/



























































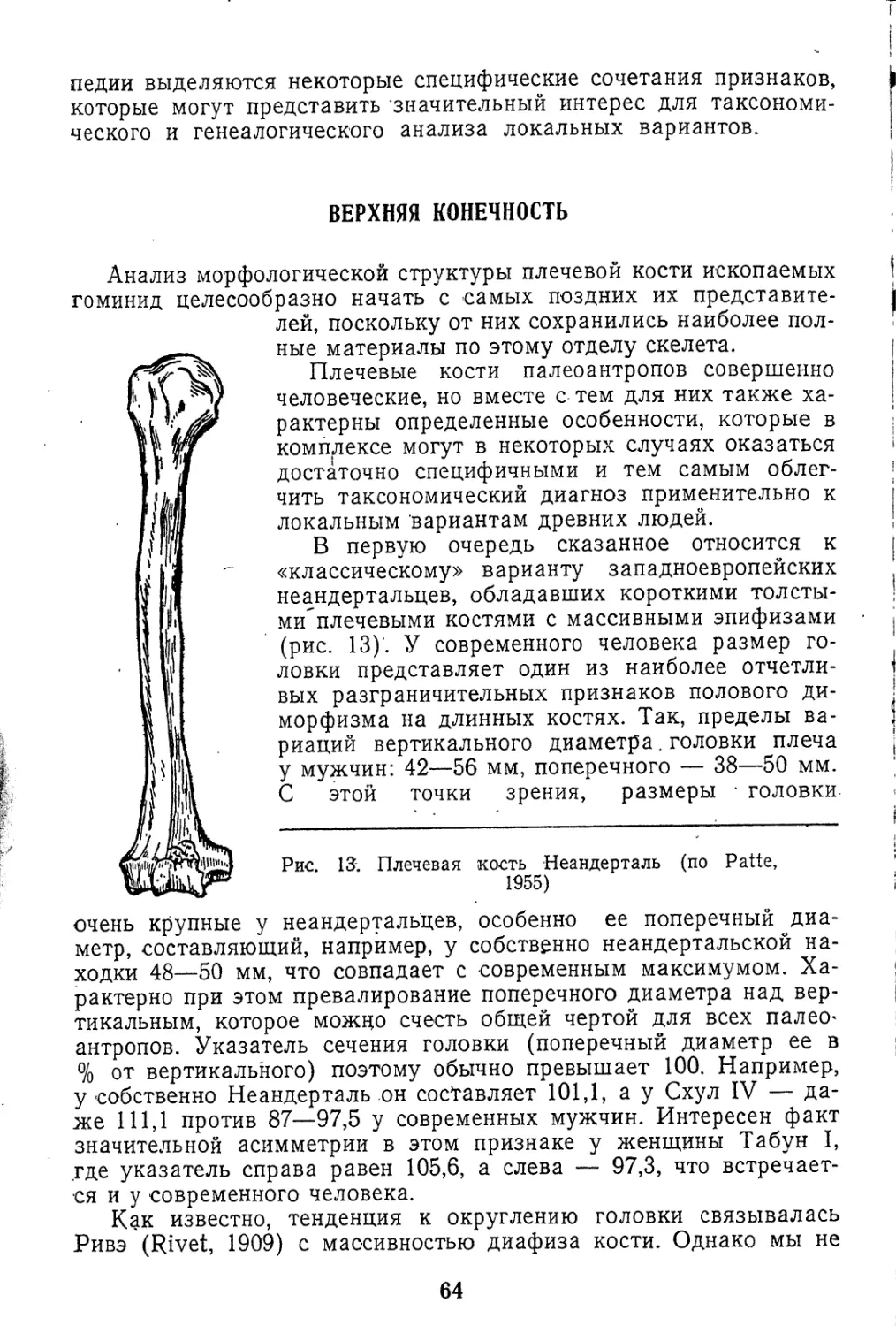


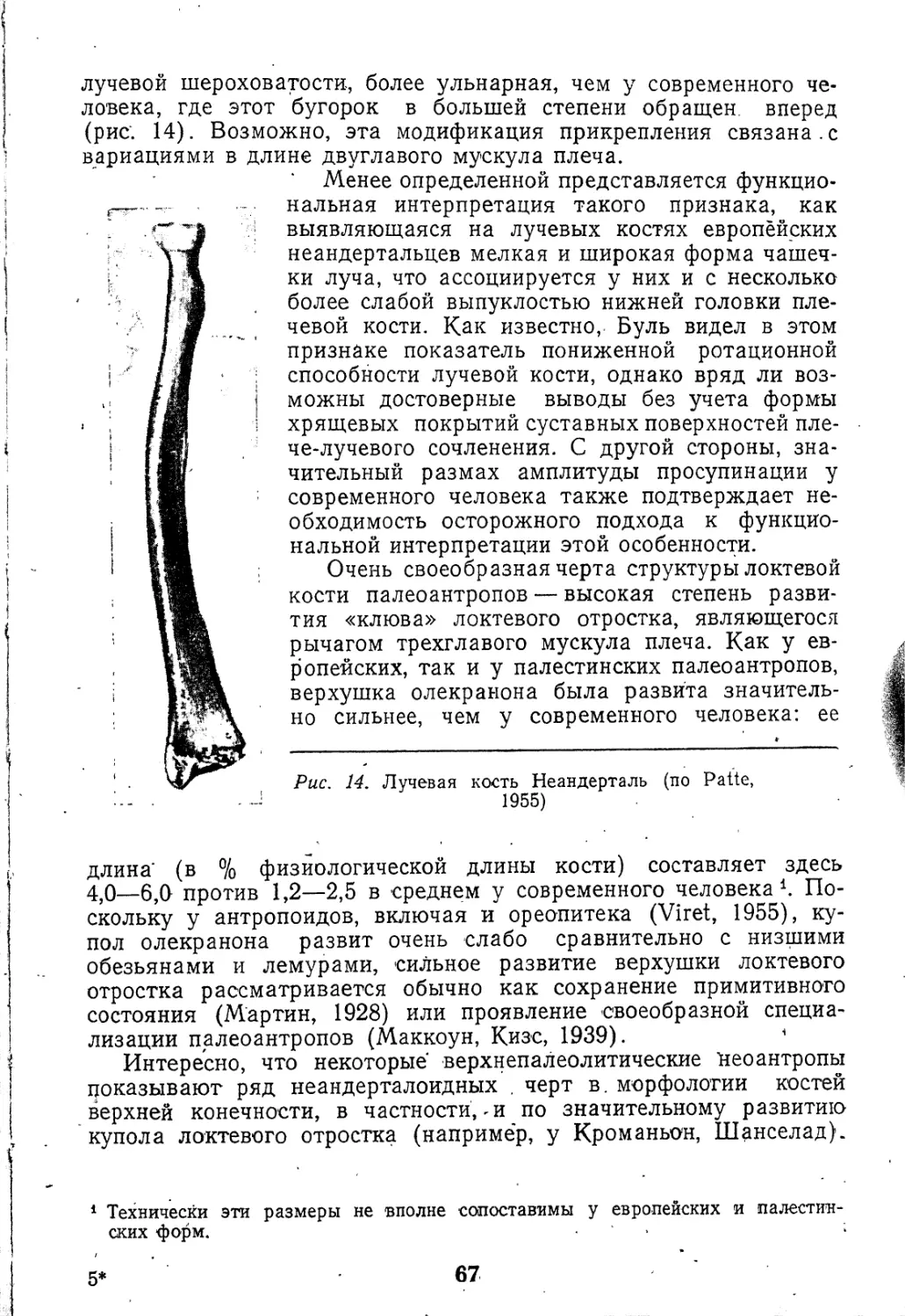





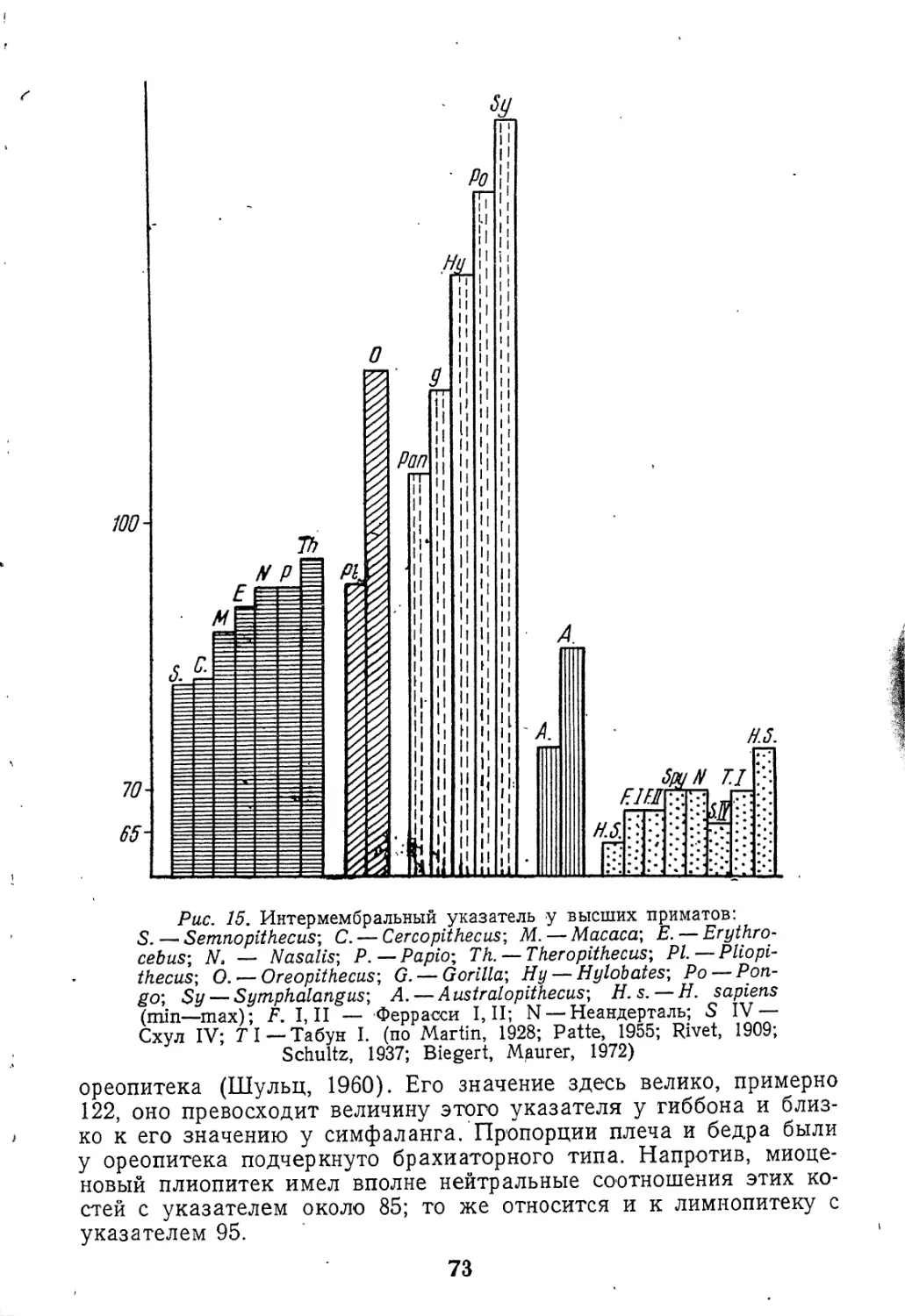












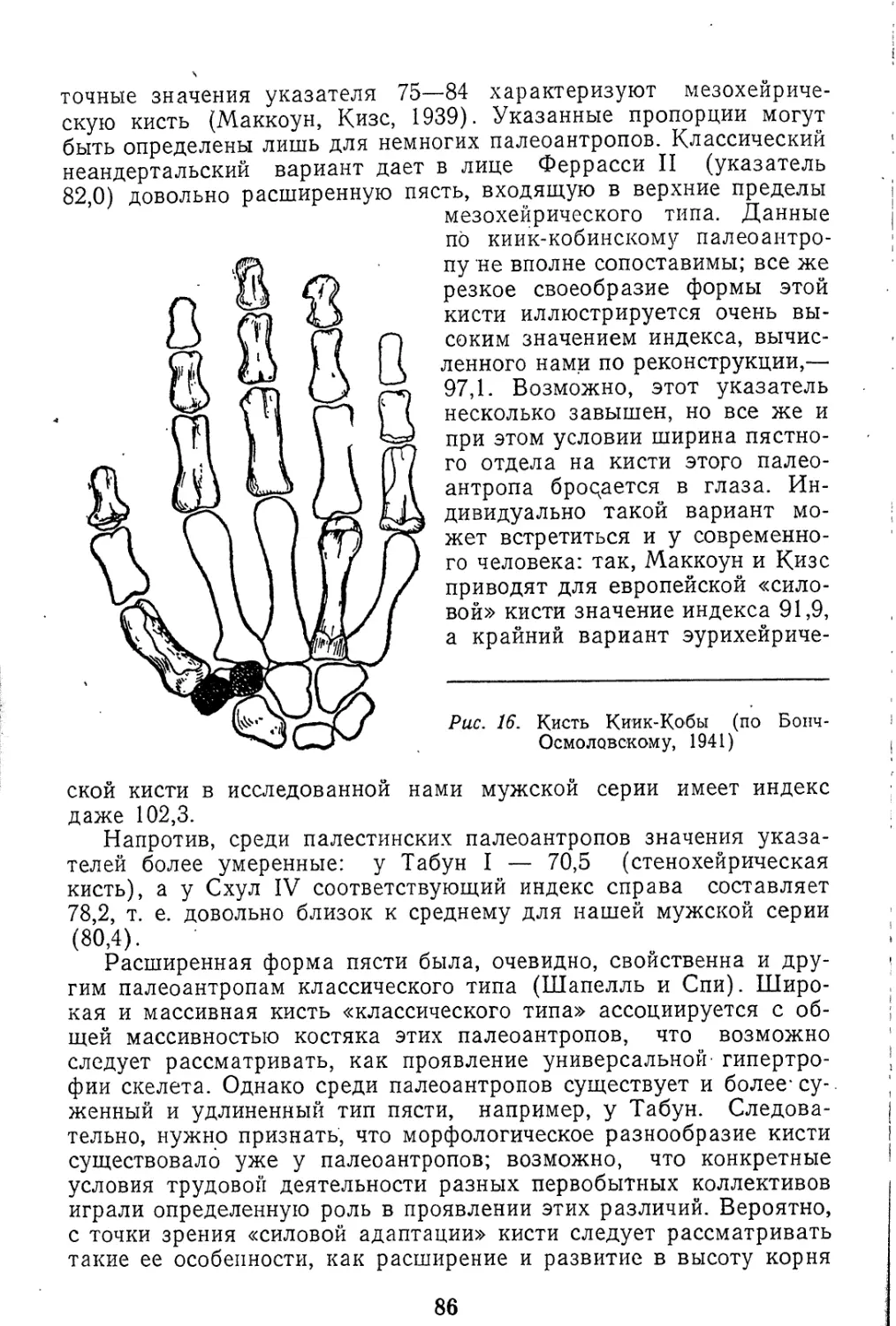



















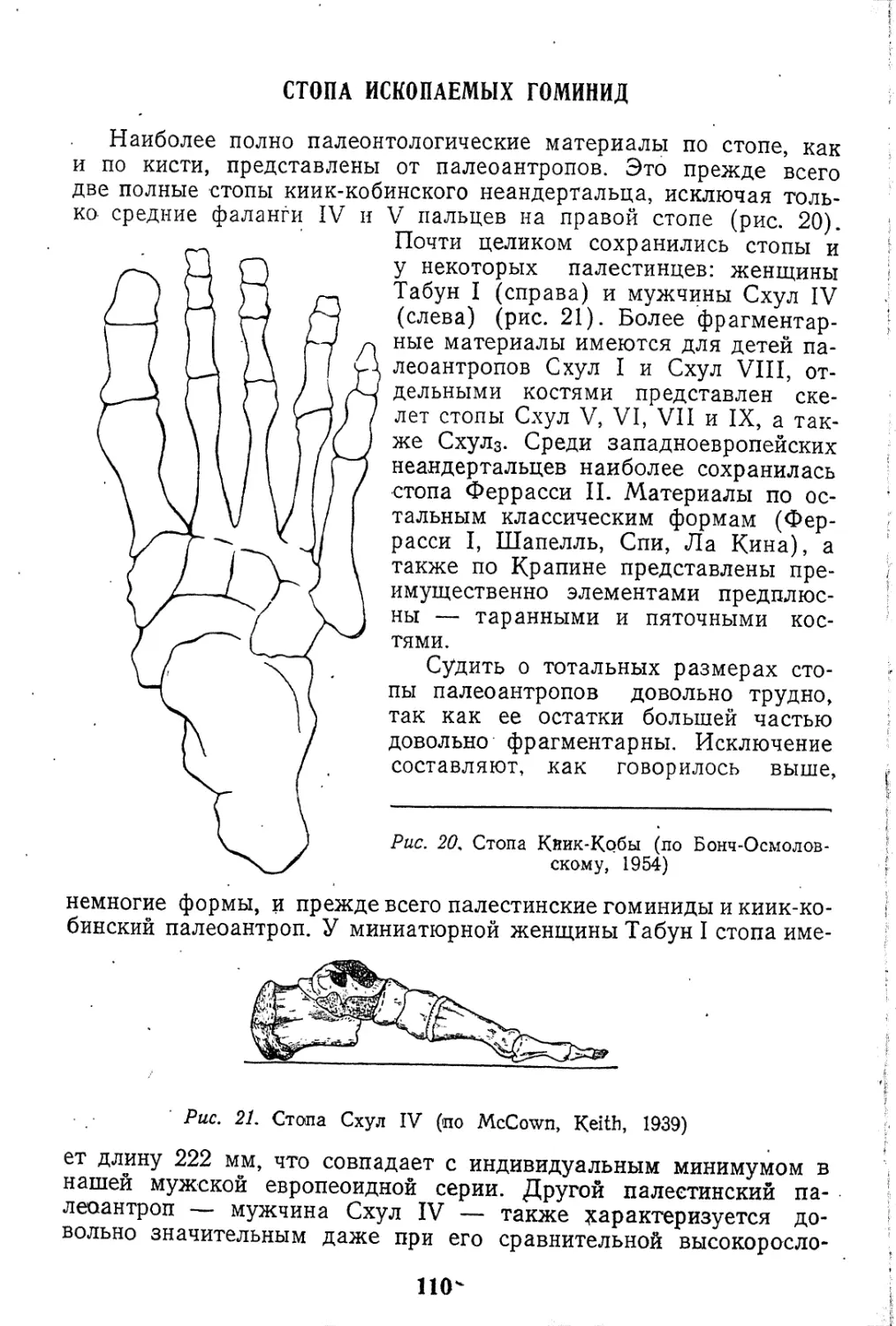


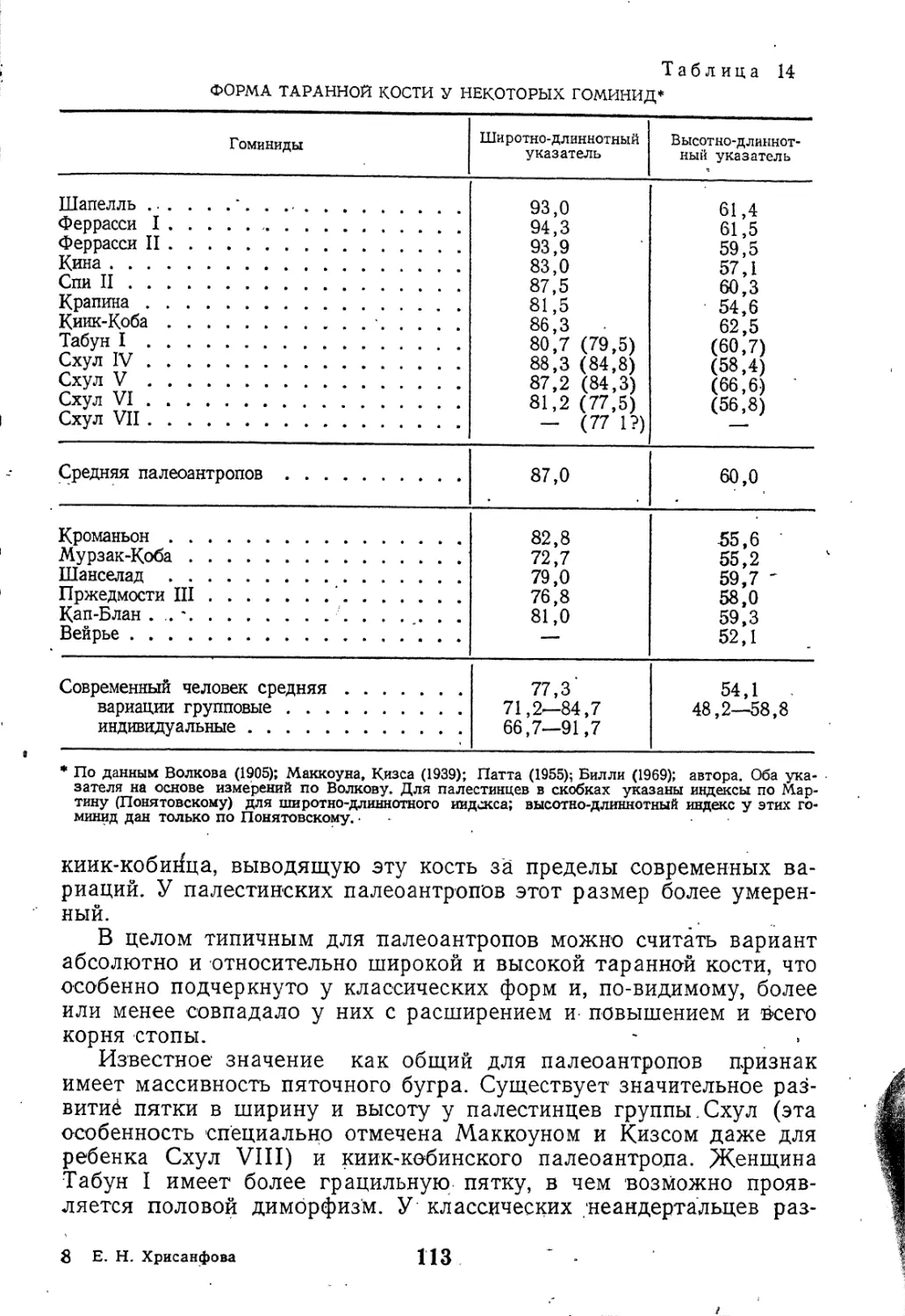

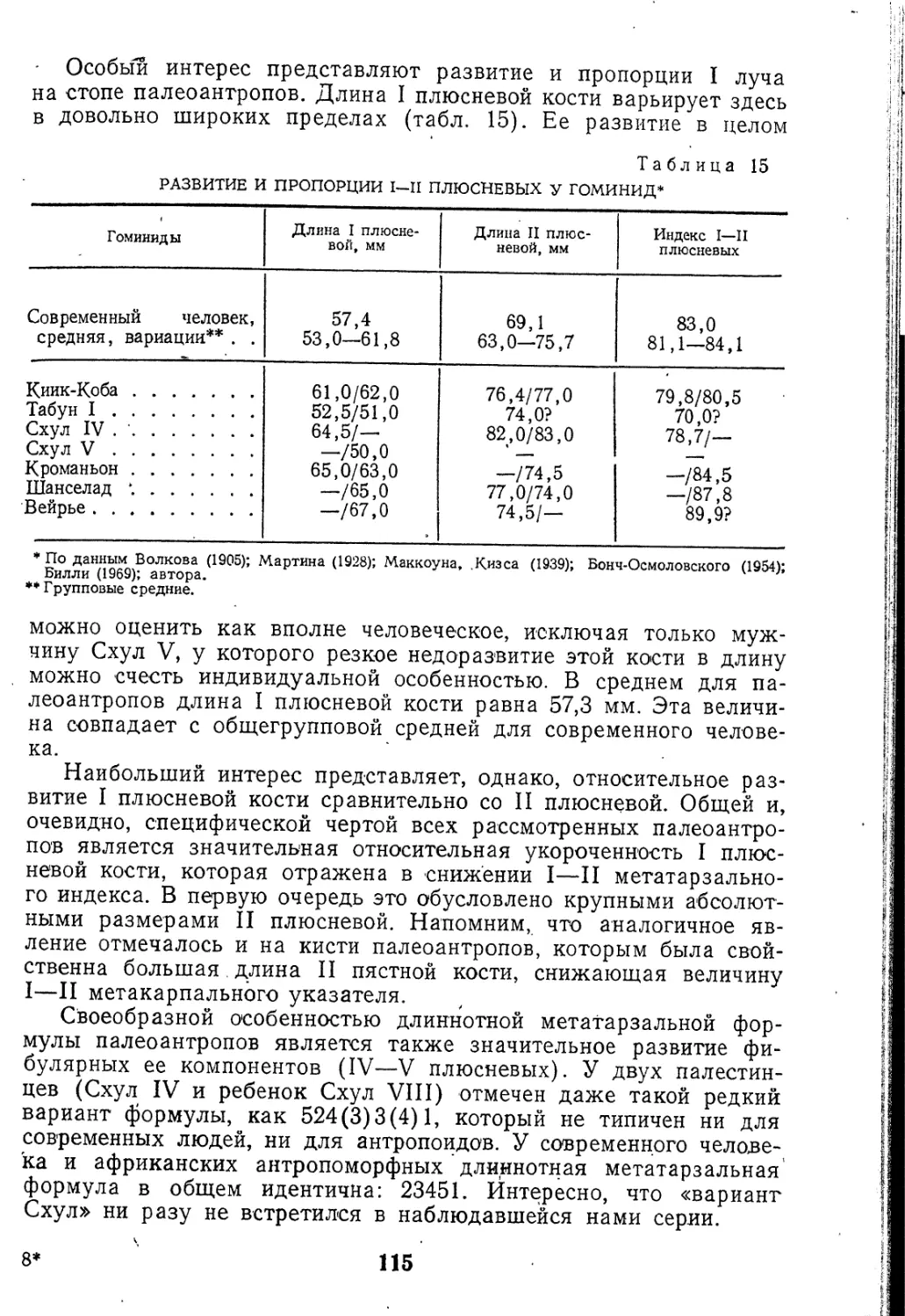






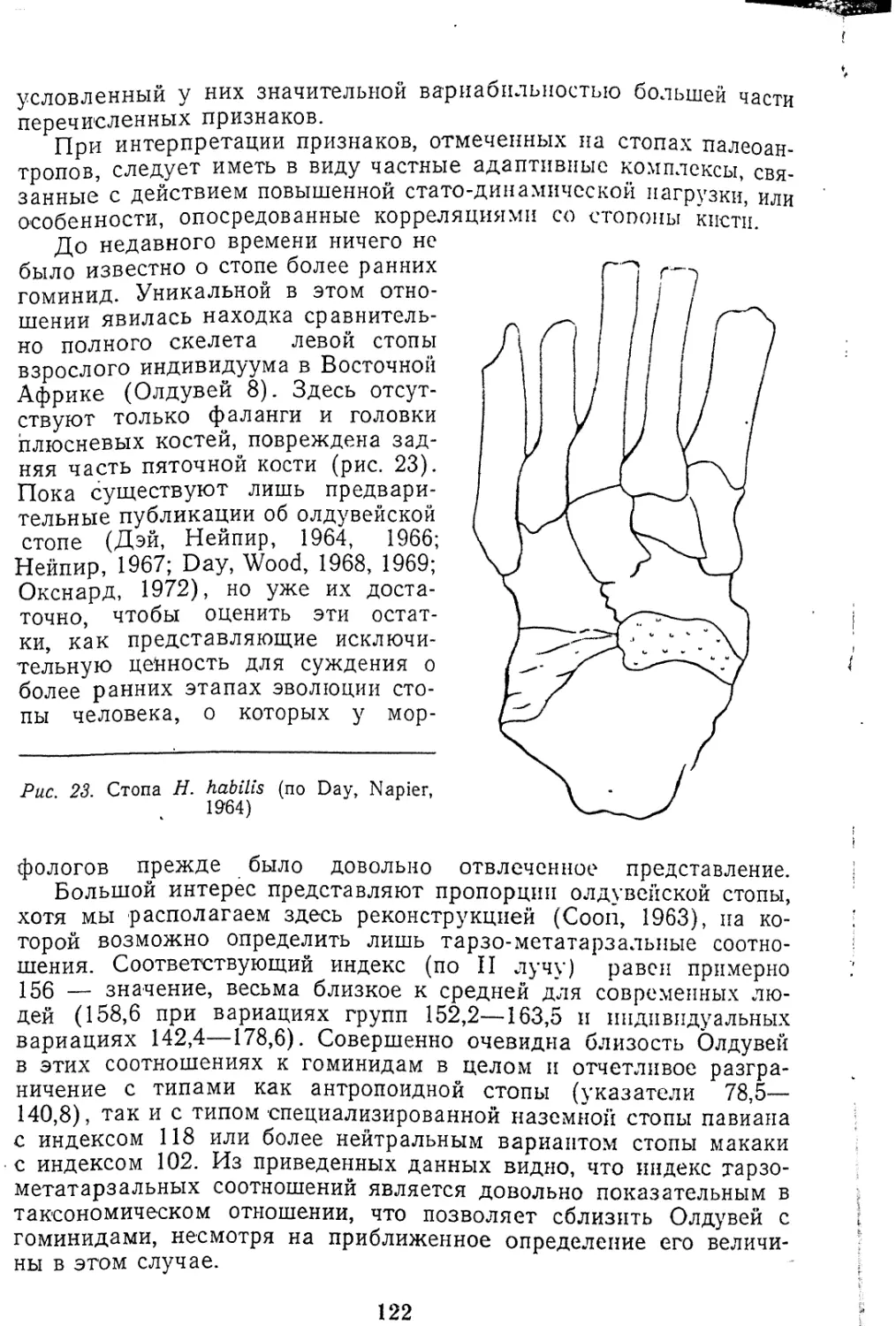



































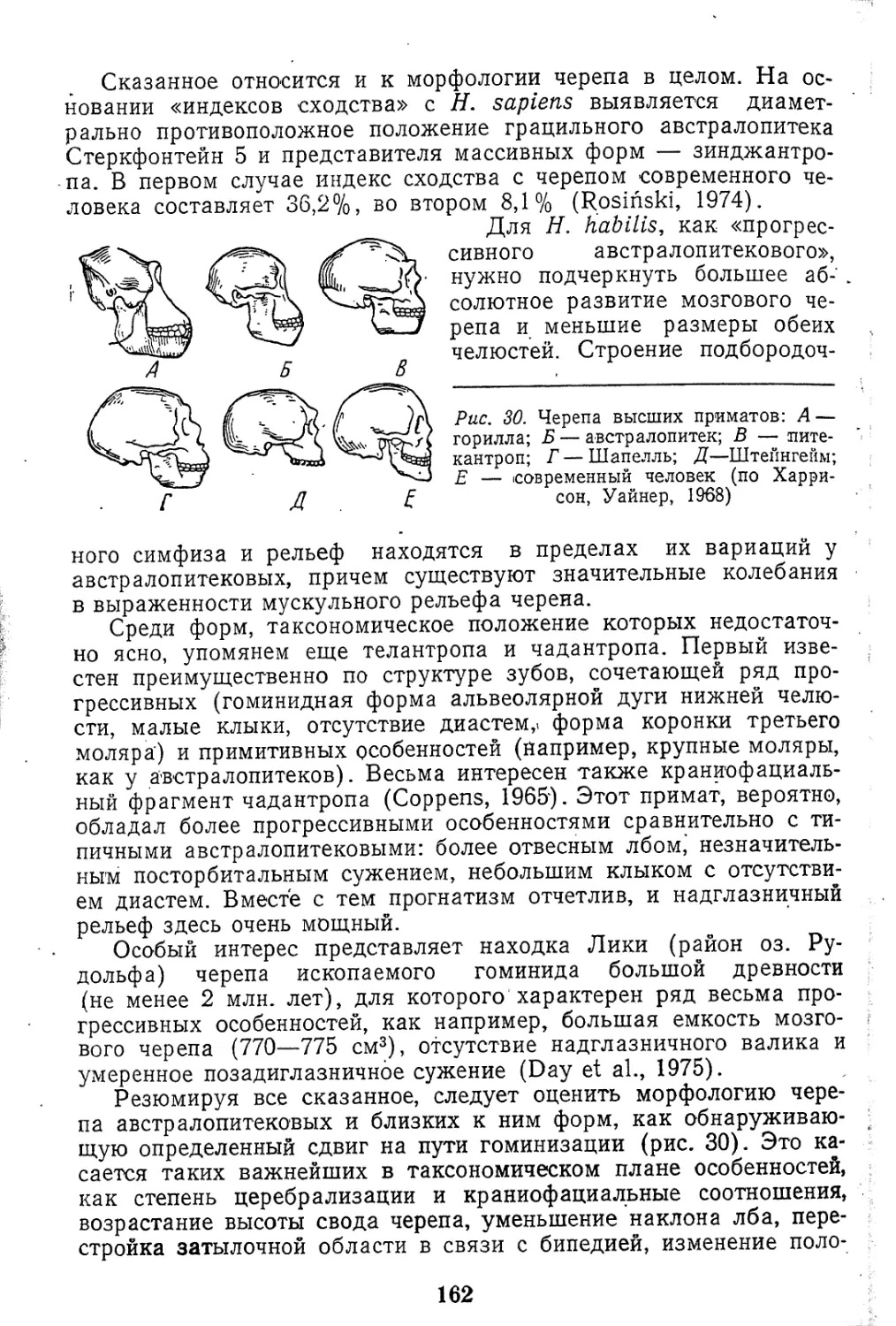









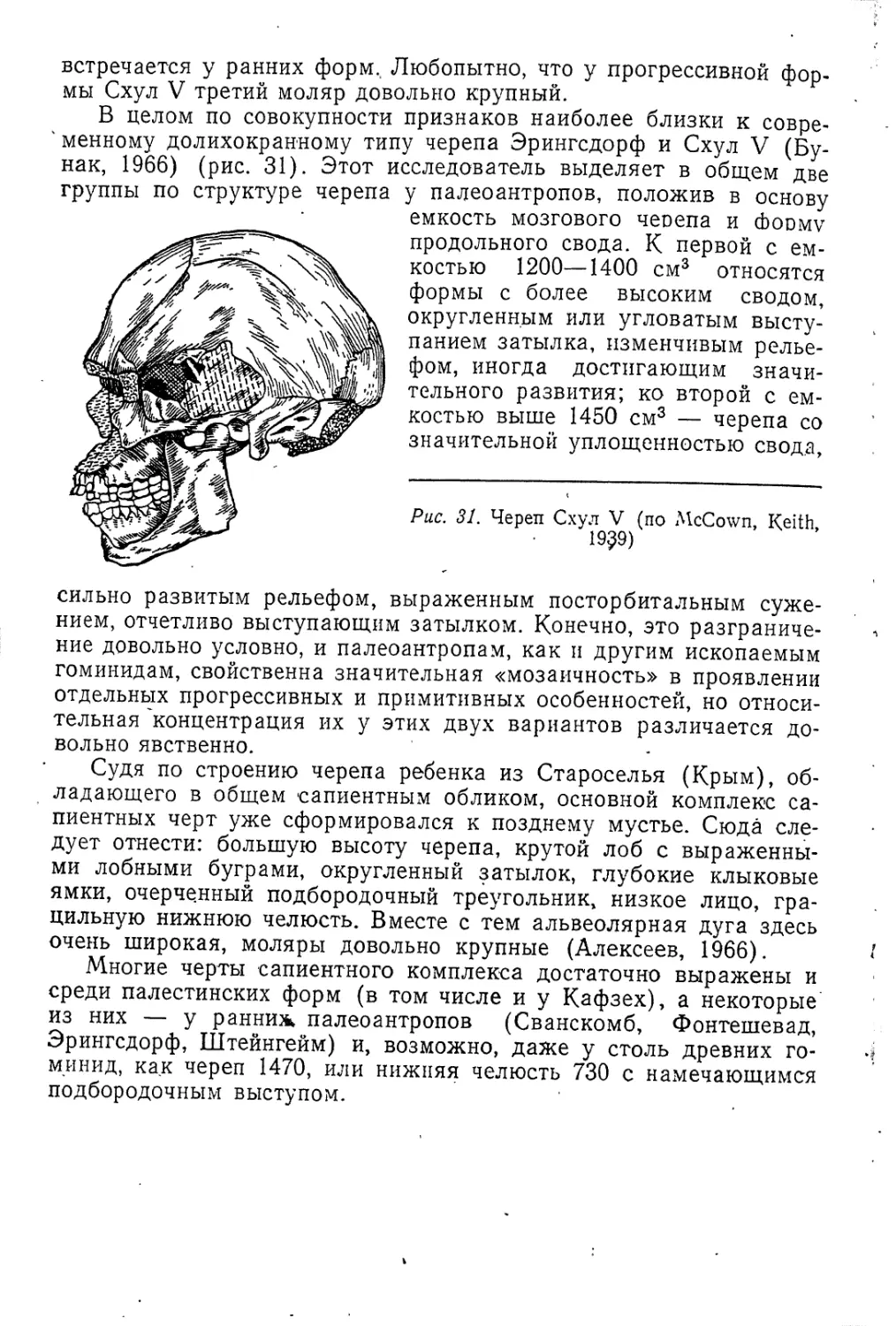














































Текст
Е. Н. Хрисанфова
ЭВОЛЮЦИОННАЯ
МОРФОЛОГИЯ
СКЕЛЕТА
ЧЕЛОВЕКА
Издательство
Московского университета
1978
УДК 571
ПЕЧАТАЕТСЯ ПО ПОСТАНОВЛЕНИЮ
РЕДАКЦИОННО-ИЗДАТЕЛЬСКОГО СОВЕТА
МОСКОВСКОГО УНИВЕРСИТЕТА
Рецензии т ы:
докт. биол. наук, проф. М. Ф. ПЕСТУ РХ,
канд. биол. наук М. И. УРЫСОН
Хрисанфова Е. Н.
Эволюционная морфология скелета человека. М.
Изд-во Моск, ун-та, 1978.
218 ст. с ил.
Монография представляет систематизированную сводку лите-
ратурных п оригинальных данных по структуре скелета совре-
менного человека и его хронологических предшественников в
аспекте антропогенеза. В ней прослежены основные тенденции
и закономерности 'морфологической эволюции гоминид, оцени-
вается сравнительная таксономическая значимость отдельных
морфологических признаков и их комплексов, обосновывается
морфологический критерий гоминид. Монография включает
также основную информацию о вариациях важнейших морфо-
логических признаков на скелете современного и ископаемого
человека.
Издание рассчитано на морфологов, палеонтологов. этно-
графов, антропологов, биологов, анатомов, а также всех лиц,
интересующихся проблемами происхождения человека и эво-
люции его физического типа.
21010—015 ,
-----------БЗ 69—24—77
077 (02)—78
© Издательство Московского университета, 1978 г«
Предисловие
Бурно развивающиеся в последнее время исследования по
палеонтологии человека привлекли значительное внимание к
проблеме его морфологической эволюции. Морфологами уже
многое сделано для того, чтобы осветить главные этапы исто-
рического становления гоминид, проследить большой путь из-
менений, происшедших в их физическом облике. Основу такого
анализа составляют, естественно, скелетные материалы. Спе-
циальная литература содержит богатый фактический материал
по остеологии человека и других приматов, однако все же
в эволюционном плане посткраниальный скелет человека осве-
щен недостаточно, и таксономия гоминид до сих пор еще
построена преимущественно на краниологическом фундаменте.
Причины такого состояния кроются отчасти в неполноте па-
леонтологических материалов по посткраниалъному скелету и
их разнокачественной представленности. Сюда следует приба-
вить значительную степень трансгрессивной изменчивости мно-
гих морфологических признаков у приматов и трудности их
таксономической оценки и функциональной интерпретации.
Понятно также, что анализ остеологических признаков с пози-
ций эволюционной морфологии предполагает предварительное
изучение разных аспектов их изменчивости у современных
людей и других приматов.
Сказанное в достаточной мере объясняет, почему пробле-
ма эволюции посткраниального скелета гоминид занимала по
существу второстепенное место в антропологической литера-
туре. Соответствующие материалы привлекались обычно как
дополнительные в плане общих эволюционных концепций
антропогенеза. Исключение составляют главным образом ди-
стальные отделы конечностей человека, по которым имеются
подробные публикации.
Основной задачей настоящей книги и было поэтому пред-
ставить формирование посткраниального скелета человека во
времени.
Перед исследователем, желающим разобраться в сущности
эволюционных преобразований на скелете гоминид и их пред-
шественников, неизбежно возникают и специальные задачи,
связанные с необходимостью учета закономерностей морфоло-
гической изменчивости, факторов, влияющих на морфогенез
скелета вообще, уточнением функциональной интерпретации.
Это потребовало привлечения обширной литературы, посвя-
щенной изучению костной системы в различных аспектах:
сравнительно-анатомическом, возрастно-половом, этно-терри-
ториальном, профессиональном, обменно-гормональном, кли-
нико-патологическом, биомеханическом и т. д. Автором исполъ-
3
зованы и некоторые данные собственных работ, выполненных
в этих же аспектах.
Не претендуя на сколько-нибудь подробный разбор су-
ществующих (и достаточно различающихся между собой)
классификационных и филогенетических схем, ограничимся
здесь лишь указанием, какие из ископаемых форм гоминид и
их вероятных предшественников мы считаем нужным включить
в наш анализ.
В соответствии с наиболее распространенной точкой зре-
ния, изложение палеонтологических данных будет построено
применительно к основным хронологическим этапам эволю-
ции гоминид, признаваемым большей частью исследователей.
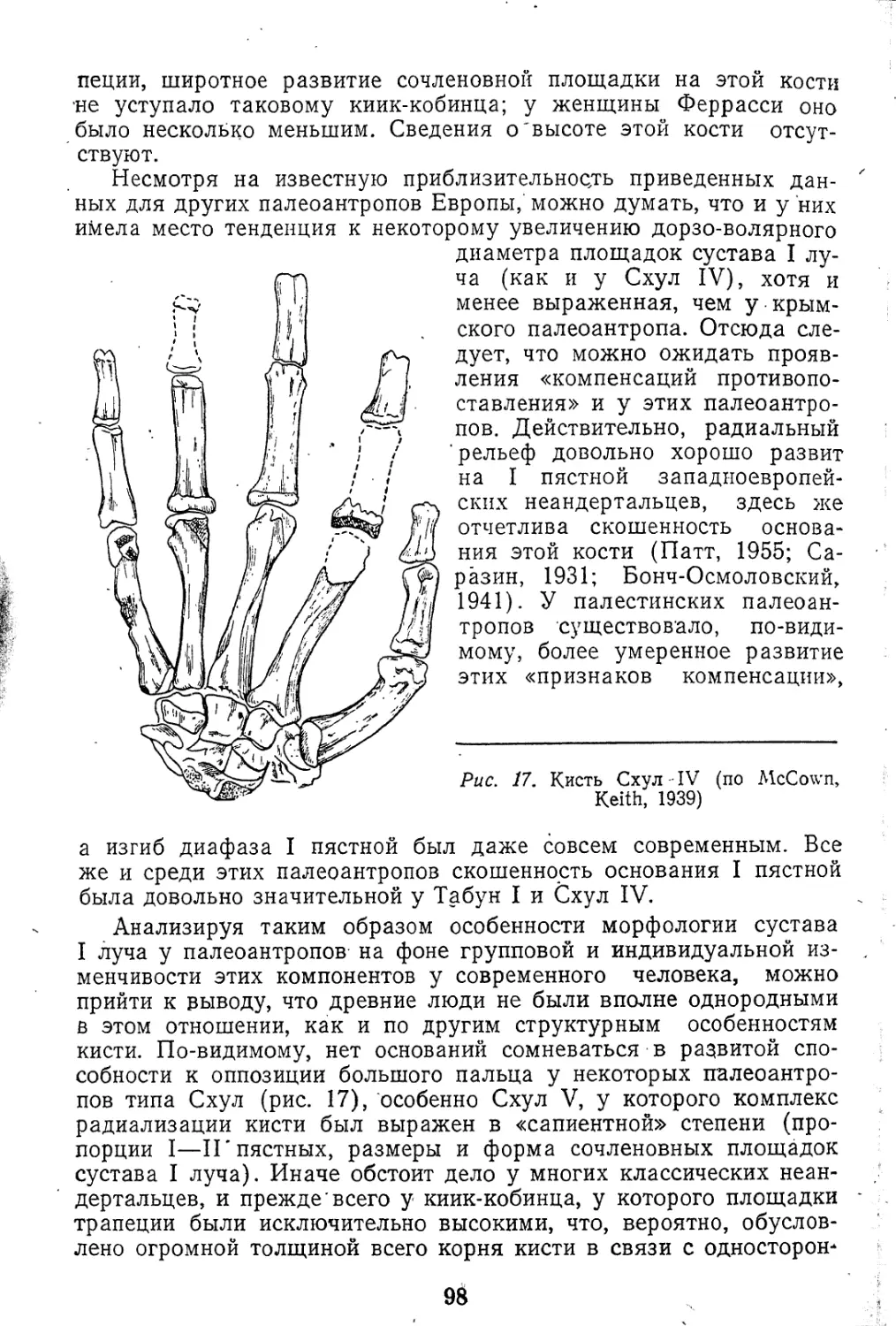
В качестве представителей древнейшего этапа филогенеза го-
минид здесь будут рассмотрены австралопитеки и близкие
к ним формы, примерный возраст которых от 4—5 до 0,7 млн. лет.
Следующая стадия эволюции гоминид, по-видимому, связы-
вается с архантропами, верхняя граница существования кото-
рых составляет примерно 300 тыс. лет, тогда как относительно
нижней существуют известные разногласия (от 700 тыс. до
1,9 млн. лет и даже ниже). К этой группе древнейших людей
относятся Широко известные находка питекантропов Явы и
Восточной Африки (Олдувей), атлантропа, гейдельбергского
и древневенгерского человека, синантропов. Наконец, более
поздний этап антропогенеза представлен многочисленным и
весьма пестрым в морфологическом отношении населением
Афревразии периода от 300 до 35 тыс. лет (палеоантропы).
Человек современного типа (неоантроп) появился, очевидно,
примерно 35—40 тыс. лет тому назад или, может быть, еще
раньше.
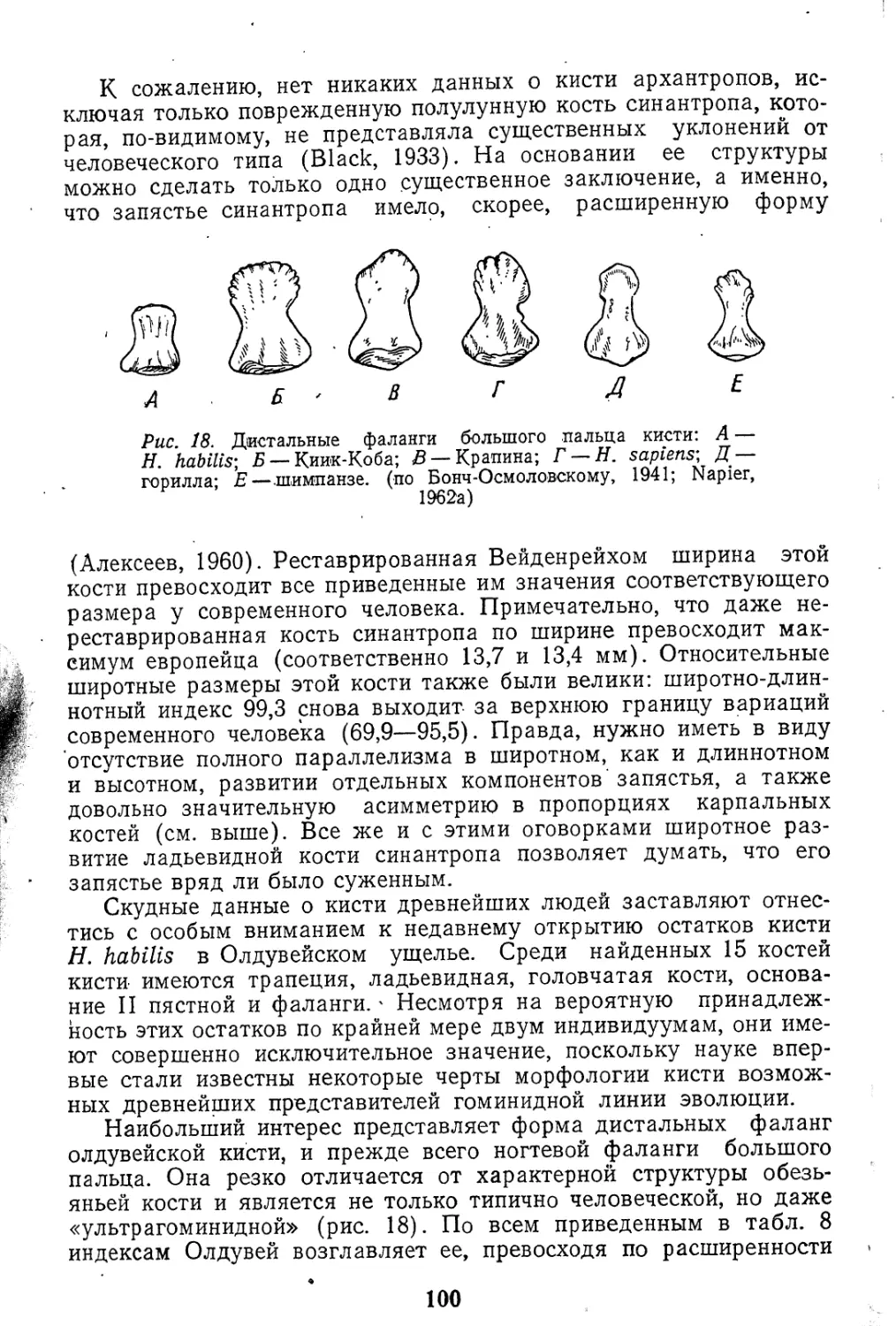
Следует подчеркнуть, что эта схема весьма условна и на
границах выделяемых «стадий» наблюдается сосуществование
гоминид. По современным данным уже в начале плейстоцена
в Африке представлены роды австралопитеков и Ното. О том
же свидетельствует и наличие «промежуточных» в морфологи-
ческом отношении форм, относительно которых не всегда легко
установить их «стадиальную» принадлежность.
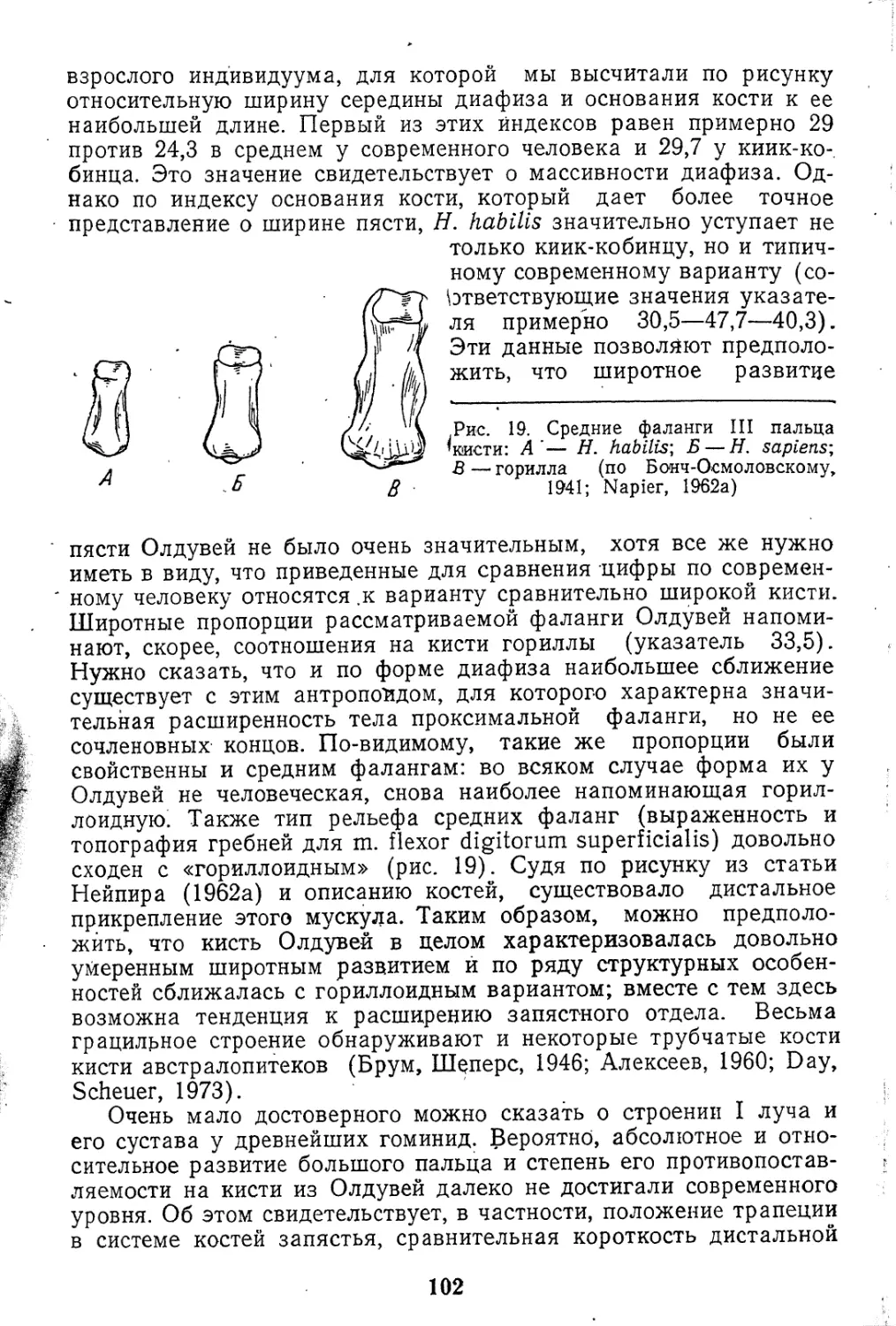
Поскольку, как уже указывалось, основной нашей задачей
был анализ палеонтологических материалов по посткраниаль-
ному скелету человека, изложение и начинается с этих отде-
лов скелета. Лучшая разработанность краниологии и одонто-
логии ископаемых гоминид (см. Бунак, 1959, 1966; Зубов,
1966, 1973) освобождает нас от необходимости систематизи-
ровать все многочисленные данные. Мы ограничимся лишь при-
веденным в конце книги кратким очерком, отражающим ее
современное состояние.
Глава I
ОСЕВОЙ СКЕЛЕТ
Эволюционная морфология осевого скелета человека разрабаты-
валась преимущественно в сравнительно-анатомическом аспекте.
Это объясняется не только скудостью и слабой сохранностью со-
ответствующих палеонтологических материалов, но и большой от-
четливостью сравнительно-анатомических рядов по многим важ-
ным структурным признакам. Совершенно особое место занимают
здесь вариации позвоночной формулы, с которыми связываются
основные теоретические концепции морфогенеза осевого скелета,
почему эти вопросы и излагаются нами в начале главы.
ПОЗВОНОЧНАЯ ФОРМУЛА
Типичная позвоночная формула человека: CyTh^LsSsCo^s,
т. е.. при общем числе сегментов 33 (34) она включает 24 до-
крестцовых позвонка. Вместе с тем в литературе описаны доволь-
но нередкие отклонения от этого модального типа, которые
обычно носят характер так называемых «компенсированных ва-
риаций», т. е. сдвигов границ между отделами позвоночника на
один сегмент. Напротив, «некомпенсированные вариации», состо-
ящие в изменении общего числа позвонков, чрезвычайно редки.
Изучение различных вариантов позвоночной формулы человека
позволяет заключить, что каждая граница может смещаться в
обоих вероятных направлениях — краниальном и каудальном..
Частота вариаций возрастает в кранио-каудальном направлении,
причем помимо «грубых» сдвигов границ между отделами, сопро-
вождающихся изменениями позвоночной формулы, существует и
обширная группа менее значительных изменений, отражающих
тенденции краниального или каудального перемещения.
Преобладающее большинство скелетов-человека имеют, 24 сег-
мента в дбкрестцовом отделе: по данным разных авторов,’это ти-
пичное для Н. sapiens «видовое число» отмечается в 85—95%.„ в
остальных случаях наблюдаются отклонения на один сегмент (23
или 25,позвонков) (табл. 1).
В большей части работ отмечается некоторая тенденция к пре-
обладанию удлиненных позвоночников (с 25 сегментами), однако
здесь, вероятно, следует принять во внимание такие обстоятель-
ства, как представительность материала (численность серии) и
его половую принадлежность. В некоторых случаях преобладание,
вариантов «удлиненного позвоночника» может связываться с пре-
имущественно мужским контингентом обследованных индивидуу-
мов, так как известна частичная зависимость этого варианта от
, пола. Так, например, в представительной серии из 1239 скелетов
5
Таблица I
ВАРИАНТЫ ЧИСЛА ДОКРЕСТЦОВЫХ ПОЗВОНКОВ У СОВРЕМЕННОГО ЧЕЛОВЕКА
Группа Автор n 25 (%) 24 (%) 23 (%)
Европеоиды Bardeen, 1904 1059 4,4 91,3 4,3
Willis, 1923 557 4,2 95,1 0,7
Lanier, 1939 100 2,0 95,0 3,0
» d Разные авторы 554 7,0 89,7 3,3
» 2 » » 416 3,4 91,6 5,1
Негры США Willis, 1923 185 4,9 93,6 1,6
» , » Lanier, 1939 100 7,0 88,0 5,0
Восточноафриканцы All brook, 1955 206 11,7 84,9 3,4
Южноафриканские банту Shore, 1930 82 5,0 95,0 0,0
Австралийцы <5 Tulsi, 1972 74 5,4 90,6 4,0
» 2 » 51 1,9 92,2 5,9
» вместе 125 4,0 91,2 4,8
Японцы (5 Hasebe, 1913; Nishi, 1928 227 4,9 91,2 4,0
» 2 95 5,3 89,5 5,3
Эскимосы Stewart, 1932 217 12,0 88,0 0,0
Европеоиды, <5 Bernstein, Peterson, 1966 647 6,8 89,0 4,2
монголоиды,
негроиды
> 2 » » 592 3,0 89,1 7,9
» вместе 1239 5,0 89,0 6,0 .
европеоидов, монголоидов и негроидов 25 докрестцовых позвонков
встречается у мужчин чаще, чем у женщин, тогда как 23 докрест-
цовых сегмента, напротив, значительно чаще наблюдаются у жен-
щин. При объединении же обоих полов частота 23 и 25 сегментных
вариантов оказывается примерно одинаковой. С другой стороны,
следует принимать во внимание и специфику скелетного материа-
ла. Так, необычная частота удлиненных туловищных позвоночни-
ков на ископаемых скелетах эскимосов, возможно, определяется
исключением из серии некоторого количества «укороченных» поз-
воночников, как предположительно неполных (Stewart, 1932).
В целом можно заключить, что обычно в тех случаях, когда су-
ществует вполне доброкачественный и полноценный в количест-
венном отношении материал, причем с примерно однозначной
представленностью обоих полов частота обоих основных вариан-
тов докрестцового позвоночника мало различается.
Вопрос о групповых вариациях пока остается открытым, но
роль половой принадлежности сказывается довольно отчетливо.
При этом оба типа (25- и 23-сегментцый) встречаются и у муж-
чин, и у женщин, но с разной частотой. Можно предположить, что
в известной мере эти вариации опосредуют и модальные тенден-
ции к половому диморфизму в длине позвоночника человека. •
Оба нижних отдела осевого скелета (крестцовый и копчико-
вый) значительно более вариабильны. Так, для крестца «типич-
ное число» позвонков 5 встречается, по данным разных авторов,
6
в 69—89% случаев; из двух основных вариантов (6- и 4-членные
крестцы) первый значительно более част. При этом здесь, как и
для докрестцового отдела, отмечена частичная связь с полом: ше-
стичленный вариант чаще встречается у мужчин, а четырехчлен-
ный — у женщин.
Изучение вариаций позвоночной формулы у современных при-
матов позволяет сделать некоторые предположения относительно
ее возможного преобразования в примато- и антропогенезе
(табл. 2).
Таблица 2
ТИПИЧНЫЕ ПОЗВОНОЧНЫЕ ФОРМУЛЫ ПРИМАТОВ*
Приматы Позвонки
шейно-грудно- поясничные докрестцовые крестцовые копчиковые
Тупайеобразные C?Thi3Le 26 3 24—26
Лемурообразные C7Th12_i5 Ц_8 26-30 3—6 6—28
Долгопятообразные C7ThlsLe 26 3 26—27
Игрунковые СДЬ^-тзЦ-? 26 3 25—32
Капуциновые C7Thi3_14L5_7 26—28 3—4,5 16—28
Коата C7Thi4L4 25 3—4 31
Церкопитековые СДЧ^з-^Ев-? 26 3 3—28
Гиббон С7ТЬ1зЬ5_4 25—24 5 3—2
Орангутан C7Th12L4 23 5—6 3
Шимпанзе C7Th13L4_3 24 6-5 3
Горилла С7ТЬ1зЬз«4 24 6—5 3
Человек C7Thi2L5 24 5 4
* По данным Радлауэра (Radlauer, 1908); Дэкворса (Duckworth, 1915); Мартина (Martin, 1928);
Вебера, (1936); Фика (Fick, 1936); Шульца (Schultz, 1961, 1963).
Тенденция в приведенном сравнительно-анатомическом ряду
прослеживается достаточно отчетливо: по направлению от низших
приматов к высшим наблюдается явственная редукция числа до-
крестцовых позвонков, затрагивающая преимущественно пояснич-
ный отдел позвоночника. Число докрестцовых сегментов, состав-
ляющее у низших приматов, к которым многие систематики при-
числяют и тупайеобразных, обычно 26—30, уменьшается до 26 у
низших обезьян и 25—24 (и даже 23) у высших узконосых при-
матов. К такому выводу пришли многие авторы, изучавшие осе-
вой скелет в сравнительно-анатомическом аспекте (Ruge, 1892;
Keith, 1903; Todd, 1922; и др.). Вместе с тем выявляется тенден-
ция к укреплению крестцового отдела: от 3 сегментов у низших
приматов и низших обезьян до 5 и даже 6—7 позвонков у высших
узконосых. Таким образом, основные преобразования позвоночной
формулы в филогенезе приматов, очевидно, были связаны с пояс-
нично-крестцовой границей, где осуществлялись редукция числа
поясничных сегментов и возрастание количества позвонков крест-
цового отдела (прежде всего «истинных» сакральных сегментов,
участвующих в сочленении с подвздошными костями).
7
Следует подчеркнуть, что эволюция позвоночной формулы в
приматогенезе имела, по-видимому, и достаточно выраженную
локомоторную опосредованность, которая выявляется на всех
уровнях сравнительно-анатомического ряда. Так, уже среди низ-
ших приматов в известной степени может быть отмечена и тен-
денция к укорочению туловищного отдела позвоночника (до 25
сегментов у некоторых собственно лемуров), а у передвигающих-
ся прыжками при выпрямленном положении тела крупных индри
наблюдается укрепление крестца (до 4 сегментов). Среди лорие-
вых формы, передвигающиеся по типу медленного лазания с об-
хватыванием, что многие исследователи считают первичным ти-
пом локомоции приматов, сохраняют примитивное высокое число
докрестцовых позвонков 29—30 (например, Perodicticus, Arctoce-
bus, Nycticebus).
Другой полюс в отряде приматов представляют высшие узко-
носые, особенно крупные антропоиды (орангутан, шимпанзе, го-
рилла), которые даже превосходят человека в редукции туловищ-
ного отдела позвоночника. Так, число докрестцовых сегментов
одинаково у человека и африканских антропоморфных (шимпанзе
и гориллы), однако снижение его до 23 сегментов происходит у
человека, как упоминалось выше, обычно не более чем в 5—6%,
тогда как у шимпанзе эта редукция наблюдается втрое чаще —
19%. Что касается орангутана, то здесь 23 докрестцовых позвон-
ка — типичное число для этого антропоида, а в 15% отменено
всего 22 сегмента. Сакрализация нижнего поясничного сегмента
особенно часта у горилл, причем, в отличие от ора'нга и шимпан-
зе, у гориллы этот сакрализованный позвонок обычно сохраняет
свою независимость. Одновременно у антропоидов и человека наб-
людается укрепление крестца с повышением числа сегментов до
пяти, а у крупных антропоидов — даже до шести-семи. В послед-
нем случае преобразование позвоночной формулы имеет, по-види-
мому, непосредственное отношение к своеобразному способу ло-
комоции этих обезьян — брахиации при которой туловище дер-
жится в довольно выпрямленном положении. С этой точки зрения
следует рассматривать и нередкое, особенно у массивных форм
антропоидов — гориллы и орангутана, появление дополнительных
подвздошно-крестцовых сочленений, наблюдающихся также у че-
ловека. По-видимому, подобный процесс укорочения туловища и
укрепления подвздошно-крестцовых сочленений мог возникать па-
раллельно или конвергентно в разных таксонах приматов. Так,
например, у «брахиаторов Нового Света» — коат (Ateles), кото-
рых по локомоции нередко сопоставляют с гиббонами, как и у
последних, наблюдается укорочение докрестцового позвоночника
до 25 сегментов (см. табл. 2). У человека перестройка позвоноч-
* Способ передвижения, при котором основную роль играют верхние конечности
(от лат. brachium — плечо, рука), испытывающие при этом более или менее
выраженное удлинение.
8
ной формулы в связи с Прямохождением в общем идет в том же
направлении, что в известной степени можно объяснить влиянием
общего фактора — повышения статической нагрузки на пояснич-
но-крестцовый отдел позвоночника при выпрямлении туловища.
Эти примеры подчеркивают роль локомоторных адаптаций в эво-
люции позвоночной формулы у приматов.
Палеонтологические данные при всей их ограниченности все
же подтверждают существование указанной тенденции в филоге-
незе приматов. В качестве примера можно привести одного из ве-
роятных миоценовых предшественников современных гиббонов —
плиопитека (Pliopithecus vindobonensis) ’, у которого было увели-
чено число поясничных и уменьшено число крестцовых позвонков,
сравнительно с современными гиббонами, что указывает на про-
исшедшую позднее редукцию в числе туловищных сегментов и
укрепление крестца в гиббоноидной линии. Строение скелета
плиопитека свидетельствует о нейтральности пропорций его ко-
нечностей и бтсутствии сколько-нибудь отчетливых признаков-
«структурной брахиации». Вместе с тем у настоящего брахиа-
тора, каким являлась другая миоценовая обезьяна — ореопи-
тек, — число поясничных позвонков уже сократилось до пяти, как
у современных гиббонов, однако отставание наблюдалось срав-
нительно с современными крупными антропоидами, у которых
обычно существует только четыре или даже три поясничных
сегмента.
К сожалению, малочисленность и очень слабая сохранность
соответствующих материалов не позволяет сделать прямого опре-
деления позвоночной формулы у непосредственных предшествен-
ников человека. От наиболее интересной в этом плане группы ав-
стралопитековых известны лишь единичные находки позвонков,
которые не дают возможности реконструировать позвоночную
формулу у этих приматов. .Можно только предположить, что, по-
скольку локомоторная обусловленность позвоночной формулы у
человека и других приматов достаточно отчетлива, формула ав-
стралопитековых, прямохождение которых может считаться до-
казанным на основании других остеологических данных, была
весьма близкой к таковой прямоходящих гоминид вообще, в том
числе и современного человека, хотя морфологическое своеобра-
зие отдельных сегментов позвоночника очевидно, имело место —
у массивных австралопитековых в большей степени, чем у гра-
цильных (Robinson, 1970). ,
Скудные данные об осевом скелете более поздних ископаемых
гоминид также не позволяют сделать никаких заключений о чис-
ленных вариациях позвоночника. Намек на легкое каудальное пе-
ремещение грудно-поясничной границы существует, правда, на
позвоночниках некоторых поздних палеоантропов — Схул V и,
1 Прямая генеалогическая связь плиопитека с гиббонами оспаривается некото-
рыми исследователями (Lewis, 1972).
9
возможно, Схул IV, у которых реберные фасетки были найдены
на поперечных отростках XI грудного позвонка; возможно су-
ществование каудального сдвига и на верхней границе грудной
клетки У'Схул I, но подобные признаки, как известно, не пред-
ставляют ничего исключительного, встречаясь и у современного
человека.
Что касается вариаций позвоночной формулы у Н. sapiens fos-
sitis, то имеющиеся в литературе данные свидетельствуют как
будто о существовании здесь той же полиморфности вариаций,
что и у Н. sapiens recens. Указания некоторых авторов на стойкое
преобладание тех или иных морфологических вариантов по отно-
шению к отдельным группам ископаемого человека (Ferembach,
1963) также не противопоставляют его Н. sapiens recens, посколь-
ку оно может встречаться и у последнего.
Резюмируя все сказанное, можно предположить, что исходная
позвоночная формула приматов была близка к той, которая наб-
людается у современных тупаий и, может быть, лемуров с первич-
ным обхватывающим типом лазанияВ ее дальнейших преобра-
зованиях существенную роль сыграли разнообразные локомотор-
ные адаптации, приведшие у высших форм узконосых приматов
к существенной перестройке пояснично-крестцовой границы.
Учитывая связь позвоночной формулы с локомоцией, можно пред-
положить, что, по крайней мере, у всех прямоходящих высших
приматов она уже приобрела типичное соотношение поясничных
и крестцовых сегментов, хотя это пока находит лишь косвенное
подтверждение. Ввиду вообще довольно раннего оформления
структурного комплекса бипедии в антропогенезе можно предпо-
лагать и достаточно большую древность позвоночной формулы
современного человека.
В специальной литературе получила широкое распространение
концепция, связывающая вышеописанную сравнительно-анатоми-
ческую тенденцию с численными вариациями позвоночника сов-
ременного человека — прежде всего в аспекте биогенетического
закона (Rosenberg, 1876, 1899, 1907). Этот автор основывался
преимущественно на онтогенетических данных (маленькая серия
из 9 эмбрионов очень раннего возраста), где существовала вы-
сокая частота удлиненных (25-сегментных) докрестцовых позво-
ночников. Розенберг считал, что в ‘раннем онтогенезе человека
происходит перемещение таза в краниальном направлении при
посредстве так называемого «копчикового архива», в который
«сдаются» нижние элементы позвоночника. Однако Бардин (Bar-
deen, 1904, 1905), изучивший более представительный материал
,по ранним эмбрионам человека (46 эмбрионов 13—50 мм длины),
пришел к противоположному результату, т. е. определил, что в те-
чение данного периода подвздошная бластема дифференцируется
1 По Шульцу (1953), предположительная исходная позвоночная формула при-
матов: С7Т/г13Ев5зСо_25.
10
в области более краниальной, чем та, .которую впоследствии за-
нимают подвздошные кости. Более поздние работы (Степанова,
1955, 1961; Попова-Латкина, 1956), проведенные на сериальных
препаратах эмбрионов человека очень ранних стадий развития
(5—50 мм длины), показали, что подвздошные кости и крестец,
по-видимому, независимы в своем развитии, и не наблюдается
следов каких бы то ни было перемещений таза по отношению к
позвоночнику. Предполагается, что причины изменчивости числа
позвонков в пояснично-крестцовой области следует искать на бо-
лее ранних стадиях, когда в каудальном отделе появляется боль-
шое количество мелких мезенхимных закладок, от которых зави-
сит длина крестцово-копчикового отдела позвоночника (Попова-
Латкина, 1965).
По-видимому, уже на очень ранних стадиях эмбриогенеза су-
ществует значительное морфологическое разнообразие в регио-
нальном расчленении позвоночника, закономерно дополняющее
аналогичную картину для более поздних возрастов. Это предпо-
ложение вполне соответствует мнению, высказанному А. Н. Се-
верцовым (1939) в итоге анализа меристических вариаций позво-
ночника некоторых рептилий, о ранней детерминации позвоночной
формулы в онто- и филогенезе (архаллаксис).
А. Н. Северцов предполагал, как известно, что меристические
вариации позвоночника имеют наследственную базу. Примени-
тельно к человеку генетическая детерминация позвоночной фор-
мулы была показана Кюнэ (Kiihne, 1932, 1936) на основании изу-
чения близнецов и семейных линий. Интересны в этой связи и
данные Уильямса (Williams, 1965), определившего наличие пере-
ходного пояснично-крестцового позвонка в ряду: мать — дочь в
трех различных семьях. Также и необычная концентрация неко-
торых морфологических вариаций позвоночника (в частности,
25-сегментных докрестцовых позвоночников), отмечающаяся иног-
да в популяциях современного человека, объясняется в отдельных
случаях высоким уровнем инбридинга (Bennett, 1972).
Первоначально было высказано предположение, что укороче-
ние позвоночника у человека обусловлено доминантным, а удли-
нение — рецессивным геном. Однако признак дополнительного
докрестцового позвонка, по-видимому, не определяется простыми
менделевскими генами, но существует известное число возможных
детерминантов меристических вариаций (Sawin, 1937; Green,
1941), причем наследуются прежде всего не аномалии позвоноч-
ника, но тенденция к краниальному или каудальному сдвигам *.
На экспрессию этих генов существенное влияние оказывает пол
(Fischel, 1906; Trotter, 1929; Danforth, 1930; Bornstein, Peterson,
1966;.Tulsi, 1972).
1 Практически этот признак уже используется в рамках полисимптоматического
диагноза сходства для антропологического доказательства отцовства (Martin,
Sailer, 1958).
11
Другая сторона концепции Розенберга связана с истолковани-
ем изменчивости позвоночной формулы на скелете современного
человека с точки зрения упомянутой филогенетической тенденции;
она "разделяется многими авторами (Ruge, 1892; Ancel, Sencert,
1902; Adolphi, 1905; Regalia, 1895; Видерсгейм, 1900; Keith, 1903;
Todd, 1922; Willis, 1923; Мартин, 1928). Применив принцип онто-
филогенетической редукции к объяснению численных вариаций
на скелете современного человека, Розенберг ввел их разграниче-
ние на «прогрессивные», идущие в направлении краниального
сдвига туловищного позвоночника и двустороннего укорочения
грудной клетки, и «регрессивные», происходящие в противопо-
ложном направлении. Ансель и Сансер (Ancel, Sencert, 1902}
обозначают эти процессы как «преувеличение» в первом и «за-
паздывание» во втором случае. При этом некоторые исследовате-
ли идут еще дальше, утверждая, что и в настоящее время в мно-
гообразной картине изменчивости осевого скелета выявляется
определенная тенденция, а именно в направлении той же филоге-
нетической редукции в длине туловищного отдела позвоночника
и грудной клетки (Видерсгейм, 1900; Бушмакин, 1913; Лехе, 1913;
Сперанский, 1925; Тонков, 1953; Жеденов, 1962). Суть этих изме-
нений они видят в постепенном краниальном перемещении крест-
ца при редукции числа позвонков и ребер и слиянии атланта с
черепом, причем указанный процесс рассматривается как про-
грессивный, связанный с выгодой сохранения равновесия при пря-
мостоянии и прямохождении.
Как известно, концепция «направленной изменчивости» была
рассмотрена А. П. Быстровым (1957), который, впрочем, ограни-
чился тем, что критиковал ее с точки зрения отрицания формооб-
разующей роли естественного отбора у современного человека.
Вместе с тем специального рассмотрения заслуживает и другая
сторона вопроса, а именно проблема коррелирующих вариаций,
т. е. соотношения региональных сдвигов на целом позвоночнике.
Действительно, рассматривая не изолированные меристические
вариации позвоночника, но сопоставляя позвоночные формулы в
целом nd литературным материалам, представляющим различные
этно-территориальные группы земного шара, можно прийти к вы-
воду о наличии определенной сопряженности региональных сдви-
гов на индивидуальных позвоночниках.
Разберем этот вопрос на примере грудной клетки, в отноше-
нии которой Розенберг и ряд исследователей его школы призна-
вали тенденцию к ее двустороннему укорочению на скелете сов-
ременного человека. Действительно, на границах грудной клетки
наблюдается достаточно тесная корреляция (Kiihne, 1936). Одна-
ко в преобладающем большинстве случаев оба полюса сдвигаются
однозначно — чаще краниально, значительно реже — каудально.
При разнонаправленных перемещениях границ грудной клетки
ее верхний полюс имеет тенденцию к краниальному, .нижний —
к каудальному сдвигу. В целом, на границах грудной клетки пре-
12
обладает однозначное перемещение («компенсация» по Адольфи)
или удлинение, но не укорочение, как предполагал Розенберг.
Компенсаторные явления на границах грудной клетки прослежи-
ваются даже при отсутствии «грубых» изменений позвоночной
формулы, при стертых тенденциях к краниальному или каудаль-
ному перемещению границ.
Так, при редукции XII пары ребер на нижней границе грудной
клетки происходят компенсаторные процессы: двенадцатый груд-
ной позвонок не вовлекается в процесс редукции. Об этом свиде-
тельствует, в частности, отрицательная корреляция между длиной
XII ребра и высотой тела XII позвонка. В выраженных случаях
компенсации редукция XII пары ребер может сопровождаться
появлением «шейных ребер» на седьмом шейном позвонке; по-
видимому, проявление той же компенсаторной тенденции можно
усмотреть и в отрицательном знаке связи между длинами XII
ребра и костального элемента VII шейного позвонка, что согла-
суется с фактами компенсации при «стертых» формах шейных ре-
бер. Другой пример «компенсаторных корреляций» — тенденция к
усилению I ребра при редукции XII; между длиннотными разме-
рами этих ребер также намечается отрицательная связь. Приме-
чательно, что при этом происходит некоторое увеличение и I груд-
ного позвонка.
Сравнение величин коэффициентов вариации в ряду ребер сви-
детельствует, что у сходных (гомодинамных) элементов скелета
величина этого параметра может служить действительным пока-
зателем редукционных процессов. Так, при почти равной в сред-
нем длине ребер 1и XII пар (соответствующие величины в нашей
серии 108,1 и 110,9 мм1 значения коэффициентов вариации раз-
личаются примерно в четыре раза — 6,55 и 26,55%). При этом
изменчивость XI пары ребер значительно ниже, чем XII
(ц=8,62%).
Поскольку гистологическая редукция является одним из про-
явлений рудиментации органа (Северцов, 1939), не лишено осно-
ваний предположение, что столь существенные отличия в в ар на-
бильности размеров XII ребра могут ассоциироваться и с опреде-
ленными отклонениями в его гистологической структуре. Для его
проверки нами были изучены препараты XII ребра в рудиментар-
ном (длина ребра меньше 5 см) и нерудиментарном состоянии, а
также I и VII ребер — общим числом 240 препаратов. В итоге
можно прийти к заключению, что максимальные различия в строе-
нии компакты проявляются на концах рудиментарного XII реб-
ра, особенно на его грудинном конце, где существуют явные при-
знаки гистологической редукции. Слой компакты редуцирован до
минимума, встречаются лишь единичные остеоны атипичного
строения и весьма варьирующей формы, число полных колец осте-
она уменьшено. Здесь отмечен максимум очень крупных остеонов
1 Серия мужских мацерированных скелетов (л=50).
