






















































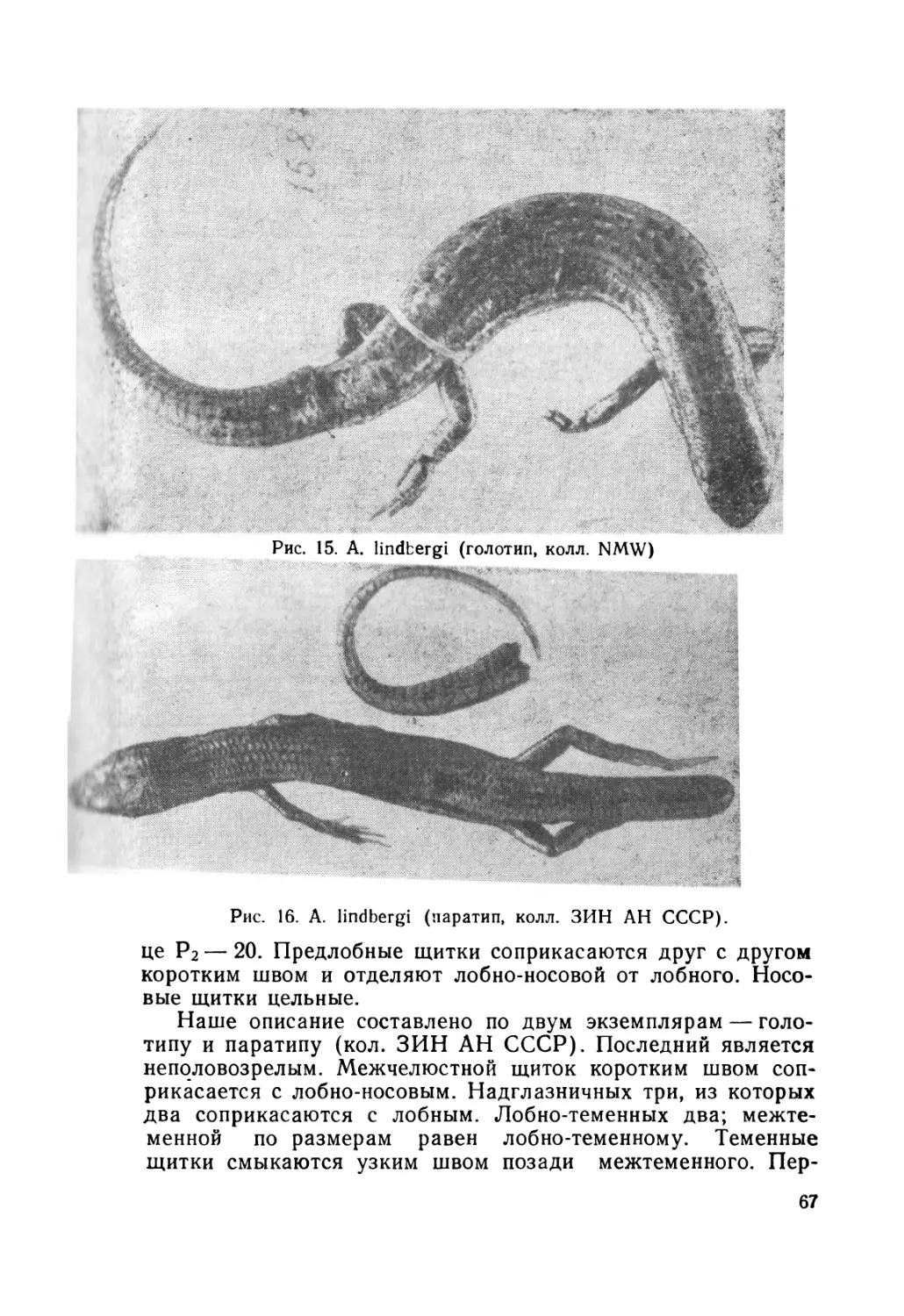


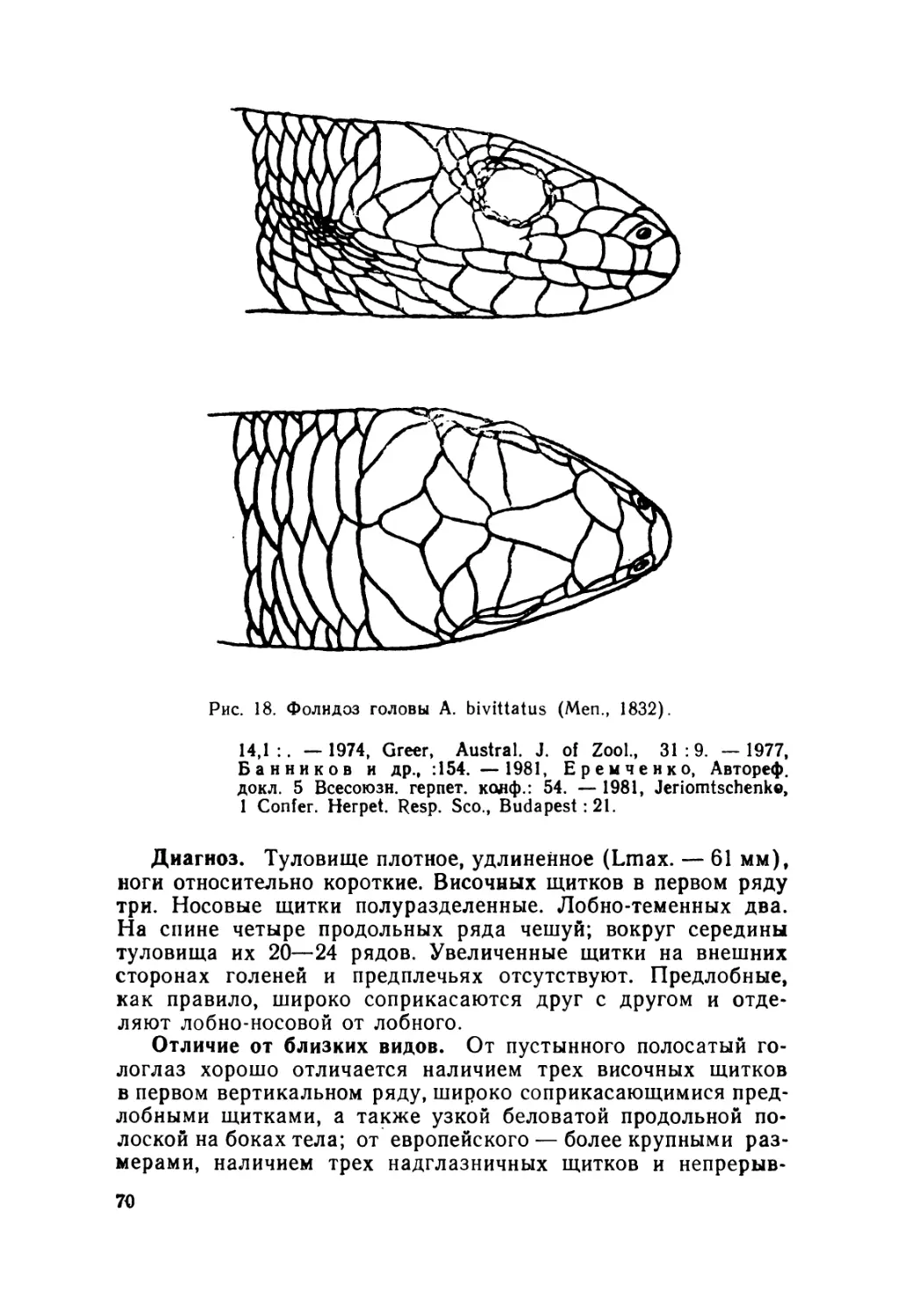


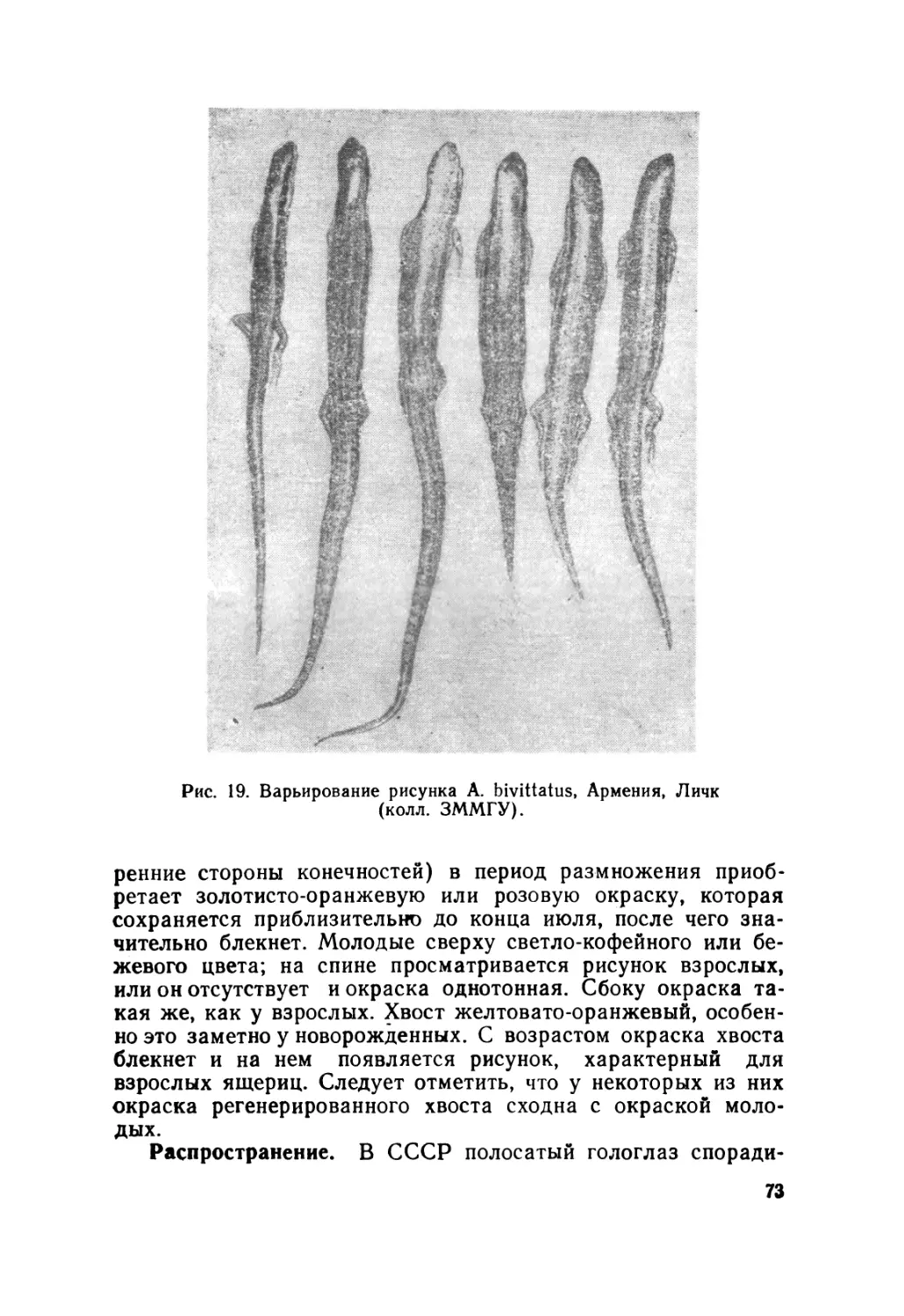


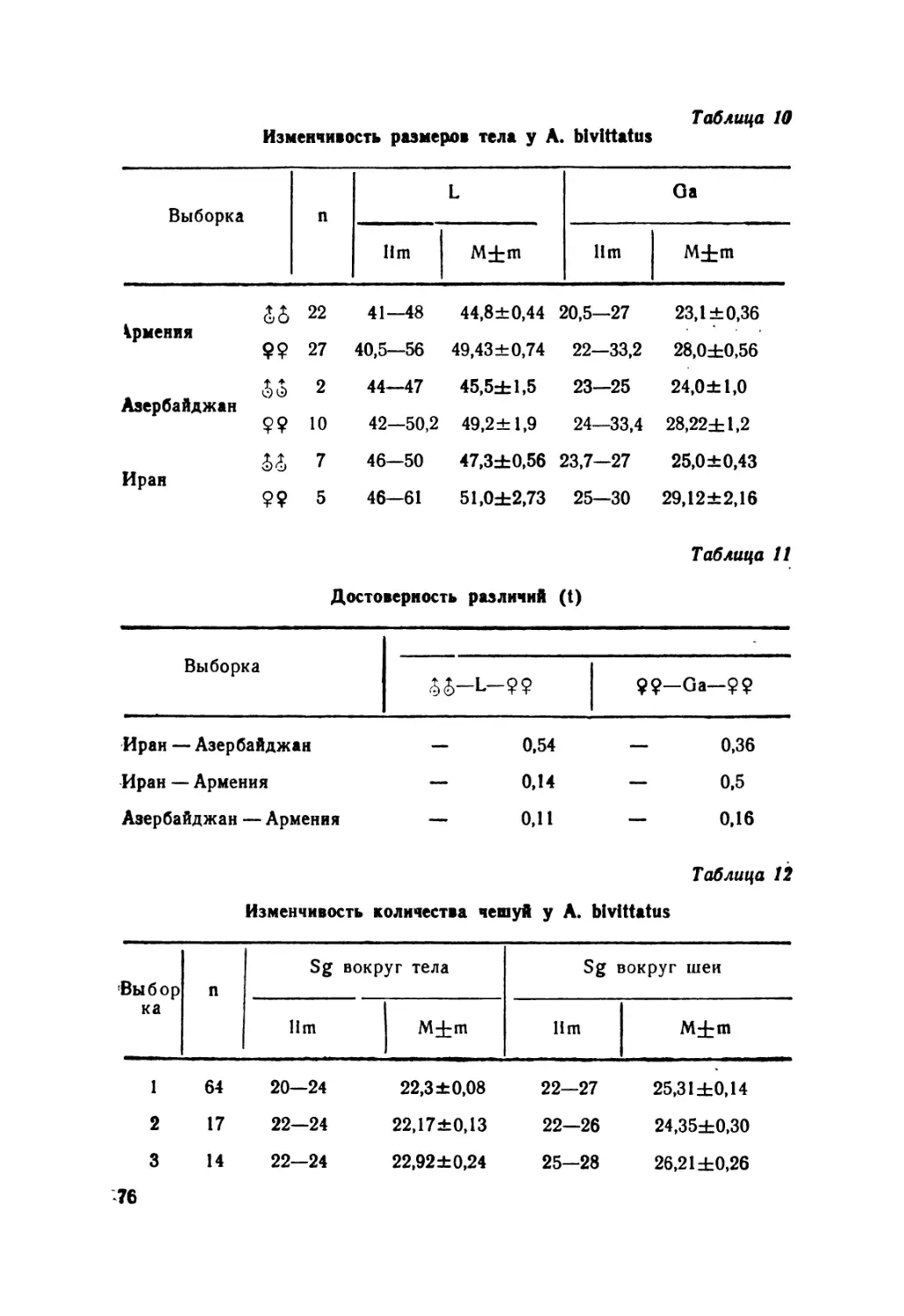

























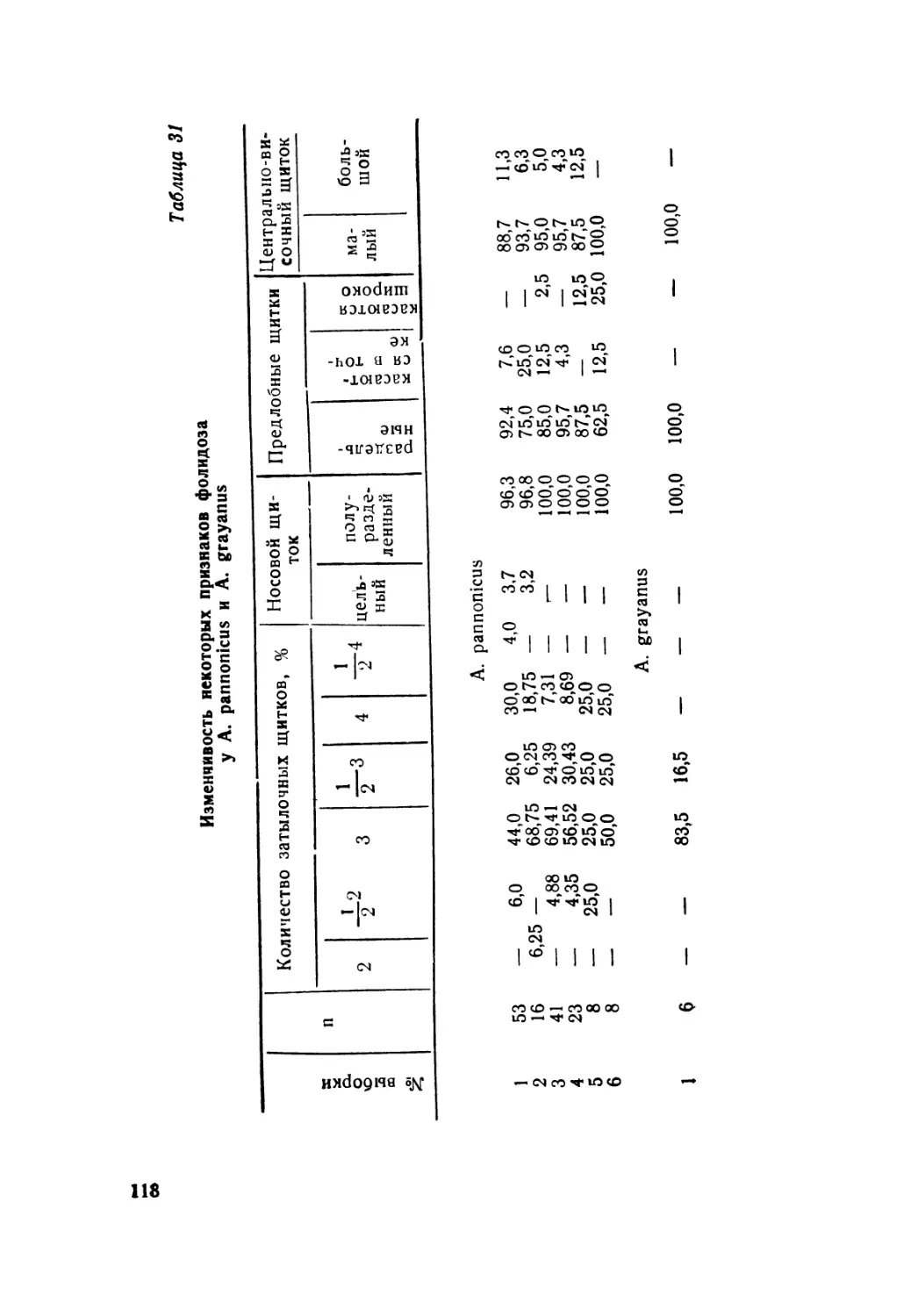











































Автор: Крёмченко В.К. Щербак Н.Н.
Теги: reptilia рептилии пресмыкающиеся герпетология биология ссср зоология фауна ящерицы географическая изменчивость
Год: 1986




АКАДЕМИЯ НАУК КИРГИЗСКОЙ ССР
ИНСТИТУТ БИОЛОГИИ
В. К. Еремченко, Н. Н. Щербак
АБЛЕФАРИДНЫЕ ЯЩЕРИЦЫ
ФАУНЫ СССР
И СОПРЕДЕЛЬНЫХ СТРАН
Издательство «Илим»
Фрунзе 1980
УДК 598.113.7(47 + 57)
< В монографии рассматривается широкий круг вопросов,
связанных с систематикой, географической изменчивостью,
филогенией и экологией палеарктических аблефаридных яще-
риц (гологлазов). Составлены определительные таблицы ис-
следованных таксонов, показаны основные закономерности
дифференциации н возможные пути их становления. Даны
сведения о редких и исчезающих видах фауны СССР.
Книга рассчитана на зоологов-систематиков, герпетологов,
специалистов в области охраны природы, заповедного дела,
музеев и преподавателей биологических факультетов вузов.
Утверждено к печати
Ученым советом Института биологии
и принято РИСО
Академии наук Киргизской ССР
Ответственный редактор канд. биол. наук Э. Д. Шукуров.
Рецензенты: докт. биол. наук, проф. И. С. Даревский
канд. биол. наук П. Л. Чиров
Издательство «Илим», р.
ВВЕДЕНИЕ
Существует мнение, что после недавней ревизии гологлазов
рода Ablepharus (puhn, 1969 а, в) вопросы систематики этих
ящериц решены удовлетворительно (Greer, 1974; Банников и
др., >977; и др.). В связи с этим наша работа была задумана
как эколого-фаунистическое исследование гологлазов, распро-
страненных в пределах Тянь-Шаня. Однако по мере накопле-
ния полевого материала, в процессе работы с коллекциями и
критического переосмысливания литературных данных у нас
правились сомнения относительно справедливости существую-
щих положений по систематике гологлазов нашей фауны и в
целом Палеарктики.
Прежде всего это касается алайского гологлаза A. alaicus,
который, по мнению некоторых западных герпетологов (Wet-
tstein, 1960; Mertens, 1965; Fuhn, 1969 а, в; Greer, 1974), пред-
ставлен лишь подвидом полосатого гологлаза A. bivittatus
alaicus (Elpat., 1901). При этом, как считали эти исследова-
тели, афганский подвид полосатого гологлаза A. b. lindbergi
Wettstein, 1960 выступает как бы связующим звеном между
номинативным подвидом и формой «alaicus».
В то же время отечественные герпетологи (Никольский,
1915; Терентьев, Чернов, 1949; Чернов, 1959; Яковлева, 1964;
Банников и др., 1977) не подвергали сомнению валидность
A. alaicus. Более того, описывали близкие к «alaicus» фор-
мы— A. kucenkoi Nik., 1902 и A. saposhnikovi Kaschtschenko,
1909, которые впоследствии были формально сведены в сино-
нимы A. alaicus (Терентьев, Чернов, 1949; Банников и др.,
1977). Все это свидетельствовало о более сложной структуре
данного вида.
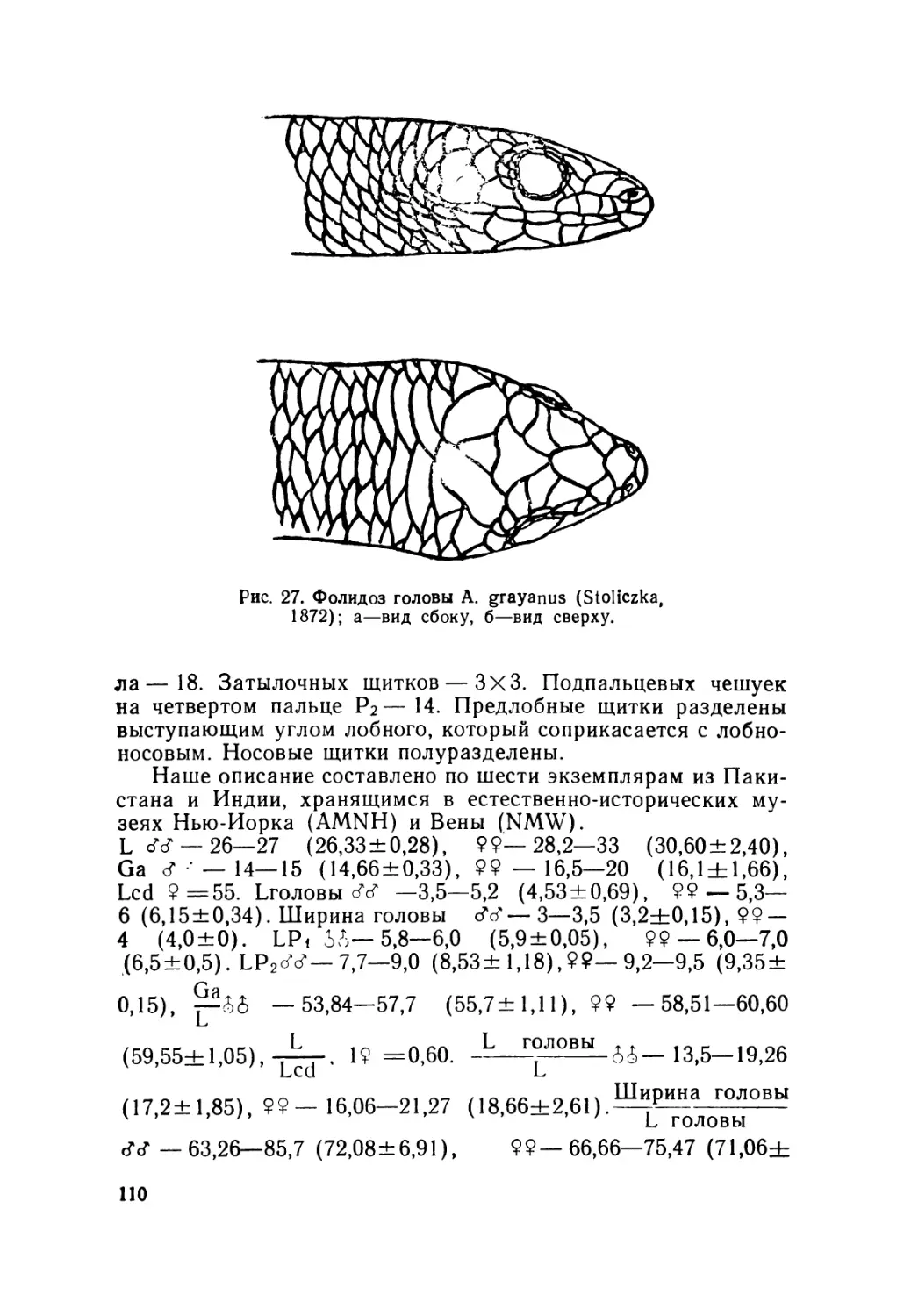
Далеко не благополучной выглядит и систематика двух
близких видов — A. pannonicus (Licht., 1823) и A. grayanus
(Stoliczka, 1872), основные ареалы которых находятся на соп-
редельных с СССР территориях в Иране, Афганистане и Се-
веро-Западной Индии. Так, одни исследователи (Smith, 1935;
Терентьев, Чернов, 1949; Mittleman, 1952; Чернов, 1959; Min-
ton, 1966; Банников и др., 1971; Саид-Алиев, 1979) признава-
ли видовую самостоятельность A. grayanus, другие (Mertens»
з
1965; Fuhn, 1969, а, в; Greer, 1974; Банников и др., 1977; То-
поркова и Степанов, 1977) рассматривали его в качестве под-
вида азиатского гологлаза Л. pannonicus grayanus (Stoliczka,
1872).
Мнение о политипичности A. pannonicus, как впрочем и
вышеупомянутого вида A. bivittatus, было основано на изуче-
нии единичных экземпляров, а также весьма ограниченном
числе использованных для этой цели признаков. Подобный
подход к систематике указанных видов определяется моно-
типической концепцией вида. В частности А. М. Никольский
(1907, 1915), работая с единичными экземплярами гологла-
зов, описал из Северного Ирана вид A. persicus и форму
A. brandtii var. brevipes, а из Семиречья—A. tenuis. Вполне
закономерным является то, что позже эти таксоны рассматри-
вали (Шнитников, 1928; Терентьев, Чернов, 1949; Fuhn, 1969 в;
Банников и др., 1977) в качестве синонимов различных видов:
первые два — A. pannonicus (Licht., 1823); последний —
A. deserti Strauch, 1868. Конкретные доказательства этого
мнения не были опубликованы, в связи с чем необходимо пе-
реисследовать указанные таксоны с учетом географической
изменчивости видов, к синонимам которых они были отне-
сены.
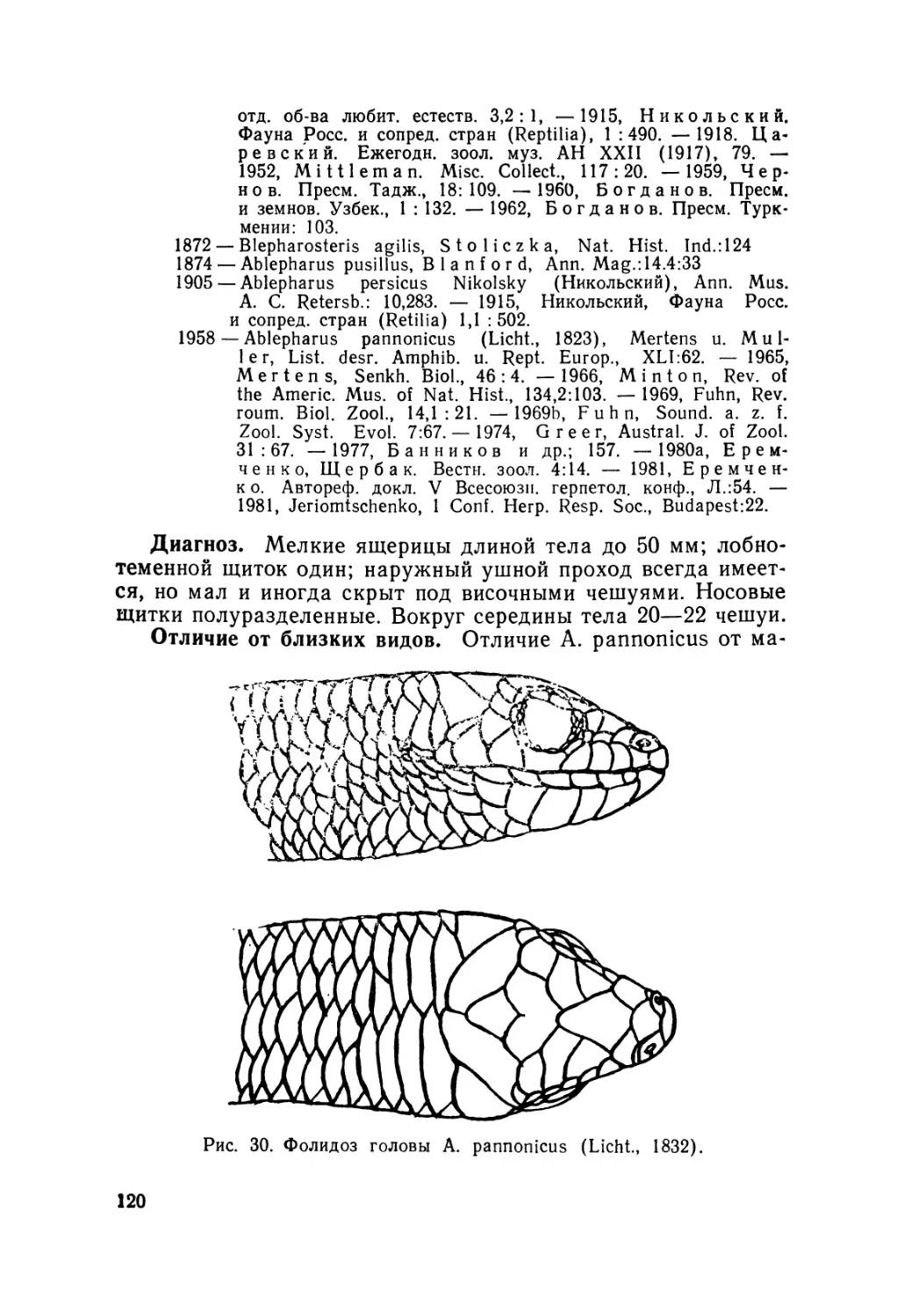
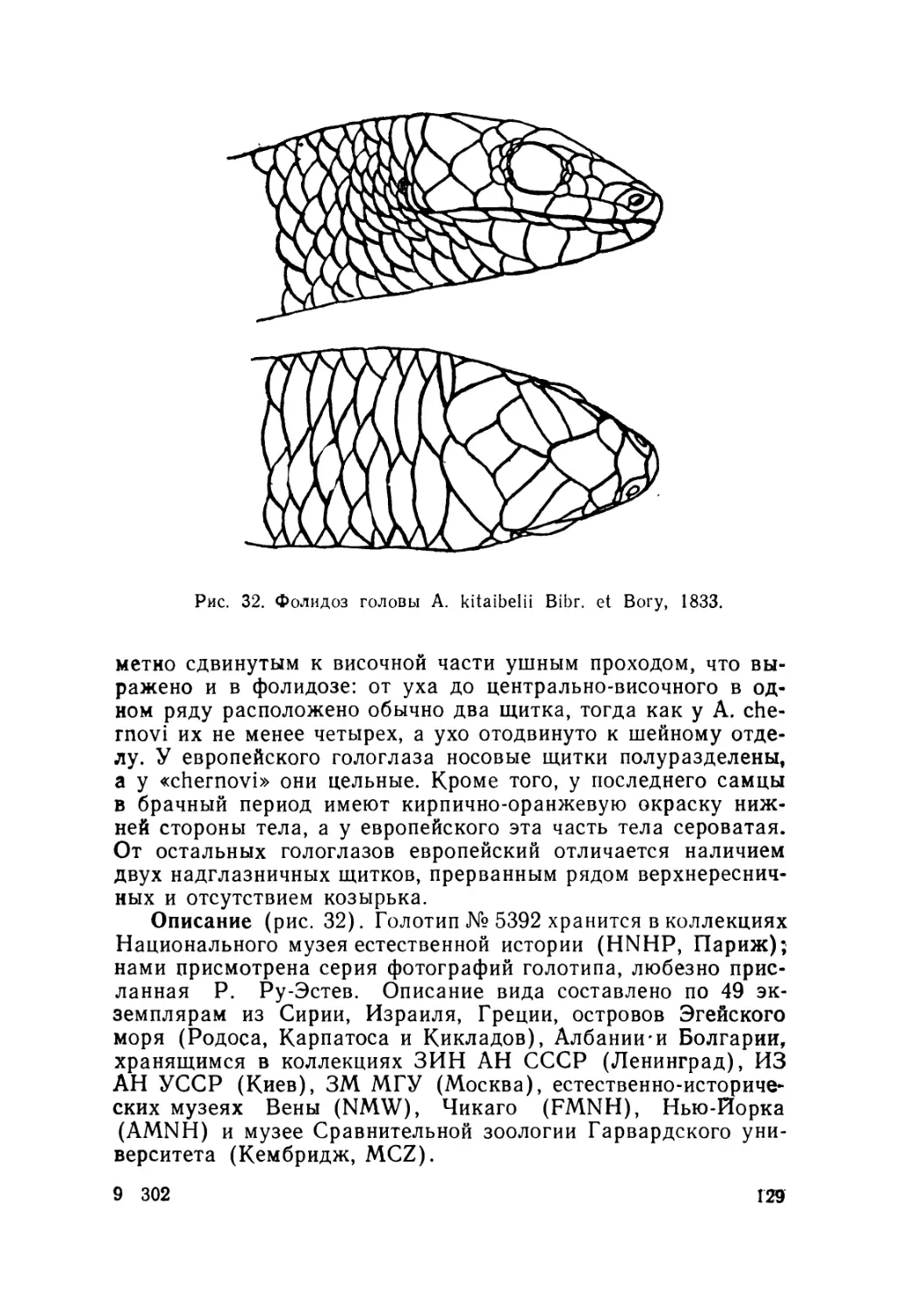
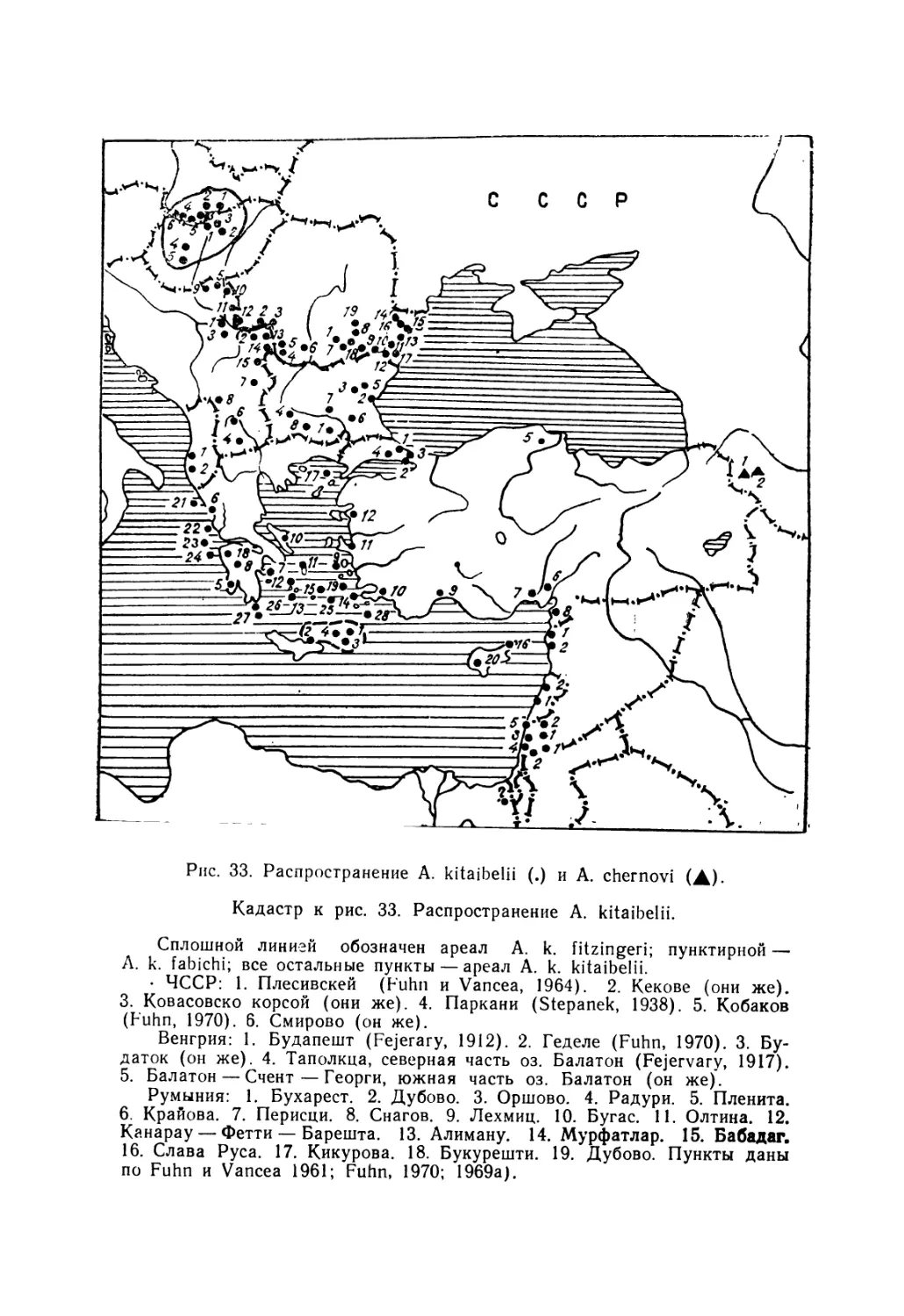
Европейский гологлаз A. kitaibelii Bibr. et Вогу, 1833—
единственный вид данной группы, внутривидовую системати-
ку которого изучали неоднократно (Stepanek, 1938; 1944; Mer-
tens, 1952; Fuhn, 1969 в, 1970; Gruber, 1981; и др.). Он был
разделен на 5 подвидов: номинативный, A. k. fabichi Stepa-
nek, 1938; А. к. fitzingeri Mertens, 1952;. А. к. stephaneki Fuhn,
1969 и А. к- chernovi (Darevsky, 1953). При этом последняя
из указанных форм ранее была описана как самостоятельный
вид (Даревский, 1953).
Используя ограниченное число признаков, невозможно
верно и в достаточной мере глубоко отразить особенности из-
менчивости и систематику одного вида, поскольку все осталь-
ные в этой группе практически не изучены. Если учесть еще,
что данная группа обладает мелкими размерами тела и к то-
му же развитым внешним сходством, то станет понятным, нас-
колько сложна эта задача. Как нам представляется, не лише-
на недостатков и систематика европейского гологлаза.
Определенную трудность вызывают некоторые разногла-
сия номенклатурного характера. Одни нггледонатеЛЙ /Маг-
tens, 1952; Fuhn, 1969 а, в; Greer, 1974; Банников И Др * 1977)
считают автором рода Ablepharu*. X ЛнмтоиштейМ (Lich-
tenstein, 1823), другие (Straiirh, 1НПН, БлнатьевсЯдо, 1901;
Smith, 1935; Minton, 1966) нагianhaioi на приоритете Л. Фи-
цингера (Fitzinger, 1824). Таким иОразом, систематика голо-
4
глазов рода Ablepharus носит лишь предварительный харак-
тер.
Скрытый образ их жизни и распространение в пустынных
и горных районах затрудняет полевые экологические иссле-
дования этих животных. Сведений по образу жизни ящериц
до сих пор было явно недостаточно.
Настоящая работа выполнена в лаборатории зоологии
позвоночных животных Института биологии АН Киргизской
ССР и Зоологическом музее Института зоологии
им. И. И. Шмальгаузена АН Украинской ССР. В основу дан-
ного труда, являющегося результатом десятилетних исследо-
ваний, положены материалы, собранные в 1973—1982 гг. во
время многочисленных экспедиций по Средней Азии, Казах-
стану и Закавказью. Кроме того, были обработаны фондовые
коллекции Зоологического Института АН СССР (Ленинград,
ЗИН), Зоологического музея Института зоологии АН УССР
(Киев, ИЗАН УССР)» Зоологического музея Московского го-
сударственного университета (Москва, ЗММГУ), Института
зоологии АН Казахской ССР (Алма-Ата, КазЗИН), Инсти-
тута зоологии и паразитологии АН Таджикской ССР (Душан-
бе ТаджИЗиП) и Института биологии АН Киргизской ССР
(Фрунзе, АН ИБ). Для камеральной обработки нам предо-
ставили свои коллекции и некоторые зарубежные хранилища:
естественно-исторические музеи Нью-Йорка (AMNH), Чика-
го (MNH), Вены (NMW) и Парижа (MNHN), Музей сравни-
тельной зоологии Гарвардского университета, Кембридж, Мас-
сачусетс (MCZ), зоологические музеи Будапешта (BZM) и
Берлина (ZMB), а также частная коллекция американского
герпетолога доктора Ш. Минтона, Индианаполис (SAM).
Проведение данного исследования было бы невозможно
без постоянной помощи наших коллег и товарищей по рабо-
те. Так, в определении компонентов питания ящериц прини-
мали участие сотрудники Института биологии АН Киргиз-
ской ССР: Л. В. Пэк, Ю. С. Тарбинский, Т. Калтаев и
С. Н. Овчинников. Паразиты, обнаруженные на этих живот-
ных, определены сотрудниками Института зоологии АН УССР
В. Г. Гущей и Г. И. Щербак. Коллекционный материал был
получен благодаря любезности И. С. Даревского, Л. Я. Бор-
кина и Н. Б. Ананьевой (Ленинград), В. Ф. Орловой и
Т. О. Александровской (Москва), С. А. Саид-Алиева и
Т. С. Сатторова (Душанбе), 3. К- Брушко (Алма-Ата), О. Де-
ли (Будапешт), Р. Г. Цвейфеля (Нью-Йорк), Р. Ф. Инжея и
А. Резетар (Чикаго), Дж. Розадо и П. Альберха (Кембридж),
И. Айзельта и М. Хепля (Вена), Ш. А. Минтона (Индиана-
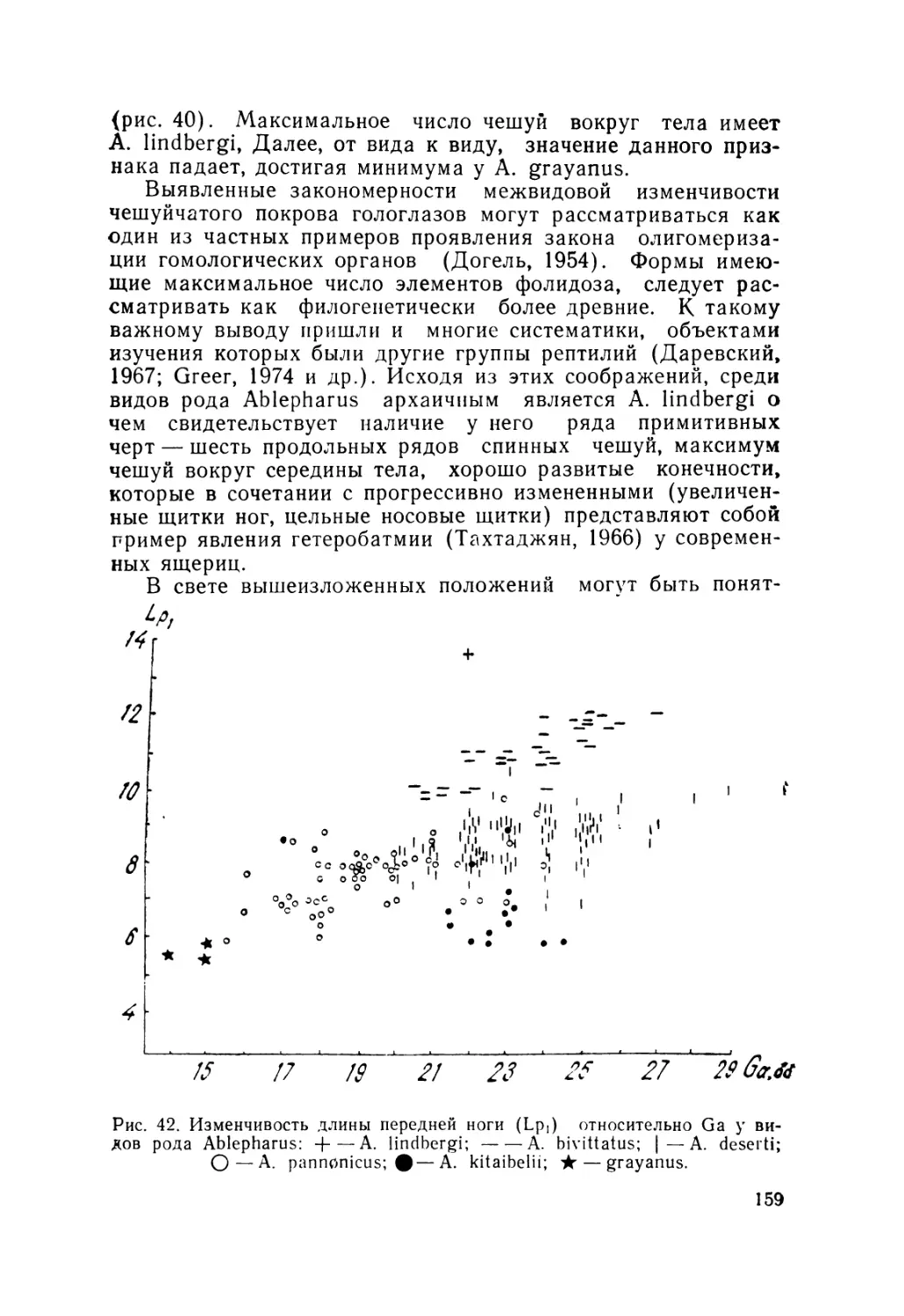
полис), Р. Ру-Эстев (Париж).
5
В статистической обработке материалов авторам оказана
помощь сотрудником Института биологии АН Кирг. ССР
В. И. Тороповой. При подготовке рукописи к печати цепные
замечания и советы мы получили от проф. И. С. Дарев-
ского и Л. Я. Боркина. Перечисленным выше лицам авторы
выражают искреннюю благодарность.
ОБЩАЯ ЧАСТЬ
КРАТКИЙ ОБЗОР ИССЛЕДОВАНИЯ
ЯЩЕРИЦ РОДА ABLEPHARUS
Родовое название Ablepharus впервые было предложено
Л. Фицингером (Fitzinger, 1824) для ящерицы из Средней
Азии (Бухары), диагноз которой, однако, был опубликован
на год раньше X. Лихтенштейном (Lichtenstein, 1823). Типо-
вым видом данного рода является азиатский гологлаз — Ab-
lepharus pannonicus (Lichtenstein, 1823) (Scincus pannonicus
Lichtenstein, 1823).
По Л. Фицингеру и X. Лихтенштейну, род Ablepharus ос-
нован на двух характерных для этой ящерицы признаках: не-
подвижные прозрачные веки и пятипалые конечности. В та-
ком виде содержание диагноза рода Ablepharus достаточно
четко выразило его отличия от ранее известного рода Scincus
Gronovius, 1767. Позже крупнейший отечественный герпето-
лог А. Штраух (Strauch, 1868), автор описания нескольких
форм гологлазов, столкнувшись с трудностью определения
э*гих ящериц, впервые провел ревизию их систематического
положения. Особое значение при этом он уделил наличию у
гологлазов пятипалых конечностей и включил в род Ablepha-
rus девять видов: A. pannonicus; A. bivittatus; A. deserti;
A. brandtii; A. nigropunctatus; A. wahlbergi; A. boutoni; А. Н-
neooccllatus и A. wotjulum. Спустя Шлет после ревизии А. Шт-
рауха известный британский герпетолог Г. Буланже (Boulen-
ger, 1887) указал на доминирующее значение в диагнозе рода
Ablepharus признака «неподвижный прозрачный диск покры-
вает глаз». Он и отнес к данному роду еще 16 видов, принад-
лежавших ранее к 10 различным родам: Lerista Bell, 1833;
Gryptoblepharus Wiegmann, 1834; Microblepharus Fitzinger,
1943 и др. Эти выводы Г. Буланже длительное время (около
90 лет) являлись руководством для многих герпетологов и к
началу 70-х годов XX в. было уже описано-около 43 видов ро-
да Ablepharus, распространенных в Евразии, Африке, Мада-
гаскаре, Австралии и Полинезии. Практически всех мелких
сцинков, имеющих ясно выраженное прозрачное «окошко» на
глазах, разные исследователи принимали за гологлазов рода
7
Ablepharus. К этому времени в фауне нашей страны и Пале-
арктики в целом было описано около 14 их различных форм:
A. kitaibelii Bibron et Bory, 1833; A. pannonicus (Lichtenstein,
1823); A. bivitlatus (Menetries, 1832); A. deserti Strauch, 1868;
A. grayanus (Stoliczka, 1872); A. brandtii Strauch, 1868; A. alai-
cus Elpatjewsky, 1901 и др.
Выдающийся русский герпетолог А. М. Никольский (1899»
1902, 1907, 1915), хороню знакомый с палеарктическими ви-
дами гологлазов, писал, что у этих ящериц «нижнего века
нет» или «век нет, вместо них один цельный прозрачный непод-
вижный кружочек, покрывающий глаз». Британский герпето-
лог М. Смит (Smith, 1935, 1937) установил, что ряд видов,
включенных в род Ablepharus, имеют веки и так называемую
палпебральную щель( palpebral slit). Скрытые под ресничной
частью фолидоза (supercilium), они долгое время оставались
неизвестны исследователям- М. Смит (Smith, 1939) высказал
предположение о полифилитическом происхождении аблефа-
ридных ящериц и счел характерным для представителей ро-
да Ablepharus полное или почти полное слияние нижнего ве-
ка с верхним. С учетом этого М. Смит (Smith, 1939) относит
к роду Ablepharus и такие виды, как A. spencery; A. lineoocel-
latus; A. egerinae и др., у которых веки несросшиеся.
Так же широко понимали объем рода Ablepharus и другие
герпетологи, поддержавшие мнение М. Смита (Parker, 1936;
De Witt, 1936; Mittleman, 1952 и др.). Однако М. Миттлман
(Mittleman, 1952), признав самостоятельность африканских
гологлазов рода Panaspis Сорре, 1868, реконструированного
ранее М. Смитом (Smith, 1935), восстанавливает и другой
род — Gryptoblepharus (Wiegmann, 1834), выделив его так-
же из рода Ablepharus. Эти выводы требовали, однако, допол-
нительных доказательств. Важным шагом в этом направле-
нии явились исследования в области краниологии сцинковых
ящериц, выполненные австралийским герпетологом Ф. Мит-
челлом (Mittchcll, 1950). В указанной работе на основании
особенностей строения некоторых частей черепа удачно клас-
сифицированы «трудные» в систематическом отношении сцин-
ки родов Tiligua и Egernia- Он же (Mittchell, 1958) придавал
диагностическое значение и другим остеологическим призна-
кам, в частности, строению конечностей.
Позднее еще большее внимание краниологии в изучении
систематики сцинковых ящериц уделяет австралийский гер-
петолог А. Грир (Greer, 1967, 1970, 1974, 1979; Greer and
Parker, 1968). Базируясь на краниологических, а также дан-
ных наружной морфологии, экологии и особенностях распрост-
ранения, он восстановил австралийский род Lerista Bell., 1833,
относимый ранее к роду Ablepharus, а также обосновал неко-
8
торые высшие таксоны, в том числе подсемейств Feylininae и
Acontinae. Параллельно с ним работал румынский герпетолог
И. Фун (Fuhn, 1969а, в, 1970, 1972 и др.). На основании кра-
ниологии и признаков наружной морфологии он показал ав-
тохтонное происхождение евразиатских гологлазов и рамки
рода Ablepharus сузил до 5 видов (pannonicus, kitaibelii, bi-
vittatus, deserti, borealis), исключив из него формы африкан-
ского и австрало-полинезийского происхождения. Выводы
И. Фуна были приняты А. Гриром (Greer, 1974) и с неболь-
шими изменениями, не затрагивающими родовые критерии,
советскими специалистами (Банников и др., 1977).
На этом можно было бы считать решенным вопрос родо-
вой таксономии Ablepharus. Однако, как показано нашими
исследованиями, признаки некоторых видов гологлазов не со-
ответствуют родовому диагнозу, отчего систематика данного
рода выглядит несовершенной. В первую очередь это касает-
ся оценки таксономического положения формы Ablepharus
nlnicus Elpatjewsky, 1901. Мнения относительно систематики
итого вида, как уже было сказано, являются глубоко противо-
речивыми..
Нс менее сложная проблема возникла в настоящее время
и и поирогпх видовой и внутривидовой систематики гологла-
зов, в также их номенклатуры. До того, как получил свое
признание род Ablepharus, М- Менетри (Menetrie,
1832) описал новый вид гологлаза из Закавказья A. bivitta-
1нн (Scincus bivittatus), объем которого был расширен не-
которыми западными герпетологами (Wettstein, 1960; Mertens,
1965; Fuhn, 1969 а, в) за счет внесения в него форм «alaicus»
н «lindbergt».
В 1833 г. Биброн и Бори (Bibron et Вогу) по материалам
из Греции описали ’гологлаза и назвали в честь венгерского
зоолога Китайбела — A. kitaibelii. Этот вид долгое время пу-
тали с A. pannonicus Lichtenstein, 1823 ' (A. pannonicus
I’llzinger, 1824). Спустя 40 лет после выхода работы Биб-
ропа и Бори Ф. Столичкой (Stoliczka, 1872) по материалам
из Западной Индии и Пакистана была описана форма «gray-
anus», которая по признаку отсутствия наружного ушного
прохода была выделена им в отдельный род Blepharpsteres-
Позднее «grayanus» был переведен (Boulengen 1887) в род
Ablepharus, а его видовая самостоятельность признана многи-
ми специалистами (Smith, 1935; Mittleman, 1952; Чернов,
1959; Minton, 1966 и др.). С другой стороны, Р. Мертенс (Mer-
tens, 1965), а вслед за ним И. Фун (Funh, 1969 а, в и др.) рас-
сматривали данный вид только в качестве подвида азиатско-
го гологлаза — A. pannonicus grayanus (Stoliczka, 1872).
Как уже отмечалось, для выяснения видового состава оте-
9
чественных гологлазов большое значение имела работа*
А. Штрауха (Strauch, 1868), в которой он описал два новых
вида — A. deserti и A. brandtii и впервые провел ревизию это-
го рода. Видовая самостоятельность A. deserti Strauch обще-
признана (Boulenger, 1887; Никольский, 1899, 1915; Mittleman,
1952, Терентьев, Чернов, 1949; Greer, 1974; Банников и др.,
1977; и др.). Статус второго вида—A. brandtii вызывает но-
менклатурные разногласия. По некоторым данным (Mertens:
und Wermuth, 1928; Mertens, 1952; Fuhn, 1969 а, в; и др.)
A. brandtii является синонимом A. pannonicus (Lichtenstein,
1823). Согласно другой точке зрения (Никольский, 1915; Чер-
нов, 1959) A. pannonicus считается синонимом A. brand-
tii й даже A. deserti.
Большое внимание вопросам систематики гологлазов уде-
лял А. М. Никольский, описавший три новых вида: A. kucen-
koi Nik., 1902, A. persicus Nik., 1907, A. tenuis Nik., 1915, а так-
же форму A. brandtii var. brevipes Nik., 1907. Все они в нас-
тоящее время сведены в синонимы: первый — A. alaicus Elpat,
1901, второй — A. deserti Strauch, 1868, третий и четвертый —
A. pannonicus (Lichtenstein, 1823) (Терентьев, Чернов,
1949; Банников и др., 1977; и др.). Форма A. saposhnikowi,
описанная в 1909 г. из восточного Тянь-Шаня В. П. Кащенко,
впоследствии также была сведена в синонимы A. alaicus
Elpat., 1901 (Терентьев, Чернов, 1949; Банников и др., 1977;
и др.), хотя, как это будет показано ниже, без достаточных
оснований.
Сравнительно недавно И. С. Даревский (1953) описал
A. chernovi по материалам из Армении. Позднее И. Фун
(Fuhn, 1969 в, 1970), проводя ревизию вида A. kitaibelii,
рассматривает A. chernovi в качестве подвида А. к.
chernovi (Darevsky, 1953). Мнение И. Фуна в настоящее вре-
мя общепринято (Greer, 1974; Банников и др., 1977; Gruber,
1981).
Внутривидовую систематику европейского гологлаза A. ki-
taibellii изучали неоднократно. В соответствии с современны-
ми взглядами этот вид состоит из пяти подвидов: A. k. kitaibelii
Bibron et Bory, 1833; A. к. fabichi Stepanek, 1938; A. k. fitzin-
geri Mertens, 1952; A. k. stepaneki Fuhn, 1969 и A. k. cherno-
vi (Darevsky, 1953) (Fuhn et Vancea, 1961; Fuhn, 1969 a, 1970;
Greer, 1974; Банников и др., 1977; Gruber, 1981).
Подводя итог обзору работ систематического плана, отме-
тим, что несмотря на некоторые положительные решения, как,
например, выделение И. Фуном евразиатских видов, в целом
систематика указанной группы до сих под еще не разработана.
Первые и весьма скудные сведения по экологии гологла-
зов мы находим у М. Менетри (Menetrie, 1832). Отдельные
10
данные по образу жизни гологлазов Ирана приводйт Н. А. За-
рудный (1898). Интересные наблюдения по экологии A. alai-
cus в условиях высокогорья Алайской долины описаны
Л. А. Андрушко (1951). Р. П. Зимина (1959), долгое время
работавшая на Иссык-Куле, также описывает некоторые чер-
ты экологии A. alaicus. Результаты экологических наблюде-
ний за A. deserti в Узбекистане опубликовала М. В. Калужи*-
на (1951) и другие (Кашкаров, 1924; Дубинин, 1954). Особо
следует отметить статью И. С. Даревского (1955), в которой
автор обсуждает пути проникновения A. deserti в долину Мур-
габа. Интересующие пас материалы о гологлазах содержатся
в статьях и монографических сводках по рептилиям Средней
Азии и Казахстана (Чернов, 1934, 1945, 1959; Параскив, 1956;
Богданов, 1960, 1962, 1965; Яковлева, 1962, 1964; Саид-Алиев,
1979). Отдельные данные о распространении, численности,
биотопическот распределении и врагах содержатся в ряде
публикаций (Шульпин, 1948; Даль, 1935; Щербак, 1959; Кар-
пенко, 1970; Макеев, 1975; Второв и Макеев, 1972; Второв и
др., 1978; Сатторов, 1977; Раджабов, 1975; Ядгаров, 1973; Ков-
iiiapi», 1966, 1981). Некоторые сведения по экологии гологла*
зов приводили в ряде работ западные герпетологи: по евро-
пНн кому юлоглазу A. kitaibelii (Hejervary, 1912, 1917; Bolkay,
1914, 1919; Wettstein, 1953, 1960; Karaman, 1939; Radovano-
vic, 1957; F?uhn et Vancea, 1961; Paculjevic, 1964, 1965; Daan,
1967; Mertens, 1968; Dzukic, 1972); по малому гологлазу —
A, gniyanus (Smith, 1935; Wettstein, 1960; Mertens, 1965; Min-
lonb 1966; Leviton and Anderson, 1970).
МАТЕРИАЛ И МЕТОДИКА
Материалом для настоящей' работы послужили сборы иг
наблюдения с 1973 по 1982 г. в различных районах Средней
А <ин, Казахстана. Закавказья. Стационарные исследования
выполняли в некоторых пунктах Иссык-Кульской котловины
(ур. Кар-Кыра), Киргизского хребта (ущ. Аламедин), Чуй-
ской долины (окрестности г. Токмак и с. Манас).
