
(44 + XY).
МЕНСТРУАЦИЯ И ОВУЛЯЦИЯ
Менструация и овуляция относятся к циклическим
процессам женского организма (рис. 26.8). В связи с тем что они
тесно связаны между собой, они будут рассматриваться
одновременно. Менструальный цикл равен в среднем 28 дням, однако
Длительность его может быть различной. Если у женщины
вообще не было месячных, то такое состояние называется первичной
аменореей; если же месячные были и прекратились, то говорят
0 вторичной аменорее. Болезненные менструации называются
Дисменореей.
32-1705
498
Глава 26
Во время месячных, т. е. первые 4—5 дней менструального
цикла (менструальная фаза), эпителий матки отторгается и
выходит с кровью, вытекающей из мелких разорванных сосудов.
Менструации предшествует снижение синтеза прогестерона в
результате дегенерации желтого тела; именно это явление и
служит причиной менструации.
Дни менструального цикла
Рис. 26.10. Содержание в плазме гормонов гипофиза (ЛГ и ФСГ), эстрадиола
и прогестерона в различные фазы менструального цикла — перед овуляцией,
в момент овуляции (14-й день) и в последующие дни. (Guyton А. С, 1977.
Basic Human Physiology.)
Во время постменструальной (фолликулярной) фазы
фолликулы растут, созревают и начинают вырабатывать эстрогены.
Продукция эстрогенов достигает максимума перед овуляцией,
что совпадает с пролиферацией эндометрия. Примерно в
середине цикла (14-й день), в конце поздней постменструальной
фазы, наступает овуляция и из разорвавшегося фолликула
образуется желтое тело, секретирующее прогестерон — гормон,
способствующий сохранению целостности эндометрия, а также
поддержанию и увеличению его толщины. С этого момента
начинается пременструальная фаза, или фаза желтого тела\ она
длится до конца 28-го дня, затем цикл повторяется.
Перед овуляцией и после нее повышается секреция
некоторых гормонов (рис. 26.8 и 26.10). Уровень ЛГ в крови начинает
возрастать примерно за 3 дня до овуляции, достигая максимума
Размножение
499
приблизительно за 16—24 ч. Этому пику Л Г предшествует
примерно на 24 ч максимум концентрации эстрогена. Полагают, что
пик содержания эстрогена служит пусковым механизмом для
резкого увеличения секреции Л Г (oihk ЛГ), которое и приводит
к овуляции. Одновременно с концентрацией ЛГ растет и
содержание ФСГ; эти два показателя достигают максимума
одновременно.
Через 3—4 дня после достижения максимума содержание
ФСГ и ЛГ резко падает и держится на низком уровне в течение
всей фазы активности желтого тела (лременструальной фазы,
или фазы желтого тела). Непосредственно после овуляции
содержание и эстрогена, и прогестерона начинает возрастать; на
21—22-й день менструального цикла уровень их достигает
наибольшей величины. Спустя 4—5 дней, непосредственно перед
очередной менструацией, концентрации эстрогена и
прогестерона возвращаются к низкому уровню.
Эстрогены вырабатываются преимущественно клетками
внутренней оболочки фолликула. К ним относятся 17 р-эстрадиол,
эстрон и эстриол. Из всех эстрогенов наиболее активен эстради-
ол, а наименее — эстриол. Секреция эстрадиола также наиболее
высока; в зависимости от стадии развития фолликула она
составляет: 1) 50 мкЕД/сутки в ранней фолликулярной фазе;
2) 400 мкЕД/сутки непосредственно перед овуляцией; 3) резко
уменьшается после овуляции.
У мужчин также вырабатываются некоторые эстрогены,
однако большинство эстрогенов, циркулирующих в крови мужчин
(40 мкЕД/сутки), образуется из андрогенов.
Последовательность событий 1П,ри менструальном цикле следующая:
1. Во время менструации и начала роста фолликула
вырабатывается ФСГ и ЛГ.
2. Растущие фолликулы секретируют эстроген, вызывающий
пролиферацию постменструального эндометрия.
3. В фолликулах происходит овогенез.
4. Примерно за 2 дня до овуляции уровень эстрогена достигает
максимума, что, возможно, приводит к пику секреции ЛГ,
вызывающего овуляцию.
5. За 16—24 ч уровни ФСГ и ЛГ достигают наибольшего
значения.
6. Через 16—24 ч происходит овуляция.
7. После овуляции образуется желтое тело, вырабатывающее
прогестерон (фаза желтого тела); шод действием этого
гормона сохраняется толщина и целостность эндометрия.
8. К середине фазы желтого тела (21-й день цикла) секреция
прогестерона и эстрогена (эстрадиола) достигает
максимума.
32*
500
Глава 26
9. Непосредственно перед очередной менструацией содержание
прогестерона и эстрогена возвращается к низкому уровню.
10. В результате отторжения эндометрия и разрыва
поверхностных сосудов наступает очередная менструация.
МЕНСТРУАЛЬНЫЙ И ЭСТРАЛЬНЫИ ЦИКЛЫ
Эстральным циклом называется -периодическое наступление
течки у животных; в этот период млекопитающие, за
исключением приматов, спариваются и размножаются. Самка
млекопитающих принимает самца лишь во время течки, или эструса,
примерно соответствующей моменту овуляции (1-й день). У
женщин же овуляция наступает на 14-й день цикла, а
менструация — на 1-й.
Период после течки называется метаэструсом; длительность
его у разных видов животных различна. Затем следует диэструс
(соответствует менструации у человека), потом проэструс (про-
лиферативная фаза), и вновь наступает эструс.
РЕГУЛЯЦИЯ ВЫРАБОТКИ ФСГ И ЛГ ПО МЕХАНИЗМУ
ОБРАТНОЙ СВЯЗИ
Высвобождение ФСГ и ЛГ из гипофиза регулируется
уровнем этих гормонов в крови и их секреции по механизму
короткой петли отрицательной обратной связи (см. предыдущий
раздел), уменьшающей выделение рилизинг-фактора гипоталамуса
для ФСГ и ЛГ.
Содержание эстрогенов и прогестерона влияет на секрецию
ФСГ и ЛГ посредством длинной петли обратной связи, изменяя
•высвобождение гормонов гипоталамуса и гипофиза (см. гл. 25).
Низкий уровень эстрогена тормозит выработку ЛГ во время
ранней стадии фолликулярной фазы менструального цикла.
Усиленную секрецию эстрогена связывают с усиленным синтезом
ЛГ и наступлением овуляции (пример положительной обратной
связи). После овуляции (в фазу желтого тела) секреция
эстрогена снижается, а прогестерона — возрастает; очевидно, это
увеличение секреции прогестерона служит главным фактором, обус-
^ ловливающим угнетение выделения ФСГ и повышение
выработки ЛГ.
ГОРМОНЫ ПОЛОВЫХ ЖЕЛЕЗ
На рис. 26.6 изображены структурные формулы эстрогенов,
тестостерона и прогестерона. К естественным эстрогенам
человека относятся 17 р-эстрадиол (Эг), эстриол (Э3) и эстрон
Размножение
501
(3i). Последние два гормона являются метаболитами наиболее
активного из эстрогенов — эстрадиола, большая часть которого
вырабатывается в фолликулах яичников. Эстрон частично
превращается в эстриол в печени. Считается, что эстриол
образуется е плаценте из дегидроэпиандростерона. При приеме внутрь
естественные эстрогены относительно неактивны, так как они
инактивируются в печени. Синтез 17 р-эстрадиола
осуществляется следующим образом:
Холестерин ► Прегненолон ► Прогестерон ►
► 17-гидроксипрогестерон * Тестостерон ►
* 19-нортестостерон *■ 17|3-эстрадиол.
Этот процесс протекает преимущественно в клетках оболочки
фолликулов яичника, однако он, возможно, имеет место и в
клетках Лейдига яичек, в клетках желтого тела и коры
надпочечников.
Мощным пероральным эстрогеном является синтетическое
производное эстрадиола — этенилэстрадиол, а также другое
синтетическое нестероидное соединение с эстрогенной
активностью— диэтилстильбэстрол, часто применяющийся в качестве
противозачаточного средства.
Прогестерон не только секретируется в больших количествах
в фолликулах (желтых телах) яичников, но и является важным
промежуточным продуктом биосинтеза стероидных гормонов,
протекающего в ряде органов. Период полураспада
прогестерона невелик; в печени он превращается в прегнандиол, который
в дальнейшем подвергается конъюгации и выводится с мочой.
ДЕЙСТВИЕ ЭСТРОГЕНОВ
Как указывалось выше, эстрогены вырабатываются
развивающейся яйцеклеткой и играют важную роль в менструальном
Цикле и процессе имплантации. Кроме того, они способствуют
повышению кровоснабжения матки и увеличивают активность
и силу сокращений ее мускулатуры. Под действием эстрогенов
Повышается связывание кальция мышцами и несколько
увеличивается задержка воды и солей в тканях. Эстрогены влияют
также на развитие молочных желез.
Действием эстрогенов обусловлено также развитие в
пубертатном периоде вторичных половых признаков. Широкие бедра
и узкие плечи у женщин—пример влияния эстрогенов на
телосложение. Они влияют также на распределение жировой ткани
и накопление ее в области молочных желез и ягодиц.
502
Глава 26
БЕРЕМЕННОСТЬ И РОДЫ
Оплодотворение яйцеклетки наступает в фаллопиевых
трубах после овуляции; при этом в яйцеклетку проникает только
один сперматозоид. Ядра яйцеклетки и сперматозоида
сливаются и образуется зигота. Затем зигота опускается по трубе в
матку и через 7!/2 дня после оплодотворения наступает
имплантация зиготы в эндометрий. Это происходит после того, как
зигота достигает стадии 200 клеток (бластоциста). Имплантация
возможна лишь в том случае, если эндометрий соответствующим
образом подготовлен к этому процессу в результате действия
эстрогенов и прогестерона. По мере внедрения бластоцисты в
эндометрий последний разрастается вокруг нее; с этого
момента бластоциста называется зародышем, или эмбрионом. В
дальнейшем образуется плацента, а затем — амнион и хорион
(рис. 26.11).
5-7 месяцев ]
Рис. 26.11. Различные стадии развития эмбриона; видна плацента и оболочки
плода.
Размножение
503
СЕКРЕЦИЯ ГОРМОНОВ В ПЕРИОД БЕРЕМЕННОСТИ
Во время беременности желтое тело под действием ХГТ
сохраняется и продолжает вырабатывать эстрогены и
прогестерон. У большинства млекопитающих удаление яичников
приводит к выкидышу; у женщин же этого не происходит, так как
после 6-й недели беременности эстрогены и прогестерон в
достаточных количествах вырабатываются плацентой.
В раннем периоде беременности секреция ХГТ низка; затем,
по мере развития плаценты, она быстро увеличивается,
достигает максимума к 8—9-й неделе, а к 16—20-й неделе резко
снижается (рис. 26.12). Уровень прогестерона в течение всего
срока беременности повышается, что указывает на
функциональную активность желтого тела. Содержание прогестерона в крови
может колебаться от 2 мкг/мл на 2-й неделе беременности до
17 мкг/мл к 40-й. В последнюю неделю значительно возрастает
уровень эстриола.
0 4 8 12 16 20 24 28 32 36 40
Недели
Рис. 26.12. Уровни гормонов в плазме при беременности (0—40 недель). (По
Catt К. J. 1971. An ABC of endocrinology.)
У женщины беременность длится около 9 месяцев или
270 дней. Сроки беременности для некоторых других видов
млекопитающих следующие: у коровы—280 дней, у овцы—148 дней,
У лошади — 337 дней, у собаки и кошки — 63 дня, у крысы —
22 дня.
Механизм начала родов не ясен. Не вызывает сомнения тот
факт, что на последней стадии беременности матка
чувствительнее по отношению к различным раздражителям. В начале
родовой деятельности матка сокращается один раз в 30 мин,
затем частота сокращений достигает 1—3 в 1 мин. Очевидно, на
504
Глава 26
родовую деятельность влияет гормон задней доли гипофиза ок-
ситоцин, хотя роды могут протекать нормально и при
пониженной секреции этого гормона или в его отсутствие.
По всей вероятности, главными факторами, приводящими к
родовой деятельности, служат рост плода и растяжение матки.
Изгнанию плода способствует также натуживание, сокращение
брюшных мышц и некоторые рефлексы. В большинстве случаев
сначала рождается головка плода, но иногда наблюдается
ягодичное предлежание.
ЛАКТАЦИЯ
Лактация включает рост и развитие молочных желез,
выработку молока и его выделение. Молочные железы человека —
это парные органы, расположенные в верхней части грудной
Молочные протоки
Молочный синус
Альвеолы
Сосок
Междольковая
перегородка
Жировая ткань
Рис. 26.13. Схема строения молочной железы взрослой женщины. Видны
альвеолы в различных стадиях развития: 1 — в покое, 2 — при беременности,
3 — в период лактации.
Размножение
505
клетки по бокам от срединной линии. У взрослой женщины
молочные железы представляют собой упругие округлые
образования, состоящие из долек; в центре каждой железы
расположен сосок (рис. 26.13). В препубертатном возрасте молочные
железы сходны у представителей обоих полов и размеры их
невелики. С наступлением половой зрелости у женщин молочные
железы начинают расти и развиваться главным образом под
действием эстрогенов и прогестерона. Эстрогены усиливают
рост и развитие долек и альвеол, в которых вырабатывается
молоко. Альвеолы сообщаются с соском посредством млечных
ходов.
После того как молочные железы с их протоками и
альвеолами полностью сформируются, начинается выработка молока
под действием гормона гипофиза пролактина. Для этого
необходимо также, чтобы секреция других гормонов, влияющих на
рост и строение тканей, была нормальной. К этим гормонам
относятся, помимо эстрогенов и прогестерона, СТГ, тироксин,
инсулин и кортикоиды. Серьезные гормональные сдвиги могут
приводить к нарушению выработки молока даже при нормальном
уровне пролактина.
Примерно с 5-го месяца беременности молочные железы
начинают увеличиваться, что связано с увеличением количества
жира, а также размеров и числа альвеол и долек (рис. 26.13).
После родов выработка молока значительно увеличивается
и прекращается лишь через несколько месяцев после отнятия
от груди. Это совпадает с уменьшением секреции эстрогенов и
прогестерона, а также ФСГ и ЛГ, и с повышением выработки
пролактина. Секреция пролактина происходит периодически с
интервалами по 20—30 мин. Она значительно колеблется даже
в течение суток. Содержание пролактина в крови небеременных
женщин составляет в среднем около 8—10 нг/мл, повышаясь
при беременности до 100—400 нг/мл. При повышении выработки
молока уровень пролактина в плазме возрастает, а при
увеличении длительности лактации — снижается.
Секреция пролактина значительно увеличивается (в 10 раз)
в начале вскармливания. Кормление грудью способствует
уменьшению выделения ФСГ и ЛГ и предупреждает или
отдаляет наступление овуляции и менструации. У женщин, не
кормящих грудью, менструальный цикл восстанавливается через
6 недель после родов.
Выделение молока. Как указывалось выше, гормон задней
Доли гипофиза окситоцин вызывает еыброс молока из млечных
Цистерн и протоков по направлению к соску, не влияя при этом
и а секрецию молока. Окситоцин выделяется в ответ на
кормление грудью или связанные с кормлением рефлекторные
воздействия, а также при раздражении молочных желез (см. гл. 25}«
506
Глава 26
ПОЛОВОЙ АКТ
Как и головка полового члена у мужчин, клитор у женщин
содержит чувствительную к раздражителям эректильную ткань.