13
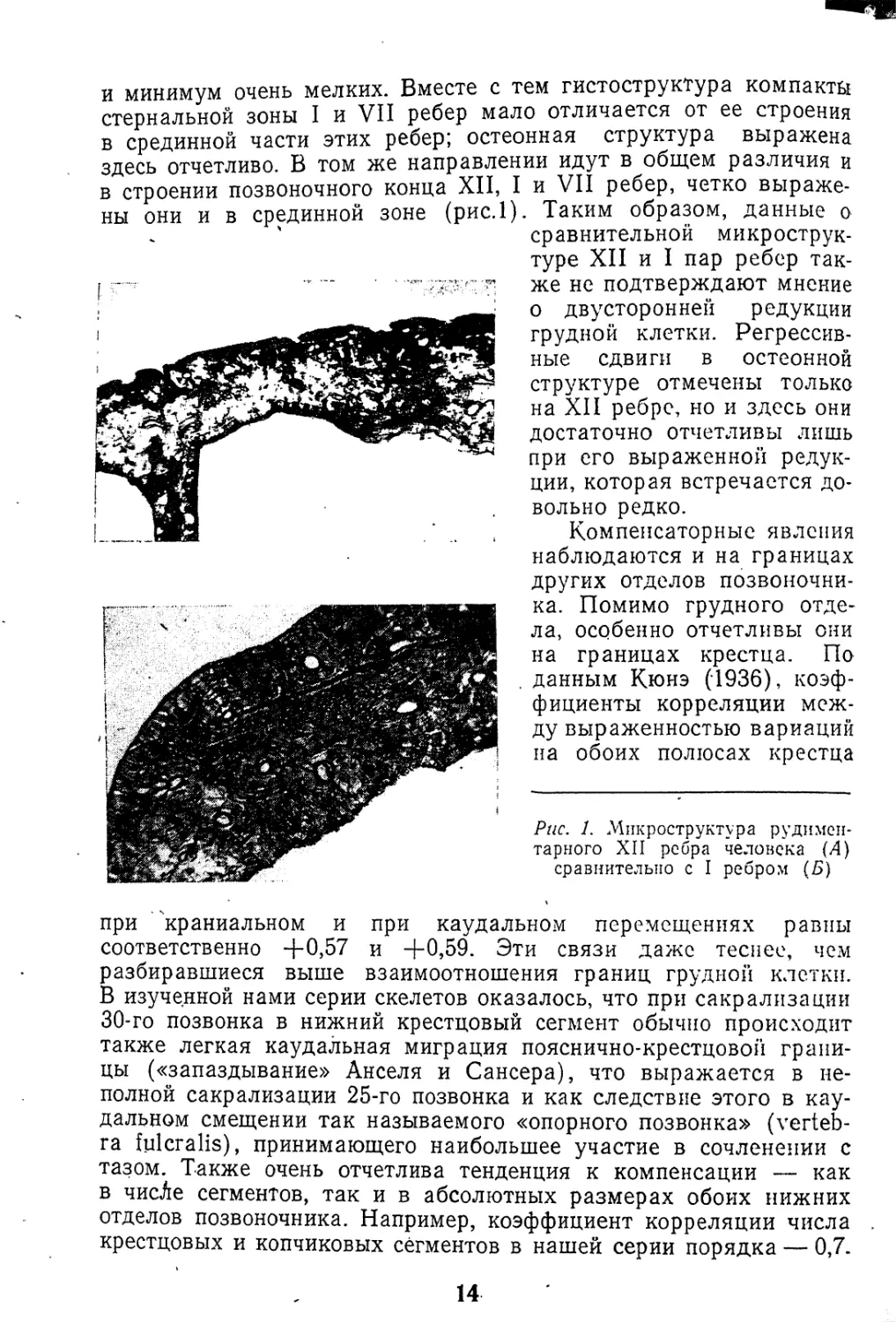
и минимум очень мелких. Вместе с тем гистоструктура компакта
стернальной зоны I и VII ребер мало отличается от ее строения
в срединной части этих ребер; остеонная структура выражена
здесь отчетливо. В том же направлении идут в общем различия и
в строении позвоночного конца XII, I и VII ребер, четко выраже-
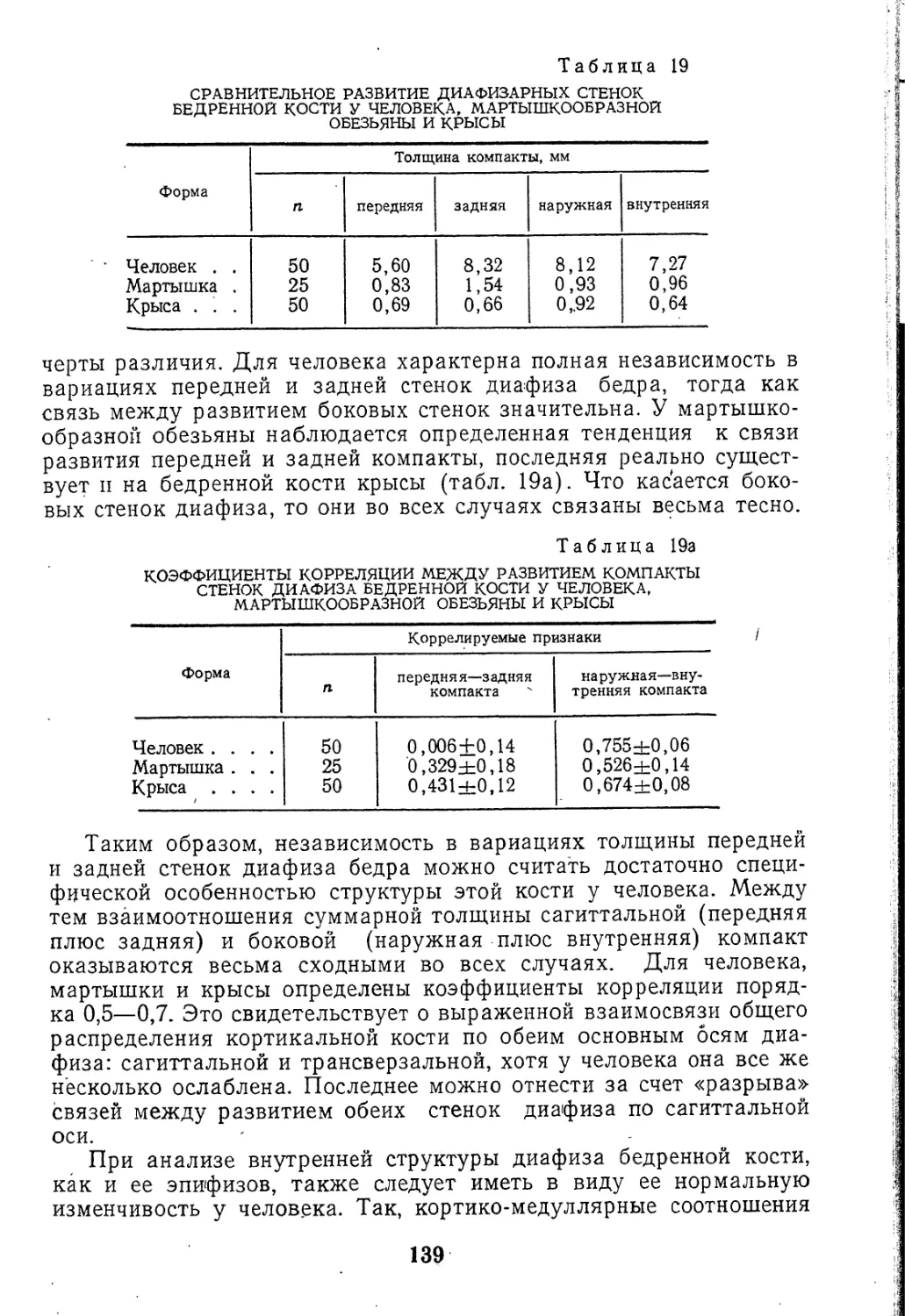
ны они и в срединной зоне (рис.1). Таким образом, данные о
сравнительной микрострук-
туре XII и I пар ребер так-
же не подтверждают мнение
о двусторонней редукции
грудной клетки. Регрессив-
ные сдвиги в остеонной
структуре отмечены только
на XII ребре, но и здесь они
достаточно отчетливы лишь
при его выраженной редук-
ции, которая встречается до-
вольно редко.
Компенсаторные явления
наблюдаются и на границах
других отделов позвоночни-
ка. Помимо грудного отде-
ла, особенно отчетливы они
на границах крестца. По
данным Кюнэ (1936), коэф-
фициенты корреляции меж-
ду выраженностью вариаций
на обоих полюсах крестца
Рис. 1. Микроструктура рудимен-
тарного XII ребра человека (4)
сравнительно с I ребром (Б)
при "краниальном и при каудальном перемещениях равны
соответственно +0,57 и +0,59. Эти связи даже теснее, чем
разбиравшиеся выше взаимоотношения границ грудной клетки.
В изученной нами серии скелетов оказалось, что при сакрализации
30-го позвонка в нижний крестцовый сегмент обычно происходит
также легкая каудальная миграция пояснично-крестцовой грани-
цы («запаздывание» Анселя и Сансера), что выражается в не-
полной сакрализации 25-го позвонка и как следствие этого в кау-
дальном смещении так называемого «опорного позвонка» (verteb-
ra fulcralis), принимающего наибольшее участие в сочленении с
тазом. Также очень отчетлива тенденция к компенсации — как
в числе сегментов, так и в абсолютных размерах обоих нижних
отделов позвоночника. Например, коэффициент корреляции числа
крестцовых и копчиковых сегментов в нашей серии порядка — 0,7.
14
Сказанное выше не позволяет видеть в изменчивости позвоноч-
ной формулы современного человека сколько-нибудь выраженной
тенденции к укорочению туловища и грудной клетки. Оба экстре-
мальных варианта докрестцового позвоночника встречаются, по-
меньшей мере, с одинаковой частотой.
Применительно к приматам модальная тенденция к компен-
сации отмечалась Шульцем (1938/1939). При межгрупповых со-
поставлениях на узконосых приматах, очевидно, что редукция •
(возрастание) числа докрестцовых сегментов не вызывают соот-
ветствующих изменений в абсолютной длине туловища. Также и
относительные размеры докрестцовых позвонков или вовсе не
связаны с их числом в соответствующем отделе (как для шейных
сегментов), или обнаруживают даже некоторую тенденцию к сни-
жению при возрастании числа позвонков (поясничный и грудной
отделы). При внутривидовых сопоставлениях у узконосых обезьян
намечается некоторое снижение относительной длины грудно-по-
ясничных позвонков при уменьшении их числа, но меньшее, чем
следовало бы ожидать теоретически (Хрисанфова, 1962).
С подобными явлениями компенсации мы встретимся и даль-
ше, при рассмотрении других отделов скелета человека.
ФОРМА ПОЗВОНОЧНИКА И ГРУДНОЙ КЛЕТКИ
ИСКОПАЕМЫХ ГОМИНИД
Палеонтологические материалы по осевому скелету представ-
лены главным образом более поздними формами гоминид — па-
леоантропами и верхнепалеолитическим Н. sapiens. Существуют
лишь единичные сегменты позвоночника от ранних предшествен-
ников человека — австралопитековых. Правда, предпринимались,
попытки реконструировать длину туловища 1 (основным компонен-
том которой является длина грудно-поясничного отдела позвоноч-
ника) у A. africanus и A. robustus (Biegert, Maurer, 1972). Расче-
ты, произведенные на основе региональных пропорций у близких
таксонов приматов, дали для длины туловища австралопитеков
соответственно 39СГ и 440 мм, что помещает их между крупными
гиббоновыми и шимпанзе среди современных гомшюидов, а из
ископаемых форм сближает с плиопитеком (387 мм) и ореопите-
ком (425 мм). Однако подобная реконструкция, основанная в
каждом случае австралопитековых всего на двух сегментах, ра-
зумеется, лишь приблизительна. Более полные сведения о тоталь-
ных размерах позвоночника и его региональном расчленении су-
ществуют для палеоантропов (табл. 3 и За).
Реконструировать примерную длину (высоту) докрестцового
отдела возможно только для двух палестинских палеоантропов.
1 Условный составной размер.
15
чи
Табл ица 3
АБСОЛЮТНЫЕ И ОТНОСИТЕЛЬНЫЕ ДЛИНЫ ОТДЕЛОВ ДОКРЕСТЦОВОГО ПОЗВОНОЧНИКА *
- НЕКОТОРЫХ ГОМИНИД*
Отделы
шейный (3-7) грудной поясничный докрестцовый |
Гоминиды абс. отн. абс. отн. абс. отн. абс. отн.
м И. sapiens Min. ... Max. Схул V <- Схул IV ? 1 Табун I (0 Шапелль 62,1 52,1— 68,4 55,5 15,0 14,1— 15,7 14,1 224,5 201,4— 243,1 225,0? 54,1 53,4— 56,0 57,3? 128,3 111,3— 143,6 112,4 119,6 132,0 30,9 29,9— 31,4 28,6 414,9 364,8— 455,1 392,9 377,0 24,5 23,3- 25,7 , 22,3 24,5
* Соответственно в мм и %; по данным разных авторов (Hasebe, 1913; McCown» Keith, 1939; La*
nier, 1939; Patte, 1955; Хрисанфова, 1962; Tulsi, 1972).
Таблица За
СООТНОШЕНИЕ НА ШЕЙНО-ГРУДНОЙ ГРАНИЦЕ
У НЕКОТОРЫХ ГОМИНИД
Ci + C. + C, | Tt-f-Ta + T,
Гоминиды мм
sapiens... ,... ... ......
' ЛПапелль................... '.
"Крапина......................
Схул V ......................
Шанселад...............' • . .
39,4—40,8
37,0
37,0
36,0
38,5
47,3—62,7
. 47.0
53,0
женщины Табун I и мужчины Схул V. Первый размер составляет
около 377 мм при росте примерно 154 см, что дает вполне совре-
менные соотношения. Указатель относительной длины докрестцо-
вого позвоночника (в % длины тела) даже совпадает здесь со
средним для современного человека. Иначе обстоит дело у вто- .
рого палеоантропа — Схул V. Этот Высокорослый мужчина с ми-
нимальной реконструированной длиной тела 176,5 см обладал
относительно весьма укороченным'позвоночником, что, по-видимо-
му, выводило его за рамки известных групповых вариаций этого
указателя у современного человека. Из табл. 3 следует, что эта
резкая укороченность докрестцового отдела создавалась преиму-
щественно за счет шейных и поясничных позвонков. В определен-
ной степени подобная тенденция была, очевидно, свойственна и
другим людям Схул — мужчине Схул IV и женщине Схул VII, а
также ребенку Схул I. Несколько иной была ситуация у класси-
ческого неандертальца Шапелль. Шейные и верхнегрудные по-
звонки шапелльца малы, но поясничный отдел приближается по
длине к среднему для современных мужчин (см. табл. 3 и За).
16
Напомним, что и Буль в своей реконструкции придал докрестцо-
вому отделу позвоночника Шапелль в целом длину, соответствую-
щую средней для современного европеоидного варианта, что осо-
бенно существенно ввиду малых размеров тела этого неандер-
тальца. О других палеоантропах (Крапина, Ла Кина) трудно ска-
зать что-либо определенное из-за отсутствия достаточно полных
материалов по осевому скелету.
Аналогичная картина наблюдается и у верхнепалеолитических
неоантропов. Одна из наиболее полных находок — Шанселад —
свидетельствует, что размеры и пропорции позвонков здесь были
вполне современными, учитывая к тому же небольшую длину тела
этого человека (160 см, по Billy, 1969). То же относится и к более
высокорослым неоантропам (например, Оберкассель, Вейрье
и др.). В то же время кроманьонец из Грота Детей (Verneau,
1906) имел, по-видимому, поразительное укорочение докрестцово-
го позвоночника относительно роста, очень большого у этого го-
минида. Эти своеобразные пропорции подчеркнуты здесь в еще
большей степени, чем у мужчины Схул V: так, длина докрестцо-
вого отдела (реконструированная, с дисками) составляла у кро-
маньонца 580,5 мм против 583,8 мм у Схул V при длине тела со-
ответственно 190 и 176,5 см.
Таким образом, можно констатировать, что среди ископаемых
гоминид среднего и верхнего палеолита существовали различные
варианты абсолютного и относительного развития докрестцового
позвоночника, как это наблюдается и среди современных популя-
ций. Особое положение занимают люди .Схул (прежде всего
Схул V), сближающиеся по исключительной укороченности ту-
ловища с кроманьонцами, что в меньшей степени было свойствен-
но и некоторым другим верхнепалеолитическим (Пржедмости) и
неолитическим популяциям. Как этого и следовало ожидать, ва-
риации туловищного позвоночника в целом не противопоставляют
палеоантропа неоантропу.
Наибольший интерес в связи с проблемой локомоции и осан-
ки представляет изучение на палеоантропологическом материале
«палеогенных изгибов» позвоночника, связанных с прямохожде-
нием, — шейного и поясничного.
Поскольку шейный лордоз обусловлен исключительно межпоз-
воночными дисками и вертикальный указатель этого отдела не
позволяет сделать никаких заключений о степени выраженно-
сти изгиба, о нем судят косвенно, преимущественно по морфо-
логии остистых отростков, а также положению затылочного отвер-
стия.
Классическое представление о крайне незначительном, если
не вовсе отсутствующем, изгибе шейного отдела позвоночника
неандертальца связано главным образом с известной реконструк-
цией Буля, на которой шапелльский человек изображен с положе-
нием головы, выдвинутым вперед и весьма отличным от современ-
ного. Эта ориентировка основана на морфологии остистых от-
2 Е. H. Хрисанфова
17
ростков: прямизна, горизонтальность и отсутствие раздвоенности
этих апофизов считаются атрибутом неполного выпрямления.
Рассмотрим два вопроса: 1. Существует ли действительная
связь между морфологией остистых отростков и шейным лордо-
зом? 2. Как отражаются вариации шейной кривизны на положе-
нии головы человека? .
Как известно, три нижних шейных позвонка шапелльского че-
ловека, как и первый грудной, имели горизонтальное положение
остистых отростков, не показывающих признаков бифуркации; то
же в общем относится и к Схул V, где известны все шейные сег-
менты. Такие особенности показывает позвоночник шимпанзе,
тогда как у современного человека апофизы коротки и сильно
наклонены назад. Эти морфологические различия связываются,
согласно распространенному мнению, с положением головы, по-
скольку остистые отростки у антропоидов функционируют как ап-
парат подвешивания для относительно тяжелого черепа, являясь
местом прикрепления особенно мощной затылочной мускулатуры.
Однако в- последнее- время этот взгляд подвергся существенному
пересмотру. По мнению некоторых авторов (Straus, Cave, 1957;
Stewart, 1962), ориентировка остистых отростков у палеоантропов
не выходит за пределы вариаций этого признака у современного
человека. Арамбур (Arambourg, 1955) приводит, например, рент-
генограмму шейного отдела позвоночника современного европей-
ца с удлиненными и выпрямленными апофизами, что не сопро-
вождалось, однако, какими-либо нарушениями в положении шеи
или головы. Подобные же факты в современных популяциях от-
мечены Саразином (Sarasin, 1916/1922), Питтаром (Piitard,
1944), Паттом (Patte, 1955). Отсутствие бифуркации остистых
отростков шейных позвонков также нередко наблюдается у совре-
менных людей, тогда как у антропоидов она может встречаться в
норме на II и III шейных сегментах, а Ле Дубль (Le Double,
1912) указывает даже на случай раздвоения апофиза пятого шей-
ного позвонка у шимпанзе. По этим причинам нельзя приписы-
вать этому признаку большую важность ни с таксономической, ни
с функциональной точки зрения
Об этом же свидетельствуют критические замечания в адрес
самой реконструкции Буля (Патт, 1955): при принятом этим ав-
тором способе ориентировки позвонков их тела у ископаемого
человека больше наклонены вперед, чем у современного, что из-
вращает сравнение и преувеличивает различия; к тому же, если
придать телам позвонков шимпанзе тот же сагиттальный размер,
что и у человека, станет очевидным относительно гораздо большее
1 Высказанное некоторыми исследователями (Capecchi, 1965) мнение, что наклон-
ное положение остистых отростков у современного человека возникло в про-
цессе адаптации к тяжелому^ труду, не очень согласуется с тем фактом, что
именно на скелетах неандертальцев наиболее выражены признаки «силовой
адаптации».
18
развитие у антропоида остистых отростков позвонков. К этому
следует еще прибавить/'что, вероятно, морфология остистых от-
ростков варьировала у палеоантропов, как это видно хотя бы из
сравнения трех последних шейных позвонков Крапины и Ла Ша-
пелль. Но даже, приняв для некоторых палеоантропов, как воз-
можно и для ископаемых неоантропов из Пржедмости, более сла-
бый шейный лордоз, мы не сможем заключить на этом основа-
нии о сколько-нибудь заметных отклонениях в положении головы
от современного типа. Действительно, по степени выраженности
изгиба шейный отдел широко варьирует у современных людей.
При рентгенографическом изучении шейных позвонков у совре-
менных европейцев, например, нередко выявлялись варианты с
прямолинейным контуром и даже кифотическим изгибом при нор-
мальном положении тела и головы (Girard, 1947). Об этом же
свидетельствуют данные Джаля, Миллера и Робертса (Juhl, Mil-
ler, Roberts, 1962), нашедших выраженный лордоз только в 60%,
тогда как в Vs всех случаев наблюдалось обратное направление
изгиба, т. е. кифоз.
Конфигурация шейного отдела позвоночника непостоянна и,
по-видимому, в значительной степени зависит от осанки индиви-
дуума. Отсюда следует, что вариации его изгиба не имеют прямой
связи с обусловленностью равновесия головы, обладающей соб-
ственным физиологическим балансом по отношению к шейному
отделу (лабиринт-окуломоториус), что позволяет ей сохранять
равновесие даже при сильном сколиозе (Cramer, 1956). Следует
подчеркнуть, что преувеличение лордоза вовсе не является бла-
гоприятным фактором ни с точки зрения статики головы, ни с
точки зрения условий физического развития. Нужно также иметь
в. виду, что при весьма сходной морфологии шейного отдела по-
звоночника различия в степени его подвижности могут быть зна-
чительными и наоборот (Тугапсе, 1958).
Обычно при обсуждении развития шейного лордоза у иско*
паемых гоминид привлекается также морфология области заты-
лочного отверстия, поскольку его положение зависит у млекопи-
тающих, в том числе и приматов, от типичной ориентировки го-
ловы (Ковешников, 1968). С этой точки зрения, основание черепа
было, по-видимому, вполне гоминидным у представителей самых
различных вариантов палеоантропов, например Схул V, Ла Фер-
расси,. Саккопасторе I, Нгандонг, Брокен-Хилл. Некоторые от-
клонения в ориентации затылочного отверстия на черепе ша-'
пелльца, возможно, объясняются деформацией основания черепа
при его реконструкции.
Следовательно, не выявляется существенных отличий в строе-
нии шейного отдела позвоночника и его ориентировке у ископае-
мых и современных гоминид. Самое большее, можно предполо-
жить проявление «компенсаторных тенденций» прежде всего у
массивных форм палеоантропов с относительно крупным лицевым
отделом черепа.
2*
19
Большой интерес представляет развитие поясничного изгиба
у ископаемых гоминид, поскольку его выраженность в значитель-
ной степени обусловливает статику, т. е. осанку индивидуума.
Здесь также в первую очередь возникает вопрсс методическо-
го порядка: насколько правомерно заключать о степени пояснич-
ного лордоза на основании морфологии поясничных позвонков.
Действительно, общие поясничные указатели значительно отлича-
ются от'костных, ибо поясничный лордоз в гораздо большей сте-
пени обусловлен межпозвоночными дисками, чем позвонками.
Одна и та же разница между вентральной и дорзальной высо-
тами тела позвонка может соответствовать разной степени изгиба
(Vallois, Lazorthes, 1942), причем компенсаторные свойства дис-
ков проявляются даже по отношению к различным популяциям и
нивелируют различия в вертикальном поясничном указателе
(Schaepelynck, 1959) *. Кроме того, следует иметь в виду, что по-
ясничный лордоз — сложное образование, состоящее из несколь-
ких компонентов, коррелирующих между собой, — при том, что
ни. один из них в отдельности не характеризует полностью его
степень ( см., например, Louis, Guerinel, Mambrini, 1965; Цивьян,
Швец, 1973). Это целиком относится и к таким остеометрическим
показателям, широко использующимся при анализе ископаемых
материалов, как вертикальный поясничный указатель и указатель
клиновидности поясничного позвонка1 2. Сказанное заставляет
отнестись с осторожностью к данным, полученным на основании
измерения позвонков, но было бы неверно, с другой стороны, не
придавать им никакого значения; так, например, при вертикаль-
ном поясничном указателе ниже 98,0 существует особо отчетли-
вая выраженность поясничного лордоза, так называемые куртора-
хидные позвоночники.
Представление о конфигурации поясничного отдела позвоноч- •
ника палерантропов также связано с реконструкцией Буля. Тела
I и III позвонков щапелльца выше сзади, чем спереди, тогда как
последний (у,'поясничный сегмент имеет противоположные соотно-
шения. У Схул IV Поясничные позвонки II и IV также характери-
зуются преобладанием дорзальных высот над вентральными, III
позвонок отличается равенством обоих размеров, и только пятый,
как и у Шапелль, выше спереди, причем разница довольно вели-
ка, составляя 4,5 мм против 5 мм в среднем у современных ев-
ропейцев. Интересно, что подобные же соотношения наблюдаются
и на пятом поясничном позвонке Схул VII, где разница равна
1 Так, по Мартину (Martin, 1928), существует постоянное преобладание вентраль-
ной высоты дисков поясничного отдела над дорзальной — в среднем более чем
на 2 см.
2 Вертикальный поясничный указатель — процентное отношение суммы задних
высот тел поясничных позвонков к сумме их передних высот; указатель клино-
ввдности — процентное отно'шение 'задней высоты тела отдельного позвонка
к его передней высоте.
20
3 мм; и даже у 4—5-летнего ребенка Схул I, у которого эта раз-
ница на последнем поясничном позвонке составляет уже 2 мм.
В целом вертикальный поясничный указатель, вычисленный
для четырех сегментов Схул IV, дает величину 97,4, т. е. вполне
современную, причем курторахидность обусловлена здесь пол-
ностью только формой пятого поясничного позвонка. Вероятно,
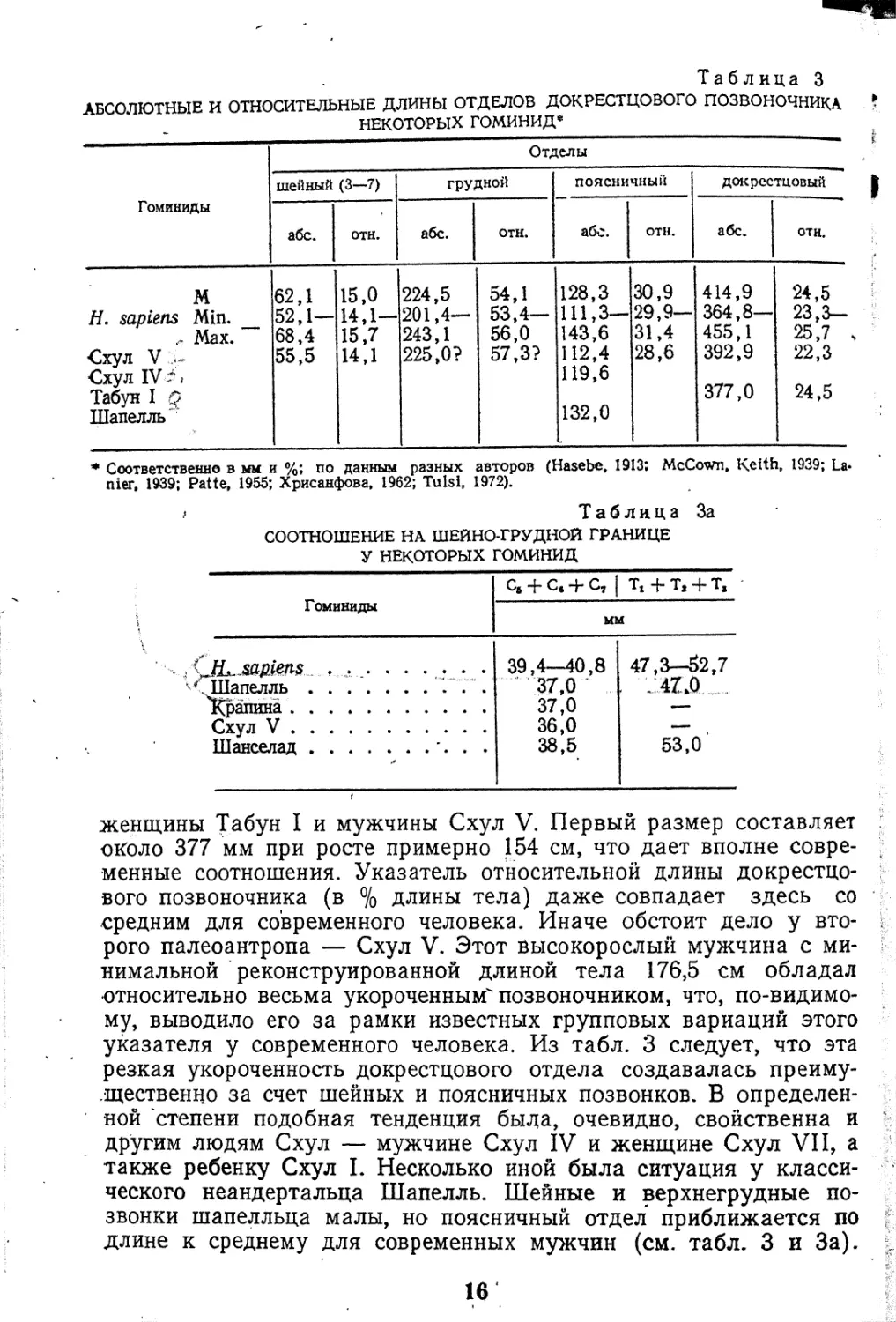
подобный тип поясничного от-
дела (переходный сегмент—пя-
тый) был свойствен почти
всем известным позвоночни-
кам палеоантропов, как пока-
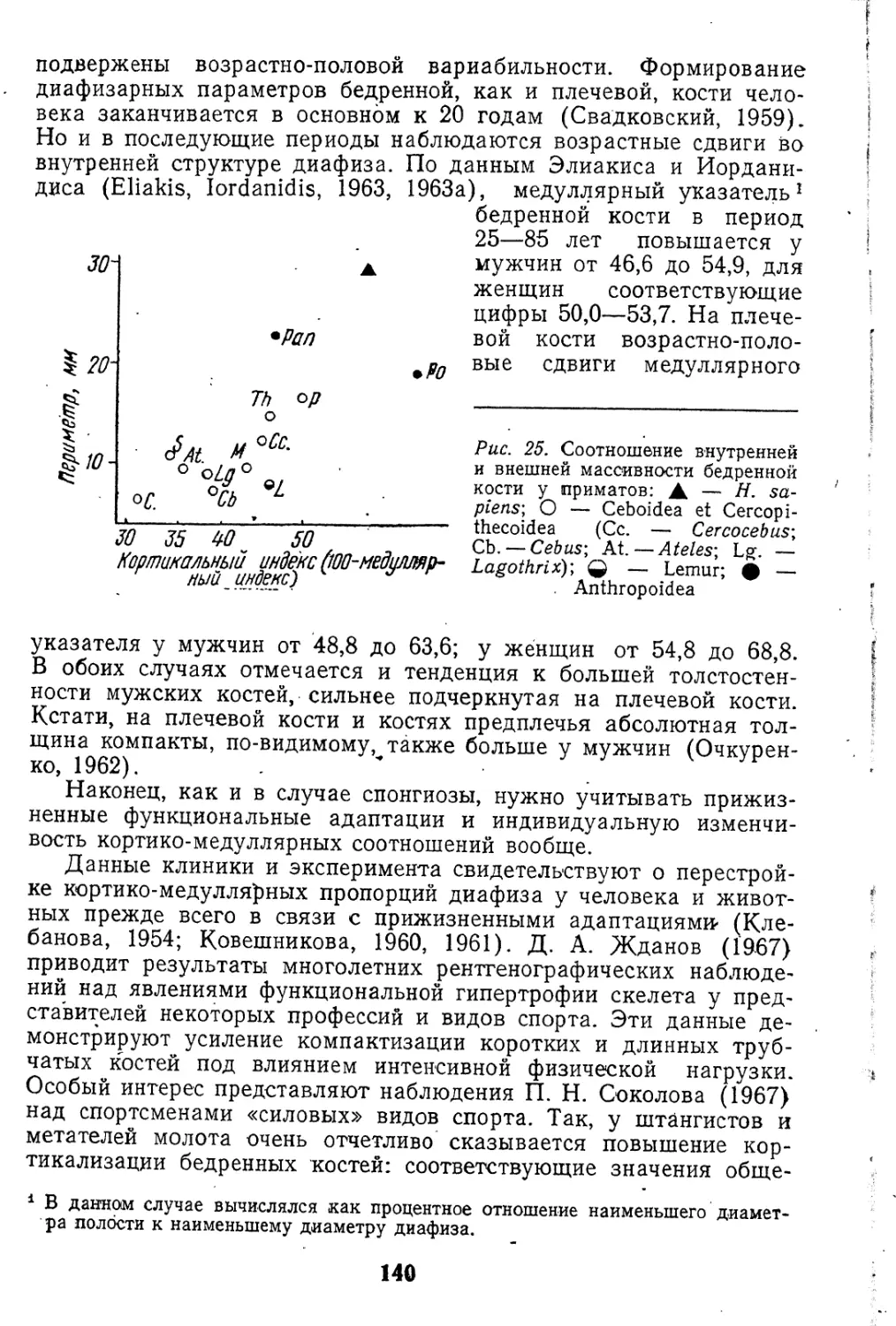
Рис. 2. Грудные (вверху) и пояснич-
ные (внизу) позвонки австралопите-
ковых. А — A. africanus-, Б — А. го-
bustus; В — Н. sapiens (по Robin-
son, 1970)
зано выше. Он достаточно широко распространен и среди верх-
непалеолитических людей, и в мезолите (Vallois,. Lazort-
hes, 1942). Однако и здесь нет сколько-нибудь заметного
противопоставления ископаемых гоминид Н. sapiens recens, у ко-
торого положение переходного позвонка достаточно изменчиво, и
подобный вариант нередок (так, в нашей европеоидной серии, на-
пример, он встретился почти в 1/з всех случаев). Кроме того, су-
ществуют различия в выраженности поясничного лордоза у сов-
ременного человека в связи с разным образом жизни. Возможно,
что он более подчеркнут при преимущественно сидячем образе
жизни, являясь в этих условиях проявлением известной стабиль-
ности, тогда как некоторая сглаженность изгиба может быть свя-
зана с большей подвижностью и гибкостью позвоночника, что
представляет, с этой точки зрения, преимущество и не примитив-
ный признак (Patte, 1955). Подчеркнем, в этой связи, что гипер-
лордоз, например, вовсе не является для человека благоприятным
признаком (Gillot, 1972). При обзоре положения переходного по-
ясничного позвонка у ископаемых гоминид нужно также иметь в
«иду, что существует, по-видимому, половой диморфизм в этом-
отношении: у женщин этот позвонок расположен выше, чем у
мужчин той же популяции (Louis, Guerinel, Mambrini, 1965;
Tulsi, 1972).
Огромный интерес представляют, конечно, структурные особен-
ности поясничных позвонков австралопитековых (рис. 2). Сущест-
вование у них поясничного лордоза предполагается по косвенным
и по прямым данным (Robinson, 1970). Так, у одного из массив-
ных австралопитеков индекс клиновидности последнего пояснич-'
ного позвонка составляет около 90,0, что указывает на достаточно
выраженное преобладание вентральной высоты и хорошо разви-
21
тый промонториум. Существование поясничного лордоза у ранних
гоминид является еще одним свидетельством их прямохождения,
а с другой стороны, подтверждает приведенную выше интерпре-
тацию некоторых особенностей поясничных позвонков у поздних
гоминид.
Известны фрагменты грудной клетки только для некоторых
палеоантропов (Шапелль, Крапина, палестинцы, Тешик-Таш);
впрочем, они не позволяют даже приблизительно определить ве-
личину грудного индекса у этих гоминид. Можно только предпо-
лагать, что форма грудной клетки приближалась у Шапелль и
Табун I к бочкообразному типу. Упомянем в связи со сказанным
о морфологии сочленовных отростков первого поясничного по-
звонка шапелльского неандертальца, которая, согласно Камерону
(Cameron, 1934) „ связывается с «гиперстеническим» телосложе-
нием. Особенно замечательная черта — большая емкость грудной
клетки у малорослой женщины Табун I, у которой, кроме того,
верхние три ребра очень быстро расширяются латерально, со-
ставляя куполообразную верхушку грудной клетки. Реберные
сегменты были длинными, особенно относительно ее роста, и
располагались наверху более горизонтально. Эти особенности на-
ряду с сильным изгибом обусловили очень большую респиратор-
ную емкость ребер, примерно вдвое превышающую этот показа-
тель у современного человека. Другим важным признаком являет-
ся значительная протяженность прикрепления межреберпых
мускулов, что наблюдается у палестинских палеоантропов, ша-
пелльца и крапинцев.
Однако тип грудной клетки, очевидно, варьировал у палеоант-
ропов, как и у современного человека. Так, другая грудная клетка
женщины палеоантропа (Схул VII), правда крайне фрагментар-
ная, имела более современное строение, близкое также к типу
мужских грудных клеток Схул. Последние составляют ряд от более
неандерталоидных соотношений у Схул V к вполне современным
у Схул IV, показывающего в некоторых признаках (например,
морфологии II ребра) даже «ультрагоминидные» особенности.
Наиболее выдающийся признак неандертальского ребра —
массивность и крепость медиального гребня, формирующего дно
реберного желобка, что способствует толщине и крепости ребра.
Очень показательна в этом отношении форма сечения неандер-
тальских ребер. Большинство известных ребер палеоантропов во-
обще мало уплощены и поэтому много толще, чем у современного
человека, особенно по отношению к высоте ребра. Это.отражается,
на форме их поперечных сечений, почти треугольных, с округлен-
ными краями. Подобный тип сечения встречается на ребрах Ша-
пелль, Неандерталь, Мустье, Крапины. Однако не все европейские
неандертальцы характеризуются такой особенностью: например,
мужчина Ла Феррасси имел сечения ребер, не столь крупные^
как шапеллец, и более уплощенные, приближаясь в этом отноше-
нии к современному человеку. Треугольное «неандертальское»
22
сечение ребер отмечено у Тешик-Таша (VII ребро). Среди палес-
тинцев в этом отношении снова существует разнообразие: напри-
мер, II ребро Схул V имеет треугольную, а гомологичный сегмент
у Схул IV — уплощенную «ультрагоминидную» форму поперечно-
го сечения. В целом верхние (грудинные) ребра палестинцев име-
ют тенденцию к округленности сечения, нижние (диафрагматичес-
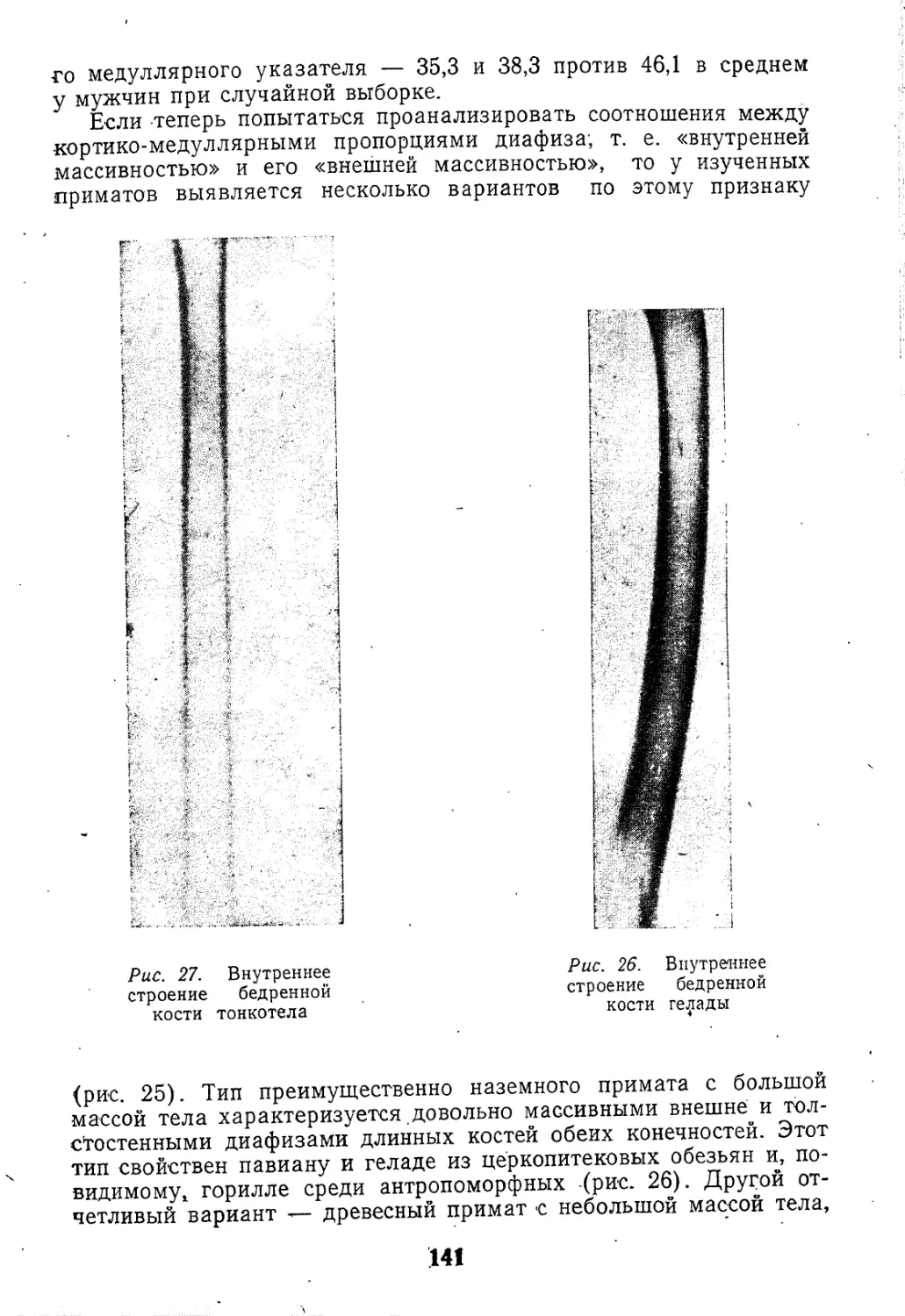
Рис. 3. Сечения ребер палеоантропов.
Стрелка обозначает положение sulcus
costalis. А — Неандерталь; Б —
Табун I; В — Схул IV; Г—Н. sa-
piens. (по McCown, Keith, 1939)
кие) — к уплощенности. Очевидно, неандертальцы Европы чаще
имели толстые узкие ребра, тогда как у палестинских палеоант-
ропов намечается начало уплощения диафрагматических "ребер,
типичного для современного человека (рис. 3). Сказанное можно
иллюстрировать цифровыми примерами: так, VII ребро Неандер-
таль имело размеры сечения в середине: 17X10,8 мм (соответст-
венно ширина и толщина), что дает указатель сечения, равный
63,5. У современного человека соответствующие величины 12,9Х
Х5,8 с указателем около 45 (Sedlin, Frost,' Villanueva, 1963).
У Табун I сечение ребра тоже округленное, но в меньшей степени
(указатель 59,5). Диафрагматические ребра Схул IV дают резко
отличные индексы: например, для VIII ребра указатель равен
всего 34, хотя оно и имеет медиальную неандертальскую выпук-
лость сечения, тогда как у крапинца соответствующий индекс со-
ставляет 6-1,7 — типично неандертальскую величину.
Толщина ребер, впрочем, значительно варьирует у современ-
ных людей, снижаясь в крайних вариантах до 2,5 мм — «ноже-
видные ребра», — что представляет противоположный неандер-
тальскому тип сечения. С другой стороны, вариация, близкая к
неандертальской, была указана Хрдличкцй (Hrdlicka, 1930) на
некоторых современных европеоидных ребрах. Что касается
верхнепалеолитического Н. sapiens, то здесь, по-видимому, также
встречались как толстые «неандертальские» ребра (например, у
Шанселад), так и уплощенные (у Комб-Капелль и кроманьонцев).
Высказывалось предположение о возможной связи неандер-
тальского типа с сильным развитием межреберных мускулов или
с общей массивностью скелета; эта особенность рано проявлялась
у палеоантропов: так, округленное сечение ребра встречено, на-
пример, уже у 4—5-летнего ребенка Схул I, массивные ребра
описаны и у 6—7-месячного ребенка из Киик-Кобы (Vlcek, 1973).
Особый интерес представляет морфология грудины, поскольку
23
эта кость в значительной мере отражает особенности строения
грудной клетки и несет на себе черты полового диморфизма.
У человека в связи с испытанным в филогенезе сокращением
числа грудинных ребер грудная кость заметно расширилась осо-
бенно в сравнении с «палочковидной» сегментированной грудиной
низших узконосых обезьян. Хотя сходный процесс расширения
грудины и ее укорочения наблюдается и у антропоидов, грудина
человека примерно в 70% случаев имеет' специфический «гоми-
нидный» тип: тело кости, узкое вверху, каудально расширяется
(Ashley, 1956). В остальных 30% случаев, по данным этого ав-
тора, наблюдаются «паноидный» и «понгоидный» варианты (соот-
ветственно узкое и широкое тело грудины с параллельными края-
ми). Своеобразный «гориллоидный» тип (очень массивная рукоят- .
ка, тело грудины с тенденцией к краниальному расширению), по-
видимому, не встречается у человека.
Единственные сведения о строении грудины у ископаемых го-
минид содержатся в монографии Макоуна и Кизса (1939). Мате-
риал весьма фрагментарен, и хотя здесь представлены все части
кости — рукоятка, тело, мечевидный отросток, но они, как прави-
ло, относятся к разным индивидуумам. Так, рукоятка грудины
известна от Схул IX, тело — от Табун I и Схул VII. Наиболее
полный фрагмент грудины принадлежит мужчине Схул IV (тело
с мечевидным отростком). Таким образом, наше, суждение о
строении грудины палестинских палеоантропов основывается
главным образом на фрагменте ее у Схул, IV-, Сравнительно с
современным типом это прежде всего относительно чрезвычайно
широкая грудина, конфигурация которой указывает на сильное
развитие диафрагматических ребер, что действительно имело ме-
сто у этого гоминида. В то же время ее форма не имеет ничего
общего с «гориллоидным типом», нет также никаких отклонений
от типичного современного варианта и по числу стернальных
ребер. •
Известно, что грудина современного человека в некоторой сте-
пени отражает половые различия: мужская грудина в среднем
абсолютно длиннее женской прежде всего за счет развития тела
грудины, так как размеры рукоятка примерно одинаковы у обоих ,
полов. Определенная длина тела грудины совпадает у мужчины i
Схул IV и женщины Табун'Г — соответственно 103 и 104 мм; по |
нйшим данным, средняя длина тела грудины у мужчин-евро- ?
пеоидов равна. 103 мм- И высокорослый мужчина Схул IV, |
и низкорослая женщина Табун I имели одинаковую длину тела
грудины, соответствующую средней. для современных мужчин.
Это свидетельствует о том, что различия между Схул и Табун по
этому признаку, вероятно, выходят за рамки половых или инди-'
видуальных. Значения широтно-длиннотного указателя у этих '
двух палеоантропов (44,6 и 33,6 у Схул IV и Табун I соответст-
венно.) снова подчеркивают полиморфизм палестинцев по строе-
нию грудной клетки. •
24
В целом можно предположить довольно раннее оформление
некоторых основных черт гоминидного комплекса (прежде всего
связанных с развитием прямохождения) на осевом скелете чело-
века' (поясничный лордоз, по-видимому, позвоночная формула).
Поздние гоминиды — палеоантропы имели вполне человеческий
тип позвоночника и грудной клетки. Вместе с тем, здесь в ряде
случаев выявляется комбинация некоторых «примитивных»
(структура шейных позвонков) и своеобразных особенностей
(укороченность шейного отдела, низкое положение переходного
поясничного позвонка, тип ребра), что позволяет говорить о не-
равномерности формирования отдельных морфологических при-
знаков осевого скелета и специфичности его структуры прежде
всего у неандертальского варианта.
/ Г л а в a II
Z ПОЯСА КОНЕЧНОСТЕЙ
ПЛЕЧЕВОЙ ПОЯС
В соответствии с локомоторными адаптациями и соотносительным •
развитием хватательной и опорной функции конечностей в срав-
нительно-анатомическом ряду приматов произошла существенная
морфофункциональная перестройка плечевого пояса. Структурные
адаптации проявляются наиболее отчетливо по отношению к важ-
нейшим локомоторным комплексам высших приматов — четверо-
ногости (проноградности), брахиации и прямохождению. Однако
оба элемента плечевого пояса весьма неравноценны, с точки зре-
ния морфофункционального и таксономического анализа. В наи-
большей степени эволюционное преобразование передней (верх-
ней) конечности в филогенезе приматов сказалось на лопатке.
Морфологическое преобразование лопатки у приматов корре-
лирует с постепенным появлением и развитием разнообразных
движений в трехосном плечевом суставе и сопутствующих движе-
, ний плечевого пояса. С увеличением аппарата переднего зубчато-
го мускула лопатка, имеющая у полуобезьян форму равнобедрен-
ного треугольника, удлиняется/и передвигается медиально, рас- 'г
полагаясь все больше и больше на задней поверхности грудной
клетки. Основные признаки филогенетической перестройки этой
кости — уменьшение морфологической длины и увеличение мор-
фологической ширины, т. е. возрастание внутреннего края, —
процесс, связанный с перемещением лопаток из бокового положе-
ния на стенке грудной клетки в дорзальное. Эта трансформация
обеспечила у человека большую свободу движений (особенно
ротационных) в плечевом суставе.
Далекие филогенетические связи прослеживаются, по-видимо-
му, и в онтогенезе человека: на очень раннем этапе эмбриогенеза
будущая лопатка имеет форму сагиттально расположенной треу-
гольной хрящевой пластинки, а по мере дальнейшего развития
она испытывает поворот во фронтальную плоскость и одновремен-
но перемещается по сагиттальной оси так, что первоначальная
краниальная ориентировка суставной впадины превращается в
г латеральную (Zawisch, 1954). Следовательно, как и данные срав-
нительной анатомии, эмбриогенетическая тенденция также свиде-
; тельствует об активном вовлечении лопатки в процесс онто-фило-
I генетических преобразований.
В отличие от лопатки, морфологической и функциональный
анализ ключицы является сложной задачей, поскольку для него
не дает опорных моментов и сравнительная анатомия приматов.
Так, не приводит к определенным результатам попытка связать
довольно существенные среди приматов различия в степени изги-
26
ба ключиц с особенностями строения некоторых мускулов, в пер-
вую очередь ключичной порции ' большого грудного мускула
(Патт, 1955): изгиб значительно варьирует даже и при отсутствии
этого мускула. Столь же мало результативными оказываются со-
поставления с образом жизни в целом (например, очень различ-
ная изогнутость кости у азиатских брахиаторов — гиббона и
орангутана). Также и по торзиону ключиц, человек в большей
мере сближается с четвероногими проноградными приматами, чем
с антропоидами (Oxnard, 1967). К этому следует прибавить, что
ключица, рано формируясь в эмбриогенезе, принадлежит к числу
«онтогенетически стабильных» костей (Corrigan, 1960); уже ее
первичная эмбриональная конфигурация весьма напоминает де-
финитивную. В картине изменчивости ключиц на передний план
выступает ее индивидуальный аспект; весьма отчетливы также и
половые различия, проявляющиеся уже при рождении (Pavlov,
P6trov, 1964).
Наибольший интерес для таксономического и функционально-
го анализа могли бы представить элементы плечевого пояса ран-
них форм гоминид и их предшественников, переходивших к дву-
ногому передвижению, как основному способу локомоции.
К сожалению, лопатки от ранних форм гоминид и прегоминид
пока неизвестны, исключая не вполне ясный по своей принадлеж-
ности фрагмент из Стеркфонтейна, приписываемый австралопи-
теку африканскому (Broom, Robinson Schepers, 1950). Впрочем,
морфологическая характеристика этого фрагмента недостаточно
достоверна, а в интерпретации его особенностей существуют раз-
ногласия (Campbell, 1966; Oxnard, 1968). Один из немногих более
достоверно определимых размерных признаков — аксиллоглено-
идальный угол — на лопатке из Стеркфонтейна составляет при-
мерно 103°. Кэмпбелл (Campbell, 1966) считает, что здесь есть
сближение с «полубрахиаторами», тогда как по Окснарду (Ox-
nard, 1968) фрагмент из Стеркфонтейна занимает по этому при-
знаку крайнее положение в ряду брахиаторов. В последнем слу-
чае следует, однако, учитывать большие вариации формы лопат-
ки-среди антропоидов (Шульц, 1963), что в немалой степени
отражается и на изменчивости формы наружного края, а следо-
вательно, и на величине обсуждаемого угла. Данные разных ав-
торов 'не вполне совпадают в отношении последнего: так, по Пат-
ту, средняя для антропоидов составляет всего 108° при очень1
сильной индивидуальной изменчивости этого признака, а у сов-
ременного человека — 132° при групповом минимуме 119,5° и
наименьшем индивидуальном значении 111°. Величина угла на ло-
патке из Стеркфонтейна с учетом возможной неточности его опре-
деления оказывается тогда не столь отклоняющейся, как на гра-
фике, приведенном Окснардом (1968), на котором все цифры
сильно сдвинуты вправо. Следует также иметь в виду, что данные
этого автора (1967) свидетельствуют о . значительно большей
близости человека по величине угла сочленовной впадины к
27
четвероногим мартышкообразным обезьянам, чем к брахиаторам
и полубрахиаторам, хотя соотношения «нропульсирующпх» й ;
«подвешивающих» мускулов весьма отличны у человека и четве-
роногого примата. Функциональная интерпретация этого признака
оказывается поэтому не вполне ясной, а принадлежность самого
фрагмента австралопитеку — пока недоказанной.
Что касается другого признака, обсуждаемого упомянутыми
выше авторами, — аксиллоспиналыюго угла, — то его определе-
ние нельзя счесть достоверным, так как на фрагменте отсутствует
медиальная часть лопаточной ости, на ориентировке которой
главным образом иосновано определение этого угла.
Наиболее полные данные о лопатках ископаемых гоминид от- \
носятся к палеоантропам, но и здесь находки большей частью до-
вольно фрагментарны. Нет определенных заключений о форме
' лопаток европейских неандертальцев, за исключением указаний'
Патта о близости лопатки Ла Феррасси к I типу Хрдлички. Су-
у i ществует также мнение, что лопатки Шапелль были короткими и
широкими. Более подробно описаны лопатки палестинцев. Общие
' размеры лопатки высокорослого мужчины Схул V, как и лопа-
точный указатель1 (63,2), входят в современные групповые гра-
дации (60,3—72,5). Очень мал указатель у Схул IV (54,7), одна-
ко подобная «скафоидная» форма лопатки встречается и у совре-
менного человека. В целом для группы Схул типичны относитель-
но малые , (по росту этих людей) и узкие лопатки, несколько на- г
поминающие по своим пропорциям кроманьонские, что согласует-
ся и с умеренным развитием ключиц у палестинских палеоантро-
пов. Общие пропорции лопаток и направление лопаточного гребня
. можно считать здесь, как и у западноевропейских неандерталь-
цев, совершенно человеческими. Это замечание важно, так как
пропорции лопатки и наклон ости связаны с локомоцией и осан-
кой.70днако нельзя не отметить, что у некоторых палеоантропов
наблюдается довольно значительный краниальный наклон ости
(Феррасси, палестинцы), маскирующий у Крапины даже лопаточ-
ную вырезку. Эту особенность, впрочем, нельзя признать прими-
, тивной, поскольку она встречается не у всех неандертальцев и
j наблюдается и у современных людей: функциональное значение
ее неясно. Насколько можно судить по сохранившимся фрагмен-
там лопаток палестинских палеоантропов, соотношение надостной
и подостной ямок варьирует; выше упоминалось, что особенно
сильное развитие подостной ямки встретилось у Схул IV (скафо-
идный тип), что соответствует индивидуальной вариации, сущест-
вующей и у современного человека.
Особый интерес представляет морфологический анализ строе-
ний наружного (аксилл яркого) края лопатки; эта наиболее проч-
ная часть костной пластинки обычно хорошо сохраняется на иско-
паемых костях и поэтому представляет лучшие возможности для
1 Морфологическая длина лопатки в % ее морфологической ширины.
- ' V 28
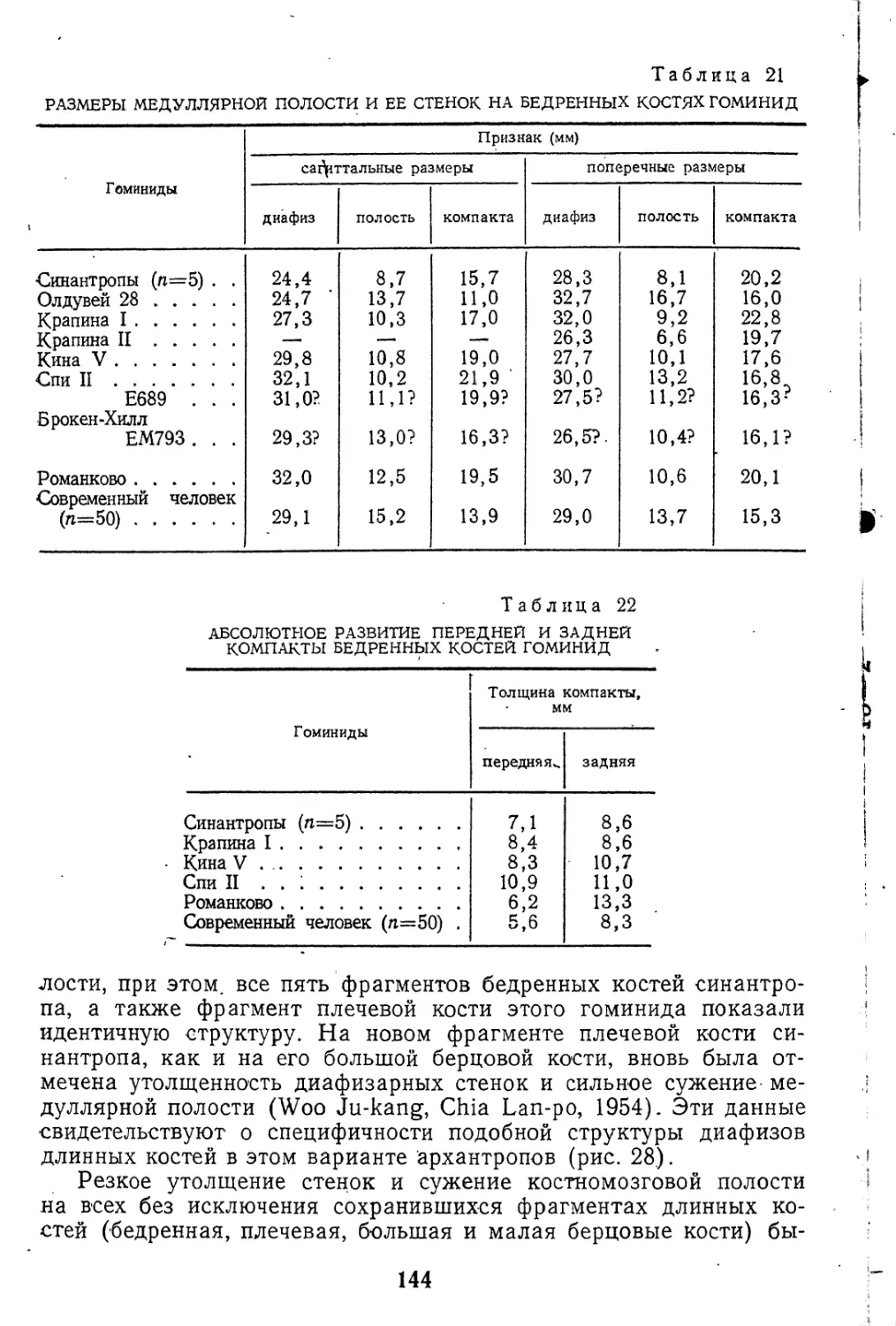
сравнительного изучения (рис. 4). На лопатках многих палеоант-
ропов, как европейских, так и палестинских,, наблюдается его осо-
бая «амфимаргинальная» форма, что выражается в равномерном
развитии обоих гребней наружного края, в результате чего здесь
может возникать дополнительная бороздка с дорзальной стороны,
описанная Маккоуном и Кизсом, как «бороздка Буля». Этому
признаку нередко придают значение главного критерия в диффе-
Рис. 4. Сечения латерального края лопатки. )/ \ I
Стрелкой обозначена дорзальная губа. А — // \ |
Н. sapiens; Б — Неандерталь (по Patte, 1955) А Б
ренциации неандертальских лопаток от современных, что вряд
ли вполне правомерно.
Строение наружного края варьирует среди самих палеоантро-
пов; подобная амфимаргинальная форма четко выражена преи-
мущественно у западноевропейских неандертальцев и непостоян-
на у крапинцев, тогда как у палестинцев существуют все стадии
перехода от более простого «дорзомаргинального типа» (Схул-
IV), свойственного также и антропоидам, к этой «ультрагоминид-
ной» формации. Последняя наиболее отчетлива на лопатке Та-
бун I. У современных людей также отмечается иногда неандер-
талоидная форма наружного края, ойа была определена на
многих лопатках из Пржедмости, у Шанселад, Оберкассель,
Кап-Блан, Вейерье. Описана также на скелете эпохи бронзы из
Пюи-дехДом (Billy, 1968). В этом признаке, очевидно, следует,
видеть позднюю гоминидную специализацию, но не черту прими-,
тивности, с точки зрения сопоставления с антропоидами, которым
свойственна значительно более простая форма края. Функцио-
нальное значение этой особенности неясно, тем более, что оба со-
седних мускула, начало которых разграничивает сверхчисленный
гребень, имеют почти тождественную функцию. Представляется
наиболее обоснованным связывать этот признак с проявлением
тенденции к общей массивности скелета, которая явственно выра-
жена в своеобразной группе западноевропейских палеоантропов.
С этой точки зрения, палестинцы снова сближаются с крапинцами.
Существуют значительные разногласия в вопросе об ориенти-
ровке суставной впадины. Так, Швальбе (Schwalbe, 1914) считал
специфической чертой неандертальской лопатки дорзальный пово-
рот суставной впадины, то же утверждал и Клаач (Ktaatsch, 1902)
для палеоантропов из Неандерталь и Мустье. Это не подтверж-
дают Валлуа (Vailois, 1946) и Хрдличка (1930), который считает,
что на неандертальских лопатках сочленовная впадина наклонена
даже несколько'слабее, чем у современного человека; по мнению
29.
Горяновича-Крамбергера (Corjanovic-Kramberger, 1906), на кра-
пинской лопатке она была направлена вперед (Haas, 1957). Эти
расхождения объясняются не только трудностями определения ука-
занного признака на фрагментарных материалах, но и индивиду-
альными вариациями, существующими, по-видимому, и у современ-
ного человека. Попытка измерить степень дорзофлексии сочленов-
ной впадины у палестинцев, предпринятая Маккоуном и Кизсом
(1939), свидетельствует все же о существовании различий между
«типичным» неандертальцем (Неандерталь) и палестинскими па-
леоантропами. У первого отмечается сильный дорзальный поворот
впадины, тогда как у палестинцев ее положение вполне современ-
ное, Возможно, что крайняя степень дорзального поворота сочле-
новной впадины у Неандерталь (угол ретроверсии 29° против 8—1 Iе
у современного человека, по Маккоуну и Кизсу) является инди-
видуальной особенностью этого гоминида.
Более характерной для 'палеоантропов чертой может считаться
форма сочленовной впадины лопатки. Она здесь относительно очень
суженная и вытянутая в высоту: гленоидальный (широтно-длин-
нотный) указатель составляет у собственно неандертальской на-
ходки 63,2, что значительно ниже группового минимума у совре-
менного человека (вариации средних: 66,9—81,8). У палестинцев
индексы, немного выше: 65,5 у Табун I и 67,7 у Схул V, что уже
близко к групповому минимуму Н. sapiens. Все же у всех этих
палеоантропов значения указателей очень низкие. По-видимому,
существуют в’ариации на лопатках крапинцев, но средняя для ше-
сти лопаток равна всего 65,0, что снова указывает на специфиче-
скую суженную форму сочленовной впадины. У верхнепалеолити-
ческого Н. sapiens индекс варьирует от 64,0 (Оберкассель) до 78,4
(Шанселад), т. е. он почти воспроизводит современные вариации,
но снова со сдвигом всего ряда влево.
Мы попытались статистически оценить различия в соотношении
сочленовной впадины лопатки и головки плеча у типичного неан-
х дертальца (Неандерталь) сравнительно с современным человеком
(табл. 4).
Максимальные различия наблюдаются таким образом для со-
отношений широтных размеров костных компонентов плечевого
сустава, эти различия реальны. Интересно, что у современного че-
ловека широтные размеры сочленовной впадины лопатки и голов-
ки плечевой кости варьируют сильнее вертикальных, что особенно
подчеркнуто для дуговых размеров, между ними наблюдаются и
более слабые связи. Вероятно, широтные размеры обоих компонен-
тов.плечевого сустава наиболее реагируют на различия в типе
суставной деятельности. -Об этом же свидетельствует и тенденция
к несколько большей вариабильности правосторонних размеров на
скелете современного" человека (Малявская, Хрисанфова, 1962).
О незначительном преобладании дисперсии для измерительных
признаков, правой лопатки свидетельствуют и данные де Море
(De Morais, 1966), также полученные для европеоидных серий.
у 30
Таблица 4
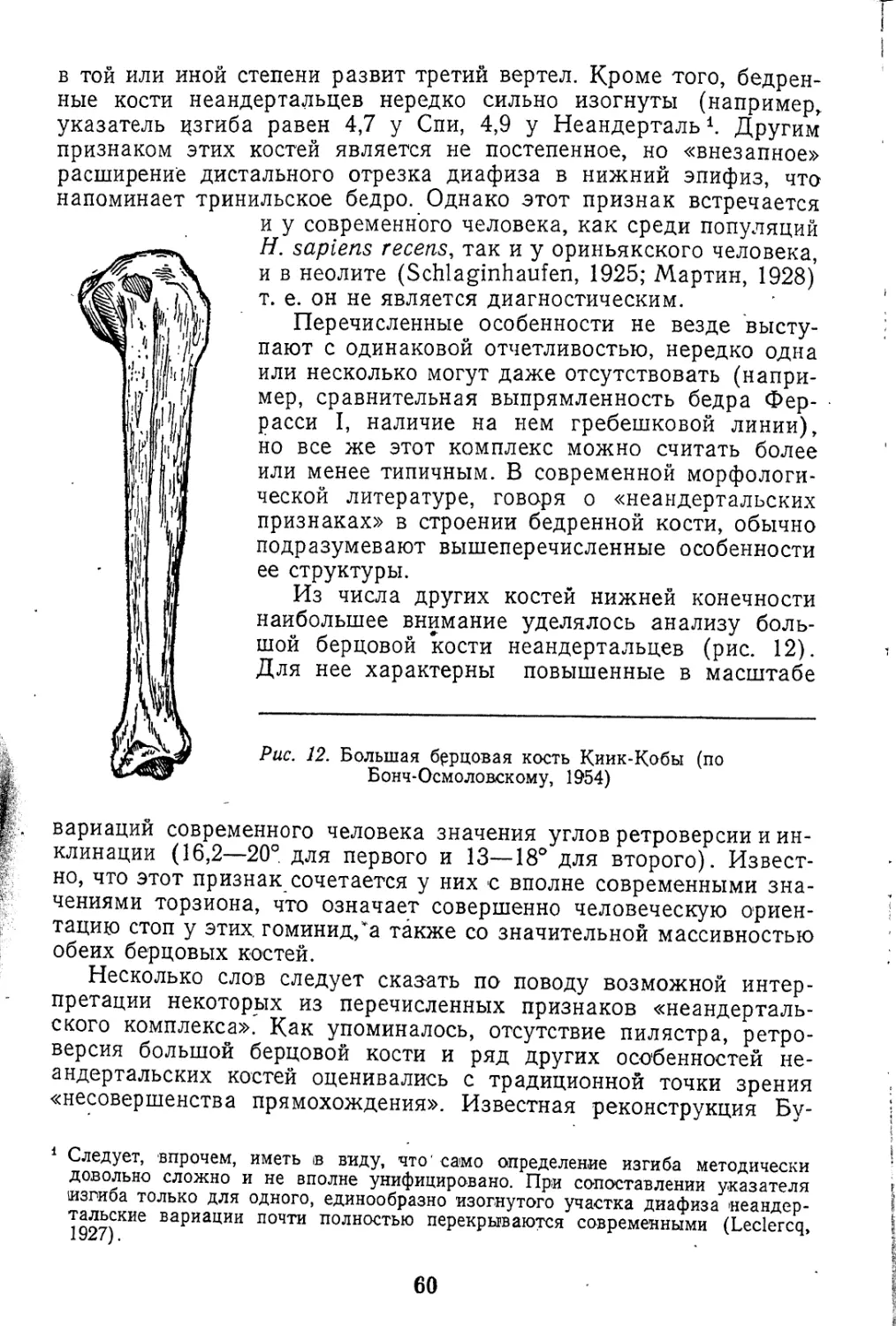
СРАВНЕНИЕ РАЗМЕРОВ КОМПОНЕНТОВ ПЛЕЧЕВОГО СУСТАВА
У НЕАНДЕРТАЛЬЦА И СОВРЕМЕННОГО ЧЕЛОВЕКА
(по уравнению регрессии)*
Коопелиоуемые признаки** X 1 У 1 У1 |ух —У t
х Y мм
Лопатка Плечевая кость
Длина суставной впади- ны Продольный диаметр головки 38*** 46,89 49*** 2,11 1,00
Ширина суставной впа- дины Поперечный диаметр головки 27*** 41,93 50*** 8,07 4,18.
Продольная дуга сус- тавной впадины Продольная дуга голов- ки 39 62,45 64 1,55 0,64
Поперечная дуга сус- тавной впадины Поперечная дуга голов- ки 28 58,67 78 19,33 7,80-
Окружность суставной впадины Обхват головки 111 144,24 152 7,76 1,43.
♦ X и у — коррелируемые признаки: у — значение признака у в современной серии при значении
признака X, соответствующем таковому у ископаемой формы (х); у1 — значение признака у у
ископаемой формы; I — критерий реальности различий.
** По правой стороне: п — 50.
*** Длина и ширина суставной впадины и головки плечевой кости Неандерталь измерялись по
муляжу, что дает для гленоидального индекса несколько большую величину, чем на подлин-
нике.
Примечательно, что на лопатках новорожденных еще не наблю-
дается подобных различий между правой и левой сторонами (Pet-
rov, Pavlov-, 1964). Следует отметить также, что, по нашим данным,
все корреляции между размерами суставной впадины и тотальны-
ми размерами лопатки выше слева.
Из табл. 4 следует, что у неандертальца меньшей по ширине
сочленовной впадине лопатки соответствует более широкая, чем у
современного человека, головка плеча, т. е. головка значительно
шире и более выпукла по отношению к сочленовной впадине у
ископаемого гоминида. . Это усиливает и без того значительную
инконгруэнтность обоих компонентов сустава, что, возможно, до-
пускало большие дуги движений у неандертальца сравнительно с
современным человеком (Малявская, Хрисанфова, 1962).
Палеоантропологические материалы по ключице представлены
синантропами и некоторыми палеоантропами («классические» не-
андертальцы, крапинцы, Тешик-Таш, палестинцы).
От более ранних.предшественников человека известен только
небольшой фрагмент ключицы из Олдувея (Олдувей 8), возмож-
но принадлежащий форме, близкой к австралопитековым (Ното
habilis). Относительно этого фрагмента в литературе также нет
полного единства мнений. Так, Нэйпир (Napier, 1965) указывает на
человеческий тип этой кости; Окснард (1968, 1969), исходя из мо-
дификации стернального конца фрагмента, предполагает, что здесь,
возможно, существовала способность к подниманию рук и подве-
шиванию тела. Определение торзиона ключицы на столь фрагмен-
31
тарном материале вряд 'ли может дать основание для подобных
широких выводов, учитывая, к тому же, что этот признак, как и
особенно форма сечения стернального конца кости, очень измен-
чивы у гоминид. Кроме того, функциональная интерпретация и в
этом случае остается недостаточно определенной, так как по тор-
зиону ключицы человек значительно ближе к четвероногим проно-
градным приматам, чем к антропоидам, хотя по соотношению сил,
действующих на плечевой сустав, он наиболее сходен с последними
(Окснард, 1967).
Большая часть известных ключиц ископаемых гоминид по сво-
им общим размерам входит в пределы современных групповых ва-
риаций. Длина ключиц значительно варьирует у Н. sapiens, состав-
ляя -116—156 мм по средним мужских серий при общей средней
143 мм (Martin —Sailer, 1958; Мартин, 1928). По-видимому, рост
не. является единственным фактором, обусловливающим эти раз-
личия, однако внутри группы отмечается реальная, хотя и невысо-
кая, связь длины ключиц с ростом. По нашим данным, получен-
ным при изучении скелетов мужчин-европейцев, между длинами
ключицы и бедра существует корреляция порядка 0,5.
Длина реставрированной ключицы синантропа составляла
145—150 мм; причем ни абсолютно, ни относительно (например, к
длине плечевой кости) синантроп не отличался здесь от современ-
ного человека, обладая сравнительно умеренной шириной плеч по
•современному масштабу.
Среди палеоантропов нет единообразия в этом отношении. Так,
некоторые классические неандертальцы Европы (собственно Неан-
дерталь, Шапелль, Феррасси) обладали необычайно длинными
ключицами, например у женщины Ла Феррасси длина этой кости
составляла 179 мм(1), т. е., далеко превышала современный груп-
повой максимум для мужчин. Большой длиной ключиц отличался
и ребенок из Тешик-Таша, превосходивший по этому признаку со-
временных детей того же возраста. При наличии у большинства
европейских форм палеоантропов сравнительно коротких плечевых
костей ключично-плечевой указатель у них оказывается чрезвычай-
но высоким, часто выходящим за верхнюю границу современных
вариаций, т. е. при сравнительно небольшом росте эти древние
люди имели очень широкие плечи. Однако не всем ископаемым
гоминидам были свойственны такие брахиморфные пропорции: на-
пример, ключицы не были особенно длинными в палеолитической
популяции крапинцев. Эта оговорка относится и к палестинским
палеоантропам, у которых вариации длиннотного развития ключиц
были велики: 135 мм (женщина Табун I) — 154 мм (мужчина
Схул V), но полностью входили в верхнюю половину современного
ряда. Поскольку палестинцы обладали большей частью значитель-
ной длиной тела, относительная ширина плеч у них (исключая
снова женщину Табун I), по-видимому, не была^ большой. Верхне-
палеолитические люди имели вполне современные значения дли-
ны ключиц (148 мм Щанселад — 156 мм мужчина Оберкассель).
32
Массивность ключиц также сильно варьировала у ископаемых
людей. Она была весьма умеренной у синантропа с указателей
массивности 23,4 и, по-видимому, крапинских палеоантропов, где
это «тонкое» строение ключиц совпадало с грацильностью длинных
костей скелета. Напротив, у классических европейских неандер-
тальцев наблюдается резкая дисгармония между грацильным
строением тонких и длинных ключиц и массивостыо трубчатых
костей конечностей. Так, указатель массивности ключицы
Неандерталь такой же, как у синантропа (23,8), т. е. довольно
близкий к минимуму современного ряда вариаций (22—27), а ин-
декс массивности плечевой кости у первого почти совпадает с мак-
симумом, у второго — с минимумом групповых вариаций Н. sa-
piens.
Такие соотношения являются специфичными для классических
палеоантропов и отличают их от современных людей, где обычно
не наблюдается столь резкая степень независимости между разви-
тием ключиц и длинных костей скелета (Беневоленская, Хрисан-
фова, 1961). Современный мужчина-европеец с длиной плечевой
или бедренной кости, как у Неандерталь, имел бы значительно бо-
лее короткую ключицу, причем различия статистически достовер-
ны (табл. 5). То же самое можно сказать.и о массивности ключиц:
при равном периметре диафиза ключица Неандерталь много длин-
нее, чем у современного человека. Из приведенных в табл. 5 дан-
Таблица 5
СРАВНЕНИЕ РАЗМЕРОВ КЛЮЧИЦЫ И ДЛИННЫХ КОСТЕЙ
У НЕАНДЕРТАЛЬЦА И СОВРЕМЕННОГО ЧЕЛОВЕКА
(по уравнению регрессии)*
Коооелио^емые поизнаки х | у | У1 | у1— У
X Y мм
Длина плечевой кости Длина ключицы 307 141,09 168 26,91
Длина бедренной кости Длина ключицы 445 144,49 168 23,51
Обхват плечевой кости Обхват ключицы 75 41,34 40 —1,34
Обхват бедренной кости Обхват ключицы 97 42,34 40 —2,34
Обхват ключицы Длина ключицы 40 145,85 168 22,15
Длина плечевой кости Обхват плечевой кости 307 64,79 75 10,21
Длина бедренной кости Обхват бедренной кости 445 ,86,55 97 10,45
* Обозначения, как в табл. 4.
ных следует также, что при длине плеча (или бедра), равной зна-
чению этих признаков у Неандерталь, современный человек имел
бы гораздо меньший обхват диафиза, т. е. его длинные кости были
бы более грацильными. Полученные данные не подтверждают
мнения Словика (Slowik, 1945), что характерная для неандерталь-
цев независимость между массивностью ключиц и других костей
скелета свойственна и современному человеку. Напротив, у послед-
него существует довольно значительная связь между массивностью
ключиц и длинных костей конечностей (порядка 0,5—0,6).
3 В. Н. Хрисанфова
33
Таким образом, сочетание гранильной ключ ины с массивными
костями конечностей можно счесть специфической чертой строения
неандертальского типа, что лишний раз подтверждает необходи-
мость использования в палеоморфологическом анализе не единич-
ных признаков, но их комплексов. Очень интересно, что «неандер-
тальский комплекс» выявляется и на скелете одного из представи-
телей верхнепалеолитического человека в Европе у Шапселад,
где указатель массивности плечевой кости был очень высок при
относительно гранильной ключице с индексом массивности 23,6.
Выше ^же говорилось, что связь длпннотного развития ключиц
с ростом невелика как при межгрушювых сопоставлениях, так и
внутри популяции. Это соответствует вариациям в пропорциях у
современных людей: широкие плечи могут встречаться у них в
комбинации с разной длиной тела. Вместе с тем между длиной
ключицы и шириной лопатки (и развитием лопаточнс^го гребня)
связь довольно тесная, что обусловлено, по-видимому, морфофунк-
циональной общностью этих признаков (Petrov, Pavlov, 1964).
Следует еще раз подчеркнуть, что основные различия в морфо-
логии ключиц современного и неандертальского человека опреде-
ляются удлиненностью этих костей в последнем случае, тогда как
периметр ключиц представляет мало различий. Эта выраженная
удлиненность ключиц при относительной малорослости и обуслов-
ливает своеобразие широтных пропорций верхней части туловища
на скелете классических неандертальцев. Иными были эти соотно-
шения в группе палестинских палеоантропов, у которых массивные
ключицы (индексы массивности: 25,2—27,0) комбинировались с
относительно грацильными плечевыми костями. Подобный вари-
ант был, по-видимому, свойствен и Тешик-Ташу (относительно мас-
сивная ключица — очень «тонкое» строение плечевой кости). Клю-
чица Табун I снова наиболее неандерталопдна в серии палестин-
ских костей.
Следовательно, у древнейших и древних людей, как и у совре-
менного человека, наблюдаются различные типы ключиц. В палео-
антропологии морфология ключиц может представить интерес, в
первую очередь, для характеристики широтных пропорций тела у
разных вариантов ископаемых гоминид: брахиморфные пропорции
классических неандертальцев и более умеренное развитие плечево-
го диаметра у палестинцев, крапинцев и, возможно, синантропов
(оговорка в последнем случае связана с фрагментарностью мате-
риалов и тем обстоятельством, что ключица и другие кости скелета
могли и не принадлежать здесь одному индивидууму).
Л
ТАЗОВЫЙ ПОЯС
‘ . *
В ряду приматов тазовый пояс играет совершенно особую роль
в качестве индикатора бипедии. Его структура является четким
критерием разграничения гоминид, как двуногих прямоходящих
34
приматов, от всех остальных форм. Причем у этих последних не
выявляется сколько-нибудь выраженной связи между строением
таза и образом жизни, и прежде всего локомоцией (Leutenegger,
1970). Вместе с тем в пределах двух основных групп приматов Ста-
рого Света—низших и высших узконосых—выступает зависимость
общей формы таза от весового фактора (Biegert, ЛГаигег, 1972).
Для человека, как двуного-
го примата, характерен следу-
ющий комплекс основных при-
знаков тазовой кости: укороче-
ние подвздошной кости вслед-
ствие дорзальной ротации под-
вздошно-крестцовых сочлене-
w-
Рис. 5. Ширина крестцовой поверх-
ности таза у высших приматов (в %
от длины туловища):О — Cercopithe-
coidea; • — Anthropoidea; Q— Ore-
opithecus; x — Australopithecus; A —
H. sapiens (no Biegert, Maurer, 1972)
tr
ty OP »Po . *g
L. /V
lob loo loo
Длина тдлобища, мм
x/4.r.
ний по отношению к тазобедренным суставам, что обусловило
и уменьшение расстояния между этими суставами, расширение
пластинки крыла подвздошной кости в вентральном и дорзальном
направлениях и укрепление ее средней части, укрепление верт-
лужной впадины и изменение ее положения, отчетливое оформле-
ние большой седалищной вырезки и обеих передних подвздошных
остей.
Рассмотрим некоторые из наиболее существенных особенностей
«комплекса бипедии» подробнее.
При сравнительно-анатомических сопоставлениях в ряду при-
матов весьма отчетливая тенденция проявляется в отношении об-
щей формы таза. Динамика ее изменения отражает два различных
процесса — расширение пластинки крыла подвздошной кости и
укорочение последней. Первый из указанных признаков не специ-
фичен для человека, так как тенденция к расширению подвздошной
кости характерна для Hominoidea, т. е. человека и антропоидов в
целом, сравнительно, например, с Cercopithecoidea (табл. 6). Го-
раздо более характерно для человека сравнительно с негоминид-
ными приматами расширение сакральной части подвздошной кости
\ (см. табл. 6 и рис. 5). Как следует из приведенных данных, человек
занимает здесь исключительное положение — с указателем, почти
вдвое превосходящим максимальные его значения в обеих группах
узконосых обезьян. Это укрепление подвздошно-крестцовых сочле-
нений ассоциируется также с расширением,пластинки крыла под-
вздошной кости в дорзальном и вентральном направлениях. Боль-,
шее дорзальное протяжение подвздошного гребня позволяет более
интенсивное прикрепление мускулов, поддерживающих туловище
з*
35
Таблица 6
НЕКОТОРЫЕ ИНДЕКСЫ ТАЗА АВСТРАЛОПИТЕКОВЫХ
В СРАВНЕНИИ С ДРУГИМИ ПРИМАТАМИ*
Индексы Огео- р it ho- cus A. afri- canus A. ro- bust us Homo sapiens 6 2 Anthropoi- dea Cercopi- thecoidea
Длина подвздошной кости (к длине туловища) . . . 33,5 25,6 (26) 26,4—25,2 31,7—37,8 22,6—27,9
Нижняя длина подвздошной кости (к длине туловища) . 16,0 11,0 (10,6) 10,5 17,9—19,5 11,5—14,7
Ширина подвздошной кости (к длине туловища) . . . 25,0 29,0 (38) 28,8 13,9-33,1 8,1—12,5
Ширина крестцовой поверх- ности (к длине туловища) . 5,7 8,0 (11,8) 11,2 5,4— 6,6 4,7— 6,4
Ширина пластинки подвздош- ной кости (к длине туло- вища) 19,3 22,0 (26,4) 17,5 10,0—25,2 2,6— 6,6
Длина седалищной кости (к длине туловища) 15,0 14,0 (15,6) 14,1 11,0—17,5 9,8—12,2
Диаметр вертлужной впадины 1 (к высоте таза) 14,8 24,6 (26,3) 26,5—24,8 13,9—18,7 12,4—12,5
Индекс большой седалищной вырезки (глубинно-широт- 1 ный) — 44,7 —' 71,1—59,0 13,5—22,2 —
♦ По данным Шульца (1969); Biegert, Maurer (1972); автора.
в выпрямленном положении, в частности М. sacrospinalis (Le Gros
Clark, 1955). Некоторыми исследователями (Snell, Donhuysen,
1968) подчеркивается важное значение также вентрального расши-
рения подвздошной кости и бокового изгиба ее среднего сегмента,
что явилось необходимым условием для соблюдения балансирова-
ния тела в фазе стояния на одной ноге при передвижении. Ука-
занные авторы видят в этой структуре одно из кардинальных раз-
личий с тазом современных антропоидов и других обезьян и при-
писывают ей решающую роль в развитии бипедии.