Мы придерживались концепции политического вида и ис-
следовали выборки из отдельных популяций. Всего было об-
работано 1260 экземпляров гологлазов всех видов фауны
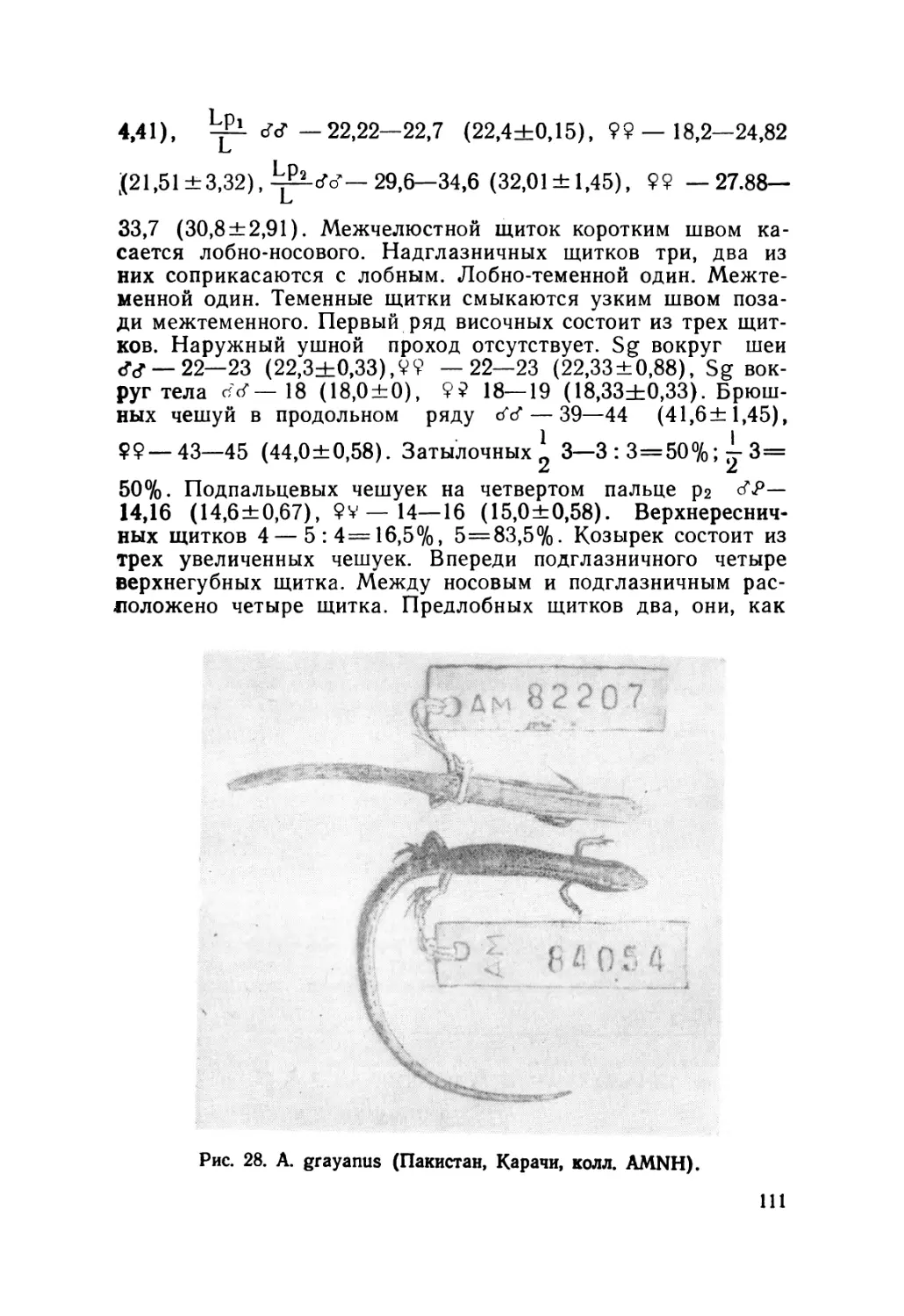
СССР и сопредельных стран: рода Ablepharus — A. deserti —
402; A. pannonicus— 151; A. grayanus —6; A. bivittatus — 95;
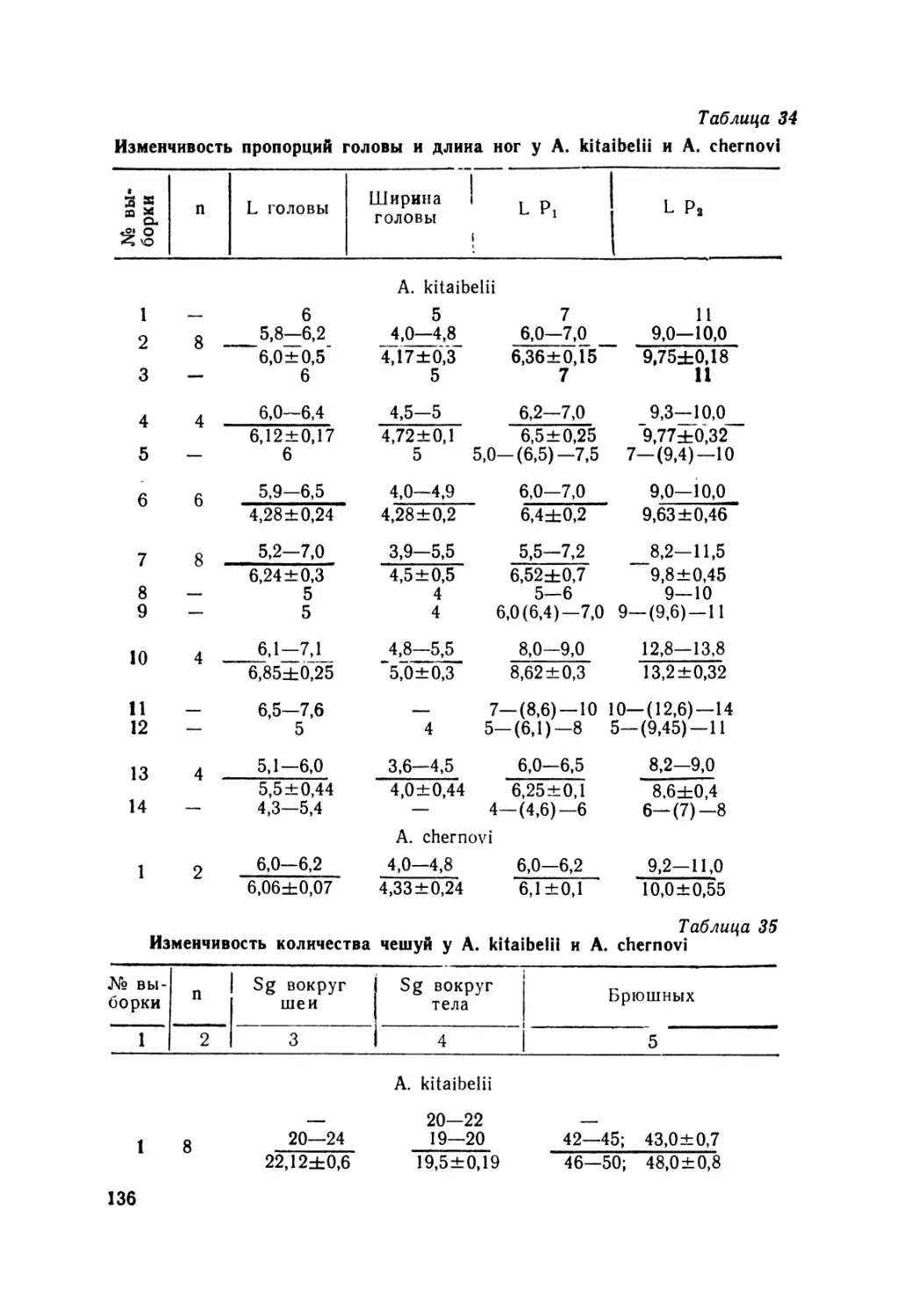
A. kitaibelii — 49; A. chernovi — 4; A. lindbergi — 2. Рода Asym-
blepharus — A. alaicus — 551. Изучены, за редким исключе-
нием, все типовые экземпляры. Кроме того, в сравнительных
целях были просмотрены родственные гологлазам сцинки po-
ll
да Scincella: S/himalayana; S. potanini; S. przsewalsky, S. la-
dacensis и некоторые другие (всего 58 экз.).
Приступая к изучению систематики и географической из-
менчивости гологлазов, прежде всего необходимо подобрать
признаки и оценить их таксономическое значение (Майр,
1971). Используя ограниченное число признаков, авторы
предшествующих исследований значительно упрощали реше-
ние вопросов систематики данной группы. Попытки найти от-
дельные диагностические признаки, не изучив достаточного
серийного материала, приводили к весьма поверхностному
суждению о признаках сходства. В один таксон нередко объе-
динялись формы, обладающие лишь внешним сходством.
Как известно (Вавилов, 1967; Шарова, 1973), низшим
близкородственным таксонам (видов, родов) часто весьма
сложно дать оценку по их сходству, поскольку оно проявляет-
ся на основе гомологических признаков.
Опыт ведущих советских специалистов-герпетологов (Да-
ревский, 1967; Яблоков, 1968, 1980; Щербак, 1974) показы-
вает, что решение указанных задач возможно популяцион-
но-морфологическим методом. Его применение позволяет
выявить границы изменчивости морфологических признаков
и на этой основе оценить их диагностическое значение. По-
добранные таким образом комплексы признаков с учетом
особенностей экологии и географического распространения
исследуемых видов используются при таксономическом анали-
зе. При выборе признаков мы обращались также к опыту сис-
тематиков, исследовавших непалеарктических аблефаридных
ящериц (Smith, 1939; Mitchell, 1950, 1958; Greer and Parker,
1967; Greer, 1970, 1974, 1979; Storr, 1973, 1976 и др.). Как бы-
ло установлено, весьма перспективным в изучении система-
тики данной группы ящериц является краниологический
метод.
Для выяснения родовой принадлежности данной группы
ящериц были подобраны следующие -признаки:
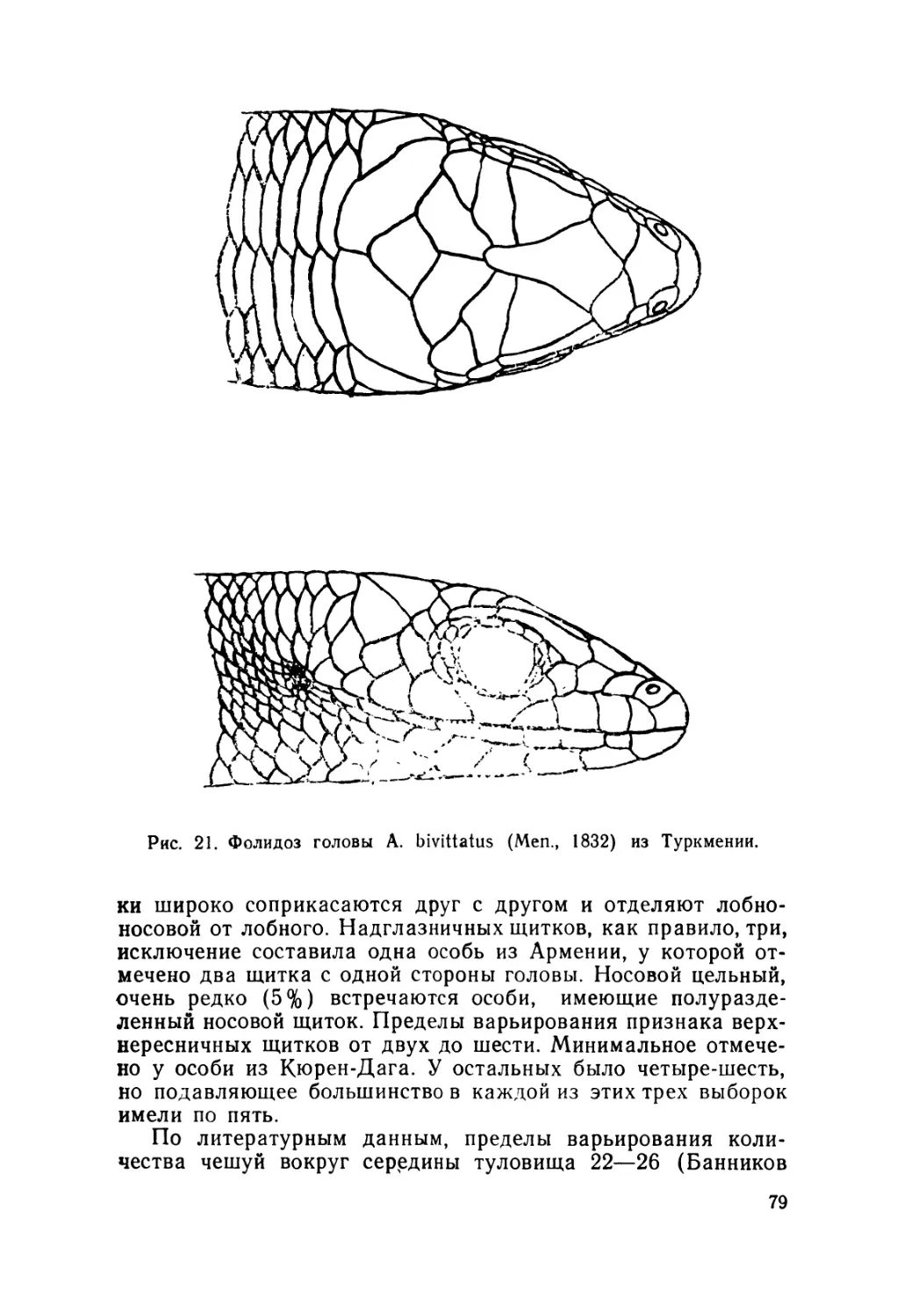
1. Наружное строение глаз. Исследовали фолидоз, окру-
жающий верхний участок глаза, и определяли степень срас-
тания верхнего и нижнего век.
2. Количество предчелюстных зубов (Premaxillare).
3. Особенности строения и расположения костей небной
рамы — эктоптеригоидов и птеригоидов — изучали у фиксиро-
ванных особей после удаления эпидермиса, покрывающего
небные кости.
4. Особенности щиткования конечностей.
5. Размеры наружного ушного прохода относительно гла-
за. Устанавливали следующим образом: голову ящерицы
слегка наклоняли набок, при этом отверстие уха становилось
12
максимально открытым. Фиксировали именно эти размеры
уха, так как нами установлено, что гологлазы улавливали
звуки, слегка повернув голову набок.
6. Характер распространения. Анализировали видовой
ареал в Палеарктике, его особенности, ландшафт, высотную
зональность и др.
7. Особенности биологии. Выделены группы ящериц по
способу размножения: яйцеживородящие или яйцекладущие.
Для составления видовых диагнозов и выявления законо-
мерностей географической изменчивости нами подобраны
следующие признаки: 1. Длина туловища — L. 2. Расстоя-
ние между передними и задними конечностями — Ga. Изме-
ряли от гленоидной впадины до ацетобулярной (Ананьева,
1977). 3. Длина нерегенерированного хвоста — Led. 4. Длина
головы — расстояние между передним краем уха и кончиком
морды. 5. Ширина головы. 6. Длина передней ноги — Lpl.
7. Длина задней ноги — Lp2. Длину передней и задней ног
измеряли от четвертого пальца до ацетобулярной или гле-
попдпой впадины соответственно. Индексы: 8. Отношение от-
резка толя, расположенного между конечностями, (Ga) к дли-
не iv 1ОПППН1 9 Отношение длины туловища к длине хвоста.
10 (h шипение длины головы к длине туловища. 11. Отноше-
нии ширины головы к длине туловища. 12. Отношение дли-
ны передней конечности к длине туловища. 13. Отношение
длины задней ноги к длине туловища. Значения индек-
сов даны в % и для удобства умножались на 100. Признаки
фолпдоза: 14. Число чешуй вокруг середины шеи.
15. Число чешуй вокруг середины туловища. 16. Число про-
дольных рядов спинных чешуй. 17. Число брюшных чешуй в
продольном ряду. 18. Число подпальцевых чешуек на чет-
вертом пальце задней ноги. 19. Число верхнересничных щит-
ков. 20. Число надглазничных щитков. 21. Число лобно-те-
менных щитков. 22. Число затылочных щитков. Подсчитыва-
ли по методике (Greer, 1974), причем первый неполный щи-
ток, часто встречающийся у гологлазов, для удобства прини-
мали за целого. 23. Число верхнегубных щитков впереди
подглазничного щитка. 24. Число щитков, расположенных
между носовым и подглазничным. Подсчитывали те, которые
касались верхнегубных щитков. 25. Число увеличенных че-
шуй верхнего «века-козырька». 26. Число височных щитков
в первом вертикальном ряду. 27. Число щитков между ухом
и центрально-височным щитком. 28. Положение предлобных
щитков относительно друг друга (раздельные, соприкасаю-
13
щиеся в одной точке, широко соприкасающиеся или между
-предлобными имеется дополнительный щиток. 29. Строе-
ние носового щитка. Он полуразделенный внизу
или цельный. 30. Расположение и форма центрально-височ-
ного щитка: он маленький и свободный сверху или относи-
тельно большой трапецевидный н сверху прижат теменным
щитком. 31. Окраска и рисунок. У ецннковых ящериц они
являются сложным элементом описания. Особое внимание
нами уделено рисунку спины ящериц, который порой выгля-
дит весьма пестрым. Наличие на спине темных и светлых
черточек и пятен и их расположение использовали в качестве
материала для построения схем рисунков. У половозрелых
особей фиксировалась брачная окраска, выраженная обычно
на нижней стороне тела. Окраска ящериц приведена по шка-
ле цветов (Бондарцев, 1954). Признаки 1, 3, 15, 18—23, 27-й
измеряли по общепринятой методике (Банников и др., 1977).
Все измерения даны в миллиметрах. Их проводили с по-
мощью циркуля, затем данные переносили па линейку и за-
писывали с точностью до 0,1 мм. Измерение мелких деталей
тела — уха, глаза, головы и просмотр фолидоза производи-
ли с помощью микроскопа МБС—9. Для указанных выше
признаков по общепринятым рекомендациям (Рокитский,
1961; Лакин, 1968) с помощью микрокалькулятора БЗ-21 и
разработанной для этой цели программы (Францевич, 1979)
подсчитаны средние значения (М), ошибка средних (ш),
квадратическое отклонение (б) и критерий достоверности
Стьюдента (t). Учеты численности ящериц проводили во вре-
мя их наибольшей активности в первую половину дня при
солнечной погоде. Учитывая неравномерность их распределе-
ния в горных условиях, мы ставили задачей выявить очаги
поселений, где производили учет ящериц на линейном марш-
руте 500 ил и Л ООО мХ4 м. Характер суточного цикла ящериц
и сезонной активности изучали по известным методикам (Да-
ревский, 1967; Щербак, 1974). Размножение ящериц A. alai-
cus, A. deserti изучали в полевых условиях, а также при их
полувольном содержании на специально отведенном для этой
цели месте (фиксировали время откладки яиц, порционность,
появление новорожденных, их количество). Линьку регистри-
ровали у добытых экземпляров, а ее характер — путем изу-
чения линных шкурок от ящериц, содержащихся в террариу-
ме, а также собранных в природе.
Данные по питанию видов основаны на материале иссле-
дований содержимого желудочно-кишечного тракта и наХэлю-
дедий за ящерицами в террариуме (A. alaicus и A. deserti).
14
РОДОВАЯ ПРИНАДЛЕЖНОСТЬ
АБЛЕФАРИДНЫХ ЯЩЕРИЦ
ФАУНЫ СССР И СОПРЕДЕЛЬНЫХ СТРАН
Первичный диагноз рода Ablepharus (Fitzinger in Lich-
tenstein, 1823) содержит два признака, характерных для
включенных сюда видов: неподвижные прозрачные веки и пя-
типалые конечности. Позже А. Штраух (Strauch, 1868), ре-
визовавший данный род, подчеркивал значение признака пя-
типалых конечностей, а Г. Буленже (Boulenger 1887) особое
значение придавал признаку «неподвижный прозрачный диск
покрывает глаз». Он и отнес к данному роду представителей
10 ранее известных родов, распространенных в Евразии, Аф-
рике, Мадагаскаре, Австралии и Полинезии. В 1901 г.
В. С. Елпатьевский описал новый вид A. alaicus, который,-
как казалось автору, отвечал приведенному родовому диаг-
нозу.
М. Смитом (Smith, 1935, 1937, 1939) было показано, что
ряд южноазиатских видов, включенных в род Ablepharus,
имеют несросшиеся веки и палпебральную щель (palpebral
slit), которые скрыты под ресничной частью фолидоза (su-
percilium) и поэтому оставались ранее незамеченными. Он
высказал предположение о полифилитическом происхожде-
нии аблефаридных ящериц и счел характерным для предста-
вителей рода Ablepharus полное или почти^полное срастание
век. Многие другие также широко понимали объем этого ро-
да (Parker, 1936; De Witt, 1936; Mittleman, 1952 и др.). Позд-
нее и Фун (Fuhn, 1969а) обнаружил палпебральное углубле-
ние у европейских видов. Он полагал, что оно является гра-
ницей срастания верхнего и нижнего века. Однако, как было
выяснено нами, палпебральное углубление имеется и в том
случае, когда верхнее и нижнее веки не срастаются, т. е. оно
существует независимо от срастания. На основании изучения
краниологических признаков И. Фун показал автохтонное
происхождение евразиатских видов и рамки рода Ablepha-
rus сузил до пяти видов — «bivittatus», «pannonicus», «deser-
ti», «kitaibelii» и «borealis», исключив из него африканские
и австрало-полинезийские формы. Это было принято А. Гри-
15
ром (Greer, 1974) и с небольшими изменениями, внесенными
Банниковым и др., 1977.
Как указывалось, некоторые зарубежные герпетологи (Mit-
tleman, 1952; Wettstein, 1960; Mertens, 1965; Fuhn, 1969a, в;
Greer, 1974) сомневались в видовой самостоятельности A. alai-
cus и рассматривали его в качестве подвида A. bivittatus.
Подобные взгляды в работах отечественных зоологов (Ни-
кольский, 1915; Параскив, 1956; Чернов, 1959; Богданов,
1960; Яковлева, 1964; Банников и др., 1977) отсутствуют.
Это обстоятельство побудило нас детально исследовать
A. alaicus. Работа привела к весьма важным выводам, свя-
занным с более широким кругом вопросов таксономии абле-
фаридных ящериц. Мы исследовали признаки, имеющие глав-
ное значение при выяснении родовой принадлежности абле-
фаридных ящериц и установили следующие особенности.
Наружное строение век. Изучение наружной морфологии
глаза показало, что в отличие от других видов у A. alaicus
нижнее и верхнее веки не сросшиеся. Внешне это почти не-
заметно, так как верхний край нижнего века обычно скрыт
под увеличенными щитками верхнего века-козырька. Разме-
ры щели между краями век, как правило, равны длине уве-
личенных щитков.
В соответствии с терминологией А. Грира (Greer, 1974)
глаза сцинковых ящериц подсемейства Lygosominae, имею-
щих полностью сросшиеся веки, утратившие таким образом
подвижность, следует называть аблефаридными, а не пол-
ностью — преаблефаридными. Следоватёльно, аблефаридная
стадия глаз свойственна всем видам данного рода, за исклю-
чением A; alaicus, у которого глаза имеют преаблефаридную
стадию, близкую к таковой у представителей родов Scincella
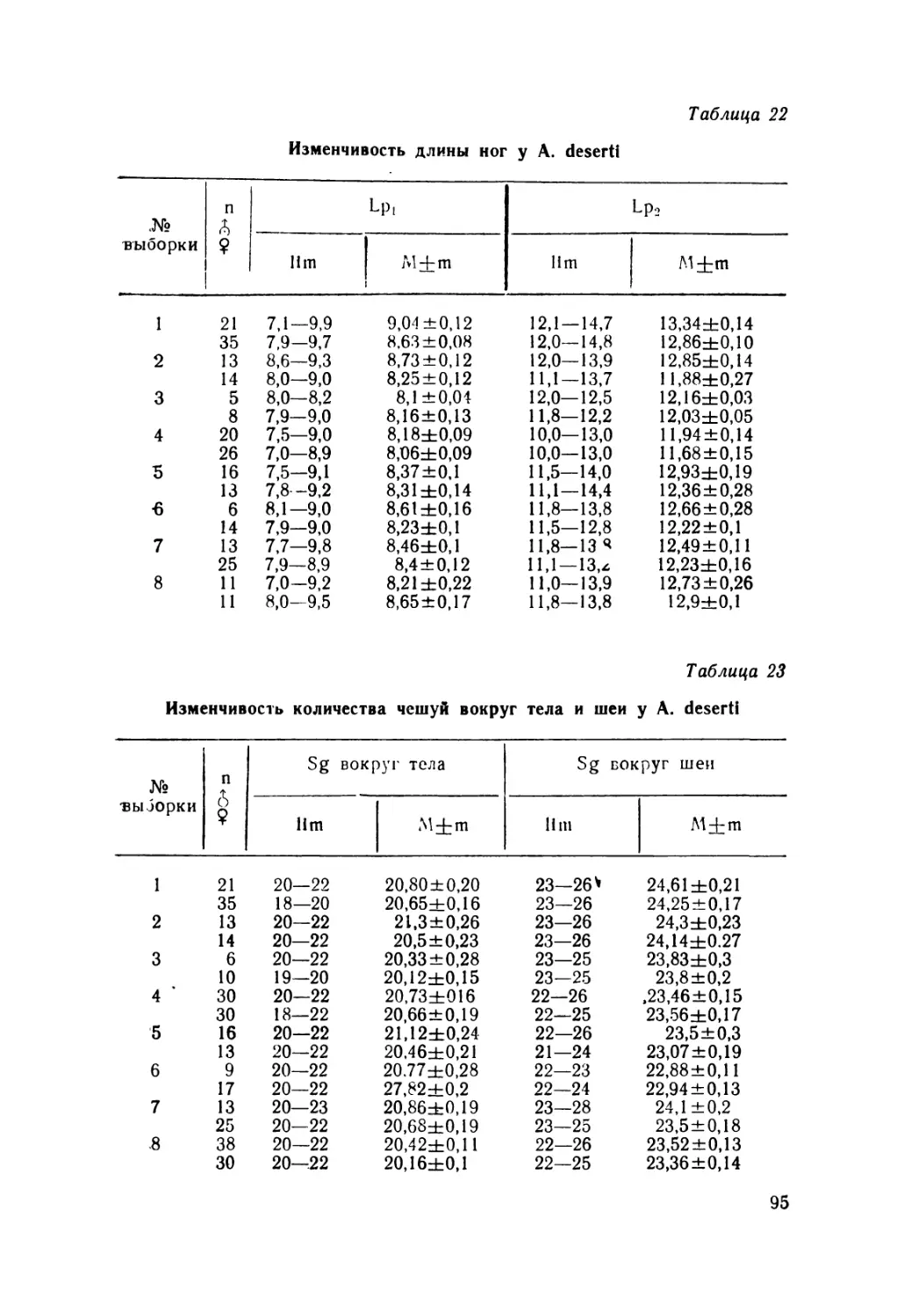
Mittleman, 1950 и Leiolopisma Dumeril et Bibron, 1839 (рис. 1).
Внешнее сходство глаз представителей рода Ablepharus и
A. alaicus возникло независимо, что подтверждается анали-
зом краниологических признаков.
Число премаксиллярных зубов. У изученных нами пред-
ставителей рода Ablepharus обнаружено по девять премак-
силлярных зубов, только у A; alaicus число равно семи. Как
известно (Greer, 1970, 1974), наличие девяти премаксилляр-
ных зубов свойственно группе лигозомных сцинков, состоя-
щей примерно из 68 видов и объединенных в 5 родов: Scin-
cella, Ablepharus, Lobulia, Lipinia и Prasinohaema. Две дру-
гие группы лигозомид, включающие около 167 видов и 17 ро-
дов, характеризуются наличием 11 предчелюстных зубов. Иск-
лючением является род Carlia, представители которого имеют
13 таких зубов. Таким образом, число премаксиллярных зу-
бов— важный таксономический признак.
16
1
Рис. I. Внешний вид глаз некоторых лигозомных сцинков: а. б. в—
преаблефаридный тип (а—Scincella lateralis, б—Leiolopisma spence-
ri, в—Asymblepharus alaicus), г, д—аблефаридный тип (г—Morethia
lineoocellata, д—Ablepharus deserti). I — глазная щель между несрос-
шпмпся верхним п нижним веками; 2—палпебральное углубление
между краем верхнего века и верхнересничными щитками.
Рис. а, б, г даны по Greer, 1974.
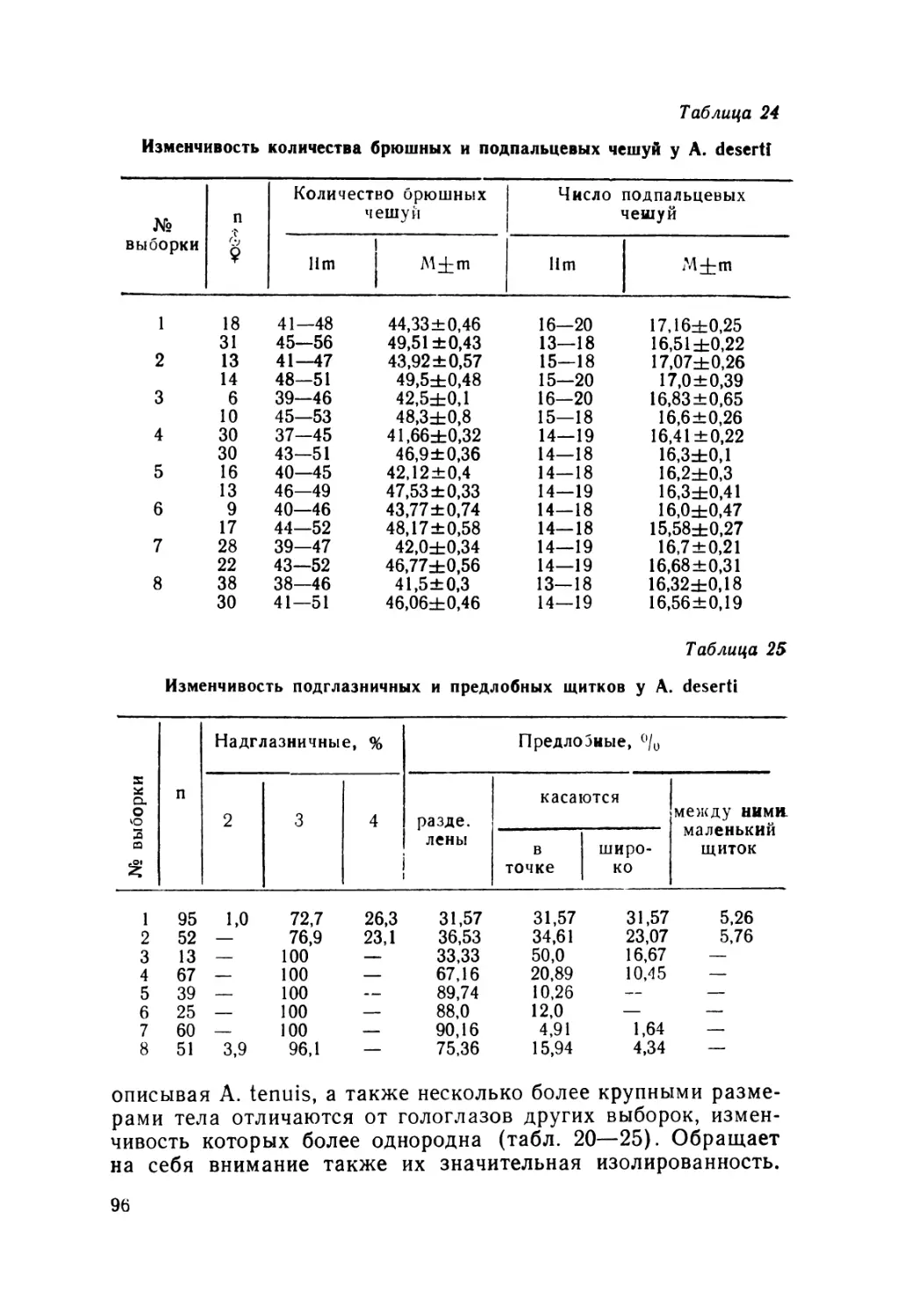
Особенности небных костей. В строении небных костей
представителей рода Ablepharus прослеживаются две тенден-
ции: а) рост птеригоидов и отставание роста эктоптеригоидов
по внутреннему краю орбит, небные кости при этом соприка-
саются (рис. 2, б, в) у A. deserti, A. pannonicus, A. kitaibelii,
A. bivittatUs и A. grayanus; б) рост эктоптеригоидов и отста-
вание роста птеригоидов по внутреннему краю орбит, небные
кости не соппикасаются (рис. 2, а), что характерно для
Рис. 2. Особенности строения костей небной рамы: a—Asymblepharus
alaicus, б —Ablepharus deserti, в — A. pannonicus, A. kitaibelii; Р —
palatinum, Pt — pterigoidum, E — ectopterigoidum.
2 302
17
A. alaicus. Следует отметить, что вторая тенденция в неко-
торой степени наметилась и у A. bivittatus, однако исключи-
тельность по последнему признаку A. alaicus вне всякого сом-
нения.
Выявленные нами особенности расположения небных кос-
тей, птсригоидов и эктоптеригоидов имеет важное таксоно-
мическое значение. Сходные с вышеописанным нами строе-
нием этих костей приводит в своей работе и Митчелл (Mit-
tchell, 1950), впервые указавший на различия этих признаков
у австралийских сцинков родов Tiligua и Egernia.
Фун (Fuhn, 1969а) включил в диагноз рода Ablepharus
следующую краниологическую характеристику: на небных
костях нет задних отростков, отделяющих птеригоиды; неб-
ные отростки птеригоидов не встречаются; изогнутые отсут-
ствуют. Однако, как позже выяснилось (Greer, 1974), это
свойственно лигозомным сцинкам пяти родов: Ablepharus,
Scincella и др.
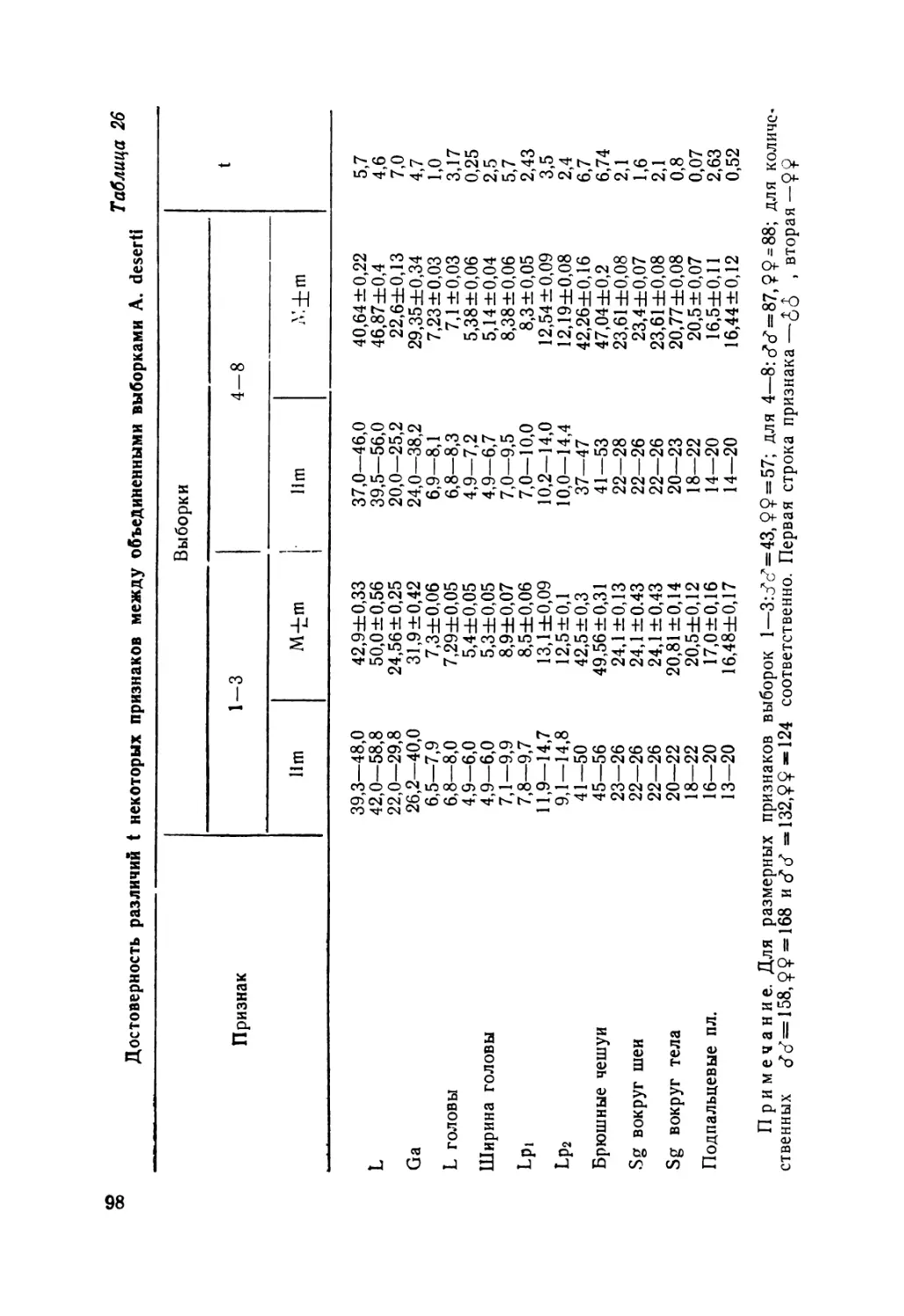
Особенности фолидоза конечностей. Таксономическая цен-
ность 9£обенностей чешуйчатого покрова конечностей у сцин-
ковых ящериц хорошо известна (Smith, 1937; Brongersma,
1942). По характеру щиткования внешней стороны пред-
плечья и голени A. alaicus и A. lindbergi резко отличается
от всех других форм, отнесенных к данному роду. Только у
этих видов имеются здесь сильно расширенные щитки
(рис. 3).
Рис. 3. Щиткование внешней стороны пред-
плечья: а—Asymblepharus, alaicus б—Able-
pharus bivittatus, в—A. deserti, г—A. pan-
nonicus, д—A. kitaibelii.
Ушное отверстие. Для A. alaicus характерно относительно
крупное ушное отверстие (его длина укладывается в диамет-
ре глаза не более четырех, трех раз), в то время как это от-
ношение у других видов составляет от 5 до 20 раз. У A. pan-
nonicus и A. chernovi отверстие уха очень мало и почти скры-
то под чешуей, а у A. grayanus оно исчезает совсем. Размер
18
Таблица 1
Сравнение родов Scincella, Ablepharus, Asymblepharus
Признак Scin- cella Ablepha- rus Asymble- pharus
Тип глаза:
преаблефаридный аблефаридный + + +
Число премаксиллярных зубов Преобладание роста: 9 9 7
эктоптеригоида — 1 +
птеригоида Положение небных костей друг к другу: + + —
касаются + + —
не касаются На предплечье и голени имеются — — +
сильно расширенные щитки Длина ушного отверстия укладывает- ся в диаметре глаза: 2 +
не более 4 раз + — +
более 7 раз пли отсутствует Особенности распространения: на запад и юго-запад от Тянь-Шаня — + —
и Памиро-Алая — + —
Тянь-Шань и Памиро-Алай на восток и юго-восток от Тянь- — — +
Шаня и Памиро-Алая Особенности размножения: + — —
яйцекладущие — + —
яйцеживородящие + — +
ушного отверстия как таксономический признак широко ис-
пользовали в работах многие герпетологи (Smith, 1935, 1939;
Чернов, 1959; Даревский, 1953; Fuhn, 1969 а, в и др., Greer»
1974).
Особенности распространения. Ареал A. alaicus располо-
жен восточнее ареалов всех других видов и не выходит за
пределы гор Тянь-Шаня и Памиро-Алая, где данная ящерица
обитает на высотах 800—4000 м над ур. м1 2 3 (Яковлева, 1964;
наши наблюдения). Ареалы всех других видов охватывают
Балканы, Переднюю Азию и доходят до гор Тянь-Шаня и Па-
миро-Алая, причем все они высоко в горы не поднимаются.
1 У A. bivittatus наблюдается рост эктоптеригоида, а небные кости
у 40% особей не соприкасаются.
2 Имеются только у A. lindberg 1
3 Здесь и далее приведена высота над уровнем моря.
19
Алайского гологлаза можно считать автохтоном указанных
горных систем, что дает возможность предположить независи-
мое происхождение алайского и всех других евразиатских
гологлазов.
Особенности размножения. A. alaicus, в отличие от всех
видов евразиатских гологлазов, является яйцеживородящим.
Эта особенность впервые была установлена В. Н. Шнитни-
ковым (1928). Выводок у A. alaicus состоит из одного-семи
детенышей (Яковлева, 1964; наши данные). Следует отметить,
что яйцеживорождение свойственно и многим ящерицам ро-
да Scincella, ареал которых, по-видимому, граничит на севе-
ре с ареалом A. alaicus. Последнее обстоятельство определи-
ло наш выбор объектов для сравнения (табл. 1).
Как следует из данных таблицы, по ряду признаков (на-
личие девяти премаксиллярных зубов и др.) роды Ablepharus
и Scincella имеют сходство, но отличаются от A. alaicus. На
обособленное положение «alaicus» указывал и известный со-
ветский герпетолог С. А. Чернов (1959), который писал:
«Филогенетические отношения и связи A. alaicus в настоящее
время неясны; во всяком случае он резко отличается от всех
остальных палеарктических видов рода...». Наши материа-
лы подтверждают эту точку зрения. По нашему мнению, вид
«alaicus», несомненно, относится к особому роду, который
характеризуется рядом примитивных черт (например, преаб-
лефаридные глаза), и связан в своем происхождении, вероят-
но, с родом Scincella Mittleman, 1952.
Новый род был назван нами Asymblepharus (от греческого
asymphio — несросшийся и blepharis — веко).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
ABLEPHARUS И ASYMBLEPARUS
2(1). Глаза аблефаридные, веки полностью сросшиеся;
зубов на премаксиллярных костях — 9... Ablepharus.
1(2). Глаза преаблефаридные, края верхнего и нижнего
века несросшиеся, скрыты под увеличенными щитками верх-
него века; зубов на премаксиллярных костях — 7... Asymb-
lepharus.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
РОД АСИМБЛЕФАРЫ —
ASYMBLEPHARUS JERIOMTSCHENKO ET SZCZERBAK,
1980
Типовой вид — Asymblepharus alaicus (Elpat., 1901).
Син.: Ablepharus (Fitzinger in Lichtenstein, 1823). Part.
Глаза преаблефаридные — края верхнего и нижнего век
несросшиеся и скрыты под увеличенными щитками верхнего
века-козырька; на предчелюстной кости (premaxillare)
семь плевродонтных зубов. Конечности пятипалые, разви-
ты относительно хорошо; на внешних сторонах предплечий и
голеней два-восемь (обычно три-пять) сильно расширенных
щитков. Наружный ушной проход хорошо выражен, его дли-
на укладывается в размерах глаза не более четырех раз; вок-
руг середины туловища расположено 24—30 продольных ря-
дов гладких чешуй. Особенность размножения — яйцеживоро-
дящие. Род включает два вида: 1. Рецентный — A. alaicus
(Elpat., 1901), широко распространенный в горных системах
Тянь-Шаня и Памиро-Алая на высотах 800—4000 м. 2. Вы-
мерший— A. borealis (Darevsky et Tchumakov, 1962), извест-
ный из нижнего плейстоцена Рудного Алтая.
АСИМБЛЕФАР АЛАЙСКИЙ —
ASYMBLEPHARUS ALAICUS (ELPAT., 1901)
Terra typica: Алай, южный склон Алайского хр. окрестности
с. Дараут-Курган (Киргизия).
1901 — Ablepharus alaicus, Е л п а тьевски й. Дневн. зоол. отд. Об-
ва любит, естествозн., 3 : 37. — 1905, Никольский. Преем,
и земн. Росс, импер., 12,1 169. Ежегодн. зоол. —1915, Ни-
кольский. Фауна Росс., Преем., 2:495, — 1928, Ш нит-
ник о в. Преем. Семиречья, 8,3:199. — 1935, Чернов.
Ящерицы (Sauria) Таджикистана. Тр. Тадж. базы АН СССР,
5:469. — 1951, Андрушко. Зоол. ж. 30,3:276. — 1949,
Терентьев и Чернов. Опред. преем, и земн. :200. —
1956, Па раски в. Преем. Казахстана: 102. — 1959. Чер-
нов. Фауна Тадж. ССР Преем., 18 : 98. — 1960, Богданов.
Земн. и преем. Фауна Узбек. ССР, 1:145. — 1964, Яков-
лева. Преем. Киргизии: 72. — 1971, Банников и др.
Земн. и преем. СССР: 141. — 1977, Банников и др.
21
:155. — 1979, Саид-Алиев. Земн. и преем. Таджикиста-
на :91.
1902 — Ablepharus kucenkoi, Никольский. Ann. Mus. Zool. Acad.
Sci. Petersb.: 7. —1908, Никольский. Ежегодн. Зоол.
музея АН, 13:340. —1915, Никольский. Фауна Росс.
Преем., 2 : 497.
1909 — Ablepharus sapozshnikovi, Кащенко. Ежегодн. Зоол. му-
зея АН, 13:126.
1927 — Ablepharus alaicus kulzchenkoi, Кашкаров. Изв. Ср. —
аз. комиссии по делам музеев и охраны памятников стари-
ны, 2 : 76.
1969 — Ablepharus bivlttatus alaicus (Elpat., 1901). Fuhn. Rev.
roum. Biol. ZooL, 14,1 : 23. 1969, в, Fuhn, Sound, a. Z. f.
Zool. Syst. u. Evolut., 7:69. — 1974, Greer, Austral. Journ.
of Zool., 31 : 9.
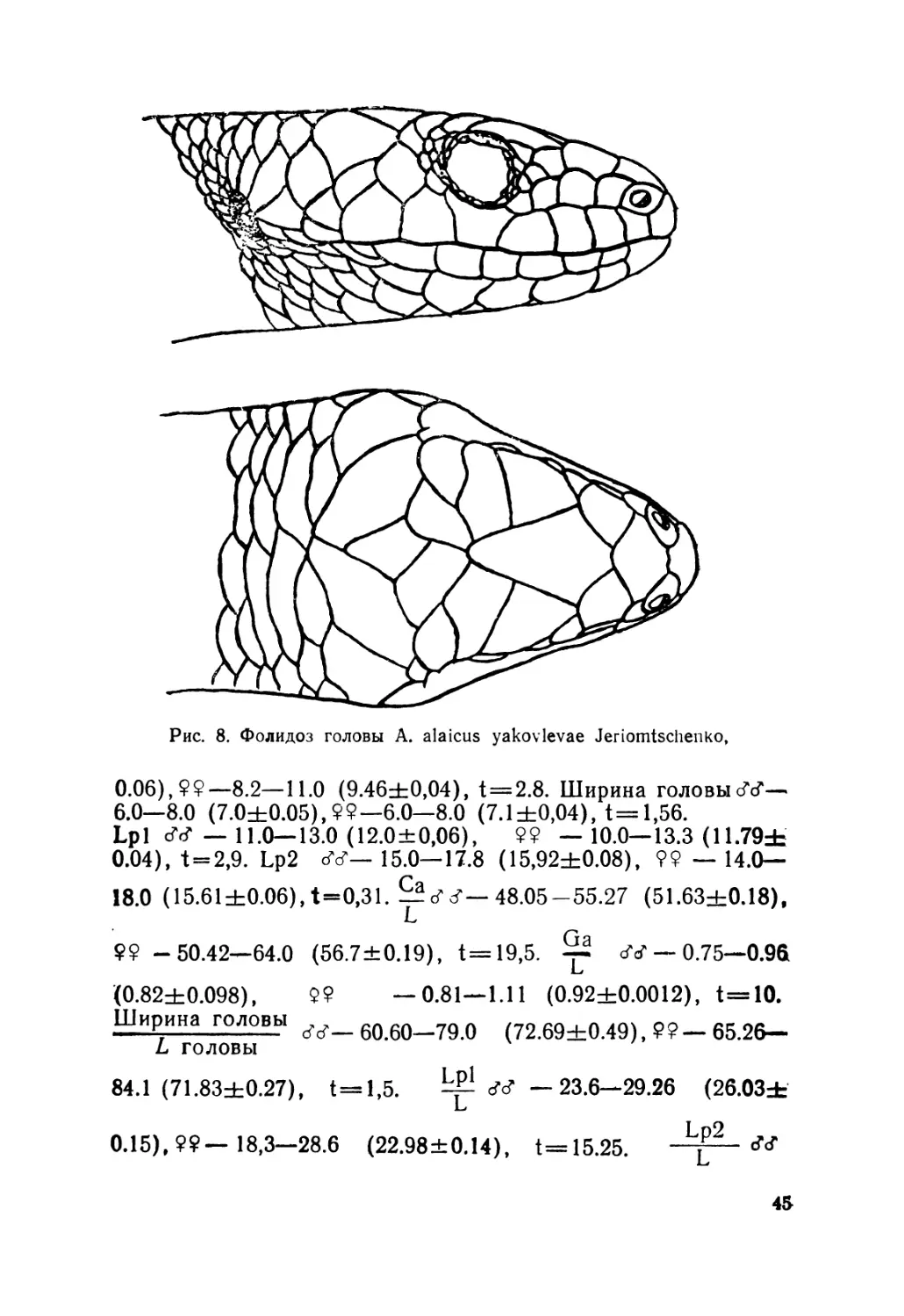
1980 — Asymblepharus alaicus (Elpat., 1901), Еремченко, Щер-
бак, Вести, зоол., 4:14. —1981, Еремченко. Автореф.
докл. 5 Всесоюзн. герпет. конф.: 54. — 1981, Jeriomtschen-
k о, 1 Conf. Негр. Resp. Soc., Budapest: 21.
Диагноз. Ящерицы длиной тела до 65 мм, имеют прерван-
ный ряд верхнересничных щитков (впереди их один-три, ча-
ще два) и хорошо развитый козырек верхнего века, состоя-
щий из трех-пяти увеличенных щитков; вокруг середины те-
ла 24-30 чешуй; на спине они расположены четырьмя продоль-
ными рядами.
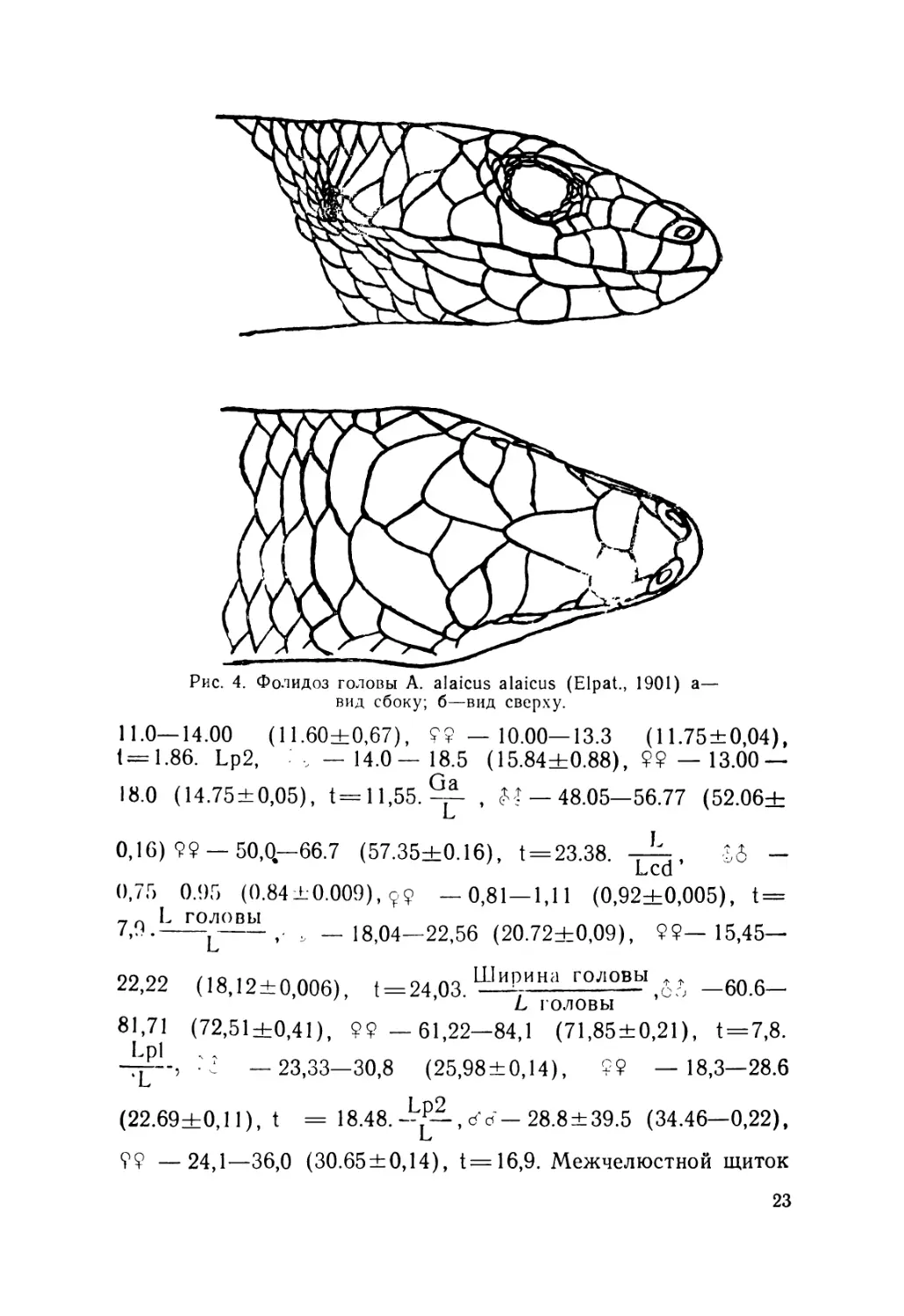
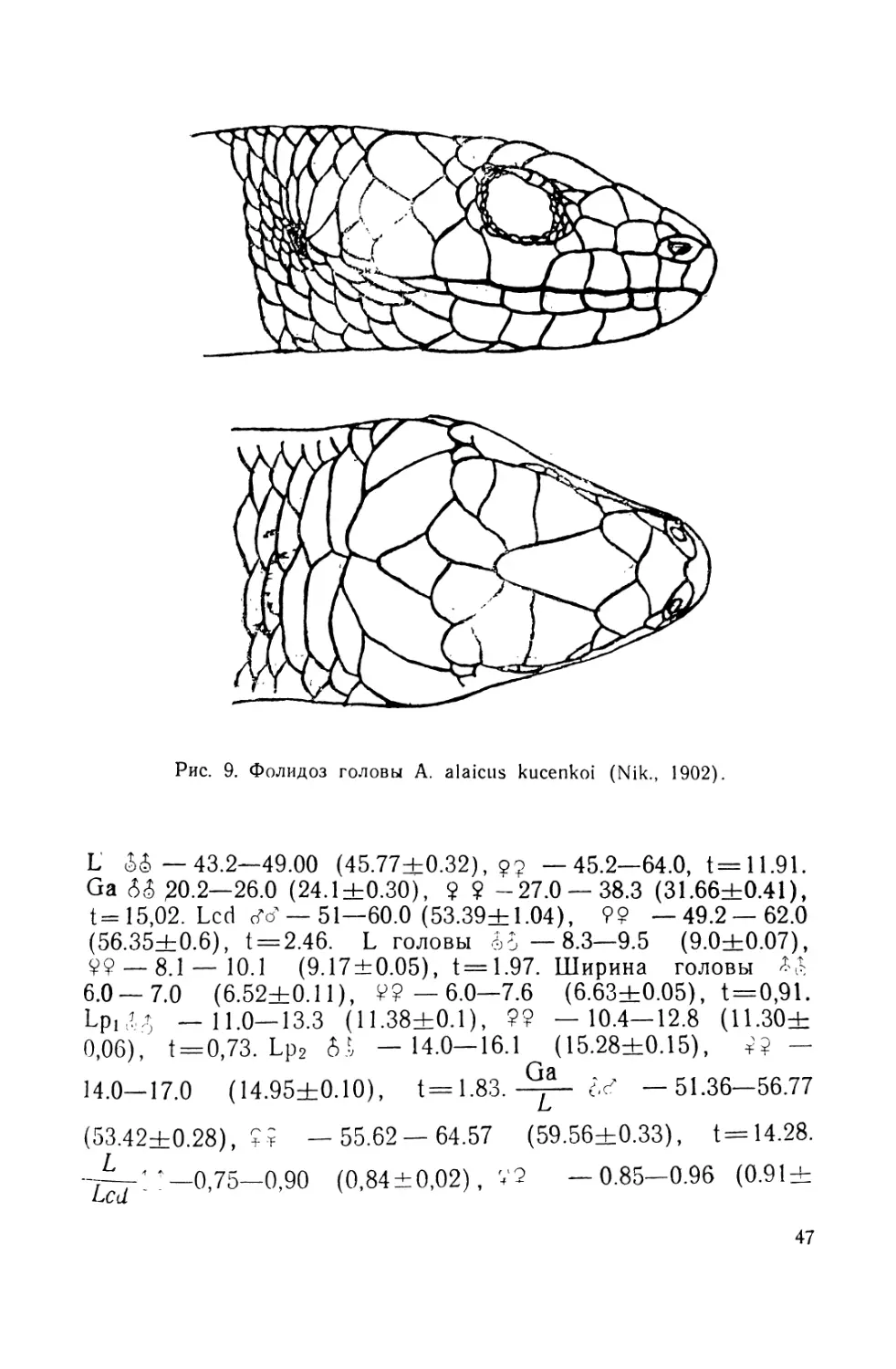
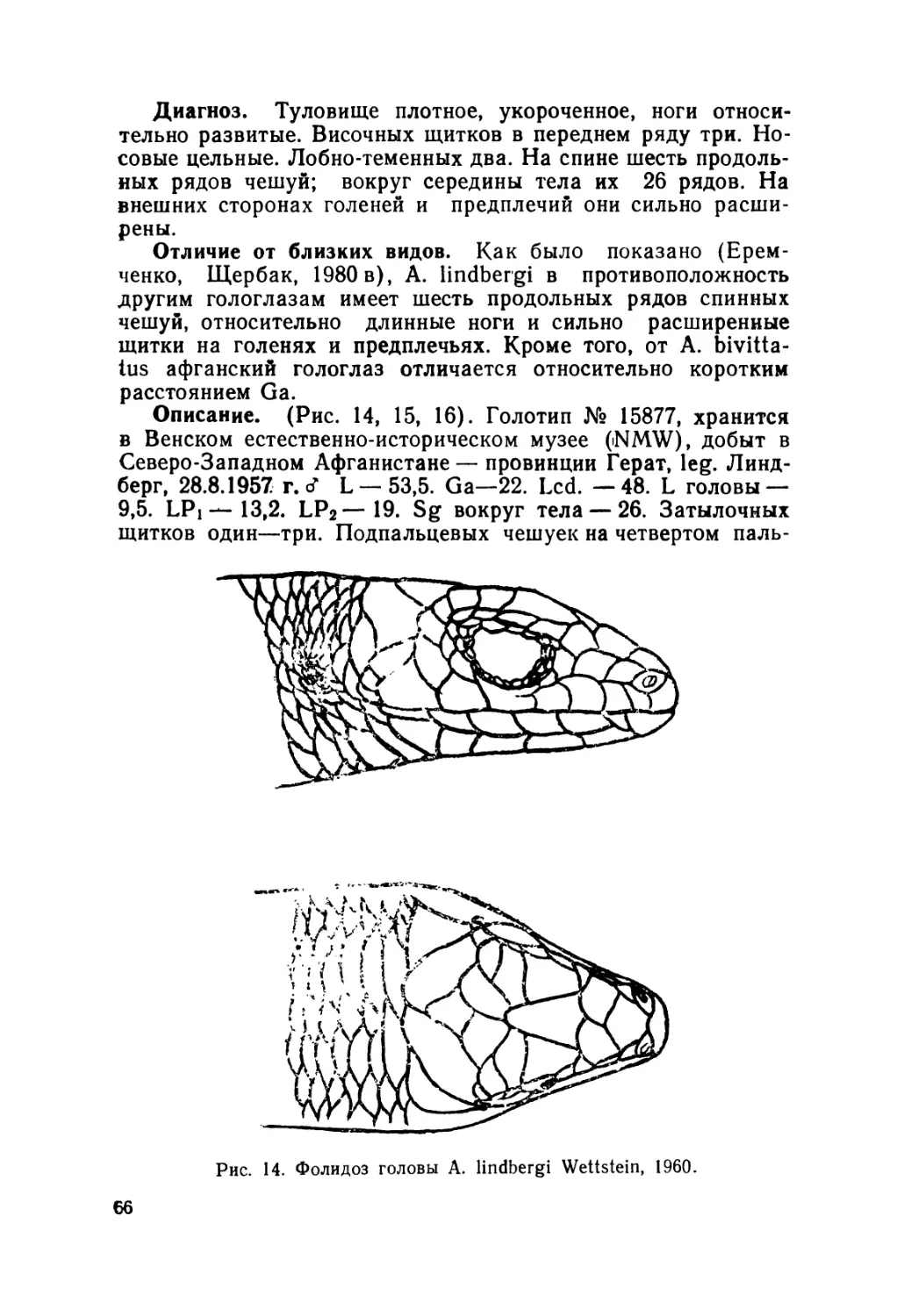
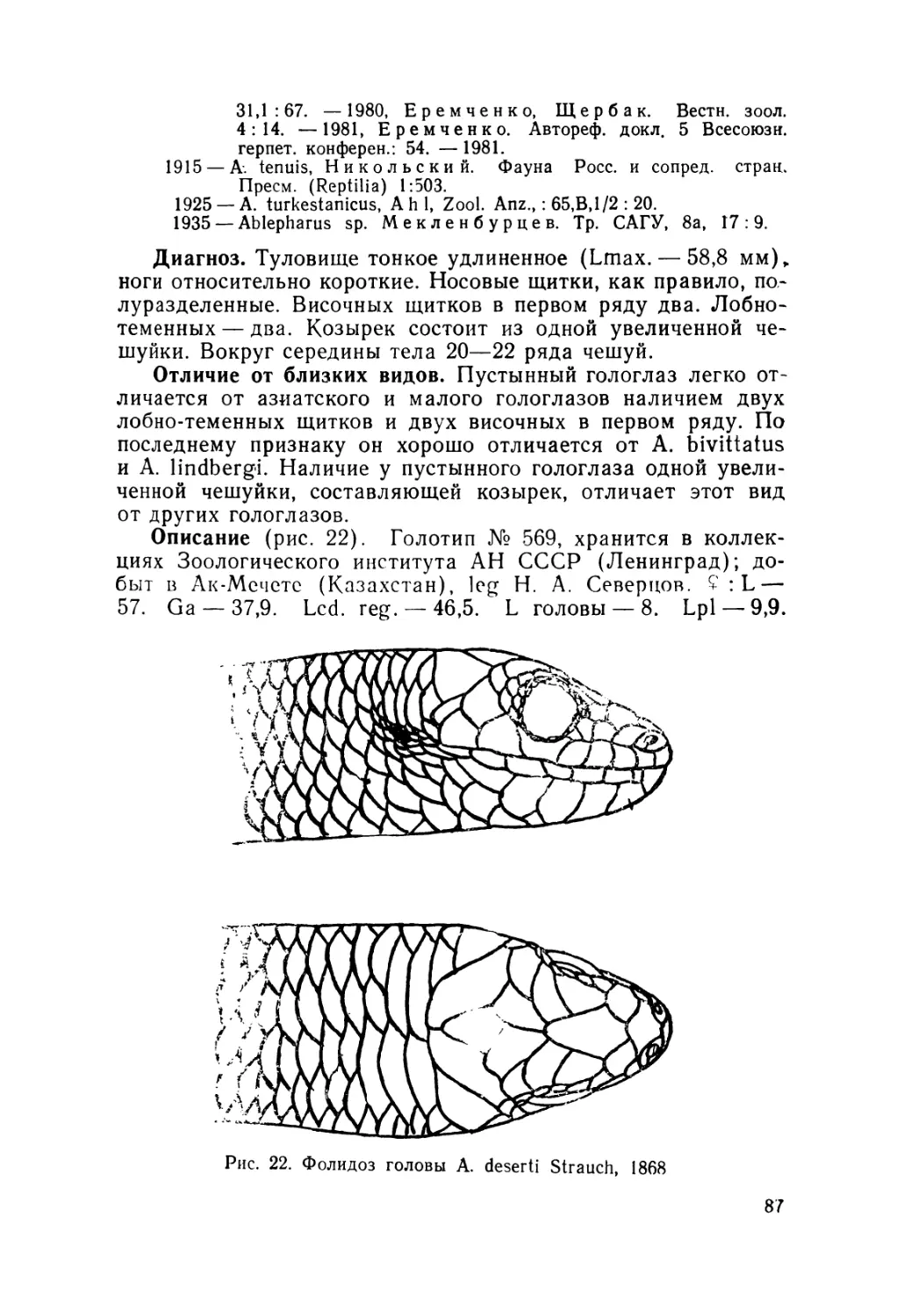
Описание (рис. 4). Лектотин № 2248 выделен нами из ти-
повой серии (всего девять экземпляров), хранящейся в зоо-
музее Московского государственного университета; добыт в
Алае на южном склоне Алайского хребта в окрестностях
с. Дараут-Курган (Киргизия),leg. Н. Н. Назаров, 1892 r.9L—
49,2. Ga — 27. L cd. reg. — 29. L головы — 9. Ширина голо-
вы— 6. Lpl — 12. Lp2—15,7. Sg — 26. Затылочных щитков
2" 1—2 Подпальцевых чешуек на четвертом пальце Р2—17.
Предлобные срослись в один крупный щиток и отделяют
лобно-носовой от лобного. Носовые щитки цельные.
Видовое описание составлено нами по экземплярам (551)
из Киргизии, Казахстана, Узбекистана, Таджикистана и
КНР, собранным авторами, а также хранящимся в коллек-
циях ЗИН АН СССР, ИЗ АН УССР, ЗМ МГУ, ИЗиП АН
КазССР, АН ИБ КиргССР и Чикагского естественно-исто-
рического музея (FMNH). L, dV —40.5—52.0 (46,0+0,20),
99— 40—64,8 (51,9=4=0,27), t= 17,56 GA. ;+ — 20.2-27.2
(23,9+0,15), 99 — 23.0—38,3 (29,9±0,21), t = 23.25. Led.
—46.0—61,2 (54,0±0,55), 99— 46.0—70.5 (55.72+0,45),
t=2.42. L головы, —8.3—11.0 (9,52±0.05), 99 —8.1 —
11 (9.4+0.03), t = 2.06. Ширина головы, бб —6.0—8.0
(6.9±0,05), 99 —7.0—8.0 (6.77+0,01), t = 2.25. Lpl, SS-
22
Рис. 4. Фолидоз головы A. alaicus alaicus (Elpat., 1901) а—
вид сбоку; б—вид сверху.
11.0—14.00 (11.60+0,67), 79 — 10.00—13.3 (11.75 + 0,04),
1 = 1.86. Lp2, , — 14.0— 18.5 (15.84+0.88), 99 — 13.00 —
18.0 (14.75 + 0,05), t=ll,55.-Д , сЧ — 48.05—56.77 (52.06±
0,16)99 — 50,0—66.7 (57.35+0.16), t = 23.38. -Д, 66 -
Led
0,75 0.95 (0.84 + 0.009), 99 —0,81—1,11 (0,92+0,005), t=
7,9.—ловы , — 18,04—22,56 (20.72+0,09), 99- 15,45—
2 2,22 (18,12 + 0,006) t = 24 03 голо^ 66 —60.6—
L головы
81,71 (72,51+0,41), 99 —61,22—84,1 (71,85 + 0,21), 1=7,8.
