/
































































































































































































































































































































































































































































































































Текст
P. J. ВАЯЬЩСТОХ, 5г.
THE GEOGRAPHICAL DISTRIBUTIONOF ANIMALS
Ф. ДАРЛИНГТОН ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ЖИВОТНЫХ Перевод с английского М. М. Мержеевской
и K П. Филонова Предисловие и редакция H. А. Гладкова ИЗДАТЕЛЬСТВО «ПРОГРЕСС»
МОСКВА 1966
° Ф
° ° ° а °
gi вава ~ °
° ° ° °
° »ее ° 3
° ° ее ае °
° в °
р
'Ъ
° ° а а~~ ° Фа Ф °
° ° а, ееje
,Э 'Iy iy ° . ° ° ° е ° °
а ° ° ° ° ° ° е /В ° °
Ф ° ° ° У
° °
~~,е
° °
° °
° °
Ф ° °
° ев
° ° УЗ'е,в
° ° 1 °
° 'с ° '~ °
° - ° ° . г ° ° °
° С..е В.
° а ° ° ° ° ° ° °
° ° I ° ° ° ° °
° °
° ° ° вв Ва р
° Ф ° °
» ° ° ° ° ° e ee
Л ° ° °
е ° ° ° ае eе ° ° ° °
° ' ° ° ° ° 1 ° ° ° Ф
° Ф ° ° ° е ° °
а
° ° ° °
° re ° ° r» ° В ° ° °
° ФВФ ° ° ° er ° ° ее В
° е ° ° ° е ° в °
° °
° ° ° Ф ° ° eae ° еа ° ° °
° ° ° ° вв ° Ф °
е ° Ф ° ° ° ° е ° ° ° ° е ° ° °
° Ф ° ° ° ° ° ° ~ ° В ° в ° ° ° ° ° ° ° ° ° ° ° ва е ° ° ° а °
° ° °
е ° е ° °
° еее °
° ае °
° 1
° ° ° ° ° ° ° фве
ф4!ва'5В
° ° ° ав
° Ф °
° °
° ° ° ° ° в Ф °
С ° ° ° ° В °
° °
° °
еа ° ° ° е °
ее » ° ° ° ° ° ' ' ° ° °
° °
° ° ° ° Ф °
° °
° ° ° ° °
° ° ° Ф ° ° ° ° °
° ° ° ° ° °
° Ф ° в ° ° еr»е ° ° °
° °
ае ае ° ° ° ° ° °
° ° ° В»в
а
Ф ° Фа»ФР
е ° °
° e eОВ ее °
Ф: . ° ° Эа
° j е ° °
° ° ° ° ° ° °
° °
° ° ° ° °
° Ф ° ° ° ° ° е °
° Ф °
° °
° ° ° ° ф °
° ° °
° ° в ° е
а ° ° °
Ф
° Э
Ф
° °
° °
а
° °
° ° °
Ф
° °
° °
° ° ° Ф °
° ° ° ° °
° °
° а ° °е 1 ° °е o ° Ф °Ф в ° °° °° е °° в °° °
O
° а ° ° °
° 1
° °
° °
° ° ° е
°, ° ° °° °° °3»
° °
в
° °
в
° °
O
° °
а °
а
° а
а ° °
° °
O
° °
° ° ° ° °
° е ° ° 11 аеа
Ф ° В ° а
° ° ° ° °
° ° ° ВВ
° \ °
е ° ° °
° ° °
° ° ф
° °
° °
° ее
° °
° °
° ° °
° °
° °
еа
° ° °
° ° ° ° °
° ° °
° ° ° °
° °
° ° ° в °
° ° ° ° °
° Ф ° ° ° ° % е ° °,в ° а ° ° е °
° ев ° е ° ° ° ° ° ° ° ° ° >
° ° °
° ° ° ° ° g ° ° ° е °
y ° ° ° в ° ° в °
° °
° ° ° ° ° °
° ° ° °е ° °ае ° °
° а
° °
° °
° � ее '~у~'~
ф
Ф
» ° в ° °
° °
° \ °
° а
° °
° °
° Ве
° °
Ъ Ф
° °
е
° ° e °
е °
° а °
° е °
° ° °
° °
° ° ° °
'„,J:4e °
ВЪ
° °
° ° ° ев ° ° в °
O ° ° °
V ав 11 ° °
° °
° ° o e °
° °
° ° ° веее ° °
° e ° ° ° °
1 ° ° ° ° °
° ° ° ° ° ееэ
° ° ° 1 ° °
° f e ° ФЭЭФ
° а~ ° °
В е °
ФВ аэ ° ° ° °
° ва ° 1 е1
° ° ° ° e °
° ° 1 ° °
r 11
° °
° ее °
ф9'. ° С
° ° °° е ° М° ° ° °° °
ав
а °
а °
ав
° ° °
° ее
° Э ° Фе
° 3,'-.'- '
.Я
° ееа °
б. ° '
.к, °
' Фе
Э) °
ев °
фе
ЭВ в
Ф Фв Фе °
° Of ~~в
е
° O
е ° °
° ееа е
° °
° °
° O j
j<<
В
Ф
° °
° ° ° ° ° °
° °
° ° Э а
° В
° ° ° ° ° ° . а
е ° ° ° е ° ° ° ° е ° ° ° ° °
в ° ° е ° ° °
е e ° ° ° ° ° ° ° ° ° ° ° ° ° ° ' ° ° ° ° °
° еe ° ° ° ° °
° ° а ° ' e ° e ° ° ° ° ° ° ° ° ° ° ° ° ' ° ° а ° ° °
°, ee4 ° ° ° ° ° °
° ° °
ее ° ее Ф ° ° ° ° je ° ° е ° ° ° е ° ° ° ° ° °
° в ° ее ° ° ° ° ° ° е
° а Ф °
° ° «А- ае 4 е ° ° °
ве ~ ° ° ° ° 1 ее
:Ж -.~".~ ° ° ° ° ° ° °
° ° ° ° ° ° ° е ° ° ° °
° ° ° ° ° ° ° ° ° °
° Ф ° В ° е ю, ° ° е
Ф ° °
Ф е ° ° ° °
O °
° ° ° ° ° ° ° ° ° ° е ° °
° е ° ° е
° ° е ° ° ° ° ° ° °
у ° ° °
в фФафД1 л ° ° ° ° ' ВФ
в Фа ° „° ~ ° ° ° ° ° ° ее~е~еевае ° ° ФВ ° ° ° °
° °
r ° °
° ° ° 1 ° ° в ° е 4 ° ° ° ° ° ° ° ° ° ° ° в ° ° е ° ° ° ° ° ° ° ° O ° Ф
° ° ° °
° ° ° » ° ° ° °
» ° ° ° Фа» ° а ф ° ° ° ° ° ° ° ° в °
° ° ° ° ° ° e ° ° ° ° ° ° ° ° в ° ° ° ° е °
° ° ° Ф ° е ° вв
° ° а ° ° ° в ° ° Ф ° o е ° ° ° ° ° е ° е а ° ° ° ° ° е ° ° ° ° ° ° ° Вв ° °
° ° ° ° ° ° ° ° ° ° ° ° ° ° Ф ° Ф ° ° ° ° ° вф °
° ° ° ° ° ° ° ° ° ° Ф ° ° ° r ° ° ° ° ° ° ° ° ° ° ° ° ° °
° ° ° ° е ° ° ° ° ° ° ° e e° ° е ° е ° P а ее ЭФ ее ° ° ° ° ° ° ° ° ° ° ° ° ° ав ° 1 ° ° ° 1 °
° ° ее ее ° Ве а ° еа аа ° ° ае ее ° ° ° ° °
° ° ° ° O ° ° 1 ° ° ° ° ° ° ° ° ° 1 °
° ° ° ее в е ° ° ° Ъе ве ~ в '4И
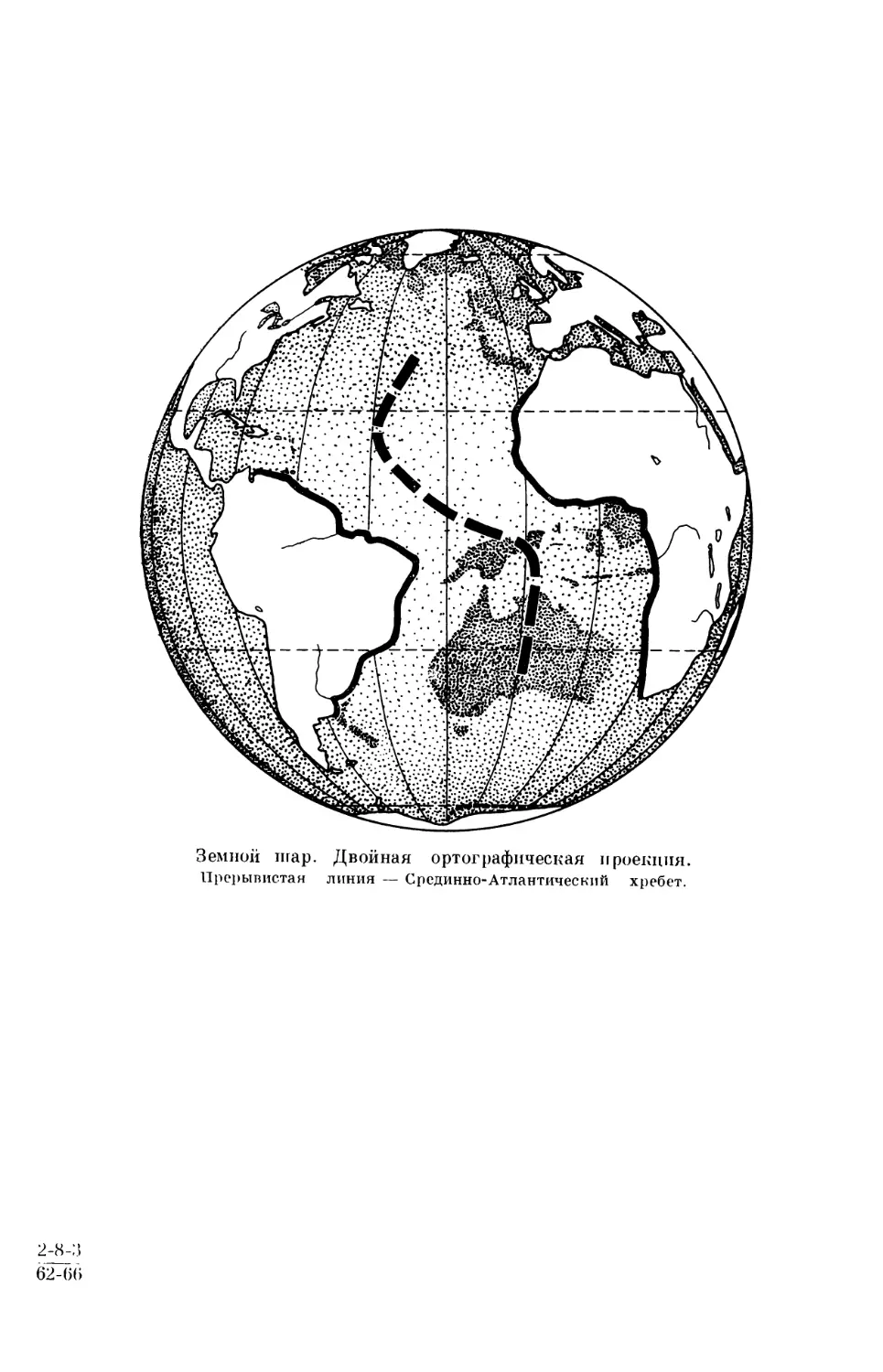
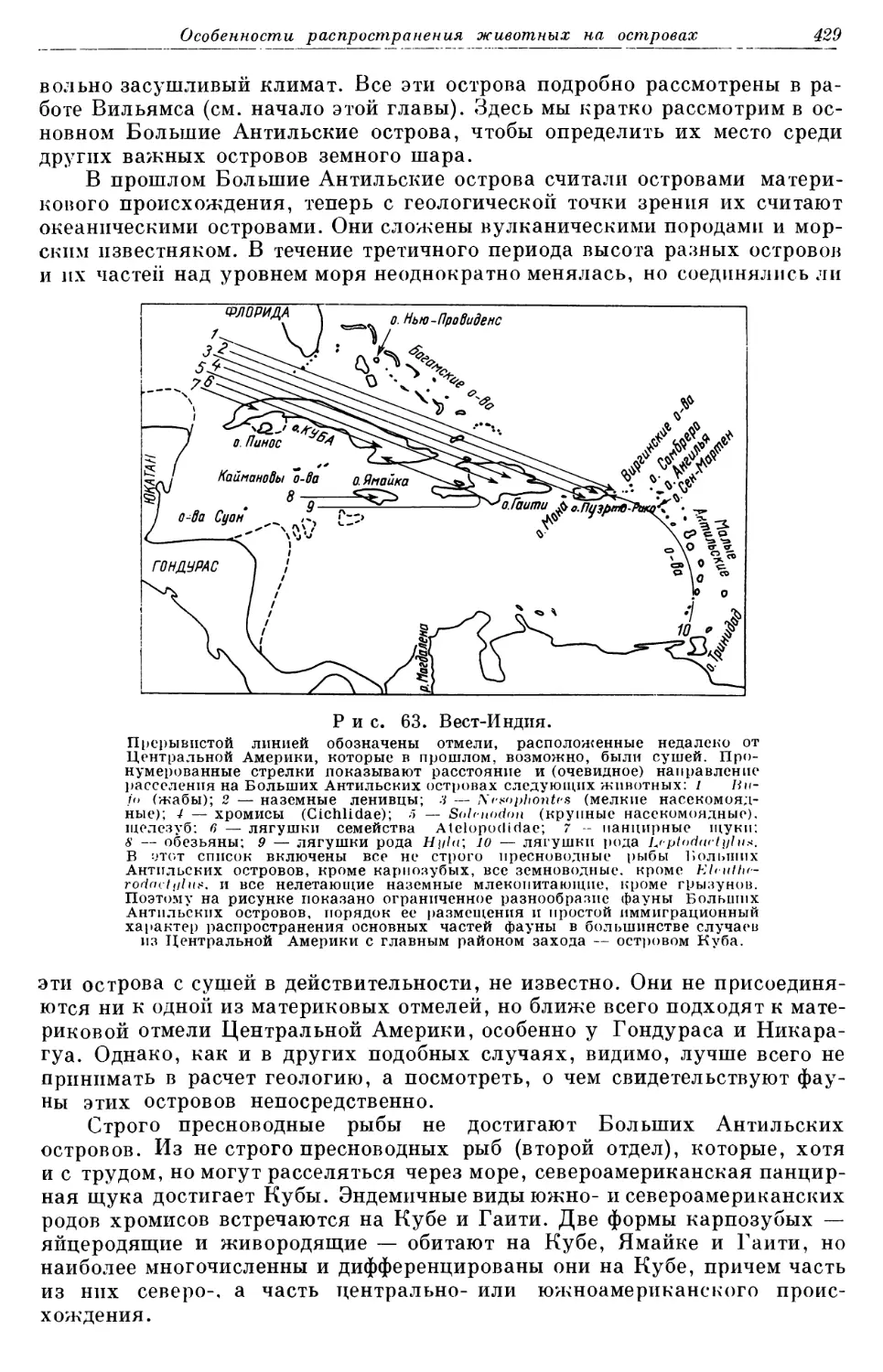
3 81NIIOlI IlI 8 P . Дав ОЙ Нв Я ОР ТО~' f)8lflIIBBCIiBH I I P08liII I M.
Прерывистая линия — Срединно-Атлантический хребет.
П Р Е Д И Г,Л О В И Е
Вопросам географического распространения животных посвящена об- ширная литература. Здесь и частные исследования — выявление отдель- ных фактов и их объяснение. И общие рассуждения — попытки отыскать основные закономерности расселения животных и объяснить всю картину их современного распространения. Фактов собрано много. Теорий выска- зано также немало. Некоторые из теорий быстро отошли в прошлое, дру- гие укрепились и, как показала проверка временем, довольно хорошо объясняют современное распространение животных. Однако осталось еще много неясных вопросов и многое предстоит еще сделать.
В период своего бурного развития, которое было обусловлено про- никновением в науку эволюционных принципов Ч. Дарвина, зоогеогра- фия, помимо решения непосредственно стоящих перед ней задач, выполня- ла и известную, можно сказать подсобную, роль в развитии естествозна- ния. На основании отдельных фактов распространения животных дела- лись смелые обобщения о прошлом облике Земли, истории континентов и морей, но многие из них в дальнейшем не были подтверждены, и в на- стоящее время зоогеографы для объяснения добытых ими фактов нередко прибегают к помощи геологов или палеогеографов. Конечно, это не иск- лючает того, что и последние в свою очередь обращаются к зоогеографам, так как многие особенности распространения животных действительно являются свидетельством прошлого.
Главная задача зоогеог рафа — выявить закономерности распрост- ранения отдельных видов животных и целых фаун и дать им причинное объяснение. Естественно, что это объяснение должно находиться в орга- ническом единстве со всеми естественноисторическими науками о Земле и ее истории. И оно будет в известной мере «служить» этим наукам, так же как они «служат» зоогеографии. Науки о Земле взаимосвязаны. Они способствуют развитию друг друга и не могут существовать сами по себе, в отрыве от других наук.
В последнее время зоогеография находится в несколько «ложном» положении. Повышенный интерес к наиболее характерным видам каждой местности (эндемикам и т. д.) приводит к тому, что в центре внимания ис- следователя оказывается только часть фауны местности, часто очень не- значительная. Другая же часть, обычно значительно большая, которая составляет «лицо фауны», остается за пределами внимания исследователя как не характерная. А между тем она есть, взаимодействует со всем био- ценозом и отражает общий характер условий существования в данной мест- ности. Как быть с данной «пренебрегаемой» частью фауны".
Впрочем, что составляет лицо фауны и что является в ней главным. это зависит от точки зрения исследователя. Во всяком случае, сейчас в зоогеографии развивается и приобретает все больше сторонников количе- ственный метод, назначение которого, в частности, определить, какие виды количественно богаче представлены на местности и тем самым являются
Пред исл овие
на ней «главными». Вероятно, зоогеографу надо не в последнюю очередь заниматься этими многочисленными членами фауны и по HHM вести райо- нирование.
Очевидно, следует говорить о двух параллельных, сопутствующих друг другу, но не «отменяющих» друг друга направлениях в нашей науке.
Интерес к количественно преобладающим видам — это подход гео- графа, который стремится выяснить компоненты ландшафта (в данном слу- чае зоологические компоненты — зооту), страны, или другой географи- ческой единицы. Подход «от эндемиков» более традиционный. Оба подходя нужны, и «зоолог» не должен упрекать «географа» за то, что последний мало интересуется видами, количественно слабо представленными в фауне. и интересуется даже не фауной, а так называемым населением. Также и «географ» не может поставить в вину «зоологу» его предпочтительное вни- мание к наиболее «характерным» видам, в частности к малочисленным эндемикам.
В большинстве зоогеографических работ общего содержания (своч- ки, руководства и т. д.), как правило, разбираются основные понятия зоогеографии: ареал и вытекающие из него представления, способы рас- селения животных и преграды, а в последние десятилетия также и эколо- гические факторы существования животных, и обычно дается описание отдельных фаун с большим или меньшим вниманием к их истории. Одно время намечалось разделение зоогеографии на две самостоятельные вет- ви: историческую зоогеографию и экологическую зоогеографию. Но это можно считать явлением временным, скорее, даже пройденным эта- пом. История не может противопоставляться экологии, тем более что история в нашем случае — это смена во времени экологических условий прошлого.
Рекомендуемая вниманию читателя книга Ф. Дарлингтона «Зоогео- графия: географическое распространение животных» построена по иному плану по сравнению со многими другими зоогеографическими сводками. Здесь не говорится о трактовке понятия «ареал», а также о теории оттес- ненных реликтов и о принципе Тэйлора. Здесь нет и разбора положений, которые иллюстрировались бы избранными примерами из разных групп животных. ф'лавное внимание автор уделяет описанию распространения и вероятного хода расселения отдельных классов всех позвоночных живот- ных — водных и наземных. Ято описание проводится исходя из определен- ных, сформулированных в предисловии, рабочих принципов автора, кото- рых он строго придерживается. Выделяя процветающие группы в каждом классе и группы угасающие, вытесняемые процветающими (при этом часто пользуясь терминами «конкуренция», «доминирование»), автор разби- рает вопросы территориальной взаимозаменяемости (викариат), который у автора обозначается как комплиментарность и т. д.
Далее Ф. Дарлингтон разбирает закономерности островного распро- странения животных и затем материкового. В последнем случае описывает- ся состав фауны зоогеографических областей, которые автор называет фаунистическими, что, пожалуй, является более правильным. Общим положениям истории фаун Ф. Дарлингтон уделяет сравнительно немного внимания. Он не склонен к гипотетическим построениям, в частности не прибегает и к теории перемещающихся материков. Большинство фак- тов современного распространения животных вполне может быть объяс- нено, как говорит Дарлингтон, если исходить из признания только незна- чительных изменений в конфигурации материков, возникновения между ними относительно небольших сухопутных связей (Берингия и др.), а так-
11ред и~. > овис
же учитывая, что возможности расселения сухопутных животных через морские преграды значительно шире, чем это обычно считается.
Книга Ф. Дарлингтона содержит обширный материал. Она отличает- гя обилием новых данных и является большим вкладом в познание основ- ньтх закономерностей географического распространения животных. ~та книга, пожалуй, самое значительное, что было создано в мировой лите- ратуре по зоогеографии за последние 15 — 20 лет. Весьма демонстративны и книге своеобразно построенные картосхемы и прозрачный глобус. Иск- лючительно полезны и интересны приложенные к первым шести главам систематические списки семейств (с указанием некоторых родов и нередко с «есьма существенными примечаниями) и литература, данная в каждой главе. К сожалению, в книге Ф. Дарлингтона мало использованасовет- ская литература. Из работ советских авторов им упомянуты лишь очень немногие (Л. С. Берг), вышедшие в переводе на английский язык.
Можно надеяться, что книга Ф. Дарлингтона будет по достоинству оценена советским читателем, интересующимся географическим распрост- ранением животных, и также преподавателями вузов.
Приверженность автора к определенным рабочим принципам приве- ла местами к некоторому схематизму изложения, так, словно автора инте- ресует не то, как происходит расселение той или иной группы животных, а скорее, как тот или иной сформулированный им рабочий принцип может быть применен по отношению к рыбам, птицам и т. д. А это не одно и то же. И вообще в изложении книги чувствуется известный схематизм, что. может. в какой-то мере зависит от того, что Ф. Дарлингтон, будучи энто- мологом, не является специалистом ни по одной из рассматриваемых им групп животных.
Автор все время пользуется рядом терминов, объяснение которых да- но в гл. 1 книги. Некоторые из этих терминов употребляются в советской литературе, хотя в ряде случаев они понимаются Дарлингтоном в несколь- ко ином. нередко расширенном и усложненном смысле, другие не упот- ребляются вовсе. Мы старались избегать по возможности сложной терми- нологии. В частности, нам удалось совсем избежать термина «комплимен- тарность», так как в болыпинстве случаев оказалось возможным говорить о викариате, взаимозамещении разных групп. Избегали мы и терминов «доминантность», «доминантный» в связи с тем, что эти термины (так же как и предыдущий) в ряде случаев упрощают и схематизируют явление. Ф. Дарлингтон часто говорит о радиации и зонации, противопоставляя их как два разных образца (pattern) распространения (или расселения) животных. Между тем вряд ли требуется особый термин, когда речь идет о зональном распространении животных. А радиация у автора в одном слу- чае соответствует тому, что принято у нас называть адаптивной радиа- цией, а в другом — представляет собой какую-то геометрическую абстрак- цию — распространение группы животных из одного центра по радиусам (насколько можно было понять, и в этом случае имеет место адаптивная радиация).
Трудности с терминологией возникали не только в этом случае. На- пример. подразделению пресноводных рыб на три раздела по их отноше- нию и солености воды (и, следовательно, по возможности пересекать мор- ские преграды) сопутствуют термины, которые могут ввести читателя в за- блуждение и будут требовать напряжения внимания при чтении. Термины «первичный», «вторичный» и «периферический» разделы пришлось заме- нить терминами «строго пресноводные», «не строго пресноводные» и «соле- выносливые» рыбы. Нельзя сказать, что это наилучший выход, но иного
(Уред исл овие
нет. Необходимо также отметить слишком вольное обращение автора со словами «лягушки» и «саламандры», когда под «лягушкой» понимаются и жабы, а в понятие «саламандр» включаются и тритоны. Фактически 'ке в данном случае речь идет об известном подразделении амфибий на бес- хвостых и хвостатых. Также пришлось избегать таких слов, как, например, «вычитание фауны» и «уменьшение фауны». Термин «переход» употреб- ляется нередко автором в очень усложненном смысле. Большие трудности не только для перевода, но и просто для понимания представляют слова Ф. Дарлингтона: «основная часть суши», «основная часть материка» и некоторые другие.
При всем том мы позволили себе и одну вольность по отношеник~ к тексту автора. В книге всюду говорится, в соответствии с терминологи- ей Уоллеса и Склэтера, о Восточной области. Между тем и советской ли- тературе эта область называется теперь Индо-Малайской. Мы произвели в книге необходимую, по нашему мнению, замену названия, кроме тех мест, когда речь идет непосредственно о работах Уоллеса или о работах (:клэ- тера.
(;дечаны и небольшие сокращения текста. Помимо некоторых ненуж- ных повторений, мы позволили себе опустить раздел, посвященньш так называемой географической истории человека, как не относящийся непо- средственно к основному содержанию книги. Небольшие сокращения сде- ланы и во введении, когда речь идет об элементарных для географа и зоогеографа понятиях.
Н. А. Гладков,профессор 6иогеографии
1Тпми,ти, Хомвсгс Б'арбор«— женимого нвт урилисюпа т р~гководилпсля
ОТ АВТОРА
О книге
Эта книга представляет основы зоогеографии. Б ней сделана попытка собрать вместе все факты, сопоставить их и рассмотреть или пересмотреть принципы географического распространения животных на всем земном шаре. Книга касается только животных суши и пресных вод и в основном позвоночных, потому что они изучены лучше всего и с точки зрения 300- географии это наиболее важная группа животных. В этих рамках автор книги пытался ответить на четыре вопроса:
Каковы основные особенности распространения животных?
Как эти особенности возникали?
Почему они возникали~
Что говорит нам распространение животных о древних землях и кли- матах?
Настоящая:книга — географическая, а не экологическая зоогеогра- фия. Это говорится не для оправдания. Сейчас моден экологический под- ход к географии животных и растений, но книги, написанные с этой точ- ки зрения, подобно замечательной «Ecological Animal Geography» (Hesse, Allee, Schmidt, 1951), по сути дела, работы экологические, а не географиче- ские. Я не собираюсь умалять значение экологии, а хочу только напомнить, что это не зоогеография. Еще Дарвин говорил, что распространение живот- ных нельзя объяснить исходя только из климата и местных физико-гео- графических условий. Например, была высказана мысль, что в Южной Африке, Южной Америке и Австралии млекопитающие объедают молодые побеги кустарников, но и млекопитающие и кустарники на каждом из этих материков различны, и экология не может объяснить эти различия. Конечно, экология может сделать большой вклад в зоогеографию, так же как и зоогеография может внести свою лепту в экологию.
Зоогеография имеет замечательную историю. Распространение расте- ний и животных в Южной Америке — первое, что натолкнуло Дарвина на мысль об эволюционном развитии, о чем и говорится в первой же фразе «Происхог1ъдения видов». Зоогеография привела также и Уоллеса, одно- временно с Дарвином, к теории эволюции. Возможно, в дальнейшем эоо- география и не приведет к таким кардинальным идеям, вроде идеи орга- нической эволюции, однако она может дать еще много нового. Распрост- ранение животных может подсказать биологам новые факты об эволюции и помочь геологам восстановить мир прошлого. И кроме того, зоогеогра- фия сама по себе интереснейшая наука.
О методике н терминологии
Зоогеография рассматривает множество своих фактов, а также при- влекает данные экологии, эволюции и геологии. Такая обширная область может быть охвачена однои книгои двумя путями: можно коснуться всех
От. автора
вопросов поверхностно при помощи избранных примеров, ограниченного рассмотрения фактов и произвольных обобщений, но это будет просто не- критический пересмотр старых взглядов с небольшими шансами на разви- тие новых; или можно осветить одни вопросы, насколько позволяет место, очень подробно, а другие — очень кратко. Я придерживался последнего метода: наиболее детально описал и рассмотрел распространение наземных и пресноводных позвоночных, а многие другие вопросы сократил до голых ссылок или просто ограничился изложением своих взглядов. Чтобы понять и критиковать мою зоогеографию, читатели должны знать точку зрения автора, например, по вопросам геологии и эволюции. Но даже и при та- ком подходе я должен был при описании позвоночных ограничивать себя: не углубляться в детали и удовлетворяться уже известными данными.
Сказанное относится и к приводимой мною литературе, которая име- ет отношение к зоогеографии и охватывает не только зоогеографические ра- боты, но также все то, что опубликовано по систематике, образу жизни и истории всех рассматриваемых животных, и многое из того, что опубли- ковано по общей географии, климатологии, экологии и эволюции. Я не устанавливал единого стандарта в подборе всей этой литературы, но пы- тался дать для каждого раздела исчерпывающие источники, а иногда толь- ко одну последнюю работу, чтобы этим открыть читателю доступ к другой литературе.
Мне также не удалось полностью осветить историю зоогеографиче- ских воззрений. Я сосредоточил свое внимание именно на изучении рас- пространения животных, а не на суждениях об их распространении и очень часто утверждал уже как факты те вещи, которые теперь кажутся оче- видными, но которые выяснились в результате большого труда многих поколений зоогеографов. Я весьма сожалею, что не всегда отдавал долж- ное тем вещам, которые этого заслуживают, но если бы я попытался про- следить историю зоогеографии более детально, очевидно, я не закончил бы эту книгу.
Определение зоогеографических терминов дано ниже, здесь же стоит упомянуть о некоторых общих словах и выражениях, являющихся люби- мыми фразами ученых — «как убедительно показывают...», «согласно на- учному мнению... », «согласно моему тщательно продуманному суждению... » и т. д.,— но которые обычно обозначают не больше чем «вероятно»; и это именно то слово, которое я буду, как правило, употреблять здесь. Я также буду говорить «я думаю» и «я предполагаю» без дальнейших обоснований.
Оо авторе
Кто может написать зоогеографию? Никто, если требуется непосред- ственное знакомство с предметом в полном его объеме. Ни один человек не может знать всех животных всех частей земного шара и всех дисциплин, связанных с зоогеографией. Но зоогеография должна быть написана, да- же если нет столь разносторонне подготовленного человека.
Как зоогеографу мне в известном отношении повезло, потому что я вырос и получил образование в городе с большими музеями; был своим человеком сначала в старом Бостонском обществе естественной истории, а позднее в Музее сравнительной зоологии в Гарварде и знал та- ких ученых, как М. Л. Ферналд, В. М. Уиллер и Томас Барбар, которые, будучи систематиками или работниками музея, также занимались препо- даванием.
11
С самого начала я глубоко интересовался (и продолжаю интересовать- ся) распространением-животных в Северной Америке и собирал там кол- лекции. Тропики я увидел впервые в 1926 г. двадцатидвухлетним студен- том-выпускником, когда провел три месяца на Гарвардской исследователь- ской станции в Соледаде, центральная Куба. Эта практика произвела на меня неизгладимое впечатление, и я возвращался в Вест-Индию четыре раза и вновь занимался коллекционированием на главных островах. Очевидно, ни одни острова, кроме этих, не считая Филиппин, не отражают так xopOIIIo зоогеографические особенности островной жизни.
В 1928 — 1929 гг. мне пришлось провести год на северном побережье Южной Америки в качестве постоянного энтомолога Колумбийского фи- лиала «Юнайтед фрут компани», удаленного от Санта-Марта в глубь мате- рика. Здесь я познал все разнообразие американских тропиков — от сухих равнин, краевых полос, болот и дождевых лесов дельты реки iVIar- далена, склонов с различной степенью облесенности, парамосов' до снеж- ных пиков Сьерра-Невада-де-Санта-Марта. В 1931 — 1932 гг., находясь с группой сотрудников Музея сравнительной зоологии в Австралии, я уви- дел все многообразие природы этой страны в разные времена года. И, нако- нец, ро время второй мировой войны в качестве энтомолога за два года я проделал путь из Австралии в Новую Гвинею, через экватор и линию Уоллеса до Филиппин и Японии.
Еще мальчиком я интересовался птицами и, если бы имел разрешение на сбор коллекций, вероятно, стал бы профессиональным орнитологом. Однако я стал энтомологом. Тем не менее я систематически собирал для музея не только насекомых и других беспозвоночных, но также лягушек и рептилий в Вест-Индии, птиц и других позвоночных в Южной Америке, а в Австралии в некоторые сезоны года занимался преимущественно кол- лекционированием млекопитающих, птиц, рептилий и амфибий. Из всех основных классов позвоночных только в отношении рыб я не могу пре- тендовать на сколько-нибудь основательное изучение их в природе, и мне пришлось немало поработать. чтобы знакомством со специальной лите- ратурой восполнить этот серьезный пробел.
hpoMe всего, необходимо добавить, что я, к счастью, систематик. Клас- сификация животных — моя профессия. Правда, я занимаюсь системати- кой жуков, а не позвоночных, но моя работа дала мне непосредственные знаю~я о том, что такое систематика, как она выявляет взаимосвязи ме- жду животными и их распространением и каковы ее слабые стороны. Зна- ния такого рода совершенно необходимы для зоогеографа.
Таким образом, мне повезло во многом, а еще одна удача увенчала все остальные. В 1932 г. я был зачислен в штат Музея сравнительной зо- ологии, где работал сначала под руководством Томаса Барбора, а затем Альфреда Ромера. Во всем мире не было места лучшего, чем этот музей, для проведения зоогеографических исследований под руководством этих уче- ных. 3ту книгу я задумал в музее почти двадцать лет назад и системати- чески работал над ней, насколько это позволяли другие занятия. У меня было время для работы, благоприятные условия, помощь и поддержка, и, возможно, лишь благодаря этому я смог выполнить поставленную задачу.
Филипп Дж. Дарлингтонмладший
' Тип растительности верхнего пояса высокогорпй экваториальных широт.— Прил. ред.
1'лаеа 1
ВВЕДЕНИЕ
В этой главе приводятся общие сведения о земном шаре, климате и растительности на нем, краткое описание геологической истории Земли, данные по зоогеографии в целом и ее некоторые конкретные аспекты, от- дельные факты о расселении животных, очень краткая история зоогеогра- фии, некоторые зоогеографические понятия и перечень методов, которыми я пользовался при написании данной работы и которые я назыгаю < рабо- чими принципами». Земной шар и карты
Зоогеографию:следует начинать с напоминания некоторых простых фактов о земном шаре. Земной шар — это сфера. Данньш факт весьма су- ществен, но часто, считая его само собой разумеющимся, о нем забы- вают. Зоогеографы должны постоянно помнить, что Земля — шар и что карты, составленные ими, должны это отражать. Р и с. 1. Карта земного шара (ЪУа11асе, 1876) в ироенции Мер-
катора. Упро|ценный контур с материковыми фаунистическими областями, 11о
Уоллесу (см. фронтиспис и рис. ' G).
Карты могут давать как правильное, так и искаженное изображение земного шара. Карты хорошо известного зоогеографа Уоллеса были весьма примитивны. Они были выполнены в проекции Меркатора (рис. 1), которая дает представление о климатических зонах, но в связи с иска- жением площади в сторону ее увеличения к полюсам не дает представле- ния об истинном положении континентов на шарообразной Земле. Дру- гие зоогеографы пользовались картами, на которых земной шар был по- хо;к на корку апельсина, или морскую звезду, или бок луковицы. Мне
Введение
кажется, что все эти причудливые проекции приносят больше вреда, чем пользы, просто потому, что они не показывают земной шар таким, какой он есть в де~~ствительности.
Глобус — лучшая карта земного шара, причем удобнее всего пользо- ваться прозрачным глобусом. В зоогеографии чаще всего пользуются картами в ортографической проекции, представляющими собой просто картину или диаграмму земного шара с бесконечно удаленной точкой зрения. Ортографическая проекция чаще всего игнорируется картогра- фами, но ни одна другая карта не позволяет достаточно хорошо опреде- лить на глаз расстояние, площадь, расположение климатических зон и обширные зоогеографические области по отношению к круглой Земле, в то время как на ортографической карте это можно увидеть и понять, причем последнее наиболее важно. Создать такую проекцию в плоскости экватора нетрудно, и зоогеографу стоит потратить время на ее изучение. То обстоятельство, что на карте может быть показана только одна полови- на земного шара в одном аспекте, в некоторых случаях можно исправить накладыванием двух карт противоположными сторонами земного шара, чтобы воссоздать, по существу, картину прозрачного глобуса (см. шмуц- титул). В этой книге выполненные мной контурные карты даны в ор- тографической проекции. Все они сделаны почти схематически, чтобы дать общее представление, а не детали, так как наиболее важна именно общая схема.
(:уша и океаны
Суша занимает только около 29~о земного шара и распределена нерав- номерно. К северу от экватора суши более чем в два раза больше, чем к югу от него, а в восточной части полушария (Старый Свет) более, чем в два ра- за больше по сравнению с западной (Новый Свет). А если Тихий океан при- нять за центр полушария и сопоставить с противоположной стороной земного шара, несимметричность проявляется еще сильнее: большая часть суши находится на противоположном полушарии и совсем незначитель- ная — в Тихом океане. Единственно вероятное объяснение этому мож- но найти в том, что Луна, согласно одной из гипотез, оторвавшись от Земли со стороны Тихого океана, забрала с собой большую часть земной коры с этой стороны.
Несмотря на небольшую площадь и неравномерное распределение, большая часть суши образует единую более или менее непрерывную си- стему. Все основные континенты (за исключением Антарктиды) соедине- ны, или почти соединены, или связаны между собой архипелагами, так что, как говорит Уоллес (Wallace, 1876, vol. 1, р. 37), вероятно, можно со- вершить путешествие по всей системе континентов, всегда имея в поле зре- ния сушу. Более того, вероятно, только 10 тыс. лет назад суша была еще более непрерывной: Азия и Северная Америка соединялись через море Беринга; острова Суматра, Ява и Калимантан (Борнео) смыкались с Азией, остров Сулавеси (Целебес) отделялся от острова Калимантан, вероятно, всего только 25 милями водного пространства; других проходов между Азией и Австралией было меньше, и они были уже, чем теперь. Схема рас- положения континентов такова, что к северу от тропиков находятся боль- шие, почти не разделенные площади суши; в пределах тропиков болыпие площади суши, отделенные друг от друга и к югу от тропиков, меньшие площади, которые разделены большими расстояниями.
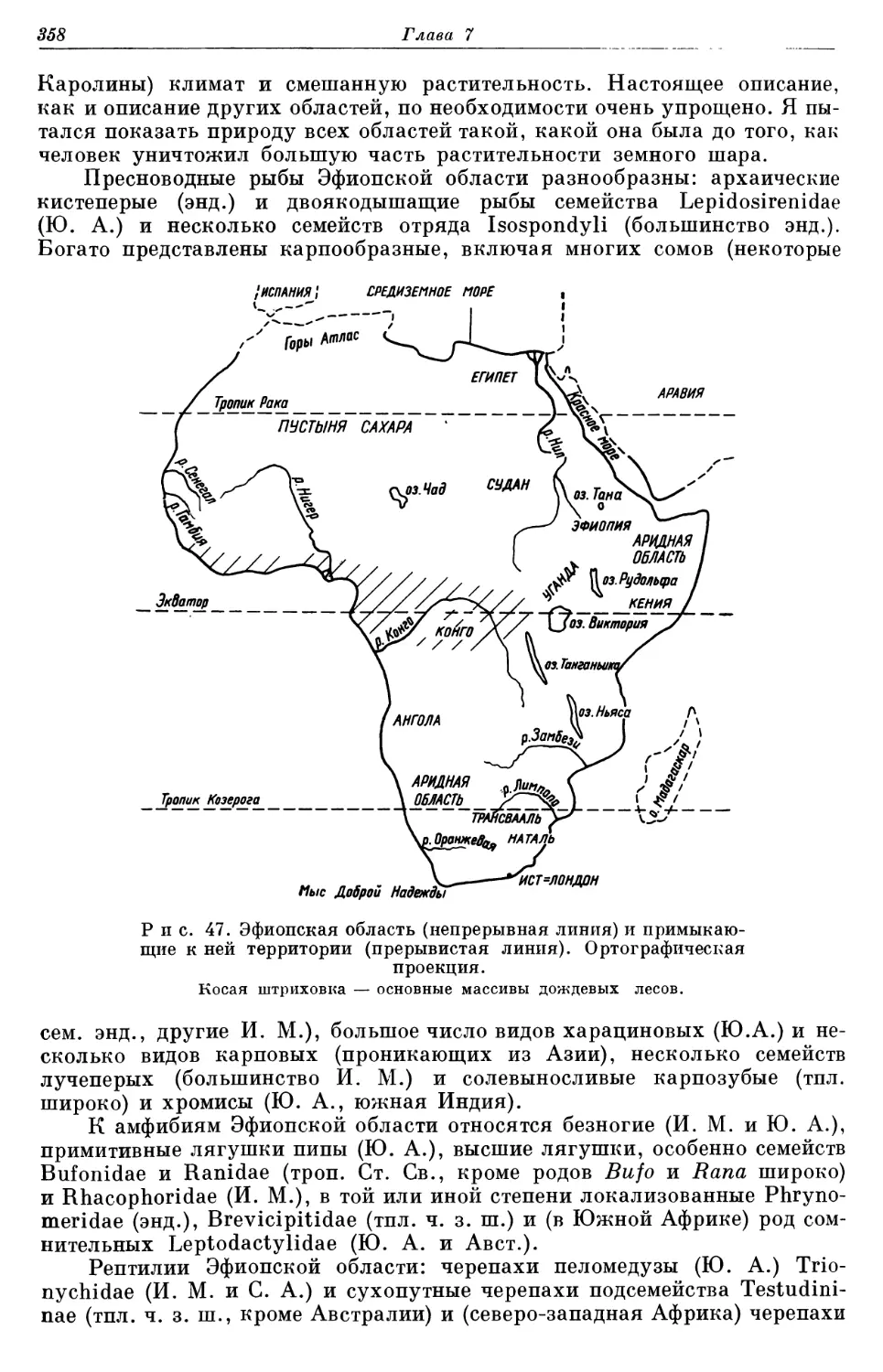
Глава 1
Влияние океанов на распространение наземных животных различно. Они выступают естественными преградами при расселении животных и в то же время смягчают климат континентов, о чем говорится ниже. Океа- нические течения иногда заносят наземных животных на острова или, воз- можно, даже переносят с одного материка на другой. Океаны и животные рассказывают нам историю Земли. Например, нахождение близкород- ственных морских животных в атлантических и тихоокеанских водах, омывающих берега Центральной Америки, свидетельствует о морском пу- ти, который недавно разобщил сушу; а ископаемые прибрежной морской фауны говорят о распределении климатов и, возможно, даже о самой суше в прошлом. Изучение дна океанов дает представление (или может его дать) о происхождении и истории материков и о древних сухопутных мостах. Следовательно, имеется достаточно оснований для того, чтобы зоогеогра- фы, интересующиеся сушей, знали кое-что и об океанах. К счастью, по океанам есть много хороших справочников (Sverdrup, Johnson, Fleming, 1942; Ekman, 1953, перевод с издания 1935 г.; Kuenen, 1950. Книга пос- леднего автора особенно полезна).
Кчимат
Температура и количество выпавших осадков — основные показатели климата. Температура поверхности Земли в основном определяется коли- чеством тепла, получаемого от Солнца, а распределение температуры по поверхности земного шара — главным образом формой Земли и ее движе- нием (рис. 2). Наибольшее количество тепла приходится на тропики. Гра-
бебернаязима
Мерноелето
Соп- ние
Р и с. 2. Движение Земли вокруг Солнца показывает, как наклон земной оси (С — Ю) в высоких широтах вызывает смену времен года (лета и зимы) (масштаб и
перспектива не выдержаны).
ницами тропической зоны являются самые северные и самые южные широ- ты, над которыми солнце находится в зените в день летнего или соответ- ственно в день зимнего солнцестояния. Эти широты простираются до "3'27' к северу и югу от экватора. В тропиках жарко в течение всего го- да: максимальные температуры, по крайней мере во влажных тропиках, ниже, чем температуры в жаркие летние дни во многих частях умеренных зон северного полушария, но тропическая жара с небольшими суточными колебаниями температуры устойчива во все времена года и днем и ночью. К югу и северу от тропиков в связи с шарообразностью Земли количест- во тепла, получаемого земной поверхностью, уменьшается, а сезонность выражается все более отчетливо. В умеренных зонах возникает смена зи- мы и лета, и по мере удаления от тропиков суровость зимы возрастает, а дальше к полюсам, пересекая северный и южньш полярньш круг, на-
Введение
блюдаются периоды, когда летом солнце никогда не заходит за горизонт, а зимой никогда не поднимается над горизонтом. Продолжительность этих периодов увеличивается по мере приближения к полюсам, достигая шести месяцев на самих полюсах. Смена времен года и суровость зимы на севере могут ограничивать распространение животных в большей степени, чем средние температуры.
B горах понижение температуры с высотой происходит не в резуль- тате того, что солнце отдает тепла меньше (на больших высотах оно греет сильнее), а потому, что воздух становится разреженнее и, следовательно, холоднее. Понижение температуры с высотой можно сравнить с пониже- нием при продвижении к северу (рис. 3). Если режим снега и льда на вы-
ечных
АРКТикА
30НА
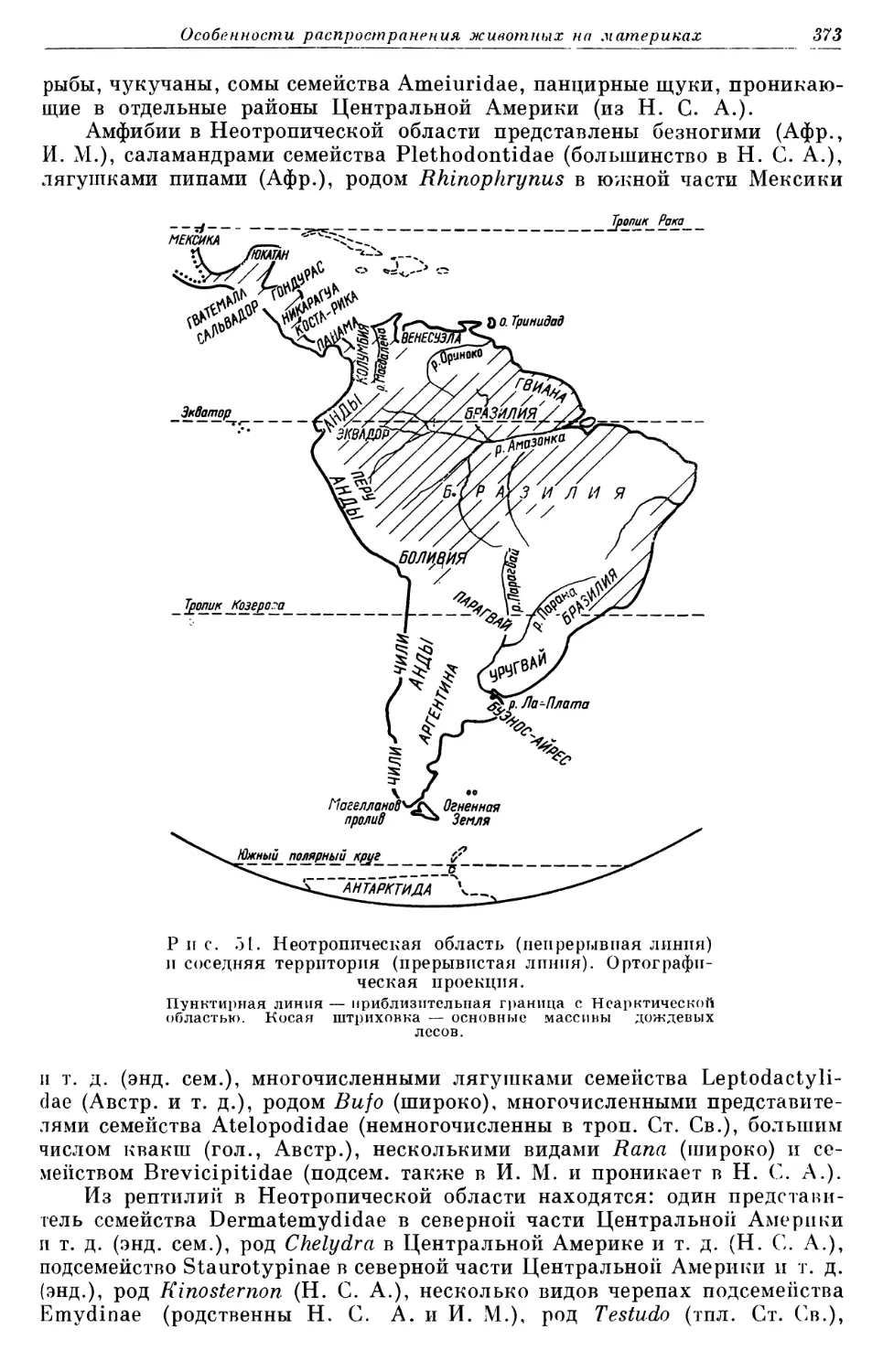
УпадСОЛИпу
ЗОНА
Я 30IYA
30НА
P и с. 3. Сравнение понижения температуры по мере продвижения к северу Земли и с изменением ее
в горных районах тропической зоны.
соте 16 000 футов в горах вблизи экватора, Карстенс-Топпен, в Новой Гви- нее (Umbgrove, 1949, Pl. IX), сравнить с подобным на уровне моря на 80' с. ш., то можно сказать, что каждые 200 футов высоты влияют на кли- мат так же, как и 1' широты в северном направлении. Это сопоставление правильно лишь отчасти. Влияние продвижения на 1" к северу по-разному будет сказываться в различных местах между экватором и полюсом. Это влияние весьма незначительно в тропиках, но оно будет неизменно увели- чиваться при прохождении через умеренные зоны, но и это не столь сущест- венно. Главное заключается в том, что характер понижения температуры с высотой и по мере продвижения на север по своей природе различен и по-разному влияет на жизнь. Зоогеографы знают об этом, но не придают данному факту должного значения. По привычке они называют последо- вательно сменяющиеся высотные зоны в горах тропиков «субтропической», «умеренной» и «арктической». Это зачастую приводит к неправильному представлению, которого, однако, можно было бы избежать, если бы зоо- географы пользовались правильными картами, показывающими распре- деление климата на шарообразной Земле. Высокогорные климаты тропи- ков — не одно и то же, что климат высоких широт, по аналогии с которым они получили свое название. Климат умеренной зоны северного полуша- рия прохладный, с хорошо выраженной сезонностью; климат на больших высотах в тропиках также прохладный, но почти неменяющийся в тече- ние всего года. На севере средняя температура 10' (;, но ее экстремальные значения могут находиться в пределах от 37,8 до — 17,8'. Высоко в го- рах тропиков средняя температура может быть такой же, но она никогда
Глава 1 не падает до нуля. Вот почему пальмы, которые так пышно растут и «уме- ренных» зонах гор тропиков, не могут существовать на большей части умеренных зон северного полушария.
Осадки также имеют зональное распределение. В основном оно опре- деляется характером атмосферного давления и атмосферной циркуляцией (рис. 4). Зоны высокого давления расположены в северном и к~жном полу- шариях за пределами тропиков. Ветер дует от зон высокого давления, унося с собой влагу и оставляя за собой пустыни. Большинство великих пустынь мира в Северной Африке, Юго-Западной Азии и юго-восточной части Северной Америки и юго-восточной Африке, Австралии и в юго-за- ПОЛЯРНАЯ 39НА
Or0 ДАВЛЕНИЯ УМЕРЕННАЯ ЗОНА НИЗКОГО ДА ВЛЕНИЯ
l ЦБТРОПИЧЕСКАЯ 30НА ВЫСОКОГО ДАВЛЕНИ
p++ Напрабление бращеиия -+
ТОРИАЛЬНАЯ AHA НИЗКОГО ДАВЛЕНИ 6ТРВПИЧЕСКАЯ 3РНА ВЫСОКОГО ДАВЛЕНИ УМЕРЕННАЯ ЗОНА НИЗКОГО ДА ВЛИПНИ~
псяяятя ЗОил
ВЫСОКОГО ДАВЛЕНИЯ Р и с. 4. Давление и зоны ветров одного 1foJIv-
шария. падкой части Южной Америки находится в зонах высокого давления или захватывает краевые области тропиков. Пустыни восточной части Цент- ральной Азии (Гоби и др.) являются исключением: сухость вызвана, поми- мо других причин, удаленностью этих районов от моря.
Ветры (пассаты), дующие из субтропических зон высокого давления к экватору и приносящие с собой влагу в виде тропических дождей, спо- собствуют произрастанию дождевых лесов. Пассаты, дующие на север и на юг в прохладные районы умеренных широт, встречают на своем пути различные воздушные потоки и характеризуются неустойчивыми осад- ками.
Основная зональность в распределении осадков видоизменяется и на- рушается в результате многих причин. На местные осадки влияет харак- тер рельефа. Обычно самые сильные дожди выпадают вблизи моря, с на- ветренной стороны и в горах. Там, где рельеф неровный и преоб.гадают вет- ры одного направления, распределение осадков может сильно варьировать на небольшом расстоянии. На северном пооережье Южноп Америки от Санта-Марты в глубь материка в течение одного дня я мог совершить про- гулку от полупустыни, где преобладают акации и гигантские кактусы, через колючие кустарники к долине с дождевыми лесами, а затем опять
Введение
вверх по склону Сьерра-Невады через листопадный лес — к вечнозеленому подоблачному лесу. Такая резкая смена растительности, отражающая местные колебания в выпадении осадков, обычна для тропиков. Во многих районах тропиков осадки имеют сезонный характер и засушливый сезон года может быть выражен очень четко: леса теряют листву, исчезает боль- шинство насекомых, а некоторые птицы в поисках кормных мест мигри- руют за сотни миль.
Влияние океанов на климат суши сказывается и на температуре и на количестве выпадающих осадков. Вообще океан смягчает экстремальные температуры на близлежащей суше, а океанические течения могут влиять на климат обширных территорий. Например, Гольфстрим несет теплые воды из Мексиканского залива по диагонали через Атлантику к Британ- ским островам и Европе, смягчая климат и позволяя растениям и живот- ным расширять свой ареал значительно дальше на север, чем на востоке Северной Америки. Я побывал, насколько это позволили сделать мест- ные дороги, на севере Квебека, в редких еловых лесах, угнетенных холо- дом, и это было почти на одной широте с южной оконечностью Англии. Таким образом, большую роль здесь играют океан и теплое течение Гольф- стрим. И, наоборот, течение Гумбольдта или Перуанское, несущее холод- ные воды с юга вдоль западных берегов Южной Америки, позволило пинг- винам достичь островов Галапагос на экваторе.
Более детально климат рассматривается в работах Кендрю (Kend- rew, 1949) и Гейгера (Geiger, 1950), которые являются кратким руковод- ством. В книге «The Climates of the Continents» Кендрю (Kendrew, 1936) рассматривает климат скорее с географической точки зрения. Очень ин- тересна книга Таннехилла «Weather around the world» (Tannehill, 1952а).
Факторы, благодаря которым климат является частично зональным, частично местным, очень сложны и существовали с момента возникновения воды и суши. Земля всегда представляла вращающуюся сферу с экватором, который получает большое количество солнечного тепла, и полюсами, ко- торым его не хватает. Таким образом, климаты всегда были в какой-то степени зональными; правда, зональные климаты временами могли быть менее дифференцированными, чем теперь. Более того, земная ось всегда была наклонена так, что, когда Земля проходит вокруг Солнца, в север- ных и южных странах происходит поочередно смена теплого и холодного сезонов, что в свою очередь увеличивает влияние зональности климата на распространение жизни на Земле. Характер взаимосвязей суши, водной поверхности и ветра очень сильно отличался в различные времена в прош- лом, так же как различные части суши меняли свои очертания и высоты, что всегда приводило к разнообразию местных климатов. В общем каждый существующий континент имеет местами влажный, а местами засушливый климат, и, вероятно, так было всегда. Геологи иногда говорят об однород- ном климате на всем земном шаре, но понимать это буквально нельзя.
Ни температура, ни количество выпадающих осадков в действительности не могли быть одинаковыми на всем земном шаре.
Растительность
Растительный покров земного шара в основном распределяется в соот- ветствии с климатом, особенно температурой и количеством выпадающих осадков, и связан с природой и историей Земли, а такн е с другими факто- рами. Распределение наиболее важных растительных формаций земного
Глава I шара показано на рис. 5. Растительность имеет зональное распространение, но растительные зоны видоизменяются, прерываются и смещаются так же, как зоны температур и осадков. Дождевые леса — самая типичная расти- тельность тропиков — распределены неравномерно. Наибольшие пло- щади этих лесов находятся в Южной Америке (долина Амазонки), в восточ- ной Индии и Западной Африке, но многочисленные небольшие участки п по- лоски этих лесов широко разбросаны по всем тропикам.
Дождевые леса высокого бонитета постепенно переходят в другH(', оолее бедные типы низинных лесов, включая тропический листопадныЙ
2 3 Ф Р ц с. 5. Распространс нпс основными т11иов рас п~тельности земного ~пара (чрезвыча11по упрощенное) отражает основные черты распро- странения жнвотных. (Е. М i 1 п е-8 е d Ь е а d, 1954, «Proc. 1 1ппеап ~ос.», 1.опс!оп, 165. р. ' 7; (The Australian Environment», Commonwe- д1[11 facie»tifie a»tl Industrial research Organization, 2»cl ed., Mel- hourn, 1950, fi<>'. ',), р. 88 — 89; «Proc. National Inst. Sei. I»gaia», 1949, fp, р.:),iЬ; l' е r ~1 о о r n et al., Plants a»d Plant Science
in Latin Ан1ег1са, AValtham, Mass., 1945.) ~~[дотность растительного покрова весьма приближенно соответствует густоте штрихов на карте. 1 — — дождевой тропический лес; 2 — темнохвой- ный лес умеренных широт; ~ — северньп~ хвойный лес; I — ре.~колесье,
зла ковники;:> — и олупустын и и 1I устыни. лес; последовательно сменяющиеся типы влажных вечнозеленых (обычно не хвойных) лесов низменностей встречаются и в горах влажных тропиков (часто на высоте примерно 12 000 футов) до тех пор, пока они не сменян>тся rðanÿHèñTûì покровом. Затем, часто на высоте 16 000 футов, травянистый покров уступает место скалам и снегу. Во многих местах тропиков встре- 'IaIoòcÿ большие площади колючих кустарников и саванны, в частности в Африке имеются значительные площади колючих кустарников и злаков- ннков. Редкая пустынная растительность покрывает обширные площадц вблизи окраин тропических областей. Далее на север, до умеренных ши- рот, простирается зона, в которой чередуются неравномерные участки ле- сов с опадающей листвой и открытые травянистые пространства, затем следует более однообразная и беспрерывная зона северных хвойных не- ~ ов и, наконец, зона арктической тундры. К югу, в умеренных широтах
Введение
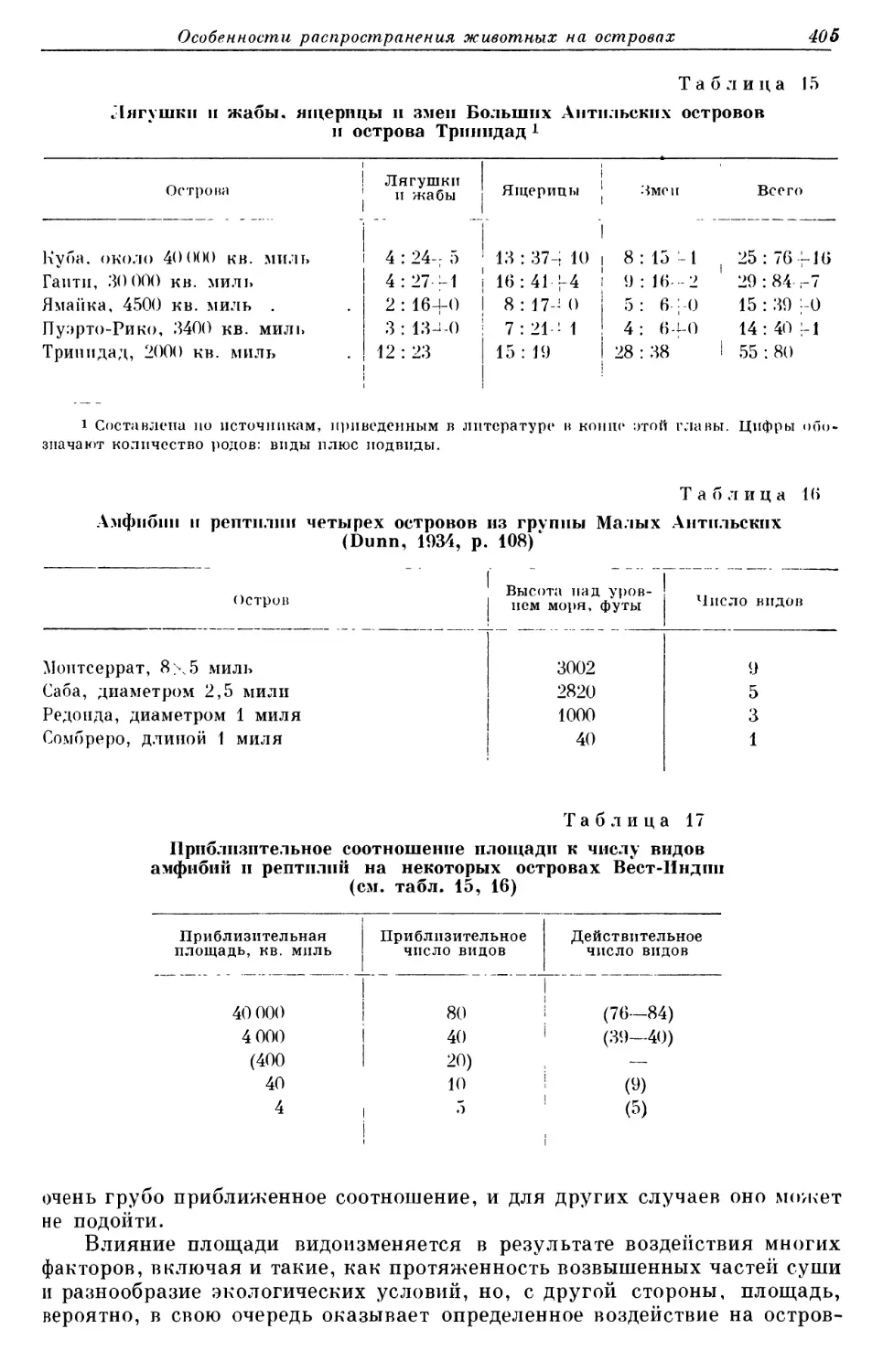
южного полушария, в Африке, Австралии и Южной Америке распростра- нена смешанная растительность.
Прежде чем закончить описание климата и растительности, необхо- димо добавить еще несколько слов о тропиках. Совершенно очевидно, что, как во времена Дарвина и Уоллеса, так и теперь, нужно побывать в тропи- ках самому, чтобы увидеть, что такое настоящая древняя, нетронутая че- ловеком флора и фауна и к чему приводит эволюция при благоприятных оостоятельствах. В умеренных зонах северного полушария нет ничего похожего на тропические леса. Вест-Индия и горы Центральной Америки полны чудес, но даже и там нет настоящих, очень древних, невероятно оогатых, величественных тропических дождевых лесов низменностей. Наилучшее место для изучения тропиков североамериканскими биолога- ми представляет собой остров Барро-Колорадо в зоне Панамского канала, где теперь расположена исследовательская биологическая станция Смит- сонианского института. Это настоящие тропики в полном смысле этого слова — близкие и доступные. Описание их дано Чэпменом (СЬаршапи, i929; 1938), но недостаточно только читать о них, их нужно увидеть и почувствовать самому. Я думаю, что любой молодой натуралист, который считает, что он может понять мир, живую
природу и эволюцию без познания тропи- Псспебнае 100�лет ков, заблуждается и причиняет себе огром- ~IПвейстоцен ный вред.
15мли.
30млн 40млд.
Гео: огпческое время
~25 млн
f80nrgy
Р д с. 6. Геологические времепа(последние ~200 [млн. лет).
Земля существует два, три, четыре
50мЛп или даже больше миллиардов лет. Низшие 60плп. растения (водоросли) появились на Земле более миллиарда лет назад, а простейшие беспозвоночные, вероятно, более чем пол- миллиарда лет назад. Древнейшие бесче- :иостные рыбоподобные позвоночные воз- никли 400 млн. лет назад, и прошло еще очень много миллионов лет до того, как на Земле появились первые земноводные, и еще много миллионов лет до того, как по-
<50млп звоночные стали настолько многочислен- ными, что находки их ископаемых, оона- руженные во многих местах земного шара, приобрели важное значение для географии. Я не знаю, насколько, в конце концов, далеко смогут заглянуть зоогеографы в
200млд, прошлое Земли, чтобы увидеть и понять характер и значение распространения животных, но я ограничусь пределами примерно 200 млн. лет, так как даже на этом отрезке времени доволь- но трудно составить достаточно ясную картину. Таким образом, в таб- лице геологических времен я обращаюсь назад через плейстоцен, третич- ный период и мезозой к перми (рис. 6). Эта таблица не нуждается в под- робном объяснении, так как она детально разобрана в работе Цейнера (Zeunev, 1950). Точная датировка таблицы также не столь важна: был ли пермский период действительно 200 млн. лет назад, или еще раньше, или
Глава 1
поз;ке — большого значения не имеет. Важна последовательность эр, пе- риодов и эпох прошлого, являющаяся шкалой, на которой все геологиче- ские и биологические события рассматриваемого времени тесно связаны друг с другом.
Зоогеография: границы, ст1иени и материал
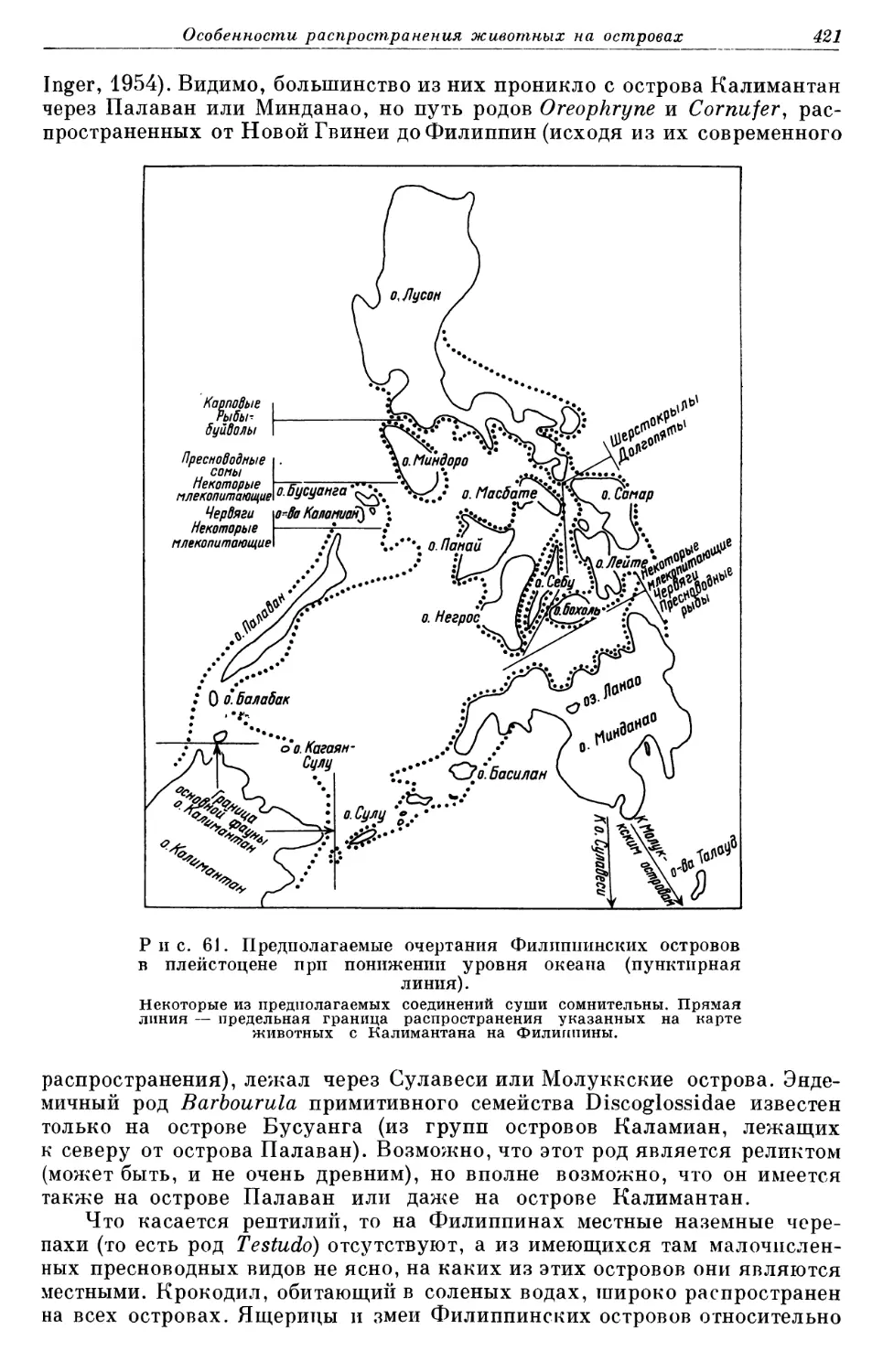
Зоогеография изучает распространение всех животных на всем зем- ном шаре. Земной шар огромен и сложен, и на нем насчитывается около миллиона различных видов животных. Поэтому, очевидно, зоогеограф дол- жен установить рамки для своей работы. Как уже говорилось, эта книга ограничивается рассмотрением вопросов распространения наземных и пресноводных позвоночных. Она также ограничивается в основном рас- смотрением общих черт распространения животных, а не локальных.
Различие между географическим и локальным распространением бы- ло сделано Уоллесом. В действительности, удобнее выделять три ступени: географическое распространение в самом широком смысле этого слова- ка всем земном шаре, распространение по областям в избранных частях земного шара и локальное распространение, принятое Уоллесом, кото- рое включает географию и экологию видов. Конечно, я буду касаться до какой-то степени всех этих ступеней, даже самой низшей (подробно все факты представлены в гл. 2 — 6), но главной моей целью было выяснить и понять основные особенности распространения животных. География ви- дов — их географическое распространение относительно друг друга и по отношению к экологии и эволюции — здесь не затрагивается. Эволю- ционные аспекты географии видов трактуются по Майру (Мауг, 1942). Вопросы экологии в книге не рассматриваются потому, что, как я уже го- ворил выше, это не зоогеография. Единственно, что с данной точки зре- ния здесь необходимо учитывать, это прежде всего ряд общих и очевид- ных утверждении: что животные для своего существования нуждаются в пространстве и пище; что наземные животные испытывают затруднения, пересекая водные пространства, а водные животные — пересекая сушу; что летучие мыши и большинство птиц могут летать, а другие позвоночные не могут и т. д. Здесь будут затрагиваться также отдельные экологические проблемы (например, отношение пресноводных рыб к соленой воде и раз- личных позвоночных к северному климату), но я постараюсь решить их анализом распространения этих животных, а не разбором книг по эколо- гии. Решение данных вопросов таким путем может оказаться полезным для экологов. Кроме того, по экологии существуют хорошие справочники (АИее, Emerson, О. Park, Т. Park, Schmidt, 1949; Hesse, Allee, Schmidt, 1951).
Материалом для настоящей работы послужили наземные и пресновод- ные позвоночные. Позвоночные составляют только 3 или 4",о всего живот- ного царства. Однако это наиболее изученные животные; они лучше всех других собраны и систематизированы и по ним имеются самые полные ископаемые данные. И по ряду других причин позвоночные более всего подходят для зоогеографической работы. Они очень разнообразны: одни живут на суше, другие - — в воде; одни могут летать, другие — не могут; одни — холоднокровные, другие — теплокровные; одни — древние, другие — нет. Такое разнообразие говорит об истории, особенностях и значении их распространения значительно больше, чем это могли бы сде- лать другие, менее варьирующие группы. Многие позвоночные являются
Введение
21
строго наземными или пресноводными животными, которые при расселе- нии с большим трудом пересекают соленые воды и, следовательно, так же, как и другие животные, свидетельствуют о соединениях суши как в на- стоящем, так и в прошлом. Однако при решении некоторых вопросов ло- кального распространения не всегда можно использовать в качестве до- казательств позвоночных, так как, по-видимому, они не остаются продол- жительное время на небольших изолированных площадях и в связи с этим не отражают истории некоторых древних островов (например, Новой Каледонии) и других небольших отдельных территорий (например, юж- ной оконечности с характерным для нее холодным климатом материка Южная Америка), как это отражают некоторые беспозвоночные.
Книга Ромера (Вошег, 1945) предварительно знакомит зоогеографа с позвоночными, их классификацией, данными по ископаемым и их эво- люциеи.
Существует пять основных классов позвоночных: рыбы, амфибии, рептилии, птицы и млекопитающие. (Рыбы в действительности образуют несколько классов, но большинство пресноводных рыб относится к клас- су Osteichthyes.) Каждый основной класс рассматривается в самостоятель- ной главе (гл. 2 — 6). Каждая глава начинается с краткого обзора класси- фикации ископаемых данных и соответствующей характеристики рас- сматриваемых животных, а затем приводится описание и обсуждение их распространения по следующим рубрикам:
Границы распространения
Зональное распространение
Радиальное распространение
Региональное распространение
Переселение и преграды распространению
Доминирование и конкуренция в связи с распространением Заключение: Особенности распространения
История расселения
Общая последовательность каждой главы следующая: описание совре- менного распространения в зависимости от климата, преград и т. д., далее идет изложение значения доминирования и конкуренции (а также эволю- ции) для распространения и, наконец, обзор основных особенностей рас- пространения и значение переселения, определившего эти особенности. Некоторые вопросы рассматриваются под отдельными заголовками, на- пример: «Что такое пресноводные рыбы?» или «Влияние географических факторов на миграции птиц». После каждой главы дается общая класси- фикация и список семейств с распространением каждого семейства. Семей- ство как систематическая единица лучше всего показывает распространение позвоночных на всем земном шаре. В списках семейств тщательно собран весь фактический материал, на котором построена книга, включены все существующие семейства наземных и пресноводных позвоночных, а отно- сительно ископаемых форм сделаны замечания в начале каждого списка.
Распространение животных — явление сложное, трудно поддающе- еся простому описанию. Многие подробности географического распростра- нения даже позвоночных неизвестны, а те, что известны, не всегда досто- верны из-за сомнительной классификации или из-за сомнений в том, что данные животные являются действительно местными на данной террито- рии, а не интродуцированными человеком. Я приложил все усилия, чтобы по возможности проще описать случаи сложных распространении, и был оеспристрастен в сомнительных местах.
Глава 1
Расселение
В зоогеографии расселение можно рассматривать с двух точек зре- ния. В одном случае речь может идти о групповом расселении, которое связано с потерей одной территории и занятием другой. Из этих двух яв- лений и складывается история расселения. В более узком смысле расселе- ние — это перемещение отдельных особей (яиц, молодых или взрослых животных), особенно их проникновение в новые места за пределы ареала.
11ри этом следует учитывать два основных факта: существует сплошное рас- селение по территориям и расселение через преграды.
Все позвоночные могут расселяться довольно быстро. Если бы благо- приятные для обитания места были непрерывными, то есть если бы не существовало препятствий или преград и т. д., то животные распростра- нились бы по всему земному шару за короткое время(большинство из них за небольшой промежуток времени в миллион лет). Таким образом, скорость расселения и расстояние сами по себе не играют существенной ро- ли в распространении позвоночных. В основном оно зависит от других причин, особенно от экологических и физических преград.
Вопрос о расселении животных через физические преграды был пред- метом горячих дискуссий зоогеографов. Часто ход рассуждений был сле- дующий: сначала формулировали закон, затем находили несколько случа- ев, которые, по-видимому, подтверждали этот закон, а затем утверждали, что он применим ко всем случаям. Так поступил даже Уоллес (Wallace, 1880, р. 235), который утверждал, что амфибии и наземные млекопитающие никогда (или едва ли когда-либо?) не достигают океанических островов че- рез преграды, которые создают соленые воды, и что, следовательно, ост- рова, на которых встречаются эти животные, должны быть материкового происхождения. Я считаю, что это самая большая ошибка, допущенная Уоллесом как зоогеографом. Но я не хочу начинать с формулировки за- кона, а хочу выяснить, как в действительности преграды влияют на рас- селение животных, а затем уже делать какие-то выводы. Однако сначала необходимо кратко остановиться на двух факторах расселения, которые способствуют наземным животным в преодолении преград: «плотах» и ветре.
«Плоты» с точки зрения зоогеографии — это расселение наземных жи- вотных по воде на плавучих предметах. Эти предметы могут быть более крупными, чем предполагает слово «плот». Когда разливающаяся река подмывает берега, обрушивает целые склоны холмов и несет выкорчеван- ные и сплетенные между собой деревья в море, то на пути могут быть зах- вачены также и многие наземные животные. Чаще всего это происходит во влажных тропиках, где ливни обычны и где имеются очень крутые, не сглаженные последним оледенением склоны холмов.
Пример образования более прочных плотов в пресных водах дал Брю- стер (Brewster, 1924, р. 190, 191), описывая гнездовья американской выпи на озере Амбейгог в Мэке и Нью-Гэмпшире. Он пишет:
«...мы набрели на плавучий остров... круглый по форме, примерно 20 ярдов в по- перечнике, покрытый частым кустарником... то там, то здесь виднелись комья гря:ц| или пучки травы. Почти в середине острова на высоту 15 или 20 футов вздымались несколько мощных молодых зеленых лиственниц. Сплетенные корни деревьев, кустар- ников и травы смешивались с гниющими растительными остатками, аллювиальными почвами и сплавным лесом, образуя огромный сырой плот, довольно плавучий, чтобы подниматься и опускаться по мере колебания воды в озере, но такой тонкий и гибкий, что большие волны, проходящие под ним, вызывали очеиь заметные волнообразные движения на его поверхности. Он был достаточно приподнят и казался относительно
Введепие
«ух~1м, пока на него не ступили, и, подобно трясине в болоте, он опустился под нашими ~~о1 ами, а вода дошла до половины колен. В то время остров был крепко прикреплен. «озможно несколькими сильными корнями, к твердой почве, находящейся примерно ;1 — 6 футами ниже, по нам сказали, что незадолго до этого он прибился сюда, плывя через за.~ивные луга от их западной части. Примерно 10 лет спустя остров снова сор- «ялся со своего якоря и пропутешествовал еще дальше на восток, пока не был оста- новлен группой пней».
В 1892 г. плот, очень похожий на описанньш Брюстером, площадью 100 футов с деревьями высотой до 30 футов, очевидно, прочно скреплен- ный корнями произрастающих на нем растений, был обнаружен близ ое- рега Северной Америки в Атлантическом океане. Как стало известно, он был отнесен и проделал путь по крайней мере в 1000 миль (Powers, 1911). Такие плоты могут образоваться и уплыть в море в любом месте земного шара. где сток из мелких озер или болот в море происходит по небольшим со спокойным течением рекам. И так как эти плоты отрываются, скорее всего, во время паводка, они, вероятно, уносят наземных животных, спа- сающихся от половодья. Мне кажется, что такие плоты могут унести поч- ти любое из наземных животных на любое расстояние, в случае если океа- нические течения, ветры и климат окажутся благоприятными.
Издавна ведущиеся дискуссии относительно плотов в основном сво- дились к вопросу, возможно ли перенесение наземных животных на пло- тах, но дело не в этом. Следует ставить другой вопрос: указывает ли пре- бывание животных, особенно на островах, на то, что эти животные дейст- вительно были перенесены на плотах? И если это так, то соответствующие плоты должны были существовать в прошлом так же, как и теперь.
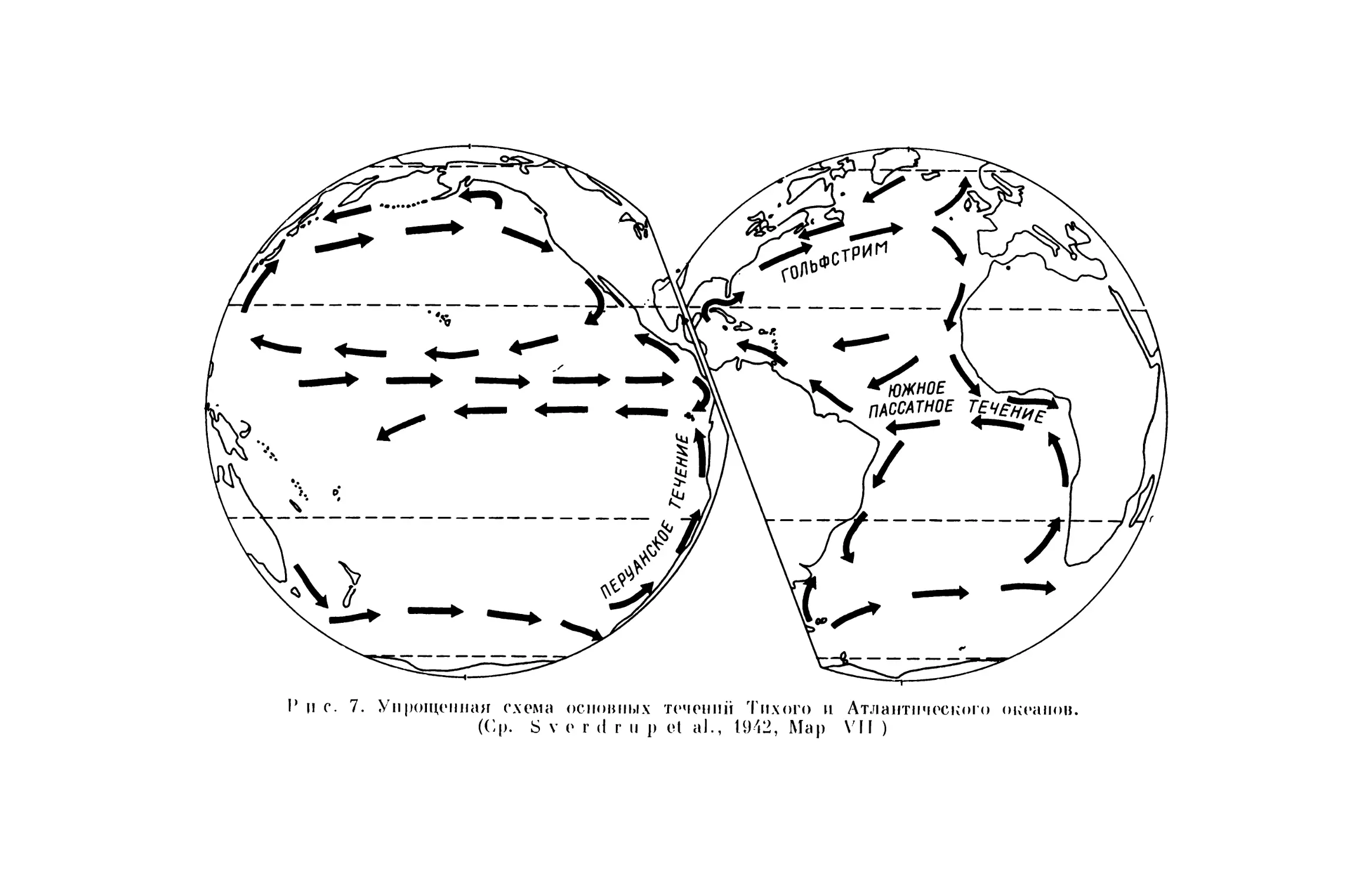
Направление движения плотов, по-видимому, отчасти зависит от те- чений океана (рис. 7). Особое внимание следует обратить на Южное пас- сатное течение Атлантического океана. Это межконтинентальное течение, которое пересекает тропическую Атлантику от Западной Африки к Юж- ной Америке. Гуппи (Guppy, 1917, р. 80), используя данные меченых дрейфующих бутылок, вычислил, что на путь из Гвинейского залива к Бра- зилии потребуется только около 12 недель. Течение несет множество пла- вучих предметов; некоторые из них выносятся из рек Нигера и Конго. Одни плавучие предметы попадают на берега Бразилии, а другие, после продолжительного плавания,— на берега Вест-Индии. Это течение имеет важное значение, так как независимо от того, приносило ли оно наземных животных или нет, оно могло принести личинки мелководных морских беспозвоночных. Шухерт (Schuchert, 1932) рассмотрел эту воз- можность и отклонил ее, но цифры, из которых он исходил, о дистанции и скорости дрейфа, по-видимому, были неправильны. Он принял расстоя- ние в 4000 миль, а оно может быть значительно меньше; самое короткое расстояние от Африки до Южной Америки менее 2000 миль. Если время. необходимое для прохождения этого расстояния (12 недель), вычисленное Гуппи, и длительность личиночной стадии (в некоторых случаях до 3 ме- сяцев), вычисленная Шухертом, правильны, в чем я не совсем уверен, то личинки некоторых морских беспозвоночных могли быть принесены Южным пассатным течением из Африки к Южной Америке.
Другим фактором расселения является ветер. Человек часто недооце- нивает силу ветра. Подобно тому как когда-то люди с трудом верили в то. что машины, более тяжелые, чем воздух, могут летать, так и сегодня неко- торые зоологи с трудом верят в то, что ветры и восходящие воздушные токи могут поднимать и нести живые организмы. Это объясняется тем, что суждения человека об этом явлении, как правило, основаны на знаком- стве с более умеренными ветрами, дующими горизонтально у поверх-
)
)> с. 7. Уирощ«инли с'кс~мл оси~виы~ та ~~агний Тихого и Атлантического ~~иж~ии~цз. (('.р. S ~ < r ~l г и р И 'il., 1М2, Мар %' II )
Введение
ности моря. Действие ветра на мелких животных зависит от соотношений поверхности тела животного и его веса. Площадь поверхности изменяется пропорционально квадрату, а вес — пропорционально кубу. Очень мел- кие животные имеют во много раз большую поверхность тела по отношению к своему весу, чем крупные, и поэтому в большей мере испытывают воз- действие ветра. Кроме того, давление и подъемная сила ветра увеличивают- ся пропорционально квадрату скорости ветра. Можно подсчитать, что воз- действие ветра со скоростью 100 миль/час на животное весом в 1 унцию (взрослая домовая мышь весит примерно 1 унцию) приблизительно в 224 ра- за (14 х 16) сильнее, чем воздействие ветра скоростью 25 миль на человека; или что влияние ветра скоростью 100 миль/час на животное длиной 1 дюйм (например, очень маленькая лягушка) в 1040 раз (65 X 16) сильнее, чем влияние ветра скоростью 25 миль/час на человека. Это грубо приближен- ные цифры, но они все же помогают составить представление о том, как сильные ветры могут воздействовать на небольших животных. Если жи- вотное, подхваченное ветром, проносится над землей, оно входит в зону обычных восходящих воздушных течений. Все эти вопросы более подроб- но рассматриваются в другой работе (Darlington, 1938). Ветер обладает силой переносить мелких животных независимо от того, летают они или нет; вопрос только в том, как часто и насколько далеко он действительно переносит их.
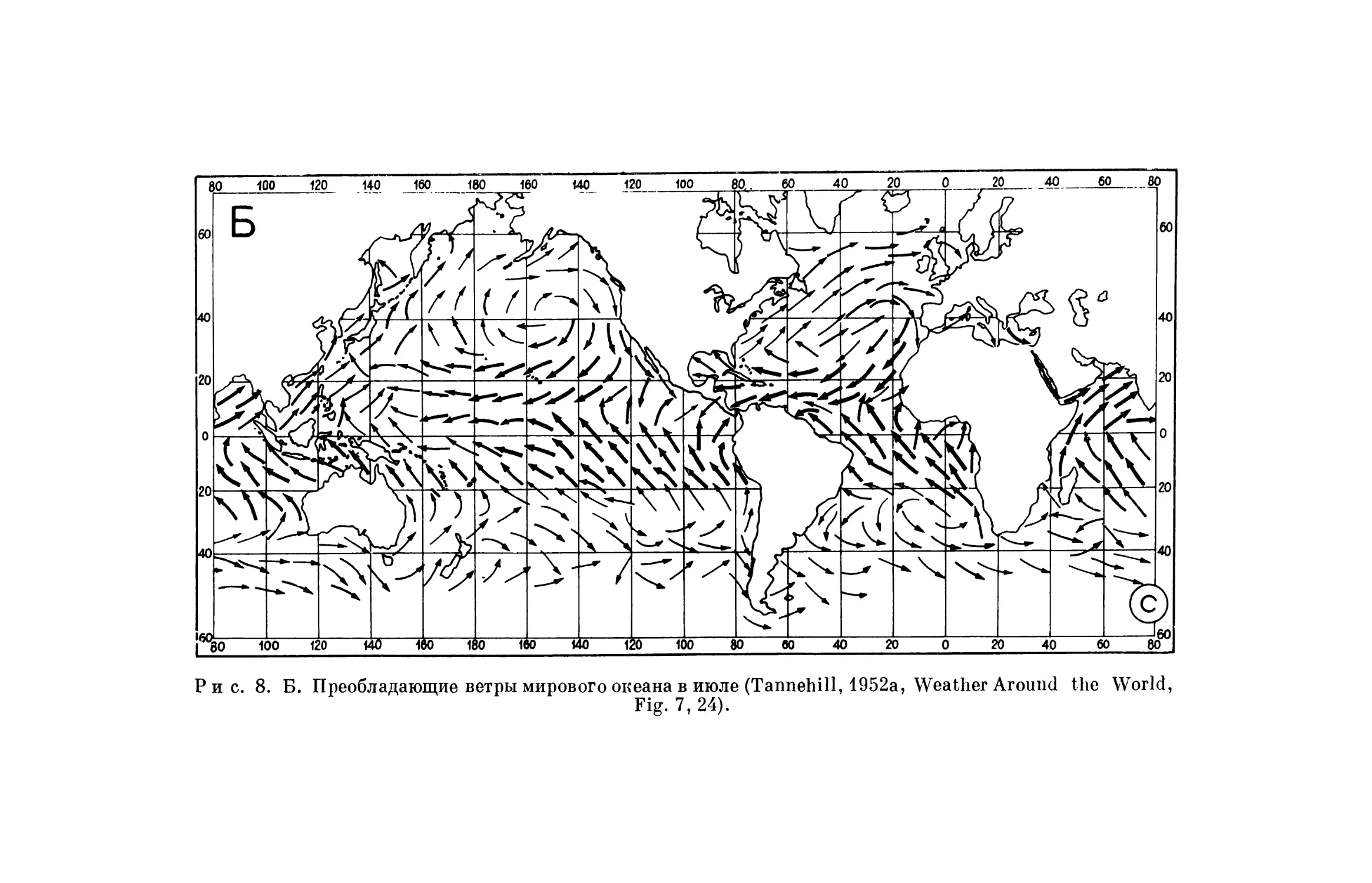
Система господствующих ветров земного шара представлена на рис. 8, но это только ветры, наблюдающиеся вблизи земной поверхности. Выше господствуют другие ветры, дующие в различных направлениях и с боль- шей силой, и эти ветры также могут уносить мелкие организмы, способные переносить холод. Такие организмы, очевидно, могут переноситься вет- рами на том или ином уровне почти на всем земном шаре. Кроме основной системы ветров, наблюдаются местные ветры, которые могут оказывать особое влияние. Торнадо и грозы, наблюдающиеся во многих частях зем- ного шара, порождают сильные восходящие движения воздуха, которые могут переносить небольшие предметы высоко над землей. Ураганы,проно- сящиеся над определенными частями Атлантического, Тихого и Индий- ского океанов (рис. 8), могут являться важным фактором в расселении жи- вотных. Эти ураганы представляют собой колоссальные вихри, движимые восходящими токами воздуха, которые могут подхватывать большое коли- чество легких предметов и переносить их на дальнее расстояние (Раг1и1 ton, 1938). Не думаю, что ветры сыграли большую роль в расселении поз- воночных, но они могли быть важным фактором в расселении более мел- ких животных — насекомых, моллюсков, а также растений. В частности, в Антарктике ветер может быть одним из возможных объяснений расселе- ния организмов.
История зоогеографии
Капитальная история зоогеографии — история взглядов человека на географическое распространение животных еще не написана, но Гадов (Gadow, 1913) дает по ней весьма полезную сводку. Я же попытаюсь дать здесь только краткий обзор.
Представления о географической истории животных в древние вре- мена были весьма примитивными. Одно из них — библейская легенда о Ноевом ковчеге, легенда о перерасселении человека и животных, как ска- зали бы современные зоогеографы, из рефугиума.
Р и с. 8 Л. Лреобладающие направления тропических ураганов.
Р и с. 8. Б. Преобладающие ветры мирового океана в июле (Tannehill, 1952а, Weather Around 1Ье World,Fig. 7, 24).
Незадолго до Дарвина столь же надуманные представления о распро- странении животных поддерживались Бюффоном и многими другими ав- торами, из которых более десяти упомянуты в работе Гадова ((adow, 1913). Некоторые из них пытались найти «центр мироздания». Такую же цель преследовал и Склетер. В своей работе, написанной в 1857 г. (опуб- ликована в 1858 г.), за два года до выхода в свет «Происхождения видов» Дарвина, он разделил земной шар в соответствии с распространением птиц на 6 областей. И все же Склетер проанализировал распространение птиц настолько хорошо, что его деление на области с небольшими изменениями сохранилось и до наших дней.
Для Дарвина и Уоллеса особенности распространения животных в не затронутых деятельностью человека частях земного шара явились одним из доказательств эволюции, а признание идеи эволюции дало тол- чок для развития современной зоогеографии. Дарвин придерживался зво- люционной точки зрения. Его две главы в «Происхождении видов» о гео- графическом распространении животных до сих пор являются одной из лучших трактовок эволюционных принципов зоогеографии. Уоллес ка- сался больше распространения современных животных. В 1876 г. он сде- лал первую и единственную попытку полно и детально показать в своей классической сводке распространение современных животных. Эти два великих, каждый в своем роде, ученых представляли две точки зрения, которые продолжают существовать среди последующих поколений зоо- географов.
После Уоллеса несколько поколений зоогеографов работало над дал ь- нейшим накоплением фактов по распространению современных животных и занималось разделением земного шара на фаунистические области. Для некоторых из них линия Уоллеса — предполагаемая граница между дву- мя фаунистическими областями — была самым главным вопросом. Одна ко нельзя умалять значение этих работ, которые в свое время обогатили зоо- географию немалым количеством фактического материала, столь необхо- димого для ее развития. Несколько подробнее этот вопрос будет рассмот- рен в гл. 7.
В то же самое время другая группа зоогеографов, можно сказать, следовала за Дарвином, очень часто не понимая его. Они пытались решить вопросы географической истории животных и для этого передвигали континенты и строили длинные материковые мосты, мало заботясь о дока- зательствах или даже вероятности этих явлений. Я не собираюсь недооце- нивать роли перемещения материков или существования мостов, если это- го требуют обстоятельства, но не следует прибегать к этим теориям оез оснований. Дарвин возмущался людьми, которые строили сухопутные мосты с такой же легкостью, с какой повар печет блины. Гадов допускал эту же ошибку, передвигая континенты без всякой причины и неооходи- мости, а Шарф, Арлдт и другие ученые совершали еще более грубые ошиб- ки в этом вопросе. Книги последних двух авторов были написаны без кри- тического подхода и изобиловали ошибками и предположениями, выда- ваемыми за факты. Эти работы и сейчас еще иногда цитируются как источ- ники «фактов» зоогеографами, которые не знают, насколько эти книги не- надежны. Мэтью (Matthew, 1915) в своей книге «(;limate and Evol»ti(ii»> проделал большую работу, чтобы доказать несостоятельность теорий дан- ных авторов. Независимо от правильности его выводов он подал хороший пример, представив доказательства и сделав попытку восстановить историю расселения животных и историю материков при помощи этих доказательств. (:интез такого рода — соединение, так сказать.
a7 (]
Введение
теорий Уоллеса и Дарвина с привлечением геологии и других наук в разумных пропорциях — должен быть целью современных зоогео- графов.
Не так давно среди зоогеографов существовало два не совсем оправ- данных направления. Одно из них пыталось сделать зоогеографию разде- .лом экологии. Хорошо, когда вводят экологию в зоогеографию, но зоо- география не есть экология. Другое возвеличивало историческую зоогео- графию — эволюцию и передвижение животных в прошлом — почти с полным пренебрежением к изучению современного «статического» распро- странения. Мне кажется, что это большая ошибка. Справедливо, что со- временное распространение животных невозможно понять, не обращаясь к прошлому, но в такой же степени справедливо и то, что прошлое не мо- жет быть понято без настоящего. Именно современный характер расселе- ния животных показывает, как в деиствительности оно связано с клима- том, преградами и т. д. Без знания этого невозможно понять, что говорит историческое распространение животных о климатах и преградах в прош- лом? Мне кажется, что в зоогеографии и настоящее и прошлое в равной сте- пени важны и зависят друг от друга, о чем мне еще придется упомянуть в гл. ).
11оследние книги по зоогеографии (на английском языке): «Plant and Animal Geography» (Newbigin, 1936); «Zoogeography of the Еапй and Inland ЖаСегз» (Beaufort, 1951) — слишком сжаты, чтобы исчерпываю- ще осветить предмет, и в основном являются компиляцией общепринятых понятий, а не пересмотром предмета в целом.
Большинство зоогеографов занималось определенными группами жи- вотных, или определенными частями земного шара, или определенными ситуациями, но в результате их объединенных усилий были охвачены исследованиями все важные стороны распространения животных.
Зоогеографпческпе понятия
;-)то не словарь незнакомых слов. Я старался их избегать, а те, кото- рые я употреблял, могут быть найдены в словарях или поняты из текста. Это в основном список знакомых слов, которые могут быть поняты непра- вильно. Я привел их здесь, потому что читатель может не знать, что эти слова нуждаются в определении, и может не посмотреть их в специальном словаре.
Автохтонный. Местный в смысле происхождения (развития) в рас- сматриваемом месте.
Преграда. (1) Физические препятствия, такие, как горные хребты, которые отдельные животные не могут преодолеть или просто не преодо- левают. (2) Климатические или экологические факторы, такие, как тем- пература или осадки, которые изменяются постепенно от места к месту и не препятствуют перемещению отдельных особей, но мешают успешному обоснованию и распространению видов.
Конкуренция. Борьба за место на земном шаре, борьба за существо- вание. В этом смысле конкуренция — это любые взаимоотношения ме- жду животными независимо от того, насколько сложны или опосредство- ванны они могут быть, независимо от того, являются ли они неблагопри- ятными или могут оказаться неблагоприятными для любого из них. Неко- торые зоологи употребляют слово «конкуренция» в узком смысле, и они могут возражать против такого толкования, но я не смог найти другого
1'.гава 1
более подходящего слова; кроме того, данный термин уже применялся (начиная с Дарвина) моими предшественниками.
Комплиментарный' (complementary). Означает встречающийся в раз- личных областях обитания, что вместе составляет весь ареал. Здесь этот термин применяется к доминирующим, экологически эквивалентным, по- видимому, конкурирующим группам животных, которые взаимно допол- няют друг друга. Эту географическую взаимосвязь я назвал комгглимен- тарной. Последнего слова в словарях нет, но я не нашел иного слова, что- оы выразить это значение. Я думаю, что это единственное слово, за иск- лючением одного-двух имен собственных, которые я дал здесь от себя.
Производный (дериват). Происшедший от чего-то другого; сравни- тельно поздний в эволюционной последовательности. Противоположное значение этого слова — примитивный.
Дифференциация. (1) Процесс возникновения различий. (2) Разница между различными животными или различными фаунами, происшедшая в результате эволюции или других процессов. Это обычное значение дан- ного слова в английском языке, применяемое в зоогеографии. Оно имеет другие специальные значения в других областях науки.
Расселение. (1) Географическое переселение групп животных в целом. (2) Перемещение отдельных особей (яиц, молодых или взрослых особей), особенно их распространение за пределами ареала.
Доминирующий. Заметно преуспевающие формы, успешно конку- рирующие (см. стр. 464). Это обычное значение данного слова в англий- ском языке, но оно имеет другое, специальное значение в генетике.
Эндемик. Ограниченный, не встречающийся нигде больше, кроме то- го места, о котором идет речь. Это слово имеет и другие значения, и его следует употреблять с осторожностью. Я бы совсем не стал его употреб- лять, за исключением очень нужных производных от него слов — энде- мизм, существование эндемических форм.
Дикий. Вторично одичавший после одомашнивания, подразумевает- ся не местны~~.
Ингиродуцированный. Введенный в фауну человеком преднамеренно или случайно.
Миграция. (1) Непериодические перемещения в определенном на- правлении (см. стр. 463). (2) Периодические сезонные кочевки, как. на- пример, у птиц (стр. 211).
Мес пный. Обитающий в естественных условиях, не интродуцирован- ньш человеком. Общий термин, включающий в себя и специальные: анде- личный, автохтонный, реликтовый. Синоним: аооригенный.
Примитивный. Первоначальный: ранний в данной эволюционной последовательности. Противоположное значение — деривапгный.
Радиацггя. (1) Возникновение многих различных и различно приспо- собившихся к среде форм от одного предка. (2) Расселение во многих на- правлениях из одного центра.
Ареал. Определенная территория, занимаемая группой животных. т1асто на картах ареалы изобрая'аются так, словно у них есть установлен- ные постоянные границы, или монолитными «блоками». Зоогеографы до- пускают ошибку, считая, что ареал, таким образом, можно переносить
' Данный термин, не встречающийся в советской зоогеографической литера- туре. очень схематизпрует явления. Поэтому далее в зависимости от контекста мы оу;~ем употреблять: взаимодополняющие, взаимопсключающие или викарпиру ющие г руп пы. — Прилс. ред.
Введение
с места, двигать и соединять вместе, подобно строительным блокам. Аре- алы — это площади, населенные живущими популяциями и они так же сложны, нестабильны и зависят от особенностей окружающей их среды, как и живущие на них популяции.
Релитп. Выжившии: растение или животное, которое (1) продолжа- ет существовать в данном месте после его вымирания или вымирания род- ственных ему форм в других местах (географический реликт) или (2) про- дочжает существовать после вымирания большей части группы (эволю- ционный, или филогенетический реликт). Употребляется также и в каче- стве прилагательного.
Специализированный. Приспособившийся к особым, ограниченным условиям жизни. Противоположное значение этого слова — неспеииали- зированный, но не примитивный. Хеспециализированный и специализи- рованный показывают отношение животного к окружающей среде. При- мшпивный и дериватный рассматривают филогенетическое взаимоотно- шение животных друг к другу. Неспециализированное 'гкивотное мог!ъет оыть дериватным, а специализированное — примитивным.
Неспециплизированный. См. спеииализированный.
Рабочие прпнципы
Ученые, изучающие географию животных и растений от Уоллеса (Wallace, 1876) до 'Кейна (Cain, 1944), пытались сформулировать свои рабочие принципы, но мне кажется, что им это не совсем удалось сде- лать, отчасти потому, что они обычно смешивали два различных вида прин- ципов. Методика работы и первоначальные предпосылки — это рабочие принципы, которые должны быть установлены до начала работы. Общие факты, выявленные в процессе работы,— это производные принципы, и их не следует окончательно формулировать до тех пор, пока работа не закончена. Например, рабочим принципом зоогеографии является или должен быть установлен следующий: животные постоянно имеют тенден- цию к размножению и расселению, умирают и теряют территорию. Невни- мание к этому рабочему принципу приводит к необоснованности выво- дов некоторых зоогеографов. С другой стороны, при зоогеографическом исследовании следует в начале работы выяснить влияние преград на рас- пространение животных, а в конце изложить установленные факты в ка- честве производных принципов. Но если принять уже как исходное поло- жение, что горы и пустыни обязательно являются важными преградами, это значит не только преувеличить их роль, но и не суметь правильно по- нять значение этих факторов в дальнейшем.
Рискуя показаться педантом, H все iree xo Iy предложить ррН рассмот- рения те рабочие принципы, которые, на мой взгляд, особенно важны для зоогеографии.
1. Сформулировагпь рабочие приниипы до начала раба пы. Зоогеогра- фия — предмет сложный, имеющий много трудностей, поэтому надо под- ходить к нему, продумав все заранее. Кроме того, читатели зоогеографии должны знать рабочие принципы автора, чтобы дать оценку его выводам.
2. По возможноспи работагпь с фактами, и не с мнениям. и и умение понять природу этих «фактов». Майерс (Myers, 1938. р. 340) замечает, что, «вообще говоря, зоогеографы делятся на три группы: на тех, которые строят мосты, тех, которые их не строят, и тех, кто является сторонника- ми перемещения материков». Это утверждение совершенно справедливо,
1'л ава
и к нему следует еще добавить, что каждая сторона упорно отстаивает свое мнение. Должна быть еще четвертая группа, и я хотел бы присоединить- ся к ней: это зоогеографы, которые берут факты, тщательно их анализи- руют и, вместо того чтобы защищать предвзятое мнение, стараются най- ти наиболее приемлемое решение.
Использование чужих мнений по вызывающим сомнение вопросам— не совсем хорошая основа для зоогеографии. Если зоогеограф опирается на авторитет мнений, это означает, что у него не хватает фактов или что он ошибается. Например, Шухерт в 1935 г. сопоставил мнения 14 квали- фицированных специалистов и нашел, что большинство из них, 13 против одного, склоняются к теории промежуточных материков и происхожде- нию животных и растений Вест-Индии на этих материках. Сейчас боль- шинство зоогеографов, изучающих животный мир Вест-Индии, выступает против этой теории. Независимо от решения этого вопроса, рассматривае- мого в гл. 8, большинство зоогеографов и в 1935 г. и в более позднее вре- мя очень сильно ошибалось. Еще раз подчеркиваю, что взгляды ученого всегда должны основываться на фактах, а не на мнениях других ученых, и никогда не следует отстаивать ошибочное мнение слишком упорно.
Сырой материал зоогеографии — это факты — сведения о том, где встре- чаются какие животные. Однако в «факты» иногда могут вкрасться ошиб- ки. Например, Огилби в 1905 г., описывая каймановую черепаху из Новой Гвинеи, отнес ее к новому роду. И более чем на протяжении 40 лет зоогео- графы считали ее нахождение там фактом. Но больше не было найдено ни одного экземпляра этого вида, а в 1947 г. Лавридж и Шрив доказали, что эта черепаха не из Новой Гвинеи и что это не новый род, а неправильно этикетированный экземпляр черепахи из Северной Америки. Итак, в кон- це концов, оказалось, что никакой каймановой черепахи в Новой Гвинее нет. Нетрудно представить, как могла произойти эта ошибка.Огилби дей- ствительно получил партию черепах из Новой Гвинеи. Возможно, груз прибыл вечером и Огилби, распаковав образцы и разложив их на сто- ле без этикеток для предварительного осмотра, положил старое чучело американской черепахи на пол, чтобы оно не мешало. Возможно, сторож. нашедший его ночью, подумал: «Опять этот рассеянный хранитель музея!»— и положил ее снова на стол к новогвинейским экземплярам, с которы- ми она и была этикетирована утром. Все это лишь плод моего воображе- ния, но из-за подобных ошибок в музеях имеются тысячи неправильно этикетированных экземпляров различных видов животных среди миллио- нов с правильными этикетками. Большинство ошибок не причиняет oco- ()hIx неприятностей, так как они или слишком очевидны, или слишком не- значительны, а большинство действительно важных находок животных в неожиданных для них местах обитания подтверждено повторными сбора- ми этих экземпляров.
Значительная трудность заключается в том, что мы еще недостаточно хорошо знаем взаимоотношения некоторых групп животных, чтобы отли- чить подлинные факты. Вот еще один наглядный пример, касающийся но- возеландской лягушки (1егоре1та; в действительности только два или три близкородственных их вида приурочены к Новой Зеландии), который должен заставить задуматься любого мыслящего зоогеографа. Сам факт. что какая-либо лягушка могла появиться в Новой Зеландии, кажется невероятным. До того как этот факт был точно установлен, Дарвин (Dar- win, 1859, 1950, р. 333) слышал о ее нахождении там и усомнился в этом. но все оказалось верным: лягушка существовала в Новой Зеландии. В 1861 г. Фитцингер описал ее внешне как наиболее близко родствен-
Иведенue
ную квакше, но в действительности это не родственные виды. Более осто- рожный систематик должен был бы говорить о сходстве, а не о родстве. Систематики (в том числе и я) слишком часто говорят «родственный» тогда, когда нужно говорить «сходный с...». В 1876 г. Уоллес, очевидно, следуя Штейндахнеру, отнес новозеландскую лягушку к семейству Вот- buratoridae, которое, по его предположению, встречается только в Европе и в умеренных широтах Южной Америки. Но оказалось, что это семей- ство было создано искусственно, так как отнесенные к нему разные лягушки фактически не были родственны друг другу. В 1911 г. Уоллес, вероятно следуя более ранней классификации Буланже, отнес новозе- ландскую лягушку к семейству Discoglossidae, в то время известному только в Европе и Китае, но и это также не было настоящим родством. В 1910 г. Буланже отнес эту лягушку, так же как и Меткалф в 1923 г., к семейству Leptodactylidae. С географической точки зрения, видимо, бо- ;le( удачно, так как свистуны многочисленны в Австралии и Южной Аме- рике, но его определение также не было верным. В 1925 г. Данн поправил ~11еткалфа, уверенно заявив, что новозеландская лягушка относится к круглоязычным Discoglossidae, но, как я уже сказал, это неверно. В конце концов, в 1931 г. Нобль после тщательного изучения анатомии лягушки пришел к заключению, что род Ieiopelma не принадлежит ни к одному из тех семейств, к которым его относили, и что это почти уникально примитивный род из двух самых примитивных родов суще- ствующих лягушек. Другои 'род, Агсарйиг [то же семейство.— Ред.~, открытый в 1889 г'.', встречается почти на другой стороне земного шара, н северо-западной части Северной Америки. Такая связь оказалась с гео- графической точки зрения еще более удивительной, чем любые из указан- ных вьипе, но, видимо, это деиствительно так.
Могут спросить, можно ли после всего этого считать положение ново- зеландской лягушки установленным в классификации и принимать за факт. Ответ таков — до времен Нобля накапливались знания по анатомии лягушек и совершенствовалась их классификация, что все время способ- ствовало уточнению положения рода Leiopelma. После работы Нобля но- вые факты, обнаруженные при изучении родов Leiopelma, Аscaphus и лягушек вообще, только подтвердили его выводы. Вероятно, в подоб- ном случае не может быть еще окончательного решения, но многие извест- ные теперь детали позволяют предположить, что роды ~.еюреЬаа и As- capllus хотя и не прямо родственны, но являются единственно уцелевши- ми родами древней группы лягушек, и поэтому их обычно относят вместе к одному семейству 1 eil>pelmidae.
Vi сожалению, родство многих животных, включая прочих лягушек, многих змей, некоторых птиц и млекопитающих (особенно некоторых пев- чих птиц и грызунов) и многих других беспозвоночных, также до сих пор еще не установлено, как и у рода Leiopelina во времена Уоллеса. До тех ш>р пока систематики не усовершенствуют классификацию, зоогеографы будут испытывать большие трудности. Даже работая с наиболее хорошо известными группами животных, зоогеографы должны помнить, что клас- сифнка1~ия несовершенна и пользоваться ею надо с осторожностью. Опыт систематика очень полезен и почти всегда необходим для зоогеогра- фа. Опытный систематик знает, какие существуют классификации и что другие систематики имеют в виду, когда говорят о родстве. Опытный си- «тематик может иногда предполагать необоснованную классификацию да- 'к11е ' 'ех ЖИВОТНЫХ hoTOpbIX он не знает, так же как Инженер может подо- зревать ошибки в конструкции моста, который cлм он не строил.
Гiaea 1
Но все это не совсем ясная сторона вопроса. Основное заключается в том, что имеющиеся в настоящее время сведения по географическим на- хождениям животных и классификации последних в достаточной степени хороши для выводов. Даже 80 лет назад, когда фактического материала было известно меньше и систематика была более примитивная, Уоллес смог, однако, сделать обоснованные общие выводы о географической встре- чаемости животных. Сегодня мы располагаем по крайней мере таким фак- тическим материалом, который позволяет нам сделать это лучше, чем он.
3. Определить и ограничить работу, котпорая должна быть сделана, и фактический материал, с которым придется работать. Необходимость установления рамок для исследования такого обширного предмета, как зоо- география, уже обсуждалась. Например, эта книга по своей цели ограни- чивается рассмотрением основных особенностей распространения нсивот- ных, а по своему материалу — наземными и пресноводными позвоночными, особенно их семействами.
4. Представишь подобранный материа.г полное пью и гпщательно от- работанным. Это зависит от опыта и желания самого автора. Таблицы, схемы и текст должны отвечать своему назначению. Контурные карты для ясности и легкости сравнения должны быть стандартными. Все рисунки должны быть составлены так, чтобы представить факты как можно полнее и нагляднее.
5. Постоянно помнить, что животпные — это живые существа, кото- рые в одних местах постоянно развиваются и размножаются, в других- расселяются, а в третьих — вымирают, создавая, таким образом, новые фаунистические комплексы. Все зоогеографы знают это, но не все прини- мают к сведению.
Распространение животных — результат сложных равновесии. В ос- нове его лежит равновесие между размножением и смертностью отдельных особей. В устойчивой популяции воспроизводство и смертность сбаланси- рованы противоположно направленными, но равными силами. Никто не скажет, что в этом равновесии важно только воспроизводство, а смертность можно игнорировать. Изменения как в степени воспроизводства, так и в степени смертности могут изменять равновесие и сдвигать его с одинако- вым успехом в любом направлении: либо к увеличению размеров попу- ляции, либо к уменьшению. Увеличение и уменьшение размеров популяции определяются занятием территории или потерей ее у вида в целом. А заня- тие территории или потеря ее многими видами приводят или к расселению, или соответственно к вымиранию родов, семейств, отрядов и целых фаун. Равновесие в конечном счете зависит от равновесия между воспро- изводством и смертностью популяции, и вся сложная система может быть сдвинута с одинаковым успехом в любом направлении: или к расселе- нию, или к сокращению даже основных групп и фаун (см. гл. 9).
Излагая «принципы» работы, зоогеографы почти всегда подчеркивают способность животных к размножению и расселению, но часто умаляют значение, а иногда просто забывают об их подверженности смертности и потерям территории. Это серьезная ошибка. Дарвин (Darwin, 1859, р. 95— 97) — один из очень немногих зоогеографов, тщательно продумавших этот вопрос. Я упоминаю о нем здесь не для того, чтобы сослаться на авто- ритетное мнение, а лишь потому, что его утверждения абсолютно верны. Он считал, что вымирание — это обычное явление, что эволюция новых форм и исчезновение старых имеют тенденцию к уравновешиванию друг друга, что есть предел числу видов животных, которые могут существо- вать, и что по мере того, как новые виды развиваются, старые должны
Введение
встречаться реже и наконец исчезать. Его рассуждения более подробны, чем те, что я привожу здесь, и заслуживают, чтобы их внимательно про- читали.
Я уже говорил об этом равновесии на уровне семейств и фаун, а не видов (Darlington, 1948, р. 1, 2). В процессе эволюции животных на Зем- ле многие фауны сменяли друг друга. Последовательность смены фаун влекла за собой, с одной стороны, эволюцию, радиацию и распространение новых доминирующих групп животных, а с другой — уменьшение и вы- мирание большинства видов ранее доминировавших групп. Как говорит Дарвин, эти два процесса взаимосвязаны. Существует какой-то предел общему числу видов на Земле. Оно, вероятно, увеличивается постепенно, по мере того как животные вступают в действительно новую среду или на- чинают вести новый образ жизни, но общее увеличение — всего лишь малая толика в сумме происходящих перемещений. Когда старая фауна исче- зает или замещается новой, то, вероятно, в общем итоге происходят лишь небольшие изменения в окружающей жизни. Таким образом, видимо, по- явление новых видов почти уравновешивается вымиранием старых; каж- дой расселяющейся доминирующей группе животных противостоит старая группа, исчезающая или уменьшающаяся до нескольких реликтов.
Это предположение не может быть доказано, но оно может быть пояс- нено. Согласно Симпсону (Simpson, 1940, р. 158), к концу плиоцена, перед тем, как Северная Америка и Южная Америка были соединены пере- шейком, там насчитывалось соответственно около 27 и 29 семейств назем- ных млекопитающих. В то время за одним или двумя исключениями об- щих семейств там не было. После соединения обоих континентов произо- шел чрезвычайно широкий обмен млекопитающими. В плейстоцене они имели 22 общих семейства и продолжалось дальнейшее перемещение и исчезновение многих семейств. Теперь, после всех перемещений, в Север- ной Америке насчитывается только 23 семейства наземных млекопитаю- щих, а в Южной Америке — снова 29. Каждый континент приобрел одни семейства, но потерял другие, общее же число семейств на каждом мате- рике осталось примерно такое же, как и раньше.
Мне кажется, что возможность постоянного распространения на зем- ном шаре новых групп животных, без почти равного количества высе- ляющихся старых групп, не большая, чем возможность появления новых поколений постоянно нарождающихся животных без вымирания старых. Поэтому я считаю, что расселение и выселение почти в одинаковой степе- ни — обычные и важные явления. Можно утверждать, что эволюция и расселение — постепенные процессы, которые продолжаются в течение дли- тельного времени, а выселение и вымирание — быстро протекающие и быстро завершающиеся процессы. Таким образом, в любое рассматривае- мое время большинство групп животных расселяется и немногие выселя- н~тся, но это не совсем точно. Если вселение одних животных происходит за счет других, то оба процесса могут протекать в равной степени медленно.
Я разбираю этот вопрос подробно, потому что он весьма важен. Пере- оценка расселения и недооценка выселения животных привели к воздви- 'кению ненужных сухопутных мостов просто потому, что создатели этих мостов не смогли предположить, что некоторые животные могли исчез- нуть с обширных территорий земного шара. Я склонен считать, что высе- ление так же обычно, как и расселение, и что только тщательный анализ ситуаций и раскрытие этих процессов могут подсказать (если можно что- либо сказать), занимают ли конкретные группы животных территории или теряют их.
36
6. Суметь npnsuëüíî подобрать и умело использовтпь «ключи» к рас- ш,ифровке исаории ге«графии животных. Лучше всего это пояснить на примере семейства пресноводных рыб Catostomidae, к которому относятся чукучаны и буйволовы рыбы (рис. 9).
(:емейство СаФояФотЫае в настоящее время представлено главным об- разом в Северной Америке: около 18 родов; 8~~ вида встречается в Амери- ке к югу до Гватемалы. Только два вида этого семейства известны в дру- гих местах. Один иэ них относится к роду С(йозЬнпиз. Около 20 видов этого рода находится в Америке, и один из них, ареал которого проходит через северную часть Северной Америки от Мэка к арктической Аляске, встречается в качестве слегка отличающегося подвида в Восточной (;иби- ри. Другой не американский вид этого семейства с тремя подвидами встре- чается в Китае и сильно отличается от других видов, образуя свой со- вершенно отдельный род Myxocyprinus.
рыб семейств»
Р >r «. Ч. 1'ж'н р(1л р»нентхе пресн<~водных
(",Моя( о ш i чае.
Обычно зоогеографы пользуются числовым подходом, как я его назы- ваю «ключами чисел». Они находят место, где теперь встречается наиболь- шее число родов или видов данной группы животных, и принимают его за место происхождения этой группы. Поступая так, они предполагают, что обычно количество родов и видов увеличивается везде примерно в оди- наковой степени. Но это может быть верным только в том случае, если большинство животных численно увеличивается и расселяется на протя- жении довольно длительного времени. Однако если выселение и вымира- ние животных примерно происходит такими же темпами, как и количест- венное увеличение и расселение, а я думаю. что это так, то числовой под- ход, применяемый без должного основания, чаще всего приведет к непра- вильныи выводам. Согласно формально применяемому числовому подхо- д~, местом происхождения семейства (;atostomidae должна быть (;еверная Америка, но это, вероятно, не так. В ряде же других случаев числа могут служить наилучшим «ключом». Если группа животных на своей тер- ритории доминирует и если она оставила многочисленные ископаемые в пределах, но не за пределами настоящего ареала группы, или если име- н>тся другие доказательства, что эта группа является доминирующей, увеличивающейся и расселяющейся, то тогда число родов и видов в раз- ных местах будет играть важную роль. Например, у доминирующего рода Cetostornus число видов указывает на американское происхождение рода и на проникновение одного вида в (:ибирь независимо от истории всего с,еме~"тства в целом.
Другой «ключ» к истории географии животных — это степень диффе- ренциации. Различия между родами и видами и наиболыпий эндемизм должны проявляться и наибольшей степени там, где семейство находи-
лось более продолжительное время, чем там, где оно только что появилось. Этот «ключ» можно противопоставить числовому. У семейства 'atostomi- dae единственный вид рода Catostomus. находящийся в Восточной Сибири, лишь немного отличается от других видов, и это, учитывая и то, что 20 видов этого рода находятся в Америке и только один в Сибири, дает воз- можность предположить1 что происходило перемещение из Америки в Си- бирь. Но представитель этого семейства в Китае сильно отличается от других, и это указывает на то, что независимо от числа родов и видов он может быть реликтом, оставшимся с тех времен, когда чукучаны были более многочисленны в Восточной Азии.
Третий «ключ» — протяженность ареала. Некоторые зоогеографы предполагают, что территория, занимаемая группой организмов, увели- чивается с возрастом группы (стр. 461). Если бы это было так, то местом происхождения семейства был бы центр его ареала. Если выселение так же обычно, как и расселение, то этот «ключ» сам по себе не представлял бы никакого интереса, но он имеет некоторую ценность в связи с другими «ключами». Тот факт, что единственный вид рода Cntostornus занимает в Си- бири сравнительно неболыиой ареал, увеличивает реальность предполо- жения, что он недавно проник из Америки.
четвертый наиболее важный «ключ» — неразрывность ареала. Аре- ал, занимаемый сибирским видом рода Catostomus, почти неразрывен с аре- алом этого рода в Америке, и это свидетельствует о том, что он достиг Си- бири недавно. Род Myxocyprinus в Китае также занимает сравнительно небольшой, но расположенный далеко от ареалов других ныне живущих представителей этого семейства, ареал, и это соответствует тому, что он является реликтом более ранней китайской фауны:atostomidae.
Пятый очень полезный «ключ» к познанию истории семейств — рас- пространение систематически близких и конкурирующих семейств. Бли- жайшими родственниками чукучановых являются карповые рыбы: кар- пы и им близко родственные. Распространение этих родственных семейств позволяет предположить, что некогда чукучаны были многочисленны в Во- сточной Азии, а возникшие от них карповые заместили там большинство
ИЗ НИХ.
Пользуясь всеми этими «ключами», вместе взятыми, можно прийти к мысли, что место происхождения чукучан — Восточная Азия, затем они переместились из Азии в Северную Америку и (за исключением рода Myxocyprinus) были замещены карповыми в Азии; что чукучаны вторич- но радиировали в Северной Америке и что один вид рода Catostornus вер- нулся в ближайшую часть Азии.
Эта история семейства Catostomidae иллюстрирует один второстепен- ный принцип распространения животных: сильно отличающиеся формы (подобно Myxocyprinus), изолированные от основного ареала своей груп- пы, особенно если они находятся внутри ареала, заселенного конкуриру- ющими группами (в данном случае Cyprinidae), или, разделенные этим ареалом, часто являются не иммигрантами, а реликтами, сохранившими- ся там, где первоначальные группы были когда-то многочисленными, но затем были замещены другими. Это дает возможность предположить даль- нейший характер распространения: если группы животных появляются в определенных местах в связи с благоприятными там условиями, то, ве- роятно, они начнут вымирать в этих же местах в связи с появлением позд- нее новых семейств, для которых эти же условия также оказываются бла- гоприятными. Другими словами, вероятно, центры эволюции и расселе- ния являются также центрами вымирания. Таков логический вывод из
Глава 1 предположения, что расселение и выселение — взаимосвязанные, почти равноценные процессы.
Я хочу немного остановиться на этом вопросе. Совместно рассмотрен- ные выше «ключи» к познанию истории географии животных имеют су- щественное значение, и я думаю, что именно они раскрыли истину о се- мействе Catostomidae. Но и они не лишены недостатков. Это можно пока- зать на другом примере. Род Езох относится к пресноводным рыбам: в не- го входят настоящие щуки, несколько видов мелких щук и щука-маски- нонг. Род состоит из шести видов: четыре приурочены к восточной части Северной Америки, один — к северо-восточной части Азии (река Амур и др.) и один вид (Е. lucius) встречается исключительно в северной части земного шара — через Северную Америку от Лабрадора и Нью-Йорка
Ъ P и с. 10. Распространеппе пресноводными рыб ро:~а sox. Прерывистой линией обозначена ириблпзитсльная южная граница
Е. Luciics в Евразии и западной части Севсрной Амсрикп. до Аляски и через Северную Азию и Европу <о Британских островов (рис. 10). На основании только этих фактов почти любой зоогеограф скажет что
7 род Езох произошел в восточной части Северной Америки (пять видов, четыре эндемичных), затем достиг Восточной Азии (два вида, один энде- мик) и, наконец, недавно достиг Европы (один вид, не эндемичный). Рас- пространение щуковых почти непрерывное, словно они расселились сов- сем недавно. Род Esox — единственный род в семействе, но современное распространение родственных семейств, особенно семейства Umbridae, соответствует североамериканскому происхождению всего отряда Нар1о- mi, к которому относится род Езох. Тем не менее имеется несколько хоро- шо сохранившихся ископаемых видов рода Esox, найденных в олигоцене и мпоцене Европы, а самый древний представитель отряда Haplomi- Palaenesox известен в эоцене Европы, в то время как в Америке ни ископае- мых рода Esox, ни каких-либо родственных этому роду рыб не известно. История географии рода Esox оказалась, во всяком случае, сложнее, чем это следует из распространения ныне живущих видов. Хотя это и противо- речит выводам, сделанным с помощью «ключей», возможно, щуковые в Европе — скорее выжившие представители выселившейся группы, чем иммигранты из Америки. Причиной выживания рода может быть его необычайная холодовыносливость: щука — одна из немногих настоящих пресноводных рыб, широко распространенных в Арктике. Это и дало ей возможность пересечь последний сухопутный мост между Сибирью и Аля- ской. и, видимо, она расселилась из Азии в Северную Америку, а не нао- борот, так что все американские щуки, очевидно. произошли из евро- азиатскои ветви.
История семейства Catostomidae насчитывает по крайней мере два пересечения этого сухопутного моста: раннее — из Азии в Америку и позд-
нее — обратно на близлежащую часть Сиоири. История семейства Esoci- dae также, вероятно, включает два переселения: одно раннее и другое позд- нее, но оба из 1;вразии в Америку.
Распространение других животных, которые расселяются быстрее, чем пресноводные рыбы, может быть результатом еще более сложных пере- мещений. Сложность географических перемещений сама по себе является причиной осторожного подхода к слишком простым решениям («ключам») при выяснении истории распространения семейств.
Несомненно, имеются другие специальные «ключи», которые в от- дельных случаях помогают раскрыть историю распространения. Напри- мер. когда ареал семейства включает территорию, недавно покрытую лед- ником, то переселение на эту территорию совершенно очевидно. Един- ственный метод, к которому я отношусь с недоверием,— это метод выяс- нения распространения примитивных форм, которые, по мнению одних ученых, показывают место происхождения семейства, а по предположе- нию других — периферию своего ареала. Возможно, они показывают од- но из двух или и то и другое (стр. 466). Если существующие представители групп животных могут быть размещены на нескольких филогенетических линиях и если все эти линии из разных направлений ведут назад к одному месту, возможно, это и будет местом происхождения группы, но просле- дить историю географии животных таким методом очень трудно и часто приводит к ошибкам.
Заканчивая, я хочу сказать, что в нескольких последних абзацах я пытался показать, что «ключи» есть только «ключи», а не доказательства и что пользоваться ими следует разумно.
Самая лучшая основа для познания истории распространения живот- ного мира — это, несомненно, ископаемые, достоверные ископаемые, най- денные в соответствующем месте, но даже ископаемые данные следует ин- терпретировать с осторожностью. Ископаемые материалы по многим жи- вотным, даже по многим позвоночным, бывают очень неполны или ошибоч- ны. Ископаемые могут очень хорошо показать эволюцию группы живот- ных и все же не показать географическую историю этой группы; одно иско- паемое может показать стадию эволюции, но, для того чтобы показать распространение в определенный период времени, требуется большое коли- чество ископаемых данных. Скудные ископаемые данные могут ввести в за- блуждение. Например, находки ископаемых попугаев в Северной Америке и Европе относятся к середине третичного периода и до плейстоцена нигде больше не найдены. Однако это не означает, что попугаи произошли на севере и до недавнего времени не водились в тропиках. Подобные не- оправданные выводы делались и в других аналогичных случаях. Во мно- гих случаях, так же как и с попугаями, распространение современных форм — значительно лучший показатель прошлого, чем бедные ископае- мые данные.
T. Седьмой и последний предлагаемый рабочий принцип зоогеогрп- фии — пьиппться предложить гипотезу в mex случаях, когда не хватает факпгов, не забывая в то же время о том, чтпо гипотеза не есть факты. Та- кая методика полезна, например, при попытке проследить географическую историю животных со скудными ископаемыми данными. Методика заклю- чается в том, чтобы разработать возможный вариант для каждого отдель- ного случая, затем суммировать эти случаи и, таким образом, найти об- щий характер распространения. Если он найден, то это усилит вероятность отдельных случаев. К данному методу я буду прибегать при описании различных классов наземных и пресноводных позвоночных: анализировать
их распространение отдельно, а затем искать общий характер распростра- нения.
Иногда сталкиваешься и с такими взглядами — что если об истории некоторых животных, например пресноводных рыб, и об их связях с сухо- путными мостами и перемещающимися материками можно только догады- ваться, но их нельзя доказать, то о них вообще не следует писать. Мне ка- жется, что такой подход указывает на отсутствие понимания научного метода. Наука развивается при помощи постановки проблемы и попытки разрешить ее. Невозможно сказать заранее, разрешима ли данная пробле- ма в данное время. Мы можем предполагать, что она не разрешима, но мы должны пытаться ее решить. Объявить же ее заранее неразрешимой — зна- чит вообще ее не решить. Тот, кто потерпел неудачу, не должен утвер- ждать, что другие не должны делать новых попыток, так как исследовать сложную проблему настолько тщательно, чтобы ничего не оставалось до- бавить нового, невозможно. Необходимо пересмотреть как саму проблему, так и ее связь с другими проблемами, и если даже все попытки окажутся неудачными, то они все-таки покажут, какая работа должна быть проде- лана для достижения цели. В этой книге я буду рассматривать систему гипотез об истории расселения животных и прошлом земного шара. Если это будет сделано мною правильно, то, я думаю, мой вклад в общее дело принесет пользу на каком-то отрезке времени. Я думаю, что предполагае- мая история будет содер>кать много истины. Но чтобы быть честным, я дол- жен добавить, что домыслы о прошлом и для меня и для других зоогеогра- фов имеют свою прелесть, и опустить их — значит пожертвовать полови- ной привлекательности этого предмета. Но делать догадки надо умело. Они должны быть осторожным изложением предположений и вероятностей, а не просто полетом фантазии и не навязчивыми идеями, отстаиваеиыми оез надлежащих основании.
Это все, что касается предлагаемых рабочих принципов. За исключе- нием пятого и шестого, они являются просто научными рабочими принци- пами в применении к зоогеографии. Вместе взятые, они гласят: решить, что ты собираешься делать; отобрать и понять фактический материал, с ко- торым собираешься работать; наметить план, как соединить все факты вместе, чтобы получить из них истину, и осторожно идти вперед.
О литературе
Библиография в конце этой и последующих глав преследует две цели: дать основные источники по используемому материалу и открыть читателю доступ к остальной литературе. (:писки литературы настолько коротки, насколько это было возможно сделать. Многие приведенные работы имеют свою литературу, которая даст возможность любознательному читателю проникнуть в далекое прошлое или в глубину сложных вопро- сов. Например, литература к этой главе не претендует на то, чтобы охва- тить полностью историю зоогеографии, но работа Гадова (Gadow, 1913) дает ряд дополнительных источников, которые раскрывают историю этого предмета. Другой пример, Г. Г. Симпсон сделал существенный вклад в:п1о- географию, но здесь даны только две его работы. Одна из них приведена потому, что на нее имеется ссылка в тексте. На другую ссылки в тексте нет, но в списке литературы она стоит потому, что это последняя работа, кото- рая открывает путь к ранним зоогеографическим исследованиям Симп- сона. (,лисок составлен в алфавитном порядке и хронологически по каждо-
Литература
му автору. Если статья помещена в журнале, указывается название жур- нала, номер тома (жирным шрифтом) и номер страницы. Цитаты в тексте отсылают к этим же спискам с указанием фамилии автора, года издания ра- боты и иногда страниц.
Vll ТЕ РАТУРА
Л llee W. (;., Етегяоп А. Е., Park О., Park Т., Schmidt Ii. Р.,
1949, Principles of animal ecology, Philadelphia — London.
В е а u f о r С I.. F. de, 1951, Zoogeography of the land and inland waters, Lo»cia»- New York.
В г e w s С е r W., 1924, The birds of the Lake Umbagog region of Мя)и1е, «Hull. AI»».
(omparative Zool.», 66, pt 1, 1 — 209.
(.' а i n S. А., 1944, Foundations of plant geography, New York — I.ondon.
(', h а р m а n F. М., 1929, Му tropical air castle, New York — London.
(.' h а р m а n F. М., 1938, Life in an air castle, New York — Еопдоп.
D а r 1 i n g t о п P. J., Jr., 1938, The origin of the fauna of the Greatet' Antilles, v ith
discussion of dispersal of animals over water and througli the а1г, «Quarterly
R eview В iol. », 13, 274 — 300.
B а r 1 i n g С о п P. J., Jr., 1948, The geographical distributic)n of соЫ-bloodecl i.c r- tebrates, «Quarterly Веыеи Biol.», 23, 1 — 26, 105 — !23.
1) а r и i n С., 1859, 1950, On the origin of species..., London (переиздано в l,)5(. г.).
[Имеется русский перевод: Ч. Д а р в и и, Происхождение видов путем есте- ственного отбора, Соч., т. 3, М.— Л., 1939.]
Е k m а n S., 1953, Zoogeography of the sea, London.
6 а d о w H., 1913, The wanderings of animals, 'ambridge,,»placid, (:arnbriclqe ~ .
Press, New York.
С е i g e r R., 1950, The climate near the ground, (.ambridge, Маяя., Иагтагд С. Press.
6 и р р у Н. В., 19l 7, Plants, seeds, and currents in tlie West Indies and Azores, Lon- don.
ц е s s e R., А 11 е е O'. ('., S c h m i d С К. P., 1,)51, Ecological;animal geograI)liy.
2nd ed., Меж York — London.
K. e n d r e w W. G., 1937, Tlie climates of the contiiients, I.о»йн~.
K e n d r e w W. G., 1949, Climatology, London, Oxford U. Press.
К и е п е и P. Н., ]950, Marine geology, New York — London.
'.]]1 а t С h e w W. D., l915, 1939, Climate and evolutioii, «Ann. ibew York Асад. ~ci.»,
2~, 171 — 318; reprinted (1939) as «Special РиЬ. Меъ York Acad. Яг1.», 1.
М а у r Е., 194', Systematics аЫ the origin of species, New York, Ccilumbia U. 1 ress.
[Имеется русский перевод: Э. M а Й р, Систематика и происхождение видов,
ИЛ, М., 1947.]
.'11 у е r s G. S., 1938, Fresh — water fishes ance West Indian 7ооцeography, «Smit)>sc)- ni an 8ep.», 1937, 339 — 364.
4 е w b i g i n М. I., '1936, Plant and animal geograpliy, Еondon.
Р о w e r s S., 1911, Floating islands, «Popular Science Mcinthly», 7,), 303 — 307.
К о m e r А. S., 1945, Vertebrate paleontology, 2пй ed., (:hicago. [Имеется ругткий
перевод: А. P о м е р, Палеонтология позвоночных, 1'011ТИ, М.— Л., 1Мс).] ~ с h u с Ь е r С (:., 193', Gondwana land bridges, «Hi]11. ('ео1. Scarc. Атег~са»,
43, 875 — 915.
с 11 и с h e r С (,, J,)35, lfistorical geology of (11е A~>tillean-(:апЬЬеап region, ~<и- York.
S c l а t e r P. L., l858, On the general geographical ЛЫributic»i of the шетЬег of
the Гlass Aves. «J. Proc. Linnean Soc. (London), Zool.», 2, I:30 — 145.
1'лави 1
S i ш р s о и 4. (., [9~0, Mammals andri 1аМ bridges, «3. Was)iir>gton Aca(l.
Sci.», 30, '! 37 — ! 63.
~ i ш р s о n ('. С., [!)53, Ечо1ийоп апй оеодгар4у, Eugene, Огерои, Oregon State
Hysteron Н igher Кducation.
<i ~ гагар П. U., J ohnson M. W., Р1епъ~пд H. if., 1042, 7l)e oceans,
X еъ' York.
'Г;~ ri и е 11 i 11 [. И., [952. Иигг~сапея, Princeton, Хек Jersey, Ргinceton CJ. Рress. Т а ~i n e h i 11 I. H.. 1!)52а, Weather around the world, 2nd ed., E'rinceton, ".сею. Jer- sey, Princetcin Г. Ргеяя.
~ ~1~ 1> о' г о ~ е ~. И. 1'., 194~, Structural history о1 the East Iisdies, London — ~eve.
York, СашЬпй~~~е Ь. Press.
9, а 11 а с е A. Н., 1876, The geographical distributio» of animals, London 2 ~ol;. W а 11 а с е А. H., I880, Island life, London.
9; а 11 а с е Л. И., 19! ), Island life, Згй ed., London.
7 е и п е r F. Е., 1050, 0ating the past, 2nd ed., London. [Имеется русскяй
вод: Ф. II, е й н е р, Цлейстоцен, Москва, ИЛ, 1963.)
Глава 2ИРЕСНОВОДНЫЕ РЫБЫ
Ни одна часть книги не стоила мне стольких усилий в начале работы и не доставила большего удовлетворения в конце, чем глава о рыбах. Я во- ооще не смог бы закончить ее оез терпеливой помощи д-ра Джорджа С. Майерса, работу которого о пресноводных рыбах и их распространении (Муегз, 1938) должен знать каждый зоогеограф, и д-ра Карла Л. Хаббса, от которого я получил сведения после опубликования моей работы по гео- графическому распространению холоднокровных позвоночных (Darling- t nn, 1948) .
Пресноводные рыбы играют исключительно важную роль в зоогео- графии. Но, являясь обитателями вод, они так же тесно связаны с мате- риками, как и другие животные. Пресноводные рыбы неизбежно привя- заны к своим бассейнам и могут попадать из одного изолированного бас- сейна в другой только при медленном изменении самой поверхности Зем- ли или чисто случайно. Поэтому они расселяются на земном шаре медлен- но H в какой-то мере сохраняют древние черты распространения. По срав- нению с большинством других холоднокровных животных (рептилий, ам- фибий и беспозвоночных) рыбы с точки зрения систематики, эволюции. современного распространения и возможностей расселения изучены до- вольно хорошо. а по сравнению с еще лучше изученными теплокровными позвоночными (млекопитающими и птицами) они, по-видимому, имеют более обобщенный характер распространения, который отчетливее пока- зывает влияние климата. Если в распространении животных существуют общие закономерности, то пресноводные рыбы должны демонстрировать их лучше, чем любые другие животные, и, действительно, они показывают об- щие особенности распространения более наглядно. Объем книги не поз- воляет сделать детальный обзор всех классов позвоночных, и поэтому для более подробного рассмотрения я выбрал именно рыб, потому что они, как уже говорилось, наиболее хорошо показывают общие особенности и принципы распространения животных.
Основная система классификации рыб, принятая здесь, вплоть до отряда, дается по Ромеру (Romer, 1945). В отдельных случаях пришлось также прибегнуть к классификации Берга (Берг, 1947). Семейства были взяты из работы Ригэна (Regan, 1929), хорошо известного своими геогра- фическими исследованиями по рыбам, но в их классификацию пришлось внести довольно много изменений и добавлений. Все эти классификации отличаются в деталях, но для зоогеографии эти различия существенного значения не имеют.
Пресноводные рыбы в отличие от морских более малочисленны. Мно- гие строго морские группы здесь даже не упоминаются, а многие ископае- мые группы, с неизвестным образом жизни или зоогеографическое значе- ние которых не выяснено, опущены, в связи с чем классификация рыб, при- веденная в конце главы, местами значительно сокращена. Для ознакомле- ния с основными чертами их эволюции и систематики читатель может оо- ратиться к учебнику Ромера (Romer, 1945).
1 лава 2
В целях зоогеографии пресноводных рыб можно сгруппировать по нескольким различным принципам. Круглоротые и хрящевые рыоы игра- ют в пресных водах сравнительно незначительную роль, в то время как все костные рыбы составляют важные пресноводные группы. Для того чтобы противопоставить примитивные формы производным, доминирую- щие — недоминирующим и раскрыть различные особенности их распро- странения, полезно провести сравнение этих групп (табл. 1). В одном слу-
Табли цд 1
Основные таксономпческне группировки пресноводных рыо
Ируглоротые и хрящевые рыоы не игра~от большой роли Иостные рыбы
Некостистые
Ii остистые
Все отряды, кроме Ostariophy~i
Os t aI'iOPhyS i
l.l. ~la () I1 QQ6 ]~i>:< l l fiI("
Otal iophy~i
чае сопоставить некостистых, не Teleostei (архаические костные рыбы) с костистыми, Teleostei (высшие костные рыбы), а в другом — единствен- ный доминирующий отряд костистых пресноводных рыб Ostariopliysi со всеми другими отрядами, объединенными нами в одну группу некарпо- образных (не Ostariophysi). Более детальное описание этих групп дается в систематическом списке cBMBI1cTB в конце главы.
Что такое пресноводные рыоы.'
Рыбы встречаются почти во всех пресных водах, драже при самых не- благоприятных климатических условиях и на самых отдаленных остро- вах. Все рыбы, которые обитают в пресных водах, в известном смысле являются пресноводными, но место, занимаемое ими в зоогеографии, ме- няется в зависимости от их солевыносливости и поведения. Это совершенно очевидно, и длительное время делались различные попытки классифици- ровать пресноводных рыб по их солевыносливости (например, Bouletiger, 1905, р. 414, 415) или объяснять некоторые детали их распространения про- хождением солевыносливых форм через моря, например, Ригэн (Rigai), 1912) в своем исследовании по рыбам и линии Уоллеса. Но лишь Майерс (Муегя, 1938) сделал действительный шаг вперед, предложив экологиче- скую классификацию, основанную просто на способности или неспосоо- ности рыб переносить соленые воды.
Рыб, которые приурочены только к пресной воде, Майерс выделил в первый отдел (primary division). Это строго пресноводные рыбы. Тех, ко- торые живут преимущественно в пресной воде, но обладают незначитель- ной (не слишком высокой) солевыносливостью, он отнес ко второму отделу (secondary division). Это не строго пресноводные рыбы. А рыбы, которые встречаются в пресных водах, но обладают высокой солевыносливостью, могут быть отнесены к отделу периферических (peripheral division). 3ro солевыносливые рыбы. Термин «периферические» заимствован не у Май- ерса, а у Николса (Nichols, 1928, р. 6 — 7). Все три выделенных отдела очень сложны, нередко не соответствуют таксономическим подразделениям, и чтобы лучше в них разобраться и более точно определить, необходимо рас- смотреть каждый отдел в отдельности.
1JРеСНОВОдНЫе РЫбЫ
h первому отделу относятся строго пресноводные и только костные рыбы. К примитивным группам костных рыб (не Teleostei) относятся аф- риканские многоперы, китайские и североамериканские веслоносы, севе- роамериканская ильная рыба и австралийские и афро-южноамериканские двоякодышащие рыбы. Многие костистые рыбы имеют разное происхожде- ние. но некоторые из них встречаются на каждом материке с подходящими местами обитания, за исключением, возможно, Австралии, где имеется только один (сомнительный) представитель костистых строго пресновод- ных рыб из семейства Osteoglossidae. В Северной Америке к костистым рыбам первого (строго пресноводного) отдела относятся хиодоны, многие карпообразные (Ostariophysi) (см. ниже), щуки, евдошки и им близкородст- венные, траутпэрчи (Percopsidae) и афредодерусы (Aphredoderidae) сол- нечные. или ушастые окуни семейства Centrarchidae, настоящие окуни, включая род Ftheostoma. Полный перечень семейств строго пресноводных (первый отдел) включен в систематический список семейств рыб в конце ГЛЯВЫ.
Около ',8 всех строго пресноводных рыб земного шара относится к од- ному отряду карпообразных (Ostariophysi). Для представителей этого от- ряда характерно наличие веберова аппарата, соединяющего плавательньш пузырь с внутренним ухом. Считают, что этот аппарат передает вибрации внутреннему уху и увеличивает чувствительность слуха. Эта способность приобретает особенно большое значение в подводных условиях (Rigan, 1922. р. 204), и этим в какой-то мере можно объяснить удивительное про- цветание карпообразных в пресных водах. Существенно при этом, что кар- поооразные преобладают преимущественно в пресных водах, а в морях их очень мало. Примерно из 40 семейств и 5000 видов карпообразных толь- ко два семейства сомов, Ariidae и Plotosidae, примерно с, 150 видами яв- ляются морскими, да и те, вероятно, произошли от пресноводных предков. К карпообразным относятся многочисленные харациновые, электрические угри. карпы, вьюны, чукучаны и сомы, из которых те или другие изоби- луют в пресных водах любого материка, кроме Австралии (5fyers, 193<' р. 3 j5).
Не строго пресноводные рыбы приурочены к пресным водам, но неко- торые из них заходят в море; и этой группе относятся только немногие кост- ные рыбы. Из примитивных некостистых (не Teleostei) сюда относятся только панцирные щуки Северной и Центральной Америки, а из кости- стык — только карпозубые и хромисы. Все эти группы в основном оби- тают в пресных водах, однако панцирные щуки проникают и встречаются в море вдоль побережья Мексиканского залива. На многих карпозубых соленая вода, по-видимому, не оказывает отрицательного воздействия. Один из них, представитель рода Mollienisia, свободно заходит в море и размножается в соленых заводях Манильской бухты, куда он был завезен случайно; известно, что представитель рода Rivulus встречается в лужах после приливов на Куракао, а некоторые представители рода Fundulus и Cgrprinodon постоянно живут в морской воде (близ побережья), за пре- делами досягаемости ими эстуариев рек. Экспедиция «Челленджера» в Атлантике обнаружила представителя рода Fundulus даже в открытом океане. Полагают, что среди карпозубых род Ар1осйег1игобладает необычной солевыносливостью, так как он был встречен на островах Индо-Малайской области за пределами района распространения большинства пресноводных рыб. Большинство хромисов может существовать в море несколько часов или несколько дней, а один вид (Tilapia), найденный в солоноватых водах Мозамбикского пролива, прожил в морской воде Нью-Йоркского аква-
риума несколько месяцев. Очевидно, многие виды второго отдела (не стро- го пресноводных рыб) могут переносить недлительное пребывание в мор- ской среде, и их распространение показывает, что некоторые из них рас- селялись именно таким образом. Часть материала из этого абзаца взята из работы Майерса (Myers, 1938, р. 345). Возможно, некоторые рыоы. ог- несеяные в настоящее время в группу строго пресноводных (первьш от- дел), могут быть перенесены во второй отдел, когда их солевыносливость будет лучше изучена. Это относится даже к немногим рыбам из отряда Ostariophysi, особенно к некоторым карповым . (Сyprinidae) и сомам, о которых будет говориться ниже.
К отделу солевыносливых рыб относятся и пресноводные, хорошо пе- реносящие соленую воду рыбы, и те, которые произошли, очевидно, от групп с высокой солевыносливостью. Солевыносливые рыбы включаю1. большое число форм (из которых здесь будут упомянуты только немногие)— одно семейство круглоротых и несколько видов акуловых рыб, одно семейство (осетры умеренной зоны северного полушария) хрящевых га- ноидов и довольно многочисленные разнообразные костистые рыбы. (;юдэ относится и несколько представителей отряда Ostariophysi, сомы семейств Ariidae и Plotosidae, которые в основном являются морскими, и только немногие из них заходят в пресные воды или стали их обитателями, осо- бенно в изолированных местах.
Солевыносливые рыбы ведут различный образ жизни. Многие из ннх мигрируют. Некоторые.,(миноги, осетры, лососи, форели и др.) разино;ка- ются в пресной воде, но большую часть жизни проводят в море, за исклю- чением нескольких видов и родов каждой из названных групп, ставших постоянными обитателями пресных вод. Другие (американские угри, га- лаксиды, некоторые бычки и т. д.) размножаются в море, но большую ~асть жизни проводят в пресных водах, и среди этих рыб некоторые формы также стали обитателями пресных вод (галаксиды и бычки, но не угри). Некоторые мигрирующие рыбы регулярно перемещаются между пресными и солеными водами (не только с целью размножения), а такие, как, напри- мер, некоторые колюшки, могут, оказывается, жить как в пресных, так и в соленых водах. Мигрирующих рыб можно классифицировать различ- ными способами по их поведению (Myers, 1949), но тонкости поведения этих рыб, важные при изучении их биологии, малосущественны с точки зрения зоогеографа. Одно из преимуществ подразделения рыб Майерсом на отделы состоит в том, что сложные вопросы их эволюции, не имеющие большого значения в географическом отношении, опущены и основное вни- мание обращено на то, до какой степени рыбы могут переносить соленую воду и расселяться через нее.
Кроме мигрирующих рыб и видов, нейтральных по отношению к со;ле- ности воды, многие рыбы, живущие в пресных водах, местами произошли, видимо, от мигрирующих или от истинно морских предков. Эти рыбы встре- чаются в пресных водах всех частей земного шара, но больше всего их в таких отдаленных местах, как, например, Австралия, где они составляют основную часть фауны пресноводных рыб, а также на отдаленных остро- вах. В Северной Америке к таким рыбам относятся: миноги, осетры, лосо- си и форели (и родственные им сиги и хариусы), которые теперь стали по- стоянными обитателями пресных вод; белые и желтые окуни (семейство морских окуней), рыба-барабанщик (морское семейство барабанщиков), налимы (семейство тресковых), некоторые бычки-подкаменщики и атерин- ки. Более подробные сведения о них приводятся нин;е в списке семейств пресноводных рыб.
Пресноводиые рыбы
Наконец, очень большое число морских рыб разных семейств под- нимается вверх по рекам или заходит время от времени (или местами) в пресные воды. Это наблюдается в любой части земного шара, и подобные примеры можно найти почти в каждой фаунистической работе по пресно- водным рыбам. Семейство Atherinidae (см. список семейств рыб) дает це- лый ряд подобных примеров. Все известные рыбы, встречающиеся и в прес- ных и соленых водах Северной и Центральной Америки, внесены в списки Гюнтером (Gunter, 1942); его список дает представление о разнообразии поведения этих рыб. Из других работ, касающихся этого вопроса, отме- тим Бредера (Breder, 1934) и Майерса (Myers, 1949, 1949а), причем послед- ний приводит дополнительную литературу.
В целом вопрос отношения солевыносливых рыб (периферический от- дел) к пресной и соленой воде слишком сложен для дальнейшего обсуж- дения здесь. Главное заключается в том, что все отнесенные к этому отде- лу рыбы либо могут проходить через моря, либо, вероятно, весьма недав- но образовались от рыб, которые могли проходить, и, таким образом. их современное распространение, возможно, отчасти результат расселения через моря.
Такое подразделение пресноводных рыб на три отдела по их соле- выносливости — простое теоретически, но очень сложное фактически— приводит к бесконечным спорам о деталях. Солевыносливость многих рыб неизвестна, а поэтому невозможно и, вероятно, всегда будет невозможно провести резкую границу между этими отделами. В настоящее время боль- шинство рыб сгруппировано по семействам на основании предположения, что все члены одного семейства обладают примерно одинаковой солевы- носливостью, что, однако, не всегда бывает так.
Многие виды семейств даже строго пресноводного отдела могут пере- носить солоноватую воду, а некоторые из них могут даже заходить в море. Например, у отряда Ostariophysi (кроме двух семейств сомов, которые фак- тически являются морскими) различные харациновые, карпы, чукучаны и пресноводные сомы заходят в солоноватые воды эстуариев (Myers, 1938, р. 344 и другие источники). Японский карп (из семейства Cyprinidae), широко распространенный в реках, часто встречается и в море (Jordaii.
1905, vol. 1, р. 256; Okada, 1955, р. 93); маленький карпик Британской Колумбии иногда встречается в море и достигает островов Ванкувер и Нельсон, в то время как строго пресноводные карповые района Британ- ской Колумбии здесь отсутствуют (Carl, Clemens, 1948, р. 77), а из Индо- Малайской области некоторые виды рода Mystus (или Macrones) семейства пресноводных сомов Bagridae, обитающие в низовьях рек, по которым под- нимаются приливы, могут заходить в море или иногда жить в соленой воде (Weber, Beaufort, 1911 — 1915, vol. 2, р. 345; Н. М. Smith, 1945). Распрост- ранение ряда других видов семейств первого отдела, то есть строго пресно- водных рыб, позволяет предположить, что они все же могли пересечь пре- граду в виде небольшой полосы океанического пространства на пути к Фи- липпинам или Австралии. Эти случаи будут рассмотрены ниже.
Что касается солевыносливости древних рыб, то палеонтологи и физио- логи сходятся на том, что общие предки всех рыб, вероятно, явили в прес- ных водах: самые ранние ископаемые рыбоподобные позвоночные найдены в отложениях пресных вод (Romer, 1945, р. 44, 45),— и вообще все рыбы имеют такие почки, которые указывают на вероятность их происхождения в пресной воде (Н. W. Smith, 1932).
Акулы, скаты и родственные им другие хрящевые рыбы, вероятно, произошли от морского предка, который вышел из пресноводной среды.
Глава 2
(: тех пор как известно их историческое прошлое, они были обитателями океана (Romer, 1945, р. 60), и их мясо, пропитанное мочой, еще одно сви- детельство приспособления к жизни в соленой воде (Н. W. Smith, 1932). Некоторые исследователи, перенося морских акул и скатов из морской во- ды в солевой раствор или пресную воду, получали один и тот же неизбеж- ньш результат — животные поглощали воду, разбухали и гибли, вероят- но. потому, что не смогли достаточно быстро приспособиться физиологиче- ски (H. W. Smith, 1932, р. 1 i). Тем не менее некоторые акулы все-таки за- ходят в пресные воды, а два вида рода Carchari~as (в основном морской род акул) живут только в пресных водах: один в озере Никарагуа, а другой в Замбези (Regan, 1929); пила-рыба обычно из моря проникает в тропиче- ские реки; некоторые скаты заходят в пресные воды, а другие стали их постоянными обитателями (Garman, 1913, р. 415 — 417). Имеются и другие данные об акулах, встречающихся или живущих в пресных водах. Эти фак- ты показывают, что исторически длительное существование в водной среде одного типа не обязательно уничтожает у рыб способность переносить другую водную среду. Они свидетельствуют также, что результаты экс- периментов, проведенных с несколькими видами рыб, могут ввести в заблужение. Опыты в лабораторных условиях, очевидно, не всегда выявляют настоящую природную выносливость даже у подопытных видов. Результаты таких экспериментов не следует слишком поспеш- но распространять на целые отряды или семейства или на их древних предков.
Древнейшие костные рыбы, вероятно, обитали в пресных водах (Bo- mer, 1945, р. 75). Многоперые и двоякодышащие, видимо, сохранили не- прерывные линии в пресных водах с момента их возникновения. Выяс- нено. что по крайней мере только одна боковая ветвь двоякодышащих рыб совершала кратковременные миграции в море в девоне (Schaeffer.
1952. р. 101). Однако предки большинства современных пресноводных рыб «какой-то отрезок времени, вероятно, прошли через море. Костистые ры- бы. возможно, возникли в море (Romer, 194,э, р. 10+). Если это так, то некоторые из них вернулись в пресные воды, а у других, возникших от морских сельдей отряда Isospondyli, развился улучшенный слуховой ап- парат, они эволюционировали и дали разнообразные формы в пресных во- дах оольших материков, образовав отряд Ostariophysi. Некоторые сомы этого отряда затем вновь возвратились в море. И наконец, некоторые иэ морских сомов вернулись в пресные воды в таких местах, как Австралия и Мадагаскар. Если это соответствует действительности, то эндемичные сомы австралийских рек находятся в конце эволюционной линии кости- стых рыб, которая бере"свое начало в пресных водах, ,заходит в море, за- тем «озвращается в пресные воды снова делает заход в море и опять воз- «ращается «пресные воды.
Все это говорит о том, что первый отдел рыб (строго пресноводных) «подразделении Майерса — это гетерогенная группа неродственных от- рядов и семейств. Несколько видов даже самых больших групп пресно- «одных рыб могут заходить в море; солевыносливость и образ жизни мно- гих видов, особенно в тропиках, не известны. Судя по предкам большин- ства строго пресноводных рыб, нельзя сказать, что эти рыбы всегда жили в пресных водах. Наоборот, многие из них, включая Ostariophysi, «ероят- но, когда-то возникли от морских рыб. Тот факт, что пресноводные рыбы да«но были приурочены к пресным водам, подтверждается прежде всего их распространением, которого мы коснемся ниже, и в меньшей степени их природой и историей их эволюции.
II ресновог)иые рабы
Майерс (Муегз. 1949а, р. 317) пишет, llo некоторые ихтиологи сом- неваются в реальности его деления. Несомненно, до некоторой степени оно искусственно. Между отделами нельзя провести отчетливую границу, и имеется множество сомнительных моментов в отношении деталей. Но это деление основано на том реальном и очень важном факте, что пресноводные рыбы различаются по своей способности переносить соленую воду, и их распространение свидетельствует об этом. Мне кажется, что выделение этих групп или других, подобных им, совершенно необходимо для пра- вильного понимания географического распространения пресноводных рыб.
Исходя из изложенного выше и практических целей, определение групп рыб, выделенных Майерсом, может быть слегка изменено следую- щим образом. Первый отдел состоит из семейств или других категорий пресноводных рыб, которые обычно не заходят в море (хотя отдельные виды некоторых групп все же составляют исключение) и которые при- урочены к пресным водам, видимо, настолько давно, что их современное распространение является результатом расселения через пресные воды. далеке ecJIII Hx отдаленные IIpeplcN, возможно, II "I;NJIH B море. Это строго пресноводные рыбы. Второй отдел состоит из семейств или других групп, которые встречаются главным образом в пресных водах, но могут заходить в море и находиться там в течение какого-то ограничен- ного времени, или которые недавно произошли от форм, ведущих себя по- добным образом. Следовательно, их современное распространение может быть, и, видимо, часто является результатом расселения вдоль береговой .линии или через узкие океанические пространства. 3УО, таким образом. не строго пресноводные рыбы. Отдел периферических, или солевыносли- вых, рыб состоит из семейств или других групп, которые хотя и сущест- вуют в пресных водах все же тесно связаны с, морем или совсем недавно вышли из моря, так что их современное распространение может быть, н часто и есть результат расселения через море.
Эти отделы удобны для изучения распространения пресноводных рыл> на всем земном шаре. Возможно, в отдельных случаях для изучения мест- ных распространений более приемлемы другие классификации, так как некоторые группы рыб, являющиеся несомненно солевыносливыми, что относится ко всем районам земного шара, в целом в определенных местах придерживаются пресных вод. В качестве примера можно взять некоторых северных лососевых, которые, хотя и произошли от мигрирующих форм, теперь приурочены к местным пресным водам и отражают историю этих oacceiiHQB, а также семеиство Atherinidae. В целом 'Iso семеиство, несом- ненно. является солевыносливым и достигло Австралии через море. Одна- ко ряд австралийских родов этого семейства, образуя подсемейство Me- lonotaeniinael теперь строго приурочен к пресным водам, и их распро- странение по направлению к Соломоновым и Молуккским островам огра- ничено солеными водами. Таким образом, создается впечатление, что это подсемейство относится к первому отделу строго пресноводных. дтот слу- чай приведен как гипотеза, так как данный факт вызывает еще некоторое сомнение (см. семейство Atherinidae в списке семейств рыо).
Границы расllpocтраненпя пресноводных рыо
Границы распространения пресноводных рыб показаны на рис. 1'l. Несколько видов из строго пресноводных рыб имею' более или менее ши- рокое циркумполярное распространение. Из некарпообразных щука (Еяох
~> и с. < I. > ранш«< р«спростране><»» «ресно»о;<и> <к рыо первого (строго ><ресно»о<ного)
»<>I><» <> (пе строг<> пресново <>«>r<>) от <<;«»> («<><>«< t<!rly )3щ>ец о> ф3><>I<>„y> 23 ф <)я3 р.
1'>~„. 1).
/ °
~lIëîшна л .1Ини Я - — г I)а н II I I3 рас I1р()странен II л ] ) ыб I! ср вого ()тдслй ll„'Ill о('H() ви < >Й (~и уны $) ыб 'дтого отдела (в восточной Индии); «рсрывиста» lllHIIH — граница расицространения рыб llсрвог0 отдела (в боль- шинстве случаев мелких ка~~иовых) в восточной И ниии; иунктирная линия -- граница расиространсния рыб
второго отдела на островах.
Пресноводные рыбы
lucius) достигает всех главных северных речных систем Евразии(кроме за- падной и северной (:кандинавии) и северных частей Аляски, дельты рек Маккензи и Андерсон, которая полностью находится за Северным полярным кругом в западной Канаде. Черная рыба (Dallia) встречается в центральнои и западной частях Аляски, достигая на севере по крайней мере полуост- рова Сьюард, в бассейнах северо-восточной оконечности Азии (район Чу- котского полуострова) и на острове Святого Лаврентия на севере Берин- гова моря. Это единственный вид из строго пресноводных рыб, наиболее приспособившийся к условиям севера и ограниченный в своем распрост- ранении арктическим местообитанием. Эта маленькая (до 8 дюймов) очень жирная рыбка исключительно хорошо переносит холод и живет на мелко- водье [мелкие речки, озера и сфагновые болота.— Ред.] в тундре. Заслу- ,кивает внимания тот факт, что эта рыбка, столь исключительно приспо- собившаяся к условиям тундры, занимает чрезвычайно небольшой ареал, главным образом в южных районах Арктики, в то время как щука, ме- нее приспособленная к северным условиям, и некоторые другие строго пресноводные рыбы имеют значительно более широкое распространение в Арктике и проникают дальше на север. В Евразии два вида окуневых до- стигают Арктики, а американский судак доходит до дельты Макензи. Пер- копсис (Percopsidae) пересекает Северный полярный круг в западной Ка- наде. Северный чукучан (Catostomus catostomus) из отряда Ostariophysi распространен циркумполярно в западной части (.'еверной Америки и Во- сточной Сибири;. несколько видов карповых (Cyprinidae) проникает в Арк- тику в Евразии, и по крайней мере один американский род этого семейства (Couesius) достигает дельты Маккензи.
Некоторые из вышеупомянутых рыб достигают настоящих арктиче- ских районов, а другие, вероятно, только проходят через большие текущие на север реки: Маккензи в Канаде, Енисей и Лена в Сибири,— но в зоне тундры, за пределами этих рек, они не встречаются. Некоторые арктиче- ские пресноводные места обитания, особенно стоячие водоемы в тундре, не заселяются ни одной рыбой из строго пресноводных, кроме даллии (в пределах ее небольшого ареала); на севере имеются места в северных частях Квебека и Лабрадора, где рыб первого отдела (строго пресноводных) нет вообще. Щука, северный чукучан и карловые рода Couesi us подни- маются по реке Джордж, на север полуострова Лабрадор до 56 — 57' с. rn., но в других частях этого полуострова строго пресноводные рыбы не изве- стны. Однако их распространению, вероятно, препятствует не климат. Скорее всего, они просто не смогли вернуться в отдаленные северо-восточ- ные бассейны, из которых в свое время были вытеснены ледниками позд- него плеистоцена.
(;олевыносливые рыбы, особенно лососевые и им близкородственные, изобилуют во многих арктических пресных водах как в границах распр<1- странения строго пресноводных рыб, так и на север от них, но на крайнем севере разнообразие их форм уменьшается. Берг (Берг, 1947) указывает, что в Евразии голец (Salvelinus, род лососевых) является самой северной пресноводной рыбой, которая достигает Новой Земли и Новосибирских островов. В Америке тот же самый род простирается еще дальше на север и был обнаружен в пресных водах по крайней мере на 8' "34' с. ш. на ост- рове Элсмира, близ конечной северной границы суши. Эти подробности по распространению пресноводных рыб в Арктике взяты в основном из ра- бот Берга (разные работы) и Виня-Эдвардса (Wynne-Edwards, 1952).
В южном направлении некоторые группы африканских строго прес- новодных рыб достигают оконечности Южной Африки, а многие другие не
l .л ава
достигают ее (более подробно см. стр. 61). Вероятнее всего, что препятстви- ем для рыб, которым не удается достичь Южной Африки, является скорее ограниченность мест обитания, чем прохладный климат. Строго пресно- водные рыбы отсутствуют (но это не связано с, климатом) на большей ча- сти Австралии. Солевыносливые формы встречаются на юге, включая Тасманию и Новую Зеландию. В Южной Америке сомы семейства Pygidii- dae доходят до 47 "30' ю. ш., примерно в 500 милях от мыса Горн (Еigen- mann, 1918, р. 269). Возможно, это самая южная рыба первого отдела на земном шаре, но солевыносливые рыбы встречаются в пресных водах на 1ех же широтах, что и в Южной Америке и на Огненной Земле.
Фауны настоящих строго пресноводных рыб в пределах своих север- ных и южных границ приурочены ко всем материкам, исключая Авс пра- лию, и к некоторым материковым островам. Ни один факт в зоогеографии не является более поразительным, чем этот, и именно он доказывает, что многие группы рыб давно были приурочены к пресным водам. Географи- ческие границы материковых фаун рыб показаны на рис. 11. Эти границы в действительности переходят очень немногие рыбы, не считая, конечно, тех, которые могут переносить соленую воду или которые завезены человеком.
И Северная и Южная Америка богаты пресноводными рыбами, а ряд кя;ноамериканских групп достигает материкового острова Тринидад, но ни одна строго пресноводная рыба не является по-настоящему местной в Вест-Индии, хотя некоторые не строго пресноводные и солевыносливые рыбы являются таковыии (Муегя, 1938, р. 356, 357). На северном острове Ньюфаундленд, хотя он и является материковым, также совершенно от- сутствуют все строго пресноводные рыбы, хотя там имеются солевыносли- вые (Frost, 1940). Строго пресноводные рыбы, вероятно, не смогли проник- нуть на этот остров со времен плейстоцена.
Африка имеет обширную фауну и древних, и современных семейств пресноводных рыб, но ни одна рыба из первого отдела не является мест- ной на Мадагаскаре, хотя несколько форм не строго пресноводных (второй отдел) и, конечно, солевыносливых рыб достигли этого острова.
Европа и Азия дали ту или иную часть своей фауны рыб (первого от- дела) материковым островам Британии (Берг, 1932, и многие другие ав- торы), Японии (Мог1, 1936, р. 10 — 14; Okada. 1955), островам Тайвань и Хайнань (Mori, 1936, р. 47 — 51) и Цейлону (Day, 1889; Munro, 1955, и ссылки в статье Хора), а большинство семейств этого отдела в Юго-Во- сточной Азии достигает островов Суматра, Ява, Калимантан. Однако давно известно (Regan, 1912), что здесь строго пресноводные рыбы резко оста- навливаются в своем распространении (рис. 11). Некоторые из них оста- навливаются недалеко от внешней оконечности острова Ява (Dammerman, 1929, р. 15, 16) и северного Калимантана, но данных об этом немного. Богатая фауна рыб Индо-Малайской области в целом хорошо представлена на острове Ява, а некоторые (очень немногие) представители этой фауны достигают даже первого острова из группы Малых Зондских. Многие ихтиологи, от Ригэна (Regan, 1912) до Бьюфорта (Beaufort, 1951, р. 180), предполагали, что значительная часть яванской фауны рыб достигает острова Бали или что на этом острове значительно больше пресноводных рыб. чем на острове Ломбок. Это предположение ошибочно. Единствен- ные строго пресноводные рыбы, которые известны на острове Бали и, ве- роятно, являются там местными, — две-три мелких карповых из родов Rasbora и Puntius; оба этих рода также встречаются на острове Ломбок, а представители рода Rasbora — также и на острове Сумбава. Пресноводные рыбы не доходят до южного конца линии Уоллеса.
ИРеСНОВОдНЫе РЫ6Ьl
Севернее многие строго пресноводные рыбы Индо-Малайской области достигают острова Калимантан, но ни одна из них не доходит до острова Сулавеси, за исключением случаев (вероятно) их интродуцирования. Очень немногие рыбы острова Калимантан достигают южной части Филип- пинских островов (Herre, 1928). В большинстве случаев это мелкие карловые, которые встречаются от острова Балабак, Палаван и островов Каламиан до Миндоро и через архипелаг Сулу до острова Минданао и на самом острове (рис. 11). Они представлены несколькими родами, но осо- бенно родами Rasbora и Puntius (Barbodes). Вдоль обоих описанных выше путей распространения локализуются некоторые эндемичные виды: в озере Ланао (остров Минданао) фауна карповых представлена по крайней мере 4 эндемичными родами и 17 видами, очевидно ведущими свое про- исхождение от одного предка (Herre, 1933). На острове Палаван и островах Каламиан, кроме карповых, имеются еще один эндемичный род и виды сомов семейства Siluridae, а на Минданао — эндемичный вид сомов из семейства Clariidae. На этом список известных местных строго пресновод- ных рыб Филиппинских островов заканчивается.
Единственные рыбы Индо-Малайской области (кроме рода хромисов на Цейлоне и полуострове Индостан), отнесенные теперь к не строго прес- новодному отделу — карпозубые (Cyprinodonts). Из них род Aplocheilus (включая Oryzias) характерен для островов, которых он достиг. Это острова Рюкю и Андаманские, где строго пресноводные рыбы отсутствуют или встречаются редко; он также встречается на островах Ломбок и Тимор и достиг острова Сулавеси, где от него развились эндемичные виды (Aurich, 1935), а, возможно, также и эндемичное семейство Adrianichthyidae (см. список семейств рыб), и, наконец, был описан еще один вид из северного Лусона (Herre, Ablan, 1934), но так как карпозубые нигде больше, кроме Филиппинских островов, неизвестны, возможно, этот вид не является местным. В связи с таким характером распространения карпозубых и из- вестным фактом, что многие из них солевыносливы, почти не остается сом- нений в том, что род Aplocheilus расселился частично через море.
Вернемся к мелким карповым. Из них два рода, Rasbora и Рипйик (Barbodes), неоднократно встречались на островах за пределами границ других строго пресноводных рыб. Оба рода пересекают линию Уоллеса на Малых Зондских островах и проходят большую часть обоих иммигра- ционных путей карповых на Филиппинские острова. Фактически один вид Puntius binotatus, который встречается в пресных водах от Сингапура че- рез острова Суматра, Ява и Калимантан, заходит за пределы границ боль- шинства пресноводных рыб в трех направлениях: к островам Бали и Лом- оок, h острову Палаван и к острову Минданао, где он является очевидным предком эндемичного карпа озера Ланао. Такой характер распростране- ния мелких карповых, аналогичный характеру распространения рода Aplocheilus, но только меньшего масштаба, позволяет предположить, что эти карповые обладают более чем обычной солевыносливостью, хотя и не столь высокой, как у рода Aplocheilus, и что они смогли пересечь узкие океанические преграды.
Выше я опустил описание некоторых строго пресноводных рыб. осо- оенно лабиринтовых — АпаЬая testudineus, и Ophicephalus striatus, а так- же сома Clari:as batrachus, которые являются обычными съедобными рыоа- ми и живут в различных пресных и солоноватых водах; они очень живучи, могут длительное время жить без воды (если их держать во вла; 'ной среде), что очень облегчает их перевозку. Эти рыбы встречаются за гра- ницами распространения большинства пресноводных рыб на Малых Зонд-
ских островах, Филиппинских, Сулавеси и др. Возможно, в эти места они были завезены человеком.
В Австралии далеко за пределами границ больших материковых фа- ун имеются две рыбы предположительно строго пресноводных семейств. Одна из них — представитель костноязычных Scleropnges leichhnrdti— большая рыба, 2 — 3 фута длиной, населяющая реки тропических частей Квинсленда и южноё части Новой Гвинеи. Другой вид, часто относимьш к тому же роду, встречается в восточной части Индо-Малайской области. Ни один другой род из современных рыб строго пресноводного отдела не может сравниться с распространением этого рода, охватывающим прост- ранство между Азией и Австралией. Со времен Ригэна (Regau, 1911) кост- ноязычных рыо считали древними, но первые известные ископаемые, от- носящиеся и третичному времени, указывают на то, что они не древнее Osta- ri0physi или некоторых других ныне живущих рыб, а их филогенетические связи не предполагают большой древности. Мы не знаем, насколько они приспособлены к соленой воде. Африканский представитель костноязыч- ных (Clupisudis) заходит в солоноватые воды. Ископаемые остатки предпо- лагаемого представителя костноязычных также найдены в морских отло- ;кениях эоцена Англии. Я не пытаюсь утверждать, что Scleropages достиг Австралии через речные системы древних соединений Земли или (частично) через море. Этот случай, несомненно, следует толковать с осторожностью.
Австралийская двоякодышащая рыба Epiceratodus forsteri, как извест- но, существует только в средних частях двух маленьких речных систем— Мэри и Бернетт — в субтропиках юго-восточного Квинсленда. Прямого родства между этой рыбой и современными двоякодышащими рыбами Аф- рики и Южной Америки нет, но она родственна ископаемым формам, кото- рые появились в триасе и встречались на болыпей части земного шара в мезозое. Это реликт семейства значительно более древнего, чем костно- язычные — единственный несомненный реликт древнего семейства пресно- водных рыб за пределами границ распространения больших материковых фаун. Но мы слишком мало знаем о солевыносливости рыб, особенно древ- них, и не можем сказать уверенно, могла ли даже эта рыба достичь Авст- ралии только через пресные воды.
В пределах общих границ распространения рыб строго пресноводно- го отдела каждая отдельная группа их еще более ограничена. Ни одно семейство не является космополитом или почти космополитом. Костно- язычные и рыбы семейства Nandidae могут быть названы тропикополита- ми (кроме тех представителей семейства Nandidae, которые не достигают Австралийской области), но их представители до какой-то степени локали- зировались в пределах тропических районов; тот факт, что они рассели- лись полностью через пресные воды, еще недостаточно достоверен. Семей- ство карповых (;yprinidae, простирающееся почти через всю тропическую и умеренную зону Евразии, Африки и Северной Америки, но не достиг- нувшее Австралийской области или Южной Америки, имеет самое широ- кое, по существу непрерывное, распространение по сравнению с любым семейством рыб строго пресноводного отдела. Несколько родов строго прес- новодных рыб, такие, как Barbus (СурппЫпе), Clnriius (сомы), Annbas и Ophicephnlus (змееголов) и ЛХагйасетбеКиг (угри), имеют меридиональное распространение через весь континент Африки и широтное — на боль- шеё части Южной Азии. Род Esox имеет почти непрерывное распростра- нение через северную Евразию и восточную часть Северной Америки. Не- сколько других родов строго пресноводных рыб обычны для Евразии и Северной Америки, но их распространение более прерывистое. Вероятно,
(/perноводные рыбы
щука (Esox luci~us), самый широко распространенный вид строго пресно- водных рыб, встречается в широтном направлении по всей северной Евра- зии и (;еверной Америке.
Зональное распросграненпе пресноводных рыо
И пределах границ, описанных выше, многие пресноводные рыбы распространяются скорее вдоль климатических зон, чем вдоль материко- вых массивов. Большие группы пресноводных семейств первого и второго отделов умеренной зоны северного полушария и тропической зоны имею1 четкие отличия. В арктической зоне и умеренной зоне южных широт отс- утствуютт не столь хорошо выраженные фауны строго пресноводных се- мейств, но имеется много солевыносливых рыб. Распространение этих рыо также связано с климатом. (."олевыносливые лососевые и им близкород- ственные доминируют в арктических пресных водах; в южном направле- нии они встречаются на большей части умеренной зоны северных широт. ио нигде не заходят в настоящие тропические воды. В тропиках бычки явlHB)TcH самыми обычными солевыносливыми рыбами, хотя они и не ти- пичные рыбы тропиков, а многие другие тропические морские семейства отчасти могут встречаться и в пресных водах. В умеренных широтах юж- ной зоны галаксиды и им родственные — это наиболее ярко выраженные зональные солевыносливые рыбы. Род Galaxias более чем с 40 видами, ко- торые живут в пресных водах или по крайней мере заходят в них, встре- чается повсеместно в умеренном климате южного полушария — в южной Австралии и Тасмании, на острове Лорд-Хау, в Новой Зеландии, на юге Южной Америки и на южной оконечности Африки (см. список семейств рыб). Миноги также имеют зональное распространение как в южном, так и в северном полушарии; в южном они заходят в пресные воды южной Ав- стралии, Тасмании, Новой Зеландии и умеренной зоны Южной Америки (в южном полушарии), но не Южной Африки.
Распространение многих морских рыб следует климатическим зонам: обширные фауны рыб побережий тропических частей земного шара во многих отношениях удивительно сходны и заметно отличаются от фаун умеренных зон. находящихся к северу и к rory от них (Муегя, 1940). Некоторые морские семейства характерны для обеих зон: северной и умеренной зоны южных широт,— но отсутствуют или бедно представ- .лены в тропиках (см. iVIyxinidae, (~adidae, (:ottidae в списке семейств рыб. а также см. НиЬЬя, 195' ), а другие характерны или для северных, или для далеких южных морей. Многие солевыносливые и морские рыбы, которые могут расселяться через моря, имеют тенденцию придерживаться еди- ных климатических зон, и этот факт говорит о том, что климат сильно воз- действует на их распространение. Очевидно, климатическая зональность оказывает существенное влияние также и на распространение настоящих пресноводных рыб, хотя это не всегда учитывается.
Чтобы проанализировать зональное распределение строго пресновод- ных рыб, лучше всего разделить их на две группы, предлагаемые в нача- ле этоп главы. К первой группе относятся все рыбы, за исключением от- ряда Ostariophysi, а ко второй — только один отряд Ostariophysi.
Распределение некарпообразных также в основном происходит в со- ответствии с климатическими зонами (рис. 12). Из них 16 семейств являют- ся тропическими: 13 встречаются в Африке, 9 — в Индо-Малайской оо- ласти (6 общие с Африкой), 3 — в Южной Америке (все 3 общие с Африкой
и ' — также с Индо-Малайской областью). Семь исключительно африкан- ских семейств относятся к древним или генерализованным отрядам и включают некоторые очень древние формы. Три семейства исключительно Индо-Малайской области, высшие колючеперые рыбы по происхождению оолее современны. Надо отметить, что в отличие от Африки в Индо-Малай- ской области нет древних пресноводных рыб. Южноамериканские семей- ства отличаются своим малым числом; тем, ITo все они встречаются также
восточная часеь
Зайадноя чосп~ьСео. Ядрили
9иеренная ~он~
Восточной Азии
Юрооа Сей. АмерикиPolyodonfidue (1)Amiidae (1)Hiodontidae (2)Umbri dae (1) — — Umbri due (1 — — — — — Umbridoe (2)Percopsidae (1) — — — — — percopsi 4ae (1)Aphredoderidae(f)Centrarchidae (1)- — — — CentrarcН4ае (40)Perci doe (100+)
Polgodontidae (1)
Perci 4ae (10) — — — — Percidae Я(„~о~ф~~~~)
"" '." ~и
— — — — граница иежду себеро уверенной — — — — и тропической раунаии
11I
~Аика Индо-Малай ская I 1 I Южн А
область I I I Apxauvec~ue фориы 111
Lepi dosirenidae (3)
Polgpteridae (f3)
Isospondgli "
111
Phroctolaemidae (1) 11
Pan tadontidoe (1)
1
11
11
Ggmnarchidae (1) 111
ПогтугЫае (150)
11
Kneriidoe (3) '1(
111
Notopteridoe (2)- — — Notopterf doe (3)
1
0steoglossidae (f) — — Osteoglossi doe ( j
.11 8ь!вшие колючеперь!е
Nandidae (f) — — — — — Йап4а1ае (4)
Pristolepidae(4) ~ I ~
Anobantidae (16) — — — Anobantfdae(25) — ~ ~ ~
Laciocephalidae (f)
Channidae (3) — — — — Channi dae (20) — — — ф
ffostocembeli dae(34) Mmfocembelidae ино~очосл.)
СИаийИигй4ае 1)
Lepidosireni dae (f)
0steofflossi due (2)
Nandi dae (2)
Р и с. 12. Зональное распространение некарпообразньтх пресновод- ных рыб первого отдела («Quarterly Кейеъ. of Biolo<zy», 23. 1948,
table2, р. 5).
Цифры в скобках означают приблизительное число видов в больших семействах. Стрелки указывают тропические семейства Индо-Малайской области, которые проникают в умеренные широты Восточной Азии. (Один вид мелких окуней из семейства Percidae, достигающий западной ~%сксики, здесь не приводится, так как он ке имесг ничего общего с более северными видами этого семейства, для которых ха1~актсрно прсрывпстос
распространение.)
в тропиках Старого Света, и своим филогенетическим разнообразием. Vi ним относятся одна группа архаических некостистых рыб, одна Ьоя- pondyli и одна группа колючеперых. Хотя эти 16 семейств образуют зо- нальную фауну, большая часть которой приурочена почти к тропикам, а остальная распространена по всему земному шару, совершенно ясно, что эта фауна на разных материках имеет коренные отличия, отражающие как различия в истории этих материков, так и в эволюции и расселении разнообразных групп рыб.
Пресноводные рыбы
Девять семейств некарпообразных пресноводных рыб первого отде; и (не считая арктической даллии) являются северо-умеренными. Все девять семейств представлены в восточной части Северной Америки (только 5 из 9 семейств находятся на западе от Скалистых гор). и все они прекрасно там локализовались (более подробно см. кихоте раздел «Пресноводные ры- бы Северной Америки»). Только 4 семейства из 9 представлены в Евразии: семейства Esocidae (рис. 10) и Percidae (рис. 13) широко распространены вдоль северной Евразии; семейства Polyodontidae и Umbridae сооп~ет- ственно представлены единственным реликтовым видом в Китае и Восточ- ной Европе.
Очень немногие из рассматриваемых рыб пересекают границу между тропической и умеренной зонами северного полушария. (Следует помнить, Р п с. 13. Распространение пресноводных рыб семейства Регс1с1ае. Лриблизительные границы трех подсемейств: 1 — Регс~11ае; 2 — ?лсн~-
p ercia ne; 3 —, theostoli i i »ae. что в настоящий момент мы ограничиваемся рассмотрением некарпообраз- ных рыб строго пресноводного отдела, рис. 12.) В Америке ни одна из этих рыб не пересекает этой границы; фактически ни одна из них не захо- дит даже в Центральную Америку. В Африке некоторые тропические се- мейства следуют по Нилу почти до устья и, таким образом, заходят в кра- евые части умеренной зоны, но в водах настоящей умеренной зоны они не встречаются. И только в Восточной Азии группы рыб умеренной и тропи- ческой зон перекрывают друг друга. В Восточной Азии из трех семейств умеренной зоны ни одно не достигает тропиков, и все они фактически яв- ляются или реликтами (Polyodontidae в Китае), или при; рочены к край- нему северу (семейство Esocidae к Амуру и севернее, семейство Pevci- dae — только севернее реки Амур). Но из 9 тропических семейств Индо- Малайскохх области три заходят в умеренную зону: семейства АпаЬа~й1- dae и Alastacembelidae достигают Северного Китая, а семейство (;hannidae (0phicephalidae) — Амура. Эти семейства не имеют эндемичных родов в умеренной зоне. Ьлиз северных границ распространения видов также очень мало эндемиков, а ареалы этих семейств в умеренной зоне значи- тельно меньше их обширных ареалов в тропиках Старого Света. Все это указывает на то, что в Восточной Азии семейства умеренной зоны отсту- пили, а тропические — продвинулись на север, в умеренную зону.
Некарпообразные строго пресноводные рыбы имеют разное происхо- ждение. Развитие двоякодышащих и многоперых рыб, вероятно, происхо- дило только в пресных водах, а развитие всех остальных основных групп, скорее всего, происходило в морях, но когда и где — не известно. Некото- рые из них, очевидно, имели сложное расселение в пресных водах, а дру-
гие, возможно, просто оставались на месте происхождения или поблизо- сти от него. Ископаемые остатки этих рыб указывают на два существен- ных факта. Во-первых, они показывают, что некоторые семейства сущест- вовали в умеренной зоне северных широт, начиная с эоцена, а в течение третичного периода они начали исчезать в Евразии и западной части (:еверной Америки и сохранились только в восточной части Северной Аме- рики; распространение некоторых уцелевших семейств (рис. 12) подтвер- ;кдает это отступание. Во-вторых, ископаемые остатки подтверждают, ITO два семейства отступили еще с больших площадей. Семейство Osteoglos- sidae независимо от его происхождения отступило из умеренной зоны север- ных широт в тропики и существовало там в течение всего третичного пе- риода. Семейство Ceratodontidae, двоякодышащие рыбы, которое встре- чалось в пресных водах почти всего земного шара в мезозое, сократилось 30 одного вида, который населяет две небольшие речки в Австралии.
Для пресноводных рыб второго отдела (не строго пресноводных), относящихся к некарпообразным костным рыбам, характерно также зо- нальное распространение, но они в какой-то степени переходят климати- ческие границы. Панцирные щуки теперь встречаются в умеренных ши- ротах восточной части Северной Америки, в Центральной Америке и на Кубе. Их ископаемые найдены как в умеренных широтах Азии и Европы, так и Северной Америки, в отложениях позднемелового и ранне-третич- ного периодов, а в тропической Индии, вероятно, в отложениях ранне- третичного периода. Следовательно, они являются древней, главным об- разом северо-умеренной группой, но, появившись однажды в тропической зоне Азии, они продвинулись в американские тропики дальше, чем любые другие северные пресноводные рыбы. Однако Южной Америки эта группа не достигла. Карпозубые характерны для тропиков всего земного шара, кроме Австралийской области, но они встречаются также к северу и к rory от тропиков в умеренных зонах, особенно в Америке. Хромисы встречают- ся главным образом в тропиках Африки и Америки, хотя несколько их видов находится в прилегающих областях с умеренно-теплым климатом, а несколько — на юге Индии и на некоторых островах (см. список се- мейств).
Радиальное распространение пресноводных рыб
Распространение отряда Ostariophysi менее зональное, чем распро- странение некарпообразных. Харациновые и болыпинство сомов в основ- ном являются тропическими рыбами, но их распространение не строго зональное. Карловые и родственные им рыбы имеют преимущественно не- зональный характер распространения, хотя у некоторых родов или групп родов (единица меньшая, чем семейство) тенденция к зональности наблю- дается. Основной характер распространения семейств, относящихся к над- семейству Cypriniformes, радиальный, а не зональный; они расселяются вдоль материковых массивов из одного главного центра распространения. Таким центром является тропическая Азия — Индо-Малайская область. Семейство Homalopteridae приурочено к этому району. Семейство Cobi- tidae является там самым многочисленным и разнообразным, но несколь- ко его видов распространено по всей Евразии и в Северной Африке. Се- мейство Catostomidae (чукучановые), которое теперь встречается главным образом в Северной Америке, не имеет радиального распространения, но оно соответствует радиальному распространению надсемейства Cyprini-
1ormes в целом. (;ypris)idae очень разнообразны в тропической Азии и довольно большое число их простирается по всем умеренным широтам Евразии; сравнительно немногие ветви этого семейства достигают Афри- ки. но некоторые из них распространились по всему Африканскому кон- тиненту, а несколько ветвей достигает Северной Америки и также распро- странилось почти по всему континенту. Таков современный характер рас- пространения надсемейства Cypriniformes. Их ископаемые данные, обоб- щенные Ромером (Romer, 1945, р.,>83), слишком скудны, но все же они по- казывают, что данное надсемейство эволюционировало внутри Индо-Ма- лайскд~й области и перемещалось за ее пределы главным образо~ в третич- ное время, что создало современный характер его распространения (рис. 14).
~CEgю~ д
У("ВР~ННаЯ ЗОИа ЕВфазиЦ
Cyprinidae — -~ Cypri ni dae
Cobitidae (
Catostomi dae
Самы сем. Sitar Иае
CoMbI сем. ВаагЫае
i Сомы сем. Sisoria'
~ 1
Сомы сом. АтешгЫае) ~-,~ф — ~ — ~ <
' Йны ceo. Ameiuridae
~ ~1 11'~
ддд д дд ддю~~д дождю
Ф4д=Иалайская
облосаь Юмжая А~ерша
~~11
Cypri ril.dae ~
° ъ °
~1~~ 1 1 1
Cyprini ае
t".obi tidae ~ Cobi Ыае
~ Нотайр8ет~ае
1ц (СаШа1отШае)
QO Cyprini formes
«9
'~~~-(~фагас[по) — — — — — Charocins 5 (4)
Ъ
Ъ
+ Gymnoti formes 4(Ф)
Сомы 6(З) ~Сомы 9(6) — — — — - Сомы У(У)
-~ Сайг&Шае
~A)~us à
СЬагас1по 2(1)
Р и с. 1 -~~. Радиация иресноводныъ рыб отряда Ostariopl>ysi: современное расп ространение и основные линии расселения сеиейств. («()uarterly Bevies of Biology», 23, 19-i8, tahle 3, р. 11.) Названия в скобках означают предполагаемое распространение этих групп в прошлом, в местах, где они в настоящее время больше не существ~кл.. Цифры означают число семейств харациновых и тропических сомов B данных областях. Цифры в скобках — число эндемичных семейст».
Таким образом, различие между характером распространения некар- нообрнзных (не Ostariophysi) строго пресноводного отдела и Ostariophy- я заключается в следующем: некарпообразные в основном имеют зональ- ное распространение (рис. 12): все семейства приурочены или почти при- урочены или и тропикам, или к умеренной зоне северного полушария; из тропических рыб два семейства встречаются по всему земному шару во всех основных тропических районах обитания пресноводных рыб, а ряд северо-умеренных семейств встречается исключительно только в север- ном полушарии или встречалось там в прошлом. Распространение карпо- образных менее зональное: они более многочисленны по количеству видов и в какой-то степени по числу семейств по сравнению с некарпообразны- ми, но ни одно семейство не приурочено к единой климатической зоне земного шара. Только некоторые группы отчасти имеют зональное распро- странение, а характер распространения самой доминирующей группы
Глпва 2
Cypriniformes — главным образом радиальный, и радиация этой группы. очевидно, шла из центра, находящегося в тропиках Юго-Восточной Азии. На протяжении длительного периода весь отряд Ostariophysi, возможно, радиировал из тропиков Старого Света (рис. 14). Из этого следует. что ес- ли южноамериканские формы произошли из Старого Света или в преде- лах Старого Света, то расселение шло из тропиков в умеренные зоны. Так это, видимо, и было.
Зональное и радиальное распространение недостаточно четко разгра- ничивается. Хотя некарпообразные в основном рыбы зонального распро- странения, некоторые из них, особенно лабиринтовые рыбы (Anahaiiti- dae и Channidae), первоначально имели радиальное распространение. И хо- тя для Ostariophysi характерно в основном радиальное распространение, некоторые из них имеют частично зональное распространение или локали- зуются в отдельных зонах. Более того, зональный характер распростране- ния некарпообразных включает много локализаций, разрывов и других показателей их отступания (ископаемые данные). А в радиальном распро- странении отряда Ostariophysi наблюдается меньше разрывов и других показателей расселения. Все это позволяет предположить, что радиальное распространение, вероятно, означает раннюю стадию в расселении доми- нирующих групп пресноводных рыб, а зональное — позднюю стадию их истории. Зональность — это своего рода локализация, и, возможно, ()Hd является ступенью, через которую проходят ранее доминирующие, но от- ступающие группы на своем пути к дальнейшему сокращению ареалы и окончательному вымиранию. Это наводит на мысль о корреляции между доминированием и характером распространения, что более подрооно, будет рассмотрено ниже.
Несомненно, не все группы, имеющие зональное распространение, должны иметь одинаковую историю. Некоторые (вероятно. лососевые'.), возможно, всегда имели зональное распространение в связи с ограничен- ностью их распространения или характером приспособляем< сти.
Региональное распространение пресноводных рьи1
Предыдущее обсуждение касалось распространения пресноводных рыб на всем земном шаре или на его отдельных болыиих территориях. Теперь кратко остановимся на особенностях их распространения на каж- дом континенте в отдельности. Карты континентов (или материковых фаунистических областей) приводятся в гл. 7.
Пресноводные рыбы Африки. Фауна пресноводных рыб Африки раз- нообразна (ее состав см. рис. 12 и 14). В дополнение к рис. 14 следует ска- зать, что карповые, харациновые и сомы многочисленны и и ироко распро- странены, хотя и не совсем равномерно, в Африке и что там известно толь- ко два местных вида из семейства (;obitidae. Основные работы по распро- странению рыб в Африке следующие: Boulenger (1905; 1909 — 1916); Pei- legrin (1912; 1921; 1933, р. 10 — 19) и iVichols и Griscom (1917, р. 739 — 752). Дополнительные специальные ссылки даны при последующем рассмотре- нии вопроса.
Наиболее богата и разнообразна фауна пресноводных рыб тропиче- ской Западной Африки от реки Сенегал до Конго (в последней — самая богатая). Все архаические и генерализованные группы пресноводных рыб Африки (рис. 12) лучше всего представлены на западе тропической зоны, а некоторые группы приурочены только к ней. Большая часть этой фау-
ны через бассейн озера Чад достигает Верхнего Нила, а остальная прости- раг'reer вниз по Нилу к его низовьям, достигая даже его устья (Boulenger, 1907). Ь[ногие рыбы Нигера, озера Чад и Нила идентичны или близкород- ственны, что свидетельствует о недавней связи между этими водными систе- мами. Рыбы реки Конго, относящиеся к тем же генерализованным груп- пам, отличны от них, очевидно в связи с тем, что они более длительное время находились в изоляции. Верховья Нила и притоки Конго в настоя- щее время в сезон ливней фактически соединяются, но это соединение, вероятно, недавнее, и, по-видимому, сравнительно немногие рыбы сумели расселиться через него. Таким образом, Конго и Замбези обменялись не- которыми видами рыб, но не фаунами в целом; в Замбези имеется срав- нительно богатая фауна рыб, за исключением многоперых и некоторых друг1rz тропических групп Западной Африки.
Реки тропической Африки, текущие на восток, кроме Замбези, срав- нительноо невелики и обладают еще более ограниченной фауной рыб, в ко- торой также отсутствуют многоперые и другие характерные западно- африканские группы. Этот пробел отчасти восполняется наличием многих видов (но немногих родов) семейства Cyprinidae, которых сравнитель- но немного в Западной Африке и которые, как полагают, достигли Вос- точной Африки с севера (из Азии) аналогично вьюновым в районе озе- ра Тана.
Ьольшие озера восточно-центральной Африки обладают высокоэн- деьшчной фауной рыб с преобладанием хромисов и несколькими другими видами рыб (Brooks, 1950). Только в озере Ньяса насчитывается около 178 вндов хромисов, из которых 174 являются эндемиками, представляя 20 эндемичных родов.
В южном направлении, к rory от реки Замбези, которая сама в какой- то степени имеет обедненную фауну (см. выше), фауна пресноводных рыб постепенно обедняется (подробности см. Barnard, 1943, 1948). Один род харациновых (очевидно, только один) достигает реки Лимпопо и в преде- лах небольшого радиуса распространяется за ее пределами. Карпозубые второго отдела (не строго пресноводные) достигают южных точек на реке Замбези, но не доходят (на значительное расстояние) до южной оконеч- ности континента. Хромисы второго отдела распространены до окрест- ностей Ист-Лондона, но фактически Капской провинции не достигают. Два cтрого пресноводных семейства сомов, Clariidae и Bagridae, достигают реки Оранжевой и заходят немного дальше (но не до самых южных бас- сейнов). В последние, представляющие собой небольшие реки южных рай- онов Африки, проникает только два настоящих пресноводных семейст- ва: A»abarrtidae и Cyprinidae. Семейство Anabantidae имеет прерывистое распространение. Тропические формы этого семейства не встречаются к югу от реки Замбези, но два эндемичных вида (возможно, образующие отдельный род Sandelra) изолированы на южной оконечности континента. Семейство (;yprinidae имеет более или менее сплошное распространение. но дало эндемичную группу видов на крайнем rore. Следует отметить. что карповые, которые появились в Африке сравнительно недавно,— един- ственные строго пресноводные рыбы, сплошной ареал которых доходит до ю;кной оконечности континента. В южноафриканских пресных водах имеется также несколько видов солевыносливых рыб, включая один вид семейства Qalaxiidae. Последний, помещенный в эндемичный подрод, обычен вдоль большей части южной границы континента и к северу по реке, Олифантс. Это варьирующий вид с двумя промежуточными форма- ми — горной и озерной. Очевидно, он не заходит в море, так же как
и некоторые австралийские и южноамериканские представители этого семе~~ства.
На (;eaepe Африки на огромной территории, занимаемой пустыней (;axapoii, существует несколько видов пресноводных рыб (Pellegrin, 1921.
1937). Пресноводные рыбы первого отдела представлены несколькими видами карповых и двумя видами сомов рода Clnri~ns. а из второго отдела— несколькими видами карпозубых и тремя родами хромисов. Все эти семейства широко распространены в оазисах Сахары, а некоторые виды, встречающиеся в поверхностных водах, были найдены также и в подзем- ных (см. ссылку к хромисам в списке семейств рыб). Эти рыбы, вероятно, являются реликтовыми с тех сравнительно недавних времен, когда климат Сахары был более влажным. Их географические связи чрезвычайно сме- шаны. По крайней мере некоторые из карповых и карпозубые являются, по-видимому, северными (средиземноморскими) формами, а сомы семей- ства,lariidae и хромисы — в основном тропическими.
И, наконец, фауна пресноводных рыб северо-западной части Африки (Pellegrin, 1921), включая горы Атлас и прибрежные районы на север от них, сильно отличается от фауны основной части континента. Малое число видов здесь объясняется, по-видимому, ограниченностью пресных вод и их сезонностью.
Из строго пресноводных рыб в этом районе имеются только карло- вые — одни с европейскими, а другие с восточными связями,— и один вид европейского вьюна. Здесь также обитает несколько не строго пресновод- ных карпозубых, в горных ручьях — форель, а вблизи побережья — ко- люшка (оба вида, несомненно, представляют северные солевыносливые семейства), угри и другие солевыносливые рыбы. Сомы, хромисы и все другие типично тропические африканские рыбы здесь отсутствуют.
Таким образом, говоря о распространении пресноводных рыб Афри- ки, надо отметить следующие особенности: богатство фауны наряду с раз- нообразием древних ветвей (от западной тропической зоны к Нилу), не сравнимое ни с одним местом на земном шаре, причем наиболее богатая фауна отмечается в реке Конго; сравнительная бедность фауны на восто- ке, отчасти компенсирующаяся карповыми рыбами северного (азиатского) происхождения; местное радиирование рыб, особенно хромисов, в боль- ших озерах; постепенное обеднение фауны на юг к Капской провинции; скудость фауны рыб (представлена несколькими видами рыб в изолиро- ванных водах) и, наконец, изоляция с незначительным переходом в се- веро-западной части континента, резко отличающейся от остальной, очень ограниченной по видовому составу фауны пресноводных рыб близкород- ственной европейской.
Эти особенности обусловлены отчасти местными климатическими ус- ловиями, отчасти размером бассейнов (чем крупнее реки, тем больше в них рыб и разнообразнее их видовой состав) и в какой-то степени историей различных речных бассейнов.
О происхождении и расселении древних групп африканских пресно- водных рыб известно мало. Более современные карповые и вьюны, оче- видно, проникли из Азии. Некоторые рыбы из группы колючеперых, ве- роятно, также азиатского происхождения (рис. 12).
Пресноводные рыбы Ееразии. Состав пресноводной фауны рыб Азии и Европы представлен на схемах 12 и 14, но они недостаточно хорошо отра- жают неравномерность распространения рыб на этом материке.
Полной сводки по распространению рыб на всем континенте Евразии нет. но по отдельным частям имеются следующие работы: Pellegrin, 19'3
1! ресн овод н ые рыбы,
(Сирия); Kosswig, 1952 (Турция); Нога, 1934 (Афганистан); Day, 1889, Нога (различные статьи — Индия и т. д.); Н. М. Smith, 1945 (Таиланд); ШеЬег, Beaufort, 1911 — 1951 (Зондские острова); Nichols, 1943 (Китай); Мог(, 1936 (Восточная Азия и Япония); Берг, 1934 (Евразия, исключая тропики), 1932 (Европа) и другие его работы, окончательно завершенные изданием трехтомника «Пресноводные рыбы Советского Союза и сопре- дельных стран», и многие другие авторы.
Основная часть тропической Азии с главными островами рифа Боль- ших Зондских островов (Суматра, Ява и Калимантан) имеет обширную фауну пресноводных рыб, которая чрезвычайно богата карповыми 11 ии олизкородственными, а также относительно богата колючеперыми рыбами, но в ней отсутствуют все архаические и почти все африканские семейства сельдеобразных. Расселение пресноводных рыб в целом относительно рав- номерно шло через восточную часть Индо-Малайской области (Индоки- тай) к островам Ява и Калимантан. Оно относится ко времени недавнего существования материкового моста между островами и континентом. Рас- пространение пресноводных рыб на островах прослеживается довольно хорошо в старых речных системах, низовья которых теперь затоплены мо- рем (рис. 59). Основная фауна Индо-Малайской области включает болыпое число разнообразных рыб низменностей, а также некоторые семейства и часть семейств, приспособленных к жизни в горных потоках. Расселение многих рыб низменностей (не всех) происходит более или менее непре- рывно в западном направлении на полуостров Индостан (Нога, 1944). Рыбы стремительйых потоков не могут широко расселяться в связи с от- сутствием в настоящее время единой системы горных ручьев, и тем не ме- нее некоторые из них встречаются в изолированных горных районах Южной Индии. Хора (Нога, 1949) зарегистрировал 15 таких видов в шести семействах на полуострове Индостан, оторванных от основного ареала тех же видов дальше на востоке. Он полагает, что эти рыбы заселили Ин- дию незадолго до плиоцена (слишком поздно для рыб этого типа, чтобы достичь Цейлона) и что они проникли в Индию через горы Сатпура и дру- гие возвышенности, в то время когда последние представляли собой почти единую систему и получали большее количество влаги. Многие рыбы встречаются в горных реках северной части (не в полуостровной) Индии и в ручьях на южных склонах Гималаев, но их происхождение (с востока) понять гораздо легче.
На северных склонах Гималаев и в других высокогорных районах Центральной Азии значительно более ограниченная фауна рыб состоит главным образом из особой группы карповых (Schizothoracinae — рас- щепобрюхие), одного рода гольцов (Nemacheilus) и одного рода сомов (Glgp- tosternum) из семейства Sisoridae. Все эти рыбы приспособлены к жизни в горных реках и, очевидно, расселялись с востока на запад, в обход Гималаев, а не с юга на север, через них (Нога, 1937, и др.). Некоторые другие рыбы проникли в центральную часть высокогорной Азии иными путями, но такого широкого распространения там не имеют.
В восточной части Азии к северу от тропиков через Китай происхо- дит постоянный обмен. Некоторые тропические семейства совсем не достигают умеренной зоны. Сомы семейства Homalopteridae и Sisoridae достигают реки Янцзы, где также встречаются рыбы умеренных широт: реликтовые веслоносы и китайский чукучан, а в верховьях рек — лосо- севые. Это рыбы одной и той же речной системы, но обитающие на различ- ных ее участках. Другие тропические семейства поднимаются еще дальше на север на различные расстояния: сомы семейств Siluridae и Bagridae
и лабиринтовые рыбы семейства Channidae достигают Амура, где они встре- чаются со многими рыбами умеренных широт, включая щуку, солевыносли- вых миног, осетров, лососевых, подкаменщиков, колюшек и налимов. Ято, несомненно, слишком упрощенное изложение. Перемещение основных групп рыб с юга на север сложно, кроме того, существует перемещение и с востока на запад или, что вероятнее всего, интердигитация (взаимное проникновение) ( ichols, 1943) китайских рыб с рыбами центральной высокогорной части Азии. Далее характер распространения еще более ус- ложняется за счет большого разнообразия семейства ('yprinidae и слож- ного перехода от южных родов к северным, происходящего внутри этого семейства. Не удивительно, что при таких обстоятельствах мнения ихтио- логов о том, где следует проводить фаунистическую границу, расходятся. Берг (1934) считал, что, поскольку на большей части Китая доминируют тропические рыбы, его надо включить в тропическую Индо-Малайскую область. Мори (Мог1, 1936) хке полагал, что только самая южная часть Ки- тая может быть отнесена к Индо-Малайской области, а основная часть- к палеарктической области (Старый Свет, умеренная зона северного по- лушария). Он проводит разделяющую границу у южного Наньшаня, не- много севернее тропика Рака. Николс (Nichols, 1943, р. 5) также проводит не очень четкую фаунистическую границу для рыб, районы распростра- нения которых находятся около границы между тропической зоной и уме- ренной зоной северного полушария. Но споры о том, где должна прохо- дить эта граница, менее важны по сравнению с тем основным фактом, что китайская фауна пресноводных рыб — огромная, сложная и переход- ная фауна, ее переходы охватывают колоссальную территорию от тропи- ков до реки Амур, находящейся на одной широте с Британской Колумбией.
В Северной Азии, к северу от Амура и к северу от центрального высо- когорья, фауна рыб представлена некоторыми карпообразными, несколькими видами вьюнов, щук, двумя видами окуневых и, особенно на крайнем се- вере. оолыпим числом лососевых и других северных солевыносливых рыб. Из последних подкаменщики (семейства Cottidae, радиировали в озе- ре Ьайкал, образовав там два эндемичных семейства: голомянки — Co- mephoridae и широколобки — Cottocomephoridae) с 9 родами и 16 или 18 видами (см. список семейств). Широко распространенная фауна pbIo даль- него севера Азии в значительной части является общей и для Северной Европы. Большая часть этих рыб достигает Британских островов, а неко- торые (меньшая часть) — Ирландии.
Далее, в Европе к югу распространено только несколько других ви- дов рыб. Большой сом Silurus glands, который в восточном направлении расселился до района Аральского моря, в западном — достиг Рейна. В Юго- Восточной Европе, особенно в реках, впадающих в Черное и Каспийское моря, имеется несколько эндемичных или реликтовых окуней (несколько родов и около 10 видов) и один вид евдошки семейства Umbridae; послед- ний приурочен к определенным частям Дуная и Днестра. В фауне евро- пейских пресноводных рыб преобладают карповые, из которых одни ши- роко распространенные рыбы, а другие являются эндемиками или реликтами, что особенно характерно для Юго-Восточной Европы (Бал- канский полуостров). В средиземноморской части Европы водится не- сколько карпозубых из второго отдела пресноводных, то есть не строго пресноводных рыб. На Балканском полуострове (в Греции) в изолирован- ных местах обитания встречается сом азиатского рода Parasilurus. Ho в Южной Европе строго пресноводных рыб первого отдела африканского происхождения нет. Небогатая европейская фауна становится еще бед-
II ресноводные рябы
бо
нее в Испании и к>жной Италии, где нет ни сомов, ни щук, ни окуневых и единственными строго пресноводными рыбами первого отдела являются несколько карповых и два вида вьюновых (относящигсся к надсемейству (;урн н11огтея). Этот пробел компенсируется различными солевыносли- вымп рыбами средиземноморской (умеренно теплой) группы.
В примыкающей к Африке юго-западной части Азии (от Малой Азии до Аравийского полуострова и Ирана включительно) видовой состав фау- ны рыб весьма ограничен в связи с неблагоприятными физико-географи- ческими условиями. Большинство из них относится к карповым, из кото- рых несколько видов (обитатели ручьев) широкораспространенного (аф- ро-азиатского) рода усачей Barbus и Gnrrn встречаются даже в изолиро- ванных водах южной Аравии (Trewavas, 1941). В Юго-Западной Азии имеется также несколько представителей вьюновых, карпозубых и незна- чительное количество других строго пресноводных рыб первого и второ- го отделов с различными взаимосвязями. Так, африканские хромисы до- стигли Сирии и Палестины и образовали там эндемичный род и несколько видов. Немногие сомы широко распространенных тропических групп встре- чаются в Сирии и в системе Евфрата и Тигра, а тропический вид семей- ства МаяФасетЬе1Ыае (колючие угри) — в Евфрате и Тигре. Большой ев- ропейский сом и щука достигают северных частей Малой Азии. Несколько восточнее, в Афганистане, и также на юге Ирана появляются различные индийские рыбы, а в северном Афганистане — форель (северная) и спе- цифические группы карповых и сомов из Центральной Азии. Как видно, засушливая юго-западная часть Азии является районом двойного перехо- да фаун рыб, с одной стороны, от африканских форм к формам Индо-Малай- ской области, а с другой — от тропических к северным. Подробности та- кого двойного перехода пока не очень хорошо изучены.
Таким образом, основные особенности распространения пресновод- ных рыб в Евразии следующие: богатство фауны, несмотря на отсутствие архаических групп, на основной части тропической Азии и современных материковых островах шельфовой зоны Больших Зондских островов (Су- матра, Ява и Калимантан); обеднение фауны и специализация в районах центрального высокогорья; переход, охватывающий многие группы на большой территории, через Восточную Азию от тропиков на север до Аму- ра; широкое распространение с востока на запад особых северных групп рыо с образованием эндемичных семейств (происходящих от солевыносли- ных групп) в озере Байкал; бедность фауны рыб в Европе, увеличиваю- щаяся в западном направлении, особенно в Испании и южной Италии, но с несколькими эндемичными или реликтовыми группами в Юго-Восточ- ной Европе; оедность фауны и двойной переход в юго-западной части Азии.
I ro касается перемещения рыб Евразии — карповых и им близкород- ств 'HHbIx и некоторых других, то, по-видимому, эти рыоы произошли и распространились на север и запад из тропиков Азии, а другие, более древ- ние северо-умеренные формы, вероятно, исчезли перед их появлением. ;)то предположение основывается на взаимосвязях и распространении са- мих Сург1п1(огтея и на их способности успешно приспосабливаться к раз- ноо(~разным климатическим условиям и местам обитания; на характере распространения всей фауны рыб в Восточной Азии, где северо-умерен- ные группы рыб, по-видимому, отступили, а тропические, включая кар- повых, распространились в северном направлении в умеренную зону; на небольшом числе других рыб в умеренных широтах Евразии и на релик- тов(1м распространении некоторых из них. Перемещения рыб в умеренных широтах Азии и Европы во время и после оледенения в плейстоцене—
Г.гава 2
это особый вопрос, который здесь детально не рассматривается, а ссылки на него приводятся в гл. 10.
Пресноводные рыбы Северной Америки. Состав фауны пресноводных рыб Северной Америки также представлен на рис. 12 и 14. По сравнению с фауной умеренных широт Евразии эта фауна характеризуется большим разнообразием некарпообразных, чем карпообразных, хотя число видов в трех семействах последних в Северной Америке велико. (Еще два южно- американских семейства отряда Ostariophysi достигают только южной краевой части Северной Америки.) Все известные пресноводные рыбы (:е- верной Америки были включены в списки (Jordan et al., 1930), но по рас- пространению рыб в Северной Америке нет ни одной общей сводки, хотя имеется ряд неплохих работ по местным фаунам (Hubbs, Lagler, 1947).
Основная часть фауны пресноводных рыб Северной Америки сосредо- точена в восточно-центральной части материка (рис. 12), в реке Миссисипи и ее притоках. Фауна более мелких рек атлантического побережья, т»- кущих на восток, беднее, что становится особенно заметным на северо- востоке. Например, в штате Нью-Гэмпшир (Carpenter, Siegler, 1947) местные пресноводные рыбы первого отдела представлены только двумя видами щук (Esox), тремя мелкими американскими окунями семейства Centrarchidae, тремя окуневыми рыбами и рыбами из отряда Ostariophysi, 14 мелкими карповыми, тремя чукучановыми и двумя видами сомов из семейства Ameiuridae. Кроме того, здесь интродуцированы щука, два чер- ных морских окуня, три других представителя семейства Centrarchida», судак и др. В Новой Шотландии (Livingstone, 1953) местных строго прес- новодных рыб еще меньше: насчитывается только один вид окуневых, во- семь карповых, один вид чукучановых, один сом и один вид (из второго отдела) карпозубых. Все 12 видов встречаются на перешейке, соединяю- щем Новую Шотландию с материком, но менее половины из них достигает южной оконечности Новой Шотландии или северного острова Кейп-Бре- тон, а в некоторых изолированных ручьях Новой Шотландии строго прес- новодные рыбы отсутствуют. Оскудение фауны рыб, наблюдаемое в севе- ро-восточном направлении, отчасти можно объяснить тем, что некоторые рыбы, особенно крупные, не смогли вернуться после отступания ледника в плейстоцене в более изолированные реки побережья.
В юго-западном и западном направлениях богатая фауна рыб Мисси- сипи быстро редеет. Из некарпообразных строго пресноводных рыб (пер- вый отдел) только семейства Centrarchidae и Percidae достигают Мексики, где южные границы их распространения примерно совпадают с границей тропиков. Однако все три североамериканских семейства отряда Ostari~>- physi заходят (значительно или незначительно) дальше южных границ отмеченных выше семейств некарпообразных. То же самое наблюдается у групп из не строго пресноводного отдела, границы которых рассматри- ваются более подробно в разделе пресноводных рыб Центральной Америки.
В западной части Соединенных Штатов Америки, на запад от конти- нентального водораздела, проходящего через Скалистые горы, известно только три местных вида строго пресноводных некарпообразных рыб, каждый из которых представляет разные восточноамериканские семей- ства, каждый образует эндемичный род и приурочен к различным бассей- нам. К ним относятся: Xovumbra hubbsi (Umbridae), обитающий в реке Чехейлис (штат Вашингтон), Columbia transmontana (Percopsidae) в реке Колумбия (штаты Орегон и Вашингтон) и Archoplites interruptus (Centrar- chidae) в бассейнах рек Сакраменто и Сан-Хоакин (Калифорния). Четвер- тый вид Bafinesquiellus pottsii, представляющий еще одно семейство (Рег-
Пресноводные рыбы
cidae), достигает рек Мексики, текущих на запад. Щуковые (ЕяосЫае) доходят до западных бассейнов только на Аляске, а траут-пэрчи (Percop- sidae) — по крайней мере до верхней части Юкона, текущего на запад. Черная рыба, или даллия (Dallia), тоже приурочена к западным бассей- нам Аляски. На запад от водораздела из отряда Ostariophysi встречаются чукучаны и ряд мелких карповых, но местные сомы, за исключением двух- трех видов в западной Мексике, отсутствуют. Южнее на западе имеется несколько карпозубых из не строго пресноводного отдела. Ряд карповых и карпозубых встречается в реликтовых водах даже в самых засушливых частях юго-запада.
К северу богатая фауна строго пресноводных рыб восточно-централь- ной части Северной Америки также постепенно обедняется. Довольно не- значительная часть ее заходит в полярные воды по реке Маккензи, а еще меньшая встречается в тундре. Строго пресноводным рыбам, видимо, со- вершенно не удается достичь некоторых отдаленных районов тундры, включая часть полуострова Лабрадор. Из строго пресноводных рыб толь- ho три вида широко распространены в Аляске и еще два заходят на Аля- ску по Юкону. К этим трем видам относятся щука, северный чукучан и черная рыба, Dallia. Все эти три вида рыб строго пресноводного отдела встречаются также в Азии, проникая через Берингов пролив, и это позво- ляет предположить, что недавно здесь был холодный материковый мост. Большинство северных пресноводных рыб Северной Америки, несомнен- но, являются солевыносливыми, они схожи с аналогичными рыбами Азии и Европы и указывают на переход или родство в обоих направлениях. На- пример, несколько видов лосося характерны для северо-западной части (;еверной Америки и северо-восточной Азии, а атлантический лосось (Sal- ino salar) — для северо-восточной части Северной Америки и Европы. Из строго пресноводных рыб первого отдела ни одна (за исключением цир- кумполярно распространенной щуки) не характерна для Северной Аме- рики и Европы, но некоторые формы восточной части Северной Америки и Европы взаимосвязаны, включая род Umbra, некоторых окуневых и кар- повых. Берг (1932) рассматривает этот вопрос более детально.
Таким образом, основные особенности распространения пресноводных рыб в Северной Америке таковы. Богатая и разнообразная фауна на во- стоке, особенно в крупнейшей речной системе Миссисипи; некоторое обед- нение ее вдоль атлантического побережья, увеличивающееся на северо- восток; постепенное обеднение, по крайней мере групп первого отдела, на юго-запад и через Мексику; ограниченность карповых, чукучанов и кар- позубых в западном направлении от водораздела материка и разбросан- ность очень немногих представителей других восточных семейств (и неко- торых солевыносливых рыб, главным образом близ побережья); распрост- ранение на север в полярную фауну немногих пресноводных рыб первого отдела и многих солевыносливых рыб, близкородственных рыбам северной Азии, в случае северо-восточных форм, близких рыбам Северной Европы.
Что касается происхождения и перемещения, то многие современные пресноводные рыбы Северной Америки, вероятно, являются частью преж- ней, третичной фауны, которая была типичной для Европы, Северной Азии и Северной Америки (Берг, 1932). Большинство некарпообразных строго пресноводных семейств этой фауны (рис. 12) вымерло на западе (;еверной Америки (они также уменьшились в числе или вымерли в Европе и особенно в Азии) и сохранилось или даже радиировало в восточной части, как, например, окуневые. В восточной части Северной Америки встречают- ся также некарпообразные, возмон но, представители сс,мейства Cegtrar-
1' fueu .'?
chidae, которые, вероятно, ведут отсюда свое начало и игракп сущест- венную роль в истории распространения рыб. Наконец, три семейства от- ряда Ostariophysi в Северной Америке являются, по-видимому, иммигран- тами из Восточной Азии. Установить точно время их проникновения труд- но: сомы семейства АтешгЫае и чукучаны, возможно, появились в ран- нетретичное время, а карловые — в миоцене. Влияние плейстоцена на рыо (;еверной Америки и Евразии рассматривается в гл. 10.
Пресноводные рыбы Южной Америки. Фауна пресноводных рыб Юж- ной Америки, с одной стороны, известна малым числом исходных автох- тонных корней, а с другой — богатством и эндемизмом некоторых групп отряда Ostariophysi. Из некарпообразных строго пресноводных рыб (рис.
12) в Южной Америке имеется только три семейства, которые насчитыва- ют там пять известных видов. Из отряда Ostariophysi (рис. 14) в Южной Америке встречаются только харациновые и электрические угри (обра- зовавшиеся от харациновых) и сомы (нет ('ypriniformes), но к ним отно- сится около 2000 видов, известных в Южной Америке, со многими энде- мичными родами и примерно 17 эндемичными семействами плюс одно се- мейство (СЬагасЫае), общее с Африкой. В Южной Америке имеется также ряд не строго пресноводных карпозубых и хромисов и некоторые солевы- носливые пресноводные рыбы. Эта фауна давно привлекала внимание их- тиологов, и основные особенности ее распространения были исследованы ,"-Эйгенманном (Ligenmann, 1909, 1910, 1927, 1942 [With Allen) etc.).
Самая богатая фауна пресноводных рыб Южной Америки сконцен- трирована в Амазонке и ее притоках. Это, несомненно, наиболее богатая по видовому составу рыб речная система земного шара. Уже сейчас насчи- тывается более 1000 видов различных амазонских рыб, но возможно, что это число удвоится, когда исследование реки будет полностью завершено (Муегя, 1947 — 1949, pt. 2, р. 13). Только одних сомов в системе Амазонки известно более 450 видов, в то время как в значительно лучше изученной всей системе реки Миссисипи их насчитывается менее 20 видов (iXIyers)! .Видовое разнообразие харациновых почти такое же, как и сомов, но пер- вые намного превосходят их по числу особеи; в Амазонке обитает несколь- ко видов электрических угрей, все три встречающиеся в Южной Америке семейства некарпообразных строго пресноводных рыб, различные карпозу- бые солевыносливого раздела, хромисы и некоторые другие солевыносливые «иды. Многие группы рыб Амазонки, но не все (например, не двоякоды- шащие рыбы) достигают Гвианы и Ориноко. Многие, но опять-таки не все, например не костноязычные и не представители семейства 1М'анд Ыае, достигают системы рек 11арагвай — Парана. Истоки этих рек олизко под- ходят к системе реки Амазонки и, возможно, даже соединяются с ней. Близ северо-западной части Южной Америки в относительно неоольшой и изолированной реке Магдалена фауна рыб обедняется (М1!ез, 1947); так, в ней отсутствуют все три семейства строго пресноводных некарпо- образных рыб. Ихтиофауна реки Магдалена насчитывает 149 известных видов: 58 харациновых, 6 электрических угрей, 67 сомов, 5 карпозубых (в двух семействах), 4 хромисов и 9 видов солевыносливых рыб в 7 семей- ствах. Реки восточного побережья Бразилии на юг от Амазонки и на север от Параны большей частью невелики, их ихтиофауна ограниченна ii род- ственна фауне Амазонки.
Узкий западный водосборный бассейн тропической части К)жной Америки, к западу от Анд, характеризуется очень неравномерным распро- странением фауны рыб. В Тихоокеанском водо 'oopHOM бассейне Колум- бии и Эквадора имеется довольно большое число строго пресноводных рыб.
Il ресноводные рыбы
Рыбы районов, расположенных по обеим сторонам этой части Анд, более сходны, чем рыбы районов, находящихся по обеим сторонам ('.калистых гор. Некоторые западные формы рыб реки Колумбии, вероятно, проник- ли с. востока, обогнув горы с севера. А рыбы речных бассейнов на запад- ных склонах горных хребтов эквадора, вероятно, произошли от амазон- г«'их, проникнув через Анды или переселив«пись сн>да еще до их образо«щ- ния (Kigenmanrr, 1927, р. 5 — 6; см. также Kige««marrn, АИе««, 1942). На юг от Перу в сухом Тихоокеанском водосборном бассейне (к западу от Анд) фауна пресноводных рыб скудная.
В высокогорной части Анд обитает ограниченная, специализирован- ная фауна горных ручьев, которая на самых больших высотах (иногда до 14 000 футов) экваториальной зоны состоит в основнои из одного р«зда харациновых (~ео«аппа) и двух родов сомов (Astr««blepus и Pygirdirum) (Чуегя, 1947 — 1949). Эта фауна прослеживается почти на всем протяжении Лнд (от их северного»онца на юг в Чили и Патагоник«) с некоторыми мест- ными изменениями в ее составе: например, род EIntcherir«r, родственньш ро- ду Pygidrurn, для некоторых районов Чили и северной Патагонии является эндемичным. На более низких высотах в экваториальной части Анд появ- ляются другие рыбы, живущие в горных реках со средней скоростью те- чения, а ниже ' ООО или 3000 футов численность и разнообразие видового состава рыб быстро увеличиваются и происходит сложны«1 переход и фау- не низменностей. Последняя как очень богата, так и чрезвычайно сло;«'на экологически (Муегя 1947 — 1949). Одна часть высокогорных Анд — - Лль- типлано (южнып 11еру), включая озеро Титикака (высота более 12 000 футов над уровнем моря), обладает эндемичным подсемейством карпозу- бых Orestiinae, состоящим из четырех родов и около О видов, из которых боль«пинство населяет само озеро Титикака (Brooks, 1950, р. 162). Кроме вышеперечисленных, из других рыб в бассейне озера Титикака встречает- ся еще только сом.
