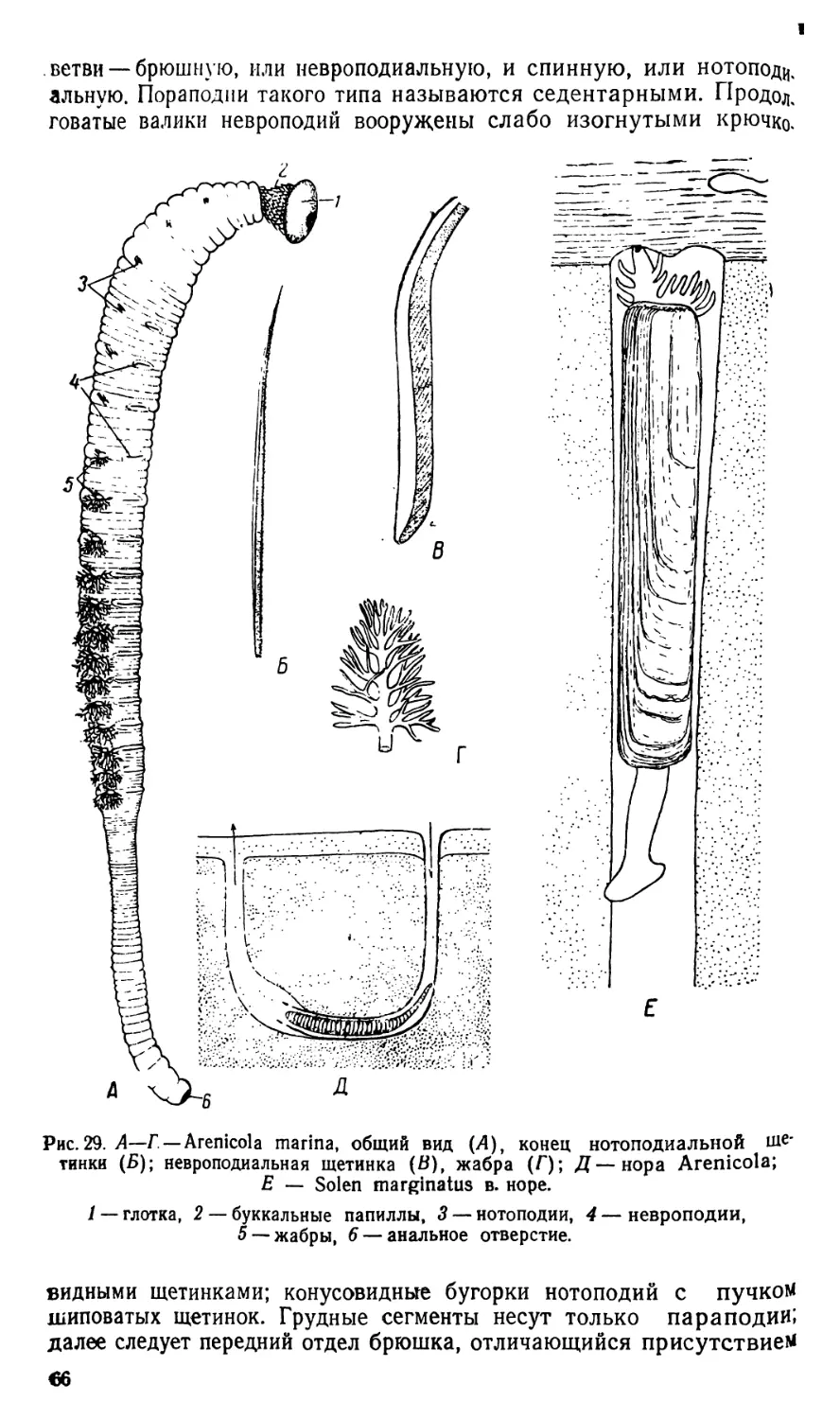
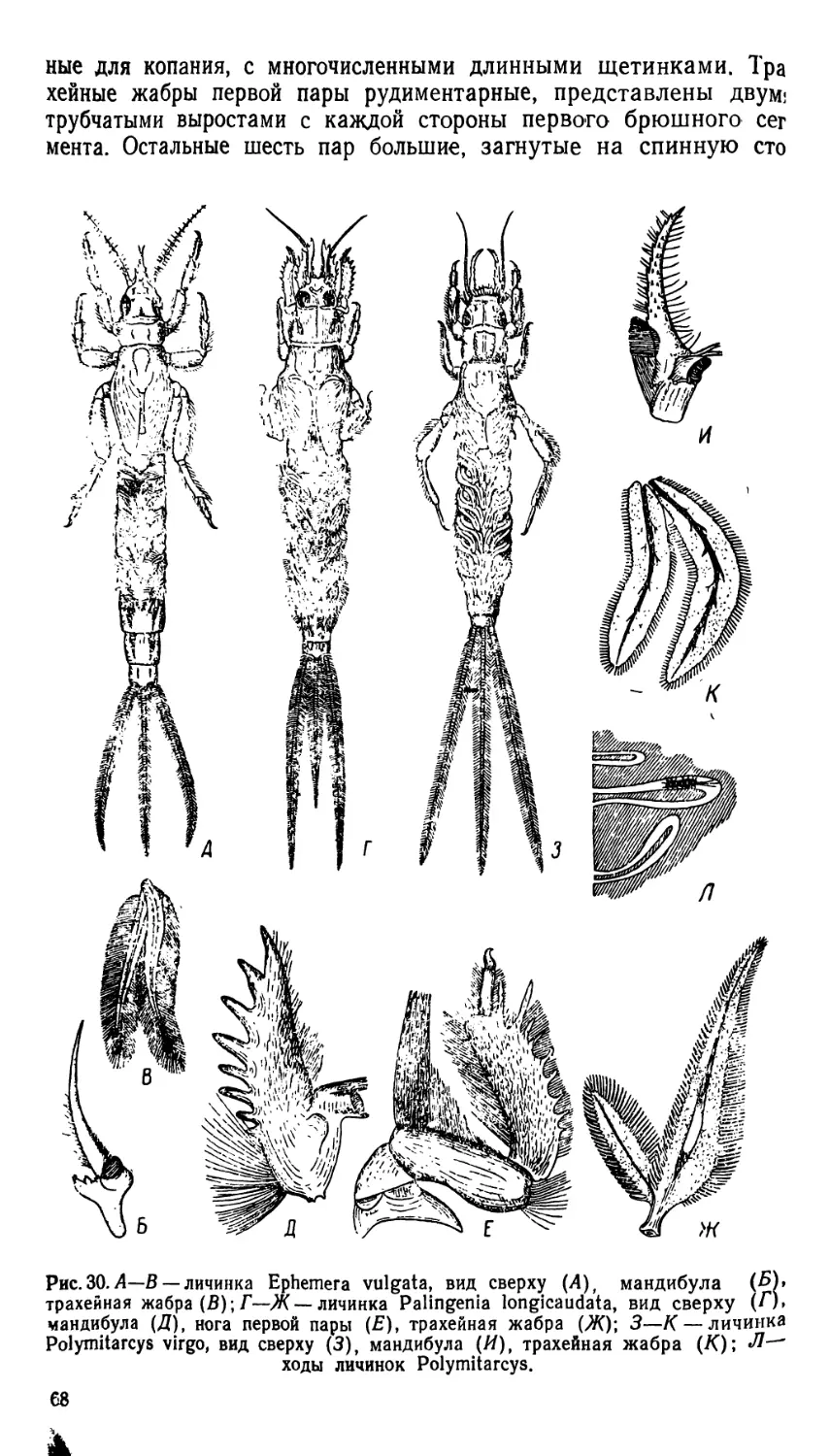
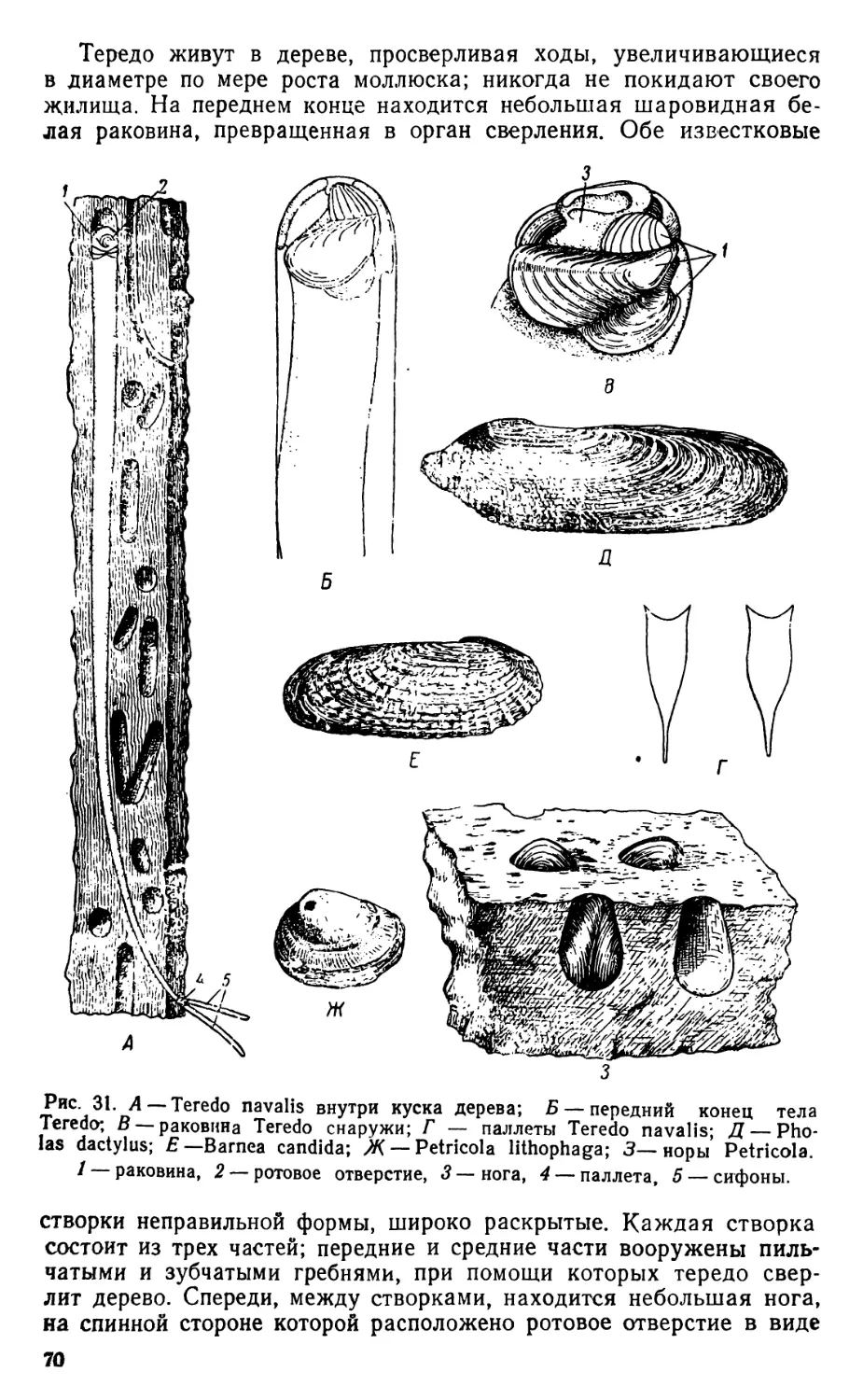
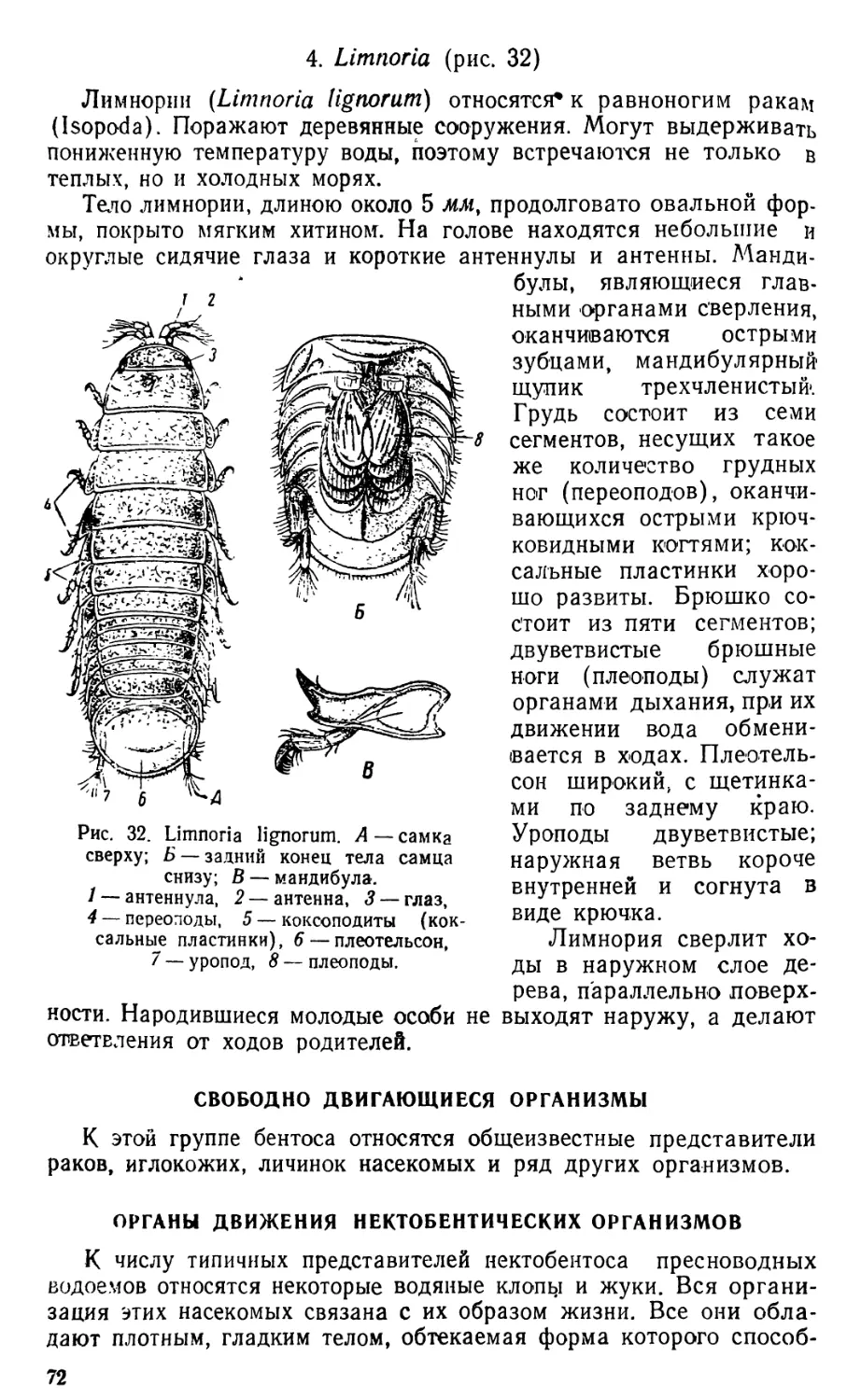
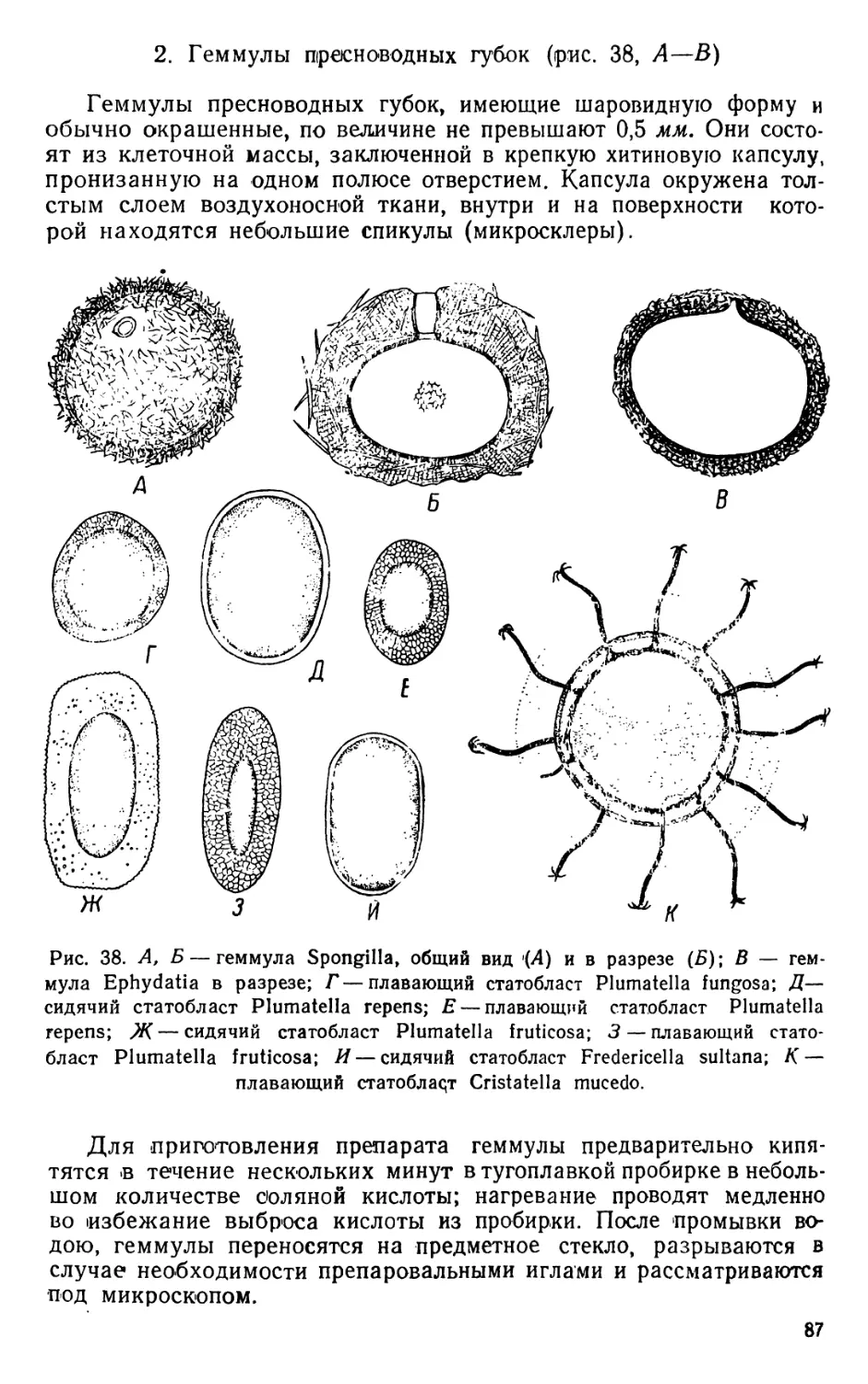
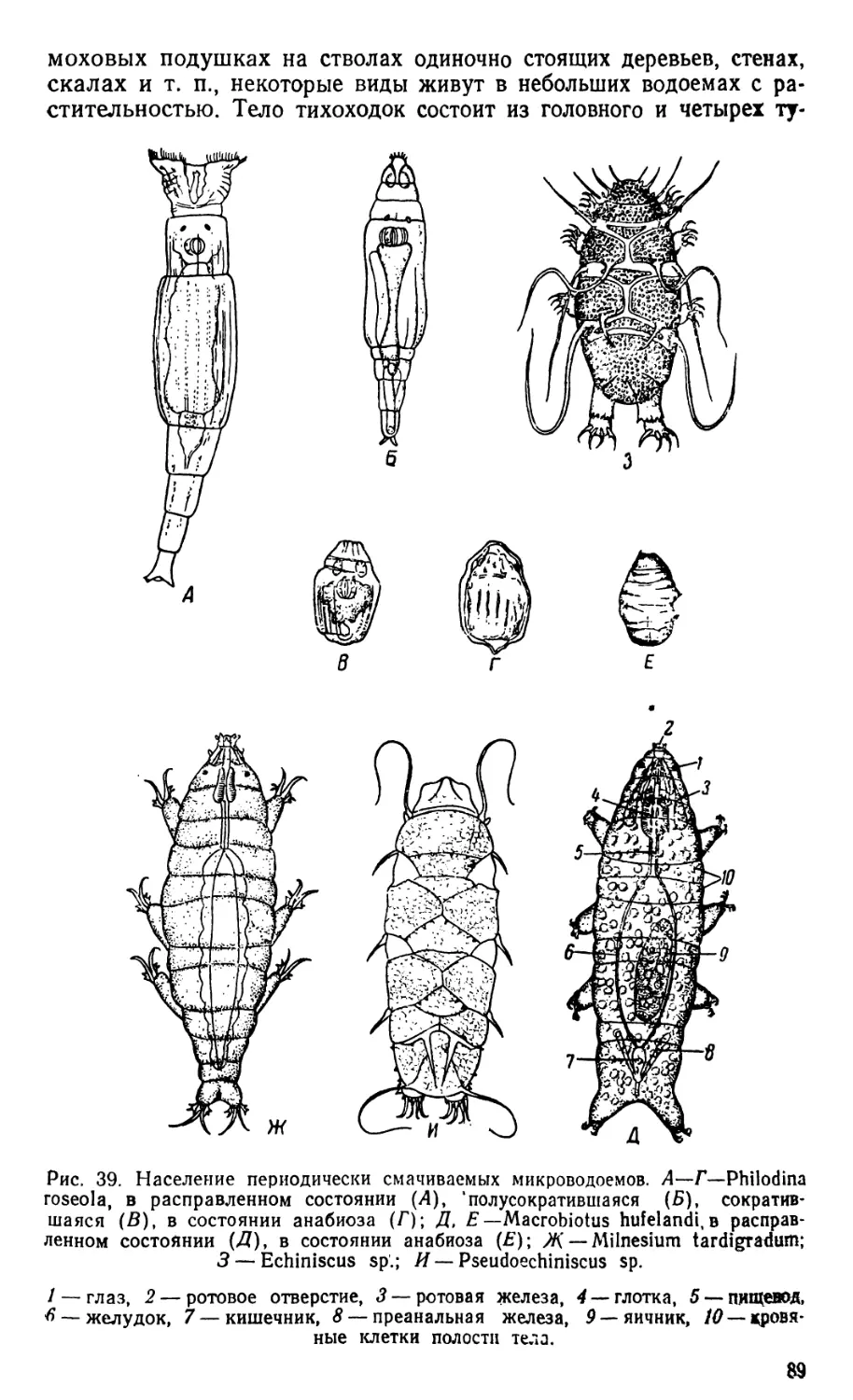
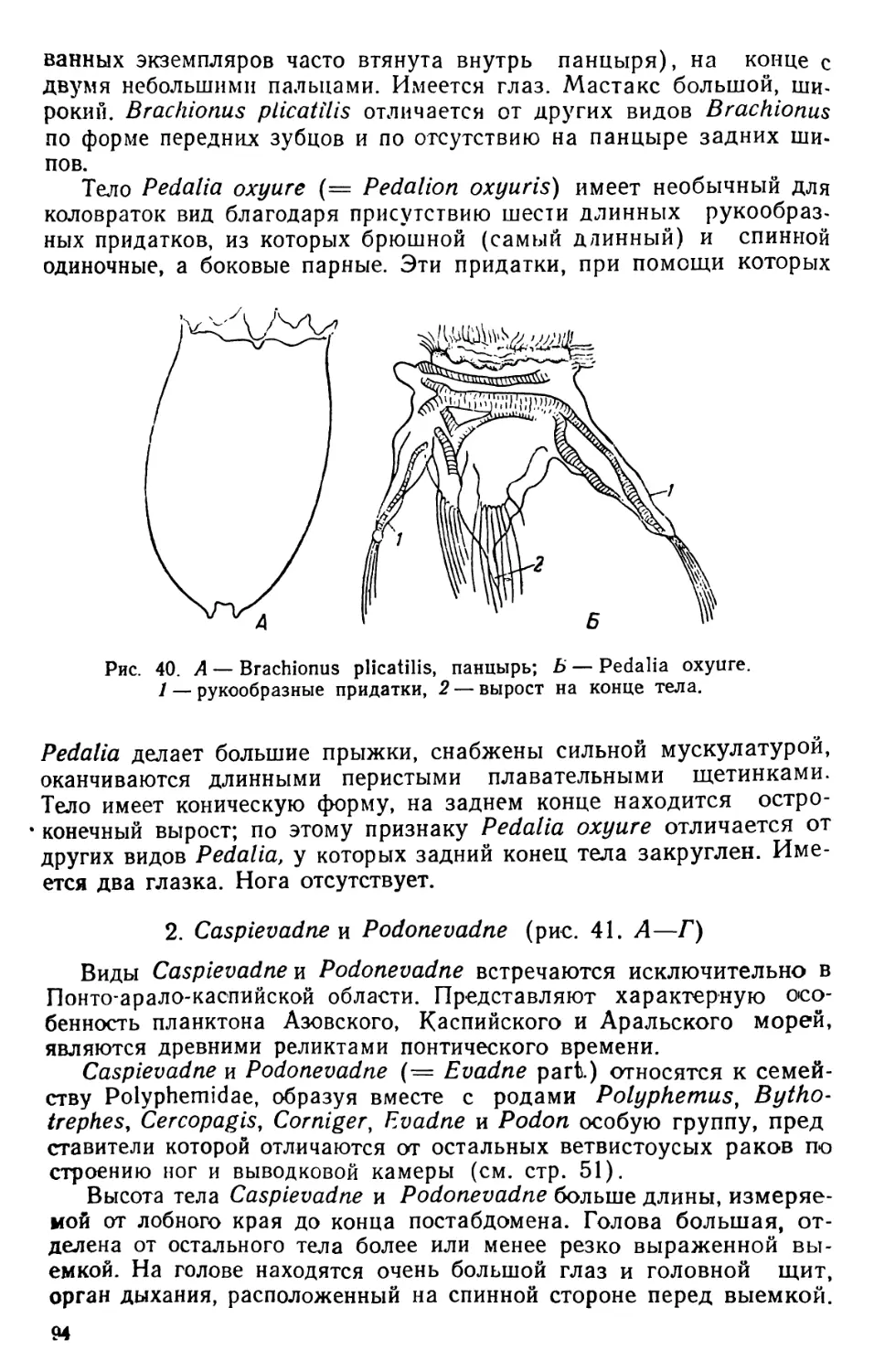
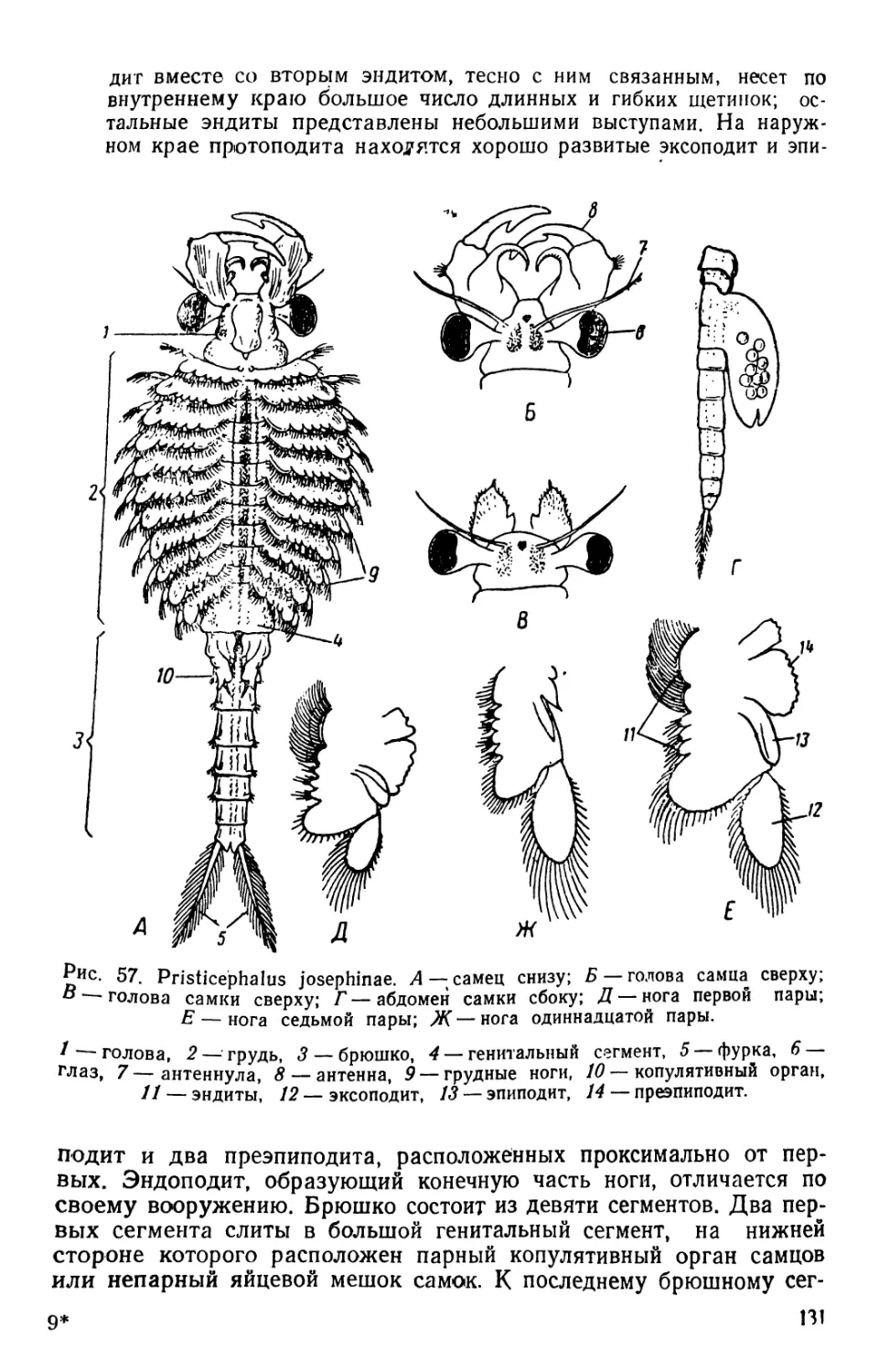
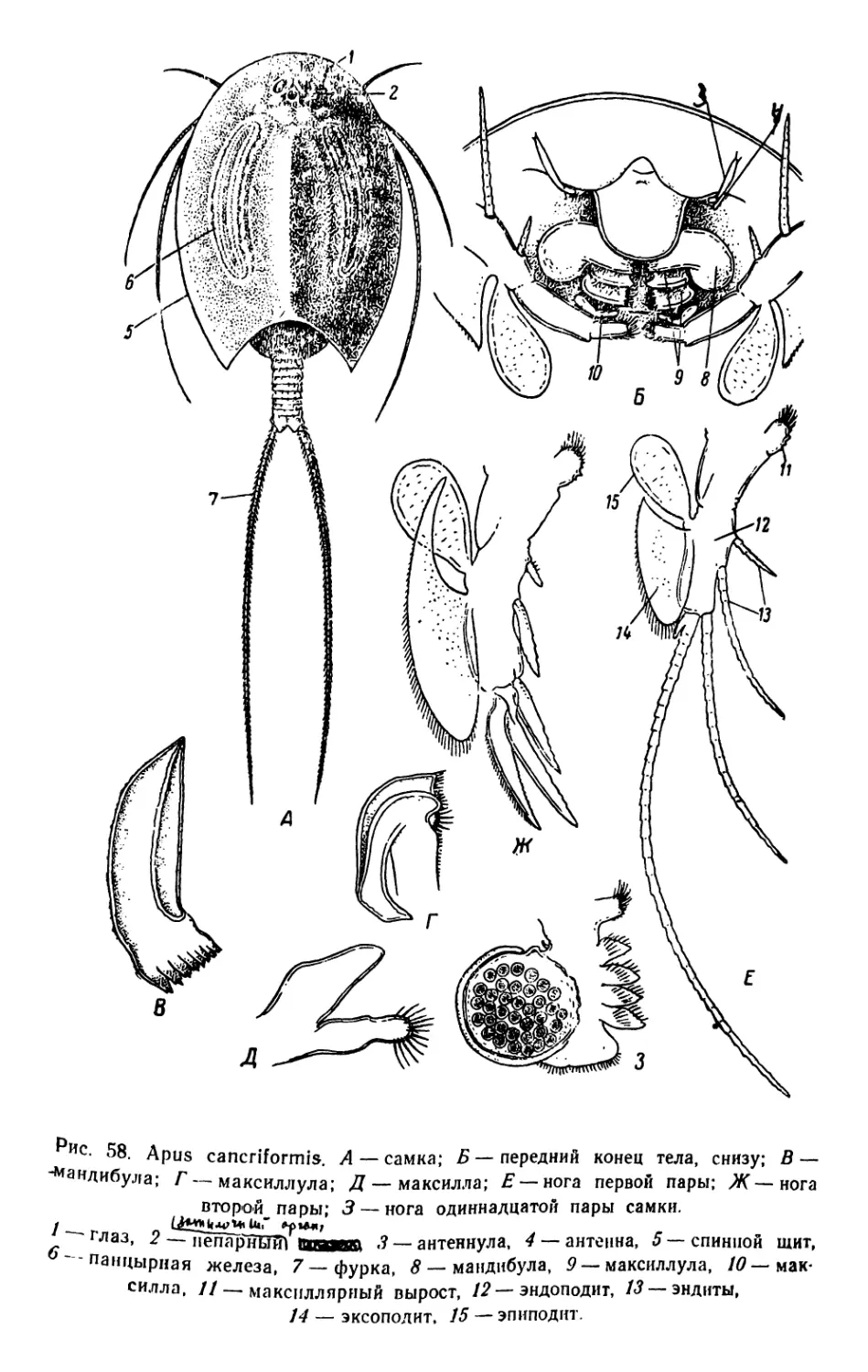
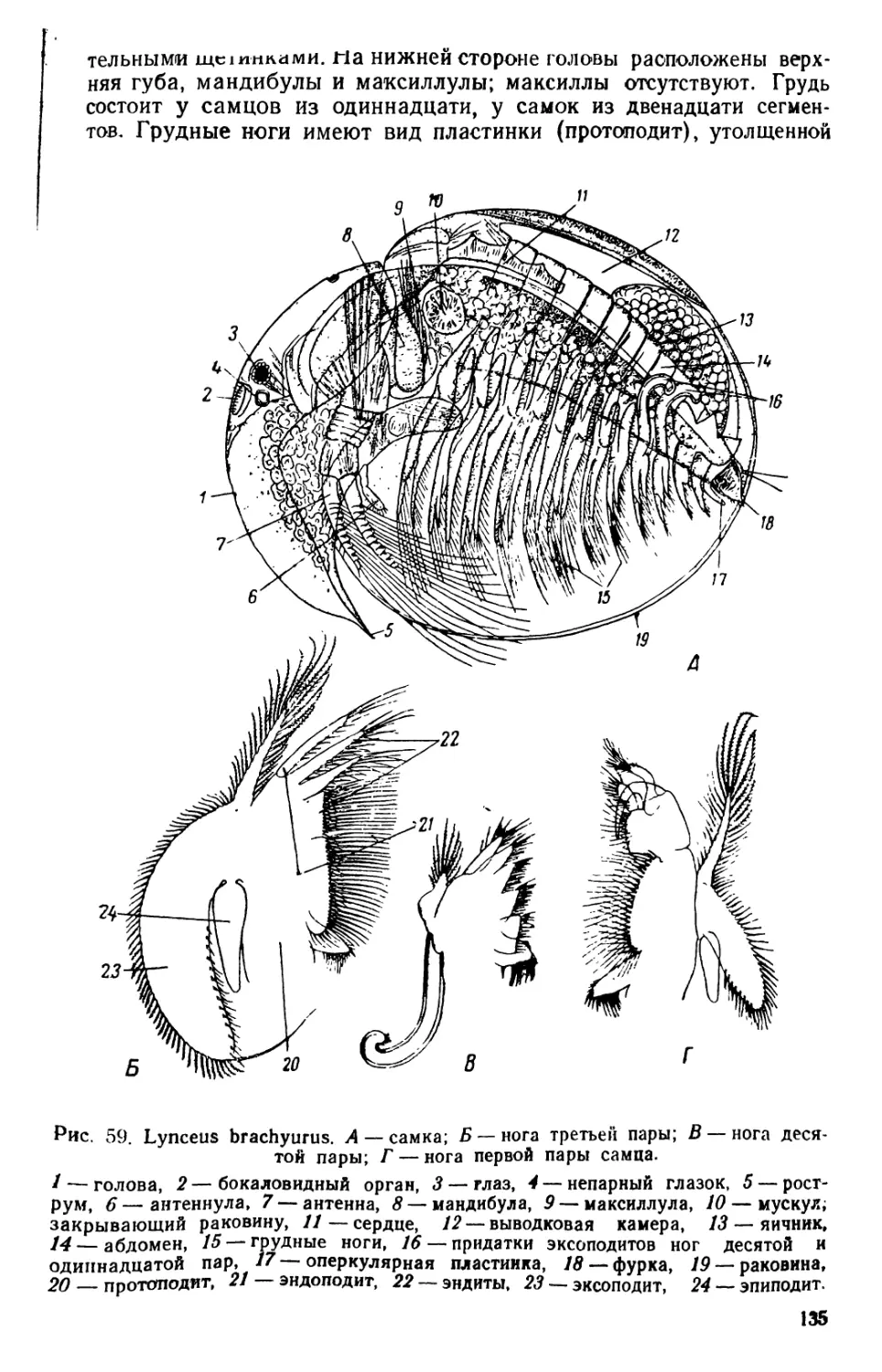
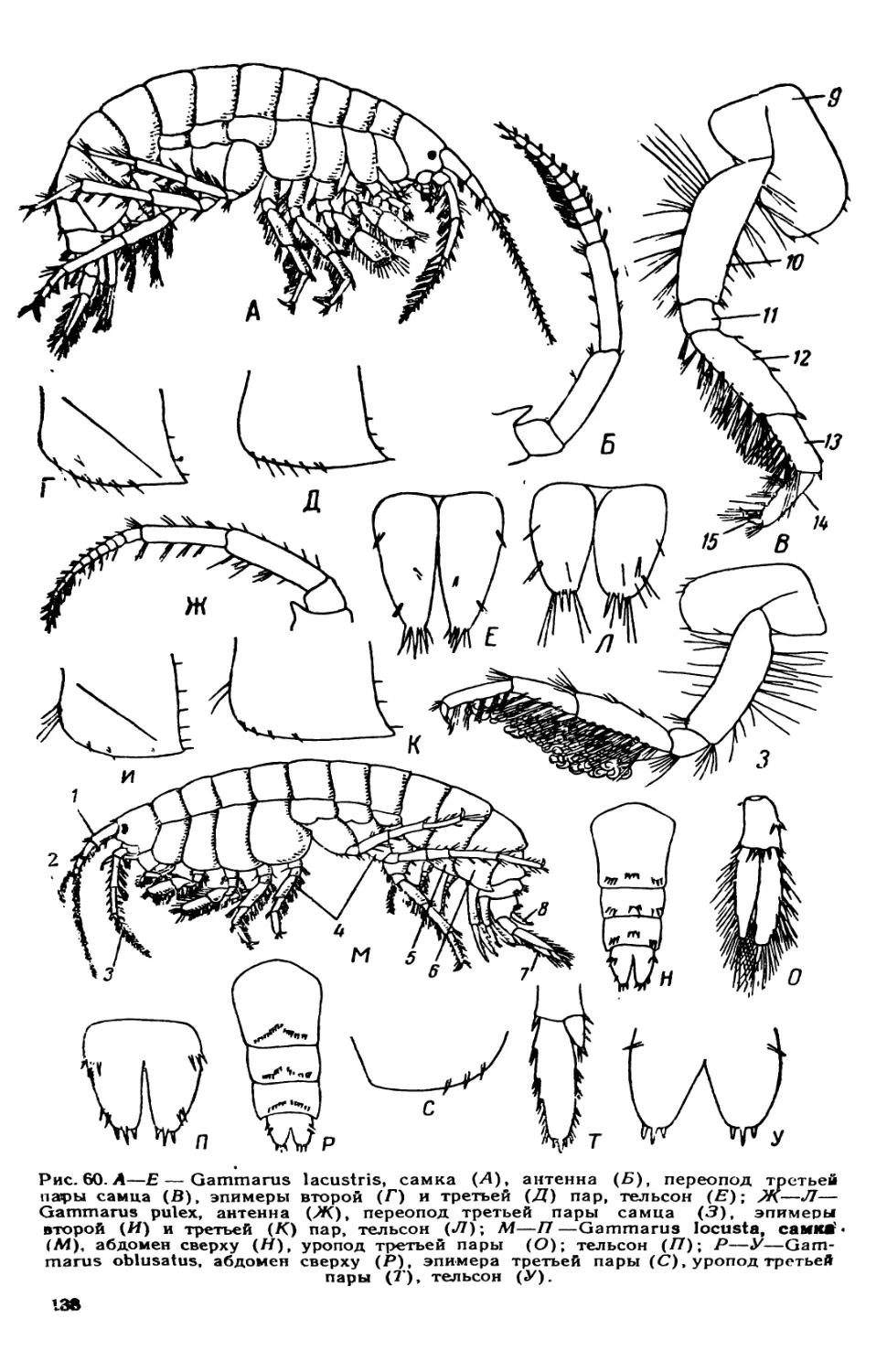
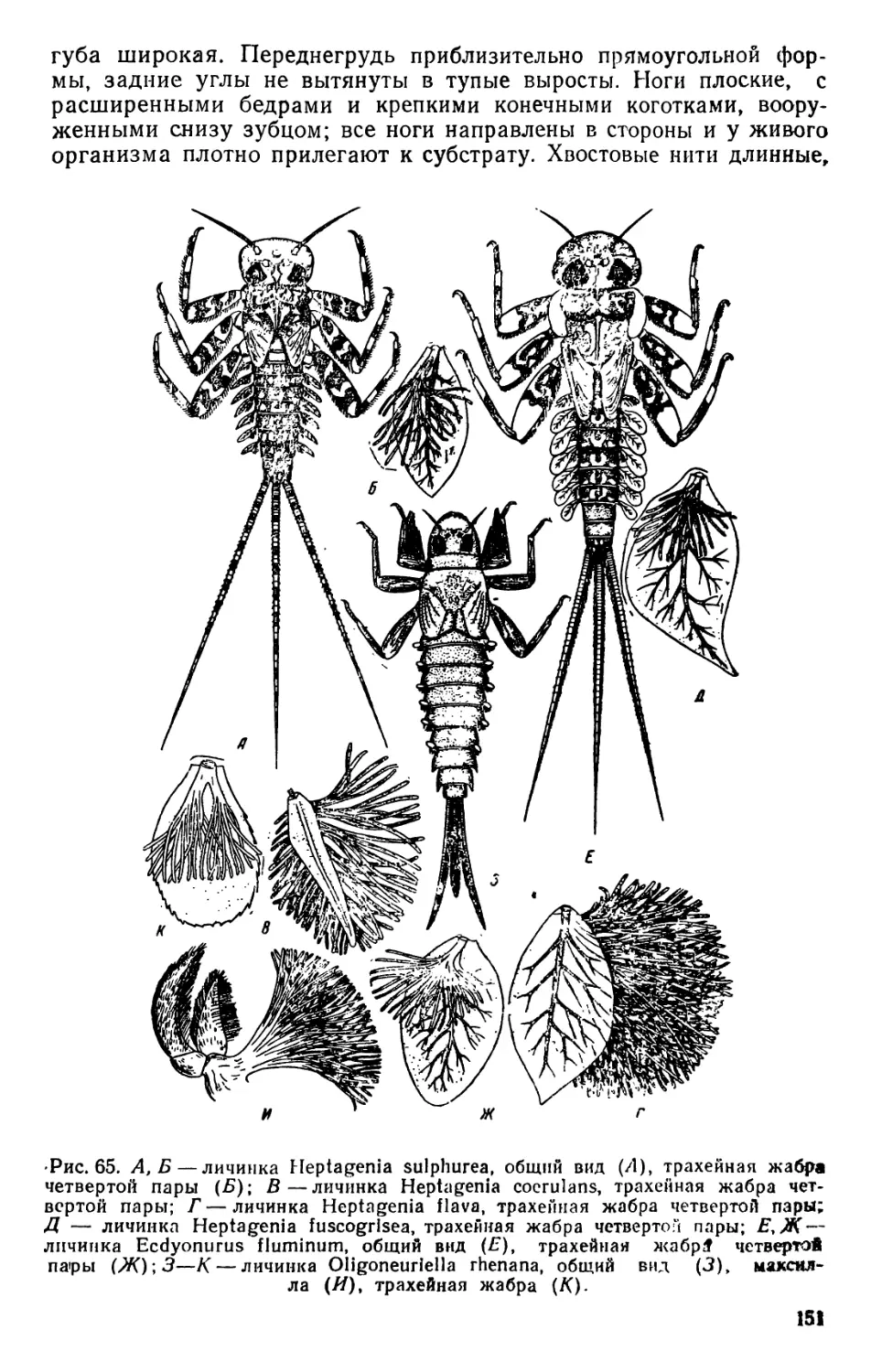
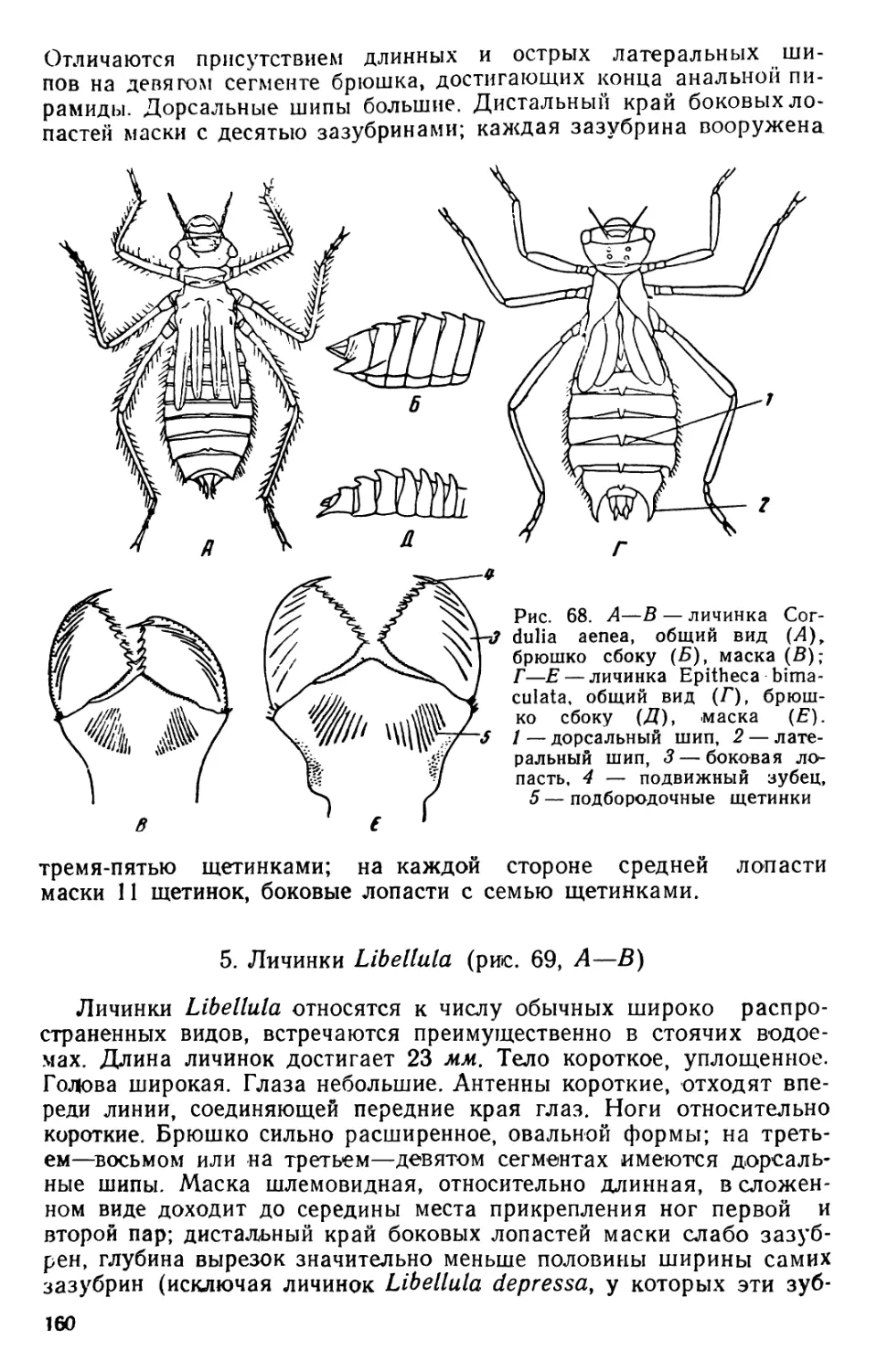
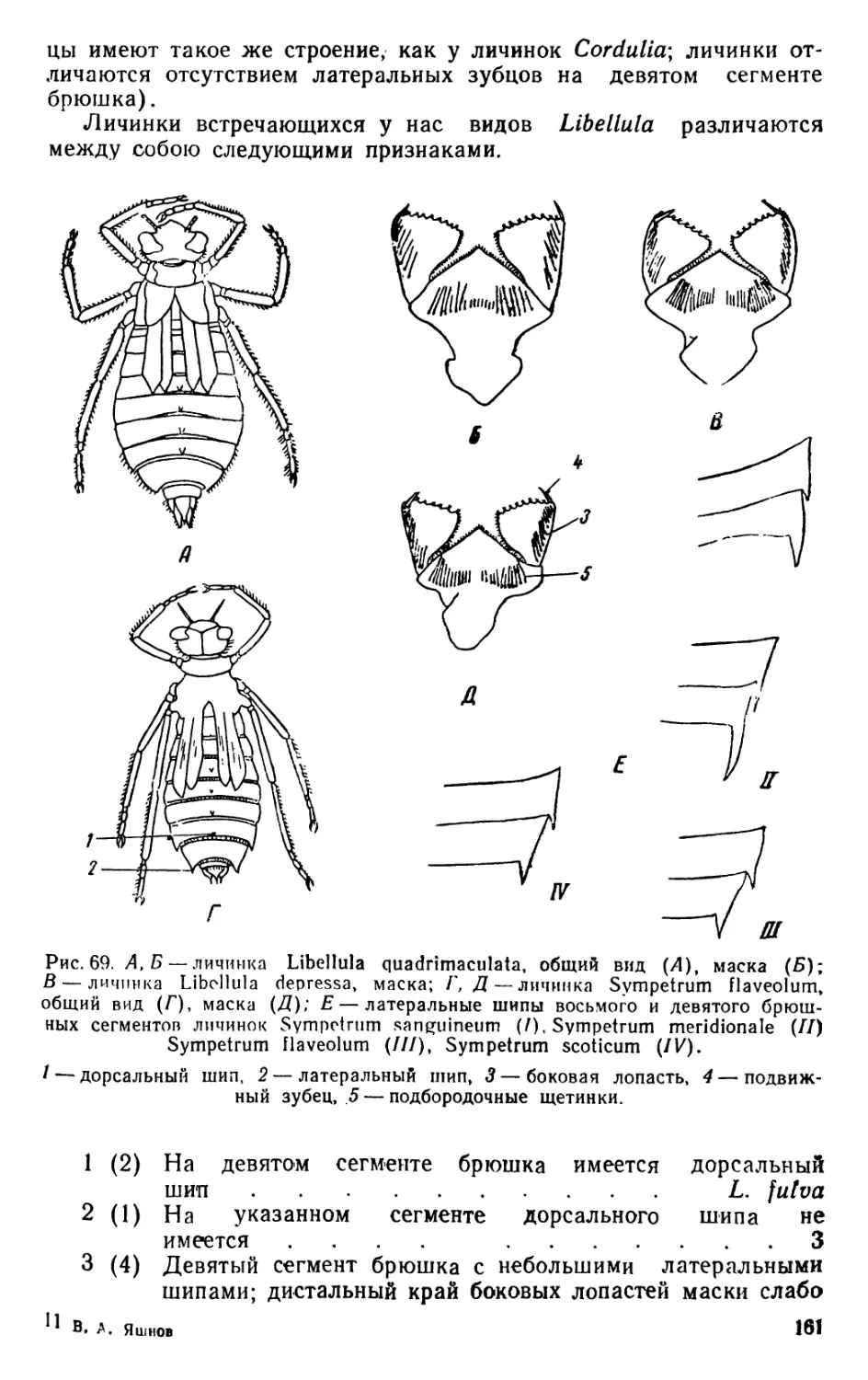
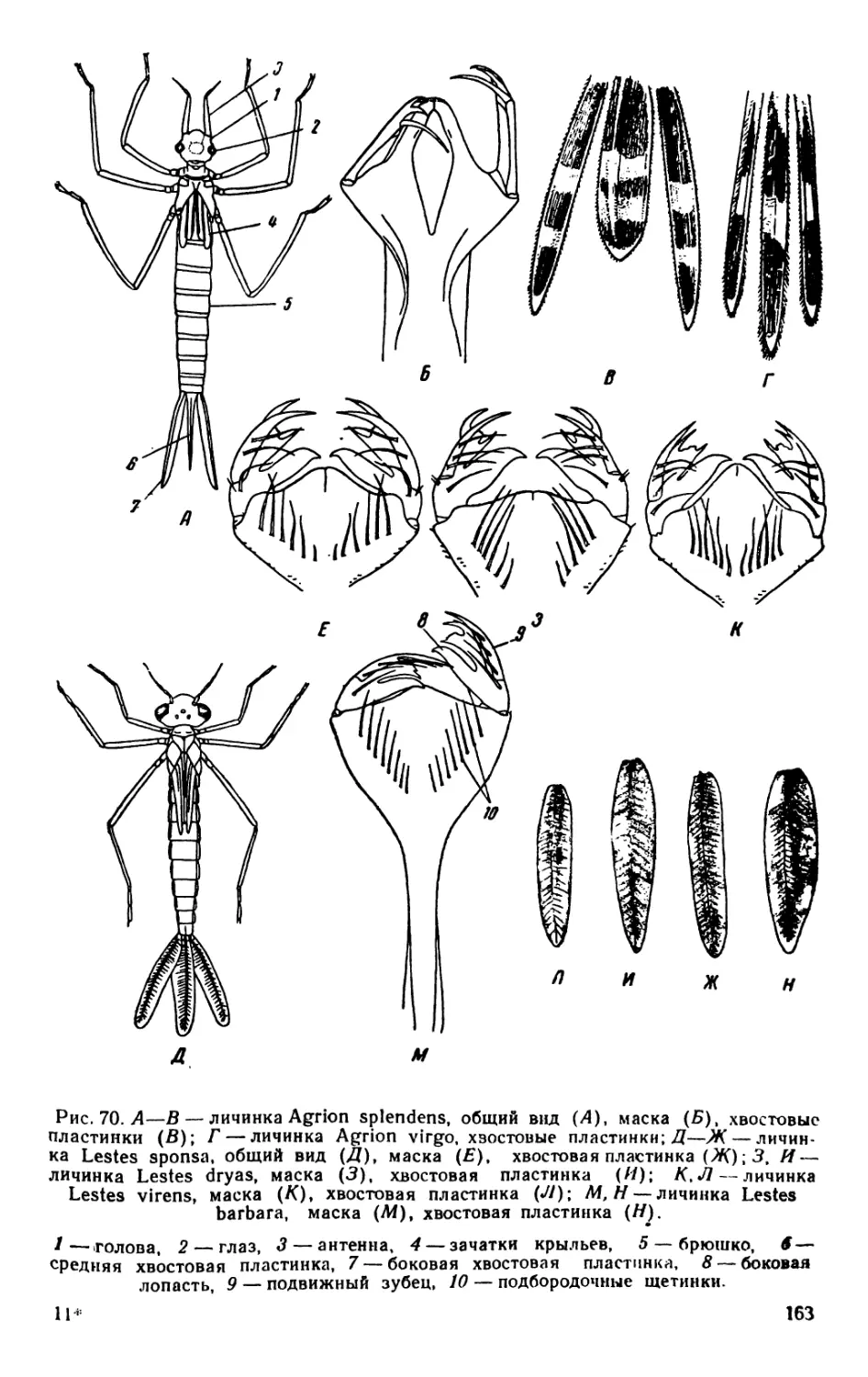
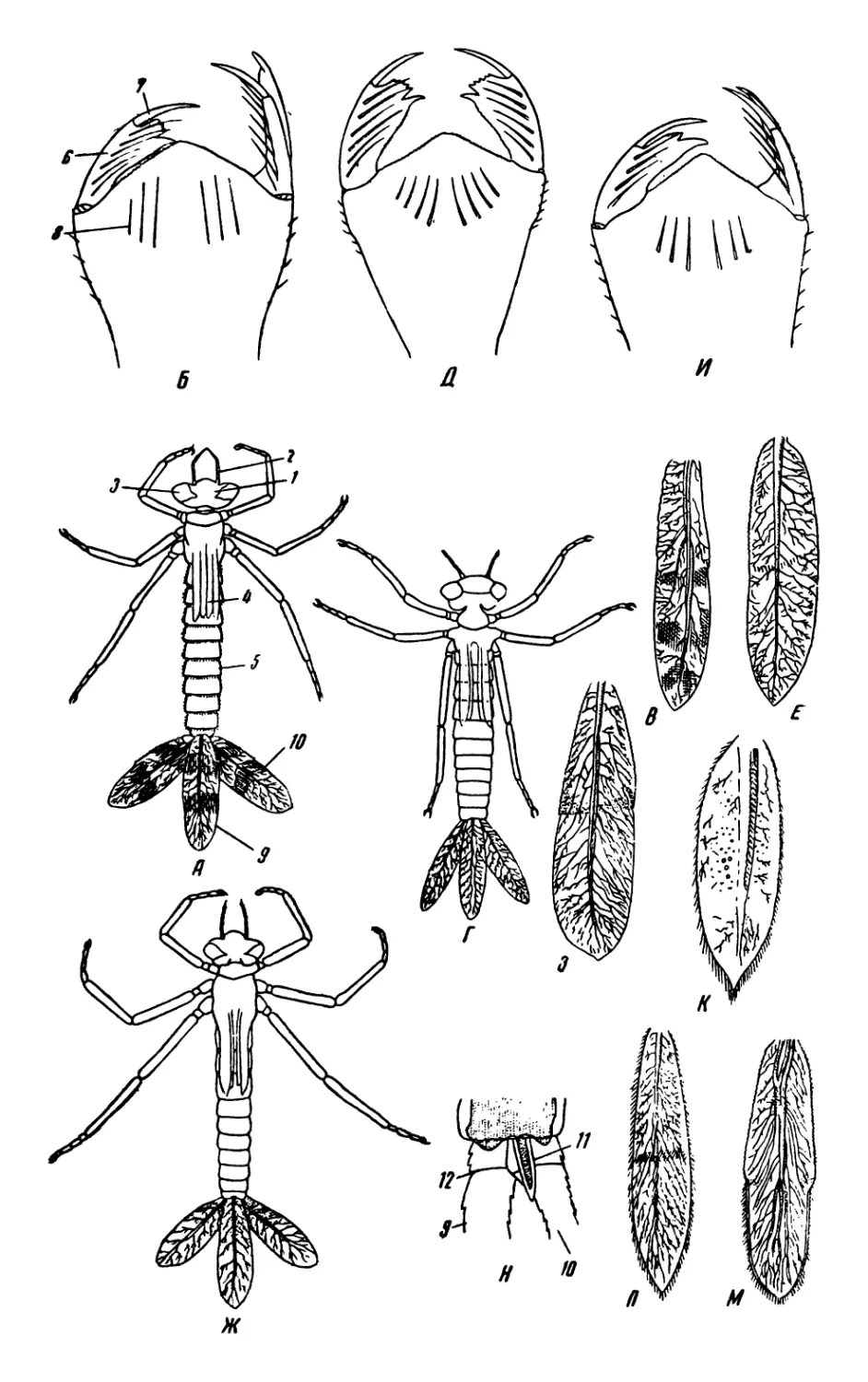
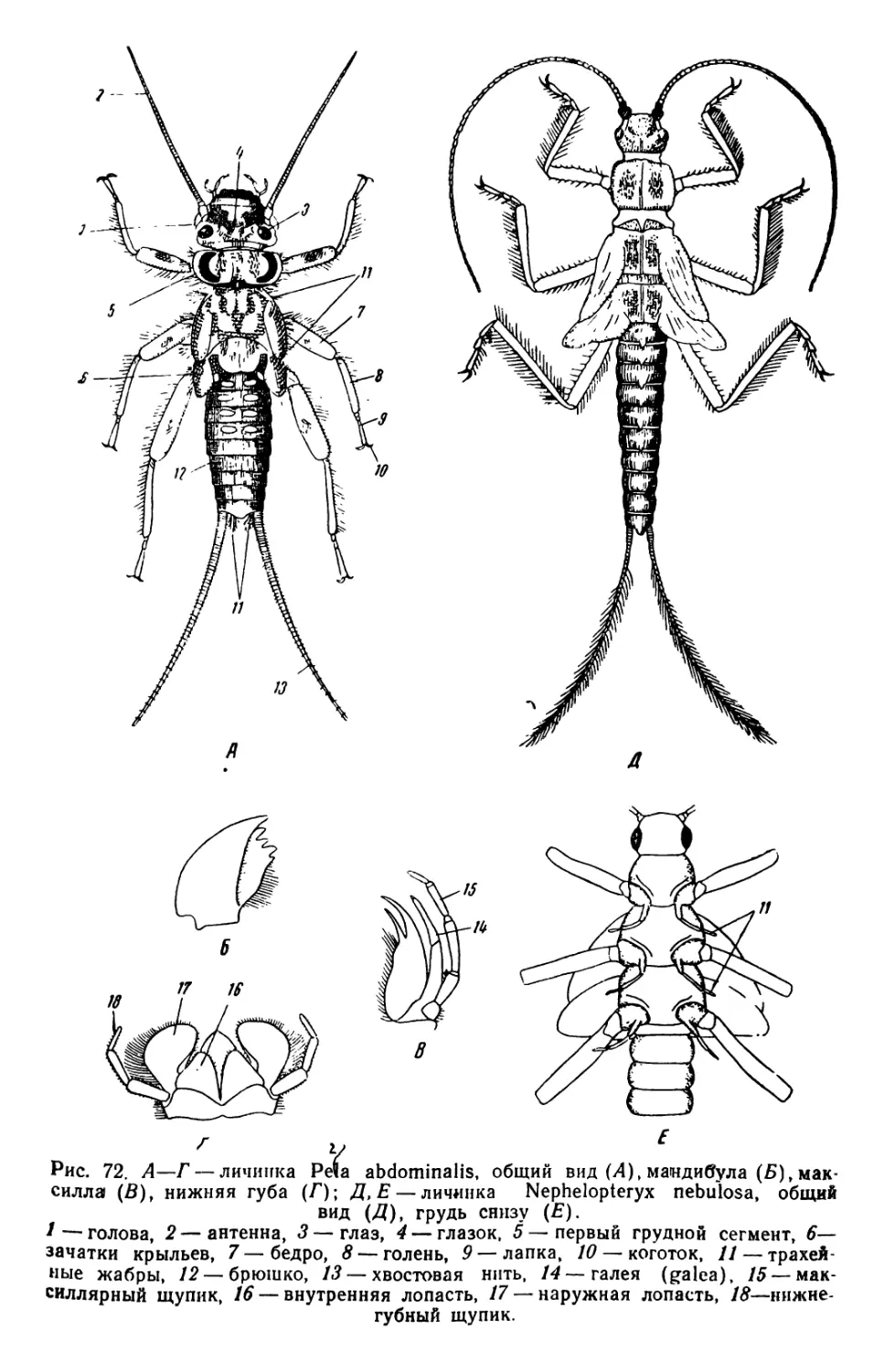
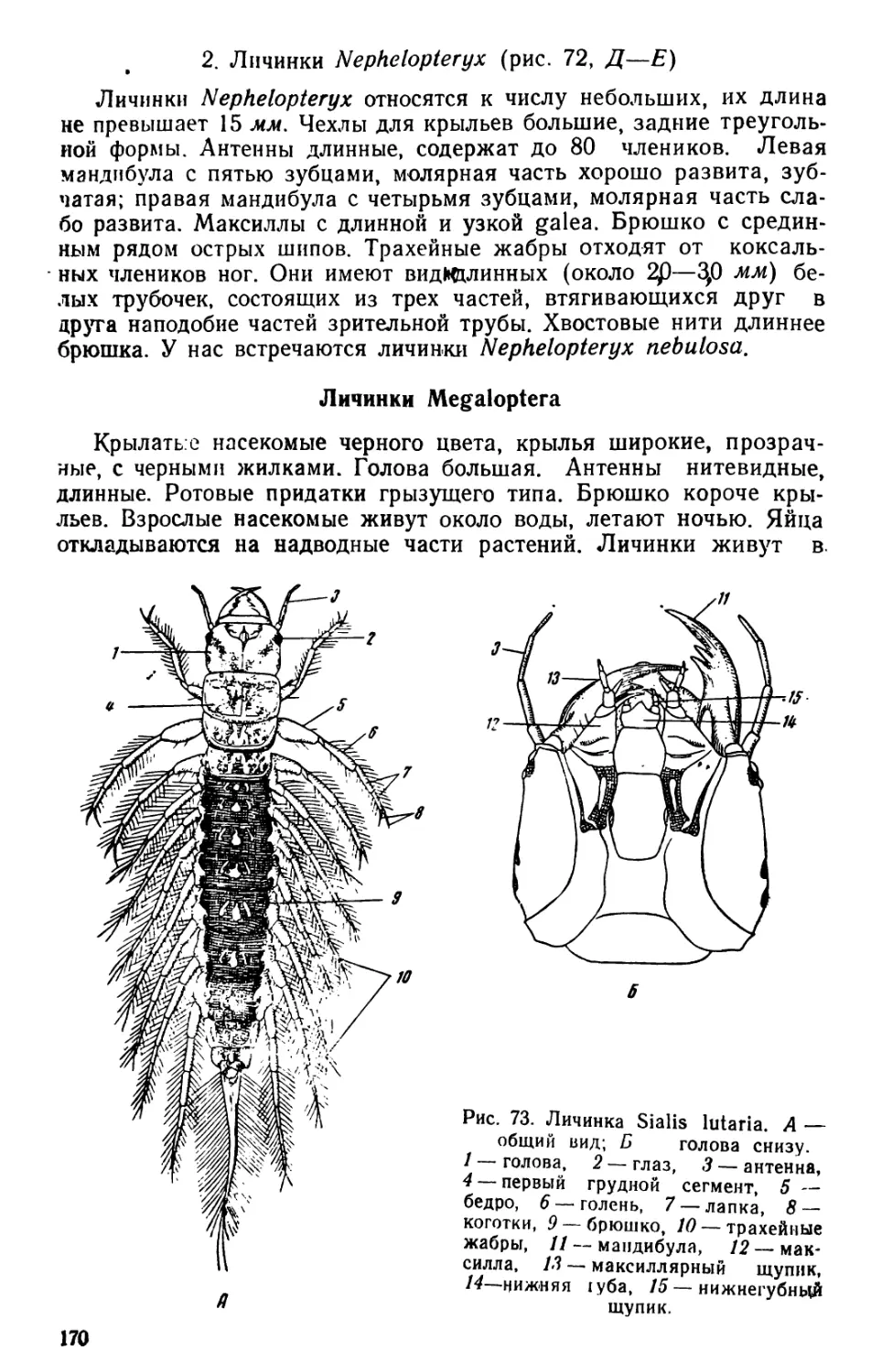
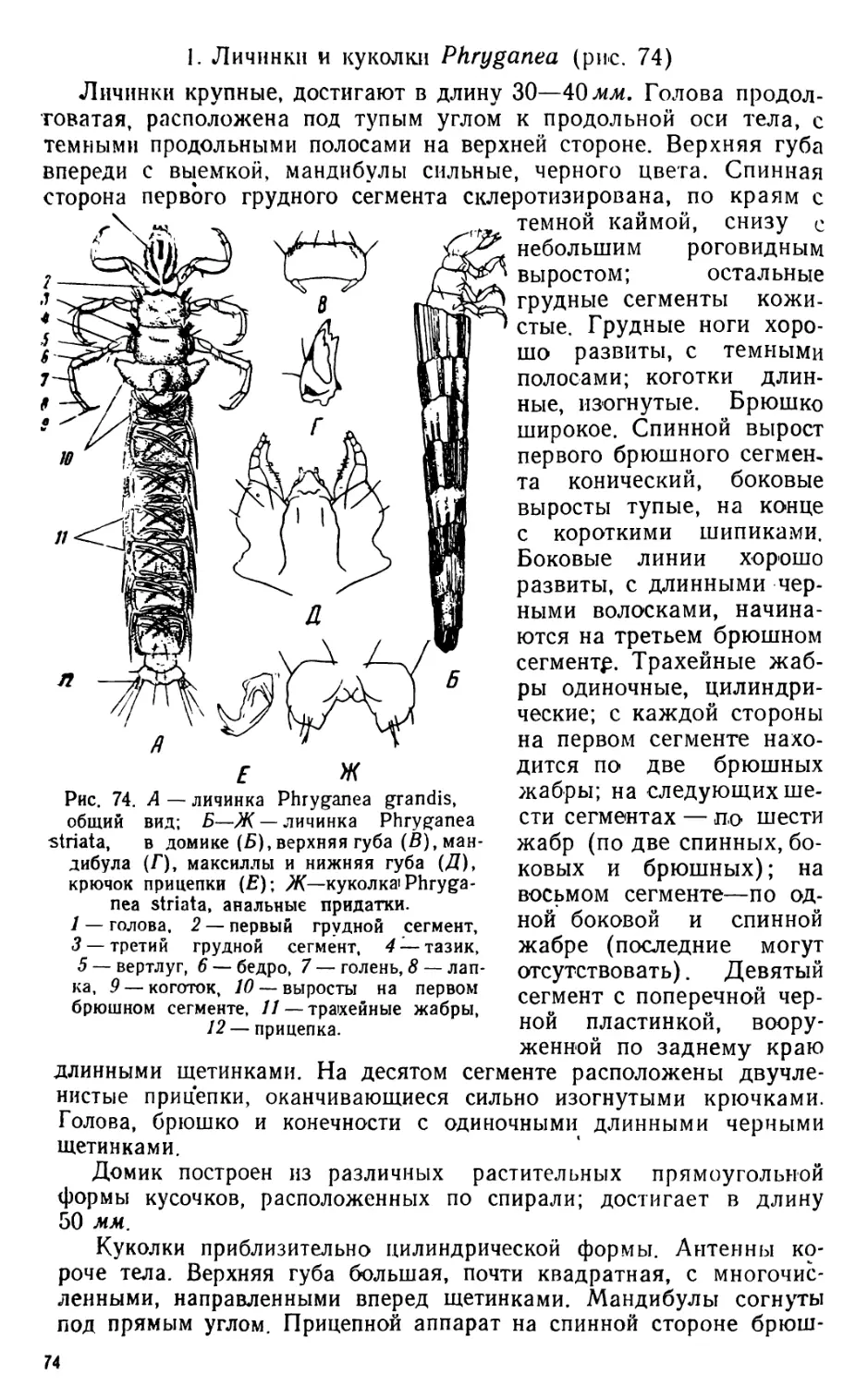
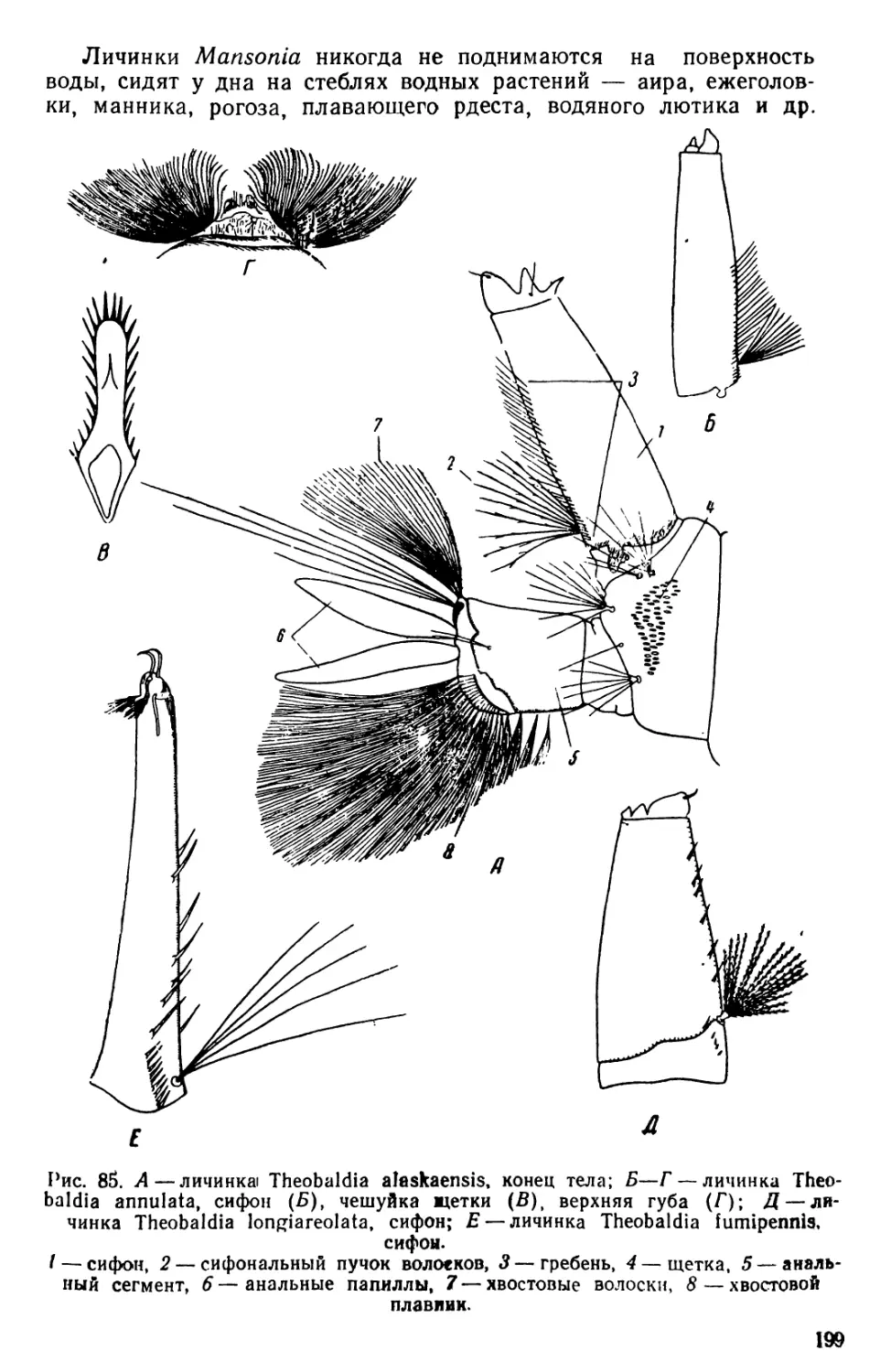
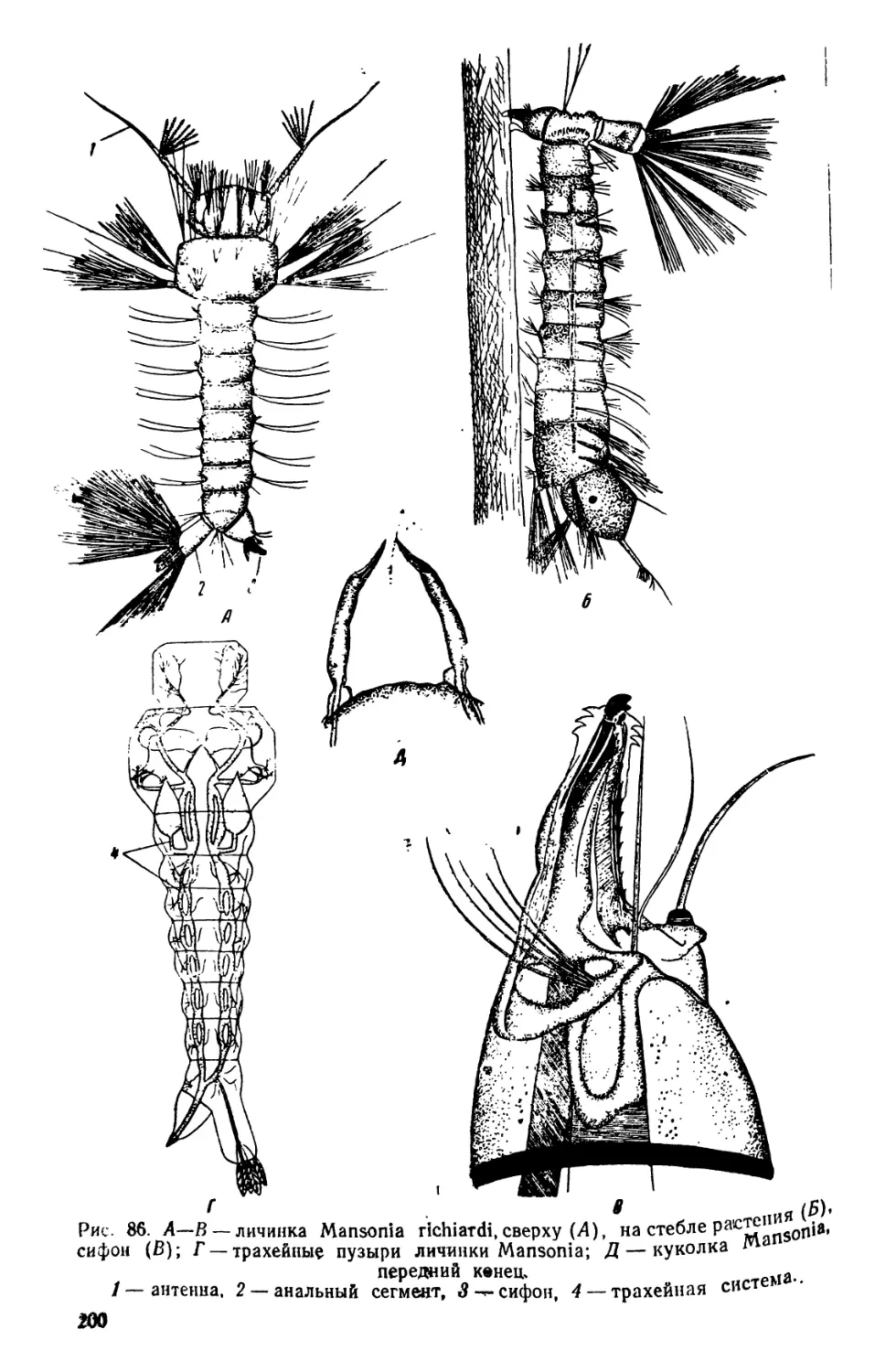
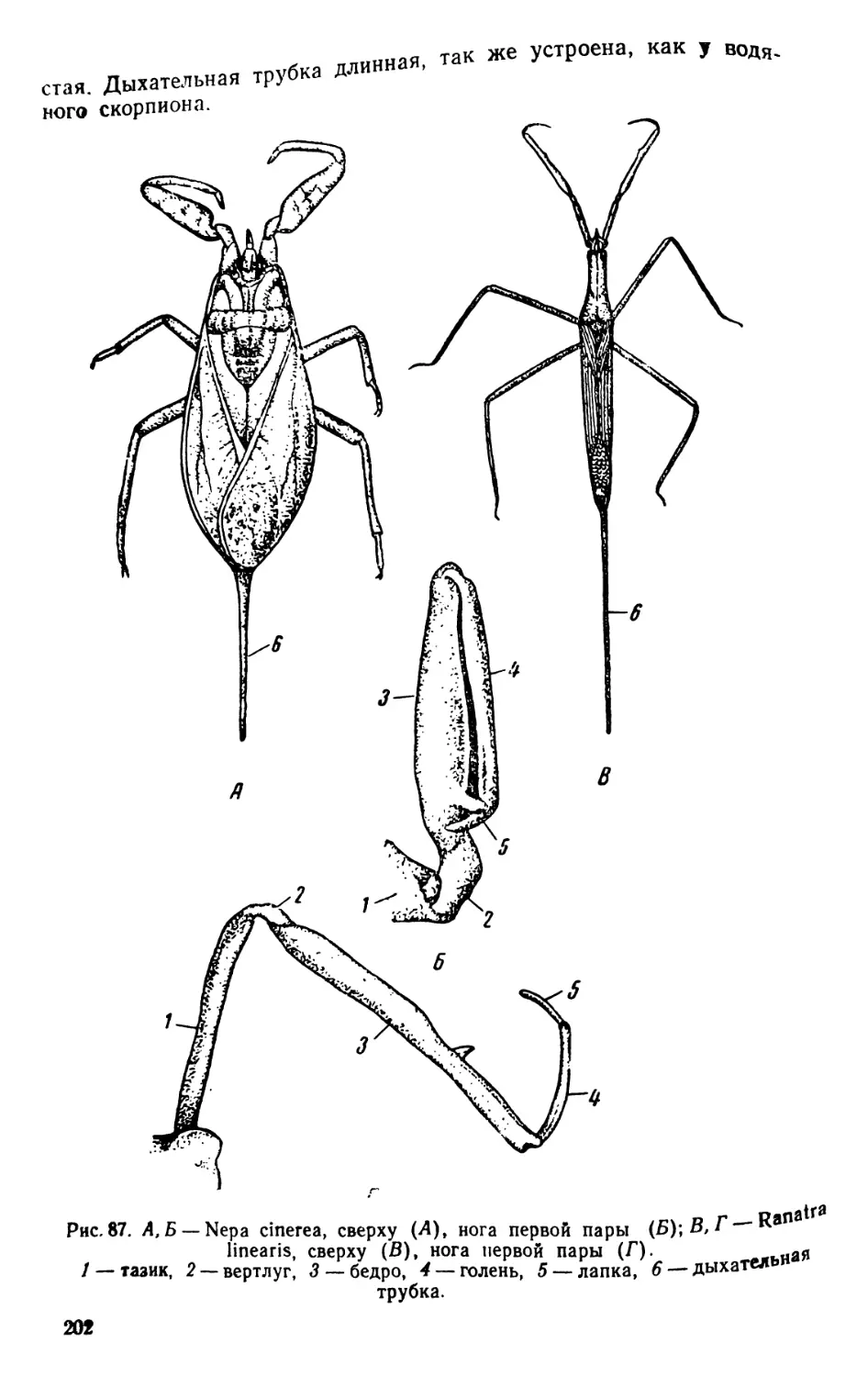
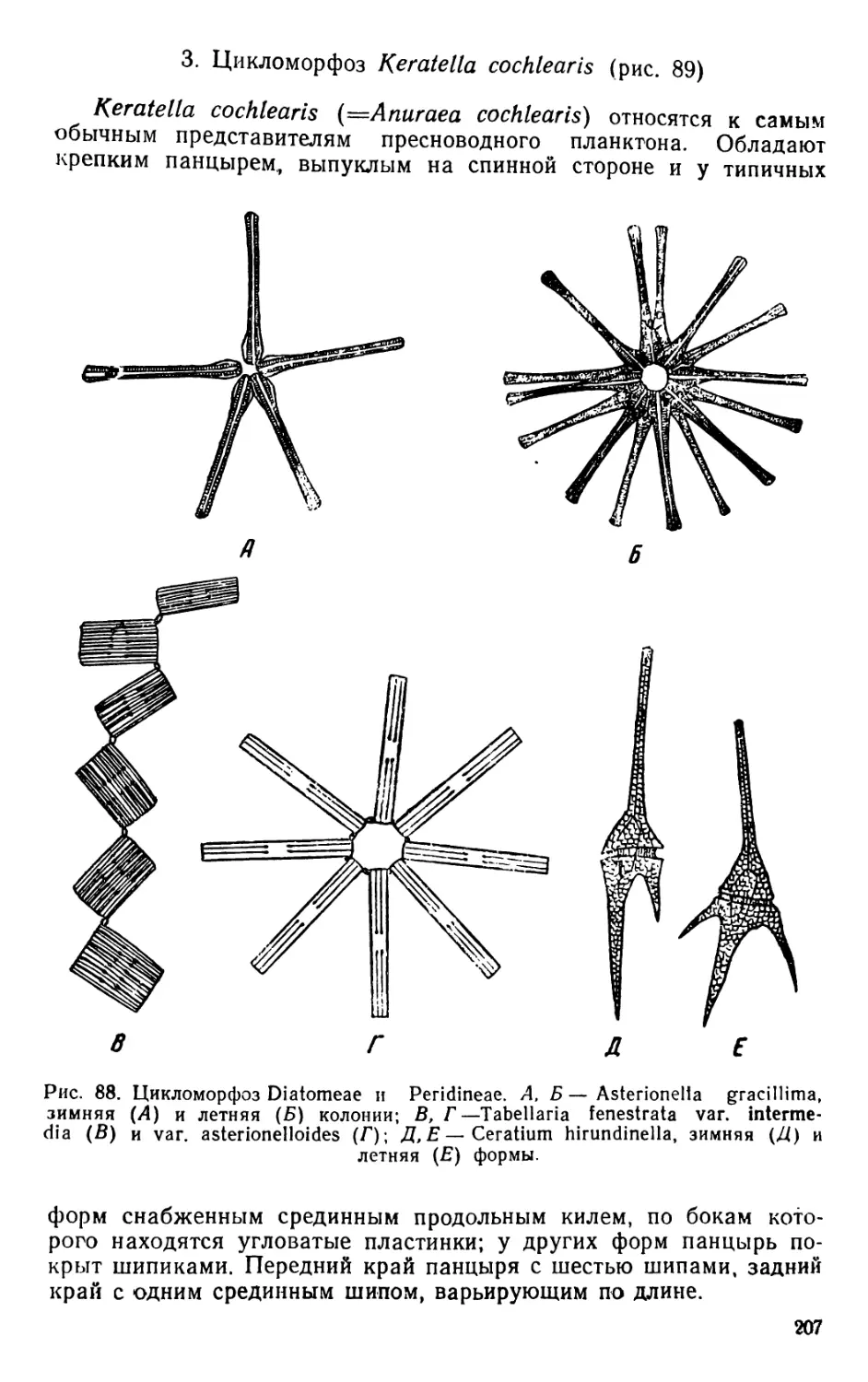
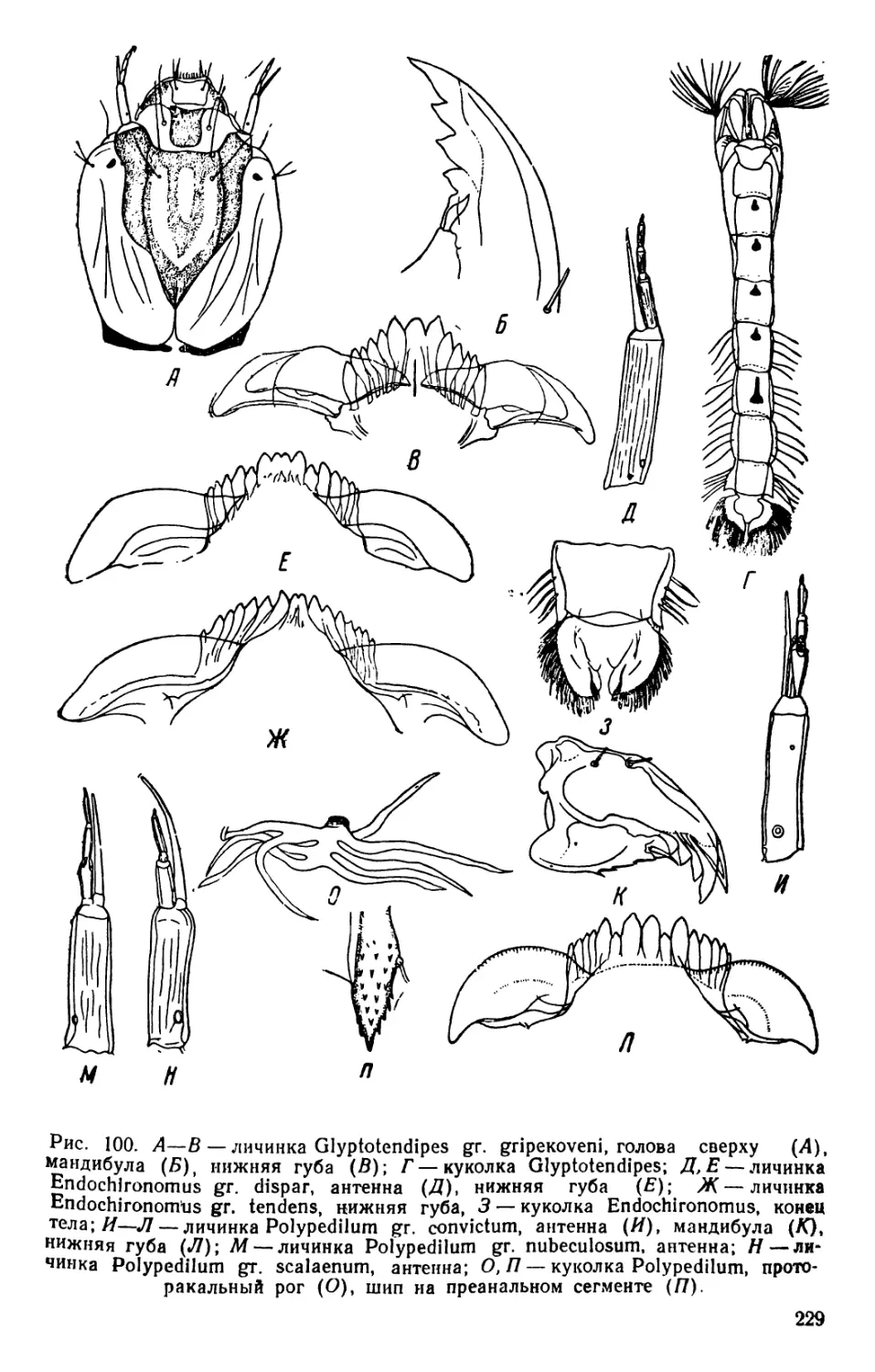
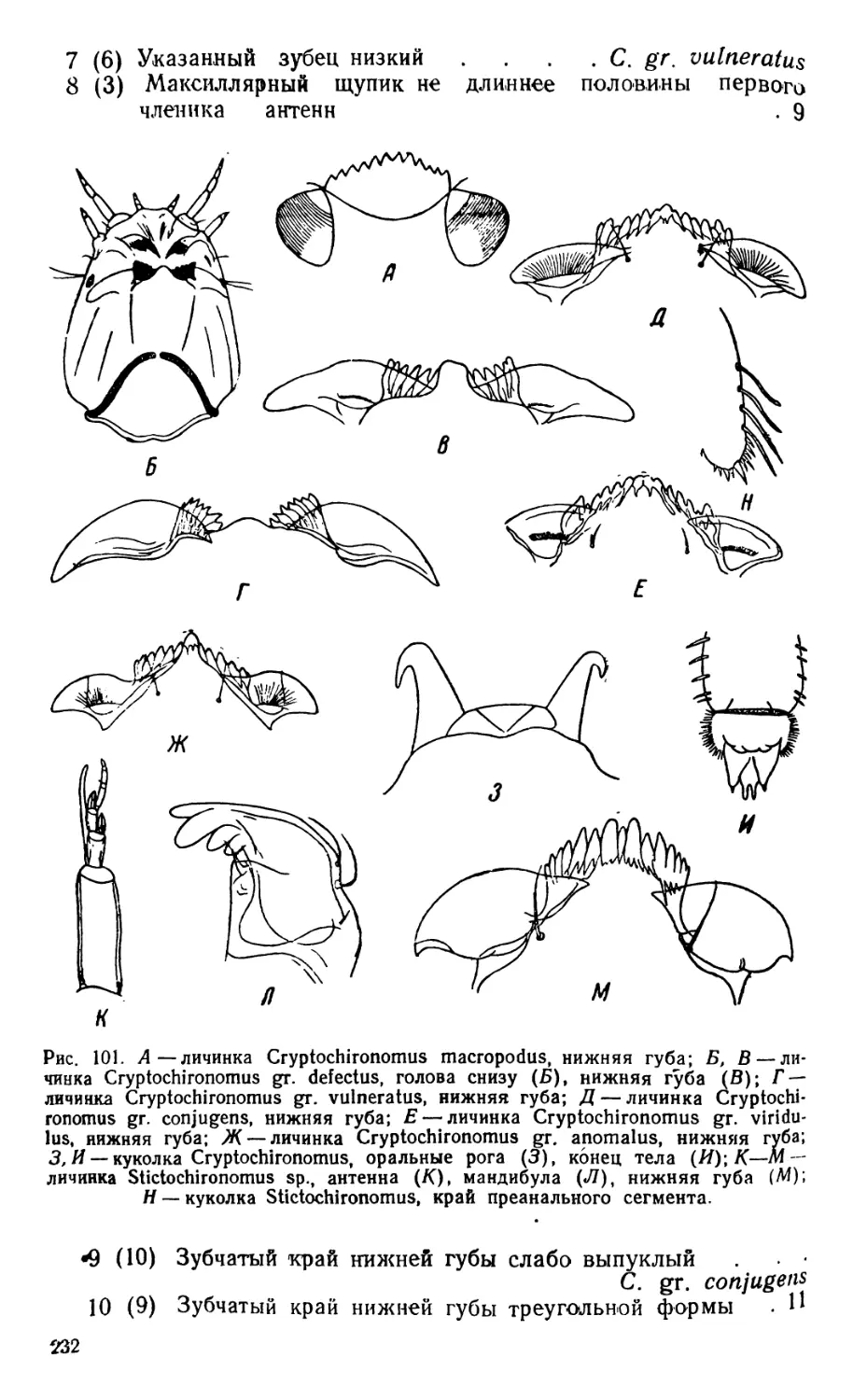
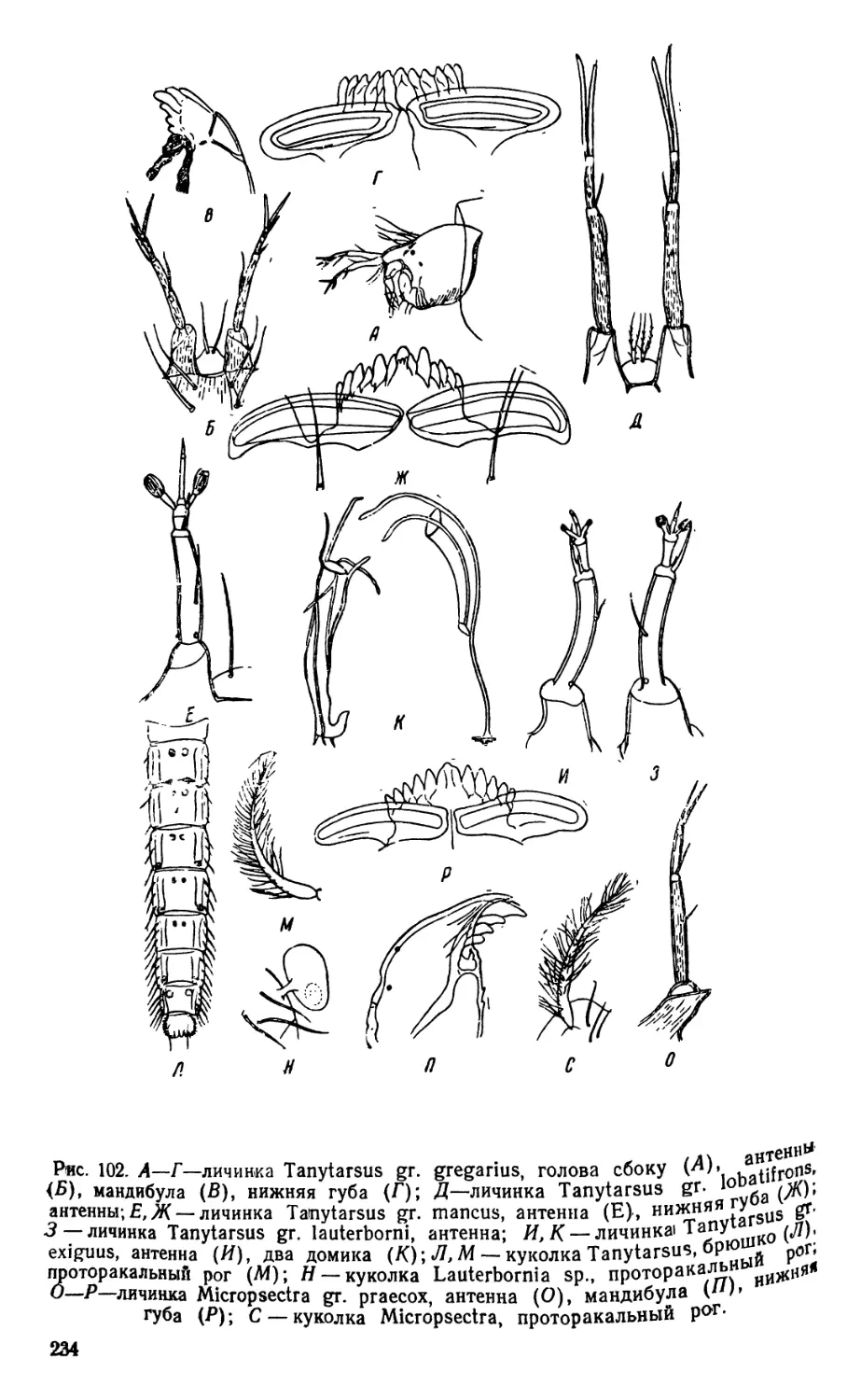
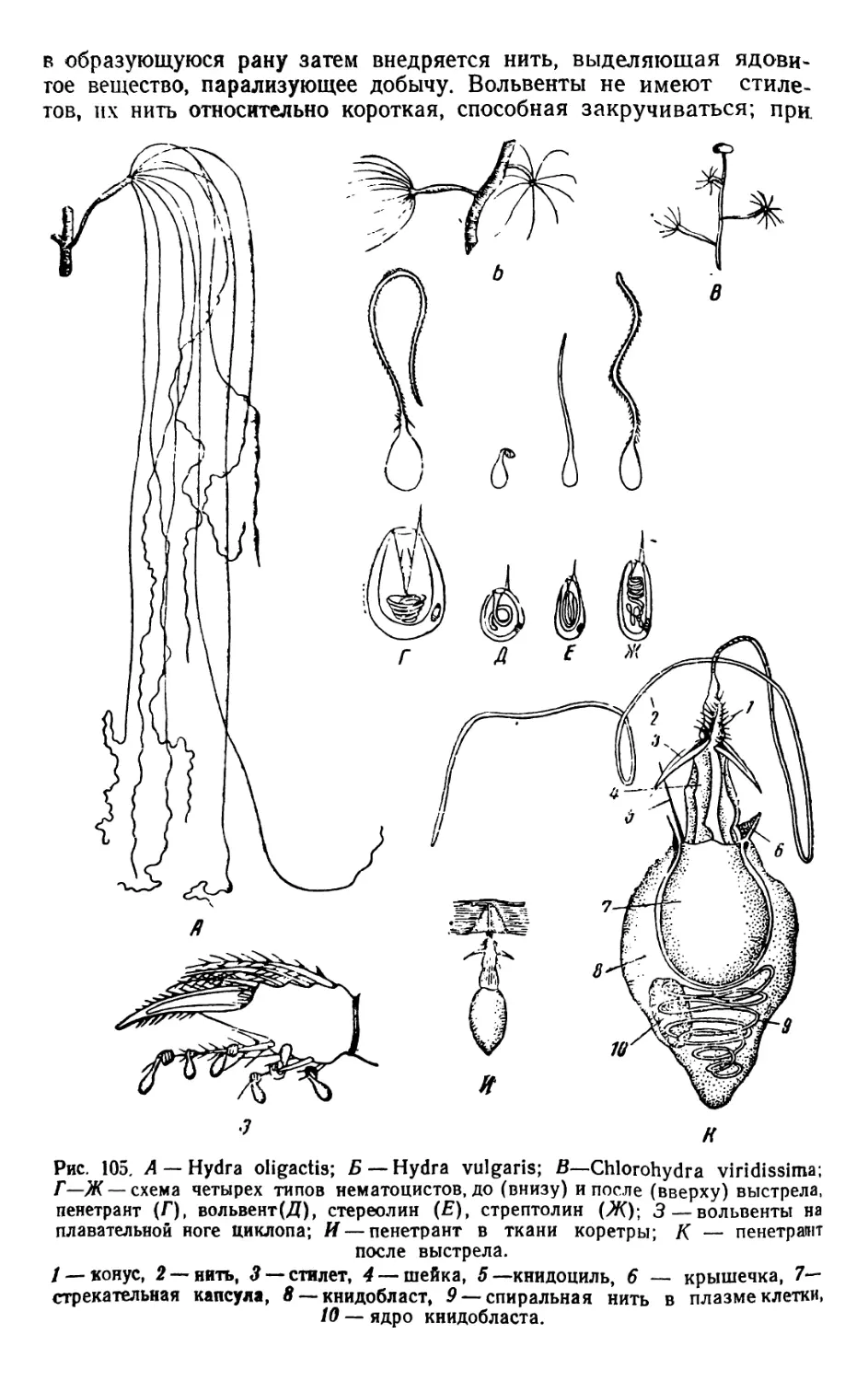
/
































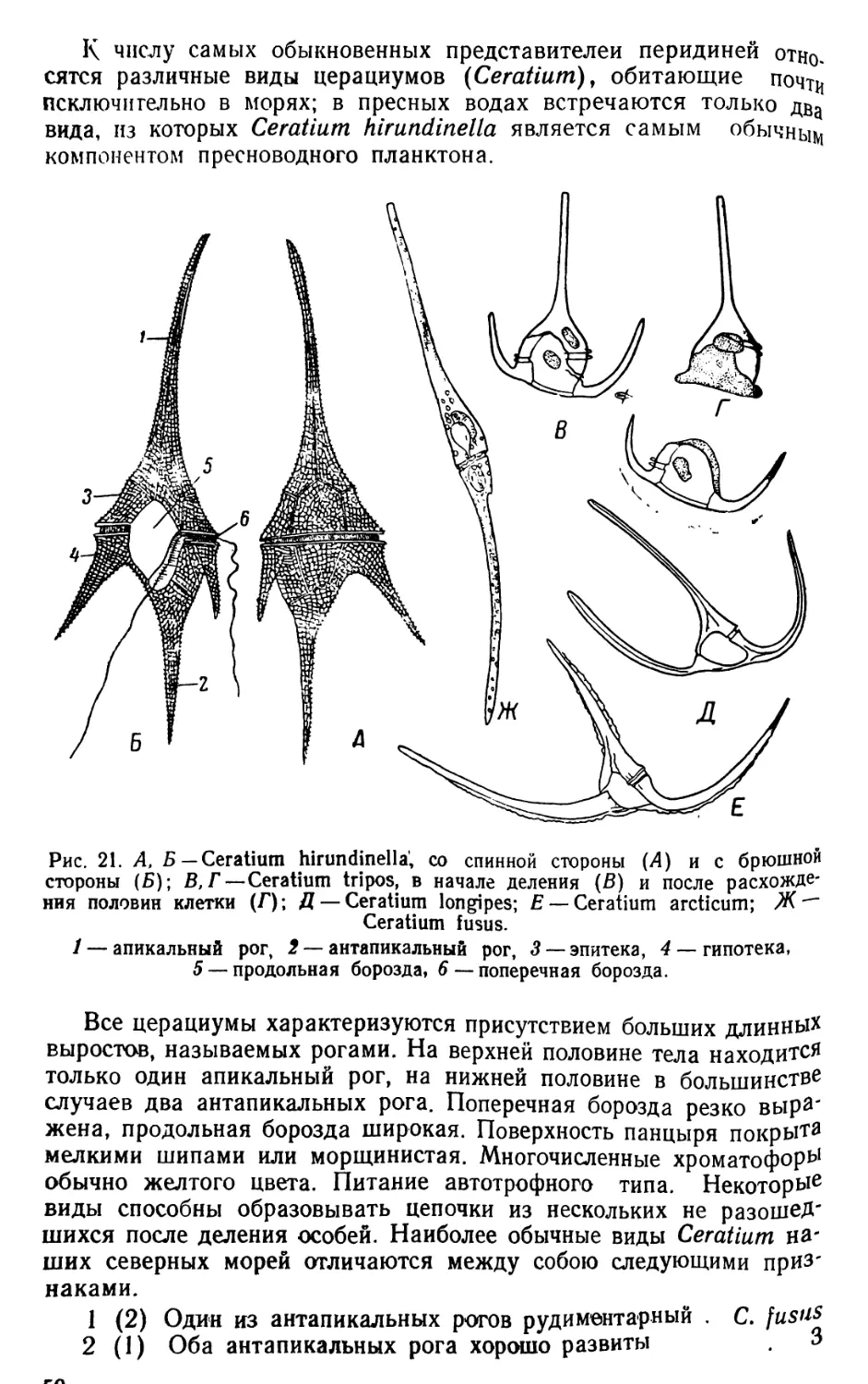

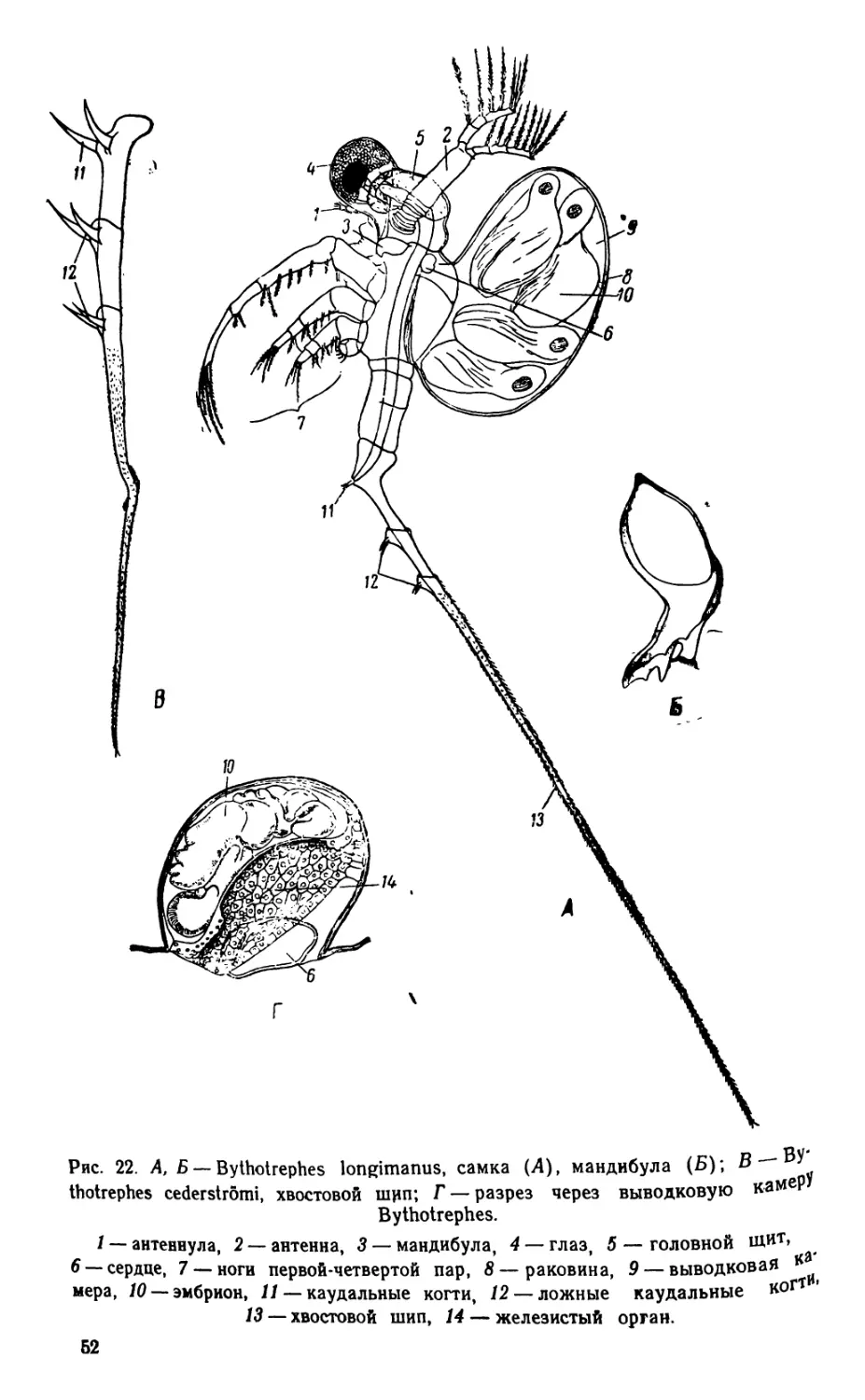














































































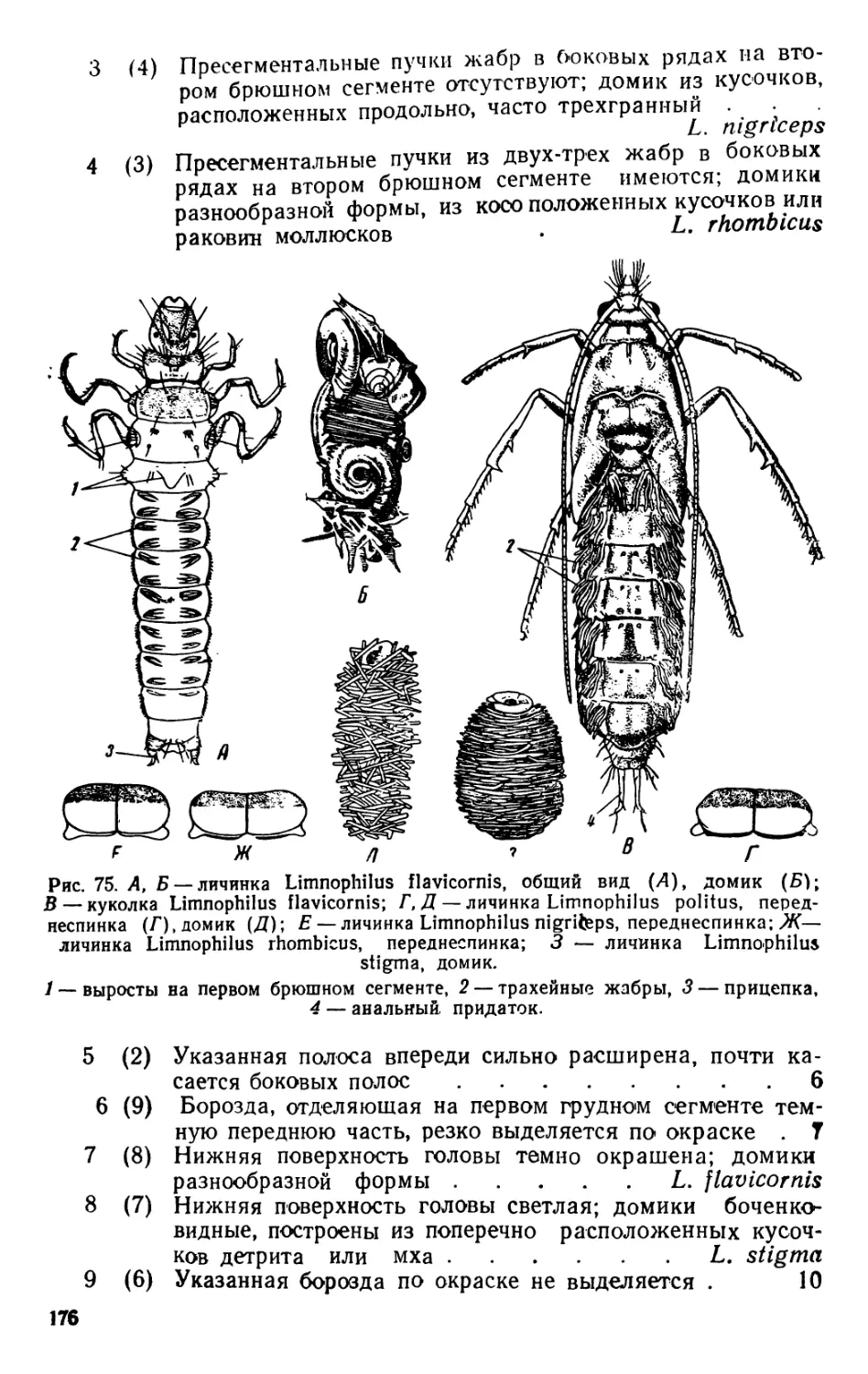
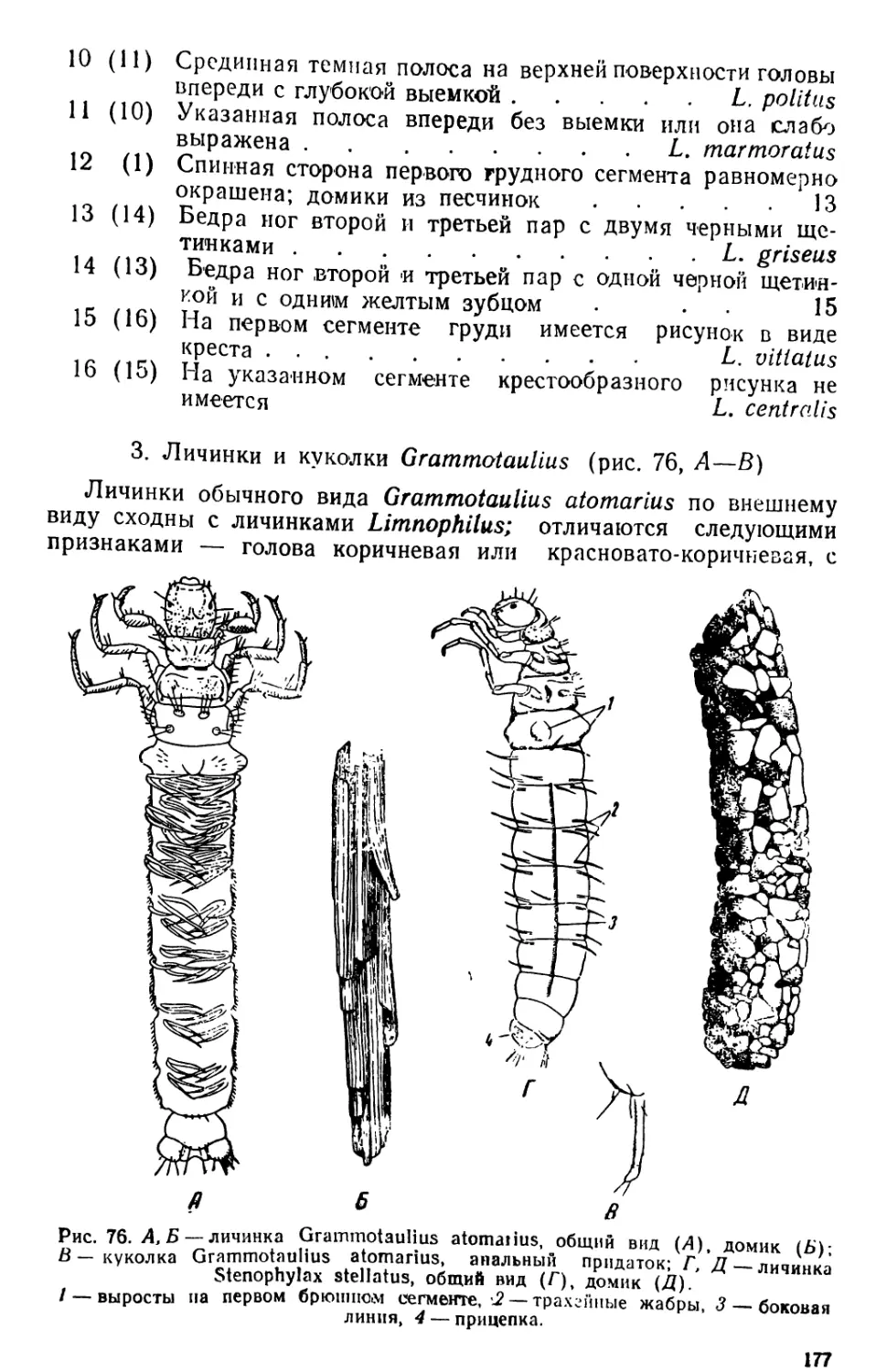
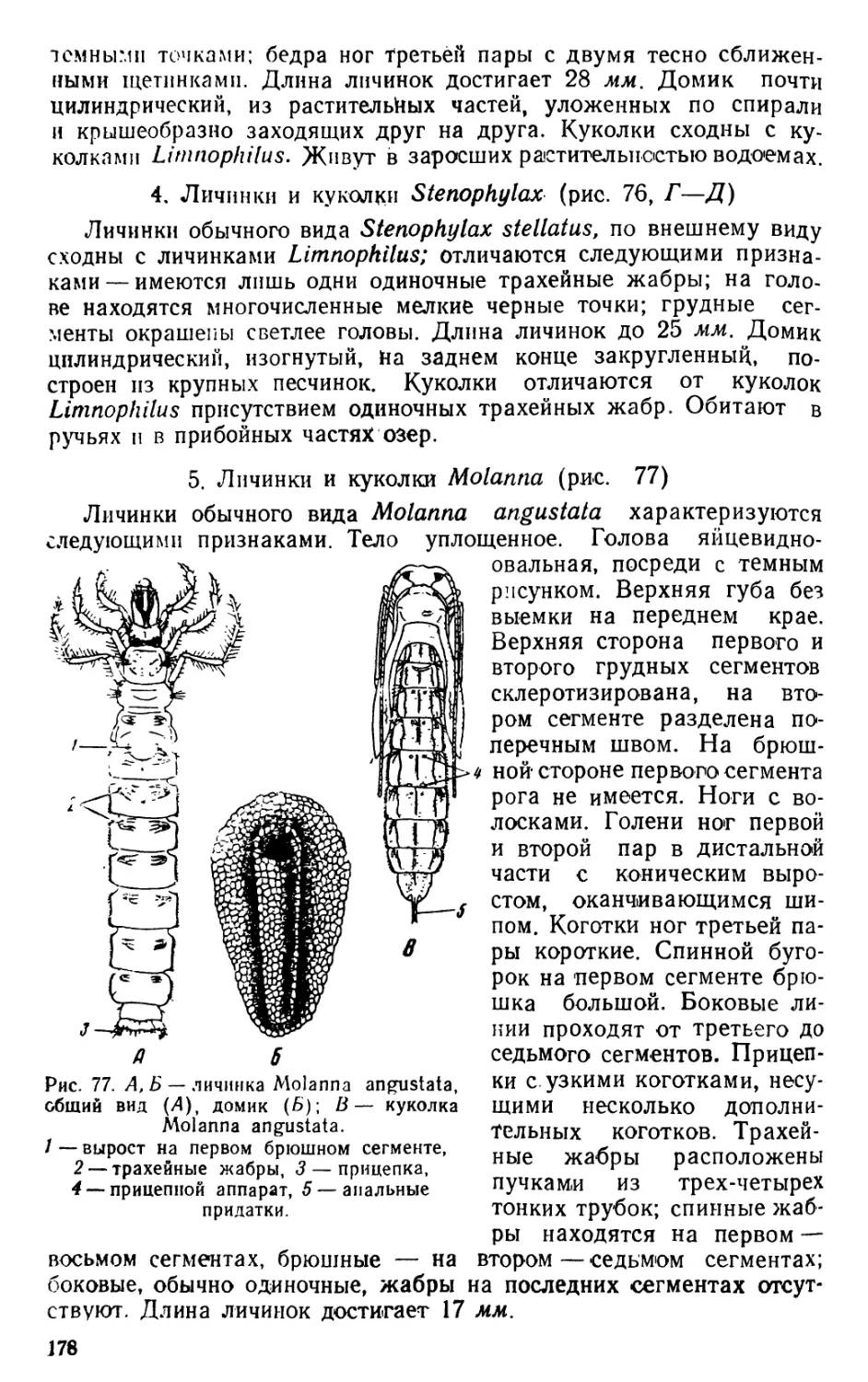
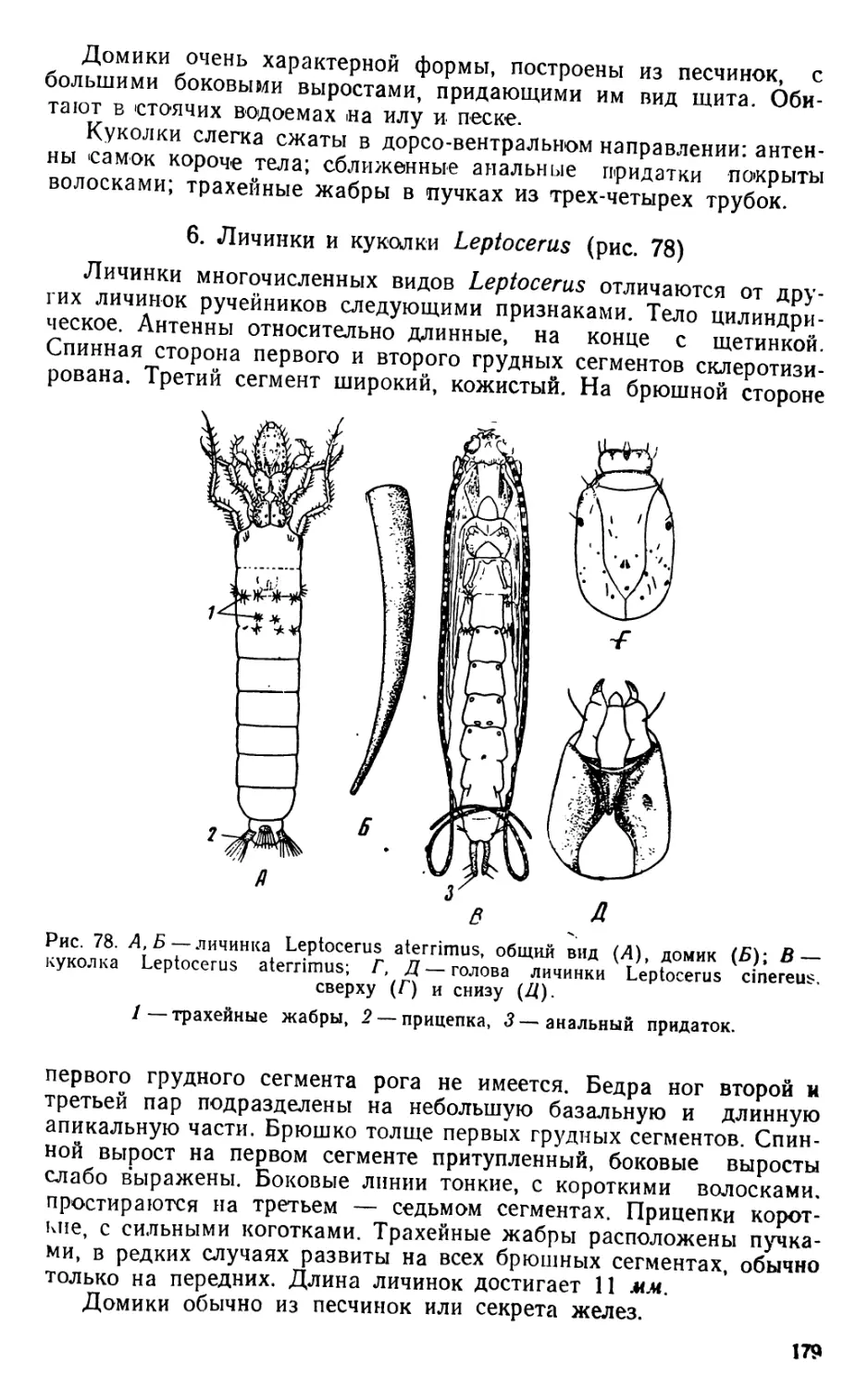






























































Автор: Яшнов В.А.
