/


















































































































































































































































































































































































































































































































































































Автор: Онипченко В.Г.
Теги: человек и окружающая среда экология человека охрана природы растения
ISBN: 978-5-396-00550-1
Год: 2014




Текст
В. Г. Онипченко
ФУНКЦИОНАЛЬНАЯ
ФИТОЦЕНОЛОГИЯ
Синэкология растений
МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ имени М. В. ЛОМОНОСОВА
Биологический факультет
В. Г. Онипченко
ФУНКЦИОНАЛЬНАЯ ФИТОЦЕНОЛОГИЯ
Синэкология растений
Издание второе, стереотипное
URSS
МОСКВА
ББК 20.1 28.5
Онипченко Владимир Гертрудович
Функциональная фитоценология: Синэкология растений. Учебное
пособие. Изд. 2-е, стер. — М.: КРАСАНД, 2014. — 576 с.
В настоящей монографии в обобщенном виде представлено современное состояние науки о растительных сообществах, при этом особое внимание уделяется функциональным аспектам их организации. С позиций результатов исследований конца XX - начала XXI века критически анализируется ряд классических теорий. В книге рассмотрены связи растений с прокариотами, грибами и животными, типы взаимоотношений между растениями, механизмы формирования состава и структуры растительных сообществ, вопросы популяционной биологии растений, продукции растительного покрова. Большой раздел посвящен различным формам динамики растительных сообществ — сезонной, флуктуационной, сукцессионной и эволюционной; изложены факторы исторической динамики растительного покрова.
Книга предназначена для научных работников — ботаников, экологов, сотрудников природоохранных организаций, а также преподавателей и студентов вузов, связанных с профессиональной подготовкой ботаников и экологов.
Рецензенты:
проф. кафедры геоботаники биологического факультета МГУ
имени М. В. Ломоносова, д-р биол. наук, член-корр. РАН В. Н. Павлов', проф. кафедры общей экологии биологического факультета МГУ имени М. В. Ломоносова, д-р биол. наук А. М. Гиляров;
доц. кафедры геоботаники биологического факультета МГУ имени М. В. Ломоносова, канд. биол. наук С. А. Баландин
Издательство «КРАСАНД». 117335, Москва, Нахимовский пр-т, 56.
Формат 60x90/16. Печ. л. 36. Зак. №1114
Отпечатано в ООО «Чебоксарская типография № 1».
428019, г. Чебоксары, пр-т И. Яковлева, 15.
ISBN 978-5-396-00550-1
© КРАСАНД, 2013
НАУЧНАЯ И УЧЕБНАЯ ЛИТЕРАТУРА
E-mail: URSS@URSS.ru Каталог изданий в Интернете: http://URSS.ru Тел./факс (многоканальный):
URSS + 7 (499) 724 25 45
Все права защищены. Никакая часть настоящей книги не может быть воспроизведена или передана в какой бы то ни было форме и какими бы то ни было средствами, будь то электронные или механические, включая фотокопирование и запись на магнитный носитель, а также размещение в Интернете, если на то нет письменного разрешения владельца.
Содержание
Предисловие......................................................... 12
Глава 1. Введение .................................................. 14
1. Основная задача и система наук о растительности.................. 15
2. Определения фитоценоза........................................... 16
3. Концепции дискретности и континуума.............................. 20
4. Представление о консорциях и консортивных связях.............. 21
5. Концепция «ключевых» видов....................................... 22
Глава 2. Растения и азотфиксирующие прокариоты........ 23
1. Симбиотическая азотфиксация...................................... 24
1.1. Бобовые и клубеньковые бактерии............................ 24
1.2. Актинориза................................................. 30
1.3. Консортивные связи с цианеями ............................. 38
1.4. Прочие случаи симбиоза .................................... 42
2. Ассоциативная азотфиксация....................................... 42
3. Несимбиотическая азотфиксация.................................... 44
Глава 3. Растения и грибы........................................... 45
1. Микосимбиотрофизм................................................ 48
1.1. Основные типы микориз...................................... 48
1.1.1. Везикулярно-арбускулярная микориза (ВАМ).............. 48
1.1.2. Эрикоидная, арбутоидная и монотропоидная микоризы .... 52
1.1.3. Чехольчатая эктомикориза ............................. 55
1.1.4. Микориза орхидных..................................... 58
1.2. Функциональное значение микориз в целом.................... 59
1.3. Возникновение микориз...................................... 62
1.4. Методы изучения в природе и использование в сельском хозяйстве.................................. 63
4
Содержание
1.5. Зависимость от экологических факторов и распространение микориз в различных природных зонах....................................... 64
1.6. Немикоризные растения............................. 66
1.7. Распространение микоризных грибов и развитие микориз.................................... 68
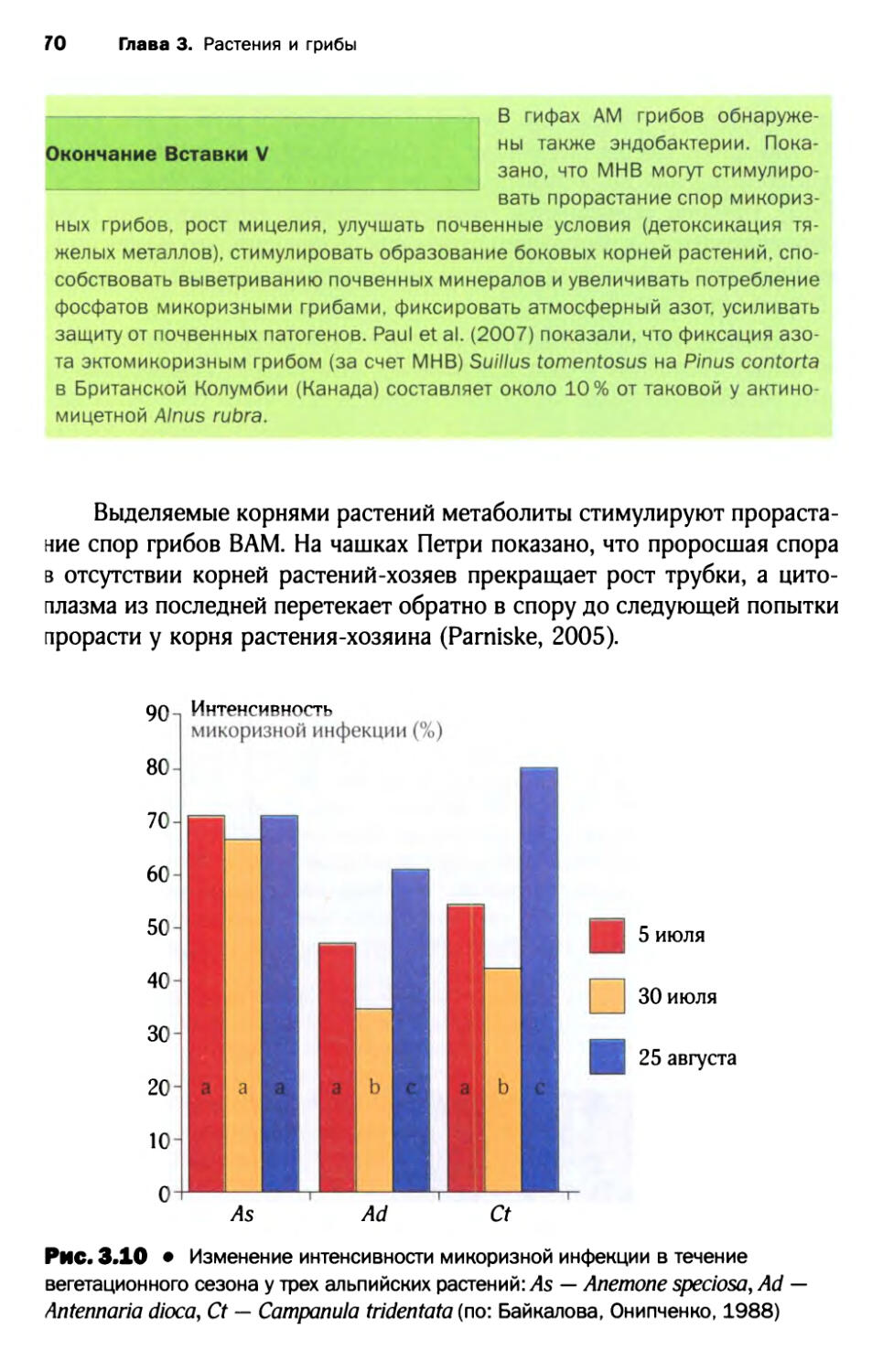
1.8. Микоризы и конкурентоспособность растений......... 71
1.9. Микоризные сети в почве........................... 72
1.10. Микогетеротрофные растения........................ 76
2. Эндофиты ............................................... 81
2.1. Распространение и систематическое положение...... 81
2.2. Функциональное значение для растений.............. 84
3. Фитопатогенные грибы.................................... 88
4. Сапротрофные грибы.................................... 95
Глава 4. Растения и животные............................... 97
1. Хищные растения ........................................ 98
2. Типы воздействия животных на растения.................. 101
3. Защитные адаптации растений от поедания ............... 103
3.1. Уход от фитофагов в пространстве и времени....... 103
3.2. Механические способы защиты ..................... 105
3.3. Привлечение животных для защиты от фитофагов..... 109
3.4. Химические средства защиты растений.............. 111
3.4.1. Токсины растений............................ 111
3.4.2. Пищевые детерренты.......................... 117
3.5. Гипотезы, объясняющие распространение защитных приспособлений у растений................... 119
3.6. Реакция растений и сообществ на повреждение фитофагами ........................... 122
4. Фитофагия и изъятие первичной продукции................ 124
5. Фитофаги и смены растительных сообществ................ 129
6. Фитофагия и флористическое богатство................... 131
7. Роющая деятельность животных........................... 134
8. Животные и опыление растений........................... 137
9. Распространение плодов и семян животными (зоохория).... 143
9.1. Беспозвоночные .................................. 144
Содержание
5
9.2. Рыбы и рептилии ................................. 147
9.3. Птицы............................................ 147
9.4. Млекопитающие.................................... 150
10. Выпас.......‘......................................... 151
10.1. Стравливание..................................... 151
10.2. Вытаптывание..................................... 154
10.3. Отложение экскрементов........................... 155
Глава 5. Типы взаимоотношений растений в фитоценозах .... 157
1. Прямые (контактные) отношения.......................... 158
1.1. Внутривидовые срастания.......................... 158
1.2. Паразитные цветковые растения ................... 159
1.3. Полупаразиты .................................... 162
1.4. Эпифиты.......................................... 169
1.5. Лианы............................................ 173
2. Трансбиотические отношения............................. 178
3. Конкуренция............................................ 178
3.1. Модель Тильмана.................................. 178
3.2. Конкуренция в одновидовых посевах. Правило и эффект Сукачева............................. 186
3.3. Закон -3/2 ...................................... 188
3.4. Конкуренция в парных посевах: схема De Wit....... 189
3.5. Влияние условий среды на исход конкуренции. Надземная и подземная конкуренция..................... 191
3.6. Адаптации растений бедных и богатых почв ........ 195
3.7. Эколого-ценотические стратегии и конкурентоспособность растений...................... 198
3.7.1. Взгляды Л. Г. Раменского.................... 198
3.7.2. Взгляды Ф.Грайма: типы стратегий растений... 199
3.7.3. Противоречия Тильман — Грайм. Интенсивность и значимость конкуренции............. 208
3.7.4. Взгляды Ю.Э. Романовского................... 212
3.8. Ауто- и синэкологические ареалы и оптимумы....... 214
3.9. Полевые эксперименты по изучению конкуренции..... 216
3.9.1. Метод фитометров............................ 216
3.9.2. Удаление отдельных особей, видов, групп..... 216
3.9.3. Корневая (траншейная) обрубка .............. 218
6
Содержание
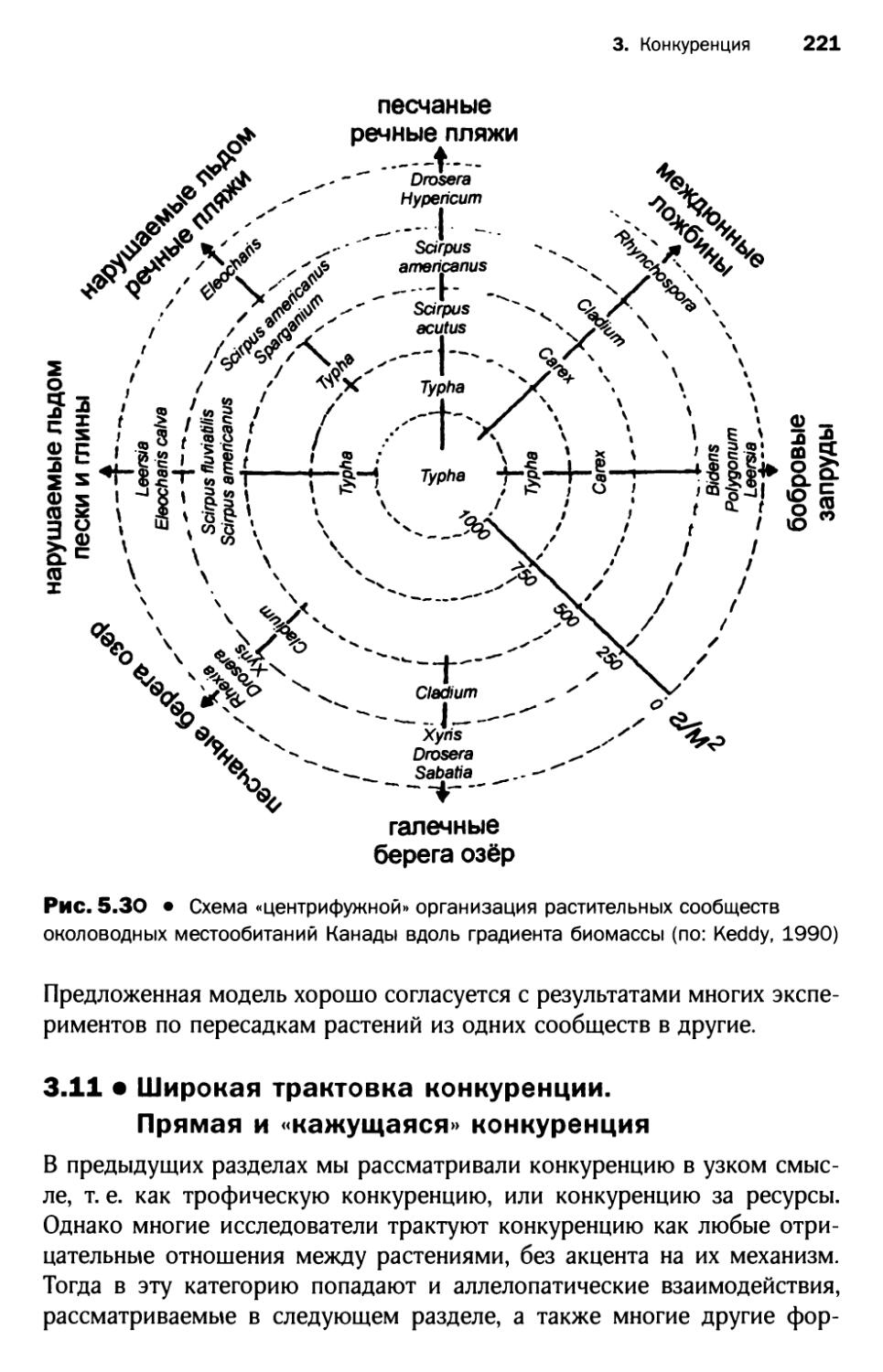
3.10. Представления Р. Keddy о конкурентной иерархии и центрифужной организации растительных сообществ................................ 220
3.11. Широкая трактовка конкуренции. Прямая и «кажущаяся» конкуренция...................... 221
4. Аллелопатия........................................... 223
5. Создание фитосреды и положительные взаимоотношения растений................................. 233
5.1. Фитогенное поле................................. 233
5.2. Изменение температуры воздуха и почвы........... 233
5.3. Водный режим.................................... 237
5.4. Изменение микрорельефа и закрепление подвижных субстратов ................ 244
5.5. Изменение солевого режима, кислотности и окислительно-восстановительного потенциала почв...................................... 245
5.6. Роль растений в изменении содержания ЭМП и органического вещества в почве..................... 248
5.7. Создание препятствий для ветра, фитофагов, привлечение опылителей............................... 253
6. Положительные отношения между растениями.............. 254
Глава 6. Состав и структура фитоценозов.................. 258
1. Состав фитоценозов.................................... 259
1.1. Свободноживущие цианеи и водоросли в составе наземных экосистем......................... 259
1.2. Лишайники....................................... 264
1.3. Мохообразные.................................... 272
2. Флористическая насыщенность и флористическая неполночленность фитоценозов............................. 275
3. Факторы поддержания флористического состава фитоценозов...................................... 281
3.1. Исторические факторы............................ 282
3.2. Современные факторы............................. 284
3.2.1. Расхождение по ресурсам и абиотическая гетерогенность среды............................... 285
3.2.2. Влияние нарушений.......................... 288
3.2.3. Влияние фитофагов.......................... 289
Содержание
7
3.2.4. Положительные взаимодействия между растениями .... 290
3.2.5. Сбалансированная конкуренция.................. 290
3.2.6. Нейтральная теория S.P. Hubbell............... 292
3.2.7. Отбор на редкость и массовость................ 294
4. Пространственная структура фитоценозов................... 295
4.1. Вертикальная структура (ярусность)................. 295
4.2. Горизонтальная структура сообществ................. 297
4.2.1. Эдафотопическая мозаичность................... 298
4.2.2. Влияние растений на горизонтальную неоднородность почв................................. 300
4.2.3. Ценобиотическая мозаичность .................. 303
4.2.4. Клональная мозаичность и разделение труда..... 306
4.2.5. Теория «масс-эффекта» и модель «карусели»..... 309
4.2.6. Возрастное состояние и «дар»-парадигма........ 312
4.2.7. Модель Тильмана: пространственная структура и колонизационная способность....................... 319
Глава 7. Онтогенез растений в фитоценозах................... 321
1. Основные понятия и термины периодизации онтогенеза...... 322
2. Латентный период (жизнеспособные диаспоры)............... 323
2.1. Размеры семян ..................................... 323
2.2. Происхождение семенных банков в почве.............. 324
2.3. Покой семян........................................ 324
2.4. Пространственная структура семенных банков......... 327
2.5. Длительность сохранения жизнеспособных семян ...... 330
2.6. Почвенные банки в различных сообществах и природных зонах...................................... 333
2.7. Стратегии растений, сукцессии и смены семенных банков................................ 334
2.8. Споры мохообразных и папоротников в почвах......... 337
3. Виргинильный период...................................... 338
3.1. Прорастание семян.................................. 338
3.2. Приживание всходов................................. 341
3.3. Длительность виргинильного периода .............. 342
3.4. Изменение свойств растений в онтогенезе............ 344
4. Генеративный период...................................... 345
4.1. Семенная продуктивность, урожай семян и репродуктивное усилие................................ 345
8
Содержание
4.2. Перерывы в цветении и вторичный покой растений.... 348
4.3. Типы регенерационных стратегий .................. 350
5. Вегетативное размножение растений. Клональные растения.... 351
6. Длительность жизни растений............................ 353
Глава 8. Биомасса и продукция фитоценозов................. 356
1. Запасы и структура фитомассы сообществ различных типов. Аллокация................................ 357
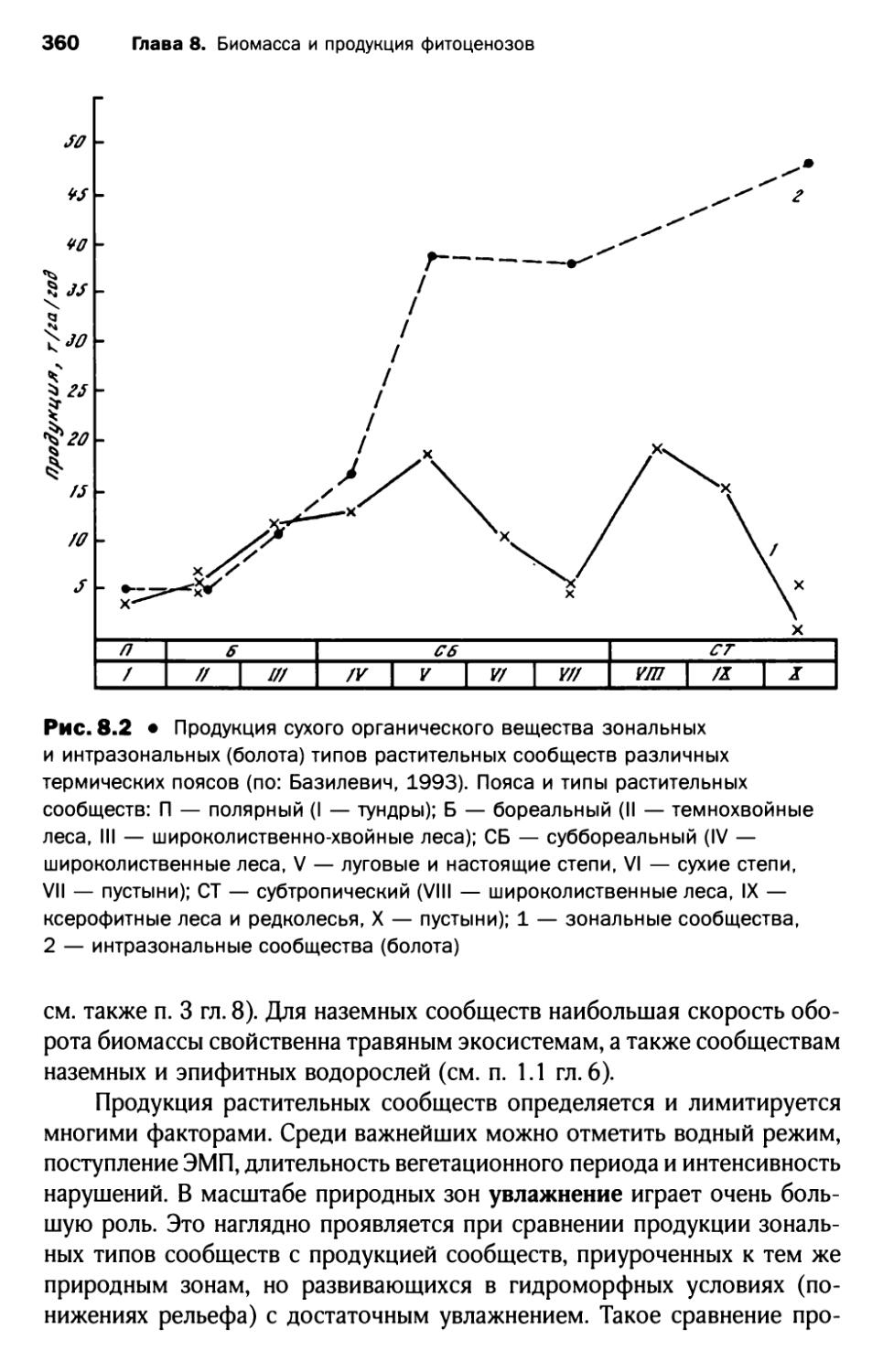
2. Продукция фитоценозов и факторы, ее лимитирующие ...... 359
3. Крупнотравье как система с максимальной продукцией...... 365
4. Энергосодержание фитомассы и энергетическая продукция фитоценозов.................... 368
Глава 9. Динамика фитоценозов............................. 370
1. Сезонная динамика...................................... 371
1.1. Температура как фактор сезонной динамики......... 371
1.2. Световой режим и сезонная динамика............... 374
1.3. Водный режим и сезонная динамика ................ 375
1.4. Снеговой режим................................... 375
1.5. Выделение фенологических периодов................ 377
1.6. Феноритмотипы и проблема вечнозелености.......... 379
2. Флуктуации (разногодичная изменчивость фитоценозов)..... 381
2.1. Экотопические флуктуации......................... 382
2.2. Антропические (антропогенные) и зоогенные флуктуации................................ 387
2.3. Фитоциклические флуктуации ...................... 387
2.4. Типы флуктуаций по степени их выраженности....... 389
2.5. Сочетание различных механизмов флуктуационной динамики в естественных фитоценозах................... 393
3. Сукцессии.............................................. 395
3.1. Определение сукцессий............................ 395
3.2. Взгляды Ф. Клементса............................. 396
3.3. Классические примеры первичных сукцессий и современные взгляды на них....................... 399
3.4. Взгляды В. Н. Сукачева на классификацию сукцессий. 408
3.5. Дальнейшее развитие представлений о климаксе в США...................................... 409
Содержание
9
3.6. Взгляды Коннела и Слатиера. Механизмы сукцессий.... 410
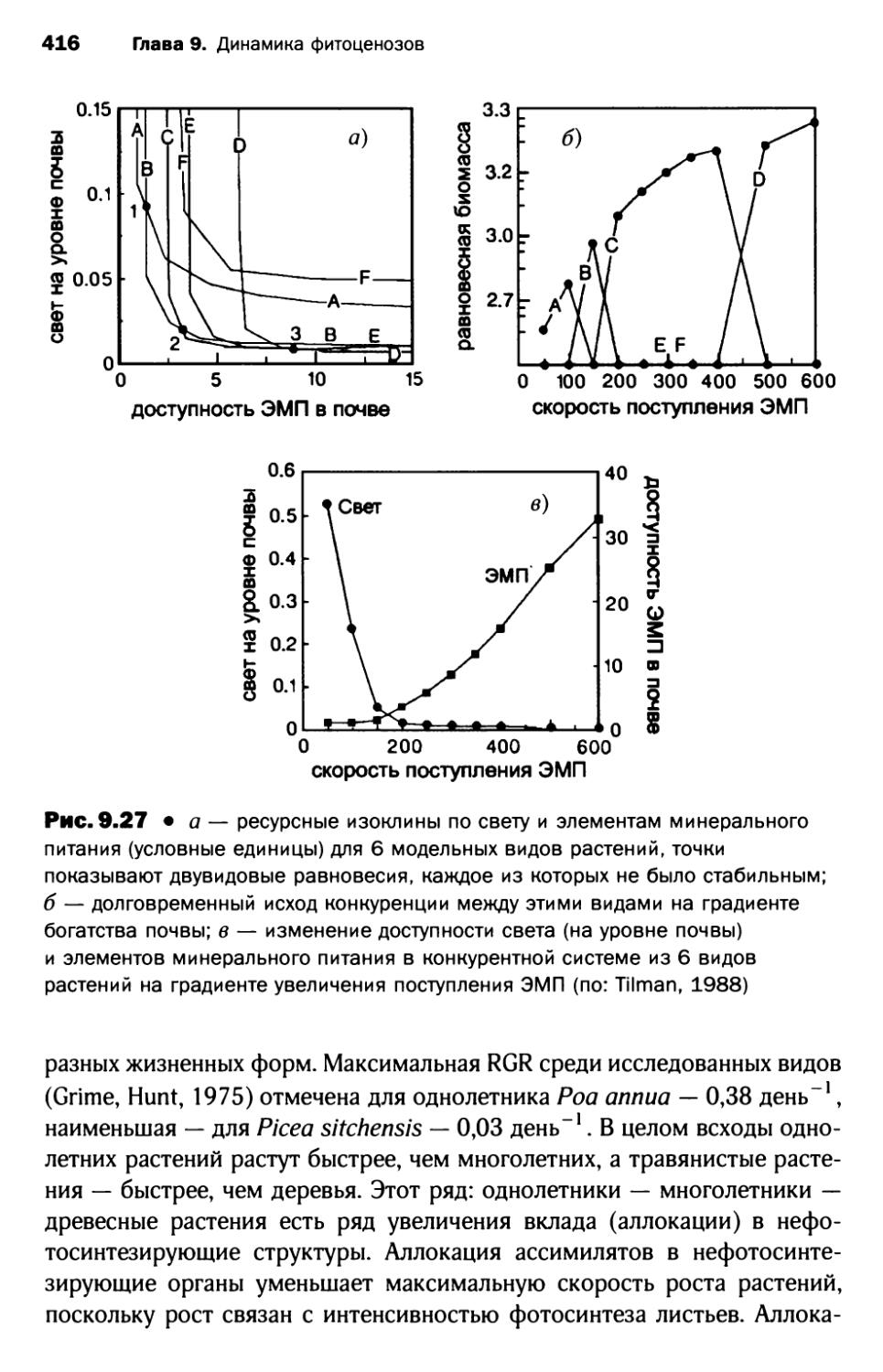
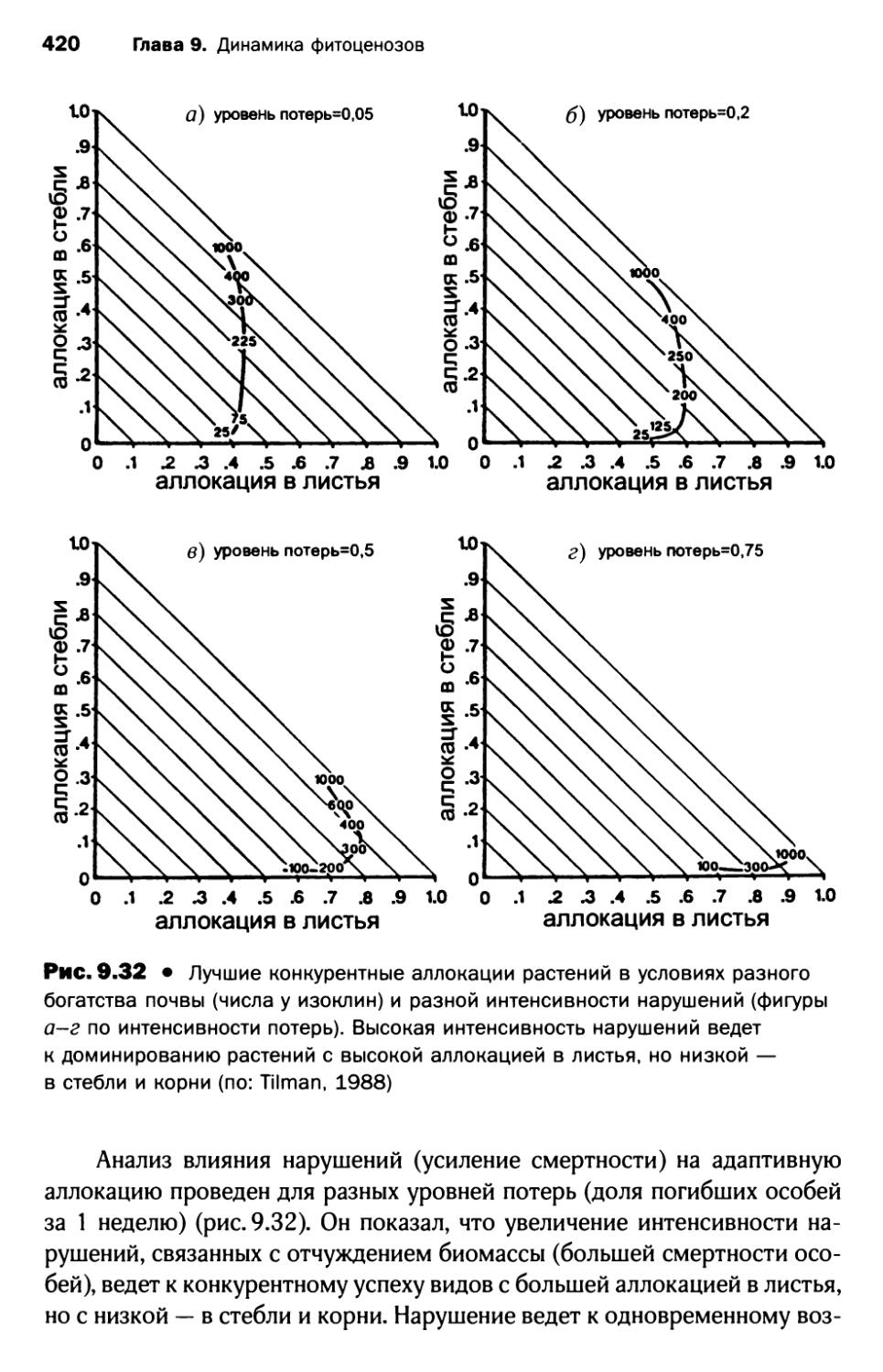
3.7. Сукцессионная теория Тильмана...................... 415
3.8. Сукцессии и стратегии растений Ф. Грайма........... 426
3.9. Факторы сукцессионнной динамики.................... 428
3.9.1. Пожары....................................... 428
3.9.2. Вырубки лесов................................ 444
3.9.3. Выпас и сенокошение.......................... 444
3.9.4. Внесение удобрений........................... 450
3.9.5. Кислотные дожди и SO2........................ 454
4. Эволюция сообществ и историческая динамика............. 455
4.1. Эволюция сообществ................................. 455
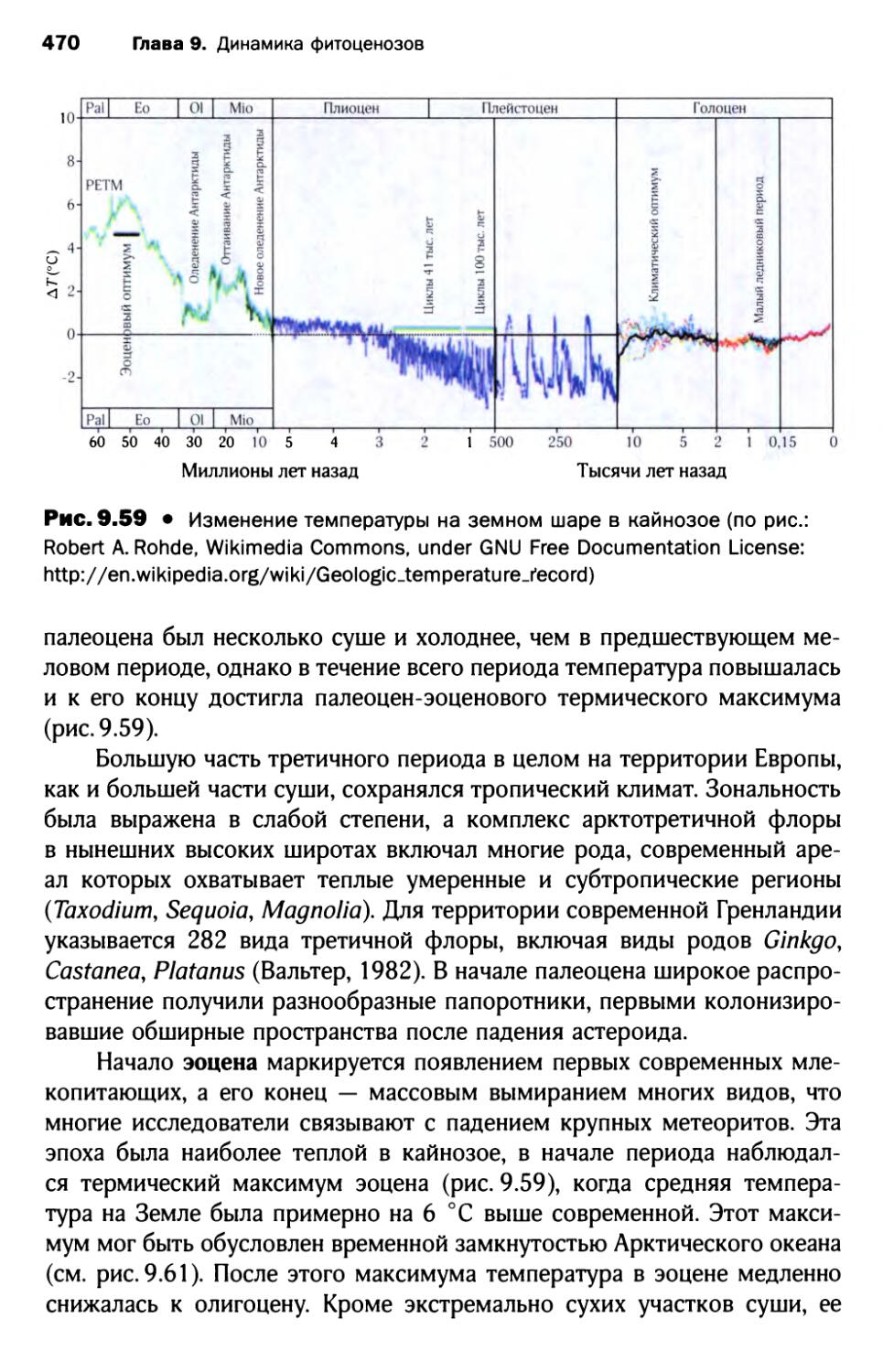
4.2. Историческая динамика в кайнозое.................. 467
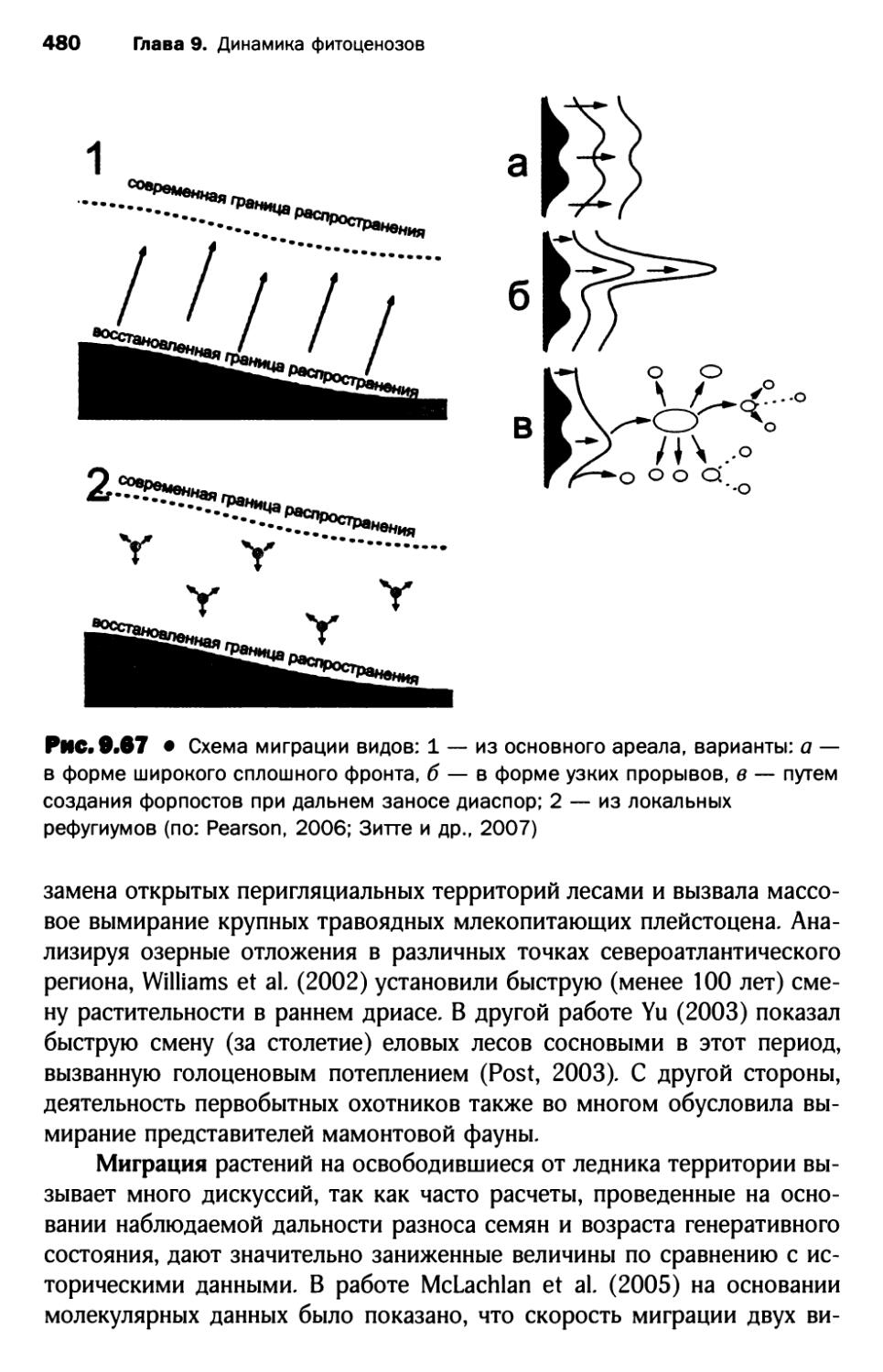
4.3. Современное изменение климата и его влияние на растительный покров.................................. 485
4.3.1. Баланс углерода и увеличение содержания СО2 в атмосфере.......................................... 485
4.3.2. Увеличение температуры....................... 490
4.3.3. Поступление окислов азота и эвтрофикация..... 492
4.3.4. Влияние увеличения поступления УФ-радиации... 493
4.3.5. Повышение концентрации озона в приземных слоях атмосферы.......................... 494
Список литературы........................................... 495
Предметный указатель........................................ 562
Посвящается светлой памяти Тихона Александровича Работнова, основателя популяционной биологии растений и Учителя многих поколений геоботаников
Предисловие
В
настоящее время в отечественной литературе отсутствуют сводки, обобщающие современное состояние функциональной фитоценологии. Фундаментальные обобщения Т. А. Работ-нова (1983, 1992) последний раз переиздавались более 20 лет
назад. В фундаментальной сводке Б. М. Миркина и Л. Г. Наумовой (1998)
вопросы функциональной организации фитоценозов рассмотрены относительно кратко. В настоящее время в мировой литературе накопился большой материал, позволяющий автору с учетом результатов собствен
ных исследований в значительной степени пересмотреть классические взгляды на функционирование наземных экосистем. Поэтому перед автором стояла сложная задача восполнить существенный пробел в современной отечественной экологической литературе. Это, естественно, нельзя было сделать с исчерпывающей полнотой, поэтому автор будет весьма благодарен за критические замечания, уточнения и дополнения к настоящей работе.
В основу книги лег анализ отечественных и зарубежных публикаций конца XX - начала XXI в., охватывающий преимущественно работы, опубликованные по 2008 г. включительно. Ясно, что выбор этих публикаций и особенно примеров, иллюстрирующих те или иные закономерности, является в значительной мере субъективным. Он основан преимущественно на работах, опубликованных в ведущих международных ботаникоэкологических журналах. Общая структура монографии с небольшими изменениями заимствована из сводки Т. А. Работнова (1983), учеником которого считает себя автор.
В книгу вошли также некоторые собственные материалы автора, его коллег и учеников, полученные в ходе 30-летнего изучения структурнофункциональной организации альпийских экосистем Тебердинского заповедника. Автор выражает глубокую благодарность всем коллегам, сделавшим критические замечания к настоящей работе, особенно В. Н. Павлову, А. М. Гилярову, С. А. Баландину, Ю. Т. Дьякову и Е. А. Игнатовой. Специальную благодарность хочу выразить Д. С. Дзыбову, предоставившему краткое описание метода восстановления степей для данной книги.
Предисловие
13
В настоящей работе использованы многие рисунки и фотографии, разрешения на публикацию которых любезно представлены их авторами и организациями, указанными в подписях к этим иллюстрациям. Все фотографии, авторство которых специально не указано в подписях к ним, принадлежат автору.
Исследования по различным аспектам функциональной фитоценологии в течение многих лет были поддержаны Российским фондом фундаментальных исследований1). Выражаю благодарность администрации Тебердинского заповедника, особенно заместителю директора Д. К. Теке-еву, за многолетнюю помощь и поддержку наших исследований. Сердечно благодарю свою жену А. А. Ахметжанову, всех своих родных, близких и друзей за всестороннюю поддержку моей работы.
1} Проекты № 93- 04- 06498,96- 04- 48420,98- 04- 49350,99- 04- 48012,02- 04- 48060, 04-04-49646, 05-04-48578, 05-04-89002, 08-04-00344, 08-04-92890, 11-04-01215.
Глава
Основная задача и система наук о растительности
Определения фитоценоза
Концепции дискретности и континуума
Представление о консорциях и консортивных связях
Концепция «ключевых» видов
Введение
1 • Основная задача и система наук о растительности
Геоботаника, или наука о растительности (vegetation science) — ботанико-экологическая наука, основной задачей которой является выявление механизмов формирования растительного покрова в различных пространственных и временных масштабах. Эта задача четко связана с фундаментальным вопросом экологии, который был сформулирован в первой половине XIX в. Heer (1835) (см. Braun-BIanquet, 1964): что определяет локальное участие и распространение видов (Stanton et al., 1994)?
Фитоценология является одной из основных наук о растительности, или геоботаники. Согласно взглядам В. В. Алехина (см. Павлов, 2004) и Л. Г. Раменского (1924), помимо фитоценологии, геоботаника включает также экологию растений, понимаемую в узком смысле, и ботаническую географию (рис. 1.1).
Рис-1.1 • Структура науки о растительности (геоботаники)
Термин «фитоценология» впервые предложил Хельмут Гаме (Н. Gams) в 1918 г., а в нашей стране впервые термин «фитоценология» использовал Л. Г. Раменский в 1925 г. (Работнов, 1995). Близкое понимание структуры науки о растительности существует в германоязычных странах (Walter, 1986), где обычно различают общую геоботанику (AHgemeine Geob-otanik), учение об ареалах растений (Floristische Geobotanik, Arealkunde), историческую геоботанику (Historische Geobotanik), экофизиологическую
16
Глава 1. Введение
геоботанику (Oekophysiologische Geobotanik) и биоценотическую геоботанику (Biozoenologische Geobotanik). В нашем пособии функциональная фитоценология целиком охватывает последнюю главу, в ней также рассмотрены некоторые вопросы общей и исторической геоботаники. Кроме того, вслед за названием основного труда Ж. Браун-Бланке («Pflanzensozi-ologie»), широко используется термин фитосоциология, который в нашей стране в советское время был отвергнут по политическим мотивам, хотя этот термин впервые ввел И. К. Пачоский в 1896 г. в работе на польском языке. Затем он независимо появился в разных странах — в России — в 1898 г., во Франции — в 1910 г., в США и Швейцарии — в 1917 г., в Швеции и Норвегии — в 1919 г. (Работнов, 1995).
В англоязычных странах обычно используют общий термин «экология растений» (plant ecology) для наук эколого-ботанического направления, часто различают ауто- и синэкологию растений, последний термин соответствует фитоценологии в понимании В. В. Алехина.
Подробную структуру науки о растительности, или геоботаники, предложили Б. М. Миркин и Л. Г. Наумова (1998) (табл. 1.1).
Основным международным научным обществом, объединяющим исследователей растительного покрова, является Международная ассоциация науки о растительности (1AVS), издающая ряд журналов. Крупнейшими национальными экологическими обществами являются Экологическое общество Америки (ESA) и Британское экологическое общество (BES).
В англо-саксонской традиции исследования ботанико-экологического направления часто подразделяют на противопоставляемые направления, такие как фитосоциология (где основное внимание уделяется выявлению полного состава растительных сообществ) и демография растений (акцентирует внимание на популяционной структуре доминирующих видов) (Silvertown, 1982). Кроме того, экология как экспериментальная наука часто необоснованно противопоставляется более описательной естественной истории (natural history), включающей зоологию, ботанику и геологию. В настоящей монографии мы, следуя отечественным традициям, будем стараться использовать знания из различных областей, помогающие выявлению природных закономерностей формирования и функционирования растительных сообществ.
2 • Определения фитоценоза
Фитоценоз, или растительное сообщество, является основным объектом изучения фитоценологии, поэтому возникает необходимость рассмотреть его определение. В отечественных учебниках часто используют определение, данное В. Н. Сукачевым (1954): «Фитоценозом, или растительным
2. Определения фитоценоза
17
Таблица 1.1 • Структура науки о растительности (по: Б. М. Миркин и Л. Г. Наумова, 1998)
Уровень организации объекта исследований Наука Основные задачи
Организменный Экофизиология Изучение процессов адаптации растений к экологическим условиям
Популяционный Популяционная биология растений Изучение популяций растений — их состава, генетического и фенетического разнообразия, динамики и пространственной структуры в связи с условиями среды
Видовой Ареалогия (ботаническая география) Изучение ареалов видов
Экология растений R-ординация и классификация видов по их отношению к градиентам факторов среды
Физиогномика (экологическая морфология растений) Изучение жизненных форм, форм роста и функциональных типов растений
Экологическая биология растений Изучение стратегий растений, экологии репродуктивного процесса (типов опыления, способов распространения плодов и семян и т. п.)
Фитоцено-тический Фитоценология Изучение факторов и моделей организации фитоценозов, их морфологии (синморфо-логии), экологии (синэкологии), синфенологии, динамики (синдинамики), продуктивности, альфа-разнообразия. Классификация фитоценозов (синтаксо-номия), география растительности
Субфитоцено-тический Фитоценология Изучение ультраценотических континуумов и мероценозов — микрогруппировок, синузий, ярусов
Инфрацено-тический Флористика (ботаническая география) Изучение флор и гамма-разнообразия растительности
Фитоценология Анализ пространственных континуумов и построение топоклинов
Симфитоценология Изучение комплексов фитоценозов разного уровня и бета-разнообразия растительности
18
Глава 1. Введение
сообществом, надо называть всякую совокупность как высших, так и низших растений, обитающих на данном однородном участке земной поверхности, с только им свойственными взаимоотношениями как между собой, так и с условиями местообитания, и поэтому создающую свою особую среду, фитосреду». Это определение, имеющее философский оттенок, не очень применимо к практическому выделению сообществ в природе, поскольку определить без специальных измерений и экспериментов, насколько своеобразны отношения между растениями, весьма затруднительно. Однако, в понимании В. Н. Сукачева, практически все сочетания растений в природе, как и искусственные посевы, могут рассматриваться как фитоценозы.
По-другому подошел к определению фитоценоза В. В. Алехин (1935): «Фитоценоз — закономерное сочетание растений, обусловленное: 1) историческим развитием, 2) экологическими условиями, 3) взаимодействием видов и обладающее: а) известной структурой, б) способностью к восстановлению и в) способностью воздействия на среду». Таким образом, по В. В. Алехину, фитоценозами могут считаться лишь самоподдержи-вающиеся, т. е. климаксовые, растительные сообщества. В этом случае при полевых исследованиях также сложно выявить, насколько изучаемая группировка растений способна к самоподдержанию. В настоящее время большинство исследователей придерживаются весьма прагматичному определению фитоценоза «как любого условно однородного контура растительности» (Миркин, Наумова, 1998), свойства которого (особенности воздействия на среду, самоподдержание и др.) могут изучаться как отдельные вопросы.
В общей экологии термин сообщество (community) чаще употребляют к совокупности совместно обитающих организмов отдельных таксонов (растений, птиц, насекомых), а термин биоценоз — к живой части экосистемы, включающей виды самых разных таксонов, но обитающие вместе на конкретной территории. Во всех случаях наличие взаимоотношений не является необходимым для выделения сообществ (Looijen, van Andel, 1999). В то же время Паркер (Parker, 2001) выдвинул концепцию сообщества индивидуумов — сообщества должны выделяться по особям (популяциям) какого-либо вида и охватывать все организмы разного таксономического положения, прямо или косвенно связанных (различными типами связей) с организмами рассматриваемого вида, т. е. сообщество — сеть взаимодействий, которая образует биотическое окружение особей. Эти взгляды весьма сходны с концепцией консортивных связей В. В. Мазинга, рассмотренной ниже.
Споры об определении растительных сообществ были обусловлены попытками выявить универсальные факторы их организации. Многочис-
2. Определения фитоценоза
19
Таблица 1.2 • Основные модели организации фитоценоза по Б. М. Миркину (Mirkin, 1994). Число плюсов отражает величину вклада факторов организации сообщества. В названиях моделей использованы буквенные символы растений, относящихся к различным стратегиям Ф. Грайма (см. п. 3.7.2 гл. 5), G-B — растения, устойчивые к отчуждению биомассы крупными фитофагами. Горизонтальные взаимоотношения — отношения между организмами одного трофического уровня, вертикальные — разных трофических уровней
Модель Вклад факторов организации Примеры
Экотоп Биотическая трансформация Горизонтальные взаимоотношения Вертикальные взаимоотношения Интенсивность нарушений Дифференциация
Абиотическая S-модель +++ Экстраординарные пустыни, водные сообщества (Lemnetea, Potametea и др.)
Биотическая S-модель +++ +++ Верховые болота (Oxycocco-Sphagnetea)
C-S-модель +++ +++ +++ Неморальные и бореальные леса (Querco-Fagetea, Vaccinio-Picetea)
G-В-модель +++ +++ Степи (Festuco-Brometea), саванны
C-R-S-модель +++ Послелесные луга (Molinio-Arrhenatheretea)
R-модель +++ Рудеральные и сегетальные сообщества
ленные исследования свидетельствуют, что роль различных факторов в формировании различных сообществ неодинакова. Это положение нашло отражение в выделении нескольких моделей организации фитоценозов, предложенных Б. М. Миркиным (Mirkin, 1994, табл. 1.2), и в концепции экологических фильтров (Belyea, Lancaster, 1999). Согласно схеме Б. М. Миркина, экотоп важен для формирования состава всех сообществ, конкуренция наиболее выражена в лесах, а формирование многих типов травяных фитоценозов обусловлено деятельностью крупных животных-фитофагов.
По схеме Belyea, Lancaster (1999) формирование видового состава сообществ из видов региональной флоры включает последовательное
20
Глава 1. Введение
Фильтр поступления диаспор
Абиотический фильтр
Биотический фильтр — взаимоотношения между организмами
Состав сообщества
Рис. 1.2 • Формирование состава растительных сообществ при последовательном действии экологических фильтров («сит») (по: Belyea, Lancaster, 1999)
действие трех факторов («сит»): 1) поступления диаспор на тот или иной участок территории, 2) отбор растений, устойчивых к локальным условиям абиотической среды, 3) формирования состава сообществ в результате биотических взаимоотношений между видами, прошедшими два предыдущих «сита» (рис. 1.2).
3 • Концепции дискретности и континуума
На ранних этапах развития геоботаники большинство исследователей придерживались концепции дискретности растительных сообществ, предполагающей наличие четких границ между ними. Однако с начала XX в. Л. Г. Раменский (1910) в нашей стране и Н. A. Gleason (1917, 1922, 1926) в США предложили альтернативный взгляд на растительный покров, который, по их мнению, является результатом взаимоотношений индивидуальных видов и среды, он непрерывен (континуален) и не образует дискретных единиц. В настоящее время эта точка зрения может рассматриваться как более соответствующая природе растительного покрова, однако она не исключает и наличие более четких границ в отдельных случаях и возможности классификации растительных сообществ по аналогиям с классификацией других континуальных природных объектов (почв, горных пород и т. п.). Сочетание континуальных и дискретных
4. Представление о консорциях и консортивных связях
21
свойств растительного покрова были изучены в специальной работе Hoagland и Collins (1997), которые провели анализ изменения состава видов растений вдоль градиента увлажнения на 42 заболоченных участках в Миннесоте (США). Авторы сравнивали распространение отдельных видов растений на этом градиенте со случайной моделью. Было выявлено, что в 10 случаях из 42 границы распространения видов были агрегированы, а моды (максимумы участия видов) были агрегированы в 19 случаях (значимые отличия от случайной модели).
4 • Представление о консорциях и консортивных связях
Термин «консорция» впервые использовал Е. Варминг (Warming, 1895). Однако детальная разработка представления о консорциях как структурно-функциональных элементах биогеоценоза (экосистемы) принадлежит В. В. Мазингу (1966). Эти представления во многом аналогичны учению
Консорты
1 - го концентра
Рис-1.3 • Структура консорции (по: Мазинг, 1966). На примере нескольких видов из консорции лугового клевера (Trifolium pratense) в 1-м концентре показан фитофаг (олень) и опылитель (шмель), гнезда которого часто разоряются ежом (2-й концентр)
22
Глава 1. Введение
о пищевых цепях и сетях, но рассматривают не только трофические и другие типы связей между организмами. В центре консорции находится детерминант — популяция автотрофного растения, которая связана с консортами прямо (консорты 1-го концентра) или через другие организмы (рис. 1.3). Выделяют несколько типов связей между организмами: трофические (поедание), топические (использование в качестве субстрата для поселения), фабрические (использование частей растений для сооружения построек, гнезд и т. п.), форические (перенос частей растений, например пыльцы, диаспор и т. п.) и др. Трофически связаны с растениями: биотрофы (фитофаги, фитопаразиты, симбиотрофы), эккрисотрофы (использующие выделения растений), сапротрофы (использующие отмершие части растений, например копытные на зимних пастбищах).
Обычно значительное число видов-фитофагов связано с отдельными видами растений, которыми они питаются. Так, более 200 видов насекомых отмечены как потребители на нескольких видах деревьев умеренной зоны. На листьях чая указывается около 80 видов, на вайях орляка — около 40 видов членистоногих (Lawton, Schroder, 1977; Linhart, 1991) (см. главу 4).
5 • Концепция «ключевых» видов
В экосистемах могут присутствовать виды, оказывающие большее влияние на ее структуру и функционирование по сравнению с их участием (например, биомассой). Такие виды, роль которых непропорционально (их массе) велика, получили название «ключевых» видов (keystone species). Впервые этот термин использовал Пейн (Paine, 1966, см. Bond, 1993) для хищников в морских сообществах. Ключевыми видами в наземных экосистемах могут быть виды разных трофических уровней (от растений до хищников), особенно сильно преобразующие среду. Примерами таких видов могут быть бобовые растения, которые симбиотически фиксируют азот, оказывая большое влияние на азотный режим почв. Как ключевые виды для зоны широколиственных лесов могут рассматриваться зубры, создающие крупные прорывы в пологе леса, а также бобры, строительная деятельность которых принципиально меняет гидрологический режим пойм малых рек, вызывая циклические смены растительных сообществ и поддерживая высокое флористическое разнообразие территории (Смирнова, 2004).
Глава
А
зот является важнейшим элементом минерального питания растений, его недостаток в почве лимитирует продукцию многих видов растений. Поэтому взаимоотношения с азотфиксирую-щими прокариотами имеют особое значение в жизни растений, так как способностью к азотфиксации обладают только прокариоты. Растения преимущественно поглощают азот из почвы в виде 1) нитратов, 2) аммония, 3) аминокислот. Последние играют очень важную роль как источник азота для многих групп растений умеренного пояса (Kielland, 1994). Эти соединения азота становятся доступны растениям в результате микробиологической деструкции органических веществ опада, а также азотфиксации. По степени связи растений с прокариотами можно выделить три основных типа азотфиксации в растительных сообществах:
1) симбиотическая — осуществляется внутри растений в специализированных структурах,
2) ассоциативная — происходит на или вблизи поверхности корней (в ризоплане или ризосфере), а также на поверхности листьев (фил-лоплане) растений,
3) несимбиотическая происходит вне прямой связи с растениями-эукариотами за счет внешних источников органики (гетеротрофная азотфиксация) или с использованием продуктов собственного фотосинтеза (у фотосинтезирующих цианобактерий).
1 • Симбиотическая азотфиксация
Для сосудистых растений в наземных экосистемах характерно 3 типа симбиотических отношений с прокариотами: ризобиальный симбиоз (бобовые и клубеньковые бактерии), актинориза и цианобактериальный симбиоз (табл. 2.1).
