/




















































































































































































































































































































































Текст
П.В. ТЕРЕНТЬЕВ
ГЕРПЕТОЛОГИЯ
ВЫСШАЯ ШКОЛА
1961
Проф. П. В. ТЕРЕНТЬЕВ
ГЕРПЕТОЛОГИЯ
УЧЕНИЕ О ЗЕМНОВОДНЫХ И ПРЕСМЫКАЮЩИХСЯ
Допущено
Министерством высшего и среднего
специального образования СССР
в качестве учебного пособия
для государственных
университетов
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
«ВЫСШАЯ ШКОЛА»
М о с к в а—1961
ПРЕДИСЛОВИЕ
В течение почти четверти века мне пришлось преподавать
герпетологию в Ленинградском университете то как самостоя-
тельный специальный курс, то как часть большого практикума.
В процессе этой работы была составлена обширная (более
12000 названий) библиография и написана рукопись в 50 печат-
ных листов. Предлагаемое пособие представляет краткое изло-
жение этого материала.
Задача предлагаемой книги дать целостный очерк фило-
генеза, частной систематики и других материалов, касаю-
щихся низших наземных позвоночных.
Предполагается, что студенты, для которых предназначено
настоящее пособие, уже знакомы с предметом, по крайней мере,
в объеме общего курса зоологии позвоночных. Такая предпо-
сылка дает возможность не останавливаться на ряде терминов
и понятий. Конечно, было бы полезно дать общий очерк разных
сторон, морфологии, экологии и распространения, но для этого
потребовалось бы значительно расширить объем этой книги.
Поэтому основное место отведено частной филогении, экологии
и географическому распространению.
Изложение сознательно асимметрично. Палеонтологические
сведения кратки. Необходимость экономии места заставила из-
лагать отличия групп в форме шведских дихотомических таблиц.
Предполагается, что студент будет использовать их не столько для
определения, сколько для внимательного изучения, причем обя-
зательно с проекцией на филогенетические схемы. Изложенные
в таблицах данные не приводятся в дальнейшем тексте. Для
сбережения места то, что явствует из рисунков, по возможности
не излагается в тексте. В первую очередь это касается филогене-
тических взаимоотношений и географического распространения.
Филогенетические схемы современных групп доведены до
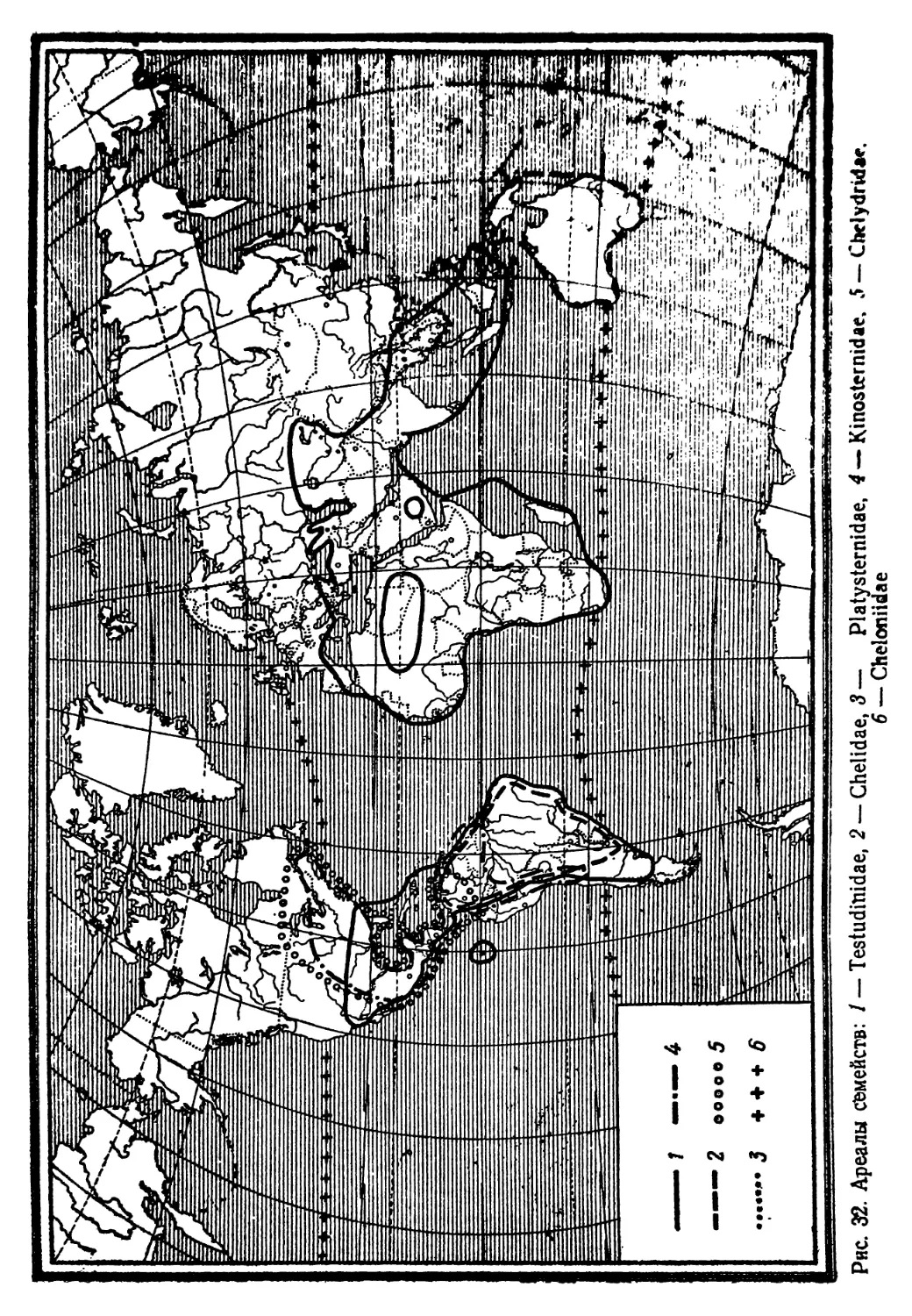
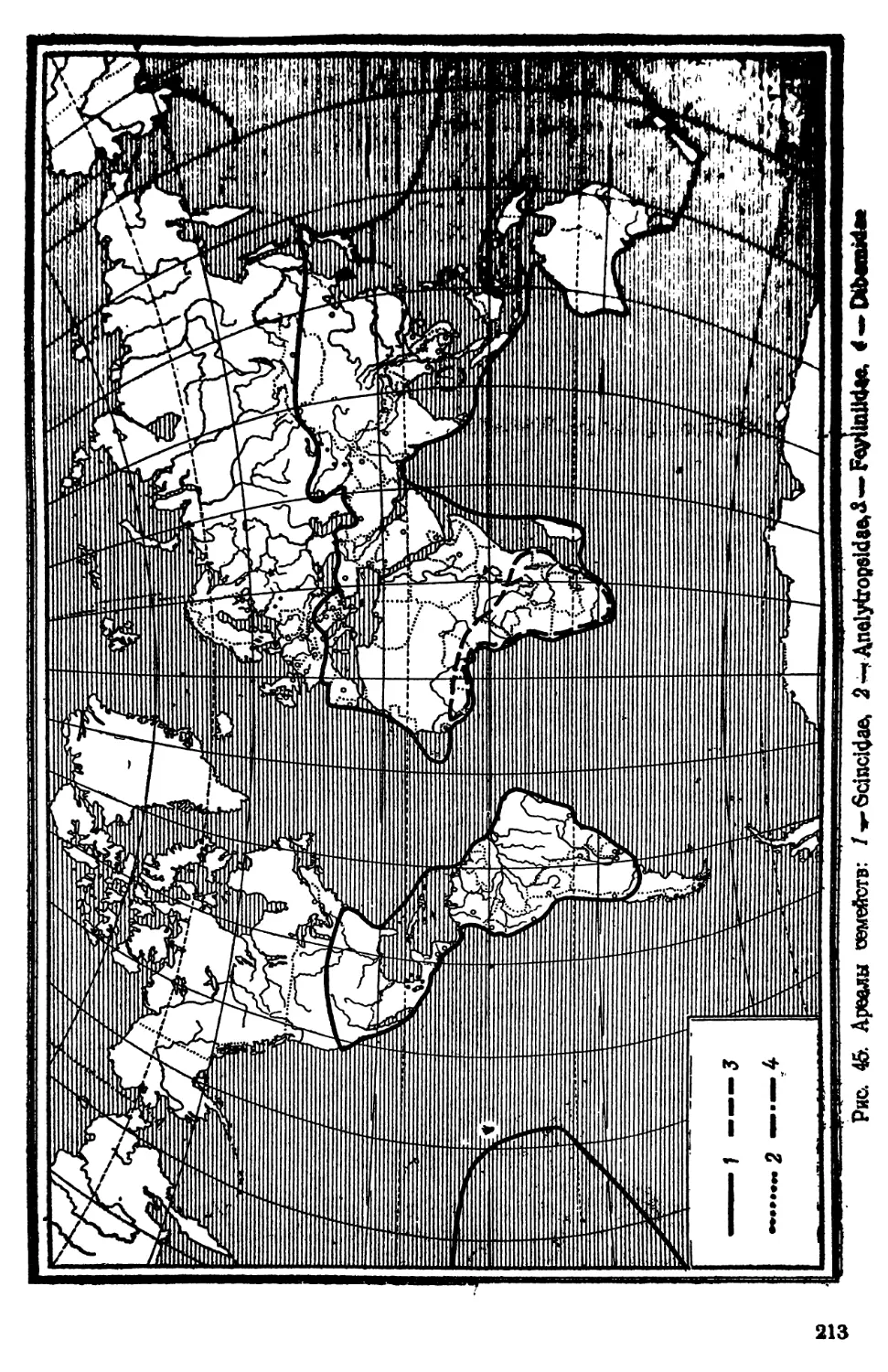
семейств, для ископаемых — до отрядов. Ареалы всех современ-
ных семейств картированы. Нумерация семейств имеет чисто
техническое значение, и ей не следует придавать особого смысла.
В библиографии указываются преимущественно сводки, в ко-
торых читатель найдет вполне исчерпывающую литературу по
3
ппппмету В перечнях иностранной литературы тома обознача-
ются жирной цифрой, а страницы — двоеточием.
Пользуюсь случаем выразить искреннюю благодарность про-
фессорам Б. С. Матвееву и А. Г. Банникову и доценту А. М. Алек-
папову за ценные указания и исправления.
Чувство глубокой признательности заставляет меня посвя-
тить эту книгу памяти моего учителя покойного профессора Мос-
ковского университета Григория Александровича Кожевникова.
Павел Терентьев
Глава первая
ИСТОРИЯ ГЕРПЕТОЛОГИИ
Ветвь зоологии, изучающая пресмыкающихся, носит назва-
ние «герпетология» (от греческого «герпетон»— ползучее жи-
вотное, змея и «логос» — слово, учение). Строго рассуждая, уче-
ние о земноводных надлежит именовать «батрахологией» (от
греческого «батрахос» — голый гад) или амфибиологией. Однако
практика показала, что специалисты, занимающиеся пресмыкаю-
щимися, обыкновенно занимаются и земноводными. Поэтому
обычно в понятие «герпетология» включают и учение о земно-
водных
Несомненно, что человек обратил внимание на змей, кроко-
дилов, ящериц, черепах, жаб и лягушек уже в самые отдаленные
времена. Причина этому — ядовитость и сила некоторых змей,
хищность^крокодилов, едкрстькожного сока жаб и своеобразная
внешность всех этих животных. Кроме того, многие тропические
формы издавна употреблялись в пищу или для поделок (кожа
крокодилов, панцири черепах и т. п.). Особенное внимание уде-
лялось змеям, часто являвшимся тотемами. С этим связывался
ряд обрядов, приводивший иногда к созданию культа змей—•
«офиолатрии»^ Интересно, что еще в 1927 г. в Пенанге существо-
вал китайский храм, где содержались многочисленные змеи.
Змеям приписывалось много чудодейственных свойств. Мысль о
том, что змеи сообщают людям понимание языка всех тварей,
возможно берет начало от галлюцинаторных представлений лю-
дей, отравленных змеиным ядом. Крокодилы и, в меньшей сте-
пени, черепахи также являлись тотемическими животными ряда
племен. Ряд земноводных получил от зоологов мифологические
имена (тритон, протей), но всеобщее внимание привлекали
жабы.
Переходя к зарождению, научной зоологии, следует вспом-
нить рассказ Гекатея Милетского (конец VI — начало V в.
до н. э.) о нильском крокодиле и попытку расшифровать миф о
Кербере как о ядовитой змее. Первым из античных авторов,
’ Автором термина следует считать, по-видимому, Я. Клейиа (1756), хо
тя он понимал этот термин оде шире, объединяя змей с червями.
5
давшим значительное количество сведений о пресмыкающихся
и земноводных, был Аристотель (384 322 гг. до н. э.). Он уже
знал о наличии у змей только одного легкого, разбирал вопрос о
жи-вородности гадюки и т. п.]К сожалению, в работах Плиния
Старшего (23—79 гг. н. э.) верные наблюдения тонут в массе
выдумок и сказок (например, утверждение, что хамелеон питает-
ся воздухом, что слюна саламандры уничтожает волосы на чело-
веческом теле и пр.). В дальнейшем удельный вес фантастики
все возрастал и в естественнонаучных сборниках средневековья
«Физиологах» (Carus, 1880) она составляла основной фон. Неко-
торое улучшение наступило в области зоологии лишь в XIII в.,
когда в Европе вновь обратили внимание на сочинения Аристо-
теля. В 1467 г. вышла, по-видимому, первая печатная книга,
касающаяся между прочим змей (Rabanus, 1467).
- Исходным пунктом, приведшим к рождению современной
зоологии, можно считать работы Конрада Геснера (1516—1565),
Гадам посвящены 2-я и 5-я книги его «Истории животных»
(1554, 1587).[Животные им описываются еще в алфавитном по-
рядке. Специально гадам посвящена книга Улисса Альдрованди
(Aldrovandus, 1640). Сказочный элемент занимает у этих авто*
ров все еще много места — «василиск», драконы и другие чу-
дища рассматриваются как реальные животные. Только в
XVIII в. такие названия как «Draco» и «Basilicus» были при-
своены реальным ящерицам. Однако отзвук сказочных драконов
дошел и до наших дней в легенде о «гигантской морской змее»,
сведения о которой проскальзывают время от времени даже в ра-
ботах XX в.
Творец понятия вида и системы животных Джон Рей (1628—
1705) различал водных и древесных лягушек, жаб, 10 видов че-
репах, 21 вид ящериц (включая крокодила) и 49 видов змей
(включая двуходковых, червяг и веретенницу).
* Создатель зоологической номенклатуры Карл Линней (1707—
1778) установил единый класс Amphibia, подразделив его на
3 отряда: Reptiles (рода Testudo с 15 видами, Draco с 2 видами,
Lacerta с 49 видами и Rana с 17 видами), Serpentes (рода Сго-
talus с 5 видами, Воа с 10 видами, Coluber с 97 видами, Anguis
с 16 видами, Amphisbaena с 2 видами и Coecilia с 2 видами) и
Nantes (некоторые аберрантные |рыбы). Выделение третьего
отряда было заблуждениемДв которое Линнежввел американский
врач Д. Гарден, принявший плавательный пузырь одной из рыб
за легкие.^Рода, принятые Линнеем, разумеется, далеко не соот-
ветствуют современным: например, все крокодилы рассматри-
вались как один из видов рода Lacerta —«Lacerta crocodilus*.
Формально первой, специально герпетологической книгой
можно считать работу Топселя (1608), но и она носит средне-
вековый характер. Фактически первым, чисто научным, специ-
альным трудом по герпетологии (в нашем, более узком смысле)
следует считать работу венского медика Лауренти, которую он
6
представил в качестве докторской диссертации (1768). Именуя
в ней оба интересующие нас класса «Пресмыкающимися», Лау-
ренти оставляет в стороне черепах, а всех прочих делит на
отряды Salientia (бесхвостые земноводные), Gradientia (наши
хвостатые земноводные и ящерицы) и Serpentia (змеи и наши
безногие земноводные и безногие ящерицы). В начале XX в.
выяснилось, что в действительности автором этой работы был
студент Винтерль, продавший Лауренти свою рукопись. Систе-
матики и сейчас всюду цитируют фамилию Лауренти, хотя по
сути он этого не заслуживает.
Несколько особняком от систематического направления свое-
го времени стоит работа художника-миниатюриста Рёзеля фон
Розенхофа (1705—1759), посвященная бесхвостым Германии
(1758). В ней собрано много верных экологических и анатоми-
ческих сведений.
Заметным шагом вперед следует считать противопоставление
лягушек, жаб, квакш и саламандр, собранных в один отряд,
всем другим отрядам (Brongniart, 1800). Дальнейший шаг
сделал П. Латрей, год спустя принявший земноводных и пре-
смыкающихся за самостоятельные классы. Однако взгляд этот
долгое время не встречал сочувствия и был принят зоологами
только после классических работ И. Мюллера (1801—1858) и
Т. Г. Гексли (1825—1895).
Большой прогресс в разработке герпетологии был достигнут
А. Дюмерилем (1774—1860), который совместно со своим асси-
стентом Ж. Биброном (1806—1848) выпустил монументальное
сочинение (1834—1854), где дал обзор, всех тогда известных га-
дов земного шара (основой служили коллекции Парижского му-
зея естественной истории). Позднее на первое место выдвинулись
зоологические коллекции Британского музея в Лондоне. Первые
описания этих коллекций были опубликованы Дж. Грэем
(1800—1881), а потом А. Гюнтером (1830—1914). Исключитель-
ное значение для развития герпетологии имели работы Дж. Бу-
ленджера = Булянже (1858—1937). Он обработал все группы зем-
новодных и пресмыкающихся в масштабе земного шара, резуль-
татом чего явилось новое издание «Каталогов» Британского
музея по этим группам животных (вышли между 1882—1896 гг.).
Кроме того, им было опубликовано множество как мелких
работ, так и крупных сводок. До наших дней не утратила своего
значения сводка Г. Гадова (1909). Крупнейшим герпетологом
Америки был Э. Коп (1840—1897). Особо следует отметить его
сводки по фауне Америки (1889—1900), содержащие многочис-
ленные анатомические сведения.
• Большой интерес представляют работы Л. Стейнегера
(1851—1943). В Американском музее естественной истории ра-
ботал Г. Нобль (1894—1940), лучшие сочинения которого были
посвящены фауне Конго и земноводным в целом (1924—1955).
Трагически погибший от укуса змеи К. Шмидт (1890—1957)
7
оставил, помимо ряда специальных работ, великолепно иллюст-
рированный обзор пресмыкающихся.
Большое значение при изучении морфологии имеют до сих
пор сводки в 4 томах лейденского профессора К. Гофмана
(1§73—1878, 1879—1890). Лучшей сводкой по фауне Германии
остается вышедшая в 1897 г. книга Б. Дюригена (1853—1924),
по Европе в целом — книга Э. Шрейбера (1912), а по мировой
фауне — соответствующие тома «Жизни животных» А. Брема.
Важное значение имеет каталог европейской фауны, выпу*
щенный Мертенсом и Л. Мюллером (1940). Классической работой
по содержанию герпетологических объектов в неволе служит
книга П. Крефта (1907). Перу Е. Циттеля принадлежит очерк
истории изучения ископаемых (1889). Описание ископаемых
дано Ф. Хюне (1956).
История отечественной герпетологии освещена в специаль-
ной статье, (Терентьев, 1957) и потому излагается здесь в сжа-
том видеЛПервый очерк нашей фауны дал П. С. Паллас (1741 —
1811) в своей «Зоографии» (1813)j Первыми учеными, занимав-
шимися герпетологией в России и выпустившими специальные
герпетологические работы, были Э. А. Эверсманн (1834) и
И. А. Криницкий (1797—1838). ^Основоположником герпетоло-
гии следует признать А. А. Штрауха (1832—1893). Большой его
заслугой явилось создание в Зоологическом музее (ныне — ин-
ституте) Академии Наук богатейшей в стране герпетологиче-
ской коллекции (при его жизни она насчитывала 10 396 номе-
ров— сейчас свыше 20 000). На основе материалов, полученных
при обработке этих коллекций, А. А. Штраух выпустил серяйб
капитальных монографий (1862, 1866, 1869, 1870, 1873).
Первый полный список отечественной герпетофауны был
составлен А. М. Никольским (1858—1942). Современником его
был Я. В. Бедряга (1854), оставивший «ряд ценных сводок (1886,
1889, 1896) и обработавший богатейшие сборы Пржеральского
(1898—1912).
Второй центр изучения отечественной герпетофауны -— Зооло-
гический музей Московского университета. Особое место зани-
мал Л. А. Лантц (1886—1953), собравший большую коллекцию
и работавший не только с консервированным, но и живым ма-
териалом.
В настоящее время советская наука располагает много-
численными кадрами герпетологов, ведущих исследовательскую
работу не только в крупных центрах, но и на периферии.
* *
♦
Линней знал 215 ныне живущих земноводных и пресмыкаю-
щихся, Дюмериль и Биброн — 846, а Буленджедр — уже 5151.
Сейчас существуют значительные разногласия, проистекающие
8
дренмуществевно из разного понимания объема вида. Однако
можно считать, что к середине XX в. герпетология знала около
6950 видов. Паллас считал, что в России обитает 11 видов зем-
новодных и 43 вида пресмыкающихся. В 1949 г. их было соответ-
ственно известно 33 и 125, причем значительного изменения этих
цифр ждать не приходится. J3 настоящее время перед герпето-
логией стоят задачи изучения внутривидовой изменчивости, уточ-
нения распространения и, главное, изучения экологии.
ЛИТЕРАТУРА
Бедряга Я. Земноводные н пресмыкающиеся. Научные результаты путе-
шествий Н. М. Пржевальского, отд. зоол., 1898—1912. т. 3.
Верндр Ф. Эттпмадиыв пресмыкающиеся Альфреда Брема, 2 тома
(Брем, Жизнь животных» перевод с 4-го немецкого издания под редак-
ЦИСй Н. М. Книповича, тт. 4, 5), 1914.
Никольский А. Пресмыкающиеся и земноводные Россй1ской империи
(Herpetotogia rossica), Зап. Академии наук. 1906, 7 серая, физ-мат.
отд., т. 17.
Никольский А. Пресмыкающиеся, «Фауна России», т. 1, 1915 (черепахи,
ящерицы), т.2, 1916 (змеи).
Никольский А. Земноводные, «Фауна России». 1918.
Терентьев П. Материалы к истории отечественной герпетологии. Труды
института истории естествознания АН СССР, 1957, т. 16, выл. 3»
стр. 97—122.
Терентьев П. и Чернов С. Определитель пресмыкающихся и земновод-
ных, изд. 3-е, «Советская наука», 1949,
Hindus I&ri duo, Bartholomaeus
Axrib'TosSnHS... opus сопсЩпмш, Bononiae, 1640.
Bastian A. Das Thier in semen Mythologischen Bedeutung, Zeitschrift fur
Ethnologic, 1869, 1: 45—66.
Boulenger G. Reptilia and Batrachia («Fauna of the British India»),
Calcutta, 1890.
Brojigniart A. Essay d’une classification naturelle des Reptiles, Bull. Soc.
Phflom.» 1800, 2: 81.
Car us V. Histoire de la Zoologie, Paris, 1880.
Cope E. The Batrachia of North America, Bull. U. S. National Museum,
No. 34. 1889.
Cope E. The Crocodilians, Lizards and Snakes of North America, Rep.
U. S. National Museum for 1898, 1900: 153—1270.
Dumdril A. et Bibron G. Erpfctologie gfendral ou histoire naturelie
complete des reptiles, Paris, 1834—1854, 9 vols.
Dfirigen B. Deutscblands Amphibien und Reptilien, Magdeburg. 1897.
Eversmann E. Lacertae Imperii Rossici variis in itineribus meis observatae,
Nouveaux M&noires de la Soc. Imp. des Naturalistes de Moscou, 1894»
3* 337 369
Gad’ow H. Amphibia and Reptiles, The Cambridge Natural History* 8* 1J®®*
Gesner C. Historiae animalium, 2 De Quadrupedibus oviparis, Ediho
I, Tiguri, 1554. , . .. _ . .. .
Gesner C. Historiae Animalium, 5 De Serpentium natura et vanisschedis et
collectaneis ejusdem compositus per J. Carronum, Editio 1, Tigun, 1587.
Hollman Ch. K. Amphibien (Bronn’s Klassen und Ordnungen). Leipsig und
Heidelberg. 1873—1878. -
Hoffmann Ch. K. Reptilien (Bronn’s Klassen und Ordnungen), Leipzig,
3 vole, 1879—1890.
9
н ы . n е ₽ PalSontologic und Phylogenie der niederen Tetrapoden. Jena, 1956
w i a i ri f T₽ntenwn hcrpctoiofji1755.
К iX Г t P Das Terrarium. Ein Handbuch der hauslichen Reptilien und
Amphibienpflege nebst Anleitung zum Bestimmen der Terrarientiere.
к r i n7c k i Tbbservationes quedam de Reptilibus indigenis, Bull de la Soc
imp des Naturalistes de Moscou. 1837 46-49 oc-
Laurenti J. Specimen medicum, exhtbens Synopsis Reptilium emendatum
cum experiments circa venena et antidote Reptilium Austriacorum. Viennae,
1768
Mertens R. und Muller L. Liste der Amphibien und Reptilien Europas
Ab ha nd I. der Senckenberg. Naturforsch. Gesellschaft, 1928. 41; 2 ed
(«Die Amphibien und Reptilien Europas»), ibidem, 1940, No 451.
Mertens R. La vie des Amphibiens et Reptiles, Paris, 1959.
Noble G. Contribution to the Herpetology of the Belgian Congo Expedition
Bull Amer. Mus. Nat. Hist, 1924, 4»: 147—347. * И
Noble G. The Biology of the Amphibia New York, 1 ed., 1931; 2 ed 1955
Oudemans A. The Great Sea-Serpent, Leiden and London, 1892
Pallas P. Zoographia Rosso-Asiatica, Petropoli, 3, 1813.
Rabanus Maurus. De Sermonum Proprietate, seu de Universe. Strassburg
1467 (книга VII имеет раздел «De serpentibus»).
Roe sei von Rosenhof A. Historia naturalis ranarum nostratium in qua
omnes earum proprietates, praesertim quae ad generationem ipsarum
pertinent fusius enarrantur, Norimbergae, 1758.
Schmidt K. Herpetology, San Francisco, Calif. Acad. Sci., 1966: 501—627.
Schmidt K. and Inger R. Living Reptiles of the World, New York, 1967.
Schreiber E. Herpetologia Europaea, 1 ed., Braunschweig, 1875; 2 ed.,
Jena, 1912.
Stejneger L. Herpetology of Japan and adjacent Territory, BuU. U. S. Na-
tion. Museum, 57, 1907.
Stejneger L. and Barbour Th. A Check List of North American
Amphibians and Reptiles, Washington, 1 ed. 1917. II ed. 1922. Ill ed. 1933.
Stejneger L. Chinese Amphibians and Reptiles in the U. S. Nat. Museum,
Proc. U. S. Nation. Museum, 66, 1925.
Strauch A. Chelonologische Studien mit besondere Beziehung auf die
Schildkrotensammlung der Kais. Akademie der Wissensch. zu St. Petersbourg,
Mem, Acad. Imp. St. Petersbourg, 1862, 7 Serie, 5, No. 7.
Strauch A. Synopsis der gegenwartig lebenden Krokodiliden, Mem. Acad.
Imp. St. Petersbourg, 1866, 10.
Strauch A. Synopsis der Viperiden, Mem. Acad. Imp. St. Petersbourg, 1869,
7 serie, 14, No. 6.
Strauch A. Revision der Salamandriden-Gattungen nebst Beschreibung
einiger neuen oder weniger bekannten Arten dieser Familie, Mem. Acad.
Imp. St Petersbourg, 1870, 7 serie 16, No. 4.
Strauch A. Die Schlangen des Russischen Reiches, Mem. Acad. Imp. St.
Petersbourg, 7 ser., 1873, 21, No. 4.
T о p s e 11 E. The Historie of Serpents; or, The second book of living creatures;
wherein is contained their divine, naturall, and moral descriptions, London,
1608 (обычно эта «вторая книга» переплетается как часть работы, вышед-
шей в 1607 г., ио известен и экземпляр, имеющий самостоятельный ти-
тульный лист и предисловие, помеченные 1608 г.).
Z i 11 е | К. Geschichte der Geologic und Palaontologie, Geschichte der
Wissenschaften in Deutschland, Neuere Zeit, 1889, 23: 827—833.
Глава вторая
ВЫХОД ЗЕМНОВОДНЫХ НА СУШУ
И ДРЕВНИЕ ЗЕМНОВОДНЫЕ
Земноводные — самый примитивным класс наземных позво-
ночных и потому вопрос о происхождении этого класса совпа-
дает с вопросом о выходе позвоночных на сушу и возникнове-
нием в филогенезе первично-наземных черепных. Для того чтобы
правильно осознать это, следует представить себе сумму разли-
чий между первично-водным («рыбаэ). и первично-наземным
(«земноводное») позвоночным.
Условия существования в водной и воздушной среде во мно-
гом различны. Эти обстоятельства нашли свое отражение в тех
изменениях, копгорые в теле позвоночных при выходе
на сушу, тйряя&в вв&тщнк из них были прямым ответом на
изменение факторов среды, а другие возникли коррелятивно.
Дыхание сменилось с жаберного на легочное. Это, в свою оче-
редь, вызвало перестройку кровеносной системы.' В дополнение
к ранее существовавшему большому кругу кровообращения
(сердце — тело — сердце) возник малый (сердце — легкие —
сердце). Последнее обстоятельство обусловило усложнение
функции и строения сердца: если у рыб через сердце проходил
только один поток венозной крови, то у земноводных через
него проходит два — венозный и артериальный. Появление двух
предсердий («трехкамерность» сердца) представляет собой мор-
фологическое отражение этого физиологического факта. Однако
несовершенство легких сделало кожное дыхание у большинства
земноводных необычайно важным, а изредка даже почти един-
ственным способом дыхания взрослых животных/] Кровяное
давление, по-видимому, изменилось мало (у лягушки 20—60 л«,
у рыб 16—120 ), но пульс несколько участился (у лягушки 40—
50 ударов в минуту, у рыб 18—30). Зато заметно изменилась
сама кровь: количество ее по отношению к весу тела возросло
(у лягушки 3—7%, у рыб 1—2%), установилась более постоян-
ная формула 1фови, увеличились размеры форменных элемен-
тов (поперечник эритроцитов земноводных 18—70 микронов, у
и
рыб 7—27), а число их уменьшилось (у квакши 700 000 эритро-
цитов на леле3 крови, у лягушки 159 000—640 000, у тритона
202 000—270 000, у протея всего 36 000, у карпа 1 900 000—
3 060 000).
Изменение преобладающего способа дыхания, в частности,
исчезновение жабр, сохраняющихся только у личинок и немногих
неотенических форм земноводных, сказалось даже на скелете.
По мнению Мензбира (1930), постепенное исчезновение став-
шего излишним жаберного скелета позволило голове приобрести
подвижность, за счет возникновения шеи. Подвижность связана
с тем, что первые позвонки не входят в состав затылочной части
черепа, как это имеет место у рыб. Связь последнего с давле-
нием плотной водной среды на передний конец тела прекрасно
продемонстрирована Дубининым (1949), установившим так-
же, что вторичное исчезновение шеи у бесхвостых земноводных
связано уже с совершенно иными обстоятельствами — с раз-
витием способности прыганья и зарывания в почву. Остатки жа-
берного скелета, меняя функцию, получают у земноводных зна-
чение подъязычного аппарата.
Эволюция разных органов совершается в филогенезе не оди-
наковыми темпами. Легкие земноводных, бывшие, по-видимому,
больше гидростатическим приспособлением, изменились медлен-
нее потребности животного в кислороде (у лягушек дыхательная
поверхность относится к поверхности кожи приблизительно как
2 3, у млекопитающих она в десятки раз больше) и потому им
на помощь пришло кожное дыхание. Последнее мыслимо только
при влажной коже, а это вызывает у земноводных прогрес-
сирующую утрату чешуй в филогенезе и усиленное развитие
слизистых желез. При этом слизистый футляр, одевающий тело
рыб, получает еще большее значение — это и механическая, и
биологическая защита. Во многих случаях слизь приоб-
ретает ядовитые свойства — защита земноводных от врагов.
Кожа становится органом водного обмена — земноводные никог-
да не пьют ртом, а пополняют запас воды погружением своего
тела в нее или «купаньем» в росе. В связи с этим кожа земновод-
ных получила особые свойства избирательной проницаемости к
жидкости, газам и другим веществам. Кроме того, она получила
и значение фоторецептора. Такая биологическая нагрузка неиз-
бежно должна вызывать быстрый износ. Можно думать, что
частые вынужденные регенерации кожи на травмированных
местах были исходным материалом, из которого развилось явле-
ние линьки. Позднее же оно попало под контроль эндокринных
факторов. Другой цикл изменений связан с переменой характе-
ра движения: парные конечности, построенные по типу плав-
ника— ichthyopterygium, уступают место наземной конечности-^-
cheiropterygium = chiropterygium, ныне в основе своей пятипалой,
но первоначально, по-видимому, бывшей семипалой. С этим
коррелятивно связаны многие изменения в скелете, мускулатуре
12
и нервах (Северцов, 1950). В частности, надо отметить исчеэно-
вение связи пояса передних конечностей с черепом и возникно-
вение прикрепления тазового пояса к позвоночнику (крестцу).
Меньшая плотность воздушной среды представляет больше воз-
можности для отклонения формы тела от строго обтекаемой,
преобладающей у рыб.
Плаванье сохраняется у большинства земноводных, но в до-
полнение к нему появляется ползанье и прыжки. В связи с по-
следними первоначальная ящерицеподобная форма тела укора-
чивается, исчезает хвост и возникает диспропорция между пе-
редними и задними конечностями (Вермель, 1931). Напротив,
потребность вторично вернуться исключительно к плаванию
выводит, в силу невозможности обратного развития наземной
конечности в плавник, на первое место змееобразные изгибания
всего тела, что имеет следствием частичное (Siren) или полное
(Aistopoda) исчезновение конечностей. Подобная безногость
оказывается полезной и при подземном образе жизни (Apoda).
Так можно понять наличие среди земноводных трех основных
типов формы тела; ящерицеобразной, бесхвостой и безногой.
Не менее интересна и перестройка органов чувств. Плотность
воды обеспечивает передачу звука непосредственно через хрящ
или кость на внутреннее ухо. С выходом на воздух, в дополнение
к нему появляется среднее ухо, развивающееся за счет исполь-
зования ненужного больше брызгальца (spiraculum), затянув-
шегося барабанной перепонкой. Из материала ушной капсулы
еще в онтогенезе земноводных отделяется звукопроводящий
аппарат, гомологичный подъязычной кости (hyomandibulare)
рыбообразных позвоночных. Все это обеспечивает довольно зна-
чительный уровень слуха, правда слуха, строго избирательного.
Возможность высыхания, загрязнения и повреждения рого-
вицы в наземных условиях привели к возникновению век и слез-
ных желез. Трудно сказать, что именно вызвало образование
внутренних носовых отверстий — хоан. Появление их было
важно и для дыхания, и для сохранения влажности слизистой
оболочки рта, которая могла слишком пострадать, если бы
дыхание шло через открытый рот. Во всяком случае хоаны пред-
ставляют характерный признак воздушно-дышащих животных.
Наряду с этим исчезли древние водные органы кожного чувства
(«боковая линия»), сохранившиеся только у личинок. Разумеет-
ся, перестройка органов чувств, как и других органов, не могла
не вызвать существенных изменений в нервной системе: передний
мозг имеет сравнительно крупные размеры, и его полушария
полностью отделяются друг от друга. Появляется настоящий
мозговой свод (архипаллиум). Средний мозг уже начинает
терять значение зрительного центра, хотя образует еще довольно
крупный отдел головного мозга. Мозжечок развит слабо. Как
примитивный признак нужно отметить то, что изгибы головного
мозга почти не выражены.
13
Стоит отметить и тот фЛКТ, что в свяли с выходом ив суЩу
впервые появляются железы ротовой полости. Правда, пома не
удалось установить пищеварительного иля убивающего действия
выделения межчелюстной и парной небной желез лягушки, ио
значение их для увлажнения и связывания (некоторая клейкость)
добычи несомненно. Можно было бы отметить еще целый ряд
любопытных связей, ио и сказанного достаточно. Важно учесть,
что земноводные благодаря уже отмеченным особенностям
своего дыхания практически не смогли полностью порвать свою
связь с водной стихией. Отсюда возникло характерное явление
метаморфоза, в течение которого комплекс «водных» признаков
личинки быстро сменяется на «наземный». Подобная смена про-
исходит, например, даже в мускулатуре, В итоге перечисленных
специализаций выработалась совокупность типичных признаков
современных земноводных, Остановимся иа главных отличитель-
ных признаках;
I) Череп сочленяется с позвоночником двумя мыщелками;
2) имеется слуховая косточка, входящая в овальное окно; 3) ко-
нечности наземного, пятипалого типа; 4) эритроциты имеют ядра,
двояковыпуклы и овальны; 5) сердце состоит из 2 предсердий м
одного желудочка; б) имеется артериальный конус, снабженный
клапанами; 7) дуги аорты симметричны; б) жабры присутствуют,
по крайней мере, иа некоторых стадиях личиночного развития;
9) почки мезонефрические, снабженные нефростомами; 10) кож-
ный орган водного чувства («боковая лиимя») присутствует во
крайней мере, на некоторых стадиях личиночного раймпнИ
II) последним черепным нервом является блуждающий; 12) не-
парные плавники если имеются, лишены поддержки хрящевых
или костных лучей; 13) истинная грудина и доходящие до нее
ребра отсутствуют; 14) отсутствует специальный копулятивный
орган; 15) онтогенез протекает без развития амниона и аллан-
тоиса.
Следует оговорить, что признак лятилалости конечности (3)
будет частично (некоторые Caudata) или полностью (Apoda) от-
сутствовать за счет вторичной редукции. Признак (5) не подхо-
дит к некоторым хвостатым (Plethodontidae), где утрата легких
ведет ко вторичному упрощению сердца, Онтогенез некоторых
немногих бесхвостых столь укорочен и видоизменен, что признаки
(б) и (10) выпадают или сильно модифицируются, Исключением
из признака (12) является Ichthyottega, Признак (14) не подхо-
дит к роду Aacaphua, у которого часть клоаки превращена в свое-
образный копулятивный орган, Если не обращать внимания иа
перечисленные исключения. то признаки (1), (2), (3) и (12) не
будут отличать земноводных от рыб. Рыбы, пресмыкающие-
ся и птицы не обладают признаком (I). Признак (4)
отделяет земноводных от млекопитающих. Признак (б) ие свой-
ственен большинству рыб и всем птицам н млекопитающим.
Таким образом, ие представляет труда дать определение сов-
14
ременным земноводным путем комбинации перечисленных приз-
наков. Абсолютно точным диагнозом их можно считать сумму
(1)и (15) признаков («Amphicondylous Anamnia»), но такое оп-
ределение не приносит пользы, если на руках имеются лишь взрос-
лые экземпляры. Поэтому в качестве практического диагноза пред-
лагается сумма (1) и (11) признаков.* позвоночные с дву-
мя затылочными мыщелками и внечерепным
подъязычным нервом. Земноводные унаследовали от
рыб слизистую кожу. Однако упрощенное отличие земноводных
от пресмыкающихся по гладкой, лишенной чешуи кожи и присут-
ствию метаморфоза, хотя и оправдывается в очень большом
числе случаев, но не приложимо к безногим и к бесхвостым с
укороченным развитием. Кроме того, характер кожи ископаемых
не всегда ясен. Столь же ненадежен и признак легочного дыхания,
ибо, с одной стороны, формально легочное дыхание имеется у
некоторых рыб (Dipnoi), а с другой стороны, не все земноводные
снабжены легкими (Plethodontidae). Поэтому характер кожи,
метаморфоз и легкие не включены в приведенный выше список.
Если определение современных земноводных, как мы видели,
не представляет непреодолимых затруднений, то совсем иначе
обстоит дело при включении в рассмотрение вымерших форм.
Признаки (1) и (11) не подходят к некоторым древним земно-
водным, а относительно многих других признаков ископаемых
форм мы не можем сказать ничего определенного.
Проблему возникновения земноводных целесообразно разбить
на три вопроса: когда вышли первично-наземные позвоночные
ва. сушу, едв был их м мак это произошло?
Отмеченная выше непрерывность переходов между рыбами
и земноводными заставляет избрать формальный признак, ко-
торый можно принять за условную границу между двумя клас-
сами. Разумнее всего принять за пограничный признак появле-
ние конечности типа cheiropterygium, ибо таковая характерна
для всех 4 классов позвоночных, объединяемых под именем
Tetrapoda=Quadrupeda. В каменноугольных слоях найдено мно-
го скелетов с хорошо развитыми конечностями подобного типа, из
чего следует, что возникновение Tetrapoda имело место раньше.
К сожалению, в девонских отложениях находки менее многочис-
ленны и часто состоят лишь из одних черепов. На помощь при-
ходит факт существования довольно многочисленных «окамене-
лых» следов пятипалых конечностей, получивших у палеонтоло-
гов общее название «ихнитов» (отсюда специальная отрасль
«ихнидология»). Впервые таковые были обнаружены в 1833 г.
в нижне-триасовых пестрых песчаниках Тюрингии. Позднее сле-
ды ископаемых были найдены как в более молодых,^ так и в
более древних слоях. Для интересующего нас в данный момент
вопроса важнее всего находка таких следов в верхне-девонских
отложениях Зап. Пенсильвании, откуда их описали под именем
Thinopus antiquus. До недавнего времени это было единственное
15
доказательство, ибо от интереснейшей пруппы девонских позво-
ночных Ichthyostega и Ichthyostegopsis, описанных Сёво-Сёдер-
бергом из Вост. Гренландии (1932), равно как и от найден-
ной в Вост. Канаде Elpistostege (1938), были известны только
черепа. Рассмотрение этих черепов позволило предположить, что
это остатки организмов, бывших связующим звеном между зем-
новодными и рыбами, но с уверенностью поставить их по ту или
иную сторону границы, разделяющей два указанные класса, было
невозможно. Только в 1952 г. установили, что животные
этой группы имели пятипалые конечности. В настоящий момент
существование земноводных в верхне-девонское время не может
вызывать никаких сомнений. Момент возникновения этого клас-
са приходится, следовательно, отодвинуть на более раннее время,
но как далеко? Решить это сейчас еще трудно. Исходя из чисто
сравнительно-анатомических сопоставлений Северцов (1926,
1950) предложил считать, что отделение предков земноводных
от рыб имело место в силуре. Но точнее, или вернее принять за
время возникновения земноводных нижний или средний девон.
Помня обычные масштабы перевода геологических периодов в
годы, следует принять древность класса земноводных примерно
в 320 000 000 лет.
Теперь можно перейти ко второму вопросу проблемы выхода
позвоночных на сушу — кто был предком земноводных? В общей
форме вопрос этот не вызывает сомнений, ибо начало им могли
дать только рыбы. Однако, по современным представлениям,
рыбы уже не могут рассматриваться как единое целое. Как изве-
стно, ныне живущие земноводные не переносят соленой воды.
С другой стороны, остатки ископаемых земноводных всегда
встречаются лишь в пресноводных отложениях (исключение
составляют захоронения на Шпицбергене, где, по-видимому, имел
место снос трупов в море.). Значит можно с большой долей ве-
роятности искать предков земноводных именно среди пресновод-
ных рыб. В девонское время в пресных водах жиЛй некоторые
аберрантные хрящевые рыбы, хрящевые ганоиды, двоякодыша-
щие и кистеперые. Приходится выбирать среди них. Стоит при
этом отметить, что костистые рыбы (Teleostei) появились только
в триасе, т. е. примерно на 130 000 000 лет позднее возникнове-
ния земноводных. Поэтому всякие попытки поисков преемствен-
ности между земноводными и костистыми рыбами должны быть
заранее отвергнуты. Организация хрящевых рыб и хрящевых
ганоидов столь своеобразна, что мысль о родстве их с земновод-
ными, по-видимому, никем не выдвигалась. Таким образом,
предками земноводных могли быть либо двоякодышащие, либо
кистеперые. Наличие воздушного дыхания у двоякодышащих
как бы гипнотизировало исследователей и потому первые эво-
люционисты безоговорочно выводили земноводных из двоякоды-
шащих. Действительно между двоякодышащими и земновод-
ными имеются общие черты и помимо воздушного дыхания:
16
аутостилический череп, хоаны, характер крыши больших
полушарий головного мозга, сходный артериальный конус
и т. п. В дальнейшем этот взгляд поддерживали, главным об-
разом, анатомы. Палеонтологи же сравнительно скоро стали на
противоположную точку зрения.
Спор этот был подробно разобран Северцовым в 1926 г
Многие морфологические особенности земноводных и двояко-
дышащих оказались, при более внимательном рассмот-
рении, в корне различными. Так у двоякодышащих имеются
лишь нижние ребра, а у земноводных — верхние или нижние.
Обычно принято думать, что эмбриональный хрящевой череп
двоякодышащих очень похож на соответственный череп личинок
хвостатых земноводных, но анализ .расположения выходных
отверстий нервов и сосудов показал, что картину, свойственную
земноводным, можно с одинаковой легкостью вывести и из хря-
щевого черепа кистеперых. Зато характер зубов дает более чет-
кое указание: большие зубы двоякодышащих, сидящие на кры-
ловидных и сошниковых костях, образовались путем слияния
целого ряда отдельных мелких зубов, чего никак нельзя ска-
зать о зубах земноводных. Совершенно невозможно вывести
череп земноводных из черепа современных двоякодышащих,
для которых характерно наличие больших непарных окостене-
ний на верхней поверхности, а также утрата предчелюстных,
челюстных и некоторых других костей.
У хвостатых земноводных хорошо сохранились вентральные
элементы жаберного скелета, сильно редуцированные у двояко-
дышащих. Некогда плавники рогозуба («бисериальный архип-
теригий») принимались за исходный материал для развития
наземной конечности и это было аргументом за происхождение
наземных позвоночных именно от двоякодышащих. Однако
теперь можно считать доказанным, что наземная конечность
развилась из эврибазального плавника рыб, пройдя стадию сте-
нобазального плавника, возникшего путем образования меж-
ду стенкой тела и осью, проходящей через метаптеригии «метап-
теригиальной выемки». Раньше считали, что носовой аппарат
двоякодышащих представляет собой переходную форму от
рыбьего к наземному типу. Северцов й его ученики показали,
что при превращении носовых ямок рыб в носовые полости на-
земных позвоночных заднее носовое отверстие осталось на дор-
зальной поверхности головы, а переднее перешло через край
ротовой щели и превратилось в хоану. Взгляд этот вполне под-
тверждается позднейшими палеонтологическими находками
древних земноводных. У двоякодышащих произошло анало-
гичное смещение, но оно пошло значительно дальше, чем у
наземных позвоночных. Плавательный пузырь двоякодышащих
располагается со спинной стороны по отношению к кишечнику,
тогда как легкое земноводных занимает брюшное положение.
Черты сходства между двоякодышащим^ и земноводными сле-
17
дует признать либо за конвергенцию, либо за древние признаки
свойственные их общим предкам. Однако некоторые исследова-
тели продолжают считать двоякодышащих предками Tetrapoda
Так полагают, что Caudata могли в отличие от прочих земновод^
ных, произойти прямо от двоякодышащих (Holmgren, 1949)
Однако взгляды эти не встречают сочувствия среди большинства
зоологов.
Итак, приходится считать установленным происхождение
наземных позвоночных от кистеперых, ибо все, что свидетель-
ствовало против родства двоякодышащих и земноводных, ука-
зывает на родство с кистеперыми. Наличие у кистеперых обоих
групп ребер позволяет вывести из этой группы рыб как Apoda,
снабженных только нижними ребрами, так и других земновод-
ных (Caudata, Salientia), у которых имеются только верхние
ребра. Черепа кистеперых и низших земноводных характери-
зуются преимущественно количественными различиями: череп
становится более плоским, постепенно .редуцируется основная
затылочная кость, уменьшение крыловидных костей компенси-
руется расширением околоклиновидной, увеличиваются боковые
затылочные, постепенно редуцируются переднеушная и верхне-
затылочная и т. п. (Watson, 1920). Различия эти возникают
столь постепенно, что, базируясь только на черепе, трудно про-
вести границу между двумя группами. Рассмотрение верхней
крыши черепа позволяет легко гомологизировать кости, причем
трудно отказать в справедливости мнению Вестолла (Westoil,
1938, 1940), о том, что наша обычная номенклатура костей не-
правильна. Однако из осторожности пока разумнее воздерг
жаться от капитальной ломки номенклатуры костей черейб
Tetrapoda. Поэтому здесь и далее будут употребляться только
традиционные названия.
Строение скелета конечностей кистеперых рыб таково, что
скелет конечностей земноводных выводится из него без всяких
натяжек. При этом имел место двойной поворот плавника и,
разумеется, перестроились пояса конечностей (Gregory and Ra-
ven, 1941). Решив вопрос о происхождении земноводных от ки-
степерых в общей форме, представляется желательным уточ-
нить от какой именно группы внутри кистеперых взяли свое
начало Tetrapoda.
По современным представлениям, подкласс Crossopterygii
распадается на две ветви: Osteolepides = Rhipidistia и Coelacanthi.
Вторая из этих ветвей была сперва пресноводной, но с триаса
ушла в море, где вторично потеряла хоаны. Такая история, а
главное морфологическое своеобразие (трехлопастный хвост,
значительная или полная редукция подкрышки, внутренней кры-
ловидной кости и т. п.) делают ее для нас неинтересной. Пред-
ков земноводных, очевидно, надо искать среди Osteolepides,
которые, в свою очередь, распадаются на три ветви: Osteolepi-
formes ( = Osteolepidae), Holoptychiiformes ( = Porolepiformes) и
18
RhiBodontlformc* (* Megalkhtby Idee). Сейчас нет единодушия а
суждении о том. от какой именно из трех названных групп про-
изошли первые Tetrapoda Олни отдают предпочтение Rhlxodonll-
formes, другие склоняются к принятию дифилетического проис
хождения Tetrapoda и считают, что HoloptychlHormc* дали
начало Apoda и Caudata, a Osteolepiformes всем прочим тем-
новодным (Huene, 1956). Пожалуй, наиболее правильно искать
корни земноводных средн наиболее примитивных Сгоячор1егуц11.
т. е. среди Osteolepiformcs.
Остается последний вопрос: как и почему осуществился выход
позвоночных на сушу? Шаблонное толкование утверждает, что
предки земноводных стали все чаще выхолить на сушу и
под влиянием новой обстановки получили все те особенности,
которые были перечислены ранее. Однако принять эту упро-
щепную точку зрения мешают некоторые факты. Прежде всего
непонятно, что именно «гнало» предков земноводных из воды на
сушу? Это не могло быть только погоней за добычей, ибо подхо-
дящей пищи тогда на суше было не больше, чем в воде. Это не
могла быть одна конкуренция или «давление» со стороны врагов,
ибо предки земноводных были достаточно крупными животными.
Нельзя ссылаться и только на ухудшение качества воды, ибо
для того, чтобы дышать воздухом, было достаточно просто под-
ниматься к поверхности воды, как это и делают многие рыбы.
По-видимому, каждый из этих факторов сыграл свою роль, но
не они явились решающим моментом. Вызывают недоумение
и некоторые морфологические факты. Так у большинства древ-
них земноводных имелись хорошо выраженные органы боковой
линии. Присутствие боковой линии типично для водных живот-
ных, так как именно она воспринимает колебания водной среды.
Не меньшую трудность представляет и покровный скелет
земноводных: представление о стегоцефалах как о животных,
закованных в чешуйчатую броню (наподобие средневековых
рыцарей), просто не верно. Лишь немногие из них (Ophiderpcton,
Kicnodon etc.) были одеты чешуей по всему телу, некоторые же
были вовсе лишены чешуи, а у большинства чешуя покрывала
только брюшную часть тела, такое расположение чешуи эколо-
гически бессмысленно у животных, ползающих по земле. Зато
оно вполне понятно, если допустить, что древние земноводные
плавали на поверхности воды: снизу можно было ждать напа-
дения, а сверху опасности не было — ведь ни птиц, ни летающих
пресмыкающихся еще не существовало. Да и с точки зрения бла-
гоприятного расположения центра тяжести у плавающего жи-
вотного брюшное расположение чешуи не представляется не-
целесообразным.
Исходя из этого, Северцов в 1926 г. выдвинул положение
(Северцов, 1950), что первые Tetrapoda были первично-водными
животными и что комплекс признаков, характеризующих земно-
водных, образовался у этих животных еще в то время, когда они
з* 19
-uwrii» в воде i ольжо -г ___
ZoTtw тип. ebrtroptery^um. легвие я т_п. лревяие эемжтод,
возможность выйти на сушу Означает ля таяо* взгляд
де^шг«»е вреахадтания и каких либо ортогенетяческж» сид
2^'^ж«о »}м**«ошмх оргаяя^аияю животного в определенном
яалравлечвв? Вовсе лет Судя по жаражтеру отложен**, в левой,
сжое время сделки земноводных жили в неглубоких водоемах,
в ч’СеЭовмвх в^эд к в х быстро перепал'
мялась разлагающимися органическими веществами к теряла
кислород В сходно* обстановке многие из рыб вынуждены
иереходятъ сперва к заглатыванию воздуха, а затем и к воз душ-
ному дыханию. Интересно, что дыхательные движения современ-
ных земноводных являются. по-видимому, производным движе-
ний жаберного аппарата рыб. Значит есть все основания пола-
гать. что легкие развились у первых земноводных в ответ на
изменения качества воды, а вовсе не в связи с попытками выйти
на сушу. Несколько труднее понять причины изменения конеч-
ностей. Неоспоримо сходство волнообразных движений тела рыб
с таковыми ряда земноводных и даже пресмыкающихся. Имен-
но эти движения создают силу, движущую тело вперед, тогда
как. конечности остаются при плаванья пассивно прижатыми к
телу (например, у тритонов). Однако переход к придонному
(бентоняческому) образу жизни обычно сильно отражается на
верных плавниках рыб. Возможно, что конечности древних зем-
новодных. образовавшиеся из парных плавников кистеперыт
давали преимущества при закапывании в грунт, имевшем место
ирн впадении в спячку во время пересыхания водоемов. Упот-
ребления плавников для копания развило их костный скелет и
мускулатуру, что пригодилось позднее, в более влажный период,
при выходе на сушу. Конечности, первоначально служившие для
закапывания в ил, могли потом пригодиться при передаляяшга
Кз пересохших водоемов в другие. Передвижение по суше долж-
но было довершить преобразование плавника в наземную конеч-
ность, разбив ее на отделы, подобно тому, как это конвергентно
имеет место у рыб. Итак, под влиянием специфических условий
жизни в мелких водоемах девонского периода в организме
кястеперых появились особенности, которые позднее позволили
этим измененным животным (первым земноводным) начать
выходить на сушу.
Почему изменились не все кястеперые, а только часть их?
Вероятно, это произошло благодаря различиям в микроклимате
и других особенностях водоемов. Изменившиеся кястеперые,
мигрируя иногда, встречались с представителями нензменнв-
шейся группы. Пока в водоеме воды было много, из этих двух
групп лучше чувствовали себя, конечно, последние, так как они
были более искусными пловцами. Если же водоем пересыхал,
то земноводные оказывались в лучшем положении, имея больше
шансов, ползая, лопасть в другой водоем, где еще сохранилась
водя- В дальнейшем, вместо немедленны* поисков ноям, древние
земноводные все дольше оставались ив берегу и, быть может,
поедали выброшенную из волы рыбу. Более крупные эемновод*
ные поедали попутно более мелких, а затем все большее висло
земноводных начало поедать насекомых, которых в те времена
было уже довольно много. Так, видимо, совершился выход поз*
воночных на сушу.
Первые остатки древних земноводных были описаны более
столетия назад из триасовых отложений Германии. На основа^
нин ископаемых, найденных в триасовых отложениях Англии/
Оуэн ввел родовое понятие Labyrinthodon Позднее Мейер рас*
ширил его до группового. Первоначально ископаемых земновод-
ных изучали вне прямой связи с современными. Заслугой
Э. Геккеля является включение лабиринтодонтов в общую си*,
стему земноводных: он подразделил этот класс, основываясь на
присутствии или отсутствии чешуи и т. п., на Phractamphibia
(Labyrinthodontia + Peromela== Apoda) и Lissamphibia (Sozura“%
Caudata 4- Anura). Такое деление нашло много сторонников,
хотя впоследствии Apoda стали относить не к Phractamphibia,
а к Lissamphibia. С точки зрения чистоты и логичности номен-
клатуры сторонникам последнего взгляда следовало бы отка-
заться от терминов Геккеля и воспользоваться терминами бра-
тьев Саразин, как более отвечающими их концепции. Последние
именовали ископаемых амфибий Archaeobatrachi, а всех прочих
земноводных объединяли в другой подкласс под именем Neo-
batrachi. Интересно, что хвостатые и бесхвостые объединялись
ими в один отряд. Аналогичной точки зрения держался и Коп,
который считал Stegocephali (вымершие формы), Urodela (хво-
статые 4- безногие) и Salientia за три равноправных подкласса.
Еще Т. Гексли избегал противопоставления «панцирных» зем-.
новодных прочим, подразделяя разбираемый класс сразу на.
4 самостоятельных группы Saurobatrachia (=Caudata), Labyrint-
hodontia, Gymnophiona (== Apoda) и Batrachia ( = Salientia). По-
следующие исследования все более показывали правоту Гексли,
и сейчас несомненно, что группа Lissamphibia или Neobatrachi
есть разнородное образование, возникшее полифилетически.
Поэтому ныне можно говорить о Stegocephali й Neobatrachi не
как о систематических единицах, а лишь как о своеобразных
эволюционных ступенях.
Поскольку факт присутствия или отсутствия чешуи не яв-
ляется решающим в систематике крупных прупп земноводных,
надо искать оснований для построения естественной систему
среди иных признаков. Важнейшим из них является строение
позвоночника, изменившееся под влиянием силы тяжести, совер-
шенно по-новому действовавшей на скелет вне воды. У кисте-
перых позвонки оставались в общем хрящевыми, с выходом же
иа сушу имел место процесс возрастающего окостенения и при-
дания большей жесткости позвоночнику. У некоторых примитив*
S1
ных земноводных отсутстеоваля цельные тела г»звов1Со,
вместо них было три костных влемента. скрепленных хря„'’
Передний (краниальный) из этих костных элементов, ней.**!
и подковообразный, получил название hypocentrum, а
задний каудальный — pleurocentrum. Первый из них назывя»
также intercentrum, но название это получило меньшее раепп
странение. Допуская разную степень развития и слияния подХл'
ных элементов, палеонтологи создали подразделение позвонкп
ископаемых земноводных на 5 типов: (1) рахитомный
понентр и плеаронентры не сращены, не равны по размещай
2) эмболомерный — гипоцентр и плевроцентры не сращена
примерно одинаковы по размерам и формируют два тела на од» ’
невральную дугу, 3) филлоспондильный — элементы п<?
звонка соединены друг с другом, но между ними сохранились
швы, 4) лепоспондильный — швов между элементами
позвонка не сохранилось и он напоминает тонкий костный бочо-
нок, охватывающий хорду, 5) стереоспондильный____
имеется целое костное тело. Эти типы были положены в основу
классификации древних земноводных. Позднее элементами
позвонка занялись сравнительные анатомы, установившие за-
кладку позвонка из 4 парных хрящевых элементов, или дуг
(arcualia): передний верхний (краниальный дорсальный) из
них был назван основным спинным (basidorsale), передний ниж-
ний (краниальный вентральный) — основным брюшным (basi-
ventrale), задний верхний (каудальный дорсальный) — меж-
спинным (interdorsale) и задний нижний (каудальный вентраль-
ный) — межбрюшным (interventrale). К сожалению, несмотря
на довольно значительное количество работ, посвященных изу-
чению позвонков, не удалось установить общности во взглядах
палеонтологов и анатомов, что объясняется многообразием воз-
можных способов слияния arcualia в целостный позвонок.
Можно различить 8 теоретических типов (рис. 1) такого слия-
ния: 1) голомерный — развиты все 4 дуги, 2) эмболомер-
ный — одинаково хорошо развиты и самостоятельны основные
брюшные и межбрюшные элементы, тогда как основные спин-
ные образуют невральную дугу, к которой в онтогенезе может
прирастать рудимент межспинных, 3) рахитомный — как
предшествующий, но основной брюшной и межбрюшной элемен-
ты разной величины (косые), 4) стереоспондильный —
крайний случай изменения рахитомного позвонка, у которого
основные брюшные элементы хорошо развиты, и он рано окосте-
невает, а межбрюшной сходит на роль вставочного хряща,
5) нотоцентральный — практически состоит только из
невральной дуги, происшедшей за счет слияния более мощно
развитого межспинного элемента и рудимента основного спин-
ного, тогда как брюшные элементы редуцированы, 6) гастро-
центральный— наиболее развиты межбрюшные,, не остав-
ляющие .места для развития основных брюшных элементов,
22
остающихся хрящевыми или окостеневающих в хвосте в форме
гемальных отростков, 7) лсевдоцеитральны* — невраль-
ная дуга, состоящая только из основных спинных элементов,
срастается с основными брюшными, а межбрюшные сведены до
роли вставочных хрящей, 8) аделоспонднльный — рано
обызвествляющаяся оболочка хорды утолщается и образует
двояковогнутое тело (сходство с акулами), к которому лрнрв-
стает невральная дуга, возникшая из основных спинных элемен-
тов. Три типа этой классификации в значительной мере совла-
дают с таковыми предшествующей. Голомерный тип встречается
только в онтогенезе. Лепоспондильному типу старой классифи-
кации в новой отвечают гастроцентральный и псевдоцентраль-
ный. Что касается до филлоспондильного типа, то Ромер (1939)
показал, что в действительности он является только онтогене-
Рис. 1. Типы слияния дуг позвонков:
/ — голоспондильный, 2 — эмболомерный, 3 — рахитомный, 4 — стереоспонднльный.
5 — нотоцентральный, 6 — гастроцентральный, 7 — псевдоцентральный, 8 — аделоспон-
днльный. Элементы позвонка: BD — основной спинной, ID — межспинной, BV — ос-
новной брюшной, IV — межбрюшной
тической стадией развития рахитомного. В последнее время
большое значение придается эмбриональному развитию позвон-
ков у современных форм. В то время как у бесхвостых позвонки
образуются преимущественно за счет дуг, причем окостенение
идет сверху вниз, у хвостатых и безногих тела позвонков возни-
кают окостенением вокруг хорды. Исходя из этого Ромер (1954)
предложил разделить всех земноводных на 2 подкласса: дугопоз-
вонковых (Apsidospondyli) и тонкопозвонковых (Lepospondyli).
По-видимому, такое решение является более правильным и прак-
тически верным, чем слишком теоретическая попытка Хюне (1956)
разделить всех позвоночных сразу на 4 ветви (Urodelidia, Bat-
rachomorphoidea, Reptiliomorphoidea, Theromorphoidea, Sauro-
morphoidea). При этом в трактовке Хюне исч-езает деление на
классы и даже такие группы, как млекопитающие и птицы совер-
шенно теряются, а земноводные разделяются между тремя пер-
выми ветвями.
Понятие класса в зоологии основывается не только на фор-
мально-морфологических (а по Хюне — только скелетных!) при-
знаках, но учитывает и физиологию, и экологию, и значение
групп в современной фауне. Эти соображения заставляют при-
нять деление Ромера.
23
в настоящем пособия земноводные резделогм (формммм^
на 9 отрядов
1 <12> Кости верхней поверхности черепа представляют со-
бой, хак правило, сомкнутую крышу, которую могут
пронизывать только ноздри, орбиты н отверстие иепар.
кого глаза; обычно имеются: верхиеэатылочная и верх,
лев и сочная кости; обыкновенно более или менее разви-
той панцирь («Archaeobatrachi»).
2(3) Сзади чешуйчатой и квадратно-скуловой имеется не-
большая предкрышка; наружные отверстия ноздрей
чаще находятся ня вентральной стороне головы.
I. Ichthyostegalia
3(2) Прекрышки нет; наружные отверстия ноздрей никое
да не бывают на вентральной поверхности головы.
4(7) Тело позвонка состоит из нескольких костных обра-
зований, скрепленных хрящей.
5 ( 6) На одну невральную дугу приходится два тела поз-
вонка (hypocentrum и pleurocentrum почти равны по
величине).
II. Embolomerl
6(5) Одной невральной дуге отвечают не равные друг
другу hypocentrum и pleurocentrum.
III. Rhachitomi
7(4) Тело позвонка целиком костное.
8(9) Зубы складчатые.
IV. Stereospondyli
9(8) Зубы простые.
10 (11) Позвонки гастроцентральные. Конечности всегда
имеются.
VI. Mlcrosauria
И (10) Позвонки пеевдоцентральные. Конечности могут от-
сутствовать.
VIII. Nectridia
12 ( 1) Кости верхней поверхности головы, как правило, не
образуют сомкнутой крыши; верхнезатылочная н верх-
невисочные кости отсутствуют; панциря нет, если же в
коже имеются чешуйки, то тело полностью лишено ко-
нечностей («Neobatrachi»).
13 (14) Конечности и их пояса полностью отсутствуют; в ко-
же могут иметься чешуйки.
24
IX. Apoda
14 (13) Всегда имеется хоть одна пара конечностей Чешуек
в коже никогда не бывает, хотя изредка встречаются
отдельные кожные окостенения
15 (16) Лобные кости самостоятельны; подвздошная кость
не вытянута в длинный отросток; всегда имеется хвост
VII. Caudata
16 (15) Каждая лобная кость в онтогенезе сливается с те*
менной соответствующей стороны; подвздошная кость
сильно вытянута вперед; хвоста у взрослых нет или он
рудиментарен.
V. Sallentla
Филогенетическое взаимоотношение отрядов изображено на
прилагаемой схеме (рис. 2). Отчетливо видны три последова*
----2
Рис. 2. Филогения отрядов земноводных. Подклассы: 1 — Apsidospondyli,
2 — Lepospondylt
2 П. В. Терентьев
25
тельных ступени: рыбы (термин «Pisces» все более понимается
сейчас как сборное обозначение), древние («Archaeobatrachi»)
и новые земноводные («Neobatrachi»). К последней ступени при-
надлежат отряды V, VII и IX. С другой стороны, первые пять
отрядов образуют подкласс дугопозвонковых (Apsidospondyli),
а последние 4 — тонкопозвонковых (Lepospondyli). Древние зем-
новодные (рис. 3) будут кратко рассмотрены в следующем раз-
деле главы, а новые составят предмет подробного рассмотрения
трех последующих глав. При этом ныне живущие отряды распо-
лагаются в традиционном порядке — безногие (Apoda), хвоста-
тые (Caudata) и бесхвостые (Salientia). Этот порядок до неко-
торой степени отражает степень древности и специализации.
Внутри ступени древних земноводных намечаются как эко-
логические типы («жизненные формы»), так и некоторые общие
филогенетические тенденции, развивающиеся во многих группах
независимо и параллельно (Watson, 1920). Так из примитивных
форм с тупой мордой (типа Pteroplax) вырабатываются, с одной
стороны, рыбоядные формы с удлиненной мордой (напр. Apha-
neramma), а с другой, — явно наземные с плоской головой и бо-
ковым расположением глаз (напр. Broiliellus и Cacops). Появля-
лись и неотенические формы, вторично приспособившиеся к вод-
ному образу жизни (напр. Dvinosaurus). Быстров (1935), изучив
структуру накладных костей, показал, что изменения в черепе
являются результатом возникновения особых зон интенсивного
роста. Действие этих зон видно как в филогенезе, так и в онто-
генезе (например, вытягивание морды Archegosaurus). Как фи-
логенетические тенденции надо отметить: 1) переход от одного
цельного или трехраздельного затылочного мыщелка, образо-
ванного основной и боковой затылочной, к двум мыщелкам, об-
разованным только боковыми затылочными (результат редук-
ции основной затылочной); 2) возникновение и увеличение меж-
крыловидных отверстий («окон»), приводящее к переходу от
широкой, плоской крыловидной кости к узкой фигурной («трех-
ветвистой») , 3) выход языкоглоточного нерва из черепа (Watson,
1920).
ПОДКЛАСС А — APSIDOSPONDYLI.
ДУГОПОЗВОНКОВЫЕ
I. ОТРЯД ICHTHYOSTEGALIA; РАБОВИДНЫЕ
Древнейшие известные земноводные, найденные в девоне и
карбоне северного полушария. Межвисочная кость отсутствует.
Заднелобная и верхневисочная отделены широким промежутком.
Семейства могут быть различены так:
1(2) Межтеменная кость непарная,
26
I
и*
Рис. 3. Представители вымерших земноводных: 1—3 — скелет и череп Ichthyos-
tega, 4—скелет Micropholis, 5 — череп Benthosuchus sushkini сверху, 6—по-
перечный разрез зуба Benthosuchus, 7 — череп Diplocaulus сверку
fchthyo«<eyto*e
2 рода, левом Гренландия.
/ I) Межтеменная кость парная.
? / 4) Верхнеушная (tabular*) подковообразная, е ш*,
3 ' боковым отверстием.
Acanthostegldae
1 род, девон Гренландии.
л ( 3) Верхнеушная без длинного свободного отростка
5(6) Межтеменные кости примерно равны лобным.
Elpistostegidae
1 род, девон Канады.
- / 5) Межтеменные много меньше лобных.
°/ 'эд Линия затылка, если смотреть сверху, со зиачятель»
7 ' впадиной в средней части. Чешуйчатая и верхиеудлц
охватывают ушной канал, замыкая его сзади.
Otocratitdae
1 род, карбон Шотландии.
8(7) Линия затылка приближается к прямой; имеется сл<
бо выраженная ушная вырезка.
Colosteidae
2 рола, карбон Сев. Америки.
II. ОТРЯД EMBOLOMERL ВСТАВЧАТОПОЗВОНКОВЫЕ
Основная затылочная и основная клиновидная кости хорошо
окостеневшие. Затылочный мыщелок одинарный или тройствен*
ный (образован основной и, обычно, боковыми затылочными).
Боковая затылочная пронизана подъязычным нервом. Верхне-
ушная кость (tabulare) еще сохраняет связь с плечевым поясом.
Крыловидные кости с большой небной частью. Меж крыловидное
окно небольшое. Зубы складчатые. Ребра двухголовчатые. С кар-
бона по пермь включительно.
Отряд объединяет следующие семейства:
1 ( 2) Межносовая кость имеется, межвисочная отсутствует;
глазница увеличена передней и задней вырезками;
верхнеушная кость с небольшим выступом; сошник ши-
рокий.
Loxommidae
3 рода, карбон Европы.
2(1) Межносовая кость отсутствует, а межвисочная имеет-
ся; глазница средних размеров; верхнеушная кость с
задним приостренным отростком; сошник удлиненный.
23
i ( 6) Ключииа в виде пластинки г пвраллельтлш перед-
ними и задними мраями.
4 ( 5) Верхнеушная кость не имеет фасетки для сочленения
с задневисочной; край крыловидной кости ие доходит
назад до сочленовной части квадратной.
Anthracoaauridae
10 родов, карбон Европы и Сев. Америки.
5(4) Верхнеушная кость соединяется суставом с задневи-
сочной, задний край квадратной ветви крыловидной
кости доходит до уровня основания черепа.
Palaeogyrinidae
1 род, карбон Шотландии.
6(3) Ключица с треугольным расширением на брюшной
стороне, скулыптурированная.
7(8) Позвонки частично рахитомные; морда сужается по-
степенно, ’И боковая граница черепа почти прямая.
Pholidogasteridae
1 род, карбон Шотландии.
8(7) Позвонки полностью эмболомерные; морда впереди
глаз заметно сужается, и боковая граница черепа во-
гнутая.
Cricotidae
2 рода, карбон и пермь Сев. Америки.
III. ОТРЯД RHACH1TOMI («TEMNOSPONDYLI»).
ДРОБНОПОЗВОНКОВЫЕ
Затылочных мыщелков чаще два. Небная часть крыловид-
ных костей средних размеров. Основная затылочная и основная
клиновидная окостенелые. Зубы с радиальными складками ден-
тина. С карбона по триас включительно. Интенсивная адаптив-
ная радиация дает большое многообразие представителей этого
отряда, сильно затрудняющее классификацию. У Dvinosauridae
вторичное приспособление к воде дало неотению. Семейства
можно пробовать различать так:
1 (12) Имеется межвисочная кость, окруженная заднелоб-
ной, заглазничной, верхневисочной и теменной.
2 (11) Затылочный мыщелок один (иногда трехраздельный) ,
обычно образованный основной и боковыми' затылоч-
ными костями («тройственный»).
3(8) Межкрыловидное отверстие маленькое, его наиболь-
шая длина содержится в наибольшей длине черепа
(считая от кончика морды до линии, соединяющей наи-
более выступающие назад части черепа) более двух
24
раз; глазницы посередине длины черепа иля я залян
его полоняне.
. । 7) Глазница посередине наибольшей длины черепе, для
4 на каждой из них содержится а наибольшей длине че
репа 3 4 с небольшим раза.
с ( 6) Череп удлиненный, его наибольшая ширина содер-
жится в наибольшей длине около полутора раж
Dendrerpetontldae
2- 5 родов, карбон Сев. Америки и Европы.
с ( 5) Череп округлый, его наибольшая ширина равна иля
° 1 чуть меньше наибольшей длины.
Pellontidae
4—5 родов, карбон Европы и Сев. Америки.
у । 4) Глазницы позади половины длины черепа, длина каждой
из них содержится в длине черепа 10—12 раз.
Edopsldae
4 рода, карбон и пермь Европы, Сев. Америки
и Южн. Африки.
g ( 3) Межкрыловидное отверстие большое, его наибольшая
длина содержится в наибольшей длине черепа не более
2 раза; глазница в передней половине черепа.
д (10) Ширина черепа немного менее его длины; ключица на
брюшной стороне расширена; вместо лобковой кости
хрящ.
Trimerorachidae
3 рода, карбон и пермь Сев. Америки и Ев-
ропы.
Ю ( 9) Ширина черепа почти равна его длине; ключицы расши-
рены слабо; лобковая кость имеется.
Dvinosauridae
2 рода, пермь Европы.
11 (2) Затылочных мыщелков два.
Cochleosauridae
I род, карбон Европы.
12 ( 1) Межвисочной кости нет.
13 (30) Межбрюшной элемент позвонков окостенелый.
14 (17) Наибольшая длина межкрыловидного отверстия содер-
жится в наибольшей длине черепа не менее трех с поло-
виной раз.
15 (16) Наибольшая ширина черепа содержится в его наиболь-
шей длине 2 и более раза.
30
16
17
18
19
20
21
22
23
24
25
26
27
28
СЭммюргиорМм
2 рода, карбон я перми Сев Америки.
(15) Ширина черепа содержится а «го длине много менее
двух раз.
Zatraehydae
5 родов, пермь Сев. Амернхя и Европы.
(14) Длина межкрыловидного отверстия содержится в длине
черепа менее трех с половиной раз.
(21) Длина межкрыловндного отверстия содержится в длине
черепа не более 2 раз.
(20) Наибольшая длина носовых костей содержится в иди*
большей длине черепа 3—4 раза.
Lysipteryglldae
2 рода, пермь Европы и Азии.
(19) Длина носовых костей содержится в длине черепа 4—
6 раз.
Mlcropholldae
1 род, триас Южн. Африки.
(18) Длина межкрыловндного отверстия содержится в длине
черепа более двух раз.
(25) Затылочных мыщелков два, если же один, то ключица
на брюшной стороне не расширена и ушная вырезка
занимает боковое положение.
(24) Глазницы взрослых в задней половине черепа; ключицы
на брюшной поверхности расширены; вместо лобковой
ко-сти хрящ.
Arch egos auridae
4 рода, пермь Европы, Азии и Южн. Америки.
(23) Глазницы на середине длины черепа; ключицы не рас-
ширены; лобковая кость имеется.
Dissorophidae
11 родов, карбон и пермь Сев. Америки и Ев-
ропы.
(22) Затылочный мыщелок один, иногда трехраздельный.
(27) Ключицы на брюшной поверхности расширены.
Actinodontldae
4 рода, пермь Европы.
(26) Ключицы не расширены.
(29) Ушная вырезка сзади открытая.
31
Eryopsldae
3 рода, пермь Европы я Сев. Америки.
29 (28) Ушная вырезка сзади замкнута или совсем отсутствует
Trema topsldae
5—6 родов, карбон и пермь Европы, Ама
и Сев. Америки.
30 (13) Межбрюшной элемент позвонков хрящевой и помеша-
ется в пространстве, оставляемом для него окостенев-
шими основными брюшными элементами (отличие от
Цельнопозвонковых!).
31 (32) Наибольшая ширина черепа содержится в его наиболь-
шей длине 1,3—1,7 раза.
Benthosuchldae
6 родов, триас Европы и Азии.
32 (31) Ширина черепа содержится в его длине 1,0—1,3 раза.
33 (34) Длина межкрыловидного отверстия содержится в на-
ибольшей длине черепа не более двух раз.
Uranocentrodontidae
3 рода, пермь и триас Южн. Африки.
34 (33) Длина межкрыловидного отверстия содержится в длине
черепа более двух раз.
35 (36) Лобковая кость замещена хрящем.
Lydekkerinidae
4 рода, триас Южн. Африки.
36 (35) Лобковая кость имеется.
37 (38) Ноздри на конце морды; морда несколько вытянута.
Rhinesuchi dae
2 рода, пермь Южн. Африки.
38 (37) Ноздри заметно сдвинуты от конца 'морды назад; морда
тупо закругленная.'
Sclerothoracidae
'2 рода, пермь Европы.
IV. ОТРЯД STEREOSPONDYLI. ЦЕЛЬНОПОЗВОНКОВЫЕ
Два затылочных мыщелка. Основная затылочная и основная
клиновидная не окостенелые. Зубы сложно-радиальноскладча-
тые. Все триасовые. Вопрос о реальности этого отряда дискуссио-
нен. Семейства можно различать так:
1 (10) Наибольшая ширина черепа всегда заметно меньше его
наибольшей длины.
32
2 ( 9) Глазницы небольшие; наибольшая длина на содержит™
6—15 раз в наибольшей длине черепа.
3(4) Голова сильно вытянута: наибольшая шяряяа черепа
содержится в его наибольшей длине 1,Я~3,5рам
Trematosauridae
7 родов, Европа, Азия и Южн. Африка
4(3) Голова короче: ширина черепа содержится IД—1,7 раз
в его длине.
5 ( 6).Глазницы в задней половине черепа.
Capltosauridae
6—7 родов, Европа, Африка и Сев. Америка.
5 ( 5) Глазница в передней половине черепа или иа середине
его длины.
7(8) Череп треугольный, с прямолинейным контуром боковых
сторон (смотреть сверху); ширина черепа в районе нозд-
рей содержится 2,9—4,7 раз в наибольшей ширине че-
репа.
Lyrocephalidae
3 рода, Шпицберген и Южн. Африка.
8(7) Череп‘спереди закругленный, иногда с несколько изог-
нутым контуром боковых сторон; ширина черепа в рай-
оне ноздрей 2,4—3,6 раза содержится в ширине черепа.
Metoposauridae
1—5 родов, Европа и Сев. Америка.
9 ( 2) Глазницы большие; длина содержится 4—5 раз в длине
черепа.
Mastodonsauridae
1 род, Европа, Азия и Южн. Африка.
10 ( 1) Ширина черепа превышает его длину.
Brachyopidae
10 родов, Европа, Азия, Африка, Австралия,
Сев. и Южн. Америка.
V. ОТРЯД SALIENTIA. БЕСХВОСТЫЕ
(см. главу 5)
ПОДКЛАСС Б — LEPOSPONDYLI. ТОНКОПОЗВОНКОВЫЕ
VI. ОТРЯД MICROSAURIA. УКОЛОВИДНЫЕ
Позвонки гастроцентральные. Небольшие или мелкие верхне-
карбоновые или нижне-пермские животные с короткой шеей, вы-
тянутым телом (видимо напоминали современную амфиуму),
33
короткими конечностями (передние с 3, а задние с 5 палыимя)
и относительно коротким хвостом. Глазницы в передне# части
черепа Затылочный мыщелок одинарный, но иногда образован
с участием боковых затылочных костей. Ребра двуголовчатые
Положение н объем группы вызывают большие споры. Некото-
рые рассматривали часть ее представителей (например, Panty-
lus) или всю группу как пресмыкающихся. Пока осторожнее
говорить всего об одном семействе Microsauridae, охватываю-
щем 7—12 родов из Европы и Сев. Америки.
VII. ОТРЯД CAUDATA. ХВОСТАТЫЕ
(см. главу 4)
VIII. ОТРЯД NECTRIDIA. ПЛАВАЮЩИЕ
Позвонки псевдоцентральные. Конечности могут отсутство-
вать. Формы приспособлены ко вторичному — водному образу
жизни. Семейства можно различить так:
1(4) Невральные дуги соединяются с телом позвонка швом;
оболочка хорды обызвествляется.