Весьма отчетлива- дифференциация гоминид от негоминидных
приматов и по относительному укорочению подвздошной кости, и
особенно ее нижней части. Из табл. 6 и рис. 6 следует, что человек
снова занимает экстремальное положение по степени относитель-
ной укороченности нижней длины подвздошной кости, т. е. по сбли-
жению между тазобедренными и подвздошно-крестцовыми су-
ставами.
Большое значение в связи с развитием прямохождения при-
дается изменению. взаимного положения подвздошной, седалищной
и лобковой костей, что наглядно отображается при сопоставлении
в сравнительно-анатомическом ряду приматов угла инклинации
крестцовой части подвздошной кости. Действительно, сакральная
поверхность тазовой кости находится примерно на одной оси с се-
36
далищной костью у низших узконосых обезьян, испытывает дор-
зальный поворот у антропоидов, который у человека заходит на-
столько далеко, что она располагается здесь почти под прямым
углом к седалищной кости. С поворотом сакральной части тазовой
кости назад и вниз развивается также хорошо очерченная большая
седалищная вырезка, что подчеркивает выступание седалищной
ости; выраженность последней определена и модификациями'
связочного аппарата (llig. sacrospinosum et sacrotuberosum). На
переднем крае подвздошной
кости следует особо отметить
оформление передне-нижней 20
подвздошной ости, связанное
с прикреплением илеофемо-
ральной связки и развитием '5'
• 10-
Рис. 6. Нижняя длина подвздошной
. Ли •Pan
НУ п •Ра V
п 90
5 й °°Д
аС.
*Aaf.
кости у высших приматов (в % ОТ „X- тс-------------------------
длины туловища). Обозначения, как гии „ MU '
на рис. '5 (по Biegert, Maurer, 1972) Длина туловища, ММ
прямого мускула. В области седалищной кости известное значение
придается укорочению ее тела, что сопровождается «восхождени-
ем» седалищного бугра с соответствующим сближением его с верт-
лужной впадиной. Впрочем, следует указать, что по отношению к
длине туловища пропорции седалищной кости человека не выделя-
ют его из группы Hominoidea.
В последнее время появились попытки новой интерпретации ге-
неза комплекса бипедии на тазовой кости человека на основании
некоторых эмбриологических данных. Так, согласно Оливье (Oli-
vier, 1962, 1967) и Мендрезу и Кобленцу (Mendrez, Coblentz, 1966),
таз человека и циноморфных обезьян обнаруживает на очень ранних
стадиях эмбриогенеза сходную конфигурацию, причем в основных
чертах она.является как бы «гоминидной» (широкая, и низкая фор-
ма крыла с отчетливой большой сеДалищной вырезкой). Отсюда
делается вывод, -что «гоминидная форма» является общим эмбрио-
нальным типом тазовой кости приматов, которые таким образом
все «преадаптированы» к прямохождению, реализующемуся лишь
у человека. Оливье высказывает даже парадоксальную (с точки
зрения классической эволюционной морфологии) мысль, что связь
между локомоцией и формой таза не есть связь причинная, хотя
он и. оставляет открытым вопрос о факторах, которые могли бы
обусловить эту зависимость. Эти данные противоречат также рас-
пространенной концепции (см., например, Krukierek, 1955) о соот-
ветствии онтогенетических преобразований формы таза человека
генеральному направлению его эволюции.
Не входя в подробное обсуждение этого вопроса, заслуживаю-
щего специальных, дальнейших исследований на более представи-
37
тельном сравнительно-эмбриологическом материале, следует все
же заметить, что расширенная форма крыла подвздошной кости
в эмбриогенезе циноморфных существует весьма короткое время, и
уже очень рано и быстро происходит ее преобразование в типич-
ном для этих форм направлении. Характерная вытянутая и сужен-
ная форма крыла наблюдается уже у зародыша макака 22 мм
длины.
В упомянутом сходстве, по-видимому, более правомерно видеть
не «преадаптацию» в смысле, близком по существу к неотенической
концепции Болька (Bolk, 1926), но известную аналогию с онтоге-
нетическими фактами, встречающимися и в других отделах скелета
и определяющимися, вероятно, общими ранними эмбриональными
признаками более широкого значения. Такова, например, свой-
ственная человеку и другим млекопитающим широкая форма за-
кладок дистальных костей конечностей, которая даже рассматри-
валась Г. А. .Бонч-Осмоловским (1941), как один из аргументов
для обоснования его ошибочной концепции о «лапообразности»
исходной формы кисти общего предка гоминид и антропоидов.
Сказанное побуждает нас отнестись с особым интересом прежде
всего к строению тазового пояса ранних гоминид и других прима-
тов, переходивших к факультативному или облигатному прямо-
хождению.
Б качестве представителя высших приматов с предполагаемыми
элементами прямохождения («факультативная бипедия») рассмот-
рим загадочную миоценовую обезьяну — ореопитека, чье положе-
ние в филогенезе (и таксономии) приматов все еще недостаточно
ясно. Как известно, особое внимание привлекла форма тазовой
кости ореопитека, на которой отмечались признаки начавшегося
расширения. По величине широтно-высотного указателя подвздош-
ной кости (80,0) ореопитек занимает как бы промежуточное поло-
жение между обоими африканскими антропоидами (66,0 у шим-
панзе, 94,0 у гориллы). По отношению к длине туловища ширина
подвздошной кости ореопитека также помещает его между шим-
панзе и гориллой, но значительно ближе к первому, а по отноше-
нию к орангутану можно говорить даже о совпадении относитель-
ной ширины подвздошной кости у этих двух форм. Если проанали-
зировать дальше пропорции подвздошной кости ореопитека, можно
подтвердить, что и по относительной ширине пластинки крыла, и
по важнейшему в аспекте бипедии признаку — расширенности
сакральной поверхности — этот примат не выходит за рамки ва-
риаций современных антропоморфных, сближаясь в наибольшей
степени с орангом (Biegert, Maurer, 1972). То же касается и отно-
сительной длины подвздошной кости, которая снова почти совпа-
дает с соответствующим индексом оранга. По весьма существен-
ному в комплексе бипедии признаку — относительной нижней дли-
не подвздошной кости — ореопитек занимает промежуточное поло-
жение между низшими и высшими узконосыми, не показывая уни-
кальной специализации человека.
38
То же нужно сказать и о другом важнейшем признаке — поло-
жении и развитии вертлужной впадины. Отношение длин седа-
лищной и подвздошной костей, определяющее положение aceta-
buli на оси тазовой кости, здесь очень низкое, что свидетельствует
о значительном смещении вертлужной впадины в каудальном на-
правлении, хотя абсолютная длина седалищной кости ореопитека
довольно велика. Очень резкие различия между ореопитеком и че-
ловеком существуют и в абсолютном, и в относительном развитии
вертлужной впадины. Особенно мала последняя по отношению к
тотальным размерам тазовой кости. Соответствующий индекс да-
леко не достигает минимума у человека. Здесь следует особо от-
метить, что развитие вертлужной впадины у человека вообще очень
стабильно: так, например, коэффициент вариации ее наибольшего
диаметра равен всего 5%, т. е. он здесь такой же, как и для всей
высоты таза, хотя последний размер превышает первый почти в
четыре раза. Относительное развитие вертлужной впадины и ее
положение также является довольно устойчивым признаком у
Н. sapiens.
По всем этим признакам различия типа' ореопитека от гоминид-
ного весьма отчетливы. По пропорциям вертлужной впадины этот
примат явно принадлежит к кругу понгидных форм. Индекс вели-
чины acetabuli (к тазовой кости) здесь фактически совпадает с та-
ковым для <5 шимпанзе (14,5 по Шульцу, 1969), Таким образом,'
морфология тазовой кости ореопитека по основным, наиболее су-
щественй’ым в аспекте бипедии особенностям, явно не гоминидная,
но понгидная. Лишь по немногим признакам можно говорить о
довольно слабо проявляющейся тенденции к промежуточному по-
ложению ореопитека: это — относительное укорочение расстояния
между вертлужной впадиной и подвздошно-крестцовым сочлене-
нием (в масштабе антропоидов), некоторое расширение подвздош-
ной кости относительно ее длины, структура передне-нижней под-
вздошной ости — признаки, интерпретирующиеся иногда как на-
чальная адаптация к двуногости (Straus, 1963). Однако значение
этих особенностей отнюдь не следует переоценивать, так как
основные черты комплекса бипедии не были выражены на тазе
ореопитека, сохранявшем в общем антропоидный облик (Biegert,
Maurer, 1972). Все же существенно, что некоторые черты этого
комплекса уже намечались у примата, жившего 10—12 млн. лет
назад.
Рассмотрим теперь с этой же точки зрения строение таза наи-
более ранних из известных сейчас гоминид — австралопитековых.
От них к настоящему времени достаточно хорошо изучены фраг-
менты тазовых костей как грацильных (Л. africanus); так и мас-
сивных форм (Л. robustus).
Как видно из табл. 6 и рис. 7, общая форма таза австралопите-
ковых вполне гоминидная, характеризующаяся прежде всего отно-
сительной укороченностью и расширенностью подвздошной кости.
Особенно бросается в глаза широкая форма пластинки подвздош-
39
ного крыла, что отчасти связано с очень заметным выступанием
области обеих передних подвздошных остей (особенно верхних) и
свидетельствует о широком туловище. Один из важнейших призна-
ков комплекса бипедии — развитие сакральной поверхности —
также совершенно не обезьяний, но вполне человеческий у А. ro-
bustus и приближающийся к человеческому у A. africanus. Воз-
можно, что при интерпретации различий между обеими форма-
ми австралопитековых следует
принять во внимание также
разницу в размерах тела, по-
скольку для этого признака
намечается положительная ал-
лометрия внутри обеих групп
катарриновых приматов (см.
рис. 5).
Рис. 7. Таз австралопитековых: А -г
плезиантроп; Б —Прометей (по Pin-
kerton, 1973)-
Сопоставление по другому важнейшему признаку комплекса би-
педии — относительной нижней длине подвздошной кости — сви-
детельствует о совершенно человеческих соотношениях у обоих
австралопитековых, занимающих по степени сближения тазобед-
' ренного и подвздошно-крестцового суставов наряду с человеком
крайнее положение в ряду узконосых приматов (см. рис. 6).
То же самое следует сказать и о таком кардинальном различии
двуногих гоминид и негоминидных приматов, как развитие и поло- .
жение вертлужной впадины. Относительные размеры ее у A. afri-
canus не только совершенно человеческие, но и совпадают со сред-
ними для современных женских тазов. То же относится, по-види-
мому, и к A. robustus', где соответствующий индекс выше и сбли-’
жается со средним для мужских тазов.
К числу признаков «комплекса бипедии», отчетливо выявляю-
щихся у австралопитековых, относятся также: извилистая форма
подвздошного гребня, ротация подвздошно-крестцового сочленения,
формирование большой седалищной вырезки. Как видно из при-
веденных данных, указатель большой седалищной вырезки A. afri-
canus совершенно'не антропоидный, но гомицидный, укладываю-
щийся в пределы индивидуальных его вариаций у Н. sapiens.
Исходя из указанного выше адаптивного к прямохождению
значения этих признаков, можно заключить, что бипедия уже
существовала у австралопитековых (Le Gros Clark, 1955; Biegert,
Maurer, 1972; Me Henry, 1975). Это не исключает, конечно, возмож- t
1 Определение у A. robustus juobh подвздошной и седалищной костей связано
со значительной реконструкцией краниальной части подвздошной кости.
40
пости некоторых различий с гоминидным типом, как например,
относительно более узкая сакральная область у A. africanus или
несколько повышенное в масштабе вариаций у Н. sapiens значение
высотно-широтного указателя подвздошной кости. С филогенетиче-
ской точки зрения весьма интересен факт вариаций в степени уко-
рочения седалищной кости у разных представителей австралопите-
ковых, отмеченный Ле Гро Кларком (Le Gros Clark, 1955). В наи-
большей степени «восхождение» седалищного бугра выражено у
австралопитека — Прометея (Д. africanus), тазовая кость которого
вообще имеет самый гоминидный вид: верхний край бугра почти
достигает уровня нижнего края вертлужной впадины. Противопо-
ложный вариант отмечен у A. robustus. Впрочем, здесь следует
иметь в виду дба обстоятельства. Во-первых, наличие четкого по-
лового диморфизма в абсолютном и относительном развитии седа-
лищной кости у человека, как и у других млекопитающих (см.
Bernstein, Crelin, 1967). При этом нужно принять во внимание так-
же и возможную роль различий в массе тела, поскольку намечает-
ся положительная аллометрия в отношении этого признака как сре-
ди гоминоидов, так и у церкопитекоидов (Biegert, iMaurer, 1972).
Во-вторых, существенно, что и современный человек по относи-
тельному развитию седалищной кости (к длине туловшДа) вовсе
не занимает крайнего положения в ряду узконосых приматов: как
среди высших, так и среди низших их представителей существуют
формы с большим относительным укорочением седалищной кости.
Вообще некоторые признаки половой дифференцировки таза
выявляются в филогенезе гоминид, по-видимому, очень рано: так,
например, по важнейшим признакам диморфизма (относительные
размеры вертлужной впадины и запирательного отверстия, форма
седалищной вырезки, седалищно-лобковый индекс) A. africanus
(плезиантроп) явно ближе к женскому типу, чем к мужскому.
Из всего сказанного вытекает, что ввиду раннего формирова-
ния прямохождения и гоминидного комплекса таза в антропогенезе
морфология последнего представляет наибольший интерес для
таксономического (и функционального) анализа ранних стадий
эволюции гоминид и их предшественников, в меньшей степени —
более поздних форм.
Из числа последних внимание морфолога должны привлечь
прежде всего архантропы. Высказывалось мнение (Zihlman, Hun-
ter, 1972), что дальнейшая эволюция австралопитековых по отно-
шению к человеку должна была сопровождаться определенными
изменениями в передней части подвздошной кости, несколько мо-
дифицировавшими характер вращательных движений в тазобед-
ренном суставе. Хотя в принципе это и не изменило локомоции
человека сравнительно с австралопитековыми, но способствовало
выработке более совершенного с биомеханической точки зрения
прямохождения, характерного для современного человека.
До недавнего времени таз древнейших людей (архантропов)
был совершенно не известен. Определенный свет на его структуру
41
пролила находка фрагмента тазовой кости в IV слое Олдувей-
ского ущелья, Олдувей 28, сделанная в сопровождении ашельских
орудий. Функциональный анализ костных остатков с несомнен-
ностью свидетельствует о хорошо развитой бипедии этого гоми-
нида, а таксономический анализ позволяет отнести находку к
Н. erectus (Day, 1971).
К числу признаков, свидетельствующих о «привычной бипедии»
этого гоминида, можно отнести: широкую форму и S-образную
изогнутость крыла подвздошной кости, отчетливое развитие перед-
не-нижней подвздошной ости, мощ-
ный «вертикальный пилястр» на
подвздошной кости (рис. 8). Очень
крупной была вертлужная впади-
на: ее вертикальный диаметр, по-
видимому, близок к крайнему вари-
анту этого размера, отмеченному у
классических неандертальцев и кро-
маньонцев. Другой важный признак
гоминидного комплекса — форма
большой ' седалищной вырезки: ин-
декс последней составляет здесь
примерно 53, т. е. свидетельству-
ет о совершенно человеческой и
С
Рис. 8. Таз Олдувей 28 (по Day, 1971)
притом подчеркнуто женской конфигурации этой области.
Особый интерес эта находка представляет и в том отношении,
что, по-видймому, вполне реально предположение о принадлежно-
сти морфологически и стратиграфически тесно связанных фрагмен-
тов тазовой кости и бедра одному индивидууму. Таким образом,
впервые представляется возможность параллельного исследования
обоих важных отделов нижней конечности архантропов — прежде
всего в! аспекте бипедии.
В итоге такого анализа оказывается, что целый ряд признаков
свидетельствует — одновременно на обеих костях — о человече-
ской шагающей походке этого гоминида. Сюда относятся: мощное
развитие вертикального пилястра на тазовой кости в сочетании с
распространенностью прикрепления среднего и малого ягодичных
мускулов на бедре. Такое сочетание указывает па хорошее разви-
тие механизма альтернативного наклона таза при шагающей поход-
ке. В этом плане следует упомянуть и о вероятной миграции перед-
не-верхней подвздошной^ости вниз, что соответствует вышеприве-
денному предположению Цильман и Хантер . (Zihlman, Hunter,
1972). Одновременно наблюдается и сильное развитие горизонталь-
ной перекладины в комбинации с хорошо выраженным прикрепле-
нием большого ягодичного мускула, что свидетельствует о мощ-
42
ном механизме экстензии тазооедренного сустава, тогда как
прикрепление прямого мускула бедра и обоих широких мускулов
показывает возможность и мощной экстензии коленного сустава.
Наконец, отчетливый рельеф в обоих пунктах прикрепления илео-
феморальной связки указывает на полное развитие опорного ме-
ханизма таза для прямохождения и.типичное для двуногого гоми-
нида расположение центра тяжести тела (Day, 1971).
Пока остается вопрос о полной идентичности этой походки с
типичной для Н. sapiens, но сам факт наличия у олдувейского
архантропа шагающей походки не подлежит особому сомнению,
тем более, что предположение о существовании шагающей поход-
ки, как известно, высказывалось уже в связи со структурой ногте-
вой фаланги большого пальца стопы Олдувей 10, имеющей возраст
около 1 млн. лет (см. ниже).
Среди палеоантропов наиболее полно представлен тазовый пояс
у палестинских форм; некоторые данные имеются и о тазовых ко-
стях неандертальцев Европы (Неандерталь, Шапелль, Крапина).
Абсолютные размеры таза (высота и ширина) как европейских,
так и переднеазиатских палеоантропов, совершенно человеческие.
Особенно относится это к широтному диаметру, который во всех
случаях входит в пределы вариаций у современного человека или.
даже превосходит максимум, как у некоторых высокорослых пале-
стинцев (Схул IV и V), где соответствующие широтные размеры
составляют 280 и 290 мм против современной средней 279 мм для
мужчин-европеоидов. То же можно сказать и о высотном диаметре,
который, по-видимому, значительно варьировал. Так, у собственно
меандертальской находки он превосходил современный групповой
максимум (225 мм против 220 мм), тогда как у Шапелль и неко-
торых палестинцев высота таза в общем близка к среднему совре-
менному варианту. Из сказанного следует, что указатель общей
формы таза везде совершенно человеческий, причем даже преде-
лы вариаций его у палеоантропов полностью совпадают с колеба-
ниями групповых средних у современного человека: 70,0
(Схул V) — 85 (Шапелль) против 70—85 по групповым средним
Н. sapiens. Особенно низкая и широкая форма таза была свой-
ственна «прогрессивному» палеоантропу Схул V. В известной мере
вариации формы таза у гоминид могут связываться с действием
некоторых экзогенных факторов: так, некоторые привычные ста-
тические позы («сидение на корточках»), привычное ношение
тяжестей, характер питания, например на развитии таза особенно
сказывается недостаток пищевого кальция, могут модифицировать
индивидуальную форму таза, если действуют длительное время в
период роста, как о том свидетельствуют многочисленные наблю-
дения. Подобная интерпретация индивидуальных различий прило-
жима и к древним людям. Напомним также, что для многих верхне-
палеолитических гоминид (Кромапьои, Пржедмости) типичны вы-
сокие значения указателя, т. е. относительно высокие и узкие по
форме тазы.
43
I
г
I
Абсолютно человеческой является и степень торзиона крестцо- '
вой части подвздошной кости по отношению к седалищной оси,
причем у классических неандертальцев (Шапелль, Неандерталь)
этот подвздошно-седалищный угол (96°) даже меньше, чем в сред- I
нем у современного, человека (109—115°), что можно рассматри-
вать как «ультрагоминидный» признак. По-видимому, и палестин-
ские палеоантропы вполне современны в этом отношении. Такие
величины угла указывают на полностью выпрямленное положение (
тела, если видеть в опускании крестцовой части подвздошной
кости относительно тазобедренного сустава результат адаптации .
к увеличению нагрузки, вызванному ходьбой в выпрямленном по-
ложении.
Абсолютное и относительное развитие вертлужной впадины у
палеоантропов также вполне человеческое и соответствует полово-
му диморфизму в этом признаке. Седалищная вырезка на тазовых
костях Шапелль и Неандерталь «ультрагоминидна», отличаясь
особенно глубокой и узкой формой'. Указанная особенность тазо-
вых костей шапелльца и неандертальца подтверждает их принад-
лежность мужским индивидуумам. Нет также существенных рас-
хождений с современными людьми и в структуре седалищной ости.
Палеоантропы разделяют, очевидно, со всеми гоминидами при-
знак извилистости подвздошного гребня; напомним, что уже у
австралопитековых она была выражена в «ультрагоминидной»
степени. На подвздошном гребне современной тазовой кости обыч-
но заметна вертикальная перекладина, спускающаяся от своей
верхней границы — бугорка гребня — к тазобедренному суставу
и являющаяся рычагом для мускулов, участвующих в балансиро-
вании туловища. Эта перекладина, отсутствующая у антропоидов, :
очень отчетлива у Олдувей 28, а также у Неандерталь и палестин- *,
ских палеоантропов, где она даже обнаруживает тенденцию к
удвоению у Схул IV.
Палеоантропы не показывают никаких отклонений от современ-
ного человеческого типа и по такому существенному признаку, как
увеличенная площадь прикрепления крестца в дорзальном и кау-
дальном направлениях. То же можно сказать и о пропорциях се- ;
далищной кости и развитии "седалищного бугра — они даже «уль-
трагоминидны» у палеоантропов.
Что касается мускульного рельефа, то Патт (1955) отмечает
современный тип ягодичных линий на подвздошной кости ша- ;
пелльца. Передне-нижняя подвздошная ость сильно выступает-у
европейских неандертальцев, что считается прогрессивным ’
признаком, поскольку указанная формация связана с разви-
тием прямого мускула бедра. Во всяком, случае, даже при самом
осторожном подходе^ к интерпретации особенностей мускульного
рельефа, нельзя найти у палеоантропов сколько-нибудь сущест- ,
.венных отклонений от современного человеческого типа. i
Тазовый пояс тесно связан также с размножением и родовым |
механизмом, здесь обычно достаточно отчетливы половые разли- )
f
I
44
чия. С этой точки зрения наибольший интерес представляет морфо-
логия малого таза и особенности строения лобковой кости. Струк-
тура лобковой кости индивидуально очень варьировала у палеоан-
тропов. Впрочем, в области лобкового симфиза нельзя найти ника-
ких отклонений от современного типа, исключая массивность и
большие размеры. Своеобразной особенностью являются очень вы-
сокие значения лобкового угла: от 80° у Схул IV и IX до 100°
у Табун I. Таким образом, маленькая женщина Табун имела вели-
чину лобкового угла, находящуюся у верхней границы вариаций
этого признака у широкобедрых европейских женщин. Также почти
все размеры конъюгат выше у Табун сравнительно с гораздо $олее
высокорослыми европейскими женщинами. Очень выражена жен-
ская, относительно широкая и мелкая форма большой седалищной
вырезки с индексом 41,7 против 59—61 у современных женщин,
велика и длина лобковой кости. Типичными являются и пропорции
вертлужной впадины (по отношению к тазовой кости): у Табун I
указатель равен 23,5 против 24,8 в среднем в современной женской
серии, тогда как у мужчин Схул IV и V и Неандерталь соответ-
ствующие значения 27—28, т. е. также близкие к средней совре-
менной мужской серии (26,5 у европейцев).
Но в группе Схул наблюдаются и некоторые черты своеобразия
в половой дифференцировке таза, например, на скелете Схул IX.
Этот гоминид характеризуется совмещением женских и мужских
признаков в строении таза: к числу первых принадлежат, напри-
мер, длина лобковой кости, размер запирательного отверстия,
площадь подлобковой дуги и лобковый угол. Поскольку по осталь-
ным признакам скелета это, очевидно, мужской индивидуум, при-
ходится предположить, как и в отношении вариаций грудины и
ключицы, своеобразную комбинацию признаков полового димор-
физма на скелетах некоторых палеоантропов.
Определенное смешение признаков полового диморфизма отме-
чается и для таза Олдувей 28. Так, по ’общей массивности и раз-
мерам вертлужной впадины он ближе к мужскому типу, а по фор-
ме большой седалищной вырезки и сочленовной поверхности под-
вздошной кости — к женскому. Следует особо подчеркнуть, что
таз Олдувей безусловно женский по наиболее" существенному из
перечисленных признаков половой дифференцировки —указателю
формы большой седалищной вырезки1.
В заключение следует упомянуть о структуре костных компо-
нентов тазобедренного сустава. Выше упоминалось, что в эволюции
гоминид значительным изменениям подверглись вертлужная впа-
дина таза и головка бедра, размеры и крепость которых увеличи-
лись, причем кульминации этот процесс достиг у палеоантропов.
В этом отношении снова выделяются европейские неандертальцы
1 Конфигурация большой седалищной вырезки .является одним из немногих остео- -
логических индикаторов пола, относительно не зависимых от общих размеров
таза и отражающих влияние локальных градиентов роста.
45
с их особенно крупными компонентами сустава, что подчеркнуто в
еще большей степени, чем на плечевом суставе. Возможно", это
увеличение вертлужной впадины и головки бедра способствовало
большей прочности сустава; обеспечивая опору и крепость соедине-
ния, большая поверхность суставных компонентов позволяет про-
являть и большую силу в движениях, хотя и менее разнообразных.
Интересно также, что по соотношениям обоих компонентов тазо-
бедренный сустав неандертальского человека, в отличие от плече-
вого, не показывает сколько-нибудь существенных отклонений от
современного типа (Малявская, Хрисанфова, 1962), причем у
Неандерталь соответствие в развитии обоих костных компонентов
сустава выражено даже более полно, чем у Н. sapiens. Сказанное
полностью относится и к палестинцам и крапинцам, у которых и
по абсолютным, и по относительным размерам суставных компо-
нентов нет отклонений от современного типа.
Почти ничего нельзя сказать о соотношении компонентов тазо-
бедренного сустава у архантропов, поскольку ни в одном случае
оба компонента не сохранились одновременно. Можно только
отметить совершенно человеческий тип и размеры головки бедрен-
ной кости питекантропов и вертлужной впадины у Олдувей. В от-
ношении синантропа, у которого разрушенные эпифизы бедренной
кости были реконструированы Вейденрейхом (Weidenreich, 1941),
сошлемся на указание этого автора, считавшего на основании неко-
торых косвенных данных (положение ягодичной шероховатости),
что синантроп был способен к полному выпрямлению ноги в тазо-
бедренном суставе.
Таким образом, морфологический анализ остатков таза поздних
гоминид подтверждает высказанное выше предположение, что
структура таза как индикатор бипедии мало специфична у этих
форм. Она во всех отношениях совершенно человеческая, причем
многие структурные особенности выражены у палеоантропов даже
в «ультрагоминидной» степени (например, ширина таза в крыльях,
величина подвздошно-седалищного угла, форма большой седалищ-
ной вырезки, пропорции седалищной кости и развитие седалищного
бугра, размеры вертлужной -впадины). Это — ожидаемый резуль-
тат, учитывая вообще очень раннее оформление комплекса бипе-
дии на тазовом поясе гоминид.
Глава III
ДЛИННЫЕ КОСТИ КОНЕЧНОСТЕЙ
В палеоантропологии и палеонтологии приматов нередко приходит-
ся иметь дело с остатками длинных костей, как преобладающим,
а часто и единственным материалом, на основании которого тог-
да делаются попытки интерпретации локомоции ископаемых форм
и даже таксономического диагноза. Трудности, встающие перед
исследователем при подобном анализе, хорошо иллюстрируются
работами Страуса (Straus, 1948) и Керна и Страуса (Kern, Stra-
us, 1949), которые в итоге сравнительного изучения фрагментов
бедренных и плечевых костей некоторых австралопитеков и совре-
менных приматов пришли к выводу о невозможности заключить
по морфологии этих компонентов скелета об особенностях локо-
моции ископаемых форм. Сходный вывод — о «нейтральности»
многих остеометрических и остеоскопических признаков длинных
костей — следует и из монографии Патта (1955), посвященной
морфологической характеристике неандертальского типа.
Основное содержание настоящей главы представляет анализ
длинных костей конечностей гоминид с точки зрения того интере-
са, который они имеют для эволюционной морфологии посткра-
ниального скелета человека. Выбор морфологических признаков,
как и в других случаях, ограничен указанным принципом и по
самому существу стоящей перед нами задачи не должен включать
все детали строения изучаемых костей. Поскольку бедренная кость
играет совершенно особую роль в анализе палеоантропологиче-
ских материалов, именно с нее мы и начнем изложение соответ-
ствующих данных.
НИЖНЯЯ КОНЕЧНОСТЬ
Как уже говорилось, морфология бедренной кости наиболее
разработана в эволюционном и сравнительном аспектах. В па-
леонтологии приматов структура бедра получила особое значение
ввиду значительной представленности этого отдела скелета среди
ископаемых форм. Вместе с тем нельзя не отметить, что оценка
признаков бедра претерпела существенную трансформацию со
временем, когда значительно пополнилась информация и об основ-
ном, палеонтологическом, и о сравнительно-анатомическом мате-
риале. Сказанное можно проиллюстрировать на примере одного
из важнейших с точки зрения комплекса бипедии признаков —
изгибе бедренной кости.
Согласно распространенной в морфологической литературе кон-
цепции, изгиб бедра обычно связывается в своем генезе с прямо-
хождением и распространением прикреплений четырехглавого мус-
кула. Первоначально его считали исключительным атрибутом пря-
47
мохождения: так, Вирхов, сопоставляя бедренную кость питекан- ;
тропа с выпрямленной бедренной костью гиббона, счел на этом )
основании слабый изгиб кости обезьяночеловека питекоидной чер- 1
той. Напротив, Фрэпон и Лоэ (Fraipont, Lohest, 1887), сравнивав-
шие бедро одного из палеоантропов (Спи) с изогнутыми костями j
африканских антропоидов, полагали на этом основании, что пите-
коидным признаком на этот раз является уже сильная изогнутость 1
кости ископаемого человека. Дальнейшее расширение сравнитель-
но-анатомического ряда с включением в него других приматов и I
млекопитающих показывает, что изгиб бедра встречается у пред- [
ставителей самых разных таксономических групп и в связи с раз- i
личными типами локомоции. Так, среди антропоидов изгиб посто-
янен на бедренных костях .африканских форм (шимпанзе, горил- \
ла) в противоположность азиатским, имеющим выпрямленные ко- !
сти. Многие виды низших узконосых обезьян имеют изогнутые 1
бедра (особенно некоторые павианы). У других млекопитающих j
выпуклый вперед изгиб встречается среди грызунов, хищных, на-
секомоядных, копытных, сумчатых. Таким образом, не выявляет-
ся исключительной связи изгиба бедра с бипедией, как и вообще I
с одним каким-нибудь определенным типом локомоции. Напри- ;
мер, в одной и той же группе животных — копытные — при оди-
наковой локомоции, специализированные бегуны, одни формы /
имеют прямое, другие — изогнутое бедро. Более того,’ значитель- J
ные вариации в степени изгиба этой кости могут существовать у
представителей одного и того же вида, а иногда даже у одного и >
того же животного бедренные кости могут сильно различаться по . !
изогнутости (такие случаи известны и среди приматов, например
у гиббона, игрунки, павиана, Патт, 1955; Швальбе, 1919/21). Сле-
дует также упомянуть о значительных колебаниях изгиба и у сов- 1
ременного человека, где максимум групповых средних индекса из- |
гиба почти вдвое превосходит минимум (Quiros, Faulhaber, Argu- !
ello, 1966; Singh, Singh, 1972).
Сказанное приводит к заключению о значительных вариациях
изгиба бедренной кости у гоминид, приматов и млекопитающих в
целом. Среди факторов, которые могли бы обусловить эту вари-
абильность, помимо генетического нужно иметь в виду и некото-
рые другие. Очевидна, например, его зависимость от конкретных
биомеханических условий: характера и величины механической на-
грузки и свойств самой костной ткани. С этой точки зрения груп-
повые различия в изгибе бедра среди современных людей в из-
вестной Мере связываются с плотностью костной ткани - (минера-
лизация, соотношение веса и объема) (Evans, 1957; Broman et al.
1958; Singh, Singh, 1972).
Следовательно, признавая специфической особенностью бед-
ренной кости человека изгиб ее диафиза, нельзя вместе с тем за-
бывать, что этот признак обнаруживает широкий диапазон измен-
чивости, вызванной самыми различными причинами. Это сообра-
жение особенно важно при анализе ископаемых материалов: оце-
48
нивая степень изгиба бедра, следует иметь в виду как масштаб
вариаций этого признака у современных гоминид и других при-
матов, так отсутствие его исключительной связи с бипедией. Ес-'
тественно, что особая осторожность в интерпретации нужна при
работе с фрагментарным материалом.
Другой важнейший признак человеческого бедра — пилястр,
который рассматривается обычно как контрфорс, возникший в ре-
зультате адаптивной реакции костной ткани на изгиб бедра (ук-
репление задней стенки), или как «преувеличение» шероховатой
линии в связи с распространением прикреплений четырехглавого
мускула бедра. Наиболее распространена компромиссная точка
зрения на генез пилястра, согласно которой его возникновение
объясняется частично статическими, частично динамическими при-
чинами, хотя разные авторы придают им различный вес. Сущест-
вуют также разногласия относительно того, какие мускулы сыг-
рали здесь главную роль. Нужно иметь в виду и сложность вза-
имоотношений ме.жду изгибом и пилястром: в некоторых случаях
последний реально выполняет роль контрфорса (например, на ра-
хитических бедрах), но можно привести и ряд примеров, когда
очень сильный изгиб не сопровождается выраженным пилястром,
как это нередко наблюдается на бедренных костях гориллы и
других приматов. По-видимому, при интерпретации этих соотно-
шений следует иметь в виду сложность генеза пилястра и множе-
ственность факторов, могущих повлиять на его развитие, как это
указывалось выше при обсуждении вариаций изгиба бедра.
В литературе распространена точка зрения на пилястр, как
на специфически человеческое образование. Например, в палеоан-
тропологии его считают одним из главных критериев при опреде-
лении принадлежности фрагментов бедренных костей гоминидам.
Напомним, в частности, что. Вейденрейх (1941) применил этот
критерий в качестве основы для отбора фрагментов диафизов бед-
ренных костей синантропа. Вместе с тем здесь снова нужно иметь
в виду, что пилястр (или структура, сходная с пилястром) может
встречаться и у некоторых животных (среди обезьян, хищных,
копытных). У антропоидов структура пилястра и шероховатой ли-
нии обычно не выражена, за редкими исключениями; у некоторых
низших узконосых обезьян, например павианов, латеральная губа
шероховатой линии развивается в отчетливый гребень; варьирует
структура задней стенки бедра и у широконосых обезьян (шеро-
ховатая линия, по-видимому, выражена у коаты и ревуна). Сле-
довательно, признавая этот признак специфичным для человека,
снова нельзя говорить об его исключительной связи с бипедией,
поскольку среди приматов он, вероятно, наиболее выражен у не-
которых проноградных мартышкообразных обезьян. Следует так-
же иметь в виду, что пилястр может отсутствовать на бедренных
костях современного человека, и почти в любой остеологической
коллекции можно найти кости со слабым развитием шероховатой
линии бедра.
4 Е. Н. Хрисанфова
49
Для количественной оценки пилястра у человека обычно при-
меняется тай называемый указатель пилястрии; т. е. - процентное
отношение сагиттального диаметра диафиза к поперечному на
уровне наибольшего развития пилястра. Соответствующие груп-
повые средние этого индекса колеблются в пределах: 100,0—129,4
против 75,4—97,1 у антропоидов. Индивидуально размах вариа-
ций, конечно, много шире и достигает, например, у населения
Баварии: 72,5—136,7 (Martin-Sailer, 1958). Изменчивость фор-
мы, диафиза бедра у человека исключительно велика (Dokladal,
1972).
Однако указатель пилястрии сам по себе далеко не всегда
адекватно отражает реальное развитие пилястра. Известны. при-
меры, когда при очень высоком указателе нельзя собственно го-
ворить о пилястре, так как кость сильно сжата с боков в вент-
ральной части;, с другой стороны, при очень слабом индексе мо-
жет наблюдаться сильное развитие пилястра, что имеет место при
резкой передне-задней уплощенности диафиза бедра. Поэтому по-
мимо указателя пилястрии необходимо давать также описатель-
ную оценку пилястра. Но в некоторых случаях и этого может ока-
заться недостаточно. Учитывая вероятную двойственность генеза
.пилястра (роль статического и динамического факторов), логич-
нее принимать во внимание развитие не только суперструктуры,
но также и всей задней стенки бедра, прежде всего его компакты,-
Примеры, иллюстрирующие роль такого «внутреннего пилястра»,
будут приведены ниже. Структура диафиза бедра отражает также
весьма существенный в гоминидном комплексе признак скручен-
ности кости — ее торзион. Этот угол определяется взаимным рас-
положением оси верхнего эпифиза (шейки) кости и касательной
к мыщелкам нижнего эпифиза; его величина связывается с дина-
микой изменений таза и действием ротирующих мускулов.
Для человека типичен «положительный торзион», когда ось
шейки проходит сзади латерально, вперед медиально. Групповые
вариации этого признака: 8—39,7°, т. е. ретрбверсия не встречает-
ся как групповой признак. Индивидуально значения угла торзио-
на колеблются более широко, от —25 до +49°, хотя отрицатель-
ные углы и .встречаются весьма редко. Этот признак имеет раз-
граничительное значение прежде всего с антропоидами, посколь-
ку у последних значительно более обычны отрицательные углы
(ретроверсия). В месте с тем антеверсия снова не является исклю-
чительной принадлежностью бедер человека и не показывает об-
лигатной связи с бипедйей, так как она свойственна многим при-
матам (низшие узконосые обезьяны, лемуры, в меньшей степени
цебусовые). . '
Пожалуй, наибольший интерес как компонент комплекса би-
педии представляет кондилодиафизарный угол — угол инклина-
ции, составленный анатомической осью кости и касательной к
плоскости обоих мыщелков («коленному базису»): у человеку в
норме ось' бедра проходит сверху медиально, вниз латерально.
50
Швальбе считал критерием гоминидности бедра значения этого
угла не выше 83° *, и в отличие от других рассмотренных выше
признаков, последующие исследования не внесли существенных
коррективов в эту интерпретацию. Действительно, хотя вариации
групповых средних и индивидуальных величин оказались несколь-
ко более широкими, чем предполагал Швальбе (обычно в преде-
лах: 78—82° для первых и 72—88° для вторых), все же этот при-
знак значительно более стабилен у человека, чем другие, класси-
ческие компоненты «комплекса бипедии». Правда, здесь следует
иметь в виду частичную трансгрессию с антропоидами (79—94°),
тогда как низшие обезьяны и полуобезьяны имеют уже значи-
тельно более высокие углы инклинации — 86,5—90,5°.
Менее важен для таксономического диагноза коллодиафизар-
ныи угол, определяющий положение шейки бедра по отношению к
диафизу. Для него характерна очень высокая групповая (121—
133°) и особенно исключительно сильная индивидуальная измен-
чивость (114—153°). В последнем случае наблюдается трансгрес-
сия с современными антропоидами (109—138°). В известной сте-
пени сказанное можно распространить и на развитие шейки бед-
ра: признак этот имеет теоретическое значение в большей мере,
чем практическое из-за своей значительной изменчивости (Оливье,
1972). Гораздо более существенно укрепление головки бедра как
компонента тазобедренного сустава.
Что касается нижнего эпифиза кости, то специфически гоми-
нидной его модификацией является, согласно Швальбе, преобла-
дание трансверзального развития обоих мыщелков над сагит-
тальным, а также относительно крупный наружный мыщелок.
В этом последнем признаке пытались видеть один из важных кри-
териев выпрямленного положения тела, поскольку между челове-
ком и антропоидами существуют различия в распределении стати-
ческой нагрузки на нижнюю конечность. Однако следует иметь в
виду; что к статике имеет отношение не вся сочленовная поверх-
ность, а только ее часть, контактирующая при полном разгибании
колена с тибиальной площадкой. При сопоставлении относитель-
ного развития этих «статических поверхностей» па обоих мыщел-
ках выяснилось, что у человека существует тенденция к их равен-
ству (средняя процентного отношения величины тибиальной по-
верхности наружного мыщелка к таковой внутреннего равна 95,3
при вариациях 80,6—108,6). Это яркий разграничительный при-
знак бедер человека и крупных антропоидов, где колебания ин-
декса 57,7—70,9. Однако снова нужно учитывать частичную транс-
грессию с проноградными циноморфными обезьянами (76,6—
96,4), хотя у последних относительное развитие наружного мы-
щелка в целом и ниже. Все же исключительная связь подобных
пропорций с бипедией вновь не подтверждается (Керн, Страус,
1 Или яе аиже 7° по другой системе отсчета.
4* 51
1949). То же самое можно сказать и о другом признаке нижнего
эпифиза кости — межмыщелковой вырезке. Ее значительное раз-
витие встречается не только у человека, но и у некоторых обезьян,
как низших, так-и высших, что, по Патту, может ассоцииро-
ваться с их способностью вытягивать ногу при «гимнастических»
движениях на деревьях. Вообще следует подчеркнуть, что ввиду
крайне сложной кинематики коленного сустава с его неконгруэнт-
ными поверхностями, тип движений в нем не может определяться
только одной формой- суставных поверхностей, но также строени-
ем и функцией связочного аппарата и способом приложения мус-
кульных сил. Поскольку к тому же для данной системы нельзя
говорить о какой-то единой определенной функции каждого из
мускулов, исходя из широкой взаимозаменяемости функций, обес-
печивающей оптимальную экономию силовых затрат (Knese,
1955), нужна большая осторожность при интерпретации воз-
можных морфологических вариаций костных компонентов этого
сустава.
Из числа признаков, характеризующих структурные особенно-
сти костей голени, наибольший интерес для комплекса бипедии
представляет наклон верхнего эпифиза большой берцовой кости:
с вариациями этого признака связывались кардинальные разли-
чия в локомоции и осанке гоминид и других приматов. Степень
заднего отклонения верхнего эпифиза выражается посредством
угла ретроверсии, составленного касательной к сочленовной по-
верхности внутреннего мыщелка и диафизарной осью; иногда упо-
требляется и в общем однозначный угол инклинации, составлен-
ный той же касательной и физиологической осью кости. Сравни-
тельно-анатомические сопоставления свидетельствуют о довольно
высоких значениях обоих этих углов у антропоидов в сравнении
с современным человеком. У последнего вариации угла ретровер-
сии в среднем 7,6—20°, инклинации — 5,3—16,5°. Однако индиви-
дуально встречаются гораздо более высокие значения этого при-
знака — до 31,5° для угла ретроверсии.
Наиболее существенный вопрос состоит в том, возможно ли во
всех случаях интерпретировать высокие значения углов ретровер-
сии как^показатель согнутого положения колен при ходьбе — на-
подобие -их положения у антропоидов? Выше упоминалось, что
признак достаточно варьирует в популяциях современного чело-
века; с другой стороны, среди современных антропоидов (шим-
панзе) встречаются значения углов, соответствующие групповому
или индивидуальному максимуму Н. sapiens. Сочленовные пло-
щадки мыщелков у прямостоящего человека, как показало рент-
генографическое изучение, могут быть ретроверсированы, тогда
как костные оси бедра и большой берцовой кости располагаются
полностью или почти полностью в одной вертикальной плоскости.
Следовательно, в большем или меньшем наклоне тибиальных пло-
щадок нельзя видеть облигатного критерия выпрямленности, по-
скольку речь идет о гоминидах.
52
Можно ли сделать отсюда вывод, что этот признак здесь ней-
трален и лишен всякого функционального значения?
Напомним, что сильная ретроверсия является фетальным при-
знаком, отсутствующим на самых ранних этапах эмбриогенеза и
достигающим наибольшего развития к концу утробной жизни. Фе-
тальная ретроверсия должна рассматриваться как следствие сог-
нутого утробного положения колен и исчезает обычно в первые
же месяцы постнатальной жизни, т. е. задолго до начала ходьбы.
Возможно, что сохранение этого признака у некоторых ископае-
мых, и современных людей во взрослом состоянии может быть свя-
зано с привычной флексией колен не при движениях, но в уравно-
вешенном положении покоя, не связанном с какими-нибудь ины-
ми напряжениями, кроме нормальных позотонических. В связи с
этим укажем на вероятное распространение среди древних людей
таких поз, как, например, встречающееся у современного человека
и широко изучающееся сейчас морфологами «сидение на корточ-
ках». При этом способность коленного и голеностопного суставов
.к гиперфункции обеспечивается рядом анатомических приспособ-
лений на эпифизах сочленяющихся костей: выпуклый сагитталь-
ный контур наружного мыщелка, сильная медиальная скошенность
тибиальной площадки, желобок четырехглавого мускула на прок-
симальном конце большой берцовой кости, сочленовная фасетка
на задней поверхности внутреннего мыщелка бедра, добавочные
контактные фасетки в области голеностопного сустава.
Другой важный с точки зрения эволюционной морфологии при-
знак — торзион большеберцовой кости; у человека он всегда по-
ложительный, и оси стоп конвергируют назад, тогда как у антро-
поидов оси сходятся вперед, что дает у них отрицательные углы
торзиона в 85%. Групповые средние колеблются здесь от —7,5°
(гиббоны) до —24° (оранги). Положительный торзион человека
является, по-видимому, примитивным признаком, встречающимся
также у проноградных низших обезьян, тогда как антропоиды
представляют здесь вторичную специализацию (Anthony, 1912).
Групповые средние у современного человека варьируют от +6°
до +33°, индивидуальные значения колеблются от 0 до +53°; от-
рицательные значения торзиона практически не встречаются (по
Патту, в 0,1%).
Типичным для формы сечения большой берцовой кости у боль-
шинства млекопитающих является преобладание передне-заднего
диаметра над поперечным или их примерное равенство. Увеличе-
ние сагиттального диаметра связывается с повышением сопротив-
ления продольному изгибу, причем у человека этот процесс со-
провождался, очевидно, ослаблением связи между обоими диамет-
рами диафиза, являющейся наиболее тесной у эурикнемичных
форм. Как показали исследования Паувельса (Pauwels, 1950),
Кнезе и др. (Knese et al., 1954), наибольший момент инерции пло-
щади диафиза большеберцовой кости находится в сагиттальной
плоскости. Вместе с тем многообразие факторов, моделирующих
53
диафиз кости, обусловливает чрезвычайно большие колебания в
величине указателя формы сечения диафиза (указателя кнемии),
варьирующего у приматов в пределах 40—90, а у современного
человека по группам от 59,3 до 74,1 и индивидуально от 50 до 90.
Таким образом, существует высокая степень трансгрессии у чело-
века с приматами в целом. Больший интерес в аспекте прямохож-
дения представляет форма переднего края большой берцовой ко-
сти — заостренная у человека и сглаженная у антропоидов.
В итоге краткого анализа основных признаков «комплекса би-
педии» на длинных костях нижней конечности человека можно
прийти к заключению, что большая часть этих признаков, взятых
в отдельности, не является исключительным атрибутом прямо-
хождения, но может иметь более широкое значение. Отсюда сле-
дует, что для целей таксономического и функционального диагно-
за следует привлекать по возможности не изолированные призна-
ки, но их комплекс. Это имеет особое значение при фрагментар-
ном .состоянии материалов и, в частности, является обязательным
для находок костей ранних гоминид и их предшественников.
Подтверждение сказанному вытекает прежде всего из струк-
турно-функционального анализа костей австралопитеков и близ-
ких к ним форм. Сейчас известны фрагменты бедренных костей
как' грацильных (Л. africanus), так и массивных (A. robustus)
их представителей. В литературе высказывались противоречивые
суждения о роли различных морфологических признаков бедра
в определении локомоции австралопитековых, для которой все же
решающее значение признается за структурой таза. Вместе с тем,
при всех расхождениях в функциональной интерпретации отдель-
ных морфологических признаков, существует, по-видимому, единое
мнение об отсутствии каких-либо «противопоказаний» в призна-
нии бипедии австралопитековых на основе строения их бедренных
костей. Остановимся на некоторых наиболее существенных момен-
тах.
Известно скептическое мнение Керна и Страуса (1949) даже
по поводу такого важного разграничительного признака, как угол
инклинации бедра Л. africanus. Он был определен здесь в 83°
(7°) и показал совпадающие значения у антропоидов в 7—8%.
В еще большей мере сглаживались различия при измерении угла
инклинации только дистального отдела диафиза, что дает боль-
шую точность при фрагментарном состоянии кости австралопите-
ка. Тогда процент совпадений повышается до 32 у горилл, т. е.,
каждое 3-е бедро этих антропоидов имеет такое же значение угла
инклинации, как и фрагмент австралопитека. Однако при исполь-
зовании для определения угла инклинации «оптической оси» диа-
физа, которой приписывается ва'жное биомеханическое значение
(Heiple; Lovejoy, 1971), для двух фрагментов бедер австралопи-
тека определены уже значения угла 14—15° против 9,4—10,5° у че-
ловека и 1,72—0,97° у гориллы и шимпанзе. Это —'абсолютно че-
ловеческие значения.
54
Столь же противоречива оценка и другого важного признака,
гоминидного комплекса — морфологии межмыщелковой вырезки.
Как известно, она в «ультрагоминидной» степени воспроизводит
обычный тип человеческой вырезки у грацильного австралопите-
ка, особенно на бедре ТМ-1513, где ее широтно-высотный указа-
тель составляет всего 47 против 42 и 50 у человека. Но, как ука-
зывалось выше, интерпретация этой структуры не совпадает:
одни авторы видят здесь адаптацию к вытягиванию ноги в ко-
ленном суставе, возможную в различных по типу локомоции груп-
Рис. 9. Контур латерального мыщел-
ка бедра некоторых приматов: А —
Н. sapiens; Б—A. africanus; В —
Pan (по Heiple, Lovejoy, 1971)
пах приматов; некоторые же считают, что на фоне других призна-
ков комплекса бипедии у австралопитека она должна прямо свя-
зываться здесь с прямохождением (поддержка латерально-меди-
альной стабилизации patellae при полной экстензии колена) (Hei-
ple, Lovejoy, 1971).
t Оценка соотносительных размеров самих мыщелков бедра сно-
ва не вполне совпадает у разных исследователей. На одном из
фрагментов бедра A. africanus пропорции были, очевидно, вполне
человеческими (Gieseler, 1959), на другом — характеристика их
расходится (Ле Гро Кларк, 1947; Керн, Страус, 1949). Можно
предположить, что эти соотношения здесь варьировали, как это
наблюдается и среди других приматов. Напомним, что существует
и значительная трансгрессия этого признака У двуногих и про-
ноградных форм приматов.
Вместе с тем контур сочленовной поверхности мыщелков, пред-
ставляющий более важный компонент «комплекса бипедии», уже
эллипсоидный и уплощенный у A. africanus, хотя и не в такой
степени, как у Н. sapiens. Важно, что и у обоих австралопитеко-
вых наиболее затронут этим изменением формы латеральный мы-
щелок (рис. 9).
В целом морфология дистальной части бедра австралопитеко-
вых не противоречит признанию бипедии этих форм, а некоторые
факты, по-видимому, даже могут рассматриваться как прямое
свидетельство продвинувшейся адаптации к прямохождению, осо-
бенно если прибавить сюда существование на бедренной костц
этих гоминид таких особенностей; как развитие шероховатой ли-
нии, увеличение головки бедра, выраженность межвертельной ли-
нии (Howell, 1962). Необходимость использования при структур-
55
ном анализе комплексов признаков особенно выступает из при-
веденных выше данных.
Известный интерес представляет анализ костей голени из I
слоя Олдувейского ущелья, относящихся, возможно, к массивному
австралопитеку. В области коленного сустава существуют некото-
рые отличия от типичных человеческих соотношений. Это относит-
ся, в частности, к прикреплению переднего большеберцового мус-
кула: округленный, а не острый, как у человека, передний край боль-
шеберцовой кости, по-видимому, ассоциируется с распространением
части упомянутого мускула на передне-медиальную поверхность ко-
сти (Davis, 1964). В литературе высказывались соображения, что
прямоходящие массивные австралопитековые Восточной и Южной
Африки обладали некоторыми структурными особенностями бед-
ра, позволяющими предположить у них меньшее развитие ме-
ханизма альтернативной поддержки таза и, в частности, сред-
него и малого ягодичных мускулов (относительно малые разме-
ры большого вертела, отсутствие L. intertrochanterica) (Day,
1969). Вполне человеческий тип имели бедренные кости очень
раннего гоминида из Кении (район оз. Рудольфа). Их отличи-
тельные особенности: малые размеры, грацильность и сравнитель-
ная прямизна и уплощенность диафиза (указатель пилястрии
.бедра 1481—88) (Day et al„ 1975).
Более представительны материалы по бедренным костям ар-
хантропов. Как у питекантропа, так и у синантропа в общем вы-’
ражен весь набор признаков, описанный выше как «комплекс би-
педии». Известно, что бедро питекантропа имело даже настолько
типичный «сапиентный» облик, что.неоднократно возникали сом-
нения в принадлежности его столь древней форме, до сих пор еще
не вполне разрешенные (Day, 1971).
Вместе с тем у этих древнейших гоминид отмечается опреде-
ленное морфологическое разнообразие, накладывающее специфи-
ческий отпечаток на общий для них «гоминидный» фон кости. По-
скольку бедренные кости синантропов и Олдувей 28 находятся во
фрагментарном состоянии, сравнительная характеристика бедер
архантропов может опираться преимущественно на признаки диа-
физа. Для синантропов, имеющих довольно массивные бедра
(прежде всего, вследствие их короткости), специфической являет-
ся уплощенная в сагиттальном направлении — платимерная фор-
ма диафиза. Особенно характерна для них платимерия на уровне
середины диафиза: указатель пилястрии равен' здесь в среднем
для пяти костей 86,3 при вариациях от 79,4 до 91,2. Подобные
значения индекса встречаются у современного человека только в
качестве индивидуальных вариаций. Вместе с тем шероховатая
линия бедра, насколько можно судить о ее развитии при весьма
фрагментарном состоянии костей, несомненно, все же существо-
вала, по крайней мере, на Femur III и Femur IV; возможно, что на
обеих этих костях возвышался пилястр (Вейденрейх, 1941).
56
Указатели пилястрип у питекантропа для пяти бедер
колеблются в пределах 90,5—103,6 при средней 97,1 *, что
уже ближе к типичным величинам этого индекса у сов-
ременного человека. Что касается формы подвертельной об-
ласти, то точное определение указателя платимерии на триниль-
ском бедре (I) затруднительно из-за значительного разрастания
костной ткани 1 2; на «лейденских бедрах» питекантропа указатели
снова обычные для современных бедер. Существуют определен-
ные различия между обоими архантропами и в структуре рельефа
верхнего конца кости, который отчасти влияет на форму диафиза
в подвертелыюй области. У синантропа описан выраженный под-
вертельный гребень, продолжающийся к основанию большого вер-
тела в виде «подвертального бугорка» (tuberculum hypotrochante-
ricum), отсутствует «гребешковая линия», идущая от проксималь-
ного разделения шероховатой линии к основанию малого вертела;
отсутствует или «стерта» и межвертельная линия. Такие особен-
ности встречаются, как будет показано ниже, на бедренных костях
и других ископаемых гоминид, тогда как у питекантропа снова
существовало более «сапиентное» строение этой области: на бед-
ре Триниль I обе линии — межвертельная и гребешковая — бы-
ли, очевидно, выражены. Можно указать и на другие различия
между бедренными костями этих двух типов архантропбв: так,
торзион на кости Триниль I равен примерно 25°, тогда как у си-
нантропа предположительно были более высокие значения этого
угла, близкие к максимуму современных вариаций: 35—36°. Вме-
сте с тем для обоих представителей древнейших людей характер-
ны слабые величины изгиба бедра (указатели 1,8—2,3) и весьма
близкие значения коллодиафизарного угла: 124° па бедре Три-
ниль I и предположительно не более 119—127° на реставрирован-
ных бедрах синантропа I и IV.
Заслуживает большого внимания фрагмент диафиза бедрен-
ной кости, открытый недавно в слое IV Олдувейского ущелья и
принадлежащий, по-видимому, архантропу (Н. erectus, Олдувей-
ский гоминид 28 Дэй, 1971) (рис. 10).
Морфологический анализ этой кости обнаруживает своеобраз-
ное сочетание признаков. С одной стороны, как-будто намечают-
ся некоторые черты сходства Олдувей с синантропом. Это касает-
ся прежде всего выраженной платимерии на подвертельном и
особенно на срединном уровне с указателем пилястрии 75,5 (!)
при наличии шероховатой линии бедра, что соответствует очень
редкой индивидуальной вариации у современного человека и поч-
ти на 11 единиц ниже, чем в среднем у синантропа. Определенное
1 По результатам переисследования этих костей Дэем (Day, 1971).
2 По-видимому, следствия оссифицирующего миозита (Рохлин, 1965). Относи-
тельно связи этой патологии со фторовой интоксикацией мнения расходятся
(Soriano, 1970; Day, Molleson, 1973).
57
сходство отмечено также по некоторым признакам общей формы
диафиза (дистальное положение наименьшей ширины, выпук-
лость медиального края). Вместе с тем есть и ряд расхождений с
синантропом — в общей длине, много большей у Олдувей 28, у
которого она такая же, как и у Триниль I, т. е. 455—456 мм, м'ас-
Рис. 10. Фрагмент
бедра Олдувей 28:
А — спереди; Б —
сзади (по Day,
1971)
Рис. 11. Бед-
ренная кость
Неандерталь
(по Patte,
1955)
сивности, меньшей у Олдувей, кортико-медуллярных пропорциях
диафиза, рельефе проксимального конца кости (отсутствующие у
синантропа межвертельная и гребешковая линия существуют на
бедре Олдувей 28, как и у питекантропа). Трудно поэтому безо-
говорочно согласиться с мнением Дэя (1971), что указанные чер-
ты сходства между синантропом и Олдувей являются аргументом
против принадлежности тринильских бедер архантропу. Безотно-
сительно к тому, что дает переисследование морфологии Триниль,
видно, что многие структурные признаки бедра достаточно варь-
ируют у" этих древнейших, как и современных людей, причем в не-
которых отношениях Олдувей'28 сближается с синантропом (на-
пример, платимерия диафиза, форма его дистального отдела), но
58
в других он далек от него и даже ближе к питекантропу (тор-
зион, рельеф проксимального конца, общие размеры). Есть и при-
знаки, общие для бедренных костей архантропов, как например,
слабый изгиб диафиза (максимум индекса изгиба 2,9 у Олдувсй).
К тому же переисследование «сапиентных» тринильских бедер с
Явы, приписываемых питекантропу (Day, Moileson, 1973), пока
все же не принесло решающих аргументов в пользу пересмотра
этой точки зрения.
Приведенные факты соответствуют представлению о полимор-
физме структуры бедра у ранних гоминид и отсутствии здесь, как
и на более поздних этапах морфологической эволюции человека,
его единообразного типа.
Последнее находит себе подтверждение и при анализе бедрен-
ных костей палеоантропов. Здесь также совершенно отчетлив го-
минидный фон «комплекса бипедии», но морфологическое разно-
образие этой кости достаточно велико. Наиболее четко выявляет-
ся комплекс специфических черт на костях «классических» неан-
дертальцев (рис. 11).
Сюда следует отнести небольшие длиннотные размеры бедрен-
ных костей (430 мм у Шапелль, 440 мм у Неандерталь, 428 мм у
Спи) при очень большом периметре, что дает высокие значения
указателей массивности. Очень велики размеры эпифизов — аб-
солютно и относительно особенно крупными были сочленовные го-
ловки, тогда как размеры шейки бедра имеют меньшее разграни-
чительное значение при сравнении неандертальских и современ-
ных костей. При этом существует явная тенденция к опущенному
положению шейки (слабые значения коллодиафизарных углов).
Средняя этого угла для неандертальцев составляет, по Булю, 116°
при вариациях от 106 до 125°. Можно говорить, следовательно, об
эксцентрическом положении неандертальцев в ряду современных
вариаций даже при индивидуальных сопоставлениях. На нижнем
эпифизе отмечается суженный и удлиненный наружный мыщелок,
уступающий по широтному развитию внутреннему. Вероятно, так-
же несколько повышенное значение кондилодиафизарного угла.
Диафиз имел тенденцию к округлению в середине: индекс пиля-
стрии в среднем 99,0 при слабом развитии шероховатой линии и
отсутствии пилястра. Известно, что в ранней палеоантропологиче-
ской литературе- этот признак использовался для обоснования
концепции о «несовершенстве» прямохождения у палеоантропа.
Однако, как уже упоминалось, сложность генеза пилястра требует
учета развития не только его наружной части (суперструктуры),
но также и привлечения данных о строении всей задней стенки
бедра в целом («контрфорс»). Тогда оказывается, что как раз у
классических неандертальцев задняя компакта бедра была осо-
бенно хорошо развита — «внутренний пилястр».
В рельефе проксимального конца кости этих палеоантропов
обычно отсутствуют межвертельная и гребешковая линии, часто
отчетливо выражена структура подвертельного гребня и ямки, и
59
в той или иной степени развит третий вертел. Кроме того, бедрен-
ные кости неандертальцев нередко сильно изогнуты (например,
указатель цзгиба равен 4,7 у Спи, 4,9 у Неандерталь *. Другим
признаком этих костей является не постепенное, но «внезапное»
расширение дистального отрезка диафиза в нижний эпифиз, что
напоминает тринильское бедро. Однако этот признак встречается
и у современного человека, как среди популяций
Н. sapiens recens, так и у ориньякского человека,
и в неолите (Schlaginhaufen, 1925; Мартин, 1928)
т. е. он не является диагностическим.
Перечисленные особенности не везде высту-
пают с одинаковой отчетливостью, нередко одна
или несколько могут даже отсутствовать (напри-
мер, сравнительная выпрямленность бедра Фер-
расси I, наличие на нем гребешковой линии),
но все же этот комплекс можно считать более
или менее типичным. В современной морфологи-
ческой литературе, говоря о «неандертальских
признаках» в строении бедренной кости, обычно
подразумевают вышеперечисленные особенности
ее структуры.
Из числа других костей нижней конечности
наибольшее внимание уделялось анализу боль-
шой берцовой кости неандертальцев (рис. 12).
Для нее характерны повышенные в масштабе
Рис. 12. Большая берцовая кость Киик-Кобы (по
Бонч-Осмоловскому, 1954)
вариаций современного человека значения углов ретроверсии иин-
клинации (16,2—20° для первого и 13—18° для второго). Извест-
но, что этот признак сочетается у них с вполне современными зна-
чениями торзиона, что означает совершенно человеческую ориен-
тацию стоп у этих, гоминид,*а также со значительной массивностью
обеих берцовых костей.
Несколько слов следует сказать по поводу возможной интер-
претации некоторых из перечисленных признаков «неандерталь-
ского комплекса». Как упоминалось, отсутствие пилястра, ретро-
версия большой берцовой кости и ряд других особенностей не-
андертальских костей оценивались с традиционной точки зрения
«несовершенства прямохождения». Известная реконструкция Бу-
1 Следует, впрочем, иметь в виду, что' само определение изгиба методически
довольно сложно и не вполне унифицировано. При сопоставлении указателя
изгиба только для одного, единообразно изогнутого участка диафиза неандер-
тальские вариации почти полностью перекрываются современными (Leclercq,
60
ля, многократно воспроизведенная последующими авторами, изо-
бражала шапелльского неандертальца с ногами, согнутыми в ко-
ленях. Выше уже указывалось на неправомерность подобного
прямолинейного решения вопроса и на вероятную роль в этио-
логии этих признаков, так называемого «комплекса положения на
корточках» или других «поз покоя», несомненно предшествовав-
ших возникновению сидячих поз (Huard, Montagne, 1953).
С этой точки зрения можно объяснить такую характерную для
костей нижней конечности многих ископаемых гоминид комбина-
цию признаков, как сильный изгиб бедра, опущенная шейка, зна-
чительная ретроверсия большой берцовой кости, поскольку экспе-
риментально и на теоретической модели было показано возраста-
ние изогнутости бедра и дорзальной инклинации проксимального
конца большой берцовой кости при повышении нагрузки, особен-
но при флексированном коленном суставе (Жданов, 1967; Kum-
mer, 1967).
Однако описанный выше «неандертальский комплекс» вовсе не
является универсальным даже в пределах Европы. Уже здесь
мы встречаемся с палеоантропами, бедренные кости которых не
воспроизводят полностью неандертальского типа. Речь идет об
отклоняющихся в своей морфологии и, очевидно, менее специа-
лизированных бедренных костях Эрингсдорф и отчасти крапин-
ских (последние, правда, находятся в весьма фрагментарном со-
стоянии). Что касается эрингсдорфского бедра, то, обладая мас-
сивностью и изгибом, оно существенно отличается от неандерталь-
ского типа в строении рельефа наличием гребешковой и межвер-
тельной линии, хорошо развитой шероховатой линией. Некоторые
крапинские бедра характеризовались, напротив, выпрямленностью
диафиза и значительными вариациями массивности, среди них
есть и весьма грацильные кости. На одном из фрагментов раз-
вита и межвертельная линия. Нужно, впрочем, иметь в виду, что
по многим из перечисленных признаков бедренной кости сущест-
вует умеренно выраженный половой диморфизм, который не всег-
да легко учесть на ископаемых объектах.
Среди палестинских палеоантропов можно выделить два типа
бедра: один — с определенным налетом неандерталоидности и
другой — более близкий к современному. К первому варианту
следует прежде всего отнести Табун I, хотя неандерталоидность
здесь весьма относйтельна. Так, бедренная кость Табун, по-види-
мому, была лишена сильного изгиба и напоминает неандерталь-
ский вариант по относительной массивности верхнего эпифиза (но
не нижнего) и слабому развитию шероховатой линии. На всех ко-
стях Табун низки значения указателя пилястрии (средняя индек-
са для Табун I, ТабуН1 и Табун £а составляет всего 90,4), что на-
поминает соотношения на костях западноевропейских неандер-
тальцев. Но этими признаками и. ограничивается сходство.
Для большинства представителей группы Схул типично до-
вольно значительное длиннотное развитие бедра при сравнитель-
61
но небольшом периметре, а следовательно, весьма умеренные и
современные значения указателей массивности. Столь же ограни-
ченным было и развитие эпифизов; нет тенденции к округлению
диафиза в середине, напротив, часта стеномерия вплоть до таких
высоких значений указателя пилястрии, как 142,5 у Схул V.
В среднем по группе Схул этот указатель равен примерно 115,
что значительно превышает соответствующую величину индекса
у Табун, как и у классических неандертальцев. Это ассоциирует-
ся также с хорошим развитием шероховатой линии и наличием
пилястра в варианте Схул. Значительно уступает классическим
неандертальцам также абсолютное и особенно относительное раз-
витие головки бедра. Описанный тип наиболее выражен на бедре
Схул IV. Но среди палестинцев существуют и отклонения от него, что
неудивительно, принимая во внимание исключительный размах ва-
риаций, .почти совпадающий для большинства остеометрических
признаков с колебаниями современных групповых средних. Так,
например, наряду с очень слабо или умеренно изогнутыми костя-
ми существуют и крайние степени изгиба на костях Схул V и осо-
бенно Схул VI. Рядом с резко выраженной стеномерией подвер-
тельной области, свойственной, например, бедру Схул V, наблю-
дается и гиперплатимерия (Схул IX). Очень мал был индекс пи-
лястрии на бедре Схулэ, и у ребенка Схул VIII, тогда как у
Схул V существовала крайняя степень,боковой уплощенности этой
кости.
Таким образом, даже из краткого перечисления некоторых ва-
риаций остеометрических признаков бедра создается картина
чрезвычайно высокой их вариабильности, на фоне которой все же
более или менее отчетливо выделяются два варианта: «сапиент-
ный», наиболее полно воплощенный в бедре Схул IV, и уклоняю-
щийся в сторону «неандерталоидности», подобно Табун. Среди лю-
дей Схул последний вариант наиболее выражен на бедре Схул IX
(абсолютная и вероятно относительная массивность верхнего эпи-
физа, большая длина шейки бедра сзади, округленность диафи-
за, отсутствие развитой шероховатой линии).
Заслуживает внимания и тот факт, что по. многим из рассмот-
ренных признаков существует определенное сближение бедрен-
ных костей Крапины с типом Схул. В этой связи следует упомя-
нуть о еще одной интересной находке бедра восточноевропейского
палеоантропа (Украина, Романково) вместе с мустьерскими ору-
диями в неперемытой ледниковой морене, датируемой днепров-
ским оледенением (Накельский, Карлов, 1965). Отличительными
особенностями этой кости являются ее значительная массивность
при довольно большой длине (470 мм) • и крайняя степень подвер-
тельной платимерии. Очень велика относительно и особенно абсо-
лютно верхняя эпифизарная ширина. Однако хорошо выражен пи-
лястр и структура нижнего эпифиза, настолько можно судить о
ней при разрушенном состоянии внутреннего мыщелка, не напоми-
нает неандертальского типа. По ряду морфологических признаков
62
(большие абсолютные размеры верхнего эпифиза, значительная
длина кости, гиперплатимерия, развитие' пилястра)- Романково
напоминает «неандерталоидную» форму Схул IX, с которой име-
ется сходство в развитии верхнего эпифиза и резкой уплощенно-
сти подвертельной области. К сожалению, эта последняя находка
очень фрагментарна, что делает дальнейшие сопоставления с ней
невозможными. Все же очевидно, что романковскому бедру свой-
ственна такая же пестрая комбинация признаков, которая отли-
чает бедренные кости Схул. При значительной фрагментарности
находок бедренных костей палеоантропов с территории Восточной
Европы представляет особый интерес этот «прогрессивный вари-
ант» европейского палеоантропа менее специализированной при-
роды, чем классические неандертальцы Европы (Хрисанфова,.
1965). '
На бедренной кости ребенка палеоантропа из Тешик-Таша ана-
лиз значительно затруднен возрастными особенностями формы
диафиза, которые, безусловно, йаскируют некоторые морфологи-
ческие различия, что относится, например, к слабой выраженно-
сти изгиба бедра и ягодцчной шероховатости. Однако знамена-
тельно, что развита шероховатая линия (Рохлин, 1949) и сущест-
вует гребешковая линия, что отличает тешик-ташца, по крайней
мере по этим признакам, от классических европейских неандер-
тальцев.
Отчетливо выделялась шероховатая линия и на бедре одного
из представителей палеоантропов Африки.— Брокен-Хилл.
В заключение следует остановиться и на некоторых вариантах
структуры бедра верхнепалеолитического человека, представляю-
щих интерес в аспекте сделанного выше обзора морфологического'
типа этой кости у палеоантропов. Речь идет в первую очередь о
бедренной кости Шанселад, в строении которой обращает на се-
бя внимание явный «неандерталоидный» налет: это короткость,
бедра (408 мм), его высокая массивность (включая диафиз и оба.
эпифиза), сильный изгиб, округленная форма середины диафиза,,
опущенная шейка. Существует также частая на бедрах палео-
литического человека комбинация подвертельной ямки и третьего-
вертела.
Вместе с тем шероховатая линия бедра хорошо развита,
и выражен пилястр. Отметим также значительную ретроверсию на
большой берцовой кости и ее значительную массивность. Многие
из перечисленных признаков встречаются на бедренных костях и
других верхнепалеолитических гоминид: например, общим для них
признаком является опущенная шейка бедра (Billy, 1969), что
снова напоминает об адаптациях к флексии колена в «позах по-
коя» (см. выше).
Краткий обзор строения бедренных костей ископаемых, гоми-
нид убеждает в том, что морфологическое разнообразие структуры
этой.кости было велико уже на самых ранних стадиях эволюции.
На общем для всех прямоходящих гомннид фоне комплекса би-
63
педии выделяются некоторые специфические сочетания признаков,
которые могут представить значительный интерес для таксономи-
ческого и генеалогического анализа локальных вариантов.
ВЕРХНЯЯ КОНЕЧНОСТЬ
Анализ морфологической структуры плечевой кости ископаемых
гоминид целесообразно начать с самых поздних их представите-
лей, поскольку от них сохранились наиболее пол-
ные материалы по этому отделу скелета.
Плечевые кости палеоантропов совершенно
человеческие, но вместе с тем для них также ха-
рактерны определенные особенности, которые в
комплексе могут в некоторых случаях оказаться
достаточно специфичными и тем самым облег-
чить таксономический диагноз применительно к
локальным вариантам древних людей.
В первую очередь сказанное относится к
«классическому» варианту западноевропейских
неандертальцев, обладавших короткими толсты-
ми'плечевыми костями с массивными эпифизами
(рис. 13). У современного человека размер го-
ловки представляет один из наиболее отчетли-
вых разграничительных признаков полового ди-
морфизма на длинных костях. Так, пределы ва-
риаций вертикального диаметра. головки плеча
у мужчин: 42—56 мм, поперечного — 38—50 мм.
С этой точки зрения, размеры • головки
очень крупные у
Рис. 13. Плечевая кость Неандерталь (по Patte,
1955)
неандертальцев, особенно ее поперечный диа-
метр, составляющий, например, у собственно неандертальской на-
ходки 48—50 мм, что совпадает с современным максимумом. Ха-
рактерно при этом превалирование поперечного диаметра над вер-
тикальным, которое можно счесть общей чертой для всех палео-
антропов. Указатель сечения головки (поперечный диаметр ее в
% от вертикального) поэтому обычно превышает 100. Например,
у собственно Неандерталь он составляет 101,1, а у Схул IV — да-
же 111,1 против 87—97,5 у современных мужчин. Интересен факт
значительной асимметрии в этом признаке у женщины Табун I,
где указатель справа равен 105,6, а слева — 97,3, что встречает-
ся и у современного человека.
Как известно, тенденция к округлению головки связывалась
Ривэ (Rivet, 1909) с массивностью диафиза кости. Однако мы не
64
нашли статистически достоверной зависимости между этими при-
знаками в серии плечевых костей европейцев (из 50 объектов),
что подтвердилось и при изучении их бедренных костей. Этот ре-
зультат кажется нам вполне согласующимся и с данными палео-
морфологии. Действительно, среди палеоантропов, например, цир-
кулярную форму головки имеют кости с самой разной степенью
массивности диафиза, что подчеркивает специфику этого призна-
ка как свойственного палеоантропам в целом (см. выше).
Диафиз плечевых костей классических неандертальцев обнару-
живает тенденцию к массивности и округлению в середине: ци-
линдрический тип его встречается у шапелльца и на собственно
неандертальской кости, а в смягченном виде — у Феррасси. Для
современного человека, как и для млекопитающих в целом, типич-
но отклонение от исходной цилиндрической формы диафиза в сто-
рону более или менее выраженного превалирования сагиттально-
го диаметра над трансверзальным (Бунак, 1956). Среди людей
Схул встречаются как округленное «неандерталоидное» сечение
диафиза (Схул IV и V), так и .уплощенное «сапиентное» (Схул
II). По-видимому, Табун занимает в этом отношении промежуточ-
ное положение, что отмечается также для Феррасси и крапин-
цев.
На нижнем эпифизе плечевой кости западноевропейских не-
андертальцев особенно выделяется своим сильным развитием
внутренний надмыщелок (Неандерталь, Шапелль, Феррасси. I,
Спи, а также Крапина). Табун I снова сближается с неандер-
тальским' типом по развитию внутреннего надмыщелка. На неко-
торых неандертальских костях имеется, далее, «супинаторный гре-
бень» в области наружного надмыщелка, что следует рассматри-
вать как индивидуальную вариацию, встречающуюся и у совре-
менного человека. Возможно, что эта особенность в какой-то ме-
ре связана с расширением области прикрепления плечелучевого
мускула и внутренней головки трехглавого мускула плеча (Патт,
1955).
Возможно также, что у западноевропейских неандертальцев
существовала тенденция к более горизонтальному положению
блока: величины кондилодиафизарного угла у классических форм
(Спи, Шапелль, Феррасси, Неандерталь) составляли 86—87°, что
обусловливает их эксцентрическое положение в ряду вариаций
современного человека (70—86°). Этот признак характерен и для
крапинцев, тогда как у палестинцев кондилодиафизарные углы
несколько ниже, что означает более косое положение блока (83—,
84°). Снова отмечается неандерталоидность Табун I со значением
угла 86°. Наконец, для неандертальского типа обычна довольно
объемистая локтевая ямка; что касается перфорации последней,
то она у ископаемых гоминид, как и современных, по-видимому,
ассоциируется с грацильностью плечевых костей (Benfer,
Me Кегп, 1966), как и с возрастными инволютивными процессами в
костной ;ткани. Среди древних людей перфорация отсутствует на
5 Е. Н. Хрисанфова
65
массивных костях Неандерталь и Шапелль и, напротив, отмечается
на более грацильных костях Ла Феррасси и Крапины — у по-
следних почти на половине всех найденных фрагментов плече- '
вых костей. Это очень высокая частота, приближающаяся к мак-
симуму у современного человека, вариации 4,1—58%.
Таким образом, по структуре плечевой кости палеоантропов,
выделяется прежде всего специализированный тип классических
неандертальцев; в группе Схул встречаются преимущественно са-
пиентные плечевые'кости, однако с определенной примесью неан-
дертальских признаков. Так, тенденция к округленности диафиза
в середине отмечена, например, у Схул IV и V, у них же абсолют-
но велики эпифизы, обширна локтевая ямка. По многим призна-
кам с классическими неандертальцами сближается женщина Та-
бун I, занимающая как бы промежуточное положение между ти-
пами Схул и неандертальским (относительная массивность верх-
него эпифиза, широтное развитие внутреннего надмыщелка, гори-
зонтальность блока, тип локтевой ямки, массивность и форма се-
чения диафиза). Следует также отметить, что по некоторым из
этих признаков крапинцы и даже Ла Феррасси отличаются от
классических форм грацильностыр и формой диафиза.
Нужно, впрочем, все время иметь в виду, что указанные у па-
леоантропов вариации не представляют ничего исключительного:
большая часть перечисленных признаков широко варьирует сре-
ди современных гоминид — индивидуально и в межгрупповом ас-
пекте (Gozdziewski,' 1962). Специфична только их комбинация
прежде всего у классических палеоантропов.
Кости предплечья также более или менее полно представлены
только у палеоантропов. Хотя и на них можно распространить вы-
вод, сделанный на основе анализа структуры плечевой' кости, — о
вполне человеческой их природе — все же здесь существуют не-
которые структурные особенности, которые в комплексе позво-
ляют высказать определенные предположения о специфическом
характере функциональной деятельности руки этих гоминид.
Значительный интерес в этом плане представляет прежде все-
го морфология шейки луча, на длину и выраженность которой
влияет положение места прикрепления дистального сухожилия
двуглавой мышцы плеча. У всех палеоантропов наблюдается ди-
стальный сдвиг этого прикрепления, обусловливающий удлинение
шейки. Индекс ее относительного''развития (в % физиологической
длины кости) равняется здесь 11,1—15,3 против 9—10 у совре-
менного человека. Возможно, что это перемещение прикрепления
сочеталось с менее обширными по амплитуде, более медленными,
но более мощными движениями, что подтверждается и сравни-
тельно-анатомическими параллелями. Патт (1955) указывает, в
частности, что самые низкие индексы длины шейки луча наблю-
даются у приматов, способных к движениям с относительно боль-
шой амплитудой (коата, гиббоны, "оранги, тонкотелы). Палеоан-
тропам свойственна, очевидно, и несколько отличная ориентировка
66
лучевой шероховатости, более ульнарная, чем у современного че-
ловека, где этот бугорок в большей степени обращен, вперед
(рис. 14). Возможно, эта модификация прикрепления связана.с
вариациями в длине двуглавого мускула плеча.
Менее определенной представляется функцио-
г-гг— нальная интерпретация такого признака, как
' выявляющаяся на лучевых костях европейских
'\~w неандертальцев мелкая и широкая форма чашеч-
I I ки лУча’ чт0 ассоциируется у них и с несколько
Г Л более слабой выпуклостью нижней головки пле-
I Ш, чевой кости. Как известно, Буль видел в этом
I я признаке показатель пониженной ротационной
i IЯ ‘ ; способности лучевой кости, однако вряд ли воз-
/Л I можны достоверные выводы без учета формы
' i г Д хрящевых покрытий суставных поверхностей пле-
j i№ че-лучевого сочленения. С другой стороны, зна-
|я№' чительный размах амплитуды просупинации у
И современного человека также подтверждает не-
|S обходимость осторожного подхода к функцио-
|Ж нальной интерпретации этой особенности.
IB Очень своеобразная черта структуры локтевой
уК кости палеоантропов — высокая степень разви-
lak тия «клюва» локтевого отростка, являющегося
уВ рычагом трехглавого мускула плеча. Как у ев-
ропейских, так и у палестинских палеоантропов,
i | Д. верхушка олекранона была развита значитель-
но сильнее, чем у современного человека: ее
Рас. 14. Лучевая кость Неандерталь (по Patte,
1955)
длина’ (в % физиологической длины кости) составляет здесь
4,0—6,0 против 1,2—2,5 в среднем у современного человека1. По-
скольку у антропоидов, включая и ореопитека (Viret, 1955), ку-
пол олекранона развит очень слабо сравнительно с низшими
обезьянами и лемурами, сильное развитие верхушки локтевого
отростка рассматривается обычно как сохранение примитивного
состояния (Мартин, 1928) или проявление своеобразной специа-
лизации палеоантропов (Маккоун, Кизс, 1939). 1
Интересно, что некоторые верхнепалеолитические неоантропы
доказывают ряд неандерталоидных . черт в. морфологии костей
верхней конечности, в частности, и по значительному развитию
купола локтевого отростка (например, у Кроманьон, Шанселад).
1 Технически эти размеры не вполне сопоставимы у европейских и палестин-
ских форм. • •
5*
67
Крайняя степень групповых и Индивидуальных вариаций на-
блюдается у нижнего края лучевой вырезки локтевой кости. Па-
леоантропы в целом обнаруживают здесь высокую степень эуро-
лении, стоящей даже на границе с гиперэуроленией с сапиентным
типом связывается реальное увеличение костного вещества в
прекороноидной области, благодаря чему трансформируется поле
прикрепления плечевого мускула. Некоторые исследователи (Мак-
коун, Кизс, 1939) видят в этих преобразованиях связь с эволюци-
ей тонких движений просупинации, более точных движений при
флексии и экстензии предплечья, поскольку главной функцио-
нальной нагрузкой на локтевую кость при ротационных движениях
луча является обеспечение ее фиксации — тоническим действием
плечевого мускула и оппонирующим — трехглавого. Возможно,
что увеличение локтевого отростка, свойственное палеоантропам,
следует понимать как модификацию, связанную с возрастанием
силы трехглавого мускула при согнутом локтевом суставе.