1.1 • Бобовые и клубеньковые бактерии
Симбионты:
Растения семейства Fabaceae (бобовые в широком смысле, включая мимозовые и цезальпиниевые), а также этот тип симбиоза обнаружен у видов рода Parasponia (Ulmaceae) (Law, Lewis, 1983). Виды рода
1. Симбиотическая азотфиксация
25
Таблица 2.1 • Сравнение основных типов симбиотических ассоциаций сосудистых растений с азотфиксирующими прокариотами (по: Law, Lewis, 1983;
Работнов, 1992; Bergman et al., 1992; Sprent, 1999; Rai et al., 2000)
Показатель Ризобиальный симбиоз Актинориза Цианобактериальный симбиоз
Прокариоты Rhizobiaceae Frankia Nostoc, редко Anabaena
Растения большинство Fabaceae s. 1., Parasponia spp. (Ulmaceae) более 200 видов двудольных, гл. об. древесных растений: Alnus (Betulaceae), Hippophae, Elaeag-nus (Elaeagnaceae), Dryas (Rosaceae), Myrica (Myricaceae), Casuarina (Casuari-пасеае) и др. папоротники (Azol 1а, 6 видов), саговники (все изученные, 150 видов), из цветковых растений — только Gunnera (Gunneraceae, 65 видов)
Локализация прокариот Чаще коротножи-вущие клубеньки на корнях, стеблевые мамиллы (Sesbania rostrata) Долгоживущие клубеньки на корнях Полости в лопастях вай (Azolla), межклетники первичной коры (саговники), внутриклеточно в гландах на стеблях и черешках листьев (Gunnera)
Максимальная интенсивность азотфиксации, кг N/ra в год 450-550 (клевер в Новой Зеландии), максимум до 680 210-225 у Alnus glutinosa 720 — Gunnera a re па ria в Новой Зеландии
Экологические предпочтения растений нейтральные или слабощелочные почвы пионерные субстраты, болотные кислые почвы хорошее обеспечение влагой (Gunnera), водные растения (Azolla)
Parasponia (всего 5) достигают в высоту 15 метров и естественно произрастают на островах Индо-Малазийского архипелага и Тихого океана. Они часто выступают пионерами на бедных азотом лавовых субстратах (Vessey et al., 2004). Большинство, но не все бобовые образуют клубеньки. Лишь около 20 % видов бобовых к началу XXI в. было исследовано на наличие клубеньков. Из исследованных видов 97 % мотыльковых, 90% мимозовых и 23 % цезальпиниевых имели клубеньки. В низинных тропических лесах Нового Света древесных бобовых больше, чем деревьев любого другого семейства, по числу стволов они составляют 12-15% (Rundel, 1989). Бобовые хорошо представлены и в более сухих тропических экосистемах, например в листопадных тропических лесах в Chamela
26
Глава 2. Растения и азотфиксирующие прокариоты
(Мексика), бобовые — самое богатое семейство, включающее 14% флоры. МсКеу (1994) полагал, что изначально бобовые эволюционировали как требовательное к азоту семейство на богатых почвах в тропических областях. В умеренных областях травянистые бобовые весьма обычны, а древесные — редки (Crews, 1999).
Бактерии Rhizobiaceae. Ранее все клубеньковые бактерии рассматривались внутри одного рода Rhizobium. В настоящее время их относятся к нескольким родам: Rhyzobium, Sinirhyzobium, Bradyrhyzobium, Mezorhyzobium, Azorhyzobium и др., общее число видов превышает 50 (Barea et al., 2005; Willems, 2006). Помимо ризобиальных симбионтов бобовых, которые относятся к а-протеобактериям, найдены /3-протеобак-терии, включая рода Ralstonia и Burkholderia, которые также образуют клубеньки на бобовых (Elliott et al., 2007; van der Heijden et al., 2008). Необходимо отметить, что в почве обитает много штаммов ризобиальных бактерий, которые не могут образовывать клубеньки на корнях бобовых и/или неэффективно фиксируют азот. У бобовых существуют (хотя и не универсальные) механизмы защиты от таких штаммов, предотвращающие неэффективный симбиоз (Sachs, Simms, 2008).
Отмечается специфичность отдельных видов и рас бактерий к видам или родам бобовых. Физиологически бактерии отличаются, как по способности образовывать клубеньки (инфекционность), так и по эффективности фиксации азота. Клубеньки формируются из корневых волосков при попадании клубеньковых бактерий из почвы. При формировании клубеньков наблюдается сложный обмен метаболитами между корнями и бактериями в почве, в том числе трансформация выделяемого корнями триптофана бактериями в индолилуксусную кислоту (гетероауксин), которая в свою очередь стимулирует рост корневых волосков перед инвазией бактерий (Goss, 1991) (рис. 2.1). Обычно клубеньки живут менее одного сезона, но у некоторых древесных растений из подсемейства цезальпиниевых (например, Campsiandra laurifolia) клубеньки многолетние (Barrios, Herrera, 1993; Sprent, 1999).
Корень бобового растения
Рис. 2.1 • Схема инициации образования клубенька у бобовых растений (по: Goss, 1991)
1. Симбиотическая азотфиксация
27
Фиксация молекулярного азота весьма энергоемкий процесс, источником энергии в биологических системах служат органические соединения. Эффективность использования органического углерода для фиксации молекулярного азота у бобовых весьма значительна. Они затрачивают на фиксацию 1 г азота 5-7 г углерода (углеводов) (Lundquist, 2005), в то время как свободноживущие азотфиксаторы затрачивают на порядок больше. В то же время у бобовых растений ощутимая часть (обычно около 10%) продуктов фотосинтеза идет на поддержку азотфиксации.
Количественные показатели азотфиксации. Максимально известные величины составляют 680 кг N/ra в год (Spehn et al., 2002), что сопоставимо с самыми высокими дозами азотных удобрений, применяемых в интенсивном растениеводстве. Для посевов клевера в Новой Зеландии отмечена фиксация до 450-550 кг N/ra в год, в Средней России посевы бобовых фиксирует около 200 кг N/ra в год (при благоприятных условиях посевы люцерны могут связывать из воздуха 271, клевера — 149, люпина — 265 кг N/ra в год) (Михневский, 1970).
Оценить интенсивность азотфиксации в поле можно по размерам и окраске клубеньков: чем они крупнее и розовее, тем эффективнее идет азотфиксация. Розовая окраска клубеньков связана с высоким содержанием в них растительных гемоглобинов, которые связывают кислород. Все симбиотические азотфиксирующие прокариоты содержат Mo-Fe нитрогеназный комплекс, работающий в условиях пониженного парциального давления кислорода. Леггемоглобины составляют около 5 % белков зрелых клубеньков (Vessey et al., 2004). В целом же гемоглобины широко встречаются у растений (не только у бобовых). Они могут временно депонировать кислород и улучшают энергетический баланс клеток при кислородном голодании (Hill, 1998).
Развитие большинства бобовых лучше происходит на нейтральных и слабощелочных почвах. Ризобиальные бактерии активны в широких пределах pH (5,0-8,5). При этом бобовые образуют клубеньки в очень широком диапазоне экологических условий, в том числе и при низких pH, в условиях затенения, при низкой доступности Р и иногда даже на богатых нитратами почвах (Sprent, 1999). С другой стороны, азотфиксация бобовых ведет к снижению pH в их ризосфере, что способствует большей доступности фосфатов, которые бобовые эффективно потребляют (Marschner, Romheld, 1983).
Клубеньки образуются не только на корнях в почве. У однолетнего тропического бобового Sesbania rostrata на стебле образуются «стеблевые мамиллы» — клубеньки верхушек придаточных корней (Spencer-Barreto et al., 1989, рис. 2.2).
28
Глава 2. Растения и азотфиксирующие прокариоты
Рис. 2.2 • Ризобиальный симбиоз: а — Parasponia andersonii (Ulmaceae) образует ризобиальные клубеньки (фото R.Taputuarai — Moorea Biocode, с любезного разрешения автора); б — стеблевые клубеньки (мамиллы) у Sesbania sp. (David Patriquin, Dalhousie University, с любезного разрешения автора); в — клубеньки на корнях люцерны (Medicago sp.) (фото Andrew Davis, Bio-imaging John Innes Centre, с любезного разрешения автора)
Как поступает фиксированный азот в почву и к другим растениям? Чаще всего это происходит при отмирании и разложении корней и побегов бобовых растений. При дефолиации (удалении части надземных побегов и листьев) клевера поступление азота к корням плевела (Lolium регеппе) увеличилось в 5 раз (Ayres et al., 2007). Таким образом, фитофаги могут играть существенную роль для транспорта азотных соединений от бобовых к небобовым растениям. В эксперименте (Bardgett, Wardle, 2003) было показано, что клеверная нематода (Heterodera trifolii), питающаяся на корнях фиксирующего азот Trifolium repens, не только увеличивает корневые выделения и микробиологическую активность в ризосфере, но и усиливает рост корней и содержание в них азота у соседствующих растений плевела (Lolium регеппе). При совместном выращивании обра
1. Симбиотическая азотфиксация
29
зующих и не образующих клубеньки растений арахиса (Arachis hypogaea) транспорт азота от первых к последним составил до 48 % содержания азота в неклубеньковых растениях. При этом не было замечено значимого снижения участия донорных растений. Аналогичный эксперимент с выращиванием арахиса совместно с сорняками (Sida spinosa, Senna obtusifolia) показал, что последние получают 30-80 % азота от арахиса за 60-дневный период выращивания. В то же время транспорт к немикоризным сорнякам (Cyperus esculentus, Amaranthus palmer!) был очень низким, что свидетельствует о передаче азота главным образом по мицелию микоризных грибов (Moyer-Henry et al., 2006).
Но бобовые используют и минеральные соединения азота в почве. Определить долю азота из разных источников можно по естественному содержанию природного стабильного изотопа азота 15N. Обычно за счет изотопного фракционирования содержание этого изотопа в минеральном почвенном азоте ниже, чем в атмосферном. Растения, получающие минеральный азот из почвы, обычно имеют более низкие концентрации 15 N
Рис. 2.3 • Внешний вид трех видов бобовых растений альпийского пояса северо-западного Кавказа и содержание 15 N (%о<5) в их листьях, показывающее интенсивную азотфиксацию у Astragalus levieri и Oxytropis kubanensis и практическое отсутствие азотфиксации у Trifolium polyphyllum
30
Глава 2. Растения и азотфиксирующие прокариоты
по сравнению с растениями, получающими азот от клубеньковых бактерий. У последних изотопный состав азота близок к атмосферному. Таким образом, по содержанию 15 N можно определить долю фиксированного азота. Например, наблюдения над 3 видами Trifolium в альпийских сообществах Скалистых гор в Колорадо и сравнение содержания 15 N в тканях этих видов при выращивании на безазотистой среде и в природе показали, что от 70 до 100 % азота эти растения получают за счет азотфиксации и лишь незначительную долю — из почвы (Bowman et al., 1996). Анализ содержания этого изотопа в альпийских бобовых северо-западного Кавказа подтвердил практическое отсутствие азотфиксации у Trifolium polyphyllum, у которого отсутствовали нормально развитые клубеньки, и активную азотфиксацию других видов (рис. 2.3).
Клубеньки, образованные другими группами бактерий, иногда встречаются на корнях небобовых растений, например клубеньки с Bacillus alopecuri на корнях Alopecurus pratensis. Они могут фиксировать азот, но интенсивность и эффективность азотфиксации в этих случаях крайне невелика и вряд ли имеет экологическое значение.
1.2 • Актинориза
Симбионты:
Растения: многолетние двудольные, чаще древесные растения (деревья, кустарники, кустарнички), а также некоторые травы — известно более 200 видов из 10 семейств (Huss-Danell, 1997). Основные семейства и рода: Betulaceae (Alnus), Elaeagnaceae (Hippophae, Elaeagnus и др.), Rosaceae (Dryas, Cercocarpus и др.), Datiscaceae (Datisca — травянистые растения), Myricaceae (Myrica и др.), Coriariaceae (Coriaria), Casuarinaceae (Casua-rina, Ceuthostoma и др.), Rhamnaceae (Adolphia, Ceanothus и др.), т. e. систематически довольно удаленные друг от друга семейства двудольных растений (рис. 2.4). Не совсем понятно, почему не все растения в рассматриваемых семействах формируют актиноризу, например почему ольха формирует актиноризу, а береза — нет. Одно из предположений, объясняющих это явление, связано с развитием арбускулярной микоризы (AM). Этот тип микоризы, способствующий лучшему получению преимущественно фосфатов, в Скандинавии встречается в семействе Betulaceae только у Alnus (Bousquet et al., 1989; Huss-Danell, 1997).
Рис- 2-4 • Актиноризные растения: а — Alnus incana (Betulaceae); 6 — Casua-rina equisetifolia (Casuarinaceae); в — Elaeagnus angustifolia (Elaeagnaceae); г — Dryas octopetala (Rosaceae, тундры северной Швеции); д — Coriaria nepalensis (Coriariaceae, Юньнань, Китай); e — Morelia (Myrica) pensylvanica (Myricaceae)
1. Симбиотическая азотфиксация
31
32
Глава 2. Растения и азотфиксирующие прокариоты
Актиномицеты: род Frankia (монотипное семейство Frankiaceae), облигатные симбионты, но имеют свободноживущую в почве стадию. Для процесса азотфиксации необходим молибден и кобальт. Отмечается довольно высокая специфичность к отдельному роду или группе родов цветковых растений.
Проникновение актиномицетов в корни растений осуществляется как через корневые волоски (Alnus, Casuarina, Myrica), так и по межклетникам (Elaeagnus, Rhamnaceae, Rosaceae). Frankia может в значительном количестве накапливаться в почве сообществ и безактиноризных растений, например под Betula nigra (Paschke, Dawson, 1992). Показано присутствие этих микроорганизмов также в гнездах птиц, как использующих почву для их постройки, так и не использующих ее. Это свидетельствует о возможности распространения Frankia птицами (Paschke, Dawson, 1993). Большинство изученных почв содержало диаспоры Frankia. Почвы без активных для Alnus штаммов Frankia отмечены только из самых северных районов Сибири, Северной Америки, подповерхностных торфяных слоев. Споры Frankia могут долго сохраняться в почве и переноситься по воздуху (Huss-Danell, 1997).
Структура клубеньков. Клубеньки актиноризы содержат центральный цилиндр (в отличие от клубеньков бобовых) и являются многолетними образованиями (рис. 2.5). Они имеют верхушечную меристему, которая продуцирует новые клетки только базипетально, т. е. не формирует корневой чехлик (Huss-Danell, 1997). За исключением родов Casuarina и Allocasuarina, актиномицеты в клубеньках формируют так называемые везикулы, в которых локализуется нитрогеназа. Ориентация и форма везикул варьирует среди растений-хозяев. В культуре Frankia формирует сферические везикулы. Структура оболочек этих везикул служит для предотвращения проникновения кислорода и зависит от степени аэрируемости почвы. В отличие от ризобиальных бактерий, Frankia обладает собственной защитой от высо-
л кого парциального давления
Рис. 2.5 • Многолетние актиноризные г х
клубеньки на корнях ольхи (Alnus incana) кислорода и поэтому способ-на фиксировать азот в сво-
1. Симбиотическая азотфиксация
33
5одноживущих (почвенных) стадиях (Huss-Danell, 1997). Для актиноризы Alnus incana + Frankia) энергетическая стоимость фиксации азота сходна : таковой для бобовых растений и составляет 4,5-5,8 г углерода орга-шческих соединений на 1 г фиксированного азота (Lundquist, 2005).
Многие актиноризные растения (но также бобовые и некоторые 1ругие), обычно не имеющие микориз, образуют так называемые «кластерные корни», специфические структуры, имеющие вид ершика (Skene, 1998) [см. Вставку /].
Вставка I. Кластерные корни
Кластерные корни (см. рис. 1.1, 1.2) развиваются у ряда растений, обитающих на очень бедных фосфором почвах. Они характер-
ны для представителей различных семейств (преимущественно Proteaceae почти все виды — около 1600 из 73 родов, Casuarinaceae, Fabaceae, Myri-
caceae, Betulaceae и др.), среди которых встречаются актиноризные растения. Эти корни улучшают фосфорное питание растений за счет одновременного выделения в среду органических кислот (подкисление), хелатирующих агентов (комплексообразователей) и фосфатаз. Выделение этих агентов происходит в пульсирующем режиме (в течение нескольких дней). Jeschke и Pate (1995)
Рис-1.1 • Кластерные корни различных растений (по: Lambers et al., 2003)
34
Глава 2. Растения и азотфиксирующие прокариоты
Окончание Вставки I
показали, что кластерные корни поглощают примерно в 10 раз больше фосфора, чем некластерные. Протейные могут расти при самых низких концентрациях фосфора в листьях (0,33 мг/г сухого листа), у других видов этот показатель обычно не ниже 1 мг/г. Протейные часто угнетаются даже при умеренном содержании фосфора в почве (Shane, Lambers, 2005).
Рис. 1.2 • Структура «кластерных корней»: а — кластерные корни (CR) образованы укороченными, густыми мелкими корешками (RO), в свою очередь в зрелости покрытыми густыми корневыми волосками (RH); б — простой кластерный корень, обычный у многих растений, за исключением видов семейства Proteaceae; в — сложный кластерный корень с редкими корешками второго порядка, изредка встречающийся у растений с простыми кластерными корнями; г — сложный кластерный корень, типичный для приземистых видов Proteaceae (по: Skene, 1998).
Интенсивность азотфиксации актиноризных растений колеблется от 10 кг N/ra в год у Myrica gale до 210-225 кг N/ra в год у Alnus gluti-nosa. Содержание в почве минеральных форм азота может регулировать уровень азотфиксации, так почвенные нитраты могут ингибировать образование актиноризных клубеньков. Засуха также сильно ингибирует нитрогеназную активность у Alnus incana (Sundstrom, Huss-Danell, 1987). Наблюдения над сезонным ходом азотфиксации у этого вида на бедных почвах в северной Швеции показали, что азотфиксация начинается вскоре после развертывания листьев в мае, возрастет в июне, поддержи
1. Симбиотическая азотфиксация
35
вается высокой с июля по август, уменьшается в сентябре и практически прекращается в октябре (Huss-Danell, 1997).
По своим экологическим особенностям актиноризные растения часто являются пионерами (Dryas octopetala — зарастание карбонатных □сыпей в тундрах и высокогорьях) и видами средних стадий первичных сукцессий (Alnus spp.), на которых происходит обогащение почвы N. Они гакже встречаются на кислых, часто заболоченных почвах. Обогащение почвы азотом, например в зарослях ольхи, ведет к интенсификациии нитрификации, с чем связано произрастание многих нитрофилов совместно с актиноризными растениями, например в черноольшатниках.
Актиноризный кустарник Myrica faya, естественно произрастающий на Канарских островах был завезен на Гавайи, где большая
часть почв была образована на вулканических породах, бедных доступными формами азота. На островах сформировалась богатая флора, содержащая много эндемичных видов, адаптированных к бедным азотом почвам. Myrica faya, семена которого распространяются птицами, быстро вытеснил многие местные виды растений с бедных лавовых почв, обогащая эти почвы азотом. Этот вид был впервые отмечен в национальном парке «Гавайские вулканы» в 1961 г., в 1966 г. его заросли занимали около 600 га, а в 1985 г. — более 12 000 га (Vitousek, 1990).
Вставка II. Завоз Myrica faya — экологическая катастрофа на Гавайях
Рис. 11.1 • Myrica faya (фото Peter Schoenfelder, с любезного разрешения автора)
36
Глава 2. Растения и азотфиксирующие прокариоты
Основной механизм обогащения почвы азотом в случае актиноризного симбиоза — это поступление N через листовой опад и отмирающие корни. Для листьев этих растений обычно характерен невысокий уровень оттока азота из отмирающих листьев (реутилизация) [см. Вставку III] азота. Например, из опадающих листьев березы отток азота составил 81 %, рябины — 77 %, а ольхи — только 19 % от содержания в вегетирующих листьях (Манаков, 1970).
— Реутилизация (ретранслокация, _ ... _ возврат) — перемещение элеме-
Вставка III. Реутилизация
нтов минерального питания из отмирающих органов растений в живые, которое позволяет обеспечить более экономное использование этих элементов за счет их внутреннего цикла.
Наиболее детально исследована у древесных растений. По наблюдениям в Испании, реутилизация азота сильнее выражена у листопадных деревьев и кустарников по сравнению с вечнозелеными. Максимальная из 19 изученных видов отмечена у Betula pubescens — 70 %, минимальная у Prunus spinosa — 23 % (del Arco et al.. 1991). Реутилизация возможна лишь до некоторой нижней пороговой концентрации. По данным Killingbeck (1996), такая минимальная возможная концентрация в отмирающих листьях деревьев и кустарников для азота составляет 0,3%, для фосфора — 0,01% сухой массы. Эффективность реутилизации может быть оценена по отношению наблюдаемой реутилизации к этим величинам.
Реутилизация наблюдается не только у древесных растений. Например. Lolium регеппе реутилизирует 30-70% азота, содержащегося во взрослых листьях перед их отмиранием (Thomas et aL, 1990). Средняя реутилизация азота у двудольных травянистых растений была ниже, чем у граминоидов. С другой стороны, весеннее отрастание побегов Festuca rubra и Agrostis capillaris на 70% и 82% соответственно обеспечивалось реутилизированным из старых листьев азотом (Bausenwein et aL, 2001).
Источником реутилизируемого азота являются протеины, а фосфора — нуклеиновые кислоты и фосфолипиды (Aerts, 1996). Механизм реутилизации азота связан с распадом белков в отмирающих листьях и синтезом аминокислот с двумя атомами азота (аспарагин и глютамин), которые служат транспортными соединениями при реутилизации (имеют минимальное отношение С : N) (Eckstein et al., 1999).
Реутилизация азота из листьев сильнее выражена у растений бедных почв. Поэтому их опад беднее азотом, что вызывает его более медленное разложение, способствуя консервации азота в ценозе. Напротив, у растений богатых почв опад, богатый азотом, разлагается быстро, и возможные потери азота из почвы при этом выше (Hobbie, 1992). Растения с более яркой листвой реутилизируют из листьев больше элементов минерального питания (ЭМП), чем растения с тусклыми листьями (Taylor. 1998; Lee et aL, 2003; Schaefer. Wilkinson. 2004), поэтому многие растения тундровой и таежной зоны имеют очень яркую осеннюю окраску (рис. 111.1).
1. Симбиотическая азотфиксация
37
Рис-111.1 • Горная лесотундра на Верхоянском хребте (Якутия) осенью: а — общий вид участка лесотундры с осенней окраской листьев кустарничков (Arctous alpina, Vaccinium uliginosum и др.); б — Alnus fruticose, имеющая актиноризу и фиксирующая азот, не обладает яркой осенней окраской; в — Salix berberifolia иг — Arctous alpinus с ярко-красный осенней окраской листьев
38
Глава 2. Растения и азотфиксирующие прокариоты
Окончание Вставки III
Реутилизация азота не всегда выгодна растениям, не испытывающим в нем недостатка. Tateno (2003) сравнил фенологию ли-
стьев двух видов листопадных деревьев — азотфиксирующей (нереутилизи-рующей азот) Alnus firma и неазотфиксирующей (реутилизирующей из листьев около 49% азота) Morus bombycis (шелковица) со сходной морозоустойчивостью листьев (-2 °C) (табл. 111.1). Растения шелковицы заканчивали вегетацию в середине октября, когда минимальные температуры воздуха были еще выше О °C. Листья шелковицы постепенно желтели и опадали. Напротив, молодые растения ольхи продолжали фотосинтезировать до середины ноября, отмирали при наступлении низких температур без специальной осенней окраски. Расчеты показали, что увеличение длительности фотосинтеза на 1 месяц у ольхи примерно в 6 раз покрывает энергетические затраты на фиксацию реутили-зированного шелковицей азота. Поэтому стратегия нереутилизировать азот с максимально возможным увеличением фотосинтетического периода адаптивна для растений с симбиотической азотфиксацией.
Таблица 111.1. Сравнение фенологии актиноризных и неактиноризных деревьев (Tateno, 2003)
Виды деревьев:
Morus bombycis
Alnus firma
Актинориза
Осенняя окраска листьев Окончание вегетации Реутилизация азота
Нет
Желтая
Середина октября 49%
Есть
Нет
Середина ноября
Нет
1.3 • Консортивные связи с цианеями
Симбионты:
Растения. Этот тип симбиоза известен в разных группах растений (диатомеи, мохообразные, водные папоротники Azolla, саговники и цветковые растения рода Gunnera) (рис. 2.6). Кроме того, цианеи часто выступают в качестве основного или дополнительного фотобионта в лишайниках, образуют симбиоз с разными группами животных (простейшие, губки, морские черви класса Echiuroidea, см. Rai, 1990).
Цианеи (цианобактерии, синезеленые водоросли). Виды рода Nos-toc, а также Anabaena azollae, таксономически близкая роду Nostoc.
Известны различные случаи симбиоза цианей с мохообразными, хотя чаще цианеи обитают на поверхности листьев мохообразных, являясь эпифитами. Например, в гиалиновых клетках сфагновых мхов (Sphagnum
1. Симбиотическая азотфиксация
39
Рис. 2.6 • Лишайники и растения, имеющие симбиотические связи с цианеями (цианобактериями): а — эпигейный (напочвенный) лишайник Peltigera aphthosa с основным фотобионтом — зелеными водорослями и дополнительным — Nostoc — в цефалодиях на поверхности таллома; б — водный папоротник AzoIla mexicana (с любезного разрешения Russ Kleinman, Courtesy of the Dale A. Zimmerman Herbarium, Western New Mexico University); в — саговник Encephalartos altensteinii (Южная Африка); г — Gunnera manicata в ботаническом саду Кунминга, Китай
riparium и S. linderbergii), где создается близкая к нейтральной среда, Nostoc фиксирует до 5 кг/га N, в то же время на сфагнумах чаще поселяются эпифитный Nostoc, имеющий высокий процент гетероцист и в 14-22 раз более интенсивно фиксирующий азот, чем свободноживу-щие колонии (Meeks, 1990). Nostoc отмечен в коробочках мхов Andreaea rupestris (Филин, Филина, 1989) и Tetraphis pellucida (Филина, Филин, 1989). У антоцеротовых цианеи заселяют специальные полости таллома. У цианей, живущих в полостях Blasia и Anthoceros, резко увеличивается
40
Глава 2. Растения и азотфиксирующие прокариоты
Рис. 2.7 • Агеотропные корни саговника Dioon spinulosum (Мексика) с цианеями на поверхности почвы
доля гетероцист (у свободноживущих 2-5 % клеток, у симбиотических — до 60%) (During, van Tooren, 1990). Симбиотические ассоциации были экспериментально ресинтезированы в лабораторных условиях.
Способность водного папоротника Azolla увеличивать плодородие почв упоминается еще в китайских словарях более 2000 лет назад. У этого папоротника в верхних лопастях вай имеются полости, заселяемые цианеями. Azolla (около 6 видов) издревле используется на рисовых полях, прибавка урожая риса составляет от 14 до 40 % за счет внесения в почву 40-80 кг N/ra в год (Moore, 1969).
Симбиоз с саговниками (порядок Cycadales с 3 семействами: Су-cadaceae — Cycas, Stangeriaceae — Stangeria, Bowenia, Zamiaceae — 8 родов, всего в порядке около 240 видов). У всех исследованных видов саговников обнаружены коралловидные корни, в межклетниках первичной коры которых поселяются цианеи. Развитие этих корней определяется генетически, они развиваются и в отсутствии симбионтов. Коралловидные корни с цианеями обладают не типичным для корней агеотроп-ным ростом (к поверхности почвы) (Lindblad, Bergman, 1990; Peterson, 1992; Vessey et al., 2004) (рис. 2.7). Во всех случаях у симбиотических цианей отмечен высокий процент гетероцист (более крупных клеток с утолщенными оболочками, в которых и происходит азотфиксация), часто также двойных или тройных, которые практически не встречаются у свободноживущих цианей. Интенсивность азотфиксации симбионтов примерно в 7 раз выше, чем у свободноживущих цианей. Однако абсолютные показатели азотфиксации невелики, например для Macrozamia в австралийских эвкалиптовых лесах отмечена интенсивность азотфиксации до 19 кг/га в год.
1. Симбиотическая азотфиксация
41
Среди цветковых растений симбиоз с цианеями отмечен лишь для видов рода Gunnera (Gunneraceae). Род назван Линнеем в честь норвежского монаха и ботаника Ernst Gunneriis (1718-1773), он насчитывает около 65 видов, распространенных в основном в Южном полушарии (Африка, Мадагаскар, Центральная и Южная Америка, Новая Зеландия, Индонезия). Ископаемые представители рода известны из меловых отложений возрастом 95 млн лет (Jarzen, 1980). В верхнем мелу и начале третичного периода род был более широко распространен и встречался на всех континентах. Размеры растений рода Gunnera колеблются от 2 см до 6 м. Все — многолетние травы, обычно корневищные, часто столонообразующие. По экологической приуроченности представители этого рода характерны для влажных лесов, заболоченных участков, влажных скал, нестабильных осыпей и т. п., почвы которых имеют низкую обеспеченность азотом и фосфором. Интересно отметить, что совместно с растениями этого рода почти не встречаются бобовые или актиноризные растения.
Цианеи (точно идентифицирован Nostoc puctiforme) поселяются в специализированных железоподобных органах (гландах) на стебле и в черешках листьев. Эти органы формируются и без присутствия цианей, поселение которых происходит сначала в выделяемой гландами слизи, а затем проникновением внутрь. Размеры железок с цианобионтом — от 2 мм до более 2 см, они увеличиваются с возрастом растения. Циано-бионт составляет около 1 % общей биомассы растения. Это единственный тип симбиоза с цианеями, когда последние поселяются в сосудистом растении внутриклеточно (Rai et al., 2000).
В стерильной культуре растения Gunnera могут развиваться и без ностока, однако в природе такие случаи неизвестны. Таким образом, этот симбиоз факультативен, но развитие растения-хозяина обычно улучшается при поселении цианей. Так, в экспериментах с Gunnera tinctoria биомасса листьев увеличивалась в 10 раз при поселении Nostoc. У симбиотических цианей резко увеличивается доля гетероцист, достигая в старых гландах 50 %. Азотфиксация идет весьма интенсивно, например для Gunnera arenaria в Новой Зеландии отмечены величины около 720 кг N/ra в год (Bergman et al., 1992). В связи с высоким содержанием защитных веществ, виды Gunnera обычно мало повреждаются фитофагами, за исключением овец и коров в местах выпаса. Двукрылые (Tipulidae, Chi-ronomidae) могут быть векторами переноса ностока, так как их личинки часто колонизируются ностоком и часто развиваются при основании побегов Gunnera во влажные периоды года (Osborne, Sprent, 2002).
Общие особенности симбиоза с цианеями. Цианобионт получает от хозяина защиту от высыхания и выедания, а в случае саговников
42
Глава 2. Растения и азотфиксирующие прокариоты
и Gunnera, возможно, также и сниженное парциальное давление кислорода, поскольку нитрогеназа функционирует в анаэробных условиях. Существует специфичность цианобионтов по отношению к хозяевам. Не все (но и не единственный!) штаммы Nostoc способны инфицировать хозяев. Инфицируя хозяев, Nostoc изменяет свои свойства. Из нитчатой водоросли он становится почти одноклеточным, размеры клеток возрастают, полифосфатные гранулы исчезают, уменьшается количество карбоксизо-сом и гликогена, утоньшается клеточная стенка. Рост клеток и их деление замедляется по сравнению с чистой культурой. Процессы фотосинтеза уменьшаются, например у симбионтов печеночников и антоцеротовых до <1-12% от нормальной интенсивности. Цианосимбионты саговников и Gunnera, даже изолированные из хозяев, не способны к фотосинтезу (Lindblad, Bergman, 1990; Bergman et al., 1992). Глютаминсинтета-за — фермент, связывающий полученный при азотфиксации аммоний у диазотрофных цианобактерий. Этот фермент блокируется различными способами у симбиотических организмов, что позволяет хозяевам получать несвязанный аммоний. Исключение составляют саговники, которые не подавляют активность глютаминсинтетазы цианей, а получают от них связанный азот, видимо, в форме глютамина или цитруллина. Дальнейший же транспорт фиксированного азота по тканям хозяина осуществляется преимущественно в форме аминокислот (Rai et al., 2000).
1.4 • Прочие случаи симбиоза
Во многих случаях в теле растения могут поселяться бактерии, не приносящие ему видимого вреда. Наиболее часто они заселяют межклетники и относятся к родам Pseudomonas, Bacillus, Azospirillum. Функции их не всегда ясны. Иногда они вырабатывают вещества, увеличивающие устойчивость растений к грибным и бактериальным патогенам. Для сахарного тростника (Saccharum officinarum) показано обитание в его тканях (листья, стебли, корни) бактерии Acetobacter diazotrophi-cus, на 80-100% обеспечивающих растение азотом, поэтому тростник в Бразилии может беспрерывно выращиваться на одних и тех же плантациях около 100 лет без внесения азотных удобрений (Boddey et al., 1991; Stone et al., 2000).
2 • Ассоциативная азотфиксация
Понятие об ассоциативной азотфиксации введено М. М. Умаровым (1984), этот тип азотфиксации происходит в ризосфере и ризоплане, а также филлоплане (на поверхности листьев) растений.
2. Ассоциативная азотфиксация
43
Таблица 2.2 • Интенсивность азотфиксации
в филлосфере растений в оптимальных условиях (по: Умаров, 1986)
Семейство Вид Азотфиксация в филлосфере (мг N2 10-4 дм2ч-1)
Asteraceae Matricaria perforata 2,15
Taraxacum officinale 3,31
Achillea millefolium 7,01
Роасеае Avena sativa 3,34
Phleum pratense 2,73
Festuca rubra 3,04
Brassicaceae Barbarea vulgaris 1,59
Ericaceae Ledum palustre 1,36
Vaccinium uliginosum 1,16
Rosaceae Malus domestica 0,68
Sorbus aucuparia 0,59
Tiliaceae Tilia cordata 0,29
Polygonaceae Rumex acetosella 0,22
Betulaceae Betula pendula 0,15
Sapindaceae Acer platanoides 0,12
Комплексы ризосферных микроорганизмов отличаются у растений разных видов. Они существенно изменяются при выращивании моно- и смешанных культур растений в эксперименте на изначально однородном почвенном фоне (Westover et al., 1997). Особенно велики показатели ассоциативной азотфиксации в ризосфере быстрорастущих С4 тропических злаков, в Бразилии у видов Paspalum отмечена фиксация до 90 кг N/ra в год.
Большинство (~70%) почвенных бактерий потенциально способны к фиксации атмосферного N2, проблема состоит в источнике энергии — доступности органических веществ. Корневые выделения могут быть одним из таких источников. По наблюдениям М. М. Умарова (1984), интенсивность азотфиксации в засеянных полях Подмосковья составляет 30-40 кг/га в год, а на полях без растений (пар) — 10-13 кг/га в год.
Ассоциативная азотфиксация идет также на поверхности листьев (филлосфере). Ее интенсивность различна у видов разных семейств. Она выше у травянистых растений (сложноцветные и злаки) и существенно ниже у многих древесных растений (табл. 2.2). В лесах умеренной зоны — 3-5 кг/га азота фиксируется в кронах, в посевах злаков в США —
44
Глава 2. Растения и азотфиксирующие прокариоты
до 10 кг N/ra в год. Для естественных ценозов эти величины значительны, хотя в условиях сильного загрязнения воздуха в Европе с осадками поступает: в Англии — около 15-20 кг аммония/га в год, максимум в Нидерландах — 40-50 кг/га в год (Pearson, Stewart, 1993). Nostocsp., обитающий на листьях зеленого мха Pleurozium schreberi, фиксирует 1,5-2 кг N/ra в год и служит важным поставщиком азота в бореальных лесах, где этот вид покрывает до 80% площади (DeLuca et al., 2002; van der Heijden et al., 2008), являясь одним из наиболее массовых видов мхов на нашей планете.
В целом на листьях большинства растений бактерий мало, они обнаруживаются только около устьиц, на каждое устьице отмечается обычно 1 -40 бактерий (Гузев и др., 1984). Корни заселены бактериями значительно плотнее. По данным Rovira и Campbell (1974), от 4 до 10% поверхности корней пшеницы покрыто бактериями. В филлосфере основную долю составляют грамотрицательные бактерии, в то время как основные деструкторы отмерших растительных тканей — грамположительные спорообразующие бактерии (Звягинцев и др., 1993).
3 • Несимбиотическая азотфиксация
Азотфиксация в значительных масштабах происходит и в почве, и на ее поверхности вне непосредственной связи с растениями. Ее можно подразделить на фототрофную, осуществляемую свободноживущими цианеями с собственным фотосинтезом, и гетеротрофную, осуществляемую почвенными бактериями с использованием внешних источников органических веществ.
Прокариоты составляют примерно половину геномного разнообразия на Земле, а их разнообразие в почвах примерно на три порядка превосходит таковое во всех других средах, вместе взятых (Curtis et al., 2002; Crawford et al., 2004). Разнообразие почвенных бактерий даже на небольших участках очень велико. С использованием молекулярных методов на экспериментальной ферме Sourhope в Шотландии бактериальное богатство почв оценивается в 0, 5 х 106 видам (Fitter, 2005). Один грамм почвы содержит 1010 —1011 бактерий, принадлежащих к 600-50 000 видов (Curtis et al., 2002; Horner-Devine et al., 2003).
В целом масштабы ассоциативной и несимбиотической азотфиксации оцениваются в 90 млн т азота в год (более 50 % суммарной азотфиксации) (Venkataraman, 1982).
В пустынях огромное значение в балансе азота имеет поверхностная корка, состоящая из цианей, лишайников и мхов. Ее разрушение (при выпасе, рекреации) может существенно снижать поступление фиксируемого азота в почву (Evans, Ehleringer, 1993).
Глава
Микосимбиотрофизм
Эндофиты
Фитопатогенные грибы
Сапротрофные грибы
Растения и грибы
-
к
концу XX в. было описано около 70 тыс. видов грибов, что, возможно, составляет лишь около 5 % от реально существующих в природе (Каратыгин, 1994). По отношению к растениям грибы выступают как: симбиотрофы (микосимбиотрофы, эн-
дофиты), паразиты, сапротрофы, эккрисотрофы (Работнов, 1992). Одним
из ярких примеров эккрисотрофии является развитие на стволах деревьев из рода Nothofagus в Новой Зеландии грибов, придающих стволам черную окраску [Вставка IV]. Многие виды дрожжевых грибов, раз
вивающихся на поверхности листьев растений, также являются эккри-сотрофами. Остальные трофические группы грибов рассмотрены ниже
в соответствующих разделах.
В наземных экосистемах разнообразие почвенных грибов обычно выше, чем фитопаразитных в надземной сфере. Например, в отдельных степных фитоценозах в Казахстане отмечено 1 -4 вида паразитных грибов и 71-82 вида почвенных сапротрофов (Биокомплексные исследования в Казахстане, 1976).
Эволюция наземных растений во многом шла под влиянием грибов. Развитие механической ткани, усиление ксилемы на базе лигноцеллюлозных соединений могло происходить в силуре-девоне как защитная реакция на фитопатогенные грибы. С другой стороны, мицелий в целом развивался как форма освоения тканевых субстратов (Каратыгин, 1993).
I_______________________________________
Вставка IV. Сообщество на коре Nothofagus: медведяная роса щитовки как источник энергии
На стволах антарктических буков (Nothofagus solandri, N. fusca) в Новой Зеландии обитают щитовки (Ultracoelostoma assimile, U. brittini, Homoptera). Они питают-
ся флоэмным соком деревьев, выделяя его богатые гексозами излишки через длинную анальную трубку на поверхность стволов («медведяная роса»). Этими
выделениями питаются многие насекомые и даже птицы, однако значительная часть выделений попадает на поверхность стволов, где ее используют ряд видов грибов, среди которых наиболее часто встречаются представители рода Capnodium (аскомицеты) и дрожжи Candida railenensis и Hanseniaspora osmophila (Serjeant et al., 2008) (рис. IV.l).
Глава 3. Растения и грибы
47
Рис. IV.1 • Сообщество на коре Nothofagus: а — анальная трубочка Ultracoelostoma с каплей «медведяной росы» (фото Garry McDonald, http://www.backpack-newzealand.com, с любезного разрешения автора); б и в — ствол Nothofagus, покрытый налетом эккрисотрофных грибов и анальными трубочками Ultracoelostoma (фото Peter Bray — Airborne Honey Ltd., с любезного разрешения автора)
48
Глава 3. Растения и грибы
1 • Микосимбиотрофизм
Исследователи по-разному определяли понятие «микориза». Allen (1991: 4) дает следующее, наиболее распространенное определение: «Микориза — это мутуалистический симбиоз между растением и грибом, локализованным в корнях или корнеподобных структурах, в котором энергетически богатые вещества перемещаются преимущественно от растения к грибу, а неорганические ресурсы — от гриба к растению». Несколько иное определение приводит Read (2003): «Микориза — структура, в которой симбиотический союз между грибом и адсорбирующим органами растения приводит к увеличению приспособленности одного или обоих партнеров».
Микосимбиотрофизм («питание при помощи микоризных грибов») широко распространен в природе. Например, по данным И. А. Селиванова (1981), из изученных 3449 видов сосудистых растений бывшего СССР образуют микоризу 2697 видов, или 78%. По базе данных Wang и Qiu (2006), включающей 3617 видов из 263 семейств, микоризу образуют 44% мохообразных, 52% папоротникообразных, 100% голосеменных и 85 % цветковых растений.
1.1 • Основные типы микориз
1.1.1 • Везикулярно-арбускулярная микориза (ВАМ)
Растения. ВАМ свойственна большинству изученных сосудистых растений земного шара (рис. 3.1), хотя в настоящее время в природе на присутствие микориз исследовано не более 10000 видов дикорастущих растений.
Факультативные ВАМ (12%)
Другие типы микориз (15%)
Немикоризные (18%)----.
ВАМ совместно с другими типами (5%)
ВАМ (50%)
Рис. 3.1 • Доля сосудистых растений с везикулярно-арбускулярными микоризами во флоре земного шара (по: Trappe, 1987; Brundrett, 2002)
1. Микосимбиотрофизм
49
Грибы. Везикулярно-арбускулярная, или арбускулярная микориза (AM) образована особой ветвью грибов — Glomeromycota (гломалевые, или гломусовые, грибы) (Schussler et al., 2001; Smith et al., 2003). Их таксономия основана преимущественно на морфологии бесполых покоящихся спор, в то время как молекулярные методы выявили большое разнообразие этих грибов. Концепция вида у этой группы грибов весьма неопределенна, большинство авторов признает около 200 видов, главными родами являются Acaulospora, Gigaspora, Glomus и др. Сообщества грибов AM устроены аналогично таковым у других организмов — немного доминирующих видов и много редких. Несколько видов Glomeromycota являются космополитами. Возникновение этой группы грибов датируется 353-452 млн лет назад, а время дивергенции близкородственных видов Glomus оценивается между 10 и 100 млн лет (Rosendahl, 2008). Специальные исследования показали, что генетическое разнообразие этих грибов необычайно велико, даже в пределах одного корня (Rodriguez et al., 2004).
Анатомо-физиологические особенности. ВАМ относится к эндомикоризам. Грибы присутствуют в первичной коре корней, где образуют арбускулы (древовидные разветвления внутри клеток, рис. 3.2) и пузыревидные везикулы. Именно по наличию таких образований рассматриваемый тип микориз и получил свое название. При этом если арбускулы образуются практически всегда, то везикулы наблюдаются значительно реже. Поэтому в современной литературе данный тип микориз часто называют арбускулярными микоризами (AM). Обычно гифа гриба проникает внутрь клеточной стенки и как бы вдавливает клеточную мембрану, увеличивая поверхность соприкосновения с ней за счет многочисленных древовидных разветвлений. Арбускулы внутри клетки живут по разным данным от 2 до 15 дней (Alexander et al., 1989; Bonfante, Perotto, 1995), затем перевариваются клеткой, которая становится восприимчивой к образованию новых арбускул.
Основное функциональное значение ВАМ состоит в обеспечение растений соединениями фосфора, которое происходит в ходе активного обмена, а не только за счет переваривания гиф, в которых одномоментно содержится лишь около 1 % фосфора, получаемого клеткой в процессе обмена. Внешние гифы грибов, отходящие в почву от корня, проникают на расстояние не менее 10-12 см от его поверхности (Li et al., 1991; Jakobsen, 1999). Монофосфаты, поглощаемые гифами грибов, трансформируются затем в полифосфатные гранулы, которые в вакуолях перемещаются по гифам с током цитоплазмы. Затем они переводятся обратно в монофосфаты и передаются через мембраны в клетку растения-хозяина
50
Глава 3. Растения и грибы
Рис. 3.2 • Арбускулы грибов в корнях растений: а — арбускулы во внутреннем слое первичной коры корня Asarum canadense; б — арбускула гриба Glomus sp. при большем увеличении (фото Mark Brundrett, с любезного разрешения автора)
(Clarkson, 1985, рис. 3.3). В некоторых работах показано также значение ВАМ в переносе NH4-ионов из удаленных источников в почве к корням (Johansen et al., 1992). В экспериментах в плошках на относительно сухих почвах ВАМ-грибы обеспечивали 7-49% поглощенного растениями азота почвы из внесенных нитратов и аммония (Hawkins et al., 2000;
1. Микосимбиотрофизм
51
Корень
Монофосфаты
Почвенные частицы
Гифы грибов
Монофосфаты
Рис. 3.3 • Схема транспорта фосфатов от почвенных агрегатов к корням растения гифами грибов ВАМ (по: Clarkson, 1985). Поглощение из почвы и передача клеткам корня осуществляется в форме монофосфатов, а транспорт по мицелию — в форме полифосфатных гранул
Johansen et al., 1994; Leake et al., 2004). В эксперименте c Plantago lanceolate и видами Glomus показано, что грибы транспортируют в растение меченый азот, но не углерод из сухого растительного материала, при этом до трети азота из этого материала транспортировалась грибами в растение (Leigh et al., 2009). В работе Maherali, Klironomos (2007, см. van der Heijden et al., 2008) показаны функциональные различия между семействами грибов, образующих ВАМ: виды Gigasporaceae увеличивает потребление фосфатов растениями, а виды Glomeraceae обеспечивает преимущественно защиту от грибных патогенов.
Мицелий гломалевых грибов плотно пронизывает почву. По наблюдениям в травяных сообществах Северной Америки длина внешних гиф возрастала в течение вегетационного сезона и достигала 111 м/см3 почвы в прерии и 81 м/см3 — на пастбище (Miller et al., 1995). В то же время длина корней составляет 0,5-50 см на куб. см почвы. Длительность жизни гиф — 5-7 дней (Friese, Allen, 1991; Jakobsen, 1999). Длина гиф внешнего мицелия у разных видов зависит от свойств корней растений: Lolium регеппе образует 14 м на 1 г почвы, но только 1 м гиф на 1 м длины корней, в то время как для Trifolium repens эти величины составили 3 м на 1 г почвы и 46 м на 1 м длины корней. Тонкие поглощающие гифы ВАМ-грибов имеют диаметр около 2 микрометров, корневые волоски 10-20 микрометров, тонкие корни 100-500 микрометров.
52
Глава 3. Растения и грибы
Скорость роста гиф ВАМ-грибов 0,3-3,3 мм в день, эктомикоризных грибов — 5-8 мм в день (Leake et al., 2004). Гифы грибов, в отличие от корневых волосков, могут проникать в микропоры почвы диаметром 10-20 микрометров, где только и сохраняется вода при иссушении почвы, поэтому микоризные растения могут быть более устойчивы к засухе (O’Keefe, Sylvia, 1992).
Этот тип микориз характерен для растений из очень широкого спектра растительных сообществ, но особенно широко представлен в травяных фитоценозах с хорошо развитой дерниной (луга, степи, прерии). Исследования van der Heijden et al. (1998) показали, что видовое разнообразие сообществ на карбонатных лугах в Европе и залежах в Северной Америке сильно зависит от разнообразия таксонов грибов, образующих ВАМ. В экспериментальных посевах смеси семян при добавлении различного числа таксонов ВАМ — грибов, индекс разнообразия и общая биомасса растений строго увеличивались с увеличением разнообразия микоризных грибов. Это увеличение шло параллельно увеличению длины гиф грибов в почве и уменьшению содержания в почве фосфатов.
1.1.2 • Эри ко и дна я, арбутоидная и монотропоидная микоризы
Растения. Эти типы микориз названы по растениям-хозяевам, для которых они характерны. Все они свойственны представителям семейства вересковых (Ericaceae), или порядка Ericales в более традиционных системах (рис. 3.4). Эрикоидную микоризу формируют представители родов Erica, Empetrum, Calluna, Vaccinium и другие, всего около 25 родов и 2000 видов кустарников и деревьев. Арбутоидную микоризу формируют растения трех родов (Arbutus, Arctostaphylos, Pyrola) (Smith et al., 2003). Монотропоидная микориза свойственна родам Monotropa, Pterospora и Sarcodes и другим бесхлорофилльным вересковым.
Грибы. Эти типы микориз образованы высшими грибами (аско- и баз-идиомицеты). Так, эрикоидную микоризу формируют дискомицеты из родства Hymenoscyphus(Pezizella)ericae и редко несовершенный гриб Oidioden-dron. Оба гриба могут присутствовать в одном корне (Monreal et al., 1999). Базидиомицеты образуют арбутоидную (например, Cortinarius, Amanita) и монотропоидную (Boletus spp. на видах рода Monotropa) микоризы.
Анатомо-физиологические особенности. Для растений с эрикоид-ной микоризой характерны тонкие, часто волосовидные корни. Грибы образуют рыхлую сеть у поверхности корня и колонизируют эпидермальные клетки. После проникновения в клетку и временного контакта отмирает клетка. В корнях вереска 80 % их объема могут занимать гифы
1. Микосимбиотрофизм
53
Рис. 3.4 • Растения, образующие эрикоидную (а — Rhododendron caucasicum; б — Diapensia lapponica) и арбутоидную (в — Pyrola rotundifolia; г — Arctous alpinus) микоризу
54
Глава 3. Растения и грибы
Рис. 3.5 • Поперечный разрез молодого корня вереска Calluna vulgaris с интенсивной колонизацией эпидермальных клеток микоризным грибом (по: Read, 1996)
грибов (Read, Stribley, 1975) (рис. 3.5). При формировании арбутоидных микориз грибы образуют хорошо развитый чехол вокруг корней, их гифы интенсивно колонизируют клетки первичной коры, формируют по межклетникам сеть Гартига. Сети Гартига (впервые описал Hartig в 1840 г.) — это сети мицелия вокруг клеток первичной коры (рис. 3.6). Отличие мо-нотропоидной микоризы от арбуто-идной состоит в том, что гифы грибов в первом случае не проникают внутрь клеточных стенок корня.
Функциональное значение. Грибы, образующие микоризы с вересковыми, способны выделять широкий спектр ферментов и использовать почвенную органику как источ
ник Р, N и Са, эти элементы могут затем передаваться растениям. Растения, образующие монотропоидную микоризу, практически лишены хлорофилла и являются микогетеротрофами (см. ниже). Растения с эрико-идными и арбутоидными микоризами часто произрастают на кислых,
Рис. 3.6 • Чехольчатая эктомикориза: а — поперечный разрез корня Picea abies — виден наружный чехол из гиф (коричневатый) и сеть Гартига — мицелиальные тяжи по межклетникам первичной коры; б — микоризные оранжевые корни сосны (фото M.Vohnik, Institute of Botany ASCR, Czech Republic, www.mykorhizy.webpark.cz, с любезного разрешения автора)
1. Микосимбиотрофизм
55
бедных элементами минерального питания, но богатых органикой почвах (например, торфах).
1.1.3 • Чехольчатая эктомикориза
Растения: преимущественно доминирующие виды деревьев лесов умеренного пояса, а также тропических и субтропических лесов. Эктомикоризу образуют представители 140 родов из семейств Betulaceae, Dipterocarpaceae, Fagaceae, Leguminosae, Myrtaceae (Eucaliptus), Pinaceae, Salicaceae. Лишь немногие кустарники и травы также могут формировать микоризу этого типа (Cistaceae, Polygonaceae, Rosaceae) (Smith et al., 2003).
Важно отметить, что одни и те же виды растений могут образовывать микоризу разных типов. Например, корни некоторых эвкалиптов формируют эктомикоризу в подстилке, а ВАМ — в нижележащих минеральных слоях почвы (Reddell, Malajczuk, 1984; Ozinga et al., 1997).
Грибы: агарикоидные (шляпочные) и гастероидные базидиомицеты, а также некоторые аскомицеты. Всего известно не менее 6000 видов эктомикоризных грибов, из них примерно 2000 видов представляют род Cortinarius (паутинник). Примерно 40 % всех агарикоидных грибов образуют микоризу. Далеко не все из них продуцируют надземные базидиомы (часть — подземные, часть ресупинатные, а некоторые, как Cenococcum geophilium, могут быть стерильными (Brundrett, 2002).
Как правило, один вид растения-хозяина образует микоризу со многими видами грибов. Так, для Великобритании отмечено 55 видов микоризных грибов для дуба, 103 — для березы и 90 — для сосны (Alexander, Watling, 1987).
Анатомо-физиологические особенности. Для этого типа микориз характерно образование наружного мицелиального чехла вокруг тонких корней, а также формирование сети Гартига (рис. 3.6). В тонких корнях бука 34-45 % их массы приходится на эктомикоризные грибы микоризный чехол может также служить хранилищем ЭМП (Harley, Smith, 1983; Ozinga et al., 1997).
Функциональное значение для растений. Основное положительное воздействие эктомикориз заключается в улучшении поглощения растениями соединения азота и фосфора. Грибы обладают мощным ферментным аппаратом и способны расщеплять различные органические соединения в почве, в слабой степени даже целлюлозу. Эктомикоризные грибы выделяют низкомолекулярные органические кислоты, которые являются активными агентами выветривания минералов. К поглощаемым грибами и передаваемым растениям катионам относятся также Са и Mg. На основании наблюдений и экспериментов в широколиственных лесах Hub
56
Глава 3. Растения и грибы
bard Brook в США показано, что растворение апатита (фосфата кальция) представляет собой ощутимый источник Са, сравнимый с поступлением Са из атмосферы и при выветривании силикатов. Кальций апатита используется преимущественно деревьями с эктомикоризами, поскольку эктомикоризные грибы могут выветривать апатит и поглощать ионы напрямую (минуя почву) (Blum et al., 2002). Эктомикоризы увеличивают поглощение железа из биотита (Watteau, Berthelin, 1994), Р из апатита (Wallander et al., 1997), кальция из кальциевых фосфатов (Lapeyrie et al., 1990) за счет выделения органических кислот (Simard et al., 2003). Эктомикоризные грибы могут также снижать токсичное действие алюминия на корни деревьев в кислых почвах (Landeweert et al., 2001). В работе Hobbie и Hobbie (2006) показано, что до 80% азота растения бореальных лесов получают через эктомикоризные грибы (van der Heijden et al., 2008).
Эктомикоризные грибы увеличивают поглощение растениями элементов минерального питания (ЭМП) за счет увеличения доступа к различным участкам почвы, изменения химических свойств в ризо- и ги-фосфере, изменении бактериальных сообществ в микоризосфере. Van Tichelen et al. (1999) показали, что удельная поглощающая способность корней (поглощение на 1 г корня за единицу времени) была в 10 раз выше для фосфатов и в 5 раз выше для аммония в случае эктомикоризных корней Pinus sylvestris по сравнению с безмикоризными. Эктомикоризы увеличивают всасывающую поверхность корней примерно в 60 раз. Поскольку корни в почве преимущественно ограничены макропорами, гифы эктомикоризных грибов делают доступными для растений микропоры в почвенных агрегатах, а это большой дополнительный источник ЭМП (Simard et al., 2003).
Длина гиф внешнего мицелия эктомикоризных грибов составляет 30-8000 м на 1 м длины корней и 3-600 м на 1 г почвы. Скорость роста гиф эктомикоризных грибов — 5-8 мм в день. Площадь, занимаемая генетами (клонами) эктомикоризных грибов, зависит от возраста леса и интенсивности нарушений и составляет более 50 м2(Anderson et al., 2001; Sawyer et al., 2003).
Эктомикоризы преобладают в бореальных и листопадных лесах умеренного пояса с хорошо развитой подстилкой (опадогенным горизонтом). Микориза развивается главным образом на бедных почвах, особенно лимитированных по азоту. Микоризные грибы сокращают цикл круговорота азота до немногих звеньев: деревья —> опад —> микоризные грибы -> деревья. Это может приводить к монополизации цикла азота эктомико-ризными грибами и доминирующим видом деревьев, что ведет к подавлению развития других растений и снижению флористического разнообразия (Francis, Read, 1994) (рис. 3.7).
1. Микосимбиотрофизм
57
Рис. 3.7 • Монополизация цикла азота эктомикоризными грибами. При разрушении опада грибами, образующими микоризу с деревьями, соединения азота через грибы поступают обратно к деревьям, другие растения практически исключаются из потребления этого азота
Многие эктомикоризные грибы (аско- и базидиомицеты) образуют подземные плодовые тела, распространение их спор при этом целиком зависит от поедающих их млекопитающих (Johnson, 1996). Происхождение подземных плодовых тел связано с условиями, затрудняющими развитие надземных: сухость почвы (например, в австралийских эвкалиптовых лесах) или промерзание верхнего слоя почвы (в субальпийских лесах).
58
Глава 3. Растения и грибы
Подземные базидиомы имеют сильный запах для привлечения млекопитающих, преимущественно мелких. Так, в лесах Австралии почти все сумчатые весом менее 3 кг используют в пищу подземные базидиомы.
Всходы и молодые древесные растения могут инфицироваться эк-томикоризными грибами от соседних взрослых растений. Эксперименты по подсадке сеянцев Quercus rubra к взрослым особям своего рода (Quercus montana), пенькам Q.montana и взрослым особям Acer rubrum показали лучший рост сеянцев в связи с лучшим развитием различных видов эктомикоризных грибов в первом случае (Dickie et al., 2002).
1.1.4 • Микориза орхидных
Растения: семейства орхидные (Orchidaceae).
Грибы: обычно базидиомицеты или их несовершенные формы, часто близкие к роду Rhizoctonia. В целом изучение микориз орхидных показало широкий спектр грибов, образующих этот тип микориз, широкое распространение по земному шару отдельных таксонов и в то же время наличие ряда специфичных связей с отдельными видами орхидей. Кроме базидиомицетов у представителей рода Epipactis выявлено несколько родов симбиотических аскомицетов (Selosse et al., 2004; Bidar-tondo et al., 2004). В ходе онтогенеза орхидные могут менять микоризные грибы, например Gastrodia elata сменяет Mycena на Armillaria (Xu, Mu, 1990). Многие орхидные имеют микоризу с представителями семейства Russulaceae (Dearnaley, 2007). Европейские орхидеи образуют симбиоз с представителями родов (анаморфы) Ceratorhiza, Moniliopsis, Epulorhiza. Соответствующие телеоморфы (половые, или совершенные стадии) относятся к родам Tulasnella, Ceratobasidium, Thanatephorus (Dijk et al., 1997).
В целом для орхидей более обычна специализация к грибам одного рода, реже нескольких родов или одного вида. У ряда орхидных, в том числе бесхлорофилльных, микоризу образуют грибы не из родства Rhizoctonia, а другие базидиомицеты. Так, у Corallorhiza trifida и Cepha-lanthera austinae — грибы из Thelephoraceae (McKendrich et al., 2000), Corallorhiza maculata и C.mertensiana — грибы из Russulaceae (Taylor, Bruns, 1997,1999). Прорастание семян многих зеленых и бесхлорофилльных орхидей наблюдается только в окрестностях материнского растения, где присутствует специфичный вид гриба (Taylor et al., 2003).
Анатомо-физиологические особенности. Для рассматриваемого типа микориз характерен паразитизм растений на грибе на ранних стадиях онтогенеза или в течение всего жизненного цикла. Гриб образует клу
1. Микосимбиотрофизм
59
бочки мицелия в клетке (пелотоны), а затем лизируется клеткой, которая может быть инфицирована несколько раз.
После прорастания семян орхидей, которое наблюдается обычно лишь в присутствии микоризных грибов, образуется подземная стадия (протокорм). В этой стадии растения находятся обычно от 4 до 20 лет, лишь затем развивая надземные побеги. В процессе развития масса протокорма увеличивается в 2000 и более раз за счет органических веществ, поставляемых грибом. Баланс отношений между грибом и орхидеей обусловлен синтезом полифенолов растением и полифенолоксидаз грибами. Концентрация доступных форм азота в почве влияет на баланс отношений гриба и растения. При высоких концентрациях азота усиливается патогенная активность грибов, что ведет к увеличению отмирания растений, особенно на ранних стадиях их развития (Dijk et al., 1997).
Функциональное значение: грибы обеспечивают растения как элементами минерального питания, так и органическим веществом, при этом те же грибы могут выступать в качестве паразитов на других видах растений. В большинстве случаев не удалось показать транспорт органики от орхидей к микоризным грибам (Smith, Read, 1997; Gebauer, Meyer, 2003). Некоторые эпифитные орхидеи во взрослом состоянии становятся немикоризными.
1.2 • Функциональное значение микориз в целом
Если в большинстве случаев (кроме микориз орхидных) микоризные грибы получают от растений органические вещества (продукты фотосинтеза), то положительное влияние грибов на растения весьма разнообразны и могут проявляться в следующих направлениях:
1. Обеспечение растений соединениями фосфора, малоподвижными в почве. Поверхность мицелия значительно больше, чем у корней на единицу биомассы. Так, Bowen (1984) показал, что энергетические затраты на построение 1 см корня эквивалентны таковым, требуемым на построение 1000 см эктомикоризных гиф (см. также: Dighton, 1990).
2. Обеспечение отдельных групп растений другими ЭМП (N, Са, Mg, S) и микроэлементами (Си, Zn, Cl, Ni), в то же время некоторые микоризные грибы могут снижать поступление в корни тяжелых металлов (Zn, Cd) и токсичность алюминия в кислых почвах. Изучение широкого спектра субарктических растений в Швеции на естественное содержание 15 N показало, что растения с микоризами разных типов используют разные источники азота. Содержание 15 N в листьях экто- и эрикоидномикоризных
60
Глава 3. Растения и грибы
растений было существенно ниже, чем у безмикоризных и ВАМ-растений, поскольку первые могут использовать азот из органических соединений в почве (Michelsen et al., 1996). Экспериментально показано, что арбускуля-рные микоризы могут ускорять разложение опада (листья злаков — почти вдвое) и увеличивать поглощение азота из опада. Растения, гифы микоризных грибов которых имели доступ к разлагающемуся опаду, поглощали втрое больше азота, чем контрольные растении (Hodge et al., 2001).
3. Обеспечение водой — повышение устойчивости к засухе.