2(3) Глазницы в передней половине черепа; верхнеушная
кость отсутствует; имеется хорошо развитое пинеальное
отверстие между теменными костями.
Adelogyrinidae
3 рода, карбон Европы и Азии.
3(2) Глазницы примерно посередине длины черепа; верхне-
ушная кость имеется; пинеальное отверстие маленькое
или совсем отсутствует.
Lysorophidae
3 рода, карбон Сев. Америки.
4(1) Невральные дуги прирастают к телу позвонка; оболочка
хорды не обызвествляется.
5(6) Конечности отсутствуют; глазницы примерно на сере-
дине длины черепа.
Aistopodidae
4 рода, карбон и пермь Европы и Сев. Аме-
рики.
6(5) Конечности имеются; глазницы чаще в передней поло-
вине черепа.
7(8) Длина межтеменных костей почти равна длине темен-
ных; заднебоковые углы черепа очень сильно вытянуты
вбок и назад.
34
[MfrtocatHMM
3 рола, яарбоя м пермь Се* Айерами и Ев
ропы.
а / 7) Длина межтеменных мосте* заметно меньше длины те-
" меннмх; ладнебокоаме углы черепа вытянуты нетал нлн
совсем не вытянуты,
а (10) Верхнеушная кость образует направленные назад вы*
росты в форме рогов.
Ceraterpetontldae
2 рода, карбон Европы и Сев. Америки.
Ю ( 9) Верхнеушная кость маленькая, часто незаметная, без
1 выростов.
Urocordylidae
4 рода, карбон и пермь Европы и Сев. Аме-
рики.
IX. ОТРЯД APODA. БЕЗНОГИЕ
(см. главу 3)
ЛИТЕРАТУРА
Быстров А. Некоторые данные о скульптуре и росте накладных костей
стегоцефалов. Архив анатомии, гистологии и эмбриологии, 1935, т. 14.
Вермель Ю. Эскизы о факторах, направляющих эволюцию, Тр. ин-та зооло-
гии Моск, университета, 1931, т. 4. вып. 3.
Дубинин В. Изменения шейного отдела позвоночника животных в фило-
м 3 генезе и онтогенезе, <Журн. общей биол.», 1949, т. 10, № 2.
Ромер А. Палеонтология позвоночных, М.—Л., 1939.
Гевеоцов А. Происхождение наземных позвоночных, Собр. соч., М.—Л.,
1950, т. 2, стр. 313—404.
Трофимов Б. Новые данные о древнейших наземных позвоночных, При-
рода, 1955, 3.
Шмальгаузен И. История происхождения амфибий. Изв. АН СССР,
1958, сер. биол. № 1.
Gregory W. and Raven. Н. Studies on the Origin and Early Evolution of
Paired Fins and Limbs, Annals New York Acad. Sci., 1941, 42: 273—360.
Holmgren N. On the Tetrapod Limb Problem-Again, Acta Zoologica, 1949,
30, No. 3 485—508.
H u e n e F. Palaontologie und Phylogenie der niederen Tetrapoden, Jena, 1956.
Jarvik E. On the Fish-like Tail in Ichthyostegid Stegoceptaiians, Meddelel-
ser om Gronland, 1952, 114, No. 12.
Menzbier M. On the Combination of Factors, to which is due the Origin and
Development of Terrest-ial Vertebrates, Comp. Rend. Acad. Sci. URSS,
1930, A, No. 22: 589—594.
Romer A. Notes on Branchiosaurs, Amer. Journ. Sci., 1939, 23: 748—761.
Romer A. Vertebrate Paleontology, II ed., Chicago, 1945.
Save-Soderbergh G. Preliminary Note on Devonian Stegocephalians
iiom E. Greenland, Meddelelser om Gronland, 1932, 94, No. 7.
Traite de Paleontologie, Paris, 1955, 5: 3—249. 275—305, 314—316.
Watson D. The Structure, Evolution and Origin of the Amphibia, Philos.
Transac. Roy. Soc. London, 1920, B, 209: 1—72.
Westoll T. Ancestry of Tetrapods, Nature, 1938, 141, No. 3559: 127—128.
Г лава третья
БЕЗНОГИЕ ЗЕМНОВОДНЫЕ
Тело у большинства видов безногих земноводных * в попереч-
ном сечении круглое и лишь у некоторых сжатое с боков (рис. 4),
Кожа образует многочисленные складки, разбивающие поверх*
ность тела животного как бы на ряд колец. В онтогенезе число
этих колец первоначально совпадает с числом позвонков, а по-
том возрастает за счет возникновения вторичных бороздок. Пер-
вичные бороздки распределены, обычно, по всему телу на равных
расстояниях и чаще образуют полное или почти замкнутое кольцо
вокруг тела. Вторичные же бороздки встречаются лишь у некото-
рых видов и помещаются чаще на заднем конце тела и, у некото-
рых видов, правильно возрастают по направлению от головы к
хвосту. Кожа богата железами. Железы эти располагаются в оп-
ределенном соотношении с «кольцами» кожи: передняя половина
каждого кольца имеет пояс тесно сидящих очень больших слизи-
стых желез, рядом с которыми встречаются и мелкие. Полагают,
что мелкие железы могут выбрызгивать едкую жидкость, вызы-
вающую истечения из носа у лиц, вскрывающих свежие экземп-
ляры. Во второй половине каждого кольца располагаются в осо-
бых сумках чешуйки, служащие защитой для желез. Таким обра-
зом, чешуйки представляют собой целиком мезодермические об-
разования, лежащие в глубоком слое кожи. Замечательна ком-
пактность и твердость черепа, вся верхняя поверхность которого
фактически представляет собой почти сплошную крышку, а лице-
вая часть прочно срослась с черепной коробкой.
У большинства родов носовая кость слита с предчелюстной.
Чешуйчатая сливается с квадратно-скуловой. Между ними и
теменной костью может находиться значительное отверстие, ана-
1 Старые исследователи (Линией, Лауренти, Литрей, Додей) относили
безногих к змеям. Дюмериль был, по-видимому, первым, указавшим в 1807 г.
на близость безногих к лягушкам и жабам. Оппель твердо установил в 1811 г.
принадлежность рассматриваемой группы к земноводным и дал ей назва-
ние «Apoda».
логичное височной ямке пресмыкающихся. Глежит мялеяыквя,
а иногда прикрывается расширенной челюстной костью Некото-
рые (Ichthyophis) имеют кольцо окостенений в склеротике глаза
Верхнеушная и переднеушная сливаются с боковыми зятылочны
ми и околоклиновидной. Ушной столбик сочленяется с квадрат
ной костью таким же образом, как у змей. Затылочных мышел
ков два. Сочленовная кость образует толстый отросток, иаправ
Рис. 4. Строение безногих: А — череп Ichthyo-
phis glutionosus сверху, Б — череп Chthonerpe-
ton indistinctum сверху, В — внешний вид
Siphonops annulatus. Г — задняя часть тела
Geotrypetes petersii, Д — голова Caecilia ten-
taculata:
1 — ноздря, 2 — глаз, 3 — щупальце, 4 — увеличенное
щупальце, 5 — кольцевые складки кожи, 6 — задне-
проходное отверстие, 7 — хвост, 8 — первичные склад-
ки, 9 — вторичные складки
ленный вверх и назад от сустава нижней челюсти. Многочислен-
ные, мелкие, загнутые назад зубы сидят в 2 ряда: наружный на
предчелюстной и челюстной кости, а внутренний — на нёбной и
сошнике. Подъязычный аппарат наиболее примитивный среди
всех современных земноводных. Позвонки псевдоцентр а льныё,
внешне двояковогнутые. Пронизывающая их хорда в середине
каждого позвонка прерывается хрящем. Межпозвоночные хрящи
имеют вид колец, пронизанных хордой. Общее число позвонков
200—300. На первом из них нет межсуставного («зубовидного»)
отростка. Ребра, при своем появлении в онтогенезе, слиты с поз-
37
вонками. позднее двуголовчатые. Они свойственны всем позвои
кам, начиная с десятого до предпоследнего включительно. Ника
ких следов конечностей или их поясов в скелете не обнаружено
Движение происходит путем змееобразных изгибаний позвоноч
ника. Никаких следов мускулатуры поясов конечностей нет.
Передний мозг более массивен чем у других современных
земноводных. Промежуточный и средний мозг не отличимы друг
от друга. Мозжечок, зрительные и слуховые нервы недоразвиты.
Переходя к органам чувств надо прежде всего отметить утрату
барабанной перепонки и среднего уха (конвергенция со змеями!).
Маленькие глаза практически бесполезны, будучи скрыты под
кожей или даже под костями. В онтогенезе внутренняя (истин-
ная) и внешняя роговица срастаются, и способность глаза к
движению утрачивается. Функция зрения заместилась усилением
обоняния. Безногие имеют на голове маленькое щупальце кони-
ческой или округлой формы, помещающееся в ямке, которую
называют кожной ноздрей, или слезной ямкой. Внутрь ямки
открывается проток большой глазной железы (гардеровой).
Одно время это образование неосновательно считали за ядовитый
аппарат. При жизни щупальце постоянно высовывается из ямки,
являясь вероятнее всего, особым органом чувств (видимо не
только осязания, но и обоняния). Пищевод и желудок незаметно
переходят друг в друга. Тонкая кишка мало извитая. Печень
очень вытянутая и косыми щелями разбита на большое число
долек.
В онтогенезе возникает в стенках глотки 7 пар жаберных
мешков, вместо 6 обычных для других земноводных. Первые
6 мешков дают почки зобной железы, разбитой у взрослого
животного на последовательные дольки. Первая жаберная щель
открыта лишь короткое время в течение эмбрионального перио-
да, а потом образует брызгальце. По бокам головы зародыша
имеются жабры либо в форме ветвистых пучков (Ichthyophis,
Hypogeophis), либо в форме пластинок (Typhlonectes, Sipho-
nops). Дыхание взрослых легочное. Легкие асимметричны: пра-
вое очень короткое, а левое вытянуто в длинный мешок. Сход-
ство со змеями подчеркивается тем, что у Typhlonectes разви-
вается трахеальное легкое. Как и у других земноводных, воздух
вбирается в ротовую полость за счет движения гортани. Ребра
в дыхании не участвуют. Сердце располагается в передней части
тела, вытянутое. У Siphonops короткий артериальный’ конус
имеет один ряд клапанов, занимающий почти всю высоту стенки
конуса. Спиральной складки нет. У Ichthyophis две легочных
артерии, а у Gegeneophis левая исчезает в онтогенезе. Лимфати-
ческая система носит сегментальный характер: у Hypogeophis
под кожей всего тела (исключая голову) располагается более
200 лимфатических сердец, помещающихся межсегментно на
границе между спинной и боковой поверхностями животного.
Почки очень длинные и узкие (ширина 1—2 мм). Имеется моче-
38
юй пузырь. Половые железы косят также сегмегтвльиый
пер, хотя короче почек. Клоака самая одета особым млткуэк
<ым футляром, при помощи которого может выпятгкатыя (
»грать роль копулятивного органа. Оплодотоореияе ачутреяяег
Некоторые яАцекладуши (fchthyoohit, Н wngeophit). другие
юрод я щи (Typhlonectes, Dermopnis) У лоследияж змбраом мне
ет несколько рядов кожных зубов, посредством которым скоблЯ1
стенки яйцевода.
Вопрос о происхождении отряда вызывал горячие споры
Первоначально предполагали, что это утратившие йога хвоста
гые, в частности, потомки Amphiumidae. Потом было указам,,
что по своему строению безногие значительно примитивнее мво-
статых и чешуя их гомологична чешуе других ископаемых зем-
новодных. Особенности строения позвоночника делают ныне
несомненным непосредственное происхождение безногих от при-
митнвных тонкопозвонковых. Остается только ответить на воп-
рос: почему и как у них исчезли конечности?
Обычно принимают, что утрата конечностей была вызвана
либо продвижением средн густой растительности, либо жизнью
в норах. Однако знание экологии земноводных делает более ве-
роятным иной взгляд. Движение ящероподобных животных
всегда содержит в себе элемент змееобразных волн. Если изгибы
тела в горизонтальной плоскости начинают преобладать, то тело
удлиняется, а конечности редуцируются. Много примеров воз-
никновения безногости имеется среди сцинковых ящериц, кото-
рые как бы «плавают» в песке. Поскольку земноводные не
живут внутри сыпучих песков, остается допустить, что безногие
потеряли свои конечности в результате возрастающего приспо-
собления к плаванию в воде. Какие причины привели к этому?
Первые девонские земноводные были первично-водными живот-
ными, но в карбоне среди них имелось уже немало типичных
наземных животных. Вскоре все подходящие «влажные» стации
оказались достаточно полно насыщены земноводными, и тогда
«давление жизни» вынудило перейти к освоению «сухих» стаций.
В результате в среднем или верхнем карбоне из земноводных
возникли пресмыкающиеся. Уже в перми последние достигли
значительного многообразия и в триасе, заполнив «сухие» ста-
ции, стали теснить своих предков — земноводных. Земноводные
дали гигантских стегоцефалов, но и они не выдержали борьбы
н отступили назад, в свою исходную стихию — появились вто-
рично-водные (даже неотенические!) формы стегоцефалов. Од-
нако адаптивная радиация пресмыкающихся получила столь
значительный размах, что они смогли дать ряд водных ветвей,
вызвавших к концу триаса вымирание даже водных земновод-
ных. Уцелели лишь немногие мелкие наземные формы (хвоста-
тые и бесхвостые), сильно изменившиеся и приспособившиеся
к совершенно специфическим стациям, да та часть водных тонко-
позвонковых стегоцефалов, которая сумела укрыться от конку-
39
Рис. 5. Географическое распространение Оеамогич
цли, перейдя яэ воды в почву. Значит, если скаэашюе сярв-
^дднво, безногие потеряли конечности в воде.
^ныне известно всего одно семейство червяг, распространен-
в тропическом поясе Америки, Африки (исключая Мада
!яскар) и Азии (рис. 5).
1 Латинское название семейства червяг пишут по-разному, но
правильнее именовать его Caecihidae. Ныне принимают 19 ролов
г 55-—56 видами:
। (24) В коже есть округлые чешуйки.
2 (21) Глаза под кожей, просвечивают.
3 (18) Нижняя челюсть с двумя рядами зубов.
4 (II) Чешуйчатая и теменная кости соприкасаются.
5 ( 6) Щупальце коническое.
Ichthyophis
2 вида, Индия, Цейлон, Суматра, Калиман-
тан (Борнео), Ява.
6(5) Щупальце не коническое.
7(8) Щупальце круглое.
Dermophis
7 видов, Америка, Африка.
8(7) Щупальце клапанообразное.
9 (10) Щупальце сзади и книзу от ноздри.
Hypogeophis
4 вида, Сейшельские о-ва, Занзибар.
10 ( 9) Щупальце под ноздрей.
СаесШа
8 видов, Южн. Америка.
11(4) Чешуйчатая и теменная кости разделены промежутком.
12 (15) Щупальце клапанообразное.
13 (14) Щупальце около глаза.
Rhinatrema
3 вида, Южн. Америка.
14 (13) Щупальце несколько ниже и сзади от ноздри.
Geotrypetes
1 вид, Зап. Африка.
15 (12} Щупальце не клапанообразное.
16 (17) Щупальце коническое.
Uraeotyphlus
3 вида, Азия, Африка.
17 (16) Щупальце круглое.
pni*N0i>
I вид. Сейшельские о-ва.
, нижняя челюсть с одним радом зубов.
;«го3 ₽' ^мое-
Cfyptop«opW«
1 вид. Сейшельские о-ва.
20 (19) Щупялы*' клвпанообраэиое.
Amphlumophls
1 вид, Южн. Америка.
21
22
23
( 2
(23‘
(22)
Глаза покрыты костями черепа.
Щупальце около ноздри.
Herpele
5 видов, Южн. Америка, Зап.
Щупальце ближе к углу рта, нежели к
Gymnophls
4 вида, Америка.
АФРика, Азия
ноздре.
24
25
26
27
( О
(32)
(29)
(28)
28
(27)
29
30
(26)
(31)
31
(30)
32
33
34
(25)
(36)
(35)
35
(34)
Чешуй нет.
Глаза под кожей.
Зубы на нижней челюсти расположены * о
Чешуйчатая и теменная кости соприКас1ю Р*Да*
Typhlonectes ТСя>
3 виде, Южн. Америка
Чешуйчатая и теменная не соприкасаются
Chtfionerpeton
2 вида, Южн. Америка.
Зубы на нижней челюсти расположены »
Чешуйчатая и теменная кости соприкаСаю°АИН ряд-
Siphonops ТСЯ*
5 видов, Южн. Америка.
Чешуйчатая и теменная кости разделены пп
Bdellophis Делены промежутком.
1 вид, Вост. Африка.
Глаза под костями черепа
Зубы на нижней челюсти ©я-ттлп™
щупальце круглое, около ноздонЫ В 2 ряда-
Gegeneophis
1 вид’ южн. Азия
Sa уХТГ’
расстояния между
42
Boulengeruta
2 вида, Вост. Африка.
36 (33) Зубы на нижней челюсти в один ряд.
Scotecomorphus
1 вид, Вост. Африка.
Род Ichtyophis, рыбозмей, включает два вида, распространен-
ные в Индии к югу от Гималаев, на Цейлоне, Суматре, Калиман-
тане и Яве. Большая часть ареала рода занята I. glutino-
sus (рис. 6). Последний достигает 38 см длины и 14 мм толщины.
Рис. 6. Представители безногих: А—самка Ichthyophis glutinosus с яйцами,
Б — яйца Siphonops annulatus, В — зародыш Siphonops annulatus в яйце;
Г—подросшая личинка Ichthyophis glutinosus; Д—зародыш Typhlonectes
compressicauda:
J — жаберное отверстие, 2 — хвостовой плавник
Кольцевые складки многочисленны (240—400). Окраска
взрослых темно-бурая или голубовато-черная с широкой ярко-
желтой продольной полосой по бокам тела. Самка откладывает
43
яйца диаметром около 6-9 мм. одетые плотной белковой обо
ломкой. Кладка прон.чнолнтся в ямку, вырытую во влажной земли
около текучей воды. Самка свертывается вокруг 2 дюжин отло
женных яиц н лежит пока не выйдут личинки. Такой hhcthhri
латиты направлен, видимо, против роюших змей и безноги»
ящериц, в изобилии живущих в почве тропиков. Яйца ие толькг,
остаются влажными, но и увеличиваются в весе, по-видимому
благодаря выделениям кожи матери. Еще внутри яйцевой обо
лочки личинка развивает нежные ветвистые жабры, орган боко-
вой линии и хвостовой плавник. Ко времени вылупления из яйца
наружные жабры исчезают, появляются жаберные щели и, по-
падая в воду, личинка начинает двигаться наподобие угря.
Однако дышит она уже легкими. После довольно длительного
пребывания в воде, личинка претерпевает метаморфоз и, в каче-
стве взрослого животного, переходит исключительно к подзем-
ному образу жизни. Если бросить взрослое животное в воду,
то оно тонет. Взрослые держатся близ плоских, сырых берегов
ручьев на глубине 30 см под дерном. Питаются земляными чер-
вями и змеями из семейств Typhlopidae и Uropeltidae.
К роду Caecilia, или настоящих червяг, принадлежит круп-
нейший представитель всего отряда — С. thompsoni, водящийся
в Колумбии на высоте до 1150 м над уровнем моря. Достигает
117 см длины.
Род Typhlonectes, помимо уже указанных признаков, отли-
чается телом, более или менее сжатым с боков. На брюхе и
вокруг заднепроходного отверстия развивается подобие приса-
сывательного диска. Длина тела около 46 см при толщине в.
19 см. Окраска от оливково-коричневой до черной, бею жизнь
проводит в воде. Живородяща. Эмбрионы, в числе 6 штук, снаб-
жены лепестковидными жаберными придатками, по-видимому,
отпадающими вско-ре после рождения.
Род Siphonops («кольчатая червяга») встречается, между
прочим, внутри муравейников.
ЛИТЕРАТУРА
Brauer A. Beitrage zur Kenntnis der Eritwicklungsgeschichte und der
Anatomie der Gymnophionen, Zool. Jahrb., Abt Anat. 1897, 10: 389, 1898,
12: 477, 1904, Suppl. 7: 381.
Kingsley J. The Systematic Position of the Caecilians. Tufts Coll. Stud.,
1902, 6: 123.
Marcus H. Beitrage zur Kenntnis der Gymnophionen, Archiv mikr. Anat
Entw. 1908, 71: 695—774, Morphol. Jahrb., 1908. 38: 590—607, 1909, 40.
105—183, Zeitschr. Anat. Entw. 1923, 69: 328—345.
Nieden F. Gymnophlona (Das Tierreich, Lief. 37), Berlin, 1913.
Parker H. Viviparous Caecilians and Amphibian Phylogeny, Nature, 1956,
178, No. 4527: 250—252.
—— Глава чпвертал
ХВОСТАТЫЕ ЗЕМНОВОДНЫЕ
Форма тела большинства представителей отряда хвостатых ’
наименее специализированная, но наиболее характерная для
земноводных вообще. Однако встречаются формы, у которых
тело столь сильно удлиненно, что напоминает угря. У таких форм
конечности оказываются рудиментарными (Amphiuma) или зад-
няя пара их совсем отсутствует (Siren). Большинство хвостатых
ползает или плавает, змееобразно изгибая тело; конечности при
плавании пассивны. Только немногие бегают (Hydromantes,
Oedipus) или даже делают прыжки (Chioglossa).
Подробное описание внутреннего строения можно здесь опу-
Иб° имеются хорошие сводки по саламандре (Francis,
1934), тритону и аксолотлю. (Воронцова и др., 1952)—живот-
ным, служащим «модельными» для всего отряда. Основная мас-
са морфологических признаков носит примитивный характер.
Кожа всегда плотно прилегает к глубжележащим образованиям.
У водных представителей, кроме переднеушной кости, может
встречаться 1—2 ушных. Околоклиновидная кость широкая.
Квадратноскуловые элементы, как правило, редуцированы до
связок. В подъязычном аппарате всегда сохраняется не менее
3 самостоятельных дуг. Позвонки псевдоцентральные, двояко-
вогнутые или задневогнутые. Число их значительно у вытянутых
в длину водных форм (98 у Amphiuma) и меньше у наземных.
Все туловищные позвонки, кроме первого, несут двухголовчатые
ребра. Грудной пояс остается в значительной мере хрящевым,
и его элементы не разделяются швами. В тазе характерно нали-
чие предлобкового хряща (epipubis), играющего большую роль
в выдавливании воздуха из легких при погружении в воду. Эле-
менты зевгоподия всегда раздельны. Автоподий не окостеневает.
1 В нынешнем объеме отряд был впервые установлен в 1807 г. Дюмери-
м В отношении латинского наименования отряда до сих пор имеются рас-
*Ле«г пения: систематики чаще употрябляют термин Caudata, а несистематнки_
Однако, как еще в 1907 г. показал Стейнегер, последнее название
результат ошибки, ставшей традицией.
45
Сходство эдпястъя я предплюсны велико. Наблюдается *
4 центральных элементов. На передней конечности от 4 к
3 пальцев, а на задней — от 5 до 2. Вся спина и верх хяоеп
покрыты одним мощным спинным мускулом (m. dorsalis truncij
Мускулатура боков и низа тела состоит из многих слоев. Полу
шария переднего мозга более вытянуты, нем у бесхвостых
Крыша среднего мозга без заметных зрительных долей. Бара-
банная перепонка и среднее ухо редуцированы. В онтогенезе
около глаза возникает особый вырост — «балансир», несущий
осязательные (? и дыхательные) функции и помогающий личин-
ке не погружаться в ил. У личинок развиты 3 пары наружных
___>
—-3
Рис. 7. Филогения семейств хвостатых. Подотряды: 1—Cryptobranchoidea,
2 — Ambystomoidea, 3 — Salamandroidea
жабр, которые иногда остаются на всю жизнь (протей, сирена).
В прежнее время признавалась группа постоянножаберных
(Perennibranchiata), но сейчас ясно, что это только сборная
группа разнородных по происхождению форм с разными степе-
нями неотении («permanent larvae»). Личинки имеют 4 пары
жаберных щелей, из которых во взрослом состоянии сохраняют-
ся у сирены 3, у протея и колодезного тритона — 2, у всех про-
чих они замыкаются. При обитании в воде гидростатическая
роль легких несомненна. В филогенезе она, видимо, была пер-
вичной. Иногда во взрослом состоянии исчезают и жабры, и лег-
кие (апневмония). Весь газообмен в таких случаях идет через
кожу и слизистую оболочку ротовой полости. Одновременно
перестраивается и кровеносная система: сердце становится как
бы функционально двукамерным. Первоначально апневмония
46
возникает у обитателей горных ручьев с богато ахрираеаююя
водой. Почка и семенник у хвостатых короче таковых у безногих.
Вся передняя половина почки несет следы дегенерашги Мо-
Рис. 8. Представители хвостатых: 1 — Cryptobranchus alleghaniensis,
2 — Amphiuma means, 3 — Typhlomolge rathbuni, 4 — Ambystoma tigrinum,
5 — Aneides lugubris, 6 — Batrachoseps attenuatus
чевой пузы<рь имеется. Отверстие клоаки обычно в форме про-
дольной щели. Количество яиц сравнительно невелико (150—
700 у аксолотля, 60—200 у тритонов, 2—5 у живородящих форм).
47
Внутри отряда наблюдается эволюция от наружного оплодотво-
рения и отсутствия спаривания (только «брачные мгрыэ) до
внутреннего оплодотворения н охвата самки самцом у некоторых
наземных форм. Сперматозоиды откладываются в особых студе-
нистых пакетах (сперматофорах), которые затем втягиваются
клоакой самки. У личинки передние ноги появляются раньше
задних. Высокая способность регенерации сохраняется на всю
жизнь.
Первые несомненные представители отряда найдены в мелу.
Палеоценовых находок не известно и только с эоцена остатки
хвостатых становятся многочисленными.
Начиная с Бонапарте (1839), стали различать в качестве
особых семейств саламандр, амфиум и сирен, выделяя еще иног-
да и протеев. Штраух предложил различать по характеру нёбных
зубов продольнозубых (Mecodontia) и поперечнозубых (Lechrio-
donta). Однако сейчас этому признаку, равно как и присутст-
вию жабр, не придают особого значения. Известные ныне семей-
ства отряда хвостатых группируются в 3 подотряда (рис. 7, 8):
1 ( 4) Оплодотворение наружное; угловая кость свободная
[подотряд (1) Cryptobranchoidea].
2 ( 3) Подвижные веки и слезная кость имеются.
1. Hynobiidae
3(2) Подвижных век и слезной кости нет.
2. Cryptobranchidae
4(1) Оплодотворение внутреннее (вторично утрачено Sireni-
dae); угловая кость срастается с предсуставной
5 ( 6) Серии нёбных зубов поперечные; никогда не* бывает
зубов на околоклиновидной кости [подотряд (2) Amby-
stomoidea]. 1 7
3. Ambystomidae
6(5) Серии нёбных зубов продольные, расходящиеся назад,
если же поперечные, то на околоклиновидной кости име-
ются зубы [подотряд (3) Salamandroidea].
7 (12) Жабры у взрослых, как правило, отсутствуют; верхне-
челюстная кость имеется.
8 (11) Веки, как правило, имеются; взрослые без жаберных
щелей.
9 (10) На отростках нёбных костей зубы в виде 2 продольных
серий; околоклиновидная кость без зубов; легкие име-
ются; позвонки задневогнутые.
4. Salamandridae
10 ( 9) Серии зубов располагаются на задней части сошника;
легких нет; позвонки двояковогнутые или, значительна
реже, задневогнутые.
46
I ( 8) Век мет; на всю жгись смряяяется ипинтими
жаберная «ель (брызг алый)
V*
2(7) Жабры сохраняются на вею жизнь. аержяечелкхгпюй
кости нет.
3 (14) Две лары конечностей; на предчелюстной хостя я ияж-
ней челюсти имеются зубы.
1. Proteidae
14 (13) Задние конечности отсутствуют; зубов нет; челюсти по-
крыты роговидными пластинками.
7. Sirenldae
Подотряд 1 — Cryptobranchoidea. Скрытожаберные
Позвонки двояковогнутые. Яйца откладываются в желатино-
подобных мешках.
1. СЕМЕЙСТВО HYNOB1IDAE. УГЛОЗУБЫЕ
Ранее, вероятно, имели сплошной ареал от Средней Азин до
Тайваня включительно. Ныне центр многообразия в Вост. Азин
(рис. 9). Отличия родов следующие:
1(6) Сошниковые зубы в виде более или менее непрерыв-
ных угловатых серий.
2(5) Сошниковые зубы ясно изогнуты под углом; хвост не
длинный и хоть немного сжат с боков; когтей нет; легкие
развиты нормально.
3(4) На ступнях есть роговые образования
Pachypalaminus
1—6 видов, Япония, Китай.
4(3) На ступнях роговых образований нет.
Hynobius
18 видов, Вост. Азия, Сибирь.
5(2) Сошниковые зубы почти поперечные; хвост длинный и
почти цилиндрический; когти личинки могут сохраняться
и у взрослых; легкие редуцированы.
Onychodactylus
2 вида, Япония, Корея, Уссурийский край.
6( 1) Сошниковые зубы в виде двух небольших разделенных
групп.
5 П- В. Терентьев 4g
Рис. 9. Ареалы семейств: /—Hynobiidae, 2 - Cryptobranchidae, X- Ainbystomidtr, J <чгггш1л<г
7 ( 8) На задних конечностях по 5 па льне»; ступни без рого
вых образований.
Ranodon
1 вид, Казахстан. Афганистан (?).
8(7) На задних конечностях по 4 пальца; ступни с роговыми
образованиями.
Batrachuperus
1 вид, Сычуань, Хами.
Род Hynobius, углозуб — центральный и, вероятно, исходный
для всего семейства. Большинство видов островные и только
4 встречаются на материке. Сибирский углозуб, Н. keyserlingii
часто рассматривался как представитель особого рода Salaman-
dreHa=Jsodactylium, характеризующегося всего 4 пальцами иа
задних ногах (четырехпалый тритон). Однако число пальцев
не может служить родовым критерием, ибо среди других видов
рода иногда попадаются особи с редуцированным пятым паль-
цем. Как исключение у сибирского вида попадаются иногда
особи всего с 3 пальцами. Сибирский углозуб распространен от
Камчатки, Курил, Сахалина и Японии по всей Сибири. В первые
десятилетия XX в' перешел через Урал в Европу (найден в Горь-
ковской области и под Сыктывкаром). По-видимому, вся жизнь,
исключая время размножения, протекает на суше. Икра заклю-
чена в колбасовидные прозрачные мешки длиной около 15 см.
Общая длина взрослых особей до 13 см. Окраска серо-коричне-
вая или буровато-серая с мелкими черными пятнышками. Осо-
бого брачного наряда нет. На Колыме были случаи нахождения
в вечной мерзлоте. В сильно затененных ручьях Уссурийского
края живет Onychodactylus (=Geomolge) fischeri, а в горных
ручьях Вост. Казахстана — Ranodon sibiricus.
2. СЕМЕЙСТВО CRYPTOBRANCHIDAE. СКРЫТОЖАБЕРНИКИ
Став целиком водными животными, выработали ряд вто-
рично-примитивных признаков. В прошлом имели значительно
более обширное распространение. Отличия родов таковы:
1(2) После метаморфоза сохраняется пара или одно жабер-
ное отверстие и 4 верхнежаберных хряща.
Cryptobranchus
1 вид, Сев. Америка.
2(1) После метаморфоза жаберных отверстий нет, а верхне-
жаберных хрящей всего 2.
Megalobatrachus
1—2 вида, Вост. Азия.
Единственный современный вид рода Cryptobranchus alleg-
hanlensis скрытожаберник (см. рис. 8), встречается в реках
51
иосточиой часта США Днем забивается вод камня Мгдлтаяь
подтает по дяу. но может и хорошо плавать Питается червя**
раками, лягушками и рыбами. От светло*аспидного до светло,
бурого цвета. обычно с темными пятнами. Общая дляил до 56 о.
Исполинская саламандра. Mcgalobatrachus japonic*» (*•*!
matimm). живет в горных ручьях Вост Китая и Япония Обмч
но держатся а темных пещерах поя нависающими берегам*
и только очень редко по ночам после сильных дождей выход*?
на берег Питаются рыбой, .темноводными и беспозвоночным*
ЯАиа откладывают в горизонтальные норы. Светло-бурого шмт*
с темными облаковидными пятнами. Общая длина до 159
Мясо употребляется в пищу и как лекарство. Остатки, опяса*.
иые в XVIII в. Шейхцером из миоцена Германии под именем
«Человек свидетель потопа*, были отнесены Кювье к хвостатый
Палеонтологи называют их Andrias scheuchzeri. Позднее был*
описаны еще 2 вида этого рода. Однако все эти формы прмиад.
лежат к роду Megalobatrachus, возможно не заслуживая даже
выделения в особый вид. Родовая самостоятельность миоцено-
вого Zajssanurus и плиоценового Plicognathus требует проверка.
Подотряд 2 — Ambystomoidea. Амбистоыовые
3. СЕМЕЙСТВО AMBYSTOMIDAE. АМБИСТОМЫ
Позвонки двояковогнутые. Известно 4—6 современных *
1 ископаемый род (Plioambystoma). Указания на нахождение вне
Сев. Америки оказались основанными на путанице этикеток.
Современные рода таковы:
1(2) Серии сошниковых зубов не прерваны посередине ал*
разделены лишь очень небольшим промежутком; слез-
ной кости нет.
Ambystoma,
12—20 видов, Сев. и Центр. Америка.
2(1) Серии сошниковых зубов разделены посередине значи-
тельным промежутком; слезная кость имеется.
3(4) Носорая кость имеется; легкие нормальные; общая для-
на взрослых особей до 27—40 см.
Dicamptodon
2 вида, запад Сев. Америки.
4(3) Носовой кости нет; легкие редуцированы; размеры
меньше.
Rhyacotriton
1 вид, запад Сев. Америки.
Род Ambystoma широко известен в литературе под названием
*Amblystoma*. Неправильность последнего названия была убе-
дительно доказана Стейнегером еще в 1907 г. Большинство
видов откладывает яйца ранней весной в воду и лишь цемно-
52
_на землю (самка закручивается вокруг них). Яйца рядл
Садов помещаются в желатянеподобных мешках, более порот
^х, чем у Угл°зубов. Наибольшей известностью пользуется
4 tigrinum (см. рис. 8); сошниковые зубы в форме изогнутой
цли ПОЧТИ прямой линии между хоанами, по бокам тела J2 бо-
ооздок и 2 ясных тарсальных бугорка. Окраска сильно измен
LBa. коричневая или черноватая, обычно, с неправильными
желтыми пятнами. Общая длина до 28 см. Самки обычно круп
нее. Описан ряд подвидов, охватывающих почти всю территорию
СД1А и северную часть Мексики. Личинки этого вида известим
под туземным названием «аксолотль» («играющий в воде»).Пер-
поначально их считали представителями постоянножаберных
и именовали Siredon pisciformis. В 1865 г. в Париже наблюдали
сперва размножение этих личиночных форм (неотения), а по-
том метаморфоз части особей. В обширной литературе, посвя-
щенной «проблеме аксолотля», указано, что причиной затяжки
личиночной стадии считали внешние условия, но позднее была
доказана эндокринная природа этого явления. Ныне превраще-
ние аксолотля в амбистому путем воздействия гормоном щито-
видной железы стало простым школьным опытом.
В аквариумах часто держат выведенных в неволе альбиносов.
В неволе размножение происходит несколько раз в год. Самки
захватывают клоакой сперматофоры, отложенные самцами.
Общее число яиц от одной самки может доходить до 500 (в не-
скольких кучках). Оптимальная температура для икрометания
от +18° до +20°. Вылупление на 14—30 день. Молодь питается
инфузориями и мелкими ракообразными. Более крупные посте-
пенно начинают поедать мотыль, дождевых червей, головастиков
и нарезанное сырое мясо. Половозрелость аксолотлей наступает
примерно через 6 месяцев при длине в 20—25 см. Экземпляры
в 30 см встречаются редко. Икрометание в условиях Мексики
2 раза в год; амбистомы живут в это время в воде наподобие
аксолотлей. В Колорадо аксолотли наиболее активны при +21°.
Днем они идут в мелкую воду, а ночью уходят в глубину. По
окончании метаморфоза скрываются в норы грызунов и другие
убежища. У. A. gracile на уровне Моря метаморфоз наступает
обычно в возрасте одного года и лишь небольшая часть личинок
оказывается неотеническими. В горах же личинки этого вида
растут медленнее, и отдельные неотенические личинки достига-
ют исключительно крупных размеров. Интересны наземные виды
рода. Так, А. орасит держится в дуплах пней и даже яйца от-
кладывает на суше. Достигающая 97 мм A. talpoideum может
закапываться в почву и рыться в ней, как крот.
Подотряд 3 — Salamandroidea, Саламандровые
Два основных ствола — настоящие и безлегочные саламанд-
ры— разошлись, видимо, очень давно (?мел). Протеи, амфиумы
и сирены, вероятно, значительно моложе (?третичные).
аз
< t.EMEH* TWO SAI AMANDPIDAE HA< ТОЯ1ШН < АЛАМАНДPh
Hl®VC THo 14 современных и 4вымерших рода Среди н>
нямсчяются нязсымяя (Лв/олил/шлв, MtrttnsItlш, ChioftfosiQ) *
водная (Neurons. Triturus etc ) группы Первая характеризуй
ся округлым хвостом, а вторая — филогенетически, вероятна
более молодая хвостом, сжатым с боков. Современные
таковы:
I ( 6) Нёбные зубы в форме S-образных кривых.
2 ( 5) Язык короткий и толстый.
3(4) Хвост не длиннее туловища с головой.
Salamandra
2 вида, Европа, Сев. Африка.
4 ( 3) Хвост заметно длиннее туловища с головой.
Mertensiella
2 вида, Закавказье, Мал. Азия.
5(2) Язык длинный, высовывающийся.
Chioglossa
1 вид, Пиренейский полуостров.
6(1) Нёбные зубы в форме Д-образной фигуры.
7 ( 8) На задних ногах по 4 пальца.
Salamandrina
1 вид, Италия.
8 ( 7) На задних ногах по 5 пальцев.
9 (12) Предчелюстная кость парная.
10 (11) Хвост примерно, равен туловищу с головой; остистые
отростки высокие и широкие.
Tylototriton
6 видов, Юго-Вост. Азия.
11 (10) Хвост длиннее туловища с головой; остистые отростки
весьма низкие, маленькие.
Pleurodeles
2 вида, Пиренейский полуостров.
12 ( 9) Предчелюстная кость непарная.
13 (14) Хвост сжат с боков лишь в задней своей половине.
Hypselotriton
1 вид, Юннань.
14 (13) Хвост сжат с боков на всем или почти на всем протя-
жении.
15 (18) Хвост длиннее тела с головой.
64
16 (17) Паротиды (заглазничные железы) хорошо заметны:
лобно-чешуйчатая дуга полная, костная.
Taricha
3 вида, запад Сев. Америки.
17 (16) Паротиды не выдаются; лобно-чешуйчатая дуга не пол-
ная.
Neu rergus
1 вид, Мал. Азия.
18 (15) Хвост равен длине тела с головой, чуть короче или
длиннее.
19 (20) Края клоакального отверстия сильно выдаются, обычно
в виде конуса или груши.
Euproctes
3 вида, Пиренеи, Сардиния, Корсика.
20 (19) Края клоакального отверстия не бывают коническими
или грушевидными.
21 (24) Паротиды хорошо выражены, выдающиеся.
22 (23) Лобно-чешуйчатая дуга полная; остистые отростки ши-
рокие и высокие.
Cynops
4 вида, Япония, Китай.
23 (22) Лобно-чешуйчатая дуга в задней части сухожилия;
остистые отростки небольшие, но высокие.
Pachytriton
I вид, Юго-Вост. Китай.
24 (21) Паротиды не выдаются, плохо или совсем не заметны.
25 (26) Остистые отростки широкие и высокие; лобно-чешуй-
чатая дуга широкая.
Notophthalmus
3 вида, восток Сев. Америки.
26 (25) Остистые отростки менее развиты; лобно-чешуйчатая
дуга иногда сухожильная, иногда костная, но не широ-
кая.
Triturus
9 видов, Европа, Зап. Сибирь, Мал. Азия,
Иран.
Наиболее известным видом настоящих саламандр (Salamand-
га) является пятнистая S. salatnandra (=S maculosa). Черные
пятна на теле перемежаются с ярко-желтыми или красноватыми.
Сильная изменчивость окраски дала повод к описанию многих
разновидностей, ценность которых сомнительна. Каммерер пы-
55
талей доказать прямое влияние окраски почвы и температуру ц
степень развития черного пигмента, но последующие ясследоммв
заставили в этом сомневаться. Яркую окраску этого вида при
нято считать за предупреждающую, ибо едкие кожные выдел*
ния животного раздражают слизистые оболочки человека j
могут вызвать смерть мелких животных. Встречается в Средни
и Южной Европе. Алжире, Марокко, Египте, Сирии и Мал. Азии
У нас — в Карпатах. Держится среди влажного мха, меж
ду корнями, в ямках, мышиных норах и развалинах. Избе
гает известковой и сухой почвы. В горах встречается на уровж
900, редко 1250 м. Зимует в земле. Питается улитками, червями
и мелкими членистоногими. Для размножения собираются я
водоемы с текучей водой, где откладывают сперматофоры. Иног.
да же спаривание происходит на суше; самец подползает под
самку. Сперма может храниться в теле самки до 3 лет. В неволе
наблюдалась откладка икры. Нормально молодь имеет жабры.
Половозрелость наступает на третий или четвертый год. Макси-
мальная длина 285 мм.
В Альпах и на Карпатах на высоте 600—3000 м живет аль-
пийская саламандра, S. atra, нормально сплошь черная. Из мно-
гих яиц, вынашиваемых самкой этого вида, окончательного
развития достигают всег.о два, причем молодые рождаются уже
без жабр. Удалось воспитать -в воде зародышей, вырезанных из
тела матери. Каммерер показал, что при температуре ниже
+ 12° беременность <S. salamandra удлиняется, а на нижней гра-
нице своего распространения S. atra иногда кладет молодых в
воду (у них на теле бывают небольшие желтые пятна). Общая
длина S. atra до 130, очень редко до 178 мм. Описано 5—12 ис-
копаемых видов.
Длиннохвостая саламандра Mertensiella, представлена у нас
в Закавказье видом М. caucasica. Держится на высоте 500—
2800 м вблизи текучей воды. Активна ночью. Схваченная за
хвост, иногда его отбрасывает. Общая длина до 19 см.
Язык Chioglossa может высовываться вперед на 2—3 см.
У иглистого тритана Pleurodeles, ребра часто протыкают кожу.
Живет в воде, годами не выходя на сушу.
Pleurodeles, совместно с Taricha, Neurergus, Euproctes, Су-
nops, Notophthalmus и Triturus, объединяет водные формы, ра-
нее входившие в обширный род тритонов Molge=Triton. Позднее
его разбили на самостоятельные рода, причем не оказалось воз-
можным сохранить старое родовое название.
Род Triturus включает большинство видов, оставшихся от
старого рода. Самцы многих из них в брачйое время имеют спин-
ной гребень, являющийся также органом дополнительного кож-
ного дыхайия. После брачной игры самка захватывает клоакой
отложенные самцом сперматофоры. Икринки откладываются по
одиночке на водные растения. Не редки неотенические личинки.
Удалось получить разнообразные межвидовые гибриды.
56
Наиболее известный вид - обыкновенный тритон, Т vulgt
ris, имеет самцов, у которых фестончатый гребень без лерерма
переходит в хвостовую оторочку. Спина оливково-зеленая ид
бурам с темными пятнышками или без них. Брюхо оранжевое
темными пятнышками. У самца в брачное время хвост с сии<
наточшрламутровой полосой. Распространен от Ирландии
франции до Зап. Сибири включительно. Описаны многочислен
ныг разновидности. В воде проводит только время размножения
переселяясь далее на сушу, где и зимует. Питается насекомыми
водяными осликами, пауками, червями и т. л. Весной (под Моск
вой обычно в апреле — мае) -самка кладет около 700 икрииок
В северных районах (Вологда) личинки перезимовывают. Об
тая длина взрослых до 8, редко до 11 см. У самцов Т. crlstatui
гребень имеет понижение, либо в области крестца совсем пре-
рывается. Окраска темнее. Самка часто с тонкой желтой линией
вдоль спины. Кожа зернистая. Распространен почти по всей
Европе, за исключением Южной Франции и Пиренейского полу-
острова. Описан ряд разновидностей. У нас покидает воду позд-
нее обыкновенного тритона. Общая длина до 15, редко 18 см.
Чрезвычайно красив Т. vittatus (Зап. Кавказ, Мал. Азия, Пале-
стина), с очень высоким, зазубренным гребнем. Напротив, сам-
цы, Т. boscai (Пиренейский полуостров), Т. italicus (Южн. Ита-
лия) и Т. montandoni (Карпаты, Чехословакия, Румыния),
вообще лишены гребня. •
5. СЕМЕЙСТВО PROTEIDAE
Вся организация .носит личиночный характер. Известно
2 современных и 1—2 ископаемых (эоцен-миоцен Германии)
рода. Единственный вид рода Proteus anguinus, европейский
протей отличается скрытыми под кожей глазами и присутствием
на передних конечностях всего 3, а на задних — 2 пальцев. При
жизни окрашен в цвет мяса с ярко-красными жабрами, а в
спирту белый. При содержании на свету кожа темнеет. Обитает
•в подземных водах Югославии. Оптимальные условия: темнота,
чистая вода и температура +10°. Вся поверхность тела — кожа —
воспринимает свет. В желудках пойманных особей находили
червей и рачков. В неволе ест улиток, червей, мелких ракообраз-
ных и нарезанное л(ясо. Может годами обходиться без пищи.
£амка кладет 12—56 яиц. При температуре ниже +15° живоро-
дящи. Общая длина до 28, редко 30 см.
Американский протей, Necturus (2 вида) имеет функциониру-
ющие глаза, пигментированную кожу и по 4 пальца на обоих
конечностях. Наиболее известный вид N. maculatus живет в на-
земных водоемах восточной половины США. Активен ночью.
Питается членистоногими, червями, рыбами и лягушками. Весной
откладывает яйца. Общая длина до 36 см.
58
6 СЕМЕЙСТВО amphumidae
Тело угревидное с 2 парами илом развитых копечиогтей
Единственный род Amphiuma представлен 2 видами в восточиЫ
половине США. А. теши (см. рис. 8) с 2 пальцами ма обоих ко
ценностях и A. tridactyla — с 3. Впрочем некоторые зоологи
ннмают это за индивидуальную изменчивость. Черного цвета <
зеленоватым отливом. Живет в болотистых водоемах со стоячей
водой и илистым дном. Может долго оставаться вне воды. Пи
тается ракообразными, моллюсками и рыбами. Спаривание в мае
а икру откладывает в августе — сентябре. Ноги у молодых отио
снтельно длиннее, чем у взрослых. Общая длина до 89, ред-
ко 100 см.
7. СЕМЕЙСТВО SIRENIDAE
Два современных рода в Сев. Америке. Представителя рода
Siren имеют по 3 жабры и по 3 брызгальца с каждой стороны.
На передних конечностях по 4 пальца. Два вида. Более крупная
S. lacertina живет в канавах и прудах юго-востока США. Серого
цвета, иногда с маленькими желтоватыми пятнами. Плавает по-
добно угрю. Обитает в неглубоких затененных местах. Иногда
вбуравливается в сырую землю. Питается моллюсками, червями,
рыбами и тритонами. Жабры молодых сперва уменьшаются, а
потом развиваются вновь («обратный метаморфоз» Копа). Об-
щая длина до 72 см.
Единственный вид другого рода Pseudobranchus striatus обла-
дает всего 1 жаброй и 1 брызгальцем с каждой стороны. На
передних конечностях по 3 пальца. Длина до 20 см. Живет в бо-
лотах Южн. Каролины, Георгии и Флориды.
8. СЕМЕЙСТВО PLETHODONTIDAE. БЕЗЛЕГОЧНЫЕ САЛАМАНДРЫ
Больше половины родой сем. безлегочных — водные живот-
ные, и, вероятно, утрата легких произошла в связи с обитанием
в горных ручьях. Сухопутные рода вторичного происхождения.
В связи с особенностями дыхания число колебаний горла очень
велико (120—180 в мин.). Наиболее наземные виды менее актив-
ны и потребляют кислорода меньше, чем водные. Перегородка
между предсердиями утрачена. Возникли, видимо, в Аппалачах
Сев. Америки, но имеют представителей и в Зап. Европе. Разде-
ляется на следующие рода:
1(4) Глаза полностью или частачно скрыты под кожей.
2(3) Жабры сохраняются на всю жизнь; язык спереди сво-
бодный.
Typhlomolge
1 вид, Техас.
3(2) Жабры с возрастом исчезают; язык спереди прикреп-
ленный.
59
TypMotrtton
2 вида, центр США
4(1) Глаза нс скрыты под кожей, н веки свободны.
5 (16) Язык грибовидный: свободный со всех сторон кружзд
прикрепленный центральной ножкой н могущий выоовы
ватъея далеко вперед.
6(9) Сошниковые зубы сливаются с сидящими на околоклв
нов и дней кости.
7(8) Предчелюстные кости разделены.
Gyrinophilus
4 вида, восточн. часть Сев. Америки.
8 ( 7) Предчелюстные кости слиты.
Pseudotriton
2 вида, восточн. часть Сев. Америки.
9(6) Сошниковые зубы отделены от сидящих на околоклино-
видной кости.
10 (П) Предчелюстные кости разделены.
Hydromantes
4 вида, Зап. Европа, Калифорния.
П (10) Предчелюстные кости слиты.
12 (13) На задних ногах по 4 пальца.
Manculus
1 вид, юг Сев. Америки.
13 (12) На задних ногах по 5 пальцев.
14 (15) Обитатели горных ручьев; теменные кости всегда пол-
ностью окостенелые; доги всегда имеются.
Eurycea
10 видов, восток и юг Сев. Америки.
15 (14) Наземные; теменные чаще не окостеневают; имеются
виды с редуцированными конечностями.
Oedipus
30 видов, Центр, и Южн. Америка.
16 ( 5) Язык прикреплен спереди по средней линии.
17 ’(20) На задних ногах по 4 пальца.
18 (19) Предчелюстные кости слиты, если же раздельны, то тело
сильно вытянутое; теменные кости не окостенелые.
Batrachoseps
6 видов, запад Сев. Америки.
19 (18) Предчелюстные кости свободны; тело нормальных про-
порций; теменные кости окостенелые.
60
Hemldactyllttm
1 вид, Сев. Л мерила.
20 (17) На задних ногах по 5 пальцев.
21 (22) Позвонки задневогнутые.
Desmognathiis
9—10 видов, восток. Сев. Америки.
22 (21) Позвонки двояковогнутые.
23 (26) Предчелюстные кости слиты.
24 ( 25) Светло-желтого или пепельного цвета с черными пятна
ми; общая длина до 12 см.
Leurognathus
1-вид, Сев. Каролина.
25 (24) Темно-пепельного» почти черного цвета с примесью жел
того; общая длина до 8 см.
Stereochilus
1 вид, от Виргинии до Георгии.
26 (23) Предчелюстные разделенные.
27 (28) На челюстных костях зубы клыкообразно увеличены.
Aneides
5 видов, Сев. Америка.
28 (27) Клыкообразных зубов нет.
29 (30) Околоклиновидные пластинки разделены.
Ensatina
2 вида, восток Сев. Америки.
30 (29) Околоклиновидные пластинки соприкасаются.
Plethodon
16—17 видов, Сев. Америка.
Представители родов Gyrinophilus, Pseudotriton, Leurognat-
hus, Desmognathus, Eurycea, Manculus и Stereochilus ведут вод-
ный образ жизни, чаще всего в горных ручьях. Лишь изредка не-
которые из них покидают воду и держатся под старыми деревья-
ми и камнями. На голове Stereochilus всю жизнь сохраняются
органы кожного чувства (боковой линии). Личинки Eurycea про-
водят в воде, видимо, 2—3 года. Наибольшая длина представи-
телей этих родов достигает 7—17 см.
Два рода приспособились к жизни в подземных водоемах.
Typhlotriton spelaeus встречается в пещерах Миссури, Оклахомы,
Канзаса и Арканзаса (Сев. Америка). Во время метаморфоза у
него исчезают не только жабры, но и почти весь пигмент, а глаза
полностью или частично закрываются срастающимися веками.
Длина до 93 мм. Typhlomolge rathbuni (см. рис. 8) добыт из ар-
61
геэмднского холода* в Сан-Маркос (Техас) с глубины 57 м. Г ла
ла полностью скрыты под кожей, пигмента нет, жабры coxpaww
ются всю жизнь. Длина до 10 см.
В Европе живут 2 вида Hydromantes, а 2 другие описаны г
Сев. Америки. Оба европейские вида долгое время были объели
нены под общим названием Spelerpes fuscus. Правильнее имемо
вать экземпляры из Сардинии Н. genei, а из Приморских Альп i
Сев. Италии —- Н. italicus. Образ жизни целиком наземный »
ночной. Живут в пещерах и тенистых местах, чаще всего в гора,
(до 1800 м). Питаются паукообразными, многоножками я насеко
мымя. Язык высовывается вперед на расстояние, почти равное по-
ловине длины животного. Общая длина взрослых 70—105 мм
Молодые рождаются весной.
Виды родов Ensatina, Hemidactylium, Plethodon, Oedipus,
Batrachoseps и Aneides (см. рис. 8) ведут наземный образ жизни
и откладывают яйца на суше. Держатся под камнями, упавшими
деревьями и в пещерах. Самки обвиваются вокруг яиц. Предста-
вители Hemidactylium имеют способность отбрасывать хвост.
Внутри рода Oedipus имеются как виды с хорошо развитыми
конечностями, так и целая серия червеобразных видов с разной
степенью редукции конечностей. Возможно, что некоторые виды
живородящи.
ЛИТЕРАТУРА
Воронцова М., Лиознер Л., Маркелова И. и Пухальская Е.
Тритон и аксолотль, М., 1952.
Шмальгаузен И. Морфология аппарата звукопередачи хвостатых амфи-
бий, Зоол. журнал, 1956, 35, № 7: 1023—1043.
Шмальгаузен И. Морфология позвоночника хвостатых амфибий, Зоол.
журнал, 1957, 36, № 11: 1717—1734.
Boulenger G. Catalogue of Batrachia Gradientia s. Caudata and Batrachia
Apoda in the Collection of the British Museum, London, 1882.
Dunn E. The Salamanders of the Family Hynobiidae, Proc. Amer. Acad, of
Arts and Sci, 1923, 58, No. 13: 445—523.
Dunn E. The Salamanders of the Family Plethodontidae, Norhampton Mass.
Smith College, 1926.
Francis E. The Anatomy of the Salamander, Oxford, 1934.
H e r r e W. Die schwanzlurche der mitteleocan Braunkohle des Geiseltales und
die Phylogeriie der Urodelen unter Einschluss der fossilen Formen, Zoologies,
1935, 87, Heft 87.
Wahleet G. Biogeographische und okologische Tatsachen zur Phylogenie
amerikanischer Schwanzlurche, Zoolog. Jahrbucher, Abt. System. 1957, 85.
Heft 3: 253—282.
Глава fttrai'
БЕСХВОСТЫЕ ЗЕМНОВОДНЫЕ«
Форма тела н многие детали внутреннего строения подавляю-
щего большинства Salientia связаны с особенностями движе-
ния — прыжками. Поэтому отсутствие этой способности (напри-
мер, Breuiceps, Vibrissaphora) следует считать вторичным. Была
экспериментально доказана положительная связь увеличения
относительной длины задних ног с размерами прыжка. У разных
видов и родов длина передних конечностей составляет от 30 до
61 % задних. При плавании передние конечности пассивны и при-
жимаются к телу, а задние двигаются назад согласованными
толчками. Такой способ плавания наряду с некоторыми другими
соображениями привели некоторых исследователей (Татаринов)
к мнению о водном происхождении диспропорциональности ко-
нечностей бесхвостых. Однако вероятнее «наземная гипотеза».
В прыжке бесхвостого земноводного, например, лягушки, следует
отличать три фазы: отскок, пребывание в воздухе («полет») и
приземление. Длительность первого и последнего не велика —
порядка 0,1 сек. Рабочий толчок сообщается телу задними конеч-
ностями. Последняя оказывается разбитой на ряд сегментов, по-
следовательно уходящих при отскоке от земли, начиная с прокси-
мальных. Наибольшее напряжение падает на первые два отдела,
что привело к укреплению бедра, а большая и малая берцовые
слились друг с другом. При отскоке более раннее отнимание от
земли одной из ног может менять направление движения («асим-
метричный отскок»). Вероятно, увеличение числа сегментов зад-
ней конечности выгодно в этом отношении, а потому у бесхвостых
и возник новый сегмент—«вторичная голень», за счет удлинения
1 В выиешнем объеме отряд был впервые установлен в 1768 г. Лауренте
вод именем Salientia. Спустя 9 лет Скололи предложил для вето термин
Ecaodata, а еще через 36 лет Филер — название Anuri. Именно эта два по-
следние термша получили в разных своих вндонзменевнях шаровое распро-
странение, особенно средн несмстематнков. Однако, как обстоятельно доказал
в 1907 г. СтеЯнегер, термин Salientia со всех точек зрения является наилуч-
0жм в потому рекомендуется для употребаеиия.
63
tibinlr и ftbularr Аналогичное опъясиспкг можно предложить ।
для удлинения четвертого пальца >адней ноги. Ра твит и* сустав
ных бугорков препятствует скольжению лапки. Значительное ув*
лнченне внутреннего пяточного бугра роющих форм, несомненно
является позднейшей специализацией. Во второй фазе прыжщ
животное подчиняется только законам свободного падения. Нт
преодоление сопротивления воздуха тратится при этом мене»
2% энергии. В «полете» тело может только вращаться вокру>
центра тяжести, причем максимальная маневренность требу»)
увеличения компактности тела. Видимо поэтому в филогении
Salientia наблюдается последовательное перемещение вперед
места крепления тазового пояса к позвоночнику; крылья под.
вздошной кости удлиняются, и роль крестцового позвонка начи-
нает выполнять все более впереди лежащий. Следствием является
укорочение позвоночника и слияние задних туловищных позвон-
ков, прошедших через стадию крестцового, с передним концом
хвостцовой кости. В первый момент приземления исключительна
роль передних конечностей, следствием чего является слияние
друг с другом лучевой и локтевой кости. Не исключена связь с
этой фазой и прогрессирующей редукции ребер, что в свою оче-
редь объясняет сохранение личиночного типа дыхания при помо-
щи колебания нижней стенки ротовой полости. Несовершенство
такого типа дыхания в наземных условиях требует хорошего
развития кожного дыхания, что возможно только при сохранении
влажности кожи. Вероятно, в этой связи, при условии необходи-
мости некоторой экономии влаги, у Salientia особенно хорошо
развиты подкожные лимфатические полости. Твердые кожные
образования представляют исключения (когти Xehopus и Нуте-
nochirus, спинные пластинки Megophrys, Nototrema, Phyllome-
dusa, Lepidobatrachus, Ceratophrys и Brachycephalus). Характер-
ную трехветвистость крыловидной кости, возможно, следует
рассматривать как результат увеличения глазного яблока,
понятного при возрастании активности животного. Конкретные
сведения по анатомии группы лучше всего получить, изучая
строение лягушки.
Долгое время самые древние остатки бесхвостых были из-
вестны из юрских отложений, но в 1936 г. Пивето установил
наличие их в нижнем триасе. Ватсон пытался рассматривать в
качестве предков всего отряда Miobatrachus ( = Amphibanius) из
карбона Сев. Америки, но сейчас это признано неправильным.
Линней относил всех представителей отряда к одному роду
Rana. В 1768 г. Лауренти принимал уже 4 рода: Rana, Bufo, Hyla,
Pipa. Фитцингер в 1826 г. делил нынешних Salientia на 4 семейст-
ва (Ranoides, Bufonoides, Bombinatoroides, Pipoides), Кювье в
1829 г. признавал всего 3 (Ranae, Hylae, Bufones). В 1831 г. Ваг-
лер впервые придал серьезное значение отсутствию языка
(Aglossa). Последующие авторы комбинировали эти деления,
дополняя их то характером кончиков пальцев (Oxydactyla и Р1а-
64
tydactyla Гюнтера), то строением зрачка (Cyclogirnidrt, Piagtog
Icnides и Orthoglenidrs Бруха), то другими признаками, Важны*
шаг вперед был достигнут в 1864 г. Копом, обратившим внима
кие на строение грудного пояса (рис. II) и форму боковых от
ростков крестцового позвонка. Если вр а новые кости соединены по
средней линии подвижно и налегают друг на друга, то такой пояс
называется «подвижногрудым» (Arcifera). Его противололож
ностью является «неподвижногрудый» тип (Firmisternla). Тот
факт, что у молодых лягушек грудной пояс сперва построен по
Рис. 11. Грудной пояс с брюшной стороны. «Подвижногрудый» тип
(/ — Bombina bombina, 2—Bufo bufo), переход в онтогенезе от под-
вижногрудого к «неподвижногрудому» (3 — взрослая и 4 — молодая
Rana temporaria), разные случаи «неподвижногрудости» (5—Cacopus
systoma, 6 — Breviceps gibbosus, 7 — Hemisus guttatum)
подвижногрудому типу, заставил считать неподвижногрудых за
более молодую группу. До первой четверти нашего века включи-
тельно всеобщим признанием пользовалась система Буленджера,
представляющая собой по сути только модификацию идей Копа.
Он принимал деление на подотряды безъязычных и явноязычных
(Phaneroglossa). К первому принадлежали только липовые, а
второй подразделялся на 2 серии: 8 семейств подвижногрудых и
3 семейства неподвижногрудых. В 1916 г. Никольс предложил
считать основой классификации строение позвоночника. Нобль
(1922) проверил его выводы, принял их, дополнил их анализом
мускулатуры и нацело перестроил систему отряда. Спустя 15 лет
Пивето дополнил ее установлением наиболее примитивного под-
отряда первичнобесхвостых. Сейчас можно дать такое деление на
12 семейств, объединяемых в 6 подотрядов (рис. 12):
65
i 21 Xeocrcu (уростиль > отсутствует; имеете* хвост, номер
живаемы* нешиогими позвонками [подотряд I Proa
noura)
1. Pretobatraehtdae
> ( I) Хвостец имеется; хвоста у взрослых мет.
i ( 4) Все туловяшные (— предкрестцовые) позвонки дю*о
вогнутые (подотряд 2 AmpnicoelaJ.
2. Uopdmldae
Рис. 12. Филогения семейств бесхвостых. Подотряды:
/ — Ргоапоига, 2 — Amphicoela, 3 — Opisthocoela,
4 — Anomocoela, 5 — Procoela, 6 — Diplasiocoela
4(3) Все или большая часть туловищных позвонков не двоя-
ковогнутые.
5(8) Ребра имеются в течение всей жизни или только в онто-
генезе; все туловищные позвонки задневогнутые [подот-
ряд 3 Opisthocoeiaj.
6(7) Туловищных позвонков не менее 8, крестцовый свобод-
ный, с расширенными поперечными отростками; у взрос-
лых имеются свободные ребра; верхняя челюсть с зуба-
ми, нижняя беззубая; язык и веки хорошо развиты;
евстахиевы трубы раздельны; грудной пояс типично под-
вижногрудый.
№.
/
я
9
10
11
12
13
14
15
16
17
18
19
>. DhceftoMMfte
( 6) Туловищных позвонков 5 7, крестимый обычно глии
шийся с хвостцом, с очень сильно расширенными попе
речными отрос псами; свободные ребра имеются лишь ня
ранних стадиях развития; обе челюсти беззубые иля
только нижняя; язык и иеки у большинства утрачивают
ся; евстахиевы трубы открываются одним общим отвер
стием; грудной пояс частично или полностью иеподвиж-
ногрудый.
4. Pipidae
( 5) Ребер никогда не бывает; туловищные позвонки чаше
передневогнутые.
(10) Крестцовый позвонок сращен с хвостцом, если же сво-
бодный, то сочленяется с хвостцом одним бугром; порт-
няжный мускул обычно не отделен от полу перепончатого
[подотряд 4 Anomocoela].
5. Pelobatidae
( 9) Крестцовый позвонок свободный, сочленяющийся с хвост-
цом двумя бугорками, очень редко сращенный с хвост-
цом; портняжный и полуперепончатый мускул самостоя-
тельны.
(18) Все туловищные и крестцовый позвонки передневогнутые
[подотряд 5 — Procoela].
(13) Между конечной и предпоследней фалангой имеется
вставочный хрящ.
8. Hylidae
(12) Фаланги без вставочных хрящей.
(17) Грудной пояс подвижногрудый.
(16) Туловищные позвонки свободны.
7. Bufonidae
(15) Седьмой-девятый туловищные позвонки срослись.
6. Palaeobatrachidae
(14) Грудной пояс полностью или частично неподвижно-
грудый.
9. Brachycephalidae
(11) Обычно первые 7 туловищных позвонков передневог-
нутые, а восьмой двояковогнутый, редко 2 первых поз-
вонка слиты или все позвонки передневогнутые; крест-
цовый позвонок спереди выпуклый [подотряд 6 Diplasi-
ocoela].
(22) Поперечные отростки крестцового позвонка цилиндриче-
ские или чуть расширенные.-
67
20 (2!) Между конечной и лрелпослслнги фалангой имеет..
вставочный хряш
II. Polypedatldae
21 (20) Вставочных хрящей нот.
10. Ranldae
22 (19) Поперечные отростки крестцового позвонка расширены
12. Brevldpitidae
Подотряд 1 — Proanoura. Первичнобесхвостые.
I. СЕМЕЙСТВО PROTOBATRACHIDAE. ДРЕВНИЕ БЕСХВОСТЫЕ
Единственным представителем семейства и подотряда в целом
является Protobatrachus massinoti (рис. 13), описанный в 1936—
1937 гг. по одному экземпляру из нижне-триасовых отложений
северной части Мадагаскара. Общая длина тела около 10 см.
Череп в основном построен уже по типу Salientia, но присутствует
небольшая заднеушная кость. Чешуйчатая кость имеет только
слабо выраженную заднюю ветвь горизонтального отростка.
Общее число позвонков около 16. Все туловищные, исключая пер-
вого, н'есут ребра. Между подвздошными костями лежали 3 поз-
вонка, видимо гомологичные хвосту. Окостенения в области
грудины отсутствовали. Лопаточно-врановые элементы были
объединены. Крылья подвздошных костей умеренно вытянутые,
а сами кости не соединены по средней линии. Задние конечности
лишь немного длиннее передних, хотя общие контуры тела уже
были лягушкообразны. Лучевая и локтевая, а также большая и
малая берцовые кости еще не слились. Предплюсневые кости
(tibiale и fibularc) несколько увеличены, но еще раздельны. Кожа
была бугорчатая, без всяких следов чещуй. Практически все
современные исследователи признает это животное низшим пред-
ставителем отряда в целом, но недавно (Griffiths, 1956) были
высказаны критические замечания, основанные на присутствии
17 окостеневших хвостовых позвонков у головастиков Megophrys
major.
Подотряд 2 — Amphicoela. Гладконогообразные
2. СЕМЕЙСТВО LIOPELMIDAE. ГЛАДКОНОГИЕ
Зубы только в верхней челюсти. Ребра сохраняются всю
жизнь. Предгрудины нет. Небольшая грудина целиком хрящевая.
Имеются рудименты хвостовых мускулов. Барабанная перепон-
ка, евстахиевы трубы и резонаторы самцов полностью редуциро-
ваны. Разорванный ареал семейства (рис. 14), представленного в
современной фауне всего 2 родами, свидетельствует о его
68
глубокой древности. Род Ascaphut — гладхоног, свракгервэуетгв
вертикально-эллиптическим зрачком и нахождением сошниковых
зубов между хоанами. Единственный вид A. truri (рис. 15) ветре*
чается в Сев. Америке (Вашингтон, Орегон, Сев Калифорния).
Рис. 13. Скелет Protobatrachus maesinoti: А — череп сверху, Б — череп
снизу, В — три первые позвонка, Г — тазовый пояс и хвостовой отдел,
•4 —правая половина грудного пояса, Е — передняя конечность, Ж—зад-
няя конечность:
/ — носовая кость. 2 — лобно-теменная, 3 — переднеушная, 4 — чешуйчатая, 5 — верх-
нечелюстная» 6 — квадратно-скуловая, 7 — квадратная» 5 — крыловидная. 9 — боковая
затылочная. 10 — кость пояском. 11 — околоклиновидная. 12 — нёбная. 13 — заднеуш-
ная. 14 — первый позвонок, 15—поперечный отрострк второго позвонка, 16 — ребра.
17 — поперечный отросток третьего позвонка» 18 — крючковидный отросток, 19 —>• под-
вздошная кость. 20— седалищная. 21 — хвостовые позвонки. 22 — крестцовые ребра,
23 — надлопатка, 24 — клейтрум. 25 — лопаточно-врановая. 26 — ключица, 27 —* головка
плечевой кости, 28 — правовая кость. 29 — лучевая кость. 30 — локтевая, 31 — бедрен-
ная, 32 — большая берцовая, 33 — малая берцовая, 34 — сесамовидные кости, 35 — боль-
шеберцовая предплюсны, 36 — малоберцовая предплюсны
где живет в воде холодных горных источников. В брачное время
самец ползает по дну потока в поисках самки. Оплодотворение
внутреннее,- роль копулятивного органа играет удлиненная часть
клоаки. Имеются непарные предлобковый и залобковый хрящи.
69
рис. 14. Ареалы семейств. / — Liopelmldue. У е*.
Кожа бугорчатая. Хорошо развиты парогмлм я сиииж, Лжомы.
складки Сверху красновато бурого цвета с черноватыми пятил
ми; снизу беловатого. Общая длина вхрослых до 50 мм Е сме-
ренный вид другого рода Ltopttma hnchitetleri из Новой Зетаная!
ведет сходный водный образ жиэии, достигает 33'
Рис. 15. Представители Liopelmidae и Pipidae; А — Ascaphus truei самец
я его клоакальный придаток, частично (Aj) и полностью выпяченный (Аз).
Б— Liopelma hochstetteri, В—Pipa pipa самка с выпяченным яйцекладом,
/'—вертикальный срез через кожу спины самки Pipa pipa, Д — Нутепо-
chirus boettgeri:
/ — крышечка, 2 — перегородка между ячейками. 3 — желточный мешок. 4 — гзроллп
что описанные из верхнеюрских отложений Montsechobatrachus
(Испания) и Eobatrachus (Вайоминг) принадлежат к этому се-
мейству.
Подотряд 3 — Opisthocoela. Жерлянкообразные
3. СЕМЕЙСТВО DISCOGLOSSIDAE. КРУГЛОЯЗЫЧНЫЕ
Известны 4 современных рода из разных мест Евразии:
1 ( 2) Пальцы передних ног соединены перепонкой.
Barbourula
1 вид, Бусуанга (Филиппинские о-ва).
2(1) Пальцы передних ног свободные.
3(6) Зрачок круглый, треугольный или сердцевидный.
4(5) Кожа гладкая; брюхо беловатое, без пятен или с корич-
невыми точками.
Discoglossus
1 вид, зап. часть Средиземноморья.
5 ( 4) Кожа бугорчатая. брюхо при жи»ни красное или желл>
с редкими черными пятнами
Bombta*
4 чида. Европа, Вост Азия
6 ( 3) Зрачок вертикальный.
Alytes
2 вида, Европа.
Первый из упомянутых родов целиком водный, два следующие
тесно связаны с водой, а последний целиком сухопутный. Род
Bombina (Bombinator), жерлянка. В Европе представлен 2 вида
ми. одни из которых с красным брюшком — В. bombina — доходи?
на восток примерно до 61° долготы, а другой с желтым брюш-
ком— В. uariegata ( = salsa) идет на восток только до Карпат.
Там, где ареалы этих видов налегают друг на друга, они разобще-
ны стационарно: желтобрюхая водится в холмистых или горис-
тых (до 1800 м) местностях, а краснобрюхая в низинах и равни-
нах (не выше 250 м). В остальном образ жизни сходен.
Значительную часть времени проводят в воде прудов, луж, канав,
ручьев и т. п. Застигнутые на суше, заворачивают лапки на спину
(«Unken — reflex»). При раздражении кожа выделяет мыльную
пену. Крик: «уу... уу...». Икрометание может растягиваться на
долгий срок (под Москвой начало его обычно в конце апреля).
Икра откладывается небольшими комками или поодиночке. Ме-
таморфоз примерно через 90 дней. Половозрелость на 3-м году.
Зимует чаще на суше. На Дальнем Востоке, в Корее и Сев. Ки-
тае живет В. orientalis, а в горах Юннаня (1800—2591 м) —.
В. maxima. Общая длина тела трех первых видов не превосходит
50 мм, а представителей четвертого доходит до 68 мм.
По внешности и образу жизни Discoglossus pictus напомина-
ет водяную лягушку.
От оредней Германии до Пиренейского полуострова включи-
тельно распространена обычная повитуха, Alytes obstetricans
( =campanisona), замещаемая в песчаных местностях Португалии,
Эстремадуры, Аррагонии и Нов. Кастилии близким видом. A. cis-
ternasii. Образ жизни обоих наземный и ночной. Хорошо копают
землю, зарывшись в которую зимуют. Во время спаривания (с
марта по сентябрь), происходящего на суше, самец наматывает
шнуры икры, переплетая их в форме восьмерки, на свои задние
ноги. На 11—17 день он сходит в воду, где и вылупляются голо-
вастики. Развитие последних требует 3—7 недель. Взрослые до-
стигают длины 50 мм. Опытами установлено, что при температу-
ре от +25° до +30° спаривание происходит в воде и икра не
наматывается.
Из миоценовых слоев Швейцарии описана Latonia, близкая к
Discoglossus. Скелеты представителей последнего рода и Alytes
найдены в миоцене Франции и Германии.
72
4. СЕМЕЙСТВО PIPIDAF ЛИПОВЫЕ
Утрату языка и другие особенности представителей семейс тв,
пытаются объяснить водным образом жизни. Специализация вс
дет к редукции век и зубов в верхней челюсти Ныне извести*
следующие <рода:
] ( 6) Концы пальцев передних ног без звездообразных при
датков; на 3 внутренних пальцах задних ног когти; Аф
рика [подсемейство Xenopinae, Когтистые лягушки).
2 ( 3) Верхние веки имеются.
Pseudhymenochlrus
1 вид, Французская Гвинея.
3 ( 2) Веки редуцированы.
4 ( 5) Верхняя челюсть с зубами.
Xenopus
5 видов, Африка к югу от Сахары.
5 ( 4) Обе челюсти без зубов.
Hymenochirus
2 вида, Зап. Африка.
6(1) Концы пальцев передних ног с кожистыми звездообраз-
ными придатками; когтей на пальцах задних ног нет;
Южн. Америка [подсемейство Pipinae, пипы собственно].
7 ( 8) Верхняя челюсть с зубами.
Protopipa
1 вид, Гвиана.
8 ( 7) Обе челюсти беззубые.
Pipa
3 вида, Гвиана, Бразилия.
Представители рода Xenopus, шпорцевой лягушки, имеют под
глазом более или менее длинное щупальце. В Южн. Африке (на
север до Анголы и Килимаджаро) живет X. laevis. Образ жизни
ее исключительно водный. Яйца откладывают группами от 1 до
4 на камни и подводные растения. В неволе икрометание продол-
жается все лето, причем одна самка откладывает до 15000 икри-
нок. Вылупляются личинки через двое суток. На третий день у
них вырастают длинные «усы». Эти головастики питаются, фильт-
руя воду через жаберные отверстия. Метаморфоз через 10—
12 недель, половозрелость на второй год. Длина взрослых до
80 мм. Из других мест Южн. Африки описано еще 4 более мелких
вида. В неволе -растут и размножаются хорошо; служат удобным
объектом для эндокринологических экспериментов (в частности,
тест беременности). В Камеруне, Габоне и Конго встречаются
чрезвычайно мелкие (длина взрослых 28—46 м,м) виды рода
Hymenochirus, у которых плавательная перепонка имеется между
73
пяльцами не только на задних, но н на передних ногах. С а мм,
крупный (до 20 см) из пилон рода Pipa—P. pipa (^americana*
surinamensis) встречается в воде каналов и луж Гвианы и Бразя
лни. и во время дождей в лесах. В углах рта у нее имеют
кожные лоскуты и ряд кожных придатков на голове. Поедае
червей и мелких рыб. Спаривание длится сутки. Не исключено
что оплодотворение внутреннее. При помощи выпячиваюшегос,
конца прямой кишки («яйцеклада») самка откладывает себе и;
спину 40—114 яиц. Вскоре вокруг каждой икринки образуете,
ячейка в 10—15 мм глубиной, закрытая крышечкой в 5—6 мл
диаметром. Очевидно имеет место поступление воды и, возможно
других веществ от матери, ибо икринка весит в начале 2,95 г, ъ
к концу развития 3,37 г. Молодые животные покидают мать уже
по окончании метаморфоза примерно на 82 день. Представителя
другого вида — Р. snethlageae, значительно мельче (до 75 мм) и
не имеют лоскута в углах рта, а Р. parva лишена всяких придат-
ков. Ископаемые представители семейства обнаружены в мелу
(Eoxenopoides) и миоцене Африки (Xenopus), а также в эоцене
Германии (Opisthocoelellus, Germanobatrachus).
Подотряд 4 — Anomocoela, Чесночницеобразные
5. СЕМЕЙСТВО PELOBATIDAE. ЧЕСНОЧНИЦЫ
Чесночницы, по-видимому, возникли в третичное цремя в
Южн. Азии, где и сейчас еще встречается самый примитивный
род Megophrys. В Европу могли попасть не ранее олигоцена
(рис. 16). Филогенез внутри семейства идет к редукции зубов и
уменьшению размеров тела. Распадаются на 3 подсемейства, из
которых Megophryinae характеризуется более или менее свобод-
ными межпозвоночными дисками (охватывает большинство ро-
дов). Подсемейство Pelobatinae отличается передневогнутыми ту-
ловищными позвонками, срастанием крестцового позвонка с
хвостцом (исключая Pelodytes) и охватывает 4 рода (Pelobates,
Scaphiopus, Macropelobates, Pelodytes). Напротив, подсем. So-
oglossinae имеет передневогнутые позвонки и свободный хвостец
(Sooglossus, Nesomantls). Современные рода различаются так:
1(8) Зрачок горизонтальный.
2(7) Зубы в верхней челюсти имеются.
3 ( 4) Межпозвоночные диски свободны. *
Nesobia
1 вид, о-ва Бунгуран (Натуна).
4(3) Межпозвоночные диски не свободны.
5 ( 6) Сошниковые зубы имеются.
Nesomantls
1 вид, Сейшельские о-ва.
6(5) Сошниковых зубов нет.
Рис. 16. Ареалы семейств: 1 — Bulonidae. 2 — Pelobatidae
Soofftottus
? яидл, Сейшельские о-ва
7 ( 2) Чглмк ти без чубов.
Ophryophryne
I вид, Бакбо (Тонкий),
я < I) Зрачок вертикальный.
9 (12) Хвостец сращен с крестцовым позвонком
10 Грудина целиком хрящевая.
Scaphlopus
6 видов, Сев. Америка, Мексика
11 (10) Часть грудины окостенелая.
Pelobates
3 вида, Европа, Марокко, Зап. Азия
12 ( 9) Хвостец свободный.
13 (14) Вся грудина хрящевая.
Leptobrachella
1 вид, Калимантан (Борнео).
14 (13) Часть грудины костная.
15 (16) Сошниковые зубы имеются.
Pelodytes
1—2 вида, Зап. Европа, Кавказ.
16 (15)
17 (22)
18 (19)
Сошниковых зубов нет.
Верхняя челюсть с зубами.
Зубы хорошо развиты; кожа более или менее гл
Megophrys а^Кая.
25 видов, Южн. Азия, Малайский архигге
Зубы развиты слабо; кожа бугорчатая. Лаг‘
Верхняя губа самца с шипиками.
19 (18)
20 (21)
Vibrissaphora
2 вида, Сычуань, Фуцзянь.
21 (22) Верхняя губа гладкая.
Scutiger
4 вида, - Гималаи.
22 (17) Зубов в верхней челюсти нет.
Aelurophryne
5 видов, Кашмир, Хами, Сычуань, Тибет.
Все виды рода Megophrys живут в горах (до 2042 л).