Многие рыбы тропической фауны низменностей, подобно некоторым африканским рыбам, проникающим по Нилу в умереннук> зону северных широт, достигают умеренной зоны южных широт по речным системам рек Парагвай — Парана, доходя до места их слияния. Но по мере продвижения на юг происходит непрерывное обеднение фауны пресноводных рыб. ко- торое начинается уже на юго-востоке Бразилии, а южнее Буэнос-Airpeca скудость фауны пресноводных рыб становится особенно очевидной. Из строго пресноводных рыб только несколько харациновых и сомов дости- гают Чили и 11атагонии. Некоторые из них близкородственны широко рас- пространенным тропическим южноамериканским формам или, очевидно, произошли от них, а один из родов сомов, видимо, самый примитивный из современных американских сомов, отнесен и самостоятельному семей- сr'r«y Diplomystidae, которое приурочено к центральным частям Чили и Лргентины. Другой сом, Nernatogenys, видимо, самьш примитивный из современных представителей семейства Pygidiidae, встречается в отдель- ных районах Чили (Eigenmann, 1927, р. 13). Географическая история этих родов неизвестна, но вполне разумно предположение, что они являются реликтами сравнительно примитивных групп, переместив«««ихся в тропи- ческую часть Южной Америки. Самая южная точ»а, котор««««дoс«««га«««« представители харациновых, например Cherry«don, живущий «««royal rrrr;»- бренностей.— 41""18' ю. пт. (Ki~enmarrrr, 1927). Выносливые сомы семей«та- ««а Pygidiidae обитак«щие в горных реках и ручьях, заходят значитесь«но дальше на юг, а род Hatcheria достигает по крайней мере «7'3««' н«. ш. (I';i- епта«««1, 1918, р. ' 69, 83). Южнее границы распространения этих рыо
встречаются только миноги, галаксиды и рыбы рода Aplocliiton, доходя- щие до южной оконечности материка. Все они являются антарктическими солевыносливыми группами, которые встречаются также в Новой Зеландии и южной Австралии и которых в Южной Америке в северном направлении становится все меньше, в то время как убывание числа тропических форм пресноводных рыб происходит к rory. Согласно Эйгенманну (Eigenmann.
1927, р. 15), антарктическая фауна рыб в целом заходит на север только до Вальдивии, один представитель рода Galaxias и Aplochiton встречается на севере до Консепсьона и только миноги достигают широты Вальпараи- со. Далее к северу антарктических рыб со стороны Южной Америки, омы- ваемой Атлантическим океаном, встречается меньше. В южном полушарии в более теплых частях умеренной зоны Южной Америки встречается не- сколько солевыносливых рыб других групп: Mugilidae, Atherinidae, Ser- ranidae.
Таким образом, наиболее характерные особенности распространения фауны пресноводных рыб в Южной Америке следующие: богатство фауны в центральной части (несмотря на бедность предковых форм), особенно в Амазонке; более бедная фауна в прилежащих речных системах к северу и rory от Амазонки и еще беднее в относительно изолированной реке Маг- далена; сравнительная бедность фауны в западных бассейнах Колумбии и Эквадора; постепенное обеднение фауны в юханом направлении, начи- ная с юго-восточной Бразилии, но прерываемое системами рек Парагвай— Парана, текущими.на юг; бедность и специализация рыб в горных реках Анд и радиация некоторых карпозубых, преимущественно в озере Тити- кака, и переход на крайнем юге к совершенно другой фауне с антаркти- ческими солевыносливыми рыбами.
Происхождение рыб Южной Америки будет рассмотрено ниже. Сов- ременное богатство фауны почти полностью обусловлено ее эволюцией и перемещением внутри континента. Между большими реками происхо- дил обширный обмен рыбами, а наибольшая из них — Амазонка до сих пор имеет самую многочисленную и самую разнообразную фауну рыб; здесь наблюдается та же общая корреляция ме~кду размером рек и числен- ностью и разнообразием рыб, которая отмечалась в Африке и Северной Америке. Видимо, вообще из Амазонки рыбы расселялись во всех возмож- ных направлениях (Eigenrnann, 1909, р. 371). Северная часть Анд не яви- лась для них существенной преградой, так как целый ряд рыб либо обог- нул, либо пересек горы. К югу от Амазонки фауна рыб меняется в основном за счет уменьшения числа видов, и это уменьшение идет до южной части района Ла-Платы, где остается всего несколько харациновых и сомов, а еще далее остаются только сомы и, наконец, строго пресноводные рыбы исчезают совсем. Эйгенманн в этих особенностях распространения пытался ~видеть свидетельство прежнего деления Южной Америки на отдельные части: основную — Архамазонскую и южную — Архиплата (Kige~rmann, 1909, р. 370) или Архиоразильскую п Архигвианскую (Eigenmann, Allen, 1942. р. 35), имеющие каждая свою независимую фауну, но я этого взгляда не разделяю. По-видимому, есть только одна основная фауна строго пресноводных рыб Южной Америки, которая радиирует из Амазон- ки. и отдельные распространившиеся за пределы бассейна Амазонки части этой фауны. которые имеют обедненный видовой состав и которые лока- лизовались или специализировались в разных местах, чего и следовало ожидать при таком разнообразии мест обитания, как в Южной Америке.
Пресноьодные рыбы Центральной А.мерики. Переходная территория Центральной Америки. хотя последняя не является ни континентом, ни
Пресноводные рь~бч
на<"гоящей фаунистической областью, настолько важна, что заслуживает детн.»» ного рассмотрения. Из более ранних работ по распространению рыо на этой территории можно назвать работы Ригэна (Regan, 1906 — 1908, 1». ~ и — ХХ Х [1). Эйгенманна (Еigenmann, 1909, р.309, 310) и Майерса (Муегз.
1938. р. 350 — 351). Все рыбы Центральной Америки. известные в то время. »»к:»в>чены в перечень Джордана и др. (Jordan et al., 1930). Пресноводные рыоы Панамы, которая является своего рода переходной территорией. расеи»»триваится Хильдебрандом (Hildebrand, 1938). Работы Буэна (Buen, 19-I(i) и Майлса (МИез, 1947) о рыбах Мексики и реки Магдалена хотя и не касаются непосредственно фауны Центральной Америки, интересны с точ- ки зрения природы и происхождения данной территории. Карта Цент- 1»н:»ьной Америки дана в гл. 7.
Фауна пресноводных рыб Центральной Америки — сильно обеднен- ная. В ней полностью отсутствуют все некарпообразные строго пресно- водные семейства (рис. 12) как Северной, так и Южной Америки. Выпа- дение этих рыб фактически начинается на континентах недалеко от самой 1~ентральной Америки. Из 9 семейств умеренных широт Северной Америки только два (Centrarchidae и Percidae) достигают Мексики. Каждое из этих семейств представлено несколькими видами в бассейне реки Рио-Гранде, но только один вид от каждого семейства проникает дальше на юг пример- но до границы тропиков: представитель семейства Centrarchidae — на восток, а представитель семейства Percidae — на запад от континенталь- ного водораздела. Из 3 семейств (всего 5 видов) некарпообразных строго пресноводных рыб Южной Америки ни одна не достигает даже реки Маг- далена или более мелких рек северо-западной части Южной Америки.
('. представителями отряда Ostariophysi дело обстоит лучше, но они немногочисленны даже в Центральной Америке и также начинают умень- шаться в числе на прилежащих участках Южной и Северной Америки. Из карпообразных Северной Америки карповые идут на юг, уменьшаясь ко- :»и»ес твенно, к реке Бальсас в южной Мексике (зона тропиков), недале- »о от перешейка Теуантепек. Чукучановые идут на юг к Гватемале, так- ;»;е количественно уменьшаясь: два североамериканских рода останавли- на»отея у северного края Мексики; третий род (ЛХохог1ота) доходит до к»жно-центральной части Мексики, и только один, четвертый, род (Car- pi»»»des) достигает Гватемалы. Сомы семейства Ameiuridae также постепен- н»» исчезают в южном направлении, и только род Ictalurus достигает Гва- темалы. Из южноамериканских представителей отряда Ostariophysi не- которое сокращение основных групп наблюдается даже в реке Магдалена. Довольно большое число их все же достигает Панамы, но лишь немногие проникают значительно дальше. Фактически очень немногие из тех кар- пообразных. которые заходят в восточную Панаму из Южной Америки, достигают даже западной Панамы. Хильдебранд (Hildebrand, 1938, р. 221.
»7) описывает 19 родов с 32 видами харациновых в восточной и централь- ной Панаме и только 5 родов и 9 видов в западной. Из 7 видов сомов се- мейства Loricariidae, из которых некоторые обычны для Центральной 11анамы, только один вид известен в западно»» части страны. Еще мены»»е »»и'»»оамериканских представителей отряда Ostariophysi проникает в 1(е»»тральную Америку, где они имеют различные границы распростра- нения. Из харациновых эндемичный род Bra»nocj»arax является эндемиком озера Никарагуа. Из южноамериканских родов Boeboides достигает Гва- темалы, Brycon — Гватемалы и Юкатана и Л styanax — притоков Рио- Гранде в Техасе. В Гватемале водится единственный вид электрического угря, который, как известно, обитает за пределами центральной Панамы.
Из пресноводных сомов Южной Америки, очевидно, только представите- ли семейства Pimelodidae, главным образом род Л/гапгйа, заходят в глубь Центральной Америки, практически достигая реки Пануко на восточном берегу Мексики. Приведенные здесь подробности, возможно. недостаточ- но полны и иногда неточны, но они хорошо показывают обеднение фауны строго пресноводных рыб в Центральной Америке. Эти рыбы не только немногочисленны, но и очень бедны здесь эндемичными родами.
Бедность строго пресноводных рыб частично восполняется неп р«- порциональным обилием не полностью пресноводных групп. Хромисы значительно многочисленнее в Центральной Америке, чем в Южной, и площадь их наиболее высокой численности довольно точно викариирует с той, которую занимают строго пресноводные рыбы: хромисы становятся многочисленными и в западной Панаме (Hildebra»d, 1938, р. 221), там, где происходит резкое обеднение фауны строго пресноводных рыб, и, хотя несколько родов хромисов достигают Мексики, их ареал там суживается к северу и только один род достигает реки Рио-Гранде. Ни один из этих родов не контактирует с фауной рыб Северной Америки. У хромисов Цент- ральной Америки наблюдается значительный эндемизм родов и вид«в. хотя большинство из них (или все), по-видимому, образовались от южно- американского рода Cichlasumrc (Regan, Муегя). Карпозубые также непро- порционально многочисленны и отличаются высокой степенью эндемизма в Центральной Америке (Муегя, 1938, р. 350 — 351). Панцирные щуки. ко- торые теперь, несомненно, являются главным образом представителями Северной Америки, проникают на юг через восточную часть Мексики и Центральную Америку до озера Никарагуа. В пресных водах Централь- ной Америки имеется также довольно большое число солевыносливых рыб. Состав фауны пресноводных рыб в более изолированных частях Централь- ной Америки примерно такой же, как и на полуострове Юкатан (H«l>t>s, 1936), который включает только один вид (с двумя подвидами) хараццно- вых и один вид (с несколькими местными подвидами) сомов семейства Pi- melodidae и не имеет никаких других строго пресноводных форм, а из рыб не строго пресноводных здесь насчитывается 13 видов карпозубых, 6 хромисов и несколько солевыносливых видов.
Таким образом, можно отметить следующие особенности распрост- ранения пресноводных рыб в Центральной Америке: большая бедность строго пресноводных групп (нет некарпообразных рыб этого раздела, мало представителей отряда Ostariophysi); уменьшение, наблюдающееся в связи с постепенным обеднением фауны числа североамериканских групп на всем протяжении от Северной Америки к Мексике (только немногие группы достигают Гватемалы, а южноамериканские группы уменьшаются по мере удаления от Южной Америки, хотя некоторые из них распростра- нены по всей Центральной Америке до Мексики и одна достигает реки Рио- Гранде); относительно большое число ряда не строго пресноводных групп, причем площадь их наиболее высокой численности не совпадает с той, ко- торую занимают строго пресноводные группы, а эндемизм среди них прояв- ляется в большей степени, чем у строго пресноводных форм.
В общем плане Центральная Америка является местом, где встречанп- ся две большие фауны рыб: северо-умеренная и тропическая (рис. 15), но переходная область этих фаун охватывает более обширную площадь, чем только Центральная Америка. Наиболее богато фауна рыб Северной Америки представлена в реке Миссисипи, и ее обеднение идет постепенно в западном и южном направлениях к Мексике и через нее до конечной гра- ницы распространения строго пресноводных форм в 1'ватемале. ~!>ау»в
1)ресноводные рыбы Южной Америки, как уже говорилось, богаче всего представлена в Ама- зонке, и обеднение ее начинается в других болыпих реках Южной Аме- рики, оно становится очевидным в реке Магдалена, особенно резко выра- ;кено в реках Панамы, хотя небольшая часть фауны распространяется че- рез Центральную Америку и достигает близлежащих участков Северной Америки. Отсутствие всех строго пресноводных рыб, за очень небольшим исключением, в Центральной Америке и частично в Мексике привело к развитию в этих районах характерной эндемичной фауны, образовав- шейся от групп не строго пресноводных рыб.
Природа фауны пресноводных рыб Центральной Америки свидетель- ствует о том. что при расселении пресноводные рыбы с трудом достигли р и с:. I.5. Переход фаун строго пресноводных рыО в Мексике
>r Центральной Америке. Прерывистые стрелки показывают проникновение двух североамерикан- ских семейств некарпообразных рыб первого отдела в Мексику; сплошные стрелкп — распространение отряда Ostariophysi из Северно~~ ll Южной Америки. Стрелки показывают приблизительное расстояние,
на которое рыбы продвинулись в переходной зоне. Центральной Америки, области довольно молодой в биологическом отно- шении, и что препятствия, затрудняющие их распространение, возникали на прилежащих участках материков Северной и Южной Америки. При дан- ных условиях, вероятно, первыми в Центральную Америку проникли соле- выносливые рыбы, распространяясь вдоль побережий или преодолевая узкие пространства соленых вод, а затем они, видимо, радиировали в изо- ляции. Строго пресноводные рыбы появились, видимо, позже, в связи с тем, что им потребовалось болыпе времени для преодоления более значитель- ных преград, встречающихся на их пути, или, возможно, они стали рас- селяться после геологических преобразований, которые уменьшили число существовавших препятствий. Следует обратить внимание на то, что боль- шинство строго пресноводных рыб достигло Центральной Америки с юга, а не с севера и что граница распространения некоторых южных групп, рас- селившихся через Центральную Америку, заходит значительно дальше, чем у любых северных групп.
Глава 2
Теперь стоит снова вернуться к не строго пресноводным рыбам, то есть ко второму отделу пресноводных рыб (по Майерсу). Последний. как мы уже указывали, состоит из групп, которые хотя и обитают в основ- ном в пресных водах, включают виды, которые заходят в море или некото- рое время могут находиться в морской воде. Эти группы встречаются на многих островах, где строго пресноводные рыбы отсутствуют. И эти же самые группы представлены непропорционально многочисленными высо- ко дифференцированными формами в Центральной Америке. Все факты, вместе взятые, показывают важные особенности расселения рыб и историю некоторых участков суши, где встречаются эти рыбы. При желании можно сомневаться в реальности подразделений Майерса, но нельзя сомневаться в их пользе для зоогеографии.
Не мешает снова взглянуть на контрастное распространение строго пресноводных рыб и рыб отряда Ostariophysi. Из некарпообразных ряд семейств, часть из них древние, распространен в Северной Америке. причем некоторые семейства довольно многочисленны. Из отряда Ostariophysi только 3 семейства находятся на основной части материка, и, вероятно, некоторые из них проникли туда сравнительно недавно. Но все семейства отряда Ostariophysi проникают на юг Северной Америки значительно дальше, чем любые некарпообразные рыбы строго пресно- водной группы. (Эти три семейства карпообразных, видимо, пришли из Азии и распространились по территории, населенной древней фауной не- карпообразных рыб в умеренных широтах Северной Америки.) Такая же особенность распространения характерна и для юга: из строго пресновод- ных рыб Южной Америки только представители Ostariophysi достигают Центральной Америки. Это одна сторона ухсе отмеченного основного разли- чия: некарпообразные на земном шаре в целом распределяются главным образом зонально, их возможности расселения ограничены, в то время как карпообразные имеют радиальное распространение и их расселение не ограничено. Это очень существенное различие.
Пресноводные рыбы Австрплии. Кроме двоякодышащих и костноязыч- ных рыб, которые уже рассматривались, в Австралии имеется много соле- выносливых пресноводных рыб. Они включены в сводный список рыб Ав- стралии (McCulloch, 1929 — 1930), а сведения о их происхождении и распро- странении даны тем же автором (McCulloch, 1927).
V Фауна рыб в пресных водах Австралии лучше всего представлена ми- ногами, галаксидами и т. д., сомами (морские семейства), окунеобразны- ми рыбами (морские семейства), бычками и атеринками. Некоторые другие морские группы представлены только одним или очень малым числом прес- новодных австралийских видов. Имеется также несколько видов угрей се- мейства Anguillidae, один широко распространенный вид глубоководных yrpeii семейства Synbranchidae и один вид семейства Amphipnoidae. И, наконец, есть несколько интродуцированных видов лососевых и кар- повых. В настоящее время некоторые виды карповых сильно размножи- ЛИСЬ.
Одни из местных пресноводных (но солевыносливых) рыб широко рас- пространены по всей Австралии, другие, преимущественно миноги, галак- сиды и т. д., имеют зональное распространение, придерживаясь более прохладной южной части континента. В отдельных случаях наблюдается локализация родов и видов.
Все пресноводные рыбы Новой Гвинеи, за исключением костноязыч- HhIx. являются солевыносливыми, и в целом фауна рыб Новой Гвинеи тесно связана с фауной Австралии. Фактически в южных реках Новой Гви-
))ресноыодные рыбы
неи водятся те же рыбы (и другие водные животные), что и в реках север- ного побережья Австралии (Whitley, 1943). Это дает возможность пред- положить (так же, как и в шельфовой зоне Больших зондских островов в Азии) существование огромной речной системы в плейстоцене, нижняя часть которой в настоящее время затоплена морем (рис. 59). Реки север- ной части Новой Гвинеи имеют родственную, но в какой-то степени и от- личающуюся фауну рыб с эндемичными родами (Whitley, 1943).
Таким образом, происхо~кдение большинства пресноводных рыб Авст- ралии не является загадкой, за исключением двоякодьппащих и костно- язычных, которые, возможно, проникли в Австралию непосредственно из моря или, преодолев его, из других мест земного шара.
Иереходные области и преграды распространению пресноводных рыб
В местах соприкосновения соседних различных фаун пресноводных рыб наблюдаются явления, дающие возможность судить о факторах, кото- рые регулируют распространение рыб. Резкой границы, отделяющей одну фауну от другой, обычно нет, но имеются зоны перехода или перекрыва- ния фаун с постепенным уменьшением элементов обеих фаун в противо- положных направлениях. Степень перехода варьирует в зависимости от рассматриваемых факторов.
Как правило, зональные климатические различия помогают лишь раз- граничить основные фауны рыб, но в одном случае зональный климат яв- ляется единственным очевидным и главным причинным фактором. Это наблюдается в Восточной Азии, где в большой переходной зоне шириной около 2000 миль, от тропиков до реки Амур, населенной многими видами рыб, встречается тропическая фауна Индо-Малайской области и уме- ренная фауна Евразии. Далее на запад, где влияние климатической зо- нальности усиливается преградами в виде гор и пустынь, происходит уже незначительное смешение фаун рыб тропической и умеренной зон; в зоне между тропической Африкой и умеренными широтами Европы, где кли- матический фактор усиливается пустыней и морем, фауна строго пресно- водных рыб не имеет точек соприкосновения, в крайнем случае это проис- ходит только внутри семейства Cyprinidae. В Америке тропическая и се- веро-умеренная фауны рыб отделяются не только климатическими зонами, но и узким перешейком, который несколько миллионов лет назад был за- топлен соленой водой, и отчасти таким препятствием, как засушливая территория на юго-западе Северной Америки. Тем не менее немногие стро- го пресноводные рыбы Южной и Северной Америки встречаются и пере- крывают ареалы друг друга в Гватемале и Мексике.
В двух других случаях зональные климатические преграды помогают разграничить фауны pbIo, которые сами по себе находятся в сходных кли- матических условиях. Фауны тропиков Африки и Азии разделяются Юго- Западной Азией, климат которой скорее не тропический, а умеренно-теп- лый и относительно засушливый. Эта преграда оказалась очень эффектив- ной, ибо тропическая фауна рыб Африки и Индо-Малайской области в це- лом сильно отличается друг от друга.
Однако несколько видов тропических рыб встречается в районе этой своеобразной преграды. Очевидно, некоторые из них расселились здесь уже в настоящее время, а некоторые в прошлом.
Своеобразной преградой между фауной умеренной зоны Азии и Се- верной Америки является арктический климат. Теперь их разделяют так-
же и соленые воды океана, но это произошло относительно недавно, и, очевидно, основной преградой все-таки стал климат. Совсем недавно, когда Азия и Северная Америка соединялись через Берингов пролив сухопутным мостом, климат здесь, видимо, был настолько холодным, тто преодолеть это препятствие могли только арктические рыбы. Однако в более древние времена, когда климат был теплее, через:этот район смогли расселиться многие виды рыб.
11устыни — это также очевидные преграды, но их «лияние H«р«с- пространение рыб не столь велико, как обычно считают. Нет такого ме- ста, где бы две различные основные фауны рыб отделялись только одной пустыней без участия других факторов. Наличие некоторых рыб в изоли- рованных водах пустынь, включая Сахару, показывает, что когда-то эти пространства были менее сухими, чем теперь, и представляли мень иун1 преграду для рыб. Очень часто пути расселения рыб огибают пустыни или проходят через них, например река Нил, которая протекает через пу- стыни Северной Африки. Таким образом, пустыни сами по себе не могут быть серьезной преградой для рыб, но в сочетании с другими факторами они приобретают важное значение.
Горы также служат преградой для распространения рыб на корот- ком отрезке времени, но их роль на протяжении более длительных перио- дов незначительна, так как животные всегда находят обходные пути или успевают пересечь их. Так, многие рыбы обогнули горы Центральной Азии с востока; различия в фаунах рыб низменностей Южной и (;еверной А:шя объясняются скорее влиянием климата, чем горных преград. В Южной Америке рыбы, расселяясь, огибали Анды или проникали через их север- ную часть. Кажется, что Скалистые горы Северной Америки для расселе- ния пресноводных рыб являются самой непреодолимой из существуп>щцх горных преград, однако карповые и чукучановые, а также и карп~луоьи широко распространены по обеим сторонам этих гор. Известно также по меньшей мере три других вида рыб, которые преодолели их на тер(эито- рии (",оединенных Штатов Америки, два вида пресноводных рыб, преодо- левших этот водораздел на крайнем севере, и еще два — на севере Мекси- ки. Тот факт, что фауны рыб на востоке и западе от (,'калистых гор столь различны, можно только отчасти объяснить наличием горных прегр«д. а отчасти ограниченностью мест обитания на западе.
Единственными наиболее эффективными преградами для пресно«~>д- ных рыб являются, по-видимому, соленые воды. Даже такие относите;~ ь- но неболыпие пространства соленых вод, которые теперь отделяют 13ест- Индию от Северной и Южной Америки, Мадагаскар от Африки и остро» (;улавеси от Калимантана, могут остановить распространение строго пресноводных рыб на длительное время. В тех редких случаях. когда пред- полагаемые строго пресноводные рыбы пересекают узкие пространств« СОЛеНЫХ ВОД, ВОЗНИКаеТ СОМНОНИР ОТНОСИТЕЛЬНО CTPIIBHN СОЛеВЫНОСЛИИОСТИ ЭТИХ ВИДОВ.
Такие преграды, как зональный климат, пустыни и горы, лишь з«- медляют темп расселения и ограничивают или обусловливают численносп и видовой состав расселяющихся рыб. Наиболее действенно сочетание д«ух видов преград, например климатической зональности и засухплп»ых областей. Но и в этом случае расселение пресноводных рыо в как~и~- то мере продолжается. Те рыбы, которым со временем удается прсо (о- .[еть даже сложные преграды, являются рыбами либо дом~~н~~рующими (например, (yprinidae), либо с особой приспособляемостью и..ш солены- носливостью (например, (;yprinodontidae).
1) ресн овод и ы е рыбы
,Доминирование и конкуренция
Тсперь обратимся к понятию «доминирование» и его связи с особен- ностями распространения пресноводных рыб.
Доминирующие животные — это преуспевающие животные. (;ледует ш>мнить, что карпообразные являются доминирующими пресноводными рыоами, значительно превосходящими по численности всех других строго пресноводных рыб, вместе взятых. Из отряда Ostariophysi семейство (;ург)- шаве и близко им родственные рыбы численно преобладают.
(;емейство (;yprinidae, несомненно, самое большое семейство пресно- водных рыб по числу родов, видов и очень часто такн'е и по количеству особей. В фаунах пресноводных рыб карповые в пределах своего ареала численно преобладают до такой степени, какой ни одни другие пресновод- ные рыбы нигде не достигают. ( ypriniformes, главным образом сами Cyp- ~iiiidae. господствуют во всей тропической и умеренной зонах Евразии.
13 !китае, например, на территории, простирающейся от краевых зон тропиков и захватывающей значительную часть умеренной зоны север- ных широт, Николс (lVichols, 1943) насчитывает 268 видов (:ург1«Ыае. <~3 вида других Сургип1огтея, 65 видов пресноводных сомов и только 15 видов некарпообразных из строго пресноводной группы (плюс 2 вида карпозубых не строго пресноводной группы и 73 вида солевыносливых рыо). В Европе Берг (1932, р. 174) насчитывает 94 вида Cyprinidae, 10 дру- гих (;ypriniformes, только 2 вида пресноводных сомов и 13 некарпообраз- ных рыб строго пресноводной группы (плюс 4 вида не строго пресновод- ных и 1' 1 вид солевыносливых рыб, из которых многие встречаются в прес- ных «одах только случайно).
(:емейство,yprinidae довольно разнообразно по своей структуре. а надсемейство Сург1п1!огтея в целом представлено еще более разнообраз- нычи формами. Структурное разнообразие не является необходимым кри- терием доминирования, но оно обычно увеличивается в том случае, если доминирующая группа радиирует.
Семейство (,"ург111Ыае обладает также очень высокой выносливость|о или болыиой способностью приспособления к различной пресноводной среде. Это семейство доминирует как в тропиках, так и в умеренной зоне северного полушария, и в этом отношении оно единственное в своем роде. Ни одно другое семейство пресноводных рыб не является столь порази- тельно преуспевающим в обеих зонах. Более того, карловые сумели рас- прос граниться через места с неблагоприятными условиями обитания, ко- торыс стали преградой почти для всех других пресноводных рыб, и достиг- ли многих относительно изолированных мест в пределах общих границ распространения семейства. В отдельных местах семейство (,yprinidae и нек«торые другие представители Сург1п11огтея являются единственными представителями строго пресноводной группы, как, например, в Испании, ю,'кной Италии, в умеренной зоне северо-западной части Африки, в юж- ной Лравии, в западных частях (:еверной Америки (чукучановые относят- ся к «адсемейству (:ypviniformes) и, возможно, в отдельных частях Южной Африки. В некоторых из этих мест карловые встречаются совместно с»ар- иозубыми, но последние относятся к не строго пресноводному разделу и являются по сравнению с карповыми значительно менее многочисленной и менее преуспевающей группой. Наличие карповых в изолированных местах с неблагоприятными условиями обитания, по-видимому, объяс- няется присущими им особыми характерными чертами, включая, воз- можно, их неболыиой размер и способность переносить не только соле-
Глава 3
ную, но, вероятно, и щелочную воду, в то время как существование в этих местах карповых объясняется общим доминированием этого семей- ства.
И, наконец, ископаемые остатки и другие сведения (см. рис. 12 и 14) позволяют предположить, что в умеренных широтах Евразии расселение семейства Cyprinidae было связано с отступанием и исчезновением некото- рых других рыб и что здесь имела место прямая конкуренция, в которой карповые одержали верх.
Итак, Cyprinidae очень многочисленны по количеству видов и числу особей и довольно разнообразны по структуре, обладают высокой вынос- ливостью (или хорошей приспособляемостью), весьма преуспевают и в тро- пическом и в умеренном климате и занимают различные места обитания. включая участки с относительно неблагоприятными условиями или изо- лированные участки. Таким образом, в целом их распространение почти не прерывается на значительной территории, охватывающей большой диа- пазон климатов и труднодоступные места, которые оказались непреодо- лимыми преградами для большинства других пресноводных рыб. Очевид- но также, что карповые конкурировали с некоторыми другими рыбами и заставили их отступить в другие места. Все эти характерные черты созда- ют картину сильно доминирующей группы, в связи с чем характер распро- странения такой группы можно назвать радиальным. Столь очевидное до- минирование этой группы дает нам в руки наиболее достоверные «ключи», которые показывают, что Cyprinidae и Cypriniformes радиировали из глав- ного центра в тропической Азии.
Весь отряд Ostariophysi, к которому относятся "ypriniformes, являет- ся доминирующим и составляет около '/з всех пресноводных рыб первично- го отдела. Ostariophysi доминируют в основном в тропической и умеренной зонах Евразии, в меньшей степени в Африке и Северной Америке и в ооль- шей — в Южной Америке, где, вероятно, имеется 1800 или '000 извест- ных видов отряда Ostariophysi и только всего 5 видов некарпообразных пресноводных рыб первого отдела (плюс значительное число пресноводных рыб второго отдела и некоторых солевыносливых рыб). Распространение всего доминирующего отряда Ostariophysi указывает на очевидную ра- диацию из тропиков Старого Света. Ostariophysi умеренной зоны северного полушария, по-видимому, появились из тропиков Старого Света, а Osta- riophysi Южной Америки явились частью первоначально очень ограни- ченной и, возможно, дериватной фауны. Однако это не нарушает положе- ния, что несомненным центром радиации этого отряда (рис. 14) были тро- пики Старого Света.
Отдельные группы отряда Ostariophysi, за исключением Cyprinifoi- mes и семейства Cyprinidae, не имеют четкого радиального распростра- нения. Харациновые и им близко родственные, хотя и многочисленны в районах своего распространения, а в Южной Америке и очень разнооб- разны, все же в основном приурочены к Африке и Америке, и именно к тропическим зонам этих стран.
Что касается сомов, то они заслуживают более детального рассмот- рения.
Сомы многочисленны, разнообразны и широко распространены (рис.
16), однако ни в одной фауне рыб они не доминируют в такой степени, как семейство Cyprinidae. Очень многочисленны сомы только в тропиках. (Даже морские семейства являются главным образом тропическими.)
В умеренной зоне северного полушария только в Китае и на востоке Северной Америки численность сомов так же велика, как и в тропиках
Иресноводпые рыбы (но нет ничего общего с разнообразием их в тропиках). Очень мало сомов в Юго-Западной Азии, только два локализованных вида имеется в Европе и ни одного в Северной Азии (севернее широты Амура). На большей части западных районов Северной Америки и на других небольших отдельных территориях, где обитают карповые (например, северо-западная часть Африки и часть Аравии), ни одного местного вида сомовых не встречает- ся. Поэтому в умеренной зоне северного полушария сомов не только мень- ше, но и распространены они там менее равномерно и менее непрерывно, Р и с. 16. Распространение строго пресноводных сомов (первый
отдел) (см. текст и список семейств в конце главы). Жирные пересекающиеся линии — основные фауны сомов; стрелки— распространение отдельных родов или небольших частей фаун, включая распространение рода 'lpptostrrnum через Центральную Азию; тонкие пересекающиеся линии — изолированные области с одним ил1т двумя видами сомов; прерывистые линии — другие границы, которых достигли п~н.сн оводные сомы, включая границы распространения вида Sil is~ us
фалья в Европе и т. д. чем карповые. Помимо того что сомы не заходят так далеко на север, как карловые, они также не заходят далеко и на юг Африки, хотя в Южной Америке в южном направлении они проникают дальше, чем любые дру- гие строго пресноводные рыбы; здесь они достигают наивысшего доми- нирования и очень разнообразны по структуре и приспособляемости.
Взаимосвязи различных семейств сомов недостаточно выяснены, и поэтому можно строить только предположения об их эволюции и рассе- лении. Так, если Ostariophysi произошли в тропиках Старого Света, то сомы, вероятно, ведут свое начало оттуда же, но Южной Америки они, вероятно, достигли давно. Вполне реально, что предки южноамерикан- ских сомов проникли в Южную Америку через север и затем там исчезли. Их современное распространение дает возможность предположить, что несколько более поздних групп сомов распространились из тропической Азии на север, но не нашли там благоприятных условий обитания. Предки семейства Ameiuridae, возможно, переместились из тропической Азии в умеренную, а затем в Северную Америку. Если это так, то позднее они вымерли в Азии и на западе Северной Америки. Предполагают, что семей- ство Ameiuridae родственно формам Старого Света, но ископаемые этого се- мейства, найденные в Восточной Азии, сомнительны. Еще более сомнитель- но такое свидетельство, как характер окраски, наводящий на мысль о су-
Глава; ?
ществовании вида этого семейства в Китае. Семейство Siluridae, возможно. распространилось довольно недавно из тропиков Индо-Малайской области в Северную Азию и Европу. Возможно, сейчас в Европе происходи' от- ступание этого семейства, так как два современных европейских вида. обитающих в изолированных местах, вероятно, являются реликтами (см. список семейств рыб). И, наконец, семейство Bagridae, достигшее Амура. и семейство Sisoridae, проникшее в Центральную Азию, по-видимому, яв- ляются еще более современными вселенцами с еще не определившейся суд ьоо~~.
В целом можно сказать, что сомы имеют частично зональное, а ча- стично радиальное распространение. Они являются главным образом тро- пическими рыбами, но несколько их групп незначительно радиировали из тропиков Азии в умеренную зону северных широт.
Из других пресноводных рыб лабиринтовые рыбы, скорее всего. име- ют радиальный характер распространения. В тропической Азии они яв- ляются самыми многочисленными и разнообразными рыбами. Границы распространения двух азиатских родов А п((Ьаз и Ophicephnlus прости- раются до Африки, и те же два рода Восточной Азии распространены на севере южной Маньчжурии и Амура соответственно. Однако лабирин- товые рыбы отсутствуют в Западной Азии и Северной Африке. и эт()т раз- рыв наводит на мысль о том, что умеренная радиация этих рыб уже закон- чилась и они начали отступать.
Для зоогеографии очень важен тот факт, чт() одни группы жив()тных более преуспевают по сравнению с другими (по крайней мере в определен- ные времена и в определенных частях земного шара) и что эти более преус- певающие группы иногда имеют особый характер распространения, ко- торый не ограничивается зональным климатом и болыпинством видов пре- град. Этот факт так же хорошо иллюстрируется на примере пресноводных рыб. особенно семейства (;yprinidae, как и на других животных.
Доминирование весьма относительно, и ()Ho всегда связано с конку- ренцией. Имек)тся различные доказательства того, что среди рыб действи- тельно происходит конкуренция. Интродукция человеком в какие-то во- ды одних рыб иногда вызывает уменьшение численности или истребление других видов рыб. 11одобное вытеснение одних рыб другими, очевидно, происходит и в природе. Распространение в некоторых водах щук отри- цательно влияет на численность других рыб, а появление чукучанов при- водит к уменьшению численности форели. В природе подобные явления возможно проследить только косвенно. История распространения пресно- водных рыб в умеренных широтах Евразии, где карповые расселились, а некоторые другие рыбы, видимо, отступили, предполагает конкуренцик) целых групп. Викариирующее распространение доминирун)щих рыб на всем земном шаре (рис. 17), особенно Cypriniformes и других, основывает- ся на их конкуренции. Видимо, та картина распространения пресновод- ных рыб, которую мы наблюдаем сейчас, есть в какой-то мере резуль.(ат конкуренции между большими группами рыб на больших территориях в течение длительного времени. Конкуренция, вероятно, была очень слож- ной в деталях и влекла за coooii как длительные, так и кратковременные адаптации.
Нелегко установить. какие особенности делают одних пресноводных на больших территориях и в различных местах обитания доминирук)щи- ми, более преуспевающими. чем другие. Все представители отряда Ostario- physi обладают аппаратом Вебера, который, видимо, обостряет слух и. возможно, является причиной доминирования этого отряда в пресных
Ijресноводцые рыбы
иодах. но не в морских. Это объяснение можно считать правдоподобным, но правильно оно или нет, сказать трудно. Более простое объяснение до- минированию карповых среди отряда Ostariophysi найти трудно. Умерен- ное доминирование лабиринтовых рыб происходит, возможно, отчасти и благодаря тому, что они обладают органом дополнительного дыхания.
Зона смешоннмо дотниро5ания
Зона некарпобыхкостистих
Зона скешанного8птниродания
рыбы)
сеоеро=умеренной
-- - — --/юница магд
Южная Аперика
боны
Харацинобые и ор.
Карпову Юые Хром сы
Адстраюия
(салеаынаслааые
рыбы)
Р п с. 17. Доминирующие группы пресноводных рыб основных областей континентов («Quarterly Review of Biology», 23, f94fl,
table 4, р. 12).
Иочему Юго-Восточная Азия была признана центром радиации карпо- ь е оторь сомов и лабиринтовых рыо — это отдечьный вопрос, на который я постараюсь ответить ниже.
Заключение: особенности распространения пресноводных рьгб
Начне с того, что рыбы встречаются почти во всех пресных вода.,- на всем земном шаре и самыми основными чертами их распространения яв- ляются ограниченность распространения и частичная взаимозаменяемость (викариирование). Распространение строго пресноводных рыо, как пра- вило. почти всегда ограничено континентами и современными материковыми островами (кроме Австралии). Пресноводные рыбы не строго пресноводной группы в какой-то степени восполняют отсутствие строго пресноводных рыб на не слишком удаленных от материков островах и в других местах (особенно в Центральной Америке), достичь которых строго пресноводным рыоам было трудно. Рыбы с высокой солевыносливостью хотя и встреча- ются почти везде, но относительно многочисленны на крайнем севере, край- нем юге, в Австралии и на удаленных от материков островах, где строго пресноводных и даже не строго пресноводных рыб мало или совсем нет.
А~щгика
Ap~auvec~ue генерали-зо5онные некорпсоорроз-Сомы алые Харац ивы~
Capri'oi doe(oooo- ~ численны, но со5рвюенные не апт~аапся розно5ра заем)
Хро~ие»
бст5ыност5ые рыбы) (к сеаеру)
ось! реп. Атешгйае Чукучано5ые
урга Ыае(ннееанасленнн но собрененные не оалн-' чаннасл разнееаразаеи) 8екарпоо5розные (есеаенна акунебые) и тропической фауна
Глава 2
В пределах границ распространения строго пресноводных рыб для их расселения характерны и некоторые другие особенности. Некарпооораз- ные имеют тенденцию к зональному распространению: большинство их семейств придерживается одной из двух основных климатических зон земного шара или локализуется в одной зоне. Многие рыбы не строго прес- новодной группы, солевыносливые и некоторые морские рыбы также име- ют зональное распространение. Для представителей отряда Ostariophysi характерно скорее радиальное, а не зональное распространение: рассе- ление по материкам Cypriniformes свидетельствует о наличии главного центра радиации в Юго-Восточной Азии, а распространение отряда Osta- riophysi в целом явно указывает на радиацию из тропиков Старого Света. Однако между зональным и радиальным характером распространения имеются промежуточные, переходные случаи.
Другая общая особенность распространения пресноводных phI()— дифференциация фаун на различных континентах. Фауна Африки пред- ставляет сложное смешение архаических и высших рыб, но с относительно немногочисленными представителями Cypriniformes. В фауне тропиче- ской Азии отсутствуют все архаические рыбы, но она богата представи- телями Cypriniformes и высшими, колючеперыми группами, особенно лабиринтовыми рыбами. В фауне умеренных широт Евразии преобладают Cypriniformes; что касается фауны пресноводных рыб Северной Америки, то она довольно разнообразна: в нее входят два семейства Cypriniformes. но они не доминируют, как это происходит в Евразии. Фауна Южной Аме- рики сильно ограничена, в ней полностью отсутствуют Cypriniforme, но имеются богатые эндемичные группы других видов отряда Ostariopliysi. Б фауне Австралии почти нет строго пресноводных рыб. Эти различия в фаунах соответствуют общему характеру распространения: в тропиче- ской и умеренной зонах Евразии находится центральная область домш(и- рования карповых (Cyprinidae); далее, на прилегающих материках Лфри- кп и Северной Америки находятся смешанные фас ны, вклю (ая некоторых представителей семейства t;ypriniformes; и, наконец, на более изолирован- ных континентах Южной Америки и Австралии отмечаются высокоэнде- мичные фауны, в которых отсутствует семейство Cypriniformes (рис. 17).
Фауны пресноводных рыб разных континентов, несмотря на большие разли(ия в составе, имеют общий характер распространения на каждом континенте. Основная часть каждой фауны концентрируется в самых боль- и(их реках и на самых больших и благоприятных для обитания участках, а постепенное оскудение ее происходит в более мелких реках, на мен (((ux и менее подходящих для обитания удаленных у|астках. Этот характер распространения ярко выражен в Африке, Евразии в целом, в Северной и Южной Америке. Фауны рыб соединяющихся материков обычно отде- ляются друг от друга участками с неблагоприятными условиями обита- ния, на которых численность рыб относительно уменьшается. Том не менее (>тдельны(, ком1тоненты каждоп фауны обычно все же проникают на эти участки и их ареалы перекрывают друг друга, образуя переходные зоны, в которых происходит постепенное уменьшение элементов каждой фауны в диаметрально противоположных направлениях.
Короче говоря, общий характер распространения пресноводных рыб оонаруживает по крайней мере пять следующих особенностеи: 1) геогра- фическая ограниченность строго пресноводных рыо и местами частичное замещение их более солевыносливыми рыбами, 2) зональность, 3) радиа- ция, 4) дифференциация материковых фаун на земном шаре, в которой ярко выражен викариат доминирующих гр~ пп, .i) концентрация кажд(и(
Пресноводные рыбы
основной фауны на больших благоприятных для обитания участках с обед- нением по краям, причем окраины прилегающих фаун перекрывают аре- алы друг друга через большинство видов преград. Эти особенности рас- пространения связаны между собой. Основные из них — ограниченность II взаимозаменяемость (викариат) содержат или влекут за собой все осталь- ные. Викариат отчасти является зональным и выражается в дифференциа- ции материковых фаун. Зональность и радиация взаимосвязаны и copeN- ствуют дифференциации материковых фаун, а концентрация основных фаун на больших участках с обеднением по краям усиливает другие осо- оенности распространения, даже если они выражены по краям не столь часто. Все это так же сложно, как сложно и распространение животных в целом.
Общий характер распространения животных имеет бесконечные мест- ные особенности распределения, создаваемые местными климатами, местны- ми преградами, местными изменениями в водоемах, местной эволюцией и исчезновением различных рыб. О некоторых таких местных особен- ностях распространения упоминалось выше, но большинство из них не является предметом обсуждения в данной книге.
О месте реликтов в распространении пресноводных рыб будет сказа- но ниже. У круглоротых и акуловых рыб в пресных водах древних релик- ТоВ нет. Современные пресноводные представители этих классов относи- тельно недавно были морскими формами. Из костных рыб все не костистые (не Teleostei) являются филогенетическими реликтами. Они немногочис- ленны: из строго пресноводного раздела можно назвать только семейство Polypteridae с двумя родами и 13 видами в Африке, веслоносов с одним ро- дом и одним видом в Китае (другой род с одним-двумя видами в восточ- но-центральной части Северной Америки), ильную рыбу (Amia calla) на востоке Северной Америки, двоякодышащих из семейства Lepidosirenidae с одним родом и тремя видами в Африке (другой род с одним видом в Юж- ной Америке) и двоякодышащих из семейства Ceratodontidae с одним ви- дом в Австралии. Из рыб не строго пресноводного раздела встречается только семейство Lepisosteidae с одним родом и четырьмя или пятью ви- дами в восточной части Северной Америки, в Центральной Америке и на острове Куба. Из этих рыб географически изолирован только один авст- ралийский вид двоякодышащих, и это единственный вид, который строго ограничен в современном распространении. Все другие рыбы, так же как и основные фауны современных пресноводных рыб, водятся — одни в тро- пической зоне, другие в умеренной зоне северного полушария, но все они оолее или менее широко распространены в пределах тех областей, в кото- рых встречаются (см. список семейств рыб). Из всех основных областей, в которых встречаются фауны строго пресноводных рыб, только в тропи- ческой Азии отсутствуют реликты названной выше группы (не Teleostei). Одни реликтовые рыбы, особенно двоякодышащие, ограждены от конку- ренции с большинством других рыб особыми адаптациями и образом жиз- ни, другие — нет. Например, ильная рыба Авиа са1аа, хотя и является единственным реликтом отряда, которьш доминировал в морях юрского периода и когда-то был широко распространен в пресных водах умеренной зоны северного полушария, существует совместно с другими крупными хищными рыбами современной богатой фауны пресноводных рыб восточной части Северной Америки, успешно с ними конкурируя. Эти факты не сов- сем соответствуют широко распространенному мнению среди биологов, что филогенетические реликты обычно встречаются н изолированных или защищенных местах.
Л'ак происходило распростр«нение пресносодных рыб? Современное распространение пресноводных рыб является в основном рсзультатом пе- ремещения самих рыб, сложного процесса их эволюции, расселения, огра- ничения различными препятствиями, отступания и исчезновения их в тех или иных частях земного шара. По особенностям распространения и на- ходкам ископаемых остатков установлено, что в геологически недавние времена одни группы рыб очень широко расселялись, а другие столь же интенсивно отступали. Соответственно прилегающие фауны рыб обычно не резко отделяются друг от друга, а перекрывают друг друга по краям. Это дает возможность предположить перемещение отдельных компонен- тов каждой фауны в противоположных направлениях.
Самым важным недавним событием в расселении пресноводных рыб является расцвет Cypriniformes в Юго-Восточной Азии и их радиация от- туда, сопровождаемая отступанием различных других рыб с большей ча- сти Евразии. Еще более важным и более ранним событием был расцвет Озфаг1орЬуз1 в пресных водах и их радиация из тропиков Старого Света. Эти два события, в основе которых так или иначе заложено зональное распро- странение, по-видимому, и составляют важнейшие черты современного распространения рыб.
Почему происходило распространение рыб~ Распространение рыб происходило благодаря самой природе рыб, природе земного шара и слож- ному взаимодействию их обеих. Это совершенно очевидно, но некоторые стороны этого явления не так легко понять.
В связи с существованием древних преград в виде соленых вод и в связи с тем, что строго пресноводные рыбы не обладают солевыносливостью, их распространение действительно ограничивается территориями материков.
Однако многие другие рыбы, живущие в пресных водах, могут пере- носить как пресную, так и соленую воду. Некоторые группы рыб, обла- дая именно такой способностью, смогли распространиться за пределы гра- ниц строго пресноводных групп, достичь пресных иод на изолированных островах и развиваться в них. Это создало основную особенность распро- странения рыб: частичное взаимозамещение строго пресноводных и не стро- го пресноводных форм.
Зональное распространение пресноводных рыб возникло в результа- те того, что целые группы рыб приспособились к какому-то одному кли- мату — тропическому, умеренному или арктическому. Насколько боль- шое значение играет сам климат, подтверждается также зональным рас- пространением многих солевыносливых и морских рыб. Зональное распро- странение строго пресноводных рыб акцентируется преградами, так как гропическая зона и умеренная зона северного полушария в большинстве случаев разграничиваются на континентах территориями засушливых областей, а в некоторых местах горами или др~ гими преградами, препят- ствующими расселению рыб. Такие преграды не существуют только в во- сточной части Азии. Распространение рыб в этой части показывает, что, несмотря на отсутствие преград, многие пресноводные рыбы все же при- держивались зонального распространения, но при этом резкой границы между фаунами не было.
Радиальное распространение пресноводных рыо происходило в связи с тем, что некоторые доминирующие группы пресноводных рыб эволю- ционировали в определенных местах, особенно в тропи хеской Азии, и распространялись из нее в разных направлениях, насколько позволяла непрерывность суши, относительно мало ограничиваясь климатическими зонами или оольшинством видов преград (исключая соленые воды).
II ðåñêîâîäíûå рыбы
Дифференциация материковых фаун рыб происходила в результате очень сложных факторов, которые вызывали расширение или сокращение ареала, выживание, эволюцию или вымирание различных рыб на различ- ных континентах. Основная особенность дифференциации — доминирова- ние семейства Cyprinidae в Евразии, смешанное доминирование на прилежащих материках и отсутствие карповых на более отдаленных материках — является результатом их расцвета и в Юго-Восточной Азии.
Концентрация основных фаун рыб на больших территориях с олаго- приятными условиями обитания дает возможность предположить связь между площадью этих территорий, численностью и природой фауны. Это положение играет важную роль в распространении животных.
В заключение можно сказать, что основная черта распространения пре- сноводных рыб определилась тремя факторами: древними преградами соленых вод, зональностью климата, природой и эволюцией самих рыб, а также площадью обитания, играющей пока еще неопределенную роль.
Что говорит распространение рыб о древних сушах и климат«х? Ископаемые данные и современный характер распространения рь б на севере от тропиков показывают, что с раннетретичного времени в умерен- ной зоне северного полушария существовала достаточно хорошо выра- женная фауна пресноводных рыб. Таким образом, можно предположить, что климатические зоны существовали по крайней мере с раннетретично- го времени и были расположены так же, как и сейчас. Это говорит и о том, что в третичном периоде Евразия и Северная Америка были соедине- ны и что климат в районе соединения был достаточно теплым, чтобы обес- печить обмен многих рыб умеренной зоны. Более тонкие подробности рас- пространения рыб рассказывают нам об изменениях климата и водных систем в плейстоцене. К этому периоду Азия и Северная Америка были все еще соединены (или соединились вновь через Берингов пролив). но климат в этом районе был холодный и это соединение пересекали аркти- ческие, а не южные рыбы.
К югу от тропиков (в умеренной зоне) распространение пресноводных рыб не свидетельствует о таких болыпих изменениях.
Строго пресноводные рыбы Южнои Африки и Южно1х Америки. оче- видно, проникли в южные части этих континентов из тропиков незави- симо друг от друга. В Южной Австралии строго пресноводных рыб нет. Таким образом, на этой группе рыб нельзя проследить связь между юж- ными оконечностями континентов. Я полагаю, что галаксиды и другие со- левыносливые рыбы, которые все же встре чаются в южном полушарии, распространились через море, хотя некоторые ученые не придерживаются этой точки зрения (например, Stokell, 1945).
По характеру распространения пресноводных рыб в пределах тропи- ков можно узнать об истории материков гораздо больше, чем по распро- странению любых других ныне живущих на этих материках животных.
Расселение пресноводных рыб не позволяет предполагать большие изменения в связях между материками Старого Света. Фауны рыб Африки п тропической Азии в целом очень различны, но они имеют около 1<> об- щих семейств строго пресноводного раздела и даже несколько оощих родов. Эти две фауны отделяются теперь довольно эффективной прегр»- дой, и, по-видимому, так было в течение длительного времени, за иск,ио I(— нием факта случайного взаимопроникновения нескольких видов рыб 'tv.— рез Юго-Западную Азию. У некоторых видов рыб, особенно у карповых, оомен через этот путь произошел недавно. 11олное отсутствие строго прес-
Глава 2
новодных рыб на Мадагаскаре указывает на то, что ранние обмены также происходили через Юго-Западную Азию, а не через прямое соединение суши. Различия в фаунах рыб Африки и Азии указывают также на боль- шие различия в истории обоих материков. Фауна Африки представляет сложное смешение архаических и более современных, в основном проник- ших из Азии, рыб. Фауна тропической Азии состоит главным образом из более современных рыб, относящихся к группам, которые, по-видимому, возникли и радиировали в тропической Азии. История формирования Аф- риканского континента, по всей вероятности, благоприятствовала сохра- нению там древних рыб, а тропической Азии — эволюции новых. Более специфические подробности распространения пресноводных рыб в этих частях земного шара раскрывают многое из истории рек и озер, но это не может быть освещено здесь с исчерпывающей полнотой. Работы Хора (несколько статей) дают интересный материал по истории пресноводных рыб и рек Индии, а работа Бьюфорта (Beeaufort, 1951) раскрывает с этой точки зрения историю некоторых озер и рек Индии, Больших Зондских островов и Африки.
Пресноводные рыбы не свидетельствуют о том, что в связях Австра- лии с другими материками произошли какие-либо значительные измене- ния. Австралийский представитель костноязычных и австралийская двоя- кодышащая рыба, очевидно, как и любые другие рыбы, могли появиться из Азии. Но, несмотря на то что распространение двоякодышащей рыбы свидетельствует о предположительном соединении Австралии с другими материками, все же одного этого примера недостаточно.
Прежде чем сделать заключение о том, что могут рассказать нам прес- новодные рыбы о древних сушах и климатах, остановимся на происхож- дении фауны рыб Южной Америки.
Происхождение пресноводных рыб Южной Америки (нехарактерный случай). Анализ фауны рыб Южной Америки вызывает болыцие трудности. Если бы было известно, каким образом пресноводные рыбы достигли Юж- ной Америки, то была бы больше известна и история всех материков с ме- лового периода. Но мы этого не знаем и можем делать только некоторые осторожные предположения. Лучше всего начать с четкого представления о положении Южной Америки на земном шаре по отношению к другим материкам (см. фронтиспис).
Фауна Южной Америки — самая большая из всех фаун пресновод- ных рыб — произошла от немногочисленных предков. Пресноводные рыбы Южной Америки первого и второго отделов относятся только к семи ос- новным группам (табл. 2).
Все семь основных групп представлены и в Африке и только часть из них — в Азии. В Азии есть также один представитель костноязычных, два рода семейства Nandidae и два рода сомов, но нет рыб из семейства Lepi- dosirenidae, нет харациновых, имеется только один род хромисов, кото- рый, очевидно, сравнительно недавно проник в южную Индию из Африки. и только несколько видов карпозубых, которые, вероятно, не являются прямо родственными южноамериканским. Более того, между южноаме- риканскими пресноводными рыбами и современными настоящими северо- американскими группами нет прямых родственных связей, за исключени- ем солевыносливых карпозубых. В целом фауна пресноводных рыб Юж- ной Америки по своим связям главным образом африканская. И эти свя- зи древние. В настоящее время в Африке и Южной Америке имеется четы- ре общих семейства строго пресноводных рыб и два семейства не строго пресноводных, но нет ни одного общего рода. Далее, важен тот факт, что,
Пресноводн.ые рыбы.
Табл пца 2
Основные гр1.плы пресноводных рыб Южной Америки С~ рого пресноводная группа, некарпообразные
1. Osteoglossidae: одно семейство, имеется также в Африке, Индо-Малайской
области, Австралххйской областхх (возиожно, солевыносливо)
. Nandidae: одно семейство, имеется также в Африке и Индо-\[алаххскохх
области
:3. Lepidosireni<lae: одххо семейство, имеется также в Афрххке Строго пресноводные Ostariophysi
Сомьх: девять семейств; другие сеиеххства в Африке, Ипдо-Малаххскохх ооластхх
5. Characiformes с происходящим от него Gymnoti form es: девя гь семейств; ха- рациновые встречаются также в Африке
Н» строго пресноводная группа
6. Cichlidae: одно семейство, также в Африке до южной Индии
7. Cyprinodontes: два главных семейства; широко распространены Cyprinodonti- dae, а доминирующие роды в Южной Амерххке и Африке являются родствен- ными Poeciliidae. Общие с Северной Америкой
несмотря на то, что все основные группы строго пресноводных рыб Южной Америки имеются и в Африке, в последней существуют группы, которых нет в Южной Америке. Это простой факт, но некоторые его детали могут вы- звать сомнение. Более подробно, но в несколько иной интерпретации он оыл изложен Майерсом (Myers, 1938, р. 348 — 349). Общая связь южно- американских рыб с африканскими была хорошо известна и более ран- ним авторам (Eigenmann, 1909, р. 363 и след.). Разнообразие фауны прес- новодных рыб Южной Америки резко контрастирует с малочисленностью ее предков. В подавляющем большинстве здесь доминируют представи- тели отряда Ostariophysi, которые насчитывают около 1800 или больше известных южноамериканских видов рыб; из них немного меньше полови- ны составляют харациновые и электрические угри и более половины — со- мы (по некоторым данным харациновые превосходят сомов по числу осо- бей). Три другие группы строго пресноводных рыб объединяют TOJIbRO пять известных в Южной Америке видов. Сюда не входят карпозубые и хромисы, представленные довольно большим числом, но менее важные в зоогеографическом отношении в связи с известной солевыносливостью некоторых из них. Заслуживает внимания и тот факт, что три семейства из строго пресноводных рыб некарпообразных Южной Америки сильно отличаются друг от друга. Двоякодышащая рыба — примитивный пред- ставитель костистых (не Teleostei). Это крупная рыба, приспособившаяся к существованию во временных водоемах. Два представителя семейства Osteoglossidae — мягкоперые костистые рыбы отряда Isospondyli — очень крупные рыбы, которые нуждаются в больших и постоянных водоемах. И два представителя семейства Nandidae — высшие колючеперые кости- стые рыбы — мелкие, могут жить в небольших водоемах. Эти различия в эволюционном уровне и экологических требованиях следует учиты- вать при рассмотрении вопроса, каким образом рыбы достигли Южной Америки.
Известная ограниченность и несбалансированность фауны пресно- водных рыб Южной Америки позволяет предполагать, что эта фауна— производная, происшедшая от нескольких иммигрантов, которые каким- то образом сумели достичь Южной Америки из Африки, или от фауны, аналогичной той, которая сейчас в целом приурочена к Африке. Ostario physi, сомы и харациновые радиировали в Южной Америке изолирован-
Глава 2
но, подобно хромисам в африканских озерах, или хромисам и «арпозубым в Центральной Америке, или некоторым другим пресноводным рыоам в иных местах, но только в большей степени. Малочисленность предков и высокая степень радиации свидетельствуют о том, что рыбы с трудом достигли Южной Америки. Вопрос заключается в том, каким образом это произошло.
Я думаю, что Ostariophysi по причине их многочисленности в Н)жной Америке и строгой приуроченности к пресным водам, а также в связи с тем, что они являются доминирующими рыбами на большей части зем- ного шара, могут послужить своеобразными «ключами» к истории проис- хождения южноамериканских пресноводных рыб, причем эти ««лючи», по вышеуказанным причинам, относительно достоверны.
Пресноводные рыбы отряда Ostariophysi предположительно могли достичь Южной Америки несколькими различными путями. Возможно, они достигли ее через море, если они обладали достаточной солевыносли- востью в древние времена, или через существовавший в те времена сухо- путный мост из Африки, или через существующие северные континенты (rye II настоящее время aTIIx рыб y~Ie Her). Если IIa amx IIpepIIo.Iu I;eHIIII сделать выбор, то он должен основываться главным образом на современ- ном распространении, так как ископаемых по рассматриваемым рыбам имеется недостаточно.
Харациновые и пресноводные сомы могли достигнуть Южной Амери- ки через море только в том случае, если они были по происхождению мор- скими рыбами или если их некоторые ранние пресноводные формы оола- дали солевыносливостью. Предположение о морском происхождении этих рыб маловероятно. Ostariophysi первоначально либо приспособились к пресной воде, либо даже произошли в ней. Расселение через Море соле- выносливых компонентов ранних пресноводных групп рыб возможн~>, но неправдоподобно. Если бы эти рыбы могли преодолевать даже небольшие моря, то они должны были бы существовать на Мадагаскаре, или, возмож- но, в Австралии, или на других изолированных островах, но их там нет, за исключением сомов семейств Ariidae и Platosidae, у которых способ- ность жить в морях возникла вторично. Весьма показательно отсутствие других представителей отряда Ostariophysi в отдаленных районах. Ведь эти рыбы, являясь доминирующими и хорошо приспособляющичисн, должны выжить на любой довольно большой, постоянной и изолированной суше, которой им удалось достичь. следующий аргумент, выдвинутыш про- тив морского расселения этих рыб,— наличие в Южной Америке других пресноводных животных, включая некоторых моллюс)ов и лягуше«, «о- торые аналогично рыбам указывают на те же связи с Африкой, однако и они вряд ли могли расселиться через море. Более вероятно, что соле- выносливые хромисы и карпозубые достигли Южной Америки, распрост- раняясь из Африки через море. Эти рыбы достигли и Мадагаскара, но не смогли проникнуть в Австралию.
Что касается непосредственного соединения материков Африки и Южной Америки в прошлом, а затем их разъединения, эта гипотеза дав- но интересовала лиц, изучающих распространение рыб. На первый взгляд зто предположение соответствует фактам. Все настоящие пресноводные рыоы Южной Америки имеют древние связи, «оторые ведут к Африке, а пресноводные рыбы относятся именно к таким животным, которые долж- ны были сохранить прежние географические связи. При более вниматель- ном взгляде картина оказывается иной. Пресноводные рыбы Южной Аме- рики не представляют всей фауны рыб Африки или какой- IIIuo части ео.
Пресноводные рыбы
В частности, они не представляют ее древнюю часть. В Южной Америке двоякодышащие рыбы имек~тся, но древних многоперых и большинства сравнительно древних африканских групп Isospondyli нет (рис. 12). Юж- ноамериканские пресноводные рыбы представляют незначительную часть фауны рыб Африки, которая, как уже отмечалось, чрезвычайно разнород- на по степени развития отдельных особей и их экологическим требовани- ям. Подобное явление — не результат перемещения материков, а, скорее всего, результат того, что несколько видов рыб, возможно разными пу1.я- ии или в разные времена, с трудом, но все же достигли Южной Америки. Более того, теории перемещения материков обычно охватывают все кон- тиненты, а не только Южную Америку и Африку, а распространение стро- го пресноводных рыб не свидетельствует о том, что такие перемещения происходили в других местах земного шара. Отсутствие строго пресновод- ных рыб на Мадагаскаре — заслуживающий внимания факт, который не- совместим с теорией перемещения материков. Мне кажется (правда, в раз- рез с мнениями некоторых зоогеографов), что распространение современных групп пресноводных рыб на земном шаре не свидетельствует о перемеще- нии материков.
Вывод таков: связи южноамериканских пресноводных рыб почти все ведут к Африке, что на первый взгляд говорит о возможности перемеще- н11я материков, но при более тщательном анализе данное предположение отпадает.
Предположение, что пресноводные рыбы достигли Южной Америки через длинныи материковый мост из Африки, можно было бы отвергнуть, ссылаясь на то, что пресноводные рыбы относятся к таким животным, ко- торые, если и могут расселяться через сухопутные мосты, то этот путь они избирают в последнюю очередь. Но все же они пересекают сухопут- ные мосты. Такой мост существует и теперь. Он соединяет Южную и (',е- верную Америку, протягиваясь от Панамского перешейка до перешейка Теуантепек. Его протяженность примерно 1300 миль, что составляет около '-", длины предполагаемого моста между Африкой и Южной Америкой. Более того, ныне существующий мост, довольно узкий и гористый, изме- нялся и разрушался на протяжении большей части третичного периода. Полное соединение (существующее и до настоящего времени) произошло в позднем плиоцене, несколько миллионов лет назад.
Современное распространение пресноводных рыб в районе этого M()- ста было рассмотрено в разделе «Пресноводные рыбы Центральной Аме- рики». Один род харациновых и один род пресноводных сомов преодоле- ли этот мост на всем его протяжении, несколько других представителей отряда Ostariophysi проделали только часть этого пути, но он не был пере- сечен ни одной строго пресноводной рыбой из некарпообразных. Однако солевыносливые рыбы (несколько видов карпозубых и один род хромисов) преодолели это препятствие также на всем его протяжении, а панцирные щуки проделали примерно половину всего пути.
К рыбам, которые пересекали этот мост в прошлом или пересекают его в настоящее время, относятся только несколько (причем очень немно- гочисленных по сравнению с их числом в Северной и Южной Америке) представителей доминирующего отряда Ostariophysi и несколько соле- выносливых рыб. Последние сильно отличаются друг от друга. Панцир- ные щуки — примитивные костные рыбы очень крупных размеров. hap- позуоые относятся к костистым рыбам (не к отряду Isospondyli), но все- таки по степени эволюционного развития они стоят ниже рыб пз семейства колючеперых; это мелкие рыбы, которые могут жить в небольших, иногда
Глава 2
временных водоемах. Хромисы, представители колючеперых,— рыбы сред- него размера. не требующие особых условий. Эти рыбы проникли через сухопутный мост и образовали фауну, которая в целом (хотя некоторые группы различаются очень сильно) аналогична фауне, из которой, пронзо- irma. видимо, фауна рыб Южной Америки: несколько доминирующих Os- tariophysi плюс несколько других рыб, сильно отличающихся по своему развитию и экологическим потребностям. Следовательно, если считать, что из Африки рыбы могли проникнуть в Южную Америку через прямой уз- кий сухопутный мост. то в силу трудности его преодоления он оказал сильное избирательное или фильтрующее действие и позволил пройти только нескольким доминирующим плюс нескольким солевыносливым группам пресноводных рыб. После этого последовала радиация домини- рующих Ostariophysi. Вышесказанное дало бы очень хорошее представле- ние о происхождении южноамериканской фауны пресноводных рыб, если оы двоякодышащая рыба, костноязычные и представители семейства Xandidae, так же как и карпозубые, обладали хотя бы небольшой соле- выносливостью. Имеется еще один путь, по которому пресноводные рыбы могли достичь Южной Америки, и еще одна более приемлемая альтерна- тива, позволяющая отказаться от гипотезы прямого моста. Возможно, сведения по другим животным помогут решить, какое объяснение наибо- лее правильно. Как уже говорилось, строго пресноводные рыбы относятся к таким животным, у которых было меньше всего возможностей пересечь сухопутный мост. Поэтому если они могли это сделать, то, естественно. должны быть и другие животные, которые преодолевали этот мост значи- тельно быстрее и в большем числе. Наличие их должно подтвердить суще- ствование предполагаемого моста.
Остается, таким образом, предположение, что пресноводные рыбы— 0stariophysi — достигли Южной Америки через север по существующим континентам, где в настоящее время их уже нет. Это могло произойти в том случае, если названные рыбы или произошли на севере и вторглись в тро- пики Старого и Нового Света или если они произошли в тропиках Старого Света и переместились через север в Южную Америку. Оба эти предпо- ложения не связаны со значительными изменениями суши. Но эволюци- онный характер и география расселения рыб различны, следовательно. различны должны быть и ископаемые. Если рыбы развивались на севере, то там должны были остаться значительные ископаемые остатки. но если они просто просочились через краевые части северных зон, то эти данные будут очень скудными.
В своей книге «Climate and Evolution» Мэтью (%1atthew,1915, р. 297— 299; '1939, р. 127 — 130) пытался доказать северное происхождение прес- новодных рыб. Но последнее не соответствует действительности. Данные Мэтью о предполагаемых доказательствах с помощью ископаемых, как показал Майерс (Муегз, 1938, р. 351 — 352), ошибочны. Едва ли найдена хотя бы одна ископаемая рыба, которая подтвердила бы взгляды Мэтью. Не северные группы современных пресноводных рыб продвигались в тро- пики, а, наоборот, видимо, тропические группы проникали в северном на- правлении. Надо учитывать, что тропические пресноводные рыбы имеют определенный характер эволюции и расселения в пределах тропиков. Он проявляется в сохранении древних рыб в Африке до настоящего вре- мени, эволюции и радиации новых доминирующих групп в тропической Азии и в первоначальной ограниченности фауны с последующей самостоя- тельной радиацией в Южной Америке.
Факты, только что приведенные в качестве аргументов против север-
17ресноводные рыбы
ного происхождения пресноводных рыб, позволяют предполагать, что ры- оы просочились через северные районы. Этот вопрос был уже рассмотрен мною в другой работе (Darlington, 1948, р. 13 — 15), но его необходимо кос- нуться и в данно~т книге.
Если тропические рыбы проникли через север при условиях, сходных с ныне существующими, то они, вероятно, распространялись через Восточ- ную Азию, мост Беринга, Северную Америку и Центральноамериканский мост в Южную Америку. Поэтому важно проследить, пользовались ли рыоы этим путем в последнее время. Некоторые, например тропические, рыбы, несомненно, начинали расселение по этому пути. В частности, лаби- ринтовые рыбы радиировали (умеренно) в тропической Азии и распростра- нилпсь в возможных пределах почти повсюду в тропиках Старого Света, а два широко распространенных тропических вида продвинулись в Во- сточной Азии в северном направлении до южной Маньчжурии и до района реки Амур (хотя других случаев расселения лабиринтовых рыб в север- HbIx широтах нет). Эти рыбы не достигли Северной Америки, но несомнен- но, что они могли бы туда проникнуть, если бы климат там был более мяг- ким, а суша не прерывалась Беринговым проливом. Карповые (Cyprinidae) по этому пути проникли значительно дальше. Начав, вероятно, с тропи- ческой Азии, они распространились через умеренные широты Евразии и Северной Америки, вторично проникнув в краевые части тропиков уже в южной Мексике. Но едва ли можно сказать, что эти рыбы просочились через север, так как они, доминируют там так me, как и в тропиках Старого Света. Однако этот случай надо отнести к особым: карповые — единствен- ные пресноводные рыбы, которые заметно преуспевают в условиях как тропического, так и северного климата.
Можно предположить, что сомы H харациновые, которые давно до- стигли Южной Америки, начали свое расселение так же, как и лаоирин- товые рыбы, а затем пересекли Северную Америку так же, как карповые, не играя, однако, доминирующей роли на севере.
(:омы являются главным образом тропическими рыбами. Некоторые современные группы их распространяются в умеренную зону северного полушария, но, очевидно, не находят в ней достаточно олагоприятных ус- ловий для обитания (стр. 79). Древние сомы, возможно, распространились в умеренной зоне северного полушария довольно давно и через нее проникли в Южную Америку, однако сведений об этом перемсще- нил нет.
Харациновые в настоящее время приурочены и Африке и Америке, а на самих материках — почти исключительно к тропикам. В тропической Азии они не встречаются, и их ископаемых там не найдено. Однако пола- гают. что встречающиеся там Cypriniformes произошли от харациновых рыо, что совместно с последующим доминированием Cypriniformes в тро- пической Азии указывает на существование там харациновых в прошлом и объясняет их исчезновение. Очевидно, когда-то эти рыбы существовали в тропической Азии, распространились к северу в Восточной Азии и через Северную Америку достигли Южной (как, вероятно, это произошло с, со- мами). Возможно, эти рыбы прямо пересекли старую северо-умеренную фауну рыб, так же как это сделали относительно недавно и три современ- ные группы североамериканских Ostariophysi (стр. 72).
Другие некарпообразные пресноводные рыбы Южной Америки, воз- можно, проникли через север так же, как сомы и харациновые, или рас- селялись какими-либо другими путями, частично через море, или прямо из Африки, или вдоль побережья.
Глава 2
Для дальнейшего обсуждения подробности распространения этих рыб существенного значения не имеют.
Если тропические рыбы Старого Света достигли Южной Америки через Восточную Азию и Северную Америку, то, следовательно, они про- исходят не от первичной африканской фауны, а от фауны, которая была когда-то общей и для Африки, и для тропической Азии и более или менее полностью сохранилась в Африке, но в результате эволюции более позд- них групп в значительной степени была вытеснена в Азии. Таким образом, особенностью распространения пресноводных рыб в тропиках Старого Света является сохранение старых групп в Африке и эволюция новых до- минирующих групп в Азии.
Учитывая все вышесказанное, можно считать образование фауны ю'к- ноамериканских пресноводных рыб от пришельцев, проникших через се- вер, вполне вероятным. Путь, по которому шло проникновение, не обяза- тельно должен был быть подобным описанному выше. Это мог быть и любой другой путь, соответствующий положению материков Евразии и ('.е- верной Америки во время расселения рыб, которое, скорее всего, пр~>исхо- дило в меловом периоде, так как изоляция Южной Америки от всех дру- гих континентов произошла примерно в начале третичного периода. Ископаемых Ostariophysi и большинства других пресноводных рыб в меловом периоде, по существу, нигде на земном шаре нет. Поэтому от- сутствие на севере в меловом периоде ископаемых харациновых и других рыб — это только частность. Недостаток существенных ископаемых дан- ных заставляет нас попытаться восстановить историю расселения рыб дру- гими путями.
Из всех различных путей, которыми пресноводные рыбы мог.чи до- стичь Южной Америки,— из моря или через него, через непосредственное соединение Африки и Южной Америки или прямым сухопутным мостом между этими материками, с севера или через север, по-видимому. T() Ibli0 два подходят для строго пресноводных форм. Первый — расселение че- рез прямой, но узкий мост из Африки; второй — через умеренную зону северного полушария (но не из нее). Оба пути можно считать «фильтром» через мосты. Фильтрация в первом случае была связана с трудностью пе- ресечения узкого сухопутного моста, а во втором — с прохладным или сезонным климатом районов, лежащих севернее тропиков (см. фронтис- пис). В том и другом случае следует предполагать, что с тех пор, ки~; ры- бы достигли Южной Америки, произошли некоторые изменения. В первом случае длинный сухопутный мост, вероятно, опустился в Атланти~;у. во втором — некоторые группы рыб, должно быть, исчезли, по крайней ме- ре на части умеренной зоны северного полушария. И в этом последнем слу- чае также можно предположить некоторые изменения суши, например существование позднемелового сухопутного соединения между Северной и Южной Америкой. Если исходить только из изучения расселения рыо, то пока трудно решить, какое из этих двух предположений наиболее пра-
ВИЛЬНО.
Что говорит распространение рыб о древних сушах и климато.т (за- ключение). Распространение пресноводных рыб свидетельствует. что про~и- лое земного шара, по-видимому, незначительно отличалось от его наст<>я- щего: изменения в расположении климатических зон или материков не- значительны, климат умеренной зоны северного полушария я третичном периоде был теплее, а в плейстоцене холоднее, чем в настоящее время, в тре- тичном периоде и плейстоцене на севере материки соединялись по крайней мере через Берингов пролив, в меловом периоде материк Южной Америки
Пресноводные рыбы
,:»н.дпнялся сухопутным мостом с Африкой или с Северной Америкой. Рас- пространение рыб не указывает на тесную связь Африки с Южной Амери- кой, а также не подтверждает предположения о перемещении каких-лиоо материков, во всяком случае в пределах времени, относящегося и суще- ствованию данных pbl0, а их датирование можно начать только с поздне- мело»»ого периода. Расселение рыб дает кое-какие сведения и оо истории раз III IHbIx материков. Можно предположить, что Африка была относи- тельно постоянным материком, а Южная Азия — непостоянным и каким- п> образом благоприятствовала эволюции новых доминирующих групп пресноводных рыб, что в Южной Америке была хорошо развита система по»»ерхностных вод и что этот материк был изолирован на протяжении длительного времени после того, как его достигли пресноводные рыбы, и что Австралия была еще более изолированной, чем Южная Америка, и»» течение еще более длительного времени.
Вполне закономерно высказать мысль, что известная нам история от- рядн Ostariophysi совпадает с историей моря Тетис — великого предка Сре- ди;»емного моря. В настоящее время оно занимает только часть его бывшей акватории, а в прошлом простиралось вдоль всей Южной Европы и Азии до Восточной Индии. Это море было непостоянным, так как оно совпадало с, частью геосинклинали, где наблюдался ярко выраженный процесс горооб- ра:»ования (в результате которого в третичном периоде образовались Аль- пы и Гималаи). Очевидно, этот район постоянно преобразовывался: изме- нялись береговые линии, возникали и исчезали преграды и мосты, из- менялись архипелаги и постоянно смещались границы между солеными и пресноводными местами обитания рыб. Эта область как раз и могла быть местом, где в меловой период предки Ostariophysi зашли в пресные воды и где начали доминировать харациновые и древние сомы. А восточная (юж- ноаз»1атская) часть района Тетис, где были более благоприятные условия или где они, возможно, наблюдались в течение более продолжительного времени, очевидно, была тем местом, где примерно в начале третичного периода харациновые дали начало,ypriniformes и откуда затем послед- ние радиировали.
1!сторпя расселения пресноводных рыо
Ниже приводится краткая частично гипотетическая история рассе- ,чен»1я современных пресноводных рыб (рис. 18).
Я предполагаю, что в меловой период существовали разнообразные f(мейства некарпообразных пресноводных рыб, самостоятельно»»о:»никшие и море, болыиинство которых расселилось зонально в соответствии с кли- матом: одни — в тропиках Старого Света, другие — в умеренной зоне север- ного полушария. (Некоторые из некарпообразных, возможно, проникли в пресные воды позднее, но здесь они не выделены.)
В течение мелового периода Ostariophysi, возникшие в пресных водах, »»о:»можно в тропиках Старого (лета, а вполне вероятно, и в районе моря 'Гст»1с, распались на харациновых и сомов. И только часть образова»»»пейся и Ст'»ром Свете тропической фауны некарпообразных рыб и некоторое чис- I(» примитивных OstBr1орЬуз1 (харациновые и сомы) достигли Южной Аме- рики. распространяясь или из Африки вдоль прямого сухопутного моста. или и; » тропиrec»;oii Азии через Се»»ерную Америку.
Позднее, примерно с начала третичного периода, предположительно от харациновых Юго-Восточной Азии (восточная часть района Тетис), раз-
Р п с. 18. Исторпя расселенпя пресно- водных рыб.
Упрощенная диаграмма, наглядно пока- ;~~ бывающая, каким образом происходило распространение современных рыб, в каких местах и каковы его результаты: светлые полосы ио диагонали — зональное распростра- нение древних фаун некарпообразных рыб на основной части тропиков Старого Света и в умеренной зоне северного полушария; расцвет примитивных рыб (харациновых II сомов) отряда Ostariophysi (светлые верти- кальные полосы) на основной части тропи- ков Старого Света; перемещение отдельных частей древней тропической фауны в Южную Америку, возмотжно, по краевым частям умеренной зоны севсрного полушария (пре- ) ~ывистые стрелки) с последующим увели- чением примитивных Oiiariopliysi в Южной Америке; радиация из тропиков Индо-5ja- лайской области доминирующего надсемей- ства Сург[ш1огше~ (сплошной круг со сплош- ными стрелками), явившаяся причиной вымирания (светлые полосы) многих более древних рыб в Индо-Малайской области, и умеренных широтах Евразии и в западно~~ части Северной Америки; выживание (чер- н ьи полосы) древней тропической фауны ~~ыб в Африке и Южной Америке и преоб- ~1азование древней северо-умеренной фауны
в западной части Северной Америки.
вились чукучановые. Последние и предки сомов семейства Ameiuridae распространялись затем через Восточную Азию и Северной Америке. Далее, очевидно, чукучановые положили начало роду карповых Cyprini- dae, которые дали бурную радиацию в Юго-Восточной Азии. расселились по всей Евразии и достигли Африки и Северной Америки. В то же самое время некоторые более высокоорганизованные колючеперые группы, осо- оенно лабиринтовые рыбы, радиировали, но в меньшей степени из Юго- Восточной Азии, и достигали Африки и умеренных широт Восточной Азии,
но дальше не распространялись. ать"нттнвоытт~нрнв и е в~ноя ттарнн~ Вновь расселившиеся доминирующие рыбы, особенно Cyprinidae, заняли место многих более древних рыб.
p~~~ggg pj's ~" 1 ~' харациновых, хромисов и большин-
ство древних пресноводных семейств А ~eш ° рыб, а в умеренных широтах Евра- зии — большинство чукучановых и несколько семейств некарпообраз- ных, особенно окуневых (в Восточ- ной Азии). Гибели древних рыб в тро- пиках и умеренных широтах Евразии, а также в западной части Северной и в Центральной Америке содейство- вали различные геологические и кли- матические изменения; таким обра- зом, в целом древняя тропическая пресноводная фауна рыб мелового периода сохранилась только в Афри- ке и частично, в производной форме, в Южной Америке. Несомненно, это всего лишь простая схема главных исторических событий, которые в действительности были очень сло к- ными.
В заключение в paccMoTpeHllbfx. нами вопросах распространения пре- сноводных рыб необходимо подчерк- нуть некоторые существенные мо- менты. Первый важный момент — то, что многие рыбы имеют зонал ь- ное распространение, ограниченное климатом. Второй — то, что некото- рые доминирующие рыбы имеют радиальное распространение, которыми не ограничивается климатом, и преодолевают как климатические, так и оольшинство других преград. Видимо, радиирование доминирующих рыо первоначально почти не оыло связано с климатом, и уже только поздне~ их распространение стало идти по зонам. Наиболее доминирующие совре- менные пресноводные рыбы, вероятно, радиировали из тропиков Старого (.вета, а умеренная зона северного полушария, возможно, послужила мо- стом, через который ряд из них проник в Южную Америку. Следует так- ;ке отметить связь между величиной рек, численностью и видовым разно- образием рыб. Самое большое число рыб находится там, где имеется наи- оольшее пространство. Этот факт может показаться слишком элементар- ным, чтобы подчеркивать его значение, однако влияние пространства играет значительную роль, и с этим нельзя не считаться.
Пресноводные рыбы
СПИСОК СЕМЕЙСТВ 11РЕСНОВОДНЫХ РЫБ И ПРИМИТИВНЫХ"РЫБООБРАЗНЫХ ПОЗВОНОЧНЫХ
Общий характер этого списка семейств и других списков в конце последующих глав изложен в первой главе. Классификация и природа пресноводных рыб рассмот- рены в начале этой главы. В приведенный ниже список включены все современные семейства пресноводных рыб первого и второго отделов, то есть строго и не строго пресноводных рыб (сокращенно 1, 2) и только наиболее важные семейства отдела соле- выносливых рыб (слв). Если бы в списки были включены все морские группы, из кото- рых один-два компонента или более живут или случайно заходят в преспые воды, то он увеличился бы еще примерно на 100 семейств. Тем не менее я старался сделать спи- сок достаточно полным, чтобы можно было представить разнообразие связей пресно- водных рыб с морскими. Ископаемые рыбы упоминаются только в тех случаях, когда они в той или иной степени раскрывают историю современных пресноводных семейст». Знак звездочка (*) перед латинским названием означает, что это вымершая груш>>>.
КЛАСС AGNATHA, примитивные, бесчелюстные позвоночные. Древххий класс, из кото>»>- го сохранился только отряд Cyclostomata (круглоротые). Этот отряд состоит из двух современных групп или подотрядов. К одному из них относятся миксины (Myxi- nidae), которые являются типично морскими обитателями и живут главным образом в прохладных водах северного и южного полушарий, изредка в тропиках (Myri»e circifro»s Garman в глубоководных частях Панамского залива). Другой подотряд состоит из следующего единственного семейства.
(cIB) Petromyzonidae, миноги. Прохладные воды северного и южного полушарий, но
не тропики: северное побережье Африки, большая часть умеренных широт Евра- зии, Япония, западная часть Северной Америки от Арктики до Цеххтральххой ча- сти Мексики (до реки Лерма и т. д., захватывая краевые части тропической зоны), восточная часть Северной Америки от Лабрадора до Флориды, включая боль- шую часть системы внутренних вод (Великие озера, большая часть бассейна реки Миссисипи и т. д.),''западная 1'ренландия и Исландия; южная Австралия, Тасма- ния, Новая Зеландия и большая часть умеренной зоны южных широт 10жной Америки (но не Южной Африки). Около 8 родов (различные северные и южные роды) и 24 видов. Ни один вид не является исконно морским, многие живут в мо- рях, но заходят в пресные воды для икрометания, а некоторые, по-видимому, оби- тают в пресных водах северного полушария, но не южного. Класс Agnatha ~~„ti~i- чает древнейших ископаемых позвоночных, ископаемые самих круглоротых неизвестны.
КЛАСС СНОМРВ1СНТНУЕ8, акулообразные рыбы, включая скатов. Многочислен и с
верхнего девона. В настоящее время и, вероятно, всегда был морским, но не- сколько видов акул заходят в реки. Полагают, что 2 вида акул рода C a rcl~ a ri n,~ (первоиачаль»о морской род акул) живут только в пресных водах: один — в озере Никарагуа, другой — в реке Замбези (Regan, 1929). Пилы-рыбы (Pris- ti dae) обычно поднимаются по тропическим рекам во многих частях земхюго шара.
Некоторые морские скаты (Trygonidae) заходят в пресные воды, а роххстххеххххое им семейство, приводимое ниже, почти всегда приурочено к пресным водам. (с;хв) Potamotrygonidae, речные скаты. Приурочены к Южной Америке, в осххоххххом
к следующим речным системам: Магдалена, Ориноко, Амазонка, Ла-плат».
Около 3 родов, 14 видов (Garman, 1913).
КЛАСС 08ТЕ1СНТНУЕЬ, костные рыбы. Возникли в девоне, вероятно в пресных водах.
С тех пор история разных групп была различной. Т4есколько современни~ не- костистых рыб, возможно, возникло и расселилось исключительно в пресны х водах. Костистые рыбы, вероятно, произошли в море в юре (B orner, 1945, р. 10 >), а многие группы возвратились в пресные воды, причем некоторые из них в сто,1ь давние времена, что в настоящее время они также строго приурочены к прессам водам. В целях изучения зоогеографических проолем»р»- митивных реликтовых некостистых рыб, противопоставив им производ~~> <Х доминирующих костистых рыб.
В тексте костистые рыоы подразделяются в свою очередь на доминирук)ци 11 отряд OstAI'lopl1vsl, который противопоставляется всем прочим костистым рыоам (табл. 1) .
Не 7cleosic.i, архаические костные рыбы: несколько неродственных групп, кот()рые
и в эволхоцххоххххх>ххх и в географическом отношении являются реликтами.
Надотряд Chonclrostei, хрящевые ганоиды
Отряд Polyp terini
(1) Polypteridae, многоперые. Западная и центральная тропххчехх'ая Лфрххка до о:и ра
Та»га»ьика и по всему Нилу (нет в озерах Викторххя или Ньяся, в восточных >»>г-
1 саеа 2
сейххах и в Южной Америке). Два рода: Polgpieius, примерно с 1' видами, знпхх- мает весь ареал семейства; угревидный род |"alamicht/hays с одним «идом, прц~ро- чеххххххч к Западной Африке, в болотистых эстуариях от дельты реки Нигер до устья реки Конго (Pellegrin, 1923a, р. 3,)). Многоперые — довольно крупххьхех хищные рыоы, обладающие частичным воздушным дыханием, что, возможно, и является причиной их выживания в районах сезонных засух (IIomer). Их иско- ххаемые (чешуя) бххли найдены в эоцепе Египта, по за ххределамхх Африки это семейство ни в настоящее время, ни как ископаемое не известно. Очевидно, оно произошло от семейства Palaeoniscoidae, которое когда-то было многочисленным н широко распространенным на всем земном шаре и имел» длительиун~ историю (с девона) расселения в пресных и соленых водах.
Отря;~ Acipenseroidei
(слв) Acipenseridae, осетровые. Ilpoxëàäèûå воды северного полушария: большая чсссть
умеренной зоны и северной Евразии (не Северной Африки), Исландия, Япония.
западное побережье Северххохх Америки от Аляски до Калифорнии (и, возможно.
часть Мексики), Гудзоххов залив и Великие озера, бассейн реки Миссисхххххх, восточное побережье Северной Америки от реки Святого Лаврентия (и, возможххо, Лабрадора) до Флориды. Около 5 родов, более 20 видов. Очень крупные рыбы, ни один вид не является полностью морским. Некоторые заходят в пресные воды только для размножения, некоторые обитают в пресных водах. Ископаемые остатки современного рода 3cipenser относятся по крайней мере к;юцену Европы п Северной Америки.
(1) Polyodontidae, веслопосы. В настоящее время локализованы в Китае и восточно-
центральной части Северной Америки; 2 современных рода и вида. Часто считают, что вид Pse phurus gladius приурочен к реке Янцзы в Центральном Китае, но в спи- сках Мори (Mori, 1936), дается также еще одно местообитание — Хванг-Хо, Север- ный Китай. Вид Polyodoii spathula встречается в системе реки Миссисипи — от реки Миссури в Южной Дакоте до Пенсильвании и Нью-Йорка (озеро Шотокуа); в юж- ном направлении — до западной части Северной Каролины, Миссисипи, Х? уизианы.
Техаса. Несколько раз этот вид был зарегистрирован в бассейне Великих озер, куда, как предполагают, он проник через каналы. Но, скорее всего, это свидетель- ство вымирания вида на севере... Чаще всего встречается в открытых водах больших заиленных рек... (Hubbs, 1.agler, 1947, р. 28 — 29). (Возможно, в Мисси- сипи имеется еще один неописанный вид веслоносов; Муегэ, 1949а, р. 119.) Веслсх- посы очень большие рыбы. Ископаемые остатки рода, найденные в Европе в верхнем мелу, в западной части Северной Америки, и верхнем мелу Монтаны (VlacAIpin, 1947) и в эоцене Ваххомххпга, позволяют предххсэлсэнхххть, что в ххропслсэм это семейство встречалось во всей умеренной зоне северного полушария.
Н а д о т р я д Н о 1 о s t е i, некостистые
Отряд Semiono(оИеа
( ) 1.ерЫоМеЫае, панцирные щуки. Восточная часть Севе1эиой Америки в «притоках
реки Миссисипи от Моххтахсьх через Великие озера (за ххск.ххочеххием озера Верх- него) до водораздела рек (: и. Лавреххтия — Шамплейн, штатов Квебек и Вермонт, спускаясь по реке Св. Лаврентия почти до соленых вод» (НиЬЬэ, Lagler, 1947, р. 31). В южном направлении достигает Флориды и на восточном псэсэереэкье до- стигает Мексики. Центральная Америка на н>г до озера Ппкарагуа (согласно схххсьмеххххоъху сообщению Хаббса, данные для 11аххамьх, хэероятнсэ, ошибочные), на Кубе. ((:ообщалось об одном современном вххде в Китае, что, хэерсэятххох являет- ся ошибкой.) '1 род, 4 — 5 видов. Крухшые рыбы, ххсэкрытх.хс» ххрочпьхм паххцирем, ведут хищный образ жизни, обитают в пресных «одах, но, как известно, и неко- торых случаях заходят в море. Вид Лерсдог1еиг 1 istoeс/си8 обычно заходит в соле- ные воды в «проходах> дельты Миссисипи; один»кземпляр был добыт Гн нтером (Gunter, 1942, р. 317) в открытом море в трех четхэертях мили от ххоберен<ья Лун- зианы. Возможно, это был экземпляр с Кубы, хотя подлинность его не coeceii установлена. Ископаемые остатки современного рода относятся к Европе (верхне- мелохэсэхх до мноцена), умереннххм хххиротам А:ши (:эоцен) и С,'eeep»oii Америке (верхнемеловой и позднее), а тнкже к ряду районов т1эсэпхх1хес кой Ин;хии (прнмерно в начале третххчсхого периода) (llora, Menon, 1952 — 195:1).
Отряд Amioidea
(1) АххххЫае, ильные рыбы. В хх;хстоясцее время приурочены к востсэчпсэ-центральной
части Северной Америки «от системы реки Мххссисххххи (за хсс клхочением озера Ву;х] в Миннесоте до бассейна рек Св. Лаврентия — Б[амплейн и Квебеке и Вермонте.
В южном направлении до Лппалачей и далес до Флориды и Техаса. И северном направлении в бассейне Атлантического океана до ooplix Каролин и рски (:аскуп- хахххха (зарегистрировахх в Коннектикуте, но, хэсэзмсэжххо, является резуххс,татом интродукцхххх}. Весь район Всликих озер не зсэ1эепхст1эххрован только хэ сяссс'псхах
Пресноводные рыбы
рек Верхнего озера, за исключением его стока в реку Сент-Мери)»(НиЬЬз, Lagler.
!947, р. 32). Один современный вид Amia calva — довольно крупная, типично хищная рыба, населяющая озера, реки и болота,— успешно и явно конкурирует ~ другими хищными рыбами такого же размера. Согласно Ромеру (Romer, 1945, р. 99 — 100), отряд Amioidea включает около 4 ископаемых семейств, которые до- минировали в юрских морях. Семейство Amioidea включает юрские и меловые роды, некоторые из них были морскими. Ископаемые вида Amia найдены в прес- »оводных отложениях в Европе (нижний эоцен до нижнего миоцена) и Северной Америки (средний эоцен и позже). Таким образом, очевидно, когда-то эта группа доминировала в морях, затем была приурочена к пресным водам северного полу- шария, а затем сократилась до единственного сохранившегося вида в восточно~~ части Северной Америки.
ПОДКЛАСС AMPHIBIOIDEA (Choanichthyes). Следующие два отряда в иастояшее
время обычно ставят в конце систематики рыб как предков амфибий, но с точки зрения зоогеографии их лучше помещать с другими примитивными, реликтовыми костными (не Teleostei) рыбами. Amphibioidea появились в девоне и были обыч- ными в пресных водах в позднем палеозое (Romer, 1955, р. 114), а некоторые из них заходили в море. В настоящее время осталось несколько представителей этих рыб, которые приводятся ниже.
Отряд Crossoplerygii, кистеперые. Как известно, в настоящее время существуют только
в океане недалеко от юго-восточного побережья Африки. Ископаемые кисте- перые были обнаружены и в других частях земного шара (девон — меловой период). Более поздних ископаемых не найдено, а современные формы были неиз- вестны. Предполагали, что этот отряд вымер в конце мезозоя. Однако современ- ный экземпляр, целакант, называемый Дж. Л. Б. Смитом (J. L. В. Smitt, 1939) J-atimeria chalumnae, был пойман 22 декабря 1938 г. в океане в нескольких милях от Ист-Лондона (Южная Африка) тралом на глубине примерно 40 морских саже- ней. Другой экземпляр был пойман в Мозамбикском проливе вблизи Мадагаскара в конце декабря 1952 г., о чем сообщалось в прессе («Time», 12 Jannuary 1953, р. 60); после этого было поймано еще несколько особей. Пока еще не ясно, все ли они являются представителями одного и того же вида. Это большие рыбы, весом свыше 100 фунтов, живущие, видимо, на умеренных глубинах, там, где оби- тают и многие другие рыбы; следовательно, они выжили не потому, что ушли в очень глубокие воды. Ранние целаканты были пресноводными рыбами, но в триасе, когда они стали обычными и разнообразными, они появились и в океа- не, где последние из них, видимо, сохранились (Romer, 1945, р. 121).
Отряд Dipnoi, двоякодышащие рыбы. (<...âïåðâûå известны из девона... В позднем па- леозое и триасе двоякодышащие рыбы достигали умеренной численности и раз- нообразия в пресноводной среде, в которой они произошли и приспособились» (R orner, 1945, р. 122). В свое время они были широко распространены, но не очень многочисленны, некоторые из них встречались даже в море (в девоне), но в настоя- ц1ее время численность их уменьшилась и распространение локализовано, о чем будет сказано ниже. Это крупные рыбы, которые имеют легкие и в случае необ- ходимости могут дышать воздухом.
(1) Сега1ойопСЫае. В настоящее время это семейство локализовано в Австралии: один
современный вид (Epiceratodus forsteri) приурочен к средним течениям двух небольших рек — Мэри и Бёрнетт — в субтропиках юго-восточного Квинсленда (Spencer, 1926). Ископаемые этого же рода, относимые к верхнему мелу, найдены в других местах восточной Австралии. Близкородственный, но более древний вымерший род *Ceratodus известен из триаса; был более или менее космополити- чен и в мезозое, по-видимому, исключительно в пресных водах.
(1)'I.epidosirenidae. Значительная часть тропической Африки и Южной Америки. 2 ро- да. Protopterus, с 3 видами, распространен в Африке от Сенегала до верховьев Нила, к югу до Конго и Замбези и в районе озер Танганьика и Ньяса (но не в рай- и»е озера Виктория). Род .Lepidosiren с 1 видом широко распространен в систе- мах рек Амазонка и Парана в Южной Америке. Как теперь известно, эти рыбы довольно обычны вдоль нижнего течения Амазонки (Myers. 1947 — 1949). По име- ющимся данным, этот род встречается в нижнем течении Параны у Ресистенсии.
Аргентина (Barrio, 1943), и ниже Сан-Педро, в дельте Параны, примерно на 34= ю. ш. (MacDonagh, 1945). Ископаемые зубные пластинки Protopterus были най- дены в Египте (олигоцен) и в Восточной Африке (нижний миоцен), а ископаемый представитель семейства Lepidosirenidae (поздний миоцен) — в верховьях доли- ны реки Магдалеиа, в северной части Южной Америки, где в настоящее время ;>ro семейство не встречается (Savage, 1951), но ископаемые, несомненно, отно- сящиеся к этому семейству, за пределами Африки и Южной Америки не най- дены.
Глава 2
Н а д о т р я д Т е 1 е о s С е i, высшие костные рыбы. Teleost,ei, очевидно, произо- шли в море в юре (Romer, 1945, р. 102 — 104) и теперь являются домиииру1опц1ми рыбами как в морях, так и в пресных водах. История их пресноводных групп различна.
()~рдд lsospondyli. Основная, самая примитивная группа современных костистых рыб
(очевидно, морского происхождения), до настоящего времени очень многочислен- ных в морях. Некоторые из морских семейств заходят в пресные воды или имеют пресноводных представителей. Например, Сlupeidae (сельдевые) в основном мор- ские рыбы, но некоторые заходят в пресные воды или поднимаются в реки ца не- рест, а ряд видов живет в пресных водах. Родственные им семейства Е1орнЬ» (тарпуны) и Chanidae — это небольшие морские семейства, обычно заходяп~ие в пресные воды, а семейство Osmeridae (корюшки} — главным образом м»рек»е, но некоторые виды нерестятся в реках, а некоторые в изолированных озерах. f'ë;~ других семейств отряда Isospondyli, перечисленных ниже, является или гла»- ным образом пресноводными, или строго пресноводными группами. Не»се~ ихтиологи согласны с подразделением этого отряда на подотряды, и не все семей- ства, отнесенные к одному подотряду, в действительности родственны друг другу. II о д о т р я д С 1 и р е о i d е а. Различные морские и солевыносливые груп1~~.~
(сельди и т. д.) и 2 небольших африканских пресноводных семейства.
(1) Kneriidae (включая Cromeria). Тропическая Африка. Ilo крайней мере 3 роля, (! ви- дов.
(1) PhractoIaemidae. Тропическая Западная Африка. 1 род и 1 вид.
П одотряд Salmonoidea
(слв) Salmonidae, лососи, форели и др. Северное полушарие от высоких арктических
широт к к~гу до прохладных вод северо-западной части Африки, холодные воль~ горных рек и ручьев Кавказа и северного Афганистана (Hora, 1934) (ио не И»дни), Западный и Северный Китай (но не на основной части собственно Китая), 1! и»- ния и горы Тайваня, северо-западная часть Мексики (Виеп, 1946, р. 87); иптро- дуцировано во многих других частях земного шара. Разнообразные роды и м»»- гочисленные виды, но ни один из них не является полностью морским; ми лги~ заходят в пресные воды на нерест, а некоторые приурочены к пресным водам. (слв) Coregonidae, сиговые. Северная Евразия и Северная Америка, в основном на тер- риториях, подвергавшихся ледниковой деятельности. Все виды нерестятсн в пресных водах и все, кроме нескольких, приурочены к ним. Однако си говы».
иногда заходят в соленые воды (Gunter, 1942, р. 313). Берг (1948 — )!)4!)} от- носит сиговых к семейству Salmonidae.
(слв) Thymallidae, хариусы. Северная часть Евразии н Северной Америки. Мал»чис- ленные виды приурочены к пресным водам. Родственно семейству Salmonidae. (слв) Galaxiidae. В прохладных пресных водах южного полушария несколько ви:~»в
заходит в море. Довольно мелкие, в ряде случаев форелеобразные рыбы песк»ль- ких родов, из которых главный — Galazias — объединяет множество видов (при- мерно от 40 до 70 в зависимости от того, насколько мелко они «дробятся»} и н>ж- ной части Австралии, Тасмании и на близлежатцих островах, на острове,1орд- Хау, в Новой Зеландии и на близлежащих и субантарктических островах, н» Огненной Земле. К северу от южной оконечности Южной Америки достигаст примерно 38' ю. ш. (в Чили), встречается на близлежащих островах и на Фолк- лендских островах и (один эндемичный подрод с двумя иитерградирующими фор- мами) в самой южной части Африки. Другие небольшие роды (существ»вы»»а отдельных из них вызывает сомнение) локализованы в пределах ареала родя Gnln- xias на Тасмании, в Новой Зеландии и в Чили. Один эндемичный род встреча»т- ся в Новой Каледонии в краевой зоне тропиков. Граница его распространения является северной границей распространения всего семейства. (Как унте дав»» было доказано, экземпляр из Индии, предположительно относимый к этому семейству, к нему не относится.) Считалось, что это семейство приурочено к прес- ным водам, ио уже давно известно, что некоторые виды заходят в море или даже размножаются там. Один вид Galaxias attenuatus, который размножается в море, встречается в пресных водах, образуя слабо отличимую расу в южной Австралии, Новой Зеландии и в южной части Южной Америки. Основные данные по этому семейству, интересному с точки зрения зоогеографии. можно найти в ряботях нижеследующих авторов: (Boulenger, 1902; Regan, 1905; Eigenmann, 1927; Scott, 1936 — 1941; Barnard, 19-~3 (африкаиские формы); Stokell. 1945), а также и ряля других, ссылки на которых приводятся в работе ('токелла. Ископаемые остатки, видимо, относящиеся к самому роду Galax~as, были найдены в пресновоии~к отложениях плиоцеиа в Новой Зеландии (Stokell, 1945).
(ели) Aplochitonidae. Юго-восточная Австралия, Тасмания, Новая Зеландия и тожияя
часть Южной Америки к югу от Консепсьона. Обитатель пресных вод, но некото-
Пресноводные рыбы
рые виды заходят или размножаются в море. Около 3 родов, малочисленные виды.
Вероятно, родственно семейству Galaxiidae (Chapman, 1944).
(с;и~) Hetropinnatidae. Южная и Западная Австралия, Тасмания и Новая Зеландия.
1 род, состоящий из нескольких близкородственных видов. Одни — обитатели пресных вод, другие — соленых, но все они в виде исключения из семейства Galaxiidae размножаются только в пресных водах (Stokell, 1941, 1949). Родст- «енные связи этого семейства неясны.
lI îäîòðÿä Osteoglossoidea
(1) Osteoglossidae, костноязычные. Эндемичные роды рассеяны по всем главным тропи- ческим областям: Clupisudis (Heterotis) — тропическая Западная Африка до вер- ховьев Нила (не ниже Асуана); один вид обычно встречается в пресных водах, но Буланже (Boulenger, 1907, р. 85), цитируя Штейндахнера, отмечает, что нередко он встречается в солоноватых водах устья реки Сенегал. Scleropageÿ отча- сти встречается в Индо-Малайской и Австралийской областях. 2 вида: один— в южном Таиланде, Индокитае, на полуострове Малакка, на островах Суматра, Калимантан, а другой — в южной части Новой Гвинеи и восточной части тропи- ческой Австралии (в реках залива Карпентария и восточного Квинсленда, дохо- :ггг на юг до реки Доусон). Случаи захода в соленые воды неизвестны (Longman, письменное сообщение). Osteoglossum и Arapaima — монотипические роды тро- пической Южной Америки, широко распространены на участке Амазонка — Гви- ана — Ориноко (точные границы не определены), но не достигают реки Магда- лена или системы реки Парана. Относится ли это семейство в действительности к основному отделу пресноводных рыб или нет и насколько оно является «древ- ним», см. текст (стр. 54). Современные формы — крупные или очень крупные рыбы, для существования которых необходимы большие реки или другие постоян- ные бассейны. Ископаемые костноязычные вымершего рода встречаются в прес- новодных отложениях в Северной Америке (эоцен Вайоминга), Индии (эоцен) (Hora, Мепоп, 1952 — 1953), на Суматре (вероятно, эоцен) и в Австралии (в Квин- сленде, вероятно, в одигоцене, встречается предположительно тот же самый род, который был найден в эоцене Северной Америки). Ископаемое, предположитель- но относимое к костноязычным, обнаружено в морских глинах эоцена Англии.
В Южной Америке ни одно ископаемое этого семейства обнаружено не было. (j) Pantodontidae, африканские пресноводные рыбы-бабочки. Тропическая Западная
Африка. 1 род, 1 вид.
(1) Мotopteridae. Основные районы тропиков Старого Света. 2 рода: род 1Votapterus
с одним видом в тропической западной Африке, от реки Гамбии до реки Конго, и 3 вида в Индо-Малайской области, от Индии до Явы и Калимантана; род Yeno- mystus с 1 видом в тропической Африке, включая верховья Нила; обитает в прес- ных, иногда солоноватых водах.
(1) Mormyridae. Тропическая Африка и весь Нил. Около 10 родов и 150 видов.
(1) Gymnarchidae. Тропическая Африка, включая верховья Нила. 1 род, 1 вид.
(1) Hiodontidae, хиодоны. Восточно-центральная часть Северной Америки, вероятно,
от верховьев системы реки Маккензи, по западным и южным притокам рек Г~ дзо- нова залива и в бассейнах рек Св. Лаврентия — Шамплейн и к югу почти через всю систему Миссисипи. 2 рода, 2 или 3 вида.
Отряд Ostariophysi. Доминирующий отряд пресноводных рыб широко представлен
во всех основных областях земного шара, но различные основные группы отряда имеют самое различное распространение. Но только те рыбы, которые достигают Мадагаскара и Австралийской области, являются вторично морскими рыбами (см. ниже). Отряд охватывает примерно 7/s современных строго пресноводных рыб (первый отдел) и насчитывает около 5000 известных видов приблизительно в 40 семействах. В этот отряд включено также 2 семейства сомов, которые являют- ся главным образом морскими, хотя произошли, вероятно, от пресноводных форм.
Некоторые рыбы этих семейств снова вернулись в пресные воды, особенно в от- даленных местах. Ригэн разделил этот отряд на 2 подотряда — на более или менее карпоподобные формы и на сомов, а первый подотряд он подразделил erne rr;r на 3 группы, называемые здесь надсемейством. В данной книге я следовал клас- сификации Ригэна, а не Николса (Nichols, 1938), так как в работе последнег~~ пр11- водится меньше данных по их распространению. В годы издания работы Рпгэна (Regan, 1922) не было известно ни одного ископаемого пресноводных семейств отряда Ostariophysi за пределами распространения их современного ареала.
В последующие годы было найдено несколько ископаемых за пределами границ ареала, но в большинстве случаев их связи и значение все еще вызывают сомне- ние. Известные ископаемые свидетельствуют о том, что неко горые специализи- рованные семейства отряда Ostariophysi существовали в р;11и1етретичном време- ни, что в свою очередь позволяет отнести возникновение и первоначальное рас-
100
Глава 2
селение этого отряда к меловому периоду (Regan, 1922, и некоторые более позд- ние авторы), хотя ископаемые остатки карпоподобных мелового периода мало- численны и в какой-то степени сомнительны.
П одотряд Cyprinoidea, карпоподобные рыбы
(Надсемейство Characiformes), харациновые. Тропические и субтропиче- ские части Африки и Америки. Около 6 семейств. В Африке — 2 семейства с О родами и 130 видами. Они распространены в тропической Западной Африке, по всему Нилу, Восточной Африке и (несколько) в субтропической части Южной Америки, но не достигают крайнего юга материка. В Амери- ке — 5 семейств (одно общее с Африкой) приблизительно со 100 родами и, вероятно, более чем с 800 известными видами. Наибольшей численности достигают в тропиках Южной Америки. Примерно 19 родов достигают Па- намы (Hildehrand, 1938), но только немногие из них проникли дальше — в тро- пическую часть Центральной Америки, и только один род достигает реки Рио- Гранде в южном Техасе и Нью-Мексико; 3 рода встречаются на юге от Буэнос- Айреса, и ни один не известен в пределах 700 — 800 миль от мыса Горн (дальше всего на юг заходит Cheirodon, который достигает 41'18' ю. ш.; Eigen- mann, 1927, р. 14). Несколько ископаемых харациновых известны в пределах сов- ременного ареала этой группы, а также небольшое число ископаемых вехрнеме- лового и раннетретичного периодов известно из Северной Америки (Нау, 1929, р. 718 — 719) и из раннетретичного периода во Франции (Piton, 1940, р. 269— 270), но (если они являются харацинами) их значение очень сомнительно. Заслу- живает внимания полное отсутствие харациновых как современных, так и ис- копаемых, в тропиках Азии. Но если Cypriniformes развились от харацинов в тропической Азии, следовательно, и харациновые должны были находиться там с давних времен. Харацины варьируют от мелких рыб до рыб среднего раз- мера с различным образом жизни и включают и хищную пиранью, и многих ак- вариумных рыб. Ниже приводится список семейств харациновых.
(1)Characidae. Африка и Америка (в пределах выше приведенных границ). Около 5 ро- дов, 70 видов в первой и около 75 родов и 600 видов в последней; один достигает Рио-Гранде по южному побережью Соединенных Штатов (представители аме- риканских семейств пересмотрены Eigenmann, Myers, 1917 — 1929).
(1)Citharinidae. Африка. Около 15 родов, 60 видов.
(1)Hemiodontidae. Америка. Около 5 родов, 60 видов.
(1)Anostomidae. Америка. Ококо 9 родов, 150 видов.
(1)Xiphostomidae, Америка. 3 рода, около 8 видов.
(1)Gasteropelecidae, летающие харацины. Америка, к северу до !!анамы и к югу до
Санта-Фе (северная Аргентина). 3 рода, 8 видов (Fraser-Вгиппег, 1950).
(Надсемейство Gymnotiformes). Электрические угри. Южная и Центральная Америка.
4 семейства, около 17 родов и 35 видов. В основном в тропиках К) жной Америки, достигая на юге Ла-Платы, к востоку от Анд, и западного побережья Эквадора и Колумбии, несколько видов достигают Панамы, но только один вид проникает дальше в Центральную Америку до Гватемалы. Угреобразные рыбы, родствен- ные харациновым и, видимо, от последних образовавшиеся. К этой группе, пересмотренной Эллисом (Ellis, 1913), относятся следующие семейства:
(1) Rhamphichthyidae, Sternarchidae, Gymnotidae и Electrophoridae
(Надсемейство Cypriniformes)
(1)Catostomidae, чукучановые и рыбы-буйволы Ictiobus. Северная Америка и Восточ- ная Азия. Около 18 родов и 84 видов распространены в Северной Америке (Jor- dan et al., 1930), проникая на юг до Гватемалы, включая рыб, распространенных в Тихоокеанском бассейне Северной Америки, вплоть до Калифорнии. Один аме- риканский вид Catostomus catostomus распространен в Восточной Сибири (слегка дифференцированный подвид). Один эндемичный род Myxocyprinus, с одним до- вольно большим, очевидно реликтовым, но специализированным видом и с Зпод- видами, изолирован в восточно-центральном Китае (Nichols, 1943, р. 58 — 61; Nelson, 1948). Распространение этого семейства рассматривалось выше (стр. 36, рнс. 9). Ископаемые данные позволяют предположить, хотя и не наверняка, существование этого семейства в эоцене Северного Китая (Nelson, 1949), а также в эоцене Северной Америки (Boulenger, 1905, р. 415; Regan, 1922, р. 206; Gosli- ne, 1944, р. 216; Darlington, 1948, р. 2), но самые ранние достоверные данные для Северной Америки, возможно, относятся к миоцену (Нау, 1929 — 1930; Romer.
1945) .
(1)Cyprinidae, карповые. На всех континентах, за искл~очением Южной Америки н Ав- стралии. Самое разнообразное и самое многочисленное семейство в Евразии, осо- бенно в Юго-Восточной Азии и Китае. Менее многочисленное по сравнению со всей фауной рыб и менее разнообразное (меньше основных ветвей) в Африке и Северной Америке, но все же широко распространенное на обоих этих материках—
11ресноводные рыбы
101
по всей Африке, до ее южной оконечности и почти по всей Северной Америке к востоку и западу от Скалистых гор и от полярных частей (на севере до дельты реки Маккензи) до Мексики (где на юг до бассейнов Пануко и Бальсасакарпо- вые встречаются в меньшем числе) (Buen, 1946). Данных о существовании этого семейства в Аляске нет, но, вероятно, оно там встречается, так как один вид С"oue- sius р1итпбеиздостигаетверховьев Юкона (Wynne-Edwards, 1952, р. 18). Солевы- носливость некоторых представителей этого семейства и их встречаемость в во- сточной Индии относительно линии Уоллеса (многие достигают ее, но очень немно- гие пересекают) рассматривались выше (стр. 53, 54). Семейство,yprinidae, ве- роятно, самое большое из всех семейств рыб, с несколькими сотнями родов и приблизительно 2000 известных видов. Ригэн разделил это семейство на 3 сле- дующих подсемейства, которые имеют почти одинаковое распространение: Cypri- ninae, отличающееся разнообразием в Юго-Восточной Азии и многочисленное в умеренных широтах Евразии и в Африке; Bariliinae, также наиболее разно- образны в Юго-Восточной Азии, несколько его представителей имеется в уме- ренных широтах Евразии и в Африке; Leuciscinae — многочисленное и в умерен- ных широтах Евразии и в Северной Америке. Последнее подсемейство единст- венное, которое (по Ригэну) встречается в Америке. Николс (Nichols, 1938) подтверждает это, но он помещает один американский род (Xotemigonus в восточ- ной и центральной частях Северной Америки) в другое, не признаваемое Ригэ- ном, подсемейство Старого Света — Abramadinae. Позднее Николс опубликовал таблицу (Nichols, 1943, р. 10) адаптаций главных групп карпообразных рыб с ука- занием их распространения. Ископаемые данные указывают на существование карповых в Европе начиная с эоцена, а в Северной Америке только с миоцена. (1)Cobitidae, вьюны. Евразия и Северная Африка. Многочисленны в Юго-Восточной
Азии до островов Ява и Калимантан. Менее многочисленны в умеренных широтах Евразии (до Испании, Португалии) и на Британских островах. Один европейский вид находится в Марокко, в палеарктической Северной Африке, и по крайней мере один эндемичный вид — в Эфиопии. Это семейство встречается главным образом в быстрых водах горных рек.
(1)Homalopteridae, вьюны. Юго-Восточная Азия, Индия и Южный Китай до островов
Ява и Калимантан. 17 родов, около 50 видов (Hora, 1932). Обитатели стремитель- ных горных потоков.
П о д о т р я д $ i 1 u r о i d е а, сомы. Пресные воды многих частей земного шара
(рис. 16), но наибольшей численности достигают в тропиках. Малочисленнее и более локализованы в умеренных зонах и отсутствуют в районах с очень zo лодным климатом. Сомы Мадагаскара, Австралии и т. д. являются солевыносли- выми пресноводными формами, развившимися от морских семейств. Из настоя- щих пресноводных сомов в Африке имеется 6 семейств и около 250 или более ви- дов, большинство из которых находится в тропической части континента. Некото- рые достигают почти самого устья Нила (Boulenger, 1907), а род Clarias встре- чается в реликтовых водах Сахары (Braestrup, 1947). В умеренных широтах Се- верной Америки сомов нет. Несколько пресноводных сомов достигает умеренно- теплых широт Южной Африки, несколько не доходят до ее южной оконечности (Barnard, 1943). По крайней мере 2 африканских вида достигают Сирии и 2 вида пресноводных сомов встречаютсяв системе Тигра и Евфрата. В тропической Азии— 9 семейств (3 общих с Африкой) пресноводных сомов. Виды многочисленны, на- чиная от Индии через Гималаи и Китай до островов Ява и Калимантан. Все се- мейства Индо-Малайской области, кроме монотипического Cranoglanididae, достигают этих островов, но ни одно из них не является местным на острове С~.— лавеси (хотя 2 ветви все же достигают южных Филиппин). Несколько азиатских тропических семейств простирается, как уже говорилось, в умеренные широты Евразии, а распространение отдельных семейств рассматривается ниже. В Се- верной Америке имеется одно эндемичное семейство Ameiuridae, которое дохо- дит на юг только до Гватемалы. В Южной Америке — 12 пресноводных семейств, которые составляют примерно половину (около 1000 видов) пресноводных рыб Южной Америки. Несколько южноамериканских семейств достигают Панамы или Панамы и Коста-Рики, нотолько одно семейство (Pimelodidae) через Центральну1о Америку проникает в Мексику. В Центральной Америке эндемичных родов мало или нет совсем. На юге (не дальше центрального Чили или центральной Арген- тины) встречается только 4 рода, но один из них Diplomystes — предположитель- ио самый примитивный из родов современных сомов, образующий отдельное се- мейство, частично локализован в умеренной зоне Южной Америки, а дру- гой Nematogenys — предположительно также наиболее примитивный из совре- менного рода семейства Pygidiidae, локализован на небольшой территории в Чи- ли. Два других семейства сомов Ariidae и Plotosidae, видимо, возникшие от прес-
102
Глава 2
новодных предков, являются морскими, за исключением тех, которые вернулись в пресные воды (особенно в изолированных местах). Эти два семейства, очевидно, не имеют прямого родства и представляют две самостоятельные инвазии из моря.
Специалисты до сих пор не пришли к единому мнению по классификации и гене- алогическим связям сомов, поэтому здесь их распространение дано в общих чер- тах, а описание некоторых семейств сведено до минимума (см. ниже). Ископаемые формы сомов известны начиная с эоцена (в отложениях и пресных, и соленых вод), но их связи еще недостаточно выяснены, и в связи с этим они не могут быть использованы для решения вопроса о происхождении и расселении семейств, ряд из которых ведет свое начало, вероятно, с мелового периода (Gosline, 1944, р. 216) .
(Пресноводные сомы Старого Света), в основном даны по Ригэну
(1)Bagridae. Многочисленное в Африке. 1 род (Macrones) представлен в Сирии (РеПе- grin, 1923), в системе Тигра и Евфрата и многочислен в Южной и Восточной Азии.
Расп ространен на юго-восток до островов Ява и Калимантан и на север (в умень- шающемся числе) до Амура. Это колоссальное семейство родственно североаме- риканскому семейству Ameiuridae (Regan) или южноамериканскому Pimelodi- dae (Myers). Один широко распространенный вид Индо-Малайской области, Macrones gulio,— частично морской (Weber, Beaufort, чо1. 2, р. 345), а другие виды того же рода в большей или меньшей степени морские (Н. М. Smith, 1945). (1)С1аг11йае, второе большое семейство. Африка, Сирия (один африканский вид) и Ин- до-Малайская область до островов Ява и Калимантан с одним эндемичным ви- дом на острове Минданао, южные Филиппины. Некоторые виды, как, например, Clarias batrackus, могут некоторое время находиться без воды и с помощью че- ловека (перенос) пересекают линию Уоллеса. Род Heterobranchus с несколькими видами в Африке и одним изолированным видом на островах Калимантан и Бан- ка в Индо-Малайской области; ископаемые этого рода найдены в Индии (Сива- лик, нижний плиоцен) (Hora, 1937а; но см. Gosline, 1944, р. 219).
(1) Schilbeidae. Африка и Индо-Малайская область.
(1)Mochochidae и Amphiliidae. Африка.
(1)Malopteruridae, электрические сомы. Тропическая Африка и весь Нил. 1 вид.
(1)Siluridae. Евразия. Многочисленно в тропической Азии. На север распространено
в уменьшающемся числе до умеренных широт Восточной Азии (до реки Амур).
Нет в северных частях реки Амур и на большей части Центральной Азии. Один очень большой вид (S i l u rus g l an is) распространен от района Аральского моря на за- пад до Рейна и один эндемичный вид азиатского рода Parasilurus изолирован в реке Ахелус (Греция). Это семейство отсутствует в Южной и Западной Европе (к западу от Рейна). В Индо-Малайской области сомы семейства Siluridae распр«- странены на юго-восток до островов Ява и Калимантан с одним эндемичным родом и видом (а не двумя, как предполагалось ранее, по письменному сообщении> Майерса) на острове Палаван, островах Каламиан и южных Филиппинах. (1)Sisoridae. Индо-Малайская область, в основном в горных ручьях. 1 род Glyptoster- пит с 2 видами (Hora, 1937) в Центральной Азии (к северу от Гималаев) и на за- пад до района Аральского моря (Берг, 1949).
(1)Amblycepidae, Pangasiidae (1 род), Chacidae (1 вид) и (;ranoglanididae (1 вид). Эти
семейства населяют всю или часть Индо-Малайской области.
(Пресноводные сомы Южной и Центральной Америки) (в основном даются по Gosline,
1945). Надо отметить, что североамериканское семейство сомов Ameiuridae (см.
ниже) также достигает Центральной Америки.
(i)Diplomystidae. Встречается повсеместно в центральной части Чили, между Сант- Яго и Вальдивией (Eigenmann, 1927, р. 13), в центральной Аргентине, в реке Рио- Негро. Один предположительно примитивный реликтовый род с двумя видами. (l)Doradidae. Южная Америка и Панама. 45 родов, 131 вид (только 1 вид достигает
Панамы) .
(1)Ageneiosidae. Южная Америка и Панама. 2 рода, 27 видов (только 1 вид достигает
Панамы).
(1)Pimelodidae. Южная и Центральная Америка на север до центральной Мексики
(река Пануко; Впеп, 1946): 56 родов, 281 вид. Несколько родов достигают Пана- мы, но род Rhamdia с многочисленными видами — основной или единствеиный род, который проникает дальше на север.
(1)<elogencidae. Частично на севере Южной Америки. 1 вид.
(1) Hypophthalmidae. Часть Южной Америки. 1 вид.
(1)Се1орз1йае. Южная Америка. 4 рода, 12 видов.
(~)Pygidiidae. Южная Америка и Панама. 27 родов, 135 видов, но только 1 род (Pitgidi- um) достигает Панамы. Это семейство обитает как в горных реках, так и в реках низменностей. На юг проникает гораздо дальше, чем любые другие строго прес-
Иресноводньсе рыбы
иоводные рыбы, достигая примерно 47'30' ю. ш. (Eigenmann, 19! 8, р. 269). ['олы этого семейства рассмотрены Майерсом (Myers, 1944).
(l)Bunocephalidae. Южная Америка. 10 родов, 31 вид.
(I)(;allichthyidae. Южная Америка и Панама. 8 родов, 50 видов (только 1 вид в 1! ани- ме) .
(1) Loricariidae, панцирные сомики. Южная Америка и прилежащие части Центральной
Америки. 49 родов, 415 видов. Ряд родов достигает Панамы и только 1 род дости- гает Коста-Рики.
(l)Astroblepidae. Южная Америка, в северной части Анд. 1 род, 37 видов, только 'I
«ид достигает Панамы.
(11ресноводные сомы умеренной зоны северного полушария) См. семейства Bagridae,
Siluridae и Sisoridae и следующие:
(1)Ameiuridae. Северная Америка к востоку от Скалистых гор. Один (или несколько)
из восточных видов пнтродуцирован на западе, и несколько видов встречается в природе в бассейнах западных частей Мексики. На юге семейство проникает и уменьшающемся числе видов до Гватемалы (предположение о существовании сома из семейства Ameiuridae в Китае не подтверждается). 10 родов, примерно 36 видов (Jordan et а1., 1930). Вымерший род Rhineastes из эоцена Северной Аме- рики и сомнительные ископаемые в Восточной Азии из плиоцена, возможно, от- носятся к этому семейству (Myers, 1938, р. 347). Ископаемые рода Ameiurus относятся к олигоцену Северной Америки.
(Сомы соленых вод и их производные)
(слз) Ariidae. Теплые моря, в прибрежных частях морей и эстуариях по всей тропиче- ской зоне и (в уменьшающемся числе) заходит в умеренную зону (например, 2 ви- да достигают мыса Код, Массачусетс). Во многих частях земного шара некоторые виды заходят в реки, и ряд видов, вероятно, обитает в пресных водах. Виды, кото- рые встречаются в пресных водах Южной и Центральной Америки, приведены Гослине (Gosline, 1945). По описанию ископаемые этого семейства относятся к эо- цену, но лучше всего считать их сомнительными.
(слн)0ойсЫЬу1йае. Южная часть Новой Гвинеи. 1 вид, в какой-то степени родствен- ный семейству Ariidae и, по-видимому, от него возникший, живет в пресных и со- лоноватых водах.
(с ia)PIotosidae. Моря Индийского и Тихого океанов. Некоторые виды заходят в реки,
другие живут в пресных водах Мадагаскара и Австралийской области. Роды "cpidoglanis и Cnidogla»is приурочены к рекам Австралии и Новой Гвинеи. Отряд Аросев. Около 20 морских семейств и семейство пресноводных угрей. (сaa)Anguillidae, пресноводные угри. Семейство имеет широкое, но прерывистое рас- пространение. Пресные воды Европы, побережья Средиземного и Черного морей, восточное побережье Африки (но не западное), Мадагаскар, Маскаренские и Сей- шельские острова, большая часть Южной и Восточной Азии, Япония, Индо- Австралийский архипелаг, большая часть Северной и Восточной Австралии, Но- вая Зеландия и многие отдаленные острова Тихого океана (но не Гавайские или острова Галапагос), восточная часть Северной Америки (но не западная) и изредка северная часть Южной Америки, от южной Гренландии и Лабрадора до Бразилии и Вест-Индии (но не на большей части Южной Америки). 1 род, око- ло 16 видов. Все размножаются в море, но взрослые особи, по крайней мере сам- ки, проводят большую часть жизни в пресных водах. Европейские и североаме- риканские виды размпожаются на площади, где их ареалы перекрывают друг друга — в западной части Атлантического океана непосредственно к северу от Вест- Индии. Полагают, что африканские виды нерестятся в Мадагаскарской котло- вине (Barnard, 1943). Места размножения большинства других видов до сих пор неизвестны. Последние сведения по этому семейству приводятся в работе Эге (Ege, 1939), который дает ссылки на раннюю работу Шмидта.
Отряд МеясЫЬт.s
П о д о т р я д Н а р 1 о m i (Еsocoidei, по Бергу, 1936).
(1)*Palaeoesocidae: Средний эоцен Германии, один хорошо сохранившийся ископаемый
род (Берг, 1936).
())1.'inbridae, евдошки..Локализованы на различных территориях в умеренной зоне
северного полушария. 2 рода: Утбга с 3 видами, один в среднем и нижнем тече- нии Дуная и в нижнем течении Днестра в Восточной Европе (Берг, 1932); один ()т Манитобы через большую часть района Великих озер до Квебека и бассейна озера Шамплейн (зарегистрирован один раз в канале Эри на водоразделе реки Мохок), на юге в центральном бассейне континента до верховьев системы реки Огайо, в озере Рилфут (северо-восточный Теннесси), северо-восточном Аркан- засе, Канзасе и, возможно, в восточной Небраске (Hubbs, Lagler, 1947, р. 74).
Третий вид встречается по атлантическому побережью, в реках и болотах иизмен-
104
Глава 2
ностей, от острова Лонг-Айленд до Северной Каролины. Род Ь'оьитога (иногда помещаемый в отдельное семейство) с одним видом в западной части Соединенных Штатов, в реке Чехейлис, штат Вашингтон. Евдошки — маленькие очень вынос- ливые рыбки, хорошо переносят неблагоприятные условия, включая недостаток кислорода и холод.
(1)Dalliidae, черная рыба. Северная Аляска и крайние точки Восточной Сибири (полу- остров Чукотка; Берг, 1949). 1 род и 1 вид, в мелких реках, озерах и болотах тундры.
(i)Esocidae, щуки. Прохладные воды северного полушария. 1 род, около 6 видов. Рас- пространение дано в гл. 1. Крупные, хищные рыбы.
П о д о т р я д С у р r i n о d о n t e s (Microcyprini): распространение основных
семейств см. Майерса (Myers, 1931). Кроме семейств, перечисленных ниже, есть еще несколько небольших семейств — Jenynsiidae, Anablepidae и Tomeuridae (Myers, 1947 — 1949, pt. 3, р. 9) в Америке и Horaichthyidae (Hubbs, 1941) в Индии, часто относимых к этому подотряду. Все они локализованы в пределах ареалов больших семейств, приведенных ниже. Карпозубые — в большинстве случаев мелкие рыбы. Одни солевыносливы, другие живут в очень мелких водоемах, включая оазисы или реликтовые воды в пустынях. Некоторых из них содержат в аквариумах или в водоемах для уничтожения комаров. Часто бывают интро- дуцированы за пределами их естественных ареалов.
(2)Amblyopsidae, пещерные рыбы Северной Америки. Юго-восточная и центра.1ыия
части Соединенных Штатов. 3 слепых рода живут в пещерах, а четвертый, зря- чий род обитает и в болотах, и в пещерах. Майерс относит это семейство к строго пресноводным рыбам, хотя нет веских оснований для того, чтобы отделять его от других семейств не строго пресноводных рыб этого подотряда.
(2)Cyprinodontidae, карпозубые. Тропические и умеренно-теплые области земного ша- ра, за исключением Австралийской области, Средиземноморье Европы, большая часть Африки, не достигая южной оконечности материка (Barnard, 1943), Мада- гаскар, Сейшельские острова, Южная Азия на север до Японии и Северного Ки- тая и на юго-восток до островов Тимор, Сулавеси, Тайвань и т. д. и, вероятно, на острове Лусон в Филиппинском архипелаге, Северная, Центральная и Южная Америка от реки Св. Лаврентия и южно-центрального побережья Калифорнии на юг до реки Ла-Плата и на некоторых Вест-Индских островах и Бермудах (распространение в основном приводится по Майерсу (Myers, 1931, р. 248) . Много- численность родов и видов, особенно в Америке, идет на убыль в Индо-Малайской области. Большинство из них обитает в пресных или солоноватых водах, но некото- рые и в море. Как правило, морские формы встречаются вблизи берегов, но экс- педиция «Челленджера» поймала представителя рода Fundulus в средней Атлан- тике, а один вид этого рода достиг Бермудских островов, где он встречает- ся в более или менее солоноватой воде, иногда в мангрове. Считают, что до- минирующий южноамериканский род этого семейства родствен африканскому, а не североамериканским формам (Myers, 1938, р. 348). Одна группа из 4 родов и около 20 известных видов, образующих подсемейство Orestiinae, приурочена к высокогорью на небольшом участке западной части Южной Америки (высота «Альтиплано», включая озеро Титикака), где эта группа претерпела заметну1о радиацию (Tchernavins, 1944; Brooks, 1950).
(2)Poeciliidae, живородящие пецилиды. Америка, «от Делавэра, Иллинойса и Аризо- ны до западной части Эквадора и северной Аргентины» (Myers, 1938, р. 348) и на некоторых островах Вест-Индии. Многочисленные роды и виды; большинство— обитатели пресных вод, но некоторые обладают значительной солевыносливостыо. (2)Goodeidae. Локализованы в центральной Мексике, большинство в системе реки Лер- ма. Многочисленность родов и видов (около 30) (Hubbs, Тигпег, 1939; Turner, 1946) — еще один замечательный пример радиации на ограниченной территории. (2)Adrianichthyidae. Остров Сулавеси в озерах Посо и Линду. 2 рода, 3 вида, по-ви- димому, образовавшиеся от солевыносливых карпозубых. Бьюфорт (Beaufort, 1934) описал ископаемые формы, возможно, относящиеся к предкам карпозубых (возраст не установлен) с острова Сулавеси.
П о д о T р я д S у п е n t о g n а С h i. Группа, состоящая примерно из 4, главным
образом морских, семейств, включая следующие:
(ezra)Hemirhamphidae, полурылы. Все тропические и некоторые моря умеренной зоны.
многие виды заходят или живут в пресных водах. Разные роды и многочисленные виды. Большинство пресноводных форм относится к живородящему подсемейству (или семейству) Dermogenyinae, которое распространено от Юго-Восточной Азии до острова Сулавеси и Филиппин и почти всегда приурочено к пресным водам (IIubbs, письменное сообщение). Один из пресноводных родов Yomorhamphus, с 2 видами, приурочен к острову Сулавеси.
Пресноводные рыбы
П о д о т р я д Т h о г а с о s t e i. Преимущественно морская группа, примерно из 8
или более семейств, включая следующие:
(слв) Gasterosteidae, колюшки. Прохладные воды северного полушария. Небольшое
семейство, включающее примерно 10 видов мелких рыб, некоторые из которых встречаются и в соленых, и в пресных водах (некоторые детали см.: Myers, 'l 930, р. 99 — 100; Wynne-Edwards, 1952, р. 22 — 23).
П о д о т р я д S а 1m о р е г с а е. Северная Америка. 2, очевидно, реликтовых
филогенетически изолированных семейства. Довольно мелкие окунеобразные рыбы. (1)Percopsidae, перкопсис (или траут-пэрчи). 2 рода и 2 вида. Percopsis omiscomaycus
в северной и восточной частях Северной Америки от «Юкона и Альберты до Гудзонова залива и Квебека и от Великих озер через систему реки Св. Лаврентия до озера Шамплейн; на юг по Атлантическому склону (обычно редко), от системы реки Гудзона до реки Потомак и от долины Миссисипи до Западной Виргинии, Кентукки, Миссури, Канзаса и Южной Дакоты (редко и локально на юге)» (Hubbs, Lagler, 1947, р. 79). Вид Columbia transmontana локализован в западной части Северной Америки в реке Колумбия, штаты Орегон и Вашингтон.
(1) Aphredoderidae, афредодерусы. Центральная и восточная части Северной Америки.
1 вид (по меньшей мере 2 подвида) распространен на юг от «юго-восточной Минне- соты, южного Висконсина, южного Мичигана, южных притоков озера Онтарио и западных склонов Аппалачских гор и их предгорий до залива Галф-Коаст [Мексиканский залив.— Ред.] и Техаса» (Hubbs, Lagler, 1947, р. 80), а также по Атлантическому склону от Нью-Йорка до Флориды. Ископаемые родыобнару- жены в западной части Соединенных Штатов (эоцен и олигоцен).
Отряд Acanthopterygii, колючеперые. Состоит более чем из 20 подотрядов (Romer)
и большого числа семейств. Более половины подотрядов и большое число семейств исконно морские или изредка встречаются в пресных водах. Около 20 других семейств главным образом морские, но обычно заходят или представлены в прес- ных водах. Несколько семейств приурочено только к пресным водам. В нижесле- дующий список включены главным образом последние и несколько морских семейств, имеющих представителей и в пресных водах.
П о д о т р я д P e r с о i d е а, Окунеобразные. Состоит примерно из 50 морских
семейств (некоторые имеют представителей в пресных водах) и также из ряда име- ющих важное значение пресноводных семейств первого и второго отделов. (слв) Serranidae, морские окуни. Большое морское семейство. Кроме множества строго
морских видов, имеется несколько видов, которые заходят в пресные воды для размножения, и несколько видов, включая североамериканского белого окуня Lepibema chrys0ps и желтого окуня Morone interrupta, которые обитают в прес- ных водах. «Нильский окунь» (Lates niloticus) и близкородственные виды — круп- ные рыбы, широко распространенные в пресных водах Африки, Южной Азии и Австралии,— относятся к этому же семейству. Более подробные сведения о севе- роамериканских представителях этого семейства см. в работе Хабса и Лаглера (Hubbs, Lagler, 1947, р. 80 — 81) .
(слв) Sciaenidae, горбылевые, барабанщики. Другое большое морское семейство, несколь- ко видов которого заходят в пресные воды, и очень немногие обитают в них.
К последним относится пресноводная рыба-барабанщик Ар1ойто1из grunniens.
Северной Америки, которая встречается от бассейнов Гудзонова залива до Гва- темалы (Hubbs, Lagler, 1947, р. 95).
(1) Centrarchidae, пресноводные ушастые окуни. Северная Америка. Несколько
родов, около 24 видов. На восточной стороне Скалистых гор, от южной Канады до Мексики. Один вид достигает реки Сото-ла-Марина, почти до тропика Рака в восточной Мексике (Buen, 1946). Один эндемичный род и вид .4 rchoplites inter- ruptus изолирован в Калифорнии в системах рек Сакраменто и Сан-Хоакин.
Несколько восточных видов интродуцировано на западе от Скалистых гор.
1) Percidae, окуневые. Европа, Северная Азия и восточная часть Северной Америки.
Нет на крайнем востоке Азии и в западных бассейнах Северной Америки, кроме одного вида (Rafinesquiellus pottsii), достигающего в Мексике рек, текущих на запад. Границы распространенияэтого семейства показаны на рис. 13. Три под- семейства. (1) Percinae встречается в Евразии и Северной Америке. Вид Perca fluviatilis встречается по всей Евразии от Британских островов до реки Колыма, но не достигает бассейнов Тихого океана на крайнем востоке Азии, а Perca fla- гезсепз (желтый окунь) — в восточной части Северной Америки на северо-западе озера Атабаска (в западной част~и Северной Америки нет за исключением интро- дуцированных видов). Третий вид рода Perca является эндемиком в районе озера Балхаш — Тарим, Центральная Азия. Это единственный эндемик семейства оку- невых в Азии. Еще 3 рода и 7 видов находятся в Южной Европе, особенно в бас- сейнах Черного и Каспийского морей, и один из видов (Acerina сегпиа) прости-
106
Глава 2
рается на запад до Англии и на восток до Колымы, достигая восточных границ вида Ретса fluriatilis. (2) Luciopercinae: Евразия и Северная Америка. Lucioperca (3 вида) распространен в Европе от района Аральского моря до Эльбы; в се- верных бассейнах и в Азии нет.Близкородственный род Stizostedion (' вида) в Се- верной Америке в пределах границ этого семейства (северная граница этих видои является северной границей семейства). (3) Ftheostominae в восточной части Северной Америки от южной Канады до Мексики. Л. pottsi достигает реки Мескиталь, пересекающей в западной Мексике тропик Рака (Виеп, 1946). Оно включает около 30 родов и 100 видов (Jordan et al., 1930). В большинстве случаев очень мелкие рыбы, живущие в быстрых водах. Можно предположить, что они за- нимают те же самые ниши в восточнойчасти Северной Америки, что и представи- тели надсемейства Cypriniformes, живущие в быстрых водах в Евразии. Иско- паемые окуневых встречаются в Европе и Северной Америке в эоцене и позднее. (1) 3andidae. Разрозненные тропические территории: 1 или 2 эндемичных рода в каж- дой из трех основных тропических областей (но не в Австралийской области).
Каждый род имеет только 1 или ' вида: в (Западной) Африке — Polycentrop- sis; в Индо-Малайской области — Nandus (Индия до острова Калимантан, но не до Явы, в пресных и солоноватых водах) и Badis (Индия до острова Калимантан и, возможно, за его пределами), в Южной Америке — Polycentrus(Tprntnpag и Гви- ана) и Мопосьг~Ьиз (Гвиана и низменность Амазонки). Довольно мелкие рыбы.
Майерс, видевший представителей всех родов, сообщает, что все роды, кроме рода Badis, несомненно, могут быть отнесены к одному семейству и что даже близко- родственный Badis смело может быть отнесен к этому же семейству.
(1) Pristolepidae. Индо-Малайская область. Распространение прерывистое: Индо- Китайская подобласть до островов Ява и Калимантан с 1 восточным видом, встре- чающимся также на изолированной территории в полуостровной части Индии и на острове Цейлон (Hora, 1944). 1 род, около 4 видов. Очень похожи на рыб семейства Nandidae.
(2) Cichlidae, хромксы. Сконцентрированы в тропиках Африки и Америки, с несколь- кими видами на прилежащих территориях. Много родов, более 600 видов. В Аф- рике большинство хромисов находится в западных и центральных тропических районах и особенно в озерах Ньяса, Танганьика и др., где образовались обшир- ные группы близких видов (Brooks, 1950; Greenwood, 1951). Но несколько видов встречается до самых низовьев Нила и даже (с несколькими видами других рыб) в изолированных или подземных водах северной части Сахары (Boulenger, 1907, р. 465, о02; Braestrup, 1947), но ни один вид не достигает крайней северо-западной части Африки к северу от Атласских гор. Несколько видов достигает крайнего юга, несколько не доходя до мыса Доброй Надежды (Barnard, 1943); несколько видов имеется в Сирии и Палестине, включая 1 эндемичный род в долине Джор- дан (Trewavas, 1942); 3 эндемичных рода находятся на Мадагаскаре и 1 эндемич- ный род (близкий мадагаскарскому) с 3 видами встречается в эстуариях рек и в реках береговых районов Цейлона и полуострова Индостан. По сообщению Бьюфорта, этот род встречается также в Таиланде, что, скорее всего, является ошибкой, так как ни один представитель этого семейства не вошел в работу Сми- та (Smith, 1945) по пресноводнымрыбам Таиланда. В Америке хромисы довольно многочисленны на большей части Южной Америки (но ни один вид не достигает более прохладных частей Чили и Аргентины) и относительно многочисленны в Центральной Америке до южной Мексики (Buen, 1946), с 1 видом, достигающим южной краевой части Техаса (предполагают, что формы Центральной Америки и Мексики образовались от южноамериканских). Широко распространенный южно- и центральноамериканский род Cicklasoma встречается также на Кубе и Гаи- ти и, как сообщалось (вероятно, это ошибка), на Ямайке. Виды этого семейства обычно встречаются в пресных или солоноватых водах, но некоторые виды встре- чаются также в сильно солепых внутренних водах и некоторые могут в течение довольно продолжительного времени жить в настоящей морской воде (Myers, 1938). Хромисы — небольшие, типично окунеобразные рыбы. Подробное описа- ние этого семейства дано в монографии Пеллегрина (Pellegrin, 1903), но в отно- шении некоторых вопросов эта работа уже устарела. Известно несколько иско- паемых хромисов, но они не представляют особой ценности для разработки воп- росов географической истории этого семейства.
П о д о т р я д В 1 е n n i о i d е а. Морская группа рыб примерно из 20 семейств,
включая следующие:
(с;1в) Brotulidae. Морское семейство, некоторые представители которого встречаются
в глубоководных частях. Единственные известные представители пресных вод- слепые формы, обнаруженные в пещерах на Кубе (2 эндемичных рода) и па Юкатане (1 эндемичный род) (Hubbs, 1938).
Пресноводные рыбы
П o;t о т р я д А п а с а n t b i n i. Другая морская группа рыб из нескольких
семейств, включая следующее:
{слв) (.'adidae, тресковые. Главным образомморское семейство, состоящее из нескольких
десятков морских видов, распространенных от арктических до антарктических морей. Немногочисленные виды, в основном глубоководные формы, встреча1отся в пределах тропиков (Hubbs, Lagler, 1947, р. 79) и l род Lotn (налимы) с 1 видом, несколькими подвидами в более холодных пресных водах северного полушария {группе-Edwards, более подробное описание). Род Lota на большей части своего ареала обитает в пресных водах, но на крайнем севере он встречается также и в море и считается мигрирующим. Еще один род этого семейства иногда заходит в реки северо-восточной части Северной Америки (Gunter, 1942, р. 314), и несколь- ко родов встречаются в пресных водах Северной Азии (Берг, 1949).
Il о д о т р я д С h а е С о d о п С о i d е а. Еще одна морская группа из нескольких
семейств, включая:
{слв) Toxotidae, брызгуны. Между Индией и Австралией — от Индии до Филиппин.
на юг до Северной Австралии и на восток до Соломоновых островов и Новых Гебрид. Один род, несколько видов «в водах литорали с низкой соленостью, в солоноватых водах эстуариев и в реках» (Weber, Beaufort).
! 1 о д о т р я д S с о г р а е п о i d е а. Группа, состоящая из 20 или больше семейств,
большинство из которых являются строго морскими, кроме двух, приведенных ниже:
(слв) Cottidae, бычки-подкаменщики. Множество родов и приблизительно 300 видов
в северных морях (и несколько в антарктических морях). Некоторые обитают в более холодных реках и озерах северного полушария (Hubbs, Lager, 1947, р. 96, Wyne-Edwards, 1952, р. 21).
{слв) СотерЬог1йае (голомянки) и Cottocomephoridae (широколобки). Почти полностью
приурочены к озеру Байкал, Сибирь (2 вида достигают Енисея, а 1 эндемик реки Лены). Их происхождение, очевидно, связано с радиацией морского предка из семейства Cottidae; 9 родов, 16 или 18 видов (Берг, 1949; Brooks, 1950).
Подотряд Gobioidea, бычки и т. д.
(слв) Gobiidae (с Eleotridae и др.). Очень большое семейство или группа семейств мел- ких рыб, которые распространены в основном вдоль побережий всех морей тро- пиков и многих морей умеренной зоны. Можно часто встретить в коралловых рифах и в водоемах, оставляемых приливом, вдоль песчаных или илистых побере- 'кий. Некоторые виды большую часть времени могут проводить вне воды, охо- тясь за насекомыми, летающими над мокрым песком или илом. Большинство других видов живет в пресных водах рек, заходя в море только в период нереста (иногда не заходят). Некоторые из пресноводных форм часто встречаются в быст- рых реках в разных частях земного шара, включая Мадагаскар, Австралию.
Вест-Индию и многие другие тропические острова, такие, как Лусон и Гавай- ские, где строго пресноводных рыб нет. Например, род Bhyacichthys, иногда выделяемый в самостоятельное семейство, известен на островах Ява, Сулавеси, Филиппинах и Соломоновых островах (все одного вида). Рыбы этого рода живут в стремительных реках, присасываясь к камням (как это делают и другие бычки.
живущие в горных потоках), а если их потревожить, плавают вокруг камней или прячутся под ними. Распространение и хорошо изученный образ жизни дру- гих бычков дают возможность предположить, что этот род может преодолевать такие препятствия, как соленые морские воды (Herre, 1927). Ряд эндемичных бычков обитает в Черном и преимущественно в Каспийском морях.
ll о д о т р я д А п а b а n t о i d е а, лабиринтовые рыбы. Подотряд пресноводных
рыб, состоящий примерно из 3 семейств. В большинстве случаев это мелкие или среднего размера рыбы, имеющие дополнительный орган дыхания.
(1) А11нЬапЯае (включая Osphronemus), анабасы, или ползуны и петушки (Bettn)
п др. Африка и Индо-Малайская область, на север до Восточной Азии. В Африк< .
вероятно, имеется только род Anabas (который встречается также и в Индо- Малайской области, хотя иногда африканские и восточные формы относят к раз- ным родам) приблизительно с 20 видами от верховьев системы Нила до Кейптауна {отсутствует в Северной Африке — тропические формы, распространяясь на юг, достигают только Замбези, 2 вида из Кейптауна образуют эндемичную географи- чески изолированную группу; Barnard, 1943). Предполагаемый западноафрикан- «кий род Micracasttkus, вероятно, является неправильно этикетированным родо~~ Betta из Индо-Малайской области (в соответствии с письменным сообщением Май- ерса). В Индо-Малайской области насчитывается примерно 11 родов (включая .4nabas) и множество видов, некоторые из которых достигают островов Ява и Ка- лимантан и некоторые виды на севере доходят до Центрального Китая или (1 вид) до южной Маньчжурии и Кореи. Один из видов Индо-Малайской области, Лп~-
108
Глава 2
has testudipieus,— обычная рыба, живущая в различных пресных и солоноватых водах, довольно регулярно переносится человеком в разные районы. Этот вид встречается за пределами выше описанных границ до Филиппин и островов Сула- веси и Хальмахера, но возможно, что в этих местах он был интродуцирован.
Предполагаемый эндемик этого семейства на Кагаян-Сулу (маленький остров на северо-востоке от Калимантана) теперь считается таким же обычныи, как и вид на острове Калимантан (Myers), и, возможно, был интродуцирован. (1) Channidae (Ophicephalidae), змееголовые. Африка и Индо-Малайская область, на
север до Восточной Азии. 2 рода, приблизительно 20 видов. Главный род Ophi- cepkalus включает около 3 видов в Африке — от Западной Африки до верховьев Нила и на юг до залива Делагоа (J. L. В. Smith, !950) и ряд видов в Индо-Ъ! алай- ской области — от Индии до островов Ява и Калимантан и далее на север в умень- шающемся числе через Китай (1 вид достигает Амура). Другой род (>..hanna), вероятно только с 1 видом, приурочен к Юго-Восточной Азии. Вид Ophicephalus г~па~иг встречается за пределами выше обозначенных границ на Малых Зондских островах, на островах Сулавеси, Хальмахера и Филиппинах, но возможно, что он был там интродуцирован. Это обычная рыба, которая может некоторое время жить без воды и часто переносится человеком.
(1) Luciocephalidae. Индо-Малайская область, от Малаккского полуострова до острова
Калимантан. 1 вид.
П о д о т р я д М и g i ! о i d е а, кефали и т. д. В основном морская группа, но не- которые кефалевые часто заходят или постоянно живут в пресных водах ( >!уегз, 1938; Schultz, 1946), а нижеследующее семейство еще более примечательно чис- лом солевыносливых пресноводных форм.
(слв) Atherinidae, атеринковые. Побережья всех морей тропиков и морей умеренной
зоны и во многих изолированных пресных водах. Некоторые роды aBëÿþòñ~ строго морскими, другие — в основном морскими, но заходят в пресные воды.
Например, ЛИеппа — широко распространенный морской род со многими видами, но в Средиаемном море вид А. caspia, проникающий в пресные воды (по побережью), образовал местные пресноводные расы, а вид А. tamarensis обнаружен в реках береговых районов и в реках Тасмании и Южной Австралии. Другие роды, которые встречаются и в пресных, и соленых водах, встречаются вдоль восточного побережья Соединенных Штатов и местами на восточном и западном побережьях тропической части Америки. Род Austromenidia обитает в прибрежных частях морей, рек и озер умеренной зоны южных широт Южной Америки на юг до Магелланова пролива. Но из 59 родов этого семейства (Jordan, Hubbs. 19!9) половина ограничена пресными или иногда солоноватыми водами. Например, в Старом Свете на Мадагаскаре имеется два эндемичных рода, 1 на острове Сула- веси и около 10 в Австралии, на Новой Гвинее и на соседних островах. В Новом Свете 1 пресноводный род приурочен к восточным прибрежным районам Соеди- ненных Штатов; 3 локализованы в Мексике (1 род из Кубы описан совсем неда»- но), 2 — в юго-восточной Бразилии и 1 — в реках Анд, Перу и Чили, встречается до высоты 8000 футов и выше. Надо о>гметить, что в большинстве случаев пресновод- ные роды этого семейства находятся в тех местах, где строго пресноводные рыбы основного первого отдела отсутствуют или относительно малочисленны. Клас- сификация Джордана и Хаббса изменена в соответствии с последней работой (например, Schultz, 1948), но основной характер распространения семейства не изменился. Подсемейство Melanotaeniinae примерно с 10 родами, иногда считает- ся приуроченным к более теплым частям Австралии, Новой Гвинеи и прилежа- щим островам того же материкового шельфа (Beaufort, 1926, р. 103; !951, р. 154— 155), но Джордан и Хаббс включают Telmatherina с острова Сулавеси в ту же самую группу. Атеринки — довольно мелкие, похожие на корюшку рыбки. (слв) Phallostethidae. Таиланд и полуостров Малакка до Калимантана и Лусона. Около
10 родов, 18 видов (Herre, 1942). Одни обитают в соленой или солоноватой воде, другие в пресной. «Крошечные и почти незаметные (прозрачные) рыбки» (Herre). П о д о т р я д О р i s t h о m i. Подотряд пресноводных рыб, состоящий из 2 се- мейств.
(1) Mastacembelidae, колючие угри. Африка до Индо-Малайской области, 2 )>од'>—
Mastacembel us со множеством видов, из которых около 30 находится в тропической Африке (но не в Северной Африке и не на большей части Нила), 1 — » системе Тигра — Евфрата, 12 — в Индо-Малайской области на н>I'<>- восток до островов Ява и Калимантан и 1 — в Китае на север до Пеtlx() (Mori, 1936); род Rhynchoi>della с 1 видом в Индо-Малайской области. ( !ти подробности в основном взяты из работы Боуленджера — (Boulenger. 1912).
(1) Chaudhuriidae. Индо-Малайская область. 1 вид, известный только в озере )!»ли
Б ирма (Regan, 1919) .
Литература
И о д о т р я д $ у n b r а п с h i i, ильные угри. Группа, состоящая из несколькихнебольших семейств, которые хотя и живут главным образом в пресных водах,цо не считаются строго пресноводными рыбами.
(слв) Synbranchidae. Тропики. Несколько родов в соленых, солоноватых и пресныхв(~дах. Род БуиЬга)>chus, обитающий в пресных и солоноватых водах и изредкав море, имеет 1 вид в тропической Западной Африке, 1 — от Индии до НовойГвинеи и Австралии (McCulloch) и 1 — в тропической Америке — от Мексикиц Ь~ бы до Бразилии. Родственные слепые рыбы были найдены в пресных водах,ио в районах, расположенных близ моря, в Западной Африке и на Юкатане.
(слв) Amphipnoidae. Индия и Бирма, а также Квинсленд, Австралия (МсСи11осЬ,)929). 1 вид в пресных и солоноватых водах.
(слв) Indostomidae. Индо-Малайская область. Известен только экземпляр из озерав Бирме. 1 вид. Систематическое положение этого семейства сомнительно (Rolin,I!) 36) .
ЛИ ТЕРА ТУРА
А u r i c h Н., 1935, Mitteilungen der Wallacea-Expedition Woltereck, X I I I, X IV,
Fische I, II, «Zool. Anzeiger», 112, 97 — 107, 161 — 177.
В а r п а r d К. Н., 1943, Revision of the indigenous freshwater fishes of the S. 9'. Саре
region, «Ann. South African Mus.» 36, 101 — 262.
В а r п а r d К. EI., 1948, Report on а collection of fisches from the Okovango River,
v ith notes on Zambesi fishes, «Апп. South African Mus.», 36, 407 — 458.
В а r r i о А., 1943, ...Lepidosiren paradoxa... [in Argentina], «Revista Argentina de
Zoogeografia», 3, 9 — 20.
В е а п f о r С L. F., de., 1926, Zoogeographie van den Indischen Archipel, Нааг)ет,
'ч etherl ands.
В е а и f о r С L. F. de., 1934, On а fossil fish from Gimpoe (central — Celebes), «Verb.
Geol.— Mijnb. Genoot. Ned. Kolonien, Geol. $ег.», 10, 180 — 181. Взято из «Zoo- logical R ecord».
В е а u f о r t Ь. F., de., 1951, Zoogeography of the land and inland waters, London- ' ew York.
В е r e L. S., 1932, Ubersicht der Verbreitung der Susswasserfische Europas, «Zoogeo- graphica» (Jena), 1, 107 — 208.
Б е р г Л. С., 1934, Разделение палеарктики на зоогеографические области на
основании распространения пресноводных рыб, «Тр. Первого Всесон~зного
географического съезда», вып. I I I, Москва.
1> е р г Л. С., 1936, Подотряд Esocoidei (Pisces) «Изв. Виол. ин-та Пермского гос.
унив.», т. Х, вып. 9 — 10.
Б е р г Л. С., 1940, Система рыб, ныне живущих и ископаемых. «Тр. Зоол. ин-та»,
вып. 2, М.— Л.
Б е р г Л. С., 1947, Географические зоны Советского Союза, Москва.
Ь е р r Л. C., 1948 — 1949, Рыбы пресных вод СССР и сопредельных стран, 4-е изд.,
ч. 1 — 3.
В о 1 i n R. Ь., 1936, The systematic position of 1т~ойогииз paradozus... «J. Washing
t on Асад. Sci.», 26, 420 — 423.
I) о и1 е n g e r G. A., 1902, The explanation of. а remarkable case of geographical
distribution a~nong fishes, «Nature», 67, 84.
В о и 1 е n g e r G. А., 1905, The distribution of African fresh-water fishes, «Ха1цге»,
72, 4) 3 — 421.
В о и 1 е n g e r G. Л., '1907, The fishes of the Nile. См. А n d e r s о n, Zo<ilogy of
Egypt, London, Ltd., 2 vols.
В о и 1 е n g e r G. А.. 1909 — 1916, Catalogue о~ the fresh-water fishes of Africa in 1Ье
British Museum, London, British Mus., 4 чо1я.
Ь о и 1 е n g e r G. A., 1912, А synopsis of the fishes of the genus Mastacenibclus,
«J. Acad. Nat. Sci. Philadelphia», Ser. 2, 15, 197 — 203.
110
Глава 2
В г а е s С г и р F. W., J 947, Remarks on faunal exchange through the Sahara, «Vi- denskabelige Medclelelser Dansk naturhistorisk Forening», 110, 1 — 15.
В г е d e r (:. М., Jr., 1934, Ecology of an oceanic fresh-water lake, Andros Island, ВаЬя- mas, with special reference to its fishes, «Zoologica», (Sci. Co»tril>. Пеи York
Zool. Soc.), 18, 57 — 88.
В г о о k s J. Ь., 19950, Speciation ~n ancient lakes, «~»charter)y Review Biol.», 2;i, 131—
176.
В и е n F., de., 1946, Ictiogeograplxia Continental hlexicana, «Bevista Soc. 'Alezicana
Hist. Nat.», 7, 87 — l 38.
С а г l С. С., С 1 е m e n s W. А., 1948, The 1геяЬ-water fishes of British Columbi
B ritish Columbia Provincial M us. H andbook Х о. 5.
С а г р е п С е г R. G., $ i e g 1 е г H. R., 1947, A sportsrnan's guide to the 1геяЬ-
water fishes of New hampshire, Concord, New НатряЫге, State Fish ancl б mme
Commission.
С Ь а p m а n %. М., 1944, ...Лр1осй|8ои "eh~a Jenyns, «J. Alorphology», 75, 149 — 165. D а m m е г m а п К. W., 1929, On the zoogeography of J ava, «Treubia», 1 I, 1 — 88. D а r 1 i n g t о п P. J., 1г. 1948, The geographical distribution, of cold-blooded ~.c rteb- rates, «Quarterly Review Biol.», 23, 1 — 26, 105 — 123.
D а у F., 1889, Fishes, См. «The fauna of E3ritish India, including Ceylon and Впгша»,
London, 2 vols.
D y m о n d J. R., V 1 а d y k о ч V. D., )9934, The clistribution and relationship о1
the salmonoid fishes of North America and North Asia, «Proc. Fifth Pacific Sci.
Congress», 5., 3741 — 3750.
E g e V., 1939, А revision of the genus Аnguilla Shaw..., «Dana Rep.», No. 16.
К i g e n m а n n С. Н., 1909, The fresh-water fishes of Patagonia and an examination
of the Archiplata — Archhelenis theoty, «Rep. Princeton U. Exp. Patagonia, 1896—
1899», 3, pt 3, 225 — 374.
Е i g е п m а n n Г,. Н., 1910, Catalogue of the fresh-ъater fishes ul' tropical a»d soiith
temperate America, «Rep. Princeton U. Ехр. Ра[agonia, 1896 — 1899», 3, pt 4,
375 — 511.
Е i g е п m а n n С. Н., 19'18, The Pygidiidae, а family of So»th American catfislies,
«Mem. Carnegie Mus.», 7, 259 — 398.
Е i g е п m а n n С. Н., 1927, The fresh-water fishes of СЫ1е. «Меп~. Ai ational Acad.
Sci.» (Washington), 22, No. 2.
Е i g e n m а n n С. Н., A 1 1 е n W. R., 1 942, Fishes of western South Лшег~с»,
Lexington, Kentucky, U. of Kentucky.
Е i g e n m а n n С. Н., М у е г s G. $., 1917 — l 929, The American Characidae,
«Мет. Мця. Comparative Zool.», 43.
Е 1 1 i s М. М., 1913, The gymnotid eels of tropical Amebicÿ, «Мет. Carnegie Мця.»,
6, 109 — 204.
F r а s е г - В г u n n e г А., ! 950, ...Gasteropelecidae, «Лип. апд Map. X atural Il isto- ry», $ег. 12, 3, 959 — 970.
F г о s С Х., 19~0, Л preliminary study of Хewfoundland trout, Xewfoundlancl I)epart- ment Ма(. Resources, «Research Ви11. 9» (Fisheries).
G а r т а n S., 1913, The Р1аа1ояйот1а (sharks, skates, and rays), «Мет. Mus. Сошря- rative Zool.», 36.
G о s l i n e W. А., 1944, Т1~е problem of the derivation of the South American «ncaa
African freshwater f jsb faunas, «Anais Асад. Brasileira де Ciencias», 16, ' 11—
мм3 е
G о s l i n е Ж. А., 1945, Сatalogo dos iiematognatos de apua-doce da Атег~са до Sul
е Central, «Boletim Mus. Nacional» [Brazil], New Ser., No 33.
G r е е n w о о d P. H., 1951, Evolution of the African cichlid fishes'. the FIaplochromis
species flock in Lake Victoria, «Хature», 167, 19 — 20.
„7итература
G u n t e r G., 1942, А list of (11е fishes of ...North and Middle America recorded from
both freshwater and sea water, «American Midland Naturalist», 28, 305 — 326. Н а у О. P., 1929 — 1930, Second bibliography and catalogue of the fossil Vertebrata
of North America, «Carnegie Inst. Washington Pub.», 390.
II e r r e A. W. С. Т, 1927, бо2нея of the Philippines апй the China Sea, Philippine
Bureau of Sci., Monograph 23.
Н е r r е А. 'А'. С. Т., 1928, True freshwater fishes of the Philippines. (."м. D i c k e r- s о n et al., Distribution of life in the Philippines, 242 — 247, Bureau of Sci., Ма- пйа, Monograph 2l.
Ji e r r e A. %. С. Т., 1933, The fishes of Lake Lanao: а problem in evolution, «Arne- rican Naturalist», 67, 154 — 162.
И е r r е А. W. С. Т., .1.942, New and little known phallostethids, with keys Со the
~епега and Philippine species, «Stanford Ichthyological Bull.», 2, 137 — 156. Н е r r е А. W. [С. Т.], А Ь 1 а и G. L., '193 ~, .1р1осЬел[из 1иопепзьз, а new Philip- pine cyprinodont, «Philippine J. Sci.», ~4, 275 — 277.
Н i 1 d e b r а n d S. F., 1938, А new catalogue of the freshwater fishes of Panama,
«Pub. Field Mus. Nat. Hist., Zool. Ser.», 22, No. 4, 2l5 — 359.
Н о r а S. Ь., 1932, Classification, bionomics and evolution of homalopterid fishes,
«Mem. Indian Mus.», 12, 263 — 330.
1I о r а S. L., 1934, Fish of Afghanistan, «J. Bombay Nat. Hist. Soc.», 36, 688—
706.
Н о r а S. Ь., 1937, Comparison of the fish-faunas of the northern and the southe vn
faces of the Great Himalayan Range, «Rec. Indian Mus.», 39, 241 — 250.
ll о r а S. Ь., 1937а, '-Geographical distribution of Indian fresh-water fishes and its
bearing on the probable land connections betwenn India and the adjacent count- ries, «(;urrent Science» (Bangalore), 5, 351 — 356.
l I о r а S. L., 1938, On the age of the Deccan trap as evidenced by fossil fish remains,
«C;urrent Science» (Bangalore), 6, 370 — 372.
Н о г а S. Ь., '1938а, "hanges in the drainage of India, as evidenced by the distribution
of freshwater fishes, «Proc. National Inst. Sci. India», 4, 395 — 409.
)! о r а S. T., !9-~4, On the Malayan affinities of the freshwater 6яЫаипа of peninsular
India..., «Proc. National Inst. Sci. India», 10, 423 — 439.
Н о г а S. Ь., 1949, Вайпд the period of migration of the socalled Malayan element in
the fauna of peninsular India, «Proc. National Inst. Sci. India», 15, 1 — 7.
И о r а S. L., М е п о п А. G. К., 1952 — 1953, Distribution of Indian fishes of the
past and 1Ье~г bearing on the geography of India, «Everyday Science», 1, 26 — 37;
2, 105 — l 13.
ll и b b s С. T.., ,1936, Fishes of the Yucatan peninsula. См. The Cenotes of Yucatan,
«Carnegia Inst. Washington Pub.», 457, 157 — 282.
I I u h b s С. L., 1938, Fishes from the caves of Yucatan, «(;arnegie Inst. Washington
Pub.», 491, 261 — 295.
ц u b b s С. T., 1941, А newfamily of fishes, «J. Bombay Nat. IIist. Soc.», 42, 446 — 447. И u b Ь s С. Х., 19,э2, Antitropical distribution of fishes and other organisms, «Proc.
Seventh Pacific Sci. Congress», 3, 324 — 329.
lt u Ь Ь s (:. I ., T а g 1 е r K. F., 1947, Fishes of the Great Lakes region, «(.:гапЬго- ok Inst. Sci., Bull.», 26.
ll u Ь b s (:. I., М i 11 е r R. R., 19-~8, Correlation between fish distribution аш1
hydrographic history in the desert basins of western United States. См. The Great.
Вамп, with етпрЬаяя on glacial and postglacial times, «13ull. 1 . Utali, Biol.
Series», 10, No. 7, 18 — 166.
If u b b s С. Т,., Т и r п е r С. L., 1939, Studies of the fishes of the order Cyprinodon- tes. 16. А revision of the Goodeidae, «Misc. Pub. Mus. 7ool.», U. Michigan, 4 1 о г d а n D. S., 1905, А guide Со the study of fishes, Хеъ Y~irk, 2 vols.
112
Глава 2
Л ordan D. S., Ivermann В. Ж., Clark El. %., 1930, (;lreck list of the
fishes... of North and Middle America... Rep. United States Commissioner of
Fisheries for 1928, pt 2.
3 о r d а n D. S., Н u b b s С. ? ., 1919, А monographic review of the family of Athe- rinidae or silversides, «Leland Stanford Junior U. РиЬ.», U. Ser. (без номера). К о s s w i g С., 1952, Die zoogeographie der turkischen Siisswasserfische, «Pub. Hydro-
biological Research Inst., Faculty Sci.», U. Istanbul, $ег. В, 1, 85 — 101.
Ь i т i n g s С о п е D. А., 1953, The freshwater fishes of Nova Scotia, «Ргос. Хоча
Scotian Inst. Sci.», 23, 1 — 90.
М а с А 1 р i n А., 1947, ...polyodontid fish from Upper 'retaceous of Montana...,
«Contrib. Mus. Paleontology, U. Michigan», 6, 167 — 234.
M c C ( и11 о с h) А. R., 1927, Fishes of Australia. CM. «Australian Encyclopaedia».
Згй ed., vol. 1, Sydney 465 — 467.
М с С u11 о с h А. R., 1929 — 1930, A check-list of the fishes recorded from Australia,
«Australian M us. (Sydney) M em.», 5.
М а с D о п а g h Е. J., 1945, Hallazgo de una I-epidosire~i рагайоха en el detta del
Parana, «Notas del Mus. de la Plata», 10, 11 — 16.
M а t t h e w W. D., 1915, 1939, Climate and evolution, «Ann. New York Асад. Sci.», 24,
171 — 318 (переиздано в 1939 г.), см. «Special РиЬ. New York Асад. Sci.», 1. M i 1 е s С., '1947, Los Peces del Rio Magdalena, Ministerio de la Economia Wacional,
Seccion de Piscicultura etc., Bogota, Colombia.
M о г i Т., 1936, Studies on the geographical distribution of freshwater fishes in eastern
Asia (частная публикация).
М и n r о 1. $. R., 1955, The marine and freshwater fishes of Ceylon, Canberra (Austra- lia), Dept. of External Affairs.
M у е г s G. $., 1930, The killifish of San Ignacio and the stickleback of San Rarnon.
Lower California, «Proc. California Acad. Sci.», Ser. 4, 19, 95 — 104.
М у е r s G. $., 1931, The primary groups of oviparous cyprinodont fishes, «Stanford
Pub., U. Ser., Biol. Sci.», 6, 241 — 254.
М у е r s G. $., 1938, Freshwater fishes and West Indian Zoogeography, «Smithsoni- an Rep.», 1937, 339 — 364.
M у е r s G. S., 1940, The fish арпа of the Pacific Ocean..., «Proc.. Sixth Pacific Sci.
Congress», 3, 201 — 210.
M у е r s G. S., 1944 [Rearrangement of Pygidiidae], «Proc. (alifornia Асад. Sci.»,
Ser 4, 23, 591 — 602.
М у е r s G. S., 1947 — 1949, The Amazon and its fishes (в '> частях) «Aquarium J.»
1947, March, р. 4 — 9 (pt 1); April, р. 13 — 20 (pt 2); Мау, р. 6 — 13 and 32 (pt 3);
July — Aug., р. 8 — 19 and 34 (pt 4); 1949, Feb., р. 52 — 61; МагсЬ, р. 76 — 85
(pt 5).
М у е г s G. S., 1949, Usage of anadromous, catadromous and allied terms for migra- tory fishes, «Copeia», 1949, 89 — 97.
М у е г s G. S., 1949а, Salt-telerance of freshwater fish groups in relation to zoogeo- graphical problems, «Bijdragen Tot de Dierkunde», 28, 315 — 322.
М у е г s G. S., 1951, Freshwater fishes and East Indian zoogeography, «Stanford Ic li- thyological Bull.», 4, 11 — 21.
M у е г s G. S., 1953, Paleogeographic significance of freshwater ЙяЬ distribt]tion in
the Pacific, «Ргос. Seventh Pacific Sci. Congress», 4, 38 — 48.
Х е 1 s о п Е. M., 1948, The... ЖеЬег~ап apparatus of the (,atostomidae..., «J. Могр1н~- logy», 83, 225 — 251.
Х е 1 s о п E. Ъ[.. 1949, The орегси1аг series of the 'atostomidae. «J. МогрЬо1оуу».
85, 559 — 568.
1М i c h о 1 s J. Т., 1928 Fishes from the White Nile... [with table of «world' s fresh- water fish Гыцпае»], «American Mus. Novitates», No. 31,).
Литература
Х i с Ь о 1 s J. Т., 1938, Classilication of сагр-like fishes, «Zoologica» (New York»).
23, 191 — 193.
3 i c h о 1 s 3. Т., 1943, The fresh-water fishes of (Ыпа, Natural Ilistory ol,entral
Asia (American Мця. Nat. Hist., New York), vol. 9.
X i c h o 1 s J. Т., G г i s c о m L, 1917, Fresh-water fishes of the (.'опто Basin
[ittcluding а discussio>i о1 African fresh-water fish faunas, р. 739 — 752], «Bull.
American Mus. Sat. Ilist.», 37, 653 — 756.
О ~ а d а Y., 1955, Fishes of Japan, Tokyo.
Р е 1 1 е р г i n 3., 1903, ...1а 1апи11е des cichlides, «Меж. Soc. Zool. France», l6, 41 — 3%) Р е 1 1 е у г i п 3., 1912, Les Poissons d eau douce d'1 Afrique et leur distributioIl g('()g- raphique, «Мст. Roc. Zool. France», 25, 63 — 83.
Р е 1 1 е g г i n J ., 1921, Les Poisso»s des eaux douces de 1'Afrique du N ord Francaise:
Магос, Algerie, Tunisie, Sahara, «Mem. Soc. Sci. Nat. Maroc», l, No. 2.
Р е11е g г i n J., 192;), Etude sur les poissons..., «Voyage Zool. d'Henri Gadeau de
Кегы11е en Syrie», 4, 5 — 37.
Р е 11 е g r i n J., 1923а, Les Poissons des eaux douces de 1'Afrique Occidentale (с1и
$епоеа1 аи Nфег), Paris.
Р e l l е р г i n J., 1933, Les Poissons des eaux douces de 51adagascar..., «Меп1. Acad.
AIalgache Tananarive», 14.
P e 1 1 е g г i n 3., 1937, Les 1 oissons du Sahara occidental, «Compte Rendu Associa- Mon Franqaise Avancement Sci.», 60th Session, 337 — 338.
Р i t о п ? ., 1940, Ра1еопФо1од~е du Gisement & ocene de Menat;, (1егтоМFer- rand.
В е g а n (.. Т., '1905,"Ë revision of the fishes of the family Galaxiidae, «Ргос. Zool
Soc. Ьопйоп», 1905, pt 2, 363 — 384.
В е g а п С. Т., 1906 —.1.)08, Pisces, Biologia (;entrala-Ашег~сапа.
R e g а и C. Т., 1911, The Classification of the teleostean fishes of the order Ostario- plrysi, «Ann. and Mag. Nat. Hist.», Ser. 8, 8, 13 — 32, 553 — 577.
К е о а n (.. Т., 1912, Discussion on Wallace s Line, «Rep... British Association Advan- cement Sci.», 1911, 433 — 435.
В р р а п С. Т., 1919, Note on Chaudhuria..., «Ann. and Nag. Nat. Hist.», Ser.,), 3,
198 — 199.
8 e g а п С. Т., 19' ', The distribution of the 6я11ея of the order Ostariophysi, «Bijdra- gen Tot de Dierkunde», 22, 203 — 208.
R е р а п С. Т., 1329, (,'ус1ояФотпайа. Selachians. Fishes. (',м. «Encyclopaedia Britan- naca», 14th ed.
R о m e r А. S., 194.'1, Vertebrate Paleontology, 2nd ed. Chicago. [Имеется русский
перевод: А. P о м е р., Палеонтология позвоночных, ГОНТИ, М.— Л., 1939.] Я а v а g e D. Е., 1951, Beport on fossil чег1еЬгаСея from the upper Magdalena Valley.
(;olombia, «Science», 114, 186 — 187.
~ c h а е f f е г В., 1952, Hates о1 evolution in the coelacanth and dipnoan fishes, «Ет o- l ution», 6, 101 — 111.
S с 11 а е f f e r В., 1952a, The evidence of the fresh-water fishes [in regard to Mesozoic
1анй connections across the South Atlantic], «Bull. American Mna. Nat. Efist.»,
99, 227 — 234.
8 с Ь и l t z L. P., ! 946, А revision... Mugilidae, «Proc. United States National М us.».
96, 377 — 395.
Я h ц 1 t z L. P., 1948, Л revision of ях subfamilies of atherine fishes..., «Ргос. United
States Хational Mus», 98, 1 — 48.
Я с о t t Е. О. (., 1936 — 19~1, Observations on fishes of the family (alaxiidae, «Рарег»
a n d P roc. H. Soc. Tasmania», 1935, 85 — 11 2; 1937, 1 ) 1 — 143; 1940, 55 — 69.
S m i С Ь Н. М., 19~5, The fresh-water fishes of Siam, or Thailand, «Hull. United Sta- tes National Mus.», 188.
1'.гава 2
S m i С h Н. W., 1932, Water regulation and its evolution in the fishes, «ОцагСег1~»
Review Biol.», 7, 1 — 26.
ь m i t h 3. Ь. В., 1939, А living fish of Mesozoic type, «Nature», 1~3, 455 — ~56.
S m i С h J. Ь. В., 1950, Two noteworthy non-marine fishes from South Africa, «Ann.
and Mag. Nat. Hist.», Ser. 12, 3, 705 — 710.
Я (р е п с е r) W. В., 1926, Ceratodus. CM. «Australian Encyc!opaedia», Згй ed., vol. I
Sydney, 248 — 250.
S t о k е 11 G., 1941, A revision of the genus Retropi»»a, «Вес. Canterbury plus.»,
4, 361 — 372.
5 t о k e 1 1 G., 1945, The systematic arrangement of the New Zealand Galaxi i~lw,
pt. 1, «Trans. апй Proc, R. $ос. New Zealand», 75, 124 — 137.
S С о k е 11 G., 1949, A fresh water smelt from the Chatham Islands, «Вес. СаМегЬигу
М ця.», э', 205 — 207.
Т c h e r п а v i n V. V., 194~, А revision of the subfamily Oresti danae, «Ргос. 7o(il.
Soc. London», l j.4, 140 — 233.
Т r e w а v а s Е., 1941, Fresh-water fishes. CM. «British. Мин. (i%at. Hist.) Ехр. Ы~и1Ь-
west Arabia 1937 — 1938», 1, No. 3, р. 7 — 15.
Т r e w а ч а s Е., 1942, The cichlid fishes of Syria and Palestine, «Ann. а~н1. Мло.
'Mat. Hist.», Ser. 11, 9, 526 — 536.
T и r п е r С. Ь., 1946, A contribution to the taxonomy and zoogeography о1 the ц(мну М
fishes, «Occasional Papers Мия. Zool.», U. Michigan, 495.
Ж e b e r M., В е а u f о r С Ь. F. de, 1911 — 1951. The fishes of СЬе Indo-A ustral i~n
Archipelago, 9 vol., Leiden), vol. 8 by Beaufort alone, vol. 9 by Beaufort a<id
W. М. Chapman).
% h 1 С 1 е у G. Р., 1943, The fishes of New Guinea, «Australian Mus. Mag.», 8, ! ~!—
144.
% у n n e — Е d w а r d s V. С., 1952, Freshwater чегСеЬгаСея of the arctic and subarctic,
«Bull. Fisheries Research Board Canada», Хо. М.
Глава 8ЗЕМНОВОДНЫЕ
Доктор Томас Барбар, мистер Артур Лавридж и мистер Бенджамин Шриф, профессор Альфред Ромер (в отношении древних земноводных), доктор Карл Шмидт и профессор Е. Р. Данн (замечания по первым вариан- там рукописи этой главы), доктор Дж. С. Майерс и доктор Пауло Ванзо- лини помогли мне в решении многих вопросов, касающихся современных ЗВМНОВ ОДНЫХ.
А. Ромер (Romer, 1945, переиздано в 1947 г.) выделяет десять отря- дов земноводных. Семь из них, примерно около сорока семейств, содержат только давно вымершие формы. В свое время некоторые из последних имели важное значение. Это были первые наземные позвоночные, домини- ровавшие до эры рептилий. К трем современным отрядам земноводных от- носятся Anura (лягушки и жабы), Urodela (саламандры)1 и Apoda (безно- гие). Эти отряды, вероятно, все древние и хорошо отграничены один от другого. Предполагаемые предки представителей отряда Anura встреча- лись до мезозоя в Пенсильвании(Вотег,1947,р.259), а собственно бесхвостые известны с юры. Самые ранние известные ископаемые формы саламандр относятся к нижнему мелу. О них нет никаких данных ни из юры, ни из триаса, но, вероятно, эти формации произошли от еще более ранних, доме- зозойских Lepospondyli (Romer, 1945, р. 161). Отряд Apoda также, оче- видно, образовался от домезозойских Lepospondyli (Romer), хотя ка- ких-либо достоверных ископаемых, относящихся к этому отряду, не из- ВеСТНО.
Классификация семейств и родов современных земноводных заставля- ет желать лучшего. Наиболее упорядочена классификация саламандр. Apoda — упрощенные безногие, роющие формы, мало известны и таксо- номически изучены недостаточно. Лягушки и жабы изучены полнее, но у них отсутствуют какие-либо наружные признаки, имеющие большое филогенетическое значение, а в эволюции и адаптации различных групп и даже в эволюции скелета (Parker, 1932) наблюдается параллелизм. Поэто- му действительное родство некоторых лягушек и жаб остается до сих пор еще под вопросом, разрешить который я, в силу недостаточной компетент- ности, не берусь и поэтому пользуюсь последней, наиболее подробной клас- сификацией современных земноводных (Nolle, 1931) с небольшими изме- нениями, сделанными Паркером (Parker, 1934; 1940).
Термины Anura, Urodela, Apoda, а также Salientia (означающий при- мерно то же самое, что и Anura) применяются уже в течение многих лет, но все же необходимо остановиться здесь на значении этих слов. Эти тер- мины часто вносят путаницу, и я стараюсь по возможности избегать их, хотя это создает другую незначительную трудность. Обобщающего слова,
' Правильнее было бы называть хвостатые, так как, кроме саламандр, сюда относятся и тритоны.— Прим. ред.
обозначающего и лягушек, и жаб вместе, нет. Употребляя слово «лягуш- ки» в широком смысле слова, то есть вклн)чая также и 'каб, я обхожу эту трудность. Это вполне естественное расширение значения термина, так как жабы не таксономическая единица, а просто лягушки с утолщенной и бородавчатой кожей.
Земноводные — холоднокровные животные. Настоящие лягушки и саламандры, а также некоторые безногие являются земноводными,кивот- ными с водной личинкой (головастики) и наземными взрослыми особя- ми. Однако в каждом отряде одни формы стали полностью водными (на протяжении всей своей жизни), а другие стали полностью наземными. У последних стадия личинки либо отсутствует, либо проходит на земле в яичной капсуле, либо переносится взрослыми особями, и, таким образом, животные проводят вск) свою жизнь вне воды. В частности, лягушки, Осо- бенно лягушки Индо-Малайской области и Вест-Индии, а также некото- рых островов, включая Сейшельские и Новую Зеландию, обладают раз- личными приспособлениями для такого рода размножения (Вагооцг, 192()).
У современных отрядов земноводных наблюдается корреляция между числом видов, таксономическим и экологическим разнообразием, про- тяженностью и разнородностью занимаемой территории. Безногие обра- зуют одно семейство с 17 родами и 73 известными видами. Во взрослом состоянии все они являются роющими или водными животными. В основном земноводные приурочены к влажным тропикам Африки, Индо-Малайской области и Америки. Современные саламандры образуют 8 семейств (око- ло 43 или более родов и около 200 видов). Большинство из них во взрос- лом состоянии наземные или водные животные (в поверхностных слоях воды), некоторые встречаются в подземных водах и незначительная часть живет на деревьях. Саламандры в основном приурочены к влажным райо- нам умеренной зоны северного полушария, некоторые проникли в тропики, но ни одна из них не встречается в умеренной зоне южного полушария. Лягушки, включая жаб, образуют около 37 семейств и подсемейств, око- ло 200 или более родов и, возможно, 2000 или даже больше известных видов. К этому отряду относятся не только земноводные, водные и наземные формы (последние встречаются во многих районах земного шара), но так- же и многие древесные формы, образовавшиеся независимо от нескольких различных семейств. Последние широко распространены в обеих основ- ных климатических зонах и могут встречаться как в местах с благоприят- ными условиями существования, так и в засушливых и других небла- гоприятных местах. Граница их распространения на север и на юг, а также на островах заходит значительно дальше, чем у безногих и са- ламандр.
Земноводные обладают ограниченной солевыносливостью. Лягушка Лапа сапсгп,ага и родственные ей виды в Индо-Малайской области живут вблизи морских побережий и у солоноватоводных эстуариев. Непродол- жительное время взрослые особи могут находиться в морской воде, оче- видно без особого вреда для себя, а их головастиков находили в норках кра- бов, даже расположенных ниже высокой линии прилива (Boulenger, 1903; Pearse, 1911; М. Smith, 1927). По другим сведениям, саламандры и лягушки, видимо случайно попавшие в соленую воду, не погибали. В со- лоноватых водах они также чувствовали себя более или менее нормально (Schmidt, 1951). Многие ученые, а также Дарвин (Darwin, 1859, переизда- но в 1950 г.), считали, что на всех земноводных контакт с соленой водой оказывает немедленное губительное действие, что не совсем верно (Schmidt,
3емноводные
117
1951). Как уже говорилось, некоторые земноводные, по крайней мере некоторые лягушки, могут переносить случайное погружение в море или действие соленых брызг, и это доказывает возможность перемещения их на плотах через узкие океанические проливы. Действительная солевынос- ливость различных земноводных при разных условиях не изучена и заслу- живает серьезного исследования. Просто же опыты, проведенные с этой целью, не дадут окончательных результатов. Расселялись ли земноводные, пересекая соленые воды, или нет, скорее покажет их распространение, чем результаты лабораторных исследований, а их распространение по сравнению с распространением пресноводных рыб. несомненно, показы- вает, что лягушки неоднократно преодолевали небольшие пространства соленых вод.
Земноводные плохо переносят потерю влаги, и в этом отношении наблюдаются видовые различия. Торсон и Свила (Thorson, Svihla, 19Я) обнаружили, что Scaphiopus (чесночницы) могут переносить больп1ун~ потерю влаги, чем Ви/о (обыкновенные жабы). 11оследние могут терять вла- ги больше, чем Ну1«(древесные лягушки), а они в свою очередь теряют влаги больше, чем Капа (обыкновенные лягушки), которые переносят потерю влаги тяжелее всех. Эти опыты проводились колько с несколькими видами. Здесь снова наблюдается болыпая разница между приспособляе- мостью и выносливостью подопытных животных и приспособляемостью и распространением этих животных в природе. В пустынях иногда встре- чаются не только представители рода Scaphiopus и Bufo, но также и рода R«na, и его производные (стр. 134) и даже представители рода Hpla (см. список семейств). Фактически различные лягушки имеют различные при- способления, для того чтобы избежать потери влаги (Buxton, 1923, р. 96). Следовательно, другой вопрос — в какой степени засушливые местности являются преградой для земноводных — следует решать по их распространению, а не априори или на основании экспери- ментов.
Распространение некоторых земноводных связано с человеком, кото- рый употреблял их для еды или привозил как диковинку или как комнат- ное животное, а также для уничтожения вредных насекомых или случай- но. Поуп и Боринг (Роре, Boring, 1940) упоминают о коммерческих пере- возках гигантской саламандры из Японии в Китай и отмечают, что опре- делить распространение этого вида в Китае Было трудно не только потому, что саламандр привозили в качестве деликатесной еды, но также и пото- му. что их держали как диковинку во дворцах и богатых домах, и это, вероятно, объясняет некоторые совершенно неожиданные данные, вклю- чая и те, которые были получены в Сямыне (Амой).
Зеленая лягушка (Rana esculenta), которая в Англии приурочена и бо- лотистым местностям графств Норфолк и Кембриджшир, очевидно, была интродуцирована там в средние века итальянскими монахами, и, таким образом, некоторые ее особи сходны с итальянскими (Boulenger, 1936, р. 307). Независимо от того, действительно ли эта лягушка была завезена в Англию так давно, она, конечно, не раз была интродуцирована там позд- нее, и история ее расселения в Англии является примером неоднократного интродуцирования, а затем после целого ряда лет исчезновения интроду- цированных колоний (M. Smith, 1951, р. 141 — 143).
Морская жаба (Bufo marinus) — местный вид на материке тропиче- ской части Америки — была интродуцирована на многих островах Вест- Индии, Бермудских и Гавайских островах, в тропической части Австра- лии, на Новой Гвинее, Соломоновых и других островах Тихого океана
Глава 8
с целью регулирования численности вредных насекомых. Можно привести много подобных примеров. Не менее важным является и тот часто не учи- тываемый факт, что мелкие лягушки перевозятся детьми всех народов. И, вероятно, самым ва'кным из всего этого является то, что некоторые лягушки, включая древесных, которые могут спрятаться на лодках в связ- ках тростника или бананов, очень часто перевозятся случайно, особенно в тропиках. И одних случаях посредничество человека выявить легко, в других — нет. В тех случаях, когда земноводные появляются в самых неожиданных местах, всегда следует предполагать вмешательство чело- века. Но это лишь частности, не имеющие ничего общего с основным характером распространения земноводных.
Границы распространения земноводных
Все северные и южные границы распространения современных зем- новодных определяются лягушками (рис. 19). Настоящие лягушки рода Лапа проникают на самый крайний север. В Европе северные границы рас- пространения вида Лапа temporaria совпадают с границами суши в Скан- динавии (примерно у 71' с. ш., то есть значительно севернее Полярного круга). Этот или родственные ему виды пересекают Полярный круг так- же в нескольких точках в Азии. Подвид лесной лягушки (Rana sylvatica) достигает Полярного круга или пересекает его на западе Северной Америки и встречается почти до северной оконечности и субарктического полуост- рова Лабрадор (на востоке). Границы распространения настоящих жаб рода Bufo в Европе подходят близко к Арктике, достигая приблизитель- но 65' с. ш., но на западе Северной Америки они достигают только 60' с. ш. и заходят немного выше 55" на полуострове Лабрадор. Древесные лягушки рода Ну1а проникают только до 57' с. ш. в Европе и, возможно, расселяются несколько севернее 52' в Восточной Сибири и примерно до 62' на западе Северной Америки и до 55' на востоке; другой род древес- ных лягушек (Pseudacris), который преимущественно обитает в болотах и на земле, а не на деревьях, на западе Северной Америки почти достигает Полярного круга.
Из саламандр сибирский углозуб (IIynobius keyserlingii) пересекает Полярный круг по направлению к Верхоянску (Dunn, 1923) — одному из самых холодных мест на земном шаре 1 (средняя январская температу.ра доходит до — 48,9' С, а абсолютный минимум до — 60,8' С, хотя средняя температура июля доходит примерно до 15' С. Этот пример подтверждает факт, отмечаемый Винн-Эдвардсом (Wynne-Edwards, 1952, р. 24), что границы распространения северных земноводных определяются скорее летними температурами, чем зимними. О нахождении других саламандр около Полярного круга ничего не известно. Они достигают примерно только 63' с. ш. в Европе и, возможно, только 59 и 53' соответственно на западе и востоке Северной Америки. Таким образом, саламандры не до- стигают северных границ родов Лапа и Bufo в Европе и Лапа и семейства Hylidae в Северной Америке (Еkman, 1922; Mertens, Мй11ег, 19 ~0; Mills, 1948; Schmidt, 1953; Wynne-Edwards, 1952; Logier, Toner, 1955; Harper, 1956 и другие источники).
' По цосдедним данным советских ученых, полюсом холода являемое» Оймякон в бассейне реки Индигирка. Средняя температура января — 50,1' С; абсолютный мини- мум — 71' С. — 17рим. ред.
л Q
4 Ц
Ж
Яо~"
ьО
'~в 'с
д ~ м
й~[ сс
0+<~
в Й
E
Х
0
Ж
в .р, я Ф~
Я ~ ~~ ~ яО:
Е~ р,
,„' о,~р,
о о
( с
3|~ в Е ~~-1' Й~
-о
й ~, с~ О = o+вв g ж й
О
~д д ~=-й 3 n,~~у
во е о~~~й
о r о с~
д W г'~~
Ф ~~
~î> й сО~~~~~ � ~~о Й ьч ',[; g g („„
° р: - ~; Q
~~Я 0~3
О
д c ~ ~ фЯ а5
М р ° ° е
~ в о ~ с~
~И 40;
в с~~ ~6»
у,до~~
~ Я, д д~о~ д
ожжо
а. в ~ Вч ~. в ~ ~Й оч gD<
с~ се<~ д ~,'Д, ~н
в ~ ~~-ж
~ ~о <z
а5 О
1 а,~.
Х
4
ОО"- 4
олово~
в
в~о
и
~ ;Я н ~.
д мй в
ьО р~
а. XZ Д«б
;.:;,-) М,й
1 о
~а.о
юИаа.
[~й
Ж g5
~М в ~~
в
ГЖ=
Глава 8
Самые северные лягушки (Лапи) достигают районов вечной мерзлоты (Harper, 1956, р. 99). Я думаю, что до сих пор не известно, каким ооразом они переносят полярную зиму. Не исключено, что для того, чтобы защи- тить себя от сильного холода, они впадают в спячку довольно глубоко ПОД ВОДОИ.
В южном направлении лягушки достигают оконечности Южной Аф- рики, острова Тасмания (на юг от Австралии), Новой Зеландии, оконеч- ности Южной Америки и Огненной Земли. Бриджес (Bridges. 1948, р. 447) упоминает о мелких еще не определенных лягушках на Огнен- ной Земле.
Границы распространения земноводных на островах также определя- ются лягушками. Саламандры приурочены к материковым областям и материковым островам, таким, как Британские, Японские, Рюкю и Taii- вань. Безногие обитают в тропиках континентов Африки, Азии и Америки и встречаются на таких материковых островах, как Цейлон, Суматра, Я ва и Калимантан, а в Америке — на острове Тринидад, но их нет на Мадагаска- ре, а также в собственно Вест-Индии, и они не пересекают линию Уоллеса. В этом отношении безногие резко ограничены, как и строго пресноводные рыбы. Один вид безногих, так же, как и несколько видов карповых рыб, достигает южных Филиппинских островов — Палаван и Минданао. Толь- ко в одном месте, на Сейшельских островах в Индийском океане, границы распространения безногих заходят значительно дальше, чем границы рас- пространения строго пресноводных рыб. В связи с отсутствием безногих на Мадагаскаре этот факт тем более интересен.
Лягушки существуют на всех пригодных для обитания материках и большинстве материковых островов, включая и те, что упоминались выше; они являются местными также и на некоторых островах неясного происхождения или в биологическом смысле океанических, включая остров Мадагаскар, Сейшельские острова, весь Индо-Австралийский архи- пелаг, включая Филиппины, Соломоновы острова и, возможно, Палау и острова Фиджи, на Новой Гвинее, в Австралии, Тасмании, Новой Зе- ландии и в Вест-Индии. Дополнительные сведения и ссылки приводятся в гл. 8. Интересно, что на острове Ньюфаундленд, хотя он и является боль- шим материковым островом, расположенным вблизи континента, местные земноводные, так же как пресноводные рыбы и рептилии, отсутствуют. Это трудно доказать, я по крайней мере не мог найти никаких сведе- ний о земноводных Ньюфаундленда, кроме данных об одной лягушке (Rnna clamitans), которая, очевидно, была завезена с грузом из Новой Шотландии (Mills, 1948, р. 7; Н. J. Squires, письменное сообщение от 24~ ок- тября 1951 г.; С. Н. Lindroth, письменное сообщение).
В пределах общих границ распространения земноводных широко распространено несколько родов, особенно лягушек. Род Лапа в широком смысле (рис. 21) распространен по всей Африке, Евразии и Северной Аме- рике и достигает северных краевых частей Австралии и северной части Южной Америки. Род Bufo (рис. 22) распространен на всех континентах, кроме Австралии. Род ala (рис. 25) — на всем земном шаре, за исключе- нием большого разрыва в Африке и в тропиках Индо-Малайской области. Но все другие роды лягушек более ограничены в своем распространении. Из саламандр род Triturus (тритоны) имеет прерывистое распростране- ние в Голарктике, а распространение всех других родов более лока- лизовано. Из безногих ни один род не известен за пределами границ одной области, и в целом из земноводных вообще ни один вид не заходит далеко за пределы границ одной области.
Зели новодные
Зональное распространение земноводных
В пределах указанных границ из трех существующих отрядов земно- водных два имеют строго зональное распространение (рис. 19).
Безногие преимущественно тропические животные. Они заходят в краевые части умеренных зон только в Восточной Азии и южной части Южной Америки, да и то на очень ограниченное расстояние.
Саламандры в основном придерживаются умеренной зоны северного полушария. Данные о находке одной саламандры в тропической части Западной Африки весьма сомнительны. Но все-таки некоторые виды сала- мандр иногда встречаются в краевых частях тропиков в Восточной Азии (см. Salamandridae в списке семейств). Саламандры семейства Ambysto- тЫае встречаются в краевых районах тропиков в Мексике, но на до- вольно больших высотах. Род Oedipus из семейства Plethodontidae или его представители распространены в тропиках Америки до Боливии: одни виды этого рода обитают в горах, другие — на равнинах тропиков. Один вид находится в экваториальной зоне в устье Амазонки (см. список семейств Plethodontidae). Хотя большинство саламандр приспособлено к жизни только в пределах небольших колебаний температур прохладно- го климата, исключения позволяют предположить, что эти животные ооладают не наследственным приспособлением к холоду, а большой потен- циальной приспособляемостью, так же как и лягушки. Следовательно, надо всегда помнить разницу между приспособлением и приспособляемо-
СТЬЮ ЖИВОТНЫХ.
Как уже говорилось, лягушки очень широко распространены, но болыпинство их находится в тропиках. Из 37 приводимых здесь семейств и подсемейств лягушек 27 приурочены к тропикам или к почти тропикам, 4 или 5 других семейств и подсемейств по своему распространению также являются, скорее, тропическими, чем умеренными (рис. 20). Из 2000 из- вестных видов, вероятно, около 80 оо — тропические, немного больше 10',о — северо-умеренные и меньше 10о/о — южно-умеренные.
Ни одна группа лягушек не приурочена целиком ко всей умеренной зоне и не распространена по ней сплошь. Подсемейство Pelobatinae почти полностью приурочено к умеренной зоне северного полушария, проникая в тропики только на Мексиканском нагорье. Оно очень немногочисленно п отсутствует на большей части Азии, хотя ископаемые этого подсемей- ства найдены в Монголии в отложениях миоцена. Другие подсемейства семейства Pelobatidae изолированы в тропиках Юго-Восточной Азии и на Сейшельских островах. Круглоязычные Discoglossidae встречаются глав- ным образом в умеренной зоне северных широт, но только в Старом Све- те. Но и там (в Европе и Восточной Азии) они имеют прерывистое распро- странение. Еще один род этого семейства изолирован на маленьком островке в тропической части Филиппин. И, наконец, род Агсарйиг (один вид), который иногда относят к самостоятельному семейству, а иногда объе- диняют с родом Leiopelma из Новой Зеландии, приурочен к холодным гор- ным ручьям северо-западной части Северной Америки. Эти три группы лягушек — единственные в ранге семейства или подсемейства — приб in- жаются к группам, типичным для умеренной зоны северного полушария. Согласно классификации Нобля (Noble, 1931), все они примитивные и все имеют прерывистое или локализованное реликтовое распространение. Некоторые другие, являясь главным образом тропическими, проникают в умеренную зону северного полушария в Восточной Азии или в восточ- ной части Северной Америки, указывая, таким образом, на возможность
а
Ф
с
а
а
М
а а
а
а
а
с
° а
а а
а Ф ° а аа
а
° а
Г
I
ф а б °
е 69 Фе
° Ф
° а а
б
° ° в
° 6 ° Э в
11 6 °
° Эв ) ° ° °
а а °
1 ° °
В ° °
° °
° а
а
° 6
° ° ° ° °
° а
6 ю 6 °
° в
Ь 6
Э ) °
r ° r °
Ф °
6 °
° а
а
° °
° в
° ° °
° °
а а
° ° ° °
° ! ° ' ° 1
а
а а
° 6 °
° °а °
1 ° а
° ° ° °
ю °
а а
° °
ю ° l
° в
661
°,
° °
° °
° ° 1
ю
° 6 ° / 6 °
° ю в °
д °
° °
° в ° ° ° 1
° ° °
6
° °
° °
а° °
д °
У ° °
3 °
1 ю ° ° °
1 °
6 °
Ю а
° ° ° '
° ° а °
У °
Ф ° °
° 1
а Э
° У
° Ф
Э ° ° )
° 1 °
в ° °
° !
° ° !
! ° ° ° °
° °
° ю ю °
° 1
ю
! ° ю
° Ю °
° ° ° °
° °
1 а
а а
а
Ф ° Ф ! °
° ° °
6 °
° ° ю °
° l
° Э
° в ю
У Ю
6 Э 6
ю
° а
Э ° ° °
а а °
! 3 1 ° 3 °
° 6 ° a ° a° ° в
° ° 1 ° в ° в
° I ! °
° a 1 ' Эi °
ю а I ° ( °
ю
а ю I
° ° Ъ ва
° ° ° ° °
° ° ° в ° °
128
Земноводные
местных перемещений в северном направлении (рис. 20): по крайней мере пять тропических родов Индо-Малайской области, кроме Лапа и Bufo, относящихся к четырем семействам, достигают Центрального Китая, а два из них заходят еще дальше на север — яванская лягушка Rhacopho- rus — до северных частей Хонсю, Япония, и узкоротая лягушка Kaloula— до Маньчжурии. В Америке три тропических рода семейства Leptodacti- lydae и один род семейства Brevicipitidae достигает Техаса и даже Южной Аризоны. Vicrohyla — второй род семейства Brevicipitidae — распростра- нен на север до Айовы, Индианы и т. д. Представители доминирую- щих, широко распространенных родов Лапа, Bufo и Ну1а завершают перечень лягушек умеренной зоны северного полушария, за исключением нескольких, проникающих только в ее южные краевые части.
В умеренной зоне южного полушария фауна лягушек еще менее зональна. Для южных континентов — Южной Америки и Австралии,— ограниченных преградами, характерны семейства Leptodactylidae и Hy- lidae, но они не строго приурочены к зонам южного полушария с умерен- ным климатом. Фактически в Америке оба семейства являются в основном тропическими. Это дает возможность предположить, что лягушки, являясь главным образом тропическими животными, часто простираются локаль- но или широким фронтом в умеренные зоны и иногда существуют там зна- чительное время, но не образуют отдельных доминирующих фаун уме- ренных зон.
Радиальное распространение земноводных
Два рода лягушек имеют ярко выраженное радиальное распростра- нение. Род Лапа (рис. 21), к которому относится большинство наших обычных. хорошо известных лягушек умеренной зоны северного полуша- рия, очень хорошо представлен в тропиках Старого Света. Этот род встре- чается по всей Африке и во всех пригодных для обитания частях Азии, достигает Мадагаскара и далее через Индо-Австралийский архипелаг Новой Гвинеи и (если в него входит род Hylarana) до северной части Ав- стралии. Он встречается по всей Северной Америке и доходит (в умень- шающемся числе), пересекая Центральную Америку, до северной половины Южной Америки (см. список семейств). Семейство Ranidae, к кото- рому относится род Лапа, за исключением самого рода, приурочено к рай- онам Старого Света с теплым климатом. Род Bufo (рис. 22), включающий настоящих жаб, встречается по всей Африке, в умеренных широтах и тро- пической части Евразии и на островах до Сулавеси и т. д. (но не в Австра- лийской области), в Северной, Центральной Америке и по всей, или почти по всей, Южной Америке и на некоторых островах Вест-Индии. Этот род относится к семейству Bufonidae, которое, за исключением рода Bufo, также целиком приурочено к тропикам Старого Света. Центр радиального распространения этих двух родов и их семейств находится на основной части тропиков Старого Света.
Два других семейства лягушек, очевидно, не имеют радиального рас- пространения. Одно из них — семейство Brevicipitidae (Microhylidae) (рис. 23), подразделяемое Паркером (Parker, 1934) на 7 подсемейств. Все подсемейства встречаются в тропиках Старого Света, и большинство из них там локализовано или имеет прерывистый ареал. В настоящее время наибольшей концентрации они достигают на Мадагаскаре, в тропической Азии и в тропической части Австралийской области. Одно из подсемейств
Ю
° °
° 1
-t
.г ' ° ' .,' .. °
° °
° ° ° °
Ъ,:.'
° °° ° °,° °° ° °, °, °
° '
е °
° °° °
1
е
°, '.' ', '.1
° ° ° Ф ' °
е° °
Фв
° °
Ъ, ° . ~Р..
° ° °
° ° ° °
~'в 9%
° ° ° ° ° в ° ° ° ô ° °
°,
° °
°,) 'в.' ° °
° °
° °
° °
° °
° ° ° °
4
Р и с. 21. Распростраиенио лягушек семейства 14апЫае.
Жирные пересеченные 11олосы — области распространения других представителей этого семейства с
))()д~)м В«)) «; (;тр(лки--радиация родя Я«)) «(сх( ма); че1)ньн; II() JI()('I:è — п риблииит('лт.HEI(', гу)ании ц f)il(;ll))l)(т~)') ненин )и) Р Вт)«» Лв( Tf)llJlllи II Н)л;пой AM( ()I)it .
° вв
° °
° °
° °
° ° ° ° °
° ° °
в ° ° ° ° ° g °
° ° ° ° ° ° °
° ° Ф ° ° ° °
° °
4.."ю.. ° ° ° ° ° ° ° ° ° ° ' ° ° °
° ° ° ° °
ю ° ° °
° °
Ф
° в ° °
° °
° ° °
° °
° ° °
° ° ° ° ° °
° ' ° ° °
° °
° ° ° 1
° ° °
в ° ° °
° ° °
° ° ° °
° °
° °
° В °
° ° ° °
° °
в °
° ° ° ° ° ° ' ° ' ° ° .в е ° ° °
° °
в ° ° ° ° ° ° ° ° 1 ° ° В ° е °
е
° °
° ° °
'Т:... ~Т4 Ф:.
° ° °
° °
° ° °
° ° ° °
в ° ° '. ° . ° ' ' °
° °
вв ° ° ° °
° ° ° ° °
° ° ° °
° °
° ° ° ° °
° ° ° ° ° ° ° ° ° ° ° °
° °
° ° °
° ° ° +
° ° °
° ° ° ° ° ° °
° °
° °
° °
° ° ° ° е
В'-
° °
° ~ ° °
'A ~л ~В,
° ( ° °
° °
° °
° Ь
° ° ° °
М° °° ° ' ' ° Р' ° ° ° °
° °
° °
I,
° Э(
° °
~ ФЮВ В'
°, ° ° °
° е, °
В
° ° В
° r ° °
° ° °
"..:М.'. ° ° . м ° ° ' . ° .
° °
° ° ° ° ' ° ° °
Ф
° °
° °
° °
° ° ° ° ° ° ° ° ° °
° ° °
° °
° е ° °° ° °° ° °~е °° °° ° ° °° ° °° ° .1 ° °° ° + ° ° ° °° ° Ф ° ° ° °° ° °
° °
° °
° ° ° ' ° е ° ,с ° ° °
° °
° °
° °
° В
° ° ° ° ° е е~ ° ° ° ° \ ° ~ ° ° °
, °,г~.
° ° ° °
° °
° ' ° 'Ф
° ° ° Ф ° в ° °
° °
° °Р ., ° °° еъ
° ° ° ° ° °
° ° °
° ° ° °
° ° °
° ° ° ° 'g
° ° ° ° ° °
° O ~pa
° ° ° ° °
° ° °
° ° ° ° °
с ° ° °
° °
° ° °° ° °° °° ° °
° Э° \° °° °° °
° °
° °
е Ф. ° °
° ° ° °
° ° ° °
-'ВВ '.
° ° ° °
° °
° °
° ° Ф ° ° ° ° °
° ° ° ° ° ' Ф
° ° °
° У
° ° В °
° °
° '" ЪВ,. .:(:В ° ° ° ° ° е ~(.
° ~Э ФВ
° ° ° ° rr
° ° ° °
° °
Р и с. 22. Распространение жаб семейства Bufonidae.Жирные пересеченные полосы — области распространения других представителей этого семейств»с родом Ии/0; стрелки — радиация рода Ви(о (схема); черные полосы — границы рода 1~><J() к:я~ядуот Африки и к востоку от Индокитая.
Ье! ° °
се~ ФВ
ФВ
° °
° ° ° °
° ° °
° °
° °
° ° °
г~ '~' °
%
° °
° °
° ° ° °
° ° °
° В
° °
°, ° ° ВЭ ° '
° ° ее
° °
' г.' .'"
° ° °
° Ю Ф~ ° ° ч
° Ф ° Ю ° ° ° , е °
I ° \ ° ° °
' I;:.' ° ° ° ° ° В Ъ"
° ° ° ° ° °
'.с . ° .
° ° ° °
.фэа.ф~ .
° °ра ° °
° °
° °
° °
° °
c-. °
° Ф
° °
° °
.г °
° Э
° Ф °
° ° Ф ° ° ° °
8
° ° ° °,. б
° °
° Г
° ° ° °
° °
° ° ° ° Ã ° ° ° °
° ° Г °
° ° Г ° ° ° ° ° Г
° ° ° ФЭ °
Ч °
° ° ° ° ° . ° °
° °
° ° Ф °
° °
° °
° °
° ° ° °
° ° ° ° ° Э ° °
° ° ° ° ° °
° ° -~с~ ~'Э~М ° ° ° °
° ° ° ° ° ° °
° °
° °
° ° °
ё~.":=° Э ° ~ °га
° ° ° ° °
° ° ° ° ° ~ ° °
«Ю»
В
Фу~
В ° В ° °
° °
° 'Г ° ° °
° °
° °
° ° ° ~
° ° г
° ° °
° °
° ° ° ° ° ° С ° ° ° 'а' ° Г»
° ° ЭЭС
° °
° ° ° а °
° а ° ° ° ~ °
г ° а ° а
° ° ° ° Г
° Э ° ° ° °
° °
° Â
° а ° Ф~ф° °
° °° °° °
ФВ° ° ° °° ° °° °
° °
° °
° ° Фа ° °: ° с".
° ' ° Г
° °
° °
%у ° °
I° ФФЭ
~1 !
° °
° Ф °
° ° °
° Э
.'..: Ъ° °
ю Г
° ав
° Г °
° Э
° а
° ° °
° ° °
° ° Г
° 4 ° °
С
° ° ° °
а.
° ° ' ° Э
Г
В
° °
° ° ФВ,
!' и с. 2'4. 1'ic)i~iIii,траиение лягушек семеиства Hrevici~>iti~lae: ттрттблттзтттельтттто граттттцттр.кпр<~стрдн«ттття 7 ттодсемецл в, иеречислеттиык в списке семейств.
(~' с
° ° °, ° °
° ° I Э ° ° °
° ° ° ° ° ° ° ° ° ° ° ° ° ° ° °
° ° ° ° ° ° ' ° а
Э,
° ° ° °
Г ° °
° ° ° ° ° °
° ° ° °
° ° ° ° ° °
° ° Ф ° ° ° °
° ° ° °
° ° ° ° ° ° °
° °
° ° ° ° ° ° °
° °
° ° ° ° ° ° ° ° Г
° ° ° °
° ° ° ° ° ° r ° ° ° ° ° °
Ф
° ° ° °
° ° °
° °
Гб
° 4ГФ ° °
'. С ф ° С эаб
° ° ° ° ГФ ° ° ° ° ° ° ° ° ° °
Г ° °
° а ° ° ° ° °
° °
° ° В Ф °
а ° °
° ~~ ° Г °
У"
ЧФ(
Зе.чноводные
Alicrollylillae, встречающееся в основном в тропической Азии; в северном направлении заходит в умеренные широты Восточной Азии и достигает Маньчжурии. Это единственное подсемейство, которое встречается в Аме- рике и является там также в основном тропическим (подробнее см. список семейств).
Второе семейство — Leptodactylidae, свистуны (рис. 24). Свистуны многочисленны в Австралии и Тасмании (некоторые достигают Новой Гвинеи), в Южной и Центральной Америке (некоторые достигают север- ных районов южной части Техаса и Аризоны). Род Не1еор1пупе, изоли- рованный в Южной Африке, возможно, относится к этому семейству, однако это недостаточно точное предположение. Современные представители семейства Leptodactylidae, за исключением рода Heleophryne, не извест- ны за пределами Австралийской области и теплых частей Америки. Од- нако ископаемая (эоцен) лягушка из Бомбея, тропическая часть Индии, отнесена к этому семейству. Жабы, которые, как считают, ведут свое нача- ло от представителей семейства Leptodactylidae, произошли в основной части тропиков Старого Света, что подтверждается существованием там в прошлом свистунов.
Распространение этих четырех семейств — Ranidae, Bufonidae, Bre- vicipitidae и Leptodactylidae — находится в определенной согласован- ности (рис. 21 — 24). Об истории каждого семейства можно сказать следую- щее: семейство Ranidae радиировало в тропиках Старого Света, откуда род Лапа расселился на большей части всего земного шара. Семейство Bufonidae и род Bufo в основном имеют сходную историю — радиацию в тропиках Старого Света и расселение рода Bufo на большей части зем- ного шара. Семейство Brevicipitidae радиировало главным образом в тро- пиках Старого Света, где теперь оно, по-видимому, отступает, и одно его подсемейство проникло через север в тропики Америки и вновь там ради- ировало. Свистуны, очевидно, когда-то встречались на основной части тропиков Старого Света, но отступили оттуда и теперь сохранились в Ав- стралии, в более теплых частях Америки и, возможно, в Южной Африке. Если рассмотреть историю всех четырех семейств вместе, мы получим менее конкретные, но более важные данные, указывающие на определенную последовательность распространения, что позволяет предположить общие черты распространения этих семейств и их эволюции. По-видимому, эти семейства развивались последовательно на основной части тропиков Старого Света, радиировали оттуда, а затем древние семейства отступили или вымерли, с одновременным расцветом там более поздних семейств. Древние же семейства сохранились главным образом на периферических или ограниченных преградами территориях. Роды Лапа и Bufo находят- ся почти на высшей точке этого цикла, семейство Brevicipitidae — не- сколько отошло от нее, а семейство Leptodactylidae отошло еще больше.
Рассмотренные четыре семейства — преимущественно земноводные или наземные. Ни одно из них в основном не ведет древесного образа жиз- ни, хотя к ним относятся дериваты древесных форм. Два основных семей- ства древесных лягушек Hylidae и Rhacophoridae имеют самостоятель- ный характер географического распространения.
Род Ну1а (рис. 25) широко распространен на всем земном шаре, за исключением большей части Африки и тропической Азии. Число видов этого рода увеличивается по мере удаления от этого разрыва: несколько видов встречается в умеренных широтах Евразии, их число увеличивается в умеренной зоне Северной Америки и в Центральной и Южной Америке, особенно 13 тропиках. Кроме того, множество изолированных видов нахо-
1' и с. 2~i. Распространешм лягушек семейств» Lepto(lactyli(lac.
А
Ф
» \,
ОHO
(P У*в
Ъ» . ° . ° ° а.--
. У:.'.: "':.;:...
° ' ° °
° ° °
Ф
;х -:.:..
Ф ° °
, ° ' ° ° ° ' ° ° °
° °
° °
° ф ° °
° °
° ~М
r °
..Ф °
° °
° °
Ф '- ° °
° ° ° °
° ° ° ° °
° ° rð ° 'а °
i+;)
° °° ° °
° °
° ° .' В Ф, °
° Ф
° ° ' ° ° ° ° °
° °
« ° ° r ° °
° °
° °
Ф
° °
a.~v ' Ф ° °
° ° °
:Ф Ъ
° °
° ° ° аа
фв
° ° °
° ° ° °
° ° ° °
аа '\
° °
° ° ° ° °
° а °,
° °
+1
° ° °
° ° ° ° °
а
° °
° °
° °
+ИНОГО
а
° ««
с °
%«
Вава '
° ° а
« °
« ° °, °
° ° ' °
«а ° ., ° Ф ° ',
g I
!
° °
° ° °
.-Г:В <ГЪ .
° ° °° °
° °° °
° °
В °
° °
° °
° ° °
° ° ° °
° °
° °
° ° ° ° ° ° °
Ф
° °
° °
° °
° °
° °
Р и с. 25. Распространение осповных семейств древеснык лягушек.
34 1исорй ог~ <1д( (жирны( ~ ~('р('сеч(нны( иолосы) и Ethyl i (Ia('. (~ ~ рерывистые линии). Цифрамп ч(но число родо11 с(',мсиства Нfli(IB('. (иомимо род» 7Туl~с) на разных T('ððèò~~риях.
0ДозЕФA
в
» °
~ °
/ °
° °
° °
° ° °
° °
° °
° °
° °
° °
° ° ° ° °
в ° ° ° ° ° °
° ° ° ° °
° в
° ° ° ° °
Ф °
° ° ° ° ° ° ° ° °
Ф 'в
° °
° ° ° ° ° ° ° ° ° °
° Ф ° ° °
° °
Ф ° ° Фа
° ° °
° °
° ° ° « ° °
° ' ° ° °
° ° ° ° ° ° ° ° °
° ° ° - ° ° ° ° ° ° Ф ВФ
° °
° ° ° ° ° ° ° ° r
° °
° °
° ° ° ° ° ° ° ° °
° °
° ° ° ° ° ° в ° ° °
° ° °
° ° ° ' ° °
° ° r ° °
,Ф' ° ° а °
° б
° аа а ° ° ° ° а ° Ф ° ° °
° ° ° r Ф
r ° ° ° ° ° ° ° ° ° °
Ф
л. ° а Фв«
,ю
° r °
V ° аа° °в° ° ° °° °° °° ° , ° ,", ° « ° °
130
дится в Новой Гвинее и Австралии. Большинство других представителей семейства Hylidae находится в Южной и Центральной Америке, где род Iiyla пополняется другими родами (см. список семейств). Такое распро- странение могло быть результатом либо первичной радиации этого семей- ства в тропической Америке и расселения оттуда рода Ну1п (если сущест- вовал открытый путь в Австралию), либо первичной радиации из какого- либо другого центра и вторичной радиации в Америке и Австралийской области. Если это расселение произошло в результате последней, ro ис- тория распространения семейства Hylidae соответствует характеру их распространения, о котором можно судить на основании современных семейств. Вероятно, квакши сначала радиировали в основной части тропи- ков Старого Света и позднее исчезли там, а род Оу1а, возможно, проник оттуда в Австралийскую область и Америку, а затем вновь радиировал уже в этих местах. Кроме того, другие представители семейства Ну1Ыае могли достичь Америки, возможно, до появления там рода Ну1а. В своем расселении квакши не связаны со сплошным лесом. Несмотря на то что бол ь- шинство видов этого рода являются древесными, некоторые из них живут на скалах или даже являются наземными видами, и немногие (сходные по своему строению с другими видами) живут в пустынях и могут пересекать пх (см. список семейств).
Исчезновение квакш в тропиках Старого (:вета имеет прямую снязь с расцветом древесных лягушек семейства Bhacophoridae (рис. 25). Это семейство многочисленно на основной части Африки, на Мадагаскаре, в тропической Азии и т. д. Представители семейства Rhacophor idae достигают Южного Китая и Японии, но по тем или иным причинам отсут- ствуют в умеренной зоне северного полушария и не достигают Австралий- ской области или Америки. Это семейство доминирует на территории, которая почти точно викариирует с территорией, занимаемой представи- телями рода Hyla. Семейство Rhacophoridae частично перекрывает ареал 1jyln в Восточной Азии и на острове Тимор и т. д. (см. список семейств). Возможно, что цикл радиации семейства Bhacophoridae с основной части тропиков (:тарого Света находится в начальной стадии, и по мере даль- нешпей радиации они выйдут за пределы своих границ и заместят род Ityln.
Гипотеза последовательного расселения групп лягушек из тропикон Старого Света излагалась мной несколько иначе в статье, помещенной в «Quarterly Review» (Darlington, 1948, р. 21). Она была основана на сран- нении распространения семейств без учета их филогенетического поло'кения.
Лягушки произошли от амфибий, у которых уже имелись ребра и хвост. Из современных лягушек (1) Leiopelmidae до сих пор имеют гно- бодные ребра и мускул, приводящий в движение хвост, то есть у них остался мускул для движения хвостом, которого у амфибий уже давно нет. ('.нободные ребра имеются и у представителей семейства (2) Discoglossi- dae, даже у взрослых особей, а у семейства Pipidae — на стадии личинки. ('.емейстно Rhinophrynidae предположительно связано с этллми семейст- вами, так как его представители имеют такие же позвонки и личинки, сх<~д- ные с личинками семейства Pipidae. (3) (:емейство Pelobatidae не имеет свободных окостенелых ребер, но обладает другими в какой-то мере прими- тивными признаками. Оставшиеся «ныси~ие» лягушки образуют дне основные группы, различающиеся строением скелета и мускулатуры: (1) f.eptodactylidae, Atelopodidae, Bufonidae и Hylidae и (5) Bariidae. Rlia- сорЬогЫае, Phrynomeridae и Brevicipitidae.
Пять выше перечисленных групп рассматриваются обычно как под- отряды (Noble, 1931; Romer, 19 л5, р. о61) и, помимо ~ же указанных, харак-
3ея иоводные
теризуются и различными другими признаками. Подотряды приводятся в списке семейств. Эти пять групп, не обязательно прямо или косвенно родственные друг другу, наглядно представляют стадии эволюции лягушек. Их распространение находится в соответствии с их местом на шкале эво- люции. Первая, самая примитивная группа, сохранилась только местами в северо-западной части Северной Америки и в Новой Зеландии. Семей- ства второй группы, хотя и не являются строгим реликтом, имеют преры- вистое или локализованное распространение, но нигде не доминируют. Третья группа также имеет прерывистое распространение и приурочена к определенным частям земного шара, но субдоминирует частично в Индо- Малайской области. Четвертая и пятая группы почти всегда космополиты и обе включают семейства, которые радиируют, и семейства, которые доми- нируют в разных местах, но четвертая группа имеет тенденцию к числен- ному преобладанию на периферических или ограниченных преградами континентах, а пятая, которую Нобль считает самой высшей, ныне числен- «о преобладает на основной территории тропиков Старого Света. Короче говоря, самые примитивные группы лягушек являются самыми локали- зованными, а последовательно более высокоорганизованные группы- оолее доминирующими. Следующая группа, стоящая рядом с высшими, доминирует на периферических территориях и высшая — главным обра- зом в тропиках Старого Света. Такая корреляция довольно хорошо сов- падает с предположением о радиации последовательно идущих групп из тропиков Старого Света. Частные случаи, не соответствующие этому пред- положению, не нарушают, на мой взгляд, основной корреляции.
Я не люблю придавать особого значения подробностям, но при рас- смотрении распространения лягушек одна деталь все же является чрез- вычайно важной. Существование очень примитивной лягушки в Новой Зеландии и полное отсутствие родственных ей лягушек в Австралии и Южной Америке позволяют выявить последовательность и замещение фаун лягушек на материках.
(: удить об истории географического распространения лягушек по ископаемым данным трудно. Indobatrachus из семейства свистунов, най- денный в эоцене Индии, возможно, является единственной ископаемой лягушкой, которая имеет важное географическое значение. Другие иско- паемые остатки указывают лишь на то, что определенные лягушки нахо- дились в определенных местах в определенное время, но в основном эти ископаемые незначительны и некоторые из них сомнительны. Например, Нобль (Noble, 1928), описывая вид рода Hyln из нижнего миоцена Европы, признает несовершенство экземпляра, но, не колеблясь, относит этот вид к роду Hyla. Паркер (Parker, 1929, 1930), всесторонне исследовав этот экземпляр, пришел к выводу, что он не является представителем семей- ства Hylidae. Однако это ископаемое до сих пор приводится в сведениях о роде Ну1п (Вотег, 1945; Darlington, 1948; Schaeffer, 1949). Как посту- пить в данном случае, я не знаю, но, несомненно, это неверные сведения. Другая ископаемая лягушка из олигоцена — миоцена Европы была отне- сена к семейству Hylidae и к существующему высокоспециализированному роду Лгггр1ггgnn codon, приуроченному в настоящее время к небольшой области в Южной Америке, но, по данным Хоффштеттера (Hofstetter, 1945), оказалось, что европейский экземпляр ископаемого принадлежит не к Атр1ггдппйойоп, а что это череп лягушки с неправильно присоединенной гг нему челюстью ящерицы! Заслуга Шеффера (Schaeffer, 1949) заключает- ся в том, что он составил сводку современных данных но ископаемым лягушкам, но я сомневаюсь, что ископаемые формы имеют столь важное зна-
Глава 8
182
чение. Я считаю, что лягушки относятся к тем животным, ископаемые которых все еще недостаточно изучены и распространение современ- ных форм которых является лучшим путеводителем по их географической истории '.
Региональное распространение земноводных
1 Меткалф (Metcalf, 1923, 1940) пытался проследить эволюцию и расселение лягушек друпиг путем — при помощи ис паразитов опалцн.,")òî так называемая «зоо- география «хозяина» паразита». Она может привлечь внимание зоологов и в некоторых случаях оказаться полезной, но пример Меткалфа был неудачным. См. Нобля (ХоЬ]е.
1925) и Данна (Dann, 1925).
В связи с тем, что объем книги не позволяет детально рассмотреть фауну земноводных в разных частях земного шара, придется ограничить- ся обзором общего состава фаун и их связей.
Земноводные умеренной зоны северного полушария уже были упомя- нуты выше: это саламандры, несколько более или менее локализованных примитивных групп лягушек, несколько видов лягушек из тропических групп, которые достигают умеренной зоны Восточной Азии или проникают в умеренные широты Северной Америки, и (доминирующие в фауне) представители трех больших родов Лапа, Bufo и Ну1а (плюс два других рода семейства Hylidae в Северной Америке). Фауны земноводных обоих полушарий умеренной зоны северных широт Евразии и Северной Аме- рики, отличаясь в деталях, в целом очень сходны между собой и распро- странены на каждом из этих континентов очень неравномерно. Это хорошо иллюстрируется следующим примером.
В пределах умеренной зоны северного полушария саламандры встре- чаются в следующих местах (табл. 3): в Европе и примыкающих к ней частях Юго-Западной Азии, Северной Африки — 3 семейства, 8 родов, около 20 видов; на большой площади в Западной и Северной Азии встре- чается только 1 вид Hynobius keyserlingii; в Восточной Азии, вклю- чая восточные Гималаи и т. д., в Китае и Японии — 3 семейства, 11 ро- дов и по крайней мере 42 вида; в западной части Северной Америки от южной части Аляски до Калифорнии включительно — 3 семейства, 10 ро- дов, 25 видов; в центральных районах западной части Северной Америки, включая Большой Бассейн, Скалистые горы и Великие равнины, только один широко распространенный вид рода Ambystoma и несколько локали- зованных, географически изолированных представителей семейств Amby- stomidae и Plethodontidae; в восточной части Северной Америки от плато Озарк, восточный Техас, и т. д. до Атлантического океана и от южной части Канады до Флориды — 7 семейств, 20 родов и около 64 видов. Как видно, саламандры концентрируются на четырех довольно отдален- ных друг от друга территориях умеренной зоны северного полушария: и Европе, Восточной Азии, западной и восточной частях Северной Амери- ки — и только в последней современные семейства представлены почти полностью. Многие семейства и роды саламандр имеют прерывистый ареал (табл. 3). а их ископаемые формы указывают на то, что некоторые из них встречались когда-то далеко за границами современного ареала: например, гигантская саламандра (Megalobctrachus) из семейства (;ryp- tobranchidae и представитель саламандровых Tylototriton теперь оба при- урочены к Восточной Азии, а их ископаемые найдены в миоцене Европы. Географические связи разных групп идут в различных возможных направ-
Beë новодные
Таблица 3
Распространение саламандр на четырех основных территориях умеренной зоны северного полушарпя
В ос точная частьСе в. Америки
Западная часть Сев. Америки
Европа
Восточная Азия
Hynobiidae (5, 26+) Cryptobranchidae
(1, 1)
('ryptobranchidae
(1, 2)
Ambystomidae
(1, 9)
Ambyst.ота (9) S alamandridae
(1, 3)
Тгкйи us (3)
Ambystomidae
(3, 6)
Ambystoma (4) Sa lamandri dae
(1,4)
Tr i t urus (4) (Notophthalmus)
Salamand ridae
(5, 15+) Triturus (3) (Not ophthalmus)
Salamandrirlae
(6, 18)
Triturus (10)
Атр111итМае (1, 2) Piet}iodontidae
(13, 38)
Plethodon (11) Aneides (1)
Plethod ontidae
(6, 15)
Р1еПо<1ои (5) Aneides (3) Hydrosnantes (2)
Р1е1 leod on t Mae (1, 1)
Hydroniantes (1) Proieidae (1, 1)
Proteidae (1, 7) Sirenidae (2, 3)
семейства (8, 20) 3 семейства (11, 42) 3 семейства (10, '5) 7 семейств (20, 64)
П р п м е ч а н и е. Перечислены все семейства и роды, которые встречаются более чем на одной из четырех территорий. Цифры в скобках после названия семейств — число родов и видов, цифры после названия рода — число видов. В некоторых случаях приводится только приближенное число.