4. Перекачка органических веществ (см. п. 1.9 гл. 3).
5. Повышение устойчивости растений-хозяев против патогенов и увеличение длительности жизни корней. В ризосфере микоризных корней уменьшается численность бактерий, резко снижается поражаемость корней фитопатогенными грибами. Эктомикоризные сеянцы Pinus Silvestris продуцируют в 2-8 раз больше летучих защитных соединений (терпены, сесквитерпены), чем неинфицированные растения (Krupa, Fries, 1971). В экспериментах с инокуляцией всходов однолетнего злака Vulpia ciliata корневым патогеном Fusarium oxysporum совместно с микоризным грибом Glomus sp. повреждаемость растений была существенно ниже, чем при инокуляции только патогеном Fusarium(oM). Предполагается, что для растений с сильно разветвленной корневой системой защитное значение ВАМ против патогенов существенно важнее, чем перенос Р (Newsham, Fitter, Watkinson, 1995). У тонких корней Pinus palustris микоризная колонизация увеличивала длительность жизни корней более, чем на 45 % (Guo et al., 2008).
Хотя микоризные растения обычно меньше поражаются почвенными патогенами, поражение надземных органов может увеличиваться. Имеющие ВАМ растения ячменя более восприимчивы к листовому фитопатогену Erysiphe graminis. Этот гриб образует на микоризных растениях в три раза больше конидий, чем на безмикоризных. Однако микоризные растения при поражении этим грибом меньше снижали число семян, число колосков и массу семян, чем пораженные безмикоризные растения (Gernns et al., 2001).
6. Увеличение устойчивости растений к стрессовым факторам. Например, Tsang и Maun (1999) показали, что микоризная инфекция увеличивает устойчивость к засолению бобового растения Strophostyles helvola.
7. Отрицательное воздействие на неспециализированных и положительное — на специализированных фитофагов. Эта закономерность была показана на примере представителей семейства Lamiaceae. В Великобритании облигатно микоризные растения этого семейства, имеют более
1. Микосимбиотрофизм
61
Рис. 3.8 • Относительные (%) изменения в росте трех пар фитофагов (в паре на каждом виде растений один — специализированный, другой — неспециалист — «генералист») при питании на растениях, имеющих микоризу по сравнению с растениями того же вида, не имевшими микоризу. На Clinopodium vulgare (а) и Origanum vulgare (б) специализированные фитофаги значимо увеличивали, а неспециализированные уменьшали рост при микоризации растений. На Stachys sylvatica (в) оба вида фитофагов увеличивали рост при микоризации растений (по: Gange et al., 2002)
высокий процент специализированных насекомых-фитофагов, чем слабомикоризные виды растения. Эксперименты с рядом видов показали, что микоризная инфекция увеличивает рост специализированных фитофагов на 30-40 %, при этом рост неспециализированных фитофагов уменьшался у Clinopodium vulgare и Origanum vulgare (рис. 3.8) (Gange et al., 2002).
62
Глава 3. Растения и грибы
8. Формирование почвенных агрегатов. Показано, что различные микоризные грибы могут способствовать образованию почвенных агрегатов в разной степени. Растущий корень может создавать давление 2 МРа и уплотнять почву вокруг себя (Goss, 1991). Образуемый грибами белок гломалин обладает склеивающими свойствами (Rilling, Mummey, 2006).
1.3 • Возникновение микориз
Палеонтологические данные девона и карбона показывают, что уже первые наземные растения (например, Rhynia) имели микоризы. Микоризы современных наземных растений возникали независимо и многократно в разных группах.
Новые палеонтологические находки свидетельствуют, что арбускуляр-ные микоризы появились 460 млн лет назад, предшествуя появлению сосудистых растений. До этого первые находки датировались ранним девоном (400 млн лет) и относились к Aglaophytes major — растению, сочетавшему признаки сосудистых растений и бриофитов. Работа Redecker et al. (2000) показывают находку близкого к Glomus таксона из пород ордовика. Эктомикоризы возникали в эволюции независимо не менее 9 раз! (Wilkinson, 2001).
В настоящее время печеночники и антоцеротовые часто образуют симбиоз с ВАМ-грибами, формирующими арбускулы в их талломе. В ряде случаев эволюция шла от мутуалистического симбиоза к контролю корнями грибного компонента и формированию бесхлорофилльных микогетеро-трофных растений (см. ниже). Плауны и селягинеллы имеют нормальную ВАМ, как и большинство папоротников. Взрослые особи Psilotum также имеют ВАМ. Подземные гаметофиты Ophioglossum и Botrichium являются микогетеротрофными ВАМ-растениями. В семействе Orchidaceae микоге-теротрофия возникала в процессе эволюции независимо около 20 раз. Развитие эктомикориз, эрикоидных и орхидных микориз, как и безмикоризных корней связано с быстрой радиацией покрытосеменных в меловом периоде. Основные ветви эволюции эктомикориз связаны с порядком Fagales и семейством Pinaceae, в остальных группах этот тип микориз возникал независимо. Корни голосеменных растений (Taxodiaceae) из нижнемеловых отложений (Австралия) имели вздутия как у современных эктомико-ризных видов хвойных (Cantrill, Douglas, 1988). Растения с эрикоидными микоризами имеют возраст не менее 80 млн лет. Толстая первичная кора и опробковение экзодермы характерны для микоризных таксонов, в то время как безмикоризные растения чаще имеют тонкие корни с большим числом корневых волосков и относительно хорошей химической защитой. Среди наиболее ранних групп безмикоризных растений — семейства Proteaceae и Restionaceae (100 млн лет назад) (Brundrett, 2002).
1. Микосимбиотрофизм
63
1.4 • Методы изучения в природе и использование в сельском хозяйстве
Микоризы интенсивно исследуются как в лабораторных, так и в полевых условиях. Для оценки степени микоризной инфекции ВАМ обычно изучают встречаемость мицелия в тонких корнях растений. И. А. Селиванов (1981) предложил оценивать интенсивность микоризации корней в баллах в зависимости от доли клеток первичной коры, заполненных мицелием, изучаемых на срезах или тотальных препаратах корней. Эта методика позволяет использовать статистические методы и сравнивать разные растения, их группы или сообщества по интенсивности микоризной инфекции.
Полевые экспериментальные исследования микориз часто направлены на оценку их роли в формировании состава и структуры растительных сообществ. Для подавления развития микориз используется стерилизация почв или применяются фунгициды (чаще всего беномил), которые, к сожалению, оказывают воздействие и на почвенные грибы, не образующие микориз. Такие эксперименты показывают изменение отношений между растениями при подавлении микоризной инфекции. Опыт с внесением фунгицидов, проведенный в высокотравной прерии США, показал, что угнетение микориз изменяет конкурентные отношения между видами (увеличивает участие Carex spp., Aster ericoides и др., уменьшает появление всходов Elymus canadensis, Koeleria cristata) и по-разному воздействует не только на разные виды, но и разные этапы онтогенеза (Hartnett et al., 1994). Пятилетний эксперимент в этой прерии показал резкое снижение биомассы доминирующих облигатно микосимбиотрофных С4 злаков и увеличение роли недоминирующих факультативно микосимбиотрофных СЗ злаков и разнотравья при подавлении микоризной инфекции беномилом (Hartnett, Wilson, 1999). Применение беномила привело также к существенному снижению продукции высокотравной прерии, что, однако, не наблюдалось, если одновременно вносили фосфорные удобрения (Bentivenga, Hetrick, 1991).
В аналогичных экспериментах с применением беномила на пустошах в Великобритании уменьшилось участие микоризных видов (Crepis capil-laris, Erodium cicutarium) и увеличилось участие безмикоризных (Arenaria serpyllifolia, Rumex acetosella) (Newsham et al., 1995). На горных лугах в Северной Греции 2-летнее подавление микориз беномилом уменьшило содержание фосфора в растениях и снизило участие более требовательных к Р видов разнотравья и большинства бобовых. Многие виды разнотравья исчезли, видовое разнообразие на экспериментальных площадках снизилось, при этом возросло участие злаков, осок и бобового Doricnium herbaceum, что свидетельствует о большом значении микориз для формирования состава изученных сообществ (Karanika et al., 2008).
64
Глава 3. Растения и грибы
В экспериментах часто сравнивают развитие растений с и без инокуляции (заражения) микоризными грибами. В этом случае обычно рассчитывают «индекс полезности микориз» по формуле:
(Мм - Мь) х 100%
Мм ’
где Мм — биомасса микоризного растения, — биомасса немикоризного растения.
Для повышения урожайности сельскохозяйственных культур часто используют искусственную инокуляцию культурных растений перед посадкой, которая может обеспечивать значительную прибавку урожая. Lekberg и Koide (2005) провели мета-анализ 290 публикаций, посвященных влиянию различной сельскохозяйственной практики на развитие микориз и урожайность растений. Микоризная колонизация увеличивалась при искусственной инокуляции (на 29 %), оставлении пашни под паром (на 20 %), снижении нарушений почвы (на 7 %). Эти мероприятия также увеличивали биомассу растений и концентрацию фосфора в побегах: инокуляция на 57 и 33 % соответственно, парование — на 55 и 24 %. Увеличение микоризной колонизации было менее вероятным на богатых фосфором почвах.
Отклик растений на микоризную инфекцию во многом зависит от дефицита фосфора в нем. Дефицит Р — разность между требуемым количеством Р (минимальное поступление Р, обеспечивающее максимальный прирост и семенную продукцию) и скоростью его поступления. Уменьшение дефицита Р возможно за счет:
1) увеличения его поглощения (более разветвленная корневая система, сильнее развиты корневые волоски). Например, виды с интенсивным развитием корневых волосков (Lolium регеппе) менее отзывчивы на микоризную инфекцию, чем виды с более грубыми, менее «опушенными» корнями (Allium, Manihot);
2) уменьшая потребности в Р, например за счет низкой скорости роста и т. п. В любом случае уменьшение Р дефицита ведет к уменьшению положительного отклика растений на микоризную инфекцию (Koide, 1991).
1.5 • Зависимость от экологических факторов и распространение микориз в различных природных зонах
Микоризы встречаются в растительных сообществах всех природных зон земного шара, но отдельные типы микориз распространены неравномерно. В тундрах (особенно влажных) относительная роль микоризных видов
1. Микосимбиотрофизм
65
может быть невелика, но здесь среди микоризных растений широкое распространение имеют виды с эрикоидной микоризой, ВАМ- и эктомикоризой. В умеренном поясе в лесной и степной зонах микоризные виды обычно преобладают над безмикоризными. В бореальных и неморальных лесах доминирующие виды деревьев образуют эктомикоризу. На лугах и в степях преобладают растения с ВАМ, как и пустынях и тропических лесах, однако в пустынях велика роль безмикоризных видов. На верховых болотах велика роль вересковых кустарничков, образующих эрикоидную микоризу, большинство же водных растений микориз не образуют.
Микоризы не формируются в наиболее экстремальных условиях существования растений, например в очень холодных областях (в полярных пустынях на равнине и в субнивальных растительных сообществах самых высоких поясов гор). В условиях сильного засоления (шоровые солончаки) микоризные растения также отсутствуют.
Развитие микориз зависит от многих экологических факторов. Лучше представлены микоризы в средних по условиям увлажнения сообществах, их развитие подавляется как в экстремально сухих, так и в переувлажненных или водных местообитаниях. Недостаточная аэрация почв сдерживает рост микоризных грибов. На микоризные грибы также неблагоприятно воздействует очень низкая или высокая кислотность почвы, а также засоление. С другой стороны, микоризы лучше представлены на бедных или средних по богатству элементами минерального питания почвах. Высокое почвенное плодородие, как и внесение удобрений (особенно фосфорных) подавляет развитие микориз.
Жизненность растений, образующих микоризу, обычно выше, чем этих же растений при отсутствии заражения микоризными грибами. В этом состоит принципиальное отличие действия микоризных и фи-топатогенных грибов. С другой стороны, микоризы лучше развиваются у здоровых растений, а некротрофные фитопатогенные грибы легче поражают ослабленные и угнетенные особи растений. Воздействие фитофагов может снижать микоризную инфекцию. Так, в экспериментах с разными видами ив, их защита от фитофагов (гуси и зайцы) повышала эктомико-ризную инфекцию на 10% в слое почвы 0-5 см, на 23% в слое 5-10 см и на 42% в слое 10-15 см (Rossow et al., 1997).
Воздействие растений-полупаразитов также может снижать интенсивность микоризной инфекции у растений-хозяев, что было показано, например, для Lolium регеппе и Rhinanthus minor (Davies, Graves, 1998). В то же время, полупаразит развивается значительно лучше, если растение-хозяин имеет микоризу, по сравнению с развитием на безмикоризном хозяине.
Интенсивность микоризных инфекций растений обычно возрастает в ходе сукцессий. Однолетники начальных стадий вторичных сукцессий
66
Глава 3. Растения и грибы
часто не микосимбиотрофны. Смена типов микориз в ходе первичных сукцессий может быть связана с доступностью пропагул микоризных грибов, что в свою очередь зависит от способов их распространения. На Гавайях показано увеличение частоты ВАМ и ее интенсивности при увеличении возраста вулканического субстрата (на молодой лаве обитают 57 % микоризных видов, на пепле — 84%, на почве — 100%) (Gemma, Koske, 1990). Грибы ВАМ могут также оказывать отрицательное воздействие на немикоризные виды растений, формируя состав сообществ в ходе сукцессий. Такие закономерности были установлены экспериментально в работах Francis и Read (1994). Рядом с потенциально микоризным растением подорожника ланцетолистного (Plantago lanceolate), отгороженным непроницаемой для корней, но проницаемой для мицелия, сеткой, выращивался потенциально микоризный золототысячник (Centaurium егу-thraea) или безмикоризная песчанка (Arenaria serpyllifolia). В отсутствие микоризных грибов песчанка развивалась лучше, чем золототысячник, а в присутствии таких грибов картина менялась на противоположную — лучше развивался золототысячник. Авторы говорят о возможном подавлении развития безмикоризных видов нарушенных местообитаний (начальных стадий сукцессий) гломалевыми грибами за счет выделения токсичных для этих растений метаболитов. Таким образом, гломалевые грибы как бы отбирают виды на ранних стадиях онтогенеза, способные образовывать ВАМ. Присутствие же в сомкнутых сообществах поздних стадий сукцессий безмикоризных видов (например, осок Carex spp.) связано с их вегетативным, а не семенным размножением (Francis, Read, 1994).
1.6 • Немикоризные растения
По систематическому положению среди цветковых растений не образуют микориз большинство представителей семейств осоковые (Сурегасеае), рестиевые (Restionaceae), ситниковые (Juncaceae), крестоцветные (Bras-sicaceae), гвоздичные (Caryophyllaceae), маревые (Chenopodiaceae), гречишные (Polygonaceae), протейные (Proteaceae). Как правило, отсутствие
Рис. 3.9 • Примеры немикоризных растений: а — крапива Urtica d/o/ca (растение богатых почв); б — телорез Stratiotes aloides (водное растение); в — чесночница Alliaria petiolata (Brassicaceae, растение содержит сильные и фунги-статические вещества); г — Protea neriifolia (Proteaceae, растение, формирующее кластерные корни); д — Halocnemum strobilaceaum (Chenopodiaceae, галофит); е — Melampyrum cristatum (полупаразит)
1. Микосимбиотрофизм
67
68
Глава 3. Растения и грибы
микориз более характерно для травянистых растений с укороченным жизненным циклом и корневой системой с обильными корневыми волосками (Baylis, 1972, 1975). Исследование встречаемости немикоризных растений в почвах скальных выходов в Швеции показали, что безмикоризные виды чаще встречались и были более обильны на более кислых почвах с более высоким содержанием обменного фосфора. Для луговых сообществ аналогичная закономерность не наблюдалась (Olsson, Tyler, 2004).
По экологическим свойствам безмикоризные растения относятся к следующим группам (Работнов, 1992, рис. 3.9):
1. Растения богатых почв (например, крапива Urtica dioica).
2. Водные растения (например, частуховые, шейхцериевые, рдестовые) и растения переувлажненных почв. В воде фосфаты находятся в растворе. В то же время водная среда неблагоприятна для развития грибов.
3. Растения, выделяющие сильные фунгистатические вещества, подавляющие развитие грибов (например, горчичные масла у крестоцветных). Живые корни видов Brassica ингибировали прорастание спор видов Glomus, этот эффект снимался (иногда частично) при добавлении лизина, аргинина и глютатиона, которые реагировали с горчичными маслами (изотиоциантами) (Schreiner, Koide, 1993а, b).
4. Растения, использующие специальные механизмы для поглощения фосфатов, такие как формирование кластерных корней [см. Вставку 1], что характерно для люпина (Lupinus albus) и многих представителей протейных.
5. Растения, развивающиеся в экстремальных условиях среды, неблагоприятных для грибов (при сильном засолении, низких температурах и т.п.).
6. Паразитные, полупаразитные и насекомоядные растения, получающие ЭМП от других растений или животных. Однако некоторые паразитные растения (Cuscuta) могут быть микоризными на ранних стадиях развития (до прикрепления к растениям-хозяевам) (Khalid, Iqbal, 1996), а виды полупаразитных мытников (Pedicularis spp.) могут иметь довольно высокую степень микоризной инфекции даже на корнях с гаусториями (Li, Guan, 2008).
1.7 • Распространение микоризных грибов
и развитие микориз
По воздуху распространяются споры аско- и базидиомицетов, формирующих чехольчатые эктомикоризы, эрикоидные микоризы и микоризы орхидных. Животными микофагами распространяются споры аско-, ба-
1. Микосимбиотрофизм
69
идиомицетов и гломалевых грибов, формирующих экто- и некоторые >А-микоризы. Споры гломалевых грибов сохраняют жизнеспособность ри прохождении по пищеварительному тракту грызунов (Mosse et al., 981). Наконец, большинство гломалевых грибов, формирующих ВАМ, |аспространяется с частичками почвы или растениями, что может оканчивать доступность их пропагул на ранних и средних стадиях сукцессии Ггарре, Luoma, 1992).
Существует тенденция среди древесных растений: ВАМ-виды обыч-о имеют съедобные плоды, а эктомикоризные виды — семена. Живот-ые, поедающие плоды, часто захватывают и источники спор эндогоно-ых грибов — растительные остатки, беспозвоночных. Например, дрозды спользуют в пищу плоды многих растений одновременно с дождевыми ервями. А синзоохорные виды деревьев распространяются животными особенно грызунами), часто поедающими плодовые тела эктомикориз-ых грибов и делающими запасы семян, что способствует совместному 'аспространению семян и спор (Pirozynski, Malloch, 1988).
-----------------—-------------------1 При образовании и функциони-Зставка V. «Бактериальные------------ровании микориз важную роль
юмощники микориз» могут играть некоторые группы
бактерий, названные «бактериальными помощниками микориз» (mycorrhiza helper bacteria, МНВ). Различают бактерии, улучшающие формирование микориз (экто и AM), и бактерии, улучшающие функционирование микоризного симбиоза. МНВ разнообразны по систематическому положению (грам-отрицательные протеобак-терии Agrobacterium, Azospirillum, Azotobacter, Burkholderia, Bradyrhizobium, Enterobacter, Pseudomonas, Klebsiella, Rhizobium, грамположительные Firmi-cutes (Bacillus, Brevibacillus, Paenibacillus), грамположительные актиномицеты (Rhodococcus, Streptomyces, Arthrobacter). Некоторые МНВ могут формировать биопленки у гиф микоризных грибов (рис. V.1).
Поглощаемые от растений углеводы микоризные грибы переводят в трегалозу, которая может использоваться бактериями при ее выделении из гиф грибов.
Рис. V.1 • «Бактериальный помощник микориз» Pseudomonas fluorescens образует биопленочную структуру на поверхности эктомикоризного гриба Laccaria bicolor in vitro (Эпифлюоресцентная фотография: по Frey-Klett, 2007)
70 Глава 3. Растения и грибы
В гифах AM грибов обнаруже-
Окончание Вставки V ны также эндобактерии. Пока-
; зано, что МНВ могут стимулиро-вать прорастание спор микоризных грибов, рост мицелия, улучшать почвенные условия (детоксикация тяжелых металлов), стимулировать образование боковых корней растений, способствовать выветриванию почвенных минералов и увеличивать потребление фосфатов микоризными грибами, фиксировать атмосферный азот, усиливать защиту от почвенных патогенов. Paul et al. (2007) показали, что фиксация азота эктомикоризным грибом (за счет МНВ) Suillus tomentosus на Pinus contorta в Британской Колумбии (Канада) составляет около 10% от таковой у актино-ми цетной Alnus rubra.
Выделяемые корнями растений метаболиты стимулируют прорастание спор грибов ВАМ. На чашках Петри показано, что проросшая спора в отсутствии корней растений-хозяев прекращает рост трубки, а цитоплазма из последней перетекает обратно в спору до следующей попытки прорасти у корня растения-хозяина (Parniske, 2005).
Рис. 3.10 • Изменение интенсивности микоризной инфекции в течение вегетационного сезона у трех альпийских растений: Hs — Anemone speciosa, Ad — Antennaria dioca, Ct — Campanula tridentata (по: Байкалова, Онипченко, 1988)
5 июля
30 июля
25 августа
1. Микосимбиотрофизм
71
Заражение микоризными грибами происходит из почвы, с семенами инфекция обычно не передается (может быть, только на покровах). Грибы инфицируют клетки корня выше точки роста, растущая меристема никогда не инфицируется (Harley, 1984). При формировании микориз большую роль играют некоторые группы почвенных бактерий, стимулирующие образование симбиотических связей между корнями и грибами («бактериальные помощники микориз», [см. Вставку V]). Заражение микоризными грибами обычно происходит на самых ранних стадиях развития проростков, при этом интенсивность микоризной инфекции может быть выше у всходов и ювенильных особей, чем у взрослых растений. Например, у альпийского одуванчика (Taraxacum stevenii) средняя интенсивность микоризной инфекции составила у всходов 50 %, у ювенильных особей — 48 %, а у взрослых вегетативных особей — 17 % (Байкалова, Онипченко, 1988). У генеративных особей может наблюдаться снижение интенсивности микоризной инфекции в связи с оттоком ассимилятов в репродуктивные структуры. В течение вегетационного сезона у многолетних растений обычно наблюдается увеличение интенсивности от весны к осени, при этом в период цветения интенсивность микоризной инфекции чаще несколько снижается (рис. 3.10). Осенью часто происходит увеличение числа везикул в корнях.
1.8 • Микоризы и конкурентоспособность растений
Улучшая поступление элементов минерального питания с помощью микоризных грибов, растения затрачивают на поддержание последних значительное количество продуктов фотосинтеза. По разным оценкам эти затраты составляют от 10 до 50% синтезированных органических веществ. Во многих работах показано, что интенсивность микоризной инфекции строго скоррелирована с количеством выделяемых корнями экссудатов (Schwab et al., 1991). Несколько меньше растения тратят на поддержку ВАМ-грибов (10-20% продуктов фотосинтеза), значительно больше (40 %) эта величина достигает у некоторых эктомикоризных видов (Smith et al., 2003). Несмотря на такие «издержки» микоризы чаще повышают конкурентоспособность растений, поэтому микоризы обычно лучше выражены у доминантов растительных сообществ (табл. 3.1).
Наличие или отсутствие микориз в экспериментах изменяет исход конкурентных отношений в экспериментальных посадках. Например, при выращивании безмикоризной солянки Salsola kali и Agropyron repens без инокуляции микоризными грибами Salsola имела конкурентное преимущество, а при инокуляции — наоборот, большую биомассу образовывал Agropyron (Allen, Allen, 1984). При совместном выращивании Lolium
72
Глава 3. Растения и грибы
Таблица 3.1 • Относительная (с учетом безмикоризных видов растений) интенсивность микоризной инфекции (%) для доминирующих и всех видов растений фитоценозов Зауральской лесостепи (по: Селиванов, 1981)
Сообщество Все растения Доминанты
Березняк 52,4 81,6
Разнотравно-ковыльная степь 52,5 62,8
Типчаково-ковыльная степь 54,1 64,4
Ковыльная степь 60,0 76,0
Полынно-типчаковая степь 74,8 86,6
регеппе и Holcus lanatus без микоризных грибов эти злаки развивались примерно одинаково, а при инокуляции грибами ВАМ преимущество получал Holcus, a Lolium угнетался. Поэтому, при применении фунгицидов может существенно изменяться структура фитоценозов (см. 1.4), так как микоризные грибы обычно более чувствительны к фунгицидам, чем паразитные. В опытах Gange и др. (1990) по применению фунгицида ипродиона на ранних стадиях сукцессий показано снижение общего проективного покрытия и флористической насыщенности сосудистых растений по сравнению с контролем (опыт 11,6±0,8, контроль 14,2±0,9 видов сосудистых растений на площадку 2,5 х 2,5 м).
Улучшение фосфорного режима растений за счет микоризных грибов способствует увеличению содержания органических соединений фосфора в семенах, что может увеличить вероятность приживания всходов. Так, семена, полученные от микоризных Abutilon theophrasti и Avena fatua, содержали больше соединений Р, чем семена от безмикоризных растений (Koide, 1991). Анализ дальнейшей судьбы проростков, образовавшихся из семян микоризных и безмикоризных растений канатника (Abutilon), показал, что проростки микоризных родителей были существенно крупнее, а их выживаемость вдвое выше, чем у проростков безмикоризных родителей (Heppell et al., 1998).
1.9 • Микоризные сети в почве
Пионерные работы, показывающие важную связь обмена веществами между растениями через микоризные грибы с флористическим разнообразием сообществ, были выполнены Grime et al. (1987). Ими был произведен посев смеси семян луговых растений в плошки с почвой без и с грибами, образующими ВАМ. Через несколько месяцев выращива
1. Микосимбиотрофизм
73
ния было показано увеличение разнообразия, числа и размеров всходов в варианте с микоризными грибами. Показан также перенос меченых 13 С органических веществ от взрослых особей доминирующего вида — овсяницы овечьей (Festuca ovina) к молодым особям других видов, но только образующим микоризу. Таким образом возникло представление о единой микоризной сети в почве, образованной гломалевыми грибами, мицелий которых, кроме того, образует анастомозы (срастания) чаще, чем мицелий зигомицетов. Перераспределение продуктов фотосинтеза между особями одного или разных видов может происходить через единую микоризную сеть, существующую в почве большинства природных фитоценозов и являющуюся важным фактором поддержания их флористического богатства.
В экспериментах по изучению внутривидовой и межвидовой конкуренции между взрослыми особями и всходами Черноголовки (Prunella vulgaris), а также взрослыми особями земляники (Fragaria vesca) было показано (Moora, Zobel, 1996), что ВАМ усиливает внутривидовую конкуренцию и ослабляет межвидовую, таким способом способствуя поддержанию флористического разнообразия сообществ.
Современные исследования показали перенос органических соединений между растениями через эктомикоризные и ВАМ-системы. Спорным и дискуссионным остается значение этого переноса для растительных сообществ и динамики экосистем. Simard et al. (1997а, b) показали двунаправленный транспорт органических соединений между сеянцами Pseudotsuga menziesii и Betula papgrifera. Псевдотсуга получала больше органики от березы в условиях затенения, чем при освещении. В экспериментах Graves et al. (1997) показан существенный транспорт меченых продуктов от Festuca ovina к другим растениям, но большая часть перемещенных продуктов оставалась в подземной части акцепторных растений. Значительный поток азота между удобренными и неудобренными растениями также показан для видов с арбускулярной микоризой (Eissenstat, 1990). Показан также перенос аммония и нитратов от азотфиксирующих растений (бобовых, актиноризных растений) к другим растениям, хотя возможен обратный перенос (от ячменя к гороху, от эвкалипта к ка-зуарине) (Dickie et al., 2002; Selosse et al., 2006). Существенный поток органических соединений от отмирающих и поврежденных растений отмечен через микоризу (Giller et al., 1991; Simard et al., 2003).
Watkins et al. (1996) показали однонаправленный транспорт органики от СЗ растения к С4 злаку (около 10% ассимилятов донорного растения перемещалось к реципиенту). Lerat et al. (2002) выявили существенный транспорт органики от травянистого растения Ergthronium americanum, рано развертывающего листья весной, к сахарному клену
74
Глава 3. Растения и грибы
Весна Лето
Рис. 3.11 • Сезонная смена направления транспорта органических веществ через микоризные сети между травянистым эфемероидом Erythronium атепсапит и сахарным кленом (Acer saccharum) (по: Lerat et al., 2002). Весной поток направлен преимущественно от эфемероида к распускающему листья клену, а во второй половине лета — от клена к переходящему в покой эфемероиду. Acer saccharum — фото Karren Weisel, TreeTopics.com, с любезного разрешения автора, Erythronium атепсапит — фото David G. Smith, delawarewildflowers.org, с любезного разрешения автора
(Acer saccharum) в весеннее время и обратный транспорт — в осеннее. Транспорт органики через эктомикоризные грибы осуществляется в форме трегалозы, маннитола и арабитола (Soderstrom et al., 1988; Simard, Durall, 2004) (рис. 3.11).
Микоризные грибы существенно увеличивают конкурентоспособность заносного сорняка Centaurea maculosa в естественных прериях
1. Микосимбиотрофизм
75
на северо-западе США. В работе Carey et al. (2004) приводятся экспериментальные доказательства, что это может быть связано с переносом продуктов фотосинтеза от доминанта прерий Festuca idahoensis к васильку через грибы ВАМ. В эксперименте биомасса Centaurea maculosa была существенно выше в варианте совместного произрастания с Festuca idahoensis при внесении микоризных грибов. В этом варианте и изотопный состав С василька был близок к таковому для Festuca idahoensis. Авторы оценивают, что транспортируемая от овсяницы органика составляет до 15 % надземной биомассы василька.
Передача углеводов между растениями может происходить по градиенту концентраций от лучше обеспеченных взрослых растений к молодым особям (Read et al., 1985; Finlay, Read, 1986; Grime et al., 1987). Микоризная сеть может также улучшать приживание всходов, например молодые дубы Quercus rubra лучше развивались вблизи других дубов, поддерживающих эктомикоризную сеть, чем вблизи Acer rubrum (Dickie et al., 2002). Транспорт органических веществ к молодым растениям, несомненно, имеет адаптивное значение и должен быть особенно важен для растений, размножающихся преимущественно семенным путем, так как в случае вегетативного размножения такой транспорт может осуществляться через другие органы (столоны, корневища и т. п.). Поэтому важность микоризной связи между взрослыми особями и их всходами должна быть наибольшей для растений, размножающихся семенным, а не вегетативным путем. Проведенные нами (Онипченко, 1996) наблюдения подтвердили эту закономерность для альпийских растений северо-западного Кавказа. Как в целом для всех исследованных видов, так и для видов в пределах отдельных растительных сообществ показана меньшая степень развития микориз у вегетативно подвижных видов (горизонтальный годичный прирост более 2 см в год) по сравнению с вегетативно мало- и неподвижными (рис. 3.12).
Механические нарушения почвы (порой, вспашка и пр.) ведут к разрушению микоризной сети, поэтому интенсивность инфекции обычно уменьшается на нарушаемых почвах. В экспериментах Jasper et al. (1989) было показано, что просто обрыв мицелия от микоризных растений не приводил к существенному уменьшению инфекции у тестовых особей клевера подземного (Trifolium subterraneum), но если почва до посадки тестового растения перемешивалась, микоризная инфекция резко снижалась (с 40-50 % в контроле до 1 % в этом эксперименте).
Почвенная микоризная сеть сильно зависит от деятельности почвенных беспозвоночных, активно потребляющих мицелий гломалевых грибов. К ним относятся амебы, нематоды, клещи и коллемболы. В экспериментах с применением селективных инсектицидов или нематодоцитов,
76
Глава 3. Растения и грибы
Рис. 3.12 » Сравнение интенсивности микоризной инфекции (%, ордината, среднее и ошибка) вегетативно-подвижных растений (разрастание более 2 см в год, белые колонки) и неподвижных/малоподвижных растений (серые колонки). АЛП — альпийские лишайниковые пустоши, ПЛ — пестроовсяницевый луг, ГКЛ — гераниево-копеечниковый луг, АК — альпийский ковер (по: Onipchenko, Zobel, 2000)
подавляющих соответствующие группы животных, показано усиление микоризной инфекции у растений (Smith, Read, 1997). В то же время в нескольких исследованиях показано, что микофаги предпочитают потреблять сапротрофные и фитопатогенные грибы микоризным (Hiol et al., 1994; Johnson, Gehring, 2007).
Воздействие грибов, образующих более специализированные к растениям-хозяевам чехольчатые эктомикоризы, может существенно отличаться от действия гломалевых грибов. Эктомикоризы чаще образуют доминанты древесного яруса лесов умеренного и тропического регионов, что усиливает их конкурентные позиции по отношению к недоминирующим видам. Этим объясняется, например, то, что в тропических лесах с очень высоким флористическим разнообразием преобладают ВАМ-виды растений, а в тропических лесах, где доминанты образуют эктомикоризу, флористическое разнообразие существенно снижено (Ozinga et al., 1997).
1.10 • Микогетеротрофные растения
Транспорт органических веществ от грибов к высшим растениям обеспечивает существование на ранних стадиях онтогенеза растений семейства орхидных, самого крупного среди цветковых растений. В то же время, многие
1. Микосимбиотрофизм
77
Рис. 3.13 • Примеры основных групп микогетеротрофных растений
(по: Leake, 1994): а — Monotropaceae — Monotropa uniflora; б — Gentianaceae — Voyria chionea; в — Petrosaviaceae — Petrosavia sp.; г — Triuridaceae — Sciaphila albescens; d — Peltophyllum luteum, мужское и женское растение; е — Triuris alata, мужское и женское растение;
ж — Burmanniaceae — Campylosiphon purpurascens; з — Burmannia tenella; и — Gymnosiphon brachycephalus; к — Trismia saulensis; л — Corsiaceae — Corsia ornate; м — Arachnites uni flora; н — Orchidaceae — Neottia nidus-avis
78
Глава 3. Растения и грибы
Рис. 3.14 • Микогетеротрофные растения лесов Европейской части России: а — Monotropa hypopitis; б — Corallorhiza trifida; в — Neottia nidus-avis; г — Epipogium aphyllum. Фото автора
1. Микосимбиотрофизм
79
зеленые лесные орхидеи используют органику, полученную от грибов. Ее доля в биомассе растений составляет, например, 7-26% у Cephalanthera rubra (100% у бесхлорофилльных мутантов этого вида), 14% у Epipactis helleborine, 27% у Listera ovata (Selosse et al., 2006). Такой смешанный тип получения органических соединений получил название «миксотро-фии». К миксотрофам, помимо орхидей, относятся многие грушанковые тенистых лесов. Для изученных видов (Orthilia secunda, Pyrola chloran-tha, Pyrola rotundifolia, Chimaphila umbellata) доля органики, полученной от грибов, составляла от 10 до 67,5 % углеродного баланса. Эта величина увеличивалась в более тенистых местообитаниях (Tedersoo et al., 2007).
От этого же транспорта полностью зависит существование целой группы цветковых растений, получившей название «микогетеротрофные растения» (Leake, 1994). Эти растения лишены зеленой окраски, но в отличие от паразитных растений, не имеют прямой связи с растениями-хозяевами. Раньше их рассматривали как «сапротрофов», питающихся органическими веществами почвы.
Всего известно около 400 видов из 87 родов, большинство из них (88 %) — однодольные растения (рис. 3.13). Основные семейства двудольных: Monotropaceae, Pyrolaceae, Polygalaceae, Gentianaceae, однодольных: Petrosaviaceae, Triuridaceae, Lacandoniaceae, Geosiridaceae, Bur-manniaceae, Corsiaceae, Orchidaceae. Среди мхов к этой группе растений может быть отнесена Buxbaumia aphylla. Безхлорофилльные гаметофит-ные стадии характерны для некоторых папоротников (заростки Botrychi-ит) и плаунов, так же как для спорофитов и гаметофитов печеночника Cryptothallus mirabilis (Read et al., 2000; Taylor et al., 2003).
Большая часть микогетеротрофных растений встречается в тенистых и влажных тропических лесах, но отдельные представители (Monotropa, Neottia, Corallorhiza, Epipogium) обитают в умеренной зоне (рис. 3.14). Все имеют очень мелкие, пылевидные семена и являются облигатными микосимбиотрофами. При этом типы микориз могут быть самыми разными: арбутоидная (у Pyrolaceae), монотропоидная (Monotropa), везикуляр-но-арбускулярная — Polygalaceae, микориза орхидных. Одни и те же виды грибов могут образовывать с разными растениями микоризы разного типа, а иногда и выступать как паразиты на других растениях (табл. 3.2).
В настоящее время показано (Simard, Durall, 2004), что микогетеротрофные растения получают органическое и минеральное питание от микоризных грибов, которые, в свою очередь, во многих (если не во всех?) случаях получают их от зеленых растений, паразитируя на них или образуя с ними микоризу. Например, McKendrick et al. (2000) показали, что питание микогетеротрофной орхидеи Corallorhiza trifida поддерживается прямым транспортом органики от близ растущих автотрофных деревьев
80
Глава 3. Растения и грибы
Таблица 3.2 • Грибы, образующие разные типы микориз и/или паразитирующие на растениях (по: Harley, 1984)
Гриб Микоризные связи Микоризные или патогенные связи
Boletus spp. Эктомикоризы сосны Монотропоидная микориза Monotropa hypopitys
Rhizoctonia solani (Thanatephorus cucumeris) Микориза орхидных (Orchis purpurella) Деструктивный паразит на многих травах
Rhizoctonia goodyerae-repentis (Ceratobasidium cornigerum) Микориза орхидных (Goodyera) Деструктивный паразит на многих травах
Armillaria mellea Микориза микогетеротрофных орхидных, монотропоидная микориза Monotropa uniflora Деструктивный паразит древесных и травянистых растений
(Betuia pendula, Salix repens) через общий микоризный гриб. Поэтому микогетеротрофные растения могут рассматриваться как эпипаразиты зеленых растений.
Микогетеротрофные эпипаразиты влияют на хозяев опосредовано, поэтому у хозяев отбор не может быть направлен против эпипаразита, не затрагивая грибной мутуализм. Микогетеротрофный симбиоз возникал в эволюции растений неоднократно: по крайней мере один раз у печеночников, один раз у папоротников, один раз у голосеменных, вероятно десять раз у однодольных и 4 раза у двудольных. В настоящее время молекулярными методами показана очень высокая специализация микогетеротрофных растений к грибным симбионтам, особенно в сравнении с микоризообразующими грибами зеленых растений (Monotropa hypopithis с родом Tricholoma). Несмотря на меньшее развитие вегетативной сферы, микогетеротрофные растения могут достигать больших размеров: высота Pterospora и Allotropa достигает 2 м, а масса Sarcodes — нескольких килограмм. Транспорт органики от грибов к растениям может осуществляться двумя путями: 1) разрушением и повторной колонизацией внутриклеточных гиф — у орхидных, 2) через временно нарушенные мембраны в контакте гриб-растение, например в форме грибного дисахарида трегалозы (Muller, Dulieu, 1998). Микогетеротрофные растения часто являются индикаторами ненарушенных старовозрастных лесов (Bidartondo, 2005). Среди этих растений многие виды являются редкими и требуют специальной охраны.
2. Эндофиты
81
2 • Эндофиты
2. 1 • Распространение и систематическое положение
Термином «эндофиты» называют организмы, живущие внутри растения. В последние десятилетия этот термин приобрел более узкий смысл, под эндофитами чаще понимают грибы, обитающие внутри растения (чаще в надземных органах) и не вызывающие у него явных патологических изменений, т. е. способные бессимптомно поселяться в видимо здоровых тканях растений. Предполагается, что эндофитизм может быть также широко распространен в природе, как и микосимбиотрофизм.
Эндофиты обычны в тканях злаков и многих двудольных растений, особенно древесных. Практически во всех случаях, когда отдельные виды деревьев изучали на присутствие эндофитов, обнаруживали несколько видов последних: у Sequoia sempervirens — 26 (Espinosa-Garcia, Langen-heim, 1990), пальмы Licuala ramsayi — И видов (Rodrigues, Samuels, 1990). Большое число эндофитных грибов отмечено в коре деревьев, они колонизируют молодую кору, а спороносят на отмерших старых ветвях. Виды отдельной группы грибов из родства Hypoxylon заселяют ксилему, их активная колонизация связана с периодами ослабленного тока при засухе. В корнях ольхи (Alnus glutinosa, A. incana) поселяются также Penicillium nodositatum, формируя клубеньки, похожие на актиноризные. Другие виды Penicillium отмечены в корнях ряда видов деревьев в Орегоне и на Sorbus в Германии. В листьях Dryas octopetala в Швейцарии найдено 23 вида эндофитов (Fischer et al., 1995), в стеблях Salicornia perennis — 31 вид (Petrini, Fischer, 1986). Из талломов лишайников также изолировано большое число эндофитных грибов (Petrini et al., 1990). Грибы обитают в тканях мохообразных и папоротников (Stone et al., 2000).
При изучении разнообразия грибов-эндофитов в листьях растений от тундр до тропиков в Северной Америке было выявлено, что эндофиты в высоких широтах характеризуются небольшим числом видов разных классов аскомицетов, в то время как эндофиты в тропиках представлены очень большим числом видов из небольшого числа классов аскомицетов. В тропиках очень много редких видов эндофитов. Практически все исследованные виды растений содержат эндофиты в своих тканях (Arnold, Lutzoni, 2007).
Интересно отметить, что видовой состав эндофитов растений-хозяев и их полупаразитов может существенно различаться. Так, из изолированных из пихты (Abies alba) и растущей на ней омелы (Viscum album) 70 видов грибов лишь 10 видов было общими (Petrini, 1996). У Sequoia sempervirens разнообразие эндофитных грибов в хвое зависело от ее возраста (хвоя живет до 12 лет). Оно возрастало с 1 года до 4-7-летнего
82
Глава 3. Растения и грибы
Рис. 3.15 • Epichloe sp. (половые спороношения) — умеренный паразит на злаках холодного климата
возраста, а затем уменьшалось в возрастном классе 8-12 лет (Espinosa-Garcia, Langenheim, 1990).
Лучше исследованы эндофиты злаков и древесных растений. В качестве эндофитов выступают обычно аскомицеты (локуласкомицеты, дискомицеты, пиреномицеты) и их несовершенные формы. Более подробно изучены эндофиты злаков. Чаще всего это спорыньевые грибы (Clavicipitaceae), как эндофиты известны представители не менее 5 родов и 30 видов на нескольких сотнях видов злаков из 80 родов.
Среди представителей семейства Clavicipitaceae встречаются облигатные паразиты насекомых, паразиты злаков и осок, паразит двудольных, эндофиты. Epichloe (рис. 3.15) и Parepichloe представляют две линии эволюции к хозяевам (первый к злакам холодного сезона, а второй — злакам теплого сезона). Переход к эндофитизму имеет следующие положительные стороны для гриба: 1) большая доступность веществ хозяина, 2) защита от высыхания, 3) защита от поверхностно обитающих насекомых — микофагов, 4) защита от паразитических грибов и конкуренции с другими микроорганизмами. Конидиальные спороношения (Neotyphodium) распространяются мухами (Phorbia), питающимися мицелием, но не переваривающими конидиеспоры (White et al., 2000).
Clay (1998) выделяет три типа связей злаков с грибами (табл. 3.3): 1) паразитический — все соцветия злаков поражены грибом и не дают семян;
2) умеренный паразитизм — часть соцветий поражено грибом, а часть дает нормальные семена;
3) мутуалистический — все соцветия дают семена, симптомов угнетения нет.
Третий тип распространен только среди злаков, произрастающих в условиях наличия холодных сезонов. Эндофиты становятся более патогенными в стрессовых для растений условиях, они более обычны в много-
2. Эндофиты
83
Таблица 3.3 • Эволюционный ряд перехода грибов от паразитизма к эндофитизму (+ означает развитие признака, — его подавление) (по: Clay, 1998)
Признак Паразиты Умеренные паразиты Эндофиты
Половое размножение гриба + + -
Половое размножение злака - ± +
Жизненность злака - + +
летних дерновинных злаках (80%), реже у однолетников (16%), очень редко у корневищных злаков (3 %).
Редко ветвящиеся гифы эндофитов растут по межклетникам параллельно длинной стороне клеток стеблей и листьев злаков, где они, вероятно, получают сахара и аминокислоты, выделяемые в апопласт. В отличие от других растений, часто инфицируемых многими эндофитными грибами одновременно, в отдельных растениях злаков обычно присутствует один штамм эндофитного гриба. Попытки заражения злаков несколькими грибами одновременно приводили к выпадению всех штаммов, кроме одного. Epichloe festucae является умеренным паразитом, чередуя половой и бесполый циклы, которые сменяются по сезонам в зависимости от размеров растения и условий среды. Поскольку выживание настоящих системных эндофитов зависит от семенной продуктивности хозяев, генотипы грибов, снижающие образование семян, элиминируются отбором. Умеренные паразиты и системные эндофиты представлены только среди СЗ злаков подсемейства Pooideae. Встречаемость эндофитов в Lolium регеппе в европейских популяция составила 62 %, хотя встречаемость внутри популяций обычно была ниже 50 %. Уровень инфекции у злаков возрастал с увеличением интенсивности выпаса и возрастом искусственных пастбищ (Clay, Schardl, 2002).
По механизмам заражения растений выделяют 2 основных типа эндофитов:
1. Структурные (системные) — инфекция передается с семенами, гифы проникают в ткани зародыша. Например, виды Acremonium могут передаваться с семенами, но гибнут при их длительном хранении, хотя семена еще сохраняют всхожесть (Vicari, Bazely, 1993). В поверхностно стерилизованных семянках подсолнечника обнаружено 98 видов различных грибов (Roberts et al., 1986; Hirsch, Braun, 1992).
2. Индуктивные — с семенами не передается, заражение происходит из среды каждый раз заново.
84
Глава 3. Растения и грибы
На примере эндофитных грибов подтверждается правило Law—Lewis (1983), которое говорит об общей закономерности подавления полового размножения и снижении систематического разнообразия в условиях благоприятной биотической среды (у эндосимбионтов) [Вставка VI].
Вставка VI. Правило Лоу-Левиса (Law, Lewis, 1983)
Сравнивая таксономическое разнообразие симбиотических организмов Law, Lewis (1983) выявили, что разнообразие эндосим-
бионтов обычно значительно ниже, чем более крупных организмов, в которых они обитают. Они предположили, что обитание внутри других организмов (более постоянной биотической среде) ведет к редукции полового процесса и снижает скорость видообразования. Эта закономерность в дальнейшем получила название «правила Лоу—Левиса». Примером могут служить симбиотические связи растений с прокариотами и грибами (см. табл. VI.1). Среди грибных компонентов микориз наибольшее разнообразие наблюдается у эк-томикоризных грибов, большая часть мицелия которых располагается в почве, а не в тканях растений.
Taunt Число видов и родов и распространение полового процесса
среди эндо- и экзобионтов в мутуалистических симбиозах
(по: Law, Lewis, 1983, с изменениями)
Симбиоз Половой процесс у: Эндобионт Экзобионт
эндобионтов родственных свободно-живущих видов число родов число видов число родов число видов
Rhizobium — растения Имеется, но ограниченно Имеется, но рас-пространенность неизвестна 7 более 50 600 17 500
Frankia — растения отсутствует 1 ? 17 более 200
Эрикоидные микоризы отсутствует Имеется, обильно 2 2 25 2000
ВАМ отсутствует Имеется, обильно 8-10 200 11000 225000
Эктомикориза имеется, обильно Имеется, обильно 200 6000 140 5000
2.2 • Функциональное значение для растений
Если для грибов польза симбиоза с растениями связана с получением от последнего органических веществ, то основное положительное влия
2. Эндофиты
85
ние эндофитов на растения, видимо, заключается в синтезе защитных веществ от фитофагов, как позвоночных, так и беспозвоночных.
Примеры: гриб Rlabdocline parkei (несовершенная стадия Meria parkei) на дуглассии Pseudotsuga menziesii инфицирует ее хвою в первый год ее жизни (хвоя живет в среднем 9 лет). На хвое этого вида часто поселяются галлообразователи — галлицы рода Contarinia, вызывающие ее отмирание. Эндофит инфицирует галлы, что резко снижает выживаемость насекомых (Carroll, 1986). Отмирание личинок Besbicus mirabilis, образующих галлы на дубах Quercus garryana, вызывается эндофитом Discula quercina, обычном в листьях этого вида. Но грибы сами не поражали личинок, а делали непригодной ткань галла для питания насекомого (Wilson, 1995).
В субарктическом горном лугу в Канаде было показано, что частота встречаемости эндофита Neotyphodium и плотность его гиф в листьях Festuca altaica существенно выше на участках близ колоний пищухи и сурков. Пищухи предпочитали потреблять свободные от эндофитов побеги овсяницы вблизи колоний (Koh, Hik, 2007).
Эндофиты злаков выделяют токсичные для травоядных метаболиты (эргот-алкалоиды), которые вызывают у скота паралич и другие нарушения нервной системы (в медицине спорынья использовалась как абортивное средство). Разные алкалоиды обладают различным действием на фитофагов. Эрготические алкалоиды действуют преимущественно на позвоночных, лолитремы также действуют на млекопитающих, перамин — на насекомых-фитофагов. Например, эндофит Lolium регеппе — Neotyphodium lolii подавляет развитие стеблевого вредителя Listronotus bonariensis на лугах в Новой Зеландии (Bultman et al., 2003). В экспериментах (Knoch et al., 1993) инфицированные и неинфицированные эндофитным грибом Neotyphodium (Acremoniurri) coenophialum семена Festuca arundinacea были предложены пустынным семеноядным муравьям в Аризоне (США). Муравьи избирательно предпочитали неинфицированные семена. В случае же сбора инфицированных семян, последние чаще выбрасывались неповрежденными в специальные «мусорные камеры», где условия для прорастания были лучше, чем на поверхности почвы.
Молекулярные данные показывают, что многие системные эндофиты злаков являются межвидовыми гибридами. Эрготический алкалоид эрговалин, лолитремы и лолины продуцируются только одним видом с половым процессом — Epichloe festucae, но очень обычны среди системных эндофитов. Формулы основных типов алкалоидов эндофитных злаков приведены на рис. 3.16. Все классы алкалоидов синтезируются грибами в чистых культурах (Clay, Schardl, 2002).
86
Глава 3. Растения и грибы
Рис. 3.16 • Примеры алкалоидов, вырабатываемых эндофитами (по: Clay, Schardl, 2002)
В то же время не во всех случаях эндофиты воздействуют отрицательно на фитофагов. В работе Gange (1996) было показано, что присутствие грибного эндофита Rhytisma асеппит в листьях Acer pseudo-platanus увеличивало численность тлей на этих листьях. Кроме того, алкалоиды эндофитных грибов могут оказывать отрицательное влияние на развитие паразитоидов насекомых-фитофагов, что было, например, показано для паразитоидов Euplectrus, обитающих на гусеницах бабочки Spodoptera fruigiperda, питавшихся побегами Festuca arundinacea, инфицированными и неинфицированными грибом Acremonium coenophialum (Bultman et al., 1997).
Другое важное свойство эндофитов — это их антогонизм к патогенным грибам и бактериям, с которым связано уменьшение поражаемости растений-хозяев.
В то же время взаимодействие эндофитов с микоризными грибами может быть отрицательным. Baker (1987) описывает отношение стеб-
2. Эндофиты
87
Таблица 3-4 • Показатели роста и выживаемости (среднее и ошибка) растений Festuca arundinacea, инфицированных и неинфицированных эндофитом Neotyphodium (Acremonium) coenophialum (по: Clay, 1990)
Показатель Инфицированные Неинфицированные
Выживаемость рамет на третий год, % 84 56
Число вегетативных побегов 15,0 ± 1,0 9,9 ± 1,5
Число соцветий 2,6 ± 0,3 1,8 ±0,3
Биомасса растений, г 5,0 ± 0,4 3,0 ± 0,5
левого долгоносика Listronotus bonariensis, листового эндофита Neotyphodium loliae, гриба ВАМ Glomus fasciculatum и многолетнего плевела Lolium регеппе. Инфекция эндофита резко снижает поражаемость плевела долгоносиком, но в присутствии микоризного гриба этот эффект подавляется. В отсутствии эндофита не наблюдалась связь между микоризной инфекцией и фитофагией (Gehring, Whitham, 1994).
Кроме защитных, эндофитные грибы, могут выполнять и другие функции, связанные с изменением обмена веществ растений хозяев. Эндофиты могут увеличивать устойчивость растений к засухе, жаре, низкой освещенности и низкой концентрации ЭМП в почве, увеличивать интенсивность фотосинтеза и роста. В отличие от других симбиозов, связанных с улучшением поглощения минеральных ресурсов, эндофитные ассоциации основаны на защите хозяина от биотического и абиотического стресса. Например, инфицированные эндофитами и неинфицирован-ные растения Роа alsodes развивались сходным образом при хорошем обеспечении водой, но при недостатке воды биомасса инфицированных растений была на 17 % выше (Kannadan, Rudgers, 2008).
В экспериментах Marks и Clay (1996) с Festuca arundinacea и грибом Neotyphodium coenophialum изучали фотосинтез инфицированных и неинфицированных растений при разных температурах. При низких температурах показатели фотосинтеза не отличались, а при высоких (+35 °C) инфицированные растения фотосинтезировали на 20-25 % интенсивнее, чем неинфицированные. С другой стороны, Е. Ю. Благовещенская (личное сообщение) выявила лучшую выживаемость инфицированных эндофитами злаков в условиях низких температур. Clay (1990) показал лучшую выживаемость и жизненность растений Festuca arundinacea инфицированных эндофитом по сравнению с неинфицированными (табл. 3.4).
88
Глава 3. Растения и грибы
3 • Фитопатогенные грибы
Более 20 000 видов грибов — паразиты растений, по крайней мере, на некоторых стадиях жизненного цикла. Известных грибных фитопатогенов на несколько порядков больше, чем бактериальных или вирусных заболеваний растений (Agrios, 1988; Mitchell et al., 2002).
Можно выделить несколько основных групп фитопатогенных грибов (рис. 3.17).
1. Некротрофные патогены, происходящие из почвы и убивающие ткани растений в ходе инфекционного процесса. Многие из них видонеспецифичны и поражают широкий круг растений-хозяев и способны питаться сапротрофно. В природе изучены преимущественно на лесообразующих древесных породах. Примеры: оомицет Phytophthora cinnamomi поразил леса юго-восточной Австралии. Он существенно изменил их структуру, поскольку ряд видов эвкалиптов (Eucalyptus baxteri, Е. radiate и др.) были более восприимчивы, чем другие (Е. syderoxilon, Е. viminalis и др.). В результате инфекции погибло около 40 % деревьев первой группы, в то время как устойчивые виды почти не пострадали. Грибная эпидемия вызвала значительное увеличение участия травянистых растений под пологом этих лесов (Weste et al., 1973). Другой, менее специализированный гриб — Armillaria 1и-teobubalina — вызвал гибель отдельных участков эвкалиптовых лесов и замену их открытыми сообществами (Kile, 1983).
2. Опухолевые, вилтовые и камбиальные заболевания. Вызываются не-кротрофными грибами, поражающими сосудистую систему и/или камбий растений и чаще всего приводящими растение к гибели. Типичные примеры — Cryphonectria parasitica (заболевание каштанов в США, поражение камбия) и голландская болезнь вязов — Ophiostoma ulmi, О. novo-ulmi. Cryphonectria parasitica, происходящая из Китая, первый раз была отмечена в Северной Америке в 1904 г., а к 1940 г. каштановые леса из Castanea dentate в США были уничтожены практически целиком на огромных площадях. Распространение Cryphonecria parasitica в США проходило в среднем со скоростью 37 км в год (Brasier, 1990; Dighton, 2003).
3. Листовые паразиты, вызывающие локальные повреждения. Эти грибы могут существенно снижать выживаемость, размножение, рост и конкурентоспособность растений, но редко приводят к полному отмиранию хозяина, так как должны питаться на живых тканях. Примерами могут служить ржавчинные грибы: Puccinia lagenophorae на Senecio vulgaris, Puccinia chondrilla на Chondrilla juncea. Развитие
3. Фитопатогенные грибы
89
этих грибов сильно зависят от погодных условий сезона и микроклиматических условий местообитания хозяев. Для них характерна высокая пространственная и временная вариабельность развития.
4. Несистемные патогены генеративных органов. Часто вызывают резкое уменьшение семенной продуктивности, но не приводят к гибели само растение. Примером может служить спорынья злаков — Clav-iceps purpurea. Перенос спор многих патогенных грибов может осу-ществляеться при опылении цветков насекомыми или ветром. Привлечение насекомых происходит грибными «псевдоцветками» или секрецией сахаров. Для передающихся половым путем инфекций характерны следующие черты: 1) плотностно-зависимая передача, 2) передача во взрослом состоянии хозяина, 3) патогенность, связанная чаще со стерильностью, но не с гибелью хозяина. Поражение головней пыльников наиболее характерно для гвоздичных, где оно отмечено более, чем у 100 видов (Thrall et al., 1993; Hood, Antonovics, 2003). Опылители могут передавать не только споры грибов, но и бактериальные и вирусные инфекции (передача вирусных инфекций с пыльцой показана для 19 вирусных инфекций растений — Mandahar, Gill, 1984), дрожжи и патогенных клещей. В то же время нектар многих растений содержит антимикробные вещества, препятствующие развитию патогенов (Antonovics, 2005).
5. Системные патогены, поражающие различные органы растений и передающиеся с семенами. Примером может служить Puccinia monoica, развивающаяся на Arabis holboellii. В этой группе часто наблюдается переход к мутуализму за счет приобретения новых функций.
Растения могут вырабатывать различные приспособления для защиты от фитопатогенных грибов. Б. М. Козо-Полянский (см. Работнов, 1992) различал 3 линии защиты от фитопатогенов: 1) внешняя — выделение фунгицидных и бактерицидных веществ в окружающую среду — эфироносные растения весьма устойчивы к грибным паразитам (Симонян, Берсегян, 1974); 2) особенности покровов (толстая кутикула, опушение, мелкие размеры устьиц), 3) внутренняя — химический состав цитоплазмы и особенности метаболизма клеток. Многие виды растений могут синтезировать фитоалексины в ответ на внедрение патогенов. Более устойчивы к грибным инфекциям галофиты и виды, содержащие алкалоиды. Многие защитные адаптации растений к грибной инфекции возникли изначально как реакции к неблагоприятным (стрессовым) условиям (толстая кутикула, опушение и т.п. — к засухе и т.п.) (Heath, 1991).
Поражаемость растений фитопатогенными грибами сильно зависит от условий среды и типа сообщества. Высокая влажность воздуха
90
Глава 3. Растения и грибы
3. Фитолатогенные грибы
91
Рис. 3.17 • Примеры фитопатогенных грибов из различных экологических групп. Некротрофные патогены: а, б — Heterobasidion annosum — корневая губка сосны и ели (фото А. В. Александровой). Опухолевые и вилтовые патогены: в, г — Cryphonecria parasitica на Castanea sativa (фото А. С. Коничева и А. А. Белова); д — Melampsorella caryophyllacearum — ржавчинный рак пихты. Листовые паразиты, вызывающие локальные повреждения: е — Puccinia роагит на листьях Petasites album. Несистемные патогены генеративных органов: ж — склероции Claviceps purpurea (фото А. В. Александровой)
92
Глава 3. Растения и грибы
усиливает развитие фитопатогенов. Так, по наблюдениям в естественных сообществах в Гвинее, (Kranz, 1990) во влажный год сильно поражались грибами 12 видов (23 % общего числа), в то время как в сухой год только 5 видов (10%). При долгом лежании снега на лугах усиливается поражение растений фитопатогенными грибам. С другой стороны, развитие фитопатогенных (как и микоризных) грибов резко подавляется в водной среде, поэтому крайне мало грибов, поражающих гидрофиты. Калийные удобрения повышают, а азотные понижают устойчивость растений к грибным заболеваниям (Работнов, 1984а).
УФ радиация резко увеличивает гибель спор грибов и подавляет развитие их мицелия, с чем, видимо, связано незначительное распространение фитопатогенных грибов в высокогорьях. Показано, что и в целом с увеличением абсолютной высоты происходит обеднение видового состава паразитных грибов и уменьшение их роли в сообществах (Hirsch, Braun, 1992).
Влияние паразитных грибов на растение-хозяина может изменяться по годам. Например, наблюдения в течение 6 лет за поражаемостью Linum marginale ржавчинным грибом Melanospora lini выявляли наличие инфекции каждый год, но только в течение 2-х лет гриб существенно снижал приспособленность пораженных растений (Burdon, 1993).
В разных типах ельников Центрально-лесного заповедника число пораженных видов травяно-кустарничкового яруса составило: в ельнике осоково-сфагновом — 4 из 19, ельнике травяном — 21 из 54, ельнике приручьевом — 26 из 41 (Частухин, 1947, см. Работнов, 1992). Таким образом, доля пораженных растений была выше в более влажных и продуктивных сообществах. В целом, в естественных сообществах патогенными грибами поражаются по разным наблюдениям от 21 до 63% (чаще 30-50%) видов растений, т.е. далеко не все виды.
Для многих фитопатогенных грибов плотный растительный покров служит препятствием распространения их спор. Strong и Levin (1979) отмечают, что чем выше растения, тем больше у него видов паразитных грибов. В среднем в США отмечено 15 видов патогенных грибов на один вид деревьев, 7 видов — на кустарник и 5 видов — на травянистое растение (Burdon, 1993).
По данным (Oakes, 1966; см. Работнов, 1992) реальные потери продукции растений из-за фитопатогенных грибов на естественных пастбищах США составили 6 %, в посевах люцерны — 18 %, в посевах лугового клевера — 35 %. Такие различия связаны как с полидоминантностью естественных сообществ, так и с длительностью сопряженной эволюции растений и патогенов, поскольку люцерна и клевер относительно недавно были завезены в США из Европы.