Самый мелкий вид М. kempii (длина 15 мм, Ассам), а самый
крупный М. carinensis (длина до 15 см). Личинки могут посред-
ством своих ротовых полей вертикально висеть в воде, держась
на пленке поверхностного натяжения. Один из видов Aelurophry-
76
nt - Afl mam ma tat (Кашмир. Хам»
наличием на каждой стпроиг груди tvwori я*
леям В свази с роющим обравом жизни РгЫыг». чет
ночница, имеет очень увеличенный. юпауеюбрвхяый пятой
ныЙ бугор, функционирующий наподобие лопаты будут*
активным ночью, ив день животное зажапмавеп в. пятжь м.м*«
довольно глубоко а почву Обычный в средней Европе • Крым^
и на Сев. Кавказе пил Р fuxcun имеет вззутый между г ним*
лоб и желтоватый паточный 6vrop. Длина до ЙО мм 8 воле
только вовремя икрометания Последнее прайс аплит пол аплой
причем икра выходит 2 шнурами, обычноелнваюшимнеа а один
Головастики могут достигать длины до 175 мм Временами кожа
издает сильный запах, напоминающий чеснок. На южных сило
нах Кавказа, в Мял. Азии и Сирин живет плосколобая форма
Р syriacus. На Пиренейском полуострове и в Южн. Франции
встречается Р. cultripes с черным пяточным бугром. Остатки из-
вестны из олигоцена Франции. В Сев. Америке наша чесночница
заменена родом Scaphiopus (^Spea). В олнгоцене Монгол ни най-
ден. Macropelobates, превышавший чесночницу в длину на 10—
20%. Описаны также многочисленные рода из олигоцеиа — плио-
цена Германии и Сев. Америки, но значение их нуждается в про-
верке. «Ильная лягушка» Pelodytes живет по окраинам водоемов;
пяточный бугор у нее небольшой. По-видимому, оба вида этого
рода (A caucasicus — горы Закавказья и Р. punctatus — низ-
менности Южн. и Зап. Европы) являются только подвидами. Из
эоцена Германии описана Propelodytes, а из миоцена Невады —
Miopelodytes. Наличие на Сейшельских островах лишь бурных
потоков воды обусловило специфику размножения Sooglossus
sechellensis (рис. 17). Взрослые особи этого вида достигают в
длину 18—19 мм. Живут в старых лесах среди влажного мха,
питаются преимущественно термитами. Крупные яйца отклады-
вают на землю, на влажные листья. Лишенные роговых челюстей
и наружных жабр головастики прикрепляются к спине самца.
Дыхание их идет через кожу, в особенности хвоста.
Подотряд 5 — Procoela. Жабообразные.
6. СЕМЕЙСТВО PALAEOBATRACHIDAE. ДРЕВНИЕ ЖАБЫ
Известны с юры до миоцена Европы включительно. Имелись
зубы в верхней челюсти и на сошнике. Вероятно были водные.
Кроме взрослых найдены также головастики. Более известен род
Palaeobatrachus, менее — Halleobatrachus.
7. СЕМЕЙСТВО BUFONIDAE. ЖАБЫ
Чрезвычайно обширная группа, центральная для всего под-
отряда, включившая и прежнее семейство Cystignathidae.
77
Рис. 17 Представители Pelobatldae и Btdonldae; / Bufo |arhoa, Я го/иин»
Meaophrya feae, 3— головастики Megophrya morrtana, 4 ЛсуМорма ы
brookil, I -Soogloaaua aechalfenah с головастиками, 6 МтлоНулаМв» «for.
•alia, 7 Crlnla taamenlftnala, Я — Myobatrachiia K'>Hldil, U l,a,*2PhW
purcillt, /0-РвешИв mlnuta, // — Caratophrya appendiculate. U ElawW
rodactylua palmatua, 13 — Rhlnophfynua doraafta, И голова Bufo fyphoniti»,
/5 f^pfodactylua longtroatrla, 16 Pahidlcola fiiacomacMlata, /7 ftoogk»
aua sechellenala, 13 - nnwn Eloala naaua, !9 M»«ophfya naauta
7Л
И «десь. подобно другим бесхвостым, прими гноим А пригни*
челюстные зубы постепенно утрачивается полностью Ра «ля
чают следующие подсемейства.
I ( 2) Остаток хорды пронишвает see по i нон к и
(I) Crinilnae
15 родов, Австралия и соседние о-ва
2(1) Позвонки имеют плотное тело, не пропитанное остатком
хорды.
3 ( 4) На верхней расширенной поверхности пальцев имеется
по паре щиткоподобных образований.
(6) Elosiinae
3 рода, Вост. Бразилия.
4(3) Таких образований на концах пальцев нет.
5 ( 6) Язык спереди свободный и может высовываться; груди-
на редуцирована.
(4) Rhinophryninae
1 род, Мексика, Гватемала.
6(5) Язык иного строения; грудина имеется.
7 (12) Поперечные отростки крестцового позвонка цилиндриче-
ские или чуть расширенные; верхняя челюсть обычно с
зубами.
8 (11) Грудина широкая, хрящевая (редко с обызвествления-
ми).
9 (10) Конечные фаланги Т-образные; зрачок вертикальный;
предгрудина редуцирована.
(2) Heleophryninae
1 род, Южн. Африка.
10' ( 9) Конечные фаланги иные, если же Т-образные, то зрачок
не вертикальный, и предгрудина хорошо развита.
(3) Pseudinae
12 родов, Центр, и Южн. Америка.
! 1 (8) Грудина узкая, всегда костная.
(7) Leptodactylinae
7 родов, Центр, и Южн. Америка.
J2 ( 7) Поперечные отростки крестцового позвонка расширен-
ные; обе челюсти без зубов.
(5) Bufoninae
5 родов, распространение почти космополити-
ческое (нет в Австралии, Новой Гвинее, По-
линезии, на Мадагаскаре и в полярных
областях).
79
//«мйежгбсгяа (1) Спямме. яястшышбсммг ямбм, ow» як
г»ж>бр»»*<х «вНМкэег пркмктиямое Огжмвиме <*»*• ytiimai»
гяк
I fЯ) Зубы в верявей челюсти есть
2 ( 5) Первый na.teu пере ли» ж «юг может при* и во пос та алеть
ся другим
ч i 41 Зрачок гориюмтальиый
6 видоа. Австралия
4(3) Зрачок вертикальный.
Mytrolysis
I вид. Сев. Австралия.
5 I 2) Первый палеи не может противопоставляться осталь-
ным.
6 < 13) Зрачок горизонтальный.
7 ( 8) Сошниковые зубы в виде 2 длинных рядов; барабанная
перепонка ясная.
Raaaster
2 вида. Австралия. Новая Гвинея.
8 ( 7) Сошниковые зубы в виде 2 коротких рядов или отсут-
ствуют; барабанная перепонка отсутствует или неясная.
9 (10) Язык округлым; поперечные отростки крестцового по-
звонка ясно расширены.
Ptiiloria
1 вид, Австралия.
10 ( 9) Язык овальный или эллиптический; поперечные отросла
чуть расширены.
11 (12) Предгрудина хорошо развита.
Crbria
8 видов, Австралия, Тасмания.
12 (11) Предгрудина маленькая.
Addotus
8 видов. Австралия.
13 ( 6) Зрачок вертикальный.
14 (19) Перепонка между пальцами задних ног хорошо развита.
15 (18) Барабанная перепонка ясная.
16 < 17) Кожа спины гладкая; грудина хрящевая.
Mixophyes
1 вид. Вост. Австралия.
17 (16) Кожа бугорчатая; грудина полуокостенелая.
80
| вил, Австралии
1Я (15) Бврвбамной перепонки нет
Helioportn
2 вида, Австралии.
19 < 14) Перепонка задних ног развита слабо или совсем отсут-
ствует.
20 (21) Сошниковых зубов нет.
Hyperolla
1 вил. Вост. Австралия.
21 (20) Сошниковые зубы имеются.
22 (23) Сошниковые зубы в виде одного непрерывного ряда; ба-
рабанная перепонка ясная.
Lechrlodus
1 вид, Новая Гвинея.
23 (22) Сошниковые зубы в виде 2 рядов, с перерывом между
ними; барабанная перепонка отсутствует или неясная.
Limnodynastes
8 видов, Австралия, Тасмания.
24 ( 1) Обе челюсти беззубые.
25 (26) Зрачок вертикальный; барабанная перепонка ясная.
Myobatrachus
1 вид, Зап. Австралия.
26 (25) Зрачок горизонтальный, барабанной перепонки пет или
неясная.
27 (28) Сошниковые зубы имеются.
Notaden
1 вид, Австралия.
28 (27) Сошниковых зубов нет.
Pseudophryne
7 видов, Австралия, Тасмания.
Наиболее примитивными родами являются, по-видимому,
Mixophyes и Lechrlodus (длина тела 60—80 мм). По всей Австра-
лии широко распространена Limnodynastes, лягушковидная жа-
ба. Длина 42—67 мм. Образ жизни напоминает нашу чесночницу.
Икрометание как у большинства австралийских Salientia, не
фиксировано календарно, а связано с дождями. Обычно с июля
до мая откладывают в воду белые пенистые комки икры. Наибо-
лее известный вид рода Chiroleptes (длина 31—82 мм)—Ch. pla-
tycephalus во время засухи зарывается, сильно раздувшись, на
глубину около 30 см. При этом большой запас воды находится
7 П. В. Терентьев
81
не ТОЛЬКО В МОЧГВОМ пулырс И В ПОДКОЖНЫХ ПОЛОСТЯХ, НО Я в Г!<)
логти теля. Легкие чрезвычайно развиты и доходят до заднее
копна тела. Большинство родов подсемейства мелкие (20- 44 сж|
животные. Crtnia laeuis откладывает яйца под землей в гладко
стенных норах. Напротив, С. signifera живет в воде. Яйца Рмц.
dophrynr, отложенные поодиночке под камнями, могут месяцами
выжидать дождя. Myobatrachus живет в термитниках. Из эоцена
Индии описан Indobatrachus, видимо, принадлежащий к этом)
же подсемейству.
Подсемейство (2) Heleophrynlnae. Горные жабы.
Имеет всего I род Heleophryne, 5 видов которого жив^т ?
Южн. Африке. Верхняя челюсть с зубами, нижняя беззубая
Длина 40—42 мм. Головастики приспособлены к жизни в горны»
потоках.
Подсемейство (3) Pseudinae. Южноамериканские жабы.
Происходят, видимо, прямо от Criniinae, охватывает значи
тельное количество родов, главнейшие из которых таковы:
1 ( 2) Обе челюсти без зубов.
Batrachophrynus
3 вида, Перу.
2(1) Верхняя челюсть с зубами. п
3(4) Грудина с широким костным стержнем; конечности, осо-
бенно пальцы, очень длинные, зрачок вертикальный.
Hylorina
3 вида, Чили.
4(3) Грудина хрящевая или частично обызвествленная; ко-
нечности не удлинены; зрачок горизонтальный.
5 (10) Плавательная перепонка вдается между метатарсаль-
ными костями 4-го и 5-го пальцев задних ног.
6(7) Первый палец передних ног может противопоставляться
остальным.
Pseudis
5 видов, Южн. Америка.
7(6) Первый палец не противопоставляется остальным.
8(9) Нижняя челюсть с зубовидными выростами.
Grypiscus
1 вид, Бразилия.
9 ( 8) Нижняя челюсть гладкая.
Cycloramphus
2 вида, Бразилия.
10 ( 5) Плавательная перепонка не вдается между метатар-
сальными костями двух последних пальцев. <
Il (14) Конечные фаланги Т-образные
12 (13) Сошниковые зубы имеются
Eleutherodactylu*
Около 200 видов. ю« Сев. Америки. Вес т
Индии. Центр, и Юж и Америка
13 (12) Сошниковых зубов нет
Syrrhophus
22 вида. Центр, и Южн. Америка.
14 (11) Конечные фаланги не Т-образные.
15 (20) Плавательная перепонка задних ног хорошо развита.
16 (17) Кожа головы срастается с черепом; глазница замкнута
костями.
Calyptocephalus
3—5 видов, Панама, Чили.
17 (16) Кожа головы не срастается с черепом; глазница не
замкнута костями:
18 (19) Язык сзади с вырезкой, сердцевидный.
Ceratophrys
16 видов, Южн. Америка.
19 (18) Язык без вырезки.
Telmatobius
10—12 видов, Южн. Америка.
20 (15) Плавательной перепонки нет или она очень маленькая.
21 (22) Язык сзади совершенно свободный.
Borborocoetes
11—17 видов, Центр, и Южн. Америка.
22 (21) Язык сзади полностью или частично прикрепленный.
Zachaenus
2 вида, Бразилия, Патагония.
Наиболее примитивны 6 водных родов с плавательными пе-
репонками и простыми конечными фалангами. Примером их
может служить Pseudis, гвианская водяная жаба, головастики
которой могут достигать длины свыше 25 см, взрослое живот-
ное— 19—69 мм. Особое место среди всего подсемейства зажи-
мает род Hylorina, имеющая удлиненные конечности. Наконец,
5 родов образуют наземную группу (плавательные перепонки
короткие или их совсем нет). Обще распространен род Borbo-
rocoetes, южноамериканская наземная жаба. Широко известна
Ceratophrys, рогатка, остатки которой найдены также в плей-
стоцене Бразилии. Длина ее достигает 20 см и более. Яркая
окраска и странные придатки многих видов имеют защитное зна-
7* 83
чеиие (когда животное сидит полузарывшись в почву сиры*
лесов). Установлен факт пожирания рогатками других бесхво-
стых и мышей. Многочисленные виды Eleut herodactylus ("Hylth
des) живут на деревьях, кустах и в траве. Обитатель Антильскях
островов Е/. martinlcensls (длина 40 мм) откладывает пакеты
яиц в лаяухи листьев. Весь метаморфоз протекает внутри ик-
ринки.
Подсемейство (4) Rhinophryninae, носатые жабы, ло-видя-
мому, происшедшие от Pseud in ае, заключает в себе всего I вид
Rhinophrynus dorsalis. Ведет роющий образ жизни. Пяточный
бугор лопатообразный. Питается термитами. Обе челюсти без-
зубые. Длина тела до 85 мм.
Подсемейство (5) Bufoninae, настоящие жабы, близко род-
ственно Criniinae. Обе челюсти беззубые. Характеристики основ-
ных родов таковы:
1 ( 8) Язык прикреплен по всей длине или свободен сзади.
2(3) Ноздри направлены прямо вверх.
Pseudobufo
2 вида, Малайский архипелаг.
3(2) Ноздри направлены несколько вбок.
4(5) Конечные фаланги Т-образные; пальцы передних ног
обычно соединены перепонкой и несут диски или пла-
стинки.
Nectophryne
13 видов, Африка, Малайя, запад Индо-Ма-
лайского архипелага.
5(4) Конечные фаланги не Т-образные; перепонок между
пальцами передних ног нет.
6(7) Предгрудина есть.
Nectophrynoi des
2 вида, Вост. Африка.
7 ( 6) Предгрудины нет.
Bufo
250 видов, почти всюду, кроме Австралии.
-'ф ( 1) Язык спереди свободный.
Wemerla
1 вид, Камерун.
Наиболее примитивный по своей морфологии род Nectophry-
noides, живородящая жаба (единственная живородящая форма
среди всего отряда). Органы внутреннего оплодотворения пока
не найдены. В теле самки наблюдали более сотни личинок, хвос-
ты которых, по-видимому, являются местом газообмена. Длина
взрослых 27 мм. Термитная жаба Werneria конвергентно схожа
84
строением языке с Rhinnphrynua. Длина 33 мм. Водяная жаба
Pseudobufo достигает 150 мм. Основная масса видов принадле
ж нт к роду Bufo, настоящих жаб. Как правило, это наземные,
сумеречные животные, сравнительно плохо плавающие и делаю-
щие лишь короткие скачки. Яйца выходят в виде шнуров. Круп-
ные виды могут нападать на позвоночных. В роде Bufo встре-
чаются виды, подобные нашим европейским, голова которых ие
имеет особых возвышений. Далее, многие виды хфактеризуются
развитием вертикальных гребней между глазами и на конце
морды. Как крайний случай надо отметить В. typhonius, гребен-
чатоголовую жабу из Южн. Америки. По-видимому, отсутствие
гребней является более древним признаком, хотя доказательств
этому пока нет. В северной и умеренной Евразии широко распро-
странена В. bufo, серая или обыкновенная жаба. Основная часть
ее ареала (Европа, Сибирь до Байкала) занята более молодой
формой В. b. bufo. В южных местностях сохранились остатки
более древней доледниковой формы: В. b. spinosus в Испании,
В. b. uerrucosissitna на Кавказе, В. b. asiaiucus на Дальнем Вос-
токе, В. b. faponicus в Японии ив В. b. gargarizans в Китае.
Все эти формы отличаются значительно большей величиной и
более сильным развитием бугорков на спине. Встречают-
ся под корнями, бревнами, в пещерах, подвалах, хлевах
и т. п. Обычно придерживается раз избранного местооби-
тания. В горах на высоте до 3048 м. Питаются жуками, мура-
вьями, пауками, многоножками, слизнями, дождевыми червями
и т. п. Икрометание под Киевом в конце марта, севернее — в
апреле — мае. Шнуры икры от 3 до 5 м. Одна самка кладет до
6840 икринок. Метаморфоз на 77—91 день. Половозрелость на
четвертый год. Длина взрослых до 200 мм. Зимует на суше. В то
время как этот вид связан с полосой лесов и не избегает тайги,
значительно меньшая (до 140 см) В- viridis, зеленая жаба, встре-
чается в сухих местностях, попадаясь даже в оазисах среди
пустыни. Ареал зеленой жабы очень широк: все Средиземно-
морье, Европейская часть СССР до 59° с. ш., Крым, Кавказ, Сред-
няя Азия, Иран, Зап. Монголия, Сибирь до Алтая. Часто попа-
дается в норах грызунов. В горах до 4572 м (Гималаи). На
Дальнем Востоке, в Монголии, Китае, Корее и Вост. Сибири за-
меняется В. raddei. В Зап. Европе и западных частях СССР
встречается В. calamita.
Из экзотических видов надо прежде всего упомянуть
широко распространенную в Пент, и Южн. Америке В. та-
rinus, ага, достигающую 200 и более мм в длину.
Кожные выделения этого вида употребляются для смачивания
стрел. В Сев. Америке обычна В. lentiginosus. В Африке не ред-
ки В. regularls, В. mauritanicus и В. superciliosus, а в Южн. Азии
широко распространена В. melanostlctus, чернорубцовая жаба.
Стоит отметить необычайную длинноиогость тушканчиковой
жабы—В. jerboa (Калимантан, Суматра, Малайский архипелаг) и
85
lepfofus (Калимантан) В ископаемом состоянии Ил
^-^стны из миопена Европы и Флориды В Герм «ют
н’ м миоценовые Rufonopxix, Eobufella и Parabufella.
сЯ fl оде f лейст по (6) Elosiinae, щиткопалые жабы, видимо. о*,
Л1И**о °’ P*eud*naf’’ встречается только в Бра тя л им &*1>к«*н
н ‘.ть с тубами, нижняя беззубая. Длина тела 23 40
1с\якт<фнстики описанных ролов таковы
। ( 4) Сошниковые зубы есть.
•’ ) 3) Ннжиятгчелюсть гладкая.
Elosia
5 видов.
* । 2) Нижняя челюсть с отростками («ложные зубы»).
Megaetosla
I вид.
4 J {) Сошниковых зубов нет.
Crossodactylus
3 вида.
Подсемейство (7) Leptodactylinae, свистуны- произошло от
Pseiidjnae. Главные рода; ' к
I (10) °- х” » “^пхней челюсти есть
2 4 9)
(8)
( 7)
( 6)
4
5
(5)
6
Зубы в -верхней челюсти есть.
Зрз.чок горизонтальный.
Сошниковые зубы имеются.
Предгруднна хрящевая.
Верхнее веко гладкое; сошниковые зубы между хоана»"-
Physalaemus
24 вида, Южн. Америка.
Верхнее веко с бугорками; сошниковые зубы сзади хоан.
Edalorhina
3 вида, Перу, Эквадор.
у ( 4) ПреДпруДина костная.
Leptodactylps
-52 вида, Центр, и Южн.. Америка.
8 ( 3) Сошниковых зубов нет.
Paludicola
23 вида, Центр, и Южн. Америка.
gi...( 2) Зрачок вертикальный.
Limnomedusa
Г вид; Уругвай.
10 ( -1) Обе.ярлюсти беззубые.
Eupemphix
7 видов, Центр, й 'Южн. Америка.
Наиболее поимнтии н рол f'htMaiurrrtu*. лр*ин он-лтий. ш*
видимому, от Bmht/rMwtfi и давший начало жем лр\>жм рл^яч
подсемейства. Представители этого и большмттяя тртгич ротов
мелкие формы <11 47 мм) Вилм ролл Lrptf>da< tutu*. сяи*т\>»н.
сильно варьируют по величине от 23 до 2<Ю «и <X,p/»i аичнм
их напоминает наших водных лягушек, хотя иляяягглышя лере
понка у них редуцирована. Самый крупный L. pcntadm tulus
свистун пятипалый, получил свое название та пвльиеподобкый
вырост на внутреннем пальце передней ноги Самки отклады-
вают икру (похожа на сбитые белки) в «гнездо» среди водяных
растений; самки других видов устраивают ямки вблизи луж.
Личинки могут переживать засуху, собравшись в комок под ли-
стьями. Длина Limnotnedusa достигает 66 мм. Виды Paludicotu.
болотниц, мельче (15—41 мм). Образ жизни полуводный. При
кваканьи Р. fuscomaculata может сильно надуваться, потом
разом уменьшаться; при этом получается крик, напоминающий
мяуканье котенка. Мелкие (1 мм) яйца складывают в январе в
виде пенистой массы, плавающей на поверхности воды. Голо-
вастики вылупляются через 18—24 часа. На (бедрах взрослых
имеется по яйцевидной кожной железе. Пяточный бугор напо-
минает таковой у чесночниц.
8. СЕМЕЙСТВО HYLIDAE. КВАКШИ
Особенности строения пальцев большинства видов стоят в
несомненной связи с древесным образом жизни. Имевшие место
попытки разбить это семейство на 2 подсемейства (Gadow—на
Amphignathodontinae и Hylinae; Noble — на Hemiphractinae и
Hyliriae) нельзя признать удачными ввиду наличия большого
числа переходов. Основные рода различаются так:
1 ( 6) Обе челюсти с зубами.
2(3) Зубы настоящие; поперечные отростки крестцового поз-
вонка сильно расширены.
Amphignathodon
1 вид, Эквадор.
3 ( 2) Зубы нижней челюсти ложные — выросты кости; попе-
речные отростки крестцового позвонка цилиндрические.
4 ( 5) «Псевдозубы» сидят также на сошнике и нёбных ко-
стях; голова в виде костного шлема.
Hemiphractus
7 видов, КЫн. Америка.
5(4) «Псевдозубы» сидят на нёбных костях и околоклино-
видной; костного шлема на голове нет.
87
I
2 ВЯЛЯ. Браавлвя. Тряямяжт
Нижняя челюсть безгубая
Верхняя челюсть с яублмя
Сошниковые я
Зрачок горизонтальный; нМные вост* * '
Dtagfena
2 вилл. Мексике. Эквадор
Ю ( 9) Зрачок ромбический; нйбиые кости без »убо«
Triprion
I вид, Юкатан.
..«пыпняя кость всегда, а сошниковые «ыгц
11(8) Околокл иновидна я
11 ' без зубов* £
!з (22) Г^’ьиы 'Ряс^^Тк^рХГтц<»^гоИ позвонка растру
< ill -стного шлема с выд"оти—
гребнями.
Ptemohyla
1 вид, Зап. Мексика.
16 (15) Голова не имеет вида каски, лишь иногда кожа прим,
стает к черепу.
17 (18) Самки не имеют на спине сумки для яиц.
Hyla
350 видов, все части света, исключая Африку
к югу от Сахары, Мадагаскар, Полинеэзд
к востоку от Соломоновых островов и по-
лярных стран.
18 (17) Самки имеют на спине сумки для яиц.
Gastrotheca
15 видов, Центр, и Южн. Америка.
19 (14) Поперечные отростки крестцового позвонка цилиндра-*
ческие или слабо расширенные.
20 (21) Конечные фаланги Т-образные.
Centrolene
2 вида, Эквадор.
21 (20) Конечные фаланги простые.
Cryptobatrachus’
22 1 вид» Гвиана.
663 Дисков, самое большое — конечные фалан-
ги несколько вздуты.
R8
23 (24) Плавательная перепонка разлита хорошо; барабаня л я
перепонка неясная.
Acrte
1 вид, Сев. Америка.
24 (23) Перепонка только при основания пальцев задних ил,
барабанная перепонка хорошо видна.
Pseudacrfe
6 видов, Сев. Америка, Перу.
25 (12) Зрачок не горизонтальный.
26 (27) Зрачок ромбический; голова с костным шлемом.
Corythomantis
1 вид, Бразилия.
27 (26) Зрачок вертикальный; голова без костного шлема.
28 (29) Язык сзади лишь немного свободный.
Nyctimantis
3 вида, Эквадор.
29 (28) Язык сзади свободный на большом протяжении.
Phyllomedusa
22 вида, Центр, и Южн. Америка.
30 ( 7) Верхняя челюсть беззубая.
Allophryne
1 вид, Южн. Америка.
Самки большинства видов откладывают икру в воду или око-
ло нее. Центральным и самым обширным родом является Hyla,
длина 17—135 мм. Особенно многочисленны в Австралии и
Папуасии, но отсутствуют в тропических частях Азии и Африки
(рис. 18). От Средиземноморья через Зап. Европу, юго-запад
Европейской части СССР, Крым, Кавказ и Мал. Азию до Ирана
включительно обитает И. arborea, обыкновенная квакша, или
древесница (длина 40—50 мм); вид распадается на ряд подвидов.
Большую часть своей жизни проводит среди листвы деревьев и
кустарников, а иногда даже широколиственных травянистых
растений. В горах до 1500 м. Особую активность проявляет в
сумерках. Может сравнительно быстро менять окраску. В воде
бывает только во время икрометания. Крик издает все лето.
Чаше зимует в иле водоемов. На Дальнем Востоке, в Японии
и Корее и Сев.-Вост. Китае живет Н. japonica. В то время как
наши виды этого рода окрашены обычно в разные оттенки зеле-
ных тонов, тропические виды иной раз поражают красотой рас-
цветки. Так австралийская Н. coerulea (длина 100 лсм) при
жизни зеленая с красными концами пальцев. Живущая там же
Н. аигеа имеет золотистый или бронзовый оттенок. Наиболее
А И Л TwAMtrrwHB
89
обычны ДЛЯ Сев. Америки Н. versicolor (сверку бугорчатая, о
_йя с темным рисунком). //. regilla (чаще агленая с продольной
полосой вдоль спины). Н. carolinensls (2 светлые продольные
полосы ня оливково-зеленой спине) и Н. pickerlngt (серо-желтая
или красновато-бурая с темным рисунком) Из многочисленны*
Рис. 19. Представители Brachycephalidae и Hylidae: 1 —r-Amphignathodon
guentheri, 2—голова Diaglena spatulata сбоку, 3 — G astro thee a marsu-
piata (часть кожи справа отрезана для показа яиц,, находящихся в сумке),
4 — Hemiphractus scutatus, 5 — Gastrotheca pygmaea (яйца вынуты из
широко раскрытой сумки), 6 — Cryptobatrachus evansi, 7 — Oreophrynella
quelchii, 8 — Dendrobates braccatus, 9 — Brachycephaltis ephippium
южно-американских видов особенно интересна И. faber (длина
80—90 мм) с темной продольной полосой на желтой или бурой
спине. Название «кузнец» дано за голос, напоминающий стук
молотка по медной пластинке. Самка Н. fabqr сооружает из ила
луж или болот кольцевидное гнездо, диаметром около 30 см и
со стенками около 10 см вышиной, предохраняющее икру от
нападения хищных водяных насекомых и т: *п. Бразильская
Г 91
H. nebuloea укрепляет комки икры, окруженные бел**
массой, на внутренней поверхности гниющих банажэвм.Т^’ч
Откладывание икры в наполненные водой пахужи листы* «ы^
дается у европейской Н. arborea.Диски на палы,», «,М(в «ч
как и других древесных Salientia, богаты лимфатая*,-..^/*^'-
странствами и слизистыми железами. Прикрепление я ’
тем сильнее, чем меньше остается воздуха между воглфдд *’>
Дисками. Особые мускулы дают возможность дискам гт>Мг<<? •
более плоскими. Обыкновенно прикрепление к субстрату ж *
также за счет кожи брюха и горла. Род Phyllomeama (р*
листовая квакша (длина 23—110 мм) откладывает икру * ,
нутые листья над стоячими водоемами, в которых личиими
должают свое развитие. Северо-американская Ас^
на 31 мм) держится по берегам стоячих вод и на листьях *
пых растений, a Pseudacris (длина 23 35 мм) живет преиму^
ственно в зарослях и на хлебных полях.
Интересна эволюция заботы о потомстве в некоторых pti3St
Hylidae. Если самки Cryptobatrachus (длина 75 мм) пр<>7
носят комок яиц на спине, то у Gastrotneca (^Nototrema) И1
спине развивается специальная сумка, причем у G. pygma^
этот выводковый мешок открывается длинной щелью, а у G,
ferum — уже только небольшим отверстием в области кресту
Большое значение имеет и величина яиц: у G. marsuplata
относительно мелкие и их много, и головастики продолжав
свое развитие в воде, у G. ovlferutn всего около 15 крупнщ
(1 см в диаметре) яиц и весь метаморфоз протекает в сумке
В ископаемом виде семейство известно начиная с миоцод
{Ну 1а в Европе).
9. СЕМЕЙСТВО BRACHYCEPHALIDAE. КОРОТКОГОЛОВЫЕ
Принято различать 3 подсемейства, которые, возможно, воз-
никли самостоятельно. Основные отличия таковы:
1 (12) Предгрудина имеется; верхняя челюсть с зубами илк
беззубая.
2 ( 7) На верхней сторонке концов пальцев кожные щитки
[подсемейство (2) Dendrobatinae. Древолазы].
3 (.6) Верхняя челюсть с зубами.
4(5) Задние ноги с плавательной перепонкой.
Hyloxalus
1 вид, Южн. Америка.
5(4) Плавательной перепонки нет.
Phyllobates
20 видов, Центр, и Южн. Америка.
6(3) Обе челюсти беззубые,
ад
Deftdretetes
10 я я доя, Центр я Южн Атрия*
10 ( 9>
Кожных щитков на пальияж мет 1подсет*етм (I)
Rhlnodermatlnae. Носатые лягушяя}
Верхняя челюсть с хубамя.
Наружный палеи передней иогн хороню разнят.
Smlnthlifaa
3 вида, Куба, Перу, Вост. Бразилия.
Наружного пальца передней ноги нет.
Geobatrachus
I вид, Колумбия.
11
8) Обе челюсти беззубые.
Rhinoderma
1 вид, Чили.
12 ( О
13
14
(18)
(15)
Предгрудины нет; обе челюсти беззубые [подсемейство
(3) Brachycephalinae, собственно Короткоголовые].
Грудина хрящевая.
Кожа спины с известковой бляшкой; пальцы, особенно
1-й и 4-й передней ноги, сильно укорочены.
Brachycephalus
1 вид, Гвиана, Бразилия, Чили.
15 (14) Известковой бляшки на спине нет; пальцы длиннее.
16 (17) Первый или 2 первых пальца задней ноги могут про-
тивопоставляться прочим.
Oreophrynella
2 вида, Гвиана.
17 (16) Пальцы задней ноги не противопоставляются друг другу.
Atelopus.
26 видов, Южн. Америка.
18 (13) Грудина костная.
Dendrophryniscus
3 вида, Бразилия, Парагвай, Аргентина.
Большинство представителей семейства — это ярко-окрашен-
ные мелкие (длина 20—-76 мм) животные, держащиеся вблизи
лесных потоков. Sminthillus откладывают яйца на землю, при-
чем каждая самка кладет, видимо, только по 1 крупному яйцу.
Напротив, у Rhinoderma darwini (длина 30 мм) яйца после опло-
дотворения помещаются в сильно растягивающийся горловой
мешок (видоизменение внутренних резонаторов) самца, где
протекает все или почти все развитие, причем на более поздних
93
стадиях головастики получают питание при п<м ред< гв<* гт*-»*-*
мешка. Самцы Dendrobates и Phyltobates переносят своих гол#»
метиков, присосавшихся к коже спины, из пересыхающих tvisnr
мои в новые. Ядовитая кожная слизь Dendrobatea употребляется
местными охотниками для отравления стрел и для прижизненная
перекраски зеленых перьев попугаев в желтые.
Подотряд 6 — Diplasiocoela. Лягушкообразные
10. СЕМЕЙСТВО RANIDAE. НАСТОЯЩИЕ ЛЯГУШКИ
Настоящие лягушки, видимо, возникли в Старом Свете, при
чем Африка явилась местом их наибольшей дифференциация
Ископаемые Rana известны начиная с эоценовых отложений Ев-
ропы, a Ooeidozyga — из равновозрастных отложений Индии
Принимаемые ныне 6 подсемейств и ряд родов в ряде случаев
соединены незаметными переходами, затрудняющими системати-
ку. Основные рода таковы:
1 ( 4) На верхней стороне конца каждого пальца по паре
кожных щитков [подсемейство (5) Petropedetinaej.
2(3) Сошниковые зубы имеются.
Pet^opedetes
5 видов, Камерун., Сиерра-Леоне.
3(2) Сошниковых зубов нет.
Arthroleptides
1 вид, Танганьика.
4(1) Концы пальцев без щитков.
5 (12) Зрачок вертикальный; Зап. Африка [подсемейство (2)
Astylosterninae].
6(7) Пальцы задних ног не связаны перепонкой.
Gampsostednyx
1 вид.
7(6) Пальцы задних ног более или менее связаны пере-
понкой.
8 ( 9) Перепонка на задних ногах развита слабо.
Astylosternus
3 вида.
9(8) Перепонка на задних ногцх связывает пальцы на 3/4.
10 (11) Конечные фаланги слегка изогнутые.
Nyctibates
1 РИД.
Н (10) Конечные фаланги 2-го и 3-го пальцев задней ноги длин-
ные и сильно изогнутые.
94
12
!3
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
ScotoMeps
I вил.
( 5) Зрачок горизонтальный, если же нет, то уго род из
Индии. . . .
(24) Сошниковых зубов нет; Африка [подсемейство (I)
ArthroleptinacJ.
(15) Грудина длинная, костная.
Arthroleptella.
1 вид.
(14) Грудина короткая, чаще хрящевая.
(21) Верхняя челюсть с зубами.
(18) Нижняя челюсть несет длинные выросты («ложные
зубы»).
Dlmorphognathus
1 вид.
(17) Нижняя челюсть гладкая.
(20) Язык сзади с глубокой вырезкой.
Phrynobatrpchus
14 видов.
(19) Язык сердцевидный.
Arthroleptis
35 видов.
(16) Обе челюсти беззубые.
(23) Окраска контрастная из черного и белого цветов; Габун.
Cardioglossa
5 видов.
(22) Окраска иная; Катанга.
Schoutedenella ’
1 ВИД.
(13) Сошниковые зубы имеются, если же нет, то это рода
из Азии или с Соломоновых островов.
(42) Концы пальцев расширены в диски или, по крайней
мере, вокруг конца фаланги имеется бороздка, отде-
ляющая расширенную часть [подсемейство (6) Cornu-
ferinae].
(27) Нижняя челюсть с выростами («ложные зубы»).
Ceratobatrachus
1 вид, Соломоновы о-ва.
(26) Нижняя челюсть гладкая.
(37) Задний конец предгрудины расщепленный.
95
30
(34)
(31)
31
32
(30)
(33)
Сошниковые зубы есть *o9C&° ромите
Переломки ми ваднмх и<ни»
ENscodefes
I вид, Соломоновы
Перепонка на задних i-
ствует.
Компы
33
(32)
Конны
соски.
- ] о*в0-
ногах слябе
пальцев немного расширены.
Гвн^'ф^нппины. Новая Гвинея. По,,
пальцев образуют хорошо выражении,
35
(29)
(36)
Comufer
12 видов, Бирма, Филиппины, Калисз*.
(Борнео), Фиджи, Соломоновы о-ва.
Сошниковых зубов нет.
Пальцы передних. ног с перепонкой.
Palmatorappia
1 вид, Соломоновы о-ва.
36 (35) Перепонки на передних ногах нет.
Batrachylodes
1 вид, Калимантан (Борнео).
47 (28) Задний конец предгрудины не расщеплен.
38 (39) Пальцы передних ног с перепонкой.
Simomantis
1 вид, Калимантан (Борнео).
39 (38) Пальцы передних ног без перепонки.
40 (41) Сошниковые зубы имеются.
Hylorana
64 вида, Южн. и Вост. Азия, Сев. Австралия.
Африка.
II (40) Сошниковых зубов нет.
Micrixalus
26 видов, Южн. и Вост. Азия, Индо-Малайскнй
архипелаг.
12 (25) Концы пальцев не расширены в диски, если же неболь-
шие диски имеются, то зрачок вертикальный или квад-
ратный.
13 (46) Грудина хрящевая [подсемейство (3) Phrynopsinae).
И (45) Голова большая; зубы удлиненные.
2 вида. Моаамбаж. Камерун
маленькая; зубы не удлинен и ые
- (44) Г°Л teptodactytodon
р 3 вида. Камерун.
гпудни»’ по К,«?АНеЙ/Ле|» У •3Р?КЛЫХ косную
, /дЭ) Г2£, [подсемейство (4) Raninee).
|б 1 Ллк горизонтальный.
. (б6} гошниковые зубы имеются.
4’ (51) поперечные отростки крестцового позвонка цялиидри
*9 (8°) веские.
Rana
Около 200—300 видов, всюду, кроме большей
части Южн. Америки, Австралии, Нов. Зе-
ландии, Полинезии и полярных стран.
Поперечные отростки крестцового позвонка слегка рас-
50 <49) ширены.
Nannophrys
2 вида, Цейлон.
„ч гошниковых зубов нет.
(48) лпч отверстиями евстахиевых труб небольшой пре-
52 (58)
Oreobatrachus
1 вид, Калимантан (Борнео).
53 (<54)
52 (68)
(55)
Altirana
1 вид, Тибет.
55 (54) Предгрудина костная.
4°ввдагУЮжн - Вост. Азия, Индо-Малайский
архипелаг.
Nannobatrachus
1 вид, Малабарский берег.
58 (57) Зрачок вертикальный.
Nyctibatrachus
4 вида, Индия.
97
К подсемейству (j) Arthroleptinae, Карликовых ля^ушяк. Яри
чядлежат маленькие африканские вилы, живущие иа яемле ,
лгсах илиi тарослях кустарника Самиы некоторых видов ЛгГЛг.,
Irpiii. Schoutcdenelia и Cardioylossa имеют очень удлиненна
3 й палеи передней ноги (рис. 20)
Рис. 20, Представители Ranidae, Polypedatidae и Brevicipitidae:
1 — Schoutedenella globosa самец, 2 — Astylosternus robustus самец,
3— Asterophrys turpicola, 4 — Polypedates dennysi, 5 — Microhyla pul-
chra, 6 — Hylophorbus bibroni, 7 — Glyphoglossus molossus
X подсемейству (2) Astylosterninae, Лесных африканских ля-
гушек относятся животные среднего размера. У некоторых ро-
дов, например, Scotobleps и Gampsosteonyx, конечные фаланги
могут прободать кожу, образуя «ложные когти», значение кото-
рых неизвестно. В брачное время самцы Astylosternus (-Tricho-
batrachus) robustus (Гвинея, Камерун) развивают на боках тела
и бедрах длинные волосовидные сосочки, кожи, являющиеся
С|Я
дополнительными opt ямами дыхания («вело яг* и лягушка»)
Последнее связано со слабым разнигисм легких
Подсемейство (3) Pfirynopainae Жабовидгш мелкие жи
вотные.
Подсемейство (4) Rani пае, Собственно лягушки Самый
обширный род — капо, лягушка, остальные рода имеют узкий
ареал н включают по I--4 видя. Одной из характерных особей
костей Rana является вырезка на заднем свободном конце языка.
Различают 7 подродов, некоторые из которых подразделяются
на секции. Наиболее примитивен подрод Rana (s. st.), возник*
шнй, по-видимому, в Азии. Характеризуется довольно массивны-
ми прямыми ключицами, нерасширенными концами пальцев,
явной барабанной перепонкой и отсутствием соприкосновения
между чешуйчатой и верхнечелюстной костями. Из принадле-
жащих к нему видов 14 водится ib Евразии, 22 в Америке, II в
Африке, 45 в Индо-Малайских странах и 1 в Полинезии.
Озерная лягушка, R. ridibunda, может считаться типичным
представителем водных видов нашей фауны. Она распростра-
нена в Средиземноморье, Европе (на север до 60° с. ш.), на Кав-
казе и в Средней Азии. Вся жизнь проходит в воде или около
нее. В горах до 2438 м. Чрезвычайно прожорлива и питается не
только беспозвоночными, но систематически заглатывает голо-
вастиков, лягушек и мальков рыб. Икрометание в Италии в
марте, под Одессой в апреле, под Москвой в середине мая.
Метаморфоз на 82—125 день. Половозрелость на 3-й год. Длина
взрослых до 170 мм. Зимует на дне водоемов. Менее крупен (до
100 мм), ярче окрашен и более строен другой вид наших водя-
ных лягушек — прудовая, R. esculenta. На восток распростра-
нена примерно до 50° долготы. На Дальнем Востоке эти виды
заменяются R. nigromaculata. Все водные лягушки, обитающие
на территории нашей страны, имеют в окраске зеленый цвет.
Самцы их снабжены наружными резонаторами (голосовые
мешки), в виде надувающихся по углам рта пузырей. Напротив,
наши наземные лягушки обычно имеют резонаторы, скрытые под
кожей («внутренние»), и окрашены в коричневые, серые или
темные тона. Кроме того, у них имеется темное «височное» пят-
но, идущее от заднего края глаза через барабанную перепонку
почти до плеча. Типичным представителем наземных лягушек
можно считать травяную лягушку, R. temporaria. Она распро-
странена от Франции до Сахалина включительно, идя на север,
в ряде случаев, до пределов континента. В Крыму, на Кавказе,
в низовьях Волги и в Сибири (между 70° и 130° долготы) отсут-
ствует. Встречается в лесах, рощах, среди кустарников и т. п.
В горах до 3048 м. Активна в сумерках. Икрометание ранней
весной, почти тотчас после пробуждения. Метаморфоз на 50—
90 день. Половозрелость на 3-й год. Длина до 100 мм. Большая
часть зимует под водой. На сырых лугах, по опушкам, полям и
т. п. держится R. terrestrls (=R. arvalis). На Кавказе встречают-
99
ся R camerani я /?. macrocnemis. От С*шф*чън по Сюлл*»<
Китая распространена R. ch&nsinensis.
Из представителей иных фаун следует упомянуть север»
риканскую лягушку-быка, R. catesbeiana (-R. nutgifin),
тельные размеры которой позволяют ей нападать даже на и.
кнх птиц. Образ жизни сходен с нашими водными лйгувмг*^
самцы лишены резонаторов. Подрод Tomopterna (4 явлл 9 Ц
рике) отличается раздвоенной при основании лредгрулиьл
Образ жизни роющий. К роющим относятся и представят*»,
подрода Pyxicephalus (I вид в Африке), у которых чешуйчат
кость соединяется с верхнечелюстной; предгрудина иельиэ»
Прямая ключица, цельная предгрудина, отсутствие барабаня^
перепонки и соединения чешуйчатой кости с челюстной харажт*
ризуют подрод Nanorana, единственный вид которого N. pletk*
(длина до 58 мм) живет в горах Тибета и Кашмира на высот»
от 365 до 4572 м. Тонкие, изогнутые и широко разделенные дру.»
от друга ключицы отличают африканские подроды Hildebrandto*
(предгрудина цельная; 7 роющих видов) и Ptyehadena (пр^
грудина, раздвоенная при основании; 16 видов). Положен»*
крупнейшего представителя современных Salientia (длинз до
326 мм) несколько неопределенно: некоторые выделяют R. go-
liath из Камеруна в самостоятельный род Gigantorana. Остро-
язычная лягушка Ooeidozyga (—Oxyglossus) ведет водный образ
жизни.
Подсемейство (5) Petropedetinae. Щиткопалые лягушки, по
строению пальцев аналогичны Elosiinae. Конечные фаланги у
них часто Т-образные.
Подсемейство (6) Cornuferinae. Дископалые лягушки. Самый
обширный род Hylorana, образом жизни напоминает Rana, хотя
головастики ряда видов живут <в горных ручьях. Некоторые си-
стематики выделяют в самостоятельный род Staurols виды, го-
ловастики которых снабжены большим присасывательным дис-
ком на брюхе.
11. СЕМЕЙСТВО POLYPEDATIDAE. ВЕСЛОНОГИЕ ЛЯГУШКИ
Большинство членов этого семейства ведут древесный образ
жизни. Основные рода:
1 (18) Верхняя челюсть с зубами.
2 (11) Зрачок горизонтальный.
3 (6) Первые 2 пальца передней конечности могут противо-
поставляться остальным.
4(5) Сошниковые зубы имеются.
Chiromantis
12 видов, Африка.
б ( 4) Сошниковых зубов нет.
юо
CMrixalns
6 видов. И ИЛИЯ. И я до-Китай
е ( 3) Пальцы не противопоставляются друг другу.
7 ( 8) Сошниковые зубы имеются.
Polypedates
120—130 видов, Южн. и Вост. Азия. Мада-
гаскар.
8(7) Сошниковых зубов нет.
9 (10) Язык сзади глубоко вырезанный.
Ixalus
18 видов, Индия.
10 ( 9) Язык сердцевидный.
Hyperolius
190 видов, Африка и Мадагаскар.
11(2) Зрачок вертикальный.
12 (13) Первые 2 пальца передней ноги могут противопола-
гаться остальным.
Leptolepis
8 видов, Африка.
13 (12) Пальцы не противополагаются друг другу.
14 (15) Диски на пальцах исчезающе мелкие.
Kassina
4 вида, Африка.
15 (14) Диски на пальцах хорошо заметны
16 (17) Сошниковые зубы имеются.
Hylambates
10—18 видов, Африка.
17 (16) Сошниковых зубов нет.
Megalixalus
29 видов, Африка и соседние острова.
18 ( 1) Обе челюсти беззубые.
Mantella
1 вид, Мадагаскар.
Наиболее примитивен обширный род Polypedates (—Rhaco-
phorus). К нему относится, между прочим, Р. nigropalmatus
(Ява, Суматра), получившая название «летающей лягушки» за
свои длинные планирующие прыжки. Недавно наблюдали
прыжки в 10—12 м также у Р, malabaricus. Особи этого рода
обычно взбивают свою икру движениями задних ног до пено-
образного состояния. Ряд видов помещает комки икры среди
toi
листвы (Р niRropalmatus. Р. leucomystax Зондский ярхип<Мвг
Р. dennysi — Китай). Р. schlegeli (Япония) помещает их в r>oi’
♦смпых норах, а самка Р reticulatus (Цейлон) приклеивав
свои 20 икринок к брюху. Интересны устройством пальнув рот»
Chiromantis, Chirixalus и Leptoiepis.
12 СЕМЕЙСТВО BREVICIPITIDAE (-ENGYSTOMATIDAE) УЗКОРОТЫ»
Несмотря на свое многообразие (14 подсемейств), это семей,
ство представляет собой довольно естественную группу, если не
считать Cacosterninae и Hemisinae (рис. 21). Основные рода
различаются так:
1 (10) Верхнечелюстные кости вытянуты вперед и встречаются
друг с другом впереди предчелюстных; Новая Гвинея
[подсемейство (5) Symphygnatinae. Сростночелюстные
жабы].
2(5) Сошниковые зубы есть.
3 ( 4) Язык с глубокой продольной бороздкой.
Xenobatrachus
6 видов.
4 ( 3) Язык без такой бороздки.
Callulops
1 вид.
5(2) Сошниковых зубов нет.
6(7) Верхнее веко с кожными придатками; на черепе про-
дольный гребень, разделяющий 2 группы височных
мышц.
Asterophrys
1 вид. I
7(6) Веко без придатков; продольного гребня на черепе нет.
8 ( 9) Концы пальцев расширены в небольшие диски. |
Mantophryne
1 вид.
9 ( 8) Концы пальцев не расширены.
Xenorhina
1 вид.
Ю ( 1) Верхнечелюстные кости не соприкасаются.
11 (12) Грудной пояс соединяется с черепом [подсемейство (13)
Hemisinae. Лягушки-поросята].
Hemisus
2 вида, Южн. и тропич. Африка.
12 (И) Грудной череп не соединяется с черепом, если же с0' ]
опыиартга тп ипшиитт и ппдпппямпппгп ХПЯШЯ Н€Т*
t'Ol
Рис. 21. Ареалы семейств: 1 — Brevicipitidae, 2 -• Polvoedatkb*
’WWFW
13
14
IS
16
17
18
19
20
21
22
23
24
25
26
27
28
29
104
(хнлиим v каждой стороны разбит на 2 куеочмл; Меда
гаскар {подсемейство (2) Rbombophrynlftae Ромбов*»
ине жабы)
(25) Зубы в верхней челюсти есть
(24) Зрачок горизонтальный.
(23) Сошниковые зубы имеются.
(22) Сошниковые зубы в виде 2 рядов.
(21) Ряды сошниковых зубов длинные.
(20) Грудина большая; ключица достигает лопатки.
Mantlpus
1 вид.
(19) Грудина маленькая; ключица не достигает лопатки.
Platyhyla
2 вида.
(18) Ряды сошниковых зубо® короткие.
Platypelis
• 2 вида.
(17) Сошниковые зубы в виде одного короткого поперечного
ряда.
Cophyla
1 вид.
(16) Сошниковых зубов нет.
Anodontohyla
1 вид.
(15) Зрачок вертикальный.
Plethodontohyla
4 вида.
(14) Обе челюсти беззубые.
(27) Ключицы имеются.
Rhombophryne
1 вид.
(26) Ключиц нет.
Stumpffia
1 вид.
(13) Сошник с каждой стороны цельный.
(30) Между последними двумя фалангами каждого пальца
по вставочному хрящу [подсемейство (8) Phrynome-
Нпае].
Phrynomerus
5 видов, Африка к югу от Сахары.
-m (2®) Вставочных хрящей нет.
Xi (36) Два первых позвонка сращены (подсемейство (I!)
э‘ Brevlclpitinae. Короткоглавы].
42 (35) Задняя нога с 5 пальцами.
^3 (34) Крестцовый позвонок сращен с хвостом.
Brevlceps
6 видов, Африка.
34 (33) Крестцовый позвонок свободный.
Spelaeophryne
1 вид, Африка.
35 (32) Задняя нога всего с 4 пальцами, из которых 2 сильно
4 редуцированы.
Didynamipus
1 вид, Камерун, Фернандо По.
«6 (31) Такого сращения нет.
37 (40) Первый палец передней ноги сильно редуцирован,
д В. Африка [подсемейство (12) Hoplophryninae].
38 (39) Ключица узкая, но длинная.
Parhoplophryne
1 вид.
39 (38) Ключица редуцирована до узелка.
Hoptophryne
2 вида.
40 (37) Первый палец передней ноги не редуцирован.
41 (60) Предгрудина имеется, если же отсутствует, то барабан-
ная перепонка не видна, и концы пальцев расширены.
42 (53) Ключицы имеются, если же отсутствуют, то нет и пред-
грудины.
43 (52) Сошники большие, охватывающие хоаны почти со всех
сторон, за исключением их наружного края [подсемейст-
во (1) Dyscophinae. Красные жабы].
44 (47) Зубы в верхней челюсти есть.
45 (46) Грудина большая.
Dyscophus
6 видов, Мадагаскар.
46 (45) Грудина в виде маленькой хрящевой пластинки.
Calluella
1 вид, Южн. Китай, Индия, Суматра.
47 (44) Обе челюсти беззубые.
48 (51) Предгрудина имеется; поперечные отростки крестцового
позвонка сильно расширены.
105
49 (50) Зрачок горизонтальный, концы пальцев рвеогврмц*
Callulina
I пил, Танганьика.
50 (49) Зрачок вертикальный; концы пальцев не paemepewi
Pseudohemlsua
I вид. Мадагаскар
51 (48) Предгрудины нет; поперечные отростки крестцового
звонка расширены слабо.
Scaphlophryne
1 вид, Мадагаскар.
52 (43) Сошники редуцированы до 2 маленьких кусочков меж.
хоанами [подсемейство (10) Melanobatrachinaej.
Melanobatrachus
1 вид, Индия.
53 (42) Ключиц нет; предгрудина имеется.
54 (57) Челюсти беззубые; конечные фаланги Т-образные (под
семейство (9) Kaloulinae, Азиатские бычьи лягушки),
55 (56) Сошники большие, простирающиеся до хоан.
Kaloula
11 видов, Южн. и Вост. Азия, Малайский ар
хипелаг.
56 (55) Сошники маленькие, не доходящие до KOHita.
Ramanella
1 вид, Индия.
57 (54) Верхняя челюсть обычно с зубами; конечные фаланги
иной формы [подсемейство (14) Cacosterninae).
58 (59) Поперечные отростки крестцового позвонка расширены
сильно.
Cacosternum
2 вида, Южн. и Вост. Африка.
59 (58) Поперечные отростки крестцового позвонка расширены
слабо.
Anhydrophryne
1 вид, Южн. Африка.
60 (41) Предгрудины нет, если же есть, то барабанная перепон-
ci /пт !£а яв9твенная, и концы пальцев не расширены.
о1 (90) Сошники большие, охватывающие хоаны со всех стопой
исключая их наружный край. н ’
106
62(69) Ключицы я предвранояые хряшя имеются (подсемеАгтяо
(3) Sphcnopnryninae).
Сошниковые зубы имеются
(64)
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
Uophryne
3 айда, Новая Гвинея
(63) Сошниковых зубов нет.
(68) Ключица достигает лопатки.
(67) Концы пальцев расширены.
Sphenophryne
11 видов, Целебес, Калимантан (Борнео), Но-
вая Гвинея, Квинсленд.
(66) Концы пальцев не расширены. (
Oxydactyla
1 вид, Новая Гвинея.
(65) Ключица не достигает лопатки.
Oreophryne
9 видов, Новая Гвинея, Ару, Молуккские
острова.
(62) Ключицы и, обычно, предврановые хрящи отсутствуют
[подсемейство (4) Cacopinae].
(73) Зубы в верхней челюсти есть.
(72) Задний конец языка загибается складчатой сумкой.
Colpoglossus
1 вид, Калимантан (Борнео).
(71) Такой сумочки нет.
Calliglutus
1 вид, Калимантан (Борнео).
(70) Зубов в верхней челюсти нет.
(75) Зубы в нижней челюсти есть.
Geriyophryne
1 вид, юго-восточный остров у Новой Гвинеи.
(74) Обе челюсти беззубые.
(89) Грудина есть.
(84) Зрачок горизонтальный.
(81) На нёбе 1 складка.
(80) Задние ноги с перепонкой между пальцами.
Cophixalus
1 вид, Новая Гвинея.
(79) Задние ноги без перепонки.
107
Ml
я»
(*м> If* нёбе 2 складня
(Ml Обе складки e «убами
О видов. Понам Гвяивя. Молумисяя,
M (R2>
Только ладная складка с дубами
CHooraplifyiw
I аил. Новая Гвинея
М (77) Зрачок не горизонтальный.
86 (86) Зрачок круглый.
Phrynomantis
4—о видов. Африка. Амбониа.
86 (85) Зрачок вертикальный.
87 (88) Небная складка одна.
CMyphofftossus
I вид, Индия.
88 (87) Нёбных складок две.
Cacopus
2 вида. Индия.
89 (76) Грудины нет.
Aphantophryne
I вид. Новая Гвинея.
90 (61) Сошники маленькие, охватывающие только передний ।
внутренний край хоаны.
9) (102) Ключица и предврановые хрящи имеются (подсемейство
(6) Kalophryninae].
92 (93) Предгрудина имеется; зрачок горизонтальный.
Kalophrynus
8 видов, Мадагаскар, Южн. Азия, Индо-мв'
лайский архипелаг.
93 (92) Предгруднны нет; зрачок не горизонтальный.
94 (95) Барабанная перепонка явственная.
Otophryne
1 вид, Гвиана.
95 (94) Барабанная перепонка не вндна.
96 (99) Нёбная складка одна.
97 (98) Пальцы передних ног с перепонкой.
10В
Nectodactylue
I вид, Бразилия.
до (97) Перепонки на передних ногах нет.
Chiasmoclels
1 вид, Парагвай.
ал ( 96) Нёбных складок две.
100 (101) Зрачок вертикальный; вокруг глаза нет окостенелого
кольца.
Hypopadius
13 видов, от Техаса до Гватемалы.
101 (100) Зрачок круглый; вокруг глаза окостенелое кольцо.
Stereocyclops
1 вид, Бразилия.
102 ( 91) Ключиц и предврановых хрящей нет [подсемейство (7)
Microhylinae. Азиатско-американские узкороты).
103 (104) Имеется тонкий предврановый хрящ.
Gastrophrynoides
1 вид, Калимантан (Борнео).
104 (ЮЗ) Предвранового хряща нет.
105 (106) Зрачок горизонтальный.
Phrynella
2 вида, Малайский архипелаг.
106 (105) Зрачок не горизонтальный.
107 (110) Зрачок круглый.
108 (109) Лопатка соединена с черепом.
Dasypops
1 вид, Бразилия.
109 (108) Лопатка не связана с черепом.
Microhyla.
29 видов, Южн. и Вост. Азия, Малайский ар-
хипелаг, обе Америки.
ПО (107) Зрачок вертикальный.
Ctenophryne
1 вид, Колумбия.
Большинство представителей 2—4 см длиной (пределы 15—
100 мм). Образ жизни древесный (Didynamipus, Mantipus, Pla-
tyhyla и др.) или роющий (Cacopus, Glyphoglossus, Breviceps,
Spelaeophryne и др.), в связи с чем на ногах иногда развиваются
лопатообразные наросты. Самки Hoplophryne кладут яйца меж-
ду листьями бананов или в старые бамбуковые стволы. Самка
109
Hemisus mar mor at us прикрывает яйца брюхом, кожа на
ром краенест и вздувается. Головастики вылупляются значит?.^
но развитыми. Многие проходят все развитие внутри я*
цевой оболочки (Xenobatrachus, Hylophorbus и др.). У Mwitopfatf
пе robusta яйца соединены четковидно. Самец охраняет отложи
ные самкой яйца, обнимая их передними лапами. Все развит»
проходит в яйце, причем органом дыхания головастика является
хвост. Интересна красная окраска представителей родов Dytcf,
ph us, Calluella и Calliglutus.
ЛИТЕРАТУРА
Банников А. и Денисова М. Очерки по биологии земноводных, ,М
1956.
Терентьев П. Лягушка, М., 1950.
Boulenger G. Catalogue of the Batrachia Salientia s. Ecaudada in the
Collection of the British Museum, London, 1882,
Boulenger G. The Tailles Batrachians of Europe, London, 2 vols. 1897—1898.
Boulenger G. Les Batraciens et principalement ceux d’Europe, Paris, 1910.
Griffiths I. Status of Protobatrachus massinoti, Nature, 1956, 177: 342—343.
Noble G. The Phylogeny of Salientia, Bull. Amer. Mus. Nat. Hits., 1922,
46: 1—87.
Piveteaux J. Anoura (in: Traite de Pal6ontologie, Paris, 1955, 5: 250—274).
Г лава шестая
ПРОИСХОЖДЕНИЕ ПРЕСМЫКАЮЩИХСЯ И
ВЫМЕРШИЕ ГРУППЫ
Пресмыкающиеся (Reptilia, от «reptare»— ползать на брю-
хе), возникшие из примитивных стегоцефалов в верхне-каменно-
угольное или нижне-пермское время, представляют собой второй
шаг позвоночных на суше, шаг, приведший к освоению сухих ста-
ций. Последнее сделалось возможным благодаря сильному раз-
витию рогового слоя кожи, что обусловило почти полное исчезно-
вение кожных желез й кожного дыхания. Переход взрослых осо-
бей к наземному об(разу жизни привел к исчезновению наруж-
ного оплодотворения и развитию как плотных, часто пропитан-
ных известью, яйцевых оболочек, так и особых зародышевых
образований—амниона, серозы, аллантоиса. Все это сопровожда-
лось другими коррелятивными изменениями, позволяющими рас-
сматривать возникновение пресмыкающихся как ароморфоз.
Подобная перестройка организации позволила пресмыкающимся
не только занять все сухие стации, но уже в пермское время на-
чать интенсивную конкуренцию с земноводными. Юра и нижний
мел представляют собой время господства пресмыкающихся
(в том числе вторично-водных), но уже к концу мела многие из
этих групп начинают клониться к упадку. Начало кайнозоя от-
мечено отсутствием господствующих групп пресмыкающихся и
адаптивной радиацией млекопитающих.
Основные отличия современных пресмыкающихся от земно-
водных— сухая кожа и отсутствие водной личинки. Более под-
робно строение пресмыкающихся можно характеризовать сле-
дующими признаками:
1) Череп сочленяется с позвоночником одним затылочным
мыщелком, образованным преимущественно основной затылочной
костью; 2) нижняя челюсть образована несколькими костями и
сочленяется с черепом при посредстве квадратной кости;
3) имеется слуховая косточка, входящая в овальное окно; 4) ко-
нечности в основе наземного, пятипалого типа, но могут внешне
превращаться в ласты или редуцироваться; 5) подъязычный
111
игра является черепным нервом; О) репря формирует неги я му
грудину; 7) подпэлошнО'Крестцовое сочленение лежит c»j
вертлужной впвдмиы. 8) поверхность теля почти совершеммо Л1
шгиа желех и покрыта чешуей или щитками. 9) температура тг«
переменная (пойкилотермные); 10) эритроциты имеет ядра
овальны; ! I) сердце состоит из 2 предсердий и одного желудгл
кв. но не вполне разделенного на две части; 12) дуги аорты да»
13) лыхаиие легочное; жабры полностью отсутствуют; 14) кожнм|
орган водного чувства («боковая линия») полностью отсутству
ет; 15) почки метанефрические; нефростомов нет; 16) веет и
имеется типичная клоака; 17) яйца меробластические; 18) опло
дотворение внутреннее, всегда с помощью копулятивного орган;
(отсутствует только у Sphenodon); 19) в онтогенезе формирует»
амнион и аллантоис.
Признаки (1). (5), (6), (7), (8), (13). (14), (15), (17) и (19|
отличают пресмыкающихся от земноводных. Признаки (9), (11)
и (12) отличают их и от птиц, и от млекопитающих, а признаки
(2), (7) и (10) особо важны при различении от млекопитающих
Интересно, что признак (6) позволил пресмыкающимся, птицам
и млекопитающим развить новый тип дыхания при посредстве
ритмических движений грудной клетки. К сожалению, строение
мягких органов, равно как и способ размножения ископаемых
остаются нам в большинстве случаев неизвестными. Поэтому
разграничение вымерших пресмыкающихся и вымерших земно-
водных подчас весьма затруднительно. Особенно ярко это высту-
пает при рассмотрении группы Seymouriamorpha. Остатки этих
юрско-пермских животных стали известны впервые в 1847 г.
(Khinosaurus из-под Ульяновска), но особое внимание на них
обратили после открытия в 1904 г. рода Seymouria (Техас). Сей-
час их различают 12 родов и разделяют на семейства Seymo-
uriidae и Lanthanosuchidae. Это были животные среднего разме-
ра (длина черепа 10—16 см) с ящерообразным телом, полуэм-
боломерными позвонками, складчатыми зубами и пятипалой
передней конечностью. Мозаичность их морфологии заставляла
исследователей относить их то к низшим пресмыкающимся
(Абель, Броили, Кэз, Виллистон, Ромер), то к земноводным
(Брум, Сушкин, Вудвард, Быстров). Ячеистая структура крыши
их черепа, сейсмосензорные каналы и, по-видимому, личиночная
стадия в развитии заставляют причислять Seymouriamorpha к
земноводным (Пивето, 1955, Хюне, 1956), но, по сути, это типич-
ная промежуточная группа, которой было даже предложено
дать специальное название «Batrachosauria» (Ефремов, 1946).
Любопытно, что Lanthanosuchus (Средняя Волга) имел височное
отверстие, конвергентное с таковым у пресмыкающихся.
Различать современные группы (черепахи, клювоголовые,
змеи, ящерицы и крокодилы) не трудно, но систематика ископае-
мых очень сложна. В 1830 г. Мейер попытался разместить иско-
паемых в системе Броньяра, поместив все известные тогда вымер-
112
шяе формы в отряд ящериц Чсрг» 5 лгт Ьлгмвиль выделял д
качестве самостоятельных отрядов ихтиозавры* и плезиозавров,
а позднее Оуэн довел число отрядов пресмыкающихся до 11 Кол
обратил внимание ня важность хяряктера строения и нрикреплг
ния квадратной кости — неподвижная она (мояммостнлия) или
подвижная (стрептостилия). В дальнейшем особое значение
получило строение височной области черепа. Древнейшие пре-
смыкающиеся имели черепа, у которых все кости черепа образо
вывали сплошную кровлю. Потом какой-то фактор вызвал особо
сильное развитие мышц (в частности, височной), двигающих
нижнюю челюсть. В связи с этим началось образование «окон»
в височной области черепа — появление подобных отверстий дает
дополнительное место сократившимся мышцам. Костные мости-
ки между этими «окнами» («ямками») или около них получили
название височных «дуг». В латинских названиях групп употреб-
ляется соответственно термин «apsis» — дуговидная арка,—
заимствованный из архитектуры. Развитие всех этих образований
шло несколькими путями. В наиболее сложном случае возникают
2 «окна» — верхнее и нижнее и, соответственно, 2 дуги — верхняя
(из заднелобной или заглазничной и чешуйчатой кости) и ниж-
няя (из скуловой и квадратноскуловой). Бывает и смешанная
дуга, состоящая из скуловой и чешуйчатой кости. Такая дуга
встречается только при одной ямке, которую принято считать
верхней, если заднелобная или заглазничная кости участвуют в
образовании дуги. В прртивном случае ямку называют
нижней. Наблюдаются случаи слияния ямок и их исчезновения
(рис. 22).
У некоторых черепах развиваются вырезки в задней части
височного отдела крыши черепа. Их не принято считать за ви-
сочные ямки («ложные»). Строение височной области черепа по-
служило основой ряда классификаций (Осборн, 1903; Виллистон,
1925) и до сих пор сохранило первостепенное значение. Для си-
стематики имеет значение характер крепления зубов: сидящие в
лунках называются текодонтными, прирастающие к вершине
челюсти — акродонтными, а прирастающие к боку челюсти —
плевродонтными. Недавно возникли сомнения в монофиле-
тичносуи класса пресмыкающихся. Настаивают даже на от-
несение ихтиозавров к земноводным. Однако такая крайняя точ-
ка зрения не может быть признана справедливой. Не понятно, в
частности, почему ихтиозавры потеряли личиночную стадию, если
их предки не были пресмыкающимися. Ныне предпочитаются
смешанные классификации. Можно принять деление пресмыка-
ющихся на 3 подкласса (рис. 23) и 18 отрядов, которые различа-
ются по следующим особенностям:
1(4) Череп массивный, без височных ямок, если же височная
ямка есть, то имеется кольцо окостенений в склеротике
глаза при наземных конечностях [подкласс А — Анар-
sida],
9 Л. В. Тероиъп ИЗ
2 ( 3) Ветви нижней челюсти свободны. имеете* яеяврвв» u
меииое отверстие; тело голое или с костимым влвгтг,
ками; пргдкреетиовых позвонков 23—26
I. CotytotBtirla
3 ( 2) Ветви иижней челюсти ерошены; теменного отвереп»
нет; тело обычно покрыто панцирем; преямреспияыь
позвонков 18.
II. Chelonla
4(1) Имеется 1—2 височных ямки или череп ажурный, яд»
тело покрыто черепахоподобным панцирем, не закрыв*
ющим, однако, крестец.
Рис. 22. Схемы типа черепа по строению височноб области. А — без дуг
и ямок (анапсидный); Б — с двумя ямками и двумя дугами (диапсядный):
В— с одной замкнутой и одной незамкнутой ямкой (диапсидный упрощен-
ный с исчезновением нижней дуги); Г — с одной ямкой и одной сме-
шанной дугой (синапсидный)
5 (18) Височная ямка одна, всегда замкнутая или ее совсем
нет [подкласс В — Synapsida]-
6 (11) Височная ямка нижняя: заглазничная или заднелобная
кость не участвуют в ограничении ее снизу.
7(8) Сухопутные — конечности ходильные; лобковая и седа-
• лищная кости чаще слиты.
III. Theromorpha
8(7) Водные — конечности плавательные; лобковая и седа-
лищная кости раздельны.
9 (10) Предчелюстные кости не касаются лобных; зубы мно-
гочисленные, длинные; глазница в последней четверти
черепа.
114
IV, AUteMwrla
10 ( 9) Предчелюстные костя яке «юте я лобнмж. зубы мелочи
еденные, конические; глятииил к яечлле второй оотоом
иы черепе.
V. ThalattOMurte
11(6) Височная ямка верхняя: ааглааиичиая или аадиелобаая
кость участвует в ограничении ее симу ялр височной
ямки совсем нет.
12 (15) Предкрестцовых позвонков 24—30.
13 (14) Обычно наземные; шея длинная; зубы конические или
вытянутые в поперечные пластинки; панциря ие бывает.
VI. Protorosaurla
14 (13) Водные; шея короткая; задние зубы обычно увеличены
и уплощены; тело часто одето панцирем.
VII. Placodontia
15 (12) Предкрестцовых позвонков 40—105.
16 (17) Имеется поперечная кость; позвонки с плоскими перед-
ними и задними поверхностями (плятицельные) или
слабо двояковогнутые; шея обычно длинная; хвост от-
носительно короткий; непарных плавников нет.
VIII. Sauropterygia
17 (16) Поперечной кости нет; позвонки явно двояковогнутые;
облик рыбовидный — короткая шея снаружи не замет-
на, хвост, длинный, имеются спинной и хвостовой плав-
ники.
IX. Ichthyosauria
18 ( 5) Височные ямки две, или 1 замкнутая и 1 незамкнутая,
или одна большая незамкнутая и череп ажурный [под-
класс С — Diapsida].
19 (34) Передние конечности не превращены в летательный ап-
парат; ветви нижней челюсти свободные.
20 (33) Квадратная кость неподвижная (монимостилия).
21 (22) Предкрестцовых позвонков много (около полусотни).
XII. Pleurosauria
22 (21) Предкрестцовых позвонков 23—34.
23 (30) Предглазничного отверстия нет, если же есть, то, брюш-
ные ребра отсутствуют.
24 (27) Ребра одноголовчатые; обычно имеются непарное.темен*
ное отверстие и ключицы.
115
\ i /
i i i i
\ | I }
SavroptenW*
plecodoatio }
V I
*tesa$auria
Chelonla
Cotylosoona
— Anapsida.
----2 I
--- 3 I
Ряс. 23. Филогеяия отрядов пресмыкающихся. Подклассы: /
2— Synapsida, 3—Diapsida
Theromorpha
25 (26) Зубы в лунках (текодонтные).
X. Eosuchia
26 (25) Зубы приросшие к вершине челюсти (акродонтные).
XL Rhynchocephalia
27 (24) Ребра двухголовчатые; теменного отверстия и ключицы
нет.
28 (29) Таз трехлучевой: лобковая кость простая; брюшные
ребра имеются; предчелюстная кость с зубами.
XV. creeodllle
29 (28) Таз, как правило, четмрехлучевой. лобковая кость раз-
делена на 2 ветви; брюшных ребер нет; лредвелюстивв
кость чаше без зубов.
XVII. Ornlthlschla
30 (23) Предглазничное отверстие и брюшные ребра имеются
31 (32) Вертлужная впадина без отверстия; непарное теменное
отверстие чаще имеется.
XIV. Thecodontia
32 (31) Вертлужная ©падина с отверстием; теменного отверстия
чаще нет.
XVI. Saurischia
33 (20) Квадратная кость подвижная (стрептостилия).
XIII. Squamata
34 (19) Последний палец передней конечности очень сильно уд-
линен и служит для прикрепления летательной перепон-
ки; ветви нижней челюсти обычно слиты.
XVIII. Pterosauria
ПОДКЛАСС A —ANAPSIDA. БЕЗДУГНЫЕ
I. ОТРЯД COTYLOSAURIA. ЦЕЛЬНОЧЕРЕПНЫЕ
Название отряда предложено Копом в 1881—89 гг. Эта груп-
па, исходная для всех прочих пресмыкающихся, взяла начало
по-видимому, от Palaeogyrinidae. Первоначально это были вод-
ные хищники (Limnoscelidae), постепенно ставшие наземными
и перешедшие к питанию насекомыми, червями и др. (Bolosauri-
dae). Некоторые виды начали употреблять и растительную пищу
(Procolophonidae), другие целиком перешли на питание водной
растительностью (Pareiasauridae), наподобие современных беге-
мотов. Размеры возросли с 1,5—1,8 м (Limnoscelis) до 3 м
(Pareiasauridae) (рис. 24). Основные семейства различаются так:
1 ( 4) Квадратная кость горизонтальная; ушной вырезки нет
[подотряд Captorhinomorpha].
2 ( 3) Лобные кости не доходят до глазницы, ибо предлобные
и заднелобные соприкасаются.
Limnoscelidae
2 рода, пермь Сев. Америки.
3 ( 2) Лобные кости доходят до глазницы.
П7
CaptodHfitdee
7- 12 родов. пгрмь Сев Америки
. . п Каалр*™** хост*» вертикальная; обычно имеется увм«
’ 1 вырезка (подотряд Diadectomorpha).
_ । Ворхнгвнсочная кость имеется; ушная вырезка хлроиц.
6 1 заметна.
п и Theromorpha:
Рис. 24. Представители Seymouriamorpha, Cotylosauna ___________pareiasaurus,
Z —скелет Seymouria baylorensis, 2 — Limnoscelis palustrts, a
11R
4— Ophlacodon, 5—череп Seymouria сверху, 6—Edaphosaurus
7 — Galechlrus, в — Keratocephalus, 9 — Cynognathus
119
• ) fr*r*M** КОСТЬ t*r*nc| ЧГШУЙЧЯТОЙ. Г.1Л*Н9Я4 'Pt**,
f, i '' чсяа ctt«jy яерхпгчглюгтплй я скуловой »or *»w <►
0К(Х^<*нгиий ж <• мл грот и же глктк, ок ня ня йй^яй»
Diadeetlde*
2-4 рода, пермь Сев Америки
-1 Теменная не клслгтся чешуйчатой, гл ангина лчраажч»^
7 < ™ снизу только скуловой костью; окостенений в скжротяю
глаза нет; между скуловой, чешуйчатой я каадратяож»
лоаымн костями небольшое окно.
Botosauridae
I род. пермь Сев. Америки.
8 < 5)
9 (1°>
Верхневисочной нет; ушная вырезка слабая или сжрм'»
другими костями.
Лобная кость не касается глазницы.
Pareiasauridae
8—11 родов, пермь — триас Европы и Африки
•л / 9) Лобная касается глазницы.
И (12) Глазница снизу ограничена скуловой и заглазничной
11 ' костями.
Procolophonidae
10—12 родов, пермь Африки и Европы, триас
Сев. и Южн. Америки.
12 (Н) Глазница снизу ограничена только скуловой.
Millerettidae
3 рода, пермь Южн. Африки.
II. ОТРЯД CHELONIA. ЧЕРЕПАХИ
(см. главу 7)
ПОДКЛАСС В — SYNAPSIDA. СЛОЖНОДУГИЕ
III. ОТРЯД THEROMORPHA. ЗВЕРОПОДОБНЫЕ
Заполняют пробел между пресмыкающимися и млекопитаю-
щими: в филогенезе этой группы идет дифференцировка зубов,
образуется вторичное нёбо, зубная кость получает исключитель-
ное значение в нижней челюсти и т. п. Возможно возникновение
теплокровности. Ближе всех к Млекопитающим стоят Ictidosau-
ria. Возникнув в верхнем карбоне, звероподобные достигли
особенного расцвета в пермское время и угасли в триасе.
Адаптивная радиация их была очень широка: по-видимому, самые
примитивные были рыбоядами (Ophiacodontidae, Eothyridae), а
большинство—настоящими хищниками (Varanosauridae, Gorgo-
nopsidae, Titanosuchidae и др.). Встречалось, однако, немало и
120
растительноядных (Edaphosaurldae, TapInocrphalM*# и др) Эяо-
логическую нишу болотных травоядных занимали Dteyno4orWM«e.
a Lystrosaurus называют иногда «пресмыкавшимся ткмемем»
У Euchambresldae подозревают наличие ядовитой железы в веря*
ней челюсти. Размеры колебались от крысы до быка и более
(Jnostrancevia 3 м длиной, Jonker la 4 м, Ophlacodon до 5 м)
Уже описаны сотни видов этого отряда. Брум допускает, что их
существовало 20000—50000. Основные пруппы различаются так:
t (16) Верхнетеменная кость имеется; затылочная поверхность
наклонена вперед; ноздри направлены в стороны (подот-
ряд Pelycosauria].
2 ( 5) Челюстной сустав примерно на уровне зубного ряда; ниж-
няя челюсть по высоте ровная, без ясного венечного от-
ростка; вставочные элементы позвоночника большие,
3(4) Череп длинный; клыки небольшие.
Ophiacodontidae
4 рода, карбон — пермь, Сев. Америка.
4(3) Череп короткий; клыки сильно увеличены.
Eothyridae
5—6 родов, пермь Европы и Сев. Америки.
5 ( 2) Челюстной сустав ниже уровня зубного ряда; нижняя
челюсть заметно‘повышается в задней половине; вста-
вочные элементы небольшие.
6(9) Тела спинных позвонков сжаты с боков, с брюшным ки-
лем; лопатка длинная.
7(8) Крестцовых позвонков 2; череп низкий.
Varanopsidae
3—7 родов, пермь Сев. Америки, ?Африки,
?Европы.
8(7) Крестцовых позвонков 3; череп высокий.
Sphenacodontidae
12—14 родов, карбон — пермь (?нижний три-
ас) Европы и Сев. Америки.
9(6) Тела спинных позвонков бочонковидные, снизу закруг-
ленные, без киля; лопатка короткая.
10 (13) Остистые отростки спинных позвонков чрезвычайно вы-
сокие.
И (12) Остисгые отростки спинных позвонков имеют попереч-
ные выросты.
Edaphosauridae
1 род, пермь — нижний триас Европы и Сев.
Америки.
12 (11) Поперечных выростов на остистых отростках нет.
121
UpeouvrldM
I род, пермь Сев Америки
13 (Ю) Остистые отростки спинных позвонков обычной, мбв«
шой длины.
14 (15) Зубы острые; ноздри обычной величины.
Nltotauridae
2—4 рода, пермь Сев. Америки.
15 (14) Зубы тупые; ноздри очень большие.
Ca&eidae
3—5 родов, пермь Сев. Америки.
16 ( 1) Верхнетеменной кости нет; затылочная поверхность вер
тикальная или наклонена назад; ноздри направлена
более или менее вверх.
17 (34) Предтеменная и заднелобная кости имеются.
18 (23) Заглазничная часть черепа короткая; вторичного нёба
нет. с
19 (22) Брюшные ребра имеются; шейных позвонков о; зуоы
однообразные [подотряд Dromasauria].
20 (21) Зубы имеются; задний угол нижней челюсти почти го-
ризонтальный.
Galechiridae
2 рода, пермь Южн. Африки.
21 (20) Зубов нет; задний угол нижней челюсти загнут книзу.
Galeopsidae
1 род, пермь Южн. Америки.
22 (19) Брюшных -ребер нет; шейных позвонков 7; увеличенные
клыки ясно отличаются от других зубов [подотряд
Gorgonopsida].
Gorgonopsidae
Более 73 родов, пермь Южн. Африки и Ев-
ропы.
23 (18) Заглазничный череп длинный; вторичное нёбо более или
менее развитое [подотряд Anomodontia].
24 (25) Один или несколько рядов зубов.
Endothlodontidae
33 рода, пермь Южн. Африки.
25 (24) Обычно имеются только клыки или зубов совсем нет.
26 (29) Череп очень короткий и широкий: ширина его иногда
больше длины.
27 (28) Теменная область крыши черепа очень широкая.
122
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
3 род*, пермь Юж я Афряяя
(27) Теменная область небольшая
GeMdtdae
1 род. пермь Шотляидяи.
(28) Ширина черепа «сегда заметно меньше его алиям
(31) Ноздря лежит близ глазницы.
Lystrosauridae.
1 род, триас Южн. Африки и Китая.
(30) Ноздря между концом морды и глазницей.
(33) Носовые кости по длине примерно равны иля меньше
лобных.
Dicynodontidae
29 родов, пермь — триас Южн. Африки, Южв.
Америки, Европы и Азии.
(32) Носовые кости заметно длиннее лобных.
Kannemeyeridae
4 рода, триас Африки, Сев. Америки и Китая.
(17) Предтеменной нет; заднелобная чаще отсутствует.
(56) Вторичного нёба нет, если же есть, то заглазничная
часть черепа короткая и присутствует квадратно-скуло-
вая кость.
(43) Челюстной сустав ниже уровня зубного ряда.
(42) Поперечное крыло крыловидной кости большое [под-
отряд Dinocephalia. Пермь].
(39) Клыков нет.
Tapinocephalidae
4 рода, Африка, Европа.
(38) Клыки имеются.
(41) Череп вздут за ноздрями и над глазницами.
Bumetidae
2 рода, Африка.
(40) Вздутий черепа нет.
Titanosuchidae
6 родов, Европа, Африка.
(37) Поперечное крыло крыловидной кости маленькое [под-
отряд Rubidginia].
Rubidginidae
3 рода, пермь Южн. Америки.
123
43 (36) Челюстной сустав не ниже уровня зубов (подотряд
rocephallft, дмроголовые].
44 (47) Зубы сзади клыков отсутствуют или плохо ре ввиты; овл
сзади нёбной кости отсутствует или маленькое.
45 (46) Непарное теменное отверстие имеется; глубокой
глазничной ямки в верхнечелюстной кости нет.
Whaitsldae
9 родов, пермь Южн. Африки.
46 (45) Непарного теменного отверстия нет; верхнечелюстная
кость имеет перед глазницей глубокую ямку.
Euchambresidae
1 род, пермь Южн. Африки.
47 (44) Зубы сзади клыков хорошо развиты, если же отсутст-
вуют, то окно сзади нёбной кости хорошо развито.
48 (55) Крыша черепа в районе теменных костей сужена.
49 (50) Височная ямка примерно равна глазнице; зубная кость
очень длинная (примерно 84% длины черепа).