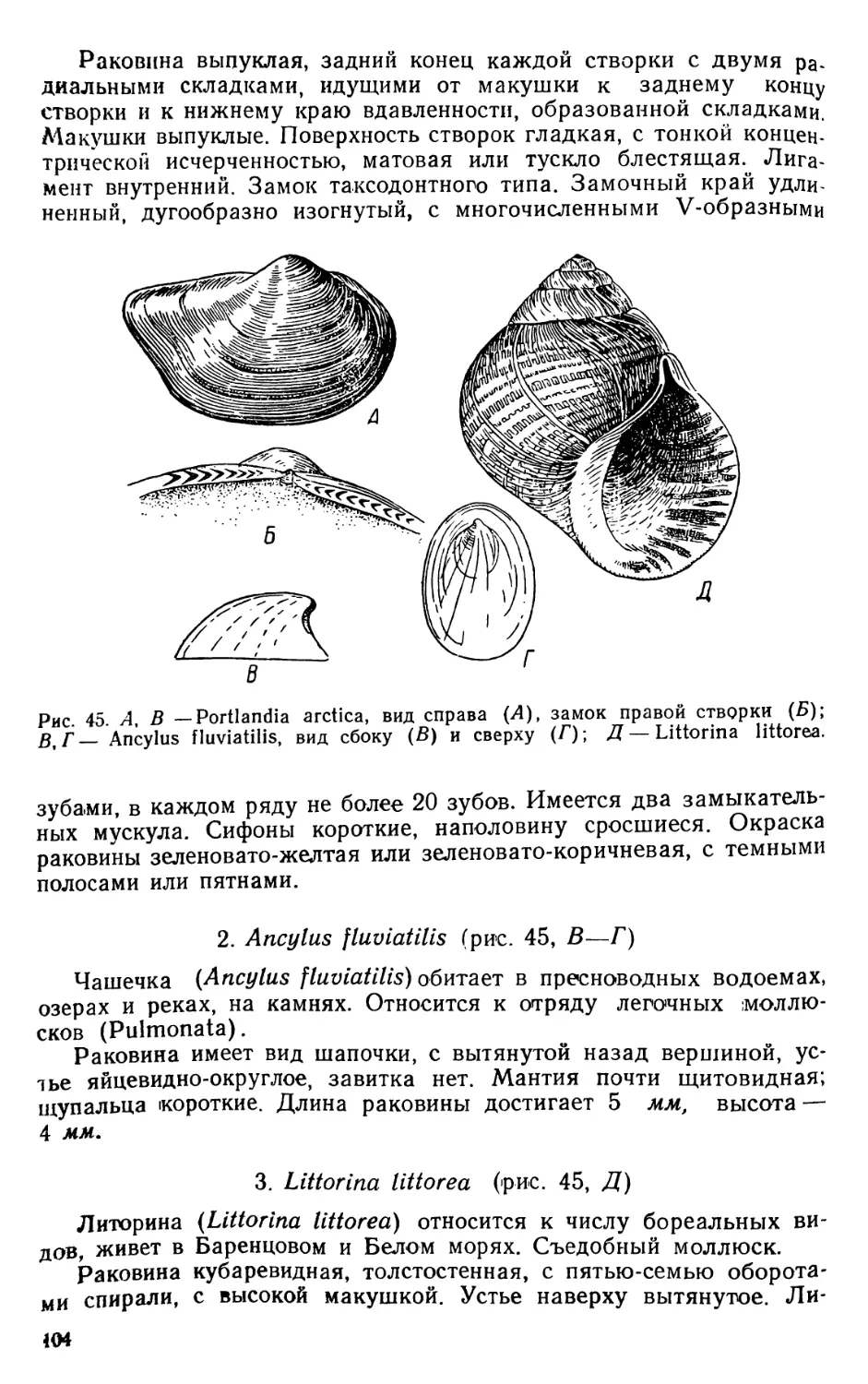
Теги: биология микробиология практикум гидробиология издательство советская наука
Год: 1952




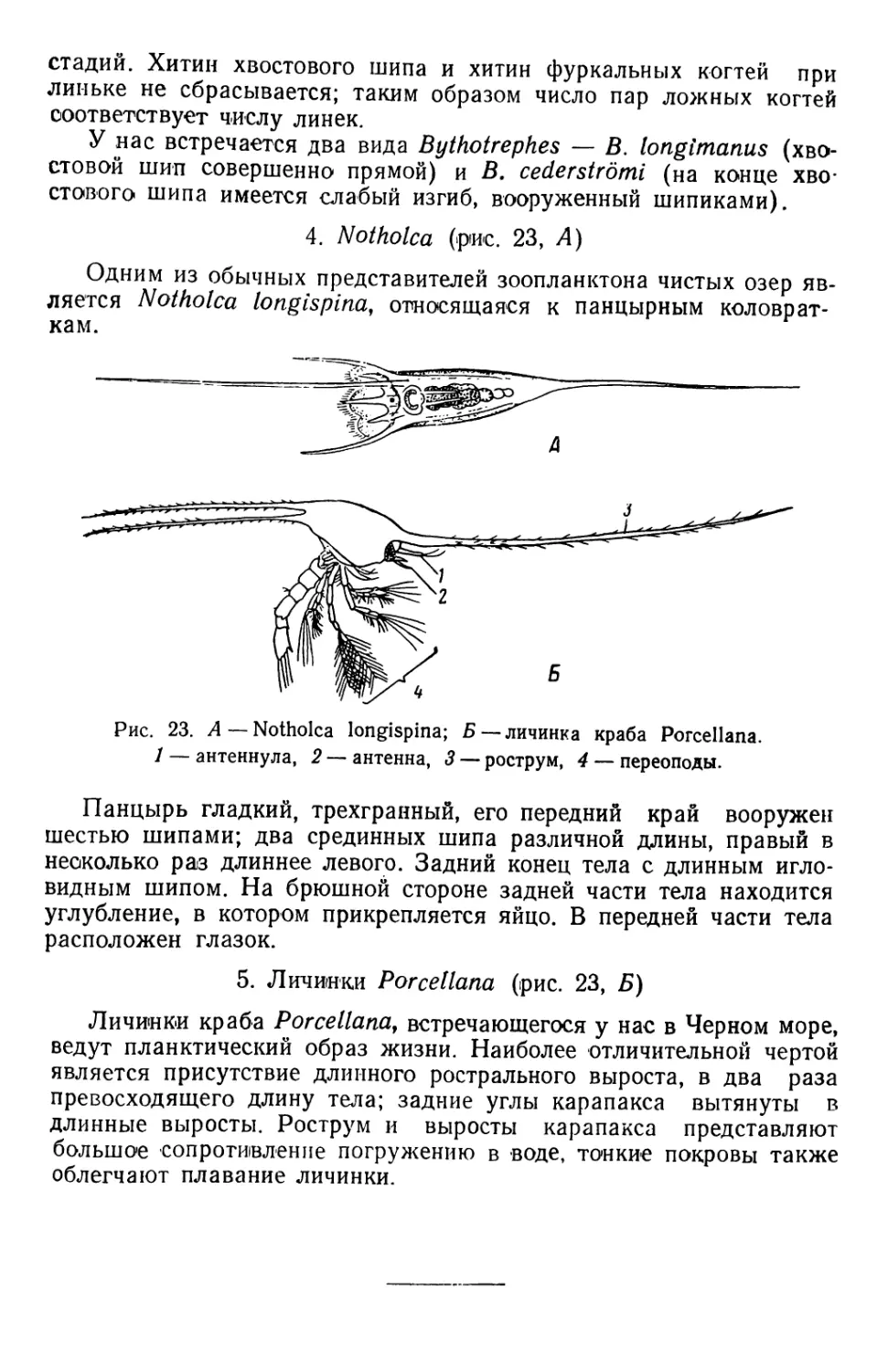
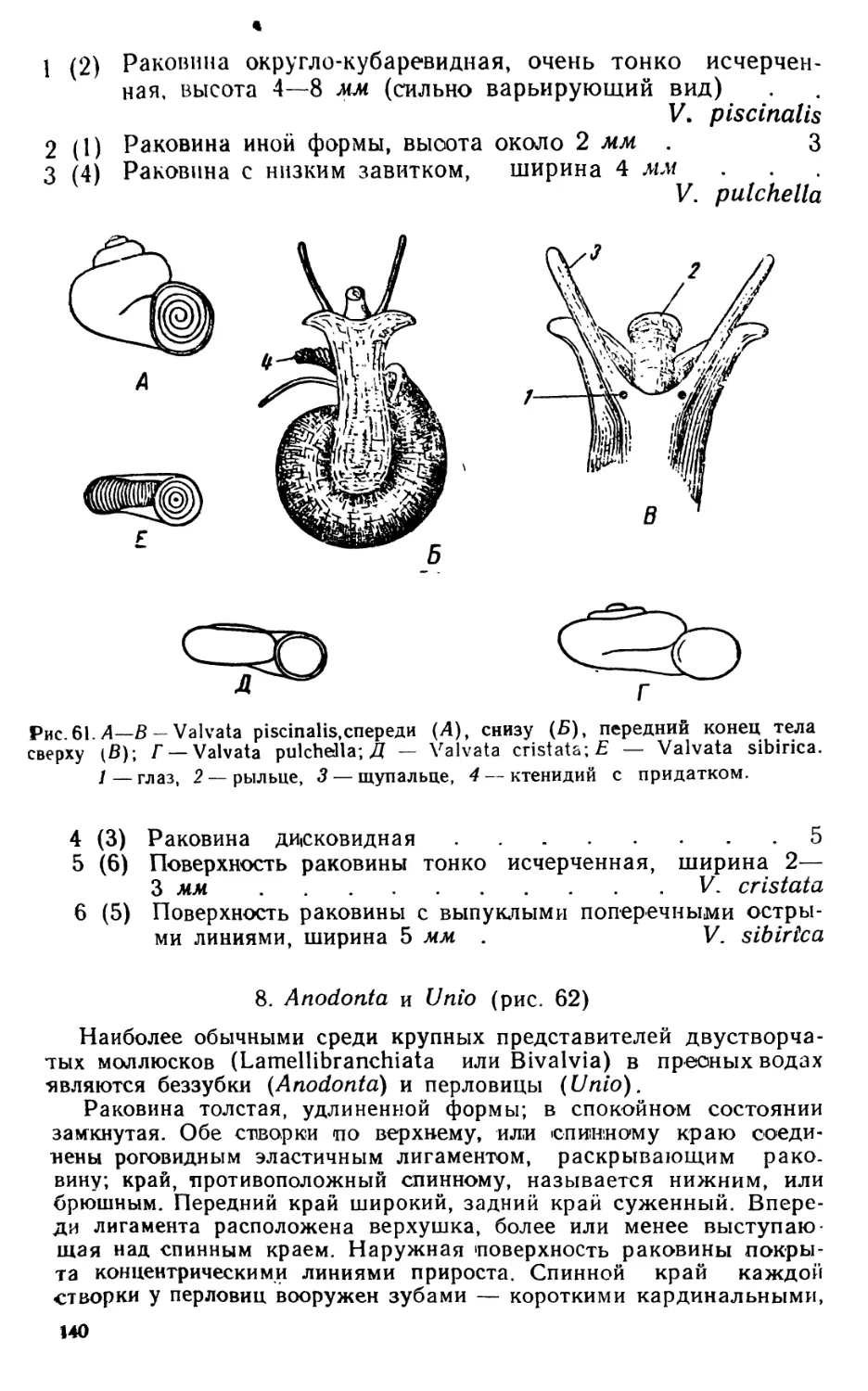
Текст
В. А. ЯШНОВ
МАЛЫЙ
ПРАКТИКУМ
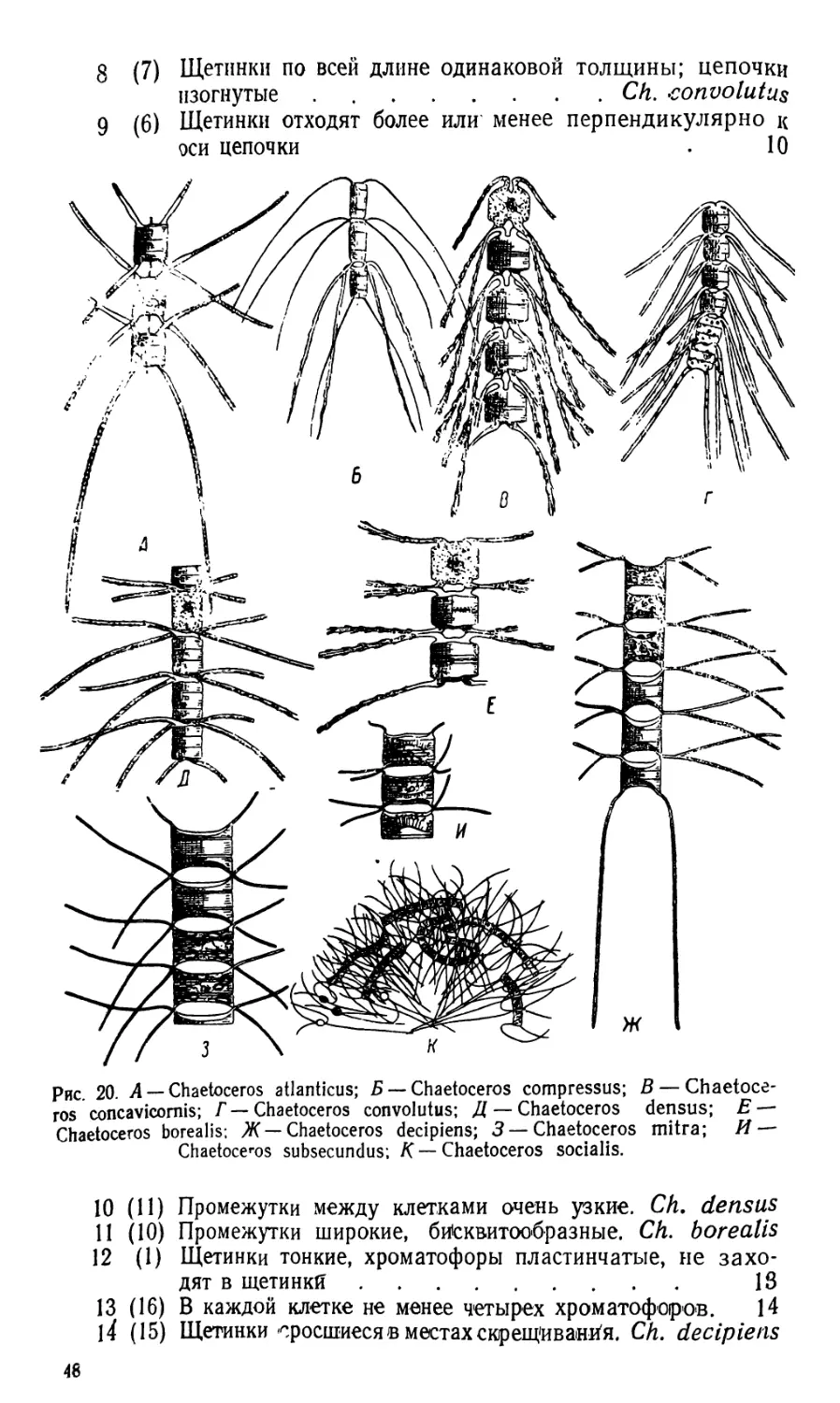
ПО
ГИДРОБИОЛОГИИ
в. А. ЯШ НОВ
МАЛЫЙ ПРАКТИКУМ
ПО ГИДРОБИОЛОГИИ
Допущено
Министерством высшего образования СССР
в качестве учебного пособия
для биолого-почвенных факультетов университетов
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО «СОВЕТСКАЯ НАУКА»
Москва — 1952
ПРЕДИСЛОВИЕ
Главная задача малого практикума по гидробиологии заклю-
чается в ознакомлении с различного рода приспособлениями, вы-
работавшимися у водных организмов под воздействием абиотиче-
ских и биотических условий среды. Практикум соответствует, та-
ким образом, той части курса общей гидробиологии, которая рас-
сматривает взаимоотношения между водными организмами и сре-
дой. В соответствии с этим материал практикума распределен на
следующие разделы: основные экологические группы водных орга-
низмов; приспособления организмов к солености воды, растворен-
ным в воде органическим веществам, активной реакции воды, ра-
створенному в воде кислороду, температурному режиму и свето-
вым условиям; пища и питание водных организмов.
Проведение практических занятий по изучению размножения
гидробионтов, а также по водным биоценозам, т. е. по тем разде-
лам, которые требуют для своей проработки большого количества
времени, целесообразнее включить в программу большого практи-
кума по гидробиологии.
Выбор материала для занятий весьма обширен, в любом во-
доеме можно найти большое число организмов, пригодных для
этой цели. В качестве примеров рассматриваются преимуществен-
но те формы, которые широко распространены и могут быть лег-
ко собраны, а также организмы, имеющие большое практическое
значение. Однако в некоторых случаях приходится обращаться к
сравнительно трудно доступному материалу; пути к его получению
будут указаны ниже.
Материал по отдельным занятиям в большинстве случаев пре-
вышает тот, который может быть рассмотрен в течение сравни-
тельно небольшого времени, выделяемого на практические заня-
тия по общему курсу гидробиологии. Это сделано, с одной сторо-
ны, для того, чтобы в случае необходимости можно было заменить
одни организмы другими, а с другой стороны, чтобы предоставить
дополнительный материал для самостоятельной работы, в особен-
ности по тем группам, которые важны в практическом отношении.
Этим объясняется, в частности, и включение в практикум сравни-
тельно большого 'материала по водным личинкам насекомых. В
общем курсе зоологии эти личинки обычно подробно не рассмат-
риваются, значение же их в жизни пресноводных водоемов весьма
велико, к тому же многие личинки насекомых важны и в практи-
3
четком отношении. Представители нектона в практикум ие вклю-
чены, так как они на занятиях по ихтиологии рассматриваются
значительно полнее, чем это можно было бы сделать в практикуме
по гидробиологии.
Морские организмы, а из пресноводных — редко встречающие-
ся, рассматриваются, естественно, в фиксированном виде; пресно-
водные организмы, в особенности те, которые легко содержатся в
аквариумах, должны изучаться в живом состоянии. В тех случаях,
когда имеется возможность делать выбор между пресноводными
и морскими формами, предпочтение отдается первым.
Само собой разумеется, что при проведении занятий нельзя ог-
раничиться только демонстрированием приспособлений организмов
к определенным условиям среды, необходимо также возможно
более полное ознакомление с их строением, без чего трудно по-
нять значение многих приспособлений. Поэтому большое внимание
уделяется изучению морфологии животных и растений, являющих-
ся объектами исследования.
Не менее существенно определение систематического положения
организмов. Совершенно не безразлично, будут ли рассматривать-
ся представители определенных видов или же родов и других выс-
ших таксономических единиц. Необходимо знать признаки, харак-
теризующие самых обычных представителей водных организмов.
Кроме того, при определении хорошо уясняются особенности раз-
личных видов. Существующие определители не всегда могут быть
использованы во время занятий, кроме того для ряда групп до
сего времени у нас не имеется доступных определителей, позво-
ляющих вести определение до вида..Поэтому в тех случаях, ког-
да это возможно, приводятся небольшие таблицы для определе-
ния самых обычных видов, составленные по наиболее бросающим-
ся в глаза признакам. Одновременно даются указания по технике
определения.
В практикум введены также некоторые работы эксперименталь-
ного характера, требующие сравнительно небольшого времени
для своего осуществления.
В общем практикум должен дать начальные сведения по строе-
нию и систематике водных организмов, приспособлению их к раз-
личным условиям среды и элементарным методам исследования.
Практикум по гидробиологии составляется впервые, на основе
только личного опыта, поэтому его следует рассматривать как
первую попытку по составлению руководства для практических
занятий по общему курсу гидробиологии. Все замечания и поже-
лания, которые будут приняты с признательностью, следует на-
правлять по адресу — Москва. Московский государственный уни-
верситет. Кафедра гидробиологии биолого-почвенного факультета.
За просмотр рукописи и ряд ценных замечаний приношу ис-
креннюю благодарность доценту Московского государственного
университета К. А. Воскресенскому.
Проф. В. ЯШНОВ
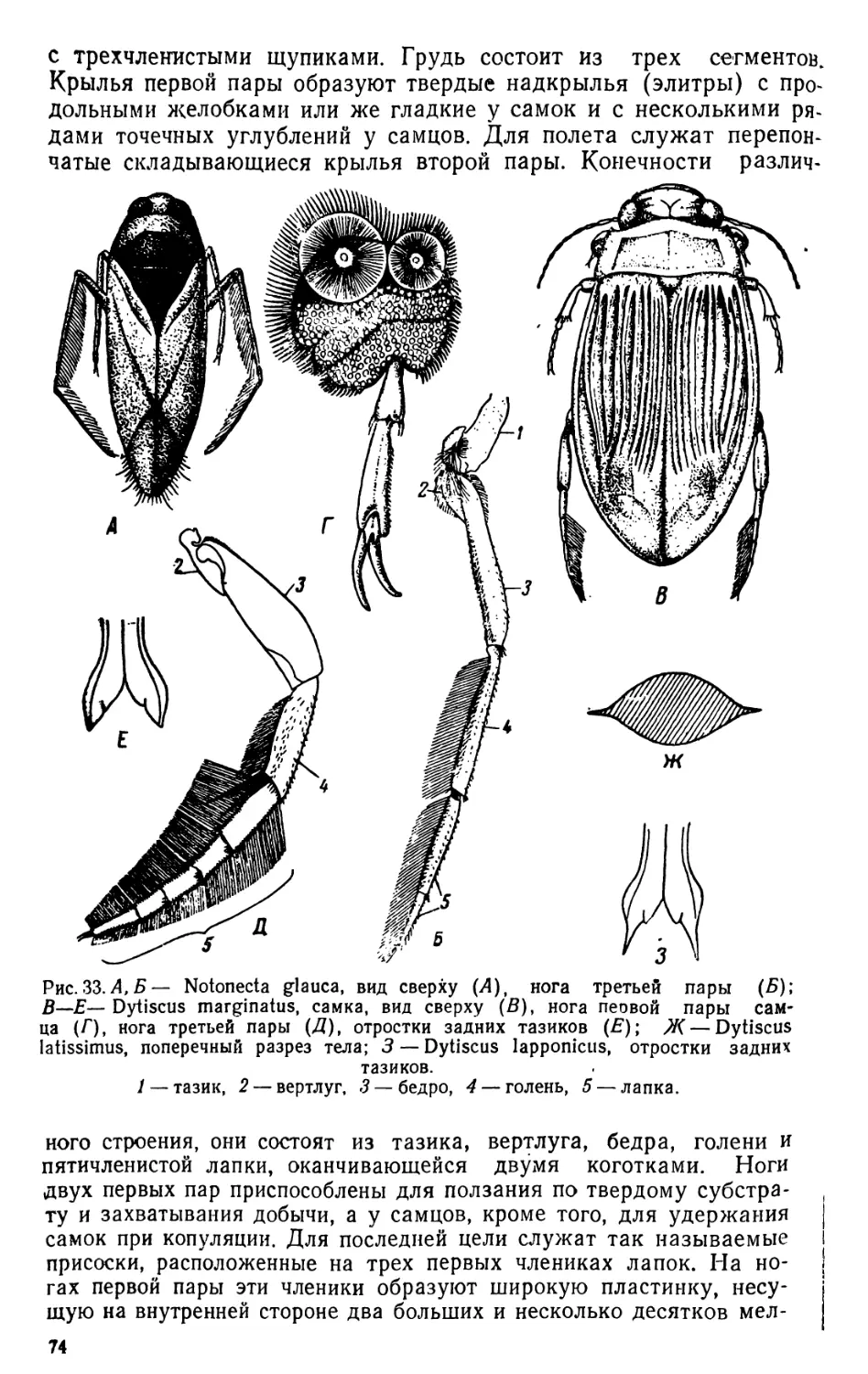
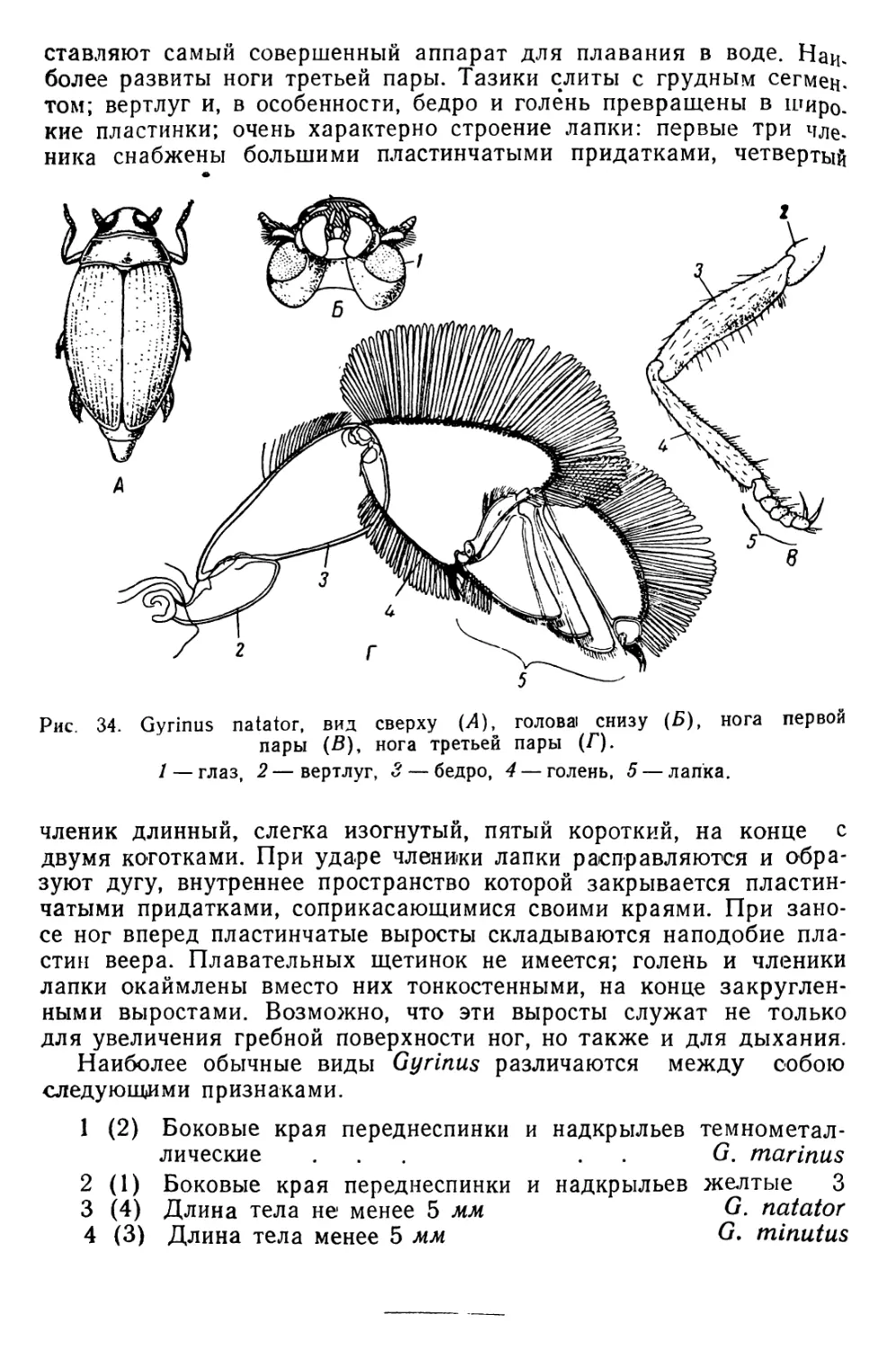
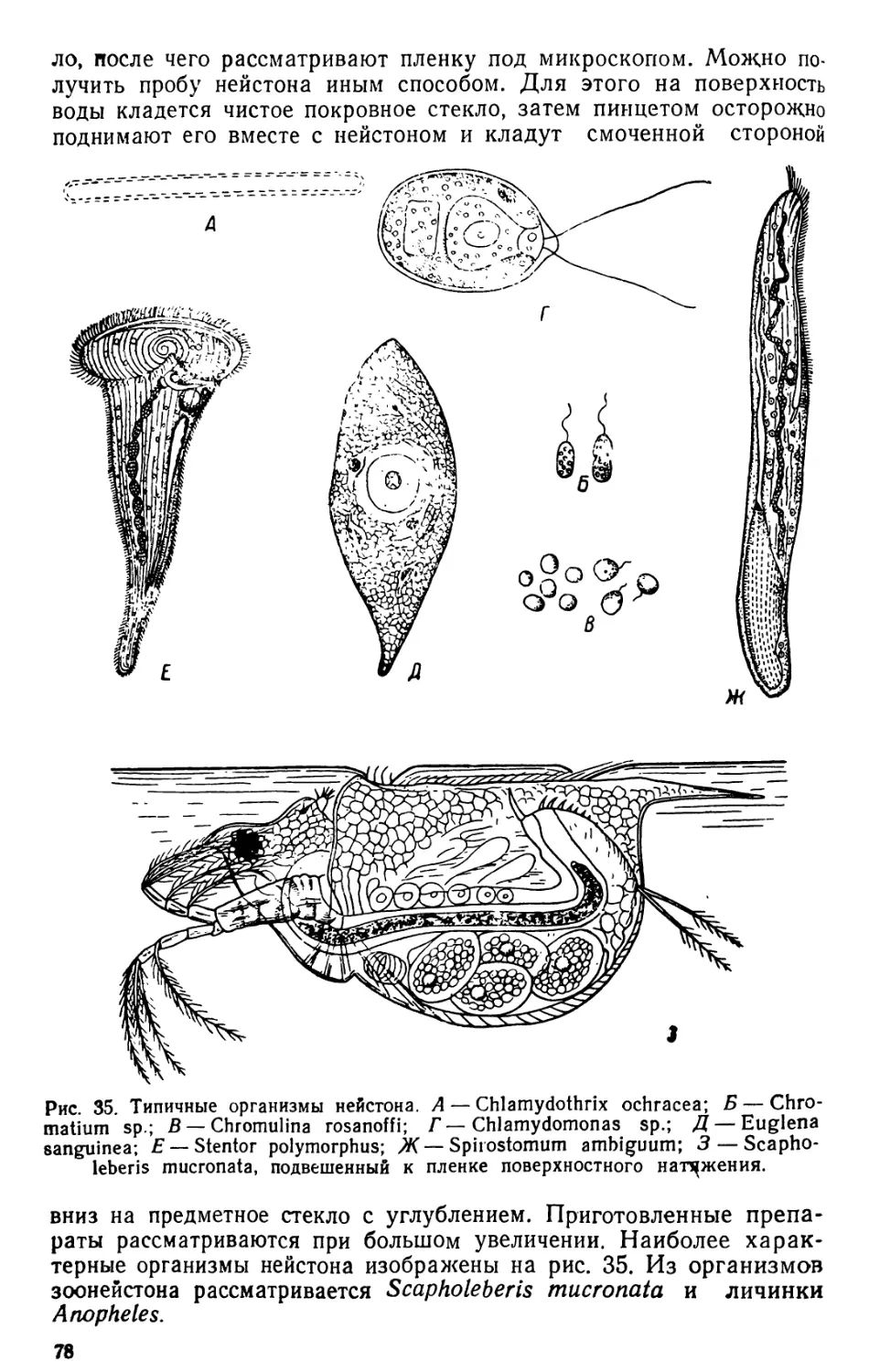
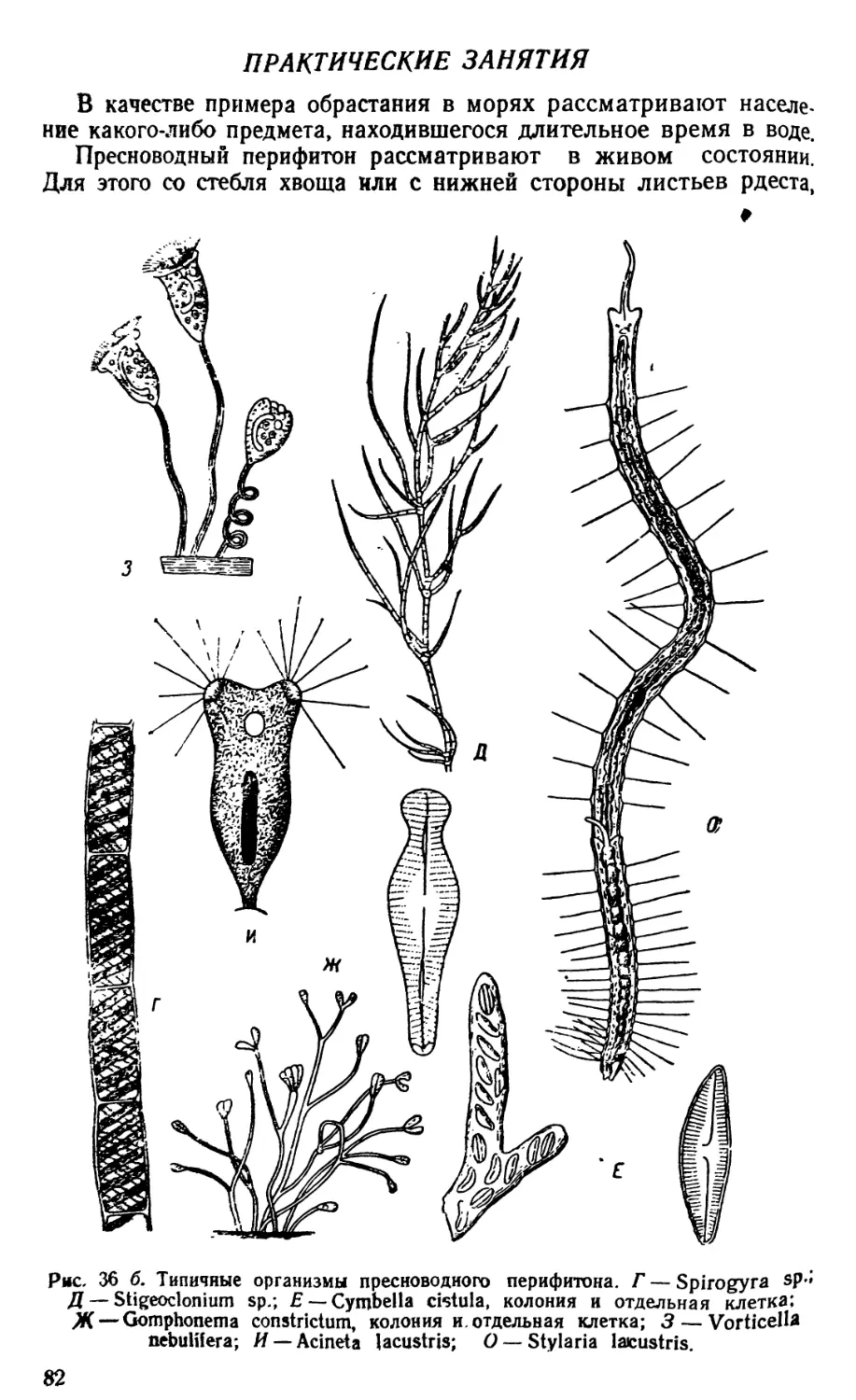
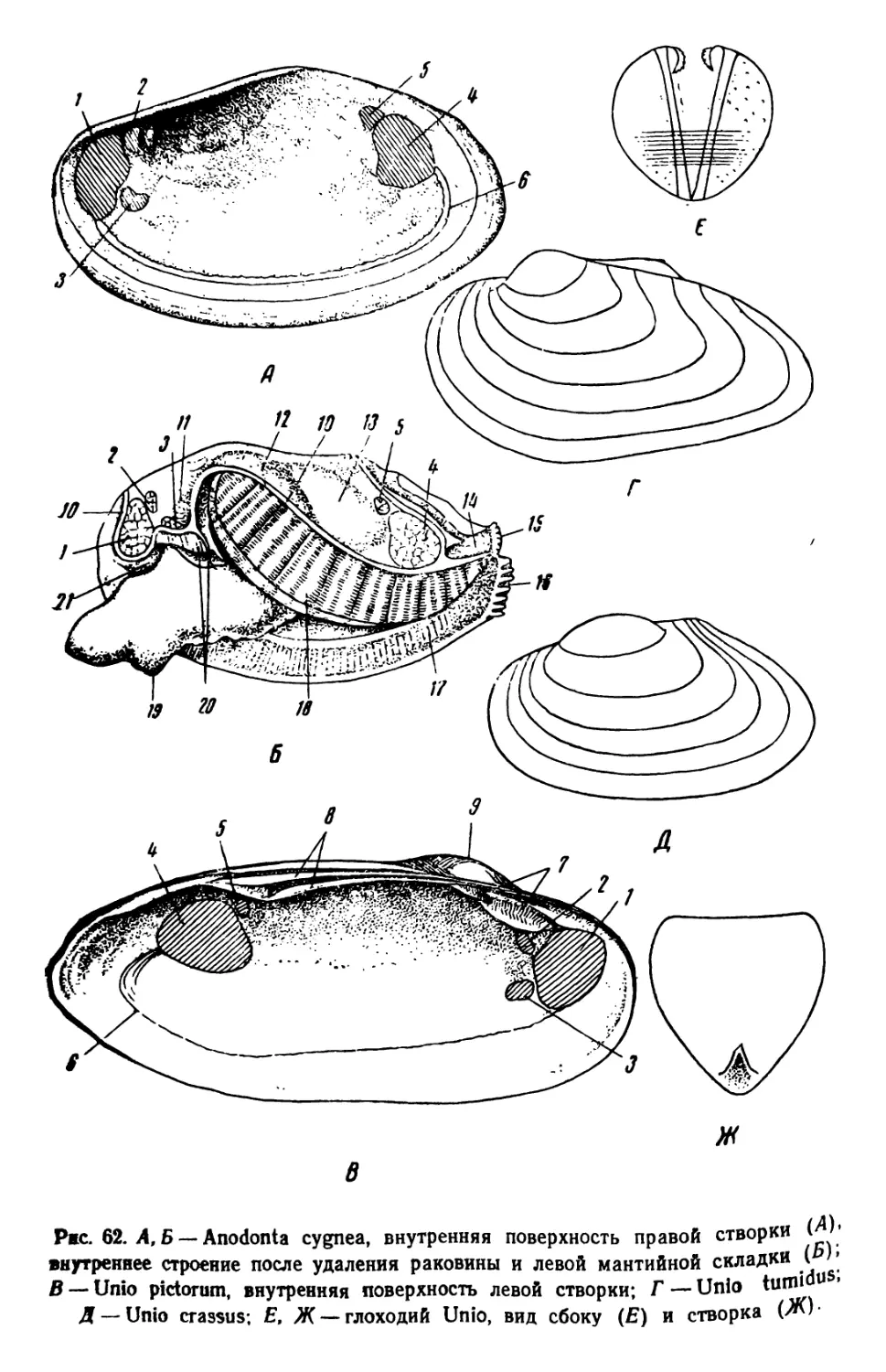
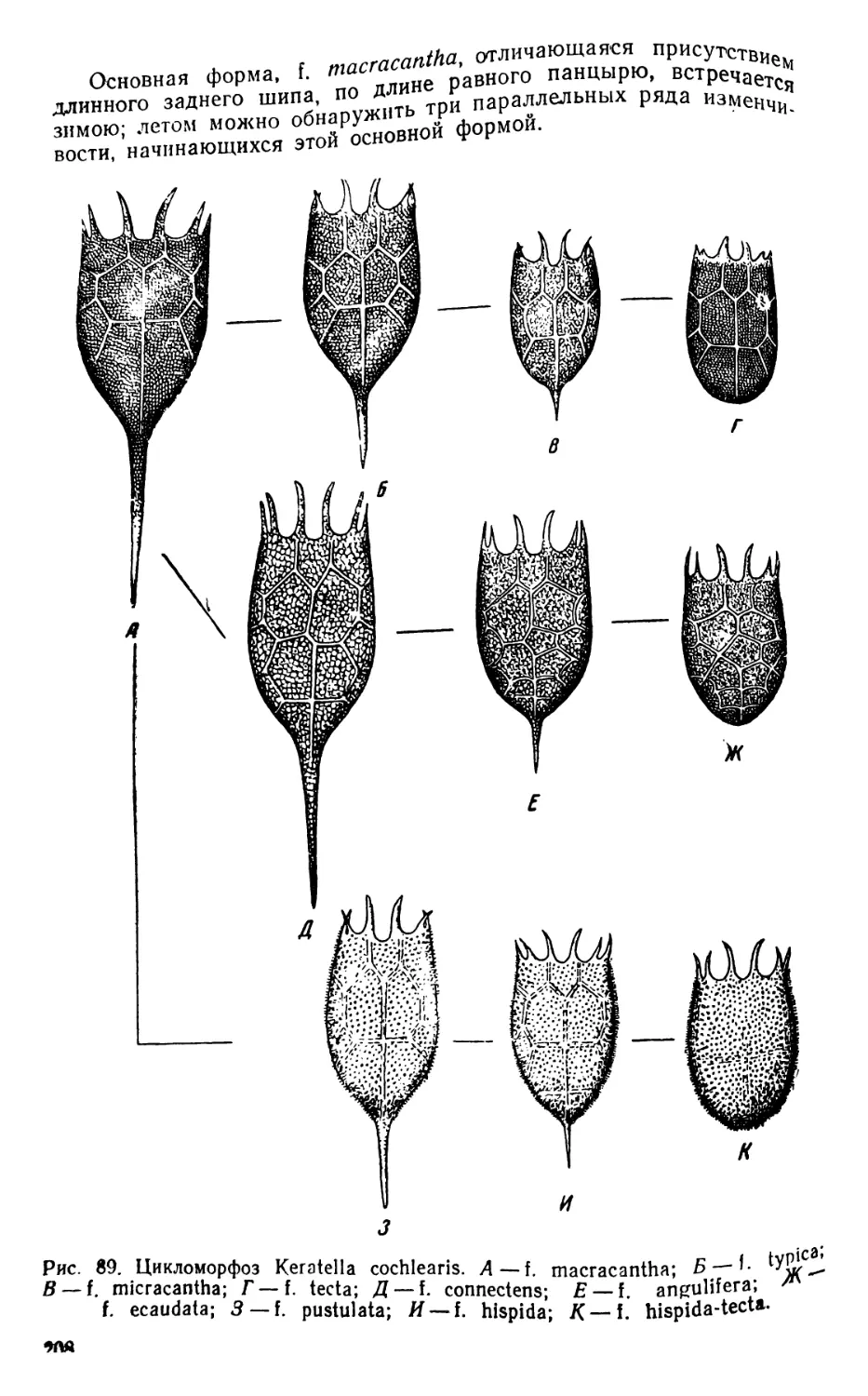
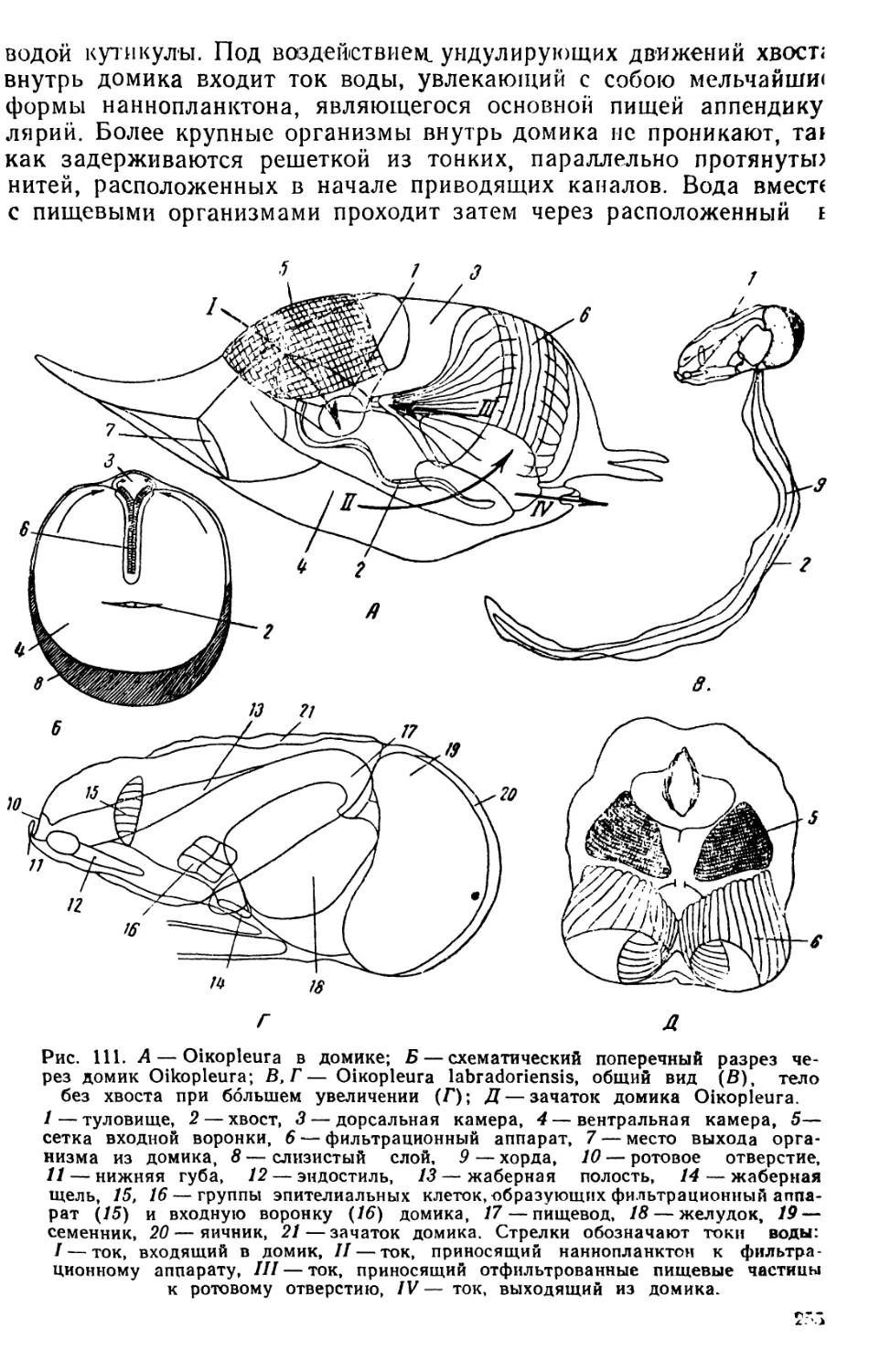
ОРГАНИЗМЫ ПЛАНКТОНА
Планктон состоит из организмов, свободно живущих в толще
воды и не способных противодействовать собственным движениям
водной среды (течениям, конвекционным токам и т. п.) из-за от-
сутствия или относительно слабого развития у них органов движе-
ния. В систематическом отношении планктон делится на раститель-
ный планктон, или фитопланктон, и животный планктон, или зоо-
планктон.
В состав планктона входят, с одной стороны, голопланктиче-
ские организмы, всю свою жизнь, включая также и период разви-
тия, проводящие вне связи с твердым субстратом, а с другой сто-
роны— меропланктические организмы, которые определенный пе-
риод своей жизни проводят на дне водоемов. К числу последних
относятся, например, планктические личинки червей, иглокожих,
моллюсков, ракообразных и других морских донных животных,
отпочковывающиеся от полипов гидроидные медузы, а также мно-
гие живущие в прибрежной области организмы, цисты и покоящи-
еся яйца которых для дальнейшего развития опускаются на дно.
В зависимости от размера организмов планктон разделяется на
следующие группы.
1. Ультрапланктон (бактерии)—величина организмов не
превышает нескольких микронов, нижняя граница за пределами
видимости.
2. Н а н н о п л а н к’тон, или карликовый планктон (мельчайшие
низшие растения и простейшие), — величина организмов измеряет-
ся микронами и десятками 'микронов; благодаря своим ничтожно
малым размерам организмы наннопланктона проходят через са-
мый густой шелковый газ, могут быть исследованы только при
применении центрифугирования или камерного метода, поэтому эта
группа организмов называется также центробежным, или камер-
ным, планктоном.
3. Микропланктон (главная часть фитопланктона, а так-
же инфузории, коловратки, мелкие ракообразные и г. п.)— вели-
чина организмов измеряется десятыми и сотыми долями милли-
метра; улавливается планктическими сетями из густого шелкового
газа или методом осаждения, поэтому называется также сетяным,
или осадочным, планктоном.
4. Мезопланктон (крупные представители фитопланктона,
главная часть зоопланктона морей) —величина организмов изме-
ряется миллиметрами; улавливается планктичеокими сетями из
редкого шелкового газа —сетяной планктон.
5. Макропланктон (высшие раки, медузы; пелагические
черви и т. п.) — величина организмов измеряется сантиметрами,
встречается исключительно в морях; улавливается большими
планктическими сетями. (
6. Мегалопланктон (многие сцифоидные медузы, крупные
сифонофоры и т. п.) — величина организмов измеряется десятка-
ми сантиметров; встречается исключительно в морях.
Отличительная особенность планктических организмов — их
способность находиться в воде во взвешенном состоянии — накла-
дывает определенный отпечаток на их строение.
Большинство видов относится к числу активных пловцов.
Удельный вес этих организмов почти всегда немного больше удель-
ного веса воды, поэтому при отсутствии движения, например в
анестезированном состоянии, они опускаются вниз; скорость погру-
жения, в зависимости от размеров организмов, измеряется милли-
метрами или сантиметрами в секунду. Веслоногие раки (Copepoda),
являющиеся самыми распространенными представителями пресно-
водного и морского планктона, двигаются, при помощи ударов своих
грудных ног; органами движения ветвистоусых раков (Cladocera)
являются сильно развитые антенны; движения коловраток (Rota-
toria) вызываются работой коловращательного аппарата, а инфу-
зорий (например, морских Tintinnoinea) — биением ресничек; пла-
вание различных медуз (Hydromedusae и Scyphomedusae) происхо-
дит при помощи сокращений колокола. Многие организмы фито-
планктона также обладают способностью к самостоятельным движе-
ниям, вызываемым обычно, как, например, у перидиней (Peridi-
пеае) и кокколитофорид (Coccolithophoridae), колебаниями плаз-
матических жгутов.
Другие планктические организмы, к числу которых относятся
среди растений диатомеи (Diatomeae) и синезеленые (Cyanophy-
сеае), а среди животных —радиолярии (Radiolaria) и форамини-
феры (Foraminifera), никаких органов движения не имеют, они
не плавают, а парят в воде. Способностью ж парению обладают
также и многие животные, имеющие органы движения, как, напри-
мер, сифонофоры (Siphonophora), аппендикулярии (Appendicula-
ria) или личинки комара коретры (Chaoborus); плавание этих ор-
ганизмов происходит преимущественно в горизонтальном направ-
лении.
Для облегчения плавания и парения в воде у планктических
организмов выработался ряд приспособлений, уменьшающих оста-
точный вес и увеличивающих сопротивление формы.
Остаточным весом называется разность между весом какого-
либо тела и весом вытесненной им воды. Чем больше остаточный
вес, тем больше скорость погружения этого тела, и обратно; при
остаточном весе, равном нулю, устанавливается безразличное рав-
новесие.
R
Приспособления к уменьшению остаточного веса планктических
организмов можно разделить на несколько групп.
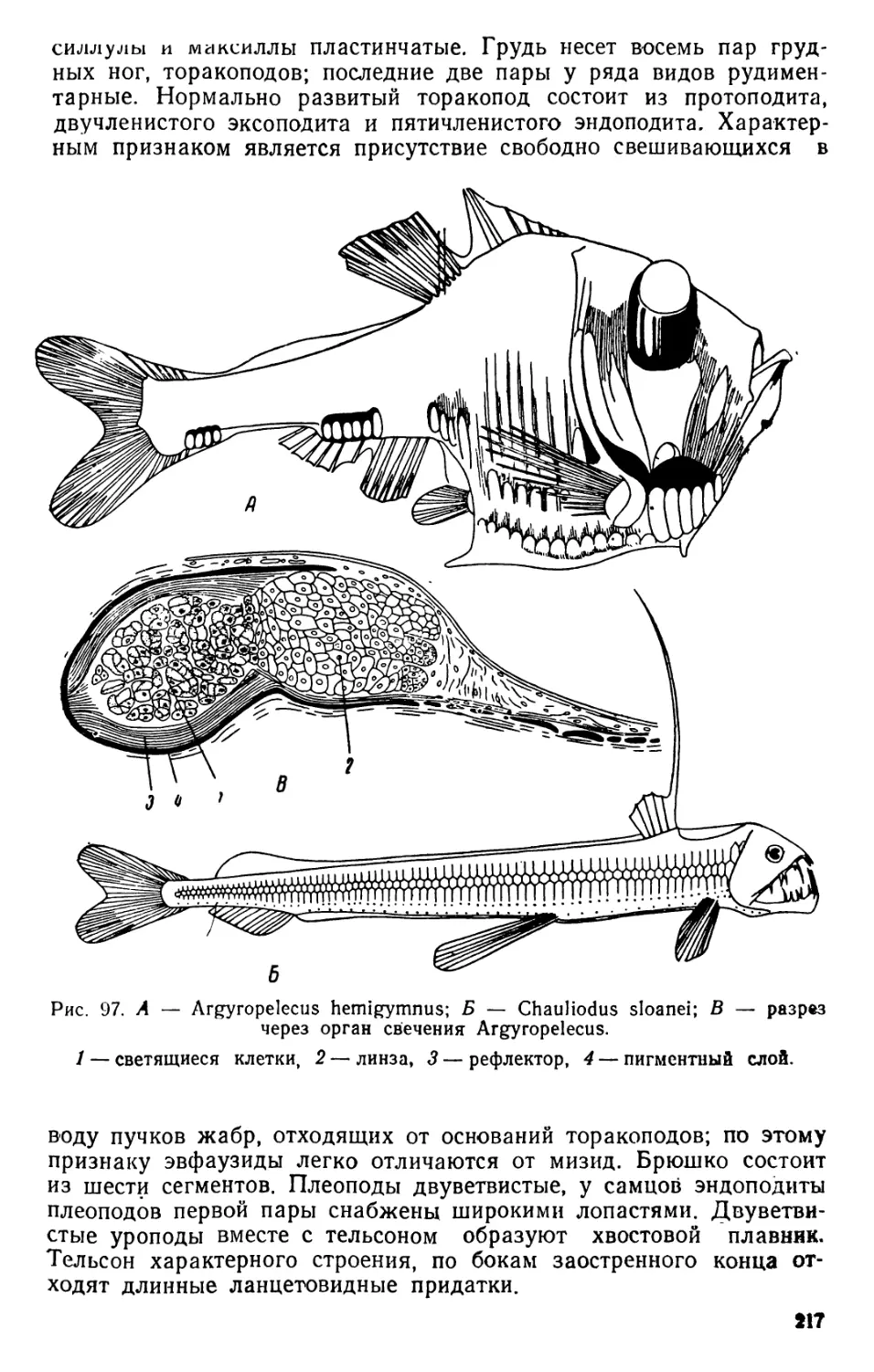
1. Редукция скелетных образований. Все планкти^
ческие организмы лишены тяжелого скелета и по этому признаку
резко отличаются от близких форм, ведущих донный образ жизни.
Планкти ческие диатомеи обладают более легким скелетом, по срав-
нению с донными видами, створки которых сильнее пропитаны
кремнеземом и значительно тяжелее. У пелагических видов ветви-
стоусых раков хитиновая раковина значительно тоньше и нежнее,
чем у видов, обитающих на дне. Пелагические высшие раки (Агп-
phipoda, Euphausiacea, Decapoda) в противоположность донным
видам никогда не обладают тяжелым, пропитанным известью пан-
цырем. То же наблюдается и у морских крылоногих (Pteropoda) и
киленогих (Heteropoda) моллюсков, характеризующихся или пол-
ным отсутствием раковины, или очень слабым ее развитием.
2. Пропитывание водою. Многие планктические расте-
ния и животные отличаются очень большим содержанием воды.
Количество ее нередко превышает 90%, что имеет большое значе-
ние для уменьшения остаточного веса, так как плазма, удельный
вес которой в среднем равен 1,05, тяжелее не только пресной, но
и морской воды (удельный вес пресной воды равен 1,00, а для мор-
ской воды колеблется, в зависимости от температуры и солености,
от 1,02 до 1,03). Пропитывание водою приводит к образованию
студенистого вещества, особенно сильно развитого у медуз и си-
фонофор, а также у пелагических моллюсков, сагитт (Chaeto-
gnatha) и ряда других животных. У пресноводных организмов ча-
ще наблюдается выделение прозрачного студенистого вещества,
покрывающего тело снаружи, как, например, у некоторых сиыезе-
леных и десмидиевых (Desmidiaceae) водорослей, а также у неко-
торых коловраток. У многих радиолярий внутри тела имеются ва-
куоли, содержащие жидкость удельно более легкую, по сравнению
с морской водой. Внутри Noctiluca (Cystoflagellata) находится
большое количество клеточного сока.
3. Жировые включения. Масляные и жировые включе-
ния являются, главным образом, резервным веществом, однако они
одновременно служат для уменьшения остаточного веса. У планк-
тических водорослей первым продуктом фотосинтеза является не
крахмал, а более легкое масло. Очень часто жировые включения
встречаются и у различных планктических животных. Нередко
внутри тела пресноводных и морских веслоногих раков можно ви-
деть большое количество жировых включений в виде капель или
даже специального жирового органа, лежащего вдоль кишечника.
Такие же включения наблюдаются у ветвистоусых раков, коловра-
ток и многих других планктических животных.
4. Газовые включения. В противоположность жидким
включениям, которые практически не сжимаются, газовые включе-
ния изменяют объем в зависимости от изменений температуры и
давления. Поэтому организмы, обладающие газовыми включени-
ями, могут при помощи этого гидростатического аппарата не только
сохранять безразличное равновесие, но, в случае необходимости,
подниматься вверх или опускаться вниз. Наиболее совершенным
с этой точки зрения является гидростатический аппарат личинок
иеристоусого комара коретры (Chlaoborus), ведущих планктиче-
ский образ жизни. У многих сифонофор имеется большой пнейма-
тофор, наполненный воздухом и сообщающийся с наружной сре-
дой каналом. Планктические синезеленые водоросли содержат в сво-
их клетках многочисленные мелкие газовые включения, с помо-
щью которых они поднимаются со дна на поверхность воды.
Сопротивление, испытываемое погружающимся в воде телом, за-
висит от его удельной поверхности и величины вертикальной проек-
ции; сопротивление, определяемое этими величинами, сокращенно
называется сопротивлением формы. Удельная поверхность, т. е.
отношение абсолютной поверхности к объему тела, меньше всего
у шара; по мере удлинения одной или двух осей тела, а также об-
разования различных выростов, удельная поверхность возрастает.
Вертикальная проекция зависит от положения плоскости тела по
отношению к вертикали. Если тело имеет пластинчатую форму, то
величина вертикальной проекции становится максимальной при та-
ком положении пластинки, когда она расположена в плоскости,
перпендикулярной к направлению действия силы тяжести.
Приспособления к увеличению сопротивления формы планктиче-
ских организмов можно подразделить на следующие группы.
1. Удлинение одной оси. Многие растительные и живот-
ные виды имеют удлиненную, палочковидную форму тела. Такой
формой обладают некоторые диатомеи, перидинеи, а среди живот-
ных — сагитты и ряд других организмов. В нормальном положении
они держатся горизонтально. Если под влиянием волнения или
иных причин это положение сменится на вертикальное, то, разу-
меется, организмы будут быстро погружаться острым концом
вниз. Однако благодаря присутствию направительных выростов
или изогнутой форме тела это падение пойдет не прямолинейно,
а по кривой, поэтому организмы вскоре займут опять горизонталь-
ное положение. Удлинение одной оси достигается также образова-
нием колоний, часто встречающихся у различных планктических
водорослей, а из животных — у сальп.
2. Удлинение двух осей. Среди планктических орга-
низмов часто встречаются дисковидные, пластинчатые формы.
Многие диатомеи и перидинеи имеют вид пластинки или образуют
яарашютообразные выросты. Уплощенная форма тела свойственна
также некоторым представителям зоопланктона — радиоляриям,
медузам, пелагическим червям (из Polychaeta), веслоногим ракам
н другим животным.
3. Образование выростов. Трудно указать какую-либо
группу планктических организмов, среди которых не имелось бы
большого количества видов, снабженных шипами, иглами, длин-
ными выростами и другими подобными образованиями. Все эти
выросты обычно рассматриваются в качестве приспособлений, уве-
личивающих сопротивление формы. Однако далеко не всегда это
соответствует действительности. Наблюдения над живыми организ-
мами показали, что, например, выросты Daphnia и Bosmina распо-
лагаются у нормально ориентированных особей в вертикальной
плоскости и служат не для уменьшения скорости погружения, а
для руления и сохранения прямолинейности движения. Выросты
десмидиевых водорослей заключены в покрывающее колонию сту-
денистое вещество, а шипы ряда радиолярий — в экстракапсуляр-
ную плазму, поэтому вряд ли могут считаться придатками, увели-
чивающими сопротивление формы. Во многих случаях, как, напри-
мер, у Chaetoceros среди диатомей, выросты служат скорее не
для уменьшения скорости погружения, а для использования сла-
бых конвекционных токов воды, поднимающих эти организмы в зо-
ну фотосинтеза; при отсутствии движения воды водоросли опуска-
ются на дно.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
ОПРЕДЕЛЕНИЕ СКОРОСТИ ПОГРУЖЕНИЯ АНЕСТЕЗИРОВАННЫХ
ПЛАНКТИЧЕСКИХ ОРГАНИЗМОВ
1. Скорость погружения дафнии (Daphnia)
Для определения скорости погружения дафнии берут высокий,
до верха наполненный водою стеклянный цилиндр и переносят на
поверхность воды пинцетом с мягкими концами (чтобы не повре-
дить животное) одну крупную, предварительно анестезированную
дафнию (Daphnia magna или D. pulex). Анестезирование ведут в
часовом стекле, прибавляя к воде по каплям однопроцентный
раствор серного эфира или слабый раствор соды или соляной ки-
слоты. Зная высоту цилиндра и время (отмечаемое по секундомеру
или секундной часовой стрелке), в течение которого дафния опус-
кается до дна сосуда, можно вычислить скорость погружения орга
низма.
2. Скорость погружения коловраток (Rotatoria)
Аналогичные наблюдения можно провести с коловратками (на-
пример, Keratella, Brachionus), а также с другими животными ми-
кропланктона. Так как эти организмы слишком малы для просле-
живания их пути простым глазом, то для отсчета времени их по-
гружения следует применять лупу большого диаметра, а сами на-
блюдения вести в узком цилиндрическом сосуде; анестезированные
организмы переносятся пипеткой.
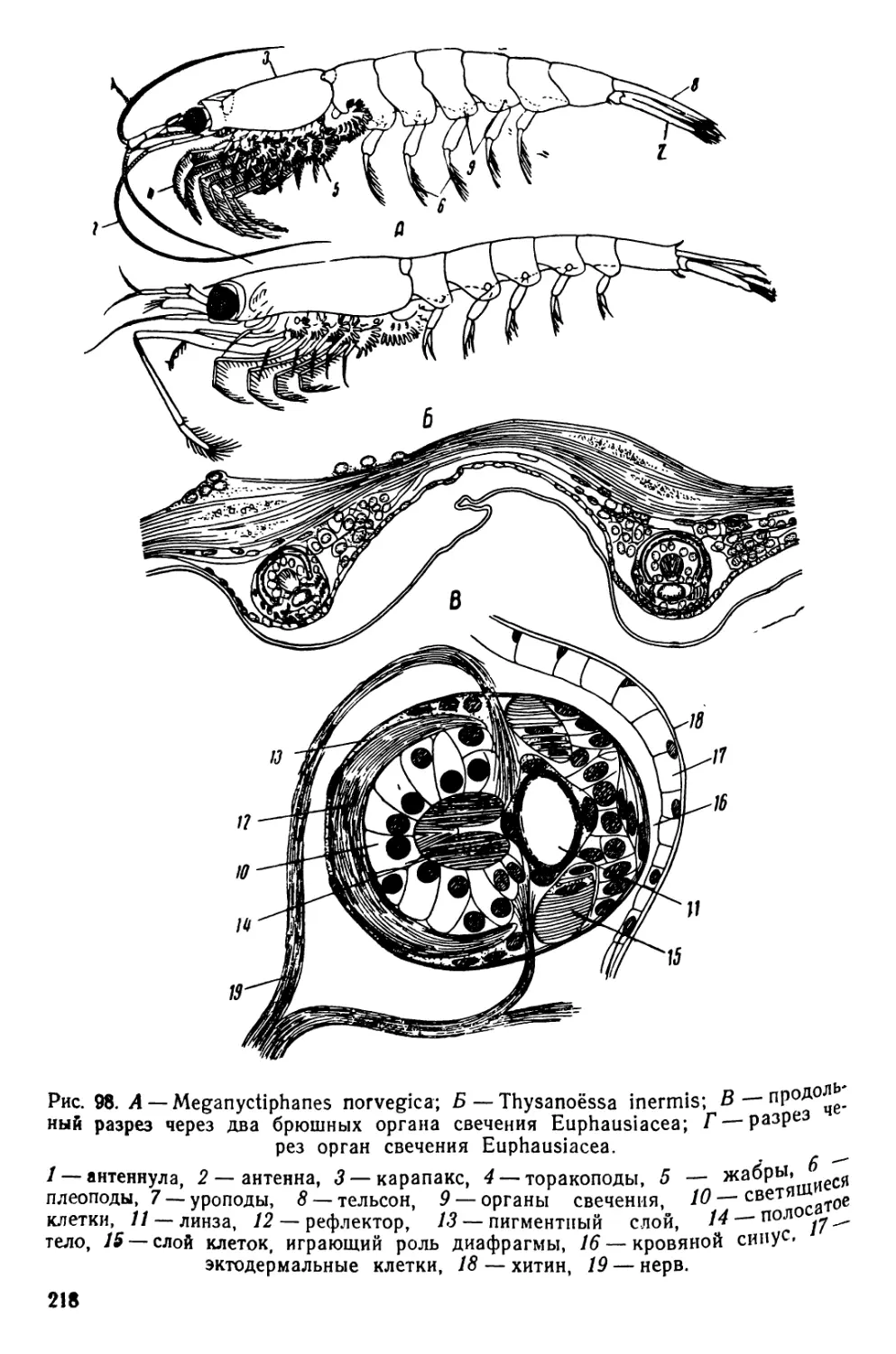
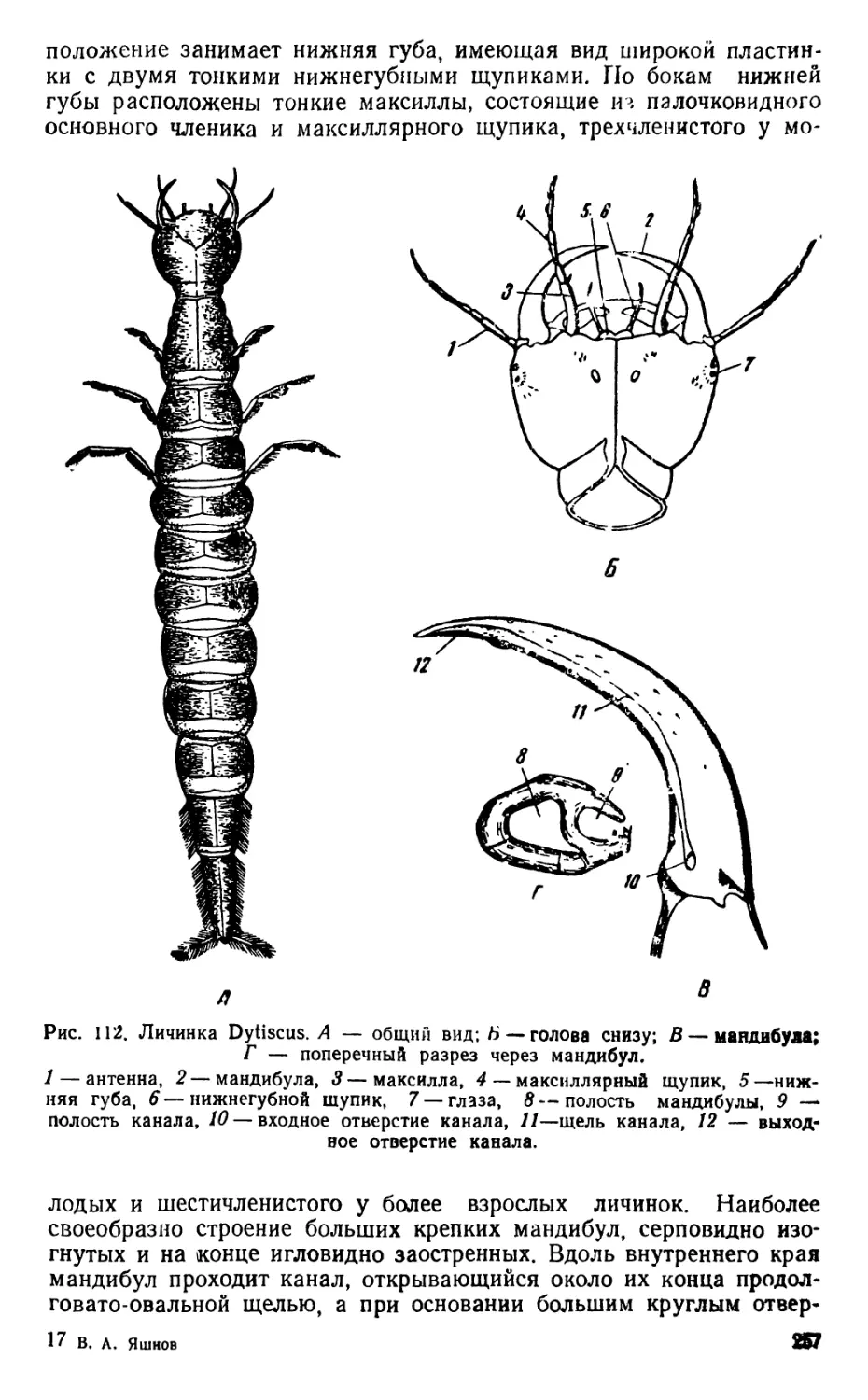
ОРГАНЫ ДВИЖЕНИЯ ПЛАНКТИЧЕСКИХ ОРГАНИЗМОВ
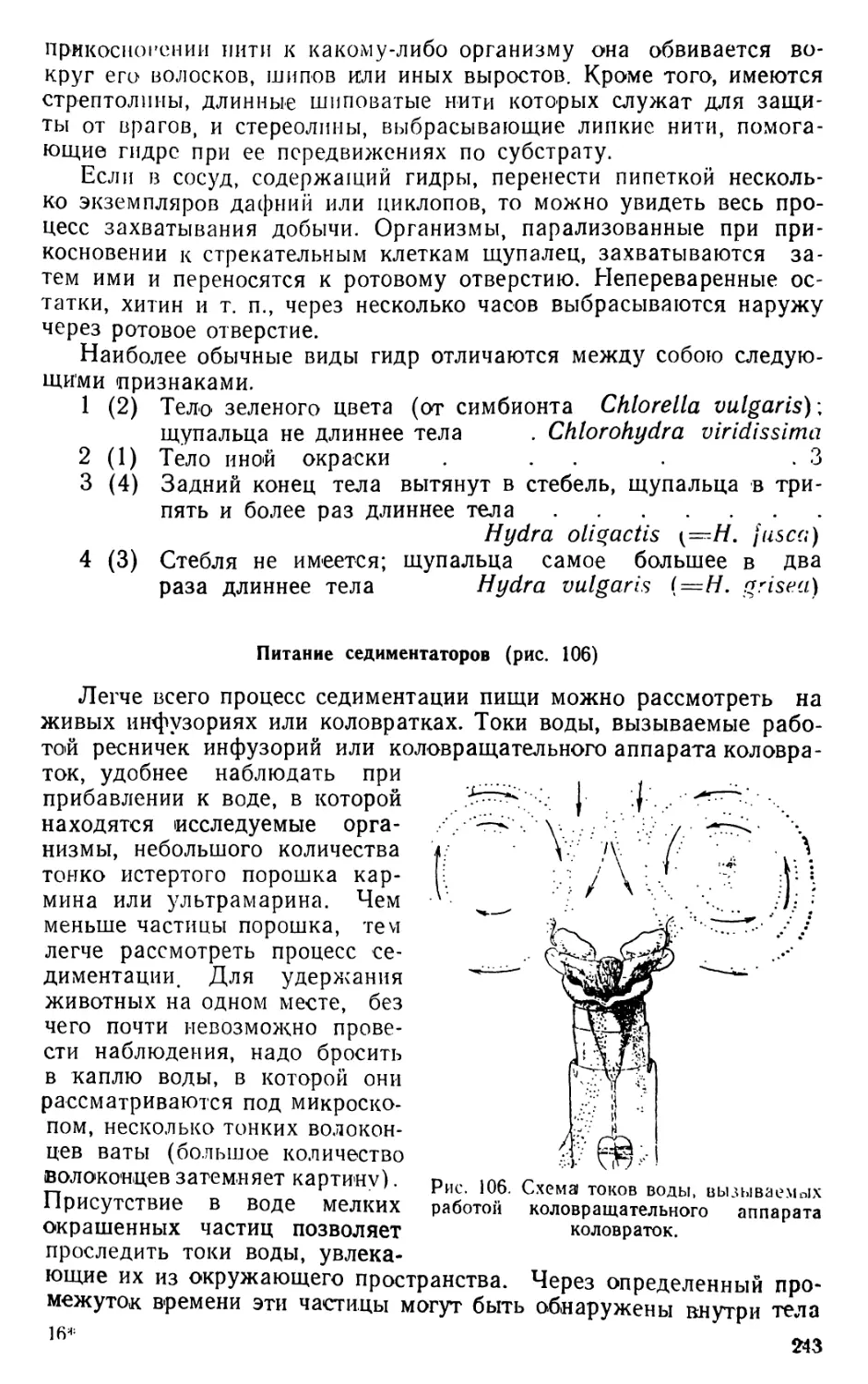
Движение при помощи грудных ног
Наблюдения ведутся над живыми диаптомусами (Diaptomus) и
циклопами (Cyclops), относящимися к отряду веслоногих раков
•(Copepoda); Diaptomus входит в состав подотряда Calanoida, а
Cyclops — подотряда Cyclopoida.
Прежде всего следует рассмотреть строение диаптомуса и
циклопа.