3. Фитопатогенные грибы
93
В Миннесоте, экспериментальное подавление грибных фитопатогенов вызывало увеличение продукции растений на 0,5-9 % (Mitchell et al., 2002). Большинство исследователей отмечает, поражаемость растений в сбалансированных сообществах незначительна.
Поражение фитопатогенными грибами существенно снижает жизненность и конкурентоспособность растений (отличие от микоризных грибов и эндофитов), их устойчивость к неблагоприятным условиям среды. Например, несистемный паразит Puccinia lagenophorae поражал в эксперименте осенние всходы Senecio vulgaris. Из 400 всходов суровую зиму пережили в контроле — 232 всхода (58%), а в опыте (пораженные грибом) — 126 (31 %) (Paul, Ayres, 1986а, b; Kranz, 1990).
Фитопатогенные грибы оказывают влияние на взаимоотношения растений с животными. Чаще пораженные растения хуже поедаются животными, но бывают и обратные ситуации. Так, поражение ржавчинными грибами (Puccinia sesselis на Allium ursinum, Coleosporium tussilaginis на Tussilago farfara) резко увеличивает поедание этих растений моллюсками (Ramsell, Paul, 1990). Бабочка Lobesia botrana быстрее развивается на винограде и яблоне, если их плоды поражены грибом Botrytis cinerea (Linhart, 1991). Седмичник Trientalis еигораеа, инфицированный головневым грибом Urocystis trientalis, предпочтительно поедается полевками в лесах Швеции (Ericson, Wennstrom, 1997). Экспериментальная изоляция части популяций седмичника от полевок привела к значительному увеличению зараженности растений этим грибом. Таким образом, фитофаги могут выступать как фактор, снижающий поражаемость растений фитопатогенными грибами.
С другой стороны, повреждение животными часто облегчает проникновение грибной инфекции внутрь растения-хозяина. Животные могут быть переносчиками грибной инфекции. Например, показан перенос паразитного гриба Phomopsis subordinaria на Plantago lanceolata долгоносиками (De Nooij, 1988). Насекомые-опылители могут переносить споры фитопатогенных грибов. Такой случай описан для переноса гриба Micro-botryum (ранее Ustilago) violacea на цветки Lychnis viscaria (Skogsmyr, 1993).
Известны также случаи, когда повреждение растений фитофагами уменьшает их поражаемость паразитными грибами, видимо, в связи с интенсификацией синтеза защитных веществ. Например, для щаве-лей (Rumex crispus, R.obtusifolius) показано уменьшение поражаемости листьев ржавчинным грибом Uromyces rumicis, если они до этого повреждались листоедом Gastrophysa viridula (аналогичный эффект получали при искусственном повреждении листьев вырезанием мелких дисков) (Hatcher et al., 1994).
94
Глава 3. Растения и грибы
Таблица 3.5 • Поражаемость грибными инфекциями трех видов травянистых растений в зависимости от возраста их популяций на островах Швеции (по: Carlsson et al., 1990)
Вид растения Возраст популяций с максимальной инфекцией, лет % пораженных растений при максимальной инфекции Возраст снижения инфекции до 10 %, лет
Valeriana sambucifolia 40-45 84 > 50
Trientalis еигораеа 50-100 45 > 400
Silene dioica 50-120 60 > 300
Поражаемость грибными паразитами зависит также от возраста популяций растений. На поднимающихся из воды островах разного возраста в Швеции изучена поражаемость фитопатогенными грибами трех видов растений (Valeriana sambucifolia, Trientalis еигораеа, Silene dioica) (Carlsson et al., 1990). Показано, что во всех случаях, наибольший процент пораженных растений наблюдался на ранних или средних фазах развития популяции (табл. 3.5). Положительная корреляция между по-ражаемостью растений и плотностью их популяций установлена только для системы Valeriana-Uromyces, где паразит имеет однолетний цикл развития. Авторы считают возможным различное объяснение полученным закономерностям: 1) зависимость поражаемости от плотности популяции, 2) эффект основателя и отбор на устойчивость, 3) изменения в среде в связи с первичной сукцессией (Carlsson et al., 1990).
Поражаемость растений грибами связана с возрастом растений и фенофазой. Для многих видов наибольшая поражаемость отмечена после цветения (Kranz, 1990).
У вегетативно-подвижных растений даже системная инфекция не всегда передается от пораженного материнского растения к вегетативным потомкам. Исследованиями В. А. Брызгаловой (1928) было показано, что системная инфекция Puccinia suaveolens проникает в горизонтальные корни бодяка Cirsium arvense на расстояние до 20-30 см, тогда как длина этих корней, дающих корневые отпрыски, часто превышает 1-2 м. Поэтому при вегетативном размножении бодяка молодые особи вегетативного происхождения не получают инфекцию от материнского растения. Эксперименты с выращиванием Lactuca sibirica, пораженного Puccinia minussensis, показали, что при удалении дочерних побегов
4. Сапротрофные грибы
95
на расстояние более 45 см от материнского растения (но сохранении связи с ним!) инфекция также не передается (Wennstrom, Ericson, 1992). Таким образом, вегетативно-подвижные растения могут «убегать» от системных фитопатогенов, в то время как вегетативно слабо подвижные растения этого сделать не могут (Wennstrom, 1994).
Фитопатогенные грибы могут быть использованы для контроля участия нежелательных (обычно интродуцированных) видов в составе сообществ. Так, в Австралию был завезен европейский сорняк Chondrilla juncea. Для борьбы с ним были испытаны грибы Puccinia chondrillina из разных географических регионов. Эффективными оказались лишь грибы из Италии, из чего исследователи заключили, что именно оттуда и был занесен этот сорняк. После заноса активного штамма гриба плотность Chondrilla уменьшилась в среднем с 200 особей/м2 до 10 особей/м2, т.е. приблизилась к типичной для Средиземноморья (Kranz, 1990).
4 • Сапротрофные грибы
В настоящее время многие микологи рассматривают сапротрофию как вторичный по отношению к паразитизму способ питания грибов. Многие виды грибов в ходе онтогенеза сменяют паразитизм на сапротрофизм. В ходе эволюции сапрофитизм мог неоднократно сменяться паразитизмом, особенно при усилении аридности климата.
В лесной подстилке 78-90% биомассы всех микроорганизмов составляют грибы, среди которых большая роль принадлежит сапротро-фам. Длина грибных гиф в 1 г сухой почвы лиственного леса — 393 м, а в расчете на 1 г гумуса — 4-8 км (Frankland, 1982; Christensen, 1989, см. Каратыгин, 1994).
Сапротрофные грибы играют большую роль как в разложении органических веществ, так и в накоплении дефицитных биогенов в процессе их иммобилизации. По данным Swift, Boddy (1984), в процессе разложения древесины происходит резкая относительная аккумуляция N, отношение С N меняется от 247 1 неразложившейся древесины до 44-175 1 разложившейся древесины, 35 1 грибного мицелия и 6 1 насекомых. Мицелий Stereum hirsutum на древесине дуба концентрирует Р в 9,4 раза, N в 8,2 раза больше по сравнению с субстратом (Frankland, 1984).
В ряде случаев грибы, развивающиеся на поверхности листьев растений, тормозят их дальнейшее разложение. Так, при повреждении тлями листьев платана (Platanus occidentalis) на них развиваются грибы, сни
96
Глава 3. Растения и грибы
жающие скорость разложения пораженных листьев по сравнению с контрольными (Choudhury, 1988; см: Facelli, Pickett, 1991).
По размерным характеристикам обычно выделяют 2 группы почвенных сапротрофных грибов: макромицеты и микромицеты. В подстилке лесов выше участие макромицетов (60% базидиомицеты), а в нижних горизонтах лесных почв и в луговых почвах выше доля микромицетов. Видовое разнообразие микромицетов в почве очень трудно установить, оно зависит от числа изученных изолятов. Так для 3 типов почв в лесах Вайоминга при изучении 1200 изолятов найдено от 185 до 286 видов грибов. В ряде работ показана положительная корреляция между видовым богатством растительных сообществ и разнообразием микромицетов.
Сапротрофные грибы также могут влиять на приживаемость всходов растений. Например, семена дерева Gmelina arborea лучше прорастали в присутствии гриба Chaetomium bostrychodes (Osonubi et al., 1990). Многие сапротрофные грибы филлопланы (поверхности листьев) являются антагонистами фитопатогенных грибов. Например, виды Alternar-ia и Cladosporium являлись эффективными ингибиторами Melampsora larici-populina на листьях тополя (Omar, Heather, 1979; Dighton, 2003).
Глава
Растения и животн
Ж
• Хищные растения
• Типы воздействия животных на растения
• Защитные адаптации растений от поедания
• Фитофагия и изъятие первичной продукции
• Фитофаги и смены растительных сообществ
• Фитофагия и флористическое богатство
• Роющая деятельность животных
• Животные и опыление растений
• Распространение плодов и семян животными (зоохория)
• Выпас
1 • Хищные растения
В настоящее время общее число видов хищных (насекомоядных) растений оценивается в 600, они встречаются в 6 подклассах цветковых растений (Ellison, Gotelli, 2001). Наиболее обычны насекомоядные растения в семействах Lentibulariaceae (Pinguicula, Utricularia), Droseraceae (Drosera, Dionaea, Aldrovanda), Nepenthaceae (Nepenthes). На территории России и соседних стран (бывшего СССР) отмечено 18 видов: Pinguicula (жирянка) — 7 видов, Utricularia (пузырчатка) — 6 видов, Drosera (росянка) — 4 вида и Aldrovanda — один вид.
Выделяют 5 типов ловушек хищных растений: 1) липучки (Drosera, Pinguicula), 2) капканы из двух захлопывающихся долей листьев (Dion-аеа, Aldrovanda), 3) кувшинчики с жидкостью (Nepenthes, Sarracenia), 4) ловушки, похожие на угря, привлекающие жертву длинными выростами и имеющие направленные назад волоски, не позволяющие выбраться наружу (Genlisea), 5) засасывающие током воды пузырьки (Utricularia) (Carow, Furst, 1990; Adlassnig et al., 2005) (рис. 4.1). Ряд видов североамериканских Sarracenia (S. flava, S.leucophylla, S.minor) выделяют летучие компоненты, характерные для цветков или плодов. Это, наряду с внешним видом ловушек, напоминающих цветки, способствует привлечению насекомых в ловушки (JQrgens et al., 2009).
По характеру местообитаний хищные растения чаще обитают в воде или на болотах, в условиях хорошего освещения, но дефицита ЭМП, особенно азота. Эти растения не встречаются в сухих или затененных местообитаниях (Ellison et al., 2003). В ряде случаев экспериментально показано, что хищные растения (например, Drosera rotundifolia) положительно реагирует на удобрение почвы N и Р (Stewart, Nilsen, 1992). С другой стороны, карликовые виды Drosera в Австралии показали положительную реакцию на подкормку насекомыми (коллемболами), но отсутствие реакции на увеличение богатства почвы, что свидетельствует о высокой специализации к насекомоядности (Karlsson, Pate, 1992а).
Н. М. Hanslin и Р. S. Karlsson (1996) в экспериментах с дрозофилами, обогащенными 15N, изучали эффективность использования азота хищными растениями (доля азота, переходящая в массу растения). Она составила для 4-х изученных видов (Drosera rotundifolia и 3 вида Pinguicula) in situ (в природных условиях) 29-41 %, а в лаборатории — 40-50%. Более эффективное усвоение азота в лабораториях авторы связывали с отсутствием ветра, сильных дождей, перегрева и других неблагоприятных факторов. Авторы отмечают, что потребление азота корнями этих растений сильно стимулируется при увеличении числа пойманных мух, при этом поглощение азота из субстрата увеличилось на большую вели-
1. Хищные растения
99
Рис. 4.1 • Хищные растения: а — Drosera rotundifolia; б, в — Genlisea margaretae с ловушками, похожими на угря, привлекающими жертву длинными выростами и имеющими направленные назад волоски, не позволяющие выбраться наружу (фото Stephane Joly, Франция, с любезного разрешения автора); г — Pinguicula vulgaris; д — Utricularia vulgaris с зимующими побегами (турионами); е — Nepenthes maxima
100
Глава 4. Растения и животные
чину, чем потенциальное содержание азота в пойманных мухах. Прямыми экспериментами показано, что более крупные особи росянок ловят больше мух, чем мелкие, а при увеличении числа пойманных мух улучшается жизненность растений (положительная обратная связь).
Преобразование листьев в специализированные органы ловли насекомых снижает эффективность их фотосинтеза, поэтому цена насекомояднос-ти — малая скорость роста. Насекомоядные растения могут обитать на богатых почвах только при низкой межвидовой конкуренции. Наблюдения над Utricularia macrorhiza показали, что интенсивность фотосинтеза у ловчих пузырей примерно вдвое ниже, чем у обычных листьев, в то время как интенсивность дыхания примерно одинакова. Следовательно, большее развитие пузырей в более бедных ЭМП водоемах ведет к уменьшению общей продукции на 20-80 %, поскольку для этого вида характерна высокая изменчивость числа пузырей на листьях в зависимости от трофности водоема (Knight, 1992). Сходные закономерности установлены Ellison, Gotteli (2002) в эксперименте по удобрению азотом участков с хищным растением Sarracenia purpurea в Северной Америке, которое привело к уменьшению размеров ловчих органов по сравнению с фотосинтезирующими. Показана также отрицательная корреляция между содержанием азота в почве разных болот и долей ловчих органов (Agrawal, 2002а).
Многие насекомоядные растения выделяют пищеварительные ферменты для переваривания добытых насекомых. Однако возможны и другие механизмы получения низкомолекулярных органических и минеральных веществ из пойманных животных. Так, в кувшинчатых листьях Sarracenia, заполненных водой, формируются своеобразные открытые экосистемы. У Sarracenia purpurea листья образуются каждые 20 дней, каждый лист (кувшинчик) содержит до 50 мл воды, в которой обитают многие бес
позвоночные. Наиболее обычная добыча у этого вида — муравьи (Cresswell, 1991). Конечным звеном пищевой цепи являются личинки комаров — Wyeomyia smithii, продукты жизнедеятельности которых и служат источниками ЭМП для растений (Ellison et al., 2003).
В пузырьках Utricularia в большом количестве обнаружены живые и мертвые клетки десмидиевых и зигнемовых водорослей из 45 родов, составляющие до 80% общей добычи пузырьков (рис. 4.2). Их доля
Рис. 4.2 • Водоросли в ловчих пузырьках Utricularia (по: Peroutka et al., 2008)
2. Типы воздействия животных на растения
101
существенно увеличивалась в более бедных элементами минерального питания водах. Предполагается, что водоросли могут составлять значительную долю в питании пузырчатки (Peroutka et al., 2008).
Насекомоядные растения — относительно молодая группа, их остатки находят начиная с палеоцена, в то время как в эоцене известно уже несколько групп насекомоядных растений.
2 • Типы воздействия животных на растения
Воздействия животных на растения можно разделить на прямые (поедание, паразитизм, устройство гнезд, механические повреждения и т. п.) и косвенные (через другие организмы — трансбиотические или через среду — трансабиотические).
Примером трансабиотических воздействий может служить перенос ЭМП животными из одних сообществ в другие. Растения близ птичьих базаров в Арктике и Антарктике получают большую часть ЭМП за счет морских экосистем, гнездящиеся птицы поддерживают высокое богатство почв. По наблюдениям В. Ю. Душкова (1980), в полупустынях Се-
Рис. 4.3 • Колония грачей на деревьях мелколистного вяза в Северном Прикаспии (Казахстан)
102
Глава 4. Растения и животные
верного Прикаспия мелколистный вяз хорошо растет в местах поселения грачей, а вне их часто засыхает (рис. 4.3). Животные могут влиять на азот-фиксацию, например выпас гусей на приморских маршах способствует развитию азотфиксирующих колоний Nostoc.
Трансбиотические воздействия животных на растения очень разнообразны. Их примерами могут быть распространение спор патогенных грибов животными (см. главу 3). Почвенные беспозвоночные могут оказывать влияние на растения через микоризные грибы (Gange, Brown, 2003). Дождевые черви благоприятствуют развитию ВАМ грибов за счет распространения их спор (они часто проходят неповрежденными через пищеварительный тракт червей и концентрируются в копролитах) и фрагментов мицелия, но могут повреждать мицелиальную сеть в почве. Вне
сение нематодоцидов на лугах вызывало увеличение колонизации корней микоризными грибами (Ingham et al., 1986). В экспериментах Klironomos и Kendrick (1995) было показано увеличение микоризной колонизации корней сахарного клена в присутствии почвенных клещей и коллембол.
Простейшие имеют большое значение для минерального питания растений, так как потребляют бактерий, иммобилизирующих ЭМП в почве. Наиболее важные потребители бактерий — голые амебы (рис. 4.4). Как
Рис.4Л • Типичное распределение амеб (Acanthamoeba castellanii) вдоль бокового корня риса (Oryza sativa), выращенного на агаре (по: Bonkowski, 2004)
и нематоды, простейшие используют химические сигналы для выявления своих жертв. В эксперименте с меченой подстилкой и наличием/отсутст-вием почвенных простейших Lolium регеппе удваивал свою биомассу и поглощал из подстилки в 2-3 раза больше азота в присутствии простейших, чем без них (Bonkowski et al., 2000). Простейшие также выделяют ростовые вещества, усиливающие рост корней, и участвуют в цепочке событий, стимулирующих рост боковых корней (Bonkowski, 2004).
Среди прямых воздейст-
вий животных на растения в природе наибольшее значение имеет фитофагия, т. е. поедание растений или их частей животными. Различные аспекты фитофагии и защиты растений от поедания будут рассмотрены в следующих разделах.
3. Защитные адаптации растений от поедания
103
3 • Защитные адаптации растений
от поедания
Можно различить две стратегии адаптации растений к воздействию фитофагов — избегание и толерантность. В первом случае растение имеет приспособления, снижающие вероятность быть съеденным, а во втором — снижает ущерб от отчуждения части биомассы (рис. 4.5). Стратегии устойчивости (избегания) и толерантности к отчуждению биомассы фитофагами не являются исключающими, они сочетаются у многих видов растений (Nunez-Farfan et al., 2007).
Избегание
Толерантность
Уход от фитофагов в пространстве и времени
н______। Механические средства защиты
Отавность
Компенсаторный фотосинтез
Привлечение животных для защиты
Усиление поглощения ресурсов
Химические средства защиты
Изменение аллокации и т.п.
Рис. 4.5 • Адаптации растений к воздействию фитофагов
3.1 • Уход от фитофагов
в пространстве и времени
К этому типу адаптаций относятся признаки, снижающие вероятность для растения быть найденным фитофагом (незаметная окраска, обитание на участках, свободных от фитофага, мимикрия с растениями, обладающими защитными приспособлениями, короткий жизненный цикл, развитие в период минимальной активности фитофагов) (Zamora et al., 1999). Например, окраска некоторых видов растений из рода Trichocaulon в Южной Африке делает их малозаметными на фоне окружающих скал и камней (рис. 4.6).
Вегетативная подвижность клевера снижает повреждаемость его корней почвенными нематодами по сравнению с вегетативно неподвижной люцерной (Работнов, 1992). Быстрое сезонное развитие, свойствен-
104
Глава 4. Растения и животные
Рис. 4-6 • Примеры адаптаций избегания фитофагов: а — скрывающая окраска Trichocaulon в пустынях Южной Африки (фото M.Werger, с любезного разрешения автора); б — массовое (не ежегодное!) цветение эфемеров в пустыне Калифорнии, США (фото Frank Кее, www.keesphotos.com, с любезного разрешения автора)
ное эфемерам и эфемероидам, не позволяет фитофагам потреблять значительную долю надземной биомассы этих растений. Сходное значение имеет чередование по годам высокой семенной продукции растений, когда в семенные годы фитофаги потребляют не все образовавшиеся семена. На примере Fraxinus excelsior в Англии было показано, что хотя в урожайные годы потребляется большая доля семян, абсолютное количество оставшихся и проросших семян в эти годы может быть существенно выше (табл. 4.1).
3. Защитные адаптации растений от поедания
105
Таблица 4.1 • Показатели семенного размножения Fraxinus excelsior в урожайный и неурожайный год (по: Gardner, 1977; van der Valk, 1992)
Показатели 1967 (неурожайный) 1969 (урожайный)
Продуцировано семян (на м2) 0,33 1193
Рассеяно семян (на м2) 0,08 998
Съедено семян (%) 25 75
Проросло семян (%) 0 2,5
Появилось всходов (на м2) 0 31,6
3.2 • Механические способы защиты
Механические способы защиты листьев против фитофагов включают: 1) шипы и колючки, 2) опушение, 3) жесткость (склерофилльность) листьев, 4) включение минералов в ткани растений (Hanley et aL, 2007) (рис. 4.7). Защитная роль этих приспособлений показана во многих экспериментах. Так, удаление шипов Acacia drepanolobium втрое увеличило потребление листьев у экспериментальных растений млекопитающими (Milewski et al., 1991). Северо-американская полевка (Microtus ochrogaster) в прериях (Gali-Muhtasib et aL, 1992), как и европейская Microtus agrestis (Massey et aL, 2007) избегают злаков, содержащих значительное количество кремнезема (фитолитов). Многие животные также избегают листья с кристаллами оксалата кальция. Экспериментальная дефолиация (удаление листьев) усиливает развитие колючек и шипов у многих видов растений (Hanley et aL, 2007). Многие из рассматриваемых приспособлений могут выполнять несколько функций, например опушение уменьшает испарение и защищает от фитофагов. При удалении волосков с листьев Verbascum thapsus они испаряли больше воды и больше повреждались фитофагами (Woodman, Fernandes, 1991) (рис. 4.7). В то же время волоски на растениях могут служить препятствием для хищных насекомых и па-разитоидов, питающихся фитофагами. Так, Encarsia formosa — эффективный паразитоид тепличной белокрылки (Trialeurodes vaporariorum) становится бессильной в волосках огурца, где белокрылка процветает (van de Merendonk, van Leuteren, 1978; Price et aL, 1980). Колючки у кактусов затеняют стебли и уменьшают нагревание тканей растений.
Фитолиты растений могут рассматриваться и как средства механической защиты и как пищевые детерренты (см. ниже), снижающие питательную ценность биомассы растений. Поедание растений с крем-
106
Глава 4. Растения и животные
Рис. 4.7 • Примеры механических средств защиты у растений: а — густое опушение листьев Verbciscum thapsus; б — ствол Chorisia speciosa, Malvaceae (Южная Америка); в — сильно колючие боковые побеги бамбука Bambusa sinospinosa (Юнань, Китай, ботанический сад Xishuangbanna);
г — южноамериканский кустарник Collecia paradoxa (Rhamnaceae)
3. Защитные адаптации растений от поедания
107
невыми фитолитами вызывает разрушение эмали зубов, язву пищевода и мочекаменную болезнь у млекопитающих (McNaughton et al., 1985).
В целом опушение уменьшает повреждаемость листьев насекомыми, а шипы и колючки — позвоночными. Ротанговые пальмы (Calamus spp.) имеют колючие стебли и влагалища листьев, необходимые для закрепления этих лиан. Таким образом, шипы и колючки могли эволюционно возникать не только как защита от фитофагов, но и иметь другие функции.
Р. Grubb (1992) проанализировал, каким группам растений свойственны колючки. Он выделил следующие группы, растения в которых отличаются повышенной уязвимостью к фитофагам (рис. 4.8):
а) Растения засушливых местообитаний, особенно в тропиках и субтропиках, где колючие растения часто доминируют (защита от повреждений животными, приводящих к потерям дефицитной воды). По исследованиям в Гане, доля колючих растений во флоре увеличивается с 1 % при количестве осадков 2000 мм до 8 % при осадках 750 мм. Интересно отметить, что некоторые высокие кактусы лишены колючек в самых верхних частях, недоступных для наземных позвоночных фитофагов.
б) Колючие растения богатых местообитаний умеренной зоны (Cirsium, Dipsacus, Rubus etc.). Эти растения занимают нарушенные местообитания, прорывы в пологе леса, площадь которых в доагрикультур-ном ландшафте была невелика, поэтому для выживания этим видам важна усиленная защита. По данным для субтропических лесов в Японии 40% видов светолюбивых видов деревьев, характерных для прорывов в пологе, обладают колючками, в то время как среди теневыносливых деревьев доля колючих лишь 4 %. Аналогичные данные для тропических лесов Папуа — Новой Гвинеи составили 28 % и 1,3% соответственно. Многие колючие кустарники умеренной зоны (Grossularia, Rosa, Crataegus) характерны для пойм рек, лесных опушек и других открытых участков, занимающих небольшие площади в лесном ландшафте.
в) Тропические однодольные (панданусы, пальмы, бромеливые) и древовидные папоротники, имеющие ограниченное число конусов нарастания (высокая уязвимость при повреждении точки роста). Например, из 209 видов пальм западной Малайзии, 80% видов и 53% родов — колючие (по: Whitmore, 1973).
г) Растения, имеющие ограниченный период для активного фотосинтеза. Среди рода Ilex колючим краем листа обладают только вечнозеленые виды, произрастающие в листопадных лесах (/. aquifolium,
108
Глава 4. Растения и животные
б)
3. Защитные адаптации растений от поедания
109
Рис. 4.8 • Экологические группы колючих растений (по: Grubb, 1992): а — растения засушливых местообитаний, Lamira echinocephala (Меловой хребет, КЧР); б — колючие растения богатых местообитаний умеренной зоны (Dipsacus laciniatus, Краснодарский край); в — тропические однодольные, имеющие ограниченное число конусов нарастания (пальма Aiphanes aculeate, Южная Америка); г — растения, имеющие ограниченный период для активного фотосинтеза (Hex aquifolium, Нидерланды); д — растения, имеющие повышенное содержание азота в листьях (Acacia karroo, Южная Африка); е — растения, защищающиеся от специфических групп фитофагов (Banksia baxteri, Австралия, фото Eric La Fountaine, University of British Columbia Botanical Garden, с любезного разрешения автора)
I.opaca), в связи с кратковременным периодом активной ассимиляции до распускания листьев деревьев.
д) Растения, имеющие повышенное содержание азота в листьях и, следовательно, более привлекательные для фитофагов по сравнению с соседями. Например, в Капской флористической области с ее очень высоким флористическим разнообразием (преимущественно за счет видов Ericaceae и Proteaceae) очень мало колючих растений. Среди последних — некоторые виды Aspalathus и Cliffortia. По сравнению с вересковыми и протейными, содержащими очень мало азота в листьях (4-10 мг/г), листья видов этих родов богаты азотом (11-20 мг/г) и защищены от фитофагов колючками.
е) Растения, защищающиеся от специфических групп фитофагов. Кустарниковые заросли в Австралии образованы многими сильно колючими видами семейств Proteaceae (не колючие на юге Африки!) и Ericaceae. Они содержат мало азота, но сумчатые фитофаги способны усваивать их ткани за счет низкой интенсивности метаболизма и специфической физиологии — способности к повторному использованию азота мочевины.
3.3 • Привлечение животных для защиты от фитофагов
Летучие органические соединения, выделяемые растениями, привлекают клещей и паразитических ос, а экстрафлоральный нектар — чаще муравьев. Число известных летучих соединений, выделяемых разными органами растений, превышает 1000. Экстрафлоральные нектарники известны в более, чем 300 родах цветковых растений (Elias, 1983; Koptur, 1992). Известно много примеров усиления выделения летучих веществ при поражении листьев фитофагами. Большая степень поражения паразитами у насекомых отмечалась на выделяющих нектар деревьях по сравнению
110
Глава 4. Растения и животные
с безнектарными. Выделения углеводов, которыми питаются муравьи, резко стимулируют последних к поиску белковой пищи (насекомых) для сбалансирования диеты (Davidson, 1997; Heil, 2008).
Луговой клоп Lygus lineolaris редко поражается паразитоидом Leio-phron pallipes на видах Oenothera, Daucus, Solidago, но поражаемость достигает 30-40% на видах Erigeron, которые выделяют большое количество нектара, привлекающего паразитоида (Streams et al., 1968; Shah-jahan, 1974; Price et al., 1980).
Бразильский кустарник Caryocar brasiliense выделениями экстра-флоральных нектарников привлекает муравьев, защищающих его (и окрестные растения) от многих групп фитофагов (Oliveira, 1997). Fonseca (1994) показал, что при удалении муравьев с мирмикофильных деревьев в амазонских лесах потеря их листвы от фитофагов увеличивается в 10 раз (Schowalter, 2006). В целом экстрафлоральные нектарники могут достигать значительных размеров (у Pithecellobium macradenium до 7,4 мм в диаметре) (Elias, 1972), желобчатые экстрафлоральные нектарники могут покрывать всю длину черешков листьев (до 15 см) у Ochroma pyramidale (O’Dowd, 1980; Marquis, Whelam, 1996).
Особи Chamaecrista nictitans (Caesalpiniaceae) с экстрафлоральны-ми нектарниками привлекали хищных пауков семейства Salticidae, потреблявшими значительное число фитофагов. Как результат, эти особи производили значительно больше семян, чем таковые без нектарников и пауков (Ruhren, Handel, 1999).
Корни Thuja occidentalis выделяют вещества, привлекающие энто-мопатогенную нематоду Heterorhabditis megidis, которая питается личинками долгоносика Otiorrhynchus sulcatus, повреждающего корни туи (Van Tol et al., 2001; van der Putten et al., 2001). В полевых условиях показано, что выживаемость хищных нематод Heterorhabditis marelatus в сухие зимы была выше в ризосфере Lupinus arboreus, чем в окружающих почвах луга, поскольку эта нематода чувствительна к высыханию (во влажные зимы этот эффект не проявлялся). Эта нематода играет важную роль в потреблении личинок моли Hepialus californicus, питающейся корнями люпина (Preisser et al., 2006).
Однако вопрос о пользе привлечения растениями хищных насекомых и паразитоидов далек от однозначного разрешения. Естественный отбор у растений и видов третьего трофического уровня вызывает конфликт интересов. Селективно растениям трудно выработать механизмы привлечения конкретных видов насекомых и при этом не приводить в активность другие, вредные для них, виды насекомых. Muller, Godfray (1999) показали, что популяции тлей контролируются всей гильдией
3. Защитные адаптации растений от поедания
111
хищных насекомых, а не отдельными видами. Гусеницы, пораженные па-разитоидами, могут потреблять большую биомассу, чем непораженные (Rahman, 1970; van der Meijden, Klinkhamer, 2000).
3.4 • Химические средства защиты растений
Защитные вещества растений можно разделить на две большие группы — токсины (ядовитые вещества, часто вызывающие летальный эффект) и пищевые детерренты (понижающие питательную ценность и усвояемость фитомассы). Защитные вещества растений часто в целом называют вторичными метаболитами.
3.4.1 • Токсины растений
Выделяют несколько групп токсинов по их химической природе и физиологическому действию (рис. 4.9).
Азотсодержащие
Цианогенные глюкозиды
Алкалоиды
Небелковые аминокислоты
Пептиды и гликопротеины
Рис.4.9 • Основные группы токсинов
Цианогенные глюкозиды при гидролизе дают синильную кислоту, основной физиологический механизм — блокировка дыхательной цепи в митохондриях. Цианогенные глюкозиды характерны, например, для некоторых рас Trifolium repens. Их содержание в растениях значительно варьирует. Отмечено увеличение доли растений, содержащих цианогенные глюкозиды с севера на юг и запад (рис. 4.10): при низких температурах повреждение растений слизнями резко снижается, особенно при весеннем росте растений, что снижает необходимость в химических способах защиты от фитофагов. Напротив, наибольшая частота цианогенеза у клевера наблюдается в странах с мягкой зимой (Великобритания, страны
112
Глава 4. Растения и животные
Рис. 4.10 • Доля растений Trifolium repens, содержащих цианогенные глюкозиды в популяциях в различных районах Европы и Передней Азии (показано черным в круговых диаграммах), и январские изотермы (по: Jones, 1972)
Средиземноморья). Эксперименты с цианогенными расами Trifolium repens и Lotus corniculatus показали, что они более устойчивы к фитофагам, но менее конкурентоспособны на плодородных почвах в отсутствии фитофагов по сравнению с нецианогенными расами (Herms, Mattson, 1992).
Алкалоиды (рис. 4.11) (например, кониин из Conium maculatum, которым был отравлен Сократ) делятся на 3 группы: настоящие (происходят из аминокислот и содержат азот в гетероцикле), псевдоалкалоиды (происходят не из аминокислот, например кофеин), протоалкалоиды (происходят из аминокислот, но содержат азот не в гетероцикле) (Bennett, Walls-grove, 1994). Примером может служить алкалоид капсаицин — основной компонент перца чилли, его вкус избирательно отпугивает млекопитаю-
3. Защитные адаптации растений от поедания
113
Кониин из Conium macula turn (Umbelli ferae)
CH2OH
Фенил
Атропин из At го pa belladonna (So lanaceae)
Соланин из Solanum tuberosum (Sotanaceae)
Стрихнин us Strychnos nuxt)omica (Loganaceae)
Рис. 4.11 • Примеры алкалоидов различных растений (по: Харборн, 1985): Глю — глюкоза, Гал — галактоза, Раф — раффиноза
щих-фитофагов, но не отпугивает ряд птиц — более эффективных распространителей семян. Видео наблюдения показали, что в дневное время только птицы питаются плодами чилли, среди которых Toxostoma curvi-rostre поедала 72% плодов. Эксперименты с несодержащими капсаицин плодами показали, что мелкие млекопитающие потребляют эти плоды, но полностью разрушают семена. Авторы делают вывод, что капсаицин избирательно отпугивает фитофагов, не способствующих распространению семян (Tewksbury, Nabhan, 2001).
Концентрация алкалоидов, как и других защитных веществ, подвержена разнонаправленному действию отбора и значительно варьирует как внутри вида растений, так и внутри популяций. Для Senecio jacobaea показана примерно 70% наследуемость концентраций алкалоидов (Vrieling et al., 1993). Van der Meijden (1996) анализирует модель влияния специализированных (для которых защитное вещество растений выступает уже как аттрактант) и неспециализированных (защитное вещество уменьшает повреждаемость растений) фитофагов на встречаемость в популяции растений с разным уровнем содержания защитных веществ. С позиций этой модели невыгодно иметь как очень высокую, так и очень низкую концентрацию защитных веществ. Полевые наблюдения с Cynoglossum officinale подтвердили эти закономерности. Специализированные фитофаги этого
114
Глава 4. Растения и животные
вида почти не встречались на растениях с низкой концентрацией защитных веществ и не подавлялись высокими концентрациями, в то время как все случаи питания неспециализированными фитофагами отмечены на листьях с низкой концентрацией защитных веществ. Аналогичные данные получены также для Brassica napus (Giamoustaris, Mithen, 1995).
Небелковые аминокислоты (известно около 300) могут встраиваться в белковые цепи, разрушая структуру последних, что приводит к летальному эффекту. Они типичны для бобовых и часто служат для защиты богатых белком семян.
Пептиды и гликопротеины как токсины нечасто встречаются у растений. К гликопротеинам относится рицин (основной токсичный компонент семян клещевины Ricinus communis), сходный с ним по строению и механизму действия абрин (характерен для бобового Abrus precatorius). Эти гликопротеины нарушают синтез белка в рибосомах. Циклопептиды являются главным токсином бледной поганки.
Сердечные глюкозиды оказывают кардиотоническое действие, проявляющееся в замедлении частоты и усилении сердечных сокращений. Примером может служить сердечные глюкозиды, содержащиеся в ваточнике (виды рода Asclepius), которым в Северной Америке питаются гусеницы бабочек-данаид (Danaus spp.), устойчивые к этим токсинам. Они накапливают эти гликозиды и становятся несъедобными для голубых соек, питающихся гусеницами. Это типичный пример использования токсинов растений насекомыми для защиты от хищников.
Сапонины — безазотистые гликозиды с поверхностно-активными (детергентными) свойствами. При попадании в кровь сапонины могут быть токсичны, поскольку вызывают гемолиз эритроцитов. Несколько различных сапонинов содержатся в листьях наперстянки (Digitalis). При поедании растений с высоким содержанием сапонинов происходит раздражение слизистой желудка и кишечника.
Терпеноиды — производные растительных углеводородов терпенов. К ним относится, например, ментол (обычный у губоцветных), пинен (входит в состав эфирных масел сосен), каротин. Физиологическое действие состоящих из терпеноидов эфирных масел зависит от химического строения их компонентов. Антимикробная активность терпеноидов возрастает в следующем порядке: углеводороды, окиси, фенолы, альдегиды, кетоны, спирты, сложные эфиры. Терпеноиды влияют на многие физиологические процессы в организмах животных: работу различных желез, сокращение сердца, изменение проницаемости клеточных мембран и т. п. Хлопчатник (Gossypium barbadense) содержит терпеноид госсипол, токсичный для ряда фитофагов, но в то же время являющийся пищевым стимулятором для опасного вредителя хлопчатника — Anthonomus grandis (Hedin et al.,
3. Защитные адаптации растений от поедания
115
1968). Дополнительно госсипол является фитоалексином, эффективным против ряда патогенных грибов (Ingham, 1972; McNaughton, 1983).
Отдельную функциональную группу составляют фототоксины, действие которых проявляется только на свету (активизируются солнечными лучами, главным образом ультрафиолетом). Известно более 200 различных фототоксинов, они найдены в 40 семействах, 32 порядках и 8 подклассах цветковых растений. Наиболее обычны фототоксины в семействах Umbelliferae и Rutaceae. Их химическая природа разнообразна (15 фитохимических классов, но все содержат либо ненасыщенные кольца, либо ацетиленовые (тройные) связи). Механизм действия фототоксинов — образование радикалов, оказывающих неспецифическое токсичное воздействие. Эти соединения эффективны не только против животных, но и вирусов, бактерий, грибов (Downum, 1992). Широко известны фототоксины некоторых борщевиков (рода Heracleum), содержащиеся в листьях Citrus фототоксины часто вызывают фотодерматиты у сборщиков плодов цитрусовых.
Как отмечено выше, животные могут использовать токсины растений для защиты от других животных. Птицы часто применяют для постройки или выстилки гнезд растения, отпугивающие эктопаразитов. Так, скворцы используют листья моркови для уменьшения поражаемости птенцов клещами (Шилов, 1997).
Чувствительность отдельных видов фитофагов к токсинам различна. Например, плоды видов рода Strychnos, содержащие сильнодействующие яды (стрихнин и бруцин), поедаются слонами и лангурами (обезьяны) (Janzen, 1983).
К отдельной функциональной группе можно отнести гормональные токсины. В регуляции линьки насекомых участвуют две группы гормонов: ювенильные и личиночные. Гормоны линьки насекомых экдизоны найдены в больших количествах у многих папоротников и голосеменных растений. Японские ученые (Takemoto et al., 1967) обнаружили, что равное количество (25 мг) экдизона содержится в 500 кг шелковичных червей, 25 г листьев тисса (Taxus baccata) и в 2,5 г корневищ Polypodium vulgare.
Ювенильный гормон насекомых в растениях был открыт К. Сла-ма (Slama, 1971) при неудачной попытке вырастить клопов-солдатиков на фильтровальной бумаге в США, где эту бумагу делали из Abies bai-samea. Впоследствии из бальзамической пихты был экстрагирован ювенильный гормон ювабион, высокое содержание которого, видимо, и обуславливает слабую поражаемость насекомыми некоторых видов голосеменных растений.
116
Глава 4. Растения и животные
Воздействие фитофагов обычно повышает содержание защитных веществ в растениях за счет индуцированного усиления их синтеза. В то же время в некоторых случаях фитофаги могут активно снижать содержание токсинов в процессе потребления биомассы. Экспериментально показано, что слюнные железы гусеницы Helicoverpa zea выделяют соединения, которые уменьшают продукцию никотина, синтезируемого табаком Nicotiana tabacum. В специальном эксперименте у части гусениц были удалены слюнные железы, продуцирующие глюкозоксидазу — основной фермент слюны этих гусениц. Выживаемость гусениц при удалении слюнных желез снижалась, а концентрация никотина в листьях была на 26 % выше, чем в листьях, на которых питались гусеницы с неудаленными железами. Было также показано, что обработка листа раствором с активной глюкозоксидазой снижала концентрацию никотина, таким же образом, как и при питании гусениц с неудаленными железами. При выращивании гусениц на растениях, обработанных глюкозоксидазой, их выживаемость была выше, чем на листьях, обработанных водой. Таким образом было показано, что глюкозоксидаза гусениц ответственна за подавление индуцированного синтеза никотина у табака (Musser et al., 2002).
Механизмы адаптаций фитофагов к защитным метаболитам растений могут быть подразделены на 3 группы (Dearing et al., 2005): избегание, регулирование и биотрансформация. Существует несколько вариантов избегания попадания или всасывания токсинов в организме животных:
1) разложение защитных веществ до поедания растений. Полевка Microtus pennsylvanicus поедает ветки хвойных деревьев через несколько дней после их сбора, когда концентрация защитных веществ существенно снижается (Roy, Bergeron, 1990). Бобр вымачивает ветви деревьев в воде несколько дней до поедания и предпочитает питаться вымоченными ветками (Muller-Schwarze et al., 2001);
2) наличие танин-связывающих протеинов в слюне;
3) регуляция всасывания в пищеварительном тракте с использованием двух молекулярных механизмов — гликопротеинный транспортер и биотрансформационные ферменты;
4) микробиологическая детоксикация в пищеварительном тракте.
Регуляция может быть связана с влиянием защитных веществ путем вызывания рвоты, их негативным воздействием через рецепторы вкуса и запаха. Биотрансформация включает различные механизмы: окисление, коньюгация (внедрение) молекул, чаще с глюкуронидами и сульфатами, что приводит к поляризации молекул и облегчает их вывод из организма с мочой (Dearing et al., 2005). Специализированные фитофаги
3. Защитные адаптации растений от поедания
117
могут осуществлять детоксикацию отдельных токсинов и использовать яды растительной природы для защиты от хищников. Для калифорнийского кузнечика Romalia microptera показана способность потреблять даже гербицид 2,4 дихлорфенол, что делает этот вид кузнечиков резко ядовитым для птиц и хищных насекомых (Колчинский, 1990)
3.4.2 • Пищевые детерренты
Пищевые детерренты не вызывают, как токсины, летальный эффект, но уменьшают питательную ценность тканей растений. К основным группам пищевых детеррентов относятся танины, терпены, каучуки, значительно уменьшают питательную ценность фитомассы также целлюлоза, лигнин и кремнезем.
Танины выступают как ингибиторы протеолитических ферментов (поэтому и используются при дублении кож). Содержание танинов в листьях дуба подвержено сезонной динамике. Гусеницы зимней пяденицы охотно поедают листья дуба весной и начале лета, но перестают кормиться на дубе в середине лета и переходят на другие деревья. Feeny (1970) установил резкое изменение в сезонном содержании танинов в листьях дуба, их концентрация возрастает от весны к осени. Высокое содержание танинов в листьях некоторых деревьев приводит к снижению скорости их разложения и требует большей массы тонких корней для поглощения необходимого количества азота из почвы. В экспериментах с тополями (Populus angustifolia, Р. fremontii) была показана наследуемая связь между высоким содержанием танинов и продукцией тонких корней (Fischer et al., 2006).
По мнению Hohchuli (1996), целлюлозные оболочки клеток растений являются наиболее универсальной защитой от насекомых-фитофагов, большинство из которых не усваивает целлюлозу, а вынуждено тратить энергию и иметь специализированные адаптации для разрушения клеточных стенок и извлечения содержимого цитоплазмы.
Кремнезем в клетках и межклетниках многих растений (особенно злаков и осок) может быть пищевым детеррентом. Он снижает пищевую ценность корма, усиливает стачиваемость зубов, а также может вызывать ряд заболеваний травоядных млекопитающих (Hochuli, 1993). Кремнезем также неблагоприятно воздействует на насекомых-фитофагов (Vicari, Bazely, 1993).
В качестве пищевых детеррентов для фитофагов могут выступать и тяжелые металлы, накапливающиеся в растениях. В экспериментах с потреблением саранчой Schistocerca gregaria ярутки Thlaspi caerulescens, содержащей разные концентрации цинка, было показано, что при возможности выбора саранча предпочитает растения с низкой концентра
118
Глава 4. Растения и животные
цией Zn, а при невозможности — потребляет растения с высокой концентрацией, но при этом рост животных был отрицательно скоррелирован с содержанием Zn в растениях (Behmer et al., 2005).
Детерренты могут по-разному действовать на различные виды животных. Так, танины оказывают более сильное отрицательное воздействие на млекопитающих, чем на насекомых (Bernays et al., 1989; Linhart, 1991).
Однако часто насекомые не могут сменить пищевое растение и вы
нуждены питаться тканями с высоким содержанием пищевых детеррен-тов. В таком случае возникает парадокс пищевых детеррентов. При питании на растениях с большим содержанием детеррентов увеличивается длительность жизни фитофага и количество потребленного растительного материала (так как пищевая ценность его ниже). Возможная
разгадка парадокса состоит в том, что даже при незначительном удлинении жизненного цикла фитофагов резко увеличивается их потребление хищниками. Это было показано в специальных экспериментах (Price et al., 1980): мексиканскую божью коровку (Epilachna varivestis) выращивали на растениях, содержащих и не содержащих танины в присутствии или отсутствии хищников (Podisus maculiventris). Как и предполагалось, без хищника более поврежденными оказывались содержащие танины растения, а с хищником — более повреждались не содержащие танин растения.
Многие защитные вещества обладают универсальным действием, уменьшая поедаемость фитофагами и подверженность грибным инфекциям. В выборке из 43 видов британской флоры была показана положи
Рис. 4.12 • Связь скорости разложения опада (ордината) с поедаемостью для 43 видов растений британской флоры (по: Grime et al., 1996)
тельная корреляция между поедаемостью растений моллюсками и скоростью разложения их опада (рис. 4.12) (Grime et al., 1996).
Вторичные метаболиты (защитные вещества), помимо защитных функций, могут выполнять и различные другие функции, а именно: привлечение опылителей (антоцианы и флавоноиды), защита от УФ излучения (фенольные смолы, флавоноиды, антоцианы), структурные функции (лигнин, танины), защита корней от кислой и восстановительной среды (танины),
3. Защитные адаптации растений от поедания
119
временное хранилище азота (алкалоиды и цианогенные глюкозиды), улучшение поглощения ЭМП (фенолы), выступать как фитогормоны и регуляторы взаимоотношений с азотфиксирующими прокариотами (флавоноиды и простые фенолы), повышать устойчивость к засухе (танины, фенольные смолы) (флавоноиды) (Herms, Mattson, 1992).
3.5 • Гипотезы, объясняющие распространение защитных приспособлений у растений
Многие исследователи пытались объяснить, почему у некоторых растений имеются защитные приспособления от фитофагов, а у других нет, у одних синтезируются токсины, у других — детерренты. Рассмотрим четыре основных гипотезы, связанных с этим вопросом.
1. Гипотеза «экспонированности» (Feeny, 1976) — растения, более представленные (экспонированные) как во времени (многолетние), так и в пространстве (обильные в ценозах) должны вкладывать больше асси-милятов в защитные органы или вещества, преимущественно детерренты, а более короткоживущие и редкие виды — использовать токсины. Защита от фитофагов более необходима «экспонируемым» растениям.
2. Гипотеза «недостатка ресурсов» (Coley et al., 1985) — растения, обитающие в условиях недостатка ресурсов, особенно бедных почв, должны вкладывать больше ассимилятов в защитные органы или вещества, преимущественно детерренты (безазотистые), а растения богатых почв могут синтезировать азотсодержащие токсины. Защита от фитофагов более необходима растениям при недостатке ресурсов. Наблюдается компромисс между стоимостью синтеза мобильных веществ (выгодно при небольшой длительности жизни листьев) и немобильных детеррен-тов (выгодно при большой длительностью жизни) (Howe, Westley, 1988).
Любые факторы, которые ограничивают рост сильнее, чем фотосинтез (умеренная засуха, умеренный недостаток ЭМП, низкие температуры) способствуют увеличению пула ассимилятов для синтеза защитных веществ. Напротив, затенение уменьшает этот пул и часто приводит к увеличению поражаемости растений фитофагами. Например, в экспериментах с искусственным затенением вереска Calluna vulgaris (пропускание тентов — 12 %, т. е. условия сменяющего верещатник березняка) в его побегах снижалась концентрация лигнина, фенолов и конденсированных танинов, что вызывало их большую поедаемость зайцами и оленями, т. е. ускоряло сукцессионную смену верещатника березняком (lason, Hester, 1993).
3. Гипотеза «универсальной» защиты (Edwards, 1989) — эволюционно более древние группы растений имеют комплекс адаптаций универсальной (neutral resistence) защиты (толстые стенки листьев, жесткое
120
Глава 4. Растения и животные
опушение, танины и т. п.). Эти приспособления помогают переживать неблагоприятные воздействия условий среды, действуют против патогенных микроорганизмов, а также животных. Напротив, у эволюционно молодых групп преобладает специфическая защита против животных (преимущественно токсины — алкалоиды, терпеноиды, глюкозиды).
4. Гипотеза уязвимости («недостатка-доступности», Grubb, 1992). Большее развитие защитных приспособлений свойственно растениям, более привлекательным для животных, но и более ограниченных недостатком ресурсов, временем вегетации и т. п. Эта гипотеза предполагает наиболее выраженную защиту как в наиболее бедных ресурсами местообитаниях, так и в богатых, площадь и/или время существования которых невелики. Гипотеза Грабба хорошо объясняет как механические средства защиты (см. выше), так и химические. Среди деревьев и высоких кустарников в Европе токсичны для человека и скота следующие группы (рис. 4.13):
а) растения низкопродуктивных местообитаний (Juniperus sabina);
б) быстрорастущие растения богатых почв в прорывах полога (Sambu-cus nigra, S. racemosa);
в) вечнозеленые виды листопадных лесов (Buxus sempervirens, Prunus laurocerasus, Rhododendron ponticum, Taxus baccata);
г) раннецветущие виды (Daphne mezereum);
д) растения с более высоким, чем у соседей, содержанием азота (бобовое Laburnum anagyroides);
Большинство ранневесенних эфемероидов также содержат ядовитые вещества (Allium ursinum, Anemone nemorosa, Arum maculatum, Colchicum autumnale, Dentaria bulbifera, Ficaria verna, Corydalis spp.).
Сходная с предыдущей гипотеза оптимальной защиты предполагает, что поскольку синтез защитных веществ требует затрат ассимилятов, растения защищают свои ткани в прямой зависимости от стоимости их возможной потери. Легко заменяемые ткани и органы нуждаются в меньшей защите, чем более ценные, трудно восстановимые.
Рис- 4.13 • Экологические группы токсичных древесных растений средней Европы (по: Grubb, 1992): а — растения низкопродуктивных местообитаний (Juniperus sabina); б — быстрорастущие растения богатых почв в прорывах полога (Sambucus nigra); в — растения с более высоким, чем у соседей, содержанием азота (бобовое Laburnum anagyroides); г — вечнозеленые виды листопадных лесов (Prunus laurocerasus); д — раннецветущие виды (Daphne mezereum)
3. Защитные адаптации растений от поедания
121
122
Глава 4. Растения и животные
3.6 • Реакция растений и сообществ на повреждение фитофагами
1) Усиление синтеза защитных веществ. Во многих работах показано усиление синтеза защитных веществ растениями в ответ на повреждение их фитофагами. Однако лишь в немногих случаях показана действительное угнетение фитофагов и уменьшение повреждения растения в ответ на дополнительный синтез защитных веществ.
Рассмотрим некоторые примеры. Синтез азотсодержащих защитных веществ, как показали эксперименты с содержанием никотина в табаке, усиливается при повреждении растений, а также в условиях более бедных почв. При недостатке азота его доля, идущая на синтез никотина, увеличивается более чем в 3 раза (Ohnmeiss, Baldwin, 1994).
При повреждении насекомыми-фитофагами хвойных растений они вырабатывают больше терпенов и меньше поражаются фитофагами в дальнейшем (Raffa, Berryman, 1983; Schultz, 1988).
McNaughton et al. (1985) показали, что содержание кремния в злаках саванн Африки (Серенгети) было значительно выше на участках с интенсивным выпасом. В специальном эксперименте было исследовано изменение содержания кремнеземистых фитолитов в двух злаках (Festuca ovina и Lolium регеппе) при различной интенсивности и форме дефолиации (механическом срезании листьев на высоте 5 см, поедании полевками и саранчой). Содержание кремнезема слабо возрастало при механических повреждениях, но увеличивалось при повторяющемся воздействии фитофагов до 400 %. В дальнейшем фитофаги меньше повреждали растения с более высокой концентрацией кремния (Massey et al., 2007). Аналогично при увеличении численности полевок и леммингов в Скандинавии относительное содержание фенолов (так называемый «защитный индекс» — отношение содержания фенолов к общему азоту) в листьях черники возросло почти вдвое в течение 2 лет (Oksanen, Oksanen, 1981).
Когда насекомые или исследователь повреждают лист березы, соседние листья этого же растения становятся менее питательными для гусениц (Haukioja, 1980; Howe, Westley, 1988). Более того, для некоторых растений показано усиление синтеза защитных веществ, если фитофагами повреждено соседнее растение. «Индукторами» этого процесса могут выступать газообразные выделения поврежденных растений, в частности, этилен (Schultz, 1988). Agrawal (2000) также приводит несколько примеров сигнальных взаимодействий между растениями при повреждении фитофагами. Сигнальными веществами могут служить жасмонат и салицилат — летучие вещества, обладающие сильным физиологическим действием. Вокруг дерева Alnus glutinosa, подвергшегося дефолиации,
3. Защитные адаптации растений от поедания
123
Рис. 4.14 • Различия в индуцированном синтезе защитных веществ у растений хлопчатника: а — без фитофагов; б — при повреждении надземными фитофагами; в — при повреждении подземными фитофагами. Интенсивность окраски листьев показывает индукцию синтеза защитных веществ (белая — нет индукции, светлая — низкая индукция, темная — высокая индукция) (по: Bezemer, van Dam, 2005)
уменьшалась поражаемость фитофагами других деревьев. Табак, росший вокруг поврежденной полыни, был более устойчив к фитофагам, чем росший вне поврежденной полыни.
Синтез защитных веществ растениями в ответ на повреждения фитофагами может повышать синтез этих веществ и у их потомков, возникших из семян. Так, потомки особей Raphanus raphanistrum, поврежденных фитофагами, были более устойчивы к фитофагам, чем потомки неповрежденных особей (Agrawal, 2002b).
Индукция синтеза защитных веществ в ответ на поражение фитофагами широко распространена. Поражение надземными фитофагами может усиливать такой синтез, влияющий и на подземных фитофагов, и наоборот. Это влияние может проявляться в разной степени. Например, у хлопчатника повреждение корней ведет к увеличению концентрации терпеноидов во всех листьях, в то время как повреждение листьев сильно увеличивает концентрацию терпеноидов только у молодых листьев (рис. 4.14). У турнепса (Brassica гара) при повреждении корней личинками двукрылых побеги выделяют специальные вещества, привлекающие паразитоидов этих личинок (Neveu et al., 2002; Bezemer, van Dam, 2005).
2) Компенсаторный рост. Его внешние причины: возрастание освещенности, обеспечения водой и ЭМП, возможны (хотя редко показаны) прямые воздействия на рост растений гормонов и ростовых веществ, продуцируемых травоядными. Например, листовые минеры задерживают старение листьев, в которых они обитают, по сравнению с незаселенными листьями (Engelbrecht, 1971; McNaughton, 1983).
При повреждении листьев фитофагами фотосинтез оставшихся листьев усиливается, а при повреждении завязей и плодов — ослабевает
124
Глава 4. Растения и животные
(меньше необходимо ассимилятов для созревающих плодов). В целом растения значительно чаще могут компенсировать потерю биомассы и продукции, но редко — семенную продуктивность после повреждения фитофагами (Crawley, 1999).
В ряде случаев отрастающие побеги после повреждений растений затем сильнее повреждались фитофагами, так как их листья имели более крупные размеры, высокое содержание азота, меньшую жесткость и плотность волосков, что было показано для тропического кустарника Conocarpus erectus (Spiller, Agrawal, 2003).
3) Изменения в распределении ресурсов (аллокации). Например, для инвазионного сорняка Centaurea maculosa в США показана высокая устойчивость и даже усиление роста побегов при поражении корневым фитофагом Agapeta zoegana. Выявлено, что хотя фитофаг снижает общее потребление азота растением на 30 %, а поглотительную способность его корней — на 50 %, но содержание азота в побегах не уменьшалось. Это объясняется большей транслокацией соединений азота из корней в побеги у поврежденных растений (Newingham et al., 2007).
4) Популяционные эффекты. Под воздействием фитофагов изменяется продолжительность жизни особей, приживание всходов и другие популяционные характеристики. Наблюдения в течение 20 лет за фиксированными особями растений в условиях выпаса и его отсутствия показали, что скорость обновления и колебания численности особей выше на выпасаемых участках. Средняя продолжительность жизни злаков увеличивается, а кустарников уменьшается при выпасе (West et al., 1979).
4 • Фитофагия и изъятие первичной продукции
Доля потребляемой фитофагами надземной продукции в естественных фитоценозах весьма различна, она колеблется от долей процентов до 70% и более. Например, в злаковниках Серенгети травоядные могут выедать до 94% надземной продукции (McNaughton et al., 1985; Zamora et al., 1999). В среднем для наземных ценозов приводится цифра изъятия 18% надземной продукции (Суг, Расе, 1993; Strauss, Agrawal, 1999).
Различные группы животных изымают разную долю продукции. Так, мелкие млекопитающие («мышевидные грызуны») в естественных фитоценозах потребляют обычно около 1,5-2% надземной продукции. Коэффициент усвоения энергии из пищи у грызунов высок и составляет около 80%. Насекомые фитофаги потребляют около 10% продукции естественных сообществ (Schultz, 2002). Наибольшее изъятие продукции
4. Фитофагия и изъятие первичной продукции
125
Таблица 4.2 • Три типа растительных сообществ с различной трофической структурой и относительной ролью фитофагии и конкуренции (по: Fretwell, 1977; Oksanen et al., 1981; Oksanen, 1990)
Продуктивность растительного сообщества
Низкая Средняя Высокая
Трофическая структура
Без существенного влияния фитофагов С фитофагами, но без хищников С хищниками
Роль фитофагов
Низкая Высокая Низкая
Роль конкуренции
Высокая Низкая Высокая
отмечено стадными саранчевыми — неспециализированными фитофагами, потребляющими почти любые зеленые ткани растений.
Но при оценке изъятия годичной продукции животными необходимо учитывать не только съеденную массу, но и травматический опад, который часто превышает потребление. Так, у мелких млекопитающих травматический опад примерно вдвое выше потребляемой биомассы (Добринский и др., 1983).
С. Д. Фретвел и Л. Оксанен предложили классификацию сообществ по их продуктивности и относительной роли фитофагии и конкуренции в их формировании (табл. 4.2) (Fretwell, 1977; Oksanen et al., 1981; Oksanen, 1990). Низкопродуктивные сообщества не могут поддерживать значительную численность фитофагов, поэтому роль фитофагии в них невелика, а конкуренция за ресурсы между растениями существенна. Сообщества со средней продукцией поддерживают популяции фитофагов, но не поддерживают следующий трофический уровень (хищников). Поэтому фитофаги регулируются доступностью корма, роль фитофагии в организации сообществ велика, а конкуренции — снижена. Высокопродуктивные сообщества уже могут поддерживать трофический уровень хищников, которые в свою очередь регулируют популяции фитофагов. Относительная роль фитофагии в организации сообществ снижается, а конкуренции между растениями — возрастает, хотя абсолютное потребление растений фитофагами может быть очень велико.
Представленные закономерности были проверены на нескольких примерах. Так, в Швеции исследовали участие, популяционную структуру
126
Глава 4. Растения и животные
60 -|
40 -
20 -
О -
БХ ХП
БП
Рис- 4-15 • Надземная биомасса черники (среднее и стандартная ошибка) на островах без хищных животных (БХ, п = 4), участках материка (с хищниками и полевками, ХП, п = 4) и на островах без полевок (БП, п = 5) летом 2003 г. Значимые различия наблюдались между вариантами БХ и остальными (по: Dahlgren et al., 2007)
и пищевую ценность побегов черники (Vaccinium myrtillus) на островах без основных потребителей (полевок Clethrionomys rufucanus), с полевками, но без хищников, и на материке с полевками и хищниками (Dahlgren et al., 2007). Биомасса черники была максимальной в отсутствии полевок, меньшей на материке (на 50 %) и самой низкой на островах с полевками без хищников (на 90%) (рис. 4.15). В лабораторном эксперименте полевки охотнее потребляли чернику с островов без полевок, которая содержала меньше защитных веществ.
Хотя рассматриваемая классификация сообществ по их продукции и роли фитофагов исходно была предложена для оценки влияния позвоночных-фитофагов, она была также экспериментально подтверждена в работе Fraser и Grime (1997) для беспозвоночных. На 6 лугах различной продуктивности неоднократно в течение вегетационного сезона применялись инсектициды и препараты против моллюсков. Урожай соответствующих лугов сравнивался с контрольным, подверженным действию фитофагов. Как показано на рис. 4.16, наибольшее увеличение урожая отмечено в средних по продуктивности лугах, т. е. там роль фитофагов наиболее значительна. Это также подтвердилось в экспериментах с пересадками, когда Urtica dioica была пересажена на среднепродуктивные луговые участки, ее повреждаемость без применения пестицидов резко возросла (рис. 4.16). Таким образом, в высокопродуктивных участках хищные насекомые эффективно сдерживают численность фитофагов.