Lycideopsidae
1 род, пермь Южн. Африки.
50 (49) Височная ямка в длину заметно больше глазницы; зуб-
ная кость менее длинная (примерно 70—79% длины
черепа).
51 (52) Нёбные кости вытянутые и спереди заостренные.
Ictidosuchidae
14 родов, пермь .Южн. Африки.
52 (51) Нёбные кости короче и спереди не заостряются.
53 (54) Длина предглазничной кости черепа содержится в его
общей длине 1,8—2,1 раза.
Pristerognathidae
31 род, пермь Южн. Африки, Европы и Китая.
54 (53) Длина предглазничной части черепа содержится в дли-
не черепа 2,4—2,5 раз.
Alopecopsldae
11 родов, пермь Южн. Африки.
55 (48) Крыша черепа в районе теменных костей широкая. ,
Scaloposauridae
26 родов, пермь Южн. Африки и Урала.
56 (35) Вторичное небо есть.
57 (58) Затылочный мыщелок один (подотряд BauriamorpnaJ.
Bauriamorphldae
11 родов, Триас Южн. Африки.
194
(57) Затылочных мыщелков два.
(70) Заглазнмчная дуга полная (подотряд Cynodonta Соба
козубые).
(60 Вторичное нёбо по средней линии несомкнутое: между
внутренними концами верхнечелюстных я нёбных костей
остается продольная щель.
Procynodontidae
8 родов, пермь—триас Южн. Африки.
JS13
63
64
(63)
Вторичное небо цельное, без щели.
Между верхнечелюстной и скуловой костями имеется
«ступень» (резкая разница в уровне при взгляде сбоку).
Зубы многовершинные, ширина их горизонтальной про-
екции не больше длины.
Chiniquodontidae
2 рода, триас Южн. Америки.
Зубы с уплощеной верхней поверхностью и бугорками
по краям, ширина их больше длины.
Transversodonti dae
4 рода, триас Южн. Америки.
65
66
(62) Между верхнечелюстной и скуловой костью «ступени»
нет.
(67) Сзади клыков зубы равномерные или убывают назад.
Cynosuchidae
12 родов, пермь—триас Южн. Африки и Ев-
ропы.
67
(66)
68 (69)
69 (68)
Сзади клыков зубы по направлению назад увеличи-
ваются.
Зубы многовершинные, ширина каждого из них в гори-
зонтальной проекции не больше его длины.
Cynognathidae
5 родов, триас Южн. Африки.
Зубы с небольшими бугорками по краям, низкие, шири-
на их больше длины.
Dtademodontldae
9 родов, пермь Европы и триас Южн. Африки.
70 (59) Заглазничная дуга отсутствует [подотряд Ictidosauria].
Ictidosauridae
Юродов, триас Южн. Африки, Европы; Сев.
Америки и Китая.
126
•V. ОТРЯД MCftOSAURIA СРДДМГЯИКРМ
nontmca прАсмыКАюшмяся иырябснят», лолпш +***
• <Цк~1,0 Д. ОбЛМК КрОКОДИЛООбрА 1ИЫЙ. <” ДЛИМЮТб
Ряс. Ж Пред[Стакители Mesoaauria, Thalattosauria и Protorosauria: I — Ме-
ЛО*аигив, W$g*ltisaurus,. ’5—* Protorosaurus, 4—5—череп Thalattosaurus
сбоку и CMpjcy, jp— нижняя челюсть Trilophosaurus сверху, 7 — Araeoscelis
ДО
м сжатым с боков хвостом (рис, 26). Косги зевгоподмя жлсюмык*
Йюрочены, но конечность в целом сонрлиила мвэемим* тмя
ежду пальцами предполагают наличие переломом Н^мотормм
ребра были бамакообраэно расширены. Всего 3 рода Держались
в пресноводных водоемах пермо-карбона Бразилии
пит) и Южн. Африки (Mesosaurus, NoUoxaurux).
V. ОТРЯД THALATTOSAURIA МОРСКИЕ ЯЩЕРЫ
Морские формы с веслообразными конечностями и кольцом
окостенений в склеротике глаза. Длина до 2 м. Два рода в верх*
нем триасе Калифорнии (Thalattosaurus, Nectosaurus).
VI. ОТРЯД PROTOROSAURIA. ДРЕВНИЕ ЯЩЕ£Ы
Группа, относительно объема и места которой существуют
большие разногласия. Большинство наземные, ящеровидные, но
Protorosauridae, видимо, были полуводными. weigeltisaurus,
вероятно, лазал. Длина от 0,3 до 2,5 м. Семейства характеризу-
ются так:
1(6) Зубы конические или цилиндрические.
2(3) Конечности очень тонкие и длинные: длина плечевой
кости больше длины черепа.
Araeoscelidae
1—2 рода, пермь Техаса.
3(2) Конечности короче. Длина плечевой кости короче черепа.
4(5) Задние кости черепа образуют зубчатый воротничок,
напоминающий по строению хамелеона; окостенений в
склеротике глаза нет.
Weigeltisauridae
1 род, пермь Европы.
5(4) Сзади черепа подобия зубчатого воротничка нет; имеет-
ся кольцо окостенений в склеротике глаза.
Protorosauridae
1 род, пермь Европы.
6(1) Зубы в виде пластинок, поперечных длине челюсти.
Trilophosauridae
1 род, пермь Техаса.
По-видимому, к этому отряду принадлежат также Broomia
и Aphelosaurus.
VII. ОТРЯД PLACODONTIA. ПЛОСКОЗУБЫЕ
Возрастающее приспособление к морскому об|разу жизни и
питанию моллюсками и плеченогими приводит к изменению яще-
127
рообрахной формы тела (МуЬмНсшшлн) • ламакгкмощмиб»,.
f/’lecodw*), и наконец. я черепа «обратную (PiarvcMy*.
dua). На тале развивается панцирь, зубы уплощаются (рис д.
Длина примерно от 0.6 до 3,5 л. Вероятно, для кладям и« аын
дили на берег наподобие современных морских мерена*, 0 гр^4
се Европы установлены следующие семейства:
i ( 2) Все зубы однообразные заостренные (подотряд ННт^
соза uroidea].
Hetvetlcosauridae
1 род, Швейцария.
2(1) Задние зубы увеличены и уплощены.
3(6) Передние зубы удлиненные; кожный скелет отсутетвуе|
или слабо развит [подотряд Placodontoidea],
4(5) Морда более приостренная; зубов 46.
Paraptacodontidae
I род, Швейцария, Ломбардия.
5(4) Морда более тупая; зубов 32.
Placodontidae
1 род, Германия.
6(3) Предчелюстная кость с мелкими зубами, либо зубов на
ней вовсе нет (? клюв); кожный скелет хорошо развит
[подотряд Cyamodontoidea].
7 (10) Череп сверху треугольный; имеется большая височная
ямка; в каждой челюсти по несколько зубов.
8(9) Конец морды с небольшими зубами.
Cyamodontidae
1 род, Польша, Германия, Франция, Швей-
цария.
9 (8) Конец морды без зубов.
Placochelydae
1 род, Венгрия, Альпы, Ломбардия.
10 ( 7) Череп сверху прямоугольный; височная ямка вторично
закрыта; в каждой челюсти всего по 1 зубу.
Henodontidae
1 род, Германия.
VIII. ОТРЯД SAUROPTERYG1A. ЗМЕЕШЕИНЫЕ ЯЩЕРЫ
Появляются в триасе. Средней величины (Nothosauridae от
0,2 до 3 м), полуводные ящерообразные формы, но в мелу дости-
гают очень крупных размеров (Elasmosaurus до 13 м). Приспо-
собление к жизни в море вызывает сильное удлинение шеи
123
5 Рис. 26. Представители Placodontia, Sauropterygia и Ichthyosauria: / — Hydrotherasaurus alexandrae. _ / FlyM
3 3—Placochelys placodonta, 4 — череп Placochelys снизу, 5 — Lariosaurus balsami, 5 6 ~ плавммк.Е^М4иу«1^Лч
эволюция контура ихтиозавров (7 —• Cymbospondylus, 8 —«• Mixosaurus, 9— Stenopterygiua, ю- наммнетв1**'
(МОЖГГ В 3 прввлсходмтъ длину ГуЛОЯИЩЯ), уЖО<ММЯМЯ» 1«»
ста к преярященнг конечностей лястм (сопровождается пииту
фаланг ней). Туловище становится бочокковидиым я лолуяагсц
по контурам, как некоторые выражаются, «змея, продетая сюит
тело черепахи». Существует подозрение на яйцежияородиот
Различаемые семейства можно объединить в 2 подотряда
I (4) Крыловидные кости соединены друг с другом продола
ным швом по всей длине и лишены отверстий (подотряд
Nothosauria; триас Европы).
2 ( 3) Височные ямки большие; не менее б крестцовых позвоч
ков.
Nothosauridae
11 родов.
3 ( 2) Височные ямки маленькие; крестцовых позвонков 3—4
Pachypleurosauridae
5 родов.
4(1) Крыловидные кости расходятся друг с другом в своей
задней части и обычно несут окна [подотряд Pleurosau-
ria].
5(6) Ноздри очень небольшие и помещаются только между
предчелюстными и верхнечелюстными костями [ветвь
Pistosauridea].
Pistosauridae
1—2 рода, триас Германии.
6(5) Ноздри помещаются между предчелюстными, верхнече-
люстными и предлобными или слёзными костями.
7 (10) Зубы разнообразные; 13—27 шейных позвонков; пле-
чевая кость равна или короче бедреной [ветвь Pliosau-
roidea].
8 ( 9) Шейные ребра двухголовчатые; зевгоподий вытянутый.
Pllosauridae
6 родов, юра — мел Европы, Азии и Австра-
лии.
9(8) Шейные ребра одноголовчатые; зевгоподий короткий.
Polycotilidae
4 рода, мел Европы и Сев. Америки.
10 ( 7) Зубы однообразные, маленькие; шейных позвонков 24—
U ' 76; плечевая кость длиннее бедреной [ветвь Plesiosauroi-
deaj.
jl (12) Шейные ребра двухголовчатые; зевгоподий вытянутый.
Plesiosauri dae
12 родов, мел Европы.
12 (II) Шейные ребре олиоголоачаты*. ютомодей жлрогжже
Elasmatatirldae
13 ролов, мел Европы. Сев Америка в Нов
Зеландии
IX. ОТРЯД ICHTHYOSAURIA РЫВОЯКИРЫ
Происхождение вызывает значительные споры. Начинав ги
среднего триаса до среднего мела, эволюция шла с многочислен
ными параллелизмами. Приспособление к жизни в море, где за
нимали экологическую нишу дельфинов, дало, прежде всего,
изменение конечности наземного типа в гиперфалангический
многолучевой, а позднее — малолучевой плавник. Изменились
контуры тела, и возникли непарные плавники. Имелось кольцо
окостенений в склеротике глаза. Звук воспринимался костями,
как у китообразных (барабанной перепонки нет, стремечко утол-
щенное). На плавниках некоторых форм обнаружены чешуйки.
Длина тела от 1 до 16 м. Жили стадами, питаясь головоногими в
рыбой. Omphalosaurus питался раковинными моллюсками. Из-
вестны беззубые формы. Доказана яйцеживородность (1—
12 эмбрионов на самку). Установлены случаи поедания молоди.
Можно принять такие группы:
1 ( 4) К промежуточной кости запястья прикрепляются 2 пя-
стных кости [подотряд Latipinnati].
2(3) Длина лучевой или локтевой кости укладывается в дли-
не плечевой 1,4—1,7 раза.
Mixosauridae
1 род, триас Европы и Сев. Америки.
3(2) Длина лучевой или локтевой кости укладывается в дли-
не плечевой 2,3—5,0 раза.
Ichthyosauridae ( = Macropterygiidae)
6 родов, юра — мел Европы, Сев. Америки
и Индии.
4 ( 1) К промежуточной кости запястия прикрепляется всего
1 пястная кость [подотряд Longipinnati].
5(6) Зубы расположены в несколько рядов, тупые или мос-
товидные.
Omphalosauridae
3 рода, триас Европы и Сев. Америки.
6(5) Зубы однородные, острые.
7(8) Седалищная и лобковая кости разделены.
Schastasauridae
10 родов, триас — юра Европы и Сев. Америки.
131
В ( 7) Седалищиэи и лооко®*® кости глиты.
Stenopteryglldae
3 родя* *°Р® — Европы.
ПОДКЛАСС С - DIAPSIDA. ДВУДУГНЫЕ
Входящие сюда отряды хорошо распределяются в две фило
генетические группы — Lepidosauria (первые 4 отряда по пряна
тому ниже порядку) и Archosauria (остальные 5). Хотя боль
шниство современных пресмыкающихся принадлежит к первой
из этих групп, но прогрессивной приходится признать вторую.
Трудно привести признаки, формально разграничивающие эти
группы. Можно однако отметить, что для Archosauria наиболее
характерны эволюционные изменения конечностей и их поясов,
в частности, тенденция развития хождения на двух ногах (бипе-
дализма). У Lepidosauria, обычно, исчезает сперва нижняя ви-
сочная дуга, а потом и «верхняя, тогда как исчезновение верхней
дуги при сохранившейся нижней характерно для птиц — потом-
ков Archosauria. У последних часто имеется предглазничное
отверстие, обычно исчезает непарное теменное отверстие, а зубы
текодонтные.
X. ОТРЯД &OSUCH1A. РАННИЕ ЯЩЕРЫ
Eosuchia — наиболее древние представители подкласса,
встречающиеся в перми Южн. Африки и Мадагаскара. Большин-
ство представителей, видимо, было наземными, но Tangasauridae,
вероятно, плавали. Длина черепа 6—8 см. Характеристика се-
мейств следующая:
1(4) Околоклиновидная кость тонкая, сильно вытянутая: пе-
редний конец доходит или заходит вперед за середину
длины всего черепа.
2(3) Нижняя височная дуга полная: теменные кости не каса-
ются чешуйчатых.
Younginidae
5 родов.
3(2) Нижняя дуга не полная; теменные касаются чешуйча-
тых.
Prolacertidae
1 род.
4(1) Околоклиновидная кость менее тонкая и длинная (по-
видимому, не выходит вперед из задней половины че-
репа).
Tangasauridae
2 рода.
132
XI. ОТРЯД Я MT MCMOC ВРЯ А МА «ЛИМИНОЛОЯЫТ
(си. главу S)
хи отмд pleurosauxia. змеевидные ямдоы
Всего одни род Pleuroaaurus из верхней юры Франция я Гер
мании. Молодые были описаны как Acrosaurus. Тело дляииве (у
взрослых 0.7—2.3 м), змеевидное, но с 4 конечностями наземного
типа (рис. 27). Насчитывают 172 позвонка, в том числе 120 хво-
стовых. Видимо, жили по берегам водоемов, хорошо плавали я
питались ракообразными.
XIII. ОТРЯД SQUAMATA. ЧЕШУЙЧАТЫЕ
Морфологические особенности и палеонтологические сведения
позволяют различить 3 подотряда:
1 ( 2) Предчелюстные кости парные, удлиненные; передние
туловищные ребра обычно двухголовчатые; триас.
(1) Prosauria. Древние ящерицы.
2(1) Предчелюстные кости, как правило, объединены в не-
парное образование, короткие; туловищные ребра всег-
да одноголовчатые; позднее триаса.
3(4) Челюстная и крыловидная кости неподвижны; обычно
передняя часть черепной коробки не окостенелая, име-
ется верхняя височная дуга, черепной столбик и 1—
2 пары конечностей; ветви нижней челюсти соединены
швом; зубы акродонтные или плевродонтные; веки сво-
бодны, если же сращены, то конечности имеются; с юры
до современности.
(2) Sauria. Ящерицы (см. главу 9)
4(3) Челюстная и крыловидная кости подвижны; передняя
часть черепной коробки окостенелая; верхней височной
дуги, черепного столбика и передних конечностей нет;
могут быть рудименты задних конечностей; ветви ниж-
ней челюсти соединены подвижно эластичной связкой;
зубы всегда акродонтные; веки сращены, и глаз при-
крыт прозрачной кожей наподобие часового стекла; с
мела до современности.
(3) Serpentes. Змеи (см. главу 10)
Объем подотряда Prosauria, так же как его наименование,
спорны. Можно принять, что в него входит всего одно семейство
Askeptosauridae, охватывающее 3 рода: ящеровидный водный
Askeptosaurus (Италия, длина 2,5 ж), Macrocnemus (Швейца-
рия, Италия, 7СССР, длина 0,2—1,5 ж), с несколько удлиненной
шеей и очень длинными голенями (?хищник, ?двуногое хожде-
133
. N (Германия, Альпы, алии* от ОД де б/> ж).
*5ЬА длинной шеей (видимо, жили по берегам, ввталяеь
Уловленными ив воды животными)
XIV, ОТРЯД THECODONT!А. ЛУНЧАТОЗУВЫЕ
Триасовая группа с разнообразной радиацией. Размеры ко
лебались от 0,1 (Scleromochlidae, Elachiatosuchidae) до 5 at
(Erythrosuchidae, Rauisuchidae и др.). Большинство имело раз*
меры, форму тела и образ жизни, как у крокодилов. Некоторые
phytosauridae питались рыбой (Mystriosuchus, Embrachosuchtu
л др.), другие (Brachysuchus, Machaeroprosopus) нападали на
сухопутных животных, приходивших на водопой. Erythrosuchidae
были крупными наземными хищниками. Scleromochlidae лазали
по деревьям и питались насекомыми. Интересна тенденция к раз*
витию двуногости у Euparkeriidae и Ornithosuchidae. На боко-
вых щитках спнны Desmatosuchus (Staganolepidae) и Typotho-
rax (Rauisuchidae) имелись крупные шипы. Неизвестных живот-
ных, оставивших в триасе следы, называемые Chirotherium, Хю-
не относит к Thecodontia. Род Belodon ныне разделен на несколь-
ко родов семейства Phytosauridae. Не следует обманываться фи-
лологическим значением слова Phytosauria, сохраняемым по
традиции — растительноядных форм в отряде не было. Основ-
ные отличия подотрядов и семейств таковы:
1 (18) Предчелюстные кости короткие; ноздри открываются в
передней части морды; основная клиновидная кость уд-
линенная; врановая кость с отверстием [подотряд (1)
Pseudosuchia].
2(7) Нижняя челюсть не имеет окна, непарное теменное от-
3 ( 6)
4 ( 5)
верстие имеется.
Межтеменная кость имеется.
Зубы акродонтные.
Proterosuchidae
5 родов, Южн. Африка и Азия.
3(4) Зубы текодонтные.
Erythrosuchidae
1 род, Южн. Африка.
6(3) Межтеменной кости нет.
Elachistosuchidae
1 род, Европа.
7 ( £) Нижняя челюсть имеет окно; непарного теменного от-
верстия нет.
8 (17) Спина покрыта панцирем, который может редуциро-
ваться до одного продольного ряда бляшек.
9 (12) Межтеменная кость имеется.
135
10 (И) крестцовых поя воинов 2. лобком* кость короче сем
лищной.
Euptrfcerlldae
2 рода, Южн. Африка, Китай.
II (10) Крестцовых позвонков 3; лобковая кость длиннее седа
лищной.
Omlthosuchldae
7 родов, Европа, Африка, Сев. Америка.
12 ( 9) Межтеменной кости нет.
13 (14) Ноздря близ конца морды и отделена от верхнечелюст*
ной кости заметным промежутком.
Sphenosuchidae
2 рода, Южн. Африка, Китай.
14 (13) Ноздря отодвинута от конца морды назад и касается
или почти касается верхнечелюстной кости.
15 (16) Спинные щитки заметно вытянуты в поперечном на-
правлении.
Staganolepidae
8 родов, Европа и Сев. Америка.
16 (15) Ширина спинных щитков примерно равна их длине.
Rauisuchidae
9 родов, Южн. и Сев. Америка, Южн. Афри-
ка и? Европа.
17 (8) На спине нет ни панциря, ни ряда отдельных щитков.
Scleromochlidae
1 род, Европа.
18 ( 1) Предчелюстные кости образуют удлиненное рыло; нозд-
ри удалены от конца морды; основная клиновидная
кость короткая; врановая кость лишена отверстия [по-
дотряд (2) Phytosauria].
Phytosauridae
14 родов, Северное полушарие.
XVI. ОТРЯД CROCODILIA. КРОКОДИЛЫ
(см. главу 11)
XVI. ОТРЯД SAUR1SCHIA. ЯЩЕРОТАЗОВЫЕ
В конце первой четверти прошлого века были описаны кости
гигантских ящеров, которых в 1841 г. Оуэн объединил под об-
щим иааванием Dinosauria (от «deinos»a*yjKac). Возникнув в
1Эб
рмлсг, оня были ЦИППД1 щу»,ЩИМй НЙН'ХЖМММ фОвМЯМЯ • »•
книи всегп wrwtofl и вымерли и конце мелк (per 7Д| Ил яр*
«женим 140 млн лет своего гушегг яояянмя owe лл4и mi
цепные формы и 40- 50 г мегом, тян и мелкие |»ели«гимо4 г .•»*
(\чж). Уже я конце прошлого пекл было помлжио, что к жАгмм*
гельнсх’ти дннозвяры представляют 2 соигршеиио гдылгтояг»^
ныч. независимо ночммкшим спряла После работ Хюн* ИЛ4 и
тхледующих лет разделение это получило всеобщее лрмтяяиме
Однако некоторые черты конвергентно сходны у обоих отрядов
Это прежде всего гигантизм многих фора/. Его ставят в свя*ь г
сильным развитием гипофиза динозавров, хотя в конечном счете
причиной этому, видимо, были взаимоотношения хищника и
жертвы. Кроме того, большинство особенно крупных динозавров
вели полуводный образ жизни. У них наблюдалось развитие
пустот в костях, и возможно, имелись воздушные мешки напо-
добие птиц. Не исключено, что существовало подобие тепло-
кровности (псевдогомотермия). Головной мозг был удивительно
мал и у некоторых (особенно Sauropoda) развивался «крест-
цовый мозг»— утолщение спинного в районе крестца, которое
могло до 20 раз превосходить объем головного.
Многие группы независимо друг от друга переходили к
ходьбе на двух ногах, что часто сопровождалось редукцией паль-
цев. Разнохарактерными гипотезами зоологи и палеонтологи
пытаются объяснить полное, почти внезапное вымирание дино-
завров в конце мела, но сама многочисленность этих попыток
свидетельствует о нашем незнании. Несомненно, что имело место
сочетание абиотических факторов с биотическими, а не действие
какой-либо одной причины. К сожалению, многие виды, рода и
группы динозавров описаны по слишком фрагментарным ос-
таткам.
Предками ящеротазовых динозавров явились видимо лунчато-
зубые, близкие к Euparkeriidae и Ornithosuchidae. В дальнейшем
наметилось разделение на двуногих хищных и вторично четве-
роногих травоядных Sauropoda. За немногими исключениями,
именно последние были крупнейшими наземными позвоночными
(длина Brontosaurus 18 м, Diplodocus 25,6 м). Пищу перетирали
с помощью заглатываемых камней («гастролиты»). Ветвь хищ-
ных очень скоро разделилась на небольших (0,5—1,5 м) Coelu-
rosauria, только в мелу давших сравнительно крупные (около
4 м) страусоподобные формы, и на более крупных (около 3 м)
Carnosauria, достигших к концу своей истории очень большой
величины (крупнейший наземный хищнйк всех времен Spino-
saurus был 12 м длиной). Предполагают, что Ornithomimidae
перешли к питанию яйцами или моллюсками. Можно различить
следующие основные группы ящеротазовых:
1 (10) Кости тонкостенные; 23 предкрестцовых позвонка; обыч-
но большая берцовая длиннее бедра [подотряд (1)
Coelurosauria].
137
ЗНСЫЛШНЛ СГЛАТ^.ил 0**11*0*09*
Рис. 28. Филоге
138
динозавров
139
2 ( Ш Зубы есть
.3(4) Передние конечности с 5 ни льнами
Podoltetaurldae
Г> родов, триас Европы я Сев. Америки
4(3) Передние конечности с 3, редко 4 пальцами.
5 ( 6) Ключица имеется; тела позвонков без полостей
Seglsauridae
1 род, юра Сев. Америки.
6(5) Ключицы нет; полости в телах позвонком имеются
7(8) Животные мелкие — длина 0,5—0,6 ж; лобковая косГ|
короче бедра; передние конечности укороченные.
Compsognathldae
2 рода, юра Европы и Сев. Америки.
8 ( 7) Животные крупнее — около 2 м; лобковая кость при
мерно равняется бедру; передняя конечность длиннее.
Coeluridae
4—8 родов, юра — мел Сев. Америки, ?Евро
пы и ?Азии.
9 (2) Зубов нет.
Omithomimidae
5 родов, мел Европы, Азии и Сев. Америки.
10 ( 1) Кости толстостенные; обычно 25 предкрестцовых поз-
вонков и большая берцовая короче бедра.
И (18) Зубы острые, сжатые с боков; череп крупный, равняется
длине 6—8 первых хвостовых позвонков; шея короткая,
состоящая из 9—10 коротких шейных позвонков [подот-
ряд (2) Carnosauria].
12 (17) Зубы загнуты назад; остистые отростки спинных позвон-
ков обычно короткие.
13 (14) Глазница большая, округлая; длина порядка 3 м.
Teratosauridae
5 родов, триас Европы, Южн. Африки и Азии.
14 (13) Глазница сравнительно небольшая, вытянутая' верти-
кально; длина в 2—3 раза больше.
15 (16) Дистальный конец седалищной кости утолщен; спинные
позвонки задневогнутые; длина порядка 6 м.
Megalosauridae
10 родов, юра — мел Европы, Азии и Сев. Аме-
рики.
140
, (16) Седалищная к<ж.г», ««мяичяяяогя б»» утодяиямя. гимн
1 иые почвойКЯ с ПЛОСКИМИ ПОЯГрЖЯОГГЯМЯ ИЛЯ Г.чегкл мл
нмюгнутые; длина порядка 6 10 л.
Tyrannosauri dae
3 родя, мел Сея Америки, Алии и Африки
17 (12) Зубы прямые, не загибающиеся назад; верхмяе лстш
тые, отростки спинных поавонкоа очень большие (0,7
IJ м).
Splnosauridae
1 род, мел Египта.
ig (|1) Зубы ложковидные или листовидные; череп относитель'
но маленький, равняется длине 2—4,5 первых хвостовых
позвонков, шея длинная, состоящая из 10—16 удлинен-
ных позвонков.
19 (22) Шейных позвонков 10, двояковогнутые; крестцовых поз
вонков 3; передние конечности заметно укорочены [под-
отряд (3) Prosauropoda].
20 (21) Мелкие (длина порядка 1—2 м); спинных позвонков 14.
Thecodontosauridae
4 рода, триас Европы, Южн. Африки, Азии,
Австралии и Сев. Америки.
21 (20) Крупные (длина порядка 8 л); спинных позвонков 15.
Plateosauridae
7 родов, триас Европы и Южн. Африки.
22 (19) Шейных позвонков 12—16, задневогнутые; крестцо-
вых 4—6; передние конечности длиннее [подотряд (4)
Sauropoda]
23 (28) Носовые отверстия черепа крупные; морда тупая или
слабо вытянутая.
24 (27) Остистые отростки спинных позвонков простые.
25 (26) Передние конечности короче задних; 5—6 крестцовых
позвонков.
Cetiosauridae
3 рода, юра Европы, Африки, Австралии и
Сев. Америки.
26 (25) Передние конечности равны или длиннее задних; 4 кре-
стцовых позвонка.
Brachiosauridae
3 рода, юра — мел Европы, Африки и Сев.
Америки.
27 (24) Остистые отростки спинных позвонков на концах раз-
двоенные.
141
Camarmanrlda*
5 ролов, юра Европы. Африки я Ов Aitrp^
2Я (23) Носовые отверстия черепа сравиитгльяо мялкм*.
вытянутая.
29 (30) Остистые отростки спинных поя во мной простые. 6 яр»^?
новых позвонков.
TltanoBanridae
в родов, юра — мел всех частей свет»
30 (29) Остистые отростки спинных позвонков на концаt ра,
двоенные; 5 крестцовых позвонков.
Dlplodocldae
I род, юра Сев. Америки.
XVII. ОТРЯД ORNITHISCHIA. ПТИЦЕТАЗОВЫЕ
Никогда не достигали полной двуногости и, видимо, даже
наиболее специализированные в этом отношении формы време-
нами пользовались для передвижения и передними конечностями.
Многие вторично перешли к передвижению на четырех ногах
(Stegosauria, Ceratopsia). В связи с этим расчленение лобковой
кости на 2 ветви испытывает различные прев|ращения и, в част-
ности, у Ceratopsia, может наблюдаться значительная редукция.
В тех случаях, когда передняя ветвь лобковой кости хорошо
развита, она служит для поддержки стенки брюха. Размеры ко*
лебались от 1 до 8 м. Все птицетазовые были растительноядными,
причем большинство имело подобие рогового клюва. Интересно
(развитие «зубной батареи» — многорядных зубов у Hadrosauri-
dae. Только эта группа была целиком водной, тогда как Iguano-
dontidae держались по берегам водоемов, а остальные были дре-
весными формами. Предполагают, что Hypsilophodontidae вели
древесный образ жизни. Многие выработали средства пассивной
защиты от хищников: панцирь, большие шипы и рога. У Ceratop-
sia конец черепа образовывал огромный костный «ошейник»,
защищавший их от хищников. Предполагают, что непарный гре-
бень, имеющийся у некоторых Hadrosauridae (Corythosaurus,
Parasaurolophus и других), имел отношение к дыханию (? запас
воздуха при нырянии). Отличия групп таковы:
1 (12) Череп сравнительно небольшой (расстояние от конца
морды до затылка содержится в длине туловища от 2
до 8 раз); большинство безрогие, парных рогов никогда
не бывает.
2(9) Костного панциря нет; шейные и передние туловищные
позвонки задневогнутые; обычно большая берцовая
длиннее бедра [подотряд (1) Ornithopoda].
3 ( 4) Сильно вздутые лобные и теменные кости образуют
бугор.
>42
PachycephafoMorldM
2—3 рода* мел Сем. Америки и ?Европы
3)
6)
4
Добные и теменные кости не вздуты.
Предчелюстные кости с зубами; длина тела 12л
Hypsllophodontldae
I—2 под а, юра — мел Европы* Сев. Америки
и Южн. Африки.
5) Предчелюстная кость без зубов; длина тела до Я м
Шейных позвонков 6—10, а крестцовых 4—6.
Iguanodontidae
6 родов, юра — мел Европы, Сев. Африки,
Азии и Сев. Америки.
8
7) Шейных позвонков 13—16, а крестцовых 7—8.
Hadrosauridae
18 родов, мел Европы, Азии, Сев. и Южн.
Америки.
9(2)
Ю (10
Имеется более или менее развитой панцирь из костных
щитков; позвонки двояковогнутые или с плоскими по-
верхностями; большая берцовая короче бедра [подотряд
(2) Stegosauria].
На нижней челюсти имеется предзубная кость; височ-
ные ямки хорошо заметны.
Stegosauridae
9 родов, юра — мел Европы, Африки и Сев.
Америки.
И (10) Предзубной кости нет; височные ямки частично или
' полностью закрыты.
Apraedentalidae
27 родов, мел Европы, Южн. Африки, Азии
и Сев. Америки.
12 ( О Череп очень большой (содержится в длине туловища
' менее 2 раз); обычно имеются рога, в том числе парные
[подотряд (3) Ceratopsia].
13 (И) Ноздря большая; непарный рог на носу и по рогу над
каждым глазом.
Ceratopsidae
13 родов, мел Сев. Америки.
14 (13) Ноздря маленькая; рогов нет или только один непарный,
15 (16) Предглазничное отверстие есть; глазница большая
(алина ее содержится в наибольшей длине черепа 7—
9 раз).
143
Protocer«top»ide*
3- 4 ролл, мгл Алии и Сем Америки
16 (15) Предглазиимного отверстия нет, глазница ««леммив
(в длине черепа примерно 12 раз).
Pachyrhlnoaaurtdae
I род, мел Сем. Америки.
XVIII. ОТРЯД PTEROSAURIA. ЛЕТУЧИЕ ЯЩЕРЫ
Величина тела колебалась от размеров воробья до очень
крупных форм (Pteranodon имел в размахе крыльев 7—8 м и
был крупнейшим летающим животным всех времен) (рис. 29)
Летательные способности постепенно совершенствовались (уко-
рочение хвоста, исчезновение зубов и тому подобное), но это
не могло исправить коренного недостатка летательного аппара-
та: разрыв перепонки выводил из строя все крыло. Видимо, это
обстоятельство привело к вымиранию летучих ящеров, не смог-
ших выдержать конкуренции с птицами. Обитали по берегам
водоемов (преимущественно морей), бросаясь планирующим
полетом за рыбой. Наверх карабкались с помощью пальцев.
Позднее приобрели способность дальних полетов и парения над
открытым морем. Возможно, что в покое подвешивались головой
вниз наподобие летучих мышей. Имелась перепонка между
пальцами задних ног, киль на грудине и горловой мешок. Пред-
полагают, что в предглазничной. впадине помещалась железа,
выделения которой смазывали крыло. Находка волосовидных
образований позволяет предположить теплокровность. Группы
различаются по следующим характерным особенностям:
1 ( 4) Зубы имеются; ноздря отграничена от предглазничной
впадины костным мостиком; хвост длинный; пястная
кость летательного пальца короче половины длины
предплечья [подотряд (1) Rhamphorhynchoidea]-
2(3) Предглазничная ямка очень большая; дистальный конец
передней ветви лобковой кости расширен.
Dimorphodonti dae
6—8 родов, юра Европы.-
3 ( 2) Предглазничная-ямка маленькая; концы лобковой ко-
сти не расширены.
Rhamphorhynchidae
1—4 рода, юра Европы, Африки и ? Сев. Аме-
рики.
4(1) Зубов мало или совсем отсутствуют: носовое отверстие
на черепе полностью или частично слито с предглазнич-
ной ямкой; хвост очень сильно укорочен; пястная кость
144
летательного пальца Длиннее, равна или немного мором
предплечья (подотряд (2) Pterodactyloldeal
( 6) Лопатка и врановая кость раздельны; спянкме позвонки
и предплюсневые кости свободные.
Pterodactylldae
5—6 родов, юра Европы и Африки.
Рис. 29. Представители
Pterosauria: 1 — Rhamphorhynchus, 2 — Pterodac-
tylus, 3— Pteranodon
146
• I 6) .Попатжа я араковая ж ость срастаются, часть гнидам» !
погвомжов прирастает друг к другу («noierlwm»); «р*
мпмальиые моста предплюсны срастаются с тягмм»
? ( 8) Зубы имеются.
Omtthochelridaa
3 рода, мел Европы.
8 ( 7) Зубов нет.
Pteranodonttdae
4 рода, мел Европы, Сев. я Южн. Америка.
ЛИТЕРАТУРА
Efremov I. On the Subclass Batrachosauria — an Intermediary Grr^
between Amphibians and Reptiles, Bull. Acad. Sci. USSR, 1946, No., 6,
Huene F. The Principles of Early Tetrapod Classification, Americ. Joum
Sci., 1954. 252. ”*
Kuhn O. Die fossilen Reptilien, Berlin, 1937.
Romer A. Osteology of Reptiles, Chicago, 1956.
S6ve-S6derberg G. Some Points of View Concerning the Evolution of
the Vertebrates and the Classification of this Group, Arkiv f. Zoolno?
1934 26 А. К»,
Traits de Palfcontologie, Paris, 1955, 5: 319—990.
Гдала седьмая ---
ЧЕРЕПАХИ
Представители этой группы столь сильно отличаются от всех
современных пресмыкающихся, что корни их приходится искать
в глубокой древности. Большинство исследователей согласно в
том, что они возникли в перми от Cotylosauria, видимо, близких
к Pareiasauridae или Diadectidae. Наибольшего расцвета черепа-
хи достигали в мезозое, а до наших дней из 26 семейств дожило
всего 12. Распространение некоторых групп было раньше значи-
тельно обширнее. Так мягкокожие черепахи занимали прежде
Европу и Сев. Америку. Бокошейные в триасе жили в Европе,
позднее были оттеснены южнее.
Основная особенность строения — панцирь, вероятно, находит-
ся в связи с наземным и роющим образом жизни древнейших
форм, который сохранен многими семействами и до настоящего
времени (Archaeochelyidae, Eunotosauridae, Miolanidae, Polyster-
nidae, Protochersidae, Testudinidae, Triassochelyidae). Напротив,
морские черепахи (Cheloniidae, Dermochelydidae, Plesiochelyidae,
Protostegidae, Thalassemyidae, Toxochelyidae), а также пресно-
водные и болотные (т. е. все остальные семейства) представляют
собой вторичное явление. Панцирь не дает абсолютной защиты,
но относительная выгода его столь велика,- что черепахи, суще-
ствующие огромный промежуток времени (порядка 180—
200 млн. лет), не приобрели существенных изменений в своей
организации. Первоначально пресмыкающиеся имели двоякого
рода костные образования на поверхности тела: более глубокие
костные пластинки — theca и более поверхностный слой костных
чешуй — epitheca. Сверху располагались сегментальные ряды ро-
говых чешуй. От такого первоначального строения у некоторых
ящериц (Scincidae, Anguidae) и крокодилов сохранилась epi-
theca, а костный панцирь черепах развился за счет theca, к кото-
рой присоединились части внутреннего костного скелета. Только
у Dermochelys поверх редуцированного костного панциря распо-
лагаются многочисленные костные пластинки, видимо, гомоло-
И* 14*
гмчные epitheca. Вполне развито* панцирь черепахи сосТа.
спинного (сигарах) и брюшного (plastron) шитов * М)
У большинства современных форм панцирь сверху П|>и
«ли дерм а льны мн роговыми пластинами, гомологичным* *И».
вой чешуе ящериц. Во избежание путаницы рекомендуй
нме элементы панциря называть пластинками, a Рого*^*'
Рис. 30. Скелеты и панцири черепах: А — продольный разрез скелета,
В — спинной щит Hardella (костные пластинки обозначены пунктиром, а ро-
говые щитки — сплошными линиями), В — грудной щит Stereogenys (костные
пластинки обозначены пунктиром,, а роговые щитки — сплошными линиями),
Г — скелет Eunotosaurus снизу, Д — череп Miolania спереди:
7 — грудной пояс, 2 — спинной щит, 3 — тазовой пояс, 4 — бедро, 5 — большая берцовая,
6—малая берцовая, 7 — брюшной щит, 8—локтевая, 9— лучевая, 10— плечевая, // —
аагривковая, 12 — остистые, 13 — надхвостовые, 14 — хвостовая. 15 — реберные, 16 — край-
ние» 17. — шейный, 18 — позвоночные, 19 — боковые, 20 — краевые, 21 — надгрудная,
22 — внутригрудная. 23 — грудная, 24 — средняя, 25 — нижняя, 26 — задняя, 27 — меж-
горловые, 20 — горловой, 29 — плечевой. 30 — грудной, 31 — брюшной, 32 — бедренной.
33 — заднепроходный
щитками (рис. 30). На спинном щите различают пластинки:
загривковую (nuch.ale), остистые (neuralia), реберные (costalia),
крайние (marginalia), надхвостовые (suprapygale), хвостовую
448
(pygale) и щитки шеянын (nuchalr), птвпночныг (vertebra hi),
боковые (costa На). краевые (marginalia) На брюшном шаге
имеются пластинки, надгрудная (eplplastron), янутригрудяая
(cntoplastron), грудная (nyoplastron), средняя (meaopl astron),
нижняя (hypoplastron), задняя (xiphiplastгоп) и шитки горло
вой (gularc), межгорловой (intergularc), плечевой (humeral*),
грудной (pectorale), брюшной (abdominal*), бедренной (femora-
1е), заднепроходный (anale), подмышечный (axillare), паховой
(inguinale) и межкраевые (intermarginale)- Считают, что над
грудная пластинка гомологична ключице, а внутригруднэя
межключице. Чем выше панцирь, тем большую статическую
нагрузку он выдерживает. Экологический смысл швов костного
панциря — гашение ударов. Каждый роговой щиток в онтогенезе
растет как самостоятельное целое, и на поверхности его возника-
ют концентрические годовые кольца. У Chrysemys весь наруж-
ный слой рогового щитка периодически сходит.
Развитие панциря вызвало неподвижность части позвоночни-
ка и редукцию многих туловищных мускулов. Грудной пояс ока-
зался скрытым под ребрами (единственный случай среди всех
позвоночных!). Невозможность дыхания реберного типа вызвала
ряд приспособлений. У большинства колебания подъязычного
аппарата создает вакуум в ротовой полости. Дыханию помогают
движения ног и шеи. У многих водных форм имеются анальные
пузыри (bursae anales) — боковые тонкостенные выросты задней
кишки около самой клоаки, стенки которых обильно снабжены
сосудами. Эти пузыри периодически наполняются водой, напоми-
ная водные легкие голотурий.
В миоцене Сивалинских холмов найдена Testudo (Colossoche-
lys) atlas, панцирь которой имел в длину 2—3 м. По-видимому,
самой крупной черепахой была Miolania, череп которой был 58 см
шириной, и, следовательно, панцирь имел в длину больше 5 м.
У крупнейших современных черепах (Dermochetys) панцирь до-
стигает в длину 2 м.
Наземные виды преимущественно растительноядные, а вод-
ные поедают различных животных: от медуз до рыб и лягушек.
Могут голодать до 6 лет; удивительно выносливы к травмам.
Самцы имеют непарный копулятивный орган. Откладывают на
суше в грунт шаровидные или овальные яйца.
В 1854 г. Штанниус противопоставил мягкокожих черепах
всем остальным. В 1870 г. Коп обратил внимание на строение
шеи, установив группы Cryptodira и Pleurodira, а также выделил
как особую группу Atheca, в противоположность которым в 1886 г.
Долло предложил назвать всех остальных черепах Thecophora.
Неясным до наших дней является положение Ваёшбае и Pleuros-
ternidae. Хюне относит их к Pleurodira, а Бергунью — к Crypto-
dira. Реальность пермских Archaeochelyidae многими зоологами
отвергается. Можно принять такие группы (рис. 31):.
1 ( 2) Зубы на предчелюстной и челюстной кости имеются; пан-
149
а«ря еше мет. ио ребра расширены (лодоярЯЖ (1) Ешн.
мог**)
I. Fwnofoi aurldae
Р Зубов на предчелюстной ж челгостмо* костях вет; а*.
2 ’ верь имеетеа («Testudinata ver а»).
рис, 31. Филогения черепах: / — Eunotosauria. 2 — Cryptodira, 3— Trio-
nycboidea. 4— Pleurodira, 5— Atheca
3 (50) Спинные позвонки и ребра соединены неподвижно и уча-
ствуют в образовании панциря; теменные кости про-
должаются вниз и соединяются с крыловидными или
отделены от них надкрыловндной.
150
, (33) Шея прячется в панцирь, иягкбяясь я •яргмядыюй
плоскости, если же не втяжная, то ионеяяоети првврвш**
им в ласты; тая нс срастается с пяяпярем.
а (30) Панцирь снабжен роговыми щитками; крыловидные кое>
тн встречаются друг с другом по средней линия, пальцы
имеют не больше * фаланг каждый (подотряд (2) Сгур-
tod i га).
6 (25) Ноги наземного или болотного типа, ио не в виде ластов
(«плавников).
7(8) На сошнике, околоклиновидной и крыловидной костях
имеются зубы.
2. Triassochelyidae
8 ( 7) Зубов нет.
g (10) Череп несет до 3 пар костных выростов, из которых
задние роговидны и длинны.
5. Miolanidae
10 ( 9) Рогов на черепе нет.
11 (24) Внутригрудная пластинка имеется.
12 (15) Брюшной щит из 11 костных пластинок.
13 (14) Череп заостренный.
4. Pleurosternidae
14 (13) Череп короткий.
3. Baenidae
15 (12) Брюшной щит из 9 костных пластинок.
16 (19) Загривковая пластинка с каждой стороны с ребровид-
ным боковым отростком, простирающимся под крайние
пластинки.
17 (18) Хвостовые позвонки передневогнутые.
9. Dermatemydidae
18 *(17) Хвостовые позвонки большей частью задневогнутые.
8. Chelydridae
19 (16) Загривная пластинка без ребровидного отростка.
20 (21) Хвостовые позвонки большей частью задневогнутые.
11. Platystemidae
21 (20) Хвостовые позвонки передневогнутые.
22 (23) Панцирь высокий; ширина каждой из костей пясти рав-
на ее длине или чуть меньше; пальцы без перепонок.
в. Testudlnidae
23 (22) Панцирь уплощенный; пястные кости удлиненные; паль-
цы обычно с перепонками.
151
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
152
7. EmydMae
(II) Внутри гр ул НОЙ ПЛЛСТИНКН ПГ1
10. Ktaostenildae
( 6) Ногя превращены в ласты.
(27) Брюшной щит не сердцевидный: расстояние от лопате»
ной вырезки до паховой около 3/з всей длины грудног
щита.
13. Protostegldae
(26) Брюшной щит сердцевидный: расстояние от лопаточной
до паховой вырезки меньше 2/з длины грудного шита.
(29) Сошник и нёбные кости образуют вторичное нёбо; фа
ланги без суставных головок.
12. Cheloniidae
(28) Вторичного нёба нет; фаланги снабжены суставными
головками.
14. Toxochelyidae
( 5) Панцирь без роговых щитков; крыловидные кости по
средней линии не встречаются друг с другом; некоторые
пальцы могут иметь более 3 фаланг [подотряд (3) Trio-
nychoidea]-
(32) На каждой лапе по 3 когтя; крайние пластинки отсут-
ствуют или рудиментарны; костный брюшной щит с боль-
шим окном.
15. Trionychidae
(31) На каждой лапе по 2 когтя; крайние пластинки имеются;
костный брюшной щит с маленьким окном или совсем
без него'.
16. Carettochelydidae
( 4) Шея изгибается в горизонтальной плоскости и уклады-
вается в панцирь сбоку, если же не прячется, то ноги не
в форме ластов; таз срастается с панцирем [подотряд (4)
Pleurodira].
(45) Брюшной щит имеет с каждой стороны по 1—2 средних
пластинки.
(44) Остистые пластинки имеются.
(37) С каждой стороны по 2 средних пластинки.
17. Proterochersldae
(36) С каждой стороны по 1 средней пластинке.
(39) Средняя пластинка большая, вытянутая продольно и за-
крывающая большую часть наружного яре* грудной *
нижней пластинок.
24. Polysternldae
«о (Зв) Средняя пластинка вытянута поперек или маленькая,
дд (41) Шейные позвонки двояковогнутые.
18. Kalloklbotlidae
«I (40) Второй шейный позвонок двояковыпуклый.
42 (43) Сошник большой; нижняя челюсть имеет ямку сзади
жевательной поверхности.
22. Bothremyidae
43 (42) Сошник меньше; такой ямки на нижней челюсти нет.
21. Pelomedusidae
44 (35) Остистых пластинок нет.
25. Eusarkiidae
45 (34) Средней пластинки нет.
46 (47) Брюшной щит <в средней части с большим окном.
19. Thalassemyidae
47 (46) Брюшной щит без окна.
48 (49) Ложной височной ямки нет.
20. Plesiochelyidae
49 (48) Имеется большая ложная-височная ямка.
23. Chelidae
50 ( 3) Внутренний скелет не связан с панцирем, состоящим
только из мозаики небольших костных пластинок.
26. Dermochelydidae
Подотряд 1 — Eunotosauria. Широкоспинные
1. СЕМЕЙСТВО EUNOTOSAURIDAE
Единственный представитель — маленький (чуть более деци-
метра) Eunotosaurus africanus из перми Южн. Африки. Верхняя
часть черепа неизвестна. Вместо рогового клюва на челюстях и
нёбе имелись небольшие зубы. На сильно выпуклой спине име-
лись кожные окостенения.
Подотряд 2 — Cryptodira. Скрытошейные
2. СЕМЕЙСТВО TRIASSOCHELYIDAE
Из триаса Германии описаны Triassochelys и Proganochelys,
(панцири 65 см длиной).
10 П. В. Терентьев 153
3 CE.MERCTSO BAENiftAE
Шесть ролов u> — »<>игна Сев Л меряв*.
4 сЕМЕПСТВО PLFl ROSTt'RNIDAfc
Олввмадцвтъ родов кз юры — мела Европы. Африки я
Амерявя
5 СЕМЕЙСТВО MIOLANIDAE. РОГАТЫЕ ЧЕРЕПАХИ
Симпсон показал, что рогатые черепахи, ранее относившее.'
к бокошейным, в действительности принадлежат к скрытошед.
ным. В мелу Аргентины и Квинсленда найдена Miolania (сама,
крупная черепаха всех времен), а в эоцене Аргентины — Сго,.
sochelys.
6 . СЕМЕЙСТВО TESTUD1N1DAE. СУХОПУТНЫЕ ЧЕРЕПАХИ (РИС. 32}
Известны, начиная с эоцена, состоят из 5 ископаемых и 7 сов-
ременных родов. Основные современные рода можно различить
так:
1 ( 6) Жевательная (альвеолярная) поверхность верхней че-
люсти без срединного гребня.
2(3) Задняя часть спинного щита подвижна относительно
передней (шов между четвертой и пятой реберными,
пластинками).
Kinixys
3 вида, Африка.
3(2) Задняя часть спинного щита неподвижно соединена с
передней.
4(5) Передняя часть брюшного щита подвижна относительно
задней.
Pyxis
1 вид, Мадагаскар.
5(4) Передняя часть брюшного щита неподвижно соединена
с задней.
Homopus
4 вида, Южн. Африка.
6(1) Жевательная поверхность верхней челюсти имеет кост-
ный киль, проходящий параллельно ее наружному и вну-
треннему краям.
7(8) На жевательной поверхности места сращения -верхних
челюстей продольный костный киль.
154
Рис. 32. Ареалы семейств: / — Testuditiidae, 2 —Chelidae, 3— Platysternidae, 4 —Kinosternidae. 5 — Chelydridae.
6 — Cheloniidae
Ctopberus
I вид, юг Сев. Америки.
8 ( 7) На жевательной поверхности места сращении ерхк«1
челюстей вместо киля находится ямка.
9 (10) Горловой щиток I; выдающаяся вперед горловая чат
брюшного щита закругленная или тупая.
Gonlochersus
1 вид, Южн. Африка.
Ю ( 9) Горловых щитков 2, если же I, то горловая часть брюш-
ного щита впереди приостренная.
Testudo
27 видов, Европа, Африка, Мадагаскар, Азия
и Южн. Америка.
У единственного вида рода Gopherus — G. polyphemus — пан-
цирь достигает 25 см длиной; распространен от Южн. Каролины
до Флориды и Техаса. Вырывает норы (3—5 м длиной), идущие
в землю под острым углом. Живет постоянно в одной норе, по-
кидая ее только в самое жаркое время. В мае — июне кладет 12—
20 шаровидных яиц.
Род Testudo (рис. 33) — основной в семействе, включает тра-
воядные и плодоядные виды, которые, однако, могут питаться
беспозвоночными, а в неволе поедают даже лягушек. Самцы
обычно меньше самок; в задней части брюшного щита имеют
впадину. Пережидают зиму или летнюю жару, зарывшись в
грунт. Звездчатая черепаха (Т. elegans) имеет весьма выпуклый
спинной щит. У старых особей середина отдельных щитков так
сильно выступает вверх, что щитки представляют собой высокие
бугры. Шейный щиток отсутствует. Спинной с темными радиаль-
ными линиями. Длина панциря до 26 см. Живет на сухой, зарос-
шей травой и кустарниками почве Индийского полуострова и
Цейлона. В дневную жару скрывается под камнями и в траве.
Наиболее активна в дождливое (время. Откладывают по 4 яйца
в ямку, глубиной в 15 см. Оба европейских вида характеризуют-
ся пятнистой окраской спинного щита, присутствием горловых
щитков, слабым развитием альвеолярного края верхней челюс-
ти и присутствием 5 когтей на передней лапе \ Средиземномор-
ская черепаха, Т. graeca, имеет на задней стороне бедра большой
бугор. Длина ее панциря до 28 см. Типичная форма распростра?
йена в Южн. Испании и в Сев. Африке. В Юго-вост. Европе, в
Мал. Азии, Палестине и Закавказье живет Т. g. ibera, а в Ира-
ке— Т. g. zarudnyi. Обитают в полупустынных и степных мест-
ностях, часто поросших кустарником, в горных лесах и в кустар-
никовых низинах близ моря. В горах до 1150 м. Айтивна днем.
’ Правила номенклатуры заставили в последние , десятилетия изменить
латинские названия, что может порою путать начинающего зоолога.
156
Рис. 33. Представители разных групп черепах: /—2 — Testudo gigantea, 3 —
Amyda gangetica, 4 — Eretmochelys imbricata, 5 — Chelus fimbriata, 6 — Chelo-
dina longicollte, 7 — Dermochelys coriacea
Спаривание в условна к СССР с апреля по начало «од
Одна самка кладет в вырытую ямку 2—5 яви (ДЛ1»1
вйка 32-42 мл) В ноябре такапываегся в землю до коми а фа
рала иля начала марта. Южно-европейская черепаха Т h/rmafi
(раньше называлась Т graeca!) лишена бугра на задней час’
беж>а и обитает в Вост. Испании, Южн. Франции, Южн. Итали
иаБалканском полуострове, Балеарах, Корсике и CapjuiMWi
Г. hermanni употребляют в пишу, хотя вполне съедобны я Други
виды.
Степная черепаха Г horsfieldii, живущая в Казахстане, сред
неазиатскнх республиках ССР, Иране, Афганистане и Севере
Зап. Пакистане, отличается от двух предшествующих наличие*
только 4 когтей на передней ноге. Длина панциря до 29 см
Держится в глинистых или песчаных степях и полупустынных
предгорьях. В Гиссарском хребте поднимается до 1200 м. В связи
с выгоранием растительности иногда наблюдается летняя спячка,
переходящая в зимнюю. Для зимовки роет норы или использует
норы млекопитающих. Спаривание в марте — июне. Кладка яз
2—5 яиц, каждое 42—57 мм длиной. Инкубация 80—100 дней.
Местами наносит вред культурным растениям и оросительным
сооружениям — роет норы в плотинах.
Особый интерес представляет группа видов, обычно именуе-
мых «гигантскими наземными черепахами». Крупные виды рас-
сматриваемого рода имели широкое распространение в третичное
время (Франция, Вайоминг). Некоторые полагают, что центром
возникновения их была Гондвана или Лемурия. В четвертичное
время они сохранились только двумя пятнами: на островах за-
падной части Индийского океана (Маскаренские, Мадагаскар,
Коморские,-Альдабра, Амирантские и Сейшельские) и на Гала-
пагосских островах в восточной части Тихого океана. Последние
получили свое наименование от испанского «galapago»=4epenaxa.
Отсутствие хищников сделало эти острова естественными запо-
ведниками, но после открытия их черепахи были подвергнуты
хищническому истреблению; что вызвало прогрессирующее вы-
мирание. Островная изоляция стимулировала дифференцировку,
каждый остров имел свою особую форму, ранее рассматривавшу-
юся как вид. Все черепахи этих островов были крупные (до
156 см и 240 кг) черные илй тёмно-окрашенные животные с длин-
ной шеей. Костный панцирь относительно утонченный. Предпо-
лагают, что некоторые особи жили до 300 лет. Можно различить
3 группы. Наличие шейного и парных горловых щитков характе-
ризует Т. gigantea (3 подвид a- jftsr Сейшельских островах и Аль-
да бре) и вымершую на Мадагаскаре* 71 grandidieri. Особи одного
из подвидов в-возрасте 150 лет имели панцири 150 см, длиной.
У второй группы отсутствуют шейные щитки при парных горло-
вых. К этой группе принадлежат Т. elephantopus (10 подвидов
на Галапагосских островах) и вымершая на Маврикии Т. sumei-
rei. Черепахи Галапагосских/островов питаются кактусами, ли-
стьями деревьев, ягодами и лятайяикямя Проходятся лежь до
6 кж. Во время спаривания самцы изляют сильный хриплый «уж
В октябре откладывают дюжину круглых яиц, каждое около 5 еж
в диаметре Третья группа, лишенная шейного щитка и имевшая
непарный горловой щиток, целиком вымерла Были описаны
6 видов (? под виде») этой группы, жившие ня островах Маврикия
в Родригец. Из них Т indica имела панцирь, достигавший ЙО см
длины.
7. СЕМЕЙСТВО EMYD1DAE. ПРЕСНОВОДНЫЕ ЧЕРЕПАХИ (ряс 34)
Известны начиная с эоцена. 4 вымерших и 25 современных
родов. Отличия главных современных родов:
1 (36) Спинной и брюшной щиты соединены неподвижно,
2 (19) Жевательная (альвеолярная) поверхность верхней че-
люсти широкая, с 1—2 срединными гребнями.
3 (10) Подмышечная и паховая подпорки хорошо развиты
(первая достигает или почти достигает первого ребра, а
вторая прирастает между пятым и шестым).
4(5) Четвертый позвоночный щиток вытянутый, накрываю-
щий 4—5 остистых пластинок.
Kachuga
6 видов, Индия, Бирма, Ассам.
5(4) Четвертый позвоночный щиток накрывает 3 остистые
пластинки.
6(7) Хоаны между глазами.
Callagur
1 вид, Малайский полуостров, Суматра и
Калимантан (Борнео).
7(6) Хоаны сзади глаз.
8(9) Жевательная поверхность верхней челюсти с 2 средин-
ными гребнями.
Batagur
1 вид, Индия, п-ов Индокитай, Малайя и Су-
матра.
9(8) Жевательная поверхность верхней челюсти с 1 средин-
ным гребнем.
Hardella
1 вид, Индия.
10 ( 3) Подмышечная и паховая подпорки короткие или уме-
ренные.
И (12) Хоаны позади глаз.
159
Ряс. 34. Ареалы семейств: 1 — Emydidae, 2— Trionychidae, 3—Pelomedusidae, 4 Dermttemydid**,
5
2 ямдя. Индия. Бярмя
>2 (Н) Хоаны между глаз.
L (|8) Паховая подпорка прирастает к пято* рябержй ямс
тикке; внутрнгрудмая пластинка впереди лляжгвогруя
кого шва.
>4 (15) Ширина задней доли брюшного шита у взрослых боиве
V» ширины спинного шита; киля на спинном мяте даже у
молодых нет.
Chrysemys
I вид, Сев. Америка.
15 (14) Ширина задней доли брюшного шита не больше */> ши-
рины спинного щита; у молодых спинной щит с продоль-
ным килем.
|6 (17) Межглазничное пространство очень узкое (примерно
равно половине горизонтального диаметра глаза).
Deirochelys
1 вид, Юго-восток США.
17 (16) Межглазничное пространство шире.
Pseudemys
9 видов, Сев. и Южн. Америка, Вест-Индия.
18 (13) Паховая подпорка прирастает между пятой и шестой
реберными пластинками; внутригрудная пластинка пе-
ресекается плечево-грудным швом.
Ocadia
1 вид, п-ов Индокитай и Южн. Китай.
19 ( 2) Жевательная поверхность верхней челюсти без гребней.
20 (29) Хоаны позади глаз.
21 (24) Паховая подпорка прирастает к пятой реберной плас-
тинке; голова покрыта сверху цельной кожей.
22 (23) Спинной щит крышеобразный.
Graptemys
6 видов, Сев. Америка.
23 (22) Спинной щит придавленный.
Malaclemys
1 вид, Сев. Америка.
24 (21) Паховая подпорка прирастает между пятой и шестой
реберными пластинками; задняя часть головы покрыта
маленькими щитками.
25 (28) Спинной щит с непрерывными гребнями.
26 (27) Боковые гребни спинного щита доходят до четвертого
бокового щитка.
161
ЛмГао» ИядокхтИ. Кит«й. Япоип
7? <М| Гю«о»ыг гргЛнн нг доходят ДО четвертого бодоме
щитка.
Malayemy* м _
1 лид. Таиланд. Малайский полуостров. Ям
28 (25) Спинной тит с прерывистыми гребнями или сериям^
бугров.
Geoclemys
1 вид, Индия.
29 (20) Хоаны между глаз.
30 (33) Задняя часть головы покрыта мелкими щитками.
3! (32) Спинной щит, особенно у молодых, с 3 продольными
килями; длина спайки спинного и брюшного щитков при.
мерно равна длине задней доли брюшного щита.
Siebenrockiella
1 вид, Таиланд, п-ов Индокитай, Малайя, Зонд-
ский архипелаг.
32 (31) Спинной щит с 1 тупым килем; длина спайки много
больше задней доли.
Orlitia
1 вид, Малайя, Суматра, Калимантан (Бор-
нео) .
33 (30) Голова сверху покрыта цельной кожей.
34 (35) Спинной щит с 1—3 продольными килями; хвост всегда
короткий.
Geoemyda
15 видов, Южн. и Юго-Вост. Азия, Центр, и
Южн. Америка.
35 (34) Спинной щит без киля или с одним тупым килем; хвост
средней длины, а у молодых длинный.
Clemmys
9 видов, Европа, Азия, Сев. Америка.
36 ( 1) Спинной и брюшной щиты соединены друг с другом
подвижно.
37 (44) Ложная височная ямка ограничена снизу костным мос-
тиком.
38 (39) Хвост у взрослых умеренный, а у молодых длинный.
Emys
2 вида, Европа, Сев. Африка, Зап. Азия, Сев.
Америка.
39 (38) Хвост всегда короткий.
40 (43) Задняя часть спииного шита зубчатая.
41 (42) Шов между грудным шитом я краевыми короче пни
между брюшными и краевыми.
Cydemys
2 вида. Юго-Вост. Азия.
42 (41) Шов между грудным щитком и краевыми длиннее нюа
между брюшным и краевым.
Notochelys
1 вид, Юго-Вост. Азия, Зондский архипелаг.
43 (40) Задняя часть спинного щита не зубчатая.
Cuora
5 видов, Юго-Вост. Азия, Зондский архипелаг.
44 (37) Ложная височная ямка снизу открытая.
Теггарепе
4 вида, Южн. Канада и США к востоку от
Кордильер.
Индийские кровельные черепахи Kachuga любят чистую воду
и питаются почти исключительно водными растениями. Длина
панциря К. tecta до 23, а К. trivittata — до 55 см. Повышенная
способность к нырянию вызвала образование как бы костяных
камер для легких. В США черепах употребляют в пищу. Пище-
вым продуктом служит и бугорчатая черепаха Malaclemys terra-
pin.. Держится она, главным образом, в соленых водоемах вос-
точных берегов Сев. Америки. Длина тела до 20 см. Хорошо
переносит холод. Кладка с апреля до июня. Одна самка дает
8—20 яиц. На специальных фермах ее откармливают крабами и
креветками.
Представителем рода водных черепах, Clemmys, в нашей фау-
не является С. caspica, панцирь которой достигает 23 см-. Распро-
странена от Балканского полуострова до Кавказа и Южн. Турк-
мении включительно. Живет в озерах, канавах, реках и других
пресноводных водоемах. Питается рыбами, лягушками, головас-
тиками и водными насекомыми. Спаривание в начале апреля.
В начале июня самка кладет 9—12 яиц, каждое длиной 39—
44 мм. Зимует, зарывшись в донный ил. Представителем рода
болотных черепах Emys у нас может служить Е. orbicularis. Пан-
цирь достигает 19 см. Распространена в южной и средней Европе,
Мал. Азии, на Кавказе и в Сев. Африке. Северная граница идет
через Белоруссию, среднюю Волгу и север Казахстана. Живет в
озерах, болотах и медленно текущих реках, а также на морских
побережьях, опресненных реками. Питается рыбой, земноводны-
ми, мягкотелыми, червями, насекомыми и изредка растениями.
Массовое спаривание в мае. С конца мая самки откладывают в
163
вырытые мми на берегу ямки по 3—12 яки, каждое длммой П
39 мм. Инкубация около 100 дней. Зимует на дне водоемов.
робчатая черепаха Теггарепе отличается значительной или ног.
ной редукцией перепонки между пальцами — перешла к ивземи*.
му образу жизнн. Питается улитками, слизнями, червями, яле
дами и грибами. В холодные ночи залезает в норы и под коряя
деревьев, там же и зимует. Длина взрослых 13—17 см. Клалц
в июне — июле. Инкубация 88—109 дней.
в. СЕМЕЙСТВО CHELYDRIDAE. КАЙМАНОВЫЕ ЧЕРЕПАХИ
Известны, начиная с эоцена, в числе 2 вымерших и 2 сояре-
менных родов. Все формы пресноводные, вредят рыболовству.
Единственный вид рода Chelydra — С. serpentina, кусающая че-
репаха, распространена от Канады до Мексики и Эквадора.
Общая длина тела достигает метра; вес до 20 кг. Очень хищная;
легко ловится на крючок. Пахнет мускусом. Употребляется в
пищу наряду с грифоновой черепахой, Macroctemys temminkii.
Последняя отличается глазами, смотрящими вбок, а не вбок и
вверх, и наличием на нижней поверхности хвоста только малень-
ких чешуй, вместо большого щитка у Chelydra. Грифоновая че-
репаха распространена от Техаса и Флориды до Миссури. Общая
длина 140 см.
9. СЕМЕЙСТВО DERMATEMYDIDAE
Известны, начиная с мела, в числе 4 ископаемых и 3 ныне
живущих родов. Различия между последними таковы.
1 ( 2) Брюшной щит широкий; подбородок без кожных при-
датков.
Dermatemys
1 вид, Вост. Мексика, Гватемала, Гондурас.
2(1) Брюшной щит маленький, крестообразный; подбородок
несет пару кожных придатков.
3(4) Брюшной щит соединен со спинным швом.
Staurotypus
2 вида, Мексика, Гондурас, Гватемала, Саль-
вадор.
4(3) Брюшной щит соединен со спинным связкой.
Claudius
1 вид, Вост. Мексика и Гондурас.
Пресноводные животные; длина панциря от 10 до 42,5 см.
10. СЕМЕЙСТВО KINOSTERNIDAE. ЗАМЫКАЮЩИЕСЯ ЧЕРЕПАХИ
Известны, начиная с плейстоцена. Два рода в Сев. и Южн.
Америке: Sthernotherus (2 вида), у которых брюшной щит ма-
164
леныснй с малоподвижными долями брюшного шита, и
поп (16 видов) с большим развитием брюшного шита, передняя
и задняя доли которого хорошо подвижны относительно средней
неподвижной части. Первый предпочитает лужи или болоте, где,
замутив воду, скрывается от врагов. Второй, лучше защищенный,
живете прозрачной воде. Длина панциря 11 —17 см. Кожа конеч-
ностей столь мешковата, что при втягивании скрывает их пол-
ностью.
II. СЕМЕЙСТВО PLATYSTERN1DAE. БОЛЬШЕГОЛОВЫЕ ЧЕРЕПАХИ
Из мела Китая описан Scutemys. Таращук нашел представи-
теля семейства в миоцене под Одессой. Единственный современ-
ный вид Platysternon tnegacephalum живет в реках Южн. Китая,
Бирмы, Тайланда и на Филиппинах. Голова большая с крючко-
видной верхней челюстью. Панцирь (длиной до 15,5 см) как бы
вдавленный. Питается рыбой.
12. СЕМЕЙСТВО CHELONIIDAE. МОРСКИЕ ЧЕРЕПАХИ
Известны, начиная с мела, в числе 4—11 вымерших и 4 совре-
менных родов. Каждый современный род включает всего по 1 ви-
ду, характеризующемуся следующими признаками:
1 ( 4) Спинной щит с окнами между реберными и краевыми
пластинками; боковых щитков 4 пары.
2 ( 3) Спинной щит у молодых с 1 продольным гребнем; обыч-
но спинные щитки не налегают друг на друга; клюв без
крючка; лапы с 1 когтем.
Chelonia mydas
Зеленая черепаха.
3 ( 2) Спинной щит у молодых с 3 продольными гребнями; у
взрослых спинные щитки черепицеобразно налегают
друг на друга; клюв с крючком; на каждой лапе по
2 когтя.
Eretmochelys imbricata
Бисса.
4(1) Спинной щит у взрослых вполне окостенелый; боковых
щитков 5 и более.
5 ( 6) С каждой стороны по 3 межкраевых щитка; ширина са-
мой широкой остистой пластинки меньше ее длины; свер-
ху окраска рыжая или коричневая.
Caretta caretta
Головастая черепаха.
6 ( 5) Межкраевых щитков 4; ширина самой широкой остистой
пластинки примерно равна ее длине; сверху окраска
оливковая или зеленовато-серая.*
165
Lepldochetys olivacea
Оливковая морская черепаха,
Живут в Атлантическом, Тихом и Индийском океан in t
исключая последний вид, в Средиземном море. Зеленая череп»,4
особенно часто встречается около острова Вознесения, в
Индии и у Москитного берега. Питается водорослями. Люб»,
спать на поверхности воды. Яйца кладет на песчаных берегах,
собирается сотнями. В Вест-Индии и Малаккском проливе |
множенне в апреле — нюне, а у берегов Зап. Африки — в сеитжЛ'
ре—-январе. Самка кладет, видимо, в несколько приемов
300—400 яиц, причем в одну ямку помещается по 75—200 яиц. I
Инкубация 42—70 дней. Длина панциря до ПО см, а вес
450 кг. Употребляется в пищу («суповая черепаха»). Бисса меиь.]
ше (панцирь до 85 см), чаще попадается в Карибском море,
около Цейлона, у Мальдивских островов и в море Сулу. Питается
рыбами и моллюсками. Преследуется ради роговых щитков. Пан-
цирь головастой черепахи достигает 105 см в длину.
Этот вид нередок в Средиземном море, а иногда попадается у
берегов Англии и Голландии. Питается ракообразными, мягко-
телыми и рыбами. Оливковая черепаха часто попадается около
Цейлона и Вест-Индии.
13. СЕМЕЙСТВО PROTOSTEGIDAE
Пять родов из меловых и третичных слоев Сев. Америки и
Европы.
14. СЕМЕЙСТВО TOXOCHELYIDAE
Семь родов в меловых слоях Сев. Америки.
Подотряд 3 — Trionychoidea. Голокожие
15. СЕМЕЙСТВО TRIONYCHIDAE. МЯГКОКОЖИЕ ЧЕРЕПАХИ
Встречаются, начиная с мела, в числе 6 вымерших и 7 совре-
менных родов. Последние различаются так:
1 ( 8) Брюшной щит без кожных клапанов; орудная и нижняя
пластинки раздельные; задний край крыловидных костей
без срединного отростка.
2(5) Глазница ближе к височной ямке, нежели к носовой.
3(4) Все реберные пластинки разделены по средней линии
остистыми.
Dogania
1 вид, Юго-Вост. Азия.
4(3) Последняя пара реберных пластинок по средней линии
соприкасается друг с другом.
1«б
Amy da
иялош, Аня*. Африяя, <>• Ам*р*«1
< ( 2) Глазница ближе « мпггнюй я мкг, нежели к жаггяпюА
* 7) Костные отверстия хоан между глазницами
PHochelvt
I вид, Южн. и К>го-Вост Азии. Новая Гвяя-я
у ( 6) Костные отверстия хоан позади гяазиин
Chitra
! вид, Индия.
g(|) БрюшноА шит с кожными бедренымн клапанами, под
которыми могут прятаться задние ноги; грудная и ниж-
няя пластинка срастаются; задний краб крыловидных
костей со срединным восходящим отростком.
9(12) Остистые пластинки образуют непрерывный ряд.
10 (Н) Крайних пластинок нет.
Cycloderma
2 вида, Зап. и Юго-Вост. Африка.
у (10) Крайние пластинки имеются.
Trionyx
1 вид, Индия и Цейлон.
12 ( 9) Остистые пластинки образуют прерывистый ряд.
Cyclanorbis
2 вида, Средняя Африка.
В прежние эпохи встречались также в Сев. Америке и Евро-
пе. Обитатели пресных водоемов. Для дыхания выставляют на по-
верхность воды конец морды (в виде хоботка). Могут, однако,
долго находиться под водой, используя для дыхания запасы воз-
духа, собираемого в кожистых пучках и ворсинках глотки. При
пересыхании водоемов переживают засуху в иле. Иногда попа-
даются в море. Активны ночью. Злобные, сильно кусающиеся
хищники (поедают рыбу, лягушек, мягкотелых). Могут нападать
на купающихся. Яйца круглые; откладывают в песок. У наиболее
крупных экземпляров длина спинного диска достигает 1,5 м.
Центральный род семейства — Amyda. У ряда видов с возрастом
вырабатывается своеобразный внутривидовой диморфизм: у
взрослых, питающихся позвоночными, сохраняются острые края
челюстей, у питающихся мягкотелыми образуются массивные
широкие челюсти, лишенные острого края. A. sinensis водится в
бассейне Амура, в Сев.-Вост. Китае и в Японии. На берег выхо-
дит, но от воды не удаляется. Спаривание 2—3 раза в год. Самка
кладет несколько десятков яиц, каждое диаметром 19—21 мм.
167
Ииюбэимя АМеЛ Полояоэре лесть яя в— 7 под Лжт«гм
13 < м длн*к»А, яге до 10 кг Мясо м «Лия с словим к ого4»иц
цгмятч-я я Японии В США употребляется я пящу A ftrot
16 СЕМКЛ< ТИО СЛЯКТТСМ HEt.VOIOAE
Пикетны, начинки с эоцена, в числе 3 яы мерши я и I гаер»
немного рода Вымершие рода обнаружены в Европе, Сео. Амери
не и Маньчжурии. Единственный современный представят»^
Carettocheltis insculpta живет я реках и иногда « солояояотм]
водах Новой Гвинеи. Длина спинного диска 46 см.
Подотряд 4 — Pleurodira. Бокошейные
17. СЕМЕЙСТВО PROTEROCHERSIDAE
Четыре рода нз триаса Европы.
18. СЕМЕЙСТВО KALLOKIBOTIIDAE
Один род из мела Европы.
19. СЕМЕЙСТВО THALASSEiMYIDAE
5—9 родов из юры — мела Европы и Сев. Америки.
20. СЕМЕЙСТВО PLESIOCHELYIDAE
5—6 родов из юры — мела Европы и Китая.
21. СЕМЕЙСТВО PELOMEDUSIDAE
Известны, начиная с эоцена, в числе 1—3 вымерших и 3 сов-
ременных родов. Последние различаются так:
1 ( 2) Квадратно-скуловая кость соединяется швом с теменной.
Podocnenis
8 видов, Южн. Америка и Мадагаскар.
2( 1) Квадратно-скуловая кость не касается теменной.
3(4) Средние пластинки вытянуты поперек и касаются друг
друга по средней линии; у взрослых передняя часть
брюшного щита подвижная.
Pelusios
5 видов, Африка, Мадагаскар, Маврикий и
Сейшельские о-ва.
4(3) Средние пластинки небольшие и не касаются друг друга
по средней линии; передняя часть брюшного щита не-
подвижная.
168
PetomeduM
I вид. Африка и Мадагаскар.
В ископаемом виде найдены и a Гяропг. Многие поедают лягу
шек, рыб. червей, мягкотелых и др. Волу покидают лишь в поряд-
ке исключения. Наиболее крупный представитель всего семейства
Podocnemis expansa (длина панциря до 77 см) живет в лесным
озерах тропиков Южн. Америки, в случае пересыхания которых
переселяется в реки. Питается преимущественно упавшими пяо
дямн. Взрослые особи, равно как и яйца, употребляются в пишу
Очень большое количество яиц перерабатывается на масло. Опло-
дотворение в воде. Кладка на Амазонке в сентябре — октябре, иа
Оринокко— в марте на песчаных отмелях в ямах до I м диамет-
ром и 45—60 см глубиной. В каждой яме 80—200 яиц. Инкубация
около 40 дней.
22. СЕМЕЙСТВО BOTHREMYIDAE
' Пять родов в мелу — эоцене Европы и Сев. Америки.
23. СЕМЕЙСТВО CHELIDAE. ЗМЕИНОШЕЙНЫЕ ЧЕРЕПАХИ
Известны, начиная с мела, в числе 3 вымерших и 10 современ-
ных родов. Главные из последних отличаются так:
1 ( 6) Шея длиннее спинной части позвоночника.
2 ( 3) На передней ноге 5 когтей.
Chelus
1 вид, Бразилия, Венесуэла,-Гвиана.
3 ( 2) На передней ноге 4 когтя.
4(5) Остистые пластинки имеются; шейный щиток не касает-
ся переднего края спинного щита, имитируя шестой по-
звоночный.
Hydromedusa
2 вида, Южн. Америка.
5(4) Остистых пластинок нет; шейный щиток касается перед-
него края спинного щита.
Chelodina
8 видов, Сев.-Вост. Австралия.
6(1) Шея короче спинной части позвоночника.
7 (12) Первый лозвоночный щиток шире второго; симфиз ниж-
ней челюсти узкий.
6 (11) Остистые пластинки имеются.
9 (10) Теменные кости сверху узкие: совместная наименьшая
ширина их укладывается в длине шва между ними 8
и более раз.
16»
Batrachemy*
I вид. Южн. Америка.
10 ( 9) Теменные кости сверху широкие; совместная и«И1ц.
шая ширина их укладывается в длине шва между
2,5— 3 раза. J "•*
Phrynops
2 вида, Южн. Америка.
11 ( 8) Остистых пластинок нет.
Platemys
4 вида, Южн. Америка.
12(7) Первый позвоночный щиток не шире второго; симфц,
нижней челюсти широкий. ’
13 (14) На верхней челюсти срединного гребня нет.
Emydura
10 видов, Новая Гвинея и Австралия.
14 (13) Срединный гребень на верхней челюсти имеется.
Elseya
1 вид, Сев. и Сев.-Вост. Австралия.
Водные хищники. Добычу не схватывают, а молниеносно втя-
гивают в себя (например, головастиков) вместе с некоторым ко-
личеством воды, потом выбрасываемой обратно. Матамата,
Chelus fimbriata, имеет нос, вытянутый в мясистую трубочку’
Глаза очень малы и сдвинуты далеко вперед. Кожа головы и шеи
образует многочисленные выросты. Длина панциря до 38 см. Жи-
вет в реках, активна ночью. Представители рода Chelodina,
напротив, активны днем. Берут пищу (черви, гусеницы, малень-
кие лягушки) быстрым боковым движением головы.
24. СЕМЕЙСТВО POLYSTERNIDAE
Два рода в олигоцене — плиоцене Европы.
25. СЕМЕЙСТВО EUSARKIIDAE
Один род в эоцене Туниса.
Подотряд 5 — Atheca, Бесщитковые черепахи
26. СЕМЕЙСТВО DERMOCHELYDIDAE. КОЖИСТЫЕ ЧЕРЕПАХИ
Известны, начиная с эоцена, в числе 2—3 вымерших и 1 сов-
ременного рода. Единственный современный вид Dermochelys со-
riacea встречается в Средиземном море, Атлантическом, Тихом и
Индийском океанах, но всюду весьма редок (с 1558 г. поймано
170
•сего около 40 экземпляров) Длина панциря до 2 л, а
ООО кг. Пуглива. Ныряет при первой тревоге и приближается к
берегу только для размножения. Питается медузами, мягкотелы-
ми, иглокожими, ракообразными, рыбами и водорослями. Ра^
ыиожаются в районах между 20° северной и 26е южной широт
Самка кладет 90—150 яиц, диаметром до 54 мм. Яйим находят
круглый год. Инкубационный период 65—70 дней.
ЛИТЕРАТУРА
В е u х Te8tu<iinata (in: Traitfc de Pal&mtologie, Paris, 1966, 5
goulenger G. Catalogue of the Chelonians, Rhynchocephalians and
Crocodiles in the British Museum, London, 1889.
fjeraniyagala P. Testudinate Evolution. Proc. Zool. Soc. London, 1930
part. IV.
Hoffmann C. Chelonii (in: Bronn’s Klassen und Ordnunden des Tierrei-
reichs, 6: 1—442, 1879).
Mertens R. und Wermuth H. Die rezenten Schildknoten, Krokodile und
Bruckenechsen, Zool. Jahrbuch (Syst.), 1955, 83, Heft 5: 323—440.
Siebenrock F. Synopsis der rezenten Schildkroten mit Berucksichtigung
dec in historischer Zeit ausgestorbenen Arten, Zool. Jahrbuch, 1909, Suppl.
10: 427—618.
ar a t so n D. Eunotosaurus africanus Seeley, and the Ancestry of the Chelonia,
Proc. Zool. Soc. London, 1914, 69.
Глава восьмая
КЛЮВОГОЛОВЫЕ
Значение единственного современного представителя эт<Я
группы было понято не сразу. В 1831 г. Грэй, имея только черт
этого животного, дал ему название «Sphaenodon», позднее им ж<
измененное на «Sphenodon». Спустя 11 лет в его руки попаи
цельный экземпляр животного, который он описал под именем
Hatteria punctata и отнес к семейству агам. Только спустя 30 лет
Грэй установил, что Sphenodon и Hatteria идентичны. Еще до
этого в 1867 г. Гюнтер показал, что сходство рассматриваемого
животного с ящерицами чисто внешнее, а по внутреннему же
строению оно стоит совершенно особняком среди всех современ-
ных пресмыкающихся и заслуживает выделения в особый отряд
Rhynchocephalia (от греческого «ринхос» — клюв и «кефалон»—
голова; указание на загибающуюся вниз предчелюстную кость).
Хотя Коп сперва сближал их со змеешейными ящерами и черепа-
хами, а потом со звероподобными, первоначальный взгляд Гюн-
тера о близости клювоголовых с чешуйчатыми оказался наиболее
правильным.
Типичный двудугный череп (рис. 35) с неподвижной квадрат-
ной костью, брюшные ребра, присутствие венозного синуса, отсут-
ствие копулятивного органа и ряд других признаков отличают со-
временного представителя группы от нынешних пресмыкающих-
ся. Эти особенности сохранились неизменными в течение пример-
но 190 млн. лет, ибо возникновение отряда в целом надо отнести
к триасу (|рис. 36). Принимая во внимание вымершие формы,,
можно различить следующие семейства:
1 ( 6) Морда не вытянутая; носовая кость парная [подотряд (1)
Rhynchocephalia vera].