9
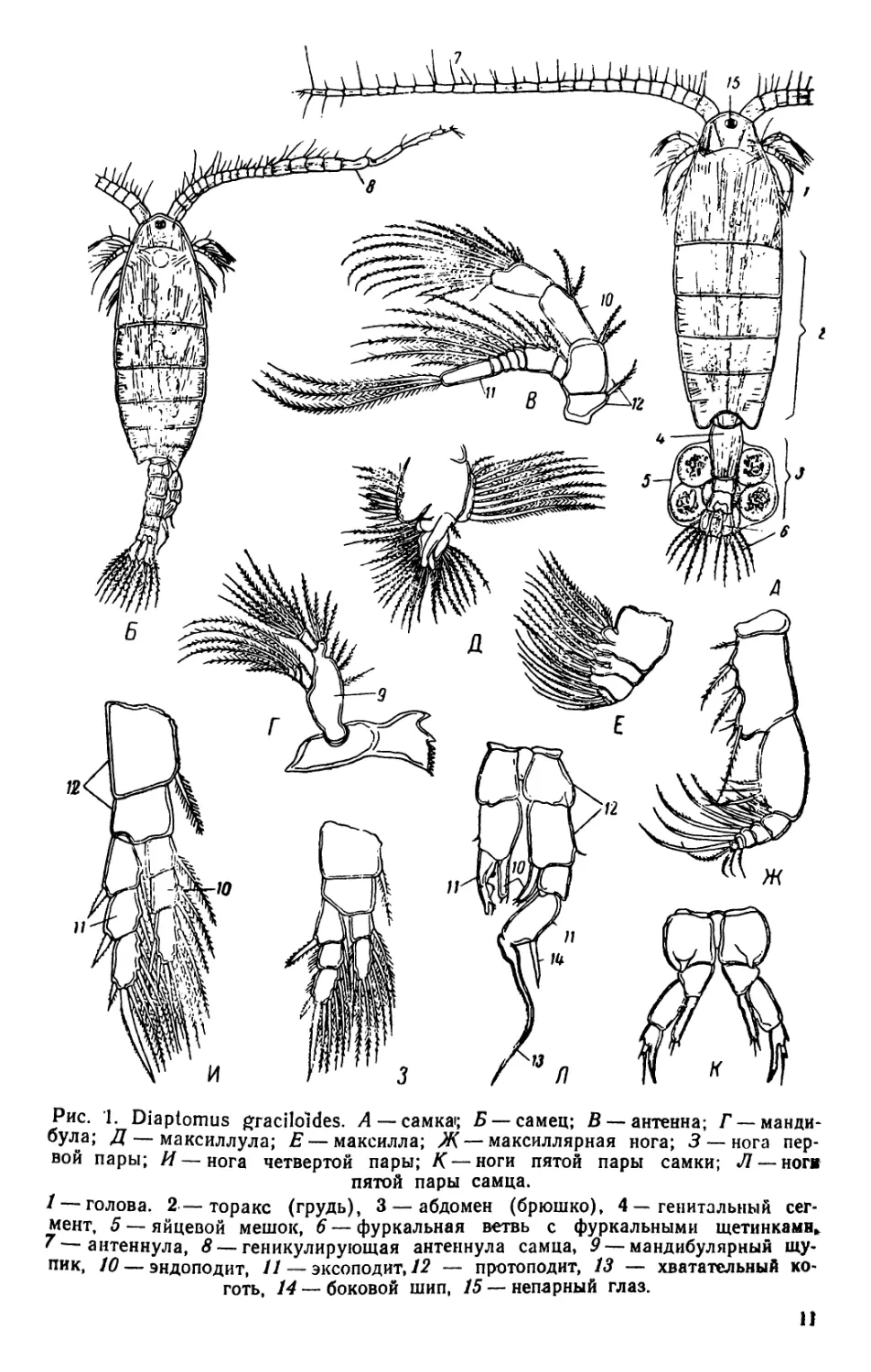
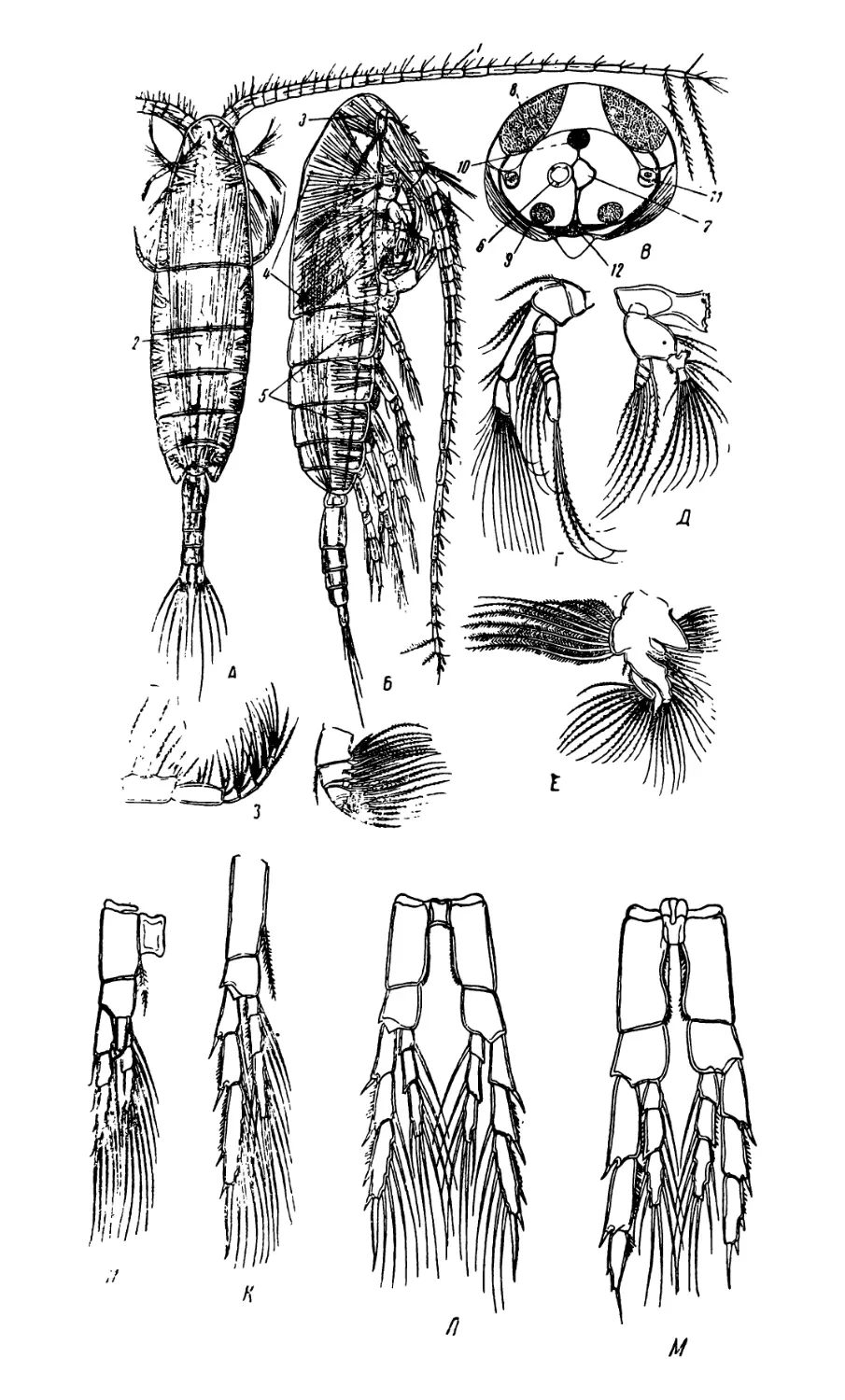
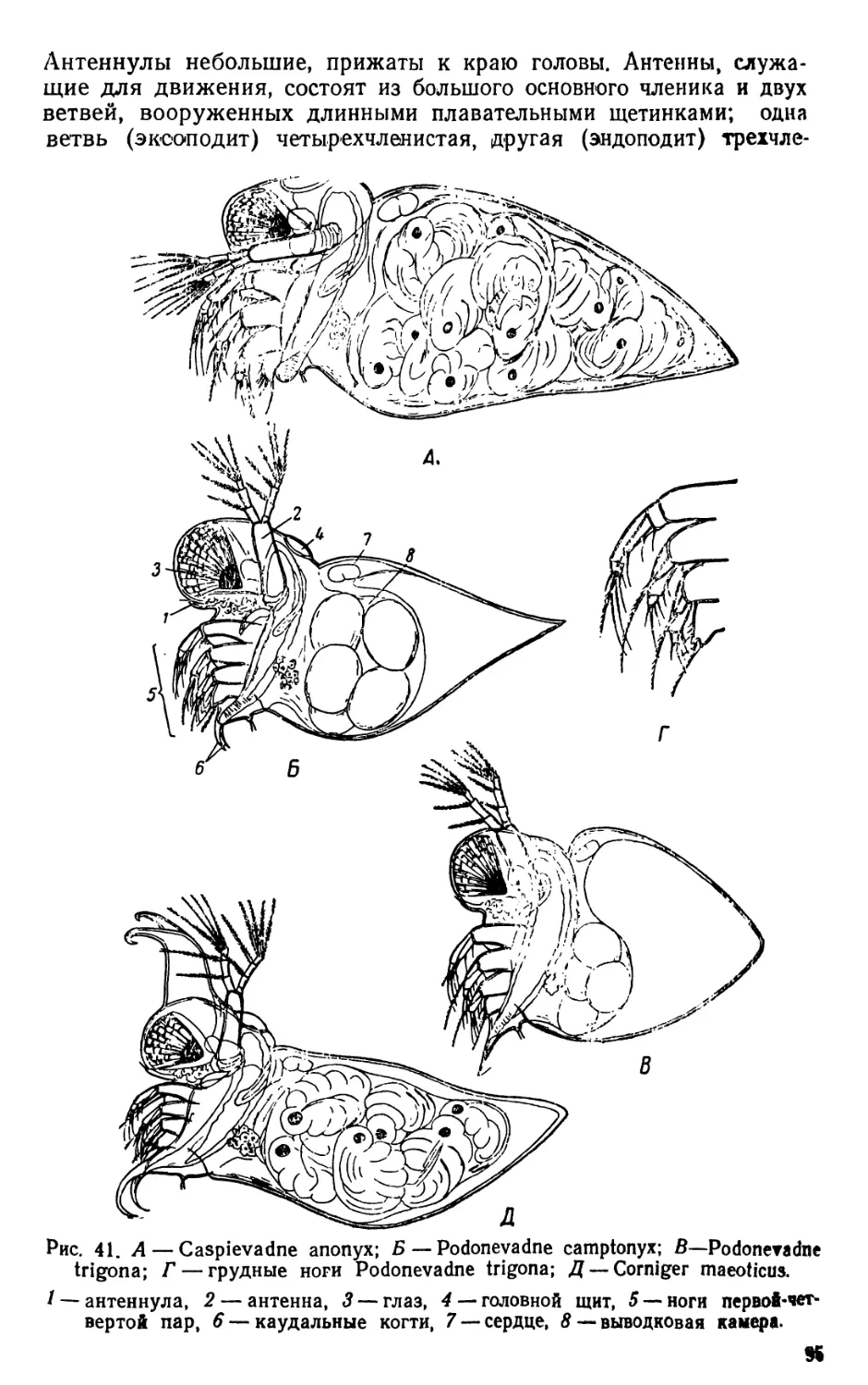
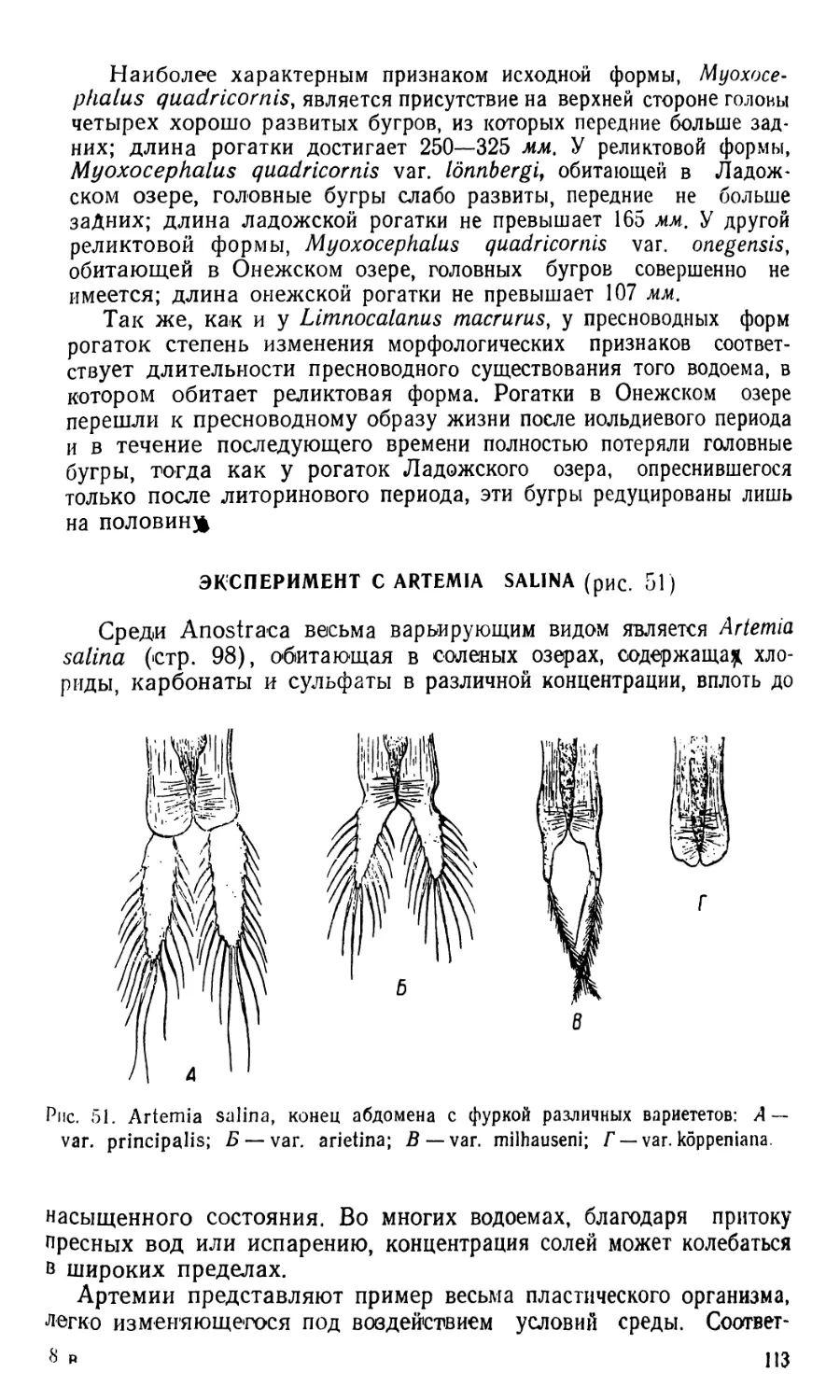
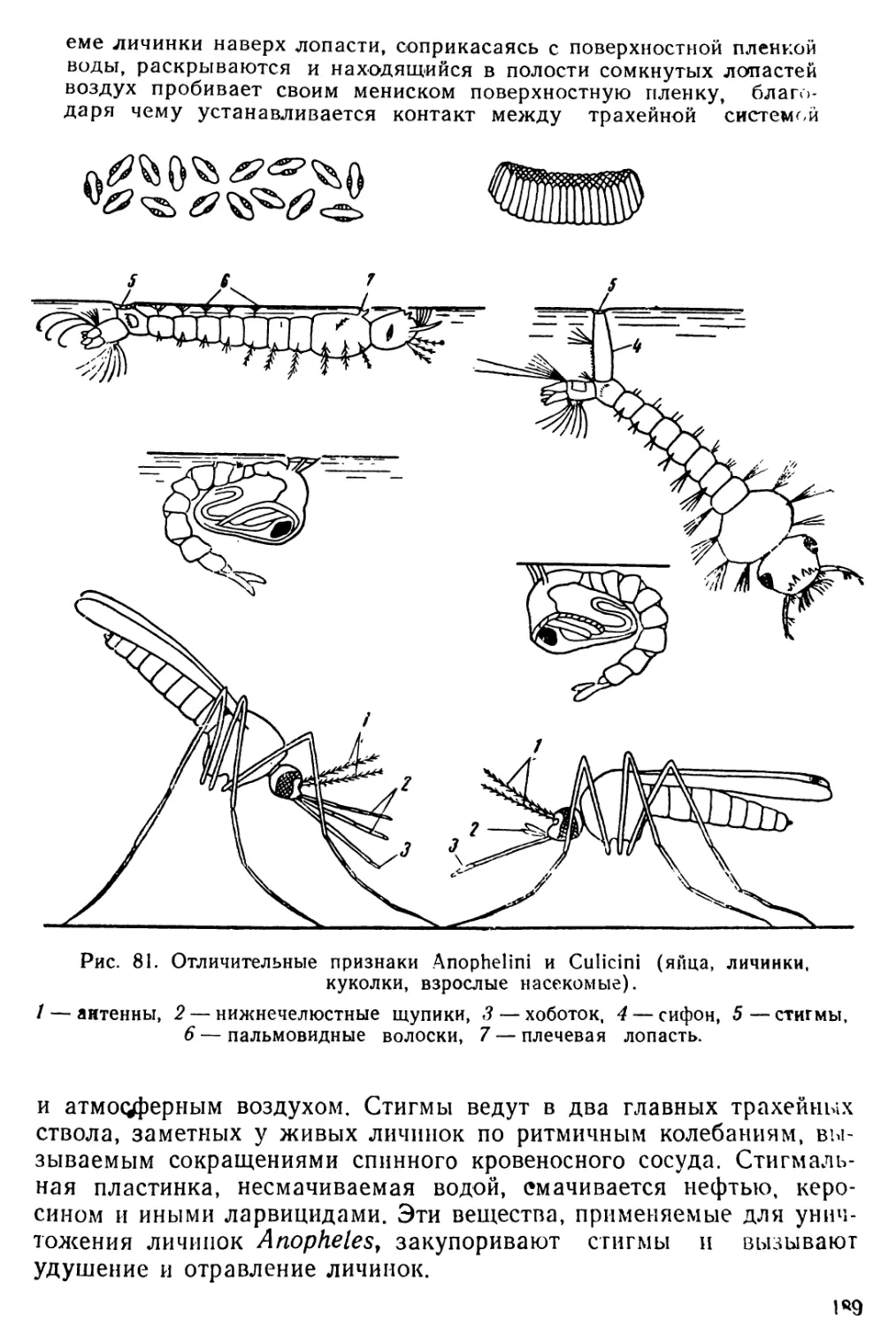
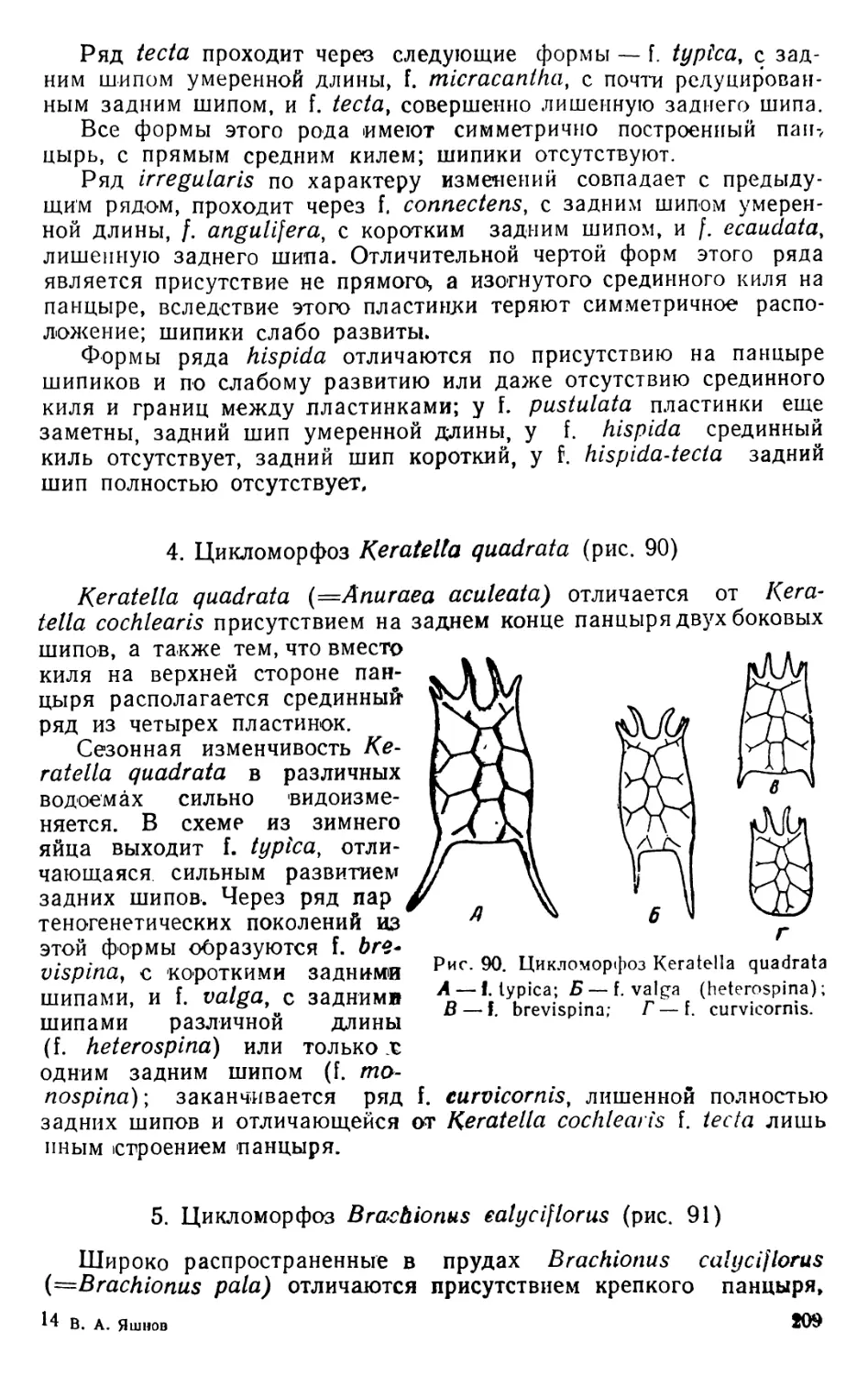
1. Diaptomus (рис. 1—2)
Тело диаптомуса резко делится на два отдела. Передний отдел
состоит из головы и четырех (у некоторых видов пяти) грудных
сегментов, плотно соединенных в компактное целое; короткий и уз-
кий задний отдел образован брюшком, в состав которого входит
три сегмента у самок и пять сегментов у самцов. Первый брюшной
сегмент называется генитальным, так как на нем открываются про-
токи органов размножения. К последнему, анальному, сегменту
брюшка причленяетсяфурка, имеющая вид двух коротких придат-
ков, называемых фуркальными ветвями; каждая фуркальпая ветвь
несет пять коротких оперенных щетинок.
На голове находится шесть пар придатков: антеннулы, антен-
ны, мандибулы, максиллулы, максиллы и максиллярные ноги; при-
сутствие последних указывает на слияние с первичными головными
сегментами первого торакомера (первичного грудного сегмента).
Антеннулы длинные, достигают конца брюшка, а у некоторых ви-
дов — фуркальных щетинок. Правая антеннула самцов превраще-
на в геникулирующую антеннулу, дистальная часть которой может
складываться наподобие перочинного ножа и зажимать при копу-
ляции придатки тела самки; находящиеся около сочленения члени-
ки вздуты и часто вооружены шипами или зубцами. Антенны дву-
ветвистые; максиллулы пластинчатые; максиллы и максиллярные
ноги одноветвистые.
Грудные ноги первых четырех пар, служащие для плавания,
состоят из двучленистого протоподита и двух ветвей, из которых
наружная ветвь, эксоподит, на всех ногах трехчленистая, а внут-
ренняя ветвь, эндоподит, на ногах первой пары двучленистая, а на
ногах трех следующих пар трехчленистая; обе ветви снабжены
длинными, перистыми плавательными щетинками, а эксоподиты,
кроме того, крепкими шипами. Ноги каждой пары соединены ме-
жду собою хитиновыми пластинками, поэтому движение их совер-
шается одновременно.
Ноги пятой пары резко отличаются по строению от остальных
ног. У самок они симметричные с одночленистыми или двучлени-
стыми эндоподитами и трехчленистыми эксоподитами. Ноги пятой
пары самцов асимметричные. Правая нога больше левой; базипо-
диты двучленистые, эндоподиты слабо развиты; эксоподит левой но-
ги двучленистый, с различными придатками; эксоподит правой
ноги большой, двучленистый, конечный членик с шипом по наруж-
ному краю и с длинным изогнутым хватательным когтем. Строе-
ние ног пятой пары характерно для каждого вида, и определение
диаптомусов ведется почти исключительно по строению ног этой
пары.
Кишечник имеет вид простой трубки, открывающейся аналь-
ным отверстием между ветвями фурки. Сердце расположено на
границе между двумя первыми грудными сегментами. Специальных
органов дыхания не имеется, газообмен происходит через тонкие
покровы тела. В передней части головы расположен небольшой не
парный глаз. Яйца вынашиваются самками в непарном яйцевод
мешке, прикрепленном к нижней стороне генитального сегмента.
10
Рис. 1. Diaptomus graciloides. А — самка; Б — самец; В —антенна; Г —манди-
була; Д — максиллула; Е — максилла; Ж— максиллярная нога; 3 — нога пер-
вой пары; И—нога четвертой пары; Д'—ноги пятой пары самки; Л —ноги
пятой пары самца.
/ — голова. 2 — торакс (грудь), 3 — абдомен (брюшко), 4 — генитальный сег-
мент, 5 — яйцевой мешок, 6 — фуркальная ветвь с фуркальными щетинками»,
'—антеннула, 8 — геникулирующая антеннула самца, 9 — мандибулярный щу-
пик, 10 — эндоподит, 11 — эксоподит, 12 — протоподит, 13 — хватательный ко-
готь, 14 — боковой шип, 15 — непарный глаз.
П
„ ябД°ме^
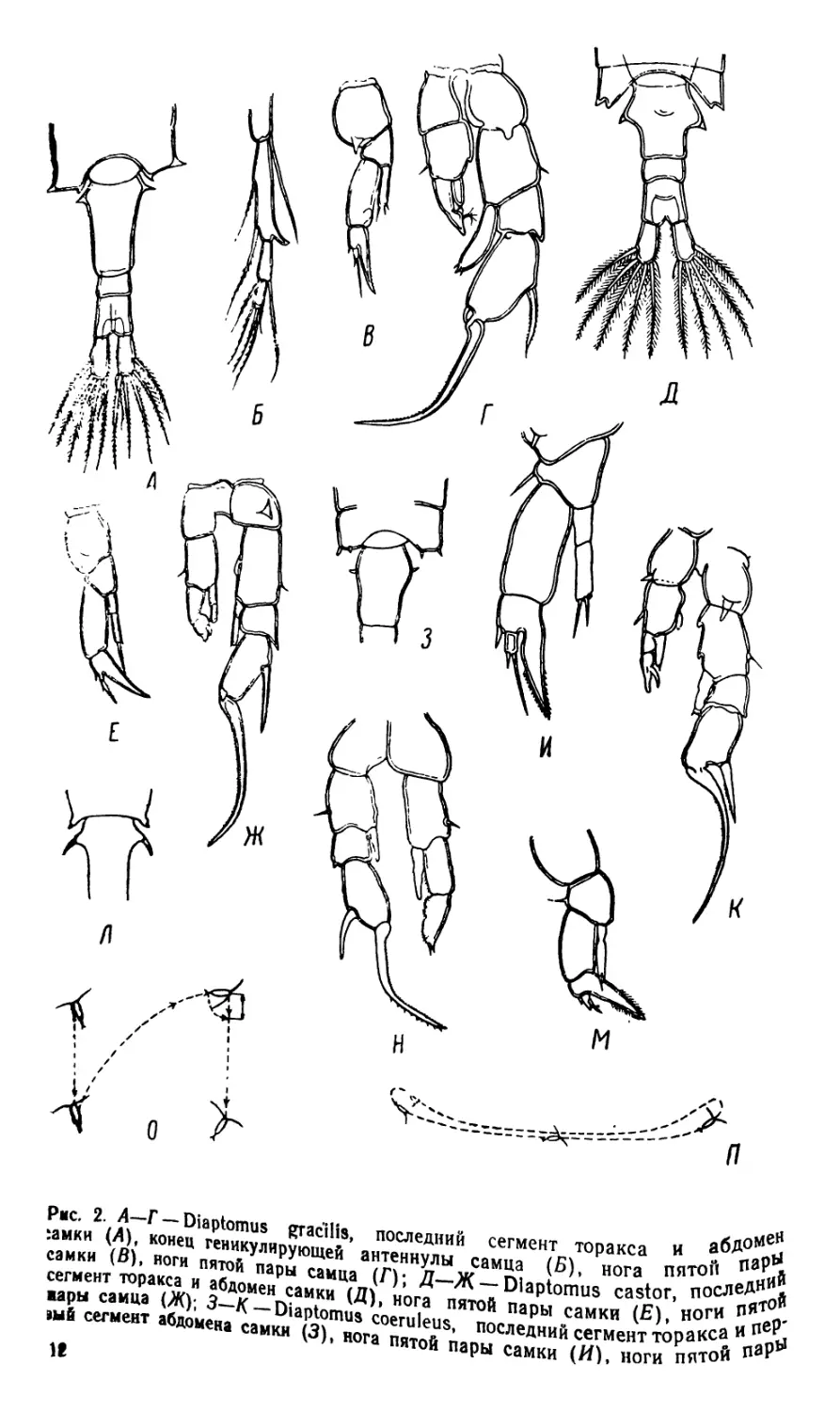
Рис, 2. А—Г — Diaptomus gracilis, последний сегмент торакса и ПЗрЧ
:амки (А), конец геникулирующей антеннулы самца (Б), нога пято
самки (В), ноги пятой пары самца (Г); Д—Ж — Diaptomus castor, по ^ofl
сегмент торакса и абдомен самки (Д), нога пятой пары самки (В), НОГ а и ЯеР
жары самца (Ж)\ 3—К-—Diaptomus coeruleus, последний сегмент торакс
вый сегмент абдомена самки (3), нога пятой пары самки (И), ноги пятой
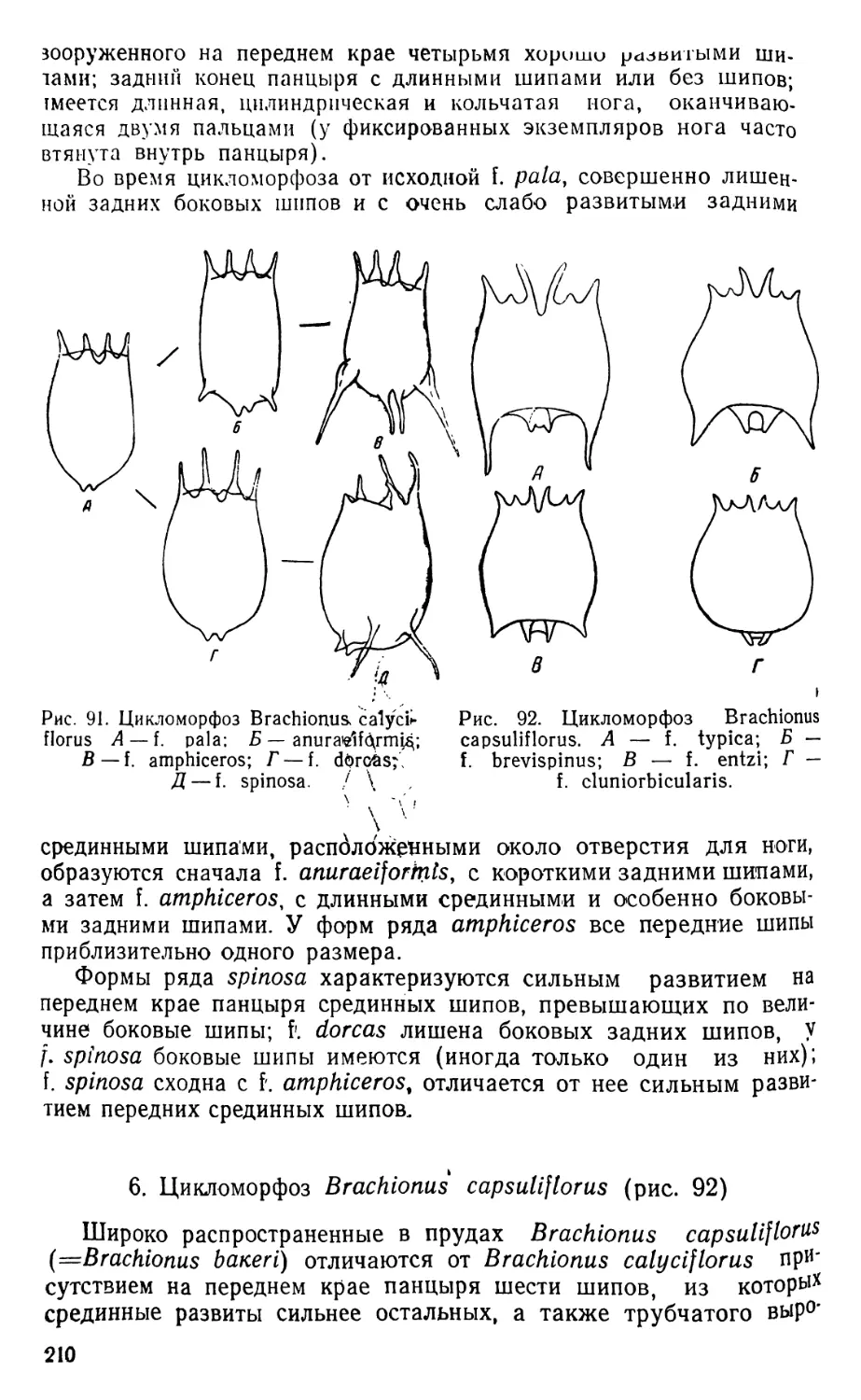
Наиболее обычные виды Diaptomus различаются между собою
следующими признаками (в основу таблицы положено строение
ног пятой пары самцов) (техника препарирования конечностей ука-
зана на стр. 259).
1 (2) Хватательный коготь на эксоподите правой ноги сильно
изогнут, при основании колбовидно вздут (пелагиаль
озер) .......................................D. graciloides
2 (1) Хватательный коготь при основании лишен воздутия. . 3
3 (4) На конечном членике эксоподита левой ноги находится
тонкий придаток с пучком волосков на конце (пелагиаль
озер).......................................D. gracilis
4 (3) Указанного придатка с волосками не имеется ->
5 (6) Боковой шип на последнем членике эксоподита правой
ноги прикреплен выше середины наружного края члени-
ка и направлен кзади (мелкие, часто пересыхающие
водоемы) ........................................D. castor
% 6 (5) Указанный шип прикреплен ниже середины наружного
края членика.............................................7
7 (8) Последний членик эксоподита левой ноги тонкий, с длин-
ным и сильно изогнутым тонким придатком (преимуще-
ственно пруды).............................D. coeruleus
8 (7) Указанный членик вздутый, на конце с двумя неболь-
шими придатками (мелкие пересыхающие водо-
емы) . . D. amblyodon
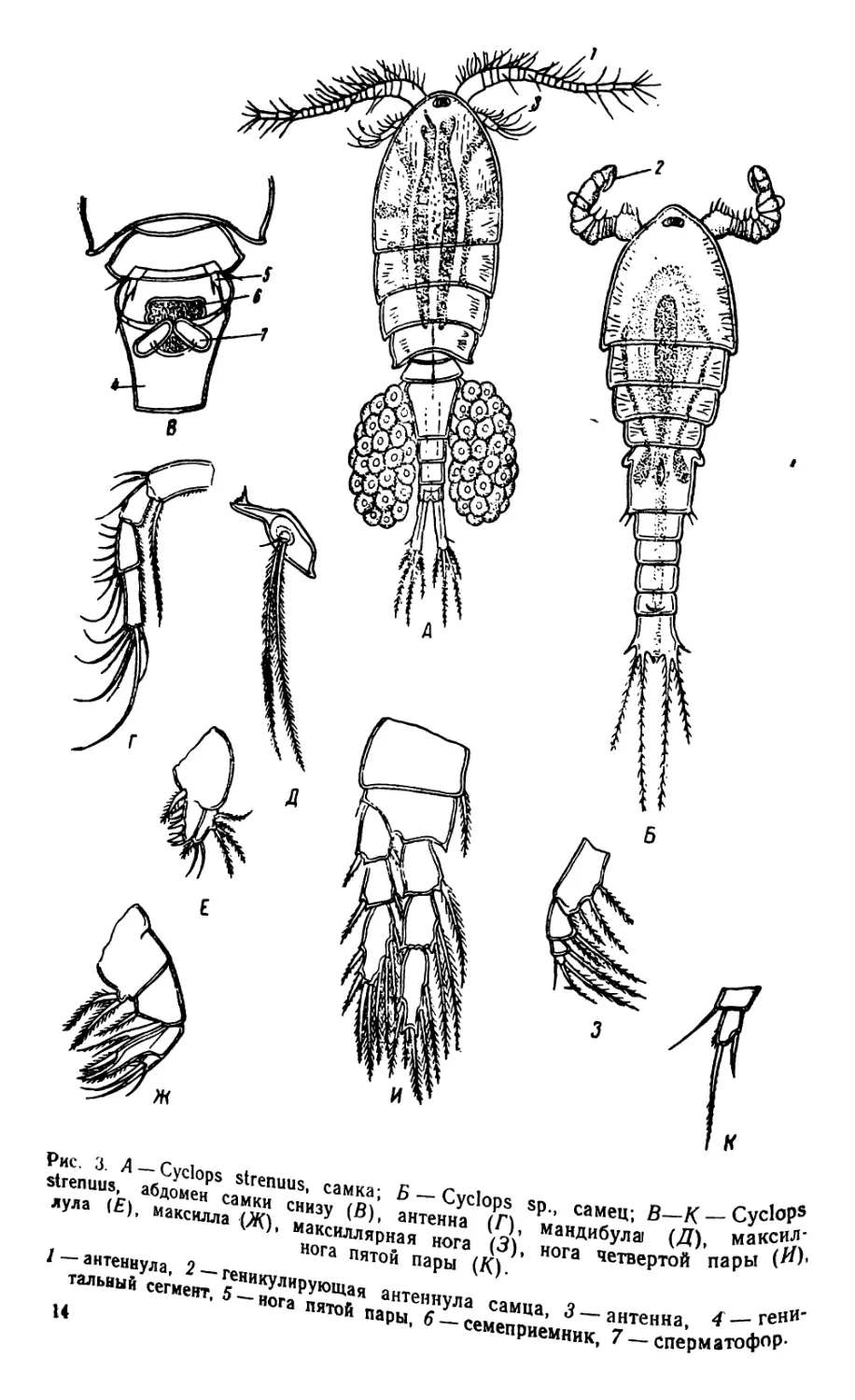
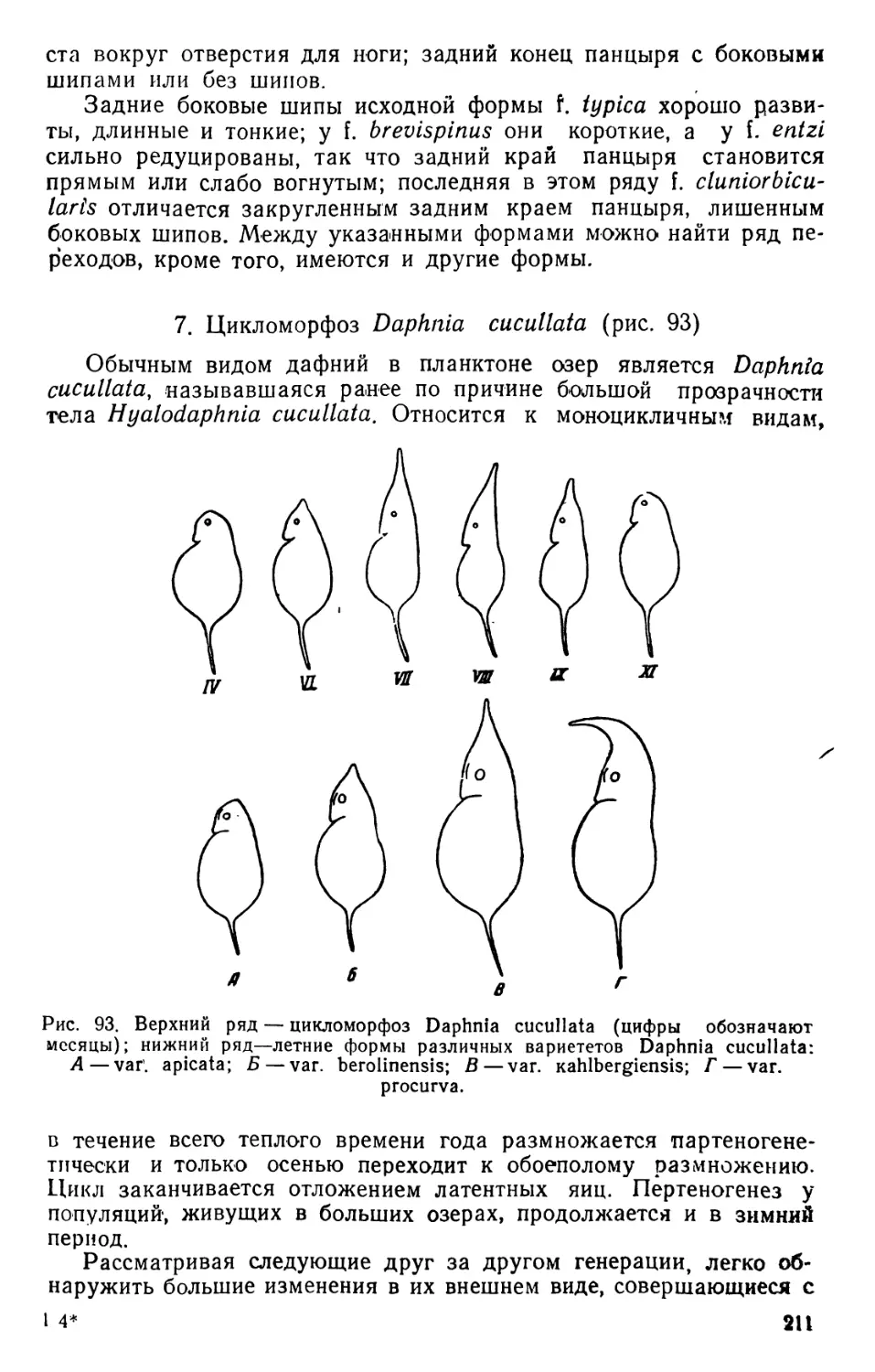
2. Cyclops (рис. 3—4)
Циклопы отличаются от диаптомусов рядом признаков. Перед-
ний отдел тела состоит из головы и трех грудных сегментов; брюшко
самок состоит из пяти сегментов. Антеннулы-короткие, не длиннее
двух третей переднего отдела тела. Обе антеннулы самцов превра-
щены в гейикулирующие органы. Антенны одноветвистые, без эк-
соподитов. Грудные ноги первых четырех пар с трехчленистыми
эндоподитами и эксоподитами. Ноги пятой пары рудиментарные,
одинакового строения у обоих полов. Сердце отсутствует. Самки
вынашивают яйца в двух яйцевых мешках, прикрепленных по бо-
кам генитального сегмента. В настоящее время прежний род Сус-
lops разделен на несколько новых родов (Cyclops s. str., Macrocyc-
lops, Eucyclops, Mesocyclops, Acanthocyclops и др.).
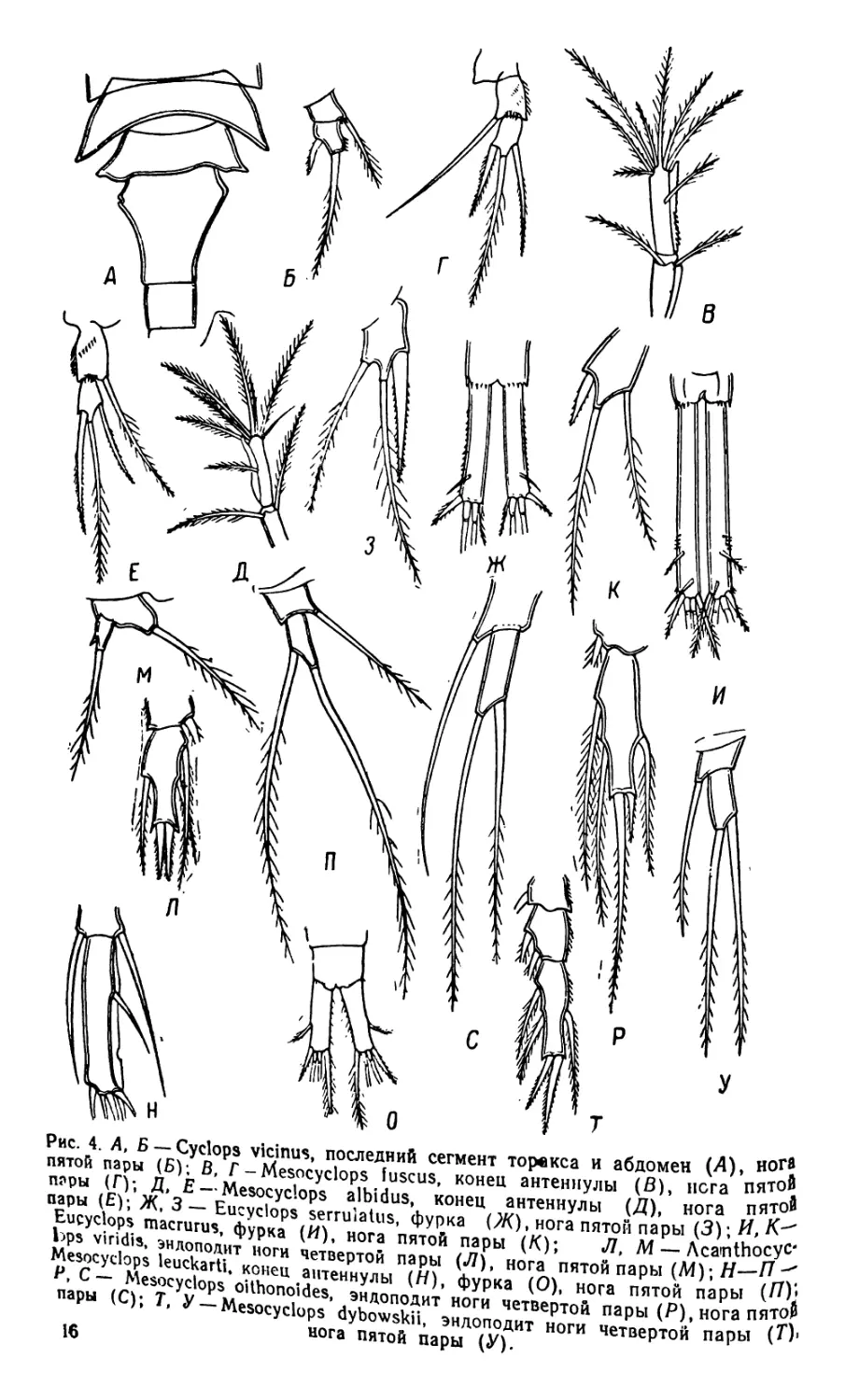
Наиболее обычные виды Cyclops различаются между собою
следующими признаками (в основу таблицы положено строение
ног пятой пары у самок).
1 (8) Последний членик ног пятой пары стремя придатками. 2
2 (5) Антеннулы 17-членистые . 3
самца (К); Л—Н — Diaptomus amblyodon, последний сегмент торакса и первый
сегмент абдомена самки (Л), нога пятой пары самки (Л4), ноги пятой пары
самца (Я); О — парение и скачок Diaptomus; П — скольжение Diaptomus.
О
Рис. 3. Д —Cyclops strenuus, самка; Б — Cyclops sp., самец; В—К —
strenuus, абдомен самки снизу (В), антенна (Г), мандибула) (Д), маКС/Ш
лула (Е), максилла (Ж), максиллярная нога (3), нога четвертой пары I
нога пятой пары (К).
тальаыйУсегмент Г^НИКуЛИруЮЩая антеннУла самца, 3 —антенна, < "^.ГД
» нога пятой пары, 6 — семеприемник 7 — сперматоф°Р’
U
о /4) Пластинка вдоль края последнего членика антеннул с
глубокими вырезами .... Mesocyclops fuscus
4 (3) Указанная пластинка гладкая . . Mesocyclops albidus
5 (2) Антеннулы 12-членистые............................6
6 (7) Внешние края фуркальных ветвей вооружены крупны-
ми шипиками.............................Eucyclops serrutatus
7 (6) Внешние края фуркальных ветвей гладкие, с одной
группой шипиков .... Eucyclops macrurus
8 (1) Указанный членик с двумя придатками .... 9
9 (14) Последний членик ног пятой пары с одним длинным и
одним коротким шипами .... 10
10 (11) Первый членик ног пятой пары широкий . . . .
Acanthocyclops viridls
11 (10) Указанный членик узкий, цилиндрический ... 12
12 (13) Боковые задние углы последнего грудного и первого
брюшного сегментов короткие . . Cyclops strenuus
13 (12) Боковые задние углы последнего грудного сегмента и
первого брюшного сегмента вытянуты в длинные за-
остренные выросты..........................Cyclops vicinus
14 (9) Указанный членик с двумя длинными шетинками . 15
15 (16) Вдоль последнего членика антеннул имеется пластинка
с глубокой вырезкой .... Mesocyclops leuckarti
16 (15) На последнем членике антеннул пластинки не
имеется ................................................17
17 (18) Последний членик эндоподита ног четвертой пары во-
оружен одним коротким и одним длинным шипами
Mesocyclops oithonoides
18 (17) Указанный членик вооружен двумя короткими, почти
равной длины шипами . Mesocyclops dybowskii
* *
Наблюдения над движением веслоногих раков производятся
следующим образом. Помещают несколько живых экземпляров
диаптомусов или циклопов в узкий, с параллельными стенками
стеклянный сосуд. После некоторого времени, когда животные успо-
коятся, наблюдают их движения через сильную большого диаметра
лупу. Траекторию движения можно зарисовать на клетчатой бу-
маге, поместив другой лист клетчатой бумаги за аквариумом.
В состоянии парения диаптомусы висят на распростертых в
стороны антеннулах, которые благодаря своей длине и большому
количеству щетинок представляют значительное сопротивление во-
де. Медленное опускание, продолжающееся у диаптомусов до тех
пор, пока организм не опустится на несколько сантиметров, сме-
няется резким' скачком, направленным косо вверх и вызванным
синхронными ударами грудных (плавательных) ног и брюшка; ан-
теннулы во время скачка прижимаются к телу и в движении .не
участвуют. К концу подъема продольная ось животного распола-
гается горизонтально. Затем антеннулы расправляются, и висящий
на них организм возвращается в исходное положение. Кроме этого
15
Рис. 4 А, Б —Cyclops vicinus, последний сегмент торвкса и абдомен И) ’ПяТОй
пятой пары (Б); В, Г-Mesocyclops fuscus, конец антеннулы (S), иста П
пары (Г); Д, Е - Mesocyclops albtdus, конец антеннулы (Д), нога п
пары (Е); Ж, 3 — Eucyclops serrulatus, фурка (Ж), нога пятой пары (
Eucyclops macrurus, фурка (И), нога пятой пары (К); Л, "7 лл v И—
bps viridis, энлополит ноги четвертой пары (Л), нога пятой пары (М);л
Mesocyclops leuckartk конец антеннулы (Я), фурка (О), нога пятой пары к Я
Р» С — Mesocyclops oithonoides, эндоподит ноги четвертой пары (Р), нога пято?
пары (С); Т, У — Mesocyclops dybowskii, эндоподит ноги четвертой пары (*)'
нога пятой пары (У).
способа движения, диаптомусы могут длительное время медленно
скользить на одном и том же уровне по спирали или по кругу.
Органами движения в этом случае являются антенны, мандибу-
лярный щупик и максиллулы, ударами которых вызывается силь-
ный ток воды; аитеннулы играют при этом роль рулей.
Циклопы являются пловцами. Брюшко их относительно длиннее,
чем у диаптомусов; антеннулы короткие. Движения циклопов,
имеющие характер «прыжков», вызываются, главным образом, уда-
рами грудных (плавательных) ног. Обе ноги каждой пары соеди-
нены между собою хитиновой пластинкой, поэтому они двигаются
одновременно; при быстром ударе ног циклопы делают порыви-
стые скачкообразные движения, которые могут быть направлены
в любую сторону. Как только плавательные ноги прекращают свою
работу, циклопы довольно быстро опускаются вниз, принимая по-
чти вертикальное положение.
Движение при помощи антенн
Наблюдения ведутся над живыми дафниями {Daphnia) и бос-
минами {Bosmina), относящимися к подотряду ветвистоу£ых раков
(Cladocera) отряда жаброногих раков (Branchiopoda).
Прежде всего следует рассмотреть строение дафнии и босмины.
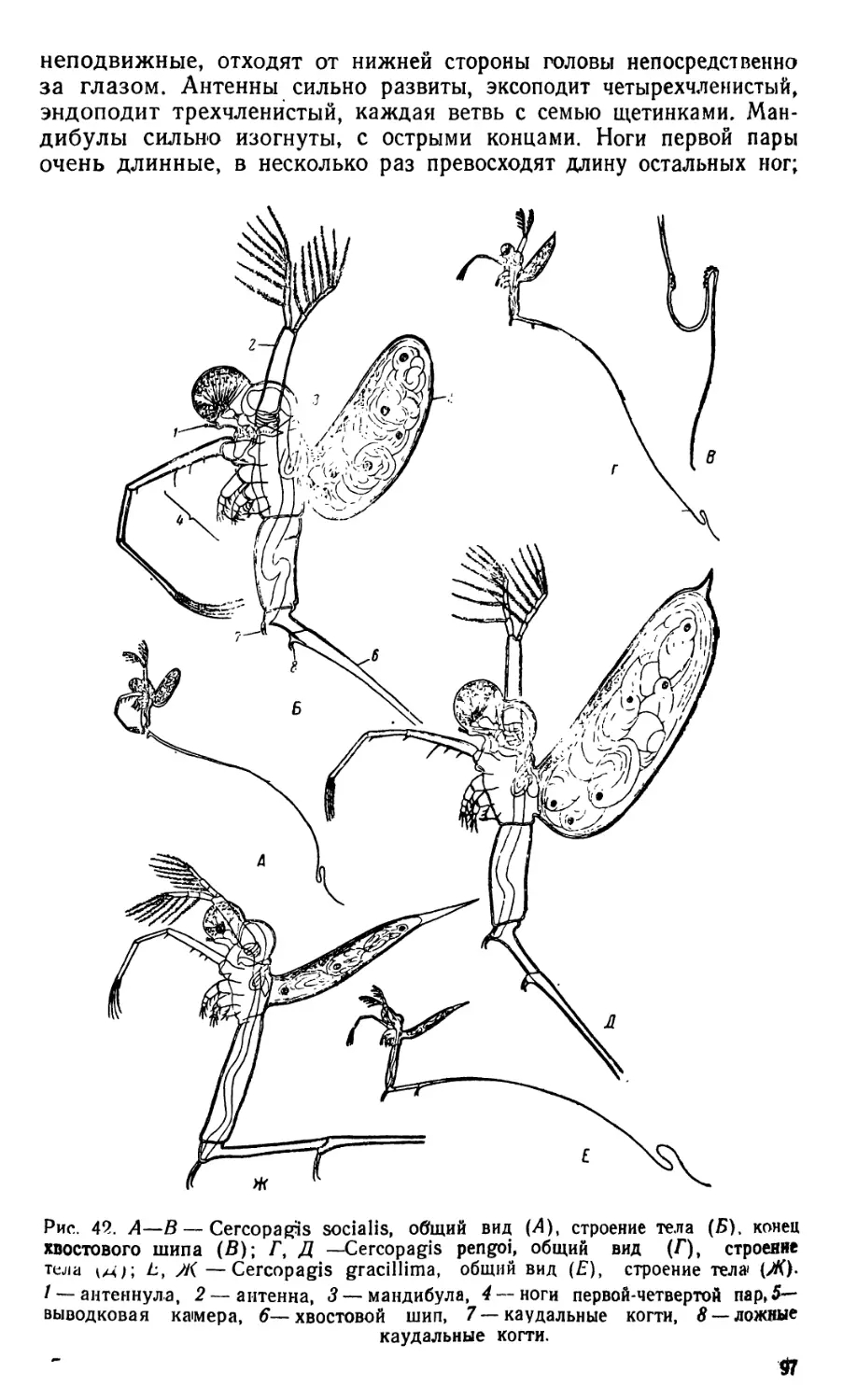
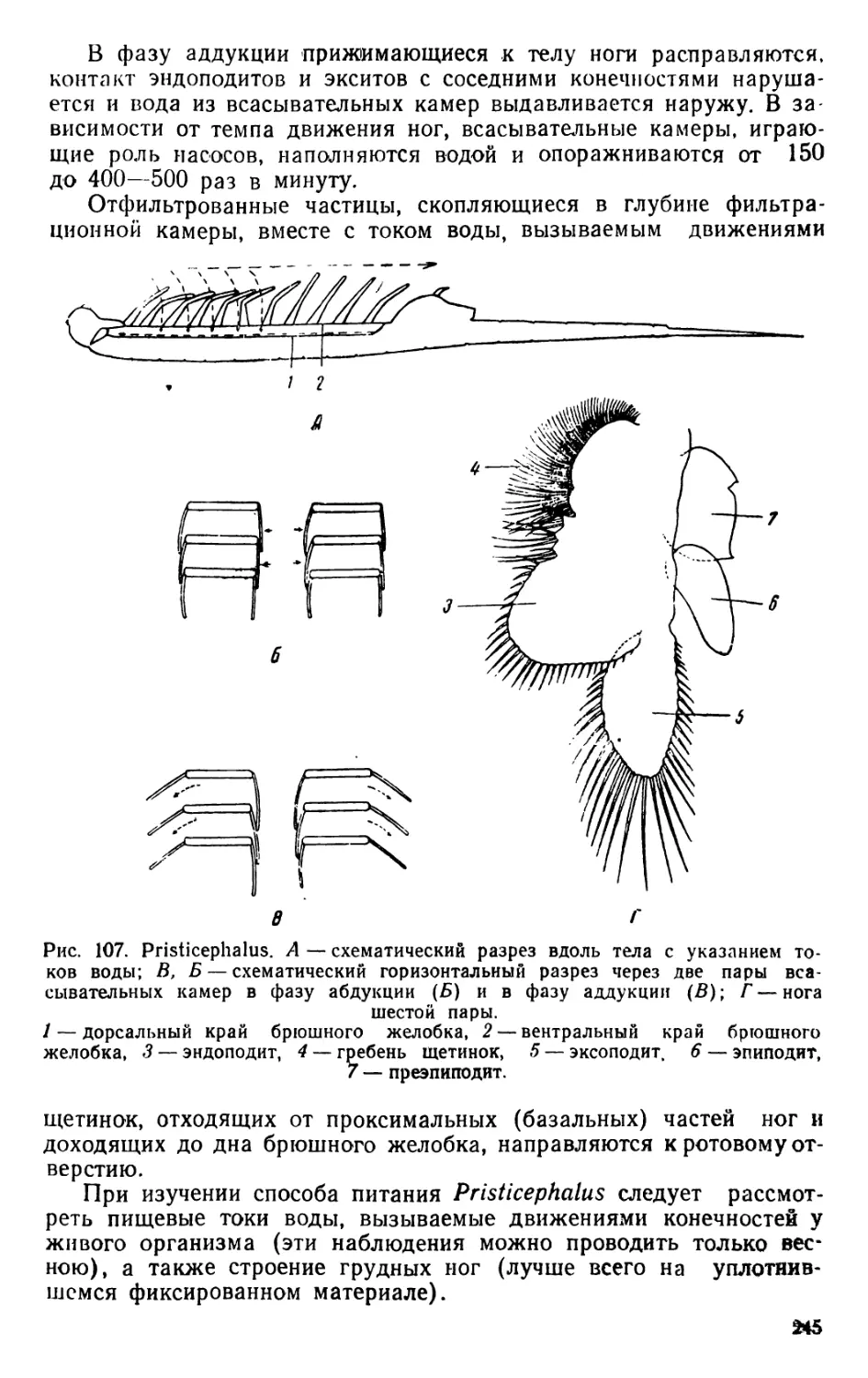
1. Daphnia (рис. 5—6 и 8)
Тело дафнии сжато с боков и все покрыто, за исключением го-
ловы, двустворчатой раковиной (карапаксом). Внутри раковины
тело помещается свободно, прикрепляясь к ней только передней ча-
стью. Между стенками створок раковины и спинной поверхностью
животного у самок расположена выводковая (зародышевая) каме-
ра. Край раковины на спинной стороне вытянут в длинный шип
(spina). Голова согнута на брюшную сторону, лобная часть вытя-
нута в направленный вниз и назад рострум. Большой срединный
глаз, образованный слиянием эмбрионально закладывающихся двух
сложных глаз, снабжен специальной мускулатурой, обусловливаю-
щей его подвижность внутри глазной камеры. У некоторых видов
дафний имеется, кроме того, небольшой непарный глазок. Короткие
антеннулы, являющиеся органами чувств, сочленены у самок непо-
движно с головой позади рострума; на конце они несут пучок чув-
ствительных щетинок.
Антенны, являющиеся единственным органом движения, хорошо
развиты, состоят из большого протоподита и двух ветвей, наруж-
ной, или эксоподита, и внутренней, или эндоподита, вооруженных
длинными оперенными плавательными щетинками. Внутри антенн
видна сильная мускулатура. Каждая антенна приводится в движе-
ние тремя абдукторами, тремя аддукторами и одним леватором; два
первых абдуктора и леватор своими бросающимися в глаза вееро-
образно расширенными концами прикреплены’к хитину спинной ча-
сти головы. Подъем антенны совершается вследствие сокращения
аддукторов и леватора, абдукторы являются их антагонистами.
Мандибулы, хорошо заметные у живых организмов по своеоб-
разным движениям, лишены щупика, на внутренней стороне с болы
2 В- А. Яшнов. 7
, 4 — рострГ
печеночный ®
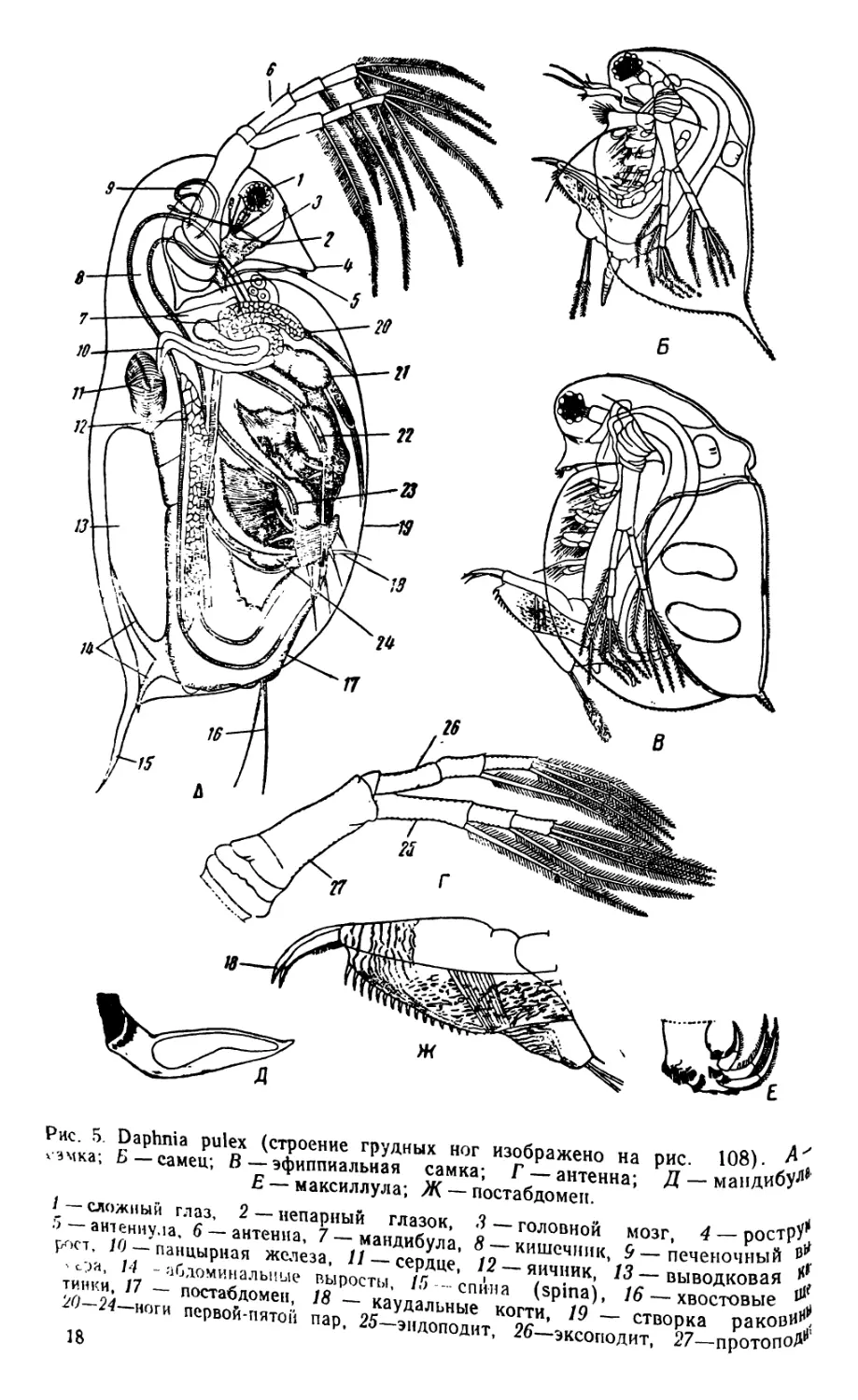
Рис. 5. Daphnia pulex (строение грудных ног изображено на рис. 108).
гэмка; Б — самец; В — эфиппиальная самка; Г — антенна; Д — мандиоу
Е — максиллула; Ж — постабдомеп.
/—сложный глаз, 2 —непарный глазок, 3—головной мозг
5— антеннула, 6 — антенна, 7 — мандибула, 8 — кишечник, 9—
рост, 10 — панцирная железа, // — сердце, /2 —яичник, 13—выводковая
. с;га, 14 -абдоминальные выросты, 15 - спина (spina), 16 — хвостовые
тинк.и, 17 — постабдомеп, 18 — каудальные когти, 19 — створка раков#*,
2)~24 ноги первой-пятой пар, 25—эндоподит, 26—эксоподит, 27—протопоп
18
шой жевательной поверхностью, состоящей из хитиновых пласти-
нок, вооруженных шипиками. Максиллулы редуцированы. Максил-
лы полностью отсутствуют. Всего имеется пять пар грудных ног,
являющихся тургорными конечностями.
На спинной стороне брюшка (абдомена) находится несколько
абдоминальных выростов, служащих для замыкания сзади вывод-
ковой камеры; позади этих выростов расположена пара оперенных
плавательных или хвостовых щетинок. Задний отдел брюшка поза-
ди хвостовых щетинок называется постабдоменом, или каудой. Он
подогнут под тело организма так, что его верхний, или спинной,
край, на котором открывается анальное отверстие и располагаются
анальные зубчики, становится как бы нижним краем; на конце
постабдомена находится фурка, образованная двумя крепкими и
слегка изогнутыми когтями, называемыми фуркальными, или кау-
дальными, когтями.
Рот, ограниченный сверху верхней губой, а с боков мандибу-
лами, ведет в короткий пищевод, переходящий в длинную среднюю
кишку, одинакового диаметра на всем своем протяжении; в перед-
ней части средней кишки находятся парные печеночные выросты,
имеющие вид коротких и изогнутых слепых придатков, располагаю-
щихся в голове.
Сердце имеет вид округлого мешка с одной парой боковых ос-
тий. Сокращения сердца совершаются с большой быстротой, при
комнатной температуре—'до 200—290 ударов в минуту, что явля-
ется пределом для животных. Кровь из сердца поступает в лакуны
тела. Осмотическое давление крови при нормальных условиях рав-
няется двум-четырем атмосферам. Это обстоятельство имеет боль-
шое значение для сохранения формы тела и упругости конечностей
(тургорные конечности).
Органами дыхания служат эпиподиты конечностей, которые
омываются токами воды, вызываемыми ритмическими движениями
ног.
В качестве органов выделения служат парные панцырные (скор-
луповые) железы, расположенные в толще створок раковины, ме-
жду наружным и внутренними слоями, и хорошо заметные после
отделения створок от тела дафнии.
Хорошо заметный головной мозг состоит из двух слившихся по-
ловин. От его передней части отходят нервы к сложному глазу, а
на нижней стороне часто находится непарный (науплиевый) глазок.
Органы размножения самок простираются по бокам кишечника
от первой пары ног до постабдомена; короткие яйцеводы открыва-
ются на спинной стороне тела в задней части выводковой камеры.
Все эмбриональное развитие молоди протекает в выводковой
камере в период между двумя линьками. Дафнии образуют парте-
ногенетические и латентные яйца. Последние заключены внутри
сильно видоизмененной и темноокрашенной верхней части створок
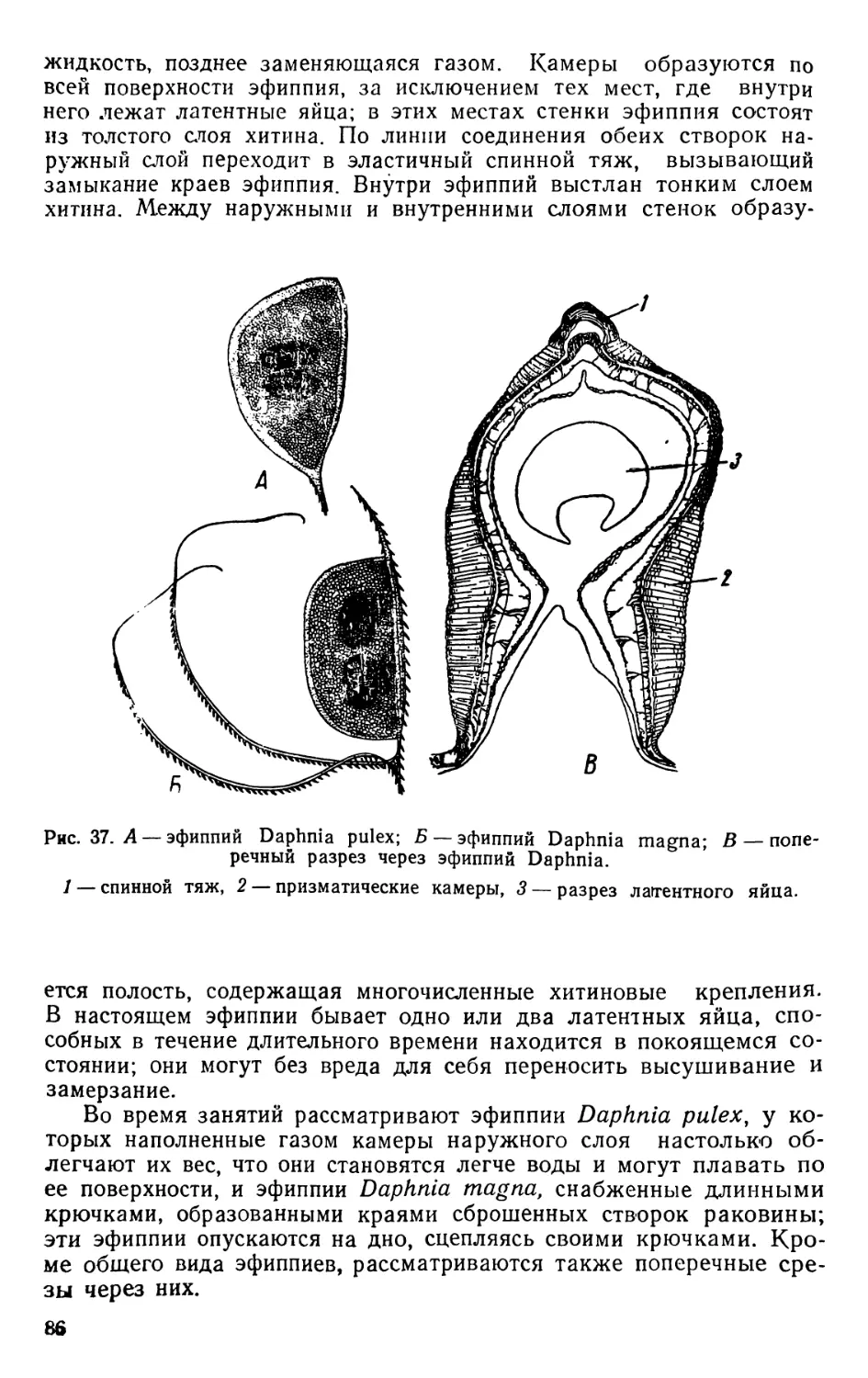
раковины, образующей так называемый эфиппий (седлышко).
Самцы дафний отличаются от самок отсутствием выводковой
камеры, наличием больших и подвижных аитеннул и иным воору-
жением пог первой пары.
19
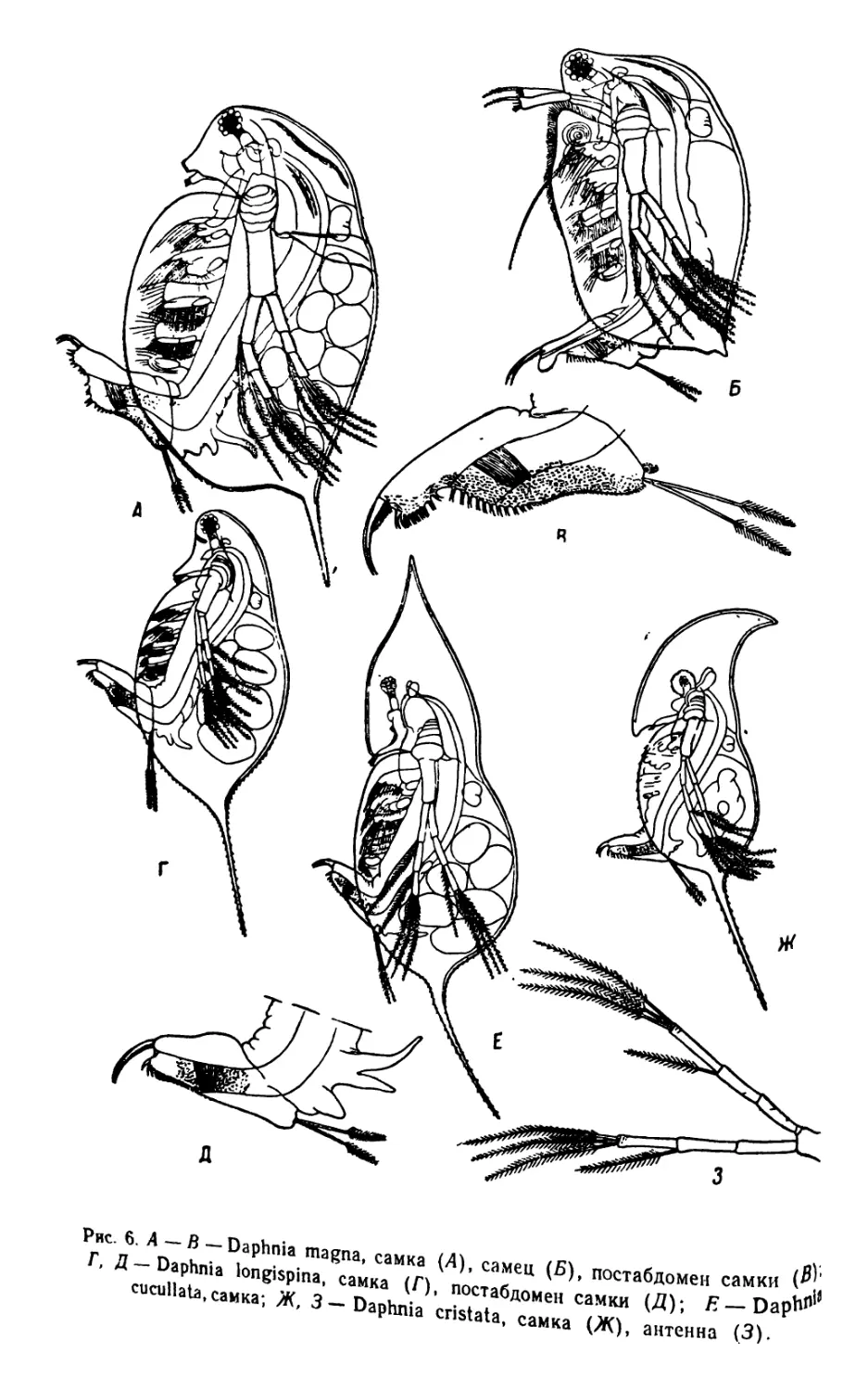
Рис. 6. А —В—Daphnia magna, самка (А), самец (Б), постабдомен самки
Г, Д —Daphnia longispina, самка (Г), постабдомен самки (Д); Б — Dap»1
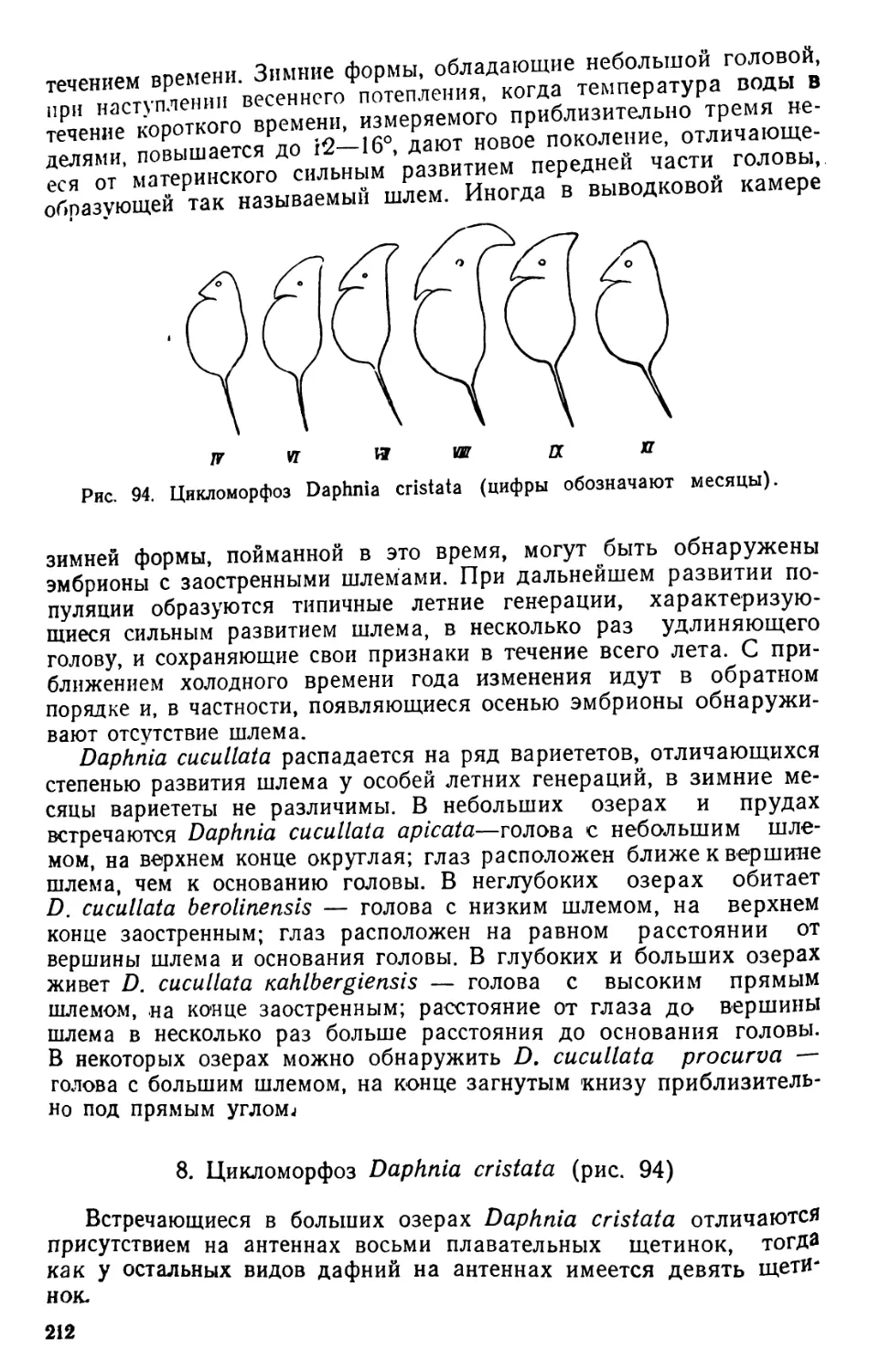
cucullata, самка; Ж, 3 — Daphnia cristata, самка (Ж), антенна (3).
Пять самых обычных видов Daphnia различаются между Ьобою
следующими признаками.
1 (4) Каудальные когти при основании вооружены крупными
шипами................................................... 2
2 (3) Верхний (спинной край) постабдомена прямой, без
выемки.................................... D. pulex
3 (2) Верхний край постабдомена с выемкой . . D. magna
4 (1) Каудальные когти при основании лишены крупных
шипов .............................................. ... 5
5 (6) Непарный глазок имеется . D. longispina
6 (5) Непарный глазок отсутствует.........................7
7 (8) Рострум острый, щетинки антеннул не заходят за
его конец..................... . ... D. cristata
8 (7) Рострум тупой, щетинки антеннул заходят за его
конец D. cucullala
2. Bosmina (рис. 7—8)
Тело босмины заключено в раковину) нижне-задние углы кото-
рой вытянуты в выросты, называемые мукро (mucro). Эти выросты
у отдельных форм различно развиты, в редких случаях полностью
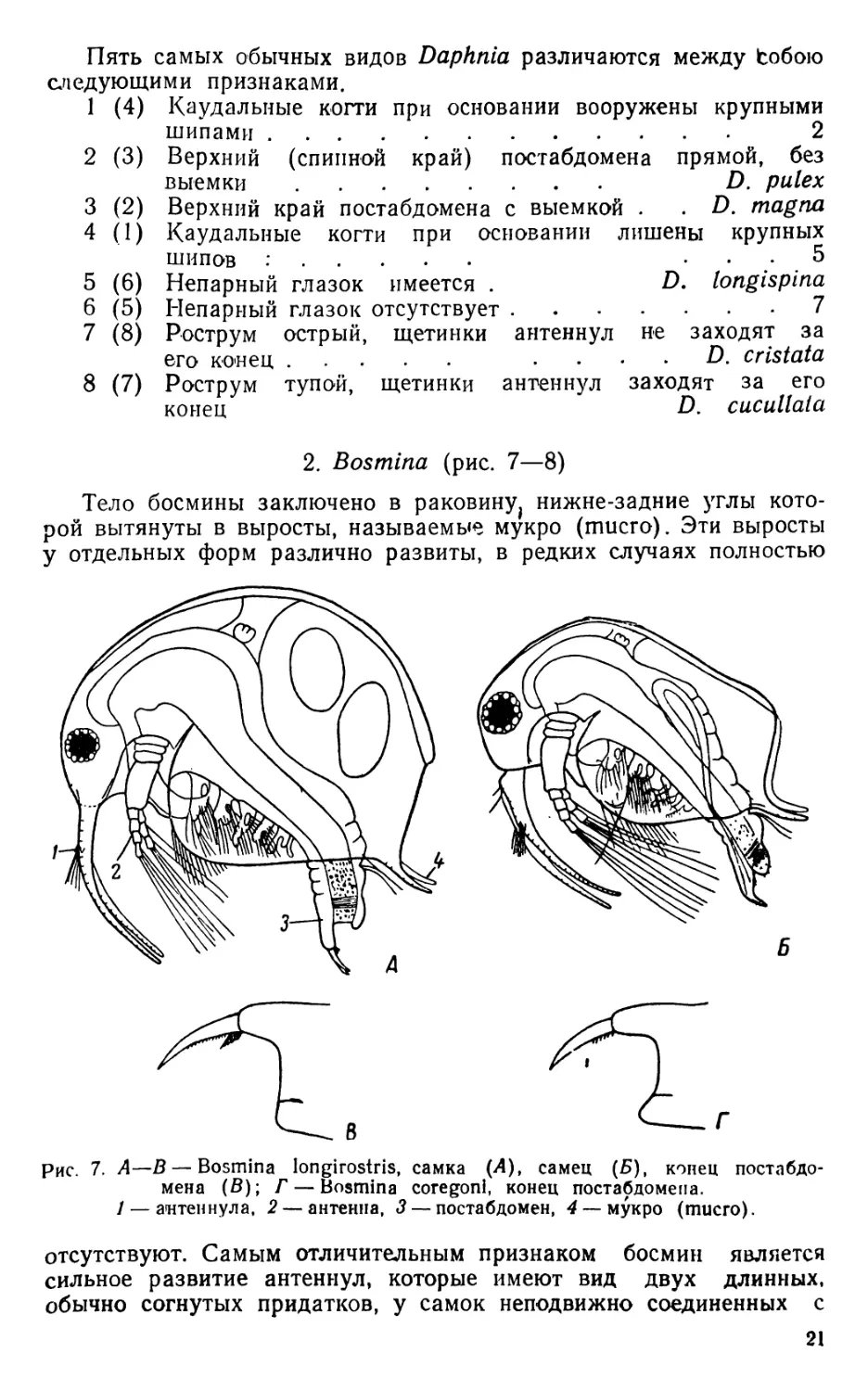
Рис. 7. А—В — Bosmina longirostris, самка (Л), самец (Б), конец постабдо-
мена (В); Г — Bosmina coregoni, конец постабдомена.
1 — антеннула, 2 — антенна, 3 — постабдомен, 4 — мукро (mucro).
отсутствуют. Самым отличительным признаком босмин является
сильное развитие антеннул, которые имеют вид двух длинных,
обычно согнутых придатков, у самок неподвижно соединенных с
21
рострумом головы, у самцов подвижных; чувствительные щетинки
расположены около середины антеннул; ряды шипиков, проходящих
поперечно, придают антеннулам вид членистости. Антенны корот.
кие, с трехчленистыми эндоподитами и четырехчленистыми эксопо-
дитамн. Грудные ноги, в числе шести пар, — различного' строения;
ноги шестой пары имеют вид пальцевидных придатков. Постабдомен
короткий, сильно сжатый с боков, на конце как бы обрубленный, на
брюшной стороне с длинным выростом, несущим большие фур.
кальные (каудальные) когти. Глаз умеренной величины, глазок от.
сутствует. Кишечник без печеночных выростов. Эфиппий неболь-
шой.
Босмины, так же как дафнии, распадаются на очень большое
количество отдельных форм, часто сильно различающихся между
собою.
В настоящее время все эти формы сведены в два вида, отлича-
ющиеся по строению каудальных когтей-
Bosmina coregoni—базальная часть каудальных когтей воору-
жена рядом мелких зубчиков, заходящих также на вырост постаб-
домена; вдоль всего края коготков находятся очень небольшие ще-
тинки.
Bosmina longirostris — базальная часть каудальных когтей во-
оружена рядом щетинок, дистально увеличивающихся в длину; вы-
ше щетинок находятся небольшие шипики.
* *
* •
Наблюдения над движением дафний и босмин производятся та-
ким же способом, как над веслоногими раками.
Движение дафнии происходит в две фазы (см. рис. 8). Во время
первой фазы дафния под действием удара антенн делает скачок
косо вверх; во время второй фазы медленно опускается вниз на
распростертых в разные стороны антеннах. Удар антеннами произ-
водится не строго вдоль продольной оси животного, а книзу и кза-
ди, вследствие этого скачок, совершаемый косо вверх, сопровожда-
ется опусканием головы и подъемом задней части тела. Центр тя-
жести дафний лежит ниже места прикрепления антенн, поэтому ор-
ганизм, опускаясь на распростертых антеннах, одновременно пово-
рачивается вокруг соединяющей их оси и возвращается в исходное
положение; этому вращению в известной степени препятствует со-
противление, оказываемое воде головой и расположенным на конце
створок шипом (spina). В зависимости от направления удара ан-
тенн, а также от длительности фазы покоя, общее направление
пути движения дафний может быть горизонтальным или же идти
под некоторым углом вверх или вниз. Образование шлема, наблю-
даемое у пелагических дафний (Daphnia cucullata, D. cristata), рас-
сматривается в качестве приспособления к сохранению горизонталь-
ного направления движения. С одной стороны, большая голова
сдвигает центр тяжести организма ближе к месту прикрепления ан-
22
тенн, с другой же стороны, большой шлем и длинная spina, ока-
зывая воде сильное сопротивление, являются стабилизаторами, пре-
пятствующими значительному изменению положения тела между
двумя ударами7 антенн.
Босмины двигаются в воде спинной стороной кверху (см. рис.
8). При ударе антенн, направляемом, в противоположность дафни-
ям, книзу и кпереди, головной конец босмины поднимается вверх,
Рис. 8. 1 — Daphnia в фазу опускания, вид сверху; 2 — фазы движения моло-
дой Daphnia; 3 — изменение курса движения Daphnia: в зависимости от дли-
тельности паузы между ударами амтенн; 4—фазы движения Bosmina (пере-
вертывания через голову, фаза III, не происходит благодаря сопротивлению,
оказываемому антеннулами).
ап — место прикрепления антенн, стрелка указывает направление толчка, вы-
зываемого ударом антенн, о, и \—действие толчка на переднюю и заднюю
части тела, g —центр тяжести, 4^ и g —действие силы тяжести на переднюю
и заднюю части тела.
а задняя часть тела опускается вниз. Центр тяжести босмпн нахо-
дится на одном уровне с местами прикрепления антенн, а иногда и
выше их, поэтому вращение организма, вызываемое силой тяже-
сти, действует в том же направлении, как и удар антенн; это дол-
жно было бы привести к перевертыванию животного через голову,
если бы не то большое сопротивление, которое оказывает воде уве-
личенная, в результате сильного развития антеннул, нижняя поло-
вина организма- По своему значению антеннулы являются рулями.
Ампутация их вызывает при плавании перевертывание организма
через голову. У некоторых босмин стабилизатором служат большие
выросты (mucro), находящиеся на конце створок раковины п вытя-
нутые вдоль продольной оси животного.
23
Движение при помощи коловращательного аппарата (рис. 9)
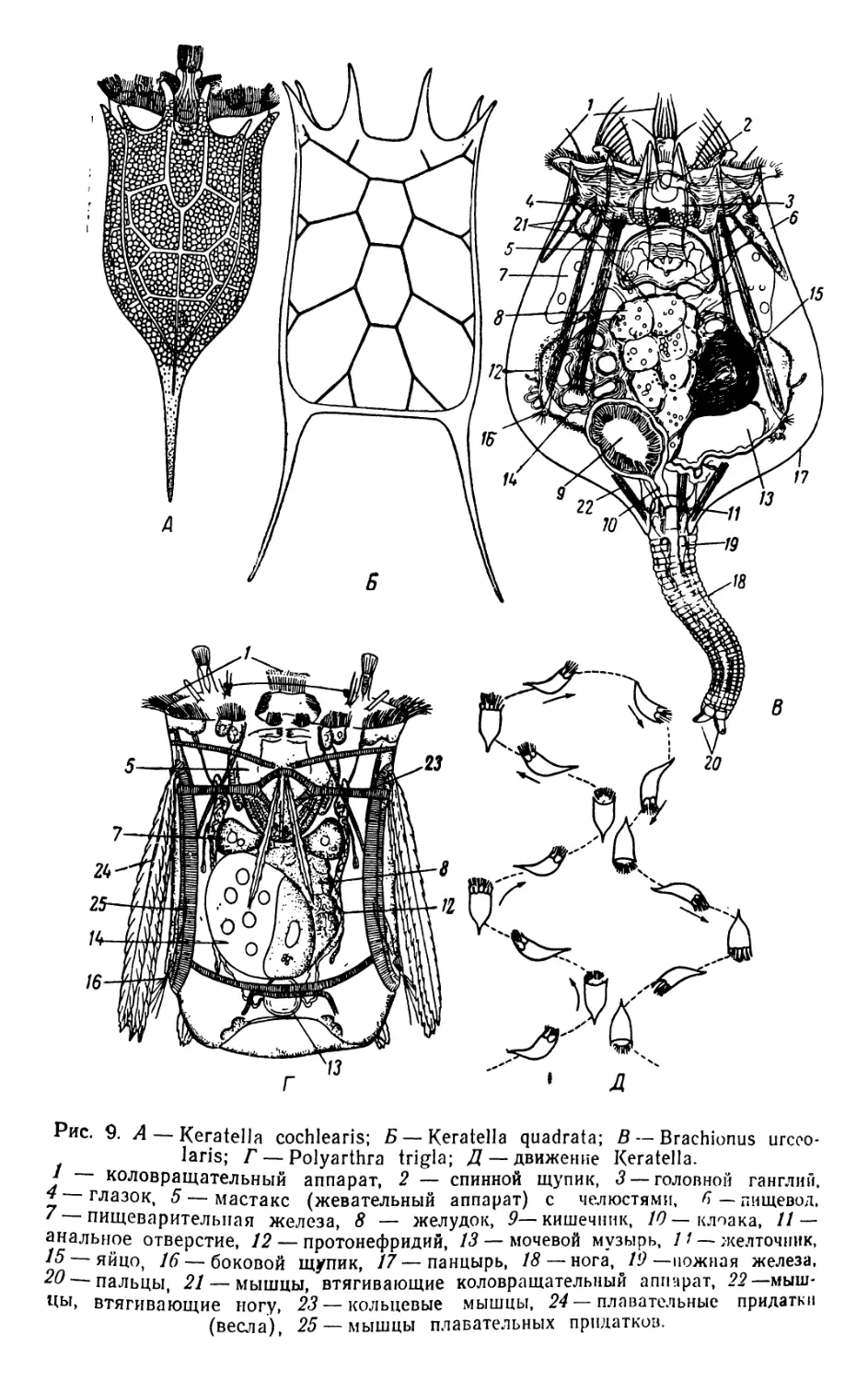
В большинстве случаев коловратки (Rotatoria) ведут свободный
образ жизни. Органом движения служит так называемый коловрд.
щательный аппарат, расположенный на переднем конце тела. Этот
орган в схеме состоит из ресничного ротового (буккального) поля,
окружающего ротовое отверстие, и ресничного циркумапикального
пояса, расположенного вокруг переднего конца головы; самая тер.
минальная часть головы (апикальное поле) лишена ресничек, несет
органы чувств. Строение коловращательного аппарата у представи-
телей различных семейств коловраток сильно варьирует. Нередко
ресничный цпркумапикальный пояс распадается на два кольца,
верхнее (trochus) и нижнее (cingulum); также может изменяться
расположение ресничек на ротовом поле.
Для изучения работы коловращательного аппарата наблюдают
живых коловраток, например Keratella (=Апигаёа) или Brachionus
под микроскопом в воде, к которой прибавлено немного гуммиара-
бика. Еще убедительнее эти наблюдения вести под микроскопом
или бинокуляром, ориентированном горизонтально; организмы рас-
сматриваются в лунке предметного стекла, покровное стекло, во
избежание скольжения, смазывается по краям ланолином или ва-
зелином.
Плавание коловраток совершается под действием ударов ресни-
чек коловращательного аппарата, главное значение для движения у
рассматриваемых видов имеют реснички ротового поля. Движение
коловраток обычно совершается по спирали. Другой тип движения,
скачкообразный, можно наблюдать у Polyarthra, снабженных по
бокам тела весловидными придатками.
ПРИСПОСОБЛЕНИЯ К ПАРЕНИЮ
Уменьшение остаточного веса
Редукция скелета
В качестве примеров рассматриваются представители отряда
крылоногих моллюсков (Pteropoda), относящегося к классу брюхо-
ногих моллюсков. Все виды Pteropoda ведут планктический образ
жизни, в связй с этим раковина у них или полностью исчезает или
сильно редуцируется. Наиболее обычными видами являются Clione
limacina и Limacina helicina, часто встречающиеся в наших север-
ных морях, нередко в массовых количествах.
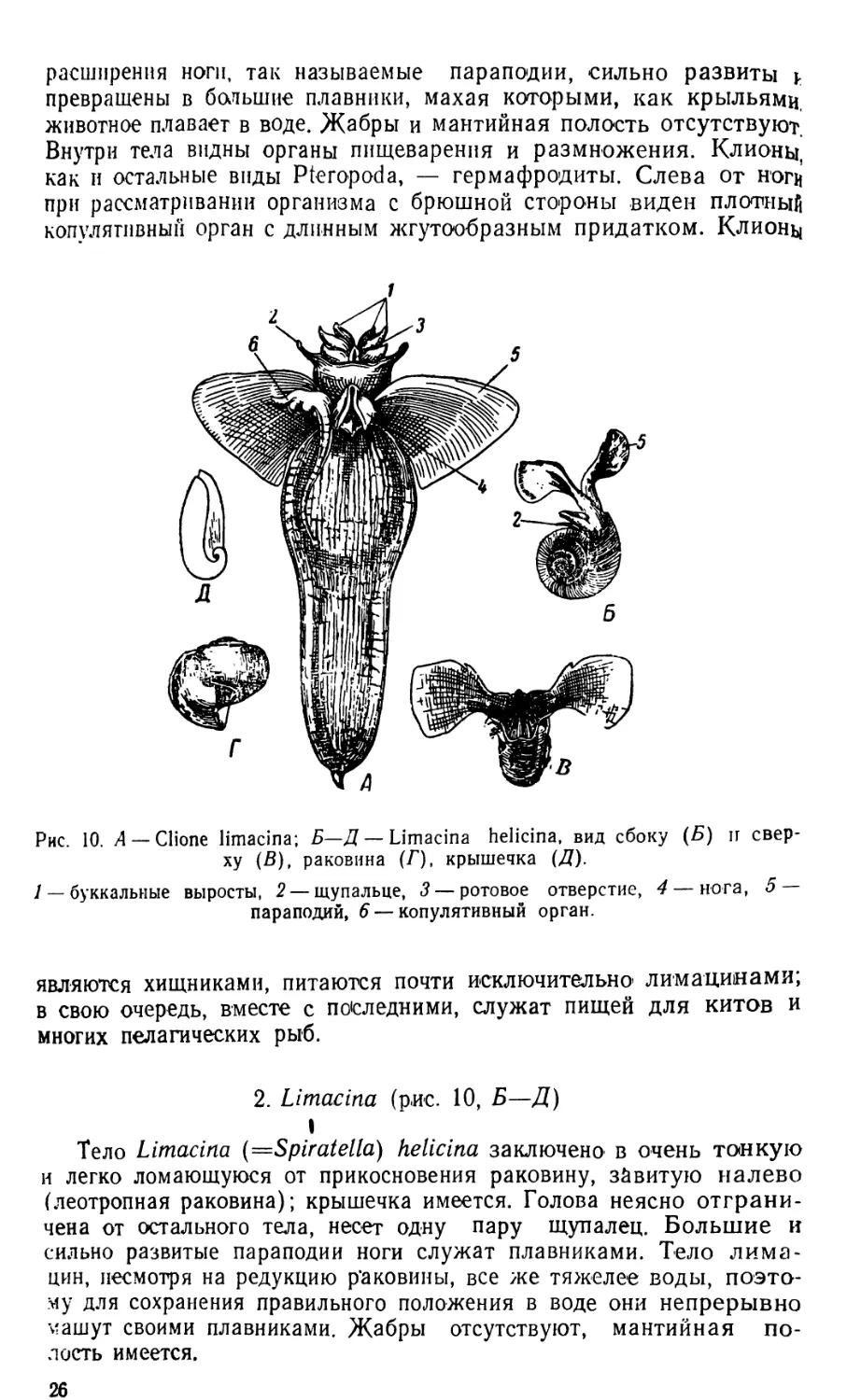
1. СНопе (рис. 10, Д)
Тело СНопе limacina лишено раковины, удлиненное, к концу
заостренное. Голова резко отделена от остального тела, несет две
пары щупалец и несколько пар буккальных выростов, расположен-
ных на вытягивающемся хоботе. На брюшной стороне, под головой,
находится рудиментарная нога, состоящая из трех частей; боковые
24
Рис. 9. А — Keratella cochlearis; Б — Keratella quadrata; В — Brachionus urcoo-
laris; Г — Polyarthra trigla; Д — движение Keratella.
I коловращательный аппарат, 2 — спинной щупик, 3— головной ганглий,
4 глазок, 5 — мастакс (жевательный аппарат) с челюстями, 6 — пищевод,
' пищеварительная железа, 8 — желудок, 9— кишечник, 10— клоака, 11 —
анальное отверстие, 12 — протонефридий, 13 — мочевой музырь, 1 / — желточник,
15— яйцо, 16 — боковой щупик, 17—панцырь, 18— нога, 19 —ножная железа,
20'—пальцы, 21 — мышцы, втягивающие коловращательный аппарат, 22—мыш-
цы, втягивающие ногу, 23 — кольцевые мышцы, 24—плавательные придатки
(весла), 25 — мышцы плавательных придатков.
расширения ноги, так называемые параподии, сильно развиты j.
превращены в большие плавники, махая которыми, как крыльямц
животное плавает в воде. Жабры и мантийная полость отсутствуют.
Внутри тела видны органы пищеварения и размножения. Клионы,
как и остальные виды Pteropoda, — гермафродиты. Слева от нощ’
при рассматривании организма с брюшной стороны виден плотный
копулятивный орган с длинным жгутообразным придатком. Клионы
Рис. 10. A — Clione limacina; Б—Д — Limacina helicina, вид сбоку (Б) и свер-
ху (В), раковина (Г), крышечка (Д).
1— буккальные выросты, 2 — щупальце, 3 — ротовое отверстие, 4 — йога, 5
параподий, 6 — копулятивный орган.