И клитор, и половой член иннервируются двигательными
парасимпатическими нервами (nervi erigentes). При раздражении
этих нервов происходит расширение артерий клитора и сужение
его вен, что приводит к эрекции клитора. Эрекция клитора
сопровождается увлажнением влагалища, обусловленным не
столько секреторной активностью бартолиниевых желез, как
считалось ранее, сколько транссудацией жидкости
непосредственно из влагалищного эпителия.
Эффект раздражения клитора и вульвы усиливается при
прикосновении к молочным железам и другим эрогенным зонам.
Психические раздражители, действующие через центральную
нервную систему, могут быть обонятельными, слуховыми и
зрительными. Когда возбуждение от действия психических и
физических раздражителей достигает наивысшей точки, наступает
юргазм. Нервные процессы, происходящие при этом у женщин
и мужчин, сходны, однако у женщин нет секреторных реакций,
подобных выбросу семенной жидкости. Кроме того, женщина
способна повторно испытывать оргазм через короткие
промежутки времени.
Оргазм наступает в результате возбуждения симпатические
двигательных волокон подчревных нервов. При половом
возбуждении и оргазме как у мужчин, так и у женщин
значительно изменяется частота дыханий и сокращений сердца и
кровяное давление. Частота сокращений сердца, составляющая в
покое около 70—80 в 1 мин, достигает 125 <при умеренном половом
возбуждении и 180 три оргазме. Кровяное давление в момент
оргазма повышается на 30—80 мм рт. ст. Влияние
гипоталамуса на половое поведение рассматривается в гл. 10.
КОНТРАЦЕПЦИЯ
В связи с тем что яйцеклетка может быть оплодотворена
лишь в течение 24 ч после овуляции, а сперматозоиды в жен
ских половых путях сохраняют жизнеспособность не более 72 ч
естественным способом предохранения от беременности являет
ся воздержание от полового сношения в течение трех дней дс
и после ожидаемого срока овуляции, т. е. 6 дней (циклические
способ). Этот способ крайне ненадежен из-за колебаний менст
руально-овариального цикла и других факторов.
Размножение
507
ГОРМОНАЛЬНЫЕ ПРОТИВОЗАЧАТОЧНЫЕ СРЕДСТВА
Вероятность зачатия у женщин может быть снижена путем
предупреждения овуляции. Для этого предложены пероральные
противозачаточные средства (синтетические гормоны). Схемы
приема этих препаратов могут быть следующими: 1) прием в
течение 20 дней комбинации эстрогена с прогестероном с
последующим пятидневным перерывом; 2) -прием только одного
прогестерона; 3) прием эстрогенов в течение 15 дней с
последующим пятидневным приемом комбинации эстрогенов с
прогестероном. Первая комбинация подавляет выработку ФСГ и ЛГ,
в результате роста фолликулов ©• яичниках \и овуляции не
происходит.
При приеме одного лишь прогестерона в больших дозах
также подавляется высвобождение ЛГ и овуляция (возможно, в
результате угнетения способности эстрогена вызывать
повышение уровня Л Г ори овуляции). Использование пероральных
противозачаточных средств почти в 100% надежно, хотя
длительное применение гормонов может привести к нежелательным
последствиям.
Прием синтетического препарата диэтилстильбэстрола после
полового сношения оказывает противозачаточное действие,
предупреждая имплантацию оплодотворенной яйцеклетки
(«перехватывающий» препарат).
Внутриматочные механические средства (ВМС), обычно
имеющие форму спирали, (вводятся в шейку матки. Они
предупреждают имплантацию (возможно, ускоряя прохождение
яйцеклетки в матке). При правильном введении они достаточно
эффективны.
ЛИТЕРАТУРА
Catt К. J. (1971). An ABC of endocrinology, Little, Brown, Boston.
Ganong W. F. (1977). Review of medical physiology, 8th ed. Lange Medical, Los
Altos, California.
Greep R. O., Koblinsky M. A. (1977). Frontiers in reproduction and fertility
control, a review of reproductive sciences and contraceptive development,
MIT Press, Cambridge.
Greep R. O. (ed.) (1977). Reproductive physiology, international review of
physiology, vol. 13, Chaps. 1, 4, and 8, University Park Press, Baltimore.
McCann S. M. (ed.) (1977). Reproductive physiology, endocrine physiology,
international review of physiology, Vol. 16, Chap. 2, University Park Press.
Baltimore.
Masters W. H., Johnson V. E. (1966). Human sexual response, Little, Brown,
Boston.
Morgan M. E. (1973). Endocrine control systems. In: Brobeck J. H. (ed.), Best
and Taylor's physiological basis of medical practice, 9th ed., Williams and
Wilkins, Baltimore.
O'Dell W. D., Moyer D. L. (1971). Physiology of reproduction, Mosby St. Louis.
508
Глава 26
Sawin С. Т. (1969). The hormones, endocrine physiology, Little, Brown, Boston.
Tepperman J. (1973). Metabolic and endocrine physiology, 3rd edn., Year Book
Medical, Chicago.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Каковы механизмы эрекции полового члена?
2. Какие нервы участвуют в осуществлении эрекции, эмиссии и
эякуляции?
3. Нарисуйте схему этапов сперматогенеза.
4. Сколько хромосом в зиготах и гаметах человека?
5. Каков объем (в мл) эякулята мужчины в норме? Сколько
сперматозоидов содержится в 1 мл эякулята?
6. За какой срок сперматозоиды переходят из влагалища в
фаллопиевы трубы?
7. Какие два основных вещества участвуют в синтезе
тестостерона?
:8. Где синтезируется тестостерон, и что влияет на его синтез?
"9. Назовите основные эффекты тестостерона.
10. Нарисуйте схему этапов овогенеза.
11. Опишите основные процессы менструального цикла в их
последовательности. Соотнесите при этом различные стадии с
днями цикла. Сопоставьте менструальный цикл с овариаль-
ным.
12. Какие гормоны гипофиза участвуют в овуляции и каким
образом? Каково участие в овуляции эстрогена и
прогестерона?
13. Где происходит оплодотворение яйцеклетки?
14. Каким образом желтое тело способствует сохранению
беременности?
15. Каков период беременности у человека?
16. Каковы механизмы родов?
17. Какие гормоны способствуют секреции и выделению
молока?
18. В чем состоит принцип методов предохранения от
беременности гормональными средствами?
19. Назовите основные эффекты эстрогенов.
Глава 27
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Щитовидная железа человека состоит из двух основных
долей, фиксированных к боковым поверхностям трахеи в верхней
части, и соединяющего их перешейка (рис. 27.1). У взрослого
человека в норме она весит около 20 г, однако размеры ее
значительно варьируют. При ряде состояний она может
увеличиваться (гипертрофия) или уменьшаться. Щитовидная железа
богато васкуляризована; иннервация ее осуществляется
симпатическими нервами.
При световой микроскопии видно, что щитовидная железа
состоит из фолликулов (рис. 27.1) диаметром около 200 мкм.
В неактивном состоянии каждый фолликул ограничен низким
однослойным кубическим элителием и содержит большое
количество коллоидного вещества — тиреоглобулина.
Во время активной секреции фолликулы щитовидной
железы становятся меньше, эпителиальные же клетки крупнее, выше
и могут приобретать призматическую форму. Содержание кол-
Рис. 27.1. Щитовидная железа человека. А. Боковые доли и перешеек.
Б. Срез железы в активном и неактивном состоянии; обратите внимание на
крупные заполненные коллоидом фолликулы с однослойными кубическими
клетками в неактивной железе и более мелкие коллоидсодержащие
фолликулы с более крупными клетками — в активной.
510
Глава 27
лоида в фолликулах ниже; наличие лакун свидетельствует о его
реабсорбции (рис. 27.1). Гипертрофия железы (увеличение
размеров ее клеток) и ее гиперплазия (увеличение числа клеток)
могут свидетельствовать о повышенной выработке тиреотропно-
го гормона гипофиза (ТТГ) и увеличенной секреции гормонов
щитовидной железы.
Вещества, блокирующие захват или связывание иода (или
и те и другие) и угнетающие синтез и выделение тироксина,
например хлораты, периодаты, перхлораты и нитраты,
увеличивают высвобождение ТТГ из гипофиза. Это в свою очередь
приводит к гипертрофии и гиперплазии щитовидной железы.
ИОДСОДЕРЖАЩИЕ СОЕДИНЕНИЯ
Этапы синтеза тироксина (Т4, или 3,5,3',5'-тетраиодтирони-
на) следующие (см. также рис. 27.2):
1. 1~ захватывается клетками щитовидной железы и
посредством активного транспорта переносится в них.
2. В клетках 1~ окисляется до I2 с помощью фермента перокси-
дазы, а затем переносится в коллоидное вещество
фолликулов, где происходит присоединение тирозиновых групп к
молекулам тиреоглобулина.
3. Тирозин присоединяет иод и образует моноиодтирозин
(МИТ).
4. МИТ присоединяет еще один атом иода, образуя дииодтиро-
зин (ДИТ).
но
CH2.CH(NH2)COOH
Тирозин
HOH^~\-CH2.CH(NH2)COOH
Моноиодтирозин
HO-<^jVcH2.CH(NH2)COOH
^Vn^^
Дииодтирозин
CH2.CH(NH2)COOH
I
Трииодтиронин (Т3)
• I
но^^о^\
-CH2.CH(NH2)COOH
I I
Тироксин (Т4)
Рис. 27.2. Этапы синтеза тироксина.
Гормоны щитовидной железы
511
5. МИТ соединяется с ДИТ, образуя 3,5,3'-трииодтиронин (Тз).
6. Две молекулы ДИТ соединяются, образуя одну молекулу Т4
или тироксина.
ВЛИЯНИЕ ИОДА НА СИНТЕЗ ГОРМОНОВ ЩИТОВИДНОЙ
ЖЕЛЕЗЫ
При недостатке иода в пище нормальный синтез тироксина
нарушается, секреция ТТГ усиливается и щитовидная железа
увеличивается (зоб). В свое время, когда иод еще не
добавлялся к поваренной соли, это заболевание было распространено в
США в области Великих озер, где поч©а бедна иодом («зобный
пояс»). Для нормального синтеза гормонов щитовидной железы
суточное потребление иода с пищей должно составлять по
меньшей мере 10 мг.
АНТИТИРЕОИДНЫЕ ПРЕПАРАТЫ
Тиокарбамиды, в том числе тиомочевина, тиоурацил и мети-
мазол, подавляют синтез тироксина и увеличивают выход ТТГ,
препятствуя присоединению иода к моноиодтирозину. Эти
препараты не блокируют захват иода, но препятствуют его
связыванию. В клинике они используются для лечения
тиреотоксикоза.
ЗОБОГЕННЫЕ ПИЩЕВЫЕ ПРОДУКТЫ
В некоторых растениях семейства крестоцветных, в том
числе капусте и репе, содержатся зобогенные или антитиреоидные
вещества. Люди, потребляющие эти овощи, должны принимать
повышенные количества иода.
СЕКРЕЦИЯ И ВЫСВОБОЖДЕНИЕ ТИРОКСИНА
Тиреоглобулин, содержащийся в коллоиде фолликулов
щитовидной железы, представляет собой высокомолекулярное
соединение, состоящее из четырех полипептидных цепей.
Тиреоглобулин связывает вещества, образующиеся в щитовидной железе —
МИТ, ДИТ, Тз и Т4. Под действием фермента протеазы эти
вещества отделяются от тиреоглобулина. Т3 и Т4 могут выделяться
в кровоток, а от МИТ и ДИТ в щитовидной железе
отщепляется иод.
Согласно общепринятой точке зрения, большая часть
циркулирующего в крови Тз образуется в результате отщепления
иода от Т4 в периферических тканях. Небольшое количество Тз
(3,3',5'-трииодтиронина) получается в результате конденсации
512
Глава 27
ДИТ и МИТ. Иодосодержащие соединения распределяются в
щитовидной железе следующим образом: МИТ—23,8%, ДИТ —
33,2 %, Т3 - 7,4 % и Т4 — 35,6 %.
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ В КРОВИ
В норме средний уровень несвязанного, или свободного,
тироксина (Т4) в крови человека составляет в среднем 3 нг/100 мл,
Т3 — 1,5 нг/100 мл. Почти весь тироксин крови (99%) находится
в связи с глобулином (60%), преальбумином (30%) и
альбумином (10%). Т3 в несколько меньшей степени связывается с
глобулином (75%), и в большей — с альбумином (25%).
Биологическая активность Т3 в несколько раз выше, чем Т4,
и его метаболический эффект проявляется быстрее. Т4 и Т3
активны лишь в свободном (несвязанном виде). Периоды их
полураспада равны примерно 7 и 3 суткам соответственно.
СЕКРЕЦИЯ И МЕТАБОЛИЗМ
Суточная секреция тироксина составляет около 80 мкг, а
Т3 — 40—50 мкг. Эти цифры колеблются в зависимости от
уровня ТТГ. Так как в крови Т3 и Т4 связаны с белками плазмы,
концентрация связанного с белком иода (СБИ) служит
надежным индикатором их содержания. В норме уровень СБИ у
человека составляет около 6 мкг/100 мл, причем большая часть его
приходится на долю Т4.
ОПРЕДЕЛЕНИЕ СКОРОСТИ СЕКРЕЦИИ
Скорость синтеза и секреции гормонов щитовидной железы
можно определить несколькими способами, в том числе: 1)
радиоиммунологическим методом, 2) измерением СБИ и 3)
определением выхода радиоактивного иода из щитовидной железы.
Радиоиммунологические методы очень чувствительны и
служат превосходным способом определения малых количеств Т3
и Т4. В клинической практике для приблизительной оценки
активности щитовидной железы используют измерение захвата
железой радиоактивного иода; скорость его захвата и
высвобождения отражает синтез и секрецию гормонов в железе.
Испытуемый принимает внутрь препарат-индикатор, содержащий
1311; затем на шею в области щитовидной железы
накладывается счетчик гамма-лучей и измеряется захват изотопа железой.
При этом определяют также захват иода в каком-либо другом
участке и вычитают его из захвата в области щитовидной
железы. Колебания захвата 1311 железой непосредственно
отражают ее активность и протекающие в ней процессы синтеза. В нор-
Гормоны щитовидной железы
513
ме через 10 ч после приема 1311 щитовидная железа захватывает
100% изотопа, а при гипотиреозе и гипертиреозе — 20 и 300%
соответственно.
Дезаминирование тироксина и трииодтиронина и отщепление
от них иода происходит во многих тканях. В печени эти
гормоны подвергаются конъюгации, образуя сульфаты и глюкурони-
ды. Метаболизм Т4 происходит преимущественно в печени и
скелетных мышцах.
ДЕЙСТВИЕ ГОРМОНОВ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
И РЕГУЛЯЦИЯ ИХ СЕКРЕЦИИ
На секрецию гормонов щитовидной железы влияют такие
факторы, как изменение температуры, физическая нагрузка и
прочие виды деятельности, а также другие гормоны. Однако
главным регулятором функции щитовидной железы служит ти-
реотрапный гормон передней доли гипофиза — ТТГ. Химическое
строение этого гормона, а также условия, влияющие на его
высвобождение из гипофиза и .выделение рилизинг-фактора
гипоталамуса, рассмотрены в гл. 25. Циркулирующие в крови
гормоны щитовидной железы влияют на уровень ТТГ по
механизму отрицательной обратной связи (см. рис. 25.6),
преимущественно на уровне гипофиза (длинная петля). При высоком
содержании Т3 и Т4 "В крови высвобождение ТТГ гипофизом
уменьшается, а при низком—увеличивается (положительная
обратная связь). Однако наряду с этими механизмами имеет
место и регуляция на уровне выработки ТТГ-РФ гипоталамусом
(короткая петля).