2(5) Костное отверстие ноздрей парное.
3(4) Седалищно-лобковое отверстие имеется; хвост немного
длиннее туловища с головой.
1. Sphenodontidae
4(3) Седалищно-лобкового отверстия нет; хвост очень длин-
ный.
Рис. 35. Представители клювоголовых: / — череп Sphenodon сбоку, 2 — реставрация скелет» СерЬв/отя
(Rhynchosauridae), 5—череп Champsosaurus, 4—внешний вид Spneaoaoa
3. ClaraxHanrtdae
r> ( 2) Костные отверстия ноздрей слиты в одно исвдриве
2. RhynchotaurldM
6(1) Морда вытянута наподобие гавиала; носовая кот ж
парная (подотряд (2) Choristodera].
4. Champaosaurldae
1. СЕМЕЙСТВО SPHENODONTIDAE
Известно, начиная с триаса Европы, Азии, Африки и Сев.
Америки, в числе 7—10 родов, из которых только 1 дожил до
наших дней. Наземные, первоначально небольшие (15—20 см
длиной) животные. Современный представитель Sphenodon рип-
ctatus, «туатера» или «туатара», общая длина достигает 75 см.
Тело сверху покрыто зернистыми чешуйками. На затылке, вдоль
спины и на хвосте гребень из треугольных сжатых пластинок.
Барабанной перепонки и обособленной^ барабанной полости нет.
Отверстие клоаки в виде поперечной щели. Хвост способен реге-
нерировать. Зубы взрослых настолько стираются (исключая
2 больших передних), что старые экземпляры кусают краями
челюсти, подобно черепахам. У молодых по 1—2 зуба на сошнике.
Окраска тусклая оливково-зеленая с мелкими белыми и более
крупными желтыми пятнами. Зрачок вертикальный. Радужина-
тёмно-бурая. До прихода европейцев водилась по всей Новой
Зеландии, но затем стала быстро вымирать в результате пресле-
дования человеком, лесных пожаров, а также от нападения крыс,
одичалых свиней, собак и кошек. До 1870 г. еще встречалась на
самом Северном острове, а в начале нынешнего века уже только
на мелких островах к востоку (в бухтах Вангарей и Пленти) и к
югу (пролив Кука) от Новой Зеландии. Несмотря на охрану за-
174
кона, туатара стала особой редкостью после второй мярояойяой
нм: цена на нее выросла с 40 руб. до 10000 руб. та мкатлвр
Правда, по данным 1949 г., на островах Карева и Стефена чягж
ее стабилизировалось. Туатара устраивает себе норы в теиистм>
ущельях или поселяется в норах буревестников Активна иочыэ
По данным Богерта, наибольшая активность животного прояв-
ляется при температуре тела +9° (пределы от +5* до ИГ)
Бегает плохо. Питается насекомыми, пауками, дождевыми червя-
ми н улитками. В неволе ест мучных червей, ящериц, лягушек,
мышей н куски мяса. Любят воду, подолгу лежат в ней и могут
хорошо плавать. При линьке отмерший эпидермис сбрасывается
кусками. С середины марта до середины августа зимует в норах.
Кладка в ноябре — декабре, вдали от жилых нор. Для кладок
вырывают особые маленькие норы, куда яйца укладывают с по-
мощью лап и рта и засыпают землей. Вход затыкают травой,
листьями или мхом. В кладке 9—17 яиц: длина около 20 мм, ши-
рина 14. Развитие длится 12—14 месяце®.
2. СЕМЕЙСТВО RHYNCHOSAURIDAE
В триасе Европы, Азии, Африки и Южн. Америки 10 родов.
Наземные. По-видимому, питались растениями (?выкапывали
корни). Длина тела 0,3—1,2 м.
3. СЕМЕЙСТВО CLARAZISAURIDAE
Два рода в триасе Швейцарии. Морские. Питались мягкоте-
лыми и ракообразными. Длина около 1 м.
4. СЕМЕЙСТВО CHAMPSOSAURIDAE
В мелу — эоцене Сев. Америки и Европы 3—4 рода. Пресно-
водные, рыбоядные животные со сжатым с боков хвостом. Длина
1,5—2,5 м.
ЛИТЕРАТУРА
Boffert Ch. The Tuatara: why is it a lone Survivor?, Sci. Monthly (New
York), 1953, 76: 163—170.
GfintherA. On the Anatomy of Hatteria, Philos. Trans., 1867, 157, No.
2; 1—34.
Schmidt K. A Visit to Karewa Island, Home of Tuatara, Fieldiana (Zool.},
1953, 34: 153—164.
Wermuth H. Briickenechsen, Aquarien und Terrarien, 1956, 3, No. 2: 56—59.
Wettstein O. Rhynchocephalia, in: Kiikenthal’s Handbuch d. Zoologie,
1936, 72: 1—235.
.... — Гклятая------I>t«
ЯЩЕРИЦЫ
Ящерицы (Sauria = Lacertilia) наиболее обширная группа сов-
ременных пресмыкающихся. Возникнув в юре, они, видимо, про-
должают дифференцироваться и в наши дни.
Передняя часть мозговой коробки ящериц не вполне окосте-
невает— теменные кости никогда не соединяются с основной
клиновидной нисходящими пластинками. Правая и левая ветвн
нижней челюсти обычно соединяются друг с другом неподвижно.
Пояса конечностей (в крайнем случае рудимент таза) имеются
даже у безногих форм. Исключая американских варанов, все
ящерицы имеют мочевой пузырь.
Большинство форм наземные животные, встречаются полувод-
ные, а в прошлом были и чисто водные. Существуют также лазя-
щие, быстро бегающие и летучие формы. Некоторые роются в
земле наподобие червей. Способность быстрого бега обычно
сочетается с удлиненным хвостом. Ящерица с оторванным хвос-
том бежит медленнее, сильнее изгибая туловище. Несение хвос-
том функции выравнивания движения при быстром беге, видимо,
объясняет укорочение его у змееобразных фор-м. Некоторые пред-
ставители агам и игуан, в особенности Chlamydosaurus, могут
бегать на задних ногах. Ящерицы, лазящие по скалам, деревьям
и др. предметам, имеют особенно хорошо развитые когти, а на
нижней поверхности пальцев у них часто имеются листовидные
выросты, обладающие присасывательной способностью. Концы
пальцев многих ящериц, живущих среди сыпучих песков, оторо-
чены по бокам бахромой роговых зубчиков, образованных увели-
ченными чешуями. Это приспособление играет не только роль
«песчаных лыж», но и помогает при рытье норки. У представите-
лей некоторых семейств наблюдается возникновение безногих
форм. В большинстве случаев передние конечности исчезают ра-
нее задних. Иногда мускулатура исчезает раньше костей конеч-
ностей, а иногда наоборот. Безногость сопровождается уменьше-
нием левого легкого. Только у двуходковых редуцировано или
полностью отсутствует правое легкое Испуганные «лм ноИуж
хенмые ящерицы могут шипеть иля фыркать, «мамин яонуж Я1
раздутых легких. Только немногие, нреимушествгмио ночные
издают писк илн чирикают. Из чувств лучше всего ранило >ре
ине. Зрачок большинства круглый. Слух развит очень неравно*
мерно, а обоняние слабо. В случаях, когда язык удлинен я раз
двоен, он играет роль органа осязания.
Большинство ящериц плотоядные животные, питающиеся раз
иообразной пищей (от червей до млекопитающих), но имеются и
растительноядные формы. У последних кишечник длиннее.
У единственной ядовитой ящерицы (Не lode г та) подъязычные
слюнные железы превращены в ядовитые. Во время размножения
между самцами происходят ожесточенные драки. Копулятивный
орган парный и строение его имеет систематическое значение.
Большинство ящериц откладывают яйца, покрытые пергаменто-
образной оболочкой, которая у гекконов со временем твердеет.
Отдельные виды разных семейств яйцеживородны. Обычно это
стоит в связи с холодным климатом или жизнью в горах. Ряд
видов сцинковых может быть отнесен к настоящим живородя-
щим, так как у них развивающийся зародыш получает питание
через кровеносные сосуды <в стенах яйцеводов. В связи с размно-
жением находятся бедренные поры (pori femorales), располага-
ющиеся у представителей ряда семейств на нижней поверхности
бедра. Каждая пора представляет собой ямку или трубковидное
образование, из которого высовывается столик ороговевших кле-
ток. Если ряд бедренных пор укорачивается до 1—2, то их назы-
вают паховыми (pori inguinales). Иногда есть ряд пор впереди
отверстия клоаки — анальные поры (pori anales), изредка -встре-
чаются и задние анальные поры (pori postanales), расположен-
ные на основании хвоста. Число и вид пор специфичен, различа-
ясь у самцов и. самок. Бедренные и паховые поры принято под-
считывать только с одной стороны, а анальные суммарно.
Кожа нормально покрыта чешуей, образованной собственно
кожей и одетой ороговелым эпидермисом. Последний, толстый и
прозрачный, периодически сбрасывается. Обычно линька идет
кусками, исключая безногие формы. Большинство ящериц спо-
собно приобретать более темную или более светлую окраску.
У некоторых появляются яркие новые цвета (хамелеоны). Пиг-
мент (черный, желтый, красный или ирризирующий) залегает в
собственной коже и, реже, в мальпигиевом слое эпидермиса.
Синие и зеленые цвета в большинстве случаев являются оптичес-
ким эффектом тонких структур. В ряде случаев чешуйки заклю-
чают окостенелые пластинки собственно кожи, называемые осте-
одермами.
Совокупность чешуй — «щиткование» (pholidosis) .играет
важную роль в систематике. Чешуя древних форм более однород-
на, но у других она сильно разнится по величине, форме и строе-
нию. Наименее изменены спинные чешуи (squamae), которые
13 П. В. Терентьев 177
могут быть гладкими, бугорчатыми, коническими идя гяжбжц
ними ребрышками («килсяатые»). Очень мелкие чешуйка ма«м
веются зернышками (granular), а крупные—щитками
Особенного развития последние достигают на голове, где лоирм
тая ими верхняя часть называется «шапочкой» (pileus). Кажлщ
щиток имеет свое название. Ня верхней поверхности головы ся*
рели назад обычно лежат щитки: межчелюстной (rontrale). кого
вые (nasalia —supranasalla), лобоносовой (fronton a sale «interna
sale), предлобные (praefrontalia), лобный (frontale), надглаз-
ничные (supraocularia), верхнересничные (supraciliaria), лоботе-
менные (frontoparietalia), межтеменной (interparietale), темен-
ные (parietalia) и затылочный (occipltale). На боковой стороне
головы могут быть щитки задненосовые (postnasalia«nasofrena-
lia), передний скуловой (frenale=loreale), задний скуловой (fre*
nooculare=°postloreale), предглазничные (praeocularia), верхне-
губные (supralabialia), подглазничный (suboculare) и заглазнич-
ные (postocularia). У некоторых форм виски покрыты довольно
крупными височными щитками (temporalia), а у других среди
более мелких образований выделяются центрально-височный
(massetericum), барабанный (tympanale) и верхневисочные
(supratemporalia). В месте соединения нижних челюстей лежит
подбородочный щиток (mentale), сзади которого находятся
нижнечелюстные (inframaxillaria = submaxillaria). Край нижних
челюстей окаймлен нижнегубными (sublabialia) щитками. У не-
которых видов шею от туловища отделяет ряд увеличенных че-
шуй—воротник (collare), впереди которого может лежать попе-
речная горловая складка (plica gularis). У многих нижняя поверх-
ность туловища покрыта увеличенными щитками, передние из ко-
торых считаются грудными, а задние — брюшными. Перед клоа-
кальной щелью часто встречается заднепроходный щиток (апа-
1е), впереди которого иногда лежат более мелкие (praeanalia)
(рис. 37).
Чешуя восстановленного после аутотомии хвоста во многих
случаях отличается от нормальной. Предполагают, что восстанов-
ленная чешуя обладает более древними признаками. Сама ауто-
томия представляет не произвольный, а рефлекторный акт: если
осторожно приклеить живую ящерицу за хвост, то она не может
отбросить часть хвоста и тем освободиться. Напротив, достаточно
небольшого болевого раздражения, как хвост отламывается кон-
вульсивными сокращениями мускулов, причем перелом происхо-
дит посредине одного из позвонков. Подвешивание трупа ящери-
цы за хвост с прикреплением дополнительного груза показывает,
что сам по себе хвост довольно прочен. Регулирующий аутотомию
нервный центр не зависит от головного мозга. Отброшенный ку-
сок хвоста может сохранять подвижность до полусуток. Новый
хвост не содержит позвонков, а лишь палочку или трубку хряща
и соединительной ткани. Способностью аутотомии восстановлен-
ная часть уже не обладает. Те ящерицы, у которых хвост играет
178
специальную роль (весло, орган прикрепления иди защятИ ля*
щены как способности аутотомии, так и регенерации. Частичное
повреждение может вызвать развитие множественных хвостов.
Понятие подотряда ящериц в современном понимании было
Рис. 37. Терминология щиткования ящериц: А — голова живородя-
щей ящерицы сверху, Б — голова живородящей ящерицы сбоку,
В — клоакальная область луговой ящерицы, Г — голова полосатой
ящерицы снизу:
/ — межчелюстной, 2 — носовые, 3 — лобоносовой, 4 — скуловые, 5 — предлоб-
ные, 6 — верхнересничные, 7 — надглазничные. 8 — лобный, 9 — лоботеменные,
10 — теменные, П — межтеменной, 12 — затылочный, 13 — передненосовой»
14 — задненосовой, 15 — передний скуловой, 16 — задний скуловой, 17 — цен-
тральновисочный, 18 — барабанный, 19 — верхнегубные, 20 — нижнечелюстные
щитки, 21 — горловая складка, 22 — воротник, 23 — бедренные поры, 24 — зад-
непроходный щиток, 25 — клоакальная щель, 26 — брюшные щитки
установлено Оуэном в 1841 г. До этого к ним то причисляли кро-
кодилов, то относили безногих ящериц к змеям. Линней разделял
всех ящериц на 2 рода: Lacerta (49 видов) и Draco (2 вида).Уже
Лауренти в 1768 г. различал гекконов, хамелеонов, игуан, васи-
лисков, драконов, агам, сцинков и других. В 1825 г. Латрей уста-
новил семейства Lacertilia, Iguana, Geckotidae, Chamaefeontidae,
13* 179
Stincidoe
t
tytropsidat f
Gerrbosnuridne
\
\
\
LnrerUda?
I
I
Mflwsauri.dM
AigLalosourLdne
AnnlelUdae Xenodauridae
4
CordylLdae^
Lanthanottdae
• Angui.dae
Dibamidae
^DolLchosadrldaB
varanLdae—^ f /
TeLLdae
I
Helodermatidae
.Eublepharldae
'Chamaeleontidw
Euposaurcdae
RgamLdae
Arretosauridae
GekkonLdae j
Xantusudae
UroplatLdae
1
2
•3
-4
Pygopodidae»
krdeosaurcdae \/
Polyglypha-
nodonUdae
i
4
i
Vwc, fta. Филогения ящериц.'. 1 — гекконообразные, 2 — игуанообраэиые, 3 — сцинкообразные. 4—асротсннниеобраяныс
Tetrapoda. Di pod я и Apode. Последуюшие автору лам
ля аналогичное деление, причем лишь Фктнингер IA2G г при
знал двухолковых за ящериц (он различал уже 16 семейств
76 ролов и 169 видов). В 1830 г. Ваг лер положил • основу деле
имя строение языка. Напротив, Дюмериль и Биброя я18.М
1845 гг. уделяли большое внимание типам зубов. Системы Кола и
Булонджера представляли собой комбинацию скелетных прятив-
ков, типов зубов и языка. Последний различал 21 семейство и
1616 видов. Капитальная переработка системы, с учетом особей
иостей мускулатуры, копулятивного аппарата, палеонтологичес-
кого материала и т. п., была произведена Кэмпом (1923). Важ-
ные дополнения к его взглядам сделали Мэкдовелл и Богерт
(1954). Сейчас можно принять такую группировку (рис. 38):
1 (12) Тела позвонков примерно одинаковой ширины иа обоих
концах, слегка перетянутые в середине; имеются подко-
вообразные интерцентры, если же их нет, то язык покрыт
сосочками, носовые кости раздельны, а обе височные
дуги отсутствуют; веки обычно слиты и неподвижны
[надсемейство (a) Gekkota].
2(3) Непарное теменное отверстие имеется.
1. Ardeosauridae
3(2) Теменного отверстия нет.
4(5) Передних конечностей нет, а задние рудиментарны.
2. Pygopodidae
5(4) Обе пары конечностей хорошо развиты.
6(9) Теменные кости раздельны.
7 ( 8) Носовые кости раздельны.
4. Gekkonidae
8(7) Носовые кости слиты.
3. Uroplatidae
9(6) Теменные кости слиты.
10 (11) Язык с сосочками; голова покрыта зернистой чешуей;
веки подвижные.
5. Eublepharidae
11 (10) Язык со складками; голова покрыта щитками; веки сли-
ты и неподвижны.
6. Xantusiidae
12 ( 1) Тела позвонков конические, без интерцентров, если же
нет, то или носовые кости слиты и веки подвижны, или
зубы верхней челюсти плевродонтные, а нижней — акра-
донтные; веки обычно свободны и подвижны.
181
13 (22) Поверхность копулятивного органа самоа вчевсг»»,
межключица обычно T-образпая или яяореобрама»
прямой поверхностный мускул почти всегда прясугстау*?
(налссмгАсгво (в) Iguania).
Н (21) Шейных позвонков 7; язык мясистый, не выбрасывав
шнйся.
15 (16) Нижний конец скуловой кости вытянут назад в доводы»»
длинный отросток (рудимент нижней височной луп).
7. Polyglyphanodontldae
16 (15) Такого отростка на скуловой кости нет.
17 (20) Зубы на верхней и нижней челюсти одного тиля; непар-
ное теменное отверстие обычно имеется.
18 (19) Зубы плевродонтные.
8. Iguanidae
19 (18) Зубы акродонтные.
10. Agamidae
20 (17) Зубы на верхней челюсти плевродонтные, на нижней —
акродонтные.
9. Arretosauridae
21 (14) Шейных позвонков 3; язык червеобразный, выбрасыва-
ющийся далеко вперед.
11. Chamaeleontidae
22 (13) Поверхность копулятивного органа с пластинками или
бахромчатая; межключица обычно крестообразная; пря-
мой поверхностный мускул всегда имеется.
23 (40) Язык с чешуйками или косыми складками; зубы редко
конические и изогнутые, заменяются путем врастания
нового зуба в полость старого; ключица, если имеется,
обычно расширена, часто с отверстием и иногда крючко-
видная [надсемейство (с) Scincomorpha].
24 (31) Чешуя черепитчатая (налегающая), округлая или ок-
ругло-шестиугольная; бедренных пор нет; имеются, по
крайней мере, рудименты брюшных ребер.
25 (26) Имеются верхняя височная и заглазничная дуги, а так-
же грудной пояс.
15. Scincidae
26 (25) Верхней височной и заглазничной дуг нет; грудной пояс
рудиментарный или отсутствует.
27 (28) Переднечелюстные кости раздельны; остеодерм нет.
12. Dibamldae
28 (27) Предчелюстные кости слиты; остеодермы имеются.
29 (30) Ноздря прорезана внутри большого рыльцевого щитка.
182
13. FeyHrHldae
&
3l
32
33
34
35
36
37
38
39
40
41
42
(29) Ноздря помещается между несколькими щитками.
14. Anelytropeldae
/^4) Спинная чешуя, если имеется, зернистая или черепитча-
' тая, а брюшная обычно квадратная и нечерепитчатая;
бедренные поры имеются; брюшные ребра встречаются
редко.
(39) Заглазничная и верхняя височная дуги имеются; тело
' покрыто чешуей.
(36) Остеодермы есть.
(35) Язык короткий, на заднем конце цельный или чуть вы*
v резанный; ключицы нерасширенные; продольной боко-
вой складки нет.
16. Cordylidae
(34) Язык довольно длинный, сзади расщепленный; ключицы
расширенные проксимально; обычно продольная склад-
ка по бокам тела есть.
17. Gerrhosauridae
(33) Остеодерм нет.
(38) Верхняя височная ямка закрыта костями черепа; перед-
ний поверхностный пучок челюстно-подъязычной мышцы
(m. mylohyoideus) и мочевой пузырь имеются.
18. Lacertidae
(37) Верхняя височная ямка открытая; переднего поверхност-
ного пучка челюстно-подъязычной мышцы и мочевого
пузыря нет.
19. Teiidae
(32) Дуги черепа отсутствуют; тело покрыто мягкой кожей,
образующей много колец, в свою очередь, разделенных
на маленькие четырехугольные щитки.
20. Amphisbaenidae
(23) Язык гладкий или с сосочками; зубы часто кониче-
ские и загнутые, заменяются путем вклинивания новых
зубов между старыми; ключица простая [надсемейство
(d) Anguimorpha].
(58) Зубы плевродонтные, если же текодонтные, то конечно-
сти укорочены и приспособлены к полуводному сущест-
вованию.
(53) Костное отверстие наружной ноздри продолжается на-
зад в форме щели, достигающей или почти достигающей
лобной кости; зубы верхней челюсти помещаются впере-
ди вертикали глазницы.
183
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
(52) Скуловая мочи, нс доходят до предлобной, едпямя »у
дсрстпА 2. ядовитых дубов нет
(45) Здглдднмчпав дуга неполная
21. Varanldae
(44) Злгладннчная дуга полная.
(49) Лобные кости раздельны; шейных позвонков 9 в бол*»
(48) Шейных позвонков 13 м более; непарное теменное m
верстис имеется.
25. Dollchosatirldae
(47) Шейных позвонков 9; непарного теменного отверстия
нет.
24. Lanthanotidae
(46) Лобные кости слиты; шейных позвонков 7.
(51) Гиперфалангии нет; таз хорошо развит; имеется крестец
из 2 позвонков; межключица якореобразная.
22. Aigialosauridae
(50) Гиперфалангия имеется; таз сильно редуцирован и
крестцового сочленения нет; межключица сильно упро-
щена или отсутствует.
23. Mosasauridae
(43) Скуловая кость доходит до предлобной; слезное отвер-
стие одно; на нижней челюсти ядовитые зубы.
, 26. Helodermatidae
(42) Костное отверстие наружной ноздри округлое или оваль-
ное; зубной ряд верхней челюсти П|род<злжается назад за
вертикаль переднего края глазницы.
(57) Височная дуга имеется; в нижней челюсти 6 костей.
(56) Скуловая кость не вытянутая; переднеушная кость свя-
зана с теменной на небольшом отрезке.
27. Anguidae
(55) Скуловая кость вытянутая и скульптурированная; перед-
неушная кость крепко соединяется с теменной.
29. Xenosauridae
(54) Височной дуги нет; в нижней челюсти 5 костей.
28. Anniellidae
(41) Зубы текодонтные(?); конечности не укорочены, назем-
ного типа.
30. Euposauridae
НАДСЕМЕЙСТВО (я) GEKKOTA. ГЕККОНООВРЛЗНЫЕ
I СЕМЕЙСТВО AROrO«AL«IDAE
Два рола мелких (череп 1,5— 2 см) ящериц в юре Гермами
и Маньчжурии.
2. СЕМЕЙСТВО PYGOPOD1DAE ЧЕШУЕНОГИ
Змееобразны, с длинным ломким хвостом. Задние коиечносп
в виде плавниковообразных придатков или совсем отсутствую!
В ископаемом состоянии не известны. Описано 8 родов с I/ им
дами. Распространение ограничено Австралией, Тасманией и Но
вой Гвинеей. Главнейшие рода различаются так:
1 (10) Теменные кости парные; голова сверху покрыта снимет
ричными щитками.
2(5) Преанальные поры имеются.
3 (4) Чешуя килеватая.
Pygopus
1 вид.
4 ( 3) Чешуя гладкая.
Cryptodelma
3 вида.
5(2) Преанальных пор нет.
6(7) Чешуя с 2 ребрышками,
Pletholax
1 вид.
7(6) Чешуя гладкая.
8(9) Имеется большой теменной щиток.
Delma
5 видов.
9(8) Теменного щитка нет.
Aprasia
1 вид.
10 ( 1) Теменная кость непарная; голова сверху покрыта мел-
кой чешуей.
Lialis
2 вида.
Новозеландский желтопузик, Pygopus lepidopus (рис. 39),
общая длина достигает 58 см. Не редок в сухих местностях Авст-
ралии и Тасмании. Активен ночью. Питается насекомыми. Неко-
торые рода (например, Aprasia) роются в почве.
12 П. В. Терентьев 185
Рис 39 Представители гекконообразных: ^~“чеРе" АэьТпЬл-У™5 иСверхУ>
J^Pvffoous feoidopus, 3 — Uroplatus fimbriatus, 4 — Ptychozoon homalo-
cephMumP 5-TarePntola mauritaniea, <5 - Eublephans maculanus, 7-Xa„.
tusia vigihs
3. СЕМЕЙСТВО UROPLATIDAE. ПЛОСКОХВОСТЫ (РИС. 40)
Единственный род Uroplatus с 5 видами на Мадагаскаре.
Наиболее известен U. fimbriatus, общая длина достигает более
25 см. Хвост его спереди сужен, а потом расширен листообразно.
Хорошо прыгает. Окрашен под цвет коры.
4. СЕМЕЙСТВО GEKKONIDAE. ЦЕПКОПАЛЫЕ ИЛИ ГЕККОНЫ
Двояковогнутость позвонков, возможно, является вторичным
явлением. Сравнительно мелкие, преимущественно ночные яще-
рицы, обычно с большими глазами; зрачок вертикальный. Лишь
немногие виды имеют настоящие подвижные веки, у остальных
186
км»
SB®
Рис. 40, Ареалы семейств; 1 — Pygopodidae, 2 — Uroplatidae, 3 — Gekkonidae, < — EublepharidM. 5 — Xantvafeda»
до» глап прикрыт, как у пней, прозрачной перепонкой, гоыплогяч
пой мигательной перепонке или нижнему веку Хвост, как пря.
вило, ломкий. Ноги всегда хорошо ратвитм Основная адаптив-
ная радиация идет по пути видоизменения пяльцев, У большим,
ствд они сравнительно короткие, часто соединенные перепонкой
и снабжены на нижней поверхности разнообразными пластин-
ками и выростами, образующими аппарат прикрепления Нали-
чие последнего позволяет бегать по вертикальным поверхностям
и даже по потолку. Отсутствие прикрепительного аппарата у
пустынных форм, вероятно, является вторичным. Кожа нежная,
тусклоокрашенная. Живут на скалах, в трещинах стен, на дере,
вьях и средн песка. Насекомоядны. Издают звуки (отсюда на-
звание «геккон»). Большинство кладет по 2 яйца и только
Naultinus elegans (Нов. Зеландия) и Р achy dactylus blbronl (Аф-
рика) яйцеживородящи. В ископаемом состоянии известны, на-
чиная с третичных отложений (Rhodanogekko) и эоцена (Cadur-
cogekko) из всех частей света. Известно около 300 современных
видов, объединяемых в 70 родов, важнейшие из которых сле-
дующие:
1 (34) Пальцы не расширены или, самое большее, кажутся рас-
ширенными только при основании.
2 (13) Пальцы снизу покрыты мелкими чешуйками.
3 ( *6) Бока пальцев с бахромой.
4(5) Низ пальцев зернистый; спинная чешуя крупная.
Teratoscinciis
4 вида, Средняя и Центр. Азия, Иран, Афга-
нистан, Белуджистан.
5(4) Низ пальцев с килеватыми чешуйками; спинная чешуя
мелкая.
Ceramodactylus
2 вида, Иран, Аравия.
6(3) Пальцы без бахромы.
7 (10) Кожа ладони и ступни вздута подушкообразно.
8(9) Когти есть.
Nephrurus
4 вида, Австралия.
9(8) Когтей нет.
Chondrodactylus
1 вид, Южн. Африка.
Ю ( 7) Подушкообразных вздутий на ладони и ступне нет.
П (12) Когти есть.
Rhynchoedura
1 вид, Австралия.
12J (И) Когтей нет.
1S8
Cetopm
I вил, Южн. Африке.
13 ( 2) Пальцы снизу покрыты поперечными пластинками
|4 (25) Концевые суставы пальцев гораздо уже, чем основной
сустав, так как более или менее сжаты с боков.
|5 (18) Коготь лежит между 3 щитками: маленьким верхним
и 2 большими нижнебоковыми.
16 (17) Веки неподвижные.
Heteronota
3 вида, Австралия.
17 (16) Веки свободные.
Aelurosaurus
3 вида, Индия.
18 (15) Коготь лежит между 2 щитками: маленьким верхним и
большим "нижним.
19 (22) Зрачок круглый.
20 (21) Хвост округлый или плоский.
Gonatodes
18 видов, тропическая Америка, Вест-Индия.
Индия.
21 (20) Хвост ясно сжатый с острым верхним краем.
Pristurus
6 видов, Сев. Африка, Юго-Зап. Азия.
22 (19) Зрачок вертикальный.
23 (24) Хвост конический и очень ломкий.
Gymnodactylis
90 видов, Юго-Вост. Европа, Азия, Полине-
зия, Австралия, тропическая Америка.
24 (23) Хвост с самого основания очень тонкий и не ломкий.
Agamura
2 вида, Иран, Белуджистан.
25 (14) Концевые суставы такой же ширины, как и основной
сустав, и не сжаты с боков.
26 (31) Пальцы задних конечностей по бокам с бахромой.
27 (28) Пальцы передних конечностей с явной бахромой зуб-
чиков.
Crossobamon
1 вид, Средн. Азия, Афганистан.
28 (27) Пальцы передних конечностей с цельным краем или не-
ясной бахромой.
189
и (.чо)
П™П>ечиые пл.стли.» нижней стороны пальнен «ил,
•<w ТЫ\ .
30 (29)
Stenodacfyfat
6 йилоч. Сев.
Поперечные пластинки
31 (26)
32 (33)
33 (32)
34 ( 1)
35 (50)
36 (37)
Африки, Юго-Зап. Азии,
«илей, почти гладкие нли"Л"5?.СТО₽2НЫ
«ли с мелкими бугорками.
Ptenopus
I вид. Южи. Африка.
Пальцы задних конечностей к
ми п^стея по бокам с цельными края-
Спинная чешуя черепицеобразная
Homonota
2 вида, Южй *
жн. Америка.
Спинная чешуя зернИстая
Alsophylax
И видов, сев . .
Пальцы расширены. ФРика, зия.
Пальцы расширены цо
Пластинки на нижней длине-
ThecadactylOs СТ°₽°Не ПаЛЬЦеВ 2 РЯДа'
2 вида, тропик
пролива. ческая Америка, о-ва Торресова
37 (36) Пластинки на нижней ет«
38 (41) Когти отсутствуют й гоРоне пальцев в 1 ряд.
39 (40) Зрачок круглый. всех пальцах.
40 (39)
Phelsuma
8 видов, Мап •
СейшельскиГГаСлКар’ Маврикий, Родригес,
и Андаманские о-ва.
Зрачок вертикальный.
Pachydactylus
10 видов. Африка
41 (38)
42 (45)
43 (44)
Когти имеются хоть на
Некоторые пальцы без когтей пальцах*
Когти имеются на 3-м и 4-м пальце.
Tarentola
6 видов, Средиземноморье, Вест-Индия.
44 (43) Когти имеются на всех пальцах, исключая первый.
Homopholis
2 вида, Южн. Африка.
190
15
16
г
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
(42) Все пальцы с когтями.
(49) Спина покрыта большими чешуйками
(48) Чешуйки спины черепицеобразиы
Geckolepla
2 вида, Мадагаскар.
(47) Чешуйки спины не налегают друг на друге.
Eurydactylus
1 вид, Новая Каледония.
(46) Спина покрыта зернистой чешуей.
Aeluronyx
2 вида, Мадагаскар.
(35) Пальцы расширены только местами.
(82) Последний конечный сустав сжатый.
(71) Сжатый концевой сустав сидит на верхушке расширен-
ной части.
(64) На нижней стороне расширенной части пальцев по 1 ря-
ду пластинок.
(61) Сжатый концевой сустав очень короткий.
(60) Первый палец не имеет сжатого конечного сустава и
когтя.
(59) Конечности снабжены ясной кожистой оторочкой.
(58) По бокам тела хорошо развитая кожистая складка.
Ptychozoon
1 вид, Малайя, Ява, Калимантан (Борнео),
Суматра.
(57) По бокам тела узкая кожистая складка.
Luperosaurus
1 вид, Филиппинские о-ва.
(56) Конечности без кожистой оторочки.
Gecko
8 видов, Китай, Япония.
(55) Первый палец имеет сжатый концевой сустав., и коготь,
как и другие пальцы.
Rhacodactylus
6 видов, Новая Каледония.
(54) Сжатый концевой сустав длинный.
(63) Концевой сустав образует угол с расширенной частью
пальца.
Hoplodactylus
5 видов, Юго-Вост. Индия, о-ва Юго-Вост.
Пацифики.
191
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
/62) Кониевой сустав лежит в одной плоскости с расшир*ч.
' ной частью.
Naultlnua
2 вида, Новая Зеландия.
/53) На нижней стороне расширенной части пальцев 2 ро*
'* плешинок.
(66) Первый палец хорошо развит, но без когтя.
Lepldodactylus
10 видов, Индия,. Австралия, Полинезия.
/65) Первый палец рудиментарный.
(70) Зрачок круглый.
(69) Коготь на первом пальце очень мал, едва заметный.
Lygodactylus
5 видов, Африка, Мадагаскар.
(68) Коготь на первом пальце большой, ясный.
Microscalabotes
1 вид, Мадагаскар.
(67) Зрачок вертикальный.
Spathoscalabotes
1 вид, Индия.
{М\ Сжатый концевой сустав сидит посредине расширенной
части пальца (как бы выходит из нее).
(73) Первый палец рудиментарный.
Perochirus
5 видов, Филиппины, Новые Гебриды, Каро-
линские о-ва.
(72) Первый палец хорошо развит.
(77) На первом пальце конечный сустав не сжат.
(76) Коготь на первом пальце есть.
Aristelliger
2 вида, Центр. Америка, Вест-Индия.
(75) Когтя на первом пальце нет.
Gehyra
10 видов, Индия, Австралия, о-ва Южн. Паци-
фики, зап. берег Мексики.
(74) Конечные суставы всех пальцев сжаты.
(79) Снизу расширенной части пальцев по 1 ряду пластинок.
Phyllopezus
1 вид, Бразилия.
(78) Снизу (расширенной части пальцев по 2 ряда пластинок.
192
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
(81) Спина покрыта крупно* чешу**; ушно* опырспы м
крытое.
Teratoleph
1 вид. Индия.
(80) Спика покрыта мелко* чешуе*; ушное отверстие открм
тое.
Hemidactylus
35 видов. Южн. Европа. Юго-Вост Азия. Аф
рика, тропическая Америка, Полинезия.
(51) Последний сустав расширенный.
(88) Расширение пальцев несет на нижней стороне гребешки.
(85) Гребешки расположены веерообразно.
Ptyodactylus
2 вида, Сев. Африка, Юго-Зап. Азия.
(84) Гребешки расположены поперечно.
(87) Задний край гребешков с тонкой бахромой.
Dactychilikion
1 вид, Южн. Африка.
(86) Задний край гребешков гладкий.
Rhoptropus
1 вид, Юго-Зап. Африка.
(83) Расширение пальцев на нижней стороне несет пла-
стинки.
(90) На каждом пальце задних конечностей по 1 такой пла-
стинке.
Sphaerodactylus
26 видов, Центр, и Южн. Америка, Вест-Ин-
дия, Флорида.
(89) На каждом пальце по 2 пластинки, разделенных про-
дольной бороздкой.
(92) На предпоследнем суставе каждого из 4 внешних паль-
цев находится пара таких же пластинок, как и на рас-
ширенном концевом суставе.
Calo dactylus
1 вид, Южн. Индия.
(91) Нерасширенные суставы всех пальцев одинаковые.
(94) Когтей нет.
Ebenaria
2 вида, Мадагаскар.
(93) Когти есть.
(98) Поперечные пластинки на нижней стороне всех пальцев
простые и очень маленькие.
193
96 (97) Расширенный ион йеной суете» нггж леям»*» с»ер1у
крупными чешуйками, зилчнтслыю отлич»ющИМ||’^а^
чешуи других густлво». ’ *>
Phyllodactytui
40 яилоп. Средиземноморье. Африка, А»,,о
лия» тропическая Америка. ™
97 (96) Расширенный концевой сустав одет сверху мелММй
чешуйками, такими же, как на остальных суставах. *
Dlplodactylus
9 видов, Австралия.
98 (95) Поперечные пластинки на нижней стороне всех пальцу
парные.
Oedura
7 видов, Австралия.
Из гекконов, живущих в пустынях, прежде всего назовем
сцинкового геккона, feratoscincus. Свое название он получил по
характеру чешуи, легко спадающей у живых. От Каспия до Ара-
ла, Казахстана, Ганьсу, Афганистана и Белуджистана включи-
тельно живет Т. scincus. Общая длина достигает 11 см. Держится
на слабо закрепленных или барханистых песках, иногда и на гли-
нистых местах. На день скрывается в вырытой им норе,
глубиной до 80 см, а иногда в норах грызунов. Может
издавать (трением хвоста) легкие шуршащие звуки. Начиная с
июня, делает 2—4 кладки по 1—3 почти круглых яиц (диаметр
15—18 мм). Поедает крупных насекомых и их личинки. Единст-
венный вид гребнепалого геккона, Crossobamon eversmanni, рас-
пространен в песках Средней Азии, Южн. Казахстана, Ирана и
Афганистана. Общая длина до 6 см. На день скрывается в норах,
чаще всего вырытых им самим. Часто влезает на кустарники и
способен перепрыгивать с ветки на ветку. Начиная с мая, не-
сколько кладок яиц, размеры яйца 11X9 мм. При поимке издает
слабый писк.
Наиболее известным представителем рода геккончиков Also-
phylax является A. pipiens, прозванный «пискливым» за способ-
ность издавать протяжный металлический звук как ночью, так и
днем. Распространен от низовий Волги до Центр. Монголии и
Алашаня, а на юг до Сев. Ирана и Сев. Афганистана. Длина тела
с головой до 43 мм. Живет в саксауловых зарослях и в глинистых
обрывах. Прячется в трещины, под камнями и в норы грызунов.
В пасмурную погоду можно встретить и днем. Обширный род го-
лопалых гекконов, Gymnodactylus представлен у нас 4 видами.
В Вост. Закавказье, Сев. Иране, Устюрте и Юго-Зап. Ср. Азии
встречается каспийский геккон, G. caspius. Длина тела с головой
до 75 мм. Живет в расселинах скал, среди развалин, в кучах
камней и в домах. Издает металлический звук. Размер яйца
|2,5 X II мм irr лдспин до Омиргяья жив^т серый геммой,
(3. rusiowl. Кроме того, в Ср Лэйн широко рмспрострмиеи 6 fed-
tschenkol, нязымемый «туркестанским», Многочисленные1 милы
тистопялых гекконов, Phyliodactylui держатся под корою де-
ревьев или под кямкями. Но Сардинии и Корсике живет Ph two
paeus. общая длина до 7 см. Размер яйна 7 8.S мм В странах
Средиземноморья обитают также Hemidactylui turcica* (общая
длина до 10 см) и Tarentola mauritanica (общая длина до 18 см).
Единственный вид рода Ptychozoon— Р. homalocephalum, дости-
гающий общей длины 20 см, живет на деревьях, к стволам кото*
рых приклеивает свои яйца. Наиболее крупный представитель
семейства Gecko stentor (тропическая Азия), общая длина пре*
вышает иногда 36 см.
5. СЕМЕЙСТВО EUBLEPHAR1DAE
Живут в земле, преимущественно в сухих местностях. Извест-
но 7 родов, из которых главные:
I ( 2) Нижняя сторона пальцев зернистая.
Hemitheconyx
1 вид. Зап. Африка.
2(1) Нижняя сторона пальцев покрыта поперечными плас-
тинками.
Eublepharis
6 видов, Южн. Азия, Центр. Америка, Мекси-
ка, Юг США.
Eublepharis macularius, достигающий 30 см, распространен
от Зап. Индии до Ирана и юга Туркмении. Активен ночью, днем
скрывается между камнями. Питается насекомыми, пауками и
скорпионами; к укусам последних, видимо, безразличен.
6. СЕМЕЙСТВО XANTUSIIDAE
Обычно относят к сцинкообразным, в настоящее время опре-
делена их принадлежность к гекконообразным. Пять родов, от-
метим главные:
1 ( 4) Спинная чешуя однородная; надглазничные Щитки име-
ются.
2 ( 3) Яоботеменные щитки большие, отделяющие межтемен-
ной от височных.
Xantusia
3 вида, Калифорния.
3(2) Лоботеменные щитки маленькие; межтеменной касается
височных.
Crlcosaura
1 вид, Куба.
195
чешуя рвач,
яя дг лязии чм*
нет.
Рис. 41. Представители вымерших
игуанообразных: 1 — Polyglyphanodon
sternbergU 2 — Arretosaurus ornatus
4 (1) Спинная
родням;
щитков
Lepldophym*
I вид, Центр Америка
Небольшие (общая длюь
10—30 см), насекомоядные и*
земные ящерицы. У Xaniutfa
vigilis обнаружено яйцеживо,
рождение.
НАДСЕМЕЙСТВО (в) IGL’A-
NIA. ИГУАНООБРАЗНЫЕ
7. СЕМЕЙСТВО
POLYGLYPHANODONTIDAE
Систематическое положе-
ние сомнительно. Зубы, сидя-
щие в лунках (?), на предче-
люстной кости конические, а
на челюстной — расширены в
поперечном направлении. Из-
вестны два рода из верхнеме-
ловых отложений Сев. Амери-
ки. Длина черепа около 8 см.
8. СЕМЕЙСТВО IGUANIDAE.
ИГУАНЫ
Достоверно известны, на-
чиная с олигоцена (Acipri-
оп) — миоцена (Tetralopho-
saurus) Сев. Америки. Начи-
ная с плиоцена, уже попа-
даются современные рода.
Многочисленны ископаемые
•находки, обнаруженные вне
Америки (С hamops, Iguana-
vus, Jguanosaurus etc), но они
сомнительны в смысле систе-
матической принадлежности.
Однако существование в на-
стоящее время представителей на Мадагаскаре и в Полинезии
указывает на широкое распространение игуан (рис. 43). Коп об-
ратил взимание «а удивительный параллелизм родов игуан и
агам, несомненный результат приспособления к сходным эко-
логическим условиям:
196
обр03 Признаки: Iguanidae:
,|СНЭНЯ' 0 Высокий СПИННОЙ И
лдеиес* хвостовой гребень, под-
1. Дг держиваемый остисты-
Иые ми отростками позвон-
ков • ..........ВазШлсщ
2) Вдоль спины и хвоста
умеренно высокий гре-
бень, не поддерживае-
мый остистыми отрост-
ками- . . fguana
3) Слабый спинной пре-
бень или совсем без
гребня, часто горловой
И. Назем- мешок . . . Anolis
ые 1) Бедренные поры име-
ются, . анальные — нет:
а) Хвост с шипами. Hoplocer-
cus . .
б) Хвост длинный, Crotaphy-
простой. tus
2) Бедренных пор нет,
анальные имеются Liolaemus
3) Бедренные и анальные
Agamldae
Н ydrosaurut
Gonyocephalus
Calo les
Uromastix
Liolepis
Agama
поры отсутствуют:
а) Ушное отверстие
явное; спинная че- Phrynoso-
шуя шиповатая. та Moloch.
б) Ушное отверстие
скрыто под кожей;
спина без высоких
шипов » Anota . Phrynocephalus
III. Полу- ‘ Amblyrhyn-
водные. chus Physignathus
Интересно, что жизнь в каменистых пустынях стимулирует
развитие шипов. Большинство насекомоядны, но некоторые
(Iguana, Brachylophus, Amblyrhynchus, Conolophus etc.) расти-
тельноядны. Некоторые виды Sceloporus и Phrynosoma яйцежи-
вородящи. По-видимому, Ctenosaura всеядна, a Leiocephalus са-
rinatus является хищником. Описано более 300 видов, группирую-
щихся в 53 рода. Основные рода имеют следующие отличия:
1 (60) Бедренных пор нет.
2(9) Пальцы расширены или сжаты.
3(8) Концевой сустав приподнят над предпоследним.
4(5) Боковые зубы с полукруглыми вершинами.
197
ChamaeleoUs
I аил. Куба.
5(4) Боковые зубы трех вершинные.
5 ( 7) Хвост может закручиваться.
Xlphocercws
2 вида, тропическая Америка.
7( 6) Хвост ие закручивающийся.
Anolis
Более 300 видов, тропическая я субтропи^
ская Америка.
8 ( 3) Концевой сустав не приподнят.
Norops
2 вида, тропическая Америка.
9(2) Пальцы не расширены.
10 (59) Зубы, сидящие на предчелюстной кости, конические.
11 (48) Чешуйки снизу пальцев явно килеватые.
12 (17) Задняя часть головы более или менее выдается.
13 (14) Горловой мешок имеется.
Corythophanes
3 вида, Центр. Америка, Эквадор.
14 (13) Горлового мешка нет.
15 (16) Пальцы задних ног без кожной оторочки; спинной гре-
бень отсутствует*
Laemanctus
4 вида, Центр. Америка.
16 (15) Пальцы задних ног с кожной оторочкой по наружному
краю; самец со спинным гребнем.
Basiliscus
4 вида, тропическая Америка.
17 (12) Задняя часть головы не выдается.
18 (35) Имеются 1—2 поперечных горловых складки или боль-
шой (заметно больше глаза) затылочный или межтемен-
ной щиток.
19 (28) Грудина без отверстия.
20 (23) Тело более или менее сжато с боков.
21 (22) Брюшных ребер и горлового мешка нет.
Ophryoessa ;
1 вид, Южн. Америка.
22 (21) Брюшные ребра, а у самцов и горловой мешок имеются.
198
23
2<
(24)
(27)
25
26
(26)
27
Eftyaltolde*
8 видов, Южн. Лмврикд.
Тело приплюснутое.
Спинной гребень имеется.
Chafarodon
1 вид, Мадагаскар.
Спинного греби я нет.
Пальцы задних йог сжаты с боков.
Hoplurus
5 видов, Мадагаскар.
Пальцы задних ног приплюснутые, по бокам зазубрен*
ные.
Pristidactylus
1 вид, Патагония.
28 (19) Грудина с отверстием.
29 (32) Хвостовая чешуя мелкая или средняя.
30 (31) Пальцы прямые.
Tropidurus
11 видов, Южн. Америка.
31 (30) Пальцы явно изогнуты в сочленениях.
Uraniscodon
2 вида, Южн. Америка.
32 (29) Хвостовые чешуи крупные и шиповатые.
33 (34) Хвост дочти цилиндрический.
Strobilurus
1 вид, Бразилия.
34 (33) Хвост плоский.
Urocentron
3 вида, Южн. Америка.
35 (18) Поперечных горловых складок нет; затылочный щиток
маленький.
36 (37) Самцы с большим горловым придатком.
Tropidodactylus
1 вид, Венесуэла, Доминика.
37 (36) Придатка на горле нет.
38 (41) Самцы с анальными порами.
39 (40) Спинная чешуя гладкая и не налегает друг на друга.
199
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
200
HekKcphelu* 3 пиля, семеро запад Южи. Ам^**
(Э») Спинная нгшу" кнлгватая и черелктчат., LtoUemti* 22 вида. Южн. Америка (южнее пва
(3S) (45) (44) qX^xBOCT0»1 образует кольца. ’* Брюшные ребра имеются. Scartlscus 1 вид, Парагвай.
(43) Брюшных ребер нет. Stenocercus 7 видов, запад Южн. Америки.
(42) (47) Чешуя, хвоста не образует колец. Брюшных ребер нет. Saccodeira 3 вида, Южн. Америка.
(46) Брюшные ребра имеются. Leiocephalus 17 видов, Вест-Индия, Южн. Америка
(П) (58) (55) (52) Чешуйки снизу пальцев гладкие или с неясным Кй„ Имеется поперечная горловая складка. ки*ем. Тело цилиндрическое или. чуть сжатое. На спине имеется небольшой гребень или ряд зубц0в Enyalius 6 видов, Южн. Америка.
(51) (54) Спинного гребня нет. Спинная чешуя разнородная. Anisolepis 1 вид, Южн. Бразилия.
(53) Спинная чешуя однородная, зернистая. Urostrophus 2 вида, Южн. Америка.
(50) (57) Тело ясно сжатое. Боковые зубы трехвершинные.
Liosaurus
1 вид, Южн. Америка.
(56) Вершины боковых зубов тупые или округлые.
5«
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
Dfplotaeimtt
1 ВИД. П1ТВГОЯИЯ.
(49) Горловой складки нет.
Ctenoblepharis
! вид, Перу.
(10) Зубы, сидящие на предчелюстной кости, трежвершвияые.
Phymaturus
1 вид, Чили.
( 1) Бедренные поры имеются.
(62) Третий и четвертый пальцы задней ноги равны.
Polychrus
3 вида, тропическая Америка.
(61) Четвертый палец задней ноги длиннее третьего.
(66) Зубы, сидящие на предчелюстной кости, равно как я бо-
ковые, трехвершинные.
(65) Хвост сильно сжат с боков; рудиментарная перепонка
между пальцами задних ног.
Amblyrhynchus
1 вид, Галапагосские о-ва.
(64) Хвост округлый, только слегка сжатый; пальцы задних
ног вполне свободные.
Conolophus
1 вид, Галапагосские о-ва.
(63) Только боковые зубы зубчатые или и они конические.
(98) Голова без шипов.
(75) Ряд бедренных пор короткий; имеется поперечная горло-
вая складка.
(72) Спинной гребень имеется.
(71) Самец с горловым мешком; хвост без шипов.
Brachylophus
1 вид, Фиджи и Тонга.
(70) Самец без горлового мешка; при основании хвоста име-
ются шипы.
Ctenosaura
4 вида, Центр. Америка.
(69) Спинного гребня нет.
(74) Спинная чешуя однородная.
Cachryx
1 вид, Юкатан.
(73) Спинная чешуя разнородная.
201
Ам,Р"*’и горловая скл.и,
U1* ПОР ДЛ««"Ы*’
__ <88) Р«* 6*‘“’,е"и Р л „ли РяД эУбио"'
75 (6В) сутСтвуст. «меетс* гребень и
78 (79) роговцдные-
слегка конически,.,
79 (78) Нос покрыт многим
7У ' щитками.
Вест-Индия.
J мешка нет. щитКИ; квост сжатый.
80 Н°аРЛв°еВр°У головы КРУ"«
2йви"а, тРопИЧеСйешуСГхвост цилиндрический,
/головы мелкая чешу
82 (81) - — сев. Амери-
1 гребня или РЯДЯ вУбц°В'
85 (86) В Sauromaluso запад Сев. Америки.
6 видо3» -рла с головой.
т заметно длиннее тела
оа /85) Хвост заме
86 Crotaphytus Америки.
б вйДов, юг я
(84) ЗатЫЛОяная^ешуяоХх Орловых складки.
| $6) ₽°ЬловЖ зазубрены.
90 190 ГвиТ, Нижняя Калифорния.
, второй горловой складки зазубрен.
/оО) абаннвя перепонка видна.
92 Н»р’*"“* КраЙ Н°ГИ бе3 ШИП°В'
»1ПН Сев, Америжм
.а коей ноги несет ряд шипов.
94 (93) Няру*н“* **
3 ?идя. Аризона, Новая Мексика.
95 (92) Б.₽.в»кн«даГ СК₽ЫТ‘-
5 видов, юг Сев. Америки.
96 (89) Хвостовая чешуя крупная.
Uta
20 видов, Сев. Америка и Мексика.
97 (88) Горловых складок нет.
Sceloporus
41 вид, Сев. и Центр. Америка.
98 (67) Задняя часть головы окаймлена костными шипами.
Phrynosoma
16 видов, Сев. Америка и Мексика.
Самый обширный род семейства—Anolis (рис. 42), включает
небольших, яркоокрашенных и быстрых древесных ящериц. За
способность быстро менять окраску их называют «хамелеонами
нового света». A. carolinensis (юго-восток США, Куба), общая
длина достигает 22 см. Верх тела блестяще-зеленый, низ серебри-
сто-белый, на горле красное утолщение. Наиболее крупные, об-
щая длина 44 см и более, A. equestris и A. edwarsii с Кубы и
Ямайки. Род Basiliscus держится обычно на деревьях или кустар-
никах около рек. Хорошо прыгает и плавает. Ест червей и гусе-
ниц. Активен днем. Яйца (13 X 20 мм) откладывают в числе 12—
18 штук в ямку или зарывают у основания древесных стволов.
Инкубация 50—55 дней. В. americanus (Панама, Коста-Рика)
достигает 80 см общей длины. Ярко-зеленая Iguana iguana дости-
гает 1,5 м длины; держится на деревьях. Мясо ее употребляется в
пищу. Напротив, Tropidurus держится среди скал и камней, а
Liolaetnus — на земле среди кустарников. Многочисленные виды
Sceloporus встречаются под корой деревьев, на заборах и в дру-
гих местах. Обитатель пустынь Phrynosoma (общая длина 9—
15 см), будучи схвачена, может выбрызгивать капли крови из
глаз. Conolophus subcristatus живет в вырытых им норах. Осо-
бенно интересна Amblyrhynchus cristatus (общая длина до
137 см), которая держится стадами на морских берегах, выплы-
вая в море за пищей, состоящей из водорослей.
9. СЕМЕЙСТВО ARRETOSAURIDAE
Единственный представитель Arretosaurus ornatus из эоцена
Монголии. Длина черепа, видимо, была около 9 см.
203
Рис. 42. Представители современных игуанообразных; 1______АпоНч г» .
(Эквадор), 2 — Moloch horridus, 3 — Chlamydosaurus kingi,
iguana, 5 — Basiliscus americanus, 6— Phrynosoma cornutum, 7_Draco /^.Uana
e —Uromastix spinicauda, 9 — Chamaeleon Chamaeleon. 10_______ ru 8ns'
leon deremensis, 11 — Brokesia nasus bhama*
204
Рис. 43. Ареалы семейств: 1 — Iguanidae, 2 — Agamidae, 3 — ChamaelewUidae, 4 t.
10 CFMBftCTBO AOAMIOAE. АГАМЫ
ЗйМеЩВЮТ ИГувМ В Стлром C.BCTC. ИчввСТИЫ ЯВЧВИВВ С 1ИЛ1
(Macrocephatosaurun ня Монголии, череп II«5 см), В Евро**
обнаружены в эоцене — олигоцеие (Ацата, Polntochaffwlto)
Ныне описано более 290 современных видов, группирующихся
35 родов (рис. 43). Большинство насекомоядны, некоторые (кв
пример, Лдо/па) питаются смешанной пишей, a Lophura, Liotepi*
н Uromastix питаются растительной пищей. Основные различия
родов следующие:
1 (48) Рот большой; зубы в верхней челюсти вертикальные.
2 (45) Резцы маленькие, конические.
3 (36) Настоящие анальные или бедренные поры отсутствуют.
4 ( 5) Удлиненные ребра поддерживают крыловидные выросты.
Draco
35 видов, Южн. и Юго-Вост. Азия, Зондский
архипелаг.
5(4) Крыловидных выростов по бокам тела нет.
6 (31) Тело не уплощенное.
7(8) На задней ноге только 4 пальца.
Sitana
1 вид, Индия, Цейлон.
8(7) На задней ноге 5 пальцев.
9 (22) Барабанная перепонка скрытая.
10 (15) Спинного гребня нет.
11 (12) Конец морды плоский, простой.
Otocryptis
2 вида, Южн. Индия, Цейлон.
iq Конец морды с придатком или бородавками.
1о (14) Конец морды с большим роговидным (выростом.
Ceratophora
3 вида, Цейлон.
14 (13) Конец морды с коническими бугорками.
Phoxophrys
1 вид, Суматра.
15 (10) Спинной гребень имеется.
меч Г0Рле или впереди груди нет складки.
17 (18) Чешуя мелкая.
Aphanlotls
1 вид, Малайские страны.
18(7) Чешуя очень крупная.
206
Cophott*
2 вила. Суматра. Цейлон.
19 (16) На горле или впереди плеч имеется складка.
20 (21) На горле имеется углообразная складна,
Lyrloceohaltts
1 вид. Цейлон.
2! (20) Косая складка проходит впереди плеч.
Japalura
6 видов, Южн. Китай.
22 ( 9) Барабанная перепонка явная.
23 (24) Конец морды заканчивается длинным сжатым отрост-
ком.
Harpesaurus
1 вид, Ява.
24 (23) Такого отростка нет.
25 (26) Поперек горла ясно выраженная складка.
Gonyocephalus _
25 видов, Вост. Индия, Австралия, Полинезия.
26 (25) Поперечной горловой складки нет или она выражена
слабо.
27 (30) Спинная чешуя по величине неравномерная.
28 (29) Самцы с горловым мешком.
Salea
2 вида, Южн. Индия.
29 (28) Горлового мешка не бывает.
Acanthosaura
9 видов, Вост. Азия.
30 (27) Спинная чешуя равномерная.
Calotes
24 вида, Индия, Малайские страны.
31 ( 6) Тело более или менее уплощенное сверху вниз.
32 (35) Барабанная перепонка явственная.
33 (34) Самцы с мозолистыми чешуями перед клоакальной
щелью, а иногда и на середине брюха.
Agama
60 видов, Южн. Евроца, Юго-Западн. Азия,
Африка.
34 (33) Мозолистых образований перед клоакальной щелью нет.
207
3 кндя, Мидия,
35
36
37
38
39
40
41
42
43
44
45
46
47
48
(32) Барабанная перепонка скрыта.
Phrynocephatas
55 видов, Юго-Вост. Европа, Цяятр,
( 3) По крайней мерс самцы имеют анальные или бедл>:
поры.
(40) Тело уплощено сверху вниз.
(39) Барабанная перепонка явная.
Amphibolurus
14 видов, Австралия.
(38) Барабанная перепонка, скрытая.
Tympanocryptis
2 вида, Австралия.
(37) Тело более или менее сжато с боков.
(42) Шея с большим плащевидным воротником.
Chlamydosaurus
1 вид, Австралия.
(41) Шея без воротника.
(44) Пальцы задних ног по бокам зубчатые.
Physignathus
7 видов, Австралия, Полинезия, Юго-Вост,
Азия.
(43) Пальцы задних ног по бокам с лопастинками.
Lophura
1 вид, Юго-Вост. Азия.
( 2) Резцы в виде 1—2 больших режущих зубов.
(47) Имеются бедренные и анальные поры.
Uromastix
7 видов, Южн. Азия, Сев. Африка.
(46) Настоящих пор нет.
Aporoscelis
2 вида, Вост. Африка.
( 1) Рот очень мал; зубы верхней челюсти горизонтальные.
Moloch
1 вид, Австрия.
Самый оригинальный род Draco: с помощью небольших
«крыльев*, расположенных по бокам туловища между передни-
ми и задними ногами, может совершать планирующие полеты на
208
расстояния до 23 м. Окраска очень яркая Наиболее гамета*
летучмА дракон, D. volant, распространенный от Малаки до Явы
включительно; общая длина более 21 см. Род Agama в яадейфау
не представлен 7 видами, нэ которых степная агама, A. cangmno-
Icnta, распространена от восточного Предкавказья до Казахстана
включительно. Длина тела с головой до 11 см. При нервном воз-
буждении, а также при повышении температуры воздуха взрос-
лые экземпляры быстро меняют окраску: у самцов появляются
ряды пятен на спине, низ и бока тела становятся темно-сииимя,
а хвост в передней половине ярко-желтым. У самок основной фон
становится голубовато-желтым, на спине выступают ржавые
пятна. Живут в саксауловых лесах, в предгорьях и на заросших
песках. Спасаясь от дневной жары, влезают на кусты. Одна —
две (?) кладки, по 5—12 яиц (18 X 10 мм) в каждой. Зимуют
в норах грызунов. В Дагестане, на восточном и центральном Кав-
казе, в Турции, Иране, Афганистане и Закаспие распространена
A. caucasica (длина тела с головой до 15 см). Круглоголовки,
Phrynocephalus, формой и окраской чешуи хорошо имитируют
грунт, на котором живут. Живущие на песках хорошо закапыва-
ются. Некоторые виды яйцеживородящи. Ушастая круглоголов-
ка, Ph. mystaceus, хорошо отличается от всех других видов рода
наличием в углах рта больших складок кожи. Будучи раздраже-
на, она приподнимается нал почвой, открывает рот, скручивает
и раскручивает хвост и подпрыгивает, чем создает устрашающий
эффект. Живет на песках от Предкавказья по всей Ср. Азии до
Ирана и Афганистана включительно. Помимо членистоногих по-
едает иногда и ящериц. Кладка из 2—6 яиц (24—25 X 12—13жж).
Длина тела с головой до 11 см. Такырная круглоголовка. Ph. he-
lioscopus (длина тела с головой до 53 жж), держится на глинистых
почвах, в степях и пустынях от центрального Закавказья до Вост.
Казахстана включительно. Самые мелкие (тело с головой до
47 мм) круглоголовки нашей фауны — хентаунская, Ph. rossikowi
(низовья Аму-Дарьи) и песчаная, Ph. interscapularis (от Каспия
до Арала). Плащеносная ящерица, Chamydosaurus kingi, инте-
ресна не только наличием большой складки кожи на шее, но и
способностью бегать на задних ногах. При этом туловище нахо-
дится в .вертикальном положении. Короткое время бежать подоб-
ным образом могут также Physignathus' и Atnphibolurus. Пред-
ставители Calotes ведут древесный образ жизни, а рода Moloch —
обитатели каменистых пустынь.
11. СЕМЕЙСТВО CHAMAELEONTIDAE. ЧЕРВЕЯЗЫЧНЫЕ
С достоверностью ископаемые неизвестны. Под вопросом на-
ходится принадлежность к этому семейству Mimeosaurus из
мела Монголии и Tinosaurus из эоцена Монголии и Сев.
Америки. Современных представителей объединяют в 3 ро-
да с 85 видами. Центр многообразия форм на Мада-
гаскаре. Большинство всю жизнь проводят на деревьях и кустар-
15 П. В- Терентьев
никах, по которым медленно передвигаются а поясняя насеном^,
и иногда мелких позвоночных. Только в порядке исключая»,
СЛшпае/дел uulgaris в Сахаре живет в вырытых им морах, a ojuti
мз видов — Brookesia — среди опавшей листвы. Тело покрыв
зернистой млн бугорчатой кожей, быстро меняющей спой нле
как под влиянием температуры и освещения, так и в зависима
стн от состояния самого животного. Барабанная перепонка отгу?
ствуст. Глаза почти совсем закрыты сросшимися веками, в сере
дине которых имеется лишь небольшое отверстие. Каждый гл а.
может двигаться независимо от другого. Череп в виде каски <
гребнями. Межключина и часто ключица отсутствуют. Язык ио
жет выбрасываться вперед на расстояние, превышающее полови-
ну тела животного. Мешкообразные выросты легких позволяли
животному раздуваться. Пальцы связаны в два пучка. Хвост
может закручиваться за ветки. Поврежденный не регенерирует.
Большинство яйцекладущие, но некоторые африканские виды
(Chamaeleon pumilis, Ch. melanocephalus etc.) яйцеживородящи.
Некоторые исследователи (Гюнтер, Буленджер, Гадов) противо-
поставляли хамелеонов всем прочим ящерицам как подотряд, но
это неправильно, ибо в этой группе только собрано воедино то,
что встречается порознь у древесных представителей Agamidae,
Iguanidae и других. Так, независимое движение глаз имеется у
Anolis и Polychrus. Изменчивость окраски велика у Anolis, Са-
lotes, Agama. Закручивающийся хвост свойствен Cophotis и
Xiphocercus. Наконец, Phyllurus (Gekkonidae) имеет сходные
конечности. Рода можно различить так:
1(2) Хвост, по меньшей мере, равен телу; чешуи на подошвах
гладкие.
Chamaeleon ( = Chamaeleo)
73 вида, Африка, Мадагаскар, Индия, Цейлон,
Южн. Испания, Мал. Азия, Аравия.
2(1) Хвост короче тела; чешуи на подошве шиповатые.
3(4) Когти простые.
Brookesia
7 видов, Мадагаскар, Носи-Бе*
4(3) Когти двувершинные.
Rhampholeon
5 видов, тропическая Африка.
Наиболее известен обыкновенный хамелеон, Chamaeleon Cha-
maeleon, распространенный по Средиземноморью, общая длина
тела до 30 см. Кладка его состоит из 25—-40 яиц, зарываемых в
землю на глубину 15 см. Инкубация 125—133 дня. Наиболее
крупный вид Ch. oustaleti (Мадагаскар) достигает почти метра.
Самцы некоторых видов, например, Ch. deremensis (Вост. Афри-
ка), имеют рога. Самый мелкий представитель семейства Bro-
okesia minima (общая длина достигает только 33 мм).
210
НАДСЕМЕЙСТВО (с) SC1NCOMORPHA
СЦИНКООБРАЗНЫЕ (р** 44».
12 СЕМЕЙСТВО DfBAMIDAE
Тело червеобразное, лишенное конечностей. Только у генное
имеются рудименты задних конечностей а виде небольших при-
датков по бокам клоакальной щели. Чешуя округляя, черспятча*
тая. Глаза под кожей. Ушного отверстия нет. Один роя с 2 вида
мм. Dibamus novat-guineae, общая длина достигает 16,5 см, най-
ден в Новой Гвинее, на Целебесе н Молуккских островах. На Ни-
кобарских островах живет D, nicobaricus. Образ жизни роюншй.
Рис. 44 Представители некоторых семейств, сцинкообразных: 1 — Trachysaurus
rugosus, 2— Egernia depressa (Юго-Зап/ Австралия), 3 — Lygosoma dorsale
(Новая Гвинея), 4 — Scincus officinalis, 5 — Acontias holomelas (Мадагаскар),
Некоторые зачисляют в это семейство также род Ophiopsiseps с
единственным видом О. nasutus (Австралия), положение которо-
го весьма спорно.
16* Ш
13, CEMBRCTBO FBYLfMlfDAE
Чер»еобр*»иое тело лишено конечностей. Глаза скрыты о
кожей. Ушное отверстие отсутствует. Описано 7 видов, объедки
емых в 3 рода. Главные отличия:
I ( 2) Число продольных чешуй вокруг тела нечетное; вперед
клоакальной щели несколько мелких щитков.
Feyllnla
I вид, Зап. Африка.
2(1) Число продольных рядов чешуй вокруг тела четное; вя«
реди клоакальной щели большой щиток.
Typhlosaurus
5 видов, Южн. Африка.
Feyllnla currorl, общая длина достигает 31 см. Из видов
Typhlosaurus наиболее крупен Т. aurantiacus (Мозамбик) —
до 17 см.
14. СЕМЕЙСТВО ANELYTROPSIDAE
Тело червеобразное. Конечностей нет. Глаза под кожей. Един-
ственный вид Anelytropsis papillosus живет в Мексике, общая
длина достигает 17 см. Образ жизни роющий.
15. СЕМЕЙСТВО SCINCIDAE. СЦИНКОВЫЕ
С достоверностью установлено существование, начиная с эоце-
на (Eutnecoides и Eumeces), в Сев. Америке (рис. 45). На Маври-
кии до XVII столетия обитал Didosaurus — самый крупный из
известных членов семейства (около метра). Описано более 700 со-
временных видов, группируемых в 32—80 родов. Такое многооб-
разие, быть может, зависит от того, что большинство видов
яйцеживородные. Образ жизни самый разнообразный, но многие
предпочитают сухой и песчаный грунт. Конечности устроены
весьма разнообразно и не редко совсем отсутствуют. Безногие
формы возникают независимо в разных родах. Хвост легко ло-
мается и регенерирует. Веки могут становиться неподвижными, а
ушное отверстие исчезать. Мелкие виды питаются преимущест-
венно насекомыми, а более крупные, хоть частично, — сочными
частями растений. Основные рода отличаются так:
I (40) Ноздря прорезана в носовом щитке, в самом заднем крае
межчелюстного или между несколькими щитками.
2 (33) Ноздря не касается межчелюстного щитка.
3 (14) Нёбные кости разделены пр средней линии нёба.
4(7) Верхненосовых щитков нет.
5(6) Непарный затылочный щиток отсутствует.
Egernia
10 видов, Австралия.
6(5) Имеется непарный затылочный щиток, касающийся меж-
теменного.
213
Ctrueia
I «мд. Соломоиоаы о-ва
4) 0rpж поисковые щитки имеются.
* lit) ИоЛД^1’ прорезана а носовом щитке или между межме-
Н ' люстным. первым верхнегубным и задненосовым, а иное
да задненосодым.
<!0) Крыловидные кости с зубами; конечности хорошо раз
$ ' виты.
Eumeces
59 видов, Сев. и ОДнтр. Америка. Азия и Сея.
Африка.
. ( 9) Крыловидные кости без зубов; конечности короткие или
' рудиментарные.
Brachymeles
4 вида, Филиппины.
И(8) Ноздря прорезана только между верхним и нижним но-
совыми или между носовым и верхненосовым.
12 (13) Конечности хорошо’развиты.
Sclncus
9 видов, Сев. Африка, Аравия, Иран.
13 (12) Конечности рудиментарны или отсутствуют.
Ophtofnorus
6 видов, Юго-Вост. Европа, Сев.-Зап. Азия.
14 ( 3) Небные кости встречаются на средней линии н^ба.
15 (30). Барабанная перепонка более или менее погружена ниже
уровня кожи или'с^рытая.
(25) Вырезка нёба заходит вперед до уровня центров глаз;
-» крыловидные кости разделены по средней линии нёба.
17 (22) Верхеносовых щитков нет.
18 (21)' Боковые зубы тупые, с округлой поверхностью или кону-
совидные, примерно одинаковые.
19 (20) Пластинки на нижней стороне пальцев подразделенные.
Trachysaurus
1 вид, Австралия.
20 (19) На нижней поверхности пальце® неразделенные попереч-
ные пластинки.
Tillqua
5 видов, Малайя, Австралия.
2J (18) Боковые зубу маленькие с закругленной вершиной, но
* на каждой стороне один из них резко выделяется своей
величиной.
214
Hemlaphserfodon
I вил, Квинсленд. Австралия.
22 (17) Всрхнеяосовые щитки имеются.
23 (24) Боковые яубы со сжатой, зазубренной вершиной.
Macroadncua
1 вид, о-ва Зеленого мыса.
24 (23) Боковые зубы конические.
Mabuya
90 видов, Африка, Мадагаскар, Южн. Азия,
Центр, и Южн. Америка, Вест-Индия.
25 (16) Вырезка нёба не доходит впереди до уровня центров
глаз; крыловидные кости соприкасаются по средней ли-
нии хоть в своей передней части.
26 (29) Веки подвижны.
27 (28) Когти не втяжные или отсутствуют.
Lygosoma
250 видов, Юго-Вост. Азия, Малайя, Австралия,
Африка, Сев. и Центр. Америка.
28 (27) Когти втяжные.
Ristella
4 вида, Индия.
29 (26) Веки сросшиеся, прозрачные, прикрывающие глаз.
Ablepharus
32 вида, Юго-Вост. Европа, Юго-Зап. Азия,
Африка, Австралия.
30 (15) Барабанная перепонка явственная и поверхностная.
31 (32) Голова в виде костного шлема, хорошо отделенного от
шеи.
Tribolonotus
8 видов, Новая Гвинея.
32 (31) Голова обычного вида.
Tropidophorus
8 видов, Юго-Вост. Азия, Калимантан (Бор-
нео), Филиппины.
33 ( 2) Ноздря прорезана в заднем крае межчелюстного щитка
или между ним и каким-либо другим.
34 (35) Нёбные кости соприкасаются.
Scelotes
13 видов, Африка, Мадагаскар.
35 (34) Нёбные кости по средней линии нёба не соприкасаются.
36 (39) Верхненосовые имеются.
37 (38) Первый верхнегубной не касается ноздри.
215
Ctateldee
12 Нилов. Юж и Европа. Африка. Югп.-и_
Азия **
за (37) Первы* верхнегубиоА касается ноздря.
Sepxtaa
10 ммдов. Южн. Африка. Мадагаскар
39 (36) Верхненосовых нет.
Chalcidoaeps
I внд, Цейлон.
40 ( I) Ноздря прорезана в межчелюстном щитке, с задямм
краем которого соединена горизонтальной щелью.
41 (42) Имеется верхнее веко.
Acontlas
10 видов, Южн. Африка, Мадагаскар, Цейлон.
42 (41) Веки отсутствуют.
Typhlacontias
1 вид, Юго-Зап. Африка.
Единственный вид рода Trachysaurus — Г. rugosus, коротко-
хвост покрыт толстой, бугристой чешуей, общая длина достигает
36 см. Держится в песчаных местностях. Питание смешанное.
Имеет место настоящая живородность, ибо 2—3 его эмбриона
лишены яйцевых оболочек. Подобное же явление свойственно од-
ному из видов исполинских гладких ящериц, Tiliqua (общая дли-
на доходит до 60 см). Обширный род МаЬиуа охватывает неболь-
ших, проворных животных, питающихся насекомыми. Некоторые
его виды яйцекладущи, но заходящая к нам в восточное Закав-
казье и на юг Туркмении и Узбекистана М. aurata (Вост. Афри-
ка, Мал. Азия, Иран) яйцеживородяща. Держится преимущест-
венно на обломках древесных пород в предгорьях, реже — на
песках. Общая длина туловища с головой до 10 см.
Необычайное обилие видов Lygosoma потребовало установле-
ния ряда подродов, которые некоторыми считаются за самостоя-
тельные рода. Часть из них имеет хорошо развитые пятипалые
конечности. Это будут Hinulia (пример L. lesuerii, Австралия),
Otosaurus (L. cumingii, Филиппины), Keneuxia (пример L. stna-
ragdinum, Индо-Малайский архипелаг и Папуасия) и Етоа
(пример L. суапигит, Малакка, Папуасия, Полинезия). Под-
род Homolepida имеет еще пятипалые, но уже короткие ноги
(пример L. ornatum, Новая Зеландия), а у остальных представи-
телей других подродов идет параллельная редукция пальцев и
конечностей. Это, несомненно, результат приспособления к сход-
ным экологическим условиям. По-видимому, большинство видов
предпочитает лесистые или вообще покрытые растительностью
местности, реже встречаясь на голой земле или среди скал.
216
C. smaragdlnum (Индо-Мвлайский архипелаг, Полинезия, Каро-
линские и Соломоновы о-ва), часто встречается на кустах; может
взбираться на высокие деревья, L. nigrum (о-ва Каролинские,
Соломоновы, Самоа, Фиджи и др.) обитает на морских побережь*
ях, при преследовании направляется прямо к воле и бежит даль-
ше по дну. L. ophloscincus (Квинсленд) полностью безнога.
Значительным распространением отличается и близкий род
гологлаз, Ablepharus. вид A. boutonl неправильно рассеян в бо-
лее жарких областях обоих полушарий. У A. branati длина тела
с головой достигает 50 мм\ встречается на юге Ср. Азии, в Иране,
Ираке, Афганистане и Зап. Индии. Держится во влажных, по-
крытых травой долинах и на горных лугах. Часто залезает в
трещины почвы и под камни. Питается насекомыми. Откладывает
3—6 яиц. A. deserti (длина тела с головой до 60 мм) живет пре-
имущественно среди ксерофильной растительности предгорий
Ср. Азии и Казахстана. Откладывает 2—6 яиц. В Таджикистане,
Киргизии и горах Казахстана живет A. alaicus, в Сев. Иране,
Азербайджане и Армении — A. bivittatus, в Зап. Индии и Таджи-
кистане — редкий у нас A. grayanus (ушное отверстие отсутству-
ет). Все упомянутые виды пятипалые, но известны виды и-с мень-
шим числом пальцев, хотя совершенно безногих нет. Распрост-
ранение длинноногих сцинков, Eumeces, также широкое.
Пожалуй, самый крупный вид — Е. algeriensis (общая
длина ' 43 см). На юге Ср. Азии, в Закавказье, Иране,
Мал. Азии и Сев. Африке -встречается Е. schneideri (тело с голо-
вой до 16,5 см). Держится в степях или полупустынях на твердой
почве. Роют норы, где проводят ночь и зимуют. Поедают насе-
комых, мягкотелых, ящериц и других мелких животных. В Южн.
Туркмению заходит из Ирака и Сев.-Зап. Индии Е. taeniolatus.
Настоящий сцинк, Scincus, живет среди сыпучих песков, внут-
ри которых Двигается с большой скоростью («песчаная рыба»
французов). S. officinalis (Сахара и пустыни около Красного
моря) общая длина достигает 21 см. Высушенный и растертый в
порошок раньше широко применялся как лекарство. Арабы упот-
ребляют в пищу. Внутри рода Chalcides, как и у Lygosoma, мож-
но подобрать ряд по редукции пальцев, сопровождаемый, как
обычно, удлинением тела. Самый крупный из современных сцин-
ков — Corucia zebrata (61 см), интересен также закручивающим-
ся хвостом. У рода Acontias из 9 видов 7 безногие. Безног и род
Typhlacontias.
16. СЕМЕЙСТВО CORDYLIDAE ( - ZONURIDAE). ПОЯСОХВОСТЫ
Быстро двигающиеся животные, питающиеся животной пи-
щей. По-видимому, все, исключая Chatnaesaura, яйцекладущи.
В эоцене Франции найден род Pseudolacerta, показывающий бы-
лое, более значительное распространение на север (см. рис. 47).
Описано 4 рода с 20 видами:
217
. цояечиостя хорошо развиты.
I ( !! Чешу» спины крупная, расположенная правильным* «*.
Я < 3 пер<яиым* рядами, с остеодермами.
Cordytat
13 видов, Южн. Африка, Мадагаскар.
Э(2»
4 (®>
Чешуя спины целиком или частично зернистая, без ост»
одер*,
доя спине округлые чешуи перемешаны с зернышками.
Pseudocordylus
1 вид, Южн. Африка.
5 (
Вся спинная чешуя однородно-зернистая.
Platysaurus
3 вида, Южн. Африка.
6(1) Конечности рудиментарны.
Chamaesaura
3 вида, Южн. Африка.
Наиболее крупный вид поясохвостов, Cordylus (—Zonurus),
видимо, С. giganteus, достигающий 40 см. Лучше известен
С cordylus (общая длина 18 см), предпочитающий крутые скло-
ны скалистых областей. Род Chamaesaura разбивают иногда на
три, так как его виды сильно разнятся степенью редукции конеч-
ностей: Ch. (Cricochalis) аепеа (общая длина 32 см), несмотря
на змеевидное тело имеет еще обе пары пятипалых конечностей,
Ch. (Ch.) anguina (52 см) имеет 2 пары конечностей, но палочко-
видных (без пальцев), а у Ch. (Mancus) macrolepis (52,5 см) со-
хранились только палочковидные рудименты задних конечностей.
17. СЕМЕЙСТВО GERRHOSAURIDAE
Подвижные наземные ящерицы с прямоугольной или ромбои-
дальной чешуей и длинным, ломким и способным к регенерации
хвостом. Все, исключая Tracheloptychus, имеют боковую продоль-
ную складку. Описаны рода:
1(6) Ноздря прорезана между двумя носовыми и первым
губным щитком; брюшные щитки образуют прямые по-
перечные серии.
2(5) Нижнее веко покрыто чешуйками.
3(4) Предлобные щитки имеются; конечности хорошо раз-
виты.
Gerrhosaurus
5 видов, тропическая и Южн. Африка.
4(3) Предлобных щитков нет; конечности укороченные или
рудиментарные.
218
TetradacMM
3 вяла, Южя. Африка.
5(2) Нижнее веко с прозрачным диском,
Cordylotaum
2 вида, Юго-Зап. Африка.
g ( I) Ноздря прорезана между двумя носовыми, ыежчелаэет-
ным и первым верхнегубным; брюшные щитки не обра-
зуют прямых поперечных серий.
7(8) Лоботеменных щитков нет; по бокам тела продольная
складка.
Zonosaunis
3 вида, Мадагаскар.
8(7) Лоботеменные имеются; боковой складки нет.
Tracheloptychus
2 вида, Мадагаскар.
Наибольших размеров достигает Gerrhosaurus validus (Южн.
и Юго-Вост. Африка), общая длина доходит до 51 ем. Более
мелкий (до 40 см) G. flavigularis живет в песках и травянистой
степи Южн. и Юго-Вост. Африки. Виды рода Tetradactylus обра-
зуют ряд последовательного уменьшения конечностей: Т. seps
пятипалый, Т. ietradactylus четырехпалый, а Т. africanus имеет
лишь незначительные нерасчлененные рудименты обоих пар
вечностей.
18. СЕМЕЙСТВО LACERTIDAE. НАСТОЯЩИЕ ЯЩЕРИЦЫ
Известны в Европе, начиная с эоцена (Eolacerta, Plesiolacer-
ta, Pseudeumeces, Dracaenosaurus, Lacerta). Некоторые (хюне)
относят плиоценового Eumecoides не к сцинковым, а к настоя-
щим ящерицам. В янтаре с берегов Балтики (олигоцен) обнару-
жена Nucras, ныне живущая только в Африке. Подвижные, сред-
ние и мелкие ящерицы с длинным, ломким и способным к реге-
нерации хвостом. Питаются животной пищей. Все, исключая
Lacerta vivipara, яйцекладущи. У группы скалистых ящериц
Кавказа возможен партеногенез. Известно 156 современных ви-
дов, разбитых на 21 род. Рода отличаются так:
1 (40) Лоботеменные щитки имеются.