лениях между четырьмя основными территориями, на которых они рас- пространены. Саламандровые являются связующим звеном между Евро- пой и Восточной Азией, Hydromantes — между Европой и западной частью Северной Америки, Proteidae — Европой и восточной частью Север- ной Америки. Некоторые представители рода Triturus — между Восточной Азией и западной частью Северной Америки, современные представители семейства Cryptobranchidae — между Восточной Азией и восточной частью Северной Америки, а некоторые представители семейства Plethodontidae и рода А mbystoma связывают западную и восточную части Северной Америки.
Когда древняя, ранее очень разнообразная и широко распространен- ная фауна сокращается и отступает на ограниченные территории, такой характер распространения называется реликтовым. Ему свойственны две основные черты. Первая — прерывистость распространения: родственные формы встречаются на сильно разобщенных территориях, а ископаемые могут быть обнаружены далеко вне границ ареалов живущих форм. Вто- рая — разнообразие географических связеи: географические связи сохра- нившихся форм очень разнообразны и смешаны (они отражают, ско- рее, процессы вымирания и сохранения, а не пути расселения). (;аламанд- ры умеренной зоны северного полушария дают представление об этом типе распространения лучше, чем любые другие животные.
1лава 8
Что касается земноводных тропической и умеренной зон южного полушария, то их лучше рассматривать вместе. О безногих следует ска- зать, что они приурочены к отдельным площадям почти всегда в границах тропиков в Западной и Восточной Африке, на Сейшельских островах, в Индо-Малайской области и в Америке и что взаимосвязи этих форм с раз- ных территорий неопределенны. Саламандры расселяются только на незна- чительное расстояние до северных краевых частей тропиков Старого (:вета, и одна их группа, широко распространенная в тропической части Север- ной Америки, образовалась на севере сравнительно недавно. Лягушки (включая жаб) изобилуют в тропиках и встречаются в ряде более или менее изолированных территорий.
Африканские лягушки, распространенные к югу от(:ахары (Noble, 1922, р. 64 — 67; 1924), относятся к 7 семействам примерно с 40 или более родами (по современной классификации, это число значительно больше). Наиболь- шей численности и разнообразия они достигают в тропических лесах Запад- ной Африки, но многие роды широко распространены по всем африкан- ским тропикам. Только роды Лапа и Bufo проникают далеко в глубь Сахары (Angel, Lhote, 1938). Pyxicephalus, зарегистрированная Гибе в центральных районах южной Сахары (Guibe, 1950), наряду с несколькими видами рода Bufo,— просто обычная лягушка рода Лапа с укороченными ногами и с усовершенствованным роющим аппаратом (Loveridge, устное сообщение). В северо-западной части Африки умеренных широт обитают европейские формы земноводных, включая по крайней мере 3 рода саламандр и такие роды лягушек, как Discoglossus, Hyla и Pelobates, а также Лапа и Bufo. В Южной Африке в районах умеренных широт имеются различные пред- ставители родов Rana и Bufo, несколько лягушек других характерных африканских групп и еще два рода, заслуживающие особого внимания. Род Heleophryne примерно с 4 видами, распространенный в Ю'кной Афри- ке в северном направлении до Трансвааля, независимо от того, относится ли он к семейству Leptodactylidae или нет (см. список семейств), является изолированным и, по-видимому, реликтовым родом. Род Breviceps с де- сятком или более видов, распространенный от Южной Африки на северо- восток до Танганьики, является главным родом африканских представи- телей семейства Brevicipitidae. Этот род почти определяет современный ареал данного семейства в Африке. В настоящее время он отступает в Юж- ную Африку аналогично роду Heleophryne.
Фауна лягушек Мадагаскара (Noble, 1931; Grandidier, Petit, 1932 и др.) сильно отличается от фауны Африки. На острове отсутствуют пины, представители семейства Bufonidae (включая род Bufo), Phrynomeri- dae и Heleophryne, обитающие на материке; настоящих лягушек мало. Семейство Rhacophoridae относительно многочисленно; два рода (Меда- lixalus и Hyperolius) — общие с Африкой, один род (Rhacophorus) — об- щий с Индо-Малайской областью, но не с Африкой, и около четырех эндемичных родов. Узкоротые лягушки еще более многочисленны (Parker, 1934). Они представлены 2 подсемействами: Cophylinae с 8 родами, при- уроченное к Мадагаскару, и подсемейство Dyscophinae, имеющее один эндемичный род (Discophus) на Мадагаскаре, но в основном приуроченное к Индо-Малайской области. На Мадагаскаре имеется также и ряд других лягушек, взаимосвязи которых недостаточно выяснены.
На Сейшельских островах насчитывается всего 5 видов лягушек (Par- ker, 1936). Один из них (Лапа rnascareniensis), возможно, интродуцирован, другой (Megalixalus seycltellensis) — четко выраженный эндемик африкан- ского и мадагаскарского рода и три оставшихся вида из двух родов оора-
3ех н овод ные
зун>т эндемичное подсемейство Sooglossinae, относящееся к семейству Ре! о hatidae.
Лягушки тропической Азии от Индии (Boulenger, 1890) до материко- вых островов Суматра, Ява и Калимантан (van Kampen, 1923) насчиты- вают по крайней мере 6 семейств и 37 (возможно, значительно больше) родов, из которых одни — общие с Африкой, а другие нет; одни широко рас- пространены в Индо-Малайской области, а другие локализованы. О ро- дах, которые достигают в Китае умеренной зоны, уже говорилось. Пример- но 12 родов распространены до Филиппин (Inger, 1954), где они составляют болыиинство лягушек названных островов, которые в свою очередь имеют одного реликтового представителя семейства Discoglossidae — Barbouru- la — и два рода, общих с Австралийской областью через острова Сулаве- си или Молуккские. Все тропические азиатские семейства (кроме семей- ства Hylidae) и по меньшей мере 9 родов встречаются за пределами остро- вов Ява и Калимантан, расселяясь по направлению к Австралийской области или в самой области (van Kampen, 1923; Mertens, 1930; Parker, 1934 и др.), но ни единого характера распространения, ни общих границ ареала у них нет. Подробности приводятся в гл. 7.
На Новой Гвинее из лягушек (Parker, 1934; Loveridge, 1948) домини- рует род Hyla (с Л'усйтуг~ег) примерно с 40 видами только на одном этом большом острове и семейство Brevicipitidae, 2 подсемейства которого с 9 родами и более чем с 50 видами большей частью приурочены к Новой Гвинее и соседним островам. На Новой Гвинее имеется также несколько представителей австралийских свистунов, один представитель рода Ла- па — настоящих лягушек и несколько других представителей семейства Hanidae из подсемейства Cornuferinae. На Молуккских островах обитают те же лягушки, что и на Новой Гвинее, но они менее разнообразны: несколько представителей рода Hyla, 2 рода семейства узкоротых лягушек и несколько представителей рода Лапа и подсемейства Cornuferinae. Это подсемейство, малочисленное на Новой Гвинее и Молуккских островах и относительно многочисленное на Соломоновых островах, где встре- чается еще только один род Hyla, является единственным подсемей- ством на островах Фиджи, где оно, возможно, не является местным (стр. 426) .
Лягушки Австралии и Тасмании (Loveridge, 1935; Parker, 1940) относятся к двум основным группам: Hyla с 24 или более австралийскими и тасманскими видами и австралийское семейство Leptodactylidae (по Ноблю, Criniinae). Последнее образует одно или два эндемичных подсе- мейства с 14 родами и 56 видами в Австралии и Тасмании, 3 родами (ни один из них не является эндемичным) и 5 видами (только 2 энде- мичных) на Новой Гвинее (ни одного за ее пределами). Вся эта группа рас- сматривается как довольно близкородственная американским свистунам (].ер1одасФу1Ыае). Кроме них, тропической части Северной Австралии достигают очень немногие новогвинейские лягушки: два близкородствен- ных вида рода Ну1агппа (или Лапа) и также два новогвинейских рода узко- ротых лягушек, представленных в северном Квинсленде 4 видами, из ко- торых 2 вида являются эндемиками.
В Новой Зеландии имеется единственный, очень примитивный род лягушек Leiopelma, о котором у>ке говорилось выше.
Фауна лягушек тропической и умеренной зон южных широт Америки (Nohle, 1931) очень богата: около 7 семейств и 60 или более родов,— точ- .ное число зависит от классификации, принятой тем или иным автором. Большая часть этой фауны достигает Центральной Америки (см. ниже),
Глава 8
а меньшая — Вест-Индии (гл. 8). О нескольких тропических родах. кото- рые достигают умеренной зоны северных широт Северной Америки, гово- рилось выше. В Южной Америке на территориях, облесенных в различ- ной степени, на равнинах и в горных местностях можно выделить различ- ные подфауны лягушек. По направлению к южной оконечности Южной Америки эта фауна сокращается, и во влажных лесах умеренной зоны, в южном Чили, наблюдается очень четкая подфауна, для которой харак- терны эндемичные роды семейств Leptodactylidae и Atelopodidae, огггсуггг- ствие всех представителей семейства Hylidae, а также отсутствие семейств Brevicipitidae, Pipidae и рода Лапа (Dunn, письменное сообщение 6 ок- тября 1946 г.).
Ранее был рассмотрен ряд более или менее отчетливо выраженных фаун лягушек тропических стран и стран умеренной зоны южного полу- шария. Теперь кратко остановимся на географических связях, которые существуют между этими фаунами.
Между лягушками Африки, Мадагаскара, Сейшельских островов и тропической Азии наблюдается смешанный характер связей. Африка и тропическая Азия имеют некоторые общие группы лягушек, но на каждом материке имеется несколько местных семейств, которых нет на другом (рис. 20). Два африканских рода (Megalixalus и Hyperolius), достигающие Мадагаскара и в одном случае Сейшельских островов (род Megalixalus), в Индо-Малайской области отсутствуют. Два других рода (Bufo, Hylara- na) встречаются и в Африке, и в Индо-Малайской области, но отсутствуют на Мадагаскаре и Сейшельских островах. Подсемейство Melanobatrachi- nae из семейства Brevicipitidae известно только в Восточной Африке и Южной Индии. Семейство Bufonidae, кроме рода Bufo, встречается в Аф- рике и в районе Малаккского полуострова, но отсутствует на Мадагаска- ре, Сейшельских островах и на основной части Индо-Малайской области. Род Bhacophorus представлен многими видами на Мадагаскаре и в Индо- Малайской области, начиная от Цейлона и Индии, и далее в восточном направлении, но отсутствует в Африке и не имеет ни одного вида на Сей- шельских островах. Подсемейство Dyscophinae из семейства Brevicipiti- dae отсутствует в Африке, один его род приурочен к Мадагаскару, нет ни одного на Сейшельских островах, на Цейлоне или на полуострове Индо- стан, два рода встречаются далее на восток в Юго-Западном Китае, Бир- ме, на полуострове Малакка, на островах Суматра и Калимантан. Семей- ство Pelobatidae отсутствует на основной части Африки и на Мадагаскаре, но представлено одним эндемичным небольшим подсемейством на Сей- шельских островах; на Цейлоне или на полуострове Индостан также нет ни одного представителя Pelobatidae, но одно подсемейство широко рас- пространено в восточной части Индо-Малайской области и еще одно— в умеренной зоне северного полушария.
В связи с усовершенствованием классификации, открытием новых видов и определением видов, интродуцированных человеком, представле- ния о распространении некоторых из вышеприведенных групп, вероятно, изменяются, но характера связей лягушек в этой части земного шара это не изменяет. В основном особенности распространения лягушек через Ин- дийский океан проявляются в разрывах и смешанных связях, которые ука- зывают, что эта фауна частично реликтовая, а существующие связи отчасти результат вымирания и выживания, а не просто расселения. При этих обстоятельствах родство некоторых мадагаскарских лягушек с лягушками Индо-Малайской области не обязательно указывает на прямое расселение в прошлом или на существование прямого материкового моста. В целом,
Земноводные
187
же фауны лягушек тропической Африки, Мадагаскара и тропической Азии удивительно различны.
От Индо-Малайской области до Новой Гвинеи и Австралии наблю- дается такое большое изменение фауны лягушек, что, за исключением рода Ну1а, имеющего чрезвычайно прерывистое распространение, ни одно из семейств тропической Азии не достигает умеренной части Австра- лии. Значительное изменение фауны происходит также между Индо-Ма- лайской областью и Новой Гвинеей, однако между ними нет единой гра- ницы распространения, это, скорее, переходная зона, в которую входят все Малые Зондские острова, острова Сулавеси и южные Филиппины (стр. 387). Другая значительная смена фаун происходит между Новой 1'ви- неей и умеренной частью Австралии, но опять-таки это происходит не у водной преграды и не у какой-либо одной границы. Около 9 видов лягу- шек (2 из рода Лапа или Hylarana, 3 из семейства Leptodactylidae, 2 из рода Hyla и 2 из семейства Brevicipitidae) встречаются и в Новой Гвинее и на прилежащей части тропической Австралии (семь из них были собра- ны мною). Это мелкие лягушки, в большинстве случаев живущие в уеди- ненных местах и вряд ли занесенные туда человеком. Они свидетельствуют о том, что между фаунами Новой Гвинеи и Австралии происходил обмен. Однако фауна лягушек Новой Гвинеи состоит главным образом из пред- ставителей семейств Hylidae и Brevicipitidae, а в Австралии в основном из представителей семейств Hylidae и Leptodactylidae, и только в умеренной части Австралии встречаются квакши и свистуны.
Фауны лягушек тропической Азии и тропической Америки очень различны. Семейства Pelobatidae, настоящих жаб, настоящих лягушек и Rhacophoridae Индо-Малайской области в тропической Америке представ- лены только родами убиквистами — Лапа и Bufo, а представители семейств тропической Америки — пины, свистуны и квакши — отсутствуют (или почти отсутствуют, как, например, род Hyla) в Индо-Малайской области. Единственные лягушки, которые, кроме тропической Азии и тропической Америки, нигде больше не встречаются (за исключением прилежащих частей умеренной зоны северного полушария),— это подсемейство М|с- rohylinae, включая род Microhyla из семейства Brevicipitidae. Семейство Atelopodidae, которое когда-то считали свойственным исключительно аме- риканским тропикам, встречается и в Индо-Малайской области, а также и в Африке, но его распространение изучено недостаточно (см. список семейств).
Фауны лягушек Африки и Южной Америки также очень различны. В африканской фауне доминируют жабы, лягушки и семейство Rhaco- phoridae (из них только роды Bufo и Лапа находятся в Южной Америке). В фауне Южной Америки доминируют свистуны, квакши и семейство Ate- lopodidae. В Африке из них известен только один род, относимый к се- мейству Leptodactylidae, что вызывает сомнение, и несколько (сомнитель- ных) представителей семейства Atelopodidae. Единственные лягушки, которые, кроме Африки и Южной Америки, больше нигде не встречаются, это пины, из которых одно подсемейство приурочено к Африке, а другое- к северным районам Южной Америки. Согласно Ноблю, пины — прими- тивные водные лягушки и их распространение соответствует распростра- нению некоторых пресноводных рыб.
Прямого родства между мадагаскарскими и американскими лягуш- ками, очевидно, нет.
Фауны лягушек Австралии и Южной Америки очень сходны, и это тем более удивительно, потому что большинство других фаун лягушек рез-
ко отличается друг от друга. В умеренной зоне Австралии обитают толь- ко квакши и свистуны, также многочисленные и в Ю'кной Америке. Хотя распространение этих групп не является чем-то особенным, вместе с тем оно не так просто, и в Южной Америке существует много других групп лягушек, отсутствующих в Австралии, а в тропической части Ав- стралийской области имеются такие группы, которых нет в Южной Аме- рике. Следует еще раз отметить, что ни одна группа лягушек Новой Зе- ландии не родственна ни одной из известных групп Австралии и Южной Америки.
В целом фауны лягушек разных частей земного шара очень различны. Широко распространенные роды Лапа и Bufo только подчеркивают замедленные темпы расселения других лягушек в последнее время. Я счи- таю, что этот факт соответствует гипотезе о том, что распространение групп доминирующих лягушек связано с локализацией и отступанием других менее доминирующих групп.
Земноводные Центральной Америки
Для земноводных Центральная Америка с Мексикой являются такой же важной переходной территорией, как и для рыб.
С севера несколько североамериканских групп земноводных достигли северных краевых частей (в некоторых случаях только северо-западной части) Мексики. Саламандры семейства Ambystomidae и чесночницы в пре- делах тропиков в южном направлении доходят до Мексиканского нагорья, но не достигают тропических низменностей. Саламандры семейства Ple- thodontidae и лягушки рода Лапа через Центральную Америку достигают северной половины Южной Америки, а род Bufo распространяется еще дальше на юг.
С юга в Центральную Америку проникает большее число земновод- ных. По Данну (Dunn, 1942), из 6 южноамериканских родов безногих 2 рода достигают Панамы (один из них распространен через Центральную Америку до тропиков Мексики). Согласно Ноблю (Noble, 1922, р. 67), из 60 южноамериканских родов лягушек 21 достигает Центральной Аме- рики, и там происходит постепенное обеднение фауны. Данн отмечает (Dunn, 1931), что не менее чем из 25 южноамериканских родов семейства Leptodactylidae только 5 родов заходят в Центральную Америку, только 3 рода распространяются до Мексики и только 1 род достигает Мексикан- ского нагорья (где он положил начало двум эндемичным родам). Из 10 юж- ноамериканских родов семейства Atelopodidae лишь 4 рода проникают в Центральную Америку и ни один не встречается к северу от Никарагуа (правда, один род имеется на Кубе). Из 13 южноамериканских родов семейства Hylidae не более 6 родов достигают Центральной Америки (еще 2 рода являются там эндемиками) и только род Hyla достигает основной части Северной Америки (есть еще 2 эндемичных рода), наконец, примерно из 8 южноамериканских родов семейства Brevicipitidae только 3 рода доходят до Центральной Америки, а 2 рода через Мексику до Соединенных Штатов.
Этого краткого и довольно поверхностного обзора вполне достаточно для того, чтобы показать в общих чертах, как фауны земноводных Север- ной и Южной Америки перекрывают друг друга с постепенным уменьше- нием отдельных элементов каждой фауны. Основная часть перехода про- исходит в Мексике и Центральной Америке, но фактически проникновение
Зем новодные
отдельных групп заходит значительно дальше, так как несколько северо- американских групп проникает в глубь Южной Америки, а несколько к1жноамериканских — в глубь Северной Америки.
1!осле сводок Нобля и Данна по фауне земноводных Мексики и Цент- ральной Америки (;мит и Тэйлор (Smith, Taylor, 19~8) довольно удачно суммировали многочисленные новые данные по земноводным Мексики, но никто не проделал подобной работы по Центральной Америке. Этого не удалось сделать и в данном труде, так как возникают трудности, отча- сти связанные с тем, что у земноводных Центральной Америки наблюдает- гя сильное дробление родов. Независимо от того, удобно ли такое дробле- ние с точки зрения систематики, для зоогеографа это сильно усложняет лело. Здесь можно принять только одно решение — сохранить старые большие роды, поскольку они естественны, а большинство новых считать подродами. Это позволит таксономистам дробить систематические едини- цы, как им необходимо, не внося путаницы в работу зоогеографов.
Один вид лягушек из Центральной Америки оказался более изоли- рованным, чем предполагали Нобль и Данн. Это Rhinophrynus dorsalis— специализированный вид роющей лягушки, в прошлом отнесенный к се- мейству Leptodactylidae, а в настоящее время вынесенный в монотипное семейство, приуроченное к тропическим частям Мексики и Гватемалы (Smith, Taylor, 1948, р. 34). Несмотря на свою специализацию, эта лягуш- ка значительно примитивнее, чем любые другие в Центральной Америке, и по степени развития близка к семейству Discoglossidae и пинам. Пред- положительно, это единственный поразительный реликт земноводных в Мексике и в Центральной Америке. К сожалению, его предки и гео- графические истоки (Северная, Центральная или Южная Америка) неизвестны.
11ереселение земноводных и преграды их распространенин>
Там, где встречаются различные фауны земноводных, обычно между ними нет резкой границы, но существует переходная область с постепен- ным уменьшением обеих фаун в противоположных направлениях.
В Восточной Азии, где основной преградой для встречающихся фаун земноводных — тропической и северо-умеренной — является климат, безногие и ряд тропических лягушек достигают восточных Гималаев и Южного Китая; 5 родов лягушек проникают на север в Центральный Китай и один из них достигает Маньчжурии, а другой — Японии; несколько северных представителей семейства Salamandridae и род Ну1а распро- страняются в южном направлении, на ограниченной части тропической Юго-Восточной Азии.
В центральной Америке среди земноводных наблюдается более силь- но выраженная переходность северных и тропических фаун, чем среди строго пресноводных рыб: она охватывает относительно большее число земноводных, и некоторые из них простираются значительно далыпе в том или ином направлении. Очевидно, земноводные перемещались через Центральную Америку чаще и проникали дальше, чем пресноводные рыбы.
Между Азией и Австралией, через линию Уоллеса, там, i<уда поч1и не проникают строго пресноводные рыбы, ареалы лягушек ~1ндо-Малай- ской и Австралийской областей перекрывают друг друга на значитель- ное расстояние, и это перекрытие идет сложным путем. Видимо, узкие
Глава 8
пространства соленой воды лягушки могут преодолевать довольно часто.
Едва ли нужно вдаваться в дальнейшие подробности. Стоит только отметить, что зональный климат, вероятно, является для земноводных такой же преградой, как и для рыб, но часть из них, так же как и рыбы, все же преодолевает эту преграду. Другие виды преград менее важны для земноводных, чем для рыб. Пустыни и горы являются существенной преградой, но это только вопрос времени. Ничто — ни климат, ни пусты- ни, ни горы, ни даже узкие пространства соленых вод — не помешало рас- селению доминирующих видов лягушек на земном шаре.
Доминирование и конкуренция в связи с распространением земноводных
Из современных земноводных численно преобладающими являются лягушки рода Лапа. Этот род представлен многими видами во всех основ- ных тропических и умеренных районах Старого Света и Северной Америки, хотя только 2 вида (фактически виды рода Hylarana) достигают Северной Австралии и только один — (северной части) Южной Америки. Некоторые виды также многочисленны. В целом же род довольно разнообразен по структуре и образу жизни и включает земноводные, водные, наземные и роющие формы. Он доминирует в двух основных климатических зонах— тропической и умеренной зонах северного полушария — и встречается в самых разнообразных стациях, включая холодные и засушливые: например, к этому роду относятся самые северные формы из всех земновод- ных, и он является одним из двух родов лягушек, широко распространен- ных в Сахаре. Таким образом, род Лапа имеет почти сплошное распростра- нение на значительной части земного шара, причем в меньшей степени, чем другие земноводные, зависит от климата и других преград. Все это, вместе взятое, дает представление о доминирующей группе с радиальным характером распространения, сравнимое в общих чертах, но не в деталях. с доминирующей группой пресноводных рыб семейства Cyprinidae.
Имеющиеся ископаемые данные недостаточны, чтобы показать, существует ли связь между радиальным распространением рода Лапа и со- кращением ареала — отступанием других лягушек. Возможно, что это и так. Распространение семейства Leptodactylidae, которое многочисленно теперь только в Австралии и теплых частях Америки, взаимно исключает площади высокой плотности рода Лапа. Расцвет семейства Ranidae, воз- можно, является причиной исчезновения семейства Leptodactylidae в ос- новных местах их расселения, причем оно сохранилось лишь на матери- ках, защищенных преградами, куда лягушки рода Лапа проникают толь- ко в настоящее время. Вытесняемые доминирующим семейством Вашйае, узкоротые лягушки, возможно, отступали другим образом. Отступание лягушек семейства Brevicipitidae — мелких, редко встречающихся в боль- шом количестве и обитающих преимущественно в скрытых местах (Par- ker, 1934, р. 1, 14),— возможно, носит как экологический, так и геогра- фический характер.
Трудно сказать, чем вызвано доминирование семейства Ranidae и рода Лапа. Очевидно, они обладают какими-то особыми качествами, кото- рые делают их по сравнению с другими лягушками наиболее приспособ- ленными к разным условиям.
Род Bu jo по характеру географической радиации и доминированию занимает только второе место среди земноводных после рода Лаиа
Земноводные 1!о сравнению с родом Лапа он менее многочислен по видам и менее раз- нообразен по структуре и образу жизни, но маркет существовать в самых различных местах обитания, включая теплые и холодные, влажные и за- сушливые.
Некоторые другие лягушки доминируют только на небольших пло- щадях или в специфических стациях. Распространение и взаимоисключе- ние доминирующих лягушек на земном шаре показано на рис. 26. Р и с. 26. Доминирующие земиоводньп семи осповшык
областей. Эта диаграмма подчеркивает взаимозамещение среди наземных груп и семейств lian i dae (жирные линии), 1.е fi t ridactyl i dae (тонкие сплошные линии), среди древесных групп ссмейства l( t>acophoridae (жирная пунктирная линия) и семейства
Hyl i dae (тонкая пунктирная линия).
0 предполагаемой конкуренции среди лягушек уже неоднократно упоминалось. Конкуренция неотъемлема от понятия «доминирование»„ «представления о замещении одних лягушек другими и понятия викари- ата» — все эти вопросы связаны друг с другом. Невозможно понять или достаточно хорошо объяснить распространение лягушек без учета конку- ренции как предпосылки. Различные группы лягушек, очевидно, чаще конкурируют на обширных территориях, чем на ограниченных, а об ис- ходе этой конкуренции можно судить по истечении длительного времени. Конкуренция — явление довольно сложное, и многие ее детали пока еще не известны. Конкуренция МоВсВТ происходить не только между группами лягушек, а также между лягушками и саламандрами, и то, что первая обладает способностью прыгать, а последняя нет, не изменяет того факта, что требования типичных форм обоих отрядов примерно одни и те же и что оба отряда имеют типично водных, иногда конкурирующих личинок.
142
Заключение: особенности распространения земноводных
Распространение земноводных, рассматриваемое в самых общих чер- тах, так же как и распространение пресноводных рыб, характеризуется по крайней мере пятью особенностями. Первая из них — ограниченность. Распространение земноводных ограничено территорией, простирающейся от точек, находящихся к северу от Полярного круга, до (или почти до) основных границ суши на юге, то есть ограничено материками и недавно возникшими материковыми (и некоторыми неясного происхождения) ост- ровами, включая Новую Зеландию. Земноводные совершенно отсутствун~т (если только они не интродуцированы) даже на пригодных для обитания, но удаленных на большое расстояние от материка океанических островах. В пределах этих границ вторая и третья особенности — зональное и р»- диальное распространение различных земноводных — у;ке были достаточно подробно рассмотрены и не вызывают каких-либо вопросов. Четвертая особенность — дифференциация материковых фаун земноводных, вопр<н сложный, но также хорошо выявлен ранее. И, наконец, пятая осанн.н- ность распространения амфибий, как и у пресноводных рыб,— концен- трация фауны с обеднением ее по краям на больших территориях с благо- приятными условиями обитания. Эта особенность хорошо иллюстрируется концентрацией саламандр на четырех основных территориях в умеренной зоне северного полушария (рис. 19), но она также может быть продемон- стрирована и на других фаунах земноводных. Для всех основных фаун характерна концентрация на больших территориях с условиями, пригод- ными для обитания. В местах, где происходит встреча различных основных фаун земноводных, наблюдается переход одних элементов фауны в дру- гую, с, перекрытием этих фаун по краям и с постепенным обеднением
как!одоп фа~уны в противоположных направлениях.
Что касается места, занимаемого реликтами в распространении, то безногие и саламандры являются архаическими и в этом смысле релик п>- вычи формами и почти полностью приурочены к основной тропической зоне материков и соответственно к умеренной зоне северного полушария. Примечателен тот факт что в Австралии или на отдаленных островах нет географически изолированных видов этих очень древних отрядов, эа исключением безногих на Сейшельских островах.
Из лягушек первые три подотряда являются филогенетическими реликтами, а из них в свою очередь только один род Leiopel~na, обитающий в Новой Зеландии, географически изолирован и находится за пределами досягаемости этих районов высшими лягушками. В данном случае можно было бы провести параллель с двоякодышащими рыбами Австралии (но в Австралии нет именно примитивных, реликтовых лягушек). Небольшая группа семейства Pelobatidae приурочена к Сейшельским островам. Дру- гие филогенетические реликты лягушек (три первых подотряда) встре- чаются на основных материках — преимущественно в тропической и уме- ренной зонах северного полушария. В тропиках Индо-Малайской области имеются представители семейств Discoglossidae и Pelobatidae, в Африке и Южной Америке — Pipidae, в тропиках Мексики и Гватемале — единст- венное семейство Rhinophrynidae и в умеренной зоне северного полуша- рия — представители семейств Leiopelmidae, Discoglossidae и Pelobati- dae. Если рассматривать фауну лягушек в целом, то в кочичественном соотношении в умеренной зоне северного полушария реликтовых групп п видов больше, чем в тропиках.
Зел~ новодньм История расселения земноводных
Вместо того чтобы обсуждать вопросы, как и почему происходило распространение земноводных и что оно нам говорит и древних материках и климатах каждый в отдельности, ниже приводится картосхема предпо- лагаемой истории расселения земноводных, из которой вытекают ответы на поставленные вопросы (рис. 27). 1' и с. 27. Схематическое изображеиие (предполагаемого) 1l()c'I( ä0âàòåëI'íîã0 расселения трек семейств,чя гушс и. I.ciupcl~»i(lac (белый hpyr с белыми стрелками), ныне сохранив- шееся только в Новой 3еландии и в изолированной стации в за- »адной части Северной Америки (двойные»ересекан~»~иеся линии); I.cubi to(lactylidae (круг со штриховыми стрелками), сохра- нившееся в основном в Австралии и Южной Америке (линии »о диагонали; семейство Ha»i(lac (черный круг с черными стрелками), которое, очевидно, заместило преды;(~ ~цне семейства на основной части земного шара и только г(»(рь достигло
Австралии и Южной Америк».
Начнем с лягушек мелового периода. В те времена предки бо:1ьшин- ства современных семейств лягушек, вероятно, находились в тропиках (:тарого Света. Возможно, были и др~гие, теперь исчезнувшие семейства в других частях земного шара, но достоверных данных об этом нет.
Вероятно, в течение мелового периода ранние Leiopelmidae, когда-то существовавшие в тропиках Старого Света, затем исчезли и сохранились только в умеренных зонах северного и южного полушарий. Позднее они остались в виде реликтовых форм в северо-западной части (:еверной Аме- рики и в Новой Зеландии. В позднем мелу из тропиков (:тарого Света в 1ожную Америку проникли водные пины. (Если пресноводные рыбы смогли проникнуть в то же самое время через умеренную зону северного полушария, то, видимо, то же самое произошло и с пинами.) Позднее пины отст~ пили с севера и из тропической Азии, сохранившись в Африке и Юж- ной Америке. Очевидно, еще позднее, в третичном периоде, круглоязыч- ные отступили из тропиков Старого Света в умеренную зону Евразии, оста- вив своих представителей на Филиппинах. А семейство Ре!оЬа1Ыае отступает и теперь в северном направлении из тропиков (:тарого (:вета. Исчезновение этих групп с основной части тропиков Старого (;вета, по-ви- димому, было вызвано развитием там высших лягушек.
Глава 8
Последовательно доминирующие семейства высших лягушек, видимо, произошли в тропиках Старого Света и радиировали оттуда на большей части или по всему земному шару. По мере расцвета в тропиках Старого Света более поздних семейств древние семейства одно за другим исчезали. Они сохранились только в периферических районах, в основном в Австра- лии и Южной Америке. Семейства Leptodactylidae, Brevicipitidae, Bufo- nidae и Ranidae, возможно, создали одну последовательность в распро- странении, а Hylidae и Rhacophoridae — другую. Но все эти семейства, кроме Rhacophoridae, достигли Америки, вероятно, через северные рай- оны. Расселение узкоротых хорошо показывает, как лягушки тропиков Старого Света смогли проникнуть через северные районы в американские тропики, но детали этого распространения очень сложны.
Что касается безногих и саламандр, то не ясно, соответствует ли их расселение предполагаемому типу последовательной радиации и заме- щения.
Можно предположить, что безногие всегда были тропическими живот- ными и проникли через северные районы из тропиков Старого Света в Но- вый, подобно некоторым лягушкам. Они или никогда не достигали Австра- лии, или, возможно, достигли ее, но позднее исчезли там, как, например, лягушки семейства Leiopelmidae.
Расселение саламандр представляет еще более сложную проблему. Были ли они приурочены к умеренной зоне северного полушария с момента их происхождения (возможно, еще до мезозоя) и до того времени, пока одна из групп (что произошло сравнительно недавно) не смогла продвинуться в глубь тропиков Америки? Или же когда-то саламандры были широко распространены в тропиках Старого Света и затем исчезли, не оставив даже реликтовых форм в изолированных горных районах или на островах (несколько саламандр, обитающих в краевых частях тропиков в Азии, являются не реликтовыми, а отдаленными видами существующих азиат- ских групп умеренной зоны)? Ни одна из этих альтернатив не является вполне удовлетворительной, но предпочтение можно отдать, скорее, вто- рой. Современная северная зональность большинства саламандр может являться лишь стадией в медленном перемещении из Старого Света в Но- вый. Конкуренция саламандр с доминирующими группами лягушек, возможно, явилась причиной их исчезновения в основной части тропиков Старого Света. А распространение современных саламандр в умеренной зоне северного полушария (табл. 3) с частыми разрывами и с наибольшей концентрацией их в восточной части Северной Америки позволяет пред- полагать их отступание из Старого Света по направлению к Новому. Если это отступание происходит и сейчас, то оно осложняется радиацией семейства Hynobiidae в Восточной Азии; другими словами, не следует счи- тать, что перемещения сложных групп, происходящие в течение длительного времени, будут простыми.
Эта предположительная история расселения объясняет распростра- нение существующих земноводных без удивительных материковых мостов или перемещающихся материков. Однако современное распростра- нение этих животных имеет мало общего с распространением их в древние времена. Современные отряды земноводных, очевидно, имеют архаичное, домезозойское происхождение, но их современное (более позднее) распро- странение, возможно, результат событий позднемелового и третичного периодов и ничего не может рассказать нам о земном шаре предшествующих времен.
3емноводные
С11ИСОК СЕМЕЙСТВ ЗЕМНОВОДНЫХ
H список включены только современиые группы земноводных и ископаемые.
которые имеют какое-то отношение к этим группам. Употребляемая классифика- ция и связи современных групп с древними и вымершими группами рассматрива- лись в начале главы. Звездочка (*) перед латинским названием означает, что эта группа вымерла.
Отряд Apoda
Caeciliiclae, безногие. Тропики Африки, Азии и Америки, Сейшельские острова в 11и- дийском океане. В Африке 6 родо~, около 17 видов: широко распространены в тро- пиках Западной и Восточной Африки (в основном разные роды), но пока еще ие найдены в центральной части Африки. На Мадагаскаре безногие отсутствук>т.
Ранее предполагали, что на Сейшельских островах имеется 2 — 3 рода безногих, но Паркер (Parker, 1941) считает, что там существует только 1 эндемичный род lgypogeopЫ8 с 6 видами, которые, по-видимому, все произошли от одного предка.
В Азии имеется 4 рода, 6 видов, распространенных от Цейлона и Индии на север к краевым частям умеренной зоны северного полушария до Сиккима (27 или 28 с. ш.) в восточных Гималаях (Boulenger, 1890) и, возможно, до южных краевых частей Китая (Роре, Boring, 1940, р. 14), в южном и восточном направлениях до островов Суматра, Ява и Калимантан (van Kampen, 1923). 1 вид достигает островов Палаван и Минданао на южных Филиппинах (Inger, 1954). В Америке Данном (Dunn, 1942) выделено 6 родов с 44 видами, распространенных от 20 с. ш. (Веракрус и Герреро в Мексике) до 35' к>. ш. (Буэнос-Айрес в Аргентине, в основном в пределах умеренной зоны южного полушария) и от уровня моря до высоты 4500 футов в Коста-Рике и до 6200 футов в Эквадоре. Все американские роды встречаются в Южной Америке: 2 рода достигают Панамы и один из двух через Центральну>о Америку проникает в тропическую часть Мексики. Безногие достигают острова Тринидад, но не собственно Вест-Индии. Широко распрост- раненных родов безногих не имеется. Все они приурочены к отдельным районам.' 10го-Восточной Азии, Сейшельским островам, африканским тропикам или тро- ицкам Америки. Данн (Dunn, 1942) указывает, что в Америке безногие обитают в тропических дождевых лесах, в тропических лесах с опадающей листвой, в тро- пических саваннах, в лесах и саваннах «умеренной» зоны и в горных подоблач- ных лесах (но в районах саванн они встречаются только в лесных полосах вдоль рек). )жизненный цикл и образ жизни большинства безногих изучены недостаточ- но. 0 некоторых имеются сводки в работах Гадова (Gadow, 1909), Нобля (Noble.
1931) и Данна (Dunn, 1942, р. 451 — 453). Часть из них земноводные со стадией водной личинки и наземными (роющими) взрослыми особями; другие — водные на протяжении всей жизни, а некоторые, включая виды, находящиеся на Сей- шельских островах, уже не имеют стадии водной личинки и стали полностью наземными. Ископаемые безногих не известны.
Отряд Urodela, с,аламандры
Hynobiidae, углозубые (азиатские наземные саламандры). Умеренные широты Азии
от Урала до Тихого океана и от краевых частей Арктики на юг до гор Юго-3а- иадной Азии (ио крайней мере до Памира и Самарканда), 10жный Китай, Тай- вань, Япония и Сахалин. 5 родов, 26 или более видов (Dunn, 1923 Liu 1950 и более ранние статьи). 1эолыиинство из этих саламандр находится в Японии и Восточной Азии. Только вид Fry)i б~ц8 й~у8егШгцй распространен по всей Сибири достигает 60' с. IIT. На Урале, пересекает Северный поляр ый круг и доходит до Верхоянска и Камчатки. Иногда углозубых неправильно называют «азиатскимц саламандрами». Они всегда имеют стадию водной личинки, а взрослые особи, хотя и являются типично наземными животными, у некоторых специализирован- ных родов также бывают водными. Оплодотворение у этого семейства внещнее, в воде, и, возможно, поэтому в отличие от многих представителей семейства ]~1е- thodîntiäàå у них сохранилась водная стадия личинки.
Это семейство, хотя его ископаемые сомнительны (миоцен Европы), оче
видно, значительно древнее (или является предком) семейства Cryptohranchidae, IcoTopoe ведет свое начало по крайней мере с верхнего олигоцена и распростр»- нено циркумполярно.
'ryptobranchidae, скрытожаберные гигантские саламандры. Восточная Лзия и восточ- ная часть Северной Америки. В Азии известен только род 1IIi gö~oböt~ö~~~ö,~ с 1 видом, распространенным во внутренних районах Китая от провинций Шань- си и Шэньси на юг и юго-запад до провинций Гуанси, Гу~~жоу, Сычуань и юго- западного Сикаиа. Другой вид или подвид находится в Японии, где приурочен
Глава 3
к южной и западной частям острова Хонсю. В Северной Америке — только род CryptobгапсЬus с 1 видом, встречающимся в западной части штата Нью-11орк ц в реке Саскуиханна, а также от системы Огайо на запад до центральной части Миссури и на юг до штатов Джорджия и Луизиана. Второй близкородстве««ый вид или подвид — в юго-восточной части реки Миссури, притоках реки Бл»к- Ривер, штаты Миссури и Арканзас. Это семейство типично водное. Азиатские формы, несмотря на гигантские размеры, живут в горных реках. Америкянские.
менее крупные, формы живут в больших реках.
Ископаемые рода Megalobatrachus найдены в Европе (миоцен), а други<
ископаемые вымерших родов этого семейства были в Европе (верхний о><игоце« и миоцен) и в Небраске (нижний плиоцен).
Ашbystomidae, различные саламандры, аксолотли. Северная Америка, от иокной
части Аляски, залива Джемса и южного Лабрадора (на север примерно до 53 ) на юг до южной части Мексиканского нагорья (краевые части тропиков, но «< низменности) и до северной части Флориды. Отсутствует частично в северной Мексике и юго-западной части Соединенных Штатов. Предполагаемое нахожде- ние одного вида в Таиланде, как указывает Нобль (Noble, 1926), явилось ошиб- кой. (Предполагаемый таиландский вид до сих пор в списках Бурре (Bourret, 1942) стоит со знаком вопроса (?), так как Бурре утверждает, что больше не было найдено ни одного экземпляра.) Основной род Ambystoma, который, взятый даже в весьма ограниченных рамках, встречается по всему ареалу семейства, вклн~чает 22 вида, из которых 10 мексиканских. Один из более северных видов (А. ti gri »um) единственный вид саламандр, который имеет более или менее сплошное раси ро- странение между основными восточными и западными областями распростран~ иия саламандр в Северной Америке (Bishop, '1943, map. 2О). А. macrodactylun< — <;<- падный вид, который встречается во внутренних частях страны до западной Моц- TaHbI, а также найден (но, возможно, не является там местным) в штате Айо«я (Bishop, map. 15). Два дополнительных монотипных рода встречаются вдоль побережья Тихого океана. Один из них (Dicamptodon ensatus) обитает также rr < v- верных районах Скалистых гор, в изолированных местностях, в штатах Айдахо и Монтана (Bishop, map. 21). Смит и Тэйлор (Smith, Taylor, '1948) выделяют в Мек- сике еще 3 рода этого семейства с 7 видами, включая неотеническую личинку яксолотль (<>iredon), однако Нобль относит эти виды к роду ~1mb<tstvma.
Все ископаемые, приписываемые семейству Ambystomidae, находятс» в
Северной Америке и относятся к верхнему мелу, плио~~ену и плейстоцеиу. Г!о- :>тому пе известно, являются лп более ранние формы подлинно скритожабер«<,<м«. ~а1а ша1ии.'Ыае, настоящие саламандры, тритоны. Все четыре основные территории
распространения саламандр находятся в Евразии и Северной Америке. В тропи- ках Западной Африки саламандр нет, однако они, песом«евно, заходят н крае<>ь« части тропиков в Восточной Азии. В Европе 6 родов, 16 видов, больптинство в Южной Европе, но род Triturus простирается на север примерно до 63 с. ш., иа восток до Урала и в Юго-Западную Азию до севериого Иран» и Палестины, а род 5alania»dry также достигает Малой Азии, Сирии и Палестины. 3 европеи- < г'rrx рода, вклточая Triturus, встречаются в северо-западной части (умере««яя зона) Африки, где они представлены 4 видами (2 э«демич«ых). 1 вид из 1!сия««« и Марокко (Р1<и>о<1е1ез u>alii) был по ошибке зарегистрирован также в Зяп«д«ой Африке (СЬаЬяпаи<< '1954). В западных и северных частях Азии за пред«;<~«< границ рода Triturus (см. вып<е) это семейство отсутствует. В Восточной Л:<ии.
ITH большей части Китая и па Японских островах, включая и остров PI()RI(), сll()- ва встречается род Triturus и 4 или более:шдемичных рода и в общей сложности примерно 1,э или более видов. Цесколько видов саламандровых «стречаются в кр»- евой зоне тропиков в Азии: T>iturus chi>«»sis — на острове Гопко«г (Роре, It<>- ri<ri>~, l9i0, р. 23), Parnmesotriton <1е1оиг1а1<', — эпдечик области Тонкин и Til<>- /о/ ito» — на плато Шаль и Ьирме, примерно на той же широте, что и Тонкин (письменное сообщение Майерся от <4,'VII 1948 г.), на rore граница его ря< «г><>- < тране«пя доходит до северного Таиланда (ВоиггеС, 1942). В Америке существует только 1 род Triturus (илп ' родственных рода, если заняться их разделе«и< «).
Одна группа из 4 видов находится на западе и распространена от и~жной ч;ити Аляски до юго-западной части Калифорнии и до прилегающей части Иижпей Ь<>- лпфорпии (предполага<от, что эта rpyrrrra родственна восточ«о<1зиятским ф<>1>м«м).
7~ругая группа из 3 видов находится на востоке, от Гяспе п Мин«есоти до Флори- ды в северо-восточной части Мексики. Семейство 3alamandridne отсутствует на обширной территории, охваты ваюшей Бо~ыпой Вассеин и Скалистые горы. Пред- ставители этого семейства, включая и род Criturus, в основном животные земно- водные, но все они различа~отся по количеству вречеии, проводимому на суше.
Ископаемые формы салям««дровых в Европе от«осят<я к:>оцену. Опи укя-
Земноводные
И7
зывают как на историю распространения современных форм, так и на европей- ское или евроазиатское происхождение семейства. Ископаемые формы Tyloto- t iton, в настоящее время обитающего в Восточной Азии, относятся к эоцену- миоцену Европы.
Amphiumidae, водные угревидные амфиумы. Юго-восток США, две формы, являющи- еся подвидами или близкородственными видами (Baker, 1947), один — от низовь- ев Миссисипи на запад до северо-восточной части Техаса и на север до ближайшей части штатов Миссури и Кентукки, а другой — на восток вдоль Мексиканского залива от Луизианы через Флориду и на север ио Атлантической прибрежной равнине до Виргинии.
Ископаемые этого семейства относятся к эоцену Северной Америки, но ев- ропейские ископаемые сомнительны.
Plethodontidae, безлегочные саламандры. Северная, Центральная и часть Южной Аме- рики. 1 вид изолирован в Европе.
В Европе имеется только 1 вид Hydromantes genet (= fuscus) с2 подвидами,
локализованными частично в горах, изредка в пещерах на юго-востоке Франции, частично в Италии и Сардинии. 2 других вида этого рода, один из которых описан Горманом и Кампом (Gorman, Camp, 1953), находятся в Калифорнии и локали- зуются на больших высотах. Герпетологи пришли к единому мнению, что европей- ский и калифорнийские виды относятся к одному роду, а изучение костного мозга и т. д. этих видов (Barret, 1947) подтвердило, что европейский вид относится к се- мейству Plethodontidae.
В Америке, к северу от Мексики, Бишоп (Bishop, 1943) выделяет 17 родов
безлегочных саламандр (включая Hydromantes) и 53 вида: 6 родов и 14 видов на западе, от нижней части Аляски до западной верхней части Мексики; 13 родов, 37 видов на востоке от залива Джемс и северного побережья залива Св. Лаврентия до северной половины Флориды, на западе до Висконсина, плато Озарк, восточ- ного Канзаса, восточной Оклахомы и центрального Техаса. Только роды Pletho- don и Asieides встречаются на обеих территориях, а в районах, расположенных между основными территориями, обитают еще 2 вида рода Plethodon (один опи- сан в работе Бишопа), изолированные в отдельных местностях в Айдахо и Нью- Мексико, и 1 вид из рода Aneides в Нью-Мексико (1 оже, 1950; Stebbins, Biemer, 1950). Вероятно, это единственные представители семейства Plethodontidae в за- падно-центральной части Северной Америки.
Безлегочных саламандр нет на большей части северной Мексики и на боль- шей части прилегающих краевых районов США. Однако южнее в Мексике, на- чиная от Наярита и южного Нуэво-Леона, они появляются вновь и встречаются по всей центральной и южной Мексике и Центральной Америке (разнообразные виды, включая горные и равнинные формы (Taylor 1944; Schmidt, 1936), и в не- которых районах Южной Америки (сравнительно мало видов) в южном направле- нии до восточного склона Анд в северной Боливии и в восточном до западной Ве- несуэлы и устья Амазонки. Нахождепие в указанных райопах вида Оесйрия paraensis в настоящее время точно установлено (Myers, Carvalho, 1945; Schmidt, lnger, 1951). Этот род был также зарегистрирован на небольших высотах у llay- ты (около 150 л над уровнем моря) на Мараньяне; у Бенжамен-Констана (око- ло 100 м над уровнем моря), на Жавари в северной Боливии примерно на высоте 200 л (Данн, письменное сообщение от 6/Х 1946 г.). Однако ранее собранные све- дения о виде западного североамериканского рода Ansatiiia из района Ла-Платы в южной части Южной Америки, а также предполагаемого рода Oedipus с Гаити и настоящее время вызывают сомнение (Myers, Сагча1Ьо, 1945). Число известных видов и подвидов рода Oedipus из Мексики и Центральной и Южной Америки и настоящее время приближается к 100. [Тэйлор (Taylor, l944) разделил род Oe- di gus, который оп называет Boli tog lossa, на 7 небольших родов, но почти все они образуют одну -естественную группу. Как и в других идентичных случаях, трудно судить о правильности подобного дробления и в целях зоогеографии хотелось бы найти пути сохранения старой классификации.]
Безлегочные саламандры ведут различный образ жизни. Многие являются
наземными формами, многие — водными, особенно в быстрых реках. Несколько видов обитает в пещерах или в подземных водах и несколько северо-умеренных п тропических видов относятся к древесным. В связи с тем, что безлегочпые сала- мандры живородящи, они не имеют водной стадии личинки.
Ископаемых семейства Plethodontidae не известно. Род Hydrorna»tes в Ев- ропе, по-видимому, реликтовый, сохранившийся с тех времен, когда представи- тели этого семейства были широко распространены в Евразит . Очевидно это семейство проникло в Южную Америку недавно, уже после с~к;I,èíåíèÿ материков в плиоцеие.
1'лава 3
Proteidae, протеи. Европа и восточная часть Северной Америки. В Европе имеется толь- ко 1 вид Proteus anguinus с 2 подвидами в подземных водах — ограниченная тер- ритория близ побережья Адриатического моря (Триест, Истрия, прибрежная часть Югославии). В Северной Америке — только 1 род Жес1игия с 7 видами, встреча- ющимися на западе от Аппалачей (от ~ожной части Канады до Луизианы и северо- западной Флориды) и на востоке от Аппалачей (от Северной Каролины до Джорд- жии). На север от Северной Каролины до нижнего течения реки Коннектикут и нескольких прибрежных реках бассейна Атлантического океана встречается цид N. maculosus, возможно, не являющийся местным. Распространение различ- ных видов см. в работе Бишопа (Bishop, 1943, map. 1).
Ископаемые указывают, что семейство Proteidae существовало в Европе
начиная с эоцена, а возможно, и с мелового периода.
Sirenidae, сирены. Юго-восточная часть Северной Америки. 2 рода, 3 вида, распрост- раненных вдоль побережья от Виргинии до Флориды и по побережью Мексикан- ского залива до северо-восточной части Мексики и на север вдоль Миссисипи до северного Иллинойса и Индианы (Bishop, 1943, maps. 54 — 56). Обитатели пру- дов, канав и болот.
Ископаемые до плейстоцеиа не известны.
Отряд Anura, лягушки (включая жаб)
Подотряд Ашрhicoela
Leiopelmidae (Ascaphidae). Северо-западная часть Северной Америки и Новая Зелан- дия. В Северной Америке имеется 1 вид Лзсау~йиг truei с 3 подвидами, встречаю- щийся от южной части Британской Колумбии (севернее 50' с. ш.) к югу до граф- ства Гумбольдта в северной Калифорнии и на восток до западной Монтаны. Жи- вет в холодных горных реках. Миттлман и Майерс (iVIittleman, Myers, '1949) при- водят, подразделяя этот вид на 2 подвида, историю открытия этой лягушки, которая была описана только в l899 г., и дают список литературы по данному виду.
В Новой Зеландии имеется только I род Leiopelma с 2 или 3 видами. L. hochstette- ,ri и L. areheyi (которые, возможно, являются разными формами одного вида) локализованы в северной части Северной Ирландии (Turbott, 1942; Stephenson, Thomas, 1945). Один или оба вида встречаются на открытых, постоянно влажных от тумана хребтах, а также в ручьях, где они откладывают яйца на земле под .камнями или корягами. Вид L. hamiltoni известен только на маленьком, в неко- торые сезоны года безводном острове Стефана, пролив Кука (АгсЬеу, 1922).
На острове Южном, где имеются вполне подходящие стации, представители семейства Leiopelmidae пока еще не известны.
Ископаемые формы, предположительно относимые к подотряду Amphico- ela, известны из верхней юры.
Подотряд Opisthocoela
Discoglossidae, круглоязычные (жерлянки и т. д.). Европа и т. д., Восточная Азия
и на одном из Филиппинских островов, 4 рода. Род Bombina имеет 2 вида в Европе- на север до южной Швеции и до 57' с. ш. в СССР, на юг до Италии, на Балканах и Северном Кавказе и еще 2 вида в Восточной Азии от Маньчжурии и т. д. до Юго- Западного Китая и Тонкина. Род Diseoglossus (1 вид) — в Южной Франции и Ис- пании, на некоторых средиземноморских островах и в Северной Африке. Род Alytes (2 вида) — в Западной Европе от Швейцарии и Западной Германии до Испании, и род Barbourula (1 вид) на острове Бусуанга в группе островов Кала- миан (на север от острова Палаван на Филиппинах) (Myers, 1943). Это самый боль- шой полностью водный (в горных ручьях) вид семейства. Другие круглоязычные имеют разнообразные формы, от наземных до водных.
Ископаемые круглоязычных относятся к миоцену Европы, но рассматри- ваемое семейство, вероятно, ведет свое начало от более древних предков.
Pipidae, пипы(шпорцевыелягушкии т. д.). Африка и Южная Америка. В Африке име- ется 3 рода: Хепориз с 5 широко распространенными к югу от Сахары видами и 2 других более локализованных африканских рода (составляющие вместе 4 или 5 видов). В Америке имеется только 1 род Pipa с 5 довольно разнообразными по своему строению видами, однако Дани (Dunn, 1948) считает, что их лучше отно- сить к одному роду. Все они распространены в северной половине Южной Америки и востоку от Анд (Dunn, 1948). Центральной Америки это семе11ство не достигает и доходит только до острова Тринидад, но не до собственно Вест-Индии. Это самое водное семейство лягушек.
Ископаемые, приписываемые этому семейству, либо не обладают большой
достоверностью, либо относятся к сравнительно недавнему «ремеии, и их трудно использовать для решения зоогеографических вопросов.
lIhinophrynidae. Тропические низменности Мексики (от Тамаулипаса со стороны
Атлантического океана и до перешейка Теуаитепек со стороны Тихого океана) иа
Зем новодние
юг до Гватемалы и Юкатана включительно. Единственный вид Bkinophrynus do rsali s. Относительно примитивной природы этой лягушки см. работу Смита и Тэйлора (Smith, Taylor, 1948). Взрослые особи име«от специальное ро«ощее пр««- способление, но сохраняют многие признаки примитивизма — не имеют ребер и т. д. Считают, что головастики этого вида сходны с головастиками пины, но Данн (Dunn, 1948, р. 7) этого взгляда не придерживается.
Ископаемые семейства Rhinophrynidae не известны.
Подотряд Апошосое1а
Pelobatidae, чесночницы. Тропическая Восточная Азия и т. д., умеренные широты
Европы и т. д., Северная Америка (нет на большей части умеренной зоны Азии) и Сейшельские острова. 3 подсемейства: (1) подсемейство Megophryinae — восточ- ная часть Индо-Малайской области (нет на Цейлоне и на полуострове Индостан) на север до Центрального Китая (Ганьсу и юго-западный Шаньси), на запад до Юго-Западного Китая (Liu, 1950) и восточных Гималаев; на юг и восток до остро- вов Ява, Бали, Калимантан и частично на Филиппинах, 7 родов, около 50 или более видов; род Медорйгуг, около 30 видов, распространен по всему или почти по всему ареалу подсемейства; другие роды локализованы в Юго-Западном Китае, на острове Калимантан и т. д.; лягушки этого подсемейства обычно являются водными, их личинки часто встречаются в горных речках. (2) Подсемейство Ре1о- batinae, жабы-чесночницы и т. д.; имеют прерывистое распространение в умере~~- ной зоне северного полушария; 2 рода в Европе и т. д. и 1 род в Северной Амери- ке; в Европе распространен род Pelobates с 3 видами — от Испании до Урал», а также Кавказ, Северный Иран, Малая Азия, Сирия, Палестина (Израиль) «« умеренная зона северной части Африки и род Pelod«ttes с 2 географически отдален- ными видами — один в 3аггадной Европе, а другой — на Кавказе; в Америке нахо- дится только род hcaphiopus с 6 — 8 видами, от южной части Британской Колумбии до Массачусетса и далее на юг до Флориды и Мексики, достигая южного края Мексиканского плато в Оахака в пределах тропиков, но не на низменностях; эти лягушки обычно живут в довольно засушливых или песчаных местах, но имеют водную личинку. '(3) Подсемейство Sooglossinae приурочено к Сейшельским островам; 2 рода, 3 вида; некоторые из них (или все) откладывают яйца на земле и затем переносят своих головастиков в воду. Однако утверждать, что эти лягушки несомненно относятся к семейству Pelobatidae, нельзя.
Из 3 подсемейств ископаемые данные имеются только о подсемействе Pe- lobatinae из миоцена Европы и раннего олигоцена Северной Америки (Zweifel, 1956) и олигоцена Монголии (границы распространения древних форм выходят далеко за пределы современного ареала этого подсемейства) (Noble, 1924а). Н«« связи и распространение современных форм позволяют предположить, что в дей- ствительности род чесночниц гораздо более древнии, чем можно судить об этол по найденным ископаемым, и что он имеет очень сложную историю (Zweifel, 1956, р. 13 — 16) .
П одотряд P roc oe 1 à
* Palaeobatrachidae. Только европейские ископаемые. Таксономические границы и пре- делы геологических времен (верхняя юра до миоцена или только миоцен) этого семейства не установлены. Возможно, это семейство имеет сложную истори~(>, и его связи с современными лягушками не известны.
Leptodactylidae (рис. 24), свистуны. Южная и Центральная Америка и т. д., A««<Ура- :«ийская область и, вероятно, Южная Африка. Ископаемые, возможно, пз Индии.
««о современных представителей этого семейства там нет (в настоящее время уста- иовлено, что головастик из Индии, похожий на Heleoph,ryne, не родствеnen свистунам — Ramaswami, 1944). В Америке даже по старой систематике насчиты- вается около 25 — 30 родов или больше (ес««и придерживаться более дробной клас с««ф««кации) и несколько сот видов. Они многочисленны по всей Южной и Цент ральной Америке. К югу несколько родов достигают леспых районов южног(> Чили или являются там эндемиками, а границы распространения одного или двух родов подходят близко или, возможно, достигают Магелланова пролива или даж~ Огненной Земли. Северные границы семейства определяются 3 родами, которые достигают Техаса и в одном случае Аризоны. Род 1.''leutherodact«tlus (личи««очная стадия отсутствует) более чем с 200 видами (возможно, некоторые следует рас- «матривать как подвиды), широко распространен в пределах рассматриваемых rpa««H«J ««доминирует в Центральной Америке и особенно в Вест-И ««;«««««, где образуетосповнуючасть фауны лягушекпоследпей. Один вид из Вест-Индии был цнтродуцировап во Флориде. Род I.eptodagtiglus также достигает ряда островов Вест-Индии. В Австралийской области Паркер (Parker, 1940) выделяет '14 родов, 58 видов, которые сконцентрированы в самой Австралии, как в тропической, так и в умеренной зоне. 3 рода достигают Тасмании и 3 рода — Иовой Гвинеи и пред--
Глави 8
ставлены там в общей сложности 5 видами, из которых только 2 вида являит(.я эндемичными. В Южной Африке находится только род Не1еор1лу»е с 4 видами, ичеющими прерывистое распространение (на горных хребтах) от Столовой го1эы (вблизи мыса Доброй ЕЕадежды) к северу до Наталя и Трансвааля и, таким обра- зом, охватывающими полностью всю умеренную зону южного полушария. Со «р»- мени выхода работы Нобля (Noble, 1931) большинство авторов рассматривает этот род как представителя семейства Leptodactylidae, но ряд авторов, включая Туа (с] и Toit, 1934), стоят на другой точке зрения. Однако все они придерживаются едино~о мнения, что южноамериканские и австралийские свистуны являются родствеи- ными группами (Parker, 1940, р. 2). По образу жизни американские свистуны p i;i- нообразны и включают многие водные, наземные и древесные формы. Австралий- ские же в основном являются наземными или земноводными. Род Не1еор!гпупе связан с горными ручьями, в которых живет его высокоспециализирован~~ая личинка.
Ископаемая (из эоцена) лягушка (*Indobatrachus), относимая к семейству
Leptodactylidae, найдена в тропической части Индии, близ Бомбея. Нобль (Nob- le, 1930) на основании целой серии, состоящей примерно из 0 остатков особей этого рода (полностью сохранившегося скелета до сих пор не найдено), считает его близкородственным роду Crinia из Австралии. Паркер (Parker, '1940, р. '10, 11), очевидно, принял его за представителя семейства Leptodactylidae, не будучи уверенным в подлинности его связей с современными формами. Вероятно, это единственная ископаемая лягушка, которая имеет большое значение для выяс- нения многих зоогеографических вопросов. Настоящие жабы, очевидно, произо- шедшие от свистунов на основной части тропиков Старого Света, указывают, таким образом, на существование там в прошлом семейства свистунов.
>ufonidae, настоящие жабы и т. д. (рис. 22). Почти космополиты иа всех главных мас- сивах суши, но отсутствуют на Мадагаскаре и в Австралийской области. Род В ufo встречается по всему ареалу семейства. Различные его виды обычны и широко распространены в каждом районе, где встречается этот род: через всю Африку до ее южной оконечности, включая изолированные оазисы в Сахаре; на север до 65' с. ш. в Скандинавии; через Евразию до Японии и на юг и восток через Индо- Малайскую ооласть до островов Ява, Бали, Ломбок, Сулавеси, Палаван и Мин- данао. В Америке примерно от 60' с. ш. через Северную, Центральную и Южнун> Америку приблизительно до Магелланова пролива и, возможно, до Огненной Зем- -ли, а также на Кубе, Гаити и в Пуэрто-Рико, Вест-Индия. Четыре других рода этого семейства приурочены к двум отдаленным ча«тям тропиков Старого Света'.
в тропической Африке (%ет»епа и Nectophry»oides) и в районе полуострова Ма- лакка (Pseudobufo и Pedostibes). Эти роды, а также род Bufo образуют одну есте- ственную группу, что подтверждается наличием у всех родов (возможно, за иск- лючением рода И'ег»епа, который не был исследован) органа Биддера (Davis.
1936). Настоящие >кабы (Bufo) имеют водную личинку и, как правило, наземных взрослых особеи, но другие роды могут включать как водные, так и древесные формы.
Ископаемые показывают, что род Bufo находился в Европе начиная с оли- гоцена и в Северной Америке с плиоцена,'но это уже второстепенные подробности истории этого рода, которыи, очевидно, радиировал из тропиков Старого Света. <telopodidae (Brachycephalidae). Южная Америка и т. д. и несколько родов в Западной
Африке и районе Малайи. В Южной Америке находится примерно 10 родов, из которых 4 рода проникли в Центральную Америку. На основной территории кон- тинента ни один из них не встречается за пределами Никарагуа (по сведениям Данна в 1931 г.), но 1 эндемичный род этого семейства имеется на Кубе. В Южном Чили встречается 1 род (Rhi»aderma). В Западной Африке — роды Мес1оЯп~»е и Didynamipus, которые, как считает Паркер (Parker, 1934, р. 5), сходны с аме- риканскими представителями этого семейства и, возможно, им родственны, а от- сутствие органа Биддера у рода Nectophryne (Davis, 1936) еще раз подтверждает это родство. В районе Малайи в настоящее время признают представителем семей- ства Atelopodidae, только вид Cacophryne borbonica (Davis, 1935, 1936), но Гриф- фитс (Griffits, 1954) считает, что он возник самостоятельно (от малайских жаб) и не имеет прямой родственной связи с другими представителями семейства Ate- lopodidae. Это семейство требует всестороннего изучения. До тех пор пока свя- зи между родами, относящимися к этому семейству, не будут хорошо изучены, оно не может иметь большого значения для зоогеографии. Большинство амери- канских представителей этого семейства — мелкие и ведут наземный образ жизни. Hvlidae, квакши, древесные лягушки и т. д. (рис. 25). Род phyla распространен почти
по всему земному шару, за исключением обширного разрыва на большей части Африки и в тропической Азии. Виды этого рода очень многочисленны в Южной
Зем и оводн ые
и Центральной Америке; несколько видов имеется на Ямайке, 1'анти и Куое. :)тот род простирается на север без разрывов, но в убываютцем числе в умеренных п~иротах Северной Америки (примерно 12 видов), достигая озера Большое 11е- нольничье и центрального Лабрадора (род квакш Pseudacris заходит еще дальше на север. почти до 11олярпого круга в западной части Северной Америки). Несколь- ко видов рода Пу1а (только один варьирующий вид?), исключая всех других пред- ставителей этого семейства, встречаются в умеренных широтах Евразии — в Ев- ропе на севере примерно до 57' и выше 52' к востоку от озера Байкал (Берг, 1947) лостигают умеренной северо-западной части Африки, Японии, островов Рюкн> и Тайвань, встречаются на юге на основной территории Азиатского континента ло границ, приведенных ниже. Квакши не встречаются или почти не встречаются на больп|ей части Африки и в тропиках Индо-Малайской области, но род Hyila снова появляется в тропической и умеренной зонах Австралийской области. Около 35 видов рода !Ху!а находится на Новой Гвинее (Loveridge, 1948), несколь- ко на Соломоновых и Молуккских островах и в восточной части Малых Зондских островов (2 на острове Тимор; ! достигает острова Омбай (Алар), Саву п Сумба— van Kampen, 1!)23) и около 24 видов (включая 2 общих с Новой Гвинеей) в Ав- с тралиии Тасмании (T,overidge, 1935). Кроме рода Hyla, большинство квакш рас- пространено в тропиках Америки. Нобль (Noble, ) 931) и Данн (Dunn, 1931) вы- деляют более десятка эндемичных родов в тропиках Америки, многие из которых приурочены к небольшим территориям. Если придерживаться более дробного де,~ения, то можно выделить еще большее число родов. Например, только в Мек- сике Смит и Тэйлор выделяют около десятка родов в дополнение к роду Hyle и еще 37 мексиканских видов (некоторые, возможно, являются подвидами) они относят к самому роду Hyla. За пределами тропиков Америки, кроме рода Hyla, имеются только род icris и Pseudacris в Северной Америке и iVyctimystess Новой Гвинее. Несмотря на такое множество и разнообразие квакш в тропиках Южной Америки, ни одна из них не достигает влажных лесов умеренной зоны в районе и>жного Чили. Это давно известный факт, еще раз подтвержденный Данном (пись- менное сообщение'от l6/Õ 1946 г.).
Несомненно, род Hyla не встречается или почти не встречается на основной части тропиков Старого Света (этот факт нуждается в дополнительной проверке). Одна из групп этого рода, распространенного в умеренной части Евразии, все-таки заходит в краевые части тропиков в Восточной Азии, достигая Тонкина, Хайнаня и Лннама (Bourret, 1942). Род Hyta также достигает Сирии, Палестины и даже н>жной Аравии (Schmidt, 1953а), но не Египта (Flower, 1933, р. 844). В Африке 1 вид рода Hyla был ошибочно описан как новый вид из Абиссинии — это ока- '~а,~ся неправильно этикетированный экземпляр обычного южноамериканского энда (Noble, 1926). Другой представитель рода Hyla был описан Алом как новьш внд из Того — тропическая часть Западной Африки близ Гвинейского залива. Отчасти это тоже оказалось случайной ошибкой, так как Нобль (Noble, 1926) установил, что это ие новый вид Hyla arborea meridinalis, а известный в Юго-За- падной Европе, Северной Африке и т. д. (Предполагаемый экземпляр из Того, вероятно, не был этикетирован там, а был просто включен в многочисленные эк- земпляры, отправляемые оттуда немецким офицером во время первой мировой войны, который делал эти сборы по пути из Германии. Поэтому и место сбора вызывает сомнение.) В Индо-Малайской области 1 вид рода Hyla был описан как экземпляр с острова Ява (Ahl, 1926) . Так как эти сведения впоследствии не прове- рялись, принимать их без подтверждения довольно рискованно. Последние дан- ные о роде Hyla с острова Негрос, Филиппины Ингер (Inger, 1954, р. 247) счи- тает также сомнительными. В таких случаях, так же как и с экземпляром из Того, вся тяжесть доказательства достоверности этих сведений ложится на тех, кт() считает, что род Hyla в действительности встречается в этих неожиданных для него местах. Это говорится, несмотря на то что предположение о нахождении ви- дов рода Hyla на острове Ява или на Филиппинах вполне бы соответствовало предположению о первоначальной радиации этого рода из тропиков Старого Света.
Большинство квакш — древесные лягушки с рядом водных, наземных II.~Ii в большей или меньшей степени роющих форм. Даже в самом роде Hyla, где боль- шинство видов древесные, есть и наземные виды (из тех, которые приспособлены к жизни на деревьях), виды, приспособившиеся к жизни среди скал, и виды, жи- вущие в пустынях. Так, Hyla rubella имеет очень широкий ареал распространения в Австралии'. от влажных частей северпого Квинсленда до засушливой западной части континента (Loveridge, 1935, р. 42). В пустынях Центральной Австралии эта лягушка выживает не благодаря структурным адаптациям или спасению от засухи в норах, а благодаря быстрому размножению и расселению во временных
Глава 3
И2
Меткалф (3Ietcalf, 1923) пытался проследить историю эволюции квакш при помощи их паразитов — опалин. Он считал, что квакши произошли в тропиках Америки, что род FIyla расселился оттуда в Северную Америку и сменил там своих паразитов или приобрел новых и перенес их в Евразию и что род Hyla пересек в южной части Тихого океана (не антар- ктический) материковый мост из ?Ожной Америки в Австралию и снова сменил паразитов или по крайней мере приобрел в Австралии новых простейших опалин от ныне вымершей предполагаемой австралийской лягушки из семейства Leiopelmidae. По этому поводу следует сделать два замечания. Во-первых. считают, что «простейшие Рго1оорайпа из подрода II». кото- рых, как предполагают, род Hyla приобрел в Австралии, встречаются также и у представите- лей семейств Discoglossidae и Pelobatidae и у некоторых других лягушек. Следовательно, род Hyta мог приобрести их почти в любом месте иа своем пути в Австралию: возможно, в Евра- зии или в Индо-малайской области, где сейчас встречаются Discoglossidae и большинство лягу- шек семейства Pelobatidae. Во-вторых, если лягушки обмениваются паразитами или приоб- ретают новых с такой легкостью, последние не представляют большой ценности при изучении истории распространения лягушек (Noble, 1925; Dunn, 1925). Выше я уже упоминал о взгля- дах Меткалфа и снова упоминаю о них, потому что совсем недавно этот вопрос был снова поп~тят Бьюфортом (Beaufort, 1951, р. 152. 166) в связи с родо~~ Ну!а.
водоемах в период дождей (Buxton, 1923, р. 96). Вид Hyla а renicolor ведет такой же образ жизни в пустынях юго-западной части Северной Америки (Bogert, Oliver, 1945). Эти примеры показывают, что «древесные лягушки» могут пересекать пусты- ни, и это в какой-то мере способствует возможности их расселения на всем зем- ном шаре.
Ископаемые данные по этому семейству не многочисленны и не достоверны
(стр. 131). Они не могут оказать существенную помо1ць при исследовании истории этого семейства. Вогатство и разнообразие семейства Hylidae и рода Hyla в тро- пиках Америки, возможно, являются результатом первичной или вторичной ра- диации. Если это результат последней, то, видимо, первичная радиация, имела место в тропиках Старого Света, как это и предполагалось (стр. 127, 131), а если это так, то не исключена возможность того, что квакши произошли от древних Bufonidae или от тех же, в настоящее время исчезнувших, свистунов из тропиков Старого Света, от которых произошли настоящие жабы (i~1оЫе, 1922, р. 71). 11о если такое объяснение происхождения семейства Bufonidae вполне логично, то для семейства Hylidae оно уже не подходит1.
П одотряд D iplasiocoela
Ranidae, настоящие лягушки и т. д. (рис. 21). Распространены почти ио всему земному
шару в пределах границ распространения лягушек; но отсутствуют на большей части Австралии и в южной, части Южной Америки. Вольшой род Лапа при- мерно с 250 видами встречается на большей части ареала этого семейства: по всей Африке, включая пригодные для обитания части Сахары, на Мадагаскаре, ~а всей Евразии от Арктики и через всю тропическую Азию, на острове Рюкю в Япо- нии, на Тайване, по всему Индо-Австралийскому архипелагу до Филиппин и Новой Гвинеи. В Америке — от арктических районов до тропической части Юж- ной Америки. По направлению к Австралии род Лапа численно убывает, но po;l,— ственные формы по-прежнему многочисленны и достигают тропических районов северной части Австралии, а также Соломоновых островов. Расселение лягушек из семейства Ranidae на островах Палау и Фиджи рассматривается в гл. 8. В Аме- рике род Лапа хорошо представлен в Северной и в некоторых районах Централь- ной Америки, но только 1 вид достигает Южной Америки (Данн, письменное сообщение). Это вид Л. palmipes, который распространен от южной Мексики через Центральную Америку и северную часть Южной Америки по крайней мере до района Амазонки. Семейство Ranidae, исключая род Лапа, приурочено к Старому Свету, а там почти приурочено к тропикам. Нобль (Noble, 1931) выделяет 6 под- семейств более чем с 30 родами. Большинство из них приурочено к Африке, но подсемейство Cornuferinae примерно с 10 родами хорошо представлено в Индо- Малайской и Австралийской областях. Классификация некоторых подсе- мейств, включая Cornuferinae, до сих пор не установлена, и их распростране- ние не заслуживает подробного обсуждения. По образу жизни большинство представителей семейства Ranidae, включая большую часть рода Лапа, явля- ются земноводными, но среди них есть водные, наземные, роющие или древесные формы.
! Ископаемые свидетельствуют о существовании рода Лапа в Евразии начи- ная с миоцена и в Северной Америке начиная с плиоцена (но это незначительные детали). Распространение современных форм ясно показывает, что возникновение настоящих лягушек приурочено к основной части тропиков Старого Света и что род Лапа распространился оттуда через северные районы земного шара, а оттуда совсем недавно в Южную Америку.
Rhacophoridae (Polypedatidae). Тропические древесные лягушки Старого Света (рис.
25) . Основная часть тропиков Старого Света и т. д. 13 родов (Noble, 1931) . 6 родов— африканских, 2 из' них достигают Мадагаскара и 1 (Megalixalus) — Сейшель-
Земноводные
ских островов; 4 рода приурочены к Мадагаскару; 2 рода — к Индо-Малайской области и последний род tacophorus (Polypedates) отсутствует в Африке, но пред- ставлен многочисленными видами на Мадагаскаре. На Сейшельских островах нет ни одного вида, но в Индо-Малайской области они довольно многочисленны.
Этот род простирается на север примерно на южную половину Китая и достигает Японии (где он встречается на острове Рюкю) и Тайваня, а также распространен на востоке и юге Индо-Австралийского архипелага до Филиппин и через них до островов Сулавеси (4 вида) и острова Тимор (2 вида), но не далее. В своей работе Бьюфорт (Beaufort, 1951, р. 70) указывает, на существование Rhacophori- dae на Новой Гвинее, что, вероятно, является ошибкой, так как ван Кампен (van Kampen, 1923) и Лавридж (Loveridge, 1948) не отмечают там ни одного вида. По образу жизни большинство представителей этого семейства являются древесными, хотя некоторые, как говорит Нобль, «возвратились на землю».
Ископаемые семейства Rhacophoridae не известны, но его разнообразие
в Африке и на Мадагаскаре и тот факт, что лягушки этого семейства не смогли достичь Австралийской области, дают возможность предположить африканское происхождение. Эти древесные лягушки и особенно сам род Rhacophorus, возмож- но, заместили род Hyla на основной части тропиков Старого Света, а род Rha- cophorus, видимо, в свою очередь был замешен в Африке родственным (дериват- пым?) родом Chiromantis и другими (Noble, 1931, р. 525).
Brevicipitidae (Microhylidae), узкоротые лягушки и т. д. Тропики (за исключением
Западной Африки) и некоторые прилежащие умеренно-теплые районы. 7 подсе- мейств (Parker, 1934). (1) Dyscophinae: 3 рода — один с 6 видами приурочен к Мадагаскару и 2 рода с 6 видами в восточной части Индо-Малайской области имеют разорванный ареал от Юго-Западного Китая (Liu, 1950) до островов Су- матра и Калимантан. (2) Cophylinae: Мадагаскар; 8 родов, 21 вид. Чрезвычайно разнообразные по форме и строению, что явилось результатом изолированного центра эволюции (Parker, 1934). (3) Asterophryine: 4 рода, 24 или более видов, большинство в Новой Гвинее, а также несколько на островах Хальмахера, Лнь- бошачжоу (Амбоина) и т. д. Майерс ( письменное сообщение от 14/VII 1948 г.) сообщает, что он обнаружил данные по Metoposti ra с островов Сулу, но они были ошибочны, так как в действительности эта лягушка была с островов Санана, Сула (Соела) и Молуккских островов. (4) Sphenophryninae: 5 родов, 36 или более видов, большинство в Новой Гвинее и т. д., 4 вида (2 эндемичных) находятся в близ- лежащих частях Австралии. Oreoph,ryne встречается не только на Новой Гвинее, а также на Молуккских островах, Сулавеси, Минданао и острове Билиран, Южных Филиппинах и вдоль Маль~х Зондских островов от Флореса до Бали. (5) Microhyli- nae. Индо-Малайская область и Америка. Microhyla (в Америке его часто называют Gastrophryne) встречается как в Америке, так и в Индо-Малайской области, с 15 видами в последней, от Цейлона и Индии до островов Ява, Бали, Калимантан, Тайвань и Рюкю (но не в Японии) и на Азиатском континенте на север по край- ней мере до Центрального Китая) и с 12 или более видами в Америке (от Брази- лии и Эквадора на север до Канзаса, юго-восточной Айовы (Klimstra, 1950), Индианы и Мэриленда). В Индо-Малайской области имеется еше 9 родов по край- ней мере с 30 видами, большинство из которых находится в пределах ареала рода Microhyla. Род Ealoula проникает еще дальше на север до центральной Маньч- журии и достигает островов Сулавеси и Малых Зондских до острова Флорес', несколько родов достигают Филиппинских островов. Еще 6 родов по меньшей ме- ре с 30 видами (включая недавно описанных и внесенных в списки Данном— Dunn, 1949) находятся в тропиках Америки, один из них (Hgpopachus) прости- рается на север до южного Техаса и еще один — на юг до Буэнос-Айреса, но ни один не достигает лесов умеренной зоны в южной части Южной Америки. (6) Brevicipitinae: Южная и Восточная Африка. Род В геисеря с 13 видами в основ- ном южноафриканский, но встречается и на севере до Танганьики; 3 осталь- ных рода с 4 видами локализованы в Таиганьике. (7) Melanobatrachinae.
Восточная Африка и IOamaa Индия. В Африке встречается 2 рода, 3 вида, все локализованы в горах Усамбара и Улугуру на Тапгаиьике. В Индии находится только 1 род Hoploph,r>g)ze с 1 видом, локализованным в горах Траваикур» и Кочина.
Большинство узкоротых — это мелкие лягушки, ведущие скрытный образ
жизни. Однако многие из них являются роющими животными и питаются мура»— ями или термитами. Некоторые наиболее активные формы живут на деревьях пли в траве.
Ископаемые этого семейства не известны. Паркер (Parker, 1934, р. 3) считает
это семейство монофплетической группой, которая географически видоизменя- лась. Ои полагает, что, возможно, оно является древпей боковой ветвью одного
Глава, 3
и того же родя, от которого произошли семейства Нап1дае и Rhacophoridae, и что, очевидно, местом его происхождения была Юго-Восточная Азия. Однако как оы там ни было, ясно, что узкоротые лягушки произошли в основном в тропиках Старого Света и что подсемейство Microhylinae достигло тропиков Америки через северные районы. Похоже, что они проделали этот путь Hp один раз. Род ЛХ~сго/ry- la, возможно, достиг Америки относительно недавно, так как виды пз Индо-М;1- ,~айской области и Америки до сих обладают больц~им сходством, а сам род пред- ставлен в Северной Америке лучше, чем в Южной (Parker, 1934, р. 14). Другие американские роды, возможно, образовались от одного или более рашп|х имми- грантов.
P1irynomeridae. Африка к югу от Сахары. 1 род, 5 или более видов. Древесные
лягушки, очевидно, образовавшиеся от узкоротых, так же как кнакши ооразо- вались от свистунов или примитивных жаб и как лягушки семейства ВЬасорЬо- ridae образовались от настоящих лягушек семейства Ranidae (Parker, ! 931, р.З).
ЛИ ТЕРЛ ТУР;1
А h 1 Е., 19'6, Мене Eidechsen иий Amphibien, «Zoologischer Anzeiger» (Leipzig), 67,
186 — [92.
А и g е 1 F., 19~7, Ъ'|е et moeurs des amphibiens, Paris, Payot (Bihliotheque Scienti- 11Ч11е) .
А n g е 1 F., Ь h о t e H., 1938, Reptiles et amphibiens du Sahara central et du Sou- dan, «Bull. СопиЫ d Etudes IIistoriques et Scientifiques de 1 Afrique Occidentale
Franqaise», 21, 345 — 384.
А r c h е у G., 1922, The habitat and life history of Liopetma hoehstetleri', «Rec. Canter- bury Mus.», 2, 59 — 71.
В а k e r С. Ь., 1947, The species of Amphiumae, «J. Tennessee Асад. Sci.», 22, 9 — 21. В а r b о и r Т., 1926, Reptiles and amphibians, Boston.
В а г r e t t 9; . С., 3 r. 19~7, EIematopoiesis in the European plethodontid... «Anatomi- cal Record» (Phi1adelphia), 98, 127 — 136.
В е а ц f о r t Ь. F., de, 1951, Zoogeography of the land and inland жМегя, London- New York.
Б е р г П. С., 1947, Географические зоны Советского Совхоза, Москва.
В i s h о р S. С., 1943, Handbook of salamanders, Ithaca, New York.
В о g е г t С. М., О 1 i v e r J. А., 1945, ...herpetofauna of Sonora, «Bull. American
М us. Nat. Н1яй.», 83, 297 — 425.
В о и 1 е n g e r Е. G., 1936, Batrachia аий Reptilia. См. Regan (ed.), «ivatural llisto- ry», London, 297 — 392.
В о и 1 е n g e r G. А., 1890, Beptilia and Batrachia. См. «Fauna of British India
including Ceylon and Burma», London.
В о и 1 е n g e r G. А., 1903, Report on the batrachians and reptiles. См. Annandale
and Hobinson, Fasciculi Malayensis, Zool., 1, 131 — 176.
В о и r r e t R., 194, Les batraciens де 1'Indochine, Institut Oceanographique de 1'In- dochine, Mem. 6.
В r i d g e s Е. L., 19~8, Uttermost part of the earth [Tierra del Fuego], London. В и х t о п P. А., 1923, Animal life in deserts..., London.
С h а Ь а п а u d Р., 1954, [On the supposed salamander larva from French Guinea],
«Bull. Inst. franqaise d'Afrique noire», Ser. А, 16, 1293 — 1294.
D а г 1 i n g С о п Р. J., J r., 1948, The geographical distribution of cold-blooded ver- tebrates, «Ouarterly Review Biol.», 23, 1 — 26, 105 — 123.
D а г w i n (;., 18о9, 1950, On the origin of species..., London, (переиздапо в 1950).
[Имеется русский перевод: Ч. Д а р в и н, Лроисхождение видов путем естест- венного отбора, Соч., т. 3, М.— Л., 1939.]
1) а v i s D. D., 193,э, А new generic and family position for Bufo borbonica, «Zool.
Ser. Field Mus. Nat. Hist.», 20, 87 — 92.
Лшпература
l~ й i i $ D. D., 1,)36. Т))е distribution of Bidder s огдж iri the Bufonidae, «Уоо1. ~ег.
Г~еИ Мия. Nat. 1l ist.», 20, 1) 5 — 125.
l)»»» Е. В., 1323, Т1и. яа1ашаш]егя of the fami]y Ilyriobiidae, «I гос. Anierican Л«ас1.
Л rts 4>с i Ф, 58, 443 — 523.
1~ и» и Е. К., 19 За, ТЬе деоргарЫса1 distribution о1 amphibialls. <American Ъ Лиг»- list», 57, l 29 — 1 36.
0 и 11 n E. R., 1925, The' host-parasite method and the distribution of 1'го~я, «Anleг~сап
~ аСигя] ist», 59, 370 — 375.
1~ а п п Е. 8., 1931, The herpetological 1аш~а of the Amerie;is, «;opeia», 193I, l(~6- -! J9. >»<»» Е. Е1., 1942, ТЬе American caecilians, «Bull. Mus. (.:omparative Хоо1.», 91,
439 — 540.
l»[ >i n Е. Н., 1948, Люепсап frogs of the family Pipidae, «Americaii M»s. Novitates»,
Хо. 1384.
I> i»»> Е. R., 1949, Notes on South American frogs of the family Microhylidae, «An~e- rican Мця. Novitates», No. 1419.
Р и Т о i t С. А., 193~, А revision of the genus Eieleuphngne, «Лпп. U. Яе11епЬоя«Ь»,
12, Sec t i on Л, iX o. 2.
Е k ш а n $., 19' 2, Р jiin;lirldeпя Utbrednings-historia pa Skandinaviska H lav<in,
Stockholm.
V 1 о w e r S. S., 1933, ...reptiles and amphibians of Egypt..., «I гос. Zool. Soc. Lon- don», 1933, 735 — 851.
(» а d о w H., )909, Amphibia and reptiles, The Cambridge Natural History, vol. 8,
London.
(т о г in а п J., (; а in р С. Ь, 19,эЗ, А new ...НцдготапКез агом (;alifornia..., «Copeia»,
1953, 39 — 43.
4 г а и d i d i e r G., P е С i С G., 1932, Zoologic de Madagascar, Paris, Soc. d Editions
(hograpliiques, Магйппея et Coloniales.
(' г i f f i С h s I., 195~, On the «otic element» in Amphibia Salientia, «Ргос. Zool. Hoc.
London», 124, 35 — 50.
(' и i Ь е J., 1,)50, Batraciens. См. Contribution а 1'etude de 1'Air, «Mem. 1'Iiist. fran- (;ais d'Afrique noire», 10, 329 — 332.
6 ii n t h e r A., 18э8, On the geographical distribution of batrachians, «Ргос. Zool.
Soc. Lond on», 26, 390 — 398.
H а r р е r F., 1956, Ampliibiaris and reptiles of the Ungava Peiiinsula, «Ргос. Biol.
Soc. Жasliington», 69, 93 — 104.
lI о f f s t e t t e r R., ) 915 [Concerning fossil r1mphignathodun], «Bull. Soc. Geologi- ц ие Ргаисе» (5), 15, (67 — 169.
1 n g e r R. F., 1950, ...amphibians of the Riu Kiu islands, «Ашег~са» Naturalist»,
84, 95 — 1! 5.
I n g e r В. F., 1954, Systematics and zoogeography of Philippine Amphibia, «Fiel- diana» (Chicago Fiat. EIist. Mus.), «Zool.», 33, 181 — 531.
К 1 i m s t r а Ж. D., 1950, Narrow-mouther toad taken in Iowa, «Kopeia», 1950, 60. I. i и <.., 1950, Amphibians of western China, «Fieldiana» (Chicago Nat. Hist. Мпя.),
(Хоо1.», 2.
L о ~ i e r E. В. S., 195~, The frogs, toads and salamanders of eastern Canada, Canada. К о g i e r Е. В. S., Т о п e r G. (;., 1955, Check-list о1 the amphibians and reptiles
of Canada and Alaska, «( ontrib. R. Ontario Mus. Zool. and Palaeontology», No. 41. Е о v e r i d ц е А., 193,э, Australian Amphibia in the Museum of Сomparative Zoology,
«Bull. Mus. 'omparative Zool.», 78, 1 — 60.
Ь о v e r i d g е А., 1945, Reptiles [and amphibians] of the Pacific world, New York. L о v e r i d g е А., 1948, Меъ Guinean reptiles and amphibians..., «Bull. Mus. Compa- rative Zool.», 101, 305 — 430.
L о ~~ е С. Н.. Jr.. 1950, ...biogeographical problems in ЛпеИе8, «Copeia», 1950, 92 — 9!>.
Глава 8
М е r t e n s R., 1930, Die Amphibien und Reptilien der Inseln Bali, Lombok, Sunibawa und Flores, «Abhandlungen Senckenbergischen Naturforschenden Gesellschaft», 42, 117 — 344.
М е r С е n s R., М й 11 е r L., 1940, Die Amphibien und Reptilien europas (Zweite Liste), «Abhandlungen Senckenbergischen Naturforschenden Gesellschaft». Abh. 451.
M e t с а 1 f М. М., 1923, The opalinid ciliate infusorians, «Bull. United State Natio- nal M us.», No. 120.
M e t с а 1 f М. М., 1940, Further studies on the opalinid ciliate infusorians ав4 tlieir hosts, «Ргос. United States National Mus.», 87, 465 — 634.
М i 11 s R. C., 1948, А check list of the reptiles and amphibians of Canada, «Herpeto- logica», 4, Supplement 2.
M i t t 1 е m а п M. В., М у е r s G. S., 1949, Geographic variation in the ribbecl frog, «Ascaphus truei. Proc. Biol. $ос. Washington», 62, 57 — 68.
М о о r e J. А., 1954, Geographic and genetic isolation in Australian Amphibia. «Ame- rican Naturalist», 88, 65 — 74.
М у е r s G. S., 1943, Rediscovery of the Philippine discoglossid frog, Ваrbou~«la tru- suang ensi s, «Copeia», 1943, 148 — 150.
М у е r s G. $., 1953, Ability of amphibians to cross sea barriers, with especial reference to Pacific, zoogeography, «Proc. Seventh Pacific Sci. Congress», 4, 19 — 27.
М у е r s G. $., С а r v а 1 h о А. Ь., de, 1945, Notes on ...Brazilian amphibians... Plata salamander, «Boletim Mus. National [Brazil], Zool.», No. 35. о b 1 е G. К., 1922, The phylogeny of the Salientia..., «Bull. American AIBA». Nat. Hist.», 46, 1 — 87.
N о b 1е G. К., 1924, ...herpetology of the Belgian Congo..., pt. 3, Amphibia [with check list of the Amphibia of Africaj, «Ви11. American Mus. Nat. Insist.», 49, 147— 347.
N о b 1е G. К., 1924а, А new spadefoot toad from the Oligocene of 4Iongolia with а summary of the evolution of the «American Mus. Novitates», iVo. 132.
N о b l e G. К., 1925, The evolution and dispersal of the frogs, «American Naturalist», 59, 265 — 271.
N о b 1 е G. К., 1926, ...remarkable cases of distribution among the Ampliibia..., «American Mus. Novitates», No. 212.
N о b 1 е G. К., 1928, Two new fossil Amphibia of zoogeographic importance from the Miocene of Europe, «American Mus. Novitates», No. 303.
N о Ь 1 е G. К., 1930, The fossil frogs of the Intertrappean beds of Bombay. india, «American M us. Novitates», No. 401.
N о Ь 1 е G. К., 1931, 1954, The biology of the Amphibia, New York (переиздано в 1954 г., см. «Dover Publications»).
О 1 i v e r 3. А., S Ь а w С. E., 1953, The amphibians апй reptiles of the Hawaiian Islands, «Zoologica» (New York), 38, 65 — 95.
P а r k e r Н. %., 1929, Two fossil frogs from the Lower Miocene of Europe, «А~ш. апд Mag. Nat. Hist.» (10), 4, 270 — 281.
P а r k e r Н. W., 1930, The status of the extinct frog, И%оба/ гас1гиз, «Апп. аЫ Мад. Nat. Hist.» (10), 6, 201 — 205.
P а r k e r H. %., 1932, Parallel modifications in the sekleton of the Amphibia +alien- tia, «Archivio Zool. Italiano», 16, 1239 — l 248.
P a r k e r Н. W., 1934, А monograph of the frogs of the family Microhylidap, London.
P а r k e r Н. W., 1936, Revised list of reptiles (excluding chelonians) and ampliibians collected in the Seychelles, «Tr. Linnaean Soc. London, Zool.» (2), 19,44'~.— 446.
P а r k e r Н. W., 1940, The Australasian frogs of the ~ам~1~ Leptoda( tylidae. «Xovi- tates Zoologicae» (London), 42, 1 — 106.
Липг е рот.ура
Р а r k e r H. 9 1941, The caecilians of the Seychelles, «Ann. and Mag'. Nat. Hist.»
(11), 7, 1 — 17.
1' е а r s е А. S., 1913, Concerning the development of frog tadpoles in sea water. «Phi- lippine J. Sci. D.» (General Biol. etc.), 6, 219 — 220.
Р о р е С. Н., В о r i n g А. М., 1940, А survey of Chinese Amphibia. «Peking Nat.
I I i st. Bull. », 15, 1 3 — 86.
Н а п1 а s w а m i Ь. $., 1944, The chondrocranium of two torrentdwelling anuran tad- poles, «J. Morphology», 74, 347 — 374.
R о ш е r А. $., 1945, Vertebrate paleontology, 2nd ed., Chicago [Имеется русский
перевод: А. P о м е р, Палеонтология позвоночных, ГОНТИ, M.— Л., 1939.] Н о ш е r А. S., 1947, Вет1еъ of the Labyrinthodontia, «Bull. Mus. Comparative Zool.»,
1 — 368.
S c h а е f f е г В., 1949, Anurans from the early Tertiary of Patagonia, «Bull. American
Мця. Nat. Hist.», 93, 41 — 68.
S c h ю i d t K. P., 1936, Guatemalan salamanders of the genus Oedipus, «Хоо1. Ser.
Field M us. Nat. Hist.», 20, 135 — 166.
S c h nx i d t K. P., 1951, Annotated bibliography of marine ecological relations of li- i.ing акр)11Ыапя, «Marine Life Occasional Papers», 1, 43 — 46.
$ с. h m i d t К. P., 1953, А checklist of North American amphibians and repti.ås, Ате- rican Soc. Ichthyologists and Herpetologists (printed by U. of Chicago Press).
Лdditions and corrections in «Copeia», 1954, 304 — 306.
$ с h m i d t К. P., 1953а, Amphibians and reptiles of Yemen, «Fieldiana» (Cliicago
%at. Hist. Mus.), «Zool.», 34, 253 — 261.
~ с h ш i d t K. Р., I n g е г R. F., 1951, Amphibians and reptiles of the Ilopkins- Branner Expedition to Brazil, «Fieldiana» ( 'hicago Nat. Hist. Mus.), «Zool.»,
31, 439 — 465.
S m i t h Н. М., Т а у l о r Е. H., 1948, An annotated checklist and key to the Amphi- bia of Mexico, «Bull. United States National Mus.», 194.
$ m i t h М. (А.), 1927, Contributions Со the herpetology of the Indo-Australian region,
«Proc. Zool. $ос. London», 1927, 199 — 225.
S m i t. h М. (А), 19.э1, The British amphibians and reptiles, London.
S t e b b i n s R. С., 1954, Amphibians and reptiles of western North America, New
York.
S t e b b i n s R. С., R i e m e r W. J.. ')950, А new species of plethodontid salaman- der from the Jemez Mountains of New Mexico, «Copeia», 1950, 73 — 80.
S t е р h e n s о n N. G., Т h о m а s Е. М., 1945, А note concerning the occurence
and life history of Leiopelma Fitzinger, «Тг. and Proc. R. Soc. New Zealand», 75,
319 — 320.
Т а у 1 о r Е. Н., 1944, The genera of plethodontid salamanders in Mexico, «U. о1 К»- sas Sci. Bull.», 30, pt. 1, 189 — 232.
Т h n r s о п Т., S v i h 1 а А., 1943, Correlations of the habitats of amphibians with
their ability to survive the loss of body water, «Ecology», 24, 374 — 381.
Т u r h о t t Е. G., 1942, The distribution of the genus Leiopelma in New Zealand...,
«Тг. and Proc. R. Soc. New Zealand», 71, 247 — 253.
van Е а m р е п P. N., 1923, The Amphibia of the Indo-Australian Archipelago, Leiden. ~' г i g b t Л. Н., ЪУ r i g h t Л. А., 1949, EIandbook of frogs and toads of the United
Садбея and Гanada, Згй ed., ILhaca, New York.
9' у п п е — Е d w а r d s V. С., 1952, Freshwater vertebrates of the агсбс аш1 subarctic.
«Вп11. Fisheries Research Board Danada», No. 94.
w e i f е 1 R. G., 1956, Two ре1оЬаСМ frogs from the tertiary of North America and
1Ье1г relationships to fossil and Recent forms, «American Mus. Novitates», No.
l 762.
Глава 4РЕПТИЛИИ
В работе над географией рептилий мне помогали лица, уже упомяну- тые в начале предшествующей главы, а также Чарльз М. Богерт, Самю- эль Б. Мак Дауэлл мл., Гарст Ундервуд и Эрнест Е. Вильямс, которые охотно отвечали на мои вопросы, сообщая оригинальные, часто еще неопубликованные сведения.
Альфред Ромер (Romer, 1945) дает очень полную общую сводку по эволюции и ископаемым рептилий, которая знакомит нас не только с самими животными, но и с географией их распространения. Основная система классификации, используемая здесь, дается по работе Ромера (Во- тег, 1956) «Остеология рептилий» («Osteology of the Reptiles»), с которой я смог ознакомиться в рукописи. Однако я буду касаться только совре- менных рептилий и их ископаемых и затем непосредственно родственных им групп и нескольких древних групп (главным образом динозавров), которые, возможно, играют существенную роль в географии животных.
Эпохой рептилий был мезозой. Современные рептилии представляют всего только несколько групп из всех тех, которые существовали в древ- нее время, и относятся к трем разным подклассам, дифференцировавшим- ся со времен триаса. Эти подклассы представлены черепахами, крокоди- лами, ящерицами и змеями с новозеландской гаттерией (Бр1гепоаоп). Чере- пахи, существующие со времен триаса, с тех пор претерпели очень мало изменений. Крокодилы также являются очень древними животными: ооо- собившись в позднем триасе, они за свою длинную историю так'ке претер- пели очень незначительные изменения. Внутри третьего подкласса в три- асе выделился отряд клювоголовых (Rhynchocep/<alia), который, очевидно в раннем и среднем мезозое, был космополитом. Ископаемые этого отряда позднее нижнего мела неизвестны, но тем не менее один представитель этого отряда (Sphenodon) сохранился до наших дней на удаленном от материков острове Новая Зеландия. Ящерицы, которые, «озможно, произошли о1 той же самой ветви, что и клювоголовые, появились в юре, а змеи, которые произошли от ящериц, существуют с мелового периода. Имеются досто«ер- ные ископаемые предки некоторых современных семейств черепах, крокодилов и им родственных, а также клювогочовых. Хорошо сохра- нившихся ископаемых ящериц и змей вполне достаточно для того, чтобы в общих чертах показать их происхождение и )BOJIloUHIo, но недостаточно для того, чтобы выяснить их географическую историю.
У современных рептилий, так же как и у амфибий, наблюдается определенная корреляция между численностью, та11сономи веским и эколо- гическим разнообразием, размерами и разнотипногтью занимаемых тер- риторий.
Древние клювоголовые с одним видом, сохранившимся до настояи~его времени, были строго ограничены во всех отношениях.
Современные крокодилы образуют одно семейство с 9 родами и 25 видами как земноводными, так и водными (но яйца откладывают на супш ).
Penmuлии
Приурочены к тропикам и к некоторым более теплым краевым частям уме- ренных зон.
Современные неморские черепахи образуют 7 семейств, 50 или более родов и около 200 или более видов. В большинстве случаев это земновод- ные или водные животные (но яйца откладывают на суше). Несколько групп семейства Testudinidae стали сухопутными. Черепахи широко распростра- нены по всем тропикам и так же широко, но не регулярно, в умеренных зонах.
Ящерицы и змеи — самые многочисленные и самые широко распрост- раненные современные рептилии. Тщательный подсчет их численности приводится в моей статье (Darlington, 1948, р. 23, 24); около 21 семейства (теперь сокращено до 18), более 300 родов и почти 3000 видов ящериц и около 11 семейств (теперь сокращено до 8), 300 родов и 2600 видов змей. Как ящерицы, так и змеи являются почти космополитами, встречающимися в самых различных климатических зонах и местах обитания. Они состав- ляют доминирующую группу современных рептилий, из которых одно семейство змей Colubridae является численно преобладающим, о чем будет сказано ниже.
Несмотря на то что ящерицы и змеи имеют очень широкое распростра- нение, их численность в разных местах различна. Ящерицы превосходят змей по числу видов в Европе, возможно, в Африке, на Новой Гвинее и в Австралии. Змеи превышают ящериц по числу видов в Южной и Восточ- ной Азии. В Северной Америке в целом обе группы примерно равны по числу видов, но в западной части Северной Америки преобладают ящерицы, а в восточной — змеи. В Южной Америке видовой состав змей более мно- гочислен на основной, тропической части материка, а ящериц — в умерен- ной зоне южных широт. На островах, за исключением некоторых (недавно возникших) островов материкового происхождения, ящерицы, как прави- ло, более многочисленны, чем змеи. Однако независимо от того, каково их соотношение по числу видов, на большей части земного шара числен- ность ящериц значительно выше, чем змей.
Экологически рептилии — настоящие наземные животные, по суще- ству, не зависящие от воды, как от окружающей их среды. Некоторые из них являются .вторично земноводными или водными, но почти все они воз- вращаются на сушу для откладывания яиц или выращивания потомства. Из современных рептилий только некоторые морские змеи размножаются в воде. Рептилии более многочисленны, чем амфибии, их наземные места обитания более разнообразны, и они могут быстрее расселяться на суше. Б связи с тем, что их кожа относительно непроницаема (Bogert, Cowles, 1947), они также более приспособлены к расселению через морские соле- ные воды, а некоторые из них могут непродолжительное время находиться в морской воде или даже жить в море. Пресной воды для питья рептилии употребляют очень мало, а некоторые из них долгое время могут обхо- диться и совсем без воды. Их покрытые скорлупой яйца намного прочнее, чем у амфибий, и могут распространяться различными способами. Все это находит свое отражение в относительно широком и сложном распростра- нении рептилий и раскрывает историю их появления на островах. Только в условиях холодного климата амфибии имеют преимущество над реп- тилиями. Рептилии — это единственные холоднокровные настоящие наземные животные. Они более требовательны по сравнению с амфиби- ями к теплу и чрезвычайно чувствительны по сравнению с птицами и млекопитающими к холоду, что также сказывается на их распростра- нении.
Глава 4
16|)
Границы распространения рептилий
Границы распространения рептилий показаны на рис. 28. Самым северным представителем является ящерица Lacerta ййрага, которая захо- дит за Северный полярный круг по крайней мере до 70' с. ш.; в Северной
Хотя рептилии являются главным образом наземными животными. современные крокодилы, а также большинство черепах — земноводные. Новозеландская гаттерия живет на суше, но обычно держится поблизости от воды. Ящерицы и змеи также в основном наземные животные, но и они разнообразны по своим адаптациям. И те и другие имеют разные формы: наземные, роющие и древесные. У ящериц наблюдается явное тяготение к относительно сухим и открытым местам, у змей — к более влажным и хорошо защищенным. Но это, скорее, только тенденция, а не правило. и здесь имеется много исключений. И ящерицы и змеи встречаются в пу- стынях, но и те и другие многочисленны и в тропических дождевых лесах. В настоящее время только некоторые ящерицы являются земноводными. и нет ни одной полностью водной формы (некоторые были водными в ме- ловом периоде), в то время как несколько групп змей ведет водный образ жизни; но ящерицы больше приспособлены к особым нишам. Ящерицы и змеи отличаются особенностями своего питания, что в свою очередь сказывается на их распространении. Большинство ящериц целиком или частично насекомоядные, а насекомые встречаются почти везде. Некоторые змеи также питаются насекомыми, но многие нуждаются в более крупной добыче, которая имеется не везде. Вероятно, относительная малочислен- ность отдельных видов змей, как правило, связана с их кормовыми потреб- ностями, но разреженность популяций может и не зависеть от пищи, а являться результатом расселения через преграды или способности попу- ляций к сохранению на небольших изолированных территориях или в ло- кализованных нишах.
Ящерицы непропорционально многочисленны на островах, и это слу- жит доказательством тому, что они обладают повышенными способностя- ми пересекать соленые воды (см. гл. 8), но это связано также и с другими факторами. Согласно вышеприведенным причинам, ящерицы, в отличие от змей, обладают лучшей способностью находить корм и сохранять свои популяции на небольших, экологически изолированных территориях. Отсутствие или малочисленность змей на отдаленных от материка островах уже само по себе благоприятно отражается на сохранении или увеличении численности ящериц.
Некоторые рептилии распространяются человеком, случайно или преднамеренно, значительно чаще, чем амфибии. Иногда существование этих ящериц настолько связано с человеком, что ряд из них называют «домашними ящерицами». В тропиках некоторых из них перевозят на боль- шие расстояния и завозят на многие отдаленные острова (см. семейство гек- конов и сцинков в списке семейств). Маленькая змейка слепун (ТурИоря) в Тихом океане и в других местах обычно расселялась с почвой перево- зимых растений, за что ее нередко называют «змеей цветочных горшков». Черепах, а также и других рептилий обычно перевозят живыми для еды или для комнатного содержания. С другой стороны, в отдельных местах чело- век стал причиной исчезновения многих рептилий, которые либо являлись съедобными, либо опасными: так, например, на некоторых островах были уничтожены гигантские черепахи, а в других местах черепахи, большие и ядовитые змеи.
Реи~пилии Европе, вероятно, до границы суши, а в Восточной Сибири почти дости- гает Полярного круга. Это единственный живородящий (яйцеживородя- щий) вид семейства Lacertidae. (Приблизительные границы распростра- нения этого вида и всех других европейских видов, перечисленных здесь, были нанесены на карту Хехтом — Hecht, 1928.) Следующее место зани- мает гадюка обыкновенная Vipera berus, которая также пересекает Север- ный полярный круг в Европе (севернее 67' с. ш.). В Сибири этот вид и его представители не доходят до Полярного круга в пределах одного-двух гра- дусов и являются также живородящим видом. Оба рода Lacerta и Vipera распространены по всей северной лесной зоне Евразии (Берг, 1947). Я не знаю, встречаются ли они севернее зоны лесов. Из северных рептилий Р и с. 28. Приблизительные северные и южные границы распро-
странения рептилий. Я — ящерицы; 3 — змеи; Ч — черепахи; Ь — крокодилы (кроме рода Л. 11ip.<t (о г); Л — приблизительный ареал рода А 1ИдаГох в Восточной
Азии и восточной части Северной Америки. третье место принадлежит яйцеродящей змее — обыкновенному ужу (Natrix natrix), которая достигает примерно 65' с. ш. в Скандинавии. Четвертое место занимает живородящая ящерица Anguis fragilis, достигающая примерно 64,5' с. ш. в Швеции (письменное сообщение проф. Линдрота. Данные о нахождении этого вида на Северном полярном круге оказались ошибочными). Пятое и шестое места занимают живородящая змея-медян- ка Coronela austriaca, которая встречается в Швеции за 63' с. ш., и прыткая ящерица Lacerta agilis, распространенная в СССР примерно до 62,5' с. ш. Последняя относится к тому же роду, что и самая северная ящерица 7л~сег 1а vivipara, но только этот вид яйцеродящий.
Самые северные рептилии в Америке — живородящие змеи рода Tkans- noplii s (подвязковые змеи, близкородственные роду Natrix) распространены (широкой полосой) по краевым частям материка, проникая до 56' с. ш. в Британской Колумбии, до 60' в Форт-Смите, Северо-Западные террито- рии, и до 49' — на востоке (Logier, Toner, 1955). Распространение других змей, включая кивородящих и яйцеродящих, ограничивается 51 или 5' на западе и 47 или 48' с. ш. на востоке. Три вида ящериц трех различных семейств достигают 50 или 51' с. ш. в Британской Колумбии: два из них- 'кивородящие, один — яйцеродящий. Самой северной ящерицей на восто- ке материка является яйцеродящий вид Еитесея fasciatus, но граница его
Глава 4
распространения доходит примерно только до 45" в южном Онтарио, а на прибрежных равнинах Атлантического океана заходит немного»»ы»»»е 42' с. ш. По атлантическому побережью (".еверной Америки севернее этой точки ящерицы не встречаются, хотя около десяти видов змей и три семейства черепах проникают дальше на север материка. (Этот факт дает основание натуралистам Новой Англии оп»ибочно считать, что ящери»»ы по своей природе не в состоянии переноситb холод.)
К югу многие ящерицы и змеи достигают южной оконечности Африки. Некоторые ящерицы и три вида змей семейства Еlapidae достигают Тас- мании. Гекконы и сцинки, но не змеи (за исключением некоторых морских змей) проникают до Новой Зеландии, включая остров Южный. В Ам(.рике два или три вида мелких ящериц из семейства Iguanidae достигают Огненной Земли. Самой южной точки — провинции (:акта-Крус в к>жной Аргентине — - достигает пористая гадюка (Trir»eresurus a)nniod(tt<>ides). Ново- зеландские гекконы (2 эндемичных рода) являк).»ся единственными изве- стными живородящими представителями своего семейства. Ново:»е..»анд- е;;ие сцинки, около 9 эндемичных видов большого рода Lygosi»r)n,, на(коль- КО ИЗВес'1'НО, Та»ъ Же hah И СЦИНКИ ДРУГИХ Мест. Я)»ЛЯЮТСЯ ЖИВОг)ОДЯЩИМИ.
(Мое утверждение в статье в (<Quarterly Review'», что новозеланские (»(ин- >(и кладут яйца, было ошибочно.) Гадк>ки рода Tri(rneresurus в x>;»;)»()ii Аргентине, по-видимому, по аналогии с другими видами этого рода. жи»»о- родящие. Способы размножения других самых к)жных рептилий мн( не известны.
Основные границы ра'просранения репти ! Опредечяю»ся в (), „()
oì "UJepèöàMè и змеями, но следует отметить пределы и других групп SPI>enodon в Новой Зеландии находится вблизи ки;ной грани) ы 1)аспро странения рептилий. Особого внимания заслуживает тот факт, »то э».)> древние рептилии сохранились в климате, который является «прг,де-»>,нь»»>» для современных рептилий. Крокодилы встречан»ся (или встречались »» п))еделах недавнего исторического времени) к c(.»«pz о» тропико)» д<> (;„ ри)»» -Мпадной Азии (Cr(>c(>dilus), вдоль нижнег() течения реки 51нц»ы в Китае (Alligator) и на Каролинских островах в»»осточной части (;еверной Америки (Alligator). В Африке и Южной Америке они проникак)т немног(, южнее тропиков. Из черепах европейская боло»ная или речная черепаха Етуз orbi(eularis (подсемейство lmydinae) в Европе распространены до ')7,о' с. ш., но ни одна из других европейских черепах не дос»игае1 да>»;~ )О . Род Trionyx (семейство Trionychidae) распространен в Восточно»» А:»ии примерно до 48= но ни одна из других азиатских черепах не >»стре<»ае»св на север дальи1е 40', а а большей ~асТН вну1р ~н 1 )1 ceaepi~bIx Р; »»()н()), Лзии черепах нет. В Америке самая северная черепаха (;/(ryse)>(ys (п(>дсе мейство Еmydinae) достигает 51" в Ьританской lio.)y»»)()nn и ш»ате ~[ани тоба, 49 или 50 севернее озера Верхнего и, возможно, '~я' вдоль (»осто»ш>г<) побережья Северной Америки. К югу черепахи достигают оконечнос»и ~фри>'»( Южной Австралии, но не Тасмании (кром(. некоторых морс»;их '»ерепах) и северной Аргентины в Южной Америке.
~!исленность рептилий начинает убывать (хотя несколько видов и д(> ( тпгают северных и южных оконечностей материков) недалеко от предел»,— )»ых границ их распространения. В Европе в северных районах Мх 1цс„.)ен- )»ocTb yìåHûïaeòñÿ значительно интенсивнее, чем у амфиоип: та)(, »; Ю;»; ной Европе насчитывается 37 родов и 89 видов рептилий и всего 14 p(>g(>)» и 39 видов амфиоий; а в Северной Европе — только 5 родов N 6 r»Bpol» реп
и 6 родов и 11 видов амфибий (Schreiber, 191' ). В Южной Америкe ооеднение фауны рептилий наблюдается к югу от тропиков. Это xopo[)»o
прослеживается на распространении змей в Аргентине, число видов кото-рых постепенно уменьшается от 55 известных вблизи тропиков в Мисьо-несе до одного вида в Санта-Крусе (табл. 4).
Табл ица 4
Постепенное обеднение фауны змей в Аргентине 1
5э
.Рио- Н е1'ро
Ч обут
(;апта-h pyc Огненная Земли~
Мисьонес
1-~орриен тес Энтре-Риос, Буэнос-Айрес Ia-Пампа
С) Qs
15
1 Провинции и территории Аргентины приводятся в наиравлспци с север» на юг. цц<~iрi.I
эбо;значакэт число Ilзвс'cTHbIx видов змей в каждой из них (Seri(.', 1936, р. 57 — 59).
Что касается границ распространения рептилий на островах, то для ящериц они почти не существуют: ящерицы многочисленны на кад;дом (в пределах возможной досягаемости материка) острове, подходящем для них в климатическом отношении, и на многих отдаленных островах, вклю- чая Маскаренские и Сейшельские в Индийском океане. Они распростра- нены также на островах западной части Тихого океана, достигая по край- ней мере островов Тонга (за островами Фиджи), в Новой Каледонии и Но- вой Зеландии, на Галапагоссах в восточной части Тихого океана. В Вест- Индии, на Бермудских и Канарских островах, острове Мадейра и остро- вах Зеленого Мыса в Атлантическом океане их численность меньше и они все, несомненно, местного происхождения. За этими пределами на еще более отдаленных островах, особенно в тропиках Тихого океана, обитает несколько видов гекконов и сцинков, но, вероятно, они были занесены сюда человеком. Более подробные сведения о встречаемости ящериц и других рептилий на островах приводятся в гл. 8.
Несколько семейств и родов рептилий распространены очень широко. Из неморских черепах семейство Testudinidae достигает всех пригодных для обитания материков, кроме Австралии. Род Testudo (сухопутные чере- пахи) встречается на большей части Африки, в теплых частях Евразии и т. д., в Южной Америке и на отдельных островах. Triongx — наиболее широко распространенный род пресноводных черепах (мягкокожистые черепахи семейства Trionychidae) встречается в Африке, Южной и Восточ- ной Азии и восточной части Северной Америки. Основной род крокодилов Crocodilus почти тропикополит. Из семейств ящериц ни одно не имеет тако- го широкого распространения, как семейства гекконов и сцинков, кото- рые, являясь тропикополитами, заходят и в некоторые умеренно-теплые страны. Самым широко распространенным родом ящериц является Lygo- яоша, встречающийся почти во всех теплых частях Старого Света, в восточ- ной части Северной Америки и в Центральной Америке. Из змей ceMeIYcT- во Colubridae (ужовые) является космополитом и не достигает только наиболее отдаленных частей Австралии. Семейство Viperidae (гадюки и т. д.) имеет также широкое распространение, исключая равнинные части Австралийской области. Семейства Typhlopidae (роющие червеподооные змеи), Boidae (удавы) и Elapidae (кобры, аспиды и др.) являются тропико- политами, но заходят и в некоторые умеренно-теплые страны. Род ТурИорг также тропикополит, Natriz (в основном полуводные водяные змеи се- мейства Со1иЬг1йае) распространен на большей части Старого Света, вклю- чая часть Австралии, и на большей части Северной Америки. Но все эти случаи являются исключениями. Несмотря на большую приспособлен-
Г.iава 4
ность некоторых рептилий к расселению и их широкое распространение. многие семейства, большинство родов и почти все виды рептилий доволь- но ограничены в своем распространении.
Зональное распространение рептилий
Крокодилы в основном являются тропическими животными. Те немно- гие виды, за исключением рода Alligator, встречающиеся в краевых частях умеренных зон, также являются тропическими и лишь заходят на ограниченные территории с умеренным климатом. Однако род Alligator приурочен и к умеренно-теплым зонам в Китае и юго-западной части Соединенных Штатов Америки.
Большинство черепах также жители тропиков. Единственный род, который встречается в умеренной зоне северного полушария, Clemmgs с 2 видами в Средиземноморской области (один из них достигает тропиков Западной Африки), 5 — в Восточной Азии (в южном направлении до кра- евых частей тропиков), 1 — на западном побережье Северной Америки 3 — — в восточной части Северной Америки. Еще один род в Европе (Еиу~) и 4 или 5 в восточной части Северной Америки приурочены к огра- ниченным территориям с умеренным климатом, но зонального распростра- нения они не имеют. В пределах умеренной зоны северного полушария черепахи многочисленны только в восточной части Северной Америки, при- чем большая их часть относится к подсемейству Emydinae, которое наибо- лее многочисленно в тропиках Азии и проникло в тропики Америки. Все семейства и многие роды северных черепах также встречаются и в тропи- ках (см. список семейств).
Большинство ящериц также тропические животные, а представители умеренной зоны северного полушария незначительно отличаются от тро- пических. Все широко распространенные ящерицы северной зоны отно- сятся к тропическим семействам, а многие и к тропическим родам. Напри- мер, род Lacerta, к которому относятся самые северные рептилии, встре- чается от полярной Евразии до тропической Азии. Некоторые роды ящериц и одно небольшое семейство (Anniellidae) локализуются в умеренных частях Евразии или Северной Америки, но каждый род, распространенный на территориях обоих материков, встречается также местами и в тропи- ках. Самым близким настоящему голарктическому роду ящериц являет- ся род Ophisaurus (семейство Anguidae), один или два вида которого оби- тают в Иго-Восточной Европе и в умеренной зоне Северной Африки и т. д.; два или три — в восточных Гималаях, Бирме, Южном Китае и на острове Тайвань; один — в восточной части Северной Америки и один вид в горах острова Калимантан, почти на экваторе. В некоторых областях умеренной зоны южного полушария также обитает несколько эндемичных родов ящериц. но зональной фауны они не образуют, то есть эти роды не обычны для умеренной зоны южных континентов.
Змеи также в основном тропические рептилии, и их фауна в умеренной зоне северного полушария не имеет четко выраженных границ. Все змеи северных районов относятся к тропическим семействам, а многие и к тро- пическим родам. Род Vipera, к которому относятся самые северные змеи, встречается также повсеместно в тропиках Африки и Азии. Настоящего голарктического рода змей не существует. Некоторые роды приурочены к умеренным частям Евразии или Северной Америки, но каждый четко выраженный род, распространенный на обоих континентах, встречается
Pe~irnuл ии
также местами в тропиках. Шмидт (Schmidt, 1946) считает, что род ОрЬеосХ- rys приурочен к Восточной Азии и восточной части Северной Америки, что фактически правильно в отношении большей части этого рода, но нельзя забывать, что он также хорошо представлен в тропиках Индокитая, а один вид встречается на Юкатане, в краевой части тропиков Америки. В уме- ренной зоне южного полушария зональной фауны змей нет.
В заключение надо отметить, что зональное распространение репти- лий выражается только в том, что они являются главным образом тропи- ческими животными. В умеренной зоне северного полушария отчетливой широко распространенной зональной фауны рептилий нет. В этой зоне рептилий значительно меньше, чем в тропической, и все они относятся к тропическим семействам, а многие и к тропическим родам. Тропические рептилии часто вторгаются в умеренную зону северного полушария, но редко в ней адаптируются, и обычно довольно короткий срок нахождения их там недостаточен для того, чтооы они стали отличными от своих тропических родственников. В умеренной зоне южного полушария также отсутствует четкая зональная фауна рептилий.
Для некоторых семейств ящериц характерен другой вид зонального распространения, который может быть назван меридиональным. Эти семейства распространены с севера на юг через все климатические зоны. Семейство Lacertidae встречается в Старом Свете от арктических областей до Южной Африки. Оно составляет основную часть фауны ящериц умерен- ной зоны Евразии, но не достигает Америки. Семейство Iguanidae в Аме- рике встречается от северных границ распространения ящериц в западной Канаде до южной оконечности Южной Америки. Оно составляет основную часть фауны ящериц умеренных широт Северной Америки, но в настоя- щее время в континентальной части Старого Света не встречается. Семей- ства Agamidae в Старом Свете и Teiidae в Новом имеют в какой-то степе- ни сходное распространение, но на севере они убывают. Несомненно, что распространение в меридиональном направлении отчасти присуще и дру- гим животным и его границы могут пересекаться и сливаться с границами других видов распространения. Существенно и то, что количественное соотношение северо-умеренной фауны ящериц, относящихся к семействам со строго меоидиональным распространением, таково, что умеренные части Евразии и Северной Америки гораздо больше отличаются фаунами ящериц, чем другими группами животных (исключая наземных перелет- ных птиц).
Радиальное распространение рептилий
Радиальное распространение у рептилий выражено менее отчетливо, чем у рыб и земноводных. Доминирующие наземные рептилии очень быст- ро расселяются и не менее быстро проходят через стадию радиального рас- селения, в связи с чем «ключи» к разгадке их истории найти очень трудно. Однако можно проследить радиальное распространение нескольких семейств и родов рептилий.
Самое большое из современных семейств рептилий — Со1иЬгЫае (ужовые) является также самым широко распространенным: ужовые мно- гочисленны во всех тропических и умеренных зонах земного шара, за исключением Австралии, где их немного и где они, очевидно, являются недавними вселенцами. С точки зрения экологии ужовые обладают необы- чайно разнообразной приспособляемостью: они могут встречаться как
166
в жарких, так и в холодных, как во влажных, так и в сухих и т. д. местах обитания. Таким образом, распространение этого семейства почти сплош- ное и охватывает такие места, которые неблагоприятны для многих дру- гих рептилий. Семейство (:olubridae, несомненно, радиировало как географически, так и в отношении эволюции. Но, характеризуя современ- ное распространение этого семейства, нельзя сказать, что оно просто радиальное, так как оно значительно сложнее. О семействе в целом можно сказать, что доминирующие, дериватные формы преимущественно встре- чаются в Африке и тропической Азии, а более малочисленные и менее раз- ноооразные формы — в умеренных зонах Евразии и (:еверной Америки. что примитивные формы встречаются на Мадагаскаре и в Южной Америке и что только недавно отмечено несколько новых вселенцев в Австралии (см. список семейств). Это один из примеров очень сложного характера радиации из основной части тропиков Старого Света. Ужовые, как доми- нирующие змеи, в Австралии замещены очень отчетливой довольно древ- ней фауной аспидовых. Это трудно объяснимая аномалия. Аспидовые, очевидно, произошли от ежовых, и поэтому, являясь предположительно более поздними по происхождению, они не должны были достичь Австра- лии первыми. Если это так, то почему же в таком случае аспидовые достигли Австралии в столь давние времена, что все их австралийские роды теперь уже являются эндемиками, а ужовые проникли туда настолько недавно, что ни один их род не стал эндемичным?
Из семейства Colubridae наиболее широко распространен род 1V«trix примерно с 80 видами. Он встречается в Африке и на Мадагаскаре, в тро- пической и умеренной зонах Евразии (приблизительно до 65' с. ш. в Скан- динавии), через острова до Северной Австралии, в восточной части Север- ной Америки (в западной части теперь их нет), на юг до Гватемалы и на Кубе. Род Thamnvphi~s (подвязковые змеи), образовавшийся от рода 1'иtrix, распространен по всей Северной Америке (на север до границы распространения змей и на юг — до Коста-Рики). В Индо-Малайской области видов рода Natrix отмечается больше, чем в любом другом месте. Это характерная особенность радиального распространения с концентра- цией на основной части тропиков Старого Света, или, точнее, в тропической Азии. По образу жизни виды рода Natrix обычно связаны с водой, но не являются полностью водными. Они обладают способностями расселения, присущими сухопутным змеям, плюс некоторыми способностями преодо- левать водные преграды, свойственными водным змеям. Род Natrix, вероятно, обладает и другими пока еще не известными особенностями. которые ставят его в эволюционном отношении на ступень выше большин- ства остальных змей и которые способствовали его радиации. Представи- тели этого рода в Старом Свете — яйцеродящие, а в Америке — живоро- дящие, а так как живорождение у рептилий иногда является адаптацией к холодному климату, возможно, что американские формы этого рода произошли от предка, проникшего из Евразии в Америку через север. (Название Natrix употребляется здесь в довольно широком смысле, так как некоторые группы видов, включая, например, тех, которые нахо- дятся на Мадагаскаре, иногда помещают в отдельные небольшие роды.)
Несколько других больших групп рептилий также имеют в какой-то степени радиальное распространение. Семейство черепах Testudinidae сосредоточено в тропической Азии: наиболее разнообразны и многочислен- ны там пресноводные черепахи подсемейства Етуо1пае, которые также (но уже в значительно меньшем количестве) достигают умеренных широт
Реижилии
1';вразии, Северной и Южной Америки и ". д. Наземные черепахи Testu- diiiiiiae, происшед~пие, видимо, от пресноводных черепах тропической Азии, в некоторых направлениях проникают еще дальше. Черепахи семейства Trionychidae наиболее разнообразны на основной части тропиков Старого Света: один род простирается в северном направлении в Восточной Азии и достигает Северной Америки, другой род достигает Новой Гвинеи. Сцинки весьма разнообразны в тропиках Старого Света: несколько видов распространены на севере и три рода достигают Америки. Гадюки наиболее разнообразны на основной части тропиков Старого Света, особенно в тро- пической Азии: несколько видов заходит на север и два азиатских рода простираются до Америки, но радиальный характер распространения .этого семейства в Америке усложняется распространением гремучих змей.
Ре гнональное распространение рептилий
Фауны рептилий двух северных областей земного шара (Евразии и Северной Америки, севернее тропиков) характеризуются скорее отрица- тельными признаками. Ни одно семейство рептилий не является здесь исключительно голарктическим. Небольшое семейство Anniellidae, лока- лизованное в юго-западной части Северной Америки, — единственное семейство, которое приурочено к умеренной зоне северного полушария, а Alliigntor является единственным исключительно голарктическим родом рептилий. (Несколько других почти голарктических родов было отмече- но выше, в зональном распространении рептилий.) Большинство домини- рующих северных рептилий относится в основном к тропическим семей- ствам, особенно пресноводные черепахи, агамовые (в Евразии) и игуано- вые (в Северной Америке), а также ужовые и гадюковые. Кроме перечис- ленных, в каждой северной области есть рептилии, ведущие свое начало от тропических или общих с тропиками (большей частью с тропиками Ста- рого Света, когда речь идет о Евразии, и отчасти с тропиками Старого Све- та и Америки, если речь идет о Северной Америке) и которые в умеренной зоне часто занимают довольно небольшую территорию. К ним относятся из черепах — каймановые черепахи в восточной части Северной Америки, род Testudo в южной Евразии, Gopherus в южной части Северной Америки и Тпопух в Восточной Азии и восточной части Северной Америки; из яще- риц — хамелеоны в южных частях Европы, гекконы — в основном в Центральной и Восточной Азии, Xontusio, в юго-западной части Северной Америки, несколько видов сцинков на больших площадях Евразии и Се- верной Америки, настоящие ящерицы на крайнем севере Евразии, один род из семейства Teiidae в Северной Америке, семейство Anguidae широко разбросано в Европе и Северной Америке, род Shinisourus — в Китае, Helodermo, — в юго-западной части Северной Америки, Vo,ronus — по южным краевым частям Евразии и разные двуходковые — в Южной Европе и т. д. и в южных частях Северной Америки; из змеи: род ТурЫорг — в юж- ной Евразии, Leptotyphlops — в юго-западной части Северной Америки, разные мелкие удавы — в Азии и в западной части Северной Америки, кобры — по южным краевым частям Азии и аспиды — в южной части Северной Америки. Из перечисленных выше рептилий в одних случаях тро- пические роды или виды простираются на север локально, в других— виды или роды, являясь эндемиками в умеренной зоне, относятся к тро- пическим семействам. Но все эти северо-умеренные группы, вместе взя- тые,— и те, которые доминируют, и те, что являются локальными или рас-
Глава 4
пространены по окраинным частям, — составляют фауны значительно оед- нее тропической.
Для рептилий умеренной зоны северного полушария характерны не только их сравнительная немногочисленность и слабое отличие от их тро- пических сородичей, но также их неравномерное распределение. Умерен- ные зоны Евразии и Северной Америки особенно отличаются своими доми- нирующими ящерицами, большинство из которых относится к разным семействам (Agamidae и Lacertidae — в Евразии, Iguanidae и Teiidae— в Северной Америке), являющимися общими для тропиков Старого и Но- вого Света соответственно. Фауны рептилий Евразии и Северной Америки отличаются многими другими деталями, часть из которых уже была отме- чена выше. Конечно, и умеренные зоны Евразии и Северная Америка име- ют некоторые общие семейства и даже роды (Cle>nmys и Trionyx; Еитесея, Lygosomo, и Ophisourus; несколько родов ужовых и гадюковых и род Alligator), но все же различия в фаунах рептилий преобладаютнад сход- ством. Вероятно, значительно отличающиеся фауны рептилий умеренной зоны Евразии и Северной Америки произошли из разных тропических областей: из тропиков Старого и Нового Света соответственно.
В пределах каждой северной области рептилии распространены неран- номерно. К северу резко снижается число видов рептилий, а с востока на запад происходит дифференциация фауны. Например, в западной части Северной Америки мало черепах и много ящериц (и много змей); в восточ- ной части — много черепах и относительно мало ящериц (и много змей). Неравномерность такого распределения происходит в результате неодно- родности климата, сухого или влажного, и усиливается еще в связи с тем, что северные рептилии, особенно ящерицы, распространены в краевых частях зоны, где они могут существовать только в исключительно благопри- ятных для них условиях. В тропиках ящерицы избегают сухих, открытых солнечных мест; многие живут во влажных лесах, а многие, избегая солн- ца, ведут ночной образ жизни. Зависимость большей части рептилий от высоких температур наблюдается исключительно в прохладном климате, что объясняется необходимостью поддерживать свою температуру в тече- ние дневной активности (Bogert, 1949) и потребностью в тепле, необходи- мом для размножения (см. стр. 469).
Короче говоря, численность рептилий умеренной зоны северного полу- шария относительно мала, и резкого отличия их от рептилий тропической зоны нет, но распределены они настолько неравномерно, что, как у;ке говорилось, между фаунами рептилий умеренных зон Евразии и Северной Америки существуют большие отличия, особенно в фаунах ящериц. Реги- ональные фауны рептилий распределены также неравномерно.
В тропической и умеренной зонах южного полушария рептилии столь многочисленны и их распространение и связи столь сложны. что прихо- дится ограничиться кратким описанием. Дополнительные сведения приво- дятся в списке семейств в конце главы.
В Африке, южнее Сахары, распространены черепахи семейств Ре!o- medusidae и Trionychidae и наземные черепахи подсемейства Тея1ий1гг пае, один вид подсемейства Emydinae (очевидно, новый вселенец, проник- ший только в северо-западную часть африканских тропиков), несколько представителей агамовых, большое число настоящих хамелеонов, а также оольшое число гекконов, сцинков и ящериц семейств Lacertidae, Сordy- lidae, род Varonus и семейство Amphisbaenidae, червеобразные змеи обоих семейств (Typhlopidae и Leptotyphlopidae), питоны, песчаный удав. (приурочен к восточной части тропической Африки), большое число у'ко-
Репшилии
169
вых, аспидовых и типичных гадюковых, крокодилы. Большинство из этих групп широко распространено в Тропической Африке, и значительная часть, но не все, проникают в Южную Африку. Ящерицы семейства Cordylidae в основном являются южноафриканскими, но проникают также и на север в тропики. Между фауной рептилий влажных лесов и более сухих открытых частей существуют различия, на которых здесь нет возможности остановиться. Все эти особенности распростра- нения рептилий в Африке рассмотрены в работе Шмидта (Schmidt, 1919 — 1923) .
На Мадагаскаре распространены черепахи семейства Pelomedusidae и подсемейства Testudininae (но отсутствуют представители семейства Тпо- пусп1йае), имеются реликтовые игуаны, большое число хамелеонов, гек- конов, сцинков, несколько видов ящериц семейства Сordylidae (только Gerrhosaurinae, а не Cordylinae), но нет агамовых, нет ящериц семейства Lacertidae, нет ящериц рода Varanus и семейства Amphisbaenidae. Здесь имеются червеобразные змеи семейства Typhlopidae (но не Leptotyplilo- pidae), эндемичное подсемейство ложноногих, но нет питонов и ужовых (включая предположительно примитивные роды), также нет представи- телей семейств Elapidae и Viperidae; встречается нильский крокодил.
По своим связям мадагаскарская фауна рептилий в основном является африканской, несмотря на то, что многие африканские семейства в ней отсутствуют и имеется много четко выраженных эндемичных родов. Очень немногие рептилии Мадагаскара прямо родственны рептилиям Индо- Малайской области. Sibynophis — единственный встретившийся мне мадага- скаро-восточный (но не африканский) род (см. семейство Colubridae в списке семейств), хотя не исключена возможность существования и дру- гих родов. Индийский хамелеон является родственным средиземноморско- му, а не мадагаскарскому виду. Маскаренский род гекконов РЬе1зита, достигающий Андаманских островов в Индо-Малайской области, скорее, островной род, чем мадагаскаро-восточный (см. список семейств).
В тропической Азии и т. д. (Индо-Малайская область) имеется один вид подсемейства Platysterninae — многочисленные пресноводные и сухо- путные черепахи и черепахи семейства Trionychidae, многочисленные ага- мовые, один вид хамелеонов, многочисленные гекконы и сцинки, несколько видов ящериц семейства Lacertidae, один вид Dibamidae, некоторые пред- ставители родов Ophisaurus, Varanus и Lanthanotus. Здесь представлены каждое семейство и почти каждое подсемейство змей, хотя некоторые из них встречаются только в краевых частях области. Есть крокодилы и га- виалы. Некоторые из этих групп широко распространены в Индо-Малай- ской области, другие локализованы: так, например, большинство прес- новодных черепах сосредоточено в восточной части области; хамелеоны приурочены к южной Индии и Цейлону; Dibamidae встречается только на островах к юго-востоку от южного Индокитая; большинство видов семейства Lacertidae находится на материке, и только один вид достигает Больших Зондских островов; род Lanthanotus встречается на острове Калимантан, а целое подсемейство Uropeltinae (роющие змеи) примерно с 43 видами приурочено частично к полуострову Индостан и Цейлону. Род агамовых Cophotis представлен одним видом на Цейлоне и одним на остро- вах Суматра и Ява. Из рода Draco один вид распространен в южной Индии (но не на Цейлоне), где он изолирован от основного ареала рода в Индо-Малайской области пространством по крайней мере в тысячу миль. В каждом роде сцинковых — Lygosoma, Dasia и Riopa — имеется по паре видов, которые распределены между южной Индией и восточной частью
По
Глава 4
Индо-Малайской области, в северной части Индии близкородственных им видов нет. Один вид рода Cylindrophis изолирован на Цейлоне, но так или иначе этот род приурочен к восточной части Индо-Малайской обла- сти и т. д. Var anus яаг,г.аког встречается на Цейлоне и в восточной части Индо-Малайской области, но отсутствует на большей части полуострова Индостан. Для этих групп характерно такое же прерывистое распростра- нение, как и для некоторых пресноводных рыб. Небольшие разрывы, вероятно происходящие в направлении, в котором наблюдается изоляция популяций в южной части Индии, встречаются среди пресноводных чере- пах и гадюковых (см. список семейств). По крайней мере у пяти видов и двух родов змей обнаруживается другой характер разрывов: они встре- чаются в Индокитае, а также на Больших Зондских островах, но отсут- ствуют на полуострове Малакка и в Таиланде (М. А. Smith, 1943, р. 25 — 26). У рода Ophisaurus наблюдается еще более широкий разрыв в этом направлении: от Бирмы и далее до острова Калимантан.
Что касается связи между африканской тропической фауной репти- ;г1ш и фауной рептилий Индо-Малайской области, то последняя имеет ряд важных семейств, большинство из которых встречается также и в дру- гих частях земного шара, а также ряд общих родов: Testudo* (сухопутные черепахи), Trionyx (водные черепахи), Agama, Мабиуа*. Lygosoma, l.acer- ta n Varanus, ТурЫорз*, Python, Boiga и 1Vatrix*, а также ряд других родов ужовых — Naja (кобры) и Vipera, Crocodylus* и другие. Боль- шинство этих родов встречается также за пределами Африки и Индо- Малайской области. Одни из них, помеченные звездочками, встречают- ся на Мадагаскаре, другие не встречаются (или встречаются, но, воз- можно, не являются там местными), и эти последние достаточно много- численны и разнообразны, чтобы можно было убедиться в том, что Африка и тропическая Азия обменялись рептилиями через материки (в то же время данных для предположения, что этот обмен произошел через Мадагаскар, очень мало). Однако различия между фаунами рептилий Африки и Индо-Малайской области показывают, что этот обмен был ограниченным. Интересно отметить некоторые удивительные различия между фаунами: в Африке водятся черепахи семейства Ре1оше- dusidae, а в Индо-Малайской области — подсемейства Emydinae; в Афри- ке многочисленны хамелеоны и ящерицы семейства Lacertidae, в то вре- мя как в Индо-Малайской области многочисленны ящерицы семейства агамовых. Из этих двух областей только в Африке распространены яще- рицы семейств Amphisbaenidae и Cordylidae, так же как и змеи семейства Leptotyphlopidae, которые едва достигают Индо-Малайской области, где (включая семейство Aniliidae, несколько подсемейств ужовых, отсут- ствующих в Африке, ямкоголовых гадюк и большинство морских змей) змей больше, чем в Африке.
В Австралийской области распространены черепахи семейства (;he- lyidae, Carettochelys (в Новой Гвинее) и Trionychidae. богато представле- ны агамовые и гекконы, имеется эндемичное семейство Pygopodidae. много сцинков, род Dibamus (в Новой Гвинее) и Varanus; большое число видов змей рода ТурИорз, ряд родов питонов, мелкие древесные удавы (в Новой Гвинее и т. д.), несколько видов ужовых, многочисленные пред- ставители семейства Elapidae (и морских змей) и крокодилы (в тропиках). Учитывая вьппеуказанные ограничения и то, что представители семейства Ps gopodidae слабо представлены на Новой Гвинее, а ужовые отсутствуют на большей части Южной и Западной Австралии, можно сказать, что пере- численные группы довольно равномерно распространены в основных
Реп~п юал ип
(71
частях Новой Гвинеи и Австралии. Однако не многим из них удается достичь Тасмании, где совсем нет черепах (кроме морских черепах в отдель- ных районах), крокодилов, а также родов Varanus, Турйорз, питонов и усовых.
Связи австралийских рептилий в большинстве случаев ведут к Индо- Малайской области. Один вид черепах семейства Trioriychidae, обитаю- щий в Новой 1'винее (если только там он действительно местный), являет- ся выходцем из Индо-Малайской области и иногда заходит в море. Один из видов крокодилов Индо-Малайской области обычно заходит в море. Из ящериц в Австралии, так же как и в Индо-Малайской области, много- численны агамовые. Роды Goniocephnlus и Physignathus встречаются толь- ко в Индо-Малайской и Австралийской областях, но их ареал разрывает- ся, так как оба рода отсутствуют на многих островах, расположенных между этими областями. Но, с другой стороны, водный род агам Lophura (FIpdrosaurus) встречается только от Филиппинских островов и острова Сулавеси до Новой Гвинеи. Род гекконов Gymnodactylus многочислен о1 Индо-Малайской до Австралийской области, но также встречается и в других местах, а ареал двух или трех других родов гекконов охваты- вает и Индо-Малайскую и Австралийскую области. Из Индо-Малайской области сцинки, особенно род Lygosoma, буквально наводнили Новую 1'винею и Австралию. Каждый из родов Trophidophorus, Ophioscincus и Rliodona распространен как в Индо-Малайской, так и Австралийской областях, но, подобно некоторым агамовым, упомянутым вьппе, отсутст- вует на некоторых Мастях архипелага, лежащего между ними. С другой стороны, Emoia и Otosaurus почти полностью или полностью приурочены к островам от Больших Зондских островов до Новой Гвинеи. Род Diba- пгия распространен от южного Индокитая до Новой Гвинеи. Вараны (род Vnrnnus), а также и червеподобные змеи (ТурЫорг) имеют в Австралии сплошное распространение (насколько позволяют границы суши). Змеи рода Cylindrophis семейства Aniliidae встречаются от Индо-Малайской области до Новой Гвинеи. Род Python, широко распространенный в тро- пиках Старого Света, простирается строго до острова Тимор, а вообще этот род достигает Новой Гвинеи и Австралии. От Индо-Малайской области до Австралии распространены следующие роды ужовых: 4 рода, ведущие только водный образ жизни (не считая морских змей), полуводный род Natrix и древесные Boiga и Ahaetulla, а также наземный род Steganophis, который распространен от Филиппинских островов до Северо-Восточной Австралии. Кроме вышеприведенных связей австралийских рептилий с Индо-Малайской областью, четкие связи их с другими частями земного шара наблюдаются лишь в одном случае: черепахи семейства Chelyidae встречаются только в Австралийской области и в Южной Америке. Све- дения о некоторых австралийских гекконах, которых считали прямо род- ственными южноафриканским или американским, не являются достовер- ными.
Переход фаун рептилий Индо-Малайской и Австралийской областей слишком сложен, чтобы вдаваться в подробности (многие детали приво- дятся в следующих работах: Rooij, 1915 — 1917, и Mertens, 1930). Выше при- ведены семейства, роды и виды, которые имеют сплошное распространение от Азии до Австралии, и групп, встречающихся в обоих областях, но от- сутствующих на островах, расположенных между ними, а также групп, которые приурочены к этим островам. Многие рептилии Индо-Малайской области приостанавливаются у линии Уоллеса, а многие другие прости- раются на различные дистанции по направлению к Австралийской обла-
172
сти, но не достигают ее. Короче говоря, большая часть фауны рептилий Индо-Малайской области простирается по направлению к Австралии с по- степенным обеднением отдельных элементов по мере увеличения расстоя- ния, но большое число групп все же достигает Новой Гвинеи, несколько меньшее — Австралии и еще меньшее — Тасмании, однако характер этого распространения усложняется у некоторых групп разрывами и эндемич- ными группами, локализованными в зоне обеднения. По-видимому, ооль- шой и очень сложный наплыв рептилий происходил из Индо-Малайской области как по направлению к Австралийской области, так и в нее. Это перемещение, видимо, наблюдалось преимущественно в одном направле- нии, но в обратном направлении было очень незначительно: такие австра- лийские группы, как черепахи семейства Chelyidae, ящерицы семейства Pygopodidae и австралийские змеи семейства Еlapidae, распространялись по направлению и Индо-Малайской области; и только несколько видов из этих групп змей распространялись на запад, достигая даже Молуккских островов.
К фауне рептилий Юя1ной Америки относятся род C//~ålódrà (в северо- западной части материка) и Einosternon, два неэндемичных рода черепах подсемейства Emydinae, род Testudo, семейства Pelomedusidae и (;lielyi- dae; большое число игуановых, гекконов и единственный не эндемичный род сцинков, многочисленные представители семейства Teiidae и А1прЫя- baenidae и некоторые представители семейства Anguidae; оба семейства червеподобных змей, один представитель семейства Aniliidae, подсемейство Boinae, многочисленные ужовые (особенно примитивные формы подсемей- ства Xenodontinae, если оно признано), аспидовые, ямкоголовые гадюки, крокодилы и кайманы. Эта фауна распространена неравномерно (но все детали привести здесь невозможно). В южном направлении она претерпе- вает постепенное обеднение, и только несколько видов ящериц семейства Iguanidae достигают Огненной Земли.
Фауна рептилий Южной Америки может быть подразделена на две основные части (табл. 21, стр. 473). Более древняя часть фауны находилась в Южной Америке по крайней мере с начала третичного периода, когда континент стал изолированным. Более современная, вероятно, возникла в позднем плиоцене, после соединения Южной Америки с Северной. Одна- ко сухопутные черепахи, очевидно, достигли Южной Америки в проме- жутках между этими двумя периодами, преодолев водные преграды тре- тичного времени (Simpson, 1943); вероятно, что и некоторые другие реп- тилии проникли в Южную Америку подобным образом.
Большинство более современных компонентов фауны рептилий Ю к- ной Америки, включая роды Chelgdro,, Einosternon, подсемейства Lmy- dinae, аспидовых, ямкоголовых гадюк и некоторые не эндемичные роды из других семейств, очевидно, достигли Южной Америки из Северной Америки или через нее из других частей земного шара. Большинство из них встречается в Северной Америке и до настоящего времени, но некото- рые уже исчезли или начали исчезать. Например, род 6еоеиуда теперь находится в тропической Азии и т. д., в Мексике, Центральной и Южной Америке, но его ископаемые относятся к Европе и умеренной зоне Север- ной Америки (см. список семейств). Далее, род Testudo сейчас распростра- нен в теплых частях Старого Света и в Южной Америке на север до Панамы (и на островах), но этот род существовал с эоцена до плейстоцена в Север- Hoii Америке. Род Sibynophis (на Мадагаскаре, в Индо-Малайской области и в тропиках Америки) и род Trimeresurus (включая Bothrops) (в Юго- Восточной Азии и тропической и субтропической частях Америки). воз-
Penmu;i ии
можно, также проникли в эти районы через Северную Америку. Един- ственный род сцинков в Южной Америке МаЬиуа встречается в тропиках как Старого, так и Нового Света. Возможно, он также расселился через север или достиг Америки через Атлантический океан, так как сцинки обладают большой способностью пересекать водные пространства. Нахожде- ние рода МиЬиуа на островах Вест-Индии и существование эндемичного вида на острове Гаити позволяет предположить, что Вест-Индия, возмож- но, была местом, куда прибыли американские представители этого рода, и что они могли достичь этого места из Африки на плоту. Некоторые роды гекконов, которые по сравнению со сцинками более легко преодолевают водные пространства, возможно, являются общими для более теплых частей Америки и Старого Света, и, возможно, они также пересекли Атлантиче- ский океан (см. список семейств, особенно род Tarentolcz).
Более древние группы южноамериканских рептилий, для которых характерен родовой эндемизм, имеют разнообразные географические свя- зи. Змеи семейства Aniliidae, ужовые подсемейства Xenoderminae и Parei- nae плюс Dipsanidae связывают фаунистически Южную (и в некоторых случаях Центральную) Америку с Индо-Малайской областью. Черепахи семейства Pelomedusidae в настоящее время являются общими для Афри- ки и Мадагаскара (Podocnemis только для Мадагаскара), а ископаемые этого семейства обнаружены в другом месте. Ящерицы семейства Amphis- baenidae и змеи семейства Leptotyphlopidae в настоящее время приуроче- ны к более теплым частям Америки и Африки и к соседним территориям, но ископаемые некоторых представителей семейства Amphisbaenidae встре- чаются и в более северных районах. Черепахи семейства Chelyidae являются общими только с Австралийской областью. Конечно, некоторые из более древних групп рептилий Южной Америки, например игуановые, до сих пор являются общими с рептилиями Северной Америки. Отмечается также и некоторое современное перерасселение родов и отдельных древних семейств. Например, один южноамериканский род семейства Teiidae проник далеко в глубь Северной Америки. И в Южной, и в Северной Америке недав- но произошел обмен несколькими видами ужовых. Из современного рас- пространения рептилий, видно, что связи южноамериканских групп просле- живаются по направлению почти к каждой части земного шара. Такое положение могло создаться только при наличии сложных расселений с последующим сложным исчезновением. Исчезновение рептилий в Северной Америке уже началось в связи с изоляцией некоторых более современных рептилий в Южной Америке (см. предыдущий абзац), а некоторые более древние группы, очевидно, были изолированы в результате вымирания рептилий на обширных территориях в течение продолжительного времени. Сложность расселения и исчезновения подтверждается современным рас- пространением игуановых и удавов (см. список семейств).
Рептилии Центральной Америки
В Центральной Америке имеется единственный сохранившийся вид черепах семейства Dermatemydidae и два рода подсемейства Staurotypi- nae (относящиеся к группам, ископаемые которых обнаружены на севере), род CI!elydrn (каймановая черепаха), Einosternon (мускусная черепаха), Pseudeznys и Geoenzydcz (пресноводные черепахи). Границы их распростра- нения простираются до отдельных районов Южной Америки, но все они, очевидно, проникли сюда с севера. В Центральной Америке отсутствуют
черепахи рода Testudu (за исключением Панамы), мягкокожистые черепа- хи (Trionyx), которые известны в Северной Америке с верхнемелового периода. Нет бокошейных черепах (Pelomedusidae и (helydae), которые известны в Южной Америке с мелового периода. В Центрально~ Америке имеется много ящериц. Некоторые игуановые и гекконы Центральной Аме- рики, по-видимому, проникли из Южной Америки, а другие, вероятно, распространились с севера (часть из них приводится ниже), и по крайней мере один род игуан радиировал в Центральной Америке и, возможно,рас- селялся оттуда. Небольшое семейство Xantusiidae почти всегда приуроче- но к юго-западной части Северной Америки и Центральной Америке. Центральноамериканское семейство Teiidae появилось недавно из Южной Америки. Сцинки Центральной Америки, включая род ЛХаЬиуа, скорсе всего, достигли ее через Атлантический огеан, и два других рода проникли с севера. В Центральной Америке, так же как в Северной и Южной, встре- чаются несколько видов семейства Anguidae. Один род семейства Xeuo- sauridae распространен от Гватемалы до южной Мексики. Родственный ему род находится в Китае. Сомнительно приписываемые этому роду ископаемые из раннетретичного периода найдены в СевернойАмерике. В Центральной Америке отсутствует семейство Amphisbaenidae, хотя раз- ные роды его встречаются в Южной Америке, в южной части Северной Аме- рики и в Вест-Индии. Из змей в Центральной Америке распространены оба семейства червеподобных змей и локализованный род семейства Aniliidae (Loxocemus), некоторые представители семейства Boidae (вероятно, родом из Южной Америки), многочисленные ужовые, из которых одни, видимо, достигли Центральной Америки из Южной, а другие из Северной Амери- ки, а также аспидовые и ямкоголовые гадюки, происшедшие на севере. Из крокодилов в Центральной Америке имеется род Crocodycus, вероятно проникший через север, и современные кайманы — из Южной Америки. Дальнейшие подробности распространения некоторых из этих групп реп- тилий приводятся ниже, а также в списке семейств.
~[тобы проанализировать фауну рептилий Центральной Америки, необходимо сказать еще несколько слов о рептилиях Мексики и примыкаю- щей к ней юго-западной части Соединенных Штатов, так как эти районы являются частью одной и той же переходно-реликтовой территории. К этой дополнительной территории приурочены род Arcelytropsi s, представители семейства Anniellidae, Helodermatidae (ядозуб) и один изолированный род семейства АтрЫзЬаеиЫае (Bipes), кроме того, на нее проникают сухопут- ные черепахи рода Gopherus.
Эта фауна рептилий Централ ьно-Американо-Мексиканская приме- чательна тремя особенностями: она переходная. разнообразная ii включает оолыиое число реликтовых форм.
В Центральной Америке (без Панамы) и в Мексике (по современной классификации) обитают представители 21 семейства плюс, еще 8 подсе- мейств рептилий (не считая распространенных в краевых зонах этих тер- риторий Trionychidae и 'rycinae) по сравнению с 18 семействами плнн 6 подсемейств (не считая встречающегося в краевой зоне Южной Америки подсемейства (;helydriiiae). Таким ооразом, Центрально-Американо-Мек- сиканская фауна богаче фауны Южной Америки, особенно ящери- ц,ами.
Из общей структуры фауны, описанной выше, ясно, что фауна репти- лий Центральной Америки и Мексики является переходной со многими североамериканскими и южноамериканскими группами, перекрывающими друг друга на разное расстояние, а в некоторых случаях и на болыш~н.
Penrnu.< ии
расстояние. 11ереход этот очень сложен, и мы не имеем возможности рас- смотреть его более детально. Однако необходимо остановиться на некото- рых группах, которые распространились с севера по всей (или на большей части) Центральной Америке, но не проникли или проникли, но на очень небольшое расстояние в Южную Америку. Род Chelgdro, (каймановая черепаха) распространен от восточной части Северной Америки через Цент- ральную до самой северо-западной части Южной Америки. Игуаны ооль- шого рода Sceloporus распространены от южной части Британской Колум- бии и к>жного Нью-Йорка до Панамы. Гекконьх рода Coleonyx, который, вероятно, представляет азиатский род Eublephnris, встречаются от нхжньхх краевых частей Соединенньхх Штатов до Панамы. Из сцинков род Eènãeñe~, «оторьххх встречается также в некоторых частях Старого Света, распрост- ранен от южного края Канады до Никарагуа, и большой род Старого Све- та Lygosooin представлен в Америке одной гр~ ппохх из нескольких видов, которые распространены от восточной части С:оединенньхх Штатов до Па- намы. Род Gerrhonotus семейства Anguidae встречается от южной части Британской Колумбии до Панамы. Из ужовых род Т!япгиор1пз (подвяэ- «овые змеи) распространен от северной границы рептилий в (:еверной Аме- рике до Коста-Рики. Широко распространенный род Elnphe достигает Коста-Рики, а род Lnrnpropeltis. встречающиххся на большей части Соединен- ных Штатов, через Центральную Америку достигает северо-западной части Южной Америки. И, наконец, род ЛдЫstrodon (ямкоголовые гадюки), встречающийся также и в Азии, распространен от штатов Айова, Масса- чусетс и т. д. до Ника,рагун. Эти рептилии, иногда встречан>щиеся на вер- шинах гор в тропиках, не являются рептилиями умеренной зоны. Боль- цхинство из них встречается на тропических низменностях Центральной Америки.
Очевидными или возможными реликтами фауны рептилий Централь- ной Америки и Ъ1ексики являются черепахи Dermatemydidae, Staurotу- pinae и Gopherus («оторый не представляет той же ветви, что и род Testu- do). Из других рептилий можно назвать Xa»tusiidae, Anelgtropsi~s, Anniei- lidae, Xen<rsnurus, FIelodennn и Bipes, а также Loxocemus. Современные реп- тилии, родственные этим рели«там, или их ископаемые (см. список семей- ства) пох'азьхвахо.х, что некоторые из этих реликтов представляют группы. которые достигли Центральной Америки и Мексики с севера. История дру- гих групп рептилий не известна, и ни один их представитель не является четким реликтом южноамериканской группы.
Переходность фауны и наличие реликтов в Центральной Америке и Мексике отчасти скоррелированы. Многие различные группы рептилихх проникают в зону перехода или перемещаются через нее на разные рас- стояния в обоих направлениях. Часть рептилий (но не все), которые рас- пространились из Южной Америки на север, останавливается в краевой зоне тропиков или вблизи (в южной Мексике). Это обычный характер распространения, при котором тропические группы достигают умеренной зоны северного полушария, но не заходят в нее. Но из многочисленных рептилий, распространявшихся на юг из Северной Америки или через нее, в Южную Америку смогли проникнуть лишь сравнительно немногие, несмотря на то, что многие из них сумели пересечь линию тропиков и рассе- лились на большей части или по всей Мексике и Центральной Америке. Некоторые реликты, по-видимому, представляют более древние группы, которые достигли Мексики и Центральной Америки с севера, но не про- никли в Ю;«ную Америку. Такой характер распространения, столь не- обычный для других позвоночных, будет рассмотрен в гл. 9.
Глава 4
1[ереходы и преграды в распространении рептилий
Там, где встречаются разные фауны рептилий, между ними нет рез- ких границ, а есть широкие зоны перехода, перекрывающие друг друга с постепенным обеднением элементов обеих фаун по краям.
В Восточной Азии, где наблюдается сложный переход между тропиче- скими и северо-умеренными рыбами и земноводными, число рептилий к северу просто уменьшается. Многие тропические рептилии Азии останавли- ваются недалеко от умеренной зоны северного полушария, другие распро- страняются на север на различные расстояния, но самостоятельной северной фауны рептилий, которая проникала бы в зону перехода с проти- воположной стороны, нет. В связи с причинами, обусловившими малочис- .ленность фауны рептилий на севере и ее отсутствие на большей части край- него севера, фауны рептилий умеренных зон Евразии и Северной Америки в настоящее время также разделены пространством, на котором отсутствует зона перехода. Однако Евразия и Северная Америка в течение третичного периода обменялись рядом рептилий (различные черепахи, ящерицы, змеи и, возможно, крокодилы).
Фауны рептилий Северной и Южной Америки перекрывают друг дру- га с постепенным обеднением их в Центральной Америке и Мексике, но некоторые элементы этих фаун заходят значительно дальше: например. североамериканские черепахи подсемейства Emydinae достигают районов Аргентины, а южноамериканские представители семейства Teiidae — окра- инных частей Британской Колумбии.
Ареалы рептилий Индо-Малайской и Австралийской областей пере- крывают друг друга в восточной части Индо-Австралийского архипелага. Распространение рептилий Индо-Малайской области захватывает в зна- чительной степени площадь перехода и даже идет через нее: некоторые реп- тилии распространены (с постепенным обеднением) через все острова и встречаются в Австралии, в то время как распространение основных реп- тилий Австралии идет только до Молуккских островов.
Приводить дальнейшие подробности здесь не столь обязательно. В об- щем очевидно, что холод является эффективной преградой для распро- странения рептилий. Даже умеренный климат является частичной пре- градой для тропических рептилий. Но ни в одной части земного шара в те- чение довольно продолжительного времени не было настолько холодного климата, чтобы навсегда остановить расселение всех рептилий. Влияние других видов преград на разных рептилий сказывается по-разному. Сухие территории являются преградой для водолюбивых рептилий, влаж- ные — для пустынных. Небольшие пространства соленых вод останавли- вают одних и не являются преградой для других. Все эти преграды задер- 'кивают расселение рептилий или останавливают распространение неко- торых из них, но ничто не смогло остановить распространение по всему земному шару таких доминирующих групп, как, например, семейство Co- lu bridae.
Доминирование и конкуренция в связи с распространением рептилий
Наиболее доминирующей группой современных рептилий является семейство Colubridae (ужовые). Только к одному этому семейству отно- сится около '/, всех современных змей. В еще большей пропорции у;ковые представлены среди змей, находящихся в обычных для них местообита- ниях, кроме роющих червеподобных и морских змей. Ужовые богаты ро-
Penmuë и и
177
дами и видами во всех основных областях земного шара, за исключением Австралии, и часто отдельные роды или виды в свою очередь также мно- гочисленны. Не будучи очень разнообразными по своему строению, ужо- вые отличаются экологическим разнообразием и представлены много- численными наземными, древесными, роющими и водными формами. Они доминируют в обеих главных климатических зонах (тропической зоне и умеренной зоне северного полушария) в разнообразных местах обита- ния. В умеренной зоне они многочисленны как на облесенных, так и на открытых пространствах и как во влажных, так и в засушливых местах. Некоторые ужовые подходят близко к северной границе рептилий в Евра- зии п определяют ее в Северной Америке. Таким образом, распространение семейства Colubridae на большей части земного шара, независимо от кли- мата и преград, по сравнению с другими рептилиями почти сплошное. Все эти факты, вместе взятые, делают это семействой особой доминирующей группой, в общих чертах сопоставляемой с семейством Cyprinidae у прес- новодных рыб или с родом Лапа у лягушек.
Ни одно семейство рептилий не может сравниться с семейством ( olu- hridae по степени доминирования на земном шаре в целом, несмотря на то, что некоторые из них доминируют на ограниченных территориях. Напри- мер, черепахи подсемейства Emydinae доминируют среди черепах на опре- деленных территориях. Они богаты родами и видами, а также числом от- дельных особей главным образом в тропической части Азии, но занимают второстепенное место в умеренных широтах восточной части Северной Америки. Их распределение даже в пределах собственных границ распро- странения неравномерно. Они разнообразны экологически: имеются раз- личные водные, земноводные и наземные формы.
Расцвет и расселение доминирующих ужовых, по-видимому, оказа- ли большое влияние через конкуренцию на других змей и, возможно, еще более значительное — на ящериц: вполне вероятно, что локализация и прерывистое распространение многих семейств ящериц частично результат конкуренции со змеями и особенно с семейством (;olubridae. Однако фак- тических данных по этому вопросу очень мало. Мы не знаем, как змеи и ящерицы были распространены до появления ужовых, и мы не знаем. где впервые появились сами ужовые и как они расселялись. Ископае- мые данные не помогают разрешить этот вопрос. В распространении ужо- вых надо отметить одну существенную деталь — территории доминирова- ния (но не ареалы) в целом семейства "olubridae и Elapidae взаимно заме- няют друг друга: Со1иЬгЫае доминируют на большей части земного шара, :lapidae — в Австралии.
Если ужовые и не заместили большинства аспидов, то они по край- ней мере ограничили их радиацию за пределы Австралии. В границах семейства ужовых у производных групп наблюдается очевидная тенденция к концентрации на оольшей части земного шара, а у примитивных — к Юж- ной Америке и на Мадагаскаре, что позволяет предположить сложную кон- куренцию и замещение групп, происходящие на большей части земного шара. Более четкие признаки замещения на большей части Старого (' лета показывает викариирующее распространение игуановых и агамовых: первые находятся в Америке, а их реликты — на Мадагаскаре и островах Фиджи, вторые — на материках Старого Света. И еще более четко выраже- но замещение бокошейных черепах, в прошлом космополитов, доминирую- щими черепахами подсемейства Emydinae в Индо-Малайской области и в умеренной зоне северного полушария (рис. 29). на что указывает распро- странение как ископаемых, так и современных форм.
1'лава 4
17'
11ри отсутствии прямых данных самым лучшим доказательством кон- куренции между разными группами животных является замещающее рас- пространение доминирующих экологически эквивалентных групп. Данные такого рода можно найти и среди рептилий. Если рептилии обнаруживаю1
1' и с. 29. Радиация черепах подсемейства Emydiuae (схематично)и рас11ространенпе боко111ейных черепах.
менее выраженную взаимозамещаемость распространения, чем пресно- водные рыбы и амфибии, то это еще не значит, что конкуренция среди реп- тилий ослаблена. 11росто они расселяются быстрее и легче пересекают пре- грады, в связи с чем викариирующий характер распространения, проис- ходящий за счет конкуренции и замещения, быстро теряется.
Заключение: особенности распространения рептилий
Общий характер распространения рептилий сводится к тем же пяти особенностям, которые были отмечены у пресноводных рыб и амфибий, но некоторые из них видоизменены или выражены слабее. Первая особен- ность — ограниченность. Распространение рептилий в направлении к се- веру более ограничено, чем у пресноводных рыб или амфибий, но менее ограничено на островах. Вторая особенность — зональное распростране- ние видоизменено, оно выражается лишь в том, что рептилии являются главным образом тропическими животными. Края тропической фауны про- стираются на север, но самостоятельной фауны рептилий и умеренной зоне северного полушария нет. Третья особенность — радиация у рептилий менее заметна или более сложна, чем у пресноводных рыб или амфибий. Если существует основной центр географической радиации рептилий, то он, веро- ятно, находится на основной части тропиков Старого Света, которая являет- ся главным центром распространения черепах семейств Testudi»idae (Ету- dir>ae плюс Testudi»inae), Trionychidae, сцинков, наиболее доминирукнцих ужовых и гадюковых змей. Четвертая особенность — дифференциация различных фаун рептилий на разных континентах очень сложна. Единого основного принципа дифференциации в фауне рептилий не существует,
Рептилии
скорее можно говорить о ряде отдельных черт, характерных для разных групп, каждая из которых охватывает большую часть или весь земной шар. Черепахи имеют свой сложный характер распространения на всем земном шаре, в котором (оставив в стороне сложность) основная дифференциация происходит между фауной Emydinae в Евразии и Северной Америке и фау- ной бокошейных черепах на южных континентах. Среди агамовых и игуа- новых, а также змей семейства Boidae основная дифференциация наОлю дается между Старым и Новым Светом. Что касается семейств:oluhridae и ilapidae, то основная дифференциация происходит между Австралией и остальной частью земного шара. Другие, более мелкие группы репти- лий, которым свойствен свой характер распространения, увеличиваюч эту сложность. Пятая особенность — концентрация на больших и благо приятных территориях с обеднением фауны по краям у рептилий выражена слабее, чем у пресноводных рыб и амфибий. Рептилии, главным образом наземные животные, почти везде встречаются в большом количестве, кроме слишком холодных мест. Они сосредоточиваются в основном в теплых рай- онах, а особые группы концентрируются местами в более отдаленных раи- онах. Например, в прохладном климате ящерицы стремятся сосредото- читься на открытых солнечных местах; черепахи — на больших площа- дях с благоприятными для них условиями, а именно Emydinae в восточной части тропической Азии и восточной части Северной Америки. Однако у большинства рептилий такая концентрация не наблюдается. Там. где разные фауны рептилий соприкасаются, обычно происходит сложный переход многих групп на обширной территории.
Реликты в общем характере распространения рептилий занимают опре- деленное место. Черепахи являются реликтовой группой. Они существуют, почти не изменяясь, со времен триаса и наибольшего разнообразия достигают в тропиках, но некоторые из них проникают довольно далеко и в умерен- ную зону северного полушария. Крокодилы также являются реликтовой группой. Они ведут свое начало с триаса и являются единственными сохранившимися архозаврами. В настоящее время обитают в тропиках или волизи тропиков. Sphenodon — единственный сохранившийся представи- тель клювоголовых. Изолирован в Новой Зеландии и является филогене- тическим и географическим реликтом, занимающим исключительное поло- жение, так как ни одно из других древних пресмыкающихся не сохрани- лось в географической изоляции. В настоящее время Австралия уже не является убежищем ни для одной более или менее архаической группы реп- тилий, хотя последний представитель древней линии черепах Meiolania (см. список семейств) сохранился там до плейстоцена.
История расселения рептилий
Современный характер распространения рептилий очень сложен, сложнее даже, чем у пресноводных рыб и амфиоии. Он сложился в резуль- тате эволюции, радиации и расселения, адаптации, конкуренции, огра- ниченности, отступаний и т. д. многих различных групп. Фактическими характер сформировавшегося распространения определяется природой самих рептилий (особенно такими фактами, что они являются главным образом наземными животными, что они предпочитают теплые районы, но иногда могут заходить и в прохладные и что некоторые из них быстро рас- селяются и легко преодолевают преграды), зональностьн~ климата и дру- гими факторами, регулирующими распространение животных на земном
1лава 4
шаре. Не мешает проследить, насколько это возможно, перемещение основных групп рептилий. Здесь будут рассматриваться только современ- ные группы и относящиеся к ним ископаемые и только основные их перемещения в течение третичного периода или незадолго до него. Мезозойское расселение динозавров кратко рассматривается в гл. 10. Исто- рия расселения, описанная здесь, частично предположительна. В других местах книги и в списке семейств приводятся детали и «ключи», на которых основана история расселения рептилий.
В течение третичного периода обыкновенные черепахи (Kmydinae) радиировали в тропической Азии и оттуда через умеренную зону северного полушария расселялись в Южную Америку. Сухопутные черепахи Tes- tudo, происшедшие от Emydinae в Азии, расселились на значительной части теплых районов земного шара, кроме Австралии. Род Testudo, вероятно, достиг Южной Америки в миоцене, подсемейство Emydinae, воз- можно, в плиоцене. Мягкокожистые черепахи (Triongx) расселились из тропиков Старого Света в Восточную Азию и Северную Америку, но это было уже более позднее перемещение этих черепах в сложном дотретич- ном расселении. И мускусные, и каймановые черепахи (Chelydridae) про- никли из Северной Америки в Южную недавно. С другой стороны, боко- шейные черепахи (по крайней мере семейство Pelomedusidae) отступили из Евразии и Северной Америки; семейство Dermatemydidae отступило из умеренной зоны северного полушария и сократилось до одного вида в Мексике и Гватемале; семейство Chelydridae отступило из умеренной части Евразии, и последний представитель семейства Meiolaniidae, сущест- вовавший в Австралии до плейстоцена, исчез.
У ящериц, очевидно, несколько раз и в сравнительно недавнее время происходил обмен между Старым и Новым Светом. Сцинки из Старого (:ве- та проникли в Америку: роды Eumeces и lygosomo, через север в Северную и Центральную Америку, а род Мабиуа, вероятно, через Атлантику от Африки до тропической Америки. Род гекконов Tarentola, возможно, пере- сек Атлантику до Вест-Индии. Только несколько других родов ящериц обычны для некоторых частей Старого и Нового Света, но и их история расселения достоверно не установлена. Агамовые радиировали на конти- нентах Старого Света и, очевидно, заменили там игуановых, но не достигли Америки. В пределах Старого Света, а также ме'кду Северной и Южной Америкой происходили и другие перемещения ящериц, но здесь они не рассматриваются. Что касается отступаний, то игуановые отступили с кон- тинентов Старого Света, а некоторые представители семейства Anguidae и Amphisbaenidae — с некоторых северных территории; Varanidae отсту- пили из Северной Америки в начале третичного периода. Это примерно все то, что представляет существенное значение в отношении географиче- ского перемещения отдельных семейств, однако современное распростра- нение ящериц в целом, со многими разрывами и локализациями, позво- ляет предположить, что в недавние времена они больше отступали, чем расселялись. Возможно, что их отступание находится в корреляции с подъе- мом семейства Colubridae.
О древнем расселении змей (Typhlopidae, Aniliidae, Boidae и т. д.) ничего не известно. (Предполагаемое замещение удавов питонами на мате- риках Старого (;вета, очевидно, является домыслом, основанным на не- правильной классификации.) Семейство ужовых ((;olubridae) распростра- нялось сложным путем на большей части земного шара. Место их про- исхождения не известно, но недавняя радиация ужовых происходила из основной части тропиков Старого Света, откуда многие различные роды
Реп~иил ии
распространились на север и достигли Америки, а несколько родов— Австралии. Направление перемещения некоторых родов не ясно, но такие из них, как Natrix, Elaphe, Coluber и Opheodrys, по-видимому, отно- сятся к тем родам, которые проникли из Старого Света в Северную Амери- ку. Семейство Elapidae произошло от ранних Colubridae, вероятно, где-то в Старом Свете. По неизвестной причине, как уже упоминалось ранее, они достигли Австралии раньше ужовых, а один их вид достиг Америки и noJIoihBJI начало аспидам. Гадюки также, вероятно, произошли от ужовых на основной части Старого Света. Ямкоголовые змеи, вероятно, возникли (от настоящих гадюк) в южной Евразии, и две или три ветви их достигли Америки, от одной из которых произошли гремучие змеи. Деталей, каса- ющихся отступания змей, немного. Некоторые группы, очевидно, отсту- пили с северных территорий. Другие (Aniliidae, Erycinae, Boidae. неко- торые подсемейства ужовых) имеют дизъюнктивный ареал, на основании чего можно считать, что были и более широкие отступания.
Настоящие крокодилы (Crocodylus) расселились в раннетретичном периоде из Старого Света в Америку, но это был уже последний этап слож- ного расселения крокодилов. Ареалы рода Alligator и современных родов гавиалов в настоящее время сократились.
История расселения водяных змей заслуживает особого внимания. Различные змеи во многих частях земного шара в той или иной степени приспособлены к водной среде, но три группы из них более приспособлены к жизни в воде, чем все остальные, благодаря наличию ноздрей с клапа- нами (М. А. Smith, 1943, р. 17 — 19) и иногда плоскому хвосту. К этим груп- пам относятся подсемейства Acrochordinae и Homalopsinae, происшед- шие от ужовых и давшие отдельную ветвь, и Hydrophiidae, или морские змеи, происшедшие от аспидовых. (Подробности распространения дан- ных групп приводятся в списке семейств.) Все эти три группы, являющиеся главными современными группами настоящих водяных змей, по-видимо- му, возникли в одном и том же месте — в тропической Азии и в Индо-Ав- стралийской области. Судя по их предкам (соответствующих ископаемых нет), они, вероятно, возникли в третичном периоде или незадолго до его начала.
Ряд доминирующих рептилий, по-видимому, переместился из Старого Света в Америку. К ним относится несколько представителей Emydinae, Testudo и Trionyx; три рода сцинков, несколько родов ужовых, один вид аспидовых, два или три вида ямкоголовых змей и крокодилы. Перемести- лись ли какие-нибудь рептилии из Америки в Старый Свет (некоторые из них могли переместиться), до сих пор остается неизвестным. Имеется также несколько групп рептилий, которые, по-видимому, отступали с не- которых частей Старого Света и сохранились в Америке: это черепахи семейства Dermatemydidae, Chelydridae, игуановые и, вероятно, примитив- ные ужовые («Xenodontinae»); но представители семейства Varanidae из Северной Америки отступали в другом направлении. Сокращение ареалов одних групп, вероятно, могло быть вызвано расширением ареала других и могло протекать в том же направлении. Хотя сказанное и не является прямым доказательством, но свидетельствует о том, что у рептилий в боль- шей степени существовала тенденция к перемещению из Старого Света в Америку, чем в обратном направлении. Более того, рептилии являются в основном тропическими животными, которые часто проникают в умерен- ную зону северного полушария, но обычно там не преуспевают. Кроме того, рептилии, по-видимому, распространились с основной территории Ста- рого Света по направлению к Австралии (см. стр. 171). Это позволяет при-
1'дава 4
нимать большую часть тропиков Старого Света, Африку плюс тропическая Азия за очевидный основной центр географической радиации рептилий, поскольку он существовал. По крайней мере имеются признаки того, что последовательные группы доминирующих рептилий возникли и распрост- ранились оттуда, по-видимому, так же, как и последовательные домини- рующие группы пресноводных рыб и лягушек.
По современным рептилиям трудно что-либо сказать о древних конти- нентах и климатах. Однако для справки можно привести несколько осо- бых случаев, остальные приводятся в гл. 10.
Черепахи семейства Pelomedusidae и ящерицы Amphisbaenidae пол- ностью или почти полностью приурочены к Африке и теплым частям Аме- рики, но по ископаемым данным можно сказать, что их расселение шло через север. Слепуны, Leptotyphlopidae, в большинстве случаев африкан- ские или американские, возможно, также расселились через север. Яще- рицы рода МиЬиуа и Тагеп1о1а, возможно, достигли Южной Америки или Вест-Индии непосредственно из Африки, но их путь мог проходить не только по суше. Не исключено, что, относясь к семействам, которые спо- собны преодолевать водные преграды, они пересекли Атлантику на
ПЛОТУ.
Черепахи семейства Chelyidae встречаются только в Южной Америке и Австралийской области и нигде больше неизвестны даже как ископаемые. В Южной Америке они почти полностью приурочены к тропикам и не до- стигают прохладных южных частей континента. В Австралийской области встречаются как в тропических, так и в умеренных районах, но не в Тас- мании и Новой Зеландии. Несмотря на то что они относятся к пресновод- ным черепахам, их распространение не совпадает с распространением прес- новодных рыб или любых других водных позвоночных. Следовательно, они расселялись из Южной Америки в Австралию другим, не совсем обыч- ным путем, возможно, что это был морской путь.
Распространение черепах семейства (;helyidae соответствует распро- странению исчезнувших черепах семейства Meiolaniidae, которые встре- чались в южной части Южной Америки примерно в начале третичного периода и в Восточной Австралии, на островах Лорд-Хау и Уолпол, лежа- щих к юго-востоку от Новой Каледонии, в плейстоцене. Это были гигант- ские сухопутные черепахи. Некоторые факты наводят на мысль, что они могли расселиться через море. Во-первых, они появились в Южной Аме- рике одновременно с ранними плацентарными млекопитающими, которые, однако, не достигли Австралии. Вполне вероятно, что черепахи пересекли преграду, которую не смогли преодолеть млекопитающие. Во-вторых, ост- рова Лорд-Хау и Уолпол (а также Новая Каледония) в биологическом смысле являются океаническими островами, на которых отсутствуют стро- го пресноводные рыбы, амфибии и наземные млекопитающие. Появление там черепах в плейстоцене почти доказательство того, что они преодолели многие сотни миль океана. И, наконец, еще одно доказательство — со- временные сухопутные черепахи (экологический дубликат Xileiolaniidae) при своем расселении пересекли океанические пространства в сотни миль.
11редположение о том, что черепахи Meiolaniidae преодолели морские пре- грады между Южной Америкой и Австралией, было высказано в свое нре- мя Мэтью ( Iatthew, 1915, р. 238), и оно до сих пор является самым лучшим объяснением их распространения.
Таким образом, во всех случаях расселения имеется один общий фак- '<ор, которы11 дает объяснение распространению разных рептилий без привлечения теорий материковых мостов или перемещения материков.
Penruuë ии
В заключение можно сказать, что современные рептилии имеют очень сложное распространение, которое крайне трудно свести и общим чертам распространения других животных. Исключением являются только чере- пахи.
СПИСОК,EMEl3.СТВ РЕИТИ.ЧИ14
В этот список включены все современные группы рептилий и только несколько и«1;онаемых групп, главным образом те, которые непосредственно связаны с совре- менными, и те, которые важны с, географической точки зрения. Классификация соответственно сокращена, но во всем остальном она дается но Ромеру (Romer, 1,)56). Знак звездочки (") перед латинским названием обозначает вымершую группу.
1К)ДК.1ЛСС ANAPSIDA
Отряд Chelonia (Testudinata), черепахи. Ромер (а также и я) придерживается клас- сификации, данной Вильямсом (Wil]iams, 1950, р. 554), с небольшими измене- ниями. Карты распространения черепах в Северной Америке приводятся в рабо- тах Kappa (Carr, 1952).
* и о д о т р я д А m p h i с, Ь е 1 у d i а, состоящий из 10 семейств, вкл1очая следую- 1цие:
'A1eiolaniidae, вымершие гигантские сухопутные черепахи, известные только в южноц
части Южной Америки (2 рода в позднемеловом периоде и в эоцеие) и в восточной части Австралии на островах Лорд-Хау и Уолпол, к юго-востоку от Новой Кале- донии (другой род в илейстоцене). Расселение этой группы рассматривается в тек- сте. Некоторые ссылки, имеющие отношение к этому семейству, можно найти в работах следу~оп(их авторов: Woodward, 190~; Matthew, 1915, р. 283; 1939, 1ó. 113, 114; Anderson, 1925; Simpson, 1938; 1943, р. 42', 423. черепахи семейства A1 eiolaniidae были последними сохранившимися представителями подотряда, появившегося в Фриасе, который доминировал и, вероятно, имел космополитное распространение в мезозое (Romer), но который (за исключением ископаемых нз илейстоцена Австралии и т. д.) не известен позднее эоцена.
ll одотряд (;ryptodira
(Надсемейство Testudinoidea)
1)ermatemydidae. Южная Мексика и северная часть Центральной Америки (по рекам
бассейна Атлантического океана, от центральной части Веракруса до Гватема- лы — Н. М. Smith, Taylor, 1950, р. 19). 1 вид. Ископаемые роды, условно отне- сенные к этому семейству, найдены в Северной Америке и Восточной Азии в позднемеловом и раннетретичном периодах.
1:lielydridae, каймановые, мускусные черепахи и др. Северная, Центральная и часть
К)жной Америки. Часто повторяемые данные о предполагаемом эндемичном роде каймановой черепахи на Новой Гвинее оказались ошибочными (Loveridge, Shre- z е, 1947). 3 подсемейства — все пресноводные. (1) Сhelydrinae; каймановые чере- пахи. Северная и Центральная Америка и т. д. 2 рода: Chelydrn (I или 2 вида)— в Соединенных Штатах к востоку от Скалистых гор, достигая в северном направ- лении южных краевых частей Канады, и в некоторых районах Мексики, Цент- ральной Америки и в северо-западной части Южной Америки (Эквадор) и ЛХасго- clemys (1 вид) — в юго-восточной части Соединенных Штатов от северной Флори- ды до центрального Техаса, в северной части долины Миссисипи до центрального >>ääHnoéña и т. д. (2) Staurotypinae. Южная Мексика и северная часть Централь- ной Америки (Веракрус до Гватемалы и Британского Гондураса) (Smith, Tay- lor, 1950, р. 26, 27). 2 рода, 3 вида. (3) Подсемейство Kinosterninae, мускусные черепахи. Восточная часть Северной Америки на север до южного Онтарио, ца н>го-запад до южной внутренней части Калифорнии, а также на н>г Мексики, Цеи- традьцая Америка и примерно северная половина Южной Америки на юг до Чако (~~'11liams). 1 или 2 рода, 12 или более видов.
Ископаемые подсемейства Chelydrinae известны из средпетретичного перцо- <;> Северной Америки и (сомнительные) Европы. Staurotypinea — из олигоцеиа Северной Америки (Williams, '1952a) и Kinosterninae — из плиоцеиа Северииой Америки. Эти, хотя и далеко не полные, ископаемые данные предполага~от радца- 11и1о семейства из Северной Америки. Вильямс считает, что это семейство суц~ест- вовало «если пе с мелового, то уж, несомненно, с самого начала третичного периода».
Тев1iп1~иЫае, обыкновенные черепахи, сухопутные и др. Сухопутные черепахи (Testu- dininae) встречаютгя в большинстве районов земного шара с теплым климатом, кроме Австралийской области. другие, главным ооразом водные по[семей-
Глава 4
ства, ограничиваются районами, указанными ниже. Существует 3 таких под- семейства.
(1) Platysterninae. Южная Бирма, Таиланд, гого-восток Индокитая, крайняя н>жняя
часть Китая и Хайнань. 1 вид, обитающий в горных реках.
(2) Emydinae, обыкновенные пресноводные черепахи (рис. 29). Тропическая и учерен- ная зоны Евразии и Америки. Большипство находится в гиропичесчогс Восточной Азии и в умеренной восточной части Северной Америки. Несколько представи- телей имеется в других частях умеренной зоны северного полушария: один достигает тропиков северо-западной части Африки и несколько находятся в Цент- ральной и Южной Америке и в Вест-Индии. В В осточной Азии и т. д. (М. А. )mltll, 1931; Роре, 1935; Bourret, 1941; Rooij, 1915) распространено примерно 17 или более родов, 38 видов. Ареал большинства из них протянулся от северной Индии и крайнего юга Китая до островов Суматра, Ява и Калимантан. Только 6 родов достигают северной 1Лндии и 1 вид (Geoemyda trijuga) — южной 1Лндии и Цейло- на, где его популяция изолирована (М. А. Smith, 1931, р. 97). В Китае к северу от южных провинций Гуанси, Гуандун и Фуцзянь известно только 4 вида (Рор», 1935). 2 вида — Clemmys mutica и Geoclemmys (или Chinem~ts) reevesii, заходят далеко на север, достигая Чжэньси, южпого Шапьдуня и Хубея соответственно, но естественные пределы второго вида неопределенны, так как его завозят в различ- ные районы для продажи. В Японии встречаются 6. )'ееге8гг и эндемичшш род Clemmys, распространенный к северу по крайней мере до Токио. Два других пред- ставителя этого подсемейства находятся на островах Рюкю и несколько — нд Тайване. На Филиппинах (Taylor, 1920) имеется 1 предположительно знлемич- ный вид в южной части острова Лейте и 2 широко распространенных вида в И н- до-Малайской области, из которых один — Cyclemys (илп Сиога) amboise<.àsis, обычно держат в качестве домашнего животного. На Филиппинах также были зарегистрированы и другие представители этого подсемейства, но эти данные недостоверны. На острове Сулавеси и других, лежащих восточнее островах нахо- дится, вероятно завезенный человеком, тот же самый вид С. amboinensis сшироким, но неравномерным распространением (Mertens, 1930, р. 335). В Юго-Западной Азии, в некоторых районах Европы и на северо-западном побережье Африки имеется 2 рода, 3 вида. Один из них (Clemmys lep rosa) распространен до западной тропической части Африки. Он был зарегистрирован в реке Гамбия (Gray. 1852) как Emys laticeps, в Порто-Ново, Дагомея (Chabaunaud, 1917) и в Эйре, южно- центральная часть южной Сахары (Angel, Lhote, 1938, р. 376). В Америке, к вос- току от Скалистых гор, в северных районах Мексики и к северу до южных крае- вых частей Канады, распространено 8 родов, 18 видов и множество подвидов (Carr). В Мексике находятся и другие виды некоторых родов. Единственная восточная форма Chrysemys picta bellii, пересекая Скалистые горы в северо-запад- ном направлении, достигает южной части Британской Колумбии и др., а также Каскадных гор и, возможно, побережья Тихого океана. Эвенден (Evenden, 1948} считает, что этот вид довольно обычный в более низких частях долины Уилламет, штат Орегон, но естественные западные границы распространения этого вида точ- но не установлены. Один вид C lemmys ma rmo rata является эндемиком на поое- режье Тихого океана от южной части Британской Колумбии до северной части Нижней Калифорнии, но его северная граница не установлена (Evenden. I 948).
В южном направлении 2 восточных рода из Северной Америки достигают тропи- ков: род наземных черепах Terrapene (коробчатая черепаха) — до южной Мек- сики, Наярита и северного Юкатана (Н. М. Smith, Taylor); водный род Pseu- demys (включающий и Trachemys, Simpson, 1943, р. 419) встречается по всей Цент- ральной Америке, частично в Вест-Индии, на большей части Южной Америки до северной Аргентины. В Южной Америке ареал этого рода прерывистый. Вильямс (Williams, 1956) указывает, что данных о встречаемости этого рода между Р>ене- суэлой и южной Бразилией нет. Из других представителей подсемейства F.mydi- nae в тропиках Америки имеется несколько видов рода Geoemyda, распространен- ных в Южной и Центральной Америке до Мексики (до южной Соноры и централь- ного Веракруса), но не далее. Этот род встречается также в тропической Азии, к северу до южных, краевых частей Китая и до островов Ртокю. Роды СЬтт~у,~ и Geoemyda — единственные из подсемейства Emydinae, которые теперь считан)те~~ общими для Старого и Нового Света. Европейские и восточные виды Сев< ршн~ Америки, в прошлом помещаемые вместе в род mys, в действптельностп не род- ственны (Williams). Болыиинство обыкновенных черепах (Fmydinae) относится к водным формам, но некоторые из них отчасти или полностьго наземггые. Р<гд Geoemyda включает и водные и наземные виды и в основном является растител ь- ноядным (М. А. Smith, 1931, р. 89, 90). Пища других черепах этого подсемейства различна. Согласно Вильямсу (Williams, 1950, р. 554), самые ранние достоверные
.Penmuëии
185
ископаемые, приписываемые этому подсемейству, относятся к палеоцену Север- ной Америки, но они известны также из эоцена Европы. Распространение совре- менных форм указывает на то, что основная радиация подсемейства происходила в Юго-Восточной Азии. Ископаемые, представляющие род Geoemydg, который теперь считается общим для тропиков Азии и Америки, известны из эоцепа Европы и олигоцена Северной Америки (Williams).
(3) Testudininae, сухопутные черепахи. Теплые районы континентов, кроме Австра- лии, и многие острова. Вильямс (Williams, 1952) выделяет 6 родов. 4 из них с несколькими видами приурочены к тропической и 10жной Африке и к 11ада- гаскару. Другой род Сор/гегия с 3 существующими видами имеет прерывистое ðac- пространение через южную часть Северной Америки, от Южной Каролины п фло- риды вдоль побережья до Техаса и северо-восточной Мексики и на ограпцчеицой территории в юго-западной части Соединенных Штатов и в северо-западной Мек- сике. В настоящее время предполагают, что этот род представляет группу (в от- личие от Testudo), которая находилась в Северной Америке по крайней мере нач11- ная с олигоцена, а также в Европе в третичпом периоде (Williams). Шестой род, Testudo (примерно с 50 современными видами, распространенными в Африке, на Мадагаскаре, в субтропических и тропических частях Евразии, в Южной Аме- рике, частично в Панаме) в Северной или Центральной Америке не встречает- ся. В Африке, особенно в южной, виды этого рода многочисленны. На Мадагаска- ре имеется всего лишь несколько видов. В историческое время гигантские формы черепах населяли более 30 островов в Индийском океане, включая острова Аль- дабра, Реюпьон, Маврикий, Родригес и Сейшельские, но из этих гигантов (чис- ло существовавших форм там не известно) только одна местная форма на одном (самом южном) из островов Альдабра сохранилась до настоящего времени. ll ев- ропейских странах Средиземноморья, на Ьалканах и в юго-западной части Азии на север до Мариуполя [ныне город Жданов.— Ред.'! (Берг, 1947), а также в paii- онах Каспийского и Аральского морей имеется 6 видов. В тропической Азиц— 7 видов, из которых 1 достигает Цейлона, 1 — крайнего юга Китая и 1 — остро- вов Суматра, Калимантан и, вероятно, Ява; восьмой вид — эндемик на островах Сулавеси и Хальмахера. В Южной Америке 2 вида.' 1 широко распространен в тропиках к востоку от Анд, на север до Панамы и 1 — в более теплых частях умеренной зоны Южной Америки. II, наконец, на островах Галапагос обитает ряд гигантских форм (некоторые теперь вымерли или редки), рассматриваемых Мертенсом (Mertens, 1934) как 13 подвидов, принадлежащих к одному виду.
Достоверных сведений об ископаемых этого подсемейства ранее эоцена не ииеет- ся (Williams). Род Testudo был широко распространен начиная с эоцена, хотя, очевидно, он никогда не достигал Австралии, а Южной Америки достиг, вероят- но, не ранее миоцена (Simpson, 1943, р. 420). В Северной Америке в настоящее время этот род отсутствует, хотя он существовал там с эоцена до плейстоцена. Его также нет и в Центральнои Америке (за исключением Панамы) и Вест-Индии, хотя некоторые виды обитали на Кубе, Мона и островах Сомбреро (Williams, 1950, р. 6; 1952, р. 552) и, по-видимому, на других островах в плейстоцене. '1ере- пахи подсемейства Testudininae, достигающие в настоящее время наибольшего разнообразия в Африке и на Мадагаскаре, вероятно, произошли от Emydinae в тропической Азии. Согласно расположению Вильямса (Williams, 1952) иско- паемых и современных видов рода Testudo в 14 подродах, можно предположить, что расселение этого рода было сложным.
(Надсемейство Chelonioidea)
Chel0niidae, морские черепахи. Теплые моря. 4 современных рода, видов мало. (Надсемейство Dermochelyoidea)
Derm0chelyidae, кожистые черепахи. Теплые моря. 1 современный род, 1 — 2 вида. (Надсемейство Carettochelyoidea)
Carett0chelyidae. Приурочено к Новой Гвинее. l вид — исключительно водны~~, «во- дится в реках и спускается в море» (Rooij). Род Carettochelys существует в Новой Гвинее с миоцена, другие довольно отдаленно родственные роды обитали в Евра- зии и Северной Америке в раннетретичном периоде.
(Надсемейство Trionychoidea)
Trionychidae, мягкокожистые черепахи. Тропическая Африка, Южная и Восточная
Азия и т. д. и восточная часть Северной Америки. 7 родов. Широко распростра- нен только 1 род Trionyz (часто называемый Ainyda). В тропической Африке- ! вид рода T rioiiyx и 2 эпдемичных рода примерно с 5 видами. T rio~~yx спу«кается по Нилу до его низовьев, изредка встречается на побережье Палестины (! !обжег, 1933, р. 753 — 754). Другой представитель рода распространен в системе Тигра и Евфрата в Юго-Западной Азии. В Юго-Восточной Азии имеется около 5 родов (включая и Тлопух) п !2 видов, большинство из которых распространено от се-
1'лава, 4
верной Индии и крайнего юга Китая до островов Суматра, Ява и Калимантан.
Один из этих видов достигает Цейлона, другой — Peloclzelyis (>ibroni (cantnrii), «часто вылавливаемый на морском побережье» (Hooij), достигает остров» Лусон, Филиппины и имеет изолированную популяцию на ! l îâîé Гвинее, н(~ ;другие сведения о мягкокожистых черепахах, существуинцих за пределами Явы и )калимантана, сомнительны. ! вид рода Тгio»yx, следуя по долинам Восточ- ной Азии, в северном направлении доходит;со реки Амур или по крайней мере до озера Ханка и реки Уссури (Берг, 1947). 1, возможно, тот же самый вид, находится в Японии. В Америке имеется только род Т лопух с 2 ви- дами и несколькими подвидами, широко распространенными в восточно-централь- ной и южной частях Соединенных Штатов Америки и достигающими южноЙ части Онтарио и северо-восточной Мексики. Этот род иптродуцировап в западном направлении в системе Хила — Колорадо. Мягкокожистые черепахи ведут иск- лючительно водный образ жизни. Ископаемые показывают, что у них была доволь- но сложная история расселения на основных материках Старого Света и в Север- ной Америке начиная с мелового периода и, что, очевидно, они никогда не дости- гали Южной Америки. Принадлежат ли разрозненные ископаемые остатки, найденные в плейстоцене северо-восточной Австралии (de Vis, 1894), мягкокожи- стым черепахам, не установлено (Williams), но если они и принадлежат этому семейству, то, возможно, относятся к виду Pelockelys bibroni (описапному выше), которыц заходит в море и до сих пор сохранился на Новой Гвинее и которы~~, вероятно, достиг Австралийской области сравнительно недавно.
П одотряд Р leurod ira, бокошейные черепахи
Pelomedusidae, пеломедузы. Тропическая и Южная Африка, Мадагаскар и Южная
Америка. 3 рода, все более или менее водные. Африканские формы были пересмот- рены Лавриджем (Loveridge, 1941). Единственный вид Pelomedusa, встречается в Африке и на Мадагаскаре. Род Pelusios имеет 5 видов в Африке. Один из них достигает островов Зеленого Мыса и Мадагаскара, Сейшельских островов и острова Маврикия. Очевидно, ни этот вид, ни вид рода I'elomedusa не отличаются друг от друга ни на Мадагаскаре, ни на других островах (Loveridge), и несомненно, что расселению этих видов отчасти способствовал человек. Из рода I'odocnemis 1 «ид распространен на Мадагаскаре и 7 — в Южной Америке (Williams, 1954), в ре- ках Амазонка, Ориноко, Магдалена и др., но ни одного вида нет в системе ре« Парагвай — Парана, а также в Центральной Америке. В Африке Podocnemis существовал только в плейстоцене, а на других материках — в более раннее вре- мя, но некоторые ископаемые, приписываемые этому роду, вероятно, к нему не относятся (Zangerl, 1947; Williams). Однако какие-то виды пеломедуз находились на «сех главных материках, кроме Австралии, в меловом или раннетретичном периоде, а в Европе и Азии они просуществовали вплоть до миоцена и плиоцена.
Есть еще старые данные, относимые к верхнемеловому периоду Новой Зеландии (Lydekker, 1899), но они основаны на сомнительных ископаемых остатках обломочного характера (Williams). С географической точки зрения эти остатки, скорее, должны принадлежать представителю семейства Ch.elyidae, чем Ре1оте- dusidae, но, с другой стороны, ископаемые остатки рода Podocp>emis были обна- ружены в морских отложениях, откуда следует, что эти черепахи заходили в море. 'he1yidae, змеиношейные черепахи. Австралийская область и Южная Америка. В As- стралии (но не на Тасмании) и Новой Гвинее и т. д. — 4 рода и около 14 видов.
В Южной Америке — 6 других родов, около 13 видов, главным образом в тро- пиках (на юг до северной части Аргентины), но не в бассейне реки Магдален» или в бассейнах западных рек (Simpson, 19~3, р. 422) в Центральной Америке нет. Все семейство — в основном водное. Ископаемые из Евразии и т. д., ранее приписываемые этому семейству, в лучшем случае сомнительные (Williams, 1953. 1954а). Достаточных доказательств, что эти черепа~и когда-либо встреча- лись за пределами Южной Америки и Австралийской области, нет. Предполагае- мый род Hyd<aspis из эоцена Индии в действительности относится к семейству Pelomedusidae (Williams, 1953).
11ОДНЛАС(.' LEPIDOSAURIA, чешуйчатые
Отряд Rhynchocephalia, хоботноголовые
Sphenodontidae. Новая Зеландия. 1 существующий вид 5pheiiodop<, гаттерия.
В прошлом эти ящерицы населяли в большом количестве оба главных остров» Новой Зеландии. В настоящее время они встречаются только па нескольких островках, расположенных к северо-востоку от заливов острова Северныи, и на островах в проливе Кука. Это ящерицеобразное пресмыкающееся, достигающее в длину 2 футов и более. В прошлом гаттерии часто посещали песчаные отмели рек, а также (как и теперь) скалистые и песчаные островки в Море. Гаттерия относится к земноводным животным, любит лежать в воде
Реты илии
и хорошо плавает. Активна главным образом ночьн> (даже в прохладные или колодные ночи); температура ее тела значительно ниже, чем у современных реп- тилий, и уровень обме»а веществ низкий (Bogert). Взрослые особи месяцами мо- гут жить без пищи или пресной воды. Яйца откладывает в песок, где они разви- »я»>тся очень медленно, примерно в течение года. (Болыиая часть этих сведений почерпнута из следующих работ: Hutton, Drummond, ) 923; Хежтап, 1877: бadow, 1909, р. 298 — 300; Сonrad, 1940; Bogert, 1953). Ископаемые показыван>т, что хоботноголовые существовали на большинстве континентов в раннем и среж- »ем мезозое, но их ископаемые позднее нижнемелового периода не известны (при- мерно 135 млн. лет назад). Поразительно, что хоботноголовые смогли сохранить- «я до настоящего времени вообще, однако то, что такое животное, как гаттерия, достигло Новой Зеландии, менее удивительно, чем то, что ее достигла лягушка. Отряд ~quanIata
Подотряд Ьа certi li а, ящерицы
(Инфраотряд Ignania)
1~~~~а»Ыае, игуаны. В Северной и Южной Америке. 2 реликтовых рода — на Мадагас- каре, 1 — на островах Фиджи и Тонга в западной части Тихого океана. В Амери- ке существует примерно 50 родов, сотни видов, большинство из которых обитает в тропиках Южной и Центральной Америки и Вест-Индии, а некоторые и в уме- ренных зонах. Из нескольких родов, распространенных в умеренных зонах(;е- верной Америки (главным образом на западе), большинство северных игуан до- стигает южной части Британской Колумбии, южной Альберты, юго-западной части Северной Дакоты, Индианы, Пенсильвании и южного Нью-Йорка. К н>гу 11о крайней мере I род l.io1aemus доходит до Огненной Земли. Места обитания «мериканских игуан очень разнообразны: пустыни, равнины, леса и джунгли.
Много наземных, древесных и несколько полуводных форм. Большие древесные игуаны встречаются в тропической Америке. Крупные наземные формы различ- ных родов встречаются как на континенте, так и в Вест-Индии и на Галапагос- ских островах. Из многочисленных родов более мелких форм древесный род А»o- lis имеет 250 или более видов и подвидов в северной части Южной Америки, Цент- ральной Америке и Вест-Индии и т. д. и 1 эндемичный вид («американский ха- мелеон») — в юго-восточной части Соединенных Штатов на север до Каролины.
Род Ь'се1орогиг более чем с 100 наземными или полудревесными видами и подви- дами (H. М. Smith, 1946, р. 179; Smith, Taylor, 1950, р. 105) распространен от южной части Британской Колумбии и южного Нью-Иорка до западной Панамы (H. М. Smith, 1939). Самая большая концентрация видов этого рода отмечается в Мексике. Ряд родов, живущих в земле, приурочен к влажным районам запад- ной или юго-западной части Соединенных Штатов и северной Мексики. Другие «мериканские роды слишком многочисленны, чтобы перечислять их здесь. На Мадагаскаре находится род Ckalarodofi, c i видом, локализованным в сухих пес- чаных районах н>го-запад»ой части Мадагаскара, и Ор1игия (Hoplurus) — при- мерно с 6 видами (это мелкие формы, напоминающие представителей американ- < ких родов, живущих в земле). Кэмп (Сатр, 1923, р. 308) подтверждает, что род (..'йа1а rodo~i действительно является представителем игуановых. На островах Фи 1- жи и Тонга встречаются I:эндемичный род и вид Brach>tlophus fasciatus, изящ- »ая игуапообразная древесная ящерица (Loveridge, 1945, р. 92).
По ископаемым (в большинстве случаев неудовлетворительные фрагменты)
можно предполагать о существовании. игуановых в Северной Америке в третич- пом периоде или до его начала, но история этого семейства остается неясной.
Ископаемые игуаны были найдены также и в Европе, но, судя ио литературе ((:amp, 1923, р. 309; Hoffstetter, 1942), этот вопрос остается открытым, хотя современное распространение семейства почти не оставляет сомнений в том, что в прошлом игуановые все-таки встречались на материках Старого Света. Agani i ~1 «е, агамы. Основные тропические и субтропические части Старого Света,
кроме Мадагаскара. Наиболее многочисленны в Индо-Малайской области. ho- :>ее 30 родов и около 280 видов (М. А. Smith, 1935, р. 132). Одни из них назем- ные, другие — древесные. К последним относится «летаюп[ий дракон» (Draco).
К северу род agama достигает южной части Греции и встречается к востоку от Кас- пийского моря. Несколько родов распространено в Центральной Азии, и самые северные представители достигают Синьцзяна и центральной Монголии.,")òîò род распространен также по всей Африке. Кроме того, в Африке встречается еще ' других локализованных рода агам. Из многочисленных родов Индо-ца,сайской области 2 встречаются также в Австралийской области, где находится еще ~~ эндемичных родов, 1 из которых достигает Тасмании. Заслуживают внимания »реалы следующих родов (все они проверены по Смиту — А. М. Smith, 1935): Goniocephalus — около '10 видов в восточной части Индо-Малайской областиот
188
Глава 4
Южного Китая до островов Ява, Калимантан и на некоторых Филиппинских островах; ни одного вида на островах Сулавеси, Церам, Хальмахера плп Малых Зондских, но около десятка на Новой Гвинее и в северо-восточной Австралии.
Pkysignathus — 1 вид в Южном Китае, восточном Таиланде, Индокитае и несколь- ко видов в Австралии, Новой Гвинее, на островах Кай и Тимор и ни одного на островах, протянувшихся между Азиеи и Новой Гвинеей. Однако некоторые дру- гие роды заполняют этот пробел. Lophura (Hydrosaurus) с 3 большими водными видами распространен от Филиппин и Сулавеси до Новой Гвинеи. Саlой я пример- но с 27 видами распространен (по островам) от тропической Азии до островов Филиппины, Сулавеси, Хальмахера, Церам и Тимор и др. и до острова Бали (но не за его пределами). Draco имеет около 40 видов, встречающихся от южных крае- вых частей Китая до Филиппин, Сулавеси и Церама, а также на Малых Зондских островах и острове Тимор и др., но не далее. 1 вид этого рода находится в южной Индии (но не на Цейлоне) и изолирован от других по меньшей мере тыгяче- мильным пространством. (.ophotis представлен только ) видом иа (:уматр~ и Яве и 1 — на Цейлоне.
Лгамы родственны игуанам, вероятно, произошли от них и, виднчо, заче- нили их на континентах Старого Света. Ископаемые включают (предположитель- но) сам род ~agama в эоцене Европы.
Chamaeleontidae, настоящие хамелеоны. Мадагаскар, Африка, Азия до южной 11ндии
и Цейлона. 4 или более родов и более 80 видов. Специализированные древесные ящерицы, образовавшиеся от агамовых. Основная часть семейства иричерно поровну делится между Мадагаскаром и всей Африкой, за исключением отдельных частей Сахары. Обычный вид Северной Африки Chamaeleo сйатае1еоп распро- странен до южной части Испании, восточных островов Средиземноморья и т. 1.
Близкородственпые формы встречаются в южной Аравии, на острове Сокотра, на Цейлоне и в Индии, в лесистых районах, расположенных к югу от равнин Ганга. Давно известно, что индийский хамелеон близкородствен осн~кновеп- ному африканскому (не мадагаскарскому!) хамелеону (Gadow, 1909, р. 579; М. А. Smith, 1935, р. 253). На Сейшельских островах находится 1 эндеипчный вид рода Chamaeleo. Его географические связи мне не известны.
«Нельзя утверждать, что ископаемые данные, несомненно, показывают,
что они относятся к хамелеонам, так как все описанные ископаемые являются фрагментами челюстей, которые в равной степени могут быть отнесены и к се- мейству Agamidae» (Сатр, 1923, р. 312).
(Инфраотряд Nyctisauria)
Gekkonidae (включая Eublepharidae, Uroplatidae), гекконы. Все тропические и неко- торые умеренно-теплые районы. Более 70 родов (М. А. Smith, 1935, р. ' 2) и около 700 видов (Loveridge, 1945, р. 67). Северные границы этого семейства определяются несколькими родами, которые достигают некоторых частей среди- земноморского побережья Европы, Балкан и далее на север Болгарии и Ерыма; родами, обитающими в пустынях — от степей Казахстана или прикаспийских пустынь, через Центральную Азию до южной Монголии; ящерицами — синан- тропами рода Gekko, достигающего Северо-Восточного Китая, Кореи и острова Хонсю, Япония. В Америке границы определяются живущим в пустыне родом Coleonyx, который из Мексики заходит в Соединенные Штаты и достигает южной Невады и юго-западной части Юты (К1аиЬег, 1945), и 3 родами, интродуиировап- ныжи в южной Флориде и Флориде-Кис (Н. М. Smith, '1946). Южные границы семейства установить труднее, но, несомненно, что гекконы достигают Южной Африки, Южной Австралии (на Тасмании они, вероятно, не местные), обоих островов Новой Зеландии, центрального Чили и Аргентины. Гекконы также широко распространены на островах в пределах тропиков. Одни виды гекконов живут в пустынях, другие — на открытых пространствах и (в тропиках) в лесистых местностях и влажных джунглях, а некоторые виды, примерно 6 родов, являются ящерицами-синантропами, близко связанными с человеческим жильем, п тор- говля ими широко распространена в тропиках. Например, Hemidactylus brooki, самый обычный синантроп Индии (М. А. Smith, (935), встречается также на не- которых частях Индо-Австралийского архипелага, на большей части Африки, и Вест-Индии и в некоторых частях Южной Америки.
Точные размеры ареалов большинства родов этого большого семейства не
могут быть здесь указаны, так как они наводняют тропики, но в умеренной зоие северного полушария мало известны. Однако некоторые из EIHx заслуживаl~)T упоминания. Довольно многочисленные сведения о геккопах Индии и Африки приводят Смит (М. А. Smith, 1935) и Лавридж (Loveridge, 1947). Род Pl gllodac- tylus, примерно с 50 или, возможно, большим числом видов, имеет самое широ- кое естественное распространение из всех родов этого семейства. но оно п~~еет
Penmu.г и и
(Ин
ф
х81
прерывистый характер: несколько видов находится на Мадагаскаре, несколько :1окализовано в тропической Африке, 1 — на островах Средиземноморья, 2— в Иране (ни одного в Индии), 1 — в Таиланде и т. д. (ни одного на Индо-Австра- лийском архипелаге), ряд видов обитает в Австралии, большое число — в запад- ной и северной частях Южной Америки и на Галапагосах, несколько видов рас- сеяно по Центральной Америке, Мексике и Вест-Индии. Некоторые виды этой группы иногда относят к отдельному роду Diplodactylus, приуроченному, как считают, к Африке, Мадагаскару и Австралии (Schmidt, 1923, р. 19), но Смит (.Ч. А. Smith, 1935, р. 80), сомневается в самостоятельности этого рода. Gymno- ilactylus (если это естественная группа) встречается от восточного Средиземно- морья и тропической Азии до Австралии и многих островов Тихого океана, в Южной и Центральной Америке. Род Tarentola приурочен к странам, гранича- ~иим со Средиземным морем (несколько видов), а также распространен до Кубы п Багамских островов в Вест-Индии (1 четко выраженный эндемичный род).
Этот род, имеющий такое необычное распространение, в последнее время подверг- ся тщательному изучению (Loveridge, 1944Ь), но некоторые исследователи до сих пор еще сомневаются, действительно ли вид из Вест-Индии вместе с другими относится к этому роду. Другие случаи родового сходства гекконов Старого ц Иового Света также вызывают сомнение: Gemidactylus, возможно, не является местным в Америке; Gonatodes был разделен (М. А. Smith, 1933); Thecadactylus также был разделен (Brongersma, 1934). Eublepharis с 3 видами распространен в засушливых юго-западных районах Азии и в Северной Индии, 1 вид — на Хай- няне и островах Рюкю. Род Со1еопух примерно с 5 видами и рядом подвидов (К1а- цЬег, 1945) распространен неравномерно от засушливых юго-западных частей Соединенных Штатов до джунглей Панамы. Возможно, эти 2 рода являются род- ственными.
Обратимся к некоторым группам, относящимся исключительно к Старому
Свету: род Diplodactylus упоминался выше; Oedura в настоящее время ограни- чен ~множеством австралийских видов', родственный или по крайней мере схо- жи12 с, ним род А/гоедига приурочен к южной части Африки (Loveridge, 1944a).
Р)к!зита (Loverdige, 1942а) примерно с 18 видами приурочен к Мадагаскару, Коморским, Сейшельским и Маскаренским островам Индийского океана, кро~~е одного мадагаскарского вида, который обитает в домах и представлен на Ацда- манских островах, предположительно, эндемичной цветовой формой, другой мада- гаскарский вид встречается вблизи восточноафриканских портовых городов. U> o- i)iatric примерно с 5 видами приурочен к Мадагаскару, и в прошлом его выделяли в самостоятельное семейство. Hoplodactylus и Naultinus приурочены к Новой :зеландии, к обоим островам — Северному и Южному; это единственные гекконд 8 собственно Новой Зеландии и единственные известные живородящие гекконь~ (.Ч. А. Smith, 1935, р. 27). Большинство остальных родов гекконов локализовано в пределах той или другой территории: Африки, Мадагаскара, Северной Африки и Юго-Западной Азии, тропической Азии и т. д., Австралийской области и тропц- ков Америки.
Ундервуд (Underwood, 1954) приводит несколько иные данные, чем указа- но вьппе. Его работа имеет важное значение и, возможно, приведет к оспователь- иой переклассификации гекконов, но она не может быть принята зоогеографами :~о тех пор, пока не вынесут своего решения другие специалисты. Между тем пред- иоложеиия Ундервуда подчеркивают, как в действительности мало известно о связях разных гекконов.
Хотя гекконы и относятся к большому широко распространенному и, ве- роятно, древнему семейству, их ископаемые не известны, кроме нескольких родов из плейстоцена и трех недавно описанных родов из третичного периода Франции.
ороси<!ае. Австралия и т. д. Около 8 родов, 14 видов (Kinghorn, 1926). Наземная
или роющая ящерица с укороченными конечностями. Все роды встречаются в Ав- стралии, но род Рцдориг достигает Тасмании, а Lialis — Новой Гвинеи и остро- «ов Лру. Это семейство, возможно, родственное гекконам, и, вероятно, так же.
к и' ц геккоиы, примитивное в общих чертах, по-видимому, не является древней реликтовой группой. Ископаемые этого семейства пе известны.
фраотряд Lep tog lossa)
r(loose urid ae. Прекрасно описанная и реконструированная ископаемая ящерица
из верхней хоры Европы. Вероятно, является предком сцинков (Romer) или гек- конов (Camp).
>tusii(lae. Юго-западная часть Северной Америки, Центральная Америка, Куба.
4 небольших рода. Аantusia (H. М. Smith, 1946, р. 499; Steh2)ins, 1954; Slevin, около 1949), 5 видов, обитает в засуп|лцвых и полузасушливых местностях штата
Глава 4
Аризона, в юго-западной части штата Юта, в южной Неваде, в некоторых районак н>жной части Калифорнии, в Нижней Калифорнии, Сопоре и на негколх,>«хх при- брежных островах. Lepidophyma и Gaigeia (Н. М. Smith, 1939a, р. 24; I!) <") c «<.- сколькими видами — в южной Мексике и Центральной Америке, па влажных тро- пических низменностях и в высокогорных зонах хвойных лесов. /.i pidophzgiiia öä юге достигает Панамы. Cricosaura с 1 видом известен только в восточно~~ части Кубы, B засушливой местности на территории протяжением несколько миль меж- ду Белигом и Кабо-Крусом.
Ископаемый вымерший род * Еирогаигиз из верхней юры Европы условя«
отнесен Ромером (Romer, 1956) к этому же семейству, но с точки зрення зоогео- графии это не имеет существенного значения.
Teiidae. Северная и Южная Америка. Около 37 родов, ' 00 видов. Все роды встрс частот( я
в Южной Америке, несколько распространено на н~г дo центральной части Чили и Аргентины. По крайней мере 6 родов, в основном СпетЫорЬогиз (Burt, !!1:11) и Л»<е<'га, встречаются также в Центральной Америке. Только род C><e»>i<doi>i>o- rus простирается до Соединенных Штатов — он достигает отдельных частей шта- тов Орегон, Айдахо, Южная Дакота, Висконсин и Мэриленд. К югу этот род достигает южной части Бразилии. В Вест-Индии широко распространеи тольк(i род 4nzeiz;a, а 2 или 3 других рода достигают Малых Антильских островов..')то наземное семейство, родственное семейству Lacertidae в Старом Свете.
Достоверные ископаемые, близкие к современному <ожноамерих'««с<1о><у
роду Tupi><ambis, известны из олигоцена и плиоцена Аргентины (Сашр, 1<>>:1, р. 316).
Scincidae (вклн>чая Anelytropsidae, Feyliniidae), сцинки. 11се тропические и суотр~>- пические районы, но более многочисленны в Старом, чем в Новом Свете. Около 40 родов и более 600 видов (М. А. Smith, 1935, р. 254). Северные границы семей- сТВВ определяются несколькими видами, которые достиган~т Южной Фра~~ции, Австрии, Румынии и районов Каспийского моря в Европе; несколькими родамц в горах центральной части Южной Азии; родом Еитесез, который достигает и р(,- винции Хубэй в Северном Китае, острова Хоккайдо, Япония, южных окраи>~- ных частей Британской Колумбии, Манитобы и Онтарио и южной части Масс»- чусетса в Америке. На юге сцинки достигают Южной Африки, Тасмании, <><>о><х островов Новой Зелаидии, центральных частей Чили и Аргентины. Здесь Ilðèèî- дится распростраиение нескольких родо» и «идов сцинков. f.yposoina в широк~о~ смысле (М. А. Smith, 1.Ë7) широко распространен в Староч (:вете п также встр»- чается в восточ<юй части Северной Америки и в Централь<и>й Америке.,'-)тот р<>д насчитывает ! 75 или более видов в Старом Свете, большинство из них расцр~н т- рапепо в районе от Южной Азии до Австралии, несколько видов в Аф)>ик<. ! х« Мадагаскаре нет ни одного вида. В Новой Зеландии примерно 9 эидемичньп видов, которые, как предполагают, представляют две разные части рода.,')то един- ственные местные яйцеживородящие новозеландские сцппки (Н. В. Fell, 'I>;<rl< s, Мс<:апп, письменное сообщение). В Америке этот род представлен только 9 <><<;ta- ми п подвидами, относящимися к 1 eiolopisma (который такнсе имеется в Индо-М<>- лайской области и т. д.), и встречается от восточной части Соедххххеххххххх Штато<> через Мексику и Центральную Америку до Панамы. 11екоторые ученые (Тну!<>г, 19;37; Stuart, 1940; Mittleman, 1950) предложили раз«ые родовые названия, «< касаясь связе~~, существующих между этими родами. Единственный вид в Соеди- ненных Штатах, приуроченный к юго-востоку, в северном направлении достигает нокххой части 11ьн>-Джерси и на западе восточного Канзаса и Техаса. В запад«о>< части Соединенных Штатов нет ни одного вида этого рода, по восточные («ъ<е<>«- капские) виды близкородственны китайским формам (Sc)<>nidt, х 046, р. I >><>).
<xm<ces (Taylor, <9:>5) примерно с 50 видами распространен в Север>>ой Африке
и Юго-Западной Азии на север до Каспийского моря и на восток до северо-запа;~- ной Индии, на изолированной территории в Восточной Азии от Индок~пая до Сех>е1>ххого Китая и Японии; в Северной и Цеххтра.<ыхохх Америке от южного к1>а>< канады до Никарагуа и на Бермудских островах, где иаходится ) четко «ыр»- женпый эпдемичпый вид. ЛХабиуа более чем с 80 видами (М. А. S>nit)>, l,)3,>, р. >58) — и Африке и на Мадагаскаре, в Ю>кпой Азии до 11овой Гвинеи (едп«гт- веппып широко распространенный вид достигает последнего острова), в 1Ож«о« и 11е<хтральпой Америке, н>жной части Мексики (па север до Колпма и 11< 1>«кру- са) и в Вест-Индии. Это единственный род семейства на материке 10>кной Лмерхх- кп. 9 видов и ряд подвидов этого рода находятся в Америке (Dunn, 1Я5). Д)>yr>« едиххстх>еххххьхе местные сципки Америки (кроме,'1 выихе уххомяпутых родов) от«о- «ятся к роду 1Veosep s, эндемичному мопотипическому роду, приуроченному к цептральнои п хожххохх частям Флориды (II. М. Smith, 1<146, р. 395, 505), и .1»<- lytropsis — к роду роющих сципков, не имеющих конечностеи, с ) «идом, црцуро-
Penniuл ии
791
ченным к южной Мексике. АЬ|ерЬагиз (или Crgptoblvpharus, или 1;'moia) bouto- u«, со многими описанными подвидами; некоторые из них, возможно, не могут быть признаны (С. Е. Burt, М. D. Burt,, 1.932, р. 514), встречается в тропическоп Африке, на Мадагаскаре, на островах Индийского океана (но не в Индии п т. д.), на Индо-Австралийском архипелаге (но не в Тасмании) и от тропических остро- вов Тихого океана до островов, расположенных близ западного побережья Эквадора и IIepy (С. Е. Burt, М. D. Burt, 1932, р. 481). Этот вид был давно зареги- стрирован в тропиках Америки, но данных о его встречаемости там в последнее время нет (М. А. Smith, 1935, р. 309). Возможно, что этот вид отчасти был распро- странен человеком. Некоторые другие сцинки родов Emoia и l.удогопга были ши- роко распространены на островах в тропиках Тихого океана ((;. F. Впг1, М. D. Burt, 1932). Несколькородов сцинков являются общимидля Юго-Восточной Азии и Австралии, но они локализованы или отсутству~от на близлежаицтх с)«т- равах: Tropidopho>us (М. А. Smith, 1935, р. 322) имеет 9 видов в !1нд>>кита~ и в прилегающих районах (ни одного на полуострове Малакка или на Сумат(э» и Яве), 5 видов на острове Калимантан, 5 — на Филиппинах (Taylor, 19 >эн) и l филиппинский вид на острове Сулавеси (ни одного между Калимантанои и I I о- вой Гвинеей), 1 на Новой Гвинее (Loveridge, (945а) и 1 вид в северо-восточной Австралии. Ophiosciucus (М. А. Smith, !93,э, р. 333) имеет 3 вида близ Индокитая и I в Австралии. ЛЬосХоиа (М. А. Smith, (937, р. 230) — 1 вид в Таиланде и "II— в Австралии. С другой стороны, некоторые роды сцинков почти всегда приуроче- ны к Индо-Австралийскому архипелагу и островам расположенным на вс)стоке: Етоэа в узком смысле (М. А. Smith, (937, р. 226) более чем с 30 видами встреч>>»т- ся от островов Ява, Калимантан и Филиппин до Новой Гвинеи и островов Тихо- го океана и с ! широко распространенным видом, достига1снцим Австралии, и Otusaurus (М. А. Smith, 1937, р. 2'(8) с 18 видами встречается от Суматры, Кали- мантана и Филиапии до Новой Гвинеи и (;оломоновых островов. И, наконец, в каждом из 3 родов 1.ygosoma, Dasia и багора имеются определенные пары видов, которые распрвделя1отся между южной Индией и восточнс)й частью И~~~о-Малай- ской области, без близкородствеппых видов в северной Индии (M. А. +n>(th.
1935, р. 15). Большинство сципков ведет наземный образ жизни и многие — рон>- щий: у некоторых и3 последних конечности укорочены или отсутствуют по|ио- стью. У ряда видов наблюдается склонность к древесному образу жизни, но нп один вид не приспособлен специально к жизни на деревьях (M. А. Smitlt, I,È,>, р. 254). Что касается размножения, многие сцинки — яйцеродящие и многие- живородящие. Н овозеландские сцинки — живородящие, большинство и: немногих сцинкс)» Америки, включая и северных, яйцеродящие, хотя в Мек- сике встречаются живородяп[ие виды рода Еиэээесея (Н. М. Smith, 1946, р. >'>()).
(; ведений об ископаемых сцинках мало, они не достоверны и поэтому не
представля>от больэпой ценности для установления географической исторпп:этог<> семейства; однако распространение современных форм не оставляет бо эьпэих сомнений в том, что эта группа радиировала в тропиках (,"тарого (",вета, особенно в Индо-Малайской и Австралийской областях, и что 3 широко распростр»- ленных рода достигли Америки самостоятельно (по крайней мере 2 из llnx север- ным путем). Каким ооразом род Мабиуа мог достичь тропиков Америки, рассмат- ривалось ранее. i%eoseps и ~4i<elytropsis, воза ожно, являются реликталш более ранних заселений.
1 >certidae, яп(ерицы. Африка (не Мадагаскар), Евразия и т. д. 22 рода, 145 индов (Bon- lenI>er, 1920 — 192(). Главным образом наземные формы. 11аиболее мпогочпгл»ээ- IIo это семейство в Африке. 1[есколько родов являются общими для:эасупиниых частей Северной Африки и Юго-Западной Азии; несколько родов находится в Пп- до-Малайской области и только [ широко распространенный вид достигает (;уматры, Явы и Калимантана. К северу несколько родов широко распространено в умеренных широтах Европы и Азии. Самый северный вид ( асе~1а л г~p~ ~'~~ встречается по всей северной Евразии — от крайнего севера Шотландии п !ап- :~андии до Амура и Сахалина, достигая 70' с. ш. в (;кандинавии. Это самьш «евериьй представитель рептилий и единственный известный живородяiций ви,l ;)того семейства.
Ископаемые показывают, что род 1 acer ta находился в Европе с эоцен»
((:amp, I!123, р. 3((э). Однако да>эпые о нахонсдении представителя нфрикнп- ского f)oäà Xucr as в балтийском янтаре, возможно, ошибочные; отпечатки предполагаемых ископаемых па копале из Восточной Африки, ээозможпо, и действительности относятся и современному африканскому виду (Loverid<~e, ( 942!э, р. 338) .
Cordylidae (Zonuridae, включая Gerrbosauridae). Африка и Мадагаскар. 2 подсемей- ства. Наземные ящерицы, встречан)1циеся главным образом в засушливых мест-
192
Глава 4
.остях. (1) Gerrhosaurinae. Африка и Мадагаскар. В Африке (Loveridge, 1!142) находятся 4 рода, 13 видов плюс еще подвиды — в основном н Южной Африке.
но род Gerrhosau~из достигает 13' с. ш. в Судане. На Мадагаскаре находятся 2 других рода примерно с 10 видами. (2) (;ordylinae. Африка. 4 рода, 23 вида п1~ос подвиды (Loveridge, 1944) — в основном в Южной Африке, но несколько видов достигают Конго и Восточной Африки и 1 вид — Эфиопии. Данные о встречае- мости этого подсемейства на Мадагаскаре ошибочны (Mocquard, 1909, р. 4). 'D ihamidae. Южный Индокитай и Никобарские острова до Филиппин и Новой Гвинеи.
) род, 3 вида. Червеобра:шые роющие ящерицы, связи которых сомните тьиы. (Инфраотряд Diploglossa)
(Надсемейство Anguoidea)
An„uidae, веретеницевые. Северная и Южная Америка, Евразия и т. д. 3 современных
подсемейства плюс I ископаемое. Большинство современных форм — наземные или роющие. (1) Diploglossinae. Тропическая Америка. 4 рода, некоторые из них локализованы в основном на главных островах Вест-Индии; 1 распространен к ~огу до Аргентины. Сомнительно родственный ископаемый род обнаружен в эо- цене — олигоцене Северной Америки. (2) * Placosaurinae. Евразия и западная часть (;еверной Америки в позднемеловом и раннетретичном периодах; довольно крупные панцирные ящерицы. (3) Gerrhonotinae. Центральная Америка и запад- ная часть Северной Америки, Евразия и т. д. 2 современных рода. Geï/гопойи распространен от 11анамы через Центральную Америку и западную часть Север- ной Америки до южной части Британской Колумбии, северного Айдахо и запад- ной Монтаны (см. Tihen, 19~9). Ophisaurus имеет I вид в западной части Евразии.