являются хищниками, питаются почти исключительно лимацинами;
в свою очередь, вместе с последними, служат пищей для китов и
многих пелагических рыб.
2. Limacina (рис. 10, Б—Д)
I
Тело Limacina (=Spiratella) helicina заключено в очень тонкую
и легко ломающуюся от прикосновения раковину, завитую налево
(леотропная раковина); крышечка имеется. Голова неясно отграни-
чена от остального тела, несет одну пару щупалец. Большие и
сильно развитые параподии ноги служат плавниками. Тело лима-
цин, несмотря на редукцию р’аковины, все же тяжелее воды, поэто-
му для сохранения правильного положения в воде они непрерывно
машут своими плавниками. Жабры отсутствуют, мантийная по-
лость имеется.
26
Пропитывание водой
В качестве примеров рассматриваются представители зеленых
водорослей (Eudorina), цистофлагеллат (Noctiluca), гидромедуз
(Bougainvillia) и ветвистоусых ражов (Holopedium), снаружи или
внутри тела которых находятся богатые водою студенистые веще-
ства.
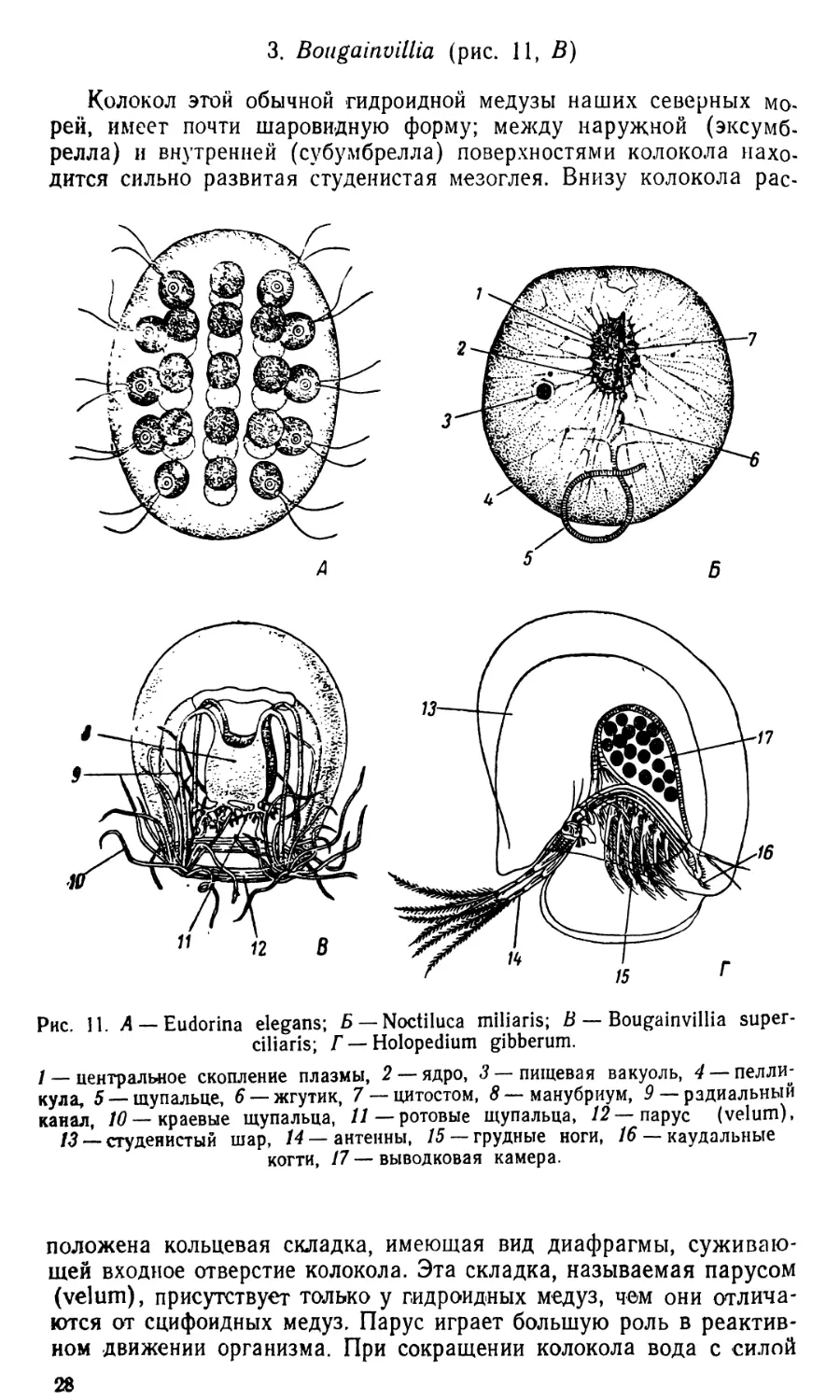
1. Eudorina (рис. 11, Л)
Эвдорина часто встречается в пресноводном планктоне, являет-
ся колониальным организмом. Относится к классу Volvocineae,
представители которых отличаются от всех остальных зеленых во-
дорослей подвижностью на вегетативной стадии. Клетки заключе-
ны, как и у других представителей семейства Volvocaceae, в студе-
нистую оболочку, имеющую эллипсоидальную форму. Колония со-
стоит из 32, реже из 16 шаровидных клеток, расположенных коль-
цеобразными рядами по периферии колонии. Каждая клетка снаб-
жена большим чашевидным хроматофором, пиреноидом, темным
глазком и двумя жгутами; под действием ударов жгутов, проходя-
щих сквозь оболочку, колонии двигаются в воде, вращаясь вокруг
своей длинной оси. Иногда можно видеть на различных стадиях
развития дочерние колонии, образующиеся в результате последова-
тельных делений материнских клеток.
Обычный вид — Eudorina elegans.
2. Noctiluda (рис. И, Б)
Ночесветка часто встречается в планктоне морей, у нас обычна
в Черном море. Свое название получила от способности светиться
под действием механического или иного какого-либо раздражения.
Тело ноктилуки приблизительно шаровидной формы, покрыто креп-
кой оболочкой. На одном полюсе находится перистом, в глубине
которого расположено щелёвидное ротовое отверстие, цитостом.
Короткий, трудно заметный жгутик отходит около цитостома, а не-
много далее — большое щупальце, по длине равное приблизительно
половине поперечника организма; параллельная исчерченность щу-
пальца зависит от параллельного расположения в нем тонких плаз-
матических тяжей. Плазма образует довольно большое скопление,
расположенное в области цитостома; от этого скопления плазмы от-
ходят по всем направлениям тонкие, между собою анастомозирую-
щие плазматические тяжи. Внутри клетки имеется большое количе-
ство клеточного сока, благодаря которому ее величина достигает
необычайных для простейших размеров, до двух миллиметров в по-
перечнике. Удельный вес ноктилук очень близок к удельному весу
воды и даже меньше его; опускание организма вызывается его сжи-
манием в результате выделения в воду клеточного сока. Ноктилу-
ки — хищники, питаются различными организмами планктона; за-
хватывание пищи совершается при помощи щупальца.
Единственный вид — Noctiluca miliaris.
27
3. Bougainvillia (рис. 11, В)
Колокол этой обычной гидроидной медузы наших северных мо-
рей, имеет почти шаровидную форму; между наружной (эксумб-
релла) и внутренней (субумбрелла) поверхностями колокола нахо-
дится сильно развитая студенистая мезоглея. Внизу колокола рас-
Рис. 11. Л —Eudorina elegans; Б — Noctiluca miliaris; В — Bougainvillia super-
ciliaris; Г — Holopedium gibberum.
/ — центральное скопление плазмы, 2 — ядро, 3 — пищевая вакуоль, 4 — пелли-
кула, 5 —щупальце, 6 — жгутик, 7 —цитостом, 8— манубриум, 9 — радиальный
канал, 10— краевые щупальца, 11—ротовые щупальца, 12 — парус (velum),
13 _ студенистый шар, 14 — антенны, /5 —грудные ноги, 16 — каудальные
когти, 17 — выводковая камера.
положена кольцевая складка, имеющая вид диафрагмы, суживаю-
щей входное отверстие колокола. Эта складка, называемая парусом
(velum), присутствует только у гидроидных медуз, чем они отлича-
ются от сцифоидных медуз. Парус играет большую роль в реактив-
ном движении организма. При сокращении колокола вода с силой
28
выбрасывается из его полости через суженное парусом отверстие,
под воздействием толчка медуза плывет вершиной колокола впе-
ред. От центра субумбреллы отходит желудок, манубриум, кубиче-
ской формы; на его конце находится ротовое отверстие, окруженное
четырьмя ротовыми щупальцами, делящимися дихотомически до
четырех — пяти раз. Выше щупалец по стенкам манубриума распо-
ложены четыре гонады. От верхнего конца манубриума отходят че-
тыре широких радиальных канала, проходящих затем в стенках ко-
локола до соединения с кольцевым каналом, расположенным по
краю колокола. Щупальца, покрытые стрекательными клетками,
собраны в четыре пучка, что является довольно редким случаем,
так как у большинства гидроидных медуз они расположены более
или менее равномерно по всему краю колокола. В каждом пучке
находится до 15 сильно сократимых щупалец, на основании кото-
рых с внутренней стороны расположены крупные глазки.
Обычный вид — Bougainvilfa. superciliaris.
4. Holopedium (рис. 11, Г)
Встречается в планктоне наших северных озер, отличается от
остальных видов ветвистоусых раков рядом существенных призна-
ков. Все тело животного заключено в большой, диаметром в не-
сколько миллиметров, совершенно прозрачный студенистый шар,
образованный разбухшими створками раковины, сброшенными при
линьках. Голова очень маленькая, без рострума. Сложный глаз не-
большой, глазок присутствует. Антеннулы очень короткие. Антенны,
являющиеся органами движения, сильно развиты, у самок одновет-
вистые, с тремя длинными плавательными щетинками, у самцов
двуветвистые. Грудных ног шесть пар. На конце тела находится
фурка, образованная двумя изогнутыми фуркальными (каудальны-
ми) когтями. Выше фурки по спинной стороне животного располо-
жены две длинные хвостовые щетинки, отходящие от конического
выроста. Выводковая камера самок высокая, с многочисленными
яйцами или зародышами.
Единственный вид — Holopedium gibberum.
Жировые включения
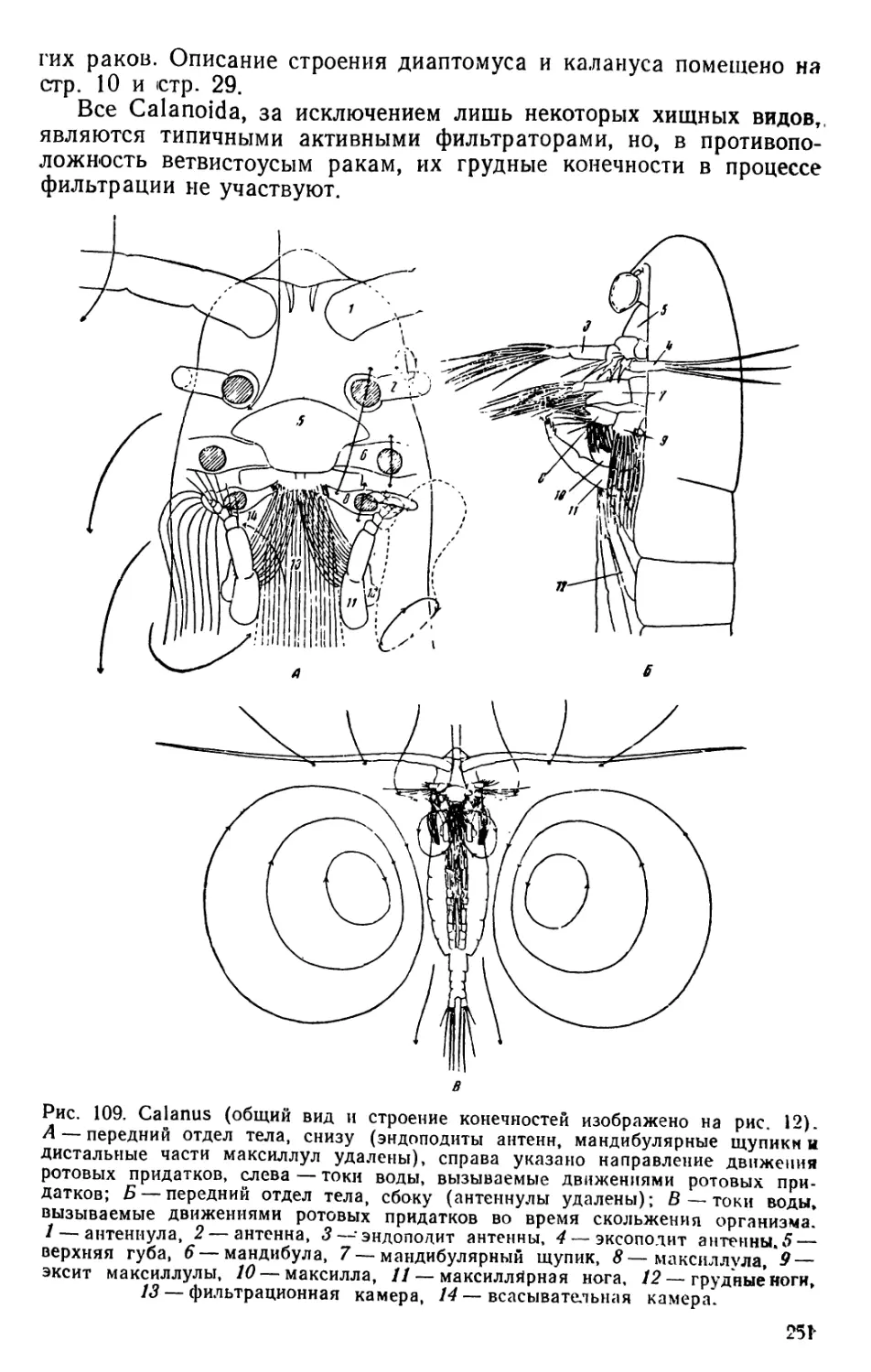
Calanus (рис. 12)
В качестве примеров рассматриваются жировые капли в теле
ветвистоусых и веслоногих раков. У некоторых свободноживущнх
видов веслоногих раков в переднем отделе тела параллельно ки-
шечнику располагается цилиндрический тонкостенный орган, напол-
ненный жировым веществом (рис. 12). Этот так называемый гид-
ростатический орган у Calanus finmarchicus, самого распространен-
ного в северных морях представителя Copepoda, проходит в перед-
ней своей части над кишечником, а затем постепенно переходит на
его левую сторону. При сильном развитии жирового вещества, лег-
29
м
ко заметного благодаря яркой окраске, гидростатический аппарат
занимает срединное положение в теле организма, оттесняя кишеч-
ник вправо. У зрелых самок его передняя часть редуцируется вслед-
ствие сильного развития расположенных в этом месте яйцеводов.
Рассмотрев расположение гидростатического органа, следует разор-
вать на предметном стекле препаровальными иглами покровы ка-
лапуса и выделить жировое вещество.
Газовые включения
В качестве наиболее обычных примеров рассматриваются воз-
душные включения синезеленых водорослей (Cyanophyceae) и ли-
чинок комаров, относящихся к подсемейству Chaoborinae семейства
Culicidae.
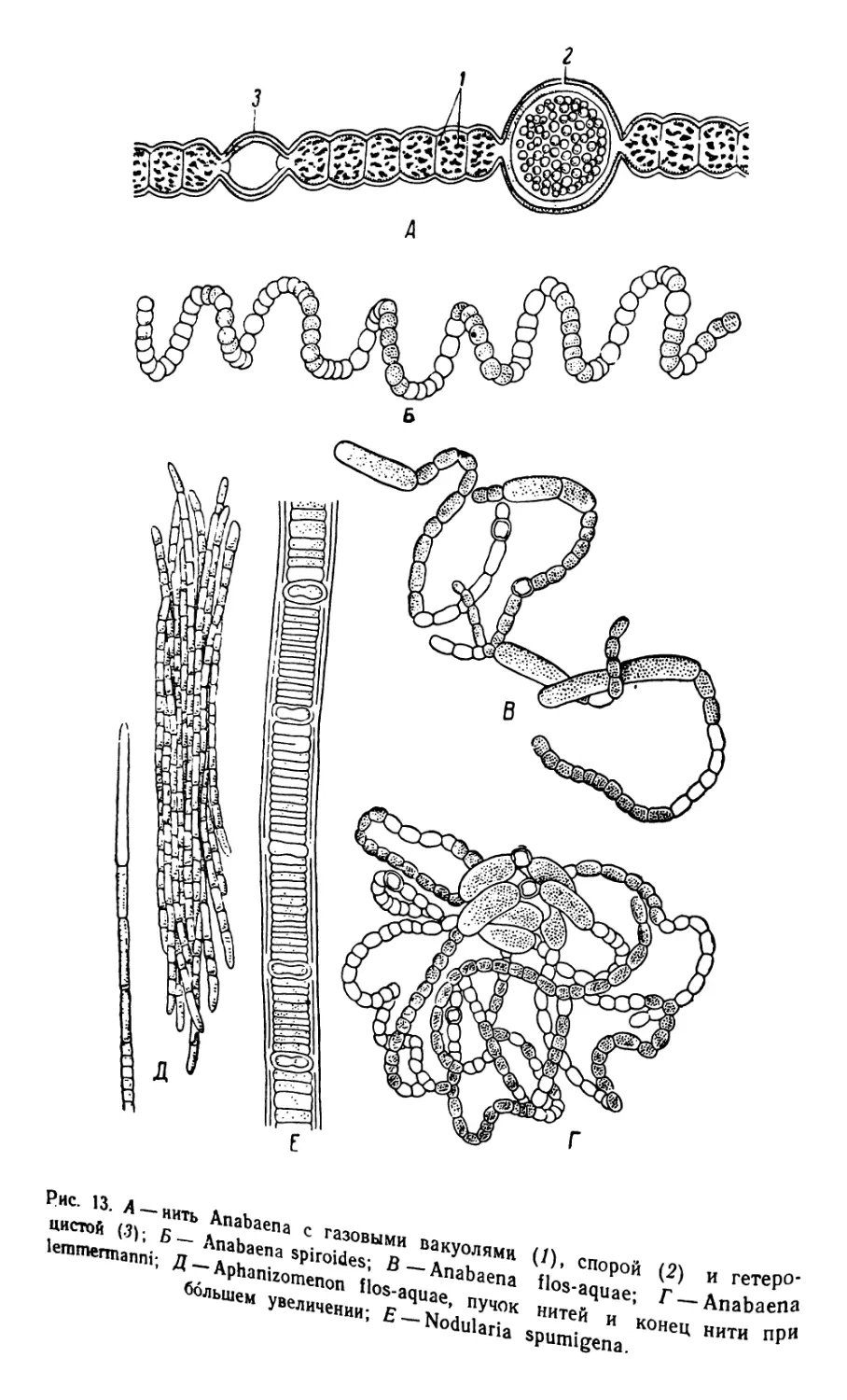
1. Anabaena, Aphanizomenon и Nodular-ia (рис. 13)
Синезеленые водоросли благодаря присутствию фикоциана и
других ассимилирующих пигментов — хлорофилла, фикоэритрина и
каротина — окрашены в разнообразные оттенки оливково-зеленого,
голубого или коричневого цветов. Характерной чертой строения
клеток является отсутствие типичного ядра и хроматофоров; прото-
пласт делится на расположенную по периферии хроматоплазму и
находящуюся внутри клетки центроплазму, которую можно считать
равнозначащей ядру клеток других растений.
Планктические синезеленые водоросли, относящиеся к семейст-
ву Nostocaceae, как, например, Anabaena, Aphanizomenon и Nadu-
laria являются главнейшими представителями фитопланктона раз-
личных пресноводных водоемов (Anabaena и Aphanizomenon) и
слабо соленых Азовского и Каспийского морей (Nodutaria); часто
встречаются в массовых количествах, вызывая летом явление цве-
тения воды. Водоросли образуют нити из многочисленных клеток,
тесно связанных между собою тончайшими плазматическими пере-
мычками (плазмодесмами); оболочки клеток тонкие. Между веге-
тативными клетками находятся особые пограничные клетки, или ге-
тероцисты, превосходящие обычно величину вегетативных клеток н
отличающиеся утолщенностью оболочки и иной окраской, что за-
висит от исчезновения ассимилирующего пигмента. Кроме гетеро-
цист в нитях находятся также покоящиеся клетки, споры, способ-
ные переносить неблагоприятные условия и прорастать в новую
нить; споры отличаются своей величиной и большей толщиной обо-
Рис. 12. Calanus finmarchicus. А — самка; Б — самец: В — поперечный раз-
рез через третий грудной сегмент; Г—антенна; Д—мандибула; Е—максилулла;
Ж — максилла; 3 — максиллярная нога; И — нога первой пары; К—нога чет-
вертой пары; Л—ноги пятой пары самки; М — ноги пятой пары самца.
1—антенпула, 2 — сердце, 3—мвпица антеннулы, 4—мышцы ротовых придат-
ков, 5—мышцы ног, 6 — кишечник, 7—гидростатический аппарат, S — спин-
ная мышца, 9— брюшная мышца, 10 — яичник, 11—яйцевод; 12— нервная
система.
Рис. 13. А —нить
цистой (3); Б— _ ^rBiQiues; L
lemmennanni; Д — Aphanizomenon ilos-aquae, пучок
большем увеличении; Е— Nna»’1--*
Anabaena с газовыми вакуолями
Anabaena spiroides; В — Anabaena
Nodularia
(/), спорой
Поз-aquae;
нитей и конец нити при
spumigena.
(2) и гетеро-
Г — Anabaena
лочки, по сравнению с вегетативными клетками. У многих видов
синезеленых водорослей можно обнаружить присутствие в клетках
многочисленных неправильной формы газовых вакуолей; благодаря
им удельный вес водорослей становится меньше удельного веса
воды. Особенно велико значение этих образований для планктиче-
ских водорослей, которые вначале развиваются у дна, а затем при
помощи газовых включений всплывают к водной поверхности. Под
микроскопом газовые вакуоли кажутся окруженными черной каем-
кой. Присутствие в них газа легко показать следующим простым
опытом. Банку, содержащую живые водоросли, наполняют до само-
го верха водою и закрывают высокой цилиндрической пробкой, не
оставляя в банке воздуха. Если затем с силой вогнать пробку в
горло банки, то в результате резко увеличивающегося давления газ
будет мгновенно вытеснен из вакуолей, и водоросли опустятся на
дно банки.
Наиболее обычными представителями синезеленых водорослей
являются различные виды Ariabaena, Aphanizomenon и Nodularia,
вызывающие цветение воды; некоторые из них поэтому несут видо-
вое название flos-aquae.
Нити Anabaerta, обычно сильно закрученные или изогнутые и за-
ключенные в слизь, содержат несколько гетероцист (пограничных
клеток), расположенных между вегетативными клетками; длина
последних равна ширине или ее превышает. Наиболее обычные ви-
ды отличаются между собою следующими признаками.
•
1 (2) Нити закручены по крутой спирали, споры округлые
или слабо изогнутые...........................A. spiroides
2 (1) Нити иной формы; споры цилиндрические, более или ме-
нее сильно изогнутые......................................3
3 (4) Нити, многократно изогнутые, образуют неправильной
формы клубок; споры разбросаны беспорядочно .
A. flos-aquae
4 (3) Нити, переплетаясь между собою, радиально отходят от
одного места; споры расположены около гетероцист
A. lemmermamu
Нити Aphanizomenon, из которых чаще всего встречается Apha-
nizomenon flos-aquae, соединены в пучки; концы нитей образованы
удлиненными бесцветными клетками; споры сильно вытянуты по
длине.
Нити1 Nodularia, из которых самым обычным видом является
Nodularia spumigena, отличаются от близких форм низкими, диско-
образными клетками; споры образуются на равном расстоянии от
гетероцист, нити прямые или вогнутые. Встречаются в солоноватых
водах.
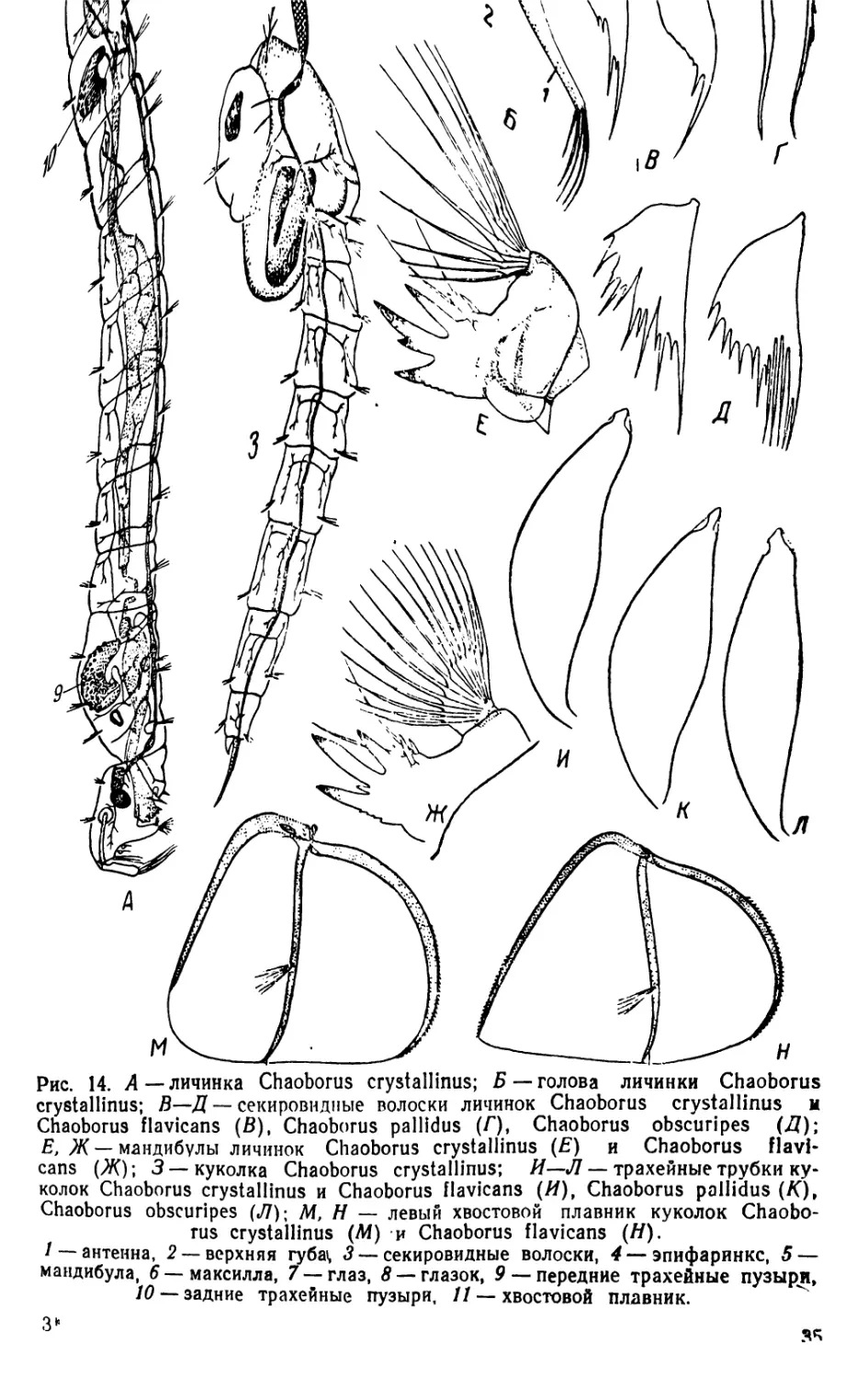
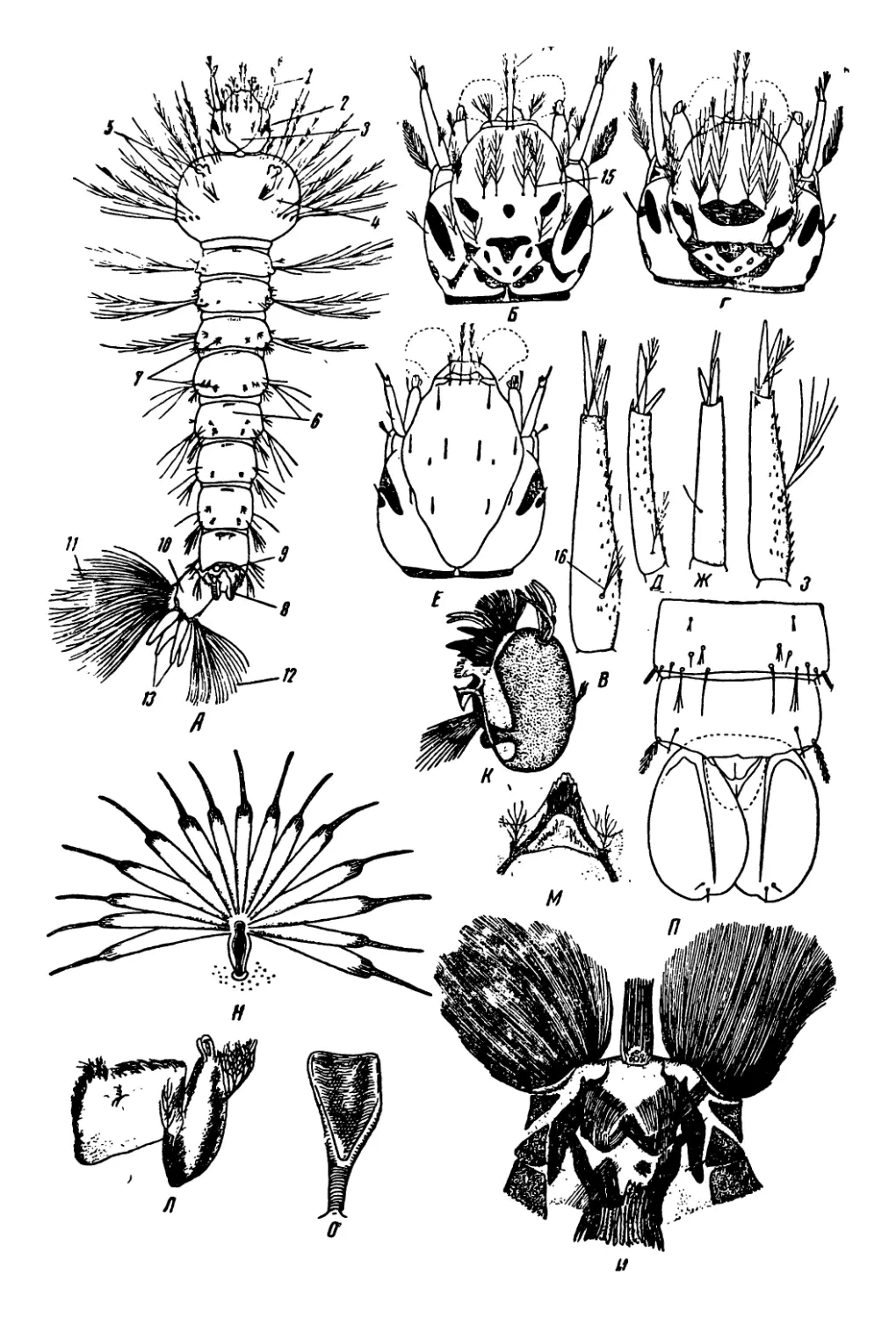
2. Личинки и куколки Chaoborus (рис. 14)
Личинки комаров подсемейства Chaoborinae отличаются от ли-
чинок других комаров присутствием двух пар, а у одного вида трех
пар, трахейных пузырей, являющихся гидростатическим аппаратом.
*3 В. A. S'liinon 33
Благодаря этому аппарату личинки держатся в воде в горизонталь-
ном положении, их удельный вес близок к удельному весу воды.
Вое личинки относятся к числу хищников, питаются дафниями,
циклопами, личинками различных комаров и т. п.
Личинки коретры (Cliaoborus, прежнее название—Corethra) яв-
ляются наиболее специализированными среди всех личинок Chao-
borinae. Тело прозрачное, бесцветное или .окрашенное в желтоватые
оттенки. Голова сильно вытянута и впереди сужена. Затылочное
отверстие широкое, это облегчает заглатывание крупной добычи.
Сложные глаза большие, позади них находятся небольшие глазки.
Антенны, тесно сближенные своими основаниями, в естественном
положении подогнуты вниз, на конце с пятью крепкими хвататель-
ными щетинками. Верхняя губа представлена длинными щетинка-
ми, отходящими от нижней поверхности головы позади оснований
антенн. Щетинки антенн и верхней губы являются органом хвата-
ния добычи. Ниже щетинок верхней губы помещаются две пластин-
ки, по одному краю зазубренные, так называемые секировндные
волоски. Большой подвижный эпифаринкс, принимавшийся ранее
за верхнюю губу, на конце, направленном к ротовому отверстию,
вооружен различного строения щетинками. Мандибулы сильно
развиты, с несколькими большими пальцевидными зубцами; на
наружном крае мандибул находится пучок длинных и уплощенных
при основании щетинок, образующих в раскрытом состоянии полу-
шаровидный хватательный веер, при помощи которого личинки
схватывают добычу для проталкивания» ее в глотку. Максиллы и
нижняя губа рудиментарные.
В грудном сегменте, лишь немного отличающемся по ширине от
брюшных сегментов, находятся два больших, хорошо заметных тра-
хейных пузыря, покрытых сверху пигментными клетками, содержа-
щими темный пигмент. Такие же трахейные пузыри расположены в
седьмом брюшном сегменте. Дыхательная система апнейстического
типа, стигмы отсутствуют, трахейные стволы слабо заметны, не на-
полнены воздухом. Дыхание кожное. На последнем сегменте тела
находится хорошо развитый хвостовой плавник, главный орган дви-
жения, и небольшие анальные папиллы.
Пищеварительная система состоит из короткой глотки, пищево-
да, расширяющегося на конце в объемистый зоб, и средней кишки,
переходящей в седьмом брюшном сегменте в заднюю кишку. На гра-
нице между зобом, занимающим весь грудной сегмент, и средней
кишкой находится ситовидная пластинка, образованная двумя ря-
дами хитиновых зубцов. Захваченная добыча, попадая в зоб, пере-
варивается под воздействием пищеварительных соков, поступающих
из средней кишки. Переваренная пища процеживается через сито-
видную пластинку, все же непереваренные хитиновые остатки уда-
ляются из зоба при его частичном выворачивании наружу. Сердце
расположено на спинной стороне личинки. Если, поместив личинку
спиной кверху, разрушить легкими надавливаниями пинцета пиг-
ментные клетки, покрывающие трахейные пузыри, то их остатки
будут отнесены кровяным током к большим перикардиальным клет-
кам, расположенным по бокам спинного кровеносного сосуда, бла-
34
Рис. 14. Д—личинка Chaoborus crystallinus; Б —голова личинки Chaoborus
crystallinus; В—Д — секировидные волоски личинок Chaoborus crystallinus и
Chaoborus flavicans (В), Chaoborus pallidus (Г), Chaoborus obscuripes (Д);
E, Ж — мандибулы личинок Chaoborus crystallinus (£) и Chaoborus flavi-
cans (Ж); 3—куколка Chaoborus crystallinus; И—Л — трахейные трубки ку-
колок Chaoborus crystallinus и Chaoborus flavicans (И), Chaoborus pallidus (K)t
Chaoborus obscuripes (Л); M, H — левый хвостовой плавник куколок Chaobo-
rus crystallinus (M) и Chaoborus flavicans (Я).
/ — антенна, 2 —верхняя губа*, 3— секировидные волоски, 4— эпифаринкс, 5 —
мандибула, 6—максилла, 7 — глаз, 8 — глазок, 9 — передние трахейные пузыри,
10 — задние трахейные пузыри, // — хвостовой плавник.
3“
годаря этому сокращения спинного сосуда становятся ясно видц.
мыми.
Личинки коретры держатся в воде в горизонтальном положении
в определенном слое воды, благодаря присутствию гидростатиче-
ского аппарата; их удельный вес почти равен удельному весу воды.
При изменении объема трахейных пузырей личинки перемещаются
в вертикальном направлении и могут опуститься на дно. Измене-
ние объема трахейных пузырей происходит вследствие диффузии
газа из крови в пузыри или, обратно, из пузырей в кровь. Некото-
рое значение для перемещений личинки имеют также покрываю-
щие пузыри пигментные клетки. Если эти клетки расширяются, то,
в результате поглощения ими проникающих в воду лучей, трахей-
ные пузыри, нагреваясь, увеличиваются в объеме, удельный вес ли-
чинки становится меньше и она поднимается кверху; сокращение
пигментных клеток приводит к увеличению удельного веса и опус-
канию личинки. Можно простым способом показать значение1 гид.
ростатического аппарата для вертикальных передвижений личинок.
Если поместить несколько экземпляров личинок коретры, до этого
длительно содержавшихся при комнатной температуре, в более теп-
лую и более холодную воду, то в первом случае личинки поднимут-
ся наверх, их пигментные клетки сожмутся, во втором случае ли-
чинки опустятся вниз, их пигментные клетки расширятся.
Личинки коретры живут в самых разнообразных водоемах, от
озер до небольших лесных луж. Встречающиеся у нас личинки от-
личаются между собою следующими признаками.
1 (2) Небольшой добавочный зубец мандибул расположен в
промежутке между основаниями среднего и наружного
зубцов; секировидные волоски такого же строения, как
у Ch. crystallinus..........................Ch. flavicans
2 (1) Небольшой добавочный зубец мандибул отходит от боко-
вой Ьтороны срединного зубца............................3
3 (4) Секировидные волоски узкие, задний край с нескольки-
ми тонкими шипами...........................Ch. ptallidus
4 (3) Секировидные волоски широкие, передний край зуб-
чатый ..................................................й
5 (6) Секировидные волоски с небольшими зубчиками по' перед-
нему краю, оканчиваются длинным и острым шипом
Ch. crystallinus (=Corethra plutnicornis)
6 (5) Секировидные волоски с многочисленными длинными и
острыми шипами по переднему краю . . Ch. obscurlpes
Прозрачное тело куколок коретры состоит* из большой голово-
груди и брюшка. От головогруди отходят две веретенообразные
трубки, замкнутые на концах и наполненные воздухом. Эти трубки,
аналогичные дыхательным трубкам куколок других комаров, вме-
сте с наполненной воздухом подкововидной полостью, расположен-
ной в нижней части головогруди, играют роль гидростатического
аппарата. Куколки держатся в воде в вертикальном положении, на
поверхность воды не поднимаются, их дыхание кожное. На восьмом
брюшном сегменте находятся две больших и гибких лопасти хво-
стового плавника, являющегося органом движения.
36
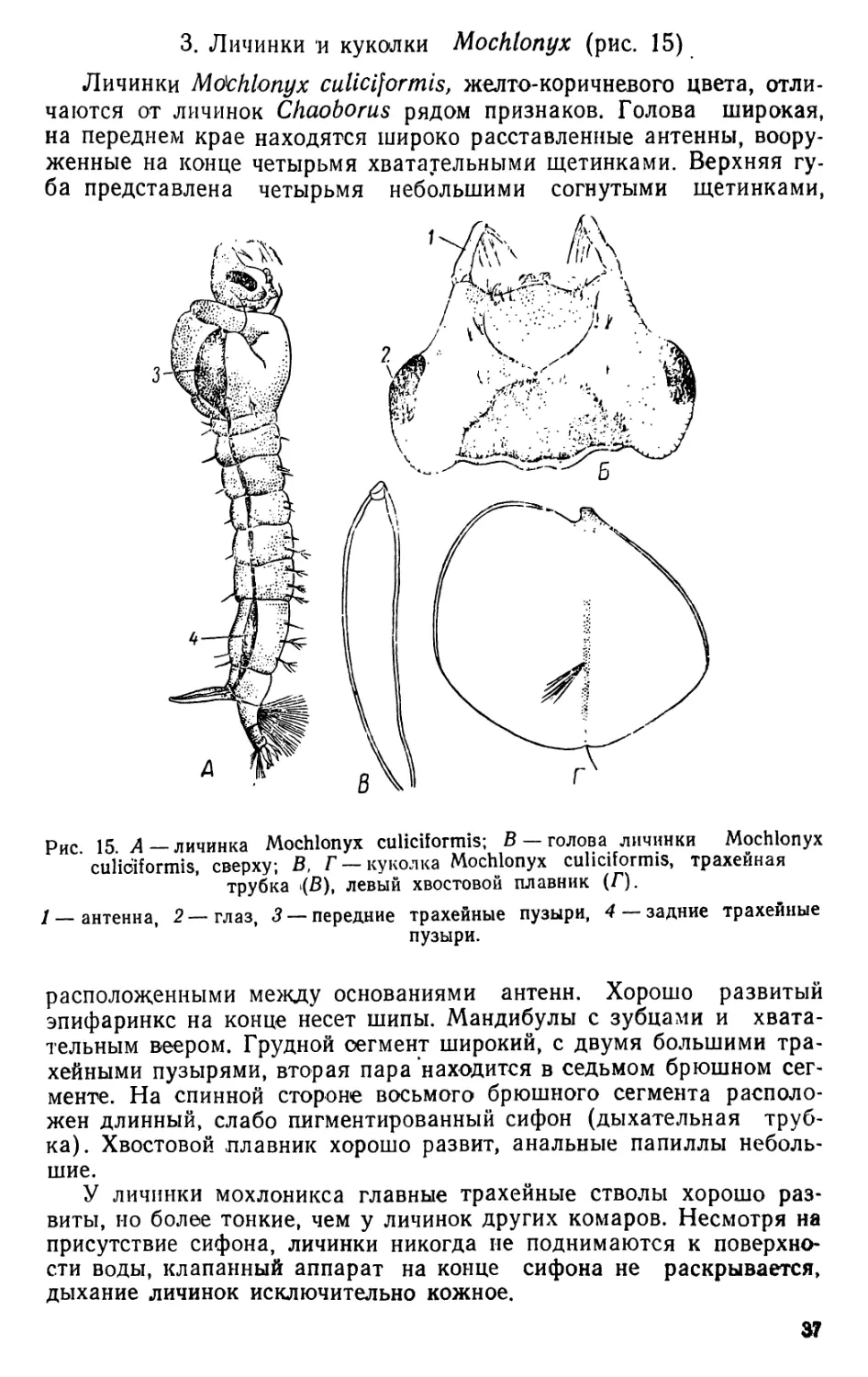
3. Личинки и куколки Mochionyx (рис. 15)
Личинки Mdchlonyx culiciformis, желто-коричневого цвета, отли-
чаются от личинок Chaoborus рядом признаков. Голова широкая,
на переднем крае находятся широко расставленные антенны, воору-
женные на конце четырьмя хватауельными щетинками. Верхняя гу-
ба представлена четырьмя небольшими согнутыми щетинками,
Рис. 15. А — личинка Mochionyx culiciformis; В — голова личинки Mochionyx
culiciformis, сверху; В, Г — куколка Mochionyx culiciformis, трахейная
трубка (В), левый хвостовой плавник (Г).
1 — антенна, 2—глаз, 3 — передние трахейные пузыри, 4 — задние трахейные
пузыри.
расположенными между основаниями антенн. Хорошо развитый
эпифаринкс на конце несет шипы. Мандибулы с зубцами и хвата-
тельным веером. Грудной сегмент широкий, с двумя большими тра-
хейными пузырями, вторая пара находится в седьмом брюшном сег-
менте. На спинной стороне восьмого брюшного сегмента располо-
жен длинный, слабо пигментированный сифон (дыхательная труб-
ка). Хвостовой плавник хорошо развит, анальные папиллы неболь-
шие.
У личинки мохлоникса главные трахейные стволы хорошо раз-
виты, ио более тонкие, чем у личинок других комаров. Несмотря на
присутствие сифона, личинки никогда ие поднимаются к поверхно-
сти воды, клапанный аппарат на конце сифона не раскрывается,
дыхание личинок исключительно кожное.
37
Личинки живут в небольших, преимущественно временных во-
доемах.
Куколки мохлоникса отличаются от куколок коретры присутст-
вием длинных, на конце открытых дыхательных трубок и жестких,
нескладывающихся лопастей хвостового плавника.
Увеличение сопротивления формы
Удлинение одной оси
В качестве примеров рассматриваются представители диатомей
(Rhizosolenia п Melosira), ветвистоусых раков (Leptodorta) и ще-
тинкочелюстных (Sagitta). Все эти организмы вытянуты по одной
оси, в воде держатся в горизонтальном положении.
1. Rhizosolenia и Melosira (рис. 16)
Диатомеи (Diatomeae), наряду с перидинеями (Peridineae) и
синезелеными водорослями (Cyanophyceae), относятся к числу са-
мых главных компонентов фитопланктона морей и пресных вод.
Оболочка диатомей состоит из двух пропитанных кремнеземом
створок, из которых меньшая (гипотека) вложена в большую (эпи-
теку), наподобие двух половин чашки Петри. Наложенные друг на
друга края створок образуют поясок. При рассматривании диато-
мей различают вид со створки и вид с пояска. Обе створки раздви-
гаются во время вегетативного деления. Дочерние клетки, сохраняя
одну створку материнской клетки, образуют новую, вторую створку,
всегда меньшего размера, по сравнению с первой. Вследствие та-
кого характера деления вегетативное размножение приводит к из-
мельчанию клеток. Когда величина молодых клеток становится рав-
ной приблизительно половине первоначальной величины, вегетатив-
ное деление останавливается, после чего начинается образование
растущих спор, ауксоспор, покрытых вначале мягкой и растягиваю-
щейся оболочкой, позволяющей им вырасти до величины исходных
клеток. По строению створок диатомеи делятся на две группы —
Centricae (Centrales), створки которых имеют радиальное строение,
и Pennatae (Pennales) с двусторонне-симметричным, дорсо-вентраль-
ным или несимметричным строением. К первой группе относятся
преимущественно планктические виды, ко второй — большинство
бентических и часть планктических видов. Плазма обычно распола-
гается тонким слоем по стенкам клетки, внутри находится клеточный
сок. Ядро имеется. Хроматофоры самой различной формы, в живом
состоянии темножелтого или коричневого цвета благодаря присут-
ствию пигментов диатомина и хлорофилла. Продуктом ассимиляции
обычно является масло.
Большинство планктических видов относится к числу парящих
организмов, выработавших различного рода приспособления к жиз-
ни во взвешенном состоянии. Скелет их значительно тоньше и лег-
че, чем у донных видов; сопротивление погружению увеличивается
благодаря образованию разнообразных выростов или удлинению
одной или двух осей.
ЯЙ
Обычными формами наших северных и южных морей являются
различные виды Rhizosolenia. Цилиндрические, слабо пропитанные
кремнеземом клетки имеют игловидную или палочковидную форму
благодаря сильному развитию в зоне пояска многочисленных про-
межуточных пластинок в виде ромбов или трапеций. Створки на
концах клетки вытянуты в
асимметричные конусы, окан-
чивающиеся обычно эксцен-
трично лежащим острием или
волоском. Клетки не могут дли-
тельное время погружаться в
вертикальном направлении, так
как благодаря асимметричному
строению концов, падение их
происходит по кривой, поэтому
через короткое время диатомеи
возвращаются в первоначаль-
ное горизонтальное положение.
Хроматофоры, мелкие; много-
численные.
Обычным видом северных
морей является Rhizosolenia
hebetata с двумя формами —
semispina (концы створок с не-
большим волоском) и hie mails
(концы створок тупые, без во-
лоска). Массовой формой Азов-
ского и Каспийского морей яв-
ляется Rhizosolenia calcar-avis.
Различные виды Melosira,
встречающиеся преимуществен-
но в планктоне пресноводных
водоемов, относятся, как и ви-
ды Rhizosolenia, к группе Сеп-
tricae. В противоположность
последним, у которых зона по-
Рис. 16. А — Rhizosolenia hebetata f.
semispina, конец клетки; Б — Rhizosole-
nia hebetata f. hiemalis; В—Rhizosolenia
calcar-avis; Г, Д — Melosira islandica.,
вид co створки (Г) и с пояска (Д).
яска сильно развита, так что
клетка получает вид очень высокого цилиндра, у Melosira высота
отдельной клетки обычно не превышает диаметра створки более,
чем в два-три раза. Клетки плотно соединены шипиками или сту-
денистой массой в длинные колонии, благодаря чему значительно
увеличивается их сопротивление погружению. Все клетки колонии
одинакового строения. Со стороны створки клетки круглые, в этом
можно убедиться, поместив изолированную клетку створкой
кверху. При рассматривании с пояска видны поры различной ве-
личины, расположенные прямолинейно или по спирали. Хромато-
форы мелкие, многочисленные.
Наиболее обычными видами Melosira, часто вызывающими цве-
тение воды, являются М. granulata, М. islandica, М. italica, отли-
чающиеся между собой мелкими деталями строения.
39
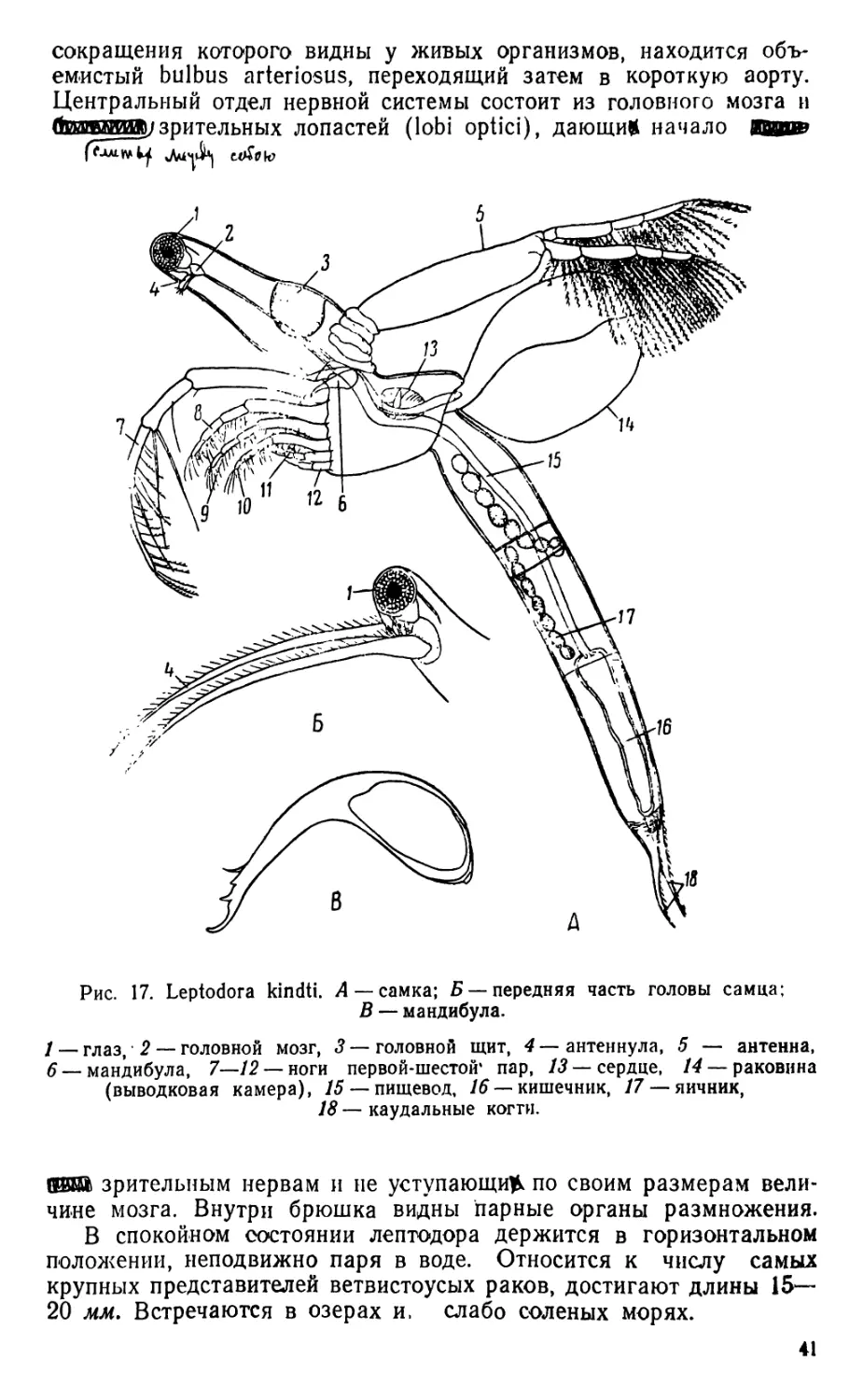
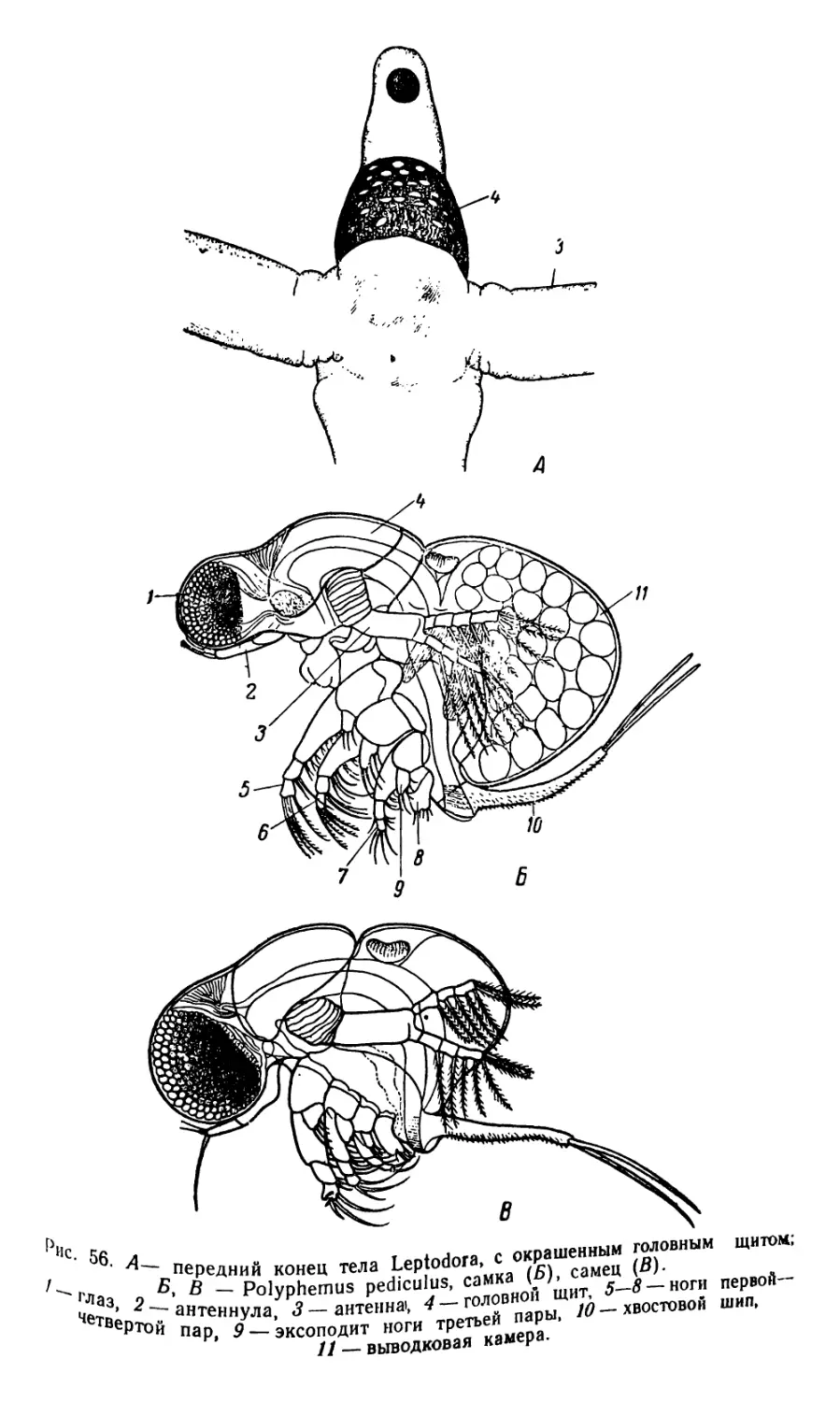
2. Leptodora (рис. Г7)
Лептодора (единственный вид Leptodora kindti) по своей орга-
низации настолько отличается от всех остальных ветвистоусых ра-
ков, что выделяется в виде единственного представителя не только
семейства Leptodoridae, на также трибы Haplopoda.
Лептодора благодаря своей исключительной прозрачности, что
послужило основанием прежнего наименования — Leptodora hyali.
па, в живом состоянии почти невидима, только движение различных
мелких частиц, увлекаемых ударами антенн, выдает ее присутствие
в воде; в фиксированном состоянии прозрачность частично теряется.
Тело сильно вытянуто в длину, почти цилиндрической формы, с
тонкими покровами, разделено довольно резко на пять сегментов.
Первый сегмент образован головой и грудью, остальные входят в
состав брюшка (абдомена). Голова сильно вытянутая, на суженном
переднем конце находится большой сложный глаз, содержащий не-
сколько сотен омматидиев; глазок отсутствует. На верхней стороне
головы, ниже середины, помещается большой округлой формы го-
ловной щит, являющийся органом дыхания, ясно отграниченный от
остальной части и состоящий из утолщенной оболочки и зернистой
или ячеистой субкутикулы. Антеннулы, расположенные позади гла-
за, у самок короткие, у самцов имеют вид длинных, слегка изогну-
тых щетинок, несущих большое число тонких хитиновых придатков
(эстетаски) — органов химического чувства (хеморецепторы). Ан-
тенны являются мощным органом движения, внутри их видна силь-
но развитая мускулатура. Они состоят из большого основного чле-
ника и двух четырехчленистых ветвей (эндоподита и эксоподита),
несущих большое число длинных плавательных щетинок. Мандибу-
лы серповидной формы, с острыми концами, приспособлены к разры-
ванию добычи. Максиллулы и максйллы отсутствуют. Верхняя и
нижняя губы хорошо развиты. На спинной стороне переднего сег-
мента находится раковина (карапакс), служащая выводковой ка-
мерой. У зрелых самок ее свободный край тесно прижат к спинной
поверхности брюшных сегментов, что обусловливает хорошую изо-
ляцию для находящихся в выводковой камере яиц. Грудные ноги
членистые, относятся к типу скелетных конечностей, встречающих-
ся среди ветвистоусых раков только у хищных видов — лептодоры и
представителей семейства Polyphemidae. Всего имеется шесть пар
ног, сближенных между собой и сдвинутых к ротовому отверстию.
Своими ногами лептодора захватывает добычу,, состоящую главным
образом из животных планктона; щипы и щетинки, находящиеся по
внутреннему краю ног, облегчают это захватывание. Эпиподиты от-
сутствуют, в связи с этим конечности теряют функцию дыхания.
Последний брюшной сегмент, соответствующий постабдомену всех
остальных представителей ветвистоусых раков, оканчивается двумя
конической формы фуркальными выростами, расположенными вдоль
продольной оси животного.
Длинный пищевод проходит почти через все брюшко и только на
его конце впадает в небольшую и широкую среднюю кишку; аналь-
ное отверстие расположено между ветвями фурки. Перед сердцем,
40
сокращения которого видны у живых организмов, находится объ-
емистый bulbus arteriosus, переходящий затем в короткую аорту.
Центральный отдел нервной системы состоит из головного мозга и
(faMWMhjзрительных лопастей (lobi optici), дающий начало пив
pAMHv Ц еЛк?
Рис. 17. Leptodora kindti. Л — самка; Б —передняя часть головы самца;
В — мандибула.
/ — глаз, 2 — головной мозг, 3 — головной щит, 4 — антеннула, 5 — антенна,
6 — мандибула, 7—12 — ноги первой-шестой* пар, 13 — сердце, 14 — раковина
(выводковая камера), /5 —пищевод, 16 — кишечник, /7 —яичник,
18 — каудальные когти.
Ш зрительным нервам и не уступающий по своим размерам вели-
чине мозга. Внутри брюшка видны парные органы размножения.
В спокойном состоянии лептодора держится в горизонтальном
положении, неподвижно паря в воде. Относится к числу самых
крупных представителей ветвистоусых раков, достигают длины 15—
20 мм. Встречаются в озерах и, слабо соленых морях.
41
3. Sagitta и Eukrohnia (рис. 18)
Стрелки, или сагитты (Sagitta), относящиеся к классу щетинко-
челюстных (Chaetognatha), являются морскими животными, веду-
щими исключительно планктический образ жизни. В живом состоя-
нии почти совершенно прозрачны, слегка просвечивают лишь мыш-
цы головы и органы размножения; в фиксированном виде живот-
ные уплотняются и теряют прозрачность.
Длинное тело делится на три отдела — головной, туловищный и
хвостовой. На голове находятся темноокрашенные парные глаза и
мощно развитый челюстной аппарат, приспособленный к хватанию
добычи. Повернув голову сагитты нижней стороной вверх, можно
рассмотреть два ряда длинных подвижных и более или менее сог-
нутых челюстных щетинок, перед которыми находятся несколько
групп небольших зубчиков, препятствующих выскальзыванию добы-
чи. Каждая челюстная щетинка, расширенная в базальной части,
где прикрепляется сильный мускул, состоит из длинного полого
стержня, в конец которого вставлен небольшой крепкий зубец. В
туловищном отделе находятся длинная средняя кишка, открываю-
щаяся на брюшной стороне, на границе туловищного и хвостового
отделов, сильно развитая продольная мускулатура, а также брюш-
ной ганглий, с отходящими от него нервами. Все сагитты герма-
фродиты. В задней части туловищного отдела расположены парные
яичники, длина которых зависит от возраста организма; яйцеводы
открываются по бокам анального отверстия. Семенники находятся в
передней части хвостового отдела, открываются по его бокам на не-
больших возвышениях. Для сохранения положения в воде служат
боковые плавники, расположенные по бокам задней части тела, и
непарный хвостовой плавник.
В состоянии покоя сагитты, находящиеся в горизонтальном по-
ложении, медленно опускаются, они немного тяжелее воды. Так как
плавники, увеличивающие поверхность тела, находятся только в его
задней части, то животное скоро начинает опускаться головою вниз.
Несколькими одиночными ударами хвоста сагитта выравнивает по-
ложение тела.
В результате молниеносно быстрых ударов хвоста, вызываемых
чередующимися сокращениями в вертикальной плоскости спинной
и брюшной продольной мускулатуры, сагитты двигаются вперед.
Быстрому движению способствует обтекаемая форма тела; голова
вместе с челюстным аппаратом закрывается гладкой кожной склад-
кой, отходящей от основания головы и имеющей вид капюшона с
небольшим круглым отверстием на брюшной стороне.
Все сагитты — хищники, питаются различными животными
планктона, преимущественно веслоногими раками, могут захваты-
вать крупную добычу, как, например, личинок сельдей.
Наиболее крупные виды сагитт, длиною до 30—40 мм, встреча-
ются в холодной воде северных морей. Sagitta elegans характери-
зуется присутствием двух пар боковых плавников, снабженных хо-
рошо заметными лучами. Eukrohnia hamata обладает одной парой
боковых плавников; лучи имеются только в их задней трети. В
Черном море встречается Sagitta euxina.
42
Ис- 18. А — Sagitta elegans; Б, В — голова Sagitta, закрытая кожной склад-
кой (Б) и со сдвинутой кожной складкой (В); Г, Д — челюстные щетинки
Sagitta elegans (Г) и Eukrohnia hamata (Д).
1— ротовое отверстие, 2 — глаз, 3 — челюстные щетинки, 4—кишечник,
5 — брюшной ганглий, 6 — боковые плавники, 7 — яичник, 8 — анальное
отверстие, 9 — семенник, 10 — хвостовой плавник.
Удлинение двух осей
В качестве примеров рассматриваются представители диатомей
(Coscinodiscus, Asterionella, Tabellaria) и зеленых водорослей
(Pedlast гит).
1. Coscinodiscus и Cyclotella (рис. 19, А—3)
Виды Coscinodiscus встречаются почти исключительно в морях,
нередко в очень больших количествах; в пресноводных водоемах
они заменяются близкими по строению видами Cyclotella.
Обычно одиночные клетки Coscinodiscus имеют вид плоских
дисков или низких цилиндров, по расположению створок в точно-
сти соответствуют чашкам Петри. При рассматривании со стороны
створки имеют вид кружков с сетчатой или ячеистой скульптурой,
образованной радиальными или тангентальными рядами кружочков
или мелких точек. Со стороны пояска имеют вид сглаженных на
углах прямоугольников, без скульптуры, часто с промежуточными
кольцами. Мелкие хроматофоры многочисленны.
Для ознакомления со скульптурой створок, без чего определе-
ние многих видов невозможно, следует применить прокаливание на
огне или проваривание в кислоте. При первом способе небольшое
количество диатомей переносится пипеткой на покровное стекло и
подсушивается. Затем помещают покровное стекло на тонкую слю-
дяную или металлическую пластинку и прокаливают на пламени
горелки в течение нескольких минут, следя за тем, чтобы края
стекла не плавились. После того, как сгорит все органическое веще-
ство, капают на препарат каплю соляной кислоты для растворения
соли и тотчас же промывают стекло в воде. После этого стекло сно-
ва подсушивается, и сухой препарат рассматривается под микроско-
пом; помещать его в канадский бальзам не следует, так как в этой
среде диатомеи невидимы благодаря одинаковым коэфициентам
преломления. Проваривание в кислоте проводится только для гру-
бых форм, для слабо пропитанных кремнеземом створок оно непри-
менимо. Само проваривание диатомей ведут обязательно в вытяж-
ном шкафу, в тугоплавких пробирках с небольшим количеством
крепкой азотной кислоты. Нагревание следует вести осторожно во
избежание выброса кислоты из пробирки. После обесцвечивания ма-
териала осадок тщательно промывается водой, и из него приготов-
ляют сухой препарат.
Наиболее обычные для наших северных морей виды Coscirtpdis-
cus отличаются между собою следующими признаками.Ц
1 (4) Скульптура створок крупноячеистая; клетки плоские2
2 (3) В центре створок находится розетка из крупных ячеек;
края створок мелкоячеистые ... С. oculus iridis
3 (2) Розетка из крупных ячеек отсутствует; края створок
радиально заштрихованы............................С. radiatus
4 (1) Скульптура створок мелкоячеистая; клетки барабано-
образные .................................................. 5
5 (6) Края створок радиально заштрихованы. С. centralis
6 (5) Края створок мелкоячеистые . С. concinnus
44
к [И
Рис. 19. А, Б — Coscinodiscus oculus iridis, вид со створки (Л) и с пояска (Б);
В, Г — Coscinodiscus radiatus, вид со створки (В) и с пояска (Г); Д —Cosci-
nodiscus centralis, часть створки; Е — Coscinodiscus concinnus, часть створки;
Ж, 3 - Cyclotel la comta, вид со створки (Ж) и с пояска (3); И —- Asterionella
Formosa, колония; К — Tabellaria fenestrata, колония; Л —Pediastrum simplex;
М — Pediastrum duplex; Н — Pediastrum angulosum, часть колонии;
О—Pediastrum boryanum.
В южных морях обычным видом является Coscinodiscus biconi-
сиз.
Пресноводные виды Cyclotella характеризуются резким делением
створок на центральную и краевую зоны; первая — гладкая или с
точками, вторая—с радиальными ребрышками; при рассматривании
с пояска видны волнистые очертания створок.
2. Asterionella (рис. 19, И)
Колониальные Asterionella, относящиеся к группе Pennatae, яв-
ляются самыми распространенными диатомеями наших пресновод-
ных водоемов. Удлиненные клетки, лишенные промежуточных пла-
стинок, соединены своими утолщенными концами в характерного
вида звездчатую колонию. Ранее считали, что они связаны также
слизистым веществом, однако последующие наблюдения этого не
подтвердили.
Два вида Asterionella различаются между собой по способу
соединения клеток в колонии. Клетки A. gracillima соприкасаются
только в одной точке, а у A. formosa — небольшими плоскостями.
3. Tabellaria (рис. 19, К)
Клетки Tabellaria fenestrata, единственного планктического вида
этого рода, соединены своими углами в зигзагообразные (var. inter-
media) или звездчатые (var. asterionelloides) колонии. При рас-
сматривании с пояска имеют вид табличек с четырьмя и более про-
межуточными пластинками, со створки — линейные, с небольши-
ми расширениями в середине и по концам.
4. Pediqstrum (рис. 19, Л—О)
Педиаструмы (Pediastrum), относящиеся к зеленым водорослям,
к классу Chlorococcineae (=Protococcineae), часто встречаются в
планктоне озер и прудов. Образуют пластинчатые колонии (ценобии)
из многогранных клеток, расположенных в одной плоскости. Число
клеток обычно, колеблется от 32 до 64, но бывает и меньше и боль-
ше. Краевые клетки отличаются от срединных присутствием одного
или нескольких выростов. Срединные клетки плотно соединены
между собою или разделены промежутками. Хроматофоры зеленого
цвета, пластинчатые, реже рассеченные, с одним пиреноидом. Раз-
множение происходит путем образования зооспор.
Наиболее обычные виды Pediastrum отличаются между собою
следующими признаками.
1 (2) Краевые клетки с одним выростом . Р. simplex
2 (1) Краевые клетки с двумя выростами . 3
3 (4) Клетки разделены промежутками . Р. duplex
4 (3) Клетки плотно соединены............................ 5
5 (6) Выросты краевых 'Клеток широко расставлены .
Р. angulosum
6 (5) Выросты краевых клеток разделены узкой выем-
кой . Р. boryanutn
46
Образование выростов
В качестве примеров рассматриваются представители диатомей
(Chaetoceros), перидиней (Cera//um), коловраток (Notholca), ветви-
стоусых раков (Bythotreph.es), десятиногих раков (личинка Porcel-
tana).
1. Chaetoceros (рис. 20)
Различные виды Chaetoceros, относящиеся к группе Centricae,
широко распространены в морях, являются массовыми формами,
вызывающими весеннюю вспышку фитопланктона.
Наиболее характерным признаком всех видов Chaetoceros яв-
ляется присутствие длинных щетинок, расположенных попарно у
краев створок; при помощи этих щетинок клетки соединены в длин-
ные колонии, имеющие вид цепочек. Обычно клетки на препарате
ложатся на сторону пояска, в этом случае они имеют вид четырех-
угольников; со стороны створки они чаще всего эллиптической фор-
мы (чтобы рассмотреть клетку со створки, надо предварительно от-
делить ее от колонии). У некоторых видов имеются промежуточные
кольца. Щетинки, значительно увеличивающие трение организма о
воду, могут быть сплошными или полыми, гладкими или с шипи-
ками; к концу обычно утончаются. Расположены они чаще всего в
одной плоскости под тупым или острым углом к длинной оси ко-
лонии, но могут быть прижатыми к ней или распростертыми в раз-
ные стороны. У некоторых видов щетинки, отходящие от концов ко-
лонии, отличаются по форме от остальных. Промежутки между
клетками могут быть самой разнообразной формы — круглые,
овальные, бисквитовидные, прямоугольные и т. д. Хроматофоры не-
большие, многочисленные, расположены обычно вдоль стенок клет-
ки, заходят у некоторых видов в полые щетинки. В некоторых слу-
чаях внутри клеток могут быть обнаружены покоящиеся споры,
покрытые плотной оболочкой, иногда с различными выростами.
Кроме покоящихся спор у Chaetoceros имеются ауксоспоры, возни-
кающие по бокам клеток.
В наших северных морях встречается больцюе количество видов
Chaetoceros. Наиболее обычные виды отличаются между собою сле-
дующими признаками.
1 (12) Щетинки толстые; хроматофоры мелкие, могут заходить
в щетинки..................................................2
2 (3) От средней части каждой створки в промежуток между
клетками отходит короткий шипик . . Ch. atlanticus
3 (2) Шипиков не имеется, а если они имеются, то не на каж-
дой створке .............................................. 4
4 (5) Щетинки двух сортов—прямые гладкие и волнистые
с зубчиками . . . Ch. compressus
5 (4) Все щетинки одинаковые ....... 6
6 (9) Щетинки прижаты к клеткам в одном направлении . 7
7 (8) Щетинки при основании немного тоньше, чем в. сред-
ней части; цепочки прямые Ch. concavicornis
47
8 (7) Щетинки по всей длине одинаковой толщины; цепочки
изогнутые..................................Ch. -convolutus
9 (6) Щетинки отходят более или менее перпендикулярно к
оси цепочки . 10
Рис. 20. А — Chaetoceros atlanticus; Б — Chaetoceros compressus; В — Chaetoce-
ros concavicomis; Г—Chaetoceros convolutus; JI — Chaetoceros densus; E —
Chaetoceros borealis; Ж — Chaetoceros decipiens; 3 — Chaetoceros mitra; И —
Chaetoceros subsecundus; К — Chaetoceros socialis.
10 (11) Промежутки между клетками очень узкие. Ch. densus
11 (10) Промежутки широкие, би1сквитообразные. Ch. borealis
12 (1) Щетинки тонкие, хроматофоры пластинчатые, не захо-
дят в щетинкй.......................................... 18
13 (16) В каждой клетке не менее четырех хроматофоров. 14
14 (15) Щетинки сросшиеся в местах скрещивания. Ch. decipiens
46
15 (14) Щетинки не сросшиеся .... Ch. mitra
16 (13) В каждой клетке один-два хроматофора 17
17 (18) Колонии прямые, без слизи ....
Ch. subsecundus (=Ch. diadema)
18 (17) Колонии сильно изогнутые, в слизи . . Ch. socialis
В южных морях наиболее обычными видами являются Сhaeto-
reros radians (мелкие клетки образуют спиральные цепочки), Ch.
curvisetus (промежутки между клетками бисквитообразные) и Ch.
socialis.
2. Ceratium (рис. 21)
Перидинеи (Peridineae, или Dinoflagellata) относятся к числу
наиболее распространенных форм фитопланктона, уступая в этом
отношении только диатомеям. Обитают преимущественно в морях,
в пресноводных водоемах встречается относительно небольшое чи-
сло видов.
Тело большинства перидиней покрыто целлюлозной оболочкой
(панцирем), состоящей из определенного числа пластинок; в более
редких случаях, как, например, у видов семейства Gymnodiniaceae,
оболочка плазматическая. Панцырь делится на верхнюю, апикаль-
ную, и нижнюю, антапикальную, половины (иначе называемые эпи-
текой и гипотекой), разделенные бороздой, опоясывающей кругом
все тело’ перидинеи. Другая, продольная, борозда проходит более
или менее перпендикулярно к поперечной. На так называемой
брюшной стороне клетки, в том месте,' где обе борозды перекрещи-
ваются, находится небольшое отверстие, через которое выходят два
жгута (на фиксированном материале не видны). Один из жгутов,
расположенный в продольной борозде и направленный назад, слу-
жит для поступательного движения, другой, лежащий в поперечной
борозде, вызывает вращение организма вокруг продольной оси.
Хроматофоры различного вида, окрашены в оттенки желтого, ко-
ричневого и бурого цветов, что зависит от присутствия пигментов—
фикопиррина, перидинина и хлорофиллина. Продуктом ассимиляции
является крахмал, быстро переходящий у морских форм в масло.
Кроме автотрофных видов, среди перидиней имеются гетеротроф-
ные, обыкновенно бесцветные, питающиеся оформленной пищей, и
миксотрофные, способные к растительному и животному способам
питания; имеется также паразиты. При вегетативном размножении
плоскость деления у многих видов проходит косо по отношению к
поперечной борозде. Каждая из дочерних клеток восстанавливает
недостающую половину, которая затем покрывается оболочкой.
Процесс деления протекает у многих перидиней в ночные часы.
Особи, только что разделившиеся, легко отличимы по неравномер-
ному развитию апикальной и антапикальной частей клетки. Кроме
того, перидинеи могут размножаться зооспорами. Некоторые виды
образуют покрытые толстой оболочкой покоящиеся споры, способ-
ные переносить неблагоприятные внешние условия. Некоторые мор-
ские виды (Ceratium, Peridinium и др.) вызывают свечение моря.
49
К числу самых обыкновенных представителен перидиней отно.
сятся различные виды церациумов (Ceratium), обитающие почти
исключительно в морях; в пресных водах встречаются только два
вида, из которых Ceratium hirundinella является самым обычным
компонентом пресноводного планктона.
Рис. 21. А, Б —Ceratium hirundinella', со спинной стороны (Л) и с брюшной
стороны (Б); В, Г—Ceratium tripos, в начале деления (В) и после расхожде-
ния половин клетки (Г); Д —Ceratium longipes; В —Ceratium arcticum;
Ceratium fusus.
1 — апикальный рог, 2 — антапикальный рог, 3— эпитека, 4 — гипотека,
5 — продольная борозда, 6 — поперечная борозда.
Все церациумы характеризуются присутствием больших длинных
выростов, называемых рогами. На верхней половине тела находится
только один апикальный рог, на нижней половине в большинстве
случаев два антапикальных рога. Поперечная борозда резко вырЯ'
жена, продольная борозда широкая. Поверхность панцыря покрыта
мелкими шипами или морщинистая. Многочисленные хроматофорь'
обычно желтого цвета. Питание автотрофного типа. Некоторые
виды способны образовывать цепочки из нескольких не разошед-
шихся после деления особей. Наиболее обычные виды Ceratium на-
ших северных морей отличаются между собою следующими приз-
наками.
1 (2) Один из антапикальных рогов рудиментарный .
2 (1) Оба антапикальных рога хорошо развиты
С. fusM
3 (4) Антапикальные рога закрытые, их концы оттянуты; апи-
кальный рог прямой....................... .С. tripos
4 (3) Антапикальные рога открытые, их концы как бы обруб-
лены; апикальный рог изогнутый 5
5 (6) Рога окаймлены пленочной оторочкой С. arcticum
6 (5) Рога без оторочки С. longipes
(Между G arcticum и С. longipes имеется ряд переходных форм).
В Черном море встречаются Ceratium tripos, С. fusus и С. furca
(антапикальные рога прямые). *
3. Bythotrephes (рис. 22)
Bythotreph.es относятся к семейству Polyphemidae. Представители
этого семейства отличаются от остальных ветвистоусых раков ря-
дом существенных признаков. Карапакс (раковина) сильно реду-
цирован, не покрывает всего тела, а защищает лишь находящуюся
внутри него выводковую камеру и по краям полностью прирастает
к спинной стороне тела. Таким образом выводковая камера, покры-
тая гиподермой, лежит под покровами тела, т. е. внутри организ-
ма, и представляет образование, совершенно отличное от выводко-
вой камеры всех остальных ветвистоусых раков, у которых она на-
ходится вне тела, между его спинной стороной и створками кара-
пакса. Гиподермальные клетки дна выводковой камеры преобразо-
ваны у всех Polyphemidae в большой железистый орган, выделяю-
щий богатый питательными веществами секрет, за счет которого
протекает развитие эмбрионов. Не менее резко отличаются предста-
вители семейства Polyphermidae по строению глаза и грудных ног.
Bythotrephes относится к числу самых крупных видов, достигает
в длину 10 мм; обитает, главным образом, в северных районах, в
озерах и небольших водоемах.
Характерным признаком Bythotrephes является присутствие очень
длинного хвостового шипа, в несколько раз превышающего длину
тела и сильно увеличивающего сопротивление погружению, так как
в нормальном положении он расположен горизонтально. Этот шип
гомологичен тому небольшому выросту, на котором у всех осталь-
ных ветвистоусых раков сидят хвостовые щетинки. Голова большая
с очень крупным сложным глазом, занимающим всю ее переднюю
часть. Глазок отсутствует. На спинной стороне головы расположен
спинной щит — орган дыхания. Антеннулы короткие, отходят от об-
щего, на конце раздвоенного основания. Антенны сильно развиты,
с трехчленистыми эндоподитами и четырехчленистыми эксоподитами,
служат органом движения. Мандибулы большие, с рудиментарным
Щупиком. Максиллулы рудиментарные. Грудные ноги, в числе че-
тырех пар, скелетного типа, состоят из' крепких, подвижных члени-
ков, приспособленных к захвату добычи Эпиподиты отсутствуют,
конечности не несут дыхательной функции. На конце тела находит-
ся фурка, образованная двумя фуркальными когтями; иногда мож-
но видеть, кроме того, еще одну или несколько пар ложных когтей,
являющихся пустыми оболочками фурки предыдущих возрастных
Рис. 22. А, Б — Bythotrephes longimanus, самка (А), мандибула (Б); В
thotrephes cederstromi, хвостовой шип; Г — разрез через выводковую к-амерУ
Bythotrephes.
1 антеннула, 2 — антенна, 3 — мандибула, 4 — глаз, 5 — головной щит,
6 сердце, 7 — ноги первой-четвертой пар, 8—раковина, 9 — выводковая
мера, 10 эмбрион, И — каудальные когти, 12 — ложные каудальные ког^’
13 хвостовой шип, 14 — железистый орган.
52
стадий. Хитин хвостового шипа и хитин фуркальных когтей при
линьке не сбрасывается; таким образом число пар ложных когтей
соответствует числу линек.
У нас встречается два вида Bythotrephes — В. longimanus (хво-
стовой шип совершенно прямой) и В. cederstromi (на конце хво-
стового шипа имеется слабый изгиб, вооруженный шипиками).
4. Notholca (рис. 23, Л)
Одним из обычных представителей зоопланктона чистых озер яв-
ляется Notholca longispina, относящаяся к панцирным коловрат-
кам.
Рис. 23. A — Notholca longispina; Б —личинка краба Porcellana.
1 — антеннула, 2 — антенна, 3 — рострум, 4 — переоподы.
Панцырь гладкий, трехгранный, его передний край вооружен
шестью шипами; два срединных шипа различной длины, правый в
несколько раз длиннее левого. Задний конец тела с длинным игло-
видным шипом. На брюшной стороне задней части тела находится
углубление, в котором прикрепляется яйцо. В передней части тела
расположен глазок.
5. Личинки Porcellana (рис. 23, Б)
Личинки краба Porcellana, встречающегося у нас в Черном море,
ведут планктический образ жизни. Наиболее отличительной чертой
является присутствие длинного рострального выроста, в два раза
превосходящего длину тела; задние углы карапакса вытянуты в
длинные выросты. Рострум и выросты карапакса представляют
большое сопротивление погружению в воде, тонкие покровы также
облегчают плавание личинки.
ОРГАНИЗМЫ БЕНТОСА
Бентос состоит из организмов, живущих на дне водоемов и не
способных продолжительное время плавать в.воде. В систематиче-
ском отношении делится на растительный бентос, или фитобентос,
и животный бентос, или зообентос.
В противоположность планктическим организмам донные живот-
ные и растения не нуждаются в облегчении своего веса, поэтому
многие из них, в особенности живущие в прибрежной области, отли-
чаются крепостью строения и нередко значительным отложением
извести. Личиночные стадии большого числа морских бентических
животных входят в состав планктона, в пресных водах это являет-
ся лишь исключением- Между бентосом и планктоном нельзя про-
вести резкой границы. Имеется довольно большое количество жи-
вотных, главным образом из ракообразных и червей, которые могут
на некоторое время подниматься в толщу воды в период размно-
жения или для добывания пищи. Те из них, которые пребывают в
воде продолжительное время, могут, другими словами, вести и бен-
тический и планктический образ жизни, относятся к группе планк-
гобентоса или бентопланктона.
Строение бентических организмов в большей степени зависит
от характера субстрата, на котором они живут, а также от осве-
щенности, силы волнения и т. п. Поэтому наблюдаются сильные от-
личия в строении близких форм, живущих на мягком грунте или на
камнях, в зоне прибоя или на больших глубинах, на полном свету
или в темноте.
По отношению к субстрату организмы бентоса делятся на сле-
дующие группы.
1. Прикрепленные организмы (сессильный бентос).
Главная масса растительного бентоса относится к числу прикреп-
ленных форм; цветковые растения укрепляются обычно на мягком
грунте при помощи корневищ, большое количество водорослей при
крепляется к твердому субстрату своими ризоидами. Среди зообен-
тоса сидячими являются губки, гидроиды, кораллы, морские лилии,
многие черви, мшанки, многие двустворчатые моллюски, усоногие
раки, асцидии и ряд других животных. Общая форма тела прикреп-
ленных животных обычно вытянутая. Очень часто они являются ко-
54
лониальными организмами, как, например, губки, гидроиды, корал-
лы и мшанки, образующие колонии путем почкования- Органы дви-
жения обычно редуцируются или изменяют свою функцию. При-
крепленные животные, несмотря на отсутствие передвижения, легко
распространяются благодаря образованию свободноплавающих ли-
чиночных стадий, переносимых течениями. Животные могут вести
сидячий образ жизни только в водной среде, так как только в ней'
они могут получать необходимую им пищу в виде приносимого во-
дою планктона или опускающегося сверху органического детрита.
Среди пресноводных животных особенно сильно развиты органы
прикрепления у реофильных форм, живущих на быстром течении.
Форма тела уплощенная, обтекаемая. Прикрепление происходит
благодаря различным присоскам, прицепкам (личинки ряда насе-
комых) .
2. Лежащие организмы. Животные, лежащие на мягком
грунте, отличаются сильно расширенным и низким телом. Много
плоских форм находится среди донных рыб, например камбаловых,
а также головоногих моллюсков. Плоской формой тела обладают
также некоторые крабы, двустворчатые моллюски, морские ежи и
другие животные, некоторые из них имеют выросты, расположенные
в одной плоскости.
3. Закапывающиеся организмы. Закапывающиеся в
грунт животные, совокупность которых называется инфауной, в про-
тивоположность эпифауне, представленной, главным образом, при-
крепленными и свободно двигающимися организмами, встречаются
во многих группах животного мира, преимущественно среди червей,
морских ежей, голотурий, брюхоногих и двустворчатых моллюсков,
плеченогих, ракообразных, личинок насекомых и ряда других групп.
Многие животные погружаются в грунт в защитных целях. Они жи-
вут в ходах или трубках, часто укрепленных какими-либо выделе-
ниями; длина ходов иногда в несколько раз превышает длину са-
мого организма. Некоторые животные свободно двигаются в грунте,
поглощая его для извлечения находящегося в нем органического ве-
щества или активно отыскивая добычу.
Закапывание в грунт вызывает ряд изменений в строении жи-
вотных. Закапывающиеся в песок неправильные морские ежи лише-
ны аристотелева фонаря, их иглы превращены в органы копания.
Раковина живущих в грунте моллюсков становится гладкой, тон-
кой, неплотно закрывающейся; хорошо развитая нога лишена бис-
сусной железы; для сообщения с внешней средой служат длинные
сифоны, нередко превышающие длину самого животного.
4. Сверлящие организмы. Сверлению подвергаются
плотные осадочные породы, скалы, сложенные из известняка, пес-
чаника, сланцев и даже гранита, а также мрамор, бетон, кирпич,
дерево и раковины моллюсков. К числу морских сверлящих орга-
низмов относятся некоторые водоросли, губки, черви, моллюски и
раки. В пресных водах наиболее обычными сверлящими животны-
ми являются личинки некоторых насекомых, минирующие листья и
стебли водных растений или делающие ходы в глинистых берегах.
55
Водоросли, а из животных губки, черви и некоторые моллюски
прокладывают ходы в известняке или в раковинах при помощи вы-
деляемой кислоты, растворяющей известь. Некоторые моллюски ме-
ханически сверлят горные породы и дерево зубцами и гребнями,
расположенными на раковине; представители бокоплавов и равно-
ногих раков сверлят дерево сильно развитыми ротовыми придатка-
ми. Сверлящие организмы обычно никогда не покидают своего жи-
лища, увеличивая его объем по мере роста, поэтому, по существу,
являются узниками. Питание происходит за счет взвешенных в воде
мелких организмов планктона и органического детрита; животные,
поражающие дерево, могут питаться древесиной. Наличие свобод-
ноплавающих личинок обусловливает широкое распространение
сверлящих организмов.
5. Свободно двигающиеся организмы (вагильный
бентос). Многие животные двигаются по дну при помощи различно
устроенных конечностей, иглокожие обладают амбулакральными
ножками, органом движения моллюсков служит нога, простейшие
двигаются при помощи ресничек или псевдоподий. Некоторые ра-
стения, как, например, донные диатомеи, также обладают способ-
ностью к движению.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
ПРИКРЕПЛЕННЫЕ ОРГАНИЗМЫ
В качестве примеров прикрепленных организмов рассматрива-
ются представители губок (Spongilla и Ephydatia), двустворчатых
моллюсков (Ostrea), усоногих раков (Balanus и Chthamalus), ли-
чинки блефароцерид (Blepharoceridae), личинки мошек (Simu-
Нит).
1. Spongilla и Ephydatia (рис. 24)
Большинство губок относится к морским животным, в пресно-
водных водоемах встречается лишь небольшое число видов. Во
взрослом состоянии губки лишены способности к передвижению, яв-
ляются типичными сидячими организмами, прикрепленными к ка-
кому-либо твердому субстрату; только некоторые морские формы
свободно лежат на дне. Среди губок встречаются одиночные и ко-
лониальные виды; последние могут быть самого различного строе-
ния— корковые, шаровидные, ветвящиеся, стебельчатые и т- п. От-
личительной особенностью всех губок является характерный запах,
по которому они легко узнаются в живом состоянии. В теле губок
находятся многочисленные полости, выстланные воротничковыми
клетками (хоаноцитами), при посредстве которых образуется ток
воды, входящий через многочисленные поры и выходящий через
крупное отверстие (устье, или оскулум). Почти все губки обладают
скелетом, который может быть известковым, кремневым, кремнеро-
говым и роговым.
гл
Рис. 24. А—Г —Spongilla lacustris, колония (Л), геммула (Б), макросклеры (5),
•‘еммульные микросклеры (Г); Д—Ж — Spongilla fragilis, геммулы (Д), макро-
•клеры (£), геммульные микросклеры (Ж); 3—К — Ephydatia fluviatilis, гем-
мула (3), макросклеры (//), амфидиски (Ю; Л Н Ephydatia mulled,
колония (Д)> мдкросклеры (^), амфидиски (Я).
Встречающиеся в пресноводных водоемах губки относятся к от-
ряду кремнероговых губок (Cornacuspongida). Тело покрыто тон-
кой дермальной мембраной, под которой расположены субдермаль-
ные полости. Через многочисленные мелкие поры в дермальной
мембране вода, вместе с кислородом и мельчайшими пищевыми ча-
стицами, входит по приводящим каналам внутрь губки, омывает
хоаноциты и проходит по отводящим каналам и протокам к оску-
лярным отверстиям, окруженным оскулярными трубками; послед-
ние у живых губок достигают длины в несколько миллиметров.
Около оскулярных отверстий видны просвечивающие сквозь дер-
мальную мембрану отводящие протоки, образующие неправильную
звездчатую фигуру.
Скелет состоит из кремневых одноосных спикул, связанных на
концах спонгином. У большинства пресноводных губок к концу лета
образуются покоящиеся стадии, геммулы, имеющие шаровидную
форму и обычно окрашенные. Весною клеточная масса выходит из
геммул и развивается в молодые губки; расположенные рядом гу-
бочки сливаются затем в общую колонию, часто окрашенную в зе-
леный цвет благодаря присутствию клеток симбиотической водоро-
сли Chlorella.
Пресноводные губки определяются по строению геммульных
микросклер. Последние представлены прямыми или изогнутыми од-
ноосными и чаще всего шиповатыми рабдами и амфидисками, напо-
минающими по форме катушку. Приготовление препарата описано
на стр. 88f *
Наиболее обычные виды пресноводных губок отличаются между
собою следующими признаками.
1 (4)
2 (3)
3 (2)
4 (1)
5 (6)
6 (5)
Геммульные микросклеры представлены рабдами (род
Spongilla) . 2
Геммулы одиночные 5. lacustris
Геммулы соединены в группы, окруженные общим слоем
воздухоносной ткани .........................S. fragilis
амфидисками;
также слабо
Е. fluviatilis
Геммульные микросклеры представлены
края дисков с зубцами (род Ephydatia)
Скелетные спикулы гладкие, встречаются
шиповатые спикулы....................
Кроме гладких скелетных спикул, имеются шиповатые или
все спикулы шиповатые Е. miilleri
2. Ostrea (рис. 25)
Устрицы (Ostrea), относящиеся к двустворчатым моллюскам,
обитают в теплых и умеренных морях.
Прикрепленный образ жизни вызвал у устриц ряд сильных из-
менений в строении. Раковина неправильная и очень изменчивая;
сложена снаружи из известковых листочков. Створки различные.
Левой, выпуклой, створкой моллюск прирастает к твердому суб-
страту, неровности которого вызывают соответствующую неровность
58
раковины; правая створка плоская; внутренняя поверхность рако-
вины выстлана фарфоровидным слоем. Замковые зубы не развиты
Нога и биссуспая железа отсутствуют. Края мантии свободные, бег
сифонов.
В Черном море часто встречается Ostrea taurica, являющаяся
объектом промысла; образует устричные банки; створки часто про-
низаны ходами губки Cliona.
Рис. 25. Ostrea taurica.
Другой менее расйространенный вид Ostrea sublamellosa отли-
чается от предыдущего по толстым многослойным створкам и по
очень широкой и длинной лигаментной площадке.
3. Balanus и Chthamalus (рис. 26)
К числу широко распространенных прикрепленных животных от-
носятся усоногие раки (Cirripedia), входящие в состав низших ра-
ков (Entomostraca). Наиболее обычными представителями являются
морские желуди, или, по-поморски, юртики (Balanidae), в массе
поселяющиеся на скалах, камнях, сваях, нередко также на донных
животных.
Тело морских желудей имеет обычно уплощенную форму и це-
ликом, вместе со всеми конечностями, заключено в состоящий из
известковых пластин домик, приросший к какому-либо твердому
субстрату.
Скелетные пластины подразделяются на две части — одни обра-
зуют крепкий домик, состоящий из непарных carina и rostrum и
двух nap lateralia, другие же, а именно парные scuta и terga, вхо-
дят в состав двускатной крышечки, operculum, подвижно соединен-
ной с домиком. Отверстие, образующееся при раздвигании обеих по-
ловин крышечки, располагается почти в горизонтальной плоскости.
Пластины домика крепко соединены между собою при помощи тон-
ких выростов, заходящих друг за друга; внутренний вырост назы-
вается ala, наружный — radius. Внутри домика находится тело ор-
ганизма, прикрепленцое передним концом к субстрату и обращен-
ное своей брюшной стороной к отверстию домика. Рудиментарные
антеннулы являются органом прикрепления. Антенны отсутствуют.
59
Рис. 26. А, Б — Balanus balanoides, домик сверху (А) и изнутри (В)» до-
Balanus balanus, домик (В), крышечка (Г), часть вскрытой пласти /^К),
мика (Д); Е—3— Balanus crenatus, домик {£), вытянутая форма домрка
крышечка (3); И —Balanus bameri, домик; К —Chthamalus stellatus,
Л — схема строения домика Balanus; М — схема строения домика China
Я —скелетные пластины домика Ba|anidae. »
/ — carina, 2 —lateralia, 3 —rostrum, 4 —scutum, 5 — tergum, 6 — alae (за
хована вертикально), 7 — radii (заштрихованы горизонтально) •
Грудные ноги, в числе шести пар, состоят из двучленистого бази-
подита и многочленистых эндоподитов и эксоподитов, несущих длин-
ные щетинки; ноги, высовываясь наружу через отверстие в домике,
образуют специальный орган для захватывания пищи, состоящей из
детрита и организмов планктона. Личиночные стадии усоногих ра-
ков, встречающиеся в весеннем планктоне в огромных количествах,
служат пищей планктоядным рыбам.
В наших северных морях встречается несколько видов Balanus,
отличающихся между собою следующими признаками.
1 (2) Основание домика мягкое, перепончатое В. balanoides
2 (1) Основание домика известковое . 3
3 (4) Пластины домика без каналов ... В. hameri
4 (3) Пластины домика пронизаны внутри вертикальными
каналами . . ... . ... 5
5 (6) Tergum с длинной, клювовидной вытянутой вершиной
В. balanus
6 (5) Tergum без клювовидно вытянутой вершины .
В. crenatus
В Черном море обычной формой побережья является Chthama-
lus stellatus. Основание домика не обизвествлено. Непарный rost-
rum с внутренними выростами (alae), по этому признаку отлича-
ются от Balanus, rostrum которых с наружными выростами (radii).
4. Личинки и куколки Blepharoceridae (рис. 27)
Блефароцериды относятся к двукрылым насекомым (Diptera, се-
мейство Blepharoceridae). Крылатые особи похожи на небольших
комаров с длинными и тонкими1 ногами. Личинки и куколки живут
исключительно в горных потоках с большой скоростью течения. У
нас встречаются в горах Кавказа, Алтая и Средней Азии.
Вся организация личинок блефароцерид хорошо приспособлена
к Ж.ИЗНИ в быстро текучих водах. Они сильно уплощены в дорсо-
вентральном направлении; сверху покрыты твердым хитином, ниж-
няя сторона, обращенная к субстрату, мягкая. Тело состоит из семи
резко отделенных сегментов. Первый, головной, образован слив-
шимися между собою всеми первичными сегментами головы и гру-
ди и первым сегментом брюшка. На голове находятся два неболь-
ших глаза и антенны, одночленистые и длинные или двучленистые
и короткие. На нижней стороне тела по средней линии расположе-
ны шесть мощно развитых присосок (одна на голове, остальные на
брюшных сегментах), при помощи которых личинки крепко прикреп-
ляются к твердому субстрату, обычно к скадам и камням, а также
могут двигаться в боковом направлении. Каждая присоска состоит
из широкого диска, посреди которого расположено полушаровидное
углубление, заполненное большим хитиновым конусом. Стенки уг-
лубления выстланы толстым слоем хитина, поэтому оно всегда со-
храняет свою форму. Периферийная часть присасывательного диска
образована мягкой кожей, легко облегающей неровности суб-
страта. Эта часть диска укреплена многочисленными гибкими хити-
новыми лучами, стилетовидной и мечевидной формы. На нижней сто-
61
Рис. 27. А, Б — личинка Liponeura
kaukasica, сверху (А) и снизу (Б);
В _ куколка Liponeura kaukasica;
Г—Е — личинка Blepharocera аг-
menica. вид снизу (Г), присоска о
(Д), поперечный разрез через при-
соску (Е).
1 — антенна, 2 — прйсоска, 3 —
анальные папиллы, 4 — трахейные
жабры, 5 — щупальцевидный прида-
ток. 6 — когтевидный придаток, 7 —
хитиновый конус, 8 — мускулы, под-
нимающие хитиновый конус, 9 — хи-
тиновые лучи, 10 — полый шип.
роне диска находятся шесть полых шипов, по которым стекает сек-
рет железистых клеток; другие железистые клетки, секрет которых
также смачивает нижнюю поверхность диска, расположены по его
краям. Прикрепление присоски происходит при сокращении силь-
ного мускула, поднимающего хитиновый конус, что вызывает обра-
зование вакуума. Крепость прикрепления зависит также от клей-
кого секрета, выделяемого железами диска. Личинки настолько
крепко присасываются, что легче их разорвать, чем оторвать от суб-
страта. По своему строению присоски блефароцерид представляют
самый совершенный аппарат такого рода среди всего животного
мира.
По краям сегментов находятся сильно хитинизированные нечле-
нистые когтевидные придатки, а у ряда видов, кроме того, щупаль-
цевидные придатки, расположенные выше первых. С нижней сто-
роны брюшных сегментов по бокам находятся пучки трахейных
жабр, состоящие из пяти-семи тонкостенных выростов; две пары
анальных папилл расположены на конце тела около анального от-
верстия.
Наиболее часто встречаются личинки Blepharocera, отличаю-
щиеся присутствием по бокам первых шести сегментов брюшка
только одних когтевидных придатков, и личинки Liponeura, у кото-
рых, кроме когтевидных придатков, имеются также щупальцевид-
ные придатки, а пучки трахейных жабр обстоят из семи выростов.
Куколки блефароцерид настолько крепко приклеиваются к суб-
страту, что всегда повреждаются при их отрывании. На переднем
конце куколок находятся пластинчатые трахейные жабры. С ниж-
ней стороны видны хорошо развитые крылья, начинающие функцио-
нировать почти сейчас же после вылупления насекомого.
5. Личинки и куколки Simulium
Описание строения личинок и кукулок Simulium помещено на
стр. 182.