2 (29) Брюшные щитки гладкие.
3 (14) Ноздря касается первого верхнегубного щитка или отде-
лена от него узкой перемычкой.
4 (13) Воротник имеется.
5 ( 8) Пальцы снизу с гладкими или бугорчатыми пластин-
ками.
6(7) Спинная чешуя мелкая, много мельче хвостовой.
7
S
9
Ю
11
13
14
15
>'6
17
18
19
20
21
22
Ltcerta
40 вяло», Европа, Сав. и Зал, Азия, Сев. я тр*
пическая Африка.
( 6) Спинная чешуя крупная, почти с хвостовую.
Alffrokkt
6 видов, Южн. Европа, тропическая Афрахд
( 5) Пальцы снизу с км лева ты ми пластинками.
(12) Пальцы без зубчиков.
(11) Носовых щитков 2; на спине 2—5 продольных рядов уае
лнченных чешуй.
Phllochortus
5 видов, Южн. Аравия, Эритрея, Сомали.
(10) Носовых щитков 3—5; вся спинная чешуя мелкая.
Latastia
7 видов, Юго-Вост. Азия, Сев. и тропическая
Африка.
( 9) Пальцы по бокам с зубчиками.
Acanthodactylus
12 видов, Пиренейский полуостров, Сев. Афри-
ка, Южн. Азия.
( 4) Воротника нет.
Psammodromus
4 вида, Юго-Зап. Европа, Сев.-Зап. Африка.
( 3). Ноздря хорошо отделена от первого верхнегубного.
(26) Нижнее веко сплошь чешуйчатое или с небольшим про-
зрачным диском.
(23) Ведренные поры есть.
(20) Спинная чешуя «рупная.
(19) Пальцы снизу с гладкими или бугорчатыми пластин-
ками.
Tropidosaura
1 вид, Южн. Африка.
(18) Пальцы снизу с килеватыми пластинками.
Ichnotropis
6 видов, тропическая и Южн. Африка.
(17) Спинная чешуя мелкая.
(22) Пальцы снизу с гладкими пластинками.
Nucras
5 видов, тропическая и Южн. Африка.
(21) Пальцы снизу с килеватыми пластинками.
Ereorias
<5а’ии?АФ?имВ0СТ 3«" «U~rF
23 (16) Бедренных пор нет.
24 (25) Воротник есть.
Macmahonla
I вид, Афганистан.
2S (24) Воротника нет.
Aporosaura
1 вид, Мосамедиш (Ангола).
26 (15) Нижнее веко с большим (прикрывающим почти весь
глаз) прозрачным диском.
27 (28) Нижнее веко подвижное.
Cabrita
2 вида, Индия, Цейлон. t
28 (27) Нижнее веко сращено с верхним.
Ophisops
5 видов, Юго-Вост. Европа, Южн. Азия, Сев.
Африка.
29 ( 2) Брюшные щитки, по крайней мере, наружные, с ясными
килями.
30 (35) Бедренных пор 10—15.
31 (32) Спинная чешуя мелкая; ноздря только между носовыми
щитками.
Gastropholis
1 вид,. Вост. Африка.
32 (31) Спинная чешуя крупная; ноздря между носовыми и пер-
вым верхнегубным.
33 (34) Носовых щитков 2.
Bedriagaia
1 вид, Центр. Африка.
34 (33) Носовых щитков 3.
Роготега
1 вид, Зап. Африка.
35 (30) Бедренных пор 1—5.
36 (37) Спинная чешуя мелкая.
Apeltoiiotus
1 вид, юг архипелага Рюкю.
37 (36) Спинная чешуя крупная.
22|
38 (39) Пальцы цилиндрические или слегка сжатые е боем
угловатые. ' *
Tachydromus
10 видов» Вост. Азия.
39 (38) Пальцы слегка приплюснутые, а последний сустав сж<т
с боков и расположен под углом»
Plafyplacoptis
1 вид, Южн. Китай, Тайвань.
40 ( I) Самостоятельные лобнотеменные щитки отсутствуют
(слились с межтеменным в один большой щиток).
' Holaspls
1 вид, тропическая Африка.
Род Lacerta известен, начиная с эоцена (L. mucronate из
Франции, близкая к современной L. agilis).. Современные виды
группируют в 6 подродов, которые различаются так:
I (10) Нижнее веко сплошь чешуйчатое.
2(7) Поперечные ряды брюшных чешуй с вырезками между
щитками.
> "3 ( 4) ЗаднеИосовых не менее 2 или 1 задненосовой и 2 перед-
них скуловых; крыловидная кость с зубами.
(1) Lacerta (s. str.)
4(3) Задненосовых 1, редко 2; зубов на крыловидной кости
нет.
5(6) Хвостовые чешуи без шипо».
(3) Zootoca
6(5) Некоторые хвостовые чешуи шиповатые.
(5) Centromastix
7(2) Задний край поперечных рядов брюшных чешуй почти I
прямолинейный. I
8(9) Брюшные щитки расположены в 6—8 продольных ря-
дов; снизу четвертого пальца задней ноги 19—36 плас-
тинок.
(4) Podarcis!
9(8) Брюшные щитки расположены в 8—20 продольных ря-
дов; снизу четвертого пальца задней ноги 24—37 плас-
тинок.
(2) Gallotia
10 ( 1) Нижнее веко с небольшим прозрачным диском.
(6) Thetla
222
Прыткая ящерица, L, agilis является, по-видимому, иаяболе*
примитивным видом рода и принадлежит к иодроду Lactrta
(s. str). Хвост у взрослых особей этого видя ие более чем а 2 ра-
за превышает длину тела с головой. Последняя доходит до II см
Самцы в брачное время зеленые, а самки бурые или коричневые,
распространена в северной и центральной Европе, в Крыму, на
Кавказе и в Сибири, доходя на восток до Саян. Предпочитает
сухие, солнечные места. Хорошо лазает. У нас пробуждается в
апреле, после чего следует линька и спаривание. Самцы дерутся
из-за самок. Кладка из 3—15 яиц, (размером 12—18 X 7—10 мм
каждое. Развитие длится около 56 дней. Ведут оседлый образ
жизни, используя норы млекопитающих или вырывая собствен-
ные. Часто самцов прыткой ящерицы принимают за особей зеле-
ной— L. viridis. Однако у последней хвост длиннее (может пре-
вышать туловище с головой в 2 с лишним раза) и межчелюстной
щиток в половине случаев явно касается ноздри (у прыткой поч-
ти всегда он не касается). Зеленая ящерица распростра-
нена в центральной и южной Европе и Юго-Зап. Азии.
Длина тела с головой может доходить до 15 см. Близ-
кими видами являются L. strigata (Кавказ, Мал.
Азия, Палестина), L. media (Кавказ) и L. parva (Армения, Мал.
Азия). В Южн. Франции, на Пиренейском полуострове и в Сев.
Африке живет очень крупная (тело с головой до 21 см) L. ocetlata
(рис. 46).
Подрод Gallotia охватывает 5 видов ящериц, водящихся на
Канарских островах. Наиболее крупная из них L. simonyi, длина
тела с головой достигает 21 см, а другие значительно меньше.
Характерным представителем подрода Zootoca является живо-
родящая ящерица A. vivipara. Длина тела с головой у этого вида
доходит до 7 см. Молодые — почти черные, а взрослые — сверху
бурого или желтоватого цвета с мелкими пятнышками. Брюхо
самцов оранжевое, а самок желтоватое или зеленоватое. Ареал
захватывает всю центральную и северную Европу и Сибирь до
Сахалина включительно, Держится в хвойных и лиственных ле-
сах, на лугах и болотах. В горах поднимается до 3 км. Может
плавать и бегать по дну водоемов, а иногда зарывается в ил.
Питается жуками, прямокрылыми, двукрылыми, улитками, пау-
ками, дождевыми червями и т. п. Появляется в Зап. Европе в
конце февраля, а у нас в марте — начале апреля, вблизи же се-
верной границы распространения— в середине июня. Примерно
через 3 месяца после спаривания рождается 2—12 молодых. Од-
нако на крайнем юге ареала (Пиренеи) эта ящерица откладывает
яйца. Общая длина новорожденных 34—42 мм. Половозрелость
наступает на 3-м году жизни. Уходит на зимовку на севере в кон-
це августа, в средних широтах — в октябре, а на атлантическом
побережье иногда остается активной -всю зиму. Родственными
формами этому виду являются L. derjugini (Грузия, Азербайджан,
Краснодарский край) и L. praticola (Кавказ, Карпаты). Подрод
223
Ряс. 48. Представители некоторых семейств сцинкообразных: / — Lacerta
ocellata, 2 — Cordylus giganteus, 3—4— Eremias spekii (Вост. Африка),
5 — Chamkesaura anguina, 6 — Gerrhosaurus flavigularis, 7 — Euchirotes ca-
naliculatus, 8 — Amphisbaena fuliginosa (тропическая Южн. Америка и Вест-
Индия), 9 — Tupinambis nigropunctata (Гвиана, Бразилия, Перу), 10 и 14 —
Lygoeoma mustelinum (Южн. Австрапия), 11 и 12 — Ly go soma afrlcanum
(Зап. Африка), 13 и 13— Lygosoma challenged (Квинсленд)*
224
Podarcis охватывает группу видов, систематика которыж трудив
в запутана. Средиземноморские представители ггого подрода то
собираются в одни вид стенкой ящерицы L muralit, то рааби
каются на несколько видов. Во всяком случае их известно около
трех десятков местных форм. У нас я Крыму, на Кавказе я в
Туркмении встречается близкий вид скалистой ящерицы, L. taxi-
cola, также, по-видимому, являющийся сборным, для него уста-
новлено уже свыше десятка разновидностей, некоторые из кото-
рых в будущем безусловно будут признаны за самостоятельные
виды. Длина тела с головой у скалистых ящериц достигает 8 см,
окраска же и щиткование варьируют в широких пределах. Дер*
жатся на скалах и глыбах корней, попадаясь также в лесу на
о18^8? деРевьев. В конце июля — начале августа откладывается
яйца. Длина новорожденных около 2,5 см. Родственными
скалистым ящерицам являются Z. taurica (Крым),£. chlorogaster
(обе последние —Закавказье и Иран). Един*
пРедставитель подрода Centromastix — L. echi-
™~1а (Зап. Африка), а подрода Thetia — L. perspicillata (Ал-
жир и Марокко).
Богато представлен в нашей фауне род Eremias, ящур-
ка. Виды этого рода, пальцы которых оторочены по бокам
зубчиками, раньше выделяли в самостоятельный род Scapteira.
иднако наличие многочисленных переходов делает такое деление
неприемлемым, равно как подвергает сомнению попытку разбить
£ОДх retnias на секции или подрода. У нас широко распростране-
на быстрая ящурка, Е. velox, от Азербайджана и Предкавка-
зья до Сев.-Зап. Китая. Пальцы у нее без зубчиков. Под-
глазничный щиток касается края рта. Молодые сверху с
продольными черными полосами, а взрослые — с темными пятна-
ми. У самцов на боках бывают голубые пятна, а у молодых низ
хвоста и голеней красные. Длина тела с головой до 8 см. Дер-
жатся на песчаных почвах, реже — на глинистых и солончаках,
обычно среди растительности. Роет неглубокие норки или прячет-
ся в норы других животных и под камнями. Часто наблюдается
2 кладки, по 4—12 яиц в каждой. Половозрелость наступает че-
рез год. Поедает жуков, муравьев, термитов, саранчевых и пауко-
образных. Отличительным признаком разноцветной ящурки Е. аг-
guia (от сев.-вост. Румынии, через Украину и Ср. Азию до
Джунгарии и Кашгарии включительно) служит подглазничный
щиток, не касающийся края рта. Сверху с неправильными чер-
ными пятнами или поперечными полосками. Длина тела с голо-
вой до 10 см. В Ср. Азии придерживается плотных почв, а в Ев-
ропе предпочитает пески. Примером .видов с зубчиками на
пальцах может служить полосатая ящурка, Е. scripta, живущая
на барханных песках Ср. Азии, Ирана и Белуджистана. Длина
тела с головой у нее доходит до 5 см. На ночь и в самые жаркие
часы дня она зарывается в песок, проделывая в нем длинные ходы.
В течение лета самка несколькораз откладывает по одному яйцу.
225
1». СЕМЕЙСТВО ТЕ It DAE АМЕРИКАНСКИЕ ВАРАНЫ (рее.
Известны, начиная с олмгоцена (современные рода). Семе*
сто обязано своим русским названием конвергентному сходству
своих крупных представителей с варанами Старого Света, ио оно
включает и более мелкие формы, напоминающие представителей
Lacertidae. Наблюдается редукция пальцев и конечностей, напо-
добие Scincidac, хотя большинство имеет хорошо развитые конеч-
ности. Голова покрыта крупными симметричными щитками, кото-
рые в отличие от таковых у Lacertidae, не срастаются с ниже
лежащими костями. Преимущественно хищники. Описано около
160 видов, (разбитых на 46 родов. Отличия главнейших родов
таковы:
1 (16) Передний носовой щиток не отделяется лобоносовым от
окружающих щитков.
2(9) Чешуйчатая часть языка цельная или со слабой выем-
кой, могущая втягиваться в основную часть языка.
3(6) Больше 20 продольных рядов брюшных щитков.
4(5) Спинная чешуя однородная; хвост не сжат или чуть
сжат с боков.
Tupinambis
3 вида, Южн. Америка, Вест-Индия.
5(4) Спинная чешуя разнородная; хвост сильно сжат с бо-
ков, с двойным продольным гребнем на верхней стороне.
Dracaena
1 вид, Южн. Америка.
6(3) Меньше 20 продольных рядов брюшных щитков.
7(8) Брюшные щитки килеватые.
Centropyx
5 видов, Южн. Америка.
8(7) Брюшные щитки гладкие.
Ameiva
27 видов, Центр, и Южн. Америка, Вест-Индия.
9 ( 2) Чешуйчатая часть языка стреловидная, двоякорасщеп-
ленная, не втяжная.
10 (15) Хвост округлый.
11 (12) Зубы сжаты в продольном направлении.
Cnemidophorus
19 видов, Сев. и Южн. Америка.
12 (11) Зубы сжаты в поперечном направлении.
1Q Uл л а пая чтит oMf'il'V Й ТТЯ.7ТЫ16В.
227
Рис. 47. Ареалы семейств: /—Cordylidae, 2—Gerrhosauridae, 3 — Lacertidae. / —T-Ji.»—
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
5 МИДОВ, 1ВПЖЯ
<I3> Hi вал»** *°riM * noWt*’
Teiwi
I жид, Южн. А мерим.
(10) Хвост сильно ежатмй с боков.
Crocodllurvs
1 вид, тропическая я Южн. Америка.
( 1) Носовой щиток отделяется от окружающих I— 2 лобс
(44) Ушное отверстие явное.
(37) Все 5 пальцев передней ноги снабжены когтями.
(34) Предлобные щитки имеются.
(21) Хвост с зазубренным гребнем, сжатый с боков,
Neusticurus
2 вида, тропическая и Южн? Америка.
(20) Хвост округлый или округло-четырехугольный.
(23) Язык покрыт косыми складками.
Apologlossus
3 вида, Эквадор, Перу.
(22) Язык покрыт чешуевидными сосочками.
(27) Спинная чешуя с ясными чешуйками.
(26) Спинные чешуи образуют только поперечные ряды.
Prionodactylus
5 видов, Бразилия.
(25) Спинные чешуи образуют правильные поперечные и про-
дольные ряды.
Cercosaura
1 вид, Бразилия.
(24) Спинная чешуя гладкая или с очень слабыми реб-
рышками.
(33) Спинная, чешуя не крупнее брюшной.
(30) Чешуя образует непрерывные ряды вокруг тела.
Anadia
4 вида, центр и -север Южн. Америки.
(29) Чешуя не образует непрерывные ряды вокруг тела.
(32) Складка воротника слабо выражена; самцы без бедрен-
ных пор.
Ceplcoptf*
2 вила, Южн. Америка.
31) Складка воротника ясная; самим имеют бедреимма
jS ' доры*
Euspondylus
4 вида, от Венесуэлы до Перу.
Спинная чешуя крупнее брюшной.
Я3 (2 ’ Argalia
1 вид, от Венесуэлы до Колумбии.
/19) Предлобные отсутствуют.
М /U) Спинная чешуя килеватая, отделенная от брюшной поя-
3» ' ' сом мелких щитков.
Oreosaurus
4 вида, север Южн. Америки.
Спинная чешуя гладкая, отделенная от брюшной
складкой.
Proctoporus
4 вида, Эквадор, Перу.
37 (18) Внутренний палец передней ноги лишен когтя или от-
сутствует.
38 (41) Веки развиты.
39 (40) Спинная чешуя узкая, шестиугольно-ланцетовидная.
Perodactylus
1 вид, Бразилия.
40 (39) Спинная чешуя округлая.
Tretioscincus
2 вида, Центр, и Южн. Америка.
41 (38) Век нет.
42 (43) • Предлобных щитков нет.
Micrablepharus
1 вид, Бразилия, Парагвай.
43 (42) Предлобные имеются.
Gymnophthalmus
4 вида, тропическая и Южн. Америка, Вест-
Индия.
44 (17) Ушное отверстие отсутствует.
45 (46) Пальцы хорошо развиты.
Heterodactylus
2 вида, Бразилия.
46 (45) Пальцы рудиментарны или отсутствуют.
47 (48) Пальцы с когтями.
229
ScotecoMtmM
2 вида, север Южн. Америк*.
4® (47) Пальиы без когтей или совсем отсутствуют
Cophiaa
5 видов, север Южн. Америки.
У представителя рода Tuplnambls, тейю— Т, tegulxln, для*
тела с головой достигает 40 см, а хвост раза в 1.5 длинна
Встречается в лесах, среди кустарников, на сахарных плантаж
ях Вест-Индии и Южн. Америки. Каждый экземпляр живет »
самостоятельно вырытой норе. Поедает мышей, лягушек, чер.
вей, насекомых, а также яйца птиц. Предполагают, что зимует
в коре, питаясь запасом плодов. Яйца откладывает внутрь тер.
мятников. Мясо и яйца тейю употребляются в пищу. Единствен,
ный вид рода Dracaena — D. guianensis, длина тела с головой
достигает 33 см. Держится в прибрежных лесных зарослях,
ежедневно затопляемых приливом. Питается моллюсками и ры-
бами. Яйца откладывает в постройки термитов. Из многочислен-
ного рода Ameiva наиболее известна A, surinatnensis, распрост-
раненная до Никарагуа, длина тела с головой до 26 см. Живет
под кустарниками, в сухой листве и среди камней. Яйца откла-
дывает в самостоятельно вырытые норы. На сухих полях и пес-
ках встречаются виды рода Cnemidophorus: С. tnurinus (Гвиана,
Тринидат; длина тела с головой до 14,5 см), С. lemniscatus (по
всей тропической Америке к востоку от Анд; длина тела с голо-
вой до 9 см) и другие. У одного из видов Oreosaurus — О. shrevei
(Тринидат) на боках тела имеются глазчатые пятна, фосфо-
рисцирующие в темноте.
20. СЕМЕЙСТВО AMPHISBAENIDAE. ДВУХОДКОВЫЕ
Известны, начиная с эоцена (Jepsibaena в Сев. Америке и
Omoiotyphlops во Франции). Найденную в олигоцёне Монголии
форму (Crythiosaurus) иногда выделяют в особое семейство. Все
современные представители имеют червеобразную форму, рою-
щие, часто попадаются в муравейниках и термитниках. Корот-
кий, толстый хвост, наличие только одного левого легкого, раз-
двоение затылочного мыщелка у некоторых родов и другие
признаки давали повод некоторым исследователям выделять дву-
ходковых даже в особый отряд. Русское название дано в связи
с тем, что вырывая узкие ходы, они могут двигаться по ним как
вперед, так и назад. Известно 108 видов, объединяемых в 13—
17 родов. Основные из них следующие:
1 (16) Зубы плевродонтные.
2(7) Имеются только передние коцечности.
3(6) Пальцев 5.
4(5) Все пальцы с когтями.
230
EodHrotet
I вид, Нижняя Калифорния
$ ( 4) Один из палшев укорочен я лишен когтя.
Bipea
1 вид, Мексика.
6 ( 3) Пальцев 3.
Hemichirotes
1 вид, Мексика.
7(2) Конечностей нет совсем.
8 (13) В районе плеча сегменты кожи более или менее увели*
чены или расположены под углом.
9 (12) Хвост цилиндрический, тупой.
Ю (11) Ноздря прорезана в межчелюстном щитке.
Lepostemon
16 видов, Южн. Америка.
11 (10) Ноздря прорезана в отдельном носовом щитке.
Monopeltis
11 видов, Вост. Африка.
12 ( 9) Хвост уплощенный, округлый.
Rhineura
1 вид, Флорида.
13 ( 8) Сегменты района плеча не отличаются от сегментов
других районов.
14 (15) Ноздря прорезана в первом верхнегубном щитке.
Blanus
4 вида, Средиземноморье.
15 (14) Ноздря прорезана в отдельном носовом щитке.
Amphisbaena
44 вида, тропики Южн. Америки и Африки.
16 ( 1) Зубы акродонтные.
17 (18) Преанальные поры есть.
Pachycalamus
1 вид, о-ва Сокотра (Аравийское море).
18 (17) Преанальных пор нет.
Trogonophis
1 вид, Юго-Вост. Африка.
Представитель рода Blanus — В. cinereus имеет на теле 110—
125, а на хвосте 22—23 кольца; общая длина до 32 см. На Пире-
331
ttefe koM полуострове и • Сев Африке живет пол темлей, яят*»,
преммущгс гвгммо ммогоиожками Наиболее к» arc тем «рум^
•ид рода Ampbisbaena ибижара. A alba (обшаи длина 9/» !Ю ш
Обитает • тропической Америке и Вест Индии Видовое латщ
г кое натваииг дано по окраске амниегших спиртовым »куеы»<Л|
ров при жизни животное желто-бурое. На поверхности мм/
встречается лишь ночью или в дождливую погоду. Обычно де|
жите я в термитниках и муравейниках, поедая хозяев Я*
ца. в числе 2—3, откладывают в земле. Представителя род
Ltposternon питаются, видимо, дождевыми червями.
НАДСЕМЕЙСТВО (d) ANGUIMORPHA.
ВЕРЕТЕНИЦЕОБРАЗНЫЕ
Первые 6 семейств этого надсемейства составляют ветвь,
именуемую Platynota (челюсти первоначально приспособлены
для захватывания добычи). Им противопоставляются следую-
щие 3 семейства, как ветвь Diploglossa (челюсти первоначально
приспособлены для раздавливания добычи). Положение послед-
него семейства (Euposauridae) сомнительно.
21. СЕМЕЙСТВО VARANIDAE. ВАРАНЫ СТАРОГО СВЕТА (рис. 48)
Известны, начиная с мела. Подразделяются на 2 подсемейст-
ва: Saniwinae (имеются нёбные зубы, заглазничная кость само-
стоятельна) и Varaninae (нёбных зубов нет, заглазничная кость
обычно слита с заднелобной). Представители первого подсемей-
ства известны из меловых отложений (Telmatosaurus—Монголия,
Palaeosaniwa, Parasaniwa — Сев. Америка), и с палеоцена по
олигоцен Сев. Америки и Европы (Saniwa). К подсемейству
Varaninae относили 8 вымерших родов из третичных отложений
Европы, Азии, Австралии и Африки, которые по новым данным
являются подродами рода Varanus, включающего также
24 современных вида (рис. 49). В плейстоцене Квинсленда встре-
чались гигантские формы (подрод Megalania — до 5—10 м дли-
ной). Некоторые виды и сейчас достигают весьма значительных
размеров. Все это подвижные хищники, большинство которых
прекрасно плавает. Мясо их во многих странах употребляется в
пищу, а кожа идет на разные мелкие поделки. Самый крупный сов-
ременный вид V. komodoensis открыт только в 1914 г. на островах
Комодо и Флорес; общая длина до 365 см (сообщения о том, что
длина их может достигать 10 м, видимо, не заслуживают дове-
рия). Этот вид исключительно наземный. Живет в выкопанных
самим норах. Питается птицами и мелкими млекопитающими.
Нередко поедают диких свиней, убитых охотниками. Более из-
вестен кабарагойа, V. salvator (от‘Цейлона до Южн. Китая и
Сев. Австралии), до 3 ле длиной. Сверху черный с желтыми пят-
232
мн, расположенными поперечными рядами. Живет близ воды
куда спасается при преследовании. Еще более связан с водой
V. niloticus (тропическая и южная Африка); держится преиму-
щественно в береговом кустарнике и среди корней подмытых
деревьев. Питается лягушками, птицами и их яйцами, ящерица*
ми, иногда даже насекомыми и улитками. Часто влезает на на*
клоненные деревья. Общая длина до 170 см. Обитающий на Фи-
липпинах, Суматре и Калимантане (Борнео) V. rudicollis (длина
около 84 еж) живет на деревьях и питается муравьями. Единст-
венный вид нашей фауны — серый варан, V. griseus, больше дру-
гих оторван от воды. Жизнь в песчаных и каменистых пустынях
обусловливает его окраску — желтовато-серую с бурыми попе-
речными полосами. Распространен от Сев.-Зап. Индии до Каспия,
Арала и Сев. Африки. Длина тела с головой до 60 см. Может
взбираться на кусты и деревья. Поедает членистоногих, грызу-
нов, яйца черепах, птиц и др. пищу. Самка зарывает в песок
от 10 до 21 яйца.
22. СЕМЕЙСТВО AIGIALOSAURIDAE
Довольно крупные, 1—2,5 м длиной, полуводные формы мела
Европы (3—5 родов). Тело было удлиненное, хвост сжатый с
боков, а конечности укороченные.
23. СЕМЕЙСТВО MOSASAURIDAE
Крупные (2—15 м) морские ящеры меловой эпохи с длинным
телом и конечностями, превращенными в плавники и лишенными
когтей. Кожа имела зернистую чешую, напоминающую современ-
ных варанов. Раньше выделялись как особый отряд Pythono-
morpha. Описаны многочисленные виды из всех частей света.
Различают 17—25 родов, которые подразделяют на 4 подсемей-
ства, каждое из которых представляет собой особый эколого-
морфологический тип. У Mesosaurinae тело было удлиненное, а
хвост расширен на конце; плавали на поверхности воды. Дер-
жавшиеся на глубоких местах Platecarpinae имели менее удли-
ненное туловище, укороченную шею и однообразный хвост. Пред-
ставители ныряющего типа (Tylosaurinae) имели более вытяну-
тую голову, уши с хрящевой защитой, сравнительно короткую
шею и достигали особенно больших размеров. Globidentinae жи-
ли в прибрежных водах и, как видно по строению зубов, пита-
лись моллюсками.
24. СЕМЕЙСТВО LANTHANOTIDAE
Единственный представитель семейства — Lanthanotus bor-
neensis (Калимантан) долгое время напрасно считался ядовитым
(рис. 50). Тело удлиненное, длина его вместе с головой пример-
но равна длине хвоста. Вдоль верхней поверхности тела тянется
233
„ некоторых семейств веретенницеобразных:
рис. 48. Представители « череп Varanus griseus снизу и сверху,
/ — Varanus salvator, 2—3 мер thiollieri, 6—7—внешний вид и
4—череп Alglaloeaur^ <&₽•!№ 8 - Adrlosaurus
234
Рис. 49. Представители некоторых семейств веретиницеобразных;
J —Lanthanotus borneensis, 2—4— внешний вид, голова сверху и снизу
Shimsaurus crocoailurus, 5— Xenosaurus rackhami 6 — Heloderma sus-
pectum, 7—Diplogloeaus etriatus (Ямайка), 8 — Anniella pulchra
235
несмолы1о р*лов бу горнов Глл1л мвлеянкие. Ушного
ж ру пи мм гол они мм и губим ж щитков нет. Короткие <#•***>»»
пятипалые Окраска сверну красновато коричяевая, еякеу ожрх
стая с коричневыми лентовидными пятнами. Обшяя длшм
около 30 гм
». СЕМЕЙСТВО DOLICHOSAURIDAE
Морские ящерицы со амеевядным телом, 60—00 см лииинЛ
с укороченными конечностями и сжатым с боной хвостом. Овясмо
4—6 родов из меловых (?верхнеюрских) отложений разных мест
Европы и Сев. Африки (Adrlosaurus, рис. 48),
26. СЕМЕЙСТВО HELODERMATIDAE. ЯДОЗУБЫ
Единственный род Heloderma известен, начиная с олигоцеиа
(Я. matthewi, Колорадо). В настоящее время о« представлен
2 видами: Н. horridum (хвост не менее 65% длины тела с голо-
вой, в окраске преобладает черный цвет, от Зап. Мексики до
Техуантепека) и Н. suspectum (хвост составляет41—55% длины
тела с головой, в окраске больше желтого, Невада, Аризона и
Новая Мексика). Общая длина первого из них может достигать
83 см, а второго — 50 см. Попытка зачислить в это семейство
Proiguana из эоцена Франции сомнительна. Тело покрыто бугор-
чатой чешуей. Ушное отверстие и пятипалые конечности хорошо
выражены. Конические зубы челюстей несут спереди и сзади
бороздку, из которых особенно глубока передняя. Железы, на-
ходящиеся под нижней челюстью, выделяют яд, убивающий мел-
ких животных, лягушек, голубей в короткий срок. Для чело-
века укусы очень болезненны. Известны смертные случаи людей.
После укуса у человека ускоряется дыхание и сердцебиение, за-
тем наступает слабость и паралич органов дыхания и сердца.
Держатся в сухих местностях, где днем прячутся в ямах у под-
ножья деревьев или под растительными остатками. Поедают пре-
имущественно мелких грызунов, яйца птиц и пресмыкающихся,
а изредка — самих пресмыкающихся. Кладка составляет от 3 до
13 яиц, хотя в самке находили при вскрытии до 30 яиц. Размеры
каждого яйца 55—75 X 23—39 мм. Инкубация около месяца.
27. СЕМЕЙСТВО ANGUIDAE. ВЕРЕТЕНИЦЕВЫЕ
Известны, начиная с мела (Isodontosaurus в Монголии,
? Peltosaurus в Сев. Америке). Описанные 9—20 родов (в том ।
числе 7 вымерших) иногда подразделяют на 4 подсемейства:
Gerrhonotinae — продольная складка по бокам тела, голова с
большими, правильными остеодермами; Glyptosaurinae — то же,
но голова покрыта многочисленными неправильными остеодер-
мами; Diploglossinae — боковой складки нет, зубы тупые или
236
237
Рис. 50. Ареалы семейств: / —Varanidae, 2 — Lanthanotidae, 3 — Helodermatidae, 4— Anguidae, 5 — АопЫШае, 6 — Xmo*
sauruidae
появятся**; Anguininae — то же, ко зубы загнуты*, клыяояяд*^
Прглстаянтелн подстмейстля Glyptmaurlnae итегтяы голый «
делена олнгоиена Европы, Лама и С* в. Америки.
яэ них достигали 2 м длиной. Внутри семейства а мелом ваш
чаегся во.арастаюшая редукция конечностей. Все фор мм «а>м
мне; питаются насекомыми, червями я моллюсками, исггйонотм
и Anguis я Анеж инородны. Современны* видов считают АО. Глм
ные современные рода семейства различаются следующая
признаками:
1(4) Вдоль боков тела имеется складка.
2(3) Имеется две пары пятипалых конечностей,
Gerrhonotus
19 видов, Сев. и Центр. Америка.
3(2) Конечностей нет совсем или сохраняются рудимент»
задней пары.
Ophisaurus
10 видов, Юго-Вост. Европа, Сев. Африка,
Азия, Сев. Америка и Мексика.
4(1) Боковой складки (нет.
5 (10) Передние конечности или их рудименты имеются.
6(7) Конечности пятипалые.
Diploglossus
16 видов, тропическая Америка.
7(6) Пальцев не больше 4.
8(9) Конечности четырехпалые.
Sauresia
1 вид, Вест-Индия.
9(8) Передние конечности рудиментарные, без пальцев, а
задние со следами 1—2 пальцев.
Ponolopus
1 вид, Вест-Индия.
10 ( 5) Передние конечности отсутствуют.
11 (12) Имеются рудименты задних конечностей; боковые зубы
конические.
Ophiodes
2 вида, Южн. Америка.
12 (11) Конечностей нет совсем; боковые зубы крючковидно-
изогнутые.
Anguis
1 вид, Европа, Зап. Азия, Сев. Африка.
Наиболее крупный вид рода Gerrhonotus — G. liocephalus
(запад Сев. Америки, Мексика), общая длина достигает 40 см.
238
«явчительно меньший (до 34 см) — G muUlcarinatut (запад Сей.
Америки до Техаса и Центр. Америка) живет преимущественно
под корой старых деревьев. Некоторые влезают даже из низкие
кустарники. Род желтопузиков, Ophisaurus в нашей фауне пред-
ставлен одним видом О. a pod us. По бокам клоакальной щели у
О. apodus имеются рудименты задних конечностей (в форме про-
долговатых сосочков). Сверху бурого цвета. Молодые оливково-
серые с темно-бурыми поперечными полосами, длина тела с го-
ловой достигает 45 см. Встречается в поросших кустарником
долинах, на лугах, полях и среди камней Юго-Вост. Европы
(у нас обычен в Крыму, на Кавказе и на юге Ср. Азии). Охотно
идет в воду и может оставаться в ней часами. Поедает улиток,
прямокрылых, жуков, ящериц и грызунов, а иногда птичьи яйца.
Кладка состоит из 10 яиц, длина яйца 32—46 мм. Единственный
вид рода Anguis — A. fragilis, веретеница встречается в наших
лесах, садах, на лугах и полянах, доходя на севере примерно до
64е с. ш. Днем прячется под камнями и пнями. На солнце гре-
ется лишь в холодные дни или ранним утром. Появляется в се-
редине марта — апреля и вскоре следует спаривание. Беремен-
ность длится 12—13 недель, после чего рождается 5—26 молодых.
Зимует в норах до 0,5 м глубиной, где собирается по 20—30 штук.
Сверху бурого, бронзового или медного цвета. Самцы с голубы-
ми или темными пятнами на спине. Молодые сверху серебристые
с узкой черной полоской, а снизу черные. Длина тела с голо-
вой до 26 см.
28. СЕМЕЙСТВО ANNIELLIDAE
Тело червеобразное, безногое. Череп приближается к змеи-
ному типу. Межглазничная перегородка, черепной столбик и че-
шуйчатая кость отсутствуют. Глаза щелеобразные. Ушного от-
верстия нет. Единственный род Anniella имеет 2 вида: A. pulchra
(побережье Тихого океана от Сан-Франциско до Сан-Диего
включительно; общая длина до 19 см) и A. texana (Техас; общая
длина до 23 см).
29. СЕМЕЙСТВО XENOSAURIDAE
Описанные 5 родов (в том числе 2 современных) подразделя-
ются на 3 подсемейства: Melanosaurinae (чешуйчатая кость от-
делена от теМенной, остеодермы касаются друг друга, только
род Melanosaurus в эоцене Сев. Америки), Shinisaurinae (то же,
но остеодермы изолированы друг от друга, Exostinus с мела
по олигоцен Сев. Америки, Necrosaurus в эоцене Европы и совре-
менный Shinisaurus в Южн. Китае), Xenosaurinae (чешуйчатая
кость касается теменной, современный Xenosaurus в Мексике и
Центр. Америке). Каждый из современных родов заключает
всего по 1—2 видам. В настоящее время встречаются очень
239
редко. Общаа длина X*пмаигиз grandts до 24 см. Примерно ,
кмх же размеров и Shinisauruc crocodiluruc (горы в ГУ|Л
Кита*).
30. СЕМЕЙСТВО EUPOSAURIDAE
Мелкие маземные ящерицы юры. Род Euposaurus (Францу
общая длина примерно до 6,5 см, Другие 3 рода сомнительны, ’•
ЛИТЕРАТУРА
Boulenger G. Catalog of the L,zards ,n the Britlsh Museum, London
ВouiVj^er^G5,Monograph of the Lacertidae, London, 2 vols. 1920—21.
Camp Ch. Classification of the Lizards, Bull. Americ. Mus. Nat. Hist., 199,
289 481
McDowell S. and Bogert Ch. The Systematic Position of к.
Lanthanotus and the Affinities of the Arjguimorphan Lizards, Bull. Amerh
Mus. Nat. Hist., 1954, 105: 1—142.
Hoffstetter R. Squamates de type moderne (in: Traite de Paleontologj,
Paris, 1955, 5: 606—640).
240
Глава десятая ’ II—
ЗМЕИ
Отсутствие конечностей «не является единственным и главным
отличием змей — в процессе эволюции ящериц безногие формы
возникали многократно. Движение ящериц с помощью рычаж-
ных конечностей изгибает тело то в одну, то в другую сторону.
Эти волнообразные изгибы выступают на первый план, если
тело вытягивается. Удлинение тела всегда вызывает укорочение,
а потом и Полное исчезновение конечностей. В последнем случае
важное значение приобретают чешуи и ребра. На просторе
изгибы тела в горизонтальной плоскости выражены очень хоро-
шо, но животное часто экономит силы, используя сцепление с
грунтом (движение гадюки по принципу «дождевого червя») или
отталкиваясь от каких-либо предметов. На сыпучем песке змее
приходится иногда забрасывать в сторону значительную часть
тела (боковое движение Cerates и Echis). Напротив, если змея
ползет в узкой норе, то продвижение вперед достигается только
за счет движения ребер и брюшных щитков («принцип улитки»).
Ползанье по ветвям представляет собой модификацию изгибов
тела при ползаньи на грунте. Некоторые змёи при плавании втя-
гивают брюшную стенку — получается особый характер попереч-
ного сечения тела, помогающий при движении в воде (рис. 51).
Своеобразные условия безногого движения предъявляют особые
требования к позвоночнику. Все позвонки передневогнутые. Чис-
ло их не менее 141, у некоторых же форм достигает
435. Каждый позвонок имеет дополнительное сочленение:
особый непарный выступ (zygosphene) и отвечающую ему впади-
НУ (zygantrum) сзади предшествующего позвонка. Обычно уже
второй-четвертый позвонок (начиная с черепа) несет ребра.
Деление позвоночника «на отделы замаскировано. Хвост не лом-
кий и не регенерирует.
Граница, отделяющая змей от ящериц, не ясна (некоторые
склонны, например, рассматривать Scolecophidia как ящериц)
и почти каждый различительный признак нуждается в оговор-
17 П. В. Терентье» 241
Рис. 51. Типы движения: А—схема движения ящерицы, Б—схема дви-
жения медянки (видно продвижение к хвосту волнообразных изгибов
/—в — ползанье гадюки, Г—использование ужом точки опоры,
Д — изменение положения поперечного сечения тела водяного ужа при
плавании, Е —ползанье эфы, Ж — ползанье по «принципу улитки»,
3 — лазанье ужа по палке
242
fX По сути, змеи представляют собой группу посгояиио-безяи
* х ящериц, получившую некоторые дополнительные особен но
<.й По-видимому, ведущую роль в изменении черепа змей игралг
приспособлен нс к заглатыванию добычи целиком Поэтому и)
первоначально стр ептос тили чес кий диапсидный череп потеря?
обе височные дуги (стал гимнокротафный) и развил некоторую
подвижность в районе между предлобными и лобными костями
(стал мезокинетическим). Характерно окостенение передней ча-
сти мозговой коробки, отсутствие непарного теменного отверстия,
межглазничной перегородки, черепного столбика, слезной и ску-
ловой костей. У большинства правая и левая половины нижней
челюсти соединены растяжимой связкой, а верхнечелюстная и
крыловидная кости подвижны. Последние особенности вырабо-
тались уже внутри подотряда змей. То, что большинство авторов
принимает за чешуйчатую кость (squamosum), другие считают
верхневисочной (supratemporale), а некоторые — даже верхне-
ушной (tabulare). В настоящем пособии принята традиционная
старая номенклатура. В частности, поперечная кость (transver-
sum) считается гомологом наружной крыловидной (ectopterygoi-
deum) нижестоящих форм. Барабанная перепонка, полость сред-
него уха, евстахиевы трубы, передние конечности, грудина и мо-
чевой пузырь у змей всегда отсутствуют.
Вопрос о происхождении змей не может считаться решенным.
Несомненно, они являются потомками ствола варанообразных
ящериц (ближе всего — Aigialosauridae) и возникли, вероятно,
в верхней юре, хотя палеонтологические остатки их найдены
только с мела. Выдвинуто 3 возможных причины исчезновения
конечностей. Чаще всего полагают, что утрата конечностей могла
идти за счет приспособления к жизни среди густого кустарника
и камней. Однако подобный взгляд не подтверждается серьезной
аргументацией. В 1940 г. Walls, основываясь на строении глаза
змей, предположил, что первые змеи возникли как роющие фор-
мы. При этом глаза частично дегенерировали, а потом, с выхо-
дом на поверхность, вновь усложнились. Такое допущение плохо
согласуется с установившимися представлениями о закономер-
ностях эволюции. Самое вероятное предположить, что змеи воз-
никли в воде, утратив ноги в результате приспособления к
плаванью. Это хорошо согласуется и с филогенетическими взаимо-
отношениями (полуводные Varanidae и Aigialosauridae и мор-
ские Mosasauridae), и с палеонтологическими материалами (все
древнейшие змеи были морские), и с неоднократно наблюдаемой
безногостью у водных форм (параллель с безногими среди зем-
новодных и угрями среди рыб). С этой точки зрения понятны и
некоторые особенности анатомического строения змей — напри-
мер, превращение большей части легкого в резервуар для воз-
духа (былой гидростатический аппарат), срастание век в про-
зрачный покров глаза (защита глаза от морской воды) и т. гг.
Вытягивание тела всегда сопровождается вытягиванием в
17*
243
длину н нарушением симметрии внутренних органов. Прямит»
ныс хмеи имеют еще 2 легких (обычно менее раэяито левог
а в дальнейшем остается только одно. Передний конец рамит
го легкого функционирует как дыхательная часть, а задний, глц
костенный, представляет собой лишь вместилище воздуха. Нек/
торне змеи (например, гадюки) имеют видоизмененное сгроеям
части задней стенки трахеи: она расширяется мешкообразна
имеет губчатое строение («трахеальное легкое»), служит дополна
тельным местом газообмена. Желудок представляет собой про
сто некоторое утолщение части пищевода, -плохо отграничен ।
лежит продольно. Печень в -форме длинной лопасти. Желчный
пузырь объемистый. Почки, семянники и яичники вытянуты
причем правые лежат впереди левых. У большинства змей яйца
имеют пергаментоо-бразную оболочку. У некоторых видов яйца
при прохождении через яйцевод склеиваются, образуя подобие
четок. Многие змеи (у нас гадюка) яйцеживородящи. Мелкие
змеи достигают половозрелости обычно на второй год, средних
размеров — на третий, а крупные удавы — на пятый. Некоторые
виды выживают в неволе до трех десятков лет.
Наиболее интересным из органов чувств является раздвоен-
ный, постоянно вытягивающийся и втягивающийся язык. В связи
с последним большинство змей имеет соответствующую вырезку
среди щитков, одевающих верхнюю челюсть. Морские змеи
высовывают только кончики языка и потому у них имеется
2 таких вырезки. Язык служит органом химического чувства: вы-
сунувшись, он прикасается к интересующему предмету, и затем,
будучи втянут, направляет 2 своих кончика в Якобсоновы орга-
ны. Жидкость, наполняющая последние, постоянно сменяется, и
потому следы вещества, принесенные языком, быстро вызывают
ощущение. В процессе принятия пищи язык втягивается в осо-
бое -влагалище и потому говорить о вкусе в обычном смысле сло-
ва нельзя. Обоняние развито хорошо, звуки воспринимаются че-
рез колебания субстрата. Немигающий взгляд змей -породил мно-
жество вздорных слухов о гипнозе и прочем. Под сросшимися ве-
ками глаз омывается слезной жидкостью. Змеи могут двигать
одним глазом, оставляя в покое другой, а так же способны сдви-
гать зрачок в разных направлениях. Перед линькой прозрачная
кожа, прикрывающая глаза, мутнеет и зрение временно ухуд-
шается. Гремучие змеи снабжены особой ямкой, расположенной
между ноздрей и глазом, воспринимающей тепловые лучи («тер-
мический глаз»).
Почти в каждой группе змей имеются роющие формы. При-
способление к -рытью («подземный паразитизм» по выражению
Копа) вызывает дегенерацию глаз, укорочение хвоста, сужение
рта, медленность движений и др. особенности. Такие группы,
как Scolecophidia, Anilidae и Uropeltidae приобрели свои особен-
ности в связи с рытьем. В качестве примеров больших или мень-
ших изменений, возникших в связи с рытьем, из других змей
244
яожно привести Егух (Старый Свет) и Charina (Сев Америка)
яз Boidae, Homalocranium (Америка) и Aparallactus (Африка)
из Qipaadomorphinae, Elaps (Америка) и Vermicella (Австралии)
нз Elapidae и Atractaspis (Африка) из Viperidae.
Все эмен питаются животной пищей, заглатывая ее целиком.
Большинство поедает позвоночных разной величины. Некоторые
(Dasypeltis, Elachistodon) питаются птичьими яйцами. Сущест-
вуют насекомоядные змеи (например, Contla) и даже питаю-
щиеся моллюсками (Amblycephalidae). Лишь немногие змеи
едят свою добычу живьем (например, Matrix), что, по-видимому,
является вторичным явлением. Как правило, жертву сначала
убивают. Древним способом убивания добычи является обма-
чивание ее несколькими кольцами тела змеи и удушение. Так
поступает большинство змей. Однако в процессе эволюции по-
явился и новый способ умерщвления добычи — яд. Не исключе-
но, что отдельные случаи выработки ядовитого аппарата имели
место среди пресмыкающихся довольно давно: подозревают, что
Euchambresia (Theromorpha) из пермских отложений Южн. Аф-
рики обладала ядовитыми железами. Однако ядовитость как
массовое явление возникает только внутри подотряда змей срав-
нительно поздно (примерно в миоцене). О змеином яде, его дей-
ствии, лечении укусов и прочем написано свыше 6000 статей и
книг. Из общего числа около 2300—2500 ныне известных видов
змей ядовитыми являются только 10—13%. На практике больше
половины этих видов столь редки, что их можно не принимать
во внимание. В Европе приходится считаться только с разными
видами гадюк. В Азии, помимо ряда весьма опасных гадюк
(Vipera, Pseudocerastes, Trimeresurus), важную роль играют
Naja, Bungarus и Ancistrodon. В Азии живет крупнейшая и самая
опасная в мире ядовитая змея Naja hannah, достигающая длины
3,6—4,6 м (по непроверенным данным даже 5,5 м). От ее укуса
человек умирает через 3 минуты. В морях Азии многочисленны
ядовитые морские змеи (особенно Hydrophis и Laticauda). В Ин-
дии, в частности, наиболее часто наблюдаются укусы людей
Naja naja, Bungarus caeruleus, Vipera russelii и Echis carinatus.
В Африке особо опасны для человека Causus rhombeatus, В it is
arietans, разные виды Naja, Dendraspis (укусы почти всегда
смертельны) и Dispholis. Около озера Танганьика, в частности,
встречается ядовитая Boulengerina (Elapidae), достигающая
1,8 м длины. Основные ядовитые змеи Сев. Америки — гремучие
(Crotalus), щитомордники (Ancistrodon) и коралловые (Elaps=
Micrurus). Ядовитые змеи Южн. Америки в основном принадле-.
жат к тем же систематическим группам, но представлены боль-
шим числом видов. На первое место претендует Bothrops =
Lachesis. Ядовитые змеи Австралии распределяются в порядке
убывающей вредности так: Acanthophis antarcticus, Notechis
scutatus, Denisonia superba, Demansia*=Diemenia и Pseudechis
ponhyriacus.
245
Трудно получить объективные денные о рдтмерах «умгим'
опасности» («офиднтм»). По-видимому, нл темном шяре ежегг/
но от укусов змей погибает 30 —40 тысяч человек, причем бол|
шня часть падает ня Азию. В Европе и ня большей части ieppj
тории СССР ядовитых амей мало. В западной Европе и прошло
веке было зарегистрировано в среднем 60, а и последнее врем
около 14 укусов людей в год.
На большей части нашей страны человек и домашние живот
ные подвергаются укусам обыкновенной и степной галюш
(смертность от 3 до 5%). Хотя, начиная с Семиречья, к гадюк
присоединяется щитомордник, в Восточной Сибири и на Дальне*
Востоке вред от ядовитых змей немногим выше. Значительна
вред от змей в Закавказье и, особенно, в Средней Азии. Наибо
лее часты укусы в ногу (55%) и в кисти рук (30%). «Молниенос-
ная» смерть ют укусов среди населения СССР очень редка. Со-
всем иначе обстоит дело в других странах. Для Индии приводит-
ся цифра в 20—25. тысяч укусов (по другим данным — даже
смертей!) в год. Можно думать, что в ряде случаев местные вла-
сти регистрировали как смерть от укуса гибель от других при-
чин. Более точные данные, видимо, несколько заниженные, сви-
детельствуют о том, что с 1940 по 1953 г. в Индии был зареги-
стрирован 1231 укус. При ©том в 690 случаях это были укусы
неядовитых змей, а плохое самочувствие укушенных объясня-
лось просто испугом. Количество укусов и смертность в Африке
велики, но меньше чем в Азии. В Сев. Америке, (Соединенных
Штатах) наблюдается в год 2—3 тысячи укусов, из которых смер-
тельный исход имел место в 10—35% случаев. В Бразилии рань-
ше в год регистрировали 19—25 тысяч укусов, а число смертных
случаев составляло 4—5 тысяч (примерно 20—21%). В 1926 г.
было отмечено 3 тысячи смертельных исходов, а в 1930 г.—меньше
100 случаев. Такое поразительное уменьшение смертности яв-
ляется результатом широкого введения лечебных сывороток.
В Австралии с 1910 по 1926 г. было зарегистрировано 195 смерт-
ных случаев.
По-видимому, даже неоднократные укусы не вызывают у че-
ловека иммунитета. Совершенно не обоснованы научно рассказы
о природном иммунитете некоторых людей.
Яд у змей вырабатывается видоизмененной верхней слюнной
железой, помещающейся сзади глаза (рис. 52). Даже введение
слюны в кровь вызывает болезненные явления, чем объясняются
очень редкие, но реальные случаи отравления от укусов неядо-
витых змей. У настоящих ядовитых змей химический состав слю-
ны сильно усложнился и потому ядовитые железы сравнительно
невелики и могут выделять одновременно всего 4—6 капель
яда. Однако имеются змеи, слабость яда которых компенсирует-
ся усиленным развитием желез (у Dollophis занимают бока
всей передней трети тела). У большинства змей зубы гладкие.
Первым приспособлением для введения яда является появление
246
Рис. 52. Ядовитый аппарат змей: А — голова гремучей змеи после удале-
ния кожи, Б — череп гремучей змеи в спокойном положении, В — череп
гремучей змеи с открытым ртом, Г—срез через влагалище ядовитого зуба
гадюки:
/ — ядовитый зуб, 2 — проток ядовитой железы, 3 — средний височный мускул,
4—ядовитая железа, 5 и /0—передний височный мускул, 6 — двубрюшный мускул»
7 — задний височный мускул, 8 — позвоночно-нижнечелюстной мускул, 9 — наружный
крыловидный мускул, // — верхнечелюстная кость, 12 — чешуйчатая кость, 13 — квад-
ратная кость, 14 — нёбная кость, 15 — поперечная кость, 15 — клиновидно-крыловид-
ный мускул, 17 — поперечный срез функционирующего ядовитого зуба» 18—20 — заме-
стители ядовитого зуба в разной степени готовности
продольной боролдкн нл передней стороне «уба. Подобные »f г.
moi у г помещаться либо и ладней части верхнечелюстной
(«задисбиролдчатые» эмен). либо в передней («переднеборб1д
чатыо). Наиболее совершенным является ядовитый зуб, у
торого края бороздки спаялись, образован трубку наподобие л '
лой иглы медицинского шприца («трубкозубые»). Вводное и
водное отверстия такого зуба помещаются на переднем верхнее
и нижнем крае (не на конце!), что высоко целесообразно согл».
совано с распределением давления тканей раны в момент укус»
Обычно имеются зубы-заместители. В наиболее специализирован
ном случае трубчатых зубов изменяется и челюстной аппарат
верхнечелюстная кость становится укороченной («вертикаль,
ной») и приобретает суставное сочленение с черепом. Верхнече-
люстная кость при помощи ряда костей сдвигается при откры-
вании рта таким образом, что ядовитый зуб занимает вертикаль-
ное положение. Напротив, при закрытом рте ядовитый зуб рас-
положен почти горизонтально. Обычный путь отравления жерт-
вы — введение порции яда в ее кровяное русло, но иногда силь-
ное выдыхание воздуха может выносить капли яда, стекающие
с зубов, на значительное расстояние (некоторые виды кобр и
Sepedon «плюют» ядом на расстояние до 3,7 м).
Попадание животных капелек яда в глаза может вызвать раз-
витие катаракты (даже гибель мелких животных). Яд представ-
ляет собой прозрачную, желтую или зеленоватую жидкость,
сложную смесь преимущественно белковых веществ, обычно кис-
лой реакции, с удельным весом 1,030—1,082. При высушивании
или охлаждении яд сохраняет свою силу годами, но нагревание до
65—420° разрушает его. Токсины змеиного яда подразделяются
на нейротоксины, геморрагины и .протеазы. Сложной природе
яда отвечает и сложная картина отравления, причем она специ-
фична для разных групп змей. Можно выделить 2 основных
типа, один из которых свойствен змеям с бороздчатыми зубами
(«ядовитые ужи»), а другой — с трубчатыми («гадюки» в широ-
ком смысле слова). Компоненты яда первой группы при элект-
розе переходят к катоду, а второй — и к катоду, и к аноду. Укус
«ядовитых ужей» действует преимущественно на нервную систе-
му и часто местные явления при нем совсем отсутствуют. Яд
морских змей по действию схож с ядом ужей — вызывает пара-
личи. Для яда гадюк характерно, напротив, прежде всего дейст-.
вне на кровь и кровеносную систему, причем местный эффект
(опухоли, отеки, кровоизлияния) обычно велик. Конкретная
картина отравления, помимо вида змеи, во многом зависит от
места укуса (легкость всасывания, особенно опасно попадание
в кровеносные сосуды), состояния змеи (после линьки ядовитее),
температуры (холод понижает) и, наконец, от вида, возраста и
массы жертвы. Для кролика летальна доза 0,01 мг яда Lachesis
atrox, 0,05 Vipera russellii, 0,15 Naja flava, 0,25 Crotalus ter-
rlficus, 0,35 Vipera aspis, 2,13 Bitis gabonica. Минимальная ле-
248
льНая доза яда Vipera ЫмНпа в мг ня I к? миноги веса жертвм
г^тавляет для карася 8, голубя 13, лягушки и мыши 15, моремой
лжинки 32, агамы 284 и ужа 1600. Яда Naja <'ютвеггтмнде
Требуется для карася 0,25, мыши 5 и ужа 40 ме. Как прмило, ядо-
витые змеи погибают от укусов других змей, в том числе и своего
эида. ио Для этого необходимы относительно большие ло-
зы яле-
Примерно половина осложнений после укуса наших змей при-
родится на долю неправильных, кустарных методов лечения.
Первая помощь при укусе состоит в немедленной изоляции уку-
шенного члена от общего кровяного русла, путем наложения
жгута или тугой повязки выше места укуса. Повязку эту не сле-
дует держать дольше 15—20 мин. и не имеет смысла наклады-
вать, если с момента укуса прошло более 5—10 мин. (I). Затем
полезно сделать надрез в месте укуса и постараться удалить по-
больше крови, содержащей наибольшую концентрацию яда.
Высасывание раны ртом таит опасность отравления через тре-
щины губ и слизистой оболочки, а с другой стороны, может ин-
фицировать рану. Кроме того, хотя действие яда, попавшего в
желудок, считается 'незначительным, но оно, по мнению некото-
рых исследователей, все ж не равно нулю. В дальнейшем рану
следует продезинфицировать и перевязать по обычным прави-
лам, а жгут или повязку снять. Прижигание раны и употребление
спиртных напитков приносит вред, а не пользу. Больного следует
как'можно скорее доставить к врачу. Активное и наиболее реаль-
ное лечебное средство — впрыскивание анти-змеиной сыворотки.
В этом направлении наибольшее значение имели работы Каль-
метта (Calmette), который в 1892 г. в Пастеровском институте
разработал технику приготовления этих сывороток.
Лошадь подвергается прививкам возрастающих доз яда в те-
чение 16 месяцев, после чего ее кровь приобретает такой имму-
нитет, что животное может перенести одновременное впрыски-
вание 80-кратной летальной дозы. Анти-змеиная сыворотка спе-
цифична по отношению к виду змеи, но в последнее время кроме
«моновалентных» сывороток стали изготовлять и «поливалент-
ные» (действующие сразу против яда змей нескольких видов).
Крупнейшим центром приготовления анти-змеиных сывороток
считается основанный в 1899 г. институт в Бутантане (около
Сан-Пауло, Бразилия). Аналогичные институты существуют в
Пенсильвании и в Бомбее. Особенно эффективны сыворотки в
течение первых часов после укуса. В отдельных случаях оказы-
ваются полезными введение под кожу больному адреналина,
камфары, внутривенное вливание физиологического раствора,
переливание крови, грелки и теплые ванны. Вопрос о пользе
применения новокаина, картизона, различных антигистаминных
препаратов и льда спорен. Были попытки использования змеино-
го яда в качестве лекарства, но чаще всего полученный терапев-
тический эффект был сомнителен.
249
Щиткование (фолидоз) змей (рис. 53) не менее дифференци-
ровано, чем у ящериц. Особое значение имеют щитки головы.
На верхней поверхности последней обычно видны межчелюстной
(rostrale), межносовые (internasalia), предлобные (praefron-
talia), лобный (frontale), теменные (parietalia) и надглазничные
(supraocularia). На боковой поверхности головы отмечают но-
Рис. 53. Терминология щиткования змей: А—голова полоза сбоку, Б—го-
лова литоринха сверху, В — голова медянки снизу, Г — спинная чешуя
полоза (черная линия показывает, как следует подсчитывать число рядов
чешуй вокруг туловища);
/__межчелюстной. 2 — носовой, 8 — межносовые, 4 — предлобный, 5 — скуло-
вой, в — предглааничный, 7 — надглазничный, 8 — заглазничные, 9 — первый
ряд височных, 10 — второй ряд височных, 11 — лобный, 12 — верхнегубные.
13 — подглазничный, 14 — теменной, 15 — подбородочный, 16 — нижнегубные,
17 — передний нижнечелюстной, 18 — задний нижнечелюстной, 19 — брюшные
сомежчелюстной (nasorostrale), 'носовые (nasalia), скуловой
(frenale), предглазничные (praeocularia), подглазничный (subo-
culare), заглазничные (postocularia), верхнегубные (suprala-
bialia) и височные (temporalia). Нижняя поверхность головы мо-
хсет нести подбородочный (mentale), нижнегубные (infralabia-
Jia), нижнечелюстные (inframaxillaria), горловые щитки (scuta
gularia) и горловую чешую (squamae gulares). У большинства
£50
мей нижняя поверхность тела покрыта увеличенными и вмтжяу
тымн поперечно пластинками, или брюшными шиткамк (vent
ra]ia), до клоакальной щели и подхвостовыми (subcxudalu)
позади нее. Последние располагаются либо в один ряд («нераз-
деленные>)« либо в 2 (<разделенные>). Количество брюшных че
шуй, равно как и рядов чешуй вокруг середины тела (не вклю-
чая брюшные!) имеет большое значение при определении. Впе-
реди клоакальной щели помещается непарный или разделенный
косым швом анальный щиток (anale).
В 1758 г. Линней включал в отряд змей, кроме признаваемых
нами за таковых (Crotalus, Boa, Coluber), также и безногих яше-
риц (Anguis, Amphisbaena) с безногими земноводными (Caecl-
На). Настоящих змей он знал около 120 видов. В современном
объеме змеи были впервые приняты Ваглером в 1830 г. Год
спустя И. Мюллер предложил различать среди змей Microsto-
mata (чешуйчатая кость в стенке черепа), Macrostomata (чешуй-
чатая кость свободна) и Peropoda (имеются рудименты таза).
В 1844—1854 гг. Дюмериль и Биброн уделили главное внимание
зубам, подразделяя змей на Opoterodonta (=Microstomata),
Aglyphodonta, Opisthoglypha, Proteroglypha и Solenoglypha.
Работники Британского музея (Gray, Gunther, Boulenger) объе-
динили гладкозубых и бороздчатозубых в группу ужей, долго
признававшуюся за семейство. В 1886 г. Коп, по сути, подтвер-
дил деление Мюллера, подразделив, однако, Angiostomata
(=Microstomata) на 3 семейства, a Eurystomata (=Macrostoma-
ta) на 9. Всеобщее признание получила система Буленджера, уста-
новленная в 1893—1896 гг. По этой системе змеи делились сразу
на 9 семейств, объединявших 1639 видов. Однако в последние
десятилетия были предприняты попытки пересмотра этой систе-
мы. Так в 1923 г. Нопча предложил именовать вымерших мело-
вых змей Cholophidia, а большинство современных объединил
под именем Alethinophidia. Предлагаемое ниже деление (рис. 54)
не может претендовать на законченность, тем более, что сведе-
ния о многих ископаемых змеях весьма фрагментарны и проти-
воречивы. В него не включено, в частности, семейство Archaeo-
phidae (эоцен), которое, видимо, в действительности должно
быть отнесено к рыбам. Нами принято следующее деление:
1(6) Зигосфены маленькие [надсемейство (a) Cholophidia].
2(5) Ребра и позвонки расширенные (пахиостоз).
3(4) Остистые отростки низкие.
1. Pachyophidae
4(3) Остистые отростки высокие.
2. Symoliophidae
5(2) Пахиостоза нет.
3. Palaeophidae
6(1) Зигосфены большие.
1в*
251
7 < 10) Pm мало йкижнмый, поперечно* ноет* яет (»»>>**
чао (Ь> ScolecophldiaJ.
Я ( 9) Вер кнг челюсти *« кость Лет зубов; ятким ч*ллеТ1
чубами; имеются рудименты лобковой и гтлялщ|^
хости.
4. LeptotyphtopidM
9(8) Верхнечелюстная кость с зубами, а нижняя челюсть
беззубая; имеется рудимент только седалищной кости.
5. Typhlopidae
10 ( 7) Рот обычно может широко растягиваться; поперечная
кость имеется [надсемейство (с) Alethinophidia],
11 (18) Предлобные кости соприкасаются с носовыми.
12 (17) Нижняя челюсть с венечной костью (coronate).
£52
|3
|4
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
(16) Имеются лвднях конечностей м чешуйчятяя
кость.
(15) Чешуйчатая кость большая, вне стенки черепа
6. Boidae
(14) Чешуйчатая кость маленькая, включенная в стенх)
черепа.
7. Anlildae
(13) Рудимента задних конечностей я чешуйчатой костя кет.
8. Uropeftidae
(12) Венечной кости нет.
9. Xenopeltidae
(11) Предлобные кости отделены от носовых.
(28) Длина верхнечелюстной кости больше ее высоты
(«горизонтальная кость»).
(27) Крыловидная кость касается квадратной или нижней
челюсти.
(22) Заднелобная кость простирается вперед вдоль всего
верхнего края глазницы.
10. Acrochordidae
(21) Заднелобная кость менее развита.
(24) Передние зубы верхнечелюстной кости гладкие.
11. Colubrldae
(23) Передние зубы верхнечелюстной кости бороздчатые.
(26) Хвост цилиндрический.
14. Elapidae
(25) Хвост сжат с боков.
13. Hydrophidae
(30) Крыловидная кость не касается квадратной или нижней
челюсти.
12. Amblycephalidae
(19) Длина верхнечелюстной кости меньше ее высоты («вер-
тикальная кость»).
(30) Между глазом и ноздрей нет ямки; верхнечелюстнай
кость без вырезки.
15. Viperidae
(29) Между глазом и ноздрей имеется ямка («термический
глаз»); верхняя часть верхнечелюстной кости с вы-
резкой. ;
16. Crotalidae
2БЗ
НАДСЕМЕЙСТВО (•) CHOI.OPHIOIA. ДРЕВНИЕ ЗМЕИ
Объем и значение спорное. Многие принимают их за формы
значите 1ьмо умлонившигся от первоначального, нам рни ней»'
местного, стола змей.
I СЕМЕЙСТВО PACHYOPHIDAE
Из нижнего мела Герцеговины описаны Pachyophis я Же-
sophis. Череп. по*вндимому, ящерицеобразный. Около 160-
180 позвонков. Имеются расхождения по вопросу о размере
вигосфенов. Длина тела около 40 см. Предполагают бентониче-
скнй образ жизни и питание червями.
2. СЕМЕЙСТВО SYMOLIOPHIDAE
Род Symoliophis из верхнего мела Египта, Португалии и
Франции. Череп не известен. Судя по размерам позвонков, долж-
ны были достигать, по крайней мере, размеров современных уда-
вов. Морские.
3. СЕМЕЙСТВО PALAEOPHIDAE
Эоценовые Palaeophis (Европа, Африка, Сев. Америка) и
Pterosphenus (Египет, Сев. Америка). Морские змеи, достигав-
шие 6—11 м длины. Некоторые сближают их с Boidae, перенося
в надсемейство Alethinophidia.
НАДСЕМЕЙСТВО (в) SCOLECOPHIDIA. ЧЕРВЕОБРАЗНЫЕ
Небольшие, роющиеся в земле формы. Глаза скрыты под
щитками. Щиткование упрощенное. Кости черепа соединены
плотно. Вероятно, сходство обоих входящих сюда семейств
лишь конвергентное. В ископаемом состоянии неизвестны, исклю-
чая Aphelophis (палеоцен Колорадо), значение которой спорно.
4. СЕМЕЙСТВО LEPTOTYPHLOPIDAE (=GLAUCONIIDAE) (рис. 55)
Внешне отличаются от представителей семейства слепунов но-
совым щитком, касающимся края рта, увеличенным анальным
щитком и немного более длинным хвостом. Всего 2 рода, один
из которых (единственный вид Anomalepis mexicana имеет
22 ряда чешуй вокруг тела) обитает в Мексике, а другой
(Leptotyphlops—Glauconia, 14 чешуй вокруг тела) представлен
примерно 46 видами в Африке, Юго-Зап. Азии и Америке. Наи-
более крупный представитель Leptotyphlops albifrons (тропиче-
ская Америка), длина тела 375 мм. а многие не достигают
100 мм. Диаметр тела заключается 36—113 раз в его общей дли-
не, а длина хвоста 8—28 раз.
254
Рис. 55. Представители червеобразных и ложноногих: 1 — череп Leptotyphlops
macrolepis сбоку, 2 — голова Leptothyphlops affinis сбоку, 3—' череп Typhiops
lumbricaiis сбоку, 4—5 — голова Helminthophis petersii сбоку и сверху,
° череп Python amethystinus, 7 — рудиментарный таз питона, 8 — рудимен-
ты задних конечностей питона снаружи, 9 — голова Eunectes murinus сбо-
ку» Ю— голова Python amethystinus, 11 — Python molurus
5. СЕМЕЙСТВО TYPHLOPIDAE. СЛЕПУНЫ
Тело с начала до конца почти одинаковой толщины. Диаметр
его содержится в общей длине 19—12£ раза. Хвост чаще очень
короткий и с небольшим шипиком на конце. Чешуя однообраз-
ная, округлая. Глаза иногда почти не видны. Развито только
правое легкое, имеющее трахеальную часть. Живут большей
частью в земле, появляясь на поверхности после дождя. Пита-
255
юГся дождеянын чгршяим. муряяьяыи. мкпгоялжкямя и
рпдпбмым Яйпгя чллушн Ок рас к л «миграция*. от ж*.тго»>ч v
яоЛ Д° чгря<»й Отличают следующие роля
I (О На тлоае крупные щитки.
2 ( 3) Нара больших предлобных шиткоа и большой
мотдря между двумя небольшими носовыми
HHmlnthophH
5 видон. Центр, и Южн. Америки
3 ( 2) Непарный предлобный и лобный шитми малы; nQrz
щиток, иногда разделенный, большой.
Typhlops
164 вида, Южн. Европа, Южн. Азия, Афьй
тропическая Америка, Австралия. ТИЧ)ч
4(1) Голова покрыта мелкой однообразной чешуей.
Typhlophis
1 вид, Бразилия, Гвиана.
Самый крупный представитель слепозмеек Typhlops humb
(Африка), общая длина тела до 775 мм. Большинство видгш
имеет длину тела от 150 до 400 мм, самый мелкий (95 мм\^
Т. reuteri (Носси Бе). От Балканского полуострова, Греции
Нижнего Египта до Таджикистана, западной Индии и Афгани*
стана распространен Т. vermicular is. У нас в Закавказье и Сред,
ней Азии он придерживается сухих, обычно каменистых мест, с
редкой растительностью. Легче всего его можно найти под кам-
нями. Питается преимущественно 'муравьями и их личинками
заползает иной раз в муравейники. По-видимому, откладывает
в мае и июне 4—8 яиц, длина каждого 12—13 мм. Длина тела
с головой до 360 мм. Хвост в 40—52 раза короче тела. Т. braminus
(Южн. Азия, Южн. Африка, острова Индийского океана) был
однажды найден в стебле сахарного тростника, причем поедал
в этот момент испражнения гусениц. Нelminthophis flavotermina-
lis (Венесуэла) завезен на остров Маврикия.
НАДСЕМЕЙСТВО (с) ALETHINOPHIDIA.
СОВРЕМЕННЫЕ ЗМЕИ
Первые 4 семейства этой группы объединяются Хофштеттером
в группу Henophidia, а остальные — в группу Caenophidia. Целе-
сообразность такого усложнения иерархии таксономической
лестницы заслуживает размышления.
6. СЕМЕЙСТВО BOIDAE. ЛОЖНОНОГИЕ (рис. 56)
Рудименты задних конечностей нередко выдаются в форме
коготков по бокам клоакальной щели. Два легких, из которых
левое несколько больше правого. Только Ungalia имеет всего
256
одно леве* легкое. Известны, начппмм и- верхнего мела. Разлвчз
ют следующие рода:
I (14) Имеется надглазничная кость; чаше 2 ряда подхвостг>
вых щитков (подсемейство Pythoninae).
2(9) Предчелюстная кость с зубами.
3 ( 8) Хвост не закручивается или закручивается очень слабо,
межчелюстной щиток без ямок или ямки неглубокие
4 ( 5) Верхнегубные щитки без ямок.
Loxocemus
I вид, Южн. Мексика, Гватемала.
5 ( 4) Верхнегубные с ямками.
6 ( 7) Ноздря между 2 носовыми щитками.
Nardoa
1 вид, Новая Гвинея, Новая Британия.
7(6) Ноздря в одном полу раз деленном носовом щитке.
Liasis
8 видов, Флорес, Тимор, Сев. Австралия, Новая
Гвинея.
8(3) Хвост может закручиваться; межчелюстной и передние
верхнегубные щитки с глубокими ямками.
Python
10 видов, Африка, Юго-Вост. Азия, Малайский
архипелаг, Ав.стралия, Новая Гвинея.
9(2) Предчелюстная кость беззубая.
10 (13) Нёбная кость с зубами.
11 (12) Губные щитки с ямками; 2 ряда подхвостовых щитков.
Chondropython
1 вид, Новая Гвинея, Ару, Майсур.
12 (11) Губные щитки без ямок; чаще 1 ряд подхвостовых.
Aspidites
3 вида, Сев. Австралия.
13 (10) Нёбная кость беззубая.
Calabaria
1 вид, Зап. Африка, Фернандо-По.
14 ( 1) Надглазничной кости нет; чаще 1 ряд подхвостовых
[подсемейство Boinae].
15 (32) Голова отличается от шеи; хвост более или менее за-
кручивающийся.
16 (21) Передние зубы очень сильно увеличены.
17 (20) Чешуя гладкая.
18 (19) Губные щитки лишены ямок или с неглубокими ямками.
258
Efricratc*
12 видов, тропическяя Америка
$ jig) Губные щитки с глубокими ямками.
Воа
5 видов, тропическая Америке, Мадагаскар
jO (|7) Чешуя с ребрышка-ми.
Enygrus
4 вида, Малуккские о-ва, Малайский архипе-
лаг, Полинезия.
21 (16) Зубы верхней и нижней челюсти постепенно убывают в
своих размерах.
22 (23) Межчелюстного щитка нет.
Trachyboa
2 вида, Южн. Америка.
23 (22) Увеличенный межчелюстной щиток имеется.
24 (27) Вокруг тела 21—29 рядов чешуй.
25 (26) Одна или. две пары предлобных щитков.
Ungalia
10 видов Вест-Индия, Гватемала, Эквадор,
Перу, Бразилия.
26 (25) Всего один большой предлобный щиток.
Ungaliophis
1 вид, Гватемала, Мексика.
27 (24) Вокруг тела 47—95 рядов чешуй.
28 (31) Чешуя гладкая.
29 (30) Носовые щитки позади межчелюстного соприкасаются.
Eunectes
2 вида, Южн. Америка.
30 (29) Носовые щитки разделены мелкими чешуйками.
Constrictor
7 видов, тропическая Америка, Мадагаскар.
31 (28) Чешуя с ребрышками.
Casarea
1 вид, остров Раунд около Маврикия.
32 (15)' Голова не отличается от шеи; хвост не закручивается
или закручивается очень слабо.
33 (34) Чешуя несет по 3—4 ребрышка.
Bolieria
1 вид, Маврикий и остров Раунд.
259
34 (33) Чешуя глзлмвя или тпльдо с одним ргбрмшюи»
35 (36) Голова помрм»« крупными пиегялми
Charina
2 аила, запад Он. Америки
36 (36) Голова покрыта мелкой чешуей.
37 (ЗЯ) Межчелюстной щиток крупный.
Егух
12 видов, Африка, Южн. и Центр. Азия
36 (37) Межчелюстной умеренной величины, заметно ^же
высоты.
Lfchanura
2 вида, запад Сев. Америки.
Все представители подсемейства питонов (Pythoninae),
ключая род Loxocemus, живут в Старом Свете. Виды рода Python
живут в лесах, кустарниках -и среди травы. При случае хорошо
плавают. Самый крупный из них сетчатый питон, Р. reticulatue
(Юго-Вост. Азия и Малайский архипелаг), в исключительных
случаях общая длина до 10 м. Он имеет 4 верхнегубных щитка
с ямками, 297—330 брюшных и 78—102 пары подхвостовых щит-
ков. Питается млекопитающими — даже такими крупными, как
поросята, антилопы или молодые олени. Поедает также птиц и
пресмыкающихся (вараны). Нападает на человека лишь случай-
но. Линька 4 раза в год. Откладывает около 96 яиц. Самка сво-
рачивается -вокруг яиц, охраняя их и, по-видимому, регулируя
температуру. Инкубация 80—82 дня. Мясо сетчатого питона,
равно как и других, употребляется, местными жителями в пищу,
а кожа идет на изготовление ме-лких вещей. Иероглифовый пи-
тон, Р. sebae (тропическая и южная Африка) достигает только
7 м. У него 2 верхнегубных щитка снабжены ямками, 269—
286 брюшных и 63—77 пар подхвостовых щитков. На Гвиней-
ском берегу он почитался и содержался в особых хижинах —хра-
мах. Чаще всего в зоологических садах попадается тигровый
питон, Р. molurus (Индия, Цейлон, Южн. Китай, Малайский по-
луостров, Ява), -редко превышающий 4 м. От предшествующего
вида он легко отличается наличием только 242—265 брюшных
щитков и 61—75 (а не 81—93) чешуями вокруг тела. В неволе хо-
рошо ест кроликов. В некоторых местах его содержат в амбарах
для истребления крыс. Молодые особи Chondropython красные с
белыми пятнами, а взрослые — зеленые. Виды родов Aspidites
(длина до 2 At) и Calabaria (до 86 см) — роющие формы.
Подсемейство удавов, Boinae представлено как в Новом, так
и в Старом Свете. Один из видов гладкогубых удавов (Epicra-
tes) — абома, Е. cenchris встречается преимущественно в лесах,
хорошо лазает по ветвям, питается грызунами, общая длина
260
i л. Напротив, стройный удав, Е. str lotus /вокруг тела 51 —
ff ляда чешуй, вместо 45—51 у предшествующего вида) держит
главным образом, «а сахарных плантациях, под крышами
f^ipux зданий и в Др. местах. Главный представитель узкобрю-
чях уДявов ^раньше — Corollas)—собакоголовый удав.
0 canina (Гвиана, Бразилия), держится на кустарниках, деревь-
ях и пнях, длина до 145 см. Питается птицами. Молодые красно-
ватые, взрослые сверху зеленые с белыми пятнами. Род настоя-
щих удавов раньше назывался Воа, а теперь — Constrictor,
Обыкновенный удав, С. constrictor, встречается в сухих лесах и
кустарниках Южн. Америки от Венесуэлы до Буэнос-Айреса.
Прячется в дуплистые стволы, под корнями деревьев, в земляные
«юры и расщелины скал. В воде попадается редко. Питается мел-
кими млекопитающими (агути, пака, крысы, мыши, редко соба-
ки) и птицами. Иногда держат в амбарах в качестве истребите-
ля крыс. Мясо идет в пищу, кожа — на изготовление сапог. Об-
щая длина до 336 см, но есть указания на экземпляры, достигаю-
щие 4 и даже 6 м. Более других видов рода любит воду
С. madagascariensis. Теснее всех членов семейства связана с во-
дой анаконда, Eunectes murinus (Гвиана, Бразилия, Сев.-Вост. Пе-
ру), которая превосходно плавает и может долго оставаться
под водой. Обычно держится на берегах рек и растущих около
них деревьях. При пересыхании водоемов наблюдались случаи
зарывания в ил. Основная пища — млекопитающие (до свинки
включительно), птица, молодые аллигаторы и рыбы. Случаи
нападения на человека чрезвычайно редки. Нормально наблю-
дается яйцеживородность (78—100 молодых), но иногда и от-
кладка яиц. Указание на анаконд длиною в 11 ж не подтверж-
дается непосредственными доказательствами. Максимальный
измеренный в нашем веке экземпляр достигал 762 см. Резиновая
змея Сев. Америки (Charind) и песчаный удав, или уДавчик
(Егух) —зарывающиеся формы. Все удавчики яйцеживородя-
щи. Западный удавчик, Е. jaculus имеет вокруг середины тела
45—54 ряда чешуй и глаза, обращенные вбок. Длина тела с го-
ловой до 75 см. Распространен от южного до восточного побе-
режья Средиземного моря, через Балканский полуостров и
Мал. Азию, до Ирана и Ирака включительно. У нас в Дагестане и
Закавказье. Обитает преимущественно в глинистых или камени-
стых пустынях или полупустынях. Во время дневной жары держит-
ся под камнями и в норах грызунов, которыми в основном и пи-
тается. В августе — сентябре рождает 18—20 молодых. Восточ-
ный удавчик, Е. tataricus (Казахстан, Ср. Азия, Зап. Китай,
Монголия, Иран, Афганистан) обитает в глинистых и лёссовых
полупустынях, в полынной степи и в песках. В связи с этим гла-
за у него обращены вбок и немного вверх. Питается ящерицами
и грызунами. Наконец, у песчаного удавчика, Е. miliaris, пред-
почитающего барханы и сыпучие пески Ср. Азии, Казахстана и
Вост. Предкавказья, глаза смотрят вверх. Обычно он зары-
261
я на лень в весок, оставляя след * виде небольшого п*т?4
ного валика. Число молодых до 10. В Индии и Белудж*'?»*
обитает Е. johnii, обшая длина до I м,
7 СЕМЕЙСТВО ANILIDAE ( - ILYSIIDAE). ВАЛЬКОВАТЫЕ ЗМЕИ
Мелкая, округлая чешуя расположена черепиисобратяо
Хвост короткий, тупой. Яйцеживородящи. В ископаемом состоя-
нии достоверно не известны. Под вопросом сюда причислят
Dinilysia (мел Патагонии) и Scytalophis (эоцен Франции). Раз-
личают рода:
1 ( 2) Предчелюстная кость с зубами; глаз под серединой
одного щитка.
Anilhis
1 вид, Южн. Америка.
2(1) Предчелюстная кость без зубов; глаз окружен щитками.
3(4) Имеется продольная подбородочная бороздка.
Cylindrophis
6 видов, Цейлон, Юго-Вост. Азия.
4(3) Подбородочной бороздки нет.
Anomachilus
1 вид, Суматра.
Единственный вид рода Anilius — коралловая сверташка,
A. scytale окрашена в ярко-красный цвет с черными кольцами
вокруг тела, общая длина до 83 см (рис. 57). Скрывается под
корнями деревьев, в норах и тому подобных местах. Питается
мелкими пресмыкающимися. Красная змея, Cylindrophis избе-
гает света, роясь под землей, общая длина до 82 см. Поедает
насекомых, червей и слепозмеек.
8. СЕМЕЙСТВО UROPELTIDAE. ЩИТКОХВОСТОВЫЕ ЗМЕИ
В ископаемом состоянии не известны. Голова маленькая, за- i
остренная. Хвост коротко усечен и покрыт на конце большим
щитком или килеватыми чешуйками. Роются в земле, упираясь
сильным хвостом. Правое легкое меньше левого. Питаются чер-
вями. Яйцеживородящи. Общая длина 17—57 см. Распростране-
ны по сырым горным лесам. Рода различаются так:
1(4) Хвост кончается большим щитком.
2(3) Межчелюстной щиток продолжается назад, разделяя
носовые.
Rhlnophis
8 видов, Южн. Индия, Цейлон.
3(2) Межчелюстной щиток иной; носовые соприкасаются.
262
UropeKis
1 вид. Цейлон.
и Хвост покрыт чешуей до самого конца.
4 ( *{ рдаз под серединой одного щитка.