Вместе с тем нет оснований считать, что повышенное развитие
олекранона могло сколько-нибудь существенно ограничивать ам-
плитуду разгибания в локтевом суставе палеоантропов, так как
последняя определяется главным образом связками, обычно бло-
кирующими разгибание локтя, не дойдя на. 15° до костного упора
(Gutierrez, 1964).
В целом в строении костных компонентов локтевого сустава
не выявляется существенных различий при сравнении палео- и
неоантропов — насколько можно судить о морфологии этого от-
дела по имеющимся материалам. Возможно, однако, что некото-
рые специфические признаки'строения этой области у палеоантро-
пов, как говорилось выше, свидетельствуют об известном ограни-
чении скорости движений в локтевом суставе при возрастании их
мощности (Патт, 1955). Более дистальное прикрепление двуглаво-
го мускула делало возможным, согласно предположению этого
автора, некоторую «разгрузку» плеча, в противоположность пред-
плечью, так что пластика этих отделов верхней конечности могла
отличаться от типичной для современного человека. При всем ска-
занном палеоантропы могли выполнять те же сгибательно-разги-
бательные и ротационные движения в локтевом суставе, хотя, воз-
можно, и с некоторыми ограничениями в смысле точности и быст-
роты.
Значительно слабее представлены длинные кости верхней ко-
нечности более ранних гоминид. От архантропов известно несколь-
ко фрагментов плечевых костей синантропа. По своей грацильно-
сти, типу сечения диафиза, углу скрученности костй (торзиону)
это также вполне человеческая кость. Как известно, торзион, оп-
ределяющийся сдвигом сочленовной оси верхнего эпифиза по отно-
шению к нижнему, является следствием преобразования грудной-
клетки и дорзального перемещения лопаток, вызванных прежде
всего развитием прямохождения у гоминид. Следовательно, в
первую очередь его и следует рассматривать в связи с этим кар-
68
динальным преобразованием локомоции, причем особенно разли-
чаются углы торзиона у проноградных приматов типа павиана и
двуногих гоминид, у которых соответствующие значения угла ко-
леблются в пределах 95—98° у первых и 129—167° у вторых. Од-
нако все же нужно иметь в виду: во-первых, что индивидуальные
колебания угла у человека'значительно выше: 113—185°; во-вто-
рых, что здесь существует частичная трансгрессия с антропоида-
ми (112—141° у последних). Средние варианты достаточно явст-
венно различаются у антропоидов 125° и человека 150—160°. Тор-
зион синантропа вполне человеческий, как и у всех палеоантро-
пов, его предположительное значение у синантропа-137°. Это, ра-
зумеется, ожидаемый результат с точки зрения древности основ-
ной локомоции, но его следует выделить, поскольку существует и
мнение, что в угле торзиона возможно видеть также показатель
развития эргомоторики у гоминид (Meitner, 1961). Из других осо-
бенностей плечевой кости синантропа можно отметить сильное
развитие дельтовидной шероховатости (Вейденрейх, 1941).
Наибольший интерес представляет морфофункциональный ана-
лиз плечевых костей, австралопитековых — и их хронологических
предшественников. Одна из наиболее полных публикаций касает-
ся структуры плечевой кости A. robustus (парантропа массивно-
го). От этой формы был известен дистальный фрагмент плеча, при
сравнительном исследовании которого выявилась значительная
однородность морфологии этого отдела плечевой кости австрало-
питека, человека и шимпанзе. В отличие от верхнего конца пле-
чевой кости, структура которого представляет достаточно отчет-
ливые различия между человеком и шимпанзе (Оливье, 1972),
нижний ее конец показал совпадение ряда измерительных
признаков, причем это относится как к развитию надмыщел-
ков, так и к собственно сочленовным поверхностям. Это об-
стоятельство не дает оснований ожидать продуктивных ре-
зультатов от сравнительно-морфологического анализа дисталь-
ного отдела плеча и при сопоставлении ископаемых и сов-
ременных форм. Действительно, анализ фрагмента плечевой
кости массивного австралопитека, проведенный Страусом для оп-
ределения положения ископаемой формы в системе приматов, по-
казал нейтральность строения этого обломка. По абсолютным раз-
мерам парантроп одинаково близок к шимпанзе и человеку, по
относительным показателям . он ближе к «среднему» шимпанзе,
но эти различия не следует преувеличивать, так как в общем зна-
чения индексов входят в пределы вариаций обеих сопоставляемых
форм.
Ле Гро Кларк считал наиболее своеобразной особенностью
плечевой кости этого австралопитека положение нижней сочленов-
ной поверхности (особенно блока) по отношению к диафизарной
оси: у человека и в еще большей степени у антропоидов блок рас-
полагается впереди относительно диафиза кости и внутреннего
надмыщелка, тогда как у парантропа он в большей мере продол-
. 69
жает диафизарную ось. Однако эти соотношения, очевидно, так-
же сильно варьируют в пределах вида. Большую или меньшую го-
ризонтальность -локтевого сустава может выразить упомянутый
выше кондилодиафизарный угол, который не поддается точному
определению у австралопитека. Страус заменил его до некоторой
степени эквивалентным «углом инклинации» (углом наклона ниж-
ней сочленовной поверхности плечевой кости к нижнему отрезку
диафиза). Сопоставление этих углов у австралопитека, шимпанзе
и человека показало нейтральность также и этого признака: зна-
чения углов у всех трех форм практически совпали (95—96°).
Другой фрагмент очень древней плечевой кости, возможно при-
надлежащий форме, близкой к австралопитековым («Канапой-
ский фрагмент» из Кении), также представляет дистальную часть
плеча. Хотя по многим признакам здесь снова констатируется
сходство с человеком и шимпанзе, но на этот раз со значи-
тельно большей близостью к человеческому типу (Patterson, Ho-
wells, 1967; Me Henry, Cocini, 1975). Это согласуется с предполо-
жением о раннем формировании гоминидного типа плечевой кости
при существовании известного морфологического разнообразия ее
у самых древних (как и более поздних) гоминид.
В этом плане представляют значительный интерес исследова-
ния морфологии длинных костей верхней конечности у высших
негоминидных приматов. Известен фрагмент плечевой кости дри-
опитека фонтанова, отличающийся своей грацильностью; подроб-
ное сравнительно-морфологическое изучение этого фрагмента по-
казало, что по большинству измерительных и описательных при-
знаков дриопитек очень близок к карликовому шимпанзе, как и
современным африканским антропоидам в целом (Pilbeam, Si-
mons, 1971), что соответствует и представлению о генеалогиче-
ской связи этих приматов. От другой интересной миоценовой
обезьяны — ореопитека — были подробно изучены фрагменты
длинных костей верхней конечности. Плечевая кость оказалась
гбминоидной по облику, что в общем характерно и для других
ископаемых миоценовых форм (Pliopithecus, Proconsul, Limnopit-
hecus; Страус, 1963)1 Результаты дискриминантного анализа трех
фрагментов локтевой кости также показали принадлежность, орео-
питека к кругу Hominoidea, причем по ряду признаков этот бра-
хиатор явно выходит за пределы вариаций церкопитековых обезь-
ян (Knussmann, 1967).
ПРОПОРЦИИ
Наибольшее значение для таксономического анализа имеет ин-
термембральный указатель, т. е..' процентное отношение суммар-
ной длины плечевой и лучевой костей к длине бедренной и боль-
шеберцовой, выражающий типичные для вида пропорции конечно-
стей. По этой причине он весьма устойчив внутри группы (коэф-
фициент вариации на скелетном материале составляет, по данным
70
Шульца (1937), 2,64%; при этом для сохранения видовых пропор-
ций наибольшее значение имеют: на верхней конечности плечевая
и лучевая, а на нижней — бедренная и большая берцовая кости;
в эксперименте также показана меньшая чувствительность луче-
вой кости сравнительно с локтевой, а большой берцовой — срав-
нительно с малой, при выключении статической функции костей
(Рогинский, 1957; Бунак, 1957).
Интермембральный указатель четко дифференцирует гоминид
от других приматов: групповые вариации его у современных лю-
дей составляют для обоих полов 64,4—74,9; индивидуальные зна-
чения колеблются в пределах 60,7—84,0 (данные разных авто-
ров) . У антропоидов средние варьируют в пределах: 106,3. (шим-
панзе) — 148,2 (гиббоны). Индивидуальные значения указателя
колеблются от 102 (шимпанзе) до 155 (гиббоны). Низшие обезь-
яны Старого Света занимают нейтральное положение со значе-
ниями указателя"78—99z
Наибольший интерес представляют, конечно, пропорции ко-
нечностей австралопитековых и близких к ним форм. Индексы,
определенные на основе реконструкций и измерений бедренной,
плечевой и лучевой костей A. africanus (плезиантроп, Прометей) и
большой берцовой кости из Олдувей, свидетельствуют о сходстве
пропорций конечностей австралопитеков с человеческими (Hel-
muth, 1968). Для интермембрального указателя приводятся, в
частности,/ значения 75,4—85,7; они сближают австралопитековых
с нейтральными по пропорциям конечностей циноморфными и
близки также к верхнему пределу вариаций индекса у современно-
го человека. Нужно иметь в виду приблизительность этих цифр.
Среди ископаемых «настоящих гоминид» более достоверные ве-
личины интермембрального указателя могут быть вычислены
только для некоторых палеоантропов. Из числа европейских форм
оба Ла Феррасси имели указатель около 68; для Спи приводится
величина 69,5 (Werth, 1928), аналогичное значение приписывает-
ся и Неандерталь. • '
У палестинских палеоантропов можно вычислить этот индекс
по данным Маккоуна и Кизса с известной степенью достоверно-
сти для Схул IV (слева), где хорошо сохранились все соответст-
вующие кости. Он равен 65,6. У женщины Табун I указатель 70,1
не столь безупречен из-за сомнительного определения длины бед-
ра, все же он может использоваться при дальнейших сопоставле-
ниях.
Следовательно, во всей группе палеоантропов известные зна-
чения интермембрального указателя колеблются от 65,6 до 70,1,
т. е. большинство их укладывается в пределы групповых сред-
них, и все величины не выходят за границы индивидуальных ва-
риаций у современного человека. Ни в каком отношении нет сбли-
жения с антропоидным типом пропорций конечностей; скорее,
можно говорить даже об «ультрагоминидности» некоторых палео-
антропов, прежде всего Схул IV с его укороченными руками.
71
У узконосых приматов в целом отмечается тенденция к поло-
жительной аллометрии в соотношениях длин конечностей и туло-
вища .при межродовых. сопоставлениях (Biegert, Maurer, 1972).
Приведенные этими авторами графики относительной длины верх-
них и нижних конечностей Hominoidea подтверждают исключи-
тельное положение наиболее специализированных форм. Таковы
гиббоны и оранги по длине верхней конечности (гиббоны и по
нижней конечности) и человек по длине нижней конечности, что
особенно • подчеркнуто на фоне генерализованной тенденции ка-
тарриновых приматов к относительному удлинению конечностей,
особенно верхней с увеличением размеров тела.
По ряду соображений, исключительная длиннорукость антро-
поидов является вторичной, что подтверждается и палеонтологи-
ческими данными. Так, дриопитек фонтанов имел сравнительно с
челюстью более короткую’ плечевую кость, чем современный шим-
панзе (Werth, 1928), а вероятный предок гиббона — плиопитек
обладал гораздо менее удлиненной конечностью, чем современ-
ный гиббон. Интермембральный указатель этого антропоида, око-
ло 94 ставит его по пропорциям конечностей рядом с современ-
ными циноморфными обезьянами (Zapfe, 1963; Biegert, Maurer,
1972). То же можно сказать и в отношении пропорций конечностей
у проконсула и лимнопитека (Ле Гро Кларк, 1950), но не орео-
питека, у которого эти соотношения соответствовали брахиатор-
ному типу (рис. 15).
Из всех этих фактов следует вывод, что предками первых го-
минид могли быть антропоиды с относительно неспециализиро-
ванными (т. е. короткими) верхними конечностями и, возможно,
менее специализированными (т. е. менее длинными) нижними ко-
нечностями; предположительные пропорции этой исходной для
гоминидной линии формы, скорее всего, были нейтральными, с
интермембральным указателем, близким к 100 (Бунак, 1957).
' Вряд ли возможно наметить какую-нибудь тенденцию в ва-
риациях интермембрального указателя у поздних ископаемых го-
минид: большинство палеоантропов скорее «ультрагоминидно»
по этому признаку сравнительно со средним современным вариан-
том, тогда как в верхнем палеолите (Пржедмости, Кроманьон)
величина указателя даже несколько больше, чем в среднем у сов-
ременных людей. Тенденция к повышению указателя в более поз-
днее время отмечена и среди современных людей на некоторых
территориях — например, при сопоставлении по этому индексу
древнего и современного населения Патагонии (Мартин, 1928).
Плечебедренный указатель также является важном диагности-
ческим критерием, дифференцирующим в первую очередь'гоминид
и антропоидов. У человека современного типа пределы его груп-
повых вариаций 68,8—72,9. У антропоидов это 101,0 у шимпанзе,
139,0 у орангутана. Индивидуальные значения могут у антропои-
дов в редких, случаях снижаться до 98 и подниматься до 147.
Большой интерес представляет определение этого индекса у
72
ореопитека (Шульц, 1960). Его значение здесь велико, примерно
122, оно превосходит величину этого указателя у гиббона и близ-
ко к его значению у симфаланга. Пропорции плеча и бедра были
у ореопитека подчеркнуто брахиаторного типа. Напротив, миоце-
новый плиопитек имел вполне нейтральные соотношения этих ко-
стей с указателем около 85; то же относится и к лимнопитеку с
указателем 95.
73
Следует подчеркнуть, что обсуждаемый индекс — единствен-
ный, на основании которого можно составить некоторое представ-
ление о пропорциях конечностей архантропов. Вейденрейх при-
водит для синантропа высокое значение указателя 79,3, выходя-
щее за современный групповой максимум. Однако, поскольку пле-
чевая и бедренная кости, возможно, не принадлежат одному ин-
дивидууму, а длина их к тому же определена при реставрации, ве-
личина индекса может оказаться не вполне достоверной. Во вся-
ком случае этот указатель надежно отграничивает синантропа от
антропоидо'в; что же касается некоторого возможного удлинения
плечевой, кости относительно бедренной, то вопрос остается от-
крытым.
Плечебедренный указатель довольно стабилен в группе- евро-
пейских неандертальцев, где он варьирует от 69,3 и 69,6 у Ла
•Феррасси I и II до- 70,4 у Неандерталь. У палестинцев соот-
ветствующие величины 68,2 и 68,9 у Схул IV и Табун I—73,2 у
Схул V.
Можно говорить о довольно устойчивой и небольшой ве-
личине этого индекса у палеоантропов, за исключением вообще
очень своеобразного по пропорциям высокорослого мужчины
Схул V, у которого, повышенное значение указателя обусловлено
очень большой длиной плечевой кости. В целом вариации плече-
бедренного индекса у палеоантропов полностью совпадают с та-
ковыми современного человека.
Последнее' утверждение применимо и к луцеберцовому указа-
телю, который варьирует у современных людей в пределах 62,6—
70,9 против 112,0—169,2 у антропоидов. У европейских палеоан-
тропов его значения также невелики — 66,4 и 65,9 у Ла Феррас-
си. У палестинцев наблюдается почти полное совпадение $ разма-
хом современных вариаций, а именно 62,7—71,6. В отличие от
плечебедренного указателя здесь существуют расхождения между
типами Схул и Табун. Максимум индекса найден у женщины Та-
бун I, минимум — у мужчины Схул IV, т. е. для первой типично
относительно длинное в сравнении с голенью предплечье, для вто-
рого — противоположные соотношения.
Значительно меньший интерес с точки зрения таксономии пред-
ставляют указатели, выражающие пропорции проксимального и
среднего сегментов каждой конечности — лучеплечевой и бедрен-
нобольшеберцовый.
Вариации первого из этих двух индексов у современного чело-
века — мужчины: 71,3-^81,5. Индивидуально указатель колеблет-
ся от 69 до 87. Из антропоидов близкие к человеческим значения
найдены у горилл (78—85). В целом для приматов характерны
более высокие величины индекса. л
Значения указателей, вычисленных разными авторами для от-
дельных форм европейских неандертальцев, несколько расходятся.
Ввиду ограниченности вполне сопоставимых материалов мы при-
ведем лишь несколько цифр для классических неандертальцев:
74
76,3 для собственно Неандерталь, 74,3 и 70,4 для Ла Феррасси
(Патт, 1955). В общем западноевропейские палеоантропы отно-
сятся к числу гоминид с относительно укороченным предплечьем.
По-видимому, у некоторых' палеоантропов группы Схул (на-
пример, Схул IV с индексом 81) предплечье было относительно
длиннее, чем у западноевропейских форм, тогда как у Схул V
(указатель 70,5) оно было исключительно .коротким, даже в боль-
шей степени, чем у представителей классического типа, к кото-
рым по этим соотношениям, очевидно, довольно близка и женщи-
на Табун I С указателем 77,3.
Удлиненное•предплечье одна из специфических особенностей
кроманьонского типа, с которым сближаются в этом отношении,
как и во многих других, некоторые представители группы Схул.
Вариации бедренноберцового указателя у современного чело-
века также частично перекрываются как внутри, так и межгруп-
повыми вариациями, его у антропоидов; пределы колебаний индек-
са у человека 77,3—86,6 и’здесь почти полностью совпадают с та-
ковыми гориллы 77—85. Подобные или близкие величины указа-
теля свойственны также шимпанзе, а у других млекопитающих
группируются вне всякой связи с таксономическим положением и
локомоцией.
У неандертальцев Европы значение индекса составляет в сред-
нем 76—79, для них типична относительно короткая голень. По-
видимому, подобные пропорции существовали и у киик-кобинца.
У палестинских палеоантропов величины указателя варьируют: он
вполне, «неандерталоидный» у женщинй Табун I (75,7) и более
высок у Схул (79,5—87,9); следовательно, палестинцам Схул свой-
ственно удлинение голени относительно бедра, сближающее их с
кроманьонцами (указатели 81—86). Однако разграничительное
значение этих пропорций нивелируется сильными индивидуальны-
ми вариациями индекса у современного человека.
Относительное возрастание широтных размеров туловища яв-.
ляется, по-видимому, параллельной филогенетической тенденцией
для человека и антропоидов. При этом, как по относительной ши-'
рине плеч, так и по относительному широтному развитию таза, на-
блюдается частичное перекрытие вариаций у человека и антропо-
морфных обезьян, что лишает соответствующие указатели особой
таксономической ценности.
У западноевропейских неандертальцев, например у Ла Феррас-
си, ключично-плечевой указатель достигал 54, что превышает сов-
ременный максимум. Однако в. группе палестинцев соотношения
этих размеров были иными; значения индекса колеблются здесь
в пределах 40—47, причем, первая из этих цифр совпадает с ми-
нимумом современных вариаций (40,1—52,1). Следовательно, па-
леоантропы в целом воспроизводят вариации современного чело-
века: для некоторых европейских форм типичны относительно ши-
рокие, а для палестинских-— узкие плечи. Умеренным было ши-
ротное развитие плеч и у синантропа, где ключично-плечевой ука-
75
затель 44,7 соответствует современной средней; впрочем,. здесь
нужно сделать ту же оговорку о его достоверности, как и в отно-
шении плечебедренного индекса.
Можно также полагать, что свойственная палестинцам тен-
денция к долихоморфии отразилась и на их относительной узко-
бедрости, хотя, возможно, и европейские неандертальцы не были
вполне однородными в отношении широтных пропорций, напри-
мер, вероятна относительно меньшая брахиморфия у некоторых
крапинцев.
Сложную задачу представляет определение длины тела у ис-
копаемых гоминид, поскольку данные, полученные по формулам
разных авторов, значительно различаются; существенный элемент
неточности привносит и необходимость пользоваться при подобных
определениях различным в качественном и количественном отно-
шении набором костей. Для таких гоминид, как австра-
лопитековые, указываются обычно малые значения длины тела,
к тому же они частично оспариваются из-за слишком фрагмен-
тарного состояния соответствующих остеологических материалов.
Ограничимся поэтому приведением лишь некоторых цифр. Так,
для Н. habilis (Олдувей 8) определен на основании длины стопы
рост в 120 см. Для грацильных австралопитеков указываются ве-
личины: 139—157 см б и 133—154 см $ (Helmuth, 1968). Воз-
можно даже, что грацильные австралопитеки могли иметь еще
меньшую длину тела (108—109 см, по расчетам Lovejoy et al.,
1970, на основе реконструкции бедренной кости). У двух массив-
ных австралопитеков определена длина тела в 146—156 см и 152—
161 см (Burns, 1971). Примерный рост раннего гоминида из Ке-
нии (F. 1481) — 156—158 см. Таким образом, среди вероятных
наиболее древних представителей гоминидной линии эволюции и
близких' к ним форм преобладали малорослые -по современному
масштабу существа.
Для архантропов также отмечаются сильные колебания вели-
чин длины тела, определенных даже на основании одной и той
же кости, как например, бедренной кости питекантропа I, рост
которого колеблется, по данным Швальбе (1919, 1921), в преде-
лах 1,59—1,77 м в зависимости от того, какая из современных по-
пуляций выбрана здесь в качестве эталона. Если для этой цели
использовать соотношения, типичные для популяции с длиной бед-
ра, равной длине его у питекантропа (455 мм), определенный
рост для этого последнего составит 1,66 м. Вейденрейх указывает
для синантропа малое значение длины тела: 1,56 м для мужчины.
Палеоантропы неоднородны в отношении длины тела: у евро-
пейских неандертальцев отмечены незначительные величины —
рост ниже среднего по современному масштабу. Средняя для муж-
чин Спи, Шапелль, Неандерталь и Феррасси I составляет 1,62 м.
Для женщины Феррасси II приводится очень малая величина:
1,48 м (Патт, 1955; Мартин, 1928). Напротив, палестинские па-
леоантропы были большей частью высокорослы, во всяком слу-
76
чае относящиеся к типу Схул. Данные, полученные Маккоуном и
Кизсом (1939), при использовании в качестве основы различных
длинных костей скелета (бедренной, большеберцовой, плечевой),
естественно, несколько расходятся. Мы ограничимся приведением
размеров, полученных по длине бедра. Длина тела у Схул V,
Схул III, Схул IV и Схул VI была не менее 1,71—1,79 м. Табун Еа,
Схул IX и Схул 7 были, по-видимому, среднего роста (1,66 м),
но и самые малорослые люди Схул превосходили по длине тела
наиболее высокорослых классических неандертальцев. Вероятны
достаточно отчетливые половые различия в этом признаке. У жен-
щины Табун I определение роста дает 1,49—1,54 м, что свиде-
тельствует, даже с учетом половых различий, о пониженной длине
тела. Больший рост имели, очевидно, женщины Схул (1,57—1,58 м
для Схул VII). Выше указывалось, что половая дифференцировка
по длине тела существовала и у палеоантропов Европы (Ла Фер-
расси) .
В целом, исключая относительно высокорослых палестинцев,
большая часть известных «формировавшихся людей» относилась
к категории среднего и ниже среднего роста, а применительно к
грацильным австралопитековым и Н. habilis, — вероятно, даже
малого роста. Впрочем, в этом признаке, кроме половых, сущест-
вуют и значительные популяционные и индивидуальные различия
у ископаемых гоминид. Например, среди архантропов встречались
как средние по росту формы (питекантроп I), так и сравнительно
высокорослые (питекантроп II с длиной тела 1,75 м, согласно
Вейденрейху). Среди палеоантропов наряду с низкорослыми за-
падноевропейскими неандертальцами классического типа выде-
ляются относительно высокорослые, хотя эти последние также не-
однородны. Например, среди палеоантропов Европы вероятно су-
ществование более высокорослого восточноевропейского варианта
(романковский человек с длиной тела не менее 1,75 м).
Следовательно, пропорции и размеры тела варьировали у ис-
копаемых гоминид, как и у современных людей. Среди палеоан-
тропов наблюдаются как высокорослые узкоплечие и узкобедрые
типы, так и выраженные брахиморфы с коренастыми короткими
конечностями и относительно крупной головой. Среди архантро-
пов в этом отношении также, очевидно, не было единообразия.
Особенно следует здесь выделить сравнительно высокорослый ва-
риант Олдувей 28, учитывая вероятную принадлежность бедрен-
ной кости женскому индивидууму. Вопрос о пропорциях австра-
лопитекбвых пока остается открытым вследствие недостаточной
достоверности данных. Но следует еще раз подчеркнуть, что все
эти вариации пропорций происходят, очевидно, на фоне гоминид-
ного интермембрального указателя, а в некоторых случаях (па-
леоантропы) речь может даже идти об «ультрагоминидности», по-
скольку это касается таких четких таксономических индикаторов,
как интермембральный и связанные с ним плечебедренный и луче-
берцовый указатели.
77
Глава IV
КИСТЬ
КИСТЬ КАК ИНДИКАТОР
ФУНКЦИОНАЛЬНЫХ АДАПТАЦИЙ
У ПРИМАТОВ И ЧЕЛОВЕКА
Среди других звеньев посткраниального скелета кисть с ее отчет-
ливо выраженным адаптивным характером эволюционных преоб-
разований занимает особое положение. В пределах отряда прима-
тов, как и у млекопитающих вообще, морфология кости обнару-
живает широкую изменчивость, теснейшим образом связанную с
функциональной активностью конечности и экологией животного
в целом..В антропогенезе весь процесс адаптации к трудовой дея-
тельности послужил основой формирования кисти нового типа,
превратившейся в специализированный орган труда. Отсюда сле-
дует высокий таксономический вес структуры кисти у гоминид.
В литературе неоднократно предпринимались попытки систе-
матизации адаптивных типов кисти у приматов. Хотя мнения
разных исследователей обычно не вполне совпадают в отношении
числа выделенных вариантов, наиболее типичные и четко очерчен-
ные структурные комплексы являются общепризнанными. Это
прежде всего типы кисти низшего примата с обхватывающим ла-
занием, проноградного примата с полуназемной, полудревесной
локомоцией (наиболее распространенный и наименее специализи-
рованный из всех), наземного примата, брахиатора и гоминидный
вариант ткисти ’. Этот перечень не претендует на полноту, так как в
него вошли преимущественно те варианты, которые имеют отно-
шение к этапам эволюции кисти приматов и человека и обсуж-
даются в связи с этой проблемой в современной литературе.
Центральным моментом эволюции кисти человека является
формирование уникального «точного» (прецизионного) зажима,
означающего развитие наиболее полного и совершенного противо-
поставления I луча.
Известно, что способность I луча кисти к противопоставлению
в той или иной степени свойственна приматам вообще и связана в
своей первоначальной обусловленности с древесным образом жиз-
ни. Из числа основных видов (способов) осуществления хвата-
тельной функции кисти у современных приматов наибольший
интерес для рассматриваемой проблемы представляют: «точный
зажим», типичный для руки человека, «силовой зажим», наблю-
дающийся у многих обезьян, но встречающийся иногда и у чело-
1 Некоторыми исследователями (Нейпир I. Р., Нейпир Р. Н., 1967) предлагается
выделять и вариант кисти «полубрахиатора», занимающий промежуточное по-
ложение между типами кисти брахиатора и четвероногого примата.
78
века, и еще более- редкий у человека «крюковидный зажим»,,
наиболее развитый на специализированной кисти брахиаторов
(Нейпир, 1956, 1965, 1967).
Каковы анатомические условия, способствующие формирова-
нию «точного зажима», который в своем развитом виде встречает-
ся, по-видимому, только у человека?
Прежде всего сюда следует отнести определенный уровень
абсолютного и особенно относительного развития I луча и его
компонентов. У человека верхушка большого пальца, как правило,
заходит несколько выше середины проксимальной фаланги указа-
тельного, тогда как у большей части остальных приматов он зна-
чительно короче. Развитие этих пропорций связано с процессом
радиализации кисти, т. е. прогрессивного развития I—II радиаль-
ных лучей, как и возрастания относительных размеров I луча
прежде всего сравнительно со II. Все понгиды обнаруживают-
резкое относительное недоразвитие Г луча, что обусловлено преи-
мущественно -удлинением центральных лучей на кисти брахиато-
ров. Специализация зашла особенно далеко у органгутана, у ко-
торого и абсолютно I луч редуцирован в размерах, тогда как
у африканских антропоидов он крупный, и снижение индексов обу-
словлено только сильным удлинением II—IV лучей. Соотношение-
1—11 лучей на кисти современного человека отличается, высокой
стабильностью: групповые вариаций индекса составляют всего-
65,3—66,1. У антропоидов соответствующие значения: 48,5—50,2.
Для индекса I—III лучей размах изменчивости несколько выше,
но также свидетельствует о высокой устойчивости этих соотноше-
ний у современного человека 60,8—64,1. У других приматов индекс-
I—III лучей варьирует в широких пределах: от 15 (коата с ее ре-
дуцированным I лучом) до 67 (потто, тонкий лори), располагаясь
в зависимости и от конкретной адаптации изученных форм прима-
тов к специфическим локомоторным функциям (Астанин, 1962).
Таким образом, человек в масштабе приматов в целом характе-
ризуется крупным абсолютным и, что особенно важно, относитель-
ным размерами I луча.
Другая анатомическая предпосылка оппозиции I луча — поло-
жение и форма костных компонентов соответствующего пястно-
запястного сустава.
Как известно, важной особенностью кисти современного чело-
века является седловидность сустава. При этом оба изгиба более
отчетливо выражены на трапеции, где сагиттальная выпуклость-
комбинируется с поперечной вогнутостью, чем на основании I пяст-
ной кости, на котором существуют противоположные соотношения.
Является ли седловидность обязательным условием противопо-
ставления? По-видимому, оно не исключено и при иных формах-
1 пястно-запястного сочленения, хцтя гораздо чаще противопостав-
ление все-таки связывается с седловидностью. Вместе с тем сущест-
вуют сравнительно-анатом'ические параллели, свидетельствующие
4 о возможности оппозиции I луча при таких типах сустава, как.
79
например, г< «ловча гый у гиббона, при котором I пястная кость
имеет чашелбра шу к* eyai;»внч:<> р-с i»,, а на трапеции рас-
положена еон.тенив-!.;'.! ! ай.: окр\глых очертаний. У остальных
антропоид» ча сустав ышичн-ы седловидной формы, причем изгиб
обеих ду \ мо/ьг‘. оьг.1 , i.iv i.pj, ;<. и м у человека. Впрочем, ин-
дивидуальна*.! и формы гусыни ссществует у антро-
поморфиь:\. ; у !»..ьшека: II.hi нашел «головча-
тый тип- у «о.а доги гиббона < HuMates 1.шоцепуя]у тогда как у
самки H'<H. .io ,<и .а ё ж: .-..a.:* К ИЛ Легкую СеДЛоВИДНОСТЬ.«Головча-
тый । ин 1 Iел I<!'v*! и описан и у Hylobates lax
(Ур, 1,-..и бр.. .иu\ iAo.ip ыкже пгмечен седловидный
1'1!!! CXVI.HI.!. Че! Я «> Ме!!е. :•. «| >.1 Л. И!' I-! И, 'Ь’Ч V ЧеДоВСКООбраЗНЫХ.
По ВИД’.’ М . :’:е чо* впервые появляется
у мбе *!.ш> : sp"r • <'.и . } ‘ <’ бь.-мх лынилняемых движений
они ш.ииыл! и . лы.и ; Ы'.о ж-фШо-м:. г И. Данилова (1965)
отмечен*!', Mb* 6ил.‘41Ьи.' <-'Орл uibix ииильян действует
сивмелнп г х лл--л;ильным, чричгм в павианы гамадрилы
широко пм.|мрок ? шч о\ч•». <йв 1 К!” । рддияльными паль-
нами кисы П‘> ;.-.<:пым b-ipi . • <B'.rb-:i. 1'С2|, и «учившего майн-
пхлчьфи\-о д.ч а .к « к и < :р;.ипр\ киг:и коркопитековых, про-
1 ни.>!>'Ч.Ь».1.-.;и<' . к *. ..и е и ;иб *.!’*• уа«витым у наземных
вид--в. ! нм --ыу-тп4 д-ю* ю-vwы топкого противопостав-
ление *. l- aii-iu vi'.’e.cs.i-й p*‘HiJ '> -ж-•! !5нр<шалось с более
проксимальным -.чделгднем л.:;!)-;.' >*• с:иб.1 :<.:.! большого пальца
и! 1 ,Т. IС А'.Ш.Ы :<:<Л...О‘ !.<*.•,;!< Й •;<!!<!!!, 10 обособления
длин нот абдукл;*.* K-’pTiv'.' Р: пнбаге.!’-! большого пальца,
большей редукки-. й :ь ш-р*"-;*! (й 1 / *ию! ириш-д-.пцей мышцы боль-
шого налы;.!.' Эю -лн.ч е ;.*у>йл;м п чд.-.ллелн*-у мартышкообраз-
ных - бе «1-ЯИ 1 еХНЬч 1 •..'• '!.’! К>-р-< 1 Ь.еГ< 1 абдуктора И ОП-
нпнирхющгй мйшшк б .!'•’.! г ' 1НЛ1-ПЛ. Тогда как другие мускулы,
облгтчл.-ицй' :г.'"'>е-; ,''.Юе’ у ч<*.*:н-н.!, у мартышкообраз-
ных I -б с . ’is VI-* р.-.Ю’.ТНЖг.'И
В ел-днл.Ч! не .•г-.л-.*' к : и!<".ых американских обезьян
нр< .’И.’ 'Но* 1; uJt.’Hi'M1 •’, . s .!? . о су; е; 4)\г г. у цеоусовых рас-
прострдден шь-лшдрг.ш-см-' .ни сустав... к|'И i.-чором существует'
ННеЙ. ’юМ у > бе ‘hHit 1 . 1 <Ь‘ ’ '> ’ И*' ' '’’И'1 i5!!i!!H 1 .1X43! ПОДВИЖ-
нчпь 1ршнш*п! лр;. л..!/..-.пи ге гшеи! и выраженного
нпш.рета |Л.ио-.! В'’д ./юм. ' ю’, шшх приматов (лему-
РП1 хж<- im.'i-a, Л1.м< -.с? !!•.'!’ р. >;’.-•-!! нротивопоставляе-
мнетй Ос, б.ю р- -ю а »: м игр *•!. очевидно, медлен-
ное и.'.-, «.»»:•• «•• г-:и «х! бх * сиь.-л’Н.-м, например. У потто и
тонког*’ и л ри ,
Пл.Ь'бный. <б б.:.л .!;рип;viиуег развитию I луча
И С!<’ !ф* iti!!*'., ш ,'СЪ<И. ч'ТИ.
У Вгрсяткых .;!лйб>.л*.-е ИрИМИ J IKHiU х !Ц *'/А*. 4ВНГеЛСН ИрНМЗТОВ —
lyiKiXH КИСТ»- чбЛиД.-и-, ЛИ!!»;- w о,М.< •!» р |!»МЧ*‘Н»1«)Й СИОСОбНОСТЬЮ
К npi>»H!U-J!iHT.iH.!e!ii!’><!>. Ю.ч’Ле.’Нгс ПЬ-Ди-Ш К К«»ИНСрГСИЦНИ И ДИ-
иерншнин и*-!» при cj я/-.sпле ра •: нблв-льных движениях
НО
L лапы в пястно-фаланговых сочленениях. Объект при этом обхваты-
' вается обеими лапами, что создает функциональный эквивалент
, хватательной руки. Такой тип кисти еще очень примитивен, однако
уже у тупаий начинает сказываться типичная для приматов спе-
циализация большого пальца (Нейпир, 1962; Данилова, 1965).
Другая важная структурная особенность, кисти человека, свя-
( занная с противопоставлением, — ладонное смещение I луча из
плоскости остальных лучей, анатомической основой которого яв-
ляется радиальное отклонение трапеции и скошенность ее ди-
i стальной сочленовной площадки. У человека существует не тыль-
' ная, как у большинства млекопитающих, а ладонная скошенность
► основания I пястной кости, которая в меньшей степени наблю-
дается и у антропоидов. Близкий к человеческому наклон площад-
ки трапеции был найден, например, у гориллы и оранга, в мень-
шей степени — у шимпанзе (Данилова, 1965). Что касается ра-
, диального отклонения трапеции, то сопоставления антропоидов с
человеком затруднены здесь иным, более проксимальным положе-
нием трапеции в запястье*.
Рассматривая проблему противопоставления, необходимо так-
же иметь в виду, что кисть человека сохраняет и псевдооппози-
цию, свойственную некоторым низшим приматам (например, тар-
зиевым), которая проявляется в увеличенной подвижности и пяст-
но-фалангового сустава большого пальца (Нейпир, 1962).
Выше перечислены только некоторые морфологические призна-
J ки, связанные с противопоставлением I луча и представляющие
для нас интерес в. плане возможной функциональной интерпрета-
! ции структурных особенностей кисти ископаемых гоминид и дру-
гих приматов. Следует, конечно, иметь в виду и широкие компен-
саторные возможности мускульно-связочного аппарата (Бонч-Ос-
моловский, 1941; Якимов, 1949; Семенов, 1950; Патт, 1955), о ко-
* торых гораздо труднее судить на основании остеологических мате-
риалов. Особую роль в эволюции кисти человека играют расшире-
ние и дифференциация корковых полей, имеющих отношение к
представительству двигательных и тактильных функций руки. При
сопоставлении структуры кисти у современных и ископаемых го-
минид нельзя не учитывать также, что более точная и тонкая
координация движений может осуществляться и за счет образова-
ния условнорефлекторных связей, без изменения структуры органа.
КИСТЬ ИСКОПАЕМЫХ ГОМИНИД
Палеонтологические материалы по кисти, как и по другим
звеньям посткраниального скелета, лучше всего представлены в
группе палеоантропов. Наиболее полные кисти известны от пале-
1 Онтогенетическая миграция сустава I луча связана у человека с радиальным
отклонением трапеции, у антропоидов — с ее действительным проксимальным
перемещением (Шульц, 19*26).
6 Е. Н. Хрисанфова
81
стинских форм, киик-кобинского палеоантропа и неандертальцев
из Ла Феррасси; в меньшей степени это относится к Шапелль и
Крапине; наконец, кисть представлена лишь отдельными костями
запястья и фалангами у Табун 3, 4 и 5; некоторыми фрагментами
пястных костей у Схул VII и Схул IX, описаны также некоторые
кости кисти ребенка Схул I.
Хотя в морфологическом отношении кисти этих гоминид не-
однородны, все же возможно выделить некоторый комплекс при-
знаков, общий- почти для всех этих представителей древних
людей.
В него входят расширенность корня кисти, мезо- или мезомакро-
карпия, относительная укороченность пальцев в связи с крупными
размерами пястных костей, общая массивность трубчатых костей
кисти, расширенность дистальных фаланг, особенно на большом
пальце, некоторое недоразвитие I пястной кости прежде всего, от-
носительно очень крупной II пястной, особенности структуры
костных компонентов сустава I луча, заключающиеся в значитель-
ной вариабильности формы и наличии в ряде случаев компенса-
торных признаков противопоставления. Рассмотрим перечислен-
ные признаки более подробно.
По общим длиннотным пропорциям (по отношению к длине
всей верхней конечности и длине тела) кисти палеоантропов нахо-
дятся в пределах современных вариаций. Следует отметить особо
укороченную относительно роста кисть вообще очень своеобраз-
ного по пропорциям конечностей высокорослого мужчины Схул V.
Возможно, этот признак можно рассматривать здесь как «кро-
маньоидный», тем более, что подобный оттенок свойствен и неко-
торым другим отделам скелета Схул V. Приведем значения отно-
сительной длины кисти (к длине тела). У современного человека
они колеблются по групповым средним в пределах 9,8—12,2 при
средней примерно И.О1. У палеоантропов эти указатели равны:
10,2 (мужчина Схул V) —11,7—11,9 (женщина Феррасси II); все
значения абсолютно человеческие. Только у киик-кобинца индекс
12,5 выходит за максимум средних; впрочем, его определение да-
леко не безупречно, так как длина кисти измерялась при рекон-
струкции, а длина тела определена на основании одной только
большеберцовой кости. Таким образом, все индексы, выражающие
относительное развитие кисти, совершенно человеческие у палео-
антропов и резко отграничивают их от антропоидов, у которых
соответствующие указатели гораздо выше, достигая 16,5 у гориллы
и даже 20,7 у орангутана.
Поскольку мы имеем дело с фрагментарными материалами,
анализ длиннотных пропорций отделов кисти целесообразно про-
водить на основе сопоставления трех компонентов: головчатой
кости, избранной как критерий развития запястья, III пястной и
III пальца (табл. 7).
1 По измерениям на живых людях.
82
Таблица 7
ДЛИННОТНЫЕ ПРОПОРЦИИ ОТДЕЛОВ КИСТИ
НЕКОТОРЫХ ИСКОПАЕМЫХ И СОВРЕМЕННЫХ ПРИМАТОВ
(в % от длины III пястной)*
Приматы Головчатая кость III палец
Шапелль 33,3
Феррасси I 33,8 135,9
Феррасси II 33,0 139,3
Табун I 34,4 136,0
Схул IV 37,5 137,5
Шанселад 39,7 139,7
Пато 36,7 —
Современный человек 31,1—38,4 ,136,8—158,5
Проконсул 33,0? 150,0?
Шимпанзе 27,0 139,3
Горилла 27,9 141,4
Орангутан 25,8 134,4
* По Булю (Boule, 1911/1913); Саразину (1932); Маккоуну, Кизсу (1939); Патту
(1955); Даниловой (1965); Нейпиру и Дэвису (Napier, Davis, 1959), данным авто-
ра.
По абсолютному развитию центрального компонента запястья
все палеоантропы снова показывают совершенно человеческие
величины: от 20,2 (Феррасси II) до 26 мм (Схул IV) при группо-
вых вариациях у современного человека: 18,3—25,9 мм. У боль-
шинства палеоантропов головчатая кость исключительно крупная,
почти достигающая современного максимума (Шапелль, Феррас-
си I, Крапина) или превосходящая его (Схул IV). Даже
наименьшие величины, определенные на кистях миниатюрных жен-
щин Табун I и Феррасси II, все же превосходят групповой мини-
мум современного человека.
. По относительному развитию'головчатой кости (в % длины
III пястной) — индексу карпии — все палеоантропы попадают в
ранг мезокарпов .(32,7—33,8, по классификации Саразина, 1932)
или даже макрокарпов. Возможно, что значения указателей для
палестинцев несколько завышены; все же, очевидно, что они обла-
дали относительно более удлиненным запястьем, чем классические
формы, принадлежа к макрокарпам (Схул IV) или находясь на
границе мезо- и макрокарпии (Табун I). В целом все индексы
карпии палеоантропов типично человеческие. Ошибка Буля, счи-
тавшего запястье шапелльца редуцированным, обусловлена толь-
ко неполнотой данных по групповым вариациям этого указателя
среди современных гоминид. Все антропоиды, напротив, обладают
редуцированным запястьем, и в этом отношении существуют отчет-
ливые различия с палеоантропами; из табл. 7 следует, что этот
признак достаточно ясно разграничивает гоминидный и антропо-
идный варианты кисти (запястья).
К сожалению, головчатая кость не сохранилась у киик-кобин-
ского палеоантропа. Для суждения о развитии запястья Бонч-Ос-
6*
83
моловский использовал здесь обе трапеции: большую и малую. >
При сравнении с соответствующими данными по современному ।
человеку оказалось, что абсолютное развитие запястья, по-видимо- I
му, не уклонялось здесь от среднего варианта; по относительному
развитию обеих трапеций запястье киик-кобинца вполне соответ-
ствовало общечеловеческой средней. Следовательно, в этом отно-
шении крымский пйлеоантроп не отличался сколько-нибудь суще-
ственно от других европейских палеоантропов.
Менее специфичен в аспекте разграничения гоминидной и ант-
ропоидной кисти индекс соотносительного развития компонентов I
.III луча (пясть—палец); Палеоантропы в целом характеризуются I
выраженной тенденцией к относительной укороченное™ пальцево- i
го отдела сравнительно с пястным; в этом отношении они прояв-
ляют сближение с европейским типом кисти (средняя индекса
для мужчин 140,9). Следовательно, кисти большинства палеоант- |
ропов были относительно короткопалыми. Нужно подчеркнуть, I
что абсолютно пальцевые отделы на кистях палеоантропов круп- ।
ные, и некоторое снижение индекса создается за счет очень силь- i
ного развития метакарпального компонента. При неполноте мате- j
риалов трудно судить, за счет каких именно компонентов пальца j
возникла эта укорененность. Судя по всему, в наименьшей степени
в этот процесс вовлекались дистальные фаланги, бывшие, по-види- '
мому, относительно длинными у большинства древних людей. По-
добные пропорции были свойственны и кроманьонцам, у которых
пальцы укорочены относительно более длинной пясти (Verneau,
1906). j
Укрепление запястья у палеоантропов заключалось не только !
в его относительном удлинении; прогрессивное развитие затронуло I
и другие размеры этого отдела кисти, прежде всего широтный.
Как известно, Бонч-Осмоловский (1941) подчеркивал чрезвычай- |
ное развитие в ширину и высоту корня кисти киик-кобинского |
человека. Общая ширина сочлененных между собой обеих трапе- j
ций (33,2 мм) значительно превосходит не только современную ।
среднюю (29,2 мм), но даже и индивидуальный максимум ;
(31,5 мм). Причем ширина каждой из этих костей в отдельности !
не выходит за предел^ современного максимума; следовательно, i
на киик-кобинской кисти существовали меньшее перекрытие сочле-
ненных костей и меньшая «сдавленность» запястья в поперечном
направлении. Другое своеобразие конструкции запястья состояло
в уменьшенном отведении трапеции. У палеоантропов из Ла Фер- j
расси больща.я трапеция была абсолютно очень широкой: для |
<5 индивидуума этот размер составлял 23,1 мм, а для $ — 20 мм, ,
что в обоих случаях близко к современному максимуму для муж- ।
ских и женских кистей. Вероятно, и относительное развитие этой I
кости в ширину было велико у всех этих палеоантропов: так, при- I
веденное Бонч-Осмоловским значение широтно-длиннотного ука- •
зателя трапеции для киик-кобинца (177,7) близко к современно- ।
му максимуму (179,1). Аналогичная тенденция к расширению ‘
84
запястья могла существовать и у Шапелль, для которого, впрочем,
нет вполне сопоставимых данных. Во всяком случае, ладьевидная
кость этого палеоантропа была очень уплощенной в проксимо-ди-
стальном направлении.
Что касается палестинцев, то широтно-длиннотный указатель
может быть вычислен только для Схул V, где достоверное значе-
ние его (слева) всего 148,1, тогда как справа индекс предположи-
тельно достигал 168,9. Столь -значительная асимметрия сама по
себе представляет большой интерес, так как лишь частично мо-
жет объясняться малыми размерами и дефектностью правой кости.
Женщина Табун I по абсолютной ширине трапеции превосходила
мужчину Схул V при огромном преимуществе последнего в росте;
это дает основание снова сблизить ее с классическим вариантом
(Феррасси). Определенную тенденцию к расширению запястья
показывает и мужчина Схул IV (по реконструкции кисти), здесь
снова обращает на себя внимание значительная асимметрия. Воз-
можно, что гораздо большая расширенность корня кисти справа
у обоих Схул свидетельствует в пользу рабочей гипертрофии орга-
на и, следовательно, о праворукости этих гоминид, подтверждаю-
щейся и другими анатомическими фактами.
Другой специфической чертой строения запястья многих палео-
антропов является значительное развитие его в высоту.. У киик-
кобинца, например, большая трапеция по высоте 23,1 мм далеко /•'
выходит за верхнюю границу, современных вариаций (13,2— Ж*
19,3 мм по групповым средним). Хотя для Феррасси и Шапелль «|
нет сопоставимых данных, но. зато по высоте других костей за- (Я|
пястья — головчатой и крючковидной — эти палеоантропы тоже .
выходят за групповой максимум. Так, средняя-высота головчатой-
кости у этих неандертальцев составляет 21,4 мм, а у крапинцев Я
даже 22,0 мм при колебаниях групповых средних у современного я
человека 16,8—20,6 мм.
У палестинцев сопоставляемые кости запястья имели меньшее '
абсолютное и относительное высотное развитие. Интересно, что
по этому признаку снова выявляется резкая асимметрия на боль-
шой трапеции Схул V: справа высотно-длиннотный указатель этой
кости составляет 112,1, слева — 88,9, что опять позволяет думать о
проявлениях рабочей гипертрофии кисти. Напомним, что по дан-
ным,- полученным на кисти современного человека, с большей
окружностью запястья связывается большая сила руки и меньшая
ее подвижность (Harcula, 1972).
Аналогичная картина выявляется и в пропорциях пясти. У со-
временного человека широтно-длиннотные . соотношения пясти
значительно варьируют: от типа узкой и длинной (стенохейриче-
ской) кисти с указателями1 ниже 75,0 до короткой и широкой
(эурихейрической) кисти с индексами от 85,0 и выше; промежу-
1 Суммарная ширина проксимальных концов II—V пястных костей в процентах
длины III пястной.
85
точные значения указателя 75—84 характеризуют мезохеириче-
скую кисть (Маккоун, Кизс, 1939). Указанные пропорции могут
быть определены лишь для немногих палеоантропов. Классический
неандертальский вариант дает в лице Феррасси II (указатель
82,0) довольно расширенную пясть, входящую в верхние пределы
мезохейрического типа. Данные
по киик-кобинскому палеоантро-
пу не вполне сопоставимы; все же
резкое своеобразие формы этой
кисти иллюстрируется очень вы-
соким значением индекса, вычис-
ленного нами по реконструкции,—
97,1. Возможно, этот указатель
несколько завышен, но все же и
при этом условии ширина пястно-
го отдела на кисти этого палео-
антропа бросается в глаза. Ин-
дивидуально такой вариант мо-
жет встретиться и у современно-
го человека: так, Маккоун и Кизс
приводят для европейской «сило-
вой» кисти значение индекса 91,9,
а крайний вариант эурихейриче-
Рис. 16. Кисть Киик-Кобы (по Бонч-
Осмоловекому, 1941)
ской кисти в исследованной нами мужской серии имеет индекс
даже 102,3.
Напротив, среди палестинских палеоантропов значения указа-
телей более умеренные: у Табун I — 70,5 (стенохейрическая
кисть), а у Схул IV соответствующий индекс справа составляет
78,2, т. е. довольно близок к среднему для нашей мужской серии
(80,4).
Расширенная форма пясти была, очевидно, свойственна и дру-
гим палеоантропам классического типа (Шапелль и Спи). Широ-
кая и массивная кисть «классического типа» ассоциируется с об-
щей массивностью костяка этих палеоантропов, что возможно
следует рассматривать, как проявление универсальной- гипертро-
фии скелета. Однако среди палеоантропов существует и более* су-
женный и удлиненный тип пясти, например, у Табун. Следова-
тельно, нужно признать, что морфологическое разнообразие кисти
существовало уже у палеоантропов; возможно, что конкретные
условия трудовой деятельности разных первобытных коллективов
играли определенную роль в проявлении этих различий. Вероятно,
с точки зрения «силовой адаптации» кисти следует рассматривать
такие ее особенности, как расширение и развитие в высоту корня
86
кисти, удлинение пясти, а также выраженный ульнарный тип кис-
ти киик-кобинца, у которого все эти проявления наиболее отчет-
ливы (рис. 16). Известно, что, по данным ряда наблюдений,
ульнаризация кисти современного человека нередко отмечается
•при повышенной нагрузке на кисть: справа чаще, чем слева, у
мужчин, чаще, чем у женщин, а у взрослых более часто по срав-
нению с детьми (Волоцкой, 1924; Астанин, 1962а; Blinkoe, 1962;
Pratesi, 1962; Lee Таи—Lung, 1964; Мацуи Рёити, Ямагута Хидэо,
1965; Исии Рэйдзиро, Охаси Хироёси, 1965; Line, Fleischmann,
1966; Мураками и др., 1966). В связи с этим высказывалось мне-
ние, что ульнарный тип кисти не является фиксированным исклю-
чительно генетически; по-видимому, оказывается возможным из-
менение основного типа кисти в ульнарном направлении прежде
всего под влиянием тяжелой физической работы (Rosler, 1957,
1957а; Wechsler, 1939). Однако у палестинских палеоантропов
можно предположить существование в некоторых случаях и ради-
ального типа кисти. Судить о пропорциях II—IV пальцев с извест-
ной долей достоверности возможно только для Схул IV, где из-
вестны проксимальная и средняя фаланги IV и средняя фаланга
II пальцев. Реконструировать на этой основе длины соответствую-
щих пальцев с достаточной точностью трудно, но в данном случае
это не имеет значения, так как именно удлинение средней фалан-
ги IV пальца и обусловливает в наибольшей степени ульнариза-
цию. Этот размер составляет у Схул IV 26 мм против 30,3 мм у
киик-кобинца, тогда как длина средней фаланги II пальца, напро-
тив, много больше у палестинца (29 мм против 24,2 мм у киик-
кобинца). Очень велика длина средней фаланги указательного
пальца и у Табун I. Уже из этого видно, что на кисти Схул IV
вряд ли мог существовать выраженный ульнарный тип, скорее
здесь можно предполагать вариант радиальной кисти, так как, в
отличие от современных приматов (в том числе, и человека), у ко-
торых IV2, как правило, длиннее (по средним), чем Щ, у Схул IV
выражено противоположное соотношение этих фаланг.
С точки зрения силовой адаптации следует рассматривать и
такие особенности кисти многих палеоантропов, как увеличение
конгруэнтности суставных поверхностей и их уплощение, столь
выраженное, например, у киик-кобинца; подобное явление неодно-
кратно отмечалось у современного человека при интенсификации
нагрузки на кисть (Гевлич, 1967; Ивкина, Супряга, 1967).
Особый интерес представляет развитие и форма ногтевых (ди-
стальных) фаланг кисти. Впервые внимание к этому признаку
привлек Бонч-Осмоловский (1941), отметивший очень большие
размеры и исключительную расширенность дистальных фаланг
большого пальца у киик-кобинского палеоантропа. Средняя длина
по 2-м фалангам равна здесь 25,2 мм; аналогичную среднюю дли-
ну имеют и три крапинские фаланги. В масштабе современных
групповых вариаций (18,6—23,5 мм) это очень большие величины,
стоящие на границе аномальной долихофалангии. У палестинских
87
палеоантропов наблюдаются менее крупные размеры: все же и
у Схул IV длина 13 соответствовала современному максимуму
(23,0 мм), хотя у Табун5 она была уже значительно ниже
(18,7 мм). По-видимому, длина дистальной фаланги очень велика
у мужчины Феррасси и значительно ниже у женщины, где она
сближалась с ее размером у ТабуНб.' Довольно высоки значения
этого признака и у кроманьонцев (23,5—24,3 мм), хотя последние
уступают -здесь некоторым палеоантропам. Особый интерес пред-
ставляет, однако, форма ногтевой фаланги киик-кобинского чело-
Таблица 8
ПРОПОРЦИИ ДИСТАЛЬНОЙ ФАЛАНГИ I ПАЛЬЦА
У НЕКОТОРЫХ ПРИМАТОВ
Приматы* Ширина ос- нования (I) Ширина, го- ловки (II) Полусумма (I + П)
(в % от наибольшей ; (ЛИНЫ)
Олдувей‘7 74,3 60,0 67,2
Киик — Коба 73,0 57,9 65,5
Крапина 63,4 51,5 57,5
Феррасси I и II — —— 55,8
(54,5—57,1)
Схул IV . 67,4 55,2 61,3
Табун5 64,2 42,8 53,5
Мурзак-Коба ...... . 64,7 42,3 53,5 .
Ментона 67,8 —.
Современный человек** . . . 63,7 43,3 53,5
(55,2—72,0) (32,4—52,0)
Горилла 62,0 33,7 47,9
Шимпанзе . . . 50,4 31,4 40,9
Орангутан 61,4 35,4 48,4
. * По данным Дэя, Нейпира (1966); Бонч-Осмоловского (1941); Маккоуна, Кизса
(1939); автора.
** В скобках приведены индивидуальные вариации.
века и некоторых других палеоантропов (табл. 8). Индекс отно-
сительной ширины головки ногтевой фаланги большого пальца
вполне соответствует «чудовищным», по определению Бонч-Осмо-
ловского, ногтям на руке этого палеоантропа. Разрыв со средней
для мужских европейских кистей, которые нередко рассматрива-
ются в качестве эталона современной «силовой кисти», составляет
примерно 15 ед. указателя! Очень велика расширенность ногтевой
фаланги и у человека из Крапины, как и у мужчины Схул IV.
В целом весь ряд вариаций индекса формы'ногтевой фаланги па-
леоантропов резко сдвинут вправо сравнительно с вариациями
групповых средних у современных людей (соответствующие циф-
ры 42,8—57,9 против 41,8—44,7). По аналогии с киик-кобинцем, у
которого все остальные фаланги были сильно расширены, подоб-
88
ную тенденцию к расширению дистальных фаланг можно предпо-
ложить и у других палеоантропов, по крайней мере у крапинцев
и Схул IV. Интересно, что гипертрофия ногтевых фаланг встре-
чается у современного человека при интенсивном силовом исполь-
зовании кисти (Привес, 1956).
Наибольший интерес представляет вопрос о развитии «точного
зажима» на кисти палеоантропов — насколько о нем можно су- *•
дить по структуре I луча и I.луче-запястного сустава, как и по
соотношению обоих радиальных лучей кисти (табл. 9).
Таблица 9
АБСОЛЮТНОЕ И ОТНОСИТЕЛЬНОЕ РАЗВИТИЕ I ПЯСТНОЙ
КОСТИ У НЕКОТОРЫХ ПРИМАТОВ*
Приматы Длина, мм Индекс I—П пястных
Человек’.средняя 44,2 -67,6
вариации / гРУт'°вые г ) индивидуальные .... 39,0—47,6 67,1—68,2
34,0—56,2 —
Шапелль • 44,5 63,5?
Феррасси I 48,5 68,4?
Феррасси II 41,1 67,1?
Киик-Коба 44,6 —--
Табун I 40,5 64,3 •
Схул IV 49,0 64,9
Схул V 46,5 69,4
Мурзак-Коба 44,2 —
Кроманьон 43,0 —
Ментона 51,3
Шанселад — 68,7
Пато ‘ ' 67,7
Австралопитек (массивный) 35,0 —
Горилла .... 47,2 53,7
Шимпанзе 42,4 50,2
Орангутан 49,1 47,6
* По-данным Саразина (1932); Маккоуна, Кизса (1939); Патта (1955); Билли
(Billy, 1969); автора.
Анализ абсолютного развития компонентов I луча у палеоан-
тропов целесообразно начать с I пястной кости, которая наиболее
полно представлена на ископаемых материалах. В сравнении с
современным человеком, древние люди не обнаруживают призна-
ков уменьшения абсолютных размеров I пястной кости. Размах
вариаций: 40,5 мм (Табун I) — 49,0 мм (Схул IV) — весьма бли-
зок к размаху современных групповых средних по этому признаку.
Средняя для приведенных в табл. 9 форм палеоантропов 44,9 мм
фактически совпадает со средней для современных серий
(44,2 мм). Также у ребенка Схул I длина I пястной кости почти
такая же, как и у современного ребенка близкого возраста.
89
По относительному развитию I пястной сравнительно со II
палеоантропы в целом, по-видимому, обнаруживают некоторое
«отставание». Указатели обычно не достигают современного груп-
пового минимума или совпадают с ним. Исключение составляют
Схул V, у которого величина индекса даже превосходит групповой
максимум, и, возможно мужчина Феррасси, значение указателя у
которого, впрочем сомнительно. Средняя 66,2 для палеоантропов
уступает современной. Учитывая недостаточную точность опреде-
ления большей части индексов у палеоантропов (реконструкция),
существенно, что и соотношения I—III пястных в общем подтверж-
дают полученный результат. У палеоантропов индекс I—III пяст-
ных колеблется от 62,6 (Шапелль) до 71,5 (Схул V), т. ,е. суще-
ствует почти та же иерархия в распределении указателей. Все эти
значения или ниже минимума групповых вариаций у современного
человека (68,3), или близки к нему. Только у двух людей Схул и,
возможно, у Феррасси I указатель соответствует среднему совре-
менному варианту или даже немного превосходит его. О пропор-
циях I пястной киик-кобинца можно судить только по ее соотноше-
нию с IV пястной. Индекс 77,2 немного не достигает группового
минимума современного человека 77,6 и значительно ниже, чем у
палестинских палеоантропов.
Следовательно, по совокупности приведенных-данных можно
говорить о тенденции к некоторому относительному «недоразви-
тию» I пястной кости у палеоантропов, хотя абсолютно она пока-
зывает у всех древних людей вполне современное развитие. Осо-
бенно подчеркнуто это «отставание» сравнительно с очень круп-
ной у палеоантропов II пястной костью. Исключение^м является
прежде всего «прогрессивный» палеоантроп Схул V, у которого
эти пропорции вполне «сапиентные».
Попытка оценить тотальные соотношения I—II лучей очень
затруднена неполнотой палеонтологических материалов, не позво-
ляющей во многих случаях достоверной реконструкции абсолют-
ных размеров большого пальца и всего I луча. Такое сопоставле-
ние может быть предпринято только в отношении довольно огра-
ниченного круга форм. Так, на наиболее сохранившихся кистях
палестинских палеоантропов более или менее достоверное восста-
новление длины большого пальца возможно для женщины Та-
бун I и мужчины Схул IV. Реконструированная длина большого
пальца Табун I составляет, по Маккоуну и Кизсу, 44,7 мм, а всего
I луча — 85,2 мм. У Схул IV известны все компоненты I луча
(дистальная фаланга левого .пальца была восстановлена). Длина
большого пальца равна здесь 58,4 мм, всего I луча — 107,4 мм.
По классическим неандертальцам есть данные для Феррасси,
относящиеся, правда, к суставной длине. Саразин приводит для
длины большого пальца Феррасси I величину 55—55,2 мм, Фер-
расси II — 43—46,5 мм (соответственно слева и справа). Тогда
суставная длина всего I луча будет равна у мужчины 101,3—
102 мм, у женщины—87,2мм (справа). Маккоун и Кизс дают для
90
длины I луча Феррасси II примерно 88 мм, не уточняя, впрочем,
о какой длине идет речь — наибольшей или суставной1.
Бонч-Осмоловский указывает значения обеих этих длин боль-
шого пальца киик-кобинца, оговариваясь, что восстановление дли-
ны большого пальца по одной только его дистальной фаланге
сомнительно. Для реконструкции длины проксимальной фаланги
этого пальца им использован вычисленный Саразином индекс про-
ксимальной фаланги к суставной длине I пястной у обоих Феррас-
си. Нами применено с этой же'целью уравнение регрессии (по
мужской европейской серии). Этот прием представляется вполне
правомерным, поскольку кисть киик-кобинца во многих отноше-
ниях сходна как раз с типом «силовой кисти» современного евро-
пейца; кроме того, при таком способе расчета можно было исполь-
зовать как основу и дистальную фалангу, и I пястную. Обе
полученные нами цифры — 31,7 и 33,3 мм — не вполне совпадают,
но вместе с тем выше, чем вычисленная Бонч-Осмоловским на ос-
нове индекса Саразина (27 мм). При примененном нами способе
определения этого размера усредненная наибольшая длина всего
I луча составила бы у киик-кобинца 102,3 (101,5—103,1 мм), а
сочленовная — примерно 100,2 (99,4—101,0 мм). В целом все при-
веденные для палеоантропов размеры вполне укладываются в пре-
делы современных вариаций.
Наибольший интерес представляют, конечно, пропорции I—II
пальцев и тотальные пропорции I—II лучей. У современного чело-
века, как и по индексу I—II пястных, существует высокая стабиль-
ность пропорций I—II пальцев: 63,5—64,5 по групповым средним
указателя. У тех палеоантропов, для которых возможно получить
сопоставимые материалы, эти индексы совершенно человеческие:
63,9 у киик-кобинца и 65,6 у Феррасси II против 63,5 в среднем в
современной мужской европейской серии. Значительное снижение
индекса отмечается только у Табун I, что обусловлено здесь ис-
ключительно большой длиной средней фаланги указательного
пальца; этот размер вызывает даже некоторое недоумение,, так
как у миниатюрной женщины палеоантропа он больше, чем на
типичной мужской кисти. Возможно поэтому, что указатель Табун
сомнителен. То же относится и к тотальным пропорциям I—II лу-
чей, которые свидетельствуют об исключительной относительной
укороченности I луча у этой формы даже на фоне вообще несколь-
ко пониженных значений указателя обоих лучей у палеоантропов.
Соответствующие величины индекса I—II лучей составляют: 60,0
у Табун I, 64,2 у Феррасси II и 64,6 у Киик-Кобы при групповых
вариациях у современного человека 65,3—66,1. Хотя все приве-
денные индексы, палеоантропов не безупречны, так как вычислены
1 При вычислении длины I луча этими авторами суммируются разные размеры:
длина I пястной приводится наибольшая, а длина проксимальной фаланги,
очевидно, сочленовная. Впрочем, судя по некоторым сопоставимым материа-
лам, разница между- обоими размерами длины большого пальца не превышает
ошибки измерения.
91
при реконструкции, полученные результаты согласуются с теми„
которые приводились выше для указателя I—II пястных.
Следует еще раз подчеркнуть, что поскольку все величины
абсолютной длины I луча у палеоантропов вполне человеческие,
причину его некоторого относительного - недоразвития следует
искать в очень крупных размерах II луча и особенно II метакар-
пального компонента. В этом отношении имеются расхождения с
укороченным вариантом I луча на кисти современного человека.
В последнем случае относительное укорочение I луча обычно было
обусловлено реальной короткостью его пальцевого компонента при
значительно меньшей изменчивости длины указательного пальца.
Основную роль в повышенной вариабильности большого пальца
играла здесь его дистальная фаланга, которая связана с разви-
тием соседних компонентов (проксимальной фаланги и пястной
кости) менее тесно, чем эти две последние между собой. Также и
связь длиннотного развития обоих отделов I луча (пальцевого и
метакарпального) у современного человека меньшая, чем на II лу-
че. Приведенные факты подчеркивают специфику соотношений
обоих радиальных лучей на кисти палеоантропов сравнительно с
«силовой кистью» неоантропа.
Большое значение в развитии противопоставления I луча и
«точного зажима» имеет также трансформация структуры костных
компонентов сустава I луча, с которой тесно связана проблема
подвижности большого пальца на кисти палеоантропов.
Как известно, в антропологической литературе на этот счет нет
единого мнения. Две крайние точки зрения связываются с имена-
ми Г. А. Бонч-Осмоловского (1941) и С. А. Семенова (1950). Пер-
вый из этих исследователей отрицал способность к выраженной
оппозиции I луча у неандертальцев Европы, тогда как второй
утверждал, что у этих гоминид не существовало ограничении в
противопоставлении I луча в силу, прежде всего развития неко-
торых компенсаторных приспособлений.
В центре этой проблемы стоит вопрос о противопоставлении
большого пальца, на кисти киик-кобинского человека, подробно
рассмотренный Бонч-Осмоловским (1941). Как известно, этот ав-
тор нашел здесь существенные отклонения от современного типа,
заключающиеся прежде всего в резкой уплощенности обеих сочле-
новных площадок. Особенно сильно сказалась эта уплощенность
на строении основания I пястной, где седловидность вовсе не вы-
ражена, а имеется простой блок полуцилиндрической формы, со- -
вершенно ровный в дорзо-волярном направлении и круто выпук-
лый в поперечном. Отсутствие седловидности Бонч-Осмоловский
считал решающим аргументом в признании ограниченности дви-
жений большого пальца киик-кобинской кисти в сагиттальной
плоскости, т. е. сгибания и разгибания, тогда как приведение и
отведение было здесь, по его мнению, не меньшим, чем у совре-
менного человека. Он основывался при этом на увеличенной кру-
тизне поперечной дуги у киик-кобинского палеоантропа: ульнарно-
92
г
радиальная- выпуклость I пястной кости была здесь выражена
.даже больше, чем у современного человека, тогда как сагитталь-
ная выпуклость почти отсутствовала на трапеции и сводилась
.лишь к легкой тыльной отогнутости дорзального края. На совре-
менной трапеции, по мнению Бонч-Осмоловского, соотношение
изгибов обратное, и сагиттальная выпуклость значительно круче
поперечной вогнутости.
Если сравнить форму сустава I луча на кисти киик-кобинского
.неандертальца и на современной мужской европейской кисти, к
которой по ряду структурных признаков тяготеет этот палеоант-
роп, окажется, что преобладание поперечного изгиба над сагит-
тальным сравнительно нередко встречается и у современного
человека. Подобный вариант соотношений обоих изгибов наблю-
дался нами не менее, чем в половине всех случаев, хотя, конечно, >
степень выраженности как сагиттального, так и поперечного изги- •
бов была здесь значительно, большей. Измерения для киик-кобин-
ца глубины сагиттальной дуги (на I пястной) дали 0, для глубины
поперечной дуги (на трапеции) — 1,5 мм (по муляжу). Подобная
степень уплощенности встречена в изученной нами серии всего в
нескольких случаях, хотя и здесь речь идет все же не о полном .
совпадении. Так, в одном случае сагиттальная дуга имела глубину
0,5 мм, поперечная — 1,5 мм. Однако при этом нужно иметь в ви-
ду весьма существенное обстоятельство: у киик-кобинца, в отличие
от современного человека, уплощенность сустава комбинируется
с огромными размерами, прежде всего высотными, обеих сочлено-
вных площадок. При сопоставлении этих размеров в изученной на- Й '
ми серии и у некоторых ископаемых гоминид можно прийти к вы- igk
воду, что исключительная высота площадки трапеции является
одним из важнейших структурных отличий киик-кобинской кисти
(табл. 10). Она далеко выходит за индивидуальный современный Я|
максимум (!), а при применении критерия среднего квадратиче-
ского отклонения оказывается, что ,этот размер у киик-кобинца 3
удаляется от средней арифметической современной мужской кис-
ти более чем на 3 о. Это колоссальные различия. По ширине *
площадки трапеции киик-кобинский палеоантроп, по-видимому,
превосходил средний современный вариант, но разница здесь ме-
нее существенна. К тому же определение широтного размера
площадки трапеции не вполне достоверно из-за повреждения уль-
нарного края кости Ч Возможно поэтому, что индекс формы соч-
1 Вероятно, ширина этой площадки у киик-кобинца не превышала 17 мм. Это
• приблизительное определение сделано нами по реконструкции, приведенной
в монографии Бонч-Осмоловского. Автор указывает на крупные размеры пло-
щадки основания I пястной, которые превосходили соответствующий размер
на площадке трапеции. Вместе с тем для всего основания I пястной дается
ширина 17,7 мм; ясно, что ширина сочленовной площадки должна быть мень-
‘ ше этого размера, а ширина площадки трапеции, по Бонч-Осмоловскому, еще
уступала ей. Полагаем поэтому, что размер 17 мм для ширины сочленовной
площадки трапеции может быть несколько завышен.
93
Таблица 10
ВАРИАЦИИ ИЗМЕРИТЕЛЬНЫХ ПРИЗНАКОВ КОСТНЫХ КОМПОНЕНТОВ
СУСТАВА I ЛУЧА У НЕКОТОРЫХ ГОМИНИД
Признак Н. sapiens,* п — 50 Киик-Коба Схул IV Л/П Схул V Л/П
Ширина площадки трапеции, мм 16,0 (13-19) 17,0? 17,8/17,5 15,5/16,2-
Высота площадки трапеции, мм 11,9 (9,3-14,5 17,0! 14,0/15,2 —/н,з
Величина** площадки трапеции, мм 27,9 (23,5—32,6) 34,0! 31,8/32,7 —/27,5
Индекс*** площадки 74,4 (60,7—94,2) 100,0! 78,7/86,9 —/69,8
Глубина**** сагиттальной дуги, мм 1,7 (0,5—3,0) о,о — —
Глубина**** поперечной дуги, мм 1,9 (0,5—3,0) 1,5 1,6/2,5 1,6/1,6
*Средняя арифметическая и минимум—максимум; данные автора.
♦♦Условно выражалась суммой обоих размеров.
♦♦♦Высота площадки в % от ее ширины.
♦♦♦♦Глубина поперечной дуги измерялась на трапеции, сагиттальной — ка I пястной кости,
т. е. в обоих случаях мы имели дело с вписывающей дугой, и сравнивали тем са-
мым аналогичные размеры. Вместе с тем, определяя всегда вписывающую дугу, мы
заведомо преуменьшаем действительную величину обоих изгибов.
леновой площадки киик-кобинца был даже выше 100. Это в еще-
большей мере подчеркивает исключительное своеобразние этого-
сустава и преобладание его развития в дорзо-волярном направле-
нии. Даже индивидуальный максимум соответствующего индекса
в современной мужской серии (94,2) все еще далек от его вели-
чины на киик-кобинской кости.
Реальность различий с современным человеком по форме
площадки трапеции была проверена статистически и по уравнению
регрессии: при одной и той же ширине площадки трапеции разни-
ца между ее высотой у киик-кобинца и на .современной кисти бо-
лее чем в пять раз превосходит частное квадратическое отклоне-
ние этого признака в современной серии. Следовательно, современ-
ная европейская трапеция, равная по ширине киик-кобинской^
имеет много меньшую высоту, и у современного человека не встре-
чается подобной «толщины» сустава, тем более в сочетании с его
уплощенностью, хотя вообще сравнительно плоские варианты не
представляют особой редкости. Напротив, имеется даже тенден-
ция к положительной связи между размерами площадки трапеции
и выраженностью обоих ее изгибов: последняя возрастает при
увеличении размеров сочленовных площадок. Применив снова
уравнение регрессии, получим, что у современного человека при
таком же, как у киик-кобинца, гипотетическом развитии суставной
площадки в высоту, глубина сагиттального изгиба была бы зна-
чительно большей, чем у палеоантропа (2,5 мм против 0,0 мм).
94
Трудно, следовательно, недооценивать наличие подобной комби-
нации признаков у палеоантропа. Она, скорее всего, должна была
затруднять движения в функционально наиболее важной сагит-
тальной плоскости, т. е. ограничивать противопоставление.
Об этом свидетельствует и целый ряд компенсаторных призна-
ков на кисти киик-кобинского палеоантропа. Таковы, например,
мощное развитие рельефа в обоих местах прикрепления ш. орро-
nens pollicis, резкая ладонная скошенность основания I пястной
и ладонный изгиб ее диафиза. Вообще говоря, во многих случаях
нужна большая осторожность при интерпретации особенностей раз-
вития мускульного рельефа, однако вряд ли можно сомневаться, что
на кисти киик-кобинского палеоантропа сильному развитию рельефа
соответствовало адекватное развитие противопоставляющей мыш-
цы большого пальца. Действительно, проксимальное прикрепление'
этого мускула — бугорок трапеции — имеет здесь вид сильно
развитого гребня, не менее чем в полтора раза длиннее нормаль-
ного, причем помимо основного бугорка появляется еще и допол-
нительный. Дистальное прикрепление мышцы представлено у ки-
ик-кобинца мощным «радиальным гребнем», отчетливо отделяю-
щимся от тела кости желобком. Этот гребень выражен лишь в
дистальной половине I пястной, как бы «приставлен» к телу кости,
что Бонч-Осмоловский считал отличительным признаком киик-ко-
бинской кисти. Правда, наличие гребня только в дистальной
половине кости нередко встречается и у современного человека,
но подобная степень его развития все же не наблюдалась нами
ни разу. О возможном компенсаторном значении этой особенности
свидетельствует также и тот факт, что и у современного человека
в обоих отмеченных нами случаях минимальной седловидности
сустава I луча описываемый рельеф достигал наибольшего раз-
вития.
Особое внимание привлекает очень сильная ладонная скошен-
ность основания I пястной киик-кобинца при слабом радиальном
отклонении трапеции и ее меньшем ладонном наклоне, т. е. при
большем приближении трапеции к плокости запястья, что, по-ви-
димому, соответствует меньшей «свернутости» большого пальца
из плоскости остальных пальцев руки. Величина наклона основа-
ния I пястной киик-кобинского человека 29° по Бонч-Осмоловско-
му или 30° по Семенову далеко выходит за верхнюю границу даже
индивидуальных современных вариаций. О компенсаторном харак-
тере этой особенности структуры киик-кобинской кисти может
свидетельствовать (в косвенной форме) и отрицательный коэффи-
циент корреляции величины этого угла с абсолютной и относи-
тельной длиной большого пальца в современной серии.
Разбирая вопрос о противопоставлении большого пальца у ки-
ик-кобинца, С. А. Семенов (1950) придает большое значение так-
же ладонному изгибу I пястной, который, по его измерениям,
составляет здесь 8 мм против 5,5—6 мм у неоантропа. Можно
вполне согласиться с мнением этого автора, что подобное развитие
95
изгиба способствовало противопоставлению большого пальца у
киик-кобинского палеоантропа.
Однако уже само существование целого комплекса «компенса-
торных механизмов» на кисти киик-кобинца свидетельствует о
структурном ограничении противопоставления, обусловленном, как
уже упоминалось, прежде всего комбинацией огромной высоты
сочленовных площадок с их уплощенностыо. При невозможности
учесть у ископаемых гоминид состояние некоторых важнейших
для функций сустава I луча элементов связочного аппарата (осо-
бенно участвующих в аксиальной ротации) окончательное сужде-
ние не может быть категорическим. Все же мы склоняемся к мне-
нию об известных ограничениях пронаторно-супинаторных движе-
ний I луча на кисти киик-кобинского палеоантропа, тем более, что
сама эволюция кисти в дальнейшем пошла не по пути усиления
компенсаторных механизмов, а, напротив, по пути их ослабления.
На сапиентной кисти все эти «признаки компенсации» значительно
менее выражены. В подтверждение этой точки зрения сошлемся
также на данные мультивариационного анализа некоторых при-
знаков неандертальской кисти, соответствующие представлению
о меньшей ловкости неандертальца сравнительно с современным
человеком (Musgrave, 1971). Об этом же, возможно, свидетельст-
вует и отсутствие особой изогнутости и перекошенности дисталь-
ной фаланги большого пальца киик-кобинца, характерной обычно
для современного человека. Этот признак связывают с особой спе-
циализацией человеческих фаланг и, в частности, с сильным раз-
витием противопоставления и осязательной способности (Бонч-
Осмоловский, 1941), хотя ему и нельзя придавать решающего зна-
чения ввиду сильной вариабильности этой структуры у современ-
ного человека.
Каково было строение I запястно-пястного сустава на кисти
других палеоантропов?
К сожалению, соответствующие материалы здесь гораздо более
ограничены. Все же можно думать, что уплощенность сустава
I луча киик-кобинского человека не была общей чертой всех па-
леоантропов. Так, например, среди известных вариантов этого
сустава только у Феррасси II он был, по-видимому, довольно
уплощенным. У мужчины Феррасси Саразин находит легкую сед-
ловидность сустава, а Буль указывает, что I пястная этого палео-
антропа имела «нормальное строение». Совершенно особый этот
сустав у Шапелль. Здесь отмечен его «головчатый тип», причем
основание I пястной кости превращено в верхнюю головку — со-
отношения, противоположные описанным на кисти гиббона. Что
касается палестинских палеоантропов, то у них, очевидно, не на-
блюдалось отклонений от современного типа. Прямое указание
Маккоуна и Кизса на этот счет существует для Схул IV, но
Поскольку какие-нибудь оговорки по отношению к другим фор-
мам отсутствуют, его можно, по-видимому, распространить и
на них.
96
Следовательно, палеоантропам свойственна значительная ва-
риабильность сустава I луча. Она 'наблюдается и среди современ-
ных людей, где существуют два основных отклонения от типичной
седловидности: в сторону ее «преувеличенной» выраженности и в
сторону ее уплощенности. Первый вариант описан Адахи (Adachi,
1905) на кисти японцев и связывается с «тонким» ремесленным
типом труда. Здесь отмечен более крутой изгиб обеих дуг сочле-
нения. Второй вариант — уплощенный — встречается как среди
европейских, так и у неевропейских народов. Выше мы ссылались
на подобные варианты в. нашей европеоидной серии, что наблюда-
лось еще Фиком (1904); Саразин нашел такой же уплощенный
тип сустава и среди представителей других групп современного
человечества. Все же, принимая во внимание сравнительную кон-
центрацию этих вариантов среди современных и древних людей,
нельзя не видеть, что последним свойственно значительно боль-
шее морфологическое разнообразие этой области.
О размерах и форме площадок сустава I луча имеются неко-
торые данные только для палестинских палеоантропов Схул IV
и V. У Схул IV площадка трапеции была велика, приближаясь к
наиболее крупной в современной серии, хотя и уступала по вели-
чине площадке на трапеции киик-кобинца. Здесь также наблю-
дается очень большая высота площадки, особенно справа, где она
превосходит современный максимум, хотя этот размер снова ниже,
чем у киик-кобинца. Ширина тоже велика, но разница менее зна-
чительная, размер в общем укладывается в пределы современных
вариаций. Однако форма площадки Схул IV уже ближе к типич-
ной для современных кистей, хотя относительное развитие ее в вы-
соту и больше, чем у среднего современного варианта (особенно
справа). Иные соотношения у «прогрессивного» палеоантропа
Схул V. Здесь оба размера площадки трапеции очень близки к
средним' для современного человека, столь же современна и ее
форма. Что касается развития изгибов, то в монографии приводят-
ся данные только для глубины поперечного изгиба, также вполне
современной для обоих людей Схул.
К сожалению, о других палеоантропах можно высказать толь-
ко весьма предположительные суждения. Ширина основания
I пястной кости Шапелль составляет 16 мм против 17,7 мм у ки-
ик-кобинца. Современный индивидуальный максимум этого разме-
ра — 18,8 мм — при средней 15,7 мм. В отличие от киик-кобинца,
шапеллец сближается, скорее, со средним современным вариан-
том. Высота основания I пястной кости Шапелль равна 16,5 мм
против 19,5 мм у крымского палеоантропа; она превосходит сов-
ременную среднюю (15,5 мм), хотя и не достигает максимума
(18,8 мм). Принимая.во внимание указания Буля и Патта, а
также Фрэпона и Лоэ для Спи о сильном развитии сочленовных
концов пястных костей у этих неандертальцев, можно думать, что
широтные и высотные размеры сочленовных площадок были здесь
достаточно большими. У Феррасси I, судя по общей ширине тра-
7 Е. Н. Хрисанфова
97
пеции, широтное развитие сочленовной площадки на этой кости
не уступало таковому киик-кобинца; у женщины Феррасси оно
было несколько меньшим. Сведения о'высоте этой кости отсут-
ствуют.