ЛЕЖАЩИЕ ОРГАНИЗМЫ
В качестве примеров рассматриваются представители двуствор-
чатых моллюсков (Pecten).
Pecten (рис. 28)
Гребешки (Pecten), обычно имеющие крупные размеры и являю-
щиеся съедобными моллюсками, относятся к числу широко распро-
страненных морских животных. Лежат свободно на грунте, могут
также проплывать некоторое расстояние, делая прыжки.
Раковина, более или менее сильно уплощенная, с многочислен-
ными ребрами, проходящими радиально; видны также концентриче-
ские линии роста. Створки у одних видов почти совершенно одина-
ковые, у других — правая створка более выпуклая, по сравнению
с левой. Замковый край прямой, образован большими выступами ра-
ковины, называемыми ушками; переднее ушко обычно больше зад-
63
него, с выемкой для выхода биссуса в правой створке, на которой
моллюск лежит на грунте. Замочные зубы отсутствуют. Внешний
роговой лигамент проходит по замковому краю; внутренний лига^
мент, соединяющий створки у вершины, находится в треугольном
углублении.
Рис.28.Л—В—Pecten islandicus, наружный вид правой створки (Л), внутрен-
нее строение после удаления правой створки (Б) и после удаления правой
складки мантии (В); Г — Pecten ponticus.
/ — заднее ушко, 2 —переднее ушко, 3 — вершина раковины (макушка), 4 —
биссусная выемка, 5 — наружный лигамент, 6 — внутренний лигамент, 7 — пе-
чень, 8 — перикардий, 9— сердце, 10 — кишечник, // — передний и задний
отделы мускула аддуктора, 12 — правая мантийная складка, 13— мантийные
мышцы, 14 — мантийные щупальца, /5 —парус, 16 — глазки, 17 — правые по-
лужабры, 18 — гонада, 19 — биссусная борозда, 20 — нога, 21 — ротовое отвер-
стие, 22 — верхняя и нижняя губы, 23 — ротовые лопасти.
64
Удалив одну из створок, для чего подрезают скальпелем боль-
шой замыкательный мускул (аддуктор), рассматривают, поместив
моллюска в воду, его внутреннее строение. Складки мантии, сво-
бодные почти на всем протяжении, срастаются между собою только
на спинной стороне. Свободный край мантии загибается внутрь об-
разуя парус, и несет многочисленные щупальца и глазки. Внутри па-
руса проходит кольцевая мышца. Парус играет главную роль в ре-
активном движении гребешка. При быстром закрывании сгворок
вода не мож,ет выйти наружу по брюшному краю раковины, так
как оба паруса, смыкаясь своими краями, препятствуют этому. По-
этому вода, находящаяся внутри мантийной полости, с силой вы-
брасывается через два отверстия, расположенные спереди и сзади
замкового края, т. е. в тех местах, где парусы не развиты. Вслед-
ствие этого моллюск делает скачок брюшным краем вперед. Попе-
ременно открывая и закрывая раковину, гребешок проплывает до-
вольно значительное расстояние. Непосредственно под внутренним
лигаментом находится печень, ниже ее с задней стороны располо-
жены сердце и прямая кишка, огибающая замыкательный мускул,
а с передней стороны — ротовое отверстие с ротовыми лопастями и
рудиментарная нога с биссусной бороздой, в которой открывается
биссусная железа. Между жабрами расположены большие гонады.
В северных морях встречаются Pecten, islandicus, красновато-ро-
зового цвета, до 80 мм в поперечнике, с многочисленными (до 35)
узкими ребрами; передние ушки немного больше задних. В Черном
море находится Pecten ponticus, желтовато-бурого или темного цве-
та, до 50 мм в поперечнике, с небольшим числом ребер (не более
14); ушки приблизительно одного размера; правая створка выпук-
лая, левая плоская.
ЗАКАПЫВАЮЩИЕСЯ ОРГАНИЗМЫ
В качестве примеров рассматриваются представители червей
(Arenicola), двустворчатых моллюсков (Solen) и личинки поденок
(Polymitarcys, Pallngenia и Ephemera).
1. Arenicola (рис. 29, А—Д)
Пескожилы (Arenicola marina), обычные представители много-
щетинковых червей (Polychaeta), живут в самой прибрежной обла-
сти морей. Строят в песке или илу U-образные норы, выстланные
внутри слизью и открытые с обоих концов; длина нор достигает не-
скольких десятков сантиметров. Заглатывают грунт, наподобие
дождевых червей; относятся к числу детритоядных животных. Вы-
брошенные из кишечника фекалии образуют на поверхности грунта
характерного вида скопления. Пескожилы часто используются в
виде наживки для лова рыбы на крючковую снасть.
Тело пескожилов с вторичной сегментацией, длина достигает
20 см и более. Простомиум гладкий, без придатков. Глотка вывора-
чивающаяся, с мелкими папиллами. Параподии, как и у других
живущих в грунте видов, резко разделены на две обособленные
65
ветви — брюшную, или невроподиальную, и спинную, или нотоподи,
альную. Пораподни такого типа называются седентарными. Продод,
говатые валики невроподий вооружены слабо изогнутыми крючко.
Рис.29. А—Г. — Arenicola marina, общий вид (А), конец нотоподиальной ше'
тинки (Б); невроподиальная щетинка (В), жабра (Г); Д—нора Arenicola;
Е — Solen marginatus в. норе.
1 — глотка, 2 — буккальные папиллы, 3 — нотоподии, 4 — невроподии,
5 —жабры, 6 — анальное отверстие.
видными щетинками; конусовидные бугорки нотоподий с пучком
шиповатых щетинок. Грудные сегменты несут только параподии;
далее следует передний отдел брюшка, отличающийся присутствием
€6
13 пар жабр, расположенных позади нотоподий. Каждая жабра
состоит из 8—14 древовидно ветвящихся выростов, связанных при
основании топкой перепонкой. Задний, варьирующий по длине, от-
дел брюшка значительно сужен и лишен пораподий и ж,абр.
2. Solen (рис. 29, Е)
Черенки (Solen), относящиеся к морским двустворчатым мол-
люскам, живут в песке, в котором строят вертикальные норы, глу-
биною до полуметра и даже до трех метров. Сечение норы оваль-
ной формы в соответствии с поперечником моллюска. Черенки
обычно живут у поверхности грунта, при опасности прячутся в но-
ру, передвигаясь в ней при помощи своей ноги.
Раковина Solen прямая, почти цилиндрическая, сильно вытяну-
тая в поперечном направлении, с параллельными спинным и брюш-
ным краями, открытая на обоих концах. Передний конец раковины
с неглубоким желобком. Створки тонкие, блестящие, с изогнутыми
под прямым углом линиями роста. Замок, редуцирован, с одним ли-
стовидным зубом на каждой створке. Лигамент наружный, удли-
ненный.
Мантия почти на всем протяжении замкнутая, за исключением
отверстия для ноги и небольшого брюшного отверстия. Сифоны ко-
роткие, соединенные, на конце бахромчатые. Нога длинная, цилин-
дрическая, на конце вздутая. Биссусной железы нет. Жабры узкие.
Черенки, встречающиеся в Черном море, относятся к виду Solen
marginatus.
3. Личинки Polymitarcys, Palingenia и Ephemera (рис. 30)
Роющие личинки поденок по своему строению резко отличаются
от личинок других видов. Они относятся к числу редко находимых,
хотя образующиеся из них крылатые насекомые в некоторых мест-
ностях иногда встречаются в очень больших количествах. Это объ-
ясняется их скрытым образом жизни. Почти всю свою жизнь рою-
щие личинки проводят в ходах, проделываемых ими в илистом грун-
те озер и речек (Ephemera) или в глинистых берегах больших рек
(Polymitarcys, Palingenia). Они строят прямые или U-образно изо-
гнутые ходы при помощи своих мощно развитых мандибулярных
зубцов, далеко выступающих за передний край головы; размельчен-
ный материал отбрасывается грудными ногами. Ходы в глинистых
берегах, местами встречающиеся на протяжении целых километров,
располагаются непосредственно под поверхностью воды; при силь-
ном изменении уровня воды личинки строят себе новые жилища.
Личинки Ephemera отличаются следующими признаками. Перед-
ний край головы продолжается за основания антенн в виде боль-
шого выступа. Антенны многочленистые, с длинными щетинками, в
особенности в нижней части. Мандибулы характерного строения —
от наружного края отходит длинный, тонкий и слегка изогнутый
зубец, далеко выступающий вперед. Максиллярный щупик трехчле-
нистый. Губной щупик двучленистый. Ноги сильные, приспособлен-
5*
67
ные для копания, с многочисленными длинными щетинками. Тра
хейные жабры первой пары рудиментарные, представлены двум;
трубчатыми выростами с каждой стороны первого брюшного сег
мента. Остальные шесть пар большие, загнутые на спинную сто
Рис.30.А—В — личинка Ephemera vulgata, вид сверху (Л), мандибула (£)»
трахейная жабра (В); Г—Ж — личинка Palingenia longicaudata, вид сверху (Г),
мандибула (Д), нога первой пары (В), трахейная жабра (Ж)\ 3—К —личинка
Polymitarcys virgo, вид сверху (3), мандибула (И), трахейная жабра (ТС); Л—
ходы личинок Polymitarcys.
68
рону брюшка. Каждая жабра состоит из двух лопастей, соединен-
ных при основании и окаймленных с обеих сторон многочислен-
ными тонкими и очень длинными выростами. Хвостовые нити короче
брюшка, вооружены волосками с обеих сторон.
Личинки двух обнаруженных у нас видов Ephemera, из которых
чаще всего встречаются личинки Е. vulgata, отличаются между со-
бою по следующим признакам.
Ephemera vulgata — головной вырост впереди с глубокой выем-
кой, его боковые края прямые; Е. danica— головной вырост впере-
ди с неглубокой выемкой, его боковые края выпуклые.
Личинки Polymitarcys отличаются от других роющих личинок
следующими признаками. Передний край головы без выступа. Ан-
тенны многочленистые, значительно длиннее головы. Мандибулы
крепкие, с большим слегка изогнутым на внутреннюю сторону зуб-
цом, несущим по наружной поверхности многочисленные зубчики.
Максиллярный и губной щупики двучленистые. Ноги первой пары
приспособлены для копания. Трахейные жабры первой пары руди-
ментарные, представлены небольшими трубочками, скрытыми под
зачатками крыльев. Трахейные жабры остальных шести пар боль-
шие, загнутые на спинную сторону брюшка. Каждая жабра имеет
вид широкой пластинки, разделенной на две лопасти, согнутые при
основании и окаймленные многочисленными тонкими короткими вы-
ростами. Хвостовые нити длиннее брюшка, с короткими волосками
на обеих сторонах. Единственным видом у нас является Polymitar-
cys virgo.
Личинки Palingenia являются наиболее специализированными
среди роющих личинок поденок. Мандибулы снабжены большими,
широкими выростами, вооруженными по наружному краю шестью-
восемью острыми зубцами. Голени ног первой пары плоские и ши-
рокие, с длинными и крепкими шипами по внешнему краю. Трахей-
ные жабры первой пары рудиментарные, остальных шести пар—
двуразделенные, перистые.
Встречающиеся у нас два вида Palingenia, Р. longicauda на за-
паде и Р. sublongicauda в центральных районах и в Сибири, в ли-
чиночном состоянии пока еще не могут быть различимы.
СВЕРЛЯЩИЕ ОРГАНИЗМЫ
В качестве примеров рассматриваются представители двуствор-
чатых моллюсков (Teredo, Pholas, Barnea, Petricola) и ракообраз-
ных (Limnoria).
1. Teredo (рис. 31, A—Г)
Тередо, или карабельный червь (Teredo), является самым опас-
ным вредителем деревянных судов и различного рода портовых
сооружений. Живет в теплых и умеренных морях; не выносит по-
ниженной солености и низких температур. У нас встречается в Чер-
ном море и дальневосточных морях.
По внешнему виду тередо совершенно не похож на двустворча-
того моллюска, поэтому на всех языках получил название червя.
Тело мягкое, червеобразное, длиною до 25 см (в Черном море).
69
Тередо живут в дереве, просверливая ходы, увеличивающиеся
в диаметре по мере роста моллюска; никогда не покидают своего
жилища. На переднем конце находится небольшая шаровидная бе-
лая раковина, превращенная в орган сверления. Обе известковые
Рис. 31. Л —Teredo navalis внутри куска дерева; Б —передний конец тела
Teredo; В — раковина Teredo снаружи; Г — паллеты Teredo navalis; Д — Pho-
las dactylus; Е—Barnea Candida; M— Petricola lithophaga; 3—норы Petricola.
1 — раковина, 2 — ротовое отверстие, 3 — нога, 4 — паллета, 5 — сифоны.
створки неправильной формы, широко раскрытые. Каждая створка
состоит из трех частей; передние и средние части вооружены пиль-
чатыми и зубчатыми гребнями, при помощи которых тередо свер-
лит дерево. Спереди, между створками, находится небольшая нога,
на спинной стороне которой расположено ротовое отверстие в виде
70
узкой поперечной щели. Биссусная железа отсутствует. На конце
длинного тела, заключающего внутренние органы животного, нахо-
дятся вводной и выводной сифоны и две известковых пластинки,
паллеты, которые защищают задний конец тела и закрывают в слу-
чае необходимости входное отверстие хода.
Тередо питаются измельченной древесиной, а также мелкими
планктическими организмами, приносимыми входящим током воды.
В Черном море встречаются два вида тередо — Teredo navalis
(паллеты глубоко вилообразно раздвоенные) и Т. utriculus (пал-
леты без выемки, листовидные). В Балтийском и Баренцевом морях
тередо встречаются лишь случайно. В дальневосточных морях оби-
тают Teredo sinensis (паллеты похожи на паллеты Т. navalis), а
также представители близкого рода BanKia (BanKia setacea), отли-
чающиеся елочковидной формой паллет.
2. Pholas и Вагпеа (рис. 31, Д—Е)
Фолады (Pholas и Вагпеа) сверлят камень, плотные осадочные
породы и дерево. В противоположность тередо, их тело целиком за-
ключено в раковину. Живут в морях. У нас. встречаются в Черном
море и дальневосточных морях.
Раковина белая, хрупкая, более или менее зияющая. Наружная
поверхность створок впереди покрыта радиальными рядами зубчи-
* ков, при помощи которых моллюск сверлит себе убежище, расши-
ряя его по мере роста. Замок без зубов и лигамента. Края мантии
сросшиеся, за исключением отверстия для ноги. Сифоны длинные,
сросшиеся, на конце бахромчатые. Нога короткая, биссусная желе-
за отсутствует.
В Черном море встречается два вида семейства Pholadidae.
Pholas dactylus характеризуется следующими признаками — рако-
вина поперечно эллиптической формы, позади округлая, а впереди
вытянута в носик; радиальные ребра расположены на переднем
конце раковины, на местах их пересечения с концентрическими
складками находятся черепицевидные зубчики. Длина раковины до
8 с,м, длина норы до 24 см; живут в известняке и мергеле.
Вагпеа Candida отличается следующими особенностями — рако-
вина почти цилиндрическая, равномерно закругленная на обоих кон-
цах; радиальные ребра, наиболее развитые на переднем конце, пе-
ресекаясь с довольно редкими концентрическими складками, обра-
зуют остроконечные треугольные зубчики. Длина раковины до 4 см,
длина норы до 14 см; живут в плотном илу, мягком мергеле и
глине.
3. Petricola (рис. 31, Ж—3)
Petricola lithophaga относится к самым обычным кзмнеточцам
Черного моря, в некоторых местах почти каждый камень, поднятый
со дна, содержит несколько норок, пустых или с моллюсками.
Раковина овальная, сильно выпуклая на переднем конце. По-
верхность створок с радиальными ребрышками. Мантия толстая.
Сифоны довольно длинные. Нога небольшая. Норки, просверленные
в известняке, неглубокие (около 2 см глубиною), колбовидные, с
коротким горлышком.
71
4. Limnoria (рис. 32)
Лимнории (Limnoria lignorum) относятся* к равноногим ракам
(Isopoda). Поражают деревянные сооружения. Могут выдерживать
пониженную температуру воды, поэтому встречаются не только в
теплых, но и холодных морях.
Тело лимнории, длиною около 5 мм, продолговато овальной фор-
мы, покрыто мягким хитином. На голове находятся небольшие и
округлые сидячие глаза и короткие антеннулы и антенны. Манди-
г 2
Рис. 32. Limnoria lignorum. А — самка
сверху; Б — задний конец тела самца
снизу; В — мандибула.
1 — антеннула, 2— антенна, 3 — глаз,
4 — переоподы, 5 — коксоподиты (кок-
сальные пластинки), 6 — плеотельсон,
7 — уропод, 8 — плеоподы.
ности. Народившиеся молодые особи не
ответвления от ходов родителей.
булы, являющиеся глав-
ными органами сверления,
оканчиваются острыми
зубцами, мандибулярный
щупик трехчленистый.
Грудь состоит из семи
сегментов, несущих такое
же количество грудных
ног (переоподов), оканчи-
вающихся острыми крюч-
ковидными когтями; кок-
сальные пластинки хоро-
шо развиты. Брюшко со-
стоит из пяти сегментов;
двуветвистые брюшные
ноги (плеоподы) служат
органами дыхания, при их
движении вода обмени-
вается в ходах. Плеотель-
сон широкий, с щетинка-
ми по заднему краю.
Уроподы двуветвистые;
наружная ветвь короче
внутренней и согнута в
виде крючка.
Лимнория сверлит хо-
ды в наружном слое де-
рева, параллельно поверх-
выходят наружу, а делают
СВОБОДНО ДВИГАЮЩИЕСЯ ОРГАНИЗМЫ
К этой группе бентоса относятся общеизвестные представители
раков, иглокожих, личинок насекомых и ряд других организмов.
ОРГАНЫ ДВИЖЕНИЯ НЕКТОБЕНТИЧЕСКИХ ОРГАНИЗМОВ
К числу типичных представителей нектобентоса пресноводных
водоемов относятся некоторые водяные клопу и жуки. Вся органи-
зация этих насекомых связана с их образом жизни. Все они обла-
дают плотным, гладким телом, обтекаемая форма которого способ-
72
ствует быстрому движению в воде. Ноги последней пары или двух
последних пар видоизменены в плавательные конечности. Они более
или менее уплощены и снабжены длинными плавательными ще-
тинками, раскрывающимися при ударе и складывающимися при
заносе ног вперед; коготки часто редуцированы.
Рассматриваются плавательные ноги водяных клопов (Notonec-
ta) и жуков (Dytiscus, Gyrinus).
1. Notonecta (рис. 33, А—Б)
Гладыши (Notonecta) относятся к числу самых обычных водя-
ных клопов, обитают в стоячих водоемах. Плавают спиною книзу.
Брюшная сторона тела плоская, с многочисленными волосками, ок-
рашена в темный цвет; спинная сторона выпуклая, у живых особей
серебристая от находящегося под крыльями тонкого слоя воздуха.
Тело легче воды, поэтому при дыхании гладыши спокойно держатся
у поверхности воды, выставляя в воздух конец брюшка. При по-
мощи хорошо развитых крыльев гладыши легко перелетают из од-
ного водоема в другой.
Тело плотное, грудные и брюшные сегменты между собой тесно
соединены. Голова с большими сложными глазами, короткими че-
тырехчленистыми антеннами и хорошо развитым чётырехчленистым
хоботком. Надкрылья лежат крышеобразно. Ноги первых двух пар
служат для прикрепления к подводным растениям и для схватыва-
ния добычи. Каждая нога состоит из тазика (соха), вертлуга
(trochanter), бедра (femur), голени (tibia) и лапки (tarsus). Ноги
третьей пары, сильно удлиненные и уплощенные, служат для плава-
ния. Лапка двучленистая, первый членик длинее второго; коготков
не имеется. Голень и лапка приблизительно равны по длине, бедро
с вертлугом немного длиннее. На голени и на обоих члениках лапки
находятся длинные плавательные щетинки, складывающиеся при
заносе ног вперед и раскрывающиеся при ударе; раскрывание и
прижимание щетинок к членикам ноги легко обнаружить, проводя
препаровальной иглой вдоль края ноги.
Два наиболее обычных вида Notonecta отличаются между собою
по цвету треугольного щитка (scutellum), находящегося между ос-
нованиями надкрыльев, черного у N. glauca и желтовато-коричне-
вого у N. lutea.
2. Dytiscus (рйс. 33, В—3)
Плавунцы (Dytiscus), наиболее крупные из водяных жуков,
Длиною 20—40 мм, являются хорошими пловцами. Окрашены в тем-
нобурые или оливково-бурые цвета, с желтыми краями передне-
спинки и надкрыльев. Тело, уплощенное в дорсо-вентральном нап-
равлении, имеет обтекаемую форму, все сегменты между собою тес-
но соединены. По бокам головы находятся большие сложные глаза
и длинные 11-членистые антенны. Ротовые придатки грызущего ти-
Па- Верхняя губа узкая, слегка подвижная, мандибулы крепкие, с
Зубцами; максиллы расчлененные, с четырехчленистыми щупиками;
1иЖ.няя губа, прикрывающая ротовое отверстие снизу, пластинчатая,
73
с трехчленистыми щупиками. Грудь состоит из трех сегментов.
Крылья первой пары образуют твердые надкрылья (элитры) с про-
дольными желобками или же гладкие у самок и с несколькими ря-
дами точечных углублений у самцов. Для полета служат перепон-
чатые складывающиеся крылья второй пары. Конечности различ-
Рис. 33. А, Б— Notonecta glauca, вид сверху (Я), нога третьей пары (Б);
В—Е— Dytiscus marginatus, самка, вид сверху (В), нога пеовой пары сам-
ца (Г), нога третьей пары (Д), отростки задних тазиков (Е); Ж— Dytiscus
latissimus, поперечный разрез тела; 3 — Dytiscus lapponicus, отростки задних
тазиков.
/ — тазик, 2 — вертлуг, 3— бедро, 4— голень, 5 — лапка.
ного строения, они состоят из тазика, вертлуга, бедра, голени и
пятичленистой лапки, оканчивающейся двумя коготками. Ноги
двух первых пар приспособлены для ползания по твердому субстра-
ту и захватывания добычи, а у самцов, кроме того, для удержания
самок при копуляции. Для последней цели служат так называемые
присоски, расположенные на трех первых члениках лапок. На но-
гах первой пары эти членики образуют широкую пластинку, несу-
щую на внутренней стороне два больших и несколько десятков мел-
74
ких присасывательных дисков, сидящих на стебельках; число при-
сасывательных дисков на ногах второй пары достигает тысячи и
более. Присасывательные диски прикрепляются к субстрату при
помощи клейкой жидкости, выделяемой ими при надавливании. Но-
ги третьей пары служат для плавания и превращены в плаватель-
ные конечности. Тазики этих ног слиты с грудным сегментом и со-
вершенно неподвижны (эта особенность строения ног свойственна
только водяным жукам семейств Dytiscidae и Gyrinidae). Бедро, го-
лень и лапка — уплощенные. На дистальном конце голени находятся
два больших шипа, нижний край с плавательными щетинками.
Лапка пятичленистая, приблизительно вдвое длиннее голени, с хо-
рошо развитыми плавательными щетинками, а на конце с двумя
небольшими коготками. Плавательные конечности двигаются в го-
ризонтальной плоскости. При заносе ног вперед членики лапки,
повернувшись вокруг своей продольной оси на 90°, разрезают воду
передним краем, плавательные щетинки складываются; при после-
дующем ударе ног о воду членики лапки поворачиваются, щетинки
широко раскрываются, благодаря этому гребная поверхность ног
сильно увеличивается.
Наиболее обычные виды Dytiscus различаются между собою
следующими признаками.
1 (2) Боковой край'надкрыльев имеет вид тонкой острой
пластинки ..................................D. latissimus
2 (1) Боковой край надкрыльев не пластинчатый . 3
3 (4) Отростки тазиков ног последней пары короткие, с выпук-
лым внутренним краем..........................D. marginalis
4 (3) Указанные отростки вытянутые, с вогнутым внутренним
краем ................................................ 5
5 (6) Отростки умеренной длины; брюшко без пятен
D. circumcinctus
6 (5) Отростки длинные и очень острые; брюшко с темными
пятнами D. lapponicus
3. Gyrinus (рис. 34)
Вертячки (Gyrinus) относятся к числу самых быстро двигаю-
щихся водяных жуков. Обычно плавают на поверхности воды, при
приближении опасности погружаются в воду. Тело, длиною 3,5—
7 мм, имеет обтекаемую форму, его покровы водою не смачивают-
ся. Окраска черно-стального цвета, блестящая. Характерной осо-
бенностью жуков семейства Gyrinidae является строение фасеточ-
ных глаз, из которых каждый разделен хитиновой полоской на две
части, так что всего получается как бы четыре глаза. Верхние ча-
сти глаз направлены вверх, нижние — вниз, поэтому глаза служат
для одновременного зрения в воздухе и воде. Антенны короткие, с
сильно расширенным третьим члеником. Ротовые придатки сходны
по строению с соответствующими придатками представителей се-
мейства Dytiscidae. Надкрылья гладкие, с продольными рядами то-
чек. Ноги первой пары обычного строения, на конце с двумя ко-
готками. Ноги второй и третьей пар сильно видоизменены, пред-
75
ставляют самый совершенный аппарат для плавания в воде. Нац.
более развиты ноги третьей пары. Тазики слиты с грудным сегмец-
том; вертлуг и, в особенности, бедро и голень превращены в широ.
кие пластинки; очень характерно строение лапки: первые три чле-
ника снабжены большими пластинчатыми придатками, четвертый
Рис. 34. Gyrinus natator, вид сверху (Л), голова снизу (Б), нога первой
пары (В), нога третьей пары (Г).
1 — глаз, 2 — вертлуг, 3 — бедро, 4 — голень, 5 — лапка.
членик длинный, слегка изогнутый, пятый короткий, на конце с
двумя коготками. При ударе членики лапки расправляются и обра-
зуют дугу, внутреннее пространство которой закрывается пластин-
чатыми придатками, соприкасающимися своими краями. При зано-
се ног вперед пластинчатые выросты складываются наподобие пла-
стин веера. Плавательных щетинок не имеется; голень и членики
лапки окаймлены вместо них тонкостенными, на конце закруглен-
ными выростами. Возможно, что эти выросты служат не только
для увеличения гребной поверхности ног, но также и для дыхания.
Наиболее обычные виды Gyrinus различаются между собою
следующими признаками.
1 (2) Боковые края переднеспинки и надкрыльев темнометал-
лические ... . . G. marinas
2 (1) Боковые края переднеспинки и надкрыльев желтые 3
3 (4) Длина тела не менее 5 мм G. natator
4 (3) Длина тела менее 5 мм G. minutus
ОРГАНИЗМЫ НЕЙСТОНА
Нейстоном называется сообщество пленки поверхностного натя-
жения воды. Для развития нейстона требуется спокойное состояние
поверхности воды, поэтому нейстические организмы всего лучше
развиваются в небольших водоемах, защищенных от ветра деревь-
ями, строениями и t п. В озерах, где волнение часто нарушает
поверхностную пленку воды, нейстон отсутствует или встречается
лишь в хорошо защищенных от ветра заливах. Некоторые нейсти-
ческие организмы развиваются только при сильном освещении, дру-
гие требуют затемнения; большинство относится к числу сапроб-
ных организмов, развивающихся при наличии органического за-
грязнения.
Массовое развитие фитонейстона вызывает характерное окраши-
вание поверхностной пленки воды, например: коричневое от железо-
бактерий Chlamydothrix (=Leptothrix)\ пурпурно-фиолетовое от
пурпурных бактерий Chromatium; золотисто-желтое от хризомонад
Chromulina rosanoffi; светлозеленое от хламидомонад Chlamydo-
monas; красное от эвгленид Euglena sanguinea. К числу организ-
мов зоонейстона относятся, например, из инфузорий Stentor poly-
morphic, Spirostomum ambiguum, из ветвистоусых раков — Sca-
pholeberis mucronaia, а из насекомых —личинки Anopheles,
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
Транспорт живого нейстона из какого-либо водоема в лабора-
торию обычно трудно выполним, поэтому на занятиях рассматри-
вают население поверхностной пленки воды, развивающееся в ак-
вариумах, вода которых предварительно была загрязнена неболь-
шим количеством органического удобрения, стимулирующего разви-
тие нейстона.
Убедившись под лупой в присутствии большого количества ней-
стических организмов, при помощи пинцета осторожно прикаса-
ются к поверхности воды кольцом, диаметром около половины сан-
тиметра, сделанным из тонкой проволоки и снабженным тремя не-
большими ножками. Подняв кольцо вместе с частью поверхностной
пленки, переворачивают его и ставят ножками на предметное стек-
77
ло, после чего рассматривают пленку под микроскопом. Можно по-
лучить пробу нейстона иным способом. Для этого на поверхность
воды кладется чистое покровное стекло, затем пинцетом осторожно
поднимают его вместе с нейстоном и кладут смоченной стороной
Рис. 35. Типичные организмы нейстона. А — Chlamydothrix ochracea; Б — Chro-
matiurn sp.; В —Chromulina rosanoffi; Г— Chlamydomonas sp.; Д — Euglena
sanguinea; E — Stentor polymorphus; Ж — Spirostomum ambiguum; 3 — Scapho-
leberis mucronata, подвешенный к пленке поверхностного натяжения.
вниз на предметное стекло с углублением. Приготовленные препа-
раты рассматриваются при большом увеличении. Наиболее харак-
терные организмы нейстона изображены на рис. 35. Из организмов
зоонейстона рассматривается Scapholeberis mucronata и личинки
Anopheles.
78
1. Scapholeberis (рис. 35, 3)
Наблюдая под бинокуляром живых Scapholeberis mucronate.
в небольшом плоском сосуде с водою, легко видеть характерное по-
ложение животного, подвешивающегося к пленке воды брюшными
краями створок. При рассматривании организма под микроскопом
следует обратить внимание на следующие особенности строения.
Голова большая, часто с более или менее хорошо развитым лоб-
ным выростом, на спинной стороне с резким перехватом. Створки
сбоку почти четырехугольной формы, с прямыми брюшными края-
ми, усаженными короткими волосками, позади вытянуты в длин-
ные выросты. Глаз хорошо развит, глазок небольшой. По осталь-
ным уризнакам Scapholeberis существенно не отличается от дру-
гих представителей ветвистоусых раков.
2. Личинки Anopheles
Описание строения личинок комаров Anopheles помещено на
стр. 188.
ОРГАНИЗМЫ ПЕРИФИТОНА
Перифитон, или оброет, покрывающий погруженные в воду рас-
тения, а также подводные части судов, сваи, тросы и т. п., встре-
чается в самых разнообразных водоемах, в морях, озерах и реках.
В состав перифитона входят многочисленные виды животных и ра-
стений, требующих' для своего развития твердого субстрата, а так-
же благоприятного кислородного режима, определенной степени
освещенности, отсутствия отрицательного воздействия грунта и т. п.
Организмы перифитона встречаются в верхних слоях воды, в прес-
новодных водоемах не опускаются ниже глубины нескольких мет-
ров.
Основную массу перифитона образуют прикрепленные организ-
мы, среди которых обитают также и свободнодвигающиеся живот-
ные.
В состав пресноводного перифитона входят преимущественно
мелкие, часто микроскопические, организмы — зеленые водоросли
{Oedogonium, Bulbochaete, Coleochaete, Spirogyra, Stigeoclonium
и многие другие виды), диатомеи {Cymbella, Gomphonema и другие
виды), а из животных — инфузории (Vorticella, Carchesium, Stentor
и другие виды), суктории {Acineta), коловратки {Rotifer, Melicerta,
Lacinularia, Limnias и другие виды) и малощетинковые черви
(Nais, Stylaria); меньшее значение имеют мшанки, губки, моллю-
ски, личинки насекомых и представители иных групп.
Значительно сильнее развит перифитон в морях, особенно тро-
пических. В его состав входят крупные водоросли — зеленые
{Cladophora, Enteromorpha, Ulvd), бурые (Ascophyllum, Fucus) и
красные {Ceramium), а из животных—гидроиды {Obelia, Laomedea,
Sertularia), мшанки {Membranipora, Retepora, Bugula и другие ви-
ды), многощетинковые черви {Spirorbis, Serpula), двустворчатые
моллюски {Mytilus, Ostrea), усоногие раки {Balanus, Lepas), обо-
лочники {Molgula, Ciona, Ascidia, Botryllus) и представители других
групп; в особенности велика роль моллюсков и усоногих раков, об-
роет которых на подводных частях судов измеряется нередко кило-
граммами и десятками килограммов на квадратный метр.
В пресноводных водоемах перифитоном питаются мальки боль-
шого числа’ рыб, а некоторые рыбы питаются им и во взрослом со-
80
стоянии. В морях, особенно тропических, перифитон имеет большое
значение для судоходства. Даже небольшое обрастание значитель-
но уменьшает скорость судна, при длительном же отсутствии доко-
вания, во время которого счищается оброет, скорость может умень-
шиться на половину. Также велика роль перифитона при коррозии
бетонных сооружений.
Рис. 36 а. Типичные организмы пресноводного перифитона. Я—Oedogonium соп-
catenatum, с антеридиями и оогонием; $ — Bulbochaete mirabilis; В —
Coleochaete setitata; Д’—Rotifer vulgaris; Л—L'imnias cerat’ophylli; M — Melicer-
ta Hngens; H — Lacinularla flosculosa.
6 В. А. Я"..... 81
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
В качестве примера обрастания в морях рассматривают населе-
ние какого-либо предмета, находившегося длительное время в воде.
Пресноводный перифитон рассматривают в живом состоянии.
Для этого со стебля хвоща или с нижней стороны листьев рдеста,
♦
Рис. 36 б. Типичные организмы пресноводного перифитона. Г — Spirogyra sp-'
Д— Stigeoclonium sp.; Е— Cymbella cistula, колония и отдельная клетка;
Ж— Gomphonema constrictum, колония и. отдельная клетка; 3 — Vorticella
nebulifera; И — Acineta lacustrjs; О — Stylaria lacustris.
82
кувшинки или иного растения острым скальпелем счищают неболь-
шое количество оброста на предметное стекло и рассматривают его
затем под микроскопом (рис. 36). Значительно лучше, не нарушая
естественного расположения организмов, рассматривать население,
развивающееся на искусственном субстрате. Для этого предметные
стекла, прикрепленные к дощечкам кнопками, покрытыми смесью
воска с парафином, некоторое время выдерживаются в воде есте-
ственного водоема или аквариума. После того, как на стеклах ра-
зовьется население перифитона, их медленно поднимают к поверх-
ности воды и, не вынимая из воды, чтобы не потерять свободнодви-
гающихся и слабо прикрепленных организмов, плавно переносят
пинцетом в небольшие кристаллизаторы. Население перифитона
лучше всего рассматривать в профиль, в этом случае можно легко
заметить ярусное расположение прикрепленных форм и нормальное
поведение животных. Для этой цели подбирается кристаллизатор,
внутренний диаметр которого равен длине предметного стекла, а
высота больше ширины последнего. Поместив в кристаллизатор
предметное стекло в вертикальном положении и отсосав пипеткой
излишнее количество воды, рассматривают перифитон при слабом
увеличении; опуская и поднимая тубус микроскопа, можно увидеть
организмы в естественном положении.
Легко поставить наблюдения над выеданием перифитона брю-
хоногими моллюсками. Для этого обросшее перифитоном предмет-
ное стекло, с помещенным на нем небольшим экземпляром Limnaea
или Planorbis, кладут в кристаллизатор в перевернутом виде на
две подпорки, сделанные из .стекла или иного материала. Под мик-
роскопом или бинокуляром можно видеть дорожку, выедаемую
моллюском в перифитоне, и работу терки.
«
ЗАЩИТНЫЕ ПРИСПОСОБЛЕНИЯ ПРИ НАСТУПЛЕНИИ
НЕБЛАГОПРИЯТНЫХ УСЛОВИЙ
.Многие пресноводные организмы, в особенности живущие во
временных пересыхающих водоемах, лужах, остающихся после
сильных дождей, таяния снега или разлива рек, способны длитель-
ный период существовать без воды. Некоторые из них при наступ-
лении неблагоприятных условий закапываются в ил, как, например,
двудышащие рыбы, проводящие засушливый период в капсулях,
построенных из слизи и донных отложений, или брюхоногие мол-
люски, закрывающие устье раковины крышечкой или затвердеваю-
щей пленкой. Другие животные, погибая сами при высыхании во-
доема, образуют стойкие покоящиеся стадии, способные длитель-
ное время находиться в сухой среде и оживающие при заполнении
высохшего водоема водою. Многие простейшие (корненожки, сол-
нечники, инфузории) при высыхании инцистируются.
К числу типичных обитателей временных водоемов относятся
крупные жаброногие раки. Отложенные яйца сохраняются у Апо-
straca в яйцевом мешке (см. рис. 57), у Notostraca в яйцевых кап-
сулях на ногах одиннадцатой пары (см. рис. 58), а у Conchostraca
в спинной части раковины (см. рис. 59); по истечении нескольких
дней они выбрасываются в воду. Находящиеся в илу яйца подвер-
гаются высыханию и промерзанию после прекращения существова-
ния водоема; весною следующего года они развиваются в новое по-
коление. Экспериментально было найдено, что нагревание высушен-
ных яйц Notostraca до 80° не препятствует их дальнейшему разви-
тию. Способность развиваться не теряется ими в течение 3—5 лет,
а иногда до 15 лет, поэтому из яиц, собранных вместе с илом со
дна высохших водоемов, можно получить, при размачивании ила в
воде, различных представителей крупных Branchiopoda, живших ра-
нее в этих водоемах.
Покоящиеся (латентные) яйца ветвистоусых раков, относящихся
к трибе Anomopoda, заключены внутри более или менее сильно ви-
доизмененной спинной части створок раковины, образующей так на-
зываемый эфиппий (седлышко). *
Латентными яйцами обладают также многие виды пресновод-
ных ракушковых раков и некоторые виды пресноводных веслоно-
гих раков, живущих в небольших водоемах. У последних фаза по-
84
коя в развитии латентных яиц наступает по достижении стадии
бластулы или гаструлы (некоторые Calanoida) или даже первой на-
уплиальной стадии (некоторые Harpacticoida). Латентные яйца, в
противоположность обыкновенным, снабжены плотной оболочкой,
под которой обычно находится еще вторая, более тонкая.
Геммулы губок, также являющиеся покоящимися стадиями, раз-
виваются осенью и служат для переживания неблагоприятного
времени года. Геммулы встречаются у большинства пресноводных
губок и лишь у некоторых морских представителей этой группы.
Шаровидное тело геммулы, покрытое хитиновой оболочкой, внутри
содержит скопление зародышевых клеток, из которых при наступле-
нии весны развивается молодая губка.
Покоящиеся стадии пресноводных мшанок, известные под наз-
ванием зимующих почек или статобластов, имеют вид чечевицеоб-
разных или шаровидных телец, окруженных плотной хитиновой
оболочкой. При наступлении благоприятного времени года стато-
бласты раскрываются и дают начало новой колонии мшанок.
Организмы, живущие в микроводоемах, в периодически смачи-
ваемых водою песке и мхе на крышах и водосточных желобах, на
коре деревьев и во мху болот, обладают способностью высыхать
полностью и впадать в анабиотическое состояние; при новом сма-
чивании водою они оживают, продолжая рост и размножаясь. К
числу этих организмов относятся некоторые коловратки, нематоды
и тихоходки, а также некоторые водоросли.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
В качестве примеров приспособления организмов к потере воды
рассматриваются эфиппии ветвистоусых раков, геммулы губок,
статобласты мшанок и представители полностью высыхающих ор-
ганизмов.
I. Эфиппии Daphnia (рий. 37)
Эфиппии представителей семейства Daphniidae являются настоя-
щими эфиппиями, в противоположность примитивным эфиппиям
представителей других семейств, у которых они соответствуют сбро-
шенным створкам раковины, отличаясь от них лишь темной пигмен-
тацией, утолщением спинного края и присутствием заполняющей их
полость рыхлой ткани, препятствующей выпадению латентных яиц.
У видов, относящихся к семейству Daphniidae, в образовании
эфиппия принимает участие только спинная часть створок ракови-
ны, всегда окрашенная в темный цвет. Эфиппий располагается на-
верху организма в виде седла, чем и объясняется его название —
седлышко.
Настоящий эфиппий характеризуется особым строением стенок.
Наружный слой состоит из высоких призматических камер, каж-
дая из которых соответствует одной клетке гиподермы, снаружи
камеры покрыты тонкой хитиновой пленкой; внутри них находится
85
жидкость, позднее заменяющаяся газом. Камеры образуются по
всей поверхности эфиппия, за исключением тех мест, где внутри
него лежат латентные яйца; в этих местах стенки эфиппия состоят
из толстого слоя хитина. По линии соединения обеих створок на-
ружный слой переходит в эластичный спинной тяж, вызывающий
замыкание краев эфиппия. Внутри эфиппий выстлан тонким слоем
хитина. Между наружными и внутренними слоями стенок образу-
Рис. 37. А — эфиппий Daphnia pulex; Б — эфиппий Daphnia magna; В — попе-
речный разрез через эфиппий Daphnia.
/ — спинной тяж, 2 призматические камеры, 3 — разрез латентного яйца.
ется полость, содержащая многочисленные хитиновые крепления.
В настоящем эфиппии бывает одно или два латентных яйца, спо-
собных в течение длительного времени находится в покоящемся со-
стоянии; они могут без вреда для себя переносить высушивание и
замерзание.
Во время занятий рассматривают эфиппии Daphnia pulex, у ко-
торых наполненные газом камеры наружного слоя настолько об-
легчают их вес, что они становятся легче воды и могут плавать по
ее поверхности, и эфиппии Daphnia magna, снабженные длинными
крючками, образованными краями сброшенных створок раковины;
эти эфиппии опускаются на дно, сцепляясь своими крючками. Кро-
ме общего вида эфиппиев, рассматриваются также поперечные сре-
зы через них.
86
2. Геммулы пресноводных губок (рис. 38, А—В)
Геммулы пресноводных губок, имеющие шаровидную форму и
обычно окрашенные, по величине не превышают 0,5 мм. Они состо-
ят из клеточной массы, заключенной в крепкую хитиновую капсулу,
пронизанную на одном полюсе отверстием. Капсула окружена тол-
стым слоем воздухоносной ткани, внутри и на поверхности кото-
рой находятся небольшие спикулы (микросклеры).
Рис. 38. А, Б — геммула Spongilla, общий вид '(4) и в разрезе (Б); В — гем-
мула Ephydatia в разрезе; Г—плавающий статобласт Plumatelia fungosa; Д—
сидячий статобласт Plumatella repens; Е — плавающий статобласт Plumatelia
repens; Ж — сидячий статобласт Plumatella fruticosa; 3 — плавающий стато-
бласт Plumatella fruticosa; И — сидячий статобласт Fredericella sultana; К —
плавающий статобласт Cristatella mucedo.
Для приготовления препарата геммулы предварительно кипя-
тятся в течение нескольких минут в тугоплавкой пробирке в неболь-
шом количестве соляной кислоты; нагревание проводят медленно
во избежание выброса кислоты из пробирки. После промывки во-
дою, геммулы переносятся на предметное стекло, разрываются в
случае необходимости препаровальными иглами и рассматриваются
под микроскопом.
87
3. Статобласты пресноводных мшанок (рис. 38, Г—К)
Статобласты пресноводных мшанок, образующиеся у большин-
ства видов при наступлении неблагоприятного периода, при высы-
хании или замерзании водоема, имеют вид чечевицеобразных или
шаровидных темных телец, покрытых плотной хитиновой оболочкой.
Статобласты развиваются в соединительнотканном тяже, фунику-
лусе, отходящем от нижнего конца желудка к стенке тела. Различа-
ются два рода статобластов — сидячие и плавающие. Первые, по-
крытые простой оболочкой, состоящей из двух половин, прикрепля-
ются к стеблям водных растений, находящимся в воде сучкам и т. п.
Плавающие статобласты окаймлены по краям кольцом мелких воз-
душных камер, помогающих им находиться на поверхности воды.
Одни виды мшанок имеют только один род статобластов, или сидя-
чих (Fredericella} или плавающих (Cristatelld), у других могут раз-
виваться и те и другие (Plumatelld); в более редких случаях стато-
бласты отсутствуют (Paludicella). На зиму колонии мшанок отми-
рают; весною перезимовавшие статобласты раскрываются и из них
выходят молодые особи, дающие начало новым колониям.
Во время занятий рассматривают статобласты различных пред-
ставителей пресноводных мшанок — широко-овальные у Plumatelia
fungosa и Plumatelia repens, узкие у Plumatella fruticosa, бобовид-
ные, или почковидные, у Fredericella sultana, круглые с двумя ря-
дами длинных якоревидных крючков, отходящих от центральной хи-
тиновой капсулы по сторонам плавательного кольца, у Cristatella
mucedo.
4. Население периодически смачиваемых
микроводоемов (рис. 39)
Среди обитателей микроводоемов наиболее часто встречаются
различные коловратки, нематоды и тихоходки. Если поместить не-
большое количество грунта, скапливающегося на крышах у водо-
сточных желобов, в чашку Петри или часовое стекло, то после сма-
чивания его водою легко наблюдать процесс оживления организ-
мов, находившихся в анабиотическом состоянии. Бесформенные ко-
мочки высохших коловраток через некоторое время расправляются
и превращаются в свободнодвигающиеся организмы; согнутые спи-
ралью нематоды вскоре после смачивания становятся способными к
движению. Такую же картину можно наблюдать, если смочить во-
дою высохший мох, собранный на крыше или стенах зданий; в этом
случае можно) найти и высохших тихоходок.
Если поместить на предметное или часовое стекло коловратку
Philodina roseola, обычную форму микроводоемов, а затем подверг-
нуть ее медленному высушиванию, то можно наблюдать, как посте-
пенно тело коловратки сокращается, пока не превратится в неболь-
шой комочек. Высушивание можно продолжить в эксикаторе с хло-
ристым кальцием. Совершенно высушенная коловратка через не-
сколько месяцев и даже лет снова оживает при прибавлении воды.
Тихоходки (Tflrdigrada), небольшие, до 1 мм длиною, организ-
мы обитают преимущественно в периодически смачиваемых водою
88
моховых подушках на стволах одиночно стоящих деревьев, стенах,
скалах и т. п., некоторые виды живут в небольших водоемах с ра-
стительностью. Тело тихоходок состоит из головного и четырех ту-
Рис. 39. Население периодически смачиваемых микроводоемов. А—Г—Philodina
roseola, в расправленном состоянии (Л), ‘полусократившаяся (Б), сократив-
шаяся (В), в состоянии анабиоза (Г); Д, Е — Macrobiotus hufelandi,в расправ-
ленном состоянии (Д), в состоянии анабиоза (£); Д'— Milnesium tardigradum;
3 — Echiniscus sp.; И — Pseudoechiniscus sp.
1— глаз, 2 — ротовое отверстие, 3 — ротовая железа, 4 — глотка, 5 — пищевод»
8 — желудок, 7 — кишечник, 8 — преанальная железа, 9— яичник, /0 — кровя-
ные клетки полости тела.
89
ловищных сегментов с четырьмя парами нерасчлененных коротких
ног; каждая нога оканчивается четырьмя коготками, у основания
которых находится железистое скопление клеток. Пищеварительная
система состоит из ротовой полости, сильно развитой мускулистой
глотки, пищевода, желудка и задней кишки. Органов дыхания и
кровеносной системы не имеется. Полость тела наполнена жидко-
стью с плавающими в ней крупными клетками. В головном сегмен-
те находится большой надглоточный ганглий. Все тихоходки раз-
дельнополы; яйца, откладываемые свободно или в сбрасываемую
при линьке шкурку, у одних видов гладкие, у других покрыты раз-
нообразными выростами.
При высыхании или замерзании тихоходки впадают в анабиоз,
оживая при каждом смачивании водою; некоторые виды образуют
при этом цисты. При высушивании тихоходок можно наблюдать
уменьшение их в объеме и втягивание ног; при медленном высуши-
вании они принимают боченковидную форму. В анабиотическом со-
стоянии тихоходки могут находится несколько лет.
Во время занятий изолированную на предметном стекле тихо-
ходку подвергают медленному высушиванию, а затем, после при-
бавления воды, наблюдают постепенное ее оживление. Быстрое вы-
сушивание действует на организмы губительным образом. На мохо-
вых подушках чаще всего встречаются различные виды Echiniscus,
спинная поверхность которых покрыта снабженным щетинками или
шипами панцырем из 10 щитков, Pseudoechiniscus, с панцырем из
14 щитков, и Milnesium tardigradum, длиною до 1,2 мм, без пан-
цыря и палочковидных включений в глотке, с околоротовыми со-
сочками и различными по длине и строению коготками. В неболь-
ших пресноводных водоемах встречаются Milnesium tardigradum
и различные виды Macrobiotus, например Macrobiotus hufelandi, у
которых на каждой ноге находится по две пары одинаковых когот-
ков, а в глотке имеются палочковидные включения; длина достига-
ет 0,7 мм.
ОРГАНИЗМЫ И СОЛЕНОСТЬ воды
Природные водоемы по количеству растворенных в воде болей
делятся на пресноводные (олигогалинные), солоноватоводные (ме-
З'огалинные), морские (полигалинные) и переосолоненные (ультра-
галинные)- Соленость океанов в среднем равна 35°/оо, а в различ-
ных морях колеблется от 17—18%о (Черное море) до 41—47%о
(Красное море). Морская вода содержит, главным образом, хло-
рид натрия, около' 89%, остальные соли играют меньшую роль.
Между водою морских и внутренних (континентальных) водоемов
существует не только количественное, но и качественное различие.
Пресноводные водоемы отличаются низкой соленостью, не более
О,5°/оо: главной составной частью солей является карбонат каль-
ция, количество которого в среднем равняется 60%. Между мор-
скими и пресноводными водоемами располагаются солоноватовод-
ные водоемы, с соленостью от О,5°/оо до 17°/оо- К числу солоновато-
водных водоемов относятся малосоленые моря, как, например, Бал-
тийское море (6—9%о), Азовское море (9—12°/оо) и Каспийское
море (не более 14°/оо), а также районы непосредственного смеши-
вания морских и пресных вод — устья рек, эстуарии, губы и прото-
ки. Вода этих водоемов является морской распресненной водой. С
другой стороны, некоторые вдающиеся в сушу существующие или
бывшие ранее морские заливы обладают, благодаря сильному мест-
ному испарению воды, повышенной соленостью, значительно пре-
вышающей соленость морской воды (ультрагалинные водоемы мор-
ского происхождения). Среди внутренних (континентальных) водо-
емов находятся стоячие соленые и переосолоненные, отличающиеся
от пресноводных по солености и по качественному составу солей—
континентальные мезогалинные (0,5—25%о), континентальные по-
лигалинные (25—1-ОО°/оо) и континентальные ультрагалинные во-
доемы (100—ЗОО°/оо и более); в этих водоемах могут находиться в
больших количествах поваренная соль (NaCl), глауберова соль
(Na2SO4), сода (Na2CO3), хлористый магний (MgCI2) и другие соли.
Организмы, не выносящие больших колебаний солености воды,
другими словами, живущие в узких солевых пределах, относятся к
стеногалинным (узкосолевым) организмам. В зависимости от при-
уроченности к определенной степени солености, стеногалинные ви-
01
Ды могут быть среди пресноводных, солоноватоводных, морских и
ультрагалинных организмов. Большинство растений й животных
пресноводных водоемов не переносит осолонения воды и, обратно,
большинство морских организмов не выдерживает более или менее
значительного понижения солености. Эти организмы относятся к
числу стеногалинных пресноводных и стеногалинных морских видов.
В противоположность этому только сравнительно небольшое число
организмов может считаться стеногалинными солоноватоводными и,
в особенности, стеногалинными ультрагалинными видами; послед-
ние, обитающие при солености воды свыше 1ОО°/оо, образуют груп-
пу галобионтов.
Те организмы, которые могут существовать при сравнительно
широких изменениях солености воды, относятся к эвригалинным
(широкосолевым) организмам. Типичными примерами эвригалинных
видов являются проходные рыбы и большое число морских лито-
ральных организмов, выдерживающих значительное понижение со-
лености, вызываемое осадками или стоком пресной воды с суши.
Пресноводные организмы, случайно заносимые в солоноватую воду
и способные в ней существовать, называются галоксенами; те из
них, которые отличаются большой эвригалинностью и могут жить и
в пресной и морской воде, относятся к галофилам.
Пребывание эвригалинных организмов в воде различной соле-
ности вызывает ряд изменений в их организации, из кототорых наи-
более легко определяются морфологические изменения. Многие эв-
ригалинные морские по происхождению организмы уменьшаются в
размерах, переходя в воду пониженной солености. Предельная ве-
личина моллюсков Балтийского моря в распресненных Финском и
Ботническом заливах в несколько раз меньше, чем в более соле-
ных западных районах моря.
Изменения солености водоемов могли произойти в течение преж-
них геологических периодов. Ранее бывшие морские водоемы или
их части в последующие эпохи мЪгли потерять, в результате текто-
нических перемещений земной коры, связь с морем и превратиться
в распресненные или пресноводные водоемы. Организмы, населяв-
шие прежние моря, должны были, в результате резкого изменения
солевого режима водоемов, или исчезнуть или, приспособившись к
изменившимся условиям среды, превратиться в обитателей распрес-
ненных или пресных вод. Такие организмы, нахождение которых в
современных распресненных или пресноводных водоемах может
быть объяснено только тем, что их исходные морские предки оби-
тали в них при иной солености, называются морскими реликтами.
Палеогеографическое прошлое наших южных морей (Черного,
Азовского и Каспийского) и Балтийского моря оказало большое
влияние на формирование населения этих водоемов, а также тех
пресноводных озер, которые в прошлом являлись частями исчез-
нувших морей.
Влияние изменения солености на организацию гидробионтов
можно показать также экспериментально. Если воспитывать, напри-
мер, Artemia salina, самого обычного животного соленых озер, в
92
воде пониженной или повышенной солености, то новые поколения
этого организма, развивающиеся при иной солености, по сравнению
с материнскими особями, будут отличаться от последних рядом
признаков.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
СТЕНОГАЛИННЫЕ ОРГАНИЗМЫ
Пресноводные организмы
Большинство растений й животных, обитающих в пресноводных
водоемах, относится к стеногалинным организмам. Некоторые груп-
пы встречаются преимущественно или исключительно в пресной во-
де— жаброногие раки, ветвистоусые раки, коловратки, малощетин-
ковые черви, водяные клещи, земноводные; среди других групп
имеются чисто пресноводные виды, например среди веслоногих ра-
ков, моллюсков, рыб и т. д.
Морские организмы
Типичными морскими стеногалинными организмами являются
кокколитофориды, радиолярии, гидроидные полипы, коралловые по-
липы, сцифоидные медузы, многощетинковые черви, крылоногие мол-
люски, киленогие моллюски, головоногие моллюски, плеченогие, ще-
тинкочелюстные, усоногие раки, эвфаузиды, ротоногие раки, игло-
кожие, оболочники и ряд других групп; большое число морских ви-
дов встречается среди диатомей, перидиней, корненожек, инфузо-
рий, губок, веслоногих раков, ракушковых раков, равноногих ра-
ков, амфипод, мизид, десятиногих раков, брюхоногих моллюсков,
двустворчатых моллюсков, мшанок, рыб и других групп.
Солоноватоводные организмы
Сравнительно небольшое количество видов можно считать ти-
пичными мезогалинными, обитающими только в солоноватоводных
водоемах. В качестве примеров рассматриваются некоторые коло-
вратки (виды Brachionus, Pedalia), и ветвистоусые раки (Podon-
evadne, Corniger, Cercopagis), обитающие в слабо соленых Азов-
ском и Каспийском морях.
1. Brachionus plicatilis и Pedalia охуиге (рис. 40)
Оба указанных вида коловраток обитают в планктоне малосоле-
ных Балтийского и Азовского морей и различных осолоненных озер.
Панцырь Brachionus plicatilis (= Br. millleri) гладкий, сравни-
тельно тонкий. На переднем конце панцыря находится шесть зуб-
цов; промежуток между средними зубцами глубже других. Основа-
ния зубцов вздутые, широкие, сами зубцы небольшие, острые. Брюш-
ной край панцыря подразделен, на четыре округлые лопасти. На
конце панцыря находится отверстие для ноги, по бокам с варьиру-
ющими по длине выростами. Нога длинная, кольчатая (у фиксиро-
93
ванных экземпляров часто втянута внутрь панциря), на конце с
двумя небольшими пальцами. Имеется глаз. Мастакс большой, ши-
рокий. Brachionus plicatilis отличается от других видов Brachionus
по форме передних зубцов и по отсутствию на панцире задних ши-
пов.
Тело Pedalia oxyure (= Pedalion oxyuris) имеет необычный для
коловраток вид благодаря присутствию шести длинных рукообраз-
ных придатков, из которых брюшной (самый длинный) и спинной
одиночные, а боковые парные. Эти придатки, при помощи которых
Рис. 40. А — Brachionus plicatilis, панпырь; Б—Pedalia oxyure.
1 — рукообразные придатки, 2 — вырост на конце тела.
Pedalia делает большие прыжки, снабжены сильной мускулатурой,
оканчиваются длинными перистыми плавательными щетинками.
Тело имеет коническую форму, на заднем конце находится остро-
• конечный вырост; по этому признаку Pedalia oxyure отличается от
других видов Pedalia, у которых задний конец тела закруглен. Име-
ется два глазка. Нога отсутствует.
2. Caspievadne и Podonevadne (рис. 41. А—Г)
Виды Caspievadne и Podonevadne встречаются исключительно в
Понто-арало-каспийской области. Представляют характерную осо-
бенность планктона Азовского, Каспийского и Аральского морей,
являются древними реликтами понтического времени.
Caspievadne и Podonevadne (= Evadne part.) относятся к семей-
ству Poiyphemidae, образуя вместе с родами Polyphemus, Bytho-
trephes, Cercopagis, Corniger, Evadne и Podon особую группу, пред
ставители которой отличаются от остальных ветвистоусых раков по
строению ног и выводковой камеры (см. стр. 51).
Высота тела Caspievadne и Podonevadne больше длины, измеряе-
мой от лобного края до конца постабдомена. Голова большая, от-
делена от остального тела более или менее резко выраженной вы-
емкой. На голове находятся очень большой глаз и головной щит,
орган дыхания, расположенный на спинной стороне перед выемкой.
94
Антеннулы небольшие, прижаты к краю головы. Антенны, служа-
щие для движения, состоят из большого основного членика и двух
ветвей, вооруженных длинными плавательными щетинками; одна
ветвь (эксоподит) четырехчленистая, другая (эндоподит) трехчле-
Рис. 41. А — Caspievadne апопух; Б—Podoneyadne camptonyx; В—Podoneradne
trigona; Г — грудные ноги Podonevadne trigona; Д — Corniger maeoticus.
I — антеннула, 2 — антенна, 3 — глаз, 4 — головной щит, 5 — ноги первой-чет-
вертой пар, 6 — каудальные когти, 7 — сердце, 8 — выводковая камера.
К
нистая. Грудных ног четыре пары, ноги последней пары меньше
других. На ногах трех первых пар хорошо заметны одночленистые
эксоподиты, на ногах последней пары они рудиментарные. Постаб-
домен небольшой, на конце он закруглен или несет два крепких ка-
удальных (фуркальных) когтя. На спинной стороне постабдомена
находится небольшой вырост, оканчивающийся двумя короткими
щетинками. Выводковая камера различной формы, в зависимости
от возраста организма, обычно более или менее треугольная, на
вершине остроконечная или закругленная.
Встречающиеся в наших южных морях виды Caspievadne и
Podonevadne отличаются между собою следующими признаками.
1 (2) Постабдомен лишен каудальных когтей .
Caspievadne апопух
2 (1) Постабдомен с каудальными когтями ... 3
3 (4) Эксоподиты всех грудных ног с двумя щетинками
Podonevadne camptonyx
4 (3) Эксоподиты ног первой пары с двумя щетинками, на ос-
тальных ногах только с одной длинной щетинкой
Podonevadne trigona
Каждый вид, в зависимости от строения выводковой камеры и
характера изгиба каудальных когтей, распадается на несколько ва-
риететов.
3. Corniger (рис. 41, Д)
Виды Corniger встречаются исключительно в Каспийском и Азов-
ском морях и в лиманах Черного моря.
Самым характерным признаком является присутствие на перед -
нем крае головы большого выроста, раздвоенного на конце. Фур-
кальные когти длинные и сильно изогнутые. Форма выводковой ка-
меры изменяется в зависимости от количества яиц. По остальным
признакам Corniger не отличается от Podonevadne.
В Каспийском море встречается Corniger maeoticus hircus—го-
ловной вырост с длинными рогообразно изогнутыми концами; в
Азовском море встречается С. maeoticus horribilis — головной вы-
рост с шипообразными концами.
4. Cercopagis (рис. 42)
Виды Cercopagis, как и Podonevadne, распространены исключи-
тельно в Понто-арало-каспийской области.
Cercopagis относятся к самым оригинальным представителям
ветвистоусых раков, произошли от сходных по строению пресновод-
ных Bythotrephes, занесенных в Каспийское море по волжской си-
стеме в прежние геологические эпохи.
Тело цилиндрическое, состоит из головы, груди, брюшка и
постабдомена. Большая голова делится перетяжкой на две части;
впереди находится большой, почти шаровидный глаз, подразделен-
ный на дорсальный и небольшой вентральный отделы; в задней ча-
сти головы находятся антеннальные мышцы. Антеннулы короткие,
«6
неподвижные, отходят от нижней стороны головы непосредственно
за глазом. Антенны сильно развиты, эксоподит четырехчленистый,
эндоподит трехчленистый, каждая ветвь с семью щетинками. Ман-
дибулы сильно изогнуты, с острыми концами. Ноги первой пары
очень длинные, в несколько раз превосходят длину остальных ног;
Рис. 42. А—В — Cercopagis socialis, общий вид (Л), строение тела (Б), конец
хвостового шипа (В); Г, Д —Cercopagis pengoi, общий вид (Г), строение
тела ; L, Ж —Cercopagis gracillima, общий вид (£), строение тел» (Ж)-
/ — антеннула, 2— антенна, 3 — мандибула, 4 — ноги первой-четвертой пар, 5—
выводковая камера, 6—хвостовой шип, 7—каудальные когти, 8 — ложные
каудальные когти.
S7
ноги последней, четвертой пары рудиментарные. Все ноги без эксо-
подитов. Наиболее характерным органом является длинный хвосто-
вой шип, гомологичный тому небольшому выросту, который у дру.
гих ветвистоусых раков расположен на спинной стороне постабдо-
мена (стр. 51). На конце хвостового шипа, в несколько раз пре-
вышающего длину тела, находится характерный двойной изгиб,
внутренние части которого вооружены острыми зубцами. Каудаль-
ные (фуркальные) когти хорошо развиты; кроме того, нередко мо-
жно видеть одну или несколько пар ложных когтей. Выводковая ка-
мера, изменяющаяся в зависимости от зрелости эмбрионов, имеет
более или менее цилиндрическую форму, на вершине закругленная
или с острым выростом. Длина тела обычно 1,5—2 мм, длина хво-
стового шипа достигает 10 мм.
Большинство видов Cercopagis обитает в Каспийском море,
Cercopagis petigoi встречается, кроме того, в Азовском и Аральском
морях.
Наиболее часто встречающиеся виды Cercopagis различаются
между собою следующими признаками.
1 (2) Выводковая камера на вершине закругленная
С. socialis
2 (1) Выводковая камера на вершине заостренная 3
3 (4) Хвостовой шип расположен под тупым углом к брюшку .
С. pengoi
4 (3) Хвостовой шип расположен под прямым углом к брюш-
ку . .С. gracillima
Ультрагалинные организмы
Число типичных ультрагалинных видов! (галобионтов) очень не-
велико. Чем больше концентрация солей в воде, тем меньшее коли-
чество видов может в ней жить. При очень большой степени кон-
центрации в соленых озерах живут из растений Dunaliella salina и
Asteromonas gracilis (оба вида относятся к Volvocaceae), а из жи-
вотных только Artemia salina.
Artemia (рис. 43)
Артемии (Artemia salina) широко распространены в различных
соленых озерах Европы, Азии и других частей света. Относятся к
подотряду Anostraca отряда Branchiopoda. Отряд Branchiopoda,
жаброногих раков, включает подотряды Anostraca (жаброноги),
Notostraca (щитни), Conchostraca и Cladocera (ветвистоусые раки).
По внешнему виду артемии сходны с Pristicephalus (стр. 130).
Тело вытянуто в длину, явно членистое. Голова небольшая; пар-
ные глаза стебельчатые; антеннулы нитевидные; антенны самок не-
большие, а у самцов превращены в хватательный орган, конечные
членики которого несут большие плоские придатки. Грудные ноги,
в числе одиннадцати пар, имеют вид широкой пластины (протопо-
дит), внутренний край которой подразделен на эндиты; по на-
ружному краю расположены эксоподит, эпиподит и преэпиподит.
М
к г я мен сверху; В —нога первой
А —самка снизу; ь-самец озер у,
пары; Г —нога шестой пары.
йс- 43. Artemia salina.
A
Ч 7^?^ 2- антенна, 3- глаз, 4- ^Х’т^-эпиподит"И//- пЙЖ
яйцевой мешок 8— фурка, 9— эксопод ,
7% подит, 12 -эндиты.
99
Брюшко образовано восемью сегментами. К последнему брюшному
сегменту причленяются две плоских ветви фурки, вооруженные по
краям щетинками. Число щетинок и размеры фурки варьируют в
зависимости от количества растворенных в воде солей.
ЭВРИГАЛИННЫЕ ОРГАНИЗМЫ
В качестве примеров рассматриваются литоральные морские
представители зеленых водорослей (Enteromorpha) и моллюсков
(Mytilus, Cardium, Муа и Масота), живущих в различных районах
Балтийского моря, а также в других морях.
1. Enteromorpha (рис. 44, Д)
Энтероморфа (Enteromorpha) относится к семейству Ulvaceae
порядка Ulothrichales. Таллом трубчатый, полый, часто ветвящийся;
сначала всегда прикреплен к субстрату узким стебельком, позднее
может оторваться и свободно плавать. Клетки округло-угловатой
формы, с одним пластинчатым хроматофором, расположены одним
слоем. Окраска обычно желтовато-зеленая или желто-зеленая.
Длина таллома достигает нескольких десятков сантиметров.
Отдельные виды энтероморфы трудно различимы. Наиболее
обычным видом является Enteromorpha intestinalis — клетки круп-
ные, расположены без определенного порядка.
Энтероморфа нередко в массе встречается в литорали морей,
может переносить сильное распреснение.
2. Mytilus (рис. 44, Б—Г)
Мидии, относящиеся к числу съедобных моллюсков, встречаются
часто в таких больших количествах, что образуют мощные скопле-
ния, называемые банками.
Раковина мидии более или менее клиновидной формы, передний
край сужен, задний расширен. Макушка сдвинута на самый перед-
ний конец. Лигмант наружный. Замочный край гладкий или с не-
большими зубовидными выростами, расположенными под макуш-
кой (дизодонтный тип замка). Имеется два замыкательных мускула,
из них передний, помещающийся под макушкой, значительно мень-
ше заднего. Нога небольшая, пальцевидная. Биссусная железа хо-
рошо развита. При помощи пучков биссуса, выходящих около ма-
кушки между створками раковины, моллюск прикрепляется к суб-
страту или к другим мидиям. Края мантии толстые, фестончатые,
позади срастаются, образуя небольшой выводной сифон. Окраска
раковины обычно черного.или иссиня-черного цвета, поверхность
гладкая.
Мидии встречаются преимущественно в литорали или в верхних
горизонтах сублиторали. В Баренцовом, Белом и Балтийском морях
обитает Mytilus edulis, в Черном море — Mytilus galloprovincialis.
Черноморская мидия отличается от атлантической съедобной мидии
высокой, почти четырехугольной формой раковины, с ясно выра-
женным спинным углом.
100
е "С- 44. Л _ р .
*аРУ*и nteromorpha intestinalis; Б-—Г—Mytilus edulis, правая ствоока
пДСгтРава (Л} ИЗНУТРИ (Я), замок левой створки (Г); Д,Е —Cardium edule,
' v» левая с ’ пРавая створка изнутри (Е)\Ж, 3~ Муа arenaria, вид справа
РУ>Ки ш/ВОрка ИЗНУТРИ (3); И—Л — Масота baltica, левая створка сна-
' '» правая створка изнутри (X), вид сверху (Л); М — Масота
calcarea, вид сверху- уй лигамент,
Це7тСйф°ны' 2“«ora, -3 "маКУШ6КоакХТТуб«- 7'ммы"*»’
^«тральные зубы, 6 — передние 6oK°®“^a, 9 - <»пеЧ^.рофор-
ечаток переднего замыкательного УнаЯ лИния, М 101
ного мускула, Ю — ман
Мидии относятся к числу типичных эвригалинных организмов.
Могут жить в распресненной воде, уменьшаясь в размерах по мере
понижения солености. Это хорошо видно на моллюсках Балтийского
моря. Мидии, встречающиеся в западном районе моря, в проливах,
при солености не менее 15°/оо, по величине не отличаются от типич-
ных морских форм, длина раковины достигает 80—100 мм. Мидии,
которые обитают в распресненных Финском и Ботническом заливах,
отличаются значительно меньшими размерами. При солености около
5°/оо длина раковины не превышает 27 мм, а при солености около
2%о не превышает 21 мм.
3. Cardium (рис. 44, Д—Е)
Сердцевидка (Cardium edule) относится, как и мидия, к эврига-
линным видам. Встречается в Баренцевом, Балтийском и во всех
наших южных морях.
Раковина крепкая, овально-сердцевидной формы. Створки вы-
пуклые, с хорошо выраженными радиальными плоскими ребрами,
покрытыми тонкими и низкими поперечными чешуйками. Макуш-
ки вздутые. Лигамент наружный. Замок гетеродонтного типа. Зам-
ковая пластинка узкая, почти прямая. В каждой створке находятся
зубы, клиновидные центральные и пластинчатые передние и зад-
ние; рядом с зубами расположены углубления, в которые при за-
крытой раковине входят зубы противоположной створки. Число зу-
бов в правой створке — два центральных, два передних и один зад-
ний; в левой створке — два центральных, один передний и один
задний. Нога удлиненная, коленчатая, гладкая. Мантия спереди от-
крытая. Сифоны короткие, при основании соединенные, на краях с
сосочками. Окраска раковины серая, с желтизной.
Размеры сердцевидок колеблются в зависимости от солености
воды, в которой они живут. В западном районе Балтийского моря
при солености свыше 15°/оо длина раковины достигает 40—44 мм,
в опресненных Финском и Ботническом заливах не превышает
22 мм при солености около 5°/оо и 18 мм при солености около 2%о.
4. Муа (рис. 44, Ж—3)
Песчаная ракушка (Муа arenaria) живет в литорали и в верх-
них горизонтах сублиторали Варенцова, Белого и Балтийского мо-
рей.
Раковина овально-яйцевидной формы, сзади зияющая. Зубов не
имеется. Лигамент внутренний, хрящевой; на левой створке при-
креплен к ложкообразному выросту (хондрофору), на правой створ-
ке к полукруглой выемке (десмодонтный тип замка). Верхний край
правой створки слегка выдается над левой створкой. Поверхность
раковины с концентрическими линиями роста, эпидермис тонкий,
шелушащийся. Сифоны очень длинные, заключены в общую обо-
лочку. Окраска раковины белая или сероватая.
102
Песчаная ракушка живет в песке, выдерживает сильное опрес-
нение. В западных районах Балтийского моря, при солености свы-
ше 15%о, раковина имеет нормальную длину, до 100 мм, в опрес-
ненных Финском и Ботническом заливах не превышает 75 мм при
солености около 5°/оо и 37 мм при солености около 2°/оо-
5. Масота (рис. 44, И—М)
Макома (Масота baltica=Tellina baltica) живет в Баренцевом
и Балтийском морях, в последнем является самым распространен-
ным видом двустворчатых моллюсков.
Раковина уплощенная, округло-треугольная, укороченная. Замок
гетеродонтного типа. В каждой створке по два центральных зуба,
задний зуб в правой створке и передний зуб в левой створке глу-
боко расщеплены. Боковых зубов не имеется. Сифоны тонкие,
длинные, разделенные. Окраска раковины внутри, а часто и снару-
жи, — розовая, фиолетовая, реже белая.
Макома балтийская выносит значительное опреснение воды. За-
ходит в предустьевые пространства рек. В западном районе Бал-
тийского моря, при солености свыше 15%о, длина раковины дости-
гает 20—22 мм, в опресненных Финском и Ботническом заливах
не превышает 17 мм при солености около 5%о и 15 мм при соле-
ности около 2%о.
Близкий вид Масота calcarda (рис. 44, М) отличается по форме
раковины (при рассматривании сверху видно, что задний конец ра-
ковины изогнут вправо) и по отпечатку, на внутренней поверхности
створок раковины мантийного (синуса — изогнутого края мантии
около сифонов (у Масота baltica отпечатки мантийного синуса на
обеих створках приблизительно одной глубины; у Масота calcarea
отпечаток на левой створке значительно глубже, чем на правой).
МОРСКИЕ РЕЛИКТЫ
Руководящие ископаемые формы
В качестве примеров руководящих ископаемых, характеризую-
щих солевой режим ранее бывших водоемов севера Европы, рас-
сматриваются моллюски Portlandia arctlca (руководящая форма
Иольдиевого моря), Ancylus fluviatilis (руководящая форма Анци-
лового озера) и Littorina littorea (руководящая форма Литорино-
вого моря). Рассматриваются не ископаемые остатки, а ныне живу-
щие организмы.
1. Portlandia arctica (рис. 45, А—Б)
Portlandia arctica, называвшаяся ранее Yoldia arctica, относится
к числу высокоарктических видов. В настоящее время обитает в мо-
рях сибирского побережья и в Белом море, преимущественно в мел-
ководной прибрежной области. Сильно изменчивый вид.
108
Раковина выпуклая, задний конец каждой створки с двумя ра-
диальными складками, идущими от макушки к заднему концу
створки и к нижнему краю вдавленности, образованной складками.
Макушки выпуклые. Поверхность створок гладкая, с тонкой концен-
трической исчерченностью, матовая или тускло блестящая. Лига-
мент внутренний. Замок таксодонтного типа. Замочный край удли-
ненный, дугообразно изогнутый, с многочисленными V-образными
Рис. 45. А, В —Portlandia arctica, вид справа (Л), замок правой створки (Б);
В, Г— Ancylus fluviatilis, вид сбоку (В) и сверху (Г); Д — Littorina littorea.
зубами, в каждом ряду не более 20 зубов. Имеется два замыкатель-
ных мускула. Сифоны короткие, наполовину сросшиеся. Окраска
раковины зеленовато-желтая или зеленовато-коричневая, с темными
полосами или пятнами.
2. Ancylus fluviatilis (рис. 45, В—Г)
Чашечка (Ancylus fluviatilis) обитает в пресноводных водоемах,
озерах и реках, на камнях. Относится к отряду легочных моллю-
сков (Pulmonata).
Раковина имеет вид шапочки, с вытянутой назад вершиной, ус-
тье яйцевидно-округлое, завитка нет. Мантия почти щитовидная;
щупальца короткие. Длина раковины достигает 5 мм, высота —
4 мм.
3. Littorina littorea (рис. 45, Д)
Литорина (Littorina littorea) относится к числу бореальных ви-
дов, живет в Баренцевом и Белом морях. Съедобный моллюск.
Раковина кубаревидная, толстостенная, с пятью-семью оборота-
ми спирали, с высокой макушкой. Устье наверху вытянутое. Ли-
104
нии роста неправильные, косо изогнутые. Крышечка роговая с дву-
мя-пятью спиральными завитками. Окраска разнообразная, часто
буроватая, черноватая, со светлыми и темными полосами. Высота
до 25 мм.
Морские реликты иольдиевого времени
В качестве примеров распространенных в пресноводных водое-
мах морских реликтов ледникового времени, а также их исходных
форм рассматриваются представители веслоногих раков — Limno-
calanus grimaldii (исходная форма) и Limnocalanus macrurus (ре-
ликтовая форма), мизид — Mysis oculata (исходная форма) и Му-
sis oculata var. relicta (реликтовая форма), равноногих раков —
Mesidothea sibirica (исходная форма) и Mesidothea entomon (ре-
ликтовая форма), амфипод — Gammaracanthus loricatus (исходная
форма) и Gammaracanthus loricatus var. lacustris (реликтовая фор-
ма) и рыб — Myoxocephalus quadricornis (исходная форма) и
Myoxocephalus quadricornis var. onegensis и var. Idnnbergi (релик-
товые формы).
1. Limnocalanus grimaldii и L. macrurus (рис. 46)
Limnocalanus относятся к подотряду Calanoida веслоногих ра-
ков. Морская исходная форма, Limnocalanus grimaldii, является
самым распространенным видом Copepoda морей сибирского побе-
режья от Карского моря до Чукотского моря, развивается в массо-
вых количествах в солоноватоводных районах; кроме того, встреча-
ется в виде реликта в Балтийском море, в которое проникла в иоль-
диевое время, и в Каспийском море, где является иммигрантом.
Пресноводная реликтовая форма, Limnocalanus macrurus, обитает
в Ладожском и Онежском озерах, во многих озерах Карелии, в
озерах Финляндии и Швеции; все нахождения лежат внутри границ
Иольдиевого моря. Limnocalanus macrurus является морским релик-
том иольдиевого времени.
По своему строению исходная и реликтовая формы очень
сходны. Тело сильно удлиненное. Передний отдел состоит из
головного и пяти грудных сегментов; брюшко у самок из трех, у
самцов из пяти сегментов. Фуркальные ветви длинные, их длина в
несколько раз превышает ширину, по наружному краю вооружены
шипиками. Антеннулы длинные, 25-членистые, у самцов правая ан-
теннула геникулирующая; антенны двуветвистые. Всего имеется
пять пар плавательных ног. Ноги пятой пары плавательные, в про-
тивоположность тому, что наблюдается у большинства других ви-
дов веслоногих раков; у самок они симметричные, с трехчленистыми
ветвями и с большим внутренним выростом на втором членике эк-
соподитов; у самцов асимметричные, с трехчленистыми эндоподи-
тами и двучленистыми эксоподитами, конечный членик левого эк-
соподита большой, с четырьмя шипами, конечный членик правого
эксоподита небольшой, с длинным крючковидным выростом.
105
Отличие между исходной и реликтовой формами заключается в
очертании спинной линии. Рассматривая обе формы сбоку, легко
заметить, что у Limnocalanus grimaldii голова низкая, профиль го-
Рис. 46. А—Д—Limnocalanus grimaldii, самка сверху (Л) и сбоку (Б), гени-
кулирующая антеннула самца (В), нога пятой пары самки (Г), ноги пятой
пары самца (Д); Е— Limnocalanus macrurus, сбоку.
ловы постепенно переходит в профиль грудных сегментов; голова
Limnocalanus macrurus высокая, с резко выраженным лобным кра-
ем, между головой и первым грудным сегментом имеется более или
106
менее ясно выраженный шейный перехват. Степень изменения этого
признака соответствует длительности пресноводного существования
того водоема, в котором обитает реликтовая форма —чем длитель-
нее пресноводная фаза, тем сильнее изменяется контур головы.
2. Му sis oculata и М, oculata van relicta (рис. 47)
Мизиды образуют отряд Mysidacea высших раков. Морская ис-
ходная форма, Mysis oculata, живет в прибрежных районах всех
Рис.47.Л—В — Mysis oculata, самка (Л), самец (Б), тельсон (В); Г -Mysis-
oculata var. relicta, тельсон.
1— антеннула, 2— антенна, 3— скафоцерит, 4— карапакс, 5 — максиллярная
нога, 6 — переоподы (торакоподы), 7 —оостегитЛ— плеоподы, 9— уропод 10—
статоцист, 11— тельсон.
ИГ
наших северных морей. Пресноводная реликтовая форма, Mysis
oculata var. relicta, встречается в Ладожском и Онежском озерах,
в озерах Карелии и Смоленской области, в озерах Швеции и север-
ной Германии, в устьях рек, впадающих в Белое море и в моря си-
бирского побережья. В Финском и Ботническом заливах Балтий-
ского моря обитает в виде реликта промежуточная форма между
типичной и пресноводной.
Тело мизиды делится на голову, грудь и брюшко. Голова несет
стебельчатые глаза, антеннулы, состоящие из трехчленистого стеб-
ля и двух длинных многочленистых жгутов, и антенны, состоящие
из трехчленистого протоподита, пластинчатого, вооруженного ще-
тинками эксоподита, называемого скафоцеритом, и многочленисто-
го эндоподита. Грудь сверху покрыта хорошо развитым карапак-
сом, срощенным впереди с грудными сегментами и лежащим сво-
бодно на остальных сегментах; сзади карапакс с выемкой. Грудные
ноги (торакоподы), в числе восьми пар, двуветвистые, с многочле-
нистыми эксоподитами; торакоподы первых двух пар превращены
в максиллярные ноги; торакоподы двух задних пор самок снабже-
ны выростами (оостегитами), образующими выводковую камеру,
содержащую у зрелых особей яйца и эмбрионы. На брюшке нахо-
дятся слабо развитые плеоподы; у самцов плеоподы четвертой пары
с длинными эксоподитами. Хвостовой плавник образован тельсо-
ном и одной парой уроподов; эндоподиты уроподов отличаются
присутствием в верхней части больших пузыревидных статоцистов
с хорошо различимым статолитом, являющимся органом равнове-
сия. Развитие протекает в выводковой камере; молодые, выходя-
щие из камеры особи в общих чертах сходны со взрослыми орга-
низмами.
Отличие между исходной и реликтовой формами заключается
главным образом в строении тельсона, а также в меньшей вели-
чине реликтовой формы. Тельсон Mysis oculata продолговатый, по
краям с 20—30 шипами с каждой стороны, вырез на конце тельсо-
на узкий и глубокий с многочисленными шипиками; длина орга-
низма колеблется от 24 до 39 мм, цвет варьирует от светлого до
бурого. У Mysis oculata var. relicta выемка на конце тельсона мел-
кая и широкая, так что ее края образуют почти прямой угол; дли-
на организма колеблется от 16 до 25 мм, тело прозрачное, с чер-
ными пятнами.
3. Mesidothea sibirica и М. entomon (рис. 48)
Морские тараканы (Mesidothea) относятся к подотряду Valvj-
fera отряда равноногих раков (Isopoda). Морская исходная форма,
Mesidothea sibirica, относится к числу высокоа1рктических видов,
живет в морях сибирского побережья от Карского моря до Чукот-
ского моря, встречается в массовых количествах в опресненных
районах. Пресноводная реликтовая форма, Mesidothea entomon,
обитает в Ладожском озере, а также в некоторых озерах Карелии,
в озерах Финляндии и Швеции, в устьях рек сибирского побережья
и Белого моря ^в некоторых реках поднимается высоко). Кроме
108
Я 6
Рис. 48. А—Mesidothea sibirica;
Б — Mesidothea entomon.
1 — антеннула, 2 — антенна, 3 — глаз,
4—коксоподит (коксальная пластинка)..
5 — плеотельсон.
того, встречается в виде реликта в распресненных районах Балтий-
ского моря, а также в Каспийском море, где является иммигрантом.
Тело Mesidothea, сильно уплощенное, делится на голову, грудь
и брюшко. Бока головы вытянуты вперед и образуют расщеплен-
ные на две лопасти отростки. Небольшие сидячие глаза находятся
на верхней стороне головы. Антеннулы слабо развиты: антенны со-
стоят из пятичленистого стебля
и сравнительно небольшого
жгута. Грудь состоит из семи
свободных сегментов, несущих
такое же количество грудных
ног (переоподов). В результате
полного слияния первого груд-
ного сегмента с головой, конеч-
ности этого сегмента превраще-
ны в максиллярные ноги, сли-
тые между собою по средней
линии. Переоподы одноветви-
стые, лишены эксоподитов.
Коксоподиты переоподов, т. е.
их первые членики, превраще-
ны в широкие и плоские кок-
сальные пластинки; коксальные
пластинки первой пары нацело
слиты с первым грудным сег-
ментом и охватывают голову
с боков; коксальные пластинки
остальных пар ясно отделены
от грудных сегментов. У самок
от переоподов отходят, направ-
ленные внутрь и усаженные по
краям щетинками, широкие вы-
росты (оостегиты), образую-
щие выводковую камеру (мар-
зупиум), в которой протекает
развитие отложенных яиц. Абдомен состоит из четырех узких сег-
ментов, три из них разделены, а последний слит с большим плео-
тельсоном (тельсон у представителей Isopoda слит с одним или
несколькими сегментами брюшка, или плеона, поэтому называется
плеотельсоном). На брюшке находится пять пар двуветвистых
брюшных ног (плеоподов), являющихся органами дыхания, и одна
пара уроподов, превращенных в своеобразного вида створки, рас-
положенные снизу плеотельсона и прикрывающие плеоподы.
Отличия между исходной и реликтовой формами заключаются в
величине и в строении плеотельсона и коксальных пластинок.
У Mesidothea sibirica коксальные пластинки ромбовидные, с ясно
заметными диагональными килями; задние углы коксальных плас-
тинок первой пары заостренные, на остальных парах направлены
острыми концами книзу. Плеотельсон ясно пятиугольной формы, с
округлым бугорком на спинной стороне у основания. Крупные ор-
10»
ганнзмы, длина достигает 95 мм. У Mesidothea entomon все Кок
сальные пластинки плоские; задние углы коксальных пластинок
первой пары закругленные, на средних парах направлены острыми
концами в стороны, а на самых задних парах — книзу. Плеотель-
сон сильно вытянутый, почти треугольной формы. На первых
трех-четырех грудных сегментах находятся спинные продольные
кили. Небольшие организмы, не превышающие в длину 60 мм
обычно меньшего размера.
4. Gammaracanthus loricatus и G. loricatus
var. lacustris (рис. 49)
Gammaracanthus относятся к отряду амфипод (Amphipoda).
Морская исходная форма, Gammaracanthus loricatus, является вы-
сокоарктическим видом, обитает по берегам Шпицбергена, Новой
Земли и Сибири. Пресноводная реликтовая форма встречается в
Ладожском и Онежском озерах, в некоторых озерах Карелии, в
озерах Швеции, в устьях рек Сибири и Белого моря (в некоторых
реках поднимается высоко); в Каспийском море встречается варие-
тет caspius.
Тело Gammaracanthus сжато с боков, делится на голову, грудь
и брюшко. Грудные и брюшные сегменты с дорсальным килем и
большими зубцами. По бокам головы находятся большие сидячие
глаза, а впереди длинный, согнутый книзу рострум. Антеннулы со-
стоят из трехчленистого стебля и длинного жгута, добавочный
жгут небольшой; антенны короче антеннул. Грудь образована се-
мью сегментами, несущими такое же количество грудных ног (пе-
реоподов). Каждый переопод состоит из семи члеников—коксопо-
дита, базиподита, исхиоподита, мероподита, карпоподита, пропо-
дита и дактилоподита. Коксоподиты превращены в большие кок-
сальные пластинки, являющиеся боковыми продолжениями грудных
сегментов. Среди переоподов резко выделяются по своему строению
переоподы двух первых пар, называемые гнатоподами, на конце ко-
торых находится ложная клешня, образованная сильно расширен-
ным шестым члеником, проподитом, и подвижным тонким и изогну-
тым дактилоподитом. К внутренней стороне коксальных пластинок
прикрепляются тонкостенные жабры, а у самок, кроме того, широ-
кие овальные пластинки с щетинками по краям (оостегиты), обра-
зующие выводковую камеру (марзупиум), в которой проходит раз-
витие отложенных яиц. Брюшко подразделено на два отдела. Пе-
редний отдел брюшка, плеон, состоит из трех сегментов, боковые
края которых расширены в большие эпимеры; каждый сегмент не-
сет пару двуветвистых плавательных ног, плеоподов. Задний от-
дел брюшка, урус, также состоит из трех сегментов; каждый сег-
мент несет пару двуветвистых прыгательных ног, уроподов. Между
уроподами последней пары помещается тельсон, имеющий вид не-
большой широкой пластинки, с выемкой по заднему краю.
Отличия между исходной и реликтовой формами заключаются в
величине и в строении рострума и эпимеров брюшных сегментов.
110
У Gammaracanthus loricatus рострум длиннее первого членика
стебля антеннул, эпимеры второго и третьего сегментов брюшка с
острыми задними углами; крупные организмы, длина достигает
Рис 49. А — Gammaracanthus loricatus; Б — Gammaracanthus loricatus var.
lacustris.
1 — голова, 2 — рострум, 3 — глаз, 4 — антеннула, 5 — добавочный жгут антен-
нулы, 6— антенна, 7—грудь, 8—14 — переоподы первой—седьмой пар, 15 —
коксоподит (коксальная пластинка), 16 — базиподит, 17 — исхиоподит, 18 — ме-
роподит, 19—карпоподит, 20 — проподит, 21 — дактилоподит, 22— плеон, 23 —
эпимера, 24—26 — плеоподы первой—третьей пар, 27 — урус, 28—30 — уроподы
первой—третьей пар, 31 — тельсон.
45—58 мм. У Gammaracanthus loricatus var. lacustris рострум ко-
роче первого членика стебля антеннул, эпимеры второго и третьего
сегментов брюшка с тупыми задними углами; величина значи-
тельно меньше.
Ill
5. Myoxocephalus quadricornis и M. quadricornis
var. onegensis и var. lonnbergi (рис. 50)
Рогатки, или четырехрогие бычки (Myoxocephalus quadri
— Cottus quadricornis), относятся к семейству Cottidae. У)сх°Гп^
морская форма, Myoxocephalus quadricornis, живет в БаренцДйа’
и Белом морях, в сибирских морях — от Карского моря д0 ц°8о,ч
Рис. 50. А — Myoxocephalus quadricornis; Б — Myoxocephalus quadricornis var.
lonnbergi; В — Myoxocephalus quadricornis var. onegensis.
ского моря, в Беринговом море, а также в устьях сибирских реК*
встречается в виде реликта в Балтийском море. Пресноводные ре-
ликтовые формы находятся в Ладожском и Онежском озерах, »
озерах Финляндии и Швеции.
ным^пгипРеТеНО°бРазное’ Голова приплющена, вооружена различ-
i uT ’ Расположенными на жаберных крышках, над гла-
вах отврпг-^УГпХ местах- С каждой стороны головы по два носо
понк^обК™*’ Верх“яя челюсть длиннее нижней. Жаберные пере-
жутка ЖаЛйЮТ свободную складку поперек межжаберного пром
ный плавник рныещели широкие. Два спинных плавника, анал
жены подобен второму спинному. Брюшные плавники сбли
112
Наиболее характерным признаком исходной формы, Myoxoce-
phalus quadricornis, является присутствие на верхней стороне головы
четырех хорошо развитых бугров, из которых передние больше зад-
них; длина рогатки достигает 250—325 мм. У реликтовой формы,
Myoxocephalus quadricornis var. lonnbergi, обитающей в Ладож-
ском озере, головные бугры слабо развиты, передние не больше
задних; длина ладожской рогатки не превышает 165 мм. У другой
реликтовой формы, Myoxocephalus quadricornis var. onegensis,
обитающей в Онежском озере, головных бугров совершенно не
имеется; длина онежской рогатки не превышает 107 мм.
Так же, как и у Limnocalanus macrurus, у пресноводных форм
рогаток степень изменения морфологических признаков соответ-
ствует длительности пресноводного существования того водоема, в
котором обитает реликтовая форма. Рогатки в Онежском озере
перешли к пресноводному образу жизни после иольдиевого периода
и в течение последующего времени полностью потеряли головные
бугры, тогда как у рогаток Ладожского озера, опреснившегося
только после литоринового периода, эти бугры редуцированы лишь
на половину
ЭКСПЕРИМЕНТ С ARTEMIA SALINA (рис. 51)
Среди Anostraca весьма варьирующим видом является Artemia
salina (стр. 98), обитающая в соленых озерах, содержащая хло-
риды, карбонаты и сульфаты в различной концентрации, вплоть до
Рис. 51. Artemia salina, конец абдомена с фуркой различных вариететов: А —
var. principalis; Б — var. arietina; В —var. milhauseni; Г — var. koppeniana.
насыщенного состояния. Во многих водоемах, благодаря притоку
пресных вод или испарению, концентрация солей может колебаться
в широких пределах.
Артемии представляют пример весьма пластического организма,
легко изменяющегося под воздействием условий среды. Соответ-
8 ” ПЭ
ственно количеству растворенных в воде солей форма тела артемин
претерпевает сильные изменения. Прежде всего варьирует об-
щая длина тела — от 20—30 мм при небольшой солености до 6—
10 мм при значительной концентрации солей; относительная длина
брюшка по отношению к длине груди возрастает параллельно уве-
личению количества растворенных солей; восьмой брюшной сег-
мент брюшка при незначительной солености более или менее ясно
разделяется на два сегмента, таким образом брюшко становится
девятичленистым, в то время как в большинстве случаев оно имеет
восемь сегментов; варьируют также по форме и размерам эпиподи-
ты ног (жаберные придатки) и выросты антенн самцов. Однако
наибольшим изменениям подвергается фурка, которая при неболь-
шой солености воды хорошо развита, явно отчленена от последнего
сегмента брюшка и несет большое число длинных щетинок (арте-
мии с таким вооружением фурки относятся к var. principalis). С
увеличением концентрации солей фурка уменьшается в размерах и
сливается с последним сегментом брюшка (var. arietina). В край-
них случаях, при очень большой концентрации солей, фурка приоб-
ретает вид небольших выростов, снабженных одной или двумя ще-
тинками (var. tnilhauseni), или же полностью отсутствует (var.
Koppeniana). Как показали экспериментальные исследования, изме-
нение концентрации солей влечет за собою соответствующие изме-
нения морфологии организмов в течение двух-трех генераций.
Так как эксперимент требует много времени, то во время заня-
тий рассматривается только несколько форм артемий, полученных
при воспитании популяции в воде различной солености — родона-
чальная форма, жившая при солености около 70—1ОО°/оо, и полу-
ченные от нее формы, воспитанные, с одной стороны, при меньшей
солености, около 20—ЗО°/оо, с другой стороны, при большей солено-
сти, около 180—2ОО%о-
ОРГАНИЗМЫ — ПОКАЗАТЕЛИ ЗАГРЯЗНЕНИЯ ВОДЫ
Сапробностью называется способность организмов существовать
в воде, загрязненной растворенными органическими веществами.
Загрязнение может быть естественным, например в результате
осеннего распада органического вещества водных растений, в массе
развившихся летом, или может явиться следствием поступления в
водоемы различных сточных вод, содержащих органические вещества.
Организмы, не выдерживающие даже следов загрязнения, обра-
зуют группу катаробионтов, или катаробов. Они живут преимуще-
ственно в водоемах, находящихся в малонаселенных местностях
или высокогорных районах, в воде, содержащей очень большое ко-
личество растворенного кислорода и лишенной свободной угольной
кислоты. Многие реликтовые организмы наших северных озер от-
носятся к этой группе.
Организмы, встречающиеся в воде, в той или иной степени за-
грязненной органическими веществами, называются сапробионтами,
или сапробами. Среди них различают четыре группы, приурочен-
ные к различной степени загрязнения; в самых загрязненных водое-
мах развиваются полисапробы, при меньшей степени загрязнения
живут а-мезосапробы и 0-мезосапробы, в практически чистых
водоемах главное значение приобретают олигосапробы.
Полисапробы обитают в водоемах полей орошения, в сточных
водах боен, кожевенных, пивоваренных и других заводов, перераба-
тывающих органические продукты. В воде этих водоемов, содержа-
щих большое количество легко разлагающихся белковых веществ,
кислород полностью отсутствует, имеется много свободной угольной
кислоты и сероводорода, часто метана; все биохимические процес-
сы несут восстановительный характер; количество бактерий дости-
гает Тотен тысяч и миллионов в одном кубическом сантиметре
воды. Сравнительно очень небольшое число видов растений и жи-
вотных может существовать при таких условиях; автотрофные ра-
стения отсутствуют; развиваются организмы, разлагающие сложные
органические вещества с образованием более простых соединений;
в больших количествах встречаются различные бактерии (Sphaero-
tilus natans, серобактерии Beggiatoa), жгутиковые (Oicomonas ти-
tabilis), инфузории (Paramaecium putrinum, Vorticella putrina),
8* 115
один вид малощетинковых червей (Tubifex tubifex при массовом
развитии) и крыски (личинки мухи Eristalis tenax).
В процессе самоочищения полисапробная зона сменяется сг-ме-
зосапробной, характеризующейся восстановительно-окислительным
характером биохимических процессов, присутствием аминокислот,
амидокислот и аммиака и полуанаэробными кислородными услови-
ями; имеется сероводород, количество бактерий попрежнему вели-
ко, не падает ниже нескольких сотен тысяч в одном кубическом
сантиметре воды. Число а-мезосапробов значительно больше чи-
сла полисапробов. Среди растений к ним относятся некоторые
грибы (Nematosporangium), синезеленые водоросли (различные
виды Oscillatoria), эвгленовые (Euglena viridis при массовом раз-
витии), зеленые водоросли (Stigeoclonium tenue), жгутиковые (раз-
личные виды Bodo), среди животных — инфузории (Paramaecium
caudatum, Spirostomum ambiguum, Stentor coeruleus, Carchesium
spectabile), водяной ослик (Asellus aquaticus при массовом разви-
тии), личинки насекомых (Chironomus plumosus при массовом раз-
витии, Sialis lutaria).
В следующей ji-мезосапробной зоне восстановительные процес-
сы сменяются окислительными; кислорода много, среди соединений
азота главное значение приобретают аммиак, нитриты и нитраты,
количество сероводорода сильно уменьшается; число бактерий не
превышает нескольких десятков тысяч в одном кубическом санти-
метре воды. К £-мезосапробам относится большое число организ-
мов — синезеленые водоросли (Aphanizomenon flos-aquae при мас-
совом развитии), диатомеи (Melosira varians при массовом разви-
тии, Navicula cuspidata), десмидиевые водоросли (Cosmariutn
botrytis), зигнемовые водоросли (Spirogyra crassa), кладофоры
(Cladophora crispata), цветковые (Elodea canadensis, Lemna minor,
Ceratophyllum demersum), инфузории (Loxodes rostrum, Paramae-
cium bursarid), губки (Ephydatia miilleri), малощетинковые черви
(Stylaria lacustris), коловратки (Asplanchna priodonta, Monostyla
lunaris, Brachionus calyciflorus, Notholca striata), мшанки (Plutna-
tella fungosa), моллюски (Limnaea stagnalis, Viviparus
viviparus), амфиподы (Gammarus pulex), ветвистоусые раки
(Daphnia magna, Daphnia pulex, Moina rectirostris, Chydorus
sphaericus)-, в ,3-мезосапробной зоне появляются рыбы (вьюн, ка-
рась, линь, карп, верховна, уклея).
Олигосапробная зона характеризуется весьма благоприятными
кислородными условиями; соединения азота представлены нитрата-
ми, сероводорода не имеется, свободная угольная кислота присут-
ствует в небольших количествах; число бактерий резко уменьшает-
ся, обычно в одном кубическом сантиметре воды содержатся Десят-
ки, самое большее сотни бактерий. К олигосапробным организмам
относится большинство растений и животных чистых водоемов. Ти-
пичными представителями этой группы являются Anabaena flos-
aquae, Anabaena spiroides (синезеленые водоросли), Melosira italica,
Tabellaria fenestrata, Asterionella formosa (диатомеи), Ceratium
hirundinella, различные виды Peridinium (перидинеи), Closterium
lunula, Staurastrum tetracerum (десмидиевые), Volvox globator,
116
Eudorina elegans (вольвоксовые), Pediastrum duplex, Botryococcus
brauni (зеленые), Lemna trisulca (цветковые), Hydra vulgaris,
Chlorohydra viridissima (гидроидные), Notholca longispina, Diurella
stylata (коловратки), Ancylus fluviatilis (брюхоногие моллюски),
Mar gar it ana margaritifera, Unio pictorum, Dreissena polymorpha
(двустворчатые моллюски), Diaptomus gracilis, Diaptomus gPaciloi-
des, Eurytemora velox (веслоногие раки), Sida crystalline, Diapha-
nosoma brachyurum, Holopedium gibberum, Daphnia longispina,
Bosmina longirostris, Bythotrephes longimanus, Leptodora Kindti
(ветвистоусые раки), личинки насекомых — Aeschna grandis, Libel-
lula depressa, Coenagrion puella, Ephemera vulgata, Polymitarcys
virgo, Phryganea striata, рыбы — стерлядь, форель, гольян, плотва,
красноперка,
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
В качестве представителей полисапробов и а-мезосапробов
рассматриваются бактерии (Sphaerotilus, Beggiatoa), синезеленые
водоросли (Oscillatoria), малощетинковые черви (Tubifex, Limno-
drilus, Lumbriculus), водяной ослик (Asellus), личинки комаров дер-
гунов (Chiroatnomus), крыска (Eristalis).