-6 —23,33—30,8 (25,98 + 0,14), 99 — 18,3—28.6
(22.69+0,11), t = 18.48.-£—,<//-28.8+39.5 (34.46—0,22),
99 —24,1—36,0 (30.65 + 0,14), t=16,9. Межчелюстной щиток
23
коротким швом касается лобно-носового. Надглазничных три
(очень редко один-два = 0,55%), два из них соприкасаются
с лобным и козырьком верхнего века. Лобно-теменных щит-
ков два. Межтеменной один, по размерам равный или нес-
колько меньше одного лобно-теменного. Теменные щитки
смыкаются узким швом позади межтеменного. Первый ряд
височных состоит из двух щитков (менее 0,5%—из трех).
На переднем крае ушного прохода расположены один-пять
(обычно три-четыре) мелких зубовидных чешуек. Sg вокруг
шеи —26—32 (28,67±0,11), —26—33 (28,45±0,07),
t= 1,68. Sg вокруг тела — 24—30 (26,53±0,09), 99 -24—
30 (26,3±0,06), t = 2,12. Брюшных чешуй в продольном ряду
—34—50 (42,83±0,22), 99 — 38—53 (45,28±0,15), t =
12,95. Затылочных 1- j5 : I - 15%,1 — 37%, 2—12,8%,
12= 18,3%, 3=36,8%, 1з =15,9%, 4=9%, J 4=1,6%,
Z Z
5= '0,2%,^5 = 0,2% • Подпальцевых чешуек на четвертом паль-
це р2е4 —15—21 (17,81±0,10), 99 — 15—23 (17,86±0,07),
t=0,41. Передних верхнересничных 1—3: 1 = 5,2%, 2=94,6%,
3=0,2%. Количество чешуй верхнего века-козырька 1—6 : 1 =
1%, 2 = 3,5%, 3 = 49.5%, 4 = 38%, 5 = 7%, 6=1%. Впереди
подглазничного обычно четыре верхнегубных щитка, редко
(0,8%) пять. Между носовым и подглазничным щитками
расположено три — пять щитков: 3—7,6%, 4—84,6%, 5—7,8%.
Предлобных щитков два, они разделены вклинивающимся
между ними передним углом лобного щитка — 65%, сопри-
касаются в одной точке— 13,7%, соприкасаются широким
швом и отделяют лобно-носовой щиток от лобного — 21,3%.
Центрально-височный щиток двух типов: 1. Малень-
кий округлый и свободный сверху —70%/, 2. Большой
трапециевидный, более или менее округлый позади и сверху
прижат нижним краем теменного щитка — 30%. Носовой щи-
ток двух типов: 1. Цельный — 75,7%; 2. Полуразде-
ленный — 24,3%. Впереди клоакальной щели два боль-
ших щитка. Хвост снизу покрыт одним рядом сильно расши-
ренных щитков. Основной фон тела сверху буроватый с олив-
ковым, бронзовым или зеленоватым оттенком. Рисунок спи-
ны трех типов: 1. Тип А — крупнопятнистый, найден у 16,0%
особей; 2. Тип Б — поперечно-полосатый, найден у 60,5% ис-
следованных экземпляров; 3. Тип В — продольно-полосатый,
найден у 23,5% особей. От ноздри через глаз и далее по бокам
туловища проходят широкие бурые полосы, испещренные свет-
24
лыми черточками и окаймленные по верхнему краю узкой
беловатой линией. Нижняя сторона тела беловатая, хвос-
та—сероватая или голубоватая. У половозрелых самцов с
конца марта по сентябрь нижняя сторона тела (кроме голо-
вы) ярко-оранжевая или кирпично-красная. У самок эта
часть тела обычно беловатая или с бледно-розовым оттен-
ком. Вместе с тем часть самок (17,5%) имеет окраску ниж-
ней стороны тела, характерную для самцов. Молодые по ри-
сунку и тональности окраски верхней стороны были сходны
с взрослыми, но нижняя сторона тела беловатая, а хвост, осо-
бенно у сеголеток, серо-голубой, синеватый или буровато-си-
ний.
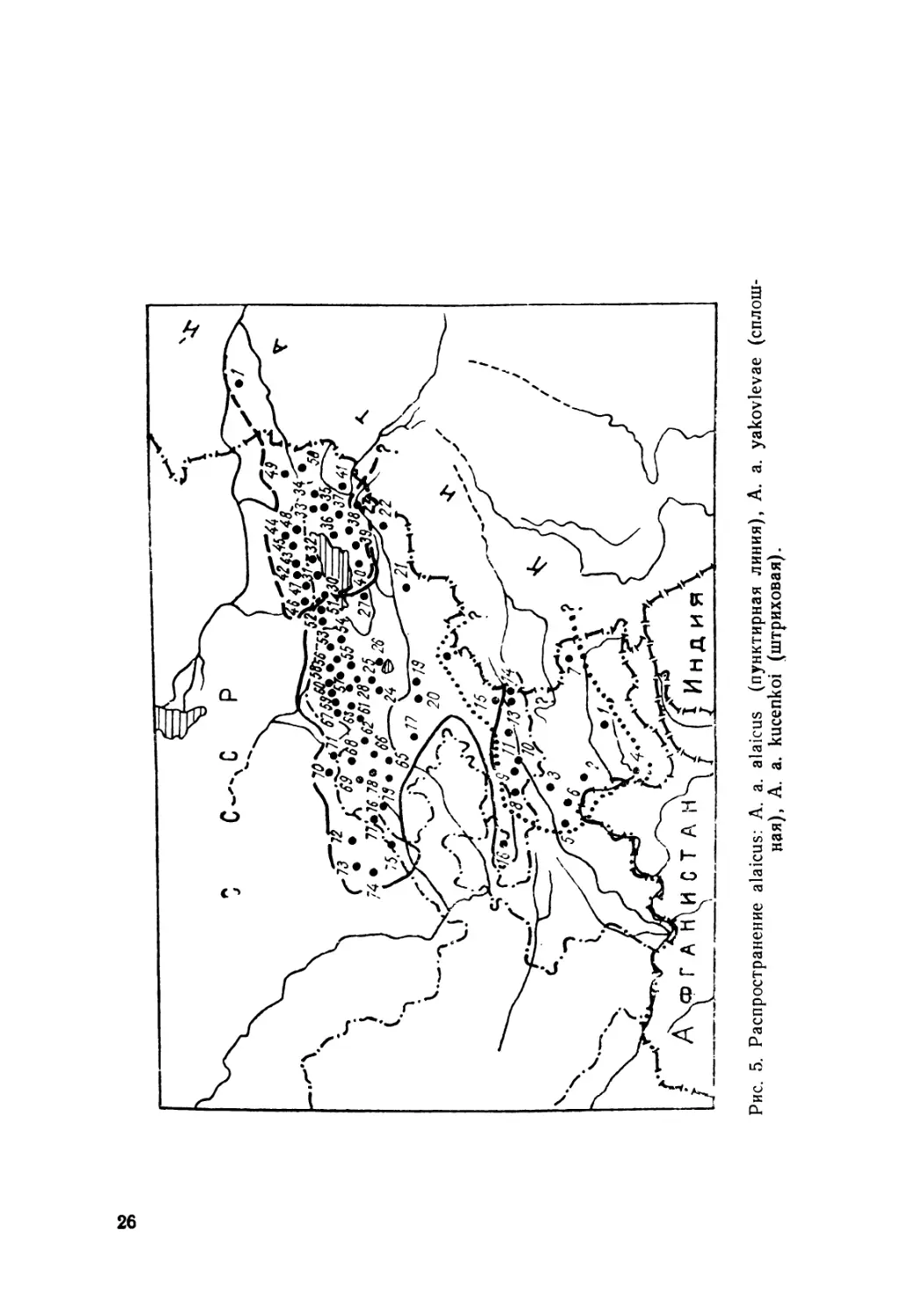
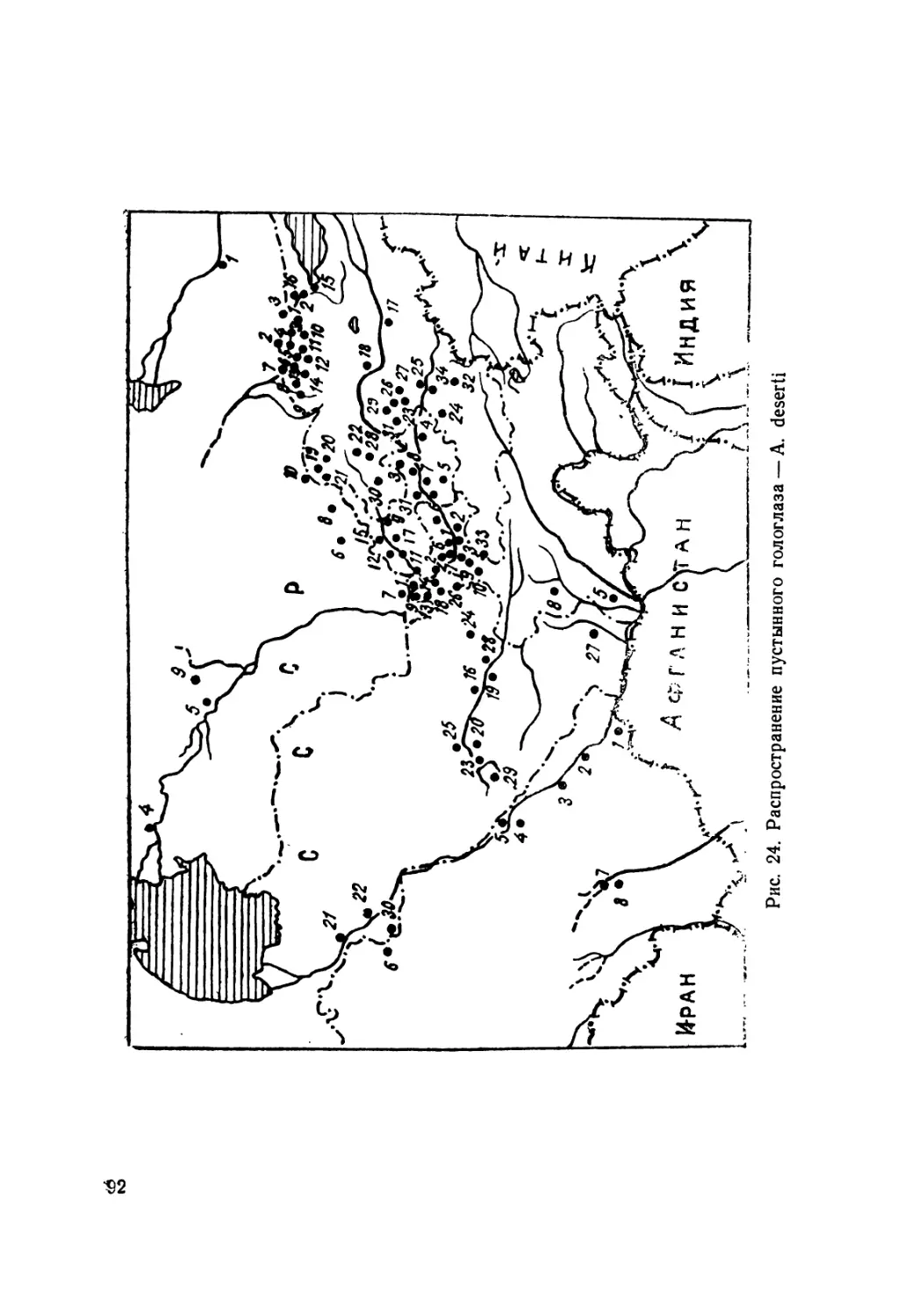
Распространение. Населяют горы системы Тянь-Шаня
и Памиро-Алая. Северная граница ареала проходит по Кир-
гизскому хребту, Заилийскому Ала-Тау и хр. Кетмень: южная
спускается к Дарвазу и оз. Яшиль-Куль (Памир), где, воз-
можно, уходит и южнее, за пределы Таджикистана — к горам
Гиндукуша; западная граница проходит по средней части
Туркестанского и Кураминского хребтов и периферии Угам-
ского и Пскемского. Наиболее восточная находка (она же и
самая северная) отмечена на территории Китая (Кульджа).
Находки асимблефара в Кульдже (коллекции ИЗ АН УССР),
а также на крайнем востоке Алайской долины (окр. с. Ир-
кештам) позволяют предполагать возможность распростра-
нения данного вида в Западном Китае в хребтах Халык-Тау,
Нарат, Боро-Хоро, Кук-Тунь, Кинг-Tay, а также Кашгарском.
В коллекциях Чикагского естественно-исторического музея
(FMNH) имеются два экземпляра асимблефара с этикеткой
A. deserti Str., добытые в Китае без точного указания места
и фамилии сборщика (см. рис. 5).
Систематика и географическая изменчивость. В. С. Ел-
патьевский (1901) обосновал описание вида Ablepharus alai-
cus рядом признаков (прерванный ряд верхнересничных щит-
ков, 26—28 чешуй вокруг середины тела, раздельные пред-
лобные щитки у части особей, относительно крупные разме-
ры тела и др.), позволяющих отличать его от ранее извест-
ного полосатого гологлаза — Ablepharus bivittatus (Men.,
1832) из Закавказья. В той же работе (Елпатьевский, 1901,
с. 38) он указывал нахождение на Памире и A. bivittatus
(№ 2856, leg., М., Воскобойников, 1898, коллекции ЗММГУ).
Изучение этого экземпляра, любезно предоставленного нам
В. Ф. Орловой, показало его принадлежность к виду Ablepha-
rus deserti Strauch, 1868. Ошибка В. С. Елпатьевского оста-
лась незамеченной. Более того, А. М. Никольский (1915) оп-
ределил экземпляры гологлазов, добытых Н. А. Зарудным в
1907 г. на Тянь-Шане (Таласском Ала-Тау, р. Аю-Тур) и в
25
to
o>
Рис. 5. Распространение alaicus: A. a. alaicus (пунктирная линия), A. a. yakovlevae (сплош-
ная), A. a. kucenkoi (штриховая).
Кадастр к рис. 5
Римские цифры — номера выборок.
КНР. 1 — Кульджа (колл. ИЗАН УССР, Киев)
СССР I—П а м и р. 1 оз. Яшиль-Куль, 3800 м; 2—Дарваз, окр. с. Софе-
Дорон, 2600 м; 3 — хр. Петра Первого, 15 км к юго-зап. от Таджикабада;
4—Хорогский Ботсад АН ТаджССР; 5—Джеканды в Каратегине; 6—Сары-
Дашт; 7 *—окр. Мургаба (Б. Памирский пост) (Чернов, 1959). II — За-
падный Алай. 8 — ур. Арча-Булак, 2531—3520 м; 9 — пер. Тенгиз-Бар;
10 — окр. с. Дараут-Курган. 2900 м; 11 — Алайская долина. III — Восточ-
ный Алай: 12—окр. с. Сары-Таш; 13—р.Кок-Су; 14—Иркештам; 15—ур.
Жошолу; 16—Туркестанский хр., ур.Кык, 2700 м. 17—Ферганский хр.,
ур.Ак-Терск. IV—Арпа: 18—р.Арпа; 19—хр.Чаар-Таш, пер.Ак-Кыз, 1290 М;
20—окр. с. Казарман; 21 —хр. Агбаши, ур. Кичи-Арча, 22—хр. Борколдой
23 — хр. Ак-Шийрак, ущ. Кур-Терез, 4000 м; V — Сон-Куль, 24 —
хр. Сон-Куль-Тоо, ур. Кара Кече: 25 — ущ. Балыкчи, 2600 м; 26 — сев. бе-
рег оз. Сон-Куль, 3050 м; 27 — верх. Кара-Куджур. VI —Восточный
Каракол: 28 — верх р. В. Каракол, 2500 м; 29 — р. В. Каракол в устье
р. Кашка-Суу, 2400 м. VII — И с с ы к - К у л ь: 30 — хр.е Кунгей Ала-Тоо,
viH. Чон-Кой-Суу; 31—ущ. Кырчин; 32—окр. с. Ананьеве; 33—междуречье
Тюп и Кар-Кыра, 2200 м; Терскей Ала-Тоо: 34—р. Ийри-Суу; 35—окр. с. Со-
ветское; 36 — окр. Пржевальска; 37 — с. Теплоключенка; 38 — сред. теч. р.
Джеты-Огуз, 2500 м; 39—ущ. р. Чон-Кызыл-Су, морены ледника Кара-Бат-
как, 3350 м (Зимина, 1959); 40 — ущ. Барскаун; 41—Сары-Джаз
в устье р. Куелю и р. Карагай; VIII — Заилийский Ала-Тау:
42—Б. Алма-Атинское ущ.; 43—М. Алма-Атинское ущ.; 44—ущ.Чим-Тургень;
45 Талгар; 46 Узун-Алы; 47—окр.Узуп-Агача; 48—Чилик; 49—верх.р.Ча-
рыи; 50 верх.р.Текес; 51 — ур.Чон-Ксмин; 52—Чу-Илийский хр., ур.Бейшеке^
1500 м; IX—Шамси: Кирг.хр. 53—ущ.Шамси, 1800 м; 54—Туюк; 55—ущ.
Иссык-Ата; X—Ал а медин: 56—окр. с. Горная Маевка, 2000 м; 57—ущ.
Аламедин, 1800 м; 58—ур. Татыр, 1800 м; 59—ущ.Ала-Арча; 60—ур.Чон-
Таш; XI—Сусамыр: 61—пер. Тюз-Ашу (южный); 62—верх. р. Сусамыр;
63 -верх. р. Зап. Каракол; 64—р. Зап. Каракол, в 1 км зап. с. Тунук; XII—
Ч и ч к а н: 65—ущ. Узун-Ахмат; 66—ущ. Чичкан; XIII—Т ю з - А ш у: 67—пер..
Тюз-Ашу (северный), 2800 м; XIV—Талас: 68—Кирг.хр., ущ. Кууганды;
69—ущ. Кара-Каин; 70—окр. с. Луговое; 71—пер. Макбал; XV—А к су-
Jl жаба гл ы: 72—заповедник Аксу-Джабаглы, ур. Чулпак; 73—окр. Чим-
кента; 74 — верх. р. Угам; 75 — Кызыл-Нура; 76 — Кураминский хр.,
пер. Камчик, 3000 м; 77 — Пскемский хр., верх. р. Кара-Корум, приток
р. Чаткал; 78 — Агуйнак; 79 — Охунь-Су-Ашу.
Пункты 1, 5, 6, 8, 9, 36, 45, 47, 71 даны по колл. ЗИН АН СССР; пунк-
ты 32. 35. 43. 44, 46, 47, 48, 49, 50, 70, 72, 73 — по колл. ЗММГУ; № 3, 4,
14 (Саид-Алиев, 1979); № 74, 78, 79 (Богданов, 1960); № 7 (Чернов, 1959);
№ 11, 16, 17, 18, 19, 20, 21, 23, 24, 27, 51, 69 77 (Яковлева, 1964), все ос-
тальные пункты — по нашим сборам (колл. ИБ АН КиргССР и ИЗ,
АН УССР).
1897 г. А. Н. Казнаковым на Памире у Сагры-Дешта (соот-
ветственно №№ 10543, 9149, коллекции ЗИН АН СССР) как
A. bivittatus. Мы, благодаря любезности, проф. И. С. Дарев-
ского, познакомились с этими экземплярами и определили их
как As. alaicus. Путаница в определении данных ящериц от-
рицательно сказывалась на выяснении таксономической при-
надлежности как «alaicus» и «bivittatus», так и «deserti». Из-
вестный исследователь сцинковых М. Миттлман (Mittleman,.
27
1952), составивший каталог ящериц подсемейства Lygosominae
мировой фауны, исключил A. alaicus из числа валидных ви-
дов, считая его, видимо, младшим синонимом A. bivittatus
(Men., 1832). Его позиция нашла позже свое логическое за-
вершение в трудах других зарубежных герпетологов (Wet-
tstein, 1960; Mertens, 1965; Fuhn, 1969, в), которые, работая
с единичными экземплярами «alaicus» и «bivittatus» и не зная
важнейших признаков, отличающих эти виды, высказывали
мнение о постепенных географических переходах между ними.
О. Веттштейн (Wettstein, 1960), а вслед за ним и другие ис-
следователи (Mertens, 1965; Fuhn, 1969а, в; Greer, 1974)
принимали за подвид полосатого гологлаза афганских голо-
глазов A. bivittatus lindbergi Wettstein, 1960. Рассматривая
указанную форму как недостающее звено, соединяющее за-
падные и BOCTQ4Hbie популяции A. bivittatus, И. Фун. (Fuhn,
1969а, в) делит данный вид на три подвида, одним из кото-
рых и явился «alaicus». Это мнение разделял и А. Грир
(Greer, 1974).
Диаметрально противоположный взгляд на систематику
«alaicus» заключался в признании его видовой самостоятель-
ности отечественными герпетологами (Никольский, 1915; Те-
рентьев, Чернов, 1949; Богданов, 1960; Яковлева, 1962, 1964;
Банников и др., 1977).
Весьма важными для изучения таксономии данного вида
представляются указания некоторых отечественных герпе-
тологов на его неоднородность. Свидетельством последнего
могут служить описание вида A. kucenkoi А. М. Никольским
(1902) и A. sapozshnikovi В. П. Кащенко (1909). Обе эти
формы происходят из Восточного Тянь-Шаня и авторы под-
черкивали их близость к виду A. alaicus. Позже П. В. Терен-
тьев и С. А. Чернов (1949) без достаточных оснований сво-
дят эти формы в синонимы A. alaicus. Мнение указанных гер-
петологов было принято многими специалистами нашей стра-
ны (Андрушко, 195J: Параскив, 1956; Чернов, 1959; Богда-
нов, 1960; Яковлева, 1962, 1964; Банников и др., 1977), а так-
же зарубежными (Fuhn, 1969а, в).
В 1927 г. Д. Н. Кашкаров без каких-либо дополнительных
исследований применяет название Ablepharus alaicus kutzhen-
ko Nik. для ящериц из Западного Тянь-Шаня.
Таким образом, существующая внутривидовая систематика
«alaicus» не имеет обоснованных решений и нуждается в реви-
зии. С целью выявления закономерностей географической из-
менчивости данного вида нами исследовано 15 выборок
(см. рис. 5), охватывающих практически весь ареал этих яще-
риц, и приведено сравнение по основным признакам фолидоза,
пропорциям тела и окраске.
28
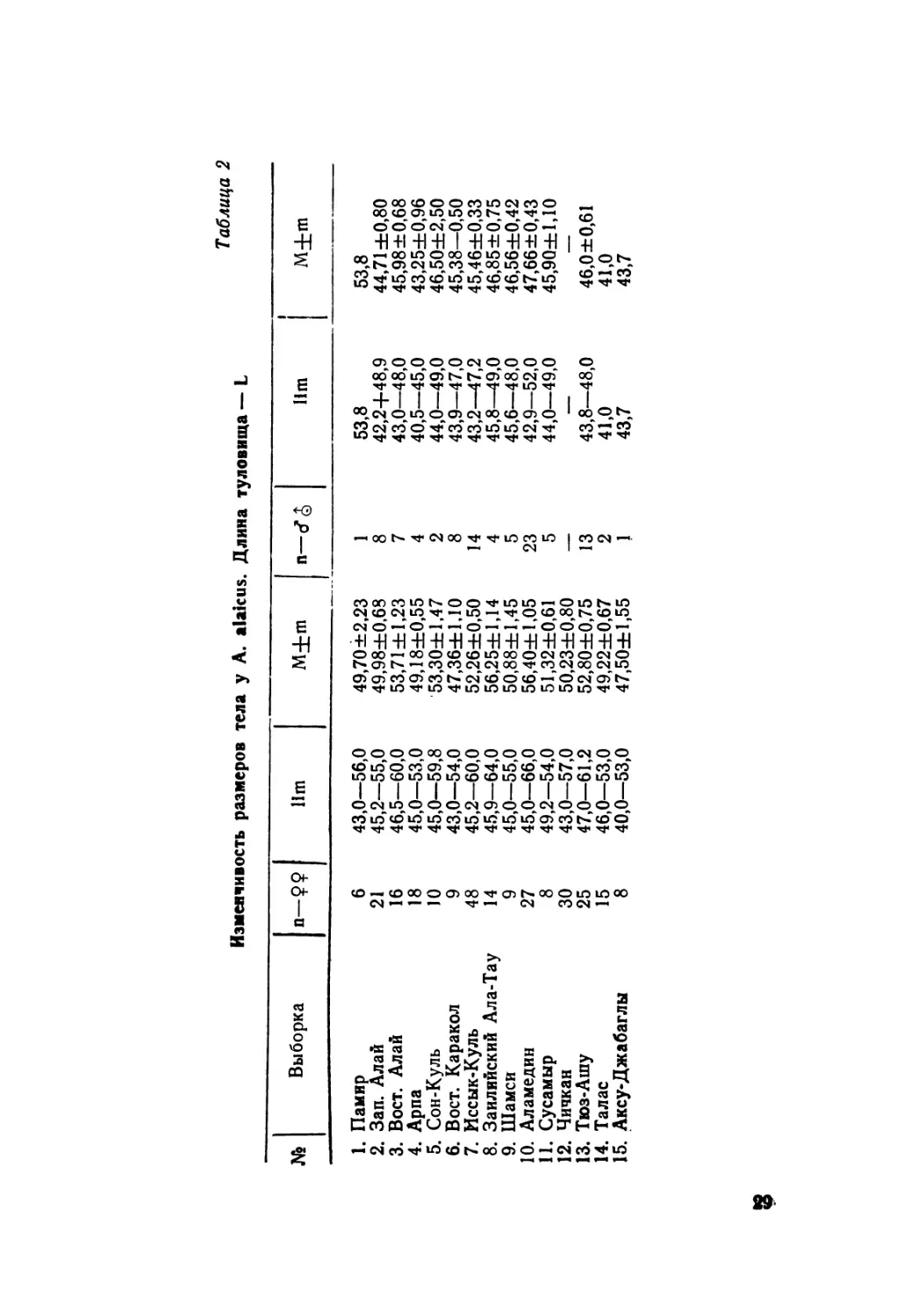
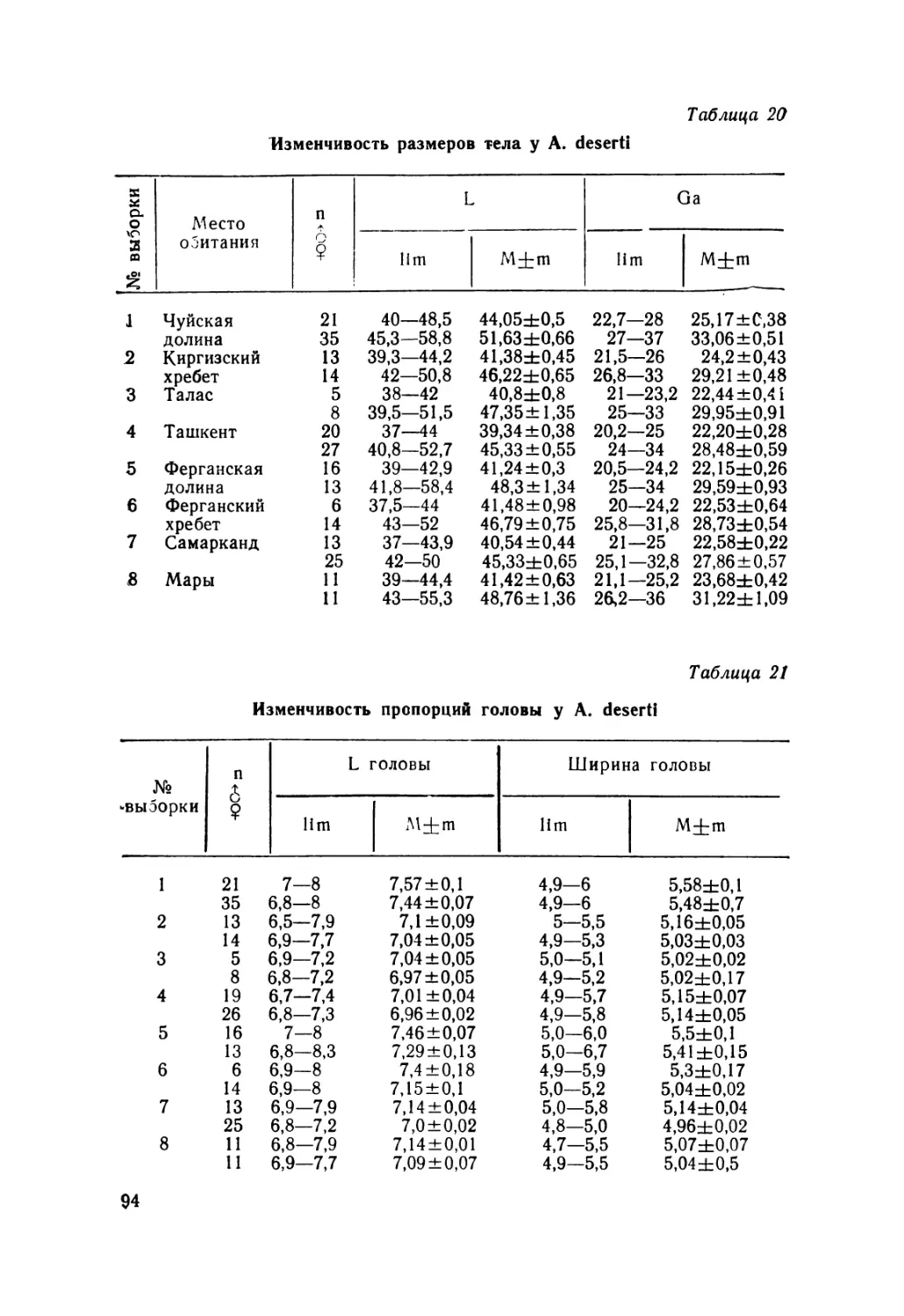
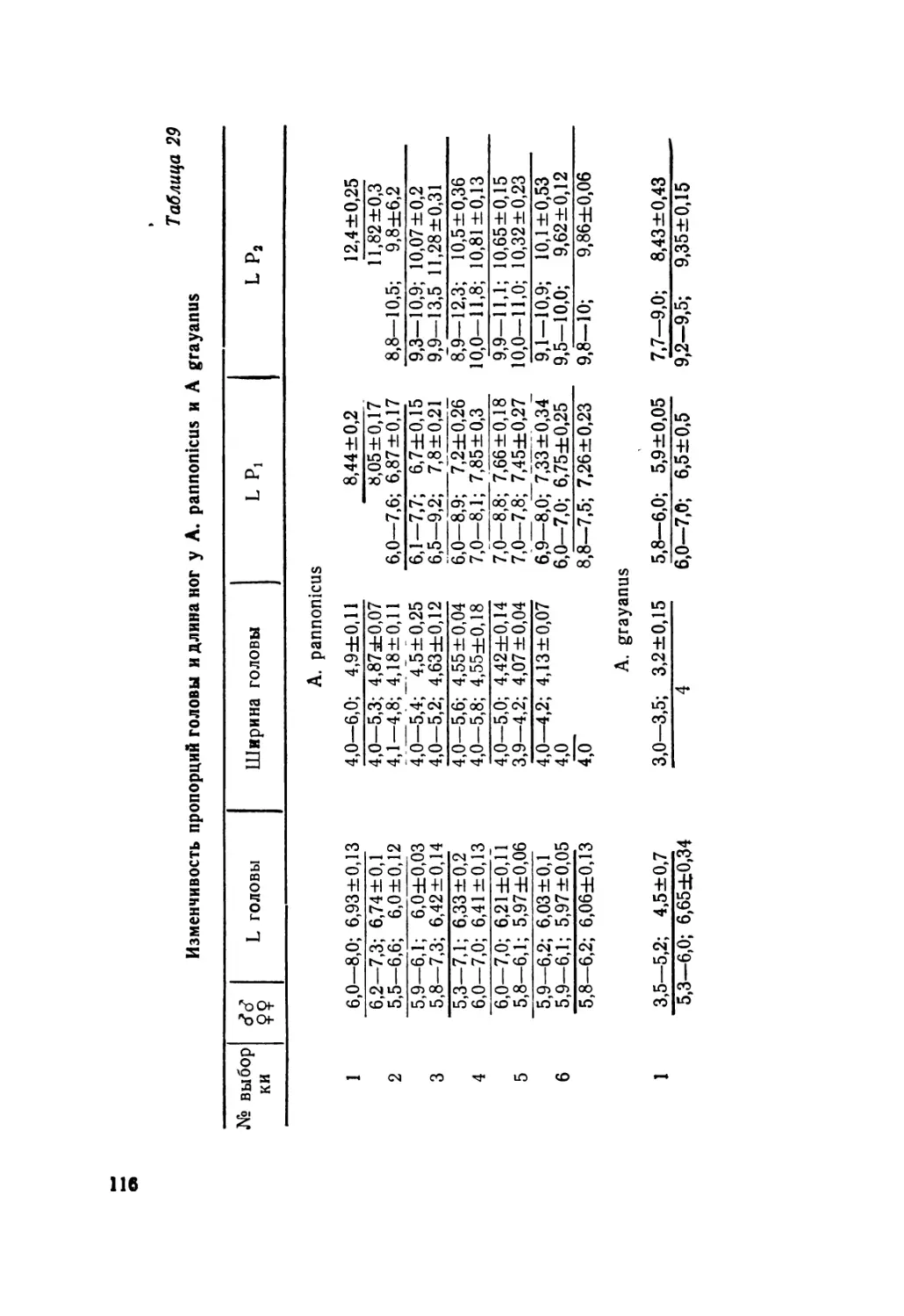
Изменчивость размеров тела у A. alaicus. Длина туловища — L
Таблица 2
№ Выборка п-?9 11m M+m n-d^ 11m i | M+m
1. Памир 6 43,0—56,0 49,70 ±2,23 1 53,8 53,8
2. Зап. Алай 21 45,2—55,0 49,98±0,68 8 42,2+48,9 44,71 ±0,80
3. Вост. Алай 16 46,5—60,0 53,71 ±1,23 7 43,0—48,0 45,98 ±0,68
4. Арпа 18 45,0—53,0 49,18±0,55 4 40,5—45,0 43,25±0,96
5. Сон-Куль 10 45,0—59,8 53,30±1,47 2 44,0—49,0 46,50±2,50
6. Вост. Каракол 9 43,0—54,0 47,36±1,10 8 43,9—47,0 45,38—0,50
7. Иссык-Куль 48 45,2—60,0 52,26±0,50 14 43,2—47,2 45,46±0,33
8. Заилийский Ала-Тау 14 45,9—64,0 56,25±1,14 4 45,8—49,0 46,85±0,75
9. Шамси 9 45,0—55,0 50,88±1,45 5 45,6—48,0 46,56±0,42
10. Аламедин 27 45,0—66,0 56,40±1,05 23 42,9—52,0 47,66±0,43
11. Сусамыр 8 49,2—54,0 51,32±0,61 5 44,0—49,0 45,90±1,10
12. Чичкан 30 43,0—57,0 50,23±0,80 — — —
13. Тюз-Ашу 25 47,0—61,2 52,80±0,75 13 43,8—48,0 46,0 ±0,61
14. Талас 15 46,0—53,0 49,22±0,67 2 41,0 41,0
15. Аксу-Джабаглы 8 40,0—53,0 47,50±1,55 1 43,7 43,7
8
оо4о5сл4^ооко —
Сл4^00К0^ОС000ЧОСл4^0Э ю I-v
№ вы-
борки
р р Ц1 О р QJ р р
О о ’cD NO о о О
I I I I I I I I
ч ч р ч ч ч ч
CD CD СП О ~<£> О СО
NONDNDNONONONONONONONONONONOND
*- СП СП NO Ч СП 4^ СО *4 4^ СП СЛ СЛ р 4*.
О О Ь (D о w О СО О КО СЛ NO О CD
I I I I I I I I I I I I I I I
OOOOOOQoOO4^GO4^OOCa0OOOOQOWW
о 7“* СП 4^ ~ О ОО с> Ч СЛ NO Ч NO ф>
00 о СИ "о ч СО NO сл ►— со ьэ о О О о
-ю
-ю
р 05 р Ч р Oi р О)
Ч СЛ СО О NO \0 СП *оо
СО сп ~ О ОС Q0 Ч О
Н- н- Н-1+ Н-1+ 1+ 1+
OoOOODoODo R
’—о ~<=> © ND 2
О 05 00 ND 00 4^ 00 СП д
о
Со
О
ОЬ
NO NO NO NO ND ND ND NO ND NO NO NO NO N0 Г\
КО О NO NO NO W 4^ NO NO ND CD ND J— CaD
bo О О СЛ NO О CD ~ О NO О 00
I I I I I I I I I I I I I
NO NO NO KO NO NO NO NO NO NO NO NO
04 4^ Ч СЛ CD СЛ СЛ CD CaD CD СЛ
CD NO 00 О О CD CD О О О О
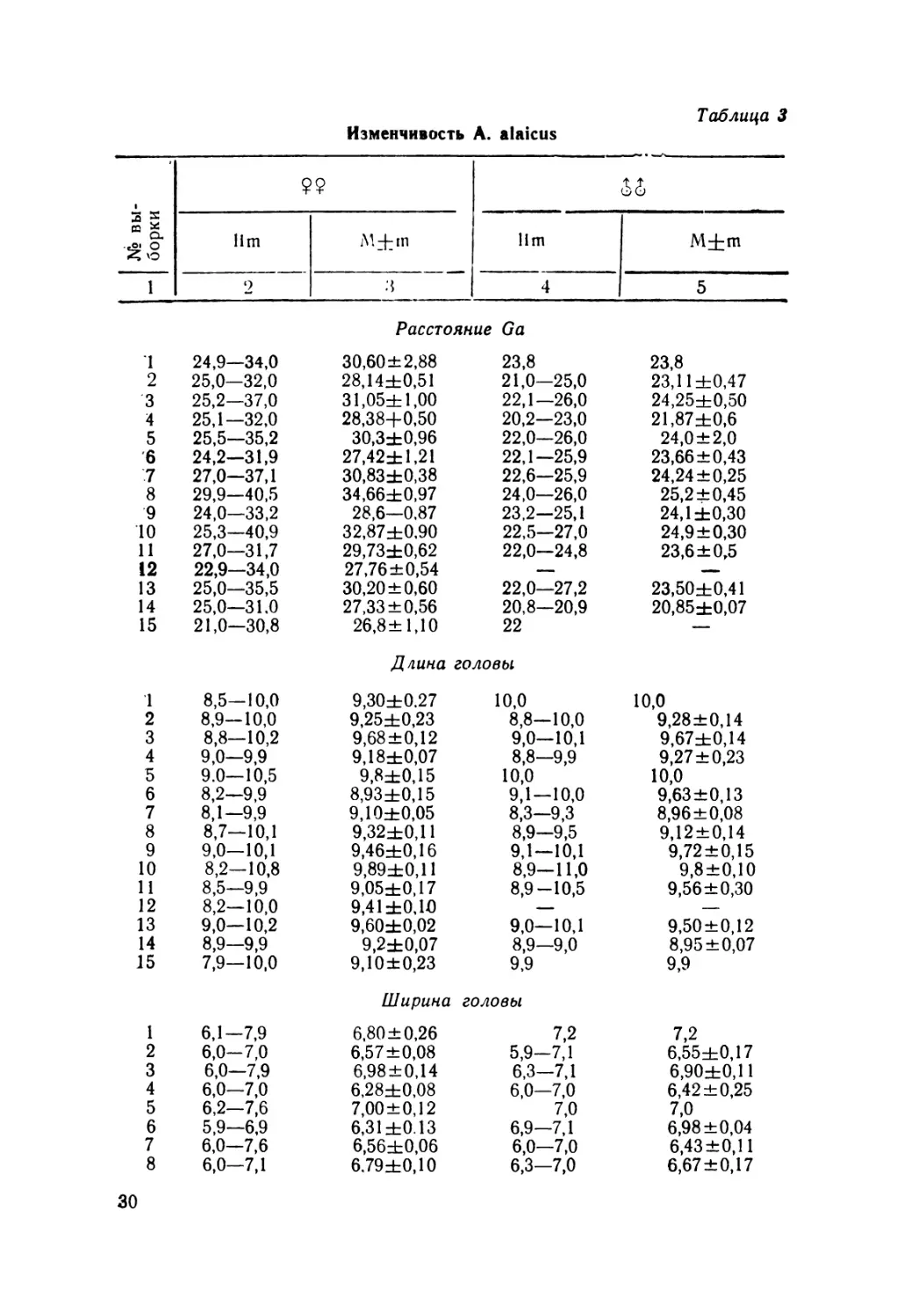
Изменчивость A. alaicus
СП — ч
1+1+
CD (£)-ь
сл 5° ч -
?“£l+l+l+
1+ 1+ 1т о о о
<—> /—ч /—1Ч - *
£> © о NO 05
1+н-!+
NO NO KONO NO NO NO KO
О W NO NO KO NO 4^ W NO 4sa OO C>3
00 СЛ -°0 S71 NO 0> 00 ND 00
, СЛ О CD CD ~ NO 4^ CD О Ч СЛ —
I H-H- I 1+ 1+ H-1+1+ 1+ 1+ H- H- H-
о о • opoppp КОО о о
о 4^ СЛ 00 О0 4^ NO Фа О СП СЛ 4^
ч •— о о СЛ СЛ W 04
Продолжение табл. 3
1 2 3 4 5
9 6,0—7,1 6,63±0,14 7,0—7,9 7,12 + 0,06
10 6,5—8,0 7,26±0,10 6,9—8,0 7,30+0,10
11 6,0—7,0 6,70±0,13 6,8—7,8 7,12+0,17
12 6,7—8,0 6,83±0,08 — —
13 6,2—7,3 6,80±0,06 6,0—7,4 6,90+0,10
14 6,0—7,6 6,70±0,1 Г 6,1 6,1
15 5,8—7,2 6,71±0,18 7,5 7,5
Длина передней ноги
1 11,1—12,9 11,90±0,13 13,0 13,0
2 11,2—12,8 12,08±0,10 11,3—13,0 12,37+0,22
3 11,0—13,5 12,43±0,18 11,0—13,9 12,20+0,37
4 11,0—12,2 11,34±0.08 11,3—11,8 11,65+0,12
5 11,6—13,3 12,77±0,16 11,6—13,0 12,30 ±0,70
6 10,2—12,5 11,56 ±0,23 12,0—12,9 12,35+0,12
7 10,4—12,0 11,26±0,05 10,9—12,0 11,32+0,10
8 10,8—12,8 11,46±0,16 11,1—12,3 11,50+0,28
9 11,0—12,9 12,16±0,25 11,5—12,2 11,94±0,11
10 11,0—13,0 11,94 ±0,10 11,0—13,0 12,03+0,11
11 11,0 12,5 1 1,58±0,21 11,1—12,0 11,76+0,17
12 10,9 12,8 11,78±0,10 — —
13 11,0—13,0 12,0±0,10 11,8—13,0 12,2+0,11
14 11,2—12,5 11,80±0,10 11,0—12,0 11,50+0,50
15 10,0—11,8 11,10±0,23 11,9 11,9
Длина задней ноги
1 14,0—17,0 15,60±0,54 17,8 17,8
2 15,1—16,8 15,96±0,07 15,2—17,8 16,21 ±0,31
3 14,5—18,0 16,34±0,25 15,0—18,5 16,35 + 0,46
4 14,0—15,9 15,06 ±0,09 15,0—16,3 15,60+0,28
5 15,8—17,9 17,0±0,21 15,8—16,9 16,35 ±0,55
6 14,0—15,8 14,92±0,18 15,7—16,9 16,06 + 0,13
7 13,9—16,0 14,80±0,07 14,0—15,8 15,10±0,15
8 14,3 16,1 15,40±0,15 15,5—16,1 15,80+0,13
9 14,8 17,1 16,02±0,25 15,1—16,6 15,96+0,25
10 15,0 16,9 15,88+0,11 15,0—17,0 15,77 + 0,14
11 14,9—16,0 15,16±0,12 15,0—16,5 15,70+0,33
12 14,4—16,8 15,58+0,10 — —
13 15,0—17,3 16,10±0,12 15,0—17,8 16,20±0,18
14 15,0—16,0 15,36+0,10 14,2—16,2 15,20± 1,00
15 13,0—16,0 14,80±0,34 16,0 16,0
Длина туловища. Известная максимальная длина тулови-
ща равна 56 мм, qq —60 мм (Банников и др., 1977). По
нашим данным —соответственно 53 мм и 66 мм. Нами также
установлено (табл. 2, 3), что асимблефары популяций север-
ной периферии Тянь-Шаня (Киргизский хребет и Заилийский
31
Ала-Тау) отличаются в целом максимальными размерами;
наоборот, минимальными обладают ящерицы из популяций
Центрального Тянь-Шаня. Последнее особенно заметно для
популяций из Арпы, Восточного Каракола и некоторых дру-
гих. Относительно мелкие размеры тела свойственны и асимб-
лефарам Западного Алая. В изменчивости длины тела мож-
но проследить наличие отдельных клин. Например, в Алай-
ской долине четкая клина с возрастанием значений проходит
с запада на восток. Аналогичное постепенное увеличение зна-
чений признака можно наблюдать и от популяции иссык-
кульской к популяции из Заилийского Ала-Тау, от Централь-
ного Тянь-Шаня к Северному. Выявленная закономерность
изменчивости размеров тела коррелирует с особенностями
климатических условий районов (главным образом с характе-
ром распределения осадков) В районах относительно боль-
шего увлажнения находятся популяции ящериц, отличающие-
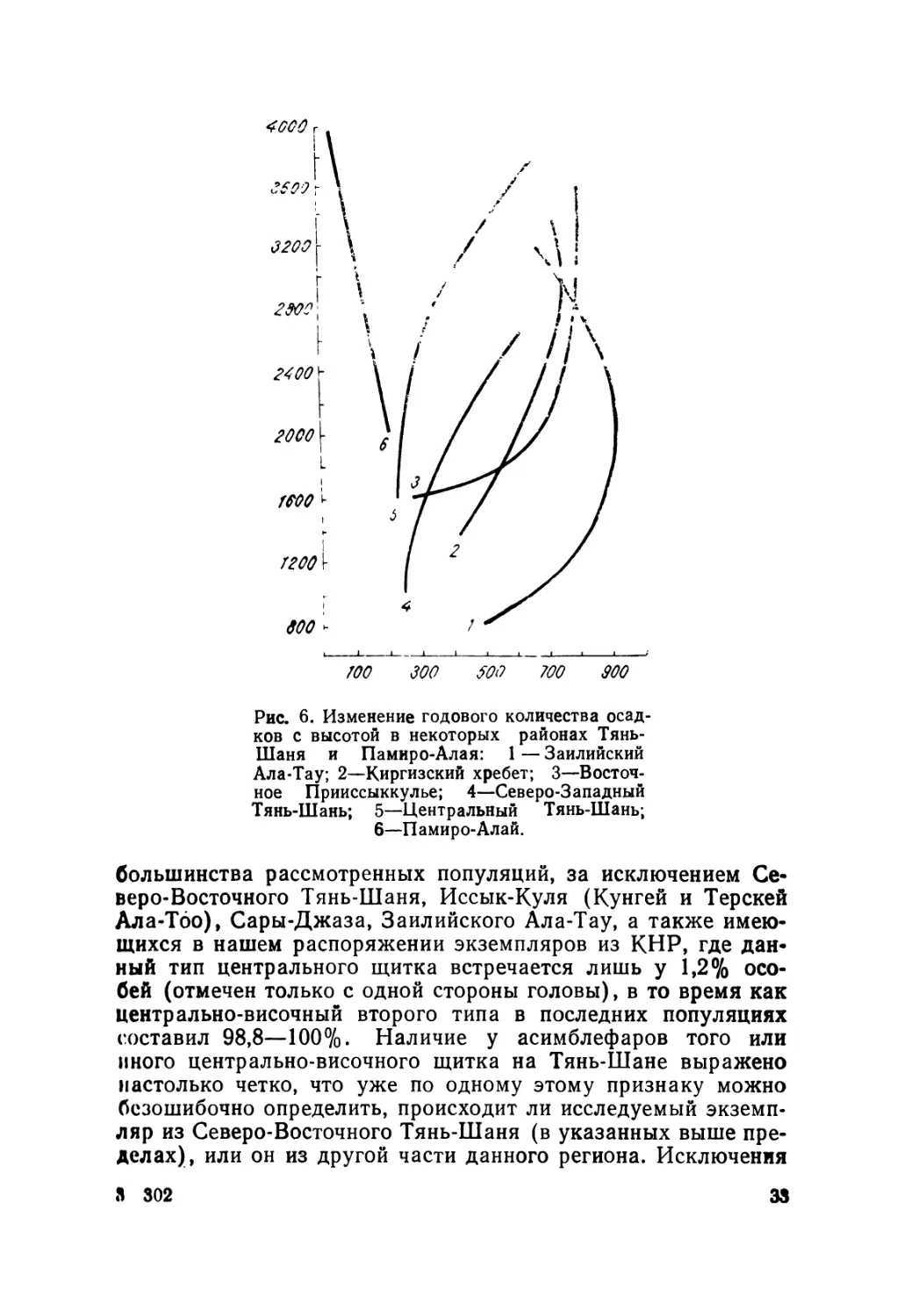
ся максимальными размерами тела (рис. 6). Наоборот, ми-
нимальные свойственны ящерицам, живущим в районах не-
большого увлажнения, т. е. внутренних районов Тянь-Шаня
и Памиро-Алая.
Все другие размерные характеристики ящериц (длина и
ширина головы, длина конечностей, хвоста) проявляют ана-
логичную изменчивость (табл. 3). Таким образом, ящерицы
северных популяций и отдельные популяции высокогорий
(оз. Сон-Куль) обладают заметно более крупным телом по
сравнению с остальными. Следует также отметить, что дли-
на конечностей по отношению к длине туловища проявляет
обратную закономерность изменчивости, т. е. с увеличением
длины туловища относительная длина конечностей уменьшает-
ся. Последнее особенно характерно для ящериц Иссык-Куля
и Заилийского Ала-Тау. На эту особенность обращал внима-
ние и А. М. Никольский (1907), описывая форму A. kucenkoi
с Иссык-Куля.
Фолидоз. Особенности щиткования головы в височной об-
ласти. Изучая височную область фолидоза A. alaicus,-мы об-
ратили внимание на форму и расположение центрально-височ-
ного щитка и выделили его два характерных типа: централь-
но-височный сверху свободный, более или менее овальной
формы и расположен на теменном щитке, закрывая его
нижний край; центрально-височный—обычно трапециевидный,
округлый позади, его верхний край находится под теменным
щитком, а нижний — под седьмым верхнегубным.
Как показали наши исследования (табл. 4), центрально-
височный щиток первого типа характерен для асимблефаров
1 Климатические особенности даны по В. Пономаренко (976).
32
4000 г
I
Рис. 6. Изменение годового количества осад-
ков с высотой в некоторых районах Тянь-
Шаня и Памиро-Алая: 1 — Заилийский
Ала-Тау; 2—Киргизский хребет; 3—Восточ-
ное Прииссыккулье; 4—Северо-Западный
Тянь-Шань; 5—Центральный Тянь-Шань;
6—Памиро-Алай.
большинства рассмотренных популяций, за исключением Се-
веро-Восточного Тянь-Шаня, Иссык-Куля (Кунгей и Терскей
Ала-Тоо), Сары-Джаза, Заилийского Ала-Тау, а также имею-
щихся в нашем распоряжении экземпляров из КНР, где дан-
ный тип центрального щитка встречается лишь у 1,2% осо-
бей (отмечен только с одной стороны головы), в то время как
центрально-височный второго типа в последних популяциях
составил 98,8—100%. Наличие у асимблефаров того или
иного центрально-височного щитка на Тянь-Шане выражено
настолько четко, что уже по одному этому признаку можно
безошибочно определить, происходит ли исследуемый экземп-
ляр из Северо-Восточного Тянь-Шаня (в указанных выше пре-
делах), или он из другой части данного региона. Исключения
Я 302
39
составляют единичные особи из южной части Центрального
Тянь-Шаня (Арпа), которые могут иметь центрально-височ-
ный щиток второго типа.
Для ящериц Памиро-Алая показательно присутствие сме-
шанных типов этого щитка. У памирских, западно-алайских
и восточно-алайских асимблефаров частота встреч централь-
но-височного щитка, характерного для особей Северо-Восточ-
ного Тянь-Шаня, составила соответственно 21, 43 и 16%, а
щитка типа 1 — 79, 57 и 84% (см. табл. 4). Среди них толь-
ко у ящериц Западного Алая соотношение частоты встреч
центрально-височного обоих типов почти равное.
Таким образом, выявленный и исследованный нами приз-
нак проявляет дискретную изменчивость, что особенно рез-
ко выражено при сравнении популяций Северо-Восточного
Тянь-Шаня. Только у асимблефаров Памиро-Алая этот приз-
нак имеет смешанный характер.
Характеристика носового щитка. Многие авторы (Терен-
тьев и Чернов, 1949; Параскив, 1956; Чернов, 1959; Богданов,
1960; Яковлева, 1964; Банников и др., 1977) отмечают, что у
данного вида ноздря прорезана в одном щитке, однако эта
характеристика не совсем точная. Нами установлено, что но-
совой щиток у асимблефаров может быть снизу полуразделен-
ный, либо цельный.
Полуразделенный щиток в значительной мере характерен
для ящериц Северо-Восточного Тянь-Шаня, в особенности
Иссык-Куля (88,7%) и в меньшей—Заилийского Ала-Тау
(52,6%) (см. табл. 4). Асимблефарам всех других популяций
Тянь-Шаня, как правило, свойствен цельный носовой щиток
(97—100%). Исключение составляют отдельные особи из Ар-
пы, 10,7% из которых обладают полуразделенным носовым
щитком. Памиро-алайские ящерицы имеют обе эти формы
носового щитка, однако частота встреч первого незначитель-
на (10—39%), причем большая доля приходится на ящериц
Восточного Алая. В целом изменчивость данного признака
обнаруживает аналогичную предыдущему признаку законо-
мерность.
Положение предлобных щитков. Результаты, полученные
нами при изучении данного признака (см. табл. 4), позволяют
разделить исследуемые выборки A. alaicus на две части: по-
пуляции Памиро-Алая и Тянь-Шаня. Памиро-алайские асимб-
лефары отличаются от тянь-шаньских резким возрастанием
(до 90%) числа особей, у которых предлобные щитки широ-
ко соприкасаются друг с другом и отделяют лобно-носовой
щиток от лобного. У асимблефаров Тянь-Шаня указанная
комбинация предлобных щитков составляет лишь 3—12,5%,
тогда как большинство особей здесь имеют разделенные пред-
34
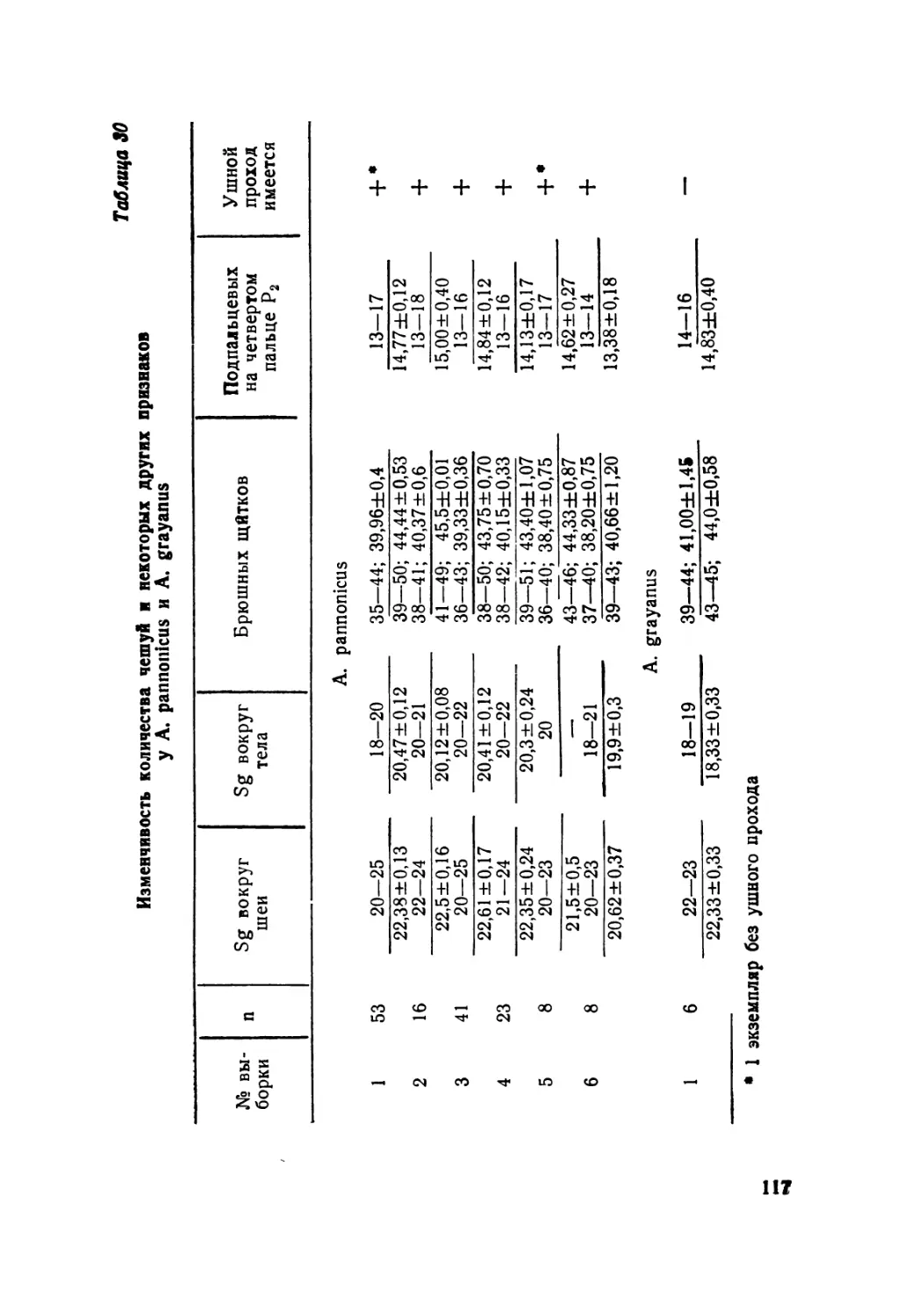
Таблица 4
Характеристика предлобных, центрально-височного и носового щитков
У A. alaicus (Elpat., 1901) из 15 выборок, %
выборки п Предложи ы е Тип централь- но-височного Носовой
разделены соприкасаются полураз- деленный цельный
в одной точке ши- роко 1 2
1 10 — 10 90 79 21 10 90
2 55 5 18,5 76,5 57 43 13 87
3 46 11 18 71 84 16 39 61
4 28 71 18,3 10,7 92,9 7,1 10,7 90,3
•5 13 92,2 — 7,6 100 100
6 36 94,5 2,5 3 100 100
7 90 83,6 6,4 10 1,2 98,8 88,7 11,3
8 38 76,3 18,4 5,3 — 100 52,6 47,4
9 10 17 54 88,2 89 5,8 5,5 6 5,5 100 98 2 2 юо’ 98
11 16 69 18,5 12,5 100 юо
12 31 77,5 19,5 3 100 100
13 45 68 27 5 97,8 2,2 3 97
14 15 18 15 89 100 5,5 5,5 100 100 ЮО 100
(68 94,5%) и предлобные, соприкасающиеся
в од-
лобные ("~ ,1,
ной точке (55—27%).
Количество затылочных щитков. По литературным дан-
ным (Банников и др., 1977), исследуемый вид имеет 2—3 па-
ры затылочных щитков. По нашим материалам (табл. 4),
1 i с
пределы варьирования признака несколько шире: 1----5 пар
2>
щитков. Установлено также, что памиро-алайские особи от-
личаются от тянь-шаньских заметно меньшим количеством
затылочных щитков (1—4), причем большинство ящериц
имеют2—— 2 щитка (до 40%), тогда как в Тянь-Шане —
£
3—уЗ. Минимум затылочных—1—в выборках Памиро-
Алая составляет 4,1—28,6%, а Тянь-Шаня всего 1,3—6,6%.
Таким образом и по этому признаку памиро-алайские асим-
блефары отличаются от тянь-шаньских.