Кроме хищников, численность фитофагов могут регулировать их паразиты и болезни. По данным для Северной Америки (Dobson, Hudson, 1986; Polis, 1999), в среднем каждая популяция позвоночных заражена 10 видами паразитических червей, а каждая особь несет в себе в среднем более 350 индивидуальных паразитов.
У разных видов растений в составе ценозов фитофаги изымают разную долю продукции. Например, по наблюдениям в мангровых зарослях на побережье Австралии (Robertson, 1991; van Andel et al., 1993), среднее повреждение развернутых листьев деревьев составляло 5 %, изменяясь
4. Фитофагия и изъятие первичной продукции
127
а)
б)
Рис.4.1в • Изменение надземной биомассы лугов разной продуктивности при применении пестицидов (а) и масса побегов крапивы (Urtica dioica) на высоко- («высокотравье») и среднепродуктивных лугах («луг») с (4-П) и без (-П) применения пестицидов (б) (по: Fraser, Grime, 1997)
от 0,3 % у Excoecaria agallocha до 35 % у Avicennia marina и Heritiera littoralis.
Но на структуру сообществ и отдельные виды растений наибольшее влияние оказывает не доля изымаемой фитомассы, а ее качественный состав и сроки изъятия. McNaughton (1983) предложил следующую функциональную классификацию фитофагов.
1. Потребители цветков, плодов и семян.
2. Потребители листьев.
3. Потребители корней.
4. Стеблевые минеры и сверлильщики.
5. Потребители соков.
6. Галлообразователи.
В отдельную группу могут быть выделены моллюски, которые на лугах оказывают очень большое влияние (непропорциональное своей биомассе), поскольку избирательно поедают всходы растений (Duthoit, 1964). Моллюски могут убивать всходы одним укусом радулы в гипокотиль (Dirso, Harper, 1980; Fenner et al., 1999). Всходы могут также поедаться многими насекомыми (Kitajima, 2007). Например, Louda (1982) показал, что исключение насекомых-семеноядов (обработка инсектицидами) у калифорнийского кустарника Haplopappus squarrosus увеличило число прижившихся всходов через год в 23 раза!
128
Глава 4. Растения и животные
Семенояды имеют большее влияние на участие растений в фитоценозах, а при потреблении вегетативных органов большое значение имеют сроки (фенологические фазы). Потребление почек и молодых побегов сильнее сказывается, чем потребление взрослых листьев и тем более ветоши. Например, Schowalter (1989) показал, что поражающая почки дерева Pseudotsuga menziesii моль Zeiraphera hesperiana потребляет биомассу листьев менее, чем на 1 %, но вызывает снижение продукции побегов и новых листьев на 13% (Schowalter, 2006). Напротив, ветошь (отмершие части растений) используется копытными на зимних пастбищах в высокогорьях, ее изъятие благоприятно сказывается на скорости биологического круговорота и продукции растительных сообществ.
Потребление цветков фитофагами широко распространено и в некоторых случаях может превосходить потребление листьев по масштабам и влиянию на растения. Например, северные олени, избирательно поедающие генеративные побеги дриады (Dryas octopetala), могут существенно снижать интенсивность семенного размножения этого вида (Cooper, Wookey, 2003). Для травянистого растения Silene latifolia отмечено повреждение цветков фитофагами у 15% всех особей в 50 популяциях в Европе (Wolfe, 2002). Поедание части цветков на растении может уменьшать роль гейтеногамии. Подобно листьям, части околоцветника могут иметь те же средства защиты от фитофагов (химические, физические барьеры, избегание во времени и пространстве, непрямые защитные механизмы). Содержание защитных веществ в цветках может быть выше, чем в листьях. Например, концентрация протеиназных ингибиторов в цветках томатов может в 400-700 раз превосходить таковую в листьях и плодах (Damle et al., 2005; McCall, Irwin, 2006).
Потребление вегетативных органов, особенно листьев, может значительно снижать семенную продуктивность растений. Фитофаги могут также воздействовать на приспособленность растений, изменяя свойства пыльцы. В экспериментах с Lobelia siphilitica (Mutikainen, Delph, 1996) после удаления 50 % листьев растения продуцировали пыльцу, скорость роста пыльцевой трубки которой была существенно ниже, чем у растений, не подвергшихся дефолиации.
Потребители соков (особенно флоэмного) больше повреждают растения за счет переноса инфекции, чем за счет изъятия ассимилятов. Потребители корней оказывают наибольшее влияние на жизненность растений в условиях лимитирования роста растений почвенными ресурсами, но имеют меньшее значение на богатых почвах (Crawley, 1999). Длительность жизни корней во многом зависит от воздействия фитофагов и патогенных грибов; по наблюдениям в миниризотронах корни чаще
5. Фитофаги и смены растительных сообществ
129
исчезают, чем отмирают и разлагаются (Johnson et al., 2001). Применение инсектицидов и фунгицидов увеличивало длительность жизни корней на 46-125 дней у персика (Wells et al., 2002а) и более чем на 500 дней у сахарного клена (Eissenstat et al., 2000; Yanai, Eissenstat, 2002).
Избирательность потребления растений и их частей часто связана с содержанием в них азота, так как ткани животных в большей степени состоят из белков (> 50 % сухой массы). Эффективность перевода растительной пищи в собственную биомассу (рост тела/количество потребленной пищи) обычно линейно возрастает с увеличением содержания азота в пище. Эта величина колеблется от 0,3 % до 58 %. Величины ниже 1 % известны для членистоногих, питающихся бедными азотом древесиной, опадом и детритом, а наибольшие значения — для животных, питающихся высококалорийной пищей с большим содержанием белков (семена, флоэмный сок, пыльца). При бедной азотом диете более выгодно иметь большие размеры тела. Эта закономерность отмечена, например, для бабочек. Самые мелкие виды (гусеницы) питаются на молодых листьях, а виды с крупными гусеницами — зрелыми, жесткими, бедными азотом листьями (Mattson, 1980).
Какие растения больше повреждаются фитофагами — ослабленные, находящиеся в условиях недостатка ресурсов или наоборот, хорошо развитые в условиях достатка ресурсов и высокого содержания белков в тканях? Было высказано две гипотезы: 1) большая уязвимость угнетенных растений, 2) большая привлекательность для фитофагов растений из благоприятных условий. Эксперименты с листовыми минерами — му-хами-агромизидами Agromyza nigripes на Holcus lanatus, выращиваемом в разных условиях, показали справедливость второй гипотезы (De Bruyn et al., 2002).
5 • Фитофаги и смены растительных сообществ
Растения, образующие сообщества разных сукцессионных стадий, часто имеют различные средства защиты от фитофагов. Виды начальных стадий сукцессий обычно образуют токсины как защитные вещества, а виды последних стадий — детерренты, что связано с разной длительностью жизни побегов. Вопрос о роли фитофагов в ускорении или замедлении сукцессий интересовал многих исследователей. Так, D. W. Davidson (1993) проанализировал 34 публикации по роли животных в сукцессиях и сделал вывод, что влияние потребителей листвы и побегов на скорость сукцессий неоднозначна (в 10 работах фитофаги ускоряли сукцессию,
130
Глава 4. Растения и животные
в 24 — замедляли). В бореальных лесах лучше поедаются виды растений средних стадий сукцессий — злаки, кустарники, пионерные деревья, т. е. фитофаги могут замедлять сукцессию на ранних стадиях и ускорять — на поздних. В отличие от потребителей вегетативных органов, семено-яды чаще предпочитают виды поздних стадий сукцессий (имеющих более крупные семена), таким образом замедляя сукцессию. В экспериментах Brown и Heske (1990) с участков пустынь были удалены зерноядные грызуны, что привело к смене летних однолетников злаками, т. е. переходу пустынь в злаковники, который ранее тормозился грызунами (Davidson, 1993). В аналогичных экспериментах по изоляции участков пустыни (Chihuahuan Desert) от птиц и грызунов показано, что такая изоляция значительно повышает участие зимних однолетников, имеющих крупные семена (Guo et al., 1995). В опытах по удалению грызунов или муравьев в той же пустыне наблюдалась тенденция к увеличению участия крупно-семянных видов при удалении грызунов и уменьшению их участия при удалении муравьев, в то время как для мелкосемянных видов тенденция была обратной (Guo, Brown, 1996).
Для начальных стадий вторичных сукцессий показана большая роль почвенных насекомых-фитофагов. Применение почвенных инсектицидов существенно изменяло структуру сообществ растений, более чем вдвое увеличивало флористическую насыщенность сообществ. В то же время применение листовых инсектицидов не вызывало таких эффектов (Brown, Gange, 1989; Gurevitch, Collins, 1994). Роль подземных беспозвоночных-фитофагов часто недооценивается, а они в экспериментах на залежах потребляли 30-40 % продукции. В то же время снижение веса надземных побегов при потреблении корней происходит скачкообразно: оно практически не проявляется при потреблении до 50-60% корней и резко проявляется при более высоких потерях. Подземные фитофаги особенно усиливают смертность всходов, в том числе проросших семян, ростки которых еще не вышли на поверхность почвы (Brown, Gange, 1991). В целом, как показали Brown и Gange (1992), в луговом сообществе применение почвенных инсектицидов ведет к доминированию двудольных, а применение листовых инсектицидов усиливает роль злаков (Wardle, van der Putten, 2002).
Роль разных групп млекопитающих на ранних стадиях вторичной сукцессии была изучена на залежи в Вирджинии (США). Поле обработали гербицидами и вспахали. В экспериментах было 4 варианта: (1) контроль, (2) исключение всех групп млекопитающих (олени, сурки, кролики и мелкие грызуны), (3) «крупные ворота» — доступ всех млекопитающих, кроме оленей, (4) «мелкие ворота» — доступ только мелких грызунов.
6. Фитофагия и флористическое богатство
131
Флористическая насыщенность через 4 года эксперимента была существенно выше при частичном исключении фитофагов (варианты 3 и 4), чем на контроле, или при полном их исключении. Покрытие трав в контроле было ниже, чем в других вариантах. Виды растений индивидуально реагировали на условия эксперимента. Так, Asterpilosus доминировал при полном исключении млекопитающих-фитофагов, a Celastrus scandens — на контроле (Bowers, 1993).
Влияние животных на участие отдельных видов растений в составе фитоценозов часто весьма значительно. Например, Haplopappus squar-rosus в прибрежных сообществах Калифорнии активно потребляют насекомые. В экспериментах с применением инсектицидов (Louda, 1982) показано, что численность всходов этого вида после обработки резко возрастает в прибрежной полосе, но не в других сообществах (Howe, Westley, 1988).
Завоз новых фитофагов может вызвать существенные изменения в структуре растительных сообществ. Например, интродукция ондатры, которая предпочтительно потребляет тростник, вызвала угнетение тростниковых зарослей и разрастание телореза (Новиков, 1976). Видимо, большое влияние на растительность оказывает и обратный процесс — исчезновение каких-либо фитофагов.
6 • Фитофагия и флористическое
богатство
Фитофаги играют большую роль в поддержании флористического богатства растительных сообществ и сосуществовании видов (Proulx, Mazumder, 1998; Olff, Ritchie, 1998). Они могут быть регуляторами участия доми-нантов, предотвращая или замедляя конкурентное исключение одних видов растений другими. Как крайние варианты этого регулирования можно рассмотреть влияние узкоспециализированных фитофагов и неспециализированных фитофагов с трофической адаптацией (избирательно потребляющих виды с большим участием). В первом случае каждый из сосуществующих в сообществе видов растений имеет своего фитофага, увеличивающего численность при возрастании участия его кормового растения. Таким образом, участие всех видов растений регулируется фитофагами, не дающими конкурентное преимущество какому-либо виду (Боголюбов, 1989; Tilman, 1982, 1986). В другом крайнем случае один неспециализированный фитофаг, обладающий способностью к трофической адаптации — избирательному поеданию видов с большим участием (доминантов), — может поддерживать сосуществование теоретически
132
Глава 4. Растения и животные
Площадь ареала растений (число квадратов со стороной 10 км)
Рис. 4.17 • Число видов насекомых-фитофагов (ордината, логарифмическая шкала), связанных с многолетними травами на Британских островах, в зависимости от площади ареалов этих трав (абсцисса, логарифмический масштаб) (по: Lawton, Schroder, 1977; Schoonhoven et al., 2005)
неограниченного числа видов растений, подавляя виды с большей конкурентоспособностью (Shmida, Ellner, 1984). Механической моделью такого фитофага может быть сенокосилка (см. о сенокошении, п. 3.9.3 гл. 9).
Естественные растительные сообщества поддерживают популяции фитофагов с различной степенью трофической специализации. Например, некоторые исследователи считали, что в тропических лесах обитают сотни видов насекомых-фитофагов, специализированных к отдельным видам деревьев. Представление об очень высоком биоразнообразии тропических насекомых была связана с переоценкой степени их специализации к питающим растениям. Более точный анализ для лесов Новой Гвинеи (Novotny et al., 2002а, Ь) показал, что из 1010 видов листогрызущих насекомых на 59 видах деревьев только < 4 % были специализированы на одном виде, большинство видов были специализированы на деревьях одного рода. Авторы делают вывод, что число видоспецифичных насекомых-фитофагов измеряется десятками, а не сотнями на один вид деревьев (Roslin, 2002).
Уровень специализации фитофагов рассмотрен в сводке Schoonhoven et al. (2005). Авторы показали, что на примерно 300000 видах сосудистых растений питается около 400 000 видов насекомых-фитофагов. Среди трофических групп фитофаги составляют 100% в отрядах Lepi-doptera, Orthoptera, Phasmida (палочники) и 90% — в отрядах Hemiptera
6. Фитофагия и флористическое богатство
133
(клопы) и Thysanoptera (трипсы). Менее 10% видов фитофагов питается на растениях более, чем 3-х семейств (Bernays, Chapman, 1978). В Великобритании 76% видов тлей — монофаги, 18% — олигофаги, и только 6 % — полифаги. Виды, питающиеся на травах, обычно более специализированы, чем виды, питающиеся на древесных растениях. Мелкие по размерам насекомые более специализированы, чем крупные. Более специализированные фитофаги предпочитают молодые и разворачивающиеся листья, а менее специализированные — зрелые листья. На дубах в Великобритании (2 вида) найдено 423 вида фитофагов, а на тисе — 6 (Kennedy, Southwood, 1984). Чем шире распространены виды растений, тем больше на них видов фитофагов (рис. 4.17) (Schoonhoven et al., 2005).
Интенсивность изъятия биомассы фитофагами также влияет на флористическую насыщенность сообществ. В многих работах показано, что наибольшее флористическое разнообразие наблюдается при средней интенсивности воздействия фитофагов. Например, Zeevalking, Fresco (1977) показали эту зависимость для диких кроликов на дюнных островах (рис. 4.18).
Анализ публикаций по воздействию фитофагов на видовое разнообразие сообществ (Proulx, Mazumder, 1998) показал, что интенсивное воздействие фитофагов снижает видовое богатство сообществ, лимитированных ЭМП, и повышает таковое для сообществ с высоким поступлением ЭМП. На 10-кратном градиенте продукции лугов в Северной Америке и Европе было изучено влияние фитофагов разных размеров на видовое разнообразие растительных сообществ. Показано, что более
Рис. 4.18 • Зависимость числа видов растений
на площадках от интенсивности изъятия биомассы кроликами в дюнных сообществах (по: Zeevalking, Fresco, 1977)
134
Глава 4. Растения и животные
крупные фитофаги (более 30 кг — копытные и др.) увеличивают разнообразие в высокопродуктивных растительных сообществах, но уменьшают — в низкопродуктивных, однако для более мелких фитофагов (например, грызунов) такой закономерности не отмечено. Меньшее влияние фитофагов в низкопродуктивных сообществах может быть связано с большей долей подземной биомассы в них (Bakker et al., 2006).
7 • Роющая деятельность животных
Изменение почвы при естественных нарушениях, вызванных роющей деятельностью животных, имеет большое значение для формирования растительных сообществ и их видового разнообразия. Среди роющих животных наибольшую роль играют млекопитающие и дождевые черви. Роющая деятельность млекопитающих особенно значимо проявляется в нелесных сообществах, где отсутствуют другие естественные убежища. Так, по данным В. В. Кучерука (1960), число входных отверстий нор на гектар в разных природных зонах Евразии составило следующие порядковые величины:
тундра тайга 1000 500
широколиственные леса луговая степь настоящие степи 1000 7500 5000
пустынные степи пустыни 2000 1500
Роющая деятельность животных оказывает как прямое (погребение растений, повреждение подземных органов), так и косвенное (через изменение почвенных свойств) воздействие на растительность. Б. Д. Абатуров (1984) выделяет следующие основные направления воздействия на почву роющей деятельности млекопитающих на аридных территориях: 1) разрыхление, улучшение аэрации, увеличение водопроницаемости
и более глубокое увлажнение;
2) рассоление почвы в местах поселений грызунов за счет лучшего промывания или, напротив, вынос на поверхность более засоленных нижних слоев почвы;
3) вынос материалов из нижних горизонтов и погребение верхних, погребение семян — важная роль в создании семенных банков;
7. Роющая деятельность животных
135
4) формирование микрорельефа с холмиками и западинами;
5) усиление подверженности ветровой и водной эрозии;
6) обогащение почвы ЭМП за счет поступления экскрементов животных;
7) формирование на разрыхленных участках залежного режима с поселением рудеральных растений.
Роющая деятельность животных способствует выносу на поверхность почвы до 10% захороненных радиоактивных элементов в местах, подвергшихся радиоактивному заражению (Криволуцкий, Покаржевский, 1996).
Нарушения, образующиеся в результате роющей деятельности (норы, выбросы почвы), создают неравномерность в пространственном распределении биомассы растений. Она обычно резко снижена на нарушенном участке, но увеличена на границе нарушения и исходного сообщества. Таким образом, изменение биомассы на пороях носит «волнообразный» характер, что было показано, например, для пороев карманного гофера (Geomys bursarius) в высокотравной прерии США. Увеличение биомассы вблизи нор и выбросов связано с лучшими условиями освещенности и поступления ЭМП на эти участки (Reichman et al., 1993).
Видовой состав растений в местах зоогенных нарушений может значительно отличаться от такового в ненарушенном сообществе. Группировки сорных растений, широко распространенные в настоящее время на обрабатываемых человеком землях, существовали задолго до появления человеческой культуры и обязаны своим происхождением роющим животным. Но в то же время многие доминанты степей возобновляются из семян только на нарушенных (в том числе и землероями) участках. В результате роющей деятельности животных в степях возникают прорывы полога («окна»), происходит угнетение плотнодерновинных злаков и разрастание корневищных злаков («залежный режим»), что особенно ярко наблюдается в степях Монголии при циклических возрастаниях численности фонового вида роющих грызунов — полевки Брандта (см. «Флуктуации», п. 2 гл. 9). На некоторых типах лугов произрастание отдельных видов может четко индицировать следы роющей деятельности животных, как, например, строгая приуроченность взрослых особей примулы весенней (Primula veris) к ходам и норам обыкновенной полевки на сухих лугах северо-западной Швейцарии (Gigon, Leutert, 1996, рис. 4.19).
В аридных злаковниках района Кару (Капская провинция) порой создаются в результате роющей деятельности трубкозубов, дикобразов и лисиц. Число всходов на открытых местах на пороях было существенно выше, чем вне их. В междюнных понижениях 96 % всех всходов росло на пороях, а выживаемость однолетников на пороях была почти в 4 раза выше, чем вне их (Dean, Milton, 1991).
136
Глава 4. Растения и животные
Рис. 4.19 • Приуроченность особей Primula veris к участкам ходов и нор обыкновенной полевки на сухих лугах Швейцарии (по: Gigon, Leutert, 1996). Обозначения: 1 — входные отверстия нор, 2 — участки пороев, 3 — действующие тропки полевок, 4 — старые тропки, 5 — генеративные особи примулы с 2 и более соцветиями, 6 — генеративные особи примулы с одним соцветием, 7 — вегетативные розетки примулы
Роющие травоядные млекопитающие представлены на разных континентах — в Северной Америке это гоферы (Geomyidae), в Африке — землекоповые (Bathyergidae), в Европе — слепышовые (Spalacidae), в Южной Америке — гребнемышиные (Ctenomyidae). Они питаются подземными органами растений, потеря которых может иметь более негативный эффект, чем потеря надземных органов. Так, О. J. Reichman и S. С. Smith (1991) показали, что потеря 25% подземной биомассы для козлобородника сомнительного (Tragopogon dubius) имеет больший эффект, чем потеря 75% листьев. Роющая активность гоферов Thomomys bottae ведет к доминированию однолетних растений, в свою очередь являющихся излюбленной пищей гоферов (Huntly, Inouye, 1988). Anderson и MacMahon (1981) показали, что карманный гофер Thomomys talpoides потребляет около 30 % первичной подземной продукции альпийского луга. Объем выбросов гоферов варьирует от 3,4 до 57,7 м3 на гектар в год (среднее — 17,8) (Smallwood, Morrison, 1999). Порой покрывают 7,5 % площади, выбросы могут занимать до 30-50 % за год на отдельных участках (Reichman, Seabloom, 2002). Подземная жизнь ведет к выработке общих адаптаций у грызунов разного систематического положения
8. Животные и опыление растений
137
(сильные мышцы для рытья, устойчивость к низкой концентрации О2 и высокой СО2, большая чувствительность к вибрациям почвы и др.). Передвижение под землей требует больших затрат энергии — чтобы вырыть 1 м ходов надо затратить в 360-3400 раз больше энергии, чем для прохождения 1 м по поверхности (Reichman, Seabloom, 2002).
Роющая деятельность дождевых червей широко распространена во всех природных зонах. Выбросы почвы (копролиты) дождевых червей обогащены ЭМП по сравнению с окружающей почвой. Например, содержание доступных форм ЭМП составляло (в частях на миллион на сухую почву) (Russell, 1973; Trudgill, 1988):
элемент Са Mg К Р N(NO3)
верхний слой почвы 1990 162 32 9 4,7
копролиты червей 2790 492 358 67 21,9
Роющая деятельность дождевых червей имеет большое значение не только для изменения структуры почвы, но для переноса семян растений между отдельными горизонтами. Особенно часто переносятся мелкие семена, например орхидных и грушанковых. Изучение переноса семян червем Lumbricus terrestris на меловых лугах в Голландии показало, что весной черви активны в верхних слоях почвы, а летом — на большей глубине (Willems, Huijsmans, 1994). Масса копролитов составила не менее 750 г/м2 в год, на поверхность выносилось 60-100 семян на м2 ежегодно. В копролитах с лугов найдены семена 26 видов растений, избирательность в поглощении семян была связана, главным образом, с их вкусом и запахом, а не размерами. Индивидуальные норы Lumbricus terrestris в почве в Англии достигают 3 м, а общая их длина до глубины 1 м на площади 1 га может достигать 888 м (Edwards, Lofty, 1977; Gange, Brown, 2003).
8 • Животные и опыление растений
Примерно 90 % цветковых растений опыляется животными (Buchmann, Nabhan, 1996) при этом 67% — насекомыми (Tepedino, 1976). Опылители могут выступать лимитирующим фактором семенной продуктивности и распространения растений. Например, Д. В. Панфилов и др. (1960) предположили, что сходный характер северной границы ареалов шмелей (Bombus), мытников (Pedicularis) и бобовых в Арктике обусловлен, тем, что распространение этих растений сдерживается опыляющими их шмелями, которые менее устойчивы к низким температурам. Применение инсектицидов в сельском хозяйстве может отрицательно сказаться
138
Глава 4. Растения и животные
8. Животные и опыление растений
139
Рис. 4.20 • Основные группы животных-опылителей: а — жуки (пример Leptura annularis); б — падальные мухи Calliphoridae, привлеченные специфическим запахом цветка Stapelia grandiflora, откладывают в него свои яйца (фото А. Бенедиктова, с любезного разрешения автора); в — муха-жужжала Bombylius sp. (фото А. Михайленко, с любезного разрешения автора); г — бражник (фото Nigel Stott, natural-japan.net, с любезного разрешения автора); д — дневные бабочки (махаон — Papilio machaon) (фото А. Михайленко, с любезного разрешения автора); е — шмель (Bombus sp.) на цветках Cirsium; ж — колибри у цветков Crocosmia sp. (Iridaceae) (фото Mila Zinkova, с любезного разрешения автора); з — летучая мышь опыляет цветки агавы (фото Charles W. Melton, www.nearfamous.com, с любезного разрешения автора)
на популяциях энтомофильных растений, произрастающих на прилегающих к обрабатываемым полям территориям. Хищники, питающиеся насекомыми-опылителями, также могут снижать семенную продуктивность перекрестно опыляющихся растений. На примере Leucanthemum vulgare показано, что сидящие в соцветиях крабовые пауки снижали семенную продуктивность соцветий за счет отрицательного воздействия на опылителей (Suttle, 2003).
Основными группами опылителей являются насекомые, птицы и летучие мыши, в отдельных районах — нелетающие млекопитающие (рис. 4.20). Хорошо документированные сведения об опылении растений нелетающими млекопитающими известны для 59 видов животных из 19 семейств и 6 отрядов. Они опыляют 85 видов растений из 19 семейств. Наиболее часто опыляются виды рода Banksia (Proteaceae) — 16 видов и Eucalyptus (Myrtaceae) — 18 видов. Поскольку многие млекопитающие — ночные животные, то цветки этих растений обычно слабо окрашены, но сильно пахнут (Carthew, Goldingay, 1997).
140
Глава 4. Растения и животные
Таблица 4-3 • Синдромы опыления цветков разными группами животных (по: Howe, Westley, 1988)
Животные Признаки цветков
открытие цвет запах форма нектар
Жуки день/ночь тусклый или белый фруктовый или амино-идный плоские или шаровидные, актиноморфные, часто с обильной пыльцой часто отсутствует
Навозные и падальные мухи день/ночь коричневый или зеленоватый вонючий плоские или глубокие, часто с ловушками, актиноморфные богат аминокислотами, если присутствует
Мухи-жуж-жалы (Вот-byliidae) день/ночь различный различный относительно глубокие, актиноморфные богат гексозами
Пчелы и шмели день/ночь различный, но не чисто красный душистый плоские или широкотрубчатые, актино- или зигоморфные богат сахарами (длиннохоботные) или гексозами (короткохоботные пчелы)
Бражники ночь белый или бледно-зеленый душистый глубокие, часто со шпорцем обильный и богат сахарами
Дневные бабочки день/ночь различный, часто розовый душистый глубокие, часто со шпорцем или актиноморфные часто богат сахарами
Птицы день яркий, часто красный нет трубчатые, подвешенные, актино- или зигоморфные обильный, богат сахарами
Летучие мыши ночь тускло-коричневый, бледный, часто зеленый затхлый плоские или глубоко-трубчатые, на ветвях и стволах, часто с обильной пыльцой, актиноморфные обильный, богат гексозами
8. Животные и опыление растений
141
Строение цветков растений, опыляемых разными группами опылителей, может существенно различаться (табл. 4.3), хотя узкая специализация к отдельным опылителям у растений встречается не очень часто. В настоящее время считается (Waser, Price, 1998), что сильная специализация эволюционно не очень выгодна. В природе основные группы опылителей выступают как «экономные покупатели на цветочном рынке», поскольку имеется противоречие между интересами животных (подешевле получить больше продуктов из цветков) и растений (произвести опыление с минимальными затратами).
Шмели имеют исходное цветовое предпочтение в двух диапазонах — 400-420 и 510-520 нм, но оно может быстро изменяться после первого опыта добычи пыльцы. Обычно в течение полета шмель посещает с вероятностью 93-98 % один предпочитаемый вид растения, но предпочтения могут меняться со временем. Переключение на другие виды происходит легче при одинаковой окраске цветков, при этом форма цветков практически не имеет значения. Чем крупнее цветок, тем меньшее время тратят шмели на его поиск (на поиск цветка 4 мм в диаметре затрачивается в 10 раз больше времени, чем на поиск цветка 28 мм в диаметре). У двудомных растений мужские цветки часто (но не всегда) выделяют больше нектара, чем женские. Симметричные цветки более привлекательны, чем цветки с нарушенной симметрией (Goulson, 2003).
Цветущее ароидное Helicodiceros muscivorus привлекает мух-опылителей трупным запахом цветков. Показано, что состав пахучих веществ очень сходен с таковым из разлагающихся трупов. Насекомые реагируют на диметил моно-, ди- и трисульфиды. Цветки этого вида остаются открытыми 2 дня, но выделяют специфический запах только в первый день (Stensmyr et al., 2002).
Нектар используется в пищу многими животными, и аминокислоты являются второй (после сахаров) группой соединений, содержащихся в нектаре. Вкус нектара обусловлен различным содержанием аминокислот. Повсеместное присутствие аминокислот в нектаре в миллимоляр-ной концентрации было показано начиная с работы (Baker, Baker, 1973). Вопрос о том, действительно ли опыляемые дневными бабочками цветки содержат больше аминокислот в нектаре, чем опыляемые птицами (которые потребляют и насекомых), окончательно не решен. Самки бабочки Lysandra bellargus предпочитают цветки с нектаром, содержащим больше аминокислот, чем самцы (Rusterholz, Erhard, 2000; Gardener, Gillman, 2002). В работе Mevi-Schuz и Erhardt (2005) показано, что бабочка Araschnia levana откладывает больше яиц при питании нектаром, богатым аминокислотами. Этот эффект исчезал, если гусеницы этой бабочки питались пищей, богатой азотом.
142
Глава 4. Растения и животные
Вставка VII. Почему так редки морские цветковые растения и насекомые?
Для объяснения этого факта выдвигались разные версии (соленость воды, конкуренция с морскими обитателями и др.). J. С. van der Hage (1996) предполо-
жил, что это связано с особенностями опыления покрытосеменных. Эффек
тивность улавливания пыльцы из среды определяется уравнением:
Е = (Рр - Р0)/п,
где Рр — плотность частиц (пыльцы), Ро — плотность среды, п — вязкость среды. Плотность морской воды и пыльцы сходны, а вязкость морской воды примерно в 50 раз больше, чем воздуха, поэтому закрепление пыльцы на рыльце очень затруднено. Опыление насекомыми в морской воде также крайне неэффективно, так как пыльца не прилипает к покровам насекомых. У настоящих водных растений пыльца имеет форму вермишели для ее лучшего улавливания.
В нектаре растений могут содержаться и защитные вещества, например алкалоиды. Эксперименты с цветками каролинского жасмина (Gelsemium sempervirens), нектар которых содержал разное количество алкалоида гелсемина, показали, что при увеличении содержания этого алкалоида цветки меньше посещаются как опылителями, так и похитителями нектара, однако это не влияло на итоговую семенную продукцию этих растений (Adler, Irwin, 2005).
Энтомофильные растения сталкиваются с дилеммой: малое количество нектара, небольшие цветки и т. п. могут не привлечь опылителей, в то время как с увеличением количества нектара, числа цветков в соцветии и т. п. увеличивается вероятность гейтеногамии, т. е. опыления внутри одной особи. Так, средний уровень гейтеногамии составил в популяции Asclepias seriaca 37 % (Pleasants, 1991), в популяции Ipomopsis aggregate — 47 % (de Jong et al., 1992). В экспериментах Hessing (1988) c Geranium caespitosum при использовании специальной краски вместо пыльцы было показано, что чем крупнее растение и больше на нем цветков, тем выше уровень гейтеногамии, достигший у крупных особей 90 % (Klinkhamer, de Jong, 1993).
Растения могут регулировать количество и состав выделяемого нектара. При опылении цветков может происходить реабсорбция нектара, которая была показана, например, для Grevillea robusta (Proteaceae) (Nicolson, 1995).
Strauss (1997) обращает внимание на необходимость совместного анализа воздействия фитофагов и опылителей на приспособленность рас
9. Распространение плодов и семян животными (зоохория)
143
тений. Было показано на примере нескольких видов растений (Raphanus raphanistrum, Nicotiana attenuate), что повреждение листьев и других вегетативных органов может приводить к ухудшению качества пыльцы, уменьшению размеров цветка и его частей. Это, в свою очередь, уменьшает эффективность опыления поврежденных особей. Такая взаимосвязь обусловлена не только сокращением ассимилирующей поверхности листьев при повреждении, но и большими затратами поврежденных растений на синтез защитных веществ.
9 • Распространение плодов и семян животными (зоохория)
По характеру прикрепления к агентам разноса диаспор выделяют следующие типы зоохории: 1) эпизоохория (на покровах), 2) эндозоохория (при прохождении через пищеварительный тракт), 3) синзоохория (распространение при поедании и запасании), подразделяющаяся на сто-матохорию (поедание частей — муравьи, птицы — прилипание к клюву и т. п.) и дисзоохорию (распространение при утере или в запасах).
Эффективность зоохории может быть оценена по следующим количественным и качественным параметрам (Schupp, 1993): количественные — число посещений (контактов) животными растений и число семян, переносимых за одно посещение; качественные — особенности обработки семян в пищеварительном тракте и качество отложений семян с позиций их возможного прорастания и приживания всходов.
Основными группами животных, распространяющих диаспоры, являются муравьи, рыбы, рептилии, птицы и млекопитающие. Главные признаки плодов и семян растений, распространяемых различными агентами, приведены в табл. 4.4.
Таблица 4.4 • Характеристика семян и плодов, распространяемых различными животными (по: Howe, Westly, 1988)
Животные Плоды
цвет запах форма «награда»
Запасающие млекопитающие коричневый слабый ароматный нераскрываю-щиеся толстостенные орехи семя
144
Глава 4. Растения и животные
Окончание таблицы 4.4
Животные Плоды
цвет запах форма «награда»
Древесные млекопитающие желтый, белый, зеленый, коричневый ароматный семена с присемян-ником или костянки, часто сложные или раскрывающиеся протеин мякоти, сахара, крахмал
Летучие мыши бледно-желтый, зеленый затхлый различные, часто висящие мякоть с липидами или крахмалом
Наземные млекопитающие часто зеленый или коричневый нет нераскрывающиеся орехи, коробочки, стручки мякоть с липидами или крахмалом
Запасающие птицы зеленый или коричневый нет округлые семена или орехи семя
Преимущественно семено-ядные птицы черный, синий, красный, зеленый, пурпурный нет крупные костянки или семена с при-семянником, часто раскрывающиеся плоды, семена более 10 мм длины мякоть с липидами или крахмалом
Частично семеноядные птицы черный, синий, красный, оранжевый, белый нет небольшие или среднего размера костянки, ягоды, семена с присемян-ником, менее 10 мм длины мякоть, часто богатая сахарами или крахмалом
Муравьи не имеет значения не различим человеком семена с присе-мянниками, менее 3 мм длины масло или крахмал в элайосомах с химическими аттрактантами
9.1 • Беспозвоночные
Среди беспозвоночных животных — агентов зоохории, наибольшее значение имеют дождевые черви, жуки и муравьи.
Дождевые черви часто переносят мелкие семена и споры. Прорастание семян и спор растений из копролитов дождевых червей может быть более успешным, чем в ненарушенной почве. Например, в экспериментах Hamilton и Lloyd (1991) было показано, что деятельность дождевого червя Lumbricus terrestris резко увеличивает численность, скорость по
9. Распространение плодов и семян животными (зоохория)
145
явления и выживаемость гаметофитов (заростков) папоротника Deparia acrostichoides в почве леса в штате Огайо. Дождевые черви могут выступать и промежуточными агентами распространения семян растений, например при поедании их дроздами (McRill, Sagar, 1973; van der Pijl, 1982).
Среди жуков важную роль в распространении семян играют жуки-копрофаги, которые закапывают семена вместе с пометом травоядных животных, что может способствовать хорошему приживанию всходов.
Мирмекохория (распространение семян муравьями) широко распространена в природе. Некоторые группы муравьев выступают дисзоохора-ми, используя в пищу семена после ферментации (Messor, Atta, Tetramo-rium). В большинстве случаев муравьи используют в пищу лишь при-семянники (элайосомы). Многие корневые паразиты и полупаразиты — мирмекохоры, так как закапывание муравьями их семян способствует прорастанию вблизи корней растений-хозяев (Cytisus, Melampyrum, Lathraea squamaria). Наибольшая роль мирмекохоров отмечена среди трав широколиственных лесов Евразии и Северной Америки (по данным Handel et al., 1981, примерно треть видов в лесах востока США имеет элайосомы), а также в кустарниковых сообществах Австралии. Из растений средней полосы России мирмекохорами являются виды рода Viola, Scilla biflora, Chelidonium majus, Melica nutans, Hepatica nobilis, Asarum europaeum (рис. 4.21). Приспособления, делающие плоды и семена доступными для муравьев — расположение цветков близ уровня почвы, полегание генеративных побегов, растрескивание плодов.
В составе австралийских склерофитных сообществ на бедных почвах встречается около 1500 видов растений, приспособленных к распространению муравьями. На изученных пробных площадках размером 0,1 га от 30 до 50 % видов, главным образом кустарников, являются мирмекохорами (Westoby, 1988). Адаптивная ценность мирмекохории в этих сообществах связана также с затаскиванием семян муравьями на некоторую глубину, где они не повреждаются частыми здесь пожарами (Willson, 1983).
В Капской области многие сотни видов растений являются мирмекохорами. Среди муравьев, распространяющих и закапывающих семена этих растений, особенно важны два («ключевых») вида Anoplolepis cus-todiens и Pheidole capensis. Агрессивный завозной (из Аргентины) вид муравьев — Iridomyrmex humilis — вытесняет оба этих вида, при этом не закапывает семян в почву. В результате ряд видов растений, например кустарник Mimetes cucullatus, резко снижает эффективность возобновления (его семена, лежащие на поверхности почвы, поедаются грызунами или уничтожаются огнем) (Bond, Slingsby, 1983, 1984; Bond, 1993).
146
Глава 4. Растения и животные
9. Распространение плодов и семян животными (зоохория)
147
Рис. 4.21 • Примеры растений-мирмекохоров: а — чистотел (Chelidonium majus); б — копытень (Asarum еигораеит); в — печеночница (Hepatica nobilis); г — перловник поникший (Melica nutans); д — пролеска (Scilla siberica); е — Mimetes fimbriifolius (у мыса Доброй Надежды, ЮАР)
В целом мирмекохория отмечена у более чем 3000 видов растений из более чем 80 семейств (Giladi, 2006). Элайосомы могут развиваться тремя разными путями, что говорит о независимом происхождении мир-мекохории в разных семействах. Биохимический состав элайосом резко отличается от состава сочных плодов, они богаты азотом и жирными кислотами, сходными по составу с таковыми в животной добыче муравьев (Hughes et al., 1993). Пять гипотез обсуждают адаптивность мир-мекохории: 1) перемещение семян муравьями защищает их от поедания фитофагами, 2) уменьшается внутривидовая конкуренция проростков, 3) транспорт семян от материнского растения, 4) перемещение семян в гнезда муравьев защищает их от огня в пирогенных сообществах, 5) гнезда муравьев — хорошие места для прорастания семян, они обогащены ЭМП (Bronstein et al., 2006).
9.2 • Рыбы и рептилии
В тропиках иногда встречаются случаи ихтио- и заурохории. Так, для Амазонии Gottsberger (1978) изучил семена 32 видов растений, поедаемых рыбами и обнаружил, что семена половины видов прошли через пищеварительный тракт неповрежденными. Это может иметь большое значение для заселения верховьев рек водными растениями с крупными семенами.
Из пресмыкающихся распространителями семян выступают черепахи, некоторые ящерицы и крокодилы. Так, например, на Галапагосских островах семена некоторых кактусов прорастают только после прохождения через пищеварительный тракт черепах, но не других видов животных (Dawson, 1962).
Интересен случай распространения семян кустарника Plocama pendula (Rubiaceae), растущего на сухих вулканических субстратах на Канарских островах. Сочные плоды этого вида поедаются всеядными ящерицами из рода Lacerta, для которых они служат источником воды. Жизнеспособные семена откладываются в трещины скал с экскрементами ящериц.
9.3 • Птицы
Птицы могут выступать как агенты эпизоохории, например, перенося семена водных и околоводных растений (Alisma, Carex, Cyperus, Glyceria, Juncus, Pleuropogon, Polygonum) с илом и почвой, что характерно для
148
Глава 4. Растения и животные
водоплавающих птиц (Ridley, 1930; van der Pijl, 1982). Такой перенос способствует быстрому заселению изолированных озер среди степной, лесной и тундровой растительности.
Многие птицы выступают агентами синзоохории. Дрозд (Turdus vis-civorus) часто отрыгивает семена омелы и чистит клюв о ветки, способствуя распространению этого растения. Семена деревьев распространяют врановые птицы — сойки, кедровки. Так, по наблюдениям Schuster (1950) в Германии в среднем каждая сойка за сезон транспортирует около 4600 желудей. Многие птицы летали с ними на расстояние 4 км, а в отдельных случаях — около 10 км. В антропогенном ландшафте вышки и линии электропередач могут способствовать появлению всходов под ними, так как на них чаще садятся птицы.
Но наиболее часто с птицами связаны различные случаи эндозоохории. Прохождение через пищеварительный тракт, как показали опыты с 14 видами растений, имеющими сочные плоды, примерно в половине случаев увеличивает всхожесть семян за счёт освобождения их от мякоти и повреждений твердых покровов (Barnea et al., 1991). В то же время у ряда видов всхожесть семян не увеличивается. Можно предположить, что прохождение через пищеварительный тракт обеспечивает распространение семян от материнского растения (Traveset, Willson, 1997). Так, у ряда видов средиземноморских растений всхожесть семян после прохождения через пищеварительный тракт птиц не изменялась, но значительно менялась скорость прорастания (как увеличивалась, так и уменьшалась) (Traverset et al., 2001). Особенно важен для растений перенос семян птицами в места, где вероятность приживания всходов выше. Например, самцы птицы Procnias tricarunculata (трехусая котинга) преимущественно распространяют семена неотропического дерева Ocotea endresiana в прорывы полога, где всходы имеют большую вероятность для приживания (Wenny, Levey, 1998; Nathan, Muller-Landau, 2000). Также при поедании многосеменных плодов (например, Morus nigra, Solanum luteum; Barnea et al., 1992) семена меньшими порциями выводятся с экскрементами, что повышает приживаемость всходов растений.
В некоторых случаях считается, что прохождение через пищеварительный тракт обязательно для прорастания семян, и растения не могут возобновляться семенным путем без соответствующих видов птиц. В качестве такого примера часто рассматривается дронт (додо). На острове Маврикий, где раньше обитал этот нелетающий голубь, было замечено отсутствие молодых деревьев Sideroxylon grandiflorum (Calvaria major) (моложе 300 лет). Считалось, что семена этого вида не прорастают без прохождения через пищеварительный тракт некоторых видов птиц,
9. Распространение плодов и семян животными (зоохория)
149
в частности дронта, полностью истребленного человеком. Однако в настоящее время облигатность этого симбиоза подвергается сомнению, так как семена могут поедаться и другими видами (черепахами), а также в ряде случаев прорастать без прохождения по пищеварительному тракту (Witmer, Cheke, 1991).
В лесах средней полосы России снижение участия можжевельника (Juniperus communis) может быть связано с уменьшением численности тетеревов, питающихся его шишкоягодами и распространяющих семена.
С другой стороны, отдельные виды растений могут играть ключевую роль в поддержании видового разнообразия многих видов птиц. Так, например, в тропических лесах Коста-Рики дерево Casearia corymbosa плодоносит в декабре, когда очень мало других видов плодоносящих деревьев, и многие виды птиц питаются плодами этого дерева, но лишь один из видов — титира (Tityra semifasciata) — является эффективным распространителем семян этого вида. Исчезновение дерева или титиры может вызвать гибель многих видов птиц тропического леса (Howe, West-ley, 1988).
Эндозоохория птицами может сочетаться с синзоохорией мелкими млекопитающими. В специальных экспериментах, проведенных в Неваде (США), грызуны растаскивали от 8 до 50 % семян из экскрементов птиц на расстоянии в среднем 2,5 м (до 12 м) и откладывали их в запасы. Весной многие семена прорастали из запасов, хотя прямого прорастания из помета птиц не наблюдалось (Vander Wall et al., 2005). Авторы подчеркивают важность такой двухфазной зоохории (птицы — перенос на дальние расстояния, грызуны — погребение в почву и места для лучшего прорастания).
Иногда сочные плоды паразитных растений поедаются птицами вместе с плодами растений хозяев и распространяются одновременно. Например, паразитный кустарник Osyris quandripartita (Santalaceae) в Испании распространяется черноголовой славкой вместе с семенами кустарников-хозяев (Herrera, 1988).
Для привлечения птиц служит обычно яркая окраска плодов или семян (красный, оранжевый, черный цвет). Типичный пример — семена бересклета. Но проблема причинной связи окраски плодов с агентами их распространения полностью не решена. Так, на вопрос «почему оранжевый цвет так часто встречается у плодов, семена которых распространяют птицы?» имеется несколько взаимно не исключающих вариантов ответа: 1) птицы наиболее чувствительны к этому цвету, 2) этот цвет создает наибольший контраст с окраской листьев, 3) птицы испытывают особую потребность в каротиноидах для поддержания остроты зрения, 4) птицы эволюционировали в направлении чувствительности к этому цвету.
150
Глава 4. Растения и животные
9.4 • Млекопитающие
Млекопитающие могут быть агентами всех 3 типов зоохории: эпизоохория встречается очень часто, синзоохория обычно связана с запасающей активностью животных, а эндозоохория бывает специализированной и неспециализированной.
Травоядные на пастбищах поглощают много семян вместе с вегетативными частями растений, часть семян проходит пищеварительный тракт, оставаясь жизнеспособной. Например, С. А. Котт (1948) обнаружил в порции помета коров (3 кг) семян: в мае — 13, июне — 61, июле — 294, августе — 1537, сентябре — 7610. Veilend et al. (2003) изучили эндозоохорное распространение семян белохвостым оленем (Odocoileus virginianus), доминирующим видом копытных на востоке Северной Америки, имеющим массу 40-100 кг. Отмечено более 70 видов семян растений в помете оленя в течение 1 года наблюдений в штате Нью-Йорк. В среднем более 30 семян прорастало из одной порции помета. Большинство семян в помете имели небольшие плоды, не имеющие специальных приспособлений к распространению. 95 % семян транспортируется оленем на расстояние более чем 100 м, 30% могут быть распространены на расстояние более 1 км (Vellend et al., 2003). Доминирующим видом в помете была Chenopodium glaucum. Наибольшая плотность семян в помете отмечена в октябре, январе и феврале. В среднем этот вид оленей распространяет около 10 семян на квадратный метр (Myers et al., 2004).
При скармливании лани (Dama dama) 25 видов растений, всхожесть сохранялась у 24. Выживаемость семян после прохождения пищеварительного тракта варьировала от 0,5 до 42 %. 50 % семян проходило пищеварительный тракт в среднем за 25 часов (диапазон 13-38 часов). Выживание семян было отрицательно скоррелировано с их массой и дисперсией линейных размерностей (т. е. сферические выживали лучше) и положительно — с длительностью жизни семян (Mouissie et al., 2005).
В несомкнутых средиземноморских лесах (dehesa) было исследовано распространение семян несколькими видами млекопитающих (кролики, олени, крупный рогатый скот) (Malo, Suarez, 1995). Помет млекопитающих содержал 6-15 всхожих семян на грамм (максимум 25-70), относящихся к 52-78 видам. Число семян в помете и их состав сильно различался по сезонам, но перекрывался по видам животных. Представленность гвоздичных (Caryophyllaceae) в помете была существенно выше, чем в сообществе. Авторы приводят следующие интегральные оценки эндозоохории: кролики могут распространять с пометом до 500 семян в день, олени до 20 000, скот — до 300 000. Всего около 350 видов или более чем 30 % местных видов может распространяться с пометом
10. Выпас
151
животных. Виды семейств Asteraceae и Роасеае, семена которых имеют специальные адаптации к распространению, реже встречаются в помете млекопитающих. В местах отложения помета крупного рогатого скота на поверхность почвы поступает около 40 семян на дм2, что примерно на порядок выше, чем содержится в почвенном банке.
Наличие твердых покровов у семян («твердосемянность»), отмеченное у ряда видов бобовых, может быть приспособлением к эндозоохории. Иногда овец используют для посева клевера (Trifolium repens), после выпаса на клеверных полях, перегоняя отары на новые места. В экспериментах, проведенных в Австралии с 10 видами бобовых и 8 видами злаков, было показано, что семена бобовых лучше сохраняются и имеют большую всхожесть при прохождении через пищеварительный тракт крупного рогатого скота (Gardener et al., 1992).
Хорошо сохраняют всхожесть семена, прошедшие через пищеварительный тракт хищных млекопитающих. Всхожесть семян борщевика из помета гризли достигает 65 %, а после зимнего промерзания помета — 85 %, аналогичные величины для семян, не прошедших через пищеварительный тракт, — 38% и 52% соответственно (Applegate et al., 1979; Шилов, 1996).
При созревании сочных зоохорных плодов в них происходят существенные биохимические изменения. Незрелые плоды обычно зеленые (малозаметные в листве) и содержат много защитных веществ, которые затем разрушаются или переносятся в другие органы. Например, содержание танинов в сухой массе зеленых бананов (Musa sapientum) достигает 17% (Janzen, 1983).
10 • Выпас
Выпас — наиболее широко распространенный и один из самых древних форм антропогенного воздействия человека. В то же время воздействие антропогенного выпаса сходно с таковым для естественного выпаса диких животных. Выделяют 3 основных направления воздействия пасущихся животных на растительные сообщества: стравливание (отчуждение биомассы), вытаптывание и отложение экскрементов.
10.1 • Стравливание
Разные виды домашних животных по-разному выедают (стравливают) травостой. Лошади и коровы обычно скусывают растения на большей высоте, чем овцы и козы, а домашние птицы часто выщипывают почки,
152
Глава 4. Растения и животные
расположенные у поверхности почвы, тем самым в наибольшей степени подавляя рост трав (Работнов, 1984а). По пищевым потребностям одно животное крупного рогатого скота соответствует примерно 8 овцам, 11 козам или 133 кроликам (Wilson, Harrington, 1984). В США за стандарт принято изъятие 12 кг сухой массы трав в день на корову (Milchunas, Lauenroth, 1993).
Флористическая насыщенность и надземная продукция растительных сообществ обычно достигают своего максимума при умеренном выпасе, несколько снижаясь при его отсутствии и резко уменьшаясь при сильном перевыпасе. Этому вопросу было посвящено много исследований в разных районах мира. Например, в Йеллоустонском национальном парке США участки с выпасом лосей и бизонов были в среднем на 47 % продуктивнее (по надземной продукции), чем на огороженных участках (Frank, McNaughton, 1993).
На основании данных по 236 наблюдениям Milchunas и Lauenroth (1993) провели анализ влияния выпаса на надземную продукцию, видовой состав, биомассу корней и богатство почвы. Различия в продукции выпасаемых и невыпасаемых сообществ уменьшались с увеличением эволюционной длительности выпаса и возрастали с увеличением надземной продукции, уровня изъятия биомассы фитофагами или года наблюдений. В выпасаемых сообществах изымалось в среднем 44 % надземной продукции.
Сравнение влияния выпаса бизонов и крупного рогатого скота на высокотравную прерию в Канзасе было исследовано в течение 10 лет (Towne et al., 2005). Доминирующие виды прерий по-разному реагировали на выпас. Покрытие Schizachyrium scoparium уменьшалось при выпасе бизонов, а покрытие Andropogon gerardii возрастало при выпасе скота. В обоих случаях выпаса увеличивалось покрытие однолетних трав, многолетних видов разнотравья и СЗ злаков, при этом покрытие однолетних трав и многолетних видов разнотравья в большей степени возрастало при выпасе бизонов.
Зависимость флористической насыщенности фитоценозов от наличия и интенсивности выпаса связана с историей территории и длительностью выпаса в регионе. Наиболее резкое снижение видового разнообразия наблюдается в сообществах, где выпас является новым фактором воздействия, — Новая Зеландия, острова южного полушария. Например, на Гавайских островах до завоза человеком не было крупных травоядных животных, и большинство аборигенных видов растений не имели защитных адаптаций против таких фитофагов. Завоз травоядных человеком вызвал исчезновение здесь многих видов растений на обширных территориях (Loope, Medeiros, 1994). В то же время в высокогорьях Альп
10. Выпас
153
с длительной историей выпаса умеренный выпас поддерживает флористически богатые сообщества. Огораживание территории от выпаса слонов в национальном парке Murchison Falls в Уганде привело к угнетению многих трав, разрастанию деревьев (особенно Acacia sieberiana) и снижению флористического разнообразия сообществ (Luken, 1990).
Chaneton и Facelli (1991, см. также Olff, Ritchie, 1998) для аргентинской пампы показали, что выпас увеличивает разнообразие растений на мелких площадках и уменьшает на больших.
В некоторых случаях выпас является необходимым условием поддержания отдельных видов растений. Так, в лишайниковых сосняках северной тайги сосна не может возобновляться при хорошо развитом лишайниковом покрове, ее всходы приживаются в местах интенсивного выпаса северных оленей. Выпас может положительно влиять на развитие геофитов, особенно рано развивающихся. Сравнение огороженных от выпаса и неогороженных участков в северном Израиле показало, что на огороженных участках плотность геофитов, особенно узколистных (Iri-daceae, Liliaceae), значительно снижается (Noy-Meir, Oron, 2001).
Выпас влияет на популяционные характеристики растений в сообществах. Так, наблюдения в течение 20 лет за фиксированными особями растений в условиях выпаса и его отсутствия показали, что скорость обновления и колебания численности особей выше на выпасаемых участках. Средняя продолжительность жизни злаков увеличивается, а кустарников уменьшается при выпасе (West et al., 1979).
Стравливание отличается от сенокошения (см. п. 3.9.3 гл. 9) большей избирательностью. Предпочтение более питательных кормов свойственно мелким копытным, более крупные животные могут использовать более грубую пищу. Например, в Серенгети: антилопа импала (вес 40-55 кг) весьма селективно выбирает злаки и побеги отдельных растений, антилопа тори (100-120 кг) питается почти исключительно различными злаками, а африканский буйвол (400-700 кг) наименее селективен и потребляет различные зеленые травы, его богатая микрофлора пищеварительного тракта разрушает многие растительные токсины (Howe, Westley, 1988).
На пастбищах часто разрастаются ядовитые или колючие непоеда-емые растения (рис. 4.22). С другой стороны, при отсутствии выпаса могут терять конкурентное преимущество и некоторые хорошо поедаемые растения. Например, Belsky (1986) на основании исследования влияния выпаса на пастбищные экосистемы Серенгети показала, что при изоляции от копытных участков огораживанием на них в течение 5 лет практически исчезает Themeda triandra — один из обычных и хорошо поедаемых копытными видов злаков (McNaughton, 1993).
154
Глава 4. Растения и животные
Рис. 4.22 • Участки с доминированием непоедаемых растений на перевыпасаемых пастбищах северо-западного Кавказа (ущелье Муху, Карачаево-Черкессия): а — Veratrum album; б — Aconitum orientate; в — Rumex alpinus; г — Digitalis ciliata
Выпас часто меняет ход сукцессий в травяных сообществах. Под влиянием выпаса в прериях США виды средних стадий сукцессий замещают позднесукцессионные виды, поскольку последние селективно поедаются (Anderson, Briske, 1995).
10.2 • Вытаптывание
Для получения одного и того же количества корма разные виды домашних животных проходят разное расстояние. Так, для альпийских лугов в Центральных Гималаях было показано, что для потребления 1 кг корма (сухая масса!) козы проходят 15,4 км, овцы 8,1 км, лошади 1,2 км, т. е. с увеличением размеров животного расстояние поиска корма сокращается (Negi et al., 1993). По данным Е. Klapp (1971), давление копыт коров составляет около 1 кг/см2, при ходьбе — 4 кг/см2. Это больше, чем у многих тракторов. Многие растения при выпасе страдают не от отчуждения биомассы, а от механического повреждения. Это особенно относится
10. Выпас
155
к растениям с крупными листьями и высоким стеблем. Сильно страдают также всходы растений. Так по наблюдениям на пастбище в штате Юта, выпас снизил за год приживаемость всходов на обнаженных участках почвы с 11,6 % (контроль без выпаса) до 0,4 % (выпасаемый участок), т. е. более чем в 20 раз (Salibi, Norton, 1987; Evans, 1998).
Выпас способствует эрозии почв как прямо (образование голых пятен почвы, ее разрушение), так и косвенно (увеличение поверхностного стока за счет уплотнения почвы). В Великобритании отмечена положительная корреляция между поголовьем овец и содержанием взвешенных частиц почвы в ручьях и речках (Mitchell, 1990; Evans, 1998). Зимний выпас часто вызывает большее разрушение почвы, чем летний. Влажная почва подвержена неупругой деформации, что часто приводит к разрушению дернового горизонта.
Уплотнение почвы при выпасе имеет различные последствия в гу-мидных и аридных регионах. В гумидных районах уплотнении снижает водопроницаемость и миграцию воды вниз по профилю, что приводит к заболачиванию. В аридных регионах аналогичное уплотнение мешает образованию внутрипочвенного запаса воды, способствует большему иссушению, усиливает капиллярный подъем влаги за счет уменьшения размеров капилляров, что часто приводит к большему засолению верхних горизонтов почвы.
Для каждого типа сообществ существует свое критическое проективное покрытие растений, ниже которого выпас приводит к резкому усилению почвенной эрозии (по: Evans, 1998). Это покрытие составляет 15 % для овсяницевого луга в Канаде (Johnston et al., 1971), 20 % для равнинных и 85% для горных лугов в Эфиопии (Coppock, 1993; Mwendere, Saleem, 1997), 50-70% для пастбищ в США (Blackburn, 1984).
10.3 • Отложение экскрементов
Копытные не переваривают 20-80 % потребленной пищи, выделяют более 90 % потребленного фосфора (на 99 % через твердые экскременты) и 65-95 % потребленного азота (на 85 % через мочу) (Ruess, 1987; Zamora et al., 1999). Таким образом, твердые экскременты обогащены азотом и фосфором, а жидкие — азотом и калием.
Локальное поступление больших доз ЭМП в местах отложений экскрементов может вызвать гибель растений, более благоприятно на развитии луговых растений сказывается поступление менее концентрированных порций навоза и мочи. Концентрация ЭМП в экскрементах зависит от количества потребленной скотом воды, поэтому хорошее обеспечение водопоями улучшает структуру лугов (Работнов, 1984а).
156
Глава 4. Растения и животные
Рис. 4.23 • Onthophagus
gazella, интродуцированный
из Африки в США для ускоренного погребения навоза домашнего скота в почву (фото Tom Murray, с любезного разрешения автора)
Потери азота (улетучивание аммиака, денитрификация) из твердых экскрементов, лежащих на поверхности почвы, весьма велики (до 80 %). Жуки-копрофаги, закапывающие навоз в почву, могут существенно снизить эти потери. Испытание 30 видов жуков на пастбищах в США показали наибольшую эффективность африканского жука-копрофага Onthophagus gazella (рис. 4.23). Закапывание навоза привело к существенному снижению потерь азота (теряется не более 5-15%). 82-88% экскрементов этот жук зарывает за неделю.
Отложение экскрементов крупного рогатого скота изменяет горизонтальную структуру пастбищ. Например, в альварах на острове Оланд (Южная Швеция) на местах отложения экскрементов часто доминируют Arenaria serpyllifolia (ее семена содержатся в значительном количестве в помете), Festuca ovina и Cerastium semidecandrum (их семена прорастают из почвенного банка под пометом). Зарастание участков с экскрементами по периферии идет за счет вегетативного размножения, а в центре — из семян. Полностью разлагаются экскременты на альварах за 6-8 лет (Dai, 2000).
Участки, удобренные жидкими экскрементами бизонов в высокотравной прерии США, значительно отличаются по флористическому составу от окружающей растительности, здесь в большей степени доминируют злаки. Бизоны предпочитают кормиться именно на таких участках. Таким образом, они поддерживают своеобразную горизонтальную структуру прерий, создавая участки, имеющие высокую кормовую ценность (Steinauer, Collins, 2001).
Глава
Типы
взаимоотношений растений
в фитоценозах
• Прямые (контактные) отношения
• Трансбиотические отношения
• Конкуренция
• Аллелопатия
• Создание фитосреды и положительные взаимоотношения растений
• Положительные отношения между растениями
п
о механизмам и агентам отношения между растениями В. Н. Сукачев (1956) предложил разделять на прямые (контактные) и непрямые (через посредников). Посредниками могут быть другие организмы (трансбиотические отношения) или среда (трансабиотические отношения). Последние можно подразделять на конкуренцию, аллелопатию и другие изменения в окружающей среде. По эффекту влияния растений одного вида на другой можно также различать отрицательные, нейтральные и положительные воздействия.
1 • Прямые (контактные) отношения
К прямым относятся как внутри- так и межвидовые отношения. Контакты между особями одного вида могут приводить к срастаниям органов, чаще наблюдаемым у корней древесных растений. Ряд специализированных жизненных форм растений связан с контактными отношениями с другими видами растений, к ним относятся паразитные и полупаразитные растения, лианы и эпифиты. Как особые случаи контактных отношений описаны охлестывания ветвями одних растений другими, повреждение (пронизывание) растущими надземными и подземными органами.
1.1 • Внутривидовые срастания
Срастания стволов и ветвей отмечены у немногих видов деревьев. Примером может служить Parrotia persica (железное дерево), произрастающее в гирканских горных лесах в Азербайджане и Иране (рис. 5.1). При длительном соприкосновении ветви этих деревьев срастаются, что может увеличивать механическую прочность древостоев.