1. Sphaerotilus (рис. 52, А—Б)
Сферотилусы (Sphaerotilus natans), относящиеся к нитчатым
бактериями (Chlamydobacteriaceae), являются одними из немногих
Д Б В г д
Рис. 52. А, Б — Sphaerotilus natans, колония (Д) и отдельные нити при боль-
шем увеличении (Б); В — Beggiatoa sp., с включениями серы; Г — Oscillatoria
princeps; Д — Oscillatoria tenuis.
бактерий, образующих большие колонии, хорошо заметные нево-
оруженным глазом.
117
Многочисленные удлиненные клетки, длиною 4—6 р и шири-
ною 2—3 р. соединены в нити, заключенные в слизистую оболоч-
ку; толщина нитей достигает 6—10р. Нити — не ветвящиеся, со-
единены в большие хлопьевидные обрастания, размером в несколь-
ко сантиметров.
Сферотилус часто встречается на сваях, досках, стеблях расте-
ний и т. п., в сильно загрязненных текучих водах. Относится к чис-
лу полисапробов, может встречаться в а мезосапробной зоне.
2. Beggiatoa (рис. 52, В)
Серные бактерии Beggiatoaceae при окислении сероводорода от-
кладывают внутри своих клеток минеральную серу в полужидком
аллотропическом видоизменении. Являются хемосинтезирующими
организмами, источником энергии служит окисление сероводорода в
cepv и серы в серную кислоту.
Beggiatoa имеют вид длинных, подвижных нитей, толщиною
2—4р; внутри клеток находятся многочисленные зерна серы.
Границы между клетками становятся видимыми только после ис-
чезновения зерен серы, что достигается путем выдерживания бакте-
рий в свободной от сероводорода воде.
Все виды Beggiatoa, из которых наиболее обычным является
Beggiatoa leptomitiformis, относятся к числу типичных полисапробов
(при нахождении вне серных источников).
3. Oscllatoria (рис. 52, Г—Д)
Oscillatoria относятся к синезеленым водорослям. Клетки сое-
динены в длинные, подвижные нити, лишенные влагалища и часто
согнутые на концах. Все клетки отдельных нитей одинакового строе,
ния, цилиндрические, в большинстве случаев короткие. Гетероцист
и спор не имеется. Окраска сине-зеленая или желто-зеленая.
Многие виды Oscillatoria относятся к числу типичных а-мезо-
сапробов. Среди последних наиболее часто встречающиеся виды от-
личаются между собою следующими признаками.
1 (2) Концы нитей ясно утончаются О. chalybea
2 (1) Нити на концах не утончаются .... .3
3 (4) Длина клеток в несколько раз больше ширины .
О. putrida
4 (3) Длина клеток меньше ширины . 5
5 (6) Длина клеток не больше четверти их ширины
О. princeps
6 (5) Длина клеток не меньше трети их ширины. . О. tenuis
4. Tubifex, Limnodrilus и Lumbriculus (рис. 53)
Тело Tubifex, Limnodrilus и Lumbriculus, относящихся к мало-
щетинковым червям (Oligochaeta), состоит из большого числа сег-
ментов, к концу уменьшающихся в размерах. Впереди находятся
предротовой (простомиум) и околоротовой (перистомиум) сегмен-
ты, лишенные щетинок. На некотором расстоянии от переднего
конца тела у зрелых особей находится поясок, clitellum, представ-
ляющий железистое утолщение кожи. Впереди пояска ’ находятся
118
тАяв “Т”Ы,“ Об“ «гХ'аг’И'
“Рган (£); Ж.З- Lumbrimlaa variegate. «««"« • Ф
поперечный разрез через кише
/ о__атриум» ’
тРубка копулятивного органа, 2 — ““Генной оболочки.
ника, 5-клетки хлорагогеннои
стенка кишеч-
предпоясковые, а позади него послепоясковые сегменты. Щетинки
являющиеся главными органами движения, расположены на каждой
сегменте, как правило, в виде четырех пучков — двух спинных и
двух брюшных. Щетинки могут быть разнообразного вида — воло.
сковидные, делящиеся на простые, перистые и пильчатые, затем
палочковидные, игольчатые и зубчатые, среди последних различа-
ются однозубчатые, двузубчатые и веерные.
Стенки тела состоят из однослойного эпителия и продольных и
кольцевых слоев мышц, образующих кожно-мышечный мешок. По-
лость целома, заполненная жидкостью, разделена поперечными пе-
регородками, диссепиментами, на метамерные участки; спинной ме-
зентерий отсутствует, а брюшной имеется только между брюшным
кровеносным сосудом и кишечником, поэтому правый и левый от-
делы целома между собою соединены. Пищеварительная система
состоит из короткой ротовой полости, глотки с сильно развитой
мускулатурой, узкого пищевода и длинной средней кишки, перехо-
дящей на конце тела в нерезко отграниченную заднюю кишку. По-
верхность средней кишки со стороны целома покрыта хлорагоген-
ной оболочкой, образованной крупными клетками, заполненными
большим количеством бурых и зеленых гранул. Клетки хлорагоген-
ной оболочки несут экскреторную функцию, кроме того, служат для
накапливания запасных питательных веществ. Кровеносная система
замкнутая, состоит из спинного и брюшного сосудов, проходящих
вдоль кишечника, и кольцевых сосудов, связанных с диссепимент-
ккми. Кроме того, имеется хорошо развитая анастомозирующая
сеть в стенках кишечника, соединенная ветвями с продольными со-
судами. Кровь красного или желтого цвета, эритроцитов нет, гемо-
глобин растворен в кровяной плазме. Специальных органов дыха-
ния не имеется.
Все малощетинковые черви являются гермафродитами. У рас-
сматриваемых видов парные мужские и женские органы размноже-
ния расположены в тех сегментах, наружные покровы которых пре-
вращены в поясок.
Систематика малощетинковых червей в значительной степени ос-
нована на строении органов размножения, поэтому определение мо-
лодых особей чрезвычайно затруднено.
В загрязненной воде часто встречаются Tubifex tubifex, Limno-
drilus udekemianus, Limnodrilus hoffmeisteri, являющиеся представи-
телями семейства Tubificidae, и Lutnbriculus variegatus, относящие-
ся к семейству Lumbriculidae. Все они являются типичными а-ме-
зосапробами. Tubifex tubifex характерен также для полисапробной
и $-мезосапробной зон.
У видов семейства Tubificidae в каждом пучке находится мно-
го щетинок; всегда имеются зубчатые щетинки, в различных пуч-
ках могут быть игольчатые и волосковидные щетинки различного
строения. Органы размножения находятся в десятом и одиннадца-
том сегментах тела. К этому семейству относятся Tubifex и Litnno-
drilus.
Tubifex tubifex характеризуется присутствием в спинных пучках
лредпоясковых сегментов двузубчатых щетинок, между зубцами ко-
120
торых находится один или несколько дополнительных зубчиков; на
послепоясковых сегментах аналогичных щетинок не имеется. Воло-
сковидные щетинки имеются. Длина тела 30—40 лги.
Виды Limnodrilus отличаются одинаковым строением двузубча-
тых щетинок на предпоясковых и послепоясковых сегментах; воло-
сковидных щетинок не имеется. Длина тела 20—60 мм. Виды отли-
чаются по строению щетинок и трубочек копулятивного органа, на-
ходящегося в одиннадцатом сегменте.
Limnodrilus udekemianus — верхний зубец двузубчатых щети-
нок длиннее нижнего; трубочка копулятивного органа короткая,
ее длина в четыре раза превышает ширину.
Limnodrilus hoffmeisteri — зубцы двузубчатых щетинок одинако-
вого размера; трубочка копулятивного органа длинная,, ее длина
превышает ширину более, чем в восемь раз.
У Lumbrlculus variegatus, единственного представителя семейства
Lumbriculidae, в каждом пучке находится по две двузубчатых ще-
тинки, верхний зубец которых короче нижнего; в виде исключения в
некоторых пучках встречаются три щетинки. Передний конец тела
часто окрашен в темнозеленый цвет. Спинной кровеносный сосуд в
средних сегментах со слепыми придатками. Органы размножения в
восьмом—двенадцатом сегментах. Количество сегментов достига-
ет 140—200 и более; длина тела 40—80 мм, толщина 1—1,5 мм.
Черви подвижные и ломкие.
5. Asellus (рис. 54)
Водяные ослики (Asellus aquaticus), относящиеся к подотряду
Asellota, являются самыми обычными представителями пресновод-
ных равноногих раков (Isopoda). Наиболее резкие отличия от рас-
смотренных ранее морских тараканов Mesidothea (стр. 108) заклю-
чаются в строении конечностей.
Уплощенное тело делится на голову, образованную слиянием
всех головных сегментов с первым торакомером, грудь, состоящую
из семи сегментов, и плеотельсон, в состав которого входят брюш-
ные сегменты и тельсон; границы первых двух брюшных сегментов
видны, остальные сегменты слиты с тельсоном без всяких следов.
По бокам головы находится по четыре простых глазка. Антеннулы
короткие, состоят из трехчленистого стебля и 9—15-членистого жгу-
та. Антенны, почти равные длине тела, образованы пятичленистым
стеблем и многочленистым жгутом. Ротовые придатки спереди
прикрыты верхней губой. Мандибулы крепкие, с большим моляр-
ным выростом и трехчленистым щупиком. Максиллулы пластинча-
тые, с двумя большими лопастями. Максиллы также пластинчатые,
с тремя короткими лопастями. Максиллярные ноги, являющиеся
конечностями слитого с головой первого торакомера, состоят из
двучленистого протоподита, первый членик которого несет большой,
окаймленный волосками эпиподит, и пятичленистого щупика (эндо-
подита). Грудных ног (переоподов) семь пар; ноги первой пары
хватательного типа, с ложной клешней. Остальные ноги ходильные,
семичленистые, состоят из коксоподита, базиподита, исхиоподита,
121
Рис. 54. Asellus aquaticus. A — самка; Б — конец тела
дибула; Г — максиллула; Д — максилла; Г
2
о ._МЯН'
1 самки снизу; я нога
Е — максиллярная нога; ' поды
первой пары самца; 3 — плеопод третьей пары самки снизу; И—пле
второй пары самца.
1 — антеннула, 2 — антенна^ 3 — глаз, 4—10 — переоподы первой—седьмой
11 — плеотельсон, 12 — уроподы, 13 — плеопод второй пары, 14 — плеопод^
тьей пары, 15 — базиподит, 16 — исхиоподит, 17 — мероподит, 18—каР”т<
19 — проподит, 20 — дактилоподит, 21 — эндоподит, 22 — эксопоД
122
мероподита, карпоподита, проподита и дактилоподита; небольшие
коксоподиты срастаются своими базальными-краями с грудными
сегментами. У самок от основания грудных ног первых четырех пар
отходят большие выросты (оостегиты), образующие выводковую
камеру (марзупиум). Брюшные конечности (плеоподы) двух пер-
вых пар различного строения в обоих полах. Плеоподы первой па-
ры у самок отсутствуют, у самцов имеются, состоят из двух члени-
ков. Плеоподы второй пары у самок небольшие, округлой
формы, с краевыми Щетинками. У самцов плеоподы второй
пары преобразованы в копулятивный орган. Расширенные эксопо-
диты плеолодов третьей пары образуют состоящую из двух поло-
вин тонкую крышечку (operculum), прикрывающую нижнюю по-
верхность плеотельсона. Приподняв крышечку, можно видеть не-
большие эндоподиты плеоподов третьей пары, а также двуветви-
стые плеоподы двух следующих пар, служащие органами дыхания.
На заднем крае плеотельсона расположены палочковидные двувет-
вистце уроподы.
Водяной ослик при массовом нахождении характеризует «-ме
зосапробную зону. Относится к числу детритофагов, питается раз
лагающимися в воде древесными листьями.
6. Личинки Chironotnus
Мотыли, личинки Chironotnus plumosus, при массовом нахожде
нии типичны для а-мезосапробной зоны; встречаются также г
полисапробной и 0-мезосапробной зонах.
Описание личинок Chironotnus plumosus помещено на стр. 225
7. Личинки Eristalis (рис. 55)
Водяные личинки мух семейства Syrphidae (журчалок), относя-
щегося к подотряду Brachycera, двукрылых насекомых, известны
под названием крысок.
Тело личинки Eristalis tenax, наиболее распространенного вида,
покрыто мягкой кутикулой белого цвета; на поверхности находятся
складки и бугорки. На голове, обычно втянутой внутрь тела, рас-
положены короткие, сближенные основаниями антенны. Ротовое от-
верстие, помещенное на нижней стороне головы, окружено двуло-
пастной складкой, способной втягиваться внутрь ротовой полости.
Находящиеся на складке шипики счищают с субстрата пищевые ча-
стицы, которые потом, при втягивании складки, переносятся в ро-
товую полость. Мандибулы рудиментарные. На брюшной стороне
личинки находится семь пар небольших ложных ножек, вооружен-
ных на конце щетинками; грудные ложные ноги отделены большим
промежутком от брюшных ног. На головном сегменте находится па-
ра небольших дыхательных трубок, снабженных на конце отвер-
стием. Задний конец тела вытянут в длинную и тонкую дыхатель-
ную трубку, превышающую в несколько раз длину самой личинки.
Дыхательная трубка состоит из трех длинных сегментов, входящих
Друг в друга наподобие частей зрительной трубы. На конце дыха-
123
1ельной трубки находятся стигмы. Около анального отверстия от-
ходят две пальцевидно ветвящиеся анальные жабры, заметные
лишь у живых особей, у фиксированных же втянутые внутрь.
Рис. 55. Личинка Eristalis tenax. 1 — антенна, 2 — ложная ножка первой пары,
3 — ложная ножка седьмой пары, 4 — дыхательная трубка.
Крыски живут в самых загрязненных водоемах, относятся к по-
лисапробам. Необходимый кислород доставляется через дыхатель-
ную трубку, которая своим концом всегда высовывается из воды.
Глубина погружения соответствует длине дыхательной трубки в
вытянутом состоянии.
ОРГАНИЗМЫ И АКТИВНАЯ РЕАКЦИЯ СРЕДЫ
Величина активной реакции (pH) обозначается отрицательным
логарифмом концентрации водородных ионов. В нейтральной воде
рН=7,0 (точнее 7,07), в кислой воде рН<^7,0, в щелочной воде
рН>7,0. В морях величина pH колеблется в пределах 3,1—8,3.
Пресноводные водоемы делятся на нейтрально-щелочные, в которых
pH изменяется от 6,0 до 10,0, и кислые, с рН<6,0; к числу послед-
них относятся богатые гуминовыми веществами сфагновые водоемы.
Высокая активная кислотность, рН<7>, зависит от присутствия
серной кислоты.
Живые организмы, выделяя при дыхании углекислый газ, изме-
няют активную реакцию воды в сторону подкисления—величина pH
понижается. При фотосинтезе углекислый газ ассимилируется —
величина pH повышается. Вследствие этого летом, в особенности в
период цветения водоема, в поверхностных слоях воды величина
pH больше, чем в нижних слоях. АсЛмиляция углекислого газа
протекает только на дневном свете, вследствие этого в водоемах
всегда наблюдаются суточные и сезонные изменения активной ре-
акции воды.
Организмы, выносящие значительные колебания активной реак-
ции воды, называются эвриионными. Те организмы, которые могут
существовать только в нейтрально-щелочных или кислых водах, от-
носятся к числу стеноионных.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
ИЗМЕНЕНИЕ АКТИВНОЙ РЕАКЦИИ ВОДЫ ПОД ВЛИЯНИЕМ
ЖИЗНЕДЕЯТЕЛЬНОСТИ РАСТЕНИЙ
В небольшой сосуд, емкостью около 100 см3, содержащий воду
нейтральной реакции, прибавляют две-три капли спиртового раство-
ра фенолфталеина (вода должна оставаться прозрачной). Затем
помещают в сосуд несколько побегов элодеи и выставляют их на
свет, например на подоконник. Через некоторое время можно ви-
деть, как на поверхности листьев образуются красные облачка,
сливающиеся затем между собой; к концу опыта вся вода приобре-
125
тает .красную окраску. Измеряя величину pH в начале и конце
эксперимента в контрольном сосуде, содержащем примерно такое же
количество ветвей элодеи, помещенных в воду без фенолфталеина,
можно определить степень изменения концентрации водородных ио-
нов.
ОПРЕДЕЛЕНИЕ ГРАНИЦ ВЫЖИВАЕМОСТИ ОРГАНИЗМОВ ПРИ
РАЗЛИЧНОЙ ВЕЛИЧИНЕ АКТИВНОЙ РЕАКЦИИ ВОДЫ
Небольшие сосуды (при работе с личинками насекомых и низ-
шими раками) или часовые стекла (при работе с коловратками)
наполняют водой из естественного водоема, и прибавлением соот-
ветствующего количества щелочи и кислоты получают ряд непре-
рывных изменений активной реакции от рН=2,0 до рН=9,0 при-
мерно через 0,5. В каждый сосуд помещают по несколько экземпля-
ров испытуемых организмов и отмечают время их выжива-
ния. Эвриионные организмы, как, например, Chironotnus plumosus,
живут долгое время при любой степени концентрации водородных
ионов. Более чувствительные организмы, живущие при рН=5,0—
9,0, как, например, дафнии и циклопы, погибают через несколько
часов при низких величинах активной реакции среды; оптимальные
величины определяются при содержании животных в опытных сосу-
дах в течение нескольких дней. Многие коловратки являются стено-
ионными организмами, могут существовать только при сравнитель-
но небольших изменениях активной реакции воды; этим, между
прочим, объясняется трудность их воспитания в экспериментальных
условиях. Например, Keratella cochlearis может жить, только при
рН=6,0—7,8; помещенные в воду с рН=9,0 погибают через не-
сколько минут.
ОРГАНЫ ДЫХАНИЯ ВОДНЫХ ОРГАНИЗМОВ
Водные организмы дышат растворенным в воде кислородом,
за исключением вторичноводных животных, сохранивших в боль-
шинстве случаев воздушный способ, дыхания. Некоторые организмы
дышат всей поверхностью тела (кожное дыхание), другие при помо-
щи специальных выростов, носящих название кровяных и трахей-
ных жабр.
Кожное дыхание встречается у большого числа водных живот-
ных, к числу которых относятся простейшие, кишечнополостные,
губки, некоторые черви (ресничные и малощетинковые черви), не-
которые ракообразные (веслоногие раки), некоторые моллюски
(часть голожаберных моллюсков, крылоногие моллюски) и некото-
рые представители других групп. У части ветвистоусых раков (Ро-
lyph£midae и Leptodora) для дыхания служит так называемый го-
ловной щит, отличающийся по строению от остальных покровов и
хорошо прокрашивающийся при применении прижизненной окраски.
Кровяные жабры имеют вид тонкостенных выростов, внутри ко-
торых находится сильно разветвленная капиллярная сеть кровенос-
ных сосудов или кровяная плазма. Кровяные жабры водных орга-
низмов, в противоположность органам дыхания наземных живот-
ных, представляют собою наружные выросты, приспособленные к
получению кислорода из окружающей воды; они могут распола-
гаться на различных участках тела. Кровяными жабрами облада-
ют многощетинковые черви, большинство ракообразных и моллюс-
ков, водные личинки некоторых насекомых, иглокожие, асцидии и
рыбы.
Трахейные жабры, свойственные водным личинкам большого
числа насекомых из отрядов поденок, стрекоз, веснянок, больше-
крылых, ручейников и двукрылых, имеют вид тонкостенных простых
или ветвящихся выростов, расположенных на самых различных ча-
стях тела и содержащих тонкие трахейные стволы или сильно вет-
вящуюся сеть трахейных капилляров. Внутри трахейных жабр на-
ходится газообразный кислород, диффундирующий из окружаю-
щей воды через тонкие покровы жабр.
Почти все вторичноводные организмы дышат атмосферным
воздухом, за исключением личинок насекомых, обладающих тра-
127
хенными жабрами или перешедших к кожному дыханию. Млеко-
питающие для вентиляции своих органов дыхания периодически
поднимаются к поверхности воды, то же делают водные насеко-
мые— жуки и клопы, а также водные личинки многих двукрылых
насекомых.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
КОЖНОЕ ДЫХАНИЕ
Рассматривается строение головного щита Leptodora или Poly-
phemus. Для этого живые организмы погружают в часовом стекле
в 0,01 %-ный раствор азотнокислого серебра; предварительно их по-
мещают на короткое время в дестиллированную воду, во' избежание
появления мути, получающейся при реагировании азотнокислого
серебра с солями воды. Спустя несколько минут под микроскопом
наблюдают потемнение головного щита, вызванное восстановле-
нием азотнокислого серебра.
Можно для той же цели воспользоваться слабым, розоватого
цвета, раствором перманганата калия, окрашивающим головной
щит в коричневые’ оттенки.
У Leptodora и Polyphemidae головной щит является единствен-
ным органом дыхания. Для сравнения рассматривают органы дыха-
ния других ветвистоусых раков, например дафнии, у которых об-
мен газов протекает в эпиподитах грудных ног. Дафнии, перенесен-
ные после промывки дестиллированной водой в 0,01—0,05 %-ный ра-
створ азотнокислого серебра, через некоторое время падают на дно
часового стекла. После этого их помещают в чистую воду, в кото-
рой они могут жить в течение продолжительного времени, позволяя
изучить строение и нормальное расположение эпиподитов грудных
ног.
Как головной щит Leptodora и Polyphemidae, так и эпиподиты
на грудных ногах других представителей ветвистоусых раков могут
быть изучены также при применении элективной прижизненной ок-
раски раствором нейтрального красного или метиленовой сини.
1. Leptodora (рис. 56, Л)
Описание строения лептодоры помещено на стр. 40. На рис. 56, Л
показана окраска головного щита раствором азотнокислого серебра.
2. Polyphemus (рис. 56, Б—В)
Полифемусы (Polyphemus pediculus) относятся к числу наи-
более широко распространенных представителей семейства Poly-
phemidae., ветвистоусых раков, встречаются в небольших водоемах,
а в озерах — среди прибрежной растительности.
Голова большая, спереди полукруглая, с очень большим глазом
на переднем конце. Головной щит широкий, с округлыми боковыми
лопастями; его поверхность утолщена и покрыта шестигранной сет-
чатостью. Голова отделена от остального тела резким перехватом.
128
^Ис.
г
головным
(В).
передний конец тела Leptodora, с
•’лаз 9 Б‘ В ~ Ро]УРЬетиз pediculus, самкаi ( Ма 5“_^_ноп|
Четвёп^ антеннУла, 3 — антенна!, 4 — голов»о Ши _ хвостовой
еРтои пар, 9 —эксоподит ноги третьей пары,
11 — выводковая камера.
щитом;
первой—
шип,
Выводковая камера, так же как и у других Polyphemidae, нахо-
дится внутри тела, но, в отличие от выводковой камеры Bythotre.
phes, прирастает к нему только по бокам. Антеннулы, прижатые к
нижней поверхности головы, слиты своими основными члениками.
Антенны сильные, с трехчленистыми эндоподитами и четырехчлени-
стыми эксоподнтами, вооруженными оперенными плавательными
щетинками. Ноги, в числе четырех пар, скелетного типа, с одночле-
нистыми эксоподнтами. Постабдомен короткий, без фуркальных
когтей. Хвостовой шип, аналогичный хвостовому шипу Bythotre-
phes, сравнительно короткий, на конце с двумя длинными хвосто-
выми щетинками. Кишечник впереди с боковыми расширениями.
Полифемусы питаются хищнически, потребляют также водоросли и
детрит. Окраска фиолетовая и коричнево-фиолетового цвета.
КРОВЯНЫЕ ЖАБРЫ
В качестве примеров рассматриваются представители многоще-
тинковых червей (Arenicola), жаброногих раков (Pristicephalus,
Apus, Lynceus), амфипод (Gammarus), равноногих раков (Asellus,
Mesidothea), брюхоногих моллюсков (Valvata) и двустворчатых
моллюсков (Anodonta и Unio).
1. Arenicola
Кустистые жабры расположены позади нотоподий на 7-м—19-м
сегментах тела. Описание арениколы помещено на стр. 65.
2. Pristicephalus (рис. 57)
Pristicephalus josephinae, являющийся одним из наиболее
обычных представителей жаброногов, относится к подотряду
Anostraca отряда Branchiopoda (жаброногих раков). По внешнему
виду сходны с Artemia salina (стр. 98). Тело, сильно вытянутое,
ясно сегментированное и совершенно лишенное карапакса, делится
на голову, челюстной отдел, грудь и брюшко. На голове находятся
парные стебельчатые глаза, нитевидные антеннулы и антенны. По-
следние у самок имеют вид сравнительно небольших выростов, а у
самцов превращены в хватательный орган, базальные членики ко-
торого несут сверху большие, сильно изогнутые придатки, а конеч-
ные членики серповидно согнуты. Челюстному отделу соответствуют
ротовые придатки — мандибулы, максиллулы и максиллы, по бо-
кам находятся парные панцырные железы, являющиеся органами
выделения. Грудь состоит из одиннадцати сегментов, несущих та-
кое же количество пар грудных ног. Грудные ноги листовидные, с
тонкими покровами. Необходимую крепость получают вследствие
большого внутреннего давления жидкости тела, поэтому называют-
ся тургорными конечностями в противоположность скелетным ко-
нечностям, крепость которых обусловливается хитином. Каждая
нога имеет вид широкой пластины (протоподит), внутренний край
которой подразделен на пять эндитов. Большой проксимальный эн-
130
диг вместе со вторым эндитом, тесно с ним связанным, несет по
внутреннему краю большое число длинных и гибких щетинок; ос-
тальные эндиты представлены небольшими выступами. На наруж-
ном крае протоподита находятся хорошо развитые эксоподит и эпи-
Рис. 57. Pristicephalus josephinae. А—-самец снизу; Б — голова самца сверху;
В голова самки сверху; Г—абдомен самки сбоку; Д — нога первой пары;
Е — нога седьмой пары; Ж—нога одиннадцатой пары.
1 — голова, 2 — грудь, 3 — брюшко, 4 — генитальный сегмент, 5 — фурка, 6 —
глаз, 7—антеннула, 8— антенна, 9 — грудные ноги, /0 — копулятивный орган,
// — эндиты, 12 — эксоподит, 13 — эпиподит, 14 — преэпиподит.
подит и два преэпиподита, расположенных проксимально от пер-
вых. Эндоподит, образующий конечную часть ноги, отличается по
своему вооружению. Брюшко состоит из девяти сегментов. Два пер-
вых сегмента слиты в большой генитальный сегмент, на нижней
стороне которого расположен парный копулятивный орган самцов
или непарный яйцевой мешок самок. К последнему брюшному сег-
9* 131
менту причленяются две ветви фурки, вооруженные по краям ще-
тинками.
Органами дыхания служат расположенные на грудных ногах
эпиподиты, постоянно омываемые токами воды, вызываемыми рит-
мичными движениями грудных конечностей. Кишечник, в передней
части с парными печеночными выростами, проходит вдоль всего
тела и открывается на последнем, анальном, сегменте брюшка.
Сердце имеет вид длинной трубки, тянущейся по спинной стороне
животного от головы до анального сегмента; в каждом сегменте,
?а исключением первого и последнего, имеется по одной парс остий.
Яичники самок расположены в абдомене; оба яйцевода впадают в
объемистый uterus, находящийся внутри яйцевого мешка.
Жаброноги обитают в мелких временных водоемах с талой во-
дой или в лужах, оставшихся после разлива реки, и вместе с их вы-
сыханием погибают. В естественном состоянии находятся в постоян-
ном движении. При плавании организма в воде, что обычно со-
вершается брюшной стороной кверху, вдоль каждого ряда конечно-
стей по направлению от головы к заднему концу тела проходят
токи воды, вызываемые быстрыми ударами наружных частей ног.
Внутренние части служат для отфильтровывания пищевых частиц
(стр. 244). При отсутствии взвешенной в воде пищи жаброноги
переворачиваются спинной стороной кверху и соскабливают щетин-
ками эндоподитов ног иловые частицы с донной растительности или
со дна' водоема. Все жаброноги относятся к числу активных филь-
траторов.
3. Apus (рис. 58)
Щитни (Apus cancriformis) относятся к подотряду Notostraca
отряда Branchiopoda (жаброногих раков).
Теле, сжатое в спинно-брюшном направлении, ясно сегментиро-
вано. Карапакс образует спинной щит, прикрывающий не только
сегменты груди, но и значительную часть брюшка. Впереди кара-
пакс сливается с головным панцырем, образуя общий щит, боковые
края которого могут сгибаться, особенно при плавании животного.
В полости карапакса, между наружным и внутренним листками,
расположены парные панцирные железы, являющиеся органами вы-
деления; их протоки открываются на максиллах. Сравнительно не-
большие, сближенные между с^бдю сложные глаза сидят на верх-
ней поверхности головы; вмшрТними расположен трудно заметный
непарный глаз. На нижней стороне головы находятся рудиментар-
ные антеннулы и антенны, верхняя губа, мандибулы, максиллулы и
максиллы. Граница между грудью и брюшком проводится с тру-
дом, так как грудные ноги по мере продвижения назад постепенно
уменьшаются в размере до полного исчезновения. Число грудных
сегментов велико. Количество пар ног превышает число торакаль-
ных сегментов, так как последние, начиная с двенадцатого, несут
по несколько пар конечностей (явление полиподии), что является
результатом слияния между собою сегментов. Ноги листовидные,
сильно уплощенные, с большими эксоподитами и эпиподитами; все
эндиты одного размера, сходного строения, за исключением первого
132 •
Рис. 58. Apus cancriformis. Я-самка; Б - нога
-мандибула; Г — максиллула; Д — максилла, гямки
второй пары; 3-нога одиннадцато., пары самки.
цмуйАуч.и.Гум,____ Янт₽ннула 4 — антенна, 5 — спиннои шит,
/-глаз, 2-пепар-нШП ВЯ® ^д„була, 9 - максиллула, «-мак-
6 - панцирная железа, 7-фурка, в-манд * «-эндиты,
силла, //-максиллярный вырост, «-эндопод
14 -эксоподит, /5-эпипод.п.
эндита, или максиллярного выроста, вооруженного по другому. По-
степенно уменьшающиеся в величину конечности в конце концов
преобразуются в небольшие пластинки. Ноги первых двух пар иного
строения. Они вытянуты в длину, сильно хитинизированы и ясно
членистые; максиллярные выросты хорошо развиты, остальные эн-
диты образуют удлиненной формы придатки, увеличивающиеся в
размерах по мере продвижения к концу конечности. На ногах пер-
вой пары три дистальных эндита превращены в длинные, многочле-
нистые жгуты, выходящие за пределы тела. У самок ноги один-
надцатой пары сильно видоизменены и превращены в выводковые
камеры, имеющие вид полусферических чашек (дистальная часть
протоподита), закрывающихся крышечкой (измененный эксоподит).
Отложенные яйца сохраняются в яйцевых капсулах в течение не-
скольких дней, а затем выбрасываются в воду. У самцов одиннад-
цатая пара ног обычного строения. Брюшные сегменты сверху с
шипиками. К последнему брюшному сегменту причленяется фурка,
имеющая вид двух длинных многочленистых и снабженных волос-
ками нитевидных придатков.
Органами дыхания служат эпиподиты, расположенные на груд-
ных ногах. В передней части средней кишки находятся сильно вет-
вящиеся печеночные выросты. Сердце, имеющее вид длинной труб-
ки, тянется по спинной стороне тела от головы до одиннадцатого
сегмента. Ветвящиеся яичники самок наполняют всю полость тела;
протоки открываются в выводковые капсулы на ногах одиннадца-
той пары.
Щитни обитают в мелких, временных водоемах с талой или дож-
девой водой. Взрослые особи могут хорошо плавать, но обычно
ползают по дну при помощи ног первых двух пар, иногда почти це-
ликом зарываются в ил. По способу получения пищи относятся к
числу активных фильтраторов (стр. 250), питаются крупными ча-
стицами детрита. Наряду с этим ведут хищнический образ жизни,
поедая живых или мертвых жаброногов (Anostiraca), икру лягушек,
малощетинковых червей и личинок Chironomidae.
4. Lynceus (рис. 59)
Линцеи (Lynceus brachyurus) относятся к подотряду Concho-
straca жаброногих раков (Branchiopoda).
Тело полностью заключено в двустворчатую раковину (кара-
пакс), почти шаровидной формы и лишенную, в противоположность
всем остальным представителям Conchcstraca, полос прироста. Тело
внутри раковины помещается свободно, прикрепляясь к ней лишь
на небольшом протяжении в передней части, около мускула, закры-
вающего створки раковины. Голова, обособленная от остального
тела слабой перетяжкой, согнута на брюшную сторону и снабжена
большим, на конце острым рострумом. Сидячие сложные глаза
сильно сближены между собою; впереди их расположен непарный
глаз и бокаловидный орган, несущий, вероятно, дыхательную функ-
цию. Антеннулы небольшие. Антенны сильные, короткие, с двумя
многочленистыми ветвями, вооруженными многочисленными плава-
134
тельными щс1 инками, ria нижней стороне головы расположены верх-
няя губа, мандибулы и максиллулы; максиллы отсутствуют. Грудь
состоит у самцов из одиннадцати, у самок из двенадцати сегмен-
тов. Грудные ноги имеют вид пластинки (протоподит), утолщенной
Рис. 59. Lynceus brachyurus. А — самка; Б — нога третьей пары; В — нога деся-
той пары; Г — нога первой пары самца.
/ — голова, 2— бокаловидный орган, 3 — глаз, 4 — непарный глазок, 5 — рост-
рум, 6—антеннула, 7 — антенна, 8— мандибула, 9 — максиллула, 10— мускул;
закрывающий раковину, 11—сердце, 12— выводковая камера, 13— яичник,
14—абдомен, 15— грудные ноги, 16 — придатки эксоподитов ног десятой и
одиннадцатой пар, 17 — оперкулярная пластинка, 18 — фурка, 19 — раковина,
20 — протоподит, 21 — эндоподит, 22 — эндиты, 23 — эксоподит, 24 — эпиподит.
135
при основании благодаря нахождению там мышечных волокон. По
внутреннему краю протоподита находятся пять эндитов, вооружен-
ных щетинками двух сортов, из которых одни, длинные и тонкие,
образуют подобие густой гребенки, а другие, короткие и сильные,
расположены вторым рядом. Проксимальный эндит, отличающийся
от других по форме и толщине хитина, называется максиллярным
выростом. Дистально от пятого эндита находится узкий и длинный
эндоподит. вооруженный щетинками, расположенными в один ряд.
Наружный край протоподита снабжен длинным эксоподитом и срав-
нительно небольшим, лишенным щетинок э^®подитом. У самок
праксимальные части эксоподитов на ногах десятой и одиннадцатой
пар вытянуты в длинные цилиндрические выросты, направленные к
спинной стороне организма и служащие для поддержания яиц, от-
кладываемых в выводковую камеру, расположенную между спин-
ной стороной тела и раковиной; у самцов эндоподиты ног первой
пары преобразованы в хватательные органы. На последнем груд-
ном сегменте вместо ног находится оперкулярная пластинка. Брюш-
ко покрыто волосками, короткое, несегментировэнное, с одной па-
рой чувствительных щетинок. Фурка рудиментарная (у других
представителей Conchostraca фурка хорошо развита и имеет вид
двух изогнутых когтей).
Органами дыхания служат эпиподиты ног, омываемые водою
при их движении. Кишечник впереди с ветвящимися печеночными
выростами. Сердце короткое, не простирается далее четвертого
грудного сегмента. Органы выделения помещаются в толще ство-
рок раковины. Яичники самок расположены вдоль тела; яйцеводы
открываются на внутренней стороне ног одиннадцатой пары. Яйца
откладываются в выводковую камеру и спустя некоторое время вы-
брасываются в воду.
Линцеи встречаются весною в пойменных водоемах, иногда в
реках. Движение совершается при помощи ударов антенн, грудные
ноги служат для отфильтровывания пищевых частиц.
5. Gammarus (рис. 60)
Гаммарусы относятся к подотряду Gammaridea отряда Amphi-
poda. В пресных водах распространены Gammarus pulex и Gamma-
rus lacustris (подрод Rivulogammarus), в северных морях обычными
литоральными видами являются Gammarus locusta (подрод Gam-
marus s. str.) и Gammarus obtusatus (подрод Marinogatnmarиs).
Тело, сжатое с боков и полностью лишенное карапакса, делится
на голову, грудь и брюшко. Голова, в состав которой входит также
первый торакомер, несет парные сидячие глаза и шесть пар при-
датков. Антеннулы состоят из трехчленистого стебля, длинного
многочленистого жгута и небольшого добавочного жгута, отходя-
щего от третьего членика стебля. Антенны состоят из длинного пя-
тичленистого стебля и сравнительно короткого жгута. Мандибулы
крепкие, с хорошо развитым молярным выростом и с трехчлени-
стым щупиком. Максиллулы и максиллы небольшие, пластинчатые.
136
Максиллярные ноги, соответствующие первому торакомеру, входя-
щему в состав головы, хорошо развиты, слиты своими основными
члениками.
Грудь (торакс или переон), состоящая из семи сегментов, несет
такое же количество пар грудных ног (переоподов), лишенных эк-
соподитов. Каждая нога образована семью члениками, называю-
щимися коксоподитом, базиподитом, исхиоподитом, мероподитом,
карпоподитом, проподитом и дактилоподитом (другие наименова-
ния: кокса, базис, исхиум, мерус, карпус, проподус и дактилус).
Коксоподиты, или коксальные пластинки, сильно расширенные и
уплощенные, являются продолжением грудных сегментов. Особенно
сильно они развиты на четырех передних сегментах, превышая вы-
соту самих сегментов; базиподиты этих ног прикрепляются не к кон-
цу коксальной пластинки, а к верхней части ее внутренней стороны.
Коксальные пластинки ног трех последних пар развиты значитель-
но слабее, базиподиты прикрепляются к их концам. Переоподы
двух первых пар, приспособленные для хватания и поэтому часто
называющиеся гнатоподами, оканчиваются ложной клешней, обра-
зованной расширенным проподитом (называемым ладонью, или
пальмой), к дистальному (пальмарному) краю которого сгибается
подвижный дактилоподит, прикрепленный к углу пальмарного края.
К внутренней стороне всех коксальных пластинок, кроме первой
пары, прикрепляются тонкостенные овальные жаберные пластинки,
содержащие внутри кровеносные лакуны. У самок от коксальных
пластинок второй — пятой пар переоподов, выше жаберных пласти-
нок, отходят широкие, окаймленные волосками хитиновые пластин-
ки (оостегиты), образующие выводковую камеру (марзупиум), в ко-
торой проходит развитие отложенных яиц. Для рассмотрения строе-
ния переоподов следует вырвать ноги второй и последней пар.
Брюшко (абдомен), по своему строению и по строению соответству-
ющих конечностей, подразделено на два отдела. Первый отдел
(плеон) состоит из трех сегментов, боковые части которых образу-
ют широкие эпимеры; отходящие от плеона плавательные конечно-
сти (плеоподы), состоят из базального членика и двух многочлени-
стых ветвей, вооруженных щетинками. Плеоподы служат не только
для плавания, но и для образования тока воды, омывающей жаб-
ры. Задний отдел абдомена (урус) состоит также из трех сегмен-
тов, несущих такое же число прыгательных ног (уроподов), дву-
ветвистых, крепких и вооруженных шипами (первые две пары),
или щетинками (последняя пара). На конце брюшка, между уропо-
дами третьей пары, помещается тельсон, имеющий вид небольшой
пластинки, глубоко подразделенной на две половины.
Характерным признаком всех видов Gamniarus является воору-
жение трех последних сегментов брюшка, несущих на спинной сто-
роне шипики.
Пресноводные виды различаются между собою следующими
признаками.
Gammarus (Rivulogammarus) pulex — задний край переоподов
третьей пары самцов с густыми пучками завитых щетинок; эпимеры
137
Рис. 60. A—Е—Gammarus lacustris, самка (А), антенна (Б), переопод третьей
пары самца (В), эпимеры второй (Г) и третьей (Д) пар, тельсон (Е); Ж—Л—
Gammarus pulex, антенна (ЛК), переопод третьей пары самца (3), эпимеры
второй (И) и третьей (К) пар, тельсон (Л); М—П—Gammarus locusta, самкяг
(М), абдомен сверху (Н), уропод третьей пары (О); тельсон (77); Р—У—Gam-
marus oblusatus, абдомен сверху (Р), эпимера третьей пары (С), уропод третьей
пары (7), тельсон (У).
138
второго брюшного сегмента с прямоугольными задними углами;
тельсон с шипами и пучками щетинок.
Gammarus (Rivulogammarus) lacustris — переоподы третьей па-
ры самцов с пучками прямых щетинок; эпимеры второго брюшного
сегмента с вытянутыми задними углами; тельсон с шипами и оди-
ночными щетинками.
Виды из северных морей различаются между собою следующими
признаками.
Gammarus (Gammarus) locusta — три последних брюшных сег-
мента с тремя группами шипиков, срединная группа содержит три-
пять шипиков, боковые по три-четыре шипика; внутренняя ветвь
(эндоподит) уроподов третьей пары почти равна длине первого чле-
ника наружной ветви (эксоподита); каждая ветвь тельсона с дву-
мя-тремя базальными, одним субапикальным и тремя апикальными
шипами.
Gammarus (Marinogamtnarus) obtusatus — три последних брюш-
ных сегмента с двумя полукруглыми рядами шипиков, в каждом
ряду по шести-девяти шипиков; эндоподит уроподов третьей пары
не длиннее трети длины первого членика эксоподита; каждая ветвь
тельсона с одним коротким базальным и тремя короткими апикаль-
ными шипами, без щетинок.
6. Asellus и Mesidothea
Строение жабр помещено на стр. 123 (Asellus aquaticus) и
стр. 109 (Mesidothea entomon).
7. Valvata (рис. 61)
Затворки (Valvata) относятся к отряду Monotocardia класса
переднежаберных (Prosobranchia) брюхоногих моллюсков.
Раковина, размеры которой не превышают 5—8 мм, имеет ку-
баревидную, коническую или дисковидную форму; в последнем слу-
чае очень сходна с катушкой (Planorbis), от которой отличается
круглым устьем и присутствием крышечки. Обороты раковины
немногочисленные. Устье круглое. Крышечка роговая, концентриче-
ская. Тело в вытянутом состоянии впереди оканчивается хоботооб-
разным рыльцем; щупальца длинные, при их основании расположе-
ны глазки; нога впереди двулопастная. Имеется один левый ктени-
дий, двоякоперистый, свободный на всем протяжении, у живого
моллюска высовывающийся из жаберной полости; рядом с жаброй
находится тонкий и длинный придаток.
Обычные виды Valvata, среди которых наиболее широко рас-
пространен Valvata plscinalis, различаются между собою следую-
щими признаками.
I — антеннула, 2 — добавочный жгут антеннулы, <3—антенна, 4— переоподы.
5 — эпимера второй пары, 6 — эпимера третьей пары, 7 - уропод третьей парни
# — тельсон, 9— коксоподит (коксальная пластинка), /0—базиподнт, 11—исхио-
подит, 12—мероподит, 13—карпоподит, 14 — проподит, /5 — да ктилопод нт.
139
I (2) Раковина округло-кубаревидная, очень тонко исчерчен-
ная, высота 4—8 мм (сильно варьирующий вид)
V. piscinalis
2 (1) Раковина иной формы, высота около 2 мм . 3
3 (4) Раковина с низким завитком, ширина 4 мм
V. pulchella
Рис.61.А—В — Valvata piscinalis.cnepeaH (А), снизу (Б), передний конец тела
сверху (в); Г — Valvata pulchella; Д — Valvata cristata;£ — Valvata sibirica.
1—глаз, 2 — рыльце, 3 — щупальце, 4 — ктенидий с придатком.
4 (3) Раковина дисковидная.................................5
5 (6) Поверхность раковины тонко исчерченная, ширина 2—
3 мм.........................................V. cristata
6 (5) Поверхность раковины с выпуклыми поперечными остры-
ми линиями, ширина 5 мм . V. sibirica
8. Anodonta и Unio (рис. 62)
Наиболее обычными среди крупных представителей двустворча-
тых моллюсков (Lamellibranchiata или Bivalvia) в пресных водах
являются беззубки (Anodonta} и перловицы (Unio).
Раковина толстая, удлиненной формы; в спокойном состоянии
замкнутая. Обе створки по верхнему, или спинному краю соеди-
нены роговидным эластичным лигаментом, раскрывающим рако-
вину; край, противоположный спинному, называется нижним, или
брюшным. Передний край широкий, задний край суженный. Впере-
ди лигамента расположена верхушка, более или менее выступаю
щая над спинным краем. Наружная поверхность раковины покры-
та концентрическими линиями прироста. Спинной край каждой
створки у перловиц вооружен зубами — короткими кардинальными,
140
находящимися около верхушки, и пластинчатыми латеральными^
лежащими вдоль спинного края; зубы обеих створок образуют за-
мок раковины, У беззубок, как показывает название, никаких зу-
бов не имеется. Наружный, конхиновый слой буро-желтого или
буро-зеленого цвета. Под ним лежит белый фарфоровидный слой,
легко обнаруживаемый при соскабливании конхинового слоя. Пер-
ламутровый слой выстилает раковину внутри; на нем видны отпе-
чатки переднего и заднего замыкательных мускулов и мантийная
линия, являющаяся следом прикрепления мантийной мышцы.
Для изучения строения тела следует удалить одну створку,
предварительно отделив от нее скальпелем мускулы-замыкатели.
После удаления створки видны складки мантии, мышцы-замыка-
тели и находящиеся рядом с ними небольшие ножные мышцы.
Сквозь полупрозрачную мантию просвечивают внутренние орга-
ны — нога, жабры, а по спинной стороне — окрашенная в серый
цвет печень, темнокоричневая перикардиальная,железа, удлиненный
перикардий и черноватая почка; утолщенным мышечным краем
складки мантии прикреплены к створкам. Сзади края мантии
сомкнуты и образуют два сифона, выше сифонов они срощены.
Жаберный (нижний) сифон большой, по краям с сосочками, аналь-
ный (верхний) сифон небольшой. Ток воды, вызываемый мерца-
нием ресничного эпителия, покрывающего жабры, поверхность ман-
тии и стенки тела, вносит в мантийную полость кислород и мелкие пи-
щевые частицы; этот ток входит через жаберный сифон. Вода, ли-
шенная пищевых частиц и кислорода и обогащенная угольной кис-
лотой, выходит наружу через анальный сифон. Входящий ток легко
обнаружить, если бросить перед сифонами находящегося в аква-
риуме моллюска небольшое количество мелких яркоокрашенных ча-
стичек, например кармина или индиго; так как эти вещества мол-
люском не потребляются, то через короткое время можно видеть их
выходящими через верхний сифон. При отгибании складки мантии
кверху, обнаруживается мантийная полость. Большая клиновидная,
лишенная подошвы нога, окрашенная в желто-оранжевый цвет, яв-
ляется единственным органом движения. В спокойном состоянии мол-
люск зарывается в грунт передним концом, выставляя наружу зад-
ний конец с сифонами. При движении нога набухает, вследствие при-
тока крови, и высовывается из раковины; укрепившись в грунте,
моллюск подтягивается затем сокращением мышц ноги. Между но-
гой и передним мускулом-замыкателем помещается широкое рото-
вое отверстие, с каждой стороны которого находится по две треу-
гольных ротовых лопасти. По бокам ноги расположены большие
коричневые жабры, поперечно и продольно исчерченные. Каждая
жабра состоит из двух полужабр, которые в свою очередь образо-
ваны двумя пластинками; нисходящие пластинки обеих полужабр
обращены друг к другу, восходящая пластинка наружной полужаб-
ры обращена к мантии, а внутренней полужабры — к телу. Веноз-
ная кровь по приносящим жаберным сосудам, расположенным в
нисходящих пластинках, вносится в жабры и окисляется в них; ар-
териальная кровь по выносящим жаберным сосудам, расположен-
ным! в восходящих пластинках, уносится из жабр. Вода входит че-
141
ж
в
(Л)
Рже. 62. А, Б — Anodonta cygnea, внутренняя поверхность правой створки i
внутреннее строение после удаления раковины и левой мантийной складки ( Ь
В — Unio pictorum, внутренняя поверхность левой створки; Г — Unio Ьпти
Д —Unio crassus; Е, Ж —глоходий Unio, вид сбоку (Е) и створка (Ж)
рез многочисленные отверстия в жабры, а из них выходит в наджа-
берные каналы, откуда поступает в анальный сифон. Следует выре-
зать небольшую часть жабры живого моллюска и рассмотреть мер-
цание ресничного эпителия при большом увеличении. В жабрах
зрелых особей находятся личинки — глохидии, дальнейшее разви-
тие которых протекает на коже или жабрах пресноводных рыб. Гло-
хидии, размером около 0,2—0,3 мм, хорошо видны в наружных
жабрах зрелых моллюсков на перегибе жаберных нитей.
Раковина беззубок (Anodonta) продолговато овальная, тонко-
стенная, коричневато-зеленого цвета. Замочные зубы отсутствуют.
У нас встречается только один (не считая дальневосточных) широко
распространенный и сильно изменчивый вид—Anodonta cygnea, до-
стигающий в длину до 160—200 мм. Живет в прудах, озерах и ре-
ках.
Раковина перловиц (Unio) — с укороченной передней частью,
толстостенная, зеленовато-коричневого или почти черного цвета.
В правой створке один кардинальный зуб и один пластинообразный
латеральный зуб; в левой створке по два кардинальных и латераль-
ных зуба. Длина раковины до 90 мм. Живут в прудах, озерах и
реках. Используются в пуговичной промышленности. Три наиболее
обычных вида Unio различаются между собою следующими призна-
ками.
1 (2) Раковина с почти параллельными верхним и нижним
краями.................................. U. pictorum
2(1) Верхний и нижний края раковины не параллельные . 3
3 (4) Задний конец раковины клиновидно заострен .
U. tumidus
4 (3) Задний конец раковины равномерно закруглен
U. crassus
ТРАХЕЙНЫЕ ЖАБРЫ
В качестве примеров рассматриваются трахейные жабры личи-
нок поденок (Ephemeroptera), стрекоз (Odonata), веснянок (Plecop-
tera), большекрылых (Megaloptera), ручейников (Trichoptera) и
некоторых двукрылых (Diptera).
1 — передний аддуктор или его отпечаток, 2 — передний ретрактор ноги или его
отпечаток, 3 — протрактор ноги или его отпечаток, 4 — задний аддуктор или его
отпечаток, 5 — задний ретрактор ноги или его отпечаток, 6 — мантийная линия.
7—кардинальные зубы, 8— латеральные зубы, 9—верхушка раковины, 10—
линия отреза левой мантийной складки, 11 — область печени, 12 — перикарди-
альная железа (кеберов орган), 13 — область перикардия, 14 — клоакальная
камера, 15 — выводящий сифон, 16 — вводящий сифон, 17 — правая мантийная
складка, 18 — левые полужабры, 19 — нога, 20 — ротовые лопасти, 21 — ротовое
отверстие.
143
Личинки Ephemeroptera
Взрослые крылатые особи отличаются следующими признаками.
Тело гладкое, без волосков. На голове находятся тонкие и обычно
короткие антенны, три небольших глазка и пара сложных глаз; по-
следние у самцов многих видов подразделяются на две части; дор-
сальные глаза отличаются от латеральных своей большей величи-
ной и деталями строения. Крылья прозрачные, блестящие; задние
крылья всегда меньше передних, у некоторых видов (например:
Cloeon, Ordella) полностью отсутствуют. Брюшко оканчивается
обычно тремя (у некоторых видов двумя) хвостовыми нитями
(cerci). Во взрослом состоянии поденки не питаются, их ротовые ор-
ганы редуцированы, кишечник наполнен воздухом, таким образом
превращен в аэростатический аппарат. Характерным признаком,
свойственным только поденкам, является наличие двух крылатых
фаз — имагинальной и субимагинальной. Яйца откладываются в
воду, только у видов Cloeon наблюдается живорождение. Все раз-
витие протекает в воде, у большинства видов личиночная жизнь
продолжается один год, личинки Ephemera и Polymitarcys живут
два года, а личинки Palingenia — три года. В течение личиночной
жизни организмы претерпевают до 20—25 линек. Из зрелых личи-
нок выходят крылатые особи на субимагинальной стадии, отличаю-
щейся от имагинальной недоразвитием ряда органов и мутностью
крыльев. Субимаго быстро линяют и превращаются, в имаго. Дли-
тельность жизни крылатых особей измеряется часами или днями. У
некоторых видов поденок (Palingenia, Polymitarcys) выход крыла-
тых особей из личинок совершается одновременно. Массовый лет
этих видов в некоторые годы достигает грандиозных размеров, в
особенности по берегам больших рек, где неоднократно наблюда-
лись огромные рои поденок на протяжении десятков и сотен кило-
метров.
Развитие поденок протекает с гемиметаморфозом. У только что
вылупившихся личинок (larvula) не имеется ни трахейных жабр,
ни зачатков крыльев, на конце тела вместо трех хвостовых нитей
находятся только две латеральные, трахеи не наполнены воздухом,
дыхание совершается через покровы тела. После первой линьки
появляются зачатки трахейных жабр и срединная хвостовая нить.
С каждой последующей линькой происходят постепенные изменения
в строении личинок — трахейные жабры достигают полного разви-
тия и начинают функционировать, появляются зачатки крыльев, ан-
тенны и хвостовые нити приобретают окончательный вид. Личинки
поденок, часто называемые нимфами, отличаются от личинок дру-
гих насекомых по присутствию на брюшных сегментах пяти-семи
пар трахейных жабр, на конце тела трех (значительно реже двух)
длинных и тонких хвостовых нитей, а на концах грудных ног толь-,
ко по одному коготку. Длина личинок поденок обычно не превы-
шает 10—15 мм, личинки Ephemera, Palingenia и Polymitarcys до-
стигают 20—30 мм.
По образу жизни, а в связи с этим и по строению, личинки по-
денок разделяются на четыре группы. К первой группе относятся
144
чичинки, живущие в небольших водоемах, богатых растительностью,
и способные к быстрому плаванию; личинки второй группы при-
способлены к жизни на илистом грунте, по которому они ползают;
третья группа включает личинок, роющих ходы в глинистых бере-
гах рек; личинки четвертой группы, обладающие уплощенным те-
лом, живут в потоках и быстро текущих реках.
Плавающие личинки
Личинки первой группы живут в прудах, различных мелких во-
доемах, болотах и в небольших речках с медленным течением,
среди зарослей растений, за которые они держатся своими длин-
ными тонкими ногами. Временами они могут быстро плавать, сги-
бая на спинную сторону брюшко вместе с хвостовыми нитями и
зачем быстро ими ударяя; хвостовые нити этих видов опушены мно-
гочисленными волосками. Среди плавающих личинок наиболее
обычными являются личинки Cloeon, Leptophlebia и Siphlonurus.
1. Личинки С1оёоп (рис. 63, А—Б)
Личинки Cloeon обладают стройным, почти цилиндрическим те-
лом, тонкими ногами и длинными хвостовыми нитями. Наверху го-
ловы находятся тонкие антенны, три глазка, образующие треуголь-
ник с направленной вперед вершиной, и парные сложные глаза. На
нижней поверхности головы находится ротовое отверстие с ротовы-
ми придатками, состоящими из верхней губы, мандибул, максилл,
нижней губы и подглоточника. Верхняя губа, расположенная впе-
реди ротового отверстия, имеет вид небольшой пластинки с выемкой
посреди переднего края. Крепкие и сильно хитинизированные ман-
дибулы вооружены с наружной стороны несколькими зубцами, а с
внутренней стороны большим молярным выростом, служащим для
перетирания пищи; в деталях строения правая и левая мандибулы
отличают|ся между собою. Максиллы имеют вид пластинки, оканчи-
вающейся зубцами, также служащими для перетирания пищи; от
наружного края максилл отходит трехчленистый (у одного вида
двучленистый) максиллярный щупик. Нижняя губа состоит из не-
парной части (подбородка, mentum) и парных внутренних и наруж-
ных лопастей, а также трехчленистых губных щупиков. Подглоточ-
ник, иначе называемый гипофаринксом, представлен небольшой,
подразделенной на три части пластинкой,, расположенной внутри у
основания нижней губы.
Грудь состоит из трех сегментов, узкой переднегруди, большой
среднегруди и небольшой заднегруди, несущих по паре грудных
ног. Зачатки передних крыльев у взрослых личинок заходят на
брюшные сегменты; задняя пара крыльев отсутствует, по этому при-
знаку личинки Cloeon отличаются от личинок большинства других
гидов. Брюшко состоит из десяти сегментов. По бокам первых семи
сегментов находятся трахейные жабры. Каждая жабра образована
двумя листками, из которых нижний больше верхнего; только жаб-
ры последней, седьмой пары представлены одиночными листками.
145
Рис. 63. А, Б — личинка Cloeon dipterum, общий вид (Д), трахейная жабра вто-
рой пары (Б); В, Г—личинка Leptophlebia vespertina, общий вид (В), трахей-
ная жабра (Г); Д — личинка Leptophlebia marginata, трахейная жабра; Е, Ж —
личинка Siphlonurus linnaeanus, общий вид (Е), трахейная жабра пятой пары
(Ж);. 3 — личинка Siphlonurus aestivalis, трахейная жабра третьей пары;
ff—Н—ротовые придатки личинки Siphlonurus aestivalis, верхняя ry6ai (Я)»
мандибула (К), максилла (Л), нижняя губа (М), подглоточник (И).
Внутри трахейных жабр легко заметить сильные ветвления трахей,
отходящих от двух трахейных стволов, расположенных по бокам
брюшка. На конце тела находятся длинные хвостовые нити, средин-
ная нить несет многочисленные волоски с обеих сторон, а боковые
нити только с внутренней стороны.
В наших водах встречаются личинки четырех .видов Cloeon,
для определения которых служит нижепомещенная таблица. Наи-
более обычным видом является Cloeon dipterum.
1 (6) Максиллярный щупик трехчленистый .... 2
2 (5) Длина антенн не превышает половину^длины тела 3
3 (4) Трахейные жабры относительно больиге, с сильным ветв-
лением трахей; верхний листок трахейных жабр первой
пары овальный, по ширине обычно составляет половину
ширины нижнего листка..........................С. dipterum
4 (3) Трахейные жабры слабо развиты, со слабым ветвлением
трахей; верхний листок трахейных жабр первой пары уз-
кий, по ширине обычно не превосходит одной трети ши-
рины нижнего листка . . С. inscriptum
5 (2) Длина антенн равна длине тела С. rufulum
6 (1) Максиллярный щупик двучленистый С. praetextum
2. Личинки Leptophlebia (рис. 63, В—Д)
Личинки Leptophlebia отличаются иным строением органов ды-
хания. Трахейные жабры первой пары двуветвистые, очень узкие.
Трахейные жабры остальных пар представлены двумя листками;
каждый листок состоит из двух частей — листовидно расширен-
ной базальной и сильно суженной конечной.
Личинки двух встречающихся у нас видов Leptophlebia разли-
чются между собою по следующим признакам.
1 (2) Базальные части трахейных жабр второй — седьмой пар
округло овальные, резко отделяются от конечных узких
выростов....................................... L. marginata
2 (1) Базальные части трахейных жабр второй—седьмой пар к
концу суживаются, более или менее постепенно переходят
в конечные узкие выросты . L. vespertina
3. Личинки Siphlonurus (.рис. 63, Е—Н)
У личинок Siphlonurus по бокам последних сегментов брюшка
находятся плоские, направленные назад шипы. Трахейные жабры
большие, листовидной формы, частично покрывающие друг друга;
/ — голова, 2 — глаз, 3 — глазок , 4 — антенна, 5 — первый грудной сегмент, 6 —
зачатки крыльев, 7— трахейные жабры, 8— бедро, 9—голень, 10 лапка, //—
коготок, 12— брюшко, 13— хвостовая нить, 14—молярный вырост, 15—мак-
силлярный щупик, 16 — внутренняя лопасть, 17 — наружная лопасть,
18 — нижнегубной щупик.
10*
147
всего имеется семь пар трахейных жабр, из которых две передние
пары состоят из двух различных по форме и размеру листков;
трахейные жабры остальных пяти пар только у одного вида пар-
ные у других же представлены одиночными листками.
Личинки трех встречающихся у нас видов Stphlonurus, из кото-
рых наиболее обычными являются личинки Siphlonurus hnnaeanus,
различаются между собою по следующим признакам.
1 (2) Трахейные жабры всех семи пар парные (на последней
паре верхний листок очень мал) ... hnnaeanus
2 (1) Трахейные»ары только двух первых пар парные, осталь-
ные пять пар представлены одиночными листками . 3
3 (4) Шипы, расположенные по бокам девятого брюшного сег-
мента,’ простираются до выгибов на десятом сегменте, от
которых отходят латеральные хвостовые нити .
S. aestivalis
4 (3) Указанные шипы не достигают выгибов на десятом
сегменте 4 S- lacustris
Ползающие личинки
Типичными представителями личинок второй группы являются
личинки Ephemerella и Ordella. Они обитают в озерах или мед-
ленно текущих реках, преимущественно на илистых грунтах; пла-
вают плохо. Тело покрыто мелкими волосками, к которым приста-
ют частицы ила, образующие оболочку вокруг личинки. Все три
хвостовых нити опушены с обеих сторон.
4. Личинки Ephemerella (рис. 64, А—Б)
Личинки Ephemerella, самым широко распространенным видом
которых является Ephemerella ignita, обладают слегка уплощен-
ным телом и небольшой головой, резко отделенной от передне-
груди. Брюшные сегменты по бокам вытянуты в направленные
назад острые шипы, задний край каждого сегмента с двумя дор-
сальными бугорками. Хвостовые нити немного короче тела. Мак-
силлярный щупик небольшой, трехчленистый. Нижняя губа боль-
шая. Трахейные жабры, в числе пяти пар, расположены дорсаль-
но по краям брюшных сегментов, от третьего до седьмого включи-
тельно. Небольшие пластинки пятой пары полностью прикрыты
пластинками четвертой пары, поэтому при наружном осмотре ли-
чинки видны лишь четыре пары трахейных жабр. Каждая жабра
состоит из большой, более или менее прямоугольно-округлой1 пла-
стинки, на нижней, вогнутой стороне которой находятся два расхо-
дящихся ряда небольших, налегающих друг на друга нежных
листочков. Эти листочки, являющиеся органами дыхания, защи-
щены от загрязнения верхними пластинками, довольно плотно
прилегающими к телу. Медленные движения личинок происходят
при помощи ног, снабженных большими и снизу зазубренными
коготками.
>48
5. Личинки Ordella (рис. 64, В—Ж)
Еще большее своеобразие в строении трахейных жабр наблю-
дается у личинок Ordella (= Caenis). Всего у них имеется шесть
пар трахейных жабр, расположенных на шести первых сегментах
брюшка. Небольшие, палочковидные жабры первой пары находят-
Рис. 64. А, Б — личинка Ephemerella ignita, общий вид (Л), трахейная жабра
(Б)\ В, Г — личинка Ordella macrura, общий вид (В), трахейная жабра (Г);
Д—Ж—переднеспинка личинок Ordella macrura (Д), Ordella horaria (В),
Ordella moestus (Ж).
ся по бокам первого брюшного сегмента. Все остальные трахей-
ные жабры расположены на спинной стороне брюшка. Большие
пластинки второй пары, соприкасающиеся своими внутренними
краями, покрывают в виде крышек трахейные жабры следующих
четырех пар. Последние имеют вид сравнительно небольших пла-
стинок, густо' усаженных по краям тонкими ветвящимися выро-
стами. Личинки Ordella живут в илу; пластинки второй пары, не
препятствуя газообмену, защищают от загрязнения находящиеся
под ними трахейные жабры.
149
Личинки наиболее часто встречающихся у нас видов Ordella
отличаются между собою по следующим признакам.
1 (2) Передний край переднеспинки по ширине почти не отли-
чается от заднего края; посреди каждой боковой поверх-
ности переднегруди находится более или менее ясное
пятно ... О. тасгига- (=0. halterata)
2 (1) Передний край переднеспинки значительно шире заднего
края......................................................3
3 (4) Передние углы переднйспинки острые; около середины пе-
реднеспинки находятся две черные точки; такие же точки
находятся и на бедрах ног у сочленения с го-
ленью ............................................О. horaria
4 (3) Передние углы переднеспинки закругленные, черных то-
чек на переднеспинке и на бедрах ног не имеет-
ся . О. moestus
Роющие личинки
Личинки третьей группы, самыми характерными представите-
лями которых являются личинки Ephemera, Palingenia и Polymitar-
cys, относятся к числу наиболее специализированных личинок по-
денок. Описание их помещено на стр. 67.
Плоские, прилегающие к субстрату личинки
Тело личинок четвертой группы уплощено в дорсо'-вентральном
направлении. Голова широкая, плоская. Глаза расположены на
верхней стороне головы. Грудные ноги и трахейные жабры направ-
лены в стороны. Хвостовые нити длинные, широко раздвинутые. Ли-
чинки очень требовательны к кислороду, поэтому живут преимуще-
ственно в быстрых потоках и реках, реже в чистых озерах. Вся ор-
ганизация личинок стоит в связи с их образом жизни. Обычно ли-
чинки держатся на нижней поверхности камней, где сила течения
слабее. Уплощенная и обтекаемая форма тела позволяет им без
труда двигаться по поверхности камня в поисках пищи.
Наиболее обычными представителями плоских личинок являют-
ся личинки Heptagenia и Ecdyonurus-, отличающиеся иным строе-
нием личинки Oligoneuriella также могут быть отнесены в эту
группу.
6. Личинки Heptagenia (рис. 65, А—Д)
Тело личинок Heptagenia сильно уплощено, сравнительно корот-
кое. Плоская голова расширена в поперечном направлении. Боль-
шие глаза и находящиеся между ними три глазка расположены на
верхней поверхности головы. Антенны короткие. Верхняя губа ши-
рокая. Мандибулы с двумя большими наружными зубцами. Мак-
силлы по наружному краю с крепкими шипиками, согнутыми на
внутреннюю сторону; максиллярный щупик двучленистый. Нижняя
ISO
губа широкая. Переднегрудь приблизительно прямоугольной фор-
мы, задние углы не вытянуты в тупые выросты. Ноги плоские, с
расширенными бедрами и крепкими конечными коготками, воору-
женными снизу зубцом; все ноги направлены в стороны и у живого
организма плотно прилегают к субстрату. Хвостовые нити длинные,
Рис. 65. А, Б—личинка Heptagenia sulphurea, общий вид (/1), трахейная жабра
четвертой пары В—личинка Heptagenia cocrulans, трахейная жабра чет-
вертой пары; Г—личинка Heptagenia flava, трахейная жабра четвертой пары;
Д — личинка Heptagenia fuscogrisea, трахейная жабра четвертой пары; Е, Ж—
личинка Ecdyonurus fluminum, общий вид (Е), трахейная жабр$ четвертой
па'ры (Ж)\3—К — личинка Oligoneuriella rhenana, общий вид (3), макенл-
ла (И), трахейная жабра (К).
151
обычно не. короче тела. Трахейные жабры расположены на первых
семи сегментах брюшка. Каждая жабра состоит из верхней пла-
стинки и пучка жаберных нитей, прикрепленных снизу к основа-
нию пластинки и содержащих тонкие трахеи.
В наших водах встречаются личинки нескольких видов Heptage-
niat для определения которых служит нижепомещенная таблица.
Наиболее обычными для средней полосы являются личинки Hepta-
genia srlphurea.
1 (4) Трахейные нити образуют сравнительно небольшие пучки;
верхний листок трахейных жабр длиннее пучка трахей-
ных нитей..............................................2
2 (3) Верхний листок трахейных жабр более или менее округ-
лой формы; хвостовые нити с чередующимися светлыми и
темными кольцами, состоящими из.двух члени-
ков Н. sulphurea
3 (2) Верхний листок трахейных жабр по форме похож на лист
сирени; хвостовые нити в полтора раза длиннее
тела.............................. Н. fuscogrisea
4 (1) Трахейные нити образуют сильно развитые пучки; верх-
ний листок трахейных жабр не длиннее пучка трахейных
нитей............................................... 5
5 (6) Верхний листок трахейных жабр узкий, языковид-
ный .......................................... coerulans
6 (5) Верхний листок трахейных жабр широкий, листовидный;
брюшко с темной срединной линией . Н. flava
7. Личинки Ecdyonurus (рис. 65, Е—Ж)
Личинки Ecdyonurus по внешнему виду очень сходны с личин-
ками Heptagenia, отличаются по строению переднегруди, задние
углы которой вытянуты назад в виде плоских округлых выростов,
лежащих по бокам среднегруди.
Личинки наиболее часто встречающихся ‘двух видов Ecdyonu-
rus отличаются между собою следующими признаками.
1 (2) Последний, десятый сегмент брюшка
лый.......................................
2(1) Последний, десятый сегмент брюшка
ный
сверху свет-
Е. fluminum
сверху те'м-
Е. venosus
8. Личинки Oligoneuriella (рис. 65, 3—К)
Тело личинок Oligoneuriella гладкое, сильно уплощенное, обте-
каемой формы. Голова спереди округлена и снабжена волосками.
Верхняя губа большая, с длинными щетинками. Максиллы с дву-
членистым широким щупиком и с большим пучком трахейных ни-
тей. Нижняя губа отличается особым строением и играет роль при-
соски. ОбЪ наружные лопасти нижней губы сильно расширены, об-
разуют диск, окруженный по периферии равномерно согнутыми
152
максиллярными и губными щупиками, покрытыми многочисленны-
ми длинными щетинками. Ноги уплощенные. Ноги первой пары с
длинными, параллельно идущими щетинками на бедрах и голенях.
Хвостовые нити короткие, не более половины или трети длины
тела. Трахейные жабры небольшие. Первая пара расположена на
нижней стороне первого брюшного сегмента, остальные шесть пар
по бокам брюшка. Каждая жабра состоит из небольшой пластинки
и небольшого пучка трахейных нитей.
Единственным у нас видом является Oligoneuriella rhenana.
Личинки Odonata
Во взрослом состоянии все стрекозы ведут воздушный образ
жизни, некоторые виды могут часами без перерыва находиться в
воздухе, являясь примером наиболее приспособленных к полету
насекомых. У многих видов во время полета происходят питание,
копуляция и откладка яиц.
Тело стрекоз стройное. На голове впереди находятся короткие
антенны, по бокам головы—сильно развитые фасеточные глаза,
между которыми располагаются в виде треугольника три неболь-
ших глазка, а внизу головы — ротовые придатки. Первый грудной
сегмент небольшой, два остальных слиты вместе в массивную
часть тела. Брюшко, состоящее из десяти сегментов, узкое и длинное.
Стрекозы относятся к насекомым с неполным превращением.
Яйца откладываются свободно в воду или ил (эксофитическая
кладка) или же в ткани растений (эндофитическая кладка). В по-
следнем случае самки погружают в воду конец брюшка или же
сами спускаются по водяным растениям под воду и надрезают яй-
цекладом ткани растения, вкладывая в каждый надрез по яйцу.
Развитие начинается сейчас же после откладки яиц. У одних видов
перезимовывают яйца, у других видов—личинки. Число личиноч-
ных стадий варьирует, обычно бывает 10—1’4 линек. Длительность
личиночной жизни колеблется от одного года (большинство
Zygoptera, Апах) до двух (Aeschna^ Cordulia) и трех лет.
Личинки стрекоз являются типичными водными организмами,
живущими в пресной воде, в виде исключения в солоноватоводных
водоемах. Тело делится на голову, грудь и брюшко. Голова непо-
движная, сросшаяся с грудью. Антенны короткие, четырех-семи-
членистые. Глаза большие, но меньшего размера, чем у крылатых осо-
бей; глазки встречаются только у некоторых видов. Верхняя губа,
мандибулы и максиллы мало отличаются по строению от соответ-
ствующих придатков взрослых организмов; нижняя губа, называе-
мая маской и служащая для добывания пищи, является для личи-
нок стрекоз самым характерным органом, легко заметным при рас-
сматривании их с брюшной стороны. Для изучения строения маски,
ее следует оттянуть пйнцетом вперед и рассматривать с внутренней
стороны. Средняя пластинка маски (подбородок, mentum) подвиж-
но соединена с другой меньшего размера пластинкой (подподборо-
док, submentum), в свою очередь подвижно-соединенной с головой;
вследствие этого маска может выдвигаться вперед и опять скла-
153
дываться. К передней части средней пластинки, называемой сред-
ней лопастью, по бокам подвижно причленены боковые лопасти,
вооруженные различного рода придатками и одним подвижным зуб-
цом. На боковых лопастях различают наружный край—от основа-
ния лопасти до подвижного зубца, дистальный край — от подвиж-
ного зубца до вершины лопасти, внутренний край — от вершины
до основания лопасти. У большинства видов на внутренней (перед-
ней) поверхности средней лопасти находятся длинные (средние
подбородочные) щетинки, обычно в виде двух сходящихся к сере-
дине лопасти рядов, такие же щетинки расположены также вдоль
наружного края боковых лопастей (боковые щетинки). Строение
маски, присутствие или отсутствие длинных щетинок, а также их
число имеют большое значение при определении личинок. Отличают
два типа масок — плоские и шлемовидные. Первые свойственны
хищным личинкам, как, например, Aeschna и Апах, хватающим
свою добычу большими подвижными зубцами боковых лопастей
маски. Плоская маска в сложенном виде закрывает только часть
нижней поверхности головы; средняя пластинка более или менее
плоская, боковые лопасти лежат вместе с ней в одной плоскости.
Шлемовидная маска, встречающаяся у личинок, живущих в илу,
как, например, Libellula и Cordulegaster, имеет вид черпака; в сло-
женном виде закрывает всю нижнюю поверхность головы, наподо-
бие забрала; широкая, треугольной формы средняя лопасть, посре-
ди вогнутая, несет на внутренней стороне два сходящихся ряда
длинных щетинок; большие, также треугольной формы, боковые л о-
пасти, довольно сильно вогнутые, несут редуцированный подвиж-
ный зубец и обычно большое число длинных, расположенных по
наружному краю щетинок; внутренний и дистальный края боковых
лопастей вооружены небольшими зубцами и щетинками. Личинки с
шлемовидной маской, открывая и закрывая ее боковые лопасти,
захватывают свою добычу вместе с илом, затем мелкие частицы
просеиваются через сито, образованное длинными средними и боко-
выми щетинками, прикрывающими сверху захваченную массу ила.
Грудь образована свободным первым и слившимися между со
бою вторым и третьим сегментами. Поверх груди лежат зачатки
крыльев, прикрывающие также передние брюшные сегменты. Брюш-
ко, состоящее из одиннадцати сегментов, из которых последний сла-
бо развит и трудно заметен, может быть гладким или вооружен-
ным дорсальными шипами, расположенными по середине сегментов,
и латеральными шипами, у многих видов хорошо развитыми по бо-
кам восьмого и девятого сегментов брюшка. Конец брюшка несет
хвостовые придатки, различного строения у представителей подот-
рядов Anisoptera и Zygoptera.
У личинок, относящихся к подотряду Anisoptera, обладающих
широким, более или менее уплощенным брюшком, на конце тела
находится так называемая анальная пирамида, образованная тремя
большими заостренными отростками—двумя церками, расположен-
ными латеро-вентрально, и одним дорсальным анальным придат-
ком; у более взрослых личинок к этому прибавляются еще два не-
больших церкоида, а у личинок самцов небольшая пластинка, при-
154
крывающая или сливающаяся с основанием дорсального придатка.
Из этих личинок выходят крупные, хорошо летающие стрекозы, с
сильно сближенными глазами и различным строением крыльев —
задние крылья при основании значительно шире передних.
У личинок, относящихся к подотряду Zygoptera и отличающихся
присутствием стройного, тонкого, цилиндрической формы брюшка,
хвостовые придатки представлены длинными и уплощенными пла-
стинками, ранее часто называвшимися хвостовыми жабрами. Все-
го имеется три хвостовых пластинки —две боковых и одна сред-
няя. При основании хвостовых пластинок находится темноокрашен-
ная полоска, по которой пластинки обламываются при автотомии.
Утраченные хвостовые пластинки восстанавливаются только при
последующих линьках, поэтому могут встретиться личинки с двумя
или одной пластинками или даже совсем без них. Вдоль каждой
хвостовой пластинки проходят один или два центральных трахей-
ных ствола, от которых отходят многочисленные трахейные ветви.
Из этих личинок выходят небольшие, медленно летающие стрекозы,
с широко расставленными глазами и одинаковым строением крыль-
ев обеих пар.
Все личинки стрекоз дышат растворенным в воде кислородом.
Трахейные жабры личинок Anisoptera находятся в прямой кишке,
являющейся задней частью кишечника. Наблюдая живую личинку
в плоской чашке, на дно которой насыпан тонким слоем мелкий пе-
сок, можно заметить, по движению песчинок, входящие в кишечник
и выходящие из него токи воды, сопровождаемые ритмичными со-
кращениями брюшка и расхождением и схождением отростков
анальной пирамиды. Частота сокращений брюшка зависит от коли-
чества растворенного в воде кислорода, чем меньше его в воде, тем
чаще и энергичнее происходят сокращения и наоборот. При вскры-
тии личинки, после удаления брюшных покровов, в задней части
брюшка видны сильно развитая мускулатура, сокращением которой
уменьшается просвет прямой кишки и вода выбрасывается нару-
жу, и многочисленные тонкие трахейные веточки, отходящие от про-
дольных трахейных стволов. В брюшке личинок стрекоз проходят
три пары продольных трахейных стволов—дорсальные, вентральные
и висцеральные, из которых дорсальные являются самыми широ-
кими и обычно окрашены. В задней части брюшка от дорсаль-
ных стволов отходят по две серии, а от висцеральных —• по одной
серии трахейных ветвей первого порядка, которые, подходя к стен-
кам прямой кишки, делятся на трахейные ветви второго порядка,
проходящие через стенки кишечника. Прямая кишка личинок Ani-
soptera расширяется перед анальным отверстием в объемистый жа-
берный (ректальный) пузырь. Если вскрыть его продольным разре-
зом, то можно увидеть многочисленные, расположенные продоль-
ными рядами тонкостенные выросты кишечного эпителия, являю-
щиеся трахейными (ректальными) жабрами. К каждой трахейной
жабре подходит веточка трахеи, распадающаяся в ней на сеть тра-
хейных капилляров. Строение трахейных жабр у личинок разных
видов различное. У личинок Cordulegaster они имеют вид длинных,
расположенных шестью продольными рядами, ундулирующих цель-
155
ных пластинок, по бокам которых находятся короткие поперечные
пластинки. У личинок Gomphus имеется шесть продольных рядов
длинных тонких папилл. У личинок Aeschna трахейные жабры, име-
ющие вид листочков, расположены шестью двойными рядами. У^ли-
чинок Апах трахейные жабры, также имеющие вид листочков, снаб-
жены многочисленными цилиндрическими короткими выростами.
У личинок Libellula, Cordulia и близких к ним видов имеется шесть
двойных рядов пластинчатых выростов.
Задний отдел кишечника личинок Anisoptera в случае необходи-
мости, как, например, бегства от врага, может служить также для
целей передвижения. Выбрасывая из кишечника сильную струю
воды, личинка делает большой скачок вперед (принцип обратного
толчка). При быстрых последовательных сокращениях и расшире-
ния^ прямой кишки личинки могут двигаться со значительной ско-
ростью. Таким образом, кишечник личинок Anisoptera не только вы-
полняет свою основную функцию при пищеварении, но является
также органом дыхания и движения.
У личинок Zygoptera жаберного пузыря и ректальных жабр не
имеется. Дорсальные продольные трахейные стволы в самой задней
части брюшка сближены между собою и входят в виде двух парал-
лельных ветвей в среднюю хвостовую пластинку, соответствующую
дорсальному анальному придатку Anisoptera. От дорсальных ство-
лов в девятом брюшном сегменте отходят ветви, входящие соответ-
ственно в правую и левую боковые хвостовые пластинки, являю-
щиеся преобразованными церками; по бокам хвостовых пластинок
расположены короткие церкоиды. От одного или двух центральных
стволов, проходящих вдоль хвостовых пластинок, отходят много-
численные трахейные ветви, только на концах переходящие в тра-
хейные капилляры.
Ранее считалось, что хвостовые пластинки личинок Zygoptera
служат для дыхания, поэтому они назывались трахейными жабра-
ми. На самом деле хвостовые пластинки, покрытые плотным хити-
ном и расположенные в вертикальной плоскости, играют роль хво-
стового плавника, при помощи которого личинки могут быстро пла-
вать. Для получения большего сопротивления воде, все пластинки
между собою складываются. При удалении хвостовых пластинок,
нормальная жизнь личинки не прекращается, замедляется лишь их
плавание. Дыхание личинок Zygoptera, так же как и личинок Ani-
soptera, совершается при помощи заднего отдела кишечника. Если
к воде, в которой находится личинка, прибавить мелкой взвеси, то
под увеличением удается рассмотреть входящий и выходяший токи
воды. У личинок большинства видов (Lestes, Coenagrion и других)
в прямой кишке проходят три продольных, резко отграниченных
складки, занимающих приблизительно две трети длины этого от-
дела кишечника и хорошо заметных снаружи кишки при вскрытии
личинки, после удаления покровов брюшка. Газообмен происходит
через богатый кровью дыхательный эпителий складок, которые мо-
гут считаться кровяными жабрами. У личинок Agrion (=^Calopte-
гух) кишечные складки снабжены большой сетью трахейных капил-
ляров, входящих в эпителиальные клетки, выстилающие внутреннюю
156
поверхность кишки. Таким образом, личинки Agrion, у которых са-
ми стенки кишечника служат как бы трахейными жабрами, зани-
мают промежуточное положение между остальными личинками
Zygoptera, дышащими кровяными жабрами, и личинками Anisop-
tera, обладающими постоянными ректальными трахейными жаб-
рами.
Личинки без хвостовых пластинок (Anisoptera)
1. Личинки Aeschna (рис. 66)
Личинки коромысла, Aeschna, живут в стоячих водоемах среди
растений и на дне. Длина личинок достигает 35—45 мм. Тело тол-
стое, плотное. Голова большая, плотно слившаяся с туловищем.
Антенны короткие, семичленистые. Сложные фасеточные глаза боль-
шие. Маска плоская, срединная лопасть на внутренней стороне без
длинных щетинок, ее передний край выпуклый, вооружен коротки-
ми волосками; боковые доли без щетинок, подвижный зубец боль-
шой; маска в сложенном виде не достигает оснований ног послед-
ней пары. По бокам первого грудного сегмента находятся парные
боковые выступы, форма и величина которых различны у отдель-
ных видов. Брюшко большое, расширенное в задней половине, свер-
ху без зубцов; боковые края шестого—девятого сегментов вытянуты
в латеральные шипы. Длина анальной пирамиды равна общей дли-
не двух последних сегментов брюшка.
Личинки наиболее распространенных у нас видов Aeschna раз-
личаются между собою следующими признаками:
1 (10) Оба боковых выступа первого грудного сегмента одина-
кового размера или передний больше заднего 2
2 (5) Оба боковых выступа острые . . . . 3
3 (4) Боковые выступы одинакового размера, выемка между
ними с прямым или тупым углом . . Ае. grandis
4 (3) Передний боковой выступ больше заднего, выемка с
острым углом . .... Ае. juncea
5 (2) Один или оба боковых выступа тупые .... 6
6 (7) Боковые выступы слабо развиты, выемка между ними
мелкая................................... Ае. affinis
7 (6) Боковые выступы хорошо развиты 8
8 (9) Выемка между боковыми выступами с прямым
углом................................ Ае. суапеа
9 (8) Указанная выемка с острым углом Ае. viridis
10 (1) Задний выступ больше переднего . • 11
11 (12) Передний выступ острый . Ае. isosceles
12 (И) Передний выступ тупой...............................13
13 (14) Передний край средней пластинки маски шире заднего
края по более, чем в два раза . ....
Ае. coerulea (=Ае. squamata)
14 (13) Передний край шире заднего края более, чем в два
раза Ае. mixta (=Ае. coluberculus)
157
Рис.66.А—В — личинка Aeschna grandis, общий вид (А), голова сбоку (£)»
анальная пирамида самца (В); Г — левые боковые выступы переднеспинки л
чинок Aeschna grandis (/), Aeschna juncea (//), Aeschna cyanea (///), Aesclin
mixta (IV), Aeschna affinis (V), Aeschna isosceles (VI), Aeshna viridis
Д—трахейная система личинки Aeschna; E — ректальный пузырь, с сетью тр
хей, личинки Aeschna; Ж— схематический поперечный разрез через ректал
яый пузырь личинки Aeschna; 3—трахейные жабры личинки Aeschna.
1 — голова, 2 — антенна, 3 — верхняя губа, 4 — глаз, 5 — переднеспинка, 6
стигма, 7 —зачатки крыльев, 8 — вертлуг, 9 — бедро, 10—голень, //—-«лап*
12—брюшко, 13 — латеральные шипы, 14 — анальная пирамида, 15 — СУ6^
тум (под под бородок), 16 — ментум (подбородок), 17—боковая лопасть, 1
2. Личинки Cordulegaster (рис. 67)
Личинки Cordulegaster annulatus живут в илу рек и ручьев.
Длина личинок достигает 43 мм. Самым характерным признаком
является строение шлемовидной маски, боковые лопасти которой по
дистальному краю вооружены большими, различного размера ост-
рыми зубцами; передний край средней лопасти посреди с раздвоен-
ным выступом.
Рис. 67. Личинка Cordulegaster annulatus. А — общий вид; Б — маска.
7 боковая лопасть, 2—подвижный зубец, 3 — подбородочные щетинки.
3. Личинки Cordulia (рис. 68, А—В)
Личинки бабки, Cordulia аепеа, единственного вида этого рода,
живут в разнообразных стоячих водоемах. Длина личинок дости-
гает 22 мм. Тело уплощенное, брюшко широкое, с небольшими дор-
сальными шипами на третьем—девятом сегментах (эти шипы все-
го сильнее развиты на средних сегментах) и с острыми латераль-
ными шипами на восьмом и девятом сегментах. Глаза небольшие.
Ноги длинные. Маска шлемовидная; дистальный край боковых ло-
пастей с девятью хорошо развитыми зазубринами, глубина вырезок
между зазубринами равна половине ширины самих зазубрин; каж-
дая зазубрина вооружена четырьмя-пятью щетинками; на каждой
стороне средней лопасти маски 13 щетинок, боковые лопасти с во-
семью щетинками. Окраска личинок серая.
4. Личинки Epitheca (рис. 68, Г—Е)
Личинки бабки, Epitheca bimaculata, единственного вида этого
рода, живут в стоячих водоемах, реже в реках. Длина личинок до-
стигает 30 мм. По внешнему виду сходны с личинками Cordulia.
подвижный зубец, /9—анальный придаток, 20 — церк, 21 — церкоид. 22— до-
бавочная пластинка (самца), 23 — дорсальный трахейный ствол, 24 вентраль-
ный трахейный ствол, 25 — висцеральный трахейный ствол, 26—ректальный
пузырь, 27 —прямая кишка, 28 —- трахейные жабры.
159
Отличаются присутствием длинных и острых латеральных ши-
пов на девятом сегменте брюшка, достигающих конца анальной пи-
рамиды. Дорсальные шипы большие. Дистальный край боковых ло-
пастей маски с десятью зазубринами; каждая зазубрина вооружена
Рис. 68. А—В — личинка Сог-
dulia аепеа, общий вид (Л)„
брюшко сбоку (Б), маска (В);
Г—Е — личинка Epitheca bima-
culata, общий вид (Г), брюш-
ко сбоку (Д), маска (Е).
1 — дорсальный шип, 2 — лате-
ральный шип, 3 — боковая ло-
пасть, 4 — подвижный зубец,
5 — подбородочные щетинки
тремя-пятью щетинками; на каждой стороне средней лопасти
маски 11 щетинок, боковые лопасти с семью щетинками.
5. Личинки Libellula (рис. 69, А—В)
Личинки Libellula относятся к числу обычных широко распро-
страненных видов, встречаются преимущественно в стоячих водое-
мах. Длина личинок достигает 23 мм. Тело короткое, уплощенное.
Голова широкая. Глаза небольшие. Антенны короткие, отходят впе-
реди линии, соединяющей передние края глаз. Ноги относительно
короткие. Брюшко сильно расширенное, овальной формы; на треть-
ем—восьмом или на третьем—девятом сегментах имеются дорсаль-
ные шипы. Маска шлемовидная, относительно длинная, в сложен-
ном виде доходит до середины места прикрепления ног первой и
второй пар; дистальный край боковых лопастей маски слабо зазуб-
рен, глубина вырезок значительно меньше половины ширины самих
зазубрин (исключая личинок Libellula depressa, у которых эти зуб-
160
цы имеют такое же строение, как у личинок Cordulia\ личинки от-
личаются отсутствием латеральных зубцов на девятом сегменте
брюшка).
«Личинки встречающихся у нас видов Libellula различаются
между собою следующими признаками.
Рис. 69. А, Б — личинка Libellula quadrimaculata, общий вид (Л), маска (Б);
В — личинка Libellula depressa, маска; Г, Д— личинка Sympetrum flaveolum,
общий вид (Г), маска (Д); Е— латеральные шипы восьмого и девятого брюш-
ных сегментов личинок Sympetrum sanguineum (/), Sympetrum meridionale (И)
Sympetrum flaveolum (///), Sympetrum scoticum (IV).
1 — дорсальный шип, 2 — латеральный шип, 3—боковая лопасть, 4 — подвиж-
ный зубец, 5 — подбородочные щетинки.
1 (2) На девятом сегменте брюшка имеется дорсальный
шитт............................................. L. futva
2 (1) На указанном сегменте дорсального шипа не
имеется .... ....................3
3 (4) Девятый сегмент брюшка с небольшими латеральными
шипами; дистальный край боковых лопастей маски слабо
И В, л. Яш нов
161
зазубрен, на этих лопастях по семи-восьми щети-
нок ........................... . L. quadrimaculata
4 (3) Девятый сегмент брюшка без латеральных шипов; ди-
стальный край указанных лопастей сильно зазубрен, на
этих лопастях по 10—12 щетинок . L. depressa
6. Личинки Sympetrum (рис. 69, Г—Е)
Личинки Sympetrum обитают в небольших, пересыхающих во-
доемах. среди растений, очень подвижные. Длина личинок дости-
гает 13—Г7 мм. По внешнему виду сходны с личинками двух пре-
дыдущих родов. Отличаются присутствием больших латеральных
шипов на девятом сегменте брюшка. Голова большая. Глаза мало
выдающиеся. Антенны отходят перед линией, соединяющей перед-
ние края глаз. На каждой стороне средней лопасти маски по 11 —
15 щетинок, на боковых лопастях по 10—12 щетинок.
Личинки наиболее легко определяемых видов Sympetrum, из ко-
торых наиболее обычным является Sympetrum flaveolum, отличают-
ся между собою следующими признаками.
1 (2) Латеральные шипы восьмого сегмента абдомена большие,
концы их заходят за середину следующего сег-
мента .............................. . 5. meridionale
2 (1) Указанные шипы короткие ... 3
3 (4) Латеральные шипы девятого сегмента абдомена изо-
гнутые .................................... S. sanguineum
4 (3) Указанные шипы прямые............................ 5
5 (6) Восьмой сегмент брюшка с дорсальным шипом 7
6 (5) Указанный сегмент без дорсального шипа . . . .
S. scoticum (-S. danae)
7 (8) Дорсальный шип восьмого сегмента абдомена короче и
слабее остальных; длина латеральных шипов девятого
сегмента абдомена не больше половины высоты самого
сегмента.....................................S.'flaveolum
8 (7) Указанный дорсальный шип большой; длина латеральных
шипов девятого сегмента приблизительно равна высоте
самого сегмента . . S. striolatum
Личинки с хвостовыми пластинками (Zygoptera)
7. Личинки Agrion (рис. 70, А—Г)
Личинки красотки, Agrion (в прежней литературе Calopteryx),
являются самыми крупными среди стрекоз этой группы, длина ли-
чинок (без хвостовых пластинок) достигает 22 мм\ встречаются в
текучей воде, среди растений. Голова приблизительно четырех-
угольная. Глаза небольшие. Антенны длинные, первый членик пре-
вышает общую длину всех остальных шести члеников более, чем
в два раза. Маска плоская, средняя лопасть с глубокой выемкой и
двумя направленными внутрь выростами, благодаря этому на кон-
це лопасти образуется ромбической формы отверстие; боковые ло-
пасти с одним подвижным и тремя неподвижными зубцами. Брюш-
162
м
Рис. 70. А—В — личинка Agrion splendens, общий вид (Л), маска (Б), хвостовые
пластинки (В); Г — личинка Agrion virgo, хвостовые пластинки; Д—Ж— личин-
ка Lestes sponsa, общий вид (Д), маска (£), хвостовая пластинка (Ж)\3, И —
личинка Lestes dryas, маска (3), хвостовая пластинка (Я); К, Л— личинка
Lestes virens, маска (К)» хвостовая пластинка («//); М,Н — личинка Lestes
barbara, маска (Af), хвостовая пластинка (Н).
/—голова, 2 — глаз, 3— антенна, 4 — зачатки крыльев, 5 — брюшко, <—
средняя хвостовая пластинка, 7 — боковая хвостовая пластинка, 8 — боковая
лопасть, 9 — подвижный зубец, 10 — подбородочные щетинки.
11* 163
ко цилиндрическое. Хвостовые пластинки длинные, с ветвлениями
трахейных капилляров; боковые пластинки трехгранные, стилето-
видные, средняя короче боковых, уплощенная. Задний отдел кишеч-
ника с складками, пронизанными ветвлениями трахейных капилля-
ров. .
Личинки двух встречающихся у нас видов Agrion различаются
между собою следующими признаками.
1 (2) Длина первого членика антенн равна ширине головы
или немного ее превосходит, боковые хвостовые пластин-
ки значительно длиннее средней; хвостовые пластинки
обычно с тремя темными пятнами ... A. splendens
2 (1) Длина указанного членика немного короче ширины голо-
вы; боковые хвостовые пластинки немного длиннее сред-
ней; хвостовые пластинки обычно с двумя темными
пятнами A. virgo
8. Личинки Lestes (рис. 70, Д—Н)
Личинки лютки, Lestes, относятся к числу характерных организ-
мов небольших пересыхающих водоемов. Тело тонкое, цилиндриче-
ское. Длина личинок (без хвостовых пластинок) достигает 14—
18 леи. Голова вытянута в поперечном направлении. Глаза большие.
Антенны семичленистые. Маска очень длинная, ложкообразная, в
сложенном виде у личинок большинства видов доходит до основа-
ния ног последней пары; расширенный конец средней лопасти на
внутренней стороне с двумя сходящимися рядами длинных щети-
нок; боковые лопасти с большим широким зубцом, отделенным глу-
бокой выемкой от остальной части, вооруженной двумя меньшими
по размеру зубцами и рядом мелких зубчиков; большой подвижный
зубец сочленен не с дистальной, как у представителей других ро-
дов, а с базальной частью боковых лопастей и снабжен двумя или
тремя длинными щетинками (ниже места прикрепления подвиж-
ного зубца имеется еще одна длинная щетинка). Хвостовые пла-
стинки, достигающие двух третей длины брюшка, темнокрашен-
ные или с темными пятнами; тонкие и прямые трахейные ветви
отходят от центрального ствола под прямым углом.
Личинки, встречающихся у нас видов Lestes, из которых самыми
обычными являются Lestes dryas и Lestes sponsa, различаются
между собою следующими признаками.
1 (2) Боковые лопасти маски с тремя длинными щетинка)ми,
из которых две расположены на подвижном зубце; хво-
стовые пластинки с тремя темными поперечными поло-
сами ................................ . L. sponsa
2 (1) Указанные лопасти с четырьмя длинными щетинками, из
которых три расположены на подвижном зубце . . 3
3 (4) Хвостовые пластинки почти по всей длине с парал-
дельными краями...................................L. virens
4 (3) Хвостовые пластинки от середины к концу постепенно
суживающиеся . 5
164
5 (6) Хвостовые пластинки узкие; на каждой стороне средней
лопасти маски по семи длинных щетинок . . L. dryas
6 (5) Хвостовые пластинки широкие; па каждой стороне сред-
ней лопасти маски по восьми длинных щетинок .
L. barbara
9. Личинки Erythromma (рис. 71, А—В)
Личинки стрелки, Erythromma, являются одними из самых рас-
пространенных личинок стрекоз; живут в разнообразных водоемах
среди растений. У нас встречается только один вид — Erythromma
najas. Длина личинок (без хвостовых пластинок) достигает 18 лш.
Голова сильно вытянута в поперечном направлении. Антенны семи-
членистые. Маска короткая, приблизительно квадратной формы, в
сложенном виде не достигает оснований ног второй пары; на каж-
дой стороне средней лопасти по три-четыре щетинки; боковые ло-
пасти с шестью щетинками; подвижный зубец без щетинок. Хвосто-
вые пластинки широкие, по внешнему краю резко подразделены
зарубкой на две половины — более плотную базальную, с зазуб-
ренным краем, и тонкую дистальную, вооруженную небольшими
волосками; темноокрашенные и древовидно ветвящиеся трахейные
ветви отходят от центрального ствола под острым углом. Личинки
зеленоватой окраски.
10. Личинки Enallagma (рис. 71, Г—Е)
Личинки стрелки, Enallagma, встречаются часто в стоячих
водоемах среди растений. Единственный вид — Enallagma cyathi-
gerum. Длина личинок (без хвостовых пластинок) около 14 мм. Го-
лова расширена в поперечном направлении. Антенны семичлени-
стые. Маска широкая, квадратной формы; на каждой стороне сред-
ней лопасти по четыре длинных щетинки; боковые лопасти с ше-
стью щетинками; подвижный зубец без щетинок. Хвостовые пла-
стинки прозрачные, на конце заостренные, с хорошо заметными
трахейными ветвями; боковые пластинки подразделены на две по-
ловины, проксимальная часть более плотная, с мелко зазубренными
краями. Личинки зеленоватой или светложелтой окраски.
1L Личинки Coenagrion (рис. 71, Ж—Н)
Личинки стрелки, Coenagrion (в прежней литературе Agrion),
встречаются преимущественно в стоячих водоемах среди растений.
Длина личинок (без хвостовых пластинок) достигает 14—17 мм.
Голова вытянута в поперечном направлении. Антенны семичлени-
стые. Маска короткая, плоская; на каждой стороне средней лопасти
по четыре, иногда по три щетинки; боковые лопасти с пятью-семью
Щетинками; подвижный зубец без щетинок. Хвостовые пластинки
широкие, обычно ясно подразделенные на две части, на конце, за
редкими исключениями, заостренные; трахейные ветви отходят от
центрального ствола под острым углом.