Несмотря на известную приблизительность приведенных дан- '
ных для других палеоантропов Европы, можно думать, что и у них
имела место тенденция к некоторому увеличению дорзо-волярного
диаметра площадок сустава I лу-
ча (как и у Схул IV), хотя и
менее выраженная, чем у крым-
ского палеоантропа. Отсюда сле-
дует, что можно ожидать прояв-
ления «компенсаций противопо-
ставления» и у этих палеоантро-
пов. Действительно, радиальный
рельеф довольно хорошо развит
на I пястной западноевропей-
ских неандертальцев, здесь же
отчетлива скошенность основа-
ния этой кости (Патт, 1955; Са-
разин, 1931; Бонч-Осмоловский,
1941). У палестинских палеоан-
тропов существовало, по-види-
мому, более умеренное развитие
этих «признаков компенсации»,
Рис. 17. Кисть Схул -IV (по McCown,
Keith, 1939)
а изгиб диафаза I пястной был даже совсем современным. Все
же и среди этих палеоантропов скошенность основания I пястной
была довольно значительной у Табун I и Схул IV.
Анализируя таким образом особенности морфологии сустава
I луча у палеоантропов на фоне групповой и индивидуальной из-
менчивости этих компонентов у современного человека, можно
прийти к выводу, что древние люди не были вполне однородными
в этом отношении, как и по другим структурным особенностям
кисти. По-видимому, нет оснований сомневаться в развитой спо-
собности к оппозиции большого пальца у некоторых палеоантро-
пов типа Схул (рис. 17), особенно Схул V, у которого комплекс
радиализации кисти был выражен в «сапиентной» степени (про-
порции I—1Г пястных, размеры и форма сочленовных площадок
сустава I луча). Иначе обстоит дело у многих классических неан-
дертальцев, и прежде всего у киик-кобинца, у которого площадки
трапеции были исключительно высокими, что, вероятно, обуслов-
лено огромной толщиной всего корня кисти в связи с односторон-
98
ней силовой адаптацией. Возможно, что и при наличии «признаков
компенсации» такая огромная высота уплощенных суставных пло-
щадок могла вызвать некоторые ограничения движений I луча
в сагиттальной плоскости.
Следует особо отметить, что один из важнейших признаков
компенсации — ладонная скошенность основания I.пястной —
обнаруживает на современной кисти отчетливую тенденцию к свя-
зи не с уплощенностью суставных площадок, но с абсолютным и
относительным развитием I луча и особенно большого пальца: от-
клонение возрастает в крайнем укороченном варианте. Возможно,
и у палеоантропов следует рассматривать эту адаптацию более
широко — в зависимости не только от степени седловидности, но
и от общих условий биомеханики сустава, ограничивающих его
подвижность. Тогда становится понятным наличие сильной меди-
альной скошенности основания I пястной у шапелльца и Табун I
с их «недоразвитыми» I лучами, но при формах сустава, по-види-
мому, обеспечивающих в обоих случаях значительную его подвиж-
ность («головчатый тип» у Шапелль и седловидный? у Табун).
Вариации в структуре компонентов сустава I луча, существующие,
как упоминалось выше, и у современного человека, были, вероят-
но, особенно выражены в период становления «сапиентного типа»
кисти: значительную роль играло при этом употребление «безру-
кояточных орудий» (Семенов, 1950). Даже у современного чело-
века структура костных компонентов сустава.1 луча весьма ла-
бильна, в эпоху же формирования кисти, в условиях еще тесной
зависимости ее морфологической эволюции от влияния средовых
факторов, разнообразие структуры кисти отражало неизбежную
закономерность развития.
В морфологии кисти древнего киик-кобинского палеоантропа,
как, по-видимому, и поздних классических неандертальцев, на пе-
редний план выдвигается односторонняя силовая специализация,
вероятно, еще преувеличенная прижизненной рабочей .гипертро-
фией. Сама многочисленность «признаков компенсации» и значи-
тельная степень их выраженности свидетельствует о недостаточ-
ном развитии здесь «точного зажима». Подобный вариант мог
сформироваться под влиянием интенсивной физической нагрузки
в условиях, мало благоприятных для развцтия тонких движений
кисти. Палестинские формы палеоантропов довольно пестры в
морфологическом отношении. Наиболее близкий к современному
вариант отмечается у Схул V; до некоторой степени с ним сбли-
жается и вариант Схул IV с его радиальным типом кисти, хотя
здесь встречаются и признаки иного порядка (относительная уко-
рененность I пястной). В еще большей степени «неандерталоид-
ность» свойственна женской кисти Табун, хотя на ней одновремен-
но отмечаются и уклонения в противоположном направлении, как
отсутствие расширения запястья, стенохейрия, укороченность уль-
нарных пястных костей, седловидность сустава I луча, умеренное
высотное развитие запястья. \
7*
99
К сожалению, нет никаких данных о кисти архантропов, ис-
ключая только поврежденную полулунную кость синантропа, кото-
рая, по-видимому, не представляла существенных уклонений от
человеческого типа (Black, 1933). На основании ее структуры
можно сделать только одно существенное заключение, а именно,
что запястье синантропа имело, скорее, расширенную форму
Рис. 18. Дистальные фаланги большого пальца кисти: А —
Н. habilis; Б — Киик-Коба; В — Крапина; Г — Н. sapiens; Д.—
горилла; Е — шимпанзе, (по Бонч-Осмоловскому, 1941; Napier,
1962а)
(Алексеев, 1960). Реставрированная Вейденрейхом ширина этой
кости превосходит все приведенные им значения соответствующего
размера у современного человека. Примечательно, что даже не-
реставрированная кость синантропа по ширине превосходит мак-
симум европейца (соответственно 13,7 и 13,4 мм). Относительные
широтные размеры этой кости также были велики: широтно-длин-
нотный индекс 99,3 снова выходит за верхнюю границу вариаций
современного человека (69,9—95,5). Правда, нужно иметь в виду
отсутствие полного параллелизма в широтном, как и длиннотном
и высотном, развитии отдельных компонентов'запястья, а также
довольно значительную асимметрию в пропорциях карпальных
костей (см. выше). Все же и с этими оговорками широтное раз-
витие ладьевидной кости синантропа позволяет думать, что его
запястье вряд ли было суженным.
Скудные данные о кисти древнейших людей заставляют отнес-
тись с особым вниманием к недавнему открытию остатков кисти
Н. habilis в Олдувейском ущелье. Среди найденных 15 костей
кисти имеются трапеция, ладьевидная, головчатая кости, основа-
ние II пястной и фаланги. - Несмотря на вероятную принадлеж-
ность этих остатков по крайней мере двум индивидуумам, они име-
ют совершенно исключительное значение, поскольку науке впер-
вые стали известны некоторые черты морфологии кисти возмож-
ных древнейших представителей гоминидной линии эволюции.
Наибольший интерес представляет форма дистальных фаланг
олдувейской кисти, и прежде всего ногтевой фаланги большого
пальца. Она резко отличается от характерной структуры обезь-
яньей кости и является не только типично человеческой, но даже
«ультрагоминидной» (рис. 18). По всем приведенным в табл. 8
индексам Олдувей возглавляет ее, превосходя по расширенности
100
ногтей даже киик-кобинского палеоантропа. Вообще ногтевая
фаланга большого пальца Олдувей представляет как бы пропор-
.ционально уменьшенную модель этой кости у крымского палеоан-
тропа. Это признак высокого таксономического -значения, свиде-
тельствующий, что древнейший олдувейский гоминид уже сделал
шаг вперед в направлении гоминизации кисти и формирования
специфически человеческих широких пальцевых подушечек. Про-
цесс расширения ногтевых фаланг затронул и другие пальцы кисти
Олдувей, хотя и не в такой мере. В этом отношении имеются от-
личия от современного человека или современной обезьяны, у ко-
торых весь ряд дистальных фаланг обнаруживает большой парал-
лелизм в расширенности головок. Так, для трех известных фаланг
Н. habilis (предположительно I—III—IV пальцев) соответствую"
щие цифры широтно-длиннотногб указателя головки составляют:
60,0—39,4—43,2; у человека аналогичные фаланги дают ряд:
41,7—42,7—40,6, а у гориллы: 33,7—35,5—32,1. Правда, принад-
лежность двух других фаланг Олдувей III—IV пальца определена
лишь приблизительно, но это не имеет-особого значения, так как
у человека и обезьяны весь ряд дистальных фаланг показывает
сходные значения индексов. Таким образом, у Н. habilis процесс
расширения ногтевых фаланг в наибольшей степени затронул
большой палец, что свидетельствует, по-видимому, об особой роли
«силового зажима» на этой стадии эволюции кисти. Известно, что
и у современного человека при таком зажиме основная нагрузка
падает на ногтевую фалангу большого пальца, что сопровождается
ее прогрессивным расширением — при условии длительности и
интенсивности действия этого фактора. Одновременно подобная
форма ногтевых фаланг Н. habilis позволяет предположить доста-
точное развитие неврологического кинэстетического аппарата уже
на этой ранней ступени антропогенеза (Нейпир, 1962). Этот автор
считает расширенность дистальных фаланг кисти одним из необ-
ходимых условий развития точного зажима, осуществляемого под
контролем осязательной рецепции.
Менее определенны другие признаки рабочей гипертрофии
кисти Н. habilis и, в частности, относящиеся к структуре запястья.
Все три элемента его были повреждены, но, по косвенным сообра-
жениям, все же можно думать, что запястье олдувейского гоми-
нида не имело выраженной суженности. Об этом свидетельствует
расширенная форма головки головчатой кости и значительная
ширина дистальной площадки трапеции, насколько можно судить
об этих пропорциях по рисункам, приведенным в статье Нейпира
(1962а).
Большой интерес представляют также широтно-длиннотные
пропорции пясти Олдувей, хотя судить о них пока возможно толь-
ко на основании косвенных данных. К-тому же на наиболее пред-
ставительной серии трубчатых костей этой кисти — средних фа-
ланг — еще не завершен процесс синостозирования. Наилучшие.
врзможности дает проксимальная фаланга (по-видимому, II па_льца)
101
взрослого индивидуума, для которой мы высчитали по рисунку
относительную ширину середины диафиза и основания кости к ее
наибольшей длине. Первый из этих индексов равен примерно 29
против 24,3 в среднем у современного человека и 29,7 у киик-ко-
бинца. Это значение свидетельствует о массивности диафиза. Од-
нако по индексу основания кости, который дает более точное
представление о ширине пясти, Н. habilis значительно уступает не
только киик-кобинцу, но и типич-
ному современному варианту (со-
ответствующие значения указате-
ля примерно 30,5—47,7—40,3).
Эти данные позволяют предполо-
жить, что широтное развитие
.Рис. 19. Средние фаланги III пальца
’кисти: А'— Н. habilis-, Б — Н. sapiens;
В — горилла (по Боич-Осмоловскому,
1941; Napier, 1962а)
пясти Олдувей не было очень значительным, хотя все же нужно
иметь в виду, что приведенные для сравнения цифры по современ-
ному человеку относятся .к варианту сравнительно широкой кисти.
Широтные пропорции рассматриваемой фаланги Олдувей напоми-
нают, скорее, соотношения на кисти гориллы (указатель 33,5).
Нужно сказать, что и по форме диафиза наибольшее сближение
существует с этим антропоидом, для которого характерна значи-
тельная расширенность тела проксимальной фаланги, но не ее
сочленовных концов. По-видимому, такие же пропорции были
свойственны и средним фалангам: во всяком случае форма их у
Олдувей не человеческая, снова наиболее напоминающая горил-
лоидную. Также тип рельефа средних фаланг (выраженность и
топография гребней для m. flexor digitorum superficialis) довольно
сходен с «гориллоидным» (рис. 19). Судя по рисунку из статьи
Нейпира (1962а) и описанию костей, существовало дистальное
прикрепление этого мускула. Таким образом, можно предполо-
жить, что кисть Олдувей в целом характеризовалась довольно
умеренным широтным развитием й по ряду структурных особен-
ностей сближалась с гориллоидным вариантом; вместе с тем здесь
возможна тенденция к расширению запястного отдела. Весьма
грацшщное строение обнаруживают и некоторые трубчатые кости
кисти австралопитеков (Брум, Шеперс, 1946; Алексеев, 1960; Day,
Scheuer, 1973).
Очень мало достоверного можно сказать о строении I луча и
его сустава у древнейших гоминид, вероятно, абсолютное и отно-
сительное развитие большого пальца и степень его противопостав-
ляемости на кисти из Олдувей далеко не достигали современного
уровня. Об этом свидетельствует, в частности, положение трапеции
в системе костей запястья, сравнительная короткость дистальной
102
фаланги большого пальца, значительное развитие рельефа в месте
прикрепления длинного сгибателя большого пальца. В косвенной
форме об этом же говорит и наличие сильной ладонной скошен-
ности на I пястной кости австралопитеков!. Вместе с тем, судя по
существующим материалам, кисти австралопитековых и близких
к ним форм (в том числе, Н. habilis) имели седловидный сустав
I луча (Нейпир, 1959; 1962). Размеры I пястной кости австрало-
питека массивного, как и головчатой кости грацильного австра-
лопитека, входили в пределы их вариаций у современных гоминид,
совпадая в обоих случаях с минимумом. По-видимому, пропорции
реконструированного II луча на олдувейской кисти, насколько о
них можно судить по очень фрагментарным материалам, явно не
антропоидные. Напомним также высказывание Нейпира (1962) о
большем, сравнительно с гориллой, относительном развитии I луча
на кисти Олдувей.
В итоге по этим сугубо предварительным данным можно за-
ключить, что кисть древнейшего гоминида еще по многим призна-
кам обезьянья с «горйллоидным» налетом, (форма и изогнутость
фаланг, вероятное полуфлексированное положение пальцев, струк-
тура латеральных гребней на средних фалангах, положение тра-
пеции, седловидность ее дистальной сочленовной площадки). Вмес-
те с тем здесь уже намечен первоначальный сдвиг на пути гоми-
низации, что прежде всего относится к форме, ногтевых фаланг,
совершенно не обезьяньей, но в некоторых 'отношениях даже
«ультрагом'инидной». Весьма вероятно, что на этом раннем этапе
развития человеческой кисти процесс прогрессивной трансформа-
ции ногтевых фаланг- мог стать одним из ведущих факторов го-
минизации и прежде всего радиализации кисти, хотя последняя
на стадии Н. habilis еще не продвинулась достаточно далеко.
Г лава V
СТОПА
СТОПА КАК ИНДИКАТОР БИПЕДИИ
У ГОМИНИД И ИХ ПРЕДШЕСТВЕННИКОВ
Стопа, как и кисть, представляет исключительный интерес для
эволюционной морфологии скелета приматов. Постоянное участие
стопы в опорной функции конечностей определило особое ее зна-
чение для гоминид, у которых именно структура стопы включает-
ся в первую очередь в «комплекс бипедии». По существу ее од-
ной уже достаточно для разграничения ортоградных и проноград-
ных форм приматов и в этом отношении стопу можно сравнить
только с тазовым поясом. Также у приматов в целом, исключая
только специализированных брахиаторов, на строении стопы в
еще большей степени, чем кисти, сказывается специализация к
определенному виду локомоции, главным органом которой явля-
ются задние конечности. При этом хорошее развитие I луча на
стопе, подобно кисти, наблюдается как среди древесных, так и
наземных форм современных приматов. В том и в другом случае
этот признак обусловлен свойственной приматам мультифункцио-
нальностью дистальных отделов конечностей и сложностью их
адаптаций. По-видимому, в филогенезе приматов особая мощность
и подвижность большого пальца стопы определяется сравнитель-
но очень ранним развитием последней, как хватательно-лазатель-
ного органа (Biegert, 1963).
Специфической чертой гоминизации стопы явилось приведение
и дальнейшее усиление I луча («тибиализация» — стопы — по ана-
логии с «радиализацией» кисти), происходившее в связи с раз-
витием прямохождения и перенесением основной нагрузки на ме-
диальный край стопы. Так, индекс соотносительного развития I—
III лучей составляет Хля человека в среднем 98 против 66—70
у африканских антропоидов (Бунак, 1954), хотя в масштабе при-
матов эти последние характеризуются довольно высоким
развитием I луча, сближаясь в этом отношении с древесными ци-
номорфными и лемурами.
Наибольший интерес в аспекте «тибиализации» стопы представ-
ляют соотношения двух ее тибиальных лучей: I и II, как и их ком-
понентов. Индекс соотносительного развития I—II плюсневых ко-
стей отграничивает человека от понгид (соответствующие сред-
ние: 82,3 у человека и 74,7—75,7 у шимпанзе и гориллы), хотя
следует отметить, что индивидуальная изменчивость этих соотно-
шений довольно велика, например минимум — максимум в изу-
ченной нами серии стоп мужчин-европейцев составляет 74,3—96,8.
По-видимому, разграничительное значение, пропорций I—II плюс-
104
невых вообще ниже, чем I—II пястных. Важной особенностью че-
ловеческой стопы является прежде всего, прогрессивное развитие
большого пальца: в отличие от кисти, где он самый короткий, на
стопе большой палец самый мощный и длинный. Характерная для
гоминидного типа стопы длиннотная пальцевая формула: 12345.
Но абсолютное развитие компонентов I луча само по себе имеет
весьма ограниченное значение для таксономического диагноза. На-
против, индекс I—II лучей явственно разграничивает человеческий
и антропоидный типы стопы: он равен в среднем 91,2 у человека
и 67,4—70,8 у африканских антропоморфных. Еще более отчетли-
во дифференцирующее значение индекса I—II пальцев: 103,1 у
человека против 60—67,1 у африканских антропоидов.
Знаменательно, что при межгрупповых сопоставлениях соот-
ношения I—II лучей на стопе современных людей варьируют боль-
ше, чем на кисти. В первую очередь это обусловлено повышен-
ной изменчивостью индекса I—II пальцев при более стабильных
соотношениях I—II плюсневых костей. Также и внутри группы
разница между максимальным и минимальным значениями указа:
теля I—II пальцев примерно вдвое выше на стопе сравнительно
с кистью. Основной причиной этих различий является повышенная
изменчивость указательного пальца на стопе, что понижает и
корреляцию между метаподиальным и фалангеальным компонен-
тами на II луче стопы сравнительно с кистью (коэффициенты кор-
реляции, соответственно, порядка 0,4 и 0,8). При фактическом
совпадении коэффициентов вариации длины больших пальцев ки-
сти и стопы (6,46 и 6,80%) на указательных пальцах разница
весьма существенна (5,32 и 8,73% соответственно).
Таким образом, в отличие от кисти, где I луч в целом варь-
ирует больше И, на стопе дело обстоит как раз наоборот. Вероят-
но, эти различия в какой-то мере опосредованы и разницей в
главной функциональной нагрузке обоих лучей на кисти и стопе
человека. В первом случае слишком «жесткие» корреляции огра-
ничили бы адаптивные возможности I луча, уменьшили необхо-
димую для него пластичность; во втором случае, напротив, опор-
ные функции стопы, связанные по преимуществу с прогрессивным
укреплением именно I луча (и его дистальной части — ногтевой
фаланги), определили его меньшую вариабильность сравнительно
со II лучом, где, возможно, пальцевой отдел частично уже претер-
певал редукцию, сказывающуюся более заметным образом на фи-
булярных пальцах стопы современного человека. Для дополни-
тельного освещения этого вопроса следует рассмотреть некоторые
особенности пальцевых отделов остальных лучей стопы.
Редукция латеральных пальцев стопы касается преимущест-
венно средних фаланг и сказывается как на размерах этих ко-
стей—«редукция длины», так и на их форме, которая становится
бездиафизарной—«редукция формы». При этом может произой-
ти даже слияние средних фаланг с соседними дистальными в. од-
ну кость с образованием бесформенной костной массы. Подобные
105
процессы наиболее выражены на IV и V лучах. Редукция средней
фаланги мизинца встречается в 96—100%, т. е. бездиафизарная
форма этой кости типична для современной _£топы. До-
вольно часто наблюдается и слияние средней и ногтевой фаланг
на мизинце стопы (по данным разных авторов, в 37—80%), при-
чем оно отмечается в самых разных популяциях современного
человека, т. е. является достаточно типичным вариантом. Про-
цесс редукции формы обычно сопровождается также и редукцией
размеров, однако при межгрупповых сопоставлениях полного па-
раллелизма не выявляется, и у европейцев, например, типичная
бездиафизарная форма фаланги сочетается с ее наибольшей в
групповом масштабе длиной *. Подобная «стабилизация» этого при-
знака прослеживается и внутри группы. Так, при слиянии сред-
ней фаланги с дистальной, что иногда рассматривается как даль-
шейшее проявление процессов редукции, оказывается, что измен-
чивость такой слившейся кости меньше, чем для суммарного раз-
мера рех же двух фаланг, но при отсутствии их слияния. Следо-
вательно, несмотря на явно рудиментарный характер средней фа-
ланги мизинца (коэффициент вариации около 22% — почти, как
у XII ребра), при слиянии ее с дистальной фалангой не наблю-
дается типичного для процессов истинной редукции дальнейшего
повышения вариабильности, но, напротив, происходит Хаже ее
значительное снижение сравнительно с тем же суммарным разме-
ром обеих неслившихся фаланг. Частично это можнсг-объяснить
явлениями компенсации прежде всего со стороны дистальной фа-
ланги. Другим проявлением относительной стабилизации мизин-
ца может быть .корреляция с развитием мизинца кисти (коэффи-
циент корреляции длин мизинцев стопы и кисти порядка 0,6).
Сказанное в значительной части применимо и к IV пальцу
стопы, также сильно затронутому редукционными процессами:
бездиафизарная форма средней фаланги этого пальца встречает-
ся в 88—100%. Значительно реже такой вариант отмечен на III
пальце — примерно в 60—70% — и особенно на II — в 20—30%.
Мы видим, что процессы редукции затрагивают все пальцы стопы,
кроме I, хотя и в разной степени; вместе с тем характер измен-
чивости II—V пальцев стопы довольно близок, что, по-видимому,
объясняется влиянием каких-то стабилизирующих механизмов.
Конечно, у современного человека невозможно связывать редук-
ционные процессы на стопе с изменением ее функций или «совер-
шенствованием прямохождения» в духе концепции Бонч-Осмолов-
ского (1943, 1954). Очевидно, что это процесс того же порядка,
что и, например, редукция XII ребер: он характеризуется повыше-
нием индивидуальной изменчивости одного из компонентов при
Стабилизирующем влиянии компенсаторных механизмов на систе-
1 Аналогичная картина наблюдается и на кисти: так, максимум длины для
средней фаланги мизинца снова отмечен для европеоидов, минимум — у
монголоидных групп, где сравнительно нередко (до 21%) встречается даже
брахимезофалангня (Cam et al. 1967).
106
му в целом. С этой точки зрения, тенденцию к слиянию обеих
дистальных фаланг V пальца следует рассматривать, скорее, как
свидетельство укрепления фибулярного края стопы, проявившего-
ся компенсаторно в связи с процессами редукции средних фаланг,
т. е. это явление слияния фаланг может иметь прямое отношение
к механизмам стабилизации мизинца стопы. При этом особо сле-
дует отметить очень раннее проявление этой тенденции в эмбрио-
генезе стопы человека и большую частоту слияния фаланг уже
на ранних стадиях утробного развития (O’Rahilly et al., I960).
Вместе с тем характеристики сравнительной вариабильности и
внутренних корреляций отделов I—V лучей стопы подчеркивают
особое положение I луча как наиболее стабилизировавшегося' из
всех на стопе современного человека (табл. 11 и 12).
Таблица 11 КОЭФФИЦИЕНТЫ ВАРИАЦИИ ДЛИНЫ I—V ПАЛЬЦЕВ СТОПЫ ЧЕЛОВЕКА Таблица 12 ВНУТРЕННИЕ КОРРЕЛЯЦИИ ОТДЕЛОВ (плюсна-палец) I—V ЛУЧЕЙ СТОПЫ ЧЕЛОВЕКА
Пальцы V ± т (у) в % Лучи Г ± т (г)
I 6,80 ±0,68 . I 0,658±0,08
II 8,73±0,87 II 0 г428±0,12
III 8,37±0,84 III 0,454±0,11
IV 10,07±1,01 IV 0,434 ±0,11
V 8,63±0,86 V 0,331±0,13
Особую роль в комплексе бипедии играет структура дисталь-
ной фаланги большого пальца стопы, представляющая признак
первостепенного таксономического значения. Расширение этой ко-
сти должно было сопровождать освоение двуногой шагающей по-
ходки, поскольку в этой области реализуется конечный этап фа-
зы контакта с опорой (субстратом) — «отталкивание». Сравни-
тельные данные по современным антропоидам показывают, что
первоначальная большая расширенность ногтевой фаланги боль-
шого пальца на стопе сравнительно с кистью является весьма
древним признаком. Вместе с тем по относительной расширенно-
сти дистальной фаланги большого пальца стопы африканские ан-
тропоморфные далеко не достигают даже индивидуального мини-
мума этого индекса у современного человека. Следует отметить
также как существенную особенность структуры этой кости у сов-
ременного человека ее уплощенность.
Один из кардинальных признаков гоминидной стопы — приве-
денное положение I луча, которое находится в тесной связи с уси-
лением его массивности, входя в упомянутый выше единый ком-
плекс тибиализации стопы. Однако у негоминидных приматов оба
эти процесса обычно разделены: у большинства из них I луч сто-
пы довольно крупный, хотя и не приведен. Исключением являет-
107
ся до некоторой степени лишь стопа взрослой горной гориллы,
передвигающейся из-за своего огромного веса почти исключитель-
но по земле: здесь большой палец проявляет тенденцию «вырав-
ниваться» по оси остальных (Abel, 1931).
При анализе палеоантропологических материалов в первую
очередь возникает вопрос: по каким признакам можно судить о
положении I луча на остеологических объектах?
Один из наиболее показательных критериев приведенности I
луча — это наличие контактных фасеток между основаниями I и
II плюсневых костей. Одновременно этот же признак позволяет
судить ио сравнительном уровне расположения тарзометатарзаль-
ных суставов обоих тибиальных лучей. Однако, как и большинст-
во других признаков, о которых будет идти речь, он не имеет
«обратного» диагностического значения. Это нужно понимать в
том смысле, что хотя наличие контактных фасеток I—II плюсне-
вых свидетельствует о приведенном положении I луча, их отсутст-
вие не говорит о противоположном. У современного человека кон-
тактные фасетки отсутствуют на каждой второй стопе.
Большой интерес представляют вариации структуры костных
компонентов сустава I луча у гоминид и негоминидных приматов.
Для человека типично отсутствие резкой медиальной скошенно-
сти дистальной сочленовной площадки внутренней клиновидной
кости, имеющейся на стопе негоминидных катарриновых прима-
тов. С этим связано и отсутствие захождения дистальной площад-
ки на внутреннюю поверхность клиновидной кости. Следует при-
бавить также уплощенность самой суставной площадки и ее су-
женную форму, обычно с одной или двумя перетяжками («почко-
видная» или в виде «песочных часов»), близкий к прямому угол
торзиона головки I плюсневой, отсутствие изгиба ее диафиза, как
особенности, характерные для человеческой стопы. Имеются дан-
ные о групповых вариациях этих признаков у современных людей.
Так, медиальная скошенность дистальной сочленовной площадки
внутренней клиновидной кости, по-видимому, более отчетлива у
некоторых популяций, не носящих обуви, с «естественной стопой»,
характеризующейся более веерообразным расположением пальцев
(Мартин, 1928); весьма вероятна в случае «естественной стопы»
также связь с характером передвижения в зависимости от ланд-
шафта (по равнинной или горной местности), что выявляется при
сопоставлении между собой некоторых популяций современных
людей (Das R., Das Bh. М., 1967). Вместе с тем на стопе совре-
менного человека в норме „не осуществляется истинное противопо-
ставление I луча, т. е. движение, когда большой палец держится
косо ниже других. Описанные в литературе факты значительной
подвижности большого пальца касаются преимущественно движе-
ний псевдооппозиции в I метатарзофалангеальном суставе и встре-
чаются в различных популяциях современного человечества. Спо-
собность эта может быть развита упражнением, при этом сущест-
вует мощное развитие абдукторов и аддукторов большого пальца.
108
Особое положение как критерий приведенности I луча всегда
занимал угол шейки таранной кости с осью блока. Однако подоб-
ная интерпретация этого признака требует большой осторожно-
сти из-за его очень высокой вариабильности, наблюдавшейся как
среди гоминид, так и у обезьян и лемуров (Бунак, 1954). То же
можно сказать и о таком косвенном показателе положения I лу-
ча, как развитие шероховатости ладьевидной кости, которое свя-
зывают с формированием в этой области медиального опорного
пункта стопы (Вейденрейх, 1922). У гоминид и других приматов
этот признак варьирует в сильной степени; высокое развитие внут-
реннего края ладьевидной кости среди обезьян встречается, на-
пример, и у преимущественно наземных павианов, и у преимуще-
ственно древесных тонкотелов. У человека этот признак также
широко варьирует, причем не наблюдается его связи со степенью
отодвинутости большого пальца. Структура шероховатости измен-
чива, что вообще характерно для элементов рельефа: по-видимо-
му, в наибольшей степени ее вариации определяются развитием
сесамоида сухожилия заднего большеберцового мускула, играю-
щего роль не только при ходьбе, но и при лазании, чем, вероятно,
и объясняется многообразие и неопределенность вариаций,
В комплекс гоминидной стопы входят также общие длиннот-
ные пропорции ее отделов. Соотношения предплюсна — плюсна —
палец (в % от длины' всей стопы) составляют у современного
человека в среднем 47,9—31,1—20,9 против 40,5—28,8—30,7 у
гориллы. Сравнительно с антропоидным типом стопы здесь на-
блюдается значительное укрепление предплюсны и укорочение
пальцевого отдела. Этот процесс, как уже упоминалось, сопро-
вождался редукционными проявлениями на пальцах стопы чело-
века. При этом произошло также ослабление внутренних связей
обоих компонентов II—V лучей. Характерное для стопы челове-
ка одновременное усиление ее проксимального и среднего отде-
лов определяется опорной функцией, тогда как на кисти запястье
и пясть связаны значительное слабее.
В заключение следует рассмотреть еще один важнейший при-
знак гоминидной стопы — развитие продольного и поперечного
сводов. Наибольшее таксономическое значение при разграниче-
нии гоминидного и антропоидного типов стопы имеет относитель-
ная высота продольного свода; меньший интерес в этом плане
представляет развитие поперечного свода, обнаруживающее час-
тичную трансгрессию с антропоидами особенно по величинам ин-
декса. Из остеологических индикаторов, косвенно указывающих
на выраженность сводов на фрагментарных палеонтологических
материалах, наибольшее' значение могут иметь: подъем пяточной
кости, ориентировка проксимальной сочленовной площадки ку-
бовидной кости, торзион головки таранной кости и связанное с
ним положение опоры таранной кости; для поперечного свода су-
щественно также и положение, и форма оснований плюсневых
костей.
109
СТОПА ИСКОПАЕМЫХ ГОМИНИД
Наиболее полно палеонтологические материалы по стопе, как
и по кисти, представлены от палеоантропов. Это прежде всего
две полные стопы киик-кобинского неандертальца, исключая толь-
ко средние фаланги IV и V пальцев на правой стопе (рис. 20).
Почти целиком сохранились стопы и
у некоторых палестинцев: женщины
Табун I (справа) и мужчины Схул IV
(слева) (рис. 21). Более фрагментар-
ные материалы имеются для детей па-
леоантропов Схул I и Схул VIII, от-
дельными костями представлен ске-
лет стопы Схул V, VI, VII и IX, а так-
же Схул3. Среди западноевропейских
неандертальцев наиболее сохранилась
стопа Феррасси II. Материалы по ос-
тальным классическим формам (Фер-
расси I, Шапелль, Спи, Ла Кина), а
также по Крапине представлены пре-
имущественно элементами предплюс-
ны — таранными и пяточными кос-
тями.
Судить о тотальных размерах сто-
пы палеоантропов довольно трудно,
так как ее остатки большей частью
довольно фрагментарны. Исключение
составляют, как говорилось выше,
Рис. 20. Стопа Киик-Кобы (по Бонч-Осмолов-
скому, 1954)
немногие формы, и прежде всего палестинские гоминиды и киик-ко-
бинский палеоантроп. У миниатюрной женщины Табун I стопа име-
Рис. 21. Стопа Схул IV (по McCown, Keith, 1939)
ет длину 222 мм, что совпадает с индивидуальным минимумом в
нашей мужской европеоидной серии. Другой палестинский па-
леоантроп — мужчина Схул IV — также характеризуется до-
вольно значительным даже при его сравнительной высокоросло-
110
сти, длиннотным развитием стопы: 256 мм против 246,9 мм для
мужчин-европейцев. Киик-кобинец также имел длинную стопу
247—248 мм, что превышает групповой максимум у современного
человека. Индексы длины'стоп (к росту) составляют: 14,6 у Та-
бун I, 15,7 у киик-кобинца и даже 16,5 у Схул IV против 14,9 в
среднем для мужчин-европейцев. В целом относительное длиннот-
•ное развитие стопы палеоантропов, можно оценить как вполне че-
ловеческое, но несколько повышенное сравнительно со средним
современным вариантом, что особенно относится к палестинцу
Схул IV. Напомним, что к аналогичному выводу можно прийти и
при оценке длиннотного развития кисти у этих палеоантропов.
Анализ общих пропорций отделов стопы возможен с известной
долей достоверности у палеоантропов Киик-Кобы, Табун I и Схул
IV. Сопоставляется абсолютное и относительное, в °/о длины всей
стопы, развитие предплюсны, а также плюсневого и пальцевого
компонентов по II лучу1.
Общей чертой для всех рассмотренных палеоантропов можно
счесть значительное развитие плюсны. Что касается предплюсны,
то она несколько укорочена сравнительно со средним современ-
ным вариантом у палестинцев, несколько удлинена у киик-кобин-
ского неандертальца. Здесь, следовательно, существует расхожде-
ние с пропорциями на кисти этих гоминид, где относительное уд-
линение запястья (мезо-макрокарпия) было как раз свойственно
палестинским формам. В генезисе этих различий известную роль
могла сыграть силовая, адаптация руки у «формирующихся лю-
дей», связанная с усилением корня кисти. Пальцевой отдел сто-
пы наиболее удлинен у Схул IV, сравнительно укорочен у киик-
кобинца (табл. 13).
Следует особо отметить сходство в длиннотных пропорциях
стопы между крымским палеоантропом (Киик-Коба) и крымским
кроманьонцем (Мурзак-Коба), хотя в последнем случае все абсо-
лютные размеры меньше. По-видимому, пропорции стопы были
вполне современными и у ребенка палеоантропа Схул VIII.
Почти ничего нельзя сказать о соотношениях отделов на сто-
пе других палеоантропов. У многих классических неандертальцев
отмечается значительное развитие главного компонента предплюс-
ны — таранной кости. Длина этой кости была довольно велика
у Шапелль, Спи II, Кина и, особенно мужчины Феррасси, мень-
ше — у женщины Феррасси, которая сближается в этом отноше-
нии с другой маленькой женщиной палеоантропа — Табун I.
Весьма умеренной й близкой к общечеловеческой средней была
1 В качестве длины..плюсны здесь принимается суставная длина. Ллдюсневой, а
в качестве длины" пальцевого отдела—сумма.длин„трех фаланг II луча..Общая
длина.. jCTonbt .получается. при' сложении . длин трех ее.. отделов. Полученная
таким образом характеристика является в известной мере условной, однако
она дает хорошие возможности для сравнений и более точна, чем оценка, осно-
ванная" на измерении длины II луча на монтированных стопах этих гоминид.
Ill
Таблица 13
ПРОПОРЦИИ ОТДЕЛОВ СТОПЫ У НЕКОТОРЫХ гоминид
(по II лучу)*
Гоминиды Относительная длина (в % длины стопы)
предплюсна плюсна | палец
Современный человек (общая средняя) Мурзак-Коба Киик-Коба Табун I Схул IV 47,9 49,1 49,8 46,2 45,5 31,1 30,2 30,2 33,8 31,6 20,9 20,7 20,1 20,0 22,9
♦ По данным Волкова (Volkov, 1905); Маккоуна и Кизса, 1939; Бонч-Осмо-
ловского, 1954; автора.
длина таранной кости у Крапины. Поскольку между длиннотным
развитием таранной кости и предплюсны в целом существует до-
стоверная связь (коэффициент корреляции порядка 0,6), можно
предполагать, что у большей части классических палеоантропов
Европы развитие всего тарзального отдела также было значи-
тельным. Об этом же свидетельствует и пример киик-кобинца, ко-
торый по морфологии стопы и кисти наиболее сближается с клас-
сическим неандертальским типом. Относительно Шапелль и Фер-
расси это предположение подтверждается и данными о развитии
пяточной кости: ее абсолютная длина близка к средней для евро-
пейцев или даже превосходит ее, как в случае крымского палео-
антропа.
Другая особенность стопы палеоантропов касается формы ее
корневого отдела, о которой обычно судят по морфологии таран-
ной кости. Общим для этих гоминид признаком является значи-
тельное расширение таранной кости: величины широтно-длиннот-
ного указателя ее особенно велики у классических форм, где они
везде намного превосходят современную среднюю (табл. 14). Ис-
ключительно велика расширенность таранной кости у Шапелль и
обоих Феррасси, но и остальные палеоантропы имеют значения
указателя, близкие к современному групповому максимуму или
превышающие его. Средняя для всех палеоантропов 87,0, почти
на 10 единиц индекса превосходит современную общую среднюю
77,3. То же можно сказать и о высотном развитии таранной ко-
сти (соответствующие средние высотно-длиннотного указателя для
палеоантропов и современного человека — 60,0 и 54,1).
В общем совпадающие результаты получены и по высотному
развитию пяточной кости: ее высотно-длиннотный указатель сно-
ва наиболее велик у классических палеоантропов. Рассматривая
другие кости предплюсны, следует обратить особое внимание на
очень большую высоту внутренней клиновидной кости на стопе
112
Таблица 14
ФОРМА ТАРАННОЙ КОСТИ У НЕКОТОРЫХ ГОМИНИД*
Гоминиды Ши ротно-дл иннотный указатель Высотно-длиннот- ный указатель
Шапелль 93,0 61,4
Феррасси I 94,3 61,5
Феррасси II 93,9 59,5
Кина 83,0 57,1
Спи II 87,5 60,3
Крапина 81,5 54,6
Киик-Коба ' 86,3 62,5
Табун I 80,7 (79,5) (60,7)
Схул IV 88,3 (84,8) (58,4)
Схул V 87,2 (84,3) (66,6) '
Схул VI 81,2 (77,5) (56,8)
Схул VII — (77 1?) —
Средняя палеоантропов 87,0 60,0
Кроманьон 82,8 55,6
Мурзак-Коба 72,7 55,2
Шанселад 79,0 59,7 ~
Пржедмости III ' 76,8 58,0
Кап-Блан . . * ....... 81,0 59,3
Вейрье — 52,1
Современный человек средняя 77,3' 54,1
вариации групповые индивидуальные 71,2—84,7 66,7—91,7 48,2—58,8
* По данным Волкова (1905); Маккоуна, Кизса (1939); Патта (1955); Билли (1969); автора. Оба ука-
зателя на основе измерений по Волкову. Для палестинцев в скобках указаны индексы по Мар-
тину (Понятовскому) для широтно-длиннотного индекса; высотно-длиннотный индекс у этих го-
минид дан только по Понятовскому. •
киик-кобийца, выводящую эту кость за пределы современных ва-
риаций. У палестинских палеоантропов этот размер более умерен-
ный.
В целом типичным для палеоантропов можно считать вариант
абсолютно и относительно широкой и высокой таранной кости, что
особенно подчеркнуто у классических форм и, по-видимому, более
или менее совпадало у них с расширением и повышением и всего
корня стопы.
Известное значение как общий для палеоантропов признак
имеет массивность пяточного бугра. Существует значительное раз-
витий пятки в ширину и высоту у палестинцев группы.Схул (эта
особенность специально отмечена Маккоуном и Кизсом даже для
ребенка Схул VIII) и киик-кебинского палеоантропа. Женщина
Табун I имеет более гр ацильную пятку, в чем возможно прояв-
ляется половой диморфизм. У классических .‘неандертальцев раз-
8 Е. Н. Хрисанфова
из
витие пяточного бугра особенно велико. Так, у мужчины Феррасси
высота пятки, измеренная от углубления свободной верхней
поверхности, по Волкову составляет 42 мм, что близко к группо-
вому максимуму у современного человека (размах вариаций 31,4—
43 мм); наименьшая ширина пятки у этого неандертальца — при-
мерно 30 мм — превосходит групповой максимум (размах сред-
них 22,3—29,5 мм по Волкову). Нужно иметь в виду возможную
зависимость этих размеров и от длины всей пяточной кости; так,
у современного человека связь высоты и ширины пятки с длиной
всей пяточной кости реальна и высока (коэффициенты корреляции
соответственно 0,74 и 0,55, Laffont, Sohier, Olivier, 1959/1960).
Большое значение в гоминидном комплексе всегда придава-
лось ориентировке пяточного бугра (большой оси задней поверх-
ности пятки) к оси голени. Маккоун и Кизс особенно подчерки-
вают гоминидность палестинцев в этом отношении, сравнительно
с классическими неандертальцами. Буль привел в своей моногра-
фии рисунок, на котором ось пятки шапелльца занимает промежу-
точное положение между ориентировкой ее у человека и антропо-
идов: она направлена здесь вверх и наружу, а не внутрь, как у
современного человека, в фибулярную, а не тибиальную сторону.
Справедливо указывалось, однако, что Буль преувеличил разли-
чия благодаря неправильной ориентировке голени, исправление
ее увеличивает разрыв между гоминидами (включая неандерталь-
цев) и антропоидами (Вейденрейх, 1922). По-видимому, неандер-
талец не выходит по этому признаку за рамки современных ва-
риаций. Действительно, ориентировка пяточного бугра варьирует
у современного человека, у которого торзион пяточной кости по
сагиттальной оси может быть как положительным, так и отрица-
тельным. У палестинских палеоантропов в ориентировке пяточно-
го бугра также не было сколько-нибудь существенных отличий
от современного человека, что наглядно иллюстрируется соответ-
ствующим рисунком из монографии Маккоуна и Кизса (1939).
То же относится и к киик-кобинцу. Таким образом, можно гово-
рить о близости ^палеоантропов в целом к неоантропу в отноше-
нии рассмотренного признака. Возможно, что существующие как
у ископаемых, так и у современных гоминид вариации можно свя-
зать в некоторых случаях с адаптацией к некоторым статическим
позам (например, «положению на корточках», см. ниже).
Более важна в смысле разграничения с антропоидами ориен-
тировка оси большой берцовой кости, которая у этих обезьян ни-
когда не бывает вертикальной, как ее изобразил Буль на упомя-
нутом выше рисунке, но очень наклонной наружу (Вейденрейх^
1922; Abel, 1931; Патт, 1955).
Отсутствие у палеоантропов принципиальной разницы с чело-
веком в ориентировке стопы по отношению к голени проявляется
и во внутренней структуре костей стопы: для таранной кости за-
падноевропейских неандертальцев это указано Паттом (1955); для
пяточной кости киик-кобинца — Бунаком (1954).
114
- Особьш интерес представляют развитие и пропорции I луча
на стопе палеоантропов. Длина I плюсневой кости варьирует здесь
в довольно широких пределах (табл. 15). Ее развитие в целом
Таблица 15
РАЗВИТИЕ И ПРОПОРЦИИ I-П ПЛЮСНЕВЫХ у гоминид*
Гоминиды Длина I плюсне- вой, мм Длина II плюс- невой, мм Индекс I—II плюсневых
Современный человек, 57,4 69,1 83,0
средняя, вариации** . . 53,0—61,8 63,0—75,7 81,1—84,1
Киик-Коба 61,0/62,0 76,4/77,0 79,8/80,5
Табун I 52,5/51,0 74,0? 70,0?
Схул IV 64,5/— 82,0/83,0 78,7/—
Схул V —/50,0 — —-
Кроманьон 65,0/63,0 —/74,5 —/84,5
Шанселад —/65,0 77,0/74,0 —/87,8
Вейрье —/67,0 74,5/— 89,9?
* По данным Волкова (1905); Мартина (1928); Маккоуна, .Кизса (1939); Бонч-Осмоловского (1954);
Билли (1969); автора.
♦♦Групповые средние.
можно оценить как вполне человеческое, исключая только муж-
чину Схул V, у которого резкое недоразвитие этой кости в длину
можно счесть индивидуальной особенностью. В среднем для па-
леоантропов длина I плюсневой кости равна 57,3 мм. Эта величи-
на совпадает с общегрупповой средней для современного челове-
ка.
Наибольший интерес представляет, однако, относительное раз-
витие I плюсневой кости сравнительно со II плюсневой. Общей и,
очевидно, специфической чертой всех рассмотренных палеоантро-
пов является значительная относительная укороченность I плюс-
невой кости, которая отражена в снижении I—II метатарзально-
го индекса. В первую очередь это обусловлено крупными абсолют-
ными размерами II плюсневой. Напомним, что аналогичное яв-
ление отмечалось и на кисти палеоантропов, которым была свой-
ственна большая длина II пястной кости, снижающая величину
I—II метакарпального указателя.
Своеобразной особенностью длиннотной метатарзальной фор-
мулы палеоантропов является также значительное развитие фи-
булярных ее компонентов (IV—V плюсневых). У двух палестин-
цев (Схул IV и ребенок Схул VIII) отмечен даже такой редкий
вариант формулы, как 524(3)3(4)1, который не типичен ни для
современных людей, ни для антропоидов. У современного челове-
ка и африканских антропоморфных длиннотная метатарзальная
формула в общем идентична: 23451. Интересно, что «вариант
Схул» ни разу не встретился в наблюдавшейся нами серии.
8*
115
Абсолютная длина второго компонента I луча — большого
пальца — может быть вычислена только для двух палеоантро-
пов: киик-кобинца и Схул IV. Оба полученных значения (53,5 мм
для первого и 59 мм для второго) свидетельствуют о вполне че-
ловеческом или даже «ультрагоминидном» его развитии; средняя
для современного человека равна 49,4 мм при групповых вариа-
циях 43,3—56,5 мм. Вероятно, вполне человеческим было разви-
тие большого пальца стопы и у палеоантропов' из Крапины и Ла
Феррасси II; по поводу стопы этой неандерталки имеется указа-
ние Буля (1946) о выступании первого пальца за уровень осталь-’
ных. Открытым остается вопрос о размерах большого пальца на
стопе Табун I и Схул V, где существовала исключительная, уко-
роченность I плюсневой кости. На современной стопе наблюдает-
ся довольно тесная корреляция длин обоих этих компонентов I
луча (порядка 0,7); на этом основании можно предположить не-
которую укороченность и большого пальца у этих двух палестин-
цев. Сказанное относится к абсолютному длиннотному развитию
и всего I луча; хотя последнее само по себе имеет весьма огра-
ниченное значение для таксономического анализа, так как у го-
риллы, например, длина этого луча даже превосходит среднюю
общечеловеческую.
Что касается пропорций I—II лучей и их компонентов, то уже
указывалось на крупные размеры II плюсневой и всего II луча у
некоторых палеоантропов, что понижает значения соответствую-,
щих индексов. Однако во всех случаях, где пропорции обоих лу-
чей могут быть определены с известной долей достоверности (Ки-
ик-Коба, Схул IV), они оказываются 'вполне человеческими.
Мы упоминали выше о значительном развитии фибулярных
метатарзальных компонентов на стопе палеоантропов. Также и
тотальные размеры IV—V лучей были велики, что объясняется
кроме упомянутого обстоятельства значительным в современном
масштабе развитием средних фаланг этих лучей (долихомезофа-
лангия). С эволюционно-морфологической точки зрения представ-
ляет большой интерес сочетание крупной средней фаланги на
стопе киик-кобинца с ее бёздиафизарной формой; «редукция фор-
мы» средней фаланги мизинца существовала, следовательно, уже
.в глубокой древности, что находится в соответствии с исключи-
тельной медлейностью редукционных процессов, следующей и Из
других палеонтологических фактов (Северцов, 1939).
Сказанное применимо и к IV пальцу: известные для палеоан-
тропов размеры средней фаланги этого пальца снова относятся
к варианту долихомезофаланги и (Киик-Коба, Схул IV), хотя тип
фаланги здесь является уже диафизарным. Интересно, что у кро-
маньонки Мурзак-Коба, обнаруживающей большое сходство с ки-
ик-кобинцем по общим пропорциям стопы, средняя фаланга IV
пальца много короче. На III пальце стопы палеоантропов длина
средней фаланги также была очень велика; в меньшей степени
это применимо ко II пальцу, хотя индивидуально здесь также мо-
116
гут встретиться очень большие размеры средней фаланги, напри-
мер, у Схул IV (рис. 22). Отметим, что у палеоантропов и на ки-
сти обычно очень велика средняя фаланга мизинца: у Схул IV
она превосходит по длине современный групповой максимум, а
у киик-кобинца близка к нему.
Приведенность I луча была выражена у древних людей во
вполне человеческой степени. У некоторых .из них существовали
контактные фасетки I—II плюсневых, что на-
блюдается далеко не на каждой современной
стопе. Они отмечены у палеоантропов группы CQ
Схул—Схул IV и Схул3. Также и дистальная Ш
площадка внутренней клиновидной кости по /Д W
положению и форме была вполне человеческой CTQ гМ
у большинства палеоантропов. Некоторые |,)1 С”/
Рис. 22. Фаланги III пальца стопы. А — Схул IV;
Б — Н. sapiens (по McCown, Keith, 1939) А Б
отклонения отмечаются у Табун I, где дистальная площадка нем-
ного заходит на медиальную сторону. Вместе с тем положение са-
мой внутренней клиновидной кости у этой женщины палеоантропа
человеческое, полностью адаптированное к бипедии. Что касается
указания Маккоуна и Кизса на сравнительно большое расстояние
между I и II плюсневыми костями, то оно варьирует и у современ-
ного человека, где, по-видимому, намечается отрицательная связь
с длиной I луча: короткий луч отклонен в среднем сильнее, чем
длинный. Вполне современна по форме и положению дистальная
сочленовная площадка на внутренней клиновидной кости Схул IV,
имеющая почковидную форму. То же относится и к детским ске-
летам Схул I и Схул VIII. У киик-кобинца скошенность этой пло-
щадки даже меньше, а ее уплощенность сильнее, чем в привлекав-
шейся для сравнения европейской серии. Также и у 'Феррасси II
тип внутренней клиновидной кости вполне человеческий; на этой
стопе in situ (частично в породе) не наблюдалось отодвинутости
большого пальца или I плюсневой кости от остального скелета
стопы.
Наконец, для палеоантропов характерно некоторое понижение
сводов стопы, хотя их выраженность находится вполне в преде-
лах вариаций этих признаков у современного человека. Непосред-
ственное определение наиболее важного в таксономическом аспек-
те продольного свода возможно у немногих формГ Только у киик-
кобинского палеоантропа произведены измерения абсолютного и
относительного развития сводов на монтированных стопах (Бонч-
Осмоловский, 1954; Бунак, 1954). К сожалению, эти материалы
не сопоставимы с данными по другим палеоантропам ввиду пол-
ного отсутствия таковых в литературе. Существуют, однако, точ-
117
ные рисунки с реконструированных стоп Табун I и Схул IV, как
и киик-кобинской стопы, на основе которых можно получить хотя
и условные, но достаточно сопоставимые данные об относительном
развитии продольного свода. Индекс высоты свода у этих палео-
антропов находится вполне в пределах его вариаций у современ-
ного человека, у которого наблюдается значительная межгруппо-
вая и внутригрупповая изменчивость этого признака. Так, индекс
относительной высоты продольного свода, определенный по сопо-
ставимым рисункам для представителей различных вариантов сто-
пы современного человека, варьирует в пределах 18—24,5. а у па-
леоантропов — от 20,8 до 25,6. Это, конечно, выборочные и при-
близительные цифры, но они вполне соответствуют предположе-
нию о значительной вариабильности этого признака как среди
современных, так и у древних людей. Бонч-Осмоловский отмечал^
правда, некоторое понижение общей высоты стопы и высоты внут-
ренней лодыжки и таранно-ладьевидного сочленения у киик-ко-
бинца, сравнительно с современным человеком. Однако это ут-
верждение справедливо лишь для сопоставлений с вариантами
стопы с относительно высоким сводом; в общечеловеческом мас-
штабе все эти признаки не обнаруживают у киик-кобинского па-
леоантропа сколько-нибудь существенных отклонений. Вполне че-
ловеческим является и такой важный в функциональном отноше-
шении признак, как наклон (подъем) пяточной кости над плос-
костью опоры. Человеческим было, как уже упоминалось, и поло-
жение оси пяточного бугра, что свидетельствует о выпрямленном
пронированном положении всей пяточной кости (Бонч-Осмолов-
ский, 1954; Бунак, 1954).
По-видимому, и развитие поперечного свода у палеоантропов
Схул (Схул IV, Схул VIII), Табун I и Киик-Кобы находилось в
пределах человеческих вариаций.
Значительно труднее судить о развитии сводов на стопе запад-
ноевропейских неандертальцев ввиду фрагментарности материа-
лов и отсутствия строгих остеологических индикаторов этого при-
знака. Представление о структуре сводов на стопе этих ископае-
мых гоминид основывается преимущественно на некоторых мор-
фологических особенностях таранной и пяточной костей, от кото-
рых имеются наиболее полные остатки.
Но ни высотное развитие таранной кости и ее блока, ни струк-
тура шейки таранной кости не являются адекватными показате-
лями развития сводов, хотя подобная точка зрения и отражена в
литературе. Большее значение как показатель сводчатости стопы
может иметь, как уже указывалось, торзион головки таранной ко-
сти, который обнаруживает определенную тенденцию к параллель-
ной соизменчивости с развитием свода на стопе современного че-
ловека. При этом нужно иметь в виду и его значительную инди-
видуальную изменчивость. Среди палеоантропов угол торзиона
наименьший у некоторых классических форм и у Табун I (28—
ЗГ). У киик-кобинца и Ла Кина он уже близок к общечеловече-
118
ской средней (36,8°) и особенно велик у некоторых представите-
лей группы Схул (37—40°).
Другой критерий выраженности свода пытались видеть в раз-
витии и положении опоры таранной кости. Действительно, разме-
ры этого отростка были' велики у палеоантропов, однако при уче-
те вообще больших размеров таранной кости они оказываются в
этом относительном выражении вполне человеческими. Также и
столь важный в оценке постановки стопы ^признак, как угол от-
клонения задней суставной площадки пяточной кости, отчетливо
разграничивающий гоминидный и антропоидный типы стопы, со-
вершенно человеческий у киик-кобинца и палестинских палеоан-
тропов (Бонч-Осмоловский, 1954; Маккоун, Кизс, 1939). По-види-
мому, и Шапелль находится здесь в пределах современных вариа-
ций (Вейденрейх, 1922).
Можно думать, что в целом развитие сводов было вполне че-
ловеческим, соответствующим двуногой локомоции у палеоантро-
пов, хотя они и не принадлежали к гоминидам с высокими сво-
дами стопы. Как и у современных людей, здесь могли существо-
вать вариации, и западноевропейские неандертальцы, возможно,
характеризовались несколько более пониженными сводами. Более
точная оценка развития сводчатости стопы палеоантропов затруд-
нительна из-за отсутствия достаточных материалов, известной про-
тиворечивости различных показателей, а также ввиду существо-
вания значительных вариаций у современного человека. Послед-
ние затрагивают различные аспекты морфологической изменчиво-
сти: возрастной, половой, индивидуальный, групповой. Сюда же
следует отнести и «функциональные вариации», играющие нема-
ловажную роль в изменчивости сводов у современного человека
и несомненно имевшие еще большее значение у ископаемых гоми-
нид. Известно, что понижение сводов стопы (ее «уплощение») у
человека связывается прежде всего с повышением нагрузки и сла-
бостью мускульно-связочного аппарата. В крайнем варианте это
приводит к возникновению плоскостопия, обязанного у современ-
ных людей преимущественно статическому фактору. Известное
значение в этиологии плоскостопия имеют и различные гормо-
нальные, ферментативные и другие обменные факторы, которые
могут существенно влиять на вязкость полимеризованных муко-
полисахаридов, являющихся основой фиброзного аппарата стопы,
как и на степень минерализованности ее костей, что наиболее яс-
но выступает в субпатологических и патологических вариантах.
С этой точки зрения достаточно объяснимо существование вари-
антов стопы с пониженными сводами среди палеоантропов, ске-
лет которых испытывал значительную статодинамическую нагруз-
ку начиная с детства, когда он особенно чувствителен к подоб-
ным воздействиям. Логично и предположение о несколько большем
понижении сводов стопы у классических неандертальцев, где си-
ловая адаптация скелета выражена в наибольшей степени.
Следы функциональных адаптаций вообще широко прослежи-
119
ваются на скелете палеоантропов. Помимо указанных структурных
особенностей, связанных с уплощением стопы, укреплением и рас-
ширением ее корня, повышением массивности всех трубчатых ко-
стей, представляют большой интерес проявления структурного ком-
плекса «положения на корточках», особенно затрагивающие об-
ласть голеностопного сустава. Выше уже упоминалось, что для
палеоантропов типична крупная таранная кость с укороченной и
сильно отведенной шейкой. Угол шейки таранной кости характе-
ризуется довольно высокими значениями, обычно превосходящи-
ми средний современный вариант, хотя изменчивость этого при-
знака как среди древних, так и современных людей достаточно
велика. Вместе с тем в условиях сформировавшейся бипедии ко-
лебания этого угла не могут иметь прямого отношения к основ-
ной локомоции. Сомнительна также его связь с дивергенцией и
подвижностью большого пальца, как это неоднократно отмеча-
лось применительно к современному человеку, у которого очень
велики также и внутрипопуляционные вариации угла.
Выше уже неоднократно упоминалось о проявлениях адапта-
ций к гиперфлексии коленного и голеностопного суставов у древ-
них и современных людей, которые связаны предположительно с
привычным употреблением некоторых статических «поз покоя».
В частности, речь шла о структурных адаптациях к «комплексу
положения на корточках», получившему в современной морфоло-
гической литературе достаточно широкое освещение (Charles,
1893; Sewell, 1904/1906; Singh, 1959; До Суан Хоп, 1964; Тгап —
Anh, Le Ngoc — Huong, 1966; Rao Prasada, 1966).
Признаки этого комплекса наблюдаются и на скелете стопы
древних людей и проявляются прежде всего в значительном раз-
витии блока, распространении его вперед и формировании особых
сочленовных фасеток. Иногда аналогичные фасетки, ^особенно с
латеральной стороны, встречаются и на нижнем конце большой
берцовой кости. Барнет (Barnett, 1954) предложил разграничивать
латеральное и медиальное распространения блока от случаев фор-
мирования отграниченных «истинных» фасеток с латеральной или
значительно реже медиальной стороны; в случае истинной фасетки
часто существует соответствующая фасетка и на нижнем конце
большой берцовой кости. Однако этот критерий «истинности» фа-
сетки далеко не всегда себя оправдывает, так как совместное су-
ществование обеих сочленовных фасеток «положения на корточ-
ках» на большой берцовой и таранной кости встречается гораздо
реже, ч'ем этого следовало бы ожидать теоретически. Это можно
отнести за счет многообразия эндо- и экзогенных факторов, уча-
ствующих в адаптивных перестройках голеностопного сустава или
влияющих вообще на его морфологию. По-видимому, различные'
фасетки возникают прежде всего в результате сочетания инвер-
сии стопы со значительной дорзальной флексией, причем фасетки
«положения на корточках» есть лишь часть всего комплекса рас-
пространения блока (Morimoto, 1960).
120
На всех таранных костях неандертальцев, изученных Булем.,
существует сверхчисленная фасетка, расположенная на шейке по
продолжению латеральной оси блока. Из описания не вполне яс-
но, идет ли речь об истинной фасетке или латеральном продол-
жении блока у разных форм. «Истинная» (в смысле Барнета) ла-
теральная фасетка «положения на корточках» найдена у Фер-
расси II, где ей соответствует фасетка и на нижнем конце боль-
шой берцовой кости. У киик-кобинца добавочные фасетки описа-
ны на обоих таранных костях, причем па правой, кости существу-
ет «истинная» фасетка «положения на корточках», которой соот-
ветствует фасетка на правой большой берцовой кости. На этой
кости выражены и другие признаки комплекса. У палестинцев,
как и у неандертальцев Западной Европы, почти вся поверхность
шейки таранной кости контактировала с большой берцовой
костью, так что трудно отграничить здесь передний край блока.
У ребенка Схул VIII есть контактная фасетка на большой берцо-
вой кости, на таранной кости также существует распространение
трохлеарной поверхности на шейку. Неоднократно подобные при-
знаки отмечались и у Н. sapiens jossilis (Ferembach, 1964/1965)..
Приведенные данные по палеоантропам свидетельствуют также
о единообразии морфологических проявлений адаптации голено-
стопного сустава к гиперфлексии, о необходимости принимать во
внимание все структурные детали, связанные с возникновением
добавочных контактных поверхностей. Не вызывает сомнения су-
ществование у палеоантропов комплекса определенных структур-
ных особенностей в области голеностопного сустава, обусловлен-
ных адаптацией к статическим позам, связанным с гиперфлекси-
ей голеностопного сустава. Эти черты нередко выступают вместе
с более значительной дивергенцией шейки таранной кости. Одно-
временно блок оказывается относительно более длинным, а го-
ловка и шейка — относительно укороченными, короткая шейка,
по-видимому, сильнее отклонена. Хотя точная функциональная ин-
терпретация признаков этого комплекса не всегда легка, весьма
вероятна его связь с привычным длительным применением стати-
ческих «поз покоя» (в том числе положения на корточках).
Перечисленные признаки строения стопы палеоантропов наи-
более отчетливо прослеживаются у классического варианта, с ко-
торым, как и пр кисти, сближается киик-кобинский неандерталец.
Большой интерес представляет и параллелизм в структуре кисти
и стопы, отчетливо выступающий у классических палеоантропов.
В обоих случаях отмечается довольно сильное развитие «корня»,
удлиненный средний и умеренно развитый пальцевой отделы, зна-
чительное широтное и высотное развитие проксимальных отделов,
исключительная расширенность дистальной фаланги большого
пальца, огромная высота площадок сустава I луча, укороченность
I плюсневой (пястной) кости относительно очень крупной II, до-
лихомезофалангия и т. д. Палестинские палеоантропы по многим
признакам стопы, как и кисти, обнаруживают полиморфизм, об-
121
условленный у них значительной вариабильностыо бозгптАГ
перечисленных признаков. ' зшеи части
При интерпретации признаков, отмеченных на стопах- -п
тропов, следует иметь в виду частные адаптивные компчклЛ0311’
занные с действием повышенной стато-динамической iiarnv^ ’ СВЯ'
особенности, опосредованные корреляциями со стопою Т. ’ ИЛИ
До недавнего времени ничего не кисти.
было известно о стопе более ранних
гоминид. Уникальной в этом отно-
шении явилась находка сравнитель-
но полного скелета левой стопы
взрослого индивидуума в Восточной
Африке (Олдувей 8). Здесь отсут-
ствуют только фаланги и головки
плюсневых костей, повреждена зад-
няя часть пяточной кости (рис. 23).
Пока существуют лишь предвари-
тельные публикации об олдувейской
стопе (Дэй, Нейпир, 1964, 1966;
Нейпир, 1967; Day, Wood, 1968, 1969;
Окснард, 1972), но уже их доста-
точно, чтобы оценить эти остат-
ки, как представляющие исключи-
тельную ценность для суждения о
более ранних этапах эволюции сто-
пы человека, о которых у мор-
Рис. 23. Стопа Н. habilis (по Day Napier
1&64) ’
фологов прежде было довольно отвлеченное представление.
Большой интерес представляют пропорции олдувейской стопы,
хотя мы располагаем здесь реконструкцией (Coon, 1963), па ко-
торой возможно определить лишь тарзо-метатарзальные соотно-
шения. Соответствующий индекс (по II лучу) равен примерно
156 — значение, весьма близкое к средней для современных лю-
дей (158,6 при вариациях групп 152,2—163,5 и индивидуальных
вариациях 142,4—178,6). Совершенно очевидна близость Олдувей
в этих соотношениях к гоминидам в целом и отчетливое разгра-
ничение с типами как антропоидной стопы (указатели 78,5—
140,8), так и с типом специализированной наземной стопы павиана
с индексом 118 или более нейтральным вариантом стопы макаки
с индексом 102. Из приведенных данных видно, что индекс тарзо-
метатарзальных соотношений является довольно показательным в
таксономическом отношении, что позволяет сблизить Олдувей с
гоминидами, несмотря на приближенное определение его величи-
ны в этом случае.
122
Прилагая метод, предложенный Шульцем (1963) для разгра-
ничения «силового» (постталярного) и «грузового» плеч рычага,
с минимальной реконструкцией к стопе Олдувей,- Дэй и Нейпир
(Day, Napier, 1964) получили здесь весьма сходные с человече-
скими внутренние пропорции «трузо'вого плеча». Это плечо вклю-
чает часть таранной кости, расположенную впереди от ее центра,
и плюсневый отдел. У человека на каждую из этих частей прихо-
дится соответственно 47 и 53%, у гориллы — 41 и 59%, т. е. у
нее существует более короткая тарзальная и более длинная ме-
татарзальная части. На олдувейской стопе пропорции промежу-
точные — 45,5 и 54,5%, но значительно ближе к типичным чело;
веческим, т. е. локомоторные адаптации в распределении сил тя-
жести и элевации стопы весьма сходны здесь с человеческими.
Форма «корневого отдела» стопы Н. habilis также может быть
оценена только приблизительно. Более определенное суждение
основывается на морфологии таранной кости; вычисление ее ши-
ротно-длиннотного указателя позволяет дать некоторые сопостав-
ления с другими гоминидами — древними и современными. Дэй
и Нейпир (1964) сообщают по этому поводу, что широтно-длин-
нотные пропорции таранной кости здесь близки к человеческим:
хотя сама кость в морфологическом отношении не напоминает
близко какой-нибудь определенный гоминидный вариант, многие
ее признаки находятся в пределах нормальной индивидуальной
изменчивости этого типа. В последующих работах (Day, Wood,
1968, 1969) своеобразие таранной кости Н. habilis (как и австра-
лопитека массивного из Кромдраай) было подтверждено данными
мультивариационного анализа.
Что касается общих широтно-длиннотных пропорций плюсне-
вого отдела стопы Олдувей, то их определение на основе рекон-
струкции недостаточно достоверно и сильно зависит от способа
монтировки: так, на рисунках, приведенных Куном (Coon, 1963)
и Нейпиром (соответственно, с реконструкцией и без реконструк-
ции), вычисление широтно-длиннотного указателя плюсны приво-
дит к очень разным результатам. В первом случае индекс почти
такой же, как на очень расширенной плюсне киик-кобинского па-
леоантропа, а во втором — соответствует пропорциям на относи-
тельно суженном плюсневом отделе стопы палестинских палео-
антропов. Вычисление указателей массивности для плюсневых
костей в отдельности свидетельствует, скорее, в пользу представ-
ления о сравнительном расширении стопу Олдувей. Так, напри-
мер, вычисленный по реконструированной длине II плюсне-
вой 1 индекс ширины основания этой кости дает величину 23,4,
т. е. совпадает с соответствующим указателем на стопе киик-ко-
бинца. Для сравнения приведем значения этого индекса на стопе
современного человека (европейца): 21,8—21,9 и палестинских па-
1 Реконструкция на основе вычисленного нами в современной серии индекса
длины этой кости без головки в % от ее сочленовной длины. .
123
леоантропов 18,0—18,2. Отсюда можно как-будто сделать вывод
о соответствии в распределении гоминид по широтному индексу
основания II плюсневой и общему широтно-длиннотному указате-
лю плюсны, а следовательно, о достаточной пригодности первого
индекса для суждения о широтном развитии плюсны на стопе Ол-
дувей. Высокая его величина у этого гоминида, как уже гово-
рилось, ‘ скорее, свидетельствует о расширенности плюсневого
отдела.
Однако окончательное решение вопроса невозможно до появ-
ления более подробных публикаций, так как многое здесь зависит
от характера самих тарзо-метатарзальных сочленений *.
По поводу других близких к Н. habilis форм упомянем мнение
Брума и Шеперса (1946), что стопа австралопитековых была, ско-
рее, суженной, хотя материалы, на основе которых сделано это
заключение, нельзя признать достаточными. По-видимому, вполне
человеческий тип имела таранная кость раннеплейстоценового
гоминида из Кении — район оз. Рудольф (Wood, 1974).
Наибольший интерес наряду с обсужденными выше пропор-
циями двух проксимальных отделов стопы представляет развитие
и положение I луча на олдувейской стопе. •
I плюсневая кость этого гоминида производит впечатление
очень массивной и крупной. Все величины индексов ее относитель-
ного развития к остальным плюсневым костям выходят даже за
верхнюю границу их вариаций у гоминид — современных и иско-
паемых. Но если и не опираться на эти цифры, как недостаточно
надежные (реконструкции), все же очень большое абсолютное и
относительное развитие I плюсневой на стопе Олдувей очевидно.
Даже значения индексов соотносительного, развития, вычисленных
без реконструкции длин I, II и III плюсневых, вполне человече-
ские, что особенно существенно, так как на I плюсневой, по-види-
мому, отсутствует пропорционально большая часть, чем на II или
III. Длиннотная метатарзальная формула Н. habilis (по рекон-
струкции) также совершенно человеческая: 23 415 и даже с «про-
грессивной» тенденцией в виде преобладания I плюсневой в
длине над V; подобный вариант был встречен в нашей серии
лишь в 2% 1 2-
Большое внимание привлекла широтная метатарзальная фор-
мула Олдувей 8. Согласно Дэю и Нейпиру (1964), III плюсневая
выделяется здесь и по длине, и по ширине, что рассматривается
или как индивидуальная вариация, или как возможное проявление
«незавершившейся» еще перестройки плюсны в связи с прямо-
1 Так, у киик-кобинца почти совпадают индексы формы плюсны, вычисленные
по суммарной ширине оснований I—V плюсневых, изолированных и на сочле-
ненной стопе, тогда как у современного человека разница между этими ин-
дексами довольно значительна.
2 Следует иметь в виду консервативность этой формулы у Hominoidea.
124
рождением. По данным этих авторов, «формула массивности»
1люсны у Олдувей 15 342 против 15 432 у современного человека
I 12345 у горной и береговой гориллы. Основное различие между
Элдувей и человеком Дэй и Нейпир видят, таким образом, в со-
отношениях III и IV плюсневых. Однако при специальном иссле-
ювании вариаций «формулы массивности» у современного человека
и антропоидов оказалось, что «типичная формула» человека
(15 432) была найдена у него лишь в 56%, а-в 44% было обна-
ружено семь различных вариантов формулы массивности, причем
в 6% она соответствовала таковой олдувейского гоминида. Ни у
одного антропоида ни разу не было встречено вариантов, наблю-
давшихся у человека или олдувейского гоминида. Отсюда выте-
кает вывод о значительной индивидуальной изменчивости «фор-
мулы массивности» у человека и отсутствии принципиальных раз-
личий по этому признаку с Н. habilis (Archibald et al., 1972).
К этому можно добавить, что при использовании для построе-
ния широтной метатарзальной формулы абсолютных, а не отно-
сительных размеров диафиза Олдувей 8 снова показывает вполне
человеческий вариант (152=34),близкий к формулекиик-кобинца.
Вообще у современного человека и по длине, и по ширине встреча-
ются оба типа соотношений III—IV плюсневых: 34 и 43; таким
образом, пропорции этих двух костей у Н. habilis не представляют
ничего исключительного. Вариант 34 по ширине диафиза особенно
характерен для палеоантропов, где он является основным. Если же
производить сопоставления по широтному развитию головки или
основания плюсневых костей, то вариант 34 вообще является всю-
ду преобладающим.
Следовательно, нельзя согласиться с выводом, что в соотноше-
нии III и IV плюсневых костей на стопе Олдувей можно видеть
незавершенность адаптаций к прямохождению. Широтная мета-
тарзальная формула в отличие от длиннотной значительно варьи-
рует у современных и ископаемых гоминид, как и высших прима-
тов в целом. Даже у одного и того же индивидуума не совпадают
широтные формулы, построенные на основе развития диафиза и
обоих сочленовных концов плюсневых костей.
Принимая во внимание значительную корреляцию между
компонентами первого луча стопы, можно предположить хорошее
развитие большого пальца на олдувейской стопе. Об этом же сви?
детельствует в косвенной форме и морфология дистальной фаланги
большого пальца стопы Олдувей 10 с отчетливыми признаками
гоминидного комплекса (рис. 24). Эта кость имеет у олдувейского
гоминида очень расширенную «ультрагоминидную» форму; вели-
чина широтно-длйннотного указателя головки здесь ‘примерно на
17 единиц превосходит среднюю для мужчин-европейцев (табл. 1-6).
Существует аналогия с «ультрагоминидной» расширенностью фа-
ланги этого пальца и на кисти Н. habilis (Олдувей 7), хотя рас-
сматриваемая фаланга стопы, по-видимому, не имеет прямой связи
с другими уже известными остатками из области FLKJ-
125
.Как один из своеобразных признаков этой фаланги, напоминаю-
щих ее структуру у киик-кобинца, отмечается преобладание шири-
Рис. 24. Дистальные фаланги боль-
шого пальца стопы: А — Олдувей 10;
Б — Киик-Коба; В — Схул IV; Г —
Н. sapiens; Д — шимпанзе (по Бонч-
Осмоловскому, 1954; McCown, Keith,
1939; Napier, 1967)
ны основания над длиной
Таблица 16
ИНДЕКСЫ ШИРОТНОГО РАЗВИТИЯ ГОЛОВКИ
НОГТЕВОЙ ФАЛАНГИ I ЛУЧА НА СТОПЕ
НЕКОТОРЫХ ГОМИНИД И ОБЕЗЬЯН*
Приматы Индекс ширины го- ловки (в % от наи- большей длины
Современный человек . .
Min—Мах. ............
Киик-Коба............
Крапина..............
Схул IV .......
Мурзак-Коба . . ’ . .
Олдувей 10...........
Шимпанзе.............
Горилла .............
54,5
45,1—80,9
68,9
57,0?
44,0?
49,8
72,1
38,1
38,1
кости: для современного че-
ловека такие пропорции фа-
ланги не характерны, а у
антропоидов относительное
сужение основания фаланги
еще сильнее, чем у человека.
С точки зрения выражен-
* По данным Маккоуна, Кизса (1939); Бонч-Осмолов-
ского (1954); Дэя, Нейпира (1966); автора.
ности комплекса бипедии
весьма существенно также
указание Дэя и Нейпира
(1966) о наличии на дис-
тальной фаланге Олдувей
10 комбинации латерального
торзиона (примерно +13°
против +16—17° у совре-
менного человека) и. валь-
гусного отклонения головки
(+2°), что, несомненно, сви-
детельствует о принадлежности этой кости двуногому гоминиду с
плантиградной шагающей походкой. По поводу вальгусного откло-
нения, впрочем, следует заметить, что если бы даже наклон был
здесь варусным, это не могло бы рассматриваться как решающий
аргумент против признания шагающей походки у Олдувей 10. Как
показало исследование Дьюднея (Dewdney, 1966), соотношение
варусного и вальгусного наклонов в популяции здоровых людей,
никогда не носивших обуви, равно 95:111, т. е. в «естественном
варианте» стопы оба типа распределяются почти поровну.
Следует также отметить, что олдувейская фаланга была плос-
кой, как у современного человека. Напомним, что характерная
уплощенность была свойственна этой кости и у киик-кобинского'
палеоантропа. Принимая во внимание очень крупные размеры и
значительную массивность I плюсневой на стопе Олдувей 8, мож-
но думать, что и она обладала расширенной ногтевой фалангой
большого .пальца.
Очень большой интерес представляет вопрос о положении I лу-
ча на стопе Н. habilis. Прежде всего следует подчеркнуть, что здесь
уже существуют контактные фасетки между основаниями I—II
плюсневых — признак, с определенностью свидетельствующий о
приведенном положении I луча. Вместе с тем высказывалось мне-
ние, что большой палец на олдувейской стопе был отведен в боль-
126
шей степени, чем у человека (Lewis, 1972). Основанием для такого
предположения служит прежде всего характер сочленовных по-
верхностей I тарзо-метатарзального сустава: выпукло-вогнутая
форма дистальной сочленовной площадки на внутренней клиновид-
ной кости. Подобный признак иногда интерпретируется как прими-
тивно-гоминоидный, сближающий эту структуру с «гориллоид-
ной». Вместе с тем нельзя не учитывать того обстоятельства, что
морфология этой области достаточно варьирует и у современного
человека, и у антропоидов (Бонч-Осмоловский, 1954). Да и вообще
структура I тарзо-метатарзального сустава весьма вариабильна у
современного человека, как это следует уже из факта отсутствия
на каждой второй стопе контактных фасеток I—II плюсневых
(Grosshans, Meyer, 1963). С этой точки зрения, наличие комбина-
ции «.примитивной» формы дистальной сочленовной площадки
клиновидной кости и «ультрагоминидного» варианта контактных
фасеток I—II плюсневых не вызывает особого удивления. Другое
заслуживающее внимания обстоятельство усматривают в несколь-
ко повышенном сравнительно с вариантом стопы современного'
человека значении угла шейки таранной кости на олдувейской
стопе. Следует, однако, напомнить, что угол этот не имеет четкой
функциональной связи со степенью отодвинутости большого паль-
ца, как об этом упоминалось выше. Кроме того, величина угла у
Олдувей совпадает со средней для палеоантропов (23°). -В извест-
ной степени сказанное распространяется и на таранную кость мас-
сивного австралопитека (парантропа), где также существовало,
повышенное сравнительно с типичным современным вариантом
значение этого угла (26°), хотя и входящее в пределы его вариа-
ций у Н. sapiens.
Относительно другого важнейшего признака прямохождения —
развития сводов — существует прямое указание о достаточной
его выраженности на олдувейской стопе (Дэй, Нецпир, .1964). По
мнению этих авторов, морфология костей дистального ряда пред-
плюсны свидетельствует и о значительной динамической поддерж-
ке поперечного свода стопы: хорошее развитие рельефа в местах
прикрепления метатарзальных связок и mm. tibialis anterior et pe-
roneus (fibularis) longus. В этом плане существенны и данные о
более высоком, чем у массивного австралопитека, развитии тор-
• зиона головки таранной кости на олдувейской стопе. Инклинация
sustentaculum tali к горизонтальной плоскости, резко разграничи-
вающая гоминидную и антропоидную стопы, в последнем случае
она много больше была мала у Н. habilis. Дэй и Нейпир пишут
о почти горизонтальном положении апофиза на пяточной кости
этого гоминида; у современного человека соответствующий угол
•равен в среднем 5° (Бунак, 1954). Таким образом, все’остеологи-
ческие индикаторы, по которым можно составить какое-то сужде-
ние о развитии сводчатости на стопе Олдувей 8, свидетельствуют
как-будто в пользу достаточной выраженности этого важнейшего,
признака гоминидного комплекса.
127
В целом предварительные результаты сравнительного морфо-
логического изучения стопы олдувейского гоминида говорят о су-
ществовании основных черт комплекса бипедии уже на этой очень
древней стадии эволюции стопы. Сюда относятся: мощное разви-
тие и приведенное положение I луча, пропорции I плюсневой, осо-
бая массивность краевых плюсневых, длиннбтная и широтная ме-
татарзальные формулы, тарзо-метатарзальные соотношения, внут-
ренние пропорции «грузового рычага» стопы, выраженность сводов.
Многие из этих признаков были развиты у Н. habilis даже в «уль-
трагоминидной» степени; не исключается полностью и возможность
такой специфически человеческой черты, как уникальная шагаю-
щая походка (Archibald et al., 1972), о чем в косвенной форме
свидетельствует структура дистальной фаланги большого пальца
Олдувей 10. Конечно, эта фаланга не имеет прямого отношения к
стопе Олдувей 8, принадлежа к более молодой геологической фор-
мации местонахождения FLKJ, однако, ее возраст тоже достаточно
древний — около 1 млн. лет; в одном слое с ней были найдены
олдувейские «чопперы». Вместе с тем, судя по некоторым данным,
среди древнейших гоминид могли существовать варианты в разви-
тии структурных адаптаций к бипедии (стопа массивного австра-
лопитека с возможно несколько повышенной подвижностью I луча
и несколько пониженными сводами; костные компоненты голено-
стопного и особенно коленного' суставов на берцовых костях из
«слоя зинджантропа»; фрагменты бедренных костей массивных
австралопитековых Восточной и Южной Африки).
ВОПРОС О ТИПЕ КИСТИ И СТОПЫ АНТРОПОИДНОГО
ПРЕДШЕСТВЕННИКА ГОМИНИД
Палеонтология высших приматов в настоящее время еще не
может с уверенностью ответить на вопрос о том, каков был тип
кисти прямого антропоидного предшественника гоминид. Решение
этой проблемы связано с целым рядом трудностей, всегда возни-
кающих там, где нет истинных филетических линий, и речь должна
идти об их реконструкции на основе различных данных косвенного
характера, нередко к тому же йротиворечащих друг дру1?у. Доста-
точно напомнить, что в качестве модельных для общего предка
•гоминид и понгид указывались самые разнообразные способы ло-
комоции и соответствующие им типы строения кисти и стопы, на-
чиная от специализированной наземной и кончая специализирован-
ной древесной.
Правда, палеонтологические открытия более позднего'времени
значительно поколебали перву!о из этих концепций, развивавшуюся
Осборном (Osborn, 1927) и особенно Бонч-Осмоловским (1941).
Представление о «лапообразной кисти» общего предка человека
и понгид,. соответствующей его наземной четвероногой локомоции,
представляется неубедительным в свете новых палеонтологических
фактов, свидетельствующих об отсутствии каких бы то ни было сле-
128
дов «лапообразности» у высших приматов миоцена и плиоцена, как
и у вероятных древнейших представителей собственно гоминидной
линии эволюции (например, Олдувей). Нельзя согласиться и с ар-
гументами, приводившимися Бонч-Осмоловским из области онто-
гении кисти (Якимов, 1949; Семенов, 1950; Алексеев, 1960), тем
более, что эмбриологические факты, относящиеся к очень ранним
этапам эмбриогенеза, обычно вообще не могут использоваться при
реконструировании облика непосредственного предка. Бонч-Осмо-
ловский принял здесь за исходные некоторые структурные особен-
ности киик-кобинской кисти, которые вовсе не воспроизводили тип
кисти древнейших прамлекопитающих, как он думал, но, напротив,
носили сугубо вторичный характер и были обусловлены резко вы-
раженной степенью «рабочей гипертрофии».