5 (Ц1 Заглазничный щиток самостоятелен.
Рис. 57. Представители некоторых семейств змей: 1 — череп Anilius scytale,
2 — череп Silybura grandis сбоку, 3—4— голова Silybura myhendrae сбоку и
хвост, 5 — череп Xenopeltis unicolor сбоку, 6 — череп Chersydrus granulatus
Ч сверху, 7—Anilius scytale, 8—Acrochordus javanicus
Plectrurus
4 вида, Южн. Индия.
7(6) Заглазничный щиток слит с надглазничным или верхне-
ресничным.
8(9) На подбородке продольная бороздка.
Melanophldium
3 вида, Южн. Индия.
9(8) Подбородочной бороздки нет.
10 (11) Увеличенная концевая чешуйка хвоста квадратная или
с 2 шипиками, расположенными на одном уровне.
Silybura
22 вида, Индия, Цейлон.
11 (10) Концевая чешуйка хвоста с двумя расположенными
друг над другом простыми или двойными шипиками.
263
Pwudoplectruru*
I пнл. Южн Индия.
12 ( 5) Глв.» окружен щитками.
Platyplectrurus
3 вида. Южн. Индия.
9. СЕМЕЙСТВО XENOPELT1DAE
В ископаемом состоянии не известны. Имеются 2 нерав
легких, которые, однако, оба функционируют. Всего одни
Xenopeltus с одним видом X. unicolor (Южн. Индия, ВиЛ'
п-ов Индокитай, Малайский полуостров и архипелаг). рОг’,и<
форма, общая длина до 1 м, с черной или бурой чешуей и вео**’
кальным зрачком. рт*
10. СЕМЕЙСТВО ACROCHORD1DAE. БОРОДАВЧАТЫЕ ЗМЕИ (рис ,
В ископаемом состоянии те известны. Чешуя совсем не нат
гает друг на друга или лишь немного черепитчатая. Вся жизС
проходит в воде. Могут часами оставаться под водой. На сущ
добровольно не выходят. Держатся в реках и около морских^
регов, уплывая в открытое -море иногда на 5—7 км. Питаются *’
основном рыбой. Яйцеживородящи. Рода различаются так: в
1 ( 4) Брюшных щитков нет [подсемейство Acrochordinae].
2 ( 3) Телю округлое или слабо сжатое; брюшной складки нет
Acrochordus
1 вид, Малайский полуостров и архипелаг, Но.
вая Гвинея.
3 ( 2) Тело и хвост сжатое; вдоль середины брюха складки
кожи.
Chersydrus
1 вид, от Южн. Индии до Новой Гвинеи вклю-
чительно.
4(1) Брюшные щитки имеются (подсемейство Xenoderminae=s
Nothopsinae).
5(6) Голова покрыта однообразными зернышками.
Xenodermus -
1 вид, Ява, Суматра, Пинанг.
6 ( 5) Голова покрыта симметричными щитками.
7 (10) Подхвостовые щитки расположены в 1 ряд.
8 ( 9) Брюшных щитков 210, подх-востовых 115.
Stolieczkala
1 вид, Юго-Вост. Индия.
9(8) Брюшных щитков 162—167, подхвостовых 43—58.
264
Рис. 58. Ареалы семейств: 1 — Anilidae, 2 — Uropeltidae, 3— Xenopeltidee, 4 — Acrochord.dae
Rmkw
I нил. Южн. Айнам.
10 ( 7) Подхаостовыс а 2 ради.
Nothopsh
2 вида, Центр. Америка.
Самый крупный представитель семейства — яванская боре,
дявчатая змея. Acrochordus javanicus. которая может превосж*,
днть 250 см. Кроме рыбы ест лягушек. Рождает 27 молодых.
Мясо в некоторых районах употребляют в пищу.
11. СЕМЕЙСТВО COLUBRIDAE. УЖЕОБРАЗНЫЕ ЗМЕИ
Еще недавно большинство систематиков принимало это се-
мейство в очень большом объеме. Его подразделяли на 8 подсе-
мейств, группировавшихся в 3 серии — гладкозубых (Agiypha),
заднебороздчатых (Opisthoglypha) и переднебороздчатых
(Proteroglypha). В настоящее время оба подсемейства последней
серии (Hydrophiinae и Elapinae) и первое подсемейство гладко-
зубых (Acrochordinae) выделены в самостоятельные семейства.
Кроме того, представляется целесообразным объединение под-
семейств Rhachiodontinae и Elachistodontinae в одно — Dasypel-
tinae. Имеются существенные подозрения, что группа Opisthog-
lypha не представляет собой единого целого, ибо, вероятно,
бороздчатость зубов возникала независимо у разных групп
Agiypha. Вопрос упорядочения систематики семейства в целом
еще остается задачей будущего. В принимаемом здесь объеме
семейство достоверно встречается в ископаемом состоянии, на-
чиная с миоцена, но можно думать, что в дейтвительности оно
древнее и возникло в Азии или Африке. Из 272 ныне живущих
родов рассмотрим основные:
1 (190) Зубы хорошо развиты.
2 (125) Все зубы гладкие (подсемейство Cblubrinae).
3 ( 38) Нижняя часть тела позвонков имеет непарные выро-
сты (hypapophyses = processus spinosi inferiores), пред-
ставленные на последних туловищных позвонках
гребнем или ясным бугром.
4(5) Зубная кость почти свободна и подвижно прикрепле-
на только к самому переднему концу сочленовой.
Sibynophis
13 видов, Мадагаскар, острова Коморо, Юго-
Вост. Азия, Центр. Америка.
5(4) Зубная кость плотно соединена с сочленовой и, если
подвижна, то в слабой степени.
6 ( 21) Зубы верхней челюсти примерно одинаковой вели-
чины.
7(8) Передние зубы нижней челюсти увеличены,
266
н 7>
-Гго уХ-«авной ми
пИ“Ьнегубные шитки ие каса,отс’' "омри.
Два "“Г™™
Чешуя с Pc6f “^^м«тся
Поедглазничиый имеется.
Зубов в верхней челюсти меньше 20.
Storeria
6 видов, Сев. и Центр. Америка.
средние
14
15
( 13) Зубов в верхней челюсти 20 и более.
(16) Зубы в верхней челюсти мелкие, числом 20—25.
Opisthotropis
8 видов, Зап. Африка, Южн. Китай, Малай-
ский архипелаг.
16 ( 15) Зубы верхней челюсти крупные, около 30 штук.
Tretanorhinus
6 видов, Центр. Америка, Антильские о-ва.
17 ( 12) Предглазничного нет и потому скуловой и предлоб-
ный достигают глаза.
Streptophorus
5 видов, Мексика, Центр. Америка.
18 ( И) Чешуя без ребрышек.
Trachischium
6 видов, Индия.
19 ( 10) Подхвостовые в один ряд.
Achalinus
5 видов, Вост. Азия.
20 ( 9) Первый верхнегубной касается ноздри.
Aspldura
5 видов, Цейлон.
21 ( 6) Зубы верхней челюсти неравной длины.
22 ( 31) Задние зубы верхней челюсти самые длинные.
23 ( 30) Имеется пара межносовых щитков.
24 ( 29) Зубы нижней челюсти почти равной величины.
25 ( 26) Чешуя в передней части, тела расположена косыми
рядами.
267
Рмчм1в*ея<м1оп
5-9 низов. Мидян. Китай. Мжлжйекчй •»>,./
лаг
26 I 25) Чешуи на всем теле расположена продольными
дани. I
27 ( 28) Верхнечелюстные зубы, в числе 18 - 40, образуют »>
прерывный ряд.
Natrbt
101 вид, почти космополит (нет в Южн. Ан»
рике, в Австралии есть только на севере).
28 ( 27) Верхнечелюстных зубов 14—22, причем 2 последних
увеличены и отделены промежутком.
Macroplsthodon
4 вида, Индия, Цейлон, Малайский архипелаг.
Китай.
29 ( 24) Передние зубы нижней челюсти много длиннее
задних.
Pseudoxyrhopus
9 видов, Мадагаскар.
30 ( 23) Межносовой щиток непарный.
Helicops
15 видов, всюду под тропиками, исключая
Австралию.
31 ( 22) Задние зубы верхней челюсти самые короткие.
32 ( 35) Зубы верхней челюсти образуют непрерывный ряд.
33 ( 34) Чешуя гладкая.
Lamprophis
4 вида, Южн. и Вост. Африка.
34 ( 33) Чешуя с ребрышками.
Gonionotophis
3 вида, тропическая Африка.
35 ( 32) Увеличенные передние-зубы верхней челюсти отделе-
ны от более мелких промежутком.
36 ( 37) Чешуя гладкая.
Boodon
И видов, тропическая и Южн. Африка.
37 ( 36) Чешуя с ребрышками.
Mehelya
15 видов, Африка.
268
, 3) Ннжияя часть тела последяиж туловищных номов*
* ' гладкая или с очень слабым килем.
( 40) Зубы с ложковидными коронками.
Iguanognathus
1 вид, Суматра.
конически® 'или веретеновидные.
/ 44) Передние зубы верхней челюсти много крутое
4 других.
42 ( 43) Два-три средних зуба верхней челюсти самые мелки*
отделенные от предшествующих и последующих от
четливым промежутком.
Dinodon
4 вида, Китай, Япония,? Гималаи.
43 ( 42) Передние зубы отделены промежутком от задних,
причем внутри каждой группы зубы уменьшаются по
направлению назад.
Lycodon
16 видов, Южн. Азия.
44 (41) Все зубы верхней челюсти примерно равной величи-
ны, задние больше или немного меньше.
45 ( 46) Средние и задние зубы верхней челюсти самые
длинные.
Stegonotus
11 видов, Моллукские и Филиппинские о-ва,
Полинезия до Самоа, Сев. Австралия.
46 ( 45) Средние зубы не увеличены.
47 (104) Задние зубы верхней челюсти несколько больше или
последние сильно увеличены.
48 ( 99) Нёбная и крыловидная кость с хорошо развитыми
зубами.
49 ( 54) Зрачок вертикально-эллиптический.
50 (51) Задние зубы верхней челюсти не увеличены.
Dryocalamus
7 видов, Индия, Цейлон, Малайский архипелаг,
Филиппины.
М ( 50) Задние зубы верхней челюсти заметно длиннее.
Z-53) Зубов в верхней челюсти 12—16; чешуя с концевыми
1 ямками.
Hypsiglena
6 видов. Сев. и Центр. Америка, Венесуэла.
z 52) Зубов в верхней челюсти 6—9; чешуя без концевых
63 I 7 ямок.
269
Я
55
49)
66)
57)
57
56
56)
61)
59
60)
60
59)
61
62
58)
63)
63
62)
64
65)
LytorhynchtM
7 пило». Ов. Африки и Звп. Алия до Сл»п
Арнзпня, Нижняя Кяляфоримя.
Зрачок круглый.
Передние зубы нижней челюсти много больше мдя*/
/Лежносовой касается ноздри.
Pseudatpls
1 вил, Южн. Африка.
Межносовой не касается ноздри.
Чешуя расположена косыми рядами, по крайней иер<
в передней части тела.
Конец морды с придатком, покрытым чешуей.
Rhynchophis
1 вид, Норвежский остров в заливе Бакбо
Конец морды -без придатка.
Leptophis
19 видов, Центр, и Южн. Америка.
Чешуя расположена продольными рядами.
Задние зубы верхней челюсти не отделены промежут-
ком от остальных.
Hypsirhynchus
1 вид, Сан-Доминго.
Задние зубы верхней челюсти отделены от предшест-
вующих промежутком.
Чешуя узкая, ланцетовидная; верхнечелюстных зу-
бов 16—20.
Uromacer
6 видов, Вест-Индия,? Бразилия.
65 ( 64) Чешуя умеренно вытянутая; верхнечелюстных зубов
11—17.
Dromicus ~
23 вида, Антильские и Галапагосские о-ва,
Чили, Перу.
66 ( 55) Передние зубы нижней челюсти примерно равны дру-
гим или немного больше.
67 ( 70)* Число рядов чешуй вокруг тела четное (10—18).
68 ( 69) Чешуя расположена продольными рядами.
Zaocys
7 видов, Центр, и Южн. Америка.
69 ( 68) Чешуя расположена косыми рядами.
270
Herpetedryas
7 видов, Центр, я Южн. Америка.
. §7) Число чешуй вокруг тела нечетное.
/О / 96) Брюшные щитки по бокам та кругленные ми с тупым
71 ' кантом.
/ g9) Зубы верхней челюсти образуют непрерывный ряж
7* ' или последние 2—3 отделены от остальных очень ко-
Fotkhm промежутком.
олова хорошо отграничена от туловиша; глаз боль*
7Э v шой или довольно большой.
74 ( 75) Чешуя расположена продольными рядами.
Coluber
43 вида, Европа, Азия, Сев. Америка, Сенегам-
бия, Сев., и Центр. Америка.
75 ( 74) Чешуя расположена косыми рядами, по крайней ме-
ре в передней части тела.
76 ( 77) Боковые чешуи равного размера со спинными.
Chlorophis
13 видов, тропическая и Южн. Африка.
77 ( 76) Боковые чешуи много короче спинных.
Thrasops
6 видов, Африка.
78 (73) Голова плохо или совсем не отграничена от туло-
вища; глаз умеренный или маленький.
79 ( 80) Межносовые слиты с предлобными.
Sympholis
1 вид, Мексика.
80 ( 79). Межносовые и предлобные раздельны.
81 ( 82) Подхвостовых один ряд.
Rhinochilus
3 вида, Сев. Америка, Мексика, Венесуэла.
82 (81) Подхвостовых два ряда.
83 ( 84) Хвост длинный, равномерно утолщенный и тупо за-
канчивающийся.
Urotheca
5 видов, Куба, Центр, и Южн. Америка.
84 ( 83) Хвост постепенно утончающийся.
85 ( 88) Верхнечелюстных зубов 12—24.
86 ( 87) Чешуя -без концевых ямок.
Rhadinaea
26 видов, Центр, и Южн. Америка.
271
я?
80
89
90
Чешу® е яонигяммн ямками
* Certmelt»
19 индо». Еиропа, Юю-Впст. Азия, Иидм
Африка. Ноям А Свет к северу от экя»уор4
Верхнечелюстных луба» Я -12.
Slmote*
35 вида». Индия. Китай, Малайский архнпела
_О1 Задние сильно увеличенные зубы верхней челюсти от.
( li} делены от остальных заметным промежутком.
( 91) Чешуя без концевых ямок.
Aporophis
8 видов, Южн. Америка.
qi ( 90) Чешуя с концевыми ямками.
до ( 95) Конец морды закругленный или тупо конический.
93 ( 94) Чешуя расположена продольными рядами.
Liophis
21 вид, Южн. Америка, Вест-Индия, юго-вос-
ток Сев. Америки.
94 ( 93) Чешуя расположена косыми рядами.
Xenodon
6 видов, тропическая часть Нового Света.
95 ( 92) Конец морды выдающийся вперед, клиновидный.
Heterodon
4 вида, Сев. Америка.
96 ( 71) Брюшные щитки с боковым ребром, похожим на киль,
и с вырезкой с каждой стороны, отвечающей этому
килю.
97 ( 98) Чешуя вдоль позвоночника не увеличена.
Philothamnus
5 видов, тропическая и Южн. Африка.
98 ( 97) Чешуя вдоль позвоночника увеличена.
Dendrophis
15 видов, Юго-Вост. Азия, Австралия.
99 ( 48) Нёбная и крыловидная кость совсем лишены зубов
или с очень небольшим числом их.
100 (101) Задние верхнечелюстные зубы отделены промежут-
ком от передних.
Arrhyton
5 видов, Куба.
272
(100)
(103)
|02
Ряд верхнечелюстных зубов непрерывный
Зрачок круглый, конги морды выдающийся вперед
тупой.
|03
104
105
106
107
108
109
ПО
111
112
113
114
Ollgodon
37 видов, Южн. Азия, Нижний Египет
(102) Зрачок приближается к вертикально-эллиптическому;
конец морды с угловатым горизонтальным краем.
Ргозутпа
11 видов, тропическая и Южн. Африка.
( 47) Задние зубы верхней челюсти примерно одинаковой
величины с другими или убывают в величине.
(112) Чешуя с концевыми ямками.
(107) Число рядов чешуй вокруг тела четное (14 или 16).
Spilotes
3 вида, Центр, и Южн. Америка.
(106) Число рядов чешуй вокруг тела нечетное.
(111) Голова хорошо отграничена от туловища.
(ПО) Задние верхнечелюстные зубы убывают в размерах;
чешуя расположена рядами.
Dendrelaphis
10 видов, Индия, Цейлон, Малайский архи-
пелаг.
(109) Верхнечелюстные зубы примерно равные; чешуя рас-
положена прямыми рядами.
Elaphe
54 вида, Европа, Азия, Сев. и тропическая
Америка.
(108) Голова совсем не отграничена от туловища или весь-
ма слабо.
Contia
28 видов, Юго-Зап. Азия, Сев. и Центр. Аме-
рика.
(105) Чешуя без концевых ямок.
(116) Верхнечелюстных зубов 15—30.
(115) Голова неясно отграничена от тела; ноздря смотрит
вбок.
Ablabes
15 видов, Юго-Вост. Азия.
115
(И4)
Голова .---- —--------
более или менее вверх.
ясно отграничена от тела; ноздря направлена
273
1|б
lie
119
120
121
122
123
124
125
126
127
128
129
130
131
274
Grayia
5 лилов. тропическая Африка
(||3) Вержнгиглюстиых лубок не более 12,
(120) Вокруг тела 13 рядов чешу А.
(1)9) Зубы нижней чел мкт и примерно одной величиям. л
' редние височные имеются.
Carpbophis
2 вида. Сев. Америка.
(118) Передние зубы нижней челюсти более крупные; в*
1 сочных нет и потому теменные щитки касаются вера
негубных.
Calamaria
70 видов, Юго-Вост. Азия, Малайский архи
пелаг.
(117) Вокруг тела 15 или 17 рядов чешуй.
(122) Передние височные имеются.
Atractus
41 вид, Центр, и Южн. Америка.
(121) Передних височных мет и потому теменные щитки
касаются верхнегубных.
(124) Ноздря располагается между 2 носовыми щитками.
Geoph is
15 видов, Центр, и Южн. Америка.
(123) Ноздря внутри одного носового щитка.
Pseudorhabdium
4 вида, Малайский архипелаг, Филиппины.
( 2) Один или более из последних верхнечелюстных зубов
бороздчатые.
(183) Ноздря на боковой стороне головы (подсемейство
Dipsadomorphinae).
(132) Нижняя часть тела последних туловищных позвонков
несет гребень или ясный бугор (гипапофиз).
(129) Зрачок круглый.
Geodipsas
5 видов, Центр. Африка, Мадагаскар.
(128) Зрачок вертикальный.
(131) Тело цилиндрическое; чешуя с ребрышками.
Langaha
4 вида, Мадагаскар.
(130) Тело более или менее сжатое; чешуя гладкая.
|32
|33
134
135
136
137
138
139
140
141
142
143
144
145
146
19*
Stenophlt
9 видов, Мадагаскар, Коморские о-ва
(|27) Нижняя часть тела последних туловишнмх позвонков
гладкая (гипапофизов нет),
(136) Верхнечелюстные зубы (не считая бороздчатых!) по
направлению назад уменьшаются.
(135) Тело цилиндрическое или слегка сжатое; подглазнич-
ного нет.
Tarbophls
10 видов, Юго-Вост. Европа, Юго-Вост. Азия,
Сев. Вост, и тропическая Африка.
(134) Тело сжатое; обычно имеется подглазничный.
Trimorphodon
4 вида, Нижняя Калифорния, Аризона, Мек-
сика, Центр. Америка.
(133) Верхнечелюстные зубы (не считая бороздчатых!)
иного типа.
(176) Гладкие верхнечелюстные зубы примерно равные или
возрастают по направлению назад.
(157) Чешуя с концевыми ямками.
(148) Зрачок вертикальный.
(141) Чешуя расположена косыми рядами.
Boiga
29 видов, тропическая Африка, Южн. Азия,
Австралия, Полинезия.
(140) Ряды чешуй не косые.
(147) Два носовых щитка.
(144) Тело сильно сжатое.
Himantodes
.7 видов, Мексика, Центр, и Южн. Америка.
(143) Тело цилиндрическое или чуть сжатое.
(146) Гладкие верхнечелюстные зубы, числом 15—18, дли-
на зубов медленно возрастает по направлению назад;
глаз круглый.
Leptodira
19 видов, тропическая и Южн. Африка, тро-
пическая часть Нового Света на севере до
Техаса.
(145) Гладкие верхнечелюстные зубы, числом 10—15, при-
мерно одинаковой длины; глаз средний или малень-
кий.
Oxyrhopus
21 вид, Центр, и Южн. Америка.
275
н7
|49
150
151
152
153
154
155
156
157
158
159
160
161
<
162
276
Носовой ткток олип. иногда полуразделеииый
Tachymenlt
4 киля, Южн Америка.
101 3p*4Oh круглый или горимштлльпый.
[’лядкнг верхнечелюстные зубы отделены от бород
(Ьм‘ чЛ1ых промежутком.
I» Гладкие верхнечелюстные зубы постепенно возраст»
(I*1' ют по направлению назад; передние зубы нижией ч»
люсти нс увеличены.
Amplorhlnus
4 вида, тропическая и Южн. Африка.
-QV Гладкие верхнечелюстные зубы примерно равные,
передние нижнечелюстные зубы увеличены.
53) Ноздря между 2 носовыми щитками; чешуя гладкая
' или с ребрышками.
Phllodryas
21 вид, Южн. Америка.
И52) Ноздря прорезана в одном, иногда разделенном, щит-
' ке; чешуя гладкая со следами продольной бороздки.
Malpolon
3 вида, Южн. Европа, Юго-Вост. Азия, Сев.
Африка.
(149) Гладкие верхнечелюстные зубы не отделены проме-
V жутком от бороздчатых.
(156) Брюшные щитки закругленные.
Oxybelis
5 видов, тропическая часть Нового Света.
(155) Брюшные щитки с килем, напоминающим шов.
Chrysopelea
3 вида, Юго-Вост. Азия.
(138) Чешуя без концевых ямок.
(175) Два ряда подхвостовых.
(164) Гладких верхнечелюстных зубов 10—15.
(163) Скуловой щиток имеется.
(162) Глаз умеренный; хвост умеренный ил и "длинный.
Erythrolauiprus
И видов, тропическая часть Нового Света,
Мексика, Техас.
(161) Глаз маленький; хвост короткий.
Scolecophis
3 вида, Центр. Америка.
(!<ИЭ) Скулового щитка мет.
Homalocranlttffl
29 видов, юг Сев. Америки. Центр Америка,
тропическая часть Южн. Америки.
м (159) Гладких всрхиегубных зубов 2—5.
(170) Нет переднего теменного и потому темениые касак»
ся верхнегубных.
1б6 (169) Нёбная кость с зубами.
167 (166) Межносового нет.
Apostolepis
14 видов, Южн. Америка.
|68 (167) Межносовой есть.
Calamelaps
7 видов, тропическая Африка.
169 (166) Нёбная кость беззубая.
Rhinocalamus
3 вида, Вост, и Южн. Африка.
170 (165) Заглазничный касается заднего теменного и потому
теменные отделены от верхнегубных.
171 (172) Предглазничных нет.
Micrelaps *
4 вида, Палестина, Сомали, Абиссиния.
172 (171) Предглазничный имеется.
173 (174) Носовой касается предглазничного, гладких верхнече-
люстных зубов 4—5.
Elapomorphus
9 видов, Южн. Америка.
174 (173) Носовой не касается предглазничного; гладких верх-
нечелюстных зубов 2—3.
Miodon
7 видов, Зап. Африка.
175 (158) Один ряд подхвостовых.
Aparallactus
22 вида, тропическая и Южн. Африка.
176 (137) Гладкие верхнечелюстные зубы достигают наиболь-
шей длины по середине ряда.
177 (178) Гладкие верхнечелюстные зубы образуют непрерыв-
ный ряд.
Taphrometopon
1 вид, Ср. Азия, Афганистан, Вост. Иран.
277
|7в
179
180
181
182
183
184
185
186
187
188
189
190
191
27в
(177) Ряд гладких верхнечелюстных рядо* имеет
(180) Зрачок круглый.
Psammophls
21 вид, Африка, Южн. Азия.
(179) Зрачок иной формы.
(182) Зрачок примерно вертикально-эллиптический.
Macroprotodon
1 вид, Сев. Африка, Юго-Зап. Европа.
(181) Зрачок горизонтальный.
Dryophis
8 видов, Юго-Вост. Азия.
(126) Ноздря помещается на верхней стороне головы я
снабжена особым клапаном [подсемейство Homalop.
sinae].
(189) Носовые щитки соприкасаются.
(186) Чешуя гладкая.
Hypsirhina
18 видов, Юго-Вост. Азия, Квинсленд, Поли-
незия.
(185) Чешуя с ребрышками.
(188) Теменные щитки развиты полностью; голова хорошо
отграничена от тела.
Homalopsis
1 вид, Юго-Вост. Азия, Малайский архипелаг.
(187) Теменные щитки более или менее разбиты на отдель-
ные чешуйки; голова слабо отграничена от тела.
Cerberus
3 вида, Юго-Вост. Азия, Сев. Австралия.
(184) Носовые щитки разделены «межносовым.
Fordonia
1 вид, Индия, Юго-Вост. Азия, Сев. Австралия,
Новая Гвинея.
( 1) Зубная система рудиментарная: только задняя часть
верхнечелюстной и зубной кости, а также нёбная
снабжены немногими зубами [подсемейство Dasypel-
tinae].
(192) На верхнечелюстной кости 2 гладких и 2 бороздча-
тых зуба; нижняя челюсть с 8 зубами; крыловидная
кость с зубами; чешуя гладкая, вокруг тела ее
15 рядов.
Elftchlstodofl
I вид, Бенгалия.
.go На верхнечелюстной хости и а нижней челюетя во
jpt V ' 3—7 гладких зубов; зубов на крыловидной кости нет,
чешуя с ребрышками, вокруг тела ее 23—27 рядов.
Dasypeitis
1 вид, Африка.
Подсемейство настоящих ужей (Colubrlnae) является самым
-мирным. Сюда принадлежит обширный род ужей. Matrix-
fjS^aonotus, некоторые виды которого, главным образом аме-
Рис. 59. Представители настоящих ужей: 1 — череп Natrix- natrix сбоку,
2—позвонок с гипапофизом (Lioheterodon), 3 — позвонок без гипапофиза
(Heterodon), 4 — Natrix natrix, 5 — Leptophis liocerus
риканские, живородящи. Различают 3—4 подрода, два из кото-
рых плохо отграничены. Подрод Natrix s. str имеет менее
30 верхнечелюстных зубов, постепенно меняющих свою длину,
и обычно разделенный анальный щиток. В нашей фауне род
представлен, прежде всего, обыкновенным ужом, N. natrix
(рис. 59), имеющим 153—193 брюшных щитка и обычно 7 верх-
негубных. Длина тела с головой до 120 см. Характерно наличие
по бокам головы по одному светлому (от белого до оранжевого)
пятну, хотя встречаются и сплошь черные особи. Распространен
в Европе (на севере до 60° с. ш.), Алжире, западной и централь-
ной Азии. Связан с водой и обычно придерживается рек, озер,
279
прудоя и других водоемов. Удаляется от воли я«чиой и
Питлгнч преймушгг।пенно чемнояодиымя. реже ммшекидим^
н рыбой В июле • пигус те я навоз, под опляшис лист* я ил* • < «
рой мох (лкллдынаст 6 30 яиц. Длинд яйпл 25 38 мм. шмгмчя
12 22. Инкубация около 2 месяце». Зимуют в норах млг*ои»
тающих и под корнями. В Закавказье, Туркмении и юго~*ямад
ной части Украины, а также на Балканском полуострове par
пространен N. natrix persa, у которого с каждой стороны те.м
идет светлая продольная полоса (видимо, более древний тая
окраски). От юго-западной Франции и Германии до Центр. Азия
включительно обитает водяной уж, N. tesselata. Он имеет 162 —
190 брюшных и обычно 8 верхнегубных щитков. С водой связая
более предыдущего. Основу пищи составляют рыбы. Кожа ис-
пользуется на галантерейные изделия.
Подрод Thamnophis, иногда выделяемый в особый род, охва-
тывает группу видов Сев. Америки и отличается неразделенным
анальным щитком, а также иным типом окраски: у него часто
встречается непарная продольная светлая полоса на спине, тог-
да как у Natrix s. str., если светлая полоса имеется, то всегда пар-
ная (светлые пятна по бокам головы обыкновенного ужа пред-
ставляют собой рудимент такой полосы). Пожалуй, самый
распространенный вид подрода Thamnophis — подвязочный уж
N. (Т.) ordinatus, многочисленные цветные разновидности кото-
рого многими принимаются за самостоятельные виды. Подрод
Amphiesma характеризуется увеличением последних 2—3 верхне-
челюстных зубов и представлен у нас тигровым ужом, N. М-)
tigrina (Дальневосточный край, Китай, Корея, Япония). Брюш-
ных щитков у него 141—170, а верхнегубных 7. Общая длина до
ПО см. Живые темно-зеленого или голубого цвета. Подрод
Macropophis имеет, в отличие от всех предшествующих, 35—
40 верхнечелюстных зубов и представлен 5 видами на Филиппи-
нах, Хайнане и в Полинезии.
Виды рода косоглазых змей. Helicops яйцеживородящи, при-
держиваются рек и болот, поедают земноводных и рыб. Пред-
ставителем' группы наземных ужей может служить полоз,
Coluber (=Zamenis). В нашей фауне он представлен группой ви-
дов, из которых наиболее интересен желтобрюх, С. jugularis,
распространенный от Синайского полуострова, Албании, Юго-
славии и Венгрии до Туркмении и Ирана. Живет в степях, полу-
пустынях, на полях и заросших песках. Питается мелкими мле-
копитающими, ящерицами, змеями и мелкими птицами. В конце
июня — июля откладывает 7—8 яиц. Длина дела с головой до
150 см. На каменистых склонах от Адриатики до Кавказа и
Туркмении включительно живет оливковый полоз, С. najadum,
в предгорьях Сев. Африки, Ср. Азии и Зап. Индии обитает крас-
нополосый полоз, С. rhodorhachis, в пустынях Ср. Азии встречает-
ся поперечнополосатый, С. karelini. Широко распространены так-
же разноцветный, С, ravergieri и пятнистый, С. tyria полозы.
280
большеглазый полоз, С. тисояия <в< я Н>ж«. Азия. я у на<* Т.р*
ясная) выделяется многими в особый рол /’////к Общая т.-ию
уела большеглазого полоза из Туркмении до 35Н см. Нлибопч
распространенный вид рода лазакнци.ч положа, [Japhr у wjp
чатыА полоз, Я. dione (от восточной Украины и I'руши ло Кчт.и
включительно). Встречается в лесах, степях и в горах. Охотна
заходит даже в морскую воду. Основная пиша - грызуны. Реже
поедает птиц, их яйца и рыбу. Откладывает 5 -16 яиц (длин;
5 см), из которых молодые выводятся примерно через месяц
В западной части нашей страны встречаются четырехполосый,
Watuorlineala, леопардовый, Е. situla, закавказский, Е. hohe-
nackeri и эскулапов, Е. longissitna полозы. На Дальнем Востоке
обитают амурский Е. schrencki, красноспинный, Е. rufodorsata
и тонкохвостый, Е. taeniura полозы.
Представителем рода Coronella у нас служит медянка,
С. austriaca (почти вся Европа, Мал. Азия, Казахстан, Иран),
обитающая в лесах среди кустарника, реже в степи. Основную
пищу ее составляют ящерицы, но поедают также мелких змей,
грызунов, землероек, птенцов и даже насекомых. Рождают по
2 15 молодых. Общая длина тела до 65 см. Наземные ужи, как
правило, душат свою добычу. Представители многих родов под-
семейства держатся «на деревьях, например, лесные ужи,
Herpetodryas. Самый крупный вид зипо, И. fuscus (Гвиана, Пе-
РУ> Бразилия, Венесуэла), общая длина до 230 см. Впрочем из-
редка они встречаются- и на земле. Хорошо плавают. Ест птен-
цов, ящериц и земноводных. Представитель рода древесных ужей,
Dendrophis— блестящий уж, D. pictus (от Гималаев до Малай-
ского архипелага включительно) живет на деревьях в холми-
стых местностях, питаясь древесными ящерицами, древесными
лягушками и мелкими птицами. Сходный образ жизни имеет
и окрашенная в зеленый цвет Leptophis. Представителем груп-
пы родов, питающихся преимущественно насекомыми и пауко-
образными, в нашей фауне служит род Contia. Эти небольшие
(длина тела с головой до 31—41 см) изящные змеи держатся
под камнями предгорий и гор. Наиболее обычны ошейниковая,
С. collaris (Вост. Кавказ, Иран, Турция) и смирная, С. modesta
(Греция, Мал. Азия, Закавказье). Насекомыми и червями пи-
таются также и карликовые змеи, Calamaria, которые держатся
под упавшими стволами, под камнями, а иногда и под землей.
Самый крупный вид 'С. occipitalis (Ява), общая длина 50 см,
а обитающей на острове Калимантан (Борнео) С. sm.it-
hii — всего 10 см. Представители рода Heterodon обладают
вытянутым вперед и вверх межчелюстным щитком, ко-
торый служит для рытья в песке. У одного из его
видов — крючковатого ужа, Н. contprtrix (Сев. Аме-
рика к востоку от Скалистых гор) при возбуждении
передняя часть тела сплющивается, становясь вдвое шире, рот
широко открывается и уж по внешнему виду походит на ядовитую
18 Р- ТсРеятьев
281
умею Питается исключительно лягушками и жябяия Зары»*»*
• чемлю 15 27 ими В нашей фауне представлены также род
I, у codon. Lytnfhyruhu* и Oligoaon.
Подсемейство ложных ужей (Dlpsadomnrphlnae) яит
ядовитые бороздчатые зубы, расположенные на жадней чаги
верхнечелюстной кости,--это так называемые «лолотрителыты
эмсиэ: укус их ядовит для мелких животных, но редко причина*
вред человеку, ибо при небольших размерах рта они просто в
достигают кожи человека задними зубами. Многие виды эты
Рис. 60. Представители ложных ужей, водяных ужей и яичных змей: 1—че-
реп Dipsadomorphus cynodon сбоку, 2 — череп Homalopsis buccata сбоку,
3 — череп Dasypeltis scabra сбоку, 4 — Dasypeltis scabra
группы душат добычу, другие — отравляют. В нашей фауне
встречается представитель рода Tarbophis—Т. fallax (Балканский
полуостров, Мал. Азия, Сирия, Иран, Закавказье), называемый
«кошачьей змеей» за вертикальный зрачок глаза. Придерживает-
ся сухих каменистых мест, иногда поросших кустарником. Ак-
тивна в темноте. Питается преимущественно ящерицами. Длина
тела вместе с головой до 70 см.
Обширный род ночных древесных змей, Boiga у нас пред-
ставлен видом В. trigonatum, заходящим в Ср. Азию из Индии.
На Кавказе и на юге-Калмыцкой АССР встречается ящеричная
змея, Malpolon monspessulanus, живущая, кроме того, в Ираке,
Иране и Средиземноморье. Держится в каменистых полупусты-
нях, питаясь ящерицами и змеями, реже мелкими млекопитаю-
щими. Единственный вид рода Taphrometopon, стрела-змея
Т. lineolatum живет в пустынях и полупустынях Казахстана,
Киргизии, в республиках Ср. Азии, Ирана, Афганистана и Монго-
лии. Питается почти исключительно ящерицами. Откладывает
2—4 яйца, длиною до 4 см. Примером древесных форм может
282
иТЬ плетневидная змея, Dryophls. Окраска почти всех в и*
♦•‘^того рода зеленвя’ Всю жизнь проводят среди листвы.
9яны днем. Питаются птицами, ящерицами и древесными ля*
А*т?ами. Наибольших размеров достигает древесная проныра,
rf^aSinus (от Гималаев до Малайского архипелага включи*
ьно). общая длина до 179 см. Сходный образ жизни ведет
1?оголовая змея, Oxybells, самый крупный ее вид—О. fulgidus
Рис. 61. Представители толстоголовых и морских змей: 1 — череп Amblycep-
halus carinatus сбоку, 2 — голова и передняя часть тела Amblycephalus monti-
coia сбоку, 3—череп Laticauda colubrina сбоку, 4—Laticauda colubrina,
5—череп Naja naja сбоку, 6 — Naja naja, 7 — передняя часть тела Elaps
fraseri сверху
(тропическая Америка), общая длина достигает 155 см, окраска
зеленая с желтой боковой линией. Особый интерес своими пара-
шютирующими прыжками представляет украшенная змея,
Chrysopelea: вытянувшись и втянув в себя часть брюха, на кото-
ром появляется продольный желобок, змея летит подобно стре-
ле. Активны днем. Питаются ящерицами (в частности Draco),
летучими мышами, грызунами и мелкими птицами. Самый
крупный вид — Ch. rhodopleuron (Молуккские острова), общая
длина достигает до 116 см.
Подсемейство водяных ложных ужей (Homalopsinae) охва-
тывает исключительно водных яйцеживородящих змей, отравля-
1в* 283
ся<>к> добычу (рыбы, лягушки, ран<»>>Лрахны*) Ьольш»
яи1оя 4ЯКЛЮЧЛГ1 рол нысом»н<хых змгй. Hupttfhina. г»
• •- ....л И bocffUffii (Сиям), обшяя длим*
иил рола llomalopiis, уляяояилиый
<ыЛГуж. И bin < at и ж трсчягк и, подобно предыдущему, я р*-
’ лях н на рисовых полях. Общая длина достигла
редставитсли родов Cerberus и f'ordonia живут ияк в
10111И я
*'.^Лр?пнмн ЯНЛ мнорой
и‘ .19 Гчинлж нный
ДО ’
ЛОЖ
кях,
105 см.
^^мейстно яичных имей, Dasypeltinae характеризуется
оедукнисй зубов, связанной с переходом и;» питание птичьими
яйцами. Нижние отростки передних позвонков прободают стен-
ки пищевода, образуя аппарат для разрезания скорлупы птичьих
яиц: при сильном сокращении мускулов скорлупа разрезается
и выбрасывается обратно через рот, а содержимое яйца прогла-
тывается. Процесс этот хорошо изучен у Dasypeltls scabra
(рис. 60), общая длина до 76 см, хуже — у Elachlstodon wester-
manni, длина до 785 мм.
12. СЕМЕЙСТВО AMBLYCEPHALJDAE. ТОЛСТОГОЛОВЫЕ ЗМЕИ
(рис. 64)
В ископаемом состоянии не установлены. Голова сидит на
тонкой шее. Глаза чаще с эллиптическим зрачком. Особенности
аубной системы определены питанием моллюсками. Образ жиз-
ни древесный. Виды Нового Света имеют только левое и трахе-
альное легкое, а виды Старого Света имеют и рудимент право-
го. Описано 6 родов с 60 видами. Главные рода различаются
так:
• / ?! Хело более или меисе сжатое.
3 ! 41 л веРхнечелюстной кости 5—6 зубов.
< 4) Один ряд подхвостовых.
Haplopeftura
1 вид, Индия, Малайский архипелаг, Филип-
пины, Молуккские о-ва.
( 3) Два ряда подхвостовых.
Amblycephalus
5 12 видов, Юго-Вост. Азия.
6 ( ?! На верхнечелюстной кости И—18 зубов.
* * Крыловидная кость с зубами.
7
6)
Leptognathus
42 вида, Центр, и Южн. Америка.
Крыловидная кость беззубая.
8 (
2&4
Dipsas
3 вида, Южн. Америка.
* Тело цилиндрическое.
Pteudopare««
I Рид, Южн. Америка.
Самый крупный представитель рода Haplvptlfura Н frorf
обшая длина до 75 см.
13. СЕМЕЙСТВО HYDROPHIDAE. МОРСКИЕ ЗМЕИ
В ископаемом состоянии не известны. Все живу» в м^ре. ис-
ключая Distira semperi (озеро Тааль на Лутоне), и лишь иногда
входит в реки во время прилива. Редко удаляются от берета
далее чем на 60 км (максимум 250 км). Поскольку хвост весло-
образно сжат с боков, верхние и нижние остистые отростки
сильно увеличены. Развито только одно легкое с дополнением в
виде трахеального легкого. Десны богато снабжены сосудами,
по-видимому, это играет роль в газообмене. Укус ядовит, иног-
да смертелен, так как на передней части верхнечелюстной кости
сидят хорошо развитые бороздчатые зубы. Живородящи. Опи-
сано И родов с 39 видами. Границы некоторых из них спорны.
Главные рода различаются так (рис. 62).
1(8) Верхнечелюстная кость не заходит вперед за передний
конец нёбной кости; брюшные щитки малы или отсут-
ствуют.
2(7) Верхнечелюстная кость длиннее поперечной кости; рас-
сматриваемой снизу, и, в дополнение к крупным ядови-
тым, несет 4—18 маленьких зубов.
3(4) Брюшных щитков нет.
Hydrus
1 вид, Индийский и Тихий океаны.
4(3) Брюшные щитки различимы, по крайней мере, в передней
части тела.
5(6) Мелкие верхнечелюстные зубы без бороздок.
Hydroph is
22 вида, от Персидского залива до Южн. Ки-
тая и Сев. Австралии.
6(5) Все верхнечелюстные зубы бороздчатые.
Distira
18 видов, от Персидского залива до Японии и
Новой Каледонии.
7(2) Верхнечелюстная кость не длиннее поперечной и несет,
в дополнение к большим ядовитым, 2—5 мелких борозд-
чатых зубов.
Enhydris
2 вида, от берегов Индии до Китайского моря
и Новой Гвинеи.
285
Рис. 62. Ареалы семейств: 1 — Colubrldae. 2— A mb j yceph a Hdee, 2 — Ну«корЫ4т
й ( i) Верхнечелюстная кость заходит вперед м пМпую;
брюшные щитки хорошо развиты.
р (10) Ноздри на верхней поверхности головы; носовые шмтяв
соприкасаются.
AipysuruB
4 вида, Малайский архипелаг я запад Тихого
океана.
( 9) Ноздри боковые; носовые щитки разделены межио
совым.
Laticauda
3 вида, восток Индийского и запад Тихого
океанов.
Регулярно в наших водах не встречаются, но мертвый экземп-
ляр Hydrus platurus однажды был найден в заливе Посьета.
Общая длина представителей этого вида может доходить до
85 см. По-видимому, активен ночью и питается головоногими мол-
люсками. Hydrophis spiralis — самый крупный вид семейства
(Индийский океан и Малайский архипелаг) — общая длина до-
ходит до 274 см. Крупных размеров (до 230 см) достигает также
ластохвост, Distira grandis (Малайский архипелаг и Сев. Авст-
ралия). Питаются рыбой. Часто держатся стаями. Интересно,
что один из видов плоскохвоста, Laticauda — L. laticauda (от
Бенгальского залива до Китайского моря и западной части Ти-
хого океана), иногда выходит на сушу.
14. СЕМЕЙСТВО ELAPIDAE. ЯДОВИТЫЕ УЖИ (рис. 63)
Древнейший представитель — Palaeonaja (миоцен Франции).
Косвенные соображения заставляют предполагать возникнове-
ние семейства в восточной Азии. Наличие бороздчатых зубов на
переднем конце верхнечелюстной кости делает их ядовитыми
даже для человека. У таких примитивных родов, как Diemenia,
верхнечелюстная кость еще длинная и несет 9—17 зубов, у Naja
эта кость уже сильно укорочена и имеет 3—5 зубов, а у Micrurus на
сравнительно короткой верхнечелюстной кости сидят только
2 ядовитых зуба. Так вырабатываются особенности, сходные с
таковыми у гадюк. Имеется одно развитое легкое и небольшой
рудимент другого. Большинство яйцеживородящи. Описано
34 рода с 192 видами. Отличия главнейших родов таковы:
1 (35) Верхнечелюстная кость без заднего отростка; нет изо-
лированных передних зубов нижней челюсти.
2 (24) Верхнечелюстная кость не выдается вперед далее
небной.
3 (23) Позвоночные чешуи не увеличены.
4 ( 7) Позади больших ядовитых зубов имеется 7—15 мелких
бороздчатых зубов.
287
Рис, 63. Ареалы семейств: 1 — Elapidae, 2 — Viperidae, J—CroUlida»
(19)
(18)
(13)
(12)
8
9
10
11
12
(11)
6) Края морды без канта; глят мелкий с вертикаль***
i ' 7 зрачком.
Psetidetaps
9 видон, Австралия, Молуккские о-м. Имм
Гвинея, Полинезия.
- / 5) Края морды образуют ясное ребро (кант); глаз сред
° ннй или крупный с круглым зрачком.
Dlemeni а ( = Demansia)
10 видов, Австралия, Новая Гвинея.
7(4) Позади больших ядовитых зубов 1—5 мелких зубов,
которые могут быть неясно бороздчатыми. ^„_Лва,,,я
Голова более или менее ясно отграничена о у
Чешуя гладкая. я„пй«цр
Подхвостовые щитки, хотя бы частично, Д
Глаз средней величины; хвост умеренный.
Pseudechis , п_»а Поли-
9 видов, Австралия, Молуккс
незия.
Глаз очень маленький; хвост короткий.
“видовая Гвинея, Соломоновы о-ва.
13 (19) Один ряд подхвостовых щитков.
14 (15) Чешуя расположена косыми рядам
Notechis
2 вида, Австралия, Тасмания.
1R Чешуя не расположена косыми рядами
(17) Брюшные щитки закругленные.
К“‘а..,,.»..
" ™> в„„„. с«—• •“р“"
Hoplocephalus
3 вида, Вост. Австралия.
Чешуя с ребрышками.
Acanthophis аохипелаг, Новая Гвинея,
2 вида, Малайский архив
Австралия.
Голова не отграничена от тела. ааМИ; хвост очень ко
Чешуя расположена косыми Р
роткий. 289
18 ( 9)
On < 8)
ElapechH
8 видок. троляческая и Юж». Афряка.
21
23
24
25
26
27
28
29
30
31
32
33
34
35
(20) Чешуя не рлсположеил косыми рядами; хвост
репный.
Boulengerina
4 вида. Центр. Африка.
( 3) Позвоночные чешуи увеличены.
Bungarus
12 видов, Юго-Вост. Азия.
( 2) Верхнечелюстная кость выдается вперед далее н$бипа
(28) Межносовой щиток касается ноздри. °"
(27) Позади больших ядовитых зубов находится оди« и.
несколько мелких.
Naja
12 видов, Африка, Южн. Азия.
(26) Мелких зубов позади больших ядовитых нет.
Sepedon
1 вид, Южн. Африка.
(25) Межносовой щиток не касается ноздри.
(30) Позади больших ядовитых зубов 1—3 маленьких зуба
Hemibungarus
3 вида, Южн. и Юго-Вост. Азия.
(29) На верхнечелюстной кости мелких зубов нет.
(34) Вокруг тела 13 рядов чешуй.
(33) Ядовитые железы не выходят за пределы головы.
Callophis
5 видов, Юго-Вост. Азия.
(32) Ядовитые железы простираются назад до конца перед-
ней трети тела.
Doliophis
4 вида, Юго-Вост. Азия, Малайский архи-
пелаг.
(31) Вокруг тела 15 рядов чешуй.
Micrurus ( = Elaps)
44 вида, Сев. и Южн. Америка.
( 1) Задняя часть верхнечелюстной кости с большим отрост-
ком, направленным назад и наружу; позади крупных
передних зубов нижней челюсти имеется значительное
беззубое пространство.
EtaidraspH
5 видов, тропическая и Южн. Африке
представителем рода Pseudechls может служить черная енц-
р porphyriacus, широко распространенная я Австралия.
•**’вет в лесах, степях и на берегах водоемов; иногда плавает.
^”н»на днем, поедает мелких млекопитающих, птиц, пресны-
ющихся и земноводных. Общая длина до 158 см и более,
наиболее крупный представитель рода Bungarus — В. flauicept
тенассерим, п-ов Индокитай, Малайя, Суматра, Калимантан,
iaa), достигает 185 см. Более известна пама, В. fasciatus (др
175 см) » распространенная от Индии и Южн. Китая до Суматры а
LBbi включительно. Окраска — желтые кольца вокруг тела на об-
дем черном фоне. Предпочитает сухие места (поля, травянистые
равнины). На день прячется. Питается мелкими млекопитаю-
щими и пресмыкающимися (особенно змеями). Известно много
случаев смерти людей от укусов этого вида. В раздраженном
состоянии большинство видов кобр (Naja=Naia) поднимает пе-
реднюю треть тела вертикально и дискообразно расширяет шею,
раздвигая в стороны передние 8 пар ребер; голова при этом
сохраняет горизонтальное положение. Наибольшей известностью
пользуется азиатская кобра, или «очковая змея», Naja n.aja=N.
tripudians. Прилагательное «очковая» обусловлено тем, что на
спинной стороне расширяющейся части тела у индийского под-
вида имеется светлый рисунок, напоминающий старинные очки.
Встречающиеся у нас на юге Туркмении, Узбекистана и Таджи-
кистана экземпляры азиатской кобры (N. п. oxiana) лишены этого
рисунка, так же как и живущие на Зондских островах (N. п.
соеса). Общий ареал вида простирается от южной Туркмении
и восточного Ирана до Южн. Китая, Тайваня, Филиппин и Зонд-
ских островов. Держится среди камней и кустарников в долинах
и ущельях, в заброшенных зданиях, в дырявых глиняных стенах
и др. Хорошо плавает. В поисках пищи может влезать на де-
ревья. Активна во вторую половину дня и в сумерках. Питается
главным образом ящерицами, лягушками, жабами и мелкими
грызунами, реже поедает мелких птиц и их яйца, а иногда даже
и рыб. В Туркмении кладет 8—12 яиц в июле — августе (в Ин-
дии 12—20). Очень ядовита и дает значительное число смертных
случаев среди людей. Яд может действовать не только при попа-
дании в кровь, но и через желудок и на глаза (развитие бель-
ма). Общая длина достигает 190 см. Еще крупнее (до 200 см)
египетская кобра, N. haie, обитающая в южной Палестине и
восточной Африке. От предшествующего вида отличается тем,
что у нее самые высокие шестой или седьмой верхнегубной щи-
ток, а не третий. Держится в полях, пустынях и горах, скрываясь
обычно в норах грызунов или в развалинах. Основную пищу
составляют грызуны, птицы, ящерицы, лягушки и жабы. Самым
крупным видом рода (до 457 см) и самой крупной ядо-
291
|INtO* A IWtoftnif ПрГМИвГЭ» МПрС»ЛГЖ»уЮ мЛ/у
JV Л^ЛПЛА- V tmnpum* От яшуж ря«**» уилыя«у*м» м
дои tn.»HM»ci< м 15 ридвми чсшуА wmpyr <*роюг«
je.ii», e.wrvto 17 25, и 19 21 четуй шшгрея ш*и, яме<го 21 V,
ДгрЖИ1СЧ В рг.1КН< леглх И ЛЖУИММ Хорошо ЛМ»Л*Т N Ч.ООГ
f|nT*vrc«t В OVHOHHUM ♦мсямн. Выли случаи, могла Крупмм* <ко
Ли пр<ч лглоплли лю fir А. В Центр. Африке жиле» <я<мбый род
водной кобры. ffoui<*ngrrina, длина которой может доходит», *с
|31 см. Укусы крупных видов роде аспидов, Micrurus — Elapt чл
сто смертельны, а укусы мелких обычно заканчиваются бляплю.
лучно. Самый крупный вид — М. surinamensis (север Южн. Ашь
р'нкн), общая длина достигает 190 см. Арлекиновая змея, М. fub
tfius (до 99 cat) распространена от восточной части Сев. Америки
до Центр. Америки включительно. Окраска из чередующихся
черных и красных или желтых колец, охватывающих тело. Дер-
жится на бататовых полях; зарывается в землю и в песок. Пи-
тается змеями, ящерицами, жуками и грызунами. Известны слу-
чаи смерти людей от укусов этого вида. Коралловая змея,
М. coraUinus (тропическая Южн. Америка и Малые Антильские
острова) достигает 79 см, окрашена в ярко-красный цвет с чер-
ными кольцами вокруг тела. Встречается главным образом в
лесах и среди кустарников. Поедает змей, двуходковых и сцинко-
вых ящериц. Людей кусает редко. Исключительно древесный
образ жизни ведет мамба, Dendraspis. Самый крупный вид этого
рода — D. antinorii (Абиссиния), общая длина достигает
269 см.
15. СЕМЕЙСТВО VIPERIDAE. ГАДЮКОВЫЕ
Видимо, возникли в Африке. Ископаемые остатки известны,
начиная с миоцена. Сзади функционирующего ядовитого зуба
помещаются 3—4 его заместителя. Внутри семейства намечается
переход от головы, покрытой крупными щитками наподобие
большинства ужей, ко вторичному распадению щитков на мел-
кие. Все, исключая Atractaspis, яйцеживородящи. Обычно нет
следа рудиментарного легкого. Описано 65 видов, объединяемых
в 10 родов, которые различаются так:
1(6) Голова покрыта большими симметричными щитками.
2(5) Зубы в нижней челюсти хорошо развиты; глаз средней
величины.
3(4) Ноздря между 2 носовыми и межносовым щитками;
зрачок круглый.
Causus
4 вида, тропическая и южная Африка.
4(3) Ноздря прорезана внутри одного щитка; зрачок верти-
кальный.
Azemlops
1 вид. Бирма.
Зубы в нижней челюсти редуцированы до 2—3; глаз
5 ( 2) маленький.
Atractaspls
26 видов, тропическая и Южн. Африка, Южн.
Аравия.
1) Все или часть щитков верха головы разбиты иа мелкие
6 ' ' щитки или чешуйки.
,14) Боковая чешуя не мельче спинной.
/13) Брюшные щитки округлые.
8 kg) Носовой щиток касается межчелюстного или отделен
' от него одним щитком.
Vipera
14 видов, Европа, Азия, Сев. и тропическая
Африка.
п / 9) Носовой щиток отделен от межносового мелкими че-
' шуйками.
11 (12) Имеется выдающийся верхненосовой щиток в форме
полумесяца.
Bitis
9 видов, Африка.
12 (Н) Верхненосового щитка нет.
Pseudocerastes
2 вида, Иран, Вазиристан.
13 ( 8) Брюшные щитки по бокам, угловатые.
Eristicophis
1 вид, Белуджистан.
14 ( 7) Боковая чешуя мельче спинной.
15 (18) Ребрышки боковой чешуи зазубренные.
16 (17) Брюшные щитки по бокам угловатые; два ряда подхво-
стовых щитков.
Cerastes
2 вида, Сев. Африка, Аравия, Палестина.
17 (16) Брюшные щитки закругленные; один ряд подхвостовых.
Echis
2 вида, Африка к северу от экватора, Южн.
Азия.
18 (15) Ребрышки боковой чешуи не зазубренные.
Atherls
4 вида, тропическая Африка.
293
Рис. 64. Представители трубчатозубых змей: 1 — череп Vipera lebetina сбоку
2 — череп Atractaspis aterrima сбоку, 3 — череп Crotalus horridus сбоку’
4 — голова Tnmeresurus flavomaculatus сбоку, 5 — голова Cerastes cornutus
6 — голова Vipera ammodytes, 7 — трещотка Crotalus (снаружи и продоль-
ный разрез), 8 — Crotalus adamanteus
Примитивная жабья гадюка, Causus, является наземной
змеей, которая держится главным образом на болотистых лугах
и близ воды. Питается бесхвостыми земноводными. Самый
крупный вид С. rhombeatus, широко распространенный по всей
тропической и Южной Африке, общая длина до 89 см. Земляная
гадюка, Atractaspis, так же примитивна, но имеет ряд особенно-
стей, вызванных роющим образом жизни: тело цилиндрическое и
голова слабо отграничена от туловища, глаза уменьшены, зубная
система сильно редуцирована, но передние ядовитые зубы верхне-
челюстной кости 'непропорционально увеличены. Причина по-
следнего неясна и некоторые даже выражают -сомнение в воз-
можности пускать в дело такие зубы. На поверхности земли
встречаются, подобно дождевым червям, после дождя. Самый
крупный вид, A. aterrima (Зап. и Центр. Африка), общая длина
до 65 см. Среди настоящих гадюк—Vipera — наиболее прими-
тивными приходится считать виды, еще не утратившие крупных
щитков на верхней поверхности головы (рис. 64). К числу тако-
294
принадлежит м обыкмовенмяя галюяа. V kw «отв а* жа
рс*ое распространение (почти вся северная и срсаяяя Гария»
гг Северного моря до Сахалина включительно) несомненно *•>
(.•леледникопого происхождения. Особенно характерна для г не
тайных лесов, болотистых мест, боров и лиственных лссоя Па
маетен мышевидными грызунами, реже ящерицами а насеяомы
ми. В августе — сентябре рождает до 18 молодым <амует под
.«емлей. часто группами. В тропиках окраска серая, буроватая иля
красновато-бурая с черным зигзагообразным ремнем вдаль спя
мы. Однако встречаются многие уклонения в окраске (напрв-
мер. сплошь черная), не имеющие систематического значения
Рыжие особи часто ошибочно именуются населением «медян-
кой». Общая длина может достигать 84 и даже как исключение
90 ел. Известны единичные смерти людей от укусов этого вида.
Заостренные верхнебоковые края морды и ноздря, прорезанная
в нижней части (а не в середине!) носового щитка, отличают
степную гадюку» V. ursini, от обыкновенной гадюки. Распростра-
нена от южной части Зап. Европы и Вост. Казахстана до Синь-
цзян-$ Йгурской авт. области включительно. По-видимому, «е за-
ходит на север далее 55° с. ш. Питается насекомыми и мелкими
грызунами» реже ящерицами. Общая длина до 62 сл. Достовер-
ных случаев смерти людей от укусов этой змеи не зарегистриро-
вано. На покрытых лесом склонах гор и субальпийских лугах
Зап. Кавказа живет близкий вид V. kaznakowi, Основной фон
тела при жизни от желтого до ярко-красного. Известны случаи
гибели людей от укусов этого вида. Примером видов, у которых
вся верхняя поверхность головы покрыта мелкой чешуей» может
служить гюрза, V. lebetina (от Сев. Африки через Палестину и
Мал. Азию до Сев.-Зап. Индии; у нас на Вост. Кавказе и на юге
Ср. Азин). Общая длина до 135 еж. Известно немало случаев
гибели людей и крупного скота от укусов этого вида. В горах
Армении, Нахичеванской АССР и в соседних районах Турции и
Ирана обитает V. raddei. Как отдельную группу можно рас-
сматривать виды, у которых кончик морды завернут кверху и
снабжен кожистым придатком. Из них на территории СССР в
каменистых местах Закавказья встречается V. ammodytes. Рога-
тая гадюка, Cerastes, имеет над каждым глазом большой покры-
тый чешуей выступ. Распространенная от Сахары до Аравии и
Палестины С. cornutus (до 75 сж) «а день зарывается в песок
(боковыми движениями тела). В Туркмению, Узбекистан и Тад-
жикистан к нам заходит один вид эфы. Echis — Е. carinatus (об-
щая длина до 75 см и более), живущий главным образом в пес-
ках. На день прячется в норы грызунов и под камни. Питается
грызунами, птицами, ящерицами и членистоногими. В августе
рождает 3—15 молодых. Известны случаи укусов, имевших
смертельный исход. Способность хвоста закручиваться отличает
древесную гадюку, Atheris. По ночам она ловит ящериц и земно-
водных.
295
(Т.МГПСТВО f.ROIAJ HJ.M: ГРЕМУЧИЕ ЗМЕИ
Сходны с предшествующим семейством н раньше входили i
него как подсемейство. По-видимому, во шикли в Ссаром Chef#
откуда мигрировали в Новый, где и достигли наикмгшегб
развития. Достоверные ископаемые остатки известны и5 миоие
на Греции и. только начиная с плейстоцена, в Сев. Америке
Эволюция внутри семейства шла параллельно по двум направ-
лениям: большие щитки верхней поверхности головы, так же как
у гадюк, распадались на мелкие, а с другой стороны, на конце
хвоста возникла трещотка («гремушка»). Последняя представ-
ляет собой видоизмененные колпачки кожи, оставшиеся после
линьки и частично вложенные друг в друга. Самым старым слу-
жит ее дистальное звено, меньшее по размерам. Новорожденная
змея обычно имеет на конце хвоста лишь одно звено. За год
прибавляется по 3—4 звена. Чаще всего встречается 5—9 звень-
ев. Трещотка, состоящая более чем из 14 звеньев, представляет
исключительное явление. Более крупные виды имеют больше
звеньев. Обитатели каменистых или заросших травой мест име-
ют меньше звеньев, чем обитающие в песчаных. Наибольшее
число звеньев наблюдается у животных, обитающих в местах,
где их меньше всего тревожат (например, на островах). Было
предложено много объяснений экологического значения тре-
щотки, «о окончательного решения «не найдено. Использование
ее входит в общий комплекс оборонительной реакции змеи: как
и многие змеи, лишенные трещотки (например, наша гадюка),
гремучие змеи подтягивают переднюю часть тела, несколько уп-
лощают и плотнее упирают о грунт заднюю часть, шипят и виб-
рируют кончиком хвоста. При этом у гремучих змей получается,
благодаря трению звеньев трещоток, своеобразный шелест, иног-
да переходящий, при большой частоте, в своеобразный свист.
Некоторые зоологи считают, что отбор развил это приспособле-
ние у предков современных гремучих змей как средство преду-
преждения крупных млекопитающих (например, бизонов). Экс-
периментально доказано, что ни одна гремучая змея не слышит
звука, издаваемого другим экземпляром. Скорость вибраций хво-
ста змей зависит от температуры среды. Описаны 91 вид, объе-
диняемые в 4—7 родов. Рода различаются так:
1(4) Верхняя поверхность головы покрыта 9 большими сим-
метричными щитками.
2 ( 3) Трещотки нет.
Ancistrodon
12 видов, берега Каспия, Азия, Сев. и Центр.
Америка.
3(2) На конце хвоста имеется трещотка.
Sistrurus
3 вида, восток Сев. Америки, Мексика.
4(1) Верх головы покрыт мелкими щитками иля чешуей.
5(6) Трещотки нет.
Lachesis
50 видов, Юго-Вост. Азия, Центр, и Южн.
Америка.
5 ( 5) На конце хвоста трещотка.
Crotatas
26 видов, от Южн. Канады и Британской Ко-
лумбии до Сев. Аргентины включительно.
В нашей фауне род щитомордника Ancistrodon (=Agkistro-
don) представлен 2 видами. Палласов щитомордник, A. halys,
имеет вокруг середины тела 23 чешуи и распространен от побе-
режья Тихого океана до низовья Волги и Азербайджана вклю-
чительно. Населяет разнообразные стации от тайги до степей и
полупустынь. Общая длина'достигает 75 см. Питается после за-
хода солнца, преимущественно грызунами. В июле — октябре
рождают 3—10. молодых. Люди от укусов этих змей обычно не
страдают. В Японии, Китае и Дальневосточном крае живет во-
сточный щитомордник, A. blomhoffi, у которого вокруг тела обыч-
но 21 чешуя. Наиболее крупным видом является китайский
A. acutus (до 150 см}. Некоторые виды яйцекладущи: A. hitna-
layanus (Гималаи на высоте 1,5—3,6 тысячи метров) и A. rhodos-
toma (Ява, Суматра, Малайский полуостров). Обширный род
Lachesis, подразделяется на 2—3 подрода или даже самостоя-
тельных рода. Бушмейстер, или сурукуку, L. mutus, отличается
от всех прочих тем, что последние подхвостовые щитки замене-
ны у него мелкой чешуей и потому его выделяют в подрод
Lachesis s. str. Это очень крупный вид, достигает 3,6, а по неко-
- торым данным даже 4,3 м. Держится на" земле в тенистых лесах
Центральной и в тропической части Южн. Америки, укусы часто
дают смертельный исход. Откладывает яйца. Другие виды, оби-
тающие в Новом Свете, объединяются в группу Bothrops. При-
зером их может служить копьеголовая змея или ярарака,
L. (В.) lanceolatus, которая живет на земле среди кустарника,
на полях, в болотах, по берегам реки в лесах тропической Амери-
ки. Общая длина доходит до 2 м. Хорошо плавает. Поедает
преимущественно крыс, а также ящериц и птиц. Рождает 20—
61 детеныша. Примером индо-малайских видов, объединяемых
в подрод Тrimeresurus, может служить древесная гадюка или
будру — пам, L.(T)t gramineus (Юго-Вост. Азия); соответствен-
но' древесному образу жизни окраска ее зеленая. Род собствен-
но гремучих змей, Crotalus, вызывает споры при разделении на
виды: их различают от 11 до 26. Наиболее обычная полосатая
дремучая змея, С. horridus (от Массачузета и Айовы до Сев.
Флориды и Техаса), общая длина достигает 188 см. Держится
между скал, в дуплах, под камнями, среди дров и в других ме-
297
стах. Питается мелкими млекопитающими, птицами и лягушка
ми. Укусы очень опасны. Крупный вид (до 267 см)—бряллим
товая. или ромбическая гремучая амея, С, adamanteus. держат*
на сырой почве по берегам пресных водоемов и моря, лоронп
плавает. Единственный представитель рода в Южн. Америке-
страшная гремучая змея, С. terrificus, придерживается сухих ка
меиистых мест н травянистых равнин. Рогатая гремучая змея
С. cerastes (пустыни Калифорнии, Невады. Аризона и Юты),
хонвергентно сходна с Cerastes тем, что у нее надглазничные
щитки образуют по рожку над каждым глазом.
ЛИТЕРАТУРА
«ян Г Яд°»итые амеи Узбекистана, Ташкент, 1950.
Л Ш их - К а Р а м я« А. Влияние ядов среднеазиатских змей на фнзиоло-
М е л * *' и патологические процессы в организме, Ташкент, 194/.
гиЧ^*"екИй Е. Ядовитые животные СССР, М.—Л., 1931.
**ав£„льев П. и Баркаган 3. К истории изучения ядовитых живот*
Перф"" Института истории естествознания АН СССР, 1957, т. 16,
кЫХ. 123—“145.
стР- * - G Catalogue of Snakes in the British Museum, London, 1—1393.
BoUJet«U 3—1896.
2—1894, о Snakes of the World> New York, XII ed., 1957.
D i t Ш a r s • patt|esnakes, Burkeley and Los Angeles. 2 vols. 1956.
Klauber • . Berner F. Giftschlangen und die Serumbehandlung der
Kraus *enbisse, Jena, 1931.
Schlange Modes of Evolution Discernible in the Taxonomy of Snakes.
Sch®idJ n 1950, 4: 79—86.
Evolution» cVnopsis der Schlangenfamilien, Mitteilungen aus Zool. Mus.
ШегПеГ 1017. 34: 191—208, Archiv fur Naturgeschichte, Abt. A, 1921, 87*
HambUJ^» ?Л2 ’ 88: 185—244, 1923, 89: 138—199, 1924, 90: 108—166, Zolf
Глава одиннадцатая
КРОКОДИЛЫ
Современные крокодилы незначительны по числу видов (все-
го 21—26), но интересны тем, что они в настоящее время единствен-
ные представители Archosauria (см. главу 6), т. е. занимают
высшее место среди всех современных пресмыкающихся. Возник-
нув в триасе из лунчатозубых (вероятно близких к Sphenosuchi-
dae), крокодилы достигли значительного расцвета в юрский и
меловой периоды. С начала третичной эпохи они быстро угасают,
причем не исключено, что процесс этот продолжается и теперь.
Исходные формы были наземными животными. Удлинение прок-
симальных костей кисти передней конечности (radiale, ulnare) и
некоторые другие особенности (таз) заставляют допустить воз-
можность кратковременной стадии двуногости, сменившейся вто-
ричной четвероногостью. Современные крокодилы при медленном
движении широко расставляют ноги, но при быстром тело их
приподнимается над почвой. В эволюции этого отряда ясно вид-
на возрастающая приспособленность к водному образу жизни.
Особенно интересно постепенное увеличение в филогении вто-
ричного твердого нёба (сперва его образуют только нёбные, а
потом и крыловидные кости), в связи с чем внутренние отвер-
стия ноздрей отодвигаются все дальше назад. Подобное строе-
ние наряду с особой складкой, замыкающей полость рта сзади,
дает возможность крокодилу оставаться под водой с открытой
пастью, выставив на воздух только кончик морды с наружны-
ми ноздрями. При этом воздух попадает прямо в гортань, минуя
ротовую полость. По-видимому, такое строение нёба надо свя-
зывать с манерой подстерегать и топить добычу. Деление на
подотряды (см. далее) отражает прежде всего этапы эволюции
нёба. Большинство крокодилов полуводные обитатели пресновод-
ных и солоноватых водоемов, но существовали и постоянные
обитатели моря: из них Teleosauridae, видимо, еще выходили на
берег, a Metriorhynchidae уже утратили эту возможность. На-
земный образ жизни Sebecidae, вероятно, был удержан ими от
сухопутных Protosuchidae. У большинства форма головы, при
299
рассмотрения сверху, напоминает треугольник. Особенно
мое и узкое рыло развивается коявергентко у ряда форм
vtalis. Tomistoma, teleorhinut, Steneosaurus etc.) в связи с рыб^
ядностью. Еще в 1766 г. Линней рассматривал всех крокодил^
Noto- Prolosu-
champ- chcdae
SLdat /
'V
Рис. 65. Филогения крокодилов: 1 — древние, 2- -средние, 3 — высшие
как один из видов рода Lacerta, но уже Лауренти в 1768 г. выде-
лил их в особый род. Выделение крокодилов в особый отряд было
сделано в 1816 г. Бленвиллем. В работах Кювье было
предложено различать гавиалов, крокодилов и аллигаторов на
правах подродов. Потом их признали за самостоятельные рода,
а позднее — семейства. Включение в отряд ископаемых форм
300
заставило Оуэна в I860 г. предложить деление крокодилов ял
Amphicoelia, Opisthocoelia я Procoelia, ио в дальнейшем пре дет а
эителн второго подотряда были признаны за лгииетазовмх ди-
нозавров. Спустя 15 лет. Т. Гексли указал иа филогенетическую
ценность положения внутренних ноздрей и установил деление на
parasuchia, Mesosuchia и Eusuchia. Первые из них теперь отно-
сятся к лунчатозубым. Подотряд Protosuchia установил в 1934 г.
Мук (рис. 65). Распространение крокодилов см. рис. 66 Отдель-
ные семейства можно различить по следующим признакам:
1 (24) Отверстия внутренних ноздрей не окружены крыловид-
ными костями; позвонки двояковогнутые или с плоски-
ми поверхностями.
2(5) Лобковая кость участвует в вертлужной впадине [под-
отряд (1) Protosuchia].
3(4) Врановая кость на обоих концах расширена.
1. Notochampsidae
4(3) Врановая кость, удлиненная и пронизанная отверстием.
2. Protosuchidae
5(2) Лобковая кость не участвует в вертлужной впадине
(подотряд (2) Mesosuchia].
6 (13) Верхняя височная ямка больше глазницы, если же рав-
на или меньше, то рыло широкое.
7 (12) Конечности не в форме плавников, с когтями.
8 (11) Носовая кость не доходит до ноздрей; обычно имеется
предглазничное отверстие.
9 (10) Морда узкая, очень длинная.
10. Teleosauridae
10 ( 9) Морда широкая, более короткая.
8. Goniopholidae
11 (8) Носовая кость доходит до наружного носового отвер-
стия; предглазничного отверстия нет.
11. Dyrosauridae
12 ( 7) Конечности в форме плавников (локтевая и лучевая, а
также некоторые кости запястья уплощены), без когтей.
12. Metriorhynchidae
13 ( 6) Верхняя височная ямка меньше глазницы.
14 (19) Расстояние от кончика морды до переднего края орби-
ты меньше максимального расстояния от -переднего края
орбиты до конца черепа.
15 (18) Между костями нижней челюсти имеется окно.
16 (17) Лобная кость не доходит до края верхней височной ям-
ки; кожный панцирь слабый или его совсем нет.
301
Рис. 66. Географическое распределение современных крокодилов: 1 — аллигаторы. J?— настомцж «рч'жол*-*»*. 3 мл
лайский гавиал, 4 — гаагский гавиал
\1
18
19
20
21
22
23
24
25
26
27
28
29
30
3. Notosucliidae
Нб) Лобная кость доходит до края вершей вясочмой лиги,
' панцирь хорошо развит.
б. Atopesaurldae
(|5) Окна в нижней челюсти нет.
7. Therlosuchidae
(14) Предглазничиая часть черепа длиннее ааглазиячмой,
(23) Череп высокий^ сжатый с боков.
(22) Поперечная кость не касается внутренних ноздрей.
4. Sebecidae
(21) Поперечная кость образует боковой край отверстия
внутренней ноздри.
5. Baurttsuchldae
(20) Череп низкий, не сжатый с боков.
9. Pholidosauridae
( 1) Отверстия внутренних ноздрей окружены крыловидны-
ми костями; позвонки передневогнутые [подотряд (3)
Eusuchia].
(26) Между поперечной скуловой и верхнечелюстной костями
большое окно.
13. Hylaeochampsidae
(25) Такого окна нет.
(28) Нижняя челюсть беззубая; боковые края черепа почти
параллельны друг другу; верхние височные ямки очень
маленькие.
14. Stomatosuchidae
(27) Нижняя челюсть с зубами; боковые края черепа спере-
ди сходятся под углом; верхние височные ямки обычно
хорошо развиты.
(30) Носовые кости доходят до переднечелюстных; верхняя
височная ямка меньше глазницы, иногда исчезает.
15. Crocodylidae
(29) Носовые кости не доходят до переднечелюстных; верх-
няя височная ямка немного больше или немного мень-
ше глазницы.
16. Gavialidae
303
Рис, 67. Представители крокодилов: 1 — скелет Protosuchus richardsoni,
2 — скелет Alligatorellus, 3 — реконструкция внешнего вида Metriorhynchus
iackeli, 4— череп Stomatosuchus inermis сверху, 5— череп Gavialis gangeticus
сверху, 6 — череп Crocodylus niloticus сверху, 7 — череп Alligator sinensis
свер/у
304
Подотряд I — Protosuchia. Дре— крокодилы
1. СЕМЕЙСТВО NOTOCHAMPS!DAE
Два рода в триасе Южя. Африки.
2. СЕМЕЙСТВО PROTOSUCHIDAE
3 триасе Сев. Америки и юре Китая 1—3 рода. Общая длина
protosuchus 80 см (рис. 67).
Подотряд 2 — Mesosuchia. Средние крокодилы
Рис. 68. Внешние части Crocodylus acutus:
/ — голова сверху, 2 — голова сбоку. 3 — кусок чешуи бока. 4 — передняя лапа сверху.
5 — клоака и задняя лапа снизу, 6 — голова снизу
3. СЕМЕЙСТВО NOTOSUCHIDAE
В меловых слоях Южн. Америки и Африки 3—5 родов. Дли-'
на черепа Notosuchus из Патагонии около 16, а африканского
Libycosuchus (иногда выделяется в особое семейство) — око-
ло 20 см.
4. СЕМЕЙСТВО SEBECIDAE
Единственный представитель — Sebecus icaeorhinus— в эоце-
не Патагонии, с длиной черепа около 48 см.
5. СЕМЕЙСТВО BAURUSUCHIDAE
Единственный представитель — Baurusuchus pachecoi — в
верхнем меле Бразилии, с длиной черепа около 38 см. Часто
объединяется с сем. Sebecidae в подотряд Sebecosuchia.
21 П. В. Терентьев 305
с семгж.пю ATOPOAAl'RtDAE
В юре Европы <овад»« м.«. г»>
22 ел) я Alligatofium (общая длина 44 см).
7. СЕМЕЙСТВО THERtOSUCHIDAF.
В юре Европы Therlosuchus — череп в см н Hannotuchut -
череп 10 см,
8. СЕМЕЙСТВО GONIOPHOLTDAE
В юре — меле Европы, Азии, Сев. и Южн. Америки 5—Н ро-
дов. Длина черепа uoniophoUs 60—70 см.
9. СЕМЕЙСТВО PHOLIDOSAURIDAE
В юре —меле Европы, Сев. Америки и Африки (?) 3—5
родов.
10. СЕМЕЙСТВО TELEOSAURIDAE
В юре Европы и Мадагаскара 8—10 родов. Известны скелеты
европейского Mystriosaurus, длина 1—5 м. В юре Steneosaurus.
11. СЕМЕЙСТВО DYROSAURIDAE
В меле — эоиене Сев. Америки и Африки 4—7 родов. Общая
длина Congosaurus 3,5 м. Иногда объединяют с Pholidosauridae.
В мелу Teleorhinus.
12. СЕМЕЙСТВО METRIORHYNCHIDAE
Пять родов в юре — меле Европы и Южн. Америки. Кольцо
окостенений в склеротике глаза. Общая длина европейских от
2 (Metriorhynchus) до 6 м (Dakosaurus).
Подотряд 3 — Eusuchia. Высшие крокодилы
13. СЕМЕЙСТВО HYLAEOCHAMPSIDAE
Единственный представитель — Hylaeochampsa vectiana из
мела Англии (череп около 12 см.)
14. СЕМЕЙСТВО STOMATOSUCHIDAE
Два рода в мелу Африки и Китая. Очень большой (2 м)
плоский череп африканского Stomatosuchus напоминал клюв
пеликана. Некоторые допускают существование также горлово-
го мешка, как у пеликана.
зов
15. СЕМЕЙСТВО CffOCOOYLtDAF
Известны, начиная с мела, на всех чвгтей света • мллеяагт*
13---46 родов, из которых 7 современных. Часто подрамеляаэтл
\ 2-—3 семейства, ио этим нарушается общий масштаб класс-»
1нкации отряда. Мощный, сжатый с боков хвост играет основ
Тую роль при плавании. На передних конечностях 5 нялыде
/обычно без переломки), на задних — 4, но сяяззмнмх плаватель
Joft перепонкой. Глаза, помимо обычных век, снабжены мига
гедьной перепонкой. Зрачок вертикальный, а радужина «елеио-
ватая. Ушные отверстия и ноздри при погружении могут плот
но зажиматься. Тело покрыто плотными, водоустойчивыми щяг-
ками, под которыми на спине помещаются костные пластинки.
Каждый щиток бока, брюха, хвоста и особенно нижней челюсти
имеет небольшую точку или ямку, к которой подходит нерв с
чувствительной почкой. Линьки нет. Поступающие в продажу
кожи крокодилов (идут на сумки, портфели, сапоги и т. п.) лише-
ны эпидермиса, удаляемого при обработке. Имеются 2 пары
мускусных желез: на горле с внутренней стороны правой и левой
половинки нижней челюсти и внутри клоакальной щели. Дея-
тельность этих желез особенно велика в период размножения.
Слюнных желез нет. В глотке имеются 2 складки, играющие роль
голосовых связок. Голос похож на кваканье, короткий лай или рев.
Все легкое губчатое и лежит свободно в грудной полости. Груд-
ная полость отделяется от брюшной мускулистой диафрагмой,
не гомологичной таковой млекопитающих (иннервируется от по-
ясничного, а не шейного сплетения). Сердце четырехкамерное, но
foramen Panizzae устанавливает связь между двумя дугами
аорты. Передняя часть желудка мускулистая. Пищеварение идет
быстро. Слепой кишки нет. Мочевой пузырь отсутствует. Клоа-
ка в виде продольной щели. Копулятивный орган — непарный
конусовидный вырост задней части клоаки с продольной борозд-
кой для спермы на спинной стороне. В скелете 24—25 предкрест-
цовых, 2 крестцовых и 35—42 хвостовых позвонка. Первый позво-
нок состоит из 4 кусков, причем верхний непарный из них счи-
тается за рудиментарный proatlas. Все шейные позвонки несут
ребра. Туловищных ребер обычно 10—13 пар. Некоторые из них
несут крючковидные отростки. Обычно 7—8 пар брюшных ребер.
Образ жизни всех видов довольно сходный. Держатся чаще
всего в тихо текущих речках, озерах и болотах. Изредка попада-
ются в солоноватой воде. Встречались как исключение в море
на расстоянии до 1300 км от берега. Максимальное время, кото-
рое могут пробыть непрерывно под водой — 6 часов. Спят на
суше, греясь под лучами солнца. Пищу поедают на суше, тут же
откладывают яйца. При пересыхании водоемов мигрируют по
суше. От засухи или зимней стужи зарываются в ил. Хватают
всякое неосторожное животное, хотя рыба, по-видимому, всегда
предпочитается. Некоторые оглушают жертву сильным ударом
21* 307
хвоста, топят, а потом разбивают ударамя о землю м камня
Наблюдается заглатывание камней. Способны голодать месяца-
мн. Опасность для людей часто преувеличивают. Время охоты
ночь и сумерки. Яйна откладывают в подобие «гнезда» в числе
20—100 штук (размеры яиц колеблются от 58X 36 до 92x58 мм);
посещаются самкой. Половозрелость на 6—10 год. В неволе
выживали до 85 лет, а на воле — вероятно больше 100. Яйца
снабжены известковой скорлупой.
Современные рода различаются так:
I (12) Носовые кости доходят до костного отверстия ноздрей,
морда от тупой до вытянутой; с каждой стороны верх*
ней челюсти 16—20 зубов.
2(9) Четвертый зуб нижней челюсти входит в ямку верхней
челюсти и при закрытом рте снаружи не виден; с каж*
дой стороны нижней челюсти 17—22 зуба [подсемейство
(1) —Alligatorinae].
3(4) Носовые отверстия разделены костной перегородкой.
Alligator
2—3 вида, США, Китай.
4(3) Костной перегородки между ноздрями нет.
5(8) Верхнее веко не вполне окостенелое; верхние височные
ямки имеются; в каждой переднечелюстной кости по
5 зубов.