встречающийся от Балкан до Средней Азии и в Северной Африке, еще 1 — в (;е- верной Африке, или 3 — в восточных Гималаях, Южном Китае, Бирме и нн острове Тайвань, 1 — на острове Калимантан (подтвержден свежим экземпля- ром в Музее сравнительной зоологии) и 1 — в восточной части Северной Америки от Висконсина и Виргинии до Техаса и Флориды. 3 ископаемых рода, приписывае- мых к этому подсемейству, были обнаружены в Европе (эоцен) и Северной Аме- рике (верхний мел и олигоцен — миоцен). (4) Anguinae. 1 современный вид, . Iiiguis fragilis, безногая европейская слепозмейка (веретеница ломкая, или медяница). Встречается примерно начиная от 64,5'с. ш. в Швеции (стр. 161) до северной части Африки, северной части Малой Азии и на Кавказе. Этот же род найден в миоцене Европы.
An»illidae. Южная половина Калифорнии и северння часть 11ижней Калифорнии.
) род, 2 вида плтос ! дополнительный подвид. Червеобразная, роющая, яйт~е- живородящая ящерица, которая, вероятно, «дегенерировала» от веретениц.
ХепояапгМае. В настоящее время локализовано в Китае и в южной части Мексики
п т. д. 1 ископаемое и 2 современных подсемейства, связи которых сомнительны.
(1) * Melanosaurinae. Северная Америка (эоцен) и? Европа (палеоцеп — эоцен). (2) Shinisaurinae. 1 вид, Skinisaurus croeodilurus, известный только в районе Яосяня.
1уанси, Китай. В работах Мак-Доуэлла и Богерта (Mcl)owell, Hogert, 1954) рас- счатриваются связи этого вида. (3) Xenosaurinae. Южная часть Мексики (нн се- »ер до Сан-Луиса-Потоси) до Гватемалы. I. род, 1 или 2 вида.
(цадсемейство Varanoidea)
Ilelo~lermatidae, ядозубы. Единственный род Не1одепаа с 2 современными «идами:
один — в южной и западной части Аризоны и в прилежащих частях штата Юта и Невада и в северо-западной Мексике, другой — в западттой и южной частях Мексики, главным образом и» побережье, от южного штата Сонора до Чиапаса (Rogert, del Сampo, 1956, р. 15). Большие красивой расцветки, действительно ядовитые ящерицы. Ископаемые этого же род» относятся к олигоцепу Колора- до.
Тагашс!ае, варановые. Теплые страны (:тарого Света. ! род VQ)clIlus причерно с '4 ви- дами и рядом подвидов (Mertens, 1 142). Вараны встречаются по всей Африке.
кроме крайней северо-западной части, но в Европе и на Мадагаскаре в настоя- 1цее время их нет. В Юго-Западной Азии распространены и» север до Каспийско- r0 моря; по всей Ипдо-Малайской области — на север до К)жттого Китая п н;1 юг и восток через Индо-Австралийский архипелаг до Филиппин, Соломоновы ~ островов, а также по всей Австралии (нет в Тасмании) и на островах западной части Тихого океана до Л!ариаиских Каролинских и .'Царша lhchlIx. Варапы- крупные плотоядные ящерицы, встречающиеся (разные инды) как и засуш;пни,ri местностях, так и во влажных джунглях. По образу жизни быватот наземньи, древесные или полуводиые. Один из полуводных видов Varaitus, «~га1о~ име(т прерывистое, ио очеиыпирокое распространение на 1[ейлоне п чл тнчно на полу- острове Иттлостан и от восточной части Индо-Малайской области до ()(тровов (;~.— лавеси и Ветар и, возможно, и до Австралии, хотя отиосителт,но последней.Черте~~с
Рептилии
«ысказы«ает сомнение. Va га~ги8 Йоиос7а.пя8 — комод~~йский гигантский «арап.
известен только на двух маленьких островах, лежащих между островами (,'yir- ба«» и Флорес, на пеб<)1~ьшой территории в западной части острова Флорес и и» Малых Зондских островах. Это самая большая современная ящерица, достигав- шая не менее 10 футов в длину.
Ископаемые рода Va rai<us известны в Европе, Индии и Л«ст рал ии. И Е вро- не — 2 или 3 вида: самые ранние в ни>кием мпоцеие (илн до него) и самые позд- ние, очевидно, просуществовавшие до неолита включительно. Гигантские виды варанов встречались в Индии в плиоцене и в Австралии в плейстоцепе: австра- ;птйские вымершие вараны рода * Megalaiiia вряд ли были крупнее грузных пред- ставителей рода Vora»us и, вероятно, достигали 15 футов в длину (Dunn, 19>7).
Другие различные ископаемые роды (отнесенные Ромером к разным подсемей- ствам) известны из верхнего мела и эоцена северного полушария, включая (:евер- ную Америку.
Lanthanotidae. Калимантан. ! вид, не ядовитый и не настолько близкородственньш
роду Heloderma, как это предполагалось раньше (Мс1>оже11, Bogert, 1954).
*Aigialosauridae, *Mosasauridae, *Dolichosauridae. 3 семейства водяных морс~;их яще- риц, которые были известны только в меловом периоде.
~ Palaeophidae. Очень мало известные формы мелового периода и эоцена, которые,
как считает Ромер (Romer, 19о6), возможно, представлены змееобразныии водя- ными варановыми ящерицами. Они известны из Европы, Северной Америки, ('.е- верной Африки и Мадагаскара.
(Инфраотряд Annu lata)
Amphislaenidae, двуходковые. Африка и т. д. (но не Мадагаскар), Южная Америка на
север до Панамы (но не большая часть Центральной Америки) и изолированные участки, разбросанные в северной краевой части Южной Америки и в районах, ~~римыка1ощих к тропической зоне. Около 24 современных родов, 150 видов. Чер- веобразные, обычно безногие роющие ящерицы. 4 подсемейства (Vanzolini, )951).
(1) Amphislaeninae. Африка и Америка. 16 родов. Род АтрИгЬаепа в Африке (1 впд в Мозамбике), Южной Америке на север до Панамы, также на Кубе, Гаити и Пуэрто-Рико. Другие роды локализованы в некоторых частях Африки (8 ро- дов, за исключением АтрШяЬаепа), в районе Средиземного моря на север до Ис- пании и Португалии, на острове Родос и в юго-западной части Азии (Bla»us); Южной Америке (3 рода); северо-западной части Мексики (!1ижняя Калифор- ния) (Bipes); на Кубе и т. д. (1 род) и на Виргинских островах (1 род). Ископаемые этого подсемейства неизвестны. (2) Rhineurinae. 2 рода в Африке, 2 — в Южной Америке и род Rhineura — во Флориде. Ископаемые рода Rhineura относятся к олигоцену западной части Соединенных Штатов, а другие — к раннему эоцену и олигоцену запада Соединенных Штатов. (3) Trogonophinae. В некоторых частях (',еверной Африки, в Аравии, Иране и на острове Сокотра (3 рода, ископаемых иет). (4) *Crythiosaurinae. Олигоцен внутренней Монголии. 1 род.
Подотряд Ophidia, змеи
(Надсемейство Typhlopaidea)
Typhlopidae, слепозмейки. Тропики и некоторые районы субтропиков. 4 рода, нри- о.1изительно 200 видов. Род ТурИорs встречается по всему ареалу семейства с многочисленными видами в Африке, па Мадагаскаре и в Южной Азии (ареал 1 вида простирается на северо-запад до Греции и района Каспийского моря).
11есколько видов находится в Гималаях и 1 — в Южном Китае (на север до юж- ного Цзянси и Фуцзяни). Также многочисленны виды на Индо-Австралийском архипелаге и в Австралии (но не в Тасмании). Относительно малочисленны они и Америке, от iожной Мексики (ин север д|о центрального Веракруса — Smith, Taylor, 1945, р. 19) до северной части Аргентины и в Вест-Индии. 3 других рода, обычно помещаемые в отдельное подсемейство, приурочены к Америке в преде- лах ареала рода Т>>р)>1оря, T>iphlops l>rami»us («змея цветочных горшков») ()chmidt, Davis, l )41, р. 91) — местный вид Индо-Малайской области, распростра- нен на всех островах почти до Новой Гвинеи и встречается также, но возможно, что он там иитродуцирован, в Южной Африке, на Мадагаскаре, на островах Индий- ского и Тихого океанов, включая 1 авайские, и в н>жной части Мексики (Smith, Т >ylor. 1, -~5, р. 19). 1!редставители:>того семейства — 1>елкие червеобразные ро~ощие змеи. Очевидно, это примитивное п древнее семейство, но представлено оно только l пли 2 ископаемыми из третичного периода Европы.
1,eptotyphlopid;>e, сзепозмейки. Африка и т. д. и теплые районы Америки. 1 род, 50
или более видов. Очень маленькие червеообразиые роннцпс змеи. В Африке их ареал простирается на юг от Сахары (нет нн Л!адагаскаре) и захватывает северо-восточную часть Африки; в Юго-Восточной Азии до северо-западной Ин- дии, в Америке от тожной части Канзаса, пустынь Аризоны и прилежащих ча-
Глава 4
стей Калифорнии, Невады и Юты до центральной Аргентины и Малых Ант«ль- ских островов и (возможно, интродуцирован) Багамских островов. Ископаемые этого семейства почти не известны.
(Надсемейство Вooidea)
Dinilysiidae. Верхний мел Южной Америки. '1 род. Вероятно, сачые древние из «сех
известных настоящих змей.
Ani1iidae (IIysiidae). Локализованы в тропической Азии и т. д. и тропической Амерх<ке.
4 условно объединенных подсемейства. (1) Loxoceminae. Центральная Америк« и южная часть Мексики (от Коста-Рики до Колимы и Цаярита), 1 род !.oxocemua.
2 вида (в прошлом считался родственным питонам Старого (.'вета). (2) Хепоре!- tinae. Восточная часть тропической Азии и т. д. от Верхней Бирмы и apoonni~nii Гуандун (Китай) до Боль<них Зондских островов, о< трова Палаван, южных ост- ровов Сулу н Сулавеси. 1 род Xenopeltis, 1 наземный ро«нций вид. (3) Aniliin,«..
Тропическая Азия н т. д. и Юхсая Америка. 3 рода (М. А. Smith, '1943, р. !)'<): Су!<«<!горЬis, 1 вид изолирован на Цейлоне и около 6 — в восточноп части Инл<>- Малайской области и на островах (от Бирмы и И«докитая до островов Ару); Л «<- тосИ~~|8, ' вида, на полуострове Малакка, на островах (;уиатр» и Калимантан, и !nilius (1lysia), ! вид, широко распространенный в северной части 1Ожной Америки к востоку от Аид. Представители этого семейства — довольно медкц~.
назем<ю-роющие змеи. (4) Uropeltinae, щитохвогтые змеи. 1Iолуостров 11нлост«н и Цейлон. 7 родов, 43 вида. Все сосредоточены на этой ограниченной территории (Ч. А. Smith, 1943, р. 61 — 94). Эти небольшие змейки, ведущие роющий onp;<з жизни, обитают в горных местностях и лесах.
~эЛгсЬаеорЫдае. В морских отложениях эоцена Европы, 1 ископаемое, oTllo(ящеес»,
скорее, к угрю, чем к змее (McDowell, Bogert, 1954, р. 66 — 67).
ВоЫае, удавовые. Все тропические и некоторые умеренные районы. Обычно выделяц>т
2 основные группы удавовых — собственно удавы с близкородственными им змеями и питоны с близкородственными им змеями, но Смит (М. А. Smitl<, I!)<:I, р. 103), Гибе (Сийе, '1949) и Мак-Доуэлл счита<от, что подразделение этого сеч« <- ства значительно более сложное. Ромер (Romer, 1,)56), следуя Мак- 1оу;>л.зг, выделяет 4 основные группы или подсемейства и. таким образом (вместе с иерем<- щением рода Loxocemus), упорядочивает распространение семейства и устр««яет оольшинство поразительных разрывов. Удавы ~I'i,tàãаскара ие являются бди:<- кородствепными удавам Америки. Фактически 1<ито«ы морфологпчески <<o:«e примитивны (1,nderwood, 1951, Мс1Эоъеll). Это семейство включает оград«ых змей, которые душат свою жертву, а также це,tlirt ряд более мелких п мсне~ заметных змей. Иитоиы Старого Свет» — <»<gepoäÿ~öèñ. а удавы IIoaoro (.'вет»— яй«ежпвородя«<ие (Schmidt, 1950, р. 82). Имеется пол«ый, хотя п несколько у< т»- реншнй список видов этого семейства (Йи11, 19:151. Ромер выделяет четыре сле- дующих подсемейства'.
(I) 1 ythoninae, питоны и т. д. Старый Свет, кроме Мадагаскара (центральноамерик;«<- сипи род J.oxocem us пере11ещен в семейство Ani llidae); 7 родов, около 18 ви- до». Calabaria c ! иебольишм наземным видом приурочен к дождевым лесаи 3аиад- Boll Африки. Руйоа —,') вида в Африке, на хог от Сахары; 2 — в Южной Азии от северной Индии и южных частей Китая на 1<>г до 1[ейлона и Индо-Австралий- ского архипелага и Филиппин, проникая почти ло Новой Гвинеи (MAR-Доу;>з;< <читает, что некоторые австралийские формы должны быть отнесены к род<,.
Python); еще ! вид — на полуострове Малакка и островах (:уматра и Калица«таи и ) вид на островах Тимор и Флорес. Остальные 5 родов и! О видов, по списку (,'~ «л ла, встреча<отся в Австралии и (или ) в Новой Гвинее, и т. д. 1 яд видов;<огтигает островов Тимор, Ветар, южной части Минданао и Соломоновых острово». В Л»- стралии некоторые питопы распространены почти повсеместно, как в за~ уш;~ ш~~,« и открытых местностях, так и во влажных лесах, и как в умеренной зоне, та lj и в тропической. В Тасмании нет ни одного вида.
(") 1!о<пае, удавы, анаконды и родственные им более мелкие формы. Тропич« кая
[Ожная Америка на юг до центральной Аргентины, включая Вест-Индию. Около 7 родов, 20 видов. Один из нпх Boa constrictor (удав обыкновенный) рагпро< т- ранен на север до Соноры и, возможно, достигает южной Аризоны (8ogert, C) liver, 1945, р. 350).
(3} Sanziniinae. Мадагаскар и т. д. 4 рода. Удавообразиый род Ь'аи <а<а с ! видом и род
.1сгац80рЫг с 2 видами встречается на Мадагаскаре. Один из видов рода . !с,.ц„,— tophis зарегистрирован из Реюпьона (Guibe, !949). Casarea и Bolyeria, по I нилу в кан<домг — удавообразные роды (по внешним признакам), известны толь- ко на острове 1'аунд близ острова Маврикий, но на самом острове Маврикий (,дин пз них (или оба) являются субфоссильными. Мак-Доуэлл (письменное сообщение) отмечает, что мадагаскарские удавы (а также некоторь|е мадагаскарские ужовые)
.Penrnuë ии
имеют нижнюю дугу на всех торакальных позвонках, что весьма необычно для змей и, возможно, является плавательным приспособлением. Это позволяет пред- полагать, что данные змеи произошли от водных предков, которые достигли Мада- гаскара водным путем. (Недавно было сделано предположение, что удавы с ост- рова Маврикий в действительности являются примитивными ужовыми.)
(4) Erycinae. Северо-восточная Африка и часть Азии, Новая Гвинея и другие острова
и западная часть Северной Америки. 4 рода. В основном мелкие змеи, ведущие скрытый образ жизни. L'ryx. наземные песчаные удавы, с 7 видами, встре- чаются в северной и восточной частях Африки до северных окраинных рай- онов Танганьики (Loveridge), по всей Юго-Западной Азии до Индии и Цейлона, на север до Балканского полуострова, на Кавказе и Урале (севернее 50' с. ш.) п по всей Центральной Азии до южной части Монголии. Епуягиз (Candoia)— очень маленький древесный удав. 3 вида в районе Новой Гвинеи, островов Бис- .марка и Соломоновых островов, 1 из этих 3 видов доходит до западной части ост- рова Сулавеси и еще 1 из 3 широко распространен на островах западной части Тихого океана до Самоа и Тонга, а возможно, и значительно дальше, но не иск- :иочено, что отчасти он был распространен человеком. 2 других наземных род» находятся в западной части Северной Америки: Charina (1 вид) встречается от Юты и центральной части Калифорнии до южной части Британской Колумбии; lлсйапига (2 вида) — в южной части Калифорнии, Нижней Калифорнии и в за- падной части Аризоны.
Ископаемые удавы — в большинстве случаев очень скудные находки—
встречаются в различных третичпых отложениях (Romer), и судить по ним о гео- графической истории этого семейства довольно трудно. Можно только сказать, что его представители встречались в течение какого-то времени в третичном периоде в Европе и Северной Америке.
(11адсемейство Colubroidea)
(;~>] «hridae, ужовые. Во всех основных частях земного шара, кроме части Австралии.
Северные границы семейства в Европе определяются яйцеродящим видом Л atпх natriz (обыкновенный уж), достигающим примерно 65' с. ш., и живородящим родом CoroïeÏa, доходящим приблизительно до тех же широт. Северные границы этого рода в Азии мне не известны. В Лмерике живородящие ужовые рода Thami<op- Ыя (обыкновенные подвязковые змеи) — самые северные рептилии, достигающие примерно 60' с. ш., в западной части Канады. К югу ужовые достигают Южной Африки и провинции Чубут в южной части Аргентины. Это огромное семей- ство, охватывающее почти 2/з всех современных змей и еще в более высокой про- порции тех, которые находятся в обычных местах обитания (за исключением глав- ным образом роющих семейств и морских змей). Оно состоит из нескольких под- семейств, но специалисты не пришли к соглашению в отношении их численности и состава. Вообще говоря, подсемейство (;olubrinae доминирует в тропиках и в умеренных широтах Евразии, Африки и Северной Америки, а подсемейство Xenodontinae (Ophiinae), если оно признается, доминирует в тропиках Америки.
Другие подсемейства более мелкие и более локализованы. Некоторые роды ужо- вых являются эндемиками на Мадагаскаре, и считается, что некоторые мадаг»- скарские формы весьма примитивны (Schmidt, 1923, р. 35). Несколько разных родов ужовых Индо-Малайской области достигают северной части Австралии (см. подсемейства) и несколько (распространяясь в южном направлении) — во- сточной части Австралии. На больших территориях Южной, Центральной и Юго- Западной Австралии и в Тасмании ужовые отсутствуют. Ромер (Romer, 1956) приводит следующие подсемейства семейства Со] ubridae плюс Xenodontinae (по Данну Ophiinae), располагая их в несколько ином порядке, чем Смит (М. А. Smith, 1943). Данн(Dunn, 1928), Поуп (Роре, 1935) и Богерт (Bogert, '(940) разделили это семейство также несколько иначе.
<1) (;olubrinae (включая Boiginae, Natricinae, Coronellinae, Sibpnophiinae). Все основ- ные части земного шара, кроме части Австралии. Более 100 родов. В Африке (8ogert, 1940), Евразии, Индо-Малайской области (М. А. Smith, 1943) и Северной Америке (Dunn, 1928) обитает много родов, в Южной Америке меньше (Dunn, 1928) и только 4 рода (этого подсемейства) достигают Австралии: Boiga, широко распространенный в тропической Африке и в Индо-Малайской области, Ahaetulla (Dendrophis) из Индо-Малайской области, 2 древесных рода, которые простирают- ся через острова к Лвстралии. Boiga имеет 1, а Айае~иПа — 2 вида в северной и восточной частях Австралии (Kinghorn, 1929; Loveridge, 1934; Glauert, 1950); Stepanotus, наземный род, известен только по нескольким видам, разбросанным на Филиппинах, Молуккских островах и Новой Гвинее, и 2 видам в Квинсленде.
Частично водный и широко распространенный по всему земному шару род Nat- nx включает 3 вида, которые достигают северной части Лвстралии, и 1 из них
Глава 4
простирается на юг в восточной части Австралии до севершш ча«ти !1ового К)ж- ного Уэльса. Классификация родов в этом подсемействе постоянно изменяется.
Ъ!ногие древние роды были раздроблены, но связи между образованными частя- 1ххх не всегда были ясны. Ниже приводятся ареалы нескольких географически интересных групп, но их нужно рассматривать с осторожностью, и следует пом- нить, что большинство родов подсемейства Сolubrinae занимает более или менее сплошные, географически ограниченные последовательные территории, и что роды, имеющие прерывистый ареал, являются исключениями. Самыи широко распространенный род ужовьхх Лахгхх (Tropidvnotus) (стр. 166). Род Ttmmnopkis, образовавшийся от Ла1пх, увеличивает его ареал в Америке. Iaphe и Colubei (в широком смысле) в основном наземные роды, которые встречаются но всему земному шару в северном полушарии, а также в тропиках Индо-Малайской области и в других местах. Род Coluber, разделенный Ингер и Кларком (Inger, С1агК, 1943), (в узком смысле), приурочен к Северной Америке и Восточной Азии. Но Богерт (письменное сообщение) сомневается в том, что азиатские виды действительно близкородственны американским. Нейсорз, в широком смысле.
имеет несколько видов, разбросанных и изолированных в тропической Африке и на Цейлоне, в Индии и Юньнани, и ряд видов в Америке — от Мексики (веро- ятно) до Аргентины. Этот род был разделен (Bogert, 1940, р. 37, 38; М. А. Smith, 1943, р. 319), но образованные в результате деления новые более мелкие роды «можно рассматривать как прямые дериваты от предковой линии рода Пейсорг, сохранившиеся как реликтовые формы, в настоящее время отделенные от ос- новного (в Америке) ареала родительской линии» (Bogert). Sibynophis (Pvlyodon- thophis, Scaphiodontophis), наземный, в основном обитахощий в горах род, рас- пространен на Коморских островах и Мадагаскаре (2 вида), в Индо-Малайской области (7 видов) от Цейлона и Индии до островов Ломбок, Калимантан, Палаван и Бусуанга на Филиппинах и на север до Шанси и Цзянси в Китае; в Центральной Америке и т. д. (9 видов и подвидов) от Атлантического склона Мексики до Па- намы и Колумбий. Богерт (Bogert, 1940) считает, что этот род следует разделить и что в результате деления образуется естественная группа рода, вероятно в ран- ге подсемейства (как это предполагает Данн — Dunn, 1928). Тэйлор и Смит (Tay- lor, Smith, 1943) уже отделили американские формы от форм Старого Света.
Opheodrys (Schmidt, Necker, 1936; Grobman, 1941) (называемый.4ЫаЬея — Booij, 1917; Eurypholis — Pope, 1935; и Liopeltis — Bourret, l936) встречается в тропи- ках и умеренных широтах Восточной Азии, включая острова Тайвань и Рюкю, в восточной части Северной Америки и на Юкатане. В Восточной Азии и т. д.
распространены 6 или большее число видов этого рода, встречающихся чаще в тропиках, чем в умеренной зоне северного полушария, и виды, ранее помещаемые в этот род, но теперь обычно выделяемые из него встречаются на Больших Зонд- ских островах. На основной части материка Азии самые северные виды достигают Шанхая и части Хубэя и т. д. В восточной части Северной Америки имеются 2 «и- да этого рода, из которых тот или другой достигает по крайней мере 50' с. ш., а на западе доходит до Вайоминга, Юты и Нью-Мексико (но не до западного ххобережья) и на юге — до северо-восточной части Мексики. Еще 1 вид изолирован в северной части Юкатана в пределах тропиков.
(2) Xenodontinae (Ophiinae). Америка и отдельные части Старого Света. 6х1 — 70 или бо- лее родов (Dunn, 1928, 1931, р. 116). Доминируют в Южной Америке и являхотся единственными представителями ужовых в Вест-Индии, за исключением водного рода iYatriz, который достиг Кубы. Несколько видов встречается в Север- ной Америке. В том случае, если это подсемейство признается (а фактически современные специалисты по змеям не признахххт его), схода могут быть вклхо- чены некоторые роды Старого Света. Мы приводим его здесь, чтобы подчеркнуть то обстоятельство, что в Южной Америке имеется большая группа относительно примитивных ужовых, представляющая особую сохранившуюся ветвь, от кото- p0ii, по-видимому, разными путями эволюционировало большинство представи- телей подсемейства Сolubrinae других частей земного шара (Bogert, I J40, р. 9).
В данном случае можно провести параллель с воробьиными птицами, у KOTopbIx огромное скопление относительно примитивных форм (Suboscines) сохранилось или эволюциопировало в Южной Америке, в то «ремя как «ысшие Oscine~ эволюционировали в других местах, в основном в Старом Свете (стр. 337 и след.). (3) Dasypeltinae (Dasypeltidae), яйцеядные змеи. Африка и Индия. 2 рода l)asytpr itis
с 1 видом, 2 подвидами (Bogert, 1940, р. 85), широко расххространено в тро- пической и Южной Африке и достигает хожной чнстхх Аравии (6нпз, устное сообщение). I: lachistodon с 1 редким видом обнаружен только в северовосточххохх части Индии. Родство этих двух родов вызывало сомнение, но М. A. Смит (М. А. Smith, 1943, р. 409) считает их родственными.
Penmuëèè
197
(4) Acrochordinae. Тропическая Азия до северной части Австралии. Два, относимых к
роду А с~.осйо rdus, водных вида: один обитает в пресных водах, другой— в эстуариях и прибрежных частях морей. Оба они распространены от основной части материка тропической Азии до северной Австралии, а вид, обитающий в соленых водах, достигает Филиппин и Соломоновых островов.
(5) Xenoderminae. Восточная часть Индо-Малайской области и тропическая Америка.
В Индо-Малайской области — 4 рода, 7 или 8 видов — от южной и восточ- ной части Китая (на север до Цзяньсу) и острова Хонсю (Япония) до Явы и Кали- мантана. В Центральной и Южной Америке — 1 или 2 других рода, виды немно- гочисленны.
(6, 7) Pareinae, Dipsadinae (Amblycephalidae), улиткоядные змеи. Восточная часть Индо-
Малайской области и тропики Америки. В Индо-Малайской области ' родя, около 16 видов, распространенных от восточных Гималаев, Южного Китая (на север до Сычуани и Фуцзяня) и Тайваня (но не на большей части Индии) до островов Ява, Калимантан и южной части Филиппин. В Америке несколько родов и около 40 или более видов, в основном в тропиках Южной Америки, до- стигая северной части Аргентины и (Sibypomorphus или только Dipsas) Централь- ной Америки и южной части Мексики до Колима и центрального Веракруса.
Некоторые авторы сомневаются в родстве азиатских и американских «Ambly- cephalidae». Ромер также относит их к самостоятельным подсемействам, но тем не менее Смит (М. А. Smith, 1943, р. 115) считает, что американские и азиатские роды являются близкородственными и, вероятно, имеют общее происхождение. (8) Jkomalopsinae. Индия, южная часть Китая, Тайвань и Филиппины до северной
части Австралии и т. д. 10 родов, более 25 видов. Все водные обитают в соленых, и в пресных водах (М. А. Smith, 1943, р. 379). 8 родов встречаются на основ- ной части континента Южной Азии. Некоторые из этих родов простираются и» Индо-Австралийский архипелаг, и 3 из них достигают северной части Австралии, 2 других рода приурочены к Новой Гвинее и северной части Австралии соответ- ственно.
Известно несколько третичных и плейстоценовых ископаемых ужовых, но
большинство из них фрагментарны, и они почти ничего не говорят о географиче- ской истории семейства.
Elapidae, кобры, аспиды, австралийские змеи и т. д. Тропические и умеренно-теплые
области земного шара. 2 подсемейства.
(1) Elapinae. В пределах ареала семейства. Примерно из 30 родов около половины при- урочено к Австралийской области. Большинство из австралийских форм — это мелкие, хотя и ядовитые, змеи, но только некоторые из них опасны и достигают значительных размеров. Поэтому разные австралийские представители семей- ства Elapidae занимают место не только кобр и гадюковых, но также и обыч- ных, не ядовитых ужовых остальной части земного шара. Ни один род этого семейства не является общим для Австралийской области и других частей земного шара. Acanthophis — австралийская шипохвостая ехидна, достигает островов Молуккских и Танимбар, прежние данные о встречаемости этого рода и» острове Калимантан, вероятно, ошибочны. Ogrnodon, приуроченный к островам Фиджи, считается близкородственным группе роющих родов Новой Гвинеи (Bo- gert, Matalus, 1945). В Африке и южной части Азии, на север по крайней мере до Прикаспия и хребта Копет-Даг на границе Ирана с Туркменией (Берг, 1947), в северной части Индии, Цзянси и Чжэцзяне и на островах Рюкю встречаются разные роды, включая кобр (нанесено на карту Богертом — Bogert, 1943, р. 312).
На Мадагаскаре представителей этого семейства нет. В Америке встречается 3 рода аспидов: Micrurus — примерно с 65 видами и подвидами (Н. М. Smith, Тау- 1ог, 1945, р. 169) от Северной Каролины и юго-западной Индианы до провинции Рио-Негро, восточные части центральной Аргентины (Serie, 1936; Роре, 1945, р. 18), но не в западной части Северной Америки или в Вест-Индии. M~c~u~o~- des — с 1 видом локализован в юго-западной части Северной Америки, Leptomii;— ru rus — с 2 видами в северной части Южной Америки.
(2) Dendroaspinae. Тропическая и Южная Африка. 1 род Dendroaspis, несколько
видов. Мамба, или древесная кобра.
*Palaeonaja — единственные ископаемые семейства Е lapidae, известные
до плейстоцена, в миоцене и плиоцене Франции. Богерт (Bogert, 1943, р. 294) сокращает это название до синонима Жауа — род современных кобр. В на<ни дни в Европе нет ни кобр, ни других представителей Elapidae, но кобры Все-таки достигают средиземноморского побережья Африки и (со стороны Азии) близко подходят к юго-восточной части Европы.
*НуйгорЬййае, морские змеи (М. А. Smith, 1926, Mertens, 1934а). Более теплые части
Индийского и Тихого океанов. 15 родов, около 50 видов, большинство из которых
1 лава 4
ооитает в тропических морях от южной части Азии до северной части Австралии и в западной части Тихого океана. Отдельные виды в северном направлении рас- сеяны до Японии и Приморского края и на юге до Тасмании и Новой Зелаидии (Phillips, 1941). Несколько видов распространено на запад до Персидского зн.1и- ва; 2 — до Мадагаскара (Mocquard) и 1 J'elamis platurus — до восточного побе- режья Африки. Последний является единственным пелагическим видом морских зме~~, который достигает западного побережья Америки, где он встречается от Эквадора на север до Калифорнийского залива; он встречается также в районе близ Гавайских и Галапагосских островов, в Атлантическом океане морских змей нет. Близкородственные морским или, очевидно, образовавшиеся от морскпк видов пресноводные формы встречаются в озере Бомбон (Таал) па Лусоне, Фи- липпины, и в озере Тунгано на острове Реннелл, Соломоновы острова (Loverid- ge, 1945, р. 170). Морские змеи родственны семейству Elapidae и ведут от него свое начало, и так же, как и змеи этого семейства, ядовиты. Мертенс считает, что, возможно, морские змеи имеют двойное происхождение — оба раза Ба терри- тории Индо-Австралии: в позднем мелу или в раннетретичном периоде. Ромер (Romer, 1956) выделяет 2 подсемейства, которые здесь не приводятся.
Viperidae, гадюки. Все континенты, кроме Австралии, и на нескольких островах.
2 подсемейства.
(1) Viperinae, настоящие гадюки. Африка и Евразия. 10 родов (М. А. Smith, 1943,
р. 478). Vipera примерно с 10 видами. Распространен в тропической и се- верной части Африки, Европе и Азии. Является местным на Индо-Австралий- ском архипелаге, где достоверно известен единственный вид (V. гигяеШ) с острова Ломбок и с острова, расположенного недалеко от острова Флорес (Mertens, 1930).
Данные с островов Суматра и Ява недостоверны (Rooij, 1917, р. 279). Ареал вида V. russelli на основной части материка Южной Азии, по-видимому, прерывистый (Роре, 1935, р. 384 — 385); этот вид встречается также на острове Тайвань. Вид Vi pe ra berus определяет северную границу семейства и всех змей, проходящую через северную Евразию. В Скандинавии этот вид достигает 67' с. ш. (как раз над Се- верным полярным кругом) и, возможно, заходит еще дальше на север в лесах Кольского полуострова (Берг, 1947). Это живородящий вид. Шесть других родов встречаются в Африке: некоторые распространены почти по всему материку, другие локализованы. На Мадагаскаре гадюки отсутствуют. 3 африканских рода простираются в прилежащие части Азии и еще 3 рода приурочены к отдельным частям Южной Азии. Никаких достоверных ископаемых этого подсемейства до миоцена не известно. Род Vipera известен из миоцена Европы, подобно роду Вitis, приуроченному в настоящее время к Африке и Аравии, но зарегистрированному из плиоцена Европы.
(2) Crotalinae, ямкоголовые гадюки, включая гремучих змей. Азия и Америка.
6 родов. Agkistrodop< встречается в Азии и Америке: в южной части Азии около 9 видов, один из которых — живородящий вид А. halys (щитомордник) — рас- пространен в Восточной Азии до 53' с. ш. и до озера Байкал, на запад до степей южных районов Советского Союза и до низовьев Волги. 1 вид встречается в Япо- нии. К югу этот род встречается на полуострове Индостан южнее 16' с. ш. (нет в северной части полуострова) и на Цейлоне, и 1 вид достигает Суматры и Явы.
В Северной Америке (Gloyd, Conant, 1943) имеется 2 вида, мокасиновая змея и водяной щитомордник, встречающиеся в юго-восточной части Соединенных Шта- тов. Мокасиновая змея заходит на север до юго-восточной Айовы и восточного Мас- сачусетса и т. д. Еще 1 вид этого рода распространен в Мексике и Центральной Америке до Никарагуа. Род Trimeresurus примерно с 22 видами (М. А. Smith, 1943, р. 502) встречается в Азии и на островах от Цейлона и Индии и на север до Гималаев и Центрального Китая (Хубэй и Чжэцзян); на островах Рюкю и к во- стоку и югу до Филиппин, островов Сулавеси, Тимор и Сангир. Ареал этого рода, очевидно, прерывается в северной части полуострова Индостан, примерно север- нее 22' с. ш. (за исключением ареала вида Т. albolabris), который встречается в се- верной части Индии и, очевидно, имеет изолированную популяцию в центральной Индии (М. A. Smith, 1943, р. 524). Этот же вид распространен на большей части Индокитая, отсутствует в Таиланде, на полуострове Малакка к ~ory от 13' с. ш., но снова встречается на Суматре, Яве и др. до острова Тимор и т. д. В Америке представителем рода Trimeresurus является Bothrops, который морфологически еще не ~ыделенв самостоятельный род (М. А. 8тйЬ, 1943, р. 502). Многие виды встречаются от восточных частей центральной Мексики (Тамаулипас — Н. Ы.
Taylor) до провинции Санта-Крус в южной Аргентине (Serie, 1936).
Род Lachesis с видом, бушмейстер, распространен от тропической части Бразилии 3,о Панамы. Гремучие змеи являются строго американскими. Основной род гре- ~~учих змей Сrotalus с 22 видами пл~ос еще подвиды лучше всего представлен в юж-
199
иой части Соединенных Штатов и в Мексике, но отдельные виды распространены иа севере до некоторых районов штата Мэн, южного Онтарио, Висконсина, Вели- ких pasnnn южной Канады и до южной части Британской Колумбии. 1 ви1 с 2 или большим числом подвидов встречается в Центральной Америке и в более засушливых частях (ио не во влажных лесах) Южной Америки, на юг до северной Аргентины. Sistrurus (карликовая гремучая змея — Gloyd, '1940, р. 76), с 2 ви- дами в восточной части Северной Америки, распространен на север до южного Онтарио, на юг до Флориды и, вероятно, в северных краевых частях Мексики и на запад до Аризоны. Третий вид изолирован на южной оконечности Мексикан- ского нагорья. Ископаемые этого подсемейства не известны до плиоцена (когда гремучие змеи находились в Северной Америке). Другие «ключи» дают основание предполагать, что это семейство возникло в Азии и что оба рода ~4gkistrodoii и Trimeresurus (Bothrops) переместились из Азии в Америку. Один из них, основной (идти более древняя ветвь), по-видимому, дал подъем гремучим змеям.
Многие, но не все гадюки и ямкоголовые змеи питаются главным образом
или частично теплокровными животными. Хотя большинство гадюк живородя- кие, в Старом Свете некоторые из них, включая ряд видов рода Agkistrodon, являются яйцеродящими (Pope, 1935, р. 387); в Америке, вероятно, все гадюки.
кроме бушмейстера, живородящие.
ПОДК,1АСС ARCHOSAURIA
Отряд (;rocodilia. 5 подотрядов, 13 семейств, из которых сохранилось только 1 семей- ство.
Croco~lylidae, крокодилы и т. д. Теплые районы земного шара. Около 8 родов, 25 су- шествуюших видов (Schmidt, 1944; слегка изменено Ингером — Inger, 1948).
Взрослые животные ведут главным образом водный образ жизни, но яйца откла- дывают на суше. В семействе выделяется 3 подсемейства.
(1) (;rocody1inae, настоящие крокодилы. 2 рода, из которых один с 2 видами приуро- чен к Западной Африке и к Верхнему Конго (Inger, 1948), а другой Crocodylus с 12 видами встречается по всем тропикам Старого Света и в северной части аме- риканских тропиков, от южной Флориды и низменностей Мексики до Эквадора, Колумбии и Ориноко (но не в Амазонке) и т. д. В каждой из основных тропиче- ских областей (Африка, Индо-Малайская область, тропическая Австралия и тро- пики Америки) имеется один широко распространенный вид рода Crocodylus, который изредка заходит в с,оленые воды, и от 1 до 3 дополнительных локализо- ванных видов, которые, вероятно, приурочены к пресным водам. В Африке к та- ким видам относится С. niloticus, распространенный в благоприятных условиях обитания на юг до мыса Доброй Надежды, на севере в некоторых частях Сахары (Angel, Lhote, 1938), спускается вниз по Нилу и спорадически заходит на побе- режье Палестины (Flower, 1933, р. 755) и в настоящее время (или в прошлом) достигает Сирии. Этот вид встречается также на Мадагаскаре (в прошлом?), на Сейшельских островах и, возможно, встречался ранее на острове Сицилия в Сре- диземном море (Flower); он часто заходит в солоноватые и даже в соленые воды (Schmidt, 1919, р. 419). В Индо-Малайской и Австралийской областях к широко встречающимся видам относится С. рогогиз, гребнистый крокодил, который рас- пространен от тропической Азии до Северной Австралии и на восток через Филип- пины и Соломоновы острова и далее. Лавридж (Loveridge, 1945, р. 41, 244) ут- верждает, что он заходит на острова Палау и Фиджи «и т. д.». Адамсон (Adamson.
1939) считает, что восточная граница крокодилов в Тихом океане неопределенна.
Он отмечает находку скелета крокодила с острова Макатеа в западных краевых частях островов Туамоту (за островами Самоа и Общества); этот вид обычно за- ходит в море. В Америке С. acutus является широко встречающимся видом, ко- торый распространен от южной Флориды, северной Мексики (Тамаулипас и Си- налоа) и т. д. до Эквадора и Колумбии, на Кубе, Гаити и Ямайке и который часто встречается в прибрежных болотах и эстуариях.
(2) Alligatorinae, аллигаторы и т. д. Настоящие аллигаторы (Alligator), 2 вида.
в настоящее время встречаются только в юго-восточной части Соединенных Штатов от долин рек Каролины и Флориды до Рио-Гранде и в низовьях до- лины реки Янцзы в Китае. Поуп (Pope, 1935, р. 66) считает, что в дальнейшем, вероятно, будет доказано, что китайские аллигаторы недавно имели обширный ареал в Восточном Китае. Более или менее родственные им кайманы, 3 рода и 9 видов, распространены в Центральной и Южной Америке в северном направле- нии до Оахаки в Мексике и к югу до системы Параны. В Вест-Индии они отсут- ствуют.
(3) Gavialinae, 2 монотипических рода рыбоядных гавиалов, локализованы в Индии
и в районе Малакки соответственно (Малайский род в действительности, возмож- но, относится к подсемейству Crocodylinae).
20о
I'.ãава 4
Обильные ископаемые этого семейства рассматриваются Ромером (Воп1ег,
1945, р. 222) следующим образом: «Все 3 1под] семейства были представлены в эоцене в северных материковых областях; крокодилы и аллигаторы были много- численны и разнообразны в этих районах в раннетретичном периоде; нередки на- ходки ископаемых на южных материках». Род 111гда8о r, который, очевидно, является единственным четко выраженным родом рептилии, строго приурочен к умеренным зонам обоих полушарий. Ископаемые этого рода известны из олигоцена Северной Америки. В Евразии ископаемые не найдены, хотя в Европе в рапнетретичном времени находились родственные роды. Мук (Mook, 1925) считает, что ископаемый Alligator, обнаруженный в нижнем миоцене Небраски, так похож на совремепный китайский вид, что он мог бы быть его прямым пред- ком. Два современных рода гавиалов теперь приурочены к разным частям Индо- Малайской области, а их ископаемые были обнаружены в Европе.
D inosaurs. Следующие 2 отряда и 6 подотрядов относятся к динозаврам. Их рас- пространение изложено в гл. 10.
~Отряд Saurischia
*Подотряд Theropoda. 3 инфраотряда, 13 семейств
*П о д о т р я д S а u r о р о d а. Brachiosauridae, Titanosauridae
Отряд Ornithisehia
*Подотряд Orthopoda. 5 с.емейств
*Подотряд Stegosauria. 1 семейство
*П одотряд Ankylosauriа. 2 семейства
~II одотряд Ceratopsia. 3 семейства
(Подкласс не определен)
*Отряд Mesosauria, мезозавры
~hfesosauridae. Южная Африка и Южная Америка, вероятно, в нижнепермском перио- де. ) род (см. стр. 514). Следует еще раз отметить, что в этой классификации реп- тилий, достаточно полной для современных семейств, опущены многие древние группы.
ЛИ ТЕРА ТУРЛ
A d а m s о п А. М., 1939, Review of the fauna of the Marquesas Islands..., «Bull.
Bernice Р. Bishop Мця.», No. 159.
A n d e r s о п С., '1925, ...Meiolania... «Rec. Australian Mus.», 14, 223 — 242.
А n g e l F., Ь h о С е Ц., 1938, Reptiles et amphibiens du Sahara central et duSahara
central et du Soudan, «Bu1l. Comite d Etudes EIistoriques et Scientifiques de
1'Afrique Occidentale Franqaise», 21, 345 — 384.
Б е р г Л. С., 1947, Географические зоны Советского Союза, 3-е изд., Москва.
В о g e r t С. М., 1940, ...arrangement of African Colubridae, «Bull. American Mus.
Nat. Hist.», 77, 1 — 107, pls.
В о g e r t С. М., 1943, Dentitional phenomena in cobras...., «Bull. American Mus.
МаС. Hist.», 81, 285 — 360.
В о g e r t С. М., 1949, Thermoregulation in reptiles..., «Evolution», 3, 195 — 211. В о g e r С С. М., 1953, The Tuatara'. why is it а lone survivor. «Scientific 5Ionthly»,
76, 163 — 170.
В о д е r t С. М., С а m р о R. М. del, 1956, The Gila Monster and its allies, «Hull.
American Mus. Nat. Hist.», 109, 1 — 238.
В о g e r t С. М., С о w 1 e s R. В., 1947, Moisture loss... in some Floridian reptiles,
«American M us. Novitates», No. 1358.
В о g e r t С. M., M а t а 1 а s В. Ь., 1945, А review of the elapid genus Ultrocalamus
of New Guinea, «American Мus. Novitates», No. 1284.
В о g e r t С. М., О 1 i v e r J. А., 1945, ...herpetofauna of Sonora, «Bull. American
Моя. Nat. Hist.», 83, 297 — 425.
В о и 1 е n g e r G. А., ']920 — 1921, Monograph of the Lacertidae, London, British ibtus.,
2 vols.
В о и r r е С R., 1936, Les serpents de 1'Indochine, Toulouse, 2 vols.
В о u r r e t R., 1941, Les tortues de 1'Indochine, Institut Oc~anograpliique de I'In- dochine, Note 38е.
»ew
В r о n g e r s m а Ь. D., 1934, Contributions to Indo-Australian herpetology, «7оо1о- gische Mededeelingen» (Leiden), 17, 161 — 251.
В u r t С. Е., 1931, ...Cnemidophorus..., «Bull. United States National Mus.», ~0.
154.
В u r t С. Е., В u r t M. D., 1932, Pacific Island amphibians ancl reptiles..., «Bull.
American Мия. Nat. Hist.», 63, 461 — 597.
(', а m р С. Ь., )923, Classification of the lizards, «Hull. American Мця. ХаФ. Hist.»,
48, 289 — 481.
(. а r 1 G. С., '1944, The reptiles of British Columbia, British Columbia I rovincial Mus.
(Victoria), Handbook No. 3.
(, а г r А., 1952, Handbook of turtles, Ithaca, New York.
(." h я b а п а u d P., 1917, ...reptiles... Afrique occidentale..., «Ви11. Atus. National
d'Hist. Nat.», 23, 83 — 105.
('. о п г а d G. М., 1940, Ву boat Со the Age of Reptiles [Sphenodon], «Natural Histo- ry» (New York), 45, 224 — 231.
D а г1 i n g С о п P. J., Jr., 1948, The geographical distribution ot' cold-blooded ver- tebrates, «Quarterly Review Biol.», 23, 1 — 26, 105 — 123.
De V i s С. W., 1894, The lesser chelonians of the Nototherian drifts, «Proc. R. Socl.
() ueensland», 10, 123 — 127.
D u и п Е. R., 1927, Notes on Vcranus komodoensiÿ, «American Мия. Novitates», No.
286.
D u n n Е. R., 1928, ...arrangement... American... "olubridae, «Bull. Antivenin Inst.
America», 2, 18 — 24.
D u n n Е. R., 1931, The herpetological fauna of the Americas, «Copeia», l93I, 106 — 119. D u n n Е. R., 1935, Notes on American Mabuyas, «Ргос. Асад. Nat. Sci. Philadel- phia», 87, 533 — 557.
Е ч е n d e n F. G., J r., 1948, Distri bution of the turtles of western Oregon, «Herpe- tologica», 4, 201 — 204.
F 1 о w e r $. S., 1933, ...reptiles and amphibians of Egypt..., «Proc. Zool. $ос. Lon- don», 1933, 735 — 851.
G а d о w Н., '1909, Amphibia and reptiles, The (:ambridge Natural History, vol. 8,
London.
G i 1 m о r e C. W., 1928, Vossil lizards of North America, «Мет. National Acad.
Sci.», 22, No. 3.
С l а и е r t L., )950, А handbook of the snakes of Western Australia, Perth, Western
Australian Naturalists'Club.
С l о у d Н. К., 1940, The rattlesnakes..., «Chicago Acad. Sci. Special Pub.», No. 4.
6 1 о у d Н. К., С о п а n t R., 1943, ...American..., «Agkistrodon. Bull. Chicago
Acad. Sci. э, 7, 147 — 170.
3 r а у J. Е., 1852, l)escription of tortoises, «Ргос. Хool. Soc. Londоп»,
20, 133 — 135.
G r о h m а п А. В., 1941, ...Opkeod~ys vernalis..., «Misc. Pub. Mus. Zool. U. Michi- gan», N о. 50.
G u i b е, J., 1949, Revision... boides de Madagascar, «Mem. Inst. Sci. Мададаясаг»,
Ser А, 3, 95 — 105.
G й n t h e r А., 1858, On the geographical distribution of reptiles [snakes], «Ргос.
Zool. Soc. London», 26, 373 — 389.
H е с h t G., 1928, Zur Kenntnis der Nordgrenzen der mitteleuropiischen Reptilien,
«Mitteilungen Zool. Mus. Berlin», 14, 501 — 595.
H о f f s t e t t e r R., 1942, Sur les restes de... Iguanidae, «Ви11. Mus. Хайопа1 d'EIist.
Nat.» (Paris) (2), 14, 233 — 240.
Н о f f s С е С С е r R., 1944 — 1945, Sur les Scincidae fossiles, «Bull. illus. NationaL
d'Íist. Nat.» (Paris) (2), 16, 547 — 553; 17, 80 — 86.
Г.сава 4
H о f 1 s С е С t. е г H., f946, ...Но1дае Йея Mascareignes..., «Bull. Мия. aviational d'I[ist.
Sat.» (Paris) (2), 18, 132 — ! 35.
Н и t t о n F. Ж., D г и m m о n d J., 1923, The animals of Xew Zealand, 4~(Ь ed.
Auckland etc.
! и ц e r R. F., 1948, The systematic status of t)ie crocodile OsEeoblepharoi~ огЬогт,
«Copeia», !948, 15 — 19.
! n g~ е r R. F., С 1 а r k P. J., ! 943, Partition of the репья Colube~, «Copeia», 1943,
l4i — i45.
K i n g h о r n J. R., l ~' 6, А brief review of the family Pygopodidae, «Rec. Australi- an Mus», 15. 40 — 64.
k i n g h о r n J. К., 1 929, Snakes of Australia, Sydney.
К 1 а ц Ь е r L. M., 1945, ...(.oleonyx... «Тг. San Diego Soc. Xat. !list.», !О,
l35 — 213.
L о д i e r Е. В. b., Т о п е r G. С., 1955, Check-list of the amphibians and reptiles
of Canada and Alaska, «Contrib. R. Ontario Mus. Zool. апд Palaeontology», No.
4l.
Ь о v e r i d g е А., 1.)34, Australian Reptiles in the Museum of Comparative Zoology,
«Bull. Mus. Сomparative Zool.», 77, 243 — 383.
L о v e r i d g е А., 1941, ...Pelomedusidae, «Bull. Mus. Comparative Zool.», 88, 467—
524.
Ь о ч е r i d g е А., ).942, ...Gerrhosauridae, «Bull. Mus. Comparative Zool.», 89, 485—
543.
Ь о v e r i d g е А., 1942а, ...Phelsuma, (<Bull. Mus. Comparative Zool.», 89, 439 — 482. Е о ъ е r i d g е А., 1942Ъ, [Reptiles of forested areas in East and Central Africa],
«Bull. Mus. Comparative Zool.», 91, 237 — 373.
L о ~ e r i d g е А., 1944, ...Cordylidae, «Bull. Mus. Comparative Zool.», 95, '1 — l).8. L о v e r i d g е А., '!944а, ...Afroedura,..., «American Mus. Novitates», No. !254. Е о v e r i d ~ е А., 1944Ь., American... Tarentola, «Copeia», 1944, 18 — 20.
L о т е r i d g е А, !94а, Reptiles of the Pacific World, New York.
L о v e r i d g е А., 194va, New scincid lizards of the genera Tropidophorus and Lygo- гота from New Guinea, «Ргос. Biol. Soc. Washington», 58, 47 — 52.
L о v e r i d g е А., !947, ...African... Gekkonidae, «Ви11. Mus. Comparative Zool.»,
98, 1 — 469.
Ь о v e r i d g е А., '1948, 'Yew Guinean reptiles..., «Bull. Mus. Comparative Zool.».
101, 305 — 430.
L о ~ е r i d g е А., S h г е v е В., 1947, The «New Guinea» snapping turtle, «Copeia».
1947, l 20 — l 23.
!, у d e k k e r R., 1889, (.atalogue of the fossil Reptilia and Amphibia in the British
Museum, pt 3, «Chelonia», London, British Mus.
М с D о w е 11 S. В., В о g e r t С. М., 1954, ...Lantkonotus..., «Bull. American
Mus. Nat. Hist.», 105, 1 — 142.
M а t t h e w W. D., 1915, 1939, Сlimate and evolution, «An.. New York Асад. Sci.»,
24, 171 — 318; reprinted (1939) as «Special pub. New York Acad. Sci.», 1.
М е r t e n s R., 1930, Die Amphibien und Reptilien der Inseln Bali, Lombok, Sum- bawa und Flores, «Abhandlungen Senckenbergischen Naturforschenden Gesell- schaft», 42, 117 — 344.
М е r t e n s R., 1934, Die Insel-Reptilien..., «Zoologica», (Stuttgart), 32, 6, Leiferung,
1 — 209.
M e r t e n s R., !934а, Verbreitung... der Seeschlangen..., «Zoogeographica» (Jena),
2, 305 — 319.
.М е r t e n s R., 1942, Die Familie der Warane (Varanidae), «Abhandlungen Sencken- bergischen Naturforschenden С esellschaft», Abh. 462, 1 — 1'1 5; 465, 117 — 233:
466., 235 — 391.
Л um е рспп у ра
М е r t е и s R., М й 11 е r 1., 1940, Die Amphibien und Reptilien Europas (zweite Liste), «Abhandlungen Senckenbergischen Naturforschungeden С esellschaft», ЛЫ1. 451.
М i 1 1 е г R. С., l 948, А checklist of the reptiles and amphibians of Canada, «Herpe- tologica», 4, Supplement 2, 1 — 15.
M i t t I e m а п М. В., 1950, The generic status of Sci incus lateralis Say, 1823, «Her- petologica», 6, 17 — 20.
М о с ц «r d F., 1909 ...reptiles ... batraciens de Madagascar, «Nouvelles Archives .'11ия. d'Elist. Nat.» (Paris), Ser. 5, 1, 1 — 110.
М о о )' (;. C., !925, Ancestry of the Alligotors, «Natural History» (New York), 25, 407 — 408.
Х е ъ ю а п А. K., 1877, N otes of the physiology and anatomy of the Tuatara, «Tr. Хеж Zealand Inst.», '10, 222 — 239.
О l i i. е г J. А., S h а w С. Е., 1953, The amphibians and reptiles of the Hawaiian Islands, «Zoologica», (New York), 38, 65 — 95.
Р h i l l i p s M. J., 1941, ...Pelamis ... in New Zealand, «Тг. and Proc. R. Soc. New Zealand», 7i, 23 — 24.
P о р е (;. H., l935, The reptiles of China, «Nat. Hist. Central Asia», IO, New York, American М us. N at. H ist.
.Р о р е С. Н., 1944 — 1945, The poisonous snakes of the New World, «Bull. New York Zool. Soc.» «Animal Kingdom»), 47, 83 — 90, 111 — '120, 143 — 152; 48, 17 — 23, 44 — 47.
В о m e r А. S., 1945, Vertebrate paleontology, 2nd ed., Chicago. [Имеется русский перевод: А. P о м е р, Палеонтология позвоночных, ГОНТИ, Москва, 1939.]
Н о m e r А. S., 1956, Osteology of the reptiles, Chicago, U. of Chicago Press.
R о о i j М. de, 191~ — 1917, The reptiles of the Indo-Australian Archipelago: vol. 1 (l 115) Lacertilia, С,helonia, Emydosauria; vol. 2 (1917) Ophidia.
S c h ш i d t Е. P., 1919 — 1923, Contributions to the herpetology of the Belgian Con- go..., «Bull. American Mus. Nat. Hist.», 39, 385 — 624; 49, 1 — 146.
S c h m i d С К. P., 1944, Crocodiles, «Fauna» (Zool. Soc. Philadelphia), 6, 67 — 72.
S c h m i d t Е. P., l946, On the zoogeography of the Holarctic Region, «Copeia», 1946, 144 — l52; slightly modified from «Lingnan Sci. J.» (Canton), 10, 1931, 441— 449.
S c h m i d t К. P., 1950, Modes of evolution discernible in the taxonomy of snakes, «Evolution», 4, 79 — 86.
S c h n> i d t K. P., 1953, А check list of North American amphibians anci reptiles, Chicago, U. of Chicago Press.
S c h т i d t К. P., D а v i s D. D., 1941, Field book of snakes of the United States and Candada, New York.
S c h m i d С К. P., N е с k e r W. Ь., 1936, The scientific name of the American Smooth Green Snake, «Herpetologica», 1, 63 — 64.
S c h r e i Ь е r Е., 1912, Herpetologia еигораеа, 2nd ed. [with Nachtrag in 1913], 3 епа.
S e r i i P., 1936, ...distribucion geografica de los ofidios argentinos, «Obra Cincuente- nario Museo de la P la ta», 2, 33 — 61.
S i m p s о n G. G., 1938, "rossockelys, Eocene horned turtle from Patagonia, «Вц11. American Мця. Nat. Hist.», 74, 221 — 254.
S i m p s о n G. G. 1943, Turtles and the origin of the fauna of Latin America, «American J. Sci.», 241, 413 — 429.
S 1е v i n J. R., 1949, Range estension of Eantusia vigilis, «Herpetologica», 5, 148.
S m i t h Н. М., 1939, The Mexican and Central American lizards of the genus Scelo- p0rus, «Zool. Ser. Field Mus. Nat. Hist.», 26, 1 — 397.
204
Глава 4
S m i t h Н. M., l939a, Notes on Mexican reptiles and amphibians, «Zool.,år. Field
Мця. Nat. Hist.», 24, 15 — 35.
S m i t h Н. М., 194, Mexican herpetological miscellany, «Proc. United States 4»Oo- nal Mus.», 92, 349 — 395.
S m i t h Н. М., 19~6, Handbook of lizards, Lizards of the United States and Сanada,
Ithaca, New York.
S m i t h H. М., Т а у 1 о г Е. H., 1945, An annotated checklist and key to t)ie sn»- kes of Mexico, «Bull. United States National Mus.», No. 187.
S m i t h Н. M., 1950, An annotated checklist and key to the reptiles of Мехico etc)u- sive of the saakes, «Bull. United States National Mus.», No. 199.
S m i t h М. (А.), 1929, Monograph of the sea-snakes, London, British Мин.
S m i t h М. (А.), 1931 — 1943, Reptilia and Amphibia. См. «Fauna of Britisli india
including (".eylon and Вигта», vol. 1 (1931) Loricata, Testudines; vol. ' (1935)
Sauria; vol. 3 (1943) Serpentes, London.
S m i t h M. (А.), 1933, Remarks on some Old world geckoes, «Rec. Indian Alus.» ((:al- cutta), 35, 9 — 19.
S m i t h М. (А.), 1937, А review of the genus Lygosoma..., «Rec. Indian IIus.», 39, 2~ 3—
234.
S т i t h М. (А.), 195l, The British amphibians and reptiles, London.
S t e b b i n s R. С., 1954, Amphibians and reptiles of western North America, New
York.
S t u а r С Ь. (:., 1940, Notes on the «Lampropkolis» group of Middle American L~]~0-
soma..., «Occasional Papers Mus. Zool. U. Michigan», No. 421.
S t u 1 1 О. G., 1935, А check list of the family Boidae, «Ргос. Boston Soc. Nat. Hist.»,
40, 387 — 408.
Т а у 1 о r Е. Н., 1920, Philippine turtles, «Philippine J. Sci.», l6, 11 — 1~4.
T а у 1 о r Е. H., 1922, The snakes of the Philippine Islands, «Philippine Bureau Sci.
Pub », N o. 16.
T а у 1 о r Е. Н., 1922а, The lizards of the Philippine Islands, «} hilippine Вигеаа
Sci. P ub. », N o. 21.
Т а у 1 о r Е. Н., 1935, ...Eumeces..., «U. of Kansas Sci. Вп11.», 23, 1 — 643.
Т а у 1 о r Е. Н., 1937, ...Leiolopisma, from Mexico..., «Copeia», )937, 5 — 1) .
Т а у l о r Е. Н., S m i t h Н. М., 1943, А review of American sibynophine sn»kes....
«U. of Kansas Sci. Bull.», 29, 301 — 338.
Т i h e n J. А., 1949, The genera of gerrhonotine lizards, «American М~Й1апд natura- list», 41, 580 — 601.
U n d e r w о о д G., ]951, On the distribution of snakes of the family Hoidae» «Ргос.
Zool. Soc. London, '120, 713 — 714.
U n d e r w о о d G., 1954, On the classification and evolution of geckos, «Ргос. Zoo).
Soc. London», ) 24, 469 — 492.
V а n z о 1 i n i P. Е., 1951, А systematic arrangement of the family Amphisbaenidае,
«Herpetologica», 7, 113 — 123.
W i 1 1 i а m s Е., 1950, ...cervical central articulations of living turtles [with а sumrn;i- ry of classification of turtlesj, «ВиИ. American Мпя. МаС. FIist.», 9 ~,
505 — 562.
Ж i l l i а m s Е., 1950а, Testudo cubensis and the evolution of Western Hemispliere
tortoises, «Bul1. American Mus. Nat. Hist.», 95, 1 — 36.
Ж i 11 i а m s Е., 1952, ...tentative arrangement of the tortoises of the юог14, «13ull.
American Мus. Nat. Нist.», 99, 541 — 560.
AV i 11 i а m s Е., 1952а, А staurotypine skull from the Oligocene of South Ii»1;~ita,
«Breviora» (Mus. Comparative Zool.), No. 2.
Ш i 1 1 i а m s Е., 1953, Fossils and the distribution of chelyid turtles, «Breviora» (Чив.
Comparative Zool.), No. 13.
Лите pa»i у рп
20 э
б' i 11 i а m s Е., '1,)54, ...the living species of ...Podoc~~emi's, «Bull. Mus. Comparative Z ool. », 11 I, 277 — 295.
'11' i 11 i а m s Е., 1954а, Fossils and the distribution of chelyid turtles. 2. Additional reputed chelyid turtles on northern continents, «Rreviora» (Mus. Comparative Z ool.), N о. 32.
M' i 11 i a m s Е., 1356, Pseudemys sc)ipta caIli'rost). from Ver)ezuela witt) а genera) survey of the scripta series, «Bull. Mus. Comparative Zool.», 1i5, 145 — 160.
б о о Й w а r d А. $., 1901, ...Miola~iia..., «Proc,. Zool. Soc. London», 1ЧО!, 16!) — 184.
J а n g e r 1 R., 1947, Redescription of Taphrosphys olssoni... Fieldia~ia (Cliica~~o ivat. llist. Mus.)..«Geology», 10, 29 — 40.
Глава ПТИЦЫ
Из всех животных наиболее хорошо изучены птицы. Известны почти все существующие виды, которых насчитывается около 8600 (Mayr, 1946a; Мауг, Amadon, 1951), не считая многих тысяч географических подвидов. Географическое распространение многих видов изучено детально. Из всех позвоночных лично мне лучше всего известны птицы, так как я на~~.щодал их в течение почти всей жизни и собрал небольшую коллекцию на севере Южной Америки и в Австралии. Большую пользу принесли мне неодно- кратные беседы с сотрудниками Музея сравнительной зоологии, а также ценные замечания по этой главе, сделанные Дж. Ван Тайном и проф. Эрн- стом Майром. С разрешения проф. Майра я пользовался его картотекой по географии птиц. Тем не менее, пользуясь такими обширными сведения- ми, я все-таки считаю, что распространение птиц понять нелегко. Совре- менный характер их распространения довольно ясен, хотя и сложен. но проследить и понять процессы, которые обусловили этот характер — эво- люцию и расселение, трудно.
Возможно, в этом и кроется причина того, что большинство американ- ских орнитологов очень мало интересуют географические связи и проис- хождение наших птиц. Фауна птиц Северной Америки непостоянна. Это временная, исторически сложившаяся группировка, часть которой бла- годаря сложным перемещениям многих видов в различных направлениях образовалась недавно и обречена отчасти на рассеивание или исчезновение, а отчасти на увеличение или беспрерывное переформирование. Крик крас- HoII танагры, или мухоловки-тиранны, ассоциируется со звуками тропиче- ского дождя в лесах Южной и Центральной Америки, откуда эти птицы проникли в Северную Америку совсем недавно, а потрескивание нашего зимородка напоминает о тропиках Старого Света. Эта птица относится к роду, происходящему из Африки и Южной Азии, и является одним из наиболее специализированных представителей семейства (в основном из Старого Света), многие виды которого питаются не рыбой, а насекомыми. ящерицами и т. д. Орлы, соколы и некоторые другие хищные птицы Север- ной Америки, очевидно, проникли в нее из Старого Света через Евразию также относительно недавно. Грифы, вероятно, появились из тропиков Америки, а представители рода Buteo (включая великолепного ямайского канюка, красноплечего, ширококрылого и другие виды канюков) входят в нашу фауну уже около 30 млн. лет. Из Европы и Азии в Северную Аме- рику проникла деревенская ласточка, хотя зимует она в настоящее время в Южной Америке, дрозд (Turdus migratorius), гаички и синицы, чижи и клесты, сойки и вороны и многие другие птицы, включая певчих. Пере- смешники, дрозды, крапивники (распространение одного вида шло через Азию и Европу), виреоны, древесницы, американские иволги (не сходные с иволгами Старого Света), кассиковые, ряд видов овсянок и большая часть других птиц проникли из тропиков Америки. Все эти детали, помимо того, что они интересны сами по себе, имеют существенное значение, ибо, чтобы
Шпицы
207
понять авифауну Америки, необходимо знать птиц всего земного шара и историю их географии.
По многим вопросам классификации птиц — какие семейства должны оыть признаны, к какому семейству относить тот или иной род, как эти семейства между собой связаны — орнитологи еще далеки от единого мне- ния. В настоящее время фактически приняты две, очень близкие друг дру- гу классификации — Уитмора (Wetmore, 1951) и Майра и Амадона (Ma- уг, Amadon, 1951). Их отличие состоит в том, что Майр и Амадон часть 1бщепринятых семейств считают только подсемействами и располагают их в другом порядке. Кроме того, Майр и Амадон классифицируют певчих птиц по группам в соответствии с предполагаемыми и возможными путями их эволюции, которые значительно лучше свидетельствуют о расселении птиц, чем целые (не разделенные на группы) семейства певчих птиц по классификации Уитмора. Поэтому здесь используется классификация Майра и Амадона.
Один отряд — воробьиные, включающий певчих птиц, является доми- нирующим на суше и охватывает более половины всех существующих видов птиц. Далее этот отряд неоднократно противопоставляется всем другим отрядам птиц — неворобьиным.
Немногочисленные остатки ископаемых птиц очень часто разрознен- ны и сомнительны, чтобы можно было судить по ним об их географических связях. Для зоогеографии наличие незначительных ископаемых данных, которым придается слишком серьезное значение, иной раз хуже, чем отсутствие этих данных вообще, и птицы хорошо иллюстрируют это поло- жение. Ни в одном случае по ископаемым остаткам нельзя точно судить о месте происхождения даже наиболее широко распространенного семей- ства птиц. Самые древние ископаемые некоторых семейств были обнаруже- ны в Европе или Северной Америке, но это, скорее, говорит о территори- альном размещении самих палеонтологов, чем о распространении птиц. Например, ископаемые попугаи были найдены в верхнем олигоцене (или миоцене) Европы и в миоцене Северной Америки и нигде больше до плей- стоцена их ископаемых обнаружено не было. Однако едва ли кто может сказать, что попугаи произошли на севере и не распространялись на юг до плейстоцена. На основании подобных односторонних географических данных не раз делались попытки определить место происхождения птиц, например уток и трогонов в Европе, крацид и журавлей-арам в Северной Америке.
В отдельных случаях ископаемые действительно наводят на мысль о roM, что определенные группы птиц всегда были приурочены к определен- ным местам: эпиорнисы — к Мадагаскару, моа — к Новой Зеландии, пинг- вины — к южным, а чистики к северным частям земного шара и т. д. В дру- гих случаях ископаемые показывают, что некоторые птицы встречались некогда за пределами их современных ареалов (обычно к северу от них): африканские страусы — в Южной Европе и Азии; фаэтоны, пеликаны, птицы-секретари, фазаны, дикие куры, цветные бекасы, попугаи, трогоны, птицы-носороги и саланганы — во Франции, Германии или Англии: фламинго — в Южной Дакоте и Орегоне; крациды и журавли-арамы, попугаи — в Небраске и т. д.; индейки — в Калифорнии, калифорний- ский кондор — во Флориде и грифы Старого Света — в Северной Амери- ке. Но изменения, происшедшие в распространении птиц (о чем свиде- тельствуют их ископаемые остатки), не имеют ничего общего с местом их происхождения и обычно еще меньше с путями их расселения. Исключе- ниями являк>тся лишь те несколько случаев, например с попугаями и тро-
Глава 5
гонами, когда ископаемые показывают, что группы птиц, которые теперь приурочены к тропическим или южным частям Старого и Нового Света, в прошлом расселялись через северные широты.
Ископаемые семейства, приводимые в списке семейств птиц в конце главы, интерполируются, по Уитмору (Wetmore, 1951), а сведения об из- вестных ископаемых современных семействе даются весьма кратко и поль- зоваться ими следует с осторожностью.
Несмотря на то что ископаемые так мало говоря" оо истории геогра- фии птиц, они все-таки дают некоторое представление об общем ходе их эволюции и о наиболее вероятном времени их расселения. Птицы поздне- юрского периода — археоптерикс и археорнис1 очень напоминают своих пресмыкающихся предков. Птицы меловой эпохи были более похожи на современных. Насколько известно, все они имели зубы, но достоверно известны только два таких семейства, относящихся к морским птицам'. Беззубые птицы, возможно, появились в конце мелового периода, а все известные птицы эоцена (включая палеоцен) — беззубые. В настоящее время некоторых из них считают, в известной мере условно, предками сов- ременных семейств. Многие ископаемые остатки птиц слишком фрагмен- тарны, чтобы по ним можно было судить о степени их родства. Уитмор (Wetmore, 1951а р. 53, 54) считает, что большинство птиц эоцена значи- тельно отличалось от современных, что и они относятся к особым вымер- шим семействам, и что большинство ныне живущих семейств возникло уже в послеэоценовое время. Но это лишь предположение. Майр нее (Mayr, 1946, р. 3) считает, что многие современные семейства ведут свое начало с более древних времен — начала третичного или еще более ран- него времени. Возможно, некоторые современные семейства неворобьи- ных птиц действительно ведут свое начало с эоцена или даже позднемело- вого периода, но из этого не следует делать вывод, что их распространение датируется этим временем. Распространение более древних групп птиц, вероятно, неоднократно изменялось в течение третичного периода. Иско- паемые формы воробьиных такх'е относят к эоцену; более ранние данные сомнительны. По-видимому, болыиая часть найденных ископаемых отно- сится к более позднему времени, чем большинство ископаемых неворобьи- ных, и многие семейства отряда воробьиных, возможно, ведут свое начало с середины третичного периода, но и это также не более, чем предположе- ние. Уитмор (Wetmore, 1951а, р. 63) считает, что роды и виды современных птиц эволюционировали главным образом в миоцене и плиоцене, а макси- мальной численности достигали в третичном периоде, после чего их чис- ленность снижалась. дто предположение, скорее, справедливо для боль- шинства неворобьиных наземных птиц. Воробьиные же, по-видимому, находятся в расцвете радиации и, вероятно, сейчас достигают максималь- Hoii численности.
Птицы — это пресмыкающиеся, у которых в процессе эволюции поя- вились перья, теплокровность и способность летать. Благодаря наличию оперения и теплокровности они переносят холод лучше, чем пресмыкаю- щиеся, что, видимо, облегчило их расселение по всему земному шару, а также отразилось на характере их распространения и в других отноше- ниях. Но совершенно очевидно, что самая важная способность птиц с точ- ки зрения зоогеографии — это способность летать.
1 Сейчас установлен~. что аркеорнис — это тоя~ «t> синош~м ар~«~итери«ca.— 71 у~ и. 1~. ред.
- Накодки гесиерорнисов и ихтпорнисов.— lip)u.ц. п~ pcв.
209
Умение летать по-разному сказывается на распространении птиц: оно дает им возможность преодолевать преграды и увеличивать возмож- ности расселения независимо от этих преград, а также совершать ежегод- ные большие миграции, которые не только усложняют распространение мигрирующих видов, H() и отражаются на общем характере распростра- Hf. ÍÈß ПТИЦ.
Тот факт, что птицы могут преодолевать преграды, не так прост, как это ка;кется. На это способны не все птицы. Очень часто перед такими пре- градами, как небольшие водные пространства, останавливаются не толь- ко нелетающие. но даже и многие летающие птицы. Например, несколько древних семейств, хорошо представленных в Южной и Центральной Аме- рике. и которым относятся тинаму. крациды, ленивки, туканы, целое над- семейство Furuarioidea (500 видов) и манакины, не могут пересечь умерен- ные водные пространства, отделяющие материк от Вест-Индии. То, что ни одна из этих птиц не достигает собственно Вест-Индии, является уди- вительным фактом в географии птиц. Некоторые другие семейства, которые также хорошо представлены в Южной и Центральной Америке, например куропатки, трогоны и котинги. достигают одного-двух островов Вест-Ин- дии, тогда как другие семейства и подсемейства, включающие таких истин- ных представителей Нового (;вета, как колибри, мухоловка-тиранна и древесницы, а также таких широко распространенных птиц, как голуби и попугаи, расселились по всей Вест-Индии.
Новая Зеландия отделястся от Австралии океаническим пространством в 100( миль. Многие австралийские семейства птиц, древние и молодые в эволюционном отношении, не смогли достичь берегов Новой Зеландии. Е ним относятся (кроме нелетающих птиц — страусов эму и казуаров) сорные куры, куропатки, козодои и близкие им виды, лирохвосты, австра- лийское семейство Atrichornithidae, медососы, нектарницы, ткачики и т. д. Некоторые другие а«стралийсгие группы,'включая попугаев и вра- новых. в Новой Зеландии представлены сильно отличающимися формами. которые, вероятно, поселились там давно. Другие, очевидно, достигли Новой Зеландии недавно, а некоторые появляются только сейчас. Vi последнии относятся австралийская сизоворонка и ласточка, которые изредка залетают в Новую Зеландию небольшими стаями (последняя была однажды обнаружена HB гнездовье); личинкоед был отмечен в Новой Зеландии по крайней мере 1 раза; австралийская белоглазка (Zosterops), случайно появившись в Новой Зеландии примерно в 1856 г., с тех пор стала там оседлым видом; большая белая цапля впервые была отмечена в Новой Зеландии приолизительно около 1865 г., а примерно с 1940 г. стала гнездиться. и сейчас она также является оседлым видом; королев- ская колпица и ржанка впервые оыли обнаружены на гнездовье в 1950 г. Ио предположениям Оливера (Oliver, 1930, 1955) и атолла (Falla, 1953), в Новой Зеландии гнездится также и австралийская лысуха. Таким обра- зом целый ряд новозеландских птиц способен преодолевать препятствия.
Из сказанного вьпие можно сделать общий вывод, что одни птицы, встречая на своем пути преграды, останавливаются перед ними, а другие, затрачивая различное «ремя и преодолевая различное расстояние, пересе-
I~ä1ÎT ИХ.
Размеры преград, которые пересекают некоторые птицы, установить не«озможно. Большинство отдаленных островов земного ~нара было достигнуто наземными птицами и заселено ими, если там были подходящие для обитания места. Например, на Гавайских островах обитает несколько наземных видов птиц как из Америки, так и из стран Старого ( вета, кото-
210
рые пересекли по крайней мере ' 000 миль океана; в южной Атлантике на островах Тристан-да-Кунья и др., вероятно самых изолированных из пригодных для обитания островов земного шара и расположенных пример- но в 000 милях от ближайшего материка (Африка), имеется 5 эндеми ~ных родов наземных птиц, представленных,) отдельными видами — двумя видами пастуlIIhoBbIx, одним дроздом и двумя видами вьюрковых. Есть осно- вание полагать, что некоторые наземные виды птиц пересекли Атланти и- ский океан между Африкой и Южной Америкой, преодолев расстояни~ немногим менее 2000 миль. Эта дистанция под силу даже таким мелким наземным птицам, как дрозды и вьюрковые, которые проникли на остров» Тристан-да-Кунья. Упускать из виду этот фагт, возможно, не следует, но тем не менее расселение большинства наземных видов птиц обычно про- исходит по материкам, а не через океаны.
Водные преграды оказывают серьезное влияние на фауну птиц. огра- ничивая число видов, отбирая их и в той или иной степени изолируя от фаун других материков. Почему одни наземные виды птиц преодолеваю1. водные преграды, а другие нет, сказать трудно. Вероятно, крупные пти~~ы имеют больше преимуществ: при прочих равных условиях они могут и о1- личие от мелких птиц покрывать гораздо большие дистанции. Быстро ле- тающие птицы, например ласточки, очевидно, также имеют преимущества. Но это только одна сторона вопроса и этого недостаточно для объяснения самой сущности этого явления. Как правило, по анатомическим особенн<>- стям птиц или по наблюдениям за их полетом трудно судить могут 1и они преодолевать водные пространства или нет. В данном случае лучшим при- мером является коростель. Вспугнув коростеля и увидев, как неохотно и с каким трудом он поднимается в воздух и пролетает всего несколько ярдов, мы сомневаемся, сможет ли он перелететь даже ~ерез небольш~ ю канаву. Однако паступп<овые птицы и болыией степени, чем другие нечор- ские птицы проникли на самые отдаленные острова океана и заселили их. Несомненно, здесь дело не только и способности перелетать через водные пространства, а так'ке и в том, что птицы, которые расселяются на новые земли, преодолевая при этом водные преграды, должны там укорениться, как, например, это происходит у пастушков даже на маленьких островах. А это в снс1ю очередь связано с рядом осложняющих обстоятельств, ljli;,~í~- чая и недостаток мест, подходящих для обитания, и конкуренцию, и при- способляемость вида.
Расселение птиц зависит от многих причин, в частности от приспособ- ляемости вида и от его способности к перемещению. Jlтицы независимо от их способности летать могут быть оседлыми или неоседлыми. Но при прочих благоприятных условиях спосооность к полету позволяет им рас- селяться с такой скоростью, что они проникаю1 во все части земного пира в пределах времени эволюции одного вида. Доказательством этому с.1у- 'кат те виды-космополиты, которые в той или иной степени распространи- лись по всему земному шару.
Быстрота, с которой некоторые виды птиц распространились по все- му земному шару, очевидно, является одной из причин, почему так труд- но проследить их историю. Чем быстрее идет расселение, тем меньше остает- ся времени для эволюции вида в течение периода. на котором происходило расселение и тем труднее проследить, как это расселение происходи:io. Допустим, что расселение предков преуспевающей группы птиц по всему земному шару произошло меньше чем за миллион лет: тогда через 10 млн. лет местонахождение видов и родов этой группы в основном будет зависеть от их эволюционных возможностей в различных местах: доступности и оо-
Шпицы
ширности территории, числа экологических ниш, количества конкурентов и т. д., а не от продолжительности времени пребывания данной группы в этих местах. Найти место происхождения и пути ее расселения обыч- ными «ключами», вероятно, будет очень трудно. Эта трудность осложняет- ся еще и тем, что очень часто расселение таких групп птиц бывает много- кратным. Несколько родов или видов птиц одной группы могут расселять- ся на всем земном шаре одновременно. Например, по крайней мере 4 рода современных хищных птиц — Circus, Accipiter, Falco и Pandion — стали почти космополитами. Род Falco распадается на подроды, из которых три в свою очередь являются космополитами или почти космополитами. Один вид рода Falco и один (единственный) вид рода Pandion почти космополи- ты. llo крайней мере полдюжины родов и два вида цапель космополиты или тропикополиты. Четыре рода семейства Rallidae космополиты или почти космополиты. Три рода или подрода ласточек космополиты или по крайней мере являются широко распространенной группой. Расселение некоторых этих групп, вероятно, было последовательным и многократным. Распро- странение цапель, пастушковых и одного семейства хищных птиц относит- ся к эоцену. Похоже, что современные космополитические роды этих групп расселились недавно и их предшественниками были ранее широко распро- страненные роды.
Короче говоря, очень часто расселение птиц проходило так быстро и так сложно, что оно не поддается анализу. ('. точки зрения зоогеографии это, пожалуй, результат одной из важнейших способностей птиц — летать, одна из основных причин (другая заключается в скудости ископаемых), почему так трудно проследить и понять историю географии птиц. На этом обсуждение этого вопроса откладывается до соответствующего раздела (см. стр. 233 и сл.), где будет сделана попытка показать, какие имеются основания для наших суждений о способах расселения птиц.
Влияние миграций птиц на их географическое распространение
Миграции птиц во многом до сих пор являются загадочным явлением. Иочему у ряда птиц в определенное время возникают определенные сти- мулы к миграциям в строго определенном направлении и как эти птицы через огромные пространства суши и воды находят свой путь — все это является еще неразрешенной загадкой, хотя к ней можно подобрать «клю- чи». Но в данном случае важно не разрешение этого вопроса, а важен общий характер миграций и их влияние на географическое распростране- ние птиц. Недавно я вновь просмотрел по этому вопросу работы Томсона (Thomson, 1926; 1949), Грискома (Griscom, 1945), Моро (Moreau, 1951), Филиппса (Phillips, 1951), Майра (Мауг, 1953) и другие источники, вклю- чая Майра и Мейсе (Mayr, Meise, 1930), которым Моро приписывает заслугу создания теоретической основы происхождения миграций.
Обычно миграции птиц рассматриваются преимущественно как пере- мещение. но их также можно рассматривать, исходя из понятия ареала. Ареал животного — это территория его обитания. На картах ареалы кажутся простыми и устоичивыми, но не следует впадать в заблуждение (как это случается с некоторыми зоогеографами), так как на деле они сложны и неустойчивы. Одна из сложностей заключается в том, что ареалы 0()Л ЫИИНСТВИ ДИКИХ ЖИВОТНЫХ В ЗИВИСИМОСТИ ОТ ИХ ЯК'1'ИВНОСТИ НеСЕОЛЫ~О изменяются. Весь ареал вида в целом обычно состоит из двух или более тер- риторий. поочередно ичи последовательно занимаемых видом. Например,
Глави 5
распределение многих видов позвоночных, которых не считают мигрирую- щими, изменяется в пределах ареала во время кормления, размножения, расселения после размножения и зимовок. Каждый из чередующихся ареа- лов, из которых состоит основной ареал вида, может до какой-то степени изменяться незаеисимо от других, причем степень изменения определяется способностями рассматриваемых животных к перемещению. Когда речь идет о животных, которые способны перемещаться быстро и на большое расстояние, отдельные части их ареалов могут быть далеко отодвинуты друг от друга. Это как раз характерно для птиц, у которых сильно выра- жены миграционные способности.
Если двойственность ареала перелетных птиц рассматривать как осо- бый случай менее заметного чередования ареалов, характерного для диких животных, тогда в некоторых отношениях явление миграции становится более понятным. Нет необходимости напоминать старый и в общем диск- редитированный взгляд на миграции птиц, как на особенное явление, вызванное у северных птиц ледниковым периодом '. Нет необходимости полагать, что перелетные виды произошли или там, где они теперь размно- жаются, или там, где они теперь зимуют. Их ареалы гнездования и зимо- вок могут изменяться в самых широких пределах, допускаемых способно- стями птиц к перемещению. Например, совершенно не обязательно счи- тать, что полярная крачка, гнездящаяся в северной части северного полушария в Арктике и зимующая в южной части южного полушария в Антарктике (рис. 30), по своему происхождению является арктической или антарктической птицей или что она внезапно начала мигрировать через тропики. Возможно, сначала это был тропический вид (несколько других видов того же рода являются тропическими). который совершал неболь- шие перелеты, гнездясь не в очень холодных частях умеренной зоны север- ного полушария и зимуя в тропиках (как это наблюдается т обыкновенной кра ~ки теперь). Впоследствии гнездовая область этого вида. возможно. сместилась в северном направлении а область зимовок — в южном. Подоб- ные двойные перемещения могут объяснить хорошо известные случаи. в которых самые северные гнездящиеся формы вида являются самыми южными зимующими, мигрирующими через ареалы менее перелетных форм. Возможно, отдельные особи птиц, гнездовая область которых сместилась к северу, сначала возвращались в места, где зимовали вместе с оседлыми птицами, но не смогли выдержать конкуренции с ними потому, что послед- ние были лучше приспособлены к местным условиям. Таким образом, область зимовок перелетных особей должна была в конце концов пере- меститься к югу. в совершенно новые районы.
Если такое толкование миграций правильно, то потенциально все птицы являются перелетными. Все имеют или могут иметь как исход- ное сезонные области обитания, причем последние могут смещаться друг от друга и любом благоприятном направлении и на любое расстояние, до- пускаемое способностями птиц к перемещению.
Иэ этого следует, что явление миграций, очевидно, происходило с дав- них времен у многих различных птиц, в разнообразных местах и при мно- Больфсон (M'01fso», 1948) предполагает, что более высокоразвитые формымигра- ций были вызваны у птиц перемещением материков. Эта мысль была довольно сенса- ционной и нашла отклик в американских журналах. Ответ на нее был помещен в жур- нале «Science» (Amadnn, 1948). Но вряд ли зта теория соответствует фактическому положению вещей. Если перемещение материков и ичело место, то это было слишко~~ давно, чтобы отразиться на современных видах птиц. В лучшем случае теория пере- мещения материков может помочь объяснить только отдельные особые случаи, которые являются лишь небольшой частицей всего явления миграций, но ие явление в цел<ос.
Пиьицы
гих различных обстоятельствах. Вероятно так;ке и то, что многие птицы, относящиеся z перелетным группам, прекращали миграции и в подходя- щих местах для обитания становились оседлыми. Так, например, произо- шло с некоторыми древесницами (Parulidae) (см. стр. 2Ы).
Теоретическая сложность миграций птиц вполне соответствует фак- тической сложности этого явления. Многие птицы совершают незначитель- ные кочевки, которые, начинаясь в одних случаях как смена местных
° ~ е ' ° э
° г
, ° ' ф
° °
° ° ° ° е
° °
ч~, ° ~ ° °
° ° ° OO °
е °
° ° ° °
° °
с
° р ее
° ° °
° °
Э
е
° °
ь1р~Э~P'Э Эф
Эе,'
° °
° °
° °
° е
.'-ъ ° ° °
Э °
е
° °
° °
° °
е °
° ° °
° °
° °
° °
° ° ° °
° ° °
° ° е
° ее
° ° ° ° ° °
° °
° ° °
° °
° ° °
е
е °
° Э ° °
° °
Э
° е °
° °
° ° ° °
° ~~ °
° °
° °
° ° ° ° °
еф ° ее ° ° °
° ° ° °
\
° °
.Эе °
° Э
Р и с. ЗО. Пути мигрмитт1 иолярнс)й крец~~ в Лг)л~пи1сс '~orокеане (схема).Крестиками обозначены фактические места зимовок, где были обнару-жены особи этого вида (Мигрйу, 1936, р. 1()9> — 1105).
кормовых участков, почти незаметно сменяются нерегулярными или регу- лярными перемещениями отдельных особей, или части популяции, илн всей популяции на несколько миль или несколько сот миль — в других случаях, и большими миграциями на тысячи и несколько тысяч миль — в третьих. Такие перемещения происходят во всех частях земного шара. Далеко не все северные птицы, которые избегают зимы, совершают миграции большой протяженности.
Моро (Могеаи, 1951) приводит еще несколько примеров миграций. Несколько видов птиц мигрируют из Северной Африки или Египта к югу на тысячу миль и более; другие виды, которые гнездятся в умеренно- теплой части Южной Африки, мигрируют на то же расстояние к се~пру, а третьи перемещаются только в пределах тропиков Африки. Эти переме- щения находятся в корреляции, скорее, с в.сажными и засушливыми вре- менами года, чем с теплыми или холодными. Рэнд (Rand, 1936, р. 300) приводит 5 видов птиц, которые гнездятся на Мадагаскаре, а «зимуют»
е ° ° ее эе
° ° à ее ° ° ° ° °
° ° ° ° ° ' °
° Э ° е ° ° °
щ.-: ° °
Э ° ° ° ° ° ° ° i i° ° °
° ° ° ', ° ° ° ° ° ° Э ° ° ° ° ° ° °
° ° ° ° ° °
° ° е ' ° ° ° ° ° ° ее ° ° Э ° °
° °
° Э~ ° ° л' ° °, ° ° ~, ° ° ° ° °
° ° ° ° ЭЭ ° °
° е е °
° Óå ÔÂ еэ ° ' ° е ° ° ° Э ° ° ° е ° ° °
° ° 9е ~ре ° Г, ° ге ° 1 ° Ъ. ° е ° ° ° + ° Э ° °
° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Э ° ° ° ° ° ° Э ° ° ° 'e ° ° ° ' ° ° Э ° °
° °
° ° °
° ° ° °
ф ° ° ° ° ° ° ° ° \ ° ° ' ° eI .
° ° °
° эее ° g °, еэ ° ° ° ° э ° ° ° ° ° Э ° ° ° ° ° ° ° ° ° ' °°, ° ° ° ° °
Ф
° ° I:"~r,
Э% ° Я
° ° ° l::;::. -::.:::',' ~,~ ~:ф;.. ° --."."'--Фг.
1ЦФ" 4ч' °
Ю.":.':."
2И
Глави 5
в Африке. По крайней мере один из них — широкорот (Eurystowus glnu- curus) достигает Конго. Другой вид (Eurystomus orientalis deignani) широ- корота Индо-Малайской области мигрирует только в пределах тропиков, преодолевая почти на 2000-мильное расстояние из северного Таиланда до островов Ява и Калимантан. Считают, что индийская популяция кукушек (Clamator )асоЬтиз pica) во внегнездовой период откочевывает еще дальше в Восточную Африку. Зеленая щурка (Merops superciliosus persicus), гнездящаяся в северо-западной Индии и т. д., зимует в центральной и юж- ной частях Африки (%1arien, 1950); другой вид щурки (Merops philippinus), близкий к индийскому, совершает более короткие миграции и в другом направлении — к южной Индии, Малайе и западной Индии; я думаю, что другие виды того же рода оседлы в Индии и других местах.
В Америке несколько видов птиц, включая мухоловку-тиранну, вире- она, козодоя, которые гнездятся на Кубе и Багамских островах и т. д., зимуют в Южной Америке. Чэпин (Chapin, 1932, р. 322 — 362) указывает на сложность миграций птиц тропической части Конго. Миграции тропи- ческих птиц вообще изучены еще недостаточно. Миграции или странство- вания некоторых морских птиц в тропики и из тропиков еще более удиви- тельны, чем миграции наземных птиц, но они здесь не рассматриваются.
Особые случаи миграций с севера на юг следует рассматривать на ос- нове тенденции к «полиареальности» и к миграциям, которые обычны для всех птиц, на основе теоретической и фактической сложности этого явле- ния. на основе многократного возникновения и многократной потери миграционных устремлений при различных обстоятельствах и с учетом многих других факторов.
Самые интенсивные миграции птиц происходят между их областями гнездования в северных частях земного шара и зимовками на крайнем юге. Характер этого перемещения, причины, его вызвавшие, его происхо- ждение и влияние на особенности распространения птиц — все должно быть принято во внимание.
Основной характер миграций с севера на юг хорошо известен, но его детали очень сложны. Многие виды птиц, которые гнездятся на севере, летят на юг различными путями. Зачастую даже особи одного вида следуют различными путями. Разные популяции некоторых видов летят на юг одним путем, а возвращаются на север другим. Перемещение не всегда про- исходит строго на север или на юг. Некоторые птицы прежде, чем они попадают на юг, летят не прямыми путями, а на восток или запад. Ряд ази- атских птиц, которые расширили свой гнездовой ареал до Аляски, возвра- щается на зимовку в страны Старого Света, и наоборот. Обыкновенная каменка, гнездовой ареал которой охватывает всю северную Евразию, рас- ширила его в Северную Америку по двум направлениям: из Европы через Гренландию в восточную часть Северной Америки на юг до северного Квебека и из Азии в Аляску, хотя американские особи до сих пор зиму- ют в странах Старого Света, а восточные летят через Европу к Западной Африке. Большинство видов из Аляски летит через Азию в меридиональном направлении к Восточной Африке (Meinertzhagen, 1954, р. 240). Восточно- сибирская форма пеночки-веснички (РЬуПозсориз trochilus) зимует в Во- сточной Африке, а другой вид этого же рода (Р. borealis) гнездится от Азии до северной Норвегии, а зимует в Таиланде и т. д., пересекая миграцион- ный путь первого вида под прямым углом (Ticehurst, 1938).
Большие различия наблюдаются также и в расстоянии, которое пре- одолевают различные мигранты, летящие с севера на юг: одни, покидая свои северные гнездовые участки, улетают на очень незначительное рас-
стояние, другие улетают в тропики или даже за их пределы. Эти различия иногда наблюдаются между близкородственными видами или даже между »с»бями одного вида. Томсон (Thomson, 1949, р. 107 — 122) и Мойр (Мауг, 1Ч. 3) в своих работах подробно останавливаются на сложностях мигра- ций птиц с севера на юг.
В умеренной зоне южного полушария миграции идут с тога на север, но они охватывают меньшее количество видов птиц, и ооль шинство из них далеко не улетает. Например, в 1или (Не11тауг, 1932, р. 25, ' 6) некоторые птицы, гнездящиеся высоко в горах в южной части Анд, зимуют в близ- лежащих долинах или перемещаются на небольшое расстояние к северу. Различные наземные птицы, которые гнездятся у южной оконечности Южной Америки, зимуют в центральной или северной части Чили и т. д. Некоторые морские птицы, включая пингвинов, совершают береговые мигра- ции. В Австралии некоторые птицы, гнездящиеся на юге, также совершан~т миграции к северу и большей частью в прохладную восточную часть мате- рика. чем в теплую западную. В Западной Австралии (Serventy, Whit- tel I, 1948, р. 5 — 6) примеров «классических» миграций наземных птиц мало. но ряд видов совершает регулярные местные, частичные миграции или же является кочующим. В Новой Зеландии наблюдаются некоторые мест- ные перемещения птиц. и два наземных вида новозеландских птиц совер- шают миграции, которые относятся к самым интересным на земном шаре. Оба вида относятся к кукушкам и покидают Новую Зеландию зимой. Один из них, Chnlcites lucidus lucidus, зимует на Соломоновых островах и т. д. (Fell, 1947), другой, Eudynamis tnitensis,— по всей Полинезии и т. д., н центральной части Тихого океана (Bogert, 1937). Вторая из этих птиц— четко выраженный вид, и характер его миграций, вероятно, установился давно. Первая же, кукушка, является только новозеландской формой вида, представленного в Тасмании и Южной Австралии другой мигри- рующей формой (зимует на Малых Зондских островах и на Новой Гвинее), а на некоторых тропических островах — оседлыми формами. Таким обра- зом, характер миграций названной новозеландской формы, возможно, недавнего происхождения. Очевидно, оба вида новозеландских кукушек совершают перелет через океан, а не через Австралию, и тем не менее воз- можно, что Австралия имела какое-то отношение к эволюции их миграций.
Основная причина миграций с севера на юг и с юга на север, вероятно. заключается в сезонных изменениях в питании птиц, хотя в некоторых слу- чаях они связаны и с другими факторами. Степень миграций различных птиц изменяется в зависимости от их пищевой специализации. Самые север- ные насекомоядные птицы, а также многие плодоядные относятся к пе- релетным, в то время как зерноядные птицы в целом являются менее пере- летными, но из этого общего правила имеется много исключений. Некото- рые северные хищные птицы относятся к перелетным (а другие — нет) или совершают миграции только спорадически. Многие северные птицы внутренних водоемов являются типично перелетными. Они много- численны на крайнем севере летом, но после сезона размножения улетают на юг, преодолевая различные дистанции. Многие птицы, гнездящиеся вблизи пресных вод. при миграциях придерживаются морских побережий, н некоторые из них, особенно кулики, покрывают огромные расстояния до умеренной зоны Южной Америки, Австралии или Новой Зеландии, где «ни обеспечены кормом в течение всей северной зимы.
Происхождение массовых миграций птиц с севера на юг, вероятно, так же сложно, как и характер этих миграций. У очень многих птиц миграции развивались с севера на юг, а некоторые перелетные формы еще
216
Ггава 5
больше усложнили это положение более поздним образованием местных оседлых форм. Являются ли птицы, мигрирующие с севера на юг, по про- исхождению южными, которые сместили свою гнездовую область к севе- ру, или северными, у которых область зимовок переместилась в южном направлении, и (в последнем случае) происходили ли миграции с севера на юг до плейстоцена или северные птицы были вынуждены мигрировать под влиянием климата плейстоцена — все это давно являлось предметом горячих споров. Вероятно, в определенных случаях все эти предположе- ния вполне реальны.
Если птицы, подобно другим животным, перемещаются для того, чтобы держаться все время в лучших условиях существования, то, вероятно, многие миграции с севера на юг начались до плейстоцена посредством сме- щения гнездовых ареалов из тропических или умеренно-теплых районов в северном направлении в целях использования обильных кормов на се- вере летом (конечно, смена времен года в северных частях земного шара происходила и до плейстоцена). Миграции большинства северных насе- комоядных птиц, видимо, начались именно таким образом. В Евразии и Северной Америке эти птицы в большинстве случаев относятся и разным семействам, которые, по-видимому, появились независимо друг от друга из тропиков Старого и Нового Света. Но у северных водных птиц мигра- ции развились в другом направлении. Эти птицы обычно гнездятся гораз- до севернее, очень часто в пределах Арктики. Они приспособились глав- ным образом к северным местам обитания, которые непригодны для насекомоядных птиц, и их миграции, возможно, явились результатом перемещений северных птиц к югу в поисках пищи во внегнездовой период. Многие из северных перелетных водных птиц Евразии и Северной Америки близкородственны, так словно они произошли от первоначального северного циркумполярного корня. Однако, какой бы ни был характер происхож- дения миграций, плейстоцен, вероятно, увеличил и число северных пере- летных птиц, и расстояние, на которое они мигрировали. Например, если северные насекомоядные и водные птицы до плейстоцена совершали толь- ко незначительные перелеты, то после него (и в течение каждого межлед- никового периода) их гнездовые ареалы, по-видимому, сместились в се- верном направлении с целью использования новых стаций и уменьшения конкуренции, а затем под давлением возрастающей численности популя- ции, ареалы их зимовок, возможно, переместились в южном направлении, расширяя, таким образом, разрывы между гнездовым ареалом и ареалом зимовок, что, как я полагаю, и произошло у полярной крачки.
Хотя миграции расширяют и ) сложняют распространение многих групп птиц, их общее влияние на распространение сказывается вполне определенным образом. Перелетные птицы не рассеиваются по земному шару беспорядочно. Правда, иногда они могут терять свой путь или появ- ляться в весьма неожиданных для них местах, но приживаются там редко.
Основное влияние миграций на общий характер распространения птиц, вероятно, заключается в увеличении связей между тропическими и севе- ро-умеренными фаунами птиц в каждом полушарии земного шара и в умень- шении этих связей между Старым и Новым Светом, особенно среди насеко- моядных птиц. В каждом полушарии земного шара в Старом и Новом Слете разные группы насекомоядных и некоторых других мелких наземных птиц, очевидно, распространились из тропиков к северу как перелетные птицы п стали оседать на севере летом, причем большинство из них благодаря привязанности к свойственным им зимовкам придерживалось пределов своего полушария. В Старом Свете luscicapinae, Sylviinae и 'Alotacill i~lac,
Шпицы представители которых многочисленны в тропиках Старого Света, распро- странились к северу как перелетные птицы, но только несколько видов славок и трясогузковых достигло Северной Америки. В Новом (.;вете семейства Tyrannidae, Vireonidae и подсемейство Parulinae хорошо пред- ставлены в тропиках, а также (в большом числе) далеко на севере как перелетные птицы, но не достигают Евразии (кроме двух видов из подсе- мейства Раги1Ыае, которые расширили свой гнездовой ареал за счет двух небольших участков Восточной Сибири). Особенности миграций этих птиц не только ограничили их распространение, но и способствовали задержа- нию обмена других насекомоядных птиц между Старым и Новым Светом. Вероятно, из всех животных, хорошо представленных на севере. нп одно не отличается так сильно в Евразии и Северной Америке, ка~; эти птицы. Среди крупных (обладающих большими способностями расселе- ния) или среди водных птиц такого влияния миграций (или по крайней. мере в такой степени) не наблюдается.
Миграции оказывают особое влияние на распространение береговых птиц. Они позволяют многим видам, которые гнездятся на крайнем севе- ре, зимовать в отдаленных районах умеренной зоны южного полушария, что, возможно, мешало эволюции значительного числа береговых птиц. гнездящихся в э roii зоне. Границы распространения птиц
Северные и южные границы распространения птиц, а также другие особенности их распространения представлены на рис. 31. 1 и с. 3i. Самые северны» и самьн южные наземные птицы и
некоторые другие детали распространения 1lTIIII,.
Абсолютной границы распространения птиц и северу нет, за исклн~че- нпем пределов суши, за которыми, как известно, кормятся и стран- ствуют морские пернатые. Но к северу разнообразие птиц уменьшается. Довольно большое число видов птиц гнездится на севере у границы леса. Многие водные птицы (гуси, утки, кулики и т. д.) и немногие наземные
(некоторые хищные, куропатки и несколько видов вьюрковых и т. д.) гнездятся еще дальше и северу, и несколько видов из них заходит даже севернее 80" с. ш., на полярные острова Северной Америки. Большинство »тих птиц зимой мигрирует на юг, но часть, включая куропаток и белую сову, часто зимует на крайнем севере. Даже некоторые чечетки (зерно- ядные вьюрковые рода .4 сссп <its), очевидно, оседлы на крайнем севере Грен- ЛЯНДИИ.
В южном направлении многие птицы достигают южных оконечностей материков; некоторые гнездятся еще южнее — на антарктических островах: императорские пингвины размножаются на льду по кроо Антарктическо- го континента, а снежный буревестник и несколько других видов трубко- носых даже проникают на незначительное расстояние в глубь Антарктики. где гнездятся на скалистых участках или на голой земле. Самыми южными птицами, которых с достаточным основанием можно считать наземными. являются белые ржанки (Chionididae, см. список семейств). Это странст- вующие, но не типично перелетные птицы. Они размножаются на антарк- тических островах и, возможно, даже по краю Антарктического конти- нен.га.
Строгих границ в распространении птиц на островах нет. Несколько видов птиц, даже несколько видов мелких наземных пернатых, достигли самых отдаленных пригодных для обитания островов земного шара. Рас- стояния или водные пространства не определяют границы распростране- ния наземных птиц, но постепенно уменьшают их разнообразие: чем даль- ше отстоят острова от материка, тем меньше встречается на них видов птиц. Например, на островах западной и центральной частей Тихого океа- на разнообразие наземных птиц, особенно певчих, уменьшается с увели- чением расстояния от континентов и больших островов Индо-Австралий- ской области (рис. 3' ), и это уменьшение происходит последовательно не п~лько на архипелагах, но в ряде случаев и на островах в пределах архи- пелагов (Mayr, 1933, р. 315). Вот еще один пример того, что при встрече птиц с преградами одни из них останавливаются, в то время как другие пересекают их, преодолевая различные расстояния.
Распространение некоторых видов и родов почти безгранично. Сокол галсан (Falco peregrinus) гнездится в различных частях всех материков и на многих островах. Скопа (Pandion haliaetus) гнездится или зимует на большей части земного шара, но в некоторых районах эта птица в основном придерживается берегов водоемов. Болыиая белая цапля (Casrnerodius «lbus) является почти космополитом и не встречается только в холодных районах. Маленькая цапля (Butorides striatus) почти тропикополит, кро- ме Северной и Центральной Америки, а также Вест-Индии, где ее заме- щает родственный вид. Каравайка (Plegadis falcinellus) почти тропикопо- лит, кроме Южной Америки, где, вероятно, обитает самостоятельный вид. Морской вуек (Charadrius alexandrinus) представлен по крайней мере на какой-либо части каждого материка. Ходулочник (Hirnantopus 3итип- tempus) встречается на всех материках, хотя его гнездовой ареал локализо- ван. Сипуха (Tyto alba) почти космополит, но отсутствует на значительной территории умеренной зоны Азии и в областях с холодным климатом. Более подробно распространение большинства этих видов дается в списке семейств. В различных частях земного шара все они делятся на подвиды. все относятся к неворобьиным, и все, кроме зуйка,— птицы средних и крупных размеров и являются либо водными, либо хищными птицами.
Кроме перечисленных выше, к родам неворобьиных наземных и прес- новодных птиц, которые являются космополитами, или тропикополитами,
или почти теми или другими, относятся еще два рода поганок, бакланы и пеликаны, являющиеся отчасти морскими птицами (PAalacloco/ax и Pe- leca»us), из хищных — Circus и Accipiter, около пяти родов цапель, дре- весные утки (Dendrocygna) и обыкновенные утки (Аnas). из пастушков —— llnlt>ls, Pr>r=ana, Gallinula и t~ulica, авдотки (Виг)»пиз). шилоклювки
° ° ° «
° ° ° °
° ве ° ° ° ° ° °°,Ф ° ° ° °,," ,°в° »Э;.'.: '
Ф °
° °
. °
° ° °
° Ф
° °
° °
° ° '
° «
° ° °
° ° °
4,
Р . ~.. y.~ QQ», . ф: /;»„
° е °
,У,
»'
° Ю > ~ф
' 'IO
у О~
° °
° ° '
Ъ
е °
° °
° r ° ~ ~° °
° Ю
~»» ~ ~ ».ф ° !
Ю
° I °
,. ч...,~И° °
° ° У
° ° ° ~Ф Ж:::;:.;~~„':Р.
М ° v,
° °
° °° ю° °° ° °
° ° ° r' r
° ° ° °
° ° г
а: ~~..::,'":::
Ф "': ° ' "."
6,
° °
° °
° ° ° ° °
»
° «
г' '.,сА, 1
ъ
° е ° ° ° . °
в.- Я~
Ъ..,
° °
'с,
° °
Ъ
~в~,~, ф»
Ф~
~б.
«~~;ф»
Ф .-т
° °
.~:-:;::~' ~.' ° ~ ° .....' .'. " ..-:.-,.'..
~ъ~
— ' ё~И,
° . '~'»
° °
° °
° е
° ° ° °
° ' ° ° ° ° ° ° ° °
",. ° ° I °
° °
Р и с. 32. Уве;и~чеш~е числа видо» наземнык нтиц с увеличе- н ис м расстроя иия от материков в западной части T Jlx ol'о
океана.
'.[иф1 ы (напримср. 'i '6/51) показывают общее число видов назем- ных птиц, превосходящее число певчих птиц (Мауг, 1933). Раз- мещение этих цифр иримерно соответствует местонахождению выше
названных островов.
(ЛесurпгоИы) — род с очень широким, но дизъюнктивным распростране- нием (кроме этого рода и рода Charadrius, некоторые другие роды и даже виды куликов охватывают миграциями и зимовками большую часть земно- го шара); обыкновенные голуби (Columba); и козодои (Caprirnulgus).
Таким образом, восемь видов и дополнительно 21 род неворобьиных птиц, приведенные выше, являются более или менее космополитами или тропикополитами. Возможно, что их насчитывается и значительно больше.
Воробьиные вообще не имеют такого широкого распространения. Ни один вид воробьиных не является полностью космополитом или тропи- кополитом. Более всех приближается к тем или другим деревенская ласточка (Hi>rundo rustica). Этот вид с несколькими подвидами встречает- ся (если считать и область зимовок, и область гнездования) на большей части земного шара за исключением Австралии, хотя изредка он достигает северного побережья последней. На большей части Африки, тропической Азии и Южной Америки этот вид все же только зимует, а не гнездится.
220
Глава э
Зональное распространение птиц
Несмотря на то, что птицы являются убиквистами и многие пернатые тропических и умеренных зон северного полушария близкородственны, все же между тропической и северо-умеренной фаунами птиц существуют поразительные отличия. Они состоят не столько в разли IIIJI семейств, сколько в самой природе и структуре обеих фаун в целом.
Среди воробьиных птиц самым широко распространенным родом, возчож- но, является род Hirundo rustic@ (деревенские ласточки и др.), который раз- множается и зимует на всех материках, кроме Южной Америки. Petro- chelidon (американская скалистая ласточка и др.), который, возможно, является подродом Hirundo, гнездится на какой-то части каждого конти- нента, но его гнездовые территории, вероятно, более ограничены и преры- висты, чем у собственно рода Hirundo. Третий род ласточек — Riip«rila широко распространен как в Старом, так и Новом Свете, но его точные границы в первом мне не известны, а во втором — гнездовой ареал н~ вклю- чает Южной Америки хотя он там и зимует. Род Апйиг (конькп из се- мейства Motacillidae), гнездится в отдельных частях каждого материка, но его распространение прерывистое, как будто при расселении эти птицы пропустили большие территории и осели только в тех местах, которые ока- зались для них особенно благоприятными. Птицы этого рода хорошие лету- ны, но их способность к полету не лучше, чем у некоторых других воробьи- ных, и особенно таких птиц, как ласточки.
Кроме четырех родов, названных выше, из воробьиных самыми широко распространенными, вероятно, являются Turdus и Corvus. Род Тиrdus хотя и не совсем космополит, но, очевидно, является наибольшим уоик- вистом среди воробьиных птиц. Он хорошо представлен гнездящимися видами на каждом материке, кроме Австралии и многих островов. и раз- личные виды этого рода встречаются в самых разнообразных экологиче- ских условиях — от открытых степных пространств в умеренных и суб- арктических районах до сплошных тропических джунглей (Ripley. 1952). Широкое распространение птиц рода Turdus, не очень крупных и обла- дающих не очень хорошими летными качествами, объясняется, вероятно, во многих случаях их хорошей приспособляемостью. Например, для нашего дрозда в Америке типичными местами обитания являются 1'ак лужайки пригородов, так и еловые леса Норт-Вуда. Род Corvus (вороны и др.) имеет представителей на каждом материке, кроме Южной Америки, и достиг Мадагаскара, Новой Зеландии (где в настоящее время исчез), некоторых островов Вест-Индии и даже Гавайских островов, где представ- лен одним эндемичным видом.
Распространение большинства видов, родов (и семейств) воробьиных птиц относительно ограничено, и этот факт, очевидно, является (хотя. быть может, и не целиком) следствием каких-либо особенностей класси- фикации, или эволюционного уровня отряда, или его геологического воз- раста. Воробьиные — относительно молодые птицы, тем не менее многие из них, несомненно, существуют довольно давно, так как они успели рас- пространиться по всему земному шару. Очевидно, у большинства из них, отчасти и потому, что в большинстве случаев это мелкие птицы, отсут- ствуют способности к быстрому расселению. Ласточки с тремя почти кос- мополитичными родами показывают, на что способны воробьиные, когда их возможности расселения увеличиваются.
Тропическая авифауна намного богаче северо-умеренной по числу семейств, родов и видов. Гриском (Griscom, 1945, р. 114) указывает, что 8%' всех видов и подвидов птиц относится к тропическим, или, иными словами, что только 150~о встречается к северу от тропиков. Голуби и попугаи изо- билуют в тропиках, но относительно немногочисленны или отсутствуют на большей части умеренной зоны северного полушария, а некоторые не- большие, но широко распространенные семейства, например ибисы, ку- кушки. козодои и зимородки, значительно лучше представлены в тропи- ках, ~ем к северу от них. К небольшим семействам, которые широко рас- пространены в тропиках, но не заходят или едва заходят в умеренную зо- ну северного полушария, относятся лапчатоноги, яканы, трогоны и боро- даткп. Многие другие семейства птиц, характерные для тропиков того или другого полушария земного шара, совсем не проникают или едва прони- кают в умеренную зону северного полушария. В Старом Свете к ним от- носятся фазаны и дикие куры (в основном Индо-Малайской области; подроонее см. список семейств), цесарки (в основном африканские), ту- рако (африканские), птицы-носороги, птицы-мыши (африканские), медо- указчики, ширококлювки, питты, бюль-бюли (последние проникают на север до Японии и т. д.), личинкоеды, тималии (несколько видов встречает- ся также в умеренных зонах Европы и один вид локализован в западной части Северной Америки), представители семейства Prionopidae (в основном африканские), артамусы (Индо-Малайская область), медососы (Индо- Малайская область и т. д.), нектарницы, белоглазки (распространены на север до Японии и т. д.) и дронго, не говоря о ткачиках и скворцах, кото- рые значительно многочисленнее в тропиках, чем на севере. В Новом Све- те к тропическим семействам относятся тинаму, шлемоносные куры, ис- полинские козодои, момоты, ленивки, якамары, туканы, муравьеловки, печники (кроме североамериканского печника), древолазы, манакины, котинги и гавайские цветочницы, не говоря о колибри и танаграх, которые очень многочисленны в тропиках Америки и сравнительно малочисленны к северу. Кроме перечисленных, целый ряд небольших семейств птиц ло- кализуется в разных местах тропиков и к северу не распространяется.
Северо-умеренная фауна наземных птиц по сравнению (; тропической, по существу, отличается меньшей численностью, а не разнообразием. В умеренной зоне северного полушария очень мало семейств или подсе- мейств птиц, характерных только или почти только для этой зоны: из на- земных птиц к ним относятся тетеревиные (Tetraoninae), завиру(ики (Pr(r- rrelli(lae, в Старом Свете только в умеренной зоне северного полушария), свиристели (Bombycillinae) и настоящие пищухи (Certhiidae). Эти четыре группы насчитывают всего 39 видов. Конечно, ряд родов и многие виды наземных птиц других семейств приурочены к умеренной зоне северного полушария и некоторые из них имеют зональное распространение, буду- чи широко представленными в северной части земного шара, но в тропиках они отсутствуют (такой зональный эндемизм встречается в рангах ниже (;емейства).
('.писок семейств или подсемейств птиц внутренних водоемов и морских птиц, которые, как гнездящиеся птицы, являются полностью или в ос- новном северными, значительно обширнее, но эти птицы более характерны для крайнего севера, чем для умеренной зоны северного полушария в це- лом. Гагары (4 вида) зимуют в пределах умеренной зоны северного полу- шария. а гнездятся значительно севернее. Некоторые группы уток и осо- бенно гусей и лебедей в большом числе гнездятся на севере, но они также представлены и в других частях земного шара. Гнездование (roJIbllIHHcTBa
песочников и им близкородственных (Scolopacinae) происходит на севере. но в связи с тем, что их миграции охватывают весь земной шар, у несколь- ких видов области гнездования разбросаны по всему земному шару. Плн- вунчики (Phalaropus, 3 вида) гнездятся на крайнем севере, но зимуют в тропических морях. Среди поморников один вид гнездится и на крайнем севере, и на крайнем юге земного шара. Гнездование трех других видок поморников приурочено только к северу. Чайки многочисленны на севе- ре, но встречаются также и в других частях земного шара. Чистиковые гнездятся только на севере (несколько видов — на юге до Нижней Кали- форнии) и зимуют в пределах умеренной зоны северного полушария.
Таким образом, основные зоны обитания птиц от тропиков к северу следующие: (1) тропики, где птицы очень многочисленны и разнообраз- ны; (2) умеренная зона северного полушария, где птицы малочисленны и менее разнообразны, чем в тропиках, но в других отношениях незначитель- но отличаются от тропических; и (3) субарктическая и арктическая зоны, в которых наземные птицы очень малочисленны и не очень разнообразны, но где наблюдается удивительная концентрация гнездящихся (в основном перелетных) водных и береговых птиц.
Строгих границ между этими зонами нет. В Африке многие тропиче- ские группы птиц останавливаются у Сахары, и лишь немногие пересе- кают ее. В Азии, особенно в восточной части, некоторые (но не все) тропи- ческие семейства птиц простираются на север через большую часть (или даже через весь) Китая и даже достигают Японии. Здесь граница между тропической и северо-умеренной фаунами птиц, вероятно, выражена менее отчетливо, чем в любом другом месте. В нескольких случаях, которые отмечены в списке семейств, восточноазиатские представители тропиче- ских семейств относятся к перелетным птицам.
В Америке многие тропические семейства птиц останавливаются у се- верной границы дождевых лесов на тропических низменностяхМексики (Мауг, 1946, р. 32), а другие семейства распространяются дальше на север на разные расстояния. Конечно, имеются в виду главным образом границы тропических групп птиц, которые придают тропикам присущее только им богатство. Некоторые другие семейства птиц распространены на боль- п1ей части земного шара независимо от границ тропиков, что делает гра- ницу их распространения расплывчатой. Далее на север самым резким ру- оежом является граница леса (Мауг, 1946, р. 30 — 31), за пределами кото- рой значительное число наземных птиц существовать не мо;кет, но даже :>ту границу переходят многие как наземные, так и водные птицы.
В умеренной зоне северного полушария сравнительная бедность се- мействами, родами и видами птиц не означает еще низкой численности особей. Не располагая точными сравнимыми количественными характе- ристиками, все же можно сказать, что хотя особей птиц на данной площа- ди в тропиках, возможно, больше, чем в умеренной зоне северного полу1па- рия, эта разница не может быть очень значительной. Если это действитель- но так, то среднее число особей одного вида в умеренной зоне северного полушария больше, чем в тропиках. Этот вопрос заслуживает более де- тального изучения.
К югу от тропиков самое болыиое изменение в фауне птиц заключает- ся в исчезновении тропических групп. Например, в Южной Америке дчя многих тропических птиц южным пределом является северная часть Ар- гентины (несколько градусов южнее фактическои линии тропиков). Дру- гие птицы проникают дальше и югу на различное расстояние, и несколь- ко видов достигает Огненной Земли. Некоторые большие тропические се-
Птицы
мейства, которые не заходят в умеренную зону северного полушария или едва заходят в нее, простираются далеко на юг. Так, попугаи многочислен- ны в южной части Австралии и Тасмании, несколько видов эндемичны в Новой Зеландии, а в Америке один вид обычен на юге до Огненной Зем- ли; печники и тапаколо хорошо представлены в умеренной зоне Южной Америки, проникая к югу до Огненной Земли, хотя в целом распростра- нение большого тропического надсемейства Furnarioidea, к которому они относятся, резко останавливается на севере — в южнои части Мексики.
Удивителен и заслуживает особого внимания тот факт, что г. умерен- ной зоне южного полушария нет зональной фауны наземных птиц и нет особых групп, общих для южных частей разных материков. Фактически между наземными птицами Южной Америки и Австралии или Новой:4- ландии прямых связей нет. Предполагаемое родство между южноамери- канскими муравьиными коньками (Conopophagidae) и новозеландскими крапивниками (Xenicidae), а также между южноамериканскими тапаколо (Rhinocryptidae) и австралийскими представителями семейства Atric<>r- nithidae о ~ень сомнительно (Мауг, Amadou, 1951, р. 12). Ю,кноамерикан- ские наяду, австралийские эму и новозеландские моа, которых когда-п> считали родственными, в настоящее время обычно рассматриваются как самостоятельные конзе ргентные группы.
h гнездящимся птицам внутренних водоемов умеренной зоны южного полушария относятся отдельные роды и виды широко распространенных семейств, которые не образуют четко зональной фауны. (:еверные кулики, зимующие в южной части Южной Америки, Австралии и Новой Зеландии. в отдельных случаях оказываются близкородственными. Они обраэунл южную зональную фауну негнездящихся птиц. Для антарктической фау- ны характерны морские гнездящиеся виды (замещаемые некоторыми не- гнездящимися): пингвины и нырковые буревестники, а также различные чайки и т. д. В основном эта фауна зональная, но частично она (включая пингвинов) следует вдоль холодных течений — течения Западных ветров и Перуанского — по западному берегу Южнои Америки, заходя далеко в тропики.
Таким образом, главные зоны обитания птиц на юге такие же, как 11 на севере но отличаются существенными деталями. Эти зоны следующие: (1) тропики с богатой фауной птиц; (2) умеренная зона южного полушария с более мало численной и разнообразной, чем в тропиках, фауной птиц (но, в сущности, не имеющей значительных отличий, кроме нескольких групп, локализованных и основном в Австралии и Новой Зеландии). В этой зоне нет строго зональных элементов, за исключением зимующих оереговых птиц, и (3) антарктическая зона, в которой преобладает глав- ным образом зональная фауна морских птиц.
Зональность в меридиональном направлении, наблюдаемая у неко- торых ящериц, проявляется и у отдельных групп птиц, особенно у мелких перелетных насекомоядных наземных птиц, которые уже были рассмот- рены ранее: в (:TapoM Свете — в основном у Aluscicapinae, Sylx11пае и 'Al<>- tacillidae, а также у Sternidae и Ploceidae, которые являются менее насе- комоядными и не строго перелетными птицами; и Америке — особенно у семейств Ту1'апнЫае, Vireonidae и Parulinae. Тиранны, или американские мухоловки (Tyraiinidae), распространены от Аляски до Огненной Земли; кассиковые (Icteridae), менее насекомоядные и не строго перелетные пти- цы, также встре иются от Аляски до Огненной Земли. Ни одна 11з этих американских птиц не встречается в Старом Свете, не считая незначитель- ных залетов в Восточную Сибирь двух видов подсемейства Pari>linage.
Глава б
Некоторые роды других семейств распространены и на севере и на юге. Например, в Америке к таким родам относятся Coccy:и8 (кукушки) и Clior- detles (козодои), причем на севере оба рода являются перелетными, а неко- торые виды распространены и на севере и на юге. Например, американ- ская пустельга (Falco sparverius) с многочисленными подвидами распро- странена от Аляски до Огненной Земли; на севере этот вид является умеренно перелетным.
Радиальное распространение птиц
Четко выраженное радиальное распространение, отражающее центры происхождения и расселение из них, у птиц почти не наблюдается. Обычно радиальное распространение имеют единичные роды или небольшие груп- пы птиц, а не целые большие семейства. Расселение последних идет таки- ми сложными путями и иногда происходит настолько быстро, что первые стадии их радиального расселения быстро завершаются, а следы вскоре стираются вследствие перерасселения, эволюции и исчезновения видов и разных местах.
Самый значительный радиальный характер распространения наблю- дается у певчих птиц (подотряд Oscines из отряда воробьиных). По числен- ности певчие птицы значительно превосходят других наземных птиц на всех материках, кроме Южной Америки. Многие их группы проникают из тропиков Старого Света на север, из Старого Света в Северную Амери- ку и (значительно меньшее число групп) из Северной Америки в Южную. Здесь имеются некоторые сложности (то есть некоторые группы вновь распространяются на север из тропиков Америки). но в целоч распростра- нение носит характер очень сложной радиации из более теплой части Ста- рого (:вета. Другие менее значительные особенности радиального распро- странения будут отмечены и рассмотрены в разделе «История расселения П ТИ Ц>>.
Региональное распространение птиц
Цтицы образуют сложную, частично перекрещивающуюся систему связей по всем> земному шару, в которой, однако, можно выделить ~иесть региональных фаун птиц. Они занимают шесть условно принятых фаунп- стических областей, которые впервые были предложены как раз именно для птиц. Это большие и сложные фауны. 1тобы не было повторений, опи- сание их здесь дается очень кратко, в общих чертах. Несколько подробнее с' руктура региональных фаун птиц рассматривается в гл. 7. Читатели. интересующиеся данным вопросом, могут обратиться к авторитетным кни- гам по региональным или местным фаунам птиц, которые упоминаются ниже в тексте и в списке литературы к этой главе. Полной современной сводки по фауне птиц всего земного шара нет. Такая книга оыла бы очень полезной, и, кроме всего прочего, она помогла бы местным орнитологам ориентироваться в авифауне любого места земного шара.
Фауны птиц двух умеренных областей — Евразии и Северной Амери- ки, исключая ее тропическую часть, характеризуются бедностью. немно- гочисленностьн> семейств, родов и видов, хотя отдельные виды могут быть и относительно более многочисленными. Бедность умеренной зоны север- ного полушария птицами уже описывалась выше, и многие тропические
lIrn и цы
22о
группы, которые гам отсутствуют, были перечислены в разделе зонально- го распространения птиц (стр. 220, 221). Кроме сходства, заключающегося и отсутствии некоторых одних и тех же представителей, обе северо-уме- ренные фауны имеют еще ряд общих положительных признаков. Для обе- их этик фаун, из тех немногих семейств и подсемейств птиц, которые при- урочены к северным частям земного шара (гагары, чистиковые, тетере- виные. свиристели и пищухи), большинство являются общими. Кроме то- го, оое фауны имеют много общих родов наземных птиц из других семейств, особенно хищных, сов, дятлов, ласточек, дроздов, корольков, синиц и т. д., вьюрков и врановых (см. список семейств), и, наконец, большая часть не только родов, но даже и видов крайне северных водных и береговых птиц является для этих фаун общей. С другой стороны, эти две области несколь- ко отличаются друг от друга. В умеренных широтах Евразии находится небольшое эндемичное семейство (завирушки), а в Северной Америке имеет- ся одно почти эндемичное семейство индеек, но они являются второсте- пенными группами. В каждой области имеется также большое число родов наземных птиц, свойственных только ей. В обеих областях обитают раз- ные группы птиц, которые являются общими соответственно с тропиками Старого и Нового Света или произошли оттуда. Это главным образом тро- пические группы, распространившиеся в прилежащие умеренные районы северного полушария; большинство из них, но не все, мигрирует на севере, и многие, но опять-таки не все, относятся к насекомоядным (о природе и истории этих групп см. стр. 216 — 223). В умеренных широтах Евразии к этим птицам относятся главным образом жаворонки, мухоловки и слав- ковые Старого Света, трясогузковые, ткачики и скворцы, а также (в мень- шем числе или только в южной части умеренной зоны северного полу- шария, или только в Восточной Азии) один или несколько видов гри- фов Старого Света, аисты и фазаны (Восточная Азия), дрофы, сизоворонки, щурки, удоды, личинкоеды (Восточная Азия), нектарницы (до Централь- ного Китая), белоглазки (Восточная Азия), иволги Старого Света и дрон- го (Восточная Азия). В Америке к тропическим птицам, которые распро- странены в северном направлении, относятся главным образом американ- ские мухоловки, или тиранны, крапивники и пересмешники, виреоны, древесницы, кардиналы, американские овсянки, кассиковые, а также (в меньшем числе) грифы Нового Света, колибри и танагры. Это далеко не полный список тропических птиц, которые заходят в пределы умерен- ной зоны северного полушария. Именно этими птицами, происшедшими от тропических, больше всего отличается фауна птиц Северной Америки от фауны Евразии.
Что касается границ, то Майр (Мауг, '1946) показывает, что фауна птиц всей Северной Америки, по существу, является переходной с включением многочисленных элементов фауны Евразии на севере, число которых уменьшается к югу, и с многочисленными элементами американской тро- пической фауны на юге, уменьшающимися к северу. Исходя из современ- ного распространения (не учитывая места происхождения и направления перемещения), фауна птиц умеренных широт Евразии является также пе- реходной с элементами североамериканской фауны, многочисленными на севере и уменьшающимися к югу, и с тропическими элементами Старого Света. многочисленными на юге и уменьшающимися к северу. 11ри таком положении границы фаун выражены не резко. Граница основной части всей тропической фауны птиц проходит примерно у тропика Рака, вблизи которой многие тропические группы птиц как в Старом, так и в Новом Свете останавливаются, а мног~~е другие группы ее пересекают. У Берин-
Глава э
гова пролива граница выражена еще менее четко (Мауг, 1946). Фауны птиц Евразии и Северной Америки образуют сложный переход на севере, хотя он ограничивается сравнительно малым числом северных наземных птиц.
Главные компоненты авифауны двух основных тропических областей Старого Света — Африки, южнее Сахары, и тропической Азии с ее сов- ременными материковыми островами (Индо-Малайская область) — пере- числяются в гл. 7. Эти фауны во многом схожи друг с другом. Обе фауны богаты родами и видами многих космополитических, зональных (тропи- ческих) семейств и семейств Старого Света, но менее богаты эндемичными семействами. В Африке из птиц эндемичных или почти эндемичных се- мейств и подсемейств, которые не встречаются в Индо-Малайской области, обитают страусы (1 вид), секретари (1), молотоглавы (1), цесарки (7), турако (19), древесные удоды (6), птицы-мыши (6), африканские речньп ласточки (Pseudochelidon — 1), кустарниковые сорокопуты (Malaconiti- nae — 42), шлемоносные сорокопуты (Prionopidae — 13), буйволовые тка- чики (3), ткачик-вдовушка (9), буйволовые птицы (Huphaginae — 2 вида), но эти группы составляют довольно небольшую часть всей африканской авифауны. К Африке приурочено TaRiEe большинство медоуказчиков, боль- шинство настоящих сорокопутов и нектарниц. В Индо-Малайской области к птицам, не представленным в Африке или представленным бедно, отно- сится большинство фазанов, совиные козодои (также и в Австралии), оольшинство питт, хохлатые или древесные стрижи (один род, распрост- раненный до Новой Гвинеи), синие птицы (семейство Irenidae), болыиин- ство тималий, артамусы (один род, главным образом австралийский) и не- которые медососы (в основном все австралийские); в Индо-Малайской об- ласти строго эндемичным является только семейство Irenidae. Лвифауны Африки и Индо-Малайской области отличаются также тем, что общие для них семейства представлены неодинаково и в каждой фауне имеется много эндемичных родов и видов. Однако сходство фаун превосходит их различие. Широко распространенные или общие семейства доминируют в обеих фаунах. Количество эндемиков и степень эндемизма в этих двух фаунах больше (причем в Африке больше, чем в Индо-Малайской области), чем в фауне умеренной зоны северного полушария, но меньше, чем в фауне птиц Австралии и Южной Америки.
Объем книги не позволяет проанализировать распространение птиц в пределах основной части Африки или Индо-Малайской области, но для понимания последующего материала следует отметить один факт. Ряд птиц южной Индии и Цейлона географически изолирован от своих ближайп~их сородичей, которые в отдельных случаях встречаются в Гималаях, Асса- ие, Бирме или Малайе; пять видов птиц, приуроченных к Цейлону, име- ют близкородственных им в Малайе (Ripley, 1949).
Между фаунами птиц Африки и Индо-Малайской области резкой гра- ницы нет. Переходной зоной между ними является Южная Аравия. До- вольно большое число тропических птиц Африки (но это лишь небольшая часть африканской фауны птиц в целом) распространено в юго-западной Аравии (Bates, 1937). Однако в юго-восточной Аравии — в Маскате и Омане — несколько видов оседлых тропических наземных птиц преиму- щественно являются индийскими, а не африканскими (Ripley, 1954, р. 245 — 246). На основной части Аравии типичные гнездящиеся птицы (раз- личные жаворонки, каменки-плясуньи, грифы, дрофы, рябки, страусы и т. д.) являются прежде всего пустынными птицами, а не африканскими или евразийскими. Их настоящая родина — засушливые районы от пу- стыни Сахары до Ирана и Центральной Азии (в некоторых случаях).
327
Эти птицы пустынь образуют своего рода переход между авифауной тро- пической Африки и умеренной зоны Евразии. Все это более подробно опи- сано в последней книге по птицам Аравии Мейнертцагена (Meinertzhagen, 1954).
Фауна птиц Индо-Малайской области сама по себе в какой-то степе- ни является переходнои между тремя окружающими ее фаунами. В своеи основе она первично африканская, и, кроме того, некоторые ее африканские элементы достигают Индии. Райпли (рукопись, с которой я имел возмож- ность ознакомиться) отмечает, что 11,50~î тропических птиц Индии, по-ви- димому, прямо родственны африканским, а не птицам восточной части Индо-Малайской области. В авифауну Индо-Малайской области входят элементы фауны умеренных широт Евразии: некоторые кулики (Scolopax). совы (Strix), дятлы (Dryocopus), поползни, синицы и др., а зимой — мно- гие северные перелетные птицы. Из австралийских элементов — несколь- ко видов сорных кур, фруктовая горлица и императорский голубь, сови- ный козодой, артамусы, медососы и, если относить к Индо-Малайской об- ласти острова Бали и Филиппинские острова, попугаи лори и какаду (см. список семейств).
В авифауне Австралийской области (Австралия, Новая Гвинея и прилегающие острова) отсутствует ряд важных семейств Индо-Малайс,кой области и часть широко распространенных семейств, включая фазанов, трогонов, бородаток, дятлов, рогоклювов, бюль-бюлей (которые дости- гают Молуккских островов), синих птиц (Irenidae) и настоящих вьюрков, но зато в значительной степени представлены обычные космополитические группы и еще в большей степени голуби, попугаи, зимородки, так же как и личинкоеды, мухоловки Старого Света, медососы и ткачики (Estrildi- uae) и т. д. К австралийской фауне относится также ряд эндемичных или почти эндемичных групп птиц: эму, казуары, сорные куры, несколько более или менее четких групп голубей и попугаев, совиный козодой, лирохво- сты, птицы семейства Atrichornithidae, австралийские славки (подсемей- ство Malurinae), медососы и несколько семейств воронообразных пти~~, включая райских птиц (см. гл. 7). Основная часть этой фауны почти по- ровну разделена между Австралией и Новой Гвинеей. Фауны птиц этих двух довольно разных частей, хотя и отличаются многими деталями, в це- лом схожи (Mayr, Serventy, 1944). Почти все установленные связи австра- лийских птиц ведут к Азии. Несколько предполагаемых африканских свя- зей. как, например, у медососов с южноафриканским родом Promeropÿ, сомнительны, а между австралийскими и южноамериканскими наземными птицами прямая связь отсутствует.
Между авифаунами Индо-Малайской и Австралийской областей нет резкой границы, но есть общая зона постепенного перехода, охватываю- щая все острова, лежащие между Явой и Калимантаном, с одной стороны, и Новой Гвинеей и Австралией — с другой. Фактически этот переход охватывает еще более обширную территорию, так как некоторые птицы Индо-Малайской области (например, птицы-носороги, хохлатые стрижи) достигают Новой Гвинеи, но не достигают Австралии, а некоторые другие (например, питты и трогоны) достигают Северной и Восточной Австралии, но не достигают остальной части материка. В другом направлении несколь- ко видов австралийских птиц, как уже было отмечено, переходят через линию Уоллеса в Индо-Малайскую область (см. гл. 7).
Южная Америка («континент птиц») имеет самую богатую фауну пер- натых. Здесь отмечается значительно большее число видов, особенно мел- ких наземных птиц, чем на любой другой равной по площади территории.
Это оогатство фауны отчасти обусловлено увеличением числа видов на раз- ных высотах различных горных хребтов (Сhapman, 1917, 1926). Часть юж- ноамериканской фауны, но не вся, достигает Центральной Америки и зна- чительно меньшая часть — Вест-Индии. В авифауне Южной Америки хорошо представлены цапли, ибисы, аисты, утки, пастушки, хищные пти- цы, совы, куропатки, кукушки, ржанки, голуби, попугаи, козодои, тро- гоны, стрижи, бородатки, дятлы, ласточки, дрозды и некоторые другие небольшие, но широко распространенные группы. В ней отсутствуют жу- равли, дрофы, трехперстки, птицы-носороги, рогоклювы и многие другие семейства Старого Света, бедно представлены зимородки. Самый большой пробел в этой фауне составляют певчие птицы (Oscines); это единственная материковая авифауна, в которой певчие птицы представлены в меньшин- стве. Однако все эти изъяны более чем компенсируются наличием эндемич- ных групп или по крайней мере наличием многочисленных групп Нового (:вета, которые часто являются таксономически изолированными и в от- дельных случаях очень богатыми по видовому составу: наяду, тинаму (33 вида), грифы Нового Света, паламедеи, крациды (38), гоацины, кариа- мы, трубачи, арамы, солнечные цапли, зобатые бегунки, исполинские козодои, гуахаро, момоты, колибри (319 видов, большинство которых на- ходится в Южной и Центральной Америке), ленивки (32), якамары (14), туканы (37), надсемейство Furnarioidea (около 500 видов, представленных как в Южной, так и в Центральной Америке), американские мухоловки (365 видов, включая североамериканские и вест-индские), манакины (59), котинги (90 видов) и траворезы. Эти группы, большинство из которых приурочено к Южной Америке или к Южной и Центральной Америке и лишь некоторые достигают Северной Америки и Вест-Индии, но ни одна не проникает в Старый Свет, насчитывают свыше 1500 видов, то есть более '/g всех известных видов птиц. Кроме того, хотя певчие птицы и пред- ставляют меньшинство в общей авифауне Южной Америки, они довольно многочисленны и в большинстве случаев относятся к группам, которые ра- диировали в американских тропиках: дрозды эндемичной группы, крапив- ники, пересмешники и компоненты главным образом американского ком- плекса: виреоны — древ есницы — гавайские цветочницы — танагры- вьюрки — кассиковые. Кроме этих, единственными певчими птицами, ко- торые достигают Южной Америки, являются жаворонки, ласточки, не- сколько видов аберрантных славок, рецентные дрозды, несколько видов оляпок, коньков, чижей и соек. Некоторые виды достигают отдельных ча- стей Центральной, но не Южной Америки: корольки, шелковистые мухо- ловки (Ptilogonatinae), свиристели (зимой), пищухи, синицы, своеобразные дубоносы (Hesperiphona vespertina), клесты и вороны.
Географические связи многих птиц Южной Америки не ясны. Ряд наиболее четких эндемичных групп, вероятно, не имеет близкого родства с какими-либо другими группами птиц. Некоторые из группы тропикополи- тов, включая попугаев, трогонов и бородаток, очевидно, являются релик- тами или по крайней мере отступающими группами, и прямой связи между птицами, которые теперь находятся в Южной Америке, и теми, которые обитают в любой другой части земного шара, вероятно, нет. Это также относится к кукушкам, большинству одноголосых воробьиных (кото- рые в настоящее время доминируют в Южной Америке) и некоторым другим широко распространенным группам. Географические связи южно- американских птиц направлены главным образом к Северной Америке или через Северную Америку Ic Евразии. Но некоторые связи идут в дру- гом направлении: так, род орлов Spizaetus встречается только от Восточ-
Пгпицы
ной Азии до Новой Гвинеи и в тропиках Америки; древесная утка (Dend- rocygna viduata) распространена в Африке, на Мадагаскаре и в Южной Америке; род камышниц Porphyrula обитает в Африке, на Мадагаскаре, а также в тропической и умеренно теплых частях Америки; род сов Ciic- саба приурочен к Африке и тропикам Америки; совы тропического амери- канского рода Lophostrix родственны африканскому роду Jubula; род карликовых дятлов встречается в Индо-Малайской области и в тропиче- ской части Америки; некоторые ласточки Южной Америки и других юж- ных континентов, возможно, родственны друг другу; аберрантные тропи- ческие американские роды славковых Ramphocaenus и Microbates родст- венны африканскому роду. Но все это редкие исключения. Прямых род- ственных связей между наземными птицами Южной Америки и Африки очень мало, и, вероятно, они совсем отсутствуют между южноамерикан- скими и австралийскими пернатыми. В целом авифауна Южной Америки значительно отличается от фаун Африки и Австралии.
Авифауна Центральной Америки (скорее, ее можно считать южно- американской, чем североамериканской) во многих отношениях является переходной. Одни южноамериканские группы отсутствуют в ней, а дру- гие распространены только частично. Некоторые североамериканские пти- цы (включая отдельных упомянутых выше певчих, которые не достигают Южной Америки) проникают в Центральную Америку. В Центральной Америке имеется также ряд своих четко эндемичных птиц. Территория перехода охватывает не только Центральную Америку, но и большую часть Северной и Южной Америки, так как многие тропические птицы Америки проникают в Северную Америку на различные расстояния (см. фауну птиц Северной Америки), а некоторые птицы Северной Америки распространяются на различные расстояния в Южную, и несколько видов достигает самой южной оконечности этого материка, например конь- ки, чижи (Spinus) и некоторые виды дятлов, не говоря уже о целом ряде перелетных птиц, особенно куликов. Но в действительности распростра- нение пернатых в Центральной Америке значительно сложнее (СгЬсот, 1942, 1950, р. 375 и след.). Природа и история авифауны Центральной Аме- рики рассматриваются в конце этой главы в связи с «тропиками Северной Америки».
В заключение о сущности и главных связях региональных фаун птиц можно сказать следующее. Фауны наземных птиц умеренных широт Ев- разии и Северной Америки являются только лишь обедненными перифери- ями прилежащих тропических фаун, в которых отсутствуют многие тропи- ческие семейства и наблюдается низкий уровень эндемизма, но имеются более определенные группы северных водных птиц. Эти две северо-умерен- ные авифауны сходны большим количеством одних видов птиц, но разли- чаются другими видами, особенно это касается мелких семейств, в основ- ном насекомоядных и кочующих наземных птиц, общих соответственно для тропиков Старого и Нового Света. Авифауны двух основных тропи- ческих областей Старого Света — Африки и Индо-Малайской области— особенно богаты представителями более или менее широко распростра- ненных семейств. Степень эндемизма в этих фаунах умеренная: в Афри- ке — несколько эндемичных семейств, а в Индо-Малайской области только одно. Обе фауны сходны в общих чертах, но различаются мн<и,ест- вом деталей. Австралийская фауна птиц менее богата, в ней отсутствунл некоторые важные семейства, но эндемичность ее выражена сильнее. Иоч- ти все установленные связи этой фауны ведут к Индо-Малайской области. Из всех фаун авифауна Южной Америки наиболее богата и наиболее
Глава .7
280
эндемична. Насколько установлено, все ее связи ведут главным обра- зом к птицам Северной Америки или через Северную Америку к Старо- му Свету.
Переселение птиц и преграды их распространению
Ломпнирование и конкуренция в связи ~ распространением птиц
Характер распространения господствующих групп птиц намного сложнее, чем других позвоночных. Каждая основная экологическая груп- па имеет свое господствующее семейство или семейства, которые часто являются космополитами. Некоторые из них упоминаются в разделе «История расселения птиц».
Совершенно очевидно, что наиболее доминирующими из современ- ных птиц являются воробьиные, численность которых далеко превосхо- дит всех остальных наземных птиц на каждом континенте: певчие воробь- иные (Oscines) доминируют на всех континентах, за исключением Южной Америки. Одноголосые воробьиные (предположительно более примитив- ные) — в Южной Америке. Воробьиные преобладают в большинстве назем- ных местообитаний от тропиков до Арктики и от влажных джунглей до пустынь. Тем не менее их доминирование имеет экологические грани- цы. Среди воробьиных имеется несколько хищных видов, однако опас- ности, что они могут занять место ястребов и сов, не существует. Среди воробьиных нет ни одного морского вида и только несколько видов свя- зано с пресными водами, не являясь при этом существенными конкурен- тами водных птиц.
Конкуренция, или взаимоисключение, вытекает из многих деталей распространения птиц, включая географический викариат Oscines и Subos- cln&s, но доказательством этого являются лишь отрицательные призна-
Переходы между фаунами птиц, происходящие в результате взаим- ного перекрывания и постепенного выклинивания элементов различных фаун и охватывающие обширные территории, сложны. Майр (Mayr, 1946) правильно утверждает, что хотя авифауна всей Северной Америки и яв- ляется переходной между Евразией и тропиками Америки, но этот переход охватывает более обширные территории, чем названные. Авифауна уме- ренных широт Евразии в значительной степени является переходной, частично переходна также и фауна Индо-Малайской области. Фактически переход между фаунами птиц тропиков Старого Света и Америки охваты- вает если не весь земной шар, то по крайней мере всю Евразию, Северную и Центральную Америку и большую часть Южной Америки. Тем не менее, как уже отмечалось выше, шесть общепринятых региональных фаун птиц являются более или менее четко выраженными. Указанное отклонение будет рассмотрено в гл. 7.
Никакие преграды не могут полностью остановить расселение птиц. Те или иные виды птиц всегда пересекают их на разные расстояния или в разные времена. Но отдельные преграды, особенно соленые воды, ока- зывают серьезное влияние на некоторые фауны птиц, что выражается в ограничении иммиграции и частичной изоляции (о чем уже говори- лось выше).
281
ки в современном распространении, так как ископаемые данные недоста- точно полноценны, чтобы показать, что отступание одних птиц было тес- но связано с расселением других.
Закл1очение: осооенности распространения итнц
В распределении птиц, несмотря на то, что их связи очень сложны, все же наблюдаются общие черты, в которых отражается пять обычных особенностей распространения.
Первая особенность — ограниченность распространения и взаимо- исключение, выражена очень слабо. Птицы не имеют абсолютных границ в распространении, но их численность и разнообразие на крайнем севере, крайнем юге и на отдаленных островах заметно уменьшаются. Взаимо- исключение ограничивается наличием многочисленных космополитиче- ских групп и обычно охватывает второстепенные группы, а не основные элементы фауны. Широкий викариат Oscines и Suboscines является иск- лючением.
Вторая особенность — зональность наземных птиц, заключается глав- ным образом в уменьшении их разнообразия от одной зоны к другой. В умеренных зонах птиц меньше, чем в тропических, и их видовой состав большей частью не столь разнообразен. Арктических птиц, по крайней мере наземных, еще меньше, и видовой состав их еще беднее. Среди аркти- ческих и антарктических водных птиц проявляются более четкие разли- чия. 11еридиональная зональность характерна для некоторых мелких наземных птиц, особенно для перелетных насекомоядных.
Третья особенность распространения птиц — радиация, у большин- ства птиц выражена нечетко, потому что расселение многих главенствую- щих групп происходило настолько быстро и сложно, что их основная радиация вскоре завершилась и следы ее не сохранились. У певчих птиц проявляются некоторые черты радиации вокруг теплой части Старого Све- та, но они очень усложнены и выражены нечетко.
Четвертая особенность — дифференциация фаун на разных конти- нентах, выражена хорошо, но очень сложна в деталях. Материковые (ре- гиональные) фауны птиц состоят из разных сочетаний космополитических, зональных и радиальных групп плюс эндемичные группы в разных про- порциях. В общем фауны умеренных широт Евразии и Северной Америки менее сложны, бедны и имеют низкую степень эндемизма. Фауны Африки и Индо-Малайской области более богаты, сложны и имеют умеренную сте- пень эндемизма. В Австралии относительно богатая фауна птиц с большим проявлением эндемизма и в Южной Америке очень богатая и высоко- эндемична я фауна.
~1то касается пятой особенности распространения — концентрации на территориях, благоприятных для обитания, и обеднения фауны по краям, то каждая основная фауна птиц сосредоточивается в наиболее благоприятных местах, а в менее благоприятных, например в Арктике и пустынях, многие ее элементы выпадают. Тем не менее довольно большое число птиц распространено почти повсеместно, даже в неблагоприятных переходных зонах. Вероятно, отчасти по этой причине, а также и потому, что многие птицы пересекают преграды и быстро расселяются, переход (область перекрытия фаун с постепенным выклиниванием отдельных эле- ментов фауны в обоих направлениях) одной фауны птиц в другую, охва- тывающий большую часть земного шара, очень сложен и обширен.
282
Глава 5
О том, какое значение имеют реликты для понимания современного распространения птиц, сказать трудно, так как в отличие от рыб и пресмы- кающихся (двоякодышащая рыба в Австралии или гаттерия. Spliettiidun. в Новой Зеландии) ни одна из современных птиц не является реликтом. Неизвестно также, какие именно птицы из современных являются самыми примитивными. Бескилевые (рис. 33) в Африке, Австралии, Южной Аме- рике, в прошлом на Мадагаскаре и в Новой Зеландии (киви до сих пор на- ходятся в Новой Зеландии) не являются примитивными реликтачи, то
Р и с. 33. Распространение нелетающих бескилевых птиц пред- полагаемых ископаемых предков (F leutherornis) и родственных
им современных летающих птиц (тинаму).
есть они, видимо, не являются остатками древней космополитической группы птиц, а скорее относятся к самостоятельным конвергентно возник- шим группам, которые произошли там, где они существуют и сейчас. Тинаму, возможно, сохранившийся представитель примитивной группы летающих птиц, от которой, возможно, возникли и некоторые бескилевые. Эти птицы приурочены к Южной и Центральной Америке, до сих пор там многочисленны и являются преуспевающими. Можно сказать, что в Юж- ной Америке примитивных птиц больше, чем в каком-либо другом месте.
История расселения птиц
Современный характер распространения птиц — результат очень слож- ного процесса эволюции, сложного, многократного и, вероятно. очень быстрого расселения, адаптации, по-видимому, конкуренции, сокращения ареалов и вымирания, что тесно связано с природными особенностями птиц, распределением климатов и наличием более или менее серьезных преград, особенно океанов. Этот процесс, очевидно, происходил отчасти самостоятельно в любой сколько-нибудь значительной экологической группе птиц. В каждой такой группе, вероятно, происходил эволюцион- ный подъем, распадение на новые формы и сложное, более или менее быст- рое расселение на большей части или на всем земном шаре следующих друг за другом доминирующих групп, а затем сокращение ареала и ис- чезновение более древних групп по мере распространения новых. Попы-
Птицы
таемся проследить некоторые детали этих явлений, что, однако. сделать не так легко.
Рассматривая разные экологические группы птиц, я предполагаю выбрать из ка'кдой группы доминирующие семейства (именно они чаще всего дают возможность «подобрать» наиболее существенные «ключи») и, если возможно, проследить их историю. (Обоснование этих выводов см. в списке семейств.) Такой метод анализа истории расселения птиц, несомненно, во многом упрощен, но это самое лучшее, что можно предло-
ЖИТЬ.
Из морских птиц чайки и крачки преобладают на всем земном шаре в целом, но их происхождение и радиация остаются, по-видимому, за пределами возможностей анализа. Чистиковые, как следует из их распро- странения, очевидно, всегда были северными птицами, а пингвины ——
ЮЖНЫМИ.
Из хищных птиц ястребиные — доминирующие дневные хищники, а совы — ночные, но в связи с тем, что эти группы древние и сложные, о первоначальном характере их распространения ничего не известно. Одна- ко некоторые подробности недавних перемещений этих птиц установлены довольно четко. Болотный лунь (Circus), два подрода Ealco, орлы Aquila,. орланы — Haliaeetus и Tyto (сипуха) попали в Северную Америку из Ста- рого Света сравнительно недавно. Но род Buteo, по-видимому, имеет севе- роамериканское происхождение. Большинство видов этого рода, включая канюков ямайского, красноплечего и ширококрылого и др., находится в Северной Амерйке (там же найдены ископаемые этого рода, относящиеся к олигоцену), шесть видов — в умеренных широтах, еще меньшее число- в Африке (один вид на Мадагаскаре) и в Южной Америке. Ни одного гнез- дящегося вида нет в Индо-Малайской области, а в Австралийской области они отсутствуют совсем. Нахождение эндемичных видов этого рода в Вест- Индии, на островах Галапагосс и Гавайских островах соответствует аме- риканскому происхождению этого рода. Анализ других родов ястреби- ных и сов мог бы раскрыть другие подробности перемещения в других ча- стях земного шара, но здесь мы имеем возможность рассмотреть переме- щения только между Старым и Новым Светом.
Из водоплавающих птиц внутренних водоемов доминирующим является семейство Anatidae. В настоящее время распространение этого семейства сложное, а первоначальный характер его расселения неизве- стен, так как оно относится к древним семействам.
Из крупных голенастых болотных и наземных птиц доминирующим является семейство Ardeidae (цапли). Это древняя группа, которая, веро- ятно, недавно вновь перераспределилась. Шесть родов и по крайней мере два вида цапель являются космополитами или почти космополитами. В этой экологической группе цапли вытесняют других птиц — журавлей, арам, дроф, пастушков и др., которых Майр и Амадон (Mayr, Amadon, 1951, р. 8) считают приходящими в упадок, а также ибисов и аистов. Боль- шинство цапель ведет водный образ жизни, но некоторые из перечислен- ных групп птиц живут вдали от воды. Однако этот факт не противоречит моей мысли, так как он является, вероятно, результатом вытеснения вслед- ствие конкуренции. Переразмещение цапель явление очень сложное. и я не пытался проанализировать его подробно. Современное проникнове- ние египетской цапли из Африки в Южную, а затем в Северную Америку прослежено Спрантом (Sprunt, 1955).
Из мелких куликов доминирующими являются ржанки (Charadrii- nae) и песочники (Scolopacinae), но их история не ясна. Ржанки рас-
Глава б
пространены широко и почти равномерно по всему земному шару. Песоч- ники гнездятся в основном на севере, но произошли ли они там, как это считает Майр (Mayr, 1946, р. 14), или сконцентрировались в этом районе позднее, возможно в течение плейстоцена, — остается под вопросом.
Пастушки (Rallidae) доминируют в экологической группе, для кото- poij характерно обитание в местах с густой растительностью вблизи воды, а некоторые из них живут на земле вдали от воды, преимущественно в тропических лесах. Распространение их перекрывает, с одной стороны, распространение куликов, а с другой — распространение куриных. Это древнее семейство со сложным распространением, которое ничего не говорит о месте его возникновения, и я не делал попытки анализировать его современные вторичные перемещения.
Из куриных птиц семейство Phasianidae доминирует на большей части (хотя и не везде) земного шара. У этого семейства явственно прояв- ляется радиация из одного главного центра — Индо-Малайской области. Настоящие фазаны, джунглевые куры и павлины почти все находятся в Индо-Малайской области, откуда предки рода A f ropavo (африканский павлин) и цесарки, возможно, проникли в Африку. Предки тетеревиных (Tetraoninae), возможно, распространились в северном направлении и за- тем циркумполярно по всему земному шару, в конечном итоге дифферен- цируясь в основном в Северной Америке. Перепела расселились по всему земному шару с вторичным центром эволюции в южной части Северной Америки. Куриные птицы обладают относительно небольшими способно- стями расселения, и их современное распространение до какой-то степени отражает их раннюю историю. Тем не менее даже история семейства Pha- sianidae, вероятно, была более сложной, чем можно предполагать, а есть и другие семейства, которые еще больше усложняют характер распростра- нения отряда,— сорные куры в Австралии и т. д., крацпды в Южной и Центрально1х Америке, которые, возможно, являются выжившими пред- ставителями одного или нескольких более ранних ответвлений куриных; примитивные, но внешне куроподобные тинаму до сих пор доминируют г Южной и Центральной Америке.
Фауна попугаев (рис. 34) почти одинаково богата по видовому соста- ву в Старом и Новом Свете, но значительно многообразнее в первом. Все формы Нового Света относятся к одному из подсемейств Старого Света. что позволяет предполагать, что попугаи произошли в Старом Свете. Но такое допущение не позволяет сделать достаточно глубокий анализ. Боль- шое разнообразие попугаев отмечается только в Австралийской области ~ на прилежащих островах. В других частях Старого Света попугаи везде
ОТНОСИТеЛЬНО МИЛОЧИСЛеННЫ И ВСЮ ОТНОСЯТСЯ К ОДНОМ~ IIOQCOMOIICТВУ, ТОМ1
же самому, что и в Америке. Говорит ли поразительное разнообразие попу- гаев в Австралийской области о происхождении их именно здесь, или оно отражает отступание некоторых групп этих птиц с других частей зем- ного шара, или местную радиацию? Ответить на этот вопрос я не могу, но. по-видимому, место происхождения попугаев не Австралийская область. За пределами этой области, в тропиках Америки, они значительно многочисленнее, чем где-либо в другом месте, но возможно, что и тропики не являются их родиной. Сомневаться сначала в австралийском, а затеи американском происхо>кдении попугаев и предполагать, что они могли произойти на какой-либо части Старого Света, где в настоящее время они Hll особенно многочисленны, ни особенно разнообразны,— значит отрицать ценность «ключей», имеющихся к этому семейству. Можно сказать, имея на то веские основания, что вообще для древних, сложных и широко
28 э
Оюицы
распространенных семейств птиц вполне достоверных «ключей» не суще- ствует. Ископаемые попугаи, найденные в Европе и Северной Америке, относятся к среднетретичному времени и до плейстоцена нигде больше неизвестны. Но это следует рассматривать как пример недостаточности ис- копаемых данных, а не как доказательство местопроисхождения этих птиц.
Голуби достигают наибольшего разнообразия в Австралийской обла- сти, в других местах Старого Света они менее разнообразны. В Аме- рике систематических групп голубей очень мало, но роды и виды много- численны. Это дает основание считать их родиной Австралийскую область.
ф °
ф
+' .
е° ° Ð
'е ..
а
е" ° °
° ° ее ~ ~° еее~
а° °° ° е °° ° °° ° ° °
Г~ °
° /,~. ° .' °
° \ °
° °
еее @ еГе„° °
'/" м rA ii+r °
ее
ъ.
ее еееЪ
°, еЛ,
Ъ
° °
Ъ
° ° ° ° ° I °
с
° °
° °
° ~ ° °
° °
° °
° е °
° ° ° ° °
° °
Ъ °
Ф °
i ie y ~ ° ° ° ° °
%
4
С '
° °
° °
«~»
° ° °
° °
° °
..:i: .. '.''
° ° °
° °
° °
° °
° ° Ъ
° ее °
° а
° ° ° е 'еЪ ее
° ° ° ° ° ' ° ° е °
° ееееае
° е еефе
е,ее
$Ф °
-ь.
ее
° Ю
° °
° °
г.'.
Ф ° ° "т''
° а
° ' ° ° ° ° r ° °
° °
Ы~
,ее g
° ° ° ;r
° ° rr
° аф
° фе
° °
° °
° ° ° °
° е °
,' '.1,
° а
° , ° %
° °
а
а ф
,Ю
° ° ,е'
Р и с. 34. Распространение попугаев.
В Австралии попугаи многочислснны и чрезвычайно разнообразны по видовому составу (жирныс перес( какнциеся полосы); в ?Ожной Америке они столь жс многочисленны, но мснсе разнообразны по видовому составу (жирные вертикальные полосы), который становится еще м(нее разнообразным в Африке, тропиках Азии и т. д. (тонки( в( ртп- кальные Il() Jlocbl). Прерывистая линия показывает 11риблизительную СОВРеМеННУ10 ССВеРНУ1О ГРаНИЦУ РаСПРОСТРаыеНИЯ ПОПУГаСВ; 11УНКТ11Рае ная линия — — 1iриблизительная граница рас11ространения вида Л1iс/гiг (c- c'01 <I1'>< t< ~ j~ 14 I I ðîøëîì; кр('гт!1ками обозначены приолизит('лЕ ные м(сто- нахо'кдения ископаемых по11угаев.
Но вряд ли история расселения голубей настолько проста. Голуои отно- сятся к древнему и, несомненно, сложному семейству, и их распростране- ние очень сходно с распространением попугаев. Эти две группы — голу- ои и попугаи — родственны, имеют одинаковую историю расселения и, возмо;кно, произошли в одном и том же месте и распространились на зем- ном шаре в одно и то же время, вместе радиировали (возможно, вторично) в Австралии 11 начали исчезать в некоторых частях земного шара примерно в одно время п по одним п тем же причинам. Возможно, они доминировали повсюду до расцвета воробьиных, и возможно, что последние теперь их вытесняют. Хотя голуби и попугаи все еще относятся к самым богатым группам птиц и до сих пор доминируют на большей части земного шара, но уже имеются два признака упадка этих групп: один из них — уход попугаев из умеренной зоны северного полушария, другой — то, что ни один род попугаев (и только один род голубей) не является общим для Старого и Нового Света. Последнее дает основание предположить, что активное расселение этих птиц прекратилось недавно. Вполне возможно, что воробьиные, являясь более активными мигрантами по сравнению с го- лубями и попугаями (хотя последние также могут совершать миграции),
286
вытесняют голубей и попугаев в умеренной зоне северного полушария. Это наглядный пример того, как перелетные птицы могут сдерживать проникновение других конкурирующих птиц в северные области и мешать взаимному обмену между Старым и Новым Светом. Нетрудно проследить, как группа птиц, приходящая в упадок, может переходить от положения, которое занимают в настоящее время попугаи, до положения трогонов, а затем в конечном итоге исчезнуть.
Columba — единственный общий род голубей для Старого и Нового Света (от этого рода произошли домашние голуби), имеет почти космополи- тическое распространение. Этот род является примером неопределенности «ключей чисел». Около 32 видов этого рода свойственны Старому Свету и около 20 — Новому; виды Старого Света более разнообразны, что наводит на мысль о их происхождении в Старом Свете. Все 20 видов Нового Света встречаются в Южной и Центральной Америке и в Вест-Индии. Один из видов Центральной Америки проникает в западную часть Северной Аме- рики, распространяясь на север до юго-западной части Канады. Не счи- тая этого случая, род Columba отсутствует на основной части Северной Америки — севернее южной Флориды. Около 14 видов этого рода нахо- дится в умеренной зоне Евразии и на прилежащих островах, 11 — на ос- новной части Африки и близлежащих островах. но на Мадагаскаре нет ни одного вида, 5 видов встречаются в тропиках Индо-Малайской области и 2 вида — в Австралийской области, но лишь один из них достигает самой Австралии (только ее восточной части) и ни один вид не достигает Новой Зеландии.
Указанное размещение числа видов позволяет предположить. что род Columba, вероятно, возник в тропиках Америки, расселился в Старый Свет через север (имеются в виду не современные виды западной части Северной Америки, а, возможно, более ранние) и с севера распространился по всему Старому Свету. Отсутствие этого рода на Мадагаскаре и во внут- ренних районах Австралии соответствует предполагаемой истории рас- селения. С другой стороны, этот род, возможно, произошел в умеренной зоне Евразии и расселялся оттуда, а затем вторично радиировал в тропи- ках Америки. Или вероятнее всего, что этот род имеет еще более cложную историю расселения.
Кукушки также имеют космополитическое распространение, но они более многочисленны в тропиках, чем в умеренной зоне северного полу- шария, и ни один род кукушек не является общим для Старого и Нового Света. Очевидно, это семейство древнее и в настоящее время приходящее в упадок. История его расселения сложна и не поддается разгадке.
Козодои — в основном насекомоядные сумеречные птицы. Они пре- терпели по крайней мере троекратную радиацию: раннюю, которая оста- вила небольшие семейства, изолированные в Индо-Малайской и Австра- лийской областях и в тропиках Америки, далее радиацию семейства Cap- rimulgidae и затем рода Caprimulgus. Центры этих радиаций мне неиз- вестны.
Ласточки и стрижи доминируют в экологической группе сильных летунов, древних насекомоядных птиц. У стрижей наиболее древний харак- тер распространения с хорошо выраженной прерывистостью и болыпой локализацией родов, но по сравнению со стрижами ласточки являются более доминирующими птицами. Три рода ласточек широко распростра- нены и, несомненно, достигли Северной Америки из Старого Света сравни- тельно недавно. Но ранняя история ласточек не очень ясна. В настоящее время основная концентрация их наблюдается в Африке и вторая — в Юж-
ной Америке, однако совершенно очевидно, что у ласточек оыло последо- вательное и многократное расселение. По современному распространению ласточек нельзя сказать, что история их расселения была простой.
Зимородки многочисленны и разнообразны в Старом Свете, и нет никаких сомнений в том, что два рода зимородков Америки возникли в Ста- ром Свете.
Дятлы доминируют в пределах своих экологических возможностей. Они многочисленны как в Америке, так и в Индо-Малайской области. Ме- нее многочисленны в умеренных широтах Евразии и Африки и отсутству- ют на Мадагаскаре и в Австралийской области. Эта особенность предпо- лагает пх расселение из Америки. но вполне возможно, что первичным
— е
Юу С1 ° Вв °
r,
° ° . ' в..'.'... ° сф .
° °
.~фй! ''..:.
° ° ' °
° ° ' °
° ° ° '; ° ° ° .'".::1.
, ° '
° ° ":В Э
е °
::.с'
° °
° 'Э ' °
е
° В r
с
° °
° ° r
ы
е ° ° ° °
° ° в~ °
° ° ° е
ч- е ° °
° °
° ° ° °
в °
° °
° °
~Ве °° .. ° °' I,
° ° ° °
° е
° °
° Э
° °
° °
с ° °
° ' °
° °
с ° °
° ° °
° ° °
с
° °
° ° е °
° °еЭ'rе° °
° т
. °...;.. ° . ° . ~':'-
° ° ° ° ° °
° ' ° °° °r° ° ° °° ° ° ВЭ° '° ° ° °Iс° Э .. ° Эе;
° °
° °
° ° ° ° ° ° ° ° ° ° ° '
° ° °
ее
Ь)
° ° :;;;.М
' °
° °
,г- в'
° ° °
все ...: в
° е° ° °. ° .'в ° '..' °° °° ° ° °° ' ° ° ве° ° ' ° ° °е °'в ' °° се ° ° ° °° ° ° ее° ° ° °Ъ° ° ° °° уВ~ е ° rrВВе
° ° \ ° ' ' °° ° е ° r ° Ю °° ° °° е ° ° °° ° ° ° ° °° °е° ° °° °°, ° °
° rеее ,° + °, ° ° ° ° ° ° ° ° °
° °
° ° ° ° ° е ° ° е °
° .,е ' Г''фВ
Р и с. 35. Распространение трогонов.H рестпком обозначено местонахождение ископаемых трогонов в Е вропе.
центром могла быть также и Индо-Малайская область, а Америка — вто- ричным. Как и у многих других групп птиц, расселение дятлов было мно- гократным (иллюстрируется широким распространением нескольких родов). и это лишний раз подтверждает, что не следует слишком доверять существующим «ключам» к распространению.
Трогоны (рис. 35) и бородатки не являются доминирующими птицами; они малочисленны и имеют прерывистое распространение. Очевидно, это угасающие группы, и поэтому едва ли современная численность и распро- странение этих птиц могут отражать их раннюю историю. Нахождение ископаемых остатков трогонов в Европе указывает на то, что они сократили свой ареал, и позволяет предполагать возможность расселения их север- ным путем. Однако отсутствие ископаемого материала из других частей земного шара оставляет вопрос о месте их происхождения открытым.
Из широко распространенных доминирующих групп остается рассмот- реть только отряд воробьиных птиц, исключая ласточек. Отряд воробьи- ных (Passeres) включает более половины всех современных птиц: примерно 5100 видов из общего числа, составляющего 8600, и он является еще бо- лее доминирующим по числу особей. Большинство птиц, которых мы где- либо встречаем, относится к воробьиным. Майр и Амадон (Мауг, Amadon, 1951) делят этот отряд на несколько подотрядов. Его действительная фило- гения. возможно, была даже еще сложнее, чем это показывает классифика-
Глава 5
ция, но лучше всего начать с деления этого отряда только на две групы: Suboscines и Oscines.
Suboscines, или ложнопевчие,— предположительно более примитив- ные. Большинство из них (всего насчитывается примерно 1100 видов). сконцентрировано в Южной и Центральной Америке. Одно из тропиче- ских семейств Америки (Тугашпйае — тиранны, или американские мухо- ловки) распространилось также по Северной Америке и по всей Вест-Ин- дии, а другое семейство (Cotingidae) достигает южной Аризоны и Ямайки. В Северной Америке на север от южной части Мексики или в Вест-Индии другие Suboscines отсутствуют, но несколько четко выраженных семейств этой группы находятся в Старом Свете: рогоклювы и питты — в тропиках Африки и Индо-Малайской области. Питты распространены до северо-во- сточной части Австралии. Филапитты встречаются на Мадагаскаре, а ново- зеландские крапивники (Xenicidae) — в Новой Зеландии.
Oscines, или певчие птицы, насчитывающие около 4000 видов, счи- таются самыми прогрессивными, преуспевающими и, возможно, из всех основных групп птиц, позднейшими в эволюционном отношении. Они раз- личны по внешнему виду, но почти одинаковы по внутреннему строеншо. Болыиинство семейств певчих птиц слабо отличается друг от друга, и многие семейства связаны промежуточными или сомнительными родами. У различных групп певчих птиц часто на разных континентах наблюдает- ся значительный параллелизм. Например, американские трупиалы и кас- сиковые (Icteridae) сходны со скворцами Старого Света (Sturnidae). хотя, вероятно, они им не родственны. Вполне можно сказать, что мы в своем безграничном невежестве еще не представляем, как развивались Oscines и каким образом различные их группы связаны между собой (Мауг, Ата- don, 1951, р. 13).
Oscines доминир~ ют на всех континентах, кроме Южной Америки, где их превосходят по численности Suboscines, примерно в соотношении трех к двум. Это оостоятельство плюс тот факт, что ни одно семейство Sul~osc nes не является общим для Старого и Нового Света и что некоторые семей- ства их в Старом Свете локализованы и, возможно, являются реликтами, в то время как ряд семейств Oscines широко распространен, позволяют предполагать, что на всем земном шаре в целом ложнопевчие воробьи- ные вытесняются певчими. Однако на этот счет могут быть и другие coo()- ражения.
Современные ложнопевчие Старого Света, возможно, не являются реликтами. Рогоклювы и питты географически не изолированы, но встре- чаются в основных районах тропиков Старого Света. На Мадагаскаре и в Новой Зеландии нет остатков древней сколько-нибудь разнообразной фауны ложнопевчих, и на каждом из этих островов имеется только одно небольшое семейство, которое является скорее автохтонным, чем реликто- вым. Но в Америке некоторые ложнопевчие (американские мухоловки) за последнее время увеличивали свой ареал, а не отступали. Совсем не обязательно считать, что все эти детали свидетельствуют о слож- ном постепенном замещении ложнопевчих птиц певчими, но они, несом- ненно, говорят о сложных изменениях в характере распространения воробьиных. Возможно, современные ложнопевчие Южной Америки не являются частью древней широко распространенной фауны, а эволю- ционировали в Ю;кной Америке от нескольких предков. В Старом Свете ложнопевчие, вероятно, никогда не были многочисленными и вскоре смешались с фауной примитивных певчих (из которых сохрани- лись австралийские лирохвосты и мелкие кустарниковые птицы семей-
Лтицы
ства Atrichornitidae). Затем в Старом Свете с эволюцией высших певчих происходило много сложных изменений, в то время как ложнопевчие эво- люционировали в Южной Америке самостоятельно. Возможно даже, ~то Suboscines возникли в Южной Америке и некоторые из них расселились на север к Аляске (так же как и американские мухоловки), а оттуда в Ста- рый Свет. Другими словами, Suboscines и Oscines могли эволюционировать в разных частях земного шара, и, возможно, каждая из названных групп давала своих вселенцев на территорию другой группы. У Oscines это про- исходило чаще, в более поздние времена и более успешно, но все же не вызывая еще общего вытеснения Suboscines из их родных мест в Южной Америке.
Когда же возникли и начали расселяться доминирующие Oscines'". Ископаемые данные (Lambrecht, 1933; Romer, 1945) только частично отли- чают на этот вопрос. Два современных семейства Oscines (Paridae и Laniidae) относятся к верхнему эоцену Европы, а несколько современных родов Oscines (Lanius, Motacilla, Sylvia, Passer) — к олигоцену Европы. Это является доказательством по крайней мере существования современ- ных семейств и родов Oscines в третичном периоде. Но может возникнуть сомнение, являются ли названные ископаемые тем, за что мы их прини- маем. Некоторые из них представлены скелетными отпечатками (а не пол- ным скелетом), некоторые — только фрагментами, и все они определены на основании только внешнего сходства с современными птицами. Их фило- генетический характер и связи не могут считаться точно установленными, поэтому они не являются достоверным свидетельством возраста певчих птиц. Эти ископаемые, несомненно, требуют критического пересмотра.
Обнаружение ископаемых Oscines в Северной Америке помогло бы установить дату их радиации, но североамериканские материалы по иско- паемым этой группы еще хуже, чем европейские. В приблизительной хроно- логии они таковы. Сомнительные воробьиные из эоцена Вайоминга- сломанная кость ноги, приписываемая Уитмором (Wetmore, 1943) древес- нице из нижнего миоцена Флориды; Palaeospiza (см. ниже) — остаток пера (первоначально описанный как мох) из миоцена Колорадо; Palae- ostruthus — верхняя челюсть, очевидно, вьюрка, близкого и Pipilo, из миоцена или плиоцена Канзаса, несколько плиоценовых и довольно большое количество плейстоценовых остатков, приписываемых суще- ствующим родам или видам Северной Америки. Palaeospiza из миоцена необычайно хорошо сохранившаяся ископаемая птица. Но почти полньш скелетный отпечаток все же не дает всего того, что надо было бы увидеть. Уитмор (Wetmore, 1925) считает, что это примитивный представитель Oscines, в некоторых отношениях приближающийся к Alesomyodi (Suho- scines), и он относит его к отдельному вымершему семейству. Таким образом, ископаемых, свидетельствующих о нахождении Oscines в Север- ной Америке до миоцена, нет, а имеющиеся миоценовые материалы
ЯВНО HOQOCTBTOЧНЫ.
В связи с тем, что по ископаемым данным нельзя установить время появления Oscines, необходимо поискать другие «ключи», которые могут быть найдены в особенностях эволюции и распространения певчих воробьи- ных. Oscines развивались бурно, и тот факт, что большинство их семейств до сих пор не очень дифференцировано, позволяет предполагать, что ска- чок и основное расселение группы произошли недавно. Если Озс~пе~ существовали и доминировали в эоцене и олигоцене, когда в северных частях земного шара было теплее, чем сейчас, многие их семейства должны оыли бы достичь Америки. Однако многие доминирующие семейства Ста-
рого Света не достигли ее или проникли туда только недавно, что дает еще больше оснований считать их происхождение более поздним. Тот факт, что некоторые Oscines (немногие изолированные ветви) были в Юж- ной Америке, вероятно, до ее соединения с Северной в позднем плиоцене. ничего не дает в отношении датировки, так как иногда Oscines пересе- кают широкие водные пространства и многие из них достигли Австралии. Все эти данные указывают на то, что, когда бы ни произошли Oscines, они радиировали и расселились не позднее, чем в среднетретичном перио- де, вероятно, главным образом в миоцене. Радиация этих птиц продолжает- ся до сих пор и, возможно, даже в настоящее время находится на высшей
ТОЧИ е.
Остается только подвести итог недавним перемещениям разных Osci- нез между Старым и Новым Светом. Ряд видов, очевидно, расселился из Старого Света в Северную Америку: жаворонок, достигший даже север- ной части Южной Америки, три вида ласточек, которые также достигли Южной Америки (как мигранты), и из них один вид образовал там гнез- дящуюся популяцию; один вид тималий; два или более видов славко- вых, пз которых один, предок комароловов (Polioptila), также не так дав- но достиг Южной Америки; четыре или более видов дроздов и др. (появив- шихся в разное время), из которых три вида достигли Южной Америки и там радиировали (Mayr, 1946, р. 19); конек, распространившийся по всей Северной и Южной Америке; один или два вида сорокопутов; воз- можно, шесть представителей группы пищухи — поползни — синицы; некоторые из них достигли Центральной Америки, но ни один не проник в Южную; вероятно, шесть родов вьюрков (подсемейство Fringilinae), из которых род Spinus также распространен в Южной Америке, и, воз- можно, четыре вида соек, ворон и др., появившихся в Северной Америке в разное время (сойки достигли так>ке и Ю>кной Америки). Если Oscines оыли первоначально птицами Старого Света, то самые отдаленные предки американских пересмешников — крапивников и группы виреоны — слав- ки — гавайские цветочницы — танагры — кассиковые должны быть доба- влены к тем представителям Oscines, которые появились из Старого Света и затем рассеялись, образовав новые формы в Америке, включая и Юж- ную. Единственные представители Oscines, у которых расселение проис- ходило, несомненно, в обратном направлении (из Северной Америки к Ста- рому Свету) — крапивники (Troglodytes). Из них один американский вид (американский обыкновенный крапивник) распространился по всей уме- ренной зоне Евразии. Некоторые дрозды, синицы, вьюрки и врановые также могли распространиться недавно из Нового Света в Старый, а пред- ки овсянковых и вьюрковых Старого Света могли расселиться в этом же направлении сравнительно недавно.
В эти списки я не включил некоторых птиц Евразии (пеночка-вес- нпчка, каменка, варакушка и желтая трясогузка), которые расширили свой гнездовой ареал до Аляски (а каменка и до северо-восточной части Северной Америки), но возвращаются на зимовку в Старый Свет, и неко- торых американских пернатых (малый дрозд, трясогузковая древесница. речная древесница), которые расширили свой гнездовой ареал до северо- восточной части Сибири, но зимуют в Америке. Подобное расширение ареала, вероятно, не имеет значения для расселения, если только зимовки птиц так же, как и гнездовой ареал, не переместятся в дальнейшем из од- ной половины земного шара в другую.
Таким образом, певчие птицы (Oscines, Passerinae), возникли в Ста- ром Свете, вероятно, претерпели быструю и сложную радиацию в средне-
247
третпчное время и продолжают радиировать до сих пор. Время от време- ни многие ответвления их достигали Северной Америки, и довольно боль- шое число проникло также в Южную Америку, где некоторые вторично радиировали и некоторые, вероятно, вернулись на север и даже в Ста- pbIl1 Свет.
Таким образом, здесь перечислены все случаи расселения, о которых имеются сведения, как воробьиных, так и неворобьиных птиц между Ста- рым Светом и Америкой. Иного, более детального списка пока не состав- лено. Надо отметить, что в приведенных мною материалах слишком мно- го сомнительных данных и в целом здесь отображена мизерная доля всех возможных перемещений птиц. Однако, исходя из всего этого, можно все-таки сделать следующее заключение. Не все эти перемещения проис- ходили в одном направлении. Несомненно, имел место и обмен: Америка не только получала, но также и что-то отдавала из своей фауны; напри- мер, видимо, роды Buteo, Columba, дятлов, почти несомненно род Troglo- dytes. и, возможно, вьюрков. Но этот обмен был неравномерным: значи- тельно больше птиц перемещалось из Старого Света в Америку, чем в об- ратном направлении. Это справедливо как в отношении сравнительно недавних перемещений, так и древних. Более того, как указывает Майр (Мауг. 1946, р. 38), целый ряд птиц Старого Света достиг как Северной Америки, так и Южной. Некоторые из них радиировали там вторично, но ни одна (или незначительное число) южноамериканская птица не достигла Старого (;вета. По-видимому, непрерывный приток птиц проис- ходил больше из Старого Света в Северную и Южную Америку, чем в обратном направлении. То же самое Симпсон выявил у млекопитающих, а я — у холоднокровных позвоночных. Это вполне возможно допустить и для птиц, но пока еще не в качестве неопровержимого факта.
Распространение птиц почти ничего не говорит нам о прошлом Зем- ли и ее климатов. (:овременное распространение птиц на материках нахо- дится в соответствии со сложным относительно недавним их расселе- нием, главным ооразом по современной суше, с некоторыми отступаниями и в исключительных случаях, возможно, с пересечением широких океа- нических пространств. (:вязи между отдельными видами наземных птиц Мадагаскара и тропической Азии, или тропической Азии и Южной Аме- рики. или Африки и Южной Америки являются редкими исключениями. и едва ли можно предположить, что какую-то роль здесь играли древние сухопутные мосты. По-видимому, ме>кду наземными птицами Южной Аме- рики и Австралии или Новой Зеландии прямой связи нет. Распростране- ние птиц кое-что говорит об истории некоторых островов, но это будет рас- смотрено в гл. 8. Полагают, однако, что по распространению птиц можно соорать более достоверные сведения об истории Северной Америки.
Некоторые орнитологи (Lonnberg, 1927; Мауг, 1946; Bond, 1948) пред- полагают, что в третичном периоде большая южная часть (;еверной Аме- рики была тропической и там находилась большая тропическая фауна птиц. которая фактически была изолирована от Южной Америки, и что Северная Америка являлась местом происхождения целых семейств аме- риканских птиц и источником большинства современных птиц Вест-Ин- дии. Если птицы действительно свидетельствуют об этом, то мы можем получить важные сведения о климате и о степени изоляции (;еверной Аме- рики в третичном периоде. Но свидетельствуют ли они ои этом
Данный вопрос был запутан из-за неточности терминов. Рассматри- вая зоогеографию третичного периода, необходимо внести четкое разгра- ничение между собственно (."еверной Америкой и Центральной Америкой,
242
Глава 5
которая, возможно, в течение третичного периода была островом или архи- пелагом (рис. 36) и, вероятно, имела самостоятельную фауну. Анализи- руя «тропическую часть Северной Америки», орнитологи зачастую не де- лали такого разграничения. Например, момотов считали тропическими птицами Северной Америки, в то время как в действительности они являются центральноамериканскими (см. ниже). Обычно употребляемый термин «Средняя Америка», допустимый в некоторых случаях, в данном случае приводит к неточности, так как он охватывает несколько отдель- ных частей суши (южную часть Северной Америки, Центральную Амери- ку и иногда Вест-Индию), история которых, вероятно, была очень раз- ЛИЧНОИ.
Р и с. 36. Схема (предполагаемая) Центральной Америкив третичном периоде.
Мне кажется, что данные, которые предполагают происхождение определенных групп птиц в тропической части Северной Америки, истол- ковывались неправильно.
Момоты, которых, как уже говорилось, предположительно считали в Северной Америке тропическими птицами, фактически имеют следующее распространение. Они насчитывают 6 родов и 8 видов. Два рода широко распространены в Южной и Центральной Америке и достигают Мексики. Каждый из этих родов включает два вида, из которых один широко рас- пространен в Южной и Центральной Америке, а другой приурочен к Цен- тральной Америке или Мексике. Возможно, это результат или происхож- дения этих птиц в Центральной Америке, или двойного проникновения в нее. Род Momotus — один из тех широко распространенных родов (а не локали- зованный реликт), который определяет северные границы маматов в север- ной части Мексики. Другой род с одним видом, главным образом южно- американский, заходит в' Центральную Америку только до Никарагуа. А остающиеся три рода, по одному виду в каждом, являются преимуще- ственно центральноамериканскими, хотя два из них пересекают перешеек Теуантепек в южной части Мексики. Но нет ни одного рода этого семей- ства, который бы встречался в Северной Америке только или главным образом севернее названного перешейка. Таким образом, в распростране- нии этого семейства нет ничего, что позволило бы предположить, что
южная часть Северной Америки (столь отличающаяся от Центральной) является местом его происхождения.
Предполагали также, что краксовые, или шлемоносные, куры по про- исхождению являются тропическими птицами Северной Америки. Они насчитывают 11 современных родов. Три рода распространены от северной части Аргентины до отдельных частей Мексики и один из этих трех (Orta- йг) достигает южного Техаса. Один род распространен от Перу до Коста- Рики; пять родов приурочено к отдельным частям Южной Америки и два рода — к отдельным районам Центральной Америки и южной части Мексики. Ископаемые рода Ortalis, относящиеся к нижнему миоцену и нижнему плиоцену Небраски и Южной Дакоты, найдены далеко на севере от современных границ семейства, но, вероятно, формы нижнего миоцена в действительности относятся не к роду Ortalis, а к вы- мершему роду, представители которого мельче, чем современные краксо- вые (Wetmore, 1951а, р. 59). То обстоятельство, что ископаемые краксо- вые указанного возраста в Южной Америке еще не найдены, вероятно, есть лишь результат неполноты палеонтологического материала. Учиты- вая современное распространение рода, нельзя сомневаться, что краксо- вые находились в Южной Америке в позднем плиоцене, задолго до соеди- нения с Центральной и Северной Америкой. Распространение краксов, по-видимому, отражает многократное расселение и дифференциацию в те- чение долгого периода в Америке в целом (не считая Вест-Индии), а не радиацию из южной части Северной Америки.
Древесницы (Parulinae) — другая группа птиц, которая предполо- жительно произошла в тропической части Северной Америки. Их распро- странение более сложное. Многие из них оседлы в Южной, Центральной Америке, Вест-Индии и южной части Северной Америки (Мексика и т. д.). Многие — перелетные: обычно они зимуют в тропиках или вблизи тропиков (часто в Центральной Америке, северной части Южной Америки или в Вест-Индии), но некоторые гнездятся на севере близ северной границы древесной растительности в Северной Америке. Одни роды этого подсе- мейства состоят только из оседлых видов в тропиках или субтропиках, другие — только из северных перелетных видов, а третьи — из тех и дру- гих: оседлых и перелетных видов в одном и том же роде. Статистически большинство существующих родов и видов является или северными пере- летными, или оседлыми на довольно небольших площадях в Мексике, или Центральной Америке, или на островах в Вест-Индии. Ни один род не приурочен к Южной Америке, но Myioborus и Basileuterus встречаются главным образом там. Если применить «ключи» чисел к истории этого семейства, эти роды следует считать первоначально южноамериканскими, которые затем распространились в Центральную Америку и Мексику. Кроме того, в Южной Америке имеются широко распространенные осед- лые виды рода Parula (к этому роду относится также северный вид Parula атепсапа), Geothlypis (к которому относится также желтогорлая дре- весница — G. trichas) и род Granatellus, общий для Центральной Америки и Мексики. Внутри рода Dendroica между перелетными и оседлыми фор- мами существуют сложные связи. К этому роду относятся многие самые северные строго перелетные американские славки. Северная перелетная желтая древесница (D. aestiva) имеет оседлые подвиды или близкород- ственные виды в Мексике и т. д., в Вест-Индии. Перелетная G. trichas имеет оседлую форму на некоторых Багамских островах. Северная пере- летная сосновая древесница (D. pinus) имеет оседлые формы на северных Багамских островах и в горах Гаити, а северная перелетная древесница
прерий (D. йгсоКог) также имеет оседлых «заместителей» (D. vitellina) на нескольких мелких островах Вест-Индии (на островах Каймановых и Суок). Другие виды рода Dendroica также являются оседлыми или кочую- щими в отдельных частях Мексики и Вест-Индии. У рода Vermi~ оп~ перелетная оранжевоголовая древесница (V. celata) образует оседлую фор- му на острове Тодос-Сантос на северо-западе, начиная от Нижней Кали- форнии. Представители несомненно перелетного вида виргинской дре- весницы (V. virginiae) являются оседлыми или кочующими в отдельных частях Мексики (Bond. 1936; Blake, 1953).
Майр и Бонд подсчитали число родов и видов подсемейства Parulinae в разных местах и пришли к заключению, что эти птицы произошли в тро- пической части Северной Америки, но такой метод анализа был бы хорош только в том случае, если бы птицы расселялись из одного места довольно простым путем, без усложнений. Но это не так. Их расселение в Америке в целом было сложным и многократным, а отдельные виды ста- новились перелетными и потом опять оседлыми, вероятно, неоднократно. Я думаю, что нельзя назвать какую-то одну территорию, которая могла бы считаться местом их происхождения. Вероятно, древесницы были широко распространены и в Северной и в Южной Америке почти с самого начала. Число родов, локализованных в Центральной Америке и Вест- Индии, вероятно, отражает не место их происхождения, а прерывистость суши в прошлом и настоящем. Число северных перелетных видов, видимо, увеличилось в плейстоцене. Многие представители подсемейства Paruli- nae (но, вероятно, не все) хорошо пересекали водные пространства. В частности, они мигрируют через водные пространства до Вест-Индии, и, видимо, водные пространства третичного времени также не задерживали их расселения. Я считаю, что это подсемейство следует вычеркнуть из списка птиц, которые, как предполагают, произошли в «тропической части Северной Америки».
Надежных подтверждений того, что какое-либо семейство птиц (не считая индеек) возникло в южной части Северной Америки или рассели- лось из нее, нет, хотя к некоторым группам, рангом ниже семейства, это положение вполне применимо. Самой достоверной иллюстрацией послед- него служат американские перепела, относимые некоторыми авторами (но не Майром и Амадоном) к самостоятельному подсемейству. Из 10 родов американских перепелов, приводимых Питерсом, 9 приурочены к южной части Северной Америки или, возможно, распространялись из нее. Деся- тый род в основном относится к Центральной Америке. Три рода достигаю1. Южной Америки, но ни один их них там не эндемичен. Очевидным цен- тром радиации этих птиц была Мексика. Но необходимо заметить, что индейки и перепела не составляют всей фауны птиц и только небольшое число других групп имеет такое же распространение, как и эти птицы.
Ископаемым птицам было придано чрезмерно большое значение в свя- зи с «тропической частью Северной Америки». Ископаемых краксов и жу- равлей семейства Aramidae (теперь в основном тропические птицы) отно- сят к среднетретичному периоду Северной Америки, но, судя по всему, они не должны были бы здесь возникнуть. Ископаемые материалы по этим птицам, да и по всем птицам вообще, слишком бедны и односторонни, что- бы по ним можно было сделать какие-то определенные выводы. О крак- сах мы уже говорили. А по ископаемым арамам можно установить не то, что они возникли в Северной Америке, а то, что это постепенно исчезаю- щая группа (теперь уменьшенная до одного вида), и эти ископаемые не подходят для анализа при помощи «ключей чисел».
Птиц,ы
Имеется другой, еще не использованный источник для решения вопро- са о тропических птицах Северной Америки. Если целые семейства птиц возникли в южной части Северной Америки и не могли преодолеть водные преграды на пути в Южную Америку, то, значит, они (как это предпола- гают теоретически) расселились из Северной Америки через Вест-Индию. Очевидно, сделать это им было трудно, и их распространение через остро- ва должно было бы иметь форму клина (рис. 37). Самое большое число их должно было находиться на Кубе, а далее, с увеличением расстояния от Северной Америки, их число должно было уменьшаться примерно так и;е, как это происходит у наземных птиц по мере увеличения расстояния от Новой Гвинеи к западной части Тихого океана (см. стр. 219, рис. 3 ). Р и с. 37. Схема «клина», который должен был бы образо- ваться (но не образовался), если бы определенные семейства птиц достигли Вест-Индии из «тропиков Северпой Америки». Цифры (например, 50/40) показывают (приблизительно) общее число (видов) наземных птиц, превосходящее число «тропических птиц
Северной Америки». Насколько я представляю, такого клина в распространении большинства птиц Вест-Индии не наблюдается. На Больших Антильских островах птиц болыпе, чем на Малых, но нельзя сказать, чтобы предполагаемых тропи- ческих птиц Северной Америки, таких, как крапивники, пересмешники, виреоны и древесницы, было пропорционально больше. Этот вопрос усложняется еще неясностью истории этих островов в третичном периоде. Перепела, правда, образуют в Вест-Индии клин; они достигли Кубы и острова Пайнс (один вид), но за пределами этих островов их нет и неиз- вестно, действительно ли они распространились из южной части Север- ной Америки.
Несомненно, что большая часть Северной Америки в третпчном периоде была теплее, чем теперь, и что птицы в южной части Северной Аме- рики были многочисленнее и в какой-то степени более изолированы от Южной Америки, чем теперь. Но, однако, сами птицы дают мало факти- ческих доказательств. А утверждение, что южная часть Северной Амери- ки имела богатую тропическую фауну птиц, изолированную от Южной Америки, и что она была самостоятельным центром эволюции важных семейств птиц, на мой взгляд, сомнительно и противоречит имеющимся данным.
Попытаемся представить положение птиц в южной части Северной и в Центральной Америке в третичном периоде. Alon выводы базируются отчасти на том, что показывают птицы, отчасти на данных пз других
Глава 5
источников: на том, что известно об истории Северной, Центральной и Юж- ной Америки третичного периода, и из характера современных фаун поз- воночных Центральной Америки и Вест-Индии.
Северная и Южная Америка, несомненно, в течение большей части третичного периода были разделены. Климат Северной Америки, конечно, был теплее, чем в настоящее время, но, вероятно, он был скорее субтропи- ческим, чем тропическим. Я думаю, что Центральная Америка была пол- ностью отрезана водой от Северной, так же как и от Южной, на протяжении большей части третичного периода. Доводом для такого предположе- ния является то, что многие из древних позвоночных Вест-Индии, по-ви- димому (судя по их распространению на островах), пришли из Централь- ной Америки. К ним относятся такие животные, которые, очевидно, не находились бы в Центральной Америке, если бы она соединялась с (;е- верной, особенно это касается дикобразоподобных грызунов. Вероятно, южная часть Северной Америки, Центральная Америка и Вест-Индия име- ли более бедные фауны птиц, чем авифауна Южной Америки, и (несмотря на водные преграды) эти обедненные фауны (кроме фауны Вест-Индии, где, очевидно, отсутствовали некоторые семейства) состояли в значительной степени из одних и тех же более или менее широко распространенных семейств птиц. Я считаю, что вся эта территория плюс соответствующие часггги Южной Америки была местом эволюции таких семейств, как кра- пивники, пересмешники и древесниг~ы, географическая история которых, как уже говорилось, была очень сложной. На острова Вест-Индии птицы попали, вероятно, из нескольких направлений (в большинстве случаев из Центральной Америки), причем расселение их было столь сложно, что простого клина уменьшающегося числа видов с удалением от Северной Америки не возникло. Краксы и некоторые другие птицы, которые не до- стигли Вест-Индии, тем не менее (как я предполагаю) были в то время широко распространены в Северной, Центральной и Юд;ной Америке. В изо- лированных местах, вероятно, было много эндемичных родов и видов, и некоторые из эндемичных родов до сих пор локализованы в южной части Северной Америки и других районах, но они свидетельствуют все же об изоляции этих семейств в прошлом, а не о месте их происхождения.
В течение третичного периода в Южной Америке должна была суще- ствовать очень богатая тропическая фауна птиц. Часть ее, но только часть. возможно, распространилась в северном направлении через водные пре- грады в Центральную Америку и южную часть Северной Америки, а еще меньшая часть достигла Вест-Индии. Другие виды птиц, вероятно, пересе- кали водные преграды в обратном направлении — от Северной Америки к Центральной и Южной, а ряд из них прямыми или окольными путями достиг также и Вест-Индии. Перемещения некоторых птиц (например, древесниц) через водные преграды, вероятно, были очень сложными. Известно, что там, где птицы сталкиваются с преградами, некоторые из них останавливаются, в то время как другие пересекают их в разное время и на разное расстояние.
Волфсон (Wolfson, 1955) пытался связать историю птиц Северной Аме- рики с теорией перемещения материков. Подобные предположения допу- стимы, но в данном случае они необоснованны. Богатство фаунистических связей и относительный эндемизм разных материковых авифаун находят- ся в корреляции с современным распределением суши, климата и преград на земном шаре. Другими словами, современное распространение птиц, очевидно, есть результат перемещения птиц по частям суши, по своим очертаниям напоминающим очертания современных материков, а не ре-
зультат перемещения материков. Перемещение материков, если оно и име- ло место, очевидно, было слишком давно, чтобы повлиять на современное распространение птиц.
С11ИСОК СЕМЕЙСТВ ПТИЦ
Ы.1АСС AVES, птицы. Этот список составлен в соответствии с классификацией Майра
и Амадона (Мауг, Amadon, 1951) по причинам, указанным выше, но подробности распространения птиц в большинстве случаев взяты из работы 11итерса (Peters.
l931 — 1951). Ископаемые семейства приводятся по Уитмору (Wetmore, 1951).
Ископаемые находки, принадлежащие современным семействам, даются с сокра- щениями по Ламбрехту (Lambrecht, 1933) и Уитмору (Wetmore, 1951a, etc.) и, как правило, проверены по Ромеру (Romer, 1945). Далее отмечается наличие или отсутствие всех африканских семейств — на Мадагаскаре, всех австралий- ских — в Новой Зеландии и всех американских семейств — в Вест-Индии.
Знак звездочки (») перед латинским названием означает, что группа вымерла.
Постоянные перемещения птиц создают особую трудность при описании их
ареалов. Так, некоторые птицы иногда залетают далеко за пределы своих обычных границ, но данные о таких залетах в этот список не включены. Многие морские птицы, гнездящиеся только на очень ограниченных участках, перемещаются в определенные сезоны года через большие океанические пространства, но и эти данные не могли быть здесь использованы. Миграции наземных птиц упоминают- ся лишь в тех случаях, когда их влияние существенно сказывается на границах или характере распространения семейств, но очень многие детали этих миграций даже наземных птиц также пришлось опустить.
:э110ДКЛАСС ARCHAEORNITHES, первоптицы
«Archaeopterygidae и »Archaeornithidae~. Европа (Бавария) в верхней юре. Каждое
семейство описано на основе одного хорошо сохранившегося скелета. Эти знаме- нитые рептилиеподобные птицы имеют большое значение для выяснения вопросов эволюции птиц и незначительное с точки зрения географии.
1!ОДКЛАСС NEORNITHES, настоящие птицы
*Отряды lchthyornithes и Hesperornithes, зубатые птицы мелового периода *Ichtyornithidae, *Apatornithidae, ~НеярегогпйЬЫае, *Enaliornithidae, *Baptorni- thidae. Меловой период Северной Америки, Европы и южной части Южной Аме- рики. Наиболее изучены только североамериканские семейства *Ichtyornithidae и *Hesperornithidae. Представителями обоих семейств были зубатые морские пти- цы. Ихтиорнисы имели сильные крылья, гесперорнисы были не летающими, а пла- вающими птицами с недоразвитыми крыльями. Остальные вышеперечисленные ископаемые птицы мелового периода недостаточно известны и помещены здесь условно. Южноамериканский род sNeogaeornis (Lambrecht, 1933, р. 258, 654) из южного Чили условноприписывается к семейству *Enaliornithidae. Все птицы мелового периода имеют скорее эволюционное, чем географическое значение. .»Отряд Caenagnathi
«(;aenagnathidae. Верхний мел Северной Америки (Альберта); единственная челюсть
без зубов, возможно принадлежащая не птице.
(Следу~ощие б отрядов — «бескилевые», Б предполагаемых неродственных групп птии,
(Mayr, Amadon, 1951) (рис. 33).]
*Отряд Aepyorni thes
":Aepyornithidae, эпиорнисы, или слоновые птицы. Мадагаскар. Вымершие птицы
плейстоцена и постплейстоцена. 2 рода, несколько видов, включая хорошо со- хранившийся ископаемый материал. Предположение о нахождении семейства в Африке основано на недостоверных остатках, и его не следует принимать во внимание (Mayr, Amadon, 1951, р. 4).
Отряд Ар teryges
Dinornithidae и Anomaloptirigidae, моа. Новая Зеландия (оба острова). Известны пз
плиоцена (и, вероятно, позднего миоцена), многочисленны в плейстоцене. Эти птицы уцелели до прибытия первых полинезийцев в Новую Зеландию. 7 родов, около 20 или более видов (АгсЬеу, 1941; Oliver, 1949). Известны по ряду более или менее целых скелетов, а также по остаткам нескольких тысяч отдельных костей.
1 Последние исследования Биба показывают, что все юрские остатки первоптиц принадлежат одному виду, и, таким образом, Arcbaeornithidae отходит в синонимы.— Прил. ред.
248
Глава 5
предполагаемое нахождение моа в Австра.сии основывается на сомнительных, очевидно неправильно определенных, остатках (Mayr, Amadou, 1951, р. 4).
ppterygidae, киви. Новая Зеландия, оба острова и остров (:тюарт. Распространены
от субтропических лесов на уровне моря до снеговой линии (Falla, 1953). ! род, 3 вида, еще 1 род и 1 вид являются субфоссильными. Предполагаемые:~а»»ые о костях, принадлежащих киви из Австралии, ошибочны (Mayr, Amado». р. 4). Отряд 8 tru thiones
"Eleutherornithidae, *Eleutheornis. Эоцен Европы (Швейцария); довольно х<>ро»н>
сохранившаяся тазовая кость. Эти семейства, вероятно, представляют пред«()вуI(7 группу современных страусов и связывают их с летающими предками (\\ vt>no- re).
Struthionidae, страусы. Почти все засушливые части (>1склн>чая густые леса) Лфрики
от гор Атлас до мыса Доброй Надежды (нет на Мадагаскаре) и (до недавнег»»ре- мени) пустыни Сирии и Аравии. Только 1 вид. Ископаемые кости, яйца и нх с кор- лупа обнаружены на территории Южной Евразии до Китая, примерно в нижнем плиоцене (Lambrecht, 1933 р. 109). Последний страус в Сирии и Аравии. псче >- нувший в 1941 г., был описан Майнертцхагеном (Meinertzhagen, 1954, р. >7'>). Отряд Gasuari
Casuariidae, казуары. Новая Гвинея и соседние острова (Ару, Церам, Салаватн,,'(>ко- би, Новая Британия) и северо-восточная часть Австралии (нет в Новой Зеландии).
1 род, 6 видов. Ископаемые (конец tibiotarsus) из плейстоцена Нового Южног» Уэльса, юг современного ареала в Австралии. В противоположность другич «руц- ным современным бескилевым эти птицы живут в густых лесах.
Dromaeidae, эму. Австралия (нет в Новой Зеландии), на Тасмании и островах !боинг
и Кенгуру вымерли. 1 род, 2 вида. Ископаемые относятся к плейстоцену Австралии. *Dromornithidae, *Dromornis, *Genyornis. Плейстоцен Австралии (но не в Новой Зе- ландии); довольно хорошо сохранившийся ископаемый материал, гигантские эму- подобные птицы.
Отряд Rheae
Rheidae, реа (американские страусы). Восточная и южная часть Южной Америки (нет
в Вест-Индии), от северной и восточной Бразилии и юго-восточной части 11еру до Магелланова пролива, 2 современных рода и вида. Ископаемые фрагментар»ь>. Отряд Gryptury
Tinamidae, тинаму. Южная и Центральная Америка, на север до некоторых районов
Мексики (2 рода) и на юг почти до южной оконечности Южной Америки: также Тринидад (нет в собственно Вест-Индии). Около 9 родов и 33 или большее число видов, из которых одни обитают в лесах, другие — в прериях. Это примитивные, внешне похожие на куропаток и перепелов птицы, которые с трудом перелетают на короткое расстояние, возможно, отдаленно родственны американским страусам (реа) (McDowell, 1948; Mayr, Amadon, 1951, р. 4). Несколько ископаемых этого семейства известны в пределах современного ареала и относятся к плиоцену. Отряд Sphenisci
Spheniscidae, пингвины. Антарктические моря к северу до берегов Южной и В осточной
Австралии (на север до южного острова Квинсленд), Новая Зеландия, Южная Африка и южная и западная части Южной Америки (к северу до Перу) и 1 энде- мичный вид на островах Галапагос, на экваторе. 6 родов, 16 видов. Ископаемые на юге Южной Америки (Патагония), в Австралии, Новой Зеландии и на острове Симор в Антарктике, вероятно, все относятся к миоцену. (Более раннее семейство *Clad0rnithidae, как считают теперь, не является родственным пингвинаи,— Simpson, 1946.) Симпсон подчеркивает отсутствие ископаемых пингвинов за пре- делами границ современного распространения этих птиц и приходит к выводу, что границы распространения семейства пингвинов в миоцене, вероятно, были примерно такими же, как и в настоящее время.
Отряд Tubinares
Di0medeidae, альбатросы. Главным образом тропические и субтропические моря н не- сколько представителей в северной части Тихого океана. 2 рода, ! 3 видов. l [о- стоянных обитателей в Северной Атлантике нет, но ископаемые остатки (~агзо- metatarsus) обнаружены в нижнем плиоцене Англии.
Pr0cellariidae, буревестники, волнорезы, глупыши. В большинстве случаев морские
птицы. 2 широко распространенных подсемейства: (1) f lydrobatinae, к»чурки.
Несколько родов, 20 видов. (2) Procellariinae, волнорезы и др. Несколько родов, 53 вида. Имеется несколько ископаемых, не представлякнцих большого знд и ция с точки зрения географии.
Pelecan0ididae, нырковые буревестники. Южные моря, на север до оерегов 10го-Во- сточной Австралии, Новая Зеландия, южная и западная части К)жной Ам«рнкц, на север до Перу. 1 род с 4 видами.
Отряд P~idicipi tides
Podicipitidae, поганки. Космополиты, включая Ъ!адагаскар, Новую Зеландию и Вест-
Индию. 5 родов (из которых Poliocephalus и Ройсерз более или менее космополи- ты). 20 видов. Главным ооразом пресноводные птицы. Ископаемые этого семей- ства относятся к верхнему эоцену (в Европе).
Отряд Gaviae
Gaviidae, гагары. Гнездятся в наиболее холодных частях северного полушария, зиму- ~от несколько южнее в пределах умеренной зоны северного полушария. 1 род, 4 вида. В основном пресноводные птицы. Несколько ископаемых остатков, отно- симых к эоцену, обнаружены в северном полушарии.
Отряд teganopodes
Phaethontidae, фаэтоны. Тропические птицы. Теплые моря. 1 род, 3 вида и l ископае- мый род, представленный достоверным материалом из нижнего эоцена Англии. Fregatidae, фрегаты. Теплые моря. 1 род, 5 видов.
Р11а1асгосогасЫае, бакланы и др. 2 подсемейства: (l) Phalacrocoracinae, бакланы. Поч- ти космополиты, включая Мадагаскар, Новую Зеландию и некоторые острова Вест-Индии. 3 рода, 30 видов. Ископаемые относятся к эоцену Северной Америки и олигоцену Европы. (2) Anhinginae, змеешейки или анхинги. Более теплые рай- оны земного шара, включая Мадагаскар и некоторые острова Вест-Индии, нет в Новой Зеландии (не считая случайных залетов). 1 или 2 вида, несколько под- видов. С географической точки зрения ископаемые не представляют интереса. ":Pelagornithidae, *Pelagornis. Миоцен Франции, плечевая кость.
Sulidae, олуши. Тропические моря и моря умеренных широт. 2 рода, 9 видов. С гео- графической точки зрения ископаемые не представляют интереса.
":Elopterigidae, *Е1орСегух, *Eostega, *Actiornis. От верхнего мела до верхнего эоцена
Европы, отдельные остатки.
Pelicanidae, пеликаны. Широко рассеяны по всему земному шару в тропических
и некоторых умеренных районах, на север до Юго-Восточной Европы и Большого Невольничьего озера в Канаде, включая Вест-Индию, но не Мадагаскар или Новую Зеландию (не считая сомнительных данных или случайно проникших туда экземпляров). 1 род, 6 видов. Имеется некоторый довольно хорошо сохранив- шийся ископаемый материал из верхнего эоцена Франции.
"'Cyphornithidae, *Cyphornis, *Palaeochenoides. Северная Америка в третичном (ве- роятно, миоцен) периоде Британской Колумбии и в миоцене Южной Каролины, ос татки костей ног.
"0dontopterygidae и *Pseudodontopterygidae, ложнозубые птицы. Первое семейство
основывается на черепе и фрагментах из нижнего эоцена Англии. Второе — на черепе неопределенного возраста, как предполагают, из Бразилии. Эти птицы «в высшей степени сомнительны по своему систематическому положению» (Wetmo- re, 1951, р. 4, 15).
*(;ladornithidae, *Cladornis, *(;ruschedula. Верхний мел южной части Южной Америки.
Остатки метатарзальных костей. Сходство этих форм и даже их родство друг с дру- гом очень сомнительны (Simpson, 1946; Wetmore, 1951, р. 2, 15).
Отряд Falcones, или Accipitres. Гетерогенная и, возможно, полифилетическая группа. Accipitridae, ястребы, орлы, грифы Старого Света и др. Космополиты, включая Мада- гаскар, Новую Зеландию, Вест-Индию и многие другие отдаленные острова.
Большое число родов, 205 видов. Многие роды локализованы, но некоторые рас- пространены широко. Род Circus (луни) космополитичен, включая Мадагаскар.
Новую Зеландию, Вест-Индию (частично) и другие острова, но на большей части Индо-Малайской области представители этого рода не гнездятся, а только зи- муют. Accipiter (тетеревятник и т. д.) почти полностью космополит, кроме Новой Зеландии; этот род встречается на Мадагаскаре, в Вест-Индии и еще на некоторых более отдаленных островах. А оийа встречается в северных частях земного шара и на юг через всю Африку (нет на Мадагаскаре); до Индии и в других частях Азии (также и в Австралии, если причислить сюда Uroaetus), в Америке на юг до цент- ральной Мексики. Этот род состоит из нескольких видов Старого Света, из кото- рых один (А. сйгугаёгог, беркут) достигает Северной Америки. Haliaeetus встре- чается на большей части Старого Света, включая Мадагаскар и Австрали~о (по не Новую Зеландию), и в Северной Америке на юг до северной части Мексики. Этот род также состоит из нескольких видов Старого Света, и один из них Il. leuco- cephatus, белоголовый орлан, населяет Северную Америку. Вuteo встречается по всей Африке, на Мадагаскаре, в прохладных районах Евразии, Северной, Центральной и Южной Америке и Вест-Индии и на многих отдаленных осТроаах, включая ГaBaiin, острова Галапагос, Хуан-Фернандес и несколько островов в Ат- лантическом океане. Представители этого рода в Индо-Малайской области не гне- здятся и зимуют не во всей, а только в части этой области и совсем не достигают
250
Глава 5
Австралийской области (см. стр. 233). Spizaetus, род орлов с песколькими видами, встречается от Южной и Восточной Азии и Японии на юг до Новой Гвинеи (но не в Австралии) и в тропиках Америки от южной Мексики до северной Аргенти- ны (но не в Вест-Индии). Ископаемые ястребиные известны из широко разбросан- ных по земному шару мест и относятся к эоцену. Buteo, очевидно, был представ- лен в олигоцене Северной Америки; грифы Старого Света («Aegypiinae»), которых теперь в Новом СВе~е нет, были обычны в Северной Америке с нижнего миоцена до плейстоцена.
Ра1сошйае, соколиные и др. Космополиты, включая Мадагаскар, Новую Зеландин)
и Вест-Индию. Различные роды, 58 видов. Большинство родов имеют более или менее ограниченный ареал, но род Falco (соколы) полностью космополитичен.
включая вышеназванные острова. Этот род является примером многократного расселения доминирующей группы. Он состоит из 10 подродов (Peters), из которых 3 почти космополиты. 1 вид (F. peregrinus, с рядом подвидов) почти космопо- лит и гнездится в отдельных частях всех материков, включая Мадагаскар и острова Фиджи, но не в Новой Зеландии или в Вест-Индии, хотя зимой этот род достигает островов последней. Известно несколько ископаемых этого семейства, относящихся к миоцену (Северной Америки). Ископаемые рода Falco относятся к миоцену Северной Америки.
Pandi0nidae, скопы. 1 вид, несколько подвидов. Гнездовой ареал и ареал зимовок
этого вида охватывают большую часть земного шара. Гнездится во всех северных частях земного шара севернее тропиков и от островов Ява, Сулавеси и Малых Зондских островов до Австралии, Тасмании и Новой Каледонии (но не в Новой Зеландии) и локально на Багамских островах (но не на основнойчасти Вест-Индии), также в Юкатане и Британском Гондурасе (Центральная Америка). Зимует частично в пределах гнездового ареала, на большей части Африки (но не на Мада- гаскаре), на части Индо-Малайской области, в Центральной и Южной Америке и в Вест-Индии. Ископаемые этого вида обнаружены в отложениях плейстоцена Флориды.
~ieocathartidae, »Neocathartes. Верхний эоцен Северной Америки (Вайоминг). «Гриф
с небольшими крыльями и сильными ногами, который, очевидно, был наземной птицей с ограниченными возможностями полета» (Wetmore, 1951, р. 5). Почти полный скелет.
Cathartidae, грифы Нового Света. Новый Свет, кроме холодных районов. В Вест-
Индии, на Кубе и Ямайке имеются только местные виды. 5 родов, 6 видов. Различ- ные ископаемые в Новом Свете относятся к олигоцену Северной Америки, вклю- чая кости калифорнийского кондора из плейстоцена восточной части Северной Америки до Флориды. Предполагали, что *Eocarthartes в среднем эоцене Герма- нии и *Plesiocathartes в верхнем эоцене или нижнем олигоцене Франции указы- вают на наличие в прошлом этого семейства в Старом Свете, но ископаемые дан- ные обоих родов фрагментарны и принадлежность их к этому семейству требует подтверждения (Wetmore; см. Mayr, Amadon, 1951).
~Teratornithidae, »Teratornis, «Cathartornis, гигантские ископаемые грифы. Плейсто- цен Калифорнии и Флориды, хорошо сохранившийся ископаемый материал. Sagittariidae, птицы-секретари. Африка (но не Мадагаскар) от Сенегала, Гамбии и Су- дана до Капской провинции. 1 современный вид и 1 ископаемый род (фрагменты) в олигоцене (и верхнем эоцене?) Франции.
Отряд Gressores
Ardeidae, цаплевые. Космополиты, включая Мадагаскар, Новую Зеландию и Вест-
Индию. 32 рода, 59 видов. Самые северные представители являются перелетными.
6 родов (Ardea, Butorides, Casmerodius, Nycticoras, Isobryckus, Botaurus, вкл|очая большую голубую цаплю, зеленую квакву, белую цаплю, квакву, малую выпь и выпь) более или менее космополиты или тропикополиты. Многие другие роды этого семейства приурочены к отдельным частям Старого или Нового Света, а не- которые еще более локализованы. Вид Casmerodius albus (5 подвидов, включая белую цаплю) почти космополит, кроме холодных районов, и достигает Мадага- скара, Новой Зеландии и более крупных островов Вест-Индии. B uto rides st hiatus (с множеством подвидов) также почти космополит, кроме холодных районов.
встречается в тропиках Африки, на Мадагаскаре, на отдаленных островах Ин- дийского океана, в Южной и Восточной Азии на север до реки Амур и Японских островов в южном направлении через острова до Северной и Восточной Австра- лии (нет в Новой Зеландии), на Таити и, возможно, на других отдаленных тро- пических островах Тихого океана, на островах Галапагос и в Южной Америке- от Панамы до Северной Аргентины. Этот вид не встречается на большей части Центральной Америки, в Вест-Индии и в Северной Америке. Здесь находится другой вид этого рода (В. virescens, зеленая кваква). Род Cochlearius (цапли чел-
251
ноклювы) с 1 видом и 3 подвидами иногда относят к самостоятельному семей- ству. Этот род встречается в тропиках Америки от Мексики до южной Бразилии (яо не в Вест-Индии). Ископаемые цапель известны из эоцена Европы и Север- ной Америки.
Threski0rnithidae, ибисы, колпицы. Более теплые районы земного шара, включая Мада- гаскар, Новую Зеландию (случайные залеты) и некоторые острова Вест-Индии.
Множество родов, 28 видов. Питерс выделяет 20 родов в этом небольшом семей- стве, из которых только один встречается и в Старом и в Новом Свете. Это род Plegadis, основные виды которого (3 подвида, включая каравайку) почти тропико- политы. Из остальных 19 родов каждый приурочен или к Старому, или к Новому Свету, а во многих случаях — к отдельным частям разных материков. Ископаемые пбисов представлены хорошо сохранившимся материалом из верхнего олигоце- на Европы и скудным из верхнего эоцена Англии.
('iconiidae, аистовые. Старый Свет (включая Мадагаскар) до Северной и Восточной
Австралии (нет в Новой Зеландии) и в Новом Свете от юго-восточной части Соеди- ненных Штатов до южной Аргентины и на некоторых островах Вест-Индии.
Северные виды Евразии совершают колоссальные миграции. Множество родов, l7 видов. Из 12 родов, выделенных Питерсом, 9 приурочены к отдельным частям Старого Света, 3 — к Новому Свету и нет ни одного рода, встречающегося как в том, так и в другом (но ископаемые рода Ciconia, приуроченного к Старому Свету.
зарегистрированы в плиоцене и плейстоцене Северной Америки). Род Balaeniceps (китоглав) с одним видом, приуроченный к ограниченной территории в Восточ- ной Африке (Белый Нил и т. д.), иногда относят к самостоятельному семейству.
Ископаемые аистов известны начиная с олигоцена (Европы и т. д.).
ЯсорЫае, молотоглавы. Вся Африка южнее Сахары, Мадагаскар и юго-восточная
Аравия. 1 вид.
Отряд Phoenicopteri
Phoenicopteridae. фламинго. В настоящее время встречается только в сильно разоб- шенных районах в Африке, на Мадагаскаре, в Южной Европе, Западной Азии до Индии и в более теплых частях Америки от южной Мексики, Вест-Индии и острова Флорида-Кис до Огненной Земли. 3 рода, 6 видов. Ископаемые фламинго представлены почти полным скелетом из верхнего олигоцена Франции, плечевой костью из верхнего эоцена Англии, вымершим родом из нижнего миоцена Север- ной Америки (Южная Дакота) и фрагментами современного рода Phoenicopterus (единственный род семейства, который встречается как в Старом, так и Ц овом Свете) из плиоцена и плейстоцена Северной Америки (на север до О регана) .
~Agnopteridae, sAgnopterus. Верхний эоцен Франции и Англии, фрагменты; в настоя- щее время ископаемые зарегистрированы также в Центральной Азии.
*Scani0rnithidae, *Scanio rnis, *Pa rasean io rnis. Верхний мел Европы, разрозненные
кости.
Отряд Anseres
*Рагапугос~йае, *Paranyroca. Нижний миоцен Северной Америки (Южная Дакота}. Anati(lac, утки, гуси, лебеди. Космополиты, включая Мадагаскар, Новую Зеландии~,
Вест-Индию. Многие северные формы являются перелетными. Много родов, 14> видов. Это семейство было пересмотрено Делакуром и Майром (Delacour, Мауг, 1945). Лебеди (которых Делакур и Майр относят к роду Cygnus) встречаются во всех северных частях земного шара (5 видов); 1 изолированный вид — в южной части Южной Америки, от Парагвая и т. д. на юг; 1 вид, черный лебедь — в Ав- стралии и Тасмании; 1, ныне исчезнувший, вид когда-то встречался в Новой 3е- ландии. Настоящие гуси (2 рода, 14 видов) являются полностью северными, кроме 1 эндемичного вида на Гавайских островах, но неродственные «гусеобразные» роды (большие длинноногие, часто наземные и травоядные формы) обитают во мно- гих других частях земного шара. Древесные утки Dead rocygna (8 видов) обитают во всех основных тропических и некоторых умеренно теплых районах, включая Мадагаскар, Австралию и Тасманию и острова западной части Тихого океана до острова Новая Каледония и островов Фиджи (нет в Новой Зеландии) и некоторые острова Вест-Индии. Ряд видов этого рода имеет очень широкое и прерывистое распространение (см. списки Питерса — Peters, Check-list, vol. 1, р. 152 — 154).
Другие утки многочисленны, имеют сложные взаимосвязи и сложное и часто ши- рокое распространение. Майр (Mayr, 1946, р. 14) отмечает некоторые виды или группы видов, которые имеют особенно широкий или дизъюнктивный ареал. Иско- паемые этого семейства известны из эоцена (Европы и Северной Америки) и (очень сомнительные) из верхнего мела (Европы). Судить о степени значения с точки зрения географии довольно многочисленных третичпых форм этого семейства трудно.
202
Глава 5
Anhimidae, паламедеи. Большая часть материка Южной Америки, нет в Центральной.
Америке и Вест-Индии. 2 рода, 3 вида.
Отряд Gal li, куриные птицы
'A1egapodiidae, сорные куры. Австралия (нет в Тасмании и Новой Зеландии), !!оворя
Гвинея и т. д., Молуккские острова, Сулавеси с широко распространенным родом МедароЖия, который достигает Соломоновых островов и некоторых (вероятно, больше в прошлом) островов западной части Тихого океана (Мариаиских островов и островов центральной 11олинезии), Малые Зондские острова (до острова Ломбок), все Филиппинские острова, мелкие острова от восточного и северного побережий Калимантана и между островами Калимантан и Ява, Никооярские острова к западу от полуострова Малакка (нет на Калимантане, Яве или Сумат- ре и ни на одной из частей азиатского материка). 7 родов, 10 видов. В Австралии различные представители этого семейства обитают в засушливых, покрытых ку- старниками местностях, а также в тропических джунглях. Заслуживающие «ни- мания ископаемые этого семейства не известны.
*Gallinul0ididae, *Gulliuulo>de~. Эоцен Вайоминга, почти полный скелет.
Сracidae, краксы, или гокко. Тропическая Америка (нет в Вест-Индии), на севср до
|ожного Техаса и на юг до северной Аргентины. l1 родов, <8 видов. Дальнейшие подробности см. текст, стр. 243.
РЬаз|апЫае, фазаны и т. д. 3 подсемейства: (1) Phasianinae, фазаны, павлины, дикие
куры, куропатки. Наибольшего разнообразия достигают в Индо-Малайской об- ласти, 1 вид локализован в Африке. Куропатки являются почти космопо:истнми.
Много родов, 165 видов. Фазаны (Delacour, 1951) преимущественно сосрсдоточе- ны в Индо-Малайской области (хорошо представлены и на тропических низмеп- ностях и в прохладных горных стациях). Формы обыкновенного фазана (Р/~а8/- «nus colchicus) распространены на запад до Кавказа и далее на север до юго-восточ- ной Сибири (не севернее 44' с. ш.). ! или несколько видов достигают или встре- чаются в Японии, на острове Тайвань, на Больших Зоидских островах (несколько родов), на острове Палаван (но на других Филиппинских островах отсутствуи>т), на Малых Зондских островах до острова Сумба (только джунглевые куры). На острове Сулавеси и других островах фазанов нет. (Предполагают, что джунгле- вые куры на Филиппинах и на Сулавеси и других островах интродуцированы.) Павлины (2 вида), которые относятся к фазановым, встречаются на большей конти- нентальной части Индо-Малайской области и на островах Цейлон и Ява (нет на (:у- матре и Калимантане и др.), и единственный, возможно им родственный, африкан- ский павлин, очень четкий монотипический род Л/>.ораго, приурочен и неболь- шой территории девственных дождевых лесов в восточной части бассейно цент- рального Конго. Дикие куры рода Са!!иг (4 вида), от которых произошли домаш- ние куры, свойственны большей части тропиков Индо-Малайской области от Цей- лона и Индии на восток до острова Ява (нет на Калимантане и др.) и до 1!алых Зондских островов. Куропатки встречаются по всему Старому Свету, вк;иочая Мадагаскар, Австралию и (в прошлом) Новую Зеландию, а также в Северной, Центральной и в тропической части Южной Америки (нет в умеренной части южного полушария Южной Америки), на Кубе и т. д. (нет на большей части Вест- Индии и др.). Американские перепела иногда рассматриваются как отзелыюе подсемейство Odontophorinae (см. текст, стр. 244). (2) i3umidinae, цесарки. Боль- шая часть Африки на север до западного Марокко, имеются необособиишиеся (возможно, интродуцированные) популяции одного вида в юго-западной Ара- вии и на Мадагаскаре. 5 родов, 7 видов. Мейнертцхаген (Meinertzhagen, 1954) по- лагает, что в Аравии цесарки интродуцированы. (3) Tetraoninae, тетеревиные. Севе- ро-умеренные и арктические районы Старого и Нового Света. ll родов, l8 видов.
1 род (Lugopus, куропатки) является циркумполярным на дальнем севере, дости- гая арктических островов Северной Азии и Северной Америки, а также Гренлан- дии и Исландии. 3 рода распространены по всей Евразии, а l — в Азии. 6 родов приурочены к Северной Америке (1 из них характерен для северных еловых ле- сов), 2 рода в настоящее время (или в прошлом) распространены по всей умерен- ной зоне Северной Америки, и 3 рода находятся в ее западной части. Такой ха рактер распространения, по-видимому, является результатом сложного расселения и эко- логической радиации, а не распространения из какого-либо единого центра.
Ископаемые показывают, что и фазаны и дикие куры когда-то населяли Европу, где теперь встречаются только домашние и интродуцированные виды, но псе.~- полагаемые ископаемые остатки фазанов в Севернои Америке сомнительны. 11с ко- паемые перепела относятся к нижнему миоцену Северной Америки; ископае- мые тетеревиных — к эоцену Северной Америки.
Aleleagridae, индейки. Восточная и южная части Северной Америки и т. д, (нет в Несг- !1идии). 2 рода, по одному виду в каждом. Обычная дикая индейка в прошлом
258
встречалась от южного Мэна, пэжного Онтарио, Южной Дакоты и т. д. до Мекси- канского нагорья. Ее современный ареал более ограничен. Сильно отличакнцийся вид — глазчатая индейка — приурочен к полуострову Юкатан, к прилежащим частям Гватемалы и Британского Гондураса. Ископаемые индеек, известные из олигоцена (Северной Америки), включают хороший ископаемыи материал вы- мершего рода из плейстоцена Техаса и Калифорнии.
Opistliocomidae, гоацины. Леса Амазонки и т. д. 1 вид. Страшная древесная птица,
связи которой сомнительны.
Отряд Cuculi
Musophagidae, турако. Африка, дожнее Сахары (нет на Мадагаскаре). 6 родов, 1,1 видов. (:uculidae, кукушки и др. Космополиты, включая Мадагаскар, Новую Зеландию и
Вест-Индию. Много родов, 128 видов. I l H один род не является общим для Старо- го и Нового Света. Большинство кукушек — тропические птицы, а те, которые гнездятся в более прохладных частях умеренных зон северного и южного полу- шария,— перелетные. Два вида, гнездящиеся в Новой Зеландии, совершают ми- грации на большое расстояние к северу через океан. Ископаемые кукушек от- носятся к нижнему олигоцену или верхнему эоцену (Франции).
Отряд Grues. «Большое число относящихся к отряду реликтовых семейств (! или 2 рода
в каждом), так же как и история его ископаемых, характеризуют этот отряд как древний и приходящий в упадок» (Mayr, Amadon, 1951, р. 8).
'ariamidae, кяриамы. Южная Америка (по не Вест-Индия) от Бразильского нагорья
до северо-западной Аргентины. 2 рода, по 1 виду в каждом.
*Bathornithidae, ~Ва~йогпis, Олигоцен (;еверной Америки.
-.:Негтояогп1И11дае, ~Herniosiornis, ~Procarioma. Нижний плиоцен Аргентины, хоро- шо сохранившиеся остатки.
1sophiidae, трубачи. Болыпая часть тропической Южной Америки (не Вест-Индии).
род, 3 вида.
*Сегаэ1ок1и!ае, *Griniioides. Нижний эоцен Северной Америки (Вайоминг), отдельные
части костей ног.