165
ж
Личинки наиболее распространенных у нас видов Coenagrion
различаются между собою следующими признаками:
1 (2) На боковых лопастях маски по семи щетинок; длина ли-
чинок (без хвостовых пластинок) достигает 16—
17 мм .........................................с. armatum
2 (1) На указанных лопастях обычно по пяти, редко по шести
щетинок; длина личинок около 14 мм.......................3
3 (4) На каждой стороне средней лопасти маски по три длин-
ных щетинки; хвостовые пластинки очень короткие, на
конце заостренные, подразделенные в дистальной трети
на две части................................С. mercuriale
4 (3) На каждой стороне указанной лопасти по четыре-пять
щетинок; хвостовые пластинки длинные, подразделенные
посреди на две части ....................................5
5 (6) Боковые хвостовые пластинки узкие (длина, по крайней
мере, в пять раз больше ширины), ланцетовидные; сред-
няя пластинка более широкая . С. hastulatum
6 (5) Все хвостовые пластинки широкие . ... 7
7 (8) Хвостовые пластинки на конце заостренные’ С. puefla
8 (7) Хвостовые пластинки на конце закругленные (на моло-
дых стадиях развития заостренные) . С. pulchellutn
Личинки Plecoptera
Крылатые особи обладают удлиненным и мягким телом. Груд-
ные сегменты резко отделены друг от друга, переднегрудь с про-
дольной бороздой. На голове находятся длинные многочленистые
антенны, два сложных глаза и три глазка. Ротовые органы недо-
развиты. Две пары прозрачных крыльев в состоянии покоя склады-
ваются на спинной стороне; задние крылья шире передних, поэтому
при их складывании образуются продольные складки. У некоторых
видов крылья укорочены. На конце тела находятся две длинные,
многочленистые хвостовые нити (cerci). Насекомые держатся пре-
имущественно около водоемов, летают плохо. Во взрослом состоя-
нии живут недолго, около месяца. Лёт начинается ранней весной.
Развитие веснянок совершается с гемиметаморфозом, продолжа-
ется от одного до трех с половиной лет. В течение этого времени
личинки многократно линяют, например личинки Perla, живущие
более трех лет, линяют до 23 раз. Личинки веснянок живут в чи-
Рис. 71. Д — В— личинка Erythromma najas, общий вид (Я), маска (Б), хвосто-
вая пластинка (В); Г—Е — личинка Enallagma cyathygerum, общий вид (Г),
маска (Д), хвостовая пластинка (Е); Ж» 3— личинка Coenagrion pulchellum,
общий вид (Ж), хвостовая пластинка (3)\ И, К —личинка Coenagrion mercu-
riale, маска (И), хвостовая пластинка (X); Л — личинка Coenagrion puella,
хвостовая пластинка; М—личинка Coenagrion armatum, хвостовая пластинка;
Н — хвостовые пластинки личинки Coenagrion сбоку.
/— голова, 2—антенна, 3—глаз, 4 — зачатки крыльев, 5 — брюшко, 6 —
боковая лопасть, 7 — подвижный зубец, 8— подбородочные щетинки, 9 — сре-
динная хвостовая пластинка, 10—боковая хвостовая пластинка, 11 — цсрконд,
12 — место облома хвостовой пластинки.
167
стой воде, богатой кислородом; загрязнение воды вызывает их ги-
бель. Обитают главным образом в текучей воде на нижней стороне
камней, преимущественно в горных местностях, в равнинных реках
и озерах встречаются сравнительно редко. Питаются они расти-
тельностью или ведут хищнический образ жизни.
Личинки веснянок отличаются от личинок других насекомых
присутствием на конце брюшка двух длинных хвостовых нитей. От
личинок поденок, некоторые из которых также имеют две хвосто-
вых нити, отличаются по строению максиллярного щупика, пяти-
членпстого у веснянок и самое большее трехчленистого у поденок,
и по присутствию двух коготков на грудных ногах. Кроме того, у
большинства видов веснянок трахейные жабры отсутствуют, дыха-
ние совершается через мягкие покровы тела; в тех случаях, когда
трахейные жабры присутствуют, они имеют вид простых или раз-
ветвленных выростов, расположенных на нижней поверхности груд-
ных сегментов и на конце брюшка, а у некоторых видов на кок-
сальных члениках ног.
1. Личинки Perla (рис. 72, А—Г)
Наиболее обычными являются личинки Perla, самки которых до-
стигают 30 мм, а самцы 20 мм. Тело уплощенное. На голове, имею-
щей приблизительно треугольную форму, находятся длинные ан-
тенны, число члеников которых доходит до 90, большие парные'
глаза и три глазка, расположенные треугольником. Ротовые органы
грызущего типа, состоят из верхней губы, мандибул, максилл и
нижней губы. Верхняя губа широкая. Мандибулы массивные, на
конце с несколькими сильными зубцами. Максиллы с двумя остры-
ми зубцами, с узким двучленистым наружным выростом (galea) и
всегда пятичленистым щупиком. Нижняя губа широкая, кожистая,
состоит из парных внутренних и наружных лопастей; щупики трех-
членистые. На внутренней стороне нижней губы расположен не-
большой языковидный подглоточник (гипофаринск). Грудные сег-
менты большие, резко отделенные друг от друга. Среднеспинка и
заднеспинка с направленными назад и вбок чехлами для крыльев.
Брюшко десятичленистое, на конце с двумя длинными и многочле-
нистыми хвостовыми нитями. Ноги уплощенные, направлены в
стороны; лапка трехчленистая, с сильно удлиненным третьим чле-
ником, оканчивающимся двумя подвижными, большими, серповидно
изогнутыми коготками. Трахейные жабры представлены шестью па-
рами плевральных жабр, расположенных по бокам груди. Каждая
плевральная жабра состоит из нескольких ветвящихся выростов, не-
сущих тонкие трахейные нити. Три пары, отличающиеся большей
величиной, находятся между сегментами груди (две первых пары)
и между последним грудным и первым брюшным сегментами (тре-
тья пара); другие три пары, меньшего размера, расположены над
основаниями ног. У некоторых видов, кроме того, имеется еще одна
пара анальных жабр, находящихся на конце брюшка между хво-
стовыми нитями.
Наиболее обычными являются личинки Perla abdominatis.
168
л у £
Рис. 72. А—Г — личинка Peta abdominalis, общий вид (Л), мандибула (Б), мак-
силла (В), нижняя губа (Г); ДБ —личинка Nephelopteryx nebulosa, общий
вид (Д), грудь снизу (Б).
1—голова, 2—антенна, 3—глаз, 4 — глазок. 5—первый грудной сегмент, 6—
зачатки крыльев, 7 — бедро, 8 — голень, 9 — лапка, 10 — коготок, 11 — трахей-
ные жабры, 12 — брюшко, 13 — хвостовая нить. 14— галея (galea), 15 — мак-
силлярный щупик, 16 — внутренняя лопасть, 17 — наружная лопасть, 18—нижне-
губный щупик.
2. Личинки Nephelopteryx (рис. 72, Д—Е)
Личинки Nephelopteryx относятся к числу небольших, их длина
не превышает 15 мм. Чехлы для крыльев большие, задние треуголь-
ной формы. Антенны длинные, содержат до 80 члеников. Левая
мандибула с пятью зубцами, молярная часть хорошо развита, зуб-
чатая; правая мандибула с четырьмя зубцами, молярная часть сла-
бо развита. Максиллы с длинной и узкой galea. Брюшко с средин-
ным рядом острых шипов. Трахейные жабры отходят от коксаль-
ных члеников ног. Они имеют видадлинных (около 2р—3,0 мм) бе-
лых трубочек, состоящих из трех частей, втягивающихся друг в
друга наподобие частей зрительной трубы. Хвостовые нити длиннее
брюшка. У нас встречаются личинки Nephelopteryx nebulosa.
Личинки Megaloptera
Крылатые насекомые черного цвета, крылья широкие, прозрач-
ные, с черными жилками. Голова большая. Антенны нитевидные,
длинные. Ротовые придатки грызущего типа. Брюшко короче кры-
льев. Взрослые насекомые живут около воды, летают ночью. Яйца
откладываются на надводные части растений. Личинки живут в.
й
Рис. 73. Личинка Sialis lutaria. А —
общий вид; Б голова снизу.
/ голова, 2 — глаз, 3 — антенна,
4 — первый грудной сегмент, 5 —
бедро, 6 —голень, 7 — лапка, 8—
коготки, 9 — брюшко, 10 — трахейные
жабры, // — мандибула, 12—мак-
силла, 13 — максиллярный щупик,
14 -нижняя iy6a, 15— нижнегубный
щупик.
170
зоде. Длительность личиночной жизни равна двум годам. За это
время личинки линяют девять раз.
Личинки Stalls (рис. 73)
Длина личинки на последних возрастных стадиях достигает
25 30 мм. Тело уплощенное. Голова почти квадратной формы, пло-
ская. Глаза небольшие. Антенны четырехчленистые. Верхняя губа
пластинчатая. Мандибулы сильно развиты, высовываются вперед,
с тремя зубцами. Максиллы с четырехчленистым щупиком. Нижняя
губа небольшая, с трехчленистыми щупиками. Каждый грудной
сегмент несет пару грудных ног, вооруженных щетинками; лапка
двучленистая, на конце с двумя коготками. Абдомен длинный, к
концу суживающийся, состоит из девяти сегментов; последний сег-
мент с длинным, суживающимся к концу отростком, опушенным с
обеих сторон волосками; внутри отростка проходят два трахейных
ствола. По бокам первых семи брюшных сегментов отходят длин-
ные пяти-шестичленистые трубчатые трахейные жабры, покрытые
волосками. В каждой трахейной жабре проходит трахейный ствол
с многочисленными ветвями. У молодых личинок трахейные жабры
короткие, одночленистые.
Личинки Stalls, единственные у нас представители отряда Mega-
loptera, встречаются в стоячих и медленно текучих водоемах с или-
стым дном — лужах, канавах, ручьях и т. п. Сильные ноги помо-
гают личинкам двигаться в илу, они могут также плавать. Личинки
являются хищниками, питаются мелкими червями, личинками раз-
личных насекомых. Весною личинки выходят на сушу и окуклива-
ются в сырых местах.
Встречающиеся у нас личинки Stalls lutarta (=S. flavilatera) и
Stalls fuliginosa по внешнему виду не различимы.
Личинки и куколки Trichoptera
Во взрослом состоянии все ручейники ведут воздушный образ
жизни, только некоторые виды, являющиеся эндемиками Байкала,
живут на поверхности воды, плавая при помощи ног.
Крылатые особи отличаются следующими признаками—голова
небольшая; антенны длинные, многочленистые; глаза обычно не-
большие; ротовые придатки грызущего типа; крылья большие, ко-
жистые, у большинства видов покрытые волосками, передние кры-
лья обычно тверже задних. Встречаются около водоемов; летают
плохо, некоторые виды могут только ползать. Развитие сопровож-
дается полным превращением.
Яйца откладываются в воду в виде студенистых кладок, содер-
жащих большое число зеленоватых или желтоватых яиц. Форма
кладок разнообразная — мешковидная, шаровидная, в виде кольца
и т. п. У ряда видов яйца приклеиваются к твердому субстрату
клеевидным веществом.
Тело личинки делится на голову, грудь и брюшко. На голове
находятся парные глаза, впереди них отходят рудиментарные ан-
171
тенны; снизу расположены ротовое отверстие и ротовые придатки—
большая верхняя губа, крепкие и изогнутые мандибулы и неболь-
шие максиллы, слившиеся своими основаниями с нижней губой.
Большие прядильные (паутинные) железы открываются на нижней
губе. Грудь состоит из трех ясно отграниченных сегментов, несу-
щих такое же количество хорошо развитых грудных ног. Спинная
сторона первого грудного сегмента всегда сильно хитинизирована,
второй сегмент кожистый или со спинным хитиновым щитом, тре-
тий сегмент обычно кожистый, реже с небольшими склеритами.
Между ногами первой пары на нижней стороне первого грудного
сегмента у личинок многих видов находится длинный вырост, на-
зываемый рогом. Второй и третий грудные сегменты у личинок
Hydropsyche и некоторых Rhyacophila снабжены хорошо развитыми
трахейными жабрами; у других личинок ручейников трахейных
жабр на грудных сегментах не имеется. Брюшко состоит из десяти
сегментов. У личинок тех видов, которые живут в домиках, на пер-
вом брюшном сегменте находится три, реже два, тугих мускули-
стых выроста, а по бокам остальных сегментов проходят боковые
линии — тонкие складки, снабженные волосками. На последнем
брюшном сегменте находятся ложные ножки (прицепки), оканчи-
вающиеся коготками. От брюшных сегментов отходят трахейные
жабры, простые или перистые, обычно одиночные, в более редких
случаях они образуют пучки. В наиболее развитом виде трахейные
жабры расположены шестью рядами —/двумя спинными, двумя бо-
ковыми и двумя брюшными — на втором — восьмом сегментах брю-
шка; по положению различают пресегментальные и постсегменталь-
ные жабры, расположенные у переднего края и, соответственно, у
заднего края сегментов. У личинок ряда видов задние трахейные
жабры могут отсутствовать, некоторые личинки лишены их полно-
стью. На заднем конце тела у некоторых личинок находятся не-
большие анальные папиллы.
По своему строению личинки ручейников делятся на две груп-
пы — комподеовидных и гусеницевидных, отличающихся между
собою также и образом жизни. Камподеовидные личинки характе-
ризуются тем, что у них продольная ось головы является продол-
жением продольной оси тела; брюшные сегменты с глубокими
перехватами, боковых линий не имеется. Среди этих личинок пре-
обладает хищный способ питания. Почти все они ведут свободный
образ жизни, за редкими исключениями домика не имеют. Личинки
некоторых живущих в реках видов (Hydropsyche, Neureclipsis и др.)
относятся к группе пассивных фильтраторов, строят л'овчие сети,
улавливающие сносимые течением планктические организмы и дет-
рит. У личинок второй группы, гусеницевидных, продольная ось го-
ловы составляет прямой или тупой угол с продольной осью тела,
перехваты между сегментами неглубокие, имеются боковые линии
и выросты на первом брюшном сегменте. Большинство личинок пи-
тается растительной пищей. Все они строят переносный, служащий
для зашиты тела домик из самого разнообразного материала, скреп-
ленного выделениями прядильных желез. Внутренняя цилиндриче-
ская полость домика выстлана паутинным слоем. Личинки держат-
172
ся в своих домиках при помощи выростов (на первом брюшном
сегменте) и находящихся на конце тела прицепок. В качестве стро-
ительного материала для постройки домика используются песчинки,
мелкие камешки, детрит, раковины моллюсков, части листьев и
стеблей растений и т. п. Личинки многих видов обладают домиками
характерного строения, у других он варьирует по форме. Домики
тех личинок, которые живут на дне, построены из тяжелого мате-
риала, песка или камешков, раковин моллюсков, пропитанных во-
дою кусочков дерева и т. п. В противоположность" этому, у личинок,
живущих среди растений около поверхности воды и способных к
непродолжительному плаванию, домики легче или лишь немного
тяжелее воды, материалом для них служат листья и стебли ра-
стений.
После нескольких, не более шести, линек личинки превращаются
в куколок, по многим признакам напоминающих взрослых насеко-
мых, с длинными антеннами, хорошо развитыми грудными ногами
и зачатками крыльев. Верхняя губа снабжена направленными квер-
ху крепкими щетинками. Обычно имеются трахейные жабры, а на
спинной стороне брюшных сегментов группы шипиков, сидящих на
твердых пластинках и образующих так называемый прицепной ап-
парат. Шипики, расположенные на третьем — седьмом сегментах,
направлены остриями назад, на пятом сегменте верхние шипики
такого же строения, как остальные, а нижние направлены остриями
вперед; на спинной стороне первого брюшного сегмента находится
вырост, вооруженный шипиками. На конце тела расположены длин-
ные палочковидные анальные придатки, несущие на себе щетинки
и шипики.
Куколки ручейников заключены в домики, которые с обеих сто-
рон закрыты крышечками, пронизанными отверстиями. Куколки
пои помощи шипиков на брюшных сегментах могут передвигаться
вверх и вниз для прочищения засорившихся отверстий крышечек
щетинками верхней губы и анальных придатков. Непрерывными
движениями брюшка куколки обменивают воду внутри домиков,
опорой при этих движениях служит вырост первого брюшного сег-
мента.
Перед вылетом насекомого куколка разрушает верхнюю крыш-
ку своими крепкими мандибулами, затем выходит из домика и вы-
ползает из воды по растениям или иным твердым предметам.
Личинки ручейников встречаются в различных водоемах, обыч-
но живут в прибрежной области, так как отличаются большой тре-
бовательностью к содержанию в воде кислорода. Встречаясь не-
редко в массовых количествах, служат пищей для многих рыб.
Гусеницевидные личинки
•К этой группе относятся самые обыкновенные личинки ручейни-
ков. Все они живут в домиках, у большинства видов имеются хо-
рошо развитые трахейные жабры.
17Т
Е Ж
Рис. 74. А — личинка Phryganea grandis,
общий вид; Б—Ж— личинка Phryganea
striata, в домике (Б),верхняя губа (В), ман-
дибула (Г), максиллы и нижняя губа (Д),
крючок прицепки (Е); Ж—куколка'Phryga-
nea striata, анальные придатки.
1 — голова, 2 — первый грудной сегмент,
3 — третий грудной сегмент, 4— тазик,
5 — вертлуг, 6 — бедро, 7 — голень, 8 — лап-
ка, 9 — коготок, 10 — выросты на первом
брюшном сегменте, 11 — трахейные жабры,
12 — прицепка.
1. Личинки и куколки Phryganea (рис. 74)
Личинки крупные, достигают в длину 30—40 .мл/. Голова продол-
говатая, расположена под тупым углом к продольной оси тела, с
темными продольными полосами на верхней стороне. Верхняя губа
впереди с выемкой, мандибулы сильные, черного цвета. Спинная
сторона первого грудного сегмента склеротизирована, по краям с
темной каймой, снизу с
небольшим роговидным
выростом; остальные
грудные сегменты кожи-
стые. Грудные ноги хоро-
шо развиты, с темными
полосами; коготки длин-
ные, изогнутые. Брюшко
широкое. Спинной вырост
первого брюшного сегмен.
та конический, боковые
выросты тупые, на конце
с короткими шипиками.
Боковые линии хорошо
развиты, с длинными чер-
ными волосками, начина-
ются на третьем брюшном
сегментр. Трахейные жаб-
ры одиночные, цилиндри-
ческие; с каждой стороны
на первом сегменте нахо-
дится по две брюшных
жабры; на следующих ше-
сти сегментах — по шести
жабр (по две спинных, бо-
ковых и брюшных); на
восьмом сегменте—по од-
ной боковой и спинной
жабре (последние могут
отсутствовать). Девятый
сегмент с поперечной чер-
ной пластинкой, воору-
женной по заднему краю
длинными щетинками. На десятом сегменте расположены двучле-
нистые прицепки, оканчивающиеся сильно изогнутыми крючками.
Голова, брюшко и конечности с одиночными длинными черными
щетинками.
Домик построен из различных растительных прямоугольной
формы кусочков, расположенных по спирали; достигает в длину
50 мм.
Куколки приблизительно цилиндрической формы. Антенны ко-
роче тела. Верхняя губа большая, почти квадратная, с многочис-
ленными, направленными вперед щетинками. Мандибулы согнуты
под прямым углом. Прицепной аппарат на спинной стороне брюш-
74
ных сегментов сильно развит. Первый членик брюшка с широким
отростком, заходящим за задний край сегмента. Анальные придат-
ки плоские, на конце с четырьмя длинными щетинками. Трахей-
ные жабры расположены подобно жабрам личинок, на первом
брюшном сегменте отсутствуют.
Личинки Phryganea живут в прибрежной растительности, пита-
ются растительной и животной пищей.
Личинки двух наиболее часто встречающихся видов Phryganea
различаются между собою следующими признаками.
У личинок Phryganea grandis в передней части восьмого брюш-
ного сегмента находятся спинные трахейные жабры; верхняя губа
позади выемки с большой светлой, иного строения прямоугольной
частью.
У личинок Phryganea striata на восьмом брюшном сегменте
спинных трахейных жабр не имеется; верхняя губа обычного строе-
ния.
2. Личинки и куколки Limnophilus (рис. 75)
Личинки многочисленных видов Limnophilus отличаются следую-
щими признаками. Тело цилиндрическое. Голова, загнутая книзу, и
первый сегмент груди по ширине значительно меньше других сег-
ментов тела. Глаза расположены приблизительно на одинаковом
расстоянии от переднего и заднего краев головы. Верхняя сторона
первого и второго грудных сегментов склеротизирована, на третьем
сегменте с тремя парами небольших склеритов. Ноги первой пары
значительно короче остальных ног. Брюшко широкое. На первом
брюшном сегменте находится три тупых выроста. Боковые линии
простираются от третьего до восьмого сегментов. Трахейные жабры
расположены пучками из двух-трех нитей; имеются, кроме того,
одиночные жабры. Последний сегмент брюшка узкий, несет короткие
прицепки с темными коготками. Длина личинок достигает 25 мм.
Домики, построенные из самого различного материала, имеют
разнообразную, часто неправильную форму; в качестве материала
применяются кусочки веточек, иглы хвойных, пластинки ряски, ра-
ковины моллюсков, детрит и т. п.
Куколки отличаются следующими признаками. Антенны не
длиннее тела. Первый брюшной сегмент с двумя боковыми лопа-
стями на заднем крае. Прицепной аппарат развит только на четвер-
том — седьмом брюшных сегментах. Трахейные жабры в пучках.
На конце тела находятся длинные палочковидные анальные при-
датки.
Личинки Limnophilus встречаются в самых различных водоемах,
в литорали озер и прудов, в весенних лужах и иных мелких водое-
мах.
Личинки наиболее обычных видов Limnophilus различаются
между собою следующими признаками.
1 (12) Передняя часть спинной поверхности первого грудного
сегмента окрашена в темный цвет............................2
2 (5) Срединная темная полоса на верхней поверхности го-
ловы резко отделена от боковых полос . 3
175
3 (4) Пресегментальные пучки жабр в боковых рядах на вто-
ром брюшном сегменте отсутствуют; домик из кусочков,
расположенных продольно, часто трехгранный •
R L. mgrtceps
4 (3) Пресегментальные пучки из двух-трех жабр в боковых
рядах на втором брюшном сегменте имеются; домики
разнообразной формы, из косо положенных кусочков или
Рис. 75. Л, Б —личинка Limnophilus flavicornis, общий вид (Л), домик (Б);
В— куколка Limnophilus flavicornis; Г,Д — личинка Limnophilus politus, перед-
неспинка (Г), домик (Д); Е — личинка Limnophilus nigriteps, переднеспинка; Ж—
личинка Limnophilus rhombicus, переднеспинка; 3 — личинка Limnophilus
stigma, домик.
/ — выросты на первом брюшном сегменте, 2 — трахейные жабры, 3 — прицепка,
4 — анальный придаток.
5 (2) Указанная полоса впереди сильно расширена, почти ка-
сается боковых полос...................................6
6 (9) Борозда, отделяющая на первом грудном сегменте тем-
ную переднюю часть, резко выделяется по окраске . Т
7 (8) Нижняя поверхность головы темно окрашена; домики
разнообразной формы..........................L. flavicornis
8 (7) Нижняя поверхность головы светлая; домики боченко-
видные, построены из поперечно расположенных кусоч-
ков детрита или мха..........................L. stigma
9 (6) Указанная борозда по окраске не выделяется . 10
176
10 (11) Срединная темная полоса на верхней поверхности головы
впереди с глубокой выемкой..........................L. politus
11 (10) Указанная полоса впереди без выемки или она слабо
выражена........................................L. marmoratus
12 (1) Спинная сторона первого грудного сегмента равномерно
окрашена; домики из песчинок...............................13
13 (14) Бедра ног второй и третьей пар с двумя черными ще-
тинками ............................................L. griseus
14 (13) Бедра ног второй -и третьей пар с одной черной щетин-
кой и с одним желтым зубцом . . 15
15 (16) На первом сегменте груди имеется рисунок в виде
креста............................................. L. vitiatus
16 (15) На указанном сегменте крестообразного рисунка не
имеется L. centralis
3. Личинки и куколки Grammotaulius (рис. 76, А—В)
Личинки обычного вида Grammotaulius atomarius по внешнему
виду сходны с личинками Limnophilus; отличаются следующими
признаками — голова коричневая или красновато-коричневая, с
Рис. 76. А, Б —- личинка Grammotaulius atomarius, общий вид (Л), домик (Б);
В — куколка Grammotaulius atomarius, анальный придаток; Г, Д— личинка
Stenophylax stellatus, общий вид (Г), домик (Д).
/ — выросты па первом брюшном сегменте, -2 — трахейные жабры, 3— боковая
линия, 4 — прицепка.
177
томными точками; бедра ног Третьей пары с двумя тесно сближен-
ными щетинками. Длина личинок достигает 28 мм. Домик почти
цилиндрический, из растительных частей, уложенных по спирали
и крышеобразно заходящих друг на друга. Куколки сходны с ку-
колками Litnnophilus. Живут в заросших растительностью водоемах.
4. Личинки и куколки Stenophylax (рис. 76, Г—Д)
Личинки обычного вида Stenophylax stellatus, по внешнему виду
сходны с личинками Litnnophilus; отличаются следующими призна-
ками— имеются лишь одни одиночные трахейные жабры; на голо-
ве находятся многочисленные мелкие черные точки; грудные сег-
менты окрашены светлее головы. Длина личинок до 25 мм. Домик
цилиндрический, изогнутый, На заднем конце закругленный, по-
строен из крупных песчинок. Куколки отличаются от куколок
Limnophilus присутствием одиночных трахейных жабр. Обитают в
ручьях п в прибойных частях озер.
5. Личинки и куколки Molanna (рис. 77)
Личинки обычного вида Molanna angustata характеризуются
следующими признаками. Тело уплощенное. Голова яйцевидно-
Рис. 77. А, Б — личинка Molanna angustata,
общий вид (Д), домик (£); В — куколка
Molanna angustata.
/ — вырост на первом брюшном сегменте,
2 — трахейные жабры, 3— прицепка,
4 — прицепной аппарат, 5 — анальные
придатки.
восьмом сегментах, брюшные — на
боковые, обычно одиночные, жабры
ствуют. Длина личинок достигает И
овальная, посреди с темным
рисунком. Верхняя губа без
выемки на переднем крае.
Верхняя сторона первого и
второго грудных сегментов
склеротизирована, на вто-
ром сегменте разделена по-
перечным швом. На брюш-
ной стороне первого сегмента
рога не имеется. Ноги с во-
лосками. Голени нот первой
и второй пар в дистальной
части с коническим выро-
стом, оканчивающимся ши-
пом. Коготки ног третьей па-
ры короткие. Спинной буго-
рок на первом сегменте брю-
шка большой. Боковые ли-
нии проходят от третьего до
седьмого сегментов. Прицеп-
ки с узкими коготками, несу-
щими несколько дополни-
тельных коготков. Трахей-
ные жабры расположены
пучками из трех-четырех
тонких трубок; спинные жаб-
ры находятся на первом —
втором —седьмом сегментах;
на последних сегментах отсут-
мм.
178
Домики очень характерной формы, построены из песчинок, с
большими боковыми выростами, придающими им вид щита. Оби-
тают в стоячих водоемах на илу и песке.
Куколки слегка сжаты в дорсо-вентральном направлении: антен-
ны самок короче тела; сближенные анальные придатки покрыты
волосками; трахейные жабры в пучках из трех-четырех трубок.
6. Личинки и куколки Leptocerus (рис. 78)
Личинки многочисленных видов Leptocerus отличаются от дру-
гих личинок ручейников следующими признаками. Тело цилиндри-
ческое. Антенны относительно длинные, на конце с щетинкой.
Спинная сторона первого и второго грудных сегментов склеротизи-
рована. Третий сегмент широкий, кожистый. На брюшной стороне
Рис. 78. А, Б — личинка Leptocerus aterrimus, общий вид (Л), домик (Б); В —
куколка Leptocerus aterrimus; Г, Д—голова личинки Leptocerus cinereus,
сверху (Г) и снизу (Д).
1—трахейные жабры, 2 — прицепка, 3—анальный придаток.
первого грудного сегмента рога не имеется. Бедра ног второй и
третьей пар подразделены на небольшую базальную и длинную
апикальную части. Брюшко толще первых грудных сегментов. Спин-
ной вырост на первом сегменте притупленный, боковые выросты
слабо выражены. Боковые линии тонкие, с короткими волосками,
простираются на третьем — седьмом сегментах. Прицепки корот-
кие, с сильными коготками. Трахейные жабры расположены пучка-
ми, в редких случаях развиты на всех брюшных сегментах, обычно
только на передних. Длина личинок достигает 11 мм.
Домики обычно из песчинок или секрета желез.
179
Куколки отличаются присутствием очень длинных антенн, закру-
ченных вокруг тела. На конце тела длинные палочковидные при-
датки. Прицепной аппарат хорошо развит. Трахейные жабры соот-
ветствуют жабрам личинок.
Личинки Leptocerus широко распространены, встречаются в озе-
рах и реках, обычно в прибрежной области.
Личинки наиболее часто встречающихся видов Leptocerus разли-
чаются между собою следующими признаками.
1 (4) Дсмпк нестроен из секрета; трахейные жабры находят-
ся на первом — восьмом сегментах.......................2
2 (3) Голова светлая; домик конический, гладкий . L. senilis
3 (2) Голова с темным рисунком...................L. fulvus
4 (1) Домик построен из песчинок или иного материала. . 5
5 (6) Трахейные жабры находятся на втором—седьмом сег-
ментах ..................................L. annulicornis
6 (5) Трахейные жабры находятся только на первом — треть,
ем сегментах . ................................7
7 (8) Голова сверху с пестрым рисунком, образованным тем-
ными пятнами и полосками.....................L. aterrimus
8 (7) Голова сверху с неявным рисунком, снизу темная, почти
черная L. cinereus
Камподеовидные личинки
В эту группу входят в большинстве случаев свободноживущие
личинки, ведущие хищный образ жизни. Личинки некоторых видов
относятся к числу пассивных фильтраторов, строят из паутины лов-
чие сети.
7. Личинки и куколки Hydropsyche (рис. 79, А—Д)
Личинки гидрспсихе (Hydropsyche) камподеовидные. Голова не-
большая, не согнута по отношению к продольной оси тела. Антенны
рудиментарные. Верхняя губа без выемки. Мандибулы сильные, с
многочисленными зубцами. Нижняя губа с рудиментарными щупи-
ками. Спинная сторона всех трех грудных сегментов склеротизиро-
вана, с одинаковыми квадратными темноокрашенными щитками, с
черными краями и пятнами. Грудные ноги короткие, крепкие, в
особенности ноги первой пары, с многочисленными шипами и ще-
тинками. Брюшко покрыто короткими темными волосками. Боковые
линии отсутствуют. Прицепки сильно развитые, двучленистые. Ба-
зальный членик длинный, темноокрашённый, на конце с пучком длин-
ных черных щетинок; второй членик очень короткий, коготок силь-
но изогнут. Трахейные жабры ветвящиеся, расположены пучками на
первом — восьмом сегментах брюшка, а также на втором и тре-
тьем грудных сегментах. На конце тела, между прицепками, рас-
положены четыре анальных папиллы. Длина личинок достигает
22 мм.
Личинки гидропсихе переносного домика не строят. Сооружают
ловчую камеру на камнях, упавших в воду сучках и т. п.; одна сто-
180
Рис. 79. А—В личинка Hydropsyche ornatula, общий вид (Д), прицепка (5),
ловчая сеть (В); Г, Д — куколка Hydropsyche ornatula, анальные придатки (Г),
домик; (Д); Ё— личинка Rhyacophila nubila; Ж— куколка Rhyacophila nubiia,
домик снизу; 3, И — личинка! Neureclipsis bimaculata, общий вид (3), ловчая
сеть (И).
1 — трахейные жабры, 2 — анальные папиллы.
181
рана камеры образована сетью из правильных прямоугольных ячеек.
Личинки питаются попавшими в сеть организмами.
Куколки в коконах, отличаются присутствием длинных, расши-
ренных анальных придатков, вооруженных щетинками; жабры раз-
ветвленные, как у личинок.
Обитают в текучей воде, реках, ручьях, иногда в огромных ко-
личествах; наиболее часто встречаются личинки Hydropsyche ото-
tula.
8. Личинки и куколки Rhyacophila (рис. 79, Е—Ж)
Личинки наиболее часто встречающегося вида Rhyacophila
nubila легко отличаются от других личинок ручейников по ряду
признаков.
Личинки камподеовидные. Голова с темными пятнами. Глаза
расположены у переднего края головы. Спинная сторона первого
грудного сегмента склеротизирована, на втором и третьем сегментах
кожистая. Грудные ноги одинаковой длины.- Прицепки хорошо раз-
витые, двучленистые; первый членик с одним длинным заостренным
и одним коротким тупым отростками; конечный коготь большой, с
добавочными шипами при основании. Трахейные жабры располо-
жены пучками по бокам первого — восьмого сегментов брюшка и
при основании ног второй и третьей пар. Домики., не строят, окрас-
ка зеленовато-фиолетовая. Длина личинок достигает 25 мм.
Куколки заключены в гладкий, красновато-бурый кокон; домик
из небольших камешков, открытый с нижней стороны, обращенной
к субстрату.
Личинки встречаются в ручьях и реках с холодной водой, пре-
имущественно в горных местностях.
9. Личинки и куколки Neureclipsis (рис. 79, 3—И)
Личинки обычного вида Neureclipsis bimaculata отличаются сле-
дующими признаками. Голова овальная, с черными пятнами. Ан-
тенны редуцированы. Глаза расположены ближе к переднему краю.
Грудные ноги короткие, коготки ног с базальными шипами. Брюш-
ко с глубокими перетяжками. Прицепки трехчленистые, последний
членик с острым когтем. Трахейные жабры отсутствуют. Длина до-
стигает 27 мм.
Личинки строят ловчие сети в виде труб, загнутых на конце.
Живут в реках с небыстрым течением.
Куколки заключены в серый кокон; домик из песчинок, камеш-
ков или частей растений.
Личинки и куколки Simuliidae (рис. 80)
Мошки относятся к двукрылым насекомым (Diptera, семейство
Simuliidae). Крылатые особи небольшого размера, не более 4—5 мм,
черного цвета. Голова с большими глазами, особенно у самцов, и
коротким и толстым хоботком. Антенны, небольшие. Грудь сильно
182
выпуклая. Ноги короткие и толстые; крылья широкие. Самцы пита-
ются соками растений, самки, кроме того, — кровью животных.
Мошки широко распространены, особенно в таежных лесах. Во
многих местах Сибири и Дальнего Востока мошки, называемые там
гнусом, являются наиболее злостными кровососами из всех двукры-
Рис. 80. А—личинки и куколки Simulium на листе растения; Б, В — личинка
Simulium ornatum, сверху (15) и сбоку (В); Г — веерообразный придаток ли-
чинки Simulium; Д, Е — куколка Simulium ornatum, в коконе (Д) и без коко-
на (Е); Ж— кокон Simulium ornatum; 3 — кокон Simulium subornatum.
/— антенна, 2 — веерообразный придаток, 3— глазок, 4 — ложная ножка. 5 —
присоска.
лых насекомых, нападают в огромных количествах на человека и
животных. Слюна мошек ядовита. В придунайских странах колум-
бацкая мошка (Simulium reptans, columbaczense) вызывает массо-
вую гибель домашнего скота. Мошки нападают на добычу только
днем.
Все развитие мошек протекает в текучей воде (рис. 80). Яйца
приклеиваются самками к листьям и стеблям растений, камням и
другим предметам, погруженным в воду или смачиваемым брыз-
гами воды.
183
Личинки обладают червеобразной формой, на заднем конце взду-
ты, Готова резко отделена of груди, сверху, на лобном склерите, с
темными пигментными пятнами, форма и расположение которых
характерны для личинок различных видов. По бокам головы рас-
положены глазки и зачатки сложных глаз. Антенны четырех— шести-
членистые. От передней части головы ниже антенн отходят большие
веерообразные придатки, состоящие из короткого двучленистого
стержня, несущего по дистальному краю 40—80 сильно изогнутых
щетинок. Веерообразные придатки, встречающиеся только у личи-
нок мошек, морфологически соответствуют боковым лопастям верх-
ней губы.
Ротовые придатки хорошо развиты. Верхняя губа имеет вид
пластинки, покрытой волосками. Мандибулы вооружены зубцами
и несколькими густыми пучками щетинок. Максиллы широкие, с
коротким, одночленистым щупиком. Между максиллами располо-
жен гипофаринкс, несущий по переднему краю короткие волоски.
Под гипофаринксом находится нижняя губа, состоящая из трех
частей; утолщенный субментум вооружен по наружному краю ще-
тинками и зубцами.
Грудь образована слившимися между собою тремя грудными
сегментами. На нижней стороне груди находится непарный кониче-
ский вырост (ложная нога), состоящий из двух члеников, большего
основного и меньшего конечного, способного втягиваться внутрь
первого; конечный членик вооружен на конце венцом радиально
расположенных крючков. Брюшко неясно сегментированное, глад-
кое, полупрозрачное. На последнем сегменте находятся присоска и
анальные папиллы. Присоска имеет вид хитинового диска, по пери-
ферии которого расположены 60—80 радиальных рядов крючков,
направленных остриями к краю диска. Прикрепление личинки к суб-
страту происходит благодаря этим крючкам, а также присасыва-
тельному действию диска, снабженного специальной мускулатурой.
Анальные папиллы (анальные жабры), расположенные над сдви-
нутым на спинную сторону анальным отверстием, имеют вид трех
простых или разветвленных выростов, могут втягиваться внутрь, не-
сут, по всей вероятности, дыхательную и осморегуляторную функ-
ции. ;
Движения личинок очень ограничены, обычно они, прикрепив-
шись к субстрату при помощи присоски, свободно полощутся в
токе воды. При передвижении личинка сначала приклеивает к суб-
страту паутинные нити, выделяемые большими расположенными
под кишечником паутинными железами, общий проток которых от-
крывается между гипофаринксом и нижней губой. Затем, прикре-
пившись к этим нитям крючками грудной ноги, личинка освобож-
дает заднюю присоску и, изогнувшись дугообразно, переносит зад-
ний отдеп тела вперед, к месту прикрепления грудной ноги. После
того, как присоска укрепится на новом месте, личинка освобождает
передний конец тела и делает следующий «шаг» вперед.
Пиша личинок состоит из мелкого детрита и организмов планк-
тона — низших водорослей, инфузорий, мелких ракообразных и
т. п. Для улавливания пищи служат веерообразные придатки. Рас-
184
крытые веера благодаря -присутствию добавочных волосков «пред-
ставляют хороший фильтрационный аппарат. Отфильтрованные пи-
щевые частицы счищаются щетинками мандибул, когда сложенные
веера попеременно вводятся в ротовую полость. По способу полу-
чения пищи личинки мошек относятся к группе пассивных фильтра-
торов, изменения в составе пищи всецело зависят от изменений в
составе сестона.
Дыхательная система личинок апнейстического типа, все стигмы
закрыты. Дыхание совершается через покровы тела, анальные жаб-
ры служат дополнительным органом дыхания.
Куколки мошек мало подвижны, заключены в темноокрашен-
ный карманообразный кокон, прикрепленный своей нижней плоской
стороной к субстрату. Стенки кокона сплетены из паутинных нитей.
По бокам куколки между головой и грудью отходят два боль-
ших сильно разветвленных выроста, расположение и число ветвей
которых характерны для куколок различных видов. Эти выросты
являются органами дыхания — трахейными жабрами, при основа-
нии они соединены с трахейными стволами куколки. Трахейные
жабры закладываются на личиночной стадии. У личинок, готовых
к окукливанию, они видны под кутикулой по бокам грудного сег-
мента в виде свернутых пучков ветвей. На спинной стороне треть-
его и четвертого брюшных сегментов куколки находятся направлен-
ные вперед шипы; такие же шипы, но направленные назад, распо-
ложены на спинной стороне конечных сегментов брюшка; несколько
шипов находится также и на брюшной стороне куколки. Все шипы
служат для удержания куколки в коконе, из которого в естествен-
ном положении высовываются вперед только трахейные жабры.
Крылатое насекомое в момент вылупления поднимается к поверхно-
сти воды в пузырьке воздуха.
Личиночные стадии мошек относятся к числу типичных реофиль-
ных организмов, обитают в реках, ручьях, водопадах, при скорости
течения не менее 0,2 м/сек.\ никогда не встречаются в стоячей воде.
Личинки очень требовательны к содержанию кислорода, не менее
90—1(5О°/о насыщения, и не выносят даже небольшого загрязнения.
Текучая вода приносит все время новые количества кислорода и
уносит немедленно все продукты обмена веществ.
Личинки мошек в некоторых реках являются существенной ча-
стью пищи рыб.
В пределах СССР встречается большое количество видов мо-
шек, особенно в Сибири и на Дальнем Востоке. Почти все они
относятся к роду Simulium. Личинки и куколки обычных видов,
встречающихся в Европейской части СССР, среди которых наиболее
распространенным является Simulium ornatum, отличаются между
собою следующими признаками.
1 (6) Последний брюшной сегмент личинок без конических
выростов; трахейные жабры куколок содержат восемь
ветвей................................................. 2
2 (5) Трахейные жабры куколок и их зачатки у личинок имеют
вид длинных и тонких выростов 3
185
3 (4) На лобном склерите личинки находятся восемь пигмент-
ных пятен; кокон куколки сплошной, без отвер-
стий .........................................5. ornatum
4 (3) На лобном склерите личинки находится пигментное пят-
но в виде буквы Н; кокон куколки на переднем крае с
многочисленными небольшими отверстиями .
S. subornatum
5 (2) Трахейные жабры куколок и их зачатки у личинок име-
ют вид толстых и коротких трубок . . S. equtnum
6 (1) Последний брюшной сегмент личинок на нижней стороне
с двумя коническими выростами; трахейные жабры ку-
колок содержат четыре ветви . 5. аигеит
ДЫХАНИЕ АТМОСФЕРНЫМ КИСЛОРОДОМ
В качестве примеров рассматриваются дыхательные трубки ли-
чинок комаров, а также некоторых клопов и личинок мух.
Личинки и куколки Culicidae
Во взрослом состоянии комары обладают стройным, хрупким те-
лом. На голове находятся сложные глаза, длинные антенны и тон-
кий и длинный хоботок, образованный верхней губой, мандибула-
ми, максиллами, подглоточником и нижней губой; на верхней сто-
роне нижней губы имеется желобок, в который вложены все ос-
тальные части хоботка. Среднегрудь, развитая значительно сильнее,
чем переднегрудь и заднегрудь, несет два прозрачных, удлиненно
овальных крыла; жужжальца отходят от заднегруди. Ноги длинные
и тонкие. Брюшко состоит из десяти сегментов, из которых два по-
следних превращены в генитальные придатки. Самцы питаются ра-
стительными соками, самки, кроме того,—кровью животных. Многие
виды комаров относятся к числу кровососов человека (Aedes, Cu-
te* и др.), некоторые виды являются переносчиками малярии
(Anopheles), японского энцефалита (Aedes, Culex), энцефаломиэ-
лита лошадей (Aedes), туляремии (Aedes, Culex, Mansonia, Ano-
pheles) .
Комары относятся к насекомым с полным превращением, все
развитие протекает в воде. Яйца откладываются поодиночке (Ano-
pheles) или склеенными в пакеты (Culex, Theobaldia, Mansonia), на
поверхность воды или на поверхность почвы на месте временных
водоемов (Aёdes). Вышедшие из яиц личинки линяют три раза, с
каждым разом увеличиваясь в размере. Личинки четвертой возраст-
ной стадии после линьки превращаются в куколки, из которых за-
тем вылупляются крылатые насекомые.
Личинки комаров живут в самых разнообразных стоячих и сла-
бо текучих водоемах, некоторые виды выдерживают загрязнение и
значительное увеличение солености воды. Длительность личиночной
жизни зависит от температурных условий, в течение лета на севере
бывает одна-две генерации, на юге до шести и более генераций.
Тело личинки делится на голову, грудь и брюшко и несет мно-
186
гочисленные, обычно перистые волоски, которые облегчают плава-
ние, вследствие увеличения сопротивления воде, а также служат
для улавливания малейших колебаний среды; при приближении
врага личинка быстро опускается вниз или резко меняет свое поло-
жение. По бокам головы расположены небольшие глазки и хорошо
развитые сложные глаза; последние появляются обычно на третьей,
а иногда на четвертой возрастной стадии. Антенны одночленистые,
с конечными щетинками и антеннальным волоском или пучком во-
лосков.
Строение ротовых придатков соответствует способу получения
пищи. Среди личинок комаров имеются фильтраторы (личинки
Anopheles, Culex, Mansonia), соскребыватели (личинки большин-
ства Аёйез, Theobaldia) и хищники (личинки Chaoborus, Mochio-
nyx), Ротовые придатки состоят из верхней губы, надглоточника,
парных мандибул, парных максилл, нижней губы и подглоточника.
Грудные сегменты большинства личинок комаров слиты между
собою. Брюшко обычно состоит из девяти сегментов; большой вось-
мой сегмент образован слившимися между собою восьмым и девя-
тым первичными сегментами.
На верхней поверхности восьмого сегмента брюшка расположе-
ны парные стигмы, ведущие в два главных трахейных ствола. У
личинок Anopheles стигмы находятся посреди стигмальной пла-
стинки непосредственно на поверхности сегмента; у большинства
остальных личинок стйгмы, окруженные стигмальной пластинкой,
расположены на вершине длинной, склеротизированной, темно пиг-
ментированной дыхательной трубки, иначе называемой сифоном.
Большинство личинок комаров дышит атмосферным воздухом, про-
рывая поверхностную пленку воды несмачивающимися стигмальной
пластинкой или концом сифона. У личинок Chaoborus стигмы отсут-
ствуют, трахейная система замкнутая.
Десятый, анальный сегмент, отогнутый на брюшную сторону,
несет различного рода щетинки, из которых наиболее развиты ще-
тинки хвостового плавника, расположенные на нижней стороне сег-
мента. Хвостовой плавник является главным органом движения ли-
чинки. Около анального отверстия находятся две пары тонкостен-
ных мешковидных анальных папилл (анальных жабр). Ранее их
рассматривали в качестве дополнительных органов дыхания, в на-
стоящее время считают, что они проницаемы для воды и являются
органами осморегуляции. Можно думать, что анальные папиллы
несут обе функции, так как установлено, что у молодых, первых
трех возрастных стадий, личинок Anopheles они служат органами
газообмена.
Куколки комаров имеют характерную форму. Головогрудь силь-
но вздута, сквозь ее прозрачные покровы просвечиваются глаза,
ротовые придатки, крылья и ноги взрослого насекомого. На спинной
стороне головогруди находятся две имеющие вид рожков трахей-
ные трубки, открытые на концах и ведущие в трахейные стволы.
Ротового отверстия не имеется. Брюшко узкое. От восьмого брюш-
ного сегмента отходят две лопасти хвостового плавника, между их
основаниями с нижней стороны виден небольшой девятый сегмент.
187
Фаза куколки длится лишь несколько дней. Куколки легче воды
благодаря присутствию большой, наполненной воздухом полости,
расположенной в нижней части головогруди. В естественном поло-
жении куколки держатся около поверхности воды, подвешиваясь к
поверхностной пленке двумя дыхательными трубками и двумя вет-
вистыми волосками, расположенными на спинной поверхности пер-
вого брюшного сегмента. Эти волоски служат также для улавли-
вания колебаний воды. Куколки комаров в противоположность ку-
колкам других насекомых, отличаются большой подвижностью.
При помощи резких движений, вызываемых быстрыми сгибаниями
и разгибаниями брюшка, они спасаются бегством от своих много-
численных врагов.
Личинки без сифона (Anopheiini)
1. Личинки и куколки Anopheles (рис. 81—82)
Типичными представителями этой группы являются личинки ма-
лярийных комаров Anopheles, относящихся к трибе Anopheiini под-
семейства Culicinae. Взрослые малярийные комары отличаются от
других комаров по строению максиллярных (нижнечелюстных)
щупиков, отходящих от основания хоботка; у самок и самцов
Anopheles щупики длинные, не короче хоботка. Личинки окрашены
обычно в зеленоватые тона. На переднем крае головы находятся
две пары простых или ветвящихся волосков (клипеальные волос-
ки); поперечный ряд из трех пар перистых, редко простых волосков
(фронтальные волоски) проходит посреди верхней поверхности го-
ловы. Антенны короткие, палочковидные, с шипиками по всей дли-
не и с простым или перистым антеннальным волоском в прокси-
мальной части; на конце антенн расположены два длинных, два ко-
ротких шипа и один простой волосок. Грудь большая, вздутая. На
верхней поверхности передней части груди по бокам находятся
чолые прозрачные двулопастные выросты, плечевые клапаны, слу-
жащие для подвешивания личинки к поверхностной пленке воды.
На спинной стороне брюшных сегментов расположены пальмовид-
ные волоски, обычно хорошо развитые на втором—седьмом сегмен-
тах. Каждый пальмовидный (звездчатый) волосок состоит из ко-
роткого стерженька, от которого под прямым углом отходят много-
численные лепестки, на конце обычно заостренные. В раскрытом
состоянии лепестки лежат на поверхности воды, удерживаемые по-
верхностным натяжением; при погружении личинки лепестки смы-
каются своими концами, захватывая пузырек воздуха, облегчающий
при последующем подъеме личинки прохождение лепестков через
поверхностную пленку воды.
Дыхательная система личинок Anopheles метапнейстического
типа. Стигмы, расположенные на восьмом сегменте брюшка, окру-
жены пятью лопастями — одним передним, двумя боковыми и дву-
мя задними, образующими стигмальную пластинку, лежащую на
спинной поверхности сегмента. При погружении личинки лопасти
стигмальной пластинки смыкаются и вследствие своей несмачивае-
мости препятствуют проникновению воды внутрь трахей. При Подъ-
еме личинки наверх лопасти, соприкасаясь с поверхностной пленкой
воды, раскрываются и находящийся в полости сомкнутых лопастей
воздух пробивает своим мениском поверхностную пленку, благо-
даря чему устанавливается контакт между трахейной системой
«СТ
Рис. 81. Отличительные признаки Anophelini и Culicini (яйца, личинки,
куколки, взрослые насекомые).
/ — антенны, 2 — нижнечелюстные щупики, ,3 — хоботок, 4 — сифон, 5 — стигмы,
6 — пальмовидные волоски, 7 — плечевая лопасть.
и атмосферным воздухом. Стигмы ведут в два главных трахейных
ствола, заметных у живых личинок по ритмичным колебаниям, вы-
зываемым сокращениями спинного кровеносного сосуда. Стигмаль-
ная пластинка, несмачиваемая водой, смачивается нефтью, керо-
сином и иными ларвицидами. Эти вещества, применяемые для унич-
тожения личинок Anopheles, закупоривают стигмы и вызывают
удушение и отравление личинок.
1»9
На последнем брюшном сегменте с нижней стороны находится
сильно развитый хвостовой плавник, а с верхней стороны—хвосто-
вые волоски, оканчивающиеся крючками, при помощи которых ли-
чинки прикрепляются к растениям, камням и другим предметам, со-
прикасающимся с поверхностью воды. Вокруг анального отверстия
расположены две пары анальных папилл.
Удельный вес личинок Anopheles больше удельного веса воды,
они удерживаются у самой поверхностной пленки силами поверх-
ностного натяжения; органами прикрепления служат плечевые кла-
паны, пальмовидные волоски и стигмальная пластинка.
Тело личинки покрыто волосками, особенно сильно развитыми
по бокам груди и трех передних сегментов брюшка. Эти длинные
перистые волоски являются стабилизаторами, удерживающими ли-
чинку в горизонтальном положении параллельно поверхности воды.
При погружении личинки вглубь, они играют роль парашюта,
уменьшающего скорость опускания.
По способу принятия пищи личинки Anopheles относятся к
группе активных фильтраторов, непрерывно отфильтровывающих
пищевые частицы, находящиеся в самом поверхностном слое воды—
мелкий детрит, бактерии, инфузории, коловратки, мелкие водорос-
ли, споры, пыльцу и т. п. Подвесившись к поверхностной пленке
воды и повернув голову на 180°, чтобы ротовое отверстие приходи-
лось бы наверху, личинки при помощи быстрых ударов (до 200
ударов в минуту) двух пучков волосков (вееров), расположенных
на верхней губе по бокам средней лопасти, вызывают в поверхно-
стном слое токи воды, направленные к ротовому отверстию.
Пищевые частицы отфильтровываются волосками, находящи-
мися на мандибулах и максиллах, и скапливаются в ротовой поло-
сти; крупные частицы раздробляются зубцами мандибул и нижней
губы. В течение суток личинка четвертой возрастной стадии про-
фильтровывает около 20 мм3 воды. Личинки питаются без выбора,
на этом основано применение для их уничтожения ядовитых пыле-
видных инсектицидов, распыляемых по поверхности воды. Личинки
Anopheles могут, кроме того, соскребывать перифитон с подводной
растительности при помощи зазубренных волосков, находящихся
на средней лопасти верхней губы.
Куколки Anopheles по внешнему виду сходны с куколками дру-
гих комаров, отличаются от последних следующими признаками.
Дыхательные трубки на головогруди конической формы, сильно
Рис. 82. А—В— личинка Anopheles maculipennis, общий вид (Д) (последний
сегмент брюшка отогнут), голова (Б), антенна (В); Г, Д — личинка Anopheles
bifurcatus, голова (Г), антенна (Д); Е, Ж — личинка Anopheles plumbeus. го-
лова (£), антенна (Ж)\ 3 — личинка Anopheles hyrcanus, антенна; И—М—
ротовые придатки личинки Anopheles, верхняя губа (/7), мандибула (К), мак-
силла (Л), нижняя губа (М)\ Н — пальмовидный волосок; О, П — куколка
Anopheles, дыхательная трубка (О), хвостовой плавник (/7).
/ — антенна, 2— глаз, 3— голова, 4 — грудь, 5—плечевая лопасть, 6—брюшко,
7—пальмовидные волоски, 8— стигмальная пластинка, 9 — стигма, 10— аналь-
ный сегмент, 11—хвостовой плавник, 12 — хвостовые волоски, 13—анальные
папиллы, 14 — клипеальный волосок, 15—фронтальный волосок, 16— антен-
нальный волосок.
191
расширяющиеся к концу. На третьем — седьмом сегментах брюшка
в задне-боковых углах расположены шипы, а на восьмом сегмен-
те — перистые волоски. Куколки различных видов Anopheles меж-
ду собою почти ничем не отличаются.
Личиночные стадии Anopheles встречаются преимущественно в
небольших стоячих водоемах, постоянных или временных, хорошо
освещенных солнцем, с зеленой растительностью, защищенных от
ветра и волнения. К числу фитофилов, обитающих в водоемах, бо-
гатых зарослями погруженных и плавающих растений, относятся
личинки наиболее опасных для человека видов — Anopheles таси-
lipennis и Anopheles pulcherrimus. В осоковых и тростниковых бо-
лотах встречаются личинки Anopheles hyrcanus. Личинки Anophe-
les btfurcatus обитают преимущественно в водоемах родникового
питания. Некоторые личинки, как, например, личинки Anopheles
plumbeus, обитают в небольших скоплениях воды, образующихся в
дуплах бука, ясеня, граба и других деревьев.
Основными переносчиками малярии являются Anopheles таси-
lipennis, A. superpictus, A. pulcherrimus-, второстепенными перенос-
чиками — A. hyrcanus, A. bifurcaius, A. plumbeus-, не являются у нас
переносчиками—A. algeriensis, A. lindesayi, A. marteri.
Наиболее распространенным видом является Anopheles таси-
lipennis (с пятью подвидами), встречающийся в Европейской части
СССР до Кольского полуострова и Белого моря, в Сибири до
таежной зоны, на Кавказе и в Средней Азии. Anopheles btfurca-
tus встречается в Европейской части СССР, Западной Сибири, на
Кавказе и в Средней Азии; Anopheles plumbeus — на Украине, Кав-
казе, в Туркмении и Таджикистане; Anopheles hyrcanus на Украи-
не, Кавказе, в Средней Азии, Алтае и на Дальнем Востоке; Ano-
pheles superpictus, A. pulcherrimus и A. algeriensis—на Кавказе и в
Средней Азии; Anopheles lindesayi и A. marteri — в Таджикистане.
Личинки Anopheles, встречающиеся в Европейской части СССР,
отличаются между собой следующими признаками.
1 (6) Фронтальные волоски длинные и перистые ... 2
2 (5) Антеннальный волосок короткий, отходит значительно
ниже середины антенны................................... 3
3 (4) Боковые клипеальные волоски древовидно разветвлен-
ные; антеннальный волосок отходит на расстоянии ’/а
длины антенны от основания ... A. maculipennis
4 (3) Боковые клипеальные волоски простые или слабо развет-
вленные; антеннальный волосок отходит от самого осно-
вания антенны...............................A. bifurcatus
5 (2) Антеннальный волосок длинный, отходит приблизительно
от середины антенны .... A. hyrcanus
6 (1) Фронтальные волоски короткие и простые A. plumbeus
Личинки с сифоном (Culicini)
К этой группе относятся личинки комаров, объединяемых в
трибу Culicini подсемейства Culicinae. Взрослые комары отлича-
ются (см. рис. 81) от малярийных комаров по строению максилляр-
!Р2
ных (нижнечелюстных) щупиков, отходящих от основания хобот-
ка, у самок Culicini щупики короткие, в несколько раз короче хо-
ботка, у самцов они длинные. К числу Culicini относятся наши са-
мые обыкновенные кровососущие комары.
Личинки Culicini отличаются от личинок Anophelini присут-
ствием длинного сифона (дыхательной трубки) на восьмом сегмен-
те брюшка. На конце сифона расположены стигмы на стигмальной
пластинке, образованной пятью клапанами—одним передним,
двумя боковыми и двумя задними. При опускании личинки вглубь
воды клапаны смыкаются, при подъеме ее к поверхностной плен-
ке клапаны раскрываются, благодаря этому устанавливается кон-
такт трахейной системы с атмосферным воздухом.
2. Личинки и куколки Culex (рис. 83)
Личинки Culex, прозрачные или окрашенные в желтовато-ко-
ричневые или зеленоватые тона, в естественном положении подве-
шиваются к поверхностной пленке воды своим длинным сифоном
и висят в воде под некоторым углом к поверхности. Все тело ли-
чинки покрыто многочисленными волосками, особенно хорошо раз-
витыми на груди; на верхней поверхности головы расположены
длинные, обычно разветвленные фронтальные волоски. Антенны от-
носительно длинные, с большим и хорошо развитым антеннальным
пучком волосков, отходящим выше середины антенны, и с длинны-
ми щетинками на конце. Грудь большая, вздутая. Брюшко относи-
тельно узкое. На спинной стороне восьмого брюшного сегмента
находится длинный сифон (дыхательная трубка), длина которого
превышает ширину у основания обычно в четыре-восемь раз.
На боковых и задней сторонах или только на одной задней стороне
сифона находится несколько пар (от четырех до восьми пар и бо-
лее) сифональных пучков. По этому признаку личинки Culex лег-
ко отличаются от сходных по внешнему виду личинок Aerfes, у ко-
торых имеется только одна пара сифональных пучков. По бокам
сифона, в его нижней части, проходят боковые гребни из неболь-
ших острых зубцов. На каждой стороне восьмого брюшного сегмен-
та, под основанием сифона, расположены так называемые щетки,
состоящие из многочисленных мелких чешуек; сверху и позади ще-
ток находятся длинные волоски. Анальный сегмент на спинной сто-
роне несет хитиновую пластинку (седло), придающую ему необхо-
димую при движении прочность; наверху анального сегмента на-
ходятся длинные хвостовые волоски, а внизу хорошо развитый хво-
стовой плавник; около анального отверстия расположены две пары
анальных папилл.
Личинки Culex питаются мелкими организмами планктона. То-
ки воды вызываются движениями двух широко расставленных пуч-
ков волосков (вееров), расположенных по краю верхней губы. Каж-
дый веер состоит из наружных волосков, более длинных и направ-
ленных вбок, и внутренних волосков, сравнительно коротких и на-
правленных вперед и внутрь. Находящаяся между двумя веерами
средняя лопасть верхней губы имеет вид широкого полукруга. При-
.4 193
Рис. 83. Л — личинка Culex pipiens, конец тела; Б — личинка Culex modestus, сифон; В — личинка Culex theileri,
сифон; Г — личинки Culex apicalis, сифон; Д — личинка Culex hortensis, сифон; Е — куколка Culex.
1 — сифон, 2 — сифональные пучки волосков, 3 —гребень, 4 — щетка, 5 —анальный сегмент, 6 — анальные
папиллы, 7 — хвостовые волоски, 8 — хвостовой плавник.
несенные током воды мелкие организмы планктона и детрит от-
фильтровываются при помощи длинных волосков мандибул и мак-
силл.
Куколки Culex отличаются от куколок Anopheles рядом призна-
ков. Дыхательные трубки цилиндрической формы. На брюшных сег-
ментах в задне-боковых углах имеются лишь одни ветвистые воло-
ски. Куколки различных видов между собою почти не отличаются.
Личинки Culex живут в самых разнообразных стоячих, обычно
небольших водоемах; некоторые виды только в чистой, другие в
той или иной мере загрязненной воде; часто встречаются в неболь-
ших лужах, канавах, бочках с водою и других временных водое-
мах.
Личинки наиболее часто встречающихся у нас видов Culex, из
которых наиболее обычным является Culex pipiens, отличаются ме-
жду собою следующими признаками.
1 (2) Сифоналъные пучки явственно расположены на боковых
сторонах сифона.............................С. pipiens
2 (1) Сифональные пучки (кроме самых верхних) располо-
жены на задней стороне сифона, зигзагообразно или в
один ряд ............................................ 3
3 (6) Длина сифона превышает его ширину у основания самое
большее в шесть раз...................................4
4 (5) Нижний сифональный пучок расположен ниже верхнего
зубца гребня сифона...........................С. modestus
5 (4) Нижний сифональный пучок расположен выше верхнего
зубца гребня сифона.........................С. theileri
6 (3) Длина сифона превышает его ширину у основания не
менее, чем в восемь раз...............................7
7 (8) Нижний сифональный пучок расположен ниже верхнего
зубца гребня сифона...........................С. hortensis
8 (7) Нижний сифональный пучок расположен выше верхнего
зубца гребня сифона С. apicalis
3. Личинки и куколки Aedes (рис. 84)
Личинки Aedes по внешнему виду более или менее сходны с
личинками Culex. Сифон относительно короткий, его длина превы-
шает ширину у основания не более, чем в три-четыре раза. На си-
фоне имеется только одна пара сифональных пучков, располо-
женных приблизительно посреди его заднего края (у некоторых
видов, кроме того, имеется несколько волосков на передней сто-
роне сифона). В нижней части сифона с каждой стороны проходит
по боковому гребню, образованному продольным рядом небольших
острых зубцов, вооруженных в свою очередь дополнительными
зубчиками. По бокам восьмого брюшного сегмента, под основанием
сифона, расположены щетки, состоящие из заостренных и направ-
ленных назад небольших чешуек. Сверху и позади щеток находятся
длинные, ветвящиеся или перистые волоски. Строение, число и рас-
положение чешуек в щетках, а также строение зубцов сифональ-
ных гребней характерны для отдельных видов, поэтому имеют боль-
шое таксономическое значение.
195
Рис. 84. А—В — личинка Aedes cinereus, общий вид (А) (два последних сегм . я
отогнуты), голова (Б), конец тела (#); Г — личинка Aedes geniculatus, cl
и восьмой брюшной сегмент; Д—личинка Aedes cataphylla, сифон; ^..ТлИЧ«сГи-
Aedes maculatus, сифон и восьмой брюшной сегмент; Ж—личинка Aedes ® jeS
cians, конец тела; 3—личинка Aёdes caspius, конец тела; И-—личинка А
communis, сифон и восьмой брюшной сегмент; К — личинка Aedes puncto »
196
По способу получения пищи личинки многих видов Aedes отно-
сятся к группе соскребывателей. Верхняя губа несет по бокам два
сближенных между собою веера; каждый веер состоит из двух
сортов волосков, наружные волоски тонкие, внутренние более тол-
стые и на конце зазубренные; при помощи внутренних волосков
личинки соскребывают пищевые частицы с поверхности водных ра-
стений или их остатков. Личинки Aedes обычно питаются на дне
водоема, для дыхания периодически поднимаются на поверхность
воды.
Куколки Aedes по внешнему виду сходны с куколками других
комаров.
Личиночные стадии Aedes жийут в небольших временных водое-
мах — в канавах, лужах, образующихся после дождя или таяния
снега, в пойменных водоемах с травою и опавшей листвой, в не-
больших скоплениях воды в дуплах деревьев или под их корнями;
некоторые виды выдерживают повышенную соленость воды. Кома-
ры Aedes являются обычными нападающими на человека кровосо-
сами.
Род Aedes содержит большое число видов. Личинки наиболее
обычных видов отличаются между собою следующими признаками.
1 (2) Фронтальные волоски расположены в один вогнутый
назад дугообразный ряд; анальные жабры лишь нем-
ного короче сифона...........................A. cinereus
2 (1) Фронтальные волоски на каждой стороне располо-
жены треугольником, средние волоски впереди внут-
ренних ........................................ 3
3 (4) Антенны без шипиков .... A. geniculatus
4 (3) Антенны покрыты шипиками, иногда слабо развитыми 5
5 (6) Дистальные зубцы бокового гребня заходят за основа-
ние сифонального пучка .... A. cataphylla
6 (5) Указанные зубцы не заходят за основание сифональ-
ного пучка............................... .. .. 7
7 (10) Перед хвостовым плавником находится четыре-шесть
отдельных коротких пучков, не связанных общим осно-
ванием; сифональный пучок отходит от середины
сифона..................................................8
8 (9) Боковые гребни содержат по 24—27 тесно сидящих
<зубцов................................A. maculatus
9 (8) Боковые гребни содержат по 17—25 черных зубцов,
расставленных на дистальном конце гребня
A. excrucians
10 (7) Перед хвостовым плавником находится не более трех
отдельных коротких пучков (если четыре, то сифональ-
ный пучок отходит выше середины сифона) 11
фон и восьмой брюшной сегмент; Л — личинка Аёдея vexans, сифон и восьмой
брюшной сегмент.
/— сифон, 2—сифональный пучок волосков, 3 — гребень, 4.— щетка, 5 —
анальный сегмент, 6 — анальные папиллы, 7 — хвостовые волоски, 8 — хвосто-
вой плавник.
197
11 (12) Анальные жабры короче девятого сегмента брюш-
ка ...........................................A. caspius
12 (11) Анальные жабры длиннее указанного сегмента . 13
13 (14) В щетках находится по 40—80 чешуек A. communis
14 (13) В щетках находится по 10—15 чешуек . . 15
15 (16) Сифональный пучок расположен посреди си-
фона .........................................A. punctor
16 (15) Сифональный пучок расположен значительно выше се-
редины сифона . A. vexans
4. Личинки и куколки Theobaldia (рис. 85)
Крупные личинки Theobaldia окрашены в темные и коричневые
цвета. Ротовые придатки приспособлены для соскребывания пище-
вых частиц с субстрата. Имеется только одна пара сифональных
пучков, расположенных у основания сифона. По этому признаку
личинки Theobaldia легко отличаются от личинок других комаров.
Личинки обитают в небольших стоячих водоемах — ямах, ко-
лодцах, канавах, в бочках и цистернах; выносят значительное за-
грязнение воды.
Личинки наиболее обычных видов Theobaldia отличаются меж-
ду собою следующими признаками:
1 (6) Длина сифона превышает ширину у основания не более,
чем в четыре раза.......................................2
2 (3) Боковые гребни состоят из четырех-семи зубцов, рас-
ставленных по всей длине сифона . Th. longiareolata
3 (2) Боковые гребни в дистальной части образованы тонкими
волосками.............................................. 4
4 (5) Чешуйки в щетках узкие, с параллельными краями
Th. alaskaensis
5 (4) Чешуйки щеток по середине суженные . Th. annulata
6 (1) Длина сифона превышает ширину у основания не менее,
чем в пять раз...........................................7
7 (8) Отдельные пучки перед хвостовым плавником постепенно
увеличиваются в длину . Th. morsitans
8 (7) Указанные пучки короткие Th. fumipennis
5. Личинки и куколки Mansonia (рис. 86)
Личинки Mansonia, слабоокрашенные, желтоватого или молоч-
но-белого цвета, по строению и образу жизни отличаются от всех
остальных личинок комаров. Голова большая, глаза развиты сла-
бо. Антенны длинные, с длинным дистальным отделом. Ротовые
придатки приспособлены к питанию организмами планктона. В зад-
ней части груди находятся два трахейных пузыря, являющихся
концевыми расширениями трахейных ветвей, отходящих от главных
трахейных стволов. Сифон характерного строения, имеет вид усе-
ченного конуса, сильно хитинизированные клапаны превращены в
буравящий аппарат. Хвостовой плавник хорошо развит.
198
Личинки Mansonia никогда не поднимаются на поверхность
воды, сидят у дна на стеблях водных растений — аира, ежеголов-
ки, манника, рогоза, плавающего рдеста, водяного лютика и др.
Рис. 85. А—личинка Theobaldia alaskaensis, конец тела; Б—Г — личинка Theo-
baldia annulata, сифон (Б), чешуйка щетки (В), верхняя губа (Г); Д — ли-
чинка Theobaldia lon^iareolata, сифон; Е — личинка Theobaldia fumipennia,
сифон.
/ — сифон, 2 — сифональный пучок волоеков, 3 — гребень, 4 — щетка, 5 — аналь-
ный сегмент, 6—анальные папиллы, 7—хвостовые волоски, 8 — хвостовой
плавник.
199
I
Рис. 86. Л—В — личинка Mansonia richiardi, сверху (А), на стебле PalCwarlsOnia’
сифон (В); Г —трахейные пузыри личинки Mansonia; Д — куколка
передний кенещ ма.,
/—антенна, 2 — анальный сегмент, 3^ сифон, 4— трахейная сист
200
Дышат воздухом, находящимся в полостях растений. При помощи
буравящего аппарата личинки прорезают ткани растения и погру-
жают стигмы в воздухоносные полости. При плавании держатся
горизонтально благодаря присутствию трахейных пузырей.
Куколки отличаются от куколок других комаров присутствием
очень длинных дыхательных трубок, заостренных на конце. Дышат
воздухом воздухоносных полостей водных растений, вбуравливаясь
в их ткани концами дыхательных трубок. При наступлении вре-
мени вылупления взрослого насекомого концы дыхательных трубок
обламываются и куколка всплывает на поверхность воды. Комары
относятся к числу кровососов.
Представители рода Mansonia распространены в тропической и
субтропической областях. У нас встречается единственный вид
Mansonia richiardi. Личинки обитают в богатых макрофитами во-
доемах, на небольшой глубине.
Водные Hemiptera
1. Nepa (рис. 87, А—Б)
Тело уплощенное, темноокрашенное. На небольшой голове на-
ходятся глаза и крепкий хоботок. Сильно развитые ноги первой
пары служат для схватывания добычи; тазик (соха) большой, на-
правленный вперед; вертлуг (trochanter) узкий; бедро (femur)
длинное, уплощенное, с желобком по внутреннему краю и высту-
пом, играющим роль защелки, в базальной части; голень (tibia)
длинная и узкая; лапка (tarsus) одночленистая; в сложенном виде
голень и лапка входят в желобок бедра. Ноги второй и третьей пар
ходильные. Брюшко состоит из восьми сегментов. Последний сег-
мент короткий, преобразован в генитальные придатки. Седьмой
сегмент образует длинную и тонкую дыхательную трубку, состоя-
щую из двух половинок; внутренняя поверхность дыхательной
трубки водою не смачивается. Раздвинув половинки дыхательной
трубки препаровальной иглой, можно увидеть стигмы седьмой па-
ры; стигмы первых шести пар не функционируют. Дыхательная
трубка личинок водяного скорпиона короткая, имеет вид желобча-
того отростка; этот желобок переходит затем в два желобка, про-
ходящих по нижней стороне брюшка к брюшным и грудным стиг-
мам. Все желобки покрыты волосками.
Водяной скорпион (Nepa cinerea) широко распространен в раз-
личных водоемах.
2. Ranatra (рис. 87, В—Г)
Тело узкое, сильно вытянутое. Глаза большие; антенны корот-
кие, трехчленистые; хоботок трехчленистый. Ноги первой пары,
прикрепленные у переднего края удлиненного первого грудного
сегмента, по своему строению сходны с соответствующими ногами
водяного скорпиона; тазик узкий и длинный, бедро с желобком в
дистальной части; голень относительно короткая; лапка одночлени-
201
стая. Дыхательная трубка длинная,
так же устроена, как у водЯ-
него скорпиона.
сверху (Л), нога первой пары (В); В, Г — Rana
Рис. 87. А, Б — Nepa cinerea,
linearis, сверху (В), нога первой пары (Г).
/ — тазик, 2 —вертлуг, 3 —бедро, 4 — голень, 5 — лапка, 6 — дыхатель
трубка.
202
Ранатра (Ranatra /means) встречается преимущественно в
центральных и южных районах, на севере — редко.
Личинки Brachycera
Многие личинки мух дышат атмосферным воздухом. Стигмы от-
крываются на последнем сегменте брюшка или на конце дыхатель-
ной трубки.
Описание строения личинок Eristalis, отличающихся присут-
ствием длинной дыхательной трубки, помещено на стр. 123.
ОРГАНИЗМЫ И ТЕМПЕРАТУРА
Температура воды в морях колеблется от —3° до +36°, в кон-
тинентальных водоемах от 0° до +45®, а в горячих источниках
почти достигает точки кипения воды. Температурный диапозон для
водных организмов, если исключить сравнительно малочисленную
группу видов, живущих в горячих источниках, не превышает таким
образом 45° — 50°, т. е. он значительно меньше, чем в воздушной
среде. В зависимости от распределения температуры верхних сло-
ев воды, простирающихся от поверхности до глубины в среднем
200 м, в Мировом океане различают пять основных температурных
областей — арктическую, бореальную, тропическую, антибореаль-
ную (потальную) и антарктическую. Вся масса воды, лежащая
ниже 1000 м, на всем протяжении от приполярных районов до эк-
ватора имеет одну и ту же температуру, около 2°—5°, и только в
арктических и антарктических областях температура воды глубин-
ных слоев опускается ниже ноля градусов (вода с отрицательной
температурой). Континентальные водоемы, в зависимости от темпе-
ратуры воды, разделяются на арктические, умеренные и тропиче-
ские.
Организмы, не выносящие больших колебаний температуры,
другими словами — живущие в узких температурных границах, на-
зываются стенотермными организмами. В зависимости от приурочен-
ности к определенной части температурной шкалы, стенотермные
организмы делятся на тепловодных и холодноводных. К числу мор-
ских тепловодных организмов относится большое количество ви-
дов животных и растений, живущих в тропической области. Холод-
новодные стенотермные организмы, обитающие нередко только в
воде с отрицательной температурой, свойственны арктической и,
соответственно, антарктической областям. Пресноводные холодно-
водные организмы встречаются на севере или в высокогорных во-
доемах, а в умеренных широтах — в глубоких слоях воды с низкой
температурой.
Те организмы, которые могут существовать при сравнительно
больших колебаниях температуры, образуют группу эвритермных
организмов. Наиболее обычными представителями этих видов в мо-
рях являются литоральные животные и растения, которые во вре-
204
мя отлива могут подвергаться сильной инсоляции. К этой группе
относится также большое количество пресноводных организмов.
Амплитуда годовых колебаний температуры воды в морях срав-
нительно невелика, в среднем не превышает десяти градусов, в кон-
тинентальных водоемах умеренных широт достигает большей вели-
чины — от ноля градусов зимою до тридцати и более градусов ле-
том. С годовыми изменениями температуры воды связаны сезонные
изменения в составе и распределении планктона и бентоса, сроки
размножения и цикличность организмов, а косвенным образом
также явление цикломорфоза, или сезонной изменчивости.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
СТЕНОТЕРМНЫЕ ОРГАНИЗМЫ
Стенотермные тепловодные организмы
В качестве примеров рассматриваются представители тропиче-
ской флоры и фауны: из синезеленых водорослей — Trichodesmium;
из сифонофор—Velella, Porpita; из альционарий — Tubipora; из
мадрепоровых кораллов — Madrepora, Fungia, Maeandra и другие
виды; из многошетинковых червей — Eunice (палоло); из весло-
ногих раков — Corycaeus, Copilia, Sapphirina; из мечехвостов —
Limulus; из двустворчатых моллюсков — Tridacna; из крылоногих
моллюсков — Creseis; из брюхоногих моллюсков — Janthina; из
киленогих моллюсков — Carinaria, Pterotrachea; из голожаберных
моллюсков — Phyllirrhoe; из головоногих моллюсков — Nautilus:
из оболочников — Salpa, Doliolum, Pyrosoma; из рыб — Exocoetus
(летучая рыба); Chaetodon, Zanclus, Ostracion, Scarus и другие
виды.
Стенотермные холодноводные организмы
В качестве примеров рассматриваются представители арктиче-
ской фауны наших северных морей: из гидромедуз — Sarsia prin-
ceps, Aeginopsis laurentii; из веслоногих раков — Calanus hyperbo-
reus; из амфипод — Themisto llbellulafAcanthostepheia malmgreni;
из равноногих раков — Mesidothea sibirica; из двустворчатых
моллюсков — Portlandia arctica; из крылоногих моллюсков —
Limacina helicina; из иглокожих—Urasterias UncKi, Icasterias
panopla, Stegophiura nodosa; из аппендикулярий — OiKopleura
vanhoffeni; из рыб— Gadus saida (сайка), Mallotus villosfus
(мойва).
ЭВРИТЕРМНЫЕ ОРГАНИЗМЫ
Эвритермные организмы специально не рассматриваются, так как
наиболее обычные представители этой группы рассматривались на
предыдущих занятиях.
СЕЗОННЫЕ ИЗМЕНЕНИЯ ПЛАНКТОНА
Массовое развитие организмов, протекающее в связи с годо-
выми изменениями температуры воды, вызывает явление цветения
водоемов. В зависимости от состава населения, вода окрашивается
«5
в различные цвета: от зеленых водорослей Eudorina, Pandorina н
Volvox — в зеленый цвет; от диатомей Asterionella, Tabellaria и
Fragilaria — в желтовато-бурый цвет; от жгутиковых Dinobryon —
в зеленоватый, Euglena — в зеленый, Synura — в коричневый,
Trachelomonas — в желто-коричневый цвета; от перидиней Cera-
tium — в желто-бурый цвет.
Рассматриваются пробы свежесобранного планктона, собран-
ные в периоды цветения воды.
ЦИКЛОМОРФОЗ
Цикломорфоз (или сезонная изменчивость) наиболее резко вы-
ражен у пресноводных планктических организмов, относящихся к
диатомеям (Asterionella, Tabellaria), перидинеям (Ceratium), коло-
враткам (Keratella, Brachionus и другие виды) и ветвистоусым ра-
кам (пелагические Daphnia и Bosmina). Это явление, причины ко-
торого еще не вполне ясны, наблюдается у организмов, имеющих
в течение года большое число генераций, получающихся в резуль-
тате деления клеток у водорослей или партеногенетического размно-
жения у животных; у литоральных организмов, а из пелагических
у тех, которые имеют только одно или два поколения в течение
года, цикломорфоз не проявляется. Наблюдая в естественных водое-
мах развитие организмов в течение года, можно видеть, как изме-
няется форма тела особей, последовательно друг за другом идущих
генераций, причем нередко эти изменения настолько резко прояв-
ляются, что, не зная генетической связи отдельных форм, легко
счесть их за самостоятельные виды. После изучения сезонной из-
менчивости пелагических коловраток и ветвистоусых раков много-
численные прежние «виды» были сведены на степень морф очень
небольшого количества видов.
1. Цикломорфоз диатомей (рис. 88, А—Г)
Рассматриваются типичные планктические диатомеи Asterio-
nella gracillima, колонии которой зимою содержат четыре-пять
коротких клеток, а летом 12—18 клеток, отличающихся большей
длиной, и Tabellaria fenestrata, с цепными колониями зимою и
звездчатыми летом.
2. Цикломорфоз перидиней (рис. 88, Д—Е)
Рассматриваются различные формы Ceratium hirundinella, об-
ладающие в холодное время года двумя антапикальными рогами
(трехрогая форма), а летом тремя антапикальными рогами (че-
тырехрогая форма).
20в
3. Цикломорфоз Keratella cochlearis (рис. 89)
Keratella cochlearis (=Anuraea cochlearis) относятся к самым
обычным представителям пресноводного планктона. Обладают
крепким панцырем,, выпуклым на спинной стороне и у типичных
Рис. 88. Цикломорфоз Diatomeae и Peridineae. А, Б — Asterionelia gracillima,
зимняя (Л) и летняя (Б) колонии; В, Г —Tabellaria fenestrata var. interme-
dia (В) и var. asterionelloides (Г); Д, Е — Ceratium hirundinella, зимняя (Д) и
летняя (В) формы.
форм снабженным срединным продольным килем, по бокам кото-
рого находятся угловатые пластинки; у других форм панцырь по-
крыт шипиками. Передний край панцыря с шестью шипами, задний
край с одним срединным шипом, варьирующим по длине.
207
„ rrnrantha. отличающаяся присутствием
Основная форма, f. ^сгй^не равного панцырю, встречает^
длинного заднего шипа, по ДЛ параллельных ряда изменчи.
зимою; летом можно обнаружу V формой.
вости начинающихся этой основн
J
Рис. 89. Цикломорфоз Keratella cochlearis. А — f. macracantha; 5 —
В — f. micracantha; Г — f. tecta; Д — f. connectens; E — f. angulifera;
f. ecaudata; 3 — f. pustulata; И — f. hispida; К — f. hispida-tect*-
МЯ
Ряд tecta проходит черев следующие формы — f. typlca, с зад-
ним шипом умеренной длины, f. micracantha, с почти редуцирован-
ным задним шипом, и f. tecta, совершенно лишенную заднего шипа.
Все формы этого рода имеют симметрично построенный пап-,
цырь, с прямым средним килем; шипики отсутствуют.
Ряд irregularis по характеру изменений совпадает с предыду-
щим рядом, проходит через f. connectens, с задним шипом умерен-
ной длины, f. angulifera, с коротким задним шипом, и f. ecaudata,
лишенную заднего шипа. Отличительной чертой форм этого ряда
является присутствие не прямого, а изогнутого срединного киля на
панцыре, вследствие этого пластинки теряют симметричное распо-
ложение; шипики слабо развиты.
Формы ряда hispida отличаются по присутствию на панцыре
шипиков и по слабому развитию или даже отсутствию срединного
киля и границ между пластинками; у f. pustulata пластинки еще
заметны, задний шип умеренной длины, у f. hispida срединный
киль отсутствует, задний шип короткий, у f. hispida-tecta задний
шип полностью отсутствует.
4. Цикломорфоз Keratella quadrata (рис. 90)
Keratella quadrata (=Anuraea aculeata) отличается от Kera-
tella cochlearis присутствием на заднем конце панцыря двух боковых
шипов, а также тем, что вместо
киля на верхней стороне пан-
цыря располагается срединный
ряд из четырех пластинок.
Сезонная изменчивость Ke-
ratella quadrata в различных
водоемах сильно видоизме-
няется. В схеме из зимнего
яйца выходит f. typlca, отли-
чающаяся сильным развитием
задних шипов. Через ряд пар
теногенетических поколений из
этой формы образуются f. bre-
vispina, с короткими задними
шипами, и f. valga, с задними
шипами различной длины
(f. heterospina) или только х
одним задним шипом (f. mo-
nospina) ; заканчивается ряд
задних шипов и отличающейся
Рис. 90. Цикломорфоз Keratella quadrata
А — f. typica; Б—f. valga (heterospina);
В — f. brevispina; Г — f. curvicornis.
f. curvicornis, лишенной полностью
от Keratella cochlearis f. tecta лишь
иным строением панцыря.
5. Цикломорфоз Bra-chionus ealyciflorus (рис. 91)
Широко распространенные в прудах Brachionus ealyciflorus
(=Brachionus pala) отличаются присутствием крепкого панцыря,
14 В. А. Яшнов 109
вооруженного на переднем крае четырьмя хороши развитыми Ши-
вами; задний конец панцыря с длинными шипами или без шипов;
шеется длинная, цилиндрическая и кольчатая нога, оканчиваю-
щаяся двумя пальцами (у фиксированных экземпляров нога часто
втянута внутрь панцыря).
Во время цикломорфоза от исходной L pala, совершенно лишен-
ной задних боковых шипов и с очень слабо развитыми задними
Рис. 91. Цикломорфоз Brachionus. calycii- Рис. 92. Цикломорфоз Brachionus
floras A— f. pala; Б — anuraeifc\rmi^; capsuliflorus. A — f. typica; Б —
В — f. amphiceros; Г — f. dbroas;. f. brevispinus; В — f. entzi; Г —
Д — f. spinosa. / \ , f. cluniorbicularis.
' x V
срединными шипами, расположенными около отверстия для ноги,
образуются сначала f. anuraeiforhzis, с короткими задними шипами,
а затем f. amphiceros, с длинными срединными и особенно боковы-
ми задними шипами. У форм ряда amphiceros все передние шипы
приблизительно одного размера.
Формы ряда spinosa характеризуются сильным развитием на
переднем крае панцыря срединных шипов, превышающих по вели-
чине боковые шипы; f1. dorcas лишена боковых задних шипов, У
f. spinosa боковые шипы имеются (иногда только один из них);
f. spinosa сходна с f. amphiceros, отличается от нее сильным разви-
тием передних срединных шипов.
6. Цикломорфоз Brachionus capsuliflorus (рис. 92)
Широко распространенные в прудах Brachionus capsuliflorus
(=Brachionus baiceri) отличаются от Brachionus calyciflorus прИ'
сутствием на переднем крае панцыря шести шипов, из которых
срединные развиты сильнее остальных, а также трубчатого выр0'
210
ста вокруг отверстия для ноги; задний конец панцыря с боковыми
шипами или без шипов.
Задние боковые шипы исходной формы f. typica хорошо разви-
ты, длинные и тонкие; у f. brevispinus они короткие, а у f. entzi
сильно редуцированы, так что задний край панцыря становится
прямым или слабо вогнутым; последняя в этом ряду f. cluniorbicu-
larts отличается закругленным задним краем панцыря, лишенным
боковых шипов. Между указанными формами можно найти ряд пе-
реходов, кроме того, имеются и другие формы.
7. Цикломорфоз Daphnia cucullata (рис. 93)
Обычным видом дафний в планктоне озер является Daphnia
cucullata, называвшаяся ранее по причине большой прозрачности
тела Hyalodaphnia cucullata. Относится к моноцикличным видам,
Рис. 93. Верхний ряд — цикломорфоз Daphnia cucullata (цифры обозначают
месяцы); нижний ряд—летние формы различных вариететов Daphnia cucullata:
А — var. apicata; Б — var. berolinensis; В—var. Kahlbergiensis; Г — var.
procurva.
в течение всего теплого времени года размножается партеногене-
тпчески и только осенью переходит к обоеполому размножению.
Цикл заканчивается отложением латентных яиц. Пертеногенез у
популяций, живущих в больших озерах, продолжается и в зимний
период.
Рассматривая следующие друг за другом генерации, легко об-
наружить большие изменения в их внешнем виде, совершающиеся с
1 4* 211
течением времени. Зимние формы, обладающие небольшой головой
ион наступлении весеннего потепления, когда температура воды в
течение короткого времени, измеряемого приблизительно тремя не-
тями повышается до 12-16°, дают новое поколение, отличающе-
еся ОТ материнского сильным развитием передней части, головы,
образующей так называемый шлем. Иногда в выводковой камере
ir Vi w чя а и
Рис. 94. Цикломорфоз Daphnia cristata (цифры обозначают месяцы).
зимней формы, пойманной в это время, могут быть обнаружены
эмбрионы с заостренными шлемами. При дальнейшем развитии по-
пуляции образуются типичные летние генерации, характеризую-
щиеся сильным развитием шлема, в несколько раз удлиняющего
голову, и сохраняющие свои признаки в течение всего лета. С при-
ближением холодного времени года изменения идут в обратном
порядке и, в частности, появляющиеся осенью эмбрионы обнаружи-
вают отсутствие шлема.
Daphnia cucullata распадается на ряд вариететов, отличающихся
степенью развития шлема у особей летних генераций, в зимние ме-
сяцы вариететы не различимы. В небольших озерах и прудах
встречаются Daphnia cucullata apicata—голова с небольшим шле-
мом, на верхнем конце округлая; глаз расположен ближе к вершине
шлема, чем к основанию головы. В неглубоких озерах обитает
D. cucullata berolinensis — голова с низким шлемом, на верхнем
конце заостренным; глаз расположен на равном расстоянии от
вершины шлема и основания головы. В глубоких и больших озерах
живет D. cucullata xahlbergiensis — голова с высоким прямым
шлемом, на конце заостренным; расстояние от глаза до вершины
шлема в несколько раз больше расстояния до основания головы.
В некоторых озерах можно обнаружить D. cucullata procurva —
голова с большим шлемом, на конце загнутым книзу приблизитель-
но под прямым углом.)
8. Цикломорфоз Daphnia cristata (рис. 94)
Встречающиеся в больших озерах Daphnia cristata отличаются
присутствием на антеннах восьми плавательных щетинок, тогда
как у остальных видов дафний на антеннах имеется девять щетй'
нок.
212
Зимние формы почти не отличимы от зимних форм других даф-
ний, голова не образует шлема. Летние формы характеризуются
развитием широкого шлема, направленного вершиной на спинную
сторону организма (Daphnia cristata cederstroemi). Между обеими
крайними формами имеется ряд переходов.
tv w vn а я
Рис. 95. Цикломорфоз Bosmina coregoni (цифры обозначают месяцы).
9. Цикломорфоз Bosmina coregoni (рис. 95)
Bosmina coregoni распадаются на большое число различных
форм, отличающихся разнообразием внешнего вида и ходом цикло-
морфоза; у некоторых форм сезонные изменения слабо развиты, у
других выражены сильно. При цикломорфозе ряда longispina лет-
ние формы отличаются от зимних меньшей длиной антеннул; в ряду
coregoni, представители которого всего чаще встречаются в наших
озерах, зимние формы обладают короткими антеннулами и срав-
нительно низкой высотой тела; у крайних летних форм антеннулы
становятся длинными, увеличивается также высота тела.
ОРГАНИЗМЫ И СВЕТ
Количество проникающего в воду света уменьшается с глуби-
ной В морях различают световую область, от поверхности воды
до глубины, в среднем, 30—80 м, полусветовую область, от нижней
границы световой до глубины, приблизительно, 300 500 м, и бес-
световую область, от этой границы до дна. Глубина проникновения
света в воду зависит от широты места, времени года и суток, ме-
теорологических факторов, а также от прозрачности воды. В при-
брежных районах моря, с менее прозрачной водой, все границы
сдвинуты кверху; еще выше они лежат в пресноводных водоемах;
в некоторых из них мощность освещенного слоя не превышает
нескольких метров, а в мутных реках измеряется лишь деци-
метрами.
Волны различной длины абсорбируются водою не одинаково.
Наиболее интенсивно поглощаются крайние части светового спек-
тра — инфракрасные и красные лучи, с одной стороны, и фиолето-
вые, с другой стороны, причем красная часть спектра поглощается
полностью самыми поверхностными слоями воды. На глубину ниже
100 м в морях и 10—20 м в пресноводных, водоемах проникают
только синие и зеленые лучи.
Все автотрофные растения могут жить, естественно, только в
верхних, освещенных слоях воды. В морях граница массового раз-
вития водорослей не опускается в среднем ниже ста метров; менее
требовательные к свету растения в тропической области могут су-
ществовать до глубины 400 м; -в пресноводных водоемах мощность
слоя, в котором развиваются водоросли, не превышает нескольких
десятков метров. В глубоких, неосвещенных слоях морей и пресно-
водных водоемов обитают только животные и бактерии.
Не менее велико значение света для животных. Многие орга-
низмы зоопланктона морей и пресноводных водоемов, находящиеся
днем на определенной более или менее значительной глубине, вече-
ром поднимаются в вышележащие слои, а затем, пробыв в этих
слоях некоторое время, опускаются вниз на ту глубину, с которой
начался подъем. Размах вертикальных суточных миграций у мор-
ских животных, живущих в низких широтах, где световые лучи
наиболее глубоко проникают в воду, достигает 500—600 м. В прес-
214
поводных водоемах размах суточных миграций не превышает не-
нсскольких десятков метров. Подъем организмов и опускание их
происходят путем активных движений, поэтому суточные верти-
кальные миграции сильнее всего выражены у животных, обладаю-
щих хорошо развитыми органами движения, как, например, гидро-
медузы, веслоногие раки, ветвистоусые раки, мизиды, сагитты и
ряд других организмов. Причины, вызывающие у них суточные пе-
редвижения, не вполне еще ясны, вероятнее всего это явление за-
висит от нескольких факторов, тем не менее первенствующая роль
света совершенно несомненна, так как всегда вертикальные ми-
грации организмов связаны с суточным ходом изменения интен-
сивности света в воде.
У многих морских организмов, рыб и беспозвоночных, живущих
преимущественно в полусветовой области, органы зрения сильно
увеличены и могут функционировать даже при том небольшом ко-
личестве света, которое проникает в эту область. У глубоководных
животных, обитающих в абсолютном мраке, глаза в той или иной
степени редуцированы или полностью отсутствуют.
Большое число морских организмов обладает способностью све-
титься. К числу их относятся многие одноклеточные животные и
растения, кишечнополостные, черви, ракообразные, головоногие
моллюски, иглокожие, оболочники и рыбы. Некоторые из них све-
тятся диффузным светом, у других имеются специальные органы
свечения. Особенно сложно последние устроены у некоторых пред-
ставителей высших раков, головоногих
моллюсков и рыб. Значение этих органов
заключается в освещении пространства,
приманивании добычи, защите от врагов
и распознавании особей одного вида.
'ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
фототропизм (рис. 96)
Некоторое количество дафний поме-
щают в наполненный водою высокий
стеклянный цилиндр и затемняют его
картонным футляром со съемной крыш-
кой. Убедившись в равномерном распре-
делении дафний в темноте по всему со-
СУДУ, что контролируется быстрым при-
подниманием футляра, снимают верхнюю
крышку и освещают цилиндр сверху при
помощи сильной электрической лампы.
При сильном освещении дафнии отрица-
тельно фототропичны, при слабом осве-
щении положительно фототропичны. Ана-
логичные наблюдения можно привести и
над другим.и свежепойманными планктиче-
^ими организмами (Scapholeberis. Poly-
p^must Bosmina),
a 6
Рис. 96. Фототропизм Daph-
nia. A — распределение в
темноте (цилиндр покрыт
футляром) ; />—распределе-
ние при наличии сильного
источника света.
215
СУТОЧНЫЕ ВЕРТИКАЛЬНЫЕ МИГРАЦИИ ПЛАНКТОНА
Просматриваются пробы планктона, собранного в морях или
больших пресноводных водоемах в различные часы суток одним и
тем же орудием лова при соблюдении одних и тех же методов
сбора. Всего удобнее для этих целей воспользоваться материалом,
получаемым при проведении горизонтальных ловов планктона в по-
верхностных слоях воды в дневные, вечерние и ночные часы. Под-
счет организмов по видам или по группам дает представление о
суточном ходе изменения плотности населения поверхностного слоя
воды. Полученные данные можно изобразить графически, отклады-
вая по абсциссе время суток, а по ординате количества организмов.
СВЕТЯЩИЕСЯ ОРГАНИЗМЫ
Просматриваются некоторые организмы, вызывающие диффуз-
ное свечение моря — Ceratium, Peridiniutn из перидиней, Noctiluca
из цистофлагеллат и Pyrosoma из сальп.
ОРГАНЫ СВЕЧЕНИЯ
Рассматриваются органы свечения рыб и Euphausiacea.
1. Светящиеся рыбы (рис. 97)
Рассматриваются расположение и строение органов свечения
глубоководных рыб, например Argyropelecus или Chauliodus.
Органы свечения рыб, глубоко погруженные в кожу и покрытые
с поверхности прозрачным эпителием, при наибольшем развитии
состоят из светящихся клеток, окруженных со стороны, обращенной
внутрь, рефлектором, покрытым слоем пигмента; с наружной сто-
роны находится линза. К светящемуся органу подходят нервы и
кровеносные сосуды.
2. Euphausiacea (рис. 98)
Эвфаузиды (Euphausiacea) относятся к высшим ракам (Malaco-
straca). Ранее их соединяли с Mysidacea под названием Schizo-
poda; однако между этими группами имеется лишь внешнее сход-
ство. По ряду признаков эвфаузиды гораздо ближе стоят к Deca-
poda, поэтому в настоящее время их ставят в системе в виде осо-
бого отряда, образующего вместе с отрядом Decapoda группу
Eucarida.
Эвфаузиды отличаются характерным строением. Карапакс на
спинной стороне срастается со всеми грудными сегментами, по бо-
кам свободный; впереди он оканчивается небольшим рострумом.
На голове находятся большие стебельчатые глаза и пять пар при-
датков. Антеннулы состоят из трехчленистого стебля и двух длин-
ных многочленистых жгутов. Антенны состоят из неясно расчленен-
ного протоподита, от дистального конца которого отходит длинный
жгут с трехчленистым стеблем (эндоподит) и чешуевидный скафо-
церит (эксоподит). Мандибулы с хорошо развитым щупиком; мак-
216
силлулы и максиллы пластинчатые. Грудь несет восемь пар груд-
ных ног, торакоподов; последние две пары у ряда видов рудимен-
тарные. Нормально развитый торакопод состоит из протоподита,
двучленистого эксоподита и пятичленистого эндоподита. Характер-
ным признаком является присутствие свободно свешивающихся в
Рис. 97. А — Argyropelecus hemigymnus; Б — Chauliodus sloanei; В — разрез
через орган свечения Argyropelecus.
1 — светящиеся клетки, 2 — линза, 3 — рефлектор, 4 — пигментный слой.
воду пучков жабр, отходящих от оснований торакоподов; по этому
признаку эвфаузиды легко отличаются от мизид. Брюшко состоит
из шести сегментов. Плеоподы двуветвистые, у самцов эндоподиты
плеоподов первой пары снабжены широкими лопастями. Двуветви-
стые уроподы вместе с тельсоном образуют хвостовой плавник.
Тельсон характерного строения, по бокам заостренного конца от-
ходят длинные ланцетовидные придатки.
Рис. 98. А — Meganyctiphanes norvegica; Б — Thysanoessa inermis; В—проД°л^
ный разрез через два брюшных органа свечения Euphausiacea; Г — разрез
рез орган свечения Euphausiacea.
/ — антеннула, 2 — антенна, 3 — карапакс, 4 — торакоподы, 5 — жабры, 6
плеоподы, 7 — уроподы, 8— тельсон, 9 — органы свечения, 10—светя ое
клетки, // — линза, 12 — рефлектор, 13 — пигментный слой, /4— поЛ°/7^
тело, 15 — слой клеток, играющий роль диафрагмы, 16 — кровяной синус.
эктодермальные клетки, 18 — хитин, 19 — нерв.
218
У всех эвфаузид имеются органы свечения, расположенные на
глазных стебельках, при основании торакоподов второй и седьмой
пар и на нижней стороне четырех первых брюшных сегментов меж-
ду основаниями плеоподов.
Органы свечения имеют приблизительно шаровидную форму.
В центре органа лежат клетки, выделяющие свет, перед ними рас-
положена линза, а позади — полушаровидный рефлектор, покрытый
пигментным слоем. Органы свечения на глазных стебельках проще
по своему строению и лишены линз.
Все эвфаузиды ведут пелагический образ жизни. Служат пищей
для рыб. Особенно большое значение имеют в питании сельдей; в
Баренцевом море не менее половины съедаемой сельдью в год
пищи падает на эвфаузид; кроме того, потребляются тюленями и
китами.
Наиболее обычные виды эвфаузид северных морей различаются
между собою следующими признаками.
Ь (2) Первый членик антеннул несет большой лопастной при-
даток .............................Meganyctiphanes norvegica
2 (1) Указанный членик без придатка.....................3
3 (4) Последний сегмент абдомена вооружен шипом, располо-
женным на спинной стороне заднего края и выступаю-
щим над тельсоном .... Thysanoessa inermts
4 (3) Последний сегмент абдомена без шипа .... 5
5 (6) На боковых краях карапакса находится по одному ши-
пику с каждой стороны; торакоподы приблизительно
одной длины.............................Thysanoessa raschi
6 (5) На краях карапакса шипиков не имеется; торакоподы
второй пары значительно длиннее остальных
Thysanoessa longicaudata
ПИТАНИЕ водных животных
По характеру воспринимаемой пищи водные животные разде-
ляются на сестофагов, питающихся сестоном, т. е. взвешенными в
воде планктоном и детритом, нектофагов, потребляющих рыб и
крупных головоногих моллюсков, бентофагов, пищей которых слу-
жат донные животные и растения, и грунтофагов, поглощающих
ил и песок вместе с находящимися в них живыми организмами и
органическими веществами.