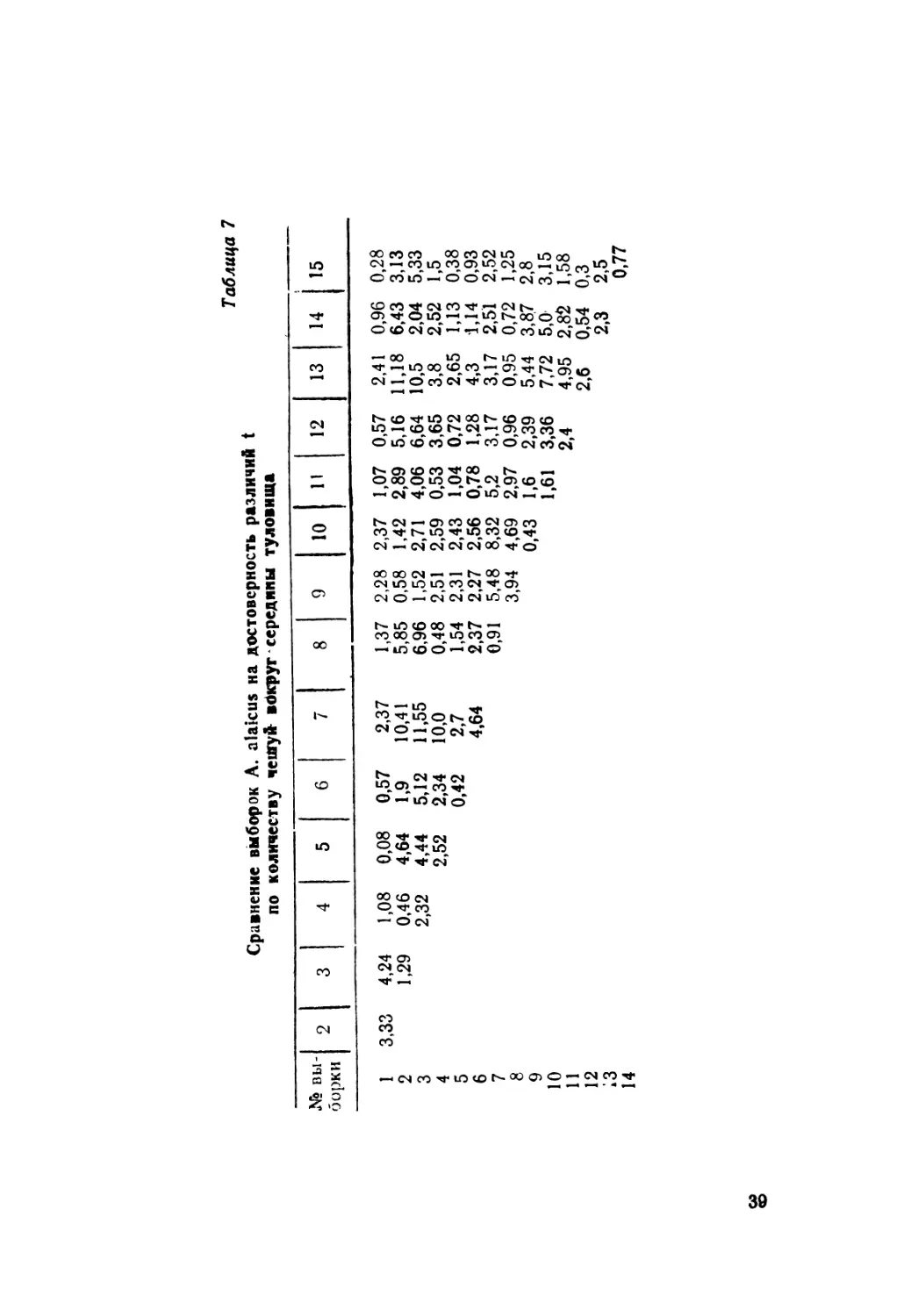
Количество чешуи вокруг середины туловища. По нашим
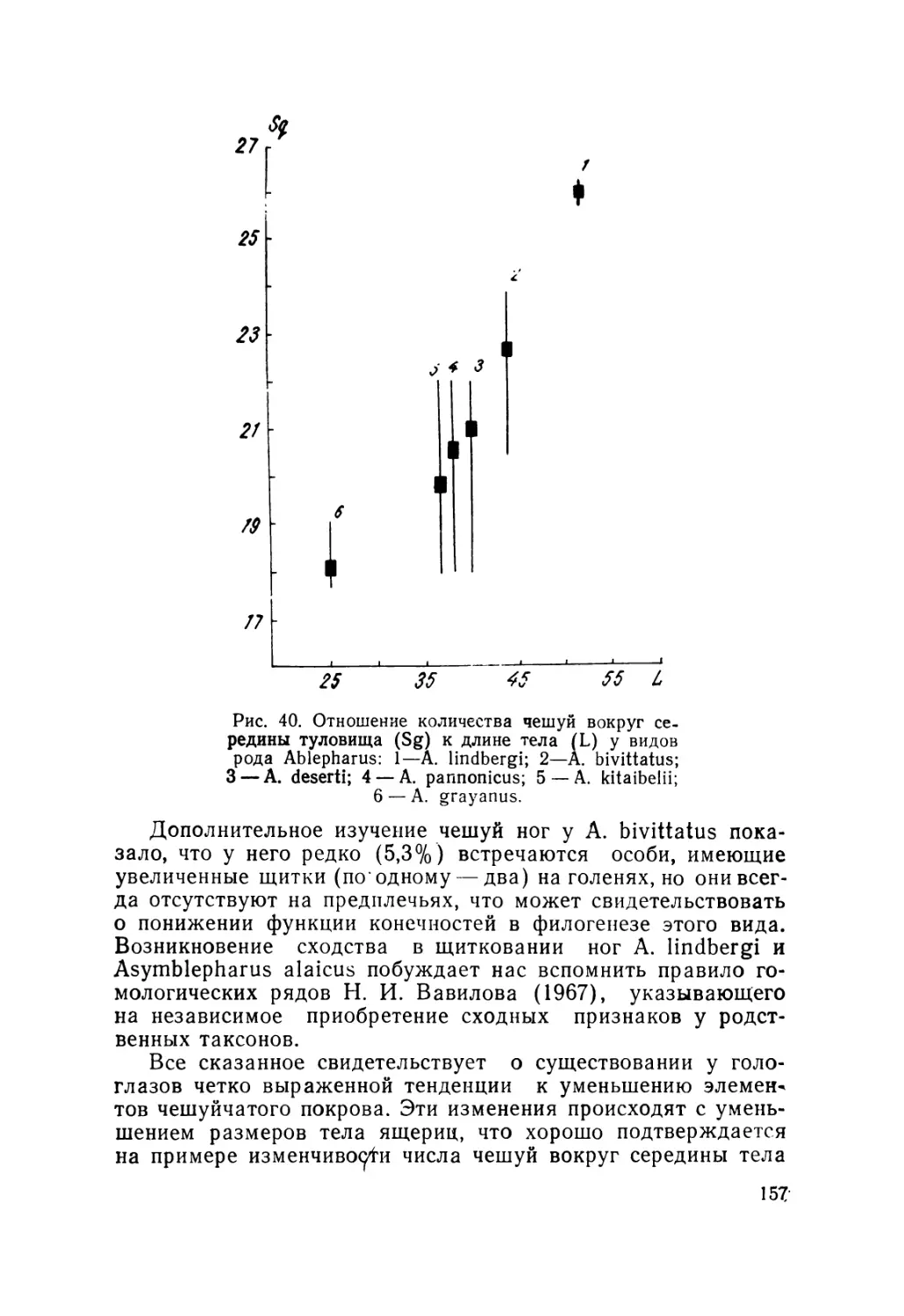
данным, пределы варьирования признака 24—30 чешуй
35
'(табл. 6), а не 26—28, как было известно из литературы
(Банников и др., 1977}. Максимальное число (30) в наших
выборках отмечено у особей Северного Тянь-Шаня (Киргиз-
ский хр., ущ. Аламедин). По средним значениям величина дан-
ного признака максимальна для ящериц Алая (27,4+0,17) и
отдельных выборок Киргизского хребта (27,2±0,28 и 27,06+
0,06) против минимального у ящериц Иссык-Куля (25,62 +
0,17), Заилийского Ала-Тау (25,82+0,21), а также Тюя-Ашу
в Киргизском хребте (25,82+0,10). Указанные различия дос-
товерны (табл. 7). Можно проследить некоторое клинальное
изменение признака в отдельных регионах: в Памиро-Алае,
например, клина идет с повышением значений от Памира к
Алаю (26,3 + 0,28; 27,7+0,17), причем в Алае она направлена
с запада на восток; переходя в Центральный Тянь-Шань
(Арпа), исчезает, но постепенно вновь образуется и идет с воз-
растанием к периферии Киргизского хребта, что заметно по
выборкам из Сон-Куля (26,33+0,19), Восточного Каракола
(26,46+0,17), Сусамыра (26,66+0,19), Шамси (27,2 + 0,28).
По направлению к Иссык-Кулю значение признака резко па-
дает (25,62±0,07) и затем вновь слабо возрастает на север-
ных склонах Заилийского Ала-Тау (25,82+0,21). Обращает
на себя внимание и наличие минимального количества чешуй
в выборке Тюя-Ашу (Киргизский хребет) по сравнению с со-
седними. Все отмеченные различия достоверны (см. табл. 7).
Количество передних верхнересничных щитков. У данного
вида может быть один-три таких щитка. В случае, когда он
один, это значит, что второй сросся с первым, либо редуци-
рован и тогда он маленький; когда верхнересничных два, то
размеры второго обычно в два-четыре раза меньше первого;
когда их три — последний, третий щиток, еще меньше второ-
го и первого.
Изучение показало (табл. 8), что ящерицы из выборок
Восточного Каракола, Иссык-Куля, Заилийского Ала-Тау,
а также Сусамыра и Таласа имеют кроме двух также и пер-
вый передний верхнересничный щиток, причем наиболее час-
то особи с одним щитком встречаются в Заилийском Ала-Тау
(40%) и Иссык-Куле (11,95%). В остальных вышеуказанных
выборках один щиток встречался довольно редко: в Восточ-
ном Караколе — 2,78%, Сусамыре — 7,14% и Таласе — 5,55%.
В последней выборке отдельные особи (5,55%) имеют по три
щитка. Во всех остальных — только по два передних верхне-
ресничных.
Количество щитков верхнего века-козырька. Изучение
признака показало, что его изменение имеет тенденцию в сто-
рону уменьшения количества этих щитков. Асимблефары Алая,
Восточного Каракола и в целом Северо-Восточного Тянь-
36
Изменчивость затылочных щитков у A. alaicus %* Таблица 6
В ы б о Р к а
Количе- ств э Памиро-Алай 1 Тянь-Шань
щитков 1 2 3 4 51 6 7 8 9 1 10 11 12 13 14 15
1 2 —
1 — — 4,1 — — — 1,3 — 6,6 2,1 — — 2,7 — —
— 5 28,6 — — — 3 3,5 1,4 — — 6 — — —
2 28,6 20 35 7,3 — 3 3,7 13,8 6,6 6,3 — 21,2 19 13,5 2,5
~г2 29,4 40 26,3 10,7 2,5 13,3 21 13,2 6,6 5,2 20 6 24,5 18 12,5
3 28 20 4 29,3 41 37,2 40 34,5 66,7 55,4 33,4 45,5 35 36,5 44
1 2 3 14 7,5 2 32 40 31 20 22 6,6 12,5 46,6 10 8,5 16 6,5
4 — 7,5 — 10,7 8,5 13,2 11 6,8 5,5 16,5 — 9 5 6,5 12
— — — — 8 2,3 — 6,2 — — — 2,3 5,3 6,5 —
5 — — — — — — — — — 2,1 — — — — —
4* — 3 -
Таблица б
Сравнение выборок A. alaicus
по количеству чешуй вокруг середины туловища
№ I выборки 1 1 Sg
п | Пт
1 8 26—28 26,3±028
2 55 26-28 27,4 ±0,17
3 46 26—29 27,7±0,17
4 28 26-28 26,7±0,29
5 13 26—28 26,33 ±0,25
6 36 24—28 26,46±017
7 90 24—26 25,62 ±0,06
8 38 24—28 25,88 ±0,21
9 15 26—29 27,2±0,28
10 54 26—30 27,06±0,06
11 16 26-28 26,66 ±0,19'
12 31 24—28 26,12±0,14
13 45 24—26 25,6±0,10
14 18 24-28 26,0 ±0,14
15 15 26—28 26,2±0^2
Шаня имеют несколько большее их число, а остальные, и осо-
бенно из выборок Сон-Куля, Аламедина, Ала-Арчи и Сусамы-
ра, характеризуются меньшим числом щитков козырька и от-
носительно большими их размерами. Анализ данных показы-
вает существование по этому признаку, хотя и не всегда ясно
выраженной клинальной изменчивости, идущей от Памиро-
Алая к северу Тянь-Шаня (за исключением Северо-Восточно-
го), точнее, от южной периферии ареала вида к северной.
Общее количество щитков между носовым и подглазнич-
ным. Их может быть три-пять. Меньшее количество (три) соп-
ровождается относительным увеличением размеров этих щит-
ков и наоборот, большее характеризуется мелкими размерами.
Как видно из приведенных данных (см. табл. 8), ящерицы
Западного и части Северного Тянь-Шаня, как правило, имеют
четыре щитка (100%), а часть ящериц Тюз-Ашу, Восточно-
го Каракола, Заилийского Ала-Тау и особенно Иссык-Куля
имеют их по три (2,77—20,87%). Вместе с тем 20% особей
из Заилийского Ала-Тау (точнее, из ущелья Талгар) имели
пять мелких щитков. В остальных выборках, начиная от Ар-
пы и далее к Памиру, ящерицы обладали четырьмя (42,85—
96,96%) и пятью (3,03—57,15%) щитками. Наибольший про-
цент встреч особей с пятью щитками отмечен на Памире —
38
Таблица 1
Сравнение выборок A. alaicus на достоверность различий t
по количеству чешуй- вдкруг середины туловища
№ вы- борки 2 3 4 5 6 7 8 9 10 11 12 13 14 15
3,33
1
2
3
4
5
6
7
8
9
10
11
12
;з
14
4,24 1,29 1,08 0,46 2,32 0,08 4,64 4,44 0,57 1,9 5,12 2,37 10,41 11,55 1,37 5,85 6,96 2,28 0,58 1,52 2,37 1,42 2,71 1,07 2,89 4,06 0,57 5,16 6,64 2,41 11,18 10,5 0,96 6,43 2,04 0,28 3,13 5,33
2,52 2,34 10,0 0,48 2,51 2,59 0,53 3,65 3,8 2,52 1,5
0,42 2,7 1,54 2,31 2,43 1,04 0,72 2,65 1,13 0,38
4,64 2,37 2,27 2,56 0,78 1,28 4,3 1,14 0,93
0,91 5,48 8,32 5,2 3,17 3,17 2,51 2,52
3,94 4,69 2,97 0,96 0,95 0,72 1,25
0,43 1,6 2,39 5,44 3,87 2,8
1,61 3,36 7,72 5,0 3,15
2,4 4,95 2,82 1,58
2,6 0,54 0,3
2,3 2,5
0,77
Таблица 8
Сравнение выборок A. alaicus по количеству передних верхнересничных
щитков и между носовым и подглазничным щитками, %
№ вы- борки п Передние верхнересничные Между носовым и подглазничным
1 2 3 3 4 5
1 10 — 100 — — 43 57
2 55 — 100 — 5.5 50,5 45
3 46 — 100 — — 97 3
4 28 — 100 — — 92,9 7,3
5 13 — 100 — — 100 —
6 36 2,8 97,2 2,7 97,3 —
7 90 12 88 — 21 79 —
8 38 40 60 — 7 73 20
9 17 — 100 —. — 100 —
10 54 — 100 — — 100 —
11 16 7 93 — — 100 —
12 31 — 100 — — 100 —
13 45 — 100 — 13 87
14 18 — 100 — — 100 —
15 15 — 93,5 6,5 — 100 —
57,15% и в Западном Алае — 47,05%, наимек [ЬШИЙ — 3,03%
у яшериц Восточного Алая и Арпы — 10,71%.
Итак, изменчивость этого признака проявляет все же кли-
вальную закономерность, которая слабо прослеживается от
Памира, Алая через Центральный Тянь-Шань к популяциям
Северо-Восточного и характеризуется уменьшением числа
щитков в процессе их олигомеризации.
Количество верхнегубных щитков впереди подглазничного.
В наших выборках изменчивость данного признака не наблю-
дается. Как исключение у единичных особей (0,8%) отмече-
но пять верхнегубных вместо обычных четырех.
Окраска и рисунок. Описания окраски и рисунка A. alai-
cus, изложенные в работах (Елпатьевский, 1901; Никольский,
1907, 1915; Кащенко, 1909; Терентьев и Чернов, 1949; Анд-
рушко, 1951; Параскив, 1956; Богданов, 1960; Яковлева, 1964;
Банников и др., 1977), весьма противоречивы. Мы считаем
целесообразным привести описания окраски и рисунка
A. alaicus некоторых вышеупомянутых авторов. В. С. Ел-
патьевский: «...спина бурая, немного светлее боковых полос,
с двумя узкими светлыми продольными полосами». А. М. Ни-
кольский: «...сверху тело оливкового цвета с двумя продоль-
ными рядами темных пятен, внутри которых помещается зе-
леновато-белый стержень». Н. Ф. Кащенко: «...цвет сверху олив-
40
ковый, с четырьмя продольными рядами черных черточек,
окаймленных по бокам белыми пятнышками». П. В. Терен-
тьев и С. А. Чернов: «...сверху буроватых тонов с оливковыми
или зеленоватыми оттенками. В середине некоторых спин-
ных чешуй светлые продольные черточки, располагающие-
ся по спине четырьмя продольными рядами, с боков они окай-
млены темно-бурым». А. Г. Банников и др.: «Посередине от-
дельных спинных чешуй светлые короткие черточки, распо-
ложенные четырьмя продольными рядами, каждая из них по
сторонам с небольшим темным пятном».
В вышеприведенных и, на наш взгляд, неудачных приме-
рах описания рисунка A. alaicus особое внимание уделено
наличию на спине отдельных светлых и темных черточек и пя-
тен, образующих общий вид. Изучая именно эти элементы
окраски ящериц, мы выделили три основных типа рисунка
верхней стороны тела (рис. 7).
Крупнопятнистый: на фоне темно-бурых пятен, по-
перечная направленность которых выражена слабо или от-
сутствует, выделяются два, реже четыре прерывистых про-
дольных ряда относительно крупных ярко-белых черточек.
Поперечно-полосатый: темно-бурые пятна, расположен-
ные на отдельных чешуях, образуют неровные попереч-
ные полоски, прерывающиеся светлыми черточками.
Продольно-полосатый: тусклые буроватые пятна и нечет-
кие светлые черточки образуют полоски продольной направ-
ленности.
Данные изменчивости рисунка показали, что первый и
второй типы характерны для асимблефаров Тянь-Шаня, при-
чем А доминирует в северо-восточной его части (Иссык-Ку-
ле— 89%, Заилийском Ала-Тау — 67%); В — тип встречает-
ся у ящериц Памиро-Алая — 84%.
В итоге анализа внутривидовой изменчивости A. alaicus
можно сказать, что отдельные важные признаки фолидоза,
пропорции тела и рисунок очень изменчивы, а многие из них
дискретны. Клинальная изменчивость признаков выражена
лучше в размерах тела и менее заметна в фолидозе. По диск-
ретным признакам (комбинации височных щитков, форме
носового щитка, положению предлобных щитков, количеству
затылочных и др.) ареал данного вида распадается на три
четко обособленных группы популяций: Памиро-Алая; Севе-
ро-Восточного Тянь-Шаня; Центрального, Юго-Западного и
части Северного (Киргизский хребет) Тянь-Шаня. Их мы
рассматриваем в качестве реальных подвидов:
1. Asymblepharus alaicus alaicus (Elpat., 1901)
2. Asymblepharus alaicus kucenkoi (Nik., 1902)
3. Asymblepharus alaicus yakovlevae Jeriomtschenko, 1983
41
Рис. 7. арьирование рисунка A. alaicus: слева направо— '—3—А. а.
alaicus; 4—6—A. a. yakovlevae (Киргизский хр.» ущ. Карабалта); 7—9—А.
a. kucenkoi (Кунгей Ала-Too. ущ. Чон-Кой-Су).
Таблица для определения подвидов A. alaicus:
1 (2) Предлобные щитки широко соприкасаются друг с дру-
гом и отделяют лобно-носовой от лобного (см. рис. 4)...
A. a. alaicus (Elpat, 1901).
2(1) Предлобные щитки обычно разделены передним углом
лобного щитка, последний соприкасается с лобно-носовым.
3(4) Центрально-височный маленький и свободный сверху
(рис. 8)... A. alaicus yakovlevae Jeriomtschenko, 1983.
4(3) Центрально-височный щиток большой трапециевидный,,
его верхний край прикрыт теменным щитком (см. рис. 9)...
A. alaicus kucenkoi (Nik., 1902).
Асимблефар алайский обыкновенный — Asymblepharus alaicus alaicua-
(Elpat., 1901).
Terra typica: видовая.
Лектотип видовой. Описание (см. рис. 4) составлено нами
по 121 экземпляру из Памиро-Алая (Киргизия и Таджи-
кистан).
L £.Л —42—49 (45,32 ± 0,57), 99 —43—60 (51,37±0,69)\
t=6,71 Ga + —21—26 (23,64 ± 6,36), 99 —25—37 (29,5±
0.56), 1=8.80 Led*-? — 46—52 (49,73± 1,88), 99 — 45—66
(55,44± 1,42), 1=2,42 L головы <Ь<Ь —8,8—10.1 (9,46±0,П),
$9—85—10,2 (9,18+0,08), t=2,06. Ширина головы —6—
7,2 (6,7±0,11, 99—6—8 (6,77 ±0,08), t=0,51, Lpl * S —11 —
14 11 lt65±0,27), 99 —11—13,5 (12,2±0,09), t=l,93. Lpa
-11—81,5 (16,23±0,27), 99—14,5±18 (16,06+0,13).
( = 0.73. 51 r,l —52.1—55.11 (52.28±0.40), — 52.5—62.74
(56.96±0.34), t=8.81. —-64.95 ( 70,43±
' ’ Lголовы
0,93), 99 —66,66—76,4 (71,20+0,45), t = 0,74. -LP- io —
25,0—30,80 (27,28+0,43), 99 —19,77—26,77 (23.48±0,26),
t=7.6. —31.25 — 39.12 (36.1±0.53), 99 —27.32—34.2&
(31.31+0.29), t=8. Sq вокруг шеи dd —26—32 (28.53+0.18),
99 — 26—31 (28.6±0.12), t=0,32. Sq вокруг тела dd — 26—
29 (27.03±0.21), 99 — 25—29 (27.07±0.15), t=0.15. Брюш-
ных в продольном ряду dd—40—47 (42.66±0.35), 99— 41 —
51 (46.7+0.31), t=8.64. Подпальцевых чешуек на четвертом
пальце Р2 — dd— 16—20 (17.55±0.16), 99 — 15—20 (17.62 +
0.16), t = 0.31. Затылочных щитков 1 —4 : 1=2%,^- 1 = 14,5%,
43
3-27%, 2=33,5%/3=14,5% ~ 3=5%, 4—3,5%. Передних
2 *
верхнересничных щитков 2(100%). Центрально-височный щи-
ток двух типов: 1. /Чаленький свободный сверху — 52%;
2. Трапециевидный большой, верхний край которого прижат
нижним краем теменного щитка — 48%• Носовой щиток двух
типов: 1. Цельный — 87%; 2. Полуразделенный—13%. Ос-
новной фон верхней стороны тела буроватый с золотистым
оттенком. Рисунок спины продольно-полосатый, найден у
84% особей, рисунок поперечно-полосатый и смешанный, най-
ден у 16% особей.
Распространение. Населяет горы Памиро-Алая (Киргизию'
и Таджикистан) на высотах 2000—4000 м. Возможно, что под-
вид имеется и на территории КНР в сходных условиях, в
частности в хребтах Кук-Тунь, Кинг-Tay и Кашгарском. На
стыке Ферганского и Алайского хребтов проходит узкая зона
интерградации этого подвида с подвидом A. a. yakovlevae
(см. рис. 5).
Асимблефар алайский Яковлевой — Asymblepharus alaicus yakov-
levae Jeriomtschenko, 1983.
Terra typica: Киргизия, Киргизский хребет, ущелье Аламе-
дин (урочище Татыр), 1500 м.
1902 — Ablepharus alaicus Nikolsky (Part.)
1927 — Ablepharus alaicus kutzenkoi Kashkarov (part.)
Голотип. Re № 14, хранится в коллекциях Института зоо-
логии АН УССР (Киев); добыт в Киргизии, Киргизском хреб-
те, ущелье Аламедин (урочище Татыр), на высоте 1500 м leg.
В. К. Еремченко, 13.8.1977. t? L — 50, Ga — 27. L cd. reg.—
41.5. L головы—10.2, Lpl — 12.5 Lp2—17. Sg вокруг тела —
30. Затылочных щитков 1—2. Подпальцевых чешуек на
четвертом пальце Р2—19. Предлобные щитки разделены пе-
редним углом лобного. Наше описание (рис. 8) составлено по
302 экземплярам из Тянь-Шаня (исключая ящериц Иссык-
Куля, Сары-Джаза, Заилийского Ала-Тау, хребта Кетмень и
Кульджи), добытых в основном автором, а также по коллек-
ции Ирины Дмитриевны Яковлевой, автора первой сводки по
герпетофауне Киргизии, в честь которой назван данный под-
вид.
L сГс? — 40.5—52.0(46.234=0.20), 99 — 40.0—64.8 (51.63±
0,35), t=12. Ga^ 20.2—27.0(24.24=0.20), 99 —23.3—38.3
(29.374=0,26), t=15.66. Led cfcT—49.0—61.2 (54.724=0,63), 99 —
46.0—70.5 (55.514=0,56), t=0.94. L гол.dV—8.8—11.0 (9.664=
44
Рис. 8. Фолидоз головы A. alaicus yakovlevae Jeriomtschenko,
0.06),99—8.2—11.0 (9.46±0,04), t=2.8. Ширина головы ЛГ—
6.0—8.0 (7.0±0.05), 99—6.0—8.0 (7.1 ±0,04), t= 1,56.
Lpl сГсГ — 11.0—13.0 (12.0±0,06), 99 — 10.0—13.3 (11.79±
0.04), t = 2,9. Lp2 dV— 15.0—17.8 (15,92±0.08), 99 — 14.0—
18.0 (15.61±0.06),t=0,31. 48.05-55.27 (51.63±0.18),
Стя
99 -50.42—64.0 (56.7±0.19), t=19,5. dV — 0.75—0.96
(0.82±0.098), 99 —0.81—1.11 (0.92±0.0012), t=10.
Ширина головы 60 60_79 0 (72.69±0.49), 99—65.26—
L головы
84.1 (71.83±0.27), t=l,5. c?d* —23.6—29.26 (26.03±
0.15), 99—18,3—28.6 (22.98±0.14), t=15.25. -^2 -
48
ояя-39 5 (34.41+0.26), 99 — 24,1—36.0 (29.0±0.2), t-
164 Sg вокруг шеисГУ — 26—31 (29.18+0.15) — 26—
Д’ Ля 8+0 09) , 1 = 2,17. Sg вокруг телалг - 24—30 (26.7±
0 13) + —24—30 (26.34 ± 0.07), 1 = 2.4. Брюшных щитков
в продольном ряду: „V 39-50 (43.48+0.28), 99 -38-53
(46.26+0.1) t = 9.36. Подпальцевых чешуек на четвертом
пальце р2 сГсГ— 16—22 (18.39 ±0.15), $+ 15 23 (18.08+
009), 1=1.8. Затылочных 1—— 5: 1 = 1.2%, 9 1=0,7%, 2 =
9;0%,J 2=12,0%, 3 = 44,0%,-1 3= 19,0%, 4=10,3%, ^4 =
3.0%, 5 = 0,4%,^- 5 = 0,2%. Передних верхнересничных 1—
3 : 1=0,7%, 2 = 99%, 3 = 0,3%. Центрально-височный, как пра-
вило (99%), маленький и свободный сверху. Носовой щиток
цельный (100%). Между носовым и подглазничным еще три-
четыре щитка: 3 = 5,5%, 4 = 94,5%. Предлобные обычно (84%)
разделены передним углом лобного щитка, который соприка-
сается с лобно-носовым, реже они соприкасаются в одной точ-
ке (10%) или широким швом (6%). Основной фон верхней
стороны тела — от оливково-бронзового до буроватого с зо-
лотистым радужным отливом. Рисунок спины с выраженной
.поперечно-полосатостью (87%).
Распространение. Населяет весь Тянь-Шань, за исклю-
•чением Северо-Восточного. Северная граница подвида —
Киргизский хребет, восточная — хребет Борколдой, южная —
стык Алайского и Ферганского хребтов (см. рис. 5).
Асимблефар алайский центрально-азиатский —
Asumblepharus alaicus kucenkoi (Nik., 1902).
Terra typica: Киргизия Иссык-Куль.
1902 — Ablepharus kucenkoi Nikolsky
1909 — Ablepharus saposzhnikovi Kastschenko
Голотип. № 9542 (A. kucenkoi) хранится в коллекциях
ЗИН АН СССР (Ленинград); добыт на берегу оз. Иссык-
Куль. А. Кащенко, 8.8.1901.? . L — 56. Ga — 34. Led — 55:
L головы — 9. Lpl — 11. Lp2— 15. Sg вокруг тела 26. Заты-
лочных 2— 2. Подпальцевых чешуек на четвертом пальце
Р2—16. Предлобные щитки разделены предельным углом
лобного. Наше описание (см. рис. 6) составлено по 128 эк-
земплярам из Иссык-Кульской котловины (хребтов Терскей и
Кунгей Ала-Тоо), бассейна р. Сары-Джаз, Заилийского Ала-
Тау, хребта Кетмень и особей из Кульджи (КНР).
46
Рис. 9. Фолидоз головы А.
alaicus kucenkoi (Nik., 1902).
L ^—43.2—49.00 (45.77+0.32), qq —45.2—64.0, t=11.91.
Ga £0.2—26.0 (24.1+0.30), $ 2 -27.0 — 38.3 (31.66+0.41),
t= 15,02. Led +9 — 51—60.0 (53.39+1.04), 29 —49.2—62.0
(56.35+0.6), t = 2.46. L головы +2 — 8.3—9.5 (9.0+0.07),
99 — 8.1 — 10.1 (9.17±0.05), t=1.97. Ширина головы
6.0— 7.0 (6.52+0.11), 99 — 6.0—7.6 (6.63+0.05), t=0,91.
ЬрьЦ —Ц.О—13.3 (11.38+0.1), 99 — 10.4—12.8 (11.30+
0,06), t=0,73. Lp2 —14.0—16.1 (15.28+0.15), 92 -
14.0—17.0 (14.95±0.10), t= 1.83. U — 51.36—56.77
(53.42+0.28), ;? — 55.62— 64.57 (59.56+0.33), t=14.28.
’-0,75-0,90 (0,84 + 0,02), '.'2 —0.85—0.96 (0.91 +
cl
47
0,006), t=3,68. Ширина головы ££ — 65.60 — 81.17 (72.21±
L головы
±1.06), 99 — 66.7—77.77 (72.27±0.44), t=0.005. -^-65 —
23.40—26.08 (24.66+0.17), 99 — 18.56—24.33 (21.26±0.17),
t= 14.46. -^1^29.37—35.11 (33.20±0.36), 99 —25.0—
32.68 (28.08±0.22), t = 12.2. Sg вокруг шеи 55 26—32 (27.95±
0.21), 99 —26—31 (27.4±0.15), t = 2.1. Sg вокруг тела 65—
24—28 (25.75±0.12) 99 —24—28 (25.60+0.07), t=l.l.
Брюшных в продольном ряду 56 —34—46 (41.63+0.42),
99 —39—53 (46.07+0.28), t=8.9. Подпальцевых чешуек на
четвертом пальце Р2 53— 15—21 (17.32±0.22), 99 — 13—
19 (17.24+0.13), t=0,32. Затылочных 1—2 4:1=0,8%,
1 1=2,6%, 2=6,0%, 2~ 2=19,0%, 3 = 38,5%, 1 3=21,3%,
4=10%,“ 4=1,8%. Передних верхнересничных— 1—2 : 1 =
21,0%, 2=79,0%. Между носовым и подглазничным три—пять
щитков: 3=16,5%, 4 = 78,0%, 5 = 5,5%. Предлобные разделе-
ны передним углом лобного — 81,0%, соприкасаются в одной
точке—10,0%, соприкасаются широко и отделяют лобно-но-
совой от лобного — 9,0%. Центрально-височный щиток, как
правило (99,2%), большой трапециевидный и сверху прижат
нижним краем теменного. Основной фон верхней стороны те-
ла буроватый с оливковым, бронзовым или зеленоватым от-
тенком. Рисунок спины крупнопятнистый — 78,0% особей; со
слабо выраженной поперечно-полосатостью — 22,0%.
Распространение. Северо-Восточный Тянь-Шань. В СССР:
Киргизия (Кунгей и Терскей Ала-Too, бассейн р. Сары-Джаз),
Юго-Восточный Казахстан (Кунгей Ала-Too, хребет Кетмень).
За пределами СССР достоверно известен из Кульджи (КНР).
В последнем районе распространен возможно шире, в частно-
сти, в китайской части хребта Кетмень, а также Боро-Хоро.
Просмотренные нами два экземпляра под названием A. de-
serti (№ 1866 (2), хранящиеся в коллекциях Чикагского ес-
тественно-исторического музея (FMNH), добытые в Китае
без указания точного места сбора, даты и фамилии сборщика,
по нашему определению, также относятся к подвиду A. a. ku-
cenkoi. Подробнее его распространение показано на рис. 5.
Стации и количественные данные. В пределах своего
ареала алайский асимблефар населяет практически все пояса
48
гор: предгорья, средне- и высокогорья, поднимаясь до линии
вечных снегов. Верхний предел распространения вида 4000 м
(хр. Акшийрак в Центральном Тянь-Шане (Яковлева, 1964).
Нами эта ящерица найдена в западной части Сусамырско-
го хребта на высоте 3300 м у нижней границы снега. В Вос-
точном Тянь-Шане Р. П. Зимина (1959) указывает ее для
каменистых скоплений морены ледника Кара-Баткак до вы-
соты 3350 м (Терскей Ала-Too). На Памире ее добыл
В. Громбчевский на высоте 3800 м в окрестности высокогор-
ного оз. Яшиль-Куль (коллекции ЗИН АН СССР). Главной
особенностью распространения алайского асимблефара яв-
ляется то, что его ареал нигде не выходит из горной зоны,
причем на севере ареала (Северный Тянь-Шань) он обитает
на высоте не менее 800 м и не спускается ниже 1500 м его
южной части (Памиро-Алай). По характеру местообитаний
О. П. Богданов (1954, 1965) отнес эту ящерицу к обитателям
горизонтальных поверхностей с твердым грунтом. Причем,
эта группа пресмыкающихся способна находиться не только
на ровных, но и на поверхностях с большим углом наклона.
По нашим наблюдениям и данным в литературе (Шнитников,
1928; Андрушко, 1951; Параскив, 1956; Зимина, 1959; Богда-
нов, 1960; Яковлева, 1964) в Киргизии, Казахстане и Узбе-
кистане асимблефары живут на склонах гор, покрытых полу-
пустынной, степной травянистой растительностью, на альпий-
ских и субальпийских лугах, по берегам многочисленных рек
и ручьев, в каменистых осыпях, у отдельных камней, выхо-
дов скал; в лесной зоне встречаются на опушках южных скло-
нов, среди кустарников или каменистых нагромождений. В
Таджикистане (Саид-Алиев, 1979) живут на склонах гор с
зарослями арчи, густой травянистой растительности, с об-
ломками скал; обитают на степных участках, по долинам рек
и ручьев. А. Г. Банников и др. (1971) отмечают этот вид для
с^дов и виноградников южных предгорий, по берегам арыков,
на поливных огородах и т. д. Однако последние указания для
A. alaicus маловероятны и относятся к другому виду — A. de-
serti Strauch. Асимблефары на юге своего ареала (Южная
Киргизия, Узбекистан, Таджикистан) не встречаются ниже
1500 м^т. е. в пределах культурной зоны.
Разнообразие местообитаний алайского асимблефара мож-
но разделить на два основных типа, в которых наблюдается
и различная численность вида: это закрытые и открытые био-
топы. К первым относятся прежде всего природные условия
Северного и Северо-Восточного Тянь-Шаня, отличающиеся от
условий других районов Тянь-Шаня и Памиро-Алая относи-
тельно большим развитием растительного покрова, присут-
ствием еловых лесов, других древесных пород и кустарников
4 302
49
Рис. 10. Стация A. alaicus: Казахстан, Заилийский
Ала-Тау.
(рис. 10, И). Ко второму типу местообитаний асимблефара
относятся биотопы, представляющие основные пространства
средне- и высокогорных степей и пустынь Центрального и
Юго-Западного Тянь-Шаня, а также Памиро-Алая, которые
отличаются от предыдущих относительно слабым развитием
растительного покрова и большей каменистостью стаций
(рис. 12). Следует отметить, что в Ферганском хребте данный
ьид ящериц встречается выше пояса леса (Яковлева, 1964).
Вопрос численности асимблефара изучен недостаточно.
В Алайской долине А. М. Андрушко (1951) отмечает этот
вид как многочисленный по левую сторону р. Кызыл-Суу, од-
нако конкретных сведений о его численности не приводит.
Отсутствуют эти данные и в других источниках (Чернов,
50
Рис. 11. A. alaicus: Киргизия, Киргизский хр., ущ. Лламсдин
1959; Саид-Алиев 1979). По нашим наблюдениям, в Алай-
ском хребте (окрестностях с. Дараут-Курган) на высоте
2500—3000 м по берегу р. Дараут среди камней, полыни и
редких кустов шиповника на 500 м маршрута 5.09 1979 г.
встречено 65 особей. Осмотр склонов на 100 м выше от пой-
мы этой речки показал низкую численность вида (на 500 м—
3 особи), но заметно большее присутствие гималайской ага-
мы— Agama himalayana (Steind.). Незначительное число
A. alaicus мы отметили в восточной части Алайской доли-
ны (3400 м) в окрестностях с. Сары-Таш. Здесь на 1 км
каменистой типчаковой степи отмечено всего три особи.
В центральном Тянь-Шане в долине р. Арпы, по наблюде-
ниям И. Д. Яковлевой (1964), в июне на маршруте 200 пог. м
найдено 46 особей, что соответствует 0,23 особи на 1 м. По
данным того же автора, в каменистых россыпях верховий
р. Каракуджур (Центральный Тянь-Шань) в мае на 9 м их
было учтено 20; в среднем по две ящерицы на 1 м. По на-
шим данным, в Восточном Караколе в устье р. Кашка-Суу
(2500—2800 м) 1 июня 1980 г. на 200 м каменистой пустыни
южной экспозиции отмечено 20, а рядом, по склону юго-вос-
точной экспозиции, в это же время — 45. В той же стации, но
выше от речки на 500 м асимблефары нами не встречены
(5 ч экскурсии), но найдены глазчатые ящурки —Eremias
51
Рис. 12. Стация A. alaicus: Киргизия, Центральный
Тянь-Шань.
multiocellata Gunther. По берегу высокогорного оз. Сон-Куль
(3050 м) на отдельном участке каменистого берега с кусти-
ками полыни, чебреца и шиповника 12.7 1978 г. на 10 м на-
ми учтено девять ящериц. На северном склоне Киргизского
хр. численность асимблефара была следующей: в ущ. Шам-
си (1600—2000 м) на остепненных лугах юго-западной экспо-
зиции в середине августа 1977 г. на маршруте в 500 м встре-
чено 27 особей; в урочище Горная Маевка (2000 м) среди
скальных обломков, кустов шиповника, полыни и др. травя-
нистой растительности соответственно на 100 м 28.9 1979 г.
было учтено 18; здесь же 28.9 1979 г. их отмечено 19; в
ущ. Ала-Арча (2000 м) среди кустов полыни и душицы на 500 м
26.5 1979 г. встречено 16; у подножья перевала Тюя-Ашу
(2600 м) по каменистому берегу р. Карабалтинки на 100 м
8.6 1979 г. — 42; здесь же, но выше на 200 м от речки по кру-
тому склону, среди щебнистых обломков скал и редкой рас-
тительности в это же время и на том же расстоянии учтено
только шесть особей; на Сусамыре 1.6 1978 г. по южному скло-
ну Киргизского хребта у подножья перевала Тюя-Ашу
(3000 м) среди отдельных камней найдено 12 особей на марш-
руте 500 м; в верховьях р. Сусамыр (сев. склон Сусамырско-
52
го хребта — 3500 м) 2.6 1978 г. на 500 м подъема в альпий-
ском поясе среди камней встречены только три ящерицы.
На Сусамыре в устье р. Западный Каракол на 100 м в
скальных обломках, кустах полыни и шиповника 16.7
1978 г. — восемь. В Таласе, на южном склоне Киргизско-
го хребта в ущ. Кууганды (2000 м), в сходной стации на
500 м 17.5 1979 г. — 12. В ущелье р. Чичкан в поясе елового
леса среди камней 7.6 1979 г. на 500 м — 48 ящериц.
В Северо-Восточном Тянь-Шане (хр. Кунгей Ала-Тоо),
в ущ. Чон-Кой-Суу (2300 м), в поясе елового леса, среди
камней, кустов караганы, шиповника и другой рас-
тительности на 500 м 17.8. 1978 г. отмечено семь особей; в
этом же хребте, но в ущ. Кырчин в сходной станции 22.6
1979 г. за 4 ч экскурсии встречено три. В Терскей Ала-Тоо,
ущелье р. Джеты-Огуз (2600 м) в поясе леса с обильной рас-
титель. эстью за 3 ч. 25.6 1980 г. учтена одна особь; одна
ящерица на 2 км сходной станции отмечена 29.6 1980 г. и в
ущелье Барскаун (2200 м). В Заилийском Ала-Тау по нашим
наблюдениям в окрестности Медео, в поясе елового леса
9.8 1978 г. за 3 ч экскурсии отмечено четыре особи. По уст-
ному сообщению сотрудника КАЗ ЗИН (Алма-Ата)
3. К. Брушко, в Заилийском Ала-Тау выше пояса леса в каме-
нистых россыпях они встречаются чаще и местами много-
численны.
Численность A. alaicus распределена очень неравномерно
и зависит от наличия подходящих станций. Максимальное
количество вида сосредоточено по каменистым берегам рек
и ручьев Центрального, Северного и Юго-Западного Тянь-
Шаня, а также Алая. Минимальная численность свойствен-
на Северо-Восточному Тянь-Шаню, а также альпийским
участкам с ограниченным числом подходящих местообита-
ний (свыше 3200 м) и крутыми склонами.
Сезонный и суточный цикл активности. Как показывают
наши наблюдения, а также данные литературы (Параскив,
1956; Яковлева, 1964), сроки выхода асимблефаров из зим-
них убежищ зависят от климатических условий района, экспо-
зиции склона, а также высоты. В Северной Киргизии наиболее
раннее появление ящерицы отмечено нами 26.3 1977 г. в
ущ. Иссык-Ата (1600 м, Киргизский хр.), в восточном При-
иссыккулье, ур. Кар-Кыра (2400 м) при стационарном иссле-
довании— только 21.4 1976 г. На западных склонах хр. Сон-
Куль-Тоо в ур. Кара-Кече (2700 м) эта ящерица добыта 27.4
1958 г. (Яковлева, 1964).
По нашим наблюдениям, самцы появляются весной рань-
ше самок. Так, среди половозрелых особей, добытых 5—10 ап-
реля 1977 г. в ур. Татыр (Киргизский хребет), самцы соста-
53
вили 80%, самки — 20%. К лету частота встреч самок резко
возрастает. Например, в ущ. Аламедин 7.7 1978 г. из 17 яще-
риц 88,3% составили самки; в ущ. Ала-Арча 26.5 1979 г. из 11
девять были самки; в ур. Чон-Таш в июне 1979 г.—из 12—10»
в Таласе (ущ. Кууганды) все 12 особей, добытых 17.5 1979 г.»
Оказались самками. В ущ. Чичкан (Юго-Западная Киргизия)
среди отловленных 7.6 1979 г 36 половозрелых ящериц сам-
ки составили 100%. На Сусамыре, по данным И. Д. Яковлевой
(1964) 16.7 1963 г., соотношение самцов и самок было 1 7,3.
О большей встрече самок в весенне-летний период можно су-
дить и по серии из 31 взрослой ящерицы, добытой М. Рафаэ-
лем в Кунгей Ала-Too (окр. с. Ананьево). Самок оказалось
здесь 83,3%. Осенью (сентябрь-октябрь), т. е. ко времени ухо-
да ящериц на зимовку, число встреч самок на поверхности сок-
ращается, а самцов резко возрастает. Так, заметное увеличе-
ние их отмечено 27.9 1978 г в ущ. Аламедин: здесь из 26 прос-
мотренных ящериц 75% самцы. В ущ. Шамси среди 21 особи,
добытой с 16 по 29.10 1974 г., на долю самцов приходилось
84,5%. Последняя встреча асимблефара (4)в Киргизском хр.
отмечена нами 3.11 1974 г. в том же ущелье. Весной, в нача-
ле апреля и мае, они активны с 10—12 до 18—19 ч. На вре-
мя продолжительной облачности (более 1 ч) ящерицы исче-
зали в убежищах и до наступления солнечной погоды не по-
являлись на поверхности. Летом, в июне—июле, они активны
в течение всего дня с 10 до 19 ч; в самый сухой сезон года,
август, заметно некоторое смещение активности к первой по-
ловине дня (10—13 ч) и ко второй (16—18 ч), с периодом
депрессии между 13—16 ч.
Отношение к температуре. Возможность существования
рептилий в горных условиях при относительно низкой темпе-
ратуре воздуха (Стрельников, 1944; Андрушко, 1951) объяс-
няют относительно высокой солнечной инсоляцией. Мы нахо-
дили активных асимблефаров в солнечные дни апреля при
5—12°, а осенью, в конце октября, при 10—15° Обнаружен-
ная нами популяция асимблефаров на побережье высокогор-
ного оз. Сон-Куль (3050 м) существует при средней июль-
ской температуре воздуха 5—10° и частых осадках в виде сне-
га и града. Большинство других районов Тянь-Шаня и Пами-
ро-Алая, где распространен этот вид, также характеризуют-
ся экстремальными условиями.
Размножение. Характерной особенностью размножения
алайского асимблефара является яйцеживорождение. Для
вида оно впервые установлено В. Н. Шнитниковым (1928).
Имеющиеся в литературе (Андрушко, 1951; Параскив, 1956;
Богданов, 1960; Яковлева, 1964; Банников и др., 1977; Саид-
Алиев, 1979) сведения о размножении даного вида отрывоч-
54
ны. По данным И. Д. Яковлевой (1964), соотношение поло-
возрелых самцов и самок, добытых в Центральном Тянь-Ша-
не с апреля по сентябрь, равнялось 1 : 2,5, т. е. самцов на
61,7% меньше самок. По данным, приводимым А. М. Андруш-
ко (1951), для Алайской долины (с 8.6 по 24.9 1933 г.) их со-
отношение равнялось 1 : 1, а вместе с неполовозрелыми сам-
цов было на 15% больше. В окрестностях Алма-Аты тоже 1:1
(Параскив, 1956). Наши материалы указывают на неравно-
мерность полового соотношения асимблефаров для различ-
ных районов. Так, в Иссык-Кульской популяции оно состави-
ло 1 : 1,35, в целом с неполовозрелыми— 1 : 1,1. В Заилий-
ском Ала-Тау — соответственно 1:1,5 у половозрелых и
1 : 1,2 — вместе с неполовозрелыми. В Киргизском хребте для
143 ящериц, добытых с марта по октябрь, соотношение поло-
возрелых составило 1 : 1,84 и 1 : 1,2—всех вместе; в Централь-
ном Тянь-Шане у 65 ящериц, добытых в июне— 1 : 2; в Юго-
Западном Тянь-Шане у добытых с мая по август — 1 : 7,5 и
в целом — 1 :3,5. Для вида соотношение половозрелых сам-
цов и самок составило 1 : 2,55, а с неполовозрелыми 1 : 1,35.
Таким образом, в половом соотношении вида наблюдается
«асто заметное преобладание самок, причем в различных по-
пуляциях оно может быть обычным— 1 : 1, с заметным пре-
обладанием самок 1 : 1,35—1,84, и, наконец, явно нарушен-
ным — 1 : 7,5 (Юго-Западная Киргизия).
Согласно И. Д. Яковлевой (1964), спаривание у асимбле-
фара в Киргизии происходит в мае-июне и, возможно, в июле,
а в Таджикистане — в конце мая — начале июня (Саид-Алиев,
1979). По нашим наблюдениям, в Киргизии оно отмечено уже
с конца последней пятидневки марта — начале апреля (Кир-
гизский хребет: ущ. Иссык-Ата, Аламедин), т. е. спустя при-
мерно неделю—полторы после их выхода из зимних убежищ.
Здесь же массовое спаривание этих ящериц отмечено нами в
первых двух декадах апреля, а на стационаре Кар-Кыра в
Восточном Прииссыккулье — с конца апреля и до середины
мая. Косвенным показателем этого в популяции Тюя-Ашу
(2600 м) является наличие в июне на теле самок следов, ос-
Рис. 13. Следы спаривания у самки A. alaicus из Тюя-Ашу, Киргизия
тавленных челюстями спаривающихся самцов. И. С. Дарев-
ский (1967) подробно изучил данный момент у скальных яще-
риц, мы отмечаем его для сцинковых ящериц (рис. 13). У 24
самок из Тюя-Ашу отметины чаще находились на передней
части брюха в области сердца (33 отметины); реже — на зад-
ней (16) и средней (3) области брюха, передних (8) и задних
ног (12), хвосте (7), голове и шее (11). Некоторые самцы
тоже не лишены отметины, что свидетельствует об агрессив-
ности их поведения, а также оборонительной реакции самок.
Наличие нескольких отметин на теле самок позволяет судить
о неоднократности попыток спаривания. Так, одна из 24 ос-
мотренных самок из Тюя-Ашу имела на теле 15 отметин, но
чаще всего их бывает 2—5. Интересен факт абсолютного от-
сутствия отметин у 32 самок, добытых на день позже в ущ.
Чичкан. Укажем, что они, в отличие от самок Тюя-Ашу из
других районов, имеют в это время яркую оранжевую окрас-
ку нижней стороны тела, аналогичную самцам. Наличие пос-
ледних в ущ. Чичкан отмечено лишь однажды 20.8 1981 г.—
два экзмпляра.
По нашим данным и материалам И. Д. Яковлевой (1964),
у асимблефаров Киргизии наибольших размеров семенники
достигают в апреле 6—8х 3.1—5.2, а также в конце сентяб-
ря— 6—7.2X3—3.4 и марта — 3.9—7X2.3—4; в остальное
время они меньших размеров: в мае — 2.8—8X1.2—2.7; в ию-
не— 2.4—5X1—3; в июле — 3—5x1.5—2.5; августе — 2.5—
5.5X2—4.5. Согласно И. Д. Яковлевой (1964), у асимблефа-
ров от спаривания до появления детенышей проходит не ме-
нее двух месяцев; автор указывает на раннее развитие
яиц у одной самки из ущ. Аламедин, добытой 2.4 1954 г.
А. М. Андрушко (1951) считает, что в Алайской долине разви-
тие эмбрионов наблюдается в течение месяца. Учитывая, что
спаривание происходит с конца марта по июнь, по нашему
мнению, время развития эмбрионов длится 1.5—3, а в отдель-
ных районах 3,5 месяцев (Киргизский хр.: ущ. Аламедин, Ала-
Арча).
По И. Д. Яковлевой (1964) и нашим данным, у эм-
брионов длиной тела 14—18 мм закладываются конечности,
глаза, хорошо выражен хвост; у зародышей длиной 22—
26 мм — фолидоз; у эмбрионов 26—30 мм — хорошо выра-
жен фолидоз и конечности; у эмбрионов 29—34 мм появ-
ляется рисунок, имеется глазная щель; при длине 39—
40 мм окраска такая же, как у взрослых ящериц, глазная щель
более или менее прикрывается краями верхнего века-козырь-
ка: эмбрион сформирован. По нашим наблюдениям и данным
литературы (Андрушко, 1951; Яковлева, 1964), молодые появ-
ляются во второй половине июля и первой половине августа.
56
В Казахстане (Угамский хребет) массовое рождение этих
ящериц отмечено в середице июня (Параскив, 1956; Банников,
1977). Молодых в выводке 1—7. Как отмечено И. Д. Яковле-
вой (1964), количество эмбрионов и их размеры зависят от
величины самок. Установлено, что максимальное число эмб-
рионов (4—7) характерно для более крупных самок ущелий
Аламедин, Ала-Арча, а минимальное (1—4)—для обитате-
лей Центрального и Юго-Западного Тянь-Шаня, которые от-
личаются более мелкими размерами тела.
Содержание беременных самок этого вида в неволе позво-
лило установить сроки рождения молодых и их размеры:
первые две новорожденные ящерицы (L = 21—22.5; Lcd =
22.5—24 мм) появились 24 июля от самки, отловленной 2.6'
1980 г. в Восточном Караколе. У самки, добытой 13.7 1981 г.
вущ. Аламедин, первый детеныш родился утром 31 июля, а
к вечеру их было пять (L = 22—23.2; Lcd=23—24.3 мм). В
этот же день у двух других самок из Восточного Каракола
было три детеныша: первые два — в середине дня, третий —
спустя 5 ч; 4 августа явились на свет два детеныша от сам-
ки из Восточного Каракола. 2 августа — один от самки, най-
денной 25.6 1981 г. в ущ. Джеты-Огуз, второй и третий отме-
чены в этот же день (L = 22—22.5; Led = 23—24 мм).
Половозрелость асимблефаров наступает на втором году
жизни при длине самок из Алайской долины 52 мм, самцов —
45 мм (Андрушко, 1951). Согласно И. Д. Яковлевой (1964), —
при длине тела самок 43 мм, самцов — 40 мм. К. П. Парас-
кив (1956) указывает, что в Заилийском Ала-Тау половозре-
лыми эти ящерицы становятся при еще меньшей длине: сам-
ки— 35 мм, самцы — 43—44 мм. По нашему мнению, мини-
мальные размеры половозрелых асимблефаров в Казахстане,
приводимые К. П. Параскивом, не точны. Нами установлено;
что половозрелыми, т. е. участвующими в размножении, самки
асимблефаров становятся при длине тела не менее 40 мм в
популяциях, где ящерицы характеризуются меньшими разме-
рами, и не менее 43—45 мм среди относительно крупных яще-
риц; соответственно самцы — 40—41 и 43—45,8 мм.
Питание. О. П. Богданов (1960), проанализировав содер-
жимое 40 желудков ящериц из Узбекистана, указывает, что
в их рационе преобладали жуки (82,6%), затем перепонча-
токрылые (37,5%), двукрылые (22,5%), равнокрылые хобот-
ные (15%)- Среди прочей пищи найдены прямокрылые
(6 экз.), тараканы (1 экз.) и блохи (1 экз.). В Казахстане
(Шнитников, 1928; Шульпин, 1948) среди объектов питания
асимблефаров отмечают большое количество тлей, мух из се-
мейства Syrphidae, бабочек, оводов, моллюсков, личинок ко-
былки, поденок, цикад, комаров, клопов. О питании асимбле-
57
фаров в Алайской долине можно судить по данным А. М. Ан-
друшко (1951), проанализировавшей 78 желудков этих яще-
риц, где также преобладали жуки (33,3%) жужелицы, черно-
телки, листоеды, слоники и двукрылые (25,6%)—ктыри, мухи
и тахины; на втором месте были перепончатокрылые — пчелы,
муравьи (17,9%), прямокрылые (саранчовые, Acrididae, Acri-
dinae, Chortnippus sp.) и клопы, в основном щитники (16,6%).
Реже встречались равнокрылые: цикадовые, тли, Jassidae
(11,5%) и чешуекрылые (огневки — 5,1%). По данным
И .Д. Яковлевой, в Центральном Тянь-Шане (Арпе) в июль-
ском питании ящериц преобладали жуки, перепончатокрылые
и клопы; осенью встречались саранчовые, уховертки, клопы-
щитники, жуки и муравьи; в пище молодых ящериц обнару-
жены в основном муравьи. О питании этого вида в Таджи-
кистане сведения отсутствуют (Саид-Алиев, 1979).
Нами исследовано содержимое желудочно-кишечного трак-
та 112 асимблефаров, добытых в Киргизии в июле-августе
(табл. 9). Установлено, что основную пищу A. alaicus состав-
ляют насекомые (95%), среди которых преобладают жуки
(58,93%) и их личинки (20,54%). Несколько уступают по
своему значению летающие: равнокрылые-хоботные представ-
лены в основном массовыми видами псиллнд (19,64%), дву-
крылые— комариками-хирономидами и мошками (17,86%).
Перепончатокрылые в большинстве наездники (14%). Далее
следуют пауки (16,07), гусеницы бабочек (10,7%) и муравьи
(8,93%). Незначительно представлены клопы (5,36%), ухо-
вертки и бабочки (по 4,46%), прямокрылые и личинки мух
(по 2,68%). Единично встречаются улитки и многоножки.
Имеются также некоторые отличия в составе корма в зави-
симости от местообитания ящериц. Так, на побережье круп-
ных водоемов (Сон-Куль) возрастает процент встречаемости
летающих насекомых и хортобионтов: двукрылые здесь сос-
тавляют 50%, а равнокрылые хоботные — 41,67%. Однако и
здесь значительную часть представляют жесткокрылые (50%)
и их личинки (20,83%). Во влажных биотопах (Сусамыр), пе-
реходных к гумидным, по сравнению с остепненными (Вос-
точный Каракол, Аламедин, Шамси, Ала-Арча) возрастает
встречаемость личинок жуков (37,5%), гусениц (29,16%);
только здесь в питании ящериц обнаружены личинки мух
(12,5%).
Стереотип поведения, убежища. С утра ящерицы прогре-
ваются под солнцем в характерной для них позе: тело вытя-
нутое и прижатое к поверхности камня, почвы или стебля
травы, оно как бы расплющивается, увеличивая площадь
обогрева, передние ноги кистями прижимают спину,
иногда задние обнимают хвост сверху. В такой позе по-
58
Таблица 9
Результаты анализа содержимого желудочно-кишечного тракта
112 экземпляров A. alaicus из Киргизии
(Ала-Арча, Аламедин, Шамси, Каракол, Сусамыр, Сон-Куль)
Объект питания Встречаемость Общее количество
эк. % экз. %
1 2 3 4 5
Aranei 18 7,73 25 6,23
Chilipoda 1 0,43 1 0,25
Insect а
Blattoptera
Blatelidae 4 1,72 4 1,0
Ortoptera
Ciliptamus sp. 3 1,29 3 0,75.
Derrrrsiptera
Anisolabis anulipes Luc. 5 2,15 5 1,25,
HomOptera
Dtelphacidae 2 0,86 2 0,5
Cicadellidae 10 4,29 15 3,74
Psillinea 8 3,43 97 24,19
Aphidinea 3 1,29 8 2,0
Hemopt era
Nfibidae
Nabis sp. 3 1,29 3 0,75
Berytidae
Berytinus sp. 2 0,86 6 1,5
Miri&ae
Pithanus sp. 1 0,43 1 0,25
Coleopter a
Carabidae
Carabus erosus Notsch. 1 0,43 1 0,25
(личинки)
Amara aenea Deg. 1 0,43 1 0,25
Harpalus amphicollis Men. 1 0,43 1 0,25
H. auxins Duft. 1 0,43 2 0,5
H. masoreides 1 0,43 2 0,5
Aphomis azureus 1 0,43 1 0,25
Прочие виды 12 5,15 16 3,99
Staphilininae (имаго) 1 0,43 1 0,2®
(личинки) 1 0,43 1 0,25
Histeridae
Hister bissexstriatus F. 1 0,43 1 0,25
Scarabaedae 4 1,72 4 1,0
Aphodius grombczewskyi
D. Koch. 1 0,43 1 0,25
A. scroba F. 1 0,43 1 0,25
Rhyssemus germanus L. 2 0,66 2 0,5
Buprestidae 1 0,43 1 0,25
Anthaxia sp. 1 0,43 1 0,25
59»
Продолжение табл. 9
1
5
Elateridae 1 0,43 1 0,25
Agriotes sgtialidus Schw. 3 1,29 4 1,0
A. meticulosus 1 0,43 1 0,25
Melanotus sp. Melyridae 1 0,43 1 0,25
Elacus sp. 4 1,73 4 1,0
Cryptophagidae 1 0,43 1 0,25
Coccinellidae 3 1,29 3 0,75
Coccinella septempunetata L. 1 0,43 1 0.25
Tenebrionidae
Oodescelis sp. 2 0,86 2 0,15
Blaps transversalis Gebl. 2 0.86 2 0.5 ‘
Opatrum sabulosHs L.
(личинки) 1 0,43 1 0,25
Прочие виды (имаго) 10 4,29 10 2,49
(личинки) 8 3,43 10 2.49
Mordeelidae
Anaspis sp. 1 0.43 1 0.25
Anthicidae
Pedilus sp. 5 2.15 6 1.49
Chrysomelidae
Luperus sericeus Jebz. 1 0,43 1 0,25.
Chaefocnema sp. 3 1,29 3 0,75
Прочие виды (имаго) Прочие семейства 8 2,15 5 1,25
(личинки) 2 0,86 2 0.5
Bruchidae
Euspermophagus sp. 1 0,43 1 0,25
Curculionidae 4 1,72 4 1,0
Polydrosus sp. 3 1,29 4 1,0
Apion sp. 5 2,15 6 1,49
Прочие виды 2 0,86 2 0,5
Прочие семейства 3 1,29 4 1.0
Hymenoptera
Tenthredinidae 1 0,43 1 0,25
Helictidae 2 0,86 2 0,5
Formicidae 6 2,57 8 2,0
Tetramorium caespitum L. 2 0,86 3 0,75
Formica picea Nyl. 1 0,43 2 0.5
Betillidae 1 0,43 1 0,25
Ichneumonidae 2 0,86 2 0,5
Braconidae 6 3,58 6 1,5
Trchoptera 1 0,43 1 0,25
Lepidoptera 1 0,43 1 0,25
Noctuidae — (имаго) 2 0,86 3 0,75
(гусеницы) 5 2,15 7 1.74
(куколки) 1 0,43 1 0,25
Прочие ночные бабочки
(имаго) 4 1,72 5 1,25
(гусеницы) 2 0,86 2 0,5
Diptera
60
Продолжение табл. 9
1 2 3 4 5
Tipulidae 1 0,43 1 0,25.
Chironomidae 8 3,43 36 8,98
Scimuliidae 6 2,57 14 3,5
Culicidae 1 0,43 1 0,25
Asilidae 1 0,43 1 0,25.
Leptogaster sp. 1 0,43 1 0,25.
Terevidae 1 0,43 1 0,25
Bibionidae Syrphidae 1 0,43 1 0,25
Eumerus sp. 1 0,43 1 0,25
Platycheirus anguatatus Zet. 1 0,43 1 0,25
Muscidae Прочие семейства 3 1.3 3 0,75
(личинки) 2 0,86 2 0,5
Gastropoda Всего: 3 234 1,3 3 398 0,75
коя ящерица находится в течение часа и более. Она меняется
с появлением движущегося насекомого. Если ящерица к
этому времени успела обогреться, она напрягается (конеч-
ности ее опущены и упираются в субстрат, голова устремле-
на вперед) и следит за движением жертвы. Мгновение, и жи-
вотное быстрым движением схватывает добычу, трясет,
трет ее о почву, затем целиком заглатывает. После этого
начинает активно разыскивать пищу или вновь возвращается
на прежнее место и принимает прежнюю позу отдыха. Насы-
тившись, долго отдыхает, время от времени передвигаясь к
тени, и таким образом регулирует температуру своего тела.
На период облачности (более часа) ящерицы исчезают под
камни, кусты травы, корни, другие убежища, но быстро появ-
ляются вновь, как только выглянет солнце. Самки по мере
развития эмбрионов значительно дольше остаются на поверх-
ности и чаще встречаются, чем самцы. По отношению к опас-
ности (приближение человека) асимблефары, как это было
замечено Р. П. Зиминой (1959) и нами в окрестностях Иссык-
Куля, затаиваются, забиваясь в дернину трав, в травянистую
ветошь, кусты шиповника, под камни или рыхлую почву под
корнями кустаников, бурьянистой растительности. Реже мы
замечали короткие перебежки этих животных, спасающихся
бегством. Чаще их наблюдали на перевале Тюя-Ашу, Восточ-
ном Караколе, Ала-Арче, Шамси и Алае. Особенно это харак-
терно для самцов и неполовозрелых особей. Затаившись в убе-
жище, спугнутая ящерица обычно вскоре появляется снова.
Интересно отметить, что асимблефары Сон-Куля, в от-
61
личие от других, совершенно не пугливы, их можно подманить
к себе шевелением пальцев рук из-за камня, они с любопыт-
ством ощупывали пальцы языком и даже устраивались в
своей излюбленной позе прямо на ладони. Возможная причи-
на столь необычного поведения сонкульских ящериц объяс-
няется отсутствием здесь змей (Яковлева, 1964; наши наблю-
дения), а один из самых опасных врагов — обыкновенная
пустельга (Cerchneis tinnunculus (L.) встречается здесь ред-
ко. Воды асимблефары не боятся и могут переплывать арыки
и небольшие ручьи (Банников и др., 1977). По нашим наблю-
дениям, в местах, где имеются колонии сурков, ящерицы мо-
гут находить убежище в их норах. Однако одним из излюб-
ленных мест укрытия служат камни и кусты травянистой рас-
тительности, под которыми они могут устраивать себе ходы,
как, например, в ущ. Аламедин. Ходы представляют собой от-
верстия диаметром 10—15 мм, идущие вниз или вбок от од-
ного камня к другому на глубину 1—40 см, причем наверху,
сразу под камнем, имеется как бы небольшая площадка 2Х
3 см с углублением 1—2 см, где ящерицу можно найти свер-
нувшуюся полукольцом.
Линька. По И. Д. Яковлевой (1964), линька аснмблефа-
ров совпадает с периодом их размножения. В Алайской доли-
не (Андрушко, 1951) этот процесс отмечен с мая по конец сен-
тября; наибольшее количество линных особей встречается в
июне, июле, августе и начале сентября. За время работы нами
было найдено и проанализировано более 240 остатков линных
шкурок, собранных с восьми ящериц, содержащихся в течение
трех лет в террариуме, а также обнаруженных в природе.
Ниже мы приводим систематизированное описание этого ма-
териала в единицах проб: линные части пилеуса — 26; подче-
люстные щитки с частью горловых чешуй—15; части туло-
вища, сошедшие кольцами (1,5—2 мм) — 56; крупные сегмен-
ты с крестцовой и плечевой области, сошедшие вместе с пе-
редних и задних конечностей — 25; отдельные чехлы с конеч-
ностей— 10; клочья шкурок со спины — 8; с брюха — 5; уз-
кие кольца шкурок туловища с разрывом в спинной — 24, и
брюшной части — 3; сегменты колец с основания хвоста — 17.
На основании вышеописанных проб частей и по визуаль-
ным наблюдениям нами установлено, что процесс линьки у
A. alaicus происходит в следующем порядке: первым ее приз-
наком у асимблефаров, как и у змей, является помутнение
глаз; к этому времени ящерица становится коричневой или
темно-бурой, радужные оттенки чешуй исчезают, затем в нес-
кольких точках тела шкурка лопается (чаще поперек спины)
я «сходит» кольцами, отдельными крупными сегментами или
клочьями. Подавляющее большинство шкурок находится в
62
вывернутом положении. Передняя часть тела обычно первой
заканчивает линьку. Таким образом, можно сказать, что про-
цесс у данных ящериц напоминает линьку у змей, но присут-
ствие у первых конечностей препятствует выворачиванию
цельной шкуры.