ВЛИЯНИЕ НА МЕТАБОЛИЗМ
Основной эффект Т3 и Т4 заключается в повышении
потребления Ог большинством органов и тканей организма — калори-
генный эффект (см. также гл. 18).
Все другие влияния этих гормонов, в том числе увеличение
всасывания пищевых продуктов, усиление распада белков,
жиров и углеводов, действие на процессы размножения, лактации
и роста, в большинстве своем являются вторичными по
отношению к калоригенному эффекту.
При повышении потребления 02 и выделения тепла
наступает распад (катаболизм) запасенных в организме белков и
Жиров, и еес тела уменьшается. Все эти явления наблюдаются
при гипертиреозе.
Под действием гормонов щитовидной железы наступает
мобилизация глюкозы, однако одновременно повышается и ее ка-
33-1705
514
Глава 27
табол,изм, так что уровень глюкозы в крови может несколько
снижаться.
Гормоны щитовидной железы влияют на обмен холестерина,
усиливая как его синтез, так и процессы, приводящие к
выделению холестерина и снижению его содержания в крови.
От уровня гормонов щитовидной железы зависит нормаль-
ный рост и развитие плода и ребенка. При снижении содержа-
Рис. 27.3. Кретинизм у ребенка 37г лет. А. До лечения. Б. После длительного
лечения гормонами щитовидной железы (тот же ребенок в возрасте 7 лет).
ния этих гормонов наблюдается кретинизм — одно из
'проявлений гипотиреоза (рис. 27.3). Гормоны щитовидной железы
ускоряют метаморфоз головастиков в нормальных лягушек.
НАРУШЕНИЕ ФУНКЦИИ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Если в результате снижения функции щитовидной железы
уменьшается потребление кислорода и скорость обменных
процессов, может развиться гипотиреоз. У детей это нарушение
сопровождается одной из форм карликовости — кретинизмом. Это
заболевание лечат введением тироксина (рис. 27.3). У взрослых
же снижение уровня гормонов щитовидной железы
сопровождается микседемой. У таких больных редкие волосы, сухая,
желтоватая и одутловатая кожа, низкий хриплый голос; основной
обмен у них снижен (рис. 27.4). Это заболевание также лечат
введением гормонов щитовидной железы.
ГИПЕРТИРЕОЗ
При повышенной секреции гормонов щитовидной железы
возрастает потребление кислорода, что может сопровождаться
такими вторичными симптомами как: 1) нервозность, 2) тре-
Гормоны щитовидной железы
515
Рис. 27.4. А. Гипофункция щитовидной железы (микседема) у взрослого до
лечения. Б. После лечения гормонами щитовидной железы. В. Гипертиреоз
(тиреотоксикоз). Обратите внимание на экзофтальм.
мор, 3) тахикардия, 4) потливость, 5) непереносимость тепла,
6) утомляемость, 7) потеря веса, 8) экзофтальм и 9) зоб.
Гипертиреоз часто наблюдается при тиреотоксикозе,
характеризующемся зобом 'и экзофтальмом (рис. 27.4). Синтез гормонов
щитовидной железы при этом возрастает в 3—10 раз по
сравнению с нормой. Гипертиреоз при этом связан с действием двух
или более гамма-глобулинов крови, стимулирующих активность
щитовидной железы.
ГИПОТИРЕОЗ
При гипофизэктомии функция щитовидной железы
снижается из-за отсутствия ее основного стимулятора ТТГ. Удаление
щитовидной железы приводит к угнетению роста и изменениям
кожи и волос. В отсутствие гормонов щитовидной железы
потребление кислорода снижено, и в результате наблюдается
тенденция к замедлению всех физиологических процессов.
ЛИТЕРАТУРА
Bentley P. J. (1976). Comparative vertebrate endocrinology, Cambridge Press,
New York.
Catt K. J. (1971). An ABC of endocrinology, Little, Brown, Boston.
Ganong W. F. (1977). Review of medical physiology, 8th edn., Lange Medical,
Los Altos, California.
McCann S. M. (ed.) (1977). Endocrine physiology II, international review of
physiology, Vol. 16, Chap. 2, University Park Press, Baltimore.
Morgan H. (1973). Endocrine control systems. In: Brobeck Jr. (ed.) Best and
Taylor's physiological basis of medical practice, 9th edn., Chap. 7, p. 29,
Williams and Wilkins, Baltimore.
33*
516
Глава 27
Геррегтап J. (1973). Metabolic and endocrine physiology, 3rd edn., Year Book
Medical, Chicago.
Woeber K. A., Braverman L. E. (1976). Thyroids. In: Ingbar S. H. (ed.), The
year in endocrinology, 1975—1976, Plenum, New York.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Каковы основные этапы синтеза тироксина (Т4) из
тирозина?
2. Как влияет ТТГ на секрецию гормонов щитовидной железы?
3. Объясните, как действуют Т3 и Т4 крови через отрицательную
обратную связь.
4. Каково процентное соотношение свободных и связанных с
белками плазмы Т3 и Т4 в крови?
5. Расскажите о биологическом действии Т3 и Т4.
6. Как влияют гормоны щитовидной железы на а) потребление
кислорода и теплопродукцию, б) обмен углеводов, в) обмен
холестерина, г) рост?
7. Что такое кретинизм? Опишите это заболевание и раскройте
его причины.
8. Что такое микседема? Опишите и объясните это заболевание.
9. Что такое тиреотоксикоз? Опишите и объясните это
заболевание.
Глава 28
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА
КАЛЬЦИЯ
Обмен кальция, и в частности содержание Са в крови и
тканях, регулируется преимущественно тремя гормонами: 1) парат-
гормоном (ПтГ); 2) кальцитонином (КТ) С-клеток щитовидной
железы и ультимобранхиальных телец и 3) дигидроксихоле-
кальциферолом (кальциферолом), образующимся в печени и
почках из витамина D и увеличивающим всасывание Са в
кишечнике.
Выделение кальцитонина и ПтГ регулируется содержанием
Са в крови и костной ткани независимо от влияний гипофиза.
СОДЕРЖАНИЕ Са В ТКАНЯХ
В организме человека содержится в среднем около 1000—
1200 г Са. Большая часть его приходится на долю костной
ткани; в крови и мягких тканях содержится 1 % общего количества
Са. В крови Са находится в двух видах — связанном с белками
плазмы (преимущественно с альбумином) и свободном (Са2+).
Связанный кальций не способен проникать через
полупроницаемые мембраны, а Са2+ проникает через них. Общий Са
плазмы состоит из обеих фракций. У человека общее
содержание Са в крови составляет 10,0 мг/100 мл (5 мэкв/л), причем
5,36 мг/100 мл приходится на долю Са2+, а 4,64 мг/100 мл на
долю связанного кальция.
Физиологически активным является свободный Са2+,
способный к диффузии. Именно он участвует в свертывании крови,
возбуждении мышц и нервов и в процессах жизнедеятельности
клеток. Все эти процессы рассматриваются в различных
разделах настоящего пособия. У человека значительное повышение
кальция плазмы выше 10 мг/100 мл является признаком гипер-
Кальциемии, а понижение — гипокальциемии.
Удаление паращитовидных желез сразу же приводит к
резкому снижению содержания кальция в крови от 10 мг/100 мл до
6—7 мг/100 мл. После введения ПтГ уровень кальция быстро
Восстанавливается. У самок птиц во время откладывания яйца
Уровень Са в крови значительно выше, чем у самцов (около
518
Глава 28
20 мг/100 мл), что связано с действием не только ПТГ, но и
женского полового гормона — эстрогена.
Кости образованы жестким органическим каркасом, в
котором отложены соли кальция (см. гл. 1). В состав этих солей,
.называемых гидроксиапатитами, входят кальций и фосфаты.
Формула гидроксиапатитов следующая: Саю (Р04) е (ОН) 2.
Большая часть кальция в костной ткани прочно связана,
однако некоторое количество его может свободно обмениваться
между этой тканью и кровью. В костях происходит постоянное
обновление кальция (отложение и высвобождение, или
вымывание). Обновление кальция за год составляет у детей 100%, а у
взрослых — от 16 до 20%.
Органический матрикс костей образован тремя основными
типами клеток — остеобластами, остеоцитами и остеокластами.
Из остеобластов образуются новые клетки, преобразующиеся в
остеоциты (поздняя стадия развития остеобласта).
Остеокласты представляют собой многоядерные клетки, разрушающие и
поглощающие костную ткань. Они играют важную роль в
высвобождении из костей Са и его выделении в кровоток.
Недавно в лейкоцитах был обнаружен мощный стимулятор
активности остеокластов, однако его физиологическое значение
еще не выяснено.
ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Обычно имеются две пары паращитовидных желез,
расположенных в области нижних и верхних полюсов долей
щитовидной железы (рис. 27.1), однако их локализация может
значительно варьировать. Это относительно небольшие образования—
6 мм в длину и 3 мм в ширину. На срезе через паращитовид-
ную железу взрослого человека видны два основных типа
клеток — главные и оксифильные. Главные клетки небольшие,
цитоплазма их прозрачна; они расположены в виде шнуров. Эти
клетки секретируют паратгормон. Оксифильные клетки у
многих видов отсутствуют. У человека их меньше, чем главных,
Функция их неизвестна.
ГОРМОН ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОН)
Паратгормон (ПтГ), образующийся в 'паращитовидных
железах, образован полипептидной цепью из 84 аминокислотных
остатков с определенной для каждого вида
последовательностью. У свиней и коров структура ПтГ сходна, однако
некоторые аминокислоты у них разные. ПтГ кур, полученный в
очищенном в(иде, по молекулярному весу .и биологической
активности сходен с ПтГ коров (молекулярный вес последнего состав-
Гормональная регуляция обмена кальция 5Ш
ляет 9500). Биологическая активность ПтГ коров почти
полностью определяется первыми 34 аминокислотами, ближайшими
к N-концу цепи. ПтГ человека, состоящий из 84 аминокислот,
сходен с ПтГ коровы.
Биологические пробы основаны на определении повышения
уровня кальция в крови под действием ПтГ. При использовании
интактных кур или куропаток пробы эти быстрей и
чувствительней, чем при использовании крыс.
Существуют также радиоиммунологические методы
определения ПтГ, однако в связи с наличием нескольких различных
по иммунореактивности типов и фрагментов этого гормона
необходимы более специфичные антитела.
ВЫСВОБОЖДЕНИЕ ПтГ И ЕГО АКТИВНОСТЬ
Высвобождение ПтГ регулируется уровнем кальция в крови
и мягких тканях. При нормальном содержании Са (около 9—
10 мг/100 мл) ПтГ секретируется в умеренных количествах, при
высоком — выделение его минимально или не происходит вовсе.
При низком уровне Са в крови секреция ПтГ максимальна
(рис. 28.1). Низкое содержание Са в крови усиливает выброс
ПтГ © кровоток из главных клеток паращитовидных желез.
Основной эффект ПтГ состолт в мобилизации кальция из костной
ткани (рис. 28.2). Паратгормон активирует остеокласты, разру-
Рис. 28.1. Факторы, регулирующие содержание кальция. При высоком уровне
кальция в крови (>10 мг%) выделяется кальцитонин, под действием
которого снижается уровень кальция в крови и увеличивается его накопление в
костной ткани. Низкое же содержание кальция в крови приводит к
выделению ПтГ, способствующего повышению уровня кальция в крови и
уменьшению количества связанного кальция (резорбция Са).
520 Глава 28
шающие костное вещество и способствующие его резорбции;
при этом в кровоток поступает свободный кальций.
Органическое вещество костной ткани распадается с образованием окси-
лролина, выделяющегося с мочой.
Под действием паратгормона активируется аденилатциклаза
мембранных рецепторов, что сопровождается усиленным
образованием цАМФ (подробнее см. гл. 25). Механизм действия
цАМФ еще не выяснен.
0,6
0,5 -Н
0,4 *-
и»° о
0,2 I
0,1 |
0
Рис. 28.2. Влияние изменений содержания кальция в плазме на уровень каль-
цитонина и ПтГ в крови. (По Catt К. J., 1971. An ABC of endocrinology.)
При разрушении костной ткани остеокластами вместе с
кальцием высвобождается фосфор в виде фосфатов. Большая
часть кальция поглощается из кровотока, однако некоторое
количество его, а также основная часть фосфатов выделяется с
мочой. Таким образом, ПтГ увеличивает реабсорбцию кальция
и снижает реабсорбцию фосфора в канальцах почек.
Под действием ПтГ увеличивается также активный
транспорт и всасывание кальция в кишечнике при условии
нормального содержания витамина D в пище. Недостаточность этого
витамина у детей может привести к гипокальциемии и
нарушению минерализации костей, что сопровождается их
размягчением и возникновением рахита.
ГИПО- И ГИПЕРПАРАТИРЕОИДИЗМ
Хирургическое удаление паращитовидных желез
сопровождается гипопаратиреоидизмом, характеризующимся
постепенным снижением уровня кальция и повышением уровня
фосфатов в крови. Это может привести к тетании — заболеванию,
проявляющемуся быстрыми нерегулярными сокращениями мышц.
Симптомы тетании исчезают при введении ПтГ или (временно)
витамина D и солей кальция.
6 7 8 9 10 11 12 13 14 15 16
Кальций плазмы, мг/100 мл
Гормональная регуляция обмена кальция
521
При опухолях паращитовидных желез может наблюдаться
повышенная секреция ПтГ (гиперпаратиреоидизм), приводящая
к гиперкальциемии, гипофосфатемии, увеличению содержания
кальция © костях и почках и образованию в последних кальций-
содержащих камней. Эти же явления возникают при введении
больших доз ПтГ.
КАЛЬЦИТОНИН
Кальцитонин (КТ) представляет собой гормон,
вырабатываемый парафолликулярными клетками (С-клетками)
щитовидной железы млекопитающих. Эти клетки в процессе
эмбриогенеза развиваются из ткани ультимобранхиальных телец; у
рептилий, птиц и рыб они находятся преимущественно в этих тельцах.
С-клетки крупные, четко различаются и могут быть
выявлены методами иммунофлуоресценции.
БИОХИМИЧЕСКАЯ АКТИВНОСТЬ
У некоторых видов животных кальцитонин был выделен и
получен в очищенном виде. Он представляет собой полипептид
из 32 аминокислотных остатков; у человека, свиньи, крупного
рогатого скота, птиц и рыб строение его сходно. Для
полноценного биологического эффекта КТ необходимы все 32
аминокислоты. У рыб (лососевых) КТ значительно более активен, чем у
млекопитающих, и обладает более высоким сродством к каль-
цитониновому рецептору.
Разработаны чувствительные биологические методы
определения КТ in vitro, при помощи которых можно обнаружить
такие малые его концентрации, как 0,5 нг/мл. Существуют также
радиоиммунологические методы определения различных типов
КТ. Кальцитонин может содержаться в кровотоке в
концентрации 50 пг/мл.
Высвобождение КТ также регулируется содержанием
кальция в плазме (рис. 28.1 и 28.2). Высокий уровень кальция
приводит к увеличению выделения КТ, под действием которого
усиливается накопление кальция в костной ткани и снижается
содержание его в крови. КТ оказывает эффект, противоположный
действию ПтГ, но не является его прямым конкурентом.