. Значительно больший интерес представляет гипотеза «нейтраль-
ного», генерализованного, типа кисти антропоидного предка, кото-
рый соответствовал его полуназемной — полудревесной локомоций;
многие исследователи видят возможную модель такого рода в кис-
ти миоценового антропоида из Восточной Африки — проконсула
(НеЬегег, 1956, 1960; Ulrich, 1955; Бунак, 1954; Майр, 1968; и др.).
Анализ некоторых особенностей кисти проконсула в общем
согласуется с таким -предположением. Относительная длина рекон-
струированной кисти проконсула в % от длины, всей верхней конеч-
ности составляла примерно 25,5, что соответствует средней для
современного человека и значительно отличается от соответствую-
щего индекса у современных антропоидов даже с нерезко выра-
женной брахиаторнрй специализацией. То же относится и к разви-
тию корня кисти. Индекс длины головчатой кости (в % к длине
III пястной) составляет у проконсула около 33 (по реставрации),
т. е. помещает его в ранг типичных человеческих соотношений.
Разница с современными антропоидами (указатели 25,8—27,9 -у
понгид) достаточно четкая.
Особый интерес представляет анализ пропорций пясти и паль-
цев (по III лучу) на кисти проконсула. Вполне достоверен здесь
только индекс развития средней фаланги (48,0), сближающий про-
консула одновременно с человеком (45,2), шимпанзе и гориллой
(49,8—47,8) с древесными циноморфными (мангобей с указателем
45,9 _и тонкотел с указателем 51,0). Определение тотальных про-
порций III- луча возможно в данном случае только по реконструк-
ции и потому не вполне достоверно. Ясно все же, что по предполо-
жительной величине соответствующего индекса проконсул (150)
снова обнаруживает определенную близость с мангобеем (144,8),
тонкотелом (156,6), а также с человеком (136,8—158,5) и отчасти
с гориллой (141,4). Такой тип пропорций пясти и пальца вполне
мог быть исходным в развитии гоминидной кисти.
Широтное развитие запястья снова обнаруживает у проконсула
известную гёнерализованность строения в смысле возможного
сближения его по этому признаку как с антропоидами, так и с
гоминидами; так, в последнем случае существует фактическое сов-
9 Е. Н. Хрисанфова
129
падение величины индекса общей формы запястья, вычисленного
для проконсула по реконструкции, с типичной для современного
человека. Сравнительно-анатомические параллели свидетельствуют
также, что пропорции кисти проконсула вполне могли существо-
вать и у раннего предкового типа (лазание с обхватыванием), что
подтверждается, например, при изучении кисти нотарктуса.
Наибольший интерес представляет абсолютное и относительное
развитие I луча на кисти проконсула. К сожалению, от него сохра-
нилась только проксимальная фаланга; естественно поэтому, что
представляющиеся наиболее существенными пропорции I пястной
и всего I луча могут быть оценены здесь сугубо предположительно.
Косвенные расчеты длины I пястной (на основе пропорций I луча
у наиболее близких к проконсулу таксонов современных обезьян)
свидетельствуют в пользу довольно хорошего развития большого
пальца и всего I луча, даже если допустить, что здесь имело место
некоторое «отставание» развития пястного компонента сравнитель-
но с пальцевым, как, по-видимому, и на других лучах кисти про-
консула. Действительно, проксимальная фаланга большого пальца
проконсула была очень крупной в масштабе размеров остальных
трубчатых костей кисти; ее длина (18 мм) совпадает с максималь-
ной у современных мартышкообразных обезьян (вариации
12,6—18,1 мм, по данным Даниловой, 1966), хотя далеко не дости-
гает величин этого признака у современных понгид (22,4—26,8 мм).
Учитывая к тому же сравнительно весьма умеренную длину
II пястной кости, можно думать, что относительное развитие I лу-
ча было довольно значительным.
Следовательно, в случае проконсула речь идет о кисти антро-
поида с умеренно выраженной тенденцией к флексии пальцев и
общими и внутренними пропорциями нейтрального циноморфного
типа, с хорошим развитием корня и большого пальца. Вместе с
тем это, безусловно, кисть лазающего примата, хотя и без следов
узкой древесной специализации. Следует особо подчеркнуть, что
подобные признаки проконсула нужно рассматривать, скорее всего,
как «модельные». Возможно, что общий предок гоминид и афри-
канских понгид уже должен был обладать такими свойственными
обеим этим группам чертами, как отсутствие самостоятельной os
centrale, уменьшение гороховидной кости запястья, седловидный
сустав и т. д., ибо возникновение их путем параллельной эволюции
все же менее вероятно. Напомним, что пока отсутствуют прямые
данные о локомоции рамапитека, признаваемого сейчас многими
за более вероятного предка гоминидной линии (Pilbeam, 1967,
1972) ’. Однако сказанное не умаляет роли этих палеонтологиче-
ских данных. Даже если подтвердится предположение о двуногой
1 В этой связи большой интерес представляет анализ двух таранных костей, при-
писываемых близкой к рамапитеку форме — Kenyapithecus africanus, который
показывает наибольшее их сходство с соответствующими костями не двуногих,
но четвероногих приматов «циноморфного типа» (Wood, 1973).
130.
локомоции рамапитека, вопрос о локомоции на предшествующей
стадии эволюции не снимается, но только переносится. Нет сомне-
ний, что достаточно нейтральный тип локомоции проконсула был
очень распространен в миоцене, когда, по мнению большинства
исследователей, произошло разделение гоминидной и понгидной
линий. Об этом свидетельствуют находки многих высших приматов
миоцена, и прежде всего плиопитеков и лимнопитеков. По целому
ряду соображений подобный тип кисти или близкий к нему вполне
мог быть исходным для развития этих.двух ветвей высших при-
матов По-видимому, то немногое, что известно о структуре стопы
проконсула, также не противоречит такому предположению. Сюда
следует отнести, например, высокое положение таранной кости на
пяточной, способность к прочной фиксации предплюсны в выпрям-
ленном положении и некоторые другие черты (Бунак, 1954). Сто-
па, как и кисть, проконсула принадлежала лазающему примату
с довольно значительным отведением большого пальца.
Менее вероятной представляется пока концепция, связывающая
тип кисти антропоидного предка гоминид с брахиаторной специа-
лизацией и сближающая его с африканскими антропоидами по
способу локомоции (Remane, 1965; Tuttle, 1969), причем крайние
сторонники «брахиаторной гипотезы» признают и вытекающее
отсюда «хождение на суставах пальцев» — «knuckle—walking» —
предка человека (Washburn, 1967; Marzke, 1971).
Как уже указывалось, крюковидная кисть брахиаторов харак-
теризуется целым комплексом специфических адаптаций, затраги-
вающих как ее скелет (например, пропорции, редукция запястья,
относительное недоразвитие I луча, структурные модификации в
области луче-запястного и пястно-фаланговых суставов и т. д.), так
и мускульно-связочный аппарат (например, укороченность сухожи-
лий флексоров пальцев и прежде всего длинного сгибателя боль-
шого пальца, общее ослабление мускулатуры большого пальца
и т. д.). Сравнительно-анатомический анализ кисти гоминид сви-
детельствует об отсутствии в ее морфологии сколько-нибудь от-
четливых следов переживания брахиаторных функций (Данилова,
1965, 1968). Применительно к стопе функциональный анализ
также показывает, что исходным типом формирования этого орга-
на у человека не могла быть стопа с определенно выраженным
приспособлением к брахиации (Бунак, 1954; Kalin, 1964). «Релик-
товые признаки» брахиаторной специализации в строении кисти,
на которые указывают некоторые авторы (например, Marzke 1971)1 2,
или не могут с уверенностью рассматриваться как реликты бра-
1 Признается'и возможность родственных связей между проконсулом и кениапи-
теком (Aguirre, 1972).
2 Нельзя не согласиться с мнением этой исследовательницы, что для аргумента-
ции брахиаторной гипотезы можно привлекать только «реликтовые признаки»,
для которых невозможно объяснение с точки зрения параллельной функцио-
нальной адаптации.
9*
131
хиаторной адаптации (например, слияние os centrale, которое сре-
ди брахиаторов типично только для африканских форм, а среди
остальных приматов встречается спорадически без отчетливой
связи с локомоцией), или же ассоциируются не столько с брахиа-
цией, как таковой, сколько с «хождением на флексированных ।
кистях» (глубокая поперечная метакарпальная связка, апоневроз i
mm. contrahentes). >
Против теории происхождения гоминид от специализированных |
брахиаторов свидетельствуют также общие'соображения о времени ।
разделения понгидной и гоминидной линий эволюции и данные па- i
деонтологии приматов. Большинство исследователей считает, что 1
адаптация к брахиаторной локомоции у предков современных ан-
тропоидов развилась сравнительно поздно — не раньше плиоцена. i
Настоящий адаптивный комплекс брахиации встречается, по-види- j
мому, только на рубеже миоцена и плиоцена (ореопитек), но даже ,
если бы возраст этой формы был более древним, в ней трудно ви- |
деть возможную модель антропоидного предшественника гоминид; !
«элементы прямохождения», которые находили у ореопитека, так- I
же значительно преувеличены, во всяком случае, применительно к
тазовому поясу и стопе, т. е. двум важнейшим индикаторам бипе- I
дни. Мы видели выше, что таз ореопитека имел вполне понгидный [
облик. Что касается стопы, то она также сохраняет в общем пите-
коидный тип по таким важным признакам, как морфология сустава
I луча и таранной кости, свидетельствующая об абдуцированном
большом пальце и значительной подвижности в голеностопном
суставе: заметное медиальное отклонение шейки таранной кости,
относительно большая ширина sustentaculum tali, выпуклость ди-
стальной сочленовной площадки внутренней клиновидной кости,
слабый торзион головки таранной кости, ее узкость, низкое поло- ;
жение и т. д. Одна из немногих особенностей пяточной кости орео- ,
питека, которая оценивалась как специфически гоминидная, — раз- ;
витие пятки — при более детальном рассмотрении также оказы- 1
вается не имеющей особого диагностического значения. Так, Стра- ।
ус (1963) приводит для индекса относительного развития пятки j
в % длины всей кости величину 40, действительно входящую в
ранг современных-человеческих вариаций, но при сравнении с ан-
тропоидами оказывается, что указатели у шимпанзе и оранга не-
сколько ниже, а у гориллы — значительно выше, чем у ореопитека
(соответствующие цифры: 35,3; 36,7 и 51,7).
В основе филетических линий находятся обычно малоспециа-
лизированные формы, и хотя обратное развитие отдельных призна- i
ков возможно, ретроспективная эволюция целого комплекса все же ’
представляется маловероятной. Напомним, что такое обратное I
•развитие должны быть испытать не тЬлько особенности кисти, но i
и ряд других брахиаторных адаптаций, достаточно широко пред- ’
ставленных во многих отделах скелета. Таким образом, по боль-
шому числу важнейших структурных особенностей предки человека
должы были испытать вначале брахиаторную специализацию, а
132
затем «вернуться» к более генерализованному типу кисти и других
отделов скелета, сближающему их в большей мере с циноморф-
ными обезьянами. Перечислим только некоторые из этих особенно-
стей: общие пропорции кисти (в % от всей верхней конечности),
развитие I луча и большого пальца, размеры локтевого отростка,
положение сочленовной впадины лопатки по отноше^ю к лате-
ральному краю, торзион ключицы, торзион бедренной и большой
берцовой костей, относительная длина подвздошной кости (в %’
длины туловища), кортико-медуллярные соотношения на плечевой—
бедренной костях (см. ниже). Все эти признаки сближают чело-
века с типом проноградного циноморфного примата в большей
степени, чем со специализированным антропоидом; обратное раз-
витие некоторых из них в более генерализованном направлении
хотя и не может быть исключено, но остается малопонятным с
точки зрения функционального параллелизма, поскольку, напри-
мер, функция проксимальных суставов верхней конечности в боль-
шей мере сближает человека с антропоморфными, чем с циноморф-
ными приматами, и такая вторичная конвергенция в данном слу-
чае не была бы .опосредована функциональной адаптацией.
Что касается концепции о «неспециализированной» («примитив-
ной», «факультативной») брахиации предка человека или «полу-
брахиации» (Нэйпир, 1967), то здесь трудно увидеть принципиаль-
ное противоречие с разбиравшейся выше гипотезой генерализован-
ного понгидного предка. Расширительное толкование термина
«брахиация», включающее в себя и «элементы брахиации», под
Которым подразумевается способность к известной свободе
движений плечевого и локтевого суставов, в том числе и к подве-
шиванию туловища, позволяет распространить ее на многих обезь-
ян, включая и таких миоценовых антропоидов, как проконсул
(Morbeck, 1975).
Более ранняя фаза в эволюции кисти — нижнетретичных пред-
шественников миоценового понгидного предка человека — связы-
вается некоторыми исследователями (Страус, 1962; Нэйпир, 1967)
с гипотетическими «вертикально цепляющимся» эоценовым лему-
роидным приматом. Подобный тип локомоции наряду с прыжками
был главным у приматов, живших примерно 50 млн. лет назад.
Его несомненным преимуществом была выраженная адаптация к
выпрямленному положению туловища, которая является одной из
характерных особенностей приматного типа и должна была су-
ществовать также у проконсула. По Биджерту (Biegert, 1963),
наиболее ранний этап эволюции кисти может быть связан с «суб-
приматным» генерализованным типом кисти — стопы тупаий.
Вопрос о способе локомоции и конкретной структуре дисталь-
ных отделов конечностей антропоидного предка гоминид не решен
окончательно, хотя наиболее распространено мнение о генерали-
зованном типе полуназемного — полудревесного антропоида с эле-
ментами свободы движений в проксимальных суставах верхней
конечности.
133
Глава VI
ВНУТРЕННЯЯ СТРУКТУРА
СКЕЛЕТА И ЕЕ РОЛЬ
В ЭВОЛЮЦИОННОЙ МОРФОЛОГИИ
ВНУТРЕННЕЕ СТРОЕНИЕ
ТРУБЧАТЫХ КОСТЕЙ ЧЕЛОВЕКА
СРАВНИТЕЛЬНО С ДРУГИМИ
СОВРЕМЕННЫМИ ПРИМАТАМИ
В морфологии человека внутренняя структура скелета затронута
в более слабой степени, чем внешняя; между тем во многих отно-
шениях она представляет не меньший интерес для проблемы его
эволюции. Особенно это относится к длинным костям нижней ко-
нечности — в первую очередь к бедренной кости, на примере ко-
торой- мы и рассмотрйм значение внутренней структуры в эволю-
ционной морфологии гоминид и других приматов.
Особый интерес в аспекте разграничения двуногих гоминид от
других приматов представляет структура верхнего эпифиза бедрен-
ной кости1. Архитектоника спонгиозы верхнего конца бедра соот-
ветствует оптимальному распределению веса при переносе центра
тяжести на бедренные кости (Lodge, 1963). Напомним, что у че-
ловека здесь различают три системы траекторий трабекул об-
ласти шейки и головки бедра, причем система I, восходящая от
так называемой «дуги Адамса», к эпифизу, связывается с давле-
нием, тогда как II и III системы условно принимаются за траек-
тории тяги.
Для человека, как прямоходящего существа, типична значи-
тельно более четкая дифференцировка костных пластинок, форми-
рующих определенные системы, чем для обезьян. Даже у антро-
поидов они развиты намного слабее, что особенно относится ко
II системе, радиирующей от медиальной стенки шейки к большому
вертелу. У орангутана и гориллы в этой области найдены только
очень немногочисленные тонкие и неправильные пластинки; в не-
сколько большем числе встречаются они у шимпанзе, но и здесь
ни по форме, ни по концентрации не напоминают человеческого
типа. Система III в наибольшей степени развита у гориллы, наи-
менее — у оранга. Среди ископаемых антропоидов рейнский пайдо-
питек, как й современный гиббон, отличается еще менее отчетли-
вой структурой спонгиозы, чем крупные антропоиды (Бунак, 1956).
У низших обезьян тензионных трабекул, по-видимому, не сущест-
вует или они выражены очень слабо (Ujwal, 1962). Поэтому тра-
1 Спонгиоза наиболее активно вовлекается *в процесс адаптивных перестроек в
. филогенезе, как и в онтогенезе, что соответствует ее большому участию в мета-
болизме кости, сравнительно с компактом (Машкара, 1970).
134
бекулярная структура верхнего эпифиза бедра может рассматри-
ваться как таксономический признак большого веса — в плане
разграничения двуногих и проноградных форм приматов.
Используя этот критерий в таксономическом анализе, нужно
иметь в виду возможные вариации структуры спонгиозы в зависи-
мости от возраста, пола, прижизненных функциональных адапта-
ций, например рабочей гипертрофии. Так, у человека период отно-
сительной стабилизации структуры верхнего эпифиза бедра охва-
тывает только возраст 25—35 лет; уже в 40—45 лёт может сти-
раться рисунок трабекул нижней поверхности’большого вертела и
центра шейки, т. е. II и III систем, — процесс, становящийся осо-
бенно интенсивным после 50 лет и более отчетливый у женщин,
чем у мужчин. Нужно учитывать и нормальную индивидуальную
вариабильность, в детерминации которой значительную роль играет
функциональный фактор. Одним из проявлений функциональной
гипертрофии костей является крупноячеистый тип губчатого веще-
ства (Привес, 1967). С этой точки зрения, представляет интерес
возможная связь вариаций трабекулярной структуры с развитием
и наклоном шейки бедренной кости у человека (Арипов, 1956).
Следует учитывать и патологию: например, при артритических за-
болеваниях тазобедренного сустава отмечены не только «грубость»
трабекул, но и нарушения в их расположении (Pietrogrande, 1953).
В последнее время немалое внимание уделяется также рентге-
номорфологическому изучению с самых различных сторон корти-
ко-медуллярных соотношений диафиза. В интересующем нас аспек-
те эволюционной морфологии следует в первую очередь оценить
эти пропорции в сравнительно-анатомическом ряду приматов
(табл. 17).
Мы изучили рентгенологически серию длинных костей некото-
рых приматов, в которую вошли представители полуобезьян и
обезьян — широконосых и узконосых, среди последних — мартыш-
кообразные и человекообразные приматы, включая человека. На
боковых и передне-задних рентгенограммах бедренных костей на
уровне середины диафиза-определялись сагиттальный и попереч-
ный медуллярные указатели. Сагиттальный медуллярный указа-
тель у изученных приматов колеблется в пределах: 40,7 орангу-
тан — 66,0 коата. Человек с указателем 49,8 занимает близкое к
экстремальному положение в этом ряду, уступая только орангу и
превосходя всех других изученных приматов.
Для поперечного медуллярного указателя соответствующие
колебания средних составляют: 41,0 орангутан — 67,0 тонкотел и
даже 72,2 мартышка. Человек (указатель 42,4) снова находится
почти-на краю этого ряда. Напомним, что подобное направление
различий, по" крайней мере у антропоморфных, выявилось уже в
исследованиях Вейденрейха (1941), вычислившего поперечные ме-
дуллярные указатели на нескольких экземплярах бедренных костей
сиаманга, шимпанзе и гориллы (65—57,7), которые в этом отно-
шении заметно отличались от оранга (41,7).
135
Таблица 17
МЕДУЛЛЯРНЫЕ УКАЗАТЕЛИ БЕДРЕННОЙ КОСТИ ПРИМАТОВ*
Приматы Медуллярные указатели
сагиттальный поперечный общий
Лемур 52,4 59,2 55,8
Капуцин . 58,9 63,6 61,3
Коата 66,0 62,6 64,3
Шерстистая обезьяна . . 64,7 59,1 61,9
Мартышка \ 65,6 72,2 68,9
Тонкотел 64,2 67,0 65,6
Мангобей . 59,5 53,0 56,3
Макака 54,1 61,9 58,0 -
Павиан - 52,3 52,3 52,3
Ге лада 54,5 57,0 55,8
Сиаманг . — 65,0** —
Орангутан 40,7 41,0 40,9
Шимпанзе 57,3 53,1 55,2
Горилла — 57,7** —.
Человек*** 49,8 42,4 46,1
* Сагиттальный и поперечный медуллярные указатели — процентное отно-
шение сагиттального (поперечного) диаметра медуллярной полости к со-
ответствующему диаметру диафиза на уровне его середины. Общий ме-
дуллярный указатель — средняя из сагиттального и поперечного индек-
сов.
**По Вейденрейху (1941).
*** Данные, полученные по измерению 100 рентгенограмм (50 боковых и 50
передне-задних) бедренных костей мужчин; изученная серия включала
2 и 3 возрастные стадии по Немешкерн и др. (Nemesk£ri et al., 1960).
Приведенные цифры относятся к середине диафиза. Форма ме-
дуллярной полости довольно изменчива как во внутри-, так и в
межгрупповом масштабе, и наибольшее сужение ее может распо-
лагаться на разных уровнях. У человека наибольшее сужение по-
лости чаще локализуется, по-видимому, в верхней половине диа-
физа, несколько выше его середины, тогда как у шимпанзе указа-
тель мало варьирует на протяжении диафиза (Вейденрейх, 1941).
Следовательно, нужно иметь в виду, что приведенные значения
медуллярного указателя могут и не отражать во всех случаях наи-
меньшее развитие полости. Однако метод определения кортико-
медуллярных пропорций на уровне середины диафиза был более
пригодным по ряду соображений, и прежде всего, поскольку нашей
основной целью было получение сопоставимых данных для палеон-
тологических материалов, обычно представленных весьма фраг-
ментарно.
В итоге анализа приведенных результатов можно прийти к за-
ключению, что в масштабе приматов гоминидам, по-видимому,
свойственна сравнительно толстостенная бедренная кость. Вместе
с тем существует тесная связь между компактизацией бедренной
кости и Гомологичного сегмента верхней конечности (табл. 18).
На плечевых костях изученных приматов сагиттальный медул-
лярный указатель дает вариации: от 50,7 павиан до 71,5 мартышка;
136
Таблица 18
МЕДУЛЛЯРНЫЕ УКАЗАТЕЛИ ПЛЕЧЕВОЙ КОСТИ У-ПРИМАТОВ
Приматы Медуллярные указатели
сагиттальный поперечный общий
Лемур 54,8 63,8 59,3
Капуцин 65,5 61,2 63,4
Коата 70,2 67,5 68,9
Шерстистая обезьяна . . 61,0 57,3 59,2
Мартышка 71,5 65,7 68,6
Тонкотел 67,4 67,0 67,2
Мангобей 52,0 59,2 55,6
Макака 57,0 64,3 60,7
Павиан " 50,7 52,4 51,6
Гелада 57,0 55,5 56,3
Орангутан 55,2. 48,7 ' 52,0
Шимпанзе 68,5 57,2 62,9
Человек* 47,0 46,6 46,8
* Данные, полученные по измерению 50 боковых и 50 передне-задних рент-
генограмм плечевой кости мужчин.
поперечный: от 48,7 орангутан до 67—67,5 тонкотел, коата. Соот-
ветствующие значения индекса для человека — 47,0 и' 46,6. В дан-
ном случае даже в большей степени, чем для бедренной кости,
снова выявляется экстремальное положение человека в ряду при-
матов.
Обращает на себя внимание фактическое совпадение общих ме-
дуллярных указателей на бедренной и плечевой костях человека
(соответствующие цифры — 46,1 и 46,8). Это ассоциируется с су-
ществованием весьма тесной связи абсолютного развития компакты
обеих этих костей; коэффициент корреляции порядка 0,8 у чело-
века. Подобную же близость, гомологичных костей верхней и ниж-
ней конечности по степени кортикализации показывает большин-
ство проноградных приматов, исключая только специализирован-
ных брахиаторов (оранг, шимпанзе, коата). (Можно предположить,
что известный параллелизм в развитии компакты длинных костей
обеих конечностей является характерной чертой проноградных
четвероногих приматов без резко выраженной специализации. При-
мечательно, что «разрыв» в относительной компактизации плечевой
и бедренной костей, существующий у всех рассмотренных брахиа-
торов, в наибольшей мере выражен у самого специализированного
из них — орангутана. .
На основании приведенных данных можно прийти к заключе-
нию и о достаточно «крепкой» структуре длинных костей гоминид
в масштабе приматов. По степени компактизации плечевой и бед-
ренной костей человек занимает экстремальное положение, усту-
пая в последнем -случае только орангу. Значительная «внутрен-
няя массивность» этих костей сочетается у человека с определен-
137
ными особенностями их остеонной структуры. На большом сравни-
тельном материале (млекопитающие разных отрядов) В. В. Бунак
и Е. А. Клебанова (1960/1961) показали, что по структуре ком-
пакты человек наиболее сходен с приматами. Вместе с тем для
него характерно такое соотношение всех структурных компонентов
компакты трубчатой кости, которое обеспечивает наибольшую ее
прочность при сравнительно небольших размерах. Если сопоста-
вить остеонную структуру на длинных костях человека и наиболее
распространенного среди высших приматов варианта четвероногой
проноградной обезьяны с умеренным развитием массы тела, то
сказанное выше можно иллюстрировать следующим примером.
Мы изучали параллельно микроструктуру плечевой кости у че-
ловека и мартышкообразной обезьяны (зеленой мартышки). Ока-
залось, что у человека существует гораздо более редкая сеть тол-
стостенных остеонов, тогда как у обезьяны она много гуще и пред-
ставлена мелкими, сравнительно тонкостенными системами. Так.
например, наружный диаметр остеона составляет для человека
и мартышки в среднем соответственно 208 и 109,5 мк; толщина
стенок остеона у первого 139,7, у второй 80,6 мк. Применительно
к человеку можно говорить, следовательно, о прогрессивном раз-
витии остеонных структур в направлении прежде всего укрепле-
ния стенок остеонных систем и увеличения их общих размеров.
Сравнительно-анатомические параллели в общем, скорее, согла-
суются со взглядом на крупный редкий тип остеонов как «силь-
ный» в биомеханическом отношении (Evans, 1958), хотя в суще-
ствующей литературе по этому вопросу и нет полного единства
мнений. По-видимому, намечается тенденция к обратной зависи-
мости между густотой расположения и размерами остеонных систем,
отмеченная у приматов и некоторых млекопитающих (Джигора,
1957). Подобная связь наблюдается и в индивидуальном масштабе:
для плечевых и бедренных костей человека она реальна и имеет
отрицательный знак с коэффициентом корреляции порядка 0,5.
К числу разграничительных признаков,, которые представляются
нам заслуживающими внимания, следует отнести и сравнительное
развитие компакты в различных участках диафиза. Мы рассмотрим
в этом плане только одну структурную особенность — распреде-
ление кортикальной кости на уровне середины диафиза бедра.
Общее расположение кортикальной кости в области середины
диафиза снова сходно у человека и четвероногого проноградного
примата (табл. 19): самая толстая стенка в обоих случаях задняя,
самая тонкая — передняя, латеральная и медиальная компакта
мало различаются по толщине. Близость этих форм по топографии
компакты подчеркивается на фоне соответствующих данных для
четвероногого млекопитающего с разнообразной моторикой, но не-
лриматного типа (крысы), у которой распределение кортикальной
кости существенно отличается.
Вместе с тем при всем общем сходстве в строении диафиза че-
ловека и мартышкообразной обезьяны между ними выявляются и
138
Таблица 19
СРАВНИТЕЛЬНОЕ РАЗВИТИЕ ДИАФИЗАРНЫХ СТЕНОК
БЕДРЕННОЙ КОСТИ У ЧЕЛОВЕКА, МАРТЫШКООБРАЗНОЙ
ОБЕЗЬЯНЫ И КРЫСЫ
Форма Толщина компакты, мм
п передняя задняя наружная внутренняя
Человек . . 50 5,60 8,32 8,12 7,27
Мартышка . 25 0,83 1,54 0,93 0,96
Крыса . . . 50 0,69 0,66 0„92 0,64
черты различия. Для человека характерна полная независимость в
вариациях передней и задней стенок диафиза бедра, тогда как
связь между развитием боковых стенок значительна. У мартышко-
образной обезьяны наблюдается определенная тенденция к связи
развития передней и задней компакты, последняя реально сущест-
вует и на бедренной кости крысы (табл. 19а). Что касается боко-
вых стенок диафиза, то они во всех случаях связаны весьма тесно.
Таблица 19а
КОЭФФИЦИЕНТЫ КОРРЕЛЯЦИИ МЕЖДУ РАЗВИТИЕМ компакты
СТЕНОК ДИАФИЗА БЕДРЕННОЙ КОСТИ У ЧЕЛОВЕКА,
МАРТЫШКООБРАЗНОЙ ОБЕЗЬЯНЫ И КРЫСЫ
Форма Коррелируемые признаки
п передня я—задняя компакта наружная—вну- тренняя компакта
Человек .... 50 0,006 +0,14 0,755±0,06
Мартышка . . . 25 0,329±0,18 0,526±0,14
Крыса .... 50 0,431±0,12 0,674±0,08
Таким образом, независимость в вариациях толщины передней
и задней стенок диафиза бедра можно считать достаточно специ-
фической особенностью структуры этой кости у человека. Между
тем взаимоотношения суммарной толщины сагиттальной (передняя
плюс задняя) и боковой (наружная плюс внутренняя) компакт
оказываются весьма сходными во всех случаях. Для человека,
мартышки и крысы определены коэффициенты корреляции поряд-
ка 0,5—0,7. Это свидетельствует о выраженной взаимосвязи общего
распределения кортикальной кости по обеим основным осям диа-
физа: сагиттальной и трансверзальной, хотя у человека она все же
несколько ослаблена. Последнее можно отнести за счет «разрыва»
связей между развитием обеих стенок диафиза по сагиттальной
оси.
При анализе внутренней структуры диафиза бедренной кости,
как и ее эпифизов, также следует иметь в виду ее нормальную
изменчивость у человека. Так, кортико-медуллярные соотношения
139
подвержены возрастно-половой вариабильности. Формирование
диафизарных параметров бедренной, как и плечевой, кости чело-
века заканчивается в основном к 20 годам (Свадковский, 1959).
Но и в последующие периоды наблюдаются возрастные сдвиги во
внутренней структуре диафиза. По данным Элиакиса и Иордани-
диса (Eliakis, lordanidis, 1963, 1963а), медуллярный указатель1
бедренной кости в период
25—85 лет повышается у
3Q-\ А мужчин от 46,6 до 54,9, для
женщин соответствующие
цифры 50,0—53,7. На плече-
•Рап вой кости возрастно-поло-
*р0 вые сдвиги медуллярного
§10-
Th ор
<ht.
of °Cb L
30 35 00 50
Кортикальный индекс (ЮО-медулляр-
ный-шВекс)
Рис. 25. Соотношение внутренней
и внешней массивности бедренной
кости у приматов: А — Н. sa-
piens; О — Ceboidea et Cercopi-
thecoidea (Cc. — Cercocebus;
Cb. — Cebus; At.—Ateles; Lg. —
Lagothrix); Q — Lemur; ф —
. Anthropoidea
указателя у мужчин от 48,8 до 63,6; у женщин от 54,8 до 68,8.
В обоих случаях отмечается и тенденция к большей толстостен-
ности мужских костей, сильнее подчеркнутая на плечевой кости.
Кстати, на плечевой кости и костях предплечья абсолютная тол-
щина компакты, по-видимому, также больше у мужчин (Очкурен-
ко, 1962).
Наконец, как и в случае спонгиозы, нужно учитывать прижиз-
ненные функциональные адаптации и индивидуальную изменчи-
вость кортико-медуллярных соотношений вообще.
Данные клиники и эксперимента свидетельствуют о перестрой-
ке кортико-медуллярных пропорций диафиза у человека и живот-
ных прежде всего в связи с прижизненными адаптациями- (Кле-
банова, 1954; Ковешникова, 1960, 1961). Д. А. Жданов (1967)
приводит результаты многолетних рентгенографических наблюде-
ний над явлениями функциональной гипертрофии скелета у пред-
ставителей некоторых профессий и видов спорта. Эти данные де-
монстрируют усиление компактизации коротких и длинных труб-
чатых костей под влиянием интенсивной физической нагрузки.
Особый интерес представляют наблюдения П. Н. Соколова (1967)
над спортсменами «силовых» видов спорта. Так, у штангистов и
метателей молота очень отчетливо сказывается повышение кор-
тикализации бедренных костей: соответствующие значения обще-
1 В данном случае вычислялся как процентное отношение наименьшего диамет-
ра полости к наименьшему диаметру диафиза.
140
го медуллярного указателя — 35,3 и 38,3 против 46,1 в среднем
у мужчин при случайной выборке.
Если теперь попытаться проанализировать соотношения между
кортико-медуллярными пропорциями диафиза; т. е. «внутренней
массивностью» и его «внешней массивностью», то у изученных
приматов выявляется несколько вариантов по этому признаку
Рис. 27. Внутреннее
строение бедренной
кости тонкотела
Рис. 26. Внутреннее
строение бедренной
кости ге^ады
(рис. 25). Тип преимущественно наземного примата с большой
массой тела характеризуется довольно массивными внешне и тол-
стостенными диафизами длинных костей обеих конечностей. Этот
тип свойствен павиану и геладе из церкопитековых обезьян и, по-
видимому» горилле среди антропоморфных (рис. 26). Другой от-
четливый вариант — древесный примат с небольшой массой тела,
141
для которого характерна комбинация внешней грацильности и от-
носительной тонкостенности длинных костей (рис. 27).
В обоих этих типах степень кортикализации диафизов плече-
вой и бедренной костей весьма близка. На графике отчетливо вы-
является особое положение орангутана, у которого существует
значительная толстостенность диафиза в сочетании с его внешней
грацильностью, что, возможно, объясняется «нетипичной» для
большинства приматов комбинацией большой массы тела с дре-
весностью. Здесь же в наибольшей мере проявляется и свойствен-
ная специализированным брахиаторам дифференциация обеих ко-
стей по «внутренней массивности», значительно меньшей на пле-
чевой кости. На рис. 25 отчетливо видно также особое положение
человека среди других изученных приматов — по сочетанию зна-
чительной внешней и внутренней массивности кости.
ВНУТРЕННЯЯ СТРУКТУРА ДЛИННЫХ КОСТЕЙ
У ИСКОПАЕМЫХ ГОМИНИД
Трабекулярная структура проксимального эпифиза бедренной
кости изучалась у некоторых архантропов и палеоантропов. По-
видимому, во всех случаях рентгенографическая картина соответ-
ствовала бипедальному типу. Прямые указания на этот счет су-
ществуют и для архантропов (питекантропа, согласно Дюбуа,
Дэй, 1971), и для палеоантропов, тад Маккаоун и Кизс свиде-
тельствуют об отсутствии сколько-нибудь существенных отклоне-
ний у палестинских палеоантропов в этом отношении. Это ожи-
даемый результат, поскольку прямохождение этих гоминид не под-
лежит сомнению. Вместе с тем как среди древнейших, так и древ-
них людей встречаются варианты с некоторыми отличиями
трабекулярной структуры от типичной для современного челове-
ка. Прежде всего это относится к синантропу, на одной из бедрен-
ных костей которого Вейденрейх (1941) описал своеобразный ва-
риант гоминидного типа. У человека I и II системы, начинаясь
от общего участка концентрации, компакты на уровне малого вер-
тела, дивергируют затем так, что между ними остается свободное
от трабекул поле — «треугольник Варда». У синантропа отмече-
на непрерывная радиация костных пластинок через все поле, т. е.
нет ни треугольника Варда,- ни отчетливой дифференциации I и
II систем. Кривая III системы более уплощенная, чем у современ-
ного человека. Трабекулярная сеть более густая и крупноячеи-
стая.
Подобный же «диффузный» тип строения спонгиозы встречается
и на бедрах некоторых классических неандертальцев,— Неандер-
таль, Спи. Интересна в этом плане также рентгенографическая
картина верхнего эпифиза бедра ребенка из Тешик—Таша (Рох-
лин, 1949). Здесь мощные группы трабекул отмечены не только
в медиальном, но и в латеральном отделах шейки бедра, что у
142
современного человека наблюдается только при наличии третье-
го вертела, а у ребенка палеоантропа встретилось независимо от
последнего. Исходя из глубокой древности функции прямохожде-
ния у гоминид, а также из параллелей с современным человеком
в отношении силовой адаптации, которая сопровождается транс-
формацией структуры трабекул в более грубоячеистую, можно
предположить у этих гоминид проявление рабочей гипертрофии,
спонгиозы *. Вопрос о возможном таксономическом значении осо-
бенностей трабекулярной структуры бедра внутри семейства го-
минид остается пока открытым из-за недостаточности палеоантро-
пологических материалов.
Более представительны последние по структуре диафизарных
отделов бедренных костей. Рассмотрим прежде всего кортико-ме-
дуллярные пропорции у различных представителей ископаемых
гоминид (табл. 20—22).
Т а б л ид а 20
МЕДУЛЛЯРНЫЕ УКАЗАТЕЛИ БЕДРЕННЫХ КОСТЕЙ ГОМИНИД*
Гоминиды Медуллярные указатели
сагиттальный поперечный общий
Синантропы (п=5) . . . 37,6 32,9 35,3
Олдувей 28 55,4? 51,0? 53,2?
Крапина I *. . 37,7 28,8 33,3
Крапина II — 25,1 —•
Кина V 36,2 36,5 36,4
Спи II 31,8 44,0 37,9
Е689 35,8? 40,7? 38 3?
Брокен-Хилл ЕМ793 .... 44,3? 39,3? 41,8?
Романково 39,4 34,2 36,8
Современный человек (п=50) 49,8 42,4 46,1
♦ Здесь и в дальнейшем использованы данные из монографии Вейденрейха
(1941) для бедренных костей синантропов, Крапины,' Ла Кина и Спи; циф-
ры для Брокен-Хилл получены на основе измерений, приведенных в статье
Кларка и др. (С1агк et al., 1968); для Олдувей 28 — вычислены по
данным из статьи Дэя (1971); для Романкова и современного человека —
по нашим данным.
Как известно, при изучении длинных костей синантропа и не-
которых палеоантропов (Крапина I и II, Ла Кина V, Спи II)
Вейденрейх (1941) обратил внимание на различия с современ-
ным человеком по соотношению компакты и медуллярной поло-
сти. Все эти ископаемые гоминиды обладали очень толстыми диа-
физарными стенками, при относительно суженной медуллярной по-
1 В отдельных случаях вероятна и связь с патологией (например, артритической.
у некоторых классических неандертальцев).
143
Таблица 21
РАЗМЕРЫ МЕДУЛЛЯРНОЙ ПОЛОСТИ И ЕЕ СТЕНОК НА БЕДРЕННЫХ КОСТЯХ ГОМИНИД
Гоминиды Признак (мм)
сагиттальные размеры поперечные размеры
диафиз полость компакта диафиз полость компакта
Синантропы (п=5) . . 24,4 8,7 15,7 28,3 8,1 20,2
Олдувей 28 24,7 ’ 13,7 11,0 32,7 16,7 16,0
Крапина I 27,3 10,3 17,0 32,0 9,2 22,8
Крапина II — — 26,3 6,6 19,7
Кина V 29,8 10,8 19,0 27,7 10,1 17,6
Спи II 32,1 10,2 21,9 ' 30,0 13,2 16,8л
Е689 . . . 31,0? 11,1? 19,9? 27,5? 11,2? 16,3?
Брокен-Хилл ЕМ793. . . 29,3? 13,0? 16,3? 26,5?. 10,4? 16,1?
Романково 32,0 12,5 19,5 30,7 10,6 20,1
Современный человек (п=50) 29,1 15,2 13,9 29,0 13,7 15,3
Таблица 22
АБСОЛЮТНОЕ РАЗВИТИЕ ПЕРЕДНЕЙ И ЗАДНЕЙ
КОМПАКТЫ БЕДРЕННЫХ КОСТЕЙ ГОМИНИД
Гоминиды Толщина компакты, мм
передняя». задняя
Синантропы (п=5) 7,1 8,6
Крапина I 8,4 8,6
Кина V 8,3 10,7
Спи II . . ; 10,9 и,о
Романково 6,2 13,3
Современный человек (п—50) . 5,6 8,3
лости, при этом, все пять фрагментов бедренных костей синантро-
па, а также фрагмент плечевой кости этого гоминида показали
идентичную структуру. На новом фрагменте плечевой кости си-
нантропа, как и на его большой берцовой кости, вновь была от-
мечена утолщенность диафизарных стенок и сильное сужение ме-
дуллярной полости (Woo Ju-kang, Chia Lan-po, 1954). Эти данные
свидетельствуют о специфичности подобной структуры диафизов
длинных костей в этом варианте архантропов (рис. 28).
Резкое утолщение стенок и сужение костномозговой полости
на всех без исключения сохранившихся фрагментах длинных ко-
стей (бедренная, плечевая, большая и малая берцовые кости) бы-
144
ло констатировано при рентгенологическом изучении скелета те-
шик-ташского палеоантропа (Рохлин, 1949).
Одна из сравнительно недавних находок палеолитического че-
ловека (вероятно, палеоантропа) из Романкова (Украина), бед-
ренная кость которого была изучена' нами рентгенографически,
также обнаруживает характерное
для этих ископаемых гоминид
соотношение компакты и полости
(Хрисанфова, 1965).
В последнее время представи-
лась также возможность исследо-
вания внутренней структуры диа-
физа на двух фрагментах бед-
Рис.'28. Внутреннее строение бедренной
кости ископаемых гоминид: А—Д—си-
нантропы; Е — Крапина; Ж— Ла Ки-
на; 3 — Спи; И — современный чело-
век (по Weidenreich, 1941)
А Б 8 Г Д
‘ ©
ренных костей (Е689 и ЕМ.793) из Брокен-Хилл (Кларк и др.,
1968). Однако цифры, приводимые этими авторами, могут быть
сопоставлены с данными наших таблиц 2Q и 21 только условно.
В обоих случаях у Брокен-Хилл приведенные цифры относятся не
строго к середине диафиза особенно на фрагменте Е689, где на-
ми усреднялись данные для дистального конца проксимального
фрагмента й проксимального конца дистального фрагмента. На
фрагменте ЕМ.793 («новом фрагменте»), представляющем сред-
нюю часть диафиза, тоже усреднялись данные для обоих его кон-
цов. Наконец, большой интерес представляют совсем недавно
опубликованные сведения о внутренней структуре диафиза бедра
архантропа из Восточной Африки (Олдувей 28) (Дэй, 1971).
Из приведенных значений медуллярных указателей бедренной
кости ископаемых людей вытекает значительная толстостенность
большинства костей. Таким образом, признак диафизарной струк-
туры, являющийся типичным для человека, был даже «преувели-
чен» у подавляющего большинства этих гоминид. Вместе с тем об-
ращают на себя внимание и некоторые интересные особенности.
Так, для современного человека характерно преобладание сагит-
тального медуллярного указателя над поперечным, т. е. лучшее
относительное развитие компакты в трансверзальном направле-
ний. Среди ископаемых гоминид такой тип внутренней структуры
середины диафиза выявляется у синантропов, Олдувей 28, Кра-
пины, Романкова, но не в специализированном варианте класси-
ческих неандертальцев, где соотношения иные. Бедренная кость
этих гоминид испытала наибольшее утолщение стенок именно в
передне-заднем направлении. И у Ла Кина V и у Спи II значения
сагиттального медуллярного указателя самые низкие :в ряду со-
10 Е. Н. Хрисанфова
145
поставляемых форм. С Кина сближается по абсолютному и отно-
сительному развитию компакты «типичное бедро» Брокен-Хилл
(Е689). На «новом фрагменте» относительное развитие компакты
более слабое в масштабе ископаемых гоминид. Хотя выше уже
говорилось о невозможности прямых сопоставлений, из приведен-
ных данных следует, что во внутренней структуре бедра нет про-
тивопоказаний для определения этого бедра, как принадлежаще-
го палеоантропу. Если принять абсолютный возраст обоих фраг-
ментов Брокен-Хилл равным 35—45 тыс. лет (Кларк и др.,
1968) \ возможность их хронологического и морфологического со-
поставления с бедренными костями* поздних палеоантропов клас-
сического типа становится довольно реальной.
В целом можно прийти к заключению, что изученные в этом
аспекте бедренные кости палеоантропов обладают специфически-
ми кортико-медуллярными соотношениями, свидетельствующими
о значительном утолщении диафизарных стенок. Что касается ар-
хантропов, то сказанное полностью распространяется на вариант
синантропа. Особняком стоит бедренная кость Олдувей 28 с ее
исключительной в‘ масштабе ископаемых гоминид тонкостённостью
диафиза. Подобная структура в сочетании с преобладанием сагит-
тального медуллярного указателя над поперечным наиболее при-
ближается к современному женскому типу, что находится в неко- i
тором противоречии с внешней морфологией бедра, не представ- I
ляющей типичного сапиентного ее варианта1 2. На тринильских
бедренных костях, приписываемых питекантропу, определение
внутренних пропорций диафиза затруднено очень сильной мине-
рализацией, по-видимому способствовавшей значительному суже-
нию медуллярной полости, характерному для этих костей.
Заслуживает внимания, что при известном сходстве в некото- i
рых признаках внешней структуры бедренных костей Олдувей 28 '
и синантропа их внутреннее строение оказалось столь различным.
Трудно дать этому факту вполне достоверную интерпретацию, по
крайней мере, до появления новых сведений о внутреннем строе-
нии бедра у архантропов. Возможно, что морфологические вари-
анты существовали здесь, как и во внешней морфологии скелета,
уже в глубокой древности.
Вместе с тем. нельзя игнорировать и тот факт, что основным
вариантом у ископаемых гоминид являлся тип бедренной кости '
с очень высокой «внутренней массивностью» диафиза^ Остановим-
ся на его характеристике несколько подробнее.
Отличительная особенность развития диафизарных стенок бед-
ра у ископаемых людей сравнительно с современными — утолще-
1 Укажем на возможную большую древность родезийского человека (не менее
il25 тыс. лет по Klein, 1973).
2 Здесь нужно иметь в виду указание Дэя о мощном развитии компакты (кор-
тикального «контрфорса») на нижней части шейки бедра: в месте разлома
ее толщина достигала 8 мм, что, согласно этому автору, значительно больше,
чем обычно у современного человека.
146
ние передней компакты относительно задней. Оно выявляется поч-
ти во всех приведенных в табл. 22 вариантах толстостенного бед-
ра-палеоантропов и архантропов и отличает их от среднего совре-
менного типа. В среднем для синантропов, палеоантропов и
современного человека относительное развитие передней компак-
ты в % от толщины задней составляет соответственно 82,6; 91,5
и 67,5.
• Следует обратить внимание, что абсолютное развитие задней
стенки компакты во всех случаях по меньшей мере не ниже сред-
него современного варианта, а у классических неандертальцев
оно даже значительно выше. У Спи II, например, разница с сов-
ременным человеком по развитию всей сагиттальной компакты
соответствует шести квадратическим отклонениям. Это позволяет
внести существенное уточнение в сложившееся представление об
исключительно слабом развитии пилястра в группе классических
неандертальцев. Факт значительного развития как раз задней
диафизарной стенки на бедренных костях этих гоминид («внут-
реннего пилястра») приобретает особое значение, так как указы-
вает на усиленную аппозицию кортикальной кости со стороны эн-
доста при отсутствии видимого внешнего1 пилястра. Очевидно, мы
сталкиваемся здесь с каким-то своеобразным способом адаптации,
причем слабость суперструктуры можно объяснить и большой мас-
сивностью бедренных костей этих гоминид, так как и у современ-
ного человека неоднократно отмечались случаи слабого развития
рельефа на массивных костях. В известной мере сказанное’ можно
отнести и к синантропу, у которого указатели пилястрии также
очень низкие, тогда как абсолютная толщина задней стенки такая
же,-как на типичном современном бедре.
В целом распределение компакты по основным, координатам
диафиза у всех ископаемых гоминид такое, же, как у современно-
го человека, т. е. задняя стенка обычно толще передней, а преоб-
ладание развития латеральной стенки над медиальной встречает-
ся почти столь же часто, как и противоположный вариант. Основ-
ные различия в структуре середины диафиза бедра «толстостен-
ного варианта» связываются помимо общего усиления
компактизации кости с относительно повышенной толщиной пе-
редней стенки диафиза.
Как показывает сопоставление суммарной толщины компакты
с размерами диафиза (суммой обоих диаметров), абсолютная кор-
тикализация в целом повышается с возрастанием внешней мас-
сивности диафиза у ископаемых гоминид (рис. 29). Особое поло-
жение занимает синантроп, у которого толстостенность диафиза
сочетается с внешней грацильностью кости. На графике отчет-
ливо выступает разница в положении синантропа, с одной сторо-
ны, Олдувей и современного человека, с другой, — а также сход-
ство в соотношении внутренней и внешней массивности диафиза
у двух последних гоминид. Следует, конечно,. иметь в виду, что
палеоантропологические материалы, изученные рентгеноморфоло-
10*
147
гически, весьма ограниченны, и расширение данных может вне-
сти известные коррективы в этот вопрос. Все же широкое распро-
странение «внутренней массивности» бедренной кости среди иско-
паемых гоминид достаточно демонстративно.
65-
во-
(
52
•Р.
•К
•Вт.
•L.K.
• Вт.
Значительно более скуд-
ными являются материалы
по другим длинным костям
конечностей. Так, для пле-
чевой кости синантропа
приводится только величина
поперечного медуллярного
указателя . (Вейденрейх,
Компакта-суммарная, мм
Рис. 29. Соотношение внутренней
и внешней массивности бедренной
кости у гоминид: ▲ — Н. sapiens-,
О — архаятропы (Олдувей 28,
синантроп); • — палеоантропы
(Брокен-Хилл, Ла Кина, Крапина,
Спи, Романково)
о/7
1941). Значение этого индекса крайне низкое (22,0) и выходит
за пределы вариаций в современной серии плечевых костей
(28,6—58,4). Указатель синантропа не достигает даже
минимума этого вариационного ряда и отстоит от средней
арифметической более чем на три квадратических отклонения.
Выше уже указывалось, что и другие длинные кости этого архан-
тропа (новый фрагмент плечевой, большая бердовая) тоже отли-
чаются значительной толстрстенностью диафиза. Также и у па-
леоантропа из Тешик-Taiiia плечевая кость имела сильно разви-
тые стенки диафиза. Это было свойственно и другим длинным
костям скелета этого ребенка палеоантропа.
Большой интерес в указанном отношении представляет вари-
ант Схул, для которого характерны многие признаки «сапиент-
ного типа». По-видимому, это подтверждается и для внутренней
структуры диафиза; о последней, мы можем судить, например, по
большой берцовой кости Схул IV (табл. 23).
Приведенные ’ данные свидетельствуют о близости внутренней
структуры диафиза большой берцовой кости Схул к среднему сов-
ременному варианту, хотя намечается явственная тенденция к
большой абсолютной и относительной компактизации этой кости
у палеоантропа. Последнее подтверждается и при сопоставлении
медуллярных указателей. Так, например, значение сагиттального
медуллярного указателя составляет у Схул IV 37,9 против .41,8 в
среднем у современного человека. Значение индекса Схул проме-
жуточное между средней и нижним лимитом современного вари-
ационного ряда (33,8—60,6). Эта величина свидетельствует о до-
вольно значительном развитии плотнрго вещества кости у этого
148
Таблица 23
СОПОСТАВЛЕНИЕ НЕКОТОРЫХ ПРИЗНАКОВ ВНУТРЕННЕЙ СТРУКТУРЫ ДИАФИЗА
БОЛЬШЕБЕРЦОВОЙ КОСТИ СХУЛ IV И СОВРЕМЕННОГО ЧЕЛОВЕКА
Признаки, мм Схул IV' Современный человек*
Сагиттальный диаметр диафиза 29,0 30,1
Сагиттальный диаметр полости 11,0 12,6
Толщина сагиттальной компакты 18,0 17,5
Толщина передней компакты 10,0 11,4
Толщина задней компакты 8,0 6,1
Поперечный диаметр диафиза 21,0 22,4
Поперечный диаметр полости "... 10,5 12,7'
Толщина боковой компакты 10,5 9,7
Толщина наружной компакты . : ........ 5,0 4,6
Толщина внутренней компакты , 5,5 5,1
* По данным рентгенографического изучения 50 больших берцовых костей современного человека,
гоминида, хотя в целом внутренняя структура диафиза здесь впол-
не современная. Подобный ее тип хорошо согласуется и с общим"'
морфологическим обликом «прогрессивного» палеоантропа Схул
IV. Хотя этот вывод не может безоговорочно распространяться на
другие палестинские формы из-за их известной морфологической
неоднородности, все же заслуживает внимания весьма умеренное
развитие компакты большеберцовой кости ребенка Схул VIII
(Маккоун, Кизс, 1939, рис. 236С).
Данные о внутреннем строении скелета верхнепалеолитическо-
го неоантропа далеко не достаточны. Возможно, что при некото-
рых сопоставлениях можно усмотреть тенденцию к прогрессивной
грацилизации внутренней структуры диафиза длинных костей у
Н. sapiens. Медуллярный указатель бедренной кости у верхне-
палеолитических людей из Чжоу-коу-тьен выше, чем у синантро-
па (Вейденрейх, 1941). Известны и другие факты такого рода:
значительной грацильностью отличались кости скелета предпо-
ложительно верхнепалеолитического человека из Салемас (Пор-
тугалия), по данным Ферембах (Ferembach, 1964/1965). Опреде-
ленный интерес представляет и изучение внутренней структуры
ископаемого бедра из Анатолии (Турция), предположительный
возраст которого составляет, по-видимому, 15—20 тыс. лет (Erd-
brink et al., 1966). Внутренние пропорции диафиза оказалось воз-
можным охарактеризовать лишь в поперечном направлении, по-
скольку в статье приведена только передне-задняя рентгенограм-
ма'кости. Значение поперечного медуллярного указателя этого
бедра (41,9) соответствует среднему для современного человека1.
1 Середина диафиза этой кости маркировалась нами при сравнении с помещен-
ной рядом идентичной рентгенограммой современной бедренной кости; получен-
ные параметры диафиза на рентгенограмме анатолийского бёдра предвари-
149
Вполне современные внутренние пропорции диафиза имеет и бед-
ренная кость восточноевропейского неоантропа из Сунгиря I и IV
(соответствующие значения общих медуллярных указателей —
45 и 48, по нашим данным).
Упомянем еще о результатах изучения внутренней структуры
бедра у средневекового и более позднего населения Великобри-
тании — в последнем случае толщина компакты была меньше
(BrottrCwell et al., 1968). Аналогичные данные получены при сопо-
ставлении бедренных костей жителей Латвии эпохи бронзы и сов-
ременного населения. Интересно, что на ископаемых костях на-
блюдается и большая густота спонгиозы шейки бедра, распрост-
раняющаяся также и на латеральный ее отдел (Дэруме, 1973).
Резюмируя изложенные выше факты о внутреннем строении
длинных костей скелета гоминид (на примере бедренной кости),
можно прийти к заключению, что оно является вероятным, разгра-
ничительным признаком прямоходящих гоминид и негоминидных
приматов. В первую очередь это относится к трабекулярной струк-
туре верхнего эпифиза бедра, которая представляет важный ком-
понент комплекса бипедии. Также и внутреннее строение диафи-
за (кортико-медуллярные соотношения) может иметь немаловаж-
ное значение в аспекте антропогенеза. Для гоминид характерна
относительно толстостенная бедренная кость со значительным раз-
витием задней компакты — «внутреннего пилястра» — и толсто-
стенными редко расположенными остеонными системами.
Все ископаемые люди, бедренные кости которых были изучены
рентгенологически, имеют вполне выраженный гомипидный тип
этой кости. Вместе с тем существуют известные локальные ва-
рианты внутренней структуры бедра как среди архантропов, так
и у палеоантропов. Наиболее распространенный тип толстостен-
ной бедренной кости с «диффузным строением» спонгиозы прок-
симального конца ее встречается среди древнейших людей у
синантропов и у некоторых палеоантропов (прежде всего классиче-
ского типа, но также и у крапинцев и среднеазиатского палеоан-
тропа из. Тешик-Таша). Трудно дать вполне достоверную функ-
циональную интерпретацию этому комплексу, хотя следы рабочей
гипертрофии прослеживаются достаточно отчетливо. Однако этот
фактор не может быть признан единственным в этиологии подоб-
ного варианта. У современного человека функциональная гипер-
трофия всегда локально детерминирована, здесь же. в ряде слу-
чаев она затрагивает все длинные кости (синантропы, Тешик-Таш,
у которого очень высокая степень кортикализации наблюдалась
даже на ключице). Видимо, следует иметь в виду и другие воз-
можные причины, в том числе состояние обменно-гормонального
статуса индивидуума (см. гл. VIII).
тельно корригировались на основе известного абсолютного размера попереч-
ного диаметра диафиза, поскольку масштаб изображения не соответствовал
натуральной величине кости.
150
С другой стороны, среди ископаемых гоминид существовали
и иные варианты внутренней структуры бедренной кости. В пер-
вую очередь это относится к восточноафриканскому архантропу
Олдувей 28, у которого выявляется близкий к сапиентному тип
кортико-медуллярных соотношений диафиза при достаточно свое-
образной внешней морфологии бедренной кости. Некоторые чер-
ты сходства с синантропом во внешней структуре бедра (исключи-
тельно высокая степень платимерии диафиза) сочетаются у Ол-
дувей с очень большими различиями во внутренней массивности
диафиза. Что касается палеоантропов, то, судя по другой важной
кости нижней конечности, большой берцовой, в группе Схул, так-
же существовал вполне сапиентный вариант ее внутренней струк-
туры. Последнее вполне логично увязывается с наличием и дру-
гих сапиентных признаков на скелете этих палеоантропов. Вопрос
о локальных вариантах внутреннего строения скелета ранних го-
минид требует дополнительного освещения как с точки зрения
пополнения пока еще немногочисленных данных, так и в аспекте
функциональной интерпретации существующих различий между
ископаемыми гоминидами. .
Глава VII
ЧЕРЕП
ОСНОВНЫЕ ЧЕРТЫ
КРАНИОФАЦИАЛЬНОГО КОМПЛЕКСА
СОВРЕМЕННОГО ЧЕЛОВЕКА
Череп человека принадлежит (наряду с тазом и стопой) к числу
отделов скелета, имеющих наибольшее значение в аспекте таксо-
номического анализа. Однако далеко не все краниологические
признаки равноценны в данном отношении. Мы приведем лишь
основные из разграничительных черт, выявляющихся при срав-
нении черепа человека с черепами современных негоминидных
приматов, и прежде всего черепом шимпанзоидного типа.
Из числа факторов, влияющих на перестройку черепа в фило-
генезе приматов и гоминид, особую роль играли: способ локомо-
ции, характер питания и прогрессивное развитие мозга и соответ-
ствующих рецепторов.
Среди краниологических признаков первостепенного таксоно-
мического значения нужно назвать прежде всего абсолютную и
относительную емкость мозговой коробки, тесно связанную с аб-
солютным и относительным развитием массы мозга. Человек за-
нимает исключительное положение среди приматов по обоим по-
казателям. Емкость мозгового черепа в среднем составляет здесь
1400 см3 для мужчин, что почти втрое превышает его размеры у
наиболее крупных современных обезьян—горилл, у которых сред-
нее значение емкости для самцов равно 500 см3. Наибольший ин-
терес представляет, однако, относительное развитие мозгового от-
дела черепа, которое можно оценивать применительно к массе
тела (в переводе на весовое выражение массы мозга) или разви-
тию лицевого отдела. В первом случае следует иметь в виду не-
пропорциональное изменение массы мозга при увеличении общих
размеров тела. Поэтому для оценки относительного развития моз-
га (степени церебрализации) при сравнении разных таксонов ис-
пользуется формула гетерогонического роста -(Дюбуа): Е=
=кхР°’5в. Коэффициенты церебрализации (к), вычисленные по
этой формуле для человека и шимпанзе, как наиболее близкой к
человеку по этим соотношениям форме среди современных при-
матов, составляют соответственно 134 и 45 г на 1 кг веса тела,
т. е. различаются примерно втрое (Quiring, 1941; Бунак, 1966).
Сопоставление развития мозгового и лицевого черепа произ-
водится разными способами. Планиметрический индекс Штраца
(отношение площадей лицевого и мозгового черепа в профиль)
дает для современного человека значения: 30 (женщины) — 42.3
(мужчины) против 90,4 у шимпанзе и 102 у орангутана. При срав-
нении объемов обоих отделов (пересчет по цифрам Штраца) со-
152
ответствующие индексы для человека и шимпанзе составляют:
212,4—337,8 и 856,7—859,5; для оранга этот показатель равен
1030 (Патт, 1955).
При сопоставлении частных краниофациальных указателей во'
всех трех направлениях роста (продольном, поперечном и высот-
ном) оказывается, что череп человека наиболее отличается от че-
репа шимпанзоидного типа в вертикальных соотношениях обоих
отделов (средняя разница 42,6 против 38,0 в длиннотном и 16,6 в
широтном направлениях). Это соответствует общим тенденциям
изменения массы мозга в филогенезе: так, по Вейденрейху
(1936), у человека сравнительно с антропоидами величина мозга
возросла в наибольшей степени в высотном и длиннотном направ-
лениях роста.
С увеличением массы мозга на черепе человека связан целый
комплекс морфологических признаков. По данным, полученным
на млекопитающих, выявляется важнейшая роль объема мозговой
массы как первичного морфогенетического фактора; содержимое
мозгового черепа играет 'при этом роль капсулярной функцио-
нальной матрицы (Moss et al., 1972). Исследования на приматах
показали, что в процессе эволюции неокортекса передняя часть
мозга принимала более сферическую форму, вследствие чего про-
изошло каудальное смещение вертекса и увеличился также кифоз
основания черепа (Hofer, 1969).
Из сказанного следует, что повышение и округление черепного
свода представляет особый интерес в плане таксономии. Для ха-
рактеристики этих черт структуры используют различные спосо-
бы. Наиболее существенны для палеокраниологии показатели от-
носительной высоты свода и прежде, всего индекс высоты калот-
ты над линией глабелла-инион (Швальбе). У современного чело-
века индекс Швальбе составляет 50—63 против максимума 37,7
у антропоидов (шимпанзе). Большое значение в последнее время
придают также указателю надглазничной высоты черепа (Rosen,
McKern, 1971), предложенному Ле Гро Кларком (1950).
В известной мере показателем цефализации является также
степень изгиба костей черепного свода — лобной, теменной и за-
тылочной. Для ее характеристики можно использовать как обыч-
ный хордо-дуговой указатель, так и высоту краниологических то-
чек продольного свода над какой-либо ориентировочной горизон-
талью или хордой. Для черепа человека характерно значительное
повышение изогнутости лобной и теменной костей и удлиненйе
последней. Одновременно происходило и выпрямление лобной ко-
сти и расширение лба. Угол назион-брегма составляет у человека
и шимпанзе соответственно 49—52,9° (индивидуально 45—60°) и
35° (индивидуально 31—41°). Нарастали размеры и менялась кон-
фигурация височной кости и происходило поднятие крыла основ-
ной кости (Delattre, Delblock, 1-972).
Сложной перестройке подверглась затылочная область черепа,
которая очень рано была затронута трансформацией, прежде всего,
153
в связи с прямохождением. В филогенезе гоминид происходила
ротация затылочной части черепа и точки астерион назад и вниз
(Delattre, 1956). В косвенной форме об этом процессе свидетель-
ствуют и наблюдения над млекопитающими при эксперименталь-
ной гидроцефалии и бипедии (Destrombes, Fenart, 1970; Riesen-
feld, 1969).
Другие преобразования затылочной области заключались в ок-
руглении задней части черепа, разрастании верхней части чешуи
затылочной кости, понижении положения точки инион. Измене-
ние положения выйной площадки в филогенезе отражает угол на-
клона ее оси (инион-опистион) к франкфуртской горизонтали: у
человека этот угол равен 35° против 65° у шимпанзе. По отноше-
нию к плоскости естественной ориентировки черепа ось пнион-ба-
зион составляет в среднем угол 30,6° у человека против 60,1° у
шимпанзе, что соответствует смещению иниона вниз и назад в фи-
логенезе (значения этого угла у неприметных млекопитающих:
92,5—135,6°, по данным Ковешникова, 1968). Подобные измене-
ния наблюдаются и в положении точки лямбда.
Особый интерес представляет трансформация положения за-
тылочного отверстия в филогенезе приматов. Угол плоскости это-
го отверстия к линии назион-базион составляет у человека 24°
(9—35°), у антропоидов — 53° (46—59°) (Вейденрейх, 1943). Ана-
логичную тенденцию можно наблюдать в сравнительно-анатоми-
ческом ряду приматов и по отношению к плоскости естественной
ориентировки головы: угол оси затылочного отверстия человека
и негоминидных приматов в целом равен соответственно 11,4° и
46—52° (Ковешников, 1968).
Другой весьма существенный признак человеческого типа че-
репа — изгиб его основания. Сфеноидальный угол у человека
варьирует в пределах: 126—150° против 147—189° у антропоидов
(Патт, 1955). Изменение взаимного положения мозгового и лице-
вого отделов черепа сопровождается также перемещением заты-
лочного отверстия к центральной вертикальной осп.
Большое внимание в эволюционной морфологии приматов всег-
да уделялось структуре надглазничного рельефа. Последнему
придается исключительное (и, возможно, даже несколько преуве-
личенное) значение в таксономии гоминид. Нет единого мнения
относительно генеза и классификации рельефа лобной кости у
приматов, но наиболее распространенные схемы (Cunnigham,
1908/1909; Бунак, 1966) учитывают как собственно структуру, ^гак
и степень ее развития. По-видимому, в филогенезе приматов все
же наблюдается постепенное усиление развития рельефа у выс-
ших их представителей. Так, сколько-нибудь выраженный рельеф
отсутствовал у эоценовых просимий, не было его и у примитив-
ных широконосых, как и у многих низших узконосых обезьян.
Слияние всех компонентов рель'ефа (глабеллярное возвышение,
надглазничные дуги, надглазничный край) с распространением
его на область скулового треугольника встречается у немногих
154
высших приматов: антропоидов Африки, а из циноморфных — у
павиана и, возможно, макаки. У остальных приматов существуют
разные комбинации элементов рельефа надглазничной области,
как и разная степень их развития, причем наиболее четкие вари-
анты отражают полное разделение всех этих компонентов или их
частичное слияние в медиальной части и распространение на
верхнеглазничный край с отграничением от области скулового
треугольника надглазничной бороздой. Можно выделить также
вариант с полным или почти полным отсутствием рельефа (Бу-
нак, 1966).
Достаточно сложен вопрос и о генетической преемственности
вариантов надглазничного рельефа: существует даже точка зре-
ния, что надбровные дуги, надглазничные дуги и надглазничный
валик (торус) не связаны между собой ни в филогенетическом,
ни в функциональном (биомеханическом) отношении (Ehara, Sei-
ler, 1970). Все же пример миоценового проконсула без выражен-
ного рельефа, скорее, говорит о вторичности торуса африканских
антропоидов.
У современного человека наиболее обычен довольно генерали-
зованный тип рельефа — отчетливое разделение надбровных дуг,
надглазничного края и скулового треугольника или распростра-
нение центральных компонентов рельефа на надглазничный край,
при наличии разграничительной верхнеглазничной борозды, от-
деляющей скуловой треугольник (I и II типы Каннингема и II—
III Бунака). В некоторых случаях отмечено отсутствие рельефа
или же, напротив, максимальное его развитие при слиянии всех
компонентов. Последний вариант описан как весьма редкий, ин-
дивидуальный, у представителей различных этно-территориаль-
ных групп современного человека (Патт, 1955). Таким образом,
все отмеченные у современного человека типы рельефа наблю-
даются и у других приматов, хотя вопрос об их полной гомологии
остается открытым. По Бунаку (1966), надбровные дуги человека
не идентичны надглазничным дугам катарриновых негоминидных
приматов, подобных павианам или макакам, отличаясь по внеш-
ней структуре костной ткани, гладкой и округлой у обезьян. Та-
кого же мнения придерживаются и Эхара и Зайлер (Ehara, Sei-
ler, 1970), связывающие возникновение надбровных дуг с дейст-
вием на периост глазничной части лобной кости мимических мышц
(прежде всего m. corrugator supercilii et m. frontalis. Однако и,
по Каннингему, . «шагреневидность» рельефа определяется на
черепе макаки резуса.
Как у человека, так и у некоторых негоминидных приматов
изменчивость надглазничного рельефа даже в однородной популя-
ции очень высока; существуют и возрастно-половые его измене-
ния, так что у одного и того же индивидуума в разные периоды
онтогенеза степень развития и даже сама структура рельефа мо-
гут существенно различаться: например, одна и. та же обезьяна
может обладать в разные периоды жизни всеми тремя типами
155
рельеф# по схеме Каннингема (Патт, 1955), что необходимо учи-
тывать и при таксономическом анализе.
Можно предполагать, что на формирование надглазничных су-
перструктур действует целый ряд эндо- и экзогенных факторов,
среди которых следует назвать генетический, эндокринный (поло-
вые гормоны, возможно, гормон роста), биомеханический (жева-
тельная мускулатура, мимические мышцы, прикрепляющиеся не-
посредственно в этой области), характер ростовых процессов в
передней части мозга и основания черепа. Поэтому вряд ли сле-
дует переоценивать роль этого признака для таксономии гоминид
и других приматов; его генезис весьма сложен и требует даль-
нейших исследований.
Не вполне выяснен также и вопрос о корреляциях между над-
глазничными суперструктурами и пневматизацией лобной кости.
Принято считать, что у приматов отсутствие развитого рельефа
обычно связано и с отсутствием лобных пазух, хотя индивидуаль-
ные варианты возможны и здесь (так, например, Патт нашел в
одном случае небольшой синус у-орангутана). Еще менее строгой
является обратная связь; у многих кйтарриновых приматов с
очень большим развитием рельефа не обнаружено лобных пазух.
Отсутствует внутренняя связь этих двух формаций и на черепе
современного человека, что характерно, по-видимому, для инди-
видуумов всех рас (Патт, 1955). Один из возможных экзогенных
факторов детерминации развития лобных пазух у человека, оче-
видно, температурный: так, популяции эскимосов Аляски из бо-
лее холодных областей характеризовались меньшей средней пло-
щадью лобных синусов, чем популяции из более теплых зон (Ко-
ertyelyessy, 1972).
Определенное значение в «сапиентном комплексе» черепа име-
ет также и развитие затылочного рельефа. Для современного че-
ловека типична его дисйерсная структура с отсутствием гребне-
видных формаций и ослаблением верхних выйных линий в лате-
ральном направлении, тогда как в центральной части развивает-
ся наружное затылочное возвышение. У антропоидов вероятным
гомологом этих образований является затылочный гребень — ме-
сто прикрепления мощного нухального мускульно-связочного ком-
плекса; однако гребень четко представлен лишь на черепах взрос-
лых самцов крупных антропоидов. У самок и молодых животных
на этом месте развивается обычно округлый торус. Сагитталь-
ный гребень никогда не наблюдается на черепе современного че-
ловека, и височные линии обычно расположены здесь довольно
низко. Наконец, известную роль в гоминидном комплексе играет
также сосцевидный отросток височной кости: его общеё развитие
и особенно выраженность свободного медиального края и лате-
рального округления также составляет специфическую черту са-
пиентного типа. ' .
Для лицевого отдела черепа современного человека характер-
но прежде всего сокращение его общих размеров. Особенно оно
156
• затронуло продольное и высотное направления роста. При сопо-
ставлении средних абсолютных разниц размеров лицевого отдела
черепа во всех трех направлениях роста у человека и крупных ан-
тропоидов максимум везде приходится на продольное направле-
ние. Укорочение лица наблюдалось и в эксперименте при выклю-
чении функции жевательных мышц, что лишний раз подтверждает
..важную роль биомеханического фактора для структуры всего ли-
цевого отдела (Dokladal, 1964; Endo, 1970).
Уменьшение лицевого черепа сочетается у человека с выражен-
ным укорочением неба и уменьшением выступания лица. Общий
лицевой угол (назион-простион к Франкфуртской горизонтали) со-
ставляет у современного человека в среднем 83,5 против 56° у
шимпанзе; индивидуальный максимум у последнего (69°) все еще
далеко не достигает минимума групповых средних человека (77°).
Примечательно, что при сходном в общем характере роста черепа
человека и приматов в целом (Petit-Maire, 1972; Duterloo, Enlow,
1970) наблюдаются некоторые существенные локальные различия.
- Так, и у человека, и у макаки максиллярный комплекс в онтоге-
незе растет вниз, однако у обезьяны передняя часть верхнечелюст-
ной дуги и премаксилла растут и вперед и выступают впереди но-
са. У человека отмечены- особенности костно-хрящевого роста но-
совой области, обусловливающие формирование характерного для
него выступающего носа на фоне уменьшения общего прогна-
тизма, а также возникновение наружной носовой ости (Enlow,
1966).
Тотальные размеры лицевого скелета у современного человека
значительно варьируют в этно-территориальном и расовом аспек-
тах, что отчасти связывается и с некоторыми конкретными адап-
тациями прежде всего к климатическим факторам (Steegmann,
1972).
Целый комплекс черт «сапиентного типа» ассоциируется ' со
структурой нижней челюсти и морфологией зубной системы. Ха-
рактерными для современного человека признаками считают уве-
личение угла нижней челюсти, уменьшение ее массивности, изме-
нение формы обеих альвеолярных дуг с большим округлением
переднего отдела, утончение (абсолютное и относительное) сагит-
тального размера альвеолярного отростка. Форма зубной дуги
обычно почти параболическая на верхней и гиперболическая на
нижней челюсти. Значительные преобразования затрагивают сим-
физеальную область: они заключаются в увеличении угла ее на-ч
—клона к альвеолярной плоскости и возникновении подбородочного
/ возвышения, хотя и широко варьирующего по степени выражен-
ности у современного человека. Далее характерно отсутствие ба-
зальной пластинки нижней челюсти; рельеф внутренней части
симфиза представлен обычно двойной подбородочной остью, в отли-
• чие от ямки на челюстях антропоидов; но, значительная изменчи-
вость рельефа этой области заставляет относиться с осторожно-
стью к этому признаку как таксономическому (Никитюк, 1966а).
5
Известный интерес представляет структура нижнечелюстного
сустава: «сапиентным» вариантом ее является глубокая сочленов-
ная ямка с малым передне-задним диаметром и развитыми соч-
леновным и позадисочленовным бугорками; увеличение глубины
нижнечелюстной ямки связано со значительным размахом движе-
ний' нижней челюсти в вертикальном направлении, что прослежи-
вается и в онтогенезе этой области у современного человека (1п-
gervall, 1972). Весьма существенно также в аспекте антропогене-
за изменение положения надостного отверстия, которое отражает
соотносительные размеры альвеолярной и базальной частей тела
нижней челюсти. Этот индекс составляет у современного человека
34,8—43,8 против 15,8 у шимпанзе (Никитюк,' 1966а), что свиде-
тельствует о гораздо более высоком положении надостного отвер-
стия у человека. Для последнего характерна также значительно
большая частота одиночных подбородочных отверстий и наличие
костного выступа — «язычка», прикрывающего начало канала
нижней челюсти.
Наконец, существует достаточно отчетливый комплекс призна-
ков, выявляющийся 'и в морфологии зубов современного человека.
Сюда относятся: редукция клыка, исчезновение диастем и специа-
лизации первого нижнего премоляра, имеющего у человека двух-
бугорковую форму, уменьшение размеров премоляров. Весьма су-
щественные специфические особенности касаются коренных зу-
бов. Здесь характерны: округленность, сглаженность жевательной
поверхности, отсутствие сильной складчатости бугорков. Далее,
существует обычно прогрессивное уменьшение размеров коронки,
в ряду моляров, «узор дриопитека» сохраняется лишь на Мь тог-
да как для остальных моляров типичен «плюс-узор». Характерны
и уменьшение размеров метаконида и увеличение энтоконида,
сглаженность рельефа передней и задней ямок, а также отсутст-
вие цингулюма и сужения талонида (Зубов, 1966; Twiesselmann.
1973).
КРАТКАЯ КРАНИОЛОГИЧЕСКАЯ
ХАРАКТЕРИСТИКА ИСКОПАЕМЫХ
ГОМИНИД
Для австралопитеков характерна сравнительно небольшая ем-
кость мозговой коробки черепа, причем усиленные поиски новых
материалов и периодическое переисследование старых вносят все
новые коррективы в прежние данные. Так, например, согласно
цифрам, приведенным Тобайясом (Tobias, 1968) для шести чере-
пов грацильных австралопитеков, средний объем мозга состав-
ляет здесь около 500/498 см3; по Холлоуэю (Holloway, 1970), со-
ответствующие определения дают для шести австралопитековых
уже размах емкости 428—485 см3, т. е. в среднем немного мень-
158
ше. Для двух массивных австралопитеков (район оз. Рудольфа)
емкость мозгового черепа несколько выше — 506—570 см3 (Hol-
loway, 1973). Однако нужно учитывать малые тотальные размеры
тела этих приматов; при вычислении коэффициентов церебрализа-
ции на основании приведенных выше данных (по формуле Дю-
буа) даже минимальный из них равен 56 г на 1 кг веса тела
т. е. значительно больший, чем у шимпанзе. Разрыв с последним
особенно велик у Н. habilis со средней емкостью мозгового черепа
около 625 см3 и коэффициентом церебрализации 76 г на 1 кг веса
тела.
Что касается краниофациальных соотношений, то почти все
авторы указывают на заметное преобладание относительного раз-
вития мозгового черепа у австралопитековых сравнительно с
вариантом шимпанзе. Правда, эти различия не всегда четки в циф-
ровом выражении: tqk, величины вертикального краниофациаль-
ного указателя составляют для мужских индивидуумов A. africa-
nus (плезиантропа) и шимпанзе соответственно 119,0 и 100,0. По-
видимому, внутри группы самих австралопитеков возможна диф-
ференциация в этом отношении, и детеныш A. africanus отличает-
ся особенно выраженным сдвигом краниофациальных пропорций
в «гоминидном» направлении сравнительно с особями современ-
ных африканских антропоидов соответствующего биологического,
возраста.
Одна из самых прогрессивных черт в организации мозгового,
черепа австралопитековых — развитие свода. Указатель надглаз-
ничной высоты калотты равен 68 у грацильного австралопитека
Стеркфонтейн V против 63—77 у современного человека и 49,8—
50,8 у шимпанзе. Сближение с человеческим типом здесь отчетли-
во. Однако, как и по другим признакам мозгового черепа, среди
австралопитековых отмечается вариабильность, и у массивных,
форм указатель значительно ближе к шимпанзоидному. Прогрес-
сивный сдвиг церебрализации отражается и в тенденции к вып-
рямлению лба прежде всего у грацильных австралопитеков. По-
добные изменения высоты черепа предполагают нарастание пло-
щади извилин мозга (Бунак, 1936).
Первостепенную важность имеет трансформация затылочной:
области в связи с изменениями локомоции и осанки и иными ус-
ловиями балансирования головы. Она затрагивает положение за-
тылочных мыщелков (более центральное), разрастание верхней:
части чешуи затылочной кости и опускание точки инион, выра-
женные у австралопитеков в степени, довольно близкой к челове- •
ческой, а также относительное сокращение выйной площадки, от-
ражающее ослабление мускульно-связочного аппарата этой обла-
сти.
1 При среднем весе тела 36 кг (Howell, .1962), а возможно, еще меньше (Lo-
vejoy et al., 1970). Н. habilis вряд ли существенно отличался по общим раз-
мерам тела от грацильных австралопитеков (если судить о нем по Олдувей 8)..
159
Коснемся, наконец, развития рельефа на черепах австралопи-
тековых. Грацильные и массивные формы, по-видимому, разли-
чаются в этом отношении; у последних рельеф сильнее. В над-
глазничной области, он обычно имеет вид торуса (III тип Кан-
нингема), хотя и несколько ослабленного в латеральных частях.
Если к тому же принять во внимание полное отсутствие следов
надглазничного рельефа на черепе детеныша A. africanus, понят-
но, что у ранних гоминид можно предположить его значительную
групповую, индивидуальную и половую изменчивость, как и в по-
пуляциях современных приматов, в том числе человека. Это под-
тверждается также морфологией надглазничной области так на-
зываемого чадантропа, имеющего исключительно мощный торус,
и отсутствием развитого надглазничного .валика на очень древнем
черепе гоминида из района оз. Рудольфа (Day et al., 1975).
В затылочной области на черепе австралопитековых отсутст-
вует формация гребневидного рельефа, в отличие от современ-
1 ных антропоидов Африки, но существует округленный валик, об-
разовавшийся из слияния верхней и высшей выйных линий. Осо-
бенно примечательно, что затылочный гребень отсутствует даже
на тех черепах,, где есть сагиттальный, хотя у современных круп-
ных антропоидов они обычно встречаются вместе. Согласно Ле
Гро Кларку (1955а), отмечается округленность внешней поверх-
ности сосцевидного отростка, что тоже рассматривается как ха-
рактерная принадлежность гоминидного типа.
В целом можно говорить, по-видимому, о тенденции к ослаб-
лению рельефа черепа у австралопитековых, имея в виду некото-
рых их грацильных представителей. Значительная индивидуаль-
ная (а возможно, и популяционная) изменчивость суперструктур,
особенно лобной кости, не является специфическим признаком
этих гоминид, но присуща приматам вообще.
Несколько слабее представлен лицевой отдел черепа австра-
лопитековых. Судя по наиболее достоверной величине лицевого
угла на черепе одного из A. africanus (плезиантропа), равной 70°,
они занимают по этому признаку примерно промежуточное поло-
жение между африканскими антропоморфными (55—56°) и совре-
менным человеком (83,5°).
Особенно характерен сдвиг в гоминидном направлении в строе-
нии носовой области. У A. africanus (плезиантропа) наблюдается
значительное выступание носовых костей за профильную линию
(назион — простион), что свойственно черепу человека и совер-
шенно нехарактерно для африканских антропоидов (Никитюк,
1966).'
Существенно также, что у некоторых из австралопитеко-
вых была развита наружная носовая ость. По-видимому, до неко-
торой" степени промежуточным было положение A. africanps (пле-
зиантропа) по форме альвеолярного отростка верхней челюсти,
относительно более широкого и укороченного, чем в шимпанзоид-
ном его варианте.
160
Что касается нижней челюсти, то ее общие размеры варьиро-
вали в связи с дифференциацией грацильных и массивных ав-
стралопитеков по развитию и массивности скелетных формаций.