Враги, паразиты. К врагам A. alaicus относят змей — Ag-
kistrodon halys (Pall.) и Elaphe dione (Pall.) (Шульпин,
1948; Яковлева, 1964; наши данные). Из птиц, по нашим наб-
людениям, этих ящериц поедает пустельга (Cerhneis tinnuncu-
lus (L.). Орнитолог А. Ф. Ковшарь неоднократно (1964, 1966,
1979, 1981) отмечал эту ящерицу в питании черногрудой
красношейки — Calliope pectoralis Sev., пестрого каменного
дрозда — Monticola saxatilis (L.). По наблюдениям в Алае
(Андрушко, 1951) асимблефаров поедает барсук (Meles ше-
les (L.). Данный список врагов является неполным и в даль-
нейшем, несомненно, будет увеличен.
На этих ящерицах нами впервые были обнаружены пара-
зиты: личинки клещей-краснотелок семейства Trombiculidae
родов Eltonella, Neotrombicula и Leptotrombidium (Централь-
ный и Юго-Западный Тянь-Шань), зараженность которыми не
составляет исключения для таких высокогорных популяций,
как Сон-Куль (3050) и Алай (3000 м). В то же время крас-
нотелки на ассимблефарах в Северо-Восточном Тянь-Шане,
Киргизском хребте и Сусамыре нами не обнаружены.
Практическое значение. Почти полное отсутствие в горах
Тянь-Шаня и Памиро-Алая настоящих ящериц (Lacerta) и са-
мое широкое распространение здесь A. alaicus указывает на
то, что они являются для этой горной системы ее важным ком-
понентом. Относительно высокая численность и широкий
спектр их питания показывают, что значение данного вида в
биоценозах гор значительно и его можно приравнивать к роли
ящериц рода Eremias других районов Средней Азии или та-
ковой скальных ящериц Кавказа.
Данный вид встречается во всех горных заповедниках Кир-
гизии, Узбекистана и Казахстана и наряду с другими живот-
ными подлежит охране.
РОД ГОЛОГЛАЗЫ — ABLEPHARUS
(FITZINGER IN LICHTENSTEIN, 1823)
Типовой вид: Ablepharus pannonicus (Lichtenstein, 1823)
Син.: 1843.— Microblepharus Fitzinger
1972.— Blepbarosteris Stoliczka
Глаза аблефаридные, нижнее веко срослось с верхним, об-
разовав прозрачную покрывающую глаз оболочку; на пред-
челостной (premaxillare) кости девять плевродонтных зубов.
Конечности пятипалые, относительно слаборазвитые; на внеш-
них сторонах предплечий и голеней обычно нет сильно увели-
ченных щитков1; наружное ушное отверстие маленькое, его
длина укладывается в диаметре глаза более пяти раз или пол-
ностью отсутствует; вокруг середины туловища 18—26 рядов
гладких чешуй. Яйцекладущие.
Род включает семь видов: гологлаз афганский — A. lind-
bergi Wettstein, 1960; гологлаз полосатый — A. bivittatus
(Men., 1832); гологлаз азиатский — A. pannonicus (Licht..
1823); гологлаз малый — A. grayanus (Stoliczka, 1872); голог-
лаз европейский— A. kitaibelii Bibr. et Вогу, 1823; гологлаз
пустынный — A. deserti Str.» 1868; гологлаз Чернова — A. cher-
novi Darevsky, 1953.
Ареал видов охватывает Евразию: Южную Европу (Чехо-
славакию, Венгрию, Румынию, Сербию, Албанию, Грецию, ост-
рова Эгейского моря), Переднюю и Среднюю Азию на вос-
ток до Западной Индии включительно. В СССР распростра-
нены в Закавказье, республиках Средней Азии и Южном
Казахстане.
Таблица для определения видов рода Ablepharus:
1(12). На спине четыре продольных ряда чешуй. Расширен-
ные щитки на предплечье и голени отсутствуют.
2(6). Лобно-теменных щитков два.
3(9). Надглазничных щитков три, два из них соприкасаются
с лобным;
1 Исключение представляет A. lindbergi Wettstein, 1960.
64
верхнересничные щитки отделяют надглазничные от
края глаза; впереди подглазничного четыре верхнегуб-
ных щитка.
4(5). Височных щитков в первом вертикальном ряду три;
вокруг середины тела 22—24 рядов чешуй; предлобные,
как правило, широко соприкасаются и отделяют лоб-
но-носовой от лобного.........................A. bivittatus.
5(4). Височных щитков в первом вертикальном ряду два;
предлобные обычно разделены передним углом лобно-
го .............................................A. deserti.
6(2). Лобно-теменной щиток один.
7(8). Ушное отверстие маленькое, но всегда имеется; вокруг
середины тела 20—22 чешуи ....................A. pannonicus
8(7). Ушное отверстие отсутствует; вокруг середины тела
18—19 чешуй ..................................A. grayanus.
9(3). Надглазничных щитков два, оба соприкасаются с лоб-
ным; ряд верхнересничных щитков прерван, и надглаз-
ничные касаются глаза; впереди надглазничного три—►
четыре верхнегубных.
10(11). Число чешуй в продольном ряду от уха до централь-
но-височного щитка не более двух; впереди подглаз-
ничного три — четыре верхнегубных, вокруг середины
тела 18—22 чешуи. Носовой щиток обычно полураз-
делен ...................................... A. kitaibelii.
11(10). Число чешуй в продольном ряду от уха до централь-
но-височного равно пяти. Впереди подглазничного
три верхнегубных, вокруг середины тела 18—19 че-
шуй. Носовой щиток цельный ..............................
.....................................A. chernovi.
12(1). На спине шесть продольных рядов чешуй. На пред-
плечье и голени имеются расширенные щитки................
.....................................A. lindbergi.
ГОЛОГЛАЗ АФГАНСКИЙ —ABLEPHARUS LINDBERGI
WETTSTEIN, 1960
Terra typica: Северо-Западный Афганистан, Герат.
Син.: 1960 — Ablepharus bivittatus lindbergi Wettstein, Zool. Anz. 165:
61;— 1965, Mertens, Senkenb. Biol., 46: 3. — 1969 a,
Fuhn, Rev. roum. Biol. Ser. Zool., 14: 1: 31.—1969 b,
Fuhn, Sounderdr. a. z. f. Zool. Syst. u. Evolut. 7 : 69. —
1970, Levi ton and Anderson, Proc. Calif. Ac Sci.,
38,10:190.— 1974, Greer, Austral. J. Zool., 34:9.- 1977,
Банников и др., : 155.
1980 — A. lindbergi Wettstein, 1960, Еремченко, Щербак,
Вести, зоол., 6:84. — 1981, Еремченко, Вопросы герп.,
Автореф. докл. V Всесоюзн. герпет. конфер., Л.: 54. —1981,
Jeriomtschenko, 1 Conf. Herpet. Resp. Soc., Buda-
pest : 21
5 302
65
Диагноз. Туловище плотное, укороченное, ноги относи-
тельно развитые. Височных щитков в переднем ряду три. Но-
совые цельные. Лобно-теменных два. На спине шесть продоль-
ных рядов чешуй; вокруг середины тела их 26 рядов. На
внешних сторонах голеней и предплечий они сильно расши-
рены.
Отличие от близких видов. Как было показано (Ерем-
ченко, Щербак, 1980 в), A. lindbergi в противоположность
другим гологлазам имеет шесть продольных рядов спинных
чешуй, относительно длинные ноги и сильно расширенные
щитки на голенях и предплечьях. Кроме того, от A. bivitta-
tus афганский гологлаз отличается относительно коротким
расстоянием Ga.
Описание. (Рис. 14, 15, 16). Голотип № 15877, хранится
в Венском естественно-историческом музее (NMW), добыт в
Северо-Западном Афганистане— провинции Герат, leg. Линд-
берг, 28.8.1957 г. L — 53,5. Ga—22. Led. —48. L головы —
9,5. LPi—13,2. LP2—19. Sg вокруг тела — 26. Затылочных
щитков один—три. Подпальцевых чешуек на четвертом паль-
Рис. 14. Фолидоз головы A. lindbergi Wettstein, 1960.
66
Рис. 16. A. lindbergi (паратип, колл. ЗИН АН СССР).
це Р2 — 20. Предлобные щитки соприкасаются друг с другом
коротким швом и отделяют лобно-носовой от лобного. Носо-
вые щитки цельные.
Наше описание составлено по двум экземплярам — голо-
типу и паратипу (кол. ЗИН АН СССР). Последний является
неполовозрелым. Межчелюстной щиток коротким швом соп-
рикасается с лобно-носовым. Надглазничных три, из которых
два соприкасаются с лобным. Лобно-теменных два; межте-
менной по размерам равен лобно-теменному. Теменные
щитки смыкаются узким швом позади межтеменного. Пер-
67
Рис. 17. Распространение A. lindbergi (+) и A. bivittatus (•).
Кадастр к рис. 17.
Распространение афганского гологлаза — A. lindbergi
Афганистан: 1. Обе (Герат), Западный Афганистан (Wettstein, 1960;
Еремченко и др., 19806). — 2. Хр. Кух-и-Баба у оз. Банд-и-Амир,
2800 м; 70 км западнее Бамиана (Еремченко и др., 19806). —3. 5 км юж-
нее Тайвара (Mertens, 1965).
вый вертикальный ряд височных состоит из трех щитков.
Ушное отверстие относительно маленькое, его размеры ук-
ладываются в размерах глаза более семи раз. На переднем
крае уха две-три мелких зубовидных чешуйки. Sg. вокруг
шеи — 32; Sg. вокруг тела — 26. Брюшных в продольном ря-
ду 41—48. Затылочных три-четыре пары. Подпальцевых че-
шуй на четвертом пальце Р2—19-20, Верхнересничных — пять.
Количество чешуй верхнего «века-козырька три-четыре. Впере-
ди подглазничного четыре верхнегубных. Между носовым и
подглазничным расположено четыре щитка. Предлобных два,
они коротким швом соприкасаются друг с другом и отделяют
лобно-носовой щиток от лобного. Центрально-височный — ма-
68
ленький и свободный сверху. Носовой — цельный. Впереди
клоакальной щели два крупных щитка. Основной фон верх-
ней стороны тела буровато-коричневый. На шее и спине по-
середине отдельных чешуй имеются беловатые черточки,
окантованные по краям черно-бурыми пятнышками, которые
в сочетании образуют слабовыраженный поперечно-полоса-
тый рисунок. По бокам туловища от скулового щитка через
глаз и далее до основания хвоста (заходит за последний) про-
ходит узкая темная полоса, испещренная черно-бурыми и
светлыми крапинками; под ней такой же ширины ярко-белая
с неровными краями, очерченная снизу бурым. Низ тела фик-
сированных экземпляров беловатый. Неполовозрелая особь
имела в общем более светлую окраску и менее выраженный
рисунок спины.
Распространение (рис. 17). Известен из горных районов
Северо-Западного Афганистана, провинции Герат (Wettstein,
1960), в хребте Кух-и-Баба (коллекции ЗИН АН СССР).
Ссылка Р. Мертенса на обитание афганского гологлаза в Се-
веро-Западном Пакистане (Пенджаб) нуждается в уточне-
нии, поскольку номера, под которыми он (Mertens, 1965) упо-
минает данный вид, указаны те же и для азиатского голо-
глаза — A. pannonicus.
Экология вида не изучена. По имеющимся сведениям
(Wettstein, 1960) и наблюдениям научного сотрудника ЗИН
АН СССР Е. Сугоняева, этот гологлаз встречается на полу-
пустынных склонах гор. Его отмечали на высоте 2926 м (Le-
viton and Anderson, 1970).
ГОЛОГЛАЗ ПОЛОСАТЫЙ —ABLEPHARUS BIVITTATUS
(MEN., 1832)
Terra typica — Талышские горы—Юго-Восточный Азер-
байджан.
Син.: 1832 — Scincus bivittatus М е n е t г i е, Catal. rais. des obj. de Zool.
1839 — 64. — Ablepharus Menetreesi, Dumer et Bibr. Er-
pet gened., 5 : 8811.
1844 — Ablepharus bivittatus Menetrie, Gray, Catal. Lis., 64.— 1867,
S t r a u c h, Melange, biol. Ac. Petersb. 6:563. — 1887, В о u-
lenger, Cat. Liz. Brit. Muz. 3:353. — 1899, Никольский,
Изв. Импер. о-ва люб. естест., антроп. и этнограф. 2,23.7:42. —
1901, Елпатиевский, дневн. зоол. отд. о-ва люб. естест.
3,1:2.
1915 — Ablepharus bivittatus (Menetr., 1832), Никольский. Фауна
Росс, и сопр. стран, Преем. (Reptilia), 1:492. — 1929, Со-
болевский, Мемуары зоол. отд. о-ва люб. естествозн.,
антроп. и этногр., 5:45. — 1939, Чернов, Зоол. сб. 1:
131. — 1949, Терентьев и Чернов, Опред. преем,
и земн.: 87. — 1952, М i 111 е m a n, Smiths. Miscell.
Coll., 117,17:22. — 1969a, Fuhn, Sounderdr. a. z. f. Zool.
Syst. n. Evol. 7,1 : 7. — 1969в, Fuhn, Rev. roum. Biol. Zool
6»
Рис. 18. Фолидоз головы A. bivittatus (Men., 1832).
14,1:. — 1974, Greer, Austral. J. of Zool., 31:9. — 1977,
Банников и др., :154. — 1981, Еремченко, Автореф.
докл. 5 Всесоюзн. герпет. конф.: 54. — 1981, Jeriomtschenke,
1 Confer. Herpet. Resp. Sco., Budapest: 21.
Диагноз. Туловище плотное, удлиненное (Lmax. — 61 мм),
ноги относительно короткие. Височных щитков в первом ряду
три. Носовые щитки полуразделенные. Лобно-теменных два.
На спине четыре продольных ряда чешуй; вокруг середины
туловища их 20—24 рядов. Увеличенные щитки на внешних
сторонах голеней и предплечьях отсутствуют. Предлобные,
как правило, широко соприкасаются друг с другом и отде-
ляют лобно-носовой от лобного.
Отличие от близких видов. От пустынного полосатый го-
логлаз хорошо отличается наличием трех височных щитков
в первом вертикальном ряду, широко соприкасающимися пред-
лобными щитками, а также узкой беловатой продольной по-
лоской на боках тела; от европейского — более крупными раз-
мерами, наличием трех надглазничных щитков и непрерыв-
70
ним рядом верхнересничных. От двух других видов — азиат-
ского и малого — двумя лобно-теменными щитками.
Описание (рис. 18). Лектотип № 565, выделен нами из
типовой серии (всего 3 экз.) коллекции ЗИН АН СССР, до-
быт в Талыше в окрестности с. Перимбель, leg. Менетри,
1830 г. $ L — 52. Ga — 30,8, Led. reg. —60. L головы — 9,5.
LPi— 10,5. LP2— 16. Sg вокруг тела — 22. Затылочных три-
четыре. Подпальцевых чешуй на четвертом пальце Ра—17.
Предлобные щитки широко соприкасаются и отделяют лоб-
но-носовой от лобного. Носовой цельный.
Наше описание составлено по 95 экземплярам из Армении,
Азербайджана, Туркмении и Ирана, хранящимся в коллек-
циях ЗИН АН СССР (Ленинград), Зоологического института
АН УССР (Киев), Зоомузея Московского Государственного
университета (Москва) и Чикагского естественно-историче-
ского музея (FMNH). •
L £6—41,0—50,0 (45,4±0,4), 9? —40,5—61 (49,55±0,7),
t=5,12. Ga 55 — 21,0—27,0 (23,62±0,31), 99 —23—33,4
(28,14±0,51), t=12,9. Led. 55 —60,0—87,7 (74,6±2,43),
59 — 63—87 (73,4±2,61), t=0,34. L головы 55—8—9,3
(6,9±0,05), 99 —7,2—10 (8,47±0,08), t=16,7. Ширина го-
ловы '5—5,8—7 (6,3±0,07), 99—5—7,2 (6,0±0,07), t=
3,03. LPi <55—8,8—12 (10,7±0,23), 99 —9—12,7 (10,5±
0,16), t=0,7. LP2 55—12,3—17,3 (15,6±0,23), 99 — 12,5—
пя
17 (15,5±0,15), t=0,36. -^55 —47,7—56,25 (51,90±0,35),
99 - 51,54-60,8 (50,5 ±0,35), t=9,18. -Д 55 — 0,52-0,66
ЬСа
(0,54±0,03), 99—0,61—0,83 (0,68±0,02), t=3,9. - ™лавь!
55—17,71—20,93 (19,6±0,14), 99— 15,63—18,83 (17,1±
0,11), t= 13,9. —И?ИНа ГОЛОВЫ55 _ 64,7—77,77 (70,8±0,68),
L головы
— 64,0—75,3 (7Ю,4±0,5), t=0,47. 5^ — 21,25—
26 ( 24,1±0,20), 99—19,28—24,51 (21,4±0,2), t=9,43.^55
— 32,27—39,3 (35,5±0,44), 99 —27,03—36,7 (31,5±0,32),
t = 7,26. Межчелюстной щиток широко соприкасается с лоб-
но-носовым. Надглазничных три, из которых два соприка-
саются с лобным. Лобно-теменных два; межтеменной по
величине равный лобно-теменному. Теменные смыкаются уз-
ким швом позади межтеменного. Первый ряд височных сос-
71
тоит из трех щитков. Ушное отверстие маленькое, его разме-
ры укладываются в размерах глаза 6—10 раз; на переднем
крае уха расположены одна—три мелких зубовидных чешуек.
Sg вокруг шеи ££—22—28 (25,37±0,11), 99—22—27
(25,18±0,17), t=0,8. Sg вокруг тела, ££ 22—24 (22,35±0,12),
9?—20—24 (22,43±0,13), t=0,45. Брюшных. ££ — 36—
45 (40,0±0,3), 99 — 39—47 (43,6±0,33), t = 8,0. Затылочных
2—6 пар: 2=1%,4 2 = 1%, 3 = 23%,-^- 3=9,6%, 4=46,8%,
-^-4 = 8,5%, 5=5,3%, -^-5=1%, 6 = 3,3%. Верхнересничных
2—6:2—1 %, 4=10,5%, 5 = 82,2%, 6 = 6,3%. Козырек сос-
тоит. из одного-пяти щитков: 1=3%, 2=6,3%, 3=70,4%,
4=14%, 5=6,3%. Верхнегубных впереди подглазничного 4—
5:4 = 98%, 5 = 2%. Между подглазничным и носовым распо-
ложено три—четыре щитка: 3 = 2%,4 = 98%. Подпальцевых че-
шуек на четвертом пальце р2 44—14—19 (16,62±0,16),
99 — 15—20 f 16,77±0,16), t = 0,68. Предлобных щитков два,
они широко соприкасаются — 99%, соприкасаются в одной
точке—1%. Центрально-височный щиток маленький и сво-
бодный сверху. Носовой цельный — 95%, редко полураздё-
ленный — 5%. Впереди клоакальной щели два больших
анальных щитка. Хвост снизу покрыт одним рядом сильно
расширенных щитков. Основной фон верхней стороны тела
фиксированных гологлазов оливково-коричневый, светло-ко-
ричневый, буроватый, зеленовато-коричневый с харак-
терным бронзовым отливом. На спине имеются два—
четыре прерывистых продольных ряда беловатых чер-
точек, окаймленных по краям черно-бурыми или темно-ко-
ричневыми пятнышками; они образуют выраженный попе-
речно-полосатый рисунок, встречающийся у 64% особей
(рис. 19, 20), у остальных (30%) на спине три узких бурова-
тых линии, проходящих по краям двух средних рядов спин-
ных чешуй; между ними могут располагаться редкие нечеткие
светлые черточки, окантованные бурыми пятнышками. 6%
особей имели однотонную окраску спины, лишенную рисун-
ка. От ноздри через глаз и далее по бокам туловища и ос-
нованию хвоста проходит широкая бурая полоса, окаймлен-
ная сверху и снизу уз'кой белой линией, выраженная в пе-
редней части туловища, а далее к хвосту она нечеткая, неред-
ко испещрена мелкими бурыми крапинками. Нижняя сторо-
на тела беловатая или у фиксированных — серовато-зелено-
ватая. Как сообщают некоторые авторы (Банников и др.,
1977; С. Ахмедов — устное сообщение), у половозрелых
самцов она (часть шеи, брюха, передняя часть хвоста и внут-
72
Рис. 19. Варьирование рисунка A. bivittatus, Армения, Личк
(колл. ЗММГУ).
ренние стороны конечностей) в период размножения приоб-
ретает золотисто-оранжевую или розовую окраску, которая
сохраняется приблизительно до конца июля, после чего зна-
чительно блекнет. Молодые сверху светло-кофейного или бе-
жевого цвета; на спине просматривается рисунок взрослых,
или он отсутствует и окраска однотонная. Сбоку окраска та-
кая же, как у взрослых. Хвост желтовато-оранжевый, особен-
но это заметно у новорожденных. С возрастом окраска хвоста
блекнет и на нем появляется рисунок, характерный для
взрослых ящериц. Следует отметить, что у некоторых из них
окраска регенерированного хвоста сходна с окраской моло-
дых.
Распространение. В СССР полосатый гологлаз споради-
73
Рис. 20. A. bivittatus (Иран,'Керманщах, колл. FMNH)
чески распространен в горах северной, центральной и юго-за-
падной частей Армении и Юго-Восточном Азербайджане до
высоты 2000 м (Алекперов, 1970). Единственный раз он был
добыт и в Юго-Западной Туркмении (хр. Кюрен-Даг). Вне
СССР этот гологлаз живет в Северном, Северо-Восточном,
Западном и Юго-Западном Иране. Его распространение в
Иране изучено недостаточно. По-видимому, здесь (горная
система Загроса) этот вид распространен более широко. Па
некоторым сведениям полосатый гологлаз, возможно, имеет-
ся и в Северо-Восточной Турции (Mertens, 1952), однако, па
мнению других (Basoglu, Baran, 1977) его там нет (см.
рис. 17).
Кадастр к рис. 17.
Распространение полосатого гологлаза — A. bivittatus
СССР. Армения: 1. Окр. Кировокана. —2. Окр. с. Спиток. —
3. Окр. с. Даштун. —4. Хосровский заповедник. —5. Окр. Анкавана. —
6. Окр. Коджерана. —7. Мергннский р-он, с. Личк. —8. Окр. с. Мегри.
Пункты 1, 3 (Чернов, 1939); 2, 7 (колл. ИЗ АН УССР); 4, 5, 6 (колл.
ЗММГУ), 8 (колл. ЗИН АН СССР).
Азербайджан: 1. С. Перимбель. —2. С. Внлянчай. —3. Окр. постов-
Аваш и Дыманского. — 4. Лерикский р-н, с. Келваз. — 5. С. Дыман. —
6. Окр. с. Джанги. —7. С. Розанове. Пункты даны по Н. И. Соболевско-
му (1929).
Туркмения: 1. Хр. Кюрен-Даг, окр. с. Даната (колл. ИЗ АН УССР).
Иран: 1. Шахруд (Никольский, 1915). —2. Карадаг, Сарма-Дария.—
3. Подножье горы Сехенд. —4. Горное плато от горы Сехенд, между
с. Картевюль и Эшпа. —5. Кушкизард, между Ширазом и Исфаханом. —
6. Табриа. —7. Казвнн. —8. Керманшах (Курдистан). Пункты 2—4’
(колл. ЗИН АН СССР), 5—7 (Fuhn, 1969 а), 8 (колл. FM NH).
Систематика и географическая изменчивость. До недав-
него времени большинство исследователей (Wettstein, 1960;
Mertens, 1965; Fuhn, 1969а, в; Leviton and Anderson, 1970^.
1974; Банников и др., 1977) принимали точку зрения, сог-
ласно которой полосатый гологлаз является видом политипи-
ческим, включающим по крайней мере три подвида: номина-
тивный, A. b. lindbergi (Wettstein, 1960) и A. b. alaicus (El-
pat., 1901). Правомочность, выделения последнего не была
признана отечественными герпетологами, поскольку видовая
самостоятельность «alaicus» с этой стороны (Банников и др.^_
1977) не вызывала сомнений. Нами показана ошибочность
включения в данный вид форм «alaicus» и «lindbergi», кото-
рые, как выяснилось, в действительности представляют со-
бой самостоятельные виды, относимые к различным родам:
Asymblepharus alaicus (Elpat., 1901) и Ablepharus lindbergi
Wettstein, 1960. Объем A. bivittatus, таким образом, был вос-
становлен в прежних рамках монотипического вида.
75-
Таблица 10
Изменчивость размеров тела у A. bivittatus
Выборка п L Оа
11m M±m 11m M±m
££ 22 41—48 44,8±0,44 20,5—27 23,1 ±0,36 Армения $9 27 40,5—56 49,43 ±0,74 22—33,2 28,0±0,56 2 44—47 45,5±1,5 23—25 24,0±1,0 Азербайджан 10 42—50,2 49,2 ±1,9 24—33,4 28,22±1,2 7 46—50 47,3±0,56 23,7—27 25,0±0,43 Иран 9? 5 46—61 51,0±2,73 25—30 29,12±2,16
Таблица И
Достоверность различий (t)
Выборка
U-L-99 99—Ga—99
Иран — Азербайджан — 0,54 — 0,36
Иран — Армения — 0,14 — 0,5
Азербайджан — Армения — 0,11 — 0,16
Таблица 12
Изменчивость количества чешуй у A. bivittatus
'Выбор ка п Sg вокруг тела Sg вокруг шеи
11m M±m lim М±ш
1 64 20—24 22,3 ±0,08 22—27 25,31 ±0,14
2 17 22—24 22,17±0,13 22—26 24,35±0,30
3 14 22—24 22,92±0,24 25-28 26,21 ±0,26
76
Таблица 16
Изменчивость количества подпальцевых пластинок
на четвертом пальце Ра у A. bivittatus
№ вы- борки п 11m M±m
1 64 15—20 16,8±0,20
2 17 15—18 16,7±0,23
3 14 16—19 16,8±0,58
Таблица 14
Достоверность различий (t)
Выборка Sg вокруг тел Sg вокруг шей Количество подпальцевых пластинок на четвертом пальце Р2
Иран — Азербайджан 2,77 5,02 0,11
Иран — Армения 2,41 3,1 0
Армения — Азербайджан 0,9 2,9 0,13
Таблица J*
Изменчивость количества чешуй у A. bivittatus
№ выборки п Нт М±т
1 и 31 36—44 40,0±0,37
1 99 30 39—47 43,5±0,42
2 8 38—45 40,0±0,83
99 9 41—47 43,2 ±0,70
3 U 9 38—43 40,4±0,68
99 5 44—47 45,0±0,55
Таблица 16
Достоверность различий по количеству
брюшных чешуй у A. bivittatus
№ вы- борки ?2
1-2 0 0,37
1-3 0,59 2,0
2-3 0,52 2,17
2 16 — — 56,5 6,0 37,5 — — — —
3 14 7,0 — 14,0 28,5 43,5 7,0 — — —
Таблица 18
Изменчивость количества верхнересничных щитков у A. bivittatus, %
№ выборки 4 5 6
1 12,3 81,5 6,2
2 6,2 87,6 6,2
3 7,1 85,8 7,1
С целью выяснения закономерностей географической из-
менчивости собственно A. bivittatus имеющийся в нашем рас-
поряжении коллекционный материал разделен на три выбор-
ки: 1. Армения (64); 2. Азербайджан (17); 3. Иран (14). Ни-
же изложены результаты изучения основных признаков
(табл. 10—18).
L туловища и Ga. Максимальные размеры туловища
(61 мм) отмечены у $ из Ирана, но расстояние Ga больше у
9? двух других выборок. Однако достоверных отличий меж-
ду ними по этим признакам нет.
Положение предлобных щитков. У всех изученных нами эк-
земпляров, за исключением Кюрендагского, предлобные щит-
78
Рис. 21. Фолидоз головы A. bivittatus (Men., 1832) из Туркмении.
ки широко соприкасаются друг с другом и отделяют лобно-
носовой от лобного. Надглазничных щитков, как правило, три,
исключение составила одна особь из Армении, у которой от-
мечено два щитка с одной стороны головы. Носовой цельный,
очень редко (5%) встречаются особи, имеющие полуразде-
ленный носовой щиток. Пределы варьирования признака верх-
нересничных щитков от двух до шести. Минимальное отмече-
но у особи из Кюрен-Дага. У остальных было четыре-шесть,
но подавляющее большинство в каждой из этих трех выборок
имели по пять.
По литературным данным, пределы варьирования коли-
чества чешуй вокруг середины туловища 22—26 (Банников
79
и др., 1977); по нашим—20—24; минимальное число (две ) их
отмечено у особи из Армении, однако в среднем гологлазы
Армении и Азербайджана несколько отличаются от иранских
меньшим числом чешуй (см. табл. 12—14). Изменчивость
брюшных щитков менее выражена.
Обращает на себя внимание наличие у большинства осо-
бей Азербайджана (56,5%) трех затылочных щитков, тогда
как в остальных выборках максимум особей имеют по четы-
ре. Изменчивость всех остальных признаков (количество под-
пальцевых пластинок, щитков между подглазничным и носо-
вым, верхнересничных) практически не выражена.
Приведенные данные показывают, что в пределах иссле-
дованного ареала популяции полосатого гологлаза не обнару-
живают прерывистой изменчивости; некоторые признаки фо-
лидоза (Sg вокруг тела, шеи, количество брюшных щитков)
проявляют клинальную изменчивость с юга на север.
На этом фоне исключительный интерес приобретает недав-
но обнаруженный экземпляр гологлаза в Юго-Западной Турк-
мении (хр. Кюрен-Даг, окрести, с. Даната, 20.04 1963, leg.
Н. Н. Щербак), который мы предварительно отнесли к A. bi-
vittatus. Данный экземпляр выявлен в коллекциях Зоомузея
ИЗ АН УССР (Киев), где он хранился под названием A. de-
serti Str. При осмотре музейной коллекции мы обратили вни-
мание на необычную для пустынного гологлаза окраску, ко-
торая при сравнении с другими видами соответствовала толь-
ко A. bivittatus и в некоторой степени — A. lindbergi. При
тщательном исследовании этого экземпляра были обнаруже-
ны и другие признаки (рис. 21), указывающие на большее
его сходство с A. bivittatus. В частности, у него имелось че-
тыре спинных ряда чешуй, три височных щитка в первом ря-
ду, цельный носовой щиток, три — четыре увеличенных щитка
верхнего века, а также пара лобно-теменных. В то же время
по другим признакам (наличию маленьких, едва соприкасаю-
щихся в одной точке предлобных щитков, двух — трех верхне-
ресничных щитков) кюрендагский экземпляр отклоняется от
A. bivittatus. Наибольшее значение при этом приобретает
признак положения предлобных щитков: у всех просмотрен-
ных нами экземпляров A. bivittatus из Ирана, Армении и
Азербайджана они хорошо развиты и, как правило, широко
соприкасаются друг с другом и отделяют лобно-носовой щи-
ток от лобного (100%). Таксономическое значение данного
признака становится еще более весомым при сравнении кюрен-
дагского экземпляра с экземпляром A. bivittatus из Шахруда
(Иран, № 6522, коллекции ЗИН АН СССР), добытым
А. М. Никольским в 1885 г. примерно в 300 км юго-западнее
с. Данаты. Гологлаз из Шахруда имеет типичные для A. bi-
80
vittatus широко соприкасающиеся предлобные щитки. Не от-
личается он от остальных особей из Ирана, Армении и Азер-
байджана и по другим признакам. Таким образом, шахруд-
ский экземпляр полосатого гологлаза вопреки ожиданию не
разделяет, а соединяет ареал данного вида, подчеркивая
своеобразие кюрендагского экземпляра.
На возможность нахождения A. bivittatus в Копет-Даге
впервые указал еще А. М. Никольский (1899). Он высказал
свое предположение, добыв в Шахруде экземпляр этого ВИ'
да, который первоначально был определен А. Штраухом как
A. deserti (Никольский, 1899). В свете этого факта небезын-
тересно отметить также и упоминание A. deserti П. Варен-
цовым (1894) для Закаспийской области. Однако, как позд-
нее отмечает А. М. Никольский (1915), последний вид ник-
то здесь не находил. Не найден он и в настоящее время (Бан-
ников и др., 1977; Шаммаков, 1974, 1981). Особого внимания
заслуживает указание Н. А. Зарудного (1897, 1898, 1904) по-
мимо азиатского гологлаза A. pannonicus, еще «какого-то го-
логлаза..., вероятно, A. bivittatus в горной стране между Ас-
хабадом и Кучаном в полосе туй и можжевельника».
Все это позволяет предполагать существование в Копет-
Дагском горном сооружении неизвестного в настоящее время
вида гологлаза, который, судя по всему, занимает промежу-
точное положение между видами A. bivittatus и A. deserti. До-
полнительные находки помогут окончательно установить его
таксономическое положение. Ниже приведено краткое описа-
ние экземпляра из Кюрен-Дага.
9L—25, Ga—13. Led — нет. L головы — 5,8. Lpi—6,3.
Lp2 — 9,1. Надглазничных щитков три, из них два соприка-
саются с лобным. Первый ряд височных состоит из трех щит-
ков. Ушное отверстие маленькое, его длина укладывается в
размерах глаза более 6 раз. На переднем крае уха располо-
жены три— четыре мелких чешуйки. Вокруг середины тулови-
ща 22 щитка. Брюшных — 45. Затылочных — три — четыре.
Верхнересничных — два и три. Козырек состоит из четырех
щитков. Впереди надглазничного четыре верхнегубных. Между
носовым и подглазничным тоже четыре щитка. Подпальце-
вых чешуй на четвертом пальце Р2-17. Предлобных два, они
маленькие и едва касаются друг друга в одной точке. Носо-
вой цельный. Основная окраска верхней стороны тела
голубовато-оливковая. На спине отдельные чешуи имеют
беловатые черточки, окаймленные по краям буроватыми пят-
нышками. Края спинной полосы ограничены беловатыми уз-
кими линиями. По бокам от скулового щитка до конца туло-
вища проходят темно-бурые полосы, отграниченные снизу бе-
ловатыми узкими полосками.
6 302
81
Стации и количественные данные. По литературным дан-
ным (Соболевский, 1929; Чернов, 1939; Банников и др., 1977;
Джафарова, 1979), полосатый гологлаз придерживается
сухих, сильно каменистых склонов, поросших колючими по-
душковидными астрагалами и другими ксерофитами, обра-
зующими растительные группировки типа фриганы. Реже они
встречаются на травянистых склонах, а также на южных
опушках лесов и кустарниковых зарослей. В Юго-Восточном
Азербайджане живет в песчано-каменистой полупустыне с
характерной сухолюбивой растительностью. На Малом Кав-
казе в пределах Азербайджана вид отмечен отдельными изо-
лированными очагами на участках, занятых астрагалами из
группы трагакантовых и кустами держи-дерева (Джафаро-
ва, 1979). По данным того же автора, в Кельбаджарском и
Лачинском районах A. bivittatus немногочислен: за 3 ч. эк-
скурсии в характерных для них биотопах можно встретить
два — четыре экземпляра, а в среднегорье Малого Кавказа на
2 км маршрута — до 8—12 особей. В Северной Армении
(Банников и др., 1977) на участке 100 м2 в начале июня учи-
тывали до 17 особей, а в Азербайджане на той площади в
конце мая—12 гологлазов. В жаркое время (июль — август)
число встреченных ящериц резко сокращается. Осенью, в
первой половине сентября, у подножья склонов среди мелких
камней можно встретить на участке 10 м2 до 37 сеголеток
(Банников и др., 1977).
Сезонная и суточная активность. После зимовки появле-
ние полосатых гологлазов отмечено в начале апреля — мая, а
уход на зимовку — в первой половине октября, причем моло-
дые исчезают значительно позже взрослых и встречаются еще
в первых числах ноября (Банников и др., 1977). Наиболь-
шую активность они проявляют в теплые солнечные часы
дня.
Линька. Коллекционные особи с признаками линьки от-
мечены нами в августе.
Размножение. Соотношение половозрелых самцов и са-
мок в нашей выборке (73 экз.) равно 1 : 1,35, т. е. самцов
42,5%, самок — 57,5%. В целом вместе с неполовозрелыми
(95 экз.) соотношение самцов и самок равно 1 : 1,05.
Исходя из данных, полученных при вскрытии 14 полово-
зрелых особей (б^с и 899), и сведений литературы (Бан-
ников и др., 1977; Джафарова, 1979) максимальные разме-
ры семенников (5X3 мм) отмечены у самцов с середины мая.
С конца июня и в августе они заметно уменьшаются. Отклад-
ка яиц начинается с последних чисел мая, но самки с яйцами
в яйцеводах встречаются до конца июня, возможно, до сере-
дины июля. Так, из восьми самок из Армении, добытых
82
.28.06 1974 г., одна имела готовые к откладке яйца размером
10x4,7, а остальные семь — лишь полусформированные раз-
мером 4—5,5X4—5,6 мм. Две самки из Ирана (г. Сехенд)
18 мая имели по 3—4 яйца размерами 6—4,5X5 мм; третья —
готовые к откладке два яйца размерами 10X5 мм. У самок,
пойманных в августе, фолликулы находились в состоянии
покоя, их размеры обычно не превышали 2 мм. Приведенные
материалы свидетельствуют о возможности у полосатого го-
логлаза повторных кладок, что отмечают и другие авторы
(Банников и др., 1977). По данным в литературе (Банников
и др., 1977) и нашим материалам, самки полосатого гологла-
за откладывают четыре, реже пять яиц размерами 9—11X5—
—5,5 мм. В августе и сентябре длина тела детенышей дости-
гает 22—38 мм. Половозрелость наступает к весне второго
года жизни при длине тела самцов 41, самок 43 мм, реже
отдельные самцы участвуют в размножении при длине тела
39—40, самки — 40,5 мм.
Питание. По литературным данным (Банников и др.,
1977), в желудках гологлазов из Армении преобладают ос-
татки мелких саранчовых, цикадок, перепончатокрылых, жу-
ков, тлей, пауков и мокриц. Найдены также крупные гу-
сеницы бабочек. Нами проанализировано содержимое 36
желудков данного вида (табл. 19), из Армении (20 экз.),
Азербайджана (7 экз.) и Ирана (9 экз.). Основу их корма
составляют жесткокрылые, в основном долгоносики (70,27%),
перепончатокрылые (43,24), из которых половина — муравьи
(21,62%), равнокрылые-хоботные преимущественно цикадки
(43,24%). Реже и в меньшем количестве встречались пауки
(38,84%), клопы (35,14%), гусеницы ночных бабочек
(21,62%), а также прямокрылые и мухи (по 10,8%). Еще ре-
же— личинки жуков и двукрылых (по 5,4%). Кроме того, в
желудках ящериц из Северного Ирана единично отмечены ос-
татки веснянок, богомолов, муравьиных львов, а у армян-
ских ящериц — улитки.
Убежища, стереотип поведения, враги. По Е. Менетри
(Соболевский, 1929), этот гологлаз иногда взбирается на низ-
кие кустарники. При преследовании прячется под камни,
кусты астрагалов, временами перебегает с одного растения
на другое, ловя насекомых (Даль, 1935). Его враги неиз-
вестны.
Охрана и практическое значение. Ввиду того, что биото-
пы полосатого гологлаза пока не затронуты деятельностью
человека, состояние популяций в Азербайджане (Лачинском
и Кельбаджарском районах) относительно стабильное (Джа-
фарова, 1979). На численность этого вида, как предполагает
вышеупомянутый автор, в отдельные годы влияют селевые
83
потоки, выкорчёвывающие кустарники, камни, под которы-
ми гологлазы находят себе убежища.
Поскольку вид является узкоареальным животным, он вне-
сен в красные книги Армянской и Азербайджанской ССР.
Мы предложили внести его в Красную книгу Туркменской
ССР. С целью сохранения полосатого гологлаза в Азербай-
джане некоторые авторы (Алекперов, Джафарова, 1979) ре*
комендуют выделить контролируемые участки на территории
Лачинского района.
Таблица 19
Результаты анализа содержимого
желудочно-кишечного тракта A. bivittatus
Объект питаиия Встречаемость Общее количество
экз. % экз. %
1 2 3 4 5
7 экземпляров из Азербайджана (с. Дыман, май)
Aranei Ortoptera 1 4,0 1 2,17
Acrididae Homoptera 1 4,0 1 2,17
Cicadellidae 5 20,0 15 32,61
Psyllinea 1 4,0 1 2,17
Hemiptera 2 8,0 2 4,35
Pentatomidae Coleoptera 1 4,0 1 2,17
Carabidae 1 4,0 1 2,17
Aphonus sp. Chrysomelidae 1 4,0 3 6,52
Alticinae Curculionidae 1 4,0 1 2,17
Ceutorzhinehinae Apininae 1 4,0 2 4,35
Apion sp. Neuroptera 3 12,0 5 10,87
Myrmeleontinae Hymenoptera Formicidae Crimatogaster sordidula 1 4,0 1 2,17
Hyl. 1 4,0 1 2,17
Monomorium sp. 1 4,0 3 6,52
Tapimona sp. 2 8,0 3 6,52
Lepidoptera 2 8,0 3 6,52
Всего 25 44
Продолжение табл. 19
1 2 3 4 5
20 экземпляров из Армении (Каджеран, Огбин, июль — август)
Aranei 10 16,95 15 20,0
Salticidae 1 1,69 1 1,33
Ortoptera Homoptera 3 5,08 3 4,0
Cicadelidae 5 8,47 8 10,67
Aphidinae 1 1,69 1 1,33
Hemiptera 4 6,78 4 5,33
Anthcoridae 2 3,4 4 5,33
Miridae 2 3,4 2 2,66
Pentatomidae Coleoptera 2 3,4 2 2,66
Meliridae 1 1,69 1 1,33
Tenebrionidae 3 5,08 2 4,0
Chrysomelidae Alticinae 1 1,69 1 1,33
Curculionidae 5 8,47 6 7,0
Mylacus sp. 1 1,69 1 1,33
Sytona sp. 1 1,69 1 1,33
Sibinia sp. 1 1,69 1 1,33
Apion sp. 6 10,16 9 14,67
Hymenoptera
Formicidae
Plagiolepsis sp. 1 1,69 1 1,33
Chalcidoidae 2 3,39 2 2,66
Jehneumonidae 1 1,69 1 1,33
Lepidoptera (бабочки) 5 8,47 5 6,66
1 1,69 1 1,33
Всего 59 72
9 экземпляров из Северного Ирана, окр. горы Сехенд, 17—18 мая
Агапеа 1 2,08 1 1,2
Plecoptera 2 4,16 3 3,61
Mantoptera 1 2,08 1 1,2
1 2,08 1 1,2
Ortoptera 1 2,08 5 6,02
Homoptera
Cicadellidae 3 6,25 5 6,02
Psillidae 3 6,25 15 18,02
Hemiptera 3 6,25 3 3,61
Coleoptera
Carabidae 1 2,08 1 1,2
Scarabaiidae 4 8,33 4 4,82
Cociineidae 1 2,08 1 1,2
Chrysomelidae 1 2,08 1 1,2
Curculionidae 2 4,16 2 2,41
Otiorzchinchinae 1 2,08 2 2,41
Ceutorchinohinae 2 4,16 2 2,41
Hymenoptera
Halictidae 1 2,08 1 1,2
85
Продолжение табл. 19
1 2 3 4 5
Formicidae
Messer sp. 1 2,08 1 1,2
Tetramorium sp. 3 6,25 11 13,25
Plagiolepis sp. 1 2,08 2 2,41
Jchneumonidae 3 6,25 3 3,61
Braconidae 1 2,08 3 3,61
Cynipoidea 1 2,08 1 1,2
Diptera 4 8,33 6 7,23
Tachinidae 1 2,08 2 2,41
Lepidoptera 5 10,42 6 7,23
Всего 48 83
ГОЛОГЛАЗ ПУСТЫННЫЙ - ABLEPHARUS DESERTI S Т RA U С Н, 1968
Terra typica: Ак-Мечеть (Казахстан).
Кариотип: 16v + Or + 12i (v+r), NF = 48; 2п = 30 (Куприянова^
1973).
1868 — Ablepharus deserti Strauch, Mel. Biol. Acad. Petersb.
6:564. — 1873, Северцов. Туркестан, животн.: 72. —
1882, Богданов. Очерки природы Хивинского оазиса:
98. — 1887, Boulenger, 3:857. — 1888, Грум-Гржи-
майло. Изв. геогр. о-ва, 23:96. — 1888, Кулагин. Изв.
импер. о-ва люб. естеств. 56:13. — 1888, Boettger, Zool.,
Jahrb. Syst. 3:218. — 1895, Варенцов. Приложение к
обз. Закасп. обл. за 1894 г. : 34, — 1899, Boettger in
R a d d е, Muz. Caucas. (Transcaspia) : 282. — 1899, Николь-
ский. Ежегод. Зоол. муз. АН; 176. — 1901, Елпатьев-
с к и й. Изв. Моск. о-ва. АН : 176. — 1908, Никольский.
Ежегодн. зоол.муз. АН. 13:340. — 1911, Никольский.
Ежегодн. зоол. муз. АН. 16:283.— 1915, Никольский.
Фауна Росс, и сопред. стран, пресмык. (Reptilia), 1 : 498. —
1928, Шнитников. Пресмык. Семиречья: 71. — 1936,
Даль, Тр. У ЗГУ, 7:1939. Андрушко и др., Вопросы
экол. и биоцен., 4 : 247. — 1948, Чернов. Преем, в кн.: Жи-
вотн. мир СССР, 2 : 142. — 1949, Терентьев и Чернов.
Опред. преем, и земн.: 87. — 1951. Калужина. Тр. биол.
поч. фак-та УЗГУ (зоол.) нов. сер. 46:88. 1952, Mi t tie-
man, Smiths. Miscell. Coll. 117, 17:24. — 1954, Дубинин.
Тр. Ин-та зоол. и паразит. АН УзССР, 3:165. —1955, Д а-
ревский. Изв. Туркм. АН:72. — 1956. Корелов. В кн.:
Природа и хоз. усл. горн, части Бостандыка : 266. — 1956,.
Параскив. Преем. Казахстана: 100. — 1959, Чернов.
Преем. Таджикистана: 112. — 1960, Богданов. Фауна
УзССР, 1:136. —1962. Богданов. Преем. Туркмении:
Ю4 —1961, Яковлева. Опред. преем. Киргизии:.
45. — 1964, Яковлева. Преем. Киргизии: 85. — 1977,
Еремченко. Торопова, Автореф. докл. 4 Всесоюзн.
герпетол. конф.: 89. — 1977, Банников и др. Опред. земн.
и преем, фауны СССР : 159. — 1969, Fuhn, Rev. roum. BioL.
Zool. 14,1:21. — 1969, в, Fuhn, Sounderdr. a. z. f. Zool.
Syst. u. Evolut 1:67. — 1974, Greer, Austral. J. of Zool.,
86
31,1:67. — 1980, Еремченко, Щербак. Вести, зоол.
4:14. —1981, Еремченко. Автореф. докл. 5 Всесоюзн.
герпет. конферен.: 54. — 1981.
1915 — A. tenuis, Никольский. Фауна Росс, и сопред. стран.
Преем. (Reptilia) 1:503.
1925 — A. turkestanicus, Ahl, Zool. Anz.,: 65,В,1/2 : 20.
1935 — Ablepharus sp. Мекленбу рцев. Tp. САГУ, 8а, 17:9.
Диагноз. Туловище тонкое удлиненное (Lmax. — 58,8 мм),
ноги относительно короткие. Носовые щитки, как правило, по-
луразделенные. Височных щитков в первом ряду два. Лобно-
теменных— два. Козырек состоит из одной увеличенной че-
шуйки. Вокруг середины тела 20—22 ряда чешуй.
Отличие от близких видов. Пустынный гологлаз легко от-
личается от азиатского и малого гологлазов наличием двух
лобно-теменных щитков и двух височных в первом ряду. По
последнему признаку он хорошо отличается от A. bivittatus
и A. lindbergi. Наличие у пустынного гологлаза одной увели-
ченной чешуйки, составляющей козырек, отличает этот вид
от других гологлазов.
Описание (рис. 22). Голотип № 569, хранится в коллек-
циях Зоологического института АН СССР (Ленинград); до-
быт в Ак-Мечетс (Казахстан), leg Н. А. Северцов. £ :L —
57. Ga — 37,9. Led. reg. — 46,5. L головы — 8. Lpl — 9,9.
Рис. 22. Фолидоз головы A. deserti Strauch, 1868
87
Lp2 — 14,2. Sg вокруг тела — 20. Затылочных g 2—3. Под-
пальцевых чешуек на четвертом пальце Р2— 17. Предлобные
соприкасаются в одной точке. Носовые полуразделенные.
Наше описание составлено по 402 экземплярам из Кирги-
зии (185), Казахстана (25), Узбекистана (141) и Туркмении
(51), хранящимся в коллекциях Зоологического института
АН СССР (Ленинград), Института зоологии АН УССР
(Киев), Зоомузея МГУ (Москва) и Института биологии АН
Киргизской ССР (Фрунзе).
L и— 37—48,5 (41,38+0,20), 99 —40,8—58,8 (48,13±
0,35), t= 16,9. Ga 20,0—29,8 (23,25±0,15), 99 — 24,0—
40,0 (30,36+0,3), t=21,8. Led —52,0—84,0 (66,84+1,36),
99 — 52,0—89,0 (67,51±1,4), t=0,34. L головы <*><Ь— 6,5—
8,0 (7,27±0,03), 99—6,8—8,3 (7,17±0,03), t=2,4. Ширина
головы 4т,—4,9—7,2 (5,4±0,04), 99 — 4,9—6,7 (5,21±0,03),
t = 3,52. LPi Sd —7,1—9,9 (8,57+0,05), 99 — 7,0—9,7
(8,4±0,04), t=ll. LP2 £*—10,2—12,7 (12,72±0,07), 99-
Ст *1
9,1—14,8 (12,34+0,07), t = 3,8. -^-££ — 49,04—62,63 (56,18±
0,23), 99 — 57,83 —68,73 (62,8±0,17),'t = 22,8.L г--^--в— <?>£>
— 14,5—19,18 (17,52+0,07), 99 — 12,84—18,14 (14,97±
0,08), t = 23,2. ££ —67,53—81,43 (73,57±
0,37), 99 — 84,28—64,1 (72,70+0,3), t= 1,8. ^££-16,35—
25,79 (20,65±0,13), 99 — 13,86—21,43 (17,6±0,12), t=16,9.
—26,77—35,15 (30,83 + 0,017), 99 —20,0—32,62
(25,72±0,18) t=20,6. Д-££ —0,53—0,69 (0,61+0,01), 99 —
LCD
0,61—0,88 (0,70±0,01), t = 6,4.
Межчелюстной щиток коротким швом касается лобно-носо-
вого. Надглазничных 2—4:2 = 0,01 °/о, 3 = 88,99%, 4= 11 %, если
надглазничных четыре, то с лобным соприкасаются три. Лобно-
теменных щитков два. Межтеменной один, по размерам пример-
но равный одному лобно-теменному или несколько меньше его.
Большие теменные смыкаются позади межтеменного. Первый
ряд височных состоит из двух щитков. Ушное отверстие ма-
ленькое, его размеры укладываются в размерах глаза не ме-
нее 8—10 раз или оно почти скрыто под чешуями. На перед-
нем крае уха имеются одна — две мелких зубовидных чешуйки,
88
которые часто бывают скрыты под височными щитками. Sg
вокруг тела U — 20—22 (20,81±0,07), 59 — 18—22 (20,5±
0,06), t = 3,4. Sg вокруг шеи — 23—26 (23,84±0,08), 98
22—26 (23,64+0,07), t = 2,0. Брюшных в продольном ряду
ео —37—50 (42,49+0,16), 99—41—56 (47,86±0,17), t=
23,3. Затылочных щитков % —5: = 0.5%, 1=0%,
-11=0,6%, 2—2,5%, *2=13,3%, 3—28,8%, *- 3—34,8%, 4 =
Ж
11,5%,-^ 4—7,6%, 5—0,3%. Подпальцевых чешуек на четвер-
том пальце Р2 <Ь6 — 13—20 (16,64±0,009), 99—13—20
(16,48±0,1), t=l,23. Верхнересничных 3—6:3= 1,6%, 4 =
12,4 %, 5 = 84,4%, 6=1,6%. Козырек верхнего «века», как пра-
вило, состоит из одной увеличенной ногтевидной чешуйки.
Верхнегубных впереди подглазничного четыре (100%). Меж-
ду подглазничным и носовым расположено три — пять щитков:
3—8%, 4—91%, 5—1%. Предлобных два, оба разделены пе-
редним углом лобного — 60,2%, соприкасаются в одной точ-
ке— 22%, соприкасаются коротким швом— 15%, или между
предлобными имеется дополнительный небольшой щиток —
2,8%. Центрально-височный обычно маленький и свободный
сверху — 98% или большой трапециевидный — 2%. Носовой,
как правило, полуразделен (99%), цельный редок— 1%. Впе-
реди клоакальной щели два больших овальных щитка. Хвост
снизу покрыт одним рядом расширенных щитков.
Основной фон верхней стороны тела сероватого, серовато-
оливкового, коричневого или буроватого цвета, часто с золо-
тистым оттенком. Однотонная окраска спины, ^не имеющая
рисунка, встречена у 11,8% исследованных особей; рисунок
в виде одной узкой буроватой линии, проходящей вдоль сред-
ней части спины, —у 3,6% особей; с тремя буроватыми про-
дольными линиями, проходящими по краям двух средних ря-
дов спинных чешуй, — 70% особей; тот же рисунок, но до-
полнительно между буроватыми линиями на середине отдель-
ных чешуй имеются светлые яркие черточки, окаймленные
по краям бурыми пятнышками, —у 5% особей; на спине пять
узких бурых линий, проходящих по краям четырех рядов
спинных’чешуй, между которыми на середине отдельных че-
шуй двух средних рядов могут быть светлые черточки,
окаймленные бурыми пятнышками — 9,6% (рис. 23). От
ноздри через глаз и далее по бокам туловища проходит бу-
рая полоска, ниже которой только на голове, шее и части ту-
ловища выражена беловатая узкая линия, на остальной час-
ти тела она отсутствует. На хвосте сбоку и сверху обычно
89
Рис. 23. Варьирование рисунка A. deserti (Таджикистан, Кайрак-Кум,
колл. ИЗАНУССР)
имеются продольные прерывистые ряды светлых черточек,
края которых окантованы темно-бурыми пятнышками. Ниж-
няя сторона тела ящериц беловатая или сероватая, низ го-
ловы и шея бывают испещрены темно-бурыми мелкими кра-
пинками. В период размножения у половозрелых самцов
брюхо, внутренние стороны конечностей и хвоста розового,
оранжевого или кирпично-красного цвета. Новорожденные
кремового или светло-коричневого цвета с золотистым оттен-
ком, хвост яркий, желтоватый или светло-оранжевый. По ме-
ре роста появляется рисунок взрослых и к осени большин-
ство молодых похожи на старших.
Распространение. Пустынный гологлаз является автох-
тоном глинистых полупустынь равнинной, а также предгорно-
пустынной зоны Средней Азии. Распространен в Южном Ка-
захстане, Киргизии, Узбекистане, северной части Таджикис-
тана и Восточной Туркмении. В северной части ареала дохо-
дит примерно до 46 параллели (Кызыл-Орда), южной — не
ниже 33-й (Мары, Сурхандарья), на запад — до 58° (Нукус),
на восток — до 77°(Йлийск). Вне СССР не найден (Банников
и др., 1977). Современный ареал пустынного гологлаза в зна-
90
чительной степени разорван; изолированные периферийные
популяции данного вида известны на значительном удалении
от основного ареала: в Южной Туркмении — по Мургабу (Да-
ревский, 1955), в Центральном Тянь-Шане — в долине р. Ат-Ба-
ши (Яковлева, 1964), Юго-Восточном Казахстане — в окрест-
ностях Илийска (Бутовский, 1964). Основной ареал пустынного
гологлаза (Киргизия, Узбекистан) в настоящее время также
не представляет единого целого и разбит на многочисленные по-
лу- или полностью изолированные поселения. Особенно замет-
но прослеживается изоляция между долинными и предгорны-
ми популяциями, например, в Северной Киргизии и Ферган-
ской долине. Высоко в горы этот гологлаз не поднимается,
но в отдельных районах он проникает на высоту 2400 м над
ур. м. (у перевала Камчик в Кураминском хр.) и около 3000 м
над ур. м. в Алайском хр., в районе Гульчи (Яковлева, 1964)
(рис. 24).
Систематика и географическая изменчивость. В настоя-
щее время, несмотря на отсутствие специальных работ, пос-
вященных внутривидовой систематике, принята точка зрения,
согласно которой A. deserti считается видом монотипическим
(Чернов, 1959; Богданов, 1960; Яковлева, 1964; Fuhn 1969а,
в; Greer 1974; Банников и др., 1977). Среди синонимов A. de-
serti эти исследователи указывают A. tenuis Nikolsky 1915 и
A. turkestanicus Ahl, 1925. Первое из этих названий дано
А. М. Никольским единственному экземпляру из Семиречья
(окрестностей Верного). Такие его признаки, как широко соп-
рикасающиеся лобные щитки и наличие четырех надглазнич-
ных заслуживают внимания и ниже будут нами рассмотрены.
Второй синоним A. turkestanicus появился благодаря А. Алю
(Ahl, 1925), который предполагал, что A. tenuis Nik. является
преокупированным (его уже применили к австралийскому
сцинку A. tenuis Broom, 1896). Вероятно, это обстоятельство,
а также отсутствие дополнительных материалов склонило
П. В. Терентьева и С. А. Чернова (1949) оба эти названия
считать синонимами A. deserti Strauch.
Возвращаясь к вопросу диагностической оценки призна-
ков A. tenuis Nik., следует рассмотреть географическую из-
менчивость A. deserti. Помимо типов нами изучены 10 выбо-
рок пустынных гологлазов, охватывающих практически весь
ареал (см. рис. 24).
В результате исследования установлено, что популя-
ции гологлазов Северного Тянь-Шаня (Чуйской долины
и обрамляющих ее хребтов Киргизского и Чу-Илийского)
по некоторым признакам (наличию четырех надглаз-
ничных (26,3%), широко соприкасающимся предлобным щит-
кам (31,57%), на которые указывал А. М. Никольский (1915),
91?
Рис. 24. Распространение пустынного гологлаза — A. deserti
Кадастр к рис. 24.
Казахстан: 1. Окр. г. Илийска (Бутовский, 1964). 2. Северо-западные-
отроги Чу-Илийского хр., 40 км на СВ от г. Фрунзе (он же). 3. Юж-
ный склон Чу-Илийского хр., ущ. Кара-Кунуз, окр. с. Ново-Александров-
ка. 4. Окр. Казалинска. 5. Кызыл-Орда (Ак-Мечеть). 6. Чимкент. 7. Окр.
Сары-Агач, р. Келес. 8. Аксу-Джабаглы. 9. Телекуль, 75 км от Петровска.
10. Джамбул (Ауле-Ата). Пункты 3,8 колл, автора (АН Кнрг. ССР, ИБ);
все остальные по колл. ЗИН АН СССР.
Киргизия: 1. Окр. г. Токмак (Еремченко и др., 1980 в). 2. Боомскоа
ущ. р. Чу в устье р. Чон-Кемин. 3. Пгт. Кант. 4. Окр. с. Норус. 5.
Окр. г. Фрунзе. 6. Окр. аэропорта «Манас». 7. Окр. с. Тюлек, р. Аксу.,
8. Окр. с. Полтавка. 9. с. Панфиловское. 10. Предгорье Киргизского хр.
ущ. Иссык-Ата, 1500 м. 11. Окр. совхоза «Ала-Тоо», 1600 м. 12. Ур. Та-
тыр, 1500 м. 13. Предгорье Киргизского хр., 5 км южнее Фрунзе. 14.
По дороге Сокулук — Белогорка. 15. Иссык-Кульская обл., Боомское ущ.к
отщелок Кок-Бак. 16. Южный склон Чу-Илийского хр., окр. с. XXII
партсъезда, ущ. р. Бейшекен. 17. Пойма р. Атбаши, 2400 м. 18. Окр.
с. Сары-Булун. 19. Долина р. Талас, 1234 м. 20. Совхоз им. ак. М. Н. Лу-
щихина, 20 км от Джамбула. 21. Таласский хр., окр. с. Кек-Сай. 22. Чат-
кальский хр., Сары-Челекский заповедник, с. Аркит, 1600 м. 23. Окр.
Джалал-Абада. 27. Ош. 25. Окр. г. Узгена. 26. Предгорье Ферганского,
хр., окр. с. Михайловка. 27. С. Ийри-Суу. 28. Ур. «Кара-Алма». 29. Арс-
ланбоб. 30. Окр. с. Узбек-Гава (Щербак, 1959). 31. Лесхоз Кызыл-Унгур.
32. Предгорье Алайского хр., ущ. Джантык. 33. Андарак. 34. Куршаб.
Пункты 1—7, 9—16, 20—21, 27, 28 сборы автора, колл. АН Кирг. ССР„
ИБ. Пункты 8, 17, 19, 24, 28, 31, 33 (Яковлева, 1964). Пункты 22, 23, 28
(колл. ИЗ АН УССР); 25, 34 (колл. ЗММГУ); 30. (колл. ЗИН АН
СССР). Выборки: I (пункты 1, 4—9, 11—14, 16), II (20, 21); VI (26, 27).
Узбекистан: 1. Окр. Орто-Токойского водохранилища. 2. Андижан. 3.
С. Балыкчи. 4. Урганча. 5. Фергана. 6. Коканд. 7. Маргелан. 8. Окр. На-
мангана. 9. Кураминский хр., пер. Камчик, 2400 м. 10. Искандар. 11.
Азат-Баш. 12. Чирчик. 13. Ташкент. 14. Чиназ. 15. По р. Ангрен у с. Той-
Тюбе. 16. Окр. с. Челек, р. Булунгур. 17. Оз. Большой Калтансыр па
р. Сыр-Дарья. 18. Хавастский р-он, окр. с. Джейлоянгар (Дубинин, 1954).
19. Самарканд. 20. Бухара. 21. Окр. Нукуса. 22. Бий-Базар. 23. Кара-
Куль (в Бухаре). 24. Джизак. 25. Окр. к. Хурам, 30 км к северу от
г. Бухара. 26. Хавает. 27. Сурхандарьинская обл., окр. ст. Шурчи. 28.
Китаб. 29. Шафрикан. 30. Хива. Пункты 13, 14, 20, 21, 23, 25, 26, 28, 30
даны по колл. ЗИН АН СССР; 2—5, 7, 24 (колл. ЗММГУ): 16, 27 (колл.
ИЗ АН УССР); 8, 9 (колл, автора; АН Кирг. ССР, ИБ); все остальные
пункты по О. П. Богданову (1960). Выборки: IV (пункты 13 и 7 из Ка-
захстана); V (6, 16 и 1 из Таджикистана); VII (14, 19).
Таджикистан: 1. Окр. с. Кайрак-Кум. правый берег р. Сыр-Дарья.
2. Предгорье Туркестанского хр., окр. с. Шахристан (Перешкольник,
1968). 3. Окр. с. Димай. 4. Аштский р-он, к. Кыр-Кудук. 5. Заповедник
«Тигровая балка». 6. Окр к. Жранбулак. 7. к. Авзыкеит. 8. Северный
скдой Гиссарского хр., к. Кызыл-Таш. 9. Северный склон Туркестанского
хр., ур. Кумбельсай, 2400 м. 10. Ура-Тюбе. Пункты 1 (колл. ИЗ АН
УССР); 3—5, 8, 10 (ЗИН АН СССР); остальные по С. А. Саид-Алиеву
(1979).
Туркмения: 1. Хатаб (Шаммаков, 1974). 2. Керки (Шаммаков, 1981).
3. С. Пальварт (Редженалыев, 1974). 4. Чардисоу (колл. ЗИН АН
СССР). 5. Фараб (Мориц, 1929). 6. С. Тахта (Шаммаков, 1974). 7. Па
дороге Майры-Байрам-Али (Даревский, 1955; Шаммаков, 1981). 8. Мары
(колл. ИЗ АН УССР).