Все нектофаги являются хищниками, среди бентофагов боль-
шинство видов также является хищниками, другие относятся к ра-
стительноядным организмам, или фитофагам.
Питание водных животных грунтом имеет такой же характер,
как и у наземных грунтофагов, примером которых могут служить
дождевые черви.
Совершенно особыми, свойственными только водным организ-
мам, являются способы питания сестофагов. Наличие в окружаю-
щей среде взвешенных пищевых частиц вызвало у них целый ряд
приспособлений для получения пищи.
К числу сестофагов относится огромное количество видов среди
водных животных, от простейших до рыб и млекопитающих. Среди
них можно выделить группу планктофагов, питающихся только од-
ними планктическими организмами без примеси детрита, а для
пресных вод, кроме того, группу нейстофагов, существующих за
счет организмов нейстона.
По способу получения пищи сестофаги делятся на ощупывате-
лей, седиментаторов, активных фильтраторов и пассивных фильтра-
торов.
Ошупыватели, представителями которых являются кишечнопо-
лостные, ловят добычу при помощи своих распростертых, как бы
ощупывающих воду щупалец, покрытых стрекательными капсулами.
Седиментаторы, как, например, инфузории и коловратки, произ-
водят своими ресничками круговорот воды, увлекающий мелкие
пищевые частицы, осаждающиеся затем на дне водяной воронки,
или же, как, например, мшанки и плеченогие, образуют ток воды,
из которого действием ресничек пищевые частицы направляются к
ротовому отверстию.
220
Активные фильтраторы, к числу которых относятся многие ра-
кообразные, водные личинки некоторых насекомых, двустворчатые
моллюски, оболочники, беззубые киты и некоторые другие орга-
низмы, вызывают движениями частей тела или ресничек токи воды,
приносящие к фильтрационному аппарату взвешенный в воде
планктон и детрит.
Пассивные фильтраторы, как, например, личинки некоторых на-
секомых, живущие в текучей воде, строят ловчие сети, улавливаю-
щие сносимые течением пищевые частицы.
Кроме монофагов, питающихся строго определенным видом
пищи, среди водных животных имеется большое количество поли-
фагов, потребляющих разнообразную пищу. Характер питания не-
редко меняется с возрастом. Многие организмы, не питающиеся во
взрослом состоянии планктоном, в личиночный период своей жизни
являются типичными планктофагами, как, например, различные
представители червей, ракообразных, моллюсков, иглокожих, рыб и
ряда других групп. Многие хищные виды, даже среди рыб, при от-
сутствии обычной пищи переходят на заменяющую или вынужден-
ную пищу; в качестве последней часто служат организмы планктона.
ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
ПИЩЕВЫЕ ОРГАНИЗМЫ
Рассматриваются пищевые организмы пресноводных промысло-
вых рыб.
Пищевыми организмами планктофагов, к числу которых отно-
сится снеток (Osmerus eperlanus), ряпушка (Coregonus albula),
проходной сиг (Coregonus lavaretus), уклейка (Alburnus lucidus),
а также мальки большинства рыб, являются, преимущественно,
ветвистоусые раки, веслоногие раки и коловратки. Некоторые
рыбы, как, например, толстолобик, или толпыга (Hypophthalmich-
thys molitrix), питаются почти исключительно фитопланктоном.
Пищевые организмы планктофагов рассматривались на преды-
дущих занятиях.
Пищевыми организмами бентоядных рыб, к числу которых от-
носится большинство пресноводных рыб, служат водные личинки
различных насекомых, например: ручейников, веснянок, стрекоз,
мух и в особенности комаров дергунов (Chironomidae), затем мало-
щетинковые черви, моллюски и ряд других организмов.
Большинство водных личинок насекомых, а также малощетин-
ковые черви, рассматривались на предыдущих занятиях. Личинки
комаров дергунов, или хирономид, являющиеся главнейшим видом
пищи многих промысловых рыб, служат также показателями раз-
личных типов водоемов. Некоторые из них не выносят загрязнения
и требуют большого количества кислорода, другие, напротив, очень
нетребовательны к кислородному режиму и могут длительное вре-
мя, измеряемое днями, жить в воде, совершенно лишенной кисло-
рода; эти личники встречаются также в загрязненных водоемах.
Таким образом, личинки хирономид могут считаться главной груп-
пой пресноводного бентоса.
±21
Личинки m куколки Chironomidae
Крылатые насекомые немного сходны с обыкновенными кома-
рами, обладают прозрачными вытянутыми крыльями и длинными
ногами; отличаются большей величиной и недоразвитием ротовых
придатков. Передние ноги у сидячего насекомого обычно вытянуты
вперед и непрерывно подергиваются. В безветренные дни в больших
количествах роятся в воздухе, толкутся над землей и водой или над
кустарниками и деревьями.
Развитие Chironomidae протекает в воде. Яйца заключены в сли-
зистые кладки, имеющие вид удлиненных мешков, лежащих на дне
или свободно плавающих в воде, или длинных шнуров, прикреплен-
ных к различным подводным предметам, или округлых комков, при-
клеенных одним концом к растениям или камням. Число яиц в
кладках достигает нескольких сотен или тысяч. Вышедшие из яиц
личинки линяют три раза, четвертая возрастная стадия превраща-
ется в куколку. Все Chironomidae зимуют на стадии личинки. В за-
висимости от возраста перезимовавших личинок окукливание про-
исходит ранней весной или в течение лета.
Личинки Chironomidae отличаются от личинок других двукры-
лых насекомых рядом признаков. Тело их обычно цилиндрической
формы. Общая длина колеблется от нескольких миллиметров до
двух-трех сантиметров. Голова сильно хитинизирована, резко отде-
лена от остальных сегментов, среди которых три первых являются
грудными, а десять следующих — брюшными сегментами. Груд-
ные сегменты у молодых личинок по внешнему виду не отличают-
ся от брюшных, только перед окукливанием они значительно уве-
личиваются по ширине.
По бокам головы находятся одна или две пары небольших ли-
чиночных глаз. Антенны короткие, в большинстве случаев пяти-
членистые; у личинок, относящихся к подсемейству Pelopiinae, мо-
гут втягиваться в каналы, расположенные в голове и хорошо за-
метные при рассматривании личинок сверху. От дистального кон-
ца первого членика антенн отходит длинная антеннальная щетин-
ка; у некоторых личинок эта щитинка срастается со вторым члени-
ком антенн; на поверхности первого членика находится один или
несколько кольцевых органов, несущих, вероятно, чувствительную
функцию. Характерными придатками антенн, свойственными толь-
ко личинкам Chironomidae, являются так называемые лаутербор-
новы органы, расположенные на дистальном конце второго или вто-
рого и третьего члеников антенн и состоящие из центральной па-
лочки, окруженной венчиком пластинок, изогнутых в виде лепест-
ков цветка; лаутерборновы органы сидят непосредственно на чле-
нике или находятся на конце стебелька. По всей вероятности они
являются органами какого-то чувства.
Ротовое отверстие расположено на нижней стороне головы. Ро-
товые придатки состоят из верхней губы, надглоточника (эпифа-
ринкса), парных мандибул, парных максилл, подглоточника (гипо-
фаринкса) и нижней губы. Верхняя губа, ограничивающая рот
сверху, имеет вид пластинки, вооруженной многочисленными ще-
222
тинками и зубцами. Надглоточник причленен снизу к верхней губе,
по его бокам у многих личинок расположены боковые лопасти верх-
ней губы (премандибулы). Сильно окрашенные мандибулы по вну-
треннему краю дистальной части вооружены несколькими сильными
зубцами. Максиллы имеют вид плоских лопастей, несущих обычно
небольшой, одночленистый щупик. Под ротовым отверстием распо-
ложена нижняя губа, представленная нижним отделом (субменту-
мом), нацело слитым, кроме переднего края, с головной капсулой.
Передний край нижней губы, обычно выпуклый или треуголь-
ной формы, вооружен темноокрашенными зубцами, число и строе-
ние которых постоянны у личинок различных видов, что исполь-
зуется в систематическом отношении. По бокам нижней губы у
личинок, относящихся к подсемейству Chironominae, находятся так
называемые паралабиальные пластинки (пластинки субментума),
низкие и широкие, с поперечной штриховкой (триба Tanytarsini),
или веерообразные, с радиальной штриховкой (триба Chironomi-
ni). Внутри ротовой полости на внутренней стороне нижней губы
расположен подглоточник, у основания которого открываются про-
токи слюнных (паутинных) желез; поверхность подглоточника во-
оружена различными шипами и зубцами.
На нижней стороне первого грудного сегмента находятся две пе-
редние ложные ножки, всегда слитые своими основаниями; ди-
стальные, более или менее разделенные, концы ножек вооружены
хитиновыми крючками. На девятом брюшном сегменте сверху рас-
положены два пучка жестких щетинок (кисточки), сидящих на
подставках, небольших у большинства личинок, реже сильно удли-
ненных (у личинок, относящихся к подсемейству Pelopiinae). По-
следний, десятый сегмент несет пару задних ложных ножек (назы-
ваемых также подталкивателями), вооруженных на концах хитино-
выми крючками, и две пары анальных папилл (анальных жабр),
находящихся около анального отверстия. Анальные папиллы в на-
стоящее время считаются органами осморегуляции. Трахейная си-
стема личинок Chironomidae замкнутая, апнейстического типа, ды-
хание совершается через покровы тела.
Все личинки Chironomidae живут на дне водоемов или среди
растений. Личинки Pelopiinae и некоторых Chironominae (напри-
мер, Cryptochironomus) ведут хищный образ жизни, нападают на
Tubificidae, мелких ракообразных и личинок Chironomidae, преиму-
щественно мелких Tanytarsini. Остальные личинки питаются детри-
том и остатками планктических организмов, скапливающихся на
дне, или же поедают мягкие части^растений и организмы перифи-
тона. .
Хищные личинки ведут свободный образ жизни. Другие, нехищ-
ные, личинки обыкновенно строят домики различного вида. У мно-
гих личинок Chironomidae домики, прикрепленные к твердому суб-
страту, строятся из паутинок, выделяемых слюнными железами ли-
чинок. Многие личинки обитают в илу, выстилая паутиной внутрен-
ние стенки ходов. Длинные трубки личинок Chironotnus откры-
ваются на поверхности грунта кратерообразными отверстиями.
Вентиляция ходов происходит благодаря колебательным движе-
223
ниям, производимым личинками. Личинки Tanytarsus живут в ило-
вых и рыхлых песчаных трубках или строят из детрита или песка
домики с килевыми нитями, простирающимися за его пределы. Не-
которые личинки строят переносные домики, напоминающие доми-
ки ручейников, из песчинок (личинки Stempellina, Zavlerid), или из
остатков водорослей (личинки Lauterborniella). Личинки многих
видов, относящихся преимущественно к подсемейству Orthocladii-
пае, минируют листья и стебли растений. Одни личинки выгрызают
борозды на поверхности листа, другие делают ходы внутри тканей
растения; личинки ритмичными движениями своего тела обновляют
воду через отверстия, расположенные на концах ходов. К осени
нередко можно видеть, как листья плавающих растений почти це-
ликом источены минами. Личинки некоторых видов Chironomidae
живут на губках, мшанках и яйцевых кладках ручейников.
Куколки Chironomidae тяжелее воды. Большинство личинок
окукливается в своих домиках, куколки хищных личинок свободно
лежат на грунте или плавают в воде. Передний вздутый отдел ку-
колки образован головогрудью. От спинной стороны головогруди
отходят зачатки крыльев, загибающихся на брюшную сторону, и
две дыхательных трубки, иначе называемые проторакальными ро-
гами. Куколки, относящиеся к подсемейству Pelopiinae, обладают
открытыми стигмами; периодически поднимаются к поверхности
воды. Проторакальные рога этих куколок большие, часто темно-
окрашенные и дистально расширяющиеся, на конце с небольшим
отверстием или продырявленной ситовидной пластинкой. Куколки
остальных Chronomidae поднимаются к поверхности воды только
ко времени вылупления крылатого насекомого. Обмен воды внутри
домика происходит благодаря колебательным движениям брюшка.
Стигмы у этих куколок отсутствуют, дыхание совершается через
покровы тела. Главное значение при газообмене имеют протора-
кальные рога, превращенные в простые или ветвящиеся тонкостен-
ные мешочки или в пучки тонких нитей, покрытых нередко воло-
сками. Брюшко куколок Chironomidae сравнительно узкое, состоит
из девяти сегментов; последний сегмент расширен в двухлопастной
плавник, снабженный по краям волосками.
Личинки Chironomidae относятся к числу самых распространен-
ных донных организмов пресных вод, встречающихся в самых раз-
нообразных водоемах, от луж и болот до больших рек и озер. Не-
которые виды обитают в морях, многие выдерживают значительное
загрязнение воды. Встречаясь в больших количествах, личинки
Chironomidae являются излюбленной пищей бентоядных рыб. Мно-
гие рыбы собирают их с поверхности грунта, другие роются в илу
и достигают глубоко зарывшихся личинок. Колебания количества
личинок Chironomidae зависят, с одной стороны, от времени вы-
лета крылатых насекомых, с другой стороны, от выедания их ры-
бами.
Личинки Chironomidae являются хорошими индикаторами раз-
личных типов водоемов. В илу прудов и эвтрофных озер встреча-
ются личинки Chironomus группы plumosus и semireductus, способ-
ные выдерживать резкое падение содержания кислорода и временно
224
даже его полное отсутствие. Олиготрофные озера заселены личин-
ками Lauterbornia, Protanypus и ряда других видов, не встречаю-
щихся при содержании кислорода ниже трех-четырех кубических
сантиметров на литр воды.
Определение личинок Chironomidae до вида возможно в сравни-
тельно небольшом числе случаев. Это зависит не только от того,
что для некоторых из них еще неизвестна имагинальная фаза, но,
главным образом, от того, что однотипные личинки близких видов
между собой ничем не отличаются, поэтому из морфологически не-
различимых личинок при воспитании получаются крылатые особи
нс одного, а нескольких видов. Если личинки всех видов какого-
либо рода морфологически неразличимы, то их называют только
одним родовым названием, например личинки Lauterbornia, Procla-
dius и т. п. Если же внутри рода имеется несколько типов личинок,
и каждому из них соответствует ряд видов, то они обозначаются
групповыми названиями, например личинки Tanytarsus группы
lauterborni, Tanytarsus группы gregarius и т. п. (в дальнейшем
слово группа будет обозначаться gr.).
Личинки с паралабиальными пластинками (Chironominae)
Личинки, объединяемые в подсемейство Chironominae, легко от-
личаются ют личинок других подсемейств присутствием паралаби-
альных пластинок; на каждой стороне головы имеется по два глаз-
ка, расположенных в поперечном направлении; подставки для ки-
сточек на девятом сегменте тела короткие. Подсемейство Chirono-
midae делится на две трибы — Chironomini и Tanytarsini.
Личинки Chironomini
К трибе Chironomini относятся личинки, отличающиеся следую-
щими признаками — антенны сидят непосредственно на головной
капсуле, только в редких случаях на очень низких цоколях; лаутер-
борновы органы обычно рудиментарные; паралабиальные пластин-
ки более или менее веерообразной формы, с радиальной штрихов-
кой; крючки на задних ложных ножках расположены венчиком.
В эту трибу входят личинки Chironomus, Glyptotendipes, Endochi-
ronomus, Polypedilum, Cryptochironomus, Stictochironomus и мно-
гих других видов.
1. Личинки и куколки Chironomus (рис. 99)
Крупные, до 20—25 мм длиною, красного цвета личинки, из-
вестные под названием мотыля, имеют червеобразную форму тела.
С каждой стороны головы находится по два глазка, расположенных
в поперечном направлении. Антенны пятичленистые; первый чле-
ник значительно длиннее всех остальных члеников, вместе взятых,
на конце с длинной антеннальной щетинкой; кольцевой орган рас-
положен ниже середины членика. Второй членик на конце с двумя
противостоящими лаутерборновыми органами. Верхняя губа с раз-
нообразными щетинками, среди которых выделяются верхнегубные
15 в. а. Яшнов. 225
^С";£!?“личинка Chironotnus gr. plumosus, общий вид (Л), голо”
“Х?/₽ТНЯ” губа ? эп«ФаРинкс (5), антенна (Г), мандибула (Аг
ший^п НИЖНЯЯ губа 3’И~ кУк°лка Chironotnus gr. plumosus, об'
щии вид (3), шип преанального сегмента (Я); /( — личинка Chironotnus &
thummi, конец тела.
боюш°КоОВ7 2~глаз’/~ антенна, 4- грудь, 5-передние ложные ножки,
ник. 10 талгтаЛЬНЫ^ сегмент»$ “-задние ложные ножки, 9 — хвостовой П*
веохнег^гпТ щ?тин“’ анальные папиллы, 12 -верхняя губа,
°®Рхн^убной гребень (край верхней губы), 14 — премандибула 15 — надгЛ0^.
эпифарингеальный гребень, 17 — эпифарангеальные крючки,
гребни, имеющие вид пластинок с мелко рассеченным нижним кра-
ем. С внутренней стороны к верхней губе причленен сложного
строения надглоточник (эпифаринкс), наверху с рядом одинаковых
зубчиков (эпифарингеальный гребень), а ниже их с двумя сходя-
щимися рядами изогнутых крючков, несущих многочисленные ши-
пы. По бокам верхней губы расположены длинные премандибулы,
оканчивающиеся двумя подвижными плоскими зубцами, принимаю-
щими участие в проталкивании пищи в глотку; кроме того, вместе
с различными другими придатками верхней губы они служат для
плетения паутинных сетей. Мандибулы сильно хитинизированы, по
внутреннему краю вооружены четырьмя темноокрашенными зубца-
ми; непосредственно под ними находится короткая и плоская ще-
тинка; в базальной части мандибул расположена большая сильно
ветвящаяся щетинка. Максиллы слабо хитинизированы, максилляр-
ный щупик небольшой. Нижняя губа сплошь зубчатая, с непарным
срединным зубцом, по сторонам которого находится по семи боко-
вых зубцов; первые боковые зубцы небольшие, образуют вместе с
срединными как бы тройной зубец; все зубцы черно-коричневого
цвета. По бокам нижней губы находятся веерообразные паралаби-
альные пластинки с радиально расходящейся тонкой штриховкой;
внутренние углы паралабиальных пластинок широко расставлены.
Тело личинок состоит из трех грудных и десяти брюшных сег-
ментов. На первом грудном сегменте находятся передние ложные
ножки, почти по всей длине слитые и вооруженные на концах хи-
тиновыми крючками. На спинной стороне девятого сегмента имеется
два пучка щетинок, отходящих от сближенных между собой неболь-
ших подставок. Десятый сегмент значительно меньше предыдущих,
несет пару задних ложных ножек, вооруженных на концах хити-
новыми крючками, расположенными венчиком, и две пары сильно
напряженных анальных папилл, окружающих анальное отверстие.
При рассматривании сверху живых личинок на восьмом брюшном
сегменте можно заметить биение сердца.
Личинки Chironomus относятся к числу самых распространенных
личинок хирономид. Одни из них живут в илу прудов и эвтрофных
озер (личинки Chironomus gr. plumosus, gr. semireductus, gr.
thummi), другие преимущественно в олиготрофных озерах (личин-
ки Chironomus gr. bathophilus, gr. salinarius), или в реках (личинки
Chironomus gr. plumosus-reductus, gr. reductus)\ личинки Chiro-
nomus gr. salinarius могут жить в соленых озерах, встречаются
также в Каспийском и Азовском морях.
Газообмен у личинок происходит через покровы тела. Некото-
рые личинки, как, например, личинки Chironomus gr. plumosus, gr.
semireductus и gr. thummi, могут выдерживать длительное отсут-
ствие кислорода, другие, как личинки Chironomus gr. bathophilus.
к нему более требовательны. Большое значение для дыхания имеет
дибула, 19 — максилла, 20 — нижняя губа,
22 — антеннальная щетинка, 23 — кольцевой
25 — пучки нитей (проторакальный рог), 26
ный сегмент.
21 паралабиальная пластинка,
орган, 24 лаутерборнов орган,
зачатки крыльев, 27 — преаналь-
содержащийся в крови гемоглобин, абсорбирующий кислород из
окружающей среды, благодаря чему повышается способность личи-
нок переносить в течение длительного времени полное или почти
полное отсутствие кислорода в воде; личинки погибают лишь при
развитии сероводорода. Красная окраска гемоглобина маскирует
зеленую окраску жирового тела, которое можно обнаружить только
тогда, когда из поврежденной живой личинки вытечет кровь. Пи-
таются личинки донным или взвешенным детритом. Строят трубки
в илу.
Личинки встречающихся у нас групп Chironotnus отличаются
между собою следующими признаками.
1 (8) По бокам седьмого брюшного сегмента находится по
одному мягкому, пальцеобразному выросту . . 2
2 (7) Восьмой брюшной сегмент с цилиндрическими отрост-
ками ................................................. 3
3 (6) Восьмой сегмент с двумя парами отростков . 4
4 (5) Отростки восьмого сегмента длиннее задних ложных
ножек..................................Ch. gr. plumosus
5 (4) Указанные отростки короче задних ложных ножек
Ch. gr. semireductus
6 (3) Восьмой сегмент с одной парой коротких отростков
Ch. gr. plumosus-reductus
7 (2) Восьмой брюшной сегмент без отростков Ch. gr. reductus
8 (1) Седьмой брюшной сегмент без боковых выростов 9
9 (12) Восьмой брюшной сегмент с двумя парами отростков
16
10 (11) Отростки восьмого сегмента обычно длиннее задних
ложных ножек, с заостренными концами .
Ch. gr. thummt
11 (10) Указанные отростки короче задних ложных ножек, с
тупыми концами..........................Ch. gr. bathopilus
12 (9) Восьмой брюшной сегмент без отростков .
Ch. gr. dalinarius
Куколки Chironotnus отличаются следующими признаками. Про-
торакальные рога превращены в пучки тонких нитей. На спинной
стороне второго брюшного сегмента находится полоса хитиновых
крючков. На преанальном сегменте задние углы с каждой стороны
вытянуты в сильный, направленный назад шип, желтого или корич-
невого цвета, сложенный из нескольких узких шипов. Анальный
сегмент превращен в двулопастную плавательную пластинку,
вооруженную по краям длинными волосками.
2. Личинки и куколки Glyptotendipes (рис. 100, А—Г)
Тело массивное, длиною до 15 мм. Антенны пятичленистые,
кольцевой орган расположен на нижней трети первого членика. Ман-
дибулы с четырьмя зубцами, внизу которых находится заостренная
листовидная щетинка. Нижняя губа с непарным срединным зубцом;
в боковых рядах по шести зубцов; первые боковые зубцы длиннее
срединного. Паралабиальные пластинки о мелко зубчатым верхним
краем.
228
Рис. 100. А—В — личинка Glyptotendipes gr. gripeKOveni, голова сверху (Д),
мандибула (Б), нижняя губа (В); Г—куколка Glyptotendipes; Д,Е— личинка
Endochlronomus gr. dispar, антенна (Д), нижняя губа (Е)\ Ж—личинка
Endochironomtis gr. tendens, нижняя губа, 3 — куколка Endochironomus, конец
тела; Я—Л — личинка Polypedilum gr. convictum, антенна (И), мандибула (К).
нижняя губа (Л); М — личинка Polypedilum gr. nubeculosum, антенна; Н—ли-
чинка Polypedilum gr. scalaenum, антенна; О,/7 — куколка Polypedilum, прото-
ракальный рог (О), шип на преанальном сегменте (/7).
229
Личинки Glyptotendipes живут в илу эвтрофных озер. Личинки
Glyptotendipes polytomus отличаются присутствием на восьмом
брюшном сегменте двух небольших отростков, у личинок Glypto-
tendipes gr. gripeKoveni этих отростков не имеется.
Куколки Glyptotendipes характеризуются присутствием на спин-
ной стороне второго — шестого брюшных сегментов темных хитино-
вых пластинок, имеющих форму эполетов. Проторакальные рога
превращены в пучки длинных нитей. Преанальный сегмент на каж-
дой стороне вооружен несколькими шипами.
3. Личинки и куколки Endochironomus (рис. 100, Д~3)
Тело стройное, длиною 10—16 мм. Антенны пятичленистые; пер-
вый членик длинный, кольцевой орган расположен у самого его
основания. Мандибулы с пятью черными заостренными зубцами, из
которых нижний слабо отделен от базальной части мандибулы.
Зубцы нижней губы и соединяющая их полоска — черного цвета;
в боковых рядах нижней губы по семи зубцов; срединный зубец це-
лый или на вершине подразделенный на две части; второй боковой
зубец ниже первого. Паралабиальные пластинки большие, с широ-
кими и тупыми наружными углами.
Личинки Endochironomus встречаются в зарослях и илу прудов
и эвтрофных озер; некоторые личинки минируют растения.
Личинки трех встречающихся у нас групп Endochironomus от-
личаются между собой следующими признаками.
1 (2) Срединный зубец нижней губы цельный, тупой . . । п .
£. gr. signaticwis
2 (1) Указанный зубец на вершине подразделен на две части . 3
3 (4) Второй боковой зубец ниже третьего . Е. gr. dispar
4 (3) В|брой боковой зубец выше третьего . Е. gr. tendens
Куколки Endochironomus отличаются от куколок других хироно-
мид следующими признаками. Шипы преанального сегмента, в чи-
сле 6—15, расположены тесно, образуя направленный назад гре-
бень. На лопастях плавательной пластинки, кроме обычных длин-
ных волосков, на внутренней стороне имеется по одному пучку ко-
ротких, темных щетинок на стебельке.
4. Личинки и куколки Polypedilum (рис. 100, И—П)
Тело стройное, длиною 6—10 мм. Кольцевой орган расположен
на нижней четверти первого членика антенн. Мандибулы с тремя
зубцами, над которыми расположен один зубец в другой плоскости
(так называемый внутренний зубец). Зубчатый край нижней губы
почти прямой, с двумя срединными зубцами; боковые ряды содер-
жат по семи зубцов; срединные и вторые боковые зубцы значи-
тельно выше остальных зубцов. Паралабиальные пластинки с ост-
рым наружным углом. Задние ложные ножки короткие.
Личинки Polypedilum живут в илу озер и в речном песке.
Личинки наиболее обычных групп Polypedilum отличаются ме-
жду собою следующими признаками.
230
1 (4) Антенны состоят из пяти хорошо различимых члеников;
антеннальная щетинка не доходит до конца антенн . 2
2 (3) Третий членик антенн короче второго; первый членик с
двумя кольцевыми органами . Р. gr. conuictum
3 (2) Указанный членик по длине равен второму; первый чле-
ник с одним кольцевым органом . Р. gr. nubeculosutn
4 (1) Только два первых членика антенн ясно различимы, ко-
нец антенны образован небольшим лаутерборновым орга-
* ном, расположенным на длинном стебельке; антенналь-
ная щетинка заходит за конец антенн . Р. gr. scalaenum
Куколки Polypedilum отличаются следующими признаками. Про-
торакальные рога состоят из небольшого числа, не более 12, круп-
ных ветвей. Преанальный сегмент с каждой стороны с толстым ши-
пом, покрытым добавочными шипиками.
5. Личинки и куколки Crytochironomus (рис. 101, А—И)
Тело стройное, длиною 5—15 мм. Легко отличаются от близких
форм по ряду признаков. Антенны длинные; антеннальная щетинка
приросла основанием ко второму или третьему членикам антенн.
Максиллярные щупики длинные, легко заметные, не короче поло-
вины первого членика антенн, а у ряда видов длиннее е-го. Ман-
дибулы серповидной формы, с сильно развитым вершинным зуб-
цом. Зубчатый край нижней губы прямой или вогнутый, реже сла-
бо выпуклый; срединный зубец в большинстве случаев значительно
шире других зубцов и более светло окрашен. Паралабиальные пла-
стинки длинные.
Личинки Cryptochironomus живут преимущественно в реках,
реже в озерах. Чувствительны к содержанию в воде кислорода.
Многие виды являются типичными псаммореофилами, живут на бы-
стром течении в песке; их тело тонкое, сильно вытянутое, приспо-
собленное к змеевидным движениям, у некоторых видов с вторич-
ной сегментацией. Личинки Cryptochironomus домиков не строят,
ведут хищный образ жизни, питаются крупной добычей, часто ли-
чинками различных Tanytarsini, которых хватают своими острыми
серповидно изогнутыми мандибулами. Личинки наиболее часто
встречающихся групп Cryptochironomus отличаются между собою
1следующими призиа»ками.
1 (2) Тело состоит из 20 сегментов С. macropodus
2 (1) Тело с обычным числом сегментов . . . ^ . . . 3
3 (8) Максиллярный щупик длиннее половины первого членика
антенн............................. • 4
4 (5) Антенны длиннее трети головы .......
Псаммореофильные виды, обитающие на сильном течении
5 (4) Антенны не длиннее трети головы..................6
6 (7) Срединный зубец нижней губы высокий, с темными боко-
выми сторонами С. gr. defectus
231
7 (6) Указанный зубец низкий
8 (3) Максиллярный щупик не длиннее
членика антенн
. С, gr. vulneratus
половины
первого
. 9
Рис. 101. А—личинка Cryptochironomus macropodus, нижняя губа; Б, В — ли-
чинка Cryptochironomus gr. defectus, голова снизу (Б), нижняя губа (В); Г —
личинка Cryptochironomus gr. vulneratus, нижняя губа; Д — личинка Cryptochi-
ronomus gr. conjugens, нижняя губа; Е— личинка Cryptochironomus gr. viridu-
lus, нижняя губа; Ж —личинка Cryptochironomus gr. anomalus, нижняя губа;
3, И — куколка Cryptochironomus, оральные рога (3), конец тела (И), К—м —
личинка Stictochironomus sp., антенна (К), мандибула (Л), нижняя губа (Л4)J
И — куколка Stictochironomus, край преанального сегмента.
«9 (10) Зубчатый край нижней губы слабо выпуклый
С. gr. conjugens
Ю (9) Зубчатый край нижней губы треугольной формы . И
232
П (12) Срединный зубец нижней губы с выемкой . . . .
С. gr. viridulus
12 (Н) Указанный зубец на конце заостренный . . . .
С. gr. anomalus
Куколки Cryptochironomus отличаются следующими признаками.
На переднем ЛраВе головогруди находятся крупные, загнутые на
концах выросты (оральные рожки). Полоса шипов на спинной сто-
роне второго брюшного сегмента посреди прервана. На преаналь-
ном сегменте боковых шипов не имеется. Плавательная пластинка
с двумя пальцевидными выростами.
6. Личинки и куколки Stictochironomus (рис. 101, К—Н)
Тело стройное, длиною 8—14 мм. Антенны шестичленистые, с
чередующимися лаутерборновыми органами, расположенными на
втором и третьем члениках; антеннальная щетинка не доходит до
конца антенн. Мандибулы с тремя зубцами, над которыми располо-
жен один зубец в другой плоскости (внутренний зубец). Посреди
зубчатого края нижней губы находится один непарный или два
небольших срединных зубца; первые боковые зубцы значительно
выше срединных или почти одной высоты с ними. Личинки Sticto-
chironomus встречаются в озерах и реках, преимущественно на
песке.
Куколки Stictochironomus отличаются следующими признаками.
По бокам пятого и шестого брюшных сегментов находится по три,
а седьмого и восьмого сегментов — по четыре длинных придатка; на
преанальном сегменте по несколько шипов, из которых средней
крупнее остальных.
Личинки Tanytarsini
К трибе Tanytarsini относятся личинки, отличающиеся следую-
щими признаками. Антенны сидят на высоких столбикообразных
хитиновых выступах головной капсулы — цоколях; лаутербориовы
органы хорошо развиты; паралабиальные пластинки узкие, сильно
вытянутые в ширину, с продольной штриховкой; крючки на задних
ложных ножках расположены подковообразно. В эту трибу входят
личинки Tanytarsus, Lauterbornia, Micropsectra и некоторых других
родов.
7. Личинки и куколки Tanytarsus (рис. 102, А—М)
Небольшие, длиною 5—7 мм, зеленоватого или красноватого
цвета, личинки Tanytarsus, напоминающие в общем личинки Chi-
ronomus, отличаются от последних рядом признаков. Тело тонкое,
по бокам брюшных сегментов (от первого до седьмого) находятся
двулопастные перистые щетинки, хорошо заметные лишь у живых
личинок. Антенны длинные, пятичленистые; цоколи гладкие, без
шипов; на первом членике антенн, кроме кольцевого органа, нахо-
дящегося у самого основания членика, имеется еще довольно длин-
ная чувствительная щетинка, расположенная около середины чле-
233
. -ч антен^
Рис. 102. Л—Г—личинка Tanytarsus gr. gregarius, голова сбоку И)’inhatifr°nS'
(5), мандибула (В), нижняя губа (Г); Д—личинка Tanytarsus gr. 10
антенны;Е,Ж — личинка Tanytarsus gr. mancus, антенна (Е), иня^^Лгзиз
3 —личинка Tanytarsus gr. lauterborni, антенна; И, К — личинка1 Tany Q (*/?)•
exiguus, антенна (Я), два домика (К); Л, М — куколка Tanytarsus, бр рог,
проторакальный рог (М); Я —куколка Lauterbornia sp., проторакаль
О—Р—личинка Micropsectra gr. ргаесох, антенна (О), мандибула v
губа (Р); С —куколка Micropsectra, проторакальный Рог-
234
ника; другая щетинка находится на его конце. Лаутерборновы ор-
ганы противостоящие, расположены на более или менее длинных
стебельках, отходящих от второго членика антенн. Мандибулы с
различным числом зубцов; щетинка под зубцами длинная, заходит
за конец мандибулы. Нижняя губа с одним одиночным или трой-
ным срединным зубцом, в боковых рядах по пяти одинаковых
или различных зубцов. Паралабиальные пластинки низкие, очень
широкие, почти соприкасающиеся своими внутренними краями, с
продольной (по отношению к длине головы) штриховкой. Задние
ложные ножки короткие, с несколькими рядами мелких крючков,
расположенных изогнутой полоской.
Личинки Tanytarsus живут в олиготрофных озерах и реках, не-
которые (личинки Tanytarsus gr. exiguus)—исключительно в быстро
текущих водах. Чувствительны к количеству растворенного в воде
кислорода. Строят прикрепленные песчаные или иловые домики,
иногда с длинными килевыми нитями. Для получения пищи личин-
ки высовывают из домика передней конец тела, удерживаясь в нем
при помощи крючков на задних ложных ножках.
Личинки наиболее обычных групп Tanytarsus отличаются между
собой следующими признаками.
1 (4) Стебельки лаутерборновых органов в три-пять раз длин-
нее общей длины трех последних члеников антенн 2
2 (3) Шетинки между цоколями антенны простые .
Т. gr. gregarius (= Eutanytarsus gr. gregarius)
3 (2) Указанные щетинки расщепленные . . . Т. gr. lobatifrons
4 (1) Стебельки лаутерборновых органов не заходят за конец
антенн .......................... ....:. 5
5 (6) Второй членик антенн короче третьего; лаутерборновы
органы крупные; личинки строят рыхлые песчаные до-
мики .........................Т. gr. mancus {= Atanytarsus)
6 (5) Указанный членик длиннее третьего; лаутерборновы ор-
ганы некрупные или мелкие.................................7
7 (8) Стебельки лаутерборновых органов короче третьего чле-
ника антенн; личинки строят песчаные или иловые до-
мики ... Г. gr. lauterborni (= Paratanytarsus)
8 (7) Стебельки лаутерборновых органов по длине равны
третьему членику антенн или длинее его; личинки строят
домики с килевыми нитями...................................
Т. gr. exiguus (= Rheotanytarsus)
Куколки Tanytarsus отличаются от куколок трибы Chironomini
присутствием простых, не разветвленных проторакальных рогов.
На спинной стороне брюшных сегментов находятся парные, а
иногда, кроме того, непарные сильно хитинизированные щитки.
Преанальный сегмент с одним или несколькими боковыми шипами.
Плавательная пла|стинка с одной или двумя парами длинных
щетинок.
Куколки различных групп отличаются по следующим призна-
кам. Куколки Tanytarsus gr. lauterborni — на брюшных сегментах,
кроме парных, имеются также непарные щитки; преанальный сег-
235
мент на каждой стороне с несколькими боковыми шипами. Кукол-
ки Т. gr. exiguus — непарных щитков не имеется; преанальиый сег-
мент на каждой стороне с одним боковым шипом. Куколки Т. gr.
gregarius — непарных щитков не имеется; преанальиый сегмент на
каждой стороне с несколькими шипами.
8. Личинки н куколки Lauterbornia (рис. 102, Н)
Личинки кроваво-красного цвета, длиною до 10 мм, по внешне-
му виду сходны с личинками Tanytarsus. Антенны сидят на цоко-
лях, на конце которых на внутреннем крае находится очень неболь-
шой, часто незаметный тупой шип. Лаутерборновы органы мелкие,
расположены на длинных стебельках, превышающих в 2—2*/2раза
общую длину трех последних члеников антенн. Личинки Lauterbor-
nia отличаются от личинок Tanytarsus gr. gregarius (сходных с
ними по сильному развитию стебельков лаутерборновых органов)
высотою цоколей, которая у личинок Lauterbornia равна приблизи-
тельно их ширине и не превышает расстояния между основаниями
цоколей, а у личинок Tanytarsus gr. gregarius значительно больше
этого расстояния. Анальные папиллы разного размера — спинные
длиннее и толще брюшных.
Личинки Lauterbornia встречаются в олиготрофных озерах и хо-
лодных ручьях. Очень требовательны к количеству растворенного
в воде кислорода.
Куколки Lauterbornia отличаются от куколок Tanytarsus строе-
нием проторакальных рогов, имеющих вид небольших, темнокорич-
невых пузырьков, на которых сидят длинные, опушенные волос-
ками отростки (эти отростки в большинстве случаев спадают). По
бокам преанального сегмента находится по 5—8 шипов.
9. Личинки и куколки Micropsectra (рис 102, О—С)
Личинки Micropsectra gr. praecox (=Eutany tarsus gr. inermipes),
розового или зеленого цвета, длиною до 15 мм, отличаются от дру-
гих личинок Tanytarsini присутствием на внутреннем крае конца
цоколей антенн ясно заметного шипа, равного приблизительно
’А высоты цоколя. Стебельки лаутерборновых органов в три-че-
тыре раза длиннее общей длины трех последних члеников антенн.
Зубцы нижней губы постепенно уменьшаются в размерах к краям.
Личинки Micropsectra живут обычно в глубоких олиготрофных
озерах.
Куколки Micropsectra отличаются следующими признаками.
Проторакальные рога удлиненные, с длинными щетинками. По бо-
кам преанального сегмента находится по группе темных шипов.
Личинки с втяжными антеннами (Pelopiinae)
Личинки, объединяемые в подсемейство Pelopiinae (=Tanypodi-
пае), резко отличаются от личинок других хирономид по ряду при-
знаков. Ведут свободный образ жизни, никогда не делают домиков
и не минируют растений. Относятся к числу типичных хищников.
236
Ротовые придатки приспособлены к захватыванию живой добычи;
зубцы нижней губы часто редуцированы, на подглоточнике имеется
зубчатая глосса. Органы осязания хорошо развиты. Задние ложные
ножки длинные, на конце с венчиком крючков.
10. Личинки и куколки Procladius (=Tanypus) (рис. 103, А—Д)
Личинки беловато-желтоватые или красноватые, с мраморным
рисунком, длиною до 11 мм, являются одними из самых распростра-
ненных среди всех Pelopiinae. Тело слегка уплощенное, с каемкой
волосков по бокам сегментов. На голове имеется одна пара глаз..
Антенны короткие (не превышают четверти длины головы), лаутер-
борновых органов не имеется. При помощи специальных мускулов,
отходящих от основания головной капсулы, антенны могут втяги-
ваться в длинные каналы, расположенные в голове. У живых личи-
нок видно, как они то втягиваются внутрь каналов, то высовывают-
ся из них; втягивание и высовывание обеих антенн может происхо-
дить одновременно идд разновременно; втянутые антенны хорошо
заметны при рассматривании головы сверху. Верхняя губа редуци-
рована, премандибул и надглоточника не имеется. Мандибулы но-
жевидные, с черным концевым зубцом. Максиллы пластинчатые, с
большим щупиком. Нижняя губа характерного вида, состоит из
мембранообразной срединной части (языка), имеющей треугольную
форму, и двух гребней, содержащих на каждой стороне по 6—8 ко-
ричнево-желтых зубцов. Паралабиальных пластинок не имеется.
Подглоточник на вершине несет узкую глоссу, состоящую из пяти
черных зубцов, из которых средний является самым низким. По бо-
кам глоссы расположены небольшие желтые чешуевидные пара-
глоссы, вооруженные по обоим краям тонкими зубцами. Передние
ложные ножки короткие, задние очень длинные, с желтыми крюч-
ками. Пучки щетинок на спинной стороне девятого брюшного сег-
мента расположены на высоких подставках, длина которых в четыре
раза больше ширины. Анальные папиллы треугольной формы, одна
пара находится около анального отверстия, другая — при основа-
нии задних ложных ножек.
Личинки Procladius живут в стоячих и текучих водах. Личинки
разных видов между собой не различаются.
Куколки Procladius, темноокрашенные с белыми пятнами, сво-
бодно плавают в воде. Проторакальные рога веретеновидные, ко-
ричневого цвета, на конце с ситовидной пластинкой. Хвостовой плав-
ник состоит из двух сближенных лопастей, каждая лопасть несет
по две длинных щетинки, дистально от которых находятся мелкие
шипнки.
11. Личинки и куколки Ablabesmyia (= Pelopia) (рис. 103, Е—Р)
Личинки, длиною 5—11 мм, цилиндрической формы, без боко-
вой каймы волосков. Голова узкая, вытянутая. Антенны длинные.,
достигают трети или половины длины головы, втяжные. Мандибулы
серповидные. Нижняя губа без зубчатых гребней, представлена
237
М Р п
. / с\ глосса
Рис. 103. А—В — личинка Procladius sp., антенна (А), нижняя губа KOflett
с параглоссами (В); Г, Д — куколка Procladius, проторакальный рог у ииЖняЯ
брюшка (Д); Е, И — личинка Ablabesmyia gr. lentiginosa, антенна Kf JI^
губа (ЛС), глосса с параглоссами (3), конец задней ложной ножки ("Ь
личинка Ablabesmyia gr. monilis, общий вид (А), конец брюшка
М—личинка Ablabesmyia gr. tetrasticta, глосса с параглоссами; Н
лишь треугольной формы мягким языком, снабженным по бокам
пузыревидными ^выростами. Глосса с пятые черно-коричневыми зу-
бцами. Пароглоссы на конце с двумя неравной длины шипами. Пе-
редние ложные ножки очень длинные? на большем протяжении
слитые между собой. Задние ложные ножки длинные, расставлен-
ные, на конце с крупными крючками. Анальные папиллы короткие,
расположены вокруг анального отверстия.
Личинки Ablabesmyia встречаются в самых разнообразных во-
доемах. Некоторые, как, например, личинки Ablabesmyia gr. fla-
vida, gr. binotata, gr. tetrasticta, gr. fulva, gr. falcigera, gr. tenuical-
car, живут в канавах, лужах и родниках. Очень чувствительны к
загрязнению (за исключением личинок Ablabesmyia gr. fulva), Ве-
дут хищный образ жизни.
Личинки наиболее часто встречающихся групп Ablabesmyia от-
личаются между собой следующими признаками.
1 (2) С каждой стороны головы по два сближенных глаза . .
A. zavreii
2 (1) С каждой стороны головы по одному глазу 3
3 (10) Все крючки задних ложных ножек простые . .4
4 (5) На каждой задней ложной ножке по два черно-коричне-
вых крючка, остальные крючки желтые . A. gr. monilis
5 (4) Все крючки задних ложных ножек желтые . 6
6 (7) Длина первого членика антенн превышает общую длину
всех остальных члеников в пять-семь раз; тело с длин-
х ними щетинками . . A. gr. lentiginosa (=costalis)
7 (6) Длина указанного членика превышает общую длину всех
остальных члеников в три-четыре раза . . 8
8 (9) Боковые зубцы мандибул почти одинаковые .
A. gr. flavida
9 (8) Боковые зубцы мандибул резко неравные . ...
A. gr. binotata (=minima)
10 (3) Некоторые крючки задних ложных ножек с одним или
несколькими шипами по внутреннему краю 11
И (14) Все крючки задних ножек желтые ... 12
12 (13) Один крючок каждой ножки с крупным шипом .
A. gr. tetrasticta
13 (12) Несколько крючков каждой ножки с острыми шипиками
A. gr. fulva
14 (11) Некоторые крючки задних ложных ножек коричневые . 15
15 (16) Глосса с равными зубцами ... A. gr. falcigera
16 (15) Срединный зубец глоссы ниже других . . . . .
A. gr. tenuicalcar
Куколки Ablabesmyia сходны с куколками других Pelopiinae,
отличаются следующими признаками. У основания проторакальных
рогов находится поперечный ряд из многих шипов (отсутствует у
куколок некоторых групп). Последние сегменты брюшка с длинны-
Ablabesmyia gr. fulva, голова с втянутыми антеннами; О—личинка Ablabesmyia
gr. tenuicalcar, крючки на задних ложных ножках; П, Р — куколка Ablabesmyia,
общий вид (/7), конец брюшка (Р).
239
ми боковыми щетинками. Хвостовой плавник сравнительно слабо
развит, с треугольными заостренными лопастями. Длинные щетин-
ки сидят около середины лопастей плавника.
Личинки без паралабиальных пластинок и без втяжных антенн (Orthoctadiinae)
Личинки, объединяемые в богатое видами подсемейство Ortho-
cladiinae, отличаются от личинок подсемейства Chironominae отсут-
ствием паралабиальных пластинок (у некоторых видов имеются па-
ралабиальные пластинки, но без штрнховатости), а от личинок под-
семейства Pelopiinae отсутствием втяжных щупалец и зубчатой
глоссы. На каждой стороне головы у большинства личинок имефтс^
по два глаза, расположенных в продольном направлении. Грудные
сегменты разделены. Длина личинок не превышает 8—10 мм; толь-
ко в редких случаях достигает 12—15 мм. Встречаются чаще всего
в чистой воде, среди растений, многие виды только в холодных ру-
чьях. Некоторые личинки минируют ткани водных растений. Пита-
ются растительной пищей.
12. Личинки и куколки Cricotopus (рис. 104)
Личинки, коричневого или зеленого цвета, длиною 5—7 мм.
Брюшные сегменты по бокам с пучками щетинок, по 6—12 щети-
нок в каждом пучке. Первый членик антенны с двумя кольцевыми
•органами. Верхняя губа с длинными премандибулами. Мандибулы
Рис. 104. Личинка Cricotopus brevipalpis. 4 — голова сбоку; Б — голова сверху;
В — антенна; Г — нижняя губа; Д — мины в листе телореза.
широкие. Нижняя губа с непарным срединным зубцом, в боковых
рядах по пяти-шести зубцов. Крючки передних ложных ножек
пильчатые, на задних ложных ножках простые. Подставки для ще-
тинок на девятом брюшном сегменте короткие.
Личинки Cricotopus живут среди растительности в озерах и ре-
ках, часто минируют листья водных растений. Среди последних наи-
более обычными являются личинки Cricotopus brevipalpis, коричне-
вого цвета, с сильно вытянутой головой и очень короткими антен-
нами, и Cricotopus gr. silvestris, зеленого цвета, с обычной формой
головы и антенн.
240
Куколки Cricotopus отличаются следующими признаками. Про-
торакальные рога тонкие. Лопасти хвостового плавника слабо раз-
виты, с тремя щетинками.
ПИЩА водных животных
Для ознакомления с составом пищи производится вскрытие раз-
личных водных животных. Выделенный из организма кишечник по-
мещают на предметное стекло, прибавляют для просветления
материала небольшое количество глицерина, выдавливают за-
тем содержимое кишечника и распределяют его при помощи препа-
ровальных игл более или менее равномерно тонким слоем по по-
верхности стекла. При вскрытии больших животных на предметное
стекло переносится только часть материала, которую следует брать
из переднего отдела пищеварительного аппарата, где пища еще не
переварилась полностью. В зависимости от размера пищевых орга-
низмов, их просмотр ведется под лупой или микроскопом; в послед-
нем случае препарат предварительно покрывают покровным стек-
лом.
Определение пищевых организмов по остаткам, иногда по от-
дельным фрагментам, требует хорошего знания их строения, по-
этому в протоколе вскрытия достаточно указать лишь большие так-
сономические группы, а также примерное количество встреченных
организмов каждой группы по трехбальной системе — мало, много,
масса.
Для изучения состава пищи вскрываются из планктофагов —
усоногие раки (Balanus, Chthamalus), асцидии (Styela и другие
виды), рыбы (атлантическая сельдь Clupea harengus, мальки раз-
личных рыб); из бентофагов—амфиподы (Gammarus и другие
виды), рыбы (пикша Melanogratnmus aeglefinus, карп Cyprinus car-
pio и другие виды); из грунтофагов — черви (Tubifex, Arenicola и
другие виды), иглокожие (Ctenodiscus и другие виды).
СПОСОБЫ ПИТАНИЯ СЕСТОФАГОВ
Питание ощупывателей (рис. 105)
Способ захватывания добычи ощупывателями проще всего рас-
смотреть на живой гидре, не получавшей некоторое время пищи. В
голодном состоянии гидра вытягивает тело и распростирает свои
щупальца в виде большой сети, свешивающейся вниз; при длине
тела, равной одному-трем сантиметрам, длина щупалец может
достигнуть десяти-двадцати сантиметров. Периодически гидра со-
кращается, а затем вытягивается в другом направлении, как бы
ощупывая окружающее пространство. Пищей гидры служат даф-
нии, циклопы, мелкие личинки насекомых, черви и мальки рыб.
Добыча, при прикосновении к щупальцам, парализуется и захваты-
вается нитями стрекательных капсул (нематоцистов). Различают
несколько типов нематоцистов. Пенетранты имеют у основания
нити три больших стилета, пробивающих в момент выстрела капсу-
лы даже твердый хитин панцыря дафнии или личинки насекомого;
16 В. А. Яшнов 241
в образующуюся рану затем внедряется нить, выделяющая ядови-
тое вещество, парализующее добычу. Вольвенты не имеют стиле»
тов, их нить относительно короткая, способная закручиваться; при
Рис. 105, Л — Hydra oligactis; Б —Hydra vulgaris; В—-Chlorohydra viridissima;
Г—Ж — схема четырех типов нематоцистов, до (внизу) и после (вверху) выстрела,
пенетрант (Г), вольвент(Д), стереолин (Е), стрептолин (Ж); 3 — вольвенты на
плавательной ноге циклопа; И — пенетрант в ткани коретры; К — пенетрамт
после выстрела.
/ — конус, 2 — нить, 3— стилет, 4 — шейка, 5—книдоциль, 6 — крышечка, 7—
стрекательная капсула, 8— книдобласт, 9 — спиральная нить в плазме клетки,
10 — ядро книдобласта.
прикосновении нити к какому-либо организму она обвивается во-
круг его волосков, шипов или иных выростов. Кроме того, имеются
стрептолпиы, длинные шиповатые нити которых служат для защи-
ты от врагов, и стереолины, выбрасывающие липкие нити, помога-
ющие гидре при ее передвижениях по субстрату.
Если в сосуд, содержащий гидры, перенести пипеткой несколь-
ко экземпляров дафний или циклопов, то можно увидеть весь про-
цесс захватывания добычи. Организмы, парализованные при при-
косновении к стрекательным клеткам щупалец, захватываются за-
тем ими и переносятся к ротовому отверстию. Непереваренные ос-
татки, хитин и т. п., через несколько часов выбрасываются наружу
через ротовое отверстие.
Наиболее обычные виды гидр отличаются между собою следую-
щими признаками.
1 (2) Тело зеленого цвета (от симбионта Chlorella vulgaris);
щупальца не длиннее тела . Chlorohydra viridissima
2 (1) Тело иной окраски . . . . . 3
3 (4) Задний конец тела вытянут в стебель, щупальца в три-
пять и более раз длиннее тела ......................... .
Hydra oligactis (-Н. jusca)
4 (3) Стебля не имеется; щупальца самое большее в два
раза длиннее тела Hydra vulgaris <=Н. grisea)
Питание седиментаторов (рис. 106)
Легче всего процесс седиментации пищи можно рассмотреть на
живых инфузориях или коловратках. Токи воды, вызываемые рабо-
той ресничек инфузорий или коловращательного аппарата коловра-
ток, удобнее наблюдать при
прибавлении к воде, в которой
находятся исследуемые орга-
низмы, небольшого количества
тонко истертого порошка кар-
мина или ультрамарина. Чем
меньше частицы порошка, тем
легче рассмотреть процесс се-
диментации. Для удержания
животных на одном месте, без
чего почти невозможно прове-
сти наблюдения, надо бросить
в каплю воды, в которой они
рассматриваются под микроско-
пом, несколько тонких волокон-
цев ваты (большое количество
волоконцев затемняет картину).
Присутствие в воде мелких
окрашенных частиц позволяет
Рис. 106. Схема токов воды, вызываемых
работой коловращательного аппарата
коловраток.
проследить токи воды, увлека-
ющие их из окружающего пространства. Через определенный про-
межуток времени эти частицы могут быть обнаружены внутри тела
животных. Так как не все коловратки поглощают без выбора взве-
шенные в воде частицы, то следует для эксперимента брать питаю-
щихся без выбора Ker atelia, Conochilus или Triarthra.
Питание активных фильтраторов
В качестве примеров активных фильтраторов рассматриваются
представители жаброногих раков (Pristicephalus, Daphnia, Apus),
веслоногих раков (Diaptomus, Calanus), двустворчатых моллюсков
(Anodonta, Unio), личинок комаров (Anopheles), аппендикулярий
(Oikopleura).
1. Pristicephalus (рис. 107)
Pristicephalus, описание строения которого помещено на стр. 130,
относится к подотряду жаброногов (Anostraca).
Процесс питания связан с движениями грудных ног, обладаю-
щих двойной функцией — наружная часть конечностей служит для
движения, внутренняя—для отфильтровывания пищевых частиц. При
плавании организма в воде вдоль каждого ряда конечностей по
направлению от головы к заднему концу тела проходят токи воды,
вызываемые быстрыми ударами ног. Эти токи увлекают также
воду, находящуюся в пространстве между обоими рядами ног; во-
да этого последнего тока, более слабого, по сравнению с первым,
всасывается вместе с находящимися в ней частицами сестона в
фильтрационную камеру. Дно последней образовано брюшным же-
лобком, идущим по нижней стороне тела, боковые же стбнки огра-
ничены гребнями щетинок, отходящими от первых двух эндитов
каждой ноги. Благодаря своей длине щетинки каждой конечности
достигают щетинок непосредственно за ней следующей соседней
ноги; вследствие этого боковые стенки фильтрационной камеры
приобретают вид сплошной решетки.
Между каждой парой уплощенных ног находятся щелевидные
пространства — всасывательные камеры, число которых на каждой
стороне равно десяти соответственно одиннадцати парам ног.
В фазу абдукции, когда ноги отходят от тела, каждая всасыва-
тельная камера с брюшной стороны закрывается эндоподитом, сги-
бающимся в сагиттальной плоскости и прижимающимся к находя-
щейся позади соседней ноге; аналогичным образом экситы ноги
(эксоподит, эпиподит и преэпиподит), сгибающиеся в фронтальной
плоскости, образуют наружную стенку камеры. Таким образом в
фазу абдукции всасывательные камеры оказываются закрытыми со
всех сторон, за исключением внутренней,-представленной, как ука-
зывалось выше, решетчатой стенкой. Последняя состоит из боль-
шого числа параллельно идущих щетинок, снабженных, в свою оче-
редь, добавочными волосками, еще более суживающими щелевид-
ные просветы между ними. При подъеме ног всасывательные ка-
меры увеличиваются в объеме, вследствие этого вода всасывается
в них, оставляя на внутренней стороне фильтра все находящиеся в
воде частицы сестона.
зм
В фазу аддукции прижимающиеся к телу ноги расправляются,
контакт эндоподитов и экситов с соседними конечностями наруша-
ется и вода из всасывательных камер выдавливается наружу. В за
висимости от темпа движения ног, всасывательные камеры, играю-
щие роль насосов, наполняются водой и опоражниваются от 150
до 400—500 раз в минуту.
Отфильтрованные частицы, скопляющиеся в глубине фильтра-
ционной камеры, вместе с током воды, вызываемым движениями
Рис. 107. Pristicephalus. А — схематический разрез вдоль тела с указанием то-
ков воды; В, Б — схематический горизонтальный разрез через две пары вса-
сывательных камер в фазу абдукции (Б) и в фазу аддукции (В); Г—нога
шестой пары.
/ — дорсальный край брюшного желобка, 2 — вентральный край брюшного
желобка, 3 — эндоподит, 4 — гребень щетинок, 5 — эксоподит. 6 — эпиподит,
7 — преэпиподит.
щетинок, отходящих от проксимальных (базальных) частей ног и
доходящих до дна брюшного желобка, направляются к ротовому от-
верстию.
При изучении способа питания Pristicephalus следует рассмот-
реть пищевые токи воды, вызываемые движениями конечностей у
живого организма (эти наблюдения можно проводить только вес-
ною), а также строение грудных ног (лучше всего на уплотнив-
шемся фиксированном материале).
2*5
2. Daphnia (рис. 108)
Большинство ветвистоусых раков (Cladocera) относится к числу
типичных активных фильтраторов. Исключение представляют лишь
представители семейства Polyphemidae и Leptodora, единственный
вид семейства Leptodoridae, ведущие хищный образ жизни. Опи-
сание строения дафнии помещено на стр. 17.
Для изучения работы фильтрационного аппарата дафнии, необ-
ходимо сначала ознакомиться с строением грудных ног, играющих
основную роль в процессе получения пищи. Рассматриваются изо-
лированные ноги, вычлененные из уплотненных фиксаций орга-
низмов.
У дафний имеется пять пар тургорных конечностей. Внутренний
край ног первой пары разделен на пять снабженных длинными ще-
тинками лопастей, из которых первые три соответствуют эндитам,
а два последних эндоподиту и эксоподиту типичной ноги жаброно-
гих раков; кроме того, имеется эпиподпт, являющийся жаберной
пластинкой. Ноги второй пары характерного строения; на удли-
ненном протоподите расположены пять эндитов, из которых прокси-
мальный (базальный) образует большой максилярный вырост; сре-
ди щетинок последнего выделяется по своей величине щетинка,
снабженная по одной сторонке рядом крепких волосков; остальные
четыре эндита слабо развиты, каждый из них несет одну или две
длинных щетинки; нога оканчивается хорошо развитым эксоподитом
с двумя длинными щетинками; по наружному краю ноги располо-
жен эпиподит. Ноги третьей пары по своему строению резко отли-
чаются от предыдущих; самым характерным признаком является
присутствие гребня щетинок, прикрепленных вдоль всего края боль-
шого первого эндита; в естественном положении этот гребень ото-
гнут под острым углом к плоскости самой ноги; остальные эндиты,
слабо выраженные и снабженные небольшим количеством щетинок,
находятся у дистального (конечного) края первого эндита, между
последним и хорошо развитым эксоподитом, несущим четыре конеч-
ных и две боковых больших щетинки; на наружной стороне ноги
находятся эпиподит и хорошо развитый преэпиподит. Ноги четвер-
той пары в общем сходны с ногами третьей пары, отличаясь, глав-
ным образом, полной редукцией дистальных эндитов и другой фор-
мой эксоподитов. Ноги последней, пятой, пары имеют иное строе-
ние: в базальной части находятся эпиподит и преэпиподит; корот-
кий эксоподит на дистальном конце несет две маленьких щетинки
и вытянут в длинную полукруглой формы щетинку; по внутреннему
краю нога лишена выростов, имеется только одна большая щетин-
ка, между нею и эксоподитом находится овальной формы дисталь-
ная часть протоподита.
Процесс питания дафнии тесно связан с движениями ног, кото-
рые потеряли полностью двигательную функцию и приспособились
к улавливанию мелких частиц сестона.
Фильтрационный аппарат дафний достигает наибольшей слож-
ности, по сравнению с аналогичными образованиями других пред-
ставителей жаброногих раков. Рассматривая живую дафнию в во-
246
де,, в которой прибавлено мелко растертого кармина или китайской
туши до получения явной окраски, можно констатировать наличие
двух токов воды — одного входящего в полость раковины через пе-
Рис. 108. Daphnia. А — фаза абдукции; Б — фаза аддукции; В — продольны!
разрез через тело дафнии; Г — токи воды, вызываемые движениями ног; Д—
яога первой лары; Е — нога второй пары; ЛК— нога третьей пары; 3 — нога
четвертой пары; И — нога пятой пары.
1 — входящий ток воды, 2 — выходящий ток воды, 3—фильтрационная камера,
4 — брюшной желобок, 5—мандибула, 6—максиллула, 7—максиллярный вы-
рост, 8 — постабдомеп, 9— эндит, 10— гребень щетинок, 11—эксоподит, 12—
эпилодит, 13 — лреэпилодит4 I—JV — ноги первой-четвертой пар..
реднюю часть щели между створками и другого выходящего нз
этой полости около постабдомена. Вносимые внутрь раковины ча-
стицы окрашенных веществ, после отфильтровывания через фильтр,
образованный щетинками грудных ног, скопляются в брюшном же-
лобке, проходящем по нижней стороне тела, и затем направляются
к ротовому отверстию. Через некоторое время после начала экспе-
римента весь кишечник заполняется тем веществом, которым дафния
кормилась. Следует отметить время поступления в кишечник пер-
вой порции вещества и время заполнения этим веществом всего
кишечника, чтобы определить так называемый коэфициент возоб-
новления.
Фильтрационная камера, находящаяся между грудными конеч-
ностями, сзади закрыта ногами пятой пары и расположенным меж-
ду ними постабдоменом. Процеживание воды через фильтр совер-
шается в результате понижения давления в всасывательных каме-
рах, вызываемого увеличением их объема в фазу абдукции. С каж-
дой стороны имеется по четыре всасывательных камеры в соответ-
ствии с пятью парами ног.
Главную роль в отфильтровывании пищи играют две последних
пары всасывательных камер, снабженных с внутренней стороны
прекрасно развитой решеткой из щетинок, отходящих от вну!рен:
них краев ног третьей и четвертой пар. Щетинки решетки идут па-
раллельно друг другу и снабжены добавочными тонкими, косо от-
ходящими волосками; благодаря этому решетки приобретают харак-
тер очень тонкого фильтра, не пропускающего через свои отверстия
даже мельчайшие организмы наннопланктона. Снаружи каждая из
всасывательных камер ограничена створкой раковины, к которой те-
сно прижаты эпиподит, большой окаймленный волосками преэпипо-
дит и две боковых щетинки эксоподита; с нижней стороны всасы-
вательная камера закрыта хорошо развитой пластинкой эксоподита,
кончающейся оперенными щетинками. Пространство между эксопо-
дитом и фильтрационной решеткой закрывается в всасывательных
камерах третьей пары щетинками рудиментарных эндитов ног тре-
тьей пары, а в четвертых всасывательных камерах — направленным
внутрь выростом эксоподитов ног четвертой пары. Со спинной сто-
роны камеры ограничены брюшной стороной тела, по бокам кото-
рого проходят плотно прижатые к створкам раковины боковые вы-
росты, отделяющие полость фильтрационного аппарата от полости
выводковой камеры.
В фазу абдукции, сопровождающуюся расширением всасыва-
тельных камер, конечности, находившиеся до того в горизонтальном
положении, начинают двигаться в сагиттальной плоскости и обра-
зуют с телом острый угол. Исключение представляют ноги пятой
пары; боковая часть этих ног, состоящая из преэпиподита, эпипо-
дита и большой, окаймленной с обеих сторон волосками, щетинки
эксоподита, сгибается в фронтальной плоскости по линии сочлене-
ния с базальной частью ноги, наподобие движущейся на петлях
двери. Ноги этой пары, находившиеся вначале в трансверсальной
плоскости, почти одновременно с движением ног четвертой пары на-
чинают отходить назад и занимают, наконец, положение, почти па-
248
раллсльное продольной оси организма. Во время этого движения,
сопровождающегося увеличением объема четвертых всасыватель-
ных камер, ноги пятой пары не прерывают контакта со створками
раковины, скользя по ним щетинками эксоподитов. В фазу аддукции
эксоподиты ног четвертой пары отделяются от ног пятой пары, вода
выдавливается из всасывательных камер, чему способствует также
движение, вызываемое ногами пятой пары, поднимающимися в это
время в свое исходное положение.
Вдоль всей фильтрационной камеры проходит брюшной жело-
бок, в котором отфильтрованные частицы сестона направляются
вперед вместе с током воды, вызываемым движениями проксималь-
ных щетинок ног третьей и четвертой пар, а также длинных щети-
нск максиллярных выростов ног второй пары; остальные щетинки
максиллярных выростов служат для сжимания скопившихся между
ними пищевых частиц в комочки и для отправки их в ротовое от-
верстие.
Темп движения конечностей дафнии в зависимости от физиоло-
гического состояния организма достигает 200—300 ударов в ми-
нуту.
Пища состоит из мелких организмов наннопланктона и соответ-
ствующей величины частиц детрита, являющегося продуктом рас-
пада остатков животных и растений.
Изучение механизма отфильтровывания дафниями пищевых ча-
стиц проводится на живом организме, помещенном в небольшом
количестве воды на предметном стекле с углублением или в часо-
вом стекле. Вся работа распадается на три этапа.
1. Обнаруживание входящего и выходящего то-
ков воды. Для рассмотрения этих токов дафнию помещают в
воду с внесенным в нее мелко истертым порошком кармина или
иного красителя, не растворяющегося в воде, или китайской туши,
содержащей частицы сажи (краска тушь для этой цели не пригод-
на). Взвесь не должна быть очень густой, иначе трудно будет ве-
сти наблюдения. При надлежащем содержании взвеси легко обна-
ружить входящий в полость раковины ток, увлекающий с собой ча-
стицы кармина или другого вещества, и выходящую около постаб-
домена из этой полости воду, совершенно их лишенную. Одновре-
менно отмечают скопление отфильтрованного материала в брюш-
ном желобке и переход первой порции его в пищевод; определяют
также время, потребное для заполнения всего кишечника прогло-
ченным веществом.
2. Определение частоты ударов ног. Свеже пой-
манную дафнию помещают в часовое стекло и определяют число
сокращений ног в минуту; применение секундомера значительно
уточняет подсчет.
3. Изучение работы фильтрационного аппара-
т а. Это является наиболее трудной частью исследования. Для рас-
смотрения фазы абдукции, при которой конечности отходят от тела
и фазы аддукции, прижимания их к телу, наблюдения следует вести
не только на лежащем боком организме, но также и на располо-
женном брюшной стороной кверху; в последнем случае легко
249
видеть поднятие и опускание грудных ног и особый характер дви-
жения ног пятой пары. Для удержания дафнии в вертикальном
положении, лучше всего держать ее в небольшом количестве воды
е несколькими волоконцами ваты; легкими движениями препаро-
вальных игл обычно без особо большого труда удается поставить
дафнию в желаемом положении.
3. Apus
Щитень (Apus), описание строения которого помещено на стр.
132, относится к подотряду щитней (Notostraca).
Строение фильтрационного аппарата у щитней в принципе сход-
но со строением его у жаброногов. По брюшной стороне распола-
гаются двумя рядами всасывательные камеры, образованные груд-
ными конечностями. В фазу абдукции каждая всасывательная ка-
мера закрывается сверху эндоподитом, а с наружной стороны при-
жимающимися к щиту эксоподитом и эпиподитом находящейся
впереди конечности. Однако в противоположность другим фильтра-
торам из отряда жаброногих раков у щитней внутренняя сторона
всасывательных камер совершенно открыта благодаря отсутствию
на ногах специальных гребней щетинок. Вследствие этого всасыва-
ющиеся вместе с водой частицы ила направляются не только в
срединную центральную камеру, достигающую у этих организмов
большой ширины в передней части, но и во все всасывательные ка-
меры. В нормальном положении голова и передняя часть щита
прижаты к поверхности дна; вызываемые деятельностью всасыва-
тельных камер токи воды увлекают с собою частицы ила, счищае-
мые с грунта зазубренными с внутренней стороны эндоподитами
ног.
Благодаря этим особенностям строения конечности щитней мо-
гут задерживать лишь крупные частицы ила, являющиеся для них
пищей; мелкие же частицы, приходящие через щетинки ног, уносят-
ся вместе с выходящими при аддукции токами воды. Отфильтро-
ванные частицы ила движениями эндитов ног передвигаются к се-
редине брюшной стороны животного и направляются затем максил-
лярными выростами ног к ротовому отверстию. В задней части тела
оба ряда конечностей, сильно сближенные между собою и называе-
мые ундуляторами, служат для удаления мелких иловых частиц,
скапливающихся в задней части фильтрационной камеры.
Являясь примером активных фильтраторов, щитни не могут
считаться сестофагами в узком смысле этого слова. Они питаются
придонным детритом, однако могут вести хищный образ жизни,
питаясь живыми или мертвыми жаброногами, икрой лягушек, чер-
вями, мелкими ракообразными и т. п.
При изучении способа питания щитней рассматривается строе-
ние грудных ног на фиксированном материале.
4. Diaptomus и Calanus (рис. 109)
Диаптомус (Diaptomus), обычный представитель зоопланктона
пресноводных водоемов, и каланус (Calanus), обитающий в наших
северных морях, относятся к подотряду Calanoida отряда веслоно-
250
гих раков. Описание строения диаптомуса и калануса пометено на
стр. 10 и стр. 29.
Все Calanoida, за исключением лишь некоторых хищных видов,,
являются типичными активными фильтраторами, но, в противопо-
ложность ветвистоусым ракам, их грудные конечности в процессе
фильтрации не участвуют.
Рис. 109. Calanus (общий вид и строение конечностей изображено на рис. 12).
А — передний отдел тела, снизу (эндоподиты антенн, мандибулярные щупики и
дистальные части максиллул удалены), справа указано направление движения
ротовых придатков, слева — токи воды, вызываемые движениями ротовых при-
датков; Б — передний отдел тела, сбоку (антеннулы удалены); В — токи воды,
вызываемые движениями ротовых придатков во время скольжения организма.
/ — антеннула, 2 — антенна, 3---эндоподит антенны, 4 — эксоподит антенны.5 —
верхняя губа, 6 — мандибула, 7 — мандибулярный щупик, 8—максиллула, 9—
эксит максиллулы, 10 — максилла, 11 — максиллйрная нога, 12 — грудные ноги,
13— фильтрационная камера, 14—всасывательная камера*.
251
Получение пищи у диаптомуса' и калануса совершается автома-
тически при процеживании воды через фильтрационный аппарат,
образованный ротовыми придатками. Антенны хорошо развиты, со-
стоят из двучленистого протоподита, двучленистого эндоподита и
многочленистого эксоподита. Ротовые придатки представлены ман-
дибулами, максиллулами, максиллами и максиллярными ногами.
Мандибулы состоят из сильно развитого первого членика, протопо-
дита, и щупика, образованного вторым члеником протоподита и
причлененными к нему эндоподитом и эксоподитом. Максиллулы
имеют вид расчлененной пластинки, снабженной на конце эндоподи-
том и эксоподитом, по внутреннему краю несколькими эндитами,
а снаружи большим экситом; последний несет длинные и парал-
лельно идущие щетинки. Максиллы состоят из протоподита и мно-
гочленистого эндоподита с эндитами, вооруженными длинными ще-
тинками. Максиллярные ноги образованы двучленистым протопо-
дитом и многочленистым эндоподитом, несущими щетинки. Кроме
того, имеются верхняя губа и нижняя губа.
Базальные части антенн, мандибул и максиллул плотно соеди-
нены друг с другом, а также с верхней и нижней губами. В проти-
воположность этому, между максиллулами и максиллами, с тесно
прижатыми к ним максиллярными ногами, находится большой про-
межуток, в котором расположены длинные и параллельно идущие
щетинки максилл, сходящиеся своими концами у ротового отвер-
стия. Полость треугольного очертания, находящаяся между стен-
ками, образованными щетинками максилл, является фильтрацион?
ной камерой. Некоторые из щетинок, а именно те, которые распо-
ложены ближе к телу организма, изогнуты в первой трети, наруж-
ные щетинки проходят прямолинейно; вследствие этого фильтра-
ционная камера в глубине, т. е. около брюшной поверхности жи-
вотного, становится более узкой, чем в наружных частях. По всей
своей длине фильтрационные щетинки несут косо отходящие во-
лоски, благодаря которым стенки фильтрационной камеры приобре-
тают характер очень тонкого фильтра, не пропускающего даже
мельчайшие организмы и детрит.
В результате частых ударов эндоподитов антенн, мандибуляр-
ных щупиков и дистальных частей максиллул образуются токи
воды, которые, с одной стороны, вызывают то скольжение живот-
ного в воде, которое уже ранее рассматривалось (стр. 17), а с дру-
гой стороны, приносят в фильтрационную камеру организмы нан-
нопланктона и мелкий детрит, служащие пищевым материалом.
Главная роль в образовании пищевого тока, несущего мелкие ча
стицы по направлению к фильтрационной камере, принадлежит ан-
теннам.
Движение ротовых придатков можно изучить только при приме-
нении киносъемки, так как частота ударов достигает у калануса
1200, а у диаптомуса 2700 раз в минуту, другими словами длитель-
ность каждого удара не превышает 1/20—1/45 секунды. Отфильтро-
ванные частицы продвигаются к ротовому отверстию крепкими ще-
тинками, расположенными на эндитах максиллул, и всасываются
затем в пищевод. Для проглатывания сравнительно крупных орга-
252
низмов, как, например, диатомей, мандибулы могут раздвигаться в
стороны. Большие частицы, не проходящие в пищевод, выбрасы-
ваются наружу, для чего щетинки максилл отходят от нижней губы.
При изучении способа питания веслоногих раков рассматривает-
ся строение ротовых частей калануса или диаптомуса. При содер-
жании живых диаптомусов в воде, с взвесью кармина или иного
вещества, не растворяющегося в воде, можно через некоторое вре-
мя обнаружить его присутствие внутри кишечника животного.
5. Anodonta и Unio (рис. 110)
Беззубка (Anodonta) и перловица (Unio), описание которых по-
мещено на стр. 140, относятся к числу самых крупных представи-
телей двустворчатых моллюсков пресноводных водоемов.
Рис. 110. Anodonta. А — вид сзади; Б — поперечный разрез через середину
тела; В — поперечный разрез через заднюю часть тела.
/ — выводной сифон, 2 — вводной сифон, 3 — спинное мантийное отверстие, 4—
брюшные края мантийных складок, 5— лигамент, 6 — прямая кишка, 7 — пери-
кардий, 8—почка, 9— наружный наджаберный канал, 10— внутренний наджа-
берный канал, 11 — наружная и внутренняя полужабры, 12 — внутрижаберный
канал, 13— перерезанные петли кишечника, 14 — нога, 15 — задний аддуктор.
Пища двустворчатых моллюсков, состоящая из мельчайших ча-
стиц сестона, вносится внутрь мантийной полости через вводной
сифон вместе с током воды, вызываемым мерцанием ресничного
эпителия покровов мантии, жабр и поверхности тела. Через много-
численные мелкие отверстия жаберных пластинок вода проходит в
внутрижаберные каналы, оставляя принесенные пищевые частицы
на стенках жабр, покрытых слизью, а затем в наджаберные кана-
лы, открывающиеся в клоаку; вода из выводного сифона вытекает
лишенной взвешенных частиц. Отфильтрованные частицы взвеси,
склеенные слизью, переносятся движениями ресничек к ротовому
Ж
отверстию. Излишнее количество их скапливается в поджаберной
полости и затем выбрасывается наружу в виде псевдоэкскремен-
тов. Процесс фильтрации происходит непрерывно, так как он связан
с получением необходимого организмам кислорода. В зависимости
от размера моллюска в час профильтровываются десятки и сотни
кубических сантиметров воды.
Поглощение взвеси наблюдается на живых моллюсках, находя-
щихся в аквариуме с чистой водой. Если пипеткой внести некоторое
количество мутной взвеси, приготовленной из глины или иного .ве-
щества, в пространстве около сифонов, то можно видеть втягивание
ее через вводной сифон, и одновременно выхожденпе чистой воды
из выводного сифона.
6. Личинки Anopheles
Личинки Anopheles относятся к числу нейстофагов, питаются
нейстическими организмами, профильтровывая их через щетинки
ротовых придатков. Описание строения личинок Anopheles поме-
щено на стр. 188.
7. Oikopleura (рис. 111)
Одними из самых своеобразно устроенных активных фильтратов
являются представители класса аппендикулярий (Appendicularia),
относящегося к оболочникам (Tunicata).
Аппендикулярии ведут планктический образ жизни. Встречаются
в морях; у нас обычными представителями планктона северных мо-
рей являются виды Oikopleura.
Тело Oikopleura, достигающее в длину 4—7 мм, удлиненно
овальной формы. От брюшной стороны отходит так называемый
хвост, являющийся органом движения. По отношению к продольной
оси тела хвост повернут на 90°, так что его левый край, в сущности,
является спинным, а правый край — брюшным. По всей длине хво-
ста посреди проходит хорда, окруженная мышцами. Длина хвоста
в несколько раз превышает длину самого организма.
На переднем конце тела помещается рот, ведущий в обширную
жаберную полость; две жаберных щели открываются наружу, на
брюшной стороне тела перед основанием хвоста. Внизу жаберной
полости находится эндостиль. Идущий косо вверх пищевод впадает
в обширный желудок; небольшая задняя кишка открывается на
брюшной стороне тела. Oikopleura—гермафродиты; у зрелых особей
позади желудка видны мужские и женские гоноды. На переднем
конце тела находятся группы железистых эпителиальных клеток,
участвующих в образовании сложно устроенного фильтрационного
аппарата.
Тело аппендикулярий частично или полностью, как, например,
у Oikopleura, заключено в студенистый, очень нежного строения
домик, легко разрушающийся при фиксации или механическом воз-
действии. Этот домик, служащий для ловли организмов наннопланк-
тона, представляет редкий среди животного мира случай образова-
ния фильтрационного аппарата из сильно разбухшей и пропитанной
254
водой кутикулы. Под воздействием ундулирующих движений хвост;
внутрь домика входит ток воды, увлекающий с собою мельчайшие
формы наннопланктона, являющегося основной пищей аппендику
лярий. Более крупные организмы внутрь домика не проникают, та»
как задерживаются решеткой из тонких, параллельно протянуты;
нитей, расположенных в начале приводящих каналов. Вода вместе
с пищевыми организмами проходит затем через расположенный е
Рис. 111. А — OiKopleura в домике; Б — схематический поперечный разрез че-
рез домик Oikopleura; В, Г— OiKopleura labradoriensis, общий вид (В), тело
без хвоста при большем увеличении (Г); Д — зачаток домика OiKopleura.
/—туловище, 2— хвост, 3— дорсальная камера, 4 — вентральная камера, 5—
сетка входной воронки, 6 — фильтрационный аппарат, 7 — место выхода орга-
низма из домика, 8 — слизистый слой, 9 — хорда, 10 — ротовое отверстие,
11 — нижняя губа, 12 — эндостиль, 13— жаберная полость, 14 — жаберная
щель, 15, 16 — группы эпителиальных клеток, образующих фильтрационный аппа-
рат (15) и входную воронку (16) домика, 17 — пищевод, 18 — желудок, 19—
семенник, 20— яичник, 21 — зачаток домика. Стрелки обозначают токи воды:
I— ток, входящий в домик, II— ток, приносящий наннопланктон к фильтра-
ционному аппарату, III — ток, приносящий отфильтрованные пищевые частицы
к ротовому отверстию, IV — ток, выходящий из домика.
задней части домика фильтрационный аппарат, очень тонкого и
сложного строения. Отверстия фильтрационной сетки настолько
малы, что задерживают даже мельчайшие организмы наннопланкто-
на, по величине не превышающие нескольких микронов. Отфильтро-
ванная пища затем всасывается аппендикулярней внутрь тела по
короткой трубке, ведущей от фильтрационного аппарата ко рту.
Так как рассмотреть функционирующий домик аппендикулярии
на фиксированном материале невозможно, то рассматривают зача-
ток нового домика, обычно облегающий передний конец тела в виде
футляра. При большом увеличении можно обнаружить решетку
входных отверстий, а на противоположном конце смятые, еще не
расправленные части фильтрационного аппарата.
Питание пассивных фильтраторов
В качестве примеров пассивных фильтпатов рассматриваются
личинки живущих в текучей воде мошек (Simulium). При помощи
перистых придатков (вееров), расположенных в передней части го-
ловы, личинки улавливают сносимый по течению сестон. Описание
строения личинок мошек помещено на стр. 182.
СПОСОБЫ ПИТАНИЯ БЕНТОФАГОВ
В качестве примеров хищных бентофагов рассматриваются ли-
чинки стрекоз (Odonata), водные клопы (Nepa, Ranatra) и личинки
жука плавунца (Dytiscus).
1. Личинки Odonata
Личинки стрекоз захватывают пишу при помощи маски, преоб-
разованной из нижней губы. Описание строения масок различных
типов помещено на стр. 153—167.
2. Nepa и Ranatra
Свою добычу водяной скоропион (Nepa) и ранатра . (Ranatra)
захватывают при помощи грудных ног первой пары. Описание
строения Nepa и Ranatra помещено на стр. 201.
3. Личинки Dytiscus (рис. 112)
Личинки плавунца (Dytiscus) являются одними из самых боль-
ших хищников среди всего населения пресноводных водоемов.
Тело личинки состоит из широкой уплощенной головы, трехчле-
нистой груди, с тремя парами грудных ног, и длинного восьмичле-
нистого брюшка, оканчивающегося двумя церками.
По бокам головы расположены по шести небольших черных
глазков. Перед ними находятся тонкие антенны, трехчленистые у
молодых личинок и шестичленистые у личинок третьей стадии. Все
ротовые придатки вытянуты в поперечном направлении, видны с
нижней стороны головы. Верхняя губа отсутствует, над ротовыми
придатками простирается передний край головной капсулы. Среднее
256
положение занимает нижняя губа, имеющая вид широкой пластин-
ки с двумя тонкими нижнегубными щупиками. По бокам нижней
губы расположены тонкие максиллы, состоящие из палочковидного
основного членика и максиллярного щупика, трехчленистого у мо-
Рис. 112. Личинка Dytiscus. Л — общий вид; Б — голова снизу; В — мандибула;
Г — поперечный разрез через мандибул.
1 — антенна, 2—мандибула, 3—максилла, 4 — максиллярный щупик, 5—ниж-
няя губа, 6 — пижнегубной щупик, 7 — глаза, 8 — полость мандибулы, 9 —
полость канала, 10 — входное отверстие канала, И—щель канала, 12 — выход-
ное отверстие канала.
лодых и шестичленистого у более взрослых личинок. Наиболее
своеобразно строение больших крепких мандибул, серповидно изо-
гнутых и на конце игловидно заостренных. Вдоль внутреннего края
мандибул проходит канал, открывающийся около их конца продол-
говато-овальной щелью, а при основании большим круглым отвер-
17 В. А. Яшков Ш
стнем, хорошо заметным при отгибании мандибул наружу. Канал
образован глубокой складкой хитина, края которой сближены меж-
ду собой. Мандибулы двигаются только в одной горизонтальной
плоскости. Ротовая щель, проходящая поперек головы, замкнута по
всей длине, за исключением самых боковых частей, в которые по-
гружены основания мандибул.
У личинок плавунцов наблюдается редкий случай внекишечного
пищеварения. Схватив добычу (личинок насекомых, головастиков,
небольших рыб и т. п.) своими мандибулами, личинка погружает
их затем глубоко в тело жертвы и по каналам изливает пищевари-
тельный сок; образовавшаяся после переваривания тканей добычи
жидкость всасывается личинкой в полость глотки через каналы в
мандибулах. Спустя некоторое время от жертвы остаются только
одни наружные покровы.
Личинки плавунцов дышат атмосферным воздухом через задние
стигмы, находящиеся на конце последнего, восьмого, сегмента. Кроме
этих стигм, имеются еще не функционирующие стигмы, расположен-
ные по бокам брюшных сегментов. Боковые края двух последних сег-
ментов брюшка вооружены рядами плавательных щетинок. Одно-
членистые церки, несущие по краям длинные плавательные щетин-
ки, при подъеме личинки на поверхность воды сгибаются под пря-
мым углом к продольной оси личинки и притягиваются капиллярным
притяжением к поверхностной пленке воды; находящиеся на конце
брюшка несмачиваемые водой стигмы прорывают пленку воды,
благодаря этому устанавливается контакт трахейной системы с ат-
мосферным воздухом.
При изучении питания личинок плавунца рассматривается строе-
ние ротовых придатков; при кормлении голодных личинок какой-
либо животной пищей можно наблюдать, как она переваривается
вне тела личинки.
ПРАКТИЧЕСКИЕ УКАЗАНИЯ
Большинство пресноводных организмов, рассматриваемых в
Практикуме по гидробиологии, относится к числу широко распро-
страненных и легко может быть собрано обычными орудиями лова.
Труднее доступны личинки Polymitarcys, Palingenia и Ephemera,
которых следует искать в глинистых берегах больших рек, и личин-
ки Mansonia, для сбора которых лучше всего вырывать водяные ра-
стения с корнями, а затем трясти корни в большом сосуде с водой.
Встречающиеся в горных реках личинки Blepharoceridae, Heptage-
nia и Ecdyonurus можно получить при содействии Северокавказской
биологической станции в Дзауджикау, а морские реликты Онеж-
ского озера при содействии Биологической станции в Петроза-
водске.
Морские организмы можно получить при содействии Севасто-
польской биологической станции Академии наук в Севастополе,
Полярного института морского рыбного хозяйства и океанографии
(Пинро) в Мурманске, Азово-Черноморского института морского
рыбного хозяйства и океанографии (Азчерниро) в Керчи, Кас-
пийского отделения Всесоюзного института морского рыбного хозяй-
ства и океанографии в Астрахани, Балтийского отделения Всесоюз-
ного института морского рыбного хозяйства и океанографии в Риге.
При фиксации тех организмов, которые не могут быть рассмот-
рены в живом состоянии, применяют спирт или формалин. Для
личинок насекомых хорошие результаты получаются при фиксации
горячей водой с последующим переносом в спирт. Для просветления
объектов применяют глицерин или, после проводки через спирты
возрастающей крепости и абсолютный спирт, гвоздичное или кедро-
вое масла.
Для начинающих некоторое затруднение представляет препари-
рование мелких объектов (Copepoda, Cladocera и т. п.). Работа
проводится следующим образом. Сначала помещают один исследуе-
мый экземпляр на предметное стекло в каплю воды, к.которой для
уменьшения хрупкости объекта, если материал фиксирован форма-
лином, прибавляют немного глицерина. Под лупой, а еще лучше под
бинокуляром, организм кладется при помощи препаровальных игл
в таком положении, чтобы были видны отделяемые придатки, затем
иглой, находящейся в левой руке, его прокалывают и прижимают к
]7* 259
предметному стеклу. После этого другой иглой осторожно надавли-
вают на основание придатков и отделяют их от тела организма.
Отделенные придатки при помощи иглы или кисточки из нескольких
волосков переносятся в капли воды, заранее приготовленные на
других предметных стеклах. Перенос облегчается, если иглу с под-
детым придатком перевернуть з воздухе на 180°, в этом случае при
прикосновении иглы к поверхности капли переносимый придаток
легко соскальзывает в воду. После этого покрывают препарат по-
кровным стеклом. Успешность препаровки мелких объектов зависит
от препаровальных игл, особенно той, которая служит для отделе-
ния придатков от тела организма. Наиболее пригодными являются
иглы, приготовленные из тонких энтомологических булавок, которые
или привязываются ниткой к деревянным основаниям или укрепля-
ются в стеклянных трубках, диаметром в 5—6 мм и длиною около
15 см; в последнем случае один конец трубки сначала оттягивается
на огне, чтобы уменьшить диаметр отверстия до !•—2 мм, затем в
этот конец помещают немного сургуча, нагревают его, пока он не
заполнит весь оттянутый конец трубки, и вставляет в еще расплав-
ленный сургуч энтомологическую булавку. Свободный конец булав-
ки должен высовываться приблизительно на 2 см. При пользовании
обычными препаровальными иглами необходимо их предварительно
отточить до необходимой тонины.
Препараты обычно рассматриваются в неокрашенном виде. В
случае необходимости, хитин окрашивается водным раствором пи-
криновой кислоты.
Для получения препарата из хитинового скелета помещают весь
организм или какую-либо часть его на сутки в крепкий раствор ед-
кой щелочи или кипятят в этом растворе до растворения мягких
частей. После промывки водой препарат проводят через спирты
возрастающей крепости и заключают в кедровое масло.
Обязательным условием является зарисовка общего вида рас-
сматриваемого организма и его отдельных придатков.
ЛИТЕРАТУРА
Липин А. Н., Пресные воды 'и их жизнь, изд. 3-е, Учпедгиз, 1950.
Павловский Е. Н. и Л е п н е в а С. Г., Очерки из жизни пресноводных
животных, Советская наука, 1948.
Большой практикум по зоологии беспозвоночных. Под ред. проф. Ю. И. П о-
л я нс к о го, Советская наука, ч. I, 1940; ч. II, 1946.
Жизнь пресных вод СССР. Под ред. проф. В. И. Жадина, Изд. Академии
наук СССР, т. I, 1940; т. II, 1949.
Определитель фауны и флоры северных морей СССР. Под ред. проф.
Н. С. Гаевской, Советская наука, 1948.
ОГЛАВЛЕНИЕ
Предисловие
Стр.
3
Организмы планктона
Определение скорости погружения анестезированных планктическнх
организмов 9
1. Скорость погружения дафнии (Daphnia) 9
2. Скорость погружения коловраток (Rotatoria)
Органы движения планктическнх организмов 9
Движение при помощи грудных ног 9
1. Diaptomus 10
2. Cyclops 13
Движение при помощи антенн 17
1. Daphnia 2. Bosmina я
Движение при помощи коловращательного аппарата . 24
Приспособления к парению 24
Уменьшение остаточного веса 24
Редукция скелета . 24
1. Clione .... • 24
2. Limacinai ... • 26
Пропитывание водой 27
1. Eudorina 27
2. Noctiluca . . • • 27
3. Bougainvillia 28
4. Holopedium . ... £19 29
Жировые включения 29
Calanus . ... 30
Газовые включения 1. Anabaena, Aphanizomenon и Nodularia . 31 оч
2. Личинки и куколки Chaoborus . 3. Личинки и куколки МосЫопук . Ок/ 37 38
Увеличение сопротивления формы . 38
Удлинение одной оси . 38
1. Rhizosolenia и Melosira . 261
<?тр.
2. Leptodora 40 J d
3. Sagitta и Eukrohnia . 42
Удлинение двух осей . • • 1. Coscinodiscus и Cyclotella . 44
2. Asterionella 46
3. Tabellaria 46
4. Pediastrum 46
Образование выростов . 47
1, Chaetoceros 47
2. Ceratium 49
3. Bythotrephes 51
4. Notholca . 53
5. Личинки Porcellana . • 53
Организмы бентоса
Прикрепленные организмы .... 54
1. Spongilla и Ephydatia 56
58
3. Balanus и Chthamalus 59
4. Личинки и куколки Blepharoceridae 61
5. Личинки и куколки Simulium . 63
Лежащие организмы 63
Pecten .... . . 63
Закапывающиеся организмы ... 65
1. Arenicola ... 65
2. Solen 67
3. Личинки Polymitarcys, Palingenia и Ephemera . 67
Сверлящие организмы .... 69
1. Teredo .... 69
2. Pholas и Bamea 71
3. Petricola .... * • • t • 71
4. Limnoria 72
Свободно двигающиеся организмы 72
Органы движения нектобентических организмов 72
1. Notonecta . 73
2. Dytiscus 73
3. Gyrinus 75
Организмы нейстона
1. Scapholeberis
2. Личинки Anopheles
Организмы перифитона
Защитные
1
2
3
4
262
приспособления при наступлении и.х а
Эфиппии Daphnia У " " неблагоиРиятных условий
Геммулы пресноводных губок . '...........................86
Статобласты пресноводных мшанок ' ' ‘ ‘ »7
селение периодически смачиваемых и ‘ 8в
иваемых микроводоемов с
Организмы и соленость воды
Стеногалинные организмы . Пресноводные организмы Морские организмы Солоноватоводные организмы Стр. 93 ’93 93 93
1. Brachionus plicatilis и Pedalia oxyure 2. Caspievadne и Podonevadne . 3. Corniger 4. Cercopagis Ультрагалинные организмы Artemi a . 93 94 96 96 98 98
Эвригалинные организмы . 1. Enteromorpha . 2. Mytilus . 3. Cardium . 4. Mya . 5. Macoma . 100 100 100 ioi 102 103
Морские реликты .103 Руководящие ископаемые формы ЮЗ 1. Portlandia arctica .... . . 103 2. Ancylus fluviatilis ... . . 104 3. Littorina littorea 104 Морские реликты иольдиевого времени . . . . 105 1. Limnocalanus grimaldii и L. macrurus . . . 105 2. Mysis oculata и M. oculata var. relicta . . . . 107 3. Mesidothea sibirica и M. entomon 108 4. Gammaracanthus loricatus и G. loricatus var. lacustris . 110 5. Myoxocephalus quadricornis и M. quadricornis var. onegen- sis и var. lonnbergi 112
Эксперимент c Artemia salina 113
Организмы — показатели загрязнения воды
1. Sphaerotilus 2. Beggiatoa . 3. Oscilatoria ... 4. Tubifex, Limnodrilus и Lumbriculus . 5. Asellus ... 6. Личинки Chironomus 7. Личинки Eristalis 117 118 118 118 . 121 123 123
Организмы и активная реакция среды
Изменение активной реакции воды под влиянием жизнедеятельности
Определений ' границ выживаемости’организмов при различной величине
активной реакции воды..............................................
* ‘ 263
Органы дыхания водных организмов
Стр.
Кбжное дыхание ... . 12Й
1. Leptodora . 2. Polyphemus 128 128
Кровяные жабры . 1. 130
1. Arenicola . . . 130
2. Pristicephalus 130
3. Apus 132
4. Lynceus . 134
5. Gammarus ... 136
6. Asellus и Mesidothea 139
7. Valvata ... . 139
8. Anodonta и Unio 140
Трахейные жабры 143
Личинки Ephemeroptera 144
Плавающие личинки . . . 145
1. Личинки С1оёоп . . . . 145
2. Личинки Leptophlebia 147
3. Личинки Siphlonurus < 147
Ползающие личинки . ... 148
4. Личинки Ephemerella .... 148
5. Личинки Ordella ...... 149
Роющие личинки 150
Плоские, прилегающие к субстрату личинки . 150
6. Личинки Heptagenia 150
7. Личинки Ecdyonurus . . 152
8. Личинки Oligoneuriella . . .? . 152
Личинки Odonata . 153
Личинки без хвостовых пластинок (Anisoptera) . 157
1. Личинки Aeschna 157
2. Личинки Cordulegaster 159
3. Личинки Cordulia . 159
4. Личинки Epitheca 159
5. Личинки Libellula 160
6. Личинки Sympetrum 162
Личинки с хвостовыми пластинками (Zygoptera) . 162
7. Личинки Agrion . 162
8. Личинки Lestes . . . 164
9. Личинки Erythromma ... 165
10. Личинки Enallagma . ... 165
11. Личинки Coenagrion . 165
Личинки Plecoptera .... .... 167
1. Личинки Perla ... .... 168
2. Личинки Nephelopteryx . .... 170
Личинки Megaloptera .... ...» , 170
Личинки Sialis , • 170
Личинки и куколки Trichoptera . .... . . 171
Гусеницевидные личинки . * . , 173
264
1. Личинки и куколки Phryganea
2. Личинки и куколки Limnophilus
3. Личинки и куколки Grammotaulius
4. Личинки и куколки Stenophylax .
5. Личинки и куколки Molanna
6. Личинки и куколки Leptocerus .
Камподеовидные личинки .
7. Личинки и куколки Hydropsyche
8. Личинки и куколки Rhyacophila .
9. Личинки и куколки Neureclipsis
Личинки и куколки Simuliidae
Дыхание атмосферным кислородом .
Личинки и куколки Culicidae
Личинки без сифона (Anophelini)
1. Личинки и куколки Anopheles
Личинки с сифоном (Culicini)
2. Личинки и куколки Culex
3. Личинки и куколки Аёбеэ
4. Личинки и куколки Theobaldia .
5. Личинки и куколки Mansonia .
Водные Hemiptera . . . .
1. Nepa . .
2. Ranatra .
Личинки Brachycera .
Организмы и температура
Стенотермные организмы ./..........................
Стенотермные тепловодные организмы .
Стенотермные холодноводные организмы .
Эвритермные организмы ...
Сезонные изменения планктона
Цикломорфоз....................................
1. Цикломорфоз диатомей .
2. Цикломорфоз перидиней .
3. Цикломорфоз Keratella cochlearis .
4. Цикломорфоз Keratella quadrata .
5. Цикломорфоз Brachionus calyciflorus
6. Цикломорфоз Brachionus capsuliflorus .
7. Цикломорфоз Daphnia cucullata .
8. Цикломорфоз Daphnia cristata .
9. Цикломорфоз Bosmina coregoni
Организмы н свет
Фототропизм ...............................
Суточные вертикальные миграции планктона .
Светящиеся организмы ..-•••
Органы свечения .
1. Светящиеся рыбы .
2. Euphausiacea
Стр.
174
175
177
178
178
179
180
180
182
182
182
186
186
188
188
192
193
195
198
198
. 201
201
. 201
203
205
205
205
205
205
206
206
206
207
209
209
210
211
212
213
215
216
216
216
216
216
'65
Питание водных животных Стр.
Пищевые организмы • ’ личинки и куколки ^^ымипластинками (Chironominae) Личинки с паралабиальными пл- 1. Личинки и куколки Chironomus 221 222 225 225
2. Личинки и куколки Glyptotendipes .... 228
3. Личинки и куколки Endochironomus 230
4.. Личинки и куколки Polypedilum 230
5. Личинки и куколки Cryptochironomus 232
6. Личинки и куколки Stictochironomus 233
7. Личинки и куколки Tanytarsus 233
8. Личинки и куколки Lauterbornia 236
9. Личинки и куколки Micropsectra 236
Личинки с втяжными антеннами (Pelopiinae) 236
10. Личинки и куколки Procladius (=Tanypus) 237
11. Личинки и куколки Ablabesmyia (=Pelopia) Личинки без паралабиальных пластинок и без втяжных антенн 237
(Orthocladiinae) 240
12. Личинки и куколки Cricotopus ... 240
Пища водных животных • 241
Способы питания сестофагов ... 241
Питание ощупывателей .... ... 241
Питание седиментаторов ... ... 243
Питание активных фильтраторов ... 244
1. Pristicephalus .... ... . 244
2. Daphnia ... . 246
3. Apus ... 250
4. Diatomus и Calanus , . 250
5. Anodonta и Unio . 253
6. Личинки Anopheles . 254
7. Oikopleura . 254
Питание пассивных фильтраторов . 256
Способы питания бентофагов . 256
1. Личинки Odonata . 256
2. Nepa и Ranatra 256
3. Личинки Dytiscus 256
Практические указания
Литература . 2*
Редактор Н. М. Передельская
Техн, редактор Л. И. Стрельникова
Подписано к печати 9/Х—1952 г.
Т 07947 Тир. 5000 экз.
Бумага 60Х921/1С=33/8 бум. л. 163/4
п. л. 18,5 изд. л.
Издательство „Советская наука".
Зак. 47 Цена 6 руб. 55 коп.
(в переплёте)
Тип. изд-ва „Советская наука",
Неглинная ул., д. 29/14. Зак. 1154