":Vogr<iidae, «Ео, гиз. Верхнии:эоцеп Азии (Внутренняя Монголия), кости пог и фраг- менты крыла.
tlrnidae. журавли. Старый Свет и Северная Америка (более северные формы — перелет- ные). Распространение в какой-то степени прерывистое: в Африке (но не на Мядя- гяскаре), в Евразии и т. д., включая тропическую Азию, но не на островах Су- мятра, Ява, Калимантан, Сулавеси и т. д. (серый журавль Grus grus встречался пи острове Ява в ледниковое время, вероятно, как мигрант,— Жейтоге, 1,)5la): локяльно оседлы на острове Лусон (но не на других Филиппинских островах} в Лвстралии, но не в Новой Зеландии; в (;еверной Америке на юг до Мексики толь- ко как мигранты, сэседлы в западной части Кубы и на острове 1!яйнс (но пе пя других островах Вест-Индии). Два американских нида относятся к широко распространенному роду Старого (лета Grus. 4 род», ! + «идов. Исконяемые относятся к эоцену (Европы и Северной Америки).
Aramidae, я!замы. Более теплые части Америки от южной Джорджии, Флорндь~, неко- торых острово» Вест-Индии и южной Мексики до восточной Аргентины. I соире- менный «ид (5 подвидов). Ископаемые роды известны из олигоценя и миоцепя ('.еверпой Америки (Южная Дакота и Небраска).
Еигурудк!ае, солнечные цапли. Большая часть (но пе вся) тропической К?жной и 1(епт- р;~льной Лмерики от центральной Бразилии до Карибского склоня Гватемалы (пет в !!ест-Индии). 1 вид (;3 подвида).
!1е11огп1!!эЫнс, лапчатопогие. Тропическая и частично нэжпяя часть Африки (пет ня
Мадагаскаре), Индо-Малайская область от северо-восточной Индии до Суматри и тропическая Америк» от неюжной Мексики до северо-восточной Лргентины и т. д.
(нет в Вест-Индии). В каждом из этих трех районов имеется по 1 зидемичпоиу роду и виду.
RhynocI)etidae, кягу. 11овая Каледония. ! вид, Довольно крупные, с хоропиэ рязвпты- мп крыльями, но не летаю1цие птицы.
Oticl idee. дрофипые. Материки Старого С1эета: Евразия (перелетные у северного кряя
яреала), Африка (не Мадагаскар) и Австралия (ие ! Тоиая Зеландия), пет пи ост- ровах между Лзией и Австралией. i! родов, 23 «ида. Ископаемые относят( я и эоцену (Европы).
Иа31~с)яе, пастушковые. Космополиты, вк;почая Мадагаскар, Новую Зеландии~, Вест- ll»<~rzc) и многие отдаленные острова. В более холодиой части умеренной зоны се- «ерного нолушапия — перелетные. 52 рода, l 32 вида. Из пастушко» роды Л«/- ~ и г и Ро ~ =а д а, из камышниц род Gn Пйг и 1а и из лысух род У'u li ca — космопол п- ты. встречмощиеся также и иа вьппеперечисленных островах. Из них только Calli»n ln 1э1 ulirn не достигают !1овой Зеландии (где, однако, много других пястуш-
Глава 5
ковых). Род Coturnicops распространен на обширных, но прерывающихся терри- ториях в Восточной и Юго-Восточной Африке (не на Мадагаскаре), Восточной Азии, Северной Америки, Мексики, Британской Гвианы и южной части К)жной Америки (нет в Вест-Индии). Porpkyrula имеет 1 вид в Африке южнее Сахары и на Мадагаскаре и 2 вида в Америке от Техаса, Южной Каролины и Вест-Индии до северной и восточной Аргентины. Другие роды этого семейства приурочены или к Старому или к Новому Свету, или (многие из них) к отдельным островам, или группам островов; многие островные роды — нелетающие. Ископаемые па- стушки были найдены в различных частях земного шара в отложениях эоцена (Европы, Азии и Северной Америки).
*Orthocnemidae, *Orthocnemis, *Elapkrocnemis. Нижний олигоцен Франции, только
кости ног, положение условное.
Меяоепайдае, пастушковые куропатки. Мадагаскар. 2 род», 3 вида. Связи сомнитель- ны, среднего размера с хорошо развитыми крыльями, но нелетающие птицы. Turnicidae, трехперстковые. 2 подсемейства: (1) Turnicinae, трехперстки. Африка (на
север до Испании), Мадагаскар, Южная Азия до Австралии (нет в Новой Зелан- дии). 2 рода 15 видов. (2) Pedionominae. Засушливые равнины внутренней части Австралии (нет в Новой Зеландии). 1 вид.
(Дополнительные ископаемые отряда Grues; по отношению к современным семей- ствам их место не определено.)
*Phororhacidae, *РЬогогЬасоз и др. Третичный (с олигоцена до плиоцена) период юж- ной части Южной Америки. Обильный ископаемый материал. Гигантские нелетаю- щие хищного образа жизни птицы. Заметки по этому семейству и двум следующим, связанным с ним, см. в работе Петерсона (Patterson, 1941).
Рйор1еги1ае, spsilopterus, sSmiliornis. Третичный период (олигоцен и миоцен) юж- ной части Южной Америки.
*>rontornithidae, *Brontornis и др. Третичный период (олигоцен и миоцен) ~ожной
части Южной Америки, обширный ископаемый материал.
*Opisthodacty1idae, 0pistkodactylus. Миоцен южной части Южной Америки, фраг- менты.
~Cunampaiidae, *Сипатра~а. Олигоцен западной Аргетины, положение сомнительное. Отряд Diatrymae. Возможно, родственный отряду Grues или происшедший от него
(Mayr, Amadon, 1951, р. 8).
eDiatrymidae, *Diatryma и др. Эоцен Северной Америки (Вайоминг, Нью-Мексикс,
Нью-Джерси) и, вероятно, Европы, гигантские нелетающие птицы.
*Gastornithidae, Са81огтз, *Remiornis, нижний эоцен Европы, разные кости.
Отряд Laro- Limicolae
Jacanidae, яканы. Большая часть континентальных тропиков и некоторые острова,
включая Мадагаскар и Австралию, но не Новую Зеландию, на севере в Америке- до южного Техаса и на некоторых островах Вест-Индии. 6 родов, 7 видов. Il ресно- водные птицы.
ТЬшосогЫае, зобатые бегунки. Западная и южная части Южной Америки, от Эква- дора до Огненной Земли. 2 рода, 4 вида. Связи этого семейства сомнительные. Chionididae, белые ржанки. Гнездятся на антарктических островах и, вероятно, также
в краевых частях антарктического материка. 1 вид достигает (но не гнездится там) Фолклендских островов побережья Огненной Земли и крайнего юга Аргентины.
Либо оседлы, либо спорадически совершают миграции. 1 род, 2 вида. Это самые южные неморские птицы (если их можно назвать неморскими) и единственные птицы с неперепончатыми ногами, достигающие Антарктики. По внешнему виду и движениям напоминают голубей, по голосу и клюву — ворон, по образу жиз- ни — болотных птиц, но воды боятся. Эти птицы драчливы и практически все- ядны (МугрЬу, 1936, р. 999 — 1006).
Dromadidae, рачьи ржанки. Побережье и некоторые острова Индийского океана, or
Восточной Африки до Индии и т. д. и на Мадагаскаре. 1 вид.
Burhinidae, авдотки. Африка (нет на Мадагаскаре). Южная Европа и Азия до Австра- лии, Тасмании, Новой Каледонии и т. д. (нет в Новой Зеландии) и тропики Аме- рики от южной Мексики до северной и западной частей Южной Америки, на a~i до южной части Перу, с 1 подвидом, локализованным на острове Гаити, Вест-IIH- дия. 3 рода, 9 видов. Довольно крупные наземные птицы, обитающие па открытых местах. Род Виг)ипиз с 7 видами занимает ареал семейства, за исключением Иидо- Австралийского архипелага, но в Австралии он имеется. Два других монотипи- ческих рода приурочены к Индо-Малайской и Индо-Австралийской областям соответственно. Ископаемый представитель этого рода относится к миоцену Европы.
~R h egminornithidae, *Rhegmino rnis. Нижний миоцен Флориды, известна только нога
этой птицы (Wetmore, 1951, р. 6).
1Tmu pbbs
*Presbyornithidae, ePresbyornis. Северная Америка (Юта) в эоцене, несколько костей
и фрагменты.
Haemat0p0didae, кулики-сороки. Имеют широкое, но прерывистое распространение:
морские побережья и часть внутренних вод, главным образом в прохладных зонах как на север, так и на юг от тропиков Старого Света. Достигают Новой Зе- ландии, но не Мадагаскара, и побережья большей части Нового Света, включая тропики и Вест-Индию. 1 род, 6 видов. Ископаемые остатки другого рода (tarso- metatarsus) относятся к нижнему миоцену Небраски.
Charadriidae, ржанковые. Космополиты. 5 подсемейств: (1) Charadriinae, ржанки.
Космополиты, включая Мадагаскар, Новую Зеландию и Вест-Индии~. 32 рода, 63 вида. Род Charadrius (типичные ржанки) гнездится по всему основному ареа- лу семейства, включая вышеназванные острова. Некоторые северные виды это-
Ф
го рода мигрируют на большие расстояния. Squatarola (тулес) и Pluvialis (золо- тистая ржанка и др.) гнездятся только в северных районах, но их миграции охва- тывают большую часть земного шара. Другие роды или являются северными, ми- грирующими на юг, или крайне южными (в Старом или в Новом Свете, но ни в том, ни в другом месте не встречаются, и в некоторых случаях мигрирующими в северном направлении. Разные локализованные роды встречаются в разных ме- стах в тропиках, а также в холодных районах и на островах. Ископаемые ржанок относятся к олигоцену Европы и миоцену Северной Америки. (2) Scolopacinae, бекасы. По своим миграциям космополиты, но большинство гнездится только в холодных частях северного полушария. 29 родов, 77 видов. Из 29 родов — 22 гнез- дятся только в арктических или умеренных районах северного полушария.
Исключение составляют следующие 7 родов: Scolopax, гнездящийся в северных ча- стях Старого Света на юг до гор Индо-Австралийского архипелага (до Новой Гвинеи); Catoptropkorus (перепончатопалый улит), гнездящийся в северной части Северной Америки и к югу вдоль атлантического побережья до Техаса, па Ьагам- ских островах и, возможно, также на Больших Антильских в Вест-Индии', Capel la (бекасы), гнездящийся в северных районах, а также в Южной и Восточной Африке, на Мадагаскаре, на острове Маврикий в Индийском океане и в отдель- ных частях Южной Америки (но не в Вест-Индии); Ckubbia, приуроченный к го- рам Южной Америки и Фолклендским островам; 1 род, приуроченный к неболь- шим островам, находящимся недалеко от Новой Зеландии, и 2 рода, приурочен- ные к отдаленным островам в тропиках Тихого океана. На большей части Африки, в тропических частях Азии и т. д., в Австралии и тропических частях Америки гнездящихся видов этого подсемейства нет. Ископаемые бекасов относятся к эо- цену (Европы и Северной Америки). (3) Phalaropinae, плавунчики. Гнездятся в северных районах, а зимуют на море в некоторых тропических и крайне южных районах. 3 рода и вида. (4) Recurvirostrinae, тилоклювки, ходулочники. Космо- политы (гнездовые территории часто локализованы), включая Мадагаскар, Но- вую Зеландию и Вест-Индию. 4 рода, 7 видов. Единственный вид (несколько под- видов) рода Himantopus (ходулочники) занимает ареал семейства. Recurvirostra (4 вида) гнездится местами во многих частях Евразии, в тропической и неюжной частях Африки и, возможно, на Мадагаскаре, в Австралии и Тасмании (но не в Новой Зеландии), в Северной Америке и на небольшой площади в горах к>жного Перу, северного Чили и т. д., в Южной Америке (но не в Вест-Индии). Два дру- гих рода приурочены: один — к высокогорным плато в Центральной Азии, дру- гой — к Австралии. Ископаемые этого подсемейства неизвестны до плейстоцена (Северной Америки). (5) Rostratulinae, цветные бекасы. Африка южнее Сахары, Египет, Мадагаскар, Южная Азия до линии Уоллеса (нет от острова Сулавесн до Новой Гвинеи), Австралия и Тасмания (нет в Новой Зеландии) — все это места распространения одного вида; другой род и вид находится в южной части Южной Америки (но не Вест-Индии); третий ископаемый род (почти полный отпечаток скелета), известен из эоцена Германии.
Glareolidae, тиркушки. Большая часть земного шара: Африка и Мадагаскар, Южная
Европа и Азия, Австралия. Некоторые (азиатские и австралийские) виды зимун>т на Индо-Австралийском архипелаге, но ни один вид там не гнездится. Набл~о- даются случайные залеты в Новую Зеландию. 6 родов, 16 видов.
Laridae, настоящие чайки и т. д. 4 подсемейства: (1) Stercorariinae, поморники.
Большой поморник (1 вид Stereo rarius squa, несколько подвидов) гнездится в аркти- ческих и антарктических и в соседних с ними районах; поморники рода Stercorarius (3 вида) — только на крайнем севере. И те, и другие зимуют во всех теплых мо- рях. Ископаемый поморник рода йеrcoгаriuã найден в плейстоцене Орегона.
(2) Larinae, чайки. Космополиты, включая Мадагаскар, Новую Зеландию и Вест- Индию. Несколько родов, 43 вида. Ископаемые относятся к эоцену (Европы и Северной Америки). (3) Sterninae, крачки. Космополиты, включая Мадагаскар,
256
Глава 5
Новую Зеландию, Вест-Индию; многие гнездятся в тропических районах, мно- гие — на отдаленных островах, но сравнительно мало в районах дальнего севера'.
некоторые северные представители — типично перелетные птицы (стр. 212): не- сколько родов, 39 видов. (4) Rynchopinse, водорезы. Побережья и реки троххи- ческой Африки (но не Мадагаскара); большие реки Индии, Бирмы и Индокитая; восточное побережье Северной Америки, по берегам и рекам Южной Америки и» юг до Магелланова пролива с отдельными представителями до Вест-Индии.
1 род, 3 близкородственпых вида.
.А1сйае, чистиковые. Гнездятся в холодных частях северного полушария (цесколько
видов в южном направлении достигают Нижней Калифорнии) и зимуют главным образом в прохладных водах морей умеренной зоны северного полушария. В тро- пиках (кроме случайных залетов) и в южном полушарии представители этих птиц отсутствуют. I,i родов, 22 вида. Крупный нелетающий вымерший вид — бескры- лая гагарка (*Pii~guinus impennis) гнездился на острове Фанк недалеко от Нью- фаундленда, в Исландии, на Фарерских островах, на островах Сент-Килда, ххв Оркиейских островах и странствовал или мигрировал на юг по обеим сторонач северной части Атлантического океана до Массачусетса, Британских островов и Дании и (случайно) до Флориды и северной Испании (Peters). Он существовал (; плейстоцена до исторического времени. Ископаемые других чистиковых (все в северном полушарии) относятся к эоцену (западной части Северной Америки).
Из них род ьЛХаисайа, плиоцен Калифорнии, — нелетающие птицы крупного размера с редуцированными крыльями, по этим признакам сходны с бескрылой гагарой (Miller, H oward, 1949) .
:Отряд Columbae
Pterncliclae, рябки. Африка и Мадагаскар, Южная Европа и Центральная и Юго-За- падная Азия на восток до Индии, в основном обитает в пустынях и степях. 2 ро- да, 16 видов. Ископаемые относятся к олигоцену (Европы).
('olumbidae, настоящие голуби и др. Космополиты, включая Мадагаскар, Новую:М-
ландию и Вест-Индию. Много родов, 289 видов. Б ольшипство находится в тропи- ках или в теплых районах. Некоторые северные виды — перелетные. Но Питерс в отличие от Майра и Амадона, которые не выделяют подсемейства, различает следующие подсемейства, возможно и не очень четко определепные, но в какой- то мере раскрывающие особенности распространения всего семейства: Treroninae.
приурочено к Старому Свету, в Африке и на Мадагаскаре, очень мцогочислешхо в Индо-Малайской и Австралийской областях и на островах западной части Тихого океана, с одним эидемичным родом па Новой Зеландии; Сolumbinae — поч- ти космополит, вк.',ххочая Мадагаскар и Вест-Ипдихо («u пе liosyro Зеландию); это единственное подсемейство голубей в Новом Свете, где оно представлено многочисленными родами и видами; 6оигигае, приурочено к Новой Гвинее и сосед- ним островам, l род, 6 видов, и Didunculinae приурочено к острову Самоа, t вид.
Род Columba, с множеством видов, от одного из которых произошли домашние голуби, встречается в диком виде на большей части земного шара (стр. 335, 336).
flu один другой род голубей не является общим для Старого и ) 1ового Света.
Ископаемых голубей очень мало, и они пе представляют иитереса и известны только начиная с миоцена (Европы).
*НарЬЫае, додо. Острова Индийского океана. 'Реюньон, Маврикий и Родригес. ' родо,
3 вида (по 1 виду на каждом острове). Все вымерли оолее 200 лет ~хазад. Фило- генетически эти птицы представляют собой очень крупных пелетаннцих голубей.
13 работе Хатчххххсона (Hutchinson, 1954) приводятся все известные данные об этххк птицах и дано их изображение, воспроизведенное по старым картинам.
Отряд Psi Нас~
.РяХ1нсп1ае попугаи (рис. 34). Тропические и умеренные районы хожихого ххо.хушнрх я,
включая Мадагаскар, Новую Зеландию и Вест-Индию, отсутствуют в Европе (хотя их ископаемые там найдены). В Азии распростра~хеиы не сенернее Гима.хаев lI южного Китая; на севере Америки в настоящее время только до северной М 'h- сики (несколько родов) с 1 видом, спорадически странствухохцххм до гор южной Ари- зоны, тогда как вымерший каролинский попугай в прошлом встречался на севе- ре до хожного Висконсина, Индианы, Огайо и штата западный Нью-11орк (Рeters).
К тогу попугаи несколько не достигают хожцой оконечности Африки ххн запад, от Книсны, некоторые формы попугаев достигахот Тасмаихххи. несколько ответвлений отряда существует в Новой Зеландии, очевидно, уже продолжительное «рсмя.
цескоц,ко «идов встречается в Чили и т. д. и по крайней мере ! вид достигает Огненной Земли. Много родов, 316 видов. 1Гитерс в отличие от Май ра и Лмадоил.
це призпающих выделения подсемейств, выделяет 6 подсемейств: Ыг~~ор1цде.
Новая Зеландня. ! вид. х~еМогхпае. Новая Зеландия и остров 11орфо:хк (п,х пос- ледххеи исчез). 1 род, 3 вида; Loriinae (лори). Австралххххская оолвхть (це 11ов хя
Птичь~
2о7
О
Зеландия), на всех Малых Зондских островах, вк;ххочая Ьали, а также Сулавеси.
Филиппины п острова западной части Тихого океана. t~ родов, много видов, осо- бенно многочисленны на )1овой Гвинее и т. д.', Micropsittinae. Новая Гвинея и т. д.
и (;оломоновьх острова. I род, разные виды; Kakatoeinae (какаду). Австралия (пет в Новой Зеландии) и Новая Гвинея и т. д. до Малых Зопдских островои.
их<.х<очая Бали (возможно, интродуцирован на этом острове), Сулавеси и Филип- пины, 5 родов, 17 видов; Psittacinae (настояхххие попугаи и т. д., икл<о хая ара).
Весь ареал семейства (кроме некоторых мелких островов), много родов и видов.
в которые входят попугаи всех частей земного шара, кроме Лвстралийской оо- ласти и отмеченных выше соседних островов. Следует добавить, что ни один род попугаев не является оощим для Старого и Нового Света, что почти «се ав«тра:ии1- скпс роды эндемичны или распространены только до Малых Зондскпх остро- вов, Сулавеси, Филиппин и др. и что многие роды этого семейства приурочены к неболыпим территориям в других частях земного шара. В Австралии находится оогатая фауна ххопугаев с разнообразным образом жизни: одни вххдьх обптапэт к<<к в густых тропических лесах, так и в разреженных лесах и зарослях кустарник».
,~ругие живут на открытых пространствах, питаясь семенами злаков и трав: несколько видов населяет полупустыни и гнездится среди скал или на земле. Иско- паемые остатки малочисленны и разрозненны: крыло и кости ног из верхнего оли- гоцена или миоцена Франции, которые предположительно относят к африканско- «у попугахо, и плечевая кость из среднего миоцена Северной Америки (Небраск<х).
очевидно, от вида, родственного каролинскому попугаю. Вероятно, попугаи ве- дут свое начало с более древних времен, но упомянутые скудные ископаемые пи- ч~го не говорят о их былом раснространении.
тряд 8 (гlgos
I'rotostrigidae, ~Protostrix. Эоцен Северной Америки. Фрагменты, очевидно, пред- ставляющпе 4 вида. Вероятно, эти виды более родственны настоящич совам, чем сппухам (Ketm<>re).
Str ipi<l;ie, совы. Космополиты, включая Мадагаскар, Иовую Зеландию и Вест-Индию.
э подсемейства: (1) gtriginae, типичные совы. Космополиты, вклхочая вышеназван- ные острова. 27 родов, ! 34 вида. Ни один род этого подсемейств» не является кос- моп<э.хххтом, но роды Охus (сплюшки), Вибо (филхх<хьх) и Glaucidium (сычикп) встречаются во всех основных частях земного шара, кроме Австралийской обла- сти. Лри этом род Otus достигает Мадагаскара и Вест-Индии и пересекает линию Уо;~леса, заходя на восток до Молуккских Островов и острова Биак недалеко от Новой Гвинеи; Вибо и Glaucidium не достигают Мадагаскара или Вест-Индии (зн псклхочением Е~убьх, где встречается Glaucidium) и не пересекают линии Уол- леса, хотя оба рода достигают Явы, Бали и Калимантана и т. д. В восточнои ча- < ти Северной Америки представителей рода Glaucidium нет. Etrix (пеясыть) встре- чается в Северной Африке и в умеренных широтах и тропиках Евразии до остро- «о~з (.уматра, Ява, Калимантан и Палавап; в Северной Америке на юг до Гонду- раса и в южной части Южной Америки от южной Бразилии и т. д. до Магеллано~~;~ пролива (по не в Вест-Ипдяи). Лгхо (ушастая и болотная совы) встречается в Аф- рике и на Мадагаскаре. по всей умеренной, но не тропической части (за иск.х«эчс- «<хе<х редких залетов зимой) Евразии и в Северной, Центральной и Южной Аме- рике и на некоторых островах Вест-ЕЛххдии. Болотная сова (Аsio flammcus) с несколькими подвххдаыхх встречается во всех северных частях земного шара (бо- лее или менее перелетный вид), на больших высотах в горах северной части Юж- ной Америки, в южной части южной Америки от южной Бразилии и т. д.на иг п па некоторых островах Вест-Индии, Фолклендских островах, островах Галана- гос, Гавайских, на острове ЕЕонаххе, на Каролинскпх островах. Acgoltus (сыч мох- «опогий и т. д.) встречается во всех северных частях .земного шара н в отдельных ч«стях Центральной и Южной Америки (нет в Вест-Индии). Лххсреа. (полярная «>и<э) H h'tu rllta. (ястребиная сова) с 1 видом в каждом встречавтся по всему зеи- ~~(~му шару в районах «райпего севера. Ряд видов рода Л'и~ох распространен от liocIoI)IoII и Южной 1; пп до Австралии, Тасмании и Новой Зеландии, и J «и;~ и западной части <Еадх<гаскнра. 1 вид рода Сlccaáa (нес<'олько подвпдои1 «,<ходится в Африке южххее Сахзрхх (но не на Мадагаскаре), и 4 вххдн — в тропп- ««х Лмернки от Мексххкхх до северо-вэсточпой Аргентины (нет и Ьест-1! пдпп), 11е следует заоывать, что есть еще 17 других родов сов, не упомянутых здел,, ~) ол ьшиттстпо из которых имеет оолее ограниченный и более или менее сп..иип- пой ареал. Ископаемые подсемейстхза Ыгхдхххае (вк:почая сухххеству«нцххй p<)..[ Либо) относятся по крайней мере к олххг<эце<ху (Е",вроххы), а род t< iz истре хает< я и отзожепххях мпоцена Северной Америки (Южная Дакота). (2) Ту1о<хша<., ан, «хх.
!1очти космополиты, «к„|ю ~ая BID;IaracIcap, Новую Зеландито Ii )~ест-Индию, от- сутствуl()T и» бвчыией части умеренной зоны Азии и в других холодных райоцах,
где обитают типичные совы. 2 рода. Ту1о с 10 видами занимает ареал подсемей- ства, но «се виды, кроме одного, приурочены к Старому Свету, где они широко рассеяны в тропиках и в умеренной зоне южного полушария; второй род с i »uzoii встречается в Индо-Малайской области. Десятый вид рода Tyto (Т. alba, сипуха) с множеством подвидов почти космополит, занимая почти весь ареал этого ~y();[a (кроме Новой Зеландии и, возможно, некоторых других островов), включая э1«- дагаскар, многие острова в Атлантическом океане и в западной части Тихог1) океана (до острова Самоа и островов Общества), Вест-Индию и острова Галнц«- гос. Это единственный представитель подсемейства в Америке. По распро- странению современных форм можно предполагать, что сипухп представляют со- бой птиц Старого Света, лишь недавно проникших в Америку (единстээепиьш вид), но '5 гигантских ископаемых вида из рода T<tto известны из плейст<э«е«« Вест-Индии (Гаити, Куба и Багамские острова — эз'еСтоге, 1<)51a, р. 62}. И< <1<э- паемые этого подсемейства до плейстоцена неизвестны.
Отряд Сарг1111и i@i
ЛероФЬе1Ыае, совиные лягушкороты. Австралийская область до острова Хальмахер»
и 11овой Каледонии (нет в Новой Зеландии). I род, 8 видов.
I odargidae, совиные козодои. Распространены диз ьюнктивно в Индо-Малайской и
Австралийской областях: 1 род, 9 видов в Индо-Малайской области до островов Я ва, Калимантан, Филиппин, другой род с 3 видами в Австралии, Тасмании, Новой Гвинее, на Соломоновых островах и др. (нет ни одного вида на Мн.зых Зондских островах, Сулавеси или Молуккских островах и Новой Зеландии). (.'аpvimulgidae, настоящие козодои. Почти космополиты, включая Мадагаскар, Австра- лию (но не Новую Зеландию) и Вест-Индию. На севере перелетные. 1!) родов, 67 видов. Большинство обитает в теплых странах. Род Caprimulgus (вкз«эчая восточноамерикапского козодоя и др.) почти космополит. Другие роды приуроче- пы к отдельным частям (;тарого или Нового Света. Ископаемые рода Сар~imul<~ti» относятся к плиоцену (Европы).
~ус6Ь1Ыае, исполинские козодои. Тропическая часть Америки от южной Мексики д1)
1]арагвая и т. д., на Ямайке и Гаити, Вест-Индии. t род, 5 видов.
)teatornithidae, жиряки. Распространены в северной части Южной Америки, вкли>чая
остров Тринидад, но не собственно Вест-Индию. 1 вид.
Отряд Troppo nes
Tr<rgoni<1ae, тр<эг<э<эьэ (рис. 35). В трех разобщенных тропических районах: тропич< < ««ээ
Африка (по не на Мадагаскаре) (2 рода, 3 вида); Индо-Малайская область (1 род,1 ~ видов) от Индии и 1(ейлона и т. д. до островов Ива, Калимантан и Филиппицскэ< х; тропическая ча<ть Америки (5 родов, эО видов) на север до южного Техаса и А1э«- зоны, на юг до северной Аргентины, а также па Кубе и Гаити в Вест-Индии. В на- стоящее время ни один род трогонов не встречается более чем в одном из трех ос- новных районов. Ископаемые — крыло и кости ног трогонов — обнаружены в олц- гоцепе (и верхнем .эоцене?) Франции. Troppo«, приуроченный в настоящее вреъ<я к Америке, зарегистрирован из миоцена Европы, но это родовое сходство все же сомнительно.
Отряд (:oraciae
«,rracii<1ae, сизоворонки. Ь<элее теплые части Старого Свет<э.,'1 подсемейства: (1) Lepl<>- somatinae, кирумбо. Мадагаскар и Коморские острова. ! вид; (' ) Brachyptevarii- nae, земляные ракши. Мадагаскар. 3 рода, о видов; (3) 1'oraciinae, типичные < и- зоворонки. Большая часть Старого Света, включая Мадагаскар и часть Лвстр»- лии,— австралийский вид изредка достигает Новой Зеландии, иногда пебо,.и,ши- ми стаями. Северо-умеренные (а также австралийские) виды — пере;«т<эые;
род», !! видов. Ископаемые — плечевая кость из верхнего эоцена или нижнего
олигоцена Франции.
Л lcediiiidae, зимородки. Космополиты, включая Мадагаскар и Вест-Индию. 11ере- летньте на севере. 14 родо», 87 видов, боль1пинство из которых находится в более теплых частях Старого (;вета. В Америку проникает только 2 рода, а именно: С< гуlе, обитающий в Африке (но не на Мадагаскаре) и в Южной Азии (rre достиг<э» Зондских островов), встречается в Северной, 1\ентральной и Южной Америке и <эн некоторых островах Вест-Индии; и СЫо ~ос~гц1е, приуроченный к более тепл и ч частям Америки от Техаса до северной Аргентины (нет в Вест-Индии). 11ек<эт<э1эь«э зимородки Старого Света питаются насекомыми, ящерицами и т. д., а не рыбой.
Оба американских рода (и некоторые другие из Старого Света) представляют crie- циализированных рыбоядшах птиц. Ископаемые этого семейства не известны ранее плейстоцеиа.
МегорЫае, щурки. Тропические и умеренно-теплые части Старого Света, включая Ъ[а- дагаскар и Австралию (но не 11овую Зеландию). Северные и некоторые тропиче- ские виды — перелетные (стр. 214). 7 родов, 25 видов.
lo>llotidae, момоты. Тропики и субтропики Америки от Мексики до северной Аргенти- ны (но не в Вест-Индии). 6 родов, 8 видов. Более подробно описаны в тексте. Todidae, тоди (плоскоклювы). Вест-Индия (Большие Антильские острова). ! pop 5
близкородственных видов.
L, pu Pidae, удодовые. Большая часть Старого Света, кроме Австралийской области
2 подсемейства: (1) Upupinae, типичные удоды. Африка, Мадагаскар, умеренная и тропическая зопы Евразии до Суматры; на севере — перелетные, ! вид, g под видами; (2) P11oeniculinae, древесные удоды. Африка южнее Сахары (но не Мада- гаскар). 2 рода, 6 видов.
Вцсего.„1дае, птицы-носороги. На большей части, но далеко не на все1~, тропиков Ста- рого Света: Африка южнее Сахары (но не Мадагаскар), ! вид, достигающий юго- западной Аравии, и Индо-Малайская область, на островах до Филиппин, Новод Гвинеи и Соломоновых островов (нет в Австралии и на Новой Зеландии), 12 ро дов, 45 видов. Ископаемые известны из эоцена Германии.
Отряд Colii
Coliidae, птицы-мыши Африка южнее Сахары (но не Мадагаскар). 2 рода, 6 видов. Отряд Macrochires
Aegialornithidae, +Aegi~lo«is. Верхний эоцеп или нижний олигоцен Франции, кости
ног и крыла; положение условное (Wetmore, 1951, р. 19).
Арой~ае (Micropodidae), стрижи. 2 подсемейства. (1) Apodinae, типичные стрижи. Поч- ти космополиты, включая Мадагаскар, Вест-Индию и (случайные залеты) Новую Зеландию; перелетные на севере, !6 родов, 76 видов. На большей части Австра- лии южнее тропиков нет гнездящихся видов стрижей, но там зимует 2 северо-во- сточных азиатских вида. Chaetura (иглохвостые стрижи) — встречаются в тропи- ках Африки (но не Мадагаскара), в некоторых частях Индо-Малайской области, в С.'еверной, Центральной и Южной Америке и на некоторых островах Вест- Индии1...1pus встречается в Африке и на Мадагаскаре, в умеренной и тропической Евразии до Явы, Калимантана и т. д. и в центральной части Анд в Южной Аме- рике, 13 Перу, северном Чили и т. д. Другие роды этого подсемейства приурочены к разным частям Старого или Нового Света. Ископаемые этого подсемейства отно- сятся к нижнему олигоцену или верхнему эоцену (Европы). Collocalia, салан- ганы. Гнезда этих птиц съедобны и идут на продажу. В настоящее время этот род широко распространен от Индо-Малайской области до северо-восточной части Австралии и по островам западной части Тихого океана и островам Индийского океана (до островов Маврикий, Реюньон и на Сейшельских). Ископаемые заре- гистрированы из миоцена Франции (tibiotarsus); (') Hemiprocninae, древесные, или хохлатые, стрижи, Индия и т. д. до Филиппин и Новой Гвинеи и т. д. (нет в Австралии и Новой Зеландии), 1 род, 3 вида.
Trocli ilidae, колибри. Новый Свет, включая Вест-Индию. Много родов, 3! 9 видов с боль- 1пим количеством подвидов. Большинство находится в тропиках, но многие тро- пические формы обитают в горах, некоторые на очень больших высотах, даже «ыше границы леса — в зоне парамос. Несколько видов встречается в умерен- ной зоне Северной Америки, главным образом на западе (1 вид достигает ого-восточной Аляски и 1 вид гнездится в восточной части Северной Америки, достигая па севере южной Канады,— северные формы мигрируют). К югу ряд «идов достигает отдельных частей Аргентины и Чили и по крайней мере l вид — Магелланова пролива.
Отряд Pici
Hucconidae, ленивки. Тропическая часть Америки от южной Мексики до северно@
Аргентины (нет в Вест-Индии). 10 родов, 32 вида.
(~albulidae, якамары. Тропическая часть Америки от южной Мексики до северо-восточ- ной Аргентины (нет в Вест-Индии). 5 родов, 14 видов.
Сapitonidae, бородатки. 3 отдельных преимущественно тропических района: Африка
от южной части Сахары и нижнего Нила к югу (но не на Мадагаскаре), Южная Азия до островов Ява, Бали, Калимантан и Филиппины и тропики Америки рт Коста-Рики до Бразилии и т. д. (но не в Вест-Индии). Разные роды, 76 видов.
Более многочисленны в Старом Свете. Ни один род этого семейства не встречает- ся более чем в одном из трех основных районов его распространения. Ревизия этого семейства была недавно сделана Райпли (Ripley, 1945).
Picidae,:tÿòëû и т. д. На оольшей части умеренной и тропической зон земпого шара,
кроме Мадагаскара и Австралийской области (до островов Сулавеси и Флорес, но не далее на восток), включая Вест-Индию. 3 подсемейства: (1) Jynginae, вер- тишейкн. Гнездятся в тропической и южной частях Африки и по всей умеренной
1 Гнездятся и на юге Азиатской части СССР от реки Васьн~ган до Курильских
островов, а также и C.åâåðo-Восточном Китае, Корее, Японии.— Прим ред.
260
Глава б
зоне Евразии и т. д. Некоторые северные формы мцгрируl<)T на юг н тропики Африки, Индии и т. д. 1 род, 2 вида. (2) Picumninae, карликовые дятлы. 4 рода: однц род с 1 видом в Западной Африке, другой род с 2 видами приурочен к Индо- Яалайской области от северной Индии и Гималаев до западной части Явы и 1ia- лимантана; Ргсипгггиз с. 1 видом (5 подвидами) распространен в Юго-Восточной Азии от Индии и т. д. до островов Суматра, Калимантан и на север до Цецтрнль- цого Китая и около 25 видов в тропиках Америки от Гондураса до северной Арген- тины (но не в Вест-Индии) и еще 1 род с 1 видом приурочен к острову Гаити в Вест- Индии. (3) Picinae, дятлы. Зацимахот весь ареал семейства. 33 рода, 179 видов, Ни один из родов ххе зншхмает всего ареала. De«d<vcopos (Drpobates, включая пушистоперогои волоснтогодятлов и т. д.) — самый широко распрострнххеххньхй род: на большой части Африки (но не в хожххой и, конечно, це на Мндага< каре), по всей Евразии ц в Индо-Малайской области до Малых Зондских островов (до острова Флорес ц т. д.), Сулавеси и Филиппин; в Северной и Центральной Aite- рике на хог до западной Панамы и Багамских островов (цо не в основной чнстц Вест- Ицдни) и на изолированной территории в южной части Южной Америки от нгго- восточпой Ьргхзххлххи и т. д. еще дальше на хог. Вус (Ъ oous, 19+7) считнет, что этот род распространился из Восточной Азии. D>.yocopus (включая шлемовпдцогодят- ла) встречается по всем умеренным широтам Евразии и по всей Индо-Мгг,хаххскохх области до островов Бали, Калимантан и Филиппин; в Северной, Центрнльххой гх 10жнохх Америке до северной Аргентины (цо не в Вест-Индии). Ptcoid< s (трехпа- ;цхе дятлы) — циркумполярцый род. Campepkilus (белоклховые дятлы) — локн- дизован в юго-восточной части Соединенных Штатов; на Кубе, в Сьерра-Мадр~ и Мексике и на крайнем юге Южной Америки, в лесном районе у Магелланова пролива. Кроме первых трех родов, перечисленных выше, все роды этого подсе- мейства приурочены к отдельным частям Старого или Нового (;вета, а болыцин- ство пз них еихе более локализовано. 11нпример, почти все роды Африки ц Ицдо- Малайской области — разные. Ископаемые дятлов (кости цог) известны цз верх- него олигоцена (Франции).
Кamphzstidae, туканы. Тропики Америки от южной Мексики до северной Лргенти- ны (но не в Вест-Индии). 5 родов, 37 видов.
1ndicatoridae, медоуказчики. Африка южххее Сахары (но не Мадагаскар) и в отдельньхх
частях Индо-Малайской области. 4 рода, 12 видов. Зто семейство no1HocTbIO афрхх- канское, но род liidicalor ичеет в своем составе е1це вид, локализованный в Яц- дии, Таиланде, и ви;{ на полуострове Малакка, на Гу~~атре п Балима цтане. Отряд Passeres, воробьиные птицы (см. текст стр. 337 и далее). Историхо этого отряда,
несмотря на то что имеется целый ряд ископаемых форм, раскрыть трудно. Ни одно ископаемое н» известно ранее эоцена, а большинство из более древни; сомнительны по своим связям. Поэтому очень немногие даже более поздние иско- паемые имеют важное географическое значение. По этим причинам искоххаеиххе в отдельных семействах отряда воробьиных пе упоминаются.
(Suboscines, ложно-певчие воробьиные)
Иодотряд Е uryl a imi
ЕххгуЬххпЫае, рогоклювы. Тропики Африки (цо ххе Мадагаскар) ( рода, г вхх,хн) ц 11,х
до-Малаиская область (6 других родов, 10 видов) от Гимнлнев (цо не от ххолуост- рова Индостан) до островов Явн. Калимантан и некоторых Филххппшх< кххх.
И одотряд Туга ап i
(Надсемейство Furnarioidea). Группа, состояхцня примерно из 500 хгххдохг, полностьн>
приуроченная к Южххой и Центральной Америке, на север только го и>'кххохх части Мексики и совершенно не представленная в Вест-Индии.
E1hiuocryptidae топоколо. Южная и чнсть Центральной Америки (нет в Вест-Иххдхххх)
на север до Коста-Рики и на юг до Огненной Земли. l I родов, 26 «идов. Еэ ольпдтн- ство обитает в горах тропической или умеренной зонь. Южной Америки.
Conopophagidae, гусенццееды. Тропики Южной Америки (но це Цецтрнльцня Ацерххка
и не Вест-Индия) ца хог до северо-восточной Аргентцны. 2 рода, 10 вцдов.
Formicariidae, муравьеловкп. Южная и Цеххтральххая Америка. Несколько род<>в
достигахот хожной Мексики (цо це Вест-Индххц). 1 ггзлххчих хе роды достххгнгот cexrep- Hoii Аргентины, но пп один род ие проникает дальи( и» аког. Много родов, «2 [ вид Furnariidae. 2 подсемейства: (1) Furn;<riinae, ххечххххкхх, хг<.я К)жххахх ц Цеххтрнльцггя Аме- рика; несколько родов хостххгангт хожной Мексики (цо не Вест-Ихх;ххххх) и «е< колько достигают Огххенххой Зеилхх и т. д.; несколько снмых хожнххх форм совершггх<>т небольшие миграции цн север (11ellinayr, 1932, р. 25) или сххускнхоття с х.ххгздохгых участков в Лидах в соседние <хо<хих1ьх нгг зимовку. Ми<гго рггххохг, 2! 2 хгхх;хов (Тнх; называемый североамериканский печник относится к древесппц»м и не «вдяется членом этого семейства); (2) Dendrocolaptinae, <хреволазьх. Южц»я и Цеххтра,хихня Лмерцка (нет в Вест-Индии), несколько родов достигнет хожной Мексххх'хх, но нн
1бэт~ ивы
261
один не заходит к хогу от центральной Аргентшхы. 1:! родов, 47 видов. [На згом настоящий список, составлеххньххх по незакоххчеххххому «Сххиску птиц Мира» Д;к..!.
Питерса, подходит к концу. Отсутствие полного, 11оследовательного и достовер- ного источника неизбежно влечет за собой в какой-то степени менее точное и ме- нее детальное рассмотрение распространения оставшихся семейств птиц. Сущест- «уют другие современные списки птиц, составленные для ооль;нинства частей земного шара (см. бххб хххографихо в коххххе глхххххх), и<э сопоставлять одни списки с другнми не так-то просто.]
(!!аххсемеххство Tyrannoidea). Главным обр хзом американская группа, по с ххес.х;ольки- мхх хорошо дифференцированными представителями, широко рассеяххххыихх в Ста- ром Свете. Семейства Старого Света, возможно, не имеют прямого родства i' ка- кими-либо определенными американскими семействами. 1!аххр и Амадон (Ма- yr, Amadon, 1951, р. 10) счххтахот, что 3 сеие!1ства Старого (.'вета, относимые к зтохх 1руппе, хотя и не близкородственны между co6o~i, все же, вероятнее всего, стоят ближе друг к другу, чем к американским формам.
Pit1idae, питты. Тропики Старого Света. 2 близкородственных вида находятся и Аф- рике (но не на Мадагаскаре); остальные встречаются от тропиков Азихх (хххх ~ с-зер :хо хожного Китая) па хог через Индо-Австралийский архххпелаг до Новой Гххшхехх, Соломоновых островов и (',еверо-Восточной Австралии (ххо ие до Новой Зел;хпдхххх).
род, 23 вида.
Р)iilepittidne, мадагаскарские питты. Мадагаскар. 2 рода, -~~ вида.
Xenicidae (Acanthisittidae), новозеландские крапивники. Новая Зе хаххдххя.,'5 рода,
4 вида. Внешне ххапомххнахот гусеницеедов (Conopophagidae) тропической Южной Лмерххки, но Майр и Амадоп (Мауг, Amadon, 1951, р. 12) счххтахот, что зто всего лишь параллелизм. h. этому семейству относится вымершая Тra гег8са lpga i- (i — единственная, очевидно, нелетающая птица из отряда воробьиных.
Tyrannidае, тнранны, или амерххкххххскххе мухоловки и т. д. 2 подсемейства: (!) Тугап- ип1ае, собственно тираш~ы. Весь Иовы11 (:вет, некоторые виды встречаются на се- ххере (летом) до центральной Аляски и т. д., а другие — на юге до Огненной Зем- ли (северные и некоторые самые хонхньхе формы мигрируют), многие достигают Вест-Индии. В Старом Свете нет ни одного вида. Много родов, 365 видов; (2) Oxy- runcinae, остроклювы. Тропическая Америка от Бразилии до Коста-Рики (нет в Вест-Индии). ! вид.
Pipridae, манакины. Южная и Центральная Америка, несколько родов, встречающихся
на севере до южной Мексики (но не в Вест-Индии), и несколько родов на хоге до северо-восточной Аргентины. Разные роды, 59 видов.
Cotingidae, котинги. Южная и Центральная Америка. Несколько родов распростране- но во внутренних районах Мексики и 1 зарегистрирован из южных окраинных частей Соединенных Штатов (южная Аризона). ! зндемичный вид на Ямаш'» является единственным представителем этих птиц в Вест-Индии. К югу несколь- ко родов достигают северной Аргентины; много родов, 90 видов.
Pl>ytotomidae, траворезы. Южная часть Южной Америки от Перу, Парагвая и т. д.
до южно-центрального Чили и Аргентины. I р~оo~д~, 3 вида.
(Oscines, певчие птицы) см. текст, стр. 238 и след.
П од о т р я д М е n u r а е, примитивные певчие птицы
~fenuridae, птицы-лиры. Юго-Восточная Австралия, от южного Квинсленда до Викто- рии (нет в Новой Зеландии). ! род, 2 вида.
Atrichornithidae, мелкие кустарниковые птицы. Австралия (но не Новая Зеландия).
! род, 2 вида', один — в северной части Нового Южиого Уэльса и южной части Квинслепда, другой (который, возможно, исчез) — в Юго-Восточной Австралии. И од отряд О seine s s. s., настоящие певчие птицы
Ylaudidae, нхаворонки. Большинсхво находится в Африке (1 вид на Мадагаскаре)
и в умеренной зоне Евразии, несколько видов — от Индо-Малайской области;~о Австралии (в Новой Зеландии местных видов нет) и 1 вид достигает Северной Шхерикхх. Этот вид — рогатый жаворонок (Otocoris alpestris) с многочисленххыьххх подвидами, встречается по всей умеренной зоне районов северного полушария как Старого, так и Нового Света и на юге Северной Америки до южной Мексики с 1 изолированным подвидом на болыпих высотах в горах Колумбии (северххая часть Юнсххой Америки). В Новом Свете других видов жаворонков нет. Совсх и не представлены жаворонки в Центральной Америке, на большей хастхх Ю'кихой Америки или в Вест-Индии. Некоторые северные формы более или менее ххере- летпые. Множество родов, 75 видов.
"'Pn)aeospizidae, Palae0spiza. Верхшп~ миоцен Ьолорадо, исклю пастельно хорошо
сохранившиеся ископаемые остатки воробьиных.
lirundinidae, ласточки. Космополиты, включая Мадагаскар и Вест-Индшо, и 1 вид
(Eiylochetidos nigrica><s) встречается в !Ховой Зеландии, куда он зн;хетает случайно
262
Глава 5
из Австралии небольшими стайками (однажды был отмечен на гнездовте). ' под- семейства: (1) ICirundininae. Занимает ареал семейства; северные формы, а также н какой-то степени и некоторые самые южные формы, перелетные птицы. Рял родов, 74 вида. По классификации Майра и Бонда (Mayr, Bond, 19~3) выделяется около 8 родов ласточек в Старом Свете и 6 «естественных групп», которые могут быть приравнены к родам, в Новом Свете; 3 рода или подрода, которые общи;~. ~я (;тарого и Нового Света,— «очевидные» иммигранты из Старого Света в Америку.
Большинство видов относятся к родам Hirundo, Riparia (деревенские и береговые ласточки), и их наибольшая площадь распространения находится в Старом (ле- те, но в каждом роде 1 евразийский вид распространен до Северной Америки или представлен там, причем североамериканские формы зимуют в Южной Аме- рике. Отдельные особи деревенских ласточек проникают далеко на юг — в рай()- ны Чили. Petrochelidon, скалистые ласточки (этот род мало отличается от роди Hi rundo). Также лучше всего представлен в Старом Свете и имеет l вид в Северной Америке, зимующий в Южной Америке. Кроме того, разные виды и подвиды этого рода ведут оседлый образ жизни в разных местах в тропической части Южной Америки. В Старом Свете наибольшей численности ласточки достигают в Афри- ке, где представлены все роды Старого Света, кроме одного (монотипический род, находящийся в Австралии), и большинство видов. Эти птицы более многочислен- ны в Южной Америке, чем в Северной, но отдельные виды встречаются на севере до Аляски и т. д. (летом) и на юге до Огненной Земли. Некоторые ласточки раз- ных южных континентов сходны и, возможно (возможно, и нет), имеют прямые связи. Если они родственны, то можно предполагать, что их предки пересекали океан, скажем, из Африки к Южной Америке, так же как иногда его пересекают и в настоящее время ласточки одного вида, направляясь из Австра- лии в Новую Зеландию; (2) Pseudochelidoninae, африканская речная ласточка.
Западная Африка (но не Мадагаскар), 1 вид.
(Бюль-бюли и птицы, им близкородственные), группировка исключительно Старого
Света.
Pycnonotidae, бюль-бюли. Вся Африка, Мадагаскар и т. д., Южная Азия на север до
Японии и Кореи и т. д. (перелетные) и острова на восток до Филиппинских и Мо- луккских (но не до Новой Гвинеи или Австралии). 13 родов, 109 видов. Описа- ние распространения родов этого семейства дается Делакуром (Ое1асоиг, '1943, р. 18).
Iren.i(lac, синие птицы и т. д. Приурочены к Индо-Малайской области. Широко распро- странены и обычны, особенно па Малайском архипелаге (Delacour, 1947). 4 рода, 14 видов.
Campephagidae, личинкоеды и т. д. Тропические и южные районы Старого Света (1 пе- релетный вид гнездится на севере до реки Амур), включая Мадагаскар, Австра- лию, Тасманию, острова Индийского и западной части Тихого океанов; нет в Новой Зеландии, кроме случайных залетов 1 австралийского вида, который был зарегистрирован по крайней мере 4 раза. Множество родов, 72 вида.
(«Примитивные насекомоядные»)
Muscicapidae. Согласно Майру и Амадону (Mayr, Amadon, 1951, р. 17 — 20, 36 — 7),
это большое космополитическое семейство, в которое входит ряд самых известных американских птиц, насчитывающее 1460 видов, то есть более чем ~/s всех совре- менных певчих птиц. Но некоторые орнитологи сомневаются в естественности этого семейства. Эта группировка, в целом самая многочисленная в Старом Све- те, особенно богата и разнообразна в Индо-Малайской и Австралийской обла- стях, но такие птицы, как пересмешники и некоторые другие, являются типич- но американскими, а крапивники — главным образом американскими. Некото- рые другие группы также представлены в Америке. Майр и Амадон подразделя- ют это семейство на 8 подсемейств: (1) Muscicapinae, типичные мухоловки (Старо- го Света). Весь Старый Свет, включая Мадагаскар, Новую Зеландию и острова.
включая даже Гавайи. В Новом Свете (если не считать Гавайи Новым Светом) нет ни одного вида. Северные формы — перелетные. Много родов, 378 видов: (2) Timaliinae, тималии и др. Теплые части Старого Света, включая Мадагаскар и Австралию (но не Новую Зеландию), с несколькими представителями в умерен- ных широтах Евразии (1 на севере до Уссурийского края) и 1 вид в западной ча- сти Северной Америки. Большое число родов, 282 вида. Наиболее многочислен- ны и разнообразны в Индо-Малайской области. Американский вид этого семей- ства — СЬатаеа, встречается вдоль побережья Тихого океана от Орегона до севе- ро-западной части Нижней Калифорнии. Делакур (Delacour, 1946), а также Майр и Амадон считают эту птицу близкородственной одному роду из Китая, но Уит- мор (Wetmore, 1951, р. 11) считает это предположение сомнительным. Подроб- ньш обзор подсемейства был сделан Делакуром (Delacour, 1946); (3) Sylviinaet,
268
н,ктоящие славки и т. д. 1[очти космополиты. большинство этих нтпц находпт- < я в Старом (.'вете. Достигают Мадагаскара II многих островов западной части Тихого океана (но я думаю, что Новой Зеландии они не достигают, см. следую- т~ее подсемейство); в Америке достигают (только Poliopttla) Ьагамских островов и Кубьг и т. д. (но пе большей части Вест-Индии). Много родов, 313 видов. Неко- торые северные формы — перелетные. Главная триба (ветвь) Sylviini приурочена к Старому Свету, кроме корольков (Леди!из) (по Унтмору, Regulidae — семей- ство, широко распространенное от умеренных широт Евразии до (;еверной Аме- рики, достигает на юге Гватемалы, но не Вест-Индии) и пеночек (Е'ЕсцЕЕозсориз).
гнездовой ареал которых доходит до западной части Аляски (но зимуют они в Юго- Восточной Азии). Вторая небольшая триба славок (если они действительно от- носятся к Sylviinae). Polioptilini приурочена к Америке. К ней относятся комаро- лоны (Polioptila) с очень небольшим числом видов, широко распространенные и умеренной и тропической зонах Америки (мигриру|от на севере), включая Кубу ir Багамские острова и, вероятно, также аберрантные тропические роды Амери- ки ЛатрЕсосапгиз и Mtcrobatr s. Однако Рэнд и Тейлор (Rand, Taylor, !953) считагот, что последние 2 рода представляют отдельную ветвь с африканскими связями. (4) KIalurinae (Acanthisidae), австралийские славки. Австралия, Новая Гвинея, 11олинезия и Новая Зеландия (Mayr, Amadon, р. 19). Около 25 родов, 85 видов; (5) Turdinae, дрозды и др. Почти космополиты (более или менее перелет- ные на севере), включая Мадагаскар, Вест-Индию, многие острова в западной части Тихого океана, Гавайские острова и остров Тристан-да-Кунья в южной 'I1cTH Атлантического океана, нет в Новой Зеландии (новозеландский «дрозд» Гц гггадra, вероятно, является аберрантным представителемподсемейства Muscicapi- nae). Большое число родов, 304 вида. Род Turdus, к которому относится, например.
американский странствующий дрозд, распространен почти по всему ареалу се- мейства, кроме нескольких отдаленных островов и фактически Австралийского материка (один представитель этого рода достигает Новой Зеландии и островов западной части Тихого океана и даже Новой Каледонии, Норфолка и острова Лорд-Хау). 11едавио обзор этого подсемейства был дан Райпли (Ripley, 195' ); (6) Afiminae, пересмешники. Ьольшая часть Нового Света (перелетные на севере).
включая Вест-Индию. Разные роды, 30 видов; (7) Troglodytinae, крапивниьн.
! лавным образом Новый Свет, особенно тропики, включая ряд островов Вест- Иидии; некоторые проникают в более холодные части Северной Америки, самые северные формы — перелетные. Типичный крапивник широко распространенно- го американского рода Troglodytes встречается в умеренной зоне Евразии. Это единственный крапивник Старого Света (Мауг, 1946, р. 7). Разные роды, 63 вида.
Ерапивник Евразии, как уже говорилось, относится, вероятно, к тому же виду, что и североамериканский обыкновенный крапивник. Распространен на запад до !1сландии, на юг до (:еверной Африки, а также на острове Тайвань (в высокогор- ных районах). Этот вид является перелетным только в самых северных частях сво- его ареала; (8) Сinclinae, оляпки. Приурочены к горным районам Евразии.
(:еверной Африки, к острову Тайвань, к западной части (не восточной) ('.еверной Америки, Центральной и Южной Америки (к Америке от Аляски до северо-западной Аргентины), но не к Вест-Индии. ! род, 5 видов.
Ргunvllidae, завирушки. Умеренные широты Евразии, Северная Африка, остров Taii- вань (в горах), и ! вид имеет изолированную колонию в юго-западной Аравии в тропическом поясе на высоте примерно выше 6000 футов (Meinertzhagen, 1954.
р. 271, 272). Это только кочующие птицы. ! род, 12 видов. Райпли (Ripley, 1952.
р. 4) считает этих птиц «чеканообразными дроздами», которые вторично стали «овсянкообразными», и предлагает отнести их к подсемейству Turdinae.
М окрасi 1lidae, трясогузки, коньки. Почти космополиты, включая Мадагаскар п
ffosym Зеландию. Малочисленны в Америке, и ни одного вида нет в Вест-Индии.
Разные роды, 48 видов. Северные формы — перелетные. Америки достигают только представители евразийского рода Мо1асИа., гнездовой ареал которого простирается до западной Аляски, но зимуют эти птицы в тропиках Азии (второй вид этого рода, возможно, также доходит до Аляски,— Bailey, 1948, р. 284), и род Anthus (коньки), который рассеян по всей Америке (не имеет сплошного распространения) — от Арктики до Огненной Земли, а также по многим местам Старого Света.
l.aniidae, сорокопуты. 2 подсемейства: (1) Laniinae, типичные сорокопуты. Африка (но
ие Мадагаскар), умеренная и тропическая зоны Евразии и острова (нерегулярно) до Филиппин, Тимора и Новой Гвинеи (но не до Австралии), Северная Америка на юг до перешейка Теуаптепек. 3 рода, 25 видов. Перелетные на севере. Два рода приурочены к Африке. Третий род Lanius встречается по всему ареалу под- семейства, пэто единственны,й род в Америке, представленный там только 2 ви-
Глава 5
дами, вероятно образовавшимися от общей евроазиатской ветви (Alayr): (2) М«- laconotinae, кустарниковые сорокопуты. Почти вся Африка на север до Марокко и Туниса. 1 вид достигает юго-западной Аравии (но не Мадагаскара). 6 родоп, -12 вида. Действительно ли эти птицы родственны настоящим сорокопутам, не ныя<- пено.
Prionopidae, лесные сорокопуты и др. 2 подсемейства. (1) Prionopinae, лесные сороко- путы: Африка южнее Сахары (но не Мадагаскар). 3 рода, 13 впдов; (2) Pityriasi- dinae. Калимантан. 1 впд. Действительно ли эта птица родственна африканским лесным сорокопутам, не выяснено.
3 <ngidae, ванги. Мадагаскар. Около 9 родов, 11» и:<о1я- ции на Мадагаскаре).
Artamidae, артамусьт, пли ласточковие сорокопуты. Австралия (и T,<cч«пни)
до Индо-Малайской области и Новой Каледонии, островов Фиджи и др. (но не в Новой Зеландии). 1 род, 10 видов. (Майр и Амадоп относят западноа<1<рика«- ский род Pseudochelidon к настоящим ласточкам).
Bombycillidae, свиристели и др. 4 подсемейства. Уитмор (ЪЪейшоге, 1951, р. 1l) <:т«- вит под сомнение их принадлежность к одному семейству. (1) Нуросо11шае.
Часть Юго-Восточной Азии, включая Аравию и, вероятно, примыкающую к ней часть Африки (нет на Мадагаскаре). 1 вид; (2) Dulinae. Остров Гаити в Вест-Ин- дии. 1 вид; (3) Ptilogonatinae, «шелковистые мухоловки». Юго-западная часть (:ое- динеиных Штатов до западной Панамы (но не в Вест-Индии). 3 рода, 4 вида; ('<) Hombycillinae, свиристели. Гнездятся в северных частях Евразии и Северн~ш Америки; зимуют несколько южнее — в Америке до Центральной Лиерикн и изредка на Больших Антильских островах и др. I род, 3 вида.
'erthiidae, пищухи. Умеренные широты Евразии и Северной Америки, на юге (.тарого
Света до Гималаев и в Америке до высокогорья Никарагуа (но не в Вест-Индии).
I род, 6 видов, из которых только 1 вид встречается в Америке.
Sittidae, поползни. Около 8 родов и 25 видов. Широко разбросаны по всему Старому.
Свету, включая Мадагаскар и Австралию (но не Новую Зеландию), и 1 из родов, Sitta, простирается до Северной Америки, имея там 4 своих вида. Весь ареал рода Sitta (настоящие поползни) охватывает северо-умеренную часть Старого Света, Индо-Малайскую область (на восток до острова Тимор), Северную Америку к югу через высокогорья Мексики и Большие Багамские острова (но не через Вест-1Лндию). Майр и Амадон рассматривают роды и подразделяют это семейство на 3 подсемейства. Но с точки зрения географии эти подсемейства не имеют боль- шого значения, так как связи отдельных родов сомнительны, а все семейство в це- лом напоминает «корзину с обрезками».
Paridae, синицы. Широко распространены в Африке (не на Мадагаскаре), в умеренных
и тропических широтах Евразии и на островах, связанных с Филиппинами и Ма- лыми Зондскими, в Северной Америке на юг до южной Мексики и Гватемалы (но не в Вест-Индии). Несколько разных родов, 64 вида. Майр и Амадоп выделя- ют 3 подсемейства: (1) Parinae (синицы и гаички). 2 или 3 рода, 46 видов; (2) Ae- githalinae (длиннохвостые синицы и др., 3 рода, несколько видов) широко рас- пространены в пределах ареала семейства, хотя границы пх распространения не совпадают; (3) Remizinae (ремезы и желтоголовые синицы). 3 рода, несколько видов. Очевидно, имеет реликтовое распространение в отдельных районах по южным окраинным частям умеренной зоны северного полушария; Юго-Восточ- ная Азия (Лети), вероятно, Гималаи (Cephalopyrus, если он относится к этому подсемейству) и ограниченная территория в юго-западной части Соединенных Шта- тов и в северной Мексике (Аипрагиг, желтоголовые синицы). Майр m Аиадон считают, что, возможно, это полифилетическое семейство и что подсемейство Aegithalinae, вероятно, образовалось самостоятельно от тималий, а подсемейство Hemizinae — от Dicaeidae из Старого Света.
(«Нектарницы Старого Света»). Группа, состоящая примерно из 400 видов, полностью
прпуроченных E Старому Свету, но даже и там почти не заходящих в умереннум зону северного полушария.
Dicaeidae, цветососы. Индо-Малайская и Австрал11йская области (но не в Новой
Зеландии). Наиболее многочисленны в Новой Гвинее и па Филиппинах, встре- чаются на востоке и на юге до (.оломоновых островов, в Австралии и Тасмании и на север и запад до Южного Китая, Индии и Цейлона (Мауг, Amadou, 1917).
Разнообразные роды, 54 вида.
~ес1аг<п1Ыае, нектарницы. Африка, южнее Сахары, ~1Ладагаскар, Южная Азия на север
:<o южной Сирии и Центрального Китая', (перелетные) и на островах до Северо- Восточной Австралии (но не Новой Зеландии). Австралийской ооласти дости- гают, однако, только несколько видов, 5 родов, ! 04 вида. Оозор этого семейства дан Делакуром (Delacour, 1914). Майр и Амадон (Мауг, Amadon, 1951, р. 26)
отмечают, что распространение этого семейства и семейства Decalidae в значи- тельной степени исключают друг друга.
.'t1eIiphagidae (Melithrepidae), медососы. - подсемейства. (I) IIelipl>aginae. Больши»- ство в Австралийской области, включая Новую 1 »инею, достиг«~от островов Ь«ли, ('.~лавеси и многих других островов в западной части Тихого океана до 1а»ай- ских островов и ]-1овой Зеландии. Много родов, 15,) видов; (2) Promeropinae.
10жная Африка. 1 вид. Является ли зта птица родственно>1»встрал111>скпм представителям этого семейства, вопрос пока еще не решенньн'. (Мау1. Аща- (lon, !951, р. 26 — 27).
Zosteropidae, белоглазки. Африка южнее Сахары, Чадагаскар n il»;to-Ма:»1»гкая
область, на север в Восточной Азии до Северного Китая (1>ере >ет»ые), 11»о»»я и острова до Австралии и Тасмании. В Новой Зеландии представлены «встр'ипй- скпч видом, который появился там неожиданно и примерно с 1856 г. стол осе;<- ,чым. Несколько родов, 80 видов. Большинство относится к роду Zowie r<iy),~. Vire»nidae, виреоны и др. Новый Свет, от внутренних районов Канады (перелет»ые»»
севере) до северной Аргентины, включая Вест-Инди>о. 11есколько родов, в»; »» »,и «беррантные Vireolanius и Сус1агй~г, 41 вид.
Drepaniidae, гавайские цветочницы. Гавайские острова до острова Лусон. 1 «зиьк
роды, 22 вида. Возможно, образовались от американских пищух-медососов или других представителей семейства Thraupidae и др. Классический пример и;:о»и- ровапной радиации. Это семейство тщательно изучено Амадонои (Лтайоп, I J5II). Thraupidae, древесницы, танагры и др. Новый Свет, 5 подсемейств: (I) Parulinae, tvc- ные певуны, или древесницы. Новый Свет, включая Вест-Индшо, на севере Север- ной Америки до границы древесной растительности (2 вида — древесная слав- ка желтоголовая и американская славка желтобрюхая — расширили свой гиез- довой ареал до небольших участков в северо-восточной Сибири, но на зимовку возвращаются в Америку) и на юг до северной Аргентины. Северные формы— перелетные. Множество родов, 109 видов. Дальнейшие подробности см. текст; (2) Coerebinae, пищухи-медососы. Южная и Центральная Америка на север до южной Мексики и на юг до северной Аргентины, вкл~очая Вест-Индию и Багам- ские острова, иногда (случайно) встречается во Флориде. Несколько родов, 3ti ви- дов; 3) Catamblyrhynchinae. В горах Южной Америки от Колумбии до 1 оли- вки. вид; (4) Thraupinae, танагры. Новый Свет, включая Вест-Индию. Боль- шинство тропических или субтропических видов, но 3 типично перелетных вида рода Piranga (также встречается в тропиках) имеют широкий гнездовойареал в Соединенных Штатах и по южным окраинным частям Канады, 2 вида на восто- ке и l — на западе. К югу разные виды танагр достигают Аргентины, но ни один пз них не заходит далеко на юг. Много родов, 196 видов; (5) Pyrrhuloxiinae, кар- диналы. Новый Свет, большинство видов распространены в тропиках, но некото- рые широко распространены и в умеренной зоне, на север до южной Канады (пере- летные) или на юг, достигая Аргентины. Множество родов, 132 вида.
Tersinidae, ласточковые танагры. Южная Америка от Колумбии и т. д. до северо-во- сточной Аргентины (но не до Вест-Индии). 1 вид.
Fringillidae, вьюрки и др. Большое, но, возможно, искусственное (полифилетическое)
семейство, широко распространенное на большей части земного шара, но отсут- ствующее на Мадагаскаре и в Австралийской области. 2 подсемейства: (1) Еп1Ьег1- zinae, овсянки. Большинство в Новом Свете от Арктики до Огненной Земли.
многочисленны в умеренной и тропической зонах (большинство видов крайнего севера в большей или меньшей степени перелетные); но род Calcarius (подорож- ники) и род Plectropkenaz (пуночки) встречаются во всех районах крайнего севе- ра Евразии и Северной Америки; ЕтЬегыа (с близкородственными родами) при- урочен к Старому Свету и широко распространен в Евразии, Африке (но не»а Мадагаскаре) и в Индии (Hartert). Много родов, 171 вид. Maiip и Амадон не от- деляют галапагосских вьюрков (Geospizinae — по некоторым авторам) от этого подсемейства; (2) Fringillinae, зяблики, KOHOIIлянки и т. д. Большинство в Старом Свете, но ни одного вида на Мадагаскаре и в Австралийской области, 'более мало- численно в Новом Свете от Арктики до Огненной Земли. Много родов, ! 22 ви;~а.
В Старом Свете сравпительно немногие представители этого подсемейства д~н ти- гают Индо-Малайской области и только 1 вид гнездится на Суматре и Яве высоко в горах, 2 вида на острове Лусон (1 из них — подвид клеста-еловика приурочен к сосновым лесам в горах северного Лусона) и 1 вид на острове Минданао: на других Филиппинских островах, на Калимантане, Сулавеси или за прете.~'о1и этих островов не встречаются. Орнитологи еще не пришли к решению, какие роды Нового Света относятся к этому подсемейству. Хеллмейр в «Списке птиц Амери- ки» («Catalogue of Birds of the Атег1сая», pt Х1, 1938) приводит 28 родов Fri»I>il- li»ae и 'arduellinae, в то время как Маир и Лма;~он объединяют эти ' подсечей-
ства и ххск~ххочахот все роды, кроме 13 (5 из которых случайные или нитродуциро- ваны в Америке). Остающиеся 8 родов, которые Майр и Ама;хон считают ххнстоя- ~цимхх Fringillinae и которые действительно относятся к американской фауне, встречаются также и в Евразии, за исключением Hespertplzozza (родствеииьш роду Гвразии) хх Loztmit z г (ириуроченному к острову Гаити). В Америке нз этих 8 ро- дов Pizzicola (шуры), Leucosticte (горные вьюрки) и Flcazzthis (чечетххи) даже зххчой не заходят далее хохкньхх краевых частей ('.оединенххьхх Штатов. Carpodacus (че- чевичник пурпурный и мексиканский) достигает южных частей Мексикиисх;оп> нагорья. Hespertphoz~a достигает Гватемалы. Spizzus (чижи) в Америке распростра- нены от Аляски до Огненной Земли с некоторыми формами, рассеянххьхыхх высоко в горах в тропиках и с некоторыми формами на тропических низмеииостях, осо- оенно в засушливых местах. I.opia (клесты) в Америке распространены от север- ной границы древесной растительности до Гватемалы с изолированной формой в горах Гаити в Вест-Индии (ср. выше филиппинские клесты). Удивительно, что клесты Гаити являются подвидом более северного из двух американских видов— белокрылого клеста, который, как правило, не гнездится к югу от гор штатов Нью-Иорк и Новой Англии и не встречается к югу от Северной Каролины даже зи- мой. Восыхой род — Loxtmitris (чиж с Гаити) — моиотипический, приурочен к го- рам Гаити, но предположительно образовавшийся от рода Spinus в результате своих ранних миграций (подобных миграциям гаитского клеста). Как видно, харак- тер распространения этих американских вьюрковых поразительно соответствует происхождению этого подсемейства в Старом Свете, перемещению 7 их родов в Америку северными путями и распространению нескольких из них на разные расстояния к югу. Но эта сравнительно простая история распространения вьюр- ковых зависит от перемещения многих тропических родов Америки, относимых Хеллмейром к подсемейству Fringillinae (плюс Carduelinae).
1с1е idae, американские иволги, американские кассиковые и др. Новый Свет от Аляски
и т. д. до Огненной Земли, включая Вест-Индию. Множество родов, 88 видов.
Хорошо представлены в обеих зонах — тропической и умеренно~~, но более или менее перелетные на севере и в некоторых случаях кочующие на крайнем ~оге. Ploceidae, ткачики. Старый Свет, включая Мадагаскар и Австралию (но не Новухо
Зеландию); нет ни одного местного вида в Новом Свете, но есть несколько интро- дуцироваиных видов (Hellmayr, 1938, pt XI, р. 1 — 4), включая домового воробья.
который в настоящее время широко распространен в Северной Америке, а также в южной части Южной Америки и является оседлой формой па Бермудских и не- которых островах Вест-Индии и т. д. Много родов, 263 вида. Майр и Амадон выделяют 5 подсемейств, имеющих следующее распространение, хотя за точность граххххц ручаться трудно: (1) Bubalornithinae, буйволовые тххачики. Африка.
2 рода, 3 вида; (2) Passerinae, воробьи. Африка, умеренная зона Евразии и Ин- до-Малайская область. Разные роды, 35 видов; (3) Ploceinae, тххпичные ткачики.
Африка, Мадагаскар и Индо-Малайская область. Разные роды, 109 видов: (4) Estrildinae, воскоклювые ткачики и т. д. Африка, Мадагаскар, Южная Азия до Австралии и т. д. Разные роды, 107 видов; (5) Viduinae, ткачики-вдовушки.
Африка: 1 род, 9 видов.
NlutххЫае, скворцы и др. Старый Свет. 2 подсемейства: ('1) 81игихххае, скворцы. Старый
Свет, включая Мадагаскар и Австралию, но в Новой Зеландии местных предста- вителей нет (иредиолагаемые скворцы Новой Зеландии теперь помещены в се- мейство Callaeidae, см. ниже); местных представителей в Новом Свете также нет, но европейский скворец был интродуцирован в Ныо-Иорк-сити в 1890 г. и теперь широко распространен по всей Северной Америке, а хохлатая майна из Южного Еитая интродуцирована и стала оседлой на небольшой территории в северо-за- падной части Северной Америки, вокруг Ванкувера. Около 24 родов (Amadon.
1943), 101 вид, северные виды перелетные или частично перелетные; (2) Bupha- ginae, буххволовьхе скворцы. Африка южнее Сахары (ио не Мадагаскар). 1 род.
2 вида.
Ori0lidae, иволги Старого Света. Тропические и умеренные районы Старого Света па
север через большую часть Европы и умеренные хпироты Азии (иерелетиые), включая Мадагаскар (только род Tylas, если он относится к этому семейству) и Австралию (ио не Новую Зеландию). 2 или более рода, 20 видов.
Dicruridae, дронго. Более теплые части Старого Света. Африка южнее Сахары; Мадага- скар, Южная Азия и на севере в Восточной Азии до реки Амур (иерелетные), на юге и востоке через Индо-Австралийский архипелаг до Северной и Восточной Австралии (перелетные). 2 рода (один — по всему ареалу семейства, другой- монотипический, приурочен к Новой Гвинее), 20 видов, много подвидов (Vaurie, 1949). Эволюция и расселение этих птиц рассмотрены Майром и Вари (Мауг, Vaurie, 1948).
(Во)1~>пы и австралийские воро~н>образные птицы). Нет определенности, что все семей- «тва, относимые к этой группе, в действительности составляют одну группу.
(.or~ i(louie, вороны, соики, сороки и т. д. Почти космополиты, вклк~чая Ma,~àracêað,
))овую Зелаидип> (только субфоссильиые) и некоторые острова Вест-Индии.
3'.! родов, ! 00 видов. Род Соггиг (вороиы и т. д.) космополитичен: это единствеп- Ill.Ill род семейства, представленный на большой части Африки, в Австралийской области и на вышеупомянутых островах (1 эндемичный вид па Гавайских остро- вах). Но оп не достигает Южной Америки, где единственными п!эедставителями вряновых являются сойки. В Америке южной границей рода Соггиг является !1икарагуа, а у соек — северная Аргентина. Роды и связи этого семейства рас- смотрены Амадоиом (Amadon, 1944).
(;racticidae, австралийские сороки. Австралия и Новая Гвинея и некоторые небольшие
острова, включая Тасманию и Лорд-Хау (но не Новую Зеландию).:3 рода, I I ви- дов (Amadon, 195!).
('rallinidae. Австралия и Новая Гвинея (но не Новая Зеландия). 3 рода, ~ вида (Лп1а- <i on, 1 950 а) .
Сallaeidae, гуйя и др. Новая Зеландия. 3 рода, 3 вида.
Ptilonorhynchidae, беседковые птицы. Австралия, Новая Гвинея и несколько мелкик
соседних островов (но не Новая Зеландия). Различные роды, 17 видов. Много- численны на Новой Гвинее, менее распространены в восточной части Австралии п очень малочисленны в Западной Австралии.
~агadisaeidae, райские птицы. Большинство на Новой Гвинее и на мелких соседних
островах. Несколько видов на Молуккских островах и несколько в Северо- В осточной Австралии, к югу, достигая только северной части Н ового Южного Уэльса (нет в Новой Зеландии). Много родов, 43 вида. Все семейство (а также и беседковые птицы) описано Айрдейлом (Iredale, 1950).
ЛИ ТЕРА ТКРЛ
Л и а d о n D., 194;), ТЬе genera of starlings and their relationships, «Ainerican Aliis.
'~ ovitates», N о. l 247.
«п а ~l о n D.. !94+, Т11е genera of Гon idae and their relationsliips, «Лпаег~сап Мм.
X ovitates», No. ] 251.
А ш а 4~ о n D., 1948, ("ontinental drift. and bird migration, «Science», !08, 705 — 707. А т а <l о n D., 19~0, ТЬе Наща11ап hoaeyreepers..., «Bull. Ainerican Mus. Nat. Elist.»,
95., 15 1 — 262.
А m а d о n D., l950a, Australian mud nest builders, «Emu», 50, t.23 — I2'7.
А m a il о n D., l951, ... "racticidae, «American Mus. Novitates», No. 1504.
@meric'in Ornithologists Union, l93l, «Check-list of North American birds», 4th ed. Еan- caster, Ра., American Ornithologists Union. 4 arious supplements in the «Auk». А г с Ь е у G., 1941, The moa..., «Bull. Aukland Inst. and Mus.», No. 1.
~ а ~ ! е у А. М., 1948, Birds of arctic Alaska, (:olorado Мця. Nat. TIist., Рopular $ег.
~ о. 8.
В а t e s G. L., 1937, Lhirds of Asar ... [part о1 Arabia]. «Ibis» (14), 1. 786 — 830.
>~ 1 а 1' е Е. 8., 1953, Birds of Mexico, 'hicago, Ь. of Chicago Press.
В о g e r t С., 1937, The d istribution and the migration of the Long-tai led Cuckoo...,
«American Mus. Ъ ovitates», No. 933.
В о п ~1 J., 193~, The distribution and origin of the Шеям Indian avifauna, «Ргос. Ame- rican Philosophical $ос.», 73, 341 — 349.
В» ~l J., 1936, Birds of the West Indies, Philadelphia, Acad. Хat. $ci.
В о п d 3., 1948, Origin of. СЬе hird fauna of the Шеям Indies, «Wilson Bull.», 60, 207—
229.
С h a p i n J. Р., '.1932, 'ГЬе birds of the Belgian Сопело, pt I, «Biill. Лшегкап Мця.
~at. Hist.», 65.
С h а р m а n, F. М., 19l7, ТЬе distribution of bird-life in,olonibi' «]ahull. Ameri- can Mus. Ха1. IIist.», 36.
С ~ a p m а n F. М., 1926, The distribution of bird-life in Eucacior, «Bull. American
.ч'ия. Nat. Iist.», 55.
268
Глава д
D е 1 а с о и r J., 1943, А revision of the genera and species of the fainily Pychonotidae
(bulbuls), «Zoologica» (Меъ York), 28, 17 — 28.
D е 1 а с о u r J., 1943а, А revision of the subfamily Estrildinae of the family Plncei- dae, «Zoologica» (New York), 28, 69 — 86.
D е 1 а с о и г J., 1944, А revision of the family Хectariniidae (sunbirds), <Zoolopica»
(Меж York), 29, 17 — 38.
D е 1 а с о u r J., 1946, Les timali in6s, «Ь Oiseau», 16, 7 — 36.
D е 1 а с о u r J., 1947, Birds of Ма1ауна, New York.
D е 1 а с о u r J., 1951, The pheasants of the world, London — New York.
D e l а с о u r J., M а у r Е., 1945, The family Anatidae, «Ш1яов Bull», 57. 3 — 55. D e l а с о u r J., 1946, Birds of the Philippines, New York.
F а l l а R. А., 1953, The Australian element in the avifauna of Хеж Zealand. «Emu»,
53, 36 — 46.
F е 11 H. В., 1947, The migration of the New Zealand Bronze Cuckoo.... «Тг. «nd Proc.
R. Soc. Хеъ Zealand», 76, 504 — 515.
G r i s с о m Ь., 1942, Origin and relationships of the faunal areas of (:entral Лшегн'a,
«Ргос. Eighth American Sci. Congress», 3 (Biol. Sci.), 425 — 430.
G r i s с о m Ь., 1945, Modern bird study, Cambridge, Mass., Нагъ агй U. Press.
G r i s с о m Ь., 1950, Distribution and origin of the birds of Mexico, «Bull. Mus. (;отр.
Z ool.», 103, 341 — 382.
Н а r t e r t Е., 1910 — 1922, Die Vogel der pal5arktischen Fauna, Berlin, Friedlander,
3 чо1я. Supplements 1932 — 1938.
Н е 11 m а у r (different parts actually by C. B. Сагу; С. Е. Hellmayr: Hellmayr and
В. Conover), 1918 — 1949, Catalogue of birds of the Americas, «Zool. Ser. Field
Мия. Nat. Hist.», 13, in 11 parts, вате of the parts in 2 or 3 numbers-actually ! 5
Ь ook- sizeditems.
Н е 11 m а у r С. Е., 1932, The birds of Chile, «Zool. Ser. Field Мця. ~аС. БЫ.», 1,). Н е n r y G. М., 1955, А guide to the birds of Ceylon, London, Oxford U. Pres.
Н и t c h i n s о n G. Е., 1954, The Dodo and the Solitaire [etc.], «American Scientist»,
300 — 305.
! r e d а ! е T., 1950, Birds of paradise and bower birds, Melbourne.
!, а с k Q., 1947, darwin's finches, London, Cambridge U. Press. [!!~бетси рУ«'кий
перевод: Д. Л э к, Дарвиновы вьюрки, ИЛ, Москва, 1949.] Ь а m b r е с h С К., 1933, «Handbuch der Palaeornithologie», Berlin.
Ь o n n b e r g Е., 1927, Some speculations on the origin of the North American orni- thic fauna, «Kungl. Svenska Vetenskapsacademiens Handlingar» (3), 4, Хо. 6. М с D о w е 11 $., 1948, The bony palate of birds, рС 1. The Palaeognathae, «А~Й»,
65, 520 — 549.
.1! а r i e n D., 1950, ~~ойея on some Asiatic МегорЫае, «J. Bombay Rat. НЫ. Soc.»,
49, 151 — 164.
Х! а у r Е., 1933, Die Vogelwelt Polynesiens, «Mitteilungen Zool. Мця. Berlin»,
306 — 323.
1! а у r E., !940, The origin and history of the bird fauna of Polynesia, «Proc. Sixth
Pacific Sci. Congress», 4, 197 — 216.
М а у r Е., 1944, Timor and the colonization of Australia by birds, «Emu», ~4, 11:~ — 1ЗО.
1! а у r Е., 1945, Birds of the Southwest Pacific, New York.
.1! а у r Е., 19~6, Eiistory of the ХогФЬ American bird fauna, «9 ilson Bull», 58,
1 — 41.
.1! я у r Е., 1946а, The number of species of birds, «Auk», 63, 64 — 69.
М а у r Е., 1953, On the origin of bird migration in the Pacific, «Ргос. ~event)i 1'aci- fic Sci. Congress», 4, 387 — 394.
1 '~ у r Е., А m а d о n D., 1947, А review of tlute Dicaeida», «Ainerica» .vtus. 3ovi- t ates», Х о. 1360.
~7цтерпт ура
269
М н у г Е., А m a d о п 1)., 3951, А classification of Recent birds, «American Мц.,
~ ovitates», Xo. 1496.
М и у г E., В n n d 3., 1943, Ъ otes on the generic classification of the swallows, Hi- r»ndinidae, «Ibis», 85, 334 — 341.
М а у г E., М е i s e W., 1930, Theoretisches zur G esch ichte des Ar ogelzuges, «D er
Vogelzuo» (I3erlin), 1, 149 — 172.
М а у r Е., S e r v e n t y D. Ь., 1944, The number of Australian bird species, «Ешц»,
<~ 4~, 33 — 40.
М а у r Е., V а u r i е (:., ! 948, Evolution in the family Dicruridae. «Kvolutior»>,
2. 238 — 265.
М е i n e r t г h а g е и В., 1954, Birds of Arabia, Edinburgh апд London.
М i 1 ] е r L., И о w а r d Н., 194!~, Т1~е flightless Pliocene bird МапсаПа, «С carnegie
Inst. Washington Pub.», No. 584, 201 — 228.
М о r е а u R. Е., J 951, The migration system in perspective, «Proc. Теп( International
Or n thnlog ical Congress», 245 — 248.
М» r р Ь у В. С., l936, Oceanic birds of South America, Niew York, American 5Ius.
1 й. Ilist., 2 ъ-оЬ.
О l i v e r O'. R. В., 1,~30, New Zealand birds, Wellington.
О 1 i т. е r Ж. В. В., 1955, Меж Zealand birds, nd ed., Wellington.
P а С t e r s о п В., 19~1, Л new phororhacoid bird..., «Geological Ser. Field Aliis. Xat.
49 54
Р е t e r s 3. L., 393'.l — 1Ч51, (: heck-list of birds of the world, C;ambridge, 1ass., На1-
~ >rd U. Press, 7 vols.
Р 11 i 11 i p s A. В., 1951, (.omplexities of migration: а review..., '<XVils<in 11ull.»,
63, 129 — 136.
а Il (1 Л. L., ]936, The distribution and habits of М'и1а„аясаг birds, «Hull. Лшсг~сап
Мiis. ~ ~t. IIist.», 72, 143 — 499.
8 а п ~l Л. Ь., Т r а у 1 о r М. А., Jr., 1953, The systematic position of tlute ~„~епега
11аиг phocaeig us and Microbates, «Auk», 70, 334 — 337.
8 i p l е у S. D., 1945, The barbets, «Aul », 63, 542 — 563.
B i р 1 е. у S. D., 1949, Avian relicts and double invasions in peninsular Iridi» ancl (:еу- ! нп, «Evolution», 3, 150 — 159.
R i р 1 е у S. D.,t 'I!I52, The thrushes, «Posti I)a» (Yale РепЬо<1у Al «s. ~;й. I I ist„),
~ о. 13.
И i р 1 с у S. D., 1,~5~, "omments on tlute biogeogr;<pliy о1' Arabia v itli pa~tie»]ar
Ыегепсс 1о birds, «J. Bombay Nat. НЫ. Soc.», 52, 241 — 248.
8 о h e r t s Л., ]940, The birds of South Africa, London, Ч'ither2iy:,Joli.nn~~b»rg~
Сentral Хev s Agency.
R о ш с с Л. S,. 19~5, Vertebrate ра1еоп(о)ору, 2nd ес1.. Iiicago. [Имеется русский
перевод: 1 о и е р Э., Палеонтология позвоночных, ГОНТИ, Мосин», 1939.] ~ < l a t e r l . j... 1858, On the general geographical distribution of ФЬе meni2iers of
1Ье class Aves, «T. of Proc. Linnean Soc. (7ool.)», 2, 130 — 145.
е г i- c n t у I). I., W й i С t e I 1 Е1. М., 19-~8. birds of Жейегп Aiistralia...,
Реггей, Ра1егяопя Press.
1 ln р . о n G. G., 1946, Fossil peag~ui»s, «Bull. American Mus. ~I at. liist.», 87, 1 — 10(~. S p r ц ~~ t Л., Jr.. 4955, ТЬе spread of the (.attle Egret, «Smithsonian Rel>()t(», ]954~,
«,9 2 ~Г1.
Т h о ш s о п Л. L., 1926, Problems of bird-migration. London.
Т h о ш s о п Л. ? ., 1949, Bird migration. Л short account, London.
Т i с е li u r s t (;. 13., 1938, А systeinatic reviews of the genus РЬу11оясорц, I,~и<1 ю,
[)~ ~ (~сД~ Цuseum
У а и r i е С., 1949, А revision of the bird fanèlò Dicruridae, «В»11. American,цц,
3 at. Hist. », 93, 199 — 342.
2УО
Глава 5
Ъ о о и s К. 11., Jr. 1947.... history of... Dendiocopos. «Limosa» (Orgaan der ИиЬ Xederlandsche Vogelkundigen), 20, 1 — 142.
~Ъ е t m о r е А., 1925, ... Palaeospiza ..., «13ц11. ious. (;omparative Zool», 67, '18:~— 193.
Ж е t m о r е А., 1943, Fossil hirds from the Tertiary deposits ol' Florida, «Ргос. ~еъ England Zool. (;lub.», 22, 59 — 68.
~~ е С m о r е А., 1,),э1, А revised classification for the birds of the world, «Smitl>sonian Misc. (;oil.», ).17, No. 4.
'A' е t m о r е А., 195la, Recent additions Со our knowledge of prehistoric birds 1J'эЗ— '.1949, «Ргос. Tentli International Ornithological Сongress», р. 51 — 74.
'A о 1 f s о п А., 1948, Bird migration and the concept of (;ontinental drift, «Scietsce», I08, 23 — 30.
W о 1 f s о п А., 1355, Origin of the i%orth American bird I'auna. critique and reinter- pretation from the standpoint of continental drift, «American iAIidland i3 аСиг «list», 53,:3'53 — 380.
Глава 6MJIEKOIIHTAIOIgHE
Таблица 5
Число видов млекопитающих Цалеарктической области и большей частптропической Азии
273
Наземные млекопитающие, кроме за1~цеобразных и грызунов Зайцеобразные (зайцы и др.)
Грызуны
Рукокрылые
Водные (преимущественно морские) млеконитавощие
27 28f }60 (j8
Эта таблица дает вполне наглядное представление о составе фауны млекопитающих земного шара, хотя от места к месту он может значитель- но варьировать. Так, например, рукокрылых относительно больше в ти- пично тропических районах. Грызунов относительно меньше в Австра- лии и на Мадагаскаре, но в некоторых областях и на ряде ост-
Классификация млекопитающих, которой я следую в данной книге, их ископаемых и современных отрядов и семейств с почти полными спи- сками родов наиболее развернуто была дана Симпсоном (Simpson, 1945). В работе Бурлье (Bourliere, 1955) можно ознакомиться с большой серией фотографий современных млекопитающих.
Млекопитающие играют важную роль в зоогеографии. Непревзой- денные ископаемые материалы позволяют воссоздать их удивительное прошлое, хотя с точки зрения географии в этих данных еще имеются боль- шие пробелы. Не столь хорошо по сравнению с птицами изучен и видовой состав млекопитающих.
По данным Майра (Mayr, 1946), число современных видов млекопи- тающих достигает 3500. Более точный подсчет невозможен. Во многих рай- онах земного шара, в основном в тропиках, еще не все виды выявлены, а в некоторых фаунистических списках дается необоснованно большое число названии.
Большая работа по систематизации млекопитающих почти всей уме- ренной и тропической зон Евразии и Северной Африки была проделана Эллерманом и Моррисон-Скоттом (Е11егтан, Morrison-Scott, 1951). Они приводят 809 видов, что составляет около '/, всех видов млекопитающих земного шара, тогда как остальные а/, приходятся на основную часть Аф- рики с Мадагаскаром, Австралию и Индо-Австралийский архипелаг и Америку. Умножая 809 на четыре, получим 3236. Это, хотя и очень гру- бо приближенное, число близко к числу, данному Майром. Из установлен- ных родов млекопитающих Симпсон (Simpson, 1945, р. 35) выделяет около 1000 современных родов и 2000 ископаемых.
В табл. 5 представлен состав северо-умеренной и тропической фаун млекопитающих (Ellerman, Morrison-Scott, 1951).
272
Глава (i
ровов они преобладают. Виды, включенные в первую строку таблицы «Наземные млекопитающие, кроме зайцеобразных и грызунов», очень раз- нообразны и сильно варьируют в разных частях земного шара. Положение грызунов по отношению к другим млекопитающим рассматривается на стр. 288.
Основной ход эволюции млекопитающих описан Симпсоном (Simpson. '19 ы). Млекопитающие, как полагают, появились в юре, но не получили су- щественного развития до конца мезозоя, так как они подавлялись господст- вовавшими тогда рептилиями. Ископаемые остатки млекопитающих, отно- сящиеся к этому времени, чрезвычайно скудны. Примерно только к началу третичного периода млекопитающие стали доминирующими животными, и затем произошла их радиация, о чем говорят более полные и более ши- роко распространенные находки ископаемых. История географии млекопи- тающих в течение третичного периода рассматривается ниже. При анализе истории млекопитающих палеоцен обычно отделяется от эоцена, по край- ней мере американскими палеонтологами. В данной главе целесообразно поступить так же, хотя в предыдущих главах такое подразделение обычно не делалось.
Считалось, что представителями самых ранних млекопитающих бы- ли однопроходные и что от них образовались сумчатые, которые в свою очередь дали плацентарных. Теперь, однако, известно, что филогения мле- копитающих не так уж проста. На очень ранних этапах своей истории млекопитающие разделяются на два или три подкласса. Сомнительно. что однопроходные являлись самой ранней группой млекопитающих, так как о существовании их когда-либо за пределами Австралии ничего не из- вестно. Поэтому, а также по некоторым другим причинам можно полагать. что они не столь уж древние и являются, возможно, вырождающимися (хотя и морфологически специализированными) потомками примитивных сумчатых. Сумчатые и плацентарные относятся к одному древнему под- классу, но входят в него не как предшествующая и следующая за ней груп- пы, а как параллельные. существовавшие одновременно задолго до конца мелового периода. Очевидно, не было такого времени, когда на всем зем- ном шаре были широко распространены только сумчатые (без плацентар- ных).
Фнлогенегттческтт млекопитающие — это рептилии, у которых раз- вился волосяной покров, появилась теплокровность, стало совершеннее воспроизводство и в конце концов прогрессировал «интеллект». Тепло- кровность позволттла млекопитающим заселить холодные области, а более высокая организация млекопитающих явилась причиной широкого и иногда быстрого расселения многих из них по всем доступным частям земного шара.
Расселение млекопитающих ограничивается главным образом пре- градами, которые образуют соленые воды. Степень способности наземных млет'оп1ттающттх преодолевать соленые воды была предметом горячих спо- ров. которые зачастую сводились просто к утверждениям «они могут» илтт «они не могут». Распространение различных наземных млекоптттаю- щих показттвает, что соленые воды действительно являются серьезным JIp('- пятствием для нттх. но что любое млекопитающее может иногда пересечь узкое водное пространство, а некоторые — даже довольно широкие пр<>- странства открытых вод. Грызуны, по-видимому, относительно хорошо преодолевают такие преграды. Они несколько раз достигали Австралии и встречаются на некоторых островах за пределами границ распростра- нения других нелетающих наземных млекопитатощих. Установить предель-
Млекопи,тан щи,е ную ширину водных пространств, которые в отдельных случаях могут пересекать некоторые грызуны, невозможно. Предки местных хомячьпх на островах Галапагос, очевидно, пересекли более 600 миль океана. Воз- можно, как предполагали и ранее, предок дикобразоподобных грызунов Южной Америки проник из Африки через Атлантический океан, вероятно, на плоту, подобном тому, который описан в гл. 1. Нельзя утверждать, что зто было действительно так, но это могло произойти именно таким путем, если принять во внимание распространение дикобразоподобных. Руко- крылые преодолевают водные преграды значительно легче. Видимо, за исключением ряда рукокрылых (причем несколько видов летучих мышей достигли даже очень отдаленных от материка островов) и нескольких вет- вей грызунов, другие плацентарные не смогли достичь Австралии без помощи человека. Границы распространения млекопитающих
Северными границами наземных млекопитающих (рис. 38) являются границы суши, за пределами которых обитают морские млекопитающие (киты, тюлени и др.). В Америке самых северных частей суши, включая Арктический архипелаг и северное побережье Гренландии, достигают заяц- беляк, гренландский копытный лемминг, волк, песец, белый медведь (в ос- новном по морскому побережью), горностай, карибу и мускусный бык. Р и с. 38. Самые северные и самые к~жнт.те наземные м.текопи-
тан)щис . Все эти животные, кроме последнего, представлены также на крайнем се- вере Евразии теми же или родственными им видами: короткохвостый гор- ностай Америки — евразийским горностаем, а американский карибу, воз- можно, тем же родом и, возможно, тем же видом, что и северный олень Евразии. Млекопитающие северного Таймыра (самая северная точка кон- тинентальной Азии), приводимые Бергом (Берг, 1947), точно соответствуют самым северным млекопитающим Америки, кроме мускусного быка, кото- рого нет на Таймыре, и одного вида лемминга, который отсутствует в Аме-
274
рике. Несколько других видов млекопитающих, включая землероек, длин- нохвостого суслика (Citellus), а также хомячьих грызунов (кроме api'ти- ческих леммингов) и куньих (кроме горностая), распространены в отдельных районах арктической тундры, но не достигают северных границ суши. Гще болыпее число млекопитающих, включая и рукокрылых (ZVIrIc>tis). достигает северной границы леса, не проникая далеко в глубь тундры.
К югу млекопитающие многочисленны вплоть до умеренно-теплых районов Африки. Ехидна, утконос, многие сумчатые, пять видов местных грызунов и несколько рукокрылых достигают умеренно-прохладного ocI- рова Тасмания. Два вида рукокрылых имеются в Новой Зеландии. Небольшое число млекопитающих обитателей лесов и равнин, включающих в том числе несколько видов хомячьих грызунов, одного дикобразоподоб- ного грызуна (Ctenomys), лисицу, выдру, гуанако (Camelidae) и летучун> мышь, достигают умеренно-холодного острова Огненная Земля. Типично лесными млекопитающими этого острова являются только три мелких вида хомячьих и лисица (Osgood, 1943, р. 31). Еще несколько видов мле- копитающих достигают или почти достигают Магелланова пролива: к нич относятся дикобразоподобные, пума, скунс и, возможно, олень. Другие млекопитающие не доходят до этого пролива (граница их распространения находится на разных от него расстояниях). Сверх ожидания, самые древние млекопитающие Южной Америки не достигают южной оконечности мате- рика, и этот факт заслуживает внимания. Южнее примерно 47' ю. ш. (30() или 400 миль от Магелланова пролива) сумчатые неизвестны, а броне- носцы, очевидно, вообще так далеко на юг не заходят.
О распространении млекопитающих на островах можно сказать, что все пригодные для обитания материковые острова населены млекопитан>- щими. хотя нередко их здесь бывает меньше, чем на прилежащих мате- риках. На древнем острове Мадагаскар, независимо от того, материкового он происхождения или нет, имеется ограниченная фауна наземных мле- копитающих, образовавшаяся от немногочисленных предков и отличаю- щаяся небольшим числом грызунов, но рукокрылые на Мадагаскаре мно- гочисленны. Эти же животные на Маскаренских и Сейшельских островах являются единственными местными наземными млекопитающими. Австра- лии, которая является древним островом-материком, достигли только один вид однопроходных и один вид сумчатых, но там нет ни одного пла- центарного млекопитающего, кроме нескольких видов мышей и ряда руко- крылых. К востоку от Австралии и Новой Гвинеи только один род древес- ных сумчатых и несколько видов грызунов достигае"С,оломоновых остро- вов. За пределами этих островов встречаются крысы и мыши, очевидно, завезенные человеком, и рукокрылые. Рукокрылые, столь многочисленные на Соломоновых островах, в восточном направлении заметно уменьша~от- ся, и пределом их распространения являются острова Самоа. На Гавай- ских островах встречается американская летучая мышь (Lnsiurus). Остро- вов Галапагос достигают одна группа мелких американских хомячьих и летучая мышь рода 1 asiurus.
В Вест-Индии имеется или имелось большое число грызунов, две группы насекомоядных, наземные ленивцы, несколько других видов разобщенных наземных млекопитающих с не совсем ясной историей нх рас- пространения и многочисленные летучие мыши. На более мелких остро- вах Вест-Индии, включая отдельные Багамские острова, Ка11мановы ост- рова и остров Суок, кроме грызунов и рукокрылых, вероятно, никогда не было местных млекопитающих. На более отдаленных островах Атлан- тического океана наземные млекопитающие отсутствуют, опять-таки за
Млеко питая щ ие
27э
исключением рукокрылых, из которых небольшая европейская летучая мышь достигла даже Азорских островов. На Бермудских островах лету- чие мыши не обитают, но изредка появляются от случая к случаю. Как видно, на островах большинство нелетающих наземных млекопитающих имеет узкие границы распространения; грызуны в какой-то степени рас- пространены более широко, а рукокрылые почти не имеют пределов рас- пространения, хотя на очень удаленных от материка островах их чпслен- НОСТЬ НОВОЛИКИ.
Большинство семейств млекопитающих распространено не по всему земному шару. Единственными космополитическими или тропикополи- тическими семействами из наземных млекопитающих (не считая человека) являются рукокрылые: семейства Vespertilionidae, Emballonuridae и Mo- lossidae. Семейство МигЫае (мыши и крысы) можно было бы считать рас- пространенным по всему земному шару, если, как это иногда и делают, отнести к нему и семейство Cricetidae. К широко распространенным на всех континентах, кроме Австралии, относятся следующие семейства назем- ных млекопитающих: Leporidae (зайцы), Sciuridae (беличьи), Canidae (собачьи), Mustelidae (куньи) и Felidae (кошачьи). Кроме названных, одно или два семейства слонов, а также семейство Equidae (лошадиные) в прошлом, возможно, имели такое же широкое распространение. Распро- странение всех других современных семейств нелетающих наземных мле- копитающих более ограничено.
Самыми широко распространенными родами наземных млекопитаю- щих, кроме Mpotis и нескольких других родов, относящихся к семейст- вам Vespertilionidae и Molossidae (см. список семейств), а также, кроме человека и интродуцированных им животных, щвляются Lutra (выдры, которые относятся к полуводным животным) и Feli s (кошки). И те и другие широко расселены на всех континентах, за исключением Австралии. Один или два рода слонов и род Equus (лошади), возможно, в прошлом имели такое же распространение. У родов Lepus (зайцы), Mustela (ласки и хорьки) и Canis (волки и др.) очень широкая климатическая амплитуда — от са- мых высоких арктических широт (северная Гренландия) до жарких paii- онов тропиков.
Некоторые виды млекопитающих занимают широкие, с резко ра:~- личными климатическими условиями ареалы. Волк (Canis lupus) распро- странен по всему северному полушарию, достигая на севере высоких арк- тических широт (северная Гренландия) и проникая на юге в Азии (полу- остров Индостан) за пределы тропиков (на несколько градусов). Возникает вопрос — относятся ли волки, которые достигают этих противополож- ных пределов, к одному и тому же виду. Эллерман и Моррисон-Скотт отвечают на него положительно. Леопард Panthera pardus распространен по всей Африке и Южной Азии до Уссурийского края. Несколько видов рода Lepus (зайцы) имеют обширные ареалы: 1. capensis встречается во всей Африке и умеренной зоне Евразии — от Кейптауна до Испании и Китая; 1. eui'opaeus — от Южной Африки до Дании, а теперь расселяется в Азию за Урал; L. tin<idus встречается в умеренных и арктических широтах Евразии и, возможно, его граница проходит так- же через всю Северную Америку (Ellerman, AIorrison-Scott). Некоторые другие виды млекопитающих являются в той или иной степени циркум- полярными. Б Америке пума (Felis concolor) распространена от холодной северной части Британской Колумбии через тропики !центральной и Южной Америки до Магелланова пролива, встречаясь как в высоких, так и в низких широтах, как в засушливых, так и во вла'кных районах.
Глава 6
Климатическая амплитуда некоторых других млекопитающих также заслуживает внимания: Tachyglossus и Ornithorhynchus (однопроходные) встречаются от умеренно-холодной Тасмании до тропической Австралии; род Didelplus (оппосумы) — от умеренных широт Америки через американ- ские тропики; Crocidura (род землероек) — от Центральной Европы и Восточной Сибири распространен на юг через Африку и тропическую часть Индо-Малайской области; Myotis (ночницы) — от северной границы древесной растительности на юг через тропики обоих полушарий. Sciurus (белки), Rattus (крысы), Equus (лошади), Sus (настоящие свиньи), Cereus и Odocoileus (олени), некоторые роды слонов и т. д., носорогов и дикие быки — все они распространены или были распространены в прошлом от холодных до тропических зон.
Зональное распространение млекопитающих
Несмотря на то что распространение некоторых родов и видов млеко питающих не ограничивается климатическими зонами (хотя очень немно- гие группы млекопитающих являются строго зональными), климат все- таки оказывает серьезное влияние на их распределение. В тропиках отмечается наибольшее разнообразие млекопитающих, уменьшающееся умеренных зон к арктическим, и только 8 видов наземных млекопитающих достигают самых северных точек любой рассматриваемой суши (стр. 273).
Тропические фауны млекопитающих не только очень богаты, но так- ~ке и очень разнообразны по своей структуре. Ни одно семейство млеко- питающих, не считаяфрдного-двух из рукокрылых, не является пантро- пическим, то есть нет ни одного семейства, встречающегося во всех тропи- ческих областях и одновременно не заходящего в какую-либо умеренную зону. Большинство семейств наземных млекопитающих, главным образом тропических, приурочено к Старому или Новому Свету. Семейство Tapi- ridae, занимающее сейчас часть Индо-Малайской области и тропическую Америку, является не столь уж существенным исключением. Приматы в целом (не считая человека) — почти пантропические животные, но ни одно их семейство в отдельности не является таковым. Отсутствие пантро- пических семейств нелетающих наземных млекопитающих хорошо отра- ;кает изоляцию Южной Америки в третичном периоде и быстроту эволю- ции млекопитающих в течение этого времени, а также тот факт, что млекопитающие Старого Света, которые недавно достигли Южной Америки, проникли туда через умеренную зону северного полушария и существуют там до сих пор. Однако из этого следует и другой, правда не столь очевид- ный факт: не было значительной тропической фауны млекопитающих, которая накапливалась бы в южной части Северной Америки в течение третичного периода. Если же такая фауна и сформировалась, то в ее состав могли бы войти тропические семейства Старого Света, проникшие через север в более ранний и более теплый период третичного времени. В плио- цене эти семейства распространились бы через американские тропики и стали бы пантропическими. Независимо от правильности этого аргумента, доказательств, подтверждающих существование четкой самостоятельной тропической фауны млекопитающих в южной части Северной Лмерики в течение третичного периода, очень мало (стр. 311).
Северо-умеренные фауны млекопитающих не только менее богаты, чем тропические, но характеризуются и относительно меньшим числом важных эндемичных групп. Основными северо-умеренными семействами и
.Цлекопитан~щ ие
277
подсемействами наземных млекопитающих являются следующие: Sorici- nae (землеройки-бурозубки), но один род этого подсемейства распростра- нен до северной части Южной Америки и, возможно, что само подсемейство не является естественным; Talpidae (кроты), но они проникают и в тропи- ки Индо-Малайской области до полуострова Малакка; Ochotonidae (пи- щухи), но их нет на большей части Европы и в восточной части Северной Америки (кроме того, это семейство состоит только из одного современно- го рода); (astoridae (бобры) с одним современным родом и двумя видами и Microtinae (полевки и т. д.), которых по сравнению с любой другой ооль- шой группой млекопитающих можно, скорее всего, назвать типичными животными умеренной зоны северного полушария; Zapodidae (прыгун- чики), маленькое семейство, приурочено к областям умеренной зоны северного полушария, но не всегда достигает границ этих областей; Aplodontidae (бобровые белки, один вид), распространены в западной части Северной Америки; Spalacidae (слепыши) — в Юго-Восточной Евро- пе и прилежащих частях Азии и в Северной Африке; Seleviniidae (семей- ство, родственное соням, один вид) — в Казахстане; Antilocapridae (ви- лороги, один современный вид) — в западной части Северной Америки. Этот список эндемичных групп той части земного шара, которая, как пола- гали (Мэтью и др.), была основным местом эволюции млекопитающих, не дает должного представления о фауне умеренной зоны, так как большин-- ство млекопитающих умеренных широт северного полушария относится к семействам, которые также широко распространены и в тропиках, глав- ным образом в тройиках Старого Света.
Я попытался проанализировать и сравнить тропическую и северо- умеренную фауны млекопитающих, чтобы более отчетливо представить влияние климата на распространение этих фаун. Были избраны фауны Юж- ной и Восточной Азии, так как в Восточной Азии тропическая и умерен- ная фауны идут широким фронтом параллельно друг другу, не встречая существенных преград, и, таким образом, климат является главным диф- ференцирующим фактором. Сначала я подсчитал виды, приводимые Эл- лерманом и Моррисон-Скоттом (Ellerman, Morrison-Scott, 1951), и рас- положил их отдельно по семействам от тропической и умеренной зон Азии к востоку, примерно начиная от 90' в. д., и если вид оказывался главным образом по одну сторону границы тропиков, то я относил его соответствен- но к той или другой фауне. Если же это был вид, широко распространен- ный по обеим сторонам, я считал его общим для обеих фаун. Затем я до- бавлял к тропической фауне виды, приводимые для Малайского архипе- лага Чейсеном (Chasen, 1940), стараясь, когда это можно было сделать, сводить количество этих видов до стандартов Эллермана и Моррисон-Скот- та. Таким образом, получившиеся списки охватывают всю тропическую Индо-Малайскую область (кроме Филиппин) и приблизительно равную умеренную — до арктической зоны в Восточной Азии. Для предупрежде- ния серьезных ошибок я сопоставил млекопитающих Цейлона (Phillips, 1935) и Британских островов (A~lathews, 1952) как образцы тропической фауны Индо-Малайской области и умеренной фауны Евразии, и нашел, что они, так же как и целые фауны, имеют примерно сходные общие характеристики. Полученные результаты приведены в табл. 6. В целях упрощения я объединил зайцеобразных с грызунами под общим назва- нием «грызущие».
На данных этой таблицы, вероятно, сказались некоторые факторы, которые неизвестны или которые нельзя подвергнуть анализу в настоя- щее время. Так, относительное число видов умеренной зоны Восточной
278
Глава 6
Та блица
(:равнение тропической фауны млекопитающих Индо-Малайской областии умеренной фауны Восточной Азии
Наземные млеко- питающие, кроме «грызущих» и руко- крылых (в широко распространенных
семействах) 1
«Грызущие» (в широко рас- пространенных семействах) 2
Рукокрылыс(Vesperti1ionidae)
Умеренная зона Восточной
Aзии: умеренная зона Китая (кроме Синьцзяна и Тибета), Монголия, Си- бирь (к востоку от 90') и т. д., Япония Британские острова
109 (85: 78оj)) 10 з (8. 80о())
100 (83: 83о~о) 19 а (18: 95%)
43 (32: 740 ) 12 (10. 83о/о)
Индо-Малайская область
(кроме Филиппин) Цейлон .
180 (85: 47о/о) 32 (18 . 56%)
135 (123: 91О/)23 (21 . 91%)
154 (59: 38оо) 28 (12: 43%)
П р и м е ч а н и е. Эти цифры взяты из фактических подсчетов видов у келлермана и Мор- рисон-Скотта (Ellerman, Morrison-Scott, 1951) и Чейсена (Chasen, 1940), но общий итог сле- дует считать приближенным.
1 Следующие «широко распространенные» (то есть широко распространенные и в тропиче- ской и в умеренных областях) за исключением «грызущих» и рукокрылых, семейства наземных позвоночных: Erinaceidae, Soricidae, Canidae, Ursidae, Mustel idaho, Ге1 и1ае, Equidae, Suidae, Cervidae, Bovidae.
'-' Следующие широко распространенные семейства «грызущих»: I.cporidac, Sciuridac, (;ricetidae, Muridae.
З В эти цифры включены ныпе исчезнувшие на Британских островах волк, медведь, кабан и бобр.
Азии может быть завышенным в связи с тем, что умеренная зона Восточ- ной Азии не отчленена от огромной территории всего материка, в то время как тропическая часть Индо-Малайской области изолирована от других тропических территорий. Однако это не распространяется на рукокрылых, для которых Индо-Малайская область не является такой уж изолирован- ной областью, чем, вероятно, отчасти и объясняется многочисленность в ней летучих мышей. При этом надо учитывать, что Чейсен дробит виды более мелко, чем Эллерман и Моррисон-Скотт, и в отдельных случаях это могло увеличить итоговые данные тропической фауны без достаточ- ных на то оснований. Моя попытка исправить эту ошибку в отношении некоторых групп, особенно рукокрылых, успехом не увенчалась. Но на данные, приводимые для Цейлона и Британских островов, это обстоятель- ство не влияет. Эти цифры соответствуют основным итоговым данным, что свидетельствует о приближенной правильности последних.
Между тропической и умеренной фаунами, приведенными в таблице, существует два основных различия. Первое — тропическая фауна богаче и содержит почти вдвое больше видов, чем умеренная. И второе — тропи- ческая фауна разнообразнее, но состоит из широко распространенных семейств и большого числа своих семейств, а умеренная фауна представлена теми же широко распространенными семействами с небольшим числом семейств, присущих только или главным образом ей. В первой колонке таб-
Л1 л еко ~г ита и щие
лицы, которая отражает основное разнообразие наземных млекопитаю- щих, десять широко распространенных семейств составляют примерно только половину тропической фауны и более чем 4'„. умеренной фауны; из рукокрылых одно широко распространенное семейство Vespertilioni- dae составляет менее '>'5 тропической фауны, но около з,, умеренной.
Грызуны и т. д. представляют собой некоторое исключение. В умерен- ной зоне Восточной Азии они сравнительно многочисленнее, чем другие млекопитающие. Возможно, это объясняется наличием обширного прост- ранства для расселения и их эволюцией на различных частях огромной тер- ритории умеренной зоны Евразии. Или, возможно, грызуны действитель- но непропорционально многочисленны в отдельных частях умеренной зоны северного полушария в связи с тем, что они хорошо приспособились переносить северную зиму, питаясь перезимовавшими семенами северных растений. А то обстоятельство, что большинство грызунов тропической Индо-Малайской области и умеренной зоны Евразии относится к одним и тем же широко распространенным семействам, возможно, означает то, что грызуны меньше подвергаются влиянию климата, чем другие млеко- питающие, или то, что они эволюционировали и расселились совсем недавно. Доминирующие мышиные Старого Света, по-видимому, действи- тельно возникли недавно: это, очевидно, относительно поздние (поздний миоцен?) производные семейства Cricetidae (Simpson, 1945). Но в других тропических районах имеется большое число строго тропических групп грызунов.
Между тропической и умеренной фаунами млекопитающих нет рез- кой границы. Не только широко распространенные семейства, но даже и некоторые преимущественно тропические семейства (например, обезьяны, виверровые), а также многие роды и некоторые виды (тигр, леопард) про- никают на разное расстояние из тропиков Индо-Малайской области в уме- ренную зону Восточной Азии. Граница между тропической фауной млеко- питающих Африки и умеренной фауной Европы выражена более четко, но здесь уже сказывается (частично) влияние преград, образуемых пусты- нями и водными пространствами. Несколько семейств и даже родов и ви- дов (например, уже упоминавшийся род Lepus), преодолев преграды, рас- пространились из тропической Африки до Европы. В Америке большинство тропических млекопитающих (большинство сумчатых, обезьян, большин- ство неполнозубых и дикобразоподобных грызунов) не достигают север- ных частей южной Мексики, но виргинский оппосум и канадский дикобраз (представители тропических семейств Америки) распространены значи- тельно дальше на север, а броненосец проник в юго-восточную часть Сое- „циненных Штатов. Многие семейства, роды и виды широко распространен- ных семейств в Америке пересекают климатические границы.
К югу настоящей зональности в распространении наземных млеко- питающих не наблюдается, но происходит обеднение тропических фаун. Лучше всего это выражено в Южной Америке. Несколько небольших се- мейств приурочено к умеренной зоне южного полушария, но самостоятель- ной южно-умеренной фауны, свойственной всем южным континентам, нет.
Радиальное распространение млекопитающих
Примеры четкой современной географической радиации семейств сре- ди млекопитающих редки, отчасти потому, что многие млекопитающие в последнее время скорее приходили в упадок, чем радиировали. Крысы
280
Глива 6
и мыши семейства Muridae имеют сложную радиацию из основной части тропиков Старого Света в умеренную зону Евразии и Австралию, но не в Америку (кроме завезенных человеком). У семейства Bovidae наблю- дается сложная радиация из основной части Старого Света в Северную Америку. Ряд других, менее важных географических радиаций отражен современным распространением других семейств и родов. Однако самым лучшим свидетельством географической радиации большинства млеко- питающих являются их ископаемые, а не современное распро- странение.
Региональное распространение млекопитающих
Фауны млекопитающих двух северных областей Евразии и внетро- пической Северной Америки очень схожи по своим как положительным, так и отрицательным признакам. Они отличаются друг от друга главным образом небольшими ограниченными в числе видов семействами и, кроме того, тем, что в умеренной зоне Евразии многочисленны мышиные гры- зуны. В Северной Америке имеется род сумчатых Didelphis (оппосумы), который из американских тропиков проникает в восточную часть Север- ной Америки, но, конечно, отсутствует в Европе и Азии. Из насекомояд- ных в обеих северных областях есть землеройки (большинство главным образом северного подсемейства) и кроты (почти полностью северное семейство). В умеренных широтах Евразии распространены также ежи (общие для Африки и Индо-Малайской области). Один род обезьян захо- дит из тропиков Старого Света на небольшую часть умеренной зоны Восточной Азии. В обеих северных областях обитают пищухи (исключи- тельно северные и ограниченные в распространении пределами северных областей) и зайцы и т. д. (широко распространенные везде).
Из грызунов в обеих областях надо отметить многочисленных белок и т. д. и хомяков (оба семейства широко распространены и в других местах, но подсемейство iVIicrotinae почти всегда приурочено именно к этим областям), несколько видов прыгунчиков (северо-умеренных) и бобров (по одному виду в каждой области). В умеренных широтах Евразии также хорошо представлены мышиные (Muridae), доминирующие по всему Старо- му Свету: слепыши, сони и тушканчики (все более или менее локализованы или экологически ограничены). В умеренных широтах Северной Америки обитают (локализованные на западе) бобровые белки, гоферы и карманчи- ковые мыши (распространенные в основном на западе, они достигают крае- вых частей американских тропиков) и канадский дикобраз (представитель тропического американского семейства).
Из хищных (Carnivora) в обеих областях представлены широко распространенные группы: волки, лисицы, медведи, куньи и т. д. (вклю- чая скунсов в Америке) и кошачьи. В Евразии встречается панда (локали- зована), в Северной Америке — енот-полоскун и др. (встречается также и в американских тропиках). Виверровые и гиена (из тропиков Старого Света) достигают некоторых теплых частей умеренной зоны Евразии.
Африканский даман достигает юго-западной части Азии. Из высших. копытных род Едииг (лошади) находится в умеренной зоне Азии (и в Афри- ке), кабаны — в умеренной зоне Евразии (и в тропиках Старого Света), пекари — в южной части Северной Америки (и в американских тропиках), верблюды локализованы в умеренных широтах Азии (и Южной Америки)., олени широко распространены в обеих северных областях (и в тропиках
Млеко питакнц ие
Индо-Малайской области и Америки); вилороги (один вид) приурочены к западной части Северной Америки. Бычьи встречаются в обеих северных областях, но в основном в Евразии (а также в Африке и тропической Евразии). Наконец, летучие мыши представлены в обеих областях много- численными видами семейства Vespertilionidae, несколькими видами семейства Molossidae и несколькими видами, относящимися главным оора- зом к тропическим семействам, которые приурочены к Старому или Но- вому Свету и простираются на ограниченную территорию умеренных частей Евразии или Северной Америки.
В двух основных тропических областях Старого Света — Африке, южнее Сахары, и Индо-Малайской области — распространено большое количество землероек (главным образом подсемейства Crocidurinae) и не- сколько видов ежей (но «волосатые ежи», или гимнуры, приурочены к восточной части Индо-Малайской области). Выдровые землеройки Pota- mogale (в Западной Африке), златокроты (большинство в Южной Африке) и слоновые землеройки обитают только в Африке, а в Индо-Малайской области распространены кроты, которые встречаются локально по области до полуострова Малакка. Шерстокрылы приурочены к восточной части Индо-Малайской области. Там же представлено три вида землероек, и в восточной части области — долгопяты. Лори — обезьяны Старого Света и обезьяны семейства Pongidae — общие для Африки и Индо-Малайской области. Панголины (семейство Manidae) также представлены и в Африке и в Индо-Малайской области. Зайцы и т. д. многочисленны и широко распространены в Африке, но в меньшей степени в тропиках Индо-Малай- ской области. Из грызунов беличьи и т. д. и мышиные многочисленны в обеих областях.
Встречающиеся в обеих областях хомяки относительно немногочислен- ны. Сони (разных семейств), бамбуковые крысы и дикобразы Старого Света распространены в обеих областях. В Африке также имеются представи- тели семейства Anomaluridae, долгоноги и три семейства предполагаемых дикобразоподобных.
Из хищных в обеих областях хорошо представлены (anidae (включая шакалов), Mustelidae, Viveridae и кошачьи. В Африке распространены гиены, один вид которых достигает Индии; в Индо-Малайской области— медведи. Трубкозубы встречаются только в Африке.
В каждой области имеется свой род и вид слонов, два рода носорогов, а также диких свиней, оленьков (более или менее локализованы) и много- численные бычьи (большинство в Африке). Но (из этих областей) даманы, род Едииг, гиппопотамы и жирафы представлены только в Африке, а та- пиры (локализованы) и олени приурочены только к Индо-Малайской области.
Оба подотряда рукокрылых многочисленны и разнообразны в обеих областях. Таким образом, фауны млекопитающих этих двух областей богаты, во многом сходны друг с другом, но имеют и отличия.
Для Мадагаскара характерна ограниченная высокоэндемичная фауна млекопитающих, которая описана в гл. 8.
Фауна млекопитающих Южной и Центральной Америки состоит из сохранившихся частей древней (третичной) эндемичной фауны и ряда новых (позднетретичных и плейстоценовых) вселенцев. В табл. 7 приво- дится примерный состав этой фауны в настоящее время, представленный родами, а не видами, в связи с тем, что видовая систематика еще не разра- ботана. Грызуны в этой фауне по численности превосходят всех других нелетающих наземных млекопитающих, но, возможно, приведенные циф-
Глава б
ры~ не совсем точны. Роды хомяков могут быть раздроблены еще мельче. чем роды других млекопитающих Южной Америки (Simpson, 1945, р. 206, 207). С другой стороны, виды хомяков, возможно, более многочисленны, чем это можно предположить по числу родов.
Таблица 7 Современные наземные млекопитающие К)жной н Центральной Америки
Приблцзительносчисло современныкродов в области
75 (42 древние)
1 1 4 2 1 40--~- 39
86+(39 древние) 53 (по возрасту не классифици- рованы)
I'pызуны
Рукокрылые (9 семейств)
1 Достигли Южной Америки в миоцене до «селения других «но«ых» семейств.
Все семейства, отмеченные в этой таблице как «новые», кроме тапиров и верблюдов, представлены также и в Северной Америке. Многие из них встречаются и в некоторых частях Старого Света. По включенным в эту таблицу млекопитающим Центральной Америки необходимо дать следую- щие пояснения. Фауна млекопитающих Центральной Америки во многих отношениях является переходной. Ряд «древних» семейств Южной Аме- рики распространен до Центральной. Несколько (но не все) родов сумча- тых Южной Америки также достигают Центральной Америки, и из них 4 рода проникают в отдельные районы Мексики, а один — в восточную часть Соединенных Штатов. Все три современных рода муравьедов распро- странены от Южной Америки до Центральной и два из них достигают южной части Мексики. Оба современных рода ленивцев простираются из Южной Америки в Центральную, достигая Никарагуа и Гондураса соответственно. Из 9 южноамериканских родов броненосцев два захо-
Сумчатые (2 семейства) (древние) Землеройки (новые)
Обезьяны (2 семейства) (древние) Пеполнозубые (3 семейства) (древние) (:обачьи (новые)
Медведь (новый)
Енотовые (новые >)
Куньи (новые)
Кошачьи (новые)
Тапиры (новые)
Пекари (новые)
Верблюды (новые)
Олени (новые)
Наземные, кроме зайцеобразных и грызунов
Зайцы (новые)
Зайцы
Белки (новые)
Гоферы (новые)
Еарманчиковые мыши (новые) омячьи (HQBblo)
Днкобразоподобные (11 семейств) (древние)
14 2 14 14 4 1 5 10
с1
1 1 1 6
Млеко и и~пак>цс ие
дят в Центральную Америку, один достигая Гондураса, а другой — Техаса и т. д. Из дикобразоподобных грызунов Южной Америки 7 семейств при- урочены к некоторым районам Южной Америки. Один из трех южноаме- риканских родов дикобразов простирается через Центральную Америку до районов Мексики (а четвертый род этого семейства — канадский дикобраз — распространен примерно от границы Мексики с Соединенными И1татами до северной границы леса). Водосвинка достигает Панамы, а два (из четырех) южноамериканских рода агути распространены через Цен- тральную Америку до южной части Мексики. Три (из 14) южноамерикан- ских рода щетинистых крыс распространены до Панамы или Никара- гуа. Однако в Центральной Америке не имеется эндемичных родов этих древних семейств. Особого внимания заслуживает то обстоятельство, что там отсутствуют эндемичные роды дикобразоподобных, хотя некоторые из них встречаются в Вест-Индии. Из «новых» семейств, помещенных в табл. 7, все, кроме медведей и верблюдов, представлены как в Централь- ной, так и в Южной Америке. Из землероек род Sorex, распространяясь е севера в тропики, достигает только Гватемалы, а род Cryptotis — север- ной части Южной Америки.
Из собачьих род Canis распространен на юг до Коста-Рики; Ггосуоп- до северной части Южной Америки. Кроме них, в Южной Америке имеют- ся эндемичные роды. В отношении енотовых интересен не факт распро- странения их в настоящее время к югу, а дифференциация их родов в Центральной и Южной Америке. Из куньих род Mustela распространен на юг через Центральную Америку и отдельные части Южной Америки до Перу. Три рода скунсов достигают Гондураса, Коста-Рики и Магеллано- ва пролива соответственно. Род t utra (выдры) доходит до Огненной Земли. И в Южной и в Центральной Америке имеются еще свои эндемичные роды. Очень широко распространены два рода семейства Felidae. Тапиры, пе- кари, верблюды и олени все первоначально достигли Южной Америки с севера, но в настоящее время распространение родов этих групп уже не отражает их прошлых перемещений к югу. Единственный род зайцев простирается через Центральную и Южную Америку до северной Арген- тины. Из «новых» грызунов Sciurus распространен на юг через Централь- ную и Южную Америку до северной Аргентины. Гоферы достигают толь- ко Панамы, а карманчиковая мышь — северной части Южной Америки.
11роанализировать распространение хомячьих я не пытался.
Этот далеко не полный и не очень подробный список вполне достато- чен для того, чтобы показать, что Центральная Америка (с южной Мек- сикой) является основной территорией перехода тропической и северо- умеренной фаун млекопитающих Америки, несмотря на то, что несколько южноамериканских групп простираются дальше в Северную Америку и одно семейство (дикобразов) Южной Америки достигает северных преде- лов древесной растительности, а некоторые североамериканские группы проникают далеко на юг в обратном направлении — через какую-то часть Южной Америки или через весь материк. Этот переход усложняется существованием эндемичных родов в Южной Америке (даже в некоторых «новых» семействах) и родов с сомнительной историей. Некоторые роды даже «новых» семейств распространились к северу, вероятно, не так ДЯ ВНО.
В Австралийской области (Австралия, Новая Гвинея и мелкие близ- лежащие острова) местные млекопитающие представлены только одно- проходными, сумчатыми, грызунами семейства Миг1с1ае и рукокрылыми и соотношениях, приведенных в табл. 8.
284
Глава 6
Таблица 8
Сводная таблица местной фауны наземных млекопитающих Австралиии Новой Гвинеи
Австралия(Тасмания и т. д.)
Ноиая Гвинея(и прилежащие острова)
семей-ства
семей-
роды
роды
виды
1
виды
Однопроходные Сумчатые
Грызуны (Muridae) Рукокрылые
2521321
21196741~
24
4
56
"0
21
П р и м е ч а и и е. Цифры взяты главным ооразом из статей Тейта (см. библиогр;~фикц Виды грызунов, в отличие от видов сумчатых, можно подвергнуть еще более дробному деле- нию. Виды рукокрылых предположительно рассматриваются Тейтом в целом. Деление родов по Тейту более дробное, чем у Симпсона.
«Дикие» собаки (динго и т. д.) и свиньи Австралии и Новой Гвинеи, вероятно, были завезены доисторическим человеком, а европейский кро- лик, заяц, лисица, так же как и крыса пасюк, черная крыса и домовая мышь, были интродуцированы. В настоящее время два последних вида широко распространены в Австралии даже в естественных местах обитания.
Из табл. 8 видно, что фауны млекопитающих Австралии и Новой Гвинеи одинаково ограничены и схожи в целом по своему составу. Они отличаются только тем, что в Австралии более многочисленны и более разнообразны сумчатые, а в Новой Гвинее имеется большее число родов грызунов, что объясняется различной историей сумчатых и грызу нов. Сумчатые находятся в Австралии давно и, очевидно, главным образом там и дифференцировали, хотя их недавние перемещения в Новую .Гви- нею и из нее были многократными. Грызуны совсем недавно появились из, Азии, и большинство их, вероятно, достигло сначала Новой Гвинеи, и они имели больше времени для эволюции здесь, чем в Австралии, хотя их перемещения между Новой Гвинеей и Австралией также были многократ- НЫМИ.
Австралийские сумчатые являются наилучшим известным примером эволюционной или адаптивной радиации в изолированном районе. Раз- личные сумчатые приспособились к самым разнообразным условиям жиз- ни, свойственным плацентарным животным вне Австралии. Мелкие сумчатые куницы похожи здесь на землероек. Это насекомоядные или преимущественно плотоядные зверьки, большинство из них бегает, но представители рода Ап1есИпотуз прыгают, как прыгунчиковые мыши. Более крупные сумчатые семейства Dasyuridae внешне схожи с куньими, росомахой или волком и ведут хищный образ жизни. Аберрантный предста- витель этого семейства ЛХугтесоЬ~иг — сумчатый мурашеед (в действитель- ности питается термитами); у него длинная морда и редуцированные зубы. Обитатель полупустынь Notoryctes, относимый к самостоятельному семейст- ву, чрезвычайно схож с кротом и живет, подобно ему, в земле. ('.умча- тый барсук бандикут (Peramelidae) — крыса- или кроликоподобное животное, живет на земле и более или менее всеяден. Различные виды встречаются в разнообразных типах местности — от дождевых лесов до
~Ъ1лекопитак~~ц ие
28о
пустынь. В основном лазающие сумчатые Phalangeridae схожи с мьппами или опоссумами. Большинство из них ведет древесный образ жизни, некоторые виды спускаются на землю и живут в сухих скалистых местах. Большинство мелких зверьков Австралии относятся частично к насеко- моядным, а частично (более крупные) к травоядным животным, но неко- торые из них специализированы. Маленький сумчатый медоед Tarsipes, имеющий длинную морду и слабые зубы, питается нектаром цветов или мягкими кормами. Dactylopsila, полосатый опоссум, имеет долотообразные резцы. которыми он пробивает мертвое дерево, и удлиненный четвертый палец. которым он вытаскивает личинки насекомых из просверленных ими ходов. У трех разных представителей семейства Phalangeridae раз- личных размеров (от маленькой мыши до небольшой кошки) развились летательные перепонки и длинный в виде пера хвост, что позволяет им планировать в воздухе наподобие летучих белок. Это исключительно необходимое приспособление, особенно в австралийских эвкалиптовых лесах, где деревья очень часто находятся на большом расстоянии друг от друга. Коала, или сумчатые медведи (Phascolarctos), — специализирован- ные представители семейства Phalangeridae. Внешне они похожи на мед- ведя. но по образу жизни больше напоминают ленивцев, хотя и живут в кронах деревьев. Коала ведут древесный образ жизни и питаются листья- ми только нескольких видов эвкалиптов. Вомбаты (Phascolomidae) — это крупные наземные сумчатые, мощные землекопы с грызущими, как у гры- зунов. зубами. Питаются растениями и их корнями. К семейству Macro- podidae относятся разнообразные мелкие кенгуровые крысы, несколько более крупные кенгуру-валлаби и еще более крупные кенгуру. Большин- ство из них являются наземными и преимущественно травоядными животными. На каждой пригодной для их обитания части Австралии встре- чается большое число разных видов кенгуру, но из них Новой Гвинеи достигают сравнительно немногие. Крупные виды кенгуру относятся к травоядным животным, которые замещают в других частях земного шара травоядных копытных. Череп кенгуру в профиль имеет чисто внешнее сход- ство с черепом лошади. Оба этих вида приспособлены к общипыванию травы, и, наконец, имеются еще древесные кенгуру Dendrolagus, вторично приспособившиеся к жизни на деревьях в дождевых лесах Северо-Восточ- ной Австралии и Новой Гвинеи. Они довольно неуклюже взбираются на деревья, но их передние конечности и когти приспособлены для лазания, задние ноги укорочены, а хвост (у обычных кенгуру жесткий и негибкий) превратился в тонкий гибкий противовес. Помимо всех перечисленных видов, в плейстоценовое или постплейстоценовое время в Австралии существовали еще более крупные кенгуру. Это были гигантские (величиной с носорога) травоядные дипротодонты и достигавшие размеров льва Tliplacoleo, которые, возможно, были хищными животными.
Однако адаптивная радиация австралийских сумчатых имеет свои пределы. Удивительно, что в Австралии нет ни одного водного вида, хотя восточная часть Австралии, так же как и Новая Гвинея, имеет хорошо развитую речную сеть. Строго водный род Platypus просуществовал в Австралии неизвестно в течение скольких веков, а сумчатое животное С!гггггпес1ез является земноводным только в Южной Америке. Ни в Австра- лии. ни в других местах нет также мелких грызущих сумчатых. Полоса- тьш опоссум и вомбаты имеют грызущие зубы, но эти животные ведут особый образ жизни и не замещают грызунов. Возможно, что в прошлом в Австралии находились и грызущие сумчатые, похожие на грызунов, и что они были замещены настоящими грызунами, которые позднее
Глава 6
достигли Австралии. Сумчатых летучих мышей в Австралии не существует, так как развитие крыльев происходит не столь быстро, и воздухом в Австралии завладели плацентарные летучие мыши.
Распространение сумчатых в границах Австралийской области сл<я,— но. Сумчатые куницы (Dasyuridae) (Tate, 1947, р. 115, 150) обитают глав- ным образом в Австралии, но из них около шести ответвлений достигли Новой Гвинеи, причем одни, очевидно, раньше, другие позже. Бандикуты (Peramelidae) (Tate, 1948а, р. 322) примерно в равной степени многочис- ленны и в Австралии, и в Новой Гвинее. Помимо древних перемещений, происходили и современные переселения этих животных в обоих направ- лениях: один вид рода Isoodon проник из Австралии в южную часть Но- вой Гвинеи, а один вид Echymipera — из Новой Гвинеи до Северо-Восточ- ной Австралии. Представители лазающих сумчатых — австралийские опоссумы (Phalangeridae) более многообразны в Австралии, чем в Новой Гвинее, но их перемещения были сложными. Помимо древних перемеще- ний, очевидно, происходило и более позднее расселение родов Petauru, (сумчатая летяга) и Eudromecia (сумчатая соня) из Австралии в Новун~ Гвинею, двух видов Phalanger (кускусы) и одного вида Dactylopsila (по- лосатый опоссум) — из Новой Гвинеи до Северо-Восточной Австралии. Вероятно, так же происходили перемещения, как древние, так и более поздние, рода Pseudocheirus (кольцехвостый кускус) (Tate, 1945а, р. 2). Кенгуровые (Macropodidae) (Tate, 1948, р. 255 — 258) — преимущественн« австралийские животные, но мелкие кенгуру валлаби довольно давно достигли Новой Гвинеи и образовали там эндемичные роды. Более круп- ные кенгуру Protemnodon agilis проникли в южную часть Новой Гвинеи недавно, а древесные кенгуру (Dendrolagus), вероятно, совершили два перемещения: одно более древнее, а другое недавнее — между Австра- лией и Новой Гвинеей в одном или в другом направлении.
Более поздние перемещения всех этих семейств, вероятно, происхо- дили через соединения суши между Австралией и Новой Гвинеей в позд- нем плейстоцене. Более ранние переселения, возможно, имели место не раныне, чем в позднетретичном периоде. В Австралии находится ряд филогенетически изолированных или специализированных сумчатых, включая два специфических семейства (сумчатые кроты и вомбаты), сум- чатого волка, сумчатого мурашееда, сумчатого медоеда, коала и три группы «летающих» сумчатых семейств Phalangeridae и т. д. На Новой 1'винее нет таких древних зндемичных, обособленно специализироваииих- ся сумчатых, и нет ни одного вида, который бы не проник из Австралии или не произошел от австралийского предка в пределах нескольких последних миллионов лет. Этот факт говорит о том, что современные сум- чатые. возможно, достигли Новой Гвинеи несколько миллионов лет назад.
Все грызуны Австралийской области относятся к семейству luridae, что нередко вводило в заблуждение некоторых зоогеографов, которые рас- сматривали их как «мышей» или просто игнорировали. В действитель- ности же австралийские мышиные не только довольно многочисленны (табл. 8), но также и довольно разнообразны. Подсемейство Hydromyii>ae почти полностью приурочено к Австралийской области. Восемь родов этого подсемейства находятся на Новой Гвинее, один из них также широко рас- пространен в Австралии и девятый род локализован в Северо-Восточной Австралии. И только два других, монотипических рода, относимых к это- му подсемейству, находятся в горах Северного Лусона на Филиппинах. Большинство представителей подсемейства Hydromyinae Австралийской области — водяные крысы, но, так же как и на Филиппинах, здесь есть
.~7леко пинг атэц ие
несколько и наземных видов. Другие австралийские мышиные все отно- сятся к подсемейству Murinae, но и они довольно разнообразны. Во всех частях Новой Гвинеи и Австралии имеются разные эндемичные роды этого подсемейства. Одни из них наземные и мыше- или крысоподобные, а другие — тушканчикообразные (прыгающие), третьи — древесные. Иэ последних некоторые достигают большой (для крыс) величины и обладаю1 в той или иной мере развитым хватательным хвостом. В Новой Гвинее и Австралии, кроме эндемичных родов, имеется много эндемичных видов и ви- довых групп, принадлежащих к доминирующему роду Старого Света Лайия.
По крайней мере 5 различных видов семейства МпгЫае достигли Авс1- ралийской области в разное время (Tate, 1951, р. 217 — 221). Предок под- семейства Hydromyinae (вероятно, наземное животное) появился на ран- ней стадии истории мышиных, скорее всего, в миоцене. Один или несколько представителей подсемейства Мигinae проникли в Австралий- скую область также довольно рано. Предок родовой группы Uromys, вероятно, появился позже. Появление рода Rattus, видимо, произошло еще позднее и было неоднократным. Очевидно, одна или несколько групп этого рода проникли через Новую Гвинею, а другая группа — через Зондские острова и остров Тимор. После их появления разные линии мышиных развивались отдельно в Новой Гвинее и Австралии, но с после- довательным обменом в обоих направлениях. Наиболее поздние из обме- нов приводятся Тейтом (Tate, 1951, р. 220). Насколько можно судить по работе Тейта (Tate, 1946), рукокрылые Австралийской области объединяют большое число разных линий, которые проникали из Азии в течение дли- тельного периода. Из более древних рукокрылых одни в какой-то мере эво- люционировали в Австралии и Новой Гвинее (но в значительно меньшей сте- пени, чем сумчатые и грызуны), другие, возможно, вернулись (распростра- няясь в западном направлении) на небольшое расстояние. Все же основной приток рукокрылых происходил в Австралийскую область, а не наоборот.
Географическая история однопроходных неизвестна. Можно пред- положить, что независимо от места происхождения их радиация в Австра- лии происходила до появления там сумчатых и что древняя австралийская фауна однопроходных была по крайней мере такой же разнообразной, как и более поздняя фауна сумчатых. Хотя род Platypus и ехидны являются последними сохранившимися представителями древней фауны, эти при- митивные (или, возможно, дегенеративные) животные более специализи- рованы, чем любые сумчатые (словно они находятся в самом конце длинных линий адаптации). Утконос — это уникальный представитель австралий- ской фауны, а ехидна — это еще более специализированное животное, чем сумчатый муравьед. Что касается более поздней истории, то если австралийские сумчатые были приурочены к собственно Австралии до позднетретичного периода (см. выше), очевидно, это также будет относить- ся и к однопроходным. В этом случае ехидны являются главным образом австралийскими животными, которые двахсды достигали Новой Гвинеи: один раз, возможно, до плейстоцена и второй раз в течение этого периода.
11ереходные области п преграды распространению млекопитающих
Между фаунами млекопитающих смежных областей всегда имеется промежуточная переходная зона, которая, однако, часто ограничивается преградами. Область перехода фаун млекопитающих между тропической Африкой и умеренной зоной Европы очень ограниченная; между тропи-
288
Глава 6
Домпнирование и конкуренция в связи ~. распространением млекопитающпх
ВойепИа — главенствующий отряд млекопитающих. Вуд (Wood, 1947, р. 154) предполагает, что по видовому составу грызуны теперь примерно в 2 раза многочисленнее всех других млекопитающих и что,
ческой и умеренной частями Восточной Азии —; и между тропической и умеренной зонами (северного полушария) Америки— также обширная и сложная, но, если можно так сказать, более односто- ронняя, то есть многие северные группы распространяются и в Южную Америку, но немногие южноамериканские группы проникают в глубь Северной Америки. Между фаунами умеренных широт Евразии и Север- ной Америки также наблюдается переход через север. Обе фауны беднеют в северном направлении. Постепенное выклинивание элементов фауны начинается севернее тропиков и увеличивается к северу до тех пор, пока обе фауны не сократятся до одинакового числа северных компонентов. Переход между фаунами млекопитающих тропической Африки и тропи- ческой Азии в настоящее время незначителен. Эти фауны имеют много оощих черт, но в настоящее время разъединены широким барьером засуш- ливой внетропической области страны, где встречаются немногие млеко- питающие тропиков, хотя в прошлом, видимо, эту область пересекали многие виды. И, наконец, между фаунами млекопитающих тропической Индо-Малайской области и Австралии переход совершается даже через водные преграды. Из млекопитающих Индо-Малайской области многие достигают линии Уоллеса: земелеройки, долгопяты, обезьяны, белки, виверровые, дикие свиньи, олени, бычьи. На острове Сулавеси обнаружен ископаемый карликовый слон. Некоторые землеройки, обезьяны, вивер- ровые, свиньи и олени достигают отдельных островов Молуккского архи- пелага, Малых Зондских островов и острова Тимор, хотя их естественные границы не совсем ясны. Грызуны семейства Muridae распространены через острова до Австралии и Тасмании. В противоположном направлении распространение большинства сумчатых ограничивается Новой Гвинеей и ее прибрежными островами. Впрочем, несколько видов достигают Мо- луккских островов, а род Phalanger (r~yc~~yc) доходит до острова Сулавеси, где эндемичные виды этого рода встречаются с эндемичными обезьянами и другими плацентарными млекопитающими, упомянутыми выше. Большая часть наземных млекопитающих совершает переход из Индо-Малайской области в Австралию, причем в обратном направлении видов переходит меньше, что относится и к рукокрылым. Многие рукокрылые Индо-Ма- лайской области распространены или в направлении к Австралии, или достигают Австралии, но только несколько австралийских видов распро- странены по направлению к Индо-Малайской области.
Никакие преграды, кроме образованных солеными водами, не пре- пятствовали распространению доминирующих групп млекопитающих на земном шаре. Некоторые нелетающие наземные млекопитающие пере- секали (в ограниченной мере) даже и соленые воды. Наиболее часто это наблюдалось у рукокрылых. Но тем не менее преграды оказывали опреде- ленное влияние на распространение млекопитающих, а небольшие пре- грады, образованные солеными водами, почти полностью (но все же не совсем) изолировали фауны млекопитающих Южной Америки и Австра- лии в течение всего или почти всего третичного периода.
289
Млекопитан тие
Заключение: особенности распространения млекопитающих
Распространение млекопитающих к северу и к югу ограничивается только границами суши. Однако большинство нелетающих наземных мле- копитающих не распространяется за пределы материков и материковых островов или островов, расположенных очень близко к материкам, хотя граница распространения грызунов заходит несколько дальше, а ряда видов рукокрылых — еще дальше. Следует отметить поразительный викариат, взаимозамещение в распространении фауны главным образом плацентарных наземных млекопитающих на основной части земного шара, смешанной фауны в Южной Америке и в основном фауны сумчатых в Ав- стралийской области.
Зональность наземных млекопитающих выражена отчетливо, но в большинстве случаев она является результатом уменьшения числа видов
возможно, они оыли также многочисленны и в течение большей части тре- тичного периода, что, пожалуй, является уже пре~ величением (см. табл. 5, 7, 8). Во всяком случае, грызуны, равные по числу видов другим нелетающим наземным млекопитающим (или превосходящие их), превосхо- дят их и по количеству особей. Rodentia — единственный (за исключением человека) отряд космополит среди нелетающих наземных млекопитающих. Разнообразные грызуны доминируют во многих местах обитания — от самых влажных лесов до самых засушливых пустынь, где только могут существовать млекопитающие. Многие из грызунов являются наземными, древесными, в той или иной степени водными, а некоторые различные их группы стали подземными. Однако доминирование грызунов имеет экологические пределы, которые напоминают пределы у воробьиных птиц. Грызуны редко бывают хищными и не являются серьезными конку- рентами хищным млекопитающим. И несмотря на то, что грызуны в какой- то степени конкурируют с некоторыми травоядными и общипывающими рас- тительность животными, они едва ли угрожают благополучию копытных в местах обитания последних. Некоторые другие группы млекопи- тающих доминируют в пределах еще более узких географических или экологических границ (например, собачьи, кошачьи, бычьи и руко- крылые).
Конкуренция — любое взаимодействие между iEHBo THhIMH, которое оказывает неблагоприятное влияние на одного из них, — очевидна среди млекопитающих. Многочисленные примеры такого влияния дала интро- дукция крыс, кроликов, мангуста, кошек и др. в новые части земного шара. Особенно очевидно значение конкуренции из истории млекопитаю- щих в Южной Америке, где за несколько миллионов лет начиная с позд- него плиоцена новые млекопитающие заместили большую часть древней южноамериканской фауны млекопитающих. В действительности основ- ная роль конкуренции проявляется в последовательной смене фаун мле- копитающих, которая, как это показывают ископаемые, происходила во многих частях земного шара. Радиация сумчатых в Австралии происхо- дит в условиях отсутствия конкуренции со стороны плацентарных, что осооо подчеркивает значение конкуренции. Это настолько ясный вопрос, что нет надобности изучать его подробно. Однако в связи с тем, что некоторые биологи все еще сомневаются в существенном значении конку- ренции для распространения животных, придется несколько позднее (стр. 464) еще раз вернуться к этой теме.
290
Глава 6
от одной зоны к другой. Северо-умеренные фауны млекопитающих, по существу, являются тропическими фаунами, из которых выпали многие элементы. А.рктическая же фауна млекопитающих фактически представ- ляет собой умеренную фауну, из которой еще в большей степени выпали отдельные, более южные компоненты. В южном направлении фауны млеко- питающих также уменьшаются за счет выклинивания видов и групп, особенно к южной (с холодным климатом) оконечности Южной Америки. Общей зональной южно-умеренной фауны млекопитающих для разных южных континентов нет.
В современном характере распространения млекопитающих радиа- ция не обнаруживается, хотя ископаемые показывают, что в прошлом многие группы млекопитающих радиировали на большей части земного шара.
Дифференциация фаун млекопитающих в шести зоогеографических областях есть результат широко развитого викариата, взаимозаменяе- мости и зональности одних групп и дальнейшей локализации других. Фауны млекопитающих двух основных тропических областей Старого Света богаты как широко распространенными, так и локализованными группами. В этих двух фаунах много общих групп, но имеются группы, которыми они различаются. Фауны млекопитающих двух умеренных областей северного полушария характеризуются главным образом бед- ностью и отсутствием многих тропических групп. Обе эти фауны, отли- чаясь в деталях, схожи в общих чертах, особенно в направлении к северу. Фауна млекопитающих Южной и Центральной Америки представляет специфическую смесь древних эндемичных групп с новыми, общими с другими частями земного шара. Австралийская фауна млекопитающих состоит главным образом из древних эндемичных семейств сумчатых и однопроходных, не считая грызунов семейства МигЫае и множества рукокрылых.
И последняя особенность распространения — концентрация на тер- риториях с благоприятными условиями обитания с уменьшением числа видов к периферии — у млекопитающих выражена умеренно. Они наи- более многочисленны в местах с более благоприятными условиями ооита- ния и малочисленны в менее благоприятных, таких, как арктические райо- ны и пустыни. Но те или иные млекопитающие встречаются на суше почти везде, даже в самых неблагоприятных зонах перехода.
Среди современных млекопитающих древнейшими реликтами, вероятно, являются однопроходные Австралии — сохранившиеся предста- вители древнего подкласса. К древнейшему современному семейству млекопитающих относится семейство сумчатых Didelphidae (американ- ские опоссумы). Представители этого семейства сохранились без сущест- венных изменений с середины верхнемелового периода (Scott, 1937, р. 746). Но современные австралийские и южноамериканские сумчатые- это не столько выжившие представители когда-то доминировавшей на земном шаре группы, сколько продукт эволюции в изоляции. Современ- ные насекомоядные являются в той или иной степени специализирован- ными сохранившимися представителями древнего отряда, который оыл предком плацентарных животных в начале третичного периода и кото- рый, возможно, такой же древний, как и первые сумчатые. Что касается сумчатых Австралии и Южной Америки, то здесь следует сделать ударе- ние не на том, что они древние плацентарные, а на том, что они продол- жительное время развивались самостоятельно, в изолированных частях земного шара.
ЛХлекопитаюи~ ие
291
История расселения млекопитающих
Несмотря на то что по фауне млекопитающих мы располагаем обиль- ными ископаемыми данными с точки зрения географии, в них все еще име- ются колоссальные пробелы. Млекопитающие возникли в триасе, однако мезозойские формы почти не известны. Увеличение количества млекопи- тающих и основной «расцвет» их приходится примерно на начало третич- ного периода, но даже ископаемые этого времени все же фрагментарны, за исключением ископаемых Европы, некоторых районов умеренной зоны Азии, западной части Северной Америки и южной части Южной Америки. Что касается наземных млекопитающих Африки, то относительно первой трети третичного периода ничего не известно, и можно только предпо- лагать, что этот материк был, вероятно, одной из самых важных частей земного шара, а начало третичного времени было самым значительным периодом в истории млекопитающих (Simpson, 1945, р. 250). Ископаемые из позднетретичных отложений Африки также немногочисленны (Hop- ivood, 1954). В тропической Азии в Бирме известно несколько ископаемых млекопитающих из верхнего эоцена. Большинство третичных млекопи- тающих Южной Азии известно из Сиваликских гор в северной Индии. Фактически эти горы расположены примерно на 6' севернее тропиков, и материалы оттуда датируются не ранее, чем миоценом. В Австралии по- ложение еще хуже. Данные о третичных млекопитающих Австралии пол- ностью отсутствуют (кроме одного, не имеющего значения для наших це- лей, сумчатого из плиоцена Тасмании). Также нет данных по ранее третич- ным млекопитающим тропиков Старого Света, а имеющиеся незначитель- ные сведения относятся к более позднему периоду. Большая роль, которую сыграли тропики Старого Света в современном распространении млеко- питающих, заставляет признать этот пробел в ископаемых данных очень существенным. Еще один пробел в ископаемых данных относится к тро- пикам Америки. Третичные млекопитающие южной части Южной Амери- ки известны исключительно хорошо, но сведений о млекопитающих тро- пической части этого материка сравнительно мало. Сейчас этот пробел начинает постепенно восполняться (Couto, 1952; Stirton, 1953). Эти про- оелы и другие недостаточно полные ископаемые сведения, особенно скуд- ные материалы по мелким млекопитающим, делают историю расселения млекопитающих до какой-то степени предположительной.
Выводы по географической истории наиболее изученных и важных групп наземных млекопитающих сделаны отчасти по ископаемым данным, отчасти на основании заключений о современном распространении (см. список семейств). Данные по истории расселения были проверены по Сим- псону (Simpson, 1945, etc.), Ромеру (Вошег, 1945) и Скотту (Scott, 1937). История расселения млекопитающих рассматривается в том же порядке, что и список семейств (Simpson), исключающий грызунов и ру- кокрылых, которые помещены в конце этого списка. Направления пере- мещений млекопитающих более детально будут рассматриваться позднее.
Место и время происхождения предков однопроходных не извест- ны. Возможно, их предками были раннемезозойские млекопитающие или звероподобные пресмыкающиеся, но вполне вероятно, что ими были позд- немезозойские примитивные вырождающиеся сумчатые. В первом случае однопроходные могли возникнуть почти в любой части земного шара, а во втором — они, возможно, были ограничены только Австралийской областью. Но как в том, так и в другом случае они, вероятно, долгое время находились в Австралии.
Распространение и вероятное расселение сумчатых показано на рис. 39. Didelphoidea обитали в Северной Америке в верхнем мелу, а в Европе — в раннетретичное время и предположительно были более или менее широко распространены в это время по всему земному шару. На большей части земного шара они встречались вместе со многими пла- центарными и, очевидно, широко не радиировали. Они радиировали, достигнув Южной Америки, несмотря на то, что там уже находились неко- торые плацентарные. Их радиация дала Южной Америке разнообразных опоссумов (Didelphoidea), некоторые из которых существуют и по сей
Ъ ю~
%
ф
° ф, ъ
° ° ° °
° % ~е ° tl
с ' ф °
° ~ °
° ° °
° ° °
° ° °
ю
° p ', °
° ~ °
г
° ° ° ° ° ° °
O
1 1
В
ю ° , °
° ° °
° ° ° °
° °
° °
° ° Я~
° у
Ю' %
° ° °
г ° °
° с
° °
° ею'
° у У °
ю
° °
е
У..Л"
мт -:.:;.
° ° ° ° ° фе Ф
° ° ° ° ° ° ° °
°,; ee ° .Ф.
е% \~ J °
° б
ю °
8. °;
° °
° °
° °
,а. А~е '° '° ° ° ° ° ° ° ° ЪЪ ° ° ° \ ° ° ° ° °° е д,,уф\ е~ еф ° ° ° f° е ° ° ° °
с ° °6'
~М. ~
° °
° ° ° ° Ъ °
° Ф ° ° ° °
Р и с. 39. Распространение сумчаты~.
Жирные пересекающиеся линии — основные области земного шара, занимаемыс сумчатыми; стрелки — распространение (вероятно, недавнее) рода 1%а1аид~ к западу от Новой Гвинеи и рода ~id' l p]ii.' к северу от тропиков Америки; крестиками обозначены места, гдс были обнарун<ены ископаемые сумчатых в позднемсловом и (или) раннетретичном периодах. Большие прерывистые стрелки — предпо- лагаемое направление расселения сумчатых.
день, различных хищных сумчатых (в настоящее время вымерших), вклю- чая сумчатого саблезубого тигра (ТЬу1асоит1из), и разнообразных "aeno- lestoidae (ценолестовые), из которых несколько неопределенных реликтов (опоссумы) до сих пор сохранились в Андах и Чили. В южной части Южной Америки ископаемые всех этих трех групп, относящиеся к палеоце- ну, являются самыми ранними. Более ранних данных о них нет. Возмож- но, все они произошли от одного Didelphoidea, который достиг Южной Америки примерно в начале третичного периода. Но не исключено, что их происхождение было еще более сложным. В Северной Америке опоссу- мы просуществовали до нижнего миоцена, затем, очевидно, отсутствовали до плейстоцена, пока род Didelphis вторично не появился здесь в плейсто- цене. [Скотт (Scott, 1937) считает, что опоссумы сохранились в течение всего третичного периода как в Северной, так и в Южной Америке и что род Didelphis произошел в Северной Америке и попал в Южную, преодо- лев водное пространство в раннем плиоцене.1
Сумчатые, конечно, достигли также Австралии и радиировали там, не конкурируя с плацентарными, кроме грызунов семейства МигЫае. Что касается вопроса, сколько сумчатых достигло Австралии и в какое время, то ископаемые данные ответа на него не дают. Возможно, что австралийские сумчатые произошли все от одного Didelphoidea или от нескольких разных предков. Насколько позволяют судить ископаемые,
Млекопитающие
предок или предки сумчатых, возможно, достигли Австралии в какой- то отрезок времени в течение позднемелового или раннетретичного периода. Радиация и современные перемещения сумчатых в пределах Австралийской области были описаны выше.
Ископаемые насекомоядных относятся к верхнему мелу Азии и Се- верной Америки, но их распространение в целом в то время не известно. Очевидно, в древнее время они никогда не достигали Австралии и не могли достичь Южной Америки (если только не считать насекомоядным род Neerolestes). Современные землеройки появились там совсем недавно. Эволюция их потомков на остальной части земного шара проходила без параллелизмов — неспециализированные протонасекомоядные, очевидно, были предками всех плацентарных млекопитающих. Сами насекомояд- ные, будучи сложной группой, имели сложное расселение на основной части земного шара. Их расселение в какой-то мере продолжалось до конца мелового периода. Тенреки, вероятно, расселились до географиче- ских границ насекомоядных в раннетретичном времени, затем пришли в упадок, сохранившись на Мадагаскаре, в Вест-Индии и Западной Африке (Potamogale). Ежовые также, вероятно, распространились до географиче- ских границ насекомоядных в начале своей истории. Они и по сей день широко распространены в Африке и Евразии (ежи и др.). На протяжении большей части третичного периода ежовые находились и в Северной Аме- рике, но затем исчезли. Землеройки (Soricoidae), ведущие свое начало с раннетретичного времени, до сих пор широко распространены в Север- ной Америке. Каждая из этих широко распространенных групп, вероятно, имеет более сложную историю, чем это показывают фактические данные. Являются ли златокроты (вероятно, образовавшиеся от Tenrecoidae) и прыгунчики реликтами в Африке и проникали ли кроты (родственные Soricoidae и существовавшие с верхнего эоцена) когда-либо далеко за пре- делы умеренной зоны северного полушария, на этот счет нет никаких данных. Однако существование этих и других локализованных и специали- зированных групп свидетельствует о CJIoiEHOH истории расселения насеко-
МОЯДНЫХ.
Шерстокрылы (Dermoptera), сократившиеся в настоящее время до одно- го специализированного рода в восточной части Индо-Малайской области, в раннетретичном периоде находились в Северной Америке. Остальная история их не известна.
Приматы, вероятно, произошли от животных, подобных тупайям, которые в настоящее время приурочены к Индо-Малайской области, а последние в свою очередь образовались от насекомоядных, близких семейству прыгунчиков Macroscelidoidae, которые ныне приурочены к Афри- ке. Примитивные приматы (лемуры и долгопяты) в раннетретичном периоде обитали в Евразии и Северной Америке (предположительно и в не- которых других частях земного шара), и, очевидно, в то время их рассе- ление было сложным. Проникла ли эта группа в Южную Америку, где от нее могли возникнуть обезьяны Нового Света (независимо от развития обезьян Старого Света), или американские обезьяны образовались от оолее «обезьяноподобных» предков, расселившихся из Старого Света позднее и не оставивших следов в Северной Америке, не известно. Ископае- мые обезьян в Южной Америке отсутствуют до миоцена, но возможно, они находились там в более ранний период, хотя в ископаемых тропиче- ских лесах следов их пребывания не найдено.
В Старом Свете лемуры или, возможно, только один вид их радииро- вали, достигнув Мадагаскара. Так или иначе, примитивные приматы
Глава 6
пришли в упадок. В Северной Америке они не могли выжить по;я;е эоцена. В Старом Свете часть из них сохранилась до настоящего времени и приурочена к более или менее ограниченным районам тропиков Старого Света. Но высшие обезьяны расселялись и перерасселялись очень слож- ным образом в Африке и Евразии в течение среднего и позднетретичного времени и в плейстоцене. Их перемещения, вероятно, в основном проис- ходили в пределах более теплых частей Старого Света, так как не извес1- но ни одного случая, когда хотя бы одна из высших обезьян достигла Аме- рики.
Неполнозубые (Еdentata) образовались, очевидно, от протонасеко- моядных. Существовали ли в Южной Америке специализированные на- секомоядные, не известно. В Северной Америке в раннетретичном периоде находились довольно примитивные неполнозубые, которые появились слишком поздно и были слишком специализированными, чтобы являться исходными формами. Строение всех южноамериканских форм показывает. что зубы, позвонки и конечности их ближайшего предка были уже до какой-то степени специализированы. Все это позволяет предполагать, что неполнозубые произошли в Северной Америке (или даже в Старом Свете) и имели там короткую историю специализации и что одна или несколько в какой-то мере специализированных форм (возможно. броненосцеобразные, но без панциря, возможно, и древесные) достигли Южной Америки, видимо, в палеоцене. Ископаемые броненосцы относятся к палеоцену IOiEHofI Америки.
Так или иначе, в Южной Америке в течение третичного периода неполнозубые дали разнообразные формы, и некоторые из них были домини- рующими. К ним относились разнообразные животные: от мелких до гигантских земляных ленивцев, броненосцев и огромных, одетых панцирем глиптодонтов, несколько видов муравьедов и древесных ленивцев. В кон- це третичного периода некоторые неполнозубые распространились из Южной в Северную Америку, где древние формы уже давно исчезли. Один из земляных ленивцев, очевидно, достиг Северной Америки в середине плиоцена, до образования центральноамериканского сухопутного моста. Этот факт, а также и то, что земляные ленивцы достигли некоторых остро- вов Вест-Индии, дает возможность предположить, что они пересекали узкие водные пространства. К позднему плиоцену или плейстоцену относят- ся различные земляные ленивцы, глиптодонты и броненосцы, которые были более или менее широко распространены в Северной Америке, но позднее исчезли (земляные ленивцы, очевидно, просуществовали в юго- западной части Северной Америки до появления человека). В настоящее время неполнозубые, сильно сократившись в числе и разнообразии, вновь приурочены к Южной и Центральной Америке, и только один броненосец распространен на севере до Техаса.
История рода Pholidota (панголины) на основной части Старого Света не известна и, вероятно, не столь важна с точки зрения зоогеографии.
Хищные претерпели ряд неоднократных и последовательных рассе- лений в основной части земного шара, включая Северную Америку, но они не достигли Австралийской области (кроме случаев распространения их человеком) и, вероятно, до верхнего миоцена (Procyonidae) или позднего плиоцена и плейстоцена (другие семейства) в Южной Америке не появ- лялись. Самые ранние хищные — исчезнувшие креодонты — появились в палеоцене и доминировали в эоцене. Их эволюция и расселение были сложными. Классификация Симпсона (Simpson, 1945) предполагает, что из них по крайней мере 10 групп перемещались в том или другом направ-
Л1лекопит nic~ig ие
2М
ленни между Старым Светом и Северной Америкой. Основное расселение наземных хищных происходило позднее (главным образом в течение среднетретичного периода и после) и носило сложный характер. Кунице- подобные (iMiacidae) в эоцене были общими для Евразии и Северной Аме- рики. Расселение собачьих (Canidae) было сложным и последовательным. Распространение современных и ископаемых форм показывает, что из них по крайней мере дюжина различных групп в разные времена пере- мещалась из Г" вразии в Северную Америку или обратно и по крайней мере три или четыре группы достигли Южной Америки. Медведи, которые филогенетически представляли больших собак с плоскими ногами, оче- видно, возникли в Евразии в течение миоцена, но никогда не достигали основной части Африки. Они перемещались между Евразией и Северной Америкой по крайней мере три-четыре раза и один раз достигли Южной .Америки. Енотовые находились в Евразии (панда и др.) и Северной Аме- рике (еноты и др.) начиная с миоцена. К верхнему миоцену (до образова- ния центральноамериканского сухопутного моста) они достигли Южной .Америки.
История расселения куньих начиная с нижнего олигоцена была слож- ной и продолжительной с очевидной радиацией в миоцене и плиоцене, которая предшествовала радиации современных форм. Было по меньшей мере девять перемещений куньих между Старым Светом и Северной Амери- кой (не обязательно все в одном направлении) и не меньше четырех про- никновений в Южную Америку. Иммигранты Южной Америки являлись предком (или предками) эндемичных родов подсемейства Alustelinae, одного или нескольких родов выдр, скунса и американского длиннохво- стого горностая. Расселение виверровых, очевидно, также было сложным. Они распространялись в течение более чем половины третичного периода на более теплой части Старого Света, достигли Мадагаскара, а также ост- рова Сулавеси и, возможно, Молуккских островов и т. д. Но достоверных данных о том, что какой-либо представитель виверровых проник в Север- ную Америку, не имеется. Гиены, происходящие от виверровых, претер- пели второстепенную радиацию в Евразии и Африке. Кошачьи (Felidae) подвергались сложной и последовательной радиации: вымершие ложно- саблезубые и саблезубые — со среднетретичного времени до плиоцена п плейстоцена соответственно, а современные кошачьи — главным образом в плиоцене и плейстоцене. Несколько представителей семейства Felidae перемещалось между Старым и Новым Светом в различные периоды, а несколько, включая представителя саблезубых и также роды Felis и Pantliera. достигли Южной Америки. Кошачьи позволяют предполагать возможность быстрого и сложного замещения последовательно главен- ствующих групп. Семейство Felidae имеет продолжительную и сложную историю распространения, но современные представители этого семейства почти все относятся к близкородственным доминирующим родам, которые расселились сравнительно недавно и повсюду заместили всех других пред- ставителей этого семейства (кроме гепарда).
Ранние копытные примерно с начала третичного периода имели слож- ную, важную с зоогеографической точки зрения, но плохо изученную исто- рию географического размещения. Они не известны в мелу, но были разнооо- разны в палеоцене. Самые обобщенные Condylarthra (архаические копыт- ные) являлись, так сказать, «предкопытными». Они были близки к общим предкам и хищных и копытных, стояли, по-видимому, немного выше про- тонасекомоядных и сильно отличались по своим размерам и характеру адап- таций. Некоторые из них были очень мелкими животными, а некоторые
Глава 6
вместо копыт имели когти и, вероятно, вели древесный образ жизни. В начале третичного периода Condylarthra, очевидно, достигли всех кон- тинентов, кроме Австралии. Если они являлись предками копытных, то, вероятно, достигнув Африки, стали там родоначальником хоботных (Proboscidea). Из них четыре-пять семейств находились в Северной Амери- ке и (или) в Европе в палеоцене и эоцене и одно из североамериканских семейств, очевидно, было представлено в Южной Америке. Шестое семейство Didolodontidae известно (но недостаточно хорошо) только в Ю к- ной Америке с верхнего палеоцена до миоцена. Возможно, представители этого семейства были сохранившимися остатками общего предка (если таковой имелся) всех третичных копытных Южной Америки (Simpson, 1945, р. 234).
В течение третичного времени копытные Южной Америки претерпели чрезвычайную эволюционную радиацию. Симпсон характеризует боль- шинство из них как протоунгулятес (первичнокопытные) и называет их архаичными. Они не предшествовали современным копытным, а вели параллельную им линию в другой части земного шара. Южноамериканские копытные (Notoungulata) охватывают четыре или пять отрядов, кроме семейства Didolontidae. Литоптерны с двумя семействами и примерно 40 родами, приводимыми в списке, были приурочены к Южной Америке с верхнего палеоцена до плиоцена и плейстоцена. Notoungulata известны по 13 семействам и около 100 родам, существовавшим в Южной Америке с палеоцена до (в некоторых случаях) плиоцена и плейстоцена, а за пре- делами Южной Америки — по единственному семейству Arctostylopidae, с двумя родами в верхнем палеоцене Монголии и в нижнем эоцене Север- ной Америки. Неизвестно, являлись ли северные Notoungulata остатками северных анцестральных ветвей или же они распространились из Южной Америки. Astrapotheria с двумя семействами и девятью родами были приурочены к Южной Америке с эоцена до миоцена. Pyrotheria с четырьмя родами находились в Южной Америке в течение эоцена и олигоцена. 3ТО были очень своеобразные животные, некоторыми чертами своего строения похожие на слонов (это считается параллелизмом) и, очевидно, не родственные никаким другим животным Южной Америки. Два дополнительных палеоценовых рода, без должного основания припи- сываемых Симпсоном к этому отряду, были впоследствии отнесены Куто (Couto, 1952) к новому отряду Xenungulata. Вероятно, это самые древние из известных южноамериканских млекопитающих. Тот факт, что такие ранние формы, по-видимому, относятся к особому отряду и в общих чер- тах не являются анцестральными протоунгулятами, лишний раз показы- вает, как мало известно о ранней истории копытных.
Откуда берут свое начало копытные Южной Америки, не известно. Если современная классификация ранних копытных действительно отражает их филогению, то, видимо, по крайней мере четыре или пять представителей их достигли Южной Америки самостоятельно: один или два Condularthra, один Notoungulata и предки Xenungulata и пиротерии, если только Notoungulata не расселялись из Южной Америки (вместо того чтобы вселяться в нее). Но это отчасти предположительная классификация. Вполне возможно, что все эндемичные группы копытных Южной Америки произошли от одного предка, по-видимому от,ondylarthra.
Копытные Южной Америки независимо от их происхождения эволю- ционировали в полной изоляции от остальных частей света. Они были исключительно разнообразны как по размерам, так и по адаптациям и по образу жизни. При этом они обладали признаками, присущими различ-
il1.ãåê0пи~иаюи~ ие
ным животным, начиная от грызунов до лошадей, верблюдов, носорогов, слонов и Chalicotheria. Впрочем, эти сравнения неточны. Некоторые копытные Южной Америки не были похожи ни на одно современное живот- ное. Как говорит Ромер (Romer, 1945, р. 392), мы не можем представить себе облик этих животных и не имеем для них общеизвестных названий. Описывая их, Ромер (Romer, 1945) и Скотт (Scott, 1937) приводят все имеющиеся в их распоряжении сведения. Возможно, эти животные суще- ствовали в исключительном разнообразии в Южной Америке на протяже- нии большей части третичного периода. Ископаемые остатки их приуро- чены в основном к южной части Южной Америки (южнее тропиков). В тро- пических лесах существовали и многие другие специализированные животные. Они достигли своего расцвета примерно в олигоцене, затем cта.~u уоывать, и к концу плейстоцена, когда современные копытные и хищные звери заняли Южную Америку, вся группа старых южноамериканских копытных полностью исчезла. Северную Америку южноамериканские копытные, очевидно, не заселяли даже временно, как это имело место среди некоторых других южноамериканских млекопитающих, и они не оставили там специализированных реликтов, как это произошло у непол- нозубых Южной Америки.
За пределами Южной Америки эволюционировало несколько разных основных линий копытных, которые расселялись с неоднократными и последовательными радиациями. В Северной Америке временно домини- ровали частично в палеоцене и эоцене Pantodonta и Uintatheriidae, кото- рые являются отдельными отрядами. Они имелись также в Евразии, где пантодонты просуществовали до середины олигоцена. Возможно, эти отряды были прямо родственными (а возможно, и нет) ранним африкан- ским хоботным и южноамериканским пиротериям. В северных районах два надсемейства непарнокопытных (Titanotheria и Chalicotheria) временно доминировали частично в эоцене и олигоцене. Chalicotheria сохранились до более позднего времени — до плейстоцена в некоторых районах Азии— и достигли Африки. Последние представители этих животных, возможно, находились в тропиках Старого Света. Большинство других основных групп копытных, которые главенствовали на основной части земного шара, сохранились до настоящего времени как реликты.
Африканский трубкозуб — единственный современный представи- тель отряда Tubulidentata, скудные и сомнительные ископаемые которого найдены в Евразии и Северной Америке, вероятно, произошел от древнего протоунгулята Condylarthra, но это лишь только предположение.
Proboscidea — слоны и др. являются «ядром» Симпсоновского надотряда Paenungulata, в который входят также Embrythopoda (Аrsinoitherium) из нижнего олигоцена Египта, водная сирена, возможно, африканские даманы (см. ниже), пантодонты (предположительно) и Uintatheriidae (см. выше) Северной Америки и Евразии и пиротерии Южной Америки. На рис. 40 схематично представлена примерная история географии хо- ботных. Их первые находки известны из верхнего эоцена и нижнего оли- гоцена Египта, но в Африке эти животные, вероятно, эволюционировали и, несомненно, имели до этого еще целый ряд более древних форм. В те- чение этого же периода Африка, возможно, отделялась от Евразии морем Тетис. Более поздняя история хоботных представляла яркую картину расселения последовательных групп из Африки или по крайней мере из основной части Старого Света. По мере того как возникали новые группы, они распространялись с удивительной быстротой, и рано или поздно ооль- шинство из них, за исключением нескольких ветвей (с миоцена до плейсто-
298
цена), проникло в Северную Америку. Число видов хоботных, которые распространились в Северную Америку, вероятно, достигает по меньшей мере дюжины, а возможно, и больше. Очевидно, происходили и некоторые обратные перемещения этих животных из Северной Америки в Евразию. но какие, не ясно. И, наконец, в позднем плиоцене и плейстоцене три группы мастодонтов и, возможно, также мамонтов достигли Южной Аме- рики. Эти краткие сведения резюмированы из работы Скотта (Scott.
1937. р. 265 — 301), который в свою очередь позаимствовал их из знамени- той монографии Осборна по хоботным. В позднетретичное время и в плей- стоцене хоботные были все еще многочисленными на всех континентах.
1, ° °
° °
е' °
' 'е ° '..'.
Ф
е1 ° ' . ° ° ' °
r~
° - °
° ° °
'ф
t ° ° .~ a ~i i° °
е'е~е е МВ '
° ° ° °
.с
° ° е ° е~
° Ф ее,
° в
° ° °
$~O
° ° е ~~Ф ° Ñ
° °
ю ° ee
Ъ ° °
1 ° °
° Э
° °
° Ю ° ' °
е
° ° е е ю
° °
Ю
е
~~ °
° ° ° ° ° O ° ° с. ° ' °
° ° r °
ъ ° ° °,
° ° °
° ° ° ° °
ъ
' е~. ~е ° е
° °
° в ° ° ° ° ° .ее °
° ° °
°,ъ ° °
ее
ее ~р
° °
° °
Ф
° у
Ю
° ° ° °
° °
° ' ° Ъ °
° ° ° ° с ° °
° е ° ° ° ° ° ° °;( °, + ° ° ° ° е ° ° ° ~ ° ° ~е ° ° °
° ° ° ° °
ее.
° ' lI °
° °
° ° °
~4я.
.с,
„ф;
° ~ е ',е елее ° ° ~е
'е
° °
1 °
° ° е °
Ф
'е ',е ° .~
° е Ь
° ° °° ° е ее° °° 1 °,, ° °s ° Ь° °° ° ° е|е° ° е,1
° Ф
° °
° ° °
° °
° °
° > ° °
° °
° °
° ° °
° °
° ° )9
° ° ° е~
е ° °
° °
е,
° ° ~ е ю Ч,
4 ° °
° ° ф ° \.
~ю
° °
'sr
в
, ° ° ° ° °
° ° ° ~r
4 ' ° ° ° °
° е
° ° ° °
° °
'ЛФ.-Р
ю
° °
° ° ° °
° ° ,r
° ° '. е ° ее ° ° °
.:.:г;.
° ° ° ° ° °
° ° ° °
° ° °
° е
° °
с
° ° ° ° ° °
'е
I °
Р и с. 40. Распространение хоботных.
веерный круг со стрелками — место происхождения и расселен пе этой группы II3 Африки; дуга — протяженность территории, на которой эволюционировали хоботные, включая южную часть Евразии; тонкие стрелки — перемещения по крайней мере две- надцати ветвей хоботных в Северной Америке, три из них дости- гают Южной Америки. Жирные линии по диагонали — приблизи- тельное распространение двух выживших видов слонов.
.кроме Австралии, и существовали как в холодном, так и в жарком кли- мате. Но с тех пор за сравнительно короткое с геологической точки зрения время они сократились до двух видов, относящихся к разным родам и приуроченных к основным районам тропиков Старого Света.
Даманы, так же как и хоботные, вероятно, возникли в Африке и, по всей видимости, эволюционировали там до появления их в нижнем оли- гоцене Египта. Когда-то они были довольно разнообразны, но в отличие от хоботных, очевидно, не распространялись за пределы Африки и района Средиземноморья. В настоящее время даманы сократились до нескольких видов. которые встречаются в Африке и в юго-западной части Азии. Некоторые современные формы являются древесными, аналогично несколь- ким, возможно раннетретичным, Pre- или Protoungulata.
Непарнокопытные (Perissodactyla) были разнообразны и многочислен- ны на протяжении большей части третичного периода и включали титано- териев и халикотериев (см. выше). В настоящее время сократились до нескольких видов лошадей и т. д., тапиров и носорогов.
Лошади с их предками и родственными видами (семейство Equidae) представлены в ископаемом состоянии или по крайней мере описаны луч- ше всех других животных. Первые примитивные лошади, вероятно, евро- азиатского происхождения, относятся к нижнему эоцену Европы и Се-
Млекопитпют,ue
299
верной Америки. Вероятно (но не абсолютно достоверно), вся основная линия их дальнейшей эволюции, которая достигла высшей точки у рода Equus, происходила в Северной Америке, хотя ограниченные вторичные линии эволюционировали и затем исчезли в Старом Свете. Начиная с эоцена и олигоцена и особенно в миоцене и плиоцене в Северной Амери- ке происходили последовательные расселения родов, из которых три рас- пространились в Евразию, а один из них — также и в Африку. Но все роды, которые достигли Старого Света, исчезли. Затем большинство из пяти или большего числа плиоценовых линий исчезли даже в Северной Америке, но одна линия сохранилась, снова радиировала и положила начало (все еще в плиоцене) роду Едииг в Северной Америке, где он дал многочисленные виды и затем широко распространился по всему земному шару. В плейстоцене Едииг встречался на всех континентах, кроме Авст- ралии, являясь в то время единственным родом семейства Equidae, существующим за пределами Южной Америки. Что касается последней, то там он сосуществовал с тремя эндемичными родами, относящимися к од- ной или нескольким более ранним ветвям (поздний плиоцен?). И наконец род E((uus и ему близкородственные в Северной и Южной Америке стали быстро исчезать, составив только несколько современных видов Equus в Евразии и Африке. Но это изложение не может отразить должным образом сложность расселения всего семейства Equidae.
Тапиры также имели относительно сложную, но не очень хорошо известную историю расселения в Европе и Северной Америке в эоцене и позднее. Сведений о том, что они достигали Африки, не имеется. Настоя- щие тапиры берут начало в олигоцене и продолжают существовать в Евро- пе, Азии и Северной Америке в плейстоцене, распространяясь в районы с, прохладным климатом. Но с тех пор их распространение ограничивает- ся лишь частью района Малайи и тропиками Америки.
Носороги были представлены разнообразными формами в Северной Америке и Евразии в эоцене и уже тогда имели многократное расселение. Носороги, в более строгом смысле, претерпели сложную эволюцию, значительно более сложную, чем у лошадей, и, предположительно, слож- ное расселение на большей части Старого Света. Вполне вероятно, что несколько разных ветвей из Старого Света достигли Северной Америки, но подробности и в некоторых случаях направления их перемещений не установлены. Ископаемые остатки носорогов в Евразии известны из среднего миоцена и плейстоцена. В Африке их относят к миоцену (но, возможно, что они были там и в более раннее время), а в Северной Аме- рике — от верхнего эоцена до какого-то периода плиоцена. В Южной Америке носороги не были известны ни в одну эпоху. В плейстоцене Евразии они существовали как в холодном, так и в теплом климате. В на- стоящее время несколько оставшихся видов отступили в тропики Старого Света.
Парнокопытные (Artiodactyla) охватывают ряд исчезнувших третич- ных групп, некоторые из которых были общими по крайней мере для Евра- зии и Северной Америки, а также несколько групп, которые пришли в упадок, и доминирующие группы (свиньи, олени и комплекс антилопы- оыки). Вообще говоря, в настоящее время парнокопытные являются доминирующими и как таковые заместили большинство непарнокопытных (Perissodactyla).
Разнообразные формы свиных (Suidae) существовали на основной части Старого Света начиная с олигоцена и миоцена. Расселяясь, они достигли острова Сулавеси, но не Америки. Пекари и др. (Tayassuidae)
800
Глава 6
были представлены отдельными подсемействами в Евразии с нижнего олигоцена до HNiEHero плиоцена. В Северной Америке они находились с нижнего олигоцена, а в Южной Америке появились в плейстоцене.
Современные африканские бегемоты (Hippopotamidae) — последние сохранившиеся представители надсемейства, в которое входили Anoplot- heriidae Европы (средний эоцен до среднего олигоцена) и Anthracotheriidae Евразии (средний эоцен до плейстоцена), Африки и Северной Америки (олигоцен и нижний миоцен). Ископаемые собственно бегемотов относятся к позднему плиоцену и плейстоцену Евразии и Африки и к плейстоцену Мадагаскара.
Верблюды и их непосредственные предки, очевидно, были приуроче- ны к Северной Америке в течение почти всей их сложной истории, с позд- него эоцена и на протяжении всего остального третичного времени, и были особенно многочисленны в Северной Америке в позднетретичное время. В Евразии, Северной Африке и Южной Америке верблюдов до плейстоцена не было. Ныне они сохранились только в некоторых частях Азии и Южной Америки, а на своей старой родине — Северной Америке исчезли.
Оленьки Tragulidae были многочисленны в Евразии (и, возможно, так ке в Африке, хотя ископаемых форм там обнаружено не было) в позд- нем эоцене и олигоцене. У семейства Hypertragulidae примерно в это же время происходила параллельная радиация в Северной Америке. Поз- же обе эти группы пришли в упадок. Североамериканские формы, очевид- но, исчезли в миоцене и раннем плиоцене. Исчезло и большинство форм Старого Света, но род ХуетогсЬиг и по сей день существует в Западнои Африке, а род Tragulus — в тропиках Индо-Малайской области. Конечно, это слишком упрощенное изложение сложной географической истории семейства Tragulidae.
Семейство оленевые (Cervidae), вероятно, произошло от ранних Tragulidae в олигоцене Евразии. Примитивная ветвь (подсемейство Palaeomerycinae) проникла в Северную Америку в миоцене, но в течение плиоцена там исчезла. Возможно, в настоящее время эта ветвь представ- лена в горах Азии кабаргой (1V1oschus), широко распространенной от Гималаев до Восточной Сибири и острова Сахалин. В дальне~1шем эволюция оленей протекала главным образом в Евразии, но специфиче- ское подсемейство Dromomerycinae радиировало в Северной Америке в миоцене и плиоцене, а затем исчезло. Основная линия эволюции оленевых на одной из стадий (подсемейство Muntiacinae) была приурочена к Евразии и представлена двумя современными родами в Азии. Завершилась она образованием подсемейств Cervinae и Odocoileinae. Первое доминируе" в Евразии и приурочено к ней, за исключением рода Cervus, распростр ненного и в Северной Америке (олень-вапити). Второе представлено в Евразии, но особенно характерно для Америки в целом, включая и Ю,к- ную. Подсемейство Odocoileinae, вероятно, произошло в Евразии и засе- лила Северную Америку в плиоцене, а Южную Америку — в плейстоцене или незадолго до плейстоцена. Роды Alee и Rangifer относятся к северно- му подсемейству Odocoileinae, которое, как считает Скотт (Scott, 1937, р. 322), проникло в плейстоцене из Евразии в Северную Америку. Основ- ной части Африки, южнее Сахары, олени, очевидно, никогда не достигали. История географии семейства Cervidae в основном излагается по Симпсону (Simpson, 1945), и независимо от того, насколько она правильна в деталях, совершенно ясно, что эволюция и расселение семейства ("ervidae были сл ожными.
Млеко и ииг аю щие
Жирафы родственны оленям и в какой-то степени замещают их в тропиках Старого Света. С миоцена до плейстоцена они были многочис- ленны в Азии (и, возможно, также в Африке?). В нижнем плиоцене несколь- ко видов достигли восточной части Европы. К настоящему времени эти животные сократились до двух видов: (обыкновенная) жирафа, обитаю- щая на открытых пространствах Африканского континента (южнее Саха- ры), и окапи, встречающийся на ограниченной территории в густых лесах Западной Африки.
Вилорогие антилопы (Antilvcapra) западной части Северной Америки являются последними выжившими представителями этого семейства,
4У Ю
°,
° е
° °
° g e
. ':мл':.м.
,''., °
° е ° °, е ° '
В
° ю
г
° ° ° °
° °
° °
° °
Ю
° рФ °
° '
° °
° l
° у °
° ~
° °
° ° °
° °
° °
° ° °
° ° °
: . :.О . .:. ":. ': ,.
° °
° °сг
° ° с
° °
° ° °
° ' ° °
° ° ° °
° Ф
° ° ° ° ee °
° °
° °
° °
е ° °
де °
° ~ 1y
° ° ° °
' ° ю
° + ° ° ° ° ° + °
° ° °
° 'l °
ю ° ~ ° ° ° ° ~ю °
° ° ° ° °
Ю ° °
° ° ° °
р ° Ú ° е~
° .' °,1 ф
° ° ~е
° ° ° ° ~ ° ° °
° ° ° ° . ~ ° - ° Р'
° ° ° °
..~:ь ~ ~а'
° °
е ° ° ° °
° ° ° ° ° °
° ° ° °
° ° °
° °
° ° °
° ° ° ° ° ° ° ° °
° ° ° !
Р и с. 41. Распространение семейства бычьих.
Эта схема показывает сложное распространение семейства бычьих. Каждая полоска представляет одну трибу в Старом Свете; черная часть полосы — встречаемость этого семейства (приблизительно) в настоящее время (в Евразии или Африке), а светлая часть — встре- чаемость его в прошлом, подтвержденная ископаемыми данными. Короткие полоски указывают крайние границы распространения этого семейства. Стрелками обозначено перемещение восьми ветвей этого семейства в Северную Америку. Горизонтальные полосы (вверху) представляют трибы Ovibovini, Впр1сарг1п~, Сар ini: полосы по диагонали в Азии — Sai„oini, Bovini, Boselapliirri, Ап1~- 1о1i~ п~; hocbre полосы 8 Африке — Bo~ini (rrosTopHo), Ar>tilopi ni (повторно), Hippotragini, Strepsicerotini, Bedurrci ni, А1се1арй~ni. Cep1ra1opЫп1, Neotraini. Стрелки, направленные к Северной Аме- рике — (ivi~>os, Во» (плейстоцен), Я~гг~«(плейстоцен), снежная коза.
снежный баран и три вида рода Biso~~.
которое радиировало в Северной Америке (в большинстве случаев на за- паде, но один вид достиг Флориды) в позднем миоцене, плиоцене и плей- стоцене. Предок этого семейства, предположительно, проник из Евразии.
Семейство Bovidae, охватывающее антилоп, баранов, коз и т. д., а также других представителей бычьих, было одним из последних наибо- лее доминирующих и широко распространенных семейств млекопитаю- щих. но тем не менее расселение этого семейства было сложным (см. ниже- следующий текст и рис. 4i). Классификация и ископаемые данные этого семейства приводятся по Симпсону.
В подсемействе Bovinae триба Strepsicerotini (6 ископаемых, два со- временных рода, включая антилопу куду и антилопу канна) находилась в Евразии с верхнего миоцена до плейстоцена, в Северной Африке была отмечена в нижнем плиоцене, а на остальной части Африки — не раньше плейстоцена. В настоящее время сохранилась только в Африке, ю'кнее Сахары. Триба Boselaphini (шестнадцать ископаемых, два современных
802
рода, включая антилопу нильгау и четырехрогую антилопу) встречалась. в миоцене и нижнем плиоцене Европы и Азии, но сохранилась только нз полуострове Индостан. Триба Bovini (восемь ископаемых, шесть совре- менных родов, включая бычьих, буйволов и т. д.), ископаемые которой известны в Евразии из среднего плиоцена, а в Африке и Северной Аме- рике — только из плейстоцена, широко распространена в пределах гео- графических границ семейства до сих пор. Два рода Bovi ni достигли Северной Америки: род Bos в плейстоцене был распространен до Аляски, но в настоящее время исчез; несколько видов рода Bison, очевидно. последовательно достигли Северной Америки. Подсемейство Cephalophinae содержит только одну трибу (три современных рода, лесные дукеры), приуроченную к Африке (с плейстоцена до настоящего времени). Подсе- мейство Hippotraginae содержит три трибы. Триба Reduncini (8 иско- паемых, 5 современных родов, включая различных африканских антилоп) относится к плиоцену и плейстоцену Азии и плейстоцену Африки, но в последней сохранилась только южнее Сахары. Триба Hippotragini (14 ископаемых, 3 современных рода, включая различных антилоп) отно- сится к нижнему плиоцену Европы, плиоцену и плейстоцену Азии и плей- стоцену Африки, но в настоящее время она представлена только в Африке и юго-западной части Азии. У трибы Alcelaphini (3 ископаемых, 5 совре- менных родов, включая бубала, гну и др.) ископаемые датируются ниж- ним плиоценом восточной части Европы и плейстоценом Азии и Африки. но в настоящее время ее представители встречаются только в Африке южнее Сахары. В подсемействе Antilopinae триба Neotragini (одно иско- паемое, 8 современных родов, включая различных африканских антилоп) известна только в Африке южнее Сахары (плейстоцен и настоящее время); триба Antilopini (8 ископаемых, 7 современных родов, антилопы и др.) относится к нижнему плиоцену Европы, плиоцену и более позднему времени Азии и плейстоцену Африки. Эта триба сохранилась и широко распространена в Азии и Африке. В подсемействе Caprinae триба Saigini (два современных рода) встречается от восточной части Европы до Цен- тральной Азии. Ископаемые обоих родов этой трибы относятся к плей- стоцену Евразии, а один из них (Saiga) и к плейстоцену Аляски. Триба Rupicaprini (3 ископаемых, 4 современных рода, включая серну и др.) находилась в Евразии с плиоцена, где широко распространена и в настоя- щее время, в основном в горах (от отдельных районов Испании, в Альпах и т. д. до Кавказских гор и от Гималаев до Маньчжурии, Японии, Тайваня и Суматры). В западной части Северной Америки эта триба представлена известной здесь с плейстоцена и существующей в наше время снежной козой (Oreamnos), но которая не является настоящей козой. Несколько сомнительная триба Ovibovini (9 ископаемых, два современных рода), ископаемые которой встречаются в ния;нем плиоцене и повторно в плец- стоцене Евразии и в плейстоцене Северной Америки. Ныне сохранилась в Восточно-Центральной Азии (Budorcas, такин, помещен в эту трибу условно) и в арктической части Северной Америки (Ovibvs, мускусный бык). Триба Caprini (5 ископаемых, 5 современных родов, включая козлов и баранов), ископаемые которых известны из плиоцена и плейстоцена Европы и Азии и из плейстоцена в Северной Африке и западной части Северной Америки, встречается в настоящее время главным образом в горных районах. Западной части Северной Америки достиг только один род этой трибы (Ovis, бараны).
Несколько пространно изложенные подробности классификации и распространения бычьих в иных случаях могут оказаться ошибочными,
Млекопитпн тие
808
а многие детали истории этого семейства вообще неизвестны (их ископае- мые данные в Африке очень неполные), но основной характер сложной эволюции и расселения этого семейства на основной части Старого Света примерно с шестью ветвями, достигающими Северной Америки, и некото- рыми группами, отступающими с обширных территорий, несомненно, изложен верно. Мускусный бык, вероятно, из Северной Америки вновь возвратился в северную Евразию, где в течение плейстоцена исчез. Бычьи не достигли Австралии и, очевидно, также и Южной Америки, хотя род Bison проник в северную часть Центральной Америки. По сложности своего расселения бычьи не являются исключением. Многие другие доминирующие семейства млекопитающих, видимо, совершали такие же сложные географические перемещения, но свидетельствующих оо этом ископаемых значительно меньше.
Первый из двух отрядов доминирующих «грызущих» млекопитающих Lagomorpha имеет наименее сложную историю. Самым древним родом «зайцев» является Eurymylus из верхнего палеоцена Азии, но связи его с, другими, более поздними зайцеобразными мне не известны. Сеноставки эволюционировали в Евразии и, очевидно, несколько раньше, приоли- зительно со среднетретичного времени, в Африке, и с этого времени начи- нается их упадок. Северной Америки (ее западной части) они, видимо, до плейстоцена достичь не смогли. Leporidae (зайцы и др.) имели ряд после- довательно сменяющихся расселений на основной части земного шара, включая Северную Америку: одного или нескольких представителей Palaeolaginae (если это естественная группа), которые оставили реликто- вые роды (не имеющие прямого родства) в Южной Африке, на островах Рюкю и в центральной Мексике; одного или нескольких из позднетретич- ных Leporinae и одного из рода Lepus, радиация которого продолжается до сих пор. Яу1ьйадиг — единственный род зайцев, который достиг Центральной и IOiEHOH Америки, до плейстоцена там неизвестен.
Грызуны очень многочисленны, но их взаимосвязи не всегда хорошо известны и понятны, а ископаемые часто недостаточны. Возможно, по сравнению с любым другим отрядом наземных млекопитающих они имеют более сло'кное расселение, но как правило, его детали малоизвест- ны и даже основные особенности в ряде случаев вызывают сомнение. Поэтому приводимое ниже изложение крайне упрощенно и отчасти пред- положительно.
Анцестральные грызуны (Aplodontoidea) появились в верхнем палео- цене Северной Америки. Их подъем относится к раннетретичному времени. Ископаемые остатки этих животных были найдены только в Северной Америке и Евразии, но, видимо, они обитали также и в Африке. Послед- ним представителем Aplodontoidea (а скорее, их наименее видоизменен- ный потомок) является семейство Aplodontidae, бобровые белки, которое находилось в Северной Америке с верхнего эоцена, и в Азии также и в нижнем плиоцене. В настоящее время оно сократилось до одного вида (Aplodontia), обитающего в западной части Северной Америки.
Беличьи и др. (Sciuroidea), очевидно, имели сложное расселение на большей части земного шара, включая Северную Америку. Южной .Аме- рики настоящие белки достигли, вероятно, сравнительно недавно.
Мешетчатые крысы и карманчиковые мыши и т. д. (Geomyoidea) существовали в Северной Америке. в основном на западе, со средне- третичного времени. До Панамы (гоферы) и северной части Южной Америки (карманчиковая мышь) они распространились, вероятно, недавно.
Глава б
[эобры, их предки и родственные им формы (Castoroidea) были много- численны в Северной Америке и Евразии с середины третичного периода до плейстоцена, но в настоящее время сократились до одного рода с одним евроазиатским и одним североамериканским видом.
Шипохвостые белки (Anomaluroidae) в настоящее время приурочены к Африке, а те, которые (предположительно) приурочены к третичному периоду Евразии и Африки, вызывают сомнение. Долгоноги (Pedetidae) находились в южнон части Африки по краиней мере с нижнего миоцена.
Первыми мьпиеподобными грызунами были хомячьи (Cricetidae), которые появились в олигоцене Евразии и Северной Америки и, вероят- но, когда-то доминировали на большей части земного шара, включая Африку. В противном случае они не смогли бы достичь Мадагаскара, где имеется по крайней мере один представитель этого семейства. Числен- ность хомячьих значительно сократилась в Африке и тропической Азии (если последние были там многочисленны), но они до сих пор доминируют в умеренных широтах Евразии и особенно в Северной Америке. Южной Америки хомячьи достигли, очевидно, в верхнем плиоцене и также стали там доминировать. Три рода являются общими для Северной и Южной Америки, и несколько дополнительных ветвей, по-видимому, достигли Южной Америки и образовали там эндемичные группы: в настоящее время около 40 родов Cricetidae являются эндемиками Центральной и Южной Америки, но какое количество первичных иммигрантов они представляют. не известно.
Второй большой комплекс мышеподобных грызунов представляет семейство Muridae, которое, вероятно, образовалось от Cricetidae где-то в тропиках Старого Света, возможно в тропической Азии, так как мыши- ные достигли Австралии, но не Мадагаскара. Очевидно, подъем мышиных произошел уже на поздней стадии истории млекопитающих, скорее всего, в конце миоцена (Simpson), но они вполне могли радиировать в теплых частях Старого Света. В тропической Азии и в Африке они заместили большинство хомячьих (если последние были там многочисленны) и не- сколько (пять?) из них достигли Австралийской области в разное время и там радиировали. Америки, за исключением случаев занесения их человеком, мышиные не достигли. Сказанное здесь — это схематич- ное изложение истории распространения, которая включала многократ- ные и последовательные расселения, специализации и локализации гру.пп, таких, как Spalacidae и Rhizomyidae, в разных частях Старо- го Света.
История расселения сонь и им родственных (Gliroidea) на основной части Старого Света не известна. Многие из них локализованы или имеют прерывистое распространение, что дает основание полагать, что эта группа приходит в упадок.
Тушканчики и т. д. (Dipodoidea) произошли от непрыгающих форм, из которых один род до сих пор существует в умеренной зоне Евразии. Прыгающие Zapodinae возникли в Евразии (один представитель обитает и в настоящее время в западной части Китая) и, вероятно, в миоцене распространились в Северную Америку. Тушканчики, ви- димо, самостоятельно возникли от них в более засушливых частях Евразии и Африки.
Дикобразоподобные образовались от одного или нескольких предков. которые жили в раннетретичное время. Их радиация в Старом Свете, в Евразии, а также в Африке (значение Африки в связи с этим в какой-то
Млекопитаь щие
ЗОВ
степени преувеличено) была незначительной. Ископаемые дикобразы Старого Света известны из олигоцена Евразии (в настоящее время встре- чаются в Индо-Малайской области и в Африке), и несомненный предста- витель Thryonomyidae найден в Индии. На Мадагаскаре дикобразы отсутствуют, и это указывает на то, что, возможно, в Африке они никогда не оыли многочисленными. В Северной Америке дикобразоподобные не известны до позднего плиоцена. Тем не менее они очень давно, неизвестно каким путем, проникли в Южную Америку. В последней их ископаемые относятся к нижнему олигоцену. Еще более ранние ископаемые остатки позволяют представить их эволюцию и предположить, что они берут начало от одного предка, который, возможно, достиг Южной Америки в позднем эоцене (% оой, 1950, р. 88). От этого предположительного предка радиировали различные крыса- и зайцеподобные формы, включая морских свинок, шиншилл, агути, нутрий и т. д., а также несколько полуводных форм (к которым относятся водосвинки), древесных дикобразов и многие другие исчезнувшие формы, некоторые из которых были очень крупными животными, что подтверждается размером их черепов, достигающих в длину 2 футов. Эта радиация достигла своего максимума в плиоцене. Охватывала ли первоначально она Центральную Америку и Вест-Индию, не известно, так как нет соответствующего палеонтологического материала. Но рано или поздно некоторые дикобразоподобные должны были достиг- нуть этих районов. В позднем плиоцене и плейстоцене отдельные дико- бразоподобные распространились из Южной Америки в Северную. Но из них до наших дней сохранился только один вид — американский дикоораз. Многие ли, встретив дикобраза в северных лесах Америки, знают, что это недавний иммигрант из тропиков Южной Америки? Водосвинки (два рода) достигли юго-восточной части Соединенных Штатов, по крайней мере Южной Каролины, в плейстоцене, но теперь они отступили как из Север- ной Америки, так и с большей части Центральной. Возможно, это было частью недавнего обширного отступания всех дикобразоподобных, за исключением наиболее доминирующих, из центральной Америки и се- верной части Южной Америки. Такое отступание подтверждается совре- менной концентрациеи этих животных в южной части Южной Америки (см. список семейств), отсутствием древних эндемичных родов в Централь- ной Америке, а также таксономической изоляцией родов Вест-Индии. Недавнее проникновение с севера недикобразоподобных грызунов и других млекопитающих в Центральную и Южную Америку довольно хорошо объясняет отступание многих дикобразоподобных к югу, а также исчезно- вение гигантских плиоценовых форм.
Каким образом дикобразоподобные достигли Южной Америки? Можно предложить довольно много вариантов решения этого вопроса. В миоцене в Южной Америке так же нео;киданно появились обезьяны. Возможно, что все же и дикобразоподобные, и обезьяны достигли Южной Америки через Северную, несмотря на отсутствие ископаемых в пределах последней. Это наиболее вероятное решение, так как если в те времена обе эти группы были лесными и древесными обитателями, то маловероятно, чтобы в Се- верной Америке были найдены их ископаемые остатки, и вероятнее всего, что и та и другая группа достигли Южной Америки через узкие водные пространства. С другой стороны, южноамериканские дикобразоподобные и обезьяны, возможно, являются продуктом параллельной эволюции и не имеют прямого родства со своими двойниками из Старого Света (Wood, 1950). Наконец, дикобразоподобные могли на плоту пересечь Атланти- ческий океан и таким ооразом попасть в Южную Америку из Африки.
Глава 6
Остановиться окончательно на каком-либо из этих предположений моя;но лишь при наличии достаточно веских доказательств.
Расселение рукокрылых еще более сложное. 'Aiegachiroptera (плодояд- ные летучие мыши, или летучие собаки) ведут начало с олигоцена или еще более раннего времени, и история их расселения в тропических частях Старого Света была сложной. Эти животные, обладая зна хительной спо- собностью к расселению, распространились на многие острова Индийско- го и западной части Тихого океанов. Но Америки они не достигли. М1сг<>- chiroptera («насекомоядные» летучие мыши, но в действительности очень разнообразные по характеру своего питания: болынинство питается насе- комыми, некоторые виды — фруктами, рыбой, а некоторые — другими рукокрылыми и т. д. или кровью), вероятно, неоднократно pacceля- лись по всему земному шару, но ископаемых, подтверждающих эти перемещения, почти нет. Поэтому сказать что-либо об их самых ранних перемещениях, невозможно. Семейство Emhalloliuridae радиировало довольно давно, вероятно в эоцене. Распространение этого семейства пре- рывистое и тропикополитное. Все роды Старого и Нового Света разные. Современные Molossidae расселились несколько позднее. Они не строго приурочены к тропикам, и один их род до сих пор является общим для Старого и Нового Света. Современные Vespertilionidae, возможно, распро- странились еще позднее: это космополитическое семейство имеет несколько общих родов для Старого и Нового Света. Доминирующий и космопо- литический род Myotis (семейство Vespertilionidae) расселился по срав- нению с другими совсем недавно. Достопримечательно, что два предста- вителя рукокрылых, достигшие Новой Зеландии, очевидно, проникли туда в разное время: один очень давно, а другой недавно.
На этом изло'кение географической истории главных групп наземных млекопитающих заканчивается. Можно добавить, что все основные груп- пы, кроме грызунов, рукокрылых и высших парнокопытных (свиньи, олени, быки и т. д.), по-видимому, начиная с миоцена или плиоцена (Simpson, 1945, р. 34) пришли в упадок и что многие крупные млекопи- тающие в течение пле~~стоцена отступали или исчезали на многих частях земного шара.
Касаясь выше истории каждой группы, я отмечал сложность рассе- ления, но, конечно, весь этот процесс в целом был еще значительно слож- нее. Расселение млекопитающих — это какой-то калейдоскоп разнообраз- ных и меняющихся в направлении, силе и скорости отдельных переме- щен и~~.
И тем не менее самые общие черты расселения млекопитающих можно представить довольно просто (рис. 42). В конце мелового периода, когда рептилии пришли в упадок, на основной части земного шара находились две группы млекопитающих, как бы уже подготовленных к радиации: протонасекомоядные (плацентарные) и Didelphoidea (сумчатые). Все со- временные млекопитающие, вероятно кроме австралийских однопроход- ных, произошли от этих двух групп. В течение третичного периода они эволюционировали в разных сочетаниях более или менее изолированно в трех основных направлениях. Протонасекомоядные дали различных плацентарных, расселение которых на основной части земного шара было очень сложным. Сумчатые достигли Австралии и радиировали там в изо- ляции. Сумчатые и отдельные плацентарные достигли Южной Америки и радиировали там вместе.
Об эндемичных фаунах Австралии и Южной Америки существует два ошибочных представления. Согласно первому, эти фауны более древние,
807
Млекопит аюи~ие
Р и с. 42. Основной характер радиации млекопи- тающих в начале третичного периода.
'Черный круг со стрелками — плацентарные млекопи- тающие; белый круг со стрелками — сумчатые; линии по диагонали — преграды раннетретичного периода.
какой-то известной целой (единой) всемирной фауны, а представляют ре- зультат проникновения отдельных частей фаун на изолированные конти- ненты через водные пространства и притом в разное время.
Два современных рода однопроходных Австралии более специали- зированы, чем любые сумчатые (стр. 287), и можно предполагать, что они являются последними реликтами фауны однопроходных, которая находилась в Австралии задолго до появления сумчатых и которая (если она существовала), вероятно, ведет свое начало с мезозоя. Сумчатые, видимо, проникли позднее, но точное время их появления не известно. Насколько можно судить (фактов, свидетельствующих, в какое время находились сумчатые в Азии и находились ли они там вообще, нет, так ;ке как нет данных и об их пребывании в Австралийской области до плио- цена), предок или предки австралийских сумчатых могли появиться в любое время от позднемелового до первой половины третичного периода an zxslI TezrSaO.
Различные грызуны семейства МигЫае появились еще позднее. ве- роятно в позднем миоцене или плиоцене и плейстоцене. Рукокрылые время от времени, несомненно, проникали в Австралию на протяжении оольшей части третичного периода. Таким образом, австралийская фауна млекопитающих, по-видимому, образовалась от очень немногих предков, которые, преодолев водные преграды, проникли сюда в разное время.
чем фауны млекопитающих остальной части земного шара. Но это не так. Очевидно, плацентарные по своему происхождению столь же древние, как и сумчатые. Обе группы существуют уже с мелового периода. В Юж- ной Америке большая эволюционная радиация млекопитающих началась примерно в то же самое время, что и на основной части земного шара, и нет оснований считать, что радиация австралийских сумчатых проис- ходила раньше. Этот процесс происходил в разных районах, но не в раз- ное время. Второе неправильное представление, сводящееся li тому, что древние млекопитающие, должно быть, достигли Австралии и Южной Америки через (разные) сухопутные мосты, весьма сомнительно. фауны млекопитающих Австралии и Южной Америки неявляются производными
306
Глава 6
Мы не располагаем прямым доказательством, откуда прибыли в Австра- лию однопроходные и сумчатые. Грызуны и рукокрылые, очевидно, яв- ляются выходцами из Азии.
История расселения млекопитающих в Южной Америке более слож- ная, но лучше документирована. Сумчатые, неполнозубые и копытные находились здесь с конца палеоцена.
Судя по этим уже в достаточной степени разнообразным или специа- лизированным формам, можно допустить, что их предки достигли Южной Америки в раннем палеоцене или даже до него. Но сколько предковых форм там находилось и все ли они появились в одно и то же время, сказать трудно. Имеется ряд доказательств, свидетельствующих о том, что эти формы были частью более разнообразной фауны, которая существовала на основной части земного шара: неполнозубые и копытные, очевидно, уже были неполнозубыми и копытными (или протоунгулята), когда они достигли Южной Америки (это подтверждается некоторыми родственными ископаемыми, особенно в Северной Америке). Но некоторые имеющие важное значение млекопитающие палеоцена, очевидно, не могли до- стичь Южной Америки, например насекомоядные и ранние плацентарные хищные.
Подводя итог, можно сказать: один или несколько видов сумчатых и неполнозубых и один или несколько видов копытных достигли (но не обязательно все в одно время) Южной Америки примерно в начале тре- тичного периода, преодолев при этом водные преграды, которые явились препятствием для других млекопитающих. На этом история расселения не закончилась. Дикобразоподобный грызун достиг Южной Америки вероятно, в позднем эоцене. Обезьяны появились в миоцене, но возможно, что они проникли сюда значительно раньше. Енотовые (Procyonidae) достигли Южной Америки в конце миоцена. рукокрылые, очевидно, по- явились в разное время, некоторые (например, Phyllostomatidae), вероят- но, в ранне- или среднетретичном периоде. Что касается ряда других млекопитающих, то не столь важно, проникли ли они в Южную Америк~- до того, как она соединилась полностью с Севернои, или нет. Яся раНННН фауна Южной Америки, по-видимому, образовалась из компонентов фау- ны основной части земного шара, и в связи с преодолением водных пре- град происходил отбор этих компонентов, хотя, возможно, в начале тре- тичного периода эти преграды были менее труднопреодолимыми, чем в более позднее время.
Что касается направления, сумчатые, неполнозубые и два-три вида копытных, по-видимому, проникли в Южную Америку из Северной. Это предположение основывается на ископаемых Северной Америки. Другая группа ранних южноамериканских копытных — пиротерии — во многом напоминает хоботных (главным образом африканских), хотя полагают. что это только параллелизм. Предки южноамериканских обезьян и дико- бразоподобных, возможно, достигли Южной Америки непосредственно из Африки или из Северной Америки, или через нее. Енотовые и все более поздние млекопитающие, достигшие Южной Америки, распростра- нились из Северной Америки. По-видимому, если говорить объективно, все известные млекопитающие, достигшие Южной Америки, могли в лю- бое время проникнуть туда из Северной Америки или из других мест. пересекая ее. Сомневаться в этом чаще всего приходится из-за отсутствия достоверных свидетельств. И все же тот факт, что все млекопитающие. которые в течение третичного периода достигли Южной Америки, про- никли туда из одной исходной «точки» — Северной Америки (через не-
Млеко питаюгцие
престанно изменяющийся архипелаг, который в позднем плиоцене пре- вратился в сухопутный мост), как я думаю, вполне соответствует имею- щимся доказательствам.
С начала третичного периода на земном шаре эволюционировала и расселялась большая фауна плацентарных млекопитающих, отдельные части которой периодически достигали Южной Америки; еще меньше элементов этой фауны и в более позднее время достигло Австралии. Ра- диация этой фауны включала расселение и повторное перерасселение по- следовательно сменяющихся ветвей многих основных групп, находящих- ся на различных эволюционных уровнях: насекомоядных, приматов, различных хищных, многих копытных, зайцеобразных, многих грызунов и рукокрылых, если говорить только об основных группах. Нет необхо- димости повторять все детали этого расселения, но надо остановиться на трех особо важных моментах: значение Африки, особенно в раннетретич- ном периоде, обмен фаунами между Старым Светом и Северной Америкой и обмен фаунами между Северной и Южной Америкой после окончатель- ного соединения этих материков.
Самые ранние известные ископаемые данные по млекопитающим Африки относятся к верхнему эоцену и нижнему олигоцену Эль-Файюма в северном Египте (точнее, несколько градусов севернее тропиков). Иско- паемые формы представляют собой смешение современных европейских и исчезнувших, очевидно, автохтонных африканских групп. Это является фактическим доказательством того, что в то время Европа и Африка были соединены. Однако африканские группы из Эль-Файюма, куда входили первые известные хоботные и даманы, а также Arsinoitherium, настолько резко отличаются от европейских, что можно предположить, что незадол- го до этого Африка некоторое время была изолированным материком. Но тогда, по-видимому, имело место еще более раннее соединение ее с Евразией или по крайней мере существовали условия, при которых в Африку проникали анцестральные ветви плацентарных млекопитающих. Все это наводит на мысль, что основная радиация плацентарных млекопи- тающих в начале третичного периода охватывала Африку, что в то время этот континент был изолирован и что некоторые млекопитающие эволю- ционировали там самостоятельно до позднего плиоцена, после чего Афри- ка вновь соединилась с Евразией. Эта гипотеза соответствет геологиче- ской истории моря Тетис, которое когда-то отделяло Африку от Евразии н через которое могли происходить неоднократные соединения суши с последующими разъединениями.
Превосходный анализ последовательно сменяющихся ископаемых фаун млекопитающих Европы, Азии и Северной Америки позволил Симпсону (Simpson, 1947, 1947а) дать подробную историю обмена млекопитающими, происходившего между Старым Светом и Северной Америкой в течение третичного периода и плейстоцена. В раннем и позднем эоцене, раннем олигоцене, позднем миоцене, со среднего до позднего плиоцена и в плей- стоцене происходил интенсивный обмен наземными млекопитающими между Евразией и Северной Америкой. В раннем и среднем миоцене и раннем плиоцене, вероятно, имел место незначительный обмен. В среднем эоцене, среднем и позднем олигоцене этот обмен был или очень незначи- тельным, или его не было совсем. Небольшое взаимопроникновение про- исходит и в настоящее время. Однако сухопутное соединение между Евразией и Северной Америкой, которое, очевидно, существовало на про- тяжении большей части третичного периода, временами прерывалось (по-ви- димому, происходило погружение): возможно, на какое-то время в палео-
Глава 6
цене ', вероятно, в среднем эоцене, в середине позднего олигоцена, на бо- лее краткий период в первой половине плиоцена и предположительно в другие эпохи, включая и современную.
Существование сухопутного моста, соединяющего через Атланти- ческий океан Европу с Северной Америкой, данными о расселении млеко- питающих до раннего эоцена не подтверждается. Но для объяснения их расселения вряд ли стоит прибегать к помощи Атлантического моста, так как оно происходило через Тихоокеанский мост (Берингия), о чем свиде- тельствуют более поздние сведения.
В раннем эоцене обмен млекопитающими охватывал многие крупные систематические группы и большие части материковых фаун, но в после- дующем усилилась тенденция к включению в обмен более мелких частей фаун и менее различающихся типов животных, принадлежащих к более низким систематическим категориям и экологическим типам, не столь чуждым для новых областей. Основным фактором отбора в данном случае был, очевидно, климат. Группы, участвующие в обмене, вероятно, явля- лись выходцами из мест с относительно прохладным климатом, но не из альпийских районов. В период между ранке- и позднетретичным време- нем отбор таких групп становился все более четко выраженным и интен- сивным, что свидетельствует о постепенном похолодании климата на се- вере на протяжении большей части третичного периода, которое закончи- лось оледенением в плейстоцене. Каждый последовательный обмен, не- сомненно, включал перемещения в обоих направлениях, но, несмотря на то, что направления передвижений многих групп остаются неясными, преобладали, видимо, перемещения из Евразии в Северную Америку.
Группы, которые не участвовали в обмене между Старым Светом и Северной Америкой, играют немаловажную роль в расселении млеко- питающих. Такие группы существовали всегда, и некоторые из них сохра- нились в течение довольно длительного времени. Начиная с эоцена или олигоцена виверровые (кошачьи хорьки, мангусты и т. д.) и Megachirop- tera (плодоядные летучие мыши) широко радиировали на основной, более теплой части Старого Света, но, насколько известно, ни один из предста- вителей этих семейств никогда не достигал Северной Америки. Начиная по крайней мере с нижнего олигоцена высшие приматы Старого Света также претерпели разнообразную радиацию на основной, более теплой части Старого Света, однако до тех пор, пока они не были занесены туда человеком, не имелось никаких сведений о том, что они когда-либо дости- гали Северной Америки. В настоящее время представители всех этих групп проникают на ограниченные территории умеренной зоны южного полу- шария, а ископаемые свидетельствуют о том, что в прошлом эти группы были распространены к северу от тропиков.
Грызуны семейства Миг1йае достигли наивысшей точки своего подъема в Старом Свете (в Америке их не было до тех пор, пока они не были зане- сены туда человеком) только в плиоцене. Менее важные группы млекопи- тающих, включающие грызунов семейства Gliroidae, настоящих свиней и жирафов, по-видимому, были приурочены к основной части Старого Света на протяжении всей их истории. В Северной Америке грызуны семейства Geomyidae (мешетчатые крысы) эволюционировали по край-
1 Симпсон не говорит об этом разъединении, но характер обмена в раннем эоцене свидетельствует о том, что оно существовало в предшествующее время и что в течение этого времени различные группы млекопитающих (после того как между ними произо- и~ел обмен в раннем палеоиене) самостоятельно эволюционировали в Евразии и Север- ной Америке.
ней мере с начала нижнего миоцена, но так и не проникли в (."тарый Свет. )водюционные самостоятельные линии енотовых и пекари в Северной Америке берут свое начало соответственно с нижнего миоцена и нижнего олигоцена. Верблнэдовые, очевидно, были приурочены к Северной Америке с позднего эоцена до плейстоцена. А вилорогие антилопы радиировали в (:еверной Америке начиная с позднего миоцена, не достигая Старого (;вета. Число строго североамериканских групп млекопитающих неве- лико. Они не имеют столь важного значения, в ряде случаев экологически более ограничены (мешетчатые крысы и вилорогие антилопы, вероятно, в основном радиировали в более засушливой западной части Северной Америки) и по сравнению с группами Старого Света обычно являются более молодыми. Все это позволяет предположить, что хотя климат северных областей в прошлом, несомненно, был более теплым, чем сейчас. однако по крайней мере с олигоцена, а возможно и раньше, климат был достаточно зональным и, вероятно, являлся преградой распространению некоторых млекопитающих, доминирующих в тропиках Старого Света. В результате этого они не смогли достичь Северной Америки, а некоторые (но значительно меньшее число) млекопитающие, которые населяли более теплые части Северной Америки, не смогли проникнуть в Старый Свет.
Об обмене млекопитающими между Северной и Южной Америкой, происшедшем довольно поздно (в плиоцене), после соединения обоих материков сухопутным мостом, говорится в работе Симпсона (Simpson, 1940. р. 158). До соединения Северной Америки с Южной в первой име- лось примерно 27, а во второй — 29 семейств наземных млекопитающих и только одно или два семейства (енотовые и, возможно, опоссумы) встре- чались на обоих материках. После установления сухопутного моста наблюдается стремительное перемещение млекопитающих и в том и в другом направлении. В плейстоцене Северная и Южная Америка имели уже 22 общих семейства и дальнейшее перемещение все еще продолжалось. Но уже наблюдались значительные отступания и исчезновения. После всех пертурбаций в Северной Америке осталось только 23 семейства, а в Южной — 29, то есть то же число, что и было раньше. В результате всех этих перемещений фауна Северной Америки несколько сакра гилась, но коренных изменений в ней не произошло. А фауна Южной Америки пре- образилась существенным образом: многие южноамериканские млекопи- тающие исчезли, а многие североамериканские выжили и стали в ней доминирующими.
Из южноамериканских млекопитающих, проникших в глубь Север- ной Америки, известны опоссумы (если только они не сохранились на се- вере с третичного периода), 3 семейства (по меньшей мере четыре разные ветви) наземных ленивцев, две ветви броненосцев и две ветви глиптодон- тов. Из дикобразоподобных грызунов известны дикобразы и два вида водосвинок. (Один из наземных ленивцев, очевидно, достиг Северной Америки незадолго до соединения материков.) Из этих млекопитающих только опоссум и дикобраз широко распространены в Северной Америке и по сей день, а броненосец достигает только Техаса. Некогда домини- ровавшие сумчатые хищные и южноамериканские копытные, очевидно, совсем не проникали в Северную Америку, а сумчатые ценолестовые, обезьяны, муравьеды, древесные ленивцы и многие группы дикобразо- подобных грызунов или не смогли достичь Северной Америки, или, на- сколько это известно, достигли только южной части Мексики.
К североамериканским млекопитающим, которые достигли Южной Америки, относятся землеройки, один род зайцев, белка, один род кар-
Глава 6
812
манчиковых мышей, некоторые ветви грызунов из семейства Cricetidae, по крайней мере три вида собачьих, один вид медведей, один-два еното- вых (один из них проник до соединения материков), четыре вида куньих, три или более кошачьих (включая вымершего саблезубого тигра), три или четыре вида слонов (в широком смысле), два вида лошадей, тапиры, пекари, верблюды и олени. Надо отметить, что в этом списке нет ни одного животного, которое могло бы происходить от самостоятельной, исчезнув- шей до того, как произошло это вселение, тропической североамерикан- ской фауны млекопитающих. К главным плейстоценовым и современным североамериканским группам, которые не достигли Южной Америки, относятся кроты, сеноставки, беличьи, кроме типичных белок, бобры,
P и с. 43. Известные направления перемещения млекопитаю-щих в третпчном периоде и плейстоцене.Длина стрелки грубо приблизительно соразмерна расстояни~о, накоторое произошло перемещение в указанном направлении.
прыгунчики (Zapodidae), вилорогие антилопы и бычьи. Все североамери- канские группы млекопитающих, достигшие Южной Америки, сохрани- лись там, за исключением слонов, лошадей и саблезубых тигров. В Се- верной Америке исчезли и тапиры и верблюды, а также слоны, лошади и саблезубые тигры. В Южной Америке некоторые из северных групп в настоящее время доминируют. Северные хищные полностью заместили там сумчатых хищных, а северные копытные — древних южноамериканских копытных. Северные грызуны, вероятно, замещают дикобразоподобных: в Южной Америке роды семейства Cricetidae уже почти достигли многообразия дикобразоподобных, оттесняя последних в южном направ- aeHlIm.
Необходимо повторить, что расселение млекопитающих шло в опре- деленных направлениях (рис. 43). Пять групп не летающих наземных млекопитающих (различные группы семейства МигЫае) сравнительно недавно переместились из Азии в Астралийскую область, но из Австралий- ской области только одна группа (Phalanger) распространилась по направ- лению к Азии и проникла только до острова Сулавеси. Это один из при- меров, показывающий, что рукокрылые значительно чаще перемещались в Астралийскую область, чем из нее. Все данные говорят о том, что пере- мещение млекопитающих происходило в большей степени из Старого Света в Северную Америку, а не наоборот. Происходившее недавно пере-
Далеко и итаюи~ ие
мещение млекопитающих происходило преимущественно из Северной Америки в Южную. А столь же недавнее перемещение из Старого Света в Южную Америку (через Северную) шло исключительно одним путем: ряд групп, возникших в Старом Свете, достигали Южной Америки, а южноамериканские группы в Старый Свет не проникали.
Ход описанных перемещений надо понимать правильно. Происходит сложный обмен, связанный обычно с многократными перемещениями в обоих направлениях, и только со временем неравноценные обмены и селективное вымирание определяют направление этого процесса в целом. Именно так надо понимать расселение млекопитающих из Старого Света в Австралию, из Старого Света в Северную Америку и из Северной Амери- ки в Южную. Так как самые богатые фауны млекопитающих принадлежат тропикам, то самым большим и единственным центром географической радиации млекопитающих, по-видимому, были основные тропические области Старого Света.
Хорошо сохранившиеся ископаемые млекопитающие позволяют пред- полагать или обосновать ряд явлений из истории нашей Земли и ее кли- мата.
Так, судя по этим данным, можно сказать, что какое-то время перед концом эоцена Африка была отрезана от Евразии, а затем эти материки время от времени соединялись.
Ископаемые млекопитающие подтверждают тот факт, что Евразия и Северная Америка начиная сначала третичного времени периодически соединялись и что главное соединение приходилось на район Берин- гова моря. Палеонтологические материалы свидетельствуют о том, что соединения через Северную Атлантику с эоцена не было вообще. Согласно ископаемым, климат Берингии в раннетретичное время был теплее, чем в настоящее время, но постепенно становился более прохладным. Даже в раннетретичное время климат был достаточно зональным и пре- пятствовал расселению ряда групп млекопитающих, доминирующих в более теплой части Старого Света, в Америку.
Изучение ископаемых млекопитающих доказывает, что почти на про- тяжении всего третичного периода Южная Америка была изолирована от всех других континентов и что полного соединения Южной Америки с каким-либо другим материком в начале третичного периода не произошло, а существующее соединение с Северной Америкой завершилось довольно поздно — в плиоцене.
Ископаемые материалы по млекопитающим свидетельствуют также о том, что в течение третичного периода между Австралией и любым дру- гим материком полного соединения не было.
Предположение о соединениях на юге между сушами Старого Света и Южной Америкой, по крайней мере в третичном периоде, едва ли совме- стимо с тем, что теперь известно о распространении современных и иско- паемых млекопитающих. Связь между Африкой и Южной Америкой в эоцене (предполагаемая), но не столь необходимая для объяснения расселе- ния на основании распространения обезьян и дикобразоподобных грызунов. нельзя признать возможной, так как она противоречит другим палеонто- логическим материалам. Очень раннее соединение в третичном периоде между Южной Америкой и Австралией, допускаемое на основании изу- чения расселения современных сумчатых и особенно на основании порази- тельного сходства между исчезнувшими южноамериканскими хищными сумчатыми (Borhyaenidae) и современными австралийскими Dasyuridae, также неправдоподобно. Предполагаемое родство вышеупомянутых групп
Глава 6
находится под большим сомнением. В раннетретичном периоде сумчатые семейства Didelphidae встречались в Европе, Северной Америке и, по- видимому, также в Азии. Симпсон считает это семейство центральной груп- пой, от которой образовались все другие сумчатые. Поэтому примерно в начале третичного времени различные представители этого семейства из Северной Америки и Азии независимо друг от друга достигли Южной Америки и Австралии, где в результате их параллельной эволюции обра- зовались (среди других групп) Borhyaenidae и Dasyuridae (Simpson, 1945, р. 171). Разнообразие неполнозубых и характерных копытных в Южной Америке, которые существовали не только на протяжении третичного периода, но также и в плейстоцене и сохранились там до на- ших дней, свидетельствует о том, что прямого сухопутного соединения в пределах соответствующего времени не было. Однако следов пребывания этих животных в Австралии ни в фауне плейстоцена, ни в современной не найдено. Что касается вопроса существования или отсутствия соеди- нений между континентами до третичного периода, то ископаемые на этот счет ничего дать не могут.
СПИСОК СЕМЕЙСТВ МЛЕКОПИТАЮЩИХ
КЛАСС MAMMALlA, млекопитающие. Это полный список известных семейств совре- менных и ископаемых млекопитающих. Таксономически этот список следует классификации Симпсона (Simpson, 1945), но приводимая классификация упро- щена в результате исключения некоторых высших категорий и отдельных под- семейств, которые с точки зрения зоогеографии не играют существенной роли.
Подробности распространения современных млекопитающих почерпнуты из мно- гих источников, наиболее важные из которых включены в библиографию этой главы. Данные по ископаемым каждого семейства заимствованы из Симпсона и сверены по Ромеру (Romer, 1945) и Скотту (Scott, 1937), а в отдельных случаях по другим источникам. Число родов, как правило, приводится и для современ- ных, и для ископаемых семейств, а число видов в большинстве случаев только для современных, но, конечно, все эти данные нельзя считать окончательными.
Симпсон отмечает, что его списки родов недостаточно полные, в связи с тем, что многие древние исчезнувшие роды не известны до настоящего времени и даже чис- ло современных видов до сих пор во многих случаях не установлено. Доплейсто- ценовыми данными, относящимися к современным родам, следует пользоваться с осторожностью. Родовые названия для ископаемых млекопитающих часто упот- ребляются в более широком смысле, чем для современных. Многие, даже имею- щие важное значение ископаемые млекопитающие до сих пор недостаточно хоро- шо известны или их положение вызывает сомнение, и даже в общепринятых свод- ках до сих пор допускается много прямых ошибок.
Палеоцен выделяется от эоцена по всему списку. Знак * перед латинским
названием означает, что эта группа исчезла.
ПОДКЛАСС PROTOTHERIA
Отряд Monotremata, яйцекладущие млекопитающие. Как в древности, так и в настоя- щее время за пределами Австралийской области не встречаются.
Tachyg1ossidae, ехидны. Австралия (включая Тасманию) и Новая Гвинея. 2 рода, 3 или
4 вида. Tachyglossus aculeatus, ехидна (несколько подвидов), широко распрост- ранена в Австралии, Тасмании н достигает юго-восточной части Новой Гвинеи.
Обитает в самых разнообразных местах, включая как дождевые леса, так и более засушливые разреженные леса, заросли кустарников и каменистые местности.