Чаще наблюдаются внутривидовые срастания корней деревьев. Ранее предполагалось, что срастания корней способствует увеличению механической прочности древостоя, однако в настоящее время показано, что при высоком насыщении корнями верхних горизонтов почвы устойчивость стволов к выпадению практически не зависит от срастания корней. Срастание способствует передаче веществ между деревьями, при этом перенос воды требует более глубокого срастания (ксилем), чем перенос пластических веществ. Stone (1974) показал, что окольцованная
1. Прямые (контактные) отношения
159
Рис. 5.1 • Стволы Parrotia persica. Стрелка показывает место срастания ветвей
красная сосна (Pinus resinosa) в посадках этого вида выжила в течение 18 лет за счет поддержки продуктами фотосинтеза из корневых систем других растений. Отрицательной стороной срастания корней может являться ускорение распространения инфекции. Межвидовые срастания очень редки, видимо, работает механизм распознавания корней. Напротив, генетически однородные посадки Pinus resinosa имеют высокую частоту срастания. Частота срастания выше при высокой плотности корневых систем, например в поверхностных горизонтах почвы при близком залегании грунтовых вод она значительно выше, чем на сухих почвах (Loehle, Jones, 1990).
1.2 • Паразитные цветковые растения
Паразитные и полупаразитные растения широко распространены в ряде семейств цветковых растений (табл. 5.1). К собственно паразитическим (голопаразиты) относятся растения, получающие как воду и ЭМП, так и органические вещества напрямую от растений-хозяев, поэтому эти
160
Глава 5. Типы взаимоотношений растений в фитоценозах
Таблица 5.1 • Семейства растений, включающие паразитные и полупаразитные таксоны с учетом стеблевого и корневого паразитизма. Первая цифра указывает общее число родов в семействе, вторая — число родов с полупаразитными или паразитными растениями, третья — число видов паразитных растений (включая полупаразиты) (по: Heide-Jorgensen, 2008)
Полупаразиты (около 4100 видов, 90%)
Olacaceae 14/14/ около 100
Schoepfiacea 1/1/30
Opiliaceae 10/10/около 30
Krameriaceae 1/1/17
Loranthaceae 73/73/ около 910
Santhalaceae 35/35/ около 430
Голопаразиты (около 390 видов, 10 %)
Cynomoriaceae 1/1/2
Lennoaceae 2/2/5
Cytinaceae 2/2/7
Mitrastemonaceae 1/1/2
Hydnoraceae 2/2/15-18 Balanophoraceae 17/17/43-44
Rafflesiaceae 3/3/20
______________105/88/ около 1800
Misodendraceae 1 /1 /8-10
Eremolepidaceae 3/3/11
Viscaceae 7/7/около 600
Lauraceae 50/1/16 (Cassytha)
Orobanchaceae__________105/17/ около 270
Apodanthaceae 3/3/23
Cuscuta reflexa
Convolvulaceae 57/1/ около 145 (Cuscuta)
Cuscuta еигораеа
растения почти не имеют собственного фотосинтеза и лишены зеленой окраски. По разным источникам их известно от 390 до 600 видов из 48-50 родов и 9 семейств. К важнейшим семействам относятся заразиховые (Orobanchaceae, около 270 видов, одно- и многолетники, рис. 5.2), повиликовые (Cuscutaceae — около 145 видов одного рода Cuscuta, часть видов этого рода могут быть отнесены к полупаразитам), Balanophoraceae (17 родов, 43-44 вида), Apodanthaceae (3 рода, 23 вида) и Rafflesiaceae (3 рода, 20 видов) (Heide-Jorgensen, 2008). Для паразитных растений характерна относительно высокая степень специализации к растениям-хозяевам.
По месту прикрепления к растениям-хозяевам паразитные растения можно разделить на корневые и стеблевые. Органы прикрепления паразитных растений — гаустории должны проникнуть во флоэму и ксилему растений-хозяев. Гаустории рассматривались как преобразованные придаточные корни, однако А. В. Жук (1997) показал, что гаустории повилик не гомологичны придаточным корням, а являются новообразованиями, возникшими в ходе специализации к паразитическому образу жизни.
Паразиты во взрослом состоянии лишены микориз, однако лучше развиваются на растениях, имеющих микоризы. Так, в экспериментах с выращиванием Cuscuta pentagona на Abutilon theophrasti было показано, что паразит плохо развивается на хозяине при отсутствии у последнего ВАМ. Колонизация корней микоризными грибами увеличивала скорость роста паразита в 3,4 раза (Sanders et al., 1993; Salonen et al., 2001).
1. Прямые (контактные) отношения
161
Рис. 5.2 • Паразитные цветковые растения флоры России: а — Orobanche grossheimii; б — Lathraea squama ria; в — Diphelypaea coccinea (фото С. Сенатора) (Orobanchaceae); г — Cuscuta europaea (Cuscutaceae)
С другой стороны, проростки паразитных растений могут иметь микоризу. Проросшие семена паразитных растений погибают в отсутствии растений-хозяев. Однако время их жизни может существенно удлиняться при развитии в них микоризных грибов. Так, в экспериментах A. N. Khalid, S. Н. Iqbal (1996) показано, что проростки Cuscuta reflexa живут при инокуляции микоризными грибами (ВАМ) (но без хозяина) 7 дней, а без грибов — 5 дней, при этом в первом случае конечная биомасса проростков была вдвое выше.
Паразитные цветковые растения обычно образуют мелкие семена, прорастание которых стимулируется корневыми выделениями растений-хозяев (для корневых паразитов). Общее число продуцируемых семян очень велико, например виды рода Aeginetia (Orobanchaceae) образуют 40000-70000 на коробочку или около 1 млн на растение (Atsatt, 1983). Dale и Egley (1971) собрали ксилемный сок 163 видов растений и показали, что из них сок 118 видов из 57 семейств стимулировал прорастание семян Striga asiatica (Press et al., 1990). Индуктором прорастания Striga
162
Глава 5. Типы взаимоотношений растений в фитоценозах
hermonthica является стригоя; синтезирован его искусственный аналог, используемый в сельском хозяйстве. Физиологическое действие стриго-ла — стимуляция вымывания из семян ингибиторов прорастания. Одним из средств борьбы с заразихой Orobanche ситапа Wallr., поражающей посевы подсолнечника, являются провокационные посевы других культур, корневые выделения которых стимулируют прорастание семян заразихи, в то время как сами растения этим видом заразихи не поражаются. Интересно отметить, что прорастание семян паразитных растений может стимулироваться теми же компонентами корневых выделений, которые стимулируют прорастание спор грибов арбускулярной микоризы (Раг-niske, 2005).
Паразиты могут изменять конкурентные отношения между видами растений, снижая жизненность своих хозяев и положительно воздействуя на другие виды. Например, S. С. Pennings и R. М. Callaway (1996) показали, что на засоленных маршах западного побережья США повилика Cuscuta salina увеличивает участие двух недоминирующих видов — Limonium cal-ifornicum и Frankenia grandifolia, поскольку поселяется преимущественно на доминанте Salicornia virginica.
В то же время в некоторых случаях паразитные растения могут уменьшать ущерб от фитофагии, таким образом благоприятно воздействуя на хозяина. Например, в работе Gomez (1994) показано, что крестоцветное Hormathophylla spinosa может лучше развиваться в присутствии паразита Cuscuta epithymum поскольку в этом случае не выедается травоядными (Zamora et al., 1999).
Под воздействием паразитных растений может изменяться морфология растений-хозяев. Так, при развитии Orobanche minor на Trifolium repens, у клевера уменьшалось отношение биомассы корней к биомассе побегов, а пораженные корни были толще до места прикрепления гаусториев паразита и тоньше — после, по сравнению с непораженными растениями (Dale, Press, 1998).
1.3 • Полупаразиты
Полупаразитные растения, в отличие от голопаразитов, имеют зеленую окраску и способны к фотосинтезу, но получают значительную часть воды и ЭМП от растений-хозяев. Но граница с голопаразитными растениями довольно условна, так как многие паразиты способны к слабому фотосинтезу (у разных видов Cuscuta отмечена разная интенсивность фотосинтеза). С другой стороны, у стеблевого полупаразита Arceuthobi-ит собственная фиксация СО 2 была существенно ниже, чем поступление органических веществ (меченых 14 С) от растения-хозяина (Hull, Leonard,
1. Прямые (контактные) отношения
163
Рис. 5.3 • Полупаразитные цветковые растения: а — Taxillus coloreas (Loranthaceae) на побегах Pinus armandii (северо-западный Юньнань, Китай); б — омела Viscum album и в — дерево, погибшее в результате массового развития омелы в своей кроне (Карачаево-Черкесия, Кавказ); г — ленец Thesium arvense (Santalaceae); представители семейства Orobanchaceae: д — хоботник — Rhynchocorys oriental is; е — мытник Pedicu laris caucasica; ж — зубчатка Odontites verna
1964a, b; Press et al., 1990). У Striga hermonthica 28 % углерода происходило от растения-хозяина (сорго) (Graves, 1995). Некоторые полупаразиты совсем не способны существовать без органических веществ, поступающих от растения-хозяина (например, виды рода Striga). Для Striga asiatica показана возможность прохождения полного цикла развития в полной темноте, но с прикреплением к растению-хозяину (Rogers, Nelson, 1962; Press, 1995).
Всего насчитывается около 4000 видов полупаразитных растений из 235 родов и 12 семейств (табл. 5.1, рис. 5.3). По жизненным формам они
164
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.4 • Оскулы Striga hermonthica, пронизывающие углубления в стенках клеток ксилемы у кукурузы (по: Dorr, 1997)
бывают одно- и многолетними травами, лианами, кустарниками и деревьями (Okoubaka aubrevillei — до 40 м высотой). Основные семейства: Orobanchaceae (рода Pedicularis, Melampyrum, Rhinanthus, Bartsia, Castilleja, Odontites, Euphrasia и другие, преимущественно корневые полупаразиты ранее относимые к Scrophulariaceae, всего 88 родов и около 1800 видов), Loranthaceae (ремнецветные) (73 рода и около 910 видов — кустарники, кустарнички, небольшие деревья, преимущественно стеблевые полупаразиты), Viscaceae (7 родов и около 600 видов, из флоры России к этому семейству относятся омела Viscum album, арцеутобиум Arceuthobium oxicedri), Santalaceae (35 родов, около 430 видов, например, ленец Thesium) (Heide-Jorgensen, 2008). Среди голосеменных растений отмечен один вид полупаразитов — Parasitaxus ustus (Podocarpaceae), растения которого облигатно срастаются с голосеменным же Falcatifoli-ит taxoides (Musselman, Press, 1995).
Гаустории большинства видов полупаразитов проникают в ксилему. Они образуют оскулы, пронизывающие стенки сосудов ксилемы (рис. 5.4). В некоторых случаях важен транспорт гормонов от растений-хозяев, например полупаразит Orthocarpus purpurascens (Scrophulariaceae) получает от кукурузы гибберлиновую кислоту. Транспорт гормонов в полупаразиты может приводить к сходной (с растением-хозяином) морфологии их листьев (Atsatt, 1983). Полупаразиты могут накапливать в своих тканях значительное количество защитных веществ, синтезированных хозяином. Например, Castilleja sulphurea накапливает алкалоиды из Lupinus argineus (Arslanian et al., 1990), a Castilleja miniata — из Lupinus argen-teus (Adler, 2002), что снижало повреждаемость различными фитофагами
1. Прямые (контактные) отношения
165
прикрепленных к люпину особей этих видов по сравнению с растениями, прикрепленными к другим хозяевам. С другой стороны, эксперименты с выращиванием Melampyrum arvense с различными по поедаемости фитофагами растениями-хозяевами не показали корреляции между их поедаемостью и поедаемостью погремка (Schadler et al., 2005).
В серии экспериментов показано, что полупаразит Rhinanthus serotinus поглощает микотоксины, вырабатываемые эндофитным грибом растения-хозяина, при этом поражаемость полупаразита фитофагами значимо снижается. В то же время при наличии полупаразита эндофит начинает оказывать негативное влияние на развитие растения-хозяина, т. е. мутуалистический симбиоз переходит в паразитический (Lehtonen et al., 2005).
Для поддержания потока воды и ЭМП от растений-хозяев полупаразиты должны иметь высокий уровень транспирации. Он особенно высок у многих видов Scrophulariaceae, которые не закрывают устьица даже ночью, а только в периоды резкого недостатка воды. Высокая транспирация в сочетании с низкой интенсивностью фотосинтеза ведет к низкой эффективности использования воды (Press et al., 1999). Показатели уровня транпирации полупаразитов находятся среди самых высоких величин, отмеченных для цветковых растений (Press et al., 1990). При этом транспирация у многолетних полупаразитов обычно ниже, чем у однолетних. Возможно, это связано с риском потерять хозяина в период засухи, который в случае многолетних полупаразитов имеет более тяжелые последствия (Graves, 1995). Многие паразитные растения могут разрегулировать устьичный контроль транспирации растения-хозяина (Seel, Press, 1994; Pennings, Callaway, 2002). Striga hermonthica содержит высокие концентрации калия в листьях, что может обуславливать открытие устьиц и ночью, и в периоды водного дефицита. Высокая транспирация этого вида приводит к существенному охлаждению его листьев (при 40 °C на 7 °C, Press et al., 1989). Применение антитранспирантов, вызывающих закрытие устьиц, снижало транспирационное охлаждение и приводило к гибели растения.
Дефолиация резко уменьшает семенную продуктивность видов Melampyrum в эксперименте, что может быть связано с важностью как собственного фотосинтеза для развития семян, так и активной транспирации для получения веществ из растений-хозяев (Lehtila, Syrjanen, 1995).
Многие виды полупаразитных растений имеют очень высокое содержание ЭМП в тканях, особенно Р и К, даже если произрастают без хозяев (наследуемый признак). Концентрация ЭМП в тканях омел (и других стеблевых полупаразитов) превышала таковую у хозяев по К в 20 раз, по Р, Mg, Мп, N, Na, Са, Fe — в 1,4 раза. Напротив, ветви растений-хозяев, пораженные паразитами, содержат меньше ЭМП, чем непораженные
166
Глава 5. Типы взаимоотношений растений в фитоценозах
ветви. Потери ЭМП для деревьев связаны также с невозможностью реутилизации из отмирающих паразитов ЭМП, в отличие от отмирающих собственных листьев и ветвей (Pate, 1995). Анализ ксилемного сока Rhinanthus minor также показал, что у растений, прикрепленных к хозяевам, резко повышается концентрация К, NO3, Н2РО4 и аминокислот по сравнению с неприкрепленными погремками (Seel, Jeschke, 1999).
Состав неструктурных углеводов может сильно отличаться у хозяев и полупаразитов, последние часто имеют высокие концентрации многоатомных спиртов. Они различаются у разных полупаразитов: для Rhinanthus характерен маннитол, для Euphrasia, Pedicularis — галактитол (Phoenix, Press, 2005).
Семена полупаразитов обычно прорастают без растений-хозяев, но у облигатных полупаразитов проростки погибают, если не встретят растений-хозяев. Экспериментально показано, что корни полупаразитов ветвятся значительно сильнее в присутствии корневых выделений потенциальных растений-хозяев (Atsatt, 1983).
Различают облигатные и факультативные полупаразиты, последние могут развиваться и без растений-хозяев. Эксперименты с 3 видами факультативных полупаразитов (Rhinanthus minor, Euphrasia frigida, Melampyrum sylvaticurri) показали, что при выращивании их в отсутствии растений-хозяев их высота уменьшалась на порядок, а биомасса — почти на 3 порядка, по сравнению с растениями, имевшими контакт с хозяевами (Seel, Press, 1993). При изучении влияния обогащения почвы на рост неприкрепленных к хозяевам особей Rhinanthus minor было показано, что лишь добавление Р увеличивает рост этого вида, в то время как N и К не оказали существенного влияния (Seel et al., 1993; Press, Seel, 1996).
Для большинства полупаразитов характерен широкий спектр растений-хозяев. Так, для Rhinanthus vernalis на лугах установлено 64 вида растений хозяев, имевших сходные физические параметры корней (Работнов, 1984а). Castilleja может паразитировать на более чем 100 видах растений из разных местообитаний, а для тропической «омелы» Dendrophthoe falcata указывается около 400 возможных хозяев (Joshi, Kothyari, 1985 и др.). В то же время известны примеры и узкой специализации.
Рост полупаразитов зависит от состава растений, к которым они прикрепились. Так, рост Rhinanthus minor различался при разных растениях-хозяевах и зависел от концентрации фосфатов в ксилемном соке (Seel, Jeschke, 1999). Marvier (1998) показал, что лучший рост Castilleja wightii наблюдался при прикреплении к двум разным растениям-хозяевам, включая бобовое растение (Lupinus arboreus и Eriophyllum stachaedifolium), хотя при прикреплении к двум бобовым растениям полупаразит содержал больше азота. Выращивание Melampyrum arvense с монокультурами
1. Прямые (контактные) отношения
167
Интенсивность фотосинтеза (микромоль СО2 на 1 г сухой массы в минуту)
О
5 10 15 20 25 30
-------1-------1------1-------1------1----
В Неприкрепленные к хозяину
ИИ Echium vulgare
—| Роа alpina
| PI an tag о lanceolata | Festuca ovina | Brachypodium pinnatum Ц Galium verum | Poa annua | Poa pratensis | Bromus commutatus | Poa nemoralis
М1ИНМНВЯВИВЯВИИНЯНВНВННМ1 T pra tense
Рис. 5.5 • Фотосинтез при световом насыщении Rhinanthus minor при прикреплении к разным хозяевам и росте без хозяев (по: Press, 1995). Малиновый — разнотравье, желтый — злаки, зеленый — бобовые
и попарными смесями 3 видов-хозяев (Medicago sativa, Lolium perenne, Linum usitatissimum) показало, что марьянник не лучше развивается с несколькими хозяевами, чем с одним. Биомасса погремка и ущерб (потери биомассы по сравнению с контролем) возрастали в следующем ряду растений-хозяев: Lolium < Linum < Medicago (Matthies, 1996). Многие физиологические показатели полупаразитов изменяются в зависимости от растения-хозяина. Например, наибольшую интенсивность фотосинтеза при световом насыщении Rhinanthus minor показал при прикреплении к Trifolium pratense, меньшую — с видами разнотравья, самую низкую — без растения-хозяина (рис. 5.5, Press, 1995). Хотя поедаемость фитофагами марьянника (Melampyrum arvense) и различалась в зависимости от растений-хозяев, но специальные эксперименты не показали прямой связи поедаемости этого вида и его растений-хозяев (Schadler et al., 2005).
Полупаразиты могут конкурировать с фитофагами за использование растений-хозяев. В эксперименте с погремком Rhinanthus serotinus, клеверной нематодой Heterodera trifolii и клевером Trifolium pratense было показано, что эта конкуренция носит односторонний характер. Присутствие погремка не сказывалось на развитии нематод, в то время как
168
Глава 5. Типы взаимоотношений растений в фитоценозах
нематода, угнетая клевер, снижала надземную биомассу и продукцию цветков погремка на 74 % и 81 % соответственно. Поэтому рост погремка в присутствии нематоды был схож с таковым при отсутствии растения-хозяина (Puustinen et al., 2001).
Полупаразиты снижают биомассу растений-хозяев, а их собственная биомасса не компенсирует этих потерь, поэтому общая биомасса и продукция сообществ при массовом развитии полупаразитов обычно снижаются. Так, по данным Davies et al. (1997) продуктивность лугов при произрастании Rhinanthus снижается от 8 до 73 %, Ameloot et al. (2005) отмечают снижение надземной продукции в среднем на 26 % (с учетом массы погремка) и на 40 % без учета его массы. При этом наиболее сильно снижается масса злаков и бобовых, а виды разнотравья часто увеличивают свое участие. Биомасса паразитирующей на сорго Striga примерно в 50 раз ниже, чем потери в биомассе сорго, вызванные этим полупаразитом (Parker, 1984; Press et al., 1990).
Полупаразиты могут снижать семенную продуктивность растений-хозяев. Например, было показано, что при прикреплении корневого полупаразита Rhinanthus serotinus к Linum usitatissimum и Brassica г ара, у растений-хозяев увеличивалась асимметрия цветков, цветки имели более мелкие размеры и реже посещались опылителями (Salonen, Lammi, 2001).
Полупаразиты способны изменять исход конкуренции между растениями, интенсивнее подавляя рост предпочитаемых хозяев. В эксперименте с совместным выращиванием Lolium регеппе и Trifolium repens, погремок Rhinanthus minor резко угнетал клевер, усиливая конкурентное преимущество плевела. Этот эффект был меньше выражен на богатой почве, где погремок образовывал значительно меньше гаусторий, чем на бедной (Gibson, Watkinson, 1991). Поэтому полупаразиты не только избирательно снижают биомассу отдельных видов растений, но и оказывают существенное влияние на структуру растительных сообществ, особенно луговых. Такое влияние было исследовано Gibson, Watkinson (1992) на лугах Великобритании. Они показали, что хотя Rhinanthus minor обитает на богатых карбонатных лугах, микроучастки с погремком имели более низкое разнообразие (оцениваемое по числу видов, индексам Симпсона и Шеннона), чем рядом расположенные без погремка. Эксперименты с удалением погремка с отдельных участков в 3 случаях из 4 вызывали увеличение и в одном — уменьшение разнообразия. Это связано с исходной структурой лугов: там где предпочитаемые погремком виды растений-хозяев не доминировали, они увеличивали свое участие при удалении погремка и общее разнообразие увеличивалось. Обратная картина наблюдалась при доминировании Koeleria macrantha — одного из основных хозяев погремка. При удалении погремка доминирование
1. Прямые (контактные) отношения
169
Koeleria усиливалось и разнообразие снижалось. Поэтому погремок может быть использован для улучшения приживания подсеянных трав при восстановлении флористически богатых лугов в Великобритании. Экспериментальный подсев погремка значимо улучшил приживаемость 7 из 10 подсеянных видов разнотравья за счет снижения биомассы доминантов и высоты травостоя (Pywell et al., 2004).
Опад полупаразитов обладает высокой зольностью и играет положительную роль в ускорении круговорота ЭМП в сообществе (Press, 1998). Так в тундровых сообществах, листья полупаразита Bartsia alpina разлагаются быстрее и теряют в 5,4-10,8 раз больше азота, чем листья кустарничков в течение 240 дней эксперимента по разложению. Смесь листьев Bartsia alpina и кустарничков разлагалась значимо быстрее, чем ожидалось из пропорции скоростей разложения отдельных фракций. Таким образом, полупаразиты могут ускорять разложение опада в субарктических сообществах. Механизм такого воздействия, видимо, связан с обогащением смеси опада азотом, стимулирующим активность деструкторов (Quested et al., 2002). Всходы тундровых растений, развивающиеся на опаде Bartsia alpina, образовывали до 50 % больше биомассы и имели вдвое выше концентрацию азота и фосфора в тканях, в сравнении с растениями, растущими на опаде других видов (Quested et al., 2003; Quested, 2008).
Полупаразиты лучше развиваются при прикреплении к растениям, имеющим микоризу. В опытах Salonen et al. (2000) Melampyrum pratense увеличивал свою биомассу и число цветков при прикреплении к микоризным саженцам сосны Pinus sylvestris по сравнению с безмикоризными. Rhinanthus serotinus существенно увеличивал свою биомассу и продуцировал больше цветков при микоризации его хозяина Trifolium pratense (Salonen et al., 2001). В то же время сами полупаразиты снижают интенсивность микоризной инфекции у растений-хозяев (Gehring, Whitham, 1992; Davies, Graves, 1998; Pennings, Callaway, 2002).
В некоторых случаях положительное влияние полупаразитов на растения-хозяев связано с привлечением животных — агентов распространения семян и плодов. Так, «омела» Phoradendron juniperinum образует привлекательные для птицы Myadestes townsendi семена, при этом птица эффективно распространяет и семена растения-хозяина — Juniperus monosperma (van Ommeren, Whitham, 2002; Press, Phoenix, 2005).
1.4 • Эпифиты
Эпифиты используют другие растения как субстрат для поселения (топические консортивные связи). Растение-хозяин носит название форофита. Различают сосудистые и несосудистые эпифиты (водоросли, лишайники,
170
Глава 5. Типы взаимоотношений растений в фитоценозах
мохообразные) (рис. 5.6). Первые преимущественно встречаются в тропиках и субтропиках, так как не переносят зимнюю засуху, поэтому основными факторами, сдерживающими распространение эпифитов в высокие широты, выступают недостаток воды и низкие температуры. Одним из немногих сосудистых эпифитов в нашей стране выступает папоротник — Polypodium vulgare, факультативно встречающийся на крупных деревьях в лесах Закавказья на высоте 10-15 метров. В горных лесах Швейцарских Альп (47° с. ш.) численность эпифитных Polypodium vulgare достигает 300 растений на гектар (Zotz, 2002, 2005; Zotz, List, 2003), но они встречаются только на покрытосеменных деревьях. Ibisch (1996) разделил эпифиты на облигатные (более 95 % особей произрастает как эпифиты), случайные (более 95 % произрастают на почве) и факультативные эпифиты (промежуточные между ними).
Вне тропиков (> 23,5° широты) эпифиты хорошо представлены в океанических областях с влажным климатом — в дождевых лесах Новой Зеландии, Чили и горных лесах Гималаев. В отличие от тропиков, среди эпифитов умеренной зоны преобладают папоротникообразные (в тропиках — орхидные). При географическом сравнении в южном полушарии на тех же широтах видовое разнообразие эпифитов значительно больше. В Новой Зеландии 2 % флоры (50 видов) — типичные эпифиты. В районе Валдивия (Южная Америка) около 10 % сосудистых растений — эпифиты. В Гималаях среди эпифитов много представителей родов Ribes, Еиопу-mus, Thalictrum, Rhododendron (Zotz, 2005).
Около 10% всех видов сосудистых растений — эпифиты. Всего насчитывается около 23 470 видов, из них большую часть составляют представители орхидных (Orchidaceae, 13951 вид эпифитов) (LOttge, 1989).
По характеру субстрата эпифиты условно разделяют на коровые, крепящиеся в мелких трещинах коры, и гумусовые, приуроченные к нишам, развилкам ветвей, небольшим дуплам, где накапливается разлагающийся опад. Специфичность эпифитов к форофитам зависит преимущественно от водоудерживающих свойств коры последних. Эти свойства, например, оказывали сильное влияние на распространение эпифитного папоротника Polypodium polypodioides, но имели меньшее значение для неукореняю-щегося эпифита Tillandsia usneoides, что было показано специальными экспериментами с пересадками эпифитов (Callaway et al., 2002)
В тропических лесах также много видов эпифилльных лишайников и печеночников (рис. 5.6 а, б). Покров из эпифиллов может уменьшить фотосинтез растения-хозяина на 20 %. Однако в ряде случаев происходит подстройка параметров фотосинтеза листьев под условия затенения, создаваемые эпифиллами. Так, эпифилльные лишайники на пальме
1. Прямые (контактные) отношения
171
Рис. 5.6 • Эпифиты и эпифиллы: а — эпифилльные печеночники на листьях пальмы в тропическом лесу (Китай); б — покров эпифиллов на листьях Lagerstroemia venusta\ в — папоротник Neottoptehs nidus (тропический Китай); г — Tillandsia usneoides; д — Tillandsia ionantha (Мексика); е — эпифитные орхидеи в диптерокарповом лесу (Юньнань, Китай)
снижали поступление света к поверхности листьев на 50-70 %, однако фотосинтез таких листьев характеризовался более низким световым порогом насыщения, а в итоге суммарная ассимиляция не отличалась от листьев без лишайников (Anthony et al., 2002). Эпифиллы адсорбируют воду и ЭМП из листьев, увеличивают вероятность заражения патогенными микроорганизмами, но могут и защищать листья от фитофагов (Coley et al., 1993). Для эпифилльного печеночника Radula flaccida показано проникновение ризоидами в листья растений-хозяев и поглощение из листьев воды и ЭМП, что свидетельствует о его полупаразитизме. Эпифилльные бриофиты усиливают общую фиксацию азота у связанных с ними цианей, действуя как губки — поддерживая высокую влажность вблизи своего таллома (During, van Tooren, 1990).
172
Глава 5. Типы взаимоотношений растений в фитоценозах
Эпифиты также небезразличны для деревьев. Они чаще оказывают отрицательное воздействие на форофит, прежде всего за счет затенения, увеличения механической нагрузки на ветви (чаще ломаются от ветра), а микоризные грибы эпифитов могут паразитировать на деревьях (особенно у орхидных).
В связи с этим ряд деревьев имеет защитные приспособления против эпифитов, например шелушащаяся или гладкая кора. По наблюдениям в бассейне Амазонки (Gottsberger, Morawetz, 1993), было выявлено, что характер коры форофита является основным фактором в распределении эпифитов наряду с возрастом дерева и плотностью растительного покрова. Ряд видов растений привлекает животных для защиты от эпифитов. Например, виды рода Cecropia привлекают листогрызущих муравьев рода Azteca, которые не трогают листья Cecropia. Муравьи обитают в полых стеблях цекропий. Кроме того, в нижней части черешка (при прикреплении к стеблю) у цекропий образуются богатые гликогеном и содержащие протеины мюллеровы тела, которые охотно поедаются муравьями (Folgarait et al., 1994). Однако эпифиты также могут привлекать животных для защиты от фитофагов. Так в полостях, имеющихся на эпи-фитной орхидее Caularthron bilamellatum, поселяются 6 видов муравьев, при этом экстрафлоральные нектарники орхидеи обеспечивают 11 -48 % пищи этих муравьев (Fisher et al., 1990).
В тропических лесах до 50 % биомассы листвы может приходиться на эпифитов. Во влажных лесах Коста-Рики опад эпифитов составляет около 50 г/м2 (сухая масса) в год. В связи с подверженностью эпифитов водному стрессу, они лучше представлены в прохладных субтропических дождевых лесах, чем в жарких тропических. Наряду с СЗ типом фотосинтеза, у эпифитов часто встречается и САМ — тип фотосинтеза, который обычен у эпифитных представителей Bromeliaceae, Orchidaceae, Cactaceae. В то же время очень редко встречаются эпифиты с С4 типом фотосинтеза. Для поглощения воды из влажного воздуха и туманов служит губчатый веламен, покрывающий корни многих орхидных; специальное опушение из зонтиковидных волосков у бромеливых и другие приспособления. Эпифитные бромелиевые могут хорошо регулировать свои ростовые процессы в зависимости от доступности воды и ЭМП, особенно в молодом состоянии. Семена бромелиевых имеют мелкие размеры, но снабжены крючковидными волосками, позволяющими им удерживаться на ветвях полога леса (Benzing, 1990), при этом большинство семян остается в пределах материнского форофита. Некоторые бромелиевые имеют небольшой запас воды в семенах, но большинство адсорбируют воду из атмосферы. Молодые растения некоторых видов,
1. Прямые (контактные) отношения
173
формирующих во взрослом состоянии водные резервуары при основании листьев, в ювенильном состоянии (1-5 лет) имеют очень густой покров из адсорбирующих волосков на тонких листьях (Whigham et al., 2008).
Элементы минерального питания эпифиты получают из осадков, стекающей по стволам воды, разлагающейся органики близ ствола, за счет ассоциативной азотфиксации, а также приноса животными. Среди эпифитов встречаются и хищные растения (Nepenthes). На неотропиче-ской (наземной) бромелии Bromelia balansae обитает прыгающий паук Psecas chapoda (Salticidae), который на 18 % обеспечивал бромелию азотом. В полевом эксперименте растения с пауками образовывали на 15 % более длинные листья, чем растения без пауков (Romero et al., 2006). Аналогичные закономерности вероятны и для эпифитных бромелиевых. Однако рост эпифитов чаще лимитирует недостаток воды, чем других ресурсов. Так, изучение роста эпифита Vriesea sanguinolenta (Bromeliaceae) в зависимости от доступности ЭМП, воды и света показало, что добавление воды усиливало рост растений всех изученных размерных групп, добавление ЭМП усиливало рост только мелких и средних по размерам растений, а увеличение поступления света (с 30 % до 60 %) снижало рост растений (Laube, Zotz, 2003). С другой стороны, эксперименты, проведенные на Гавайях показали, что внесение фосфорных, но не азотных удобрений в почву тропического леса вызывало резкое увеличение участия и видового богатства различных эпифитов. Особенно резко увеличивали свое участие азотфиксирующие лишайники (Benner, Vitousek, 2007).
Сосудистые эпифиты — относительно молодая группа, ископаемые формы эпифитов известны со среднего эоцена.
1.5 • Лианы
Лианы рассматриваются как «структурные паразиты», так как используют другие виды как опору, не вкладывают большую долю ассимилятов в опорные органы. Общее число видов лиан трудно оценить, поскольку границы между лианами и нелианами нечеткие. Считается, что лианы представлены примерно в половине семейств цветковых растений. Наиболее богаты лианами семейства: Convolvulaceae, Cucurbitaceae, Menis-permaceae.
Лианы обычно составляют около 25 % стеблей и столько же процентов видов во многих тропических лесах (Gentry, 1991; Appanah et al., 1992). Ряд тропических лесов беден лианами (< 10 % видов), в то время как в долине Амазонки 44 % древесных видов — лианы, их богатство достигает 51 вид на гектар (Schnitzer, Bongers, 2002). Площадь кронового пространства, занимаемого одной лианой, может достигать 400-600 м2
174
Глава 5. Типы взаимоотношений растений в фитоценозах
(Кузнецов, 2004). Поэтому лианы представляют собой важный компонент низинных амазонских лесов. Крупные лианы (стебель более 10 см в диаметре) составляют менее 5 % общего числа стеблей лиан, но более 80 % биомассы лиан, достигших уровня хорошего освещения. Оборот поколений у лиан идет примерно втрое быстрее, чем у деревьев, при этом вероятность отмирания дерева с лианами примерно в три раза выше, чем дерева без лиан (Phillips et al., 2005).
Встречаемость лиан на деревьях разных видов была исследована на 7 площадках по 1 га в амазонских лесах (Перу) на основании анализа 3675 стволов (van der Heijden et al., 2008). Половина стволов имела, по крайней мере, по одной лиане. Из 41 вида наиболее массовых деревьев, 5 видов были значимо чаще, а 3 вида значимо реже поражены лианами, чем ожидалось при случайном распределении лиан. Наибольшее влияние на встречаемость лиан имел характер коры. Виды деревьев, отличающиеся быстрым ростом, большей высотой, древесиной низкой плотности, длинными листьями и длинными неветвящимися участками побегов, в меньшей степени поражались лианами (van der Heijden et al., 2008).
По типу прикрепления различают 4 группы лиан (рис. 5.7).
1. Цепляющиеся шипами или крючками.
2. Прикрепляющиеся усиками.
3. С корнями или гаусториями в качестве органов прикрепления.
4. Вьющиеся (с обвивающим стеблем).
В ходе вторичных сукцессий в тропических лесах сменяются лианы с разными механизмами лазания — численность лиан с усиками уменьшается, а вьющихся видов — возрастает (DeWalt et al., 2000), но в целом после нарушений роль лиан возрастает. При выпадении деревьев от ветра примерно 90 % взрослых лиан выживает и продолжает расти (Putz, 1984). В тропическом лесоводстве лианы рассматриваются как своеобразные сорняки, быстро заполняющие вырубки и посадки древесных растений. В ходе сукцессий наибольшее развитие лиан в целом характерно для средних стадий, они тормозят наступление заключительных стадий.
По экологическим особенностям лианы обычно характерны для затененных местообитаний, но богатых азотом почв. Лианы в целом чувствительны к морозам, поскольку замерзание воды в их проводящих тканях губительно действует на многие виды. Из этого правила есть исключения, например виды рода Vitis (виноград) выносят низкие зимние температуры, осушая свои сосуды до наступления морозов (Sperry et al., 1987; Schnitzer, Bongers, 2002).
1. Прямые (контактные) отношения
175
В средней полосе России обычными видами лиан являются Calyste-gia sepium, Humulus lupulus, а также занесенный из Северной Америки Echinocystis lobata. Но более 90 % видов лиан характерны для тропиков. Многие заходят в субтропики, например Smilax excelsa делала непроходимыми колхидские леса.
Большинство лиан синтезируют фитотоксины для защиты от фитофагов.
Во многих исследованиях показано, что даже при небольшом участии лианы уменьшают рост, размножение и выживаемость деревьев. В ряде случаев это связано не столько с затенением или механическим воздействием, сколько с подземной конкуренцией за азот (Dillenburg et al., 1993, 1995). Большее отрицательное влияние лианы оказывают на медленнорастущие теневыносливые деревья, поэтому они могут непрямо улучшать условия для быстрорастущих пионерных видов. Например, в лесу Коста-Рики из 142 стволов взрослых пионерных деревьев лишь один был оплетен лианой, в то время как от 50 до 97 % стволов теневыносливых видов с диаметром ствола более 70 см были оплетены лианами (Clark, Clark, 1990). Лианы имеют очень глубокую корневую систему, обеспечивающую поступление значительных объемов воды и высокую транспирацию по сравнению с деревьями (Schnitzer, Bongers, 2002).
В тропических лесах биомасса лиан достигает от 2 до 20 т сухого вещества на гектар (1-6% надземной фитомассы, но при этом это 25-30% биомассы листвы, которая составляет 10-30% общего индекса листовой поверхности — ИЛП). Основная морфологическая особенность лиан — сильная аллокация (распределение) ассимилятов в листья, отсюда — высокая относительная скорость роста (Hegarty, 1989).
Семена лиан обычно среднего (10-100 мг) веса, они могут прорастать в темноте, но приживаемость всходов выше в местах прорыва полога («окнах»).
Травянистые лианы изменяют свою морфологию в зависимости от наличия или отсутствия поддерживающих растений. При наличии опоры лианы имеют большую длину стебля и аллокацию в главный стебель. Лианы, выращенные экспериментально без опор имеют более высокое отношение надземной биомассы к подземной, а в надземной биомассе возрастает доля боковых стеблей, что может быть связано с «поисковым» поведением (Den Dubbelden, Oosterbeek, 1995).
Некоторые виды растений могут менять жизненную форму (дерево-лиана) в зависимости от условий среды. Например, Toxicodendron diversilobum может расти как кустарник до 4 м высоты и 15-20 см в диаметре ствола или как лиана до 30 м высоты и диаметре ствола до 15 см (Gartner, 1991а, Ь).
176
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис- 5-7 • Лианы: а — Hedera helix на стволе бука (Западное Закавказье, Россия); б — Ipomoea purpurea (родина — тропическая Америка); в — Smilax excelsa — деревянистая лиана, делавшая непроходимыми леса Колхидской низменности; г — опушка леса с плотным покровом хмеля — Humulus lupulus (Северный Кавказ); д — ротанговая пальма Calamus sp.
1. Прямые (контактные) отношения
177
е — шипы на побегах ротанговой пальмы; ж — Monstera deliciosa, прикрепляющаяся с помощью корней (з); и — Ficus altissima на ранних стадиях развивается как лиана, а затем формирует гигантский полый «стебель» (к — вид изнутри) из многочисленных сросшихся стеблей (тропические леса в провинции Юньнань, Китай)
178
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.8 • Фотоснимок верхних частей крон деревьев в тропическом лесу. Кроны форофитов разделены свободным пространством, а кроны лиан (указаны черными стрелками) находятся внутри крон форофитов (по: Rowe, Speck, 2005)
При виде сверху в тропическом лесу лианы вплотную контактируют с форофитами, а кроны форофитов обычно разделены свободным пространством (рис. 5.8.) (Rowe, Speck, 2005).
2 • Трансбиотические отношения
Агентами трансбиотических отношений между растениями служат различные организмы (прокариоты, грибы, животные). Например, микоризные грибы связывают разные виды растений, в том числе автотрофные с мико-гетеротрофными (см. п. 1.9 гл. 3). Животные также часто выступают агентами таких отношений. Классические примеры в этом случае представляют случаи совместного произрастания ядовитых и неядовитых видов растений. В Северной Швеции Hjalten et al. (1993) обнаружили, что Betula pubescens сильно страдает от фитофагов при произрастании вблизи хорошо поедаемых Sorbus аисирапа и Populus tremula, но почти не страдает вблизи малопоедаемой Alnus incana. Аналогично в Шотландии поедаемость вереска копытными сильно зависела от окружающих растений (сильнее при соседстве с Agrostis/Festuca, чем с Nardus или Molinia) (Palmer et al., 2003).
3 • Конкуренция
3.1 • Модель Тильмана
Ресурс определяется Д. Тильманом (Tilman, 1982) как любое вещество или фактор, потребляемый организмами, увеличение доступности которого ведет к увеличению скорости роста организма или популяции. Таким образом, все экологические факторы можно разделить на ресурсные (для растений элементы минерального питания, свет, вода, иногда
3. Конкуренция
179
углекислый газ, кислород и др.) и нересурсные (например, температура, соленость, кислотность почвы и т.п.). Ф.Грайм (Grime, 1979) определяет конкуренцию как «тенденцию соседних растений использовать один и тот же квант света, ион элементов минерального питания, молекулу воды или объем пространства», иными словами конкуренция — это «взаимоотнятие ресурсов». Это определение конкуренции мы и будем использовать в дальнейшем, хотя во многих работах конкуренция понимается в более широком смысле, как любые взаимоотрицательные взаимодействия между видами без детализации их механизма. Большинство почвенных ресурсов, прежде всего элементов минерального питания, для растений являются незаменимыми, т. е. увеличение доступности одного из ресурсов не увеличивает рост растений при недостатке других, согласно классическому закону Либиха (Von Liebig, 1840).
Ключевым понятием теории Тильмана выступает представление о пороговой концентрации ресурса (Я*), которое определяется как минимальная концентрация ресурса, при которой еще возможно существование популяции данного вида (скорость прироста равна скорости отмирания). При конкуренции за один и тот же ресурс в стационарной среде побеждает вид, имеющий самую низкую пороговую концентрацию. Он снижает доступность ресурса в среде до этой концентрации, когда другие виды уже не могут длительно существовать.
Пороговая концентрация ресурса для вида не является постоянной величиной, она может зависеть от других факторов. Например, диатомовые водоросли Synedra ulma и Asterionella formosa конкурируют за SiO2 в среде при его недостатке. Я* в этом случае зависит от температуры: при t > 20 °C Synedra требует меньше кремния для своего роста и вытесняет Asterionella, а при t < 20 °C — наоборот. В этом случае периодическая смена температуры способствует сосуществованию обоих видов.
Измерения пороговых концентраций трудноосуществимы в почве. В водных культурах Braakhekke (1980) исследовал конкуренцию между подорожником ланцетным (Plantago lanceolata) и нивяником (Leucan-themum vulgare) за ионы кальция. Plantago мог снижать концентрацию Са2+ в растворе до 0,4 м-экв/л, a Chrysanthemum только до 0,8 м-экв/л. Эксперименты подтвердили, что подорожник является более сильным конкурентом за Са, чем нивяник.
Значительно более сложная картина возникает при конкуренции за 2 незаменимых ресурса. Здесь Тильман делает предположение, что скорость потребления ресурса у растений положительно связана с его пороговой концентрацией, т. е. вид потребляет больше того ресурса, по которому у него выше пороговая концентрация. В этом случае возможно стабильное сосуществование двух видов в стационарной среде.
180
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.9 • Графическое представление конкуренции двух видов за два незаменимых ресурса (Rl, R2) в стационарной среде. А и В — изоклины пороговых концентраций ресурсов этих видов. С — векторы потребления ресурсов. Области диаграммы: 1 — уровень поступления (концентрации) ресурсов не достаточен для существования ни одного вида, 2 и 3 — области конкурентного преимущества вида А, 5 и 6 — области конкурентного преимущества вида В, 4 — область устойчивого сосуществования видов.
Устойчивое сосуществование возможно лишь в случае пересечения изоклин пороговых концентраций видов и потреблении ими ресурсов со скоростями, пропорциональными пороговым концентрациям (т. е. ресурс с большей пороговой концентрацией потребляется с большей скоростью) (по: Tilman, 1982)
При этом один из видов должен иметь более низкую Я* по одному ресурсу, а другой вид — по другому ресурсу (рйс. 5.9).
Примером такой ситуации в природе может быть сосуществование растений с разными типами микориз на вересковых пустошах в Голландии — злака Molinia caerulea (ВАМ-растение) и вереска Calluna vulgaris (с эрикоид-ной микоризой). Более низкая пороговая концентрацией по почвенным фосфатам характерна для молинии, а по азоту — у вереска (Aerts, 2003).
Картина конкуренции за два незаменимых ресурса принципиально меняется, если мы рассмотрим не стационарную, а пространственно гетерогенную среду (рис. 5.10). Если в большой (теоретически неограниченной) группе видов отдельные виды различаются в пропорциях потребления двух ресурсов, т. е. имеют разные отношения R\ R?, то в области лимитирования обоими ресурсами они могут устойчиво сосуществовать. В этой области каждый вид как бы находит те микрозоны, где он наиболее конкурентоспособен, т. е. в них пропорции в поступлении ресурсов наиболее соответствуют его физиологическим потребностям.
Анализ рассматриваемой модели привел автора к представлению об одновершинной зависимости видового богатства растительных сообществ от богатства почв (рис. 5.11). Из рассматриваемой зависимости вытекают несколько следствий, объясняющих организацию растительных сообществ на почвах разного богатства:
1) кривая «число видов — богатство ресурсов» имеет максимум в области относительно бедных местообитаний;
2) восходящая часть кривой должна быть круче нисходящей, т. е. увеличение видового богатства на бедных почвах происходит быстрее, чем его снижение на очень богатых;
3. Конкуренция
181
Рис. 5.10 • Графическое представление конкуренции 7 видов растений за два незаменимых ресурса в гетерогенной среде. Виды различаются в пропорциях потребления ресурсов. Области диаграммы, отмеченные разными буквами, показывают возможность сосуществования соответствующих видов (т. е. а + Ъ — область сосуществования видов а и Ъ). Внутри круга 1 могут сосуществовать все 7 видов, а внутри круга 3 — в условиях более богатых почв — только два вида. Видовой состав местообитания зависит от относительной скорости поступления двух ресурсов. В условиях их примерно равного поступления доминируют виды d и е, при обилии первого ресурса и недостатке второго — виды f и д, а при обилии второго ресурса и недостатке первого — виды а и b (по: Tilman, 1982)
3) сообщества, находящиеся вблизи пика видового разнообразия, должны иметь много содоминантов, в то время как в более богатых ресурсами местообитаниях с низким видовым богатством доминировать могут лишь немногие виды, большинство остальных должно быть редким;
4) для любого уровня богатства ресурсов возрастание пространственной гетерогенности должно приводить к возрастанию видового богатства, что особенно ярко проявляется в бедных местообитаниях;
182
Глава 5. Типы взаимоотношений растений в фитоценозах
5) уровень поступления конкретных ресурсов должен определять, какие именно виды будут доминировать в сообществе.
Рассматриваемая зависимость флористического богатства от богатства почв ЭМП известна для разных типов местообитаний и может иметь различное объяснение (см. Stevens, Carson, 1999; Grime, 2001; Partel, Zobel, 2007). Трудности определения богатства почвы в естественных сообществах связаны с необходимостью оценки не концентраций ЭМП питания, а скорости их поступления в результате минерализации органики и других процессов. Tilman (1982) предложил оценивать богатство почвы по продукции ненарушенных растительных сообществ. Используя эти оценки, характер связи между числом видов и продукцией сообществ, можно хорошо продемонстрировать на примере сообществ переувлажненных и околоводных местообитаний таежной зоны. Здесь наименее продуктивны и флористически бедны, хотя и полидоминантны, олиготрофные болота (восходящая часть кривой рис. 5.11), наиболее флористически богаты некоторые низинные болота, обладающие средней продукцией (максимум на этой кривой), а наиболее продуктивны моно-доминантные тростниковые (Phragmites australis) заросли, очень бедные по видовому составу сосудистых растений (нисходящая часть кривой рис. 5.11).
Tilman (1982) отмечает также, что самые флористически насыщенные сообщества земного шара приурочены к бедным почвам (финбош в Южной Африке, кустарниковые пустоши Австралии). Снижение флористической насыщенности при равномерном внесении удобрений также показано рядом исследователей. Один из прямых экспериментов был проведен в Великобритании (Willis, Yemm, 1961; Willis, 1963): образцы дерна были взяты на сырых пастбищах и перенесены в вегетационный домик. Первоначально они содержали в среднем по 22 вида растений на квадратный метр. Затем эти образцы в течение 39 недель поливали либо водой, либо раствором с комплексными удобрениями. В конце эксперимента в первом варианте осталось в среднем 21 вид, а во втором — только 5 видов. Видимо, увеличение биомассы растений при внесении удобрений усилило конкуренцию за свет, которая и привела к доминированию более высокорослых видов растений и выпадению многих видов, растения которых имели меньшие размеры.
К сожалению, мы практически не знаем реального соотношения при потреблении ЭМП различными видами растений в природе. Судить об этом можно предположительно на основании экспериментов с внесением различных доз ЭМП. Отдельные лабораторные исследования также показывают различия между оптимальными для роста соотношениями
3. Конкуренция
183
Рис. 5.11 • Зависимость видового богатства растительных сообществ от богатства почвы ЭМП (обобщение результаты имитационного моделирования для 40 видов растений, ресурсная диаграмма построена аналогично рис. 5.10, виды различаются пропорцией потребления ресурсов, богатство почвы в условных единицах увеличивается при движении из нижнего левого в верхний правый угол диаграммы, пространственная дисперсия
уровня поступления ресурсов принята постоянной для разных областей диаграммы) (по: Tilman, 1982)
Рис. 5.12 • Оптимальные соотношения азота и фосфора в среде для роста всходов различных видов Dactylorhiza (по: Dijk et al., 1997)
ЭМП даже для близких видов растений. Например, при выращивании в культуре нескольких видов пальчатокоренников (Dactylorhiza spp.) было найдено различное оптимальное соотношение N Р в среде без микоризного гриба (рис. 5.12, см. Dijk et al., 1997).
184
Глава 5. Типы взаимоотношений растений в фитоценозах
Таблица 5.2 • Сравнительная конкурентоспособность 6 видов луговых злаков при лимитировании различными ресурсами (по: Bradshaw et al., 1960а, б, 1964). Сокращения названий видов: Ас — Agrostis canina, /4s — Agrostis stolonifera, At — Agrostis tenuis, Cc — Cynosurus cristatus, Lp — Lolium perenne, Ns — Nardus stricta
Конкурентоспособность Ресурсы
N Са P свет
Лучшая Ns At Cc Lp
Сс Ns Ns /4s
t At Lp Ac At
Ас /4s As Cc
Lp Ac At Ac
Худшая Hs Cc Ns
По экспериментальным данным Bradshaw et al. (1960a, b, 1964 и др.), 6 обычных видов луговых злаков резко различаются по конкурентоспособности за различные ресурсы (табл. 5.2). Так, белоус Nardus stricta может расти при самых низких концентрациях доступных соединений азота в почве (т. е. согласно Тильману является лучшим конкурентом за азот), а гребенник Cynosurus cristatus лучший конкурент за почвенные фосфаты и т. п.
Другим экспериментальным подтверждением разного соотношения скоростей поглощения почвенных ресурсов могут быть наблюдения над корнями 60-летних деревьев Acer saccharum, Pinus resinosa и Picea abies. Для Pinus resinosa показано наиболее высокое отношение скорости поглощения минерального азота к скорости поглощения фосфатов (4,3), а для Picea abies — минимальное (1,9). Таким образом, в оптимальных условиях ель поглощает относительно больше фосфора, чем сосна (McFarlane, Yanai, 2006).
Для наземных растений наиболее важными ЭМП являются N и Р. По результатам многолетних опытов с внесением различных удобрений на Ротамстедской станции в Англии, Тильман построил несколько ресурсных изоклин для луговых растений (рис. 5.13). При низком отношении N Р доминирует чина луговая Lathyrus pratensis, симбиотически фиксирующая азот, при среднем — различные злаки, а при высоком — кульбаба щетинистая Leontodon hispidus, полевица обыкновенная Agrostis vulgaris и щавель кислый Rumex acetosa. Доминирование отдельных видов злаков
3. Конкуренция
185
Holcus
Dactyl is Lathyrus
Festuca
Agrostis
Rumex
Рис. 5.13 • Гипотетические относительные позиции 6 видов растений лугов Ротамстедской опытной станции в осях доступности азота и фосфора при высоком уровне поступления света и калия (по: Tilman, 1982)
при полном минеральном удобрении (среднее отношение N Р) зависело от кислотности почвы: на самых кислых почвах (pH менее 4,0) доминировал бухарник Holcus lanatus, на менее кислых (pH 4,0-4,5) — лисохвост Alopecurus pratensis, при более высоких значениях pH — райграс Arrhen-atherum elatius.
Модель Тильмана базируется на конкуренции в условиях стабильной среды при постоянном уровне поступления ресурсов. Однако характер поступления ресурсов в сообщества обычно не носит стационарный характер, а подвержен существенным погодным, сезонным, разногодичным и прочим изменениям. Поэтому эта модель имеет ограниченное приме
186
Глава 5. Типы взаимоотношений растений в фитоценозах
нение для природных систем, но, безусловно, весьма важна методологически, поскольку показывает важную роль почвенной гетерогенности в сосуществовании видов и дает направления для дальнейших исследований геохимической специализации растений в природе.
В рассматриваемой модели Тильмана характер поглощения сходен для разных ресурсов. Однако надо отметить принципиальное отличие конкуренции за свет и почвенные ресурсы. Конкуренция за ЭМП преимущественно симметрична (растения поглощают их приблизительно пропорционально своей подземной массе), в то время как за свет — несимметрична. Поскольку свет поступает направленно сверху, всегда более высокие растения будут получать непропорционально больше этого ресурса (так называемая «очередность поглощения»). Пластичность растений (способность изменять свою морфологию, усиливать рост в направлении источника ресурсов) несколько уменьшает несимметричность конкуренции за свет. Эксперименты, проведенные с одновидовыми посевами растений с генетическими дефектами рецепции отношения «красный : дальний красный свет» (они не могли пластично реагировать на источник света), показали резкое усиление асимметричности конкуренции у таких растений по сравнению с нормальными (Ballare et al., 1994; Bal-lare, Scopel, 1997; Schwinning, Weiner, 1998).
Возможна, хотя и маловероятна ситуация, когда в сообщество внедряется супервид (суперконкурент), изоклины которого лежат ниже и левее изоклин других видов, т. е. у этого вида более низкие пороговые концентрации по всем рассматриваемым ресурсам. В таком случае возможно вытеснение этим видом всех остальных. Однако дальнейшее развитие связано с поддерживаемой отбором специализацией экотипов внутри этого вида и дроблением супервида на много мелких специализированных по ресурсам видов, так как такая специализация ведет к более полному использованию ресурсов среды.
3.2 • Конкуренция в одновидовых посевах.
Правило и эффект Сукачева
В сельскохозяйственной практике и лесоводстве явление конкуренции известно очень давно. В отличие от животных, где случаи конкуренции требуют доказательства в каждом конкретном случае, произрастающие рядом растения обычно конкурируют друг с другом, так как поглощают общие ресурсы. Это не исключает наличие других форм отношений между видами, как и различную интенсивность конкуренции в разных случаях.
3. Конкуренция
187
Классическим примером внутривидовой конкуренции может служить явление изреживания густых посевов. Д. А. Сабинин (1948) приводит следующие показатели доли отмерших особей пшеницы при разных нормах высева:
Нормы высева семян (кг/га) % отмерших особей
85 9
130 25
170 51
В практике сельского и лесного хозяйства давно было замечено, что с увеличением плодородия почвы возрастает масса отдельных растений, при этом уменьшается их численность. Например, в экспериментах, проведенных В. Н. Сукачевым с посадками Tripleurospermum inodorum на неудобренную и удобренную почву, доля отмерших особей в первом случае составила 5,8 %, а во втором — 25 %. Таким образом, наблюдался кажущийся парадокс — с увеличением доступности ресурсов увеличивается смертность особей, т. е. возрастает интенсивность конкуренции. Это явление получило название эффекта Сукачева. Объяснение этого парадокса заключается в переключении конкуренции за ЭМП на бедных почвах на конкуренцию за свет на богатых. Как отмечено выше, конкуренция за свет носит несимметричный характер, поэтому смертность растений увеличивается. Растения, в отличие от большинства животных, могут иметь различающиеся на порядки размеры взрослых особей, а реакция на увеличение доступности ресурсов обычно проявляется не в увеличении численности популяции, а в увеличении биомассы и размеров особей, что может быть сопряжено с уменьшением их численности.
В. Н. Сукачев изучал также поведение разных растений при загущении и вывел закономерность, получившую в дальнейшем название «правило Сукачева». Он показал, что обычно однолетники в условиях загущения ускоряют, а многолетники замедляют цикл своего развития. Такие различия связаны с разными стратегиями развития при недостатке ресурсов. Однолетники быстро переходят к цветению, стремясь дать хоть небольшое число семян. Многолетники же имеют возможность ждать улучшения поступления ресурсов в будущем, замедляя свое развитие. Но развитие некоторых растений, например кукурузы, не согласуется с этим правилом.
Наиболее детально развитие растений при загущении изучали японские ученые. Nagashima et al. (1995; Weiner, 1995) провели эксперимен
188
Глава 5. Типы взаимоотношений растений в фитоценозах
ты по выращиванию Chenopodium album с разной плотностью (400, 800 и 3600 растений на м2), наблюдая за судьбой каждой особи. Они показали, что в густых посевах растения разделились на две категории: те кто прекратил свой рост («подавленные») и те, кто продолжал его. Вторых во всех вариантах опыта сохранялось около 100 на м2 Наблюдался также верхний предел численности выживших подавленных растений.
3.3 • Закон —3/2
Эта закономерность впервые обсуждалась в работе Yoda et al. (1963), она количественно описывает зависимость размеров растений от плотности: масса растения в густых посадках пропорциональна плотности посадки растений в степени -3/2 (см. Harper, 1977; Марков, 1986):
М = с • <Г3/2,
где М — масса отдельного растения, d — плотность растений (число особей на единицу площади), с — константа.
Это эмпирический закон, полученный на основании многих экспериментальных данных. Его можно легко вывести, используя теорию размерностей. Пусть L — линейный размер, S — площадь, М — масса отдельного растения, d — число особей на единицу площади, с — константа. Тогда:
М ~ L3, S~L2, L~ Sl/2 ~ М1/3,
М~53/2, S~\/d = d~l, М ~S3/2~(d“I)3/2~d’3/2,
заменяя знак пропорциональности на константу, М = с • d-3/2
Если мы построим график зависимости массы растений от плотности посадок в логарифмическом масштабе, он будет представлять собой прямую, тангенс угла наклона которой к оси абсцисс равен -3/2. Примером, подтверждающим эту зависимость, могут служить данные по изменению среднего объема дерева (объем пропорционален массе) при изреживании посадок Pinus ponderosa (White, 1980, рис. 5.14). Необходимо подчеркнуть, что рассматриваемый закон описывает только ситуации с ограниченной доступностью ресурсов, т. е. при конкуренции обычно в загущенных посевах и посадках. При низкой плотности посадок, когда конкуренция не наблюдается, масса растений не зависит от плотности популяций (см. рис. 5.15).
3. Конкуренция
189
Рис. 5.14 • Связь среднего объема дерева Pinus ponderosa с плотностью древостоя (масштаб логарифмический). График описывает изреживание древостоя с возрастом (возраст показан цифрами в кружках, годы) (по: White, 1980)
Рис. 5.15 • Самоизреживание в 4-х популяциях Lolium регеппе различной начальной плотности посева. Н1-Н5 — последовательные сроки учета массы растений. Масса отдельных растений и плотность популяции приведены в логарифмическом масштабе (по: Kays, Harper, 1974). Генета — особь семенного происхождения
3.4 • Конкуренция в парных посевах: схема De Wit
Одним из методов изучения межвидовой конкуренции является метод экспериментов с замещением (схема De Wit). Схема этого эксперимента включает одновидовые посевы каждого вида и смешанные посевы в разных пропорциях (рис. 5.16), в минимальном варианте смешанный посев 50 % одного вида и 50 % другого. Для оценки характера отношений между видами используется показатель RYT (relative yield total) — суммарный относительный урожай = относительный урожай А + относительный урожай В. Относительный урожай каждого вида равен абсолютному урожаю в смеси, деленному на его урожай в монокультуре (см. рис. 5.16). Конкурентное преимущество одного вида перед другим определяется
190
Глава 5. Типы взаимоотношений растений в фитоценозах
Доля вида В 0.00 0.25 0.50 0.75 1.00
Рис. 5.1в • Диаграмма замещающих серий посевов видов А и В при отсутствии взаимодействий между ними. Урожай представлен в относительной шкале: относительный урожай вида А = урожай А в смеси урожай А в монокультуре; относительный урожай вида В = урожай В в смеси урожай В в монокультуре; RYT (relative yield total) — суммарный относительный урожай = относительный урожай А + относительный урожай В (по: Silvertown, 1982)
по форме кривых урожая соответствующего вида (выпуклость говорит о конкурентном преимуществе, а вогнутость — о конкурентном подавлении). Суммарный относительный урожай показывает характер отношений между видами и возможность использования большего количества ресурсов смешанными посевами (рис. 5.17):
RYT = 1 Виды используют сходные ресурсы, расхождение по нишам не наблюдается.
RYT > 1 Внутривидовая конкуренция сильнее межвидовой, наблюдается частичное расхождение по ресурсам, виды могут оказывать благоприятное воздействие друг на друга.
RYT < 1 Наблюдается взаимное подавление видов друг другом.