93
Таблица 20
Изменчивость размеров тела у A. deserti
№ выборки Место обитания п 9 L Ga
11m M±m lim М^т
1 Чуйская 21 40—48,5 44,05±0,5 22,7—28 25,17±С,38
долина 35 45,3—58,8 51,63±0,66 27—37 33,06±0,51
2 Киргизский 13 39,3—44,2 41,38±0,45 21,5—26 24,2 ±0,43
хребет 14 42—50,8 46,22±0,65 26,8—33 29,21 ±0,48
3 Талас 5 38—42 40,8±0,8 21—23,2 22,44±0,41
8 39,5—51,5 47,35± 1,35 25—33 29,95±0,91
4 Ташкент 20 37—44 39,34 ±0,38 20,2—25 22,20±0,28
27 40,8—52,7 45,33 ±0,55 24—34 28,48±0,59
5 Ферганская 16 39—42,9 41,24 ±0,3 20,5—24,2 22,15±0,26
долина 13 41,8—58,4 48,3 ±1,34 25—34 29,59±0,93
6 Ферганский 6 37,5—44 41,48±0,98 20—24,2 22,53±0,64
хребет 14 43—52 46,79 ±0,75 25,8—31,8 28,73±0,54
7 Самарканд 13 37—43,9 40,54 ±0,44 21—25 22,58±0,22
25 42—50 45,33±0,65 25,1—32,8 27,86±0,57
3 Мары 11 39—44,4 41,42 ±0,63 21,1—25,2 23,68±0,42
11 43—55,3 48,76 ±1,36 26,2—36 31,22±1,09
Таблица 21
Изменчивость пропорций головы у A. deserti
№ -выборки п 6 9 L головы Ширина головы
Пт М±т 11т М^т
1 21 7—8 7,57 ±0,1 4,9—6 5,58±0,1
35 6,8—8 7,44±0,07 4,9—6 5,48±0,7
2 13 6,5—7,9 7,1 ±0,09 5—5,5 5,16±0,05
14 6,9—7,7 7,04 ±0,05 4,9—5,3 5,03±0,03
3 5 6,9—7,2 7,04 ±0,05 5,0—5,1 5,02±0,02
8 6,8—7,2 6,97 ±0,05 4,9—5,2 5,02±0,17
4 19 6,7—7,4 7,01 ±0,04 4,9—5,7 5,15±0,07
26 6,8—7,3 6,96 ±0,02 4,9—5,8 5,14±0,05
5 16 7—8 7,46±0,07 5,0—6,0 5,5±0,1
13 6,8—8,3 7,29±0,13 5,0—6,7 5,41±0,15
6 6 6,9—8 7,4±0,18 4,9—5,9 5,3±0,17
14 6,9—8 7,15±0,1 5,0-5,2 5,04±0,02
7 13 6,9—7,9 7,14 ±0,04 5,0—5,8 5,14±0,04
25 6,8—7,2 7,0±0,02 4,8—5,0 4,96±0,02
8 11 6,8—7,9 7,14±0,01 4,7—5,5 5,07±0,07
11 6,9—7,7 7,09 ±0,07 4,9—5,5 5,04±0,5
94
Таблица 22
Изменчивость длины ног у A. deserti
№ выборки п А 9 Lp, Ьр2
lim | М i m । 11m M±m
1 21 7,1—9,9 9,04 ±0,12 12,1 — 14,7 13,34±0,14
35 7,9—9,7 8,63 ±0,08 12,0—14,8 12,86±0,10
2 13 8,6—9,3 8,73±0,12 12,0—13,9 12,85±0,14
14 8,0—9,0 8,25±0,12 11,1—13,7 11,88±0,27
3 5 8,0—8,2 8,1 ±0,04 12,0—12,5 12,16±0,03
8 7,9-9,0 8,16±0,13 11,8—12,2 12,03±0,05
4 20 7,5—9,0 8,18±0,09 10,0—13,0 11,94±0,14
26 7,0—8,9 8,06±0,09 10,0—13,0 11,68±0,15
Ъ 16 7,5—9,1 8,37±0,1 11,5—14,0 12,93±0,19
13 7,8—9,2 8,31±0,14 11,1 — 14,4 12,36±0,28
•6 6 8,1—9,0 8,61±0,16 11,8—13,8 12,66±0,28
14 7,9—9,0 8,23±0,1 11,5—12,8 12,22±0,1
7 13 7,7—9,8 8,46=1=0,1 11,8—13 * 12,49±0,11
25 7,9—8,9 8,4±0,12 11,1 —13,Z 12,23±0,16
8 11 7,0-9,2 8,21 ±0,22 11,0—13,9 12,73 ±0,26
11 8,0-9,5 8,65±0,17 11,8—13,8 12,9=1=0,1
Таблица 23
Изменчивость количества чешуй вокруг тела и шеи
у A. deserti
№ выборки п Л 9 Sg вокруг тела Sg вокруг шеи
lim 11m M±m
1 21 20—22 20,80±0,20 23—26* 24,61±0,21
35 18—20 20,65±0,16 23—26 24,25 ±0,17
2 13 20—22 21,3±0,26 23—26 24,3±0,23
14 20—22 20,5 ±0,23 23—26 24,14±0,27
3 6 20—22 20,33 ±0,28 23—25 23,83±0,3
10 19—20 20,12±0,15 23—25 23,8 ±0,2
4 ’ 30 20-22 20,73=1=016 22—26 ,23,46 ±0,15
30 18—22 20,66±0,19 22—25 23,56±0,17
5 16 20—22 21,12±0,24 22—26 23,5±0,3
13 20—22 20,46=4=0,21 21—24 23,07±0,19
6 9 20—22 20.77±0,28 22—23 22,88±0,11
17 20—22 27,82±0,2 22—24 22,94 ±0,13
7 13 20—23 20,86±0,19 23—28 24,1 ±0,2
25 20—22 20,68=4=0,19 23—25 23,5 ±0,18
.8 38 20—22 20,42±0,11 22—26 23,52±0,13
30 20—22 20,16=4=0,1 22—25 23,36±0,14
95
Таблица 24
Изменчивость количества брюшных и подпальцевых чешуй у A. deserti
№ выборки п ? Количество брюшных чешуй Число подпальцевых чешуй
Мт MJz01 Мт М^т
1 18 41—48 44,33±0,46 16—20 17,16±0,25
31 45-56 49,51 ±0,43 13—18 16,51±0,22
2 13 41—47 43,92 ±0,57 15—18 17,07±0,26
14 48—51 49,5±0,48 15—20 17,0 ±0,39
3 6 39—46 42,5±0,1 16—20 16,83±0,65
10 45—53 48,3±0,8 15—18 16,6±0,26
4 30 37—45 41,66±0,32 14—19 16,41 ±0,22
30 43—51 46,9±0,36 14—18 16,3±0,1
5 16 40—45 42,12±0,4 14—18 16,2±0,3
13 46—49 47,53 ±0,33 14—19 16,3±0,41
6 9 40—46 43,77±0,74 14—18 16,0±0,47
17 44—52 48,17±0,58 14—18 15,58±0,27
7 28 39—47 42,0±0,34 14—19 16,7±0,21
22 43—52 46,77±0,56 14—19 16,68 ±0,31
8 38 38—46 41,5±0,3 13—18 16,32±0,18
30 41—51 46,06±0,46 14—19 16,56±0,19
Таблица 25
Изменчивость подглазничных и предлобных щитков у A. deserti
№ выборки п Надглазничные, % Предлобные, %
2 3 4 । разде. лены касаются . между ними, маленький щиток
в ши ро- точке ко
1 95 1,0 72,7 26,3 31,57 31,57 31,57 5,26
2 52 — 76,9 23,1 36,53 34,61 23,07 5,76
3 13 — 100 — 33,33 50,0 16,67 —
4 67 - 100 — 67,16 20,89 10,45 —
5 39 — 100 89,74 10,26 — —
6 25 — 100 — 88,0 12,0 — —
7 60 — 100 — 90,16 4,91 1,64 —
8 51 3,9 96,1 — 75,36 15,94 4,34 —
описывая A. tenuis, а также несколько более крупными разме-
рами тела отличаются от гологлазов других выборок, измен-
чивость которых более однородна (табл. 20—25). Обращает
на себя внимание также их значительная изолированность.
96
В целом различия (t) между последними и гологлазами всех
остальных выборок во многих случаях достоверны (табл. 26).
Вышеуказанное, несомненно, подчеркивает своеобразие
северо-тянь-шаньских пустынных гологлазов, которые, как
мы считаем, находятся в начальной стадии формообразо-
вания.
Стации и количественные данные. В Узбекистане, по наб-
людениям О. П. Богданова (1960, 1965), пустынные гологла-
зы придерживаются мест с повышенной влажностью — оро-
шаемых земель, где они живут по обочинам полей среди за-
рослей травянистой растительности, в садах, полезащитных
полосах, тугаях, по берегам водоемов и в предгорьях, по бе-
регам ручьев в зоне арчи. Чаще всего встречаются на лессо-
вых и глинистых почвах, реже — галечнике по берегам рек,
иногда на засоленных почвах с густой травянистой раститель-
ностью; эти гологлдзы наиболее многочислены на рав-
нинах и орошаемых землях, где местами можно встре-
тить весною за 1 ч. экскурсии 50—60 особей. В Ферган-
ской долине, по нашим наблюдениям, в г. Коканде пустын-
ный гологлаз живет по берегам арыков у глинобитных дува-
лов, густо заросших пыреем, бородачевником. Нами встрече-
но 14 июня 1977 г. на 20 шагов восемь ящериц. В этом же
городе на участке 14 м2 захламленного палисадника среди
засыпанных глиной бревен, кустов шиповника и сухих листьев
карагача 16 июня 1977 г. учтено 16 гологлазов различного
возраста. В Каракалпакии, восточнее Нукуса, на протяжении
350 м О. Утемисов (1973) учел 61 гологлаза. Этот же автор
(Утемисов, 1980) в целом для низовий Аму-Дарьи приводит на
1 км 14 особей в апреле и 15 в ноябре; здесь пустынный го-
логлаз живет по берегам арыков, в садах, виноградниках, ле-
сонасаждениях, люцерновых полях и неосвоенных глинистых
участках. В составе герпетофауны Приаральских Кара-Ку-
мов В. С. Лобачев и др. (1973) отмечает пустынного голо-
глаза как редкий вид, но конкретных сведений о его числен-
ности не приводят. На примыкающих к Голодной степи зем-
лях поймы Сыр-Дарьи по учетам О. П. Богданова (1965) за
однодневную экскурсию можно найти до 43 особей, а в оази
сах Язьяванской степи — до 82.
Значительно реже пустынные гологлазы попадаются в
предгорьях и низкогорьях Зеравшана (Богданов, 1960), где,
по данным С. К. Даля (1935), на 1 га отмечено только
0,13 особей. По нашим наблюдениям, на северном склоне Ку-
раминского хребта (перевал Камчик) на высоте 2400 м го-
логлазы обитают на каменистых склонах гор, поросших ши-
повником и разреженной травянистой растительностью, где
на 500 м 17.06 1977 г. их было встречено девять особей.
7 302
97
оо Таблица 26
Достоверность различий t некоторых признаков между объединенными выборками A. deserti
Признак Выборки t
1-3 4-8
lim M-tm । | lim ?/.±m i
39,3—48,0 42,9±0,33 37,0—46,0 40,64 ±0,22 5,7 L 42,0—58,8 50,0+0,56 39,5—56,0 46,87±0,4 4,6 22,0—29,8 24,56+0,25 20,0—25,2 22,6±0,13 7,0 Ga 26,2—40,0 31,9 ±0,42 24,0—38,2 29,35±0,34 4,7 6,5—7,9 7,3±0,06 6,9—8,1 7,23±0,03 1,0 L головы 6,8—8,0 7,29±0,05 6,8—8,3 7,1 ±0,03 3,17 4,9—6,0 5,4±0,05 4,9—7,2 5,38±0,06 0,25 Ширина головы 4,9—6,0 5,3±0,05 4,9—6,7 5,14±0,04 2,5 7,1—9,9 8,9±0,07 7,0—9,5 8,38 ±0,06 5,7 Lpi 7,8—9,7 8,5±0,06 7,0—10,0 8,3±0,05 2,43 11,9—14,7 13,1±0,09 10,2—14,0 12,54±0,09 3,5 Lp2 9,1—14,8 12,5±0,1 10,0—14,4 12,19±0,08 2,4 41—50 42,5 ±0,3 37—47 42,26+0,16 6,7 Брюшные чешуи 45—56 49,56±0,31 41—53 47,04±0,2 6,74 23—26 24,1 ±0,13 22—28 23,61 ±0,08 2,1 .Sg вокруг шеи 22—26 24,1 ±0.43 22—26 23,4±0,07 1,6 22—26 24,1 ±0,43 22—26 23,61 ±0,08 2,1 Sg вокруг тела 20—22 20,81 ±0,14 20—23 20,77±0,08 0,8 18—22 20,5±0,12 18—22 20,5 ±0,07 0,07 Подпальцевые пл. 16—20 17,0±0,16 14—20 16,5±0,11 2,63 13—20 16,48±0,17 14—20 16,44 ±0,12 0,52
Примечание. Для размерных признаков выборок 1— 3:JV=43, ?? = 57; для 4—8: с?сГ=87, ? 9-88; для количс^
ственных №=158, 99 =168 и dV = 132,99 = 124 соответственно. Первая строка признака—•££ , вторая—99
В Таджикистане, по данным С. А. Саид-Алиева (1979),
Пустынный гологлаз придерживается равнин, предгорий и гор
с разнообразной травянистой растительностью, каменистых
склонов гор до зоны арчевников; в ущельях — обломков гра-
нитов с редкой растительностью, тугаев, поросших турангой,
джидой, кияком, солодкой и эрмантусом (рис. 25), на зале-
жах и среди посевов хлопчатника — водосборных сооруже-
ний, густо покрытых сорной растительностью. Согласно его
данным, на 12 км маршрута по правому берегу Сыр-Дарьи
(о кр. кишлаков Яван и Коккурак) 23.03. 1958 г. (с 9 до
14 ч.) отмечено 14 гологлазов, а 26—30 марта здесь же на
расстоянии 17—18 км— 15—20 гологлазов.
По учетам Т. Сатторова (1977), в окрестностях Ура-Тюбе
и Канибадама на 50 м2 приусадебного участка встречено
19 гологлазов. В Туркмении, в Мургабском оазисе (окрест-
ности Мары), в октябре по берегам арыков, покрытых выго-
ревшей растительностью, и по окраинам виноградников на
протяжении 10—15 м И. С. Даревским (1955) учтено 10—12
пустынных гологлазов. В Северной Туркмении, по данным
С. Шаммакова (1974), в окрестностях поселка Тахта (в
20 км западнее головного сооружения Каракумского канала)
в плодовом саду по берегу небольшого арыка на протяжении
Рис. 25. Стация A. deserti: Таджикистан, предгорье Гиссарского хр.
99
Рис. 26. Стация A. deserti: Киргизия, предгорье Киргизского хр.
40 м в течение 40 мин встречено 55 особей, а неподалеку в
люцерновом поле за 10 мин. — 16.
По нашим наблюдениям и данным литературы (Яковле-
ва, 1962, 1964; Щербак, 1959), местообитанием гологлазам
служат в Чуйской и Таласской долинах глинистые обрывы
или бугры карасучной зоны (рис. 26), берега речек, каналов;
в предгорьях — каменистые участки; на юге в Ошской облас-
ти они обитают в орехово-плодовом лесу, среди камней по
берегам речек, в зарослях фисташки, боярышника и арчи.
По нашим наблюдениям, в Чуйской долине численность ви-
да на различных ее участках и в предгорьях Киргизского и
Заилийского хребтов неравномерна. В долинной части, в ок-
рестностях Токмакского заказника, в глинистых оврагах, по
краям полей на 1 км маршрута встречается до 27 особей вес-
ной, в апреле— мае, и до 49 осенью, в сентябре — октябре (То-
ропова, Еремченко, 1980). В окрестностях г. Фрунзе, по берегам
осушительных дрен, заросших различной сорной раститель-
ностью, на 20 шагов 26.08 1977 г. встречено до 8—10 сеголет-
ков, а на 500 м — 7 взрослых гологлазов. В районе с. Манас
между полями в балке, заросшей бородачевником, березкой,
полынью и другой растительностью, на протяжении 500 м мы
отмечали в сентябре до 20 особей, а весной, в апреле — до
12. В 40 км западнее Манаса, в окрестностях с. Тюлек, на
глинистом берегу р. Ак-Суу, поросшем солодкой, редкими
100
кустами шиповника и разреженной травянистой раститель-
ностью, на 500 м 29—30.06 1977 г. отмечено семь — восемь осо-
бей, а рядом, в 4—5 км по берегу р. Молтовар, среди густо-
го травостоя, на краю пшеничного поля на 500 м 15—16 ию-
ня учтено 18—20 гологлазов. Иная их численность наблюда-
лась в предгорьях: в Чу-Илийских горах в 30 км северо-вос-
точнее г. Токмака, в ущ. Бейшекен, на высоте около 1000 м,
на склоне ЮВ экспозиции на участке 250 м2 скальных облом-
ков и редких кустов шиповника найдено шесть гологлазов, а
ниже по берегу ручья среди пышного разнотравья и крупных
камней на линии 150 шагов—15. В низких предгорьях
(1100 м) Киргизского хребта, в окрестностях Фрунзе, среди
камней, кустов шиповника, магалебской вишни и злаковой
растительности на 500 м в середине марта можно встретить
до 10—12 гологлазов. В условиях Таласской долины пустын-
ный гологлаз живет по склонам многочисленных балок, густо
поросших луго-степной растительностью, где по нашим уче-
там, проведенным 18.06 1978 г., гологлазы были многочислен-
ны: на 500 м маршрута — до 40 экземпляров. В предгорьях
Ферганского хребта, в окрестностях с. Ийри-Суу (1800 м), на
пологих склонах, покрытых степной растительностью (боро-
дачевник), по берегу ручья па 250 м 30—31.08 1979 г. встре-
чалось до 38 гологлазов, из них 30 — сеголетки. Здесь же на
участке, занятом подсолнечником, численность сеголеток была
еще выше: на 100 м — 36 особей.
Сезонный и суточный цикл активности. На юге Казахстана
этот гологлаз выходит из зимних убежищ в первой декаде
марта (Параскив, 1956). В Киргизии первые гологлазы появ-
ляются в конце первой — начале второй декады марта, а мас-
совый выход — с середины марта (Яковлева, 1964, наши дан-
ные). В окрестностях г. Ош сроки такие же, а первые осо-
би появляются здесь на 5—6 дней раньше (Яковлева, 1964).
В это же время они встречаются в Таласской долине, в пер-
вой половине марта — в Таджикистане (Саид-Алиев, 1979).
В окрестностях Самарканда в теплые зимы гологлазы появля-
ются в феврале (Калужина, 1951). Под Ташкентом в годы с
ранней весной этих ящериц наблюдали в первой половине
февраля; массовый выход — в начале марта, иногда в середи-
не февраля; единичные гологлазы здесь могут появляться и
среди зимы — 5. 12 1945 г., 18. 01 1948 г. (Богданов, 1960).
На зимовку в южных районах Казахстана гологлазы ухо-
дят в октябре — начале ноября (Параскив, 1956). По нашим
наблюдениям и данным И. Д. Яковлевой (1964), в Киргизии
их уход на зимовку происходит в конце сентября — середине
октября, но в годы с теплой осенью эти сроки сдвигаются
к концу октября. Активных ящериц в окрестностях г. Токма-
101
ка мы встречали 12.11 1973 г., а единичные добыты 7, 19 и
21.12 1973 г. В Узбекистане основная масса популяции ухо-
дит на зимовку в конце октября, а в ноябре здесь наблюдали
только единичных особей (Богданов, 1960). В Таджикиста-
не этот гологлаз активен до октября (Саид-Алиев, 1979), в
Туркмении встречается в октябре еще в большом количестве
(Даревский, 1955).
Суточный цикл активности пустынного гологлаза зависит
от сезона. Так, в феврале в Узбекистане их наблюдали (Бог-
данов, 1960) на поверхности почвы с 12 до 18 ч, в марте —
с 10 ч 40 мин до 18 ч 20 мин; в апреле ящерицы появляются
рано утром и остаются на поверхности до 10 ч 30 мин; в те-
чение всего лета, с середины мая до конца августа, на равни-
не гологлазы активны преимущественно в утренние (9 ч
30 мин— 12 ч) и вечерние часы (17 ч— 19 ч 35 мин). В это
«ремя в середине дня даже в густой траве в тени полеза-
щитных полос учитывали только единичных особей. По наб-
людениям того же автора (Богданов, 1960), в горах, в арчо-
вых лесах Шахимардана в июне гологлазов отмечали чаще
в полуденные часы (13 ч 15 мин— 15 ч 40 мин), что, по мне-
нию этого автора, связано с относительно низкой здесь тем-
пературой воздуха. В сентябре гологлазы активны в течение
всего дня, в октябре — только во второй половине (с 13 ч
30 мин до 15 ч) как в солнечную, так и в пасмурную погоду.
В Киргизии (Чуйская долина), по нашим наблюдениям,
в суточной активности пустынного гологлаза в середине сен-
тября наблюдается еще два пика: в первой половине дня в
11 ч, затем наступает резкий спад к 13 ч и медленный подъем
к 16 ч, после чего идет снова спад. Последние ящерицы были
отмечены здесь до 19 ч 30 мин. Только в первой половине
октября активность гологлазов в Киргизии сдвигается к се-
редине дня. Нами замечено также, что сеголетки и молодые
появляются на поверхности раньше взрослых, а вечером ухо-
дят позже их. Данные, полученные нами в июле в предгорьях
Киргизского хребта, показывают, что утром гологлазы появ-
ляются с 8 ч 30 мин, находясь на поверхности до 12 ч, пос-
ле чего исчезают и вновь появляются с 19 ч, находясь на по-
верхности до 21 ч, т. е. до сумерек.
Отношение к температуре. Данный вопрос подробно ис-
следован в Узбекистане О. П. Богдановым (1960). По его наб-
людениям, температура воздуха в момент выхода гологла-
зов в феврале обычно бывает 16° и выше, а почвы 13—20°.
Ящерицы выходят в солнечные дни и в дни с небольшой об-
лачностью, а с наступлением дождливой или пасмурной по-
годы неделями могут не появляться на поверхности до оче-
редного потепления. В марте гологлазы покидают убежища
102
и при более низкой температуре воздуха 12—19°, почвы
16—23°. Активность гологлазов в солнечные дни при такой
температуре воздуха О. П. Богданов (1960) объясняет за
счет относительно хорошо прогретой почвы и инсоляции, бла-
годаря которой температура тела ящериц становится выше
температуры воздуха и даже почвы. В апреле, при низкой
температуре воздуха 9°, почвы 11,5° активны преимуще-
ственно самцы. К осени как в солнечную, так и в пасмурную
погоду ящерицы активны во второй половине дня при тем-
пературе воздуха 16,5° и выше (Богданов, 1960).
Размножение. Яйцекладущий вид. Соотношение полов
взрослых гологлазов, отловленных в Северной Киргизии с
марта по ноябрь : 99 =1 : 1,46; в Узбекистане— 1 : 1,32.
В целом для вида составленное по коллекционным материа-
лам соотношение полов равно 1 : 1,4. По нашим наблюдениям
и данным И. Д. Яковлевой (1964), в Киргизии у самцов пус-
тынного гологлаза увеличение гонад происходит осенью. Мак-
симальных размеров 5X3,7 мм семенники достигают в мар-
те. Спаривание начинается в середине апреля. Первые спа-
ривающиеся гологлазы в Чуйской долине отмечены нами
19.04 1977 г. Половозрелые самки, добытые в марте (14 осо-
бей) и первых декадах апреля (6 особей), еще не имели раз-
вивающихся фолликулов; их размеры в это время обычно не
превышали 2 мм. Но отдельные самки, добытые здесь же
23 апреля, уже имели фолликулы размером 3 мм. Исключе-
нием является одна, отловленная 27. 03 1977 г. в предгорьях
Киргизского хребта, с единственным и почти готовым к от-
кладке яйцом размером 8X4 мм, тогда как остальные фол-
ликулы у нее не превышали 1 мм. Данный случай, вероятно,
можно объяснить задержкой яйцекладки еще с осени. Спа-
ривающихся гологлазов в Чуйской долине мы наблюдали до
конца мая, а отдельных особей — в первых числах итбня. В
Узбекистане и Таджикистане спаривание у пустынного голог-
лаза происходит в апреле (Калужина, 1951; Саид-Алиев,
1979), а по другим данным (Богданов, 1960) — вплоть до кон-
ца июня. Как отмечает последний автор, самки, добытые в
окрестностях Ташкента в марте — апреле, так же, как и в Кир-
гизии, еще не имели развивающихся фолликулов, а семенни-
ки самцов в это время достигали размеров 4X2,5 мм. Одна-
ко в Таджикистане, как следует из приведенных С. А. Саид-
Алиевым материалов (1979), фолликулы в начальной стадии
развития (3—3,3 мм) отмечены у самки, добытой 19. 03 1962 г.
Согласно исследованиям в Узбекистане (Богданов, 1960),
развитие фолликулов у самок пустынного гологлаза продол-
жается месяц. По нашим данным, в Киргизии самки с готовы-
ми к откладке яйцами размерами 9—11,8X4,3—5,5 мм ветре-
103
чаются с третьей декады мая, но в основном в июне—июле
и первой декаде августа. Как исключение следует отметить
самок, добытых в Чуйской долине 2.09 1978 г. и 15.09 1977 г.,
имевших по одному готовому к откладке яйцу размерами
10X5 и 10X5,9 мм соответственно. Столь поздняя находка
яиц у пустынного гологлаза в Северной Киргизии свидетель-
ствует в пользу высказанного нами выше предположения о
возможных случаях длительной задержки неотложенных яиц
на позднеосенний и зимний периоды.
В Узбекистане наиболее ранние встречи самок, в яйцеводах
которых обнаружены готовые к откладе яйца, относятся к по-
следней декаде мая: нами 24.05 1964 г. в Сурхан-Дарьинской
долине добыта самка с шестью готовыми к откладе яйцами
размерами 9—10,5X4,5—5 мм, а О. П. Богданов (1960) 29. 05
1954 г. у Чиназа добыл самку, имевшую три яйца размерами
11X5,5 мм. Как показывают наблюдения того же автора (Бог-
данов, 1960), в Узбекистане в течение всего июня и июля встре-
чаются самки, имеющие развивающиеся фолликулы или гото-
вые к откладке яйца. По мнению данного автора, у пустынного
гологлаза в Узбекистане несколько кладок. По нашим мате-
риалам, в Киргизии у этих ящериц обычно одна, но возмож-
но и две кладки. Так, самки, добытые в Чуйской долине 27—
30.06 1977—1978 гг., имели готовые к откладке яйца размера-
ми 8—10,3X4,3—5 мм; отловленные здесь же 27—29 июля
имели также сформированные яйца размерами 9—11,8Х
5—5,8 мм. В приферганском районе Южной Киргизии пять
половозрелых самок, добытых 4—5.06 1979 г., имели почти
готовые к откладке яйца (8,8—10X4,5—5 мм), причем одна
из них, судя по состоянию яйцеводов, часть яиц уже отло-
жила, а размер последнего оставшегося 10X5 мм. Кос-
венным показателем наличия у пустынного гологлаза в Кир-
гизии растянутого периода яйцекладки, а также его возмож-
ного повтора является и то, что выход молодых здесь проис-
ходит с первой декады июля по первую декаду сентября.
В Таджикистане самок с готовыми к откладке яйцами до-
бывали в июле (Саид-Алиев, 1979). О размножении пустын-
ного гологлаза в Туркмении можно судить из данных С. Ша-
макова (1974). В третьей декаде мая у добытых им трех са-
мок (47—54 мм) найдены готовые к откладке яйца размером
8X5 мм. У самки из нашей коллекции, добытой в окрестнос-
ти Мары 2.05 1977 г., обнаружены 4 яйца размером 4,3—5 мм;
а добытой в августе в окрестностях Ташауза, 4,3X8,5 мм.
Размеры готовых к откладке яиц у гологлазов из Узбекиста-
на по О. П. Богданову (1960)— 9—11X4,5—5,5 мм; в Кир-
гизии (наши данные) —8—13X4,5—5,5 мм. По наблюдениям
М. В. Калужиной (1951), ящерицы откладывают яйца в вы-
104
копанные ими ямки глубиной 7—8 см. Кладку яиц находили
в пневой трухе в тугайном лесу по Сыр-Дарье (Салихбаев,
1956). В окрестностях Ташкента О. П. Богданов (1960) в ще-
ли глинистого обрывчика обнаружил кладку из двух яиц, а
рядом, в 10 см от них, еще одно. Нашими наблюдениями в
г. Коканде установлено, что гологлазы водно место могут от*
кладывать до И яиц, причем среди них мы находили и яйца
серого геккона, заметно выделяющиеся несколько иной фор-
мой и твердой известковой оболочкой. Здесь по берегу ары-
ка под кирпичным дувалом в углублении 2 см 14.06 1977 г.
найдены лежащие вместе 13 яиц (два из них были серого гек-
кона), а в 5 см от этого места находились еще четыре. Обе
эти кладки обнаружены под рыхлым слоем почвы среди кор-
ней бородачевника на глубине 1,5—2 см. Количество готовых
к откладке яиц у гологлазов из Киргизии одно — пять, обычно
четыре, а в Узбекистане два — восемь (наши данные; Яковле-
ва, 1964; Богданов, 1960). Некоторые авторы (Калужина,
1951) наблюдали, что самки пустынных гологлазов из четы-
рех созревших яиц откладывают только два, а через семь —
восемь дней еще два. Отловленная нами в окрестностях
г. Токмака 14 июля самка в течение ночи отложила три яйца,
а днем еще одно.
Инкубационный период по данным из Узбекистана (Калу-
жина, 1951; Богданов, 1960) длится 45 дней. В Киргизии, по
нашим данным, самка, добытая 25.05 1982 г. в окрестностях
с. Курдай (Казахстан), в террариуме 8—10 июля отложила
четыре яйца, из которых детеныши появились спустя 17—
18 дней. Данный факт указывает на возможную более про-
должительную задержку откладки яиц у гологлазов Север-
ного Тянь-Шаня. Согласно Богданову (I960), здесь первые
новорожденные гологлазы встречаются с 8 июля до конца
сентября, а единичные особи — ив октябре. Новорожденные
имеют длину туловища с головой 20 мм, хвоста — 26 мм. В
этот период молодые в количественном отношении преобла-
дают над взрослыми. По другим данным (Дубинин, 1954),
в Хавастском районе Ташкентской области молодые встре-
чаются в августе — сентябре с размером туловища с головой
15—20 мм, хвоста— 18—25 мм. О. П. Богданов (I960, 1962)
указывает, что даже незначительная разница в высоте
влияет на сроки размножения гологлазов: например, в Якка-
баге, расположенном на высоте 800 м, 17 июля попадались
самки с яйцами, но в соседнем кишлаке Икзак (700 м) тогда
же было обнаружено много новорожденных, в то время как
у Яккабага первый экземпляр добыт только 19 июля.
В Казахстане (Голодной степи) первая кладка появляет-
ся во второй декаде мая; в Северном Приаралье — в начале
июня (Параскив, 1956).
Таблица 27
Результаты анализа содержимого
желудочно-кишечного тракта 33 экз. A. deserti из Чуйской долины
(март — июнь)
Встречаемость Оощее количество
О >ъект питания экз. % экз. %
Opiliones 2 2,99 2 2,67
Aranei 8 11,94 9 12,0
Licisidae 3 4,48 3 4,0
Disderidae 1 1,49 1 1,33
Salticidae 2 2,99 3 4,0
Thomisidae 1 1,49 1 1,33
Teridiidae 2 2,99 2 2,67
Thysanura Ortoptera 1 1,49 1 1,33
Grylidae 2 2,99 2 2,67
Acrididae 3 4,48 3 4,0
Nomoptera Hemiptera 5 7,46 8 10,67
Miridae 2 2,99 2 2,67
Reduviidae 1 1,49 1 1,33
Coleoptera 4 5,97 ;5 6,67
Staphilinidae 1 1,49 1 1,33
Scarabaeidae 1 1,49 1 1,33
Cryptophagidae 2 2,99 2 2,77
Anticidae 1 1,49 1 1,33
Buprestidae 1 1,49 1 1,33
Bruchidae 1 1,49 1 1,33
Chrysomelidae 2 2,99 2 2,67
Longtarsus sp. 3 4,48 3 4,0
Phillothreta sp. 1 1,49 1 1,33
Curculionidae 1 1,49 1 1,33
Hymenoptera Lepidoptera 2 4 2,99 5,97 2 5 2,67 6,67
Noctuidae 5 7,46 6 8,0
Psichidae 1 1,49 1 1,33
Diptera 1 1,49 1 1,33
Asilidae 1 1,49 1 1,33
Simulidae 1 1,49 1 1,33
Tipulidae 1 1,49 1 1,33
Всего 67 75
О. П. Богданов (1960, 1962), исследуя темпы роста пус-
тынных гологлазов в окрестности Ташкента, указывает, что
полугодовой прирост молодых (с 8 апреля по 4 октября) сос-
тавляет 9—14 мм. Индивидуальный темп роста различный.
В Чуйской долине Киргизии с 8.09 по 6.10 1977 г. нами поме-
чено 35 сеголеток; из них в том же году 30 сентября добы'1'
106
один экземпляр, имевший 19 сентября длину тела 23,9 мм,
хвоста — 31 мм. За 17 дней длина тела увеличилась на 7 мм,
а хвоста—17 мм. Продолжительность жизни большинства
особей гологлазов в Узбекистане обычно равна году (Богда-
нов, 1960).
Линька. В Киргизии наблюдается в апреле — июне, авгус-
те — сентябре. На юге Узбекистана, Туркмении линные особи
встречаются в мае. Первая линька июльских сеголетков из-
Киргизии отмечена в конце сентября — октябре, но в основ-
ном они линяют весной следующего года, спустя 2,5—3 неде-
ли после выхода из зимних убежищ.
Питание. По наблюдениям О. П. Богданова (1960), в
Узбекистане пустынный гологлаз начинает питаться в конце
февраля. Данный автор проанализировал состав пищи у
19 гологлазов, добытых 21—28 февраля: первое место по час-
тоте встречаемости занимают насекомые (63,6%), на втором —
пауки (47,37%). Из насекомых преобладали жуки. Прочая
пища представлена моллюсками, многоножками, встречена
фаланга.
По нашим материалам (табл. 27), на основании содержа-
ния желудков 33 экз. из Чуйской долины питание гологлазов
состоит из пауков и насекомых. Пауки встречены у
32,56% ящериц; жуки, среди которых преобладают мелкие
долгоносики, —27,9%; гусеницы также составили 27,9%
встреч, равнокрылые хоботные— 18,6, а прямокрылые, клопы,
перепончатокрылые, мухи, личинки жуков — по 11,63%. В
незначительном количестве встречались мокрицы, сенокосцы,
муравьи, личинки двукрылых — по 4,65%; редко — сетчато-
крылые и щетинохвостки. В одном из желудков взрослой
ящерицы найден хвост молодого гологлаза. Встречается
также и линная шкура гологлазов.
Стереотип поведения, убежища. По нашим наблюдениям
и данным литературы (Чернов, 1948; Калужина, 1951; Пара-
скив, 1956; Богданов, 1960; Яковлева, 1964; Банников и др.,
1977), пустынные гологлазы при опасности характерными
змееобразными движениями стремятся скрыться в густой тра-
ве, кустарнике, под камнями, в расщелинах почвы, норах мел-
ких грызунов, копнах сена, лесной подстилке полезащитных
полос. Достигнув безопасного укрытия, затаиваются. Отыс-
кивая корм, ящерицы взбираются на камни, медленно, без-
помощи задних, а иногда и передних конечностей, ползают
в траве или, затаившись, выжидают добычу. Интересное
охотничье поведение у пустынного гологлаза нам приходи-
лось наблюдать в условиях террариума: ящерица при напа-
дении на сравнительно крупного паука из семейства Sparas-
sidae производила отвлекающие жертву быстрые движения
107
кончиком хвоста, в результате чего паук развернулся к хвосту
ящерицы, а она тем временем, медленно продвинув свою го-
лову, схватила паука за открытое брюшко. После этого
ящерица терла паука о камни, а затем медленно проглаты-
вала, как бы вдавливая его в широко раскрытый рот. При-
мечательно, что ундуляция хвоста — результат возбужден-
ного состояния — у пустынного гологлаза составляет один
из характерных элементов его демонстративного поведения
в брачный период, при встрече партнера или соперника — осо-
би противоположного пола или врага (фаланга и др.).
От убежищ пустынные гологлазы далеко не отходят; взрос-
лые обычно не далее 2—7, реже Юм, а сеголетки — не более
0,5—3 м. В мае — июне их можно видеть греющимися на
солнце или в вечерние часы на теплых камнях в характер-
ной позе: передние конечности ящерицы укладывают на спи-
не, а задними обхватывают хвост. По нашим наблюдениям,
сеголетки держатся вблизи густой травянистой раститель-
ности, обычно это заросли борадачевника, частично выгорев-
шие к этому времени. При опасности они скрываются здесь
же или в трещинах почвы, пространстве между корней. По
обочинам полей мы находили сеголетков под обрывками бу-
мажных мешков из-под удобрения, брикетами сена и т. д.
Враги и паразиты. В качестве объекта питания пустын-
ный гологлаз в Узбекистане отмечен у стрелы-змеи, узор-
чатого полоза, чернолобого сорокопута и ушастого ежа (Са-
лихбаев, 1956). В Ташкенте наблюдали охоту за пустынными
гологлазами длиннохвостого сорокопута (Богданов, 1960). Этот
гологлаз также найден в желудке разноцветного полоза (Рад-
жабов, 1975). Р. Н. Мекленбурцев (1956) обнаружил 22 го-
логлаза в желудке обыкновенной пустельги, добытой в фев-
рале. В Таджикистане пустынных гологлазов находили в же-
лудке стрелы-змеи (Саид-Алиев, 1979), в Киргизии (наши
данные) — в желудке степной гадюки и узорчатого полоза.
Среди птиц опасным врагом пустынного гологлаза является
жулан.
А. Овезмухамедов (1974), вскрывший 73 пустынных голо-
глаза из Туркмении, отмечает, что 13% заражены кокцидия-
ми Eimeria beyeri Ovesmuchamedov, 1977. Наличие в крови
этого гологлаза паразитов, относящихся к Anaplasma, обна-
ружили Красовский и др. (1968). Из гельминтов у пустынно-
го гологлаза из Туркмении найдены нематоды Thubunae
Schukurovi Annaev (Аннаев, 1973). На гологлазах, добытых
в июне 1977 г. в г. Коканде (Ферганская долина), были об-
наружены протонимфы клещей семейства Dermanyssidae Ко-
lencebi, 1859 рода Ornithonysus Sambon, 1928 — обычных оби-
тателей грызунов и птиц.
108
Практическое значение. Пустынный гологлаз — важный
компонент полупустынных, а также антропогенных биоцено-
зов Средней Азии. Являясь обычными, а местами многочис-
ленными, эти ящерицы в антропогенном ландшафте играют
заметную роль: с одной стороны, служат прокормителями и
распространителями некоторых клещей, с другой — уничто-
жают ряд вредных беспозвоночных (гусениц, жуков, клещей
и др.).
ГОЛОГЛАЗ МАЛЫЙ-ABLEPHARUS GRAYANUS (STOL., 1872)
TERRA ТУР1СА: С-3 ИНДИЯ, KAR.
1872 — Blepharosteris grayanus Stoliczka, Proc. Asiat. Soc. Bengal.: 74
1887 — Ablepharus grayanus Stoliczka, Boulenger, Cat. Liz.
Brit. Mus. 3:352. — 1935, Smith, Fauna Brit. Ind.:311.
1949 — Ablepharus grayanus (Stol., 1872), Терентьев, Чер-
нов. Опред. преем, и земн. СССР, 183. — 1952, Mittlemarr
Misc. Collect., 117:20. — 1966, Minton, Bull. Amer. Mus.
Nat. Hist.: 134,2:120. — 1970, Leviton and Anderson,
Proc. Calif. Academy, 38,10:190. — 1981, Еремченко.
Автореф. докл. V Всесоюзн. герпетол. конф.: 54. — 1981,
Jeriomtschenko, 1 Conf. Негр. Resp. Soc. Budapest: 2L
1969 — Ablepharus pannonicus grayanus (Stoliczka, 1872), Fuhn, Rev.
roum. Biol. Zool., 14,1:21. —1969b, Fuhn, Sound, a. z. f. Zoof.
Syst. u. Evol. 7:67. — 1974, Greer, Austral. J. of ZooL
31.115—122. — 1977, Банников и др. Опред. преем, и
земн. фауны СССР: 157. — 1977, Топоркова, Сте-
панов. Герпетол. сб., 74:115.
Диагноз. Самая мелкая ящерица рода (Lmax. —33 мм).
Лобно-теменной щиток один. Наружный ушной проход отсут-
ствует. Носовые щитки полуразделенные. Вокруг середины те-
ла 18—19 (обычно 18) чешуй.
Отличия от близких видов. Малый гологлаз отличается от
азиатского — A. pannonicus — отсутствием наружного ушного
прохода, окраской тела, мелкими размерами и меньшим коли-
чеством чешуй вокруг середины тела. От европейского и дру-
гих— теми же признаками, а также наличием одного лобно-
теменного щитка.
Описание (рис. 27). Голотип, по сообщению А. Левитона и
С. Андерсона (Leviton and Anderson, 1970), хранится в Каль-
кутте. Нам, несмотря на запрос, он не был представлен для
просмотра. В своей работе И. Фун (Fuhn, 1969, в) назвал го-
лотипом экземпляр из коллекции Венского естественно-исто-
рического музея № 10234, добытый Ф. Столичкой 11.16 1874 в
Северо-Восточном Каче. Данный экземпляр, однако, правиль-
но называть синтипом. Благодаря любезности профессора
И. Айзельта, мы имели возможность ознакомиться с этим эк-
земпляром. Синтип. 9 L — 26. Ga — 14. Led — нет. L головы —
4,9. Ширина головы — 3,1. LP i — 5,9. LP 2 — 9. Sg вокруг те-
109
Рис. 27. Фолидоз головы A. grayanus (Stoliczka,
1872); а—вид сбоку, б—вид сверху.
ла—18. Затылочных щитков — 3X3. Подпальцевых чешуек
на четвертом пальце Р2— 14. Предлобные щитки разделены
выступающим углом лобного, который соприкасается с лобно-
носовым. Носовые щитки полуразделены.
Наше описание составлено по шести экземплярам из Паки-
стана и Индии, хранящимся в естественно-исторических му-
зеях Нью-Йорка (AMNH) и Вены (NMW).
L 6*с? — 26—27 (26,33±0,28), 99—28,2—33 (30,60±2,40),
Ga <? ’ — 14—15 (14,66±0,33), 99 — 16,5—20 (16,14=1,66),
Led 9=55. ЬголовысГсГ —3,5—5,2 (4,53±0,69), 99 — 5,3—
6 (6,15±0,34). Ширина головы cTcf—3—3,5 (3,2±0,15), 99-
4 (4,0±0). LP, 33— 5,8—6,0 (5,9±0,05), 99 — 6,0—7,0
(6,5±0,5). LP26V—7,7—9,0 (8,53± 1,18), 99—9,2—9,5 (9,35±
0,15), ^56 —53,84—57,7 (55,7± 1,11), 99 —58,51—60,60
(59,55+1,05),-гЦ-. 19 =0,60. А..г°ловы-_^_ 13,5—19,26
Led L
(17,2±1,85), 99— 16,06—21,27 (18,66+2,61) Ш»рина головы
' ’ ’ ” L головы
dV — 63,26—85,7 (72,08 ± 6,91), 9 9 — 66,66—75,47 (71,06±
4,41), гг —22,22—22,7 (22,4±0,15), 9? — 18,2—24,82
(21,51+3,32), д^-dV—29,6—34,6 (32,01 ±1,45), 99 —27.88—
33,7 (30,8±2,91). Межчелюстной щиток коротким швом ка-
сается лобно-носового. Надглазничных щитков три, два из
них соприкасаются с лобным. Лобно-теменной один. Межте-
менной один. Теменные щитки смыкаются узким швом поза-
ди межтеменного. Первый ряд височных состоит из трех щит-
ков. Наружный ушной проход отсутствует. Sg вокруг шеи
<?<? —22—23 (22,3±0,33),99 — 22—23 (22,33±0,88), Sg вок-
руг тела 6V—18 (18,0±0), 99 18—19 (18,33±0,33). Брюш-
ных чешуй в продольном ряду dd— 39—44 (41,6 ±1,45),
99 — 43—45 (44,0+0,58). Затылочных!, 3—3 : 3=50%; 3=
50%. Подпальцевых чешуек на четвертом пальце рг
14,16 (14,6±0,67), —14—16 (15,0±0,58). Верхнереснич-
ных щитков 4—5:4=16,5%, 5=83,5%. Козырек состоит из
трех увеличенных чешуек. Впереди подглазничного четыре
верхнегубных щитка. Между носовым и подглазничным рас-
положено четыре щитка. Предлобных щитков два, они, как
Рис. 28. A. grayanus (Пакистан, Карачи, колл. AMNH).
111
правило (100%), разделены передним углом лобного щитка.
Центрально-височный щиток маленький и свободный свер-
ху (100%). Носовой полуразделен (100%). Впереди клоакаль-
ной щели два больших щитка. Хвост снизу покрыт одним ря-
дом расширенных щитков. Основной фон верхней стороны те-
ла светло-оливковый с металлическим оттенком, на спине на
двух средних рядах чешуй заметно выделяется продольная
полоса оливкового или золотисто-оливкового цвета, оканто-
ванная с каждой стороны мелкими прерывистыми черточками
темно-бурого цвета, которые на передней части тела могут
быть соединены в сплошные линии: по краям спины прохо-
дят относительно широкие продольные полосы основного фо-
на (рис. 28).
Рис. 29. Распространение A. pannonicus (•) и A. grayanus ( о ).
Кадастр к рис. 29.
СССР. Узбекистан: 1. Предгорья Туркестанского хр окр. к. Алма<
чи (Дубинин, 1954). 2. Зааминский р-он, окр. к. Талы (Богданов, 1960).
112
3. Окр. Самарканда, у к. Миранкули (колл. ЗИН АН СССР). 4. Южный
склон хр. Нуратау, пер. Капчигай (Карпенко, 1970). 5. Окр. с. Красно-
гвардейское (колл. ИЗ АН УССР). 6. Окр. к. Яккабаг (Богданов, 1960).
7. Самаркандская обл. с. Челек, берег р. Булунгур (колл. ИЗ АН
УССР). 8. Сурхандарьинская обл., в 6 км выше к. Кызыл-Алма (Богда-
нов, 1960). 9. Бассейн р. Сурхандарьи, с. Арал-Тайгамбар (Ядгаров,
1973, 1974). 10. Термез (Царевский, 1918). 11. Ак-Мечеть в Бабатаге
(Никольский, 1915)
Таджикистан: 1. Окр. Гиссара, г. Ходжа-Оби Гарм. 3. Окр. Душанбе.
4. Гиссарская долина, к. Шульмак. 5. Северный склон Гиссарского хр., у
к. Кызыл-Таш. 6. Правый берег р. Хингоу, южный склон хр. Петра Пер-
вого, к. Саят. 7. Южный склон Гиссарского хр., ущ. Кондара. 8. Данга-
ринский р-он, ур. Хозратишо. 9. Заповедник «Рамит». 10. Заповедник
«Тигровая балка». 11. Дарваз, пер. Софе-Дорон, 2700 м (колл. ИЗ АН
УССР). 12. 20 км южнее Душанбе. 13. Могол-Тау. 14. Южный склон
Вахшского хр., берег р. Шуроб-Дарья. 15. Окр. Дангара. 16. Окр. с. Му-
минабад. 17. Предгорья хр. Бабатаг, окр. с. Таша-Хур. 18. Предгорья хр.
Бабатаг, окр. с. Долон. 19. Предгорья Гиссарского хр., окр. с. Пуш-
тимиепа. 20. Нимичи-Боло. 21. Ворух. 22. Окр. с. Обикинк, 23. Джили-
куль (Чернов, 1959). Пункты 1 —10, 12, 20 даны по колл. ЗИН АН
СССР; все остальные по Саид-Алиеву, 1979.
Туркмения: 1. Сарыгядык, родник Башмугур (Шаммаков, 1981); родник
Узынакыр (Шаммаков, 1981). 2. С. Даната (колл. ИЗ АН УССР). 3. Окр.
оз. Малое Делимы (Шаммаков, 1981). 4. С. Яртыкала на р. Чандыр. 5.
Ущ. Еддере (он же). 6. С. Санарбахар (Никольский, 1915). 7. Кызыл-Ар-
ват. 8. Фирюза, ур. Чаек (Шкафф, 1916; Богданов, 1962). 9. Душак (колл.
ИЗ АН УССР). 10. Хр. Гязгядык (Богданов, 1962; Шаммаков, 1981). 11.
Ур. Акарчешме (они же). 12. Поселок Моргуновский (Гептнер, 1945; Бог-
данов, 1962). 13. Иолотань (Богданов, 1964, 1962). Станция Имамбаба. 15.
Карабиль (Щербина, 1966). 16. Ишметнест (Шаммаков, 1981). 17. Посе-
лок Карлюк, хр. Кугитанг (Шестоперов, 1936; Богданов, 1962; Шербак,
1979; Шаммаков, 1981). 18. Перевал Гаудан (колл. ЗИН АН СССР).
Азербайджан: 1. О. Обливной в Каспийском море (Ахмедов, 1976,
1977). 2. Северный Кобустан, р. Сумгаит, с. Джанги (колл. ИЗ АН УСС).
3. Северный Кобустан, среднее течение р. Сумгаит-Чай (Чегодаев, 1981).
Грузия: 1. Вашловский заповедник (Бакрадзе, Даревский, 1973).
Иран: 1. Окр. Шираза, заповедник «Бализ». 2. Шахруд. 3. Карун. 4.
Дех-и-Диз. 5. Сия-Мансур. 6. Бак-Эмищир. 7. 10 км вниз по р. Корун от
Зиарета-Хадыр. 8. Керош. 9. Хорасан (колл. FMNH). Точки 1—8 даны по
колл. ЗИН АН СССР; Никольский, 1915).
Ирак: 1. Багдад. 2. Сирсиак (Serseuk). 3. Ирак-Аджели. 4. Афак. 5.
Кишарма. 6. Сик-аш-Шуак. 7. Амара. Пункты 1—2 даны колл. FMNH и
MCZ; 3—5 —по колл. MCZ; 6—7 (Fuhn, 1969а).
Сирия: 1. Халеб (Алеппо) колл. MCZ
Афганистан. 1. Пагман, 25 км севернее Кабула. 2. Обех, с-з Афгани-
стан. 3. Еспишеварз, 30 км с-з Тагаб Ихан. 4. Масджед-Шоуб. 5. Кабул.
6. Гильменд. 7. Обех, Герат. 8. Балиган. 9. Калайи-Нау. 10. Новый Гиль-
менд. 11. Шахрак. Пункты 1—3 (Mertens, 1965); 4—по колл. NMW; 5, 10
(колл. ЗИН АН СССР); 6—9 (Fuhn, 1969а): 11 (колл. NMW).
Пакистан: 1. Гауджвар (Пенджаб) (Smith, 1935).
Кадастр к рис. 29. Распространение A. grayanus.
Пакистан: 1. Карачи. 2. 3,5 км северо-восточнее Хаб-Хоук (НаЬ-*
Chowki). 3. О. Чурма в Аравийском море. 4. 4,5 мили севернее г. Бела. 5.
5 миль севернее Утхал. 6. 4,5 мили западнее Дабеджи. 7. Ташта 8 Сиби
9. Даду. Пункты 1, 2, 3, 6—9 даны по колл. AMNH; 4,5 (Minton 1966)
Индия: 1. Кач (колл. NMW).
8 302
113
Распространение. Ареал A. grayanns к настоящему вре-
мени выяснен недостаточно. По М. Смиту (Smith, 1935) и
Ш. Минтону (Minton, 1966) он охватывает северо-западную
Индию и Южный Пакистан. В колленщиях Венского есте-
ственно-исторического музея хранится экземпляр (№ 16276)
из Афганистана (Masgjed — Tchoubi), определенный Линд-
хольмом как A. grayanus. По нашему определению этот эк-
земпляр относится к A. pannonicus. Мы считаем также, что
в СССР (Таджикистане) A. grayanus отсутствует. Достовер-
ные точки находок малого гологлаза показаны на рис. 29.
Систематика и географическая изменчивость. Автор видо-
вого описания «grayanus» (Stoliczka, 1872) по признаку от-
сутствия у этих ящериц ушного отверстия поместил данный
вид в отдельный род Blepharosteris. Позже Г. Буланже (Вои-
lenger, 1887) признал видовую самостоятельность «grayanus»
и перевел его в род Ablepharus. Данный вывод был принят
многими герпетологами (Smith, 1935- Терентьев и Чернов,
1949; Mittleman, 1952; Чернов, 1959; Minton, 1966; Leviton and
Anderson, 1970 и др.). С другой стороны, видовую самостоя-
тельность A. grayanus нс признавали и рассматривали его в
качестве восточного подвида азиатского гологлаза — A. pan-
nonicus grayanus (Stol.) (Mertens, 1965; Fuhn, 1969a, в;
Greer, 1974; Банников и др., 1977; Топоркова и Степанов,
1977). И. Фун (Fuhn, 1969в), в частности, предполагал, что
между номинативной формой «pannonicus» и «grayanus» су-
ществует зона интерградации, проходящая в Таджикистане,
Восточном Афганистане, Белуджистане. Согласно вышеупо-
мянутому исследователю, эти ящерицы в зоне интеградации
(Белуджистан) имеют 20—22 ряда чешуй вокруг середины
тела вместо обычных для вида 18—20 чешуй. Однако Ш. Мин-
тон (Minton, 1966), исследовавший серию из 16 экземпляров
A. grayanus, добытых в Пакистане, отмечает для этих ящериц
18 и очень редко 17 и 19 чешуй вокруг середины тела. У прос-
мотренных нами шести экземпляров A. grayanus из Пакиста-
на из коллекции естественно-исторического музея Нью-Йор-
ка (AMNH) также обнаружено по 18 чешуй; только один из
них имел 19. Ш. Минтон (Minton, 1966) предполагал также,
что пакистанские гологлазы из Пенджаба могут быть от-
несены к виду A. pannonicus. Поскольку Ш. Минтон не изу-
чал этих ящериц специально, проблема систематики
пакистанских, а также гологлазов из Таджикистана осталась
нерешенной. Отметим, чго И. Фун (Fuhn, 1969в), ревизуя оба
эти вида, имел в своем распоряжении только по три экземп-
ляра, среди которых был и добытый в Нимичи-Боло (Таджи-
кистан). Благодаря любезности профессора И. С. Даревского
мы ознакомились с экземплярами нз Нимичи-Боло (коллек-
114
Таблица 28
Изменчивость размеров тела и хвоста у A. pannonicus и A. graynus
№ вы- борки Место п 99 1 L Ga Led
1 т 21 1 Таджикистан уу- 2 Узбекистан — 7 3 Туркмения 1^- 4 Азербайджан, 10 Грузия 9 4 5 Иран -у 6 Ирак ± и з 1 Пакистан — 32,0—43,3 37,0—50,0 32—35,3 A. pannonicus 38,65±0,83 16,0—24,0; 19,42±0,5 42,71 ±1,1 22,0—30,0; 25,95 ±0,64 32,87±0,44 16,0—19,0; 17,74 ±0,36 _ 50,8—65,0; 58,13 ±2,9 55,0—66,0; 59,25±2,3 52,0
35,5—41,5 32,0—42,5 39,0 ±0,71 39,3±0,84 21,0—27,0; 24,28±0,72 16,8—23,0; 18,84±0,5 50,0 43,8—59,0; 51,96±3,21 46,0—57,0; 55,4 ±2,77 56,5—71,0; 61,12±3,4 43,5—57,0; 50,25±6,7
32,0—46,0 31,5—37,7 35,0—44,0 30,0—34,2 37,9± 1,34 35,3±0,6 39,6±0,95 32,1 ±0,93 20,0—27,5; 22,77 ±0,81 18,0—22,0; 20,03 ±0,46 21,2—29,0; 24,48±0,9 16,0—19,8; 17,75±0,8
37,0—41,0 20,5—34,0 39,66 ± 1,33 32,0±0,08 22,8—26,0; 24,1±0,Э7 15,5—20,0; 17,62 ±0,94 37,0
37,0—40,0; 26,0—27,0: 27,2—33,0: , 38,33±0,88 20,0—22,3; 22,1 ±1,16 A. grayanus ; 33 ±0,34 14,0—15; 14,66±0,33 ; 30,6 ±1,30 16,5—20,0; 18,25±1,66 49,0 55дГ
Таблица 29
Изменчивость пропорций головы и длина ног у A. pannonicus и A grayanus
№ выбор ки 99 L головы Ширина головы L Р, L Р3
A. pannonicus
1 6,0—8,0; 6,93 ±0,13
6,2—7,3; 6,74±0,1
2 5,5—6,6; 6,0±0,12
5,9—6,1; 6,0±0,03
3 5,8—7,3; 6,42±0,14
5,3—7,1; 6,33 ±0,2
4 6,0—7,0; 6,41 ±0,13
6,0—7,0; 6,21±0,11
5 5,8—6,1; 5,97±0,06
5,9—6,2; 6,03 ±0,1
6 5,9—6,1; 5,97±0,05
5,8—6,2; 6,06±0,13
4,0—6,0; 4,9±0,11
4,0—5,3; 4,87±0,07
4,1 — 4,8;_4,18±0,11
4,0—5,4; 4,5 ±0,25
4,0—5,2; 4,63±0,12
4,0—5,6; 4,55±0,04
4,0—5,8; 4,55±0,18
4,0—5,0; 4,42±0,14
3,9—4,2; 4,07 ±0,04
4,0—4,2; 4,13±0,07
4,0
4,0
8,44 ±0,2
8,05 ±0,17
6,0—7,6; 6,87 ±0,17
6,1—7,7; 6,7±0,15
6,5—9,2; 7,8 ±0,21
6,0—8,9; 7,2±0,26
7,0—8,1; 7,85±0,3
7,0—8,8; 7,66±0,18
7,0—7,8£_7,45±0,27_
6,9—8,0; 7,33 ±0,34
6,0—7,0; 6,75±0,25
8,8—7,5; 7,26±0,23
12,4±0,25
11,82±0,3
8,8—10,5; 9,8±6,2
9,3—10,9; 10,07±0,2
9,9—13,5 11,28±0,31
8,9—12,3; 10,5±0,36
10,0—11,8; 10,81 ±0,13
9,9—11,1; 10,65±0,15
10,0—11,0; 10,32±0,23
9,1—10,9; 10,1 ±0,53
9,5—10,0; 9,62 ±0,12
9,8—10; 9,86±0,06
1 3,5—5,2; 4,5±0,7
5,3—6,0; 6,65±0,34
A. grayanus
3,0—3,5; 3,2 ±0,15 5,8—6,0; 5,9 ±0,05
4 6,0—7,0; 6,5±0,5
7,7—9,0; 8,43 ±0,43
9,2—9,5; 9,35 ±0,15
Таблица 30
Изменчивость количества чешуй и некоторых других признаков
у A. pannonicus и A. grayanus
№ вы- борки п Sg вокруг шеи Sg вокруг тела Брюшных щйтков Подпальцевых на четвертом пальце Р2 Ушной проход имеется
A, pannonicus
1 53 20—25 18—20 _ 35—44; 39,96±0,4 13—17 +
2 16 22,38±0,13 22—24 20,47±0,12 20—21 39—50; 44,44 ±0,53 38—41; 40,37±0,6 14,77±0,12 13—18 +
3 41 22,5 ±0,16 20—25 20,12±0,08 20—22 41—49; 45,5±0,01 36-43; 39,33±0,36 15,00 ±0,40 13—16 +
4 23 22,61 ±0,17 21—24 20,41 ±0,12 20—22 38—50; 43,75 ±0,70 38—42; 40,15±0,33 14,84±0,12 13—16 +
5 8 22,35±0,24 20—23 20,3 ±0,24 20 39—51; 43,40±1,07 36—40; 38,40 ±0,75 14,13±0,17 13—17 +
6 8 21,5±0,5 20—23 18—21 43—46; 44,33±0,87 37—40; 38,20±0,75 14,62 ±0,27 13—14 +
1 6 20,62 ±0,37 22—23 19,9 ±0,3 18—19 39—43; 40,66±1,20 A. grayanus 39—44; 41,00±l,4S 13,38±0,18 14—16
22,33±0,33 18,33 ±0,33 43—45; 44,0±0,58 14,83±0,40
* 1 экземпляр без ушного прохода
Таблица 31
Изменчивость некоторых признаков фолидоэа
у A. pannonicus и A. grayanus
№ выборки I п Количество затылочных щитков, % Носовой щи- ток Предлобные щитки Центрально-ви- сочный щиток
2 —2 , 2 3 1 “3 4 1 -4 цель- ный полу- разде- ленный раздель- ные 1 касают- ся в точ- , ке касаются широко ма- лый боль- шой
A. pannonicus
1 2 53 16 — 6,0 6,25 — 44,0 68,75 26,0 6,25 30,0 4,0 18,75 — 3,7 3,2 96,3 96,8 92,4 75,0 7,6 25,0 88,7 93,7 11,3 6,3
3 41 — 4,88 69,41 24,39 7,31 — ! 100,0 85,0 12,5 2,5 95,0 5,0
4 23 — 4,35 56,52 30,43 8,69 — — 100,0 95,7 4,3 — 95,7 4,3
5 8 — 25,0 25,0 25,0 25,0 — — 100,0 87,5 — 12,5 87,5 12,5
6 8 — — 50,0 25,0 25,0 — — A. grayanus 100,0 62,5 12,5 25,0 100,0
1 6 — — 83,5 16,5 — — — 100,0 100,0 — — 100,0 —
ции ЗИН АН СССР). Среди них по признаку полного отсут-
ствия наружного ушного прохода к определению «grayanus»
подходит только один из них (№ 14075), добытый А. Н. Каз-
наковым в 1873 г. А № 18873, определенный как A. panno-
nicus grayanus (Топоркова и Степанов, 1977), на самом де-
ле не отличается от остальных A. pannonicus, так как ушной
проход у этой особи скрыт под чешуями, а не отсутствует*
как утверждают эти авторы. В очерке, посвященном A. pan-
nonicus, мы указываем, что особи с подобным строением уха
не являются исключением для этого вида и в единичных слу-
чаях встречаются в Таджикистане, Азербайджане и Иране. Воз-
вращаясь к экземпляру из Нимичи-Боло (№ 14075), мы счи-
таем, что полное зарастание у него наружного ушного прохо-
да является исключением, поскольку у семи других экземп-
ляров, добытых здесь же А. Н. Казнаковым (№ 9055 и
14084 из коллекции ЗИН АН СССР), ушной проход идентичен
A. pannonicus (№ 18873) и по другими признакам (20 чешуй...
вокруг тела и др.) они не отличаются от особей этого вида
и экземпляра из Нимичи-Боло № 14075. Принимая во внима-
ние вышесказанное, мы считаем, что в Таджикистане малый
гологлаз отсутствует. Изменчивость собственно A. grayanus
у экземпляров, исследованных нами, практически не выра-
жена.
Данные экологии. По сообщению Ф. Столички (Stoliczka,
1872), малый гологлаз добыт им в «песчаных местах между
кустами травы». По некоторым данным (Minton, 1966), эти
ящерицы населяют засушливые каменистые пустыни, город-
ские сады, оазисы, берега островов, встречаются также в жи-
лых постройках; всюду ведут скрытый образ жизни, адаптив-
ные черты которого нашли выражение в очень мелких разме-
рах тела, светлой его окраски, полном отсутствии наружного
ушного прохода. Минтон (Minton, 1966) указывает, что в Па-
кистане период размножения вида растянут, молодые с дли-
ной тела 13—16 мм появляются в течение апреля и в авгус-
те. Другие данные по экологии вида отсутствуют.
ГОЛОГЛАЗ АЗИАТСКИЙ —ABLEPHARUS PANNONICUS
(LICHT., 1823)
Terra typica: Узбекистан (Бухара)
1823 — Scincus pannonicus, Lichtenstein, Verzeich. der Doub.
des Zool. Muz. der Konigl. Univ, zu Berlin; 103
1852—Ablepharus pannonicus, Brandt it Lehman, Reise : 133
1868 — Ablepharus brandti, S t r a u c h, Mel. Biol. Acad. Petersb.