но не пустыни. В местах с более прохладным климатом она впадает в спячку.
Несмотря на то что это очень примитивное млекопитающее с очень специализиро- ванпым питанием, оно довольно обычно, и его следует считать все еще преуспеваю- щим и в некоторых отношениях приспособленным животным. Близкородствен- ный род Zaglossus (Proechidna) с 2 или 3 видами (несколькими подвидами) при- урочен к Новой Гвинее (и до западной части острова Салавати) и широко распро- странен там, особенно в горных районах. Ископаемые Tachyglossus известны из плейстоценовых слоев Австралии.
0rnith0rhynchidae, утконосы. Широко распространены в Восточной Австралии от
низменностей полуострова Кейп-Йорк (примерно в 20 милях к югу от Куктау- на,— Tate, 1952, р. 577) и далее в тропиках Северного Квинсленда, доходя иа
Млеко питаюо щие
юге до Виктории и Тасмании, а иа западе — до реки Ла1пхарт в Северном Квин- сленде. Встречаются также в краевых частях Южной Австралии. 1 род (Ornitho- rliynchus или Platypus) и 1 современный вид (с несколькими подвидами). Утко- посы до сих пор обычны во многих местах. «Утконосы встречаются от прозрач- ных стремительных горных ручьев с ледяной водой на высоте 5000 футов на горе Еосцюшко до теплых и довольно мутных рек побережья Евинсленда и от боль- 1них озер до мелких водоемчиков» (Troughton, 1951, р. 3). Ископаемые, от- носящиеся к современному роду, известны из плейстоцена Восточной Ав- стралии.
эПОДКЛАСС ALLOTHERIA
»Отряд Multituberculata. Древний отряд, состоящий из 3 или 4 семейств. Сомнитель- ные млекопитающие eTritylodontidae известны из Европы (верхний триас- средняя юра), Китая (верхний триас) и Южной Африки (верхний триас). Семей- с тва *Plagiaulacidae, *Ptilodontidae и *Taeniolabididae в общей сложности с 27 ро- дами (в большинстве случаев фрагментарные ископаемые остатки) относятся к различному времени — от верхней юры до нижнего эоцена Европы, Азии и Се- верной Америки. Подкласс и отряд сомнительные
»М1сгос1ерйЫае, Европа (верхний триас). 2 рода. Ископаемые остатки — отдельные
зубы.
Иодкласс сомнительный
*Отряд Triconodonta
*Triconodontidae. Европа и Северная Америка (юра и сомнительные находки в верх- нем триасе). 8 родов.
ПОДКЛАСС THER IA
»Отряд Pantotheria. 4 семейства (eAmphitheriidae, ePaurodontiidae, eDryolestidae,
*Docodontidae), известные из юры Европы или Европы и Северной Америки (ePa- ~irodontidae также и из Восточной Африки). Всего насчитывается 22 рода.
»Отряд Symmetrodonta. 2 семейства (eSpalacotheriidae, *Amphidontidae) из верхней
юры Европы и (или) Северной Америки. 5 родов. Этот отряд известен недо- статочно хорошо.
(Инфракласс Metatheria)
Отряд Marsupialia, сумчатые. Из 6 надсемейств этого отряда, выделяемых Симпсоном,
нервые три: Didelphoidea (находящееся в Америке и включающее всех древних североамериканских и европейских ископаемых сумчатых), eBorhyaenoidea (Юж- ная Америка) и Dasyuroidea (Австралийская область),— очевидно, близкород- ственные. Остальные 3 надсемейства: Perameloidea (Австралийская область), (:aenolestoidea (Южная Америка) и Phalangeroidea (Австралийская область до острова Сулавеси и т. д.) — филогенетически несколько более изолиро- ванпые.
(Нндсемейство Didelphoidea)
0Ме1рЬЫае, американские опоссумы. В настоящее время приурочены к южной, цент- ральной и восточной частям Северной Америки. 12 современных родов (включая C;'aluromysiops — Sanborn, 1951) и приблизительно 80 видов (более половины относится к роду Маrmosa). Все роды встречаются в Южной Америке: некоторые нз них достигают Центральной Америки, а 4 — отдельных частей Мексики (один из четырех Didelpkis проникает еще дальше — в восточную часть Северной Аме- рики). Самый северный вид — виргинский опоссум (D. virginiana, который в Юж- ной и Центральной Америке замещен D. marsupialis), распространен в большин- стве районов восточной части Соединенных Штатов. Он был обнаружен на севере в южном Нью-Гэмпшире, южном Онтарио, центральном Мичигане и южном Ви- сконсине. Это обычное, агрессивное животное, которое в последние годы расши- рило свой ареал к северу. Виргинский опоссум был интродуцирован в районе Калифорнии и в отдельных районах штатов Орегон и Вашингтон, к югу род l>i~delphis достигает центральной Аргентины, Lutreolus проникает на юг несколь- ко дальше до территории Чубут, Магтояа — до южно-центрального Чили (Валь- дивия). Dromiciops известен только на острове Чилоэ и на небольшой близлежа- щей территории материка (в Чили). Votodelpkis известен только на территории, примыкающей к заливу Сан-Хорге в Южной Аргентине, примерно у 47' ю. ш.,— самая южная точка расселения любых сумчатых. Насколько известно, на самом юге Южной Америки, в 300 — 400 милях от указанного места, сумчатых нет. Иско- паемые показывают, что опоссумы были многочисленны (2 вымерших подсемей- ства, 11 родов) в Северной Америке в верхнемеловом периоде, что подсемейство Didelphinae (4 вымерших рода) существовало там в первой половине третичного периода (до нижнего миоцена), что один из североамериканских родов находил- ся также с эоцена до миоцена в Европе и что опоссумы встречались и, возможно, были также многочисленны (хотя ископаемые данные весьма скудные) в Южной
Глава 6
Америке на протяжении третичного периода. Куто (Couto, l952a) считает, что, возможно, они достигли Южной Америки в позднемеловом времени. В Севернои Америке после нижнего миоцена сумчатые не известны до появления рода Didet- phis в плейстоцене.
*Caroloameghiniidae, Южная Америка (нижний эоцен). 1 род.
е(Надсемейство Borhyaenoidea)
*Borhyaenidae, спарассодонты, сумчатые хищные. Южная Америка (с палеоцена до
плиоцена) . 21 род.
е(Надсемейство Dasyuroidea)
Dasyuridae, сумчатые мыши, сумчатые куницы и т. д. Австралийская область. Все
9 родов, приведенные Симпсоном, встречаются или встречались в Австралии.
Несколько родов достигают Тасмании. Роды РЬазсо~а1е, Sminthopsis (сумчатые мыши) и Dasyurus (сумчатые куницы) распространены до Новой Гвинеи (п на ряде небольших близлежащих островов) и островов Ару (но не островов Кай).
Однако в действительности характер географических связей в пределах семей- ства гораздо сложнее. Более дробная классификация Тейта (20 родов, 45 видов) включает несколько примитивных родов, приуроченных к Новой Гвинее и т. д., и предполагает полдюжины или более обменов различных ветвей между Австра- лией и Новой Гвинеей (Tate, 1947, table 4): Sarcophilus (сумчатый дьявол) и Thy- lacinus (сумчатый волк) приурочены к Тасмании, а их ископаемые, так же как и ископаемые более широко распространенных родов Phascagale и Dasyurus, отно- сятся к плейстоцену Австралии. Myrmecobius (сумчатые муравьеды, или мураше- еды, 2 вида или подвида), который еще совсем недавно встречался на большей ча- сти Южной Австралии, теперь строго приурочен к небольшой территории в севе- ро-западной части Южной Австралии и к прилежащей части Западной Австра- лии.
Хо10ryctidae, сумчатые кроты. Австралия. 1 род, 2 вида. Широко распространены
в южной и центральной частях Австралии. «Песчаные холмы, покрытые Acaceia апеига и низкорослыми солончаковыми кустарниками, или равнины из красного песка, поросшие спинифексом и акацией, являются самыми подходящими ме- стами обитания для этих животных» (Troughton, 1951, р. 57). К западу распрост- ранены (непрерывно?) до того места, где Большая Песчаная пустыня доходит до побережья Западной Австралии.
(Надсемейство Perameloidea)
Peramelidae, сумчатые барсуки (бандикуты). Австралийская область до острова Церам.
Симпсон (Simpson, 1945) выделяет 5 родов, Тейт (Tate, 1948а) — 8 родов с 19 ви- дами. Это семейство хорошо представлено в Австралии и Новой Гвинее (Tate, 1948а). Два рода достигают Тасмании, один (Echymipera) — Новой Британии и Новой Ирландии, островов Ару, Кай и Мисол. Один монотипический род при- урочен к острову Церам и был известен до сих пор только по типовым экземпля- рам с высоты 6000 футов в горах Манусела. Ископаемые нескольких современных родов известны из плейстоцена Австралии.
(Надсемейство Caenolestoidea)
Caen0lestidae, ценолестовые. Южная Америка. 3 рода, встречающиеся в Андах от гра- ницы Венесуэлы и Колумбии до Эквадора (Caenolestes, 4 вида) и южного Перу (Orolestes, 1 вид) на высоте до 14 000 футов, а также изолированно в провинции Ланкухэ, включая остров Чилоэ, южно-центральное Чили (Rhyncholestes, 1 вид), где он обитает в густых лесах на сравнительно низких высотах. Ископаемые (14 вымерших родов) показывают, что это семейство существовало в Южной Америке с нижнего эоцена и что в прошлом оно было довольно многочисленным и разнообразным.
*Polydolopidae. Южная Америка (палеоцен и эоцен). 4 рода.
(Надсемейство Phalangeroidea)
Phalangeridae, австралийские лазающие сумчатые и т. д. Австралийская область до
островов Тимор, Сулавеси и Соломоновых. 14 современных или недавно вымер- ших рода и около 44 видов. Большинство родов австралийские: 4 достигают Тас- мании, несколько — Новой Гвинеи и 4 рода приурочены только к Новой Гвинее.
Распространение некоторых родов сложное. Род Phalanger (кускусы) определя- ет западные и восточные границы сумчатых на Индо-Австралийском архипелаге.
Это в основном иовогвинейский род (2 вида этого рода распространены до полу- острова Кейп-Йорк в Австралии), но он встречается и на западе (острова Таиор, Ветар, Сулавеси) и на востоке (самые отдаленные острова из группы Соломоновых).
На острове Сулавеси имеется два четких эндемичных вида этого рода, которые самостоятельно возникли в Новой Гвинее (Tate, 1945). Petaurus breuiceps распро- странен от Северной Австралии и Новой Гвинеи до Новой Британии, островов Ару и Кай и (на Молуккских островах) Мисол, Бачан, острове Хальмахера и Терна-
37леко питающ ие
И7
те (Tate, 1945 Ь). Другие представители данного семейства в этом направлении за пределами Новой Гвинеи, а также близлежащих прибрежных островов (Сала- вати и т. д.) и островов Ару не встречаются. Несколько австралийских родов имеют очень ограниченный ареал: Таrsipes, медоед, приурочен к южной части Западной Австралии. Phascola,rctos, коала, в прошлом был распространен н» лесистых территориях от южного Квинсленда (19'30' ю. ш.) через Новый Южный Уэльс и Викторию до (в последнее время) юго-восточной части Южной Австра- лии и (в плейстоцене) в южной части Западной Австралии, но в настоящее вре- мя ареал и численность этого рода сильно сократились (Нагрег, 1945, Troughton, 1951). Австралийский лисий кузу (Trichosurus) был иптродуцирован на Новой Зеландии, размножился там в необычайном количестве и в настоящеее время широко распространен на обоих главных островах и на острове Стюарт (Kodzi- cki, 1950). Ископаемые этого семейства, представляющие ряд современных и ряд исчезнувших родов, известны из плейстоцена Австралии, а *Wy~iyardi< — и~ плиоцена Тасмании.
~Thylacoleonidae, Австралия (плейстоцен). 1 род. Очень крупные животные, «дости- гавшие размеров льва» (Romer). Образ жизни не известен.
Phascolomidae, вомбаты. Восточная и Южная Австралия и Тасмания. 2 современнык
рода, 2 или несколько видов. В прошлом были многочисленны на севере до цент- рального Квинсленда и на западе до восточных краевых частей Западной Австра- лии (и до юго-западной части в плейстоцене), на островах Бассова пролива, а также в Тасмании. В настоящее время ареал их сильно сократился. Оба совре- менных рода и два исчезнувших относятся к плейстоцену Австралии (Tate, 1951a).
:Macropodidae, кенгуру, валлаби, кенгуровые крысы и др. Австралийская область.
14 (по Симпсону) или 18 (по Тейту — Tate, 1948) современных родов, около 46 видов (по Тейту), большое число подвидов. Кенгуру и более крупные кенгуру- валлаби (Масгориг, Protemnodon и др.) приурочены к Австралии (включая Тас- манию и некоторые прибрежные острова), за исключением вида Protemnodou ад~- lis из северо-восточной части Австралии, который достигает южной частиИовой Гвинеи и островов Тробриан. Другие, в большинстве случаев мелкие формы много- численны и разнообразны в Австралии (несколько достигают Тасмании), оолее малочисленны и менее разнообразны в Новой Гвинее, несмотря на то, что там встречается 8 видовых групп, из которых одни являются общими с Австралией.
а другие типичны только для Новой Гвинеи. Некоторые из них встречаются в го- рах на высоте 10 000 футов и более. Более детально распространение этого семей- ства описано Тейтом (Tate, 1948, table 2). Thylogale распространен от Австра- лии и Новой Гвинеи до островов Ару и Кай, а также до Новой Британии и Ново11 Ирландии. Dorcopsis достигает островов Мисол, Вайгео и Салавати (к западу от Новой Гвинеи). Древесные кенгуру Dendrolagus, вторично приспособившиеся к жизни на деревьях, с 3 видами, встречаются в дождевых лесах Новой Гвинеи и северного Квинсленда (в южном направлении до нижнего течения реки Гербер- та), и возможно, что этот род достигает островов Ару и Kai, хотя данные, свиде- тельствующие об этом, «весьма неполные и неудовлетворительные» (Rothschild, Dollman, 1936, р. 477). Ископаемые кенгуровые, представляющие различные современные роды и 6 вымерших родов (включающих по сравнению с современ- ными более крупных кенгуру), относятся к плейстоцену Австралии.
*Diprotodontidae. Австралия, включая Тасманию. 6 ископаемых родов (плейстоцен).
Наземные сумчатые, из которых по крайней мере один вид достигал размеров большого носорога.
(Инфракласс Eutheria, плацентарные млекопитающие)
Отряд 1пвесlivora, насекомоядные. 8 надсемейств (Tupaioidea, тупайи, см. отряд при- матов).
(»Надсемейство Deltatheridioidea)
*Deltatheridiidae. Азия (верхний мел) и Северная Америка (палеоцен и эоцен). 7 родов. (Надсемейство Tenrecoidea)
*Palaeoryctidae. Северная Америка (средний палеоцен). 1 род.
Solenodontidae, щелезубы. Существуют только на Кубе и Гаити в Вест-Индии. 1 род
(Solenodon) с 1 видом в настоящее время локализованным в восточной части Кубы (тот же самый или несколько отличающийся вид в прошлом встречался также в центральной и западной частях Кубы,— Aguayo, 1950) и 1 видом — на острове Гаити. 3 ископаемых рода обнаружены в Северной Америке (олигоцен).
Tenrecidae, тенреки. Мадагаскар (род Tenrec распространен также на Коморских
островах). 10 (по Симпсону) или 13 (по Аллену) родов, 30 видов (G. М. Allen.
1939). Экологически разнообразные, по-видимому, в результате радиации на Ма- дагаскаре..Ископаемые относятся к плейстоцену Мадагаскара.
818
Глава б
Potamogalidae, выдровые землеройки. Западная Африка (от Камеруна до Лнго1хх
и Конго) . l водххьхй вид.
(11адсемейство Chrysochloroidea)
Сhrysocliloridae, златокроты. Африка, большей частью в Юн'ххохх Африке, но расххро< т- ранены и к северу до южного Камеруна, Уганды и т. д. 4 или болес совремеииы~ рода, 29 видов (по Аллену) и еще 1 род из плейстоцена Южной Лфрики.
(Надсемейство Erinaceoidea)
*Zalambdalestidae, Азия (верхний мел). 1 род.
eLeptictidae. Северная Америка (с верхнемелового периода до олигоцепа) и Еххх1оин
(верхний палеоцен и эоцен). 11 или более родов.
EIinaceidae, ежи и т. д. В настоящее время приурочены к основной части Старого
Света (но не к Австралийской области). 2 подсемейства: (1) Echinosoricinae, во- лосатые ежи, или гимнуры. Восточная часть Индо-Ъ!алайской области от Юго-з»- падного Китая до островов Ява, Калимантан и Минданао. 4 рода, 4 вида, 12 иско- паемых родов в Европе и Северной Америке, частично из эоцена — верхнего мио- цена (Европы) или нижнего плиоцена (Северной Америки). В результате ххедахх- них исследований Симпсона было совершенно неожиданно обнаружено, что раз- нообразные ежи были представлены в Северной Америке на протяжении больше11 части третичного периода; (2) Erinaceinae, настоящие, или колючие ежи. Африки (но не Мадагаскар), Евразия к северу до Шотландии, центральной Швеции, цент- ральной Сибири и Амурской области и к югу до Индии (прерывистое распро- странение в северной и южной Индии, на Цейлоне нет совсем), а также в Цент- ральном Китае (нет в Японии, возможно, хотя в списках они приводятся, ххет цхх Тайване и в Индокитае и т. д.). 5 родов, около 15 видов. Несколько ископаемых родов из Европы (начиная с олигоцена) и 1 сомнительный род из Северной Аме- рики (верхний миоцен).
*Dimylidae, Европа (верхний олигоцен до верхнего миоцехха). 4 рода.
(Надсемейство Macroscelidoidea)
Масгозсе1ЫЫае, сгхоновые землеройки, Африка, включая Северную Африку (но не
Мадагаскар). 5 или более родов, около 40 приведенных видов (G. М. Allen, 1И9). (Надсемейство Soricoidea)
*Nyctitheriidae. Северная Америка (средний палеоцен до среднего эоцена) и, вероятно,
Европа (ххихкний эоцехх). 5 недостаточно хорошо известных родов.
Soricidae, землероххки. 3 подсемейства [Эллермахх и Моррисон-Скотт (Ellerman, Alorri- яоп-Scott,, l95l) сомневаются в их достоверности]: (l) Soricinae. Евразия и Север- ная Америка, в основном умеренная зона северного полушария — на север до Арктики, на юг до центральной Испании, южной Италии, Палестины, оче- видно, Кашмира (но не на полуострове Индостан), северная Бирма, Тонкин хх Тайвань, Америка — на юг через Центральную Америку до высоких гор Колумбии, Эквадора и западной Венесуэлы. 8 современных родов, 18 евроазиат- ских и неопределенное число американских видов. Род Ьогех широко распро- странен в Евразии и Северной Америке (на хог до Гватемалы). Cryptotis встречается на большей (восточной) части Соединенных Штатов, в Мексике, в Центральной Америке и на юге северной части Южной Америки до вышеук х- занных границ. Это единственное известное насекомоядиое, которое когда-либо достигало Южной Америки (если только сомнительный род +1Vecrolestes не отно- сится к насекомоядным). Ископаемые (несколько родов), относимые к этому под- семейству, ведут к верхнему эоцену Европы и нижнему олигоцецу Северной Аме- рики; (2) Crocidurinae. Старый (:вет, кроме Австралийской области, Афрххкхх, Мадагаскар (?), большая часть Европы, Азия на север до отдельных раххонон ('ибири, на югдоЦейлона и островов Сулавеси, Молуккских(?), Кай (?), Малых Зондских до острова Тимор (?) и Филиппин (?). l2 родов, многочисленные виды.
Сгос~с(ига и Suncus, как полагают, населяют и Коморские острова, а 5ипсиг.— также и Мадагаскар. Один из этих родов или оба, возможно, имеются и на других островах, которые упоминались выше под знаком вопроса. В связи с тем, что оба эти рода включают виды, которые более или менее связаны с человеком, их естественные границы сомнительны. Ископаемые 2 современ- ных родов относятся к верхпему миоцену и нижнему плиоцену Евразии. 1 исчезнувший род приводится из верхнего миоцена Северной Америки, ио то, что его относят к этому подсемейству с точки зрения географии, может выз- вать сомнение; (3) Scutisoricinae. Восточно-центральная Африка (прилежащие части восточного Конго и Уганды). 1 специализированный род, 2 вида.
Talpidae, кроты. Умеренная зона Евразии (к хогу по направленихо к тропикам) и Север- ная Америка. На севере они достигают южной Швеции и, очевидно, пересекают Полярный круг в Азии и Америке, доходя до юго-западнои Британской Колумбии и южного Лабрадора. В южном направлении распространены до Испании, Ита-
ЛХлекопитаюи~ ие
лии и Сицилии, Малой Азии и Кавказа, Непала, полуострова Малакка и Тайва- ня, Нижней Калифорнии, северо-восточной части Мексики и Флориды. 17 (или меньшее число) современных родов, 20 видов. Ни один род не является общим д;1я Старого и Нового Света. Самые древние ископаемые кроты известны из верхнего эоцена Европы и верхнего олигоцена Северной Америки.
*XpsophontidBe. Вест-Индия (Куба, Гаити, Пуэрто-Рико); субфосс11льно в плейсто- цене. I род, 5 или 6 видов. В настоящее время не встречается. Возможно, пред- ставители этого семейства исчезли, но, возможно, и существуют до сих пор в 1()- рах Гаити.
(*Надсемейство Pantolestoidea)
*Pantolestidae. Северная Америка (средний палеоцен до среднего эоцена) и Enpi»ia
(нижний эоцен). 2 подсемейства, 6 родов.
(эНадсемейство 'AIixodectoidea)
*A1ixodectidae, Северная Америка (средний палеоцен до верхнего эоцена). 8 род~в. (Семейства и роды насекомоядных, занимающие неопределенное место)
*Picrodontidae, Apheliscidae, *Necrolistidae и другие, не вошедшие в список, искона»- мые роды. В большинстве случаев это раннетретичные формы из Северной Аме- рики или Евразии, но *Neerolestes известен из Южной Америки (нижний миоцен).
Это специфическое животное, которое, по всей вероятности, было насекомояд- ным (но могло им и не быть), ставит в тупик эволюционистов и зоогеографов (SiIllð- son, 1945, р. 178).
Отряд Dermop tera
*Plagiomenidae. Северная Америка (верхний палеоцен и нижний эоцен). ' рода. Симп- сон (Simpson, 1945, р. 179) подтверждает возможное родство этих ископаемых со следующим семейством.
<'.ynocephalidae (Galeopithecidae), шерстокрылы. (Тенассерим и Вьетнам) до островов
Ява, Калимантан и юго-восточной части филиппинского архипелага. ! род, 2 вида.
Отряд Chiroptera, рукокрылые. (11оследние сведения о рукокрылых были любезно
представлены мне доктором Купманом.)
П одотряд iW ega chir op tera
Pteropidae, плодоядные летучие мыши, летучие лисицы. Тропические и субтропи- ческие страны Старого Света', на север до Кипра, Сирии, Индии, Сиккима, побе- режья Южного Китая и островов Рюкю; на юг через всю Африку (кроме Сахары и северо-западной части) и до северной и восточной частей Австралии (один вид изредка мигрирует на юг, доходя до краевых районов Виктории); на многих ост- ровах Индийского и Тихого океанов, достигая на востоке островов Самоа. 21 род, около 200 (или меньше) видов. Pteropus (летучие лисицы), с мнохсеством видов.
Встречаются от Индии до Австралии, на многих островах западной части Тихого океана и по всем островам Индийского океана. Несколько видов находится на Мадагаскаре, и единственный вид встречается на островах Пемба и Мафия, при- мерно в 40 и 20 милях от побережья Африки (Swynnerton, Hayman, 1951). 'дти огромные сильные летучие мыши легко могли бы достичь материка Африки, но этого не произошло. Большие плодоядные летучие мыши Африки относятся к дру- гому роду. Меда1од1огяия, обитающий в лесах Западной Африки, относится к группе цветоядных форм (Macroglossinae) и приурочен к территории Бирмы и да- лее на юг и восток Индо-Австралийского архипелага. Единственное ископаем~ю этого семейства из олигоцена Италии приводится Симпсоном.
Подотряд М i crochiroptera
(Надсемейство Emballonuroidea)
Hhinopomatidae, ланцетоносы. Северо-восточная Африка (на запад до некоторых
районов Сахары и на юг до озера Рудольфа) и Южная Азия (Аравия, Пале- стина), Иран, полуостров Индостан до Таиланда и Суматры). 1 род, около 4 видов.
Ф
Emballonuridae, свободнохвостые летучие мыши и др. Тропики Старого и Нового Ñâå- та, включая большую часть Австралии, на север до Египта, Ирака, Индии и Юньнани и в Америке на север до Мексики. 8 родов, ни один из Ij'OTopbIx не является общим для Старого и Н ового Света, множество видов. Род Ет Ьайогги ~.а имеет несколько видов, встречающихся от Таиланда до Новой Гвинеи и Соломоновых островов и на запад до Самоа, и 1 изолированный вид на Мада- гаскаре (родственный род Coleura встречается в восточной части Африки, южной Аравии и на Сейшельских островах). Единственное ископаемое, отно- симое Симпсоном к этому семейству, известно из верхнего эоцена или нижпего олицегона Европы.
Xoctilionidae, зайцеротые летучие мыши (включая рыбоядных). Тропики и субтропики
Америки на север до южной Мексики и Вест-Индии. I род, несколько видов.
Х'лава 6
(Надсемейство Rhinolophoidea)
Nycteridae, никтериды. Тропическая Африка на север до Корфу (Греция), 11алести- ны и Аравии, на Мадагаскаре и юго-востоке Азии от Тенассерима до островов Ява, Калимантан и Тимор. 1 род, 20 с лишним (или меньше) африканских видов и 1 малайский вид.
Megadermatidae, мегадермы и т. д. Районы тропической Африки, включая Абиссинию;
Индия и южная часть Китая до островов Ява, Сулавеси, Тернате и Филиппины и северная половина (тропическая) Австралии (в ископаемом виде или субфос- сильно в Южной Австралии). 3 (или больше) современных рода, все в большей или в меньшей степени локализованы, 5 видов, 2 ископаемых рода в Европе (верх- ний эоцен или нижний олигоцен и средний миоцен).
Blsinolophidae, подковоносы. Тропические и субтропические страны Старого Света на
север до Англии и Центральной Европы, на юге СССР, северная Индия, Корея и Япония; на юг через всю Африку и от Азии до восточной части Австралии (до южной части Нового Южного Уэльса). 1 современный род, много видов.
Ископаемые современного рода обнаружены в эоцене Европы, а 1 исчезнувшего рода — в верхнем олигоцене.
Hipposideridae, ложные подковоносы. Тропические и субтропические страны Старого
Света, на север до Северной Африки (но не до Европы), Ирак, Иран, Индия и южная часть Китая, на юг через большую часть Африки (представлены на Мада- гаскаре) и от Азии до Австралии. 6 современных родов, множество видов. 1 иско- паемый род в Европе (верхний эоцен или нижний олигоцен и, возможно, миоцен). (Н адсемейств о Phyl lost omatoid ea)
Phyllostomatidae, листоносы и т. д. Тропики и субтропики Америки. 35 (по Симпсону),
50 или более (по Аллену,— G. М. Allen, 1939а, р. 196) родов, много видов. Многие роды достигают Мексики, но только 4 рода распространены до южных краевых районов США. Большое число родов находится в Вест-Индии и несколько — н» Багамских островах. Ни один род не достигает Флориды, кроме случайно попав- ших туда особей. Ископаемые этого семейства известны из северных частей Юж- ной Америки, а также из миоцена Флориды.
D esmodontidae, настоящие летучие мыши-вампиры. Тропики Америки, центральная
Мексика до центральных районов Чили и отдельных частей Аргентины, на остро- ве Тринидад (но не в собственно Вест-Индии). 2 или 3 рода, 3 вида.
(Надсемейство Vespertilionoidea)
Natalidae, длинноногие летучие мыши. Тропики Америки на север до Мексики, Вест-
Индия. 1 род, несколько видов.
Furipteridae, дымчатые летучие мыши . Значительная часть тропиков Южной Америки.
1 или 2 рода, несколько видов.
7hryopteridae, дискокрылые летучие мыши. Часть тропиков Америки от восточной
Бразилии и Эквадора на север до Гондураса. 1 род, 2 или 3 вида.
Myzopodidae, липконогие летучие мыши, Мадагаскар. 1 вид. Аллен (G. М. Allen, 1939а,
р. 123) предполагает, что, возможно, это семейство является (отдаленно) родст- венным семейству Thryopteridae.
3'espertilionidae, обыкновенные летучие мыши и т. д. Космополиты распростране- ны от северной границы древесной растительности до Южной Африки, Тасмании и Новой Зеландии, на острове Огненная Земля и на многих других островах. ~> или более родов, множество видов. Myotis (ночницы) — фактически род космопо- лит. От северной границы лесов в Евразии и Северной Америке до Южной Афри- ки, большей части Австралии (но, очевидно, не доходит до крайнего юго-во- стока и Тасмании) и, «возможно, до Магелланова пролива» (Osgood, 1943) (но, очевидно, нет в Новой Гвинее, Новой Зеландии или на большей части Вест-Ин- дии). Это самый широко распространенный род рукокрылых. Его ископаемые относятся к среднему олигоцену Европы. По мнению Симпсона (Simpson, 1945), ~us встречаются в Старом и Новом Свете. Остальные роды приурочены к какому- либо одному из них. Некоторые роды достигают отдаленных островов.' например, Chalinolobus — Новой Зеландии (из Австралии), Lasiurus — Гавайских и ост- ровов Галапагос (из Америки) и Pipistrelus — Азорских (из Европы или Север- ной Африки). Из немногих известных ископаемых родов этого семейства самые древние известны из верхнего эоцена или нижнего олигоцена (Европы).
%Iystacinidae (Mystacopidae), короткохвостые новозеландские летучие мыши. Новая
Зеландия. 1 вид, который проникает далеко на юг Новой Зеландии до острова Стюарт. В настоящее время этот вид встречается редко, но, возможно, что он до сих пор существует на некоторых прибрежных островах (Powell, 1947).
Molossidae, бульдоговые летучие мыши. Тропики и субтропики земного шара. На север
до южной части Европы и Азии (до Кореи в Восточной Азии), в южных и запад-
Млекоп итп,автои~ ие
ных районах Соединенных Штатов (на западе до Британской Колумбии). 6 или более родоь, небольшое число видов. Только 1 род Tabarida (складчатогубы) (как полагает и Симпсон) встречается в Старом и Новом Свете. Ископаемые этого рода относятся к олигоцену (Европы).
(Неопределенное надсемейство)
~Arcliaeonycteridae *Palaeochiropterygidae и 2 рода. Европа (средний эоцен до ниж- него олигоцена).
Отряд 1'rimates (приматы), вклн>чая человека и всех его посленасегомоядных предков. ll о д о т р я д Р r о s i m i i, примитивные приматы (полуобезьяны или лемуры). (Налсемейство Tupai обреа)
~Anapalidae, Монголия (нижний олигоцен). 1 прекрасно сохранившийся рол.
7upai idae, т~ пайи. Индо-Малайская область, от юго-западной части Китая н полуост- ров» Индостан до островов Ява, Бали, Калимантан и южных Филиппин. 6 ро- дов, небольшое число видов.
(((адгемейство Lemuroidea). (Классифицируется по Симпсопу, Simpson, 1945). Хилл
(Hill, 1953, р. 317) дает несколько иную классификацию.
эР1ея idapidae. Европа и Северная Америка (палеоцен и эоцен). 5 родов.
эАЙарп1ае. Европа и Северная Америка (эоцен). 9 родов.
Lvmuridae, настоящие лемуры. Мадагаскар (2 вида распространены на некоторых
1~оморских островах). 6 современных родов, 15 видов. Ископаемые некоторых сов- ременных и 4 исчезнувших родов известны из плейстоцена или постплейстоцепа :1! адагаскара. Симпсон подразделяет современные и исчезнувшие формы на 5 под- семейст».
1Ыгн1ас, ипдрп. Мадагаскар,,' рода, 4 вида. Ископаемые современных родов и 3 ис- че;шувшпх рода известны из плейстоцепа или постплейстоцена Мадагаскара. (Надсемейство Daubentonioidea)
Daubentoniidae, ай-ай. Мадагаскар. j вид.
(Инфраотряд Lorisiformes)
1.orisidae, потто и лори. Тропическая Африка и Индо-Малайская область. 2 подсемей- ства: (1) Lorisinae. 1'оды l ictocebus и Perodicticus (потто), по одному виду в каж- дом, обитают в лесах Западной Африки, а последний распространен на восток но изолированным лесным участкам до Мау-Форест, Кения. f.oris (1 вид, тонкий лори) населяет ю'кную часть Индии и Цейлон. iVycticebus (толстый лори), 1 или 2 вида, встречается от северо-восточной Индии и Ассама до островов Ява и Кали- мантан, возможно, за исключением острова Минданао (Hill, 1953, р. 169, 170).
11скопаемые этого рода относятся к плиоцену Индии; (2) Galaginae. Болыиая часть Африки, южнее Сахары, за исключением самых южных частей материк» (И111, 1953, р. 219, ' 20, 237, 238). 3 рода, 5 видов.
(Инфраотряд Tarsii formes)
*Anaptomorphidae. Северная Америка (средний палеоцен до среднего эоцена), Европа
(эоцен) и, возможно, Азия (олигоцен). 5 подсемейств, 26 родов.
T~rsiidae, долгопяты. Суматра, Калимантан, Сулавеси и юго-восточные Филиппины
(Минданао, Бохоль, Лейте, Самар). 1 род, 3 вида. Распространение этого семейства показано на картах, составленных Хиллом (Hill, 1955, р. 112, 1 l3).
((;омпптельные примитивные приматы)
~Apatemyidae, *(;arp0lestidae и множество сомнительных родов. Северная Америка
и Евразия в раннетретичном периоде (палеоцен и эоцен).
Подотряд Antlsropoidea, высшие приматы.
(Надгемейство Ceboidea), американские обезьяны.
'ebidae. ревуны. Тропики Америки. 1 2 родов, небольшое число видов. Все роды встре- ча~отся в Южной Америке и по крайней мере 5 родов распространено в отдельных частях или по всей Центральной Америке. 2 рода достигают также Веракруса в южной части Мексики. К тогу по крайней мере 2 рода достигают лесных районо» северной Лргептииы. Ископаемые нескольких современных родов относятся к плейстоцепу, а исчезнувших — к миоцену Южной Америки. Ни на одном из островов Вест-Ипдии обезьяиы обиаружены не были, но 1 исчезнувший ро1 существовал на Ямайке в ылейстоцепе или даже в более позднее время, и какой- то вид обезьян, возможно, встречался на Гаити (Ш111атя, Koopman, 1952).
(:allit)>ricidae, игрупки. Тропики Америки от южной Бразилии до Панамы. 2 или
более рода, несколько видов.
(11адсеме11ство 'ercopithecoidea)
(:ere(~pitl~ecidae, обезьяны Старого Света. Тропические и субтропические страны (',та- рого Света, кроме Австралийской области. j 6 современных родов, множество видов. Северные границы обезьян определяются распространением рода ЛХасаса (чакак11, включая макаку-резус) и Марокко и Алжире (иптродуцпрован на Гпб-
ралтаре), северной Индии, Южном Китае и далее хха север до долиххы 11н>х:».х (интродуцпрован дальше на север>) и на острове Хоххсхо. Два других рода tt>c~tt- гают юго-западной части Китая. К югу и востоку эти обезьяны распространены по всей Африке (нет на Мадагаскаре), Индии, на Цейлоне, на островах Ява и 1>»;tn tt на других Малых Зондских островах до острова Тимор (только Масаса i>us), на Сулавеси (Macaca и специфический род СупорШгеси~), на Тайване (тольtt> ЛХасаса) и tta Филиппинах до острова Лусон — только Масаса i rus. Род Масаса (см.
выше) распространен от Северной Африки до границ распространеххххя ot>e:>ьян в И>хдо-Малайской области. (:вященный бабуин ((;omopithrcus) распрострапе» от северо-восточной части Африки до хожной Apatttttt. Как вид>хо, ни один ро х обезьян не является общим для Африки и Азии, но Pal>io (павианы) и Cerevt>ithe- cus (обыкновенные африканские обезьяны), в настоящее время оба приуроченные к Африке, ххзвестпы в ископаемом состоянии из нижнего плейстоцена и с1>елнего плиоцена Индии соответственно. Другой пример — Colobus, род тропической Африки, представитель подсемейства Colobinae (тонкотелые обезьяны), ххьхпе обхх- тает только в Индо-Малайской области, а его ископаемые обнаружены и пл ноще- не Европы. Известно еще несколько других ископаемых обезьян Старого (:иет», самые древние из которых относятся к олигоцену Египта. Масаса irus, как и'- «естно, была интродуцирована па некоторых островах, и нельзя быть уверенным, что на удалепных от центра архипелага Малых Зондских островах, на остров~' Тимор и Филиппинах она является действительно местным видом. Для ря..х»:>тих островов приводятся эндемичные подвиды, но это пе является доказате.и.«твом того, что эти животные также являхотся там местными (си. стр. 390).
(11адсемеххство IIominoidea)
-:.!'arapithecidae. Египет (нижний олигоцен). 1 род.
! ongidae, человекообразные обезьяны. Частично Африка и восточная часть Индо->1н- лайской области. Два современных подсемейства: (1) Flylobatinae, rttt>oott> t.
Юго-Восточная Азия, от северо-восточной Индии и западной Юньнани до остро- вов Суматра, Ява, Калимантан и, возможно, (интродуцировано?) на архи>хе1»ге (".улу. 1 или несколько родов, 7 видов и 4 ископаемых рода (илн более) х> Лфрх>- ке (нижний олигоцен и нижний миоцен) и в Центральной Европе (средни t t>tttot[ett и нижний плиоцен); (2) Ponginae, большие человекообразные обезьяны. 3 1>о;х», 3 или более вида. Po>tto (Simitt) pygmaeus, opatttyxatt (2 слегка отлича»ttttt~- ('я подви [a.) ~ встречается то1ько tta ( ) м»тре хх Ка'[хх>хсхttj liter Iхо B хtхх>хь>>е!>t >[лс!tt стоцене этот род обитал в Индии. Рап, шимпанзе (1 или 2 вида), встречается и 3»- п»дной и 1хент1>»лх ххохх Африке от Сьерра-Леоне и Камеруна до район» lit>«t »; на востоке в лесшях местностях до озер Альберта и Виктория. Gor i11à, гориллы (' вида или подвида). Одна форма распространена в Западной Африке от хог»- восточной Нигерии почти до реки Конго (но не пересекая ее), а другая — в 1(е»т- ральной Африке, узкая полоса горных лесов, тянущихся между Коххго и Угг»>- ;j,oii, непосредственно к югу от экватора до Барака в северо-восточной части озе- ра Таххгаххьика (G. М. Allen, 1939, р. 177). Четвертью пскопаемьш род *Gitta»to- pithecus из плейстоцепа Китая относится к этому же подсемейст«у. Симпсоном выделено еще 2 ископаемых подсемейства: (3) Вгуо1>х(1хесхпае. Европа, Африки и И>хдия (мххох(е>х и плиоцен). 8 родов. Возможно, это гетерогенное подсеисйст»», пре;~ставляющее стадию эволн ции, которая предшествовала или вела к совр»- менным человекообразным обезьянам ('onginae) (Simpson); (4)* Austral<>i>itho- cinae. Южная Африка (плейстоцен). 1;le Грос Кларк относит это подсемейств» к семейству llominidae, см. гл. 11). Поъхимо перечисленных выше, Сххмххсо» х>1>н- «одит < возможных представителей семейства Pongidae с пеопределепньвш род- ст~зениыми связями», ископаемые которых найдены в верхнем эоцене Бирмы и нижнем миоцепе В осточной Африкхх.
I lotuittidae, люди. Космополиты. 1 современный род и вид. Ископаемые (неопре:хе.>с»- ххое число родов и видов) в Африке, Евразии и tta острове Ява (пнейст»>нlt) -::Отряд Т i! lodonlia
~Tillotheriidae. Северная Америка (верхпий палеоцен до среднего эоцепа) и F.t>1>t>ï >
(нижний эоцеп). 4 вида.
:~Отряд Таеп1одоп (а
-:Stylinodot>tidae. Север>хая Америка (палеоцен и эоцен). 7 родов.
Отряд Eclentat;s, неполнозубые. Очевидно, всегда был приурочен к Повому (л«ту. *.tletaelteirontyidae. Северная Америка (ttepxttttii ххалеоцен до среднего эоце>х»). ' ро- да. *3Ietacheiromys известен по почти полному скелету. (; тTоoч~к~~и j: з~р~еe~н~IиIя ~зоогео- графин относятся к самым важным ххз всех ископаемых млекопит»>опхххх. Они свидетельствуют о том, что в самом начале третичного периода неполнозуоьт» обитали в Северной Америке, откуда они, возможно, проникли н Южнун~ Америку.
Млеко питакяцие
«Epoicotheriidae. Северная Америка (нижний олигоцен). 2 рода.
*(11адсемейство 3fegalonychoidea), наземные ленивцы.
eAIegalonychidae. Южная Америка (сомнительно в верхнем эоцене и несомненно в оли- гоцене до плейстоцеиа). Северная Америка (средний плиоцен до плейстоцена) и Вест-Индия (Куба, Гаити, Пуэрто-Рико) (плейстоцен). 26 родов. По крайней мере 2 разные ветви этого семейства достигли Северной Америки в плиоцене п плейстоцене. *Меда1опух проник на север до Северо-Западных территории Канады (Stock, Richards, 1949).
е>>1egatheriidae. Южная Америка (нижний миоцен до илейстоцеиа) и Северная Америка
(илейстоцеи). 8 родов.
elfylodontidae. Южная Америка (олигоцен или нххжний миоцеи до плейстоцена) и (:»- верная Америка (плейстоцен). 13 родов.
(1 1адсемейство t1yrmecophagoidea)
hlyrrneeophagicfae, муравьеды. Тропики Америки. 3 современных рода, 3 или бодее
вида. Все роды встречаются и в Южной, и в Центральной Америке: 2 ви х» достигают северных границ семейства в южиой части Мексики, а 2 — южных гр» ниц в северной части Аргентины. 5 или большее число ископаемых родов обна ружены в Южной Америке (миоцеп и плиоцен).
(Падсемейство Bradypodoidea)
Бха»)уройЫае, ленивцы. Тропики Америки. 2 рода, несколько видов. Bradypus (трех- пальххх ленивец) распространен от северной Аргентины до Гондураса. Сlго1оерхг8 (двупалый ленивец) — через севернухо часть Южной Америки и в Центральиу>»> Америку до Никарагуа. Ископаемые ленивцев неизвестны.
(1 lадсемейство Dasypodoidea)
Dasypodidae, броненосцы. Южная и Центральная Америка и на север в южно-цент- ральную часть Соединенных Штатов. 9 современных родов, небольшое число видов. Все роды встречаются в Южной Америке и только 2 рода достигают Цент- ральной Америки: Сабаггоиг достигает Гондураса и Dasypus распространен да:и,- ше через Центральную Америку и Мексику до Техаса и недавно расширил свой ареал до смежных частей Оклахомы и Луизианы и, возможно, дальше на восто~; до районов, примыкающих к Мексиканскому заливу. В южном направлении несколько родов, включая l>asypus, доходят до пампас центральной Аргентины, а Za»dyus достигает Чубута. Многочисленные ископаемые броненосцев (21 род) известны из третичного периода и плейстоцехха Южной Америки (самые древххххе относятся к палеоцеиу) . Лаяурия проник в хожиухо часть Северной Авхерики в плей- стоцеие (еслп ие раньше), а очень крупный *Holmcsina находился в это же время во Флориде.
*Ре1йер1х>1Ыае. Южная Америка (олигоцен до? нижнего плиоцена). 5 родов.
(*11адсемейстно Ь1у1>(odontoid åà)
~Glyptodontidae, глиптодонты. Юясиая Америка (верхний эоце>х до илейстоцеиа)
и (.'евериая Америка (верхний ххлиоцехх и илейстоцеи). 37 родов. Ilo крайней мере 2 различны» ветви достигли Северной Америки.
Отряд Phol Ыо(а
.">1»>хх>»1»е, ящеры, п»иголины. Значительная часть Африки х»>жнее Сахарьх (хха Ior не
;халее реки Орннжевохх и Зулулеида) и Индо-Малайская область от полуострохх» Иххдостахх и Цейлона, (:иккима и Южного Китая (Юиьиань и Фуцзянь и на север ;х»> Цзянсу) до островов Ява, Бали, Калимах>тахх, Палаван и др. 1 род с 3 или -х африк»нскххми и 3 индо-малайскими видами. Достоверные ископаемые этого р»>,ха известны из плейстоцена Азии и сомнительные — из Европы.
(;»»х»ххте хьпый представитель Pholidota. Европа (олххго>хе>х и мноцеи). 3 рода.
Отряд Lag<oinorpha, зайцеобразные.
*Eurymylidae. Азия (верхний палеоцен). 1 род.
Oc)iotonidae, пищухи, или сеноставки. Умеренная зона Евразии и западная часть С»—
верной Америки. I род, около ! 2 евроазиатских и 2 североамериканских вид».
В Евразии пищухи встречаются от восточной части СССР (на остальной части Европы в настоящее время их иет) на север до арктического побережья, на во- сток до Анадыря и северной Японии (остров Хоккайдо) и на юг до Копет-Даг», северной Индии и северной Бирмы. Большинство видов распространено в гора~ н>жньхх районов Центральной Азии. В Северной Америке пищухи распростра>х»- ны относительно локально и прерывисто от южных районов центральной Лляскц до ххеитральиой части Калифорнии и северной части Ньхо-Мексико (в больш>хп- стве случаев в горах). Ископаемые современного рода Ockoto»a относятся к илио- цену и илейстоцену Евразии (когда-то этот род был распространен и» запад до Ьрххтаххских островов) и в плейстоцене (:еверной Америки. Пскопаемьхе другххх исчезнувших родов известны из Евразии (верхний олигоце>х) и из >ого-западной части Африки (нижиий миоцеи).
Глава 6
LерогЫае, зайцы и кролики. Африка (нет на Мадагаскаре), Евразия, Северная, Цент- ральная и Южная Америка. 9 современных родов, множество видов. Н» север t.epus (зайцы) распространены до побережья Ледовитого океана в Евразии и Северной Америке, а также до арктического архипелага и северного побережь» Гренландии. На юге зайцы достигают южной оконечности Африки, Индии, Ilei' лона, Таиланда. 1 эндемичный род изолирован на Суматре (очевидно, 1 вид зайцев с Цейлона был интродуцирован в западную часть острова Ява). В Америке зайцы доходят до северной Аргентины. В Центральной и ЮжнойАмерике представлен единственный род этого семейства Sylvilagus (кролик), который распространен от южной окраины Канады до северной Аргентины. Роды Pronolagus (Южная Африка на север;lo Кении, обычно в скалистых местах), Pentalagus (приуро- чен к островам Рюкю, к югу от Японии) и Лотего1адиь (известен только нв скло- нах средней части гор Попокатепетль и Истаксиуатль в центральной Мексике) дредположительно все являются реликтами в прошлом многочисленного прими- тивного подсемейства Palaeolaginae (10 ископаемых родов), которое датируется верхним эоценом Северной Америки. Мне не известно, так же как и Jlacy (i) ice, ]92',)), относится ли род Eesolagus (изолированный на Суматре) к этому подсе- мейству. Современные кролики (ископаемые приписываются современному роду подсемейства Leporinae) известны из плиоцепа (Европы и сомнительно Северной Америки). Европейский дикий кролик Огус~о1а1,иs был интродуцирован в Австра- лии с неолагоприятными последствиями дчя человека. Ряд других представителей :~того семейства был интродуцирован довольно далеко за преде,чами их естест- венных ареалов.
Отря;~ Rodentia, грызуны. Каждый отряд млекопитающих таит в себе целый ряд неяс- ных или загадочных вопросов, связанных с гаксономическими проблемами, кото- рые в настоящее время еще не разрешены. Но ни один из отрядов ие может в этом отношении сравниться с отрядом грызунов. Большинство грызунов — животные мелкие, многие из пих редко встречаются и мало изучены, связи их в результат~ конвергенции, дивергенции и возникающих параллелизмов часто запута ниь~, а ископаемые данные (по сравнению с данными по другим млекоиитанпцим) чрезвычайно неудовлетворительны (взято из Симпсопн,— Simpson, 1945, р. 197).
Все это создает неясности и трудности при разрешении вопросов распростра- нения и истории расселения грызунов. Тем ие менее грызуны с точки зрения зоогеографии важны главным образом в связи с тем, что встреча1отся в Австра.зпп и на многих островах.
( Н адсемейств о Ap 1od onto id ea)
э1зс11уготуЫае (включая eParamyidae), анцестральпые грызуны. C,'åâåðíaÿ Америка
(верхний иалеоцен до нижнего миоцена), Европа (эоцен) и Азия (олигоцен), 23 рода.
Apl0dontidae, бобровые белки. 1 современный род и вид в западной части Северной
Америки, от южной части Британской Колумбии (на север до Хопа и района Чил- ливак-Сумас) на юг по крайней мере до округа Моно в горах Сьерра-Невада, Калифорния, и в разобщенных местностях близ побере>кья почти до Сан-Фран- циско (до графства Марин). 5 ископаемых родов в Америке (верхний эоцен) и 1— в Азии (Монголия, нижний илиоцен).
eMylagaulidae, Северная Америка (пижний миоцеп до среднего илиоцепа). (i родов. *Protoptychidae, Северная Америка (верхний эоцеи). 1 род.
*Eomyidae, Северная Америка (верхний эоцеи до с редпего олигоцепа) и Европа
(верхний эоцен до нижнего миоцеиа). 6 родов.
(Надсемейство Sciuroidea)
Sciuridae, беличьи. Многочисленны и разнообразны в Африке, умеренной и тропической
зоне Евразии и Северной Америке. Более малочисленны и менее разнообразны на большей части Центральной и Южной Америки. Отсутствуют на Мадагаскаре и в Австралийской области. подсемейства: (~ ) Sciurinae, земляные и древесные белки.
Занимает ареал семейства. 30 или более родов, много видов. К северу большин- ство северных беличьих останавливается у границы древесной растительности, по род vitellus (суслики и американские земляные белки) распространен в отдель- ных районах арктической тундры, в Азии (Бер|, 1947) и и Северной Америке, а род Marmota (байбаки и лесные сурки) обычно распространен выше липин дре- весной растительности в горах. К югу представители этого подсемейства достигатот Африки, 11ндии и Цейлона, островов Ява, Бали, Калимантан, Сулавеси, а так- же южных и восточных Филиппин (Палаван, Минданао, Лейте и Самар), а в Аме- рике — северной Аргентины. Самый широко распространенный род Sciu гцг распространен в умеренных широтах Евразии, а также в Северной, 11ептрвльной и Южной Америке. 3 рода: упомянутые выше vitellus, Магтойа и Ец1апгы8, сиоирскиц и западноамерпканский бурундуки — встречаются в отде,чыцдх
Млеко пит а~ощ,ие
частях Евразии и Северной Америки. Все другие роды этого семейства приурочены или к Старому или к Новому Свету; большинство родов Евразии и Африки раз- ные. В работе Симпсона не приводятся исчезнувшие ископаемые роды белок, но ископаемые рода Sciurus (в широком смысле) относятся к миоцену Европы и Се- верной Америки, а некоторые другие современные роды ведут по крайней мере к плиоцену; (2) Petauristinae, летяги. Евразия и Северная Америка от лесных мест- ностей в арктических районах на юг до Индии и Цейлона, Явы и Калимантана п некоторых островов южных Филиппин, а также до отдельных территорий в Мек- сике, Гватемале и Гондурасе. 11 родов в Евразии, 1 — в Америке, небольшое число видов. Ископаемые до плейстоцена (Северной Америки) не известны. (Надсемейство Geomyoidea)
Geomyidae, мешетчатые крысы, или гоферы. Северная и Центр»льпая Америка от рав- нин Альберты, Саскачевана и т. д. (к северу — только до восточных районов шта- та Джорджия) до Панамы. 9 или меньше современных родов, множество видов.
Ископаемые (7 или более родов в 2 подсемействах) из Северной Америки начи- ная с нижнего миоцена или раньше.
eteromyidae, карманчиковые мьпии, кенгуровые мыши и крысы. Западная часть (л- верной Америки (на север до южной Канады и на восток по крайней мере до Д»- коты и Техаса), Центральная Америка и северная часть Южной Америки (в рай-- онах Колумбии, Эквадора и Венесуэлы и на острове Тринидад). 5 современны х родов, множество видов. Ископаемые (9 родов) относятся к нижнему мио1(ену (или к более раннему времени) Северной Америки.
(Надсемейство (;astoroidea)
(;astoridae, бобры (с их предками и вымершими родственными им формами, большин- ство из которых, возможно, не походило на бобров по образу жизни). Умерен- ная зона Евразии и Северной Америки. l современный род, l евроазиатский и l слабоотличающийся америкаиский вид, разные подвиды. Евроазиатские боб- ры в прошлом были распространены на большей части умеренной зоны Евразии и Азии, но теперь строго приурочены к отдельным районам Скандинавии, Фран- ции (на Роне и ее притоках), среднему течению Эльбы и ее притокам), восточной Польши и отдельным районам СССР через Северный Урал, к верхнему Енисею (исчезнувший?) и в северной Монголии (более детальное описание см.
Harper, 1945). Американский бобр в прошлом встречался почти по всей Север- ной Америке от Аляски и Лабрадора (северная граница леса) до реки Рио-Гран- де и северной Флориды. Одно время был истреблен на большей части этого ареа- ла, но затем вновь интродуцирован во многих местах, особенно на востоке и даже недалеко от Нью-Йорка (G. М. Allen, 1942). Ископаемые современного рода Ca- stor относятся к плиоцену. Из 13 других исчезнувших родов этого семейства наи- более древние относятся к нижнему олигоцену Северной Америки, верхнему ол ц- гоцену Европы и верхнему миоцену Азии.
*ЕпгуротуЫае. Северная Америка (олигоцен). 1 род.
(Надсемейство Anomaluroidea)
Anomaluridae, шипохвостые белки. Тропическая Африка. 3 рода, несколько видов.
Связи этого семейства сомнительны, и ископаемые, иногда относимые к этому семейству, не достоверны.
(Сомнительные Anomal uroidea)
*Pseudosciuridae. Европа (верхний эоцен до среднего олигоцена). 3 рода.
Theridomyidae. Европа (верхний эоцен до среднего олигоцена). 6 родов, 9 сомнитель- ных родов, приписываемых семейству из Африки и Азии (олигоцеп и нижний миоцен).
Pedetidae, долгоноги. Южная Африка, на север до Кении. 1 современный род, 2 вида
и еще 1 ископаемый род в Юго-Западной Африке (нижний миоцен).
(Надсемейство Iuroidea)
Cricetidae, хомяки и полевки. Африка, Мадагаскар, большая часть Евразии, (;e»ер- ная, Центральная и Южная Америка; к северу до границы суши, на островах ile- довитого океана, северная часть Гренландии (лемминги); к югу до южной оконеч- ности Африки, Индия и Цейлон (только Gerbillinae), северная Бирма п Юго- Восточный Китай (нет в Индокитайской подобласти), Тайвань, южная оконеч- ность Южной Америки и Огненная Земля. 5 подсемейств: (1) 'ricetinae, хомяки Старого Света, большинство мышей и крыс Нового Света и т. д. Хомяки (триба ('riceti») распространены по всей умеренной зоне Евразии (5 современных родов) п изолированы в Южной Африке (l род). Ископаемые (несколько родов) дати- руются нижним плиоценом Евразии. ] покоры (Myospalacini) встреч»1отся в Цент- ральной и Восточной Сибири и в Северном и Центральном Китае (1 род, 5 видов).
Ископаемые (l род) относятся к верхнему миоцену Азии. Все другие современ- ные Cricetinae образуют трибу Hesperomyini состоящую пример~~о из 50 родов
826
Глава 6
и многочисленных видов, приуроченных к Новому Свету (около 40 родов- к 11еитральной и Южной Америке), включая рисовых, хлопковых, дреиесггых крыс, саранчовую мышь, хомячка и др., которые встречаются от районов Аляски и, [абрадора до Огненной Земли (только немногие достигают этих крайних точек), с отдельными формами на островах Галапагос, Ямайке и Малых Антильских островах. Ископаемые (несколько родов) относятся к нижнему плиоцену Север- ной Америки. 3 ископаемые трибы с 12 родами и также различные сомнительные ископаемые роды этого подсемейства встречаются в олигоцене и позднее в Европе и Северной Америке; (2) Nesomyinae, Мадагаскар. 7 современных родов, 12 видов.
Ископаемые нескольких современных родов и 1 исчезнувшего рода обнару- жены в виде ископаемых или субфоссильно на Мадагаскаре (плейстоцеи).
Эллерман (E1lerman, 1940 — 1949) относит эти роды к подсемействам, Симпсон (Simpson, 1945, р. 86, 207), рассматривал их вместе. Не имея ио »тому вопросу собственного мнения, я следовал классификации Симпсогга.
Согласно классификации Эллермана, «Nesomyinae» состоит примерно из г разных ветвей, которые самостоятельно достигли Мадагаскара; (3) Lophi<>- myinae. Тропическая Африка. 1 род, около 11 видов;(4) Microtinae, лемминги, полевки и т. д. 3 трибы. Лемминги (триба Lemmini) встречаются от самых высо- ких арктических широт Евразии и Северной Америки на юг до Алтая и районов Канзаса и Виргинии. 4 современных рода, несколько видов: Dicrostonyx и I emmus распространены в Евразии и Северной Америке. Ископаемые этой трибы отно- сятся к верхнему плиоцену (Северной Америки). Некоторые северные лемминги г овершают значительные периодические массовые эмиграции (Elton, 1942).
Е трибе Microtini относится доминирующий род Microtus (обычные полевки) примерно с 260 видами и подвидами, которые распространены через Еврази~о и Северную Америку от Арктики на юг до Северной Африки (Киреиаика}, 11алестииы, Ирана, Гималаев, Восточного Китая (на север до Янцзы), возможно, Тайваня, южной Мексики и Гватемалы. Большинство из 15 других современных родов этой трибы распространено в пределах ареала рода Micro- tus, но некоторые в северной Индии, северной Бирме и Юго-В осточном Китае, простираются несколько дальше. Кроме рода Мicrotus, общими для Евразии и Северной Америки являются следугощие роды этой трибы: Сlethrionomys (рыжие полевки) в умеренно-холодных и арктических районах обоих полушарий, Pity- пгггв (сосновые полевки) на сильно разобщенных территориях в континенталь- нои части Европы (до Малой Азии и Кавказа) и в восгггочггои части Соединенных Штатов и восточно-центральной Мексике, а Лали rus — от центральной части ССС Р до Монголии и во внутренних областях западной части Северной Америки. Севе- роамериканская мускусная крыса (Ondatra) — специализированный представи- тель этой трибы. Была интродуцирована в Европе и Азии. Ископаемые M~cro- tini, охватывающие некоторые современные и i l ископаемых родов, ведут к верх- нему миоцену Северной Америки и к плиоцену Евразии. Слепушонки (Ellobiini) встречаются в Евразии от южной части СССР и Малой Азии до Монголии. 1 род, 4 вида и другой ископаемый род в Европе (верхний плиоцен); (5) Gerbillinae, песчанки и т. д. По всей Африке в благоприятных для обитания местах (но не в густых лесах Западной Африки) и от Юго-Западной Азии до Кавказа, в сосед- иих районах СССР, Центральной Азии до Монголии и т. д., на полуострове Индостан и Цейлоне. 10 родов, множество видов. Ископаемые современного рода Gerbillus относятся к плиоцену Азии.
Spa]acidae, слепыши. От Юго-Восточной Европы (Венгрия и т. д.) до районов Каспия,
Сирии, Израиля, Египта (ОАР) и Ливии. 1 род, 2 или 3 вида, 2 ископаемых родгг в Европе (плиоцен).
Rhizomyidae, ризомииды и т. д. Восточная тропическая часть Африки на север до Эфио- пии (1 род, несколько видов) и от восточных Гималаев и Южного Китая до (;у- матры (2 рода, 4 вида) и 5 ископаемых родов в Европе и Азии (верхний олигоцеи и т. д.) и Северной Африке (плейстоцен).
>>uridae, настоящие мыши и крысы. Местные по всему Старому Свету, вероятно, кромс
Мадагаскара. Интродуцированные в Новом Свете. 6 подсемейств: (1) Murinae. Ста- рый Свет. 68 современных родов, сотни видов. Многочисленны по всей Африке (ио, вероятно, нет ни одного местного вида на Мадагаскаре), многочисленны в умерен- ной и тропической зонах Евразии и на расположенных близ материка островах.
Несколько представителей достигает Арктики, но ни один из них там широко не распространен или не является особенно для нее характерным. Многие распрост- ранены через Индо-Австралийский архипелаг до Австралии и Тасмании и на восток до Филиппин и Соломоновых островов. Мелкие крысы группы Battus concolor-exul0ns, которые в известной мере являются домовыми, широко распро- странены далее на восток в Тихом океане (возможно, занесены ранними полине-
Млекопитп~тцие
827
знйцами) до Гавайских, Маркизских островов и Новой Зеландии. 2 вида крыс и 1 вид мышей были разнесены по всему земному шару в результате торговой деятельности человека. Aattus rattus (черпая крыса), очевидно, иестньхй вид тро- пххческой Азии, а Rattus norvegicus (крыса-пасхок), очевидно, — умеренной зоны Азии. В настоящее время оба вида широко распрострапе»ы почти по всему земному шару, но черная крыса предпочитает более теплые, а пасюк — более прохладные районы, и частные особенности их распространеия зависят не толь- ко от климата, но также отчасти и от конкуренций друг с другом и с другими местными животными в различных местах. Mus musculus (домовая мьппь) пред- ставлена живущими в природе формами в Южной Европе, Северной Африке и Центральной Азии до Маньчжурии и Японии, но формы «комменсалххсты», несом- ненно, распространены по всему земному шару. Aattus (более 550 выде хеиххых ,")ëëåðìaHoè форм) чаще каких-либо других родов распространеххы в природе по всему ареалу семейства. д~~демичпые видовые группы этого рода встречаются во всех частях Старого Света в границах ареала семейства (Е1!егшап, 1941, р. 149— I.50). Большинство других родов приурочено к той или другой фауххистическохх ~)бласти Старого Света или к какой-либо части одной области или к неоольшой площади, а многие — к отдельным островам или группам островов»а террито- рии Индо-Малайской и Австралийской областей. Ископаемые этого большого подсемейства (только 5 ископаемых родов) известны лишь из плиоцена (Евразии} и из плеххстоцена; (') Dendromurinae, африканские древесные мыши. Африка х' хогу от Сахары. 6 родов, более 50 видов; (3) Otomyinae, ушастые крысы. Африка южнее Сахары. ! род, 11 видов; (4) Phloemyinae, гигантские фихиппххнские кры- сы. Индо-Малайская и Австралийская области от Ассама, Бирмы и Юньнани до островов Ява, Калимантан, Сулавеси и некоторых Филиппинских, а также па Новой Гвинее, Новой Британии и прилежащих мелких островах и, возможно, lf8 острове Флорес. 6 современных родов, более 20 видов и 1 ископаемый род на острове Тимор (плеххстоцен?). Но принадлежность всех этих родов к этому под- семейству сомнительна; (5) Rhynchomyinae известны только с горы Дата, северный Лусон, Филиххпинские острова. 1 вид; (6) Hydromyinae, австралийские водяные мыши. Австралия и Тасмания, Новая Гвинея, Новая Британия и на близлежащих мелких островах (Ару и Кай). 9 родов, большинство в Новой Гви- »ее, включает как наземные, так и полуводные и специализированные водные формы. 13 видов (Tate, 1951). На северном Лусопе, Филиппинские острова, нахо- дится 2 не водных рода, 2 вида. Связи с австралийскими формами до некоторой степени сомнительные.
(}1адсемейство Gliroidea)
С'l}ridae (Muscardinidae), сони. ' подсемейства: (!) Glirinae. Умеренная зона Евразихх
и т. д. (прерывистое распространение), большая часть Европы, Северная Африка и Юго-Западная и Центральная Азия (5 родов, 6 видов), Япония (1 род и 1 вид} и ископаемые (4 рода) в Европе начиная с миоцена или раньше; (2) Graphiurinae, Африка от южной части Сахарьх до южной оконечности материка. 1 род, 39 видов.
let;~canthomyidae, иглистые сони. Локализованы в тропической Азии. 1 род и l вид
в южной части полуострова Индостан и другой род и вид в Юго-В осто чпом Китае и северном Индокитае.
Se}vviniidae, селевинии. Западная часть Центральной Азии (на запад от озера Бал- хаш). '! вид.
(11алсемейство D ipodoidea)
/ародiclae, прыгунчики и др. Умеренная зона Евразии и Северной Америки. 2 подсе- мейства: (1) Sicistinae (мышовки, морфологически не приспособлены для пры- ганья). Некоторые районы Скандинавии и Цеххтральххой Европы через Централь- ную Азию до Западного Китая, Маньчжурии и острова Сахалин. 1 современныЙ род, 6 видов и 2 ископаемых рода в Евразии (олигоцен и нижний плиоцеп); (2} Харойх1ае, прыгупчики. Западный Китай (Ганьсу и Сычуань, ! четкий вид, иног- да относимый к самостоятельному роду, а иногда включаемый в американский род) и Северная Америка от районов Аляски и Лабрадора на юг до Калифорнии, Нью-Мексико и южных Аппалачей (2 близкородственных рода, несколько видов).
1 ископаемый род из плиоцепа Северной Америки.
}}ipodidae, тушканчики. Засушливые части Северной Африки и умеренной зоны Евра- зии от Сахары, Аравии и южной части СССР через юго-западную и Центральххун> Азихо до Монголии. !2 современных родов (в 3 подсемействах), небольшое число видов. Это семейство отмечается большим разнообразием в Азии, (чем в Африке).
4 ископаемых рода в Азии (верхний миоцен).
П о д о т р я д Н у s t r i с о m о r p h а. Большинство остальных семейств грызу- нов является дикобразоподобными или считаются таковыми. Большая часть их
828
Ггава 6
находится в Южной или Центральной Америке или в Вест-Индии и только одно — на основной части Северной Америки. Некоторые группы Старого Светл (главным образом африканские), возможно, имеют связи с американскими груп- пами, но Симпсон (Simpson, 1945, р. 210 и след.) считает этот факт сомнительным, а Вуд (Wood, 1950) рассматривает сходство как результат параллелизма, а не родственных связей (стр. 304 — 305).
(Надсемейство Iiystricoidea)
Hystricidae, дикобразы Старого Света. Теплые страны Старого Света (но не Австрялий- ская область), на север до Марокко, Италии (интродуцирован?), Сирии, Турк- мении и Южного Китая (южнее Янцзы) и на юг через большую часть Африки (но не Мадагаскар), полуостров Индостан, Цейлон, Яву, Малые Зондские острова до островов Сумбава (и, возможно, Флореса), Калимантан, Палаван и на ю>кны~ Филиппинах. 5 родов, около 18 видов. Дикобразы Старого Света — наземные животные (Нового Света — древесные). Ископаемые дикобразы включчн>т 'l ис- чезнувший род и датируются олигоценом Европы.
(Надсемейство Erethizontoidea)
Erethiz0ntidae, дикобразы Нового Света. В Южной Америке находятся ' локализован- ных монотииических рода, не считая Coendou (иесколько видов), которьш встре- чается на юге до северной части Аргентины и иа севере до Центральной Аме- рики, захватывая часть Мексики. В Северной Америке Еге~lигоп (! или " видя) распространен от северной границы леса на юг примерно до границы с Мексикой на западе и по крайней мере до южной Пенсильвании на востоке. 7 ископаемых родов из Южной Америки относятся к олигоцену и более позднему времени.
Eretkizo~p появился в Северной Америке в верхнем плиоцене.
(Надсемейство Cavioidea)
*Cephalomyidae. Южная Америка (олигоцен). 2 рода.
*Eocardiidae. Южная Америка (верхний олигоцен до нижнего миоцена). 5 родов. Caviidae, морские свинки и т. д. Южная Америка, особенно ее южная часть от Пата-
гонии на север до 15' ю. ш. (только род Сала, настоящие морские свинки), про- стирается до северной части Южной Америки. 5 родов, небольшое число видов.
Кроме того, 9 ископаемых родов из Южной Америки (плиоцен).
Hydrochoeridae, водосвинки. Тропики Ю>кной Америки (на юг до дельты Параны в Ар- гентине) и Панама. 1 современный род, 2 вида (каиибары самые крупные совре- менные грызуны, ведущие полуводный образ жизни). 10 ископаемых родов (в 2 подсемействах) из плиоцена и илейстоцена !Ожиой Америки. 2 рода этого семей- ства распространились в юго-восточную часть Северной Америки в плейстоцене, достигнув Южной Каролины.
Din0myidae, диномииды, или длиннохвостые паки. Западная часть Южной Америки
(цеитральная Колумбия к центральному Перу и т. д.). ! вид.
*Heptaxodontidae, вымершие гигантские крысы. Южная Америка (миоцен и илиоцен,
19 родов в 3 подсемействах) и Вест-Индия (илейстоцен и почти современное чет- вертое подсемейство, 7 родов, включая eQuemesia из Ямайки, Гаити, Пуэрто- Рико, Ангуиллы и Сен-Мартена, но нет ни одного рода из Кубы).
Dasyproctidae, агути. Южная и Центральная Америка на юг до южной Бразилии и се- верной Аргентины и на север (2 рода) в южную Мексику. Род I>asyprocta дости- гает также некоторых Малых Антильских островов. 4 рода, небольшое число видов. В ископаемом состоянии не известны.
(Надсемейство Chinchilloidea)
СЫпсЫ11Ыае, вискаши и шиншиллы. Западная и южная части Южной Америки — OT
Перу, Боливии и северной Аргентины до или почти до Магелланова пролива.
3 современных рода, около 7 видов, 7 ископаемых родов из Южной Америки (олигоцен и позже). Настоящие шиншиллы (Chinchilla) встречаются от прибреж- ных возвышенностей северного Чили до высокогорий Анд Перу, Боливии и севе- ро-западной Аргентины, но их численность сильно сократилась в результате охот- ничьего промысла (G. М. Allen, 1942, р. 389 и след.).
(Надсемейство Octodontoidea)
'apr0myidae, нутриевые и др. Большие Антильские острова в Вест-Индии и некоторые
районы Южной Америки. 4 или 5 современных родов. Вестиндские формы (см.
и11же), включая и исчезнувшие, приводятся согласно классификации Вильямся
(Williams, 1951, более поздние неопубликованные сведения; Williams, Reynolds,
Koopman, !953). Сарготуя (4 современных вида) встречается на Кубе и острове Иайнс, и ! ископаемый вид — иа Каймановых островах. Geocapromys (3 совре- менных вида) — на Багамских островах, Ямайке и острове Суон; ископаемые- на Кубе, а также на Ямайке и Багамских островах. Исчезнувший род *Иехоlо- codon (! вид) на Гаити является родственным 2 предыдущим родам. Р1а~ьойо~~йа (2 современных и 2 исчезнувших вида) на Гаити представляет другую группу
Млекопитающие
829
рода, к которой относится также eApbaetreus (1 вид) на Гаити и eIsolobodon (eithy- odontia) (1 вид) — на Гаити, Моне, Пуэрто-Рико и Виргинских островах (возмож- но, занесены доисторическим человеком). Все исчезнувшие вестиндские формы относятся к плейстоцену или постплейстоцену. Procapromys (1 вид) предположи- тельно с северного побережья Южной Америки (центральная часть горных при- брежных районов Венесуэлы), как полагают, представляет материковую ветвь, от которой произошел вестиндский род Capromys, но достоверными фактами это подтверждено не было. Эллерман (Ellerman, 1940) не видел представителя это- го рода. Возможно, это был экземпляр молодого животного вестиндского рода Capromys с неправильно указанной местностью. Myocastor (! вид), полуводный зверек нутрия, обитает в южной части Южной Америки от крайнего юга Брази- лии до (или почти до) Магелланова пролива. Этот род был интродуцпровап в н>ж- но-центральной и (локально) в северо-западной частях Соединенных Штатов, а также во Франции. Муоса81ог значительно отличается от современных вестинд- ских нутриевых, но, очевидно, связан с ними ископаемыми формами (Simps<)n).
7 ископаемых родов семейства приводятся из нижнего миоцена — плиоценд Южной Америки.
0ctodontidae, осьмизубые. Южная часть Южной Америки от южного Перу, Боливии
и южной Бразилии далеко на юг, но, очевидно, не достигая Магелланова проли- ва. 5 родов, 8 видов и еще 3 ископаемых рода в Южной Америке (верхний олиго- цен и плиоцен) и ! вид (eAlterodon) — на Ямайке (плейстоцен).
ИепотуЫае, туко-туко. Южная часть Южной Америки от южного И еру, Боливии
и южной Бразилии до Огненной Земли. 1 современный род, множество ви;~о» и подвидов, 4 ископаемых рода в плиоцене Южной Америки.
ЛЬгосопидае, крысы-шиншиллы. Юго-Западная часть Южной Америки от южной Боли- вии до центрального Чили. 1 род, несколько видов. 1 ископаемый род в Южной Америке (плиоцен) .
Echimyidae, щетинистые крысы. Тропики Южной и частично Центральной Америки.
14 современных родов, небольшое число видов. Все роды встречаются в некоторых частях тропиков Южной Америки, несколько родов заходят на юг до южной Бразилии и т. д., 2 — в Центральную Америку до Никарагуа и по крайней мере еще 1 достигает Панамы. I;ckimys зарегистрирован с острова Мартиника на Малых Антильских островах. Около 7 ископаемых родов обнаружено в Южной Аме- рике (верхний олигоцен и нижний миоцен) и 4 — на Больших Антильских ост- ровах (плейстоцен или постплейстоцен): эВоготуг (2 вида) — на Кубе; eBroto- mys (2 вида) — на Гаити, *Ноторзотув (1 вид) и *Heteropsomys (1 вид) — в Пу- эрто-Рико. Proeckimys, который в настоящее время встречается от южной Брази- лии до Никарагуа, представлен ископаемыми (вероятно, плейстоценовыми) из Пуэрто-Рико (Williams, Koopman, 1951).
Т)1гуопотуЫае, тростниковые крысы. Большая часть Африки южнее Сахары. 1 род,
около 7 видов и 1 ископаемый род (*Paraulacodus,— Hinton, !933) в северной Индии (плиоцен). Принадлежность к этому семейству 3 других ископаемых родов из Африки (миоцен и плиоцен) вызывает сомнение.
Petromyidae, горные мыши. Юго-западная Африка (от Намакваленда до юэкной Анго- лы). ! род, '! или 2 вида, несколько подвидов.
(Сомнительные II ystri corn orpha)
(Надсемейство Bathyergoidea)
Bathyergidae, пескорои. Африка южнее Сахары, 5 родов, небольшое число видов, 1 иско- паемыи род в плейстоцене Южной Африки.
(Надсемейство Ctenodactyloidea)
!.'tenodactylidae, гунди, или гребпепалые крысы. Северная Африка от Сенегала и Марок- ко (включая прибрежную территорию Палеарктики) до Сомали (Ellerman).
4 рода, около 8 видов, 1 из родов (Pectinator), представленный в плиоцене север- пой Индии (Hinton, 1933),— сомнительный.
Отряд Cetacea, китообразные. Преимущественно морской (все океаны), но некоторые
дельфины (обыкновенные дельфины) заходят или живут в пресных водах (напри- мер, виды морского рода Sotalia обитают в Амазонке), а семейство Platanistirlae почти всегда приурочено к Амазонке. Строго морские семейства этого отряда здесь опущены.
Platanistidae, речные дельфины. 4 монотипических рода: (!) Индия (реки Инд, Гнпг
и Брахмапутра); (2) Китай (озеро Тангтин в бассейне Янцзы); (3) в системе рек Амазонки и Ориноко в Южной Америке; и (4) Ла-Плата (в эстуарии, но главным образом в пресных водах) и в прибрежных (соленых) водах прилегающих частей Южной Америки. Ни один из существующих членов этого семейства не является полностью морским, но современные пресноводные роды являются реликтами мор- ского семейства (миоцен и плиоцен), которые были замещены в море.
Глава 6
Отряд Garnivora, плацентарные хищные
ll одотряд (;reodonta
*Arctocyonidae, *5Iesonychidae, *0xyaenidae, э Н уаеподопФМ ае, *Сгео[агя М ае.
l3 общей сложности 68 родов из Северной Америки и Евразии (многочисленны в»»- леоцене и эоцене, малочисленнее в олигоцене и т. д.) и Африки (известны»ем»»- гие роды из нижнего олигоцена и нижнего миоцена).
1( од отряд F issiped a
eMi;>cidae. Северная Америка (средний палеоцеи до верхнего эоцена) и Евразия (иерх- ний эоцен и, возможно, нижний олигоцен), 12 родов.
Са»>чае, лисицы, волки, собаки. Космополиты, но местных форм на Мадагаскаре или
в Вест-Индии и т. д. и, вероятно, в Австралийской области нет. 'l2 современиы.~ родов, небольшое число видов. К северу лисицы и волки достигают границ мате'— риков, арктических островов, северного побережья Гренландии и т. д. h тогу разные представители этого семейства достигают оконечностей материков Африки и Австралии (но за пределами Явы и в этом направлении они не яв >яи>тся мест- ными), Огненной Земли и Фолклендских островов. Настоящие лисицы (Vulpes) встречаются в некоторых частях Африки, через всю умеренную зону Евразии и на юг до полуострова Индостан и северного Индокитая, на большей части Север- ной Америки, к югу примерно до границы Соединениых Штатов с Мексикой.
Лисицы Мексики, Центральной и Южной Америки относятся к другому роду.
Vulpes ги/ре8, красная европейская лисица. Распространена через всю умереннуи) зону северного полушария Евразии на юг до Северной Африки, Аравии, северо- западной Индии и северного Индокитая. На севере заходит в некоторые аркти- ческие районы (но не в глубь тундры). Красную североамериканскую лисицу иногда считают только ее подвидом, но чаще рассматривают как родственный вид европейской. 11есец (Alopez lagopus) встречается в заполярье в обоих полушариях от Норвегии через северную Евразию и Северную Америку до Гренландии, Ис- ландии и т. д. Волки и им близкородственные (Canis, включая шакалов, койо- тов, настоящих собак и т. д.) встречаются в естественных условиях почти по всей Африке, Евразии на юг до Цейлона и Сиама и в Северной и Центральной Америке до Коста-Рики, но не в Южной Америке (плейстоценовые ископаемые Южной Аме- рики, приписываемые роду Canis, строго говоря, к нему не относятся,— Simpson).
Волк (Canis lupus), в прошлом широко распространенный по всей Евразии, вклн>- чая Британские острова, в настоящее время исчез в за.падной части Европы, кро- ме некоторых отдельных районов Португалии, Испании, Италии, Сицилии, Шве- ции и Норвегии. Западная граница восточных волков в связи с их перекочевками постоянно изменяется, но они достигают Польши, Чехословакии, некоторых рай- онов Балкан и до сих пор встречаются на большей части Северной и Центральной Азии, достигая на востоке Японии (если он там еще не исчез) и в южном направ- лении до Аравии по крайней мере до Дхарвара в Индии (проникая на несколько градусов в пределы тропиков), а также в некоторых частях Китая. В прошлом формы этого же вида встречались почти по всей Северной Америке, от высоких арктических широт до плоскогорья Мексики, и до сих пор обычны на крайнем севере. В других местах Северной Америки они приурочены к нескольким изо- лированным диким местностям. Домашние собаки, вероятно, произошли от вол- ка, по-видимому, в Евразии в доисторические времена и были затем распростра- нены человеком по всему земному шару. Предки австралийских динго и «диких» собак на Новой Гвинее, по-видимому, проникли сюда вместе с доисторическим человеком из Азии. Lycao» (гиеновая собака) широко распространена в Африке, а Сиоп (красный волк) — в Южной и Восточной Азии на север до Амурской области и на юг до Индии и Явы. Speotkos (кустарниковая собака) встречается в тропиках Южной Америки от Эквадора и Гвианы до Парагвая. Это единственный сохранившийся член подсемейства Simocyoninae, которое было хорошо представ- лено в Северной Америке и Европе в среднетретичном периоде. Другие ископаемые представляющие более 50 исчезнувших родов и включающие несколько исчез- нувших подсемейств, показывают сложность эволюции семейства Сanidae, начи- пая с верхнего эоцена Северной Америки и Европы.
()rsidae, медведи. Евразия, Северная и Южная Америка. 6 родов (1 полярный, 1 евро- азиатский и североамериканский, 3 азиатских и 1 южноамериканский), возмож- но, только 7 или 8 четких современных видов. Белый медведь (Thalarctos) встре- чается на арктическом побережье, на островах и во льдах Евразии и Северной Америки. В Старом Свете распространение медведей к югу очень прерывистое— северная Испания (предположительно в прошлом, и Северная Африка), отдель- ные части Италии и Греции, Сирия и т. д., Иран, Индия и Цейлон, Суматра и Ка- лимантан. В Северной Америке черный медведь (U. americanus), с различными цветовыми фазами, встречается на севере по всему материку и на юге в ме-
Млеко т~тагогц ие
лах, пригодных для его обитания, до Флориды и северной Мексики. Гризли (Г. h<>rribilis) свойствен западной части материка от Аляски до Скалистых гор и иа юг до Ныо-Мексико. Этот вид или родственные ему виды в прошлом достига- Ill Калифорнии и северной части Мексики. Большой бурый медведь (U. midde><- <lо> ffi) приурочен к южному побережью Аляски и прилежащим островам. В Цент- рнльной Америке медведей нет. В Южной Америке нндский медведь (Tremarc- /оя) обитает на облесеиных склонах Анд в западной Венесуэле, Колумбии, Эквадоре, llepy и Боливии. Ископаемые медведи (8 родов) из»естны только и: > ( реднего миоцена Европы и нижнего плиоцена Северной Америки. ' или 3 ископа- емых рода были общими для Евразии и Северной Америки.
Procy<>nidae, еноты, панды. Северная, Центральная и Южная Америка и Восточная
Азия. ' подсемейства: (1) Procyoninae, еноты. Северная и Южная Америка. 6 сов- 1>емеиных пл>ос 8 ископаемых родов. Procyon (еноты, 2 вида) рнсгрост)>анен !)т южных краевых частей Канады до северной Аргентины. Североаме~~иканские виды достигают ! 1а намы, а 1 южноамериканский вид — Коста-Рики. Изолирован- ><ые популяции находятся или находились (интродуцированные?) на острове Ныо- 1!ровиденс (Багамские острова), Гваделупе и Барбадосе (Малые Антильские ост- рова). 5 других родов этого семейства встречаются в тропиках Ю>кной и (или) I!ентрйльной Америки (но не в Вест-Индии). Из них 3 рода достигают Мексики и 2 рода (из 3) проникают в юго-западную часть Соединенных Штатов. Ископае- мые этого подсемейства относятся к Северной Америке начиная с нижнего миоце- на и к Южной Америке с, верхнего миоцена до образования современного соеди- нения материков; (2) Ailurinae, панды. Локализованы в Восточной Азии. Крас- ння паида (Ailurus) в Юго-Западном Китае (Юнинань и Сычуань), в северной Бир- ме, Сиккиме, Непале. Гигантская папда (Ailuopoda) — в центральной и западной ('.ычуани в бамбуковых лесах па высоте до 12 000 футов. 2 других ископаемых рода из верхнего миоцена и нижнего плиоцена Европы и нижнего плиоцена Азии. Mustelidae, куньи. Весь земной шар, кроме Мадагаскара и Австралийской области, на
север до высоких арктических широт и на юг до мыса Доброй Надежды, южная Индия и (только выдры) на островах Цейлон, Ява, Калимантан, Палаван и т. д.
и до южной части Южной Америки и (только выдры) на Огненной Земле. 6 под- семейств: (J) Mustelinae, куньи. Почти весь ареал семейства, но нет па Цейлоне и Огненной Земле..12 родов, неболыиое число видов. Mustela (разпые мелкие куньи, норка и т. д.) распространены через умеренную и арктическуп> зоны Евра- зии и Северной Америки и на юг до Северной Африки, северной Индии, Явы и Калимантана, возможно, на острове Палаван и части Южной Америки (см.
«>1же). М. ermi»ea — горностай — общий вид для умеренно-холодных и арктиче- ских частей Евразии и Северной Америки. Имеются еще близкородствепные пары видов этого рода, свойственные обоим названным территориям. M. frenata— длиннохвостый горностай распространен от южной части Канады до Венесуэлы, Колумбии, Эквадора и Боливии. Другой вид этого рода является эпдемиком в рай- оне Амазонки. Род ЛХа>1ея (несколько видов, соболь, илька) и вид Gulo gulo (росомаха) являются общими для Евразии и Северной Америки, включая арктн- >еские районы. Другие роды этого подсемейства приурочены к отдельным частям Евразии, или Африки, или Южной Америки; в некоторых случаях проникают нн север через Центральную Америку до >ожной Мексики. Ископаемые этого подсе- мейства многочисленны (21 род, некоторые из них дивергентные) и ведут к ниж- iieMy олигоцену Евразии и Северной Америки. Из ископаемых родов 3 — общие для Евразии и Северной Америки, а распространение современных форм указы- вает еще по крайней мере иа 6 перемещений между этими материками; (2) Ме11>- i 0rinae. l современный вид МеШгога capensi's, медоед, который встречается на <>алычей части Африки (на юг до Капской провинции) и в юго-западных частях Азии до западной и северной Индии и Непала. Ископаемые этого рода найдены в среднем плиоцене Азии, а другого — в нижнем плиоцене Азии и Северной Америки; (3) <lelinae, барсуки. Умеренная зона Евразии (5 родов) на юг до >1алестины, Ирана (нет в Индии) и через Индокитай до островов Ява, Кали- мантан и Палаван, западная часть Северной Америки (другой род, 1 вид) от н>ж- ной Канады до северной Мексики. Ископаемые (5 родов) в Евразии (начиная с миоцепа) и в Северной Америке — (с верхнего плиоцена); (4) Mephitinae, скуп- сы. Северная и Южная Америка. 3 рода, небольшое число видов. Mephitis встре- чается от холодных районов умеренной зоны Канады до Гондураса. Spiilogale— от южной Британской Колумбии и северной Виргинии до северной части Коста- Рики. Сопера1и8 — от юго-западной части Соединенных Штатов до Магелланова пролива. Ископаемые этого подсемейства (6 родов) относятся к Евразии (верх- ний миоцен и плиоцен), а также к Северной Америке (плиоцен и позже) и Южной Америке (плейстоцен); (5) *Leptaretinae. Северная Америка (миоцен и нижний
Глава 6
плиоцен). 3 рода; (6) Lutrinae, выдры. Почти всегда до границ распространения семейства, кроме самых холодных арктических районов. 7 родов. Lutra (настоя- щие выдры, разные виды) имеет почти такое же распространение, как и подсемей- ство, но не достигает Филиппинских островов (другой род достигает острова Палаван). Другие роды приурочены к Африке, Южной Азии и Южной Америке, кроме калана (Enhyd ra), который встречается на побережье северной части Тихого океана от Берингова моря на юг до южной Калифорнии и (по крайней мере в про- шлом) до северной Японии (Хоккайдо). Из 11 ископаемых родов выдровых самый древний относится к верхнему олигоцену (Европы).
'1'Ь.еггМае, циветты, мангусты и др. Теплые страны Старого Света. 36 современных
родов, много видов. Ареал семейства охватывает Африку, Мадагаскар, более теплые части Евразии на север до Испании и Франции, Палестину, Иран, Афганистан, северную Индию и Южный Китай (род Paguma распространен |ггг север до Пекина), на юг и восток до Цейлона, Малых Зондских островов, до остро- ва Тимор (?), Сулавеси, Молуккских (?) островов Ару и Кай (?) и Филиппин (но на удаленных от материка островах он, возможно, не является местньв|).
На юго-востоке Macrogalidia, типичен для острова Сулавеси, Viverrа tanpalunga (малайская циветта) — для южной части Молуккских островов, а Paracloxurus kermapkroditus (пальмовая куница) — для Молуккских островов, Ару и Кай и Малых Зондских островов до острова Тимор, но естественные границы этих видов вызывают сомнение. На Мадагаскаре находится 7 эндемичных родов вивер- ровых (включая Cryptoproctn, иногда относимого к семейству Felidae), которые делятся Симпсоном между 3 подсемействами и трибами. Из этой классификации следует, что эти роды произошли не от одной пришедшей с материка ветви, а от нескольких, что окончательно еще не установлено (Simpson). Возможно, что все мадагаскарские формы имеют одного предка. Бирманская форма индийского ман- густа (Herpestes auropunctatus birmanicus) является иитродуцированной и широко акклиматизировавшейся формой в Вест-Индии. Ископаемые виверровые (7 ро- дов) относятся к верхнему эоцепу или нижнему олигоцену (Европы). Достовер- ных данных о том, что виверровые когда-либо достигали Австралии и материков Северной и Южной Америки, не имеется.
Hyaenidae, гиеновые. Африка (включая Северную Африку) и Юго-Западная Азия нгг
север до западных берегов Каспийского моря и на восток до Индии (но не до Цей- лона). 3 рода (включая Proteles, гривистый волк), 4 вида. Все виды африканские, кроме Нуаегга Йуаепа, который распространен от Северной и Восточной Африки до Азин (см. границы, указанные выше). Ископаемые гиеновых (3 исчезнувших, а также 2 современных рода) относятся к Европе и Азии (плиоцен и плейстоцен), а самое древнее ископаемое этого семейства находится в Азии (верхний миоцен приписывается современному африканскому роду). Ископаемые этого семейства в Африке, очевидно, неизвестны.
Felidae, кошачьи. Весь земной шар, кроме Австралийской области и Мада бхаскара
(Cryptoprocta помещен в семейство Viverridae) и некоторых других островов.
3 современных рода, много видов. Felis в широком смысле охватывает всех мел- ких кошек и пуму и определяет все географические границы этого семейства, встречаясь в естественных условиях по всей Африке, Евразии на север по крайней мере до облесенных частей Арктики и на юг и восток до Индии и Цейлона, остро- вов Ява, Бали (но. вероятно, не до Тимора), Калимантана и некоторых Филиппин- ских островов (но не до Лусона) и в Америке от отдельных арктических районов до Магелланова пролива (но не в Вест-Индии). Самый северный вид Felis lynx, рысь, распространен через облесенные части умеренно-холодной и арктической Евразии и Северной Америки. Р. concolor, пума, встречается от северной части Британской Колумбии на восток до южного Квебека и на юг до Mareëëàíîâë пролива в самых разнообразных местах обитания: леса арктической и умеренной зоны, тропические дождевые леса, пересеченные местности, более или менее открытые равнины и полупустыни и в районах, лежащих на уровне моря, до «ысо- ких гор. Домашняя кошка, возможно, происшедшая от двух предков: от 1'. sil- cestris (европейская лесная кошка) и F. lybica (африканская дикая кошка), бы:гн широко распространена по всем частям земного шара и в некоторых отдаленных местах снова стала дикой (или «одичавшей»). Род Pantliera состоит из 6 видов.
РаггИгега leo, лев, в прошлом встречался по всей Африке, кроме лесов Западной Африки и абсолютных пустынь, и в Юго-Западной Азии до некоторых райош>и Индии. В настоящее время в Африке встречается только южнее Сахары на юг |о Юго-Западной Африки и Трансвааля или, возможно, немного дальше, а в Азии он сохранился (находится под охраной) только в Катхиаваре и в Гирских лесах северо-западной Индии. В Марокко лев встречался до 1920 г., в Алжире и Туни- се — примерно до 1891 г. и в Капской провинции (Южная Африка) — примерно
Млекоиитт< щие
.i(i l8,i8 г. В Юго-Западной Азии, возможпо, несколько львов существун~т до сих пор в Ираке и Иране, но эти данные не достоверны. В пределах исторического «ремени львы встречались также в Аравии, Палестине, Сирии и Малой Л:ши. Они, очевидно, существовали в Греции примерно до 80 — 100 годов и. э.
Таким образом, вся известная история расселения этого вида — это его отступа- ~~не перед человеком иа протяжении почти 2000 лет. РапОмт tigris, тигр, встре- чается в Азии — на западе через Иран почти .~о Кавказа; па севере до Аральского моря и до Маньчжурии и среднего Амура II3, востоке и далее иа I()l' до оольшей ч»сти Индии (по не до 1дейлопа) и Юго-Восточной Азии, а также дв ()cTporkoe Сумат- р», Ява и Бали. Panthera pardus, леопард, встречается в Африке к северу и к игу от Сахары (до южной части материка) и через Южнуи Лзхпо и» север до Ii»âêàçà и Амурской области в Приморском крае и на юг до Цейлон», пс>луостро- в» .Чалакка и острова Ява. В Азии тякже встречается Panthera ~iebulosa,;tò,ò÷ ю- тьш леопард, от Южного Китая до С."уматры и Калимантана и PolltA('иа ипс~а„ снсжны1! барс, обитающий в высокогорных районах Центральной Азии. Panthe- ra опса, ягуар, распространен в Америке на север (в прошлом) по крайней мере до горных районов восточной Аризоны на север до Большого каньона и южных районов западной части Нью-Мексики, а также (вдоль побережья) до централь- ного Техаса (G. М. Allen, 1942, р. 253) и на юг (в прошлом) почти до южной око- нечности Южной Америки (по крайней мере до верховьев реки Санта-Крус в П»- т»гонии). В настоящее время доходит только до Чака и Мпсьонес на крайнем севе- ре Аргентины (СаЬгега, Yepes, !940). Последний род и вид современных кошачь- их -Icinonyx jubatus, гепард, который встречается на большей части Африки, вк;иочая Северную Африку; на юге (в прошлом) до Капской провинции (но теперь т»м исчез) и в Азии на север до южной части Туркмении и на восток (по крайней мере в прошлом) до северной и центральной Индии. Предполагаемый второй вид этого рода, локализованный в Родезии, сомнительный. Ископаемые этого родя известны из верхнего плиоцена Европы. Ископаемые кошачьих разпообразны.
11з существующего подсемейства Felinae 3 современных рода и несколько иско- в»емых относятся примерно к плиоцену Евразии. В Африке это подсемейство ранее плейстоцеиа неизвестно, но, вероятно, встречалось там и в более ранне~ время. Feli s и Pan the ra — единственные роды этого подсемейства, которьн достигли Америки, до плейстоцена там неизвестны. Известны еще 3 исчезнувших подсемейства кошачьих. *Proailurinae (4 или 5 родов) находилось в Евразии в среднетретичном периоде. eiVimravinae, ложносаблезубые (9 родов) — в Евр»- зпп, Африке и Северной Америке в разные времена среднетретичного периода. *Pseudaelurus — в Африке и в нижнем миоцене, в Европе на протя- же нии миоцена и в Северной Америке с верхнего миоцена до среднего плио- цена. *Machairodontinae, саблезубые (10 родов) находились в Евразии и Север- ной Америке с нижнего олигоцена до плейстоцена, в Африке в нижнем плиоцене (и, вероятно, в другие времена) и в Южной Америке в плейстоцепе. *l'.'usmilus существовали в Европе на протяжении олигоцена и в Северной Америке в по о- ~~ем олигоцене. *ЛХасйа~годиг — в Европе в верхнем миоцене и в Европе, Лзип, Африке и Северной Америке в нижнем плиоцене. эЯт!!одоп — в Северной п Южной Америке в плейстоцене.
I I о д о т р я д 1 i n n i р е d i а, ластоногие. Большинство морей, главным образом
вдоль побережий. Особенно многочисленны в холодных водах Арктики и Антарк- тики. 3 современных семейства (в настоящем списке не приводятся), 16 совре- менных родов. Преимущественно морской род РЬоса иногда, так же как и неко- торые другие тюлени, заходит в пресные воды и образует эндемичные подвиды в пресных озерах Финляндии, в северо-восточной части СССР, в озере Байкал, и» Баффпповой Земле и в Канаде в озере Тюленьем, Унгава (Wynne-ЕЙиards, 1952), а также в Каспийском море. Ископаемые тюлени известны из миоцеи». ![одотряд РгоСungulаСа
*Отряд,ondylarthra, древние копытные или предкопытные. *Hyopsodontidae, ~Phe<>»- codontidae, *Periptychidae, *Meniscotheriidae,? *Tricuspiodontidae (5 семейств, 33 рода). Северная Америка (большинство) и Европа (несколько) (палеоцен н;и~- цен). Кута (Couto, 1952) полагает, что еАятИЬиоосКиагШа из палеоцена Южной Лмерики относится к семейству Efyopsodontidae, а не Didolodontidae.
~ Didolodontidae. Южная Америка (палеоцен и эоцен), 7 или 8 родов. ')то семейство помещено Симпсоном (Simpson, 1945, р. 124, 235) между предыдущими семействами, но здесь оно выделено для того, чтобы подчеркнуть его географп- чс'скую изоляцию в Южной Америке.
*Отряд Xenungu lata, вымершие копытные Южной Америки
*C»rorlniidae. Южная Америка (палеоцеп). I или большее число родов (Сouto, 4952).
Одни из древиешпих известных млекопитающих Южной Америки.
*Отряд 1 itopterna, вымершие копытные Южной Америки
эРгохегосххегххсхае. Южная Америка (палеоцен до нижнего плиоцена), '3 рода.
эМасгяххс1хепххйае. Южная Америка (палеоцен до п;хеххстоцена). I.5 или оолее
родов.
*Отряд Xotoungulata, вымершие, главным образом южноамериканские, конытные. *Arctostylopidae. Азия (верхний палеоцен) и Северная Америка (нххжнхххх эоцен). ' ро- да. Симпсон не выделяет этого семейства, но здесь оно приводится для того, что- бы подчеркнуть его встречаемость за пределами Южной Америки.
*Иеиг1сояЬогп11с1ае, *Xiotostylopidae, *ОИ6еИСЬомаяidae, *Аrchaeopithecidae,
*Archaeohyracidae, *Isotemnidae, *El omalodotheri i dae, *Leontiniidae, *Xotohi p- pidae, эТохос1оп1хххяе, einteratheriidae, *Mesotheriidae, eliegetotheriidae (15 се- мейств, разные подотряды и подсемейства, около 100 родов). Южная Лххерикя (палеоцен до плейстоцена, но каждое семейство существовало только на ххротяже- пии какого-то определенного отрезка времени).
~Отряд Astrapotheria, вымершие южноамериканские копытные.
eTrigonostyIopidae. Южная Америка (палеоцен и эоцен). 1 род.
*Astrapotheriidae. Южная Америка (эоцен до миоцена). 7 родов в 2 хходсемействххх. Отряд Tubu lidentata
Orycteropodidae, трубкозубые. Большая часть Африки к хогу от Сахары. 1 род, воз- можно, только 1 современный вид, различные подвиды. Ископаемые этого рода обнаружены в плиоцене Евразии (Франция, Греция, Иран, Индия). Предполагае- мые трубкозубые известны из олигоцеиа и миоцена Европы. В нижнем эоцене Северной Америки обнаружен сомнительный представитель трубко- зубых.
Н адотряд P aenungulata
эОтряд Pantodonta
~(;oryphodontidae, *Barylambdidae, *Pantolambdodontidae. Объединенный ареал, Се- верная Америка (средний палеоцен до нижнего эоцена) и Евразия (эоцен до ерехх- него олигоцена). 9 родов.
*Отряд Dinocerata
*Uintatheriidae. Северная Америка и Азия (верхний пя;хеоцехх до верхнего эоценя).
8 родов.
Отряд Pyrotheria
*Pyrotheriirlae. Юя ная Америка (зо1хехх до олнгоцена). 4 рода. 11зо:хххрованныхх от- ряд неизвестного происхождения. 1! алеоценовый род. *(:а госУпга предположитель- но был отнесен си~да (;пмпсоном, но в настоящее «ремя помещен в другой отряд eXenungulata (см. выше).
Отрял Probo~cidea
': loeritheriidae. Африка (Египет) (верхний эоцен и нижний о1ххгоцен). 1 род.
«ЬотрЬо1Ьегххс(ае (eBunomastadontidae), мастодонты. Африка (нижнпй олигоцеих до
миоцена), Евразия (миоцен до плейстоцена), Северная Америка (верхний мххххце~х до плейстоцена) и Южная Америка (плейстоцен). 14 или большее число видов, из.
которых 5 были общими для Старого Света и Северной Аъхерики (в разные периоды времеххи в верхнем миоцене и плиоцене) и 2 родя оьх;ххх общими для Север~хой и Южиой Америки.
~Маттххйхс1ае, мастодонты, Африка (нижххий миоцен), Евразия (миоцен и плиоцеп),
Северххая Америка (средний миоцен до плейстоцена). ! род.
Еlephantidae, слоны. современных рода и вида. Еlephas maximus, индийский слои,
встречается в местах, пригодных для обитания, от подножий Гималаев в северной Индии до Цейлона и через Ассам до полуострова Малакка, а также на Суматре (интродуцирован на острове Калимантан). 1.оходопха и/ricana, африканский слон, распространен в по~ходящих для него местах обитания через значительную часть Афрнки к хогу от Сахары. В прошлом достигал самого хххха, но теперь в Южной Африке сохранился только в отдельных специально охраняемых местах. В древ- ние времена он встречался также в Северной Африке. Так, в плейстоцене этот род был распространен до Европы и Азии. Другой род «Матти&ив, вк;ххх> хяя мамонтов, встречался в Африке, Евразии и Северной и, возможно, Южной Аме- рике в плейстоцене, а карликовый слон *Arckidiskodo», относимый Симпсоиом к роду Мапипибгиб, в плейстоцене находился на острове Сулавеси, по южиую сторону линии Уоллеса (Нооххег, 1951). Еще 2 рода, помещаемых Симпсоном в отдельное подсемейство, находились в Евразии и поздиетретичиое «ре',ця и плейстоцепе.
*Deinotheriidae. Африка и Евразия (миоцен до плейстоцена). l род.
*Barytheriidae. Африка (Египет) (верхний эоцен). 1 род.
*Отр~~д Еmbrithopoda
«Arsinoitheriidae. Африка (Египет) (нижний олигонец), 1 род.
.11лекопитпн щ ие
Отряд lie гасоий.а
! rocaviidae, даманы. Африка и близлежащая часть Азии. 3 современных рода, несколь- ко видов. Все роды и виды африканские, 1 африканский вид Procavia capensis распространен до Азии в местах с подходящими условиями обитания от мыса Доброй Надежды до (',еверной Африки, Аравии, Палестины, Синайского полуострова и Сирии. 4 ископаемых рода известны из Африки (олнгоцен и миоцен) и Греции (плиоцен).
~Geniohyidae. Африка (Египет) (нижний олигоцен). 4 рода
~Myohyracidae. Африка (нижний миоцен). 2 рода
Отряд Sirenia, сирены, морские коровы
*Prorastomidae, Вест-Индия (Ямайка) (эоцен). 1 род.
:>РгоФоз>гепЫае. Северная Африка и Европа (эоцеи). 1 род.
l)ugongidae, дп>гони. Современный вид Dugon gdugon встречается (за иск.'>н>чепчем тех
мест, где он в настоящее время истреблен) в море вдоль побережья от Бо<.точной Африки и т. д. на восток до островов Рюкю и на юг через Индо-Австралийский архипелаг до Северной Австралии. Стеллерова морская корова 3ТуйгосЬсигл1аз stelleri, очевидно, была распространена только у островов Беринга и Медного (со стороны азиатского материка), где ее обнаружили в 1741 г. 1>еринг и Стел- лер. Сто лет спустя она была уже истреблена в результате охоты. Воз- можно, что когда-то ареал этого животного был значительно шире. Многочи- сленные и разнообразные ископаемые дюгони относятся к,>оцену (Африки и Европы).
Тrichechidae, ламантины. Прибрежные части морей и рек (пригодных для обитания),
большая часть района Карибского моря, включая Флориду. В реках Амазонка и Ориноко в Южной Америке и в низовьях рек тропической Зап»д»ой Африки.
l род, 3 вида. Ископаемые до плейстоцена неизвестны.
«Desmostylidae. В районе северной части Тихого океана (верхний о;>игоцен и мио- цен). 2 рода.
Ц адотряд Mesa xoni а
Отряд Perissodact> la, непарнокопытиые
(Надсемейство Equoidea)
*Palaeotheriidae. Евразия (эоцеп и нижний олигоцен). 7 родов.
Equidae, лошади и др. В настоящее время в естественных условиях с уществуют только
в отдельных частях Азии и Африки. 1 род, 7 видов. Лошади в строгом смысле (! ди- кий вид) считаются истинно дикими и не сильно смешанными с домаш»>хчи лошадьми в настоящее время только в восточных частях Центральной Азии. 0»агр, или азиатский дикий осел (не настоящий осел), встречается в Центральной и Юго- Западной Азии от Монголии и Тибета до Сирии. Настоящий дикий осел встречает- ся в Северной и Восточной Африке к югу до Сомали. Зебры (4 вида) встречаются н Южной и Восточной Африке на север до Эфиопии и Сомали. Большинство су- ществующих диких представителей этого семейства с появлением человека начало отступать (Нагрег, 1945), и 1 вид зебр уже исчез. Домашние ослы и лошади оыли распространены человеком по всему земному шару, в результате чего «о многих местах лошади вновь одичали. Родоначальником домашнего осла является дикий африканский вид (Л'quus asin.us) из Северной Африки или восточной части Среди- земноморской области, существовавший там, вероятно, 4000 — 3000 лет до и. э.
или еще раньше. Домашняя лошадь (Е. caballus) скретцивалась с настоящими дикими формами лошадей и да>ке с ныне существующей дикой лошадью Центр»ль- пой Азии (которая считается отдельным видом или подвидом pr=e»>alskii), и до сих IIOp еще не известно, где именно в Евразии в последнее время встречались несмешанные дикие лошади. Дикие лошади действительно когда-то встречалпгь на значительной части Европы, а также и Азии, и, вероятно, в некоторых частях Европы они существовали и в исторические времена. Возможно, что лошадь «пер- вые была одомашнена уже в 3000 г. до н. э. где-то на основной территории Азии или Восточной Европы, хотя, судя по историческим сведениям, она появилась позднее и Юго-Западной Азии и Северной Африке. Ископаемые лошади, вклн>чня их предков и им родственных (3 подсемейства, 18 ископаемых родов), вероятно, описаны лучше всех животных. История этого семейства рассматривалась вьиие (стр. 298 — 299). Более подробные сведения о лошадях и пх истории при»олятся в популярной книге Симпсона (Simpson, 195>).
(еНадсемейство Brontotherioidea)
:>ВгопФоШег>Ыае, титанотерии. Северная Америка (нижний эоцен до ни>к»его олигоце- на) и Евразия (верхний эоцен до среднего олигоцена). 39 родов, большинство из которых находилось в Северной Америке, но по крайней мере 2 разные группы проникли в Евразию.
( 1[адсемейство С>>а1>со(Ьег>оЫеа)
I'.лава 6
э;J>alicotheriidae, когтистые копытные. Евразия (эоцен до плеххстоцена), вероятно.
также Африка (миоцен, плейстоцен? и предположительно другое время) и Север- ная Америка (верхний эоцен до среднего миоцена). !3 родов.
(Надсемейство Tapiroidea)
*Isectolophidae. Северная Америка (нижний и средний эоцен) и Азия (верхний эоцен).
4 рода.
*llelaletidae. Северххая Америка (нижний эоцен до среднего олигоцена) и Азия (ве)>х- хххххх эоцехх и нижний олнгоцен). 1О или более родов.
*Lophiodontidae. Евразия (зацеп). 5 или более родов.
Tapiridae, тапиры. Е'аспространены на Суматре и полуострове Малакка до 18' с. ш.
(! вид) и в тропиках Америки от южной Мексики (Веракрус) до северной Арген- тины (3 вида: l центральпоамериканский, 1 широко распространен в Южной Америке и i — в Андах Колумбии, Эквадоре и Перу,— Hershkovitz, l954, р. 493, 495). >>се эти виды могут быть соединены в ! род, хотя иногда их делят на
и даже 3 рода. Ископаемые тапиров (включая несколько исчезнувших родов)
встречахотся в Северной Америке (нижний эоцен до плеххстоцехха) и в Европе (ниж- ний о1>хгоцех> до плейстоцена). Ископаемые современного рода Tapi rus (в шххрокохх смысле) известны из Европы и Азии (плиоцен и плейстоцен) и Северной и Южххохх Лмерххки (плейстоцеп). Насколько известно, тапиры никогда не хостххг»лхх Лфрики.
((1адсемейство Rhinocerotoidea)
э)!угас>ху~х!ае. Северххая Америка и, возможно, Азия (эоцен). 4 рода.
*i lyracodontidae. Северная Америка (средний эоцен до верхнего олигоцена) и, возмож- но, Азия (эоцен). 4 американских и 3 сомнительных азиатских рода.
*.Ëøónodontidaå. Северная Америка и Евразия (верхний эоцен до среднего о.хххгоцехха
п нижнего миоцена соответственно). 6 родов.
]<i>inocerotidae, носороги и им родственные. Существуют в Африке южнее Сахары (2 ро- ;ха, 2 вида) и в тропической Азии (2 рода, 3 вида). Один из азиатских родов Dice~ о rhi~ius встречается от северо-восточной Индии и т. д. до Суматры и Калиман- тана (по не до Явы). Другой род Rhinoceros, который в прошлом был распростра- нен на большей части Индии (но не на Цейлоне) до Суматры и Явы (но не до Кали- мантана), в настоящее время исчез, за исключением ряда местностей. Ископаемые ххосорогов и им родственных (песколько подсемейств, около 30 исчезнувших ро- дов) встречались в Европе (средний эоцен — плейстоцен) и в Северной Америке (верхний эоцеп — нижний плиоцен). В Азии и Африке неизвестны до верхнего эоцена и миоцена соответственно, но, возможно, существовали там в более ран- нее время. Современный род Индо-Малайской области Dicerorkinus, или по край- ней мере его предковая линия находился в Европе, а также и Азии с верхнего олигоцена до плейстоцена.
П адотряд Рaraxonia
Отряд Ar tiodac ta! а, парнокопытные
*Dichobunidae. Северная Америка (эоцен) и Европа (эоцен до среднего олигоцена).
J 5 или более родов.
*(:hoeropotamidae. Северная Америка (эоцен), Евразия (средххий эоцехх до среднего оли- гоцена) . 7 родов.
*СеЬосЬоегИае. Европа (средххихх эоцен до нижнего олигоцепа). 3 рода.
eLeptochoeridae. Северная Америка (олигоцен). 2 рода.
~I:ntelodontidae. Северная Америка (верхний эоцен до нижнего миоцена) и Евразия
(олигоцен и, возможно, верхний эоцен). 4 рода.
(! [адсемеххство Suoidea)
'~uidae, свиньи. Все пригодные д.хя обитания части Африки (также один, быть может,
пришлый впд па Мадагаскаре), умеренная и тропическая зоны Евразии и т. д.
5 современных родов, несколько видов. Настоящие свиньи (Sus) в настоящее время распространены на север примерно до 58' с. ш. в Восточной Европе и до Лму!>скохх области в Восточной Азии и на юг и восток до Индии и Цейлона, а также на Яве, Ка;хим>хита>хе, Малых Зопдских островах до острова Тимор (.), Сулавеси, Ь!олуккских, !! овой 1иинее, островах Ару и Кай, Бисмарка, Соломоновых и Фи- липшша». Но в отношении расселения свиней в юго-восточном направлении труд- но сказать, где оии являются действительно местными, а где они были распрост- ра>хен>,х ххервох>ыт>хыч человеком. На острове Сулавеси (ххъхеется эндемххчххьххх род Ваб> > ияяа, а также ххлейстоцехховые ископаемые рода 5'us) и, возможно, на ост- рове Ь~~ру они, несомиенио, местные. Но далее на восток — в центральных частях «рхипелаг» Малых Зондских островов, на острове Тимор или на самых дальш~» от центра Филиппииски» островах — свиньи, вероятно, не являются местными.
Домашние свиньи произошли от двух предков: от дикого каба~и (Sus sc>.op>>è) Европы хх Сервер>хохх Африки и от другой формы этого же вида в Восточ~>ой Лзхххх.
Млекопи~пин щие
11скопаемые семействн Suidae (17 родов) известны только из основной части Стн- рого Света, самые древние относятся к олигоцену (Европы).
1';~yasuidae, пекари. Находятся только в Америке. 1 род 2 вида, из которых один рас- пространен от южного Техаса, Нью-Мексико и Аризоны (в прошлом на север до А ркаизаса) до северной (и в прошлом центральной) Аргентины, а другой- от южной Мексики до северной Аргентины. Предполагаемые представители это- го семейства существовали в Евразии (отдельное иодсемеиство, 3 рода, ниж»»й »лигоцен до нижнего плиоцеиа), а более достоверные — в (.'евериои Америке (h ископаемых родов, нижний олигоцен и позднее) и в Южной Америке (только плейстоцен, 2 вымерших рода, один из них северо-американский).
(11»;(семейство Anthracotherioidea)
*А»о!>1о(>>ег>>чае. Европа (средиий эоцен до среднего олигоцена). 7 родов.
~Anti>racotI>eriidae. Евразия (средний эоцеидо плейстоцена), Африка (олигоцен и ниж- ний миоцен), Северння Америка (олигоцен и нижний миоцен). 23 рода.
Н >рроро1атЫае, бегемоты, или гиппопотамы. (:уществуют только в Африке. 2 рода,
по 1 виду в каждом. Большой бегемот (Hippopotamus) в прошлом встречался по всей Африке, там, где имелись подходящие для его обитания реки и водоемы, к се- веру до дельты Нила (ио не в Сахаре). Этот род существовал также в Европе и Азии в плейстоцене. Карликовый бегемот (Сйое>ориг) локализован у северного берега Гвинейского залива. Этот род приводится в виде ископаемых (плейстоцен) или субфоссильио на Мадагаскаре и островах (;редиземноморья, но Симпсон (Ятряо», 11145, р. 148) считает, что некоторые из этих ископаемых, возможно, являются просто конвергентными мелкими формами. Мадагаскарский карликовый беге- мот был весьма обыкновенным животным во времена, ныне вымерших эпиорнис» и гигантских лемуров (White, 1930). Самые древние из известных гиппопотамов относятся к среднему плиоцену Африки (Hippopatamus) и Евразии (1 исчезнувший род).
(*11ндсемейство Cainot 1>eri(>idea).
*(:ainotheriidae. Европа (верхиий эоцен до нижнего миоцеиа). 6 родов.
(*l- ндсемейство Merycoidodontoidea)
Agriochoeridae. Севериня Америка (верхний эоцеи до среднего миоцена). 4 рода. *!>1erycoidodontidae. (:еверная Америка (верхний эоцен до среднего миоценн).
Hесколько подсемейств, 24 рода.
Il одотряд Tylopod a
*Xiphodontidae. Европа (средний эоцен до нижнего олигоцена). 3 рода.
(:нте1Ыае, верблюдовые. В настоящее время существуют в природе только в Централь- ной Азии и в западной и южной частях Южной Америки. 2 рода. Род Camelus с, 2 видами относится к Старому Свету. Двугорбый верблюд (С. bactria»us)— единственный до сих иор (вероятно) дикий вид верблюдов, встречается в пусты- нях Центральной Азии. В других местах он существует либо в домашнем, либо в одичавшем состоянии. Одногорбый верблюд С. d romeda rius — обычныи одо- мншненный верблн>д (;еверной Африки и т. д. сомнительного происхождения, в диком состоянии не известен. Южноамериканский род (1.ama) в широком поня- тии охватывает 2 диких вида (гуанако и викунью) и 2 одомашненные формы (.>ама и альпака). ООъеди»енный ареал диких видов (которые перекрывнн>т;ц>уг друга, но все же»е совпадают) проходит от южного Эквадора и Перу на >ог через Лнды до Патагопии и по более низким высотам до Огненной Земли. Ископае- м>.>е этого семействн (иесколько подсемейств, 24 или большее число ископаемых родов) известны из (.'еиерной Америки (с верхнего эоцена до плейстоцена). В Ев- разии и Южной Америке они появились в плейстоцене.
(*11 ндсемейство Amph imeryco i d ea)
~A»>pl>imerycidae. Евроин (верхний;юцеи до иижиего олигоцена). 2 рода.
(*11ндсемейство Ilypertraguloidea)
~lIypertragu1idae. Северная Америка (верхний:>оцен до ни>кнего миоцена) и Л:»>я
(верхний эоцен). 11 или более родов.
*Рrotoceratidae. Северная Америтсн (нижний олигоцен до нижнего плио>(е»н>.
pода.
(11»деемейство Тгнни!оЫен)
*(elocidae. Евразия (верхинй эоцеи до нижнего олигоцена). 10 родов.
Tray>>lidae, оленьки. Районы Индо-Малайской области и Западная Африка. 2 рода.
В Индо-Малайской области представлен род Trapulus с 2 видами, распростра- 1I(.'ннымп от южной Ьирмы до островов Ява, Калимантан и Балабак, юго-запа,1ная часть Филиппин, и третьим видом на полуострове Индостан и Цейлоне. В Знпндиой Африке род Нуетоясйиг с 1 видом распространен от Гамбии до восточного Конго (в лесах). Ископаемые этого семейства (2 рода) известны из верхнего миоцеин kI частично из плиоцеиа Европы и Азии.
Глава 6
(Надсемейство Cervoidea)
(;ervidae, олени. Евразия и т. д., северо-западная часть Африки, Северная, Централь- ная и Южная Америка. 17 современных родов, небольшое число видов. К северу род Rangifer широко распространен в полярной тундре (так же, как и в леснстых местностях) Евразии и Северной Америки, включая арктический архипелаг и се- верное побережье Гренландии. К югу местные олени достигают северо-за1ыдиой части Африки (1 вид), Ыалой Азии и Ирана, Индии и Цейлона, островов Яви, Бали и Калимантан и, возможно, Малых Зондских, Тимора, ('5'.савеси, Молукк- ских и Филиппин (но, возможно, в некоторые места они были завезены челове- ком) и почти до самон южной части Южной Америки. Rangifer — циркумпозяр- ный род, и все его формы (северный олень и карибу), возможно, относятся и од- ному виду. Alee, лось, встречается в лесах холодных северных районов Евразии и Северной Америки. Формы Старого и Нового Света иногда рассматривают«я как один вид. Род Сел иг является общим для умеренных широт обоих полушарий.
благородный олень (С. elepkus), распространенный по всей умеренной зоне Евра- зии (и в (.'еверной Африке), и олень-вапити (С. саггагКггггия), который в п1>огилом встречался па большей части умеренной зоны Северной Америки, иногда рассм»- триваются как один вид. Американский род Odocoileus (впргинский, или белохво- стый, олень и чернохвостый олень) распространен от Канады до Амазонки и! ге- ру. Из 13 других современных родов этого семейства 8 находится в Азии (' I>o;«> достигают Европы), включая примитивные и производггые формы. В 'e»el>i«>ii Америке нет ни одного специфического рода. В Южноп Америке находится 5 1>о,гг>гг (1 род распространен до Центральной Америки и южной Мексики). Все они <>т- носятся к американской трибе Odocoilei»i. Ископаемые этого семейства (:3-~ 1>г>дгг) известны из Евразии (нижний олигоцен), Северной Америки (нп>кн>1й миг>цегг) и Южной Америки (только плейстоцеп).
(г-1адсемейство Giraffoidea)
ei agomerycidae. Евразия и Африка (миоцен), 3 рода.
Giraffidae, жирафы и др. Африка. 2 современных рода и вида, представляюцгие l>а:—
ные подсемейства. Gi raf fa camelopardalis, обьпсновенная жирафа, встреч>>ется в пригодных для обитания местах (но не в густых лесах) на большей части Африки.
к rory от Сахары. Okapia уohnsto»i, окапи, приурочены к густым дождевым лесам района верхнего Конго. Ископаемые этого семейства (I;> родов, представлян>гг(их 2 современных п 1 ископаемое подсемейство) в оспг>вггом известны пз Азии (MH(>- цен до плейстоцеиа). Несколько групп нзвестггы пз нижнего пл1гоцена Восточггг>гг Европы; ггесколько ископаемых было обнаружено и Африке. Ископаемые этого «емейства предположительно относимые к плеистоцену Северной Америки, пе «ы- зывают доверия.
(Надсемейство Bovoidea)
Antil0capridae, вилорогие антилопы. Западная часть Северной Америки. Один со«ре- менный вид c4»tilocapra ате~гсаиа, вилорог, в прошлом был распространен от прерий южной части Канады (от Манитобы до Альберты) до районов Техаса, Мек- сиканского нагорья и Нижней Калифорнии. В настоящее время ареал этого вида значительно сократился. Ископаемые показыва IoT, что вилороги были мно- гочисленны (12 исчезнувших родов в 2 подсемействах) в западной части (.'енег>- ной Америки в среднем миоцене и что они встречались на восток до Флори,~ы « плиоцене (*Hexameryz, White, 1941), но о нахождении их в других местах;(г>- стоверных данных не имеется.
lIovidae, быки, антилопы, бараны и т. д. Африка, умеренная и тропическая:и>гги
Евразии и Северной Америки. 5 подсемейств, 13 триб, 54 современных род» (в связи с трудностями, возникающими при классификации этого семействгг, и последней имеется еще очень много сомнительных мест). Более полное опггггг- ние см. выше, стр. 301 — 303.
Самым северным представителем этого семейства в Евразии является Oiis
саггайепвгв, снежный баран, который встречается в районах северной Сибири (гггг запад почти до устья Енисея), а также в западной части (:еверной Америки. В Аме- рике самый северный представитель этого семейства — мускусный бык, Оп%оя, типично арктическое животное, которое встречается (или недавно встречалось) от самой северной точки Аляски до арктических островов и северного побережья Гренландии и которое также встречалось в северной части Евразии в плейсто- цене. К югу естественные границы семейства Bovidae вклгочают всю Африку (но не Мадагаскар), Индию (но, вероятно, не Цейлон, хотя там встречаются в оди- чавшем виде буйволы Bubalus), Яву (на Бали местных видов, вероятно нет), Калимантан и Сулавеси. Один эндемичный вид буйволов (Bul>ulus) изолирован на острове Миндоро, Филиппины. Все юго-восточные границы на Индо-Австра- лийском архипелаге определяк>тся бычьими (Bovini). (',ерао (Сapricornis) дости-
~7итерат ура
гает Суматры, а также Тайваня. Другие представители этого семейства не дости- гают даже Индокитайской подобласти Азии, возможно, за исключением ее север- ного края. В Америке они встречаются и, насколько известно, встречались на юге только до Мексики и в северных краевых частях Центральной Америки. Ископае- мые этого семейства (включая 86 ископаемых родов) известны в основном из Ста- рого Света (несколько в миоцене, но большинство в плиоцене и плейстоцене (см. текст, стр. 301 — 303). Род Вison имеет прерывистое реликтовое распространение.
1i этому роду относится ряд плейстоценовых и, вероятно, постплейстоценовых видов, в прошлом, видимо, широко распространенных и умерепных широтах Евразии и почти по всей Северной Америке. Один евроазиатский вид (Bison ооиаяи8, зубр) встречался в прошлом и был распространен в Европе на восток, вероятно, до Центральной Азии и до реки Лены в Сибири, но его численность неуклонно сокращалась. Последние «дикие» (но охраняемые) стада на Кавказе и в Беловежской пуще исчезли в 20-х или 30-х годах нашего столетия, хотя этот вид до сих пор существует в зоопарках 1. Американский бизон (Biso» bison) в прошлом был распространен от равнин и некоторых облесенных местностей Канады до Мексиканского нагорья и от Орегона по крайней мере до западной части Нью-1Чорка (и до полуострова Кейп-Код в плейстоцене) и Джорджии. Последний дшсий бизон был убит в западной Пенсильвании в конце 1800 г., а последние бизоны других районов восточной части — приблизительно в 1825 г. Гибель бизонов на западе Америки — это часть американской истории. К 1900 г. остава- .лось только два стада диких бизонов, находящихся под защитой: одно небольшое стадо в Йеллоустонском национальном парке и стадо лесных бизонов в районе Атабаски, штат Альберта, Канада. В настоящее время в результате охраны чис- ленность американских бизонов снова увеличилась и небольшие стада широко разбросаны по стране. Домашний скот, козы, бараны — все берут свое начало от евроазиатских видов. Безгорбый домашний скот Европы, а в настоящее время и большинства других районов умеренной зоны, произошел от туров (*Bos pr~- mige»iuã), которые когда-то были широко распространены по всей Европе, но исчезли (как дикие животные) в начале XVII века. Горбатый скот тропиков, вероятно, произошел от разных тропических азиатских предков. Домашние буи- иолы произошли от современного вида Bubalus bubalus. Впервые он был одомаш- пен в Индии, затем в других местах Индо-Малайской области, а также в других районах на запад до Египта и других теплых районах земного шара. Домашняя коза, вероятно, произошла от современного вида Сарга Ыгсиг Юго-Западной Азии и островов восточной части Средиземноморья. Домашние бараны, возможно, име- l0T смешанное происхождение. Частично они возникли от современных муфлонов (Огis musimon) из Сардинии и Корсики и архаров (Ovis аттоп) из Азии. Неко- торые из этих животных (по крайней мере безгорбый скот, бараны и козы) были впервые одомашнены примерно 8000 лет назад, когда начало развиваться сель- ское хозяйство в Юго-Западной Азии.
ЛИТЕРА Т5'Р 1
А р и а у о С. G., l,)50, Observaciones sobre algunos inamiferos cubanos extinguidos,
«Boletin Insist. Г~а1. Soc. Felipe Роеу», 1, 12) — 13~~.
А 11 е п G. М., ]938 — 1940, The mammals of (;hina and Мопдо11а, 'iew York, Ameri- can Mus. Mat. Insist., 2 го1я.
Л 1 ] е n G. М., l939, А checklist of African maminals, «Hull. Мпя. Comparative
Zool.», 83.
А 1 1 е n G. М., 1939a, Bats, Сambridge, lass., 1;Iarvard U. Press.
А 1 1 е п С. М., 1942, Extinct and гаЫяЫпд mammals of the Western Hemisphere
with marine species of all oceans, American Committee for International ЖЫ
Life Protection (New York), «Special РиЬ.», Хо. 1 l.
А 1 1 е n J. А., 1878, The geographical distribution of the Mammalia..., «Bull. United
States Geological and Geographical Survey of the Territories», 4, 3lЗ — 377. Ii е р г,1. (:., 1947, Географические зоны Советского (:о|оза, 3-е изд., Москва.
1 В настоящее время в Беловежской пуще и на Северном J.~ÿâêîçå, а такж~в некоторых других местах имеется около 500 чистокровных:)y61)(113.— 1Хрп.и. р~~.
340
Глава 6
В о u r 1 i e r e F., 1955, Mammals of the world, New York.
8 и r t. 43'. Н., 194.), Present distribution and affinities of Mexican mammals, «Ann.
Association American Geographers», 39, 211 — 218.
ф1 и r t W. II., 1952, А field guide to the mammals [of America north of Mexico),
Boston.
(.' а b r e r а А., Ъ е р е s,j., 1940, Mamiferos sud-americanos..., Tucuman and Вщ- nos Aires, Argentina.
С а r t e r Т. D., H i 11 3. Е., Т а < е G. Н. Н., 1945, Мапппа1я о1 the Pacifi<:
world, ii ew Y orL .
С h а s e n F. Х., !940, A handlist of Malaysian inammals, «Вц11. ЯаИ1ец Мцц >
(Singapore), Х о. 1;>.
С о и Ф о (.. de P., 1952, Fossil mammals from the beginning of the;enozoic in grazi]
Сопду1аг(11га, Litopterna, Xenungulata, and Astrapotheria, «Bull. American Mus
Х аС. 1-[1яС. », 99, 355 — 394.
С p u ) p (.". de P., 195 а, Fossil mammals from the beginning of the (;enozoic in Brazil.
AIarsupialia: Polydolopidae and Borhyaenidae, «American Mus. Novitates», Г~о.
1 559.
(; о ы ф о С. йе P., 1952b, Fossil mammals from the beginning of the (;enozoic in Brazil.
Alarsupialia: Didelphidae, «American Mus. Novitates», No. 1д67.
]> i с е L. В., 1929, The phylogeny of the Leporidae..., «J. Mammalogy», I0, 340 — 344. Е 1 l e r m a n 3 . R., 1940 — 1949, The families and genera of living rodents, London,
British Mus, ') vols.
Е1lerman J..R., Мorrison-Scott Т. ('. S., 1951, 'hecklist of ра1ае~~
tic and Indian mammals..., I,ondon, British Mus.
Ellerman J. В., Morrison-Scott Т. С. S., Hayman B. W., 1953,
$puth African Mammals 17,э8 — 195'1: а reclassification, London, British Mus. E 1 ф o n (;. ! 942, Voles, mice, and lemmings: problems in population dynamics, Nez- York. Oxford U. Press.
С о о d w i n G. С., 19~6, Mammals of Costa Rica, «Hull. American Mus. >rat. pjst.>,
87, 271 — 474.
H а r p e r F., 1945, Extinct and vanishing mammals of the Old World, Атпег1сап Conl- mittee for International Wild Life Protection (New York), «Special pub» No Ii e r s h k о г i t z P., 1954, ...tapirs ... American species, «Ргос. United States Natio- nal Mus.», 103, 465 — 496.
Н i 11 %. (;. О., l953 — 195э, Primates. Comparative anatomy and taxonomy, ъ-оl. [—
Strepsirliini [lemurs etc.] (1953); vol. II — Haplorhini: Tarsioidea (1955), Edin- burgh, Edinburgh U. Press.
H i n ф p n M. А. C., 1933, ...rodents from Indian Tertiary deposits, <Апп. Мар. Nat.
Hist.», ( I Î), 12, 620 — 622.
Н о о i j e r 1). А., 1951, Pygmy elephant and giant tortoise, «Sci. Monthly», 72, 3 — Я. Н о р д~ о о d А. Т., 1954, Notes on the recent and fossil mammalian faunas of Africa.
«Ргос. Linnean $ос., London», 165, 46 — 49.
L а и r i е Е. М. О., EI i 11 3. Е., 1954, List of land mammals of New Guinea, (;p- lebes and adjacent islands 1758 — 1952, London, British Mus.
L y d e k k e r 8., 1896, А geographical history of mammals. Cambriclge, (;ambri<]g<
Press, Г~ ew York.
М а t t h e w W. D., 1915, 1939, (;limate and evolution, «Ann. New York Acad. Sci.»,
24, 171 — 318; reprinted (1939) as «Special Publ. New York Acad. Sci.», I.
M а t t h e w s Ь. Н., 1952, British mammals, I,ondon.
М а у r Е., 1946, The number of species of birds, «Auk», 63, 64 — 69.
М i 11е r С. $., Jr., К е 11 о g g H., 1955, [.1яФ of North American Recen[ inammals,
«Bull. United States National Mus.», No. 205.
М u r r а у А., 1866, The geographical distributi on of mammals, Londpn.
Литература
0 s g o o d %. ц., 1943, The mammals of Chile, «Zool. Ser. Field Mes. Nat. Iiist.», 30. P h i 11 i p s W. W. Л., 1935, ...mammals of Сeylon, (;еу1оп, Colombo Mus., 1.on- don.
P о w е 11 А. W. В., J947, Native animals of New Zaeland. Auckland Л1ия. handbook
Z oo1.
Beynolds T. Е., Koopman К. F., Williams L'. L'., 1)53, Acavefau- nule from western Puerto R ico with а discussion of the genus )solobodo», «Brevi ога»
(Мия. Comparative Zool.), No. 12.
8 о Ь е r t s А., 1951, The mamma]s о1 South Africa, Soutli А1'г~с», (;entral Xows
Agency.
8 о m e r А. S., 1945, Vertebrate paleontology, nd ed., (,hicago, [Имеется русский
перевод: А. P о м е р, Палеонтология позвоночных, ГОНТИ, Москва, l!)39].
8 о t h s c h i 1 d Lord, D о 11 m а n G., 1936, The genius Dr~id~olagus, «Trans. lоо1.
Soc. London», 21, 477 — 548.
S а и b о r n (',. (.'., 1951, Two new mammals from southern Реги, «Fieldiana» (СЬн:зло
Хай. !kist. Mus.), «Zool.», 3l, 473 — 477.
S с 1 а t e r P. ?., 1875, The geographical distribution of mammals, «3lanchester S(.;i.
Lectures», Ser. 5 — 6, 202 — 2! 9.
5 с 1 а t е г W. L., $ с 1 а t e r P. L., !890, The geography of mammals, London—
Trench.
S с о t С W. В., 1937, А )history of land mammals in the ШеяФегп lIemispl>ere, ге~.ised
ed. Мew York.
S i m p s о n G. С., 1937, The beginning of the age of mammals, «Biological Reviews
Cambridge Philosophical $ос.», 12, 1 — 47.
S i m p s о n G. G., 1940, Mammals and land bridges, «J. ЪЪаяЫпдСоп Асад. Sci.»,
30, 137 — ! 63.
S i m p s о n G. G., 19~3, Mammals ' and the nature of continents, «American J. Sci»,
241, 1 — 31.
S i m p s о n G. G., 1945, The principles of classification and а сlassification of mammals,
«Bull. American М us. Nat. Н ist. », 85.
S i m p s о n G. G., 1947, Holarctic mammalian faunas and continental relationships
during the Cenozoic, «Bull. Geological Soc. America», 58, 613 — 688.
S i m p s о n G. G., 1947а, Evolution, interchange, and resemblance of the North Ame- rican and Eurasian Cenozoic, mammalian fauna, «Evolution», 1, 218 — 220.
S i rn p s о n G. G., 195!, fiorses..., New York, Oxford U. Press.
S t i r t о п H. А., 1953, Vertebrate paleontology аЫ continental stratigraphy in
(;o1ombia, «Bull. Geological $oc. America», 64, 603 — 622.
S t о с k С., Н i c h а r d s Н. G., 1949, А Megalo»yx tootle from the Northwest Ter- ritories, (,anada, «Science», 110, 709 — 7'10.
~ ъ у n n e r С о n G. H., H а у m а п В. W., 1951, A c)ieck list of the land nxarnmals
of the Tangaayika Territory and the Zanzibar Protectorate, «J. East African '~'at.
list. Soc.», 20, 274 — 392.
Т а t e G. Н. Н., 1945, The marsupial genus Phalap>ger, «American Мпя. Novitates»,
Хо. 1283.
Т а С е G. Ц. Н., 1945а, The marsupial genys Ргеидос/геггик..., «American Mus. Xovi- tates», ГЧо. 1287.
Т а С е G. Н. Н., 1945Ь, Notes on the squirrel-like and mouse-like possums.... «Лин- rican 51us. Novitates», No. 1305.
T а t e G. II. ц., 1946, Geographical distribution of the bats in the Australasiat~ Ar- chipelago, «American Mus. Novitates», No. 1323.
Т а t e G. Н. 3l., ~947, Dasyuridae..., «Hull. American Мия. Nat. fIist.», 88, 97—
156.
T a t e G. Il. Н., 1947а, Мап1та1я of eastern Asia, New Ъ ог1г.
842
Глава 6
Т а t e G. Н. II., 1948, ...Macropodidae..., «Bull. American Mus. 1аС. Ilist.», 9].
233 — 352.
T а t e G. Н. Н., 1948а, ...Peramelidae..., «Bull. American Mus. XaC. IIist >,,)2,
3l3 — 346.
Т а t e G. Н. Н., 1951, Т1~е rodents of Australia and New Guinea, «Bull. Лтег~саи
Mus. Nàt. kIist.», 97, 183 — 430.
Т а t e G. Н. Н., 1951а, The wombats..., «American Mus. Novitates», No. 1525.
Т а t e G. Н. Н., 1952, Матта1я of Cape York Peninsula..., «ВцН. American
М us. N at. Н Ы », 98, 563 — 616.
T а у l о r Е. Н., 1934, Philippine land mammals, Philippine Bureau Sci. ЫопоцтярЬ
30.
Т r о u g h t о п Е., 195l, Furred animals of Australia, 4 th ed., Sydney.
Ъ h i t е Е. 1., 1930, Fossil hunting in Madagascar, «Nat. Hist. Mag.» (British Museum),
2, 209 — 235.
W h i t e Т. Е., 1941, Additions Со the fauna of the Florida Pliocene, «Ргос. New Eng- land Zool. Club», 18, 67 — 70.
M i 11 i а m s Е. Е., К о о р m а п К. F., 1951, А new fossil rodent from Puert~i
Rico, «American Mus. Novitates», No. 1515.
M' i 1 l i а m s Е. Е., К о о р ш а п К. F., 1952, West Indian fossil monkeys, «Ame- rican Mus. Novitates», No. 1546.
M о d z i c k i Е. А., 1950, Introduced mammals of New Zealand..., Wellington,
New Zealand, Department of Scientific and Industrial Research.
% о о d А. E., 1947, Rodents — а study in evolution, «Evolution», 1, 154 — 162. % о о d А. Е., 1950, Porcupines, paleogeography and parallelism, «Evolution», 4,
87 — 98.
% у n n е — Е d w а r d s V. С., 1952, Freshwater гегСеЬгаСея of the arctic and subarctic,
«Bull. Fisheries Research Board Canada», No. 94.
Глава 7
ОСОБЕННОСТИ РАСПРОСТРАНЕНИЯ ЖИВОТНЫХНА МАТЕРИКАХ И ФАУНИСТИЧЕСКИЕ ОБЛАСТИ
Цель этой главы, во-первых, показать общие особенности распро- странения, присущие всем пяти классам наземных и пресноводных позво- ночных, которые были рассмотрены в предшествующих главах, и, во- вторых, как можно проще и естественнее разделить земной шар на фауни- стические области.
Распространению позвоночных присущи несколько основных осо- бенностей. Пять таких особенностей уже были отмечены выше. Рассматри- вая их, надо помнить, что они не обособлены, а как бы накладываются друг на друга и взаимодействуют так, что их результирующая дает одно сложное целое.
Первую, самую главную, из пяти основных особенностей распростра- нения наземных и пресноводных позвоночных можно охарактеризовать как ограниченность. Позвоночные всех классов в своем распространении достигают каких-то конечных пределов или число их видов уменьшается к северу (и в какой-то степени к югу), в высоких (холодных) широтах и в направлении от континентов к островам. Так, пресноводные рыбы в Арктике распространены далеко на север, а строго пресноводные рыбы почти всегда ограничены пределами больших материков и близко распо- ложенных материковых островов. Амфибии, распространяясь к северу до арктических областей, не достигают северных границ распростране- ния рыб, в то время как на островах они проникают значительно дальше, чем строго пресноводные рыбы. Рептилии в большинстве северных райо- нов не доходят (на довольно значительное расстояние) до северных гра- ниц амфибий, но на островах проникают дальше их. Птицы проникают далеко и в северные районы, и на острова и практически не имеют конеч- ных границ распространения ни в том, ни в другом направлении. Но надо отметить, что птицы, как и все другие позвоночные на севере и на отда- ленных островах ограничены в видовом составе. Млекопитающие в своем распространении также проникают далеко на север, но (за исключением рукокрылых) на островах они ограничены примерно так же, как и амфибии.
Изучение северных границ распространения различных классов позвоночных дает основание предполагать, что эволюция следующих друг за другом холоднокровных групп (рыб, амфибий и рептилий), по-видимому, сопровождалась понижением их холодостойкости (или увеличением по- требностей в тепле) и соответственно отступанием из холодных северных районов до тех пор, пока теплокровность не дала возможности птицам и млекопитающим снова распространяться к северу. Ограниченность распространения различных классов животных на островах позволяет с уверенностью сказать, что она зависит от способностей животных к расселению, а не от их геологического возраста.
В пределах своих границ различные классы позвоночных проявляют различную степень незонального замещения одних групп другими, что
Глава 7
можно назвать дальнейшей ограниченностью в распространении различ- ных доминирующих отрядов или семейств в разных частях земного шара. У строго пресноводных рыб наблюдается полная замена, или полный викариат, которая охватывает большие части фаун между доминирующими карпообразными Евразии и доминирующими некарпообразными Юл;но11 Америки, а фауны Северной Америки и Африки являются (не касаясь в данном случае Австралии) смешанными. У амфибий заменяемость групп выражена не столь полно и охватывает только небольшие части фаун. Однако она более сложна, если говорить о доминирующем семействе Banidae на большей части земного шара и 1 ерСойас(у1Ыае в Южной Аме- рике и Австралии, а также древесных лягушек семейства Rhacophovidae в Африке и Индо-Малайской области и представителях семейства Hylidae, распространенных повсеместно. У рептилий наблюдается еще менее пол- ный, но более усложненный викариат, важнейшими чертами которого является доминирование черепах подсемейства Emydinae в Евразии и Северной Америке, бокошейных черепах повсеместно, ужовых змей на большей части земного шара и аспидовых в Австралии.
Из всех позвоночных наименее полная и в то же время самая сложная взаимозаменяемость в распространении проявляется у птиц, хотя Oscllla~ (певчие птицы), доминирующие на большей части земного шара и Suho~- cines, доминирующие в Южной Америке друг друга частично замещают. У наземных млекопитающих наиболее полный викариат наблюдается между почти всей фауной плацентарных основной части земного шара и главным образом фауной сумчатых Австралии. Южноамериканская фауна млекопи- тающих является все же смешанной. Все это представлено весьма упрощенно и до какой-то степени субъективно. Но на основании этого можно пред- полагать, что степень незональной заменяемости обратно пропорцио- нальна способности расселения животных. У позвоночных, для которых суша является весьма серьезным препятствием при расселении (рыбы) или которым не дают возможности расселяться такие преграды, как про- странства соленых вод между материками (пресноводные рыбы и наземные млекопитающие), наблюдается наиболее полная локализация и географи- ческая заменяемость доминирующих групп. Вероятно, это явление можно считать само собой разумеющимся, но все-таки оно заслуживает более детального рассмотрения. Совершенно очевидно, что позвоночные рассе- лялись на земном шаре активно, а не в качестве пассивного груза на пе- ремещающихся материках.
Вторая особенность распространения — зонильносигь, включающая зональную заменяемость. Лучше всего она проявляется у пресноводных рыб: большинство семейств некарпообразных приурочено или к тропикам, или к умеренной зоне северного полушария, а некоторые из них распро- странены (в той или иной зоне) по всему земному шару. Некоторые кар- пообразные и ряд других групп рыб, включая и солевыносливых, имеют также зональное распределение. У амфибий большинство семейств при- урочено главным образом к тропикам, и только некоторые из тропических групп проникают в умеренную зону северного полушария, но саламандры и несколько небольших групп лягушек, как правило, являются северо- умеренными. Большинство семейств рептилий тоже приурочено к тропи- кам. Некоторые из них простираются в северном направлении на разные расстояния, но хорошо выраженных зональных северо-умеренных групп рептилий нет. Основной особенностью зонального распространения наземных птиц и млекопитающих является также концентрация их в тро- пиках и распространение только некоторых тропических семейств к севе-
ру, хотя эти животные проникают на север дальше и в большем числе, чем рептилии. Как мы видим, зональное распределение рыб влечет за сооой значительную зональную заменяемость. У амфибий она проявляет- ся в меньшей степени, а у рептилий, наземных птиц и млекопитающих —- в еще меньшей. Северо-умеренные фауны этих классов образовались главным образом за счет выклинивания более теплолюбивых групп, а не за счет образования новых. Очевидно, многие важные группы рыб и и меньшем числе некоторые группы амфибий приобрели наследственную приспособленность к умеренному климату северных широт. Многие группы птиц и млекопитающих могут переносить холодный климат, но специально приспособленных к этому климату групп у них не возникает. Рептилии не переносят холодного климата и также не выработали при- способленных к нему групп.
Различие между западными и восточными группами (или наоборот 1) не имеет существенного значения в распространении большинства позво- ночных. Оно проявляется лучше всего у некоторых ящериц и у некоторых мелких перелетных, особенно у насекомоядных наземных птиц. В обоих случаях различия наблюдаются главным образом у тропических семейств, которые расселялись к северу в той или другой половине земного шара (или в Старом, или в Новом Свете), не заходя на противоположное полу- шарие. То, что эти животные не пересекают границ полушарий земного шара, по всей видимости, объясняется различными причинами: ящерицы, вероятно, не проникли достаточно далеко на север, а у птиц это, видимо, связано с привычкой возврата на зимовку в том или ином полушарии.
Третья особенность распространения — географическая радиация— выражена наиболее полно у пресноводных рыб (например, отряд Osla- riophysi и внутри отряда собственно карповых), в меньшей степени — у амфибий (например, семейство Ranidae) и еще в меньшей — у рептилий (например, ужовые), птиц (певчие птицы) и млекопитающих (мышиные и бычьи), у которых радиальное распространение или проявляется менее четко, или встречается лишь у групп, хотя в целом и важных, но не играю- щих столь большой роли, как Ostariophysi или Cypriniformes среди прес- новодных рыб. Самые определенные черты географической радиации. отражающие распространение больших доминирующих групп из опреде- ленных центров, проявляются у тех животных, которые расселяются на земном шаре медленнее других, и, таким образом, их эволюция протекала в наибольший отрезок времени, оставляя следы длительного пребывания этих животных в различных местах.
Центр наиболее выраженной радиации каждого класса обычно находится на основной части тропиков Старого Света: центр радиации Cypriniformes — в тропической Азии, лягушек семейства Ranidae— в тропиках Старого Света, змей прогрессирующего семейства Colubri- dae — вероятно, в тропической Азии, певчих птиц — в теплых странах Старого Света, грызунов семейства МигЫае и семейства Bovidae— в более теплых районах Евразии и Африки.
Четвертая особенность — дифференциация фаун на различных кон- тинентах — у каждого класса выражена по-разному. У пресноводных рыб эта особенность проявляется в следующих основных чертах: разно- образие и выживание древних тропических семейств в Африке, численное преобладание более современных представителей Cypriniformes в Евразии, разнообразие и выживание древних северо-умеренных семейств в Север- ной Америке, эволюционная радиация в изоляции в Южной Америке, сосредоточение солевыносливых групп и сохранение одного очень древ-
Глава 7
него реликта в Австралии. Более сложные основные черты дифференциа- ции земноводных включают ограничение распространения безногих в Африке, тропической Азии и Ю",кной Америке, концентрацию большин- ства саламандр на северных континентах и сложную взаимозаменяемость доминирующих лягушек. Дифференциация фаун рептилий на материках еще сложнее: каждая группа имеет свою, присущую только ей, дифферен- циацию: у черепах она (причем довольно сложная) одна, у крокодилов— другая, у ящериц — третья, у больших удавов — четвертая и у мелких доминирующих змей — пятая. У птиц основные черты дифференциации (исключительно сложные в деталях) сводятся к разнообразию их на основной части тропиков Старого Света, уменьшению к северу и дифферен- циации в Австралии и Южной Америке, причем южноамериканская фауна является самой богатой и самой дифференцированной. У млекопитающих наблюдается такое же разнообразие в тропиках Старого Света, уменьшаю- щееся к северу, дифференциация в Австралии и Южной Америке, но австралийская фауна млекопитающих более дифференцированная, чем южноамериканская.
Пятая и последняя особенность — кониентраиия на самых больших и благоприятных для обитания территориях каждого континента и уменьшение числа форм и групп в менее благоприятных местах, часто по периферии — лучше всего выражена у пресноводных рыб, в меньшей сте- пени у амфибий и в еще меньшей у рептилий, птиц и млекопитающих, которые являются настоящими наземными животными и в том или ином числе встречаются почти повсюду (но все же эти животные наиболее много- численны на самых больших и самых благоприятных для обитания терри- ториях). Этот факт, как уже говорилось выше (стр. 93, 94), может пока- заться слишком очевидным, чтобы на нем останавливаться, нельзя забывать о не столь простом и не столь очевидном, но важном факторе, как влияние на распространение животных пространства (протяженности благоприят- ной территории).
Заключение: осооенности распространения позвоночных
Итак, наземные и пресноводные позвоночные наиболее разнообразны и многочисленны на самых больших и самых благоприятных для их оби- тания территориях земного шара — в тропиках континентов, и их число сокращается к северу (и к югу), а также за пределами больших материков. Границы их распространения варьируют в зависимости от природы каж- дого класса и его способности к расселению. Каждый класс в пределах своих границ проявляет незональную взаимозаменяемость, которая выражена в какой-то мере обратно пропорционально способности к рас- селению. Все классы имеют зональное распределение: зональность пресно- водных рыб включает значительную зональную заменяемость (викариат), основанную на дифференциации как северо-умеренных, так и тропиче- ских групп. У амфибий она проявляется в меньшей степени, чем у рыб, у рептилий, птиц и млекопитающих — в еще меньшей, и зональность этих классов выражается главным образом в обеднении фаун от тропиков к северу. Радиальное распространение проявляется у всех классов прибли- зительно в обратной пропорции степени их расселения. Дифференциация фаун на разных континентах у разных классов различная. И на каждом континенте у каждого класса наблюдается также концентрация на боль- ших и благоприятных территориях и обеднение класса по периферии.
Особенности расирострпнения животных нп .чптгериках
847
Прерывистое распространение
Прерывистое распространение, то есть наличие одних и тех же или родственных животных на более или менее разобщенных территориях, довольно обычное явление в распространении позвоночных. Некоторые зоогеографы, особенно сторонники возведения мостов или перемещения материков, преувеличивали или неправильно понимали особые случаи прерывистого распространения. Здесь мы рассмотрим это явление в более общих чертах и постараемся представить его в перспективе.
Разрывы в распространении животных могут образоваться тремя путями: когда, преодолев водные пространства, наземные животные дости- гают океанических островов; когда в результате затопления суши отре- зается часть фауны на материковых островах (или в результате перемеще- ния ма териков) и когда происходит исчезновение животных на части территории (то есть частичное вымирание), которое разъединяет перво- начал ь ное сплошное распространение.
Отдельные случаи не играют существенной роли. Например, преры- вистое распространение амфибий семейства Leiopelmidae в северо-западной части Северной Америки и в Новой Зеландии — это особый случай, который с точки зрения зоогеографии не столь значителен (хотя по ряду других причин нахождение рода Leiopelma в Новой Зеландии имеет важ- ное значение). Прерывистое распространение имеет ряд общих особенно- стей (рис. 44). Распространение большинства животных, о которых гово- рится в связи с прерывистым распространением, дается более подробно в списках семейств в конце глав 2 — 6.
Как уже отмечалось, эти особенности проявляются очень сложно: все они взаимно накладываются и видоизменяют друг друга, неизбежно обра- зуя в целом едины~~ сложныи комплекс.
Этот сложный характер распространения, очевидно, определился в результате чрезвычайно разнообразного сочетания факторов и процес- сов живой и неживой природы. Наиболее важным фактором последней является протяженность территории и наличие благоприятных условий существования, зональность климата и преграды, которые образуют пространства соленых вод между материками. К факторам живой природы относятся специфические потребности, выносливость и способность к рас- селению различных позвоночных. Эти процессы охватывают:эволюцию различных доминирующих позвоночных в разные времена и в разных местах (но особенно в тропиках Старого Света) и их расселение разл11ч- ными путями по всему земному шару.
Это заключение дает возможность представить самые главные фак- торы в распространении животных и увидеть, что они не соответствуют тем. которые обычно предполагаются. Многие зоогеографы недооценивают зональности климата и еще в большей степени — влияние территории на рас- пространение животных. Я же считаю, что распространение позвоночных на земном шаре находится в довольно сложной зависимости от современ- ных особенностей рельефа, климата и преград, а последнее в свою очередь приводит к мысли, что современные позвоночные расселились в результате самостоятельных перемещений на земном шаре (который и во время этих расселений мало чем отличался от современного), а не посредством пере- мещающихся материков или сухопутных мостов. На этих предваритель- ных выводах мы остановимся более детально в последующих главах.
В умеренной зоне северного полушария наблюдаются ооширные раз- рывы в распространении животных. Многие группы встречаются в уме- ренных широтах Евразии и Северной Америки, но не в смежных арктиче- ских областях. В пределах этой общей для распространения животных особенности имеется много частных случаев. Например, многие пары родов и видов растений и животных встречаются в Восточной Азии и восточной части Северной Америки, но не в промежуточных областях. Из растений можно назвать тюльпанное дерево, магнолию, сассафрасы (американ- ский лавр), хамамелис, симплокарпус и многие другие растения (Feriiald, Р и с. 44. Схема, отражающая некоторые особенности иреры-
«истого распространенпя животнык. 1 — умеренные широты Евразии и Северной Америки; ' — - Восточ- ная Азия и восточная часть Северной Америки; .5 — тропики Старого и Нового Света; 4 — тропики Африки и Азии; . — 3«па~~юця Африка и тропическая Азия; 6 — Африка и тропическая Лосю ч)<п~л Азия; 7 — Мадагаскар и тропическая Азия; 8 — южная Индия или Цейлон и тропическая Восточная Азия; 9 — тропическая Азия и тропическая Америка; 10 — Африка и Южная Америка; 11 — Ма- дагаскар и Южная Америка; 1з — Австралийская область и КЭжная Америка; 1'5 — южная часть Австралии (Новая Зеландия) и уме-
ренная зона Южной Америки. 1929; Li, 1952), а из животных — веслоносов, саламандр семейства 4гур- t0branchidae, аллигаторов, некоторых мелких сцинков и змей рода Opheodrys. Некоторые особенности распространения в умеренной зоне северного полушария подчеркиваются расселением саламандр (стр. '132). Ряд других особенностей прерывистого распространения на севере был хорошо описан в работах ботаников (например, Fernald, 1929). Большин- ство разрывов в умеренной зоне северного полушария, очевидно, можн<i ооъяснить также отступанием животных с промежуточных территорип.
Еще более широкие разрывы наблюдаются в тропиках. А[нагие груп- пы животных встречаются в тропиках Старого Света и тропиках Америки, но не в проме."куточных умеренных районах северного полушария. Многие животные встречаются и в тропической Африке, и в тропической Азии, но опять-таки не в районах умеренных широт Евразии, которые находятся между ними. Некоторые случаи разрывов в распространении животных между тропиками Старого и Нового Света прослеживаются на ископаемых (результат расселения через север и последующего вымирания там этих животных). Это, вероятно, наиболее правдоподобное объяснение всех случаев (разрывов), по крайней мере среди современных позвоночных,
за исключением тех, когда животные, возможно, пересекали море. В ооль- шинстве случаев разрывы между Африкой и тропической Азией, несомнен- но, являются результатом расселения через Евразию (северные тропики), а затем последующего отступания с ее территории. Мадагаскар, очевидно, не лежал на пути расселения современных афро-азиатских позвоночных, за исключением нескольких, которые, возможно, пересекли море. В пре- делах тропиков отмечаются также особые, если можно так сказать, внут- ренние черты прерывистости. Одни группы животных (например, лягушки подсемейства МусгоЬу11иае, змеи семейства Aniliidae и некоторые из се- мейства Colubridae, один род орлов, один род дятлов и тапиры) встре- чаются только (или главным образом) в тропической Азии и тропической Америке (но не в Африке). Другие (несколько групп рыб, пины, черепахи пеломедузы, ящерицы семейства Amphisbaenidae, змейки семейства Leptotyphlopidae и несколько видов птиц) встречаются только (или глав- ным образом) в Африке и тропической Америке (но не в тропической Азии). Третьи (по крайней мере черепахи рода Podocnemëç и игуаны) встречаются только на Мадагаскаре и в Америке (но не в Африке, хотя один представи- тель игуановых находится на островах Фиджи). Отдельные группы (например, фазаны, оленьки) распределены между Западной Африкой и тропической Азией, а другие (например, некоторые рыбы, рептилии и птицы, см. стр. 62, 170, 226) — между южной Индией и восточной частью Индо-Малайской области, третьи (некоторые змеи, стр. 170) — между Индокитаем и Большими Зондскими островами. Такой характер распро- странения, как и ряд других подобных случаев, возник, очевидно, в ре- зультате отступания с промежуточных территорий.
К югу от тропиков разрывы становятся еще обширнее. Лягушки семейства Leptodactylidae, черепахи семейства (;helyidae и сумчатые встре- чаются только (или в большинстве случаев) в Южной Америке и Австра- лии. Но, несмотря на то, что эти животные проникают в умеренную зону южного полушария, по характеру распространения в целом они все же являются тропическими, а не южно-умеренными и в Новой Зеландии не встречаются. Ископаемые и другие доказательства позволяют предпо- ложить, что расселение лягушек и сумчатых происходило через основ- ную часть земного шара, а расселение в Австралию черепах семейства СЛе1у1йае, возможно, шло непосредственно через море.
Некоторые группы солевыносливых рыб (например, галаксиды) и наземных позвоночных, а также некоторых растений имеют зональное «антарктическое» распространение: они являются общими для холодных самых южных районов Южной Америки и Австралии, многие из них встре- чаются в Новой Зеландии, а некоторые также и на южной оконечности Африки. Ни одно пз наземных или строго пресноводных позвоночных не имеет такого распространения. К этому немаловажному факту мы вернем- ся в гл. 10.
Большинство перечисленных выше разрывов, по-видимому, является результатом отступания или вымирания многих животных на обширных территориях — весьма обычными процессами в географии животных.
Фаунистические области
Разделение земного шара на фаунистические области и описание региональных фаун позвоночных и их взаимосязей должно быть сделано прежде всего в пределах границ материков и близлежащих материковых
Глава 7
островов. Далее (в гл. 8) будут рассмотрены отдаленные острова и связь островных фаун с материками.
Различие между материками и островами весьма условно. Австра- лия — материковый остров, и на отдельных частях ее фауны влияние изо- ляции отразилось в значительно большей мере, чем, например, на фауне Мадагаскара. На фауне Южной Америки, еще не в столь отдаленные вре- мена бывшей материковым островом, до сих пор сказывается влияние изо- ляции. Однако уже вошло в обычай, а также и более удобно рассматри- вать Африку, Северную и Южную Америку и Австралию как континенты. Вместе с островами, которые в недалеком геологическом прошлом были соединены с ними, они образуют одну почти непрерывную систему суши (см. фронтиспис), очень неправильную по своим очертаниям, асимметрич- но расположенную на круглом земном шаре и неравномерно пересечен- ную горами, пустынями и узкими водными пространствами. Эту сушу мы разделим теперь на области в соответствии с распространением на не11
ЖИВОТНЫХ.
В наши дни понятие «фаунистические области» очень часто неправиль- но истолковывается и понимается, а значение их недооценивается. Прежде всего, среди некоторых зоогеографов утвердилось мнение, что современ- ное распространение животных не имеет значения или по крайней мере, что современные «статические» особенности распределения и фаунистиче- ские области сами по себе не заслуживают особого внимания и только прошлое фаун или некоторые моменты, связанные с ним — эволюция и перемещения животных, — в действительности имеют значение для зоогео- графии, как уже было отмечено в гл. 1, (такой подход, на мой взгляд, является глубоко ошибочным). Только в настоящее время можно увидеть, каким образом на самом деле распространены животные и как их распро- странение связано с пространством, климатом, преградами и другими аоио- тическими факторами, а также с растительным покровом и друг с другом. Если обратиться даже к не столь далекому прошлому, то все равно нельзя увидеть действительное положение вещей, и выводы в большинстве слу- чаев можно сделать только при сравнении с настоящим временем. Поэтому, чтобы понять прошлое, необходимо знать настоящее. Конечно, справедливо и то, что, не зная прошлого времени, нельзя понять и настоящее. Как прошлое, так и настоящее одинаково важны для зоогеографии, и одно нельзя понять без другого. Тем не менее логически допустимо и фактиче- ски иногда необходимо для анализа распространения животных прини- мать во внимание исключительно их распределение в настоящее время так, как будто оно всегда было таким, статическим, и уже затем привлекать элементы эволюции и перемещений животных и в прошлом (см. гл. 9, 10).
Многое из того, что будет сказано о фаунистических областях вообще, оыло сформулировано уже Уоллесом (Wallace, 1876). Он указал на значе- ние выделения фаунистических областей и описал их природу. Он сам признавал несовершенство и ограниченность такого деления, подчеркивая в то же время его сущность и положительные стороны. Этот простой и здравый подход разрешил большинство возникших недоразумений и явился ответом на критику этого деления, которого придерживаются даже в на- стоящее время. Уоллес считает, что фаунистические области предназна- чены для того, чтобы представить основные черты распространения совре- менных животных, и что, хотя причины, приведшие к современному рас- пространению, заложены в прошлом, любая попытка при выделении областей объединить прошлое с настоящим должна привести к путанице. С этим нельзя не согласиться. Тем не менее некоторые зоогеографы при
Особенности распроспьранения животных на материках
выделении фаунистических областей все еще пытаются объединить гео- логическую историю и третичные взаимосвязи животных с их современ- ньп| распространением.
Некоторые возражения по поводу выделения фаунистических областей оыли сделаны Данном (Dunn, 1922), который, между прочим, писал: «...зоогеографические царства не представляют ничего, кроме больших массивов суши с границами, которые соответствуют физико-географиче- ским барьерам. Между такими понятиями, как «голарктическая фауна» и «голарктическая область», если подходить с точки зрения философии, существует огромная разница. В первом случае мы говорим о зоологиче- ских проблемах, исходя из зоологии, во втором — о географических проб- лемах, исходя из мифологии». Изложенная Данном мысль, что фаунисти- ческие области — это нереальные, искусственные понятия, вызывает возражения. Фаунистические области не соответствуют точно материкам. Действительно, области вообще отделяются преградами, но не теми, о ко- торых говорит Данн. Единой системы преград нет. На земном шаре много самых разнообразных преград, и то, что одни преграды возникли раньше, чем другие, и разделили основные фауны и фаунистические области,— это не произвольное допущение, а очень существенный факт, не сразу выявленный зоогеографами. Что касается «философской» разницы между фаунами и областями, то фаунистические области — это территории, занимаемые фаунами в настоящее время. Разница, конечно, имеется, но если фауны реальны, то также реальны и не являются мифом и области.
Основная трудность в отношении фаунистических областей заклю- чается не в философском толковании этого вопроса, а в его практическом решении. Она возникает в результате сложного распространения живот- ных. Едва ли найдутся какие-либо две группы животных, распространение которых было бы совершенно одинаковым, и различные основные группы (например, разные классы позвоночных), которые в отдельных случаях имели бы совершенно различное распространение. Это разнообразие в распространении трудно свести в простую систему. Трудность настолько велика, что она породила сомнение в реальности самих областей. Поиски фаунистических областей, как говорил Гадов, — иллюзия. Другие утвер- ждали, что фаунистических областей нет, или их слишком много, или если они есть, то для различных групп животных они разные; но из всех этих высказываний нет ни одного вообще сколько-нибудь подходящего и удач- ного. Такое отношение к данному вопросу показывает отсутствие понима- ния природы и назначения этих областей.
Необходимо понять, что система фаунистических областей отражает осредненный результат распространения, это наиболее удачное среднее из всех разнообразных сочетаний распространения различных животных, которое могло быть разработано зоогеографами. Фаунистическая область не является каким-то единым образцом, которому бы точно соответствова- ли многие различные группы животных. Уоллес знал это. Правда, он преувеличивал четкость границ некоторых областей, но он понимал, что фаунистические области это результат разного распространения разных ЖИВОТНЫХ.
Таким образом, система фаунистических областей является средним всех типов распространения самых различных животных, у каждого из которых имеются свои особенности распределения. Значит, ecmb средний тип распрострпнения. животных. Он вполне реален и является очень важ- ным. Он показывает, как животные в своем распространении приспосабли- ваются к среде, которая их окружает, и насколько сильно влияют на их
Глава 7
распространение климат и преграды. Я думаю, что уже по одной этой причине (помимо многих других) естественная система фаунистических оОластей заслуживает того, чтобы она была создана.
Система фаунистических областей — это, можно сказать, стандартный зоогеографический эталон, наподобие измерительной линейки, при помо- щи которой может быть измерено, описано и сравнена распространение различных животных, определены их особые черты, а также вскрыты важные факты, касающиеся этих животных и их истории. Такой стандарт- ный зоогеографический эталон полезен даже в том случае, если ему соот- ветствует неболыиое число групп животных. Отказываться от применения стандартного типа распространения животных лишь по этой причине оснований не больше, чем отказываться от метра лишь потому, что в при- роде немногие предметы точно соответствуют его полной длине. Откло- нения от стандарта должны быть, и они также говорят о многом. Например, если определенная группа животных достигает Австралийской области, но проявляет там меньшую дифференциацию, чем «стандартная фауна», то это позволяет предположить недавнее расселение этой группы и нали- чие у нее способности преодолевать водные пространства.
В более общих чертах, если группа животных особенно хорошо пред- ставлена в районах, ограниченных климатическими условиями, а в райо- нах, ограниченных преградами, как это наблюдается у многих пресновод- ных рыб, она дифференцирована сильнее, чем обычно, или отсутствует. то это свидетельствует о наличии существенных связей животных с кли- матом и преградами, образуемыми солеными водами. Система фаунисти- ческих областей для того, чтобы быть полезной в качестве стандарта, долж- на стать общепринятой, она не должна изменяться, кроме тех случаев, когда для этого имеются достаточные основания, но и это должно делаться по общему соглашению. Если специалист находит, что распространение животных, которых он выбрал для изучения, не соответствует эталону, он не должен отказываться от него и создавать новыи, подходящии для данного случая. Специалист должен принять общий стандарт, установить, чем его группа животных отличается от этого стандарта и попытаться раскрыть значение и смысл этих отклонений. Уоллес хорошо понимал это. Тем не менее некоторые зоогеографы до сих пор недопонимают этого и все еще предлагают совершенно необоснованные варианты изменения стандартной системы фаунистических областей. Следует еще раз повторить, что хотя стандартную систему областей в большинстве случаев следует принимать как довольно произвольную, она по своей природе (в отличие от метра) все же не является чем-то отвлеченным, а вполне реальна. Эта система фаунистических областей является самой правильной и простой схемой распространения животных, которую удалось составить зоогео- графам.
Фаунистические области имеют и другое, менее важное назначение. Они помогают зоологам устанавливать в разных частях земного шара естественные границы региональных фаун. Что касается названия этих областей, то они являются вполне приемлемыми. Например, название «Индо-Малайская область» проще и точнее, а потому более приемлемо, чем «тропическая Азия и некоторые близлежащие материковые острова».
Из сказанного должно быть ясно, что фаунистические области не со- ответствуют однородным группировкам животных, которые равномерно распространены в пределах общих границ области. Каждая региональная фауна более или менее концентрируется в благоприятных для обитания местах, варьирует по составу в разных местах и, вступая в контакт с
Особенности распространения живогпных па л птерикпх
прилежащими фаунами, образует сложный переход. Тем не менее, несмотря на различия, животные, находящиеся в разных частях одной фаунисти- ческой области, в целом в большей степени связаны между собои, чем г животными других областей.
Можно надеяться, что приведенные аргументы вполне достаточны для уяснения, что такое фаунистические области и региональные фауны, а также для того, чтобы убедить скептиков в реальности и пользе системы фаунистических областей. Если и остаются какие-нибудь сомнения, то самое лучшее, что можно предложить для дальнейшего убеждения,— зто еще раз ооратиться к работе Уоллеса (Жа11асе, 1876). Критики фауни- стических областей, возможно, будут удивлены, увидев, как хорошо раз- бирался в этом вопросе Уоллес.
В настоящее время общепринято шесть основных фаунистических областей, которые впервые были предложены для птиц Склэтером в науч- ном докладе, прочитанном перед Обществом Линнея в 1857 г. и опубли- кованном в 1858 г. Подразделение Склэтера было принято Уоллесом (Wallace, 1876) как для позвоночных в целом, так и для некоторых беспозво- ночных. Уоллес подробно объясняет причины выделения этих конкретных областей. Существовали и другие варианты подразделения областей, но области, предлагаемые Склэтером, по-видимому, лучше всего отражают существо дела и были наиболее целесообразны (табл. 9). Области Склэтера и названия этих областей остались до наших дней с незначительными изменениями границ и некоторыми упрощениями в названиях, не считая того, что склэтеровская «Индийская область» стала называться по Уол- лесу «Восточной областью».
Табли ца Ч
Схема фауиистических областей (по Склэтеру) 1
Сrenti o Palaeogeana
I. Палеарктическая область. умеренная зона Евразии и северо-западная
часть Африки к северу от гор Атласа
II. Эфиопская, или Западная палеотропическая область: Африка (кромс
северной части) и южная часть Аравийского полуострова
Индийская, или Средняя палеотропическая область: тропическая Азия
и близлежащие материковые острова
IV. Австралттйская, или Восточная палеотропическая область: Новая Гвинея,
Австралия и Тасмания
Сrent~î Neogeonn
] 1сарктичсская, или Североамериканская область: Северная Америка на юг
до центральной Л!сксики, включая Грепландии~.
VI. Неотропическая, или К)жноамерпканская область: К1жиая и Центральная
Америка и южная часть л1ексики.
> Названия и объем областей (кроме некоторык островов) даются по Склэтеру.
Опираясь на склэтеровскую систему областей, более поздние авторы производили различную перегруппировку. Склэтер сгруппировал области Старого Света, противопоставив им области Нового Света в так называе- мые (он искал центры возникновения) «Creatio Palaeogeana» и «Creatio Хеоцеапа», но это расположение нельзя признать естественным. Гексли (Huxley, 1868) объединил вместе четыре области, которые охватывают Африку, Евразию и Северную Америку в «Арктогею», а Южную Америку
Глава 7
и Австралию отнес к «Нотогее». Это более удачный вариант. но все же фауны Южной Америки и Австралии сильно отличаются не только от фаун основной части земного шара, но и одна от другой. Уоллес (A allace.
1876), очевидно, придавал большее значение самостоятельным областям, чем их группировкам, и он свел в единую таблицу систему Гексли и Склэтера. Наконец, Бленфорд (Blanford, 1890) предложил группировку областей, которая еще лучше отражает истинные взаимосвязи шести региональных фаун.
Бленфорд разделил земной шар на три основные группы: Арктогеи
(основная часть земного шара с четырьмя областями), «Южноамерикан- ская область» и Австралийская область. Неизвестному автору (1893) осталось назвать Южноамериканскую и ABCTpaлийскую области соответ- ственно Неогея и Нотогея, а Лидеккер (Lydekker, 1896) назвал эти три группы «царствами». Однако желательно было бы изменить еще одно название. «Арктогея», к сожалению, — название, вводящее в заОлужде- ние, ибо сюда относится территория, которая охватывает основную часть тропиков Старого Света и умеренную зону северного полушария с аркти- ческой периферией. Я предлагаю называть ее «Мегагеей», что означает большая часть земного шара. Исправленная, общепринятая система фауни- стических областей (континентов) приводится в табл. 10. Обычно употреб- ляется еще один дополнительный термин. Хейлприн (Heilprin. 1887) объединил две северные области Палеарктику и Неарктику в Голарктиче- скую область. Здесь они не объединены, но их можно называть голаркти- ческими областями, и животные, которые встречаются в обеих, часто на- зываются голарктическими.
Таблица 1()
Принятая система материковых фаунистических областед
Царство Л1егагея (Арктогея): основная часть земного ~пара
>.',-)фиоиская область: Африка (кроме северо-западной части) и южная часть
Аравии
Il. Яндо->lалайская область'. тропическая Азия с близлежащими материко- выми островами
III. 11алеарктическая область: Евразия к северу от тропиков, с северо-запалио~1
частью Африки
IV. Цеарктическая область: Северная Америка, кроме тродическод час i ц
Мексики
Царство Неогея
Ч. Неотропическая область: Южная и Центральная Америка с sponge «ci<ni'
частьто Мексики
Царство Нотогея
УI. Австралийская область. 'Австралия с Новой Гвинеей n T. д.
Несомненно, это довольно краткое изложение истории взглядов на фаунистические области. Более подробные сведения приводятся в работах следующих авторов: Wallace, 1876; Blanford, 1890; Lydekker, 1911; (.-;а- дов., 1913; Scrivenor (идр.) 1943; Schmidt, 1954, и др. История фаунисти ве- ских областей является лишь одной частью истории зоогеографии, кото- рая была рассмотрена несколько подробнее во введении.
Система фаунистических областей, приведенная в табл. 10, показы вает относительную определенность разных областей, вероятно, так же,
Особенности распространения животных на материках
как любая другая простая система, но, чтобы подчеркнуть природу этих областей, их можно сгруппировать по-другому (табл. 11). Две северные области (Палеарктика и Неарктика) отделяются от тропиков не какими- либо непреодолимыми преградами, а главным образом зональным клима- том, в связи с чем их фауны дифференцированы и ограничены. Это об- ласти, ограниченные климатическими условиями.
Таолица 11
Группировка фауиистических областей,
подчеркивающая их природные особенности
Области, ограниченные климатом
1. Палеарктика 2. Неарктика
Основные области тропиков Старого Света 3. Индо-Малайская область 4. Эфиопская область
Области, ограниченные преградами 5. Неотропическая область 6. Австралийская область
л Iluzeaprmuuecwus НваратаческаяОсиная часвь ароткс6~тарого ОюзаЭернспская Иняс=даяайсмир ОераниченнияареградокцЯспрвпическаяОграниченная
Р и с. 45. Схема распределенияфаунистических областей.
Специфичность фаун Австралийской и Неотропической областей является следствием прошлых и настоящих преград, образованных соле-ными водами, которые изолировали и в некоторых отношениях ограничили зти фауны. Таким образом, это области,
ограниченные преградами. Фауны двух Итси, юграниченные
su~amon остальных областей — основных областей
тропиков Старого Света (Эфиопской и
Индо-Малайской) — в меньшей степени
по сравнению с остальными, но все же
ограничены климатом и преградами, ибо
совсем неограниченных областей не суще- ствует. Эфиопская и Индо-Малайская
лреграаапи области — это центральные наименее огра- А5сщрддууснеея ниченные области. Остальные асимме- трично располагаются вокруг них ради- ально, имея более сильную, все увеличи- вающуюся ограниченность (рис. 45). Это и
есть основной средний тип современного распространения животных и несколько похожая на него радиация из основной части тропиков Старого Света также является, очевидно, одной из основных особенно-стей расселения животных (см. гл. 9).
Шесть материковых фаунистических областей, представленных на прилагаемой карте (рис. 46), показаны более детально на отдельных кар-тах, помещенных ниже (рис. 47 — 52). Рассматривая каждую область в отдельности, сначала остановимся на том, какие материки или отдельные части материков входят в ее состав и каковы общие черты ее климата и рас-тительного покрова, а затем уже перейдем к составу и связям фауны позвоночных этой области. После этого сделаем краткий обзор ее границ и основных особенностей распространения фауны в пределах данной области. Там, где суша не имеет каких-либо значительных разрывов, установить точные географические границы довольно трудно, да это и
/'л ава 7 не столь существенно. Основная роль здесь принадлежит переходным зонам, их природе и протяженности и прежде всего составу самих фаун, их природе и взаимосвязям.
Дать всесторонне исчерпывающий обзор региональных фаун на не- скольких страницах невозможно. Уоллес (Wallace, 1876) посвятил около 500 страниц «обзору главных форм жизни животных в нескольких оола- стях и подобластях». Он назвал это «зоологической географией» в проти- воположность «географической зоологии», которая занимается вопросами распространения животных с точки зрения таксономических групп (гл.2 — 6), а не областей. Обзор региональных фаун, сделанный Уоллесом, до сих Р и с. 46. Шесть материковых фаунистических областей. Ьосая штриховка — — приблизительные границы и ..~оны перехода. пор является лучшим из всех имеющихся работ, хотя он несколько уста- рел в деталях. В настоящее время испытывается острая необходимость в новой, достоверной, подробной и тщательно разработанной региональной или фаунистической зоогеографии.
Дать статистический анализ региональных фаун на нескольких стра- ницах также невозможно. При точном статистическом методе придется иметь дело не только с семействами, но также и с родами, а возможно, и видами. Потребуется описать не только состав и взаимосвязи каждой фауны, но и переходы между ними. Проще всего это сделать для отдель- ных частей фаун, то есть для фауны лесов, саванн, пустынь, пресных вод и т. д. Но при таком подходе неизбежно возникнет целый ряд других трудностей.
Поскольку в данной работе не представилось возможности сделать всесторонний обзор региональных фаун или провести статистический анализ. то пришлось ограничиться одними списками семейств. Недо- статки этого метода совершенно очевидны, но другого выхода не было. При желании со многими подробностями можно ознакомиться в списках семейств. помещенных в конце гл. 2 — 6. Краткое описание региональных фаун, которое приводится здесь, позволит читателю получить некоторое представление об этих фаунах в целом.
При описании региональных фаун и их статистическом анализе нужно избегать одной из обычных ошибок — смешивать прошлое с настоящим. При описании связей и переходов между современными фаунами, живот- ных следует классифицировать строго в соответствии с их современным
Особенности распространения животных. на материках
распространением, а не по их происхождению в прошлом. Например, пекари в настоящее время обитают главным образом в Южной и Централь- ной Америке и лишь заходят в южную часть Северной Америки. Таким образом, они являются элементом Неотропической области на периферии неарктической фауны, и это является неоспоримым фактом, даже если они проникли из Северной Америки в Южную сравнительно недавно. Можно также делать анализ фаун, исходя из происхождения и распространения животных в прошлом, но к этому следует прибегать только в отдельных случаях.
В нижеприводимых фаунистических списках для обозначения глав- ных (не обязательно абсолютно всех) случаев встречаемости различных групп животных за пределами рассматриваемой региональной фауны употребляются следующие сокращения:
энд. — зндемичный, обозначает группу животных, приуроченную и рас'- сматриваемой области
з. ш. — весь земной шар или значительная часть его включая по край- ней мере части всех материковых фаунистических областей широко — значительная часть земного шара, но не весь
тпл. ч. з. ш. — теплые части всех материковых фаунистических областей троп. — тропики, включая по крайней мере часть всех тропических
фаунистических областей
Ст. Св. — Старый Свет
тпл. Ст. Св. — теплые части Старого Света
троп. Ст. Св. — тропики Старого Света
Ам. — вся Америка, Новый Свет
тпл. Ам. — теплые части Америки
троп. Ам. — тропики Америки
Лфр. — Эфиопская Африка
И. М. — Индо-Малайская область (некоторые группы, обозначенные
таким образом, простираются в умеренные широты Азии) Гол. — Голарктическая область
Пал. — Палеарктическая область
Н. С. А. — Неарктическая Северная Америка
Ю. А. — Южная Америка (некоторые группы простираются в Централь- ную Америку и т. д.)
Авст. — Австралийская область
Эфиопская область
Эфиопская область (рис. 47) охватывает Африку оез ее северо-запад- Hoi' части и (в зависимости от взглядов исследователя) южную часть Аравии. Часто в границы этой области включают и Мадагаскар, но я предпочитаю рассматривать его отдельно. Ниже нахождение или отсут- ствие тех или иных групп животных на Мадагаскаре упоминается только в нескольких исключительных случаях.
Эфиопская область — это в основном тропическая область. В эква- ториальной части Западной Африки располагается большой массив дожде- вых лесов, и многочисленные небольшие участки этих лесов рзз~>росаны по всему континенту, преимущественно в обособленных горах Восточной Африки. Значительная часть материка покрыта сухим скрабом или са- ваннами, переходящими в пустыни на севере и юго-западе. Южная око- нечность Африки имеет умеренно-теплый (соответствует широте Южной
Глава 7
Каролины) климат и смешанную растительность. Настоящее описание, как и описание других областей, по необходимости очень упрощено. Я пы- тался показать природу всех областей такой, какой она была до того, как человек уничтожил большую часть растительности земного шара.
Пресноводные рыбы Эфиопской области разнообразны: архаические кистеперые (энд.) и двоякодышащие рыбы семейства Lepidosirenidae (Ю. А.) и несколько семейств отряда Isospondyli (большинство энд.). Богато представлены карпообразные, включая многих сомов (некоторые
I'ÈÑÏÀIIÈß ~ t
СРЕДИЗЕМНОЕ MOPF
Иыс Яобреи Иадеж3ы
р и с. 47. Эфиопская область (непрерывная линяя) и примыкаю- щие к ней территории (прерывистая линия). Ортографическая
проекция.
Еосая штриховка — основные массивы дождевых лесов.
сем. энд., другие И. М.), большое число видов харациновых (Ю.А.) и не- сколько видов карповых (проникающих из Азии), несколько семейств лучеперых (большинство И. М.) и солевыносливые карпозубые (тпл. широко) и хромисы (Ю. А., южная Индия).
К амфибиям Эфиопской области относятся безногие (И. M. и Ю. А.), примитивные лягушки пипы (Ю. А.), высшие лягушки, особенно семейств Bufonidae и Ranidae (троп. Ст. Св., кроме родов Bufo и Лапа широко) и Rhacophoridae (И. М.), в той или иной степени локализованные Phryno- meridae (энд.), Brevicipitidae (тпл. ч. з. ш.) и (в Южной Африке) род сом- нительных Leptodactylidae (Ю. А. и Авст.).
Рептилии Эфиопской области: черепахи пеломедузы (Ю. А.) Trio- nychidae (И. М. и С. А.) и сухопутные черепахи подсемейства Testudini- nae (тпл. ч. з. ш., кроме Австралии) и (северо-западная Африка) черепахи
Особенности распространения животных на материках
подсемейства Emydinae (проникающие из Европы); крокодилы (троп.); много гекконов (троп.), несколько видов агамовых (Ст. Св.), хамелеоны (также на Мадагаскаре), много сцинков (з. ш.) несколько представителей Yeyliniidae (энд.), Сerrhosauridae (Мадагаскар), ящерицы семейства !.acertidae (Евразия), Amphisbaenidae (тпл. Ам.), род Varanus (тпл. Ст. Св.) и Cordylidae (энд.), роды ТурЫорз (троп.) и Leptotyphlops (тпл. Ам.), питоны (тпл. Ст. Св.), песчаные удавы (также род в Пал.), большое число видов ужовых (з. ш.), один или несколько видов семейства Dasypeltidae (В. 0.), аспидовые (тпл. з. ш.) и настоящие гадюки (Евразия).
Птицы Эфиопской области слишком многочисленны, чтобы приводить их подробно. Они включают многих представителей широко распростра- ненных семейств. К ним относятся: хищные птицы (включая грифов Ста- рого Света), совы, цапли, аисты, кукушки, козодои, зимородки, жаворон- ки, ласточки, сорокопуты, дрозды, вьюрки и многие другие птицы. Из семейств Старого Света (или главным образом Старого Света) — дрофы, сизоворонки, щурки, личинкоеды, мухоловки Старого Света и славки, трясогузковые, ткачики, скворцы, иволги, дронго и др. Из тропических в це- лом или тропических семейств Старого Света — трогоны (мало), птицы-носо- роги, бородатки, большинство медоуказчиков, питты (мало), бюль-бюли и большинство нектарниц (попугаи и голуби представлены небольшим числом). Кроме этих, имеется несколько приведенных ниже семейств, относящихся исключительно к Эфиопской области. Конечно, этот список далеко не полный. Из 67 семейств птиц, встречающихся на суше и в прес- ных водах Эфиопской области, 53 встречаются на всем земном шаре, или на его большей части, или на значительной части Старого Света, или в теплых частях Старого Света. Три семейства (медоуказчики, рогоклювы и бюль-бюли) являются общими только (или главным образом) с Индо- Малайской областью. Пять семейств (рачьи ржанки, рябки, удоды, сомнительно, свиристели и, сомнительно, медоед) имеют особые или сомни- тельные связи, и только шесть семейств являются исключительно эфиоп- скими. Из этих семейств, относящихся только (или почти только) к Эфиоп- ской области, три представлены одним видом каждый (страусы, птицы- секретари и молотоглавы), а остальные — различным числом видов: турако (19 видов), птицы-мыши (6) и лесные сорокопуты (13 видов, при- чем сомнительный член этого семейства находится на острове Калиман- тан). Кроме того, ряд подсемейств является только (или почти только) эфиопскими: цесарки (7 видов), древесные удоды (6 видов), кустарниковые сорокопуты (42 вида), ткачики (3 вида), вдовушки (9 видов) и буйволовы птицы (2 вида). И хотя эта географическая классификация семейств птиц в какой-то степени является произвольной, она показывает нам, с одной стороны, разнообразие птиц Эфиопской области, а, с другой — немного- численность эндемичных эфиопских семейств и подсемейств, которые в общей сложности насчитывают только 110 видов.
Связи птиц Эфиопской области легче проследить на родах, чем на семействах. Многие роды являются эндемичными; многие встречаются также и в Индо-Малайской области или имеют там им родственных, а некоторые проникают в умеренные широты Евразии (но многие из них являются перелетными). Число птиц, имеющих иные связи, невелико.
Эфиопская область богата крупными копытными, но с точки зрения зоогеографии важнее некоторые другие, более мелкие млекопитающие. Из млекопитающих в Эфиопской области представлены: выдровые земле- ройки (энд. сем.), златокроты (энд. искл.), е;ки (Евразия), слоновые земле- ройки (почти энд.) и землеройки (широко), лори (И. М.), обезьяны Ста-
860
Глава 7
рого Света (И. М.), два рода больших человекообразных обезьян (И. М.), панголины (И. М.), собачьи (широко), куньи (широко), виверровые (И. М. и т. д.), гиены (средняя часть Азии) и кошачьи (широко), трубкозубы (энд. отряд), один вид слонов (И. М.) даманы (почти энд.), лошади (Пал.). два вида носорогов (И. М.), свиньи (Евразия), два вида бегемотов (энд.). один вид оленьков (И. М.), два вида жираф (энд.), многочисленные бычьи (Евразия и несколько в Н.С.А.), кролики (широко); многочисленные грызуны: беличьи (широко), иглохвостые белки (энд.), долгоноги (энд. сем.), несколько представителей хомячьих (широко), бамбуксвые крысы (И. М.), многочисленные мышиные (Ст. Св., но некоторые подсем. энд. Афр.), сони (Пал.), тушканчики в Сахаре и т. д. (Пал.), дикобразы Старого Света (И. М. и т. д.) и еще 4 семейства, вероятно, дикобразоподооных (сем. энд., но некоторые, очевидно, родственные сем. Ю. А.), плодоядные летучие мыши (тпл. Ст. Св.) и 8 семейств насекомоядных летучих мышеи: Rhinopomatidae (сев.-вос. Афр. и Южная Азия), Emballonuridae (троп.), 3ycteridae (И. М. и т. д.), 5legadermatidae (тпл. Ст. Св.), два семейства подковоносных летучих мышей (тпл. Ст. Св.), Vespertilionidae (з. 1и.) и Мо1ояяЫае (тпл. ч. з. ш ) ..
Эта фауна млекопитающих представляет разнообразное смешение оолее или менее широко распространенных семейств, семейств, оощих с Индо-Малайской областью, эндемичных семейств и нескольких семейсти с другими связями. Эфиопская фауна имеет значительно более тесные (чем это здесь показано) связи с фауной Индо-Малайской области. так как многие роды и даже виды млекопитающих Эфиопской области встре- чаются также и в Индо-Малайской области или имеют там близкородст- венных им представителей.
В заключение следует сказать, что пресноводные рыбы Эфиопской области представляют очень цельную группировку, которая имеет довольно ясные, но отдаленные связи с Южной Америкой, более близкие связи с Индо-Малайской областью и важными эндемичными группами. Амфибии и рептилии Эфиопской области менее характерны для нее, но также имеют некоторые южноамериканские связи (лягушки пипы, черепа- хи пеломедузы, ящерицы семейства Amphisbaenidae, слепозмейки Lepto- typhlopidae) и еще более тесные связи с Индо-Малайской областью (оолее тесные, чем это отражает список семейств, так как многие роды амфибий Эфиопской области встречаются также в Индо-Малайской области или имеют там близкородственные роды) и некоторые эндемичные группы. Птицы Эфиопской области еще менее характерны. По своим основным свя- зям они являются строго индо-малайскими и имеют относительно неболь- шое число эндемичных групп. И, наконец, млекопитающие этой области представляют смешанную группировку, довольно сильно тяготеющую к Индо-Малайской области и имеющую ряд эндемичных групп.
Фауна позвоночных Эфиопской области в целом (если только можно вообще представить себе такой сложный комплекс как нечто единое) более всего сходна с фауной Индо-Малайской области. Обе фауны приоли- зительно одинаковы по размеру, примерно одинаковы по структуре (в меньшей степени по рыбам), имеют много общих семейств, очень много родов и некоторые общие виды. В определенной своей части фауна Эфиопской области имеет также тесные связи с Палеарктикой, образованные глав- ным образом за счет распространения тропических групп к северу, Bh Ji()- чая и многих перелетных птиц. Некоторые позвоночные Эфиопской об- ласти имеют очень отдаленные родственные связи с южноамериканскими позвоночными: к ним относится несколько важных групп рыб, ряд амфи-
Особенност,и распространения животных на лагпериках
бий и рептилий и крайне небольшое число птиц и млекопитающих. х1ря- мых связей между позвоночными Эфиопской и Австралийской областей почти не наблюдается. В фауне Эфиопской области имеются также важные эндемичные группы, особенно рыб и млекопитающих, у которых нет современных родственных связей за пределами Африки.
Границы Эфиопской области, кроме северной, проходят по океану. Большинство позвоночных этой области встречается только к югу от Са- хары или проникает на север, строго придерживаясь Нила. Небольшая северная часть Африки (к северу от Атласских гор), состоящая из север- ных частей Марокко и Алжира, имеет обедненную фауну, которая по своим связям в основном является европейской. Фауна простирающихся между этими двумя районами обширных пустынь северной части Африки очень ограничена, специализирована и имеет смешанные, частично (большин- ство) эфиопские и частично евразийские связи. В данном случае не столь важно, проходит ли северная граница области у Атласских гор (где ее провел Склэтер), или через центральную часть Сахары, у северной гра- ницы тропиков (где ее установили Уоллес и большинство более поздних зоогеографов), или Сахара в целом рассматривается как зона перехода. Обедненная фауна позвоночных юго-западной Аравии по своим связям также является эфиопской. Эту часть Аравии либо можно включить в Эфиопскую область, либо ее можно рассматривать как часть сложной зоны перехода.
Самая богатая и самая разнообразная фауна в пределах Эфиопской области находится в тропических районах с наиболее благоприятными условиями обитания. К северу от тропиков по направлению к Сахаре происходит сильное обеднение всей фауны в целом, а к югу от тропиков (к Южной Африке) обеднение фауны проявляется в меньшей степени и отмечается некоторая дифференциация. Кроме того, частичная дифферен- циация фауны наблюдается во влажных лесах Западной Африки и в райо- нах разреженных лесов и саванн Восточной Африки, хотя обе эти фауны перекрывают друг друга и имеют сложные взаимосвязи. В соответствии с этими различиями Уоллес разделил материковую Эфиопскую область на три подобласти, соответствующие Западной, Восточной и Южной Африке. Я не придерживаюсь этого чисто формального подразделения, но, о чем уже говорилось выше, не имею возможности остановиться здесь на более детальном анализе распространения животных в отдельных частях Эфиопской области, которое очень различно у различных групп животных.
Индо-Малайская область
Индо-Малайская область (рис. 48), по существу, охватывает тропи- ческую Азию и близлежащие материковые острова, включая Сулавеси, Суматру, Яву, Калимантан, Тайвань и, возможно, Филиппины.
Граница этой преимущественно тропическохх области на несколько градусов заходит на север от тропиков, особенно в северной Индии. В восточнохх части области, от северо-восточной части Индии, Бирмы. Индокитая и т. д. до Больших Зондскххх островов, находятся большие массивы дождевых лесов, местами прерываемых открытыми более сухими участками. На север от Бирмы до Юго-Восточного Китая и восточных 1ималаев простираются высокие горы с богатой и разнообразной расти- тельностью. На западе лежит основная часть полуострова Индостан
862
Глава 7
с относительно сухими и открытыми пространствами, ограниченными на юго-западном побережье полосой невысоких гор, покрытых влажно-тропи- ческим лесом. С северо-востока на юго-запад Индию пересекает цепь невы- соких. изредка прерывающихся гор (Сатпура), которая, очевидно, служи- ла в прошлом путем расселения по всей стране животных более влажных районов (Hora, 1953). Гималаи становятся более засушливыми и менее благоприятными для жизни в западном направлении.
Для фауны пресноводных рыб Индо-Малайской области характерно отсутствие архаических групп. Другие рыбы представлены: одним видом
4РЯДНАЯ ОбМАС7Ь
ай Ваиь
с
Тропик 7~ака
абаи
8 „
~о~о~ ~ю . -;
l
'7 у
, /
~ф/
)Q °
1~ °
аСуЭайси
Г1~
1 ~ )
аГ
г
Згдатор
OP" ~~~4 О.
P и с. 48. Индо-Малайская область (непрерывная линия) п примыкающие к ней территории (прерывистая линия). Орто- графическая проекция.
Пунктирная линия — примерные региональные границы. Косая
штриховка — основные массивы дождевых лесов.
костноязычных (некоторые сем. Афр. и простираются в Пал., другие энд.). необыкновенно многочисленными и разнообразными карпообразны- ми, включая карповых (Пал. Н. C. А., Афр.), семейством Cobitidae (Пал. и т. д.) и Homalopteridae (энд.), несколькими видами семейства Nandidae (Афр. Ю. А.), Pristolepidae (энд.), Anabantidae (большинство, но один род распространен до Афр.), Channidae (Афр.), единственным представителем семейства Luciocephalidae (энд.), Mastacembelidae (Афр.) и единственным представителем Chaudhuriidae (искл. энд.), несколькими видами карпо- зубых (тпл., широко) и одним родом хромисов (родствен. Афр., Мада- гаскар, на полуострове Индостан и Цейлоне).
Амфибии Индо-Малайской области представлены безногими (Афр., Ю. А.), несколькими видами саламандр в северном Индокитае (из Пал.), несколькими локализованными представителями круглоязычных (Пал.), лягушками семейства Pelobatidae (Гол.), Buffonidae (широко), семейством Ranidae (широко) и Rhacophoridae (Афр. и т. д.), родом Hpla в северном Индокитае и т. д. (проникли из Пал.) Brevicipitidae (тпл. ч. з. ш.).
Из рептилий характерны: многочисленные черепахи подсемейства Emydinae (Пал. и Ам.), единственный представитель подсемейства Platysterninae (энд.), Trionychidae (Афр. и C. А.), сухопутные черепахи семейства Testudininae (тпл. ч. з. ш., кроме Авст.), крокодилы и гавиалы (энд.), многочисленные гекконы (тпл. ч. з. ш.), агамовые (Ст. Св.), один
вид хамелеонов в южной Индии и на Цейлоне (родственные связи ведут к Средиземноморской обл.), многочисленные сцинки (з. ш.), несколько пред- ставителей Dibamidae (распространенных до Новой Гвинеи), несколько представителей семейства Lacertidae (Афр., Пал.), род Varanus (тпл. Ст. Св.), локализованные семейства Anguidae (Пал., Ам.) и Lanthanotus на острове Калимантан (энд.), многочисленные змеи ТурИоря (тпл. ч. з. ш.) и Leptotpphlops (краевые части Афр. и т. д.), локализованное семейство Uropeltidae (энд.), несколько видов семейства Aniliidae (троп. Ам.), один представитель семейства Xenopeltidae (энд.), питоны (тпл. Ст. Св.), песчаные удавы (в краевых частях, Пал. и т. д.), богато представленные ужовые [подсемейства Dipsadinae (троп. Ам.), Xenoderminae (Ю. А.), Сolubrinae (з. ш.), водное подсемейство Acrochordinae (Авст.) и Homalop- sinae (Ам.)]; представитель семейства Dasypeltidae (Афр.), аспидовые (тпл. ч. з. ш.), морские змеи (область теплых вод Индо-Пацифики), настоящие гадюки (Афр. и Пал.) и ямкоголовые гадюки (Пал. и Ам.).
Птицы Индо-Малайской области включают почти все широко распро- страненные семейства Старого Света, семейства, свойственные тропикам, и тропические семейства Старого Света, встречающиеся в Эфиопской обла- сти, но соотношение различных птиц здесь несколько иное; здесь мень- ше представителей одних групп (например, сорокопутов, удодов, медо- указчиков, нектарниц и вьюрков) и больше других (например, дятлов, питт, тималий и вороновых). Много фазановых (в Африке только один вид) и голубей, но попугаи (так же, как и в Африке) представлены неболь- шим числом. Из 66 семейств птиц, приуроченных к суше и пресным водам, 53 более или менее широко распространены повсеместно, 3 — общие толь- ко (или главным образом) с Африкой, 4 семейства с особыми или сомни- тельными географическими связями, 5 — общие только (или главным образом) с Австралийской областью (сорные куры, лягушкоротые козо- дои, артамусы, нектарницы и медососы, последние в Индо-Малайской области не проникают за пределы острова Бали), и только одно семейство этой области является эндемичным — Irenidae — с 4 родами и 14 видами. Такое отчасти произвольное географическое подразделение показывает общую природу авифауны Индо-Малайской области. Она очень схожа с фауной Эфиопской области, разнообразна, но число групп, свойственных только этой области, еще меньше, чем в первой, есть группы, имеющие австралийские связи. Необходимо отметить большое число эндемичных родов, а также многочисленность птиц, общих (или родственных) с Афри- кой и с Палеарктикой Евразии (многие из них являются перелетными). Птицы, общие с Австралией или с другими связями, немногочисленны.
К млекопитающим Индо-Малайской области относятся: ежи (настоя- щие и волосатые) (первые в Афр. и Пал., последние энд.), землеройки (широко), кроты (местные, проникают из Пал.), шерстокрылы (энд.), тупайи (энд.), лори (афр.), долгопяты (энд.), обезьяны Старого Света (афр.) и человекообразные обезьяны (Афр.), панголины (Афр.), собачьи (широ- ко), медведи (Пал. и Ам.), куньи (широко), виверровые (Афр. и т. д.), один вид гиен (Афр.), кошачьи, один вид слонов (Афр.), один вид тапиров (троп. Ам.), два вида носорогов (Афр.), кабаны (Афр. Пал.), оленек (Афр.), олени (Пал. и Ам.), бычьи (Афр. и Пал. и несколько в Н. С. А.), зайцы (широко); многочисленные грызуны: белки, летяги (Пал. и Н. С. А.), несколько видов хомячьих (широко), бамбуковые крысы (Афр.), много- численные мышиные (Ст. Св.), семейство Platacanthomyidae (энд.) и дико- бразы Старого Света (Афр.); плодоядные летучие мыши и 8 семейств насе- комоядных (те же, что и в Афр.). Мы видим, что фауна млекопитающих
Ггава 7
Индо-Малайской области представляет собой смешение оолее или менее широко распространенных семейств, семейств и родов, общих с Эфиопской областью, эндемичных семейств, нескольких групп мышиных грызунов и множества родов и видов рукокрылых, общих с Австралийской областью, и очень немногих групп с другими связями.
В заключение отметим, что пресноводные рыбы Индо-Малайской оола- сти представляют богатую и доминирующую группировку, для которой характерно отсутствие архаических групп и преобладание карпообразных. Амфибии и рептилии лишь отчасти сходны с фауной амфибий и рептилий Эфиопской области. Из существующих различий заслуживают внимания следующие: отсутствие пип и наличие лягушек семейства Pelobatidae. отсутствие черепах пеломедуз и наличие представителей подсемейства Emydinae, наличие гавиалов, отсутствие (идти очень небольшое число) хаме- леонов и ящерицы из семейств Gerrhosauridae, Amphisbaenidae, (ordylidae, необычная малочисленность змей семейства Leptotyphlopidae, но наличие представителей семейств Uropeltidae, Ilysiidae, ужовых и ямкоголоных гадюк. Для Африки вообще характерно большее разнообразие ящери~~, а для Индо-Малайской области — змей. Птицы по своим связям являются строго эфиопскими, со многими палеарктическими и отдельными австра- лийскими связями и с небольшим числом важных эндемичных групп. Ь[ле- копптающие Индо-Малайской области вообще в значительной степени являются эфиопскими, но включают также ряд групп с другими связями и некоторые эндемичные группы.
Таким образом, фауна позвоночных Индо-Малайской области, несмо- тря на отмеченные выше различия, в целом весьма сходна с фауной Эфиопской области. Отдельные части фауны Индо-Малайской области имеют связи и с Палеарктикой, в основном через Восточную Азию. Несколько видов позвоночных Индо-Малайской области, главным ооразом некоторые рептилии, птицы и рукокрылые, родственны австралийским. И очень немногие (отдельные виды лягушек, змей, птиц, а также тапиры) имеют родственные виды в американских тропиках. В фауне Индо-Малай- ской области мало групп позвоночных, приуроченных только к этой оола- сти, то есть эндемичных меньше, чем в любой другой тропической регио- нальной фауне. Итак, можно предположить, одно из двух: или Индо-Ма- лайская область была центром, из которого позвоночные расселялис ь в другие области, или она являлась главным «перекрестком» в расселе- нии, или тем и другим. (В действительности она, видимо, для одних позво- ночных, например карповых рыб, являлась центром расселения, а для других, например для птиц, — перекрестком).
Границы Индо-Малайской области выражены нечетко, в связи с тем. что ее фауна образует сложные переходы в нескольких направлениях. Засушливая часть северо-западной Индии и «спорная территория» (Wal- lace) за ее пределами имеет обедненную фауну, в которой наблюдаются переходы как с востока на запад, так и с юга на север (стр. 65). Где в этом районе проходят границы области, вероятно, не столь существенно. Несколько дальше на восток 1'ималаи образуют естественную границу, которая проходит на несколько градусов севернее тропиков. Этот горный барьер не является основной причиной дифференциации региональных фаун Индо-Малайской области и Палеарктики,— она, очевидно, кроется в зональности климата, но и горы и засушливая часть страны, простира- ющаяся к западу и к северу от них, все-таки в какой-то мере служат пре- градой, которая не допускает сильного проникновения отдельных элемен- тов Индо-Малайской фауны на север от тропиков, а также значительного
Особенности распространения животнь~х на латериках
смешения их с элементами северной фауны. Далее к востоку, в горах Ь11рмы, Юго-Западного Китая и вдоль побережья Китая наблюдается очень обширный и сложный переход, в связи с чем установить границу области здесь трудно. Обычно ее проводят через Южный Китай вблизи тропика Рака, где многие тропические группы животных достигают своих северных пределов. На юге и на востоке в ее границы входят Цейлон и Taii- вань (имеющие в какой-то степени обедненную материковую фауну), а также Суматра, Ява и Калимантан (с почти полностью представленной фауной Индо-Малайской области). В ее границы также могут быть вклю- чены и Филиппины, хотя фауна этих островов фактически периферийного типа — с сильным и постепенным обеднением по мере удаления от острова Калимантан (гл. 8). За пределами островов Ява и Калимантан фауна Индо- э[алайской области образует обширный и сложный переход с австралий- ской (более подробное описание дано в конце настоящей главы).
В пределах Индо-Малайской области с востока на запад и с севера на юг отмечается дифференциация фауны. Наиболее богатая и самая разно- образная фауна находится в более влажной восточной части области (от Южного Китая и т. д. до островов Ява и Калимантан), где находится боль- шинство лягушек Индо-Малайской области из семейства Pelobatidae, черепах подсемейства Emydinae и фазанов. Менее богатая и несколько отличающаяся от первой фауна простирается в западном направлении к Индии. Частичная дифференциация подфаун отмечается на востоке области (северной подфауны — в Южном Китае, Индокитае, Бирме, Таи- ланде, и южной подфауны — на полуострове Малакка, Суматре, Яве, Калимантане и т. д.) и на западе (северной подфауны — на основной более засушливой части Индии, и южной подфауны — в южной Индии и на Цейлоне, причем в последней представлены изолированные малайские элементы и даже несколько мадагаскарских). Вдоль этих линий Уоллес разделил Восточную область на четыре подобласти, назвав их (Wallace, 1876) Индо-Китайской, Индо-Малайской, Индийской и Цейлонской. Формально здесь эти подобласти не признаются, но названия «Индо-Ки- тайская подобласть» и «Малайская подобласть» (по Уоллесу — «Индо- Малайская») иногда употребляются.
Иалеарктическая область
Палеарктическая область (рис. 49), по существу, охватывает Евра- зию севернее тропиков и небольшую северную часть Африки.
!!алеарктика — это область умеренной зоны северного полушария с арктической периферией. В более влажных частях Восточной Азии, включая умеренную зону Китая и Японии, произрастают прекрасные лиственные леса. Большая часть внутренних районов Азии представляет собой засушливые открытые территории, простирающиеся через Юго- Западную Азию до Северной Африки, которые преимущественно в север- ных краевых частях переходят в травянистые степи. В Европе снова тянутся массивы лиственных или смешанных лесов, но видово~~ состав деревьев здесь значительно беднее, чем в лесах Восточной Азии. '1ерез Северную Азию и Европу простирается обширная зона хвойных лесов, а севернее лежат просторы тундры.
Пресноводные рыбы в Палеарктике представлены: веслоносами, рас- пространенными в Китае (Н. С. А.). двумя видами восточноазиатских чуку- чанов (Н. С. А.), большим количеством карповых (И. М., Афр., Н. С. А.),
Глава 7
семейством Cobitidae (И. М. и т. д.), несколькими локализованными видами сомов (сем. И. М.), одним представителем семейства Umbridae в Ду- нае и т. д. (Н. С. А.), родом Dallia в Восточной Сибири (Аляска), щуками (Н. С. А.), некоторыми окуневыми (Н. С. А.) и очень немногочисленными представителями семейств Anabantidae, Channidae и Alastacembelidae в Восточной Азии (проникающими из И. М.), несколькими видами карпо- зубых, распространенными к югу (широко, тпл.). Из этого списка только семейство карповых является многочисленным и доминирующим по всей
и-о6
1
/
Арадиа ЭФИОПСКАЯ 06Я. ~
x!
сю
1
1
1
1
1
1
1
1
Ъ
Г Ъ
Ъ ~J
' ~~л~,- �~ ~(~~
i 0EJr
° -~
I
Ъ /
г
I
г
г
r
r
г
г
/
I
1
I
)
Ъ
I
P и с. 49. Палеарктпческая область (непрерывная линия) и примыкающие к ней тер-ритории (прерывистая линия). Ортографическая проекция.
области, а недостаток других строго пресноводных рыб отчасти воспол- няется солевыносливыми рыбами, особенно лососевыми (Н. С. А.), распро- страненными в северном направлении.
Амфибии в Палеарктике представлены саламандрами (Н. С. А.), круглоязычными (локализованными в И. М.), представителями семейства Pelobatidae (Н. С. А., И. М.), родами Bufo (широко), Ну1а (широко), многочисленными видами рода Лапа (широко) и несколькими видами рода Bhacopkorus, семейством Brevicipitidae в Восточной Азии (проникаю- щим из И. М.).
К рептилиям, более или менее широко представленным в Палеаркти- ческой области, относятся черепахи подсемейства Emydinae (И. М., Н. С. А.), сцинки (широко), ящерицы семейства Lacertidae (Афр., М. И.), Anguidae (Ам., локально в И. М.), большое количество ужовых (широко), настоящие гадюки (Афр. И. М.) и ямкоголовые гадюки в Азии (И. М., Ам.), а также рептилии, которые встречаются по южным краевым частям Пале- арктики или Восточной Азии: черепахи рода Testudo (тпл., широко), Trionpx в Восточной Азии (Афр., И. М., Н. С. А.), аллигатор в Китае (Н. С. А.), гекконы (тпл. ч. з. ш.), агамовые (тпл. Ст. Св.), один вид хамелео- нов (Афр., и т. д.), представители семейства Amphisbaenidae (Афр., тпл. Ам.), роды Varanus (тпл. Ст. Св.), Shinisaurus в Китае (энд. сем.), ТурЫорз (тпл. ч. з. ш.), Leptotpphlops (Афр., тпл. Ам.) и песчаные удавы (Афр.).
Авифауна Палеарктики включает представителей 53 семейств назем- ных и пресноводных птиц. Это число незначительно уступает числу этих семейств в Африке или в Индо-Малайской области, но надо учитывать, что некоторые из этих птиц встречаются только по периферии области или име-
Особенности распроспгранения животных на материках
ют локальное распространение. Примерно только 37 семейств более или менее широко распространены в Палеарктике, а некоторые из них в ооль- шинстве случаев являются в этой области перелетными. К ним относятся (но это не полный список) поганки и гагары, соколы, цапли, аисты, утки, перепела и куропатки, кукушки, пастушки, ржанки, голуби, совы, козо- дои, зимородки, стрижи, дятлы, жаворонки, ласточки, дрозды и мухо- ловки Старого Света, славки, трясогузковые, синицы, вьюрки и ткачики, скворцы, врановые. Большинство семейств, широко распространенных в Палеарктике, имеет такое же широкое распространение и в других местах на большей части земного шара или Старого Света, но гагары и пи- щухи (как и чистиковые) относятся к голарктическим птицам. Куропаток ц свиристелей также можно назвать голарктическими птицами, но зави- рушки (один род и 12 видов) являются птицами, приуроченными исключи- тельно к Палеарктике. Кроме 37 широко распространенных семейств. по крайней мере летом в этой области имеется еще 10 семейств (пеликаны, ибисы, фламинго, трехперстки, авдотки, тиркушки, рябки, щурки и удо- ды), приуроченных только к периферии области (к теплым южным крае- вым частям) или локализованных в Палеарктике. Кроме этого, еще пять тропических семейств Старого Света (бюль-бюли, личинкоеды, нектарни- цы, белоглазки и дронго) представлены в Палеарктике только мигрирую- щими или главным образом мигрирующими формами, которые проникают на территорию Палеарктики в Восточной Азии. И, наконец, еще одно семейство представлено двумя древесницами, которые расширили свой гнез- довой ареал в Восточную Сибирь. Такая географическая классификация семейств отчасти произвольна (четкого отличия между семействами, кото- рые «широко» распространены, и теми, которые встречаются «по перифе- рии» Палеарктики, нет), но она довольно хорошо показывает, что птицы Палеарктики менее разнообразны, чем птицы Эфиопской и Индо-Малай- ской областей, хотя в остальном они имеют незначительные отличия. Связи птиц Палеарктики лучше прослеживаются на родах, чем на семей- ствах: многие роды (включая ряд перелетных на севере) общие с родами птиц тропиков Старого Света, многие роды и даже многие северные виды общие с родами и видами Северной Америки, но некоторые роды типичны только для Палеарктики.
Основными млекопитающими Палеарктики являются ежи (Афр. И. M.), бурозубки (широко, но сомнительное подсемейство Soricinae— голарктическое), кроты (Н. С. А.), собачьи (широко), медведи (И. М., Ам.), панда (энд. подсем., локализованное в Юго-Западном Китае, но сем. представлено также в Ам.), куньи (широко), кошачьи (широко), лошади (Афр.), свиньи (Афр., И. М.), локализованный вид верблюдов (Ю. А.), олени (И. М., Ам.), бычьи (Афр. И. М., Н. С. А.), пищухи (Н. С. А.), зайцы (широко). белки (широко) и летяги (И. М., Н. (;. А.), бобры (Н. С. А.), многочисленные хомячьи (широко), локализованные кротовые крысы (поч- ти энд.), богато представленные мышиные (Ст. Св.), сони (энд. подсем., но сем. Афр.), локализованный представитель селевиний (энд.), прыгун- чики (С. А.) и тушканчики (Сев. Афр.), рукокрылые — в основном семей- ство Vespertilionidae (з. ш.). Помимо этих, целый ряд других семейств достигает Палеарктики и на незначительное расстояние проникает в перифе- рические районы: слоновые землеройки в Северной Африке (Афр.), пан- голины в Восточном Китае (И. М., Афр.), обезьяны Старого Света в Север- ной Африке и некоторых частях Китая и Японии (Афр., И. М.), виверро- вые (Афр., И. М.), гиена и даман в некоторых частях Юго-Западной Азии (проникающие из Афр.), грызуны семейства Ctenodactylidae в Северной
868
Глава 7
Африке, плодоядные летучие мыши и несколько семейств насекомоядных (из тропиков). Роды млекопитающих Палеарктики — частично эндемич- ные. частично общие с тропиками Старого Света (особенно с Индо-Чалай- ской областью), некоторые — общие с Северной Америкой. Связи с послед- ней особенно сильно выражены на севере.
В заключение отметим, что пресноводные рыбы в 11алеарктике пред- ставлены карповыми (проникающими из Индо-Малайской области), несколькими в основном локализованными видами других строго пресно- водных семейств (первый отдел, многие с североамериканскими связями) и солевыносливыми рыбами (многие в северных частях с североамерикан- скими связями). К амфибиям Палеарктики относятся саламандры (Н. С. А.), лягушки трех широко распространенных доминирующих родов (Лапа, Bufo, Hyla) и более или менее локализованные представители четырех других семейств лягушек, связи которых ведут к Индо-Малайской оола- cTlt и (или) к Северной Америке. Рептилий в Палеарктике мало, связи большинства из них (но не всех) ведут к тропикам Африки и (или) Индо- Малайской области. Птицы Палеарктики менее разнообразны, чем в тро- ttttt;ax, и большинство из них (на уровне родов или видов) имеет связи с тропиками Старого Света или Северной Америки. Палеарктические мле- копитающие не отличаются большим разнообразием, и большинство их связей (на уровне родов и видов) ведет к тропикам Старого Света (особен- но к Индо-Малайской области) или к Северной Америке.
В целом фауна позвоночных Палеарктики значительно менее богатая по сравнению с фаунами Эфиопской и Индо-Малайской областей. Исходя из существующих связей, можно считать, что это фауна переходная, пред- ставляющая сложное смешение тропических групп Старого Света и севе- роамериканских групп (причем первые — наиболее многочисленны к tory, а вторые — к северу) и имеющая небольшое число эндемичных палеарк- тических групп.
На юге Палеарктика граничит с Эфиопской и Индо-Малайской обла- стями, а на севере ее граница совпадает с границей суши. На западе она охватывает Британские острова и, условно, Исландию, хотя последняя имеет фрагментарную фауну; на востоке она включает Японию и прохо- дит по Берингову проливу.
В пределах Палеарктики самая богатая фауна приурочена к более теплым районам с достаточным количеством осадков. К северу происходит постепенное обеднение фауны позвоночных, и на большей части Арктики она сокращается до пресноводных рыб (в основном солевыносливых), одного рода лягушек (Лапа) (полное отсутствие рептилий), нескольких наземных птиц (число птиц пресных водоемов, главным образом перелет- ных. значительно больше) и нескольких видов млекопитающих (8 видов), которые обитают в самых высоких широтах континента. Сокращение и спе- циализация фауны происходят также в сухих и горных районах. Неко- торая дифференциация наблюдается с востока на запад (причем на юге оольше, чем на севере): фауна позвоночных в благоприятных для обитания районах Китая богатая (это самая богатая часть Палеарктики) и включает много элементов Индо-Малайской области; фауна Средиземноморской обла- сти также довольно богатая, но в ней преобладают элементы Эфиопской области. Палеарктика была разделена Уоллесом на 4 подобласти: Сибир- скую, Европейскую, Маньчжурскую и Средиземноморскую. Но две пер- вые выражены нечетко, а другие не заслуживают признания, хотя «Сре- диземноморская подобласть» является вполне приемлемым термином для описания распространения некоторых животных.
0«обенности риспространения животных на л~атериках Неарктпческая ооласть
Неарктическая область включает Севернук1 Америку севернее тро- пиков (рис. 50).
Как и Палеарктика, Неарктика — область у.меренной зоны северного полушария с арктической периферией. Основной растительный покров большей части восточных районов Северной Америки в средних широтах— лиственные или смешанные леса. На западе в центральной части конти- нента простираются обширные прерии, а далее на запад засушливые части
дия
° ° ° ° Р и с. 50. !Хеарктическая область (непрерывная линия) и соседняя территория (прерывистая линия). Ортографиче-
ская проекция. Пунктирная л1тния — приблизитсльнан граница с Неотр(1ничсской
оГ~ластьн>. страны сменяются горами с узкими полосками смешанных или хвойных лесов. К северу, как и в 1".вразии, обширная зона хвойных лесов сменяе"ся тундрой.
Ii неарктическим пресноводным рыбам первого отдела относятся один- цва вида веслоносов (Китай), ильная рыба (энд., отряд), несколько видов хиодонов (энд.), чукучаны (Вост. Азия), большое число мелких карповых (И. М.. Пал., Афр.), сомы семейства Ameiuridae (почтп энд.), несколько представителей семейства Umbridae (Вост. Европа), род Dallia в Аляске (Вост. Споирь), род Esox (Пал.), несколько траутпэрчей и семейство Aphre- doderidae (оба энд.), морские окуни (энд. сем.), многочисленные окуневые (Пал.. одно энд. подсем. Н. С. А.). Из пресноводных рыб второго отдела (не строго пресноводных) надо отметить панцирных щук (также Центр. Ам.) и карпозубых (тпл., широко). Кроме этих рыб, здесь встречаются стро- го пресноводные харациновые и хромисы, достигающие реки ~Рио-Гранде (проникают из Ю. А.), а также многочисленные солевыносливые рыоы, особенно северные лососевые (Пал.).
Амфибии в Неарктике представлены саламандрами (Ilaë.), локали- зованным единичным представителем семейства Leiopelmidae (Новая Зеландия), Pelobatidae (также подсем. в Европе), родами Bufo (широко), Пу1а и т. д. (широко) и Лапа (широко), несколькими видами семейств I.ерСос1асйу1и1ае и Brevicipitidae (проникающими из Ю. А.).
870
1'.ю ава 7
К неарктическим рептилиям относятся каймановая и мускусная дедде- пахи (проникают в некоторые части Ю. А.), черепахи подсемейства Е туй пае (Евразия и троп. Ам.), эндемичный род сухопутных черепах на юге (пдддсем. тпл., широко) и род Тпопух (Афр., И. М., Вост. Азия), один вид аллига- торов на юго-востоке (Китай) и один вид крокодддлодд в южной часпд Ф.1дд- рддды (тпл. ч. з. ш.), гекконы на юго-западе (тпл. ч. з. ш.), многочисленные игуаны (троп. Ам. и т. д.), Heloderu~a (энд. сем.), представители семеддспддд >antusiidae на юго-западе (Центр. Ам. и т. д.), сцинд'и (шддроко), ящерддддь~ семейства Teiidae (троп. Ам.), Ampl>isbaenidae на юге (Ю. А. и ". д.. Лфр.), представители семейства Anguidae (троп. Ам., несколько в Пал. дд И. дд.) и Anniellidae на юго-западе (энд.), Leptotyphlops (троп. Ам., Афр.), не- сколько мелких представителей подсемейства Hoinae на западе (тад;,ке подсем. в дастях Афр., Азии и т. д.), большое число ужовых (з. ш.), аспи- довые (троп. Ам., сем. тпл. ч. з. ш.) и ямкоголовые гадюки (умерен. и трои. Азия, троп. Ам.).
Наземные птицы и птицы пресных водоемов Неарктики вклнд даддд г представителей примерно 49 семейств. Из них около 39 семейств более или менее широко распространены по всей Неарктике или в более теплых рай- онах этой ооласти, по крайней мере летом. К ним относятся: поганки, гагары, пеликаны (локально), хищные (3 сем.) и грифы Нового Света (троп. Ам.), цапли, утки и т. д., перепела и куропатки, индейки (почтдд энд.), д;укушки, журавли, пастушки, ржанки и песо днддки, чайки, гол уби, совы (2 сем.), козодои, зимородки, стрижи, колибри (троп. Ам.), дятлы. мухоловки-тиранны (троп. Ам.), жаворонки (один вид), ласточки, дрозды. пересмешники (подсем. троп. Ам.) и крапивники (подсем. большинство п троп. Ам.), трясогузковые (коньки), сорокопуты, свиристели, пищухи, поползни, синицы и гаички, виреоны (троп. Ам.), американские славки, танагры, кардиналы (сем. троп. Ам.), вьюрки, кассддддд (троп. Ам.), вороны и сойки. Этот список показывает, что авифауна Неарктики представляет собой смешение многих широко распространенных и некоторых доминдд- рующих семейств Нового Света («троп. Ам.»), а также ряда голарктических семейств и что только одно семейство (индейки) является исключительно ддли почти исключительно неарктическим. Представители следующих 10 семейств заходят в южные краевые части Неарктической области 1I3 тропиков, достигая (или почти достигая) южной части Соединенных Штатов. К ним относятся: ибисы, один вид аистов, один вид фламддн- го, один вид краксов, арамы, яканы, попугаи, один вид трогонов, момоты (только до северной Мексики) и один вид котинг. Некоторые роды неари- тддческих птиц (включая многих перелетных) являются общими с Цент- ральной и Южной Америкой, некоторые (особенно к северу) — с 1!алд- арктической областью и некоторые являются эндемиками.
Из млекопитающих в Неарктике представлены опоссумы (пронддкан>- щие ддз 10. А.), землеройки (дпддроко, подсем., также Пал.) и кроты, оро- неносдды в Техасе и т. д. (проникает из Ю. А.), собачьи (широко), медведи (Евразия, Ю. А.), еноты и т. д. (троп. Ам., родственны пандам в Азии), куньи (широко) и кошачьи (широко), пекари на юге (проникают из Ю. А.), олени (Евразия, троп. Ам.), вилороги на западе (энд.) и несколько вддд<в ддччьддх (11ал., И. М., Афр.), сеноставки на западе (Пал.) и крол дд ки (дддддроко), бобровые белки на западе (энд. сем.), белки (широко) и летяги (Евразия и т. д.), гоферы и карманчиковые мьдддддд в основном на западе (проникают в Центральную Америку и северную часть Южной Америки соответственно), бобры (Пал.), многочисленные хомякообразные (широко), прыгунчики (Пал.) и дикобраз (троп. Ам.), рукокрылые семейства Vesper-
871
tilionidae (широко), на юге несколько видов калифорнийских листоносов (троп. Ам.), длинноногие летучие мыши (троп. Ам.) и бульдоговые летучио мыши (тпл. ч.з.ш.). Эта фауна представляет собой смешение семейств, общих, с одной стороны, с Палеарктической областью (и другимичастя- ми Старого Света), а с другой — с американскими тропиками и небольцщм числом важных эндемичных групп животных.
Роды млекопитающих Неарктики подчеркивак>т те же смешанные свя- зи, hol орые в основном проявляются по направлению к Палеарктике, осо бенно к северу. [Сроме того, в некоторых группах, особенно у грызупон, имеется множество эндемичных родов.
Б заключение надо отметить следующее. Пресноводные рыбы Неарк- тики многочисленны и характерны для этой области, а большинство семейств является исключительно неарктическими или общими с Палеарк- пп;oi' Доминирующие амфибии Неарктики относятся к тои ".ке группе, что и амфибии Палеарктики (саламандры, роды Лапа, Bufo, Hpla), а пред- ставители семейства Pelobatidae родственны палеарктическим (несколь- ко видов лягушек Неарктики имеют южноамериканские или новозеланд- ские связи). Рептилии Неарктики представляют смешение палеарктиче скпх (или Старого Света) и тропических американских групп (к последним особенно относятся ящерицы) с немногими важными эндемичными неарк- тическими группами. Птицы Неарктики представляют смешение пале арктических (Старого Света) и тропических американских групп: первые становятся более многочисленными к северу, а вторые — к югу, хотя мно- гие виды, зимующие в тропиках Америки, гнездятся далеко на севере. У млекопитающих Неарктики также происходит смешение (относительно большего числа) палеарктических групп или групп Старого Света и (от носительно меньше) тропических американских групп с небольшим чис- лом важных эндемичных неарктических групп.
В целом фауна позвоночных Неарктики, как и Палеарктики, значи- тельно менее богатая, чем тропические региональные фауны, и является в основном переходной фауной со смешанными палеарктическими и тро- пическими американскими группами (первые более многочисленны к севе- ру, последние — к югу) и с небольшим числом важных неарктических групп, за исключением пресноводных рыб.
На севере неарктическая область охватывает остров Ньюфаундленд, Арктический архипелаг и Гренландию (в Гренландии все млекопитающие американские, но среди птиц есть ряд европейских). На юге фауна Неарктики образует сложный переход с неотропической фауной, о чем буде г сказано ниже, и граница ее обычно устанавливается в южной Мексике, где прохладные Мексиканские нагорья сменяются тропическими низмен- ностями, которые для многих тропических позвоночных являются север- ным пределом.
В пределах Неарктической области наиболее разнообразная фауна позвоночных сосредоточена в южных районах (которые варьируют для разных классов) с наиболее благоприятными условиями обитания, и она постепенно (но неравномерно) уменьшается к северу до тех пор, пока не сократится до обедненной северной фауны, очень сходной с фауной llале- арктики. В Неарктике также наблюдается дифференциация с востока на запад, причем в одних группах более сильная, чем в других: например, большинство пресноводных рыб и черепах концентрируется к востоку от (,'качистых гор; саламандры более богато представлены на востоке, чем на западе, но образуют изолированную западную подфяуну (рис. 19); фауна змей относительно богаче на востоке, а ящериц — на западе; неко-
1 лаыи 7
872
торые семейства млекопитающих представлены только или главным обра- зом на западе (вилороги, сеноставки, бобровые белки, гоферы, карманчи- ковые мыши). Но птицы в целом, доминирующие млекопитающие, кроме грызунов, и некоторые семейства и роды других классов проявляют значительно меньшую дифференциацию с востока на запад.