6(7) Верхнее веко плоское; передний край глазницы доходит
до вертикали 9 или 10 зуба верхней челюсти.
Melanosuchus
1 вид, Бразилия, Перу.
7 ( 6) Верхнее веко бугорчатое; передний край глазницы не
доходит до вертикали 10 зуба верхней челюсти.
Caiman
2—4 вида, Центр, и Южн. Америка.
8(5) Верхнее веко полностью окостенелое; верхние височные
ямки вторичного отсутствуют; на каждой предчелюстной
кости по 4 зуба.
Palaeosuchus
2 вида, Южн. Америка.
9(2) Четвертый зуб нижней челюсти входит в вырезку верх-
ней челюстй и виден снаружи при закрытом рте; с каж-
дой стороны нижней челюсти 14—15 зубов [подсемейст-
во (2) Crocodylinae].
10 (11) Носовые отверстия разделены костной перегородкой.
Osteolaemus
1—2 вида, Зап. и Центр. Африка.
зов
и ('°’
|2 ( О
Костной перегородки йежду гюхдрямя нет
Crocodyta
11—12 валов, Авия, Африм, Авгървлвя, Ном*
Гвинея, Сев., Центр, н Южн Америка
Носовые кости не доходят до костного отверстия
рей; морда очень длинная; с каждой стороны верхней
челюсти по 20—21 зубу [подсемейство (3) Tomitto
minae).
Tomistoma
1 вид, Малайский полуостров, Суматра, Калн*
мантан (Борнео).
Приведенные подсемейства, особенно первые два, многими
рассматриваются как самостоятельные семейства.
и В неволе чаще всего держат щучьего аллигатора, Alligator
ffiississippiensis (юго-восточная часть США), который отличается
от A. sinensis (Янцзы) большими размерами (общая длина до
4 6 ас вместо 1,8 л<) и наличием перепонки между пальцами
передних ног. Основная пища — рыба. На людей почти не напа-
дает. В брачный сезон (конец мая—.начало июня) самцы окра-
шены ярче и дерутся из-за самок. Гнездо имеет вид ямки (глубина
до 91 см, диаметр до 247 см), устланной опавшими листья-
ми и ветками. Число яиц в кладке 20—60. Яйца белые, эллипти-
ческие, 65—85 X 38—50 мм, средний вес 81 г. Самым распростра-
ненным видом рода Caiman — С. crocodilus (=С. sclerops),
представленный 3 подвидами в южной части Центр. Америки и в
северной и средней части Южн. Америки. Максимальная длина
до 2,6 м. Размеры яйца 63 X 38 X 40 мм, вес в среднем 53 г.
рода Melanosuchus (наибольшая длина 4,5 м) и Palaeosuchus
(наибольшая длина 1,4 м) часто включаются в род Caiman. Внут-
ри самого обширного в семействе рода Crocodylus наблюдается
значительное многообразие в форме головы. Такие виды, как
С. catapkractus (Зап. и Центр. Африка), С. johnstonii (Сев. Авст-
ралия) и С. intermedius (Оринокко), обладают длинной мордой,
превосходящей, по крайней мере, -в 2 раза свою длину при
основании. Морды средней длины свойственны С. acutus (Эква-
дор, Колумбия, Венесуэла, Центр. Америка, Вест-Индия, Фло-
рида), С. siamensis (Сиам, Камбоджа, Ява), С. niloticus (Афри-
ка, Сирия, Мадагаскар), С. porosus (Индия, Цейлон, Южн. Ки-
тай, Индо-Малайский архипелаг, Сев. Австралия, Соломоновы
острова) и С. novae-guineae (Нов. Гвинея, Филиппины). Самая
короткая морда (длина ее превосходит ширину не более как в
1,5 раза) у С. palustris (Индия, Цейлон, Бирма. Малайский по-
луостров и архипелаг), С. rhombifer (Куба) и С. moreletii (Гвате-
мала, Гондурас). Последний вид самый мелкий (до 1,5 ж), тогда
как С. porosus и С. niloticus достигают 10 м. Наибольшей из-
вестностью пользуется С. niloticus, держится в реках, озерах,
309
болотах и лужах В норме пятяятгя рыбой, птицами, тм*«м«
крысами, ио совершает кяпаяеммя и на домашних жияотима зо
верблюда включительно. Нередко нападает на людей (тобем^
в Судане) Вырывает около воды норы длиной я 9—12 м, вход *
которые находится под водой. Дяе-три отдушины доставляю*
в «ору воздух. Кладке из 20—48 яиц. размером 55 90 /
X 40 — 52 мм и весом около НО а. Они укладываются в яму s*i
0,5 м глубины и засыпаются песком. Инкубация около 90 дней
Эмбрион начинает раввиваться еще до кладки. Ня носу молодых
имеется «яйцевой зуб», высотой в 0.5—0,8 мм. Мясо и яйиа этого
вида идут в пищу. Почитался древними египтянами (до нас до-
шло много мумий). В устьях рек держится С. рог os us, част
уплывающий в море. Опасен для человека. Малайский гавиал.
Tomis toma schlegeli, длина достигает 4,7 м; питается преимуще-
ственно рыбой.
16. СЕМЕЙСТВО GAVIALIDAE
Единственный род Gavialis известен в ряде нидов, начиная с
миоцена Индии. В плиоцене Сиваликских холмов встречаются
остатки экземпляров, имевших длину до 15—18 м, выделяемые
иногда в особый род Rhamphosuchus. Большинство имеет очень
длинную и узкую морду, но в Белуджистане найдены остатки ко-
роткорылого Gavialis breviceps. Единственный современный вид
G, gangeticus живет в бассейнах рек Инда, Ганга, Брахмапутры,
Коладан и др. Длина достигает 6,6 м. Откладывает в песок до
40 яиц (85—90 X 65—70 мм). Питается почти исключительно ры-
бой; для человека не опасен.
ЛИТЕРАТУРА
Boulenger G. Catalogue of the Chelonians, Rhynchocephalians and
Crocodiles in the British Museum, London, 1889.
Colbert E., Cowles R. and BogertC. Temperature tolerances in the
american alligator and the bearing on the habits, evolution and extinction
of the Dinosauria, Bull. Amer. Mus. Nat. Hist., 1949, 86.
Hoffman C. Eidechsen und Wasserechsen (in: Bronn’s Klassen und Ordnun-
gen des Thierreichs, Leipzig, 1884, 6: 443—1399),
Kalin J. Crocodilia, «Traite de Paleontologies-, Paris, 1955, 5: 695—784.
Mo ok C. The evolution and classification of the Crocodilia, Journ! Geology,
1934, 42: 295—304.
Wermuth H. Systematik der rezenten Krokodile, Mitt. zool. Mus. Berlin,
1953, 29, No. 2: 375—514.
Wermuth H. Internationaler Naturschutz fur Krokodile, Aquarien und
Terrarien, 1956, 3, No. 8: 248—250.
Werner F. Loricata (Das Tierreich, Lief. 62), 1933.
Wettstein O. Crocodilia «Handbuch'der Zoologies, Berlin, 1954, 7, No., 4*
321—424.
указатель русский
260
45
»га 172, 176, 179, 196, 206
степная 209
аГ^отль 45, 47, 53
вКСи?атор щучий 309
аЛЛ «гаторы 300, 302, 309
аЛЛ«перов А. М. 4
Афанди 6
JSS « 6
^Л0< 63
k’aS«en 114
h«Se6]3
^хипаллиум 13
архиптеригии 17
аспид 292
•аутотомия
Б
178, 179
46
А. Г. 4, ПО
3. С. 298
балансир
Банников
Бар каган е
батрахология 5
Бедряга Я. В. 8, 9
бездугные II/
безногие 13, 23, 26, 35, 36, 37, 39,
40, 43, 47, 251
безъязычные 65
Бергунью Ф. 149
бесхвостые 7, 13, 23, 26, 33, 36,
39, 63, 66
бесхвостые древние 68
Биброн Ж. 1, 8, 181, 251
бисса 166
Бленвиль 113
Богерт Ч. 181
боковая линяя 13, 14, 19
бокошейные 168
Бонапарте Ш. 48
Брем А. 8
Броили Ф. 112
Броньяр 112
Брум Р. 112, 121
Брух 65
будру-пам 297
Вуленджер Дж. 7, 8, 65, 181, 210,
Булянже Дж. 7
Бутантан 249
бушмейстер 297
Быстров А. П. 26, 35, 112
В
Ваглер 64, 181, 251
Варан серый 233
вараны американские 176, 226
— Старого Света 226, 232
Ватсон 6
веретеницеобразные 180, 232, 234,
235
веретеница 6, 239
веретеницевые 236
Вермель Ю. 13, 35
Вернер Ф. 9
Вестолл 18
Виллистон 112, 113
Вйнтерль 7
Воронцова №. 45, 62
воротник 178
вставчатопозвонковые 28
Вудвард 112
Г
гавиал ганагский 302
— малайский 302, 310
гавиалы 310
Гадов Г. 7, 210
гадюка древесная 295, 297
— жабья 294
311
глдяжа ычвлямя 294
— настоящая 294
— обыкновенная 295
- рогатая 295
степная 246, 295
гадюковые 292
Г ардек Д. в
гастролиты 137
ГекатеА Милетский 5
Геккель 3. 21
геккон голопалый 194
гребиепялый 194
— каспийский 194
— листопалый 195
— серый 195
геккон сцинкоаый 194
— туркестанский 195
гекконообразные 180, 185, 186, 195
геккончик 194
гекконы 177, 179, 186, 188, 194
Гексли Т. 2, 7, 21, 301
герпетология 5, 6
Геснер К. 6
гипапофиз 279
гипоцентр 22
гладкозубые 266
гладконог 68
гладконогие 68
гладконогообразные 68
глаз термический 244
голень вторичная 63
гологлаз 217
голокожие 166
Гофман К. 8
Грей Дж. 7, 172
гремучая змея 245, 296, 297
Гюнтер А. 7, 65, .172, 210
гюрза 295
д
двудугные 132
двуногие 135
двуходковые 6, 176, 181, 230
Денисова М. 110
диапсидный череп 114
динозавры 301
Доден 36
Долло Л. 149
дракон летучий 209
древние земноводные 11, 15
— бесхвостые 68
древесница 89
древолазы 92
дробнопозвонковые 29
Дубинин В. Б. 12, 35
Жгопозвонновые 23, 26
омериль А. 7, 8, 36, 45, 181, 251
Дюриген Б. 8
ехидиа чераая 291
Гфреыоа И. A 112
Ж
жаба ага 85
водям я гайанская да а.
гребенчатоголовая дд
живородящая 84
- велеиая 85
— обыкновенная 85
— серая 85
— чернорубцовая 85
— тушканчиковая 85
жабовидки 99
жабообразные 77
жабы 5, 6, 7, 77
— австралийские 80
— горные 82
— древние 77
— красные 105
— настоящие 84, 85
жабы носатые 84
— ромбовидные 104
— сростночелюстные 102
— щиткопалые 86
— южноамериканские 82
желтобрюх 280
желтопузик 185, 239
жерлянкообразные 71
жерлянка желтобрюхая 72
— краснобрюхая 72
3
заднебороздчатые 266
звероголовые 124
звероподобные 120
зернышки 178
зипо 281
змеи 5 , 6, 112, 133, 179, 241 оло
244, 245, 247, 251, 263 ’
— бородавчатые 264, 266
— вальковатые 262
— высоконосые 284
— гремучие 245, 247, 296, 297
— древние 252, 254
— морские 6, 245, 248, 254, 283 285
— ужеобразные 266 ’ 0
— современные 252, 256
— толстоголовые 283, 284
— червеобразные 252
— щиткохвостые 262
— яичные 284
змея арлекиновая 292
— бриллиантовая 298
— древесная 282, 283
змея карликовая 281
312
„опьеголомя 297 »<,£*«>••» «6. 2« У»гл«»«я 280 крШяЯьЯ о?л КРясЯаЯ ^82 оа~ л^гоого-"09®* 283 - Sea* 291 - плетевидная. 293 пезин°вая 28 "" ромбическая 298 стрела 282 окрашенная 283 ящеричная 282 ^йы акродонтные 113 эУ плевродонтные 113 йг-ряг к 5*»И Ж. 84, 300 К»мл 1в| w Л Лвитц Л. А. 8 ластохвост 287 Лятрей П. 7, 179 леткло**™ 38« 63, 64, 179 300
текодонтные 113 И Литрей 36
кижара 232 *гуанообразные 180, 196, 204 Жаны 176, 179, 196, 206 ;хнйДология 15 ихниты 15 йшунин Г. 298 ихтиозавры 113 ложноногие 255, 256 лунчатозубые 135, 137 299 301 лягушка бык 100 ’ WI — водяная 72, 99 — волосатая 99 — ильная 77 — летающая 101 — озерная 99
к — остроязычная 100 — прудовая 99
кабарагойа 232 Кальметт 249 Каммерер 55, 56 киакша кузнец 91 листовая 92 Д_ обыкновенная 89 квакши 7, 12, 87, 89, 92 кербер 5 кистеперые 18 Клейн Я- 5 клювоголовые 112, 133, 172, 173, 174 кобра азиатская 291 — водяная 292 — египетская 291 — королевская 292 4 кобры 291 Кожевников Г. А. 4 контия ошейниковая 291 — смирная 281 Коп Э. 7, 21, 59, 65, 113, 117, 149, 172, 181, 196, 251 короткоглавы 105 короткоголовые 92 Крефт П. 8 Кривицкий И. А. 8 крокодилы 5, 6,112, 136, 147, 179, 299, 300, 301, 302, 304 — высшие 300 — древние 300, 305 — средние 300, 305 круглоголовка песчаная 209 — травяная 99 — шпорцевая 73 лягушки веслоногие 100 — водные 87 — дископалые 100 — карликовые 98 • лесные африканские 98 — наземные 99 — настоящие 94 — носатые 93 — поросята 102 — собственно 99 — щиткопалые 100 лягушкообразные 94 М мамба 292 Матвеев Б. С. 4 Маркелова И. 62 Матамата 170 медянка 242, 281, 295 Мейер 21, 112 Мелик-Карамян А. 298 Мензбир М. А. 12 Мертенс Р. 8 метаморфоз обратный 59 монимостилия 113 Мюллер И. 7, 251 Мюллер Л. 8 Мэкдовелл 181
20 П. В- Терентьев
313
н
неотения 53
иеподвнжногрудые 65
Ни колье 66
Никольский А. М. В. »
Нобль Г. 7. 66
Нопча 251
О
On пел ь 96
Осборн 113
Оуэн 21, 113, 136, 179, 301
офидизм 246
офиол атрия 5
П
Павловский Е. Н. 298
Паллас П. 8, 9
пама 291
первичнобесхвостые 65, 68
переднебороздчатые 266
Перфильев П. 298
Пивето 64 , 65, 112
пнпоцые 73
питон иероглифовый 260
— сетчатый 260
— тигровый 260
питоны 255, 260
плавающие 34
пластинка внутригрудная 149
— грудная 149
— загривковая 148
— задняя 149
— надгрудная 149
— нижняя 149
— средняя 149
— хвостовая 148
пластинки крайние 148
— надхвостовые 148
— остистые 148
— реберные 148
плевроцентр 22
Плиний Старший 6
плоскозубые 127
плоскохвост 186, 287
плоскохвосты 287
повитуха 72
подвижногрудый 65
позвонок аделоспондильный 23
— гастроцентральный 22, 23
— голомерный 22, 23
позвонок голоспондильный 23
— лепоспондильный 22, 23
—• нотоцентральный 22, 23
— псевдоцентральный 23
— рахитомный 22, 23
— стереоспондильный 22, 23
ломоигж фялл«киожд>1>.
ямболомеримй 22, у 22,
лолоч амурский 261
большеглазый 261
закавказский 28I
красноспинный 281
краснополосмй 280
—• лазающий 281
леопардовый 281
— оливковый 280
- поперечнополосатый 280
— пятнистый 280
— разноцветный 280
— тонкохвостый 281
— узорчатый 281
— четырехполосый 281
— эскулапов 281
попереннозубые 48
поры анальные 177
— бедренные 177, 179
— паховые 177
постоянножаберные 53
поясохвосты 217, 218
Пржевальский 8
пресмыкающий тюлень 121
продольнозубые 48
проныра древесная 283
протей 12, 4б, 48, 53, 58
протей американский 58
— европейский 58
птицетазовые 142, 300
пузыри анальные 149
Пухальская Е. 62
Р
регенерация 179
Рёзель фон Розен кор 7
Рей Дж. 6
рогатка 83, 84
Ромер А. 23, 35, 112
рыбовидные 26
рыбозмей 43
рыбоящеры 131
С
саламандра 6, 7, 45, 48, 59
— исполинская 52
саламандровые 53
саламандры безлегочные 59
— настоящие 54, 55
Саразин 21
сверташка коралловая 262
свистун пятипалый 86, 87
свистун 86, 87
Северцов А. Н. 13 16, 17, 19 5
Сёве-Сёдерберг 1 б
сирена 46, 48, 53
складка горловая 178, 179
314
„(О, ''р'п с**^жвв<Р""’'" «К М СР^оЖвберим* <9 С‘РЛ?Ы 23< 233 233 сЛ5жН<,ЛУГне 120 <“Лл^со*Увые 123 $дар»“ is” 9 ^да.л^48> 52'вз <1£птостклмя ИЗ Кукуку 297 9 Йин П. П. 112 с^нк 179 ctt длинноногий 217 настоящий 217 Шинковые 177, 212 гпннкообразные 180, 211, 224 сывоРотки антизмеиные 246, 249 м» «и 2«1 g-Maw га, **иыг ?R| ?!£"** 2*7 2ва ужы ***’’ 27V и" W >’«»роти, ГпТ“° »*•••«<»«• 1<Я Уаолопидные 33 *УС змей 245, 24Й. 248, 249. 292, 2Чв Ф •Физиолог» 6 Фитцингер 64. |8| Фишер 63 X
Т —“с™™? £ового Света 203 VHA рого. Света 6, 177, 179 210
Таращук 165 Татаринов 63 тейЮ __ — „ Терентьев П. В. 8, 9, НО тонкопозвонковые 23 Топсел 6 тритоны 5, 12, 20, 45, 46, 47, 51, 56, 58 Трофимов Б. 35 трубчатозубые 294 туатара 174, 175 туатера 174 хвостатые 7^17, 23. 26, 34 . 39. 45? хоаны 13, 17, 18 Хюне Ф. 8, 23, 112, 135, 137. 149,219 Ц цельнопозвонковые 32 цельночерепные 117 цепкопалые 186 Циттель Е. 8
У Ч
углозуб 51 углозубые 49 удав гладкогубый 260 обыкновенный 261 .— песчаный 261 — собакоголовый 261 .— стройный 261 — узкобрюхий 261 удавчик 261 — восточный 261 — западный 261 — песчаный 261 удавы 260, 261 уж блестящий 281 — водяной 280 — крючковатый 281 — наземный 280, 281 — обыкновенный 279 — подвязочный 280 Червеобразные 254, 255 червеязычные 209 червяга кольчатая 44 червяги 6,41 черепа типы 114 черепаха болотная 163 — бугорчатая 163 — водная 163 — гигантская 158 — головастая 165, 166 черепаха грифоновая 164 — зеленая 166 — звездчатая 156 — коробчатая 164 — кровельная 163 — кусающая 164 — оливковая морская 166 — средиземноморская 156 — степная 158 — суповая 166
20*
315
«армии Я, Л, 7. Ilf, 147, *Й8. ***»
158. 163. 179
бвсщитковые 170
— боЛЬШТ-ГОЛОВЫ» <65. И"*
— голокож V* 166
-- аамыкккмцивсч 164
— эмепмошейиыч 100
— каймановы? 104
— кожистые 170
— морские 185
— мягкокожие 186
— пресноводные 159
— рогатые 154
— сухопутные 154
Чернов С. А. 9
чесночняцеобравные 74
чесночницы /4, 77, 81, 87
чешуеногя 185
чешуи спинные 177
чешуйчатые 133, 172
чешуя горловая 250
Ш
шапочка 178
Шейхцер 52
шнрокосиннные 153
Шмальгаузен И. И. 35, 62
Шмидт К- 7
Шрейбер Э. 8
Штанниус 149
Штраух А. А. 8, 48
Щ
щитки боковые 149
— брюшные 178, 179, 251
— верхневисочные 178
— верхнегубные 178, 179, 250,260
— верхневисочные 178, 179
— височные 178, 250
— горловые 250
— грудные 178
— заглазничные 178, 250
— задненосовые 178, 179
— краевые 149
— лоботеменные 178, 179
— межкраевые 149
— межносовые 250
— надглазничные 178, 179, 250
— нижнегубные 178, 250.
— нижнечелюстные 178, 179, 250
— носовые 178, 179, 250
— подхвостовые 251, 260
— позвоночные 1-49
— предглазничные 178> 250
— предлобные 178, 179, 250
— теменные 178,. 179, 250
щиткование 177, 179
щиток анальный 251, 254
— барабанный 178, 179
ПИПЧШ ОНДрВММОП I4Q
бркмавой 149
горлевой 149
грудкой 149
аадягпрохпдяый ц»
затылочный 178. it® " Г
лобный 178, I?», 2&0
лобоиосовой 178, (у®
— ыежгорловой 149
— межтемеямой 178, 179
— межчелюстной 17в, |у»
— иосомежчелюстмой 25л '
паховой 149
— плечевой 149
— подбородочный 178, 250
— подглазничный 178, 250
— подмышечный 149
— скуловой 178, 179, 250
— центрально-височный 17я ..
- шейный 149 ’ ,7«
щитомордник 245, 246, 297
— восточный 297
— китайский 297
— палласов 297
Э
Эверсманн Э. А. 8
эфа 242, 295
Я
явноязычные 65
яд змеиный 245, 246, 248
ядозубы 236
ярарака 297
ящерица гладкая исполинская oi£
— древесная 203 я 2,е
— живородящая 177, 179. 223
— зеленая 223
— плащеносная 209
— прыткая 223
— скалистая 219, 225
— стенная 225
ящерицы 5, 6, 7, 112, 113, 133 147
176,178, 179,- 180, 181, 186 210
241,242 ’ Z10’
— безногие 7, 44, 251
— древние 132
— настоящие 219
— скалистые 219
ящеротазовые 136, 137
ящеры древние 127, 133
— змеевидные 133
— змеешейные 128, 172
— летучие 144
— морские 127
— ранние 132
ящурка 225
— быстрая 225
— полосатая 225
— разноцветная 225
St6
УКАЗАТЕЛЬ
ЛАТИиския
ffihes ~273
Ablepharus
Ablepharus ,
Ablepharus bivlttatus 217
Ablepharus brandti 217
Ablepharus
Ablephariis
Ablepharus _
Acanfhodactylus 220
Acanthophis 289
Acanthophis antarcticus 24ч
Acanthosaura 207
Acanthostegidae 28
Achalinus 267
Acipri°n 196
Acontias 216, 217
Acontias holomelas 211
Acris 89, 92 11
Acrochordidae 252, 253 9fizt
Acrochordinae 264, 266 ’ °4’
Acrochordus 264
Acrochordus javanicus 263 ora
Acrosaurus 133 2«6
Actinodontidae 31
Adelogyrinidae 34
Adelotus 80
Adriosaurus 234, 236
Aeluronyx 191
Aelurophryne 76, 77
Aelurophryne mammatus 77
Ael urosaur us 189 ‘
Agama 197, 206, 207 20Q
— caucasica 209 ’
215. 217
aiaicus 217
bouton! 217
desert! 217
grayanus 217
/«laopnylax 190. 194
Alsophylsx piplens 194
Altirana 97
Alytes 72
campanlsona 72
— cisternasi 72
— obstetricans 72
• -
265
210
— sanguinolenta 209
Agamidae 180, 182, 197, 205, 206
210
Agamura 189
Agkistrodon 297
Aglossa 64
Agiypha 266
Aglyphodonta 251
Aigialosauridae 180, 184, 233, 243
Aigialosaurus 234
Aipysurus 287'
Aistopoda 13
Aistopodidae 34
Aldrovandus 6,- 9
Alethinophidia 251, 252, 254 256
Algiroides 220
Alligator 308
— mississippiensis 309
— sinensis 304, 309
AHigatorellus 304, 306
Ailigatorinae 308
Alligatorium 306
Amblycephalus carinatus 283
— monticola 283
Amblyrhynchus 197, 201
— cristatus 203
Amblystorna 52
Ambystoma 52
— gracile 53
— opacum 53
— talpoideum 53
— tigrinum 47, 53
Ambystomidae 46, 48, 50, 52
Ambystomoidea 46, 48, 52
Ameiva 226, 230
— surinamensis 230
Amphibamus 64
Amphibia 6
Amphibolurus 208, 209
Amphicoela 66, 68
Amphicoelia 301
Amphiesma 280
— tigrina 280
Amphignathodon 87
— guentheri 91
Amphignathodontinae 87
Amphisbaena 6, 231, 232, 251
Amphislaena alba 232
— fuliginosa 224 .
Amphisbaenidae 180,
Amphiuma 45, 59
— means 47, 59
Amphiuma tridactyla
Amphiumidae 39, 46,
Amphiumophis 42
Amphodus 88
Amplorhinus 276
Amyda 167
— gangetica 157
— ferox 168
Amyda sinensis 167
Anadia 228
analae 149, 178, 251
Anapsida 113, 116, 117
Ancistrodon 245, 296, 297
—7 acutus 297
— blomhoffi 297
— halys 297
181,
59
49,
205, 230
57, 59
317
himalayanu* JK»7
rhodostoma W7
Andria* >cheuch«»rl W
Anetdrs Ы, 6'2
luguhris 47
Anelytropsidae 180, 183, 212. 213
Anetitropsls paplllosu* 212
Anglo'toniala 251
Anguidae 147. 180. 184 . 286. 237
Anguirnorpha 183. 232
Angininae 238
Anguis 6 . 238 , 239. 251
-- fragilis 239
Anhydrophryne 106
Ankylosaurus 138
Anilidae 244. 252. 253, 262, 265
Antlius 262
— scytale 262. 263
Anisolepis 200
Anniella pulchra 235, 239
_____ 239
Anniellidae 180, 184, 237, 239
Anodontohyla 104
Anolis 197, 198, 203, 210
— carolinensis 203
— edwarsii 203
— equestris 203
— fasciatus 204
Anomachilus 262
Anomalepis mexicana 254
Anomocoela 66, 67, 74
Anomodontia 122
Anota 197
Anthracosauridae 29
Anura 21
Anuri 63
Aparallactus 245, 277
Apdtosaurus 139
Apeltono,tus 221
Aphaneramma 26
Aphaniotis 206
Aphantophryne 108.
Aphelophis 254
Aphelosaurus 127
Apoda 13, 14, 18, 19. 21, 24, 26,
35, 36, 181
Apologlossus 228
Aporophis 272
Aporosaura 221
Aporoscelis 208
Apostolepis 277
Apraedentalidae 143
Aprasia 185
Apsis 113
Apsidospondyli 23, 25, 26
Araeoscelidae 127
Araeoscelis 126
Archaeobatrachi 21, 24, 25, 26
Archaeochelyidae 147, 149 *
Archaeophidae 251
ArrbegcMawnda»
Arrhegntauru» 26
Arthoaaurla 13?,
Arcilera 65
arcus! I a 22
Ardecnaur Id ar I Mo. igt le.
Ardeotaurut 186 '
Argalis 229
Arlsteiliger 192
Arretosauridae 180. igg
Arretosaurus ornafus f‘<Mt «
Arrhyton 272 2<M
Arthroleptella 95
Arthroleptides 94
Arthroleptinae 95. 98
Arthroleptis 95, 98
Ascaphus 14, 68
Ascaphus truei 69, 71
Askeptosauridae 133
Askeptosaurus 133
Askeptosaurus italicus 134
Aspidites 258, 261
Aspidura 267
Asterophrys 102
Asterophrys turpicola 98
Astylosterninae 94, 98
Astylosternus 94, 98
Astylosternus robustus 98
Atelopus 93
Atheca 149, 150, 170
Atheris 293, 295
Atoposauridae 300, 306
Atoposaurus 303, 306
Atractaspis 245, 292, 293, 294
Atractaspis aterrama 294
Atractus 274
axillare 149
Azemiops 293
В
Ваёшёае 149, 150, 151, 154
Barbour Th. 10
Barbourula 71
basidorsale 22
Basiliscus 6, 196, 198, 203
— americanus 203, 204
basiventrale 22
Bastian A. 9
Batagur 159
Batrachemys 170
Batrachia 21
Batrachomorphoidea .23
Batrachophrynus 82
Batrachosauria 112
Batrachoseps 60, 62
Batrachoseps attenuatus 47
Batrachuperus 51
Batrachylodes 96
Bauriamorpha 124
Bauriamorphidae 124
318
B>UfU«uchidet ЛОО. ЭОЛ, ЭМ
u*utu*uchu* pacheroi ЭМ
Bdellophh 4/
Qflodon i «s
BrnthoAuehldM 32
Benth«*uchu4 smthkini 27
gergounioux F 171
Bibron G. 0
Bip»» 231
Bin* 239
arietans 245
gabonlca 248
Blanus 231
cinereus 231
Boa 6. 251. 259, 261
___ canina 261
Bogert Ch. 175, 240, 310
Boidae 252, 253, 254 , 256
Boinae 258. 260
Boiga 275, 282
— trigonatum 282
Bolleria 259
Bolosaurldae 117, 120
Bombina 72
Bombina bombina 65, 72
— maxima 72
— oriental is 72
— salsa 72
Bombina variegata 72
Bombinator 72
Bombinatoroides 64
Boodon 268
Borborocoetes 83, 87
Bothremyidae 150, 153, 169
Bothrops 245, 297
— lanceolatus 297
Boulenger G. 9, 62, 110, 171, 240,
251, 298, 310
Boulengerina 245, 290, 292
Boulengerula 43
Brachiosauridae 141
Brachycephalidae 66, 67, 90, 91, 92
Brachycephalinae 93
Brachycephalus 64, 93
Brachycephalus ephippium 91
Brachylophus 197, 201
Brachymeles 214
Brachyopidae 33
Brach ysuchus 135
Brauer A. 44
Brevicipitidae 66, 68, 98, 102, 103
Brevicipitinae 105
Breviceps 63, 105, 109
Breviceps gibbosus 65
Broiliellus 26
Brongniart A. 7, 9
Brongniart A. 7, 9
Brontosaurus 137
Brookesia 210
RfookvsM minima
na«tf« JIM
Bfiwmia 127
Яц!«» 64. 84. 85. M
bdfo M. 85
rahmifa ЯЛ
<arg«ri/an« R5
japonicu» 85
jerboa 78, 85
- lentlginoMJ* 85
leptopus 86
~ marinus 85
mauritanicus 85
melanostictus 85
— raddei 85
— regularis 85
— spinosus 85
— superci 1 iosus 85
— typhonius 78, 85
— verrucosissima 85
— viridis 85
Bufones 64
Bufonidae 66, 67, 75, 77, 78
Bufoninae 79, 84
Bufonoides 64
Bufonopsis 86
Bungarus 245, 290, 291
— caeruleus 245
— fasciatus 291
Bungarus flaviceps 291
Burnetidae 123
bursae anales 149
C
Cabrita 221
Cachryx 201
Cacopinae 107
Cacops 26
Cacopus 108, 109
Cacopus systoma 65
Cacosterninae 102, 105
Cacostesnurp 106
Cadurcogekko 188
Caecilia 41, 44, 251
— tentaculata 37
— thompsoni 44
Caeciliidae 41
Caenophidia 256
Caiman 308, 309
— crocodilus 309
— sclerops 309
Calabaria 258, 260
Calamaria 274, 281
— occipitalis 281
— smithii 281
Calamelaps 277
Callagur 159
Calliglutus 107, 110
Callisaurus 203
319
Cal Io ph I» 290
Callurlla 105. HO
Callullna 105
Callulopa 102
Cal mat le 249
Calodaetyius 193
Calote.i 197 . 207 . 210
Cal yptocepha I its 03
Camarosaurldae М2
Camp Ch. 240
Camptosaurus 138
Capi tosauridae 33
Captorhinidae 118
Captorhinomorpha 117
carapax 148
Cardioglossa 95, 98
Caret ta caretta 165
Carettochelydidae 150, 152, 160, 168
Carettochelys insculpta 168
Carnosauria 137, 138, 140
Carphophis 274
Carus V. 6, 9
Casarea 259
Caseidae 122
Caudata 14. 18, 19, 21, 25, 26, 34 , 45
Causus 292, 294
— rhombeatus 245, 294
Centrolene 88
Centromastix 222, 225
Centropyx 226
Cephalonia 173
Ceramodactylus 188
Cerastes 241, 293, 295, 298
— cornu tus 294, 295
Ceraterpetontidae 35
Ceratobatrachus 95
Ceratophora 206
Ceratophrys 64, 8*3
— appendiculata 78
Ceratopsia 138, 142, 143
Ceratopsidae 143
Cerberus 278, 284
Cercosaura 228
Cetiosauridae 141
Cetiosaurus 139
Chalarodon 199
Chalcides 216, 217
Chalcidoseps 216
Chamaeleolis 198
Chamaeleo 210
Chamaeleon 210
— Chamaeleon 204, 210
— deremensis 204, 210
— melanocephalus 210
— oustaleti 210
— pumilis 210
Chamaeleontidae 179, 180, 182,205,
209
Chamaesaura 217, 218
— aenea 218
1,1)1яммвп ausoina 9i* _
macrolepla 218 •
Cha mope 198
Champioaauridae 174, • »«
Champaoaaurue 173
Charaala 208
Charina 245. 280. 261
Chaamatosaurus vanhoeMni
cheiropteryfium 12, 15, 20
Chelldae 150, 153. 155, iaq
Chelodina 169., 170
— long! coll la 157
Chelonia 114, 116, 120
— mydas 165
Cheloniidae 147, 150, 152 it>
Chelus 169 ’ ,&5> 16$
— fimbriata 157, 170
Chelydra serpentina 164
Cherydridae 150, 151, 155 lft
Chenoprosopidae 31 » *04
Chersydrus 264
— granulatus 263
Chiasmocleis 109
Chinemys 162
Chiniquodontidae 125
Chioglossa 45, 54, 56
Chirixalus 101,-102
Chiroleptes 80, 81
— platycephalus 81
Chiromantis 100, 102
chiropterygium 12
Chirotherium 135
Chitra 167
Chjamydosaurus kingi 176, 204, 208
Chlotophis 271
Choerophryne 108.
Cholophidia 251, 254
Chondrodactylus 188
Chondropython 258, 260
Chrysemys 149, 161
Chrysopelea 276, 283
— rhodopleuron 283
Chthonerpeton 42’
— indistinctum 37
Cistecephalidae 123
Clarazisauridae 174, 175
Claudius 164
Clemmys 162, 163
— caspica 163
Cnemidophorus 226, 230
— lemniscatus 230
— murinus 230
Cochleosauridae 30
Coecilia 6
Coelacanthi 18
Coeluridae 140
Coelurosauria 137, 138
Colbert E. 310
collare 178
320
CoIopus 189
Colossodidy* atlas 149
Coiosteldae 28
Colpoglossus 107
Coluber 6, 251. 271, 280
_ fugularis 280
karelinl 280
~~ mucosus 281
najadum 280
Coluber ravergierl 280
_ rhodorhachis 280
__ tyria 280
Colubridae 252, 253, 266, 286
Colubrlnae 266, 279
Compsognathidae 140
Congosaurus 306
Conolophus 197, 201
__ subcristatus 203
Constrictor 259, 261
__constrictor 261
__ madagascariensis 261
Contia 245, 273, 281
__ col laris 281
modest a 281
Cope E. 9
Cophias 230
Cophixalus 107
Cophotis 207, 210
Cop.hyla 104
Copiula 108
Corallus 261
Coradylidae 180, 183, 217, 227
Cordylosaurus 219
Cordylus 218
— cordylus 218
— giganteus 218, 224
Cornu fer 96
Cormiferinae 95, 100
Coronella 272, 281
— austriaca 281
Corucia 214
— zebrata 217
Corythomantis 89
Corythophanes 198
Corythosaurus 138, 142
costalia 148, 149
Cotylosauria 114, 116, 117, 118,
Cowles R. 310
Cricochalis 218
Cricosaura 195
Cricotidae 29
Crinia 80
— laevis 82
— signifera 82
— tasmaniensis 78
Crinilnae 79, 80, 82, 84
Crocodilia 116, 117, 136
Crocodilurus 228
Crocodylidae 300, 303, 307
Crocodylinae 308
147
Cracodylua 300
— aeulws 306. 309
- ‘•taphrerfui ЭО9
- intermedin» 309
- John»ton 1| 309
— more let t(l 309
- niloticus 304 , 309
— nova* guinea* 309
palustris 309
— porosus 309, 310
— rhombifer 309
— si a mentis 309
Crossobamon 189
— eversmannl 194
Crossochelys 154
Crossodactyius 86
Crossopterygii 18, 19
Crotahdae 252, 253. 288, 296
Crotalus 6, 245, 251, 297
— adamanteus 294 , 298
— ceraster 298
— horridus 248, 294 , 297
— terriflcus 298
Crotaphytus 197, 202
Cryptobatrachus 88, 92
— evansi 91
Cryptobranchidae 46, 48, 50, 51
Cryptobranchoidea 46, 48, 49
Cryptobranchus 51
— alleghaniensis 47, 51
Cryptodelma 185
Cryptodira 149, 150, 151, 153
Cryptopsophis 42
Crypthiosaurus 230
Ctenoblepharis 201
Ctenophryne 109
Ctenosaura 197, 201
Cuora 163
Cyamodontidae 128
Cyamodontoidea 128
Cyclanorbis 16T
Cyclemys 163
Cycloderma 167
Cycloglenides 65
Cycloramphus 82
Cyclura 202
Cylindrophis 262
Cymbospomdylus 129
Cynodonta 125
Cynognathidae 125
Cynognathus 119
Cynops 55
Cynosuchidae 125
Cystlgnathidae 77
D
Dactychilikion 193
Dakosaurus 306
Dasypeltinae 266, 278, 284
Dasypeltis 245, 279
321
Da*ype!<<* *cabr* №. УМ
Pasyptrps |(M>
Drirochelys IA1
Prime InS
Пгтеимя 245. 289
Dendraspis 245. 291. 2W
anti nor it 292
Drndrrlsphl* 273
Dendrerpetuntidee 30
Dendrnba 11 пае 92
Dendrobates 93. 94
braccatus 91
Dendrophls 272, 261
pictus 281
Dendrophryniscus 93
Denisonia 289
— superba 245
Deraniyagala P 171
Dermatemydidae 150, 151, 160, 164
Dermatemys 164
Dermochelydidae 147, 150, 153, 170
Dermochelys 147, 149
Dermochelys coriacea 157, 170
Dermophis 39, 41
Desmatosuchus 135
— haplocerus 134
Desmognathus 61
Diadectidae 120, 147
Diadectomorpha 118
Diademodontidae 125
Diaglena 88
— spatulata 91
Diapsida 115, 116, 132
Dibamidae 180, 182, 211, 213
Dibamus nicobaricus 211
— novae-guineae 211
Dicamptodon 52
Dicrodon 228
Dicynodontidae 121, 123
Didosaurus 212
Didynamipus 105, -109
Diemenia 245, 287, 289
Dimorphodontidae 144
Dimorphognathus 95
Dinilysia 262
Dinocephalia 123
Dinodon 269
Dinosaur! a 136
Diplasiocoela 66, 67, 94
Diplocaulidae 35
Diplocaulus 27
Diplodactylus 194
Diplodocidae 142
Diplodocus 137
Diploglossa 232
Diploglossinae 236
Diploglossus 238
— striatus 235
Diplolaemus 201
Diplosaurus 202
329
Dipnoi 15
Dtp»win |Й|
Iлp^sdorwrphinae щ
Ihntadcnnoepbu* eynn&M, 76}
Dip**-» 284 n ***
Di»m deles ‘>8
Disrogloasldae 68, 67, -
Di»cog|n*su* 71 *
pictus 72
Dhpholb 245
Dtssorophidae 31
Distira 285
grandh 287
- semper! 285
Ditmars R 298
Dogania 166
DoTichosauridae 180, 184 22л
Doliophls 246, 290
Dracaena 226, 230
— guianensis 230
Dracaenosaurus 219
Draco 6, 179, 206, 208, 283
— volans 204, 209
Dromasauria 122
Dromicodryas 267
Dromicus 270
Dryocalamus 269
Dryop his 278, 283
— prasinus 283
Dumeril A.. 9
Dunn E. 62
Durigen В. 9
Dvinosauridae 29, 30
Dvinosaurus 26
Dyrosauridae 300, 301, 306
Dyscophinae 105
Dyscophus 105, 110
E
Ebenaria 193
Ecaudata 63
Echis 241, 293, 295
— carinatus 245, 295
Ecpleopus 229
Edalorhina 86
Edaphosauridae 121
Edaphosaurus 119
Edopsidae 30
Efremov I. 146
Egernia 212
— depressa 211
Elachistodon 245, 279
Elachistodon westermanni 284
Elachistodontinae 266
Elachistosuchidae 135
Elapechis 290
Elaphe 273, 281
— dione 281
— hohenackeri 281
— longissima 281
MW241
—. schrencki 281
__ sttuia 281
taeniura 281
Elapldae 245, 252. 253, 287 288
Efapmae 266 ’
Elapomorphus 277
Elaps 245. 290. 292
__ fraseri 283
Elasmosauridae 131
Elasmosaurus 128
Elentherodactylus 83, 84
__martinicensis 84
__ palmatus 78
Elosia 86
__ nasus 78
Elosiinae 79, 86, 100
Elpistostege 16
Elpistostegidae 28
Elseya 170
Embolomeri 24, 25, 28
Embrachosuchus 135
Emoa 216
Emydidae 150, 152, 159, 160
Emydura 170
Emys 162, 163
— orbicularis 163
Endothiodontidae 122
Engystomatidae 102
Enhydris 285
Enyalioides 199
Ensatina 61, 62
entoplastron 149
Enyalius 200
Enygrus 259
Eobatrachus 71
Eobufella 86
Eolacerta 219
Eosuchia 116, 132, 134
Eothyridae 120, 121
Eoxenopoides 74
Epicrates 259, 260
— cenchris 260
— striatus 261
epiplastron '149
epipubis 45
epitheca 147, 148
Erem i as 221, 225
— arguta 225
— spekii 224
— scripta 225
— velox 225
Eretmocbelys imbricata 157, 165
Eristicophis 293
Eryonsidae 31
Erytnnriamprus 276
Erythrosuchidae 135
Eryx 245, 260, 261
— jaculus 261
261, 282
mlliarl* 261
UtarictM 261
180, |8|, |87,
bublepharis 196
F.J£acu.,erlu*
tuchambreaia 245
F»S«arn4brMldae ,2’-
Euchirotes 231
canallculatus 224
Eumeces 212, 214, 217
— algeriensis 217*
— schneideri 217
— taeniolatus 217
Eumecoides 212, 219
Eunectes 259
195
— murinus 255, 261
Eunotosauria 150, 153
Eunotosauridae 147, 150, 153
Eunotosaurus 148
— africanus 153, 171
Euparkeriidae 135, 136, 137
Eupemphix 86
Euposauridae 180, 184, 232, 240
Euposaurus 240
— thiol lieri 234
Euproctes 55
Eurycea 60, 61
Eurydactylus 191
Eurypterygius 129
Eurystomata 251
Eusarkiidae 150, 153, 170
Euspondylus 229
Eusuchia 301, 303, 306
Eversmann E. 9
Exostinus 239
F
femorale 149
Feylinia 212
-r- currori 212
Feyliniidae 180, 183, 212, 213
fibulare 64, 68
Fimbrios 266
Firmisternia 65
foramen Panizzae 307
Fordonia 278, 284
Francis E. 45, 62
frenale 178, 250
frenoculare 178
frontale 178, 250
frontonasale 178
frontoparietalia 178
G
Gadow H. 9, 87
Galechiridae 122
Galechirus 119
Galeopsidae 122
323
Galtolla VK. ГП
GampwMteonyM ’M, 00
Qa*trt*pho)<* ft I
OantroaliryfwitAM IM
Qaitrolhfea M 99
mafauplata 91. 99
ovifrrwm »»2
pvginna 91. 99
Gavin It dec 300. 303. 310
Gavial!» 300. 310
brevierf»* 310
gangeticus 304. 3|0
Gecko 191
stentor 195
Geckolepis 191
Geckotidae 179
Gegeneophls 38, 42
Gehvra 192
Geikidae 123
Gekkonidae 180. 181, 186, 187, 210
Gekkota 181. 185
Genyophryne 107
Geobatracnus 93
Geoclemys 162
Geodipsas 274
Geoemyda 162
Geomolge 51
Geophis 274
Geoirypetes 41
— petersii 37
Germanobatrachus 74
Gerrhonotinae 236
Gerrhonotus 238
Gerrhonotus liocephalus 238
— multicarinatus 239
Gerrhosauridae 180, 183, 218, 227
Gerrhosaurus 218
— flavigularis 224
— validus 219
Gesner C. 9
Gigantorana 100
Glauconia 254
Glauconiidae 254
Globidentinae 233
Glyphoglossus 108, 109
— molossus 98
Glyptosaurinae 236, 238
Gonatodes 189
Goniochersus 156
Gonionotophis 268
Goniopholidae 300, 301, 306
Goniopholis 306
Gonyocephalus 197, 207
Gopherus 156
— polyphemus 156
Gorgonopsida 122
Gorgonopsidae 120, 122
granulae 178
gradientia 7
Graptemys 161
324
S-ay J voi
rayta 274
tjregwry W l0, 38
Grint th* I M. 110
Crypt «cue 82
gulare 149
SOniber a 178, 251
ymnndacfylwa 189. 194
Gymnodactylu* caapiua 194
Gymnodaclylua fed when koi |од
Gymnodactylua rusanwi 19Л
Gymnophiona 21
Gymnophh 42
Gyrinophiiua 60, 61
Gymnophthalmus 229
H
Hadrosaur! dae 142, 143
Hadrosaurus 138
Halleobatrachus 77
Haplopeltura 284, 285
Haplopeltura boa 285
Hardella 148, 159
Harpesaurus 207
Hatteria punctata 172
Heleophryne 82
— purcelli 78
Heleophryninae 79, 82
Helicops 268, 280
Helioporus 81
Helminthophis 256
— flavoterminalis 256
— petersii 256
Helocephalus 200
Heloderma 177, 235, 236
— matthewi 236
— suspectum 236
Helodermatidae 180, 184, 236, 237
Helveticosauridae 128
Helveticosauroidea 128
Helveticosaurus 128
Hemibungarus 290
Hemichirotes 231
Hemidactylium 61, 62
Hemidactylus 193
Hemidactulus turcicus 195
Hemiphractinae 87-
Hemiphractus 87
— scutatus 91
Hemisinae 102
Hemisus 102
— guttatum 65
— marmoratus 110
Hemisphaeriodon 215
Hemitheconyx 195
Henodontidea 128
Henodus 128
Henophidia 256
Herpele 42
Hefpetodfyw 271. Ml
fuscus 281
Herre W. 62
Heterodactylus 229
Heterodon 272. 279, 281
__ con tor tri x 281
Heteronota 189
Hildebrandtia 100
Himantodes 275
Hinulia 216
Hoffman Ch. 9, 171, 310
Hoffstetter R. 240
Holaspis 222
Holbrook)a 203
Holmgren N. 18, 35
Holoptychiiformes 18, 19
Homalocranium 245, 277
Homalopsinae 283
Homalopsis 278, 284
buccata 282, 284
Homolepida 216
Homonota 190
Homop holis 190
Homopus 154
Hopl°cepbalus 289
Hoplocercus 197, 202
Hoplodactylus 191
Hoplophryninae 105
Hoplophryne 105, 109
Hoplurus 199
Huene F. 10, 19, 35, 146
humerale 149
Hydromantes 45, 60, 62
— genei 62
— italicus 62
Hydromedusa 169
Hydrophidae 252, 253, 285 '
Hydrophiinae 266 ’
Hydrophis 245, 285
— spiralis 287
Hydrosaurus 197
Hydrotherasaurus alexandrae 129
Hydrus 285
— platurus 287
Hyla 64, 88, 89, 92
Hyla arborea 89, 92
— aurea 89
— carolinensis 91
— coerulea 89
— faber 91
— japonica 89
— nebulosa 92
— pickeringi 91
— regilia 91
— versicolor 91
Hylae 64
Hylaeochampsa vectiana 306
Hylaeochampsidae 300, 303, 306
Hylambates 101
Hylidae 66, 67, 87, 90, 91, 92
286
Hyliftae 87
H у lode» 84
Hylophorbu» 108. HO
- bfbronl 98
Hylorana 96, 100
Hylorina 82, 83
Hyloxalu* 92
Hymenochlrus 64 , 73. 74
— boettgeri 71
Hynoblidae 46, 48, 49, 50
Hynobius 49, 51
— keyserlingii 51
hyomandibulare 13
hyoplastron 149
hypapohyses 266
Hyperolia 81
Hyperolius 101
hypocentrum 22, 24
Hypogeophis 38, 39, 41
Hypopacnus 109
hypoplastron 149
Hypselotriton 54
Hypsiglena 269
Hypsilophodontidae 142, 143
Hypsirhina 278, 284
— bocourtii 284
Hypsirhynchus 270
I
Ichnotropis 220
Ichthyophis 37, 38, 39, 41, 43
— glutinosis 37, 43
ichthyopterigium 12
Ichthyosauria 115, 116, 129, 131
Ichthyosauridae 28, 131
Ichthyostega 14, 16, 27
Ichthyostegalia 24 , 25, 26
Ichthyostegidae 28
Ichthyostegopsis 16
Ictodosauria 120, 125
Ictidosauridae 124, 125
Ictidosuchidae 124
Iguana 197, 202
— iguana 203, 204
Iguanavus 196
Iguania 179, 182, 196
Iguanidae 180, 182, 196, 197, 205,
210
Iguanodon 138
Iguanodontidae 142, 143
Iguanognathus 269
Iguanosaurus 196
Ilysiidae 262
Indobatrachus 82
infralabialla 250
inframaxillaria 178
inguinale 149
Inostrancevia 121
intercentrum 22
325
mlrntorMlr 22
mtrrgulare 149
intermit ginale 149
internasalr I7B
internasalla 260
Interparietal* 178
intervenirale 22
isodactyhum 61
Isodontoaaurus 236
Ixaltis 101
J
Japalura 207
Jarvik E. 35
Jepsibaena 230
Jonkeria 121
К
Kachuga 159, 163
— tecta 163
trivittata 163
Kalin J. 310
Kallokibotiidae 150, 153, 168
Kalopbryninae 108
Kalophrynus 108
Kaloula 106
Kaloulinde 106
Kannemeyeridae 123
Kassina 101
Keneuxia 216
Keratocephalus 119
Kingsley J. 44
Kinixys 154
Kinosternidae 150, 1'52, 155, 164
Kinosternon 165
Klauber L. 298
Klein P. 10
Kraus R. 298
Kreft P. 10
Krinicki J. 10
Kuhn O. 146
L
Labyrinthodon 21
Labyrinthodontia 21
Lacerta 6, 179, 219, 220, 222, 223,
300
— agilis 222, 223
— brandti 225
— chlorogaster 225
— crocodilus 6
— derjugini 223
— echinata 225
— media 223
— mucronata 222
— mural is 225
— ocellata 223, 224
— parva 223
326
I arena per «p toll ala 226
pratkola J23
•at lent a T26
- aimnnyi 223
strigata 223
- t auric a 225
- vlridla 223
vivipara 219, 223
I.acertidae 180, 183, 219, 228
Lacertllia 176, 179 a
Lachesis 245, 297
— atrox 248
— gramineus 297
— lanceolatus 297
— mutus 297
Laemanctus 198
Lamprophis 268
Langaha 274
Lanthanosuchidae 112
Lanthanosuchus 112
Lanthanotidae 180, 184, 233, 237
Lanthanotus borneensis 233, 235
Lariosaurus balsami 129
Latastia 220
Laticauda 245, 287
— colubrina 283
— laticauda 287
Latipinnati 131
Latonia 72
Laurenti J. 10
Lechriodonta 48
Lechriodus 81
Leiocephalus 197, 200
— carinatus 197
Lepidochelys olivacea 166
Lepidobatrachus 64
Lepidodactylus 192
Lepidophyma 196
Lepidosauria 132
Lepospondyli 23, 25, 26, 33
Leposternon 232, 231
Leptobrachella 76
Leptodactylinae 79, 86
Leptodactylodon 97
Leptodactylus 86, 87
— longirostris 78
— pentadactylus 87
Leptodira 275
Leptognathus 284
Leptolepis 101, 102
Leptophis 270, 281
— liocerus 279
Leptotyphlopidae 252, 254
Leptotyphlops 254
— affinis 255
— albifrons 254
— macrolepis 255
Leurognathus 61
Lialis 185
Liasis 258
Ijbycosuchus 305 pchanura 240 Limnodynastes 81 dorsalis 78 Limnomedusa 86. 87 Limnoscelidae 117 Limnoscelis ||7 __ palustris Ц8 Lioneterodon 279 Liolaemus 197. 200, 208 Liolepis 197, 206 Liopelmidae 66, 68, 70. 71 Liopelma hochstetteri 71 Liophis 272 Liophryne 107 Liosaurus 200 Lissamphibia 21 Longipinnati 131 Lophura 206, 208 loreale 178 Loxocemus 258, 260 Loxommidae 28 Lupeosauridae 122 Luperosaurus 191 Lycideopsidae 124 Lycodon 269, 282 Lydekkerinidae 32 Lygodactylus 192 Lygosoma 215, 216, 217 — africanum 224 — challenger! 224 — cumingii 216 — cyanurum 216 — dorsale 211 — lesuerii 216 — mustelinum 224 — nigrum 217 Lygosoma ophioscincus 217 — ornatum 216 — smaragdinum 216, 217 Lyriocephalus 207 Lyrocephalidae 33 Lysipterygiidae 31 Lysorophidae 34 Lystrosauridae 123 Lystrosaurus 121 Lytorhynchus 270, 282 MacfoprMmfaff 77g MseropterygiMae |3| Macrnainrus 21S Macrottomafs 2ЛI “slseletnye |6| Mslarlemya terapin 163 Malayemys 162 Malpolon 267 — monspeasuianus 282 Manculus 60, 61 Mancus 218 Manfella 101 Mantipus 104. 109 Mantophryne 102 — robusta 110 Marcus H. 44 marginalia 148. 149 massetericum 178 Mastodonsauridae 33 McDowell S. 240 Mecodojitia 48 Megaelosia 86 Megalania 232 Megalichthyidae 19 Megalixalus 101 Megalobatrachus 51 — japonicus 52 — maximus 52 Megalosauridae 140 Megalosaurus 139 Megophryinae 74 Megophrys 64, 74, 76 — carinensis 76 — feae 78 — kempii 76 — major 68 — montana 78 — nasuta 78 Mehelya 268 Melanobatrachinae 106 Melanobatrachus 106 Melanophidium 263 Melanosaurinae 239 Melanosaurus 239 Melanosuchus 308, 309 mentale 178, 250 Menzbier M. A. 35 Mertens R. 10, 171 л
M Mertensiella 54, 56 ’ — caucasica 56
Mabuya 215, 216 — aurata 216 Machaeroprosopus 135 Macmahonia 221 Macrocephalosaurus 206 Macroclemys temminkii 164 Macrocnemus 133 Mocropelobates 74, 77 Macropisthodon 268 Macropophis 280 Mesophis 254 mesoplastron 149 Mesosauria 115, 116, 126 Mesosaurinae 233 Mesosaurus 126, 127 Mesosuchia 301, 305 Metopoceros 201 Metoposauridae 33 Metriorhynchidae 299, 300, 301, 306 Metriorhynchus 306
327
MefrfocWunrhN* leekdl •‘ИМ
Mlcrablepharu* И9
Ml err I sot 277
Mlcriialus 96
MIcroh у la 109
ptilchra 98
Mictuhyllnae 109
Micropechit 289
Mlcropholidae 31
Mlcropbolis 27
Mlcrosauria 24 . 25. 33
Microscalabotes 192
Microstomata 251
Micrurus 245. 287. 290, 292
Micrurus corallinus 292
— fulvius 292
— surinamensis 292
Millerettidae 120
Mimeosaurus 209
Mlobatrachus 64
Miodon 277
Miolania 148. 149. 154
Miolanidae 147. 150. 151. 154
Miopelodytes 77
Mixophyes 80, 81
Mixosauridae 131
Mixosaurus 129
Molge 56
Moloch 197, 208
— horridus 204
Monoclonius 138
Monopeltis 231
Montsechobatrachus 71
Mook C. 310
Mosasauridae 180, 184. 233, 243
Morenia 161
Muller L. 10
Myobatrachus 81, 82
— gouldii 78
Mystriosaurus 134, 306
Mystriosuchus 135
Mytrolysis 80
N
Naia 291
Naja 245, 249, 287, 290, 291
— bungarus 292
— caeca 291
— flava 248
— haie 291
Naja hannah 245, 292
— naja 245, 283, 291
— oxiana 291
— tripudians 291
Nannobatrachus 97
Nannophrys 97
Nannopterygius 129
Nanorana pleskei 100
Nannosuchus 306
Nantes 306
328
Nardoa 2M
nasalia 176. 350
naaotrmalta 178
naaorostral* 250
Natrix 245. 268. 279
- natrix 279, 280
ordlnatua 280
— persa 280
— teuelata 280
— tigrina 280
Naultinus 192
— elegans 188
Nacrosaurus 239
— elegans 188
Necrosaurus 239
Nectodactylus 109
Nectophryne 84
Nectophrynoides 84
Nectosaurus 127
Nectridia 24, 25, 34
Necturus 58
— maculatus 58
Neobatrachi 21, 24, 25, 26
Nephrurus 188
Nesobia 74
Nesomantis 74
neuralia 148
Neurergus 54, 55, 56
Neusticurus 228
Nieden F. 44
Nitosauridae 122
Noble G. 10, 87, 110
Norops 198
Notaden 81
notarium 146
Notechis 289
— scutatus 245
Noteosaurus 127
Nothopsinae 264
Nothopsis 266
Nothosauria 130
Nothosauridae 128, 130
Notochampsidae-300, 301, 305
Notochelys 163
Notophthalmus 55, 56
Notosuchidae 300, 303, 305
Notosuchus 305
Nototrema 64, 92
nuchale 148, 149
Nucras 219, 220
Nyctibates 94
Nyctibatrachus 97
Nyctimantis 89
О
Ocadia 161
occipitale 178
Oedipus 45, 60, 62
Oedura 194
Oligodon 237, 282
Oft>ph«losauridae 131 Omphab«eurt>« 131 Onychodactylus 49 fischerl 51 Ooeidotyga 94 . 97. 100 Ophiacodon 119, 121 Ophiacodontidae 120, 121 Ophiderpeton 19 Ophlodes 238 Qphiomorus 214 Ophiopsiseps nasufus 211 Qphisaurus 238. 239 apodus 239 Ophisops 221 Ophryoessa 198 Ophryophryne 76 Opisthocoela 66, 71 Opisthocoelellus 74 Opisthocoelia 301 Opisthoglypha 251, 266 Opisthotropis 267 Opoterodonta 251 Oreobatrachus 97 Oreophrynella 93 — quelchii 91 Oreophryne 107 Oreosaurus 229, 230 — shrevi 230 Orlitia 162 Ornithoeheiridae 146 Orthoglenides 65 Ornitholestes 139 Ornithominridae 137, 140 Ornithomimus 139 Ornithopoda 138, 142 Ornithosuchidae 135, 136, 137 Ornitischia 116, 117, 142 Osteolaemus 308 Osteolepidae 18 Osteolepides 18 Osteolepi formes 18, 19, 25 Otocratiidae 28 Otocryptis 206 Otophryne 108 Otosaurus 216 Oudemans A. 10 Oviraptor 139 Oxybelis 276, 283 — fulgidus 283 Oxydactyla 64, 107 Oxyglossus 100 Oxyrhopus 275 Parhyopbn 254 PachypalaminM* 49 P«fbypl»uroa»Mrid*« IV) ParhyrtiifttMaurida* 144 Pachytnton 55 Palaeobatraehidw 66. 67, Tf Palaeobalrachua 77 Palaeochamaelro 206 Palacogynnidac 29, 117 Palaeonaia 287 Palaeophidae 251. 252, 254 Palaepnia 254 Palaeosaniwa 232 Paleosaurus 139 Paleosuchus 308. 309 Pallas P. 10 Palmatorappia 96 Paludicola 86, 87 — fuscomaculata 78, 87 Parabufella 86 Paraplacodontidae 128 Parasaniwa 232 Parasaurolophus 142 Parasuchia 301 Pareiasauridae 117, 120, 147 Pareiasaurus 118 Parhop lophryne 105 parietalia 178, 250 Parker H. 44 pectorale 149 Peliontidae 30 Pelobates 74, 76, 77 — cultripes 77 — fusciis 77 — syriacus 77 Pelobatidae 66, 67, 74, 78 Pelobatinae 74, 75 Pelochelys 167 Pelodytes 74, 76, 77 — caucasicus 77 — punctatus 77 Pelomedusa 169 Pelomedusidae 150, 153, 160, 168 Peltosaurus 236 Pelusios 168 Pelycosauria 121 Perennibranchiata 46 permanent larvae 46 Perochirus 192 Perodactylus 229 Peromela 21 Peropoda 251
P Petropedetes 94 Petropedetinae 94, 100 Petrosaurus 202
Pachycalamus 231 Pachycephalosauridae 143 Pachydactylus 190 — bibroni 188 Pachyophidae 251, 252, 254 Phaneroglossa 65 Phelsuma 190 Philochortus 220 Philocryphus 81 Philodryas 276
829
PhHotia «О
l*hllofh«mnu« 272
PholidoffattcHdae 29
Pholidoaaurldae 300. 303, 306
pholldmi* |77
Phoxophrys 206
Phractamphibla 21
Phrynella 109
Phrynobatrachus 95
Phrynoccphalus 197, 208, 209
-- helioscopus 209
— in terscapular Is 209
— mystaceus 209
— rossikowi 209
Phrynomantis 108
Phrynomerinae 104
Phrynomerus 104
Phrynops 170
Phrynopsinae 96, 99
Phrynopsis 97
Phrynosoma 197, 203
— cornutum 204
Phyllobates 92, 94
Phyllodactylus 194, 195
— europaeus 195
Phyllomedusa 64, 89, 92
Phyllopezus 192
Phyllurus 210
Phymaturus 201
Physalaemus 86, 87
Physignathus 197, 208, 209
Phytosauria 136
Phytosauridae 135, 136
pileus 178
Pipa 64 , 73, 74
— americana 74
— pipa 71, 74
— parva 74
— snethlageae 74
— surinamensis 74
Pipidae 66, 67, 70, 71, 73
Pipoides 64
Pisces 25
Pistosauridae 130
Pistosauroidea 130
Piveteaux J. 110
Placochelydae 128
Placochelys 128, 129
— placodonta 129
Placodontia 115, 116, 127, 129
Placodontidae 128
Placodontoidea 128
Placodus 128, 129
Plagioglenides 65
plastron 148
Platecarpinae 233
Platemys 170
Plateosauridae 141
Platepsaurus 139
Platydactyla 64
330
Platyhyia H>«, luv
Plafymantla 96
platynota 232
Platypeli» 104
Platyplacopua 222
Platyplectrurua 264
Platyxaurus 218
Platyaternidae 150. 151, jgg
Platysternon megacephaium
Plectrurus 263 40
Plesiochelyidae 147, 150, 153 ix«
Plesiolacerta 219 * °®
Plesiosauri dae 130 л
Plesiosauroidea 130
Plethodon 61, 62
Plethodontidae 14, 15. 46, 49 л? «-
Plethodontohyla 104 *
Pletholax 185
pleurocentrum 22, 24
Pleurodeles 54, 56
Pleurodira 149, 150, 152, 168
Pleurosauria 115, 116, 133, 134
Pleurosaurus 133, 134
Pleurosternidae 149, 150, 151 ic.
plica gularis 178 ’4
Plicognathus 52
Plioambystoma 52
Pliosauridae 130
Pliosauroidea 130
Podarcis 222, 225
Podocnemis 168
Podocnemis expansa 169
Podokesauridae 140
Pol ychrus 201, 210
Polycotilidae 130
Polyglyphanodon sternbergi 196
Polyglyphanodontidae 180, 182 юл
Polypedates 101 0
— dennysi 98, 102
— leucomystax 102
— malabaricus 101
— nigropalmatus 101, 102
— reticulatus 102
— Schlegel i 102
Polypedatidae 66, 68, 98, 100, ЮЗ
Polysternidae 147, 150, 153, 170
Ponolopus 238
pori anales 177
pori femorales 177
— inguinales 177
— postanales 177
Porolepiformes 18'
Poromera 221
postloreale 178
postnasalia 178
postocularia 178, 250
praeanalia 178
praefrontalia 178, 250
praeocularia 178, 250
Praslinia 42
pHonodactylus 228
priateroffnathldae 124
prhtfdactylua 199
prist urus 189
Proanoura 66, 68
proatlas 307
processus spinosi Infertorea 266
procoeia 6o, 67, 77
procoella 301
procolophonldae 117, 120
proctoporus 229
procynodontidae 12S
proganochelys 153
prolacertldae 132
propelodytes 77
prosaur i a 133
prosauropoda 139, 141
prosymna 237
proteidae 46, 49, 57, 58
proterochersidae 150, 152, 168
proteroglypha 251, 266
Proterosuchidae 135
Proteus anguinus 50
protobatrachidae 66, 68
Protob atrachus 68
— massinoti 68, 69, 110
protoceratops 138
Protoceratopsidae 144
Protopipa 73
Protorosauria 115, 116, 126,
Protorosauridae 127
Protorosaurus 126
Protostegidae 147, 150, 152,
Protosuchia 301, 305
Protosuchidae 299, 300, 301,
Protosuchus 305
— richardsoni 304
Psammodromus 220
Psammophis 278
Pseudacris 89, 92
Pseudaspis 270
Pseudechis 289, 291
— porphyriacus 245, 291
Pseudelaps 289
Pseudeumeces 219
Pseudemys 161
Pseudinae 79, 82, 84, 86
Pseudis 82, 83
— minuta 78
Pseudobranchus striatus 59
Pseudobufo 84, 85
Pseudocerastes 245, 293
Pseudocordylus 218
Pseudohemisus 106
Pseudohymenochirus 73
Pseudolacerta 217
Pseudopareas 285
Pseudophryne 81, 82
Pseudoplectrurus 264
Paeudorhabdium 274
Paeudwuchia 138, I.W
Pseudotriton 60, 61
Pseudoienodon 286
Pseudoxyrhoeus 266
Ptenopus 190
Pieranodon 144, 145
Ptrranodontidae 146
Pternohyla 88
Pterodactyl Idae 145
Pterodactyloldea M5
Pterodactylus 145
Pteroplax 26
Pterosaurla 116, 117, 144, 145
Pterosphenus 254
Ptyas 281
Ptychadena 100
Ptychozoon 191, 195
— homalocephalum 186, 195
Ptyodactylus 193
pygale 149
Pygopodidae 180, 181, 185, 187
Pygopus 185
— lepidopus 185, 186
Python 258, 260
Python amethystinus 255
— molurus 255, 260
— reticulatus 260
— sebae 260
Pythoninae 258, 260
127 Pythonomorpha 233
Pyxicephalus 100
Pyxis 154
166
305 Q
Quadrupeda 15
R
Rabanus Maurus 6, 10
Ramanella 106
Rana 6, 64, 94, 97, 99, 100
— arvalis 99
— camerani 100
Rana catesbeiana 100
— chensinensis 100
— esculenta 99
— goliath 100
— macrocnemis 100
— mugiens 100
— nigromaculata 99
— ridibunda 99
— temporaria 65, 99
— terrestris 99
Ranae 64
Ranaster 80
Ranidae 66, 68 , 90 , 94 , 98
Raninae 97, 99
Ranodon 51
Ranodon sibiricus 51
331
R«noid«t 64
Raui«i>ehldae 135, |эд
Reven Н. Щ, м
RepHH* 6, 25, 111
Repli humor phoidea 23
Rhachlodontinae S66
Rhachitomi 24, 25, 29
Rhacodactylua 191
Rharophorus 101
Rhadinaea 271
Rhanipholeon 210
Rhamphorhynchidae 144
Rhamphorhynchoidea 144
Rhamphorhynchus 145
Rhamphasuchus 310
Rhinatrema 41
Rhinesuchidae 32
Rhineura 231
Rhinocalanus 277
Rhinochilus 271
Rhinoderma 93
— darwini 93
Rhinodermatinae 93
Rhinophis 262
Rhinophryninae 79, 84
Rhinophrynus dorsalis 78, 84, 85
Rhinosaurus 112
Rhipidistia 18
Rhizodonti formes 19
Rhodanogekko 188
Rhombophryne 104
Rhombophryninae 104
Rhoptropus 193
Rhyacotriton 52
Rhynchocephalia 116, 133, 172
Rhynchoedura 188
Rhynchophis 270
Rhynchosauridae 173, 174, 175
Ricnodon 19
Ristella 215
Roesel von Rosenhof A. 10
Romer A. 35, 146
rostrale 178,. 250
Rubidginia 123
Rubidginidae 123
S
Saccodeira 200
Salamandra 54, 55
— atra 56
— maculosa 55
— salamandra 55, 56
Salamandrella 51
Salamandridae 46, 48, 54, 57
Salamandrina 54
Salamandroidea 46, 48, 53
207
Salientia 7, 18, 21, 25, 26, 33, 63,
64, 92, 100
332
Safliwa 232
Saniwfnar 232
Saufhia 73Л
Saur la 133, 176
Saurnchla 116. 117, 135
Saurobatrachia 21
Sauromalua 202
Sauromorpholdea 23
Sauropoda 137, 139, Ml
Sauropterygia 115, 116, |9fl
Slve — Soderbergh 0. 35, 144 ***
Scaloposauridae 124
Scaphiophryne 106
Scaphiopus 74, 76, 77
Scaphiopus hoi brook i I 78
Scapteira 225
Scartiscus 200
Scelidosaurus 138
Sceloporus 203
Sceloporus 197, 203
Scelotes 215
Schastasauridae 131
Schmidt K. 10, 175, 298
Schoutedenelia 95, 98
— globosa 98
Schreiber E. 10
Scincidae 147, 180, 182, 212. 91»
226
Scincomorpha 182, 211
Scincus 214, 217
— officinalis 211, 217
Sceromochlidae 135, 136
Scleromochlus taylori 134
Sclerothoracidae 32
Scolecomorphus 43
Scolecophidia 241, 244, 252, 254
Scolecophis 276
Scoiecosaurus 230
Scotobleps 95, 98
scuta gularia 250
scutae 178
Scutemys 165
Scu tiger 76
Scytalophis 262
Sebecidae 299, 300, 303, 305
Sebecosuchia 305
Sebecus icaeorhinus 305
Segisauridae 140
Sepedon 248, 290
Sepsina 216
Serpentes 6, 133, 180
Serpent! a 7
Seymouria 112, 119
— baylorensis 118
Seymouriamorpha 112, 118
Seymuriidae 112
Shinisaurinae 239
Shinisaurus 235, 239
— crocodilurus 240
Sibynophis 266
myhendrae 263
s momantls 96
< motes 272
S phonops 38, 42, 44
XZannulatus 37, 43
ciredon pisciformis 53
S ren 13. 45, 59
XI lacertina 59
Sirenidae 46, 48, 49, 50, 59
Sistrurus 296
Sitana 20o
Siiiinthillus 93
Solenoglypha 251
Sooglossinae 74
Sooglossus 74, 76
__sechellensis 77, 78
Sozura 21
Spathoscalabotes 192
Spea 77
Spelaeophryne 105, Ю9
Spelerpes fuscus 62
Sphaenodon 172
Sphaerodactylus 193
Sphenacodontidae 121, 172
Sphenodon 112, 172, 173
Sphenodon punctatus 174
Sphenodontidae 174
Sphenophryne 107
Sphenophryninae 107
Sphenosuchidae 136, 299
Spilotes 273
Spinosauridae 140
Spinosaurus 137
spiraculum 13
squamae 177
.— gulares 250
Squamata 116, 117, 133
Staganolepidae 135, 136
Staurois 100
Staurotypus 164
Stegocephali 21
Stegonotus 269
Stegosauria 138, 142, 143
Stegosauridae 143
Stegosaurus 138
Stejneger L. 10
Steneosaurus 300
Stenocercus 200
Stenodactylus 190
Stenophis 275
Stenopterygiidae 132
Stenopterygius 129
Stereochilus 61
Stereocyclops 109
Stereogen ys 148
Stereospondyli 24, 25, 32
c hernothfu.
--.viurnUl
Storerla 267
Stomatoeuchus inermis 304
Strauch A. 10
Strep tophorue 267
Strooilurus 199
Stumpffia 104
Styracosaurus 138
subcaudalia 251
sublabialia 178
submaxillaria 178
suboculare 178, 250
supraciliaria 178
supralabialia 178, 250
supranasalia 178
supraocularia 178, 250
suprapygale 148
supratemporalia 178
Symoliopnidae 251, 252, 254
Symoliophis 254
Sympholis 271
Symphygnatinae 102
Synapsida 114, 116, 120
Syrrhophus 83
T
tabulare 28
Tachydromus 222
Tachymenis 276
Tamnophis ordinatus 280
Tangasauridae 132
Tanystropheus 135
— longobardicus 134
Taphrometopon 277, 282
Taphrometopon lineolatum 282
Tapinocephalidae 121, 123
Tarbophis 275, 282
— fallax 282
Tarentola 190
— mauritanica 186, 195
Tan ch a 55, 56
Teiidae 180, 183, 226, 227
Teius 228
Teleorhinus 300
Teleosauridae 299, 300, 301, 306
Telmatobius 83
Telmatosaurus 232
Temnospondyli 29
temporalia 178, 250
Teratolepis 193
Teratosauridae 140
Teratoscincus 188, 194
— scincus 194
Terrapene 163, 164
Testudinidae 147, 150, 151, 154,
1 RK
333
Traludo б, I Ml
•Ila* 149
rlfg»n« IM
elephanlnpu* (ЯЙ
< if tn te • 157. IM
grave* IM, IM
grandidiert IM
nermann! |58
hondieldii 158
- ibera 156
Indic* 159
su metre! 158
— zarudnyi 156
Tetradactylus 219
— africanus 219
— seps 219
— tetradactylus 219
Tetralop hosaurus 196
Tetrapoda 15. 18. 19. 181
Thalassemyidae 147, 150, 153, 168
Thalattosaurla 115, 116, 126, 127
Thalattosaurus 126, 127
Thamnophis 280
theca 147
Thecadactylus 190
Thecodontia 116, 117, 134, 135
Thecodontosauridae 141
Thecodontosaurus 139
Thecophora 149
Theriosuchidae 300, 303, 306
Theriosuchus 306
Therocephalia 124
Theromorpha 114, 116, 117, 120,
245
Theromorphoidea 23
Thetia 222, 225
Thinopus antiquus 15
Thrasops 271
tibiale 64, 68
Tiliqua 214, 216
Ti nosaurus 209
Titanosauridae 142
Titanosuchidae 120, 123
Tomistoma 300, 309
— schlegeli 310
Tomop terna 100
Topsell E. 10
Toxocheiyidae 147, 150, 152, 166
Tracheloptychus 218, 219
Trachischium 267
Trachyboa 259
Trachysaurus 214, 216
Trachysaurus rugosus 211, 216
Transversodontidae 125
Trematopsidae 32
Trematosauridae 33
Tretanorhinus 267
Tretioscincus 229
Triassochelyicae 147, 150, 151, 153
Trissochelys 153
TflbofonottH 219
Tflwatop» IM
Trfchobafrachut <M
Tfil<»phoa*urid*e 127
Trllopho«*urue 126
Trimereauru* 245, 297
— Havomaculatus 294
— gremineus 297
Trlmerorachidae 30
Trimorphodon 275
Trinychidae 150, 152. 160, 1ЗД
Trinychoidea 150, 152, 166
Trionyx 167
Triprion 88
Triton 56
Triturus 54. 55, 56
— boscai 58
— cristatus 58
— italicus 58
— montandoni 58
— vittatus 58
— vulgaris 58
Trogonophis 231
Tropidodactylus 199
Tropidonotus 279
Tropidophorus 215
Tropidosaura 220
Tropidurus 199, 203
Tuatara 175
Tupinambis 226, 230
— nigropunctata 224
Tupinambis teguixin 230
Tylosaurinae 233
Tylosaurus dyspelor 234
Tylototriton 54
tympanale 178
Tympanocryptis 208
Typhlacontias 216, 217
Typhlomoge 59
— rathbuni 47, 61
Typhionectes 38, 39, 42, 44
— compressicauda 43
Typhlophis 256
Typhlopidae 44, 252, 255
Typhlops 256
— braminus 256
— humbo 256
— lumbricalis 255
— reuteri 256
— vermicularis 256
Typhlosaurus 212
— aurantiacus 212
Typhlotriton 60
— spelaeus 61
Typothorax 135
Tyrannosauri dae 141
Tyrannosaurus 139
U
Uma 203
Ungalia 256, 259
334
Ungaliophls 259
Unken — reflex 72
Uraeotyphlus 41
Uranocentrodontidae 32
Uraniscodon 199
Urocentron 199
Urocordylldae 35
Urodela 21. 45
Urodelidia 23
Uromacor 270
Uromastix 197, 206, 208
— spinicauda 204
Uropeltldae 44, 244, 252, 253, 262,
265
Uropeltis 263
Uroplatidae 180, 181, 186, 187
Uroplatus fimbriatus 186
Urostrophus 200
Urotheca 271
Uta 203
V
Varanidae 180, 184, 232,
Varaninae 232
Varanopsidae 121
Varanosauridae 120
Varanus 232
— griseus 233, 234
— rudicollis 233
— komodoensis 232
— Salvator 232, 234
— niloticus 233
ventral i a 25t
Vermicella 245
vertebral i a 149
Vibrissaphora 63
Vipera 245, 293, 294
— ammodytes 294, 295
— asp is 248
— berus 295
— kaznakowi 295
— lebetina 249, 294 , 295
— raddei 295
— russeli 245, 248
— ursini 295 ,
Viperidae 245, 252, 253,
W
Wahleet G. 62
Walls g. 243
237, 243
288, 292
Wataon D. 18, 26. 35. 171
Welgeithauridae 127
Weigeltlaaurua 126
Wermuth H. 171. 175. 310
Werner F. 298, 310
Werneria 84
Wastoll T. 18, 35
Wettstein O. 175, 310
Whaitsldae 124
X
Xantusia 195
— vigilis 186, 196
Xantusiidae 180, 181, 187, 195
Xenobatrachus 102, 110
Xenodermidae 264
Xenodermus 264
Xenodon 272
Xenopeltidae 252, 253, 264, 265
Xenopeltis unicolor 263, 264
Xenopus 64, 73
— laevis 73
Xenorhina 102
Xenosauridae 180, 184, 239
Xenosaurinae 237, 239
Xenosaurus 239
— rackhami 235, 240
xiphiplastron 149
Xiphocercus 198, 210
¥
Younginia capensis 134
Youginidae 132
Z
Zachaenus 83
Zajssanurus 52
Zamenis 280
Zaocys 270
Zatrachydae 31
Zittel K. 10
Zonosaurus 219
Zonuridae 217
Zonurus 218
Zootoca 222, 223
zygantrum 241
zygosphene 241
ОГЛАВЛЕНИЕ
Првднсллнн»
/ лом первая
История герпетологии
Глава вторая
Выход земноводных на сушу и дрспиие земноводные
Глава третья
Безногие земноводные Ч
Г лава четвертая
Хвостатые земноводные 45
Глава пятая Бесхвостые земноводные 63
Глава шестая Происхождение пресмыкающихся и вымершие группы 111
Глава седьмая Черепахи 147
Глава восьмая Клювоголовые 172
Глава девятая Ящерицы 176
Глава десятая Змеи 241
Глава одиннадцатая Крокодилы 299
Указатели 311
Павел Викторович Терентьев
ГЕРПЕТОЛОГИЯ
Редактор А. Г. Банников
Редактор издательства К. Г. Парсаданова
Технический редактор Л. А. Григорчук
Корректор Г. П. Меркулова
Сдано в набор 6/V-1960 г. Подписано к печати 30/XI-1960 г. Бумага 6OX92l/ie.
21 4- 2 цв. вкл. печ. л. 20,в©1 уч.-изд. л. Тираж 2450 экз. Т—(15205. Заказ 454.
Цена 86 коп.
Издательство «Высшая школа». Москва, Б-62, Подсосенский пер., д. 20.
Полиграфический комбинат Ярославского совнархоза,
г. Ярославль, ул. Свободы, 9*7.
Замеченные опечатки
Стра- ница Строка Напечатано Следует читать
14 5—6 снизу •не будут будут
26 6 снизу Рабовидные Рыбовидные
139 4 снизу Prosa и города Prosauropoda
245 1 снизу ponhyriacus porphyriacus
248 22 сверху животных капелек
капелек
284 19 сверху рис. 64 рис. 61
295 8 сверху В тропиках Типичная
Заказ 454.
Таблица I Представители земноводных:
1—2— Triturus vitiatus (самец и самка), 3 Bombina bombina, Ceratophrys
2ornuta,5- Atelopus variiis, 6— Salamandra salamandra, 7—Hyla arborea (самец
г надутым резонатором). 8- Bufo viridis, 9—Rana esculenta (самец с надутыми
резонаторами).
Таблица II, Представители пресмыкающихся:
/— Chrysemys picta, 2—Mabuya bibroni, 3—Natrix ordinatus var. sirtalis,
1—Vipera kaznakowi, 5—Testudo elegans, 6—7—Ceratophora aspera (самка и
самец), 8—9—Lacerta agilis (самка и самец) 10—Micrurus fulvius, 11—Lache-
sis gramineus.