Влияние КТ на почки и кишечник не выявлено, хотя он, как и ПтГ,
снижает уровень фосфатов в крови. Кальцитонин действует
непосредственно на клетки костной ткани и, по-видимому, угнетает
активный перенос кальция из этих клеток в кровоток.
Возможно, действие его опосредовано цАМФ. За сутки у человека
высвобождается около 0,5 мг КТ. Его физиологическая роль
окончательно не установлена, и значение его в регуляции обмена
Кальция костной ткани подлежит дальнейшему исследованию.
522
Глава 28
ЛИТЕРАТУРА
Catt К. J. (1971). An ABC of endocrinology, Little, Brown, Boston.
Ganong W. F. (1977). Review of medical physiology, 8th edn., Lange Medical,
Los Altos, California.
Morgan H. E. (1973). Endocrine control systems. In: Brobeck R. (ed.), Best and
Taylor's physiological basis of medical practice, 9th edn., Williams and
Wilkins, Baltimore.
Queener S. F., Bell N. H. (1975). Calcitonin, a general survey, Metabolism 24,
555.
Raisz L. G., Mundy G. R., Dietrich J. W., Canalis E. M. (1977). Hormonal
regulation of mineral metabolism. In: McCann S. M. (ed.), International review
of physiology; endocrine physiology II, Vol. 16, University Park Press,
Baltimore.
Tepperman J. (1973). Metabolic and endocrine physiology, 2nd edn., Year Book
Medical, Chicago.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Каково содержание кальция в организме человека? Какая
часть его приходится на долю крови и мягких тканей?
2. Какие существуют фракции кальция в крови, и каково
содержание этих фракций?
3. Назовите гормоны, участвующие в регуляции содержания
кальция в крови и костной ткани.
4. Составьте схему взаимоотношения между паратгормоном
(ПтГ) и кальцитонином, с одной стороны, и уровнем
кальция IB крови и костной ткани — с другой.
5. Какие клетки отвечают за резорбцию костной ткани? Чем
они активируются?
6. Где синтезируется кальцитонин у млекопитающих, рептилий
и птиц?
7. Какова химическая природа кальцитонина и ПтГ?
{Глава 29
ГОРМОНЫ НАДПОЧЕЧНИКОВ
ш
МАКРО- И МИКРОСТРУКТУРА НАДПОЧЕЧНИКОВ
У человека надпочечники представляют собой парные
пирамидальные образования, расположенные на верхних полюсах
Щпочек. Длина их колеблется от 2,5 до 5 см, однако их размеры
могут значительно варьировать в зависимости от действия
различных раздражителей, стимулирующих высвобождение АКТГ,
влияющего на надпочечники.
Надпочечники состоят из наружного отдела мезодермально-
го происхождения — коры и внутреннего отдела, гомологичного
симпатическим ганглиям и развивающегося из нервной ткани —
^мозгового слоя. Мозговой слой надпочечников иннервируется
| прега.нглионарными симпатическими нервами, при раздражении
' которых выбрасываются гормоны мозгового слоя — адреналин и
рнорадреналин.
Соотношение «обоих отделов у разных видов различно,
однако у большинства млекопитающих корковый слой более развит,
Л чем мозговой. В коре надпочечников происходит синтез и выде-
| ление кортикостероидов.
I У большинства млекопитающих надпочечники состоят из
^различных типов клеток, расположенных ib трех слоях, или зо-
|нах (рис. 29.1). Эти зоны (от наружной поверхности к внутрен-
|- ней) следующие: 1) клубочковая, 2) пучковая, 3) сетчатая.
| Клетки клубочковой зоны богаты митохондриями, но бедны ли-
■§пидами. В пучковой зоне находятся призматические клетки с
Цбольшим содержанием липидов. В сетчатой зоне расположены
^плоские клетки. У человека в 1-й зоне образуется альдостерон,
|а во 2-й и 3-й кортизол. В 1-й зоне могут образовываться новые
^клетки, пополняющие как саму эту зону, так и две остальные.
|-.На функцию 2-й и 3-й зон влияет АКТГ; активность
клубочковой зоны в какой-то мере независима от этого гормона.
% И корковый и мозговой слои надпочечников богато васкуля-
Цризированы и пронизаны венозными синусами, в которые посту-
йпают гормоны.
i
ЖГОРМОНЫ КОРЫ НАДПОЧЕЧНИКОВ
f Из надпочечников было выделено много различных стероид-
рйых гормонов, однако важнейшими из них (и секретируемыми
|Э физиологически активных количествах) являются: 1) минера-
524
Глава 29
Капсула
Клубочковая
зона
Пучковая
зона
Сетчатая
зона
Мозговое
вещество
Рис. 29.1. Зоны коры надпочечников и прилегающее к ней мозговое вещество
на срезе.
ло'кортикоид альдостерон, участвующий преимущественно в
регуляции обмена натрия и калия, и 2) глюкокортикоиды корти-
зол и кортикостерон, влияющие главным образом на обмен
глюкозы. Структура и этапы синтеза этих гормонов
представлены на рис. 29.2.
Существуют две основные структурные группы кортикосте-
роидов: 1) стероиды типа С-21 с 21 атомом углерода и двуугле-
Гормоны надпочечников
525
| родной боковой цепочкой
1 в положении 17 (D-коль-
цо) и 2) стероиды типа
С-19 с 19 атомами
углерода и кето- или гидрок-
сильной группировкой в
положении 17 (кетосте-
роиды). Прогестерон и
дегидроэпиандросте р о н
являются соответственно
| стероидами типа С-21 и
С-19; последний обладает
андрогенной активностью.
Общим
предшественником кортикостероидов и
половых гормонов
служит холестерин; сначала
в митохондриях клеток
коры надпочечников он
превращается в прегнено-
лон, из которого в
дальнейшем образуется
прогестерон.
Прогестерон
занимает ключевое положение
в синтезе как
кортикостероидов, так и андроге-
нов и эстрогенов (см.
гл. 25). Под действием
ферментов 17 а-гидрокси-
лазы, 21-гидроксилазы и
11 р-гидроксилазы он
последовательно
превращается в 17а-гидроксипроге-
стерон, 17 а-гидроксикор-
тикостерон и кортизол.
11 а-гидроксилаза и 21-
гидроксилаза
встречаются во всех зонах коры
|Рис. 29.2. Гормоны коры
надпочечников. А. Основное
стероидное ядро; пронумерованы
|атомы углерода и обозначены
р^Кольца. Б. Главные пути
синтеза альдостерона, кортизола и
|:Кортикостерона.
^1з А с
сн2он л5=Прегне- сн3
с=0 ноло,н с = 0
Дезоксикортикостерон Прогестерон СНз
СН20н |
17-(..гидроксипрогестеронСн2он
Кортикостерон С—°
А--он
Альдостерон
17-п-гидроксидезо-сн2он
ксикортикостерон ^ _Q
Кортизол
526
Глава 29
надпочечников, а 17а-гидроксилаза — только в пучковой и
сетчатой зонах.
Альдостерон вырабатывается только в клубочковой зоне
непосредственно из прогестерона и кортикостерона под
действием фермента 18-альдолазы; этот фермент также имеется только
в клубочковой зоне (см. рис. 29.2).
У крыс, мышей и птиц главным кортикостероидом является
кортикостерон, а у коров, овец, обезьян и человека — кортизон.
Лишь 12% кортикостероидов, синтезируемых в организме
человека, приходится на долю кортикостерона.
ВЫДЕЛЕНИЕ АКТГ И КОРТИКОСТЕРОИДОВ
Как указывалось в гл. 25, на выделение АКТГ из гипофиза
влияет АКТГ-рилизинг-фактор гипоталамуса. Под действием
АКТГ из надпочечников выбрасываются все кортикостероиды,
кроме альдосте.рона, секреция которого регулируется главным
образом компонентами ренин-ангиотензиновой системы,
активирующей клубочковую зону. Высвобождение альдостерона
надпочечниками может также усиливаться под действием гормонов
задней доли гипофиза типа вазопрессина.
глюкокортикоиды
Секреция кортизола, кортикостерона и АКТГ подвержена
суточным колебаниям: о,на максимальна рано утром (6.00) и
минимальна вечером. Внедрение и использование
чувствительных радиоиммунологичееких методов, позволяющих частый
(каждые 10 мин) забор проб, дало возможность установить, что
секреция АКТГ и кортизола происходит не непрерывно, а
периодически— в виде кратковременных «залпов» по нескольку ми-
Таблица 29.1. Секреция и приблизительные концентрации кортизола и АКТГ
Время суток, ч
Темное 21
Темное 24
Светлое 6
Светлое 12
Светлое 18
Кортизол, мкг/100 мл
5,0
5,2
15,0
9,4
5,3
АКТГ, пг/мл
30,0
33,0
60,0
44,0
28,0
нут (табл. 29.1). Такой |ритм секреции АКТГ связывают с
активностью супраоптических ядер гипоталамуса, в которых
высвобождается АКТГ-РФ.
Гормоны надпочечников 527
! СТРЕСС И АКТГ
*' Различные типы стресса, в том числе физический и
эмоциональный, сопровождаются выделением АКТГ, возбуждением
i секреции кортизола в надпочечниках и могут приводить к увели-
\ чению размеров (гипертрофии) последних. Под действием
стресса и АКТГ понижается содержание в надпочечниках
аскорбиновой кислоты и холестерина.
МЕХАНИЗМ ОБРАТНОЙ СВЯЗИ
Связь между уровнями кортизола и АКТГ в плазме не
относится к числу жестких или чувствительных. В пользу этого
свидетельствуют следующие факты: 1) содержание кортизола в-
плазме может уменьшиться до нуля без сопутствующего увели-
. чения выделения АКТГ; 2) секреция АКТГ наблюдается и при
повышенном или нормальном уровне кортизола; 3) у крыс, у
которых главным кортикостероидом является кортикостерон,.
промежуток времени между снижением содержания этого
гормона в плазме в результате удаления надпочечников и
повышением уровня АКТГ достаточно велик.
Все эти данные свидетельствуют о том, что прямой и
жесткой обратной связи между содержанием кортизола и кортико-
стерона в крови и выделением АКТГ, возможно, нет. В
определенных пределах это справедливо, однако секреция АКТГ
происходит при низком уровне кортизола в крови и уменьшается
при высоком. Считается, что пороговые значения содержания
кортизола в крови стимулируют выделение неиросекретов в
центральной нервной системе (гипоталамус). Очевидно, это
явление служит основным фактором регуляции выделения АКТГ;
при этом возможно либо непосредственное влияние на
высвобождение АКТГ гипофизом, либо непрямое, путем выделения
АКТГ-РФ из гипоталамуса и срединного возвышения. Точные
механизмы этого влияния дискутируются (подробнее об
АКТГ-РФ см. гл.25).
СЕКРЕЦИЯ АЛЬДОСТЕРОНА
Секреция альдостерона в меньшей степени зависит от
АКТГ, чем выработка глюкортикоидов. Для того чтобы вызвать
•выделение альдостерона, необходимы большие дозы АКТГ, и
, Даже при высоком уровне последнего секреция альдостерона
, Усиливается кратковременно. К факторам, влияющим на
выработку альдостерона, относятся (помимо АКТГ): 1) увеличенное
Потребление калия, 2) низкое потребление натрия, 3)
обезвоживание, 4) сужение аорты и 5) вертикальное (положение тела.
В последнем случае, так же как и при ограничении потребления
528
Глава 29
натрия и увеличения содержания в плазме калия, повышенная
секреция альдостерона обусловлена усиленной выработкой
ренина. Секреция альдостерона и глюкокортикоидов
увеличивается при стрессе и кровопотере.
РЕНИН—АНГИОТЕНЗИНОВАЯ СИСТЕМА
Синтез и выделение альдостерона регулируются главным
образом ренин-ангиотензиновой системой (рис. 29.3). Ренин
представляет собой белок с протеолитической активностью,
вырабатывающийся клетками юкстагломерулярного аппарата
почек. Под действием выделившегося почками ренина из а-глобу-
PftHMH S
1
Ангиотензиноген
(из печени)
\
Ангиотензин 1
\
.Превращающий
фермент
Ф
Ангиотензин II У
Юкстагломеруляр-
ный аппарат почек
К
Надпочечники |
Клубочковая
зона
Снижение
кровяного давления в
сосудах почек i
А "'
Снижение об'ема
внеклеточного
пространства
\
Снижение уровня
Na4,
обезвоживание |
т
Альдостерон
Рис. 29.3. Регуляция секреции альдостерона ренин — ангиотензиновой
системой (см. текст). Выработка альдостерона увеличивается при снижении
содержания натрия, обезвоживании, снижении объема интерстициального
пространства и уменьшении артериального давления. Изменения этих параметроз
в обратном направлении приводят к уменьшению выделения ренина и
альдостерона.
лина образуется ангиотензин I—предшественник октапептида
ангиотензина II; у человека ангиотензин II усиливает секрецию
альдостерона, но не кортикостерона. Синтез альдостерона
регулируется по механизму обратной связи (рис. 29.3). Секреция
альдостерона наступает при небольших сдвигах уровня Kf B
плазме (1 мэкв/л) и пр'и значительно больших изменениях
содержания Na+.
Базальная секреция альдостерона в норме колеблется от 50
до 200 мкг/сутки, а скорость его выделения составляет примерно
Гормоны надпочечников
529
2—16 мкг/сутки. Уровень альдостерона в плазме крови человека
в норме варьирует от 3 до 15 нг/100 мл; большая часть
альдостерона плазмы связана с белком, однако некоторое количество
обнаруживается в свободном состоянии. Физиологической
активностью обладают свободный альдостерон и кортизол.
ДЕЙСТВИЕ ГЛЮКОКОРТИКОИДОВ
У человека глюкокортикоиды, и в частности кортизол,
влияют на углеводный обмен, стимулируя синтез и отложение
гликогена и уменьшая его утилизацию; в результате уровень
сахара в крови повышается. Глюкокортикоиды вызывают также
отщепление аминокислот от белков скелетных мышц и
способствуют превращению этих аминокислот в углеводы (глюконеоге-
нез). Таким образом, под действием глюкокортикоидов белки
превращаются в углеводы и запасаются главным образом в
виде гликогена. Глюкокортикоиды влияют также на обмен ли*
пидов, увеличивая мобилизацию жирных кислот из
периферических жировых депо и угнетая их образование в печени.
Кортизол способствует распаду жиров под действием других
гормонов типа тироксина или катехоламинов (см. также гл. 22).
Кортизол способствует также задержке натрия, однако
эффект этот выражен в значительно меньшей степени, чем у
альдостерона. Под действием кортизола увеличиваются почечный
кровоток и клубочковая фильтрация. Кортизол может также
вызывать атрофию лимфоидной ткани, лимфопению и снижение
числа эозинофилов. Больные с недостаточным синтезом
глюкокортикоидов более подвержены инфекционным и
воспалительным заболеваниям, и реакция их на стресс нарушена.
МИНЕРАЛОКОРТИКОИДЫ
Альдостерон действует главным образом на почки, усиливая
реабсорбцию и задержку Na+ в дистальных канальцах, а
также выделение К+. Реабсорбция Na+ происходит также из пота,
слюны и желудочного сока; в этих случаях ионы Na+
диффундируют в окружающие эпителиальные клетки и активно
переносятся в интерстициальную жидкость. Механизм действия
альдостерона сходен с механизмом действия кортизола; имеется
специфический белковый рецепторный комплекс, мигрирующий к
ядру и стимулирующий ДНК-зависимую РНК и синтез белков
(см. гл. 22).