Уоллес разделил Неарктическую область на четыре подобласти: северную, простирающуюся через континент, IoiKHee ее — восточную, западно-центральную (Скалистые горы и т. д.) и побережье Тихого океана («Калифорнийскую»). Эти подобласти, очевидно, более или менее соответ- ствуют разным климатическим и растительным зонам, и их фауны в ка- кой-то степени различны. Вопрос, следует ли придерживаться такого деления, является не столь важным, но слишком сложным для обсуждения его в данной книге. Мерриэм (iMerriam, etc, 1892) поделил Северную Аме- рику на более сложную систему «зон жизни», основанную на температур- ных факторах. В отдельных случаях применение этих зон возможно, но они нечетко отражают фаунистические подобласти.
11еотроппческая ооласть
К Неотропической области (рис. 51) относятся Южная и Центральная Америка, тропические низменности Мексики с островом Тринидад и, очевидно, .собственно Вест-Индия, но фауна последней ограничена и име- ет некоторые переходные черты и носит, скорее, сложный характер фау- ны периферии, чем части континентальной фауны.
Эта область в основном расположена в тропиках, но ю,кная часть Южной Америки находится в умеренной зоне, достигая широт, эквивалент- ных широтам Лабрадора или Англии, и имеет средние между этими двумя районами температуры. К долине Амазонки приурочены обширные про- странства, занятые дождевыми лесами, отдельные массивы которых, так же как и участки более сухолюбивых лесов, разбросаны повсюду в других частях Южной Америки и в Центральной Америке. В тропиках простираются также обширные площади саванн и степей, причем на юге степи проникают в Аргентину. Во многих районах, особенно в западной части Южной Аме- рики, имеются пустыни и полупустыни. Распределение растительности в Южной Америке очень сложное, а в Центральной Америке — еще более сложное в связи с тем, что различные типы растительности (влажные и сухолюбивые леса, растительность саванн, полупустынь и гор) сосре- доточены на сравнительно небольших площадях. Леса и степи, прости- рающиеся в меридиональном направлении в северной части Южной Аме- рики, защищены Андами, но к югу климат в горах становится более засушливым. Далеко на юге в западной части Южной Америки, в южном Чили и в западной части Огненной Земли произрастают влажные буковые леса умеренной зоны к>жного полушария, аналогичные лесам Новой Зеландии, южной части Австралии и Тасмании.
Пресноводные рыбы Неотропической области (стр. 68 и след.) исключи- тельно многочисленны, но относятся к немногим основным группам. Домини- рующими являются некоторые представители отряда карпообразных: хара- циновые (Афр.), электрические угри (энд.) и сомы (энд. сем.). Имеется так- же двоякодышащая рыба (чешуйчатник) (так'ке Афр. сем.), два предста- вителя костноязычных (прерывистое распространение, троп.), два пред- ставителя семейства Nandidae (Афр., И. М.), ряд не строго пресноводных карпозубых (тпл., широко), хромисы (Афр.), некоторые солевыносливые
рыбы, чукучаны, сомы семейства Ameiuridae, панцирные щуки, проникаю- щие в отдельные районы Центральной Америки (из Н. С. А.).
Амфибии в Неотропической области представлены безногими (Афр., И. Х[.), саламандрами семейства Plethodontidae (большинство в Н. С. А.), лягушками пипами (Афр.), родом Rhinophrpnus в южной части Мексики
попик 0ока
9С
ф~ ~~ф „
ФФВ09 491 9Ф Ф Ф
с~~ Змдатор
T J» ~попик Козгр ~ р.Ла Плат фр +о~
с~~
1 Иагвллано3
дроли Ю P и с. 51. Неотропическая область (непрерывная линия) и соседняя территория (прерывистая линия). Ортографи-
ческая проекция. Пунктирная линия — нриблизительная граница с Нсарктической of) JIéñTbK). НосВН шт~) IxoBha — ocHoBHI I(.' массllHbl JoiHQQBblx
лесов. и т. д. (энд. сем.), многочисленными лягушками семейства Leptodactyli- dae (Австр. и т. д.), родом Bufo (широко), многочисленными представите- лями семейства Atelopodidae (немногочисленны в троп. Ст. Св.), болыиим числом квакш (гол., Австр.), несколькими видами Лапа (широко) и се- мейством Brevicipitidae (подсем. также в И. М. и проникает в Н. (.. А.).
Из рептилий в Неотропической области находятся: один представи- тель семейства Dermatemydidae в северной части Центральной Америки и т. д. (энд. сем.), род Chelydrct в Центральной Америке и т. д. (Н. С. А.), подсемейство Staurotypinae в северной части Центральной Америки и т. д. (энд.), род Einosternon, (Н. С. А.), несколько видов черепах подсемейства Emydinae (родственны Н. С. А. и И. М.), род Testudo (тпл. Ст. Св.),
87-1,
Глава У
черепахи пеломедузы (Афр.), Сhelyidae (Австр.) и боко|иейные черепахи, крокодилы (троп.) ll кайманы (энд.), гекконы (тп.r. ч. з. ш.), многочислен- ные игуановые (Н. С. А. и т. д.), представители семейства Xantusiidae
в H eHTpa JrbHorr AMepHI(e lr г. p. (TBRrrie B южнои части Н. С. А.), несколько
видов сцинков (з. ш.), один представитель семейства Elytropsidae в 1ож- ной части Мексики (энд. сем.), многочисленные представители семейстlr Teiidae (Н. С. А.), Amphisbaenidae (Афр. и т. д.), Anguidae (Н. С. А., 1';вразия и т. д.), род Xenosaurus в южной Мексике и т. д. (энд. сем.), змеи рода Tpphlops (троп. и т. д.), Leptotpphlops (Афр., южн. часть Н. С. А.), один-два вида Aniliidae (Австр.), удавы и т. д. (подсем. проникает в ю'и- нун> часть Н. С. А.), многочисленные ужовые (з. ш.), коралловые аспиды (Н. (;. А., тпл. ч. з. ш.) и ямкоголовые гадюки (С. Н. А., Азия).
Неотропическая фауна птиц чрезвычайно богата и своеобразна. Из 67 семейств птиц, представленных на суше или в пресных водоемах этой области, примерно половина широко распространена и в других местах земного шара. Несколько семейств имеют особые или сомнительные свя- зи и почти половина является исключительно неотропической или по край- Heir мере приурочена к Новому Свету (более подробно см. стр. 227 и далее).
Из млекопитающих в Неотропической области представлены опоссу- мы (почти энд., но один вид проникает в Н. С. А.) и ценолестовые сумча- тые (энд.), несколько землероек в Центральной Америке и т. д. (проника- ют в Н. С. А.), обезьяны (два энд. сем.), муравьеды (энд.), ленивцы (энд., броненосцы (почти энд., один вид проникает в южную часть Н. С. А.), собачьи (широко), один вид медведя (Н. С. А., Евразия), енотовые (Н. С. А.), куньи (широко) и кошки (широко), тапиры (И. М.), пекари (проникают в Н. С. А), верблюды (Пал.) и олени (Н. С. А., Евразия), зайцы (широко), белки (широко), гоферы и карманчиковые мьппи в Центральной Америке и т. д. (и те и другие проникают из Н. С. А.), большое число хомячьих (Н. 0. А., части Ст. Св.), дикобразы Нового Света (Н. С. А.) и еще 10 се- мейств (все энд.) дикобразоподобных грызунов, 9 семейств насекомояд- ных рукокрылых: Lmballonuridae (троп. и т. д.), Noctilionidae (энд.), многочисленные представители Phyllostomatidae (проникают в южные части Н. С. А.), настоящие вампиры (энд.), Natalidae (почти энд.), Furip- teridae (энд.), дискокрылые летучие мыши (энд.), Vespertilionidae (з. ш.) и lolossidae (тпл. ч. з. ш.). Эта фауна млекопитающих приводится в табл. 7 и рассматривается на стр. 28i и далее.
В заключение следует отметить, что пресноводные рыбы Неотропиче- ской области очень многочисленны и высокоэндемичны (большинство семейств является исключительно неотропическими), но имеют общие африканские связи. Фауны амфибий и рептилий смешанные: некоторые ~руппы являются главным образом тропическими и субтропическими (например, безногие, Testudo, крокодилы, ТурИорг), некоторые имеют африканские связи (например, лягушки пипы, черепахи пеломедузы, яще- рицы Amphisbaenidae, змеи рода Leptobtyphlops), некоторые имеют связ t с Индо-Малайской областью (подсемейство лягушек семейства Brevicipi- tidae, род черепах подсемейства Emydinae и Aniliidae, отдельные группы змей подсемейства Colubridae); некоторые — с Австралией (лягушки семейства Leptodactylidae и хилы, черепахи 0helyidae) и ряд групп имеет североамериканские связи (саламандры семейства Plethodontidae, игуа- новые, ящерицы семейства Teiidae и многие роды других семейств). Ави- фауна этой области чрезвычайно богата, но очень многие ее представители относятся к исключительно неотропическим семействам, хотя некоторые другие имеют широкое распространение. Млекопитающие Неотропиче-
Особенности распространения животных нп лап~ерикax
ской области представляют смесь крайне своеобразных эндемичных нео- тропических семейств и семейств, общих с другими частями земного шара, обычно включая и Северную Америку.
Короче говоря (обратимся на миг к прошлому), вся современная фау- на позвоночных Неотропической области представляет собой смесь сохра- нившихся частей древней эндемичной фауны третичного времени, гео- графические связи которой теперь стали разнообразными (по-видимому, они определяются, скорее, вымиранием и выживанием животных, а не направлением их расселения), и новой фауны, проникшей через Северную Америку. Древняя фауна включает всех строго пресноводных рыб (кроме нескольких недавних вселенцев в северную часть Центральной Америки), возможно, большинство амфибий и большинство рептилий Южной Аме- рики, но относительно немногих рептилий Центральной Америки, очень многих птиц (но отличия между древними и новыми группами птиц выра- жены нечетко) и примерно половину млекопитающих.
Северная граница Неотропической области является южной грани- цей Неарктики.
Основной характер распространения позвоночных в пределах Неотро- пической области следующий: чрезвычайное богатство фауны в основной тропической части Юн,ной Америки, уменьшение и смешение с дополни- тельными североамериканскими элементами и некоторая дифференциа- ция фауны дождевых лесов, степей — памп, пустынь и гор. Уоллес в Нео- тропической области выделял три подобласти: «Бразильскую» (основная южноамериканская), «Мексиканскую» (центральноамериканская) и «Чи- лийскую» (южноамериканская), которые в данной работе официально не признаются.
Австралийская область
К Австралийской области (рис. 52) относятся Австралия, Новая Гви- нея, Тасманпя и некоторые более мелкие острова, имеющие (обедненную) материковую фауну.
Австралийская область расположена частично в тропиках и частично .и умеренной зоне. Новая Гвинея целиком находится в пределах тропиков. Значительная часть острова покрыта дождевыми лесами, но есть также растительность и открытых пространств, и высоких гор, а на южном побережье — участки таких же, как и в Австралии, открытых эвкалиптовых лесов. В северной тропической части Австралии также произрастает раз- нообразная растительность, которая в северном Квинсленде представлена рядом изолированных участков пышных дождевых лесов. Далее, к югу, восточная и юго-восточная части Австралии, находящиеся в умеренной зоне, довольно хорошо увлажнены и покрыты занимающими значительную площадь открытыми эвкалиптовыми лесами и более влажными густыми лесами в горах. Большая часть внутренней Австралии — засушливая, а юго-восточная часть получает достаточное количество влаги, но с во- стока она отрезается пустынями. На юго-западе материка произрастают прекрасные леса, состоящие из колоссальных деревьев. Климат Тасмании умеренно-прохладный, по широте эквивалентный климату Нью-Йорка или Бостона, но зима в этих местах не такая холодная и отчасти более влажная. Остров покрыт лесами.
Пресноводные рыбы в Австралии представлены двоякодышащей рыбой (энд. сем.), одним представителем костноязычных (И. М. и т. д.) (pac-
1'лава. 7
пространение этих обоих видов очень ограничено) и различным»~ со.»евы- носливыми рыбами.
Амфибии Австралийской ооласти представлены только лягушками семейства Leptodactylidae (Ю. А. и т. д.), Hylidae (Ю. А. и т. д.), несколь- кими видами семейства Ranidae (широко) и Brevicipitidae (тпл. ч. ». ш., но подсем. почти энд.).
К рептилиям Австралии относятся черепахи Сигейосlге/уя (энд. сем.), '1'rionychidae (виды также в И. М.) в Новой Гвинее и черепахи семейства
о. gauaeo о. паааиаа о. Саладати
/
~о. Mucus+
~ ° I ° ра э
О g la К
:;,ЗОИ 90ЛЯЕС~
д~,'~~ р е==ъ
Ф 0=0
Типур "
..Иптор
b
+o~
° ~ ». ~~,
гу~
ф~ " б~
4.'.. ~
c+
ф
о. Н05ая
~, КслсЬ:..
1~~,
дТ Опик Козерога
l~
1
/ ~g
/
Я /
/ Ф
о. НоД~я Земля
o. Tuc~geug
р и с. 52. Австралийская оОласть (непрерывная линия) и прилежащая территория (прерывистая линия). Орто- графическая проекция.
Пунктирные линии — приблизительные границы Австралийской и Индо-Малайской областей. Косая штриховка — основны» мас- сивы дождевых лесов.
Сhelyidae (Ю. А.), крокодилы (троп.), многочисленные гекконы (тил ч. з. ш.), агамовые (тпл. Ст. Св.), многочисленные сцинки (з. ш.), на Новой Гвинее род Dibamus (И. М.), Varanus (тпл. Ст. Св.) и семейство Pygopodi- dae (энд.), род Typhlops (троп. и т. д.), возможно, один представитель семейства Aniliidae на Новой Гвинее (И. М. и т. д.), питоны (тпл. Ст. Св.) и большое число змей семейства Elapidae (тпл. ч. з. ш.).
— Наземные птицы и птицы пресных водоемов Австралийской о~>ласти охватывают представителей примерно 58 семейств. Из них 44 семейства более или менее широко распространены в других местах земного шара или в тропиках, или все они (или значительная часть их) встречаются в Старом Свете. Два небольших семейства (совиные козодои и артамусы) являются общими для Австралийской и Индо-Малайской областей. Остав- шиеся 12 семейств принадлежат исключительно Австралии или встречают- ся главным образом там: казуары (6 видов), эму (2), сорные куры (до пери-
Особенности распроспгринения животных па латериках
377
ферии И. М.) (10), совиные козодои (также на некоторых островах ) (8), лирохвосты (2), птицы семейства Atrichornitidae (2), нектарницы (прони- кают в И. М.), медососы (распространены также на многих островах и сом- нительно в Ю. Афр.) (159), Cracticidae (11), семейство Grallinidae (~), шалашниковые птицы (17), райские птицы (43 вида). В общей сложности эти семейства насчитывают 318 видов, включая несколько зндемичных австралийских семейств. К австралийской области приурочена значитель- ная часть голубей и попугаев, включая важные эндемичные австралийские группы, и многочисленные «австралийские славки» (51alurinae) (также По- линезия и т. д.) (85 видов).
К местным наземным млекопитающим Австралийской области отно- сятся несколько однопроходных (энд. подкласс), многочисленные сумча- тые (6 энд. сем.), довольно разнообразные грызуны семейства Мигк1ае (Ст. Св.), плодоядные летучие мыши (тпл. Ст. Св.) и 6 семейств насекоио- ядных рукокрылых (все встречаются и в других местах з. ш.). Фауна мле- копитающих более подробно рассматривалась на стр. 284 и далее.
В заключение отметим, что фауна позвоночных Австралийской обла- сти состоит только из одного-двух видов строго пресноводных групп рыо, один из которых является поразительным реликтом, и из солевыносливых рыб, 4 семейств лягушек, из которых ни одно не является ни примитив- ным, ни эндемичным, хотя два подсемейства из семейства Brevicipitidae почти эндемичные; рептилии представлены не столь разнообразно, без при- митивных семейств, но с несколькими эндемичными семействами; фауна птиц довольно разнообразна, и представители ее в достаточном количестве входят в состав эндемичных семейств; фауна млекопитающих Австралии очень ограниченная, с поразительно примитивными и эндемичными эле- ментами.
Представив опять на мгновение далекое прошлое, можно сказать, что австралийская фауна позвоночных состоит из нескольких очень древних эндемичных групп, которые сохранились или радиировали в Австралии в течение длительного времени, и накопившихся здесь групп солевыносли- вых (но, возмо;кно, не строго пресноводных) рыб и лягушек, рептилий и птиц, грызунов и рукокрылых. Двоякодышащая рыба, однопроходные и специфические надсемейства сумчатых являются строго эндемичными. Среди австралийских амфибий, рептилий и птиц нет реликтовых или эндемичных групп, аналогичных выше перечисленным. Если бы некоторые из этих животных находились в Австралии, как и сумчатые, в течение столь же продолжительного времени, то их радиация не достигла бы такой же степени. Современные животные, родственные некоторым более древним австралийским позвоночным (некоторым лягушкам, черепахам и сумча- тым), находятся главным образом в (тропиках) Южной Америке. В боль- шинстве случаев это, очевидно, результат отступания или вымирания животных на основной части земного шара. У строго пресноводных рыб, наземных рептилий и птиц, по-видимому, особых связей между Австралией II Южной Америкой не наблюдается. Австралийская фауна позвоночных в целом почти не имеет непосредственных связей и с африканской фауной. Многие из более современных позвоночных Австралии (лягушки семейст- ва Ranidae, многие рептилии, птицы, грызуны и рукокрылые) до сих пор имеют родственные связи с соответствующими представителями тропиче- ской Азии.
Северо-западная граница Австралийской области рассматривается ниже как «зона Уоллеса». Входят ли в состав Австралийской области ост- рова, расположенные на восток от Австралии и Новой Гвинеи, или нет,
1'лава 7
толкуется различно. В данной книге эти острова рассматриваются отдель- но (гл. 8) как периферия.
В пределах Австралийской области между Австралией и Новой Гви- неей имеются некоторые фаунистические различия. Из ННх отметим сле- дующие: только в Австралии имеется двоякодьппащая рыба; в Австралии богаче представлено семейство Leptadoctylidae, в Новой Гвинее— семейство Brecivipitidae, в Австралии распространено большое число яще- риц семейства Pygopodidae, в Новой Гвинее — змей семейства Colubri- dae, в Австралии обитают эму, в Новой Гвинее — преимущественно казуары, в Австралии водятся лирохвосты и мелкие кустарниковые птицы семейства Atrichornitidae, в Новой Гвинее — преимущественно райские птицы, к Австралии приурочены утконосы и имеется значительно боль- mee число (более специализированных) сумчатых, а в Новой Гвинее— относительно большее число грызунов. Но эти детали не столь существен- ны. В целом фауны позвоночных Австралии и Новой Гвинеи очень сходны (ср. млекопитающих, стр. 284, и птиц, стр. 227). Они имеют много общих групп, даже много общих родов и видов, и между ними отмечается обшир- ный переход. Различные австралийские животные проникают в районы открытых эвкалиптовых лесов в южной части Новой Гвинеи, а новогви- нейские животные проникают в районы дождевых лесов северо-восточной Австралии. В пределах самой Австралии наблюдается дальнейшая диффе- ренциация фауны с севера на юг, а также дифференциация ее между более влажной восточной частью, обширным центральным и засушливым запад- ным районом и умеренно влажной частью континента. Однако, так же как и в других случаях, нет необходимости делить эту область на подоб- ласти.
Переходы между региональными фаунами
Там, где региональные фауны встречаются, или там, где между ними имеется лишь незначительная преграда, происходит переход этих фаун: элементы одной фауны перекрываются элементами другой с постепенным уменьшением числа видов и групп в обоих направлениях.
Переход фаун — явление очень сложное (рис. 53). Две смежные региональные фауны обычно имеют некоторые семейства, роды и виды, которые являются общими или из которых одни в основном встречаются в одной области, но проникают в отдельные районы другой (это строго переходные элементы), а другие встречаются только в одной из двух. У се- мейств, общих для обеих областей, могут быть переходные роды, а в об- щих родах — переходные виды. Семейство, которое совершает переход в одном направлении, может иметь роды или виды, переходящие в другом направлении. Разные переходные группы могут распространяться на раз- личное расстояние. Как правило, положение усложняется наличием сом- нительных случаев.
Для детального анализа переходов необходимо тщательное изучение и статистика. Самое большее, что можно здесь сделать,— это описать пере- ходы региональных фаун в общих чертах и показать, какие части основ- ных рассматриваемых фаун — большие или меньшие — они охватывают, относятся ли перекрывающие элементы различных фаун к разным семей- ствам или только к разным родам или видам, охватывает ли перекрывание широкие или узкие зоны и в каком направлении из двух наблюдается наибольшее перекрывание. Эти переходы будут описаны, исходя только
из современного распространения животных, без учета направлений рас- селения в прошлом (хотя не исключено, что в некоторых случаях будут даны ссылки на прошлое).
Тропические фауны позвоночных Эфиопской и Индо-Малайской обла- стей имеют много общих семейств, родов и даже видов, а также много и таких, которые встречаются в одной области, но отсутствуют в другой; число строго переходных семейств, родов и видов очень незначительно.
ФАУНА Б
ФАУНА А
3~0емочные сепеаспйа
05ща ceneucmda
Р и с. 53. Схема перехода двух прилежащих регнональных
фаун (упрощенная).
Еаждая фауна состоит из эндемичных, переходных и общих се- мейств. В переходные и общие семейства входят эндемичные, пере- ходные и общие роды. Эндемичные семейства и роды определяют границу (прерывистая линия) между областями. Фауна А больше, чем фауна Б, и определяет направление перехода, то есть перехо.-~
в основном п~~оисходит по направлению ~< фауне Б.
Несколько африканских семейств, родов и видов проникают (в ряде слу- чаев дизъюнктивно) через Юго-Западную Азию до ближайшей части Индо- Малайской области (например, хамелеоны, некоторые птицы, гиена и лев). Несколько меньшее число групп Индо-Малайской области распростране- но до ближайшей части Африки (например, рыбы семейства Cobitidae). Кроме того, дополнительный, но очень ограниченный переход происходит и Юго-Западной Азии (описан отдельно для рыб, стр. 65, и птиц, стр. 226). Этот переход охватывает очень небольшие части основных фаун Эфиопской и Индо-Малайской областей и происходит как на уровне родов и видов. так и на уровне семейств. Зона перехода довольно широкая, но располо- жена в основном в засушливой местности и занимает площадь значительно большую по сравнению с площадью, образованной пределами границ основных фаун. Направление перехода неясное: или африканские позвоноч- ные в большей степени проникали в Индо-Малайскую область, или, наобо- рот, позвоночные Индо-Малайской области — в Африку. На Мадагаска- ре, который является своего рода зоной перехода (гл. 8), также наблюдает- ся смешение многих африканских позвоночных и (значительно меньше) позвоночных Индо-Малайской области, но проникшие на Мадагаскар
880
африканские группы очень редко проникают далее в Индо-Малай- скую область (исключение представляют солевыносливые хромисы),. а группы Индо-Малайской области, которые достигают Мадагаскара, ред- ко проникают в Африку. Поэтому мадагаскарская фауна состоит из жи- вотных, проникших на Мадагаскар по двум направлениям (причем в боль- шинстве случаев из Африки), а не является результатом обмена между Африкой и Индией через Мадагаскар.
Теперь обратимся к прошлому. Тот факт, что фауны Эфиопской и Индо-Малайской областей имеют много общего, несмотря на очень неболь- шое количество имеющихся переходов, позволяет предположить, что в прошлом происходил значительно больший обмен позвоночными, чем в сравнительно недавнее время. Подтверждением того, что обмен происхо- дил действительно через Юго-Западную Азию, служит наличие ограничен- ного перехода фаун Африки и Индо-Малайской области вдоль этого пути в настоящее время, в то время как через Мадагаскар этот обмен, по-види- мому, не осуществлялся вообще.
Тропическая эфиопская и умеренная палеарктическая фауны наряду с большим числом общих групп позвоночных имеют немало и таких групп, которые не являются общими и присущи главным образом Эфиопской области. Однако между этими областями переход ограничен, хотя он проис- ходит и в Сахаре, и в районе Средиземноморья. Он осуществляется глав- ным образом за счет распространения эфиопских (тропических) семейств, родов и видов по южной периферии Палеарктики: например, рыбы семей- ства Cyprinodontidae, черепахи рода Testudo, гекконы, хамелеоны, ящери- цы семейства Amphisbaenidae и т. д. Из птиц различные эфиопские (тро- пические) группы проникают на различное расстояние в южную часть Европы, где они могут быть как оседлыми, так и перелетными. Из млеко- питающих — слоновые землеройки и макаки (в палеарктической Север- ной Африке), дикобраз, гиена, виверровые, даман и ряд летучих мышей. Проникновение палеарктических групп на периферию Эфиопской области происходит реже: например, один представитель подсемейства Етуй1пае, возмо~кно, заходит в западную часть тропической Африки, род llIicrotus (полевки) проникает на незначительную часть непалеарктической или пере- ходной Северной Африки. Переход между фаунами позвоночных Эфиоп- ской и Палеарктической областей вовлекает сравнительно небольшие ча- сти основных фаун. Он происходит как на уровне родов и видов, так и на уровне семейств. Зона перехода этих фаун небольшая, и значительная часть ее находится в засушливых районах между границами основных фаун. Направление перехода главным образом северное: значительно боль- шее число эфиопских (тропических) позвоночных проникает на периферию Палеарктической области, чем в обратном направлении.
Между тропической фауной Индо-Малайской области и умеренной фау- ной Палеарктики в Восточной Азии наблюдается обширный переход. охва- тывающий все классы позвоночных. Ряд семейств, родов и видов пресно- водных рыб Индо-Малайской области проникает в умеренную зону Китая, а отдельные экземпляры достигают Амура. Несколько семейств и родов лягушек Индо-Малайской области проникает в умеренную зону Китая. Род Kaloula достигает Маньчжурии, Rhacophorus — северного Хонсн>, Япония. В обратном направлении несколько саламандр Палеарктики и род Hyla заходят к югу на периферию Индо-Малайской области (на срав- нительно небольшое расстояние). Многие группы рептилий Индо-Малайской области простираются на различные расстояния в умеренную зону Азии. То a*e самое наблюдается у многих, как у оседлых, так и перелетных,
Особенностями распространекия животных нп екатеринах
атиц Индо-Малайской области. Многие семейства и более мелкие группы млекопитающих Индо-Малайской области проникают на разные расстоя- ния в умеренную зону Азии, и только некоторые палеарктические предста- вители заходят в тропики этой области: например, серао (представляющий палеарктическую трибу семейства Bovidae) распространен до Суматры, крот (представляющий голарктическое семейство) — до полуострова Малакка, волк (голарктический вид) — до тропической части Индии. Пере- ход осуществляется на уровне родов, видов, а также семейств и охваты- вает широкую зону в благоприятных для этого местах. Переход происхо- дит главным образом в северном направлении, и значительно большее чис- .ло позвоночных Индо-Малайской области проникает в Палеарктику, чем наоборот.
Переходы фаун позвоночных обеих областей — Эфиопской и Индо- Малайской, с фауной Палеарктики образуются преимущественно за счет распространения тропических групп к северу, что отражает общее обед- нение фауны позвоночных к северу от тропиков, а также и тот факт, что .палеарктическая фауна во многих отношениях не намного богаче обед- ненной тропической фауны периферии.
Переход между умеренной фауной Палеарктики и умеренной фау- ной Неарктики — обширный и происходит через весь север, среди всех классов позвоночных. Северные части обеих фаун во многом сходны и да- .же имеют множество общих видов. Но к югу общая северная переходная подфауна постепенно становится все более характерной палеарктической и неарктической фауной. Этот переход захватывает значительные части обеих фаун. В основном он происходит на уровне родов и видов, а не се- мейств. Переходом охватывается широкая зона, большая часть которой располо;кена в весьма неблагоприятном холодном районе. Направление :перехода неопределенное: исходя из современного распространения, труд- но определить, проникает ли болыпе палеарктических групп в Неаркти- ку или наоборот.
Вообще палеарктическая и неарктическая фауны сами по себе во мно- гих отношениях являются переходными между фаунами тропиков Старого и Нового Света.
Рассмотренные выше переходы находятся в пределах Мегагеи, основ- ной части земного шара. Переходы, происходящие между фаунами Мега- геи п фаунами областей, ограниченных преградами, особенно примеча- тельны тем, что эти фауны имеют болыпие различия и заслуживают более детального разбора.
Зона перехода: Центральная Америка — Мексика
Фауны позвоночных Неарктической и Неотропической областей наряду с тем, что они имеют большое число общих семейств, родов и не- которых видов, во многом сильно отличаются друг от друга. Между эти- ми фаУнами происходит переход, в основном в Мексике и Централь- ной Америке (рис. 54), в какой-то степенн охватывающий еще оолее обтпир- ную зону.
Из пресноводных рыб некоторые неарктические семейства строго прес- :новодных рыб не достигают даже реки Рио-Гранде; американские окуни и окуневые (семейства РегсЫае) проникают до тропика Рака в Мексике; карповые доходят до реки Бальсас в южной ~Ъ[ексике; чукучаны и сомы
достигают Гватемалы H3 HeoTpozn feet;px семейст„
рыб первого отдела одни не достигают даже Центральной Америки, дру- гие не проникают за пределы Панамы. Самой северной границей угрей семейства Gymnotidae служит 1'ватемала. Пресноводные сомы 10жной Америки достигают реки Пануко на восточном побере;кье Мексики, являющейся их северным пределом, Рио-1'ранде является северным преде- JIoM харациновых. Переход охватывает только очень незначительные части неарктической и неотропической фаун строго пресноводных рыб, ко <р- рые представлены разными семействами, а смешение одних элементов фау- ны с другими происходит только в пределах территории от Рио-Гранде Р с. 5'~. Зона перехода Мексика — Центральная Америка. Ж ирна я I lóí êòèð Hà ÿ линия — приблизительная граница м ен',-1 у
Неарктической и Неотропичсской областям и. до Гватемалы (см. рис. 15). Семейства не строго пресноводных рыб (второй отдел) перекрывают друг друга на более обширной площади: неарктиче- ские панцирные щуки простираются к югу до озера Никарагуа, неотропи- ческие хромисы — и северу до Рио-Гранде, а карпозубые проникают настол ько далеко в Северную Америку, что их мотка считать скорее общим семейством, чем переходным.
Переходы амфибий захватывают значительные части обеих фаун и за- нимают более обширную зону, но в основном они происходят на уровне семейства. Некоторые неарктические семейства (особенно отдельные семей- ства саламандр) проникают на юг только до Техаса или до северных ча- стей Мексики. Саламандры семейства Ambystomidae и лягушки семейства Pelobatidae пересекают тропик Рака на Мексиканском нагорье, но не до- стигают низменностей Мексики, а саламандры семейства Plethodont.idae и лягушки рода Лапа распространены через Центральную Америку в се- верные части Южной Америки. Жабы рода Bufo имеют настолько широкое распространение как в Южной, так и в Северной Америке, что, исходя из современного распространения, их скорее можно считать общими, а не переходными. Из неотропических семейств пипы не достигают Центральной Америки; лягушки семейства Atelopodidae распространены по крайней
мере до Никарагуа (и Кубы); безногие простираются до южной Мексики; лягушки семейства Leptodactylidae — до Техаса и южной Аризоны, а ля- гушки семейства Brevicipitidae — до южной Айовы, Индианы и Мэрилен- да. Лягушки хилы и род Hpla настолько широко распространены в Север- ной Америке, что их, вероятно, лучше считать общими. Такой характер перехода в фауне амфибий на уровне семейств осложняется переходными родами некоторых семейств. локализованными родами и да;ке локализо- ванным семейством лягушек (Bhinophrynidae) в районах ]~ентральной Лмерики и Мексики.
Среди рептилий переход осуществляется еще сложнее и занимает ши- рокую зону. Для этого перехода характерно, что тенденция h участию в нем проявляется в большей степени у родов, а так'ке и у семейств. Из че- репах неарктические каймановые черепахи и черепахи рода Kinosternr>n через Центральную Америку проникают в отдельные районы Южной Аме- рики, но южноамериканские семейства черепах (Pelomedusidae, Chelyi- dae) не достигают даже Центральной Америки. По современному распро- странению семейство Testunidae (включая подсемейство Emydinae) является общим для северных и южных частей, в которых представлены переходные роды: с севера Terrapene проникает на юг до южной Мексики, а Pseudemps — через Центральную и значительную часть Южной Америки (вероятно, этот род следует рассматривать скорее как общий, а не как переходный); с юга род Testudo простирается на север до llaHBMhI, а Geoe- uyHa — через Центральную Америку до некоторых районов Мексики. Из крокодилов неарктический аллигатор в южном направлении не дохо- дит даже до Рио-Гранде, но неотропические кайманы и один вид крокоди- лов проникают на север в отдельные районы Мексики и (только вид кро- кодилов) до южной Флориды. В Америке основные семейства ящериц, в большинстве случаев тропические, простираются на различное расстоя- ние к северу, но характер распространения родов очень сложный. Одни роды, в основном неарктические, проникают в Центральную Америку и т. д. как северные переходные элементы, а другие, наоборот, распростра- нены в северном направлении из Южной Америки. Из змей в Л!ексике и Центральной Америке к северным переходным элементам относятся в ог- новном роды ужовых и ямкоголовых гадюк. Из неотропических змей род ТурЫорз достигает южной Мексики, Leptotyphlrips — юго-западной части Соединенных Штатов. Многие южноамериканские роды ужовых проникакп на различные расстояния в Центральную Америку и южную часть Северной Америки. Дополнительные сведения о сложном переходе фаун рептилий в Центральной Америке и Мексике приводятся в гл. 4. Неожиданно боль- шое число рептилий Северной Америки проникает в Центральную, но не достигает Южной Америки. Этот переход усложняется наличием ряда локализованных, эндемичных или реликтовых родов и даже семейств реп- тилий в Мексике и Центральной Америке (см. гл. 4).
У птиц переход неарктической и неотропической фаун очень обшир- ный и сложный, в котором участвует много родов и видов, а также семейств, и он охватывает не только Центральную Америку и Мексику, но и значи- тельную часть Северной и Южной Америки. Некоторые, но сравнительно немногие, неарктические группы проникают в Неотропическую область в период гнездования: например, индейки — до Гватемалы, корольки. пищухи, синицы, дубоносы рода FIesperiphona, клесты и вороны — до районов Центральной, но не Южной Америки, рогатые жаворонки — до северной части Южной Америки II т. д., но самыми заметными неарктиче- скими переходными элементами являются многие роды и виды, которые
Глава 7
(независимо от их географического происхождения) в настоящее время гнездятся в Неарктике, а зимуют по периферии Неотропической области. Многие, но далеко не все семейства богатой авифауны Ю,кной Америки распространены через Центральную Америку к северу. Большое число таких важных групп, как, например, тинаму, ленивки, якамары, туканы. надсемейство Furnarioidea, манакины, пищухи-медососы и многие мелкие семейства, а также многие роды и виды достигают южной части Мексики. которая является их северной границей. Многие другие неотропические группы проникают на различные расстояния в умеренную зону Северной Америки: например, момоты достигают северной части Мексики; крак- сы, яканы, попугаи, трогоны и котинги — южного Техаса и (или) Ари- зоны' ибисы и арами — юго-восточной части Соединенных Штатов; грифы Нового Света проникают еще дальше на север; колибри — еще север- нее. Н связи с этим возникает вопрос, какие группы, если исходить из с<нз- ременного распространения, надо считать переходными, а какие — общи- мп. Эти особенности перехода семейств в значительной степени усили- ваются и усложняются некоторыми переходными родами и видами.
У млекопитающих также отмечается обширный и сложный переход современных неарктической и неотропической фаун, который образуется в результате перекрывания ареалов семейств, родов и видов. В основном оН охватывает территорию южной части Мексики и Центральной Амери- ки. но отдельные неарктические элементы проникают на разные расстояния в Ю,кную Америку, а некоторые неотропические (меньшее число) — в от- дельные районы умеренной зоны Северной Америки. Подробности этого перехода фаун млекопитающих приведены в гл. 6.
Приведенный выше обзор является не более чем весьма поверхност- ной схемой перехода фаун позвоночных Неарктической и Неотропической областей. Этот переход никогда не рассматривался зоогеографами подроб- но. и ему никогда не уделялось такого внимания, какое было оказано JII[HIIH Уоллеса или «Зоне Уоллеса», хотя, по-видимому, он имеет не мене~ важное значение. Не имея возможности рассмотреть этот переход более детально, я все же приведу в заключение несколько общих положений. которые, конечно, не являются окончательными выводами и подлежат даль- нешпему обсуждению.
Переход неарктической и неотропической фаун происходит в Мекси- ке и Центральной Америке и охватывает все классы позвоночных, но у раз- ных классов он осуществляется по-разному. Самый простой переход. включающий меньше всего элементов, происходит на высшем уровне (в ос- новном на уровне разных семейств) и охватывает самые узкие зоны у тех классов, которые расселяются медленнее всех. У строго пресноводных рыо этот переход осуществляется несколькими элементами; все неарктические и неотропические компоненты относятся к разным семействам, и зона перекрытия простирается только от Рио-Гранде до Гватемалы. В противо- положность строго пресноводным рыбам переход неаркп~ческих и неотро- пических птиц образуется за счет большого числа элементов (за исключе- нием некоторых семейств, в основном только роды и виды) — зона пере- крывания охватывает значительную часть Северной и Южной Америки. Сложность, уровень и ширина перехода у других классов позвоночных находятся между этими двумя крайностями. Как и следовало о, кидать, все это показывает в целом, что степень выраженности перехода пропор- циональна способностям расселения различных животных.
Таким образом, исходя из современного распространения, переход фаун позвоночных умеренной зоны Северной Америки и тропической зо-
0(обеыыости распространения живопгы~)ьх на материках
ны хОжной Лиерики можно рассматривать как сложное перекрывание ареа- лов, оои~ей зоне. Теперь, исходя из современного распространения, попы- таемся резюмировать направление перехода через гр«нхщу между фауна- ми Неарктцческой и Неотропической об.ластей.
Фауна Центральной Америки в большей степени неотропическая, чем неарктическая, что можно сказать о всех классах позвоночных, за исклхоченххеы рептилий. Это, очевидно, можно объяснить тем, что Цент- ра хьная Америка расположена в тропической части земного шара. Многие ххз ххеотропххчесх'ххх групп простираются до тропических низмеххностей 1[ексххки, но не дальше. Главным образом это те группы позвоночных, распространенце которых определяют общепринятые границы межлу Неотропххческой и Неарктической областями в южной части Мексики, прхх- мсрно на линии между тропиками и умереннохх зоной северного полуххха- рия. С другой стороны, фауна Мексиканского нагорья — в основном неарктххчесх;ля, и отдельные неарктические группы достигают ю;кных пре- делов в южной части Мексики вблххзи северной границы неотропи хеских групп. Теперь возникает «опрос, проникают ли переходные группы через границу в обоих направлениях на одно и то же расстояние или же в одном хх,хправленхххх они заходят значительно дальше. чем в другом.
Небольшое число строго пресноводных рыб только пересекает грани- цу II в том. 11 в другом направлении, но ни одна из них не заходит дальхце другой. Несколько групп амфибии тах1же проникает через границу и в том. и в другом направлении примерно на равное расстояние. Из рептилий многххе пересекают границу как в северном, так и в южном направлении, но число неарктических групп, проникающих в Центральную (но не в 10;х'ную) Америку, оказывается неожиданно большим. Группы непере- летных птиц проникают в большем числе на север, чем на юг, но эта раз- ница отчасти восполняется за счет мигрантов, которые гнездятся на севе- ре, а зимуют на юге в краевых частях тропиков.
Однако среди млекопитающих наблюдается обратное явление: боль- шое число неарктических групп проникает через границу в Неотропцче- скую область и очень незначительное число неотропических млекопитаю- щххх. за исключением р~ кокрылых, проникает далеко на север.
Различия в направлении перехода через границу неарктической и нео- тропической фаун у разных классов позвоночных дают основание судить о различиях в относительном доминировании. Пресноводные рыбы и ам- фиохххх распространяются через региональные границы примерно одинако- во как на север, так и на юг, что позволяет предположить, что степень до- минирования этих животных в неарктической и неотропической фаунах црххмерно одинаковая. Все млекопитающие (кроме рукокрылых) в большей степени распространены к югу, чем к северу. Очевидно, неарктическая фауна млекопитающих является доминирующей в связи с тем, что многие «неархтическххе» млекопитающие в действительности являются домини- рующими группами Мегагеи, в то время как части южноамериканской фау- ньх на протяжении всего третичного периода были изолированы. Репти- лхххх также больше распространены к югу, чем к северу (по крайней мере до Центральной Америки), что опять-таки происходит, вероятно, отчасти из-за того, что некоторые неарктические рептилии представляют в дейст- вительности доминирующие группы Мегагеи.
Среди птиц наблюдается большее проникновение на север, чем на хог. Неотроппческая фауна птиц кажется относительно доминирующей. вххдххио, потому, что это очень большая фауна и что она была в меньхххехх степени изолирована (благодаря способностям птиц летать). чем остальная
886
Гл.ава 7
фауна позвоночных Южной Америки. Таким образом, в неарктической и неотропической фаунах степень доминирования рыб и амфибий пример- но одинаковая. В фауне Неарктики доминируют некоторые рептилии, а также и млекопитающие, а в неотропической фауне — птицы. Конечно, необходимо учитывать, что это предварительные и крайне упрощенные ВЫВОДЫ.
Линия Уоллеса, линия Вебера и «Зона Уоллеса»
Фауны Индо-Малайской и Австралийской областей чрезвычайно отли- чаются друг от друга. Долгое время внимание зоогеографов было при- ковано к Индо-Австралийскому архипелагу (рпс. 55), где встречаются
Р и с. 55. Острова «Зоны Уоллеса» (непрерывная линия) пприлежащие и ним острова (прерывистая л11ния).Штриховая линия — линия Уоллеса, линия Вебера (линия основногофаунистического баланса) и гоан~т~~ы Австралийской области.
эти две фауны, и «линия Уоллеса», то есть предполагаемая граница между фаунами Индо-Малайской и Австралийской областей, была в центре внимания многих поколений зоогеографов после Уоллеса.
Эта линия, впервые проведенная Уоллесом (Wallace, 1860), прохо- дила между островами Бали и Ломбок, Калимантан и Сулавеси, между. ~1>илиппинскими островами, островами Сангихе и Талауд (и Молуккски- ми). Особенно подчеркивалась разница между фаунами островов Бали и Ломоок, которые отделяются друг от друга глубоким, но нешироким, все- го лишь около 15 миль в ширину, проливом. Позднее зоогеографы оспари- вали положение этой линии, особенно ее северной части. Так, Гексли, который назвал эту линию именем Уоллеса, считал, что она должна разде- лять Филиппины, оставляя остров Палаван в Индо-Малайской области, а остальные Филиппинские острова — в Австралийской. Еще позднее
Особенности распространения животных на .чатер иках
88Т
ставилось под сомнение даже наличие какои-нибудь определеннои грани- цы между фаунами Индо-Малайской и Австралийской областей. Майр (Чауг, 1944) излагает в общих чертах историю этих точек зрения. В дан- ной работе трудно уделить этому вопросу то внимание, которого он за- служивает, но я постараюсь показать, каким образом основные классы позвоночных распределяются между Индо-Малайской и Австралийской областями и какие альтернативы предполагаются их распространением в отношении региональных границ или зон перехода. Филиппинские осг- рова в данном случае рассматриваться не будут в связи с тем, что они рас- положены по одну сторону основной линии перехода и их лучше разби- рать как периферийный район (стр. 420 — 425), хотя фауна их до неко- торой степени является переходной.
В Индо-Малайской области богатая фауна строго пресноводных рыб достигает островов Суматра, Ява и Калимантан, которые являются пределом в распространении большинства этих рыб. Из многих строго прес- новодных групп, обитающих на Калимантане, острова Сулавеси дости- гают только два-три вида, занесенные человеком. На острове Ява отме- чается увеличение фауны рыб Индо-Малайской области с запада на восток. Строго пресноводные рыбы, кроме нескольких видов мелких карпо- вых, не достигают Малых Зондских островов, если не говорить о несколь- ких видах, опять-таки, видимо, занесенных человеком. На острове Бали сколько-нибудь значительные представители фауны рыб острова Ява (стр. 52) отсутствуют. Этот факт очень неохотно принимается ихтиолога- ми. Единственные строго пресноводные рыбы на острове Бали — это два- три местных вида мелких карпов родов Puntius и Rasbora. Оба рода встре- чаются также на острове Ломбок, а Rasbora проникает до острова Сумба- ва. Карпозубые (не строго пресноводный отдел) и связанные с ними фор- мы достигают острова Сулавеси, и, вероятно, солевыносливый род карпо- зубых Aplocheilus проникает также до островов Ломбок и Тимор. На другом конце архипелага, в Австралийской области, почти полностью отсут- ствуют строго пресноводные рыбы, а находящиеся там один или два их представителя (двоякодышащая рыба и костноязычные) локализованы. Однако несомненно, что некоторые мелкие австралийские рыбы подсе- мейства Melanotaeniinae с точки зрения зоогеографии имеют важное зна- чение. Они произошли от морских или солевыносливых предков, но те- перь приурочены к пресным водам. Эти рыбы встречаются от более теплой части Австралии до Новой Гвинеи, островов Ару и Вайгео (Beaufort, 1926, р. 103), а также, очевидно, они приурочены к границам Сахульско- го шельфа. Однако и их распространение, и солевыносливость еще недо- статочно изучены.
Из земноводных безногие к востоку от островов Ява и Калимантан (не считая Филиппинских) не известны. Из лягушек Индо-Малайской об- ласти род Megophrys (семейство Pelobatidae) и Microhyla (семейство Bre- vicipitidae) распространены до острова Бали, Bufo — до Сулавеси, Бали и Ломбока, Ealoula (семейство Brevicipitidae) — до острова Сулавеси и вдоль Малых Зондских островов до острова Флорес, Rhacophorus — до (:улавеси и острова Тимор и лягушки семейства Ranidae, включая род Лапа в широком смысле,— до Новой Гвинеи, Соломоновых островов и Се- верной Австралии. Из лягушек Австралийской области представители семейства Leptodactylidae достигают Новой Гвинеи и островов Ару, под- семейство Asterophryinae (семейство Brevicipitidae) достигают Молуккских островов, австралийские формы рода Hyla в увеличивающемся числе рас- пространены до Молуккских островов, острова Тимор и до восточной части
Глава 7
Таблица 12Процентное соотношение западных (Индо-Малайская область).и восточных (Австралийская область) рептилий, распространенныхвдоль Малых Зондских островов до островов Кай и Ару(Mertens, 1934, р. 41)
Западные репти-лии, %
Восточные реп-тилии, %
Острова
93,9 85
87,2 78,1 75
65,2 40,6 32.5 22,2
6,1 15
12,8
' 1,9
25
34,8 5!), 4 67,5 77,8
Бали
Ломбок Сумбана Флорес Алор
Ветар
Танимбар Кай
Ару
Малых Зондских, до острова Сумба, и один род Oreaphryne (подсемейство Sphenophryinae, семейство Brevicipitidae) распространен от Новой Гви- неи до Молуккских островов, Сулавеси (некоторых Филиппинских) и Ма- лых Зондских (от острова Флорес до острова Бали включительно). Распро- странение родов и особенно видов лягушек в действительности значитель- но сложнее, чем представлено здесь, и, возможно, некоторые приведенные детали ошибочны, но общий характер их распространения, несомненно, п авильный. Заслуживает внимания тот факт, что десять одних и тех же видов лягушек находятся на Калимантане и Сулавеси, на противополож- ных сторонах линии Уоллеса, хотя некоторые из них, возможно, были за- несены человеком. Например, Паркер (Parker, 1934, р. 85 — 86) считает, что Ealoula pulehra, возможно, был интродуцирован на острове Сулавеси, но полагает, что другой вид Индо-Малайской области этого же рода (Ka- loula baleata) на этом острове является местным.
Распределение рептилий между Индо-Малайской и Австралийской областями слишком сложное, чтобы вдаваться в его детальное описание. Примеры семейств, родов и даже видов, имеющих, насколько это позво- ляют границы суши, сплошное распространение от Азии до Австралии, приводились в гл. 4. Кроме того, там также упоминались и роды, которые, встречаясь в обеих областях, в то же время отсутствуют на некоторых ост- ровах, расположенных между ними, а также роды, приуроченные к этим островам. Многие группы рептилий Индо-Малайской области не пересе- кают линии Уоллеса, а многие другие простираются на различные расстоя- ния по направлению к Австралийской области, но не достигают ее. Коро- че говоря, значительная часть (но все же только часть) фауны рептилий Индо-Малайской области распространена до Австралии с постепенным ее обеднением, увеличивающимся с увеличением расстояния от Индо-Малай- ской области, и с большим числом групп, которые достигают Новой Гвинеи, и значительно меньшим числом, которые достигают Австралии. Однако этот характер распространения усложняется общими группами, в неко- торых случаях разрывами в распространении и родами, локализованными в пределах зоны перехода. С другой стороны, австралийских групп реп- тилий, которые распространены до Индо-Малайской области, сравнитель- но мало. Только несколько австралийских змей (семейство Elapidae)
Особеиностаи распространения животных на материках
Таблица 13
Процентное соотношение западных и восточных птицна Малых Зондских островах (Mayr, 1944)
1Западные птицы,О/
Восточные птицы,
О/
/
Острова
87
72,5 68
63
57,5
И
27,5 32
3(
42,5
1эали
Ломбок Сумбава Флорес Алор
Многие семейства, роды и виды млекопитающих Индо-Малайской обла- сти достигают островов Калимантан и Ява, и только немногие проникают дальше на восток. К калимантанским (Индо-Малайская область) группам нелетающих наземных млекопитающих (кроме грызунов семейства luri- dae), которые достигают острова Сулавеси, относятся землеройки (Cro- cidura и Suncus), долгопяты, обезьяны (ЛХасаса и эндемичнып род Cyno- pithecus), дикобразы (Hystriz), белки, виверровые (4 рода, из которых один эндемик), свиньи (включая эндемичный род Babirussa), олени (Cer- vus) и бычьи (эндемик Апоа). В плейстоцене на этом острове находился «ар- ликовый слон (Hooijer, 1951).
Из вышеперечисленных млекопитающих острова Сулавесп некоторые проникают еще дальше на восток, достигая отдельных Молуккских ост-
проникают на запад даже до Молуккских островов. Расселение родов широко распространенных семейств ящериц — Gekkonidae, Agamidae, Scincidae — здесь не анализируется, но, судя по данным из работы Руиджа (Rooij, 1915), можно предположить, что австралийско-новогвиней- ские элементы обычно не проникают далеко на запад.
В табл. 12 приводится постепенное изменение фауны западных (Ин- до-Малайской области) и восточных (Австралийской области) рептилий вдоль Малых Зондских островов и т. д. до островов Кай и Ару.
Распространение птиц на границе Индо-Малайской и Австралийской областей еще более сложное, чем у рептилий. Среди птиц этих областей много общих и много переходных групп (семейств, родов и видов), а также много таких, которые встречаются только в одной из областей. Переход их очень сложен и охватывает очень широкую зону, так как некоторые птицы Индо-Малайской области проникают до Новой Гвинеи или до севе- ро-восточной части Австралии, а некоторые (меньшее число) австралий- ские птицы через линию Уоллеса заходят в краевые части Индо-Малай- ской области (стр. 227, 228). Останавливаться на всех этих подробностях здесь не придется. Некоторые из них приводятся Майром (Мауг, 1944) и другими авторами, на которых он ссылается. Орнитологи подчеркивают разницу в составе авифауны островов Бали и Ломбок. В процентном выражении они составляют только 14,5оо, а почти з/ птиц острова Ломбок все же относятся к Индо-Малайской области. При этом на острове Бали доминирующие, самые заметные виды в большинстве случаев являются представителями Индо-Малайской области, тогда как на острове Лом- бок — Австралийской. Процентное соотношение западных и восточных птиц вдоль Малых Зондских островов от острова Бали до группы Алор приводится в табл. 13.
1л ава 7
ровов. Это — — землеройки, обезьяны рода Cynopitliecus (на острове Качан. очевидно, интродуцированы), два вида виверровых (~'иerr« tong«lun«« и f'nradoxurus Iiermaphroditus), ИаЬгизза (только до острова hypy), свиньи и олени. Но все эти млекопитающие либо совсем не дифференцированы, либо слабо дифференцированы на Молуккских островах, и некоторые из них (возможно, даже и все) были там интродуцированы. Paradoxurus hermapIcroditus зарегистрирован даже на островах Кай и Ару, но, видичо. он был там интродуцирован.
На острове Ява отсутствуют некоторые млекопитающие Индо-~Цалай- ской области, встречающиеся на острове Калимантан. Это было известно еще и Уоллесу, а также видно из сравнения списков млекопитающих Калимантана и Явы, составленных Картером, Хиллом и Тейтом (Саг[ег. Hill, Tate. 1945). Эти авторы приводят отдельные списки млекопитающих и для многих других островов Индо-Австралийского архипелага, несмот- ря на то, что некоторые из них недостаточно точные. Тем не менее на ост- рове Ява до сих пор находится довольно богатая фауна млекопитан~- .щих. Многие группы млекопитающих, которые достигают Явы, не дости- гают острова Бали. Например, ежи гимнуры (Gymnures), шерстокрылы. лори, гиббоны, летяги (4 рода), некоторые другие роды грызунов, азиат- ский красный волк (Сиоп), куньи (мелкие куньи, барсуки и выдры), лео- пард, носорог, оленьки и, вероятно, дикие буйволы. На острове Бали нет млекопитающих, общих с основной частью фауны острова Ява (см. списки семейств, Carter, Hill, Tate). К млекопитающим, которые распространены до острова Бали (но не до острова Ломбок), относятся тупайя, два вида обезьян, панголин, два вида белок, леопардовая кошка и тигр. На восток от острова Бали (вдоль Малых Зондских островов) проникает очень незна- чительное число нелетающих наземных млекопитающих: дикобраз (FFyst- пт) распространен до островов Ломбок, Сумбава и, возможно, Флорес. а землеройки (Crocidura и Suncus), крабоядные обезьяны, виверра (Par«- doxurus hermaphroditus), свиньи и олени рода Cerrus встречаются на от- дельных или на всех Малых Зондских островах до острова Тимор. Этот список млекопитающих, достигающих острова Тимор, весьма сходен со списком млекопитающих Индо-Малайской области, достигающих Молукк- ских островов.
Возникает вопрос, не являются ли некоторые из млекопитающих этих островов местными, ибо на этот счет существуют серьезные сомнения. Дикобразы, мясо которых напоминает свинину и употребляется в пищу. по-видимому, ввозились на эти острова. Что же касается землероек, то по крайней мере одна из них (Suncus) часто живет в домах и так же, как и домовая мышь, может перевозиться на местных лодках (Carter, Hill, Ta- te). Ручные обезьяны и другие животные часто завозятся в другие места бродячими малайцами (Wallace), особенно часто перевозятся крабоядные обезьяны (Уоллес, в частности, указывает на нахождение этой обезьяны вблизи острова Тимор,— Wallace, 1869, vol. 1, р. 294). Некоторых виверр перевозят с целью получения на продажу мускуса, и весьма возможно, что после того, как мускус взят, их отпускают на волю (Wallace, 1869, vol. 2, р. 1+О). Свиньи в этих районах являются основной пищей. Оленей часто приручают и держат как домашних животных из-за их мяса, и возможно, что жители отдаленных островов (Wallace) интродуцируют с этой целью. Предполагают, что некоторые из этих млекопитающих на Молуккских или Малых Зондских островах являются определенными подвидами, но сомнительно все же, насколько они определены, а если они и являются выделившимися подвидами, то должны ли они быть местными. Некоторые
891
мачмологи разделяют эти сомнения. Так, Лорье и Хилл (Laurie. Hill.
195'~. р. 86). перечисляя эндемичные подвиды свиней с островов Флорес и 1'имор, считают их «одичавшими» потомками свиней. перевозимых жите- лями с острова на остров. Я не претендую на знание, какие именно виды и где являются местными, но я думаю, что вряд ли гакпе-либо млекопитаю- ьцие Индо-Малайской ооласти, за исключением грызунов семейства Aluri- dae н рукокрылых, обнаруженных к востоку от островов Сулавеси и Бали, яиляк~тся де~~ствительно местными.
Из австралийских млекопитающих лишь немногие распространены зна иггельно западнее Новой Гвинеи. Однопроходные (ехидна) достигают острова Салавати, недалеко от острова Бёрдхед.
11з сумчатых на островах Ару можно назвать по крайней мере следую- щих три вида: Dasyuridae, один вид бандикута, несколько представителей семейства Phalangeridae, валлаби рода Thylog ale и, возможно, кустар- никовый кенгуру. На островах Кай находятся по крайней мере бандпкут. кускус (Pit<clanger), сумчатая сахарная летяга (Petaurus), род Thylogale и, возможно, кустарниковый кенгуру. Pseudocheirus достигает острова (:алавати, валлаби рода Dorcopsis — острова Вайгео и других островов. лежащих недалеко от западной оконечности Новой Гвинеи, так же как и роды кускус, Petaurus и, возможно, некоторые другие сумчатые. Бандикут и иаллаби рода Dorcopsis проникают до острова Мисол. Единственные сум- чатые. которые (насколько мне известно) встречаются за пределами этих островов,— это бандикут на острове Церам (эндемичный род), сумчатая сахарная летяга (Petaurus), достигающая острова Мисол и северной части Молуккских островов (острова Хальмахера, Тернате и Бачан), род кус- куе, распространенньш через Молуккские острова до острова Сулавеси (на последнем находятся два эндемичных вида) и до островов Ветар и Ти- мор. Я не знаю, являются ли эти сумчатые интродуцированными на неко- торых из этих островов, но род кускус на запад от острова Сулавеси, несомненно, местны~~.
Вышеприведенные сведения могут быть неполными или отчасти неточными. Кроме того, остается сомнение и относительно видов — где и какие из них являются местными, а где интродуцированными. Но в целом характер их распространения в своей основе, несомненно, дан правильно.
Грызуны семейства (МигЫае) усложняют этот простой характер рас- пространения. По своему происхождению они относятся к элементам Индо- Малайской области, проникшим в австралийскую фауну, но по современ- ному распространению являются общими и включают как австралийские. так и восточные группы. Подсемейство мышиных Hydromyinae почти полностью приурочено и Австралийской области и проникает на запад только до островов Ару, Кай и Вайгео (Hpdromps — Tate, 1951, р. 232). не считая двух сомнительно родственных (реликтов?) родов, находя- щихся в северной части острова Лусон (Филиппины). Род Rattus— общий для Индо-Малайской и Австралийской областей. Распространение и связи других мышиных между Индо-Малайской и Австралийской областя- ми слишком сложны, а в некоторых случаях и слишком сомнительны. что- бы Ilx здесь рассматривать, но, видимо, местные мышиные встречаются на всех пригодных для обитания островах зоны перехода.
Рукокрылые чрезвычайно многочисленны на Индо-Австралийском архипелаге и имеют крайне сложный характер распространения. Я не пытаюсь дать здесь подробный анализ. Из работы Тейта (Tate, 1946) ясно, что многие группы рукокрылых являются общими, а другие встре-
Г.гава 7
чаются в одной области, но отсутствуют в другой. Многие роды и виды Индо-Малайской области простираются к Австралии, а некоторые (но таких меньше) австралийские — к Индо-Малайской области на различ- ное расстояние. Некоторые роды и виды в пределах зоны перехода отно- сятся к эндемичным.
В целом о млекопитающих можно сказать, что большая часть основ- ной фауны млекопитающих Индо-Малайской области, кроме мышиных и рукокрылых, в своем распространении задерживается на островах Калимантан и Ява и небольшие части этой фауны достигают островов (.ули- веси и Бали и очень незначительное число (или ни один из их представи- телей) не достигает Молуккских или ближних к ним Малых Зондских остро- вов. Основная фауна млекопитающих Австралии не проникает дальше Новой Гвинеи, но небольшие части этой фауны достигают островов Лру, Кай и др., а еще меньшая часть — некоторых Молуккских остроин~, и только один род — кускус — достигает островов Сулавесп и Тимор. М:те- копитающие Индо-Малайской и Австралийской областей перекрыван>т друг друга на небольшой площади. В том случае, если виды Индо-Малай- ской области на Молуккских островах или острове Тимор пнтродуциро- ванные, то естественное перекрытие ареалов происходит только на осг- розе Сулавеси, где два вида кускусов встречаются с небольшим числом, несомненно, местных млекопитающих Индо-Малайской области. Сомни- тельные отдельные случаи значения не имеют. Существенным фактом является и то, что, кроме мышиных и рукокрылых, на Молуккских и ближних к ним Малых Зондских островах число млекопитающих незначи- тельно и в большинстве случаев они слабо дифференцированы и, вероятно, проникли сюда недавно. На этих островах отсутствует древняя фауна наземных млекопитающих, за исключением мышиных. Возникает вопрос, почему? По-видимому, потому, что наиболее многочисленными и наиболее эндемичными (древними?) животными там являются мышиные, а еще более многочисленными — рукокрылые.
В заключение о переходе позвоночных Индо-Малайской и Австралий- ской областей можно сказать, что у пресноводных рыб переход не осу- ществляется: строго пресноводные рыбы Индо-Малайской и Австралий- ской областей не встречаются друг с другом совсем. Немышиные назем- ные млекопитающие образуют едва заметный переход. Если землеро1и,и, виверры, свиньи и олени на Молуккских или на Олижнпх к ним Малых Зондских островах являются местными, то тогда можно считать. что небольшая часть фауны млекопитающих Индо-Малайской области. про- никая на значительное расстояние через линию Уоллеса, встречает и пере- крывает еще меньшую часть австралийской фауны, проникающую на Малаккские острова, на остров Тимор, а ТВЕНОВ на Сулавесн. Если же эти млекопитающие Индо-Малайской области не являются местными на Молуккских и ближних к ним Малых Зондских островах (а этот факт весьма сомнителен) и если принять во внимание то, что говорили Уоллес и другие ученые о некоторых видах на отдельных из рассматриваемых островов, то тогда немышиные наземные плацентарные Индо-Малайской области и сумчатые Австралии встречаются только на острове Сулавеси и переход наблюдается исключительно у мышиных и рукокрылых.
У амфибий значительное перекрывание ареалов групп (в болыпин- стве случаев разных семейств) Индо-Малайской и Австралийской обла- стей отмечается по крайней мере от островов Бали п Сулавеси до остро- вов Тимор, Новой Гвинеи и до северной части Австралии. У рептилий и птиц перекрывание ареалов не только некоторых семейств, но также
Особенност,и распространения животных на л~атериках
и многих родов и видов происходит на еще более обширной площадн- от внутренних частей периферии Индо-Малайской области до северной 11 восточной частей Австралии.
Сложность, уровень и ширина зоны перехода позвоночных Индо- Малайской и Австралийской областей варьируют в зависимости от того, какими возможностями располагают различные классы при преодолении преград, образуемых солеными водами. Мне кажется, что этот очевид- ный и очень важный факт вытекает из предыдущего заключения. Более того, по крайней мере на некоторой части зоны перехода — на Молукк- ских и ближних к ним Малых Зондских островах — степень эндемизма различных позвоночных, по-видимому, изменяется в зависимости от воз- можностей этих животных пересекать водные пространства. Самые строгие эндемики принадлежат к животным, обладающим самыми высокими спо- собностями пересекать водные пространства. Наземные млекопитающие (кроме мышиных) на этих островах удивительно слабо дифференцирова- ны. Среди лягушек нет эндемичных родов и мало эндемичных видов (на- сколько об этом можно судить по работе Ван Кампена — van Kampen, 1923). У рептилий, птиц, мышиных и рукокрылых четких эндемикои, видимо, имеется больше. Это сравнение в какой-то мере субъективно, но относительно низкого уровня эндемизма немышиных наземных млекопи- тающих на Молуккских и ближних и ним Малых Зондских островах не может быть никаких сомнении. Даже в других классах, хотя они имеют более высокий уровень эндемизма, заметно отсутствие действительно четких, реликтовых позвоночных. Что касается направления перехода, то у всех групп, у которых оно все же выражено (амфибии, рептилпи, птицы и рукокрылые1, видимо, существует тенденция большего распро- странения групп Индо-Малайской области к Австралии, чем наоборот.
Я сделал попытку отобразить это явление, исходя исключительно из современного распространения, но из этого вытекают некоторые поло- жения, относящиеся к прошлому. Из состава и распространения переход- ной фауны следует, что острова, расположенные между Калимантаном и Явой, с одной стороны, и Новой Гвинеей и Австралией — с другой, засе- лялись и перезаселялись позвоночными животными относительно недавно за счет расселения через водные пространства и что расселение животных в большей степени шло со стороны Индо-Малайской области, чем со сто- роны Австралии.
Фауны позвоночных Индо-Малайской и Австралийской областей отделяются общей зоной, где происходит обеднение фаун и переход. Обед- нение — очень важное свойство перехода. Отсутствие строго пресновод- ных рыб и очень бедный состав наземных млекопитающих в зоне пере- хода так же существенны, как постепенное изменение состава наземных млекопитающих от фауны Индо-Малайской области до Австралийской по мере движения с запада на восток у других классов.
Границы этой зоны обеднения — перехода довольно хорошо опре- делены. Западная граница — линия между островами Калимантан и Сула- веси и между Явой и Малыми Зондскими островами. Это, по существу, линия Уоллеса, то есть линия, которая, как полагал Уоллес, отделяет фауны Индо-Малайской и Австралийской областей, но которая фактиче- ски отделяет почти в полном составе фауну Индо-Малайской области от на редкость обедненной фауны очевидного перехода. Здесь возникает един- ственный вопрос, куда относить остров Бали — к Яве или к другим Малым Зондскпм островам. Следуя обычному подразделению, я, несмотря на ограниченность фауны этого острова, выражающуюся особенно в от-
1лави Т
сутствии большинства пресноводных рыб и многих млекопитающих. отнес остров Бали к Яве, то есть в Индо-Малайскую область.
Восточная граница зоны ооеднения — перехода находится непосред- ственно к западу от Австралии и Новой 1'винеи, но она включает некото- рые небольшие близлежащие острова: Ару (возможно, и острова Han), Салавати, Батанга, Вайгео (возможно, и остров Мисол), расположенные непосредственно к западу от Бёрдхеда (Новая Гвинея). На островах, не взятых в скобки, находится почти чисто новогвинейская, правда обед- ненная, фауна (но, видимо, это обеднение находится в пропорциональной зависимости от размеров и природы этих островов). На островах Кай и острове Мисол отсутствуют некоторые довольно значительные группы, которые распространены на ряде других выше перечисленных островов: сюда относятся рыбы подсемейства Melanotaeniinae и некоторые млеко- питающие. Следует ли включать эти острова в собственно Австралийскую область или в зону обеднения — перехода, — дело вкуса.
Следуя Майру (Мауг, 1944), я отнес остров Мисол к Новой Гвинее, а острова Кай — и зоне перехода.
Между этими границами фауна позвоночных постепенно меняется с запада на восток от преимущественно фауны Индо-Малайской области до преимущественно австралийской фауны, хотя дать точное описание всего перехода в целом невозможно. Фауна острова Сулавеси по крайней мере на з/, Индо-Малайская (Мауг, 1944, р. 3), и элементы Индо-Малай- ской области, хотя и в сокращенном числе, очевидно, все еще преобла- дают в (более обедненных) фаунах островов Талауд и Сула. Но на острове Буру большинство составляют австралийские (новогвинейские) элементы, и они становятся подавляющими на острове Церам и в северной части Молуккских островов. Вдоль линии Малых Зондских островов переход более постепенный, и он лучше подтвержден фактическими данными. Фауна позвоночных острова Бали примерно на 90 оо состоит из элементов Индо-Малайской области. На группе островов Алор эта цифра постепенно падает примерно до 60 или 70оо. Но элементы фауны Индо-Малайской области на острове Тимор и даже островах Танимбар или по крайней мере на западных островах этой группы все еще составляют большинство. хотя некоторыми группами фауна этих островов очень бедна. На основа- нии этих данных можно провести границу, отделяющую острова с преоб- ладанием животных Индо-Малайской области от островов с преоблада- нием животных Австралийской области. Это линия фаунистического баланса, или линия Вебера (рис. 55). Более детально этот вопрос рассма- тривается Майром (Мауг, 1944, р. 10 — 12), который отмечает имеющую- ся до сих пор трудность в установлении южного конца этой линии. Майр отмечает, что линия Вебера находится ближе к австралийской стороне зоны перехода, чем к Индо-Малайской области. Это соответствует тому, что уже было сказано выше: в зоне перехода преобладают группы живот- ных Индо-Малайской области, а не Австралийской.
В определении границ Индо-Малайской и Австралийской областей зоогеографы могут придерживаться одного из двух приемлемых направ- лений. В первом случае — восточная граница Индо-Малайской области может быть проведена по линии Уоллеса или близ нее, а западная граница Австралийской области — сразу же к западу от Новой 1'винеи (включая некоторые мелкие острова, перечисленные выше). В этом случае площадь. расположенную между ними, можно рассматривать как отдельную зону обеднения — перехода, а не как часть одной из этих областей. Эта пло- щадь была названа «Зоной Уоллеса» (Dickerson et al.. 1928, р. 101), и это
вполне приемлемый и полезный термин и любом случае. Во втором случае — общую границу между Индо-Л!алайской и Австралийской обла- стямп можно провести вдоль линии Вебера или близ нее. Как в том. так и в другом случае границы имеют свои преимущества и недостатки. Какая из этих двух границ будет выбрана, большого значения не имеет. если понятно основное положение. Возможно, что признание зоны обед- нения — перехода, «Зоны Уоллеса», част возможность припти к опре— деленному решению.
Заключение о фаунпстпческих переходах
Общая природа, относительная четкость и структура региональных фаун и фаунистических областей уже были рассмотрены (см. также табл.
10. 11 и рис. 45), но необходимо сказать еще несколько слов о том, как эти области отделяются.
(',межные региональные фауны всегда соединяются зоной перехода. я этп зоны ооычно расположены в местах, которые или недосягаемы, или неблагоприятны для некоторых животных, и поэтому переходные подфау- ны. как правило, бывают обедненными. В результате этого смежные региональные фауны отделяются, как я ее называю, зоной обеднения— перехода. В большинстве случаев такие зоны образуют частичную прегра- ду между региональными фаунами. Аравия и прилежащая субтропиче- ская Юго-Западная Азия, образуя зону обеднения — перехода, отчасти являются преградой между тропическими фаунами Индо-Малайской и Эфиопской областей, а пустыня Сахара — крайней зоной обеднения— перехода и частичной преградой между фаунами Эфиопской области и Палеарктики. Арктика образует зону обеднения — перехода и частич- нук> преграду между фаунами Палеарктики и Неарктики: здесь в значи- тельно большей степени, чем водные пространства, обмен животных через район Берингова пролива в последнее время ограничил климат.
Центральная Америка и Мексика представляют зону обеднения— перехода и частичную преграду между фаунами Неарктики и Неотропп- ческой области. Влияние современной преграды проявляется отчасти в труднодоступности и ограниченности территории Центральной Амери- ки, а отчасти в засушливости отдельных районов Мексики. Острова и вод- ные пространства («Зоны Уоллеса») образуют зону обеднения — перехода и частичной преграды между фаунами Индо-Малайской и Австралийской областей. Только в Восточной Азии, между фаунами Индо-Малайской области и Палеарктики, переход осуществляется в полном объеме без про- межуточного обеднения.
Местные факторы, являющиеся в настоящее время причиной обедне- ния фаун и частичной преградой между региональными фаунами, в од- них случаях были теми же, что и факторы, вызвавшие или определившие дифференциацию фаун в прошлом, а в других — иными. В Юго-Западной Азии современные местные факторы, вероятно, были самыми эффектив- ными: до какой-то степени засушливый и не вполне тропический климат. вероятно (так же, как в настоящее время), разделил в прошлом эфиоп- скук> и индо-малайскую фауны и вызвал их дифференциацию. Холодный климат, иногда дополняемый такой преградой, как Берингов пролив, оче- видно, являлся основным барьером между фаунами Палеарктики и Не- арктики в прошлом так же, как и теперь. В «Зоне Уоллеса» водные пре- грады были в какой-то степени сходны с современными, что, очевид-
Глава 7
но, и определило значительную дифференциацию фаун Индо-Малайской и Австралийской областей. Но дифференциация фаун Эфиопской и Пале- арктической областей вызвана в основном не такой преградой, как лежа- щая между ними пустыня Сахара, а разницей климатов. Сахара — очень молодая пустыня, которая в прошлом играла роль преграды в значитель- но меньшей степени, чем в настоящее время. Ископаемые показывают. что в прошлом многие тропические животные Эфиопской области вполне мог- ли достичь Европы, но большинство из них не сохранилось там, по-види- мому, из-за не подходящего для них умеренного климата.
В Восточной Азии серьезную преграду между фаунами Индо-Малай- ской и Палеарктической областей образуют Гималаи, но горы представ- ляют менее серьезное препятствие, чем это кажется. К востоку от них происходит встреча двух фаун. Различия фаун Индо-Малайской области и Палеарктики, очевидно, надо искать прежде всего в зональном климате, а не в преградах, образуемых горами. В Центральной Америке сущест- вующие преграды опять-таки не являлись основной причиной дифферен- циации фаун (неарктической и неотропической) в прошлом. Основными преградами были водные пространства третичного времени. Даже в на- стоящее время трудности, возникающие при расселении животных через Центральную Америку, вероятно, не являются основной причиной, кото- рая обеспечивает отличия неарктической и неотропической фаун, воз- можно, за исключением рыб. За последние несколько миллионов лет обе фауны чрезвычайно смешались. По моему мнению, они остаются различ- ными лишь потому, что Северная и Южная Америка расположены в раз- личных климатических зонах.
Не анализируя этот вопрос дальше, можно сказать, что все основные региональные фауны отделяются друг от друга или приблизительно про- веденной линией между тропиками и умеренной зоной северного полуша- рия, или солеными водами. Из этого следует, что зональный климат и пре- грады, образуемые солеными водами,— это самые важные факторы нежи- вой природы, которые вызывают дифференциацию региональных фаун или способствуют ей. Другие преграды — пустыни и горные хребты— являются второстепенными, хотя они могут помочь уточнить фаунистиче- ские границы или немного сместить их. Зоогеографы часто считают пусты- ню Сахару и Гималаи главными преградами. Конечно, этот факт отрицать нельзя, но распространение животных через них или вокруг них как в прошлом, так и в настоящем показывает, что на протяжении длитель- ного периода они играли менее важную роль, чем это кажется на первый взгляд.
Что касается направления переходов, то между фаунами Эфиопской и Индо-Малайской областей оно неясное: или африканские группы в боль- шей степени проникают в Индо-Малайскую область, или наоборот. Между фаунами Эфиопской и Индо-Малайской областей, с одной стороны, и фауной Палеарктики — с другой, осуществляется переход в северном направлении: значительно большее число тропических групп проникает в умеренную зону северного полушария, чем наоборот. Переход между фау- нами Индо-Малайской и Австралийской областей направлен к Австралии. Направление перехода между современными фаунами Палеарктики и Не- арктики неясное. Между неарктической и неотропической фаунами напра- вление перехода смешанное: среди рептилий и млекопитающих значи- тельно больше северных групп проникает на периферию Неотропической области, чем неотропических групп на север, но среди птиц число тропи- ческих групп, проникающих на север, видимо, больше.
397
Границы региональных фаун обычно определяются группами, типич- ными для той или другой фауны, распространение которых приостанавли- вается примерно у общей границы (рис. 53). Например, в Восточной Азии северная граница Индо-Малайской области определяется многими тропи- ческюш группами, которые останавливаются в Южном Китае, хотя мно- гие другие тропические группы даже пересекают эту границу к северу. В Америке северная граница Неотропической области определяется мно- гими непереходными группами, которые останавливаются на тропиче- ских низменностях Мексики, хотя некоторые группы проникают гораздо дальше. Между Азией и Австралией граница основных фаун Индо-Малай- ской и Австралийской областей выражена очень хорошо, несмотря на то, что между этими границами имеется обширная зона перехода.
Несомненно, все вы1пеизложенное, как и многое другое в этой книге, знаштельно упрощено, но это самое лучшее, что можно оыло сделать в пределах объема книги и времени.
ЛИ ТЕРЛ ТУРА
А и ~> и у m и и, 18.13, The Xearctic Region and it» mammal», «Xatural ~с1епсе»,
3. 288 — 292.
В е а u f о r t Ь. F., de, ) 926, Zoogeographie van den Indischen Archipel, faarlem. В 1 а и 1'. о r d 9'. Т., 1890....Anniversary adress..., «Proc. Annual Мее1~ир Geologi- са1 $ос. L(~ndon», 1890, 13 — 80.
( а r t e r Т. ])., II i l 1 J. Е., Т а t е СНН., 1945, Матта1я о1 1Ье I acific
~~.огЫ, Xew York.
T) i c k e r s о n R. E., et al, 19 8, Distribution of life in the Philippines, I )iililipine
Bureau Sci. Monograph 21.
I> и и п Е. R., 1922, А suggestion to zoogeographers, «Science», 56, 336 — 338.
F е г 11 а l d М. Ь., 192), Some relationships of the floras of the Northern Hemisphere,
«Proc. International 'ongress Plant Sci.», 2, 1487 — 1507.
6 a <) ~ w Н., 1913. The wanderings of animals, Cambridge — New York.
H e i 1 р r i n А., 1887, The geographical and geological distribution of animals, Хеж
York .
Н о о I j e r D. А., 1951, Pygmy elephant and giant tortoise, «Sci. Monthly», 72, 3 — 8. Н о r a $. Ь., 1952, Recent advances in fish geography of India, «J. Bombay Nat. Hist.
~'oc. », 51, 170 — 188.
Н о r а S. L., 1953, The Satpura hypothesis, «Science Progress», 4I, 245 — 255.
14 u ~ l е у Т. И., 1868, On the classification and distribution о1 the Л1~сЯо)опгогр/гае
and VIeteromorphae, «Proc. Zool. Soc. London», 1868, 294 — З.l9.
L а и r i е Е. M. О., Н i 11 J. Е., 1954, List of land mammals of 3чеъ Guinea, Се1е- )1ея, апЛ adjacent islands..., London, British Mus.
t. i Il.-L., 195', Floristic relationships Ьейъeen eastern Asia and eastern Г~orth Ameri- ca, «Tr. American Philosophical Soc.», New Ser., 42, 369 — 420.
I., у rl t. l; k e r R., '[896. А geographical )history of mammals, 'ambridge, England,
(:an>bridge U. Press.
1, (у rl е 1, 1' е r) R., 1911, Zoological distribution, «Encyclopaedia Britannica», '11th
еd., 28, 1002 — 1О! 8.
М а у 1 Е., 1944, Wallace' s Line in the light of recent zoogeographic studies», «~>uar- terly Rei.iew Biol.», !',), 1 — ]4.
51 е г r i а m (:. Н., 189', ТЬе geographic, distributic)n of life in Ъ огай Лшег~са...,
«Ргос. Biol. Roc. Washington». 7, 1 — 64.
М е г t e n s R.. 1!134, Die Insel-Reptilien..., «Zoologica» (Stuttgart), 32, G. Lieferung,
1 — 209.
898
Гыва 7
Е' а r k e r Н. Ш., 1934, А monograph of the frogs of the family XJicrohylidae, Т оисЬи.
British Mus.
R о о i j Х. de, 1915, The reptiles of tire Indo-Australiari ЛгсЫре1а<~о, pt 1, Loire» S c h m i d t К. P., 1954, Faunal realms, regions, and ргойпсея, «Оиаг1ег1у Вей~а.
В iol. », 29, 322 — 331.
8 с 1 а t e r 1. Ь., 1858, On the general geo гарЫса1 distribution of 111е ппешЬег~ iaaf
the class Aves, «J. of Proc. Linnean Soc. (Zool.), 2, 130 — 145.
$ c r i v е п о r J. В., et al, 1943, Л discussion on 1Ье biogeographic с1л.isiоri of Мн
Indo-Australian Archipelago..., «Proc. Linnean $oc. Еondon», 15ЙЬ '.еянои, 1 2"0—
l65.
Т а t e G. Н. ц., 19-.~6, Geographical distribution of the bats in the Australaiari Ar- chipelago, «American Моя. Novitates», No. 1323.
Т а t e G. Н. Н., 1951, The rodents of Australia and ~еж Guinea, «Bull. Ашег~снп
Моя. i%at. IIist.», 97, 183 — 430.
ъ.ап Е а ill р е и P. И., 1923, The АтрЫЬ1а of the Indo-Australian ЛгсЫре1а„~о. Lei- den.
9, а 11 а с е А. Н., 1860, Оп the zoological деograIiliy of 1Ье Ма1ау ЛгсЫре1;~<~о, «J.
of Proc. Linnean $oc. London», 4, 172 — 184.
М, а l l а с е Л. H., l863, The Malay Archipelago... London, 2 vols.
9 а l 1 а с е А. Н., l876, ТЬ» geographical distribution of апппа1я, London, ' vols.
1'лава 8ОСОБ ЕННОС'П1 РАСПРОСТРАНЕНИЯ ЖИВОТНЫХ НА ОСТРОВАХ
В этой главе рассматриваются позвоночные животные островов. их природа, распространение и значение. Книга Уоллеса (Wallace, 1880, и более поздние издания) о жизни животных на островах до сих пор является классическим трудом, хотя изучение островных животных, как считают в настоящее время, фактически началось в более раннее время с исследований Дарвином птиц и зверей островов Галапагос. Кроме этого, следует указать на интереснейшие статьи Галика (Gulick, 1932) о биологи- ческих особенностях океанических островов и Мертенса (A1ertens, 1934) о рептилиях островов земного шара. Получить более специальные сведе- ния можно из отдельных работ, приводимых ниже. Наиболее важной из них является работа Перкинса «Fauna Hawaiiensis» (Perkins, 1913), с пре- дисловием к которой должны ознакомиться все зоогеографы
Я предполагал начать эту главу специальным обзором позвоночных Вест-Индии (островов, которые я знаю лучше остальных), используя их в качестве иллюстрации принципов распространения животных на остро- вах, но затем мне пришлось изменить свои намерения. По растительному и животному миру Вест-Индии небольшой группой специалистов весной 1954 г. был сделан ряд докладов в Гарвардском университете. В итоге все эти работы были подготовлены к печати Эрнестом Вильямсом, и вряд ли оы мне удалось сделать более лучший обзор фауны Вест-Индии. Поэтому я дал весьма краткий обзор Вест-Индских островов, не выделяя их спе- циально.
Сначала предварительно рассмотрим природу островов и островных фаун, останавливаясь на особо важных для выяснения основных вопро- сов моментах, после чего перейдем к фауне позвоночных главных островов и архипелагов земного шара, а затем попытаемся сделать краткий анализ
И ВЫВОДЫ.
Исследуя острова и островные фауны, приходится сталкиваться со многими трудностями.
Географы до сих пор еще не пришли к единому мнению относительно площади некоторых островов и протяженности водных пространств, отде- ляющих эти острова.
Геологи сомневаются или расходятся во мнениях по вопросу истории возникновения некоторых островов, и в ряде случаев им свойственны очень серьезные заблуждения. Зоогеографам следует пользоваться геологиче- скими данными с осторожностью и для разрешения вопросов природы и истории островных фаун прежде всего опираться на зоологические дока- зательства.
Однако мнения зоогеографов по тем или иным вопросам также неред- ко расходятся, а зоологические данные во многих случаях бывают непол- ными или сомнительными, так как фауны многих островов до сих пор изучены еще недостаточно хорошо и, что еще хуже, ископаемые
Глава 8
островных животных обычно весьма скудные и в оольшинстве случаен к временам более ранним, чем плейстоцен, не относятся.
Систематика ряда островных животных не вызывает доверия. Те фор- мы, которые считают определенными, иногда оказываются нечеткими, и наоборот. Некоторые систематики выделяют островные популяции как «виды» по самым незначительным признакам. Однако их вина здесь не столь велика. Часто они действуют из самых наилучших побуждений, поскольку надо составить представление о фауне того или иного острова. но, имея под руками недостаточно полный материал, в своих отчетах дают систематическое положение животных так, как они его предстанляют. Подлинность некоторых ocTpoBHbIx видов, TRI' же как их предки и.III свя- зи, нередко вызывает немало сомнений.
Далее могут вызывать сомнение II различные виды животных, случай- но или преднамеренно интродуцированные на многих островах как древ- ним, так и современным человеком. Если на острове животное является энцемиком или достаточно четким видом, то можно считать, что оно мест- ное, но во многих сомнительных случаях, когда островные формы недиф- ференцированы или слабо дифференцированы, есть основание предпола- гать, что они интродуцированы. В таких случаях перед зоогеографом встают две альтернативы: или рассматривать в качестве местных Тс виды ;кивотных, которые, возможно, являются интродуцированными, или те виды, которые, возможно, являются местными, принимать за интродуци- рованные. Оба эти варианта в одинаковой мере неприемлемы. В таких случаях лучше всего воздержаться от какого-либо решения, но анализ островных фаун остается не менее сложным в связи с тем, что сомни- тельные виды должны быть упомянуты, а причины, по которым они отнесены к сомнительным, объяснены. Некоторые млекопитающие Индо- Малайской области, которые встречаются на Молуккских, ближних к ним Малых Зондских (стр. 389 и 390) и Филиппинских островах, африканский речной кабан на Мадагаскаре, лягушки на островах Фиджи и еноты ряда мелких островов Вест-Индии — вот несколько из многочисленных при- меров, иллюстрирующих вышесказанное.
Некоторые виды позвоночных, особенно из ящериц и крыс, очень широко распространены на удаленных океанических островах, куда они часто завозятся на лодках. Можно считать, что те качества, которые поз- воляют им попадать на удаленные острова в груженых лодках и т. д., дела- ют особенно возможной теорию о прибытии их на плотах. Это позволяет предполагать, что названные животные могли достигать самых отдаленных островов естественным путем. Но в ряде случаев данный вопрос остается спорным. Я думаю, что таким образом вполне можно объяснить, как рас- селились эти животные — естественным путем или ~~ыли занесены челове- ком. Если они пересекли огромные океанические пространства естествен- ным путем, значит, они так делали в течение многих миллионов лет, и поэтому определенные эндемичные формы этих животных, включая энде- мичные роды, а также и виды, должны находиться на отдаленных остро- вах. Однако если они были занесены человеком, то их существование там исчисляется всего лишь несколькими тысячами лет и соответственно сте- пень их эндемизма должна быть низкой, хотя (по недостаточным выводам или из-за отсутствия информации) систематики могут рассматривать отдельные популяции островов как «подвиды» или как слегка определив- шиеся виды.
Даже в тех случаях, когда островная фауна достаточно известна, все еще может оставаться неясным, о чем говорит нам эта фауна, почему
одни 'кивотные находятся на»том острове, а другие отсутствуют. Напри- мер, на Британских островах нет древесной лягушки Ну1а arborea, xoгя этот нцд имеется на близлежащем материке. Бьюфорт (Beaufort, 1951, р. 5~. 55) считает, что этот вид не мог достичь Британских островов с вре- мен плейстоцена, а Малколм Смит (Malcolm Smith, 1951, р. 17) у.гвержда- ет, по эта лягушка была интродуцирована в Англии, но не сохранилась там. I'то же помешало проникновению этой лягушки в Англию — водные преграды постплейстоцена или для нее оказался неблагоприятным клима~ Англии '?
11рп попытке заняться проблемами жизни животных на островах зоо- географы постоянно сталкиваются с такими трудностями и сомнитель- ными случаями. Избегая ненужных споров о деталях, они могут продол- кать. насколько это возможно, попытку раскрыть принципы и особенности распространения островных животных.
Как уже было отмечено выше, обычно принимаемое различие между материками и островами очень произвольно. Все материки и острова могут быть расположены в виде одного очень неравномерного ряда соответствен- но уменьшающейся площади и (или) увеличивающейся изоляции, хотя, несомненно, эти два положения взаимосвязаны только отчасти. Даже самые большие материки некоторое время были изолированы: Африка и Ев- разия. возможно, отделялись друг от друга в раннетретичном периоде, и если это действительно так, то, вероятно, изоляция в чем-то ограни- чила фауны обоих материков. Северная Америка в настоящее время отре- зана от Старого Света. Меньшие по своим размерам материки — Южная Америка и Австралия — были еще более изолированными. И эта цепь продолжается далее через все крупные острова наподобие Мадагаскара до самых мелких. На всех сушах как с наибольшей, так и с наимень- шей площадью имеются фауны, которые в какой-то степени ограничены изоляцией от других суш, хотя в полной мере влияние этой изоляции ска- зывается лишь на самых мелких и самых изолированных островах. llo этой причине основные принципы распространения животных на островах должны быть теми же, что и на материках. Это более, чем философская концепция. Материки и острова земного шара образуют одну сложную пр~.— рыван>щуюся систему суши, на которой животные расселились определен- ным образом. и, чтобы представить характер распространения живот- ных и пх расселение в целом, необходимо к уже известным сведениям о животных материков добавить 'fo, что выяснено об островных животных.
В книге «Жизнь островов» Уоллес проводит резкое различие (или за- имствует его у Дарвина) между материковыми и океаническими островами.
Материковые острова он определяет как отделившиеся части континентов.
состоящие из комплекса континентальных пород и всегда заселенные наземными млекопитающими и амфибиями, а океанические острова — как возникшие в океане и состоящие из вулканических пород и кораллового известняка острова, на которых отсутствуют наземные млекопитающие и амфибии. С точки зрения геологии такое различие, возможно, и правиль- но, хотя и происхождение и геологическое строение этих островов значи- тельно оолее сложное, чем представлял Ъ оллес, но с точки зрения зоо- ,чогии;это неверно. Несомненно, одни фауны островов являются чисто мате- риковыми, представляя, очевидно, части материковых фаун, недавно отрезанные затоплением промежуточной суп и, а другие — чисто с>кеани- ческ~ош, но между этими двумя типами фаун имеется такое множество смешанных, промежуточных и сомнительных случаев. что классифици- ровать фауны островов просто на эти две категории неправильно.
Глава 8
Несмотря на то что различие между материками и островами весьма произвольно и что зоогеографически острова не могут быть строго разделе- ны на материковые и океанические, все же искусственная классификация островов приносит некоторую пользу. Для удобства я выделил три услов- ные категории: (1) явно материковые современные острова с более или менее обедненной материковой фауной, (2) периферийные архипелаги, не просто современные материковые, а лежащие вблизи материков и полу- чающие фауны с их окраинных частей, и (3) отдельные острова или изо- лированные архипелаги, не попадающие ни под одну из двух перечислен- ных категорий и так или иначе отличающиеся от них (сюда входят и такие большие острова, как Мадагаскар, и самые мелкие океанические). Острова, которые будут рассмотрены отдельно, классифицируются именно в этом порядке (см. табл. 14 и рис. 56).
Таблица 14
Произвольная классификация рассматриваемых островов
Современные материковые острова
Тропические
Суматра, Ява, Калимантан
Новая Гвинея
Цейлон
Тайвань
'Гринидад
Н отропические
Британские
Японские
11ьгофаундленд, Гренландия
Тасмания
Огненная Земля, Фолклендские Периферийные архипелаги (все тропические)
Филиппины
Острова западной части Тихого океана
В ес.т-И ндия
Другие острова и архипелаги (классифицируемые не
(;улавеси
Мадагаскар, Маскаренские, Сейшельские
11овая Каледония, .'1орд-Хау, Норфолк
1I овая Зеландия
Гавайские
Галапагос
Острова Атлантического океана
Антарктика
по климатическим признакам)
Для каждого из этих островов и архипелагов я приведу некоторые сведения об их местоположении, площади, природе и, возможно, истории, а также данные о составе и связях фауны позвоночных этих островов. Что касается периферийных архипелагов, то я также попытаюсь выяснить особенности распространения животных в пределах этих архипелагов.
Термины, содержание которых раскрыто в гл. 1, здесь следует при- менять с особой осторожностью: местный (туземный), зндемичный (иск- лючигггельный, специфический), пегггохтонный, реликтовый, интросгуциро- еанныи.
Р и с. 56. Карта островов, рассматриваемых в тексте,
Распространение животных на островах зависит 11т многих фак п1ров, в том числе от природы и способности различных животных к расселеншо. Все:это кратко рассматривалось для каждого класса позвоночных в гл. 2 — 6. Iso касается других внешних факторов, влияющих на распро- странение животных на островах, то некоторые из них, вполне очевидные и важные, настолько сложны и трудны для оценки. что я даже не пыта10ГЕ> определить их точно, а просто отмечу по ходу дела. К ним относятся экологические условия (наличие благоприятных условий и их разнообразие) разных островов; направление ветров и течений в прошлом и в настоящее время; природа ближайшего материка (древней или новой суши, эколо- г11чески благоприятной, имеющей болыпук1 или небольшую фауну, и реки с быстрым течением и т. д.); влияние конкуренции или ее сокращение.
Однако я хочу рассмотреть два других фактора, не столь очевидные, но которые оказывают очень существенное влияние на распространение и особенности размещения животных на островах. Этими факторамц являются площадь и расстояние (от материков).
Влияние ограниченности территории
Влияние ограниченности территории на количественный и качест- венный состав;кивотного населения островов имеет большое значение. Это видно из: сравнения фаун очень большого острова, с площадью в не- скол ько тысяч квадратных миль, и очень маленького, с площадью все- го 1 — 2 акра. Если история обоих островов одинакова, то есть если оба они были отделены от одного и того же материка недавно, то на большом острове материковая фауна будет представлена почти в полном составе.
сохранением чуть ли не всего состава наземных млекопитающих, но нн острове с небольшой площадью фауна будет очень ограниченной, а назем- ные млекопитающие могут быть совсем не представлены или представлены только 1 — 2 видами мышей. Таким образом, ограниченность территории мелких островов почти полностьк1 исключает всю фауну млекопитающих.
Ограничивающее влияние территории не проявляется внезапно. Определенного размера острова, обеспечивающего полный состав мате- риковои фауны и служащего как бы критерием, ниже которого внезапно начинается ее резкое сокращение, не существует. Такая зависимость увеличивается постепенно, по мере уменыпения размера острова. Факти- чески эти изменения начинаются еще на материках. На всех массивах суши — от самых больших до самых маленьких — имеются фауны, кото- рые ограничены территорией, пригодной для обитания, правда, влияние :1 г1>го ограничения очень сложное и видоизменяется под воздействием мно- гих других факторов. Фактически площадь земного шара в целом, видимо, очень сложным образом лимитирует число существующих на нем живот- ных (см. стр. 34, 35).
Имеющиеся в нашем распоряжении цифры позволяют представить более конкретно, как площадь ограничивает число некоторых животных на отдельных островах. Табл. 15 показывает число видов амфибий и реп- тилий, обитающих на четырех островах, входящих в состав Ьольпи1х Антильских островов, а табл. 16 — распределение числа этих 'ке живот- ных на четырех Малых Антильских островах. Анализ этих 1.аОлиц позв~1- ляет предположить, что в пределах размеров указанных 11ы1пе островов (кроме двух самых мелких) с изменением их площади в 1() раз видовой состав фауны рептилий и амфибий уменыпается 11 2 раза (тай 1. 17), но это
Особенности рас~гространения животных на островах
Таблица 15
,1ягушки и жабы. ящерицы и змеи Больших Антильских островови острова Триипдад1
I
Лягушки 1
и жабы
!
Ящерицы
!
1
Всего
Острою
:1мси
4:24-; 5 4: 27--'1
25: 76-'-16 29: 84-;-7
8: 15 '-1 !): 16-- '
13: 37-; 10 16: 41- -'4 8: 17--' ()
15: 39 -'-0 14: 4(1 -,'-1 55: 8()
2: 16~О 3: 13-'4)
6- -',-0
6-I-О 38
4:
7: 21--' 1
12: 23
15: 19
28:
1 сост'явлена llo источникам, приведенным в литературе в кони(;)той главы. Цифры I)()()- зиачают количество родов: виды плюс подвиды.
Таблица 16Ам(~)пбпп п рептилии четырех островов нз группы Малых Антильских(Dunn, 1934, р. 108)
! Высот» пад уров- нем моря, футы
Остров
Число видов
Таблица 17
Прпб;шзпте1ьное соотношение и.1ощадп к числу видовамфнОий и рептилий на некоторых островах Вест-Индии(см. табл. 15, 16)
I
I
Приблизительное
число видов
Действительноечисло видов
Приблизительная площадь, кв. миль
(76 — 84) (3!) — 40)
(о)
(5)
очень грубо приближенное соотношение, и для других случаев оно может не подойти.
Влияние площади видоизменяется в результате воздействия многих факторов, включая и такие, как протяхенность возвышенных частей суши и разнообразие зкологических условий, но, с другой стороны, площадь, вероятно, в свою очередь оказывает определенное воздействие на остров-
hy()a. около 4() ()()() кв. ми;1ь Гаити, 30 О()0 кв. миль Ямайка, 4500 кв. миль Пуэрто-Рико, 340() кв. миль Трииидад, 2()0() кв. миль
Моитсеррат, 8:-,5 миль Саоа, диаметром 2,5 мили Редонда, диаметром 1 миля Сомбреро, длиной 1 миля
40 00()4 000(400404
8~)
40 20) 10
3002
2820
100040
()
5 3 1
Глава 8
ных животных. В отдельных случаях ограниченность площади может влиять на размеры отдельных животных (но это влияние весьма ело;1;- ное,— Mertens, 1934, р. 120 — 123) и, вероятно, также на структуру попу- ляций и характер эволюции.
Влияние расстояния, иммиграционный п реликтовый характер распространения
Влияние расстояния — ширины водных пространств, отделяющих острова,— на состав и особенности распространения островных фаун также очень существенно. Чтобы понять это, необходимо располагать некоторыми сведениями о расселении наземных животных через водные пространства (Darlington, 1938).
Распространение наземных животных через водные пространства очень часто объясняется «случайным расселением» или «беспорядочным рассе- лением», но это не вполне приемлемые термины. Предпочтительно употреб- лять более нейтральный термин «расселение через водные пространства».
Расселение отдельных особей наземных животных через водные прост- ранства в значительной степени случайно (такие же случайности могут возникнуть и при расселении по суше), но с течением времени, когда на- чинает действовать статистическая вероятность, определяется, какие группы ~кивотных пересекают водные пространства чаще всего и каких островов онй преимущественно достигают. Если даже принять, что у рас- сматриваемой особи вида имеется только один шанс из миллиона преодо- леть данное водное пространство, то из многих миллионов шансов отдель- ные особи почти наверняка пересекут это пространство. А если учитывать прочие равные обстоятельства, то, скорее всего, они смогут преодолеть узкие водные пространства, а не широкие. Это ясно по крайней мере в общих чертах.
Влияние протяженности водных пространств на успешное расселе- ние не только обратно пропорционально расстоянию или площади расстоя- ния, оно зависит от величины потерь и степени смертности во время рас- селения. Во время расселения через соленые воды смертность большин- ства наземных животных, по-видимому, бывает высокой и образует гео- метрическую прогрессию: если при пересечении широкого пространства протяжением 100 миль из тысячи особей выживает одна, то из другой тысячи. оставшейся на второй сотне миль, выживает опять только одна особь и т. д. В таком случае, если из тысячи имеется только один шанс, при кото- ром рассматриваемая группа животных достигнет островов, отделенных 100 милями открытого морского пространства, то тогда из миллиона так- же будет только один шанс, что этот представитель достигнет острова. расположенного в 200 милях от места расселения, и только один из мил- лиарда, что он достигнет острова, отделенного 300 милями открытогс морского пространства.
По своей природе это явление усложняется биологическими факто- рами и разницей в природе и возрасте различных островов. направлением ветров, течений и многими другими факторами. Но в основном главную роль играет расстояние. Если другие факторы более или менее адекватны. то животные, расселяющиеся с материка на архипелаг, обычно сначала попадают на ближайшие острова, а затем уже распространяются на дру- гие острова через наиболее узкие морские проливы. В результате этого создается определенная последовательность, родственных форм, встре-
Особенности распространения животных на остпровах
чающихся на ряде островов, около которых проходят пути иммиграции. Это илмиграиионный характер распространения.
Существует и другой характер распространения, который заключается в следующем: когда-то группа животных была хорошо представлена на архипелаге, но затем начала численно уменьшаться и наконец на неко- T0pbIx островах исчезла совсем. Это могло произойти в том случае, если архипелаг был отделен от материка и на нем сначала существовала мате- риковая фауна, значительная часть которой позднее исчезла. В э=ом слу- чае у выживших элементов, вероятно, не будет наблюдаться прежняя пос- ледовательность в распространении на соседних островах, они встреча- ются на островах нерегулярно — отчасти на самых больших и самых ола- гOIIplIH THbIx для обитания островах независимо от их местонахождения и отчасти в результате случайного расселения. Это так называемый простой
Омииграционный xgpapmep расселения
~в
~еликаобый шракаер расселения
П
П
Р и с. 57. Сравнительная диаграмма простого имми- грационного характера распространения. возник- шего в результате расселения нескольких групп животных с материка на ряд островов, удаленных от него на различное расстояние, и реликтового характера распространения — результата частичного исчезновения древней фауны, общей для всех этих
островов в прошлом.
реликтовый характер распространения. Сравнение простого иммиграци- онного и реликтового характеров распространения показано на рис. 57.
Если новые иммигранты расселились на архипелаге, уже занятом реликтовой фауной, или если прежняя иммиграционная фауна частично исчезла в результате каких-то процессов (кроме замещения животными с основного материка), например в результате частичного затопления остро- вов, в этом случае возможно смешение иммиграционного и реликтового распространения. Замещение животными с материка образует отличный от других тип распространения, который описан ниже.
Иммиграционный характер распространения с течением времени видоизменяется, и эти изменения зависят от степени расселения рассматри- ваемых животных. Если степепь расселения низкая и если даже ближай- ших островов достигает небольшое число ветвей и еще меньшее их число проникает на более удаленные острова, то степень вымирания у эти х животных может быть очень небольшой. В этом случае более разнообраз- ная фауна и, по существу (по мере изменения материковой фауны). исключительно реликтовые группы должны сосредоточиться на ближайших к материку островах, а на более удаленных — менее разнообразные фау- ны (происшедшие от меньшего числа предков) без реликтовых групп. представленных на ближайших к материку островах. Но если степень рас- селения высокая, то ближайшие острова могут быть перенаселены и новые вселенцы могут заместить некоторых более древних животных. В этом слу- чае на ближайших и на более удаленных от материка островах должны
быть одинаковые по разнообразию фауны (пропорциональной площади острова), но на более дальних островах, вероятно, будут сосредоточены :эндемичные реликты — последние остатки групп, которые оыли замещ~,— ны на ближайших островах, так же как и на материке. На одном и том же архипелаге результат расселения различных групп животных будет различен. При соответствующих условиях большинство наземных мле- копитающих и амфибий, которые с трудом пересекают соленые воды. дожк- но сосредоточиваться главным образом на ближайших островах архипе.эа- га, где будут находиться и их эндемичные реликты. Другие животные.
НИЗКАЯ СТЕПЕНЬ РДССЕЛЕНИЯ ~атерик &ью~ий
ош о
л";ельни 'оброненные имми~ранты~ более ранние иммигрантымелякам, общие оля обоихостр Ш~~~еликтм, лриурочент~е к6лакнеиу осаро5у
и ю ю ю ю И Ю Ю Ю ю ю ю в И ю Ю
ВЫСОКАЯ СТЕПЕНЬ РАОСЕПЕНИЯ
~о5рененные
иикигранты более ранние
индигранты йликты, ойцие &я обоих
mmpodod' ~еликты, приуроченные
к 8альнену осводу
ю ю в а ю ю ю
ю ю в Ю
И ю и И ю И
Р и с. 58. Диаграмча, показывающая влияпие сте- ЦОНИ РЯССРЛИП[Я )ТЯ ZaPaz<~eP РЯССРЛРН[[Я ПО ГПСТЮ[Р
двух островов.
Бажды11 остров ограничивается (в связи с илощадь1о) фау- ной из п1ссти единиц. В первом случае степень расселения низкая, ближн11й остров не был перенаселен, а дальний остров ~«~лучил далеко не полную фауну; когда группа на материке исче:3ла (прерывистая линия), исключитель- ные реликты cTBJIII встречаться на ближнем острове. Во втором случае, когда степень расселения была высокой, ближний остров был перенаселен и некоторые более дрсв- ние группы исчезли там так же, как и на материке. а дальний остров получил полную фауну и на н 'N ста111
встречаться эндемичные реликты.
расселяющиеся более свободно, должны претерпеть большее замещение на ближайших островах, а их эндемичные реликты будут находиться на оолее удаленных
Это мо''но проиллюсрировать на Hpoc1ohl примере двух одина;() островов, расположенных один за другим (рис. 58). В этом случае у живот- ных с низкой степенью расселения наиболее разнообразная фауна и боль- шинство эндемичных реликтов должно быть на ближайшем острове. У жи- вотных с высокой степенью расселения на обоих островах должна быть примерно одинаковая по разнообразию фауна, но большинство эндемич- ных реликтов будет находиться на дальнем острове. Возможно, есть и Td- кие животные, у которых степень расселения и замещения п1эомеп;уточ- ная, что дает приблизительно равное число эндемичных реликтов на каж- дом острове. В этом случае иммиграционный характер распространения может походить на реликтовый. Но если:эти острова действительно оди- наковы по площади и местам обитания и если на них представлено доста- точное число ветвей, то вряд ли реликтовый характер будет походить на иммиграционный.
Несомненно, что различные особенности распространения в резуль- тате перерасселения древних животных и некоторых других причин будут
409
Особенносвги. распростгранения животных на острова~
более или менее видоизменяться. Наиболее вероятны осложнения среди самых активно расселяющихся животных. Животные с незначительными возможностями расселения, признаком чего является простота их имми- грационного характера расселения и встречаемость локализованных реликтов-эндемиков, главным образом на ближайших к материку островах. должны быть самыми надежными индикаторами прошлого.
Рассматривая площади островов и ширину проливов между ними, следует учитывать как события прошлого, так и настоящего, осооенно колебания уровня моря, которые происходили в плейстоцене. На протяже- нии всего плейстоцена, в период ледникового максимума, уровень моря, вероятно, был везде по крайней мере на 100 метров ниже, чем в настоящее время. Такое пони"кение увеличивало площадь многих островов и сокра- щало ширину многих проливов. Фактически оно должно было привести к соединению ряда островов, таких, как Суматра, Ява и Калимантан, с материком или друг с другом. С другой стороны, до плейстоцена и. ноз- ъ1ожно, в периоды межледниковья, когда ледники достигали незначител ь ных размеров или их не было совсем, уровень моря, вероятно, был на 20-- .:10 метров выше, чем теперь. Это, видимо, сокращало размеры одних ост- ровов, полностью затопило другие (например, некоторые из Багамских остро- вов) и увеличило ширину отдельных проливов между этилии островами.
Все только что сказанное об иммиграционных и реликтовых, смешан- ных и видоизмененных особенностях распространения носит предваритель- ный и предположительный характер. Остается убедиться, встречается ли предполагаемый нами характер распространения на самом деле.
Далее приводится краткое описание островных фаун. Дополнитель- ные сведения о некоторых из них приводятся по другим работам (см. спи- сок литературы), а многие животные островов рассматриваются более под- робно в списках семейств.
Материковые острова
Сулгагггра, К«лиманиг«н, Яви. Большие острова Суматра, Калимантан н Ява (рис. 48) расположены близ юго-восточной части Азии. 11лощадь Суматры приблизительно равна 167 тыс. кв. миль, Калимантана— 290 тыс. и Явы — 49 тыс. кв. миль. Это тропические острова с богатой расти- тельностью, с разнообразными экологическими условиями и с высокими горными хребтами. С точки зрения геологии — это древние и сложные по своей структуре острова (Umbgrove, 19 ~9, р. 35 и след.), но с точки зре- ния зоогеографии — это современные материковые острова. Они отделе- ны друг от друга и от Азии мелководными проливами. глубина которых большей частью не превы|пает 50 метров (рис. 59). Все эти острова в плей- стоцене, а возможно, и в более позднее время были соединены как между собой, так и с материком: согласно Бьюфорту (Beaufort, 1951, р. <4). полуостров Малакка и острова Суматра и Ява соединялись в пределах ран- них исторических времен.
Фауна этих островов почти полностью материковая, и многие ее виды весьма сходны с материковыми видами. В этой фауне хорошо представле- ны пресноводные рыбы, амфибии, рептилии, птицы и млекопитающие. Более того, значительная часть фауны в основном распространена на всех трех островах, хотя в ее распределении отмечаются отдельные неравномер- ности. Перечислить здесь всю фауну в целом невозможно, но некоторые существенные детали распространения рыб и млекопитан~щнх заслужива-
410
Гливо, 8
ют внимания. Причем особое значение придается пресноводным рыбам и наземным млекопитающим, так как они более, чем другие животные, сви- детельствуют о прошлом Земли.
Распространение пресноводных рыб не соответствует современной суше. Между рыбами Суматры и западной части Калимантана больше черт сходства, чем между рыбами западного и восточного Калимантана. Это объясняется тем (и распространение рыб отчасти подтверждает это), что когда эти острова соединялись между собой и с материком, то огромная площадь суши, которую они образовывали, дренировалась несколькими
° Ф' °
° Э
° Э
у
1
/i
'1~ О г
1
1
Р и с. 59.
3 J) '~ипил а га
П редполагаемые очертанпя И ндо-Австралпйско1 о в плейстоцене при понижении уровня океана
(пунктирная линия).
Затопление Зондского шельфа в прошлом ноказано ире~)ывистой линией, соединяющей существующие реки (упрощено, по Кюнену — Kue- lien, 1950). Затопление Сахульского шельфа представлено схематически
(подробности предполагаемые).
отдельными реками. Из них отметим особенно одну, текущую на север, между Суматрой (и материком) и Калимантаном, и другую, текущую на восток, там где теперь находятся острова Ява и Калимантан (рис. 59). Поэтому эти рыбы и до настоящего времени распространены в соответ- ствии со старыми речными системами. Однако история этих рек и насе- ляющих их рыб, очевидно, не так уж проста. На Яве больше пресновод- ных рыб, общих с Суматрой, чем с Калимантаном, словно обмен между Явой и Калимантаном был прерван значительно раньше. Более того, на Яве больше рыб, общих с рыбами реки Махакам в восточном Калиманта- не (см. рис. 59), чем с южным Калимантаном, что, очевидно, можно объ- яснить только изменением характера стока или сукцессией в фауне рыб (Dammerman, 1929, р. 17; Beaufort, р. 24, 25).
Богатая фауна млекопитающих Индо-Малайской области представ- лена на всех трех островах, но в ней уже проявляются такие черты, кото- рые, видимо, означают начало влияния изоляции и ограниченности тер- ритории. Наименьшее количество млекопитающих находится на самом небольшом по величине острове Ява (табл. 18). (Но на Суматре, хотя этот остров по размерам меньше, чем Калимантан, больше групп млекопитаю- щих, чем на последнем,— влияние расстояния от материка или времени?)
Особениост и распространения ж ивот пых на ост ровит.
Таблица 18
Сравнение фаун млекопитающих островов Суматра, Яваи Калимантан 1
1Калимантан1Ява(около 29 тыс. ~ (около 49 тыс.HB. миль) ' hB. ми ~ь)
Суматра(около 167тыс. кв. миль)
1 наземные млекопитаю-
05
зо
47 «8
щие. единицы
;:летучие мыши, роды
1 Цифры взяты из списков Картера, Хилла и Тейта (Саг(ег, Hil l. Та1е, 1945). Эти списки составлены частично из видов, а частично из родов (в тех r руппа к. в которых виды были не полностью определены). Таким образом, при- «еденныс цифры характеризуют не число видов или родов, а условное среднее (единицы). Животные, которые, вероятно, были интродуцированы на островах— слон на Калимантане, заяц — на Яве, на этик островах не учитывались.
Ьолее того, на разных островах, видимо, происходило вымирание, что приводило к началу образования реликтового типа распространения. Из хорошо известных крупных млекопитающих леопард встречается (из трех этих островов) в настоящее время только на Яве; носатая обезьяна приурочена исключительно к Калимантану; бантенг встречается только на Калимантане и Яве; орангутан, медведь и двурогий носорог — на Су- матре и Калимантане; красный волк, тигр, однорогий носорог — на Су- матре и Яве; слон, тапир и серао — на Суматре (более подробно см. Dam- merman, 1929, р. 4 — 7, 22 — 24). Некоторые из этих видов встречаются так- же на азиатском материке, а другие приурочены к островам. Ряд крупных млекопитающих, которых теперь на Яве нет, существовали там в плейсто- цене. К ним относятся — орангутан, несколько крупных кошачьих, гие- на, медведи. несколько родов слонов, род Hippopotamus, тапиры, антило- пы и более одного вида бычьих (Koenigswald, 1939). Таким образом, совершенно ясно, что эта фауна первоначально не была такой обедненной и что она сократилась. Однако некоторые детали распространения и обед- нения. пожалуй, можно отнести еще к тому времени, когда этот остров не был изолирован, а ограниченность территории может влиять на полу- острова так же, как и на острова. В целом этот вопрос слишком сложен для обсуждения на страницах этой книги. Даммерман (Dammerman, 1929) дает дополнительные сведения и проводит сравнение фауны Явы с фауна- ми других островов.
Новая Гьинея. Географические связи Новой Гвинеи с Австралией те же самые, что и у Суматры и других островов с Азией. Она расположена на материковой отмели Австралии и отделяется от материка узким и не- глубоким проливом (а в плейстоцене вообще соединялась с ним). Площадь острова немногим больше 300 тыс. кв. миль (самый большой, пригодный для обитания животных остров земного шара). Это тропический остров с богатой растительностью, с разнообразными экологическими условиями и очень высокими горами.
Фауна позвоночных Новой Гвинеи уже рассматривалась в предыду- щей главе как часть фауны Австралийской области. В Новой Гвинее, как и в Австралии. отсутствуют или очень бедно представлены строго пресно- водные рыбы. Здесь имеются те же семейства лягушек, что и в Австралии.
Несмотря на то что большинство позвоночных Цейлона является индийскими или родственно индийским, отдел ьные представители(например, ящерицы и змеи, стр. 169, и некоторые птицы, стр. 2' 6) имеют самые близко- родственные связи с аналогичными представителями Индо-Китайской и Малайской подобластей. Это географические реликты, однако на Цейлоне такие реликты, вероятно, не более многочисленны, чем в южной части Индии.
Тпйван ь. Остров Тайвань находится на материковой отмели, примерно и 90 милях от пооережья Китая, в краевой зоне тропиков. Его площадь рав- на приблизительно 14 тыс. кв. миль. Это остров с высокими горами и бо- гатыми разнообразными экологическими условиями. Г, материком он, оче- видно. был соединен недавно, так как его фауна в основном такая же, как и в Южном Китае, но менее разнообразная.
Фауна Тайваня довольно хорошо представлена всеми основными классами позвоночных. Здесь следует остановиться на рыбах и млекопи- тающих. Из строго пресноводных рыб на Тайване имеются по крайней мере 42 вида карповых и карпообразных, 8 видов сомов и 4 вида лабиринто- вых рыб (Мог1, 1936). Эти рыбы очень хорошо представляют доминирун>- щие группы пресноводных рыб Индо-Малайской области. Из млекопитан~- щих Картер, Хилл и Тейт приводят 5 родов насекомоядных, включая кро- та, 1 род обезьян, 11 родов хищных, включая азиатского черного медведя и леопарда (Рейз nebulosn), панголина, 5 родов копытных, включая сви- ней, 3 оленей и серао — единственного местного представителя бычьих, 1 род зайцев. целый ряд грызунов и рукокрылых. Фауна млекопитак>щих Тайваня несколько беднее фауны Цейлона и также характеризуется отсут- ствием большинства крупных млекопитающих, включая и большинство бычьих. Можно добавить, что авифауна этого острова приводится в рабо- те Хачисуки и Удагавы (Hachisuka, Udagawa, 1950 — 1951), которые так- 'ке дают описание самого острова и его растительности. Птицы Тайваня многочисленны, но их численность здесь меньше, чем на сравниваемых пло- щадях в тропиках азиатского материка.
Тринидпд. Этот остров находится недалеко от северного побережья Ю'кной Африки, вблизи Венесуэлы. Несмотря на то что площадь его менее 2 тыс. кв. миль, это довольно разнообразный по природным условиям тропический остров с, обширными низменностями и горами, достигающими примерно 4000 футов высоты. Тринидад расположен на материковой отме- ли и отделяется от материка проливом шириной около 10 миль и глубиной не более 15 метров. Очевидно, этот остров еще совсем недавно соединялся с материком. (: географической точки зрения этот остров иногда рассма- тривают как один из островов Вест-Индии, но с точки зрения зоогеографии он сильно отличается от нее, так как здесь представлена умеренно богатая материковая фауна всех основных классов позвоночных и многие виды очень сходны с материковыми.
ресноводные рыбы на острове Тринидад (Eigenmann, 1909, р. 327, 328, и другие авторы, кончая Ингером — Inger, 1949) представлены разно- видностями харациновых и пресноводных сомов, которые доминирун~т в IOêi;voii Америке, но не достигают (выражаясь зоогеографическ11) настоящих островов Вест-Индии. Амфибии и рептилии этого острова более многочисленны и по родовому составу более разнообразны, чем в собствен- но Вест-Индии (табл. 15), и вклн~чают по сравнению с фауной материка относительно большее число змей и меныпее — ящериц. Птицы Тринидада значительно многочисленнее птиц собственно Вест-Индии: Бонд (Bond.
1936, р. ХХ) пишет, что если птиц Тринидада и соседнего, меныиего по
Глава 8
~Ф
ф/
+%%/ /
g Ъ Ъ
Ъ
размерам, острова Тобаго добавить к авифауне Вест-Индии, то они почти удвоили бы число родов и прибавили бы к ним 14 неотропических семейсти, по тем или иным причинам не представленных в Вест-Индии.
К местным млекопитающим Тринидада (Vesey-FitzGerald, 1936) отно- сятся по крайней мере 4 вида опоссумов, 2 вида обезьян, 2 — муравье- дов, 1 — броненосцев, 1 представитель кошачьих (оцелот), 2 вида куньих, 1 — енотовых, 1 вид оленей, пекари, 1 вид белок, 9 — мышиных, 6— дикобразоподобных грызунов и 34 вида рукокрылых. Для такого малень- кого острова это очень богатая фауна. Возможно, что этот список непол- ный, но отсутствие в нем некоторых крупных млекопитающих, включая ягуара, пуму и тапира, имеет существенное значение.
Британские острова. Они состоят (рис. 60) из двух основных остро- вов, расположенных рядом друг с другом у побережья Европы: Велико- британии, занимающей около 89 тыс. кв. миль, и Ирландии — около 32 тыс. кв. миль. Климат этих остро- вов умеренно-холодный, но его суро- p~ > е. Расеи ф~ ~ вость смягчается теплым Северо-Атлан- тическим течением. Эти острова с низ- кими горами и небольшими низменно- ~/
с, ф/ стями имеют (в пределах климата) ~/ / разнообразные экологические условия.
Британские острова расположены на . Скомер -"
Ф материковой отмели и не так давно ~в~ „,~~~" рО ~ф,ф соединялись с материком. Вероятно, eel 4 Великобритания отделилась от мате- рика примерно около 7 тыс. лет назад, р и с. 60. Британские острова. а Ирландия отделилась от Великобри- прерывистая линия — приблизительная тании, очевидно, несколько раньше. Из
основных работ по позвоночным Бри- в иле~стоцене представляла сушу. танских островов следует упомянуть
список позвоночных (Hinton et. al., 1935), амфибий и рептилий (M. Smith, 1951), млекопитающих, вклю- чая главу о происхождении фауны млекопитающих Британских остро- вов (МаИЬепя, 1952) и естественную историю Ирландии (Praeter, 1950).
Фауна Британских островов охватывает представителей всех основ- ных классов позвоночных. Все виды являются европейскими или близко- родственны им, и вся фауна в целом, по крайней мере наземных позвоноч- ных, представлена в Великобритании и только часть ее — в Ирландии. Из строго пресноводных рыб в Великобритании распространены щуки (которые проникают в Ирландию), 11 видов карповых (из них пять захо- дят в Ирландию), два вида бычков (один проникает в Ирландию), два вида окуневых (один вид проникает в Ирландию). Из амфибий в Британии встре- чается три вида саламандр (Triturus — тритоны), два вида жаб, один мест- ный и, вероятно, два интродуцированных вида лягушек (Лапа). Но из шести местных видов земноводных только один (саламандра) встречается в Ирлан- дии. Из рептилий имеется три вида ящериц (Aïgèãs /ragilisи два вида рода Lacerta) и три вида змей (включая европейскую гадюку), но из этих шести местных видов рептилий только один (Lacerta vivi para) распространен в Ирландии. В Ирландии отсутствуют змеи и, вероятно, местные лягушки (обыкновенная лягушка, а также, возможно, и жаба Bubo calamita, коло- ния которой локализована в юго-западной части Ирландии, вероятно, интродуцированы). Черепахи в настоящее время отсутствуют как в Вели-
Особенности распространения животных на островах
кобритании, так и в Ирландии. Птицы Британских островов слишком многочисленны, чтобы рассматривать их здесь подробно.
Из млекопитающих на острове Великобритания распространены еж, 4 вида землероек и 1 вид крота (Ирландии достигают только еж и 1 вид землероек); красная лисица (также в Ирландии), 6 видов куньих (также в Ирландии), дикая кошка (в Ирландии отсутствует); 2 местных вида оле- ней (1 в Ирландии), 3 местных вида зайцев и кроликов (2 встречаются в Ирландии в естественном состоянии), 11 видов грызунов и 12 видов рукокрылых (некоторые из них встречаются в Ирландии). Кроме этих животных, в Великобритании в прошлом водились волки, бурые медведи, дикие кабаны и бобры. Первые 3 вида встречались также и в Ирландии.
В течение плейстоцена позвоночные Британских островов претерпе- ли очень сложную историю. Последовательно сменяющиеся фауны дости- гали островов (которые в то время были частью материка) в ледниковую и межледниковую эпохи, затем сокращались или исчезали. Мэтьюс (iMat- thews, 1952, р. 364 и след.) проследил отдельные моменты из истории мле- копитающих. Однако, за исключением нескольких субарктических релик- тов, современная фауна является совершенно новой, полностью состоящей из постплейстоценовых иммигрантов. Часть (только часть) постплейсто- ценовой фауны Западной Европы достигла Великобритании и часть (толь- ко часть) фауны последней достигла Ирландии, которая была изолирована раньше, чем Великобритания. Таким образом, основной характер распро- странения на этих островах можно представить как простой иммиграци- онный. Однако в деталях это распространение значительно сложнее, чем это представлено. К северу фауна Британских островов становится более малочисленнои, а многие виды в то?ю или инои степени — локализованными.
Если бы эти острова остались изолированными и сохранили свою фауну, в то время как фауна материка была уже изменена, то на Велико- британии — острове, ближайшем к материку,— находились бы эндемич- ные реликты, включая такие, как безногая ящерица семейства Anguidae и крот. В Ирландии нет наземных позвоночных, которые не были бы пред- ставлены также и на острове Великобритания. Однако среди мелких гры- зунов происходили отдельные местные замещения. Две формы европей- ской рыжей полевки Clethrionomys glareolus, очевидно, последовательно заселили Великобританию. Группа С. nageri, видимо, вселилась первой. Эта группа уже не смогла достигнуть Ирландии, но она заняла маленькие островки, расположенные в непосредственной близости от западного побе- режья Великобритании, которые в то время, возможно, еще были соедине- ны с основным островом. Собственно группа С. glareolus вселилась позд- нее. Она полностью заместила группу С. nageri на острове Великобрита- ния, но ее представители до сих пор сохранились на мелких островах западного побережья — Малл, Расей и Скомер. Мэтьюс, основываясь на распространении и сведениях об ископаемых, допускает и другие случаи замещения среди полевок (рода ЛХгсго~иг) в Великобритании. Несколько этих примеров показывают, как последовательные иммиграции могут замещать реликты на наиболее и наименее досягаемых островах архи- пелага.
Япония. Японский архипелаг представляет собой цепь островов, рас- положенных параллельно восточному побережью Азии. Самыи большои остров, Хонсю, занимает примерно 87 тыс. кв. миль. лежащий к югу от него остров Кюсю — около 16 тыс. кв. миль и Сикоку — около 7 тыс. кв. миль. Этп три острова образуют основную часть, иногда называемую соо- ственно Японией. Остров Хоккайдо, площадью около 30 тыс. кв. миль,
1'.гава 8
расположен на севере. Климат этого острова более холодный, и его фауна более бедная, почти чисто палеарктическая. Острова собственно Японии имеют умеренно-теплый климат и разнообразные экологические условия. Здесь есть как низменности, так и высокие горные хребты. Эти острова сое- динены с материковой отмелью Азиатского континента и, видимо, в плей- стоцене были соединены с последним. Это предположение вполне подтвер- ждается фауной этих островов.
Строго пресноводные рыбы собственно Японии представлены сорока с лишним видами карповых и тремя видами сомов. Эти рыбы представляют смесь североазиатских групп (особенно в южном направлении) с южно- китайскими и с некоторыми эндемичными родами карповых (Mori, 1936. р. 10 — 14; Okada, 1955). Из амфибий в Японии находятся саламандры (3 се- мейства) и лягушки (3 семейства, 5 родов и около 10 видов в собственно Японии,— Okada, 1931, р. 8). Амфибии также представляют смесь север- ных, южных и эндемичных видов. Рептилии Японии представлены энде- мичными черепахами: подсемействами Emydinae и неэндемичным родом Triionyx, несколькими видами гекконов, сцинков и ящериц семейства La- certidae, некоторыми видами ужовых и ямкоголовой гадюкой (Agkistro- don). Птицы Японии многочисленны. Млекопитающие в Японии представ- лены 6 родами насекомоядных, включая крота, обезьяной макакой, тре- мя видами собачьих, двумя видами медведей и шестью видами куньих. диким кабаном, оленем, серао, тремя видами зайцев и т. д., десятью рода- ми грызунов и 11 родами рукокрылых, включая Pteropus (летающие лисицы).
Основная часть Японии по площади примерно равна Великобритании, и Японские острова можно отчасти сравнивать с Британскими, но надо учитывать, что климат Японии теплее и эти острова не были покрыты лед- ником, и поэтому фауна Японии более разнообразная, древняя (более диф- ференцированная) и смешанная по своим географическим связям. Диффе- ренциация фауны в какой-то степени, вероятно, предшествовала разъеди- нению островов с сушей, так как горы Японии, видимо, до этого представ- ляли изолированные стации. Смешанные связи (северные и южные) япон- ской фауны дают основание предположить изменения в распространении некоторых групп млекопитающих с тех пор, как сформировалась остров- ная фауна.
Ньюфпундленд и Гренландия. Остров Ньюфаундленд расположен около северо-восточной части Северной Америки и залива Св. Лаврен- тия. Он занимает площадь примерно 42 тыс. кв. миль. Несмотря на то что этот остров лежит южнее Британских островов, климат его более холодный — от умеренно-холодного до субарктического. Остров покрыт об- ширными хвойными лесами и тундрой. Он соединен с материковой отмелью Северной Америки со стороны Лабрадора, где в настоящее время отделяет- ся от материка 10-километровым проливом. В плейстоценовый период этот остров в какой-то мере был покрыт ледниками, но соединялся ли он затем сушей с материком, до сих пор не известно. Для фауны острова это соеди- нение, вероятно, не имело существенного значения. Можно предположить, что наземные млекопитающие проникли на остров по льду, так как Нью- фаундленда достигло больше крупных, чем мелких млекопитающих.
Фауна Ньюфаундленда очень ограниченная. В ней отсутствун>т стро- го пресноводные рыбы и нет, вероятно, местных амфибий (была интродуци- рована по крайней мере одна лягушка) и рептилий. Птицы многочисленны. К местным млекопитающим (Bangs, 1913) относятся волк, черный мед- ведь, 5 видов более мелких хищных (белый медведь и песец изредка прони-
кают сюда с дрейфующими льдами), карибу, полярный заяц, бобр, ондатра, полевки (1lIicrotus) и один вид рукокрылых. Однако многие млекопитаю- щие. встречающиеся на полуострове Лабрадор (включая 16 родов и под- родов, перечисленных Бангсом), отсутствуют на острове Ньюфаундленд: здесь нет лося, землероек, дикоораза, белок и только одна мышь (кроме рукокрылых) является единственным мелким местным млекопитающим, если только она де~~ствительно является местно~~.
Гренландия (Clark, 1943) находится на далеком севере и расположена ближе к Северному полюсу, чем любая другая суша. Это — колоссаль- ный остров площадью примерно 840 тыс. кв. миль, но вся территория, за исключением приблизительно '130 тыс. кв. миль, в настоящее время покры- та льдом. Обитаемы только некоторые окраинные части острова. С гео- логической точки зрения это материковый остров, но фауна современных позвоночных, очевидно, достигла этого острова недавно, через льды, а не через сушу. В Гренландии, конечно, нет строго пресноводных рыб, амфи- оий и рептилий. Очень мало гнездящихся наземных птиц (Salomonsen, Gitz-1 ohansen, 1950 — 1951): тундряная куропатка (американская форма), два вида соколов, белохвостый орлан (евроазиатский), болотная сова (гнездование этого вида вызывает сомнение), белая сова, гренландская каменка(евроазиатская), дрозд-рябинник (евроазиатский), два вида коньков (американский и евроазиатский), белая трясогузка (евроазиатская), чечетка (два подвида), лапландский подорожник, пуночка и ворон. Птицы, особо не оговоренные в этом списке, относятся к циркумполярным видам. Я не пытался определять связи гренландских популяций. Дрозд-рябин- ник — это североевропейский дрозд (Turdus), вероятно, ставший оседлым в южной Гренландии после трансатлантического «нашествия» в январе 1937 г. (Salomonsen, 1951). Еще целый ряд наземных птиц был зарегистри- рован как случайно залетные, но многие водные птицы гнездятся в Грен- ландии. Из наземных млекопитающих в Гренландии обитают песец, волк, белый медведь, горностай, карибу, мускусный бык, полярный заяц, лем- минг. Все они выходцы из арктических районов Северной Америки.
Тасмания. Площадь этого острова, расположенного к югу от Австра- лии. равна примерно 26 тыс. кв. миль. Климат умеренно-прохладный, эко- логические условия довольно разнообразны, имеются невысокие горные хребты. Этот остров находится на материковой отмели Австралии и, как показывает фауна, еще недавно соединялся сушей с материком. Позвоноч- ные Тасмании приводятся в работе Лорда и Скотта (Lord, Scott, 1924).
На Тасмании, так же как и на большей части Австралии, отсутствуют строго пресноводные рыбы, но распространены многочисленные галакси- ды. Из лягушек острова достигает три рода семейства Leptodactylidae и род Ху1а, а из рептилий — несколько видов ящериц и три вида змей семейства Elapidae, но нет ни одного вида черепах. Птицы многочисленны. Из млекопитающих на Тасмании представлены ехидна и утконос, разные виды сумчатых (Dasyuridae, бандикут, Phalangeridae, вомбат, кенгуру и
д.), несколько родов мышиных грызунов, включая род Hydromys и 6 видов рукокрылых. Эти животные или относятся к австралийским видам, илп родственны им, или это виды, которые когда-то встречались в Австра- лии. Самые поразительные из них — сумчатый волк и сумчатый дьявол (хищные сумчатые довольно крупного размера). которые теперь в Австра- лии не встречаются. но пх ископаемые обнаружены в плецстоценовых отложениях этого материка.
Огненная Земля и Фолклендские острова. Огненная Земля, располо- женная у южной оконечности Южной Америки, отделяется от материка
418
очень узким проливом. Основной остров занимает площадь примерно и 18 тыс. кв. миль, а мелкие прилежащие острова увеличивают {ннцу{о площадь на юг от Магелланова пролива примерно до 27 тыс. кв. миль. !ми- ма~ умеренно-холодный, варьирующий от места к месту от влажного д« засушливого. Таким образом, на островах имеются и лесные, и открытые места обитания.
На Огненной Земле представлены те же рыбы, что и на южной ок«неч- ности материка: солевыносливые формы, включая галаксид. Амфибии, которые достигают этого острова, представлены только одной-двумя небольшими лягушками, а рептилии — только двумя-тремя мелкими ящери- цами из семейства игуановых. К наземным птицам относятся по крайней мере хищные, совы, попугай, дятел, мухоловки, кассиковые и еще несколь- ко видов других певчих птиц, но на Огненной Земле отсутствуют многие семейства, характерные для тропических дождевых лесов, и, очевидно, также некоторые птицы южных районов 1Ожной Америки: например, реа, тинаму и муравьеловки. Южные границы распространения этих птиц определяются климатом, а не водными преградами, хотя для таких птиц, как реа, вода служит преградой. Млекопитающие этого острова представ- лены лисицей, двумя видами выдры, ламой, несколькими хомячьими гры- зунами, одним видом дикобраза (Ctemomys) и рукокрылыми семейства Vespertilionidae. Но здесь отсутствуют сумчатые, неполнозубые или обезь- яны и обитает, как сказано, только один вид дикобразов. Насколько мне известно, в фауне позвоночных Огненной Земли нет животных, которые могли бы считаться важными реликтами.
Фолклендские острова находятся примерно в 300 милях к востоку {д
южной оконечности Южной Америки. Площадь двух основных островов составляет примерно 2500 и 2000 кв. миль. Высшая точка не превышает 2000 футов над уровнем моря. Климат островов умеренно-прохладный, экологическая ограниченность выражается в отсутствии древесной расти- тельности — для островов характерна только низкорослая растительность.
Здесь нет строго пресноводных рыб, амфибий и рептилий. Из птиц имеются хищные, совы и несколько видов мелких наземных и водных птиц. Един- ственное существующее (или существовавшее), несомненно местное назем- ное млекопитающее,— это огромный эндемичный вид лисицы, родствен- ный южноамериканским лисицам (Dusicyon). Дарвин (Darwin, 1839) «предполагал, что на этих островах есть полевки», но, насколько мне известно, его предположение не подтвердилось. 11редставители семействн бычьих, лошади, свиньи, зайцы, кролики, мышиные родов Rattus и .Ииг оыли интродуцированы. Эндемичный вид лисицы исчез примерно в 1875 г. Очевидно, эта лисица питалась птицами или тюленями и другими береговы- ми животными. Как она достигла Фолклендских островов — не известя{{. Эти острова находятся на материковой отмели и, возмо>кно, в прошлом сое- динялись сушей с Южной Америкой, однако странно, что этим соединением воспользовалась лишь лисица. Видимо, небольшая площадь и ограничен- ность мест обитания на островах ограничили фауну, а лисица уничтожила всех мелких млекопитающих, которые туда попадали. Вероятно, это была предпоследняя стадия исчезновения фауны млекопитающих на слишком маленьком и слишком неблагоприятном для обитания острове. Или вп{>л- не возмох'но, что в плейстоцене лисица достигла Фолклендских островов на дрейфующих льдах (так же, как песец достиг Исландии) и до сих пор изредка проникает на остров Ньюфаундленд.
Заключение о материковых островах. Рассмотренные выше материко- вые острова разнообразны, но их фауны имеют определенные характерные
Особенности рас)гроснгранения ж'ивотных на островах
419
общие черты. Некоторые моменты очевидны или только предполагаются, другие же, возможно, более трудно определимы.
Состав фаун современных материковых островов, конечно, не огра- ничивается только теми животными, которые обладают способностями пересекать водные пространства. Эти фауны, насколько это допускают климатические преграды, включают обычно представителей всех основных классов позвоночных. Но некоторые из них ограничены скорее климатом плейстоцена, чем современным.
Расстояние (ширина проливов) не является существенным фактором для расселения фауны материковых островов. Цейлон находится в 30 ми- лях от азиатского континента, а Тайвань — в 90 милях, но на обоих ост- ровах имеются довольно сходные фауны. В тех случаях, когда, как кажет- ся, имеет значение расстояние, на самом деле действуют другие факторы. Например, тот факт, что фауна Ирландии беднее фауны Великобритании, вероятно, связан не просто с удаленностью этого острова от материка, а с тем, что Ирландия была изолирована раньше, чем Великобритания, а также с тем, что в экологическом отношении это менее разнообразный остров.
Такой совершенно очевидный или по крайней мере предполагаемыи фактор, как влияние размеров площади на ограничение и изменение фаун материковых островов, уже неоднократно рассматривался выше. По-види- мому, на всех, включая и большие, островах наблюдается общая тенден- ция к исчезновению из островных фаун крупных млекопитающих. Влияние территории увеличивается со временем, и, возможно, пройдет довольно много времени, прежде чем ограниченность территории в полной мере скажется на таких больших островах, как Суматра, Калимантан и Ява. Значительно быстрее действие этого фактора проявляется на островах меньшего размера и с менее разнообразными природными условиями, таких. например, как Фолклендские.
Действие фактора времени на фауны материковых островов в отдель- ных случаях очевидно. Сравнение очень молодой фауны Британских ост- ровов с древней японской фауной позволяет предположить, что время при- вело не только к исчезновению некоторых крупных млекопитающих в Япо- нии, но, по мере изменения фауны на материке, увеличило эндемизм и раз- нообразие связей фауны острова.
Влияние климатической зональности на фауны материковых островов в основном сказывается так же, как и на фауне материков: чем холоднее климат, тем меньше разнообразие фауны. Суровый климат может ограни- чивать состав фауны самым неожиданным образом. Например, на острове Огненная Земля, имеющем холодный климат, не только отсутствует боль- шинство амфибий и рептилий, но даже и строго избранная фауна млеко- питающих, состоящая почти полностью из представителей групп, которые. распространяясь с севера, достигли Южной Америки сравнительно недав- но. Единственным древним южноамериканским млекопитающим на этом острове является один дикобразоподобный грызун. Иначе говоря, вся древ- няя часть характерной фауны млекопитающих КЬкной Америки здесь отсутствует. При других обстоятельствах подобное положение может вызвать сомнение — действительно ли островная фауна формируется из элементов фауны близлежащего материка.
Что касается характера распространения на материковых архи- пелагах, то Великобритания и Ирландия образуют серию из двух остро- вов, из которых дальний остров был менее досягаемым, чем остров, нахо- дящийся вблизи от материка, и начиная с плейстоцена наземные позвоноч-
Глаыа 8
420
ные этих островов имели довольно простой иммиграционный характер рас- пространения. Если эти острова были постоянно изолированны и если фауна материка изменялась, то основные эндемичные реликты должны нахо- диться на ближнем острове (Великобритания). Однако даже в этом случае некоторые подробности распространения мелких млекопитающих позво- ляют предполагать начало замещения фауны на ближнем острове с релик- тами, изолированными на мелких прибрежных островах, расположенных к западу от Великобритании.
Суматра, Калимантан и Ява показывают, что частичное исчезновение первоначально полного состава материковой фауны, видимо, является началом реликтового характера распространения на этих островах. Все это вполне согласуется с теоретическими рассуждениями, приводимыми в на- чале этои главы.
Периферийные архипелаги
Филиппинские острова. Филиппинские острова (рис. 61) расположены между Южным Китаем и Большими Зондскими островами, но в зоогеогра- фическом отношении это периферийный архипелаг, простирающийся на северо-восток от Калимантана почти на тысячу миль. Наиболее крупный остров Лусон, с площадью примерно 40 тыс. кв. миль, является в то же время главным''и наиболее удаленным от центра островом этого архипелага. Остров Минданао, с площадью около 35 тыс. кв. миль, находится на юго- восточном конце архипелага. Палаван, ближайший к Калимантану ост- ров, занимает площадь всего около 4500 кв. миль. Другие острова, распо- ложенные на незначительном расстоянии друг от друга между Минданао и Лусоном, имеют площадь примерно 4000 — 5000 кв. миль. Это тропиче- ские острова с разнообразными экологическими условиями. Некоторые из них имеют высокие горы. Филиппины расположены по краю материковой платформы Азии и имеют сложный геологический комплекс, но большей частью эти острова вулканического происхождения («Encyclopaedia Brit- tannica», 14 edition). Более подробно история Филиппинских островов рассматривается Ингером. Однако фауна позвоночных, по-видимому, не отражает всей длинной или сложной истории островов. Природа и распро- странение фауны Филиппин даны в работе Диккерсона (Dickerson et. al., 1928), но по некоторым вопросам она уже устарела.
Единственные пресноводные рыбы первого отдела на Филиппинских островах (рис. 11) — несколько мелких карповых, которые встречаются по всему острову Палаван до Миндоро и (в другом направлении) на остро- ве Минданао; один вид сомов обитает на острове Палаван и островах Каламиан, а другой вид — на острове Минданао. Не только на острове Иалаван, где много пресноводных рыб Калимантана, но также и на цент- ральных и на внешних Филиппинских островах нет местных строго прес- новодных рыб. А те немногочисленные мелкие формы, которые обитают на южных Филиппинах, вероятно, обладают некоторой солевыносли- Н ОСТЬЮ.
Амфибии Филиппинских островов (I»ger, 1954) сравнительно много- численны. Один широко распространенный вид безногих Индо-Малайской области достигает островов Палаван и Минданао. Лягушки встречаются на всех пригодных для обитания островах, но разные семейства и роды имеют различные границы распространения, так что и на Филиппинах они проникают на различные расстояния (Dickerson et. al., 1954, р. 219,
Особенности распространения животных на островах
421
Inger, 1954). Видимо, большинство из них проникло с острова Калимантан через Палаван или Минданао, но путь родов ОтеорЬтупе и Cornufer, рас- пространенных от Новой Гвинеи до Филиппин (исходя из их современного
Р и с. 61. Предполагаемые очертания Филиппинских островов в плейстоцене при понижении уровня океана (пунктирная
линия).
Некоторые из предполагаемых соединений суши сомнительны. Прямая линия — предельная граница распространения указанных на карте
животных с Калимантана на Филиппины.
распространения), лежал через Сулавеси или Молуккские острова. Энде- мичный род Barbourula примитивного семейства Discoglossidae известен только на острове Бусуанга (из групп островов Каламиан, лежащих к северу от острова Палаван). Возмо'кно, что этот род является реликтом (может быть, и не очень древним), но вполне возможно, что он имеется также на острове Палаван или дахсе на острове Калимантан.
Что касается рептилий, то на Филиппинах местные наземные чере- пахи (то есть род Testudo) отсутствуют, а из имеющихся там малочислен- ных пресноводных видов не ясно, на каких из этих островов они являются местными. Крокодил, обитающий в соленых водах, широко распространен на всех островах. Ящерицы и змеи Филиппинских островов относительно
Глава 8
многочисленны, и некоторые из них также имеют широкое распростране- ние, но распределение большинства рептилий соответствует распределе- нию амфибий — так, словно их распространение шло с острова Калимантан на Филиппины, где они проникали на разные расстояния (Dickerson et. al., 1928, р. 226, 232, 238). Исключением является род Hydrosaurus— крупные полуводные ящерицы семейства агамовых, который встречается в Новой Гвинее, на Молуккских островах, Сулавеси и на Филиппинах до острова Лусон (но не Палаван?), а также род наземных ужовых змей Steganotus, известный в северо-восточной части Австралии, Новой Гвинее, на Молуккских островах и в восточной части Филиппин до острова Лусон.
Птицы Филиппин многочисленны (Ре1асоиг, Мауг, 1946). Распрост- ранение большинства из них шло с острова Калимантан или через него, а также через Палаван, острова Сулу и Минданао. Несколько видов достигло Филиппин из Азии через море. А несколько видов, включая сор- ных кур, какаду, лори, некоторых голубей и ряд других, распространены таким образом, словно они распространялись из Австралийской обла- сти, с Молуккских островов или с острова Сулавеси. На восточных Филип- пинах (от Минданао до Лусона) особенно много эндемичных видов и даже имеются эндемичные роды, но (как я думаю) отсутствуют удивительные реликтовые и нелетающие птицы. На островах Палаван, Каламиан и др. еще в большей пропорции по сравнению с другими Филиппинскими остро- вами представлены птицы Калимантана.
Млекопитающие Филиппинских островов рассматриваются Тейло- ром (Taylor, 1934), а также кратко Картером, Хиллом и Тейтом (Carter, Hill, Tate, 1945, р. 216, 217). Дополнительные сведения были сообщены мне д-ром Купманом. Исходя из этих данных, надо отметить относительное разнообразие мелких форм млекопитающих, особенно грызунов и руко- крылых, и малочисленность крупных представителей этого класса. Совер- шенно очевидно, что большинство млекопитающих, кроле мышиных гры- зунов, недавно иммигрировало с острова Калимантан. Многие из них рас- пространены не дальше Палавана и других островов и (или) Минданао, и лишь очень немногие достигают Лусона. Из насекомоядных один энде- мичный род (Podogymnura) обитает на Минданао, а землеройки таких широко распространенных родов, как Crocidura и Suncus, расселились на Филиппинах повсеместно. Шерстокрылы достигают островов Басилан, Мин- данао, Бохоль, Лейте и Самар. Один род тупайи представлен на острове Палаван, а другой (Urogale) является эндемиком на острове Минданао. Долгопяты достигают островов Минданао, Бохоль, Лейте и Самар, лори— Минданао. Крабоядная обезьяна, очевидно, распространена на всех основных островах и на некоторых из них, включая Палаван и Лусон, является единственным представителем отряда приматов (за исключением человека). Род Manis достигает острова Палаван. На Филиппинах нет мест- ных представителей семейства собачьих. Единственный местный предста- витель семейства кошачьих — леопардовая кошка (небольшая обыкно- венная дикая кошка Южной Азии) обитает на островах Палаван, Кала- миан, Панай, Негрос и Себу. Выдра достигает острова Палаван, а барсук— острова Палаван и островов Каламиан включительно. Представители виверровых бинтуронг и мангуст достигают острова Палаван (мангуст также проникает и на острова Каламиан). Два других рода виверровых (Viverra и Paradozurus) достигают всех главных Филиппинских островов. Из немышиных грызунов три рода беличьих распространены на разных южных островах, но ни один из них не достигает острова Лусон; дикобраз
Особенно«ти распространения животных на ocntposax
проникает на острова Палаван и Каламиан. Из копытных оленек достига- ет по крайней мере острова Балабак (непосредственно к югу от Палавана), небольшой бык эндемичен на острове Миндоро, и только свиньи и неболь- IUoII олень (Сеговия) широко распространены на всех Филиппинских остро- вах. Плодоядные и насекомоядные рукокрылые многочисленны на всех островах. Родственные связи большинства из них ведут к Индо-Малайской области, а также в меньшей степени к Сулавеси, Ыолуккским островам, Австралийской области и др. (Tate, 1946, р. 4 — 7).
Из всех млекопитающих, перечисленных выше, землеройки родов Crocidura и Suncus, крабоядная обезьяна (Macaka irus), виверровые Virerra и Paradoxurus, свиньи и олени — единственные животные, которые достигают острова Лусон. Эта группа очень сходна с группой, достигаю- щей Молуккских и удаленных Малых Зондских островов. Большинство этих животных или почти все они так или иначе связаны с человеком, .и я думаю, что некоторые из них или, возможно, все были интродуцирова- ны на острове Лусон, так как если предположить, что эти животные дей- ствительно местные, то весьма удивительно, что они не образовали ни од- .ного четкого эндемичного рода.
Распространение мышиных грызунов самое разнообразное. Они мно- гочисленны на всех Филиппинских островах. Я не буду пытаться детально анализировать их распространение, но следует подчеркнуть, что эндемич- ные роды мышиных широко рассеяны на Филиппинах (кроме острова .Палаван) и особенно многочисленны на северном острове Лусон. Диккерсон и др. приводят из работы Томаса сведения о наличии 6 новых родов мыши- ных на острове Лусон: «...совершенно новое соотношение, которое, оче- видно, не имело себе равного в истории сборов млекопитающих». Неко- торые роды настолько специфичны, что их связи вызывают сомнения, но отдельные связи ведут к Сулавеси и Австралии. Например, два рода, находящихся в горах северной части Лусона, вероятно, родственны подсе- мейству Hydromyinae из Австралийской области.
Обычные объяснения распространения млекопитающих — через сухопутные мосты или прибытие их из разных мест на плотах, сопровождаю- щееся отчасти преодолением широких водных пространств,— не очень убедительны для данного случая. По-видимому, лучшим объяснением является то, что млекопитающие достигли Филиппинских островов и рассе- лились через все острова только (или преимущественно) в одном направле- нии (на северо-восток от Калимантана) через ряд проливов, что мышиные расселились раньше или быстрее других наземных млекопитающих, достиг- нув острова Лусон также раньше других, и что с тех пор прошло доста- точно много времени для эволюции родов мышиных на острове Лусон и для замещения некоторых более древних групп мышиных на Зондских островах с последовательно меняющимися особенностями распространения в вос- точном направлении. Мышиные (грызуны) значительно чаще, чем боль- шинство других млекопитающих, пересекают водные пространства. Толь- ко они одни из нелетающих наземных плацентарных достигли Австралии. Некоторые из тех же самых ветвей, которые достигли Австралии, воз- можно, также проникли и на Филиппины. Если все это действительно про- исходило так, то на острове Лусон, очевидно, существовала когда-то фау- на млекопитающих, которая состояла только из мышиных грызунов и руко- крылых, если только там не находились (случайно) реликтовые сумчатые.
Из этого объяснения вытекают определенные факты, относящиеся к истории Филиппин: эти острова никогда не были легко достижимыми, за исключением, вероятно, очень непродолжительного времени в плейсто-
цене, возможно, в прошлом на протяжении значительного периода своей истории они были значительно меньше по площади и более изолированны, чем в настоящее время. Симпсон предполагает (Simpson, 1945, р. 205), что если у мышиных не было подъема до позднего миоцена, то, возможно, последний и явился временем начала существования современной фауны Филиппинских островов. Остается только посмотреть, не идет ли вразрез с вышесказанным геологическая история этих островов. История распро- странения позвоночных Филиппин в целом, видимо, соответствует истории этих островов. Прежде всего бросается в глаза отсутствие реликтовых позвоночных, за исключением мышиных, особенно на внешних островах Филиппинского архипелага. Во всех классах позвоночных наблюдается некоторый эндемизм, проявляющийся в основном среди видов, а не среди родов, и в большинстве случаев на ближайших островах, а не на удален- ных. Обращает на себя также внимание недавняя, охватывающая почти всю фауну позвоночных иммиграция, которая большей частью шла с Калимантана. Те немногочисленные случаи, когда связи позвоночных Филиппин ведут к острову Сулавеси или к Австралийской области (по крайней мере у двух видов лягушек, у одного вида ящериц и змей и у не- скольких видов птиц и летучих мышей, а также у некоторых мышиных), в какой-то степени могут быть объяснены смещением основных расселяю- щихся групп к востоку, то есть их исчезновением на Калимантане и т. д. Возможно, только в нескольких исключительных случаях (если такие слу- чаи имеются) прямое расселение происходило через водные пространства от Сулавеси или Молуккских островов к Филиппинам.
Остается невыясненным, были ли Филиппины соединены с Калиман- таном и каким образом. Контуры морского дна (рис. 61) позволяют пред- положить, что остров Палаван и острова Каламиан соединялись с Калиман- таном, а остров Минданао не соединялся, однако на Палаване материковая фауна, свидетельствующая об этом соединении, отсутствует. Несколько видов (очень мало!) пресноводных рыб проникли на оба острова — и Па- лаван и Минданао, причем на первый остров проникло не большее число видов, чем на второй, а карповые распространены за пределы острова Палаван до Миндоро, то есть за границы предполагаемого сухопутного моста.
С Калимантана на Минданао, а также на Палаван расселилось довольно большое число разнообразных амфибий. Оба острова оыли достигнуты млекопитающими, из которых основные группы, имеющиеся на одном из островов, отсутствовали на другом. Млекопитающие острова Палаван, возможно, были дифференцированы в меньшей степени по сра»- нению с млекопитающими острова Минданао. На Палаван, очевидно, про- никло больше хищных млекопитающих, а на Минданао — нехищных (и больше древесных?). Все это говорит о том, что млекопитающие острова Минданао (по сравнению с млекопитающими Палавана) более древнего возраста и что, возможно, достижение ими Филиппин не представляло для них большой трудности. Эндемичный буйвол на острове Миндоро находится за пределами предполагаемого сухопутного моста между Калимантаном, Палаваном и островами Каламиан. Следовательно, самые существенные части фауны Филиппин недостаточно ясно указывают на существование (или границы) сухопутного моста, соединяющего Филиппины с островами Палаван. Остров Палаван, очевидно, был несколько (но в незначительной степени) доступнее острова Минданао. Поэтому трудно сказать, можно ли объяснить отсутствие некоторых элементов фауны на острове Палаван срав- нительно небольшой площадью этого острова.
Особенности распространения животных на островах
Относятся ли все Филиппинские острова к Индо-Малайской области или только остров Палаван или ни один из них не входит в эту область— не столь существенно, если ясна ситуация в целом. Это периферийные острова с периферийной и постепенно обедненной фауной позвоночных, относящихся главным образом к Индо-Малайской области.
Ocmpoea западной тропической части Тихого океана. Здесь будут рас- смотрены как единый периферийный архипелаг все острова западной тро- пической части Тихого океана, расположенные к востоку от Австралии и Новой Гвинеи и далее до пределов досягаемости IIx наземными позво- ночными (рис. 62). Основная цель — установить основной характер рас-
Р и с. 62. Главные острова вхого
Вертикальныс линии — границыГвинеи) в
тропической западной части Ти-океана.
распространения животных (с Новой Тихом оксане.
пространения животных на этой площади в целом, а не особенности фаун отдельных островов. Большинство островов этой территории — неболь- шие, разделенные широкими водными пространствами, но некоторые их группы, хотя и отделены друг от друга широкими проливами, образуют ряд беспорядочно раскиданных мелких островков к востоку от Новой Гви- неи: острова Бисмарка, Соломоновы, Санта-Крус, Новые Гебриды, Фид- жи, Тонга и Самоа. Многие острова Тихого океана представляют собой невысокие коралловые атоллы, но некоторые из них, входящие в каждую из упомянутых выше групп, относительно высоки и более разнообразны экологически. Геологическое происхождение этих островов до какой-то степени разное. За пределами (с внешней стороны) андезитовой линии (рис. 62) все острова сложены из базальтовых вулканических горных пород (плюс коралловые известняки). С другой (внутренней стороны) этой линии горные породы более сложные, но многие острова все-таки вулканическо- го происхождения, и я не думаю, что эта линия может иметь какое-либо отношение к современным границам распространения позвоночных. Ни один из этих островов не соединен с материковой отмелью. Даже острова Бисмарка и Соломоновы отделяются от нее и друг от друга узким проли-
ВоМ. С точки зрения зоогеографии все эти острова следует рассматривать как острова неизвестного происхождения, но их фауны говорят сами за себя.
На этих островах отсутствуют строго пресноводные рыбы. 11редстави- тели подсемейства lelanotaeniinae, предположительно вторично ставшие пресноводными к востоку от Новой Гвинеи, не известны.
Лягушки из семейства Leptodactylidae и Brevicipitidae (первые бед- но, а последние широко представлены на Новой Гвинее) не достигают даже архипелага Бисмарка или Соломоновых островов. Род Hyla и лягуш- ки семейства Ranidae (особенно Cornuferinae) достигают островов Бисмар- ка и Соломоновых: на первых обитают 2 вида рода Hgla и 8 родов (из них 3 эндемичных) с 14 видами семейства Ranidae (Brown, 1952), а также интродуцированный вид Bufo marinus. За пределами этих границ лягуш- ки не известны ни на островах Санта-Крус, ни на Новых Гебридах и др.. но 2 вида семейства Ranidae встречаются на островах Фиджи. Это виды. относящиеся к родам Cornufer и Plat@mantis. Оба рода встречаются также на Соломоновых и других островах. Виды, обнаруженные на островах Фиджи, считаются эндемичными (Brown, Myers, 1949; Brown, 1952), но они не очень характерны. Сотпи~ет vitiensis с островов Фиджи отличает- ся от С. дирруг с Соломоновых островов только тем, что они мельче по раз- меру и имеют маленькие пальцевые диски (Brown, 1952, р. 32). На мой взгляд, лягушки с островов Фиджи находятся на слишком удаленных от материка островах Тихого океана, слишком слабо дифференцированы, чрезвычайно мало известны и слишком тесно связаны с человеком (насе- ление островов Фиджи, употребляя этих лягушек в качестве еды, по-ви- димому, переносит их с острова на остров), чтобы их можно было прини- мать за достоверно местные виды. Вид рода Cornufer (pelewensis) встречает- ся на островах Палау, но, по сообщению Ингера (письмо от 16 марта 1955 г.), при сравнении экземпляров их трудно отличить от C. rubristriatus c Новой Гвинеи. Возможно, что этот вид был интродуцирован на островах Палау.
Черепахи (кроме морских), очевидно, дальше на восток от Новой Гвинеи не проникают (но в плейстоцене представитель семейства Meiola- niidae находился на острове Уолпол, юго-восточнее Новой Каледонии). Границы распространения обитающего в соленых водах крокодила про- стираются в Тихом океане очень далеко и точно не определены. Несколь- ко групп змей достигает Соломоновых островов, но очень немногие прони- кают дальше. Небольшой удав Gandoia (Enygrus) проникает на острова Тонга и Самоа, но возможно, что на эти удаленные от материка острова он был завезен человеком. Это древесный вид, который любит прятаться в гу- стых кронах деревьев. На Новой Гвинее подобный экземпляр того же рода свалился сверху в мой сачок для ловли насекомых. Этот удав может быть случайно перенесен в связках бананов или тростника. Змеи рода ТурИорз также случайно могут переноситься человеком, поэтому их дальние гра- ницы не ясны. Маленький аспид рода Ogmodon, эндемичный на островах Фиджи и родственный новогвинейскому, вероятно, определяет внешнюю границу распространения подлинно местных змей. В Новой Каледонии змеи, очевидно, не встречаются (Mertens, 1934, р. 39), несмотря на несколь- ко ошибочных сообщений о нахождении их там. Не встречаются они и в Новой Зеландии.
По сравнению со змеями ящерицы распространены гораздо дальше. Местные эндемичные ящерицы встречаются в увеличивающемся числе на восток по крайней мере до островов Тонга, за пределами Фиджи (но, оче-
видно. не на Самоа), на Новой Каледонии и Новой Зеландии. Большин- ство пз них относится к гекконам и сцинкам, но один превосходно выраженный реликтовый и эндемичный род игуановых приурочен к остро- вам Фиджи и Тонга. Некоторые гекконы и сцинки проникают еще дальше, встречаясь почти на каждом из островов, через всю тропическую часть Тихого океана, почти до берегов Южной Америки. Эндемичные виды и под- виды этих ящериц были описаны для некоторых самых отдаленных остро- вов, и обычно считают, что они расселились через океан естественным пу- тем на плавучих предметах. Однако на наиболее удаленных островах эти ящерицы фактически очень слабо дифференцированы. Например, на Га- вайских островах (Oliver, Shaw, 1953) ни один вид ящериц в действитель- ности не является эндемичным. Гавайские острова по происхождению достаточно древние и имеют высокоэндемичную фауну насекомых, назе- мных моллюсков и птиц. Если ящерицы действительно расселялись через Тихий океан естественным путем, то почему же ни одна из них не достигла Гавайских или других удаленных от материка островов в относительно давние времена и не образовала там эндемиков хотя бы низкого систематического ранга?
Из птиц казуары достигают Новой Британии (если только эти птицы там не интродуцированы), но не Соломоновых островов. Другие наземные птицы довольно многочисленны на Соломоновых островах, но более мало- численны, чем на Новой Гвинее. Но здесь не надо забывать о том факте, что Соломоновы острова имеют меньшую площадь. К востоку от Соломо- новых островов число птиц уменьшается пропорционально расстоянию от материка (рис. 32), но, однако, фауна птиц изменяется также в связи с величиной и природой различных островов. Несколько полинезийских птиц (Старый Свет) наряду с некоторыми американскими достигли Гавай- ских островов. На острове Пасхи наземные птицы в настоящее время отсутствуют, и неизвестно, существовали ли они когда-нибудь там вообще. К наземным птицам Старого Света, которые проникли далеко на вос- ток через Тихий океан, относятся (в порядке их численности) голуби, мухоловки Старого Света, попугаи, медососы и другие. К группам, которым не удалось проникнуть столь далеко, — какаду, щурки, сизоворонки, рогоклювы, питты, дронго, нектарницы и цветососы (Мауг, 1939).
Из млекопитающих, кроме рукокрылых, только небольшая часть до- вольно богатой новогвинейской фауны сумчатых и грызунов распростра- нена на восток. На острове Новая Британия обитает только бандикут, два вида родов Pl~alanger и Petaurus, валлаби (Thylogale) и около 4 родов крыс, включая Hydromys и Rattus, но отсутствуют однопроходные и сумчатые хищные. На острове Новая Ирландия встречаются бандикут, два вида рода РЬаКапдег, валлаби и род Rattus (и другие крысы?). Только род Phalan- ~er и около трех родов крыс распространены до Соломоновых островов. Ряд крыс на этих островах образует очень четкие эндемичные виды, а возможно, даже и роды. Вероятно, в этом направлении Соломоновы острова являются границей местных нелетающих наземных млекопитаю- щих. Однако мыши и крысы, связанные с человеком, встречаются даже на самых удаленных островах тропической части Тихого океана. Кроме видов комиенсалистов, распространенных на всем земном шаре, представители групп Индо-Малайской области Rattus concolor или exulans, которые часто живут в домах местных жителей, встречаются на востоке до Гавайских и Маркизских островов, а также на юге до Новой Зеландии. Слабодиффе- ренцированные формы этой группы предположительно являются энде-
миками на удаленных островах, но маловероятно, что это местные формы.
Плодоядные и насекомоядные рукокрылые Старого Света распростра- нены на восток до острова Самоа, но резко уменьшаются в числе по мере увеличения расстояния от материка. Некоторые насекомоядные рукокры- лые достигают также Каролинских и Маршалловых островов, Новой Viaae- донии и Новой Зеландии. Островов Галапагос и Гавайских достигает толь- ко американский род Lasiurus. На островах Общества, Туамоту и др. руко- крылые, очевидно, не известны.
Восточными границами распространения разных групп позвоночных вдоль основных направлений расселения в тропиках Тихого океана являются, по-видимому, следующие: строго пресноводные рыбы и немор- ские черепахи восточнее Новой Гвинеи не проникают; лягушки и нелетаю- щие наземные млекопитающие достигают Соломоновых островов; змеи —- островов Фиджи; ящерицы — островов Фиджи и Тонга; рукокрылые- Самоа; наземные птицы — самых дальних островов Туамоту. Эти границы, очевидно, пропорциональны возможным способностям расселения у раз- ных животных. Единственный удивительный реликт в фауне позвоночных наиболее удаленных островов — ящерица из семейства игуановых на островах Фиджи, которая может быть и не очень древней. Остальные позво- ночные, хотя среди них и имеются эндемики, образуют хорошо выражен- ный иммиграционный характер распространения на островах. Итак, мы имеем здесь расселение через водные пространства на периферийные архипелаги, уменьшающееся пропорционально расстоянию от материка, а не выживание остатков фаун на древних материковых массивах.
Некоторые лягушки, змеи, ящерицы и крысы встречаются за предела- ми вышеуказанных границ, а отдельные из этих животных на удаленных островах предположительно являются местными эндемичными формами. Возможно, что из этих эндемиков некоторые действительно являются таковыми. Но они не могут не вызывать сомнений в связи с тем, что все они находятся слишком далеко от материков, очень слабо дифферен- цированы, в ряде случаев малоизвестны и чрезвычайно тесно связаны с человеком. Все эти виды, либо употребляющиеся человеком в пищу, либо живущие в его жилищах, вероятно, переносятся им на другие острова случайно. В течение нескольких последних тысячелетий все эти животные или некоторые из них переносились человеком за пределы их естественных границ. До какой степени эти островные популяции дейст- вительно дифференцированы и что эта дифференциация означает, в боль- шинстве случаев все еще не выяснено.
Вест-ХХндия. Вест-Индия (рис. 63) расположена к востоку от Цент- ральной Америки, между Северной и Южной Америкой, в основном в пре- делах краевых районов тропиков. К самым большим и, вероятно, к самым древним и богатым по природным условиям из всех островов относятся четыре острова из группы Больших Антильских: Куба — площадью около 40 тыс. кв. миль, Гаити (Гаити и Сан-Доминго) — около 30 тыс. кв. миль, но с большими высотами, чем на Кубе, Ямайка — около 4500 и Пуэрто- Рико — площадью около 3400 кв. миль. Малые Антильские острова об- разуют неравномерную цепь островов между Большими Антильскими островами и островом Тринидад (который фаунистически представляет часть Южной Америки). Эти относительно небольшие по своим размерам острова имеют разнообразные экологические условия. Расположенные к северу от Больших Антильских и к востоку от Флориды, Багамские острова невелики и имеют очень небольшую высоту над уровнем моря и до-
Особенности распространен ия животных на, островах вольно засушливый климат. Все эти острова подробно рассмотрены в ра- боте Вильямса (см. начало этой главы). Здесь мы кратко рассмотрим в ос- новном Большие Антильские острова, чтобы определить их место среди других важных островов земного шара.
В прошлом Большие Антильские острова считали островами матери- кового происхождения, теперь с геологической точки зрения их считают океаническими островами. Они сложены вулканическими породами и мор- ским известняком. В течение третичного периода высота разных островов и их частей над уровнем моря неоднократно менялась, но соединялись ли
Р и с. 63. Вест-Индия. Прсрывистой линией обозначены отмели, расположенные недалеко от Центральной Америки, которые в прошлом. возможно, были сушей. Про- нумерованные стрелки показывают расстояние и (очевидное) направление расселения на Больших Антильских островах следующих животных: 1 И«-
(жабы); 2 — наземные ленивцы; .'~ — — V r м~р)~от Ь 8 (мелкие насскомояд- ные); 4 — хромисы (Cichlidae); 'i — Яо! ~одоп (крупныс насскомоядные). щелсзуб: 6 — лягушки семейства A t elopu <I i dae; 7 — панцирные щуки: 8 — обезьяны; 9 — лягушки рода Ну!«., 10 — лягушки рода (. р[ М (у!и;. В ' тот список включены все нс строго 1~ресноводныс рыбы 1'ольпи1х Антильских островов, кроме карпозубых, все земноводные. кроме Е( i(tt«- ro~l~~c t«lii;. n все нелетающие наземные млекопитающие, кроме грызунов. Поэто~1~ на рисунке показано ограниченное разнообразие фауны Больци1х Антильских островов, порядок ес размсщения и простой иммиграционный характер распространения основных частей фауны в большинстве случаев
113 Центральной Америки с главным районом захода — ocT!)()BoM Куба. эти острова с сушей в действительности, не известно. Они не присоединя- ются ни к одной из материковых отмелей, но ближе всего подходят к мате- риковои отмели Центральной Америки, особенно у Гондураса и Никара- гуа. Однако, как и в других подобных случаях, видимо, лучше всего не принимать в расчет геологию, а посмотреть, о чем свидетельствуют фау- ны этих островов непосредственно.
CTp0r0 пресноводные рыбы не достигают Больших Антильских островов. Из не строго пресноводных рыб (второй отдел), которые, хотя и с трудом, но могут расселяться через море, североамериканская панцир- ная щука достигает Кубы. Эндемичные виды южно- и североамериканских родов хромисов встречаются на Кубе и Гаити. Две формы карпозубых- яйцеродящие и живородящие — обитают на Кубе, Ямайке и 1'аити, но наиболее многочисленны и дифференцированы они на Кубе, причем часть из них северо-. а часть центрально- или южноамериканского проис- хождения.
Из амфибий ни безногие (главным образом тропические), ни саламанд- ры (главным образом северные) не достигают Вест-Индии. Здесь имее1л~ лишь пять родов лягушек. Единственный род (тропический американскиц) Eleutherodactylus составляет основную часть фауны лягушек этих островов. Проследить историю этой фауны я не имею возможности. Тропический аме- риканский род Leptodactylus достигает Малых Антильских островов, lly- эрто-Рико и Гаити (но не Кубы или Ямайки), а род Hyla достигает Ямайки (четыре вида), Гаити (четыре) и Кубы (один вид), но не Пуэрто-Рико. Род Ilufo проникает на Кубу (три вида), Гаити (один), Пуэрто-Рико (один) и на Виргинские острова (один вид), но не на Ямайку. И, наконец. род Sminthillus (один впд) встречается только на Кубе, где он распространен на небольших высотах. Очевидно, это эндемичный род тропического аме- риканского семейства Atelopodidae, не известного на материке к северу от Коста-Рики. Фауна амфибий Больших Антильских островов очень ограниченная. Она образована от небольшой части материковой фауны, и ее распределение на островах строго закономерно (рис. 63).
Рептилии Вест-Индии представлены несколькими видами черепах, дву- мя видами крокодилов (но не аллигатором или каймаком), некоторыми зме- ями и множеством ящериц. Единственные местные неморские черепа~и, которые являются не четко выраженными видами подсемейства Emydi- nae пресноводного рода Pseudemys и встречаются в настоящее время на «сех Больших Антильских, Каймановых и некоторых Багамских остро- вах, близкородственны видам Центральной Америки. Сухопутные чере- пахи (Testudo) известны в субфоссильном состоянии на островах Куба, Мона (между Гаити и Пуэрто-Рико) и Сомбреро и встречались также и на других островах. Обычный американский крокодил достигает Кубы, Гаити и Ямайки; более крупный вид крокодилов является эндемпком на Кубе и островах Пайн. Перечисление многочисленных ящериц и змей, обитаю- щих на этих островах, займет слишком много времени, но краткую характеристику их дать необходимо. На материке и материковом острове Тринидад змеи численно превосходят ящериц, а в собственно Вест-Индии, наоборот, ящерицы значительно преобладают над змеями (табл. 15, стр. 405). В Вест-Индии отсутствуют ядовитые змеи, за исключением Bot- hrops atrox (возможно, интродуцирована) на некоторых Малых Антильских островах. Многие ящерицы распространены широко и последовательно, а другие локализованы. Небольшое семейство Xantusiidae Центральной Америки и юго-западной части Северной Америки представлено в Вест-Ин- дии эндемичным родом и видом, которые локализованы в восточной части Кубы. Роющие ящерицы семейства Amphisbaenidae, в настоящее время отсутствующие на большей части Центральной Америки, представлены в Вест-Индии тремя родами, включая Cadea, эндемика Кубы и остро- вов Пайн, который представляет древнюю северную группу этого се- меиСтва.
Птицы в Вест-Индии довольно многочисленны, но многие материко- вые семейства, включая ряд доминирующих семейств Южной Америки, которые достигают острова Тринидад, отсутствуют в собственно Вест- Индии. Многие виды, некоторые роды и одно не очень четко выраженное небольшое семейство Todidae являются эндемиками Вест-Индии, но древ- них реликтов и нелетающих птиц здесь нет, если только считать, что субфоссильный пастушок на Пуэрто-Рико был нелетающим (насколько в настоящее время известно, пастушок Цапата на Кубе может летать). Раз- личные наземные птицы, очевидно, достигли Вест-Индии из разных мест: из Северной, Центральной и Южной Америки. Я полагаю, что большин-
Особенности распро<тц~анен ия животных и а островах
ство оседлых наземных птиц на Больших Антильских островах. является, вероятно, вселенцами Центральной Америки, а эндемичное семейство Todl- dae близкородственно первоначально центральноамериканским момотам. Многие перелетные наземные птицы, гнездящиеся в Северной Америке, зимуют в Вест-Индии, и некоторые из них (особенно некоторые древес- ницы) дали на этих островах оседлые формы. Дальнейшие подробности распространения этих птиц и характеристика всех американских семейств, которые не достигают Вест-Индии, приведены в списке семейств птиц и конце гл.
Наземных млекопитающих в Вест-Индии, за исключением многочис- ленных рукокрылых, очень мало, и большинство из них в настоящее вре- мя исчезло. Все известные ископаемые, а также современные животные приводятся ниже. Самые древние ископаемые относятся к плейстоцену (возможно, к более позднему). Таким образом, и ископаемые и современ- ные виды образуют единую фауну. В Вест-Индии имеются две группы насекомоядных. К одной относится Solenodon, известный только на Кубе и Гаити, который строго локализован и существует до наших дней на обо- их островах. Родственные ему формы относятся к олигоцену Северной Аме- рики. К другой — Nesophontes, встречающийся субфоссильно на Кубе, 1'аити и Пуэрто-Рико. Эта группа представляет другую древнюю ветвь, которая, по-видимому, также проникла из Северной Америки, так как насекомоядные достигли Южной Америки в сравнительно недавнее время. На Ямайке обнаружены ископаемые эндемичного рода обезьян. Одна или две ветви наземных ленивцев достигли Вест-Индии и образовали там эндемичный род. Если и было две ветви, то одна достигла Кубы, Гаити и Пуэрто-Рико, а другая, очевидно,— только Кубы и, возможно, 1'аити. Обе ветви по происхождению являются, несомненно, южноамерикански- ми. Из других наземных млекопитающих Больших Антильских островов, кроме рукокрылых, известны в основном дикобразоподобные грызуны, к которым относятся представители 4 семейств, по-видимому, все проис- шедшие в Южной Америке. Современные, нутриевые, так же как и исчез- нувшие (4 современных и 3 вымерших рода Вест-Индии), наиболее раз- нообразны на Кубе и Гаити, но единственный род достигал и достигает до сих пор Ямайки, Пуэрто-Рико (интродуцирован), Каймановых островов, острова Суок и некоторых Багамских. Цепкохвостые щетинистые крысы (пять родов; все исчезли по крайней мере на островах Вест-Индии) встре- чались на Кубе, Гаити и Пуэрто-Рико. Heptaxodontidae (семь вымерших родов) были распространены на Ямайке, Гаити, 11уэрто-Рико, Ангилье и Сен-Мартене, к востоку от Пуэрто-Рико. Осьмизубые (один исчезнувший эндемичный род) находились только на Ямайке. Распространение дикобра- зоподобных в Вест-Индии несколько более детально приводится в списке семейств млекопитающих в конце гл. 6.
Связи некоторых этих животных до сих пор вызывают сомнение. Из недикобразоподобных грызунов, очевидно, единственными местными животными в Вест-Индии являются достигшие Ямайки, а также некоторых Малых Антильских островов рисовые крысы (Oryzomys и др.), которые по происхождению относятся (в Америке) к северному семейству Crice- tidae. Кроме этих животных, два вида опоссумов, один вид броненосцев, агути и еноты обитают на некоторых Малых Антильских островах, а енот встречается и на острове Нью-Провиденс из группы Багамских остро- вов, но эти животные слабодифференцированы, и некоторые из них, воз- можно, были интродуцированы. Как мы видим, нелетающие наземные млекопитающие Вест-Индии очень ограниченны, крупные формы отсутст-
Глава 8
вуют (предкп наземных ленивцев Вест-Индии, очевидно, были неболь- шими животными); хищных, кроме (сомнительных) местных енотов на нескольких мелких островах, нет, отсутствуют и копытные. Более того, они имеют последовательный характер распространения. На Боль- ших Антильских островах пять или шесть основных групп (две группы насекомоядных и одна или две группы наземных ленивцев и две группы грызунов) распространены таким образом, будто они проникли в Вест-Ин- дию через Кубу и распространились до Гаити, а в отдельных случаях и на другие острова. Распространение следующих четырех групп (группа обезь- ян и три группы грызунов) таково, словно они проникли в Вест-Индию через Ямайку и только одна из них (из групп грызунов) проникла дальше. Грызуны по сравнению с другими млекопитающими Больших Антильских островов дифференцированы (или были дифференцированы) в большей степени на удаленных островах: с одним эндемичным родом на острове Пуэрто-Рико и одним (Amblyrhiza — очень крупные грызуны) даже на островах Ангилья и Сен-Мартен.
В целом вест-индская фауна позвоночных состоит из разных классов соответственно их способностям к расселению через соленые воды. Строго пресноводных рыб здесь нет, но слабо солевыносливые встречаются на отдельных островах. Амфибии представлены всего несколькими родами, хотя один из них дал обширную радиацию на этих островах. Неморских черепах очень мало, а многочисленные ящерицы превосходят змей. Пти- цы и рукокрылые многочисленны. Нелетающие наземные млекопитающие малочисленны, невелики по своим размерам и представлены в основном грызунами. Местные хищные (по крайней мере на больших островах) отсутствуют.
Основную часть этой фауны составляют группы, для которых, вероят- но, расселение через соленые воды представляет большую трудность. Эти группы распространены на Больших Антильских островах в строго последовательном порядке, который, видимо, отражает иммиграционный характер распространения, словно расселение большинства животных, достигших этих островов, шло с запада, из Центральной Америки, в большей степени через Кубу, а не через Ямайку. Не строго пресноводные рыбы (второй отдел) достигают наивысшей численности на Кубе. На Гаити и Ямайке их меньше и на Пуэрто-Рико нет совсем. Из лягушек, не считая не анализируемый род Eleutherodoctylus, два рода распространились, видимо, через Кубу или на Кубе, один — через Ямайку и один, возможно,— через Малые Антильские острова к Пуэрто-Рико и Гаити. Из нелетающих наземных млекопитающих пять или шесть групп, видимо, проникли через Еуоу, а четыре — через Ямайку.
Эта фауна представляет собой смешение групп, возникших в Северной и южной Америке, но и северные и южные группы, очевидно, достигали Больших Антильских островов одним путем. Куба, видимо, была конеч- ным пунктом захода для южных хромисов и северных панцирных щук. для южных лягушек семейства Atelopodidae и для северных жаб рода Bu- fo, для южных наземных ленивцев, некоторых дикобразоподобных грызу- нов и для северных насекомоядных. Ямайка, очевидно, была конечным пунктом захода для хомячьих, происшедших на севере (рисовые крысы). и для некоторых южных групп.
Большое значение имеет в Вест-Индии распространение самых четких реликтов-эндемиков позвоночных. На Кубе обитают лягушка семейства Atelopodidae, ящерица семейства Xantusiidae и род Cadea, к Ямайке при- урочены (или были приурочены) единственная обезьяна и грызун из се-
0собен ности распространения животных ла ост ровах
мейства осьмизубых. Каждый из этих видов, очевидно, представляет отдельную довольно древнюю ветвь, вселившуюся в Вест-Индию и неизвест- ную в каких-либо других местах этих островов. Они, видимо, сохранились на тех островах, которые явились конечным пунктом вселения и далее которого онн не распространялись. На Гаити нет аналогичных эндемичных групп животных, несмотря на то, что этот остров значительно больше Ямай- ки. С другой стороны, он не столь велик, как Куба, однако на нем имеется больше возвышенностеи и, возможно, он мог бы copepæà Tü идентичную фауну (см. табл. 15). Тот факт, что на Гаити, очевидно, нет реликтово-эн- демичных позвоночных из групп, не представленных на других островах, позволяет предположить, что этот остров не был важным конечным пунк- том для наземных позвоночных и что расселение этих животных в Вест- Индии было довольно простым. Если бы оно было сложным с последователь- но сменяющимися вторжениями и замещениями, то на Гаити находи- лось бы большее число реликтово-эндемичных животных. Нахождение реликтов-эндемиков на Кубе и Ямайке является важным фактом, но их отсутствие на Гаити, возможно, еще более существенно, так как этот ост- ров должен был быть внутренним резерватом древней фауны, если только там таковая находилась.
Резюмируя вышесказанное, отметим, что разные виды позвоночных в Вест-Индии представлены пропорционально их возможным способно- стям пересекать соленые воды. Важнейшие группы (те, которые испыты- вают наибольшие трудности при преодолении соленых вод) очень мало- численны и распространены на Больших Антильских островах, проявляя, видимо, иммиграционный характер распространения, так словно боль- шинство из них проникло с запада через Кубу или Ямайку. Фауна Боль- ших Антильских островов представляет смешение элементов фаун Север- HoI1 и Южной Америки, которые, видимо, достигли этих островов одними и теми же, а не разными путями. Реликты-эндемики встречаются на Кубе и Ямайке, но не на Гаити, что опять-таки дает основание предположить, что и hyoa и Ямайка были конечными пунктами расселения, что в свою оче- редь позволяет считать (так же, как и последовательность в расселении) расселение большинства важных групп позвоночных на Больших Антиль- ских островах относительно простым с незначительным числом последова- тельных смен и замещений, хотя, возможно, некоторые из них и имели место. Это тип довольно простого последовательного иммиграционного характера распространения с реликтами-эндемиками на ближайших к мате- рику (Центральной Америке) островах, который, скорее всего, даст надеж- ные ключи к прошлому (стр. 408).
llpocTbIM объяснением сказанному может служить то, что назем- ные позвоночные достигли Больших Антильских островов через соленые воды, что современные и наиболее поздние ископаемые формы, распро- страняясь первоначально, сосредоточивались главным образом вдоль первоначальных путей иммиграции и что большинство основных групп на Больших Антильских островах проникло сюда из Центральной Америки через Кубу или (реже) через Ямайку в то время, когда фауна Центральной Америки представляла смешение элементов фаун Северной и Южной Аме-
~)И lilt.
По этому поводу следует сделать еще два замечания. Симпсон (Simp- soii. 1356, р. 13) в последней работе по зоогеографип наземных млекопи- тающих Вест-Индии отмечает, что у животных, переносимых по воздуху и на плотах. разные способы расселения (что и соответствует действитель- ности), и полагаст (однако это совершенно не обязательно), что они
Глива 8
достигли Больших Антильских островов, расселяясь из разных мест. Насколько я понимаю, характер распространения на островах беспозвоноч- ных, предположительно переносимых по воздуху (например, крылатых жуков семейства Carabidae, изучением которых я занимаюсь), в о«но»е такой же, как и характер распространения позвоночных, переносимых на плотах. Трудно поверить, что эти типы распространения оставались теми же самыми, если животные достигли этих островов действительно раз- ными путями. Другое замечание. Можно допустить, что Гольфстрим дол- жен был мешать проникновению животных, переносимых течением из Центральной Америки в Вест-Индию, и мог оказаться, несмотря на ооль- шее расстояние, благоприятным для переноса животных из Южнои Ам<- рики. Но это вполне вероятное допущение не подтверждается фактами. Чрезвычайно важен тот факт, что не строго пресноводные рыбы (второй отдел), достигшие этих островов, по-видимому, в основном проникли из Центральной и Северной Америки, преодолев Гольфстрим. В связи с тем. что больпшнство из этих рыб мелкие и не могут проплывать большое ра«- стояние, они почти пассивно переносятся поверхностными течениями. Если течения действительно благоприятствовали перенесению животных из северной части Южной Америки, а не Центральной, то в Вест-Индии долж- на была бы находиться главным образом южноамериканская фауна рыо, чего не существует на самом деле. К другххвх переносимым водой животным. которые, видимо, достигли Болыпих Антильских островов из Центральной Америки, относятся пресноводные черепахи (Ш11хатя, 1956) и предок эндемичного кубинского крокодила, родственного центральноаыериканс I;<>- му виду. Единственным приемлемым объяснением всему этому, мне кажется, слу;х'ит то, что животные, переносххмьхь по <>озд~ ху и водой (>хи II<>- чая и переносимых течением), достиглхх Больших Антильских ocrp<»«><>. расселяясь из одного места, в больхххххнсr»e случаев из Центральной Лмерххкхх.
Все это наводит на мысль о сходстве характер» распростр»кения фауи позвоночных Филиппинских островов и Вест-Индии. Каждая фауна отри- жает довольно простой иммиграционный тип распространения с блххжай- шего материка на острова и через них. В каждой фауне ххмехотся четкие реликты-эндемики, главным образом на ближайших и материку остроиах. Но в каждом отдельном случае у грызунов на уд»ленных островах диффе- ренциация проявляется в болыией степени, хеч у других нелетахощ><х наземных позвоночных. 11охоже, что этхх особенности распространеH IIH вызваны одххиьххх п теми же процессами: расселением через водные II)><>r т- р»нства главным образом в одном направлении — за пределы ближайшего II« <.0÷íïr'à заселения, в первом случае — от Калх>мантахха и во втором от Центральной Америки. Допустххвхо и более сложное объяснение осооен- иостей распространения животных в Вест-Индии, но и настоящее время, мне кажется, вполне прххемлеио объяснение, прииеденное вьпие. 8<>ai«>,<,— rr<>, вопрос о происхождении фауны Ео:хьхххххх Антильских островов оудл разрешен тогда, когда будут лучше изучены геологическая ист<>ри>< Центральной Америки и ее ископаемая. фаун». Но в определенные ххьрхходьх, возможно, и в настоящее время, которое является одним из этих ххерхдодох>, современное (илп очень недавнее) распх>остраненххе;х;ххво < ных с><ххдетельствует об определенных моментах прошлого лучик. хеы геологllfl илхх палеонтология. История Центральной A~repr<r«< недостаточно xop<>r»<> изучена. Северная и 10жная Америка, несомненно, на протяженхххх ооль- meir части третичного периода были отделены друг от друга, и геологиче- ские данные свидетельствуют о наличии водного п~эс~.~ива у южно~~ оконеч-
Особенности распространен ия жливот ных на островах
ности Центральной Америки, который отделял ее от Южной Америки. Но была ли Центральная Америка отделена от Северной Америки и была ли она отделена вообще, еще не известно. Структура фауны позвоночных Больших Антильских островов и особенно ее распространение свидетель- ствуют о том, что Центральная Америка отделялась от Северной и что по крайней мере на протяжении какой-то части третичного периода это был остров, а не полуостров, что там находилась смешанная фауна северных и южных мелких млекопитающих и других позвоночных и что большин- ство позвоночных Больших Антильских островов пришло именно оттуда.
Заключение. Филиппины. острова западной части Тихого океана, Вест-Индия — это главные архипелаги земного шара, которые, не являясь материковыми, все же расположены близко от материков и получают пе- риферийную материковую фауну. Это довольно обширные архипелаги, имеющие достаточное число сходных островов, чтобы можно было увидеть аналогичные особенности распространения животных. К этому списку можно добавить архипелаг «Зоны Уоллеса» (гл. 7), который расположен между материковыми отмелями азиатского и австралийского континентов и получает периферийные фауны с обоих континентов.
На всех основных Филиппинских, ближайших островах западной части Тихого океана (Бисмарка и Соломоновых), Вест-Индии и островах «Зоны Уоллеса» обитают те или иные нелетающие наземные позвоночные и лягушки, что. по мнению Уоллеса, является критерием материковых островов. Ряд зоогеографов до сих пор считает некоторые из этих островов материковыми островами и объясняет состав фаун последних как резуль- тат соединений их с сушей. Однако на всех этих островах разные классы позвоночных представлены в пропорции, приблизительно соответствующей их возможным способностям пересекать соленые воды: строго пресновод- ные рыбы на этих островах отсутствуют, кроме очень немногих в южной части Филиппинских и западной части Малых Зондских островов; амфи- бии и нелетающие наземные млекопитающие в основных ветвях ограничен- ны. а грызуны составляют более чем обычную долю среди млекопитающих; среди рептиллй непропорционально многочисленны ящерицы; многочпс- ленны рукокрылые и птицы. Еще более важно то, что основные части каж- дой фауны распространены на архипелагах последовательно, убывая вдоль первоначальных путей иммиграции, и наличие в пх составе древних релик- тов является исключением, а не правилом. Большинство реликтов и ярко выраженных эндемиков встречается на островах, Олпжапшпх к материку, кроме некоторых животных, особенно грызунов, обладающих наилучши- ми способностями пересекать соленые воды, у которых самые четкие реликты или эндемики находятся на более удаленных от материка островах. Эндемичные роды или очень четкие группы грызунов встречаются или встречались не только на дальних Филиппинских или дальних островах Вест-Индии (Пуэрто-Рико и др.), но также и на Соломоновых островах (граница распространения наземных млекопитающих на островах в запад- ной части Тихого океана). на островах Церам и Флорес (в обедненной «Зоне Уоллеса»).
Все это свидетельствует о том, что расселение позвоночных с материков на периферийные архипелаги через водные пространства происходит в зна- чительно более широких масштабах и в некоторых случаях на энач11тель- но большее расстояние. чем предполагал Уоллес. Это"факт в,ажен сам по себе, независимо от того. что говорит он об истории эпох островов п их фаунах.
Другие острова п архипелаги
Сулавеси. Этот остров находится в Индо-Австралийском архипелаге, недалеко от Калимантана, за линией Уоллеса. Его площадь равна пример- но 70 тыс. кв. миль. Это тропический, с разнообразными экологическими условиями и, очевидно, древний остров. Он отделяется от Калимантана Макассарским проливом, ширина которого в настоящее время достигает 65 миль, а самое узкое и глубокое место имеет ширину примерно 25 миль (Umbgrove, 1949, fig. 4). Во времена плейстоцена этот пролив, вероятно, был не шире, чем сейчас. Ранее остров Сулавеси уже рассматривался как часть «Зоны Уоллеса».
На Сулавеси отсутствуют строго пресноводные рыбы, но имеется несколько видов карпозубых второго отдела. Из амфибий отсутствуют без- ногие, но многочисленны лягушки, из которых большинство является пред- ставителями Индо-Малайской области, а род ОтеорЬтупе — представите- лем Австралийской области. Предполагают, что десять видов лягушек являются недифференцированными видами Индо-Малайской области, но не- которые из них, возможно, были интродуцированы (стр. 387). Рептилии на этом острове многочисленны и по своим связям в большинстве случаев относятся к Индо-Малайской области. К ним принадлежат черепахи рода Testudo. В плейстоцене на Сулавеси находились более крупные предста- вители этого рода (Hooijer, 1951). Также многочисленны на этом острове птицы. Их фауна представляет смесь групп (больше) Индо-Малайской и (меньше) Австралийской областей. Из млекопитающих на Сулавеси оби- тают два вида сумчатых, представляющих две группы рода кускус, земле- ройки (Crocidura и Suncus), род Tarsius, обезьяны эндемичного рода и один эндемичный вид рода 11IIacacа, четыре рода виверр (один эндемичный), один эндемичный род свиней (Babirussa), один небольшой буйвол (Апоа), обыкновенные свиньи и олени, три рода беличьих (с несколькими эндемич- ными видами), енот и различные мышиные, включая пять эндемичных родов, и рукокрылые. Все эти млекопитающие, кроме рода Phalanger и, воз- можно, некоторых мышиных и рукокрылых,— выходцы из Индо-Малай- ской области. На Сулавеси были обнаружены некоторые плейстоценовые ископаемые млекопитающих. Большинство из них относится к группам, которые до сих пор представлены в современной фауне, за исключением карликового слона, достигающего высоты около 6 футов (Ноо11ег).
Эта фауна замечательна полным отсутствием рыб строго пресновод- ных семейств и в то же время наличием нескольких крупных млекопи- тающих, включая и исчезнувший вид слона. Каким образом более круп- ные млекопитающие достигли этого острова — это до сих пор является спорным вопросом. Бьюфор (Beaufort, 1951, р. 177, 178) и Хуиджер счи- тают, что они проникли на этот остров по сухопутному мосту через Филип- пины, однако на острове Минданао следов пребывания основных групп млекопитающих нет, как нет на нем и материковой фауны позвоночных. Более того, на самом острове Сулавеси также отсутствует материковая фауна позвоночных. Я думаю, что в течение плейстоцена наиболее круп- ные млекопитающие этого острова, включая и слона, пересекли 25-миль- ную преграду, образованную Макассарскии проливом.
Мадагаскар, Маскпренские и Сейше..ьские островп. Мадагаскар (рис.
64) расположен к востоку от Африки, на южнои окраине тропиков. Его пло- щадь составляет примерно 240 тыс. кв. миль. Это древний остров с разно- образными геологическими и экологическими условиями, вероятно в прош- лом более облесенньтй, чем в настоящее время. Его центральная часть
Особенносвг и распространения животных на островах представляет собой древнее гранитное плато высотой 3000 — 5000 ôóòîâ над уровнем моря, с разбросанными на нем вулканами, достигающими высоты более 9000 футов. Более низкие береговые части острова, особенно на западе, состоят из осадочных пород триаса, юры, мелового и третичного периодов. Отложения самого позднего времени находятся недалеко от моря, что дает основание предположить, что по крайней мере с триаса между Африкой и островом существовал пролив. В настоящее время ~н достигает более 1000 м глубины и 100 км или более ширины на всем его протяжении (iHillot. 1952, р. 17). Весьма сомнительно, что Мадагаскар имел сухопутное
Р и с. 64. Мадагаскар п острова Индийского океана. соединение с Африкой или Азией, а если оно и было, время его существова- ния неизвестно. Впрочем, судя по современному составу фауны, нет необхо- димости прибегать к подобному соединению. Однако в мезозое на Мадага- скаре существовали динозавры, и если их нахождение там следует объяс- нять соединением с сушей, значит, оно было (см. гл. 10). Ширина самого узкого места в проливе между Мадагаскаром и Африкой в настоящее время равна примерно 260 милям, но во времена плейстоцена, вероятно, он был несколько уже. Вполне возможно, что некоторые животные достигли Мада- гаскара через Коморские острова, хотя и там имеются не менее широкие проливы. Рэнд (Rand, 1936) дает прекрасное описание Мадагаскара, а также авифауны этого острова, а Миллот (Millot, 1952) приводит крат- кий обзор его фауны в целом.
На Мадагаскаре отсутствуют строго пресноводные рыбы, но встре- чается несколько, вероятно проникших из Африки, представителей хромисов (второй отдел — не строго пресноводные рыбы) и карпозубых, а также различные солевыносливые рыбы.
Из амфибий на Мадагаскаре нет безногих и некоторых материковых семейств лягушек, но есть другие семейства, в основном Rhacophoridae и Brevicipitidae. BoJIbIIIHHCTBo из 150 с лишним видов и некоторых родов лягушек, а также 1 подсемейство из семейства Brevicipitidae (8 родов и 21 вид которого, очевидно, эволюционировали на Мадагаскаре от одного предка) являются эндемиками. Одни мадагаскарские лягушки родствен- ны африканским, а другие — тропическим лягушкам азиатского конти- нента (более подробно см. стр. 134).
На Мадагаскаре отсутствуют многие важные материковые группы рептилий, включая черепах семейств Trionychidae и Emydinae; ящериц- агамовых семейств Lacertidae, Amphisbaenidoe и Cordylidae rt рода V«<-
Глава 8
nus; червеподобных змей семейства Leptotyphlopidae, питонов, аспидов и гадюк. Так же как и на крупных островах Вест-Индии, на Мадагаскаре нет ядовитых змей. Но зато здесь хорошо представлены другие рептилии: пресноводные черепахи пеломедузы, сухопутные черепахи (род Testudu и ему родственные); ящерицы, включая множество гекконов, релик- товых игуан, многочисленных хамелеонов, сцинков и несколько предста- вителей семейства Gerrhosauridae; змеи, включая род ТурЫорз, специа- лизированные удавы и ужовые. На Мадагаскаре имеется ряд эндемичных родов рептилий и один род гекконов (Uroplates примерно с 5 видами), иногда рассматриваемый как эндемичное семейство. По своим связям большинство мадагаскарских рептилий — африканские, но связи неко- торых из нпх ведут к Индо-Малайской области, а пресноводные черепахи пеломедузы рода Podocnemis и (наземные) игуаны, очевидно, имеют близ- кородственных современных рептилий в Америке.
На Мадагаскаре находится очень определенная фауна птиц (Rand.
1936), к которой относится примерно 51 семейство. Наиболее многочислен- ны 1айки, цапли, утки, пастушки, хищные птицы, кукушки, славковые и ванги. Эта фауна состоит из большой эндемичной части, значительной африканской группы птиц и небольшой части фауны птиц Индо-Малай- ской области. В ней представлено несколько полностью эндемичных се- мейств и подсемейств: вымершие гигантские эпиорнисы (два рода, несколь- ко видов); нелетающие пастушкообразные 51esoenatidae (пастушковые куропатки) — два рода, три вида, два подсемейства сизоворонок (с че- тырьмя родами и шестью видами); филапитты (два рода, четыре вида) и ванги (девять родов, 11 видов). На Мадагаскаре представлены и многие африканские семейства птиц, но многие из них и отсутствуют: птицы-сек- ретари, страусы, цесарки (кроме одного вида, который, возможно, был интродуцирован), бананоеды, журавли, древесные удоды, птицы-носоро- ги, птицы-мыши, бородастики, дятлы, медоуказчики, рогоклювы, питты, африканскне береговые ласточки (Pseudochelidonidae), настоящие соро- копуты, лесные сорокопуты (Prionopidae), вьюр1овыс, буйволовые тка- чики, вдовушки, буйволовые птицы и др. Таким образом, авифауна Мадагаскара значительно беднее африканской (более подробно см. спи- сок семейств птиц в гл. 5). Из элементов Индо-Малайской области в мада- гаскарской авифауне представлено 4 тропических азиатских рода, которых нет в Африке, и несколько других видов птиц с очевидными связями с Ин- до-Малайской областью (Rand, р. 298).
Фауна млекопитающих Мадагаскара очень ограниченная высокоэн- демичная. Она состоит из 1 эндемичного семейства насекомоядных (Tenre- сМае) с 10 или более родами и, вероятно, с 30 видами, 3 эндемичных се- мейств лемуров примерно с 10 современными родами и 20 видами, которые, возможно, все произошли от одного предка; одного предположительно эндемичного подсемейства грызунов (Cricetinae) примерно с 7 родами и 12 видами и около 7 эндемичных родов с 12 видами виверровых, имеющих одного или нескольких предков. Известны также ископаемые и субфоссиль- ные формы этих групп, а также карликовый гиппопотам, самое крупное млекопитающее, которое, как известно, достигало Мадагаскара. Карлико- вые гиппопотамы были многочисленны в плейстоцене, но они не были сход- ны с современными гиппопотамами, живущими на африканском материке. На Мадагаскаре распространены также, вероятно, интродуцированные, один-два вида землероек, индийская виверра, крысы, мыши, африканские речные кабаны рода Potamochoerus, ископаемые которого не встречаются в плейстоценовых отложениях, где были найдены многочисленные остатки
Особен но«т и расвростпранен ия w ивотных на островах
карликовых гиппопотамов. И, наконец, на Мадагаскаре много рукокрылых с африканскими и индо-малайскими связями. К последним относится не- сколько групп рода Pteropus.
Мадагаскарская фауна позвоночных сформировалась за счет живот- ных. проникших через соленые воды, на что указывает как отсутствие стро- го пресноводных рыб, так и характер остальной фауны, особенно наземных млекопитающих, которые, очевидно, эволюционировали от нескольких мелк»х предков. Малочисленность грызунов — противоречивая деталь, но. возможно, это является чистой случайностью. Достопримечательно от- сутствие в фауне Мадагаскара дикобразоподобных. Большинство позво- ночных Мадагаскара, очевидно, проникло из Африки и в значительно меньшей степени (по крайней мере некоторые птицы и рукокрылые)— из тропиков Азии. Некоторые мадагаскарские лягушки и рептилии име- вт связи с Индо-Малайской областью, что еще не говорит о их проис- хождении и в этои области. То, что yã11å было сказано о лягушках (стр.
136, 137), относится и ко многим другим позвоночным: их распространению в Индийском океане и через него присуще большое число разрывов и сме- шанных связей, которые указывают на то, что эта фауна отчасти является реликтовой и что современные географические связи в какой-то мере есть результат вымирания и выживания, а не прямого расселения. Связи не- которых мадагаскарских рептилий с тропическими рептилиями Америки— ;ITo результат еще более обширного вымирания на еще более обширной
п:[ощади.
Маскаренскпе острова — небольшие острова, расположенные в Ин- дийском океане к востоку от Мадагаскара. Остров Реюньон находится примерно в 400 милях от Мадагаскара. Его площадь равна почти 1000 кв. м»ль, высота над уровнем моря достигает 10 тыс. футов. Остров Маври- кий находится более чем в 100 милях к северо-востоку от острова Реюньон и более чем в 500 милях от Мадагаскара. Он занимает площадь 700 кв. миль с предельными высотами до 2500 футов. Остров Родригес лежит при- мерно в 350 милях к востоку- от острова Маврикий, его площадь составляет 40 кв. миль, а высота — 1300 футов.
Это тропические острова вулканического, океанического происхожде- ния. Они получают довольно большое количество осадков и первоначаль- но были покрыты лесами, сильно разреженными в настоящее время.
На Маскаренских островах отсутствуют строго пресноводные рыбы, нмфибии и наземные млекопитающие, кроме рукокрылых. Однако репти- л»и этих островов заслуживают особого внимания. На всех островах существовали ныне вымершие гигантские наземные черепахи (Testudo). (Африкано-мадагаскарская пеломедуза, зарегистрированная на острове Маврикий, вероятно, является интродуцированным видом.) Современные, или субфоссильные, змеи известны только на острове Маврикий и на ма- леньком острове Раунд — два эндемичных рода удавов (или, возможно, примитивных ужовых). Встречались ли они на других Маскаренских ост- ровах, не известно. Из местных ящериц на островах Реюньон и Маврикий об»тают гекконы и сцинки, а на острове Родригес — только один большой эндемичный род гекконон PItelsuina.
Также особенно интересны (или были примечательны) птицы Маска- ренских островов. Ни одна часть земного шара не имеет более необычай- ную авифауну, и нигде в другом месте уничтожение местных птиц не происходило столь быстро и столь опустошающе. Хачисука (Hachisuka, 1953, р. 6, 7) приводит список известных современных и исчезнувших мест- ных птиц трех островов (из перечисленных птиц одна или две приводятся
на основании старых, не совсем точных отчетов путешественников, и воз- можно, эти данные вымышленны). К птицам Маскаренских островов. кото- рые заслуживают особого внимания, следует отнести додо на острове Маврикий и ему родственных на других островах: пустынника (Pe=o- р/гарг solitarius) на острове Реюньон и «белого додо» на острове Родригес. Все эти птицьх были уничтожены человеком двести или более лет назад. Они произошли от голубей, кормящихся на земле и потерявших способ- ность к полету, предположительно после того, как ими были достигнуты различные острова. К другим исчезнувшим птицам Маскаренскххх ост- ровов относится один (или несколько) вид нелетающих пастушков, встре- чавшихся на каждом острове; предположительно нелетающая цапля на острове Родригес, почти нелетающая цапля с острова Реюньон и пред- положительно нелетающий голубь с острова Маврикий. На Маскаренских островах обитали или обитают также различные водные птицы, хищные пти- цы, совы, несколько видов голубей, несколько видов попугаев и немного- численные разнообразные воробьиные птицы. Огромные плодоядные руко- крылые (Pteropus) встречаются на всех островах, и один впд насекомояд- ной летучей мыши (Scotophilus) зарегистрирован на острове Маврикий.
Сейшельские острова — это группа, состоящая примерно из 45 ост- ровов, расположенных примерно в 600 милях к северо-востоку от 1хада- гаскара; они отстоят несколько дальше других островов от берега Африки и значительно дальше от Индии. Самый большой и высокий остров Маэ занимает площадь примерно 55 кв. миль, высота его достигает примерно 3000 футов. Общая площадь Сейшельских островов менее 150 кв. миль, но все эти острова охвачены 100-метровой изобатой и окружены отмехьхо, площадь которой занимает около 12 тыс. кв. миль. Обширные отмели близ этих островов позволяют предположить, что современные острова представляют собой остатки обширного невысокого архипелага. Основная порода, из которой сложен остров,— древние граниты, аналогичные грани- там центрального плато Мадагаскара. Сейшельские острова — тропххче- ские, получающие достаточное количество влаги — были когда-то покрыты густыми лесами, но в настоящее время большинство лесов сведено. hpaт- кий обзор фауны Сейшельских островов, описанной в отчетах экспедпцхххх Перси Сладена, дан в работе Гардинера (Gardiner, 1936).
На Сейшельских островах отсутствуют строго пресноводные рыбы, но есть амфибии. Безногие представлены одним эндемичным родом с ше- стью видами, происшедшими, видимо, от одного предка, который, по мне- нию Паркера (Parker, 1941), скорее был африканским, чем индийским, но на Мадагаскаре безногие не встречаются. Лягушек этих островов, представленных эндемичным подсемейством (два рода, три вида), относят с некоторым сомнением к Pelobatidae — семейству, неизвестному ни в Африке, ни на полуострове Индостан, но встречающемуся в восточной части Индо-Малайской области и в умеренной зоне северного полушария. На Сейшельских островах находятся также эндемичный вид африкано-ма- дагаскарской лягушки рода Megalizalus (семейство Rhacophoridae) и, возможно, интродуцированный вид Лапа. Особенно надо отметить отсут- сгапе рода Rhacophorus, встречающегося в Индо-Малаххскохх области и на Мадагаскаре, и лягушек семейства Brevicipitidae. столь многочислен- ных на Мадагаскаре.
К рептилиям Сейшельских островов относятся ныне вымершие ги- гантские сухопутные черепахи (Testudo) и один недифференцированный вид пресноводной черепахи пеломедузы (вероятно, интродуцированный), несколько видов ящериц (гекконы, сцинки и один вид хамелеонов), одни
Особен ности распросиг ранения животных на острова;х
нз которых африканского происхождения, а другие происходят, оче- видно, из Индо-Малайской области; два представителя ужовых змей (один род эндемичный).
Авифауна Сейшельских островов представлена 14 эндемичными видами наземных птиц (Vezey-Fitz Gerald, Betts, 1940), большинство иэ которых существует и в настоящее время. Одни из них по своим связям— африканские, а другие — индо-малайские. К ним относятся небольшой сокол, два вида голубей, два вида попугаев и воробьиные птицы. На. Сей- шельских островах нет и. насколько известно, никогда не оыло нелета- ющих птиц. Здесь нет также нелетающих наземных млекопитающих. но встречаются рукокрылые: плодоядные (Pteropus) и насекомоядные (два вида рода Coleura), у которых выражена филогенетическая близость с этими же родами Индо-Малайской области.
Фауны небольших, удаленных от материка Маскаренских и Сейшель- ских островов очень ограниченные, но имеют самые непредвиденные отли- чия. На Маскаренских островах нет амфибий, но есть или было несколь- ко групп нелетающих птиц. На Сейшельских островах имеются эндемич- ные, довольно древние группы амфибий, но отсутствуют нелетающие пти- цы. Эти отличия позволяют предполагать, что история этих островов была также резко отлична. Маскаренские острова, вероятно, всегда были уда- ленными от материка клочками суши и приобрели крайне ограниченную фауну исключительно из тех животных, которые легко пересекают водные пространства. Сейшельские острова, возможно, являются остатками оо- ширного архипелага более крупных островов, которые находились до- статочно близко от Африки или Индии и, таким образом, имели возмож- ность получить довольно большую фауну позвоночных (ее части, очевид- но, проникли не через Мадагаскар), большая часть из которой вымерла, когда происходило сокращение площади этих островов. В эту фауну, воз- можно, входили наземные млекопитающие, которые, вероятно, задержи- вали эволюцию нелетающих птиц. Эта история образования фауны, соот- ветствующая геологической природе Сейшельских островов, не нуждается в привлечении теории полного соединения этих островов с какой-либо другой сушей.
Новая Каледония, остров Лорд-Хау и Норфолк. Новая Каледония, расположенная в краевой зоне тропиков, находится примерно в 800 ми- лях к востоку от Австралии и несколько ближе к Новым Гебридам. Пло- щадь этого острова равна примерно 6500 кв. миль, высота гор свыше.
5000 футов. Это древний остров с высокоэндемичной фауной беспозвоноч- ных, с разнообразными экологическими условиями и сложным геологиче- ским строением. Фауна позвоночных здесь очень ограниченна. На острове отсутствуют строго пресноводные рыбы. Из солевыносливых рыб наиболее примечательными являются галаксиды — самые северные представители этого семейства. Здесь нет амфибий и местных млекопитающих, кроме руко- крылых, нет неморских черепах, хотя в плейстоцене на острове Уолпол. к юго-востоку от Новой Каледонии, встречался представитель семейства Ме1о1ап11йае. Также отсутствуют здесь змеи (51ertens, 1934, р. 39), не- смотря на неоднократные ошибочные случаи их регистрации. Таким обра- зом, единственными местными по происхождению нелетающими назем- ными позвоночными Новой Каледонии являются только ящерицы, а из них только гекконы (включая один эндемичный род) и сцинки. Птицы Новой Каледонии (Mayr, 1945, р. 149 — 175) насчитывают около 68 местных наземных и пресноводных видов, из которых 16 видов являются эндеин- камп или общими только с островами Лоялти. и, кроме того. имеется.
Глава 8
несколько эндемичных родов и одно эндемичное семейство. К местным пти- цам относятся хищные, совы, трехперстки, пастушки, кагу, голуби, попу- гаи. кукушки, совиный козодой, козодой, стрижи, зимородок и 24 вида певчих птиц. Эндемичное семейство Rhinochetidae состоит только из од- ного вида — кагу, довольно крупной журавлеобразной птицы, у которой хорошо развиты крылья, хотя она и не летает. Новокаледонский вид пастушка (Trircholimnas), возможно, тоже стал нелетающим. Из млекопи- тающих в Новой Каледонии распространены, видимо, интродуцирован- ные крысы и встречаются плодоядные (Pteropus) и насекомоядные руко- крылые. В этой фауне позвоночных отсутствуют какие-либо животные. в отношении которых можно было бы предполагать, что они, несомненно, достигли Новой Каледонии, преодолев водное пространство.
Лорд-Хау — небольшой вулканический остров, расположенный примерно в 300 милях к востоку от Австралии и примерно в 2 раза дальше h югу (и несколько к западу) от Новой Каледонии, с площадью 5 кв. миль, с наибольшей высотой, достигающей примерно 3000 футов. Значительная часть острова покрыта лесами. Местная фауна позвоночных ограничена, очевидно, эндемичным видом сцинков (один вид рода Lygosorna или Lei- ulupisrna) и неэндемичным гекконом (последний, вероятно, занесен чело- веком), несколькими видами наземных птиц и летучей мышью (Scoto- philus). В плейстоцене на острове находились черепахи семейства Ме1о1а- niidae. В прошлом птицы острова (Hindwood, 1940) включали около 15 местных, оседлых наземных видов (в настоящее время восемь видов пз них исчезли): один или два вида голубей, один попугай, пастушок и ка- мышница (очевидно, оба нелетающие), сова, насекомоядный вид зимород- ка и 8 видов воробьиных. Из всех этих видов два или три проникли, види- мо, из Австралии, три — из Новой Каледонии, один — пз Новой Зелан- дии. Географическое происхождение остальных не ясно. Другие назем- ные птицы и некоторые птицы пресных водоемов регулярно или периоди- чески посещают этот остров, и (как это обычно бывает, на удаленных от материка островах) здесь гнездятся различные морские птицы.
Норфолк — небольшой вулканический остров, расположенный при- мерно в 800 милях к востоку от Австралии, примерно на полпути между Новой Каледонией и Новой Зеландией, с площадью, равной приблизи- тельно 13 кв. миль, и высотой 1000 футов. Остров имеет благоприятные климатические условия. Растительный мир Норфолка сходен с раститель- ным царством Новозеландии и включает норфолкскую островную сосну, прекрасные араукарии. Местных нелетающих наземных позвоночных на э1.ом острове нет, а один или два широко распространенных вида гекко- нов. вероятно, были интродуцированы. Наземные птицы острова ( lathews.
1928 — 1936) представлены одним эндемичным видом голубей (исчезнув- ший), двумя видами попугаев (один из которых исчез), одной совой, насе- комоядньв| видом зимородка и 9 видами воробьиных, пз которых боль- шинство или все эндемичные. На этом острове, где ранее должны были. несомненно, обитать пастушковые, нет ни одного достоверно известного пастушка, если не считать одного зарегистрированного здесь и отнесен- ного к сомнительному, вида средней величины.
Новая Зеландия. Новая Зеландия расположена приблизительно в 1000 милях к юго-востоку от Австралии. Площадь двух главных остро- вов составляе" в целом более 100 тыс. кв. миль. Это древние с точки зре- ния геологии острова с высокими горами и разнообразными экологически- ми условиями и умеренным климатом. Список позвоночных Новой Зелан- дии и их описание даются Хаттоноъх и Драммондом (Hutton, Drummorid.
Особе ннотг и распространен ия ж иволог ных и а островах
1923). Другой список приводится Пауэллом (Powell, 1949, с ссылками на другие работы). Некоторые из этих позвоночных рассматриваются в дру- гих главах книги или более подробно приводятся в списках семейств.
В Новой Зеландии отсутствуют строго пресноводные рыбы, но есть солевыносливые, включая галаксид. Амфибии представлены лягушкой реликтового рода Ае~оре1та (два-три вида), которая сохранилась только в Новой Зеландии, в то время как родственные ей виды с большей части земного шара уже исчезли. Из рептилий к Новой Зеландии приурочена ящерица гаттерия (Sphenodon), являющаяся еще более удивительным реликтом, последним выжившим представителем отряда, который, судя по ископаемым, исчез 100 млн. лет назад. Однако это совсем не озна- чает. что гаттерия (местное название туатера) должна была находиться п Новой Зеландии именно с тех времен. В Новой Зеландии распростране- ны также гекконы (два эндемичных рода, единственные яйце-живородя- щие гекконы) II сцинки (эндемичные, яйце-живородящие виды рода Аудо- soma), но ни змей, ни черепах, кроме морских, нет. Для Новой Зеландии характерна уникальная группировка нелетающих птиц: огромные вымер- <иие моа (два семейства), киви, разнообразные, совершенно не летающие пастушки, нелетающий вымерший гусь, нелетающий попугай и, возможно (если эта птица была нелетающей), единственный нелетающий представи- тель воробьиных — Traversia. Из летающих птиц в Новой Зеландии отметим небольшие эндемичные семейства Xenicidae и Са11аеЫае, эндемич- ные роды и виды других семейств (включая хищных и сов, исчезнувший вид куропатки. кукушек, попугаев, голубей, один вид зимородка и раз- личных воробьиных), и некоторые неэндемичные роды и виды, включаю- щие и современных вселенцев, и случайно залетных птиц из Австралии (стр. 209). Из местных наземных млекопитающих в Новой Зеландии рас- пространено только два вида насекомоядных рукокрылых: один относя- щийся к древнему эндемичному семейству (Alystacinidae), а другой вид рода A<alinol~~bus, вероятно, недавно проникший из Австралии.
Чтобы объяснить состав современной фауны позвоночных Новой '~елаыдии, нет необходимости прибегать к теориям сухопутных соединений с материком. Лягушки иногда пересекают по крайней мере небольшие пространства соленых вод, и нет установленной ширины водных проли- вов, которую могут пересекать, возможно, отдельные их виды, в особен- ности такие. как Leiopelrna, способные развиваться на суше. Гаттерпя (Sphenodon) должна обладать хорошими, возможно, такими же, как у не- которых ящериц, способностями преодолевать водные пространства. В фа- уне позвоночных Новой Зеландии в целом проявляются все степени энде- мизма — от эндемичного отряда (Sphenodon) до незндемичных видов (птиц). Это вполне соответствует истории всей фауны, элементы которой постепенно накапливались, проникая на остров через широкие водные пространства.
Гавайские ocrr<po<,a. Гавайи — архипелаг в центральной части Тихо- ro океана, протяженностью примерно 1500 миль, находится примерно в -0<<0 милях от Северной Америки и еще дальше от материков Старого Свет«. Цепочки островов протянулись по направлению к Азии и Австра- л<ш, но между некоторыми из них имеются широкие проливы. Самый боль- u<oii остров — Гавайи — занимает площадь приблизительно 4000 кп. миль. Эти вулканические острова с высокими горными хребтами окруже- ны большими гл~ бинами. Приурочены они к северной окраине тропиков, но благодаря значительным размерам площади имеют богатые, разнооб- разные экологические условия. В связи с тем, что эти острова довольно
1дава 8
древние, на них смогла развиться высокоэндемичная фауна оеспозвоноч- ных и птиц, для чего, видимо, понадобилось не так уже много миллио- нов лет. Основные острова возникли, очевидно, не ранее плиоцена. (:пи- ски гавайской фауны были даны в прекрасной для своего времени работе «Fauna Hawaiiensis» (1899 — 1913), Циммерман (Zimmerman. 1948) приво- дит новейшие общие сведения о природе, истории, флоре н фауне этих островов.
Таолпца 19
Наземные и пресноводные птицы Гавайских островов(кроме острова Лейсан) '
Знделичное семейство
(1) Dr~ paniidae (вероятио, из Америки)
Эндемичпые роды
(2) 11астушок (неизвестного происхождения) (3) Два рода медососов (от австрало-азиатского предка) (4) Дрозд (от американского предка)
(4) Мухоловка Старого Света (от полинезийского lip(äêà) (4) Гусь (от американского предка)
Энделичные виды
(5) Ворона (вероятно, из Северной Америки) (5) Хищник (из Америки)
(5) Утка (вероятно, из Северной Америки)
9ндемичные подвиды
(6) Болотная сова (вероятно, из Северной Америки) (6) Ходулочник (из Америки)
(6) К амышница (и з Америк и)
(6) Лысуха (из Америки)
Хеэи деличный представитель (7) Кваква (из Америки)
1 Эта таблица, несколько измененная, заимствована из работы Майра (Мауг. 1943, р.
45, 46). 'четырнадцать отдельных вторжений перечисляются в порядке относительно очевидного возраста (цифры 1 — 7).
На Гавайских островах нет строго пресноводных рыо, местных амфи- бий и рептилий. Там распространены только птицы и рукокрылые, причем последние представлены видом американского рода Lasi urus. Встречан>щие- ся на этих островах восемь видов лягушек и жаб, одна пресноводная чере- паха, девять видов ящериц и один вид наземных змей (Oliver, Shaw, 1953) интродуцированы (одна лягушка и шесть ящериц ошибочно были описаны как новые формы с Гавайских островов). Из птиц семейство Drepaniidae (гавайские цветочницы) достигло уровня слаб одифференци рованного семейства с 9 родами и 22 хорошо выраженными видами, приуроченными к Гавайским островам, лежащим к северо-западу от Лейсана. Возможно, они произошли от американских пищух-медососов, питающихся некта- ром. но на островах у них развились нектара-, насекомо- плода- или зер- ноядные линии, а на разных островах основные линии привели к образо-
«аннхен различных географических форм. дто ярки1х пример адаптивной и географической радиации в изоляции, детально описанный Амадоном (Ama(ton, 1950), который дает также краткий обзор других местных наземных и пресноводных птиц. Maiip (Мауг, 1943) рассматривает их с точ- ки зрения зоогеографии. Он считает, что на собственно Гавайских остро- вах было 14 отдельных вторжений птиц с образованием 7 групп с раз- .личным уровнем эндемизма (табл. 19). Несколько эндемичных родов дали на разных островах отдельные виды. Кроме форм, перечисленных в табл. 19, на острове Лейсан находятся (или находились) еще один энде- мичньш род пастушков (возможно, родственный гавайскому) и один энде- мичный вид камышовки Старого Света (Acrocephalus). Кроме оседлых форм. на этих островах встречаются мигрирующие или странствующие птицы: золотистая ржанка и некоторые другие неморские виды. Авифау- на Гавайских островов является классическим примером океаническо11 фауны, образовавшейся от нескольких предков, которые постепенно накапливались, проникая на острова из разных мест (большинство, очевид- но, 1I3 Америки), и малочисленность которых отчасти компенсировалась радиацией некоторых древних ветвей. Гавайские и лейсанские пастушки .стали нелетающими, но, кроме них, на Гавайских островах других неле- тающими птиц нет.
Галппагос. Галапагосские острова расположены на экваторе, при- мерно в 600 милях к западу от Южной Америки и более чем в 700 милях к юго-западу от Центральной Америки. К ним относятся 5 больших и 10 более мелких островов, общей площадью около 2870 кв. миль. Остров .Албемарл занимает около 1650 кв. миль общей площади. Несколько более крупных островов поднимаются над уровнем моря на высоту 2000— 5000 футов. В геологическом отношении это вулканические океанические острова, отделенные от материка большими глубинами. Они имеют довольно засушливый климат, но склоны более высоких гор облесены.
На Галапагосах отсутствуют строго пресноводные рыбы и амфибии. Из рептилий здесь распространены гигантские сухопутные черепахи (около 13 рас одного эндемичного вида рода Testudo), две крупные игуа- ны (ооразующие два родственных эндемичных рода), более мелкие игуаны (эндемичные виды южноамериканского рода Tropidurus), гекконы (широко распространенного рода Phyllodactylus) и из змей — у~ковые (эндемпчные виды тропического американского рода Leimadophis, воз- можно, представляющие две разные материковые ветви этого рода).
Из наземных птиц на Галапагосах доминируют дарвиновы вьюрки, иногда относимые к подсемейству Geospizinae. Эта группа, за исключени- ем одного рода и одного вида, эндемичных на острове Кокос, который рас- положен между Галапагосами и Центральной Америкой, состоит из четы- рех или более родов и 12 видов, приуроченных к островам Галапагос. И эта естественная группа, которая произошла от одного предка (пре- док неизвестен, но вполне возможно, что это был американский выо- рок). радиировала на Галапагосах, подобно гавайским цветочницам на Гавайских островах, но менее широко. Для представителей этой группы характерно разнообразное питание: есть виды, кормящиеся на земле, рас гительноядные, насекомоядные, добывающие пищу в стволах деревьев, как дятлы, кактусоядные виды и виды, схожие по добыванию корма со славками, а в соответствии с характером их питания изменяется и форма клюва этих птиц. Но они различаются не только в связи с характером их питания, а также и географически — от острова к острову. Их разнооб- разие — это одна из деталей. которая натолкнула Дарвина на мысль об
эволюции животных. Лэк (Lack, 1947) в своей работе приводит все извест- ные в настоящее время данные об этих птицах и дает ссылки на посвящен- ные им более специальные работы.
Из других галапагосских птиц следует отметить эндем11чный род пере- смешников (Nesomimus) с отличающимися видами или подвидами на раз- ных островах, один эндемичны11 род голубей (Л esopelin), эндемичного канюка (Buteo), два вида сов, пастушка, ласточку, трех американских мухоловок (тирани), один эндемичный подвид золотистокрылой древесни- цы, не отличимой от южноамериканского вида кукушки. деревенскую ласточку и рисового трупиала, которые более или менее регулярно посе- щают эти острова, а также различных болотных и водоплавающих птиц. На островах Галапагос обитают баклан Гарриса (нелетающи1х эндемичный род) и пингвины, но других нелетающих наземных птиц здесь нет.
Из млекопитающих к Галапагосам приурочены мелкие эндемичные грызуны (семейство Cricetidae), включая род (Nesory=omys), которы11 иног- да считают эндемичным, но Симпсон относит всех их к роду Oryzonlgzs-- тому же самому роду, который проник и на Ямайку. Там тоже встречанп- ся рукокрылые рода АагКигиг, который проник и на Гавайские остроиа.
Вся фауна позвоночных 1'алапагосских островов, очевидно, океа1111- ческая. образовавшаяся так же, как и фауна Гавайских островов, то есть от нескольких предков, которые постепенно концентрировались на этих островах и которые частично компенсировали cRoIO мало 1исленнос ть радиацией некоторых групп. К таким предкам относятся одна назем1111н черепаха, три или большее число видов ящериц, одна, возможно две, змеи. дюжина nJln большее число наземных llTnq (nuen B anpy тол~;о ocean:11i11' формы), один грызун (СпсеЙЫае) и одна летучая мышь. Очевидно, нее они являются выходцами пз Америки, но трудно си;азать, откуда приш:1о большинство пз них — из Южно11 или из Центра.1ьно11 Америки. Эти ос1- рова находятся недалеко от Южно11 Америки, но остров Кокос может с:1у ,кигь переходной ступенью по направленихо к Центральной Америке, кро- ме того, плавучие предметы достигают островов Галапагос чаще гсег<~ из тех же мест. Агассиз (Agassiz, 1892, р. 59, 60, 70 и слсд.) обнаружил, что течения вблизи Панамы иногда достигают скорости 75 м11ль в донь. перенося массу плавучих предметов к островам Кокос и Га 1апагос. Таким образом, данные этого автора оказались не в пользу сторонников теории сухопутных соединений с этими островами. В отношении фауны островон Галапагос высказывалось предположение, что некоторые позвоночные этих островов, включая игуановых, являются родственнымн вестиндским формам и произошли от общего предка (предположительно) из Централь- ной Америки третичного периода. Несомненно, эта интересная версн11 заслуживает дальнейшего изучения.
Остроаи Атланпшческого океанп (рис. 56). Канарские, Мадейра и Азорские острова расположены севернее тропиков на расстоянии, посте- пенно увеличивающемся от Северо-Западной Лфрик11 и Юго-Западной Европы. Ближа11п1ие острова этих трех групп находятся примерно в 60, 350 и 800 милях от материка, а острова Зеленого Мыса, ле'ка1цне в преде- лах тропиков,— примерно в 300 милях от Африки. Это неоольшие (самый большой из них занимает площадь всего несколько сот квадратных миль) осТро11а вулканического происхождения, окр1 женныь ооль11111ми Mopchli- ми глубинами. Здесь нет строго пресноводных рыб, местных амфибий, эндемичных черепах. Но на островах Зеленого Мыса встре 1ается, возмо к- но интродуцированный, такой широко распространенны11 африкaHchllil вид, как Pelusios subniger, а на Канарских островах в плейстоцене нахо-
Особенности распростпранен ия ж ивотных н а островах
дился представитель рода Testudo, нет змей и наземных млекопитающих, за исключением рукокрылых. Поэтому ящерицы — это единственные местные нелетающие наземные позвоночные, количество видов, родов и т. д. зависит и от расстояния от материка, и от климата: на Канарских островах встречаются эндемичные представители трех средиземноморско- африканских родов, на острове Мадейра — одного рода, на Азорских нет ни одного представителя и на (тропических) островах Зеленого Мыса имеется около четырех или более африканских ветвей. Некоторые назем- ные птицы Старого Света, так же как и летучие мыши, достигают всех этих островов: представитель рода Pipistrellus даже является эндемиком на Азорских островах.