6:566.— 1873, Северное. Турк. жив.: 72—1887, Bou-
le n g e г, Cat. Liz. Brit. Muz. 111:351. — 1888, Кулагин.
Изв. M. об-ва люб. естест. VI, 2:13. — 1899, Boettger
in Rad de Muz. Caucas. :282. — 1899, Никольский.
Herpet. turan., 41. —1901, Ел пат ьевский. Дневн. зоол.
119
отд. об-ва любит, естеств. 3,2:1, —1915, Никольский.
Фауна Росс, и сопред. стран (Reptilia), 1 :490. — 1918. Ца-
ре веки й. Ежегодн. зоол. муз. АН XXII (1917), 79. —
1952, Mittleman. Misc. Collect., 117:20. — 1959, Чер-
нов. Преем. Тадж., 18:109. — 1960, Богданов. Преем,
и земнов. Узбек., 1 : 132. — 1962, Богданов. Преем. Турк-
мении: 103.
1872 — Blepharosteris agilis, Stoliczka, Nat. Hist. Ind.:124
1874 — Ablepharus pusillus, Blanford, Ann. Mag.: 14.4:33
1905 — Ablepharus persicus Nikolsky (Никольский), Ann. Mus.
A. C. Retersb.: 10,283. — 1915, Никольский, Фауна Росс,
и сопред. стран (Retilia) 1,1 : 502.
1958 — Ablepharus pannonicus (Licht., 1823), Mertens u. Mul-
ler, List. desr. Amphib. u. Rept. Europ., XLL62. — 1965,
Mertens, Senkh. BioL, 46:4. — 1966, Minton, Rev. of
the Americ. Mus. of Nat. Hist., 134,2:103. — 1969, Fuhn, Rev.
roum. Biol. Zool., 14,1:21. — 1969b, Fuhn, Sound, a. z. f.
Zool. Syst. Evol. 7:67.— 1974, Greer, Austral. J. of Zool.
31:67. — 1977, Банников и др.; 157. — 1980a, Ерем-
ченко, Щербак. Вести, зоол. 4:14. — 1981, Еремчен-
ко. Автореф. докл. V Всесоюзн. герпетол. конф., Л.:54. —
1981, Jeriomtschenko, 1 Conf. Негр. Resp. Soc., Budapest:22.
Диагноз. Мелкие ящерицы длиной тела до 50 мм; лобно-
теменной щиток один; наружный ушной проход всегда имеет-
ся, но мал и иногда скрыт под височными чешуями. Носовые
щитки полуразделенные. Вокруг середины тела 20—22 чешуи.
Отличие от близких видов. Отличие A. pannonicus от ма-
Рис. 30. Фолидоз головы A. pannonicus (Licht., 1832).
120
лого гологлаза приведено нами в очерке о последнем (с. 114).
От всех остальных гологлазов выделяется присутствием од-
ного лобно-теменного щитка.
Описание. Голотип утерян. Описание вида (рис. 30) сос-
тавлено нами по 151 экземпляру из Ирака (8), Ирана (8),
Афганистана (2) и СССР (Туркмении (41), Узбекистана
(16), Таджикистана (53), Грузии (2), Азербайджана (21),
хранящихся в коллекциях ЗИН АН СССР (Ленинград), ИЗ
АН УССР (Киев), ЗМ МГУ (Москва), ИЗИП АН Тадж. ССР
!(Душанбе) и естественно-исторических музеях Вены (NMW)
и Нью-Йорка (AMNH).
L66 —30—45,2 (36,05±0,51), 99 — 32,00—50,0 (40,2±0,59),
t=5,28. Ga $6 16—24 (19,49±0,26), 99 — 20,0—30,0 (24,38±
0,39), t= 10,22. Led 66 —52,0—71 (57,22+2,07), 99 —43,5—
66 (55,37± 1,85), t=0,66. L головы 66 — 5,8—7,9 (6,53±0,08),
99 — 5,3—7,3 (6,36±0,07), t=l,5. Ширина головы, 66 —
3,9—6 (4,63±0,07), 99—4—5,9 (4,60+0,07), t=0,29.
LP1 66—5,0—9,8 (7,83 ±0,12), 99 — 6,0—9,0 7,46+0,11),
t=2,33. LP2 rf'd” —8,8—14 (11,7+0,19), 99 — 8,9—13,8
Ля
(10,85±0,18), t=3,25. ++66 — 0,52—0,79 (0,64+0,018), 99-
L-/
0,59—0,82 (0,69+0,0151), t = 2,12. ^66—50,0—60,60
(54,07+0,37)99— 56,4—71,35 (60,77±0,49), t=10,89. -~
66— 15,48—20,0 (17,80+0,12), 99— 14,32—17,95 (15,80±
0,14), t= 10,67. -Шири-Н-- гол-- 66 — 64,3—82,85 (71,ll±0,5),
99—64,3—96,72 (72,32—1,0), t= 1,05. tL’dV — 15—25,64
(20,99+3,16), 9?— 13,55—21,94 (18,29+0,27), t = 5,0. Ц^-бб
— 18,38—35,9 (30+0,52) 99 — 20,22—31,23 (25,93±
0,37), t = 6,57.
Межчелюстной щиток коротким швом соприкасается с лоб-
но-носовым. Надглазничных щитков три, из которых два соп-
рикасаются с лобным, очень редко (2%) надглазничных два.
Лобно-теменной щиток один. Теменные смыкаются позади
межтеменного узким швом. Межтеменной один. Первый ряд
височных состоит из трех щитков (редко 0,5%—Два щитка).
Наружный ушной проход маленький, его размеры уклады-
ваются в размерах глаза не менее 10 раз или он скрыт под
височными чешуями. Sg вокруг шеи 45— 20—25 (22,5±
121
0,15), ?9 —20—25 (22,2+0,13), t=O,33. Sg вокруг тела
— 19—22 (20,4±0,08), 99 — 18—22 (20,2±0,1), t=l,56.
Брюшных в продольном ряду — 35—43 (39,67±0,20).
9?—38—51 (44,05±0,35), t=10,7. Затылочных щитков 2—
4:2=1,2%, 3 = 58,1%, 1 3 = 22,9%, 4= 16% 4 = 0,8%.
2 2 2,
Подпальцевых чешуек на четвертом пальце Р2 S6— 13—18
(14,58+0,12), 99—13—18 (14,71±0,11), t = 0,12. Верхне-
ресничных 3—5:3 = 8%, 4 = 28,6%, 5 = 63,4%. Козырек верх-
него века состоит из одного — пяти увеличенных чешуек: 1 —
2,6%, 2—2,6%, 3—88,3%, 4—11%, 5—2,6%. Впереди подглаз-
ничного три— четыре: 3—7,8%. хЧежду носовым и подглазнич-
ным расположено три—четыре щитка: 3—7,8%, 4—92,2%.
Предлобных щитков два, они разделены передним углом лоб-
ного— 86,3%, соприкасаются в одной точке—11,2%, или
соприкасаются широко и отделяют лобно-носовой щиток от
лобного — 2,5%. Центрально-височный щиток двух типов: ма-
ленький и свободный сверху — 92%; большой, трапециевид-
ный и его верхний край прижат теменным щитков— 8,0%. Но-
совой щиток двух типов: полуразделенный — 98,2%; цель-
ный— 0,8%. Впереди клоакальной щели два больших щитка.
Хвост снизу покрыт одним рядом расширенных щитков. Ос-
новной фон верхней стороны тела фиксированных экземпля-
ров буроватый. Однотонная окраска спины лишена рисунка,
встречена у 24% особей. Рисунок в виде трех темных узких
продольных линий, проходящих по краям средней пары че-
шуй, на некоторых из них имелись светлые черточки, окан-
тованные по краям бурыми пятнышками, встречены у 65,5%
особей. Наконец, аналогичный рисунок, но с узкими темны-
ми линиями, проходящими по краям всех спинных чешуй,
имелся у 10,5% особей. От ноздри через глаз и далее по бо-
кам туловища тянется бурая полоса, имеющая светлые пят-
нышки. Тело снизу сероватое или светло-палевое, иногда с
зеленоватым оттенком. У взрослых в период размножения
нижняя поверхность брюха, хвоста и конечностей кирпично-
оранжевого, оранжевого или золотисто-оранжевого цвета; сам-
ки окрашены менее ярко, имеют слабые оттенки оранжевого
тона.
Распространение. Среди видов данного рода азиатский
гологлаз занимает наибольшую область распространения, ох-
ватывающую Сирию, Ирак, Иран, Афганистан, Пакистан,
достигая границ Северо-Западной Индии. В СССР живет в
Южном Таджикистане, Узбекистане и Южной Туркмении.
Изолированные популяции этого гологлаза недавно обнару-
жены в Восточном Закавказье, на юго-востоке Грузии и в
122
Азербайджане (о-ва Жилой и Обливной). В горах встречается:
до высоты 2800 м. В нашей стране данный вид занимает преи-
мущественно предгорье, поднимается до 2800 м (Дарваз, кол.
ИЗ АН УССР).
Систематика и географическая изменчивость. Номенкла-
турный статус A. pannonicus ( = А. brandti) был принят срав-
нительно недавно (Fuhn, 1969а, в; Greer, 1974; Банников и
др., 1977) благодаря исследованиям Р. Мертенса (Mertens,
1952). Об отсутствии единого мнения на вопрос систематики
A. pannonicus и A. grayanus было отмечено в очерке о пос-
леднем. Описанные по единственным экземплярам из Ирана
формы A. persicus Nik., 1907 и A. brandti var. brevipes Nik.,.
1907 и позже (Шнитников, 1928; Чернов, 1959), сведенные в
синонимы A. brandti Str., 1868 ( = pannonicus Licht., 1823),
свидетельствовали о необходимости внутривидовой ревизии
азиатского гологлаза. Выше показано, что форма «grayanus»,
характеризуясь дискретными признаками, имеющими важное
таксономическое значение (отсутствие наружного уха, мини-
мум чешуй вокруг середины тела и некоторыми другими), не-
сомненно, представляет самостоятельный вид—A. grayanus
(Stol., 1872).
С целью изучения внутривидовой систематики собственно
A. pannonicus нами исследована его географическая измен-
чивость из шести выборок: Таджикистана (53), Узбекистана
(16), Туркмении (4), Азербайджана и Грузии (23), Ирана
(8), Ирака (8). В результате показано (табл. 29, 30), что из-
менчивость таких признаков, как размеры тела, количество че-
шуй вокруг тела, шеи и число подпальцевых пластинок имеет
клинальный характер, идущий с увеличением значений от по-
пуляции Ирака через Иран к популяциям Таджикистана.
Между крайними популяциями (например, Ирака и Таджи-
кистана) по указанным выше признакам выявлены достовер-
ные отличия (табл. 32). По другим признакам (количеству за-
тылочных щитков, строению носового щитка, расположению
предлобных щитков и форме центрально-височного щитка)
изменчивость выражена весьма слабо. Однако следует отме-
тить, что в Таджикистане чаще встречаются особи с боль-
шим центрально-височным щитком (11,3%) и единичны
с цельным носовым (3,7%). У таджикских гологлазов
относительно более открытое ушное отверстие. Наряду с
этим здесь же отмечены особи (45%), у которых ушной про-
ход наполовину покрыт височными щитками. Надо сказать,
что эти особенности щиткования уха часто трудно раграни-
чить, особенно это касается популяций гологлазов Узбекис-
тана, Ирана, Ирака, а также Западного Копетдага. У особей
Азербайджана и Грузии ушное отверстие редуцировано наи-
123;
Таблица 32
Достоверность различия t между выборками A. pannonicus
Выборка L сГсГ 99 Sg вокруг шеи Sg вокруг тела Брюшные Подпаль- цевые пластинки
Таджикистан —
Узбекистан 4,98 3,04 0,63 2,5 0,57 0,93 0,53
—»— Туркмения —»— Азербайджан, 2,83 2,88 1,1 0,36 1,14 0,8 0,13
Грузия 3,27 2,27 0,1 0,65 0,36 0,87 3,04
—»— Иран 5,3 1,83 1,72 3,9 1,83 0,11 0,51
—»— Ирак Узбекистан — 5,5 3,12 3,87 1,8 2,06 2,88 6,37
Туркмения —»— Азербайджан, 2,6 0,73 0,48 2,07 1,5 1,43 0,15
Грузия 3,34 1,61 0,5 0,72 0,32 1,43 2,07
—»— Иран 0,75 0,44 1,92 1,5 2,07 0,88 0,80
—»— Ирак Туркмения — Азербайджан, 0,81 0,6 4,7 0,71 2,26 3,1 3,68
Грузия 0 1,02 0,87 0,4 1,67 0,27 0,93
—»— Иран 2,57 0,93 2,1 3,26 1,12 0,53 0,31
—»— Ирак Азербайджан, 2,55 0,26 4,85 1,57 1,06 2,22 6,63
Грузия — Иран 2,92 0,05 1,6 1,25 2,13 0,68 1,53
—»— Ирак 2,87 0,98 3,84 0,5 2,38 1,7 3
Иран — Ирак 0,07 0,83 1,42 0,3 0,19 2,48 1,07
более сильно, но всегда имется (Бакрадзе, Даревский, 1973;
наши данные). Вспомним, что аналогичная тенденция редук-
ции уха отмечалась также и у другого вида гологлаза, оби-
тающего в Закавказье, —A. chernovi Darevsky, 1953. Как ска-
зано в очерке о A. grayanus, среди азиатских гологлазов спо-
радически встречаются лишенные наружного ушного прохо-
да (экземпляры из Нимичи-Боло (Таджикистан) и из Сар-
гата (Иран): № 14075 и 9249 соответственно: кол. ЗИН АН
СССР), но по другим признакам, например, колйчеству чешуй
вокруг тела и шеи, они не отличаются от типичных A. pan-
nonicus. Изучение типов A. persicus Nik., 1907 и A. brandti
var. brevipes Nik., 1907 из Ирана, описанных А. М. Николь-
ским (1907), подтвердило мнение Н. И. Шнитникова (1928)
и С. А. Чернова (1959) о полной их идентичности азиатско-
му гологлазу.
Подводя итог анализа изменчивости A. pannonicus, можно
наметить отдельные, находящиеся в стадии становления, оча-
ги формообразования — в Таджикистане, Азербайджане и
Грузии и, возможно, в Ираке.
124
Рис. 31. Стация A. pannonicus, Туркмения, Бадхыз.
Стации и количественные данные. Суммируя данные ли-
тературы (Чернов, 1959; Богданов, 1960, 1962; Карпенко,
1970; Второв, Макеев, 1972, Ядгаров, 1973; Второв, Макеев,
Бажанский, 1978; Саид-Алиев, 1979;) можно сказать, что, в
Средней Азии этот гологлаз придерживается мест с более или
менее густым травянистым покровом, тугайных зарослей по
берегам рек, склонов ущелий, а в горах — каменистых участ-
ков лугового типа и арчевников; в культурной зоне встреча-
ется по берегам арыков, на пустырях и огородах, местами —
среди деревьев и кустарников, в фисташковых редколесьях и
лессовых холмах с эффермерной растительностью (рис. 31).
Вид сосредоточен в основном в полосе предгорий, оазисах. В
Закавказье (Грузия — Вашловский заповедник) азиатский го-
логлаз живет в местах, покрытых зарослями бородача, где
за 2 ч. экскурсии можно встретить шесть экземпляров (Бак-
радзе, Пицхелаури, 1973; Бакрадзе, Даревский, 1973). В
Узбекистане (хр. Нуратау) этот вид во все сезоны редок.
Здесь он поселяется отдельными очагами; на участке 700—
800 м (высота 1500—1650 м) южного склона перевала Капча-
гай в апреле с 10 до 14 ч встречено 10 особей — в среднем
25 особей на 11 га=1,35 г/га (Карпенко, 1970). В бассейне
Сурхан-Дарьи данный вид вместе с девятью другими пресмы-
кающимися составил 8,2% по встречаемости (Ядгаров, 1973).
125
В Бадхызе в фисташковом редколесье (включая искусствен-
ные посадки) с эфемеровой и эфемероидноп раститель-
ностью на маршруте протяженностью 75 км данный
вид в пересчете на 1 га составил 0,9 особей, а его био-
масса— 0,5 г/га (Второв и др., 1978). По данным этих же ав-
торов, в Бадхызе встречаемость азиатских гологлазов среди
шести видов рептилий равнялась 16,7%. В Центральном Ко-
петдаге они приурочены в основном к высотам 500—800 м,
где их численность в мае составляет 26—32 на 1 га (Макеев
и др., 1974, 1975, 1983). В Таджикистане по некоторым дан-
ным (Саид-Алиев, 1979) в адырах на расстоянии 13—15 км
можно встретить 25—35 особей (тугаи низовьев р. Кызыл-Су),
а в другом месте (долина р. Бюрган) на 16 км — 26. В зак-
лючение отметим, что среди исследований, проведенных по
этому виду, наиболее точны учеты В. П. Карпенко (1970),
поскольку, как мы считаем, основная задача установления
численности гологлазов сводится к выявлению отдельных их
очагов в соответствующих биотопах, поэтому подсчеты их
на больших расстояниях нецелесообразны. Данные о числен-
ности и биотопической приуроченности вида вне СССР отсут-
ствуют.
Сезонная и суточная активность, отношение к температуре.
По наблюдениям в Узбекистане (Богданов, 1960), выход азиат-
ских гологлазов из зимних убежищ отмечен 20 февраля, когда
еще местами лежит снег. В это время их наблюдали только
во второй половине дня. В марте и апреле они встречались в
полуденные часы, а в июне отмечены с 7 ч 50 мин до
9 ч 30 мин утра и с 18 ч 37 мин до 20 ч 6 мин вечера; в сен-
тябре отмечены в 15 ч. В Нуратау на высоте 1500—1650 м
первая ящерица встречена 28 марта в 11 ч при температуре
воздуха 7,3° (температура приземного слоя воздуха на 1—
2 см от поверхности почвы была равна 27,2°), а температура
в норе равнялась 8,6° (Карпенко, 1970). В Таджикистане эти
гологлазы также появляются в марте, в апреле они активны
в течение всего дня, в мае и июне — с 9—10 до 14 ч; в пред-
горьях и горах появляются в И—12 ч и встречаются до 17 ч,
а в долине — с 7—8 до 19 ч (Саид-Алиев, 1979).
Убежища. По наблюдениям в Нуратау (Карпенко, 1970),
ящерицы скрываются под камнями, где может быть норка с
входом 8X10 мм. В Таджикистане убежищем гологлазам
служат камни (Чернов, 1959).
Размножение. Спаривание азиатских гологлазов в Таджи-
кистане происходит в конце марта — начале апреля (Саид-
Алиев, 1979), откладка яиц в окрестности Душанбе — в конце
апреля — начале мая. Уже в июне-июле в Гиссарской долине
беременные самки не встречаются (Чернов, 1959). По наблю-
126
дениям того же автора (Чернов, 1959) у гологлазов, живущих
на высоте 2200—2300 м, откладка яиц происходит на 1,2 2 ме-
сяца позже, чем в долине; здесь самки с готовыми к отклпдке
яйцами встречаются даже в июле. Данный факт С. Л. Чернов
объясняет более поздним наступлением весны в горах и пред-
полагает, что у 99 живущих в горах, происходит задержки
яиц в яйцеводах. Количество откладываемых яиц в кладке не
превышает шести, обычно трех — четырех (Чернов, 1959),
В Узбекистане в июле 99 с готовыми к откладке яйцами
(размером 8—9X4 мм) добывали с 12 по 20 июня на высоте
800 м (окр. Яккабада); в это же время добыты самки с яич-
никами в состоянии покоя (Богданов, 1960). В Иране, как по-
мечает С. А. Чернов (1959), откладка яиц происходит еще
раньше, в середине апреля самки по всем признакам яйца
уже отложили (район Дех-и-Диз). Молодые в Таджикнсоапе
появляются в июле (Чернов, 1959), в Узбекистане массовый
выход сеголеток отмечен в середине июля (Богданов, I960).
По исследованию В. П. Карпенко (1970), в Узбекистане coi-
ревание половых продуктов у9? азиатского гологлаза начи-
нается с конца марта до середины мая. Половозрелые самки
здесь имеют длину тела 35—40 мм; откладка яиц происходит
не раньше второй половины мая. Сопоставляй свои данные
и данные О. П. Богданова (1960), В. П. Карпенко (1070)
считает, что у азиатских гологлазов Узбекистана возможны
Zt.Be — три кладки.
В Туркмении по нашим материалам (колл. ИЗ АН УССР)
у 99, добытых в Кугитанге, 26 мая имелись яйца размером
8—9X4—5 мм; количество яиц у каждой самки было три. У
двух самок, добытых в 30 км южнее Пул-и-Хартуна 4 апрели,
обнаружено также по три яйца размером 5,1—8X3,5—4 мм,
а у самки, добытой 17 мая под Ашхабадом (г. Душак), най-
дены 4 яйца размером 8—9X4—4,3 мм. Таким образом, от-
кладка яиц у азиатских гологлазов Копет-Дага происходит
в мае.
Питание. В Узбекистане питание гологлаза изучал
О. П. Богданов (1960). По данным этого автора, анализ 16 же-
лудков гологлазов, добытых в июне, показал, что основное
место в рационе занимали насекомые (80% встречаемос-
ти), из которых больше всего было жуков — 60% и пауков —
40% встречаемости. Он отмечает также разницу в составе
пищи молодых и взрослых гологлазов, заключающуюся в
преобладании в пище молодых мелких насекомых, в то вре-
мя как пища взрослых более разнообразна. По материялнм
В. П. Карпенко (1970), в Нуратау весенняя пища гологлазов
представлена в основном личинками насекомых и очень мел-
ких паукообразных, многоножек, из насекомых преобладали
137
жуки (чернотелки, щелкуны и их личинки); среди прочей пи-
щи были бабочки и их гусеницы, личинки прямокрылых, ци-
кады, мухи, тли, мокрицы, сколопендра, пауки и скорпион.
Размеры заглатываемой пищи гологлазов из Нуратау могут
быть 12,3X0,3 мм. С. А. Чернов (1959) указывает, что в теп-
лые зимние дни эти гологлазы тоже питаются.
Враги. Среди врагов азиатского гологлаза в Таджики-
стане отмечены змеи: краснополосый полоз, поперечно-поло-
сатый волкозуб (Саид-Алиев, 1979), а в Узбекистане — стре-
ла-змея (Богданов, 1960). По сведениям из Ирана, на них
нападают змеи-зериг (Горелов, Щербак, 1966). В Азербай-
джане на этих ящерицах найдены эктопаразиты Souronussus
sararum Ond. (Гаджиев и др., 1982).
Практическое значение, охрана. Судя по питанию, азиат-
ские гологлазы осуществляют роль регулятора численности
преимущественно мелких беспозвоночных и их личинок, поэ-
тому являются полезными животными. Данный вид мы пред-
ложили внести в Красную книгу Азербайджанской ССР и Гру-
зинской ССР.
ГОЛОГЛАЗ ЕВРОПЕЙСКИЙ -
ABLEPHARUS KITAIBELII BIBRON ЕТ ВОИУ, 1833
Terra typica: о. Пилос, Средиземное море (Греция).
1833 — Ablepharus kitaibelii Bibron in Вог у, Exped Sci. Moreer
31:69. —1952, Mertens, Zool. Anz. Leipzig, 194:49 —
1970, Fuhn, Acta Soc. Zool. Bohem., 34,9:17. —1977, Бан-
ников и др.; 152. — 1981, Gruber, in Handbuch : 292.
1839 — Riopa ruppllii, Gray, Ann. Mag. Nat. Hist., London 1,2:332
1894 — Ablepharus festane Peracca, BouL Mus. Zool. Torino,
9,167:8.
1938 — Ablepharus pannonicus fabichi, Stepanek, Sborn. narodn.
Mus. Praze, 1 B, ZooL:l
1944 — Ablepharus kitaibelii fabichi, Stepanek, Vestn. Cesk.
Zool. SpoL Praze, 9:123
1952 — Ablepharus kitaibelii fitzingeri, Mertens, Zook Anz. 149,
1—2:48.
1970 — Ablepharus kitaibelii stepaneki, Fuhn, Acta Soc. Zool.
Bohem., 34.9:17.
1970 — Ablepharus kitaibelii chernovi (Darevsky, 1953), Fuhn,
Acta Soc. Zool. Bohem., 34,9:19.— 1977, Банников и др.:
153.— 1981, Gruber, in Handbuch... 1:301.
Диагноз. Туловище короткое, стройное (L max. —47 мм).
Надглазничных щитков два, ряд верхнересничных щитков
прерван и второй надглазничный касается глаза; лобно-те-
менных два. Височных щитков в первом вертикальном ряду
два. Между ухом и центрально-височным щитком располо-
жено в одном ряду не более двух чешуй. Носовые щитки
полуразделенные.
Отличия от близких видов. Европейский гологлаз наибо-
лее похож на гологлаза Чернова, но отличается от него за-
128
Рис. 32. Фолидоз головы A. kitaibelii Bibr. et Bory, 1833.
метно сдвинутым к височной части ушным проходом, что вы-
ражено и в фолидозе: от уха до центрально-височного в од-
ном ряду расположено обычно два щитка, тогда как у A. che-
rnovi их не менее четырех, а ухо отодвинуто к шейному отде-
лу. У европейского гологлаза носовые щитки полуразделены,
а у «chernovi» они цельные. Кроме того, у последнего самцы
в брачный период имеют кирпично-оранжевую окраску ниж-
ней стороны тела, а у европейского эта часть тела сероватая.
От остальных гологлазов европейский отличается наличием
двух надглазничных щитков, прерванным рядом верхнереснич-
ных и отсутствием козырька.
Описание (рис. 32). Голотип № 5392 хранится в коллекциях
Национального музея естественной истории (HNHP, Париж);
нами присмотрена серия фотографий голотипа, любезно прис-
ланная Р. Ру-Эстев. Описание вида составлено по 49 эк-
земплярам из Сирии, Израиля, Греции, островов Эгейского
моря (Родоса, Карпатоса и Кикладов), Албании-и Болгарии,
хранящимся в коллекциях ЗИН АН СССР (Ленинград), ИЗ
АН УССР (Киев), ЗМ МГУ (Москва), естественно-историче-
ских музеях Вены (NMW), Чикаго (FMNH), Нью-Йорка
(AMNH) и музее Сравнительной зоологии Гарвардского уни-
верситета (Кембридж, MCZ).
9 302
129
L jj— 36,0—41,0 (38,78±0,2), 99 —40,0—47,0 (44,6±0,58),
t=3,7. Ga 65 —21,5—24,5 (22,7+0,24), 9? —25,0—32,0
(28,37±0,51), t= 10,12. Led eV — 69,0—73,0 (71, ±2,0), 92 —
53—72 (63,5±3,7), t*=l,8. L головы — 5,7—7,1 (6,l±0,09),
?9—5,2—7,0 (6,18+0,11), t=0,57. Ширина головы —
4,0—5,0 (4,2+0,09), 99—3,9—6,0 (4,5+0,1), t = 2,3. Lp! oV —
6,0—9,0 (6,7±0,25), 99 — 5,5—8,5 (6,5±0,16), t=0,25. Lp2
d-rf —8,2—13,2 (10±0,4), 99—8,2—11,5 (9,83+0,26), t =
Ga
0,36. —p—e’er — 56,1—60,81 (58,7±0,44), 99 — 58,7—68,08
(63,65±0,55), t=7,l.r^&—0,56,99—0,60—0,87 (0,57±0,ll).
Lea
Ь головы 1463—173! (15,7+0,18), 99 — 12,76—14,9
L-*
(13,88±0,19), t=0,77. гол’ г d-64,5—78,7 (68,9 +
x L головы
1,1), 99 —66,1—83,33 (72,73±1,3), t=2,25.
14,63—21,95 (17,4±0,34), 99 — 11,7—18,78 (14,6±0,6), t =
4,12. U-20,0—33,68 (25,89—0,98), 99 — 19,15—28,26
(22,07—0,53), t=3,44. Межчелюстной щиток коротким швом
касается лобно-носового. Надглазничных два. Лобно-темен-
ных 1—2: 1 = 2,4%, 2 — 97,6%. Межтеменной один, по раз-
мерам примерно равен лобно-теменному. Большие теменные
щитки смыкаются позади межтеменного узким швом. Первый
вертикальный ряд височных состоит из двух щитков. Централь-
но-височный двух типов: 1. Маленький свободный сверху —
16,3%. 2. Большой трапециевидный — 83,7%. Ушной проход
маленький, его размеры укладываются в размерах глаза не
менее 10 раз. На переднем крае уха имеется одна-две мел-
ких зубовидных чешуйки. Между ухом и центрально-височ-
ным включительно расположено два щитка.
Sg вокруг шеи, 66 — 19—23 (21,60+0,3), 9 9— 19—23
(21,3+0,23), t=0,8. Sg вокруг тела, 66 —18-22 (19,6 +
0,22), 99 — 18—20 (19,41+0,17), t = 0,68. Брюшных в про-
дольном ряду: 66— 38—45 (42,7±0,6), 99 —45—52 (48,2+
0,43), t = 7,53. Подпальцевых чешуек на четвертом пальце р2
66 — 10-15 (11,7+0,37), 99 —9—14 (11,66+0,3), t = 0,85.
1 1 1
Затылочных щитков^ —5 : т? =2%, 2 = 9,8% — 2 = 2%; 3 =
130
41,2%;у- 3=6%; 4=35%; - 4=2%; 5=2%. Козырек отсут-
ствует, а верхний край глаза прикрыт вторым верхнереснич-
ным и вторым надглазничным щитками. Верхнегубных впе-
реди подглазничного 3—4:3 = 80%, 4 = 20%. Передних верх-
нересничных 2—4:2 = 93,7%; 3 = 4,2%, 4 = 2,1%. Между под-
глазничным и носовым щитками расположено три — четыре
щитка: 3 = 3%, 4 = 97%. Предлобных щитков два, они разде-
лены передним углом лобного — 75%, соприкасаются в од-
ной точке — 20%, соприкасаются широко — 5%. Носовой щи-
ток обычно полуразделен — 97%, но встречается и цельный —
3%. Впереди клоакальной щели два больших анальных щит-
ка. Хвост снизу покрыт одним рядом слабо расширенных
щитков.
Основной фон верхней части тела светло- или темно-брон-
зового цвета с золотистым отливом. Верхняя сторона тела
однотонная, без рисунка — 25%, имеет четыре ряда продоль-
ных прерывистых мелких темно-бурых черточек— 54%; имеет
три — четыре ряда светлых черточек, окантованных по краям
темно-бурыми пятнышками—12,5%; на верхней стороне те-
ла шесть рядов продольных прерывистых темных черточек —
8,5%. Боковые полосы начинаются широкой темной височ-
ной полосой, идущей от ноздри и продолжающейся назад че-
рез глаз вдоль боков туловища и передней части хвоста; иног-
да вдоль этой полосы проступают узкие светлые линии. Брю-
хо у самцов в период размножения сероватое.
Распространение (рис. 33). В Южной Европе (Чехослова-
кии, Венгрии, Румынии, Югославии, Болгарии, Албании, Гре-
ции), включая многочисленные острова Эгейского моря, Ма-
лую Азию, Западную и Северо-Западную Турцию, Сирию,
Ирак, Израиль, Иорданию, Кипр, Синай.
Систематика и географическая изменчивость. До недавне-
го времени систематики (Stepanek, 1938; Mertens u. Muller,
1940; Stepanek, 1944; Mertens, 1952; Mertens, u. Wermuth,
1960; Fuhn u. Vancea, 1961) различали три подвида европей-
ского гологлаза: A. k. fitzingeri Mertens, 1952, охватываю-
щий все северные — словацкие, венгерские, румынские и бол-
гарские популяции; номинативный— все южные, включая Си-
най, Иорданию, Израиль, Сирию, Турцию с многочисленными
островами Эгейского моря, за исключением о. Карпатос, Мик-
ронис, Касос и Арматиа, которые населяют подвид A. k. fa-
bichi Stepanek, 1938.
Согласно О. Щепанеку (Stepanek, 1944), указанные под-
виды отличаются друг от друга внешним видом, длиной ко-
нечностей и четвертого пальца задней ноги, числом перед-
131
Рис. 33. Распространение A. kitaibelii (.) и A. chernovi (Д).
Кадастр к рис. 33. Распространение A. kitaibelii.
Сплошной линией обозначен ареал A. k. fitzingeri; пунктирной —
A. k. fabichi; все остальные пункты — ареал A. k. kitaibelii.
• ЧССР: 1. Плесивскей (Fuhn и Vancea, 1964). 2. Кекове (они же).
3. Ковасовско корсой (они же). 4. Паркани (Stepanek, 1938). 5. Кобаков
(Fuhn, 1970). 6. Смирово (он же).
Венгрия: 1. Будапешт (Fejerary, 1912). 2. Геделе (Fuhn, 1970). 3. Бу-
даток (он же). 4. Таполкца, северная часть оз. Балатон (Fejervary, 1917).
5. Балатон — Счент—Георги, южная часть оз. Балатон (он же).
Румыния: 1. Бухарест. 2. Дубово. 3. Оршово. 4. Радури. 5. Пленита.
6. Крайова. 7. Перисци. 8. Снагов. 9. Лехмиц. 10. Бугае. 11. Олтина. 12.
Канарау— Фетти — Барешта. 13. Алиману. 14. Мурфатлар. 15. Бабадаг.
16. Слава Руса. 17. Кикурова. 18. Букурешти. 19. Дубово. Пункты даны
по Fuhn и Vancea 1961; Fuhn, 1970; 1969а).
Болгария: 1. Гора Каракуз (колл. ИЗ АН УССР). 2. Приморско (колл.
ЗИН АН СССР). 3. Котел. 4. Странджа Планина, окр. Софии. 5. Бела-
градик. 6. Лакитник. 7. Димитрово. 8. Родопы. Пункты даны по Fuhn,
1970.
Югославия: 1. Белград. 2. Звезда. 3. Авала. 4. Козьяк. 5. Палик. 6.
Скопье. 7. Брестоваука. 8. Банья. 9. Фруска Гора. 10. Белград. 11. Бружа.
12. Паска. 13. Подульево. 14. Пристина. 15. Преурен. 16. Парацин (Dzukic,
1972). Все остальные пункты даны по Fuhn, 1970; Paculjevlc, 1956.
Албания: 1. Пояни (Kopstein und Wettstein, 1921; колл. NMW). 2. Ав-
лона (Fuhn, 1970).
Греция: 1. Карпатос. 2. Микониз. 3. Касос. 4. Атматиа. 5. Пилос. 6.
Превеза. 7. Парос. 8. Олимпиа. 9. Никария. 10. Зубея. 11. Тинос. 12. Се-
рифос. 13. Милос. 14. Кос. 15. Хантус. 16. Кипр. 17. Саматраки. 18. Пе-
лопонес, Месенае. 19. Самос. 20. Менелитос. 21. Корфу. 22. Левкоз. 23.
Кефаления. 24. Закинтос. 25. Аморгос. 26. Китира. 27. Антикитира. 28. Ро-
дос. Пункты 1—4, 6—8, 10—17, 19, 21—28 (Mertens, 1968) (Fuhn, 1970;
Gruber, Dieter, 1977; Gruber, 1981). 4. (Mertens und Muller, 1928); 9 колл.
MCZ); 18 (колл. AMNH) и 32 (колл. NMW).
Турция: 1. Стамбул. 2. Полонеско. 3. Босфор. 4. Рукупермек. 5. Кас-
тамону. 6. Харбин, Хатай. 7. Адана. 8. Аманус. 9. Анталиа. 10. Будрум.
11. Измир. 12. Сома. Пункты цаны по Fuhn, 1970.
Ливан: 1. Ампум (Fuhn, 1970). 2. Бейрут (он же).
Сирия: 1. Джарнуз (Fuhn, 1970). 2. Аманис (он же).
Израиль: 1. Тиверия. 2. Хулда. 3. Изод Хама-Ала. 4. Орен. 5. Сорока-
Яффа. Пункты даны по Fuhn, 1970.
Иордания: 1. Дана (Fuhn, 1970). 2. Иерусалим (колл. ЗИН АН СССР;
NMW).
Арабская Республика Египет: Синайский п-ов (Магх, 1956; Schmidt
and Marx, 1956; колл. MCZ), пункты не указаны. Выборки: II (пункты I)';
IV (1, 2); VI (1); VII (12, 15, 28); X (1, 2); XIII (1, 2).
них верхнегубных щитков и чешуй вокруг середины тулови-
ща, а также особенностями окраски. Позднее О. Веттштейн
(Wettstein, 1953) заметил, что различия между «kitaibelii» и
cfitjzingeri» очень незначительны. В то же время он высказал
мнение, что популяция о-ва Родос отличается якобы собствен-
ной дифференциацией и представляет «Rasse in statu nas-
cendi».
Ревизия вида, предпринятая румынским герпетологом
И. Фуном (Fuhn, 1969а, 1970), показала валидность формы
«fitzingeri», а популяциям гологлазов Болгарии и Румынии
придан статус подвида A. k. stepaneki, Fuhn, 1969. Он же
(Fuhn, 1969а, 1970), не обнаружив существенной разницы меж-
ду A. kitaibelii и A. chernovi, ранее описанным И. С. Дарев-
ским (1953) из Армении, рассматривает последний в каче*
стве подвида европейского гологлаза. Указанная выше точка
зрения до настоящего времени была общепринятой (Greer,
1974; Банников и др., 1977; Gruber, 1981).
133
Наше знакомство с коллекционным материалом (изучено
53 экземпляра этих ящериц из Сирии, Израиля и Иордании
(8), Кикладов (9), Карпатоса и Микронис (4), Родоса (4),
Албании (10), Болгарии (6), Венгрии (8) и СССР (Арме-
нии— 4), а также переосмысливание литературных данных
убеждают нас в том, что формы «stepaneki» и «kitaibelii»
связаны между собой в значительной степени клинально
(табл. 18). Как видно, по основным признакам (количество
чешуй вокруг тела, число верхнегубных щитков, подпальце-
вых чешуй и др.) эти формы практически не отличаются.
Предположение И. Фуна (Fuhn, 1969а, 1970), что гологлазы
Югославии и Албании относятся к форме «stepaneki», на на-
шем материале (исследовано 10 особей из Албании) не подт-
верждается, они идентичны номинативной форме. Послед-
няя — A. k. kitaibelii — заметно выделяется от остальных при-
сутствием малого центрально-височного щитка (у особей Си-
рии и Израиля). Форма «fabichi» достоверно отличается боль-
шим числом подпальцевых чешуек четвертого пальца. Это
связано с более развитыми конечностями, что отмечалось и
ранее (Stepanek, 1938, 1944; Fuhn, 1969в, 1970).
Северная форма «fitzingeri» выделяется заметным возрас-
танием числа особей (до 50%, наши данные), имеющих впе-
реди подглазничного четыре верхнегубных щитка. В то же
время по данному признаку форма «fitzingeri» близка к «fabi-
chi», на что указывал и И. Фун (Fuhn, 1970). Мы отмечаем
присутствие четырех верхнегубных щитков и у особей номи-
нативной формы (Сирии и Израиля— 12,5%).
Окраска у просмотренных нами особей варьировала. Осо-
би с окраской «fitzingeri» отмечены и на островах Карпатос,
Киклады, а также в Болгарии. Здесь же (кроме о. Карпатос)
встречались особи однотонной окраски или с слабовыражен-
ным рисунком в виде мелких черточек. Рисунок из мелких
темных черточек, идущих в четыре ряда вдоль спины, отме-
чен наиболее часто (54%). В целом изменчивость большин-
ства признаков этих четырех подвидов носит клинальный ха-
рактер, лишь немногие из них дискретны.
Что касается пятого подвида — A. k. chernovi (Darevsky,
1953), то отдельные таксономически важные признаки резко
выделяют его среди вышеназванных подвидов. В частности,
особого внимания заслуживает указанная для этих ящериц
И. С. Даревским (1953) значительная редукция наружного
ушного прохода. Мы, в свою очередь, изучая фолидоз голо-
вы, обнаружили у особей «chernovi» в одном ряду от цент-
рально-височного до переднего края ушного отверстия четы-
ре— пять щитков, а у всех остальных форм только два таких
щитка; кроме того, ушной проход у «chernovi» всегда сдви-
134
Таблица 33
Изменчивость размеров тела у A. kitaibelii и A. chernovl *
№ вы- борки Место L Ga
A. kitaibelii
1. ЧССР — 47 —
2. Венгрия 4 37,0—39,0; 38,0±0,41 21,5—23,0; 22,25±0,32‘
4 40,0—47,0; 42,5± 1,5 25,0—30,0; 26,75±1,11
3. » — 43—(48)—51 —
4. Болгария 3 37,0—41,0; 39,1 ±0,3 22,5—24,5; 23,5±0,5
3 47,5-47,0; 45,25±0,6 26,0—31,0; 28,8±0,9
5. Румыния — 33—(39,7)—47 —
35—(45,7)—51
6. Албания 3 37,0—41,0; 35,53±2,6 _ 20,0—24,5; 23,6±0,6
3 44,0—47,0; 45,33±0,88 26,0—31,0; 30,3± 1,3"
7. Родос и 1 36 21,5
Киклады 7 40,0—47,0; 44,7±0,65 22,5—32,0; 29,2±0,6
8. Атика — _32—(35,4)—38 —
— 38—(41,6)—47 —
9. Делос — 32,5-(35,1)—42 —
— 41 —(44)—47 —
10. Карнатос и 2 38,0—41,0; 39,5±1,5 22,0—23,0; 21,5 ±0,5
Микронис 2 46,0—47,0; 46,5±0,5 28,0—29,0; 28,5±0,5
11. Микронис — 34—(39,7)—46 —
— 42— (46)—48 —
12. Кипр — 35—(39)—45 —
43—(43)—44 —
13. Сирия Из- 2 33,0—41,0; 37,0±4,0 20,0—23,0; 22,0 ±1,0
раиль, Иор- 41,0—46,0; 43,5±2,5 23,0—27,0; 25,0 ±2,01
дания
14. Израиль — 27—(29)—30 —
32— (34)—42 . — A. chernovi
1. СССР — — —
(Армения) 3 42,0—53,0; 46,66±3,33 26,5—36,0; 31,16+2,82
3 36,0^15,0 —
* Примечание. Ссылки на данные см. в табл. 36.
135
Таблица 34
Изменчивость пропорций головы и длина ног у A. kitaibelii и A. chernovi
3 s ® о. п L головы Ширина головы 1 L Р, L Ра
«о. о Ю 1
A. kitaibelii
1 — 6 5 7 11
2 8 - 5,8—6,2 4,0—4,8_ 6,0-7,0 _ 9,0—10,0
6,0 ±0,5 4,17±0,3 6,36±0,15 9,75±0,18
3 — 6 5 7 11
4 4 _ 6,0—6,4 4,5-5 6,2—7,0 _9,3—10,0__
6,12±0,17 4,72±0,1 6,5 ±0,25 9,77±0,32
5 — 6 5 5,0-(6,5)—7,5 7—(9,4)—10
6 6 5,9-6,5 4,0—4,9 6,0—7,0 9,0—10,0
4,28±0,24 4,28±0,2 6,4±0,2 9,63 ±0,46
7 8 - 5,2—7,0 3,9—5,5 5,5—7,2 __8,2—11,5
6,24 ±0,3 4,5±0,5 6,52±0,7 9,8±0,45
8 — 5 4 5—6 9—10
9 — 5 4 6,0 (6,4)—7,0 9—(9,6) —И
10 4 6,1—7,1_ 4^8—5^ 8,0—9,0 12,8—13,8
6,85±0,25 5,0±0,3 8,62±0,3 13,2 ±0,32
11 — 6,5—7,6 — 7—(8,6)—10 10—(12,6)—14
12 — 5 4 5—(6,1)— 8 5—(9,45)—11
13 4 _ 5,1—6,0 3,6—4,5 6,0-6,5 8,2—9,0
5,5±0,44 4,0±0,44 6,25±0,1 8,6±0,4
14 — 4,3—5,4 — 4—(4,6)—6 6- (7) —8
A. chernovi
1 2 6,0—6,2 4,0—4,8 6,0—6,2 9,2—11,0
6,06±0,07 4,33±0,24 6,1 ±0,1 10,0 ±0,55
Таблица 35
Изменчивость количества чешуй у A. kitaibelii и A. chernovi
№ вы- борки п Sg вокруг шеи Sg вокруг тела Брюшных
1 2 3 1 1 4 5
1 8 20—24 22,12±0,6 A. kitaibelii 20—22 19—20 19,5±0,19 42—45; 43,0 ±0,7 46—50; 48,0 ±0,8
136
1 2 3 4 5
3 — 18,6 — 3-4
4 6 20—23 20—22 46—51
21,66 ±0,5 20,33 ±0,33 47,25±1,3
5 — — 19-(20)—22 —
6 10 20—22 — 18—20 38—45; 41,5 ±0,9
21,2±0,2 19,1 ±0,46 44—51; 48,0 ±1,6
7 13 20—23 20 44
22—0,3 46—52; 48,75±0,54
8 — — 19—(20)—22 —
9 — — 19—(20)—22 —
10 4 22—24 20 38-41; 39,5±1,5
23,0 ±0,5 42—46; 44,0±2,0
11 — — 20 —
12 — — 18—(18,1)—20 —
13 8 19—22 17—20 —
20,56±0,43 18,25±0,3 48—49; 48,5 ±0,5
14 — — 18 —
A. chernovi
1 з 20—21 18 _ —
20,6 ±0,3 50—56; 52,75 ±1,25
2 4 — 18 —
Таблица 36
Изменчивость количества верхнегубных щитков
и некоторых других признаков у A. kitaibelii и A. chernovi
№ вы- борки п Верхнегуэных Подпальцевых Примечание
1 2 3 4 5
A. kitaibelii
1 — 4 — Fuhn, 1970
2 8 3 (50%) _ 10—13_ 4 (50%) 11,25 ±0,32 Наши данные
137
1 2 3 4 5
3 — — — Fuhn, 1970
4 6 3 11—13 11,66±0,33 Наши данные
5 — 3 — Fuhn, 1970
6 10 3 10—12 10,7±0,2 Наши данные
7 13 3 9—14 12,55±0,4 —>—
8 — 3 — Fuhn, 1970
9 — 3 — —>—
10 4 3 (33,3%) 4 (66,7%) 15 Наши данные Наши данные
11 — 3 (80%) 3/4 (20%) 17—18 Fuhn, 1970
12 — 3 — —>—
13 8 з (87,5%) 4 (12,5%) 11—14 12,5-0,32 Наши данные
14 — 3 14 A. chernovi Fuhn, 1970
1 3 1 экз. «2/3 2 экз.«3 11—14 12,5±0,64 Наши данные
2 4 3 — » Даревский, 1953
нут ближе к шее, а у всех остальных, наоборот, к височной
области. Среди других признаков «chernovi» заслуживает
внимание большее число брюшных щитков, а также несколько
иные пропорции тела. Наконец, эти ящерицы заметно отли-
чаются от других подвидов наличием у самцов в брачный пе-
риод яркой оранжевой окраски брюха, горла, нижних сторон
конечностей и части хвоста (Даревский, 1953), в то время как
у самцов A. kitaibelii эти части тела сероватые.
138
Таблица 37
^Изменчивость некоторых признаков фолидоза у A. kitaibelii и A. chernovi
Выборка
п Число затылочных щитков Носовой щиток
1 2 2 J-2 2 3 1 3 2 4 ±4 Г 5 6 цельный полураз- 1 делен |
A. kitaibelii
Венгрия 8 — 1 — 4 — 3 — - — — 8 6 2 — 2 6 8 —
Болгария 6 — — — 3 — 5 — — — — 6 4 1 1 — 6 6 —
Албания 10 1 1 — 4 — 2 — — — 2 8 7 3 — 1 9 10 —
Родос и Киклады 13 — 1 1 6 3 2 — — — 2 12 7 3 3 13 13 —
Карпатос и Мик он ИС 4 — 2 11 — 4 3 . 1 2 2 4
Сирия и Израиль 8 — — — 4 — 4 — — — — 8 4 3 1 5 3 8 —
A. chernovi
СССР
(Армения) 3 — — — — — — — j 3
з - - - 3-3
Таким образом, валидными подвидами европейского го-
логлаза являются номинативный, A. k. fabichi А. к. fit-
zingeri. Популяции гологлазов Румынии, Албании и Бол-
гарии находятся в зоне интеграции между номинатив-
ным подвидов и «fitzingeri», поэтому валидность фор-
мы «stepaneki» вызывает сомнение. Форма «fabichi» по
некоторым признакам, встречающимся у отдельных особей
(четыре верхнегубных, сплошной ряд верхнересничных, нали-
чие трех височных в первом вертикальном ряду) и имеющим
явно анцестральный характер, является реликтовой. Фор-
му А. к. chernovi на основании ряда признаков (четыре щит-
ка в одном продольном ряду от уха до центрально-височно-
го, сильно редуцированный ушной проход, максимальное ко-
личество затылочных щитков, цельные носовые и др.) сле-
дует восстановить в ранг самостоятельного вида — A. cher-
novi Darevsky, 1953.
Таблица для определения подвидов A. kitaibelii
1(2) Ноги длинные (Lpi — 9 мм, Ьр2—13,8 мм); на четвер-
том пальце задней ноги обычно 15—18 подпальцевых чешуек...
...............................................А. к. fabichi
2(3) Ноги относительно короткие (Lpi — 7,2 мм, Lp2—
11,5 мм); на четвертом пальце задней ноги не более 15 че-
шуек.
3(4) Передних верхнегубных чаще четыре (63%). На спине
четыре ряда беловатых черточек, окантованных по краям
темными пятнышками................................А. к. fitzingeri
4(5) Передних верхнегубных обычно три. Тело стройное, ок-
раска однородная без рисунка или в виде нечетных преры-
вистых темных черточек, идущих в два — четыре продольных
ряда..............................................А. к. kitaibelii
Гологлаз европейский обыкновенный—Ablepharus k. kitaibelii
Bib г. et Вог у, 1833
Terra typica: видовая.
1839— Riopa ruppllii Gray, Ann. Mag. Hist., London, 1,2:332 (Ter-
ra typica: Arabia Potrae? (Halbinsel Sinai).
1894— Ablepharus fostae Peracca, Boul. Mus. Zool. Torino, 9, 1967:8
Terra typica: Dscherasch und Es-Salt, Transjordanien.
1940 — Ablepharus k.kitaibelii, Mertens u. Muller, Part., Abh.
Senkenb. Naturf. Ges. Frankfurt am Main, 451:47
Голотип № 5392 (см. A. kitaibelii, рис. 34). Наше описа-
ние составлено по 37 экземплярам из Сирии и Израиля (8),
Греции — о. Киклады и Родос (13), Албании (10), Болгарии
(6), а также данным ряда авторов (Stepanek, 1938, 1944;
Fuhn et Vancea, 1961; Fuhn, 1969b, 1970; Gruber, 1981).
140
Рис. 34. A. kitaibelii (голотип, колл. MNHP)
L 66 — 36,0—41,0 (38,7±0,71), 99 —40,0—47,0 (44,6±0,6),
t=6,45. Gj Ы — 21,5—24,5 (22,7±0,97), 99 — 25,0—31,5
(28,37±0,58), t=8,4. Led <^—69,0—73,0 (71,0±1,8), 99—
63,0—71,0 (63,4±2,6), t=2,4. L головы 66—57—63 (6,1=Е;
0,07),99 — 5,2—7,0 (6,18±0,14), t=l,5. Ширина головы сГ<?—
— 4,0—5,0 (4,2±0,07), 99'—3,9—5,0 (4,5±0,12), t=3,l.
Lpi 66 —6,0—8,0 (6,7±0,20) 99 — 5,5—8,5 (6,5+0,15),
t=0,08. Lp2 <<6 —8,2—13,2 (10,0±0,24), 99 —8,2—11,5
Г?я
(9,8±0,27), t=l,5.^66 —56,1—60,8 (58,7±0,55), 99 —
58,7—67,0 (67,0±0,71), t=8,0.-j-t— 66 — 56, 99—0,60—0,87
(0,57±0,11). —--Л7— 14,63—17,31 (15,7±0,17), 99 —
12,76—14,9 (13,88+0,22), L = 5,7. ШиРин^ головы^ —64,5—
78,7 (68,9±1,23), 99 —59,0—83,33 (72,7±1,19), t=3,0.
U — 14,63—21,95 (17,4±0,56), 99 — 11,7—18,8 (14,60±-
«О
0,33), t=3,4.-^- 20,0—33,68 (25,9±0,7), 99—19,15—
141
"28,26 (22,07±0,5), t = 2,6. Sg вокруг шеи ??— 19—24 (21,3±
о,36), <5><Ь — 19—23 (21,3±0,25), t=0. Sg вокруг тела 99—
18—22 (19,6±0,31), 99 — 18—20 (19,41 ±0,19), t=0,5. Брюш-
ных щитков в продольном ряд7 <Ь&—38—49 (43,15±0,92),
99 —44—52 (48,2±0,48), t=4,95. Подпальцевых чешуек на
четвертом пальце р, c'rf —10—15 (11,7±0,33) 99 — 9—15
(11,66±0,3), t=0,3. Затылочных' — 4:^ = 1%; 2= 10,8%;-2=
1%; 3 = 51,7%, д- 3 = 3%; 4 = 32,5%. Верхнегубных 3—4:3 =
87,5%; 4= 12,5%. Передних верхнересничных 2—4:2=94,6%;
3 = 2,7%, 4 = 2,7%. Центрально-височные двух типов: малень-
кий— 16,2%, большой трапециевидный — 83,8%. Носовые
щитки полураздслеиные 83,8%, цельные—16,2%. Между но-
еовым и подглазничным расположено три—четыре щитка:
3 = 89%, 4=11%. Предлобные разделены передним углом
лобного щитка — 62%; соприкасаются в точке — 27% или
соприкасаются широко—11%.
Рис. 35. Варьирование рисунка. A. kitaibelii (Албания, колл. NMW)
142
Рис. 36. A. kitaibelii (Греция, Киклады, колл. NMW).
Основной фон верхней стороны тела светло-коричневый
с золотистым оттенком. На спине обычно имеется четыре ря-
да мелких темных черточек— 59,9% или спина без рисунка —
32,5%. Реже — 8% встречается рисунок из шести рядов тем-
ных черточек (популяция Албании) (рис. 35, 36).
Распространение (см. рис. 33). Ближний Восток, вклю-
чая Иорданию, Синай, Израиль, Ирак, Турцию (Западную),
Кипр, Грецию с многочисленными островами в Эгейском мо-
ре (кроме о. Микронис, Карпатос, Касос и Арматиа), Югосла-
вию, Албанию, а также Румынию. В последних четырех
странах подвид, видимо, интерградирует с формой fit-
zingeri».
Гологлаз европейский северный — Ablepharus k. fitzmgeri Mertens, 1952
Terra typica: Ofen (Budapest, Ungarn)
1824— Ablepharus pannonicus Fitzinger (non Scincus pannonicus
Licht., 1823), Verb. Ges. Nat. Berlin, 1:298
1952 — Ablepharus k. fitzingeri Mertens (nomen novum Pro Ablep-
harus pannonicus Fitzinger, 1824), Zool. Anz., Leipzig,
194:49
Голотип хранится в Зенкенбергском музее (NMFS). Наше
описание составлено по восьми экземплярам из Венгрии, а
также по литературным данным (Mertens, 1952; Stepanek,
1938, 1944; Fuhn et Vancea, 1961; Fuhn, 1969b, 1970; Gruber,
1981).
LSS — 37,0—39,0 (38,0±0,41), 40,0—47,0 (42,5+1,55),
t = 2,8. Ga SS— 21,5—23,0 (22,25+0,32), 99 —25,0—30,0
(26,75+1,1), t = 3,9. Led SS—нет. L головы SS—6,0—6,1
(6,05 + 0,05), — 5,8—6,0 (5,85+0,04), t=3,12. Ширина го-
ловы —4,0—4,1 (4,0+0,03), 99 — 4,0—4,8 (4,2±0,2)»
t = l. Lpi SS — 6,0—6,8 (6,37±0,17), 99 — 6,0—6,2 (6,1±
0,06). t=l,5. Lp2 SS — 9,5—10,0 (9,87+0,11), 99—9,0—
Gn
10,0 (9,37±0,23), t=2. -p &6 — 57,7—60,52 (58,55±0,66),
99—61,9—63,83 (62,9±0,43), t = 5,5. L. r2-^' dj— 15,38—
16,48 (15,9+0,23), ?? — 12,76—14,5 (13,8+0,37), t=4,8. Ши-
рина головы^ _ 65,6—67,2 (66,5±0,33), 99 — 58,9—80,0
(69,2±4,3), t=0,62. U — 15,8—18,37 (16,8±0,6), J5
13,2—15,5 (14,4±0,47), t=3,l. && —25,0—27,0—27:0
144
(26,0+0,43), 99 — 20,2—25,0 (22,14±2,0), t=l,0. Sg вокруг
шеи££ —20—24 (22,25±0,84), 99—20—22 (21,25+0,47),
t=l,0. Sg вокруг тела — 19—20 (19,75±0,25), 99 —19—
20 (19,5±0,3), t = 3,8. Брюшных щитков в продольном ряду
££—42—45 (43,0+0,7), 99 —45—50 (47,75+1,0), t=3,8.
Подпальцевых чешуй на четвертом пальце р2 ££— 10—13
(11,25±0,63), 99 — 11—12 (11,5+0,3), t=0,36. Затылочных
щитков 2—4:2=12,5%; 3 = 50%, 4 = 37,0%. Передних верх-
нересничных два. Центрально-височные обычно маленькие.
Носовые щитки полуразделенные. Между носовыми и под-
глазничным расположено три — четыре щитка. Предлобные
разделены — 75%, соприкасаются в одной точке — 25%.
Основной фон верхней стороны тела темно-бронзовый. На
спине обычно имеются четыре ряда продольных светлых чер-
точек, окантованных по сторонам темными пятнышками.
Распространение (см. рис. 33). Чехословакия, Венгрия. В
Венгрии, возможно, интерградирует с номинативной формой.
Гологлаз европейский реликтовый — Ablepharus k. fabichi S t е р ап е к, 1938
Terra typica: о. Микронис, Греция.
1938 — Ablepharus pannonicus fabichi, Stepanck, Sborn. Narodn.
Mus., 1 : 7
1944 — Ablepharus kitaibelii fabichi Stepanek, Vestn. Cesk. Zool.
SpoL, Praze, 9 : 123
Голотип № 18426 хранится в Венском естественно-истори<
ческом музее (NMW), leg. Rebel et Sturany, 1904, о. Микро-
нис: 9 L — 47. Ga — 29,0. Led — 72,0. L головы — 7,1. Lpi —
9,0. Lp2—13,8. Sg вокруг тела — 20. Затылочных 3X4. Под-
пальцевых чешуек на четвертом пальце р2—15. Предлоб-
ные щитки соприкасаются узким швом. Описание подвида
составлено по четырем экземплярам: о. Карпатос (3) и Мик-
ронис (1), а также литературным данным (Stepanek, 1938,
1944; Fuhn, 1969в, 1970; Gruber, 1981). L ££ — 38,0—41,0
(39,5+1,5), 99 — 46,0—47,0 (46,5+0,5), Ga ££— 22,0-23,0
22,5±0,5), 99—28,0—29,0 (28,5±0,5): Led ££ — 69; 99—
71—72 (71,5+0,5). L головы ££—6,1—7,1 (6,6±0,5), 99-
7,0—7,1 (7,05+0,05). Ширина головы ££—4,8—5,0 (4,9±0,l),
99 —5,0—5,5 (5,25 + 0,25). Lpi ££-8,09—9,0 (8,5+0,5),
99—8,5—9,0 (8,75+0,25). Lp2 ££ — 12,8—13,2 (13,0+0,2),
99—13,0—13,8 (13,4±0,4). 66 —56,1—58,2 (57,0 + 0,5).
99—60,18—60,87 (60,5±0,4). Д: J6—59,4; 99—64,8—
Led
rr n m nr\ L ГОЛОВЫ
65,3 (65,0+0,25). ------— 16,0—17,3 (16,65+0,65),
10 302
145
V$>— 13,26—15,1 (14,2±0,8). —ирина головы _ 12>2_
12,63 (12,4±0,2), ?9—10,9—11,7 (11,3±0,4). —
21,05—21,95 (21,5+0,4), ??—18,5—19,0 (18,75±0,25).-^-
44—32,2—33,7 (32,94±0,8), ?? —28,2—29,3 (28,75±0,5).
Sg вокруг шеи — 22—23 (22,5±0,5), 9?—23—24 (23,5±
0,5). Sg вокруг тела —20, $9 —20. Брюшных щитков
в продольном ряду, —39—41 (40±1,0), 99 —42—46
(44,0±2,0). Подпальцевых чешуй на четвертом пальце рг 56
’—15, 99—15. Затылочных щитков 4—5:4—2;^- 4—1; 5—
1 (экз.). Передних верхнересничных 2—4:2—2; 3—1; 4—1
(у последнего экземпляра верхнересничные составляют пол-
ный ряд). Центрально-височный малый и свободный свер-
ху— 2, большой трапециевидный — 2 экз. Носовой щиток
полуразделенный. Между носовым и подглазничным распо-
ложены четыре щитка. Предлобные разделены (у 3 экз.), со-
прикасаются широко—1. Основной фон буроватый. На спи-
не четыре продольных ряда светлых черточек, окантованных
по краям темными пятнышками; из них крайние более ярко
выражены (рис. 37).
Распространение (см. рис. 33). Острова южной перифери-
ческой части Эгейского моря — Микронис, Карпатос, Касос
и Арматиа.
Стации и количественные данные. В северных областях
распространения, в ЧССР, Венгрии и Румынии A. kitaibelii
занимает теплые южные склоны, встречаясь на опушках дуб-
рав, в опавшей листве, под камнями или в траве, на сухих лугах
с известняками (Fejervary 1912, 1917; Fuhn et Vancea, 1961;
Rotter, 1952; Muller, 1971; Gruber, 1974; Petzold, 1978). В
Болгарии встречается в местах с пышной растительностью,
избегает гор (Buresch u. Zonkov, 1934); в Югославии — на
почвах, заросших папоротниками, на границе каштановых ле-
сов (Dimorsky, 1959); в Центральной Сербии они распростра-
нены на высоте 300—800 м (Pasuljvic, 1964) на открытых
травянистых местах, покрытых кустарником (Fejervary, 1912;
Radovanovic, 1957); в Албании — на морском берегу в песке
(Fuhn, 1970 in Siebenrock, 1922); в Греции (Fuhn, 1970) и на
островах Эгейского моря (Wettstein, 1953) — почти только
в местах, заросших травой. В Израиле (Fuhn, 1970, по устно-
му сообщению Н. Hoofien) этот гологлаз живет повсюду, где
есть укрытия — под камнями, скалами и т. д., которые соз-
146
Рисе. 37. A. kitaibelii (Греция, о. Карпатос,
колл. NMW)
дают подходящий микроклимат, где процветает пышная од-
нолетняя растительность. На средиземноморской низменности
ящерица обитает в дубравах, в искусственных лесополосах,
а также по сухим руслам рек; гологлаз редок или отсутствует
в густых тенистых лесах типа Magnus, в местах, лишенных
147
камней, на песчаных дюнах в пустынных местах, а -также на
культивируемых почвах, где удалены камни.
Сезонная и суточная активность. По наблюдениям У. Гру-
бера (Gruber, 1974) на одном из островов Эгейского моря яЩе"
рицы проявляли наибольшую активность в течение 1—3 ч
после восхода солнца и 1—2 ч после его захода. Активных
ящериц наблюдали также и в сумерках (Fuhn, 1970). Различ-
ные периоды активности ящериц отмечал Г. Пасюлевич (Ра-
suljevic, 1964) в Югославии; здесь наблюдаются два пика ак-
тивности, причем их вершины резко отделяются друг от дру-
га летом, а весной и осенью сдвигаются в один. Ж- Роттер
(Potter, 1952) предполагал, что летом эти ящерицы какой-то
промежуток времени вообще не проявляют активности.