Trenbath (1974) проанализировал результаты 572 экспериментов по замещению, проведенных на различных растениях, и сравнил их для случая смешанных посевов 50: 50. Большинство экспериментов проведено для сорных растений и их смесей с культурными. Общая ста-
3. Конкуренция
191
Рис. 5.17 • Диаграммы замещения, показывающие 4 возможных варианта взаимодействий между видами. Урожай представлен в относительной шкале: а — каждый вид в смешанном посеве имеет относительно более высокий урожай, чем в монокультуре, RYT смеси 50 50 > 1; б — урожай вида А относительно больше в смешанном посеве, а вида В — пропорционален таковому в монокультуре, максимальный RYT > 1; в — вид А в смеси представлен относительно лучше (лучший конкурент), а вид В — хуже, чем в монокультуре, RYT может быть равен 1, несколько больше или меньше 1; г — оба вида развиваются лучше в монокультуре, чем в смеси (взаимное подавление видов), RYT смеси 50 : 50 < 1 (по: Silvertown, 1982)
тистика результатов следующая: RYT 0,9 в 14% случаев, RYT » 1 в 66 %, RYT 1,1 в 22 %. Таким образом, среди культурных и сорных растений редко наблюдается расхождение в потреблении ресурсов, а внутривидовая конкуренция чаще сопоставима с межвидовой. В случае, если RYT > 1, следует различать трансгрессивное (урожай в смеси больше, чем максимальный урожай любого из видов в монокультуре) и нетрансгрессивное (урожай в смеси больше, чем средневзвешенный из монокультур, но меньше, чем наиболее урожайная монокультура) превышение урожая смешанного посева над монокультурой (Trenbath, 1974; Vandermeer, 1989; Lanta, Leps, 2007).
3.5 • Влияние условий среды на исход конкуренции. Надземная и подземная конкуренция
Нересурсные факторы среды могут оказывать влияние на исход конкуренции. Примером может служить конкуренция между овсом и ячменем на почвах разной кислотности (рис. 5.18). При pH 6,4 ячмень более конкурентоспособен, а на более кислых почвах побеждает овес.
Богатство почвы ЭМП также может влиять на исход конкуренции между двумя видами. Например, при конкуренции между ячменем и райграсом на бедных почвах более конкурентоспособен ячмень, на бога-
192
Глава 5. Типы взаимоотношений растений в фитоценозах
pH 6.4 pH 4.0 рн 3.7
Рис. 5.18 • Серия замещающих посевов овса и ячменя на почвах с разной кислотностью. Урожай выражен в млн зерен на гектар (по: De Wit, 1960)
• Овес о Ячмень
тых — райграс (Cocks, 1974). Причиной различной конкурентоспособности растений по отношению к ЭМП может быть различная интенсивность потерь биогенов при отмирании растений или отчуждении биомассы. Так в экспериментах с Festuca rubra и Arrhenatherum elatius при внесении удобрений райграс вытеснял овсяницу, а на бедных почвах — наоборот. При этом конкурентное вытеснение райграса происходило значительно быстрее в случае отчуждения надземной биомассы (имитация сенокошения), так как в этом случае потери азота с отчуждаемой биомассой были значительно выше у райграса, чем у овсяницы (Berendse et al., 1992). При сравнении уровня совокупных потерь азота (отмирание, вымывание, выделение) у 6 видов злаков было показано, что типичные злаки бедных почв теряют меньше азота по сравнению со злаками богатых почв (Vazquez de Aldana et al., 1996), что и обеспечивает конкурентоспособность первой группы в условиях дефицита ЭМП.
3. Конкуренция
193
Рис. 5.19 • Эксперимент по изучению побеговой и корневой конкуренции между хондриллой (Chonrilla juncea) и клевером (Trifolium subterranieum), в присутствии или отсутствии фитопатогенного ржавчинного гриба, поражающего хондриллу: а — схема эксперимента; б — биомасса хондриллы в разных вариантах эксперимента (в процентах к таковой в монокультуре хондриллы без ржавчинного гриба) (по: Groves, Williams, 1975)
Кроме богатства и кислотности почв на конкурентоспособность растений могут влиять многие другие факторы (температура, уровень увлажнения, присутствие консортов и т. п.). Например, избирательное поедание бизонами злаков на отдельных участках прерий приводит к лучшему развитию некоторых видов разнотравья (Ambrosia psilostachya, Vernonia baldwinii), что связано с увеличением доступности воды для этих видов (уменьшение транспирирующей поверхности злаков) (Fahnestock, Knapp, 1994).
При изучении конкуренции обычно отдельно рассматривают конкуренцию за почвенные ресурсы (воду и ЭМП) (корневая, или подземная конкуренция) и конкуренцию за свет (побеговая, или надземная конкуренция). Изоляция корневых систем или побегов позволяет оценить их относительный вклад. Такие эксперименты были проведены, например, с парными посевами хондриллы (Chonrilla juncea) и клевера (Trifolium subterranieum), в присутствии или отсутствии фитопатогенного ржавчинного гриба хондриллы (рис. 5.19, Groves, Williams, 1975). В этом случае надземная конкуренция оказалась более значительной, она снижала био
194
Глава 5. Типы взаимоотношений растений в фитоценозах
массу хондриллы до 47 % от контроля без ржавчинного гриба и до 21 % — в присутствии ржавчинного гриба. Аналогичные величины для случая с совместной конкуренцией за свет и почвенные ресурсы составили 31 % и 6 %. Таким образом, этот пример показывает, что фитопатогенные грибы, как и другие консорты, могут влиять на исход конкуренции между растениями. Если в монокультуре ржавчина снижала биомассу хондриллы примерно вдвое, то при конкуренции с клевером — в пять раз.
Конкуренции за свет и почвенные ресурсы взаимосвязаны. Затенение уменьшает способность растений поглощать почвенные ресурсы. Например, в экспериментах Jackson, Caldwell (1992) затенение дерновинного злака Agropyron desertorum вызвало неспособность растений поглощать фосфаты из локально обогащенных участков, в то время как незатененные растения эффективно использовали эти фосфаты.
В обзоре Wilson (1988), основанном на анализе 23 экспериментов с разделением корневой и побеговой конкуренции, показано, что корневая конкуренция обычно более интенсивна, чем побеговая. В экспериментах по конкуренции Agropyron desertorum и Pseudoroegneria spicata конкурентное преимущество первого вида было, видимо, обусловлено лучшим ростом его корней при низких температурах в начале сезона, а также более тонкими корнями (Eissenstat, Caldwell, 1988а, b). В гетерогенной среде растения сокращают рост корней на бедных участках и увеличивают — на богатых. Как корневые выделения, так и непосредственные контакты между корнями, сигнализирующие о наличии соседей, могут изменять характер роста и ветвления корней. Тонкое узнавание корней возможно как на видовом уровне, так и на уровне отдельных генотипов, что было показано на примере Miscanthus sinensis — корни резко уменьшали рост лишь в присутствии корней растений своего генотипа. Анализ пространственного распределения корней Larrea tridentata в природе выявил весьма неоднородную структуру со значительно меньшим перекрытием у разных особей, чем предполагалось по модели круговых систем (Brisson, Reynolds, 1994), что обусловлено взаимодействием корней соседних растений. В целом возможны две противоположные картины взаимодействия корней соседних растений — 1) корневое избегание, приводящее к образованию корневых территорий отдельных особей, 2) корневое перемешивание, когда корни лучше растут близ корней других особей (видов). Оба механизма связаны с распознаванием корней соседей. При рассмотрении корневой конкуренции необходимо различать 2 контрастные ситуации, в которых разные свойства корней дают конкурентное преимущество: 1) конкуренция за свободный от корней богатый ЭМП объем почвы (здесь важна скорость роста) и 2) конкуренция в заполненном корнями объеме почвы с истощающимися ресурсами
3. Конкуренция
195
(здесь важны низкая скорость оборота, способность потреблять ЭМП при низких концентрациях и т. п.). Примером разной конкурентоспособности может служить пара видов Festuca rubra — Anthoxanthum odoratum. В первый год совместного посева Festuca имела конкурентное преимущество за счет более крупной и густой корневой системы, а во второй год в гетерогенной среде она сравнялась по биомассе с Anthoxanthum, который имел более высокую физиологическую пластичность корней и эффективнее потреблял ЭМП при их медленном поступлении (Fransen et al., 2001, de Kroon et al., 2003).
В присутствии конкурентов может изменяться вертикальное распределение корней, что показано, например, для Haplopappus ericoides в присутствии инвазионного Carpobrotus edulis (D’Antonio, Mahall, 1991). Аналогичную закономерность указывает F. Berendse (1981, 1983) для Anthoxanthum odoratum и Plantago lanceolata (в присутствии подорожника душистый колосок имеет более поверхностные корни (Hutchings, John, 2003).
Однако воздействие корней и побегов соседних растений не всегда связано только с поглощением ресурсов. McConnaughay и Bazzaz (1992) провели эксперименты с механическими аналогами корней (проволочные конструкции разной плотности) и побегов (стеклянные трубки, не уменьшающие световой поток). Они показали, что сама фрагментация пространства «корнями» и «побегами» отрицательно влияет на рост растений. Это отрицательное воздействие не проявлялось при высоком уровне обеспеченности ЭМП (но не водой!) в случае «искусственных корневых систем» и светом — в случае «искусственных побегов».
З.в • Адаптации растений бедных и богатых почв
Как было отмечено выше, богатство почв ЭМП оказывает большое влияние на структуру сообществ и характер конкурентных отношений, поэтому минеральное питание является важнейшим фактором дифференциации видов растений (Grime et al., 1997). Адаптации растений к бедным и богатым почвам принципиально различаются (Grime, 1979; Chapin, 1980). Для растений бедных почв характерны следующие свойства.
1. Низкая скорость поглощения ЭМП корнями и низкая пороговая концентрация ресурсов R* (см. 3.1).
2. Малая относительная скорость роста (RGR — relative growth rate).
3. Относительно большая биомасса корней.
4. Высокая степень микоризной инфекции.
5. Длительная жизнь корней и листьев.
196
Глава 5. Типы взаимоотношений растений в фитоценозах
6. Избыточное потребление («luxury consumption», потребление ЭМП из почвы в зависимости от их доступности, а не текущей потребности в них для роста и развития), часто более высокая концентрация ЭМП в тканях. Поэтому растения бедных почв, оказавшиеся в условиях высокой доступности ЭМП часто страдают от «автотоксикации», накоплении ряда соединений (например, нитратов) в избыточных (токсичных) для своих тканей концентрациях.
7. Низкая съедобность тканей для фитофагов в связи с высоким содержанием защитных веществ и соответственно низкая скорость разложения опада.
С другой стороны, для растений богатых почв характерны следующие свойства.
1. Высокая поглощающая способность корней, их быстрый рост на обогащенных участках почвы.
2. Значительная скорость роста.
3. Высокая эффективность фотосинтеза.
4. Низкие запасы углеводов в тканях.
5. Высокая дыхательная активность корней.
6. Меньшая продолжительность жизни и быстрое обновление корней и листьев.
7. Быстрая скорость разложения опада.
Ряд экспериментов был проведен для выявления роли этих адаптаций в конкурентоспособности и доминировании отдельных видов растений в различных условиях. Так, на вершинах холмов в Онтарио (Канада) — бедных местообитаниях — была изучена связь степени развития отмеченных выше признаков растений бедных почв и участия видов в сложении сообществ (Reader, 1998). Было показано, что более обильные виды имели: 1) более высокое отношение «корни:побеги», 2) более высокую степень микоризной инфекции, 3) меньшую биомассу отдельных побегов, 4) более низкую относительную скорость роста (RGR), 5) более низкую поедаемость листьев фитофагами. Три независимых параметра (масса побегов, микоризная инфекция и поедаемость листьев) объясняли 99 % варьирования участия изученных видов в составе фитоценоза (рис. 5.20).
Адаптации растений к произрастанию на бедных или богатых почвах могут быть связаны с их происхождением. Уровень нитратредукции, отражающий способность использовать нитраты как источник связанного азота, у растений, относящихся к семействам Proteaceae, Epacridaceae, Ericaceae и Myrtaceae, оказался существенно более низким, чем у растений из семейств Amaranthaceae, Chenopodiaceae, Moraceae, Polygonaceae,
3. Конкуренция
197
Рис. 5.20 • Связь
между относительным участием видов и их свойствами для 7 видов растений бедных почв на вершинах холмов в провинции Онтарио (Канада). Виды растений: Cl — Chrysanthemum leucanthemum, De — Daucus carota, Hf — Hieracium floribundum, Hp — Hypericum perforatum, Ml — Medicago lupulina, Pc — Poa compressa, Sc — Soiidago canadensis (no: Reader, 1998)
Относительная скорость роста (неделя *1)
Индекс поедаемости листьев (недель до потери первого листа )
198
Глава 5. Типы взаимоотношений растений в фитоценозах
Urticaceae, происхождение которых было связано с более богатыми местообитаниями (Stewart, Schmidt, 1999).
Интересные наблюдения проведены для высокогорных растений в горах Колорадо (Theodose et al., 1996). Авторы исследовали корреляцию между участием вида в сообществе (как показатель его конкурентоспособности в данных условиях) со следующими показателями: 1) скоростью поглощения минер, азота (по метке 15 N), 2) биомассой целого растения 3) отношением биомассы «корни:побеги», 4) содержанием азота в тканях. Было показано, что скорость поглощения азота выше у малочисленных видов (т. е. отрицательно скоррелирована с участием видов в сообществе), а признаки 2 и 3 положительно коррелировали с участием вида. Содержание азота в тканях было ниже у доминантов, а самым высоким — у двух редких видов, т. е. доминанты более эффективно используют поглощенный азот на построение своей биомассы.
При лимитировании продукции ЭМП конкурентоспособность видов может зависеть от запаса этих элементов в Тканях растений в начале вегетационного периода. Так, по наблюдениям Н. А. Журавлевой (1994) в луговом фитоценозе виды с большим участием в составе ценоза имеют в начале вегетации более высокое содержание N и Р (стартовые позиции), но в степном сообществе такая связь отсутствовала, что, по мнению автора, свидетельствует о невыраженное™ конкуренции за эти ресурсы при недостатке влаги в степях.
Как изменяется структура сообщества при обогащении почвы ЭМП (внесение удобрений)? Обычный эффект — это увеличение продукции при снижении видового разнообразия (усиление конкуренции за свет). Однако Gurevitch и Unnasch (1989) показали для залежей в США, что если при внесении ЭМП удалить главного доминанта (Dactylis glomerata), то видовое богатство даже увеличивается как за счет обычных, так и относительно редких видов. Аналогично искусственное подавление роста доминирующих растений некоторыми химикатами вызывало увеличение видового разнообразия (Yemm, Willis, 1962), как и воздействие фитофагов или нарушения.
3.7 • Эколого-ценотические стратегии и конкурентоспособность растений
3.7.1 • Взгляды Л, Г. Раменского
Л. Г. Раменский (1935, 1938) предложил различать 3 группы растений (фитоценотипы) по их поведению в составе сообществ. Виоленты «энергично развиваясь... захватывают территорию и удерживают ее за собой,
3. Конкуренция
199
подавляя, заглушая соперников энергией жизнедеятельности и полнотой использования среды». Патиенты «в борьбе за существование... берут не энергией жизнеспособности и роста, а своей выносливостью к крайне суровым условиям, постоянным или временным». Эксплеренты «имеют очень низкую конкурентную мощность, но зато способны очень быстро захватывать освобождающиеся территории, заполняя промежутки между более сильными растениями; также легко они и вытесняются последними». К сожалению, в своих дальнейших трудах Л. Г. Раменский детальнее не разрабатывал эти представления, которые 40 лет спустя были независимо и с других позиций описаны Ф. Граймом (Grime, 1977).
3.7.2 • Взгляды Ф. Грайма: типы стратегий растений
С иных позиций рассматривает конкуренцию и сосуществование видов Ф. Грайм (Grime, 1979, 2001). Он рассматривает функциональную классификацию видов растений как проявление их стратегий. Под стратегиями (синоним — функциональные типы) Ф. Грайм понимает «группы сходных или аналогичных, генетически определенных признаков, которые часто повторяются среди видов или популяций растений и вызывают у них проявляющееся сходство в экологии» (Grime, 2001). Выделение стратегий возможно на разных уровнях иерархии растительного покрова: глобальном, региональном, локальном и внутрипопуляционном (рис. 5.21). Небольшое число первичных типов глобального уровня должно охватывать, по мнению автора, все растения, а также быть применимым и к животным. Для выделения типов стратегий на глобальном уровне необходимо использовать небольшое число признаков, легко определяемых для большинства растений. Например, Westoby (1998) предложил 3 таких признака: удельная листовая поверхность (поверхность 1 г листа), высота полога и вес семени. Кроме того, адаптации растений могут
Рис. 5.21 • Иерархия функциональных типов (стратегий) растений (по: Grime, 2001)
200
Глава 5. Типы взаимоотношений растений в фитоценозах
Таблица 5.3 • Классификация местообитаний и характерных для них типов стратегий растений в зависимости от интенсивности нарушений и стресса (по: Grime, 1979)
Стресс
слабый сильный
Нарушения слабые С — конкуренты S — стресс-толеранты
сильные R — рудералы Нет жизнеспособных стратегий
существенно различаться на разных стадиях жизненного цикла. В классификации, изложенной ниже, Ф. Грайм рассматривает адаптации взрослых растений.
Ф. Грайм выделяет два важнейших фактора организации растительного покрова и дифференциации видов растений, которые непосредственно связаны с почвенными условиями, — это нарушения и стресс. Под стрессом он понимал «набор явлений, которые ограничивают фотосинтетическую продукцию, таких как недостаток света, воды и элементов минерального питания или субоптимальные температуры» (Grime, 2001: 7). Нарушения же «связаны с частичным или полным разрушением биомассы растений и возникают в результате деятельности фитофагов, патогенов, человека (вытаптывание, кошение, вспашка) или в результате действия ветра, мороза, засухи, почвенной эрозии и огня» (Grime, 2001: 8). В зависимости от сочетания этих факторов выделяется 4 типа местообитаний и 3 типа растений (конкуренты, стресс-толеранты и ру-дералы), поскольку, по мнению автора, выработка приспособлений для существования одновременно в условиях сильного стресса и нарушений эволюционно вряд ли возможна (табл. 5.3).
Для выделения основных типов стратегий травянистых растений Ф. Грайм использовал два показателя — относительную скорость роста (RGR — скорость прироста биомассы на единицу имеющейся биомассы) и морфологический индекс, отражающий размерные характеристики растений и мощность накопления опада под ними (см. рис. 5.22). Самый высокий морфологический индекс для растений окрестностей Шеффилда (Англия) отмечен у Epilobium hirsutum и Chamerion angustifolium (рис. 5.22). Включение в морфологический индекс показателя накопления опада связано с тем, что это один из важных механизмов доминирования видов конкурентной стратегии, поскольку через опад трудно пробиться всходам мелкосемянных и медленно растущих видов. Листовой опад деревьев оказывает большое влияние на развитие травянистых
3. Конкуренция
201
растений под пологом леса. Эксперименты с выращиванием лесных травянистых растений в присутствии или отсутствии искусственного опада (Sydes, Grime, 1981а, b) показали разную реакцию видов на его наличие и приводили к различной структуре травяного покрова. Наиболее устойчивыми к опаду видами оказались Lamiastrum galeobdolon и Ну-acinthoides non-scripta. Для определения типов стратегий травянистых растений Ф. Грайм предложил использовать и специальный ключ, представленный на рис. 5.23.
Растения конкурентной стратегии (С) произрастают и доминируют на богатых почвах с невысоким уровнем нарушений. Для них характерны следующие особенности:
а) наличие запасающих органов, позволяющих быстро развертываться листьям,
б) значительная высота растений, а также разрастание вширь побегов, в) полное развертывание листьев в наиболее благоприятный для фотосинтеза период,
г) высокая относительная скорость роста,
д) быстрый ростовой отклик побегов или корней на обогащение ресурсами отдельных участков пространства, показанный экспериментально в ряде исследований автора и его коллег,
е) быстрое отрастание после частичной дефолиации фитофагами, ж) невысокий вклад ассимилятов в защитные вещества и структуры. По-
следнее свойство хорошо согласуется с представлениями Oksanen et al. (1981) о меньшей роли фитофагов в высокопродуктивных сообществах, где основными регуляторами численности этих животных выступают виды следующего трофического уровня (хищники и паразиты) (см. п. 4 гл. 4).
Местообитания стресс-толерантов Ф. Грайм разделяет на 4 группы: арктоальпийские и аридные, где стресс «навязывается» внешней средой; тенистые, где стресс (затенение) обусловлен деятельностью растений; бедные элементами минерального питания (ЭМП), что может быть связано как с характером исходного субстрата, так и с деятельностью других растений или обоими факторами. Но во всех местообитаниях стресс-толерантов действие различных неблагоприятных факторов (низкие температуры, засуха, затенение) сопряжено с ведущим фактором — недостатком ЭМП. В отличие от видов конкурентной стратегии, пластично реагирующих на локальное (пространственное) обогащение ресурсами,
202
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.22 • Треугольная ординация травянистых растений окрестностей Шеффилда (светлые кружки — однолетники, темные — многолетники) по морфологическому индексу (см. текст) и относительной скорости роста — RGR (по: Grime, 1974). Морфологический индекс рассчитан по формуле:
М — (а + b + с)/2, где а — максимальная высота расположения листьев (баллы 1 — менее 12 см, 2 — 12-25 см, 3 — 25-37 см, 4 — 37-50 см, 5 — 50-62 см, 6 — 62-75 см, 7 — 75-87 см, 8 — 87-100 см, 9 — 100-112 см, 10 — более 112 см); b — горизонтальное простирание (баллы 0 — мелкие терофиты, 1 — крупные терофиты, 2 — многолетники с компактными неветвистыми корневищами или формирующие небольшие (до 10 см в диаметре) дерновинки, 3 — многолетники с системой корневищ или дерновинные растения с диаметром 10-25 см, 4 — многолетники диаметром 26-100 см, 5 — многолетники с диаметром более 1 м); с — максимальная аккумуляция опада (баллы 0 — опад (ветошь) отсутствует, 1 — тонкий не сплошной покров ветоши, 2 — тонкий сплошной покров, 3 — мощность опада до 1 см , 4 — до 5 см, 5 — более 5 см). Виды растений: Ас — Agrostis canina, Ае — Arrhenatherum elatius, Ад — Alopecurus geniculatus, Ah — Arabis hirsute, Am — Achillea millefolium, Ao — Anthoxanthum odoratum, Ap — Aira praecox, Apr — Alopecurus pratensis, Ar — Agropyron repens,
3. Конкуренция
203
— Agrostis stolon if era, Ase — Arenaria serpy Hifolia, At — Agrostis capi Haris, Вт — Briza media, Bs — Brachypodium sylvaticum, Bst — Bromus sterilis, Bt — Bidens tripartite, Ca — Chamaenerion angustifolium, Cal — Chenopodium album, Cc — Cynosurus cristatus, Cf — Carex flacca, Cfl — Cardamine flexuosa, Cfo — Cerastium fontanum, Cn — Centaurea nigra, Cp — Carex panacea, Cpr — Cardamine pratensis, Cro — Campanula rotundifolia, Cri — Catapodium rigidum, Cv — Clinopodium vulgare, Cvu — Cirsium vulgare, De — Deschampsia cespitosa, Df — Deschampsia flexuosa, Dg — Dactyli's glomerata, Dm — Draba muralis, Do — Dry as octopetala, Dp — Digitalis purpurea, Eh — Epilobium hirsutum, Fg — Festuca gigantea, Fo — Festuca ovina, Fr — Festuca rubra, Fu — Filipendula и I man a, Ga — Galium aparine, Gf — Glyceria fluitans, Gp — Galium palustre, Gr — Geranium robertianum, Gu — Geum urbanum, Gv — Galium verum, Hn — Helianthemum nummularium, Hl — Holcus lanatus, Hm — Holcus mollis, Hmu — Hordeum murinum, Hp — Helictotrichon pratense, Js — Juncus squarrosus, Km — Koeleria macrantha, Lc — Lotus corniculatus, Lea — Luzula campestris, Lh — Leontodon hispidus, Lp — Lolium perenne, Me — Milium effusum, Ml — Medicago lupulina, Md — Matricaria discoidea, Mn — Melica nutans, Ms — Myosotis sylvatica, Ns — Nardus stricta, Ov — Origanum vulgare, Pa — Poa annua, Pav — Polygonum aviculare, Pc — Polygonum convolvulus, Pe — Potentilla erecta, Pl — Plantago lanceolata, Pm — Plantago major, Pp — Poa pratensis, Ppe — Polygonum persicaria, Ps — Poterium sanguisorba, Pt — Poa trivialis, Pv — Prunella vulgaris, Ra — Rumex acetosa, Rac — Rumex acetosella, Ro — Rumex obtusifolius, Rr — Ranunculus repens, Sa — Sedum acre, Sal — Sesleria albicans, Sc — Scabiosa columbaria, Sd — Sieglingia decumbens, Sdi — Silene dioica, Sj — Senecio jacobaea, Sm — Stellaria media, Sp — Succisa pratensis, Ss — Senecio squalidus, Sv — Senecio vulgaris, Td — Thymus drucei, Tf — Tussilago farfara, Tm — Trifolium medium, To — Taraxacum officinalis, Tr — Trifolium repens, Ts — Teucrium scorodonia, Ud — Urtica dioica, Va — Veronica arvensis, Vr — Viola riviniana, Ze — Bromus erectus
стресс-толеранты хорошо адаптированы к временной пульсации доступности ресурсов, которая, по мнению автора, характерна для бедных местообитаний. Так, в экспериментах (Campbell, Grime, 1989а) с Festuca ovina (стресс-толерант) и Arrhenatherum elatius (конкурент) в условиях периодического поступления ЭМП первый вид имел более высокую относительную скорость роста при продолжительности периода обогащения ЭМП менее 10 часов, а второй вид — при более длительных периодах поступления этих почвенных ресурсов. Характерными чертами стресс-толерантов являются также отсутствие резко выраженной сезонной динамики биомассы побегов, вечнозеленость, низкая относительная скорость роста, нечастое цветение, низкая морфологическая пластичность.
Рудералы (виды рудеральной стратегии) приурочены к местообитаниям со значительным уровнем нарушений, которые, в отличие от стресса,
Цветение незадержанное
Потенциально быстрорастущие
Двулетники (монокарпики}
Весенние геофиты
Побеги не фрагментируются быстро
C-R
Потенциально крупные и быстрорастущие
Быстрое разрастание и фрагментация побегов
Небольшие и медленнорастущие
Небольшие и медленнорастущие
Однолетники
Побеги высокие и с интенсивным боковым разрастанием
Побеги короткие или ползучие
СП
3
Побеги высокие и/или с интенсивным разрастанием вширь Медленный оборот листьев
Побеги короткие, без сильного бокового разрастания
Многолетники
Быстрый оборот листьев
Не весенние геофит
204 Глава 5. Типы взаимоотношений растений в фитоценозах
3. Конкуренция
205
Рис. 5.23 • Дихотомический ключ для определения типов стратегий травянистых растений, используемый для классификации положения маркерных видов в ординационном треугольнике (вверху справа). С — конкуренты, S — стресс-толеранты, R — рудералы, C-R — конкуренты-рудералы, S-R — стресс-толеранты-рудералы, C-S — стресс-толеранты-конкуренты, C-S-R — сочетание всех трех стратегий (Grime et al., 1988) (примеры см. в тексте)
вызывают гибель растений или их частей, т. е. повреждают уже сформировавшуюся биомассу. К нарушениям автор относит и сезонное развертывание листьев деревьев, вызывающее массовое отмирание надземных частей эфемероидов в листопадных лесах. К типичным рудералам относятся также многие растения побережий (морских и пресных водоемов), полевые сорняки, растения вытаптываемых участков, пустынные однолетники и эфемероидные послепожарные растения. Они характеризуются небольшой длительностью жизни (одно- и малолетники) и очень высокой морфологической пластичностью, способностью образовывать карликовые формы с одним цветком в условиях стресса.
Таким образом, виды трех основных стратегий отличаются фундаментально реакцией на стресс: рудералы обеспечивают хотя бы минимальное образование семян, конкуренты усиливают поглощение ресурсов, а стресс-толеранты обеспечивают лучшую сохранность уже поглощенных ресурсов.
Помимо выделения 3-х первичных стратегий, автор определяет 4 переходных группы растений, относимые им ко вторичным стратегиям (CR, SR, CS, CSR).
К конкурентам-рудералам автор относит некоторые однолетние травы (Galium aparine, Impatiens grandulifera), многолетние монокар-пики (Anthriscus sylvestris, Cirsium vulgare, Arctium spp.), рудеральные многолетние травы (Ranunculus repens, Cirsium arvense, Tussilago far-fara, Agrostis stolonifera).
К стресс-толерантам-рудералам относятся небольшие однолетники и малолетники (например, арктическая Koenigia islandica, пустынные эфемеры), геофиты (Scilla verna, Orchis mascula, Anemone nemorosa), многие мохообразные (Racomitrium lanuginosum, Dicranum fuscescens).
К стресс-толерантам-конкурентам автор относит виды среднепродуктивных сообществ с низким уровнем нарушений, например многолетние травы — Brachypodium pinnatum, Bromopsis erecta, Scirpus sylvaticus, Juncus effusus, Mercurialis perennis, многие древесные растения (Quercus, Castanea, Fag us) и листопадные кустарники и кустарнички (Euonymus еи-ropaeus, Vaccinium myrtillus).
206
Глава 5. Типы взаимоотношений растений в фитоценозах
Центральное положение в ординационном треугольнике занимают виды, обладающие чертами всех трех стратегий (CSR). К ним отнесено 6 групп растений: небольшие дерновинные злаки (Anthoxanthum odoratum), небольшие розеточные глубоко укореняющиеся двудольные (Leontodon hispidus, Silene nutans), мелкие столонообразующие растения (Fragaria vesca, Ajuga reptans), короткокорневищные двудольные (Prunella vulgaris, Potentilla erecta), бобовые (Anthgllis vulneraria, Trifolium medium), мелкие осоки и ситниковые (Carex cargophgllea, Luzula campestris).
Проверка рассматриваемой CSR модели осуществлена многими исследованиями автора и его коллег. Особенное значение имеет программа изучения различных характеристик (экологических, физиологических и биохимических) значительного числа тестируемых видов растений британской флоры в стандартных условиях («Integrated Screening Programme»). Многие исследованные параметры оказались тесно скоррелированы друг с другом. Многомерная ординация 43 видов показала, что ось наибольшего варьирования связана с признаками, характеризующими минеральное питание растений. Это подтверждает мнение автора, что дифференциация по минеральному питанию является одной из важнейших для исследованных видов, относящихся к разным стратегиям. Третья ось наибольшего варьирования, полученная при ординации методом главных компонент, отражала дифференциацию по условиям нарушений. Таким образом, проекции видов на первую и третью ось хорошо отражали дифференциацию видов по 3 рассматриваемым стратегиям (Grime et al., 1997) (рис. 5.24).
В другом эксперименте (Campbell, Grime, 1992) авторы создали искусственный посев 7 видов растений, представляющих различные стратегии, на площадках, в которых искусственно создавалась пространственная неоднородность почвы — в одном направлении изменялся уровень богатства почвы, а в другом (перпендикулярном) — уровень нарушений (как на рис. 9.55, см. п. 4.1 гл. 9). В чистых посевах большинство видов лучше произрастало на участках богатых почв при невысоком уровне нарушений, а в смешанных происходила дифференциация видов, они чаще встречались на участках, отражающих их «стратегические» свойства.
При кратковременном стрессовом воздействии можно выделить два аспекта устойчивости: «стойкость» (resistance) — способность сохранять свою биомассу после воздействия и «эластичность» (resilience) — способность восстанавливать свою биомассу после воздействия (Westman, 1978). Специальными экспериментами с воздействием засухи, мороза и выжигания на участки дернины лугов в Англии было показано, что
3. Конкуренция
207
Рис. 5.24 • Положение видов британской флоры в проекциях на 1 и 3 ось наибольшего варьирования большого числа эколого-морфологических признаков с наложением на треугольник стратегий Ф. Грайма (по: Grime et al., 1997). Виды растений: Agrostis capillaris, Anisantha sterilis, Anthoxanthum odoratum, Anthriscus sylvestris, Arabidopsis thaliana, Arrhenatherum elatius, Brachypodium pinnatum, Briza media, Bromopsis erecta, Campanula rotundifolia, Carex flacca, Catapodium rigidum, Centaurea scabiosa, Cerastium fontanum, Chanerion angustifolium, Chenopodium album, Conyza canadensis, Dactylis glomerata, Dryas octopetala, Epilobium hirsutum, Eriophorum vaginatum, Festuca ovina, Festuca rubra, Galium aparine, Helianthemum nummularium, Helianthus annuus, Helictotrichon pratense, Holcus lanatus, Koeleria macrantha, Leontodon hispidus, Lolium perenne, Lotus corniculatus, Origanum vulgare, Pilosella officinarum, Plantago lanceolata, Poa annua, Poa trivialis, Rumex acetosella, Thymus polytrichus, Urtica dioica, Zea mays
208
Глава 5. Типы взаимоотношений растений в фитоценозах
стойкость хорошо коррелирует со свойствами S-стратегии, а эластичность — с R. Таким образом, признаки S и R стратегий могут служить для предсказания реакции отдельных видов и сообществ на экстремальные воздействия (MacGillivray et al., 19956).
Представления о стратегиях растений, разработанные Ф. Граймом, во многом аналогичны взглядам Л. Г. Раменского. Можно проследить хотя и неполную аналогию между виолентами и конкурентами, патиентами и стресс-толерантами, эксплерентами и рудералами.
3.7.3 • Противоречия Тильман — Грайм. Интенсивность и значимость конкуренции
Рассмотренные выше взгляды на ресурсную конкуренцию Д. Тильмана и Ф.Грайма противоречат друг другу по нескольким вопросам (см. табл. 5.4).
Ф. Грайм определяет конкуренцию как способность к поглощению ресурсов (capacity for resource capture). Напротив, Д. Тильман считает, что конкурентный успех — это способность к снижению концентрации ресурсов до низкого уровня и способность переносить этот низкий уровень доступности ресурса. По мнению Ф. Грайма, в условиях высокой конкуренции виды конкурентной стратегии должны иметь хорошо развитый аппарат поглощения как подземных ресурсов, так и света. Он считает конкурентную иерархию видов (их относительную конкурентоспособность) независимой от продуктивности (т. е. богатства почв) вели-
Таблица 5.4 • Основные расхождения в теоретических представлениях Грайма и Тильмана (по: Grace, 1991)
Ф. Грайм Д. Тильман
Основное свойство доминантов с наибольшей конкурентоспособностью Доминирование в благоприятных условиях — результат способности к поглощению большого количества ресурсов Доминирование — результат способности поглощать ресурсы и снижать уровень их доступности
Связь между способностью поглощать разные ресурсы Способности видов к поглощению различных ресурсов положительно скоррелированы Способности видов к поглощению различных ресурсов отрицательно скоррелированы
Роль конкуренции в различных по продукции сообществах Конкуренция менее интенсивна в непродуктивных условиях Конкуренция важна как в продуктивных, так и в непродуктивных условиях
3. Конкуренция
209
чиной, в то время как D. Tilman (1982) постулирует обратное. Хотя оба автора признают очевидную смену доминирования при переходе от низкопродуктивных к высокопродуктивным сообществам, у них остаются расхождения относительно механизма этой смены. Д. Тильман считает таким механизмом смену ресурсов, за которые происходит конкуренция (почвенные ресурсы в бедных местообитаниях, свет — в богатых). Ф. Грайм же рассматривает эту смену как результат ослабления конкуренции на бедных почвах и усиления здесь вклада других селективных механизмов, таких как засуха или воздействие фитофагов. На основании ряда наблюдений он заключает, что доминирование медленнорастущих видов на бедных почвах связано не с высокой способностью их корней поглощать элементы минерального питания, а с их лучшим удержанием и защитой от потерь, которые не характерны для видов конкурентной стратегии, в отличие от стресс-толерантов.
Необходимо подчеркнуть различия между значимостью («важностью») конкуренции и ее интенсивностью (Weldon, Slauson, 1986). Интенсивность конкуренции выражается в степени уменьшения численности или плотности популяции растений из-за присутствия соседей. Значимость конкуренции — относительное уменьшение численности или плотности популяции растений за счет конкуренции в сравнении с уменьшением численности или плотности, вызванным другими причинами (фитофаги, нарушения и т. п.).
Для оценки интенсивности и важности конкуренции может быть применен регрессионный и дисперсионный анализ. При регрессионном анализе (например, зависимость размеров растений от расстояния до соседа) интенсивность конкуренции оценивается по наклону линии регрессии. Важность конкуренции в данном случае может быть оценена по доли дисперсии, описываемой данной линией (квадрат коэффициента корреляции между эмпирическими данными и регрессионными). При удалении соседей важность конкуренции может быть оценена по доли дисперсии, связанной с воздействием фактора удаления (Weldon, Slauson, 1986). Интересную методику оценки роли конкуренции в формировании растительных сообществ предложила D.E. Goldberg (1994) [см. Вставку VIII]. Ее методика основана на сравнении структуры сообщества из нескольких видов с так называемым «нулевым» сообществом, где биомасса отдельных видов прямо пропорциональна их биомассе в монокультуре. Чем сильнее реальное сообщество отличается от «нулевого», тем важнее конкуренция для его формирования. Для высокогорных сообществ северо-западного Кавказа, изученных по этой методике (Onipchenko et al., 2009), наибольшая роль конкуренции отмечена в высокопродуктивных
210
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.25 • Связь продукции сообществ альпийского пояса северо-западного Кавказа (столбики) с ролью конкуренции в их формировании (показатель PD ю методике D. Е. Goldberg, 1994, линия). Сообщества: АЛП — альпийские тишайниковые пустоши, АН — альпийские ковры, ПЛ — пестроовсяницевые iyra, ГКЛ — гераниево-копеечниковые луга (по: Onipchenko et al., 2009)
гераниево-копеечниковых лугах, а наименьшая — в средних по продукции пестроовсяницевых лугах (рис. 5.25). Низкопродуктивные сообщества показали промежуточные величины роли конкуренции, оцененной по величине PD [см. Вставку VIII].
D. Е. Goldberg (1994) предложила ис-Вставка VIII. Методика D. Е. Goldberg пользовать посевы монокультур (или оценки роли конкуренции эксперименты с удалением всех ви-
в формировании сообществ дов, кроме одного) для оценки струк-
----------------------------------- туры так называемого «нулевого» сообщества. в котором биомасса отдельных видов прямо пропорциональна их биомассе в монокультуре.
Ожидаемое участие вида в «нулевом» сообществе: RY)m = Yim/^2 где Yim — финальное участие (обилие) г-го вида в монокультуре.
Реальное участие вида в сообществе (смеси видов): RYix = Yix/^ Yix, где: Ylx — финальное участие (обилие) г-го вида в сообществе.
Далее вычисляется абсолютное (D,) и относительное (PDj) отклонение участия вида в ценозе и монокультуре:
Di = RYIX - RYim, PDi = (RYix - RYim)/RYim
и для всего сообщества — D и PD:
d=^\d,\, pd = ^\pd,\.
3. Конкуренция
211
Эти показатели оценивают только относительное изменение участия видов в составе ценозов по сравнению с теоретическим «нулевым» сообществом, т. е. при пропорциональном изменении участия всех видов по сравнению с монокультурой эти показатели (D и PD) равны 0. Расчет этих показателей для экспериментов Campbell, Grime (1992) показал, что они минимальны на среднем уровне нарушений и богатства почвы.
Р. Keddy et al. (1998) предложили различать конкурентный эффект вида (способность подавлять соседей) и конкурентный отклик (устойчивость к конкурентному воздействию соседей). Наблюдения авторов показали, что эти два показателя не скоррелированы друг с другом.
Анализ публикаций результатов экспериментов по конкуренции, проведенный J. Gurevitch с соавторами (1992), показал, что для наземных растений эффект конкуренции был несколько (незначимо) выше для бедных местообитаний (тундры, пустыни), чем для более богатых (прерии, луга, залежи), что свидетельствует в пользу теории Д. Тильмана.
Вопрос о роли конкуренции в бедных местообитаниях рассмотрен в специальном исследовании D. Goldberg и A. Novoplavsky (1997). Они показали, что при наличии флуктуаций в доступности почвенных ресурсов («пульсов» и «межпульсных интервалов», рис. 5.26) конкуренция не имеет важного значения (прав Грайм), если выполняются следующие условия:
R
У \ Межпульсный / I /
г I интервал / 1 J
Пупьг^^-. t
Рис. 5.26 • Схематическое изображение динамики пульсирующего ресурса (R). Пунктирная линия — пороговая концентрация
1) «межпульсные интервалы» определяются целиком абиотически, поглощение ресурсов растениями не оказывает влияние на их частоту, длительность или интенсивность,
2) выживаемость растений в «межпульсные интервалы» не зависит или отрицательно скоррелирована с количеством поглощенных ресурсов в период «пульсов» (например, более крупные растения сильнее страдают от засухи и т. п.).
Если хотя бы одно из этих условий не выполняется, конкуренция в бедных местообитаниях имеет место, что подтверждает точку зрения Тильмана.
212
Глава 5. Типы взаимоотношений растений в фитоценозах
P.J. Grubb (1994) рассматривает два варианта корневой конкуренции на бедных почвах: в плошках (изначально почва лишена корней) и в ценозах, где почва плотно пронизана корнями. В первом случае конкурентное преимущество имеют виды, обладающие высокой скоростью роста корней и быстрее занимающие свободное пространство. Во втором случае — виды с наиболее низкими пороговыми концентрациями почвенных ресурсов. В целом, видимо, обе теории объясняют различные стороны конкурентных отношений и применимы к различным природным и искусственным сообществам.
3.7.4 • Взгляды Ю. Э. Романовского
Ю. Э. Романовский сделал попытку выявить соответствие между 2- (г-и К-стратеги, см., например, Пианка, 1981) и 3-компонентными (Раменский, Грайм) классификациями стратегий, с одной стороны, и взглядами на конкурентоспособность Тильмана и Грайма, с другой (Романовский, 1989а, б, 1998).
Достигнуть успеха в конкуренции за ресурсы можно двумя основными путями: 1) снижая пороговую концентрацию ресурса (Я*) или 2) быстро реагируя на увеличение доступности ресурса в среде увеличением скорости его потребления. В первом случае мы имеем дело с К-стратегией, а во втором — с r-стратегией. Таким образом, к К-стратегам могут быть отнесены патиенты Л. Г. Раменского, т. е. растения способные выживать
Рис. 5.27 • Флуктуации ресурсов и популяционные стратегии (по: Ю. Э. Романовскому): a — К-стратеги (патиенты, стресс-толеранты) увеличивают потребление ресурсов за счет снижения пороговой концентрации; б — r-стратеги быстро потребляют ресурсы в период их обилия за счет высокой
скорости потребления. Они переживают период недостатка ресурсов (голубая стрелка) в латентном (покоящемся) состоянии (эксплеренты, рудералы), либо в толерантном (активном) состоянии (виоленты, конкуренты)
3. Конкуренция
213
Таблица 5.5 • Первичные компоненты стратегии жизненного цикла (по: Романовский, 1989, 1998). + означает отбор на увеличение, - на уменьшение значений того или иного признака, 0 — отсутствие однонаправленного отбора
К- и г-стратегии К г
CSR-стратегии S С R
Индивидуальные черты
Относительный вклад энергии в размножение (репродуктивное усилие) 0 -
Относительный вклад энергии в 1 потомка («цена потомка») + - 0
Размер взрослых особей - + 0
Время достижения половозрелости (длительность прегенеративного периода) 0 -
Длительность жизненного цикла особи 0 + -
Популяционные черты
Пороговая концентрация ресурсов - + +
Рождаемость - + +
при постоянно низкой доступности ресурса и всегда имеющие конкурентное преимущество в стационарных условиях (Абросов и др., 1982; Tilman, 1982). r-стратеги, в свою очередь, обладают способностью с большей скоростью потреблять ресурсы в период их высокой доступности и переживать периоды с низкой доступностью ресурсов либо в латентной (покоящейся) стадии — эксплеренты, либо в толерантной (устойчивой, обычно взрослой) стадии — виоленты (рис. 5.27). Виоленты получают конкурентное преимущество в условиях высокой предсказуемости изменения уровня доступности ресурсов. В начале пика поступления ресурса им не требуется время для развития из покоящегося состояния, в отличие от эксплерентов. Поэтому виоленты (С-стратеги) получают возможность в благоприятные периоды быстро снижать доступность ресурсов для других видов, таким образом проявляя свойства эдификаторов (средо-образователей). С другой стороны, к низкой предсказуемости поступления ресурсов лучше адаптированы эксплеренты (рудералы), типичными представителями которых могут служить однолетники в некоторых пустынях, где осадки выпадают нерегулярно, раз в несколько лет.
Виды, относящиеся, к разным типам стратегий, характеризуются различными наборами индивидуальных и популяционных свойств (табл. 5.5).
214
Глава 5. Типы взаимоотношений растений в фитоценозах
Так, виоленты характеризуются низким репродуктивным усилием (долей генеративных органов в биомассе), низкой ценой потомка (доля одного семени от биомассы растения), большим размером взрослых особей, высокой рождаемостью и высокой пороговой концентрацией ресурсов.
В целом взгляды Ю. Э. Романовского согласуются с описанными выше представлениями Д. Тильмана, Ф. Грайма и других исследователей. Необходимо подчеркнуть два аспекта его теории.
1. Говорить о существовании в природе в чистом виде виолентов, патиен-тов и эксплерентов не имеет смысла, а, поскольку основные стратегии характеризуют направления отбора и проявляются в виде комплекса адаптаций, целесообразнее рассматривать большую или меньшую (по отношению к другим видам, популяциям, расам и т. п.) степень выраженности патиентности, виолентности или эксплерентности (Работнов, 19856). На вид не стоит вешать «ярлык», который может получиться при определении типов стратегий по ключам (типа рис. 5.21). И у однолетников, и, например, у деревьев, но в разной степени выражены свойства всех стратегий. Betula pendula по отношению к Picea abies обладает более выраженными чертами эксплерентности, а по отношению к Роа annua — чертами виолентности. Дифференциация по параметрам популяционных стратегий происходит (конечно, в разной степени) у растений различных, в том числе и экстремальных, местообитаний, на что также обращал внимание Б. А. Юрцев (1986).
2. Типы стратегий выделяются на основе популяционных свойств, которые, несомненно, могут быть связаны с морфологическими адаптациями. Но морфологические признаки при выделении стратегий «вторичны», а их значимость для характеристики популяционного поведения должна быть отдельно доказана, в то время как большая часть работ по выделению типов стратегий у растений опирается на морфологические признаки. Специфика растений как прикрепленных пластичных организмов заключается в том, что при изменении доступности ресурсов численность популяций может меняться разнонаправленно, и более надежными популяционными характеристиками могут быть биомасса, встречаемость и другие геоботанические характеристики.
3.8 • Ауто- и синэкологические ареалы и оптимумы
То, что конкуренция оказывает сильное влияние как на распространение растений в природе, так и на их участие в составе сообществ, было ясно исследователям XIX в. Действие абиотических факторов на виды часто преломляется через конкурентные отношения в сообществах.
3. Конкуренция
215
На одни и те же воздействия вид реагирует по-разному в чистых посевах и в многовидовых фитоценозах. Еще в 1883 г., подводя итог 20-летних опытов с внесением удобрений, Lawes с сотрудниками писали: «Растения не обязательно существуют там, где почвенные и климатические условия наиболее благоприятны для них, но лишь в тех местах, где они могут произрастать при конкуренции с другими видами. Отсутствие положительной реакции вида на удобрения при совместном произрастании с другими видами не означает, что он не будет положительно реагировать в одновидовом посеве» (цит. по: Работнов, 1995).
А. П. Шенников (1942) провел ряд экспериментов по пересадкам участков дернины из лугов одних типов в другие. Он показал, что, например, типчак (Festuca sulcata) хорошо разрастается при пересадке из лугово-степных сообществ на влажные луга (то есть является не «сухолюбом», а «сухотерпцем»). А. П. Шенников ввел представление об экологическом (потенциальном) и фитоценотическом (реальном, в условиях конкуренции) ареале вида (диапазоне экологических условий, в котором вид может существовать, не путать с географическими ареалами!). Позднее Н. ЕПеп-berg (1953, 1954) пришел к аналогичным заключениям в экспериментах с луговыми и болотными растениями и выделил физиологический (потенциальный) и экологический (реальный) ареал вида. Во избежание путаницы лучше говорить об ауто- и синэкологических ареалах и оп-тимумах. Эти понятия аналогичны представлениям о фундаментальной и реализованной экологической нише Хатчинсона (Hutchinson, 1957).
При полевых исследованиях необходимо учитывать возможности сдвигов экологических ареалов за счет межвидовых взаимодействий. Классическим примером является сосна (Pinus sylvestris), которая в культуре лучше растет на средних по увлажнению почвах, в то время как в природе чаще встречается либо на сухих (пески, скалы), либо на переувлажненных (болота) почвах, что обычно объясняется конкурентным вытеснением елью со средних по увлажнению участков. Однако не всегда только конкуренция является причиной смещения синэкологического оптимума по сравнению с аутэкологическим. Так, например, посадки сосны на средних по условиям увлажнения почвах в северной Скандинавии (в местах, обычно занятых ельниками) довольно сильно страдают от деструктивных патогенных грибов, таких как, Phacidium infestans (Burdon, 1993).
Эксперименты с пересадками часто помогают выявлению причин присутствия/отсутствия отдельных видов в составе сообществ. Ряд таких экспериментов провел A. Gigon (1987) для выявления причин различий флористического состава сообществ высокогорных лугов Альп, развитых на силикатных и карбонатных почвообразующих породах. Эти и многие
216
Глава 5. Типы взаимоотношений растений в фитоценозах
другие эксперименты позволили A. Gigon (1987) сделать вывод, что «наибольшая часть видов, приуроченных к менее экстремальным местообитаниям [более продуктивным], не встречается в более экстремальных из-за [неблагоприятного действия] абиотических факторов. С другой стороны, наибольшая часть видов, ограниченных в распространении более экстремальными местообитаниями, не встречается на менее экстремальных (более богатых) из-за конкуренции». Поэтому ауто- и синэкологические ареалы у видов виолентной (конкурентной) стратегии часто сходны, в то время как у патиентов (стресс-толерантов) они значительно различаются.
3.9 • Полевые эксперименты по изучению конкуренции
3.9.1 • Метод фитометров
Ф. Клементсом (см. Работнов, 1987) был разработан метод фитометров, состоящий в том, что тестовые растения какого-либо вида подсаживали в сообщества с различной плотностью, биомассой и т. п., после чего по их состоянию можно было судить о различиях или сходстве условий произрастания в изучаемых сообществах, а также делать выводы о взаимоотношениях между растениями. В ранних работах в качестве фитометров часто использовали культурные растения, например подсолнечник или редис. Позже стали использовать дикорастущие виды. Примером такого исследования может быть работа Т. Herben et al. (2007) на нескольких горных лугах в Чехии, в которой авторы использовали метод подсадок растений-фитометров (Anthoxanthum alpinum, Festuca rubra) для изучения свойств окружающих сообществ (надземная или подземная масса соседей, видовой состав соседей), вызывающих большее конкурентное подавление роста тестовых растений. Они показали, что рост фитометров был больше связан с массой корней соседей и видовым составом соседей, а надземная масса не оказывала существенного влияния. Наиболее значимые отрицательные связи получены для Anthoxanthum alpinum с массой корней соседей в слое 0-3 см, выявлена отрицательная связь Festuca rubra с участием в окружении этого же вида.
3.9.2 • Удаление отдельных особей, видов, групп
Изучение конкуренции в естественных сообществах часто проводят методом удаления отдельных особей, видов или групп растений с наблюдением над реакцией оставшихся компонентов сообществ. Такие эксперименты проведены во многих типах сообществ, но особенно часто — в травяных и кустарниковых фитоценозах. Наименьшее нарушающее воздействие на почву оказывает многократное срезание надземных побегов (метод истощения) (McLellan et al., 1995).
3. Конкуренция
217
Примером может служить изучение роли доминирующих видов на альпийских лишайниковых пустошах северо-западного Кавказа (Aksenova et al., 1998). В каждом из вариантов эксперимента был удален один из 5 доминирующих видов (Anemone speciosa,Antennaria dioica, Festuca ovina, Tri folium polyphyllum, Carex spp.). Отмечено приблизительно равное число случаев положительной и отрицательной реакции на удаление среди исследуемых видов. Результаты эксперимента показали, что среди доминирующих видов исследуемой альпийской пустоши преобладают конкурентные отношения (рис. 5.28 а). Кроме того, при удалении
Рис. 5.28 • Результаты 13 летнего эксперимента с удалением отдельных доминирующих видов на альпийской лишайниковой пустоши: а — отрицательные связи (положительная реакция на удаление) и б — положительные связи (отрицательная реакция на удаление) между видами альпийских пустошей. Толщина стрелок соответствует числу методов, которыми подтверждается рассматриваемая реакция. Стрелки направлены от реагирующего вида к удаляемому. Серые кружки обозначают доминанты, которые удаляли в эксперименте (по: Aksenova et al., 1998)
218
Глава 5. Типы взаимоотношений растений в фитоценозах
как Festuca ovina, так и Carex spp. резко увеличивал свою численность злак Helictotrichon versicolor. Поэтому в целом злаки и осоки в изучаемом сообществе, видимо, представляют собой единую гильдию видов, более тесно конкурирующих между собой, чем с другими видами. С другой стороны, при удалении доминирующих видов многие недоминирующие уменьшали свое участие, что говорит о положительном влиянии доми-нантов на эти виды (рис. 5.28 б, см. п. 6 гл. 5). Возможны следующие причины такой реакции (Fowler, 1981; Callaway, 1995): 1) неблагоприятные изменения в абиотической среде, вызванные удалением доминирующих видов; 2) усиление конкуренции со стороны других видов, которая ранее сдерживалась удаленным видом; 3) удаление или ослабление основных растений-хозяев, что может иметь место в случае полупаразита Euphrasia ossica, основными хозяевами которого, видимо, служат злаки и осоки (Ларин и др., 1950-1956; Seel, Press, 1994).
Анализируя в целом результаты этого и многих других экспериментов с удалением отдельных видов из состава естественных сообществ (Fowler, 1981; del Moral, 1983; Moore, 1982; Keddy, 1989; Moen, 1989 и др.) можно сделать ряд выводов:
1. Наряду со случаями, когда по результатам опытов можно было выделить группы более тесно конкурирующих видов (гильдии), иногда наблюдалась «диффузная» конкуренция (термин MacArthur, 1972), когда на удаление отдельного вида большинство оставшихся реагировало небольшим увеличением своего участия.
2. Отношения между видами в сообществе могут быть несимметричными. Если из сообщества удаляется вид А, а его нишу занимает вид В, то при удалении вида В его ниша может быть занята другими видами, отличными от А.
3. Может наблюдаться уменьшение участия некоторых видов при удалении других, что свидетельствует о большей роли положительных отношений между растениями по сравнению с конкурентными.
4. Общая первичная продукция сообщества после удаления отдельных видов не уменьшалась, а в ряде случаев даже возрастала.
3-9-3 • Корневая (траншейная) обрубка
При изучении корневой конкуренции видов деревьев с другими растениями часто используется метод траншейной обрубки, который впервые применил Fricke (1904). Lutz (1945; Aarssen, Epp, 1990) сообщает об изменении в травяном покрове леса в течение 21 года после обрубки корней. В первые годы число видов и общее покрытие на экспериментальных
3. Конкуренция
219
□ Vaccinium myrtiilus
□ Oxalis acetosella
Hylocomium splendens
Rubus idaeus
Рис. 5.29 • Изменение проективного покрытия
(ось ординат) четырех видов нижних ярусов в зеленомошном ельнике после обрубки корней ели в эксперименте
В. Г. Карпова (1969)
площадках существенно возрастало, а в дальнейшем флористическое разнообразие снижалось при увеличивающемся покрытии трав.
Большой интерес представляют классические эксперименты В. Г. Карпова (1969) по изучению конкуренции в еловых лесах Центрально-черноземного заповедника. После обрубки корней ели на площадках в 1 м2 произошло резкое изменение состава и структуры напочвенного покрова: почти исчезли зеленые мхи, резко (до 90 %) увеличилось покрытие кислицы (Oxalis acetosella), а на четвертый год на площадках появилась малина (Rubus idaeus) (рис. 5.29). На пятый год эксперимента наблюдалось разрастание кустов малины, а кислица снизила свое участие. Эти изменения были связаны с тем, что удаление корневой конкуренции со стороны ели вызвало бурное развитие кислицы, которая, в свою очередь, затенила мхи. Развитие более высокорослой малины и конкуренция за свет и привели в дальнейшем к разреживанию покрова кислицы. Эти результаты свидетельствуют о большой роли корневой конкуренции со стороны ели преимущественно за воду и минеральные формы азота в формировании нижних ярусов еловых лесов. С другой стороны, ряд видов, например черника Vaccinium myrtillus, практически не изменила своего участия после обрубки корней ели.
220
Глава 5. Типы взаимоотношений растений в фитоценозах
Опыты с траншейной обрубкой произведены в различных типах леса, иногда они сопровождались оценкой изменения доступности ресурсов. Так, эксперименты в лесах из каменного дуба (Quercus ilex L.) близ Барселоны показали, что влажность почвы, содержание и вынос нитратного азота и интенсивность минерализации соединений азота резко возрастали на участках, подвергшихся обрубке, по сравнению с ненарушенным лесом. Концентрация азота в лизиметрических водах выросла с 0,18 до 11,3 мг/л на экспериментальных участках, а содержание аммонийного азота возросло с 24 до 68 мг/кг почвы, в то время как на контрольных участках оно упало в течение сезона до 9 мг/кг почвы (Bonilla, Roda, 1990).
3.10 • Представления Р. Keddy о конкурентной иерархии и центрифужной организации растительных сообществ
Несмотря на частые случаи ассиметричной конкуренции, на основании экспериментов с замещениями (по схеме de Wit, см. п. 3.4 гл. 5), удалением отдельных видов в природе и других часто можно составить ряды конкурентоспособности видов для отдельных сообществ, т. е. определить конкурентную иерархию видов. Например, F. Clements (1933) обобщил результаты многих экспериментов, проведенных в прериях США, и сделал заключение, что более высокорослые злаки имеют конкурентное преимущество перед низкорослыми.
На примере растительных сообществ влажных лугов и болот Keddy (1990) предложил теорию центрифужной организации сообществ (рис. 5.30): имеется центральное, наиболее продуктивное (сердцевинное, core) местообитание, которое предпочли бы все (большинство) видов данного ряда сообществ, и серия менее продуктивных периферических местообитаний, к которым адаптированы немногие виды. Конкуренция наиболее интенсивна в центре, где немногие основные доминанты вытесняют прочие виды на периферию. Удаление доминирующих видов из центрального местообитания ведет к замещению их видами из ближайших периферийных сообществ.
Keddy, MacLellan (1990) также рассматривают структуру лесов ряда регионов США с позиций предложенной модели. В центре располагались наиболее теневыносливые виды региона, а высокое разнообразие поддерживалось за счет нарушений (gap-динамики). При удалении видов деревьев их место также занимали ближайшие соседи с более «периферийных» участков, а при удалении периферийных видов, лесные сообщества сменялись травяными и кустарниковыми (Keddy, MacLellan, 1990).
3. Конкуренция
221
песчаные речные пляжи
Typha
Я
Sdrpus acutus
Sdrpus americanus
Drosera Hypericum
Е
Typha
Cladfum
Xyris Drosera Sabatia
/У /<№/
ф л s ф <Q 3
3
галечные берега озёр
Рис. 5.30 • Схема «центрифужной» организация растительных сообществ околоводных местообитаний Канады вдоль градиента биомассы (по: Keddy, 1990)
Предложенная модель хорошо согласуется с результатами многих экспериментов по пересадкам растений из одних сообществ в другие.
3.11 • Широкая трактовка конкуренции.
Прямая и «кажущаяся» конкуренция
В предыдущих разделах мы рассматривали конкуренцию в узком смысле, т. е. как трофическую конкуренцию, или конкуренцию за ресурсы. Однако многие исследователи трактуют конкуренцию как любые отрицательные отношения между растениями, без акцента на их механизм. Тогда в эту категорию попадают и аллелопатические взаимодействия, рассматриваемые в следующем разделе, а также многие другие фор-
222
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.31 • Различные случаи отрицательных отношений между растениями. Е — естественные враги (фитофаги, паразиты, патогены); Pl, Р2, РЗ — виды растений; R — лимитирующие ресурсы (свет, вода, ЭМП и т.д.). Сплошные линии — прямые взаимодействия; пунктирные — опосредованные взаимодействия; полые стрелки — позитивный эффект; черные стрелки — негативный эффект. Случаи 1 и 2 Коннелл рассматривает как прямую конкуренцию, 3 и 4 — как «кажущуюся». В случае 4 «кажущаяся» конкуренция имеет место между Р1 и РЗ (по: Connell, 1990)
мы отношений. Например, Коннелл (Connell, 1990), используя широкое определение конкуренции, приводит схему возможных механизмов таких отношений (рис. 5.31). Прямые (контактные) отрицательные связи и ресурсную конкуренцию (случаи 1 и 2 на рис. 5.31) он рассматривает как прямую (direct) конкуренцию, а отрицательные трансбиотические (через организмы-посредники, например общие фитофаги или растения-конкуренты) отношения — как «кажущуюся» (apparent competition) (случаи 3 и 4 на рис. 5.31, см. п. 2 гл. 5).