Нарушение функции коры надпочечников
Атрофия или другие повреждения коры надпочечников
приводят к аддисоновой болезни (первичной надпочечниковой
недостаточности), при которой снижен синтез альдостерона и кор-
3*-1705
530
Глава 29
тизола. Дефицит этих гормонов, и прежде всего альдостероиа,
приводит к снижению содержания Na+ в крови и тканях (гипо-
натриемии) и повышению уровня К+ (гиперкалиемии).
Уменьшение количества солей сопровождается потерей тканевой
жидкости и обезвоживанием. Падение же уровня кортизола
приводит к слабости, сонливости, ослаблению реакции на стресс и к
гипогликемии. В результате пониженной выработки кортизола
по механизму отрицательной обратной связи увеличивается
секреция гипофизом АКТГ (вторичная надпочечниковая
недостаточность); при этом
возрастает также
выработка р-МСГ, что приводит
к отложению в коже
(преимущественно па
открытых местах,
например локтях) пигмента
меланина. Люди,
страдающие адднеоыовой
болезнью, нуждаются ь
длительном лечении аль-
достероном и кортизо-
лом.
Примером хронически
повышенного синтеза
кортизола, обусловленно-
го усиленной секрецией
АКТГ, может служить
болезнь, или синдром, Кц-
шинга. Природа
нарушений функций гипофиза
при этом заболевании до
конца не выяснена. Для
больных с синдромом Ку-
шинга характерно
лунообразное лицо, исхудание
конечностей и ожирение
с отложением жира в
области талии, лица, шеи
и туловища (рис. 29.4).
Обращают на себя
внимание багровые полосе
(стрии) на коже живого
и бедер. У таких больных
понижена толерантность
к углеводам; они
восприимчивы к инфекциям п
Рис. 29.4. Синдром
12 лет.
Кушинга у девочки
Гормоны надпочечников
531
1'при наличии наследственного предрасположения страдают са-
"' харным диабетом. Повышенная выработка кортизола приводит
| к задержке натрия и гипертонии.
Чрезмерная секреция альдостерона, обусловленная опухолью
I клубочковой зоны надпочечников, приводит к гипокалиемии и
w задержке Na+. Такое состояние известно под названием
синдрома Кона. К его проявлениям относятся слабость, повышение
| артериального давления, судороги и полиурия.
МОЗГОВОЕ ВЕЩЕСТВО НАДПОЧЕЧНИКОВ
И КАТЕХОЛАМИНЫ
К моменту рождения мозговое вещество надпочечников
представлено примитивными симпатическими нервными клетка-
I
I
ми. Эти клетки быстро
дифференцируются в зрелые хро-
маффинные. Венозная кровь,
оттекающая от коры
надпочечников, проходит через
мозговое вещество и обогащается
катехоламинами.
СИНТЕЗ И ВЫДЕЛЕНИЕ
КАТЕХОЛАМИНОВ
Главным гормоном
мозгового слоя надпочечников
является адреналин; норадреналин
вырабатывается в меньших
количествах. Этапы синтеза этих
;, гормонов изображены на
I рис. 29.5. Эти этапы
следующие (приведены субстраты
||.и ферменты): 1 Тирозин +
h + Тирозингидроксилаза —ь
*:*-*2) ДОФА + ДОФА-декарб-
\ оксилаза —►• 3) Дофамин +
+ р-Гидроксилаза—*4)
Норадреналин + Фенилэтанол-
амин - N - метилтрансфераза
(ФНМТ)—>5) Адреналин.
Синтезированные катехол-
амины запасаются в гранулах
Клеток мозгового слоя. Они
высвобождаются из гранул
и°Д действием
соответствующих стимулов, в том числе
"34*
но
с-с—он
I Тирозин
NH,
Тирозингмдрсксигаза
3, 4-диги-
j—Он дроксифени-
лаланин
2 (ДОФА)
ДОФАдекарбоксигаза
Дофамин
Дофамин-бета-гид роксилаза
Норадреналин
Адреналин
Рис. 29.5. Этапы синтеза норадрена-
лина и адреналина (см. текст).
532
Глава 29
при возбуждении, раздражении симпатических нервов,
физической нагрузке, охлаждении, действии гистамина и глюка-
гона.
ОБМЕН И СЕКРЕЦИЯ КАТЕХОЛАМИНОВ
Метаболизм адреналина (Адр) и норадреналина (НА)
происходит под действием двух ферментов; моноаминоксидазы
(МАО) и катехол-о-метилтрансферазы (КОМТ). В крови они
подвергаются о-метилированию, и некоторые продукты их
метаболизма выделяются с мочой. Неэкскретирующиеся
метаболиты катехоламинов окисляются до З-метокси-4-гидроксимин-
дальной (|ванилилминдальной) кислоты. Распад НА и Адр
происходит быстро: ©се промежуточные продукты находятся в
крови лишь IB течение нескольких секунд.
Примерно 80% катехоламинов, секретируемых
надпочечниками человека, приходится на долю адреналина. Большая часть
норадреналина под действием метилтрансферазы превращается
в надпочечниках в адреналин. Содержание адреналина в крови
человека равно примерно 30 пг/мл, причем адреналин поступает
в кровь преимущественно из надпочечников; уровень же
норадреналина значительно (в 5—7 раз) выше и составляет около
200 пг/мл. Большая часть норадреналина выделяется в
кровоток из симпатических нервов при их раздражении (см. также
гл. 10).
Многие факторы приводят к выбросу из надпочечников как
норадреналина, так и адреналина, однако некоторые
раздражители (например, гипоксия и асфиксия) вызывают
преимущественно выброс норадреналина. Выброс норадреналина
усиливается также в состоянии привычного эмоционального стресса; при
Таблица 29.2. Физиологические эффекты адреналина и норадреналина
Эффект
Артериальное давление
Частота сокращений сердца
Периферическое сопротивление
Теплопродукция
Мобилизация жирных кислот
Гликогенолиз и гипергликемия
Сокращение гладких мышц
сосудов
Моторика желудка
Моторика кишечника
Потовые железы
Норадреналин
+ + + +
+ + +
+ + + +
+ + +
+ + +
+
+ + +
—
—
Выделение i
Адреналин
+ +
+ +
+ +
+ + + +
+ +
+ + +
+ ИЛИ —
юта
Гормоны надпочечников
533
непредвиденных же обстоятельствах повышается выделение
адреналина.
Повышенная секреция адреналина надпочечниками является
одним из механизмов реакции в экстремальных ситуациях; она
подготавливает организм к внезапной физической нагрузке
(реакции бегства или нападения). Выработка катехоламинов,
оказывающих калоригенный эффект, происходит также при охлаж-
дениип ознобе или перегревании.
Эффекты адреналина и норадреналина рассмотрены в
нескольких главах данной книги (главным образом в гл. 14 и 15).
Некоторые из этих эффектов представлены в табл. 29.2.
ЛИТЕРАТУРА
Bentley P. J. (1976). Comparative vertebrate endocrinology, Cambridge
University Press, New York.
Biglieri E. G. (1976). Aldosterone. In: Ingbar S. H. (ed.), The year in
endocrinology, 1975—1976, Plenum, New York.
Brodish A., Lymangrover J. R. (1977). Hypothalamic pituitary adrenocortical
system. In: McCann S. M. (ed.), International review of physiology,
endocrine, physiology II. University Park, Press, Baltimore.
Catt K. /. (1971). An ABC of Endocrinology, Little, Brown, Boston.
Ganong W. F. (1977). Review of medical physiology, 8th edn., Lange Medical,
Los Altos, California.
Morgan H. E. (1973). Endocrine control systems. In: Brobeck J. R. (ed.), Best
and Taylors, physiological bases of medical practice, 9th edn., Williams and
Wilkins, Baltimore.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Назовите основные гормоны, вырабатываемые корой и
мозговым веществом надпочечников у человека.
2. На выделение каких гормонов преимущественно влияет
АКТГ гипофиза?
3. Расскажите о взаимосвязи между секрецией АКТГ и
содержанием кортикостероидов в крови.
4. Что такое ренин — ангиотензиновая система? Как она влияет
на секрецию альдостерона?
5. Каково участие холестерина и прогестерона в синтезе
кортикостероидов?
6. Что является непосредственным предшественником
альдостерона?
7. В какой зоне коры надпочечников образуется альдостерон?
8. Каковы главные эффекты 1) глюкокортикоидов, 2) минера-
локортикоидов?
9. Назовите заболевания, возникающие при недостатке и
избытке гормонов коры надпочечников.
10. Где образуются адреналин и норадреналин?
И. Перечислите несколько эффектов адреналина и
норадреналина.
Глава 30
ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
Эндокринную функцию поджелудочной железы выполняют
клетки островков Лангерганса; они вырабатывают два гормона,
играющие важнейшую роль в регуляции углеводного обмена —
инсулин и глюкагон.
Сахарный диабет — заболевание, связанное с недостаточным
синтезом инсулина поджелудочной железой, — был известен уже
в 1500 г. до н.э. В менее отдаленные времена (1647 г.)
сахарный диабет был подробно описан английским анатомом и
врачом Томасом Уиллисом, указавшим, что моча при этой болезни
имеет сладкий вкус. В дальнейшем у больных диабетом было
обнаружено повышенное содержание сахара в крови
(гипергликемия) и наличие его в моче (гликозурия).
В 1889 г. Меринг и Минковский показали, что удаление
поджелудочной железы вызывает у собак сахарный диабет, а в
1900 г. американский патоморфолог Юджин Опи указал на
возможную роль островковых клеток. В 1921 г. Фредерик Бантанг
и Чарлз Бест получили из поджелудочной железы вещество,
введение которого предупреждало развитие сахарного диабета у
собак с удаленной поджелудочной железой; в дальнейшем их
работа была удостоена Нобелевской премии. В 1922 г.
введением экстрактов поджелудочной железы была спасена жизнь
ребенка, страдавшего сахарным диабетом. Инсулин — активное
начало этих экстрактов — был выделен и получен в
кристаллическом виде в 1926 г. Аминокислотная последовательность его
цепи была установлена в 1955 г., а синтез осуществлен в 1963 г.
Открытие причины сахарного диабета и способа его
эффективного лечения остается одной из знаменательных вех в истории
медицины.
АНАТОМИЯ
Поджелудочная железа расположена в пределах петли
двенадцатиперстной кишки (рис. 20.1). В экзокринном отделе
поджелудочной железы, составляющем основную ее часть,
вырабатываются панкреатический сок и ферменты (см. гл. 21). Остров-
ковая ткань (эндокринный отдел), составляющая 1—2% общей
Гормоны поджелудочной железы
535
массы поджелудочной железы, образована тремя типами клеток:
альфа-, бета- и дельта-клетками, окруженными маленькими
ацинозным-и (экзокринными) клетками (рис. 30.1). На долю
альфа-клеток, вырабатывающих глюкагон, приходится около
20% всех островковых клеток; секретирующие инсулин бета-
клетки составляют 65—75%. Дельта-клетки содержат такие
Экзокринные структуры
Альфа-клетка
Рис. 30.1. Срез поджелудочной железы человека. Видны альфа- и
бета-клетки островков Лангерганса, окруженные ацинарными экзокриннкмл клетками.
Обратите внимание на наличие кровеносных сосудов между ацинарными
клетками.
гормоны, как гастрин и соматостатин. Кислые краски
прокрашивают цитоплазму альфа-клеток в красно-розовый цвет; цито-
плазматические гранулы бета-клеток под действием основных
красителей принимают пурпурно-синие тона. В гранулах бета-
клеток содержится инсулин, а в гранулах альфа-клеток —
глюкагон.
ИНСУЛИН
Инсулин представляет собой полипептид, состоящий из двух
отдельных аминокислотных цепей (А и Б), связанных дисуль-
фидными мостиками. Структура инсулина у большинства
животных сходна, хотя состав и последовательность аминокислот
у раз>ных видо'В могут быть различны. Цепь А состоит из 21, а
536
Глава 30
цепь Б из 30 аминокислот (рис. 30.2). Инсулин человека (мол.
вес 5734) отличается от инсулина других млеко/питающих типом
и последовательностью аминокислот в положениях 8, 9 и 10
цепи А и в положении 30 цепи Б.
Инсулин синтезируется в эндоплазматическом ретикулуме
бета-клеток и хранится в гранулах. Эти гранулы перемещаются
к клеточной мембране и затем высвобождаются из клеток
(преимущественно путем экзоцитоза) (см. гл. 2). Сначала
синтезируется так называемый проинсулин — вещество с длинной
одиночной цепью; в дальнейшем из него образуются две цепи.
A Gly-lle-Val Glu-Gln-Cys Cys Thr-Ser lle-Cys-Ser-Leu-Tyr-Gln-Leu-Glu-Asn-Tyr-Cys-Asn
1 2 3 4 5 6 | 8 9 10 11 12 13 14 15 16 17 18 19 / 21
S S
I /
S S
I / .
r- Phe-Val-Asn-GIn His-Leu Cys-Gly Ser His-Leu Val-GluAla-Leu-Tyr-Leu Val-Cys Gly-Glu Arg Gly-Phe Phe Туг Thr Pro Lys Thr
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Рис. 30.2. Строение инсулина человека: две полипептидные цепи (А и Б),
соединенные дисульфидными мостиками (S—S).
Метаболизм инсулина может происходить почти во всех
тканях, однако главным образом (более чем на 80%) этот гормон
распадается в печени и почках. У человека период полураспада
инсулина составляет от 10 до 25 мин; у более мелких животных
он намного короче.
СЕКРЕЦИЯ
Инсулин обычно изготовляется и отпускается таким образом,
что в 1 мг препарата содержится 24 ед. действия (ЕД или
ИЕ — интернациональная единица: 1 ИЕ = 40 мкг инсулина).
У человека в сутки секретируется примерно от 25 до 50 ЕД, т. е.
1—2 мг этого гормона. При этом инсулин выделяется в
поджелудочную вену; около половины его разрушается в печени и не
поступает в систему большого круга. На секрецию инсулина
влияет ряд факторов. Скорость секреции инсулина частично
зависит от скорости его распада. В бета-клетках имеются две
фракции инсулина: преформированная, реагирующая быстрым
выбросом инсулина в ответ на различные стимулы, и резервная,
в которую входит инсулин, синтезируемый для поддержания
преформированной фракции в условиях базальной секреции и
при длительно действующих стимулах.
Гормоны поджелудочной железы
537
ГЛАВНЫЕ ФАКТОРЫ, ВЛИЯЮЩИЕ НА ВЫДЕЛЕНИЕ ИНСУЛИНА
Факторы эти следующие: 1) уровень глюкозы в крови,
особенно ее повышенное содержание (гипергликемия); 2)
содержание аминокислот (при повышенном содержании секреция
инсулина возрастает); 3) возбуждение альфа-адренорецепторов под
действием адреналина и норадреналина (при этом выделение
инсулина снижается из-за уменьшения образования цАМФ);
возбуждение бета-адренорецепторов адреналином (при этом
Островок
Лангерганса
Панкреозимин
/ Альфа-клетка
Секретин
Глюкагон
кишечника
Глюкоза
аминокислоты
> Глюкагон
> Инсулиьо
Адреналин
Толбутамид
Возбуждение /3-адренорецепторов
Рис. 30.3. Факторы, влияющие на выработку инсулина и глюкагона. (По
Catt К. J., 1971. An ABC of endocrinology.)