Размножение. Спаривание происходит сразу после выхо-
да с зимовки — в конце февраля — начале марта (Fejervary,
1912), чаще всего в апреле или мае (Rotter, 1952). Кладка
состоит из двух — четырех яиц (Radovanovic, 1957; Rotter, 1952;
Petzold, 1978); яйца белые, эллиптической формы, 7—10 мм
(Radovanovic, 1957). Сроки откладки яиц растянуты — с кон-
ца июня до начала августа. Наблюдения в террариуме (Rot-
ter, 1952) показали, что самки гологлазов яйца зарывали в
рыхлую влажную почву на глубину 5—7 мм; во время отк-
ладки яйца имели длину 7,1—7,2 мм, а спустя 21 день увели-
чились до 11 мм, на 27-й день — до 12,1 мм и на 37-й день —
до 13,8 мм; вылупление молодых длиной тела 33 мм происхо-
дило на 65-й день.
Питание. По данным ряда авторов (Fejervary, 1912; Ra-
dovanovic, 1957; Rotter, 1952), питаются гологлазы мелкими
насекомыми и их личинками (настоящие моли, тли), а также
мелкими пауками (прежде всего ликозиды), сенокосцами
(опилионы) и дождевыми червями. По наблюдениям в тер-
рариуме (Rotter, 1952; Petzold, 1978), ящерицы поедали мел-
ких личинок мучного хрущака, сверчков, мух, мошек и их ли-
чинок, пауков. Жажду утоляли капельками воды (Fejervary,
1912; Radovanovic, 1957). В четырех желудках гологлазов из
Болгарии нами найдены сенокосец (Opiliones), жуки-блошки
(Altilinae), гусеницы совок (Nostuidae), остатки пауков (Ага-
пса) и перепончатокрылых (Hymenoptera). У одной ящерицы
из Израиля обнаружены остатки клещей и насекомых.
Стереотип поведения, убежища. Убежищами ящериц слу-
жат густой кустарник, травянистая растительность, опавшие
листья и т. д.; очень важно наличие в биотопах камней, под
которыми они часто прячутся (Fejervary, 1917; Radovanovic,
1957; Gruber, 1974, 1981). Из-за слаборазвитых конечностей
перемещение на поверхности дополняется извивающимися
движениями тела, при этом передние лапки работают интен-
] 48
сивнее задних (Radovanovic, 1957). Только чтб вылупившиеся
ящерицы для передвижения пользуются исключительно нога-
ми (Radovanovic, 1957).
Враги. Некоторые авторы (Gruber, 1974; Gruber u. Fuchs,
1977; Gruber, 1981) отмечают, что на островах Эгейского моря
A. kitaibelii уходит от степной ящерицы Podarcis erhardii. Вы-
воды указанных авторов сводятся к тому, что «сильные» попу-
ляции A. kitaibelii встречаются на тех островах, где отсут-
ствует степная ящерица, и наоборот, там, где она есть (на-
пример, остров Парос и др.), A. kitaibelii практически не по-
падаются. Среди видов, преследующих A. kitaibelii, отмечают
зеленую ящерицу (Lacerta viridis), змей (Elaphe longissima
и Coronella austriaca), а также сорокопута-жулана (Lanius
collurio) и других дневных хищных птиц, а среди млекопи-
тающих— насекомоядных и куньих (Gruber, 1981).
Практическое значение. Как все гологлазы, A. kitaibelii —
полезное животное и повсеместно нуждается в охране.
ГОЛОГЛАЗ ЧЕРНОВА — ABLEPHARUS CHERNOVI D A R EV S К У, 1953
Terra typica: среднее течение р. Роздан, Армения
1970 — Ablepharus kitaibelii chernovi (Dar., 1953), Fuhn, Acta Soc.
Zool. Bohcm. — 34:9. —1977, Банников и др.: 153.
Диагноз. Ящерицы с тонким удлиненным телом. Над-
глазничных щитков два; ряд верхнересничных щитков прерван
и второй надглазничный касается глаза; лобно-теменных
два. Между ухом и центрально-височным щитком в одном ря-
ду расположено обычно четыре чешуи. Носовые щитки цель-
ные.
Отличие от близких видов. Гологлаз Чернова наиболее
походит на европейского, но отличается от него тем, что
имеет не менее четырех чешуй, расположенных в одном ряду
между ухом и центрально-височным щитком, а также цель-
ными носовыми щитками. Кроме того, у / ’ A. chernovi в
брачный период окраска нижней стороны тела кирпично-оран-
жевая. От остальных видов рода отличается теми же приз-
наками, что и европейский гологлаз.
Описание (рис. 38) Голотип № 2810, колл. ЗМ МГУ, до-
быт И. С. Даревским. сГ : L — 42. Ga — 26,5. Led. reg. —41,5.
L головы — 6,2. Ширина головы — 4. Lpi — 6,2. Lp2—10.
Sg вокруг тела — 18. Затылочных щитков 6X4. Подпальцевых
чешуй на четвертом пальце—14. Носовой щиток цельный. На-
ше описание составлено по четырем экземплярам из Армении,
хранящимся в ЗИН АН СССР (Ленинград) и ЗМ МГУ (Моск-
ва), а также данным литературы (Даревский, 1953).
L (все 9?) — 42,0—53,0 (46,66+3,3), Ga — 26,5—36,0. Led.—
149
Рис. 38. Фолидоз головы A. chernovi Darevsky,
63 (31,16±2,82), L головы — 6,0. Ширина головы — 4,0—4,8
(4,33±0,24). Lpi — 6,0—6,2 (6,13± 0,06). Lp2 — 9,2—11,0
(10+0,52) -2-— 63,1—68,88 (66,63+1,8).^ =71,43. L Г°ЛО~
вы— 11,32—14,76 (13,13+1.0). Ширина _ 7,0-9,52
(8,0+0,74),11,32—14,76 (13,3+0,83), — 20,44—
23,81 (21,66± 1,07). Межчелюстной щиток несколько выдается
к лобному и соприкасается с ним узким швом. Надглазнич-
ных два, оба они соприкасаются с лобным. Лобно-теменных
два. Межтеменной один. Теменные щитки смыкаются узким
швом позади межтеменного. Первый вертикальный ряд височ-
ных состоит из двух щитков. Наружный ушной проход замет-
но сдвинут к шее и сильно редуцирован, его размеры
укладываются в размерах глаза более 15 раз или он скрыт
пэд височными щитками. Между ухом и центрально-височным
щитком в продольном ряду расположено четыре щитка. So
вокруг шеи — 20—22 (21,0±0,57). Sg- вокруг чела-- у всех
18. Брюшных в продольном ряду — 50 -56 (52,75 ± 1,25), за-
150
Рис. 39. Л. chernovi, (Армения, <олл. ЗММГУ)
тылочных 4—6 пар: 4X5— 1 экз.; 5X5— 1 экз.; 6X4—2 экз.
Подпальцевых чешуек на четвертом пальце р2— 11 —14
(12,5 г 0,64). Козырек отсутствует, а верхний край глаза прик-
рыт вторым верхнересничным щитком и краем первого над-
глазничного. Впереди подглазничного два — три верхнегубных:
у 1 экз — 2—3, у остальных — 3. Между носовым и подглаз-
ничным расположено четыре щитка. Предлобные разделены
передним утлом лобного. Носовой щиток цельный. Впереди
клоакальной щели два больших анальных щитка. Хвост снизу
покрыт одним рядом сильно расширенных щитков. Основной
фон верхней стороны тела светло- или темно-бронзового цве-
151
та с характерным золотистым отливом. На верхней стороне
туловища проходят светлые и темные прерывистые линии
(рис. 39). Темные боковые полосы проходят от ноздри через
глаз и далее вдоль тела и хвоста. Иногда вдоль этой полосы
проступают узкие светлые прерывистые линии. Брюхо у сам-
цов в период размножения яркое, кирпично-оранжевое, при-
чем окраска эта частично переходит на горло, нижнюю сторо-
ну конечностей и основания хвоста. У самок весной оно свет-
ло-оранжевое (значительно светлее, чем у самцов), а в
остальное время серое со светлыми рябинами. У спиртовых
экземпляров окраска нижней стороны тела выцветает и ста-
новится зеленовато-белой.
Распространение. В СССР встречается только в Цент-
ральной Армении, в правобережных предгорьях долины сред-
него течения р. Роздан (см. рис. 33). Возможно, что данный
вид в сходных условиях живет в сопредельной Турции.
Стации и количественные данные. В Армении по наблюде-
ниям И. С. Даревского (1953) стации этого гологлаза пред-
ставляют собой сильно каменистую разнотравно-злаковую
горно-ксерофитную степь; гологлаз придерживается пологих
склонов среди многочисленных мелких камней, под которыми
проводит большую часть времени, изредка появляясь на по-
верхности. В апреле — мае на площади 100 м2 под камнями
за час можно отловить две, максимум три — пять ящериц.
Сезонная и суточная активность. По данным того же ав-
тора (Даревский, 1953), гологлаз после зимовки появляется
в конце марта; в жаркие сухие летние месяцы (июль — ав-
густ) он скрывается в глубокие убежища и выходит на по-
верхность лишь в сентябре, после обильных дождей.
Размножение. У самки, добытой И. С. Даревским (1953)
19 июля, были обнаружены три готовых к откладке яйца поч-
ти круглой формы размером 4,8—5,0 мм. Молодые начинают
появляться, вероятно, не позднее конца августа.
Питание. По некоторым данным (Банников и др., 1977),
в их желудках найдены мелкие перепончато-крылые, в том
числе муравьи, двукрылые, тли, мелкие жуки и пауки.
Стереотип поведения, убежища. По наблюдениям И. С. Да-
ревскогб (1953), она при передвижении почти не поль-
зуется конечностями и напоминает маленькую змейку; под
камнями ящерицу наблюдали свернутой кольцом. Спугнутый
гологлаз быстро устремляется к соседнему камню, под кото-
рым обычно принимает описанное положение.
Практическое значение. Данный вид включен в Красную
книгу СССР и Красную книгу Армянской ССР.
152
ТАКСОНОМИЧЕСКИЙ АНАЛИЗ
В этой главе проведен анализ диагностических признают,
выявлены основные закономерности их распределения среди
таксонов; сделана попытка объяснить возможное адаптивное
значение признаков и дана классификация таксонов.
Ablepharus — Asymblepharus. Все виды, включенные нами
в род Ablepharus, имеют аблефаридную, т. е. эволюционно Сю
лее продвинутую стадию глаз, что соответствует основному
содержанию первичного (Lichtenstein, 1823; Fit/дпцег, 1824)
и современного (Fuhn, 1969; Банников и др., 1977; Е|Н’МЧШ11’
ко, Щербак, 1980) диагноза рода Ahlvpluirun. 11р<>гииополож*
ным и определяющим признаком роди Aayinblcphiruih КПК
установлено нами, является наличие у вида <alalcus> доабле-
фаридной или плезиоморфпой стадии глаз.
Выявив эти важные родовые различия, мы подчеркиваем,
что глаза ящериц этих групп имеют много сходных черт. Од-
на из них — присутствие хорошо развитого прозрачного «окош-
ка». При этом у Asymblepharus последнее представляет еще
вполне самостоятельное нижнее веко, в то время как у видов
рода Ablepharus это качественно новое образование, сходное
с змеевидной или гекконовидной глазными перепонками? Со-
вершенно очевидно, что это внешнее сходство глаз возникло
для одной и той же цели — защиты их от повреждений. Ука-
зывая на ведущее значение характеристики глаз в диагности-
ке рода Ablepharus, некоторые исследователи (Greer, 1974)
справедливо полагали, что этот род происходит от сценцел-
лоподобных предков путем эволюции аблефаридных глаз. От-
сюда может создаться впечатление, что представители этого
рода ведут свое происхождение от Asymblepharus, однако
это допущение опровергается анализом краниологических
признаков.
Как уже указывалось, костные элементы небной рамы у
этих групп ящериц имеют противолопожные тенденции фор-
мирования, причем, эволюционно более продвинутыми у видов
рода Ablepharus являются птеригоиды и палатинум, а у
153
Asymblepharus — эктоптеригоиды и редуцированное до семи
число премаксиллярных зубов. Функциональное значение
указанных краниологических изменений в эволюции данных
групп недостаточно ясно, но уже сейчас можно предполагать,
что недоразвитие палатинума у A.alaicus составляет резерв
черепного кинетизма, повлекший за собой значительную ре-
дукцию зубной системы.
Независимость происхождения исследуемых групп ящериц
косвенно подтверждается и образованием исключительно
сходных глаз у африканских (Afroablepharus и Panaspis)
и австралийских (Menetia и Proablepharus) родов аблефа-
ридных сцинков. Наше мнение о независимости происхожде-
ния родов Ablepharus и Asymblepharus вытекает и из анали-
за их распространения. В частности, их ареалы, и в том чис-
ле рода Scincella, обнаруживают несовпадение. Нами показа-
но, что некоторые представители рода Ablepharus (deserti и
pannonicus) проникли в Среднюю Азию значительно позднее,
когда род Asymblepharus уже не только сформировался, но и
пошел по пути развития конвергентного сходства с родом
Ablepharus.
Ablepharus. По числу височных щитков в первом верти-
кальном ряду выделяется группа видов, имеющая их три:
A.lindbergi, A.bivittatus, A. pannonicus и A.grayanus. Два ви-
сочных щитка свойственны периферическим видам: средне-
азиатскому A. deserti, закавказскому A. chernovi и европей-
скому A. kitaibelii. В качестве аицестрального признака три
височных щитка отмечены у единичных особей A. deserti
(1,4%) и A. kitaibelii (3,4%. С другой стороны, два височ-
ных редко встречаются и у A. pannonicus (2%). Анализ чис-
ла щитков одного продольного ряда, расположенных между
ухом и централь ю-височным включительно, показал, что боль-
шинство их (4—5) свойственно всем видам, за исключением А.
kitaibelii, у которого число щитков редуцировано до двух. Этой
особенностью A. kitaibelii резко отличается от A. chernovi, у
которого значительной редукции подверглись лишь разме-
ры уха.
У видов данного рода прослеживается четко выраженная
тенденция к редукции уха. Относительно самое крупное уш-
ное отверстие имеет A. lindbergi, затем его размеры постепен-
но уменьшаются в порядке: A. bivittatus, A. deserti, A. kitai-
belii, A. pannonicus, A. chernovi, и, наконец, наружное ухо
полностью исчезает у A. grayanus. У видов A. kitaibelii и
A. pannonicus размеры уха примерно одинаковые, но у пер-
вого из них произошла значительная редукция фолидоза ви-
сочной области. Что касается A. chernovi, то у него щитки
этого участка головы сходны с A. pannonicus, однако ухо
154
сильно редуцировано (Даревский, 1953; наши данные). Вы-
явленные изменения в строении уха гологлазов имеют, не«
сомненно, приспособительное значение. Есть указания (Olson,
1966; Mertens, 1971; Смирнов, 1982), что редукция уха у яще-
риц возникает в связи с приспособлением последних к роюще-
му образу жизни. При этом, как отмечают некоторые авторы
(Gans, Wever, 1972), полная редукция уха способствует раз-
витию у ящериц сейсмослуха, при котором чувствительность
восприятия воздушных звуков снижается, но повышается
чувствительность к восприятию колебаний, распространяю-
щихся в субстрате. Последняя особенность слуха, несомненно,
развита у A. grayanus и намечена у A. chernovi.
Признаками сходства A. grayanus и A. pannonicus явля-
ется наличие у них одного большого лобно-теменного щитка.
У всех остальных видов эти щитки маленькие и парные. При-
соединяясь к мнению ряда авторов (Camp, 1923; Robinson,
1967; Greer, 1974), мы также считаем, что парные лобно-те-
менные щитки представляют исходное состояние, а слитные
в один большой — эволюционно продвинутое. Нами отмече-
но, что единичные особи A. kitaibelii (1,7%) и один экзем-
пляр A. chernovi имели эти щитки слитные. С другой сторо-
ны, у отдельных особей A. pannonicus (1,3%) обнаружены
парные лобно-теменные щитки (анцестральный признак).
Анализ числа надглазничных щитков вновь подчеркивает
сходство и своеобразие периферических видов A chernovi и
A. kitaibelii: им свойственны два щитка вместо трех, обычных
для всех остальных видов. Также отмечено, что два надглаз-
ничных щитка очень редко встречаются и у A. pannonicus
(2%) в западной периферии ареала (Азербайджан, Грузия).
A. deserti — единственный вид, отдельные особи которого
(6,5%) на северо-восточной окраине ареала имеют четыре
надглазничных щитка (анцестральный признак). Напомним,
чю столько же надглазничных характерно для многих видов
рода Scihcella и др.
Аналогичная закономерность проявляется и в изменчи-
вости верхнересничных щитков. Полный ряд характерен для
большинства видов, кроме A. chernovi и A. kitaibelii: у них
эти щитки редуцированы и не составляют полного ряда. Как
исключение отмечено присутствие у единичных особей A. ki-
taibelii (0. Родос) полного (1 особь) или частично редуциро-
ванного (2 особи) ряда верхнересничных щитков. Значитель-
ная редукция верхнересничных щитков, как и других струк-
тур фолидоза головы (надглазничных, верхнегубных и др.),
у этих видов, вероятно, происходила вследствие их приспо:
собления к роющему образу жизни. Параллельное сходство
строения этих щитков отмечено и у вида рода Asymblepharus.
155
Подобное строение независимо возникало и у некоторых дру-
гих сцинцид, например, Menetia greyii Gray, 1845 (Storr,
1976).
Последствия скрытого образа жизни гологлазов прояви-
лись и на других элементах фолидоза головы. В частности, у
периферических видов A. deserti, A. chernovi и A. kitaibelii
произошла редукция фолидоза, окружающего верхний учас-
ток глаза. Так, у первого из них козырек состоит из одного
слабо увеличенного щитка. Полная редукция козырька свой-
ственна A. chernovi и A. kitaibelii. У всех остальных видов он
составляет обычно три — четыре щитка и редко один — два: у
A. bivittatus (4,2%) и у A. pannonicus (2,6%). Как нам пред-
ставляется, тенденция к редукции данной части фолидоза
проявляется у гологлазов вследствие образования аблефа-
ридныхглаз. Присоединяясь к мнению некоторых исследовате-
лей (Walls, 1942, 1967; Даревский, 1961), мы также считаем,
что преобразование элементов глаза у видов обоих исследуе-
мых родов происходило благодаря приспособлению этих яще-
риц к скрытому, роющему образу жизни, обитанию в пыль-
ном, сыпучем грунте, густой траве и колючих кустарниках.
Сравнение носового щитка показало, что цельные щитки
характерны только для двух видов: A. lindbergi и A. cherno-
vi. Они являются, несомненно, производными от полуразде-
ленного щитка, свойственного всем остальным видам рода
Ablepharus. Образование цельного носового щитка наметилось
и у A. bivittatus (5%). Единичные (менее 2%) особи с таким
щитком встречаются и у других видов данного рода.
Наличие четырех рядов спинных чешуй характерно для
всех видов рода Ablepharus, за исключением A. lindbergi, ко-
торому, как было установлено нами, свойственно шесть рядов
чешуй. Большее число спинных чешуй A. lindbergi — приз-
нак, подчеркивающий сходство этого вида с некоторыми ви-
дами рода Scincella.
Мелкие однообразные чешуи на внешних сторонах пред-
плечий и голеней свойственны всем видам рода Ablepharus,
за исключением A. lindbergi, имеющего здесь сильно увели-
ченные щитки. Как выяснено, мелкие однородные щитки свой-
ственны видам Scincella и Sphenomorphus, бывшим в нашем
распоряжении. Можно предположить, что преобразование че-
шуйчатого покрова ног у видов рода Ablepharus происходи-
ло путем редукции числа чешуй без увеличения их размеров
или с одновременным увеличением. Возникновение послед-
них у A. lindbergi, вероятно, было связано с повышением
функционального значения лап (при рытье). В пользу этого
предположения говорит тот факт, что конечности A. lindbergi
развиты относительно лучше, чем у остальных видов.
156
27
Рис. 40. Отношение количества чешуй вокруг се-
редины туловища (Sg) к длине тела (L) у видов
рода Ablepharus: 1—A. lindbergi; 2—A. bivittatus;
3 — A. deserti; 4 — A. pannonicus; 5 — A. kitaibelii;
6 — A. grayanus.
Дополнительное изучение чешуй ног у A. bivittatus пока-
зало, что у него редко (5,3%) встречаются особи, имеющие
увеличенные щитки (по’одному — два) на голенях, но они всег-
да отсутствуют на предплечьях, что может свидетельствовать
о понижении функции конечностей в филогенезе этого вида.
Возникновение сходства в щитковании ног A. lindbergi и
Asymblepharus alaicus побуждает нас вспомнить правило го-
мологических рядов Н. И. Вавилова (1967), указывающего
на независимое приобретение сходных признаков у родст-
венных таксонов.
Все сказанное свидетельствует о существовании у голо-
глазов четко выраженной тенденции к уменьшению элемен-
тов чешуйчатого покрова. Эти изменения происходят с умень-
шением размеров тела ящериц, что хорошо подтверждается
на примере изменчивости числа чешуй вокруг середины тела
157
Рис. 41. Особенности распространения, родственные от-
ношения и вероятные пути происхождения видов рода
Ablepharus.
Вид и его ареал:
1. A. lindbergi +
2. A. bivittatus —
3. A. deserti |
4. A. pannonicus Q
5. A. grayanus ★
6. A. kitaibelii
7. A. chernovi А
АП —продольных рядов спинных чешуй — 6; — 4
БП —височных в первом вертикальном ряду — 3; И-2
ВП — височных в продольном ряду обычно — 5; — 3
ГП —лобно-теменных щитков — 2; —1
ДО —носовой щиток полуразделен; — цельный
ЕП —щитки на внешних сторонах предплечий и —сильно увелччен-
голеней мелкие; ные
ЖО — надглазничных щитков—3; — 2
ЗП —верхнересничных щитков полный ряд; — ряд прерван
ИП—предлобные обычно широко соприкасаются; — разделены
КП —наружный ушной проход имеется — нет
ЛП —увеличенных чешуй верхнего века не менее 3; —1 или отсутствует
МП —малый центрально-височный щиток имеется; —нет
158
(рис. 40). Максимальное число чешуй вокруг тела имеет
A. lindbergi, Далее, от вида к виду, значение данного приз-
нака падает, достигая минимума у A. grayanus.
Выявленные закономерности межвидовой изменчивости
чешуйчатого покрова гологлазов могут рассматриваться как
один из частных примеров проявления закона олигомериза-
ции гомологических органов (Догель, 1954). Формы имею-
щие максимальное число элементов фолидоза, следует рас-
сматривать как филогенетически более древние. К такому
важному выводу пришли и многие систематики, объектами
изучения которых были другие группы рептилий (Даревский,
1967; Greer, 1974 и др.). Исходя из этих соображений, среди
видов рода Ablepharus архаичным является A. lindbergi о
чем свидетельствует наличие у него ряда примитивных
черт — шесть продольных рядов спинных чешуй, максимум
чешуй вокруг середины тела, хорошо развитые конечности,
которые в сочетании с прогрессивно измененными (увеличен-
ные щитки ног, цельные носовые щитки) представляют собой
пример явления гетеробатмии (Тахтаджян, 1966) у современ-
ных ящериц.
В свете вышеизложенных положений могут быть понят-
15 17 19 21 25 55 27 2$ ба.Л
Рис. 42. Изменчивость длины передней ноги (Lpi) относительно Ga у ви-
дов рода Ablepharus: Н-A. lindbergi;---A. bivittatus; |—A. deserti;
О—A. pannonicus;®—A. kitaibelii; ★ — grayanus.
159
ны и особенности признаковой дифференциации других ви-
дов. Исходные и преобразованные признаки у разных таксо-
нов распределяются по правилу гомологических рядов
Н. И. Вавилова (рис. 41). Приведенные материалы прежде
всего свидетельствуют о закономерностях географического
видообразования, в результате которого происходят викари-
рующие виды.
Серьезным и едва ли не самым важным свидетельством
того, что становление этих видов протекало в различных эко-
логических условиях, служит различие пропорций тела яще-
риц. Известно (Smith, 1939; Северцов, 1949 Mitchell, 1958),
что у многих сцинковых и в том числе аблефаридных ящериц
отмечена тенденция к редукции ног, происходящая с одно-
временным удлинением отрезка тела, заключенного между
передней и задней парой ног — Ga. У видов рода Ablepharus
(рис. 42) эта тенденция прослеживается, начиная от A. lind-
bergi, имеющего наиболее пропорционально развитые ноги.
У последующих видов (A. bivittatus, A. deserti, A. pannoni-
cus, A. kitaibelii, A. grayanus) между размерами ног и тела
возникает диспропорция, которая возрастает в перечисленном
здесь порядке. Некоторая стабилизация пропорций, выражен-
ная более широким диапазоном изменчивости, отмечена у
A. pannonicus, поскольку данный вид имеет и обширный
ареал, где условия жизни более разнообразны.
Рис. 43. Филогенетические отношения родов Scincella, Ablepharus и Asym-
blepharus.
160
Выявленная тенденция к диспропорции тела гологлазов
является следствием приспособления этих ящериц к змееоб-
разному способу передвижения, который выражен тем ярче,
чем длиннее тело и короче ноги. Наблюдения наши и других
авторов (Чернов, 1948; Даревский, 1953; Rabovanovic, 1957)
показывают, что такие виды гологлазов, как A. deserti,
A. chernovi и A. kitaibelii при передвижении часто не исполь-
зуют ног. Можно также считать, что A. lindbergi в этом от-
ношении— наименее специализированный вид.
Подводя итог сказанному, мы пришли к следующему зак-
лючению: архаичным видом рода Ablepharus, сочетающим в
себе признаки видов двух других родов (Scincella и Asym-
blepharus), является A. lindbergi. Наиболее близок к нему
A. bivittatus. Вместе эти два вида составляют предковую
ветвь рода Ablepharus, становление которого происходило в
переходной Ирано-Афганской провинции (зоогеографическое
деление Палеарктики дано по Н. Н. Щербаку, 1981 (Szczer-
bak, 1982). От группы «lindbergi — bivittatus» в разное вре-
мя дивергировали A. pannonicus и A. deserti. В результате
волн инвазии популяций A. pannonicus и последующей их
дизньюкции на юго-востоке ареала обособился A. grayanus,
а на западе — A. kitaibelii и параллельно последнему —
A. chernovi (рис. 43).
11 302
ЛИТЕРАТУРА
Алекперов А. М. Материалы по вертикальному распространению
и ландшафтному распределению герпетофауны Азербайджанской ССР. —
Уч. зап. Азерб. ун-та,сер. биол. наук, 1970, № 2, с. 57—65.
Алекперов А. М., Джафарова С. К. Узкоареальные и мало-
численные виды пресмыкающихся Азербайджана, состояние и перспекти-
вы их охраны. —Тр. Азерб. ун-та, серия биол. наук. Баку, 1979, № 3,
с. 27—30.
Ананьева Н. Б. Морфометрический анализ пропорций конечностей
пяти симпатрических видов ящурок (Eremias, Sauria) южного Прибалха-
шья. — В кн.: Герпетологический сборник. Тр. Зоол. ин-та АН СССР, т. 74.
Л. ,1977, с. 3—13.
Андрушко А. М. Земноводные и пресмыкающиеся в высокогорных
условиях Алайской долины (Памир). —Зоол. ж., т. 30, вып. 3. 1951,
с. 274—278.
Ан наев Л. Новая нематода. —Thubunaea schukurovi sp. nov. (Phy-
lopteridae) от ящериц Туркмении. — Изв. АН СССР, сер. биол. наук. 1973,
№ 1, 72—75.
Ахмедов М. Нахождение азиатского гологлаза (Ablepharus pan-
nonicus Fitzinger) на острове Обливном в Каспийском море. — Изв. Азерб.
Сер. биол.наук, 1976, № 2, с. 56.
Ахмедов М. Азиатский гологлаз на острове Жилой Урнус, Кас-
пийское море. — В кн.: Вопросы герпетологии: Автореф. докл. 4 Всесоюзн.
герпетол. конф. Л.: Наука, 1977, с. 19.
Бакрадзе М. А., Даревский И. С. Азиатский гологлаз Ab-
lepharus pannonicus (Sauria, Scinsidae) на Кавказе и вероятные зоогеогра-
фические обоснования этой находки. — Зоол. ж., 1973, т. 52, вып. 2,
с. 464—466.
Банников А. Г., Даревский И. С., Рустамов А. К. Зем-
новодные и пресмыкающиеся СССР. —В кн.: Справочники-определители
географа и путешественника. М.: Мысль, 1971.
Банников А. Г., Даревский’И. С., Ищенко В. Г., Руста-
мов А. К., Щ е р б а к Н. Н. Определитель земноводных и пресмыкаю-
щихся фауны СССР. М.: Просвещение, 1977.
Богданов О. П. К распространению представителей семейства сцин-
ковых в Бадыхызе и долине Мургаба. —Изв. АН ТССР, 1954, № 2,
с. 75—76.
Богданов О. П. Фауна Узбекской ССР, ч. 1. Земноводные и пре-
смыкающиеся. Ташкент: Изд-во АН УзССР, 1960. 254 с.
Богданов О. П. Пресмыкающиеся Туркмении. Ашхабад: Изд-во
АН ТССР, 1962, 235 с.
Богданов О. П. Экология пресмыкающихся Средней Азии. Таш-
кент: Наука, 1965, 258 с.
Богданов М. Н. Очерки природы Хивинского оазиса и пустыни
Кызыл-Кум. — В кн.: Описание Хивинского похода 1873 г., составленное
под ред. ген.-лейт. В. Н. Троицкого. Ташкент, 1882, 155 с.
162
Бондарцев А. С. Шкала цветов. М.-Л.: Изд-во АН СССР, 1954,
27 с.
Б у т о в с ки й П. М. К распространению и экологии пустынного голо-
глаза. Тр. Ин-та зоол. (АН Каз. ССР), т. 24, 1964, с. 218—219.
Вавилов Н. И. Закон гомологических рядов в наследственной из-
менчивости. — В кн.:Линнеевский вид как система. Л.: Наука, 1967, 92 с.
Варенцов П. А. Наблюдения над позвоночными и списки живот-
ных, найденных в 1980—1892 гг. —В кн.: Приложение к обзору Закас-
пийской области за 1892 г. Фауна Закаспийской области. Ашхабад, 1894»
38 с.
Второв П. А., Макеев В. М. Осенний герпетологический аспект
хребта Бабадаг и долины реки Кафирниган (юго-заиадный Таджикистан).
Зоол. ж., 1972, т. 51, вып. 10, с. 1591 —1593.
Второв П.П., Макеев В. М., Божанский А. Т. Рептилии
фисташников Бадхыза в осенний период. —В кн.: Охрана и рациональное
использование рептилий. М.: Изд-во МГУ, 1978, е. 57—58.
Г а дж не в А. Т., Мустафаева 3. А., Джафарова С. Г.
Эктопаразиты ящериц (Sauria, Reptilia) Азербайджана. —В кн.: Парази-
тологические исследования в Азербайджане. Баку: Элм, 1982, с. 155—161.
Гептнер В. Г. Гады Бадхыза. —Уч. зап. МГУ, Биология, вып. 83.
М., 1945, с. 95—126.
Горелов Ю. К., Щербак Н. Н. Новая находка песчаной змеи
(Psammophis schokari Forsk.) на территории СССР. — Зоол. ж., 1966,
т. XV, вып. 3, с. 466—468.
Даль С. К изучению фауны наземных позвоночных Зеравшанского
и Туркестанского хребтов. —Тр. Узб. гос. ун-та, вып. 7. Самарканд, 1936.
Даревский И. С. Новый вид гологлаза из Армянской ССР (Ab-
lepharus chernovi sp. nov. (Reptilia, Sauria). — Бюлл. Моск, об-ва испыт.
природы, т. 58, вып. 2. 1953, с. 39—41.
Даревский И. С. Нахождение пустынного гологлаза Ablepharus
deserti Strauch в долине Мургаб и вероятные зоогеографические обосно-
вания этой находки. —АН Туркм. ССР. Ашхабад, 1955, № 2, с. 72—74.
Даревский И. С. Об эволюции срастания век у ящериц. — Докл.
АН СССР, 1961, № 6, с. 1477—1480.
Даревский И. С., Чумаков И. С. Новый плейстоценовый вид
ящерицы Ablepharus из Рудного Алтая. — Палеонтологический ж., 1962,
№ 1, с. 127—130.
Даревский И. С. Скальные ящерицы Кавказа. Л.: Наука, 1967,
213 с.
Джафарова С. К. Новые сведения о нахождении полосатого го-
логлаза на Малом Кавказе. — Научн. тр. Сер. биол. наук Азерб. ун-та.
1979 ,№ 2, с. 64.
Догель В. А. Олигомеризация гомологических органов как один из
главных путей эволюции животных. Л., 1954, с. 368.
Дубинин В. Б. Эколого-фаунистический очерк земноводных и пре-
смыкающихся Хавастского района Ташкентской области УзССР. —Тр. Ин-
та зоол. и паразит., т. 3, Ташкент: Изд-во АН УзССР, 1954, с. 159—170.
Елпатьевский В. Genus Ablepharus Fitz в коллекциях Зоологи-
ческого музея Московского университета.— Дневн. зоол. отд.об-ва любит,
естествозн., антропологии и этнограф., 1901, т. 3, №2, с. 37—39.
Еремченко В. К., Торопова В. И. Изменчивость пустынного
гологлаза (Ablepharus deserti Str.) в Северной Киргизии. — В кн.: Вопро-
сы герпетологии. Автореф. докл. 4 Всесоюзн. герпетол. конф. Л.: Наука,
1977, с. 89—90.
Еременко В. К., Щербак Н. Н. О родовой принадлежности аб-
лефаридных ящериц фауны СССР (Reptilia, Sauria, Scincidae). —Вести,
зоол., 1980а, № 4, с. 10—15.
163
Еремченко В. К., Щербак Н. Н. Новый вид гологлаза — Ablep-
harus lindbergi st. n. (Reptilia, Sauria, Scincidae) из Афганистана. Вести,
зоол., 19806, № 6, с. 84—86.
Еремченко В. К. Систематика евразиатских лигозомных сцинков
родов: Asymblepharus Jeriomtschenko et Szczerbak, 1980 и Ablepharus (Fit-
zinger in Lichtenstein, 1823) (Sauria, Scincidae). —В кн.: Вопросы герпе-
тологии: Автореф. докл. 5-й Всесоюзн. герпетол. конф. Л.: Наука, 1981,
с.54—55.
Еремченко В. К. Распространение и географическая изменчивость
Asymblepharus alaicus (Sauria, Scincidae). —Вести, зоол., 1983, № 3,
с. 35—42.
3 а р у д н ы й Н. А. Заметки о чешуйчатых и голых гадах из северо-
восточной Персии. —Ежегодн. зоол. муз. АН, 1897, т. II, № 3, с. 349—361.
3 а р у д н ы й Н. А. Маршрут Н. А. Зарудного по Восточной Персии в
1898 г. —Ежегодн. зоол. муз. АН, 1898, т. 3, № 3—4, с. V—XII.
3 а р у д н ы й Н. А. Маршрут путешествия по Западной Персии в
1903—1904 гг. —Ежегодн. зоол. муз. АН, 1904, т. 9.
Зимина Р. П. О пресмыкающихся Иссык-Кульской котловины. —
Тр. Ин-та географии АН СССР, вып. 75, 1959, с. 156—167.
Калужина М. Морфология и биология отряда ящериц Зеравшан-
ской долины. — Тр. Узб. гос. ун-та им. А. Навои. Нов. сер. зоол., 1951,
вып. 46, с. 75—97.
Карпенко В. П. Ящерицы. —В кн.: Экология позвоночных живот-
ных хребта Нуратау. Ташкент: Фан, 1970, с. 12—34.
К а ш к а р о в Д. Н. Заметки о фауне позвоночных Чимгана. — Изв.
Турк-го отд. Русек, геогр. об-ва, Ташкент, т. 17, 1924, с. 45—53.
КашкаровД. Н. Результаты экспедиции Главного Средне-Азиатского
музея в районе озера Сары-Челек. — Изв. среднеаз. комит. по делам му-
зеев и охраны памяти, старины, искусства и природы, ч. 1, Ташкент, 1927,
с. 76—84.
Кащенко Н. Ф. Гады, собранные среднеазиатскими экспедициями
проф. Сапожникова В. В. в 1902 и 1906 гг. —Ежегодн. Зоол. музея АН
СССР, вып. 14, 1909, с. 119—129.
Ковшарь А. Ф. Птицы высокогорья западной части Таласского
Алатау (Тянь-Шань). —Тр. АН КазССР, т. 24, Алма-Ата, 1964, с. 121 —144.
Ковшарь А. Ф. Птицы Таласского Алатау. —Тр. заповеди. Казах-
стана, т. 1, 1966, 434 с.
КовшарьА. Ф. Певчие птицы в субвысокогорье Тянь-Шаня. —
Алма-Ата, 1979, 311 с.
Ковшарь А. Ф. Особенности размножения птиц в субвысокогорье —
Peuliarities of the birds reproduction in subalpine: на материале Passerifor-
mes в Тянь-Шане. — Алма-Ата: Наука, 1981, с. 260.
КореловМ. Н. Фауна позвоночных Бостандыкского района. —
В кн.: природа и хозяйственные условия горной части Бостандыка. Изв.
АН Каз. ССР. Алма-Ата, 1956, с. 259—325.
Кулагин Н. М. Списки и описание коллекций земноводных и пре-
смыкающихся зоологического музея Московского университета. — Изв.
Моск, об-ва любит, естествозн., антропологии и этнографии, т. 56, вып. 2.
М„ 1888, 39 с.
Куприянова Л. А. Кариологическая характеристика двух видов
ящериц семейства Scincidae — Цитология, 1973, т. 15, № 9, с. 1135—1142.
Лакин Г. Ф. Биометрия. М.: Высшая школа, 1968, 284 с.
Лобачев В. С., Чугунов Ю. Д., Чуканина И. Н. Особен-
ности герпетофауны северного Приаралья. — В кн.: Вопросы герпетоло-
гии: Автореф. 3 Всесоюзн. герпет. конф. Л., 1973, с. 116—118.
Майр Э. Принципы зоологической систематики. М.: Мир, 1971, 454 с.
Макеев В. М. Численность рептилий в Пашхурдской котловине и
164
на восточных склонах Кугитанга. —Зоол. ж., 1974, т. 53, вып. 9. с. 1428—
1430.
Макеев В. М. Численность активных рептилий в центральном Ба-
батаге весной. —Зоол. ж., 1975, т. 54, вып. 7, с. 1105—1107.
Макеев В. М., Божанский А. Т., Хомустенко Ю. Д. Рас-
пределение и численность рептилий в Центральном Копет-Даге. — Зоол.
ж., 1983, т. 62, вып. 7, с. 1122—1125.
Мекленбурцев Р. Н. Материалы по питанию некоторых хищ-
ных птиц Средней Азии. —Тр. Иа-та зоол. и паразитол. АН УзССР, т. 5,
1956, с. 165—171.
Мориц Л. Пресмыкающиеся Туркменистана и сопредельной Персии.
— Туркменоведение, 1929, №№4, 6—7, с. 17—35.
Никольский А. М. Пресмыкающиеся, амфибии и рыбы второго
путешествия Н. А. Зарудного в Персию в 1898 г. —Ежегодн. зоол. муз.
АН, 1899, т. 15.
Никольский А. М. Пресмыкающиеся (Reptilia). Chelonia и Sa-
uria— В кн.: Фауна России и сопредельных стран, Петроград: Изд-во АН,
1915, т. I, 532 с.
Овезмухаммедов А. Новый вид кокцидии Eimeria beyeri sp. n.
из пустынного гологлаза (Ablepharus deserti Strauch, 1868) в Туркмении. —
Изв. АН ТССР, 1974, №4.
Параскив К. П. Пресмыкающиеся Казахстана. — Изд. АН
Каз. ССР, Алма-Ата, 1956, 227 с.
ПерешкольникС. Л. Заметки по герпетофауне северного Тад-
жикистана. — В кн.: Герпетология Средней Азии. Ташкент: Фан, 1968.
Пономаренко В. Н. Атмосферные осадки Киргизии. Л.: Гидро-
метиздат, 1976, 134 с.
Раджабов Б. Питание разноцветного полоза (Coluber ravergieri
Menetries) в Узбекистане. — В кн.: Экология и биология животных Уз-
бекистана, ч. 2. Ташкент: Фан, 1975, с. 79—82.
Рокитский П. Ф. Биологическая статистика. Минск.: Высшая шко-
ла, 1961, 327 с.
Саид-Алиев С. А. Земноводные и пресмыкающиеся Яванской доли-
ны Таджикистана. — В кн.: Вопросы герпетологии: Автореф. докл. 4 Все-
союз. герпетол. конф. Л., 1977, с. 187.
Саид-Алиев С. А. Земноводные и пресмыкающиеся Таджикистана.
Душанбе: Дониш, 1979, 145 с.
Салихбаев X. С. Позвоночные животные защитных лесных на-
саждений Узбекистана. — Тр. Ин-та зоологии и паразитологии АН УзССР,
Ташкент, 1966, вып. 8, с. 131—193.
Сатторов Т. С. Ящерицы антропогенного ландшафта Северного
Таджикистана. —В кн.: Вопросы герпетологии: Автореф. докл. 4 Всесоюзн-
герпетол. конф. Л., 1977, с. 188—189.
Северцов Н. А. Вертикальное и горизонтальное распространение
туркестанских животных. — Изв. о-ва любит, естествозн., антропол. и эт-
ногр., 1873, т. 8, вып. 2, 157 с.
Северцов А. Н. Морфологические закономерности эволюции. —
Собр. сочин. М.—Л.: Изд-во АН СССР, 1949, 536 с.
Смирнов С. В. Среднее ухо ящериц: его строение, функция и пути
эволюции. — В кн.: Морфофункциональные преобразования позвоночных
в процессе освоения суши. М.: Наука, 1982, с. 88—102.
Соболевский Н. И. Герпетофауна Талыша и Ленкоранской низ-
менности. Опыт зоогеографической монографии. — В кн. Мем. зоол. отд.
сб-ва любит, естествен., антропол. и этнограф., 1929, вып. 5, с. 109.
Стрельников И. Д. Значение солнечной радиации в экологии вы-
сокогорных рептилий. —Зоол. ж., 1944, т. 23, вып. 5, с. 250—256.
165
Та хт а дж я н А. Л. Система и филогения цветковых растений.
М. — Л., 1966, 611 с.
Терентьев П. В., Чернов С. А. Определитель пресмыкающих-
ся и земноводных. М.: Госиздат, 1949, 339 с.
Топоркова Л. Я., Степанов Л. Н. Новая находка малого го-
логлаза на юго-западе Таджикистана. —В кн.: Герпетологический сбор-
ник. Тр. зоол. Ин-та АН СССР, т. 74, 1977, с. 115.
Торопова В. И., Еремченко В. К. Видовой состав и числен-
ность фауны пресмыкающихся в окрестностях г. Токмак. —В кн.: Геог-
рафия растительного и животного мира Киргизии (Известия КГО), Фрун-
зе: Илим, 1980, с. 94—96.
Утемисов О. Ящерицы культурного ландшафта Каракалпакии. —
В кн.: Вопросы герпетологии: Автореф. докл. 3-й Всесоюзн. конф., Л.,
1973, с. 184—185.
Утемисов О. Пустынный гологлаз (Ablepharus deserti Str.) в ни-
зовьях Аму-Дарьи. — Вести. Каракалпакского фил. АН УзССР, 1980, № 3,
с. 47—49.
Францевич Л. И. Обработка результатов биологических экспери-
ментов на микро ЭВМ «Электроника БЗ—21». —В кн.: Программы и
программир. —Киев: Наукова думка, 1979, 91 с.
Царевский С. Ф. Пресмыкающиеся и земноводные, собранные
В. Я. Лаздиным в южной и восточной Бухаре и в северо-восточной части
Закаспийской области летом 1915 г. —Ежег. зоол. муз. АН, 1918, т. 2
(1917), с. 79—90.
Чегодаев А. Е. О новых местонахождениях азиатского гологлаза
(Ablepharus pannonicus) в Закавказье. —Вести, зоол., 1981, №2, с. 71—72.
Чернов С. А. Пресмыкающиеся Туркмении. —Тр. совещан. по изуч.
произвол, сил АН ТССР, сер. Туркм., вып. 6, 1934, с. 255—289.
Чернов С. А. Ящерицы (Sauria) Таджикистана. —Тр. Тадж. базы
АН СССР, вып. 5., Сталинабад, 1935, с. 469—475.
Чернов С. А. Герпетологическая фауна Армянской ССР и Нахиче-
ванской АССР. — Зоол. сборн. Арм. фил. АН СССР, Ереван, вып. 3. 1939,
с. 79—194.
Чернов С. А. Эколого-фаунистический очерк пресмыкающихся и
земноводных Гиссарской долины. —Тр. Тадж. фил. АН СССР, т. 14. Ста-
линабад, 1945, с. 53—68.
Чернов С. А. Пресмыкающиеся. — В кн.: Животный мир СССР,
т. 2. Сталинабад, 1948, с. 127—161.
Чернов С. А. Пресмыкающиеся. Фауна Таджикской ССР. —Тр. Ин-
та зоол. и паразитол. АН Тадж. ССР, т. 48. Сталинабад, 1959, 203 с.
Шаммаков С. Нахождение пустынного гологлаза (Ablepharus de-
serti Strauch) в Северной Туркмении. — Изв АН ТССР. Сер. биол., 1974,
№ 3, с. 82—83.
Шаммаков С. Пресмыкающиеся равнинного Туркменистана. — Аш-
хабад: Ылым, 1981, 310 с.
Шарова И. X. Жизненные формы и значения конвергенций и па-
раллелизмов в их классификации. —Ж. общ. биол., 1973, т. 34, вып. 4,
с. 563—570.
Шестоперов Л. Е. Фаунистические обследования окрестностей
Теджена. — Бюл. Туркм. зоол. станции, 1936, № 1, с. 175—190.
Ш к а ф ф Б. А. Зоологическая экскурсия в Закаспийскую область. —
Изв. кавказ. отд. РГО. Ашхабад, 1916, т. 24, № 2, с. 154—181.
Шнитников В. Н. Пресмыкающиеся Семиречья. — Тр. общ. изуч.
Казахстана. Кызыл-Орда, т. 8, вып. 3, 1928, 85 с.
Шульпин Л. М. Материалы по млекопитающим и гадам Талас-
ского Ала-Тау. — Изв. АН Каз. ССР, сер. зоол., вып. 7. 1948, №51, с. 65—
83.
166
Щербак Н. Н. Пустынный гологлаз в террариуме. — Природа, 1959.
№ 2, с. 115—116.
Щербак Н. Н' Ящурки Палеарктики. Киев: Наукова думка, 1974,
296 с.
Щербак Н. Н.Новые находки ящериц и змей на территории Сред-
ней Азии. —Вести, зоол., 1979, № 1, с. 69—70.
Щербак Н. Н. Основы герпетогеографического районирования тер-
ритории СССР. — В кн.: Вопросы герпетологии: Автореф. докл. 5-й Все-
осюзн. конф. Л.: Наука, 1981, с. 157—158.
Щербина Е. И. К экологии лисицы Карабиля и Обручевской сте-
пи (Юго-восточная Туркмения). Изв. АН ТССР, сер. биол. наук, 1966, №3,
с. 66—74.
Яблоков А. В. Популяционная морфология животных. —Зоол. ж.,
1968, т. 47, № 12, с. 1749—1765.
Яблоков А. В. Популяционная морфология как новая ветвь эво-
люционной морфологии. — В кн.: Морфологические аспекты эволюции. М.:
Наука, 1980, с. 65—73.
Ядгаров Т. Состав, распределение и численность пресмыкающихся
острова Арал-Пайгамбар (бассейн реки Сурхандарьи). —В кн.: Вопросы
герпетологии: Автореф. 3-й Всесоюзн. герпетол. конф. Л., 1973, с. 229—230.
Ядгаров Т. О питании змей острова Арал-Пайгамбар (река Сурхан-
дарья). — Узб. биол. ж., 1974, № 4, с. 43—44.
Яковлева И. Д. Определитель пресмыкающихся Киргизии. Фрунзе:
Изд. АН Кирг. ССР, 1962, с. 111.
Яковлева И. Д. Пресмыкающиеся Киргизии. Фрунзе: Изд. АН
Кирг. ССР, 1964, с. 272.
Ahl D. Е. Herpetologische Notizcn. Zool. Anzcig. 1925. Bd. 65, N 1 —
2, p. 18—20.
В a s о g 1 u M., Baran I. Tiirkiye sGriingenleri. The Reptiles of
Turkey. Bornova—Ismir, 1977, p. 1, 272 p.
Beschkov A. A. Beitrag zur Zoogeographischen Unter-Suchung in
Bulgarien.— I zu zool. Inst. Muz. Sofia. 1961, N 10, p. 373—380.
Blanford W. T. Descriptions of new Reptiles and Amphibia from
Persia and Baluchistan. — Ann. Mag. Nat. Hist., 1874, ser. 4, vol. 14, N 31,
p. 453—455.
Boettger O. Herpetologische Miszellen Ber. Senk. Naturf. Ges.
Frankfurt am Main, 1889, p. 267—316.
В о 1 k a у S. Uber einen Fundirt des Ablepharus pannonicus Fitz, in
Ungarn. — Zool. Anz., Bd. 43, 1914, s. 499—500.
В о 1 k a у S. Addition to the gerpetology of the western balkanic pe-
ninsula (Prin. herpetol. zap. diejela balk. Palud.). — Glasn. Zcm. Mus.
Bosn. Herced. Sarajewo, vol. 31, 1919, p. 1—38.
В or у S. V. Exped. Scient de Moree. Zool. Rept., 1833, p. 1. 11, f.
4, 69 p.
В ou le n ger G. A. Catalogue of the Lizards in the British Museum.
London, 1887, vol. 3. — 557 p.
Brongersma L. D. Notes on Scincoid Lizards. — Zool. Mededeel.,
1942, vol. 24, p. 125—152.
Buresch I., Zonkov I. Untersuchungen ner die Verbreitung der
Reptilien und Amph\jbien in Bulgarien und auf der Balkanhalbinsel. Schil-
dkrdten (Testudinata) und Eidechsen (Sauria), Mitt. K. Naturw. Inst. Sofia,
1934, Heft. s. 106—108.
Camp C. L. Classification of the lizards. Bull. Am. Mas. Nat. Hist,
vol. 48, art 11, 1923, p. 289—481.
D a a n S. New record localities of Aegean amphibians and reptiles
Brit. J. Herpetol., 1967, N4, p. 311—312.
De Witt G. F. Description d‘unr espece nouvelle du genre Ablepha-
167
rus provenant
p. 29—33.
du Congo Beige. —Rev. Zool. Bot. Afrig, 1936, t. 29, N
4,
Dim or sky A. Beitrag zur Herpetofauna Mazedonies. Fragm. Balcan-
Mus. Mac. Sci. Nat., Sopge, 1959, N 3—1, s. 1—4.
Dzukis G. A. Contribution to the herpetofauna of Serbia. —Bull.
Mus. Hist. Nat. Belgrade, fasc. 29, (B), 1972, p. 105—110.
Fejervary G. Uber Ablepharus pannonicus Fitz.— Zool. (Syst).
Geogr. Biol.), Jena, 1912, s. 547—574.
Fejervary G. Nene Angaben fiber die geographische Verbretung des
Ablepharus pannonicus Fitz, in Ungarn. — Verh. Zool. Bot. Ges. Wien, 1917
s. 161—167.
Fitzinger L. Verh. Ges. naturf. Frende Berlin, 1824, Part 1, 297 s.
Ser., 1974, N 31, p. 1 — 16.
Fuhn I. E. Revision and Redefinition of Genus Ablepharus Lichten-
stein, 1823 (Reptilia, Scinicidae). — Rev. Roum. De biologie ser. De zool.,
1969a, T. 14, N 1, p. 23—41.
Fuhn I. E. The «polyphyletic» origin of the Genus Ablepharus (Rep-
tilia, Scincidae): a case of parrallel evolution — Sounderdruck aus z. f.
Zool. Systematik u. Evolutionsforschung ,1969b. N 7, p. 67—76.
Fuhn I. E. Uber die Unterarten von Ablepharus kitaibelii Bibron et
Bory De Vincent, 1833 (Sauria, Scincidae). — Vestn. Ceskosl. cpa lec. zool.,
1970, Bd. 34, N 1, s. 9—17.
Fuhn I. E. Revision du phulum forestier du genre Panaspis Cope
(Reptilia, scincidae, Lugosominae). — Rev. Roum. Biol. 1972, 17, p. 257—71.
Greer A. E. A philogenetic subdivision of Australian srins. — Rec.
Austral. Mus., 1979, vol. 32, NN 7-10, p. 339—371.
Gruber U. Zur Taxonomie und Okologie der Reptilien von der. In-
sel. Antikythira. — Salamandra, Frankfurt am Main, 1974, N 10, s. 31—41.
Gruber U., Dieter F. Die Herpetofauna des Paros-Archipls (Aga-
is). — Salamandra, Frankfurt am Main, 1977a, N 2, s. 60—70.
Gruber U., Fuchs G. Die Herpetofauna des Paros-Archipels
(Zentral-Agais . — Salamandra, Frankfurt am Main, 1977b, N 13, s. 60—77.
Gruber U. Scincidae. In: Handbuch dcr Reptilien und Amphibien
Europas. C. by Academische Verlagsgesellschaft, Weisbaden, 1981, Bd. 1,
s. 292—307.
Grum-Grshim ailo C., Bericht fiber neine Reise in das ostliche
Buchara.— Mem. sur. les Lepidopteres, 1887, vol. 3, p. 357—402.
Jeriomtschenko V. K. (USSR). Systematics of the Lygosomid
lisards of the Palearctic region (Reptilia, Sauria, Scincidae).— 1 Conf.
Негр. Resp. Soc., Budapest, 1981, p. 21.
К a r a m a n St. Uber des Verbreitung der Reptilien in Jugoslawien. —
Ann. Mus. Serb. Merid. Skopje, Bd. 1, N 1, 1939, s. 20.
К op stein F., Wettstein O. Reptilien und Amphibien aus Albanien.—
Verh. Zool. —Bot. Ges., Wien, 1921, Bd. 70, s. 387—457.
Leviton A. and AnrersonS. E. The Amphibians and Reptiles of
Afghanistan. — Proceding of the California Academy of Sciences, San Fran-
cisko, 1970, vol, 38, N 10, p. 163—206.
Lichtenstein H. D. Naturhistorische Anhang zu Eversmans s
Reise von Orenburg nach Buchara, Berlin. 1823, 1—8. 150 s.
Marx H. Keys to the Lizards and Snakes of Egipt., Cairo, 1956, N 3,
P Me ne tries E. Catalogue raisonne des objets de Zool. recneil-
lis dans un voyage an Caucase et juagu aux frontieres actuelles de la Per-
se. St. Petersbourg, 1832. .
Gans C. We ver E. G. The Eear and hearing in Amphisbaena (Pe-
ptilia). — J. Exp. Zool., 1972, vol., 179, N 1, p. 17—34.
168
Gray J. E. Catologue of ... lisards. In ... the British Museum, Lon-
don, 1845, 289 p.
Greer A. E. A new generic arrangement for some Australien Scin-
coid Lizard. Breviora, 1967a, N 267, p. 1 — 19.
Greer A. E. The generic relationships of the scincid lizard genus
Eumeces. — Breviora, 1967b, N 276, p. 1—9.
Greer A. E. A subfamilias classification of scincid Lizards. — Bull.
Mus. Comp. Zool., 1970, vol. 139, N 3, p. 151—184.
Greer A. E. The generic relationships of the scincid lizard genus
Leiolopisma and its relatives. — Austral. J. Zool. Suppl. Ser 31, p. 1—67.
Mertens R. und M й 11 e г L. Liste der Amphibien und Reptilien
Europas. — Abhandl. Senckenberg. naturfor. Ges., 1928, Bd. 41,-62 s.
Mertens R. und Muller L. Die Amphibien und Reptilien Europas
(Zweite Liste nach dem Stand vom 1 Januar, 1940). — Abhandl. Senchen-
oerg. naturfor. Ges., 1940,-56 s.
Mertens R. Uber den Glattechsen-Namen Ablepharus pannonicus.—
Zool. Anz., 1952a, Bd. 149, N 1—2, s. 48—50.
Mertens R. Amphibien und Reptilien aus der Tiirkei. — Instanbul
Universitesi Fen Fakultesi Mecmuasi, 1952b. t. 17, fase. 1, s. 41—75.
Mertens R. und Wermuth H. Die Amphibien und Reptilien Europas
(Dritte Liste, nach dem Stand vom 1, Lenuar, 1960), Frankfurt-am-Main,
1960, 264 s.
Mertens R. Bemerkungen uber einige Eidechsen aus Afganistan.—
Senckennd. bioL, 1965, Bd. 46, N 1, s. 1—4.
Mertens R. Nachtrage zur Reptilien fauna der insel Korfu. — Sen-
ckenberg. biol., 1968, Bd. 49, N 1—4, s. 173—180.
Mertens R. Die Riickbildung des Tympanum bei Reptilien und ihre
Bezienund zut Lebensweise. — Senckenberg. biol., 1971, Bd. 52., s. 177—191.
Minton S. A. A contribution to the herpetology of West Pakistan.—
Bull. Mus. Nat. Hist, 1966, vol. 134, art. 2, p. 27—184
Mitchell F. J. The scincid Genera Egernia and Tiligua (Lacerti-
lia), — Record of the South Austral. Mus., Pretoria, 1950, vol. 9, p. 275—
309.
Mitchell F. J. Adaptive convergence in australien Reptiles. — The
Austral. Mus. Magas., 1958, June 15, p. 314—317.
Mittleman M. B. A generic synopsis of the Lisards of the subfa-
mely Lygosominae. — Smiths. Miceli. Coll., 1952, vol. 117, p. 1—35.
Muller L. Bemerkungen zur Okologie tschechoslowakischer Ablepha-
rus kitaibelii. — Aguaterra, Biberist, 1971, N 8—3, s 27—28.
Olson E. C. The middle ear-morphological types in amphibians and
reptiles. — Amer. Zool., 1966, vol. 6, N 3, p. 399—419.
Paculjevic G. Tages und Saisonaktivitat von Ablepharus kitaibelii
(Bibgon et Bory) in Jugoslawien.— Glasn. Mus. Beograd, 1964. Bd. 20
(B), s. 311—314. . . ..
Paculjevic G. Nene Fundorten von Ablepharus kitaibelii (Bibron
et Bory) in Jugoslawien. — Glans. Mus. Beograd, 1956, Bd. 20 (B), s.
315—317.
Parker H. W. Dr. Karl Jordan's expedition to South-West Africa
and Angola: Herpetologicae collections. — Novitates Zoologicae, London,
1936, N 40, p. 115—146. l
Petzold H. G< Ablepharus kitaibelii fitzingeri Mertens, 1952.—
Aguar. Terrar., 1978, N 25, s. 359.
Radovanovic M. Zur Kenntnis der Herpetofauna des Balkans —
Leipzig, 1957, Bd. 159. s. 130—137.
Robinson P. The evolution of the lacertilia. Problemes Actuelis^ de
Paleontologie (Evolution des Vertebres). — Coll. Int. cent. natn. Rech. Sci.,
1967, vol. 163, p. 355—407.
169
Rotter J. Biologische Beobachtungen an der nordlichen Johannisech-
se, Ablepharus kitaibelii fitzingeri Mertens, 1952 (Sauria, Scincidae). — Der
Zool. Sarten, Leipzig, 1952, N 26, s. 312—318.
Schmidt K. and Marx H. The Herpetology of Sinai. — Fildiana zo-
ology, Chicago Nat. Hist. Mus., 1956, vol. 39, N 4, p. 21—40.
Smith M. A. Fauna of British India. Reptilia and Amphibia. London,
1935, vol. 2 (Sauria), 1—440 p.
Smith M. A. A review of the genus Ligosoma (Scinicidae, Repti-
lia) and ist allies. — Rec. Indian Mus., Calcutta, 1937, vol. 39, part 3,
p. 213—234.
Smith M. A. Evolutionary changes in the eye coverings of certain
lizards. — Proc. Linn. Soc., London, 1939, p. 190—191.
Stepanek 0. Einc nene geographische Rasse von Ablepharus panon-
nicus Fitzinger. — Sborn. Narodn. Mus. v. Praze, 1938, Bd. 1 (В), с. 1 —10.
Stepanek O. Zur Herpetologie Griechenlands. — Vest. Ceskc Spol.
zool. v Praze, 1944, Bd. 9, s. 123—147.
Stoliczka F. Notes on reptiles collected by Sugreon E. Day in
Sind.— In: Proc. Asiatic. Soc. Bengal, 1872, 85 p.
S t о r r. G. M. The genus Ctenotus (Lacertilia, Scincidae) in the Wes-
tern Australia, 1973, vol. 56, N. p. 86—93.
S t о г r G. M. The genus Menetia (Lacertilia, Scincidae) in Western
Australia. — Rec. West. Austral. Mus., 1976, N 4, 2, p. 189—200.
Strauch A. A. Uber die Arten der Eidechsengattung Ablepharus Fit-
zinger.— Mem. biol. Acad. Sci., Petersb., 1868, Bd. 6, s. 533—570.
Szczerbak N. N. Grundziige einer herpetoloographischen Gliederun
zinger. — Mem. biol. Acad. Sci., Petersb., 1868, Bd. 6, s. 533—570.
Walls G. L. The Vertebrate Eye and its Adaptive Radiation. — Gran-
drook Inst, of Sci. Bull., 1942, N 19 p. 1—71.
Walls G. L. The Vertebrate Eye and its Adaptive Radiation. — New
York-Lendon: Hafner Publ. Co., 1967, 785 p.
Wettstein O. Herpetologia Aegaea.— S. B. Osterr. Acad. Wiss.
math. — naturw., 1953, Bd. 162, N 160, s. 427—448.
Wettstein O. Lacertilia aus Afghanistan. Contribution a 1‘etude de
la faune d'Afghanistan. — Zool. Anz., 1960, Bd. 165, Heft. 1—2, p. 61—62.
ОГЛАВЛЕНИЕ
Введение 5
Общая часть
Краткий обзор исследования ящериц рода Ablepharus 7
Материал и методика ......................................... 11
Родовая принадлежность аблефаридных ящериц фауны
СССР и сопредельных стран......................... 15
Систематическая часть
Род Асимблефары — Asymblepharus Jeriomtschenko et
Szczerbak, 1980 ...................... 21
Асимблефар алайский Asymblepharus alaicus (Elpa-
tjewsky, 1901)........................................ 21
Род Гологлазы — Ablepharus (Eitzinger in Lichtenstein, 1823) . 64
Гологлаз афганский — A. lindbergi Wettstein, 1960 . 65
Гологлаз полосатый — A. bivittatus (Men., 1832) 69
Гологлаз пустынный — A. deserti Strauch, 1868 86
Гологлаз малый — A. grayanus (Stoliczka, 1872) 109
Гологлаз азиатский — A. pannonicus (Licht., 1823) . 119
Гологлаз европейский — A. kitaibelii Bibron et
Bory, 1833 128
Гологлаз Чернова — A. chernovi Darevsky, 1953 149
Таксономический анализ 153
Литература 162
Валерий Константинович Еремченко,
Николай Николаевич Щербак
АБЛЕФАРИДНЫЕ ЯЩЕРИЦЫ ФАУНЫ СССР
И СОПРЕДЕЛЬНЫХ СТРАН
Редактор издательства Н. Е. Федорищева
Обложка художника Н. А. Кожегуловой
Технический редактор Р. Р. Хусаинова
ИБ № 1298
Сдано в набор 25.11.85. Подписано к печати 21.07.86. Д — 00783.
Формат 60X90V16- Бумага тип. № 1. Высокая печать. Литературная
гарнитура. Объем 10,75 п. л., 10,6 уч.-изд. л. Тираж 1000 экз.
Заказ 302. Цена 1 руб.4О коп.
Издательство Академии наук Киргизской ССР,
720071, Фрунзе, Ленинский проспект, 265 а
Типография Академии наук Киргизской ССР,
720001, Фрунзе, ул. Пушкина, 144
ЗАМЕЧЕННЫЕ ОПЕЧАТКИ
Стр. Строка Напечатано Следует читать
11 9 снизу политического политипического
12 4 сверху изменчивости гологла- зов, прежде изменчивости, прежде
45 1 сверху Jeriomtschenko, Jeriomtschenko, 1983
51 рис. 11 A. alaicus: Стация A. alaicus:
109 10 сверху TERRA TYPICA: С-3 ИНДИЯ, KAR Terra typica: С-3 Индия, Кач
153 10 снизу сценцеллоподобных сцинцеллоподобных