Вместе с тем характерной общей чертой этих форм является из-
менение формы нижней челюсти сравнительно с понгидным типом,
что выражается в относительном укорочении и расширении тела
кости. .Австралопитеки занимают промежуточное положение меж-
ду человеком и понгидами по форме альвеолярной дуги: она отно-
сительно расширена и укорочена, при этом укорочение сопровож-
дается здесь редукцией прежде всего передней части дуги и ее
округлением. Характерно обычно одиночное, как у современного
человека, подбородочное отверстие, которое и по положению не
отклоняется существенно от современного варианта. Передняя
стенка подбородочного симфиза, лишенного базальной пластинки
антропоидов, почти отвесная; рельеф внутренней части симфиза
весьма изменчив, чаще более или менее развита ямка, но может
существовать и ость. Сочленовная ямка для нижней челюсти еще
не имела специфического антропинного характера.
Весьма характерен комплекс гоминидных признаков в строении
собственно зубной системы австралопитековых. Для них типично
сочетание небольшого клыка при отсутствии диастемы и соответ-
ствующей специализации первого нижнего премоляра с крупными
коренными зубами. Окклюзионная поверхность последних также
имеет гоминидный облик, с невысокими сглаженными бугорками
и отсутствием складчатости и суженности талонида. Однако су-
ществует примитивное соотношение размеров в ряду моляров
(М1<М2<МЗ); на всех нижних молярах отчетлив узор дриопите-
ка, метаконид но размерам примерно равен протокониду, исклю-
чительно велик гипоконус (Зубов, 1966). В развитии зубной си-
стемы снова проявляется дифференциация массивных и грациль-
ных форм. У первых особенно «дисгармонично» соотношение пе-
реднего и заднего отделов зубной дуги, что означает сочетание
очень крупных моляров с малыми по размерам клыками и резца-
ми, отмечены признаки тавродонтизма. Предполагается, .что в
этом отношении массивные формы более специализированы, чем
грацильные, что может быть связано с их преимущественной рас-
тительноядностью. У грацильных же форм обнаружены и соответ-
ствуют их вероятной большей древности некоторые примитивные
признаки, например, остатки цингулюма, квадратная форма верх-
них моляров, почти параллельные ряды заклыковых зубов, очень
крупные размеры гипоконуса. По мнению некоторых исследова-
телей, (Robinson, Steudel, 1973), грацильные австралопитеки го-
раздо ближе по морфологии зубной системы к Н. erectus, чем к
массивным, формам *.
1 Существует .и мнение об отсутствии резких различий между гранильными и
массивными формами по структуре зубов, как и по количеству растительной
пищи в их рационе (Koenigswald, • 1973; Wallace, 1973).
11х/2 Е. Н. Хрисанфова 161
Сказанное относится и к морфологии черепа в целом. На ос-
новании «индексов сходства» с Н. sapiens выявляется диамет-
рально противоположное положение грацильного австралопитека
Стеркфонтейн 5 и представителя массивных форм — зинджантро-
па. В первом случае индекс сходства с черепом современного че-
ловека составляет 36,2%, во втором 8,1% (Rosinski, 1974).
Для Н. habilis, как «прогрес-
сивного австралопитекового»,
нужно подчеркнуть большее аб-
солютное развитие мозгового че-
репа и меньшие размеры обеих
челюстей. Строение подбородоч-
Рис. 30. Черепа высших приматов: А —
горилла; Б — австралопитек; В — пите- ’
кантроп; Г—Шапелль; Д—Штейнгейм;
Е — современный человек (по Харри-
сон, Уайнер, 1968)
ного симфиза и рельеф находятся в пределах их вариаций у
австралопитековых, причем существуют значительные колебания
в выраженности мускульного рельефа черепа.
Среди форм, таксономическое положение которых недостаточ-
но ясно, упомянем еще телантропа и чадантропа. Первый изве-
стен преимущественно по структуре зубов, сочетающей ряд про-
грессивных (гоминидная форма альвеолярной дуги нижней челю-
сти, малые клыки, отсутствие диастем,. форма коронки третьего
моляра) и примитивных особенностей (например, крупные моляры,
как у австралопитеков). Весьма интересен также краниофациаль-
ный фрагмент чадантропа (Coppens, 1965). Этот примат, вероятно,
обладал более прогрессивными особенностями сравнительно с ти-
пичными австралопитековыми: более отвесным лбом,' незначитель-
ным посторбитальным сужением, небольшим клыком с отсутстви-
ем диастем. Вместе с тем прогнатизм отчетлив, и надглазничный
рельеф здесь очень мощный.
Особый интерес представляет находка Лики (район оз. Ру-
дольфа) черепа ископаемого гоминида большой древности
(не менее 2 млн. лет), для которого'характерен ряд весьма про-
грессивных особенностей, как например, большая емкость мозго-
вого черепа (770—775 см3), отсутствие надглазничного валика и
умеренное позадиглазничнбе сужение (Day et al., 1975).
Резюмируя все сказанное, следует оценить морфологию чере-
па австралопитековых и близких к ним форм, как обнаруживаю- •
щую определенный сдвиг на пути гоминизации (рис. 30). Это ка-
сается таких важнейших в таксономическом плане особенностей,
как степень церебрализации и краниофациальные соотношения,
возрастание высоты свода черепа, уменьшение наклона лба, пере-
стройка затылочной области в связи с бипедией, изменение поло-
162
жения затылочного отверстия, некоторая тенденция к ослаблению
рельефа, отсутствие затылочных гребневидных формаций, струк-
тура сосцевидного отростка, перестройка в гоминидном направле-
нии носовой области, изменение формы обеих альвеолярных дуг,
укорочение и округление их передних отделов, прямолинейный
контур симфиза нижней челюсти, редукция базальной «обезьяньей
полки», ряд особенностей в строении зубов.
Если теперь обратиться к более ранней стадии эволюции, воз-
можно, уже собственно гоминидного ствола (рамапитеки, кениа-
питеки), то можно отметить существование некоторых из числа
перечисленных признаков еще в миоцене. К ним принадлежат, по-
видимому, малые клыки, отсутствие диастем, изменение формы
альвеолярной дуги; указывается даже на специальное сходство
коренных зубов австрало- и рамапитеков (Pilbeam, 1966)
Суждение о емкости мозгового черепа архантропов можно со-
ставить на основании довольно представительных материалов.
Существуют данные как о более, ранних формах этих гоминид
(яванские питекантропы, лантьянский гоминид из Кунваньлиня),
так и о наиболее поздних представителях данной группы — си-
нантропах. В первом случае вариации емкости 775—975 см3; во
втором — 915—1225 см3, т. е. наблюдается некоторое повышение
величины признака с частичной трансгрессией обоих рядов. Учи-
тывая емкость мозговой коробки Н. habilis, можно предположить,
что hiatus между австралопитековыми и древнейшими так назы-
ваемыми «настоящими гоминидами» не был столь значительным,
как это предполагалось, поскольку минимум для архантропов
(Олдувей 12) составляет, по данным .Холлоуэя (1973), 727 см32.
В еще большей мере это подтверждается сопоставлением коэф-
фициентов церебрализации. Для яванских питекантропов значе-
ние «к» почти такое же, как у Н. habilis — 79 г на 1 кг веса те-
ла; у синантропов был сделан уже значительный шаг вперед в этом
отношении — величина коэффициента равна здесь 105 г, т. е. поч-
ти в 1,5 раза выше, чем у ранних и более примитивных архантро-
пов Явы. Эта величина может оказаться еще и преуменьшенной,
так как расчеты по формуле Дюбуа для обоих групп архантро-
пов производились на основе одного и того же значения веса
тела—60 кг, что могло вызвать некоторое занижение его у мало-
рослых синантропов. Во всяком случае приведенная для послед-
них цифра никак не является искусственно ^завышенной и, следо-
вательно, свидетельствует о прогрессивном сдвиге этого показа-
теля у поздних архантропов. В этом убеждают также расчеты
емкости черепа для человека из Вертешсёллеш, обладавшего, по-
1 Не все- морфологи рассматривают эти особенности как характерные для пред-
ковой формы гоминид. Нужно иметь ® виду и гипотезу о существО1Вании
сходных долговременных тенденций в эволюции зубной системы дивергирую-
щих родственных групп высших приматов (Frisch, 1965; Skaryd, 1971).
2 См. также стр. 164 о новой находке Лики в районе оз. Рудольфа.
111/!»*
видимому, очень крупным в масштабе архантропов мозгом Но
церебро-соматические соотношения не были еще вполне человече-
скими даже у синантропов, о чем можно судить на основании рас-
четов по внутривидовой («онтогенетической») формуле — £=
=кхР°’25 (Бунак, 1966).
Поскольку более или менее полно оба отдела черепа сохрани-
лись лишь у синантропов, только здесь можно оценить краниофи-
циальные соотношения. Общий указатель пропорций обоих отде-
лов 2 составляет 154 против 228 в среднем у современного чело-
века (мужская европеоидная серия); при этом основное различие
с последним отмечается в общем развитии лицевого отдела чере-
па, гораздо более крупного у синантропов. По нашим данным,
наибольшая величина абсолютной разницы в размерах лица си-
нантропа и современного человека (по средним) приходится на
продольные размеры — она более чем вдвое превышает здесь со-
ответствующую среднюю разность для высотных и широтных диа-
метров лица.
Что касается абсолютных размеров мозгового черепа, то ос-
новное различие с современным человеком выражается в значи-
тельно меньшем у синантропа высотном его развитии, что и об-
условливает здесь в первую очередь меньшую его емкость.
Сопоставление вертикального краниофациального указателя
синантропа и современного человека также подчеркивает отмечен-
ное выше сравнительное «недоразвитие» высотного диаметра моз-
гового черепа (индекс синантропа —"67 — выходит за верхнюю
границу групповых средних у современного человека).
Показатели развития свода свидетельствуют о значительной
его уплощенности в целом по группе архантропов. Все известные
значения индекса Швальбе здесь обычно ниже 40,0. Особенно
мал указатель у одного из древнейших архантропов — питекант-
ропа IV, где он составляет всего 24,6; у тринильских питекантро-
пов I и II индекс равен в среднем 35,4 (33,3—37,4); в группе синан-
тропов отмечены самые высокие его значения (38,5 в среднем при
индивидуальных вариациях: 34,8—41,2). Все эти величины в общем
ниже, чем у австралопитековых (Бунак, 1966), что объясняется
значительным «отставанием» высотного развития черепа сравни-
тельно с двумя другими его диаметрами при возрастании его об-
щих размеров у архантропов.
Если судить об округленности продольного свода черепа по
степени изгиба теменных костей, то соответствующий хордо-дуго-
вой указатель составит у питекантропов' 98, синантропов — 94,1,
современного человека — 89,4. Здесь снова подчеркнут меньший
1 Упомянем в связи с этим о концепции, определяющей для древневенгерского
архантропа место в начале прогрессивного ствола, ведущего к человеку из
Сванскомба (Thoma, 1966; Wolpoft, 1971).
2 Продольный диаметр X поперечный диаметр X высотный диаметр {Ьа—Ьг) %
Длина основания лица X скуловая ширина X полная высота лица
164
изгиб теменных костей у архантропов в целом и прогрессивный
сдвиг более поздних форм (синантропы), сравнительно с более
ранними (питекантропы). Об исключительной уплощенности сво-
да в области теменных костей на черепе олдувейского питекан-
тропа сообщает Хеберер (Heberer, 1963); низким сводом обладал
также очень древний лантьянский гоминид из Кунваньлиня (Aig-
ner, Laughlin, 1973).
Угол наклона лба (назион — брегма) у питекантропов Явы
не больше, чем у A. africanus: примерно 44° у плезиантропа, 45,5°
у питекантропов I и II и даже 35° у питекантропа IV. Но по сте-
пени посторбитальной суженности (наименьшая ширина лба в %
наибольшей ширины черепа) питекантропы I—II (средняя ука-
зателя 60,9) и синантропы (средняя 61,2) лишь немного уступают
групповому минимуму (62).
Наклон выйной площадки' и топография затылочного отверстия
даже у древнейших гоминид близки к современным вариациям
этих признаков, но положение иниона и, следовательно, степень
разрастания верхней чешуи отличаются в сторону большей при-
митивности. 'Отмечена исключительно большая высота затылочной
кости у Вертешселлеш.
Рельеф мозгового черепа архантропов отличается некоторыми
весьма своеобразными особенностями. Для всех них характерно
прежде всего значительное развитие надглазничных суперструк-
тур в виде сплошного торуса (III вариант Каннингема). Очевид-
но, нет тенденции к перерывам (дисперсному расположению)
рельефа 1. Наиболее мощная лобная суперструктура с резко обо-
значенным посторбитальным желобом отмечена пока на черепе
олдувейского питекантропа, но здесь не наблюдается утолщения
торуса в латеральной части. На затылочной кости обычно выра-
жен валик, не представляющий, однако, гомолога затылочного
гребня антропоидов. Как известно, он был наиболее развит на че-
' репе питекантропа IV, но является неподр аз деленным и у Вёр-
тешселлеш. Нельзя вывести строгой зависимости и между разви-
тием лобной и затылочной суперструктур, так как на черепе ол-
дувейского питекантропа затылочный валик был как раз выражен
в слабой степени. Сагиттальный гребень отсутствует у архантро-
пов,. хотя нужно иметь в виду его возможный рудимент у пите-
кантропа IV, где отмечено выступание отдельных участков кости.
Бунак (1966) указывает на неполную гомологию и этой структуры
с гребнем африканских антропоидов. Сосцевидные отростки также
варьируют по степени своего развития, не обнаруживая настояще-
го сапиентного типа. Любопытно, что и здесь вновь отсутствует
сколько-нибудь' выраженная корреляция с развитием других ком-
понентов рельефа; следует, конечно, иметь в виду и половой ди-
морфизм в этих признаках.
1 См. у Патта (1955) относительно лобного рельефа питекантропа.
11 Е. Н. Хрисанфова 165
Одной из отличительных черт черепа архантропов, очевидно,
являлась оченй большая толщина костей свода: соответствующие
значения для лобной кости составляют, например, 8,0 мм у пите-
кантропов и 9,3 мм у синантропов против 6,1 мм у современного
человека, причем эти различия зависят главным образом от раз-
вития диплоэ (Бунак, 1966). Напомним, что у олдувейского пи-
текантропа указывается огромная толщина лобной кости — 18 мм
(!). Очень толстыми стенками черепа обладали также лантьян-
ский гоминид и человек из Вертешселлеш. Примечательно, что,
как и в случае повышенной внутренней массивности трубчатых
костей конечностей у ископаемых гоминид (см. выше), этот при-
знак отнюдь не является «питекоидным» в смысле сближения пре-
жде всего с современными антропоидами.
Уже указывалось, что,, судя по группе синантропов, древней-
шие люди обладали более крупным абсолютно и относительно ли-
цевым отделом черепа; при этом, основные различия с неоантро-
пами касаются продольного развития лица. Вместе с тем по та-
кому важному в таксономическом отношении признаку, как угол
вертикальной профилировки (общелицевой угол), синантроп
(84,5°) не только попадает в размах значений этого признака у
современного человека, но и почти совпадает с «типичным» чело-
веческим вариантом (83,5°). Из числа других признаков лицевого
скелета для архантропов (синантропа) характерны крупные глаз-
ницы, большая абсолютная и относительная ширина носа, челове-
ческое положение носовых костей, что, впрочем, отмечено еще у
австралопитековых. Однако наружная носовая ость, по-видимому,
выражена здесь не более отчетливо, чем у самых ранних гоминид.
Также отсутствует и явственная клыковая ямка.
В соответствии с еще значительным удлинением основания ли-
ца нёбо архантропов очень длинное и суженное. Интересно отме-
тить также, что лобная, основная и верхнечелюстная пазухи у
синантропов были развиты слабо сравнительно с питекантропами,
что соответствует меньшей, чем у последних, пневматизации чере-
па (Никитюк, 1966). Выше уже указывалось, что эти вариации у
гоминид, возможно, связаны в числе других и с климатическим
фактором. Архантропам, включая также питекантропа IV и
олдувейского питекантропа, была свойственна глубокая, но еще
не вполне сапиентная, форма сочленовной ямки для нижней че-
люсти.
Нижние челюсти архантропов представлены довольно полно:
питекантропы, синантропы, лантьянский гоминид из Ченьчиаво,
атлантроп, гейдельбергский человек. Общими их признаками мож-
жно считать массивность, большую ширину, отсутствие подборо-
дочного выступа. Угол симфизеального профиля архантропов со-
ставляет вариационный ряд, не обнаруживающий трансгрессии с
современным (соответственно 59—70° и 73—94°). Как и следо-
вало ожидать, различия отражаются на длине нижней челюсти,
высоте и ширине ее ветви. Впрочем, в отношении последнего при-
166
знака существуют вариации и среди самих архантропов. То же
можно сказать применительно к рельефу внутренней поверхности
симфиза: подбородочные ости обнаружены, например, у синантро-
пов' и отсутствуют на гейдельбергской челюсти. По-видимому, су-
ществуют вариации и в рельефе наружной поверхности симфиза:
так отмечаются зачатки подбородочного выступа на одной из че-
люстей синантропа. Изменчива и структуры подборочного от-
верстия — напомним о необычной частоте его множественного ва-
рианта у синантропов.
Общими особенностями зубной системы архантропов являют-
ся: близкий к человеческому тип альвеолярной дуги, крупные раз-
меры зубов, частота наличия «узора дриопитека», тенденция к
уменьшению размеров третьего моляра, образованию «плюс-узо-
ра» на М2 и М3, а также к редукции цингулюма, некоторая на-
клонность к редукции бугорков моляров и к слиянию их корней.
Для этих гоминид была характерна известная «мозаичность» в
распределении прогрессивных и примитивных признаков. Так, наи-
более примитивная зубная система питекантропа IV с наличием
диастемы и несколько повышенного развития клыка характери-
зуется вместе с тем тенденцией к редукции М3. Полное отсутст-
вие М3 наблюдается на лантьянской челюсти, также имеющей
очень древний возраст. Вместе с тем на М] и М2 синантропа очень
крупный метаконид, суженный талонид, есть цингулюм, а на мо-
лочных молярах — даже пароконид (Зубов, 1966). Зубы человека
из Вертешселлеш в общем сходны с зубами синантропа, хотя и
не вполне им тождественны (Thoma, 1966).
Известные остатки древних людей — палеоантропов — в хро-
нологическом отношении целесообразно разделить, на ранние (до-
вюрмские) и более поздние — вюрмские. К первым относятся из
числа более или менее полных находок — Сванскомб, Фонтеше-
. вад, Штейнгейм, Эрингсдорф, Саккопасторе, Крапина, Галилея;
ко вторым — классические европейские формы, палеоантропов—не-
андертальцы, палестинцы, а также некоторые другие азиатские
(Кафзех, Шанидар, Тешик-Таш, Нгандонг) и африканские наход-
ки (Брокен-Хилл, Эяси, Танжер, Темара, Джебель Ирхуд, Дире-
дава).
Представляется трудной задачей дать в сжатом изложении
общую краниологическую характеристику столь разнообразной в
морфологическом отношении группе. Для целей настоящего иссле-
дования здесь имеют значение две стороны: во-первых, выяснение
вопроса о примерном времени появления конкретных признаков
«сапиентного комплекса» у древних людей и, во-вторых, возмож-
ность разграничения отдельных вариантов палеоантропов по отно-’
сительной концентрации «примитивных» и «сапиентных» призна-
ков.
Придерживаясь того же порядка изложения, начнем характе-
ристику черепа палеоантропов с оценки абсолютного и относитель-
ного развития его мозгового отдела (рис. 30).
И* . 167
Емкость мозгового черепа варьировала у древних людей в пре-
делах: 1035 см3 — 1700 см3 при средней в 1350 см3. Здесь снова
наблюдается, частичная трансгрессия с соответствующим рядом
архантропов. В то же время можно говорить уже о совершенно
человеческом («сапиентном») развитии мозгового отдела черепа—
его емкости при существенно иной форме эндокрана (Кочеткова,
1973). Даже средние емкости почти совпадают у палеоантропов
и современных людей. Это свидетельствует и о том, что на этой
стадии морфологической эволюции гоминид были достигнуты
вполне человеческие церебросоматические отношения.
Если теперь попытаться оценить развитие мозгового отдела
черепа сравнительно с лицевым, то общий указатель этих пропор-
ций составит для палеоантропов около 183, что определяет их про-
межуточное положение между архантропами и современными
людьми, ближе к первым. Существует дифференциация морфоло-
гических вариантов древних людей по этому признаку, так для ран-
них форм (Штейнгейм, Гибралтар) модифицированный индекс
краниофациальных соотношений 1 равен примерно 284, тогда как
у поздних, классических форм (Шапелль, Мустье) и у Брокен-
Хилл он составляет в среднем около 250. Наиболее высок этот
указатель у людей Схул, где его величина, усредненная для Схул
IV, V и IX, равна 314 против 413 у современного человека. Если
сравнить между собой указатели ранних и более поздних прогрес-
сивных форм, выявляется таким образом определенный сдвиг в
сторону возрастания краниофациального указателя. Это сопостав-
ление подчеркивает и качественную специфику краниофациаль-
ных ^отношений в типе классических неандертальцев. Интерес-
но, что различия в этом признаке между Палеоантропами и сов-
ременными людьми выявлялись, по-видимому, уже в детстве: так,
значение индекса1 2 у Тешик-Таш сравнительно с современными
детьми того же возраста —• соответственно 103,0 и 132,5.
По сравнению с архантропами (синантропами) мозговой череп
палеоантропов увеличился главным образом в высотном направ-
лении; общие же размеры лицевого отдела различаются у этих
двух групп весьма незначительно. Не существует и сколько-ни-
будь строгой корреляции между возрастанием абсолютных раз-.
меров мозгового черепа и повышением свода. Индекс Швальбе
варьирует у палеоантропов в целом в пределах 39,2—52,6 против
50—63 у современного человека. Причем как раз формы с абсо-
лютно меньшим развитием мозга (ранние палеоантропы) обнару-
живают обычно более высокий свод черепа и большее выпрямле-
ние лба сравнительно с более поздними специализированными
неандертальцами Европы. Упомянем в этой связи о значительной
(в масштабе ранних палеоантропов) абсолютной высоте черепа
1 Модификация указателя с заменой полной высоты лица на верхнюю.
2 Модификация того же указателя с заменой длины основания лица на длину
неба.
168
Сванскомб и повышении свода у позднего «прогрессивного» па-
леоантропа Схул V (индекс Швальбе 52,6).
Другой показатель развития продольного свода — степень
изгиба его костей — снова определяет промежуточное поло-
жение палеоантропов в целом между архантропами и неоантро-
пами.
Промежуточной является и структура лобной кости. Угол на-
клона лба варьирует в пределах 39—51°, вновь обнаруживая наи-
большие значения у ранних прогрессивных форм, сравнительно с
поздними специализированными (соответственно 47—51° и 39—
50°). В отличие от архантропов, индекс посторбитального сужения
лба уже входит в пределы вариаций этого признака у современ-
ного человека, хотя строение этой области в целом здесь весьма
специфично благодаря суперструктурам лобной кости.'
У древних людей, как правило, развивается мощный -сплош-
ной наглазничный рельеф (торус). Обычно он понижается в ла-
теральном направлении, но может и повышаться (например, у
Шанидар, Брокен-Хилл), или его толщина существенно не ме-
няется (Крапина). Думается, что эти различия несколько переоце-
нены в таксономическом плане (Дак называемые «континенталь-
ные вариации»), ибо, например, среди азиатских форм у Мала
и Нгандонг отмечаются несходные типы. В большинстве случаев
рельеф очень мощный, в том числе и у прогрессивных форм, но в
некоторых случаях существует тенденция к его ослаблению и пре-
рывистости (например, наличие надорбитальной борозды у Схул
IV, Штейнгейм, Крапины С), а возможно, и отсутствию (Фонте-
шевад). Лобные пазухи варьируют по степени выраженности, но
в целом, видимо, развиты слабее, чем у современных людей. Та-
ким образом, у палеоантропов, как и у современных людей, на-
блюдаются различные варианты надглазничного рельефа, но с
резко выраженным преобладанием торуса (III типа по Каннин-
гему, IV — по Бунаку) над остальными.
Выше уже говорилось о вполне человеческом положении вый-
ной площадки у архантропов. То же имеет значение и для палео-
антропов. Приведем для иллюстрации значение угла наклона вый-
ной площадки у Штейнгейм; он равен здесь 34° против 35° у сов-
ременного человека.
Однако, как и у архантропов, еще. не вполне достигнут сапи-
ентный статус по степени разрастания верхней части затылочной
чешуи: для ископаемых гоминид типично более высокое положе-
ние точки инион. По степени «поворота» окципитальной части,
отражающей происходившее в филогенезе гоминид «вращение»
дистального отдела черепа вокруг вестибулярной оси, прогрессив-
ные палеоантропы снова занимают промежуточное положение
между современным человеком и классическими формами с очень
малой трансгрессией признака между двумя последними. Инте-
ресно, что череп из Гибралтара приближается здесь к типу древ-
них палеоантропов, для которых характерна более округленная
169
конфигурация затылочного отдела с близким к современному по-
ложением иниона и обычно хорошо выраженным затылочным ва-
ликом (Сванскомб, Фонтешевад, Эрингсдорф, Штейнгейм, Крапи-
на, Саккопасторе) (Fuste, 1957; Алексеев, 1966). У классических
форм существует малый поворот затылочной кости и малый пере-
гиб основания черепа, что рассматривается как вторичная черта
специализации, но не как сохранение примитивного состояния.
Пяткообразное выступание затылка сочетается здесь с высоким
положением иниона, слабым рельефом нухальных линий. Отме-
чается, что «шиньонообразный» затылок поздних неандерталь-
цев— морфологически иная структура, чем встречающийся иногда
у неоантропа (включая кроманьонцев) внешне сходный вариант,
так как в последнем случае речь идет только о наличии надламбдо-
видного теменно-затылочного вдавления (Ducros, 1967). Особую
конфигурацию имеет затылочная область у некоторых «тропиче-
ских» палеоантропов (Брокен-Хилл, Нгандонг, отчасти Салданья).
У них существует сильный перегиб затылочной кости с почти пло-
ской нухальной площадкой и «гребневидным» затылочным вали-
ком. В некоторых случаях, как и у архантропов, отмечены взду-
тия по сагиттальной линии — следы сагиттального «гребня» (Бу-
нак, 1966). Нужно отметить и отсутствие строгой корреляции меж-
ду развитием отдельных элементов рельефа. У классических не-
андертальцев при сильной выраженности надглазничного торуса
затылочный валик обычно невелик, а верхние выйные’линии сла-
бы или вовсе не развиты.
В целом у палеоантропов еще не полностью выражен сапиент-
ный комплекс сосцевидного отростка. Лучшее его развитие от-
мечено у Эрингсдорф и Штейнгейм, вполне современен он у
Схул VI. Таким образом, хорошее развитие этого элемента рель-
ефа чаще встречается на «прогрессивных» черепах. Однако здесь,
как и в случае архантропов, есть исключения — крупные сосце-
видные отростки у Нгандонг и Брокен-Хилл.
' Для палеоантропов, как и древнейших гоминид, типична боль-
шая толщина костей черепа. В среднем толщина лобной чешуи
составляет здесь 7,8 мм (Бунак, 1966). Не наблюдается диффе-
ренциации по этому признаку в каком-нибудь определенном отно-
шении: очень толстые кости черепа были и у «прогрессивных»
форм (Сванскомб, Схул), и у более «примитивных» (Эяси, Сал-
данья). Велика толщина костей свода и у ребенка из Тешик-
Таша.
Характерной особенностью лицевого отдела черепа палеоан-
тропов, как и архантропов, являются его большие размеры: сред-
няя величина разницы с современным человеком по продольному
размеру лица (длине его основания) у них лишь немного меньше,
чем у синантропов, а средняя разница по высотному развитию ли-
цевого отдела — полной и верхней высоте лица — даже больше.
Но здесь, как и у более ранних гоминид, уже полностью преобла-
дает человеческая вертикальная профилировка лицевого отдела
170
(ортомезогнатия или даже гиперортогнатия, как у Шанидар I с
общелицевым углом 97°
Характерной чертой строения лицевого черепа палеоантропов
является, по-видимому, отсутствие клыковых ямок, хотя имеются
исключения среди наиболее прогрессивных форм (Саккбпасторе,
а также Староселье).
Как и у архантропов, обычны крупные орбиты и большие раз-
меры носа (особенно широтные — хамэ- или гиперхамэриния).
Но в отдельных случаях наблюдается «сапиентная» тенденция и
в структуре этих областей. Так, верхняя высота лица меньше, а
орбиты ниже в группе Схул, узкие носовые кости были у Схул V.
Обычно в той или иной степени выражена носовая ость. Сильное
выступание носовых костей наблюдается не у всех палеоантро-
пов (например, Брокен-Хилл),,но строение носовой области, как
и у более ранних форм гоминид, в целом вполне человеческое. Ти-
пичны также большие размеры нёба, крупные и массивные нижние
челюсти. В этом отношении палеоантропы, возможно, занимают
промежуточное положение между архантропами и неоантропами.
Угол симфизеального, профиля' палеоантропов явно распо-
лагается между аналогичными показателями у архантропов и сов-
ременных людей. Подбородочный выступ обычно отсутствует, ис-
ключая некоторые ранние и поздние прогрессивные формы (сфор-
мированный выступ у Схул V, а также Схул IV и II; зачаток вы-
ступа у Крапины). Рельеф внутренней поверхности симфиза варь-
ирует, т. е. подбородочная ость существует не на всех челюстях;
ее нет у Темара, она неотчетлива у Рабат. Подбородочное отвер-
стие также варьирует: встречаются как ординарные (Схул), так
и двойные (Табун I) и даже множественные варианты (Ла Кина).
Изменчива и форма ветви; выше говорилось о существовании та-
ких вариаций и у архантропов. Что касается сочленовной ямки
для нижней челюсти, то сапиентный комплекс здесь также не-
полный; отмечается значительное развитие сочленовных отрост-
ков в поперечном направлении (Никитюк, 1966).
Зубы палеоантропов в целом мало отличаются от зубов неоан^
тропов, речь идет лишь о разной концентрации примитивных и
прогрессивных признаков (Зубов, 1966). У палеоантропов зубы в
общем крупнее, может существовать примитивное соотношение
развития моляров — отсутствие тенденции к редукции третьих
моляров, менее выражена также редукция бугорков, отчетливее
складчатость. Сравнительно нередко наблюдается «узор дриопи-
тека», описан цингулюм, как у некоторых архантропов, часто на-
блюдается тавродонтизм. Как и в других системах, отмечаются
иногда и «ультрагоминидные» особенности: большая, чем у сов-
ременного человека, редукция задних моляров, тенденция к сра-
станию корней, квадратная форма нижних моляров, что 'чаще
4 Отмечается, впрочем, что при вестибулярной ориентировке черепа, неандер-
тальцев характеризуются выступанием лицевого отдела (Fenart, Heim, 1968) <
171
встречается у ранних форм. Любопытно, что у прогрессивной фор-
мы Схул V третий моляр довольно крупный.
В целом по совокупности признаков наиболее близки к совре-
менному долихокранному типу черепа Эрингсдорф и Схул V (Бу-
нак, 1966) (рис. 31). Этот исследователь выделяет в общем две
у палеоантропов, положив в основу
емкость мозгового чеоепа и сЬоому
продольного свода. К первой с ем-
костью 1200—1400 см3 относятся
формы с более высоким сводом,
округленным или угловатым высту-
панием затылка, изменчивым релье-
фом, иногда достигающим значи-
тельного развития; ко второй с ем-
костью выше 1450 см3 — черепа со
значительной уплощенностью свода,
группы по структуре черепа
Рис. 31. Череп Схул V (по McCown, Keith,
19?9)
сильно развитым рельефом, выраженным посторбитальным суже-
нием, отчетливо выступающим затылком. Конечно, это разграниче-
ние довольно условно, и палеоантропам, как и другим ископаемым
гоминидам, свойственна значительная «мозаичность» в проявлении
отдельных прогрессивных и примитивных особенностей, но относи-
тельная концентрация их у этих двух вариантов различается до-
вольно явственно.
Судя по строению черепа ребенка из Староселья (Крым), об-
ладающего в общем сапиентным обликом, основной комплекс са-
пиентных черт уже сформировался к позднему мустье. Сюда сле-
дует отнести: большую высоту черепа, крутой лоб с выраженны-
ми лобными буграми, округленный затылок, глубокие клыковые
ямки, очерченный подбородочный треугольник, низкое лицо, гра-
цильную нижнюю челюсть. Вместе с тем альвеолярная дуга здесь
очень широкая, моляры довольно крупные (Алексеев, 1966).
Многие черты сапиентного комплекса достаточно выражены и
среди палестинских форм (в том числе и у Кафзех), а некоторые
из них — у ранниж палеоантропов (Сванскомб, Фонтешевад,
Эрингсдорф, Штейнгейм) и, возможно, даже у столь древних го-
минид, как череп 1470, или нижняя челюсть 730 с намечающимся
подбородочным выступом.
Глава VIII
ФАКТОРЫ, ВЛИЯЮЩИЕ
НА СТРУКТУРУ СКЕЛЕТА,
И ПРОБЛЕМА РЕКОНСТРУКЦИИ
ОТДЕЛЬНЫХ СТОРОН
ПАЛЕОЭКОЛОГИИ ГОМИНИД
ПО ОСТЕОЛОГИЧЕСКИМ ДАННЫМ
На структуру скелета, как внешнюю, так и внутреннюю, дейст-
вуют многочисленные факторы эндо- и экзогенной природы. По-
видимому, грубая форма кости детерминирована генетически, тогда
как многие специфические детали строения могут зависеть и от
реакции костной ткани на функциональные воздействия (Murray,
Selby, 1930; Felts, 1959). Как и другие ткани, сохраняющие зави-
симый тип развития, костная ткань нуждается для реализации
генетически детерминированной структуры в комплексе определен-
ных факторов, среди которых особенно большую роль играет соб-
ственно функциональный.
Интерпретация формы кости как результата функциональной
адаптации имеет первостепенное значение в эволюционной антро-
пологии и палеоэкологии, позволяя восстановить образ жизни ис-
копаемых форм (Куммер, 1965).
Очевидно, в .наибольшей степени затрагивается воздействием
функционального, фактора широтный рост костей, что нашло от-
ражение в целом ряде исследований, посвященных проявлениям
«функциональной (рабочей) гипертрофии» на скелете человека
(Привес, 1956, 1967; -Жданов, 1967; Гевлич, 1967; Ивкина, Супряга,
1967; Соколов, 1967; Машкара, 1970; Никитюк, Бевзюк, 1970).
В итоге этих исследований во многих случаях выявляется до-
статочно отчетливая картина морфологических преобразований
при разных видах стато-динамической нагрузки в связи с различ-
ными типами трудовой деятельности или спортивной специализа-
ции.-Они охватывают практически все отделы скелета, но наиболее
существенно отражаются на структуре его «рабочих отделов»
(кисти и стопы). Данные ряда работ выявили зависимость адап-
тивной реакции от характера и интенсивности нагрузки, длитель-
ности ее действия, состояния эпифизарных центров окостенения
(если речь идет о растущем организме), уровня (порога) возбу-
димости скелетной ткани. Проиллюстрируем сказанное несколь-
кими примерами.
Дифференциация рентгеновской картины в зависимости от ха-
рактера нагрузки показана в наблюдениях П. Н. Соколова (1967)
и экспериментах Е. А. Клебановой (1962): усиленное отложение
173
периостальной кости ассоциируется прежде всего с действием ди-
намической нагрузки, тогда как аппозиция эндостальной кости
связана с возрастанием устойчивости структуры к сжатию и проис-
ходит под действием гравиостатических сил.
Дифференциация структурных преобразований в зависимости
от места приложения нагрузки иллюстрируется следующими, выб-
ранными из большого числа других примерами: расширение го-
ловки III пястной кости у боксеров (Астанин, 1951); увеличение
ширины большой берцовой кости на «толчковой ноге» у прыгунов
(Крылова, 1967); клиновидность позвонков и изменение формы
латерального края лопатки у штангистов; гипертрофия дельтовид-
ной шероховатости и изменение формы диафиза плечевой кости у
пловцов; резкое утолщение лучевой, шероховатости у гребцов;
«талия» на диафизах плюсневых костей у работников сидячего
труда и «веретеновидность» диафиза у артистов балета.
Особенно наглядный пример такого рода приводит П. Н. Со-
колов (1967): в губчатом веществе костей стопы спортсменов
(прежде всего таранной) выявлено 8 линейных систем трабекул,
ориентированных по направлению и количеству в соответствии с
особенностями различных видов спорта.
Зависимость картины гипертрофии скелета от интенсивности
нагрузки также демонстрируется в наблюдениях П. Н. Соколова
над спортсменами: размеры эпифизов длинных костей были про-
порциональными нагрузке соответствующих суставов.
Наконец, связь проявлений рабочей гипертрофии с длительно-
стью действия функционального фактора выявлялась'в многолет-
них продольных наблюдениях М. Г. Привеса и сотр. (1967).
Однако характер. адаптивной реакции сам зависит от ряда
эндо- и экзогенных факторов, и прежде всего от индивидуального
•порога возбудимости данной ткани, чем и объясняются наблюдаю-
щиеся иногда расхождения в результатах экспериментов и данных
клиники (Никитюк, Бевзюк, 1970); здесь нужно учитывать также
возраст, пол,- состояние нейрогормональной системы и т. д.
При всем том существуют достаточно определенные проявле-
ния рабочей гипертрофии как на макро-, так и на микроскопиче-
ском уровне.
Функциональная адаптация затрагивает прежде всего спонгио-
зу, что выражается в увеличении плотности перекладин и разви-
тии крупноячеистого типа кости. В компакте эти процессы прояв-
ляются в ее утолщении (при сужении костномозговой полости) и
повышении остеонизации. Широтный рост затрагивается в значи-
тельно большей степени, чем длиннотный. Все эти преобразования
в целом ведут к повышению прочности кости (Клебанова, 1954;
Машкара, 1970; и др.).
Выше приводились преимущественно данные динамических на-
блюдений. Примечательно, однако, что сходный характер влияния
биомеханического фактора обнаруживается и при межиндивиду-
альных и даже межгрупповых сопоставлениях у приматов.
174
Приведем в качестве примера исследование влияния статиче-
ского фактора (веса тела) на внешнюю и внутреннюю структуру
длинных костей конечностей — плечевой у бедренной — у четве-
роногого узконосого примата (табл. 24).
Таблица 24
КОРРЕЛЯЦИЯ ВЕСА И ДИАФИЗАРНЫХ ПАРАМЕТРОВ У МАРТЫШКИ*
Признак, коррелируемый с весом Г ± пг (г)
плечевая кость бедренная кость
Толщина сагиттальной компакты 0,613±0,16 0,343±0,18
Сагиттальный диаметр диафиза 0,511±0,18 0,255+0,19
Толщина боковой компакты 0,470+0,19 0,077+0,20
Поперечный диаметр диафиза 0,326±0,22 0,097±0,20
Сагиттальный медуллярный указатель —0,448+0,20 —0,043+0,20
Поперечный медуллярный указатель -0,231+0,24 —0,037±0,20
Общий медуллярный указатель —0,312±0,23 —0,048+0,20
Периметр диафиза . . . ’ 0,475±0,19 0,217+0,19
Указатель сечения** —0,246±0,23 —0,317±0,18
♦ Данные автора, полученные на однородной по возрасту и полу серии Cercopithecus sabaeus.
♦* Процентное отношение поперечного диаметра середины диафиза к сагиттальному...
Как следует из данных табл. 24, основная корреляция опреде-
ляется между весом и абсолютным развитием передне-задней
(сагиттальной) компакты. Все остальные связи существуют в виде
тенденций. Все же очевидно, что намечается достаточно явствен-
ная тенденция связи массы тела и с относительным развитием
•сагиттальной компакты, а также — с сагиттальным диаметром
диафиза. Последнее проявляется и в отрицательном знаке коэф-
фициента корреляции веса тела с указателем сечения на обеих
костях: т. е. при большей статической нагрузке существует тен-
денция к относительному возрастанию толщины диафиза, прежде
всего в передне-заднем направлении. Все указанные взаимоотно-
шения у мартышки выражены более отчетливо на плечевой кости.
Полученные результаты позволяют подчеркнуть и при межинди-
видуальных сопоставлениях особое значение двух признаков диа-
•физа в отношении связи со статическим фактором, а именно тол-
щины компакты и периметра. Причем определяющей и здесь
является связь с «внутренним» диафизарным параметром — тол-
щиной сагиттальной компакты, возрастание которой, очевидно,
влечет за собой увеличение всего сагиттального диаметра диафи-
за — как абсолютно, так и относительно к поперечному его раз-
меру '. Оба основных признака, проявляющих тенденцию к связи
1 Корреляции между сагиттальными размерами диафиза и компакты для пле-
чевой и бедренной костей обезьяны порядка 0,8 и 0,6 (соответственно); между
весом тела и развитием медуллярной полости' реальная связь здесь не вы-
явлена. ’ .
175
со статическим фактором (йериметр и толщина компакты), обна-
руживают определенную взаимную компенсацию при вычислении
парциальных корреляций. Так, при исключении влияния различий
в периметре связь , между весом и медуллярным указателем воз-
растает. То же происходит и при исключении влияния вариаций
в медуллярном указателе, соответственно повышается связь веса
тела с периметром.
Подобное направление внутригрупповых корреляций не являет-
ся, по-видимому, присущим только генерализованному типу про-
ноградного примата, но' отражает более универсальные законо-
мерности соотношений веса и структуры диафиза у четвероногих
млекопитающих. Об этом свидетельствуют, в частности, резуль-
таты параллельных исследований тех же взаимоотношений у дру-
гого четвероногого млекопитающего с весьма разнообразной мото-
рикой (крысы). В этом случае основная корреляция также
существует с развитием сагиттальной компакты и сагиттальным
диаметром диафиза: значения коэффициентов корреляции веса и
сагиттальной компакты составляют здесь для плечевой и бедрен-
ной костей соответственно 0,249±0,14 и 0,537+0,11; для веса и
сагиттального диаметра диафиза ’ 0,375±0,13 и 0,334+0,13. Для
периметров обеих костей коэффициенты корреляции соответствен-
но 0,206±0,15 и 0,413+0,13. Однако в данном случае корреляции
обычно выше на бедре;
Полученные результаты согласуются с данными экспериментов,
проведенных на бипедальных крысах (Saville, Smith, 1966). Эти
авторы нашли, что плотность и прочность бедренной кости яв-
ляются функцией веса тела; на этом основании высказывается
предположение о существенном влиянии этого фактора на скелет
животного. О зависимости прочности бедренной кости от веса
тела сообщают и Тер с соавт. (Ther et al., 1963).
Что касается межгрупповых сопоставлений веса тела и струк-
туры диафиза у приматов, в том числе .и гоминид, то они ограни-
чены преимущественно сведениями о внешней структуре костей.
Шульц (1953) установил на приматах, что абсолютная толщина
длинных костей зависит прежде всего от веса. В. В. Бунак (1967)
также подчеркивает важную роль веса в определении изменчиво-
сти поперечных размеров кости у млекопитающих в целом. По
отношению к различным группам современного человечества по-
добные сопоставления сделаны Г. Ф. Дебецом (1967), в работе
которого выводится исключительно высокий коэффициент меж-
групповой корреляции (порядка 0,9) при сравнении массы тела и
периметров трубчатых костей у групп, различающихся по обоим,
этим признакам.
Выше уже указывалось, что при параллельном анализе внеш-
ней и внутренней структуры нами получены результаты, свиде-
тельствующие, что в пределах групп как низших, так и высших
катарриновых приматов наиболее массивные по внешней (пери-
метр) и внутренней (компакта) структуре кости имели самые
176
крупные формы приматов. Хотя подобная связь и не проявляется
в каждом конкретном случае, общая тенденция в пределах каждой
из этих групп достаточно отчетлива (см. рис. 25).
Нами уже упоминалось также о широкой распространенности
среди ископаемых гоминид вариантов абсолютно и относительно
толстостенных длинных костей. При этом подобная структура
диафиза может существовать как на внешне грацильных, так и на
массивных костях. Поскольку усиленное отложение эндостальной
кости, по-видимому, связано прежде всего с возрастанием устой-
чивости структуры к сжатию и происходит под действием гравио-
статических сил, можно предположить, что длинные кости синан-
тропов и многих палеоантропов имели больший момент сопротив-
ления действию компрессии, чем у современного человека с такими
же внешними параметрами диафиза. Можно полагать, далее, что
по крайней мере у некоторых древнейших и древних людей био-
механический фактор (в частности, повышенная стато-динамиче-
ская нагрузка на скелет особенно в период его роста) сыграл
существенную роль в определении их специфической внутренней
структуры. Напомним, что Е. А. Клебановой (1954) была доказана
возможность значительного утолщения диафизарных стенок длин-
ных костей при стабильном периметре у молодых животных при
усиленной механической нагрузке. По уравнению регрессии можно
показать, что существуют реальные различия в развитии компак-
ты при одинаковом периметре у современного человека и синан-
тропа, как и некоторых палеоантропов. Однако, очевидно, что
одного только биомеханического фактора недостаточно для интер-
претации особенностей внутренней структуры длинных костей этих
ископаемых гоминид. Универсальная гипертрофия костяка синан-
тропов и палеоантропов, проявляющаяся уже в детском возрасте
(Тещик-Таш), не совпадает полностью с локализованной и воз-
растно детерминированной рабочей гипертрофией костей совре-
менного человека. В генезе этого признака структуры скелета ис-
копаемых гоминид представляется также весьма вероятной роль
и других факторов, например обменно-гормонального.
Целый ряд адаптаций на скелете гоминид связан с локомоцией
в широком и в более узком понимании этого термина. Дело в том,
что помимо вопроса об основной локомоции (прямохождении)
большой интерес в ’плане антропогенеза представляет и сам ха-
рактер походки, а также другие второстепенные виды локомоции,
как бегание или лазание на четвереньках, несомненно игравшие
роль факторов выживаемости среди охотников-собирателей древно-
сти -(Watanabe, 1971).
В качестве иллюстрации сошлемся на опыт анализа локомо-
торных адаптаций у ранних гоминид на примере тазового пояса
австр'алопитековых. В литературе высказывались соображения об
отсутствии у некоторых из ранних гоминид уникальной шагающей
походки человека, хотя их двуногость и может считаться доказан-
ной (Нейпир, 1967). Этот автор приписывает австралопитековым
177
род «рысцы» — способ передвижения, который требовал непропор-
ционального расхода энергии, так что австралопитеки не могли
проходить больших расстояний. Одним из следствий этой физио-
логически неэффективной походки мог быть переход к высокока-
лорийной пище (преимущественно мясной диете). С функциональ-
ной стороны отсутствие шагающей походки у австралопитеков
связывается с различиями в таком фундаментальном элементе
локомоции, как способность переднего комплекса mm. glutei
medius et minimus к внутренней ротации (Цильман, Хантер, 1972).
С морфологической стороны эти авторы усматривают одно из
основных различий между очень похожими тазовыми костями
^встралопитека-прометея и человека в структуре области пе-
редне-верхней подвздошной ости и изгибе подвздошной кости.
Предполагается, что момент внутренней ротации австралопитека
вдвое превосходил человеческий; в дальнейшей эволюции происхо-
дило его постепенное уменьшение, сопровождавшее миграцию пе-
редне-верхней подвздошной ости вниз и медиально; это облег-
чало внутреннюю ротацию и не требовало непропорциональной
затраты мускульной энергии. Адаптации внутренней структуры
тазовой кости также указывают на усиление у этого австралопи-
тека переднего глутеального комплекса. Таким образом, можно
предположить, что по крайней мере у некоторых австралопитеко-
вых система стабилизации и внутренней ротации, обеспечивающая
эффективность передвижения на дальние расстояния, была еще
недостаточно развита; в дальнейшем происходил отбор в направ-
лении совершенствования механизмов более эффективной ходьбы,
что имело весьма существенное значение и для орудийной деятель-
ности ранних гоминид, лучшей координации таза и нижней конеч-
ности с туловищем и верхней конечностью.
Напротив, судя по строению ногтевой фаланги большого паль-
ца стопы олдувейского гоминида (Олдувей 10) с ее латеральным
торзионом и вальгусным наклоном головки, в данном случае речь
должна идти о настоящей шагающей походке («ходьба с пятки на
палец») (Дэй, Нейпир, 1966), дающей ходоку энергетические преи-
мущества при преодолении с минимальным усилием больших
пространств.
Другой существенный момент касается пересмотра функцио-
нальной интерпретации многих структурных признаков локомотор-
ного аппарата, которые раньше без достаточных оснований свя-
зывались просто с «несовершенством прямохождения» даже у
таких сравнительно поздних гоминид, как палеоантропы. Это от-
носится к значительной дорзальной инклинации верхнего эпифиза
большой берцовой кости или сглаженности свода стопы некоторых
палеоантропов. Вообще многие из признаков, которые отражают
адаптации к привычной флексии коленного и голеностопного су-
ставов, можно объяснить длительным употреблением некоторых
статических «поз покоя», предшествовавших возникновению совре-
менных сидячих поз и широко распространенных в палеолите.
178
Напомним о структурном комплексе «положения на корточках»,
в который входят помимо вышеупомянутых такие особенности ске-
лета, как опущенная шейка бедра, углубление межмыщелковой
ямки на нижнем эпифизе этой кости, объясняющееся давлением
задней крестовидной связки при флексии коленного сустава, и
целый ряд адаптаций в области голеностопного сустава, особенно
«фасетки положения на корточках» на таранной, а иногда и боль-
шой берцовой костях, переднее распространение блока таранной
кости, значительная дивергенция ее шейки и т. д. Распространение
некоторых черт этого структурного комплекса на скелетах палео-
антропов свидетельствует о достаточно широко практикуемых
древними людьми различных «позах покоя».
Исключительный интерес представляет изучение адаптивных
реакций скелета кисти, как рабочего органа верхней конечности,
в свете ее эргомоторики у гоминид. Известно, что при применении
так называемого «силового захвата», когда нагрузка падает преи-
мущественно на пясть и пальцы, а ее максимум испытывают ног-
тевые фаланги, на кисти современного человека можно наблюдать
постепенное расширение дистальных фаланг, особенно на большом
пальце. Достаточно вероятно, что в эволюции руки подобный «си-
ловой захват» предшествовал точному («прецизионному») зажиму,
свойственному руке современного человека (Нейпир, 1965/1967).
С этим предположением морфологически очень хорошо увязы-
вается форма ногтевых фаланг на кисти Олдувей 7 (см. выше).
Здесь существует высокая степень расширения всего ряда ди-
стальных фаланг, особенно выраженного на большом пальце.
Напомним, что в небольшом эксперименте Нейпир показал, что
примитивное рубящее орудие олдувейской культуры можно изго-
товить, применив тот самый силовой захват, который и структурно
ассоциируется именно со свойственной Н. habilis «ультраго'минид-
ной» формой дистальных фаланг. Подчеркнем' еще раз, что ни
одна современная или ископаемая обезьяна не имеет подобного
типа ногтевых фаланг, так что диагностический вес этого критерия
очень высок. К сожалению, нам пока неизвестны вариации этой
структуры у австралопитековых в целом, что имело бы весьма
существенное значение для проблемы первоначального, становле-
ния трудовой функции и ее дифференциации у ранних гоминид.
При наличии очень широкой нормы реакции костной ткани и, сле-
довательно, значительной фенотипической изменчивости трудно
разграничить здесь генотипический и паратипический компо-
ненты, но наиболее вероятно, что наследственно детермини-
рованная структура ногтевых фаланг оказалась «преувеличенной»
прижизненной рабочей гипертрофией, как это имело место, по-
видимому, и у киик-кобинского палеоантропа и нередко наблю-
дается у современного человека. Из сказанного вытекает очень
существенное обстоятельство, а именно, что форму ногтевых фа-
ланг Н. habilis следует оценивать как индикатор примитивной
трудовой деятельности, связанной с применением силового зах-
179
вата, у ранних гоминид, а следовательно, и как необходимый ком-
понент их комплексного морфологического критерия.
Другой пример представляет кисть киик-кобинского палеоан-
тропа. Известно, что повышенная механическая нагрузка на кисть
современного человека приводит к развитию ряда адаптаций, к
числу которых относятся: уплощение сочленовных поверхностей
прежде всего в области корня кисти и увеличение их конгруэнт-
ности, усиление массивности трубчатых костей кисти, утолщение
компакты, крупноячеистый тип губчатого вещества, по-видимому,
возможная ульнаризация кисти, расширение ногтевых фаланг (Ив-
кина, Супряга, 1967; Гевлич, 1967; Wechsler, 1939). Напомним, что
многие из этих особенностей были характерны для кисти крымско-
го палеоантропа и логично связываются здесь с ее «силовой» спе-
циализацией; следует отметить и общее сходство кисти киик-ко-
бинца с ее «силовым» вариантом у современного человека.
Другая существенная сторона изучения структурных особенно-
стей скелета в палеоантропологии состоит в возможности косвен-
ной оценки на основании некоторых остеологических показателей
систем регуляции взаимоотношений организма со средой. Послед-
няя основывается на том доказанном факте, что ряд физиологи-
ческих состояний организма отражается на структуре скелета.
Речь идет прежде всего о некоторых звеньях системы нейрогор-
мональной регуляции. Как известно, работами ряда исследовате-
лей (Рохлин, 1936, 1965; Silinkova — Malkova 1963; Bugyi, 1963;
и др.) показана возможность такой косвенной оценки гормональ-
ного статуса по остеологическим материалам. Особое положение
в системе нейрогормональной регуляции занимает гипоталамо-ги-
пофизарное ее звено. По некоторым данным (Patsch, 1963), раз-
меры гипофиза в известной степени могут служить критерием
состояния гипофизарно-адреналовой системы. С другой стороны,-
определена существенная корреляция между размерами (объе-
мами) гипофиза и турецкого седла на черепе человека (0,6 в сред-
нем у мужчин и’женщин, по данным Пикулева и др., 1970), что
позволяет в известной степени использовать критерий развития
турецкого седла при определении функционального состояния ги-
пер- и гипопитуитаризма. Конечно, нужно всегда иметь в виду
'сугубую условность и приблизительность такой характеристики;
все же с этой точки зрения заслуживают внимания некоторые
возможные проявления «акромегалойдного акцента» у ископаемых
гоминид. Представляется интересной такая комбинация признаков
на скелетах некоторых палеоантропов, как значительная длина
черепа, сильное развитие надглазничного рельефа, массивность
нижней челюсти, выступание лицевого отдела, длина остистых
отростков верхних прзвонков, гипертрофия апофизарных бугорков
шейно-грудных позвонков, утолщение трубчатых костей и особен^
но макрофалангия дистальных фаланг. Известно, что особо круп-
ные размеры гипофизарной ямки предполагались у ’некоторых
«явантропов» из Нгандонга (Вейденрейх, 1943), обладавших
180
большими размерами черепа и массивностью черепных костей.
Палеоэндокринологические исследования Буги (Bugyi, 1963,
1964) позволили охарактеризовать по остеологическим данным и
функциональное состояние щитовидной железы на примере древ-
него населения Венгрии.
Большой интерес в аспекте палеоэндокринологии проставляют
также данные, свидетельствующие о возможности разграничения
на скелете состояний повышенной (пониженной) секреции поло-
вых стероидов. Последнее основывается на весьма существенном
влиянии, оказываемом половыми гормонами на процессы роста и
количество костной массы. Известно, что при их избыточной сек-
реции значительно возрастает толщина кортикальной кости. В дет-
ской эндокринологической клинике гиперсекреция эстрогенов и
андрогенов (преждевременное половое’развитие, адрено-гениталь-
ный синдром) влечет за собой сдвиг индекса кортикализации
трубчатых костей на несколько лет вперед. Но и в условиях фи-
зиологической нормы повышенный уровень секреции половых сте-
роидов ассоциируется с усиленной кортикализацией; об этом же
свидетельствуют и отчетливые результаты некоторых эксперимен-
тов, показавших, что эстрогены, например, прямо стимулируют
именно эндостальную аппозицию костной ткани, вызывая абсолют-
ное и относительное сужение медуллярного пространства (Wein-
mann, Sicher, 1955). Напротив, при гипогонадизме отмечаются ос-
теопоротические явления вплоть до развития генерализованного
остеопороза (Старкова и др., 1969). Кроме того, неоднократно
демонстрировалось (в клинике и эксперименте) тормозящее влия-
ние половых стероидов на линейный рост. В этой связи интересны
структурные особенности скелета некоторых ископаемых гоминид,
у которых обращает на себя внимание сочетание высокой степени
кортикализации костей скелета, принимающей нередко генерали-
зованный характер, с пониженной длиной тела. Представляется
возможным участие в формировании этой своеобразной структуры
длинных костей скелета помимо биомеханического обменно-гор-
монального фактора, и прежде всего половых стероидов, поскольку
последние обладают, по-видимому, единообразным характером
действия на остеобластические процессы у человека и млекопи-
тающих вообще. Причем у- сравнительно высокорослых палеоан-
тропов типа Схул, видимо, не существовало этой выраженной
внутренней массивности костяка, судя по строению большой берцо-
вой кости Схул IV. На этом же скелете, как и у некоторых других
людей Схул, можно отметить малоотчетливую картину полового
диморфизма (грудина, таз).
Конечно, специфика строения скелета ископаемых гоминид
отнюдь не сводится только к влиянию «эндокринной формулы»,
или вообще какого-то одного фактора, но недооценивать ее роль
у ископаемых (как и современных) гоминид было бы столь же
неверно, как и переоценивать ее. Дополнительные трудности выяв-
ления остеологических индикаторов эндокринных функций на па-
181
леоантропологических материалах' хорошо понятны и состоят
прежде всего в отсутствии достаточно представительного «фона»
для межиндивидуальных сопоставлений и вытекающей отсюда
необходимости в ряде случаев иметь дело одновременно с межин-
дивидуальными и с межгрупповыми вариациями интересующих
нас структурных признаков, что предполагает возможность и дру-
гих факторов их детерминации.
Весьма существенно для палеоантропологии изучение и неко-
v торых особенностей скелета, рассматриваемых обычно в качестве
«генетических индикаторов». Выше были отмечены некоторые из
них. Особый интерес представляют с этой точки зрения некоторые
черты внутренней структуры, а также проявления брахи- (долихо)
мезофалангии, поскольку для большинства палеоантропов были
как раз характерны удлиненные средние фаланги кисти и стопы.
Здесь заслуживает внимания предположение о возможном селек-
тивном преимуществе редукции размеров тела, с которой у чело-
. века достоверно связана брахимезофалангия, в определенных ус-
ловиях существования (Garn et al., 1972).
Существенный интерес в этом аспекте представляют и данные
о влиянии белковой недостаточности и голодания на структуру
скелета (Ahuja, 1969; Riesenfeld, 1967, 1970), причем особую роль
приобретает дифференциальный рост, обусловленный голоданием
в утробном периоде. Вообще исследования о влиянии характера
питания на структуру скелета позволяют во многих случаях выс-
казать определенные предположения о диететике древних гоминид
и их образе жизни. Так, можно упомянуть о связи внутренней
структуры костей с особенностями питания (Garn et al., 1964), о
влиянии некоторых растительных пигментов на окрашивание ко-
- стей (например, данные об окрашивании костей 'ализарином из
корней марены, что связывается с верой в магические свойства
этого растения, по Steckoll et al., 1971). Укажем также на интер-
претацию рахитических проявлений на скелетах некоторых вюрм-
ских неандертальцев с точки зрения недостаточности экзогенного
витамина Д (отсутствие в диете рыбных продуктов) и крайне огра-
. ниченных возможностей синтеза эндогенного витамина из-за слабой
инсоляции в зоне обитания этих древних людей (Jvanhoe, 1970).
Аспект палеопатологии представляет большие возможности при
реконструкции как некоторых сторон жизнедеятельности ископае-
мых гоминид, так и общественных взаимоотношений в первобыт-
ных коллективах. Эти исследования не только позволяют судить
о распространении в древности целого ряда заболеваний, в симп-
томатику которых входят черты костной патологии, но в некото-
рых случаях изучение их географического распространения может
быть связано и с определенными характеристиками внешней среды,
(недостаточность или избыток некоторых химических элементов).
На археологическом материале нередко отмечаются и раз-
личия в возрастно-половой динамике внутренней структуры ске-
лета сравнительно с современными людьми (Armelagos, 1972;
182 .
Mielke et al., 1972), что следует учитывать при сопоставлениях.
- Костная патология может пролить определенный свет и на
характер взаимоотношений древнейших и древних людей и их
предшественников. Интересны в этом отношении данные о повреж-
дениях костей черепа и посткраниального скелета австралопитеков
из различных местонахождений Южной Африки (Brain, 1972).
Автор приходит к выводу, что эти костные дефекты большей ча-
стью являются посмертными, некоторые из них обословлены дей-
ствием природных агентов в связи с условиями залегания костей.
Результаты этого анализа не дают оснований объяснять отмечен-
ные повреждения на костях этих приматов кровавыми столкнове-
ниями среди них в силу якобы присущей им агрессивности. Напро-
тив, неоднократно показана по отношению к древним людям воз-
можность многолетнего существования инвалидов, нуждавшихся
в заботе и уходе со стороны своих соплеменников, даже в очень
отдаленные времена (Рохлин, 1967; Sandison, 1969). Можно упо-
мянуть также серию работ Дастюга (Dastugue, 1967/1968) о
кроманьонцах, где автор подвергает сомнению традиционную
точку-зрения об особой жесткости первобытного человека, осно-
вываясь как раз на характере костной патологии. Большой интерес
представляют также доказательства исключительной древности
хирургии; например, ампутация мизинцев кисти у женщины Мур-
зак-Коба, многократно описанные трепанации черепа (Рохлин,
1967; Бобин, 1967).
В нашу задачу не входила характеристика собственно костной
патологии, которой посвящено несколько специальных исследова-
ний (Рохлин, 1965). Упомянем только о широком распространении
в древности заболеваний суставов, что наблюдалось и у шапелльца,
страдавшего артритом; последнее не позволяет считать его вполне
типичным представителем этого варианта палеоантропов (Brauer,
1972).
Один из оригинальных аспектов интерпретации палеоантро-
пологических материалов отражен в серии «палеоакушерских ис-
следований» (Malinas, 1970; Leutenegger, 1972). На основании
геометрической реконструкции женского таза австралопитека (про-
метен) предполагается существование расхождений в расположе-
нии некоторых' органов репродуктивной системы и вытекающее
отсюда меньшее число мертворождений' у ранних гоминид. Боль-
шая легкость родов у австралопитековых доказывается и на ос-
нове изучения соотношений между размерами новорожденных
форм и таза взрослых женских индивидуумов. Эти положения
весьма существенны ввиду особой роли дифференциальной выжи-
ваемости на самых ранних этапах антропогенеза.
Конечно, в этом кратком очерке затронуты далеко не все
стороны проблемы. Приведены лишь отдельные примеры, иллю-
стрирующие возможности использования остеологических мате-
риалов при изучении взаимоотношений человека и среды.
183
Глава IX
НЕКОТОРЫЕ ЧЕРТЫ
МОРФОЛОГИЧЕСКОЙ
ЭВОЛЮЦИИ И ПРОБЛЕМА
МОРФОЛОГИЧЕСКОГО
КРИТЕРИЯ ГОМИНИД
В заключительной части книги мы рассмотрим некоторые аспекты
морфологической эволюции человека, существенные как для по-
нимания ее общего характера, так и при рассмотрении проблемы
морфологического критерия гоминид.
ФОРМИРОВАНИЕ ОСНОВНЫХ
МОРФОЛОГИЧЕСКИХ КОМПЛЕКСОВ
НА СКЕЛЕТЕ ЧЕЛОВЕКА
Один из наиболее существенных аспектов морфологической
эволюции человека связан с изучением темпов формирования важ-
нейших гоминидных комплексов. Прямые определения «филети-
ческой скорости» (Симпсон, 1948, 1949) пока мало применимы
для гоминид, по крайней мере, при настоящем состоянии рекон-
струкции их филетической линии. Как известно, классическим
примером подобного определения филетической скорости среди
млекопитающих являются Equidae и Chalicotheriidae, средняя ско-
рость эволюции которых равна примерно 0,18 и 0,17 рода в 1 млн.
лет соответственно. Из приматов предположительная скорость эво-
люции в линии гиббонов составляет 0,1 рода в 1 млн. лет.
Для гоминид подобные расчеты могут быть только сугубо
приблизительными, поскольку существуют значительные разногла-
сия как в определении времени дивергенции гоминидной и пон-
гидной линий от общего ствола, так и в реконструкции собственно
гоминидной линии эволюции. В отношении первого вопроса мож-
но указать на диапазон мн'ений, охватывающих огромный времен-
ной период, с конца олигоцена и до плиоцена. В отношении
второй дискуссионной проблемы сошлемся на крайние точки зре-
ния Робинсона (Robinson, 1965), включающего в род Ното все
формы, начиная с грацильных австралопитеков и кончая Н. sa-
piens, и Маккоуна и Кизса (1939), выделявших три рода только
среди так называемых «настоящих гоминид» (питекантропы, па-
леоантропы, неоантропы — Ното).. Если условно принять доста-
точно распространенную точку зрения об отделении гоминидной
линии эволюции в раннем миоцене и существовании на пути ее
184
развития не менее трех родов (рамапитек— австралопитек — че-
ловек), то примерная скорость филетической эволюции за рас-
сматриваемый период составит не менее чем 0,1 рода в 1 млн. лет,
т. с. она окажется вполне сопоставимой со скоростью эволюции
гиббонов и в какой-то степени и других млекопитающих. При оби-
лии локальных вариантов гоминид, между которыми существовали
географические и хронологические различия, и неполноте палеон-
тологической летописи по отношению к вероятным наиболее ран-
ним представителям гоминидной линии более определенные рас-
четы, к сожалению, пока не представляются возможными, но и
полученный сугубо приблизительный результат свидетельствует о
достаточно интенсивной эволюции (Koenigswald, 1963; Бунак,
1966а). ' • '
Большинство работ, в которых затронут вопрос о темпах эво-
люции гоминид, преимущественное внимание уделяет скорости
изменений морфологических признаков. Обычно исследования та-
ких «морфологических скоростей» выполняются на наиболее изу-
ченных краниологических материалах (Wiercinski, 1956; Бодян-
ский, 1957; Рогинский, 1957а; Алексеев, 1968). Однако краниоло-
гические материалы отражают, конечно, только один из аспектов
морфологической эволюции гоминид, связанный с преобразования-
ми черепа и отчасти мозга; недостаточно освещенными остаются
ее другие важнейшие стороны — эволюция локомоции и транс-
формация руки в связи с трудовой деятельностью «формирующих-
ся людей».
Между тем сравнительные темпы формирования этих трех
оснрвных «комплексов гоминизации» представляют исключитель-
ный интерес для проблемы морфологического критерия гоминид,
поскольку процесс антропогенеза, по-видимому, как раз дает
иллюстрацию неравномерности развития различных систем ске-
лета. 'Рассмотрим с этой точки зрения классическую триаду ве-
дущих морфологических комплексов на скелете — прямохождения,
трудовой руки и краниофациальную систему.
Наиболее ранние и полные преобразования скелета в гоминид-
ной линии эволюции связаны с возникновением и развитием пря-
мохождения. Из предыдущего обзора палеонтологических материа-
лов по посткраниальному скелету следует, что «комплекс пря-
мохождения» в своих наиболее существенных характеристиках
был уже сформирован по меньшей мере 2—3 млн. лет тому назад,
а может быть, и значительно раньше.
Адаптация скелета к двуногому передвижению в выпрямлен-
ном положении выявляется в достаточной степени у австралопи-
тековых и близких к ним форм (Н. habilis). Сюда относятся: на
тазе — изменение общей конфигурации, расширение пластинки
крыла в дорзальном и вентральном направлениях, дорзальная
инклинация крестцовой части подвздошной кости, расширение
сакральной сочленовной поверхности, уменьшение нижней длины
подвздошной кости, укрепление вертлужной впадины, формиро-
12'Е. Н. Хрисанфова 185 ,
вание большой седалищной вырезки, как и обеих подвздошных
передних остей; на стопе — в первую очередь весь комплекс «ти-
биализации», т. е. приведение и укрепление первого луча, массив-
ность I плюсневой, пропорции плеч рычагов стопы, связанные с
распределением стато-динамической нагрузки, укрепление сводов.
К аналогичным результатам приводит и сравнительно-морфоло-
гический анализ длинных костей нижней конечности, хотя призна-
ки комплекса бипедии выражены здесь менее отчетливо в силу
большей нейтральности строения этого отдела скелета. Наконец,
и на осевом скелете, судя по строению поясничных позвонков
некоторых австралопитековых, черты адаптации к прямохождению
проявляются уже вполне определенно.
Все более поздние гоминиды (архантропы, палеоантропы) пол-
ностью разделяют с неоантропами структуру скелета по основному
комплексу локомоции, вопреки распространенным взглядам о по-
степенном формировании прямохождения на поздних этапах антро-
погенеза и неполном выпрямлении тела у древнейших и древних
людей.
Раннее оформление этого морфологического комплекса на ске-
лете гоминид согласуется также с косвенными данными по палео-
морфологии других высших приматов. Так, например, при пон-
гидном общем облике посткраниальный скелет ореопитека все же
свидетельствует о возможности появления некоторых черт адап-
тации к бипедии еще 10—12 млн. лет назад. В миоцене сущест-
вовали высшие приматы со сравнительно нейтральным строением
локомоторного комплекса, который вполне может рассматриваться
в качестве «модельного варианта» для развития как в бипедаль-
ном, так и в брахиаторном направлениях.
Значительно слабее представлены палеоантропологические ма-
териалы по кисти. Если снова обратиться к австралопитековым
и близким к ним формам, следует прежде всего указать на зна-
чительную примитивность структуры олдувейской кисти. Мор-
фологически рука Н. habilis еще не была человеческой, но в целом
питекоидной, с «гориллоидным» налетом. Сюда нужно отнести
полуфлексированное положение пальцев, значительную расширен-
ность диафизов проксимальных и средних фаланг, топографию и
выраженность гребней для прикрепления поверхностного сгибателя
пальцев, возможно, положение трапеции среди других костей за-
пястья и морфологию головчатой кости (Levis, 1973). Таким об-
разом, насколько можно судить о структуре этой кисти по имею-
щимся неполным и разнокачественным данным, она лишь слегка
затронута процессом гоминизации: вероятно, близкие к человече-
ским пропорции пясти и пальцев, уменьшение изогнутости фаланг,
возможное относительное удлинение I луча и некоторое его сме-
щение из плоскости остальных сравнительно с соответствующими
соотношениями на кисти гориллы. На этом фоне особенно выде-
ляется «ультрагоминидная» структура ногтевых фаланг Н. habilis.
Этапы эволюции гоминидной кисти представлены чрезвычайно
186
скудно. Пока неизвестна ее структура у архантропов. Что касается
палеоантропов, то по крайней мере у поздних прогрессивных форм
последних (Схул V) можно говорить о вполне современном ее типе
(степень радиализации, структура сустава I луча). Поскольку у
более ранних представителей древних людей (Киик-Коба), по-ви-
димому, наблюдалось еще некоторое «отставание» в развитии этих
важнейших черт гоминидного комплекса кисти, даже учитывая
вероятность вторичной «силовой» специализации, надо думать все
же, что кисть архантропов обладала еще менее совершенной струк-
турой по «комплексу радиализации», что следует также из косвен-
ных (археологических) данных (Krantz Crover, I960). Следова-
тельно, можно предположить, что дефинитивный тип человеческой
кисти сложился на высшем этапе антропогенеза, у прогрессивных
форм палеоантропов. Некоторые исследователи (Нейпир, 1965/
1967) приурочивают появление «точного зажима», означающего
развитие вполне человеческого противопоставления большого
• пальца, только к верхнему палеолиту.
Что касается других компонентов верхней конечности, то раз-
личия между современными и ископаемыми гоминидами в боль-
шей мере сказываются на строении сочленовных концов обеих ко-
стей предплечья. Выше уже упоминалось, что некоторые морфо-
логические особенности лучевой и локтевой костей позволяют
предположить совершенствование тонких движений просупинации
даже на сравнительно поздних ступенях антропогенеза (Макко-
-ун, Кизс, 1939).
Из сказанного следует, что развитие типичной человеческой Цр
руки, по-видимому, происходило более замедленными темпами. .wjk
чем формирование комплекса прямохождения, хотя и в несомнен- Яв
ной связи с последним. Если структурные адаптации к прямохож- яИ
дению вполне выражены на скелете нижней конечности австрало- яН
питековых, и прежде всего у Н. habilis, то кисть этого последнего W
еще мало напоминала человеческую. Даже у большей части па- Ц
леоантропов, возможно, не было завершено формирование дефи- 3
нитивных пропорций I—II лучей, как и формы сустава I луча. ,t£
Сказанное выше справедливо и для краниофациального ком-
плекса. У австралопитековых и Н. habilis существуют лишь неко-
торые начальные сдвиги на пути гоминизации в структуре мозго-
вого и лицевого отделов черепа — увеличение степени церебрали-
зации, перестройка основания черепа, и прежде всего его заты-
лочного отдел а,,уменьшение выступания лица, трансформация но-
совой области, изменение формы зубной дуги, особенно в ее пе-
редней части, уменьшение клыков, исчезновение диастем и специа-
лизации первого нижнего премоляра. По-видимому, гораздо поз-
же сформировались такие черты сапиентного комплекса черепа,
как редукция лицевого отдела и связанное с ней преобразование
краниофациальных пропорций, грацилизация нижней челюсти и
возникновение подбородка человеческого типа. Насколько можно
судить по существующим сейчас данным, подобные сдвиги про-
12*
187
изошли у прогрессивных палеоантропов (Схул V, Кафзех, особен-
но Староселье) или даже неоантропов. Это построение имеет от-
части провизорный характер, так как во многих случаях наибо-
лее существенные краниологические материалы, в том числе, по
ранним «прогрессивным» формам гоминид, достаточно фрагмен-
тарны или принадлежат очень молодым индивидуумам. Здесь за-
служивают особого внимания предварительные данные о некото-
рых прогрессивных признаках на черепе очень древнего гоминида
(череп'1470); если они подтвердятся, это может дать повод к су-
щественному пересмотру концепций, касающихся ранних гоминид.
Неравномерность морфологической эволюции проявляется и в
пределах основных комплексов при сравнении скоростей эволюции
отдельных признаков между собой. На черепе многих гоминид она
отражается в «запаздывании» увеличения высоты свода сравни-
тельно с возрастанием массы мозга. Другой пример касается опе-
режения редукцией зубной системы процесса грацилизации самих
челюстей. Регрессивная эволюция зубной системы началась, по-
видимому, еще на прегоминидной стадии антропогенеза (рамапи-
тек), т. е. значительно раньше, чем процесс уменьшения массив-
ности челюстей, несоответствие, особенно отчетливо проявившееся
позже у гейдельбергского человека, архантропа по морфологии
нижней челюсти и палеоантропа по структуре зубов (Зубов;
1966). В основных чертах зубная система человеческого типа сфор-
мировалась уже у палеоантропов, опередив преобразование всего
лицевого скелета и его тотальных пропорций с мозговым отделом
черепа.
Ярким примером неравномерности морфологической эволюции
в Пределах «комплекса руки» является формирование расширен-
ной «ультрагоминидной» структуры ногтевых фаланг уже на очень
ранней стадии антропогенеза, причем в наибольшей степени за-
трагивался этим преобразованием большой палец, вероятно, в си-
лу его особой роли в эргомоторике кисти вообще и «силовом за-
жиме» в частности. Наконец, выше упоминалось уже и о возмож-
ном «отставании» эволюции некоторых структур подвздошной ко-
сти сравнительно с другими компонентами комплекса бипедии, на
тазе ранних гоминид.
Другая черта ранних этапов морфологической эволюции, чело-
века состоит в ее выраженном «функциональном характере», что
в наибольшей степени проявляется по отношению к рабочим отде-
лам конечностей. Особое значение это имеет применительно к
структуре кисти, поскольку для проблемы морфологического кри-
терия гоминид весьма существен любой ее сдвиг в направлении
гоминизации, т. е. адаптации к трудовой функции.
Существуют, как известно, многочисленные «параллелизмы»
онто- и филогенетического развития в морфологии кисти гоминид.
Для человека специфичны: укрепление ее корня, расширение всех
отделов и особенно карпальных элементов, относительное укоро-
чение пальцев, Широкая форма ногтевых фаланг. Все это, конеч-
188
но, наследственно детерминированные признаки на трудовой , ки-
сти современного человека, но и в пределах онтогенеза, при ин-
тенсификации нагрузки на кисть могут наблюдаться изменения в,
аналогичном направлении, причем локализация адаптивных реак-
ций в скелетной ткани проявляет тесную зависимость от конкрет-
ных условий биомеханики кисти (см. выше). Локальные адаптив-
ные реакции в скелетной ткани кисти наблюдаются также в онто-
и филогенезе других приматов (Астанин, 1968). Известно, что и
для скелета в целом очень характерна «функциональная форма»
костей как во внешней, так и во внутренней их структуре. Все по-
добные признаки детерминированы генетически, но в то же время
в высокой степени способны к «целесообразной перестройке» в
онтогенезе при изменении конкретных условий функционирования.
Одно из возможных объяснений подобных параллелизмов с по-
зиций современной биологии связывается с концепцией стабили-
зирующего отбора (Шмальгаузен, 1969). Речь идет о возможной
наследственной стабилизации локальных адаптивных реакций че-
рез систему естественного отбора путем замены эргонтическйх
’корреляций морфогенетическими. Действительно, характер адап-
тивной реакции на костной ткани, как ткани с частично зависи-
мым типом развития, во многом определяется силой, местом при-
ложения и временем действия внешнего раздражителя. В про-
цессе стабилизирующего отбора, согласно предположению
И. И. Шмальтаузена, могла происходить замена внешнего фак-
тора развития внутренними, прежде всего со стороны нейроэндо-
кринной регуляции. В некоторых случаях, как указывает этот ав-
тор, подобные процессы, происходящие обычно в пределах преж-
ней адаптивной нормы, могут послужить основой для развития
новых дифференцировок, определяя, таким образом, будущее на-
правление естественного отбора.
Применяя эту концепцию' к конкретным палеоантропологиче-
ским материалам по ранним гоминидам, следует в первую очередь
обратить внимание на уже неоднократно упоминавшуюся струк-
туру ногтевых фаланг кисти у Олдувей 7. Всё эти кости принад-
. лежат одному молодому индивидууму и являются типично чело-
веческими, причем особо гипертрофированную форму имеет ног-
тевая фаланга большого пальца, испытавшая наибольшую струк-
турную перестройку и «опередившая» в этом отношении осталь-
ные фаланги. Известно, что изменение формы фаланг в анало-
гичном направлении может наблюдаться и при прижизненном
рентгенологическом изучении их на кисти современного человека,
если последняя подвергается интенсивному «силовому» использо-
ванию (Привес, 1956). Вместе с тем, согласно предположению
Нейпира (1962), именно «силовой захват» был наиболее древним
в процессе трудовой адаптации кисти. Выше уже указывалось на
трудность разграничения на олдувейской кисти генотипического и
паратипического компонентов; наиболее вероятно, что наследст-
венная структура дистальных фаланг оказалась здесь еще «пре-
189
увеличенной» прижизненной рабочей гипертрофией, как это наб-
людается у современного человека и, по-видимому, имело место у
киик-кобинского палеоантропа. Но как бы ни интерпретировать
эту структуру, ясно, что форма ногтевых фаланг Н. habilis может
оцениваться как индикатор примитивной трудовой деятельности,
связанной с-применением «силового захвата».
С точки зрения сравнительных темпов морфологической эво-
люции сама эта -способность костной ткани давать в отдельных
случаях отчетливый морфогенетический эффект в течение сравни-
тельно короткого времени могла иметь важное значение в струк-
турной перестройке таких сугубо адаптивных комплексов, как си-
стемы локомоции и руки, хотя подобный путь эволюции скелета,
конечно, не был единственным и, вероятно, не являлся даже глав-
ным.
Подобная способность к стабилизации локальных адаптивных
реакций, проявлявшая себя прежде всего на уровне скелетной тка-
ни, делает эту последнюю .наиболее пригодным морфологическим
индикатором для суждения о степени адаптации к трудовой дея-
тельности ранних гоминид. Мы подчеркиваем здесь это обстоя-
тельство, поскольку иногда в качестве основного морфологическо-
го критерия семейства гоминид используется структура мозга
(эндокрана), которая рассматривается или кцк макроморфологи-
ческое выражение того «комплекса мозга», который был необхо-
дим как минимальная основа для начальной трудовой деятельно-
сти, или же как «отпечаток» систематического изготовления ору-
дий в строении мозга *.
Между тем интерпретация структуры эндокрана по ряду при-
чин значительно более сложна и менее достоверна. Даже если
признавать возможность эволюции через индивидуальное приспо-
собление и для нервной ткани, основываясь на ее общей способ-
ности к функциональной гипертрофии (Шмальгаузен, 1969), все
же некоторые прямые и косвенные данные позволяют предполо-
жить, что подобные преобразования проявлялись здесь макро-
скопически далеко не сразу, тем более, что синхронность измене-
ний микро- и макроструктуры мозга в онтогенезе и филогенезе,
по-видимому, недостаточно отчетлива. Хотя прямые аналогии с
развитием мозга у предшественников человека и невозможны
(учитывая большую сложность и, вероятно, рано проявлявшееся
в эволюции качественное своеобразие этого процесса), все же сле-
дует отметить, что и в сравнительно-неврологическом ряду прима-
1 В нашу задачу отнюдь не озходило освещение эволюции самого мозга человека,
являющейся самостоятельной проблемой антропогенеза (см., например, ориги-
нальное исследование В. И. Кочетковой (1973), представляющее также наибо-
лее полную в мировой литературе сводку данных по эндокранам гоминид и их
предшественников и содержащее подробное изложение истории проблемы); на-
ми этот вопрос затрагивается лишь в той степени, которая необходима в
связи с обоснованием нашего подхода к выделению морфологического крите-
рия гоминид.
190
тов изменения в макроструктуре мозга следуют за глубокими пре-
образованиями в его тонком строении (Поляков, 1949- Шевчен-
ко, 1968).