увеличивается высвобождение инсулина и образование цАМФ);
5) глюкагон (увеличивает выделение инсулина как в
результате непосредственного влияния на бета-клетки, так и путем
повышения содержания глюкозы в крови, что в свою очередь
приводит к усиленной выработке инсулина); 6) другие гормоны
(секретин и холсцистокинин) и 7) блуждающие нервы (у
некоторых видов животных). Некоторые из этих факторов
изображены на рис. 30.3.
ГЛЮКОЗА
Глюкоза — наиболее мощный фактор, влияющий на
выделение инсулина. В ответ на прием глюкозы быстро (за 1—2 мин)
•выбрасывается преформированная фракция инсулина; затем
содержание этого гормона снижается и через 30—60 мин вновь
повышается и в дальнейшем удерживается на постоянном
уровне. Очевидно, эта фаза секреции обусловлена новосинтезируе-
мым инсулином. После приема глюкозы уродень инсулина в
плазме может возрасти от 20 до 50—150 мкЕД/мл.
538
Глава 30
ДЕЙСТВИЕ ИНСУЛИНА
Действие инсулина сводится к следующим основным
процессам: снижению содержания сахара в крови в результате
повышения поглощения и утилизации глюкозы тканями и усилению
синтеза следующих веществ: 1) гликогена в мышцах и печени,
2) липидов в жировой ткани и печени, 3) белка, РНК и ДНК в
клетках.
ПОГЛОЩЕНИЕ И УТИЛИЗАЦИЯ ГЛЮКОЗЫ
Уровень глюкозы в крови отражает регуляторные влияния
инсулина. В норме содержание глюкозы в крови человека после
8—24-часового голодания составляет 70—120 мг/100 мл.
Значительное повышение уровня глюкозы за пределы верхней
границы указывает на диабет. После приема лищи (особенно если
она богата углеводами) содержание глюкозы в крови быстро
Диабет
Опухоль островковой части
Часы
Рис. 30.4. Тест на толерантность к глюкозе в норме, у больного диабетом и
больного с опухолью поджелудочной железы (гиперинсулинизм). (По
Guyton А. С, 1971. Textbook of medical physiology, Saunders, Philadelphia.)
возрастает и возвращается к прежнему уровню через 2 ч. Это
происходит потому, что потребление глюкозы приводит к
выделению инсулина, и в результате содержание сахара в крови
снижается. Тест на толерантность к глюкозе, при котором
испытуемый принимает известное количество глюкозы, позволяет
определить характер активности инсулина. На рис. 30.4
представлены результаты этого теста у здорового человека и у больного
диабетом. Видно, что у больных диабетом (в отличие от
здоровых) выработка инсулина недостаточна для того, чтобы через
\ Гормоны поджелудочной железы 539
«—, :
1,5—2 ч после приема глюкозы уровень ее существенно снизился.
Инсулин повышает поглощение глюкозы мышцами
(гладкими, скелетными и сердечными), жировой тканью, гипофизом,
молочными железами и лейкоцитами. Глюкоза может
поглощаться и другими тканями, однако в головном мозгу,
канальцах поч£к, эритроцитах и пищеварительном тракте~пропесс по-
глощения глюкоаы-.Д£_регулируется инсулином.
Хотя инсулин повышает поглощение и утилизацию глюкозы
периферическими тканями, он не влияет непосредственно на ее
перенос »в клетках печени; в то же время именно печень
захватывает глюкозу из крови, превращает ее в гликоген (гликогенез) и
запасает. Когда периферические ткани нуждаются в глюкозе, то
под действием инсулина гликоген распадается (гликогенолиз) и
образующаяся в результате этого глюкоза выделяется в
кровоток.
Еще одним важным путем образования глюкозы в печени
является глюконеогенез. Этот путь включается при снижении
запасов углеводов в организме (например, при голодании); при*
этом глюкоза образуется из белков и жиров. До 60% всех
аминокислот организма могут быть превращены в углеводы. YlojX
действием глюкокортикоидов и тироксина происходит
мобилизация аминокислот, которые в дальнейшем могут дезаминировать-
ся в печени и служить субстратом для образования глюкозы.
Более подробно обмен белков, углеводов и жиров изложен в
гл. 22.
САХАРНЫЙ ДИАБЕТ
Сахарный диабет (Diabetes mellitus)—это заболевание,
характеризующееся усиленным выделением мочи (diabetes),
приобретающий сладкий привкус (mellitus). Причиной его является
пониженная выработка инсулина островковыми клетками.
Возможно, что существует наследственное предрасположение к
сахарному диабету. Заболевание встречается как в молодом, так
и в зрелом возрасте. Ювенильный диабет наблюдается в
молодом возрасте; он обычно протекает тяжелее и требует
проведения срочной заместительной терапии инсулином. Диабет
взрослых можно лечить как инсулином, так и препаратами,
усиливающими его выделение, например производным сульфонилмо-
чевины толбутамидом. Диабет легкой степени можно лечить
диетой.
У человека сахарный диабет (инсулиновая недостаточность)
характеризуется следующими основными симптомами: 1)
повышенное содержание сахара в крови (гипергликемия), 2)
усиленное выведение сахара с мочой (глюкозурия), 3) повышенный
диурез, 4) потребление большого количества воды (полидип-
540
Глава 30
I
сия), 5) потеря веса и мышечная слабость, 6) усиленный распад
белка и липидов и 7) кетоз и ацидоз.
В результате инсулиновой недостаточности угнетается
нормальное поглощение и утилизация тканями глюкозы —
основного источника энергии в организме. Это нарушение углеводного
обмена влияет на метаболизм липидов и белков: скорость их
распада увеличивается вследствие необходимости в
дополнительных источниках энергии.
Повышается также высвобождение глюкозы из печени, что
способствует гипергликемии, глюкозурии, обезвоживанию и
увеличению осмолярности мочи; это в свою очередь приводит к
увеличенному потреблению воды (полидипсия).
Все эти нарушения могут быть устранены введением
инсулина.
У многих видов животных производилось удаление
поджелудочной железы. У большинства животных это приводит к
тяжелому диабету, однако у птиц при панкреатэктомии либо
возникает диабет легкой степени, либо вовсе не появляется никаких
симптомов. Это заставляет думать или о наличии внепанкреати-
ческих источниках инсулина, или об участии других гормонов.
Такая точка зрения подкрепляется данными, согласно которым
некоторые вещества типа стрептозотоцина у птиц (в отличие от
млекопитающих) не разрушают бета-клеток островков Лангер-
ганса и не приводят к сахарному диабету.
ИЗБЫТОЧНАЯ СЕКРЕЦИЯ ИНСУЛИНА
При избытке инсулина возникает гипогликемия (содержание
сахара может падать до 20—35 мг/100 мл). Это состояние
сопровождается повышенной секрецией гормонов коры
надпочечников, способствующих мобилизации глюкозы и компенсации
гипогликемии. Пониженное содержание сахара в крови иногда
приводит к слабости, головокружению, чувству голода,
спутанному сознанию и в конечном счете судорогам и коме. Избыток
инсулина наблюдается у больных с опухолью эндокринного
отдела поджелудочной железы; сахар крови у них может падать
до 20 мг/100 мл.
ГЛЮКАГОН: СЕКРЕЦИЯ И МЕТАБОЛИЗМ
Глюкагон — продукт альфа-клеток поджелудочной железы,
представляет собой полипептид, образованный
последовательностью из 29 аминокислот (рис. 30.5). Структура глюкагона у
человека и свиньи одинакова. Молекулярный вес глюкагона
составляет 3485. У птиц (кур) глюкагон также состоит из 29 ами-
\ Гормоны поджелудочной железы 641
нокисдот, однако он отличается от глюкагона человека тем, что
в положении 28 у него находится вместо аспарагина серии.
Скорость секреции глюкагона у человека очень мала по
сравнению с дозами,использовавшимися при изучении влияний
этого гомона на метаболизм. Согласно данным, полученным
при применении радиоиммунологических методов, базальная
суточная секреция глюкагона составляет 100—150 мкг.
Возможно, что эти цифры завышены в связи с тем, что иммунореактив-
ность этого гормона выше, чем его биологическая активность.
Секреция глюкагона увеличивается под влиянием аминокислот,
глюкокортикоидов, стресса, инфекции, физической нагрузки, га-
стрина, холецистокинина и инсулина. К факторам, снижающим
His—Ser—Gin—Gly—Thr—Phe—Thr—Ser—Asp—Туг—Ser—Lys—Туг—Leu—Asq—Ser—Arg—Arg—
12 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 17
—Bla—Gin—Asp | Phe—Val—Gin—Trp—Leu— Met—Asn—Thr
19 20 21 22 23 24 25 26 27 28 29
Рис. 30.5. Строение глюкагона человека.
секрецию, относятся глюкоза, секретин, свободные жирные
кислоты и кетоны.
Период полураспада глюкагона равен 5—10 мин; он
подвергается катаболизму во многих тканях, прежде всего в почках,
© отличие от инсулина, распадающегося преимущественно в
печени.
ДЕЙСТВИЕ ГЛЮКАГОНА
Основной эффект глюкагона заключается в увеличении
уровня глюкозы в крови. Это происходит в результате того, что глю-
кагон активирует аденилатциклазу печени, что приводит к
увеличению синтеза цАМФ. Увеличенный синтез цАМФ
сопровождается распадом гликогена (гликогенолизом) и
высвобождением глюкозы из печени (но не из мышц). В печени под
действием глюкагона усиливается глюконеогенез из аминокислот.
Во многих исследованиях, посвященных глюкагону,
использовались очень высокие нефизиологичные (фармакологические)
дозы этого гормона, оказывающие отчетливое действие на
обмен углеводов. Однако в последних работах (Sherwin, Felig)
ценность подобных наблюдений подвергается сомнению и
ставится вопрос о том, играет ли глюкагон в физиологических
дозах какую-либо важную роль в регуляции уровня глюкозы (го-
меостазе). У человека физиологические дозы глюкагона
приводят к незначительному и кратковременному повышению
содержания глюкозы в плазме.
542
Глава 30
I
Последние данные заставляют также предположить, ч/о при
сахарном диабете гипергликемия обусловлена недостатком
инсулина, избыточная же секреция глюкагона не оказывает на нее
существенного влияния. Эта точка зрения в настоящее время
дискутируется. /
ЛИТЕРАТУРА
Catt К. J. (1971). An ABC of endocrinology, Little, Brown, Boston.
Ganong W. F. (1977). Review of medical physiology, 8th edn., Lange Medical,
Los Altos, California.
Guyton A. C. (1977). Basic human physiology, 2nd edn. Chap. 25, Saunders,
Philadelphia.
Hazelwood R. L. (1976). Carbohydrate metabolism (Chap. 11) and Pancreas
(Chap. 21). In: Sturkie P. D. (ed.), Avian physiology, 3rd edn., Springer
Verlag, New York.
Sherwin R.t Fellg P. (1977). Glucagon physiology in health and disease. In:
McCann S. M. (ed.), International review of physiology-endocrine
physiology II, Vol. 16, University Park Press, Baltimore.
Террегтап J. (1973). Metabolic and endocrine physiology, 3rd edn., Year Book
Medical, Chicago.
Turner С D., Bagnara J. T. (1976). General endocrinology, 6th edn., Saunders,
Philadelphia.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1. Что открыли Фр. Бантинг и Ч. Бест?
2. Какие клетки поджелудочной железы вырабатывают
инсулин? Глюкагон?
3. Инсулин представляет собой полипептид из двух
аминокислотных цепей. Сколько аминокислот содержат эти цепи?
4. Назовите основной фактор, стимулирующий ©ыработку
инсулина?
5. Каков главный эффект инсулина?
6. Назовите основные симптомы сахарного диабета и
расскажите о них.
7. Что такое: а) гликогенолиз; б) гликогенез, 3) глюконеоге-
нез?
8. Какое нарушение возникает при избытке инсулина?
Объясните это нарушение.
9. Глюкагон представляет собой линейный полипептид.
Сколько аминокислот он содержит?
10. Каков главный эффект глюкагона?