Бермудские острова находятся севернее тропиков, приблизительно в 600 милях от Северной Америки. В настоящее время площадь этого ост- рова составляет только около 19 кв. миль, но в прошлом она была знач»- тельно больше (Sayles, 1931). Это вулканические острова, окруженные глубокими водами. На них отсутствуют строго пресноводные рыбы, но в мангровых болотах встречаются солевыносливые карпозубые. Из мест- ных нелетающих наземных позвоночных на Бермудах распространены только ящерицы сцинки (четкий эндемичный вид рода Еи~песез), являю- щиеся выходцами из Северной Америки, а также две вестиндские лягушки. гигантская жаба из тропиков Америки и вестиндская ящерица (Du»nr Сonant, 1937), которые были, так же как и обыкновенные крысы и мьппи. интродуцированы. Семь наземных видов птиц (флоридская камышница. земляной голуоь, ворона, кошачий пересмешник, синяя птица ЯаШа. белоглазый виреон и кардинал) являются местными и оседлыми, и все от- носятся к североамериканским, а не вестиндским видам (Bond, 1936, р. XV). Многие другие наземные птицы мигрируют или случайно залетают на эти острова. Летучие мыши появляются здесь периодически, но, оче- видно, не являются оседлыми.
К более удаленным от материка островам Атлантического океана относятся: тропический остров Вознесения, расположенный примеры« в 1000 милях от Африки, с площадью 38 кв. миль, остров Св. Елены. :re~~~arnrrii примерно в 1200 Mrr.rHx от Африки с площадью ~7 r;n. миль, и в умеренной зоне южного полушария группа островов Тристан-да-Кунья. находящаяся почти на расстоянии 2000 миль к югу от оконечности Южной Африки и еще дальше от Южной Америки. Самый большой остров этой группы занимает площадь около 16 кв. миль. Это все океанические остро- ва вулканического происхождения, находящиеся в глубоководных 1астях океана. Единственные наземные позвоночные, представленные на этих островах — немногочисленные птицы. На острове Вознесения существовал вымерший ныне нелетающий пастушок, готорый не име~т названия и известен только по старинным рисункам и описанию. На острове Св. Елены имеется эндемичный вид ржанки (Cha rudra us), родственный африканскому виду. До того как на этих двух островах были вырублены леса (M allace, 1880, р. 283), возможно, там встречались и другие наземньи птицы, но данные о них отсутствуют. На островах Тристан-да-Кунья. хотя они удалены от материка на значительное расстояние и имеют мень- шую площадь и более прохладный климат, встречаются два неродствен- ных эндемичных рода нелетающих пастушков, один эндемпчный род дроз- да (iVesocichla), как с ситает Райпли (Ripley, 1952, р. 16, 17), американского происхождения, и два неродственных эндемичных рода вьюрков, из ко- торых один ('Vesospi=.à) с двумя видами, различающимися по размеру клю- ва, обитает на островах собственно Тристан-да-Кунья, другой (Лоией~а)
1дава 8
древние, на них смогла развиться высокоэндемичная фауна оеспозвоноч- ных и птиц, для чего, видимо, понадобилось не так уже много миллио- нов лет. Основные острова возникли, очевидно, не ранее плиоцена. 1'пи- ски гавайской фауны были даны в прекрасной для своего времени работе «Fauna Hawaiiensis» (1899 — 1913), Циммерман (Zimmerman. 1948) приво- дит новейшие общие сведения о природе, истории, флоре п фауне этих островов.
Таолцца 19
Наземные п пресноводные птицы Гавайских островов(кроме острова Лейсан) '
Эндехичное семейство
(1) Dr<;paniidae (вероятно, из Америки)
Э~tä~å.òþíûå роди
(2) 11астушок (неизвестного происхождения) (3) Два рода медососов (от австрало-азиатского предка) (4) Дрозд (от американского предка)
(4) Мухоловка Старого Света (от полинезийского проулка) (4) Гусь (от америк аиск or о предк а)
Энделичные виды
(5) Ворона (вероятно, из Северной Америки) (5) Хищник (из Америки)
(5) Утка (вероятно, из Северной Америки)
Энделичные подвиды
(6) Болотная сова (вероятно, из Северной Америки) (6) Ходулочник (из Америки)
(6) Камышница (из Америки)
(6) Лысуха (из Америки)
Хеэн деличный представитель (7) Кваква (из Америки)
1 Эта таблица, несколько измененная, заимствована из работы Майра (3Iayr. 1943, р.
45, 46). ~четырнадцать отдельных вторжений перечисляются в порядке относительно очеьид~~ого возраста (цифры 1 — 7).
На Гавайских островах нет строго пресноводных рыо, местных амфи- бий и рептилий. Там распространены только птицы и рукокрылые, причем последние представлены видом американского рода Lasiurus. Встречающие- ся на этих островах восемь видов лягушек и жаб, одна пресноводная чере- паха, девять видов ящериц и один вид наземных змей (Oliver. Sham, 1953) интродуцированы (одна лягушка и шесть ящериц ошибочно были описаны как новые формы с Гавайских островов). Из птиц семейство Ргерап1Ыае (гавайские цветочницы) достигло уровня слабодифференци рованного семейства с 9 родами и 22 хорошо выраженными видами, приуроченными к Гавайским островам, лежащим к северо-западу от Лейсана. Возможно, они произошли от американских пищух-медососов, питающихся некта- ром. но на островах у них развплнсь нектара-, насекомо- плода- или зер- ноядные линии, а на разных островах основные линии привели к образо-
единственные неплавающие птицы, которые достигли края антарктиче- ского континента. Здесь имеется 4 вида антарктических тюленей, но толь- ко тк~лень Уэдделла встречается круглогодично у южной границы своего ареала — вдоль антарктического побережья. Это самьш южный предста- витель из всех млекопитающих земного шара.
Фауна Антарктиды в настоящее время крайне ограниченна в связи с суровостью климата. Но это очень древний массив суши, и очевидно, и прошлом климат его был теплее, чем теперь. Остатки угольных пластов, ископаемые остатки деревьев и Glossopteris показывают, что там должна была быть значительная растительность и, видимо, наземная фауна. Что это была за фауна — типа фаун очень крупных, но удаленных от матери- ка островов или значительно большая фауна, пока не известно, но может быть, что когда-нибудь ответ на это будет дан ископаемыми находками. Между тем связи Антарктиды с другими материками в древние времена 11 та роль, которую она возможно~ играла в распространении жизни очень волнующие и важные вопросы.
Заключение о распространении животных на островах
На одних островах (например, Огненная Земля, Антарктида) фауны позвоночных в настоящее время ограничивает и видоизменяет суровый климат. На других состав или возраст современных фаун ограничили так же, как и ограничивает их и современный климат, ледники плейсто- цена (например, Ньюфаундленд, Британские острова в сравнении с Япо- нией). Некоторые группы животных, особенно рептилии, совсем не встре- чаются на островах с очень холодным климатом, но так же, как и на мате- риках, достигают северных и южных границ в океанах (Mertens, 1934, р. 5; карта северных и южных границ островных рептилий). С другой стороны, ряд животных, особенно крупных и среднего размера млекопитающих, проникли на отдельные острова с холодным климатом через ледниковые мосты или на плавучих льдах (например, на остров Ньюфаундленд. Гренландию, Исландию). 11оэтому острова с холодным климатом надо рассматривать как особые случаи, с резко отличающимися от обычных условиями жизни на островах.
Влияние площади и расстояния от материка на фауны островов везде очевидно и играет немаловажную роль, хотя переложить это влияние на язык математики нелегко. В результате этого влияния образуются или видоизменяются все особенности распространения, рассматриваемые ниже.
Самый важный факт в отношении позвоночных животных островов— 3T() то, что большинство из них, по-видимому, является относительно недавними иммигрантами, а не древними реликтами.
На всех крупных периферийных архипелагах (Филиппины, острова западной части Тихого океана, Вест-Индия и «Зона Уоллеса») главной осо- бенностью распространения является последовательная иммиграция, так словно фауны — это результат накопления животных, распространенных вдоль путей, которыми они расселялись на острова. На этих островах не оыло обнаружено реликтового характера расселения. Реликтовыи харак- тер расселения должен иметь место на архипелагах (как это, очевидно, произошло на островах Суматра, Калимантан и Ява) в том случае, если материковые фауны уменьшаются до небольших частей. (:охранившиеся части дол;кны распространяться неравномерно, причем в их расселении отчасти играют роль крупные или более благоприятные для обитания
450
Глава 8
острова, независимо от их местоположения, и отчасти от способности животных к расселению. Реликты могут встречаться также и при иммигра- ционном характере расселения, но в этом случае они обычно концентри- руются или на ближайших к материку, или на более удаленных островах архипелага пропорционально расстоянию и степени расселения. Нагляд- ные примеры были приведены для Филиппин и Вест-Индии. Но над<в отметить как очень важный факт, что реликтовьш zaparmep распростране- ния на этих или каких-либо других древних архипелагах обнаружен не был.
На островах встречается удивительно мало древних реликтовых позвоночных. По степени их реликтовости перечислим наиболее интерес- ных. Например, первое место здесь занимают древний реликт гаттерия (Sphenodon). Новозеландская лягушка лиопельма и двоякод~ышащая рыба Австралии (остров, который для строго пресноводных рыб является очень изолированным) — также чрезвычайно древние реликты. Хотя это и не типичные примеры, но самое важное в отношении данных реликтов, воз- можно, то, что им подобных больше нет, то есть других таких же древних и изолированных реликтов (из позвоночных) на других островах. Следую- щее место в этом списке могут занимать щелезуб и Vesophontes Вест-Ин- дии, а рядом с ними, вероятно, тенреки и лемуры Мадагаскара, хотя они относятся, скорее всего, к олигоцену или миоцену, но не к более ранним (на этих островах) периодам. Podocnemis и ~гуановые на Мадагаскаре и игуановые на островах Фиджи — это изолированные географические реликты, но ойи не обязательно должны быть очень древними, то есть, возможно, они были изолированы в течение непродолжительного времени. Имеются факты, аналогичные приводимым, и о других ihBBoTHbIx. но в большинстве случаев, вероятно, тоже не очень древних, и проводить линию между реликтовыми и просто автохтонными формами становится все труднее.
Насколько известно, ни на одном древнем острове нет фауны, состоящей главным образом из древних реликтов. Такие острова, как Мадагаскар п Сулавеси, на которых, как иногда считают, имеются реликтовые матери- ковые фауны, мне кажется, аккумулировали свои современные фауны из частей фауны, проникших через водные пространства. Даже островные материки — Австралия и (в третичном периоде) Южная Америка,— и< и- мо'кно, не имели реликтовых фаун и накапливали то, что получали через водные пространства, а разнообразием своих фаун они обязаны эволю- ции. (Тем не менее Австралия и Южная Америка были соединены с дру- гими материками еще раньше, в мезозое.) Фауны всех островов, кроме современных материковых, по-видимому, являются и основном реэулыа- том аккумуляции и местной эволюции, а не результатом изоляции элементов древних материковых фаун. Если из этого положения и есть исключения, то это относится к Новой Зеландии, но даже и там очень ~ra;z« настоящих древних позвоночных. Скорее всего, это древние иммигранты, а не элементы древней фауны. Тот факт, что ни один древний остров ш имеет фауны, которая является несомненным остатком древней MBTBpHi' вой фауны, нуждается в пояснении.
эпохи (времена начала аккумуляции и эволюции) более древних островных фаун позвоночных не известны, но их время можно предполо- жить, сделав ряд сравнений. Если геологи не ошибаются, Гавайские ост- рова и их фауна не древнее плиоцена. Менее дифференцированная фауна островов Галапагос, вероятно, моложе. Фауна Филиппин, видимо, не древнее гавайской. Большинство филиппинских позвоночных дифферен-
Особенности распростпранен,ия живо~иных нп островах
цировано слабо. Высокодифференцированными являются только мыши- ные грызуны, относящиеся к семейству, которое, очевидно, не давало подъема до конца миоцена. Современная фауна «Зоны Уоллеса», видимо, также довольно молодая, без высокодифференцированных форм или энде- мичных реликтов. Таким образом, фауны всех этих островов, возможно, произошли в позднетретичном периоде и плейстоцене, и это время можно считать возрастом большинства фаун позвоночных других островов, кро- ме Новой Зеландии, Мадагаскара, вероятно, Новой Каледонии и Боль- ши х Антильских островов.
Новая Зеландия — древний остров, на котором Sphenodon и Leiopel- uu представляют древние группы животных. Не совсем ясно, когда они достигли Новой Зеландии, и не исключено, что они первоначально у'ке побывали в Австралии. Странным кажется и тот факт, что они не радии- ровали на Новой Зеландии; особенно род Leiopelma, являющийся един- ственным представителем лягушек на этом, очевидно, наиболее подходя- щем для их обитания острове, должен был бы радиировать там, но, на- сколько мне известно, от него образовались всего лишь два или три слабо отличающихся вида. В Новой Зеландии также встречаются эндемичные роды ящериц и эндемичные семейства птиц. Некоторые из них радиирова- ли, но сколько времени потребовалось для их эволюции, не известно. Это, очевидно, очень древняя (среди островных) фауна, но ничто не помогает установить ее точное летосчисление. Возможно, что она полностью тре- тичная, но, возможно, Sphenodon и Leiopel~na относятся к еще более древ- НИМ ЖИВОТНЫМ.
Фауна Мадагаскара, несомненно, древняя, высокодифференцирован- ная, со множеством реликтов и значительной радиацией некоторых эндемичных групп. Например, тенреки (аналогичные Solenodon на Боль- ших Антильских островах) дали 10 или более родов. С другой стороны, фауна Мадагаскара, видимо, менее древняя, чем фауна Австралии и Юж- ной Америки третичного времени. Возможно, что она начала аккумули- роваться и эволюционировать примерно в олигоцене.
Новая Каледония предположительно также древний остров, но его фауна позвоночных явно не является древней. В ее составе отсутствуют древние реликты, но имеются эндемичные, по-видимому автохтонные, роды гекконов и птиц и одно эндемичное семейство птиц, радиация которых не была значительной. Эндемичное семейство птиц Rhynochetidae состоит пз одного вида; возможно, это (но не обязательно древний) реликт. Эта фауна не может быть датирована, и нет никаких очевидных причин, по которым ее можно было бы отнести к более раннему времени, чем середина третичного периода.
Возраст фауны Больших Антильских островов также неясен. Solenn- don и Nesophontes — древние насекомоядные животные. У первого имеются родственные представители, относящиеся к олигоцену. Но совсем не обя- зательно считать, что эти животные должны были достигнуть Антильских островов именно в это время. Они не радиировали там (см. тенреки на Ма- дагаскаре) и, возможно, проникли на эти острова недавно, после времен- ного пребывания в Центральной Америке. В целом фауна Больших Антильских островов, видимо, не очень древняя. Возможно, она относится h миоцену или даже к более позднему периоду.
Суммировав все вышесказанное, можно предположить, что (не счи- тая Австралии и Южной Америки) самая древняя существующая остров- ная фауна позвоночных находится на Новой Зеландии и что она начала накапливаться в раннетретичное время, если не раньше, что мадагаскар-
Глава 8
ская фауна занимает следующее место и берет свое начало (современная фауна) примерно с олигоцена. Далее, возможно, идут фауны Новой Каледонии и Больших Антильских островов, которые относятся к средне- третичному времени, а фауны позвоночных всех остальных островои являются более современными и относятся к позднетретичному времени или плейстоцену. Приведенные эпохи для современных фаун позвоночных предположительны и не соответствуют возрасту островов, на которых они находятся. Некоторые острова, очевидно, значительно древнее, чем их фауны. Например, Мадагаскар существовал в мезозое, но его современ- ная фауна, по крайней мере какая-то часть ее, которая может быть дати- рована, не мезозойская.
Встречаемость нелетающих птиц соответствует этой предполо'китель- ной датировке. Гигантские нелетающие птицы с полностью утраченными или почти утраченными крыльями имели время для эволюции на Новой Зеландии (где нелетающих птиц было больше, чем на любом другом ост- рове) и на Мадагаскаре, но они не эволюционировали на других изоли- рованных островах. Незначительно видоизмененные нелетающие птицы эволюционировали на некоторых других островах, особенно на Маскарен- ских (где других позвоночных слишком мало, чтобы датировать эту фау- ну), но их очень мало и они лишь слегка видоизменены даже на Новой Каледонии и Больших Антильских островах, если они только на последних встречались. Пастушки на многих островах также утратили способность к полету, на что, очевидно, потребовался весьма непродолжительный период времени
Распределение беспозвоночных отчасти также связано с этой дати- ровкой. Встречаемость и географические связи многих беспозвоночных еще недостаточно изучены, и делать обобщения о них по литературе рискованно, но можно сказать, что жуки СагаЬЫае, которых я изучаю,— «ысоко дифференцированные насекомые и, очевидно, являются древними на Мадагаскаре, Новой Зеландии и Новой Каледонии и в меньшей сте- пени дифференцированы и, вероятно, менее древние на большинстве дру- гих островов, включая Большие Антильские, Филиппины и Гавайские острова. Многие роды этих жуков приводятся в списках для 1авайских островов как эндемичные, и это как раз пример того, насколько ошибоч- ными могут быть опубликованные «факты», так как многие из предполагае- мых родов на Гавайях искусственны, и все они, видимо, являются слегка видоизмененными формами современных материковых ветвей, в то время как некоторые из родов СагаЬЫае Мадагаскара, Новой Зеландии и Новой Каледонии выражены очень четко. Таким образом, жуки СагаЬЫае в этом отношении соответствуют позвоночным и позволяют предположить, что фауна Новой Каледонии — древняя, а Больших Антильских островов— относительно молодая. Следует добавить, что одно это семейство жуков содержит большее число видов, чем все позвоночные, взятые вместе (но этот вопрос изучается очень немногими специалистами), и что оно хорошо представлено на островах.
Остановимся еще на одном доказательстве, имеющем отношение к природе и происхождению островных фаун,— на соотношении различных позвоночных на островах и границах, которых они достигают.
Строго пресноводные рыбы в очень незначительной степени проникают за пределы материков и современных материковых островов. Они дости- гают (очень немногие из них, возможно, обладающие незначительной соле- выносливостью) только ближних Филиппинских и ближних Малых Зонд- ских островов.
Особен ностп и распространения ж ивотн ых на островах
Отдельные амфибии и наземные млекопитающие проникли на многие не слишком удаленные острова. Их границы в ряде мест совпадают, но наземные млекопитающие (а не амфибии) достигают Галапагосских ост- ровов, и амфибии (но не млекопитающие) достигают Сейшельских остро- вов и, возможно, островов Фиджи и Новой Зеландии.
Из наземных млекопитающих крупные животные редко встречаются за границами материковых фаун. Карликовый слон, обитавший в прош- лом на острове Сулавеси, небольшие буйволы на Сулавеси и Миндоро и карликовый гиппопотам, распространенный в прошлом на Мадагаскаре,— это самые крупные наземные млекопитающие, которые, видимо, пересек- ли значительные водные пространства, причем, кроме полуводного гип- попотама, они, вероятно, преодолели лишь очень узкие полоски воды. Более мелкие животные, особенно грызуны, проникали на острова, а так- же и на более удаленные от материка острова значительно чаще. Только грызуны являются древними эндемиками на самых удаленных от центра Филиппинского архипелага островах. Грызуны являются также един- ственными наземными плацентарными, которые достигли Арстралии. но и в Австралию удалось проникнуть лишь нескольким ветвям. Нескольк<~ родов грызунов и только один род других наземных млекопитающих (Pha- 1<тцег) достигли Соломоновых островов. На Больших Антильских остро- вах грызуны численно превосходят всех других наземных млекопитан>- щих, и они единственные из наземных млекопитающих, которые достиг- ли некоторых более мелких и более изолированных островов Вест-Индии (Каймановых, острова Суон и некоторых Багамских) и проникли (одна ветвь) на острова Галапагос.
Рептилии на островах значительно превосходят амфибий и наземных млекопитающих, но из них одни преуспевают сильнее, чем другие. Прес- новодных черепах на островах удивительно мало, и они очень слабо диф- ференцированы. Сухопутные черепахи, очень часто крупные, примеча- тельны тем, что они в настоящее время встречаются на островах Галапагос и на островах Индийского океана, а в прошлом обитали на некоторых Вест- индских и Канарских островах. В ллейстоцене черепахи семейства Ме1о- 1aniidae находились на острове Лорд-Хау и близ Новой Каледонии. Змеи в большинстве случаев распространены на тех же островах, что и совре- менные черепахи (кроме Канарских и, возможно, некоторых Маскарен- ских островов), и в Тихом океане проникают значительно дальше чере- пах — через все Филиппины до Австралии и на восток до островов Фиджи. Более того, змей на острова проникло значительно больше, чем черепах. Из всех нелетающих наземных позвоночных на островах ящерицы самые многочисленные и распространились дальше всех: до островов Тонга (за островами Фиджи) в западной части Тихого океана. Они одни пз нелетаю- щих наземных позвоночных достигли Новой Каледонии и острова Лорд- Хау. Две ветви ящериц по крайней мере достигли Новой Зеландии и Tpll ветви — островов Галапагос (против одной — черепах и одной или, воз- можно, двух — змей). Ящерицы многочисленны и в Вест-Индии (табл. 15). и только они одни из нелетающих наземных позвоночных достигли остро- вов Мадейра, Зеленого Мыса и Бермудских. Но на самых удаленных ост- ровах Тихого и Атлантического океанов да",ке они, вероятно, не являются местными животными.
Рукокрылые встречаются на некоторых подходящих для обитания островах (Гавайские и Азорские) за пределами распространения ящериц.
Наземные птицы достигли всех или почти всех пригодных для обита- ния островов даже в самых отдаленных частях океанов. Правда, в настоя-
Глава 8
щее время они не встречаются на островах Г!асхи и Вознесения, но воз- можно, что они существовали там в прошлом. Встречаемость их на Гавай- ских островах и островах Тристан-да-Кунья показывает, что даже мелкие наземные птицы в отдельных случаях достигают океанических островов. удаленных на 2000 миль и более от материков.
Последовательность встречаемости основных групп позвоночных на островах показана в таол. 20.
Таблица 20
Основные группы наземных п пресноводных позвоночныхв порядке их встречаемости на островах
Строго пресноводные рыбы: очень мало; самые широкие пространства соленых вод. которые они, вероятно, пересекают, достигли всего нсскольких миль
Н аземные млекопитающие Крупные млекопитающие: очень мало; самые широкие водные пространства. пересекаемые ими, видимо, достигали 25 миль (слон и т. д. на Сулавеси в плейстоцене), только полуводный гиппопотам проник на Мадага- скар через более широкий пролив Ъ?елкие млекопитающие, кроме грызунов: немногочисленны; самый широкий пролив, пересекаемый пми, возможно, достигал 200 миль (до Мадагаскара в прошлом) Грызуны: относительно многочисленны; самый широкий пролив, преодолеваемый пми, возможно, достигал 600 миль или более (до островов Галапагос)
Амфибии: основные ветви немногочисленны, но отдельные радиировали; самый широ- кий пролив, преодолеваемый ими, возможно, достигал 500 (до Сейшельских островов.в прошлом) или 1000 миль (до Новой Зеландии, если только в прошлом не было большой разницы в расположении суши)
Рептилии
Пресноводные черепахи: мало; самое широкое расстояние, которое они преодо- левалп, возможно, 200 миль (в прошлом до Мадагаскара), но черепахи
семейства Chelyi~lae в прошлом достигали Австралии, возможно, из К)жной
Америки
Наземные черепахи: мало, но хорошо различимые виды; самое широкое рас- стояние, которое они преодолевали, возможно, достигало 600 миль или более
(до островов Галапагос)
Змеи: в некоторой степени более многочисленны; самое шпрокое расстояш1~.
которое они были способны преодолеть, вероятно, достигало 600 миль ил~
более (до островов Галапагос)
Ящерицы: многочисленны; самое широкое преодолеваемое ими расстояни ..
вероятно, достигало 1000 миль (до Новой Зеландии) Рукокрылые: мало на отдаленных островах (экологически ограничены); самое боль- шое расстояние, которое они смогли преодолеть, — по крайней мер(
2000 миль (от Америки до Гавайских островов) Наземные птицы: относительно многочисленны; самое большое расстояние, преодоле- ваемое ими, достигало 2000 миль или более (от Америки до Гавайски~
островов и островов Тристан-да-Кунья)
Короче говоря, число разных групп позвоночных, представленных на островах, и границы, которых они достигают, пропорциональны их воз- можным способностям пересекать соленые воды, а не геологическому воз- расту этих животных. Так, например, лягушки — очень древние живот- ные (их ископаемые относятся к юре), и некоторые из них преуспевают даже на мелких островах, если они туда попадают (лягушки, интродуциро- ванные на Гавайских и Бермудских островах), но на удаленных островах. кроме Новой Зеландии, они не встречаются. Ящерицы также относятся к древней группе, но не возраст определяет их многочисленность на остро- вах, так как большинство из встречающихся на удаленных островах являются не древними, а близкородственными современным материковым формам. Из млекопитающих на островах чаще всего встречаются опять- таки не самые древние, а грызуны, некоторые из которых (МигЫае) име-
Особенност и распространения животных на островах
нл относительно недавнее происхождение, и тем не менее они достигли бс>лышего числа островов, чем этого можно было оы ожидать.
В целом этот характер распространения на островах непосредственно определяется способностями пересекать водные пространства, а не их «озрастом. Исключением, видимо, является только новозеландская лягушка лиопельма.
Встречаемость различных животных на островах определяется и дру- гими факторами. Для пресноводных рыб и пресноводных черепах на неко- торых островах мало места. Крупные млекопитающие также не смогли бы существовать на небольших островах, даже если бы они туда и попали. Грызуны, которые питаются семенами и зернами п потребляют очень мало пресной воды, вероятно, лучше. чем большинство млекопитающих, приспо- соблены к жизни на таких островах, а также и к тому, чтобы дооираться ло них. Змеи, вероятно, менее, чем ящерицы, приспособлены к сущест- «о«анпю на мелких, экологически ограниченных островах (стр. 160). И ру- кокрылые по сравнению с птицами, очевидно, менее приспособлены к ост- ровной жизни. Но эти факторы создают только второстепенные предпо- сылки для изменения основного характера распространения, который преимущественно определяется расстоянием от материков и способностя- ми животных пересекать водные просторы.
В заключение надо сказать: иммиграционные особенности фаун пери- ферийных архипелагов. малочисленность настоящих древних реликтов на островах, очевидно, довольно молодой возраст большинства островных фаун и тот факт, что число позвоночных, встречающихся на островах, про- порционально их способностям преодолевать соленые воды, а не пропор- ционально пх геологическому возрасту,— все это приводит к тому же «ыводу, что островные фауны позвоночных (кроме фауны современных мате- риковых островов) в основном аккумулятивные, образовавшиеся в резуль- тате проникновения животных через водные пространства, а не являются сохранившимися частицами древних материковых фаун. Очевидно, при- ток позвоночных происходил с материков на острова земного шара, при- чем детали этого проникновения зависят от ширины водного пространства, размеров, природы островов и природы (но не геологического возраста) различных позвоночных. Обратный приток, по-видимому, был очень незначительным. Распространение немногих позвоночных дает основание предполагать. что они произошли на островах и вторглись на материки. Основной факт расселения животных — это общее направленное переме- п1ение от материков к островам.
Возникает вопрос, почему на островах так мало древних позвоноч- ных? Вероятно, отчасти потому, что многие острова не являются очень древними. Удаленные вулканические, океанические острова обычно существуют довольно непродол~кительное время, после чего они разру- шаются и погружаются в морские пучины. Но отдельные острова довольно древние — Мадагаскар существовал еще в мезозое, в то время там были динозавры. Но Новая Зеландия и, возможно, Новая Каледония также очень древние острова. Так почему же на этих островах не сохранилась мезозойская фауна? Даже на Новой Зеландии мезозойские позвоночные отсутствуют независимо от того, с какого времени — с мезозоя или с ка- кого-либо другого периода — там находились Leiopelma и Sphenodon. Видимо, отчасти это можно объяснить тем, что на островах животные не сохраняются продолжительное время. Вероятно, они вымирают с тече- нием времени если не в результате конкуренции с новопришельцами, то в связи с ухудшением условий существования на небольших изолирован-
Глава 8
ных площадях. Возможно, это также основной факт, связанный с направ- лением перемещений позвоночных от материков на острова и отчасти объясняющий его.
Значение эволюционной радиации, которая происходила на разных островах, трудно оценить. Здесь играют роль как время, так и другие факторы, включая эволюционные возможности (ответ на этот вопрос дан>т гавайские цветочницы Drepaniidae, которые радиировали на Гавайских островах в сравнительно короткое время), площадь и, вероятно, наслед- ственные качества. Насекомоядные радиировали на Больших Антильских островах так незначительно (по сравнению с Мадагаскаром), что это, не- сомненно, позволяет предположить ограниченность временем. Радиация ящериц и птиц на Новой Каледонии по сравнению с Новой Зеландией предполагает значительно меньшую ограниченность временем. Но поче- му же Sphenodon и Leiopelma не радиировали на Новой Зеландии, как это имело место у некоторых ящериц и птиц, включая гигантских нелетающих птиц? Радиировали ли они когда-либо вообще и являются ли последними сохранившимися представителями групп, которые исчезают, вероятн<>, даже последними выжившими представителями мезозойской фауны на Новой Зеландии? Вырождались ли они генетически? Может ли ослабле- ние наследственных качеств, по существу, приостановить радиацию ост- ровных животных, а затем привести их к почти автоматическому исчез- новению? Все эти вопросы будут довольно тщательно рассмотрены в сле- дующей главе.
ЛХХT EPA ТУРА
Л g а s s i z А., 1892, Reports on the dredging operations... Со the Galapagos, «Bull. Mus. Comparative Zool.», 23, 1 — 89.
A m а d о n D., 1950, The Hawaiian honeycreepers..., «Bull. American 3[us. 4at. Hist.», 95, 151 — 262.
В а n g s О., 1913, The land mammals of Newfoundland, «Bull. Мця. Сomparativv Zool», 54, 507 — 516.
В а r b о u r Т., 1937, Third list of Antillean reptiles and amphibians, «Bull. Мия. Comparative Zool.», 82, 75 — 166.
В е а и f о r С L. F., de, 195i, Zoogeography of the land and inland water, Londoii- New York.
В о n d J., 1936, Birds of the Жеай Indies, Philadelphia, Acad. Хай. Sci.
В r о w n W. С., 1952, The amphibians of the Solomon Islands, «Bull. Мця. Сои1ра1а- tive Zool.», 107, 1 — 64.
В r о w n %. С. М у е r s, G. S., 1949, ...new frog... Solomon Islands, with note» on ... Fijian frog fauna, «American Mus. Novitates», Ko. 1-«18.
С а r t e r Т. D., Н i 11 J. Е., Т а t e G. Н. Н., 1%5. %mammals of the Р;lucific 'б' ог1й, New York.
С l а r k А. Н., 1943, Iceland and Greenland, Smithsonian Inst, %аг Backgrouri~l Studies No. 15.
D а m m e r m а п К. W., 1929, On the zoogeography of 1аъ.а, «Treubia», 11, 1 — 88.
D а m m e r m а п К. W., J948, The fauna of Krakatau l88:3 — 1!)3;3, «Konir>klijl'~. iveder1andsche Akademie ЪЪейепасЬарреи», Verhandelingen (Tweede Sectie). 44, 1 — 594.
D а r 1 i n g t о п P. J., Jr., 1938, The origin of the fauna of tlute Greater Antilles, witj~ discussion of dispersal of animals over water and through the air, «Quarterly Review Biol.», 13, 274 — 300.
0 а r w i n С., 1839, 1952, Journal of researches... Н.М.S. l)eagle, London, reprinted 1952, New York — London.
JI ume pamy ðà
D е 1 а с о и r J., М а у г E., 1946, Birds of the Philippines, Яew York.
D e r а n i у а g а l a, P. E. P., 1949, Some vertebrate animals о1 Ceylon, vol. 1,
(:eylon, Сolombo.
D i c k e r s о n R. Е. et al., JO 8, Distribution of life ю СЬе Philippines, Philippine
Bureau Sci. Monograph 2l.
D u n n Е. R., 1934, Physiography and herpetology in the Ееяяег Antilles, «Copeia»,
1934, 105 — J 11.
D u n n E. R., C о п а n t R., 1937, The herpetological fauna of Bermnda, «ИегреСо- logica», 1, 78 — 80.
Е i g е и m а п n (.". Н., 1909, The fresh-water fishes of Patagonia..., «Hep. U. P>iti- ceton Ехр. Patagonia, 1896 — 1899», 3, рС 3, 225 — 374.
G а r d i n e r J. S., 1936, "oncluding remarks on thk distribution of СЬе land an~i
marine fauna [of the Seychelles etc.] ...«Тг. 1лппеап Soc. London», Ser. 2, 19, '~17—
464.
G и 1 i c k А., 1932, Biological peculiarities of oceanic islands, «Quarterly Ве~ iew
Biol.», 7, 405 — 427.
Н а «11 i s u k а М., 1953, The D odo and kindred birds, London.
Н а с h i s u k а М., U d а g а w а Т., 1950 — 1951, Contributions Со the ornithology y
of Formosa, «Quarterly 3. Taiwan Mus.», 3, 187 — 280; 4, 1 — 180.
Н i n d w о о d К. Л., 1940, The birds of Lord Iowe Island, «Emu», 40, l — 86.
H i n t о п М. А. C. et al., 4 Х35, List of British vertebrates, I ondon, British]
Мця.
Н о о i j e r D. А., 195I, I ygmy elephant and giant, СогЬиье, «Sci. МопСЫу», 72, 'l — 8. Н и С t о п F. В., D r u m m о n d J., 1923, The animals of 3ew Zealand, Auckland
etc.
I n g e r R. F., 19-.~9, .'ЧоСея on а collection of fresh-water fishes агою Trinidad, «Cop~ia»,
1949, 300.
1 п g е г R F 1914, ...Philippine Amphibia, Fieldiana (Chicago Vat. llist. illus.)
«Zool.», 33, 181 — 531.
К o e n i g s w a l d (:. Н. R. von, 1939, Das Pleistociin Javas, «Quartiir», 2, 28 — 5:>. K и е п е п P. Н., 1950, Marine geology, New York — London.
L а с k D., 1947, Darwin's finches, Cambridge, England, Cambridge ll. Press. [Имеет- ся русский перевод: Д. Л э к, Дарвиновы вьюрки, ИЛ, Москва, 1949.] ? i n d s е у А. А., 1940, Recent advances in antarctic bio-geography, «Quarterly
Review Biol.», 15, 456 — 465.
L о r d С. E., S с о t t Н. Н., 1924, А synopsis of the vertebrate animals of Таяма- nia, Ii ob art, Oldham.
М а t h e w s G. iVI., 1928 — 1936, The birds of Norfolk and Lord I[owe Islands... v;ith
supplement, London.
М а t t h e w s L. Н., 1952, British Iammals, London.
М а у r Е., 1939, The origin and the history of the bird fauna of Е olynesia, «Ргос. ЯхйЬ.
Pacific Sci. Congress», 4, 197 — 216.
М а у r E., 1943, The zoogeographic position of the Нажайап Islands, «Condor», 45,
45 — 48.
М а у r Е., 1945, Birds of the Southwest Pacific, iVew York.
М а у r Е., А m а d о n D., 195], А classification of Recent birds, «American l[~s.
Хovitates», No. 1496.
'11 е r t e n s R., 1934, Die Insel-Reptilien ...«Zoologica» (Stuttgart), 32, 6, Ые1еги11
l — 209.
М i 11 о t J., 1952,? а faune malgache et 1е mythe gondwanien, «Мйп. Inst. Sci. 51ada- gascar», Ser. А, 7, 1 — 36.
М о r i Т., 1936, Studies on the ~~еоцгарЫса1 distribution of freshwater fishes in eastern
Asia (частная публикация).
М и n r о I. S. В., 1955, The marine and fresh water fishes of Ceylon, (:мпЬехта (Austra- lia), Department of External Affairs.
О k а d а Y., 19: 1, The tailless batrachians о1 the Japanese Empire, Tokyo, Insperial
Agricultural Experiment Station.
О k а d а Y., 1955, Fishes of Japan, Tokyo.
О l i v e r J. А., S h а w С. E., 1953, The amphibians and reptiles of Мн; Hawaiian
Islands, «Zoologica» (Меж York), 38, 65 — 95.
Р а r k e r Н. W., 1941, The caecilians of the Seychelles, «Ann. and Alag. Nat. Hist.»
(l l), 7, 1 — 17.
P e r k i n s R. C. Ь. et, al., 1899 — 19j.3, Fauna Iiawaiiensis, Cainbricfge, Englan(l.
Cambridge U. Press, 3 vols.
Р о w е 11 А. %. В., 1949, Native animals of New Zealand, Auckland Мця. Handbook
Zool.
P r а е g e r R. L., 1950, Natural history of Ireland..., London.
В а n d А. L., !936, The distribution and habits of i>fadagascar birds, «ВиП. American
Mus. Nat. Hist.», 72, 143 — 499.
R а n d А. L., 1955, The origin of the land birds of Tristan da Cunha. Fieldiana (Ии- садо Nat. IIist. Mus.), «Zool.», 37, 139 — 166.
R i р 1 е у S. D., 1952, The thrushes, «Postilla» (Yale Peabody Mus. Май. Hist.), No. 13. S а 1 о m о n s e n F., 1948, The distribution of birds and the recent climatic change
in the A orth Atlantic area, «Dansk Ornithologisk Forenings Tidsskrift», 42,
85 — 99.
S а 1 о m о n s e n F., 1950 — 195l, The birds of Greenland (3 parts), Copenhagen. S а l о m о n s e n F., 1951, The immigration and breeding of the Fieldfare... in Green- land, «Ргос. Tenth International Ornithological Congress», 515 — 526.
S а у l e s R. W., 1931, Bermuda during the Ice Age, «Ргос. American Асад. АгЬ
and Sci.», 66, 381 — 467.
S c h u c h e r t С., 1935, Historical geology of the Antillean-Caribbean region, iiev;
York.
S i m p s о n G. G., 19-.~5, The principles of classification and а classification of mammal,
«Bull. American Mus. Nat. Hist.», 85.
S i m p s о n G. G., 1956, Zoogeography of ЖеяС Indian land mammals, «American
Mus. Novitates», No. 1759.
S m i t h М. (А.), 1951, The British amphibians and reptiles, London.
Т а t e G. Н. Н., 1946, Geographical distribution of the bats in the Australasiaï
Archipelago, «American Mus. Novitates», No. 1323.
Т,а у l о r Е. Н., 1934, Philippine land mammals, Philippine Bureau Sci. МоаоцтарЬ 30. Т i m m e r m а n n G., 1938 — 1949, Die Vogel Islands [Birds of Iceland], «Visinda- felag Islendinga» (Soc. Sci. Islandica), 21, 24 and 28 (total of 524 pages питЬге- res consecutively).
U ш Ь g r о ъ е J. Н. F., 1949, Structural history of the East Indies, Cambridge, Eng- land, Cambridge U. Press.
V e s е у — F i t z G e r а 1 d D., 1936, Trinidad mammals, «Tropical Agriculture»,
13, 161 — 165.
Vesey-FitzGerald D., Betts F. N., 1940, The birds of the Seychelles...,
«I b е» (14), 4, 480 — 504.
Ю а 11 а с е А. R., 1880, Island life, London.
W h i t l е у G. P., 1943, The fishes of New Guinea, «Australian Мця. 5Iag.», 8, 141—
144.
W i l l i а m s E., 1956, Pseudemys... ачФЬ а ~епега1 survey of the scripta series, «Bull.
Mus. Comparative Zool.», 115, 145 — 160.
Z i m m e r m а п E. C., 1948, Insects of Hawaii, vol. 3, Introduction, Honolulu,
of Надай Press.
Глава 9
ИЗМЕНЕН11Е PACliPOCTPAHEHIM ЖИВОТНЫХ: ЗНАЧЕНИЕПЛОЩАДИ, КЛИМАТА Ц ЭВОЛЮЦИИ
Предыдущие главы касались главным образом современных особен- ностей распространения животных, и, несмотря на то что перемещения, которые обусловили эти особенности, уже упоминались выше, наше вни- мание будет вновь сосредоточено на них, причем метод анализа будет уже иным. До сих пор мы выявляли факты и делали выводы, которые, хотя l1 были отчасти предположительными, все же вытекали непосредственно из этих фактов. В настоящей главе выводы предыдущих глав будут сведены вместе и расширены до одной общей гипотезы относительно природы и характера расселения животных и факторов, которые регулируют это расселение.
Основной характер расселения (предварительный)
Основной характер расселения вытекает из анализа и сравнения рас- пространения и достоверной истории пяти основных классов позвоночных (гл. 2 — 6), а также из общего характера распространения всех позвоноч- ных на материках и островах (гл. 7 и 8).
У современных доминирующих пресноводных рыб наиболее важными моментами, которые привели к расселению, по-видимому, были расцвет Ostariophysi в пресных водах (очевидно, на основной части тропиков Ста- рого Света) и их радиация оттуда, а позднее расцвет Cypriniformes в Юго- Восточной Азии и их расселение оттуда, сопровождаемое отступанием некоторых других рыб. У современных доминирующих амфибий рас- селение было. видимо, определено радиацией последовательно домини- рующих групп (лягушек) с основной части тропиков Старого Света, сопровождаемой оттеснением более древних групп.
Расселение рептилий было сложным, и проследить его пути довольно трудно. Однако можно сказать, что рептилии — главным образом тропи- ческие животные, которые вторгались в умеренную зону северного полу- шария, но редко могли находиться там длительное время или интенсивно эволюционировать. Более того, они, видимо, чаще перемещались из Ста- рого Света в Америку и из Азии в Австралию, чем в обратном направлении. Это дает основание предполагать, что главный центр расселения рептилий находился на основной части тропиков Старого Света и что их последо- вательно сменяющиеся группы проявляли тенденцию расселяться оттуда, замещая в некоторых случаях более древние группы.
Расселение птиц, очевидно, было еще более сложным. В каждой основной экологической группе птиц последовательно доминирующие вет- ви, вероятно, испытывали расцвет и распространялись по всему земному шару, в то время как соответственно более древние ветви отступали или вымирали, но большинство этих деталей осталось для нас неизвестным. Тем не менее то, что было сказано о рептилиях, в такой же мере относит- ся и к птицам: они достигают наибольшего разнообразия в тропиках и лишь незначительной дифференциации в умеренной зоне северного полу-
шария. Перемещались они, видимо, также значительно чаще из Старого Света в Северную и Южную Америку и из Азии в Австралию, а не в об- ратном направлении. Таким образом, для птиц очевидным главным цент- ром расселения является основная часть тропиков Старого Света.
Расселение млекопитающих также было сложным, но его детали изу- чены лучше. Начиная с раннетретичного времени между Старым Светом и Северной Америкой происходил постоянный и сложный обмен млеко- питающими, но преобладающим было направление от (:тарого Света к (:е- верной Америке. С позднего плиоцена между Северной и Южной Лчери- кой также осуществлялся сложный обмен, который преимущественно шел от Северной Америки к ЮжноХх. Некоторые млекопитающие перемещались из Азии в Австралихо, но в обратном направлении на значххтельххое рас- стояние проникли лишь немногие. Наибольшей численности и разнооо- разия млекопитающие также достигают в тропиках, но в умеренной зоне северного полушария они слабо дифференцированы. Поэтому главньш центр расселения млекопитающих, видимо, находился главным образом на основной части тропиков Старого Света. Ископаемые млекопитающие хорошо иллюстрируют последовательность в замещении древних групп новыми, расселяющимися доминирующими группами.
На больших, благоприятных для существования repptt~opttttx 1'оити- нентальных тропиков фауны наземных и пресноводных позвоночных наиболее богаты и наиболее разнообразны, но в северном ti южном направ- лении, а так>ке вне пределов материков их состав сокращается H cTBH(l- вится беднее."Две основные региональные фауны Старого Света (Эфиоп- ская и Индо-Малайская) — центральные, а другие обедненные материко- вые фауны располагаются вокруг них (гл. 7). Кроме того, по-видимому, происходило широкое перемещение позвоночных с материков на острова земного шара (гл. 8).
Все это приводит к одному общему выводу. Перемещение позвоночных на всем земном шаре в обоих направлених происходило очень сложным образом. Но результирующая этих перемещений, видимо, имела опре- деленную направленность от самых обширных и самых благоприятных для обитания территорий, особенно с основной части тропиков Старого Света, к меньшим по размерам и менее благоприятным для обитания территориям. Таков, очевидно, основной характер расселения.
Ареалы, популяции и зоогеографххческххе перемещения
Ареалы растений и животных — это площади, занимаемые популяци- ями. Изменения в распространении влекут за собой изменения в популя- циях. Можно много говорить о том, что происходит в сложных популя- циях, но мы остановимся здесь только на обобщениях (неизбежно слиш- ком простых), которые кажутся нам очевидными.
Популяции подвержены колебаниям численности, которые являются результатом изменений соотношения рождаемости и смертности. При ро- сте популяции у особей, ее составляющих, появляется тенденция к рас- селению, а при ее уменьшении — тенденция к потере занимаемого участка.
Зоогеографы иногда рассматривают ареалы как площади. ограничен- ные со всех сторон физическими преградами, которые препятствуют рас- пространению животных, что, однако, верно только в отдельных случа- ях: например, в отношении водных животных, обитающих в озерах, окруженных со всех сторон сушей, и наземных животных на островах. Чаще
Из.ченение расироспгранения экивотных
всего площади. занимаемые популяциями, ограничивак)тся такими фак- торами, как количество выпадающих осадков и температура, которые постепенно изменяются от места к месту и из года в год. В таких случаях занимаемые площади определяются не границами, которых не могут пере- сечь отдельные осоои, и даже не границами, за пределами которых условия существования неблагоприятны, а равновесием, охватывающим всю попу- ЛЯЦИК).
Т!лощадь, занимаемая популяцией, обычно не однородна: в одних местах она более благоприятная, чем в других, и отношение роп;дае- мости к смертности часто варьирует в соответствии с этой неоднородно- стью. Вероятно, самые благоприятные условия находятся вблизи центра этой площади, и, видимо, есть неблагоприятная зона периферии, где смерт- ность превышает рождаемость, но здесь популяция поддерживается за счет преооладания рождаемости вблизи центра площади, откуда отдельные осо- би постепенно смещаются к периферии. В этом случае фактические гра- ницы ареала определяются не пределами благоприятных участков, а по- стоянным колебанием соотношения между тенденцией к распространению у центра ареала и тенденцией к уменьшению по периферии. Это создает очень чувствительный механизм распространения и сокращения популя- ций. Любое изменение или в самой популяции, или в окружающих условиях, которое увеличивает рождаемость по отношению к смертности, вызывает расселение. Любое изменение или в популяции, или в окружаю- щих условиях, которое уменьшает это соотношение, вызывает сокращение, то есть вымирание, идущее от периферии ареала к центру. Таким образом, популяции должны изменяться не только по числу особей. но и по зани- маемой площади, распространяясь в благоприятные периоды и сокращаясь в неблагоприятные; расселяясь при улучшениях в самих популяциях и уменьшаясь, когда в результате конкуренции эти улучшения исче- зают. Такая пульсация популяций по периферии ареала наблюдается у многих групп животных.
f аспространение всех групп растений и животных — видов. родов, высших групп или целых флор и фаун — представляет сумму площадей, занимаемых популяциями, а перемещения самых простых и самых слож- ных групп в своей основе остаются одинаковыми. Все перемещения про- исходят главным образом благодаря расселению или сокращению попу- ляции примерно таким образом, как это было уже описано.
Перемещения отдельных особей животных в пределах популяций, по существу, настолько произвольны, что их можно сравнить с движением молекул газа. Эти перемещения могут принимать различные направле- ния (передвижение молекул также может быть направленным), например под влиянием ветров или течений или в связи с необходимостью переплы- вать реки или лететь навстречу заходящему солнцу. Но такие воздействия, видимо, не столь существенны. Обычно популяции перемещаются за счет случайного распространения отдельных особей, которые выживают в но- вых благоприятных для обитания местах, но погибают в неблагоприят- НЫХ.
Циклы расселения
При самом простом характере расселения группы растений и живот- ~ных неуклонно распространяются от места своего происхождения во всех возможных направлениях. В этом случае возраст каждой группы пропор-
Глава 9
ционален занимаемой площади, а местом происхождения является центр этой площади. По существу, это та же самая гипотеза «возраста и пло- щади», которая была предложена ботаником Виллисом п подверглась жестокой критике другого ботаника Фернальда (Fernald, 1926. со ссылками на более ранние работы Виллиса). Однако Виллис (Willis, 1949) до сих пор, правда в несколько измененной форме, придерживается этой гипо- тезы. Как говорит Фернальд, тенденция растений и животных и расселе- нию с течением времени — это трюизм, не представляющий ничего нового. Однако нельзя считать правильным, что равномерное распространение. идущее во всех направлениях, является основным процессом, который обеспечил современное размещение животных. Отступание было также важным фактором. Те позвоночные, после которых сохранились наиоолее достоверные ископаемые данные, в свое время прошли через циклы рассе- ления и отступания, и такие циклы вообще, видимо, были присущи и животным и растениям.
Отдельные группы животных часто проходят через указанные цик- лы. Группа возникает, затем происходит ее расцвет (доминирование),
Р и с. 66. С~ема последовательной сменыпростых циклов с перемещением в одномнаправлении.Каждый круг соответствует циклу, и редста в-ленному в предыдущей схеме.
Р и с. 65. С~ема простого циклараспи1рения и сокращения ареалас перемещением.Смс няющиеся стадии цикла цмлотпоследовательную нумерацию.
она становится разноооразной и расселяется, далее приходит в упадо<; и отступает, а затем исчезает. Эти циклы эволюции и перемещения срав- нивались с циклами жизни отдельных особей животных так, будт-о виды и высшие группы животных прошли через периоды молодости, зрелосгц и старости, но, как указывает Симпсон (Simpson, р. 187, l949), эта «нал<>- гия сомнительна. Симпсон (Simpson, 1940) описал некоторые из предпо- лагаемых и наблюдаемых циклов, которые изменяют распространение животных. В большинстве случаев они были прослежены по ископаемым. Приведем описание, отчасти заимствованное из работы Симпсона и отча сти пополненное новыми сведениями.
Самый простой цикл — он же является одним из тех, которые состав- ляют более сложные циклы,— состоит сначала из расселения, а затем сокращения одной группы животных (рис. 6,з). Этой группой може1 бь<ть одна популяция или любая высшая группа, рассматриваемая как едини- ца. Если площадь, занимаемая после сокращения популяции, бут~е" отличаться от первоначальной, значит — произошло перемещение.
Последовательность или продолжение простых циклов, прогресси- рующее в одном направлении (рис. 66), иногда называют «миграцией», хотя она не имеет ничего общего с сезонными миграциями, например с се- зонными миграциями птиц. Отдельные особи проявляют тенденцию к про- извольному расселению (правда, эта произвольность изменяется под влия-
Изменение распространения хсивотных
кием преград и направления), но новые участки занимают только те осоои, кото- рым удается переместиться на благоприят- ные территории, другие же погибают. Однажды занятый участок теряется не в результате преднамеренного оставления этого участка, а в результате вымирания занявшей его группы животных.
Цикл расселения и сокращения груп- пы может усложняться различными пу- тями. Когда группа сокращается, она может разделиться, и ее ареал становится прерывистым (рис. 67). Большинство типов разрывов (стр. 347 — 349), очевидно, обра- зовалось следующим образом. Если рас- селяющаяся группа встречает на своем
Р и с. 67. Схема расширения и сокращения ареалов, заканчива- ющихся в случае отсутствия не- прерывности расселения.
Р и с. 69. Схема расширения и сокра- щения ареала при пересечении северных областей и с последующттм отступанием
па юг.
Р и с. 68. Схема расширения и сокра- щения ареала в случае преодоления
преграды.
пути преграду, которую преодолевает с трудом, то за этой преградой может возникнуть самостоятельный цикл. Это может произойти в том случае, если компонент расселяющейся группы случайно попадает через водное пространство на новую сушу (рис. 68). Или это может произойти в том случае, если процветающая группа из тропиков проникает довольно далеко на север и какой-то член этой группы благодаря своей холодо- стойкости пересекает северный сухопутный мост между Старым и Новым Светом.
Циклы перемещения бывают во всех группах животных и расте- ний. Ископаемые свидетельствуют о том, что они происходили у многих групп, но только у некоторых групп, а именно у тех, которые имеют достаточно полные ископаемые, можно проследить эти циклы детально. К одной из таких групп относятся мастодонты. Мастодонты, оч»- видно, произошли в олигоцене в Африке и расселялись на основной части Старого Света в позднем миоцене до тех пор, пока не распространились довольно далеко к северу и смогли попасть в Америку. В последующее время они стали вымирать, исчезнув сначала в Старом Свете, а затем и в Америке. Этот цикл изображен Симпсоном (Simpson. 19~0, р. 143) в виде схемы.
Циклы перемещения отдельных групп животных часто очень сложны и состоят из множества второстепенных циклов со сложной серией продви-
Ггава 9
жений, отступаний и новых продвижений. Эта сложность усиливается, когда последовательно сменяющиеся доминирующие и конкурирующие группы животных проходят через последовательные циклы, причем каж- дая группа стремится заместить предшествующую доминирующую. Ile- ремещения же целых фаун еще сложнее. Когда, например, большая часть фауны млекопитающих Северной Америки в позднем плиоцене и плейсто- цене переместилась (распространилась) в Южную Америку, это перемеще- ние началось как очень сложный обмен со многими передвижениями в обо- их направлениях, и только в процессе времени последовательные оомены и избирательная смертность дали направление этому процессу в целом (стр. 311).
,[омпнирование, конкуренция и замещение
Основная история расселения позвоночных — это история циклов перемещения последовательно сменяющихся доминирующих групп.
Доминирующие группы относительно многочисленны по числу осо- оей, как правило, многочисленны по видам, нередко (в конечном счете) разнообразны по своим адаптациям и часто (в конечном счете) имеют широкое и сплошное распространение как в районах с неблагоприятными. так и с благоприятными условиями существования. Наиболее доминирую- щие из современных групп наземных и пресноводных позвоночных — карпо- вые рыбы, лягушки рода Лапа, ужовые змеи, воробьиные птицы и грызу- ны (см. о доминирующих группах в гл. 2 — 6) и, конечно, человек. Домини- рование, по-видимому, отражает основные особенности группы, которые способствуют ее расцвету и ведут к эволюции. Связь доминирования с эво- люцией рассматривается в данной главе ниже.
Доминирование — это показатель успеха в конкуренции. Конкурен- ция здесь определяется как любое взаимодействие между организмами, которое неблагоприятно отражается на одном из них. Поскольку некото- рые биологи, видимо, сомневаются в существенной важности такого явления, как конкуренция, стоит остановиться, чтобы доказать это. Отдель- ные доказательства для различных классов позвоночных приводились в соответствующих местах в гл. 2 — 6. Они показывают результаты интро- дукции доминирующих животных в изолированные фауны, где интроду- цированные формы часто конкурируют с местными и вытесняют их, что происходит либо когда две фауны встречаются в естественных условиях, как это произошло с фаунами Северной и Южной Америки несколько мил- лионов лет назад, когда после периода смешения и конкуренции многие южноамериканские группы были замещены североамериканскими, либо когда конкуренция ослаблена, как это имело место у млекопитающих Австралии, в результате чего радиировали сумчатые. Они показывают так- же многие случаи замещения одной группы другой, очевидно, конкури- рующей, что наглядно иллюстрируется ископаемыми, например частич- ное замещение некоторых более древних пресноводных рыб карпо- выми, бокошейных черепах — пресноводными (Emydinae) и целый ряд других случаев замещения, о которых можно судить, исходя из ископае- мых млекопитающих.
Эти особые случаи играют важную роль, но наиболее существенным является, так сказать, баланс современных и древних фаун. Можно уви- деть, что на протяжении всей известной истории позвоночных (в том слу- чае, если ископаемые их достаточно достоверны) земной шар в целом и все его основные части были населены фауной позвоночных, которая
И зльенение распространения эьсивотпных
або
была довольно постоянной по своим размерам и по адаптивным типам. Ни земной шар в целом, ни одна из его основных частей не были в одну эпоху перенаселены животными, а в другую — опустошены, и ни одна экологическая ниша не была не заполненной в течение длительного вре- мени. Всегда существовали (за исключением, вероятно, очень коротких периодов времени) в допустимых пропорциях травоядные и хищные, круп- ные и мелкие формы и разнообразные второстепенные адаптивные группы. У современных фаун проявляется тот же баланс. Фауна каждого материка соответствует площади и климату этого материка, и в каждой основной фауне в определенных пропорциях представлены травоядные, хищные и другие животные. Это не может быть делом случая. Что-то сдерживает размеры и структуру фаун позвоночных в определенных границах, несмотря на постоянные изменения и последовательную сменяемость отдельных филогенетических групп. Это может быть только результатом конкуренции, а, для того чтобы такое положение сохранялось, конкурен- ция дол~кна быть основной, всегда имеющейся в наличии силой.
Расселение доминирующих групп животных и отступание менее до- минирующих и менее конкурентоспособных групп взаимосвязаны слож- ным образом.
Расселяющиеся группы — это доминирующие группы. Способность расселяться является одной из характерных черт и доказательством доми- нирования. Доминирующие животные распространяются потому, что качества, которые делают их доминирующими, дают возможность прони- кать на новые территории и новые стации, приспосабливаться к ним и вытеснять там других животных.
Нормально любое животное воспроизводит лишних особей, которые постоянно или периодически вызывают сильное миграционное давление. Это давление может быть выражено как приблизительно соответствующее размеру избытка или пропорциональное особям, которые погибают, не найдя мест для обитания (такое представление, конечно, слишком упро- щенно, но это было вызвано необходимостью). Появление доминирующего конкурента, вызывающего сокращение и исчезновение прежних видов, усиливает миграционное давление среди последних. Если прежние виды не смогли расселиться до появления последующих, то вряд ли им удастся это сделать впоследствии.
Доминирующие расселяющиеся животные вряд ли «пропустят вперед себя» менее конкурентоспособных. Скорее всего они, вероятно, будут по- давлять их и способствовать их исчезновению.
Расселение доминирующих животных и отступание более слабых и менее конкурентоспособных не обязательно происходит в одном и том же направлении. Если доминирующая группа расселяется через однородную благоприятную территорию, то более слабая конкурирующая группа будет отступать в том направлении, в котором она замещается вселенцем. Однако если доминирующая группа расселяется с благоприятной терри- тории на менее благоприятную, например из тропиков, где вполне достаточно места для многих видов животных, в Арктику, где могут обитать лишь немногие, то неконкурентоспособная группа может отступать в про- тивоположном направлении, будучи замещенной в тех районах, где под- ходящих мест обитания недостаточно, и выживая в тех местах, где таких мест оольше.
Хто касается примитивных форм, то здесь имеются разногласия. Одни полагают, что примитивные формы остаются в первоначальных цент- рах расселения распространяющихся групп животных, другие — что онп
вытесняются к периферии или сохраняются там. Но ооа эти взгляда, веро- ятно, упрощают то, что происходит в действительности. Существует дв» вида периферийных территорий: те, что относятся к перифериям из-з» неблагоприятных условий существования (по сравнению с центрами расселения), и те, которые являются перифериями в результате своей удаленности от этих центров. Например, Арктика для большинства жи- вотных является неблагоприятной периферией, а Австралия — удаленной.
Проследить историю географического расселения групп животных н»» примитивным формам и интерпретировать их распространение доволым» трудно, но иногда возможно. (Однако предполагать, что большинспи» недоминирующих или примитивных форм должно находиться на отдален- ных перифериях, значит практически допускать кустарный метод иссле- дования.)
Говоря об особенностях расселения позвоночных и и том месте, г'г»тг»- рое занимают архаические реликты (вы г'ившие представители очень при- митивных древних групп), необходимо подвести краткий итог сказанно- му ранее (см. гл. 2 — 6). Из строго пресноводных рыб к архаическим ре гик- там относятся костно-ганоидные рыбы: многоперые в Африке, веслоносы в Китае и в восточно-центральной части Северной Америки, ильная рыб» в восточной части Северной Америки, двоякодышащая рыба (цератг»д). изолированная в Австралии, и двоякодьпиащие рыбы семейства ?.ерЫоя- renidae в Африке и Южной Америке. Из амфибий к ним относятся безнг»- гие и разные, тоже своего рода реликты, виды саламандр, встречающиеся главным образом в тропиках и умеренной зоне северного полушария соответственно; три самых примитивных подотряда лягушек, обитающих главным образом в тропиках (за исключением Австралии) и в умеренной зоне северного полушария. Только род Легоре1ггггг в Новой Зеландии гео- графически поразительно изолирован и уравновешивается таким же при- митивным родом А8сар/гиг в Северной Америке. Из рептилий своего рода реликтами являются разные виды черепах и крокодилов, которые встре- чаются главным образом в тропиках, проникая (особенно черепахи) в некоторые районы с умеренным климатом. Но нн одно другое архаиче- ское пресмыкающееся не имеет такой изоляции, как Sphenodon, который изолирован в Новой Зеландии.
У птиц истинно архаических реликтов нет, и если на какой- ги~н» части земного шара примитивных птиц находится больше, чем в других местах так это в Южной Америке. Из млекопитающих самыми архаически- ми реликтами, вероятно, следует считать австралийских однопроходных. Современные сумчатые также являются своего рода реликтами, но совсем не в том смысле, в котором предполагалось когда-то (стр. 272), и они встре- чаются главным образом в Австралийской области и Южной Амернг,е, а один вид проникает в умеренную зону Северной Америки. Однако н» основной части земного шара некоторые плацентарные представлены, воз- можно, такими же древними ветвями, как австралийские г1 южно»ме- риканские сумчатые.
Вообще архаические реликты проявляют тенденцию находиться глав- ным образом в двух местах в общей системе размещения позвоночных. Незначительное (меньшее, чем обычно предполагают) их число встречается почти в полной географической изоляции, за пределами границ распро- странения большинства более поздних конкурентов. Однако большинство »х встречается на больших, с благоприятными условиями и более или менее постоянных территориях, которые были центрами эволюции и расге- ления: пресноводные рыбы и амфибии — в тропиках и умеренной зоне
Изменен ие распространен ия хси воп~ ных
северного полушария, рептилии — в тропиках, птицы и млекопитающие— чаще в тропиках, чем в других местах. Единственное место, где архаиче- ские реликты позвоночных встречаются редко или никогда не встречают- ся,— это неблагоприятные или непостоянные периферийные территории, изолированные неполностью. Например, для умеренной зоны северного полушария, которая является периферией для рептилий, не характерны какие-либо особые архаические реликты. Нет особых архаических амфибий или рептилий и в Австралии, которая, вероятно, является для них периферией, хотя и менее изолированной, чем для пресноводных рыб и мле- «опитающих.
Наиболее поразительные реликты, последние остатки древних семейств и отрядов, сохраняются вследствие различных причин. Одни— благодаря чрезвычайной географической изоляции, другие, видимо,— в результате адаптаций, которые отчасти изолируют их от конкурентов: африканские и южноамериканские двоякодышащие рыбы могут дышать воздухом и жить в сезонных водоемах или в водах с недостатком кисло- рода; лягушки рода АгсарЬиг обитают в холодных быстрых ручьях, недо- сягаемых для большинства других североамериканских лягушек. Однако некоторые архаические реликты успешно конкурируют с современными животными: африканские многоперые рыбы, хотя они в какой-то степени могут дышать воздухом, живут, свободно конкурируя с другими пресно- водными рыбами Африки; ильная рыба, единственный реликт отряда, доминировавшего в rope, успешно конкурирует с другими крупными хищны- ми рыбами в восточной части Северной Америки; североамериканский опос- сум (прими~пивное сумчатое, независимо от того, является он реликтом или нет) успешно конкурирует с плацентарными млекопитающими и скорее осваивает новые участки, чем теряет свои. Все эти представители архаи- ческих или изолированных групп сохранили прежнее или вторично при- обрели современное доминирование.
Расселение и климат
Многое из того, что будет излагаться далее, заимствовано из моей ста- тьи о холоднокровных позвоночных (Darlington, 1948) и дополнено мате- риалом и по птицам и млекопитающим. Вопрос, который предстоит рас- смотреть,— это зависимость расселения от климата и особенно от основ- ных климатических зон (тропической и умеренной зоны северного полу- шария).
Из работы Мэтью (iblatthew, 1915, р. 172, 173; 1932, р. 3) следует вывод, который может быть сокращен до двух основных предположении: (1) что умеренная зона северного полушария вследствие разнообразия сво- его климата была основным центром эволюции и расселения наземных позвоночных и (2) что для объяснения распространения позвоночных не тре- буется прибегать к теории каких-либо великих изменений в географии зем- ного шара или теории возведения особых сухопутных мостов. Работа Мэтью сыграла большую роль. Она помогла поставить историческую зоо- географию на научную основу и дала возможность противопоставить ее научные положения (но не остановить!) тенденциям отдельных зоогеогра- фов к нереальным гипотезам. Однако первое из его двух основных утвер- ждений я считаю ошибочным.
Пресноводные рыбы, амфибии, рептилии, птицы и млекопитающие— видимо, все расселялись из тропиков в умеренную зону с«неяркого полу-
Глава 9
шария, а не наоборот. Ископаемые показывают, что в каждом классе есть группы, которые находились на севере и отступали оттуда, но это не озна- чает, что они там произошли. Принимать направление отступания за на- правление происхождения и расселения — весьма распространенная ошибка. Холоднокровные рептилии и теплокровные птицы и млекопитаю- щие являются самыми многочисленными и разнообразными животными в тропиках. Различные группы их простираются в умеренную зону север- ного полушария на разные расстояния, но очень немногие из них приуро- чены к:1той зоне. Такое положение едва ли совместимо с представлением о северной части земного шара как основного театра действий эволюции позвоночных или главного центра их расселения.
Умеренный климат северного полушария характеризуется двумя особенностями: более низкой по сравнению с тропиками средней темпера- турой и сменой теплых и холодных сезонов. Ограничивающее влияние этих факторов на позвоночных сложнее: северные границы разных животных могут определяться средней температурой — летней или зимней, а тем- пература может оказывать влияние на рождаемость, на молодых или взрослых животных, на их корм или на конкурентов. Разные позвоноч- ные достигают разных северных границ не на определенных изотермах и без определенного порядка (рис. 70). Очевидно, ограничивающее влия- ние климата нужно искать не в простом воздействии частных факторов на отдельные виды, а в общих корреляциях.
Расположение северных границ, образование самостоятельных севе- ро-умеренных фаун и распространение филогенетических реликтов у раз- ных классов позвоночных определенным образом взаимосвязано. Пресно- водные рыбы заходят далеко на север и многочисленны в умеренном кли- мате северного полушария. Они образовали самостоятельную северо-уме- ренную фауну, для которой характерны особые семейства и даже отряды, ы реликты древних групп этого класса встречаются в умеренной зоне северного полушария так же, как и в тропиках и в Австралии. Амфибии достигают Арктики и имеют более или менее циркумполярное распростра- нение, а также довольно многочисленны в более холодных частях умерен- ной зоны северного полушария. Они образовали самостоятельную северо- умеренную фауну, для которой характерны саламандры и несколько более или менее четких групп лягушек. Архаические реликты амфибий рассеяны в умеренной зоне так же, как в тропиках и в Новой Зеландии. Реп- тилии в большинстве районов земного шара значительно не доходят до Арктики и относительно малочисленны в более холодных частях умерен- ной зоны северного полушария. Они не образовали самостоятельной севе- ро-умеренной фауны, и у них нет архаических реликтов, характерных для
:> Т()И ЗОНЫ.
Птицы и млекопитающие в свою очередь заходят далеко на север и го- раздо многочисленнее, чем рептилии, в холодных частях умеренной зоны северного полушария, но они не образовали самостоятельной северо-уме- ренной фауны. Архаические формы этих животных находятся главным образом в тропиках или на южных материках, ограниченных преградам1~.
успех холоднокровных позвоночных на севере определяется их ме- стом на эволюционной шкале. В холодных районах в большей степени по сравнению с другими преуспевают низшие формы рыб, в несколько мень- шей амфибии и в еще меньшей, чем обе предыдущие группы,— рептилии. :~то имеет некоторое отношение к воспроизводству и местам обитания У рыо самый примитивный способ размножения, который, очевидно, мень- ше всего зависит от климатических условий. У амфибий характер размно-
Изменение распространения животных
жения и развития более сложный, но у некоторых из них личиночный период сокращен и развитие успевает завершиться за довольно короткое полярное лето. Потребности амфибий в тепле очень ограниченные. Рептилии обладают еще более высокоорганизованным способом размножения, при котором отрицательное воздействие низких температур сказывается в большей мере. Многие северные ящерицы и змеи, включая самых север- ных, стали яйцеживородящими: самка откладывает яйца и «следит за солнцем», пока не вылупятся детеныши. Это скорее приспособление к по- лучению достаточного количества тепла, чем к перенесению холода, и оно
Пресне ~ейные
1
— ры ы
I
p — Лягушки
~ f r — Салапандры
— Змеи
>— - <epengy g ~~ ~ ( ' I-Ящерицы lr 1< < ~ ~КраюШ~~
/
ПреснЯодные
РЫОЫ
Лягушко- Яи~ерицы — ~
1
Ъеи — ~~ ~
1 1аяаианфы—
'l 4epenaxu — 3 ~ ( ц
1
Пресного оные
Ф
,— Яйгушки- >
~~ ~~~алаианбры~ ~
/// /-3Meu -1!
/гЯи~ерицы -,~ ~ ц
Ц-Черепах~-~~ ~ I Ц д / Ярокодилы~ ~
/// ц li ~~
/
Р и с. 70. Схема северных границ распространения групп холод- нокровных позвоночных («Quarterly Review of Biology», 23, 1948,
Fig. 3, р. 106).
подчеркивает тот факт, что рептилии не в состоянии размножаться в таких холодных районах, где размножаются амфибии. Что касается мест оби- тания, то вода — это среда, предохраняющая от холода, и типично вод- ные животные, например такие, как рыбы, в холодном климате имеют преимущества, которые только отчасти разделяются амфибиями, а боль- шинство рептилий лишено их совсем.
При возникновении теплокровности положение изменяется. Тепло- кровность птиц и млекопитающих дала им возможность существовать на севере. Но, вероятно, в связи с тем, что теплокровность делает животных относительно независимыми от внешних температур, у них не образова- лось групп, специально приспособившихся к низким температурам и по- тому приуроченных к умеренной зоне северного полушария, как это про- изошло у рыб и амфибий.
Суммируя все это, можно отметить, что, видимо, многие рыбы и неко- торые амфибии переносят холод или сезонные изменения климата или пол- ностью приспособились к ним и что рептилии не обладают ни тем, ни дру- гим, а птицы и млекопитающие переносят и холод и смену времен года, но не приспособились к ним еще в полной мере.
Самые северные наземные позвоночные относятся главным образом к процветающим доминирующим, а не специально приспособившимся к холоду группам. Это очень важный факт, который, однако, часто недо- статочно принимается во внимание. Лягушки, относящиеся к доминирую- щему роду Вапа, распространенному в тропическом и умеренном поясах,— самые северные представители земноводных. Несущественно отличаясь от родственных им тропических представителей, они проникают на север дальше, чем любые приспособленные к суровым условиям саламандры. Самые северные ящерицы и змеи принадлежат к родам I-acerta и Vipera,
470
Глава 9
которые широко распространены в тропиках Старого Света. Большинство северных наземных птиц относится к широко распространенным домини- рующим семействам: дневные хищники, совы, вьюрки и (менее домини- рующие) фазановые. Из самых северных наземных млекопитающих можно назвать зайцев, грызунов, волков и лисиц, медведей, мелких куньих, оле- ней, бычьих.
Все эти животные, конечно, приспособлены к арктическим условиям. Например, у самых северных лягушек рода Лапа ускорилось их развитие. а самые северные ящерицы и змеи стали яйце-живородящими и могут быст- ро развиваться, используя тепло солнца. Это позволяет им закончить раз- множение в течение короткого полярного лета. Но эти адаптации, по-ви- димому, имеют второстепенное значение. Другие второстепенные адапта- ции к существованию в местах с холодным климатом — укорачивание вы- ступающих частей тела — сокращают потери тепла и уменьшают возмож- ности обмерзания (правило Аллена), а у теплокровных форм — увеличение размеров тела, что также сокращает потери тепла (правило Бергмана).
На примере рода Лапа можно проиллюстрировать тот путь, который проходят доминирующие группы позвоночных в течение циклов рассе- ления через север. У различных североамериканских видов этого рода разный темп развития, который ограничивает их продвижение к северу (Moore). Только один вид этого рода — Лапа sylvatica — (или его форма) может развиваться (завершить стадию головастика) в течение короткого полярного лета,.и это единственный американский вид, который заходит в Арктику. Родственный ему вид Лапа temporaria проникает в арктиче- ские районы в Евразии. Представитель этой группы рода Лапа, очевидно, пересек северный сухопутный мост между Евразией и Северной Америкой сравнительно недавно. Это не первый представитель рода Лапа, который достигает Америки, но именно он позволяет увидеть, каким образом более ранние представители этого рода проникли на этот материк. Лапа 61- vatica, Rann temporaria — это единственные виды рода Лапа, которые приспособлены к пересечению моста Беринга, если таковой существовал. Если бы эти виды исчезли, то среди современных представителей рода Rana ни один вид не имел бы адаптаций для пересечения этого арктического соединения; но этот род обладал и будет еще обладать такими приспособ- лениями. Надо помнить о разнице между имеющейся адаптацией и спо- собностью к адаптации. Экологи имеют дело с адаптациями, но они не мо- гут определить способность к адаптациям, и это одна из причин, почему экология не может дать полного ответа на вопросы, поставленные зоогео- графами. Род Лапа (стр. 123), очевидно, произошел в тропиках Старого Све- та., стал там разнообразным и распространялся к северу до тех пор, пока у одного из видов не появились необходимые второстепенные адаптации (включая сокращение личиночной фазы развития), которые позволили им достичь крайнего севера и попасть в Северную Америку.
Многие другие группы позвоночных, вероятно, прошли через тот же самый цикл: расцвет в тропиках и расселение к северу до тех пор, пока тот или иной приспосабляющийся вид не достиг крайнего севера и не перешел из одной половины земного шара в другун~. Таким образом, обмен живот- ными между Старым и Новым Светом происходит у небольшого числа видов северных фаун, но облегчается тем фактом, что самые северные виды обычно относятся к доминирующим, хорошо приспособляющимся группам.
Отдельные представители всех классов позвоночных во время цикла наибольшего доминирования и самого интенсивного расселения, видимо.
Изменение распространения зсивоигных
могут проходить через области как с теплым, так и с холодным климатом без существенных адаптаций как к тому, так и к другому, но после этого разные классы ведут себя по-разному. Многие пресноводные рыбы и неко- торые амфибии постепенно переходят от радиального характера распро- странения к зональному путем адаптации различных ветвей к тропикам или умеренной зоне северного полушария соответственно. Но рептилии, птицы и млекопитающие, очевидно, после радиации обычно отступают сно- ва в тропики без зональной дифференциации. С рептилиями это происхо- дит, вероятно, потому, что они не приспособлены к длительному сущест- иованию в холодном климате. Но, несмотря на то что птицы и млекопитаю- щие преуспевают в зонах с холодным климатом, они не становятся приспо- собленными к этим зонам, вероятно, потому, что северо-умеренные формы в основном не дифференцированы и не ограничены экологическими пре- градами от новых, конкурирующих групп, расселяющихся из тропиков.
Расселение между Старым и Новым Светом
В каждом классе позвоночных насчитывается больше таких групп, которые переместились из Старого Света в Северную и Южную Америку, а не в обратном направлении. Ограниченность ' и своеобразная непропор- циональность фауны пресноводных рыб Южной Америки позволяют пред- положить, что она произошла от нескольких иммигрантов, переместив- шихся из тропиков Старого Света в Южную Америку, а сомы семейства Ameiuridae, чукучаны и карповые, очевидно, проникли из Старого Света в (.'еверную Америку.
Лягушки родов Bufo и Лапа и семейство Brevicipitidae, очевидно, переместились из Старого Света в Северную и Южную Америку, и распро- странение некоторых других лягушек (Leptodactylidae и ala) также отражает раннее расселение из тропиков Старого Света через Северную в Южную Америку (и отдельно в Австралию), за которым последовало вымирание этих видов на всей или значительной части Старого Света. Черепахи подсемейства Emydinae, род Testudo, Crocodylus, сцинки, несколько родов ужовых, аспидовых и ямкоголовых гадюк, по-видимому, все переместились из Старого Света в Северную и Южную Америку. У птиц перемещения происходили, скорее, из Старого Света в Северную Аме- рику, а не в обратном направлении. Многие группы Старого Света достиг- ли Южной Америки, в то время как ни одна южноамериканская птица не проникла (или проникли очень немногие) в Старый Свет. Многие раз- личные группы млекопитающих произошли на основной части Старого Све- та и переместились в Северную и Южную Америку, в то время как значи- тельно меньшее число североамериканских и, вероятно, южноамерикан- ских млекопитающих проникло в Старый Свет (см. подробнее гл. 2 — 6).
Отдельные перечисленные выше факты, как я уже писал (Darlington, 1%8. р. 107), ведут в основном в одном направлении. Они дают основание предполагать, что при происходивших сложных обменах в расселении по- звоночных наблюдается определенная направленность от Старого Света к (:еверной и Южной Америке. Эта направленность проявляется оолее отчетливо прп анализе всей фауны, особенно фауны Южной Америки. Южная Амернка — это часть суши, имеющая огромное значение, так как это
' Напомним, что «ограниченность» у Дарлингтона вовсе не означает «бедность».
Фауна рыб Южной Америки исключительно богата и своеобразна.— Приль. ред.
Глава 9
основная часть тропиков Нового Света, которая противопоставляется тро- пикам Старого Света, а тропики, как известно, являются колоссальным источником и, несомненно, основным центром расселения позвоночных.
На протяжении большей части третичного периода Южная Америка оыла островом. Только в начале третичного (если это произошло именно в данное время) и вторично ближе к концу этого периода этот материк o~i;r соединен с остальной частью земного шара, что способствовало осуществле- нию обширного обмена наземными животными. Это подтверждается исто- рией млекопитающих и другими доказательствами.
Следует помнить, что пресноводные рыбы Южной Америки (см. гл. 2, табл 2) содержат много эндемичных семейств, но все отряды и основные ветви в пределах отрядов находятся в Старом Свете. В нем имеются так- же важные ветви пресноводных рыб, которых нет в Южной Америке. Это дает основание предполагать, что часть фауны пресноводных рыб Старого Света переместилась в Южную Америку довольно давно (в том случае, если эти рыбы проникли через пресные воды), вероятно, до начала третич- ного периода.
В Южной Америке современные семейства амфибий и рептилий рас падаются на две довольно четко выраженные группы (табл. 21; Schmidt, 1943, р. 252). Одна группа, отличающаяся высоким родовым эндемизмом, возможно, ведет свое начало со времени раннего соединения Южной Аме- рики с остальной частью земного шара. Другая группа с незначительным родовым эндемизмом или совсем без него, вероятно, ведет свое начало со времени более позднего соединения. Ранняя фауна амфибий и рептилий Южной Америки состоит из 19 семейств, из которых все, кроме семейства Teiidae, представленного в восточном полушарии близкородственным семейством Lacertidae, встречаются также и в Старом Свете. Но в Старом Свете есть еще ряд семейств, древних и разнообразных, которые не пред- ставлены в ранней фауне Южной Америки: из лягушек — Ascaphidae, Discoglossidae, Pelobatidae (все древние) и Bufonidae, Ranidae, Rhaco- phoridae (по крайней мере разнообразные), из черепах — Тг1опусЬЫае (древнее и разнообразное семейство) и из ящериц — Agamidae, Chamae- leonatidae, Scincidae и Varanidae. К этому списку могут быть добавлены другие, более мелкие или более локализованные семейства, характерные для Старого Света.
Поздняя фауна амфибий и рептилий Южной Америки включает 10 семейств, которые не были представлены в ранней фауне. Из этих 1О се- мейств 9 существуют в настоящее время или представлены ископаемыми в Старом Свете, где, однако, имеются современные семейства, которые в современной фауне Южной Америки не представлены. Итак, мы видим, что распространение амфибий и рептилий совпадает с характером распро- странения рыб, но только на уровне семейств, а не высших групп. Ни одно (или очень немногие) семейство амфибий или рептилий неспецифично для Южной Америки. Большинство южноамериканских семейств пред- ставлено в Старом Свете, кроме того, там имеются важные семейства. кото- рые отсутствуют в Южной Америке. Это относится как к ранней, так и современной фауне амфибий и рептилий Южной Америки и позволяет предположить два периода перемещений, в каждом из которых часть раз- нообразной фауны Старого Света достигала Южной Америки.
Что касается птиц, то многие из них, возможно, достигли Южной Аме- рики через водные пространства третичного времени. Видимо, отчасти по этой причине, а отчасти и потому, что в других частях земного и~ара фауны птиц быстро менялись, а отчасти также потому, что ископаемые птиц
Изменение распростпранения экивотных
Таблица 21
Семейства амфибий н рептилий Южной Америки
Находятсяв умереннойзонеСевернойАмерики
Достигли
Семейства
Находятся в Старом Свете
I(1 IРаНО 1 ПОЗДНО1
Безногие (.aeci l i Ыае
Х
Саламандры
P let hodon t i dae
(только Oedipus) Лягушки
Pipidae
Leptodactyl Ûàå
1
I
I
l
' Европа
О 10
Х Х
Bufoniclae
(только Bufo) Л1el(~podi dae
Х 0
Х
Х
Hylidae
Х
Ranidae (только Rana)
Brevicipitidae
Ерокодилы
Crocodi lidae
Черепахи
Chel ydr i nae
(только Ckelttdra)
Kinosterninae
(только Eittosternon)
Emydinae (Geoemttda и
Pseud emys)
Test udininae
(только Testudo)
Pelomedusi dae
Х
Х
0 Евразия
!
Х Х
Х(Ископае-мые)0
Х
1
I
1
Мадагаскар, Фид ки ~ Широко
; (Lacertidae)
Х 10 Х
Х Х Х
Х Х
Am pl>i sbaeni(l ae
Х
СЬе1уЫае
Ящерицы
Iguanidae
|ekkonidae
Teiidae
Scincidae
(только Мабиуа) Anguidae (2 рода)
' Африка, Нида-Малайская,
область
1 1
Африка
Австралийская область, южная Африка (иско- паемые в Индии)
Широко
Индо-Малайская область? 1
Африка?
Умеренная зона Евразии и
Австралийская область Широко
>>
(Ископаемые в Европе?)
1
Ха ~ Широко
1 Африка (ископаемые в
1
других местах)
Австралийская область
Широко
Умеренная зона северного
полушария и Индо-Ма- лайская область Африка и т. д.
Глава 9
Таблиц» 21
НахОдятСяв умереннойзонеСевернойАмерики
Достигли
Семейства
Находятся в Старом Свете
рано
поздно
'Ь~еи
Typh lopidae
(.ept ot yph1opi <lac
Xni li idae
ВоЫае
Со1иЬг1йае (сев. шир.)
Elapidae
Viperidae (3 рода)
Х Х
Широко
Африка и т. д.
Индо-Малайская область Широко
9
Х Х Х
Х
Условные об означения: О —; Х —; Ю — семейства. которые достигают только южных краевых частей умеренной зоны Северной Америки; а — про- никли в миоцене.
Площадь, климат п эволюция
Основной характер расселения, предварительно изложенный в нача- ле этой главы, теперь тщательно разработан II подтвержден путем анализа природы зоогеографических перемещений, предполагаемых и наблюдаемых циклов расселения, роли доминирования, конкуренции и замещения, свя- зи расселения с основными климатическими зонами и направления рас- селения между Старым Светом и Южной Америкой. Все это привело к не- которым повторениям, но тем не менее предварительные выводы, касаю- щиеся расселения, были подкреплены и подтверждены доказательствами.
очень скудны, я не могу разделить фауну южноамериканских птиц на ста- рую и новую таким же образом, как и фауну других позвоночных. Я толь- ко повторю то, что было сказано выше, и то, что подчеркнул Майр (Мауг, 1946, р. 38): многие птицы Старого Света достигли Южной Америки и не- которые радиировали там, в то время как из южноамериканских птиц Старого Света достигли лишь немногие (или не достигла ни одна из них).
Фауна млекопитающих Южной Америки по сравнению с фауной амфибий и рептилий еще более четко разделяется на древнюю и новую (табл. 7). Древняя фауна, очевидно, является производной только части фауны млекопитающих, которая существовала на основной части земного шара в начале третичного периода. Новая фауна представляет собой толь- ко часть современной фауны основной части земного шара.
Из всего этого мне хотелось бы сделать только один вывод. До нача- ла третичного периода или в начале его иммигранты, представлявшие не- которые, но не все основные ветви существовавших в то время позвоноч- ных, так или иначе достигли Южной Америки. А в конце третичного пе- риода дополнительные иммигранты, представлявшие отдельные, но не все ныне существующие ветви позвоночных, снова проникли в Южную Аме- рику. В обоих случаях расселение позвоночных в пределах тропиков или между тропиками Старого и Нового Света, видимо, в основном шло в на- правлении из Старого Света в Южную Америку.
1(зменение распространения мсивотпых
Доминирующие наземные и пресноводные позвоночные, по-видимому, в большинстве случаев имели расцвет на обширной, благоприятной для обитания площади тропиков Старого Света и переместились (распростра- нились) на менее благоприятные в этом отношении площади умеренной .зоны северного полушария и на меньшие по размеру площади Южной Аме- рики. В этом как раз и заключается суть характера расселения, который включает перемещение (расселение) в менее благоприятные по своим при- родным условиям районы (например, Арктику) и на меньшие площади (острова). Эволюционная теория может предположить причины расселения животных именно в этих направлениях.
Теоретически моя;но выделить три этапа эволюции, хотя фактически они, вероятно, смешаны и могут быть дополнены второстепенными про- цессами. Все три этапа имеют один и тот же главный принцип возникнове- ния и распространения некоторых мутаций в пределах популяции.
Первый этап эволюции — дифференциация вида — процесс, в котором сохранение и распространение мутаций может быть отчасти произвольным, независимым от отбора. Он может протекать чрезвычайно быстро в отно- сительно небольших изолированных популяциях. На этом этапе эволю- ции может происходить дифференциация видов на островах даже в том случае, если островные популяции теряют, а не приобретают доминиро- вание.
Второй этап эволюции — «д«пт«щия к специфическим окружающим условиям. Этот этап не произвольный. Те мутации, которые в определен- ных условиях оказываются благоприятными, закрепляются отбором. Отбор накапливает благоприятные мутации, и степень адаптированности популяции при прочих равных условиях находится в связи с размером популяций. Более значительная по размерам популяция имеет больше шансов дать начало каким-либо отдельным накапливаемым отбором мута- циям. Размер популяции зависит отчасти от площади и непрерывности среди. которую она населяет. Из этого следует. что адаптации возникают быстрее на 00IIIHpHBIx территориях с более или менее однородными усло-
ВИЯМИ.
Третий этап эволюции — общ«я адапт«сия. Он охватывает все мор- фологические и функциональные усовершенствования, которые повышают жизненные возможности одних животных по сравнению с другими в раз- HbIx средах, позволяют им быстрее реагировать на окружак>щие условия, или ставят их на более высокий уровень интеллектуального развития. или способствуют более эффективному размножению. Это адаптация к оо- щим окружающим условиям земного шара, и она должна приводить к об- щему доминированик>, преуспеванию на больших площадях и во многих специфических окружающих условиях. Общие адаптации, как и адап- тации к специфическим окружающим условиям, быстрее всего возникают и самых больших популяциях, которые предположительно должны сущес г- вовать на самых больших площадях, где общие условия жизни являются наири:1ее благоприятными. Для большинства позвоночных таким местом, вероятно, явились тропики Старого Света.
Говоря о тропиках Старого Света, мы имеем в виду и площадь, и кли- мат. Огромная площадь тропиков Старого Света дает возможность суще- ствовать большим популяциям. Благоприятный устойчивый климат тро- пиков также благоприятствует иногда существованию больших популя- ций и, вероятно, позволяет развиться максимальному числу поколений, по крайней мере холоднокровных позвоночных. В результате увеличивает- ся число особей, являющихся материалом для процесса эволюции.
Глава 9
Несомненно, что в действительности все это не так просто. Ллапти- рование может происходить очень быстро не только у больших популя- ций, но также и у тех, которые подвергаются сильным колебаниям численности или образуют множество мелких, частично изолированных микропопуляций, скрещивающихся только случайно. Положение осло;кня- ется еще и тем, что мы не знаем, где в действительности находятся самые большие популяции. Очень часто натуралисты утверждают, что старые устойчивые тропики населены громадным числом видов с небольшими раз- реженными популяциями. Мой собственный опыт пятилетнего пребывания в тропиках дает основание считать, что это правильно, но не во всех слу- чаях. В тропиках существует множество редких видов, но наряду с ними существуют и обычные виды.. Большие популяции встречаются также п за пределами тропиков, в основном на новых или периферийных территори- ях. Некоторые виды позвоночных весьма обычны на больших площадях. например, умеренной зоны северного полушария. Возможно, самые быст- рые адаптации происходят, когда животные распространяются на новые большие и благоприятные для их существования площади (см. нижс влия- ние теплокровности).
Фактические особенности распространения животных показьшаю1, что у наиболее доминирующих позвоночных (карповые рыбы, род Яапа, воробьиные птицы, мышиные грызуны и др.) расцвет происходит, види- мо, в тропиках, после чего они распространяются в умеренную зону се- верного полушария, а не наоборот (как это предполагал Мэтью), и это по- ложение относится как к теплокровным, так и к холоднокровным группам животных. Я думаю, что этот зоогеографический факт может явиться вкладом в эволюционную теорию: доминирующие позвоночные о6ычно эволюционируют в тропиках, а не в умеренной зоне северного полушария.
Далее, если общая адаптация протекает в результате ряда последо- вательных изменений видов, обладающих известным превосходством, то она должна протекать быстрее всего там, где виды многочисленны или где имеются большие популяции. Общее число особей всех видов в фауне в целом, видимо, определяется числом тех адаптаций, которые возникли в результате эволюции. А общая масса и разнообразие жизни на больших благоприятных территориях могут увеличивать и ускорять адаптацию. Идея эволюции доминирующих кивотных на больших, плотно заселенных территориях не нова. Она высказывалась еще Дарвином, и я лишь повто- рил эту мысль, применяя понятия современной генетики.
Специальная и общая адаптация, которые приводят к доминирова- нию, различаются только по степени. Связь между ними может быть про- иллюстрирована на следующих примерах. Различные змеи во многих ча- стях земного шара являются полуводными, но три группы из них более приспособлены к жизни в воде, чем другие (гл. 4). К этим группам от- носятся подсемейства Acrochordinae и Homalopsinae, змеи пресных и соле- ных вод, возникшие от семейства Colubridae и семейства морских змей Hydrophiidae, которое произошло от семейства Elapidae. Все три группы произошли обособленно, но все они приурочены или концентрируются в одном и том же месте: в тропиках Индо-Малайской области и на остро- вах, расположенных к югу и востоку от нее. Карповые рыбы и, вероятно, пресноводные черепахи подсемейства Emydinae вряд ли приобрели осо- оые адаптации к водной среде в Индо-Малайской области, но они стали там разнообразными и, очевидно, доминирующими, что позволило и дало им возможность распространиться очень широко. Эти три группы змей, вероятно,— единственные наземные рептилии, которые в течение третич-
Изменение распространения хсивотных
47Т
ного периода стали полностью водными, а карповые — единственные прес- новодные рыбы и Emydinae — единственные пресноводные черепахи, которые в течение третичного периода стали доминирующими и получили широкое распространение. Возможно, к этому списку водных животных следует добавить морских сомов, которые дали подъем в тропиках Индо- Малайской области. Из них одно семейство Plotosidae приурочено к райо- ну Индийского и Тихого океанов, в то время как другое семейство Ariidae встречается в теплых морях всего земного шара. Эти сомы — единствен- ные представители Ostariophysi и, вероятно, единственные рыбы, которые недавно успешно переместились из пресных вод в морские.
Соответствие ключей истории всех этих групп животных позволяет предположить, что места обитания в пресных водах и эстуариях в течение третичного периода в тропиках Индо-Малайской области были очень обширными и были населены большими популяциями многих животных, из которых отдельные группы (змеи) особенно приспособились к жизни в воде. в то время как другие (рыбы и черепахи), будучи уже водными или земноводными, достигли общего доминирования, которое позволило им распространиться в другие части земного шара или зайти в море. Это поло- жение подтверждается и геологическими свидетельствами. Мелкое, с изме- няющ11мся уровнем море Тетис занимало значительную часть тропической Азии в течение значительного времени третичного периода и, видимо, охватывало обширнейшую площадь водных стаций и являлось обширной зоной перехода между пресными и солеными водами.
Остановимся на особом случае, заслуживающем краткого разбора: что произошло бы теоретически, если бы группа холоднокровных позвоночных стала бы теплокровной, как это произошло с появлением млекопитающих и птиц. Они стали бы относительно независимыми от температуры, легко пересекали границы климатических зон и относительно преуспевали бы в местах с холодным климатом. Площадь, на которой про- исходила бы эволюция доминирующих групп, должна была бы увеличить- ся к северу и охватывала бы, кроме тропиков, и доступные части умерен- ной зоны северного полушария. Теплокровность способствовала бы эво- люцип некоторых процессов. Например, у рептилий потребность в тепле для размножения выше, чем у амфибий, а для размножения плацентар- ных млекопитающих необходимо еще большее тепло плюс регулирование температуры. Но самое существенное значение теплокровности заклю- чается, возможно, в том, что проникновение в области с холодным кли- матом и уменьшение роли местных климатических преград расширяет площадь, на которой могут эволюционировать виды и высшие группы животных, увеличивает размер популяций и способствует общей адаптации. Если это так, то, возможно, теплокровность позволила млекопитающим не только выработать усовершенствованный способ размножения, но и оыстро стать во всех отношениях более высокоорганизованными живот-
НЫМИ.
Способность к полету — еще один, но совершенно иного порядка фак- тор. который сводит до минимума роль преград, увеличивает размер попу- .ляций и. вероятно, способствует общей адаптации. Полет сам по себе имеет явные прямые преимущества, и он косвенно способствует эволюции дру- гих прогрессивных черт, что также может играть существенную роль в до- минировании насекомых и птиц.
Теперь можно вернуться к вопросу о связи площади и климата с раз- мером фаун, эволюцией и направлением расселения.
Ограниченность площади приводит к ограничению числа видов, а на
Глава 9
небольших площадях также уменьшается и размер популяций. Вначале это не имело ничего общего с эволюцией. Размер любого земного массива, независимо от того, материк это или остров, ограничивает число животных, которые обитают на нем. Этот факт обсуждался в гл. 8. Можно предполо- жить, каким образом площадь ограничивает число животных. Одни животные, например крупные млекопитающие, просто не могут удовлетво- рить свои потребности на небольшой площади. У других, вероятно, не вме- щается достаточное число особей, чтобы выдержать колебания численности. Третьим, очевидно, не хватает места для перемещений. В ряде случаев животные сохраняются только благодаря перемещениям и в том и другом направлении на очень больших площадях. Например, лошади (Equi- dae), очевидно, произошли в Старом Свете и в настоящее времясущест- вуют (в диком виде) только там. Но они не сохранились бы там, если бы не имели возможности перемещаться на дополнительные площади в Север- ной Америке и из нее. Кроме того, между различными животными возни- кает конкуренция, и конкурентные отношения, на небольших площадях возможно, проявляются в более упрощенном и непосредственном виде, чем на обширных. Таким образом, ограниченность площади может ока- зывать далеко ведущее воздействие, приводящее к сокращению фаун еще до того, как начнет проявляться действие факторов эволюции.
С течением времени начинает проявляться действие эволюции. Самые полноценные особи и самые полноценные виды проявляют тенденцию к эво- люционированию на больших площадях, где и особи и виды являются самыми многочисленными, в то время как эволюция (доминирующих форм) на небольших площадях задерживается. В конце концов доминирующие животные, эволюционирующие на больших площадях, расселяются на меньшие и замещают там менее конкурентоспособных животных. Такой вид перемещения и расселения доминирующих групп с больших площа- дей определяет направление распространения животных в целом.
Если бы эволюция протекала равномерными этапами или если бы она состояла только из одного этапа, тогда обмен фаун, по-видимому, проис- ходил бы в соответствии с их размером: в два раза большая фауна отдавала бы меньшей фауне в два раза больше ветвей, и распространение шло бы от большей фауны к меньшей, а не наоборот. Но эволюция обычно охваты- вает несколько этапов. Происходящий процесс сменяющихся благоприят- ных изменений (адаптаций) напоминает подъем по лестнице. Отдел ьные элементы небольшой фауны могут начать перемещение, как только какие- либо элементы большой фауны начнут расселение. Но при каждом переме- щении преимущество будет у большой фауны, которая прошла болыие этапов развития, и в результате длительного подъема компоненты большой фауны должны иметь несомненные преимущества и должны стать подав- ляюще доминирующими. Очевидно, это так и происходит. Животные мате- риков, по всей вероятности, почти всегда доминируют над животными ост- ровов, и расселение в большинстве случаев идет от материков к островам. Но даже и в этом случае передвижения в обратном направлении теорети- чески возможны и иногда, видимо, происходят, хотя со статистической точки зрения они совершенно невероятны.
Эволюция на очень небольших площадях имеет незначительный эффект и может вызвать прямое деградирование. Мелкие изолированные популяции могут ослабляться до тех пор, пока незначительные колебания численности не приведут их к почти полному автоматическому вымиранию.
Ограниченность распространения, вызываемая климатом, вероятно, подобно ограниченности площади, оказывает такое же влияние общего
Изменение распространения экивотных характера. Такие места, как Арктика и пустыни, неблагоприятные в кли- матическом отношении, населены малым числом видов животных, и фау- ны, ограниченные климатом, в эволюционном отношении, вероятно, находятся в таком же невыгодном положении, как и фауны, ограниченные площадью. Более того, они не изолированы географически и, видимо, кикты иалсгс материка
Остребные иечеэну6- — — — шие реликтедые
группы бвяь шва" /Уаяый дстроЮ
материк патерик (рауна иэ Зл
(рауна иэ Ял е3иниц) (рауна иэ 7л ейиниЩ Ииниц) Р п с. 71. Упрощенная схема связи площади (или кли-
мата) с направлением расселения животных. А — большой материк (или материк с благоприятным клима- том); В — малый материк (или с менее благоприятным клима- том) и С — остров (или очень неблагоприятная в климатиче- ском отношении территория). Площадь или климат позволяют находиться на материке А — фауне, состоящей из 9 7l, единиц, В — из 7 и единиц и С — из 3 п единиц. Последовательно доминирующие группы животных эволюционируют на мате- рике А, потому что он самый большой (или наиболее подходя- щий по климату), распространяются к материку В и в неко- торых случаях, в конце концов, достигают острова (.'. Когда в результате эволюции новых групп фа уна материка А пре- вышает пределы 9 п единиц, то начинается вымирание древ- них групп, и когда расселение новых групп и материку В и острову (.' увеличивает их фауны выше 7 в и 3 м единиц, то там также происходит вымирание. В этом процессе отме- частся отставание. В любое рассматриваемое время некоторые процветающие доминирующие группы будут все еще приуро- чены к материку А (вершина колонны), а другие достигнут В, но не С, а некоторые из более древних г~~упп, которые вымерли на материке А, возможно, сохранятся как реликты на В, и (или) на острове С. Основной процесс усложняется соотно- иением обратных перемещений и бесконечными индивидуаль- ными особенностями эволюции и расселения различных групп. постоянно наводняются пришельцами из мест с более благоприятным кли- матом. Видимо, именно это и произошло в Арктике, где ограниченная фа~- на наземных позвоночных теперь почти полностью состоит из представи- телей широко распространенных доминирующих семейств.
Сущность природы направленных перемещений животных с больших площадей на малые и с более благоприятных на менее благоприятные пред- ставлена на рис. 71. Основной характер расселения
Основной характер расселения позвоночных заключается в эволю- ции последовательно сменяющихся доминирующих групп на больших благоприятных площадях основной части тропиков Старого Света и в рас- пространении их на меньшие площади с наименее олагоприятными кли-
Глава 9
матическими условиями с последующим их замещением. Распространение может происходить по трем основным путям (рис. 72).
Наиболее короткий путь ведет в умеренную зону Южной Африки. Изменения, происходящие вдоль этого пути,— от большей площади к меньшей и от более благоприятного климата к менее благоприятному. Для расселения в этом направлении значительных препятствий, кроме климата, нет, и, хотя фауна Южной Африки значительно беднее тропи- ческой, многие тропические африканские позвоночные всех классов про- никли в умеренную зону южного полушария в Африке на различное рас- стояние, а некоторые из них достигли Капской провинции.
новь!и СВЕ1
СТАРЫИ СВЕ7
1~~ фропикод ~~~ие ~ ~ I ~ 'аг~ф .OObl — — — -~ ~аДop<н~~ ~�~�~о~рб
УМЕРЕННАЯ ЗОНАСЕВЕРНОГО ПОЛУШАРИЯ
ТРДПИКЯ
Южная Аррика
УМЕРЕННАЯ ЗОНАЮЖНОГО ПОЛУШАРИЯ
Умеренная зона Южной Америки
Но ая 3еяон3ия
72. Схема основного характера расселения («Quarterly Review of Biology», 23, 1948, Рф. 4, р. /f2)
Эта схема показывает предположительное происхождение домини- рующих групп на основной части тропиков Старого Света и расселе- ние их тремя основными путями по направлению к меньшим террито- риям с менее благоприятными климатическими условиями. Маленькие стрелки означают предполагаемое влияние площади и климата на
направление и расселение в переломных местах.
Большинство южноафриканских позвоночных слабодифференцирова- но. Самым четким из них, возмон'но, является изолированный род лягушек семейства Leptodactylidae. Лягушки подсемейства Brevicipitinae приуро- чены к Восточной и Южной Африке и, видимо, отступают на юг, так как у восточноафриканских форм отмечен прерывистый реликтовый ареал. Черепахи семейства Testudinidae и ящерицы Cordylidae (Zonuridae) гораздо разнообразнее в умеренной, а не в тропической Африке, но к степ- ным и пустынным местообитаниям они могут приспособляться лучше, чем к умеренному климату южного полушария. Из птиц ни одно семейство не приурочено Ic Южной Африке. Самым изолированным и эндемичным видом там, возможно, я~ляется сомнительный вид щурки (Promerops). Также ни одно семейство млекопитающих не приурочено полностью к Юж- ной Африке, и хотя златокрот и два небольших семейства грызунов (Pede- tidae и Petromyidae) встречаются в основном в этом районе, это иск- лючение. Большинство позвоночных Южной Африки все-таки близко- родственно тропическим животным Африки.
Фауна Южной Африки в целом, очевидно, сформировалась в ре-
Ilз,1аенение pncnpocmрпнения мсивотиых
зуль.гате перемещения к югу некоторой части тропической фауны Африки. Замещения, видимо, происходили и в тропиках и в более южных частях, но отсутствие непосредственных преград помешало сохранению многих реликтов в Южной Африке: обмен южноафриканской фауны с основной фауной Африки произошел, очевидно, несколько позже.
Второй путь прослеживается из тропиков Азии к Австралии и Новой Зеландии. Этот путь в южном полушарии опять-таки ведет от большеЙ площади к меныпей и от более благоприятного климата к менее благо- приятному. Главной преградой для распространения по этому пути являются пространства соленых вод.
Распространение большинства пресноводных рыб останавливается у островов Ява и Калимантан, у края материковой отмели азиатского кон- тинента. Двоякодышащие рыоы (Ceratodontidae), возможно,— единст- венные строго пресноводные рыбы, достигшие Австралии, но в те далекие времена, вероятно, даже они не нуждались для расселения в сплошной протяженности пресных вод.
Лягушки проникали в Австралию неоднократно, а в Новую Зелан- дию — один раз, но, очевидно, этот процесс сопровождался большими труд- ностями и имел большие интервалы. Семейство, которое достигло Новой Зеландии, более не существует ни в Австралии, ни в Индо-Малайской области. Лягушки семейства Leptodactylidae и Ну/а, которые в свое вре- мя достигли Австралии, также не встречаются более в Индо-Малайской области (кроме рода Ну~а, который заходит в северные окраинные части этой области). Лягушки семейства»evicipitidae, достигшие Новой Гви- неи и тропической Австралии, имели достаточно времени, чтобы стать здесь эндемичными подсемействами. Только распространение лягушек семей- ства Ranidae позволяет предположить, что они достигли Австралийской области не так давно.
Из рептилий Sphenodon достиг Новой Зеландии и сохранился там в изоляции, в то время как родственные ему формы с остальных частей земного шара исчезли. Данных о том, что неморские черепахи проникали когда-то в Новую Зеландию, нет, и, по-видимому, они с трудом достигают даже Австралии. Два семейства черепах Carettochelyidae и "helyidae, которые в Австралийской области, несомненно, являются местными, в настоящее время в Азии отсутствуют. В последней были обнаружены ископаемые, которые отдаленно родственны роду Carettochelys, но черепа- хи семейства Chelyidae, очевидно, там никогда не находились. 11редста- витель семейства Trionychidae проник в Новую Гвинею, возможно, отно- сительно недавно. Наземный род Testudo достиг только Молуккских ост- ровов. Ящерицы из Индо-Малайской области, видимо, проникали в Авст- ралию довольно часто и довольно легко, а гекконы и сцинки достигли даже Новой Зеландии. Несколько наземных, а также водных видов змей расселились из Индо-Малайской области в Австралию сравнительно не- давно, но Новой Зеландии змеи (кроме указанных видов морских змеи)
Не ДОСТИГЛИ.
Л1ногие птицы проникли в Австралию и в значительно меньшем чис- ле — в Новую Зеландию. Очевидно. они накапливались там на протяже- нии значительного времени, так как одни из них более дифференцирован- ны (по сравнению с. азиатскими птицами), чем другие.
Из млекопитающих, вероятно, один предок однопроходных, по s'paii- ней мере однн вид сумчатых, вероятно, пять мышиных грызунов и большое число рукокрылых достигли Австралии, но в Новую Зеландию проникло только два вида рукокрылых.
l.gaea 9
Все наиболее современные из этих позвоночных расселились вдоль дуги Азия — Австралия — Новая Зеландия. Более ранние, возможно, рас- селялись этим же путем. Достоверных свидетельств о том, что какие-либо из этих животных расселялись по какому-нибудь другому направлению, нет, если только не считать черепах семейства Chelyidae (и вымерших черепах семейства Ые1о1ап11йае), которые расселились из Южной Амери- ки через море. В Австралии и Новой Зеландии степень дифференциации в разных классах позвоночных приблизительно пропорциональна способ- ностям этих животных преодолевать соленые воды. Животные с минималь- ными способностями пересекать водные пространства, но которые достиг- ли этих изолированных районов, вообще задерживаются в развитии по сравнению с изменяющейся фауной Азии или же в их изоляции наблн>- даются отклонения.
Третий путь расселения проходит из тропиков Старого Света по дуге через умеренные широты Евразии и Северной Америки к тропикам Цент- ральной и Ю' ной Амер ки. О' тропиков Старого Света к умереннь1 широтам Евразии климат изменяется от более благоприятного к менее бла- гоприятному, и многие позвоночные перемещаются от больших по разме- рам площадей к меньшим. От умеренных широт Евразии к Северной Аме рике расселение осуществляется от большей площади к меньшей, но оез существенных изменений климата. Из Северной Америки к Центрально~1 опять наблюдается переход от большей площади к меньшей, но от менее благоприятного климата к более благоприятному. А от Центральной Аме рики к Южной распространение идет от меньшей площади к болыце11 и без серьезных изменений климата. Как видно, теоретические возможно- сти, определяющие направление расселения, уменьшаются вдоль это11 дуги и к концу ее идут в обратном порядке. Но животные, доминирующп~ в тропиках Старого Света, по мере расселения остаются доминирующими п в умеренной зоне северного полушария, и в Центральной, и Южно11 Америке.
Можно предположить, что различные позвоночные по мере расселе- ния вдоль этой дуги от тропиков Старого Света через умеренную зону северного полушария в американские тропики должны вести себя по-раз- ному. Строго пресноводные рыбы перемещаются из одного бассейна в дру- гой медленно, и их распространение задерживается относительно узкими преградами соленых вод. Они хорошо переносят холод, занимают на севе- ре большие площади и образуют обширные северные фауны, которые могут задерживать расселение остальных, кроме самых доминирующих более поздних тропических групп.
Амфибии могут расселяться быстрее и пересекают узкие пространст- ва соленых вод несколько легче. Они достаточно холодостойки и образунп довольно четкие северные фауны. Рептилии способны расселяться еще быст- рее и сравнительно легко преодолевают узкие океанические простран- ства. Они менее холодостойки, на севере занимают меньшие площади и не образуют отдельных северных фаун, способных препятствовать рассе- лению более поздних тропических групп.
Быстрее всех расселяются птицы и (некоторые из них) с легкостьн> преодолевают преграды. Они хорошо переносят холод, но не образук>т имеющих важное значение отдельных северо-умеренных фаун, хотя миг- рир~ющие группы, возможно, имеют тенденцию задерживать расселение других групп через север. Млекопитающие расселяются быстро по суше, но они не могут преодолевать преграды соленых вод с такой легкостью, как птицы. Они хорошо переносят холод и не образуют существенных
Из.ченение рпспрпстрпнения экивотпных
самостоятельных северо-умеренных фаун. Эти различия должны найти свое отражение в фаунах, которые концентрируются к концу этой дуги— в Центральной и Южной Америке.
Фауна позвоночных Центральной Америки описана класс за классом в соответствующих разделах в гл. 2 — 6. Переход неарктической и нео- тропической фаун (включая Центральную Америку) рассматривался в гл.
7 с учетом только современного распространения, а не перемещений в прошлом. Поэтому теперь перейдем к обзору фауны Центральной Америки и тех перемещений, в результате которых эта фауна образовалась.
Фауна позвоночных Центральной Америки существенно отличается or фауны Южной Америки. В Южной Америке многочисленны только стро- го пресноводные рыбы (но лишь немногие из них распространились дале- ко в глубь Центральной Америки), а североамериканских рыб там очень мало. В Центральной Америке отсутствуют важные эндемичные группы рыб, за исключением солевыносливых. Амфибий в Центральной Амери- ке меньше, чем в Южной, и эта фауна в основном является переходной. Ряд южноамериканских амфибий и очень небольшое число североамерикан- ских проникли в Центральную Америку, но важных эндемичных групп там мало. Зато рептилии в Центральной Америке представлены 21 семей- ством с 8 подсемействами против 18 семейств и 6 подсемейств в Южной Америке (гл. 4). Некоторые семейства и многие роды или приурочены только к Центральной Америке, или встречаются там, но не в Южной Америке, и многие из них, видимо, проникли в Центральную Америку с, севера. Таким образом, из холоднокровных в Центральной Америке мало строго пресноводных рыб, но много солевыносливых; небольшое число амфибий, которое в какой-то мере пропорционально площади и кли- мату Центральной Америки и ее положению между Северной и Южной Америкой. Но число рептилий непропорционально велико. Большинство рыб и амфибий, видимо, проникло в Центральную Америку из Южной, но относительно большее число рептилий, по-видимому, распространилось с севера. Я попытаюсь объяснить этот контраст среди холоднокровных позвоночных, прежде чем перейду к теплокровным.
Центральная Америка в течение какой-то части третичного периода, возможно, была островом или системой островов, отделенных от Север- ной и Южной Америки. Если это так, то этим и объясняется численное превосходство солевыносливых рыб. Отчасти этим же можно объяснить разнообразие рептилий, так как из последних многие проникли через узкие преграды, образуемые солеными водами. Но иммиграция через соле- ные воды остается несколько непонятной. Те немногие амфибии, которые проникли в Центральную Америку сравнительно недавно с севера, так- ;ке проникли и в Южную, но из большого числа рептилий лишь некоторые распространились в Южную Америку. Видимо, амфибии, которые рассе- ляются медленнее и могут дольше задерживаться преградами и которые смогли образовать северные фауны, достигали тропиков Америки только изредка, но всегда как доминирующие группы; в то время как рептилии, которые расселяются быстрее и менее задерживаются преградами, по край- ней мере пространствами соленых вод, и которые не образуют «своих» фаун на севере, проникали через север сравнительно часто, но с перемен- ным доминированием. Большое число рептилий проникло на небольшую площадь Центральной Америки и сохранилось там, но значительно мень- шее число их смогло распространиться на обширные площади тропиков К)жной Америки.
Птиц и млекопитающих, так же как и других позвоночных, в Цент-
484 ральной Америке меньше, чем в Южной. Исходя из современного распро- странения, большинство птиц и млекопитающих по своим связям является южноамериканскими, североамериканских меньше. Мало в Центральной Америке и существенных эндемичных групп птиц и млекопитающих. Од- нако, исходя из перемещений в (сравнительно недавнем) прошлом, отно- сительно большее число птиц, видимо, переместилось из Южной Амери- ки в Центральную и Северную, в то время как млекопитающих относитель- но больше переместилось в обратном направлении — из Северной Америки и Центральн~ю и Ю,кн1н1. Похоже, что в Центральной Америке в боль- Р и с. 73. Трн основнык пути расселения на карте земного
шара, выполненной в двойной ортографпческой проекции. 1ией или меньшей степени преооладают южноамериканские птицы и севе- роамериканские млекопитающие. Возможно, как уже говорилось (стр. 385), это объясняется тем, что птицы, обладающие способностью к полету, были менее изолированы, чем другие позвоночные Южной Америки, и мень- ше 11спытывали неблагоприятное влияние ограниченности площади. И пти- цы и млекопитающие, возможно, расселялись так быстро и проникали в Центральную Америку так часто (значительно чаще, чем даже рептилии), что современные доминирующие группы почти полностью заместили древ- ние центральноамериканские группы, в то время как рептилии сконцен- трировались таким образом, что многие иммигранты прибавились к древ- ней фауне, но еще не заместили ее.
Другими словами (излагая это, как обычно, по возможности проще), если рыбы и амфибии расселялись слишком медленно, а птицы и млеко- питающие — слишком быстро, то расселение рептилий, возможно, шло
именно в таком темпе, который необходим для концентрации древних и новых форм в Центральной Америке без значительного замещения. Независимо от того, правильно ли это толкование или нет, такое положение в Центральной Америке существует: непропорциональное разнообразие рептилий, происшедших главным образом из Северной Америки, и это каким-то образом должно быть объяснено.
Содержание нескольких последних абзацев, видимо, вполне соответ- ствует предположению, выраженному на рис. 72.
Воздействие площади и климата на направление расселения (рис.
71, 7' ) нуждается в дополнительном анализе. Площадь и климат сами по себе еще не определяют направления перемещения. Они воздействуют не прямо, а через эволюцию. У доминирующих животных проявляется тен- денция эволюционировать на самых больших и самых благоприятных тер-- риториях, а затем распространяться на территории, меньшие по пло- щади и менее благоприятные по условиям существования. Там, где п;и>- щади не слишком отличаются друг от друга, обычно происходит не пере- мещение в одном направлении, а обмен с множеством перемещений в сюо- их направлениях, и только с течением времени преобладают перемещения с больших площадей на меньшие. По мере распространения новых доми- нирующих групп у последних проявляется тенденция к замещению ран- них групп, а эти замещения, вероятно, чаще всего (но не всегда) проис- ходят от центров расселения наружу. Расселение отдельных групп. воз- можно, очень сложное, а расселение в целом еще сложнее. Расселение в целом — это сумма перемещений и передвижений и в том и в другом на- правлении, распространения, конкуренции, вымирания и замещения раз- нообразных растений и животных на всей многообразной поверхности земного шара. Оно калейдоскопично.
Основной характер расселения, описанный здесь, сложился из ана- лиза многих фактов и в основном (возможно, не всегда) изложен правиль- но. Но тем не менее я представляю его только как гипотезу. Это, на мой взгляд, наиболее удачное допущение, которое можно сделать и которое может служить основой для дальнейших исследований.
Три основных пути расселения животных на земном шаре показаны на двойной ортографической карте (рис. 73).
.7 И Т Е Р .-1 Т V Р Л
D a r l i n p t о n P. J., Jr, 1948, The geographical distributiorr of cold-blooded ver- tebrates, «Quarterly Review Biol.», 23, 1 — 26, 105 — 123.
F e r n а 1 d М. L., 1926, The antiquity and dispersal of vascular plants, «Quarterly
Beview Bio].», 1, 212 — '45.
M а t t h e w W. D., l,>i;"~, l939,;limate and evolution, «Апп. New York Асад. Sci.»,
24, 171 — 318; reprinted (1939) as «Special Р»Ь1. New York Асад. Sci.», 1.
.11 а у r К., 1946, History of the North American bird fauna, «Wilson Bull.», 58, 3 — 4!. S с Ь ш i д С K. P., 1943, Coroljary and commentary for;limate and Evolution. «Ame- rican Midland Naturalist», 30, 241 — 253.
S i m p s о n G. G., 1940, Mammals and land bridges, «3. Washington Acad. Sci.»,
30, t 37 — I 63.
) i m p s o п С. G., 1949, The meaning of evolution, New Haven, Yale U. Press. S i m p s o r) G ('., 39,зЗ, Evolution and geography, Eugene, Oregea, Oregon St~to
System of IIigher Education.
0' i 11 i s J. (;., ! 0~9, The birth and spread of plants, Geneva, (;onseriatoire et Jardin
botaniques de la Ville.
Глиеа 10ПРОШЛОЕ В СВЕТЕ ЗООГЕОГРАФИИ
Я крайне обязан проф. Ромеру и Вильямсу за многие полезные и кри- тические замечания, высказанные ими по этои главе, и должен сказать, что они ни в коей мере не ответственны ни за те ошибки, которые, возмож- но, в неи еще остались, ни за сделанные мною выводы.
Сведения об истории поверхности земного шара дает нам главным образом геология. Зоогеография в этом отношении стоит на втором месте. Она может добавить лишь некоторые детали к общей картине, вырисовы- вающейся на основе геологических данных, но в ряде случаев эти добав- ления не представляют большого интереса для геологов. Наличи е или отсутствие сухопутного соединения между двумя материками, столь су щест- венное для зоогеографов, обычно является для геологов весьма незначи- тельной деталью, которая указывает на небольшую разницу в уровне моря на ограниченной территории в течение короткого отрезка времени. Но иногда, в тех случаях когда зоогеография воссоздает общую картину при- роды и взаимосвязей материков в прошлом, ее значение возрастает. Гео- логам трудно разобраться в зоогеографии, в связи с чем зоогеографам лучше самим давать ответы на возникающие вопросы, так как это тре- бует глубокого понимания распространения животных как в прошлом. так и в настоящее время, а главное — понимания связей между различ- ными фаунами и пространством, климатом и преградами. Эти связи довольно сложные, однако некоторые обобщения сделать можно.
К особым случаям следует относиться весьма осторожно. Например. распространение некоторых идентичных или близкородственных видов или родов почти не зависит от расстояния, климата или преград. Леопард и африканский заяц' (Lepus cupensis) распространены от Южной Африки до Маньчжурии и т. д., несмотря на то что в целом фауны млекопитающих Южной Африки и Маньчжурии сильно между собой различаются. Пума распространена от северной части Британской Колумбии до южной части Южной Америки, хотя фауны млекопитающих этих районов в целом так- же существенно отличны. Обычные крысы (Rattus) в Австралии являются местными, несмотря на изоляцию этого материка и дифференциацию его фауны млекопитающих в целом. Род Galuxius встречается в пресных водах Южной Африки, южной части Австралии, Новой Зеландии и южной части Южной Америки, однако распространение его единственное в своем роде среди позвоночных. Ископаемые этого рода, к сожалению, отсутст- вуют, но известно, что современные представители его заходят в море. И все же, несмотря на это, некоторые ученые предполагают, что данный род должен был расселяться через сухопутные соединения, существовавшие в южном полушарии (я не придерживаюсь этой точки зрения). Эти особые случаи значительны сами по себе, но представляют меньший интерес, чем
' Согласно работам советских авторов, африканский заяц так далеко к северу. как указывает Дарлингтон, не заходит. В СССР распространен близкий к L. eape~r- sis — толай (L. tolai)i. Прим. рес).
Прошлое в свете зоогеографии
487
фауны в целом. Касаясь прошлого, основное внимание необходимо сосре- доточить на фаунах в целом, а не на частных случаях.
Существующее положение позволяет предполагать, что если в прош- лом две фауны, относящиеся к одному времени, были в целом сходны и генетически связаны, то они занимали климатически однородные, смеж- ные области. Если генетические связи охватывали основные части обеих фаун, но не были сходны в деталях и если связи касались скорее родст- венных родов, а не общих родов и видов, то возможно, что эти фауны были значительно удалены друг от друга и связаны длинным и не прямым пу- тем. Там, где фауны отличаются друг от друга, эти различия по своей при- роде могут быть весьма существенными. Если две фауны были сходны в це- лом, но не связаны в деталях, то, вероятно, они разделялись преградой. Если две фауны отличались друг от друга по своему объему или по харак- теру адаптаций, но в какой-то степени были близкородственными, то. вероятно, они занимали территории, разные в климатическом отношении, но соединяющиеся между собой. Подобные обобщения легче делать, чем применять. Различия же между большинством древних фаун, не извест- ных в достаточно полной степени, могут объясняться различиями во вре- мени.
Размещение современных животных дает не только принципы, кото- рые помогают объяснить их распространение в прошлом, но которые сви- детельствуют также и о самом прошлом. Например, современные австра- лийские млекопитающие убедительно свидетельствуют о том, что Австра- лия. по крайней мере с начала третичного периода, была изолирована от остальной части земного шара, хотя все значительные ископаемые млеко- питающие из Австралии относятся к периоду, имевшему место не ранее плейстоцена (правда, очевидность этого доказательства зависит и от иско- паемых млекопитающих в других частях земного шара). Некото- рые современные млекопитающие являются индикаторами более отда- ленного прошлого в большей степени, чем другие. Однако важен не возраст самих животных, а время их распространения. Группа животных, ведущая свое начало с мезозоя, возможно, неоднократно перерасселялась, и ее современное размещение может быть очень недавним без каких-либо следов раснространения в мезозое. Вообще, время распространения, види- мо, обратно пропорционально степени расселения и возможностям животных пересекать преграды. Строго пресноводные рыбы расселяются по материкам медленнее, чем любые другие позвоночные, и их современ- ное распространение все еще отражает географию мелового периода и мо- жет дать нам кое-какие данные о расположении суши в этом периоде. Но вряд ли по распространению каких-либо современных животных можно судить о более древних временах.
Значительная часть тех сведений, которые дает нам зоогеография о прошлом, основывается на распространении животных в древности (по ископаемым остаткам), но судить о нем следует по их современному распространению. Из различных групп позвоночных на протяжении все- го третичного периода наибольшая роль принадлежала млекопитающим. Начиная с позднего триаса и, вероятно, на протяжении большей части мезозоя существенную роль играли динозавры, а в нижнем триасе — дру- гие рептилии (в основном, возможно, Therapsidae) и стереоспондильные амфибии. В перми, а возможно, даже и раньше некоторые амфибии и реп- тилии представляют кое-какие «ключи», по которым можно судить о том, каким был земной шар в то время, но эти сведения очень скудны и с боль- ~пни трудом поддаются расшифровке.
Выводы зоогеографа о прошлом могут заранее определяться его взгля- дами. Если зоогеограф исходит из того, что распространение животных постоянно и сохраняет древние черты географического размещения на сот- ни миллионов лет, то представить прошлое пока;кется ему весьма легким делом: он, вероятно, прибегнет к возведению больших сухопутных мостов илп к теории перемещения материков. Если же он, так же как и я, считает. что животные обладают большими возможностями перемещения и что они постоянно расселялись (распространялись и отступали) на всем земном шаре, то он увидит, что в этом случае выяснить их прошлое значительно труднее. По калейдоскопическому перемещению животных трудно пред- ставить, как материки соединялись в прошлом, а также перемещались ли они вообще. Я не собираюсь отвергать другие точки зрения, но они должны быть четко выражены. Описывая историческую зоогеографию, следует изложить свою точку зрения о средствах и степени расселения животных, о связи расселения с их отступанием и исчезновением, 0 вре- мени и значении особенностей распространения.
Для целей зоогеографии прошлое может быть разделено на три от- резка времени (рис. 6). Плейстоцен и постплейстоцен — последний мил- лион лет или меньше — самое близкое к нам время, которое довольно хорошо известно, и его можно коснуться здесь очень кратко. Перед ним идет третичный период (примерно 60 млн. лет или около этого). Зоогеографн- ческие данные этого периода имеют наиболее существенное значение в свя- зи с тем, что геологические данные о третичном периоде еще очень непол- ные, а распространение животных, особенно млекопитающих, даст убеди- тельные свидетельства об истории и взаимосвязях материков в то время. Этому периоду предшествуют мезозой и домезозой, охватывающий еще более продолжительный отрезок времени, представление о котором остается весьма туманным. О том, каково было положение материков в то время, ничего не известно. Если они когда-либо перемещались, то их перемещение могло продолжаться и в мезозое, и, возможно, в будущем зоогеография, поможет выяснить истинное положение материков. Для каждого из этих трех периодов — плейстоцена и постплейстоцена, третичного, мезозоя и домезозоя — я постараюсь сначала дать описание положения земного шара так, как это представляют геологи, а затем привести те сведения. которыми располагает зоогеография.
Плейстоцен и постплейстоцен
11лейстоцен был эпохой или эпохами ледников. Известно четыре по- следовательных материковых оледенения в северных частях земного шара, отделяемых межледниковыми периодами, когда происходило отступание ледников. Южные границы северных ледников представлены на рис. 74. Первые плейстоценовые ледяные щиты, вероятно, появились в большин- стве случаев миллион лет назад. Последние материковые ледники, соглас- но датировке С,4, начали отступать только около 10 тыс. лет назад (Libby, 1956). [Я не пытался точно установить время начала и конца плейстоце- на. В последние годы подсчеты времени геологами, с началп отступания последнего материкового ледника в восточной части Северной Америки, сократились примерно с 25 тыс. до 10 тыс. лет. Таким ооразом, возможно. прошло всего лишь 10 тыс. лет с тех пор, как районы Бостона или Чикаго находились подо льдом. Значительно труднее установить начало плейсто- цена, чем его конец, и любая дата должна считаться условной].
Прошлое и свете гоогеографиа
llр чины плей оценового оледенения неи:~ве'ны, но 11редпо:(),,;е ний много. Флинт (Flint, 1947) критикует некоторые из них, которые кажутся наименее достоверными. Он считает, что оледенение произошло, вероятно, в результате колебаний (уменьшения поступающей на Землю) солнечной радиации и поднятия уровня суши в ряде мест на севере. Когда количество тепла, поступавшего от Солнца, было минимальным, на воз- вышенных частях Северной Европы, северо-восточной части Северной Лме- рики и в других местах начинал накапливаться лед, которьш затем рас- пространялся из этих районов в различных, в том числе и в ю;кном, направлениях. Биогеография может сделать свой вклад в эту гипотезу. В течение значительной части третичного периода отмечалось неуклонное смещение северных растительных зон к югу, очевидно, вызываемое Р и с. 74. Приблизительные южные границы се«ерных материко-
вых льдов в плейстоцене. На север от этих границ некоторые площади не были иокрыты ледником
(в Восточной Сибири, на Аляске и в других местах). постепенным охлаждением северного климата (Barghoorn, 1953, р. 217). Более того, анализируя обмен млекопитающими через мост Беринга, Симп- сон обнаружил, что на протяжении большей части третичного периода, оче- видно в результате постепенного усиления климатического барьера (гл. 6),. постепенно увеличивается селективность. Это позволяет предположить, что влияние солнечного фактора — охлаждения северных частей земного шара — проявлялось также постепенно, а не сразу'. В результате обще- го поднятия уровня суши, которое произошло в позднетретичном периоде и плейстоцене, материки стали (и остались такими до настоящего време- ни) выше, чем были на протяжении всей геологической истории.
Северное оледенение явилось одним из величайших проявлений холодного климата плейстоцена. Его влияние сказалось на всем земном ша- ре. На севере, а также и в тропиках и в умеренной зоне южного полушария увеличились ледники всех высокогорных районов. Увеличение ледников 1 7Т
Дж. С. Симпсон утверждал, что теоретически невозможно объяснить климат ледникового периода похолоданием на севере и что плейстоценовые ледяные щиты появились в результате увеличения тепла. И до сих пор среди дискутирующих по это~~~' вопросу геологов существуют и противннки (Flint, 1947), п сторонники (например, Shapley, 1953) этого положения. Но мне кажется, что независимо от того, возможно или невозможно это теоретически, распространение растений и животных в течение плейстоцена и после него ясно показывает, что льды образовались после проди.~- жительного прохладного периода, когда северные части земного шара имели хол(~;(- йый климат. Я излагаю это положение для дальнейших ссылок. Зоогеографы должны уважать другие науки, но не переоценивать пх значение и не умалять важность свжй науки. Ниже в этой главе мне придется коснуться, например, «опроса перемеще- нпя материков, который некоторые геологи стараются поставить на теоретическую основу. Тогда я обращусь к этой сноске, чтобы подчеркнуть еще раз, что зоогеограф обязан давать собственную оценку положению и не чувствовать себя слишком связан- ным с теоретически невозможными положениями других авторов.
490
Глава 10
повлекло за собой понижение снеговой линии и спускание ледников. а это дает основание предполагать, что во времена плейстоцена температура повсеместно была на 4 — 8' С ниже современной (Flint, 1947). Вероятно, хотя и не совсем достоверно, что понижение температуры происходило везде синхронно (Flint, 1947; выводы Флинта ставились под сомнение, хотя мне они кажутся вполне обоснованными).
Во времена оледенения, очевидно, на значительной части земного шара, помимо понижения температуры, увеличилось количество выпадаю- щих осадков. Это не означает, что на всем земном шаре были сильные дож- ди, просто влажные районы стали еще более влажными, а засушливые— менее засушливыми, чем в настоящее время. Очевидно, произошло сме- щение зон с умеренным количеством осадков по направлению к тропикам, в субтропические зоны пустынь. Кроме того, увеличился размер озер в юго-западной части Северной Америки, Юго-Западной Азии, Восточ- ной Африке, Южной Америке (озеро Титикака) и в Австралии, и те районы в юго-западной части Северной Америки, Северной Африки (Сахара) и в Австралии, которые ныне являются засушливыми, возможно, были луч- ше обводнены по сравнению с настоящим временем, в большей степени покрыты растительностью и представляли меньшую преграду для непу-
СТЫННЫХ ЖИВОТНЫХ.
В межледниковые периоды климат на большей части земного шара, очевидно, временами был теплее и суше, чем в настоящее время.
В течение плейстоцена происходили колебания уровня моря. Поте- ря массы воды, покрывшей часть суши в виде льда, вызвала понижение уровня Мирового океана на всем земном шаре порядка 100 — 120 м в пе- риод максимального распространения ледников и 70 — 80 м в течение пос- ледней подвижки ледников (Kuenen, 1950, р. 537). Таким образом, матери- ки и острова, которые отделялись друг от друга небольшими глубинами, соединились. С другой стороны, до плейстоцена и, возможно, в течение межледниковых периодов, когда на суше не было ледников или ими была покрыта лишь незначительная ее часть (когда Антарктида и Гренландия не были покрыты, как в настоящее время, ледяными шапками), уровень моря был выше, чем теперь, на 20 — 50 м (Kuenen, 1950, р. 539) и низкие краевые части материков, а также низкие острова были затоплены. Каз- нен считает, что, хотя этими цифрами следует пользоваться осмотритель- но, возможность ошибки не превышает 50о/о.
Как правило, постплейстоцен, или современную эпоху, всегда отде- ляют от собственно плейстоцена, но это выделение искусственное. Пост- плейстоцен, относящийся к современной эпохе, который, возможно, не что иное, как еще один межледниковый период, возможно, завершится другим ледниковым периодом. Ледяные шапки до сих пор покрывают Антарктиду и Гренландию. В течение нескольких тысяч лет со времени пос- леднего оледенения климат земного шара подвергался изменениям, ста- новясь в отдельные периоды несколько теплее, чем сейчас («климатиче- ский оптимум», — Zeuner, 1950), а в другие — несколько холоднее («малый ледниковый период»). В настоящее время, по-видимому, снова про- исходит потепление. Причины этих колебаний нам не известны, и мы не можем предсказать, что произойдет в будущем.
Наши знания о распространении и перемещениях животных в тече- ние плейстоцена и после него обширны. Дивей (Deevey, 1949) уделяет большое внимание этому вопросу, привлекая как свои данные, так и спе- циальные источники, чтобы дать представление о биогеографии плейсто- цена (и постплейстоцена). Тинеман (Thienemann, 1950) отводит в своей
ilpouiëîå в свете зоогеографу и работе еще оольше места истории плейстоцена, правда разбирая ее только лишь на примере пресноводной фауны Европы. В своей большой работе Холдхаус (Holdhaus et al., 1954) целый том посвящает влиянию леднико- вой эпохи на фауну Европы. Диллон (Dillon, 1956) высказывает некоторые интересные предположения о постплейстоценовых зонах жизни Северной Америки. Наши знания базируются отчасти на ископаемых материалах, отчасти на современном распространении животных и отчасти на геоло- .гических и оотанических данных, а именно на серии [MeihJIepHIIKoBbIx.— Ред. J отложений в озерах и болотах пыльцы, которые показывают смену растительности, а следовательно, и климата. Эти отложения еще одно доказательство того. что во время образования ледяных щитов климат Р и с. 75. Схематическое изображение нопуля~~т~т~. соъранпвшикся в восточной части Северной Амерш'и
в процессе наступания п отступания ледников. A — первоначальное распространение (жирный прямоуголь- н»к) субарктических видов; В — распространение тех же видов после наступания ледников с популяцией, изолированной в рефугиуме на Гаспе; С — распространение после отступания ледников с реликтовой популяцией в горах штатов Нью-Иорк
и Новая Англия. севера был холодным. Разрабатывая историю плейстоцена, ботаники и зоологи, опираясь в основном на геологию, в свою очередь делают вклад и в геологию, и в ботанику, и в зоологию.
В северных частях умеренной зоны северного полушария, включая значительную часть Европы и Северной Америки, плейстоцен был эпо- xoII катаклизмов. Плейстоцен изменил не только детали, но и всю основу распространения растений и животных. В незатронутых ледником частях земного шара размещение организмов отражает географические особен- ности, возникшие в этом месте в результате эволюции и измененные пере- мещениями. Что касается северных фаун и флор, то они в результате четырехкратного наступания и отступания ледников смешивались п пере- мешивались, в результате чего характер эволюции затушевывался.
Перемещения северных животных в течение плейстоцена и в после- дующие периоды были необыкновенно сложными, но в них проявлялась тенденция к отражению основных принципов движения вперед и обратно, или распространения и отступания, что часто приводило к географической прерывистости.
В течение ледниковых эпох арктические животные первыми переме- стились к югу, и затем, после того как ледники отступили, снова вернулись на север, оставляя иногда популяции, переместившиеся вместо севера высоко в горы, где они становились изолированными и сохранялись на боль- 1иих высотах как «заброшенные» арктические реликты (рис. 75). Нахож- leIIIi» арктических растений и животных, изолированных высоко в горах Европы (они встречаются также в Азии и Северной Америке), было давно
Глава 10
известно и объяснялось, правда, слишком упрощенно в деталях, в работе Эдварда Форбса (примерно в 1846 г., издание которой я не видел), а позд- нее Дарвином (Darwin, 1859). С другой стороны, очень часто по мере насту- пания ледников непокрытые льдом площади оставались позади ледяного фронта, и животные, которые двигались от ледника в южном направле- нии, иногда оставляли оторванные популяции в рефугиумах, отрезанных ледником. Когда происходило вторичное отступание ледников, некото- рые из этих популяций распространились из своих убежищ и смешались сложным образом с другими, вновь идущими на север южными популя- ЦИЯМИ.
Сложное распространение, перемещение и смешение, которые после- довали за отступанием последнего ледника, продолжаются до сих пор и являются предметом серьезного изучения для зоогеографов северных районов земного шара. Вполне естественно, что прежде всего внимание должно быть сосредоточено на перемещении северных животных в течение плейстоцена и в последующее время. Но постплейстоценовая перегруп- пировка северных животных для зоогеографии имеет не большее значе- ние, чем линия Уоллеса. Чрезмерное внимание к ней может только отвлечь зоогеографов от более важных вопросов. Зоогеографию cледует начинать с понимания особенностей земного шара и процессов, происходя- щих на нем, а также совокупного влияния эволюции на распространение животных. Приспособление же северных животных к современным изме- нениям климата следует воспринимать как кратковременное, играющее второстепенную роль изменение. Если этому придается слишком большое значение, то это приводит к преувеличению влияния местных экологиче- ских условий и временных адаптаций, а также умаляет важность создав- шихся ситуаций на земном шаре в целом.
В южных частях умеренной зоны северного полушария. охваченных. оледенением, влияние плейстоцена на распространение животных было еще более существенным. Температурные изменения, которые были связаны с изменениями режима осадков, сопровождались перемещением животных и югу и северу, а также к западу и востоку. В течение плювиальных ледниковых периодов происходит расширение лесных площадей, а в за- сушливые межледниковые периоды — расширение степных и пустынных частей земного шара. Временами, например, степи Западной Азии, оче- видно, простирались через Центральную Европу, а прерии (степи) запад- но-центральной части Северной Америки заходили значительно дальше„ чем в настоящее время, на восток. [Увеличение площади открытых прост- ранств, по-видимому, произошло в результате последовательной смены растительности, вызванной отступанием ледника, а не вследствие засуш- ливости климата.] Различные животные распространялись и отступали со своих мест обитания, а отдельные из них в конце концов попали в ооо- собленные места обитания (Schmidt, 1938).
Особенно влияние плейстоцена сказалось на рыбах и других пресно- водных животных умеренной зоны северного полушария. Лед подпружи- вал или изменял течения рек, позволяя, таким образом, рыбам переходить 113 одной речной системы в другую, а также выпахал тысячи впадин, кото- рые после отступания ледника превратились в новые озера. Далее к югу цикличность осадков вызвала расширение, а затем уменьшение площади озер, что сначала способствовало расселению рыб, а позже привело к их изоляции в ряде водоемов. Современное распространение рыб часто является свидетельством протяженности и соединений озер и рек в плейсто- цене. Расселение рыб Великого Бассейна в юго-западной части Соединен-
/1 роуилое в свези г зоогеогра j uu
ных Штатов (Hubb', Aliller et аl., резюмировано по Deevey., 19 ~9, р. 1395— 1'~00) является одной из иллюстраций к вышесказанному.
Влияние плейстоцена на распространение животных сказалось и в других частях земного шара, хотя там оно было значительно слабее. llериодическое сокращение преград на всем земном шаре в целом способ- ствовало расселению животных. Ледниково-плювиальные эпохи благо- приятно сказались на лесных и влаголюбивых животных, а более засупн- ,и1вые межледниковые периоды способствовали расселению пустынных и степных животных. В такие времена засушливые площади, например про- стирающиеся между юго-западными частями Северной и 10жной Аме- рики через тропики, видимо, были более непрерывными (или почти непрерывными), чем в настоящее время, что допускало взаимообмен меж- ду пустынными растениями и насекомыми, встречающимися в этих ме- стах. Периодические понижения уровня моря позволили материковым фаунам проникнуть на многие острова (гл. 8), что в свою очередь является подтверждением понижения уровня моря (которое, однако, происходило только до определенного уровня). Понижение уровня моря до 1000 м. очевидно, привело бы к соединению Кубы с Северной Америкой и острова Сулавеси — с островом Калимантан и Азией, что вызвало бы приток целых материковых фаун, но этого не произошло. Мост, образовавшийся в результате понижения уровня Мирового океана, соединил Азию и Се- верную Америку через Берингов пролив. Это соединение, очевидно, пред- ставляло широкую равнину, которая продолжалась как береговая равнина вдоль южных краевых частей Азии и Аляски (Flint). Климат этого сое- динения, вероятно, находился под влиянием теплого течения Куро-Сиво и примерно был таким же, как в настоящее время на Алеутских островах. Возможно, что территория этого соединения была покрыта высокими тра- вами, подобными, вероятно, тем, которые растут сейчас на полуострове Аля- ска. Многие крупные и мелкие млекопитающие могли пересечь и пересе- кали этот мост. Возможно, что человек впервые достиг Америки именно :эпи~ путем в позднем плеистоцене.
В течение ледниковых периодов в пределах тропиков даже на низмен- ностях температуры, вероятно, несколько понизились, а количество осад- ков возросло. Площадь озер увеличилась, по крайней мере в некоторых местах, также видимо имелась тенденция к расширению площади тропи- ческих лесов и к сокращению территории степей и пустынь. Совершенно иной характер носили изменения, происходившие, вероятно, на протя- жении более теплых сухих межледниковых периодов. Влияние плейсто- цена на тропические фауны, сказавшееся в основном только в краевых частях земного шара, в целом не было очень значительным. Современные тропические фауны низменностей — обширные и сложные, с многочис- ленными видами животных, хорошо приспособившимися друг и другу и к окружающим их растениям. Это, очевидно, древние, не затронутые катя клизмами плейстоцена фауны.
Как результат особого влияния событий плейстоцена следует отме- тить постепенное отступание и исчезновение крупных млекопитающих, которое продолжается и до настоящего времени. Человек, эволюциони- ровавший в основном в течение плейстоцена (но не обязательно благодаря ему), возможно, не несет главной ответственности за этот процесс, но он способствовал ему.
Другое воздействие плейстоцена, возможно, сказалось на увеличении как объема, так и протяженности миграций птиц с севера на юг (стр. 216).
Делая заключение о плейстоцене, я несколько перефразирую и до-
Глпвп 10
полню краткий обзор, данный Флинтом (Flint, 1947). Если м~>жно было бы показать в виде фильма упрощенный и чрезвычайно уско- ренный ход событий, то картина начиналась бы (в третичном периоде) с постепенного поднятия суши и появления высоких горных цепей ш> многих частях земного шара и (если Флинт прав) с постепенного умень- шения интенсивности излучения и колебаний солнечной радиации. В нача- ле плейстоцена, когда эти процессы получили значительное развитие, колебания солнечной радиации начали вызывать таяние щитов матери- ковых льдов не только на возвышенностях севера, но также и повсюду в районах горного оледенения. Эти циклические процессы сопровожда- лись увеличением и уменьшением осадков, смещением зон осадков в за- сушливые субтропики, а затем повсеместным увеличением и сокращением дождевых озер. В то же самое время уровень моря постепенно то опускал- ся, то поднимался, то создавая, то разрушая сухопутные соединения через пролив Беринга и со многими островами. В соответствии с этими цик- лами неорганического характера происходило общее, но нерегулярное перемещение растений и животных. По мере изменения климата целые северные флоры и фауны переместились к югу и северу, и по мере их пере- мещения менялся состав и внутренние особенности распространения. Многие растения и животные распространялись и отступали в связи с, рас- ширением и отступанием их специфических мест обитания, и во время этих перемещений отдельные представители оставались на горных вершинах или в свободных от ледников рефугиумах во многих влажных или засуш- ливых местах земного шара. В других частях земного шара некоторые растения и животные, проникнув через временные сухопутные мосты на острова, позже оказались там отрезанными. Заметно уменьшилось число крупных животных, увеличились ежегодные перемещения птиц с севера на юг, а человек к концу плейстоцена достиг почти всех благоприятных для обитания частей земного шара.
Третичный период
Третичный период охватывает примерно 60 млн. лет (больше или меньше) до плейстоцена. Это основная часть кайнозойской эры, которая подразделяется на пять отделов (рис. 6). С этим периодом, интересовавшим меня длительное время как зоогеографа, я ознакомился по соответствую- щим работам Данбара (Dunbar, 1949) и Ромера (Romer, 1945), а также по другим работам, часть из которых упоминается ниже.
На протяжении третичного периода конфигурация материков претер- пела лишь незначительные изменения, вызванные в большинстве случаец наступанием или отступанием морей в их краевых частях, но их поверхность была затронута существенными изменениями. На третичный период при- ходится формирование и переформирование всех великих горных систем земного шара. В это время происходило и общее поднятие суши, которое в конце концов привело материки к их необычайно высокому плейстоце- новому и современному уровню. В некоторых частях земного шара полу- чила распространение сильная вулканическая деятельность. Непосред- ственное влияние всех этих процессов на материковых животных вызывае ~ сомнение, но совершенно очевидно, что островные животные подверглись их воздействию. Острова некоторых архипелагов возникали и разрушались так же, как горы на материках, а там, где между материками были распо- ложены архипелаги, вероятно, образовывались и неоднократно разруша-
Прошлое в свете зоогеогрпфии
лись сухопутные соединения. Влияние всех этих процессов на животных, очевидно, зачастую было значительным, и нередко распространение животных является лучшим свидетельством событий тех времен.
В течение третичного периода Африка, очевидно, была наиболее постоянным материком, хотя по крайней мере восточная часть этого мате- рика была подвержена вулканической деятельности.
Отдельные части Евразии были очень неустойчивыми. В течение мезо- зоя и значительной части третичного периода через всю Южную Европу и Азию простиралось море Тетис. Возможно, временами Европа сокраща- лась до архипелага. Сложные процессы формирования Альп и Гималаев в течение третичного периода привели к поднятию их из моря Тетис. Их подъем вызвал сложные географические изменения, включая, видимо, воз- никновение и разрушение сухопутных соединений между Евразией и Аф- рикой, Азией и современным полуостровом Индостан.
В течение третичного периода материк Северная Америка был до- вольно постоянным по своим очертаниям, и только мелководные моря пере- крывали те части материка, где теперь находятся юго-восточные береговые равнины и низменности долины Миссисипи. Но поверхность этого матери- ка изменилась. Современные Аппалачи поднялись и подверглись разломам в основном на протяжении третичного периода, а горы западной части Северной Америки — сложным преобразованиям.
В Южной Америке Анды по своим современным формам представляют главным образом третичные горы, и своих современных высот они достигли не раньше позднетретичного периода и плейстоцена. Центральная Америка была районом вулканической деятельности, и там происходили местные изменения уровня суши.
Австралия, очевидно, была относительно постоянным материком, но острова между Азией и Австралией имели очень сложную третичную исто рию (Umbgrove, 1949).
Климат в третичное время, по крайней мере на север от тропиков, был значительно теплее, чем в настоящее время, но вплоть до плейстоцена происходило постепенное его похолодание. Те леса, которые теперь счи- таются типичными для умеренных широт, в раннетретичное время прости- рались на север в Арктику, а леса тропического и субтропического типа заходили далеко в умеренную зону северного полушария (Barghoorn, 1953). В течение третичного периода северные растительные зоны посте- пенно перемещались к югу. Так как климат даже в раннетретичное время был зональным, то и эти зоны были расположены так же, как и в настоя- щее время (Chancy, 1940; Barghoorn, 1953). Как указывает Баргхорн, вряд ли в средних широтах существовали подлинно тропические условия. Скорее всего отсутствие зимних морозов позволило тропическим расте- ниям продвинуться к северу. Тропические леса, которые простирались к северу, по-видимому, были беднее и не столь разнообразны по сравнению с собственно тропиками.
Пышное развитие травянистой растительности, происшедшее в уме- ренной зоне северного полушария в среднетретичном периоде (Barghoorn, 1953), возможно, являлось ответом на увеличивающуюся суровость север- ных зим. До этого трав везде было мало, а в тропиках их мало и до сих пор. Их появление в большом количестве на севере, вероятно, повлияло на эволюцию и распространение животных и, видимо (из позвоночных), особенно на зерноядных птиц и грызунов.
Краткий обзор предполагаемой H фактической истории позвоноч- ных третичного времени был дан по классам в соответствующих главах
(гл. 2 — 6). Ромер (Нотег, 1945, р. 546 и след.) рассматривает ее по последо- вательным эпохам. Вообще говоря, современное размещение позвоночных. в основном, по-видимому, является результатом третичных (или более поздних) перемещений.
Зоогеографические доказательства, воспроизводящие общую картину земного шара в третичном периоде, соответствуют геологическим и бота- ническим свидетельствам. Например. и пресноводные рыбы (гл. 2) и мле- копитающие (гл. 6) отражают зональность климата в течение третичного периода и постепенное похолодание его на севере. Еще более значительны в этом отношении ботанические свидетельства.
Относительная стабильность материка Африки и неустойчивость .Ю;кной Азии отражаются в распространении современных пресноводных рыб — древних и разнообразных групп в Африке и доминирующих более современных — в тропической Азии (гл. 2). Эти фаунистические различия позволяют предположить, что обмен рыб между Африкой и Южной Азией в течение третичного периода был затруднительным. Раннетретичные дан- ные по млекопитающим в этих частях земного шара весьма скудные, но млекопитающие из Эль-Файюма (в позднем эоцене и раннем олигоцене Кгипта), представляющие смесь очень четких, ранее известных африкан- ских групп и соответствующих им современных европейских, позволяют допустить, что в то время Африка и Евразия соединялись, а ранее на про- тяжении значительного времени были отделены друг от друга, хотя пред- положительно до этого они были соединены. Более поздние данные по мле- копитающим указывают на то, что Африка и Евразия были соединены по крайней мере большую часть времени на протяжении остальной части тре- тичного периода.
Некоторые авторы допускают в третичном периоде общее соединение Африки, Мадагаскара и Индии. Они назвали этот гипотетический материк «Лемурией». Но данное предположение может быть опровергнуто убеди- тельными доказательствами: например, отсутствием строго пресноводных рыб на Мадагаскаре и характером связей между другими, играющими зна- чительную роль животными. Даже лемуры, в честь которых была названа эта суша, свидетельствуют обратное: лемуры Африки и тропической Азии имеют прямое родство, а лемуры Мадагаскара относятся к очень четким самостоятельным эволюционным боковым линиям (рис. 76). Некоторые связи. наблюдающиеся между позвоночными Мадагаскара и Индии, явля- ются исключениями, которые лучше всего могут быть объяснены исчез- новением животных в Африке или их расселением через море, например некоторые птицы и летучие мыши (гл. 8). Следовательно, в третичном периоде Лемурия не существовала.
Ископаемые данные по млекопитающим третичного периода ясно показывают время и место сухопутного соединения между Евразией и Се- верной Америкой. Очевидно, эти соединения через Берингов пролив неоднократно возникали и разрушались (время, в течение которого прои~- ходили эти процессы, см. гл. 6). Но ископаемые млекопитающие не ука- зывают на существование каких-либо сухопутных соединений (через Атлантику) между Европой и Северной Америкой. Имеющиеся данные вплоть до раннего эоцена — неясные, но они и не требуют возведения Атлантического моста, а свидетельствуют о соединении в Северной Пацифи- ке. После раннего эоцена, в более поздние эпохи, имеющиеся свидетел~- ства явно располагают в пользу единственного моста в Северной Пацифике ((;импсон, статьи, цит. в гл. 6).
Об истории Южной Америки в третичный период можно судить по
прошлое в creme зоогеогрпфии
распространению ископаемых II современных позвоночных. Нахождение в ю;кной части Южной Америки в самом начале третичного периода не толь- ко сумчатых, но также и плацентарных млекопитаюших, некоторые из которых родственны североамериканским, указывает (если в этом отноше- нии были какие-либо сомнения), что этот материк в то время представлял спло1иную сушу. Современное распространение строго пресноводных рыб в Южной Америке, которые в основном сосредоточены в системе Ама- зонки (гл. 2), а также, я думаю, и ископаемые млекопитающие, оонаруженные в последовательно сменяющиеся эпохи, указывают, ч|о Южная Америка на протяжении всего третичного периода представляла единый неразделенный материк. Ископаемые данные по млекопитающим
° °° ° °
° ° °
° В
° °
° °
° °
1'
Ф.
° е'
° °
° ° ° - ° ° °
° °
° ° ° °
° °
ее ° ° ° ° \ ° °
° ° ° ° ° ° ° °
° ее
° В е ° Ъ ° ~
° Ъ ° ° e ее ° ° ~В
°,с ° ° ° ° °
° ° а\а
f ° ° ° ° ° ° ° °
° В °
° ВВ ° ° ° в
° ° ° ° °
° ° ° r+e g
° ° \В
° »
° °
еа
Юу a ° еа
е °
° °
° ° аае ° °
° ° е °
° ° е ° °
ее ° ° ° ° °
е ° ° ° ° ° ° °
° ° ° ° ° °
° ° ° ° °
° ° ° °,е
е ° ° °
ее
° е ° ° °
aaе te ° е ° ° ° ° аа ° ° ° ° ° ° ее фе ° ° °
° ° ° ° ° °
° е ° r+ ° ° е °
И ° ° 9 ° ' ° °
° е ° )gap
° ° °
° ° ее у ° ° °
е ° ° °
° °
° °
e;~.-- ° а
~е
° °
° ° ° ° ° ° ° ° °
е юа
е') °
° е ~
° еа °
° В~е °
Ф °
° а °
° аеф
° °
'Ь ВВ4
° ее ° е °
° «'е'
° ° ее ее
° ее ° °
е",е;
° °
° °
° °
° ° ° °
° °
° ° ° в °
° ° °
° ° °
° °
° ° ° ° ° ° °
О
° ° ° °
° ° ° °
° ° °
° в
в
° ° ° °
° ° ° ° ra ° ° в °
° ° °
° °
'Ф° ° aP a °° °
° ° ° ° ° ° °
1
0 °, Ф. ° ° °
l ."Ф
е ii °
° ° ° ° ° °
° ° °
° ° ° °
° ° ° °
° ° °
° ° ° ° °
° °
° ° ° е ° ° ° ' ° °
° ° в ° ° в ° ° °
° ° ° ° ° ° ° ° ° ° °
° ° °
° ° е °
° ° ае ° ° ° е ° ° ° ° °
° °
° ° ° ° ° ° ° ° ° ° ° ° ° ° °
° ° °
° ° ° ° ° ° ° ае
° °
е
° °
Ф
° ° ° ° ° ° ее ав
° ° a~a
\Вф ° °
%~Щ
° в е ° е ° e °
° ° ° °
В °
е ° ° ° ° в ° ° a ° °
° ° °
° ° °
° ° ° ° ° ° ° e ° °
в 'ъ"
е
° °
° ° ° °
° ° ° ° °
° ° ° ° ° °
е ° ° е °
° ~, ° ° e
° ° ° ° е °
е ° °
° ° ° е °
° °
° ° ° а °
° r °
° ° °
еа ° е ° а
° в ° ° ° ю °
° ° ° ° ° °
° ° ° ° ° ° ° ° ° ° °
° ° ° ° r ° ° ° ° ° ° ° ' ° ° ее ° ° еее е
° °
a ° ° ° ° ° ° ° ° ее
° °
° ° ° ° е ° °
° ° ° ° °
е ° ° а
° В е еа ° ° ° ° ° ° ° a ° ° °
° ° °
° ееа ° ee ВВ е еа ° r ° ° е ° ° е ° ° ее ° ° ева
° ° a
° ° е е °
° °
° В ° ° ee ° ° ° ° ° ° .
° ° еа
ве °
° °
° е
° °
в
° °
° ° °
° ° ° a
е ° °
° ° °
° ° ° r ° ° ° a a° °
° °
° ° ° ° ав,
° в
° ° е °
ф ra a a°
° °
е аа
° ° °
° +a gа ° r ° ° ° ° ° е ° В ° ° ° ° а °
° ° ° ° ° ° ° ° ° ° ° ° ° Q ° ° ° ° °,а ' еаееаае
° е ° а °
еа °
° ° ° еее ° ° °
° °
;~~а ее r ° е ° ° °
° ° °
° е ае а е
° в °
те
° °
° ° ° ° °
° ° ° ° ° ° °
° ° ° °
°, ° ° ° ° ° ° ° ° ° °
° ° ° ° ° ° °
° ° °
° ° е а
° ° ° ° °
° ° ° ° ° °
° °
° ° ° е ° ее' °
° ° ° ° ° ° ° ° °
° ° ° ° ° ° ° ° ° ° ° ° а ° ° ° ° e ° °
е ее ° ° ° ° ° °
° ° ° ° ° °
° ° ° е ° ° ° °
II Се'.
р и z. 76. 1еографическпе связи. лемуров.
Прерывистой стрелкой показаны те связи, которые должны были бы суще- ствовать (но не существуют), если бы лемуры расселились через «Леаму- рию». (:плошной стрелкой II()HaaaHbI существун~щие связи между лемурами Африки и Индо-Малайской области и теми, которьн не имеют с вязей с Мада- гаскаром.
уоедительно показывают, что Южная Америка на протяжении болыией части третичного периода была изолирована от других материков и что она присоединилась к Северной Америке через Центральную Америку в позд- нем плиоцене, когда происходил обширный обмен млекопитающих (гл. 6).
Каким образом в начале третичного периода Южная Америка была связана с другими материками — это еще вопрос не совсем ясный. Древ- нейшие третичные млекопитающие Южной Америки, очевидно, предста»- ляли ограниченную, избранную группировку, которая, возможно, достиг- ла Южной Америки, преодолев узкие водные пространства, а не через сплошную сушу (гл. 6). Одни из этих животных проникли из Северной Америки, другие не оставили четких следов направления, откуда они по- явились. Некоторые позвоночные достигли Южной Америки еще в третич- ный период до возникновения полного сухопутного соединения: дикобра- зоподобные грызуны и обезьяны появились, возможно, в позднем эоцене и олигоцене соответственно, но направление, по которому они проникли, не определено; черепахи Testudo и енотовые, очевидно, появились в мио- цене, причем енотовые пришли сюда по крайней мере из Северной Америки. Все это объяснимо в том случае, если Южная Америка на протяжении тре- тичного периода занимала по отношению к Северной Америке то же поло-
Глава 10
жение, что и в настоящее время, и если она соединялась с Северной Америкой посредством изменяющегося архипелага или не полностью соеди- ненного с ней перешейка, который являлся путем иммиграции для неко- торых или для всех раннетретичных млекопитающих Южной Америки и который позднее позволял время от времени проникать им в Южную Аме- рику путем «перепрыгивания через острова» и в конце концов в позднем плиоцене превратился в настоящий сухопутный мост. Эта отчасти пред- положительная история сама по себе соответствует фактам, которых она касается. Нахождение достоверных ископаемых в соответствующем месте, возможно, покажет, правильно это или нет. Если новые данные подтвер- дят предположение о том, что дикобразоподобные и, возможно, один или два других вида млекопитающих проникли в Южную Америку непосред- ственно из Африки, то я склонен считать, что они достигли ее на плотах, так как этому благоприятствует проходящее через Атлантику Южное пас- сатное течение и существуют адекватные плоты.
Была ли Центральная Америка в течение большей части третичного периода полуостровом Северной Америки, или отдельным островом, или архипелагом, не известно, но в конце концов это также будет установле- но с помощью ископаемых. Между тем природа фауны Вест-Индии, зна- чительная часть которой, очевидно, возникла в течение третичного перио- да от центральноамериканской, как мне кажется, позволяет предполагать. что Центральная Америка продолжительное время была отрезана как от Северной, так и от Южной Америки. Я считаю, что в течение этого времени в Центральной Америке накопилась и эволюционировала ограниченная высокоэндемичная, недоминирующая островная фауна смешанного про- исхождения — из Северной и Южной Америки,— большая часть которой (после того как отдельные ее части достигли Вест-Индии) при возникнове- нии сухопутного соединения распространилась за свои границы и была уничтожена материковыми животными. Мне кажется, что современное распространение птиц (гл. 5) и история млекопитающих в прошлом (гл. 6) свидетельствуют (в большинстве случаев отрицательно) о том, что в тече- ние третичного периода присоединения какой-либо тропической территории к Северной Америке не было. И это также, по-видимому, позволяет пред- положить, что Центральная Америка большую часть времени была изо- лирована. Однако этот вопрос остается до сих пор открытым.
Ископаемых третичных позвоночных Австралии почти совсем нет. Тем не менее отсутствие там в настоящее время местных плацен- тарных млекопитающих, кроме рукокрылых и довольно современных гры- зунов, говорит о том, что на протяжении третичного периода этот материк был отделен от всех других материков водным пространством. Современ- ная фауна позвоночных Австралии «наслаивалась» сложным образом. В ее состав входят эндемичный подкласс (однопроходных), эндемичные надсемейства, семейства и подсемейства, эндемичные роды широко распро- страненных семейств, эндемичные группы видов и виды широкораспро- страненных родов и еще менее дифференцированные и недифференцирован- ные формы. Это позволяет предположить, что на протяжении третичного периода позвоночные в небольшом числе постоянно проникали в Австра- лию через острова. Насколько можно судить по имеющимся данным, все позвоночные, которые достигли Австралии в течение третичного периода, пришли из Азии, за исключением некоторых галаксид и, возможно, че- репах семейств Ме1о1ап11йае и Chelyidae, которые, видимо, проникли сюда через море из Южной Америки. Это свидетельствует о том, что Австралия по отношению к Азии с островами, расположенными между этими двумя
Прогнутое в свете зоогеографии
материками, в третичный период занимала такое же положение, как и в настоящее время.
Фауна позвоночных Новой Гвинеи (по крайней мере млекопитающих) по сравнению с австралийской, по-видимому, менее древняя (гл. 6), а фау- на островов, расположенных между Новой Гвинеей и материковой отмелью Азии, еще моложе (гл. 7), причем в состав последней, распростра- нение которой до сих пор показывает очевидные пути иммиграции, вхо- дит несколько поразительно дифференцированных форм. Это соответству- ет геологическому непостоянству этих островов в течение третичного периода и их более древним фаунам, которые были в этот период уничто- жены и замещены, и, таким образом, данные о раннетретичных перемеще- ниях через эти острова к Австралии исчезли.
Что касается сухопутных мостов и перемещающихся материков, то распространение позвоночных не подтверждает наличия таковых в тре- тичном периоде. «Ключом» к этому служит Южная Америка, так как она связана с большинством гипотез относительно сухопутных мостов или пере- мещающихся материков. Позвоночные, особенно млекопитающие, свиде- тельствуют о том, что с начала третичного периода (если было именно это время), когда возникло соединение с Северной Америкой, Южная Америка не имела никаких сухопутных соединений нп с одним другим материком. Изоляция Южной Америки в третичном периоде — очевидный факт. Палеогеографы, которые отвергают его, просто умаляют силу своих соб- ственных теорий. Какое положение занимал этот материк в самом начале третичного периода, не совсем ясно, но если даже Южная Америка и име- ла какое-либо соединение в то время, то, вероятнее всего, с Северной Аме- рикой, а не с Африкой. Если и существовало какое-либо соединение с Австралией, то оно, очевидно, относится к дотретичному времени, то есть оно было еще до того, как плацентарные животные проникли в Южную Америку.
Распространение позвоночных присовокупляет некоторые сведения к тому, что уже известно о климатах третичного периода. Ископаемые флоры показывают, что Северная Америка в третичном периоде была теп- лее, чем в настоящее время, но, очевидно, она не была полностью тропи- ческой (стр.495 — 496). Ископаемые данные млекопитающих позволяют сде- лать те же предположения. В третичном периоде, по-видимому, не было резкого различия между умеренной п тропической фаунами Северной Аме- рики, и когда в конце третичного периода произошел обмен между млеко- питающими Северной и Южной Америки, то никакой третьей «тропиче- ской фауны Северной Америки» не появилось (стр. 311, 312). Если в Цент- ральной Америке и существовала эндемичная фауна млекопитающих, то, очевидно, она была слишком небольшой и слишком малодеятельной, чтобы играть значительную роль в образовании современных материковых фаун. Я думаю, что орнитологи преувеличили как тропические условия Север- ной Америки в третичном периоде, так и роль «тропиков Северной Амери- ки» как центра эволюции и расселения.
Все вышесказанное о Северной Америке в целом, возможно, правиль- но в отношении умеренных областей. В третичном периоде их климат был теплее, чем в настоящее время, но они, вероятно, не были полностью тро- п пческими.
Представить климат тропиков на протяжении третичного периода труднее. Если умеренный климат был более мягким благодаря дополни- тельному теплу, получаемому от солнца, то влияние последнего должно было сказаться также и на тропической части, но в чем именно проявилось
500
Глава 10
его влияние, мне трудно сказать. Возможно, облачный покров или атмо- сферная циркуляция держали температуру в пределах нормы. Этот воп- рос заслуживает тщательного анализа в целом. Очень богатые современ- ные тропические флоры и фауны земного шара составляют путем сложных взаимосвязей единое целое и, по-видимому, продолжительное время не претерпевали никаких существенных изменений — это все, что можно сказать о них сеичас.
Если все известные и предполагаемые сведения о земном шаре тре- тичного периода можно было бы показать при помощи ускоренной кино- съемки, то на ней были бы и пропуски, и неясные места, но тем не менее синхронность всех процессов сохранилась бы.
Даже в начале третичного периода большинство материков можно было узнать по их очертаниям, и их расположение на земном шаре было примерно таким же, как и сейчас. Азия и Северная Америка были соеди- нены на севере. Северная и Южная Америка почти или полностью соеди- нялпсь. Между Азией и Австралией лежал архипелаг. В течение третич- ного периода материки меняли форму своей поверхности, но не свое поло- жение (или меняли его в очень незначительной степени), а соединения меж- ду ними изменялись только в деталях, которые в геологическом отноше- нии не играют значительной роли, хотя по своему влиянию на расселение животных в ряде случаев они являлись существенными. В первой трети третичного периода Африка и Евразия, вероятно, разъединялись и вновь соединялись. Мост Беринга неоднократно разрушался и возникал вновь! Северная и Южная Америка были разъединены (если до этого между ними существовало соединение) в начале третичного периода и вновь соеди- нены в конце его. Архипелаг между Азией и Австралией изменялся, но сплошного сухопутного моста образовано не было.
Даже в начале третичного периода климат был зональным и распо- ложение зон было примерно такое же, как и теперь; причем последнее вы- зывало заметную зональность растительности. Субтропические леса про- никали далеко в глубь умеренной зоны, но они были там более редкими, чем леса в настоящих тропиках. На крайнем севере, где климат был всег- да строго зональным, находились небольшие площади лиственных лесов. В течение третичного периода, по мере того как климат становился более прохладным, северные зоны растительности медленно перемещались к тро- пикам, а основные перемещения усложнялись второстепенными переме- щениями флор и их частей: перемещения с востока на запад, расширенис ареалов и их сокращение, эволюция и исчезновения.
В связи с изменениями суши, климата и растительности происходили и перемещения животных, бесконечно сложные в деталях, но образую- щие несколько основных особенностей. В пределах основной части Старо- го Света происходило последовательное расселение и перерасселение последовательно доминирующих групп, у которых проявлялась тенден- ция к распространению на север от тропиков (гл. 9). Независимо от вре- мени существования соединения между Азией и Северной Америкой между животными этих материков происходил обмен. Однако даже в ран- нетретичное время зональность климата ограничивала этот обмен, задер- живая строго тропические группы, и по мере того, как северный климат становился все более холодным, все меньшие и меньшие части фаун участ- вовали в этом обмене. Перемещения животных происходили в обоих на- правлениях, но всегда преимущественно из Азии к Америке, а не в обрат- ном направлении. В Новом Свете часть фауны Северной Америки достигла Южной Америки (через сушу?) в начале третичного периода. Позднее сл~-
Прошлое в свете зоогеогрофим
чайные животные пересекали водные пространства, а в конце третичного периода уже многие животные перемещалнсь по новому сухопутному мосту в обоих направлениях, хотя этот обмен закончился большим вторжением животных с основной части земного шара в Южную Америку. Через изменяющиеся водные пространства проникает случайный иммигрант в Австралию. Наземные животные постоянно или время от времени про- .никали на другие острова в числе, более или менее пропорциональном расстоянию от материка. Фактически картина покажет нам постоянное распространение животных с материков на острова, напоминающие брыз- ги от фонтана, большая часть которых падает поблизости, но по мере уда- ления от него их становится все меньше и меньше.
Г[родолжптельность третичного периода в 60 млн. лет — это время, достаточное для значительной эволюции, различных замещений и исчез- новений как частей единой географической картины. Распределение и перераспределение современных животных происходило на основной части земного шара, а в Южной Америке и Австралии радпировали большие самостоятельные фауны. Это единственные наиболее выдающиеся особен- ности обмена.
Мезозой и дом езозой
Мезозойская эра охватывает около (несколько больше или меньше) 120 млн. лет до третичного периода и делится на три периода: триасовый, к>рский и меловой (рис. 6).
Согласно Данбару (Dunbar, 1949), мезозойская эра началась с обще- го выхода из-под воды материков и широкого распространения засушли- вого климата (в триасе). В юре суша начинает опускаться и климат стано- вится влажнее. Что касается температур, они были более низкими в ранней rope и более высокими в поздней. В меловом периоде уровень опускания материков был очень низким и они интенсивно затоплялись мелковод- ными морями. Северная Америка была временно разделена на две части морем, которое простиралось к северу от Мексиканского залива. С конца мелового периода снова происходит поднятие суши и образование системы Скалистых гор и Анд, хотя в третичном периоде они обе претерпели даль- нейшие преобразования. Климат мелового периода был вначале более хо- лодным, а затем потеплел. В Гренландии и (или) Аляске появились фиговые деревья, древовидные папоротники, саговники и пальмы. Но эти расте- ния, произрастающие в некоторых районах умеренной зоны и сейчас, не обязательно являются свидетелями тропических условий.
Я не знаю. насколько реальны эти предположения относительно цик- лов, происходящих на всем земном шаре, в общем, возможно, они и ре- .альны. Тем не менее следует помнить, что геологические перевороты не являн>тся простым процессом и что на земном шаре никогда не может быть однообразного распределения ни температуры, ни осадков.
К растениям раннего мезозоя относятся хвойные, древовидные папо- ротники, саговниковые и другие древние формы растений. Временами, особенно в среднем мезозое, древние флоры были удивительно однообраз- ны и имели широкое распространение. Широколиственные деревья (покры- тосемянные) появились в раннемеловом периоде и вскоре стали повсемест- но доминирующими.
Мезозой оыл веком рептилий. Наиболее хорошо известными и наибо- лее заметными. а потому играющими значительную роль животными
Глава 10
были динозавры. В мезозое окончили свое существование несколько дрен- них групп рептилий и появились крокодилы, черепахи, ящерицы и змеи. Рептилии господствовали и в море, и в воздухе, и на суше. В течение мезо- зоя другие позвоночные — рыбы и амфибии — претерпели сложнув~ эво- люцию и замещение. Птицы появились в юре, а первые млекопитаю- щие — в триасе. Но большинство из этих мезозойских групп слишком несущественны или слишком мало известны, чтобы иметь серьезное зоо- географическое значение.
Современные позвоночные, за исключением, возможно, некоторых. пресноводных рыб, дают нам очень мало сведений о географии мезозоя. В большинстве случаев их распространение относится к третичному или более позднему времени. Поэтому зоогеографические представления о ме- зозое должны строиться главным образом на ископаемых данных. Но для правильного толкования ископаемые данные следует сравнивать с дан- ными о современном распространении животных, что даже в этом случае дело не легкое.
Экология древних животных и растений, возможно, отличалась от экологии современных животных. Эта вероятность увеличивается отда- ленностью времен. В сравнительно недавние времена, в третпчном перио- де, значение отдельных видов может вызывать сомнение, но значение целых фаун и флор должно быть вполне определенным. Например, нахо'кде- ние в течение третичного периода на дальнем севере не просто единичных растений, а субтропических растительных сообществ — вполне приемлемое доказательство теплого климата (Barghoorn, 1953). Но для более древних времен подобные доказательства становятся гораздо более сомнительными. Обширные группы растений и животных, возможно, изменили свою вынос- ливость. Например, солевыносливость пресноводных рыб в мезозое. оче- видно, значительно отличалась от солевыносливости современных рыб (гл. 2). Отношение рептилий к климату в мезозое, возможно, также было совершенно другим по сравнению с настоящим временем, коренным оора- зом отличающимся, а не просто иным в силу видовой адаптации. На это предположение наводит нас гаттерия. Она обитает в умеренно-холодном климате Новой Зеландии, не имея каких-либо очевидных особых приспо- соблений, чтобы переносить холод или сохранять тепло. Она не стала яйцеживородящей, как современные ящерицы, гекконы и сцпнки, обитаю- щие в Новой Зеландии, и даже не ведет дневного образа жизни и активна только ночью, когда наступает прохлада, крайне ощутимая даже для оде- того человека (Bogert, 1953, р. 169). Возможно, гаттерия и ее мезозойские современники были менее чувствительны к холоду, чем современные реп- тилии. Еще одним примером иного отношения к климату в прошлом является американский аллигатор, который обладает «удивительной» холо- достойкостью (Colbert et al., 1946) и мог бы обитать далеко на севере от своих современных границ, если бы температура являлась единственным ограничивающим фактором. Но в настоящее время имеется еще один фак- тор, которого не было в мезозое, — наличие большого числа теплокровных животных. При отсутствии сильных теплокровных конкурентов в холод- ных северных районах, возможно, существовали разнообразные репти- лии, и целые фауны их в холодное время, вероятно, просто временно пре- кращали свою активность. Но расцвет теплокровных животных, активных в холодную погоду, сделал существование рептилий невоз- можным.
Ввиду чрезвычайной давности времени (продолжительность мезозоя в два раза больше третичного периода), сложности морфологии земного
Прошлое в cseme зоогеографиц
шара в то время, и как всегда, сложности эволюции и расселения разных животных, небольшого количества достоверных сведений и трудности интерпретации этих данных, я не буду останавливаться на зоогеографии мезозоя очень детально, а удовлетворюсь ответом только на один вопрос (правда, попытаюсь найти ответ на этот вопрос несколькими различными путями): была ли география земного шара — основное размещение мате- риков и климатов в мезозое и до него — по существу такои же, как и в настоящее время, или она была совершенно иной?
Динозавры
0 географии земного шара в мезозое можно судить, скорее всего, по распространению динозавров, чем каких-либо других животных, но не следует ожидать, что они помогут воссоздать довольно полную картину этого периода. Классификация и описание динозавров приводятся в рабо- те Ромера (Romer, 1945) и в более популярной, но вполне достоверной работе Кольберта ( "olbert, 1945), который любезно согласился прочитать данный раздел моей книги.
Динозавры были многочисленны и разнообразны. Их относят к двум отрядам, не более родственным друг другу, чем крокодилам, птерозаврам и птицам (Romer), а внутри каждого отряда имеется много различных эво- люционных линий. Наиболее хорошо известные динозавры имели гигант- ские размеры, но существовали и небольшие динозавры, а некоторые достигали размеров кролика и даже меньших размеров. Самые древние динозавры были плотоядными животными. Некоторые из них такими и оста- лись, но другие, включая самых крупных, стали травоядными. Вероятнее всего. динозавры были холоднокровными животными, но на протяжении большей части своей истории они не имели теплокровных конкурентов илц по крайней мере серьезных конкурентов. Возможно, что они проника- ли в районы с прохладным, но не очень холодным климатом. Большинство динозавров вели наземный образ жизни, но некоторые перешли к земновод- ному. если даже не полностью водному образу жизни. Расселялись они, по-видимому, главным ооразом по суше, но, как и другие животные суши, вероятно, обладали какими-то возможностями пересекать узкие водные преграды.
Динозавры подразделяются на следующие основные группы: Отряд Saurischia (ящеротазовые).
Подотряд Theropoda (звероногие).
Подотряд Sauropoda (ящероногие).
Отряд Ornithischia (птпцетазовые).
Четыре подотряда (см. в списке рептилий, гл. 4).
Звероногие — наземные и плотоядные динозавры, — которые появи- лись первыми и вскоре стали широко распространенными животными, астана.ц1сь таковыми до конца истории динозавров. Они достигли всех основных частей земного шара, включая Южную Америку, Австралию и. очевидно, Мадагаскар.
Ящероногие — огромные земноводные травоядные динозавры, оче- видно. приспособившиеся к питанию только мягкими растительными кор- мами. В среднем мезозое были, вероятно, космополитами, так как их остатки известны из Северной Америки, Европы, Африки, Мадагаскара, Южной Америки и Австралии. Позднее, в меловом периоде, в северных властях земного шара они встречались, видимо, редко, но были широко
Ггава 10
распространены на юге — в Африке, на Мадагаскаре, в Индии и Южной Америке.
Птицетазовые также были огромными травоядными динозаврами. Большинство из них вело наземный образ жизни (хотя некоторые стали земноводными) и приспособилось к питанию грубыми растительными кор- мами. Особенно многочисленны они были в меловом периоде в северных частях земного шара — в Евразии и Северной Америке,— хотя некоторые из них известны и в других районах в более раннее время. Одна ориги- нальная группа этих динозавров Ceratopsia, или рогатые динозавры, вероятно, возникла в Азии, но позднее эволюционировала главным образом в западной части (",еверной Америки (возможно, также и в Азии), будучи изолированной от восточной части Северной Америки морским барьсро:ч ((;olbert, 1948).
Короче говоря, плотоядные звероногие (Theropoda) были широко рас- пространены на протяжении всей истории существования динозавров, в то время как травоядные птицетазовые и ящероногие (Ornithischia, Sau- ropoda), независимо от их ранней истории, в меловом периоде имели зональ- ное распространение: наземные птицетазовые, питающиеся грубым расти- тельным кормом, были распространены главным образом на севере, а зем- новодные ящероногие, питающиеся мягким растительным кормом,— пре- имущественно далее к югу. Часто такое размещение толковалось по- разному.
Лалл (Lull, "l910) дал наилучший анализ распространения динозав- ров, но он сделал его еще до того, как стало известно достаточное количест- во фактов. Объясняя те факты, которые были ему известны, он полагал, что в позднем мезозое было два полностью отделенных друг от друга основных материка — северный и южньш — и что птицетазовые были всегда при- урочены к северному материку (Северная Америка плюс большая часть Евразии), ящероногие встречались на южном материке (Индия, Мадагас- кар плюс Африка и Южная Америка), а звероногие, расселившиеся в бо- лее ранний период времени, встречались на обоих материках. Его пред- положение было опровергнуто более поздними данными, установившими, что птицетазовые и ящероногие не были полностью географически изо- лированы.
Гипотеза Хюена (Ниепе, 1923, различные статьи совместно с Матле- ем) не допускала полного разделения северных и южных фаун в меловом периоде. Наоборот, она предполагала наличие связи между некоторыми меловыми позвоночными Северной и Южной Америки, а также Европы и Индии. Но Хюен предполагал наличие длинных сухопутных мости» через южные океаны от Мадагаскара через Индию, Австралию и Антаркти- ческий континент до Южной Америки и считал, что ящероногие мелового периода расселились этим путем. Самым лучшим критическим ответом н» эту точку зрения является известное распространение рассматриваемых динозавров. Хюен выделяет три рода: Antarctosaurus, известньш из Ин- дии и Южной Америки, Laplatasaurus, известный из Восточной Африки, Мадагаскара, Индии и Южной Америки, и Titanosaurus — из Англии, Центральной Европы, Мадагаскара, Индии и Южной Америки. Если»то размещение соответствует действительности (значительная часть иско- паемого материала, на котором оно основывается, фрагментарна), то оно скорее свидетельствует о неполноте географических данных, а не о рас- селении каким-либо определенным путем. Нет свидетельств, что именно эти динозавры когда-то встречались в Австралии (хотя возможно, что когда-то они там были). ( леды пребывания каких-либо динозавров и и»
111«««.<ое в суете <оог«<грс< /<««
Ант11р«т11чес«ом континенте также отсутствуют, и ни один из них не изве- стен также в Новой Зеландии.
Мэтью (Alatthew, 1915, р. 275 — 279) считал. что распространение ди- нозавров соответствовало расселению с севера (1) и по материкам (2), име- вщио1 примерно такое же расположение, как и в настоящее время. 11ервый пу н «т мне кажется сомнительным. Динозавры, TBH же как и друг11е холод- но«1зс1вные позвоночные, возможно, эволюционировали скорее в теплых, чем v. умеренно-холодных районах, и скорее всего расселялись через cellep. а не с севера. Но второй пункт сомнений не вызывает. Распростране- ние динозавров и то немногое, что о них известно, можно довольно хоро- шо объяснить их расселением по материкам, расположение климатиче- < «их:,oн которых напоминало современные. по крайней мере в меловом периоде. В это время большинство птицетазовых встречалось в умеренной зоне северного полушария, а значительная часть ящероногих отступили в тропики.
Нахождение динозавров на Мадагаскаре стоит под вопросом. Хотя там оыли зарегистрированы остатки предположительно наземных звероно- гих, мне не известна an их природа, ни их значение. На Мадагаскаре встре- чалос1 два рода ящероног11х (см. выше). Это были в той или иной степени водные животные. Некоторые их остатки были обнаружены в отложениях м1-..1и водных морей. и это позволяет предполагать, что они иногда обита- .,111 в мелководных эстуариях, бродили по дну и питались водными расте- н11ямн, поднимая голову на длинной шее над водой для дыхания. Ни одно из ~ о» ременных;животных, за исключением отдаленно напоминающего его гиппопотама. не ведет подобного образа жизни. Гиппопотамы не име- ют специальных приспособлений к плаванию, но могут хорошо плавать. что. вероятно, относится и к ящероногим. В плейстоцене гиппопотам достиг Мадагаскара из Африки, преодолев, видимо, Мозамбикский пролив. I'еологические данные свидетельствуют о существовании этого пролива с триаса. и я не стану отвергать это ради создания сухопутного соедине- ния. которое было бы необходимо для объяснения распространения яще- рс>ногпх, если оы они пришли из Африк11. Если же они проникли из Ин- ди11. то они могли расселяться через цепь архипелагов, защищенных мелко- водными морями (которые предполагаются целым рядом современных отме- лей). Мне кажется, 1то нахождение ящероногих на Мадагаскаре не дока- зывает полного соединения его с сушей. Роды ящероногих, встречавшиеся на Л!адагаскаре, были очень широко распространены по всему земному шару (см. выше), чего нельзя ожидать от крупных, строго пресноводных животных даже в том случае, если и существовали сухопутные соединения.
1] регноводные животные перемещаются по суше очень медленно, если толь- «о им не присуща какая-то особая способность к расселению. Хюен счи- тал, что широкое распространение ящероногих динозавров являлось дока- за11л ьством сухопутных соединений. Я полагаю, что широкое распро- странение этих динозавров почти доказывает, что они в какой-то мере могли pacceляться через море.
Енсе, что известно о распространении динозавров, является подтверж- дением одного основного положения: во время их существования земной шар не был разделен на два огромных массива суши, полностью изоли- рованных друг от друга. Это немаловажный факт. так как некоторые ученые лродолжа1от утверждать, что суша в позднем мезозое была разделена имен- но таким ооразом. Более того. распространение динозавров, насколько известно, вполне соответствует расселению по всей системе суши, которая незначительно отличалась от настоящей и имела, по крайней мере в мело-
о06
Глава 10
вом периоде, климатические зоны. расположенные так же, как и в н»стоя- щее время. Если распространение динозавров не свидетельствует о том, что поверхность земного шара в то время была сходна с современной, то по крайней мере оно не доказывает и обратного.
Все динозавры исчезли, очевидно внезапно и повсюду, в конце мело- вого периода. Причина их исчезновения неизвестна и в данном с,<у <»<. не столь существенна. Исчезнувших динозавров заместили мле ti<) II tl- тающие.
Мезозойские сухопутные мосты
В течение третичного периода происходило непрерывное или пери<>- дпческое расселение наземных позвоночных (хотя и не всегда по сп.1о<и- ноп суше) между Евразией и Африкой, Азпеп и (:евернои Америк<tit, Северной и Южной Америкой и из Азии к Австралии. Оно шло, видимо, именно этими, а не какими-либо другими межконтинентальными путями. Предположительно динозавры и другие наземные животные в течение мезозоя пользовались также только этими путями, но в связи с. тем, t < tt и» этот счет нет полных данных, следует принимать во внимание и дру- гие пути.
Сухопутное соединение через Северную Атлантику, посредством кото- рого осуществлялось расселение между Европой и северо-восточной частью Северной Америки, очень часто принималось, в основном ботаника- ми, без достаточных доказательств. Я не располагаю какими-либо убеди- тельными доказательствами an за, ни против существования сухопутного соединения в течение мезозоя или раннетретичного периода. но данные о распространении млекопитающих говорят о том, по позднее нижнего эоцена этого соединения не было (стр. 310).
В третичном периоде через Мадагаскар в связи с отсутствием оошир- ного сухопутного моста (достоверных данных о его существовании в ме:«>- зое также не имеется) путей расселения не было (стр. ~96). Но Мадагаскар существовал в этот период и имел фауну, включая динозавров, пз готорой не сохранилось ни одно животное. Современная фауна позвоночных по происхождению, вероятно, полностью постмезозойская. Если какие-<« ее части и являются древними, то они не могут быть теперь датированы. <,ущественной особенностью мадагаскарской фауны является отсутт t t<tt<.: строго пресноводных рыб, позволяющее предположить, по Мадаг»гк»р очень продолжительное время был отрезан от всех материков. Для р»спр«- странения динозавров, достигших Мадагаскара в мезозое, возможп«. Ilt' требовалось наличия сплошной суши. Геологические свидетельства «<у- ществованпп Мозамбикского пролива с начала трпаса не подтвер'«д»ют мезозо1хского соединения Мадагаскара с Африкой.
~!ежду Африкой и Южной Америкой, по крайней мере с начал» <р<- <ичного периода, также не было непосредственного соединеяпя. Но было .ш оно в мезозое?
Иэ всех современных животных пресноводные рыбы играют наи<«>:~ <— шую роль в характеристике географии мезозоя. Пресноводная ихти<>ф»у- на Южною1 Америки, рассматриваемая в гл. 2, образовалась от песк«1<- кпх предков, необычайно смешанных по своему эволюционному yp<t«ttt<> и экологическим требованиям, которые, возможно, проникли херез узкий сухопутный мост (но не через широкое соединение) из Африки в мело«<>ч пцэиоде, но которые также могли проникнуть через имеющие теплый li;llt-
Проггг.гое в свете зоогеогрофин
507
мат краевые части северных материков. Кроме рыб Южной Америки, и другие пресноводные животные, включая лягушек пцп и черепах пеломе- дуз, а также некоторых пресноводных моллюсков (Pilsbry, 1911; Pilsbry, Вег1иаегФ, 1927, р. 598 — 601), имеют африканские связи. Ископаемые боль- llIHHcTBa этих животных, за исключением пеломедуз, за пределами их современных ареалов обнаружены не были. Эти животные не дают ответа на вопрос о путях расселения в мезозое, но они позволяют предположить, откуда можно его получить. Если такое большое число пресноводных животных пересекло сухопутное соединение между Африкой и Южной Аме- рикой, то еще большее число наземных животных должны были сделать тг> же самое. (Сплошная протяженность пресных вод невозможна без сплоги- ной протяженности суши, и наземные животные расселяются через сухо- путные соединения быстрее, чем пресноводные. Если это положение нуж- дается в доказательствах, то его могут подтвердить недавние перемещения животных через Центральную Америку.) Распределение наземных групп животных по земному шару могло по сравнению с пресноводными груп- пами подвергаться более быстрым изменениям и не оставить следов древ- них связей, которые сохранились у пресноводных групп, но ископаемые должны показать перемещения наземных животных между Африкой и Южной Америкой в местах перехода из одной зоны в другую. Если временем расселения современных африкано-южноамериканских пресно- водных животных являлся меловой период (как это, видимо, и было), то у наземных животных этого периода должны были особенно четко прая»- ляться африкана-южноамериканские связи. Но в действительности этого не происходило. Рептилии мелового периода Южной Америки, например, олизкородственны по крайней мере североамериканским в такой же сте- пени, как и африканским, и, по-видимому, нет причин, которые в то время могли бы помешать рептилиям, достигшим Южной Америки, проникнуть через Северную Америку (Colbert, Romer; см. работу — Mayer et al., 1952). Насколько можно судить, в настоящее время достоверные свидетельства оо африкано-южноамериканском соединении в меловом периоде отсут- ствуют, но этот вопрос еще не решен окончательно. Существовало ли это соединение еще раньше в мезозое, будет рассмотрено ниже, в разделе пере- мещения материков.
Что касается сухопутного соединения ме'кду Южной Америкой гг Австралией через антарктический континент или Новую Зеландию (см. фронтиспис), то в третичном периоде такого соединения не было (стр. 499), и данные по позвоночным, видимо, не подтверждают его ни в один из дру- гих периодов (Simpson, 1940). Ни сухопутные, ни строго пресноводны<; позвоночные, видимо, не пересекалц антарктическ1гй мост. Пресноводные рыбы Южной Америки и Австралии совершенно разные, за исключением некоторых (галаксиды и т. д.), которые, вероятно, расселились через море. Лягушки семейств Leptodactylidae и Hylidae, которые в настоящее »ремя встречаются главным образом в Южной Америке и Австралии, но <псутствуют в Новой Зеландии, очевидно, расселились через основную часть земного шара (гл. 3). Черепахи семейст» Meiolaniidae и Chelyidae, если и достигли Австралии из Южной Америки, то они попали туда только через море. Бескилевые птицы Южной Америки, Австралии и Новой Зеландии (а так>г'е Африки и Мадагаскара), вероятно, являются конвергентными, а не родственными группами (см. список семейств птиц). Когда-то сумча- тые считались лучшим доказательством существования антарктического моста. Но, как теперь установлено, они достигли Ю;г,тгг>й Америки ц Ав- стралг1ц самостоятельно, и развитие специализации у нцх происходило
.Глпви И
параллельно (стр. 292). Однаго распространение некоторых растений и на- секомых, включая отдельных жуков семейства (;arabidae, свидетельствует о тех фактах, которые, видимо, являются следствием непосредственных связей между южной частью Южной Америки и Южной Австралией, а за- частую и Новой Зеландией. На вопрос, почему же позвоночные могут спп- детельствовать об одном, а растения и насекомые о другом, можно дал. разные ответы.
Во-первых, растения и насекомые, возможно, расселились еще до поз- воночных путем (не обязательно через сухопутный мост), который ко вре- мени распространения современных позвоночных перестал быть доступ- ным. Что-то наподобие этого могло произойти даже в том случае, если никакого моста никогда и не было. Насекомые могли расселяться через антарктические водные пространства, когда климат был относи.ельно мяг- ким, но со временем он мог ухудшиться и препятствовать расселению со- временных наземных птиц, распространение которых произошло недавно и не подтверждает обмена между Южной Америкой и Австралией (гл. 5). Но объяснения, основанные на разнице во времени, в других случаях, по-видимому, не подходят. Отдельные растения (например, южный бук, ."1othojagus) не столь уж древнего происхождения, и нет основании счи тать. что в Южнои Америке они более древние, чем, скажем, некоторые лягушки и пресноводные рыбы.
Во-вторых, распространение различных организмов через Антаркти- ку, возможно, связано с неодинаковыми способностями их к расселени1о. Некоторые растения и насекомые пересекают соленые воды легче, чем наземные позвоночные, что и подтверждается нахождением их на отда- ленных океанических островах. Следовательно, они смогли пересечь водные пространства между Южной Америкой и Австралией, чего не смогли сделать позвоночные, а некоторые из них могли быть перенесены ветром (гл. 1).
В-третьих, возможно, различные организмы расселялись одними и теми же путями, но после расселения вели себя по-разному. Некоторые растения и насекомые существовали длительное время на небольших тер- риториях с холодным климатом; некоторые из них, очевидно, продолжи- тельное время находились на Огненной Земле и южной оконечности Юж- ной Америки. У позвоночных этого не отмечалось. Среди наземных или пресноеодных позеоночных, приуроченных к южной оконечности Южной Америки, дреених реликтов нет. Самые древние существующие млеко- питающие Южной Америки — сумчатые и однопроходные — теперь не доходят даже до южной оконечности этого материка (гл. 6), хотя начиная с раннетретичного периода они находились в южной части Южной Амери- ки. Такое же положение сложилось и в Австралии. Определенные растения и насекомые, очевидно, длительное время были изолированы на Тасма- нии и в южных краевых частях Австралии, но древние реликтовые позво- ночные, приуроченные к этим районам, отсутствуют. Сумчатый волк и с~ мчатый дьявол имеют слишком недавнее происхождение, чтобы можно было на них ссылаться..Видимо, в таких холодных и отдаленных местах позвоночные не могут являться свидетельством древней истории, как некоторые растения и беспозвоночные. Что касается сухопутного соедине- ния между Южной Америкой и Австралией, то млекопитающие указывают лишь на то, что его не было в третичном периоде. Австралия в то время не была соединена сушей ни с одним материком (стр. 500). Но возможно, что до третичного периода между южной частью Южной Америки и An стралией такое соединение существовало и что некоторые позвоночные,
II pozzz.ã îå в свеггг е зоогеогр аф и и
так же как растения и беспозвоночные, могли выжить на остатках этого соединения.
Я не знаю, имелось ли сухопутное соединение через Антарктику или нг т, но могу сказать, насколько это было возможным. Очевидно, в той части земного шара расселение животных было таким же сложным, как и повсюду. Некоторые «антарктические» растения и беспозвоночные живот- ные, как и большинство южноамериканских и австралийских позвоноч- ных, очевидно, расселились через основную часть земного шара и затем вымирали в своих прерывистых южных ареалах. Некоторые другие растения и беспозвоночные, очевидно, расселились через Антарктику, ког- да климат по сравнению с настоящим временем был более мягким, нгг я считаю, что это расселение происходичо через водные пространства, а не через сплошную сушу. Сомнительно, что какие-либо строго наземные или пресноводные позвоночные пересекали эти пространства, но, вероят- но, некоторые солевыносливые рыбы и более или менее водные черепахи преодолевали их. Я полагаю, что из современных так называемых «антарктических» растений и насекомых некоторые расселялись одним путем, а другие — другим, и могу грубо допустить, кто из них пользовал- ся тем или иным путем. Насекомые, которые встречаются в Южной Аф- рике, а также в южной части Южной Америки и Австралии, но не в Но- вой Зеландии, расселились через основную часть земного шара и выми- рали по пути своего современного распространения. Насегомые, которые теперь встречаются в Новой Зеландии, а также в южной части Южной Америки и Австралии, но не в Южной Африке, как я полагаю, рассели- лнсь непосредственно через южные водные пространства, но ни одни из строго наземных или пресноводных позвоночных не распространены таким образом и ни одна из южноамериканских и австралийских групп не представлена в Новой Зеландии. Насекомые, которые не подходят нп под один из этих типов распространения, вызывают сомнение.
Окончательный ответ на вопрос об антарктических сухопутных мостах, очевидно, будет получен после исследований антарктическorii материка. В настоящее время там уже обнаружены ископаемые растения и ~голь. Дальнейшее нахождение ископаемых в конце концов, вероятно, покажет, имела ли Антарктида флору и фауну, которые проникли на этот материк через водные пространства, или они могли появиться там через сухопутные соединения.
Перемещающиеся материки
Я затрудняюсь ответить на вопрос, всегда ли материки оставались на своих местах или смещались на значительное расстояние по отношенин~ друг к другу и оси земного шара, но все;ке постараюсь внести в него какую-то ясность. Интересуясь этим предметом, я в течение многих лет при- нимал участие в двух симпозиумах по этому вопросу: в одном официаль- ном совещании Американской ассоциации содействия развитию науки в Нью-Йорке (28 и 29 декабря 1949 г.) (Mayer et а1., 1952) и в другом— неофициальном в Гарварде (13 декабря 1951 г.), организованном д-ром Льюэллином Прайсом. Таким образом, то, о чем я буду писать, отнюдь не является поспешным выводом.
Теория перемещения материков, заслуживающая особого вниъхания со стороны зоогеографов, принадлежит Вегенеру или Тэйлору и Вегенеру. В несколько измененном виде она преподносится де Туа и другими. В ра-
Глава l0
510
ооте Майра (Л!аут et al., 1952) приводится список литературы по этому воп- росу, а несколько дополнительных ведущих источников упоминаются в ли- тературе к данной главе. Мы не будем касаться истории этой теории.Тео- рия, как она представлена сейчас, заключается в том, что когда-то матери- ки образовывали единую земную массу, которая раскололась на части. и ее глыбы — современные материки,— перемещаясь, заняли современ- ное положение (рис. 77). Все это происходило незадолго до мезозоя и на
ТРА- ИЯ
P,п с. 77. Перемещение материков, по Бегеперу (упро- щенное и перенесенное на полушария, ортографпческая проекция), происходящее вокруг Африки у экватора.
всем протяжении этого периода. Иногда считают, что перемещение мате- риков продолжалось и в третичном периоде, но мне кажется, что такое предположение ничем не обосновано.
Обсуждение вопроса перемещения материков иногда заканчивают фразой: «Эта теория создает больше проблем, чем разрешает их». Воз- можно, это действительно так, но главное заключается не в этом. Вопрос состоит в том, происходило ли в действительности перемещение мате- риков?
Ничто в зоогеографии не вызвало столько споров или обсуждений, как идея о перемещении материков. Я пытался выяснить, нельзя лп по сов- ременному распространению животных найти какие-нибудь действитель- ные следы перемещения материков, но обнаружить их не смог. Распрост- ранение животных в настоящее время в своей основе представляет резуль- тат перемещения самих животных, а не передвижения материков. Раз- личные виды животных распространялись и отступали разными путями. образуя, таким образом, бесконечное разнообразие перемещений, кото- рые, видимо, определяются эволюционным возрастом животных, их воз- можностями расселения, реакцией на климат и взаимоотношениями друг с другом и т. д., и характер распространения животных в целом соответ- ствует современному, а не вегенеровскому положению материков, а так- же климатам и преградам земного шара, существующим в настоящее вре- мя. Особые случаи, подчеркиваемые сторонниками теории Вегенера, мне
.1Трош.лое в свете .юогеогра /шн
ка;кутся просто исключениями. Например, особые случаи миграций птиц л оглнсуются с идеей перемещения материков, но миграции в целом не мо- гут быть ооъяснены перемещением последних, а перемещение не может достаточно убедительно объяснить даже исключительные случаи (гл. 5).
Если даже перемещение материков и происходило, то, видимо, оно про- исходило очень давно, и современное распространение животных и рас- тений. вероятно, не может отразить его в связи с тем, что последние име- ют слишком недавнее происхождение. Тем не менее сторонники Вегенера гтаран~тся это сделать. Их метод заключается в попытке связать распро- странение современных растений и животных с гипотетическими очерта- ниями суши, по Вегенеру, и, когда это соответствие найдено, объявить его как свидетельство существования древних суш. Например, если груп- па 'tillвотных встречается теперь только в Африке и Ьразилии, то они утверждают, что она ведет свое начало от африкана-бразильского материка и является свидетельством существования этого материка. В основе это- го метода лежит допущение, как правило, не приводимое сторонниками Вегенера к точной формулировке: что распространение животных более постоянно, чем суши, что перемещение животных происходит в меньшей степени, чем перемещение континентов. Примеры такого рода зоогеогра- фических построений даны в книге Джиннеля (Jeannel, 19~~2) о происхо- ждении наземных фаун. Из этого метода вытекают свои выводы. Если жи1.»тные не перемещались, то, следовательно, перемещались материки. Но животные перемещались. Они распространялись и отступали иногда на оольшое расстояние за сравнительно короткий промежуток времени, и их современное распространение определяется еще многими факторами. помимо очертаний древних суш.
Если какие-либо из современных животных и должны были отражать влияние перемещения материков, так это пресноводные рыбы, но они его не отражают (стр. 88). Природа фауны рыб Южной Америки (свидетельст- ву1»щая оо образовании узкого сухопутного соединения из Африки, но не оо общем соединении этих материков) и отсутствие строго пресноводных рыб на Мадагаскаре являются свидетельством против перемещений мате- риков, если рыбы вообще могут служить свидетельством.
Настоящее доказательство справедливости или несостоятельности теории Вегенера о перемещении материков исходит из геологии и палеон- 1»логии, особенно из распространения растений и животных в прошлом, о котором можно судить по их ископаемым. Поэтому я делаю попытку оценить некоторые геологические и палеонтологические доказательства с точки зрения зоогеографа. Я буду неоднократно обращаться к материку Южная Америка, который по своим очертаниям, положению и другим признакам наиболее интересен в этом отношении.
Одним из первых аргументов в пользу перемещения материков были ttx очертания, особенно соответствие конфигурации Африки и Южной Аме- рики. которые выглядят так, словно когда-то они составляли единый мас- сив суши. Действительно, у них имеется три зигзагообразные линии. которые удивительно соответствуют друг другу: западный край Африки. восточный край Америки и Срединно-Атлантический хребет (см. фронтис- пис). Мне кажется, что совпадение этих трех линий, очевидно, свидетель- ствует о каких-то имеющих место событиях, но не обязательно подразуме- вать в данном случае перемещение материков по Вегенеру. Задолго до Тэйлора и Вегенера Джордж Дарвин (сын Чарльза Дарвина) и Фишер выдвинули предположение, что Луна оторвалась от Земли со стороны Тихого океана, забрав с собой значительную часть земной коры. Фишер
ý12
Глава 10
предполагал, что тогда остальная кора разломилась на части. которые в свою очередь дробились, по мере того как Земля изменяла свои очер- тания, 1I стали современными материками. Это тоже теория перемещения материков, но не по Вегенеру. Это произошло, ес.~и это действительно име- ло место, на очень раннем этапе истории Земли, задолго до того, как на не~1 появилась жизнь, и даже до того, как появилась вода в жидьоч состоянии. Вероятно, существуюти другие вполне приемлемые oRÿñíåíèí соответствия конфигурации материков. Существует и объяснение такого совпадения простой случайностью.
Южная Африка и южная часть Южной Америки имеют не толым сходное очертание береговых линий, но они сходны и стратиграфи 1ески так, будто у них была общая история до мезозоя и и течение всего этого периода. Степень и значение этого сходства оспариваются (например, Caster, РииЬа1', см. работу — Мауг et al., 1952). Возможно, если оое террито- рии всегда находились (как и в настоящее время) в одной и той же s;ëèìa- тической зоне, это объяснимо сходством их климата.
(:тратиграфическое сходство проявляется также в нахождении ана- логичных морских ископаемых фаун беспозвоночных в соответствующих отложениях мелководных морей Южной Африки и Южной Америк~~. Значение этого сходства также вызвало споры. Эти фауны обитали глан- ным образом в мелких прибрежных морях, но личинки этик животных, возможно, расселились через глубоководные пространства: так как име- ются случаи пересечения современными мелководными позвоночными до- вольно широких водных пространств в Тихом океане (Dunbar. см. pavoò~ Мауг et al., 1952). И в настоящее время личинки животных могут перено- ситься Южно-Пассатным течением Атлантического океана из Африки к Южной Америке (гл. 1), и ареал расселения таких животных, завися- щий от продолжительности личиночной стадии, возможно, в прошлом оыл более обширным. Сходство мелководных фаун беспозвоночных Южной Африки и Южной Америки в прошлом, по-видимому, дает больше осноганий предполагать непрерывность водных пространств и сходство климатов (возможно, зональность климата), чем близкое расположение этих материков.
Пожалуй, самым сильным геологическим аргументом в пользу пере- мещения материков является оледенение, которое происходило там, где сейчас находится южное полушарие и часть тропиков, примерно в начале перми, периоде, предшествующем мезозою. Раннепермскпе ледниковые отложения (в прошлом считались пермско-каменноугольные) встречаются в Южной Африке к северу по направлению к тропикам (свидетельства не одного, а нескольких ледников с отдельными центрами движения). на южном Мадагаскаре, в южной части Южной Америки, в основном в пределах тропиков, в Южной Австралии и (к северу от экватора) в Индии. В других местах, по-видимому, ледников в это время было сравнительно мало. Вегенер считает, что территории, покрытые ледником, образовывали единый материк Гондвану, который в то время находился у Южного полюса или поблизости от него. Однако возможны и другие объяснения. Еолман ((:o- leman, 1926) после сравнения недавних и древних ледниковых эпох не усмотрел необходимости привлекать теорию перемещения материков для объяснения существования древних ледниковых эпох. Однако он не установил истинных причин оледенения и предположил нечто неопре- деленное — «сочетание» астрономических, геологических и атмосферных условий. Приемлемо ли в действительности это объяснение, до сих пор является вопросом. Ответ на него можно получить, если более детально
1/ро«<.<о< и свете зоогеографии
ознакомиться с периодом плейстоценового оледенения. Есть некоторые основания полагать, что климат, который всегда был в какой-то степени зональным, в течение плейстоцена, возможно, был строго зональным (Bar- lioorn, см. работу — Shapley et al., 1953, р. 248). Если это так, то уменьше- ние солнечной радиации иногда могло вызывать оощее похолодание кли- мата на всем земном шаре. Даже в плейстоцене ледяные щиты п Европе и Северной Америке находились в 2000 или 2,эОО миль от Северного полюса, а 1ожный край ледника в восточной части Северной Америки находился оли'ке r;:ýêìàòîðó, чем к полюсу. Если похолодание ие было строг«зо- нальным, то щиты могли образоваться в любой климатической зоне тач. где уровень суши был достаточно высоким и выпадение осадков было чост»- 1'ОЧН О ООИЛЬНЫМ.
( основными пермскими ледниковыми покровами связана определен- ная флора Glossopteris и родственных ей растений. Если у этих растений оыла ограниченная возможность расселения, то пх встречаемость была оы веским доказательством того, что территории, покрытые ледником. сое- динялись. Однако семена этих растений или их споры не были достоверно определены. Двукрылые споры, подходящие для переноса ветром, в неко- торых местах связывают с Glossopteris, и если они действительно являются спорами растений, то естественно, что последние могли расселятьсяна огромные расстояния.
Если Африка и Южная Америка были непосредственно соединсны в мезозое, то это должно подтверждаться ископаемыми позвоночными. но последние недостаточно хорошо известны, а те, что известны, трудны для толкования. В меловом периоде позвоночные Южной Америки в значи- тельной степени были сходны с позвоночными Северной Америки и Афри- ки (стр. 507). Данные юры слишком малочисленны, для того чтооы иметь существенное значение. Однако из более раннего периода, триаса, позво- ночные известны несколько лучше. Кольберт и Ромер (см. работу Мауг et al., 1952) дают интересный анализ известных сведений о земноводных и рептилиях триаса Африки и Южной Америки. Они установили, во-пер- вых, что в течение триаса в целом большинство семейств амфибий и реп- тилий Африки и Южной Америки встречались также и в других частях земного шара и, во-вторых, что фаунистические связи между Африкой п Южной Америкой были несколько теснее (Colbert,) а в среднем триасе, очевидно, значительно теснее (Romer), чем связи между Южной и Север- ной Америкой. Однако последний вывод основывается на скудных данных. Наземные позвоночные среднего триаса Северной Америки в основном известны по опгпечаткам ископаемых в единственном ярусе.
Как показывают в настоящее время факты, гипотеза о достижении среднетриасовыми позвоночными Южной Америки из Африки через непосредственное соединение вполне приемлема, но для ее подтверждения нужны дополнительные факты. Эти факты должны быть подвергнуты ана- лизу на основе данных о всем земном шаре. В триасе на протяжении какого-то отрезка времени наземные позвоночные, очевидно, имели доступ на все материки. Брахиоподы имели космополитическое распространение, триасовые дицинодонты и примитивные динозавры известны из Африки. Европы, Азии и Северной и Южной Америки, проколофониды и псевдо- зухии — из Африки, Европы и Северной и Южной Америки, ринхозау- рия — из Африки, Европы, Азии и Южной Америки, и только цинодонты в триасе были приурочены (насколько это известно) к Африке и Южной Америке (Colbert, см. работу — Мауг et al., 1952, р. 240). Существовала ли такая общая доступность материков в среднем триасе? Вообще, было ли
какое-нибудь сухопутное соединение между Северной Америкой и осталь- ной частью земного шара в то время? Если это так, то почему среднетриа- совые семейства Африки и Южной Америки не были представлены в Север- ной Америке, если только действительно они не были там представлены'? Как зоогеограф, я должен сказать, что ооъяснить это пологкение одним расстоянием недостаточно. Все обыкновенные наземные позвоночные могут расселиться на земном шаре (даже не прямыми путями) за короткое время, если только ничто не останавливает их. Была ли Северная Америка и среднем триасе отрезана от других частей земного шара, если не водой. то экологическими условиями или зональностью климата?
Таким образом, прежде чем толковать с уверенностью о значени11 свя- з11 Африки с Южной Америкой, неооходимо охватить в целом детали кар- тины всего земного шара, а не только взаимосвязи Южной Америки с Аф- рикой и Северной Америкой.
Распространение позвоночных до мезозоя дает весьма слабое пред- ставление о состоянии земного шара. Олсон (Olson, 1955) считает, что незадолго до пермского периода распространение рептилий могло происхо- дить непрерывно как в Старом, так и Новом Свете. Однако в ранней перми обмен, очевидно, приостановился и неродственные группы рептилий образо- вали самостоятельные, но параллельные адаптивные типы в Старом и Но- вом Свете. Если это предположение правильно (Олсон утверждает, что его объяснение основано на «определенных предположениях»), то оно поз- воляет допустить скорее домезозойское положение отдельных материков с чередующимися соединениями, чем их непрерывность, как это считал Вегенер. Подтверждение свидетельству такого рода следует искать в отда- ленном прошлом путем сравнения фаун, а не выделением отдельных слу- чаев.
Примерно в начале перми в Южной Африке и Южной Америке встре- чались мезозавры, но больше нигде ни в какое время они известны не были. Это были небольшие (примерно 1 ярд в длину) активные водные рептилии, которые жили в пресной воде или, возможно, иногда в мелководных морях (Caster, см. работу — Мауг et al., 1952). Иногда считают, что онине могли пересекать соленые воды, но, сравнивая их с современными крокодила- мн, я думаю, что они могли заходить в море, однако установить предель- ную ширину водного пространства, которое они могли бы пересекать, не- 11озможно. Если мезозавры могли воспользоваться сухопутным соедине- нием между Африкой и Южной Америкой и распространиться черезпрес- ные воды, то и наземные животные должны были бы пересечь его также, и если будут обнаружены их ископаемые остатки, то именно они докажут существование этого соединения, если только оно вообще существовало. А пока мезозавры, вероятно, вызывают больше споров, чем дают возмож- ность прийти к какому-либо решению.
Если в заключение показать историю земного шара начиная с перми (более 200 млн. лет назад) при помощи фильма и дать только те факты, которые действительно известны, мы получим не последовательную карти- ну, а просто небольшие яркие кадры на широком экране. В первых кад- рах мы увидим материковые ледники, незнакомые растения, древние от- ряды амфибий и ранних рептилий. Они будут видны лишь одно мгновенье то тут, то там и не позволят представить нам ясной картины земного шара того периода. Позднее исчезнет ледниковый покров, а растения и живот- ные будут снова и снова изменяться. В течение перми и раннего мезозоя некоторые кадры будут свидетельствовать о том, что между Африкой и Юж- ной Америкой было больше общего, чем в настоящее время, но эта картина
11рож 1ое в
>»>»» qr>r>;enaprrrjirr>r
тоже исчезнет до того, как ее значение станет для нас. ясным. В триасе поя- вятся динозавры, которых мы вскоре увидим и на всех континентах, но мы не увидим. каким образом они туда попали. Наконец, но не раньше мелового периода, кадры дадут возможность представить земной шар как единое целое и уже будут вырисовываться основные черты распреде- ления суши и климатов, в какой-то степени все еще фрагментарные, но в целом, очевидно, такие же, как и в настоящее время. На этом земном шаре внезапно повсюду появятся современные растения и до конца мело- вого периода в умеренной зоне северного полушария среди кленов, дубон и буков будут видны силуэты динозавров. Травоядные динозавры распро- с транялись в соответствии с климатическими зонами, и когда эта часть филь- ма подойдет к концу, в ее кадрах все еще будут появляться динозавры, но г началом следующей части о третичном периоде они все исчезнут и мы увидим картины расцвета млекопитающих, занявших их место.
Заключение
То, что произошло за последние 200 млн. лет и что зоогеография, а также некоторые данные других наук, помогла нам установить, было уже кратко представлено в трех частях воображаемого фильма. Теперь я хочу представить историю земного шара другим путем, который. видимо, соответствует требованиям зоогеографии.
Распространение животных и другие свидетельства показывают, что 60 млн. лет назад, считая от настоящего времени, на протяжении третич- ного периода, несмотря на отклонения частного порядка, больших изме- нений в основных чертах расположения материков и климатических зон не было.
H отношении периода, предшествующего третичному, появляется сомнение, увеличивающееся по мере отдаленности времени. В меловом пери> де распространение позвоночных все еще позволяет предполагать, что расположение суш и климатических зон было таким же, как и в на- стоящее время. но в более древние периоды появляются признаки того, что расположение южных материков возможно оыло иным. 3То допуще- ние основывается на соответствующих очертаниях Африки и Южной Аме- рики. одинаковой стратиграфии этих материков, на сходстве их ископае- мых прибрежных фаун беспозвоночных и подтверждается пермским оле- денением южных материков и Индии, связанной с ним глоссоптериевой флорой и сходством позвоночных Южной Африки и Южной Америки в ран- нем мезозое. Каждый пунк" этих свидетельств может быть принят или подвергнут сомнениям, но как бы то ни было, сомнения относительно рас- положения суши в южной половине земного шара вполне реальны, хотя не следует их переоценивать. Допустим, что данные о распространении позвоночных могли бы позволить судить о прошлом Земли более уверенно. Тогда они могли бы показать, что когда-то суша была уже настолько сил ь- но разделена, что, например, динозавры эволюционировали на одном ма- терике, в то время как млекопитающие эволюционировали на другом. Они могли бы показать (при наличии ископаемых в Антарктиде или Новой Зеландии), что динозавры прошли именно этим путем. Идентичностью наземных фаун в целом они могли показать вероятную связь Африки и Южной Америки. Но по крайней мере пока все это не подтверждается. lIo имеющимся данным, несмотря на все сомнения, распространение животных может быть увязано с очертаниями суши и климатом, подобным
о16
1'.гава 10
современным. Фактически зональность климата прхх современном распо- логкенхххх материков може1' слугхъить наилучшим объяснением некоторых положений, выдвигаемых сторонниками Вегенера, включая сходство стра- тиграфических данных, оощие прибрежные морские фауны хх1'х;ххой Африки и связи между тропххческими животными на разных матерхх1,';хх- в любые периоды.
Несомненно, земной шар изменился в деталях. На месте ххезозоххскпх оолот образовались высокие горы. Там, где оыли мелководные моря, «оз- никли засушливые пространства, и наоборот. Изменились очертания материков и их поверхность. Прилежащие материки соедххня.хххсь и разъеди- нялись и вновь соединялись. Но это все второстепенные детали. Вопрос заключается в том, было ли основное расположение суши и клххххатон в древнейп1ххе времена, по существу, таким же, как и в настоящее время„ или оно существенно отличалось от современного. Я полагаю. что. «ероят- но, основное расположение материков было такое же, как и теперь. «ххе связи как с положительными, так и с отрицательнымхх данными и» этот сче r.
Эту главу я закончу гипотетической историей материков, которая не зависимо от ее достоверности, по-видимому, будет вполне соответство«ать теории распространения животных.
Я полагаю, что Луна оторвалась от полушария Паххххфххх'хх на очень ранней стадии истории Земли (очевидно, это до сих пор имеющая ход;де- ние, но не «модная» теория происхождения Луны), и тогда в резулыате разрывов и перемещений в короткое время произошло образование мате- риков (гипотеза Дарвина — Фишера), которые с тех пор сохранили свое сходство и приблизительное расположение.
Я полагаю, что все основные материки определеххньих ооразом после- довательно соединялись в большинстве случаев через очень узкие водные пространства. Любой материк мог быть отрезан на какое-то время от дру- гих, и животные, находящиеся на нем, могли (но только временно) эво- люционировать самостоятельно. Два материка могли быть отрезаны в од- но и то же время и, даже если они не были соединены между собой. могли сохранить одинаковые фауны, в то время как фауна остальной части зем- ного шара претерпевала изменения. Нечто подобное этому произошло в третичном периоде в Южной Америке и Австралии. В этом случае фауны обоих материков в целом были очень различными, хотя отдельные компо- ненты этих фаун (например, некоторые лягушки и сумчатые) оыли одина- ковыми. В случае более древних периодов два временно изолированных материка, возможно, имели более сходные фауны.
Климат моего гипотетического земного шара, несомненно, опре- делялся главным образом радиацией Солнца. Он всегда был зональным в результате конфигурации и движений Земли и всегда изменялся локаль- но с абсолютной и относительной высотой суши и направлением ветра. И зональность и локализация климата всегда оказывали определенное влияние на распространение животных. Строгая приуроченность неко- торых групп животных к отдельным климатическим зонам на разных мате- риках иногда напоминает воздействие сухопутных мостов.
Определенное количество тепла, получаемого от Солнца (я полагаю). определялось несколькими факторами, включая и радиацию Солнца и структуру и состав атмосферы. Понижения температуры в разные вре- мена, возможно, вызывались различными причинами и имели разный исход. В результате этого понижения температуры, возможно, иногда наступало общее охлаждение поверхности Земли с относительно слабой
517
дифференциацией климатических зон и ледниками, образующимися в воз- вышенных частях поверхности Земли на всех широтах, а иногда более зональным охлаждением и ледниками только в более холодных зонах. (.'те- пень дифференциации климатических зон, возможно, также варьировала с образованием и разрушением сухопутных соединений между материками и изменениями океанических циркуляций.
Таким образом, земной шар, имеющий постоянные общие очертания материков и климат, которые, однако, бесконечно изменяются в деталях. соответствует тому, что известно о распространении животных.
ДИТЕРА ТУРА
В а r g h о о r n Е. S., 1953, Evidence of climatic change in the geologic record of plant
life. См. S h а р 1 е у et al., «Climatic change», 235 — 248.
В о g e r t С. М., 'l953, The Tuatara: ъ.~~у is it а lone survivor? «Sci. Monthly», 76,
164 — 170.
С Ь а п е у R. '5"., !940, Tertiary forests and continental history, «Bull. Geological
Soc. America», 51, 469 — 488.
С о 1 Ь е r t Е. Н., 1945, The dinosaur book, American Mus. Nat. Hist., Man апд Х»- ture Pub., Handbook 14.
С о l Ь е r t Е. Н., 1948, Evolution of the horned dinosaurs, «Evolution», 2, l45 — !63. Со1bert Е. Н., Cowles R. В., В ogert С. М., 1946, Temperature tole- rances in the American Alligator..., «Bull. American Mus. Nat. Hist.», 86, 3' 7 — 37-~. С о l e ш а п Л. P., 1926, Ice ages recent and ancient, New York.
D а r 1 i n g t. о п P. J., Jr., 1949, Beetles and continents, «Quarterly Вечерев Biol.»,
24, 342 — 345.
D а r w i n С., 1859, 1950, On the origin of species... London (переиздано в 1950).
[Имеется русский перевод: Ч. Д а р в и н, Происхождение видов путем есте- ственного отбора. Соч., т. 3, М.— Л., 1939.]
0 е е v е у Е. S., Jr., 1949, Biogeography of the Pleistocene, «Bu]l. Geol. Sop. Arneri- ca», 60, 1315 — 1416.
D i l 1 о п Ь. S., 1956, Wisconsin climate and Ие zones in Пorth America, «Ясience»,
123, 167 — 176.
D u n Ь а r С. О., 1949, Historical geology, Меж York.
Й~ Т о i t А. Ь., С о w р е r R е е d F. R., 1927, А geological comparison of South
America with South America with South Africa, Carnegie Inst., Washington, Pub.
381.
F 1 i n t R. F 1947, Glacial geology and the Pleistocene Epoch, !Vew York. [Имеется
русский перевод: P. Ф л и н т, Ледники и палеогеография плейстоцена,
ИЛ, Москва, 1963.]
Н о 1 <1 h а u s Е., 1954, Die Spuren der Eiszeit in der Tierwelt Europas, Abhandlungen
Zool.-Botanischen Gesellschaft, Wien, !8.
Н и е п е F. von, М а t 1 е у С. А., 1933, Tire Cretaceous Saurischia and Ornithi- schia of the Central Provinces of India, «Мел. Geological Survey India,
Palaeontologia India», New Бег., 21, No. 1.
J e» n e l R., 1942, Ьа genese des faunes terrestres..., Paris, Presses Universitaires
il e France.
Е и е п е п P. Н., 1950, Marine geology, New York.
L i Ь Ь у O' F., 1956, Radiocarbon dating. «American Scientist», 44, .)8 — 112.
Е u l! R. S., 1910, Dinosaurian distribution. «American J. Sci.» (4), 29, 1 — 39.
М а t t h e w O. D., 1915, 1939, Climate and evolution, «Апп. New York Асад. Sci.»,
24, 17l — 318; reprinted (1939) as «Special Pub. New York Acad. Sci.», 1.
Глава lO
М а у r Е. et al., 1952, The problem of land connections across ФЬе South Atlantic, with special reference to the Mesozoic, «Ви11. American Mus. ~а1. НЫ.», 9!). 79 — 258.
О 1 s о п E. С., 1955, Parallelism in the evolution of the Permian reptilian fa]]nas о1 the Old and New Worlds, Fieldiana (Chicago Nat. Hist. Mus.), «Zool.», 37, 381 — ~0 I .
P i 1 s Ь r у Н. А., 1911, Non-marine МоИпяса of Patagonia, «Вер. Princeton L. Exp. Patagonia, 1896 — 1899», 3, pt 5, 513 — 633.
P i 1s Ь r у Н. А., В е q u а е r С J., 1927, The aquatic mollusks of the Belgiari Congo..., «Bull. American Mus. Nat. Hist.», 53, 69 — 602.
R о т е r А. S., 1945, Vertebrate paleontology, Chicago. [Имеется русский перевод: А. P о м е р, Палеонтология позвоночных, ГОНТИ, М.— Л., 1939.]
S c h m i d t К. P., 1938, herpetological evidence for the postglacial eastward ех4еп- sion of the steppe in North America, «Ecology», 19, 396 — 407.
S h а р l е у Н. et al., 1953, Climatic change..., (:ambridge, Mass., Iiarvard U. Press.
S i m p s о п С. G., 19~0, Antarctica as а faunal migration route, «Proc. Sixtli Paci- fic Sci. Congress», 2, 755 — 768.
T h i о п е m а n n А., 1950, Verbreitungsgeschichte der Siisswassertierwelt Europas. «Die Binnegewasser» (Stuttgart), 18.
U m b g r о ч е J. Н. F., 1949, Structural history of tlute East Indies,:ambridge, Ena- land, Cambridge U. Press.
W e g е п е г А. 1915, Die Enstehung der Kontinente uud Ozeane, Brunswicl<, Friedr.
W e g е п е г А., 1924, The origin of continents and oceans [translated from Згд German ed.], London. [Имеется русский перевод: А. В е г е н е р, Возникновение мате- риков и океанов, М.— ' Л., 1925.]
Z е и п е r F. E., 1950, Dating the past..., 2nd ed., London. [Имеется русский пере- вод: Ф. Ц е й н е р, Плейстоцен, ИЛ, Москва, 1963.]
ОГЛАВЛЕНИЕ
12
Глаыа .2. Иведени(
ночных
Глава. 4 Рептилии
Список семейств рептилий
З4З
1.гцва Х. Особенности расирострап~ ння животных и» островал........'эЖ1
илощади,
Гл(иа 10. Проп1лое в свете Зоогеографн»
Предислови» От автора
Глава 2. Пресноводные рыбы Список семейств пресноводных рыб и примитивных рыбооорлзшлк позпо-
1'лава 3. Земноводные Сиис ок семейств земноводиь~
Глава б. Птицы Список семеиств птиц
1'.га «а б. Мл~ к olj итающиг Список семейств млекопитающих
1'лава 7. Особенности раснространения жх1вотных на материках и фауннстн- ческие области
1' gaea 9. Изменение распростраиеиил животных: зиачеиие к;ив~ата и эволюции
115 i45
158 183
206 247
271 314
Ф. Дарлингтон ЗООГЕОГ РАФИК
Редактор П. M. Максил~ова Художник В. Кузякова
Худон'ественный редактор Л. ШкановТехнические редакторы С. При данцева и Г. Еп л инц в. а
Сдано в производство 1/III 1966 г. Подписано к печати 18/VIII 1966 г. 1-'ума1а 70:<1081'16161/4 бум. л. 45,5 печ. л. Уч.-изд. л. 45,79. Изд. М 11/2602. Цена 3 ~). 48 к.Заказ 213
Издательство «Ilp0rpecc» Комитета по печати при Совете Ъ|инистро» СССРМосква Г-21, Зубовский бульвар, 21
Московская типография ¹ 16 Главполиграфпрома Комитета по печатипри Совете Министров СССР. Ъ|осква, Трехпрудный пер., 9