Как бы ни решать .сложный и не рассматривающийся здесь воп-
рос о факторах эволюции структуры мозга у ранних
гоминид, уже сам факт широкого распространения в прош-
лом форм типа Н. habilis с развитым прямохождением, прими-
тивной трудовой деятельностью, затронувшей уже морфологию
кисти, но с понгидным по внешнему облику мозгом (Кочеткова,
1973), свидетельствует о трудности использования формы эндо-
крана, как критерия гоминизации. Следует иметь в виду и боль-
шие потенциальные возможности мозга антропоидных предков
гоминид с их уже хорошо развитой манипуляторной деятель-
ностью; мозг таких существ, очевидно, обладал достаточно широ-
кими проспективными функциями (Кац, 1973). Палеоневрологиче-
ские данные не позволяют, конечно, выявить тонкую структуру
мозга и, в частности, то богатство межнейронных связей, обуслов-
ленное индивидуальным опытом, обучением и освоением культур-
ной среды, которое явилось важной предпосылкой возникновения
индивидуального сознания и отвлеченного мышления (Холлоуэй,
1966; Ruffle, 1973). Между тем даже в отношении «предсказаний»
функции нейронной сети по ее тонкой структуре функциональная
интерпретация, по-видимому, возможна только для сравнительно
простых формаций (Braitenberg, 1972).
В связи со сказанным, представляется пока недостаточно обо-
снованным прямолинейное истолкование сходства в конфигурации
мозга (эндокрана) современного специализированного антропои-
да (шимпанзе) и двуногого гоминида, уже выделывавшего ору-
дия' (Н. habilis), как их тождества и в отношении тонкой струк-
туры мозга, интерпретируемой к тому же в категориях психоло-
гии и поведения. Известно, что даже в оценке уровня умственного
развития более поздних гоминид по особенностям их эндокранов
в литературе существуют значительные разногласия между раз-
ными исследователями (Overhage, 1962; Stgslicka-Mydlarska,
1971).
Поскольку концепция «мозгового рубикона» всегда привлека-
ла внимание антропологов, мы рассмотрим этот вопрос и в аспек-
те изменчивости морфологических признаков у ранних гоминид.
В антропологической литературе нашла отражение точка зре-
ния о «прерывистом» характере морфологической эволюции моз-
га в линии гомцнид и, в частности, об отсутствии трансгрессивной
изменчивости такого количественного признака, как емкость моз-
говой полости черепа (Кочеткова, 1973).
Речь идет прежде всего о различиях в емкости мозгового от-
дела черепа между ранними гоминидами, с одной стороны, и так
называемыми «настоящими гоминидами» (архантропами) —с дру-
гой. На первый взгляд, разница действительно довольно велика:
450—500 см3 в среднем у австралопитековых — около 625 см3 у
191
H. habilis—855 см3 у яванских питекантропов. Однако, как говори-
лось выше, при подобных сопоставлениях следует учитывать
большие различия в тотальных размерах тела сравниваемых
форм. Так, Н. habilis, как и гранильные австралопитековые, при-
надлежал к числу мелких форм высших приматов. Напомним,
что длина тела взрослого индивидуума Олдувей 8, вероятно, не
превышала 120 см, тогда как питекантропы Явы, по меньшей ме-
ре, приближались к среднему варианту современного человека.
Столь существенные различия делают фактически несопоста-
вимыми данные по абсолютному развитию мозга у этих форм.
При определении же степени церебрализации, оказывается, что
вычисленные на основе формулы гетеротопического роста коэф-
фициенты церебрализации Н. habilis и яванских питекантропов
очень близки (76- и 79 г на 1 кг веса тела) и вместе с тбм намно-
го ниже величины этого показателя у Я. sapiens (134 г на 1 кг
веса тела)А
Другая сторона рассматриваемого вопроса связана с количе-
ственной характеристикой изменчивости массы мозга на ранних
этапах антропогенеза; предполагается ее резкое возрастание уже
при переходе от австралопитековых к архантропам и, далее, к
современному человеку (Кочеткова, 1973).
Как известно, при сопоставлении вариабильности идентичных
признаков на черепе и скелете человека и других млекопитающих
среднее квадратическое отклонение обычно почти строго следует
по своей величине за средней арифметической (Рогинский, 1959).
Эта закономерность прослеживается и у ископаемых гоминид, не-
смотря на недостаточную представительность материалов, и толь-
ко квадратическое отклонение объема мозга Н. habilis оказывается
явно заниженным и выпадает из приведенного в табл. 25 ряда.
« Таблица 25
ПОКАЗАТЕЛИ РАЗНООБРАЗИЯ ОБЪЕМА ЭНДОКРАНА У РАННИХ ГОМИНИД
И НЕКОТОРЫХ АНТРОПОИДОВ’
Приматы п М, см8 S, см8 V, %
Австралопитековые 8 464- 44,7 9,6
Гориллы <5 400 534,8 55,3 10,3
Н. habilis 4 625 34 5,4
Питекантропы Явы 6 855 83 9,7
Синантропы 5 1043 112,5 10,8
* По данным Ф. Тобайяса (1968); Р. Холлоуэя (1970, 1973); В. И. Кочетковой (1973).
1 Возможно, что и по абсолютной величине массы мозга вариационные ряды
Н. habilis и архантропов могли перекрываться: максимум у первого составляет
650—657 см3 (Олдувей 13 и 7), а минимум у второго — 727 см3 (Олдувей 12,
Холлоуэй, 1973). Следует иметь в виду и упоминавшееся выше сообщение
об открытии очень древнего черепа с емкостью мозга не менее 750 см3.
192
Это, по-видимому, результат особенно резкой недостаточности
(как в количественном, так и в качественном отношении) палеон-
тологического материала в этом случае.
Сравнение коэффициентов вариации показывает, что они в
общем представляют собой величины одного порядка (исключе-
ние составляет, как уже упоминалось, только Н. habilis). Не на-
блюдается каких-либо принципиальных различий в характере из-
менчивости при переходе от австралопитековых к архантропам.
Нет существенных расхождений и с величиной коэффициента ва-
риации массы мозга у современного человека (7—7,5%; по Буна-
ку, 1941). Тем самым не подтверждается мнение о резко отлич-
ном характере изменчивости этого признака у неоантропа сравни-
тельно с его предшественниками.
Иного результата трудно было бы и ожидать. Напомним, что
представление об особенно высокой изменчивости у человека
уже неоднократно опровергалось разными исследователями
(Шульц, 1930; Pearl, 1940; Симпсон, 1948; Kmissmann, 1970; Ро-
гинский, 1959). Существенная особенность'изменчивости идентич-
ных признаков в различных (в том числе в хронологическом отно-
шении) группах состоит в ее выраженном единообразии у всех
млекопитающих. По данным Перла (Pearl, 1940), сопоставившего
несколько десятков коэффициентов вариации в нормальных попу-
ляциях человека, все их распределение дает картину изменчиво-
сти, вполне типичную для соответствующих признаков более мед- >
ленно эволюционирующих млекопитающих. Вероятно, этот 'факт
определенного единообразия изменчивости во всем ряду млеко-
питающих может найти свое объяснение с позиций стабилизирую-
щего отбора, поддерживающего нормальную величину и изменчи-
вость признака, подобно тому, как трансгрессивный характер ва-
риаций истолковывается с точки зрения самого механизма морфо-
логической эволюции, основанного на естественном отборе пре-
имущественно малых мутаций.
Другой пример такого рода можно привести для частных и бо-
лее" существенных с точки зрения «мозгового рубикона» характе-
ристик структуры эндокрана ранних гоминид..
Особый интерес здесь представляет количественная характери-
стика тех областей мозга, с прогрессивным развитием которых
связывается наиболее демонстративное нарушение «принципа по-
степенности» при переходе от австралопитековых к архантропам
(Кочеткова, 1973). Речь идет прежде всего об области, примерно
соответствующей триангулярной части нижней лобной извилины,
которая характеризуется количественным развитием задней части
нижней лобной дуги. В этом случае также методически правиль-
нее, принимая во внимание большую разницу в размерах тела со-
поставляемых форм; сравнивать относительные характеристики
развития этой области. Соответствующие индексы задней части
нижней лобной дуги (в % ко всей этой дуге) для шимпанзе, ав-
стралопитеков и архантропов — 50,46 и 57, т. е. показатели отно-
193
•сительного развития этого размера эндокрана не подтверждают
наличия резкого «разрыва» при переходе к архантропам. Еще бо-
лее демонстративный результат получается, если рассматривать
австралопитековых дифференцированно, принимая во внимание,
что из двух сопоставляемых здесь, форм, австралопитек массив-
ный (зинджантроп) и Олдувей 7, только первая может предоста-
вить достаточно доброкачественные исходные данные. Тогда ока-
зывается, что индекс задней части нижней лобной дуги массив-
ного австралопитека совпадает со средним для архантропов (57)
•более того, даже по абсолютной величине рассматриваемого при-
знака обнаруживается постепенность его возрастания в ряду:
шимпанзе — .массивный австралопитек — архантропы (соответ-
ственно 28,37 и 45 мм). Что касается Олдувей 7, лобная доля ко-
торого была полностью реконструирована, соотношения здесь яв-
но неестественны, так как индекс задней части нижней лобной
дуги у этой прогрессивной формы примерно в 1,5 раза ниже, чем
у шимпанзе. Эти факты не укладываются в предполагаемую схе-
му функциональной интерпретации этого признака. Сказанное
справедливо и для другого важнейшего структурного признака
эндокрана — развития области нижней теменной доли, в котором
также усматривается проявление скачкообразности при переходе
от австралопитековых к архантропам. Не говоря уже о том, что
абсолютные различия по развитию нижней теменной дуги между
шимпанзе и австралопитеками значительно превосходят таковые
между австралопитеками и архантропами, относительная харак-
теристика развития этой области, по индексу ко всей срединной
дуге не свидетельствует о разрыве постепенности (соответствую-
щие значения индекса для массивного австралопитека, Олдувей
7 и питекантропа 1 : 36,30 и 31) 1 2. Приведенные факты лишний раз
демонстрируют большие трудности на пути характеристики даже
важнейших структурных особенностей эндокранов.
Хотя все эти расчеты исходят у ископаемых форм из весьма
приблизительных характеристик, полученный результат соответст-
вует общей закономерности, наблюдающейся для морфологиче-
ских признаков во всех хорошо изученных филетических рядах
других млекопитающих, как и органических форм вообще.
Диалектическое единство прерывистого и непрерывного в эво-
люции предполагает возникновение нового качества в результа-
те постепенного и непрерывного накопления мелких количествен-
ных изменений. В палеонтологической летописи, отчасти по самой
ее природе, конечно, существуют многочисленные разрывы. Од-
нако в более или менее прямолинейных филетических линиях,
представляющих эволюцию видов и родов, новый признак обычно
развивается постепенно. Отмечают, что по мере дальнейших па-
леонтологических открытий эти генеалогические линии становят-
1 По данным графика 23 (Кочеткова, 1973).
2 По данным графика 34 (там же).
194
ся более непрерывными, откуда следует, что речь должна идти
о выборках из некогда непрерывного ряда последовательных форм
(Симпсон, 1948). Многие конкретные палеонтологические мате-
риалы свидетельствуют, что прогрессивная эволюция морфологи-
ческих признаков обычно идет через серию мелких сдвигов, транс-
грессирующих в ряду последовательных популяций (Mattew,
1926; Osborn, 1936, 1942; Северцов, 1939; Симпсон, 1948; Шмаль-
гаузен, 1969). Такая картина изменчивости, очевидно, соответст-
вует самому механизму эволюции, строящемуся’ главным образом
на основе накопления и комбинирования малых мутаций под кон-
тролем естественного отбора.
Нельзя отрицать в некоторых случаях и вероятность возникно-
вения одной крупной доминантной мутации, как это, возможно,
произошло с редукцией боковых пальцев у Equidae. Но и тогда
речь идет об эволюции единичного признака, независимой от бо-
лее постепенных изменений в других системах организма (Симп-
сон, 1948). С точки зрения таксономии «разрывность»; обуслов-
ленная неполнотой палеонтологических данных, представляет оп-
ределенное практическое преимущество, так как облегчает клас-
сификацию. Но нельзя не считаться с тем фактом, что по мере по-
полнения данных «разрывы» сглаживаются, а таксономические
'категории становятся менее определенными (Харрисон, Уайнер,
1968). Эти трудности велики уже сейчас и для гоминид (Н. habi-
lis, чадантроп, морфологическая неоднородность архантропов).
Нет сомнений, что при прочих равных условиях трансгрессив-
ный-характер изменчивости количественных морфологических при-
знаков проявляется тем отчетливее, чем ближе рассматриваемый
сравнительно-анатомический ряд к истинному филетическому ря-
ду. Можно предположить, исходя из общих механизмов эволю-
ции, как и по аналогии с филетическими линиями других млеко-
питающих, что в истинной филетической линии был бы вообще не-
возможен столь резкий разрыв в тотальных размерах тела между
последовательными формами, как тот, что существует, например,
между Н. habilis и архантропами Явы. Возможно, что само уве-
личение размеров тела, прослеживающееся почти как общая за-
кономерность в разных филетических линиях млекопитающих, бы-
ло благоприятным фактом с точки зрения преимущественно, «си-
лового» направления адаптации на ранних этапах антропогенеза,
в значительной степени определявшегося высокой интенсивностью
чисто механической нагрузки при использовании безрукояточных
орудий (Семенов, 1964). Вероятно, на самом раннем этапе раз-
вития каменных орудий дальнейший прогресс был связан с неко-
торым возрастанием величины тела, с которым в известной степе-
ни было сопряжено и увеличение мозга. Этот процесс, предпола-
гающий перестройку системы наиболее консервативных морфоге-
нетических корреляций, мог быть только постепенным. Популя-
ции, эволюционирующие таким путем, всегда хорошо приспособ-
лены в отношении твеличины тела, их оптимум всегда .находится
195
в пределах нормальной амплитуды вариаций, и постоянная асим-
метрия в интенсивности стабилизирующего отбора обусловливает
подобное медленное увеличение средних размеров (Симпсон,
1948).
НЕКОТОРЫЕ ПРЕДПОСЫЛКИ К ОБОСНОВАНИЮ
МОРФОЛОГИЧЕСКОГО КРИТЕРИЯ ГОМИНИД
Выше были рассмотрены некоторые черты морфологической
эволюции гоминид, существенные для проблемы их таксономиче-
ского критерия. Как известно, сам вопрос о принципах определе-
ния этого критерия очень сложен и дискуссионен, и в литературе
нашли отражение весьма различные подходы к его решению.
В нашу задачу не входило освещение этой проблемы во всем объ-
еме, и мы коснемся ее главным образом в аспекте рассмотренных
выше особенностей собственно морфологической эволюции го-
минид.
Хорошо известно, что современная систематика приматов зна-
чительно расширяет свой арсенал за счет привлечения, кроме
традиционных, в том числе морфологических критериев, также не-
которых биохимических признаков, особенно тех из них, которые
выявляются разными методами анализа белков; потенциально эти
особенности включаются в число таксономических и используют-
ся для качественного и количественного анализа филогении.
В приматологии помимо исследований в аспекте иммуногене-
тики широкое применение нашло изучение отдельных химических
компонентов и макромолекул (аминокислоты, пептиды, сопостав-
ление аминокислотного состава гемоглобина и других макромоле-
кул, например СТГ, инсулина) и даже молекулярная гибридиза-
ция ДНК- В целом' данные сравнительно-биохимического изучения
человека и других приматов подтвердили общую схему филогене-
тических и таксономических взаимоотношений в отряде приматов,
построенную на. основании фактов из других областей исследова-
ний, как и особую близость человека и африканских антропоидов.
Вместе с тем необходима известная осторожность при попыт-
ках использования данных сравнительной биохимии для уточне-
ния положения более мелких таксонов или времени эволюции от-
дельных ветвей. .Так, например, применительно к приматам уста-
новлена малая дивергенция гемоглобина и белков сыворотки кро-
ви шимпанзе и человека после расхождения этих форм и перехо-
да человека в новую адаптивную зону (Sarich, Wilson, 1967). По
данным этих авторов, древность общей ветви человека и антро-
поидов после отделения гиббонов составляет 10 млн. лет, а воз-
раст отделения человеческой ветви — 5 млн. лет. Эти расчеты по-
ка не находят подтверждения в фактах палеонтологии приматов,
поскольку линии эволюции гоминид и понгид, по-видимому,.были
достаточно дифференцированы уже в верхнем миоцене — рама-
196
питек и дриопитеки (Leakey, 1970; Pilbeam, 1972). Иммунологиче-
ские данные не согласуются с представлением о дриопитеках, как
предках понгид (Buettner-Janusch, 1969). Следует иметь в виду
и некоторые расхождения в результатах, полученных при исполь-
зовании биохимических методов по отношению к современным
приматам (Chedd, 1967; Rempe, 1968).
Не совпадали, по-видимому, и темпы эволюции биохимических
и морфологических признаков: первые были более равномерными
в различных линиях приматов (Sarich, Wilson, 1967). Применение
данных биохимии в «вертикальной», т. е. палеонтологической си-
стематике приматов вообще может быть только опосредованным,
хотя по мере накопления новых ф'актов значение их будет
возрастать, в том числе и в аспекте дифференциации роди раз-
личных биохимических показателей для таксонов разного ранга.
Из всего сказанного вытекает, что подход к выделению таксо-
номического критерия гоминид должен быть преимущественно
морфологическим, так как основной материал палеонтолога — ва-
риации фенотипов.
Но из этого не следует, что определение морфологического
критерия также не сопряжено с трудностями. Достаточно указать
на существующие в литературе разногласия: от полного отрица-
ния «морфологической грани» между первыми гоминидами и их
антропоидными предшественниками до включения в семейство
гоминид только крупных архантропов с высоким развитием веду-
щей системы «органов труда» (материалы симпозиума «О проб-
леме грани между животным и человеком» на VII МКАЭН, 1968,
ил.и симпозиума «Возникновение рода Ното и. его эволюция»,
1972). Первая точка зрения (Урысон, 1972) отражает генеалоги-
ческий подход, близкий по существу к позиции кладнзма, посколь-
ку речь идет о включении в семейство гоминид всех двуногих
высших приматов с момента разделения линий эволюции гоминид
и понгид. Однако эта концепция пока не подкрепляется в
достаточной мере палеонтологическими данными, так как подлин-
ный способ локомоции рамапитеков, признаваемых сторонниками
этого взгляда за первых двуногих гоминид, остается неизвестным.
Согласно другому крайнему мнению (см., например, Зубов, 1964,
1972), в «адаптивную зону» гоминид правомерно включается фак-
тор адаптации к трудовой деятельности, однако, делая акцент на
«отпечаток» этой специфической для гоминид функции только на
достаточно уже продвинувшемся этапе эволюции (высокое разви-
тие и сложная дифференциация рук и головного мозга), тем са-
мым из семейства гоминид исключают представителей самой ран-
ней и длительной фазы трудовой адаптации.
Прежде чем перейти к изложению собственной точки зрения
на морфологический критерий гоминид, следует подчеркнуть, что
любая концепция, построенная преимущественно на палеонтоло-
гических данных, т. е, относящаяся к установлению «вертикаль-
ных границ», может оказаться провизорной и имеет, скорее,' ха-
197
рактер таксономической гипотезы; как известно, даже неонтологи
неоднократно пересматривают таксономические группировки, в
большей мере относится это к палеонтологическим таксонам,
где всегда возможны коррективы, вносимые последующими откры-
тиями.
Резюмируя вышесказанное, при выделении морфологического
критерия семейства гоминид мы будем исходить из следующих
предпосылок.
Необходимость основываться в первую очередь на морфологи-
ческих признаках, отражающих переход предков человека в но-
вую адаптивную зону, что означает в данном случае прямохожде-
ние плюс систематическое изготовление примитивных орудий.
Вытекающая отсюда комплексность морфологического крите-
рия.
Постепенность формирования морфологических комплексов,
проявления которой будут возрастать по мере пополнения наших
палеонтологических познаний и приближения реконструируемого
сравнительно-анатомического ряда к истинной филетической линии
гоминид.
Неравномерность морфологической эволюции, особенно высту-
пающая на ее ранних стадиях. Помимо различной интенсивности
селективного давления, здесь может играть роль и дифференци-
альная способность систем организма к достаточно быстрым и
макроморфологически уловимым и отчетливым преобразованиям.
Полагаем поэтому, что с морфологической стороны решающи-
ми фактами принадлежности высшего примата к семейству гоми-
нид нужно считать следующие.
Исходя из постепенности формирования большей части морфо-
логических признаков и различий в темпах эволюции важнейших
гоминидных комплексов, за основу следует принять комплекс би-
педии, как хронологически самый ранний из всех. Оформление это-
го комплекса, связанное с возникновением прямохождения, было
самым ранним из узловых моментов антропогенеза, что соответст-
вует известной концепции Ф. Энгельса; комплекс прямохождения
уже сложился в главных своих чертах, по крайней мере, в ниж-
нем вилла-франке. Переход к прямохождению высокоразвитого
примата означал приобретение принципиально нового адаптивно-
го плато, допускающего воздействие на организм и новой систе-
мы селективных факторов (Biegert, 1963). Предполагается даже,
что этот переход мог быть обусловлен коренной хромосомной пе-
рестройкой — «хромосомный рубикон гоминизацйи» (Ruffie,
1973).
Вместе с тем существующие сейчас палеонтологические факты
позволяют думать, что прямохождение только потому и стало
ароморфозом, что оно было тесно связано с трудовой деятель-
ностью, являясь для нее не только важнейшей предпосылкой, но
в какой-то мере и ее следствием. Можно с большой долей досто-
верности предположить, что такой переход к систематическому
198
изготовлению даже самых примитивных орудий не прошел бес-
следно и для «рабочего отдела» руки. Об этом свидетельствует
человеческая форма ногтевых фаланг на олдувейской кисти, т. е.
остеологический критерий гоминизации в определенной степени
уже существовал на кисти Н. habilis. Напомним и о высоком
уровне церебрализации этого гоминида
Принимая во внимание приведенные выше теоретические сооб-
ражения и палеонтологические факты, полагаем, что помимо сфор-
мировавшегося в своих основных характеристиках комплекса бй-
педии морфологический критерий гоминид должен включать таю
же начальную адаптацию кисти к трудовой функции, вероятно,,
«силового» направления и достаточно высокий уровень церебрали-
за'ции, превосходящий таковой у наиболее развитых антропоидов,
современности. При таком подходе из семейства гоминид не ис-
ключается самая ранняя и длительная фаза трудовой адаптации. '
С фактической стороны это положение подтверждается палеопри-
матологическими находками последнего времени, и прежде всего,,
открытием Н. habilis, у которого перечисленные выше компонен-
ты морфологического критерия встречались в сочетании с наибо-
лее ранними и примитивными из известных сейчас каменных ору-
дий. Таким образом, Н. habilis является одним из древнейших из
известных палеонтологии достоверных представителей семейства
Hominidae.
Подобное заключение, по-видимому, наиболее согласуется с
интерпретацией добытых к настоящему времени фактов палеон-
тологии человека. Судя по предварительным данным некоторых
появившихся в последнее время публикаций, продолжается тен-
денция к удревнению нижней границы таксона Hominidae. Это,,
прежде всего, замечательные открытия гоминид в Восточной Аф-
рике — череп 1470 и сопутствующие., ему остатки посткраниаль-
ного скелета в Кении; находки в Эфиопии и Танзании, абсолют-
ный возраст которых превышает 2 и даже 3 млн. лет (Day et al.,.
1975; Koenigswald, 1976; Johanson, Taieb, 1976).
1 Большой интерес представляют значительные размеры и прогрессивные черты
неврологической организации очень древнего эндокрана. 1470, по данным пред-
варительной публикации Холлоуэя (1974).
Литература
Алексеев В. П. Некоторые вопросы развития кисти в процессе антропоге- I
неза. «Тр. Ин-та этнографии», н. с., 1960, т. 50. .
Алексеев В. П. Гомцниды второй половины среднего и начала верхнего '
плейстоцена Европы. В сб.: «Ископаемые гоминиды и происхождение чело-
века». М., «Наука», 1966. j
Алексеев В. П. О скорости эволюции в пределах семейства гоминид. Тр. j
«VII Междунар. конгр. антропол. и этнографических наук 1964», т. 3. М., |
«Наука», 1968. )
Арипов У. А. О внутренней архитектуре верхней трети бедренной кости че-
ловека и ее значении в хирургии. «Сб. научи. тр. Самаркандск. мед. ин-та», I
1956, №41. *
Астанин Л. П. О строении костей кисти боксера. «Изв. АН СССР», 1951, ;
вып» 35. '
Астанин Л. П. Пропорции кисти приматов. «Вопр. антропол.», 1962, вып. 10. '•
Астанин Л. П. К вопросу о возрастных изменениях пропорций человеческой . >
кисти. «Архив анатомии, гистол. и эмбриол.», 1962а, т. 43, № II. j
Астанин Л. П. Пропорции кисти человека и приматов. «Тр. VII Междунар. I
конгр. антропол. и этнографических наук, 1964», т. 3. М., «Наука», 1968. |
Беноволенская Ю. Д., Хрисанфова Е. Н. К сравнительной характе- j
ристике ключицы и трубчатых костей неандертальского и современного че- 1
" ловека. «Вопр. антропол.», 1961, вып. 6.
Бобин В. В. Палеопатологические находки последних лет в Крыму и на Се-
верном Кавказе. «Тр. VII Междунар. конгр. антропол. и этнографических
наук, 1964», т. 2. М., «Наука», 4967. к
Бонч-Осмоловский Г. А. Кисть ископаемого человека из“ грота Киик- г
Коба. В сб.: «Палеолит Крыма», вып. 2. М.—Л., Изд-во АН СССР, 1941. i
Бонч-Осмоловский Г. А. О типах эволюции стопы человека. «Природа», 1
1943, № 2. ~ '
Бонч-Осмоловский Г. А. Скелет стопы и голени ископаемого человека J-
из грота Киик-Коба. В сб.: «Палеолит Крыма», вып. 3. М.—Л., Изд-во
АН СССР, 1964. к
Бунак В. В. Макроструктура головного мозга в период роста. В сб.:. «Ана-
томические и гистоструктурные особенности детского возраста». М., Йзд-во г
Наркомпроса РСФСР, 1936. *
Бунак В. В. Современное состояние проблемы эволюции стопы у предков че-
ловека. В кн. Г. А. Бонч-Осмоловского «Скелет стопы и голени ископаемого
человека из грота Киик-коба». В сб.: «Палеолит Крыма», вып. 3. М.—Л., !
Изд-во АН СССР, 1954. j
Б у н а к В. В. О механизме приспособительных изменении труочатых костей. i
В сб.: «Тр. конфер. О-ва анатомов, гистол., эмбриол.». М.—Л., 1956. j
Бунак В. В. Изменение относительной длины сегментов скелета конечностей ’
в период роста. «Изв.. АН СССР», 1957, т. 84. г
Бунак В. В. Череп человека и стадии его формирования у ископаемых лю- >
дей и современных рас. «Тр. Ин-та этнографии», п. с., 1959, т. 49. *
Бунак В. В. Мозговая коробка. В сб.: «Ископаемые гоминиды и происхож-
дение человека». М., «Наука», 1966.
Бунак В. В. Краткий обзор таксономических и филетических схем гоминид. " ’
В сб.: -«Ископаемые гоминиды и происхождение человека». М., «Наука», :
1966а. - ч ।
Бунак В. В. Массивность скелета человека в сравнительном освещении. «Вопр. •
антропол.», 1967, вып. 26.
Бунак В. В., Клебанова Е. А. Строение компактного, вещества трубчатой !
кости млекопитающих в сравнительном освещении. «Тр. VI Всес. съезда |
анатомов, гистол. и эмбриол., 1958», т. I. Харьков, 1960/1961.
200
Б ушмакин Н. Д. Аномалии позвоночника с точки зрения его эволюции.
«Казанский мед. журн.», 1913, т. 13.
Быстров А. П. Прошлое, настоящее, будущее человека. Л., Медгиз, 1957.
Вебер М. Приматы. М.—Л., Биомедгиз, 1936.
Видерсгейм Р. Строение человека со сравнительно-анатомической точки зре-
ния. СПб., 1900.
Волочком М. О двух формах человеческой кисти. РАЖ, 1924, 13, № З1—4.
Волянский Н. Некоторые проблемы механизма и факторов эволюции. «Сов.
этнография», 1957, № 6.
Гевлич Е. Д. Морфофункциональ'ные особенности суставов плечевого пояса
и верхней конечности спортсменов. «Тр. VII Междунар. конгр. антропол. и
этнографических наук, 1964», т. 2. М., «Наука», 1967.
Данилова Е. И. Эволюция руки. Киев, «Наукова думка», 1965.
Данилова Е. И. Кисть и стопа гоминид и близких к ним форм. В сб.: «Ис-
копаемые гоминиды и происхождение человека». М., «Наука», 1966.
Данилова Е. И. Морфологические особенности кисти древних гоминид в
связи с некоторыми вопросами антропогенеза. «Тр. VII Междунар. конгр.
' антропол. и этнографических наук, 1964», т. 3. М., «Наука», 1968.
Дарвин Ч. Происхождение человека и половой отбор. Собр. соч., т. 5. М.,
Изд-во АН СССР, 1953.
Д,е б е ц Г. Ф. Опыт определения веса живых людей по размерам длинных кос-
тей. «Тр. VII Междунар. конгр. антропол. и этнографических наук, 1964»,
т. 2. М., «Наука», 1967.
Джигора С. Т. Изучение морфологии гаверсовых каналов при определении
видовой принадлежности костей. «Врачебное дело», 1957, № il.
До С у а н X о п. Строение стопы у вьетнамцев. «Архив анат., гистол. и эм-
бриол.», 1964, т. 46, №6.
Дэрумс В. Я. О некоторых морфологических признаках бедренных костей
у древних жителей Прибалтики. «Сб. Тр. научн. о-ва ’судебн. медиков
ЛитССР», 1973, № 3.
Жданов Д. А. Функционально-морфологические основы формирования скеле-
та. «Тр. VII Междунар. конгр. антропол. и этнографических наук, 1964»,
т. 2. М., «Наука», 1967.
Жеденов В. Н. Сравнительная анатомия приматов. М., «Высшая школа», 1962.
Зубов А. А. О систематике австралопитековых «Вопр. антрополог.»; 1964,
вып. 117.
Зубов А. А. Зубная система. В сб.: «Ископаемые гоминиды и происхождение
человека». М., «Наука», 1966.
Зубов А. А. Систематические критерии рода Ното и его эволюция. «Вопр.
антропол.», 1972, вып. 41.
3 у б о в А. А. Этническая одонтология. М., «Наука», 1973.
Ивкина К- Л., Супряга Е. Л. Динамика адаптивных изменений в строе-
нии кисти гимнаста. В сб.: «Мат-лы 8-й научн. конфер. по возрасти. • мор-
фол., физиол. и биохимии, 1967», ч. 1. М., «Просвещение», 1967.
Исии Рэйдзиро, Охае и Хироёси. Исследование длины пальцев у жи-
телей района Нагасаки, сев. область Сацума. «J. Kumamoto Med. Soc.», 1965,
vol. 39, № 12.
Кац А. И. Употребление и «изготовление орудий» приматами (шимпанзе и
низшие обезьяны). В сб.: симпозиума «Биология и акклиматизация обезьян».
М., «Наука», 1973.
Клебанова Е. А. Влияние усиленной мышечной деятельности на кости ко-
нечностей молодых животных. «Изв. Ин-та им. Лесгафта», 1954, № 2,6.
Клебанова Е. А. Изменение в костной системе у растущих животных
под влиянием физической нагрузки. «Тр. 1-й научн. конфер. по возрасти,
морфол. и физиол.». М., 1954а.
Клебанова Е. А. Микроскопическое строение компактного вещества длинных
костей конечностей у представителей семейства тушканчиков. «Зоол. журн.»,
1962, т. 12, вып. 3.
Ковещникова А. К. Приспособительные морфологические’изменения -орга-
нов двигательного аппарата в связи с возрастом'и функциональными на-
13 Е. Н. Хрисанфова
201
грузками у человека и животных. «Тр. VI Всес. съезда анат., гистол. и
эмбриол. 51958», т. I. Харьков, 1960, 1961.
Кочеткова В. И. Палеоневрология. Изд-во МГУ, 1973.
Крылова Н. В. Изменения костно-суставного аппарата человека под влия-
нием спортивной гимнастики, легкой атлетики и велосипедного спорта. «Тр. ‘
VII Междунар. конгр. антропол. и этнографических наук, 1964», т. 2. М.,
«Наука», 1967.
Куммер Б. Развитие и вариации формы длинных костей в зависимости от
механического воздействия. «Архив анат., гистол. и эмбриол.», 1965, т. 49
№ 7.
Лехе В. Человек, его происхождение и эволюционное развитие. М., 1913.
М а й р Э. Зоологический вид и эволюция. М., «Наука», 1968.
Майр Э. Принципы зоологической систематики. М., «Мир», 1971.
Малявская О. А., Хрисанфова Е. Н. К морфологии плечевого и тазо-
бедренного суставов современного и неандертальского человека. «Вопр. ан-
тропол.», 1962, выл. 12. •
Мацуи Рёити, Ям агуто Хидэо. Исследование длины пальцев жи-
телей района Мимаки, юго-западная часть префектуры Иё. «J. Kumamoto
Med. Soc.», 1965, vol. 39, № 11.
M а ш к a p а К- И. Влияние физического труда на строение скелета нижних ко-
нечностей. «Архив анат., гистол. и эмбриол.», 1970, т. 59, № 11.
Мураками Фумио, Мацуда Айто, Тигои Ясухиро. О длине
пальцев рук у жителей Амакусасимосима, префектура Хиго. «J. Kumamoto
Med. So с.», 1966, vol. 40, № 12.
Н а к е л ь с к и й С. К., Карлов Н. Н. Бедренная кость палеолитического че-
ловека из Романкова. «Вопр. антропол.», 1965, вып. 20.
Никитюк Б. А. Лицевой скелет, верхняя его часть. В сб.: «Ископаемые го-
миниды и происхождение человека». М., «Наука», 1966.
Никитюк Б. А. Нижняя челюсть. В сб.: «Ископаемые гоминиды и проис-
- хождение человека». М., «Наука», 1966а.
Никитюк Б. А., Бевз юк В. В. Влияние механического фактора на изме-
нение величины костей кисти за один год у молодых спортсменов. «Вопр.
антропол.», 1971, вып. 38.
Очкуренко А. М. Размеры длинных костей верхних конечностей и костно-
мозгового канала ца разных уровнях. В сб.: «Вопр. современной хирур-
гии». Киев, Госмедиздат УССР, 11962.
Пикулев Л. А., Герасимов С. М., Черемисин В. М., Шипи-
лов В. М. Соотношение между объемом гипофиза и объемом турецкого
седла. «Архив анат., гистол. и эмбриол.», 1970, т. 59, № 9.
Поляков Г. И. Структурная организация коры большого мозга человека по
данным развития ее в онтогенезе. В кн.: «Цитоархитектоника коры большого
> мозга человека». М., «Медгиз», 1949.
Попова-Латкина Н. В. Новые материалы по развитию органов в эмбрио-
нальном периоде у человека. «Проблемы современной эмбриологии». Л., 1956.
Попова-Латкина Н. В. О механизмах возникновения врожденных ано- '
малий и уродств. ДАН СССР, 1965, т. 161, № 2.
Привес М. Г. Влияние -некоторых видов труда и спорта на строение скеле-
та. В сб.: «Проблемы функц. морфологии двигат. аппарата». М., «Медгиз»,
4956.
Привес М. Г. Строение скелета людей различных профессий. «Тр. VII Меж-
дунар. конгр. антропол. и этнографических наук 1964», т. 2. М., «Наука»,
Рогинский Я. Я. Об устойчивости характерного для вида типа пропорций
тела. «Зоол. журн.», 1957, т. 36, № 1.
Рогинский Я- Я- Об этапах и темпах эволюции гоминид. «Сов. этнография»,
1957а, вып. 6.
Рогинский Я. Я. О некоторых результатах применения количественного ме-
тода к изучению морфологической изменчивости. «Архив анат., гистол. и
эмбриол.», 1959, т. 36, № -1.
202
Рохлин Д. Г. Скелет кисти и дистального отдела предплечья. «Рентгеноло-
гия и радиология». М.—Л., Биомедгиз, 1936.
Рохлин Д. Г. Некоторые данные рентгенологического исследования детского
скелета из грота Тешик-Таш, Южный Узбекистан. В сб.: «Тешик-Таш. Палео-
литический человек». Изд-во МГУ, 1949.
Рохлин Д. Г. Болезни древних людей. М., Изд-во АН СССР, 1965.
Рохлин Д. Г. Древность патологических процессов на ископаемых костях
человека в СССР. «Тр. VII Междунар. конгр. антропол. и этнографических
наук 1964», т. 2. М., «Наука», 1967.
СвадковскийБ. С. О росте диафизов трубчатых костей у человека. «Докл
АПН РСФСР», 1959, № 4.
Северцов А. Н. Морфологические закономерности эволюции. М.—Л., Изд-во
АН СССР, 1939.
Семенове. А. О противопоставлении большого пальца руки неандертальца.
«Кр. сообщ. Ин-та этнографии АН СССР», 1950, вып. 11.
С.е м е н о в С. А. Очерк развития материальной культуры и хозяйства палео-
лита. В сб.: «У истоков человечества». Изд-во МГУ, 1964.
Симпозиум «О проблеме грани между животным и человеком». «Тр. VII Меж-
дунар. конгр. антропол. и этнографических наук, 1964», т. 3. М., «Наука»,
11968.
Симпозиум «Возникновение рода Ното и его эволюция», «Вопр. антропол.»,
1972, выл. 41.
Симпсон Дж. Г. Темпы и формы эволюции. М., Госиздат, 1948.
Соколов П. Н. Структурные и функциональные особенности костной систе-
мы конечностей человека. «Тр. VII Междунар. конгр. антропол. и этногра-
фических наук, 1964», т. 2. М., «Наука», 1967.
Сперанский А. Д. Происхождение spina bifida occulta в крестцовом отделе
позвоночного столба человека. «Вести, хирургии», 1925, № 13.
Старкова Н. Т., Бухман А. И., Харитонов Е. И. Состояние костно-
суставной системы при мужском гипогонадизме. «Пробл. эндокринологии»,
1969, № 3.
Степанова М. П. Развитие скелета таза в эмбриональном периоде у чело-
века. Автореф. канд. дисс. Волгоград, 1955.
'Степанова М. П. Развитие тазовых костей в эмбриональном периоде у че-
ловека. В сб.: «Вопросы эмбриологии человека», вып. 2. Астрахань, 1961.
Тонков В. Н. Учебник анатомии человека, ч,. 1. М., Медгиз, 1953.’
У р ы с о н М. И. Выступление на симпозиуме «Возникновение рода Ното и его
эволюция». «Вопр. антропол.», (1972, выл. 41.
Харрисон Дж., Уайнер Дж. Биология человека. М., «Мир», 1968.
Хрисан-фюва Е. Н. Эволюция соотношений* мозгового и лицевого отделов
черепа в семействе Hominidae. «Сов. антропология», 1958, вып. 3.
Хрисанфова Е. Н. Теоретические вопросы изменчивости позвоночника и
грудной кДетки человека.-Изд-во МГУ, 1962.
Хрисанфова Е. Н. Бедренная кость палеоантропа из Романкова. «Вопр.
антропол.», 1965, вып. 20.
Хрисанфова Е. Н. Скелет туловища и конечностей. В сб.: «Ископаемые го-
миниды и происхождение человека». М., «Наука», 1966.
Хрисанфова Е. Н. Эволюция структуры длинных костей человека., М.,
«Наука», 1967.
Хрисанфова Е. Н. Внутренняя структура длинных костей и ее значение для
эволюционной .морфологии человека. «Архив анат., гистол. и эмбриол.»»
4967а, т. 53, № >11.
Хрисанфова Е. Н. Анализ морфологической изменчивости посткраниального
скелета человека с точки зрения этапов его формирования. «Тр. VII Меж-
дунар. конгр. антропол. и этнографических наук, 1964», т. 2. М., «Наука»,
1,9676.
X р и с а и ф о в а Е. Н. Сравнительная изменчивость внешней и внутренней струк-
туры диафизов трубчатых костей приматов. «Тр. VII Всес. съезд анат., гис-
тол. и эмбриол., 1966». Тбилиси, Изд-во АН ГССР, 1969.
13*
203
Хрисанфова Е. Н. Значение морфологии посткраниального скелета для
таксономии гоминид. В сб.: «Неотектоника, новейшие отложения и человек».
Изд-во МГУ, 1969а.
Хрисанфова Е. Н. Некоторые черты морфологической эволюции дисталь-
ных отделов конечностей человека. В сб.: «.Человек. Эволюция и внутри-
видовая дифференциация». М., «Наука», 1972.
Хрисанфова Е. Н. Адаптации скелета ранних гоминид в связи с некоторы-
ми факторами среды. В сб.: «Первобытный человек и природная среда». М.,
«Наука», 4974.
Цивьян Я. Л., Швец А. И. Особенности пояснично-крестцового лордоза че-
ловека. «Архив анат., гистол.,-и эмбриол.», 1973, т. 64, № 5.
Шевченко Ю. Г. Основные направления эволюции мозга приматов. «Тр.
VII Междунар. конгр. антропол. и этнографических наук, 1964», т. 3. М.,
«Наука», 1968.
Шмальгаузен И. И. Проблемы дарвинизма. М., «Наука», 1969.
К. Маркс и Ф. Энгельс. Соч., т. 20, М, 1961.
Якимов В. П. О /противопоставлении большого пальца руки. «Журн. общей
биол.», 1949, т. 10, № 3.
Abel О. Die Stellung des Menschen im Rahmen der Wirbeltiere. Jena, 1931.
Adachi B. Die Handknochen der Japaner. «Mitteil. der Med. Fak. Univ. Tokyo»,
4905, Bd. 6, No. 4.
A d о 1 p h i H. «Uber die Variationen des Brustkorbes und der Wirbelsaule der
Menschen. «Morphol. Jahrb.», 1905, Nr. 33.
Aguirre E. Les rapports phyletiques de ramapithecus et de kenyapithecus et
1’origine des hominides. «Anthropologic», 1972, t. 76, No. 5—6.
Ahuja M. Normal variation in the density of selected human bones in North
India. «J. Bone and Joint Surg.»,T969, Bd. 51, Nr. 4.
Allbrook D. V. The East African vertebral column. «Amer. J. Phys. Anthro-
pol.», 1955, vol. 13.
An cel P., Scncert L Des variations des segments vertebrocostaux. «Biblio-
graphic anat.», 1902, Bd. 10.
Anthony R. Contribution a J’etude morphologique des caracteres d’adaptation
a la vie arboricole chez les Vertebres. «Ann. des Sc. nat. (Zoologie)», 1912,
Bd. 9, ser. 15.
Ar a mb ou rg С. M. Sur I’attitude en station verticale des Neanderthaliens.
«С. r. Acad. Sci. Casablanca», 1955, t. 240, No. 7.
•Archibald J. D., Lovejoy С. О., H e i p I e K. G. Implications of relative
robusticity in the Olduvai' metatarsus. «Amer. J. Phys. Anthropol.», 1972,
vol. 37, No. 1.
A i g n e r J. S., Laughlin W. S. The dating of Lantian man and his signifi-
cance for analyzing trends in human evolution. «Amer. J. Phys. Anthropol.»,
1973, vol. 39, No. 1.
Armelagos G. J., Mielke J. H., Owen K« H., Van G erven D. P.,
Dewey J. R., Mahler P. E. Bone growth and development in prehistoric
populations from Sudanese Nubia. «J. Hum. Evol.», 1972, vol. 1, No. 1.
Ashley G. T. A comparison of human and anthropoid mesosterna. «Amer.
J. Phys. Anthropol.», 1956, vol. 14, No. 3.
Bard.een Ch. R. Numerical vertebral variation in the human adult and embrye.
«Anat. Anz.», 1904, vol. 25.
Bardeen Ch. R. The development of the thoracic vertebrae in man. «Amer.
J. Anat.», 1905, No. 4.
Barnett С. H. Squatting facets on the europeans talus. «J. Anat. Lond.», 1954,
vol. 88, No. 4.
В e n f e r R. A., McKern Th. W. The correlation of bone robusticity with the
perforation of the coronoid-olecranon septum in the humerus of man. «Amer.
J. Phys. Anthropol.», 1966, vol. 24, No. 2.
Bennett K. A. Lumbo-sacral malformations and spina bifida occulta in a
group of protohistoric Modes Indians. «Amer. J. Phys. Anthropol.», 1972,
vol. 36, No. 3.
204
Bernstein P., С r e 1 i n E. S. Bony pelvic sexual dimorphism in the rat.
«Anat. Rec.», 1967, vol. 157, No. 3.
Biegert J. The evaluation of characteristics of the skull, hands and feet for
.primate taxonomy, «Classific and human evolut.», Chicago, Aldine Publ
Co., 1963.
Biegert J., Maurer R. Rumpfskelette, Allometrien und Korperproportionen
bei catarrhinen Primaten. «Folia primatol.», 1972, Bd. 17, Nr. 1—2.
Billy G. Etude anthropologique d’un squelette du bronze ancien decouvert a
Ludesse (Puy-de-Ddme) «Bull. Soc. prehistor. frang.», 1968, t. 65, No. 7.
В i 11 у G. La squelette post-cranien de 1’homme de Chancelade. «Anthropoiogie»,
1969, t. 73, No. 3—4.
Black D. Sinanthropus skeletal remains with a note on other North China fossil
hominids. In:* «Fossil man in China», ser. a, 1933, vol. 11.
В 1 i n с о e H, Sighificant hand types in women according to relative lengths of
fingers. «Amer. J. Phys. Anthropol.», 1962, vol. 20, No. 1.
В о 1 к L. Das Problem des Menschwerdung. Vortrag auf der XXX Versammlung
der anatomischer Gesellschaft zu Freiburg. Jena, 1926.
Bornstein Ph. E., P e t e r s о n R. R. Numerical variation of the presacral
vertebral column in three population groups in North America. «Amer. J.
Phys. Anthropol.», 1966, vol. 25, No. 2.
Boule M. L’homme fossile de la Chapelle-aux-Saints. «Ann. paleontol. hum.»,
1911/1913, t. 9, No. 6—8.
Brain С. K. An attempt to reconstruct the behaviour of australopithecines.
«Zool. afr.», 1972, vol. 7, No. 4.
Braitenberg V. Remarks on the texture of brains. «Int. Cent. Theor. Phys.
Int. Atom. Energy, Agency»,’1972, vol. 83, No. 3—5.
Brauer G. Ging der Neanderthaler vorniibergeneigt? «Naturwiss. Rdsch.», 1972,
Bd. 25, Nr. 5.
Broman G. E., Trotter M., Peterson R. R. The density of selected bones
of the human skeleton. «Amer. J. Phys. Anthropol.», 1958, vol. 16.
Broom R., J. T. Robinson, G. W. H. Schepers. Sterkfontein ape-man
Plesianthropus. «Transvaal Museum Memoirs», 1950, No. 4.
Broom R., Schepers G. W. H. The South African ape-man; The Australo-
pithecinae. «Transvaal Museum Memoirs», 1946, No. 2.
В г о t h w e 11 D. R., ,M о 11 e s о n T., M e t r e w e 11 i C. In: «Skeletal Biology of
Earlier Human Populations» (edit, by Brothwell D. R.); Pergamon, Oxford,
1968.
Buettner-Janusch J. The nature and future of physical anthropology.
«Trans. N.Y. Acad. Sci.», 1969, vol. 3'1, No. 2.
Bugyi B. Zur Methodik einer Palaoendocrinologie des Menschen. «Homo», 1963,
* Nr. 13.
Burris P. E. New determination of australopithecine height. «Nature», 1971,
Bd. 232, Nr. 5309.
Burton F. D. An analysis of the muscular limitation on opposability in seven
species of Cercopithecinae. «Amer. J. Phys. Anthropol.», 1972, vol. 36, No. 2.
Cameron J. The skeleton of british neolithic man etc. London, Williams and
Norgate, 1934.
Campbell B. G. Human Evolution: An Introduction to Man’s Adaptations. Al-
dine Comp., Chicago, 1966.
Campbell B. G. Human Evolution: An Introduction to Man’s Adaptations.
London, Heinemann Educ. Books, 1967.
Capecchi V. Sul significate della disposizione dei process! spinosi cervicali
nella razza di neandertal. «Atti Soc. pelorit. sci. fis. mat. e natur.» 1965,
vol. 11, No. 1, 2.
Charles R. H. The influence of function, as exemplified in the Morphology of
the lowel extremity of the Panjabi. «J. Anat. Lond.», 1893, vol. 28.
C h e d d G. How close are we to the apes? «New Scientist», 1967, vol; 36, No. 575.
С 1 а г к J. D., В г о t h w e 11 D. R., P о w e r s R., О а к 1 e у К. P. Rhodesian man;
notes on a new femur fragment. «Man», 1968, vol. 3, No. L
205
Coon C. S. The origin of races. N. Y., 1963.
Cop pens Yv. L’hominien du Tchad. «С. r. Acad, sci.», 1965, vol. 260, No. 10.
Corrigan G. E. The neonatal clavicle. «Biol, neonatorum», 1960, vol. 2, No.2.
Cramer H. Zur Wirbelsaulendynamik. «Dtsch. orthop. Ges. 45 Kongr. 1955».
Stuttgart, 1956.
Cunningham D. J. The evolution of the eyebrown region of the forehead,
etc. «Transactions of the Royal Soc. of Edinburgh», 1908/1909, vol. 46, No. 11.
Danforth С. H. Numerical variation, and homologies in vertebrae. «Amer. J.
Phys. Anthropol.», 1930, No. 14.
D a s R., Das Bh. M.’ A study of the Hira foot. «Man in India», 1967, vol. 47,
No. 2.
Dastugue J. Pathologic des homines fossiles de I’abri de Cro-Magnon. «Anthro-
pologic», 1967/1968, t. 71, No. 5-6.
Davis P. R. -Hominid fossils from Bed. I, Olduvai Gorge, Tanganyika.-«Na-
ture», 1964, No. 4923.
Day M. H. Femoral fragment a robust australopithecine from Olduvai Gorge,
Tanzania. «Nature», 1969, vol. 221, No. 5177.
Day M. H. Postcranial remains of Homo erectus from Bed IV Olduvai Gorge,
Tanzania. «Nature», 1971, vol. 232, No. 5310.
D а у M. H., Leakey R. E. F., Walker Л. C., Wood B. A. New Hominids
from East Rudolf, Kenya. «Amer. J. Phys. Anthropol.» 1975, vol. 42, N 3.
Day M. H., Mol 1cson T. I. The Trinil femora. «J. Hum. Evol.»T London, 1973.
Day M. H., Napier J. R. Hominid fossils from Bed I Olduvai Gorge, Tanga-
nyika. Fossil foot bones. «Nature», 1964, vol. 201, No. 4923.
D ay M. H.> N a p i er J. R. A hominid toe bone from Bed I» Olduvai Gorge, Tan-
ganyika. «Nature», 1966, vol. 211, No. 5052.
D а у M. H., Scheuer J. L. SKW 14147: a new hominid metacarpal from Swart-
krans. «J. Hum. Evo!.», 1973, vol. 2, No. 6.
Day M. H„ W о о d B. A. Functional affinities of the Olduvai Hominid 8 talus.
«Man», 1968, vol. 3.
Day M. H., Wood B. A. Hominoid tali from East Africa. «Nature», 1969, •
vol. 222, No. 5193.
D e 1 a 11 r e A. F. R. L’hominisation du crane est-clle terminee? «С. r. Acad, sci.»,
1956, t. 243, No. 4.
Delattre A., Deblock R. La hauteur de la ptere chez 1’homme et les pri-
mates. «Bull. Assos. anat.», 1972, No. 154.
DesiombesP., Fen art R. Modifications craniennes chez le rat rendu hydro-
eephale. «Bull. Assoc, anat.», 1970, No. 147.'
Dewdney J. С. H. Some parameters of the footprint of the New Ireland adult
male. «Papua and N. Guinea Med. J.», 1966, vol. 9, No. 45
D о к 1 a d a I M. Versuch einer experimentellen Beeinflussung der Schadelform bei
Ratten. «Scripta med.», 1964, vol. 37, No. 4.
D о к I a d a 1 M. Significance of study of the cross section shape of the shaft
of the femur for the identification of skeletal fragments. «Folia Morphol.»
(CSSR), 1972, vol. 20, No. 3.
D u с к w о r t h W. L. H., M. A., H. D. Morphology and Anthropology. A Hand-
book for Studentes. Cambridge, 1915, vol. 1.
Duer os A. Le chignon occipital, niesure sur le squelette. «Anthropologic», 1967,
vol. 71, No. 1—2.
Du ter loo H. S., Enlow D. H. A comparative study of cranial growth in
Homo and Macaca. «Amer. J. Anat.», 1970, vol. 127, No. 4.
E h a r a A., S e i I e r R. Die Strukturen der Oberaugenregion bei den Primaten,
Deutungen und Definitionen. «Z. Morphol. una Anthropol.» 1970, Bd. 62,
Nr. 1.
E I i a к i s E. С., I о r d a n i d i s P. J. La determination de Page par Findice
medullaire des os longs.- «Ann. med. legale», 1963, t. 43, No. 6.
E 1 i a к i s E. С., I о r d a n i d i s P. J. The determination of the individual sex
studying the medullary of the long bones. «Практика тис академиас Афинон»,
1963а, No. 38.
206
Enlow D. H. A comparative study of facial growth in Homo and Macaca.
«Amer. J. Phys. Anthropol.», 1966, vol. 24, No. 3.
ErdbrinkD. P., Tacoma J., Visser H. A fossil human bone from Anato-
lia. «Proc, konikl. nederl. akad. wet.», 1966, C69, No. 4.
Evans F. G. Stress and strain in bones. Thomas Springfield, 1957.
Evans F. G. Relations between the microscopic structure and tensile, strenght of
human bone. «Acta anat.», 1958, vol. 35, No. 4.
Felts W. J. L. Transplantation* studies of factors in skeletal organogenesis.
«Amer. J. Phys. Anthropol.», 1959, No. 17.
Fenart R., Heim J.-L. La posture des neanderthaliens d’apres 1’orientation
vestibulaire du crane. «С. r. Acad, sci.», '1968, D267, No. 23.
Ferembach D. Frequency of spina bifida occulta in prehistoric human skele-
tons. «Nature», 1963, vol. 199, No. 4888.
Ferembach D. Les ossements humains ge Salemas (Portugal). «Comuns Serv.
geol. Portugal», 1964/1965, No. 48.
Fick R. Handbuch der Anatomic und Mechanik der Gelenke. Jena, 1904.
Fick R. Untersuchung an der Wirbelsaule von 30 Gibbons und fiber die Wir-
belzahlen der Menschenaffen. «Sitzungsber. Akad. Wiss.,Berlin», 1936, Bd. 17.
Fisch el A. Untersuchungen fiber die Wirbelsaule und den Brustkorb des Men-
schen. «Anat. Hefte», 1906, Bd. 31.
F r a i p о n t Ch., L о h e s t. La race humaine de Neanderthal ou de Canstadt, en
Belgique. «Arch. Biol.», 1887, No. 7.
Frisch J. E. Trends in the evolution of the hominoid dentition. «Bibl. Prim.»,
• Fasc. 3, Basel, 1965.
F u s t ё M. Algunas observationes sobre la orientation de la region occipital en
craneos neandertalenses. «Cursillos у conf. Inst, investig. «Lucas Mallada»,
1957, No. 4.
G a r n S. M. e t al., 1964. Цит. по об.: «The Skeletal Biology of Earlier Human
Populations» (edit, by Brothwell D. R.). Oxford, Pergamon Press, 1968.
Garn S. M., N a g у J. M., PoznanskiA. K., McCann M. B. Size reduction
associated with brachymesophalangia-5; a possible selective advantage. «Amer.
J. Phys. Anthropol.», 1972, vol. 37, No. 2.
G i e s e 1 e r W, Die Fossilgeschichte des Menschen. In: «Die Evolution der Or-
ganismen», 11 Aufl. Stuttgart, 1959.
Gil lot C. Confrontation entre certaines varietes de lombalgies et les dispositifs
anatomiques rachidiens «Ann. med. phys.», 1972, t. 15, No. 2.
Girard L. Port habituel de la tete et fonction vestibulaire. «Mammalia», 1947,
No. 10.
Green E. L. Genetic and non genetic factors which influence the tipe of the
skeleton in an inbread strain of mice. «Genetics», 1941, No. 26.
G or j a n о v i c - Kr a mb er ger K. Der diluviale Mensch von Krapina in Kroa-
tien. Wiesbaden, 1906. , .
Gozdziewski St. Analiza mi^dzyodmianowa kosci ramiennych. «Przegl.
antropol.», 1962, t. 28, No. 1.
Grosshans E.,'Meyer P. Morphologie et variations des facettes articulaires
des bases metatarsiens. «Arch. anat. histol. et embryol.», 1963, vol. 46, No. 1—4.
Gutierrez L. S. A contribution to the study of the limiting factors of elbow
extension. «Acta anat.», 1964, vol. 56, No. 1—2.
Haas N. Critica criteriului «Schwalbe» in determinares scapulei de Homo pri-
migenius. «Probl. de Anthropol.», 1957, No. 3.
Harcula K. Zroznicowania cech morfologicznych i funkcjonalnych reki ludzkiej •
w rozwoju osobniczym. «Rocz. nauk. WSWF Krakowie», 1972, No. 11.
Hasebe K." Die Wirbelsaule der Japaner. «Z. Morphol. und Anthropol.», 1913,
Bd. 15. . . л v
Heberer G. Die Fossilgeschichte der Hominoidea. In: «Ptimatologia. Handbuch
der Primatenkunde», harasusg. von H. Hofer, A. H. Schultz, D. Starck. Ba-
sel _____jq, у. 1956 - ’
Heberer G. Darwin’s Urteil uber die abstammungsgeschichtliche Herkunft des
Menschen und heutige palaoanthropologische Forschung. In: «Hundert Jahre
Evolutionsforschung», herausg von G. Heberer. Stuttgart, 1960.
207
Mielke J. Н.» Armelagos G, J., Van Ger ven D. P. Trabecular invo-
lution in femoral heads of a prehistoric (X-group) population from Sudanese
Nubia. «Amer. J. Phys. Anthropol.», 1972, vol. 36, No. 1.
De Mora is M. H. Estude antropologico da omoplata nos portugueses. «Contribs.
estudo ant'ropol. portug.», 1966, t. 8, No. 2.
Morbeck M. E. Dryopithecus africanus forelimb. «J. Hum. Evol.», 1975, vol. 4
No. 1.
Morimoto J. Additional note on the formation of the so-called squatting fa-
cets on the talus. «Med. J. Shinshu Univ.», 1960, No. 5.
Moss At L., Meehan M. A., Salentijn L. Transformative and translative
growth processes in neurocranial development of the rat. «Acta anat.», 1972,
vol. 81, No. 2.
Murray P. D., Selby D. Intrinsic and extrinsic factors in the primary develop-
ment of the skeleton. «Roux Arch. Entw. Meeh.», 1930, No. 122.
Musgrave J. H. How dextrous was Neanderthal man? «Nature», 1971, vol. 233,
No. 5321.
Napier J. R. The prehensile movements of the human hand. «J. Bone and
Surg.», 1956, vol. 38, No. 13.
Napier J. R. Fossil metacarpals from Swartkrans. Brit. Museum Natur. History,
London, 1959, No. 17-.
Napier J. R. Fossil hand bones from Olduvai Gorge. «Nature», 1962, vol. 196,
No. 4853.
Napier J. R. Evolution of the human hand. «Proc. Roy. Instn. Gr. Brit.», 1965,
1967, vol. 40, No. 6.
N ap ier J. R. The antiquity of human walking. «Sci. Amer.», 1967, vol. 216, No.4.
Napier J. R., Davis P. R. The fore-limb skeleton and associated remains sof
Proconsulus africanus. «Brit. Museum Natur. History. Fossil Mammals of Af-
rica», 1959, No. 16.
Napier J. R., Napier P. H. A Handbook of Living Primates. N.Y., Acad.
Press, 1967..
N ernes ke r i J., Harsanyi L., Acsadi G. Methoden zur Diagnose des
Lebensalters von Skelettkunden. «Anthropol. Anz.», I960, Bd. 24, Nr. L
N i s h i K. Beitrage zum Studium der Prasacralwirbel bei verschiedenen Men-
schenrassen. «Fol. Anat. Jap.», 1928, Nr. 6.
Olivier G. Les caracteres primitifs du squelette appendiculaire de 1’embryon
humain. «Bull, et mem. Soc. anthropol. Paris», 1962, t. 3, No. 3.
Olivier G. Embryologie comparee de Tos coxal chez les Primates. «Z. Morphol.
und Anthropol.», 1967, t. 58, No. 3<
Olivier G. Quelques caracteres specifiquement humains des os des membres;
application a la paleontologie. «Ser. med.», 1972, t. 45, No. 1—2.
Osborn H. F. Recent discoveries relating to the origin and antiquity of Man.
«Proc. Amer. Phil. Soc.», 1927, No. 66.
Osborn H. F. Proboscidea. 2 vols, 1936, 1942. N.Y., Amer. Mus. Press.
Overhage P. Zur Frage einer Evolution der Menschheit wahrend des Eiszeital-
ters. II Teil. «Acta biotheoret.», 1962, Bd. 16, Nr. 1—2.
Oxnard Ch. E. The functional morphology of the primate shoulder as revealed
by comparative anatomical, osteometric and discriminant function techniques.
«Amer. J. Phys. Anthropol.», 1967, vol. 26, No. 2.
Oxnard Ch. E. A note on the fragmentary Sterkfontein scapula. «Amer. J.
Phys. Anthropol.», 1968, vol. 28, No. 2.
Oxnard Ch. E. Evolution of the human shoulder; some possible pathways.
«Amer. J. Phys. Anthropol.», 1969, vol. 30, No. 3.
Oxnard Ch. E. Some African fossil foot bones. «Amer. J. Phys. Anthropol.»,
1972, vol. 37, No. I.
P a 11 e E. Les N6anderthaliens. Paris, 1955.
Patterson B., Howells W. W. Hominid humeral fragment from early
Pleistocene of northwestern Kenya. «Sci.», 1967, vol. 156, No. 3771.
Patsch J. Die Sella turcica als. Konstitutionskriterium. «Wiener med. Wo-
chenschr.», 1963, No. 113.
210
Pauwels F. Die Bedeutung der Muskelkrafte'fur die Regelung der Beanspru-
chung des Rohrenknochens wahrend der Bewegung der Glieder. «Z. Anat. und
Entwicklungsgesch.», 1950, Nr. 115.
Pavlov Sp., Petrov Iv. Caracteristique morphometrique de la clavicule chez
les nouveau-nes «Folia med.», 1964, t. 6, No. 3.
Pearl R. Introduction to medical biometry and statistics. 3 ed. Philadelphia a.
London, 1940.
Petit-Maire N. Evolution trends and comparative ontogenesis in Primate
cranium. «J. Hum. Evol.», 1972, vol. 1, No. 1.
Petrov Iv„ Pavlov Sp. Caracteristique morphometrique de 1’omoplate chez
les nouveau-nes. «Folia med.», 1964, t. 6, No. 4.
Pietrogrande V. Considerazioni sulle variazioni morfologiche indotte nelle
anche artrosiche dall’intervento di enervazione articolare. «Ortoped. e trauma-
tol. appar. motore»,1953, t. 21, No. 1.
P i 1 b e a m D. Man’s earlies ancestors. «Sci. J.», 1967, vol. 3, No. 2.
P i 1 b e a nr D. Adaptive response of hominids to their environment as ascertained
by fossil evidence. «Social Biol.», 1972, vol. 19, No. 2.
P i 1 b e a m D„ Simons E. Humerus of Dryopithecus from Saint Gaudens
France. «Nature», 1971, vol. 229, No. 5284.
Pittard E. Les populations jaunes de 1’Afrique. «Z. Troppenwiss. und Trop-
penmed.», 1944, vol. separe, I, nos 2, 3 et 4.
P i v e t e a u J. Les decouvertes d’Oldoway (Tanganyika) et le probleme des ori-
gines humaines. «Nature», Sci. progr.», 1964, -'No. 3356.
P r a t e s i R. Osservazioni sulla lunghezza relativa delle dita della mano. «Arch,
antropol. e etnol.», 1962, No.- 92.
Q u i r i n g D. The scale of being according to the power formula. «Growth», 1941,
vol. 5, No. 3.
Quiros P. F., F a u 1 h a b e r J., Arguello L. E. Estudio comparative de la
curvature femoral anterior en cuanto a sexo у raza en el mestizo de Mesoame-
rica. «Rev. Fac. med. Univ, autonomo Mexico», 1966, t. 8, No. 12.
Radlauer C. Beitrage zur Anthropologie des Kreuzbeines. «Z. Anat und En-
- ticklungsgesch.», 1908, Bd. 38, Nr. 3.
O’ R a h i 11 у R., Gardner E., Gray D. The skeletal development of the
foot. «Clinical orthopaedics», 1960, No. 16.
RaoP. D. Prasad a. Squatting facets on <the talus and tibia in Australian ’
aborigines. «Archaeol. and Phys. Anthropol. Oceania», 1966, vol. 1, No. 1. ’
Regalia E.‘ Sulla causa generale delle anomalie numeriche del rachide. «Arch.
Anthropol.», 1895, No. 25.
R e m a n e A. Die Geschichte des Menschenaffen. «Menschl. Abstammungslehre».
Stuttgart, Herausg. v. G. Heberer, 1965. \
Rempe U. Die Reco^istruktion von Stammbaumen aufgrund von Immunprazipi-
tationen. Dargestellt am Beispiel von Hominoiden. «Z. Morphol.' und Anthro-
pol.», 1968, Bd. 60, Nr. 1.
Riesenfeld A. Biodynamics of head form and cranio-facial relationships.
«Homo», 1967, vol. 18, No. 4.
Riesenfeld A. Head balance and brachycephalization. «Homo», 1969, vol. 20,
No. 2.
Riesenfeld A. On some «racial» features in rats, dogs and men. «Homo»,
1970, vol. 21, No. 3.
Rivet P. Recherches anthropologiques sur la Basse-Californee. «Extr. J. Soc.
amer. Paris», 1909, No. 6. , .
Robinson J. T. Homo «habilis» and australopithecines. «Nature», 1965,vol.205.
No. 4967.
Robinson ’J. T. Two new early hominid vertebrae from Swartkrans. «Nature»,
1970, vol. 225, No. 5239.
Robinson J. T., Steudel K. Multivariate discriminant analysis of dental
data bearing on early hominid affinities. «J. Hum. Evol.», 1973, vol. 2, No. 6.
Rosen S. I., McKern Th. W. Several cranial indices and their relevance to
fossil man. «Amer. J. Phys. Anthropol.», 1971, vol. 35, No. 1.
Rosenberg E. Ober die Entwicklung der Wirbelsaule und das Centrale carpi
des Menschen. «Morphol. Jahrb.», 1876, Nr. 1.
Rosenberg E, Ober eine primitive Form der Wirbelsaule des Menschen. «Mor-
phol. Jahrb.», 1899, Nr. 27.
Rosenberg E. Bemcrkungen uber den Modus des Zustandeiikommens der Re-
gionen an der Wirbelsaule des Menschen. «Morphol. Jahrb.», 1907, Nr. 36.
Ros in ski F. Budowa morphologiczna czaszki Australopithecinae w swietle niek-
torych wskaznikow ilorazowych. «Acta Univ. Bratisl.», 1.
Rosler H. D. Fingerlangenproportionen und Handarbeit. «Int. Z. angew. Phy-
siol.», 1957, Bd. 16, Nr. 434.
Rosler H. D. Zum Alterswandel der Fingerlangenproportionen. «Homo», 1957a,
No. 881.
Ruf fie J. Les fronticres chromosomiques de I’hominisation. «С. r. Acad, sci.»,
1973, t. 276, No. 11.
Ruge G. Der Verkiirzungsprocess am Rumpfe der Halbaffen. «Morphol. Jahrb.»,
1892, Bd. 18, Nr. 2.
S a b e n R. Contribution a Fetude du crane et en particular de 1'os temporal des
geants acromegales. «Bull, et mem. Soc. anthropol. Paris», 1964, t. 6, No. 2.
S ar az in F. Anthropologic der Neu-Caledonier und Loyalty-Insulaner. Berlin,
Kreidal, 1916/1922.
S ar azin F. Die Variationcn im Bau des Handskeletts verschiedener Menschen-
formen. «Z. Morphol.», 1932, Bd. 30, Nr. 1—2.
Sarich V. M., Wilson A. C. Rates of albumin evolution in Primates. «Proc.
Nat. Acad. Sci. USA», 1967, No. 58.
Saville P. D., Smith P. M. Relation between axial and appendicular skele-
tal calcium and body weight in the rat. «Anal. Rec.», 1966, vol. 156, No. 4.
Sawin P. B. Preliminary studies of hereditary variation in the axial skeleton
of the rabbit. «Anat. Rec.», 1937, vol. 69.
Schaepelynck J. Note sur la morphologic de la colonne lombaire. «Bull.
Assoc. Anat.», 1959, No. 104.
Schlaginhaufen O. Die menschlichen Skelettreste aus der Steinzeit des
Wauwilersees (Luzern) und ihre Stellung zu den anderen anthropologischen
Funden aus der Steinzeit. «Zurich—Munich—Leipzig», 1925.
Schultz A. H. Fetal growth of Alan and other Primates. «Quart. Rev. of Biol.»,
1926, vol. 1, No. 4.
Schultz A. H. Human variability compared with the variability of other pri-
mates. «Anat. Rec.», 1930, No. 45.
Schultz A. H. The relative length of the regions of the spinal column in old
world primates. «Amer. J. Phys. Anthropol.», 1938/39, No. 24.
Schultz A. H. The relative thickness of the long bones and the vertebrae in
Primates. «Amer. J. Phys. Anthropol.», 1953, No. 11.
Schultz A. H. Einige Beobachtungen und Masse am Skelett von Oreopithecus.
«Z. Morphol. und Anthropol.», 1960, Nr. 50.
Schultz A. H. (Hofer H., Starck D.). Primatologia. Handbuch d. Primaten-
kunde, Bd. 4, Lief. 5, Basel —N.Y., 196L
Schultz A.,H. Age changes, sex differences and variability as factors in the
classification of primates. In: «Classific. and human evolution», Chicago, AL
dine Publ. Co., 1963.
Schultz A. H. Die recenten Hominoidea. In: «Menschl. Abstammungslehre»,
herausg, v. G. Heberer, Stuttgart, 1965.
Schultz A. H. Observations on the acetabulum of primates. «Folia primatol.»,
1969, vol. 11, No. 3.
Schwalbe G. Kritische Besprechung von Boule’s Work: «L’Hommc fossile de
La Chapelle—aux—Saints», mit eigenen Untersuchungen. «Z. Morphol. und
. Anthropol.», 1914, Bd. 16, Nr. 3.
Schwalbe G. Studien uber das Femur von Pithecanthropus erectus Dubois.
«Z. Morphol. und* Anthropol.», 1919/21, Nr. 3.
S e d 1 i n E. D., F г о s t H. M., Villanueva A. R. Variations in rate of human
osteon formation. «Canad. J. Biochem. and Physiol.», 1963, vol. 41, No. 1.
SewellS. A study of the astragalus. «J. Anat.», 1904/6, No. 38, 40.
212
Shore L. R. Abnormalities of the vertebral column in a series os skeletons of
Bantu natives. «J. Anat.», 1930, vol. 64.
Silinkova -Malkova E. The skeletal system and endocrine glands. «Ra-
diol. diagnost.», 1963, vol. 4, No. 6.
Simpson G. Rates of evolution in animals. In: «Genetics Palaeontology and
Evolution.», N.Y., 1949.
Singh I. Squatting facets on the talus and tibia in Indians. «J. Anat. Lond.»,
1959, vol. 93.
Singh S. P., Singh Sh. A study of anterior femoral curvature in indian
subjects. «Acta anat.», 1972, vol. 83, No. 3.
Skary d S. M. Trends in the evolution of the pongid dentition. «Amer. J. Phys.
Anthropol.», 1971, vol. 35, No. 2.
SI о wi к F. Vergleichende morphologische Untersuchungen an der Clavicula des
Menschen und anderen Primaten. «These Sci.», Zurich, 1945.
Snell C., Donhuysen H. The pelvis in the bipedalism of primates. «Amer.
J. Phys. Anthropol.»’ 1968, vol. 28, No. 3.
Soriano M. The fluoric origin of the bone lesion in the Pithecanthropus erec-
tus femur. «Amer. J. Phys. Anthropol.», 1970, vol. 32, No. 1.
S t e с к о 11 S. H., G о f f e r Z., N a t h a n H., H a a s N. Red-stained human bo-
nes from Qumran4«Isr. J. Med. Sci.», 1971, vol. 7, No. 11.
Steegmann A. T.'Cold response, body form and cranio—-facial shape in two
racial groups of Hawaii. «Amer. J. Phys. Anthropol.», 1972, vol. 37, No. 2.
St^slicka-Mydlarska W. Odlewy wn§trz czaszkowych kopalnych i
wspolczcsnych hominidae. «Prz. antropol.», 1971, t. 37, No. 1.
S t e w a r t T. D. The vertebral column of Eskimo. «Amer. J. Phys. Anthropol.»,
1932, No. 17.
Stewart T. D. Neanderthal cervical vertebrae with special attention to the
Shanidar Neanderthals from Iraq. «Biblioth. primatol.», 1962, No. 1.
Straus W. The humerus of Paranthropus robustus. «Amer. J. Phys. Anthropol.»,
1948, vol. 6, No. 3.
Straus W. Fossil evidence of the evolution of the erect bipedale posture.
«Clin. Orthop.», 1962, No. 25.
Straus W. The classification of Oreopithecus. In: «Classific. and human evolut.»,
Chicago, Aldine Publ. Co., 1963.
Straus W., Cave A. Pathology and the posture of Neanderthal man. «Quart.
Rev. Biol.», 1957, vol. 32, No. 4.
T h e r L., S c h г a mm H., V о g eГ G. Uber die antagonistische Wirkung von
Trijodthyronin und Progesteron auf den Prednisoloneffekt am Epiphysenknor-
pel. «Acta endocrinol.», 1963, Bd. 42, Nr. 1.
Thoma A. L’occipital de I’homme mindelien de vertesszollos. «Anthropol.», 1966,
t. 70, No. 5—6.
T о b i as P. V. Cranial capacity in anthropoid apes, australopithecus and Homo
habilis &. «S. Afric. J. Sci.», 1968, vol. 64, No. 2.
Todd T. W, Numerical significance in the thoraco-lumbar vertebrae of the
Mammalia. «Anat. Rec.», 1922, No. 24.
Tran.- An h, Le Ngoc Huong. L’astragale du Vietnamidn et les facettes
d’accroupissement. «Bull, et mem. Soc. anthropol. Paris», 1966, t. 9, No. 3.
Trotter M. The vertebral column in whites and in american negroes. «Amer.
J. Phys. Anthropol.», 1929, No. 13.
T u 1 s i R. S. Vertebral column on the australian aborigine: selected morpholo-
gical and metrical features. «Z. Morphol. und Anthropol.», 1972, Bd. 64, Nr. 2.
Tuttle R. Knuckle-walking and the problems of human origine. «Sci.», 1969,
Nr. 166.
Twiesselmann F. Evolution des dimensions et de la forme de la mandibule,
du palais et des dents de I’homme. «Ann. paleontol. Vertebr.», 1973, t. 59.
No. 2.
Tyrance H. J. Relationship of extreme body types to ranges of flexibility. «Res.
Quart. Amer. Assoc. Health», 1958, yol. 29, No. 3.
Ujwal Z. S. The internal architecture of femur. «J. Anat. Soc. IndU», 1962,
vol. 11, No. 2.
213
Ulrich H. Die Vorfahren des heutigen Menschen. «Wiss. und Fortsch. 1955,
Bd. 5, Nr: 7.
Uzzell Th., Pilbeam D. Phyletic divergence dates of hominoid primates: a
comparison of fossil and molecular data. «Evolution» 1971, vol. 25, No. 4.
Vallois H. Nouvelles recherches sur 1’homme fossile de Chancelade. «Anthro-
pol.», 1946, No. 50.
Vallois H„ LazorthesG. Indices lombaires et indice lombaire total. «Bull,
mem. Soc. anthropol. Paris», 1942, No. 3.
VandermeerschB. Le Crane 1470. «Recherche», 1973. t. 4, No. 38.
Verneau R. Les grottes de Grimaldi. «Anthropol.», 1906, No. 17.
Viret J. A propos de I’Oreopitheque. «Mammalia», 1955, t. 19, No. 2.
Vlcek E. Postcranial skeleton of a Neanderthal child from Kiik-Koba, USSR,
«J. Hum. Evol.», 1973, vol. 2, No. 6.
Volkov Th. Variations squelettiques du pied chez les primates et dans les
races humaines. «Bull, et mem. Soc. Anthropol. Paris», 1905, No. 4—5.
Wallace J. A. Tooth chipping in the australopithecines. «Nature», 1973, vol. 244>
No. 5411.
Washburn S. L. Behavoir and the origin of man. «Proc. Roy. AnthropoL
• Inst.», 1967, No. 21—24.
Watanabe H. Running, creeping and climbing: a new ecological’and evolutio-
nary perspective on human locomotion. «Mankind», 1971, vol. 8, No. 1.
Wechsler W. Anthropologische Untersuchungen der Handform mit emem
familienkundlichen Beitrag. «Arch. Jul. Klaus-Stifb, 1939, Bd. 14, Nr. 139.
Weidenreich F. Der Menschenfuss. «Z. Morphol. und Anthropol.», 1922, Nr.22.
Weidenreich F. Observations of the form and proportions of the endocranial
cast of Sinanthropus pekinensis & «Palaeontologia Sinica», 1936, No. 7.
Weidenreich F. The extremity bones of Sinanthropus pekinensis. «Palaeonto-
logia Sinica», n. s., 1941, No. 5.
Weidenreich F.- The Skull of Sinanthropus pekinensis. «Palaeontologia Si-
nica», 1943, No. 10.
Weinmann J. P., SicherH. Bone and Bones. C. v. Mosby Co, St. Louis,
1955.
Werth E. Der fossile Mensch. Berlin, Borntraeger, 1928.
Wiercinski A. Evolutionary rate of craniometric traits in Hominidae. «Acta
Palaeont/Polonica», 1956, vol. 1, No. 3.
Williams P. C. The lumbosacral vertebral column. N.Y., McGraw-Hill Book
. Co, 1965.
W i 11 i s T. A. The thoracolumbal column in white and negro stocks. «Anat. Rec.»,
1923, No. 26.
Wolpoff M. H. Is Vertesszollos II an occipital of European Homo erectus?
«'Nature», 1971, vol. 232, No. 5312.
Wood B. A. Locomotor affinities of Hominoid tali from Kenya. «Nature», 1973* •
vol. 246, No. 5427. ' -
Wood B. A. Evidence on the locomotor pattern of Homo from early pleistocene
of Kenya. «Nature», 1974, vol. 251, No. 5471.
Woo Ju-kang, Chia,Lan-po. New discoveries about Sinanthropus pekinen-
sis in Choukoutien. «Sci. Sinica», 1954, vol. 3, No. 3.
Zapfe H. Fossile Menschenaffen im Wiener Becken. «Natur und Museum», 1963„
Bd. 93, Nr. 10.
Z a w i s c h C. Die Morpho-und Histogenese der menschlichen Scapula. «Acta
anat.», 1954, Bd. 22, Nr. 4.
Z i h 1 m a n A. L., H u n t e r W. S. A biomechanical interpretation of the pelvis
of Australopithecus. «Folia Primatol.», 1972, vol. 18, No. 1—2.
Оглавление
Предисловие.................................................... 3-
Глава I. ОСЕВОЙ СКЕЛЕТ.......................................... 5
Позвоночная формула . 5
Форма позвоночника и грудной клетки ископаемых гоминид ... 15
Глава II. ПОЯСА КОНЕЧНОСТЕЙ ........................................26
Плечевой пояс.............................................26,
Тазовый пояс...................................................34
Глава III. ДЛИННЫЕ КОСТИ КОНЕЧНОСТЕЙ................................47
Нижняя конечность.............................................47'
Верхняя конечность.............................................64
Пропорции......................................................70
Глава IV. КИСТЬ.............................................. 78.
Кисть как индикатор функциональных адаптаций у приматов и
человека..................................................78.
Кисть ископаемых гоминид . 81
Глава V. СТОПА ....................................................104
Стопа как индикатор бипедии у гоминид и их предшественников . 104
Стопа ископаемых гоминид......................................110*
Вопрос о типе кисти и стопы антропоидного предшественника го-
минид ................................................. 128.
Глава VI. ВНУТРЕННЯЯ СТРУКТУРА СКЕЛЕТА И ЕЕ РОЛЬ В
ЭВОЛЮЦИОННОЙ МОРФОЛОГИИ..................................‘ . 134
Внутреннее строение трубчатых костей человека сравнительно с
другими современными приматами............................134
Внутренняя структура длинных костей у ископаемых гоминид . . 142'
Глава VII. ЧЕРЕП..............................................1521
Основные черты краниофациального комплекса современного чело-
века .................................................... 152
Краткая краниологическая характеристика ископаемых гоминид . 158,
Глава VIII. ФАКТОРЫ, ВЛИЯЮЩИЕ НА СТРУКТУРУ СКЕЛЕТА,
И ПРОБЛЕМА РЕКОНСТРУКЦИИ ОТДЕЛЬНЫХ СТОРОН ПА-
ЛЕОЭКОЛОГИИ ГОМИНИД ПО ОСТЕОЛОГИЧЕСКИМ ДАННЫМ 173;
Глава IX. НЕКОТОРЫЕ ЧЕРТЫ МОРФОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
И ПРОБЛЕМА МОРФОЛОГИЧЕСКОГО КРИТЕРИЯ ГОМИНИД' 184
Формирование основных морфологических комплексов на скелете
человека.............................................184
Некоторые предпосылки к обоснованию морфологического крите-
рия гоминид..........................................196
Литература.....................................290
ЕЛЕНА НИКОЛАЕВНА ХРИСАНФОВА
«Эволюционная морфология
скелета человека»
Зав. редакцией II. ?Л. Г л а ; к о в а
Редактор II. .М. Горелик
Художественный редактор Н. Ф. 3 к: н и в
Переплет художника В. С. К а ? а к о в и ч л
Технический редактор 3. С. К о н д р а :п о в а
Корректоры С. С. М а л у р с к л я, Л. С К л о ч к о в а
Сдано в набор S X Г.Гб г Подписано к 7-1 1’Г". г /I ;>рл Формат Je
Бумага тип. № л. Усл. иеч. л Г.<> Уч.-и а л. ГС- Па V .гл: Зак. "‘.к Тираж Кип -,кз.
Цена 2 р. Йо к.
Издательство Московского уннверси:« га. Москва. К \л Ггрцена. 5 7.
Типография Изд на МГУ. Мо.мы, ЛашнсаФ горы'