11. Имеет ли глюкагон такое же значение для регуляции
углеводного обмена, что и инсулин? Обсудите этот вопрос.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аденозинтрифосфат см. АДФ
Адреналин 531,. См. также Катехол-
амины, Гормоны надпочечников
Адренэргические нейроны 192, 261
АДФ, роль в метаболизме 404, 408,
413, 416
Акклиматизация 362
Аккомодация 112
АКТГ, действие 473, 523, 526, 527
— строение 477
— уровень в гипофизе и крови 526
Альдостерон 458, 527
— синтез и выделение 528
Амилаза см. Ферменты
желудочно-кишечного тракта
Аминокислоты, всасывание 398
— недостаточность 432, 433
— незаменимые 428
— обмен 421, 423
Ампула 134
Анаболизм 401, 409
Андрогены, действие 490
— синтез и структура 489, 490
Анемия 230
Аномалии рефракции 114
Аносмия 139
Антидиуретический гормон 460, 482
Антикодон 420
Аортальные тельца 262, 263, 321
Аргинин-вазопрессин 481, 482
Артерии 29, 31, 239—241
Астигм атизм 115—116
Атаксия 167
Атетоз 169
АТФ 35, 44, 326, 404, 408—413, 416,
421, 422
Ацетилхолин в нервной системе 82,
84, 192
Базальные ганглии 65, 67, 168
Базилярная (основная) мембрана
126—128
Базофиллы 236, 237
Барабанная перепонка 125
Белки, всасывание 398
— крови 225
— переваривание 396—398
— синтез 421—424
Беременность 502
— гормоны 503
Блуждающий нерв 61, 189, 192
Боуменова капсула 440
Буферы крови 46, 316, 465
Вазопрессин, выброс 482
— действие 460, 482
— структура 482
— уровень в гипофизе 481
крови 481
Вазотоцин, уровень в гипофизе 481
крови 481
Венозный возврат 249
Вены 30, 31, 239—241
Вестибулярная система 131
Вестибулярные рецепторы 132, 133
Витамины 429, 432, 433
Вкусовая система 136—138
— сосочки 136—138
Вкусовые ощущения 136
— почки 136
— пути 138
Влагалище, анатомия 491, 496
— влияние гормонов 496, 506
Внутреннее тепло 343
Водорода ион, концентрация 45—47
Водянистая влага 109
Волосковые клетки 127, 128, 132, 133
Временная суммация 88
Всасывание аминокислот и белков
398
— воды 398, 399
— липидов 398
— минеральных веществ 398, 399
— углеводов 397
Вторичные половые признаки у
женщин 501
мужчин 490
Высокоэнергетические фосфаты 408
Гастрин 388
Гексозомонофосфатный шунт 414
544
Предметный указатель
Гематоэнцефалический барьер 251
Гемоглобин 235
— связывание Ог 236, 312
— типы 235
Гемодинамика 241
Гемолиз 230
Гендерсона — Гассельбаха уравнение
47, 315
Генетический код 422
Гипергликемия 539
Гиперметропия 114
Гипертермия 363
Гипертиреоз 514
Гиперфагия 198
Гипоксия 323
Гипоталамо-гипофизарная система,
нейросекреция 474
Гипоталамус 199
— анатомия 199, 472, 473
— регуляция вегетативных функций
196
— рилизинг-факторы 473, 475
— центр голода и насыщения 198,
375
— центры вегетативных функций
197—199
— ядра 197, 482
Гипотиреоз 515
Гипофиз, гормоны 472
— нейросекреция 474
механизм обратной связи 475,
500
— задняя доля 199, 472, 475
гормоны 481
— передняя доля (аденогипофиз)
199, 472, 475
гормоны 472, 476, 477, 479,
480
типы клеток 476
Гистампп 388
Глаз 107—123
— роговица 109, ПО
— сетчатка, функции 109, 116
— строение 109
— фоторецепторы 107, 116
— хрусталик 109
Гликогенолиз 539
Гликолиз 410—413
Глотание 375
Глюкагон, действие 541
— секреция и метаболизм 540
Глюкозурия 539
Глюкоза, обмен 409—411, 413—416
— поглощение и утилизация 538
— транспорт 43, 44
— уровень в крови 537, 538
Глюкокортикоиды 526
Глюкокортикоиды
— действие 529
Глюконеогенез 539
Гольджи аппарат 35
— сухожильные органы 14^
Гомеостаз 214
Гомеотермные 341
Гормоны, биологические ^робы 471
— коры надпочечников 523
выделение 526 '
действие 529
типы 523, 524
уровень в крови 523, 527
— механизм действия 469
— мозгового вещества
надпочечников 531. См. также Катехоламины
— определение 471
— рецепторы 469
Двигательная концевая пластинка 82
— кора 155, 156, 173
Двигательные ядра 161
Дерматомы 57
Дефекация 385
Диабет 539
Диализ 41
Диафрагма 299, 301 ,
Дисметрия 168
Диффузия 38, 39, 41, 221
ДНК 33, 34, 418—420
Доннана равновесие 38
Дрожь 357
Дыхание, ацидоз и алкалоз 316
— газовые законы 310
— газообмен 294, 310
— гипоксия 323
— диафрагма 299, 301
— диффузия газов 311, 315, 316
— дыхательные мышцы 300
— дыхательный воздух 302
— жизненная емкость 303
— кривая диссоциации оксигемогло-
бина 313
— легочное кровообращение 308
— механизм дыхательных движений
304
— минутный объем 303
— объем воздуха в легких 302
— остаточный воздух 303
— регуляция 318
— — в поддержании
кислотно-щелочного равновесия 47, 315
— связывание 02 гемоглобином
крови 236, 312
— содержание газов в организме 295
— транспорт газов 312—314
л
Предметные указатель
545
Дыхательная система, анатомия 294
воздухоносные пути 295
— «• легкие 298
плевра 299
Евнухоидизм 490
Еда 374
Жажда, нервная регуляция 198
— потребление воды 221, 375
Жевание 375
Железы, типы 13—15
Желтуха 395
Желудок, анатомия 369, 377, 386, 396
— гормоны 379
— моторика 377
— переваривание 396
— секреция 386, 389
Желудочно-кишечный тракт 368
анатомия 368—369
кровоток 373, 374
переваривание 396
ферменты 396
Желудочный сок 386, 387
кислотность 387, 396
состав 387
Желчные кислоты, соли 395
Желчный пузырь 395
Желчь 393
— состав 394
Женские половые органы, гормоны
498, 499
Жидкие среды, вода 215
давление 218, 251
образование и передвижение
216, 217
состав 215
Жирные кислоты 398, 427
основные (незаменимые) 428
Жиры, метаболизм 413, 417, 427
Задний мозг 63
Запоминание 185
Зародышевые листки 12
Зрачковый рефлекс ПО
Зрачок ПО
—оптическая система 109—111
— фокусное расстояние и диоптрии
112
Зрение, аккомодация 112
— дефекты 114, 115
Зрение, рефракция 111, 112
— формирование изображения ПО
— цветовое 118
— центральные зрительные пути 121
35—1705
Зрительная кора 122
— система 109
Инсулин 534, 535
— влияние на гликоген в тканях 538
глюкозу крови 538
жировую ткань 538
мышцы 539
печень 538, 539
— избыток 540
— недостаток 539
— секреция 536
— синтез 536
— структура 535, 536
Интенционный тремор 167
Инулин 446, 452
Иод, влияние на синтез гормонов
щитовидной железы 511
Иодопсин 118
Калории 325
Кальций, обмен в костях 518
влияние кальцитонина 521
— уровень в крови 519
Кальцитонин, биохимическая
активность 521
— влияние на обмен кальция 521
— секреция 521
— уровень в тканях 517, 521
Канальцевая секреция 456
Капаситация 489
Капилляры 216, 218, 241
Каротидные тельца 321
Катехоламины 531
— обмен и секреция 532
Кислородный долг 336
Кислотно-щелочное равновесие 45
роль дыхания 315
Кишечник, моторика 380, 382
— нервы 370—372
— переваривание 397
— секреция 391
— строение 380
Клетка 9, 32
— гены и хромосомы 33
— митоз 33
— строение 32—36
Клеточная мембрана 36, 37
— физиология 32
Клиренс, методика 445
Клубочки 440. См. также Почки
Клубочковая фильтрация 444, 446—
448
ауторегуляция 449
Кодон 420, 422
546
Предметный указатель
Кожа 11, 14
Колбочки 117
— пигменты 117—118
— функция 117, 118
— центральной ямки 116—117
Конечный мозг 65
Контрацепция 506
Кора головного мозга, активация
176, 179
ассоциативные области 174
вкусовая система 136
зрительная доля 66, 123
лимбическая система 65, 67
обонятельные нервы 67
соматосенсорная 106
ЭЭГ 176
— надпочечников, функции 529
Коричневый жир 360
Короткова тоны 255
Кортиев орган 127
Кортикостерон см. Гормоны коры
надпочечников
Костная система 21—23
Красное ядро 64, 160
Креатин 408
Креатинфосфат 408
Кребса цикл 413
Кровеносные сосуды, анатомия 31,
239—241
кровяное давление 251, 309
Кровообращение 239
— легочное 308
— саморегуляция 266
— физические законы 241
Кровоток в органах и тканях 251,
252
почках 252, 444, 446, 449
— распределение 251
— скорость 243
Кровь, антикоагулянты 233
— вязкость 226, 227
— гематокрит 226
— гемоглобин 235
— группы 234
— объем 214, 226, 227
— осмотическое давление 218
— распределение 246
— свертывание 232, 233
— удельный вес 226, 227
— физические свойства 226
— функции 225
Кровяное давление 251
в венах 249, 250
капиллярах 218, 309
влияние возраста 258
дыхания 259
поведения 259
Кровяное давление влияние
рефлексов 263
а- и (3-рецепторов 265
сердечного выброса 259, 263
силы тяжести и положения
тела 250, 258
физической нагрузки 248
частоты сокращений сердца
259, 263
внутрижелудочковое 271, 272
диастолическое 253, 254
измерение 254, 255
колебания 254
нормальное 257
половые различия 257
пульсовое 254
регуляция 259, 261—265
систолическое 253, 254
Кровяные пластинки (тромбоциты)
227, 228, 232
Круглое окно 127
Лактация 504
Лапласа закон 245
Легкие, объем 302
— растяжимость 304
— рецепторы 320
Лейкемия (лейкоз) 237
Лейкопения 237
Лейкоцитоз 237
Лейкоциты 236
— функции 237
— число 227, 237
Лизосомы 35
Лимбическая кора 209
— система 199
Лимфа, образование 222
Лимфатические узлы 223
Лимфоциты 228, 237
Липазы 396, 397
Липиды, всасывание 398
— переваривание 397, 398
— типы 427
— триглицериды 427
Лихорадочное состояние 339, 365
Лютеинизирующий гормон 473, 499
в гипофизе и в крови 498, 505
действие 479, 499
нейросекреция, механизм
обратной связи 475, 500
Мамиллярные тела 64
Матка, анатомия 491, 496
— во время менструального цикла
498
Предметный указатель
547
Маточка 131, 132
Медиаторы 82
— вегетативной нервной системы 192
— возбуждающие и тормозные 90
Медленные движения 160
Меланоцитстимулирующий гормон
(МСГ) 473, 479
Мембрана, поры 37
— проницаемость 37—39
— транспорт веществ 44
— модель 37
Мембранные потенциалы 45, 69—73
измерение 71
ионные основы 73
Менструальный цикл 500
Менструация 497
Метаболизм, интенсивность 330
— промежуточный 416
Метаболические пути 413
Мешочек 131, 132
Минералокортикоиды 529
Минутный объем 246
Миозиновые нити 205, 207
Миопия 115
Митохондрии 35
Мозг, анатомия 55—67
— базальные ганглии 65, 67, 168
— миндалина 67, 201, 202
— мозолистое тело 66, 175
— ножки 64
Мозжечок 63
— анатомия 163
— ножки 63
— повреждения 167
— функции 164
— ядра 163—165
Мозолистое тело 66, 175
Моноциты 228, 237
у-мотонейроны 149
Моча, концентрация 459—460
— образование 466
— состав 466
Мочевой пузырь 466
иннервация 467
Мочеиспускаш1е 466
Мышца(ы) быстрые и медленные 211
— гладкие 19, 212
— сердечная 19, 212
— скелетные 19, 24, 204
— тетанус 209, 212
— утомление 212
Мышечная система 24
— ткань 19, 24—28, 204
Мышечное веретено 147
— сокращение, механика 206
Мышечный тонус 211
Надпочечники 523
— анатомия и гистология 523
— размер 523
Нейрогипофиз 199
Нейроны 21
— определение и типы 49
Нервная система, автономная 188—
190
аксоны и дендриты 49
дегенерация и регенерация 49
иннервация скелетных мышц
82
клетки глии 51
медиаторы 82
мембранный потенциал 70
миелиновая оболочка 52, 79
нейромедиаторы 50, 82, 83
парасимпатическая 192, 193.
периферическая 55
потенциал действия 74
проведение импульса 49, 50,
77—79
рефрактерность 75
сенсорная информация,
кодирование 96
симпатическая 190
синаптическая передача 51, 82,
85
строение 49, 52, 55
черепные нервы 60—61
— ткань 21
Нервно-мышечное соединение 82—85
■ восстановительные процессы
84
Нервный импульс, возникновение 75
проведение 77
Нервы смешанные 80
Нернста уравнение 73
Норадреналин 531. См. также Кате-
холамины, Гормоны надпочечников
Нуклеиновые кислоты 417—424
генетический код 422
Нуклеотиды 422
Обонятельная луковица 139, 140
— система 138, 139
Обонятельные пути 139, 140
Оболочки 11,13
Образование тепла, измерение 330
Овальное окно 126
Овогенез 493, 497
Овуляция 497
— лютеинизирующий гормон 498,
499
— механизм 497—499
— прогестерон 498, 499
35*
548
Предметный указатель
Овуляция, фолликулостимулирующии
гормон 499
— эстроген 499
Окситоцин 483
— влияние на роды 483
— строение 482
— уровень в крови 483
Оплодотворение 488, 489
Опсин 118
Осмос 41—42
Осмотические растворы 44
Осмотическое давление 42
Острота зрения 113
Палочки 117
Память 184
Панкреатические полипептиды 393
Панкреатический сок 391
состав 391
ферменты 391, 396
Панкреатэктомия 540
Парааминогиппурат 447
Паратгормон, влияние на кости 519—
520
функции почек 520
— роль в мобилизации кальция 520
— строение 518
Пищеварительные железы, анатомия
и гистология 518
гипер- и гипотиреоз 520
Паркинсона болезнь 170
Пентозомонофосфатный шунт 414
Пепсин 396
Переваривание 396
Передний мозг 66
Перилимфа 126
Печень, анатомия 393
— функции 393
Пиноцитоз 44
Пирамидная система 155, 157, 158
Пиримидины 419, 422
Питание 425
— минеральные соли 429, 430
— населения 434
пищевые программы 437
Питательные вещества, образование
энергии 425
суточное потребление 437
Пищевод 368
— анатомия 368
— нервная регуляция 368—371
— сократительные движения 375
Пищевые вещества, недостаточность
432, 433
• потребность и суточные нормы
435
Плазма крови 214, 215
Пневмотаксический центр 320
Поджелудочная железа, анатомия
391, 534
— секреция 391
типы клеток 535
Пойкилотермные животные 341
Половое поведение 486
нервная регуляция 199
Половой акт 506
— член 485, 486
Половые гормоны мужские 489
строение 493, 500
— органы женские 494
мужские 485
Постсинаптические реакции 87
Потенциал действия 74—79
возникновение 84, 95
восстановление ионных
градиентов 77
фазы 74, 75
Потоотделение 355
Потребление пищи, центр голода 198,
375
насыщения 198, 375
Почка 439
— анатомия 439—444
— кислотно-щелочное равновесие 464
— клиренс 446
— кровообращение 252, 444, 446, 449
— образование ренина 463
— простагландиновая система 463
— противоточный умножитель 460
— реабсорбция воды 458
— фильтрация 444, 446—448
Пресинаптическое торможение 89
Прогестерон, влияние на
высвобождение лютеинизирующего гормона
498—499
овуляцию 499
— секреция и выброс 498, 505
— строение 493
— уровень в крови 499
Продолговатый мозг 59
анатомия 59, 61
вегетативные функции 196
кортикоспинальные пути 59
слуховая система 62
улитковые ядра 62
центры дыхания и
кровообращения 63
Пролактин 473, 505
— в гипофизе и крови 505
— механизм действия 505
Промежуточный мозг, анатомия 64
гипоталамус 60, 65
таламус 64, 65, 103—106
Предметный указатель
549
Промежуточный мозг, эпиталамус 64
Пространственная суммация 88
Противоточный умножитель 460
Протромбиновое время 233
Пуазейля уравнение 242
Пульсовая волна 254
Пурины 419, 422
Пятно (macula) 132
Радужка ПО
Рвота 380
Реабсорбция глюкозы 454
— механизм 452
— электролитов 453
Резус-фактор 235
Рейнольдса число 244
Ренин 463
Ренин-ангиотензиновая система 462,
528
Ретикулярная формация 61, 160, 162
Ретиналь 118, 119
Рефлекс(ы) моносинаптические 144
— отдергивания 150
-^ перекрестный разгибательный 151
— полисинаптический 150—152
— растяжения 144
— сгибательный 150
— сухожильный 149
— чесательный 151
Рефрактерность мышц 209, 212
— нерва 75
Рефракция 111
Рецепторный белок 84
Рецепторы 93
,— болевые 99, 195
'— адаптация 96, 97
— давления (прессорецепторы) 98,
262, 263
— соматосенсорные 98
а- и Р-рецепторы 265
Рецепторный потенциал 94
Реципрокная иннервация 152
Рибосомы 34, 423
; Ритм сердца 276
— — регуляция 277
, РНК 34, 419
I Родопсин 118
Роды 483, 502
Рост 435
\ Сальтаторное проведение 79
I Саркоплазматический ретикулум 204
I Свертывание крови 232, 233
Семенная жидкость,
оплодотворяющая способность 489
хранение 489
Сенсорная информация, кодирование
и адаптация 96
Сердечный выброс 246
влияние 248, 249, 274
распределение крови 251
— цикл 270
Сердце, анатомия 267
— деполяризация 281
— диастола 272
— коэффициент полезного действия
(к. п. д.) 276
— проводящая система 281
— работа 276
— реполяризация 281
— систола 270
— сокращения, саморегуляция 275
— тоны 272
— частота сокращений 263
— электрическая ось 287, 288
Сетчатка 109, 117
— строение 115
Синапсы межнейронные 85
— электрические 90
Система кровообращения 31, 239,
241
Системы живого организма 9
«Скользящих нитей» теория 208
Скотопсин 118
Следы памяти 186
Слуховая кора 131
— система 124
нервы 130
ощущение звука 124, 131
различение частоты звука 129,
131
чувствительность 125
Слуховые косточки 125
— рецепторы 63, 127, 128, 131, 132
Слюнные железы, секреция 385
Снеллена таблицы 114
Соединительная ткань 13
Соматосенсорная кора 106
Соматосенсорные пути 59, 100—106
Соматотропный гормон (СТГ) 473
механизм действия 481
строение 478
уровень в крови и гипофизе
479
Сон 177, 180—182
— центр 183
Сосудодвигательные нервы и частота
сокращений сердца 263, 279
— центры 263, 278
Сперматогенез 487, 493, 497
550
Предметный указатель
Сперматозоиды, анатомия 488
— оплодотворяющая способность 489
— число 488
Специфическое динамическое
действие пищи 338
Спинной мозг, организация 143
соматосенсорные пути 59,
100—106
строение 57, 143, 190
Спинномозговые рефлексы 142
Средний мозг 64
восходящие и нисходящие
пути 64
зрительная область 64
слуховая область 64
строение 64
четверохолмие 64
Старлинга гипотеза 218, 309
Стекловидное тело 109
Таламус 121, 122, 138, 173, 175
Температура воздуха,
термонейтральная зона 350
— среды 341, 349, 363
— тела 342, 343
измерение 342
ректальная 344, 364
тепловой баланс 345
Температурные рецепторы 350
Тепловой баланс 345
— удар 364
Теплообмен, коэффициент передачи
тепла 349
— путем излучения 245
испарения 348, 354
конвекции 347
теплопроводности 346
Теплообразование и акклиматизация
363
Теплопродукция и возраст 339
лихорадочное состояние 339,
366
температура окружающей
среды 338
— центры 352, 353
Термогенез 360
Терморегуляция 351
— адаптация к экстремальной темпе*
ратуре 362
— гетеротермных животных 341, 362
— механизм 355, 357, 360
— роль мозга и гипоталамуса 352,
353
— тепловая одышка 355
— центр 352
Тестостерон 489, 490
— действие 490
— синтез 489
Тиреотропный гормон (ТТГ) 473,511,
512
строение 477
Тироксин в крови 511, 512
— влияние на метаболизм 513
рост 514
— секреция 511—514
— синтез 511
— структурная формула 510
Ткани тела И—21
Транспорт активный и пассивный 40,
44
Тромбоциты 232
Тропомиозин 205
Тропонин 205
Т-трубочки 204—206
Углеводы, всасывание 397
— обмен 409—415
влияние инсулина 539, 542
Улитка 125, 126
Ультимобранхиальные тельца, анато
мия 517, 521
Ухо 124
— внутреннее 126
— наружное 125
— среднее 125
Фагоцитоз 44
Фаллопиевы трубы 502
Ферменты желудочно-кишечного
тракта 396
— ингибиторы 405
— природа и активность 403, 414—
416
Фика уравнение 39, 246, 247
Фильтрация 38, 218, 444, 454
Фолликулостимулирующий гормок
(ФСГ) 473, 499
в гипофизе и в крови 476, 490
действие 479, 491
строение 477
Форменные элементы крови 225, 227,
228. См. также Эритроциты,
Лейкоциты
Фосфорилирование окислительное
415, 416
Фотопигменты 118, 119
Фоторецепторы 116—117
— потенциалы 117
— фотохимические свойства 118
Франка — Старлинга закон 274
Предметный указатель
551
Хеморецепторы 262, 263, 321
Холестерин см. Липиды
Холинэргические нейроны 192, 261
Хрящевая ткань 17
Центр апноэ 319
Центральная нервная система 55—67
— ямка 116
Цикл лимонной кислоты 414. См.
также Цикл трикарбоновых кислот
Цикл трикарбоновых кислот 413
Цитоплазма 34
Цитохромная система 416
Черная субстанция 168
Шванновские клетки 51
Щелевые контакты 90
Щитовидная железа, анатомия 509,
510
гормоны, влияние на гипофиз
513
ТТГ-стимуляция 473, 511, 512
Экзоцитоз 44
Экстрапирамидная система 159—163
Электрокардиограмма (ЭКГ) 272,
282
— СА-блокада 292
— зубцы и интервалы 286
— нормальная 282
— отведения 282
— патологические изменения 289
— потенциал действия 281
— узлы 281, 292
— во время сна 177, 181
Электроэнцефалограмма (ЭЭГ),
нейрофизиологические основы 177
— ритмы 176
Эмбриональное развитие, стадии 502
Эмиссия 485
Эмоциональное поведение 199
Эндолимфа 126
Эндоплазматический ретикулум 34
Энергетический обмен 326
анаэробный 326, 337, 409
аэробный 326, 337, 409
во время сна 339
■ единицы измерения 325
интенсивность 333
основной 333, 334
— эквивалент пищи 329, 333, 425
Энергия 406
— образование 326, 406
— потребление 437
— превращение и использование 326,
406, 409
Эозинофилы 236, 237
Эпителий 11
Эритропоэз 229
Эритроциты 227, 228, 230
— гематокрит 226
— гемоглобин 230
— образование 228, 229
— при анемии 230, 231
— продолжительность жизни 230
— прочность 230
— состав 230
— размер и число 231
Эрекция и эякуляция 485
Эстроген и вторичные половые
признаки 501
— механизм действия 501
— секреция 499, 501
— уровень в крови 499
Ядерная мембрана 33
Ядро клетки 32
Яички 485
— гормоны 489
Яичники, анатомия 494
Яйцеклетка, созревание 495
ОГЛАВЛЕНИЕ
Предисловие редактора перевода 5
Предисловие . . 7
Список авторов . 8
Глава 1. Общий план строения тканей, органов и опорно-двигательного
аппарата 9
Терминология 10
Ткани 11
Анатомия 21
Вопросы для повторения 31
Глава 2. Общая и клеточная физиология . 32
Строение клетки 32
Клеточная мембрана 36
Кислотно-щелочное равновесие .... 45
Вопросы для повторения 48
Глава 3. Организация нервной системы 49
Нейрон 49
Анатомическая организация нервной системы 55
Вопросы для повторения 68
Глава 4. Физиология нейронов 69
Мембранный потенциал 70
Потенциал действия 74
Синаптическая передача 82
Вопросы для повторения 91
Глава 5. Рецепция. Соматическая чувствительность . . .93
Рецепторы .93
Кодирование сенсорной информации. Адаптация ... . 96
Соматосенсорная система . 9S
Центральные соматосенсорные пути . 100
Вопросы для повторения .108
Глава 6. Органы чувств 1°9
Зрительная система Ю9
Фоторецепция И 6
Слуховая система 1-4
Вестибулярная система 131
Вкусовая система 136
Обонятельная система 138
Вопросы для повторения 141
Оглавление 553
Глава 7. Спинномозговые рефлексы ... 142
Виды рефлексов . .142
Вопросы для повторения ... . .153
Глава 8. Центральная регуляция движений . 155
Двигательные функции коры ... 155
Мозжечок 163
Базальные ганглии 168
Вопросы для повторения 170
Глава 9. Высшие функции коры головного мозга . . . 172
Организация коры 172
Ассоциативные области коры 174
Активация коры головного мозга. Сознание 176
Сон 180
Память 184
Вопросы для повторения 187
Глава 10. Нервная регуляция функций внутренних органов . . 188
Центры регуляции функций 188
Автономная нервная система 190
Центры регуляции вегетативных функций 196
Лимбическая система ч 199
Вопросы для повторения 203
Глава 11. Мышцы 204
Скелетные мышцы 204
Механика мышечного сокращения 206
Сердечная и гладкие мышцы 212
Вопросы для повторения 213
Глава 12. Жидкие среды организма. Кровообращение в капиллярах.
Лимфообращение 213
Измерение объема жидкостных пространств организма 213
Состаз жидких сред организма 215
Фильтрация и гипотеза Старлинга 218
Фильтрация и образование тканевой жидкости 219
Образование лимфы 222
Вопросы для повторения 224
Глава 13. Кровь 225
Физические свойства крови 226
Форменные элементы крови 227
Группы крови 234
Гемоглобин 235
Лейкоциты 236
Вопросы для повторения 238
Глава 14. Кровообращение 239
Общие представления 239
Физические законы движения крови (гемодинамика) 241
Сердечный выброс и распределение крови 246
554
Оглавление
Кровяное давление 251
Нормальные показатели артериального давления .... 257
Регуляция артериального давления 259
Вопросы для повторения 266
Глава 15. Сердце как насос 267
Анатомические сведения 267
Сердечный цикл 270
Сила сокращений 273
Гомеометрическая саморегуляция силы сокращения 275
Ритм сердца 276
Вопросы для повторения 280
Глава 16. Электрокардиография ,281
Проводящая система сердца . 281
Деполяризация и реполяризация .281
Электрокардиограмма . 282
Патологические изменения ЭКГ . 289
Вопросы для повторения . 293
Глава 17. Дыхание 294
Анатомия 294
Легочная вентиляция 301
Механика дыхательных движений 304
Легочное кровообращение . 308
Легочное дыхание ". 310
Транспорт дыхательных газов 312
Насыщение тканей кислородом 316
Регуляция дыхания 318
Вопросы для повторения 324
Гл ава 18. Энергетический обмен 325
Определения и единицы измерения 325
Превращение и использование энергии 326
Энергетический эквивалент пищи 329
Определение интенсивности метаболизма (обмена) 330
Основной обмен ..... 333
Вопросы для повторения 340
Глава 19. Терморегуляция 341
Температура тела и ее измерение 342
Тепловой баланс 345
Система терморегуляции 351
Терморегуляция и мышечная работа 360
Физиологическая адаптация и экстремальная температура . . . 362
Лихорадочное состояние 365
Вопросы для повторения 366
Глава 20. Моторика желудочно-кишечного тракта ....... 368
Элементы анатомии 368
Еда и питье 374
Жевание и глотание 375
Сократительные движения пищевода 375
Оглавление
555
Моторика желудка
Моторика кишечника
Моторика толстого кишечника
Вопросы для повторения
Глава 21. Секреция, переваривание, всасывание
Слюнные железы
Желудочная секреция
Панкреатическая и кишечная секреция
Печеночная и желчная секреция
Переваривание
Всасывание
Вопросы для повторения ....
Глава 22. Промежуточный обмен
Анаболизм и катаболизм ....
Энергия
Анаэробный и аэробный катаболизм .
Метаболизм углеводов, жиров и белков
Нуклеиновые кислоты .....
Синтез белка
Вопросы для повторения ....
377
380
382
384
385
385
386
391
393
396
397
399
401
401
406
409
413
417
421
424
Глава 23. Питание 425
Жиры, углеводы и белки 425
Минеральные соли и витамины 429
Питание населения 434
Потребность в пищевых веществах и рекомендуемые суточные нормы 435
Общие соображения 436
Вопросы для повторения 438
Глава 24. Почка . . . 439
Функциональная анатомия 439
Нефрон 440
Клубочковая фильтрация 444
Канальцевая реабсорбция 450
Канальцевая секреция 456
Реабсорбция воды 458
Противоточный умножитель 460
Почечные гормоны 462
Почки и кислотно-щелочное равновесие 464
Образование мочи 466
Вопросы для повторения 467
Глава 25. Эндокринология. Функции гипофиза и гипоталамуса 469
Механизм действия гормонов 469
Определение содержания гормонов 4/1
Гормоны гипоталамуса и гипофиза 4/^
Рилизинг-факторы гипоталамуса 4Л5
Гормоны аденогипофиза **
Гормоны задней доли гипофиза ... *°'
Вопросы для повторения ...
556 Оглавление
Глава 26. Размножение 485
Эрекция, эякуляция и эмиссия 485
Сперматогенез и сперматозоиды 487
Мужские половые гормоны 486
Репродуктивная функция у женщин 490
Анатомия женских половых органов 494
Овогенез и сперматогенез 497
Менструация и овуляция 497
Гормоны половых желез 500
Беременность и роды 502
Лактация 504
Половой акт 506
Контрацепция 506
Вопросы для повторения 508
Глава 27. Гормоны щитовидной железы .... 509
Иодсодержащие соединения 510
Секреция и высвобождение тироксина 511
Секреция и метаболизм 512
Действие гормонов щитовидной железы и регуляция их секреции . 513
Нарушение функции щитовидной железы 514
Вопросы для повторения 516
Глава 28. Гормональная регуляция обмена кальция . . . 517
Содержание Са в тканях 517
Паращитовидные железы 518
Кальцитонин 521
Вопросы для повторения 522
Гл ава 29. Гормоны надпочечников 523
Макро- и микроструктура надпочечников 523
Гормоны коры надпочечников 523
Мозговое вещество надпочечников и катехоламины 531
Вопросы для повторения 533
Глава 30. Гормоны поджелудочной железы 534
Анатомия 534
Инсулин 535
Поглощение и утилизация глюкозы 538
Сахарный диабет 539
Глюкагон: секреция и метаболизм 540
Вопросы для повторения 542
Предметный указатель 543
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении,
качестве перевода и другие просим присылать по адресу:
129820, Москва, И-110, ГСП
1-й Рижский пер., д. 2
издательство «Мир»
ОСНОВЫ ФИЗИОЛОГИИ
Ст. научный редактор Е. А. Яновская
Мл. научный редактор Р. Ф. Куликова
Художник В. Е. Карпов
Художественный редактор Л. М. Кузнецова
Технический редактор Т. А. Максимова
Корректор Н. В. Андреева
ИБ № 3723
Сдано в набор 24.11.83. Подписано к печати 04.04.84. Формат 60x90Vie.
Бумага типографская № 2. Гарнитура литературная. Печать высокая.
Объем бум. л. 17,50 Усл. печ. л. 35. Уч.-изд. л. 33,77. Изд. № 4/2708.
Усл. кр. отт. 35. Тираж 15.000 экз. Заказ 1705. Цена 2 р. 70 к.
ИЗДАТЕЛЬСТВО «МИР»,
129820, Москва, И-110, ГСП, 1-й Рижский пер., 2.
Московская типография № 11 Союзполиграфпрома при Государственном
комитете СССР по делам издательств, полиграфии и книжной торговли.
Москва, 113105, Нагатинская ул., д. 1.
ИЗДАТЕЛЬСТВО «МИР»
Выйдут в свет в 1984 году
ОСНОВЫ СЕНСОРНОЙ ФИЗИОЛОГИИ: Пер. с англ.
/Под ред. Р. Шмидта, 23 л., 3 р. 80 к.
Книга специалистов из ФРГ, выдержавшая 3 издания на
немецком языке и 2 на английском, посвященная одному из самых
интересных разделов нейрофизиологии. Рассмотрены общие
вопросы сенсорной физиологии, строение и функции соматовисце-
ральной, зрительной, слуховой, вкусовой, обонятельной систем,
механизмы, лежащие в основе чувств равновесия, голода и
жажды. Материал изложен на вполне современном уровне, компактно
и четко.
Предназначена для студентов и аспирантов университетов и
медицинских институтов, а также для специалистов з области
нейрофизиологии.
МОЗГ: Сб. статей. Пер. с англ. — 2-е изд., 20 л.,
1 р. 70 к.
Первое издание этой книги выпущено издательством «Мир»
в 1982 г. Книга представляет собой специальный номер журнала
«Сайентифик Америки», посвященный головному мозгу, в
котором принимали участие такие известные ученые, как Э. Кэндел,
Э. Эвартс, лауреаты Нобелевской премии Ф. Крик, Д. Хьюбел,
Т. Визель и другие. Рассмотрены проблема познаваемости
головного мозга как «машины» мышления, методы изучения его
деятельности, организация нейрона и мозга в целом, механизмы
зрения и движения, химия и развитие мозга, специализация мозга
человека, нарушение функций мозга.
Предназначена для биологов всех специальностей, для
врачей, химиков, физиков, кибернетиков, для студентов и
аспирантов.