Примером 3-го случая могут служить отношения между кустарником Gutierrezia sarothae и многолетней травой Machaeranthera canescens в сухих злаковниках Мексики. Всходы Machaeranthera встречаются под кустами Gutierrezia, но никогда не приживаются там, что могло бы быть объяснено конкурентными отношениями между этими видами. Однако опытами с изоляцией всходов Machaeranthera от поедания прямокрылым Hesperotettis viridis (который также поедает кустарник) показано, что приживание всходов лимитируется этим фитофагом, а не прямой конкуренцией с кустарником (Connell, 1990).
Поэтому необходимо быть весьма осторожным при интерпретации результатов различных полевых экспериментов по конкуренции (например, с удалением отдельных растений, траншейной обрубкой и т.п.), так как при экспериментальном воздействии часто меняется численность фитофагов и происходят другие изменения в ценозе, которые могут вызвать сходные с конкуренцией эффекты.
4. Аллелопатия
223
Кроме того, ряд авторов (Aphalo, Ballare, 1995) предостерегают от упрощенного понимания конкуренции как процесса, связанного только с недостатком ресурсов. Растения обладают способностью воспринимать и информационные сигналы, связанные с возможной нехваткой ресурсов или стрессом в будущем (листопад при укорочении длины дня), а также реагировать на присутствие соседей, поскольку отраженный от них боковой свет имеет низкое соотношение «красный : дальний красный свет», что вызывает удлинение стебля как опережающую реакцию на возможное затенение.
4 • Аллелопатия
Основателем учения об аллелопатии считается австрийский ученый Ханс Молиш (Molish, 1937), который опубликовал работу «Der Einfluss einer Pflanze auf die andere — Allelopathie» (влияние одного растения на другие — аллелопатия). X. Молиш изучал рост растений, накрытых стеклянным куполом, под который помещали также созревающие яблоки. В результате рост многих растений уменьшался, что было связано с выделяющимся из фруктов этиленом. В прямом переводе термин «аллелопатия» означает «взаимострадание», что не совсем точно отражает характер отношений между растениями при выделении различных метаболитов. Т. А. Работнов (1992) дал следующее определение: аллелопатия — влияние одних организмов на другие через специфические метаболиты, не имеющие для них значение как источник минерального или углеродного питания.
Хотя аллелопатические эффекты определяются обычно как воздействия одних растений на другие непосредственно через метаболиты, в природе более часто имеют место воздействия метаболитов на биотические и абиотические почвенные процессы, которые в свою очередь влияют на другие растения. Inderjit и Weiner (2001) предложили более общую классификацию аллелопатических влияний:
1. Аллелопатия в узком смысле. Растение А продуцирует вещество X, которое воздействует на растение В.
11. Непрямые аллелопатические эффекты.
А) Непрямая аллелопатия:
1) Аллелопатия, связанная с разложением веществ. Растение А продуцирует вещество X, которое разлагается или трансформируется в вещество У, которое влияет на растение В.
2) Индуцированная аллелопатия: Растение А продуцирует вещество X, которое индуцирует организм D на синтез вещества Z, которое влияет на растение В.
224
Глава 5. Типы взаимоотношений растений в фитоценозах
В) Непрямая токсичность. Вещество X взаимодействует с почвой и вызывает образование вещества Z (не являющегося продуктом распада X), которое воздействует на растение В.
С) Непрямые средовые эффекты. Вещество X вызывает изменения в почвенной среде, которые влияют на режим ЭМП в почве, что приводит к уменьшению ростовых характеристик растения В без токсичных эффектов. Пример последнего: полифенолы из опада Pinus muricata в карликовых лесах Северной Калифорнии ингибируют нитрификацию и влияют на минерализацию органического азота (Northup et al., 1995; Paavolainen et al., 1998). Этот эффект снимается при внесении удобрений. Есть данные о аллелопатическом подавлении микоризных грибов актиномицетами (Friedman et al., 1989).
По мнению Inderjit и Weiner (2001), широко понимаемая аллелопатия имеет большое влияние на структуру сообществ, хотя, видимо, и меньшее, чем ресурсная конкуренция. Прямые химические взаимодействия между растениями имеют значительно меньшее распространение, чем опосредованные почвенной средой.
Химические выделения растений происходят из надземных и подземных органов. Различают также активное выделение, связанное с процессами жизнедеятельности самого растения, и пассивное, вызванное агентами внешней среды (вымывание, травматическое выделение и т. п.). Очень распространено пассивное выделение из листьев — вымывание осадками как собственных веществ, так и продуктов обмена эпифилльных сапротрофов. Так, в лесу из Quercus petraea при годовом количестве осадков 1700 мм в год вымывается около 90 кг/га различных веществ, при этом большая часть их — из старых листьев (68 кг) (Carlisle et al., 1966). Вымывание из созревающих плодов может значительно снижать урожай. Так, по наблюдениям Н. Г. Холодного (1957), урожай зерновых культур может снижаться на 20-30 % если во время созревания зерновок стоит дождливая погода.
Надземные органы растений выделяют также много летучих органических веществ. Основные компоненты этих выделений — изопрен и монотерпены. Окисление этих веществ в присутствии NO ведет к образованию ощутимых количеств озона, что отмечается в ряде городов. Различные виды выделяют разное количество летучих веществ. Большие выделения изопрена характерны для видов рода Quercus (Sharkey, Lerdau, 1999).
Корневые выделения очень разнообразны. Выделяют 3 основные группы корневых выделений (Marschner, 1995; Uren, 2001): 1) низкомолекулярные органические соединения (сахара, органические кислоты, аминокислоты, фенольные соединения и др.), 2) высокомолекулярные
4. Аллелопатия
225
Таблица 5.6 • Органические соединения, выделяемые корнями растений (по: Uren, 2001)
Группы соединений Соединения
Сахара и полисахариды Арабиноза, фруктоза, галактоза, глюкоза, мальтоза, манноза, слизи различного состава, олигосахариды, раффиноза, рамноза, рибоза, сахароза, ксилоза
Аминокислоты а-аланин, /3-аланин, 7-аминомасляная кислота, аргинин, аспарагин, аспарагиновая кислота, цитруллин, цистатионин, цистеин, цистин, деоксимугеновая кислота, 3-эпигидроксимугеновая кислота, глутамин, глутаминовая кислота, глицин, гомосерин, изолейцин, лейцин, лизин, метионин, мугеновая кислота, орнитин, фенилаланин, пролин, серин, треонин, триптофан, тирозин, валин
Органические кислоты Уксусная, аконитовая, аскорбиновая, бензойная, масляная, кофейная, лимонная, р-кумаровая, феруловая, фумаровая, глутаровая, гликолевая, глиоксиловая, яблочная, малоновая, щавелевоуксусная, щавелевая, р-гидроксибензойная, пропионовая, янтарная, сиреневая, D-винная, валериановая, ванилиновая
Жирные кислоты Линолевая, линоленовая, олеиновая, пальмитиновая, стеариновая
Стерины (= стеролы) Каместерин, холестерин, ситостерин, стигмастерин
Факторы роста р-аминобензойная кислота, биотин, холин, N-метилникотиновая кислота, никотиновая кислота, пантотеновая кислота, витамины В1 (тиамин), В2 (рибофлавин), и В6 (пиридоксин)
Ферменты Амилаза, инвертаза, пероксидаза, фосфатаза, полигалактуроназа, протеаза
Флавононы, азотистые основания и нуклеозиды Аденин, флавонон, гуанин, уридин/цитидин
Прочие разнообразные соединения Ауксины, скополетин, гидроксициановая кислота, глюкозиды, растворимые белки, соединения-восстановители, этанол, глицинбетаин, инозитол и мио-инозитол-подобные соединения, А1-индуцируемые полипептиды, дигидрохинон, сорголеон
гелеобразующие вещества, формирующие затем муцигель, 3) шелушащиеся клетки и продукты их разрушения. В целом корневые выделения очень разнообразны (табл. 5.6), они включают сахара и полисахариды,
226
Глава 5. Типы взаимоотношений растений в фитоценозах
Минеральные вещества не толь-Вставка IX. Корневые выделения Ко поглощаются, но выделяются и обмен веществ у растений корнями растений. Растения, как
и все живые организмы, характеризуются постоянно идущим обменом веществ, поэтому для каждого элемента можно рассчитать время его нахождения в организме растения. Одну из первых попыток оценить время оборота (оно рассчитывается как содержание элемента в растении, деленное на скорость его поглощения или выделения) предприняли А. Н. Тюрюканов и др. (1968), которые выращивали два растения пшеницы, корни которых развивались в трех сосудах с полным питательным раствором (см. рисунок IX-1). Часть элементов минерального питания в первом растворе была представлена радиоактивными изотопами, что позволяло оценить скорость поглощения элементов в первом сосуде, скорость выделения корнями во втором сосуде, скорость поглощения вторым растением из второго сосуда и выделения в третий. Для всех исследованных элементов время оборота изученных ЭМП не превышало двух недель, оно составило для фосфора (32Р) — 5,8 дней, кальция 45Са — 1,2 дня, калия (аналог Rb) — 7,0 дней. Конечно, эти величины получены в физиологическом эксперименте, где ЭМП не лимитировали рост растения, поэтому аналогичные величины для растений, развивающихся в естественных условиях, должны быть существенно выше, но имеющиеся в настоящее время исследования базируются преимущественно на скоростях отмирания органов и не учитывают оборот корневых выделений.
Рис. IX.1 • Схема эксперимента по изучению обмена веществ у пшеницы (по: Тюрюканов и др., 1968)
аминокислоты, органические кислоты, жирные кислоты, стиролы, ростовые факторы, ферменты, флавоноиды, нуклеотиды и многие другие соединения. Неорганические корневые выделения немногочисленны (ионы, вода, электроны), но имеют очень большое значение [Вставка IX].
4. Аллелопатия
227
@ Ф Й Ф
« « « « «
ffi $ ф @
« > « • «
® 9 < &
< « • « •
9 ® 0 &
Г>
Ф Ф ф Ф>
*® е ® Ф»
* ы >*
а)
в)
Рис- 5.32 • Прорастание семян куколя (Agrostemma githago) в присутствии соплодий свеклы: а — на фильтровальной бумаге; б — на фильтровальной бумаге над тонким слоем почвы; в — на фильтровальной бумаге над тонким слоем активированного угля (по: Grummer, 1955)
По данным Biondini et al. (1988), у злаков прерии корневые выделения, рассчитанные по углероду, составляют 8-17% от нетто-продукции, с корневыми выделениями поступает около 11 г/м2 азота в год. В стерильной среде выделения по С составляют лишь около 60 % от таковых на естественном микробиологическом фоне. В целом же в разных случаях корневые выделения составляют от 1 до 30 % продукции (в среднем около 10%) (Uren, 2001).
Во второй половине XX в. было проведено большое число исследований, преимущественно лабораторных, показывающих отрицательное влияние вытяжек, смывов, настоев и т. п. одних растений на рост, развитие и прорастание семян других (Райс, 1978). Ключевыми для анализа роли аллелопатии можно считать эксперименты Грюммера (Grummer, 1955). Он проращивал семена куколя (Agrostemma githago) в чашках Петри. В присутствии соплодий свеклы на фильтровальной бумаге всхожесть семян была очень низкой (12%), что объяснялось ингибирующим влиянием веществ, выделяемых соплодиями свеклы. Если под слой фильтровальной бумаги был добавлен тонкий слой почвы, всхожесть увеличилась до 88 %, а в случае тонкого слоя активированного угля составила 100 % (рис. 5.32). Таким образом, было показано, что адсорбция органических выделений почвой или активированным углем резко снижает их негативное воздействие на растения. Поэтому результаты чисто лабораторных экспериментов практически не имеют значения для оценки роли аллелопатии в природе.
С аллелопатией связывают так называемую проблему «почвоутомления», или «токсикоза почв», когда при длительном выращивании одной
228
Глава 5. Типы взаимоотношений растений в фитоценозах
культуры, сильно снижается ее урожай (Работнов, 1992). Это явление имеет различные механизмы и может быть связано с односторонним обеднением почвы монокультурами, накоплением токсичных метаболитов, а также просто увеличением численности патогенных микроорганизмов и фитофагов, имеющее место и в природе при первичных сукцессиях на песчаных дюнах (van der Putten, Peters, 1997; van der Putten, 1997). Отрицательное влияние взрослых растений на молодые растения своего же вида, получившее название эффекта Janzen-Connell, также может быть связано с накоплением токсичных веществ и/или патогенов в почве. В экспериментах Petermann et al. (2008) изучали развитие 24 видов европейских травянистых растений на почве, взятой из-под предварительно выращенных монокультур этих видов. Отрицательное воздействие на растения тех видов, которые произрастали на соответствующей почве ранее, в дальнейшем наиболее резко проявлялось в смешанном посеве (в условиях конкуренции). С использованием различных методов обработки почвы (гамма-облучение, активированный уголь, фунгициды, удобрение) показано, что именно почвенные патогены вызывают отрицательное влияние почвы из-под монокультур того или иного вида на особи этого же вида. Гамма-радиация снимала этот отрицательный эффект полностью. Удобрение частично снимало эффект, поэтому он может быть в какой-то мере связан и с обеднением почвы. Активированный уголь производил незначительный эффект, что свидетельствует о небольшой роли аллелопатии в «почвоутомлении». Авторы считают, что рассмотренный отрицательный эффект влияния почвы при длительном выращивании того или иного вида на особи этого вида является важным и широко распространенным в природе механизмом, способствующим поддержанию флористического разнообразия сообществ (Petermann et al., 2008).
В ряде случаев горизонтальную структуру сообществ пытались объяснить с позиций аллелопатического влияния. Так, Muller (1966) в Калифорнии описал концентрические участки голой земли вокруг кустов Salvia leucophylla, произрастающих среди однолетних злаковников (рис. 5.33 из Райса, 1978). Проведя химические анализы почв, он не обнаружил разницы между участками почв под шалфеем, голых пятен и злаковника, из чего был сделан вывод об отсутствии конкуренции. Автор объяснил эту структуру действием летучих веществ, выделяемых шалфеем, и показал, что выделения листьев в сосудах тормозят прорастание семян злаков. Эта работа может служить примером методически несовершенных исследований, поскольку роль конкуренции реально не оценивалась.
Как классические случаи аллелопатического воздействия описаны примеры угнетения травяного покрова под пологом интродуцированных видов деревьев (Bode, 1958; del Moral, Muller, 1970; Работнов, 1992). Леса
4. Аллелопатия
229
Рис. 5.33 • Участки Salvia leucophylla с опоясывающей зоной голой почвы (А, В) и окружающим сообществом однолетних злаков (С, главным образом Bromus rigidus и Avena fatua), рост которых подавлен близ кустов шалфея. Подавление роста злаков приписывалось аллелопатическому влиянию летучих выделений шалфея, однако, роль корневой конкуренции не была исследована (рис. из Райс, 1978)
из эвкалиптов в Средиземноморье и Калифорнии но О почти лишены травяного покрова, что связывают 11
с выделением фенольных соединений из опада ли- р || |1
стьев. Но в Австралии эвкалиптовые сообщества — одни из самых флористически богатых сообществ ||
земного шара. Аналогичная картина наблюдается О
и с грецким орехом (Juglans fallax). В его листьях рИСв 5.34 • юглон содержится гидроюглон, разлагающийся в почве
до юглона (рис. 5.34), обладающего мощным аллелопатическим воздействием. Однако в местах естественного распространения грецкого ореха (например, в орехоплодных лесах Южной Киргизии) хорошо развит травяной покров. Т. А. Работнов (1992) объясняет это тем, что в результате длительной сопряженной эволюции на родине подбираются устойчивые виды растений и комплексы микроорганизмов, способные разлагать ал-лелопатически активные вещества. Аллелопатия ярко проявляется лишь в случае встреч эволюционно несопряженных видов растений.
230
Глава 5. Типы взаимоотношений растений в фитоценозах
Травянистые интродуценты также могут оказывать аллелопатическое воздействие на аборигенные растения. Так, в экспериментах W. М. Ridenour, R. М. Callaway (2001) занесенный в Северную Америку из Средиземноморья сорняк — василек пятнистый (Centaurea maculosa) оказывал неблагоприятное аллелопатическое воздействие на аборигенный доминирующий вид овсяницы — Festuca idahoensis, которое снималось при добавлении активированного угля. Centaurea maculosa выделяет катехин, который также ингибирует прорастание семян многих видов растений. Активированный уголь, внесенный на почву вблизи особей Centaurea maculosa, увеличил в эксперименте численность проростков растений на 78 % через 25 дней после посева (Perry et al., 2005). Однако даже при снятии аллелопатического эффекта василек имел конкурентное преимущество при выращивании в парных культурах с овсяницей. Таким образом, аллелопатический эффект может быть сопряжен с конкуренцией за ресурсы. При дальнейших исследованиях (Callaway et al., 2005) была показана сопряженная эволюция трав, произрастающих совместно с Centaurea maculosa. Особи, происходящие от таких растений, показывали лучшую выживаемость всходов в присутствии Centaurea maculosa или его корневых выделений.
Другой интересный пример комплексного изменения свойств почв заносными видами растений в широколиственном лесу в штате Нью-Джерси приводят Kourtev et al. (2002). Здесь было показано резкое изменение структуры ризосферной микрофлоры под двумя заносными в Америке японскими растениями — Berbens thunbergii и Microstegium vimineum в сравнении с ризосферой местных видов Vaccinium spp. Свойства почвы под заносными видами существенно изменялись — pH, мощность органического горизонта, интенсивность нитрификации. Структура микробных комплексов существенно различалась под местными и экзотическими растениями, последние включали больше бактерий и меньше грибов.
Для экспериментального разделения аллелопатического и конкурентного влияния одного вида растений на другое Fuerst, Putnam (1983) предложили по аналогии с известной в микробиологии триадой Герли-Коха следующие общие принципы. Для доказательства аллелопатии необходимо: 1) установить факт снижения тех или иных ростовых характеристик одного вида растений в присутствии особей другого, 2) выделить, идентифицировать и (желательно) синтезировать предполагаемый аллелопатический агент — вещество, 3) экспериментально показать действие этого агента в отсутствии других растений в естественных концентрациях и на естественном микробиологическом фоне.
Возможны и другие методические приемы для разделения эффектов конкуренции и аллелопатии. Так, Nilsson (1994) изучила влияние
4. Аллелопатия
231
Рис. 5.35 • Аллелопатическое и конкурентное воздействие шикши (Empetrum hermaphroditum) на молодые растения сосны (Pinus syivestris). Средние величины и ошибка среднего для длины побегов (а), сухой массы побегов (б), общей сухой массы растения (в) и сухой массы корней (г) сосны в условиях конкуренции и аллелопатического воздействия со стороны шикши (контроль), снятия аллелопатического воздействия с помощью активированного угля (уголь), снятия корневой конкуренции (обрубка) и снятия аллелопатии и конкуренции одновременно (обрубка + уголь). Синие столбики — в условиях эксперимента в теплице, желтые — в полевом эксперименте. Варианты, отличающиеся значимо на уровне р < 0,05, отмечены неперекрывающимися буквами над столбиками (по: Nilsson, 1994)
Empetrum hermaphroditum на рост сеянцев сосны (Pinus syivestris). Проведено 2 серии экспериментов: в природе и в лаборатории, в каждом случае в 4 вариантах: контроль (отсутствие воздействия), поверхностное внесение активированного угля (500 г/м2, удаление аллелопатических веществ из опада и подстилки), корневая обрубка вокруг сеянцев при помощи трубы (снятие корневой конкуренции) и сочетание применения угля с корневой обрубкой. Опыт продолжался в течение 3-х лет. Во всех случаях хуже всего развивались сеянцы сосны в контроле, лучше всего — при одновременном снятии конкуренции и аллелопатии (рис. 5.35). Кон
232
Глава 5. Типы взаимоотношений растений в фитоценозах
куренция несколько сильнее подавляла развитие сосны, чем аллелопатия, но различия часто были незначимы. Возможный механизм аллелопатического влияния шикши — подавление развития эктомикоризного для сосны гриба Paxillus involutus, что было показано в ранней работе (Nilsson et al., 1993).
Хотя многие исследователи аллелопатии рассматривает непосредственное воздействие органических веществ как механизм отрицательного влияния одних растений на другие, ряд экспериментов показывает, что этот механизм может быть принципиально другим. Вносимый в почву органический субстрат вызывает развитие микроорганизмов, сопутствующее потребление ими элементов минерального питания из почвы и снижение доступности этих элементов для растений. Так, в экспериментах по влиянию вытяжек листьев тундровых кустарничков и березы на рост трех видов однодольных растений (Michelsen et al., 1995) было показано, что внесение этих вытяжек на стерильную почву не оказывает таких неблагоприятных воздействий на растения, как в случае нестерильной почвы. В последнем случае усиливалось микробиологическое дыхание почвы и содержание в ней грибов. Авторы считают, что богатые органикой растворы вызывают усиление развития микроорганизмов, увеличение иммобилизации ЭМП и ужесточение конкуренции между растениями и почвенной микрофлорой за ЭМП. Видимо, этот механизм широко распространен в природе.
Аллелопатия может выступать фактором гомеостаза в фитоценозах. Многие виды клональных растений не способны возобновляться семенным путем в собственных клонах. Так, прорастание семян рогоза может тормозиться выделениями взрослых растений (Работнов, 1992).
Интересным аспектом аллелопатических отношений является выделение пыльцой специальных веществ при попадании на рыльце пестика и другие органы растений. Эти процессы изучены для небольшого числа видов растений. Пыльца ряда растений может обладать ингибирующим действием по отношению к репродуктивным и вегетативным функциям соседних видов. В конце 80-х гг. прошлого века было показано, что пыльца кукурузы (в Мексике, действие через почву при отложении на ее поверхности) и Parthenium hysterophorus (Asteraceae, в Индии, действие при отложении на поверхности листьев) может вызывать хлороз соседних растений разных видов. Пыльца видов Hieracium и Phleum pratense снижает репродуктивный успех у конкурирующих видов. Пыльца тимофеевки действует преимущественно на злаки (Bromus inermis, Danthonia spp., Elytrigia repens), а ястребинок — на представителей сложноцветных (Achillea millefolium, Cirsium arvense, Sonchus arvensis), при этом аутотоксикация отсутствует. Видимо, пыльца этих видов при попадании
5. Создание фитосреды и положительные взаимоотношения растений
233
на рыльца других растений тормозит прорастание пыльцевых трубок. У пыльцы кукурузы основной аллелопатический агент — фенилуксусная кислота, она действует только на двудольных. В ряде случаев микробная трансформация выделений пыльцы также имеет место (Murphy, 2002).
5 • Создание фитосреды и положительные взаимоотношения растений
5.1 • Фитогенное поле
Растения, самим фактом произрастания на каком-либо участке и в про-цессе своей жизнедеятельности изменяют не только ресурсные факторы внешней среды. Для описания этих изменений применяли различные термины. Часто говорят о фитогенном поле (Уранов, 1965) или сфере средообразующего воздействия (Голубев, 1980). Часто трудно вычленить в чистом виде конкурентные и другие средоизменяющие воздействия. Интенсивность воздействия на среду обычно убывает при удалении от места укоренения растений. Например, для многолетнего злака прерий США Bouteloua gracilis показано, что эффективность использования воды значительно выше при ее добавлении в центральную часть корневой системы, по сравнению с периферической (Hook, Lauenroth, 1994).
5.2 • Изменение температуры воздуха и почвы
Температурный режим приземного воздуха и почвы сильно зависит от растительного покрова и структуры побегов отдельных видов растений, имеющих различную густоту, высоту, площадь и ориентацию листьев. В целом покров растений снижает амплитуду колебаний температуры поверхности почвы, что смягчает проявление морозных явлений и жары. Пространственная гетерогенность растительности создает, особенно в ясные дни, пеструю мозаику температур самих растений и почвы. Наиболее резкие перепады температуры почвы наблюдаются на лесных опушках, один из примеров изменения температуры разных субстратов в весенний день в Голландии по работе Stoutjesdijk и Barkman (1992) представлен на рис. 5.36. Самые сильные перепады температур наблюдаются при высокой пористости материалов и их низкой теплопроводности (затруднен отвод тепла). Поэтому даже в мартовский день температура ветоши овсяницы (Festuca ovina) достигала +64 °C. С другой стороны, при температуре воздуха около +12 °C почва с северной стороны от куста можжевельника Juniperus communis имела отрицательную темпера-
234
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.36 • Температура различных поверхностей и почвы на глубине 4 и 9 см на лесной опушке с кустами можжевельника в ясный день 3 марта 1976 г. (полдень). Температура воздуха на высоте 1 м составляла +11,8 °C. Величины солнечной радиации (Вт/м2) показаны на 3 поверхности (горизонтальную, вертикальную и перпендикулярную солнечным лучам) (по: Stoutjesdijk, 1977; Stoutjesdijk, Barkman, 1992)
туру, а в нескольких метрах на южном склоне, покрытом лишайниками, прогрелась до 18 °C.
Растения, способные пересыхать (в умеренной зоне это преимущественно мохообразные и лишайники), в сухом состоянии имеют высокую пористость и нагреваются значительно сильнее, чем сосудистые растения. Такие виды, как Polytrichum piliferum и Cladonia pyxidata в сухом состоянии выносят нагревание до 90-100°, в то время как всходы сосудистых растений в таких условиях погибают.
Микромозаичное распределение температуры свойственно и поверхности почвы в луговых сообществах (рис. 5.37, Stoutjesdijk, Barkman, 1992)
Снижение амплитуды колебаний температуры в сомкнутых сообществах по сравнению с открытыми участками может иметь большое значение для жизнедеятельности многих видов растений. Так, эксперименты, проведенные 3. Киквидзе (Kikvidze, 1996) на субальпийских лугах Кавказа, показали, что изоляция отдельных растений путем удаления их соседей в радиусе 1 м приводит к сильному нагреву и суммарно уменьшает эффективность фотосинтеза, например для Trifolium ambiguum (табл. 5.7).
5. Создание фитосреды и положительные взаимоотношения растений
235
’С 50
40-
30
20
Рис. 5.37 • Температура поверхности почвы и растений на сухом дюнном лугу. Сплошные линии — температура при высоком стоянии солнца (интенсивность суммарной радиации 839 Вт/м2), прерывистые — при низком стоянии солнца (419 Вт/м2). Стрелками показан угол падения солнечных лучей (по: Stoutjesdijk, 1966; Stoutjesdijk, Barkman, 1992)
Таблица 5.7 • Нетто-фотосинтез (мг СО2 на м2 в час) Trifolium ambiguum на субальпийском лугу Центрального Кавказа в естественных условиях и при удалении соседних растений (по: Kikvidze, 1996)
Растения на лугу Изолированные растения
Солнечная погода 18 12
Облачная погода 6,4 8,6
Итог с учетом числа солнечных и облачных дней 13 10
Защита от перегрева и иссушения очень важна для молодых кактусов, имеющих поверхностную корневую систему и закрытые днем устьица (САМ фотосинтез). Многие исследователи указывают, что приживание
236
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.38 • Колоновидный кактус Cephalocereus hoppenstedtii в пустыни Tehuacan (Мексика). Молодые кактусы приживаются в тени кустарников (фото М. Werger)
всходов ряда видов кактусов может осуществляться только при некотором затенении со стороны растений-«нянь». Это показано, например, для колоновидных кактусов Neobuxbaumia tetetzo и Cephalocereus hoppenstedtii (рис. 5.38), приживание всходов которых происходило лишь в тени кустарников, где температура почвы была на 16° ниже, чем на открытой поверхности почвы (Valiente-Banuet et al., 1991). Максимальная температура на поверхности стебля Ferocactus acanthoides была на 11° ниже под покровом растений-нянь, чем на открытом месте (Nobel, 1984).
Приживаемость всходов и молодых растений в аридных и высокогорных регионах также зависит от экранирования от прямого солнечного света. В высокогорьях Анд (осадки 200-300 мм, дневная амплитуда температур более 40°) был показан эффект нянек (Festuca orthophyIla на сложноцветное Parastrephia lucida) и пространственная ассоциированность этих видов (Kieler, Lambrinos, 2005). Но изменение температурного режима одними растениями может и негативно сказываться на других. В экспериментах с выращиванием проростков Eucalyptus pauciflora в окружении злаков или их ветоши в сравнении с голой почвой (Ball et al.,
5. Создание фитосреды и положительные взаимоотношения растений
237
2002) было показано, что именно более низкие температуры почвы под злаками или ветошью тормозят рост эвкалипта, сокращая длительность вегетационного периода для него.
Огромное влияние на температурный режим почвы оказывает моховой покров и подстилка (опадогенный горизонт), формирование и свойства которой обусловлены деятельностью различных видов растений. В большинстве случаев подстилка выступает в качестве теплоизоляционного материала, препятствующего теплообмену между почвой и атмосферой. При экспериментальном удалении подстилки на лугах обычно увеличивается скорость разложения органических веществ в почве в связи с увеличением температуры. На лугах с густым войлоком растения весной развиваются медленнее и реже цветут в связи с более низкими температурами почвы. A. Watt (1956, 1970) показал, что опад (ветошь) орляка (Pteridium aquilinum) играет защитную роль для молодых вай. При его удалении молодые вайи сильно повреждаются морозами в условиях океанического климата (Facelli, Pickett, 1991). Под участками с более мощной подстилкой или моховым покровом наблюдается подъем уровня вечной мерзлоты в континентальных регионах.
Опад различных видов растений по-разному меняет условия среды и воздействует на другие растения. В экспериментах с опадом трав и Quercus alba в Нью-Джерси (США) показано, что опад дуба по сравнению с опадом трав оказывает более сильное воздействие на многие виды травянистых растений (Facelli, Pickett, 1991). Изучение влияние опада Роа pratensis на прорастание семян и приживание всходов луговых трав показало, что опад на 26-41 % уменьшает всхожесть семян и на 95-100% уменьшает приживаемость всходов изученных видов растений (Bosy, Reader, 1995). На залежи с доминированием Solidago были проведены опыты с удалением и добавлением опада, который в этом сообществе образуется в значительном количестве (660-860 г/м2). Эти воздействия не изменяли существенно общую биомассу растений, но удаление опада приводило к значительному увеличению видового разнообразия (Carson, Peterson, 1990).
5.3 • Водный режим
Воздействие растений на водный режим их местообитаний весьма существенно и разнообразно. Рассмотрим основные механизмы такого воздействия, используя классификацию Т. А. Работнова (1992).
а) Перехват атмосферных осадков
Растения своими надземными побегами могут задерживать атмосферные осадки, которые в таком случае либо при испарении не достигают
238
Глава 5. Типы взаимоотношений растений в фитоценозах
Таблица 5.8 • Виды, предпочитающие различные участки увлажнения в ельниках (по: Stoutjesdijk, Barkman, 1992)
Тип леса Виды приствольных повышений Виды межкроновых участков
Сырые ельники Ptilium crista-castrensis, Vaccini urn uliginosum Polytrichum commune Sphagnum girgensohnii Equisetum sylvaticum
Влажные ельники Vaccinium myrtillus Hyiocomium spiendens Ptilium crista-castrensis, Vaccinium uliginosum
Сухие ельники Vaccinium vitis-idaea Pleurozium schreberi Vaccinium myrtillus Hyiocomium spiendens
поверхности почвы, либо стекают по стволам, создавая высокую гетерогенность верхних горизонтов почвы по уровню поступления воды, а с нею и ряда биогенных элементов. По данным В. Лархера (1978), в Европе хвойные деревья перехватывают 3-6 мм осадков, лиственные — 0,5-2 мм, верещатники и луга — 1-2 мм, сфагновые мхи — 15 мм. В среднем за год в Западной Европе в хвойных лесах перехватывается до 30%, а в лиственных — до 20% осадков. Из дождя в 10 мм кронами задерживают Pinus cembra 74 %, Juniperus communis 70 %, Picea abies 45 %, Fagus siivatica 37 %, Quercus robur 32 %. В тропических лесах дождь мощностью в 100 мм может быть необходим, чтобы вызвать сток по стволам.
Сток по стволам деревьев зависит от архитектуры их кроны, особенно наклона ветвей. Так по данным В. Н. Мины (1970), за 4 летних месяца средний сток по стволам у ели (горизонтальное расположение ветвей) составил около 5-6 л на дерево, а у сосны и березы, с их более вертикально расположенными ветвями, — 15-22 л. Этот сток имеет очень важное значение для развития напочвенного, особенно мохового, покрова в таежных лесах. В сосняках приствольные повышения получают обычно больше, а в ельниках — меньше воды, чем межкроновые пространства. Поэтому в зависимости от типа леса разные группы растений предпочитают приствольные или межкроновые участки (табл. 5.8). При уменьшении общего увлажнения виды приствольных повышений перемещаются в межкроновые участки.
Минеральное питание мхов часто осуществляется преимущественно за счет веществ, содержащихся в стекающей по стволам воде. Это было показано, например, для Hyiocomium spiendens (Tamm, 1950). Кроме того,
5. Создание фитосреды и положительные взаимоотношения растений
239
листья мохообразных обладают высокой адсорбционной способностью поглощения катионов, примерно в 100 раз более интенсивно поглощая катионы, чем анионы. Таким образом, влияние мохообразных на просачивающуюся по их куртинкам воду аналогично действию катионитов — в этой воде снижается концентрация катионов и возрастает Н+, т. е. идет ее подкисление, что не может не сказаться на свойствах почвы под ними (см. п. 1.3 гл. 6). Но и виды сосудистых растений нижних ярусов могут перехватывать листьями элементы минерального питания, содержащиеся в осадках, прошедших сквозь крону. Так, в дубовых лесах из Quercus robur, пролесник (Mercurialis perennis) поглощал листьями около 50% Са, Mg, К, Na, содержащихся в осадках, прошедших через крону дуба (Andersson, 1991 и др.).
б) Осаждение воды из туманов
В ряде регионов с незначительным поступлением осадков с дождями или снегом, например на юге Тихоокеанского побережья США, существование лесов в условиях аридного климата обусловлено осаждением воды из туманов (за счет усиления конденсации). По наблюдениям в Перу, где на открытом месте осадки составляли в среднем 168 мм в год, под пологом казуарины (растение с редуцированными чешуевидными листьями) годовое поступление осадков равнялось 488 мм, а под пологом эвкалипта (с большой поверхностью листьев) достигло 676 мм за счет перехвата влаги, содержащейся в туманах (Вальтер, 1968).
Осаждение воды из туманов наблюдается также в горах близ верхней границы леса. Например, на Северном Кавказе одиночно стоящие на хребтах сосны вызывали ощутимый дождь под их кронами во время прохождения облаков через хребет, в то время как осаждение воды травянистыми растениями было менее значительным.
Количественная оценка перехвата воды из туманов проведена в сообществах крупнокочкарных злаков (Chionochloa macro) в Новой Зеландии. Было показано, что покров из этих растений, имеющих многочисленные длинные и тонкие листья, улавливает и переводит во внутрипоч-венный сток дополнительно около 120 мм осадков за год (Holdsworth, Mark, 1990). С другой стороны, посадка сосен (Pinus radiata) уменьшила поступление воды во внутрипочвенный сток на 41 % после 22 лет (Fahey, Jackson, 1997). Выжигание злаковников ведет к снижению поступления воды в грунтовые воды на 32% на второй год и на 19% в третий (Mark, Dickinson, 2008)
Martorell и Ezcurra (2007) изучали поглощение воды из туманов алюминиевыми моделями с разной шириной листьев и сравнивали полученные данные с наблюдаемыми у разных видов рода Tillandsia. Поглощение
240
Глава 5. Типы взаимоотношений растений в фитоценозах
воды увеличивалось с увеличением площади листьев, но эффективность (поглощение на единицу площади) была выше при узкой форме листа. Для растений, поглощающих влагу из туманов предложен специальный термин «небулофиты». Общий признак небулофитов — узкие листья или аналогичные структуры (например, талломы лишайников). Узкие листья имеют меньшую толщину пограничного слоя, что способствует аккумуляции влаги из туманов, где частицы воды имеют размеры около 20 микрометров. Отмечено также, что узколистность растений повышается в высокогорьях (Martorell, Ezcurra, 2007).
в) Сокращение поверхностного стока, предотвращение эрозии, снижение скорости течения воды в поймах
Сосудистые растения могут снижать поверхностный сток за счет создания механических препятствий, поглощения воды корнями, увеличения водопроницаемости почв за счет развития корневых систем и другими путями. Но и почвенные водоросли могут играть существенную роль в закреплении почв, особенно в аридных условиях. Так, длина нитей водорослей составила для почв Гиссарского хребта 88 м/см3 почвы, на песках — 65 м/см3 Закрепление поверхности почвы может происходить и за счет выделения агрегирующих веществ (см. Штина, 1991).
В условиях заливания поемными водами растения снижают скорость их течения. Кусты ив (Salix spp.) таким образом способствуют осаждению песка в прирусловой части, где скорость течения велика, а надземные органы луговых растений увеличивают отложения наилка в центральной и притеррасной частях пойм.
г) Снижение уровня почвенно-грунтовых вод
В условиях высокого стояния почвенно-грунтовых вод, корни отдельных растений могут не только изменять окислительно-восстановительный потенциал в ризосфере (см. ниже — п. 5.5 гл. 5), но и локально снижать уровень воды. Это отмечено для крупных растений, испаряющих много воды в условиях низкой влагопроницаемости почв. Такое снижение может способствовать и локальному улучшению аэрации вблизи растения. Примером может служить дерновинный злак заболоченных лугов Molinia caerulea, вблизи дерновин которой несколько снижается уровень почвенно-грунтовых вод и значительно лучше произрастает мелкоуко-реняющийся Erica tetralix, не выносящий поверхностного стояния воды (Berendse, Aerts, 1984).
В аридных районах корни растений-фреатофитов достигают уровня грунтовых вод на большой глубине. Максимально известная глубина
5. Создание фитосреды и положительные взаимоотношения растений
241
проникновения живых корней — 68 м у древесного вида Boscia albitrun-са в Центральном Калахари, а глубина более 40 метров показана для 8 видов деревьев (см. Canadell et al., 1996). Даже очень небольшая доля глубоких корней может существенно улучшить водный режим растений во время засухи. Так, Gregori et al. (1978; Canadell et al., 1996) показали, что у озимой пшеницы 3 % корней, проникших на глубину более 1 м, в период засухи обеспечивали около 20 % транспирации.
д) Уменьшение испарения с поверхности почвы за счет затенения
Затенение растениями поверхности почвы резко снижает физическое испарение. В аридных условиях такое затенение способствует более долгому сохранению почвенной влаги и повышает пестроту почвенного покрова, обусловленную влиянием отдельных растений. Так, Joffre и Ram-bal (1988) в редколесьях Испании отмечали значительную «задержку» в потерях воды почвой под дубами (Quercus rotundifolia, Q.robur) в сравнении с открытыми участками, что позволяет произрастать под кронами дубов многим видам растений, отсутствующим на открытых участках.
Затенение поверхности почвы растениями влияет на сопряженные режимы трех важнейших факторов: света, температуры и увлажнения. Примером их совместного влияния может служить приуроченность многих мохообразных к северным сторонам стволов деревьев и окружающей почвы. Stoutjesdijk и Barkman (1992) провели эксперименты с Dicranum scoparium для выяснения вопроса, почему мхи лучше развиваются на затененных северных участках. Одна куртина мха была разделена на 4 части, которые поместили с восточной, южной, западной и северной стороны от куста можжевельника. На северной стороне приход радиации в ясный день составил лишь 66 % от такового на южной стороне, однако фотосинтез мхов с северной стороны продолжался существенно дольше, так как они медленнее высыхали. Эффективные суммы радиации (время до высыхания х интенсивность радиации) находились для 4 экспозиций в следующем соотношении: 288:253: 281 :836 (В: Ю: 3: С).
е) Изменение снегового режима
Снижая скорость ветра, растения, особенно древесные, могут вызывать локальное задержание снега, наблюдаемое, например, по опушкам лесов. Ярким примером влияния растений на снеговой режим могут служить перераспределение снега близ кустов можжевельника (Juniperus communis) на сухих лугах (альварах) в южной Швеции (рис. 5.39, Rosen, 1982). Мощность снега с подветренной стороны существенно выше, он стаивает примерно на 2 недели позже, поддерживая значительную влажность почвы в течение нескольких недель.
242
Глава 5. Типы взаимоотношений растений в фитоценозах
а)
б)
Рис. 5.39 • Снежный покров на альварах о. Оланд (Швеция) зимой (а) и весной (б) вблизи кустов можжевельника (по: Rosen, 1982)
5. Создание фитосреды и положительные взаимоотношения растений
243
ж) Гидравлический лифт — подъем корнями растений воды из нижних горизонтов почвы в верхние
Растения способны не только поглощать влагу корнями, но и выделять ее в более сухие горизонты почвы. Это явление широко распространено в аридных регионах. Впервые оно было описано для полыни в североамериканских кустарниковых степях (Artemisia tridentata), которая своими глубокими корнями способствует подъему влаги из нижних горизонтов в верхние, где эта влага становится доступной для других видов растений, что было показано с применением стабильных изотопов (Caldwell, 1990).
Аналогичные результаты были получены в лесах из сахарного клена (Acer saccharum), в которых виды растений нижних ярусов получали за счет «гидравлического лифта» клена от 3 до 60 % воды (Dawson, 1993). Каждую ночь взрослое дерево сахарного клена может перекачивать из нижних горизонтов почвы в верхние около 100 л воды (Emerman, Dawson, 1996). Одно дерево зонтичной акации (Acacia tortilis) в Африке поднимает за ночь от 70 до 235 л воды (Ludwig et al., 2003). В целом роль гидравлического лифта в эвапотранспирации природных сообществ оценивается в среднем от 14% до 33% (Wan et al., 1993; Richards, Caldwell, 1987), в отдельных случаях может быть существенно выше (50-100%) (Liste, White, 2008).
Гидравлический лифт описан более, чем у 90 видов растений разных жизненных форм, но, видимо, распространен значительно шире (Caldwell et al., 1998; Ryel, 2004; Liste, White, 2008). Поступление воды из корней в более сухие горизонты почвы осуществляется по градиенту концентраций у обычных растений ночью (рис. 5.40), когда резко уменьшается или прекращается транспирация. Но у растений с САМ — типом фотосинтеза и ночным открытием устьиц может наблюдаться обратная картина, когда поступление воды в почву происходит днем (если не очень высоко испарение). Если блокировать транспирацию СЗ растений днем (поместить побеги в полиэтилен), то увеличение содержания воды в верхних горизонтах почвы наблюдается и в дневные часы (см.: Horton, Hart, 1998). Напротив, кратковременное освещение растений ночью ведет к открытию ими устьиц и, соответственно, уменьшению или прекращению поступления воды в почву за счет гидравлического лифта. Выделение воды в почву способствует и лучшему поглощению ЭМП растениями из верхних, более сухих горизонтов почвы, усиливает микробиологическую активность в них, выравнивает влажность почвы. Оно приводит к увеличению длительности жизни тонких корней. Проникновение корней на большие глубины через сухие горизонты почвы также может быть связано с выде-
244
Глава 5. Типы взаимоотношений растений в фитоценозах
Рис. 5.40 • Схема действия «гидравлического лифта» растений. Стрелками показано направление движения воды. Днем при открытых устьицах основной ток воды из корней расходуется на транспирацию, а ночью при закрытых устьицах вода выделяется в более сухие поверхностные горизонты почвы (по: Canadell et al., 1996; Horton, Hart, 1998)
лением ими влаги, поглощенной в более влажных горизонтах, поскольку перемещение воды корнями осуществляется в различных направлениях от влажной почвы к сухой.
Перераспределение почвенной влаги может осуществляться корнями растений и после отмирания или удаления надземных частей, что было показано на примере Bromus tectorum (Leffler et al., 2005).
5.4 • Изменение микрорельефа и закрепление подвижных субстратов
При своем произрастании растения изменяют микрорельеф. Около стволов деревьев возникают приствольные повышения. Плотнодерновинные растения, аккумулируя приносимые ветром частицы и откладывая большое количество отмерших органов (ветоши), образуют возвышения, как в аридных, так и в гумидных условиях. Крупнодерновинные злаки и осо
5. Создание фитосреды и положительные взаимоотношения растений
245
ки могут образовывать большие кочки. Кочкарные ландшафты особенно свойственны луговым сообществам южного полушария («тассоки»), где они образованы видами рода Chionochloa. Число кочек может достигать 40 000 на гектар, они затрудняют поверхностный сток и усиливают конденсацию воды из туманов (см. выше), обеспечивая значительное поступление воды для внутрипочвенного стока.
Большое значение имеет также закрепление корнями растений подвижных субстратов, наблюдаемое при зарастании речных отмелей, осыпей в горах, солифлюкционных наплывов и т. п.
Изменяя скорость ветра, растения могут способствовать осаждению переносимого им материала, изменяя гранулометрический состав почвы в месте своего произрастания. Примером таких изменений может служить последовательность смены видов в сухих днищах озер пустыни Mojave в США. Прутняк (Kochia californica) поселяется в трещинах очень тонкодисперсной почвы (такырная корка растрескивания) и способствует аккумуляции более крупных почвенных частиц, переносимых сильными ветрами. В результате образуются небольшие холмики, которые может колонизировать лебеда Atriplex torreyi (Vasek, Lund, 1980). Лебеда, в свою очередь, способствует еще большей аккумуляции песчаных частиц и поселению других видов (Atriplex confertifolia, Haplopappus acradenius). В ходе этих смен наблюдаются существенные изменения гранулометрического состава почвы и микрорельефа.
5.5 • Изменение солевого режима, кислотности
и окислительно-восстановительного потенциала почв
В условиях аридных областей растения могут влиять на содержание солей в верхних горизонтах почвы двояко — увеличивая или уменьшая их засоление. Увеличение засоления происходит в случае высокого содержания солей в опаде растений, что наблюдается, например, у саксаула (Работнов, 1992). В большинстве случаев растения способствуют рассолению почвы в местах своего произрастания. Механизмы такого влияния могут быть различными. Например, затенение кронами растений ведет к уменьшению испарения с поверхности почвы, что снижает капиллярный подъем влаги и отложение солей. Вблизи от таких растений могут произрастать менее солеустойчивые виды растений, что, например, было показано для засоленных маршей в Новой Англии (Bertness, 1991; Bertness, Shumway, 1993). В этих условиях ситник Juncus gerardi был приурочен к зарослям Distichlis spicata. Экспериментально было показано благоприятное влияние этого растения на рост ситника в условиях
246
Глава 5. Типы взаимоотношений растений в фитоценозах
сильной засоленности. В то же время при искусственной промывке почвы это влияние исчезало, и ситник предпочитал участки без Distichlis spicata.
Другим механизмом локального снижения засоления почвы под отдельными растениями может быть отмеченное ранее снегозадержание вследствие снижения скорости ветра. Задержка снега увеличивает поступление воды в почву и способствует вымыванию солей из верхних горизонтов почвы в нижние.
Ряд видов растений может существенно подкислять почву при своем произрастании. Это подкисление связано с различными механизмами. Как уже было отмечено, листья мохообразных обладают свойствами катионитов, обменивая катионы в стекающей по ним воде на Н+. Поэтому под куртинками мхов происходит локальное подкисление почвы. При сплошном развитии мохового покрова (верховые болота, моховой ярус таежных лесов) такое подкисление захватывает значительные территории.
Основной механизм изменения pH за счет деятельности корней — освобождение Н+ или ОН- для компенсации ионного дисбаланса, получающегося при поглощении тех или иных ионов ЭМП корнями. Кроме того, органические анионы могут выделяться корнями. Корневое дыхание также может вносить вклад в изменение кислотности ризосферы. Поглощение нитратов и фосфатов ведет к соответствующему выделению ОН- а аммония — Н+ Кончики корней обычно выделяют Н+ а базальные части — ОН- (Hinsinger et al., 2003). Симбиотическая азотфиксация бобовых сопровождается высвобождением протонов и выходом их в почву. Локальное снижение pH в ризосфере способствует большей доступности фосфатов, которые бобовые эффективно потребляют (Marschner, Romheld, 1983). Сильно подкисляют почву и «кластерные корни» (см. п. 1.2 гл. 2, Вставка 1).
Подкисление почвы происходит и при воздействии продуктов разложения опада, что хорошо известно для хвойных деревьев. Так, в ходе сукцессий на карбонатных породах, освободившихся от ледника на Аляске, pH почвы в климаксовом лесу под ситхинской елью (Picea sitchensis) достигает величины 5 при начальном значении около 8 (Burrows, 1990). Способствует подкислению почвы и опад вересковых кустарничков. Значительное снижение pH наблюдается под участками вереска на мелах (Grubb et al., 1969; Trudgill, 1988, рис. 5.41). При длительном произрастании рододендроновых стлаников (Rhododendron ferrugineum) на доломитах в Швейцарских Альпах под ними формируется мощный органогенный горизонт (рис. 5.42), кислотность которого существенно выше (на 4 порядка!), чем у подстилающей породы.
5. Создание фитосреды и положительные взаимоотношения растений 247
Рис. 5.41 • pH подстилки, почвы и подстилающих меловых пород на пустоши под кустом вереска (Calluna vulgaris), угнетающего Erica cinerea. Вертикальный масштаб для почвенного профиля в 2,5 раз больше горизонтального. Высота кустов вереска — 55 см (по: Grubb et al., 1969; Trudgill, 1988)
горизонт pH (H2O)
см 3456789
органическое вещество, %
Рис. 5.42 • Почвенный профиль, pH и содержание органического вещества в почве под участком рододендрона (Rhododendron ferrugineum) в Швейцарских Альпах близ Давоса (по данным Н. Sticher, личное сообщение)
Большое значение имеет влияние отдельных растений на окислительно-восстановительный потенциал почв в условиях высокого увлажнения почв и недостаточного поступления кислорода. Растения, имеющие в корнях или корневищах аэренхиму (воздухопроводящую ткань), поставляют кислород в ризосферу, давая возможность произрастать в их окружении растениям, не адаптированным к недостатку кислорода в почве.
248
Глава 5. Типы взаимоотношений растений в фитоценозах
Так, на засоленных маршах восточного побережья США Hacker и Bert-ness (1995) установили, что в ризосфере ситника Juncus maritimus возрастает окислительно-восстановительный потенциал, что делает возможным произрастание рядом с ситником многолетника Iva frutescens, который обычно характерен для более аэрируемых почв. Однако проведенные эксперименты по имитациям воздействия ситника (увеличение аэрации, уменьшение засоления) показали, что главный положительный эффект наблюдается за счет уменьшения засоления под ситником, аэрация же имеет несколько меньшее значение (Hacker, Bertness, 1995). Этот же вид ситника на заболоченных лугах Голландии ассоциирован с Plantago coronopus и Samolus valerandi (Schat, van Beckhoven, 1991), что, видимо, связано с улучшением аэрации почв и окислением сульфидов вокруг корней ситника.
Наиболее ярко роль изменения окислительно-восстановительного потенциала почв показана в экспериментах с выращиванием рогоза (Ту-pha latifolia) в недренируемых горшках. Содержание кислорода в почве с рогозом было в 4 раза выше, чем в горшках без рогоза. Это различие ярко проявлялось при низких температурах (11-12 °C), в то время как при более высокой температуре (18-20 °C) оно нивелировались. В этом эксперименте не имеющие аэренхимы ива Salix exigua и незабудка Myosotis laxa развивались при низких температурах значительно лучше в присутствии рогоза, чем без него. В зарослях рогоза в природе Myosotis laxa, произраставшая вблизи рогоза, была крупнее и производила больше семян, чем особи, изолированные от рогоза (Callaway, King, 1996)
5.6 • Роль растений в изменении содержания ЭМП и органического вещества в почве
Растения могут влиять на содержание элементов минерального питания и органического вещества, с одной стороны, уменьшая их содержание за счет поглощения ЭМП и ускорения разложения почвенной органики (последнее показано, например, в экспериментах по влиянию живых корней растений на скорость разложения отмерших корней — van der Krift et al., 2002), а с другой, — увеличивая их содержание за счет поступления с опадом, корневыми выделениями, усиления выветривания первичных минералов и торможения разложения органики при выделении антимикробных метаболитов. Реальные воздействия растений зависят от балансов этих противоположных процессов.
Растения в процессе своей жизнедеятельности изменяют распределение химических элементов по почвенному профилю. Так, при посадках сосны на дюнах в Аргентине Na и К были изначально равномерно рас-
5. Создание фитосреды и положительные взаимоотношения растений
249
обменный К (смоль/кг)
обменный Na (смоль/кг)
Рис. 5.43 • Распределение по почвенному профилю обменных калия и натрия в исходных дюнах (слева) и после 55 лет выращивания плантаций сосны Pinus pinaster в прибрежной зоне Аргентины. Показаны средние значения и ошибки среднего (п = 5) (по: Jobbagy, Jackson, 2004)
пределены по почвенному профилю, но через несколько десятилетий выращивания К (интенсивно вовлекаемый в биологический круговорот) был значимо сконцентрирован в верхних горизонтах почвы, a Na — в более глубоких (рис. 5.43). В глобальном аспекте концентрация К в верхних горизонтах почвы без растений была бы на половину или на треть меньше, чем с растениями. Аккумуляция кремния в почве ряда тропических регионов целиком обусловлена деятельностью растений (в верхних горизонтах он присутствует только в форме остатков фитолитов — опального кремнезема) (Meunier et al., 1999). Посадка эвкалиптов на лугах с почвами, бедными Мп, привела к увеличению содержания его обменных форм в верхних горизонтах почвы (листья Eucalyptus cama/du/ensis содержали
250
Глава 5. Типы взаимоотношений растений в фитоценозах
в 15-25 раз больше марганца, чем листья травянистых растений). Листья широколистных деревьев содержат больше Са и имеют в 5 раз более высокое отношение Са К, чем листья трав, поэтому анализ данных департамента земледелия США показал, что распределение Са по профилю в таких лесах более мелкое (больше в верхних горизонтах), чем в травяных сообществах (рис. 5.44) (Jobbagy, Jackson, 2004).
Растения могут влиять на азотный режим почвы различными способами — поглощая доступные формы азота, стимулируя азотфиксацию или воздействуя на интенсивность микробиологической минерализации азота. Например, в экспериментах van der Krift и Berendse (2001) исследовалось влияние разных видов растений на интенсивность минерализации азота и нитри-течение 4-х лет. Авторы выявили сильное
Рис. 5.44 • Усредненные данные по распределению Са по профилю почвы в лесах и травяных сообществах в США (вертикальная шкала — глубина почвы, см, горизонтальная шкала — доля кальция от общего пула в 1-метровом слое почвы) (по: Jobbagy, Jackson, 2004)
фикации в монокультурах в
влияние вида растений на интенсивность минерализации и нитрификации. В среднем виды богатых почв (см. п. 3.6 гл. 5) усиливают минерализацию и нитрификацию сильнее, чем виды бедных почв. Растения также могут обогащать почву азотом в случае симбиотической или ассоциативной азотфиксации (см. главу 2).
Воздействие разных видов растений на свойства почв были исследованы в эксперименте с монокультурами альпийских растений (Макаров и др., 1999; Onipchenko et al., 2001). В них изучены изменения ряда ха
рактеристик исходно однородной почвенной смеси значительным числом видов растений после 6-летнего выращивания в близких к естественным условиям высокогорий.
Отдельные виды альпийских растений, использованные в экспери
менте, по-разному изменяли кислотность почвы и концентрацию элементов минерального питания в сравнении с контрольным вариантом (почва без растений). Большинство исследованных видов, кроме Hedysarum саи-casicum, снижало содержание нитратного и аммонийного азота, что подтверждает представления о лимитирующей роли доступных форм азота во многих альпийских сообществах. Hedysarum caucasicum повысил содержание N-NH4 выше уровня его исходной концентрации в субстрате, что, безусловно, связано с проявлением симбиотической азотфиксации. Наиболее низкая концентрация нитратного азота получена в почве под
5. Создание фитосреды и положительные взаимоотношения растений
251
0,12 0,11
0,1 0,09 0,08 0,07 0,06 0,05
Рис. 5.45 • Концентрация отдельных форм ЭМП в почве под монокультурами 17 видов альпийских растений, начальном (CI) и конечном (СЕ) контроле. Виды расположены в порядке возрастания концентрации. Значимые различие (р < 0,05) показаны разными буквами. Все величины, кроме нитратов и нитритов, в мг/100 г почвы, нитраты и нитриты — в мкг/100 г почвы (по: Onipchenko et al., 2001). Виды растений: /4s — Anemone speciosa, Ad — Antennaria dioica, Ct — Campanula tridentata, Cs — Carex sempervirens, Си — Carex umbrosa, Fo — Festuca ovina, Fv — Festuca varia, Gg — Geranium gymnocauion, He — Hedysarum caucasicum, Lh — Leontodon hispidus, Me — Matricaria caucasica, Ns — Nardus stricta, Pa — Phleum alpinum, Sc — Scorzonera сапа, Sp — Sibbaldia procumbens, Ts — Taraxacum stevenii, Tp — Trifolium polyphyllum
двумя видами растений семейства сложноцветных — Matricaria caucasica и Leontodon hispidus (рис. 5.45). Carex sempervirens, Taraxacum stevenii и Sibbaldia procumbens значимо уменьшили концентрацию обменного аммония в сравнении с контролем. Эффективное использование азота
252
Глава 5. Типы взаимоотношений растений в фитоценозах
именно альпийскими осоками отмечено нами и при внесении азотных удобрений (Ахметжанова, Онипченко, 2005; Soudzilovskaia, Onipchenko, 2005). Содержание подвижного фосфора было низким под 4 видами злаков (Festuca ovina, Nardus stricta, Festuca varia, Phleum alpinum) и Anemone speciosa, как и в конечном контроле эксперимента. Значимо более высокие величины содержания подвижного фосфора отмечены под Hedysarum caucasicum и Leontodon hispidus (рис. 5.45).
Результаты этого эксперимента показывают также значительную степень специализации растений отдельных семейств по их влиянию на свойства почвы. Бобовые уменьшают содержание катионов (Са, Mg и К) и увеличивают содержание азота в связи с азотфиксацией. Осоковые (Cyperaceae: Carex spp.) имели тенденцию снижать содержание аммония, в то время как сложноцветные (Asteraceae) и осоковые существенно снижали концентрацию нитратов. Эти результаты подтверждают предпочтительное потребление нитратов аммонию у представителей семейства сложноцветных (de Graaf et al., 1998). Среди исследованных семейств злаки снижали концентрацию подвижного Р в почве в наибольшей степени.
С этими проблемами связан один из классических вопросов геоботаники — вопрос о взаимосвязи почв и растений. Что первично — почвенная пространственная неоднородность или она создается в результате воздействия разных видов растений? Один из вариантов решения этого вопроса экспериментальным путем предложили Reynolds et al. (1997). Сухие сообщества с доминированием однолетников в Калифорнии явились удобным объектом для постановки эксперимента. Эти сообщества характеризуются неоднородной горизонтальной структурой, в их составе доминируют 3 вида однолетников, часто образующих монодоминантные группировки — Calycadenia multiglandulosum (Asteraceae), Lasthenia cal-ifornica (Asteraceae), Plantago erecta (Plantaginaceae). Участки под этими группировками значительно различались по ряду свойств почвы. Calycadenia занимала участки с более глубокими почвами, a Lasthenia — участки с самыми маломощными, но и самыми влажными почвами. Исследователи удалили растения с небольших площадок внутри каждой группировки, а затем посеяли семена удаленного или других видов, создав таким образом монокультуры на местах произрастания своего или чужого вида. Было показано, что такие свойства почвы, как почвенная влажность, мощность, почвенный микробный азот, отношение С/N были связаны с начальной неоднородностью почвы. В то же время различия в содержании почвенных нитратов и интенсивности минерализации азота обусловлены деятельностью самих растений и изменялись в течение одного сезона при выращивании «чужого вида» (рис. 5.46).
5. Создание фитосреды и положительные взаимоотношения растений
253
а)
35
30
25
20
15
10
5
0
Р P->-L L L-*-P С С-*Р
б)
0.5
ас
Ьс
а а
о Ф
Р P->L L L^P С С^-Р
Вариант эксперимента
Участки с доминированием
Q Plantago Lasthenia □ Calycadenia
Рис. 5.46 • Изменение выхода нитратов (а) и влажность почвы (б) на участках с однолетними растениями. Р — Plantago еrecta, L — Lasthenia californica, С — Calycadenia multiglandulosum, P —> L — посадка Lasthenia на участке Plantago, L -> P посадка Plantago на участке Lasthenia, C —> P посадка Plantago на участке Calycadenia. Неповторяющиеся буквы отмечают значимые различия (р < 0,05). Посев Plantago на участки других видов резко снижал поступление нитратов в почву (по: Reynolds et al., 1997)
5.7 • Создание препятствий для ветра, фитофагов, привлечение опылителей
Многие растения, особенно древесные, создают значительные физические преграды, уменьшающие скорость ветра внутри сообществ. Например, по данным Молчанова (1968), при скорости ветра на открытом
254
Глава 5. Типы взаимоотношений растений в фитоценозах
месте 7 м/сек (100%), она составляла в чистом сосняке 3,8 м/сек (54%), а в сосняке с подростом ели — 1,3 м/сек (19%).
Для растений засоленных маршей в США проведены эксперименты по влиянию сомкнутых сообществ на морфологию и выживаемость отдельных видов растений по сравнению с произрастанием на открытых участках. Особи 4 изученных видов растений в тесно сомкнутом сообществе были выше, тоньше и образовывали меньшую надземную биомассу по сравнению с особями на открытых участках. При удалении соседей большинство таких растений полегало, что свидетельствует об использовании окружающих растений как опор в тесно сомкнутых сообществах (Harley, Bertness, 1996).
Сильный ветер может отрицательно сказываться на росте растений. Russel! и Grace (1979) выращивали Lolium регеппе и Festuca arundinacea в ветровом туннеле при 4 скоростях ветра (от 1,1 до 10 м/сек) в течение 14 дней. С увеличением скорости ветра ростовые процессы замедлялись, размеры растений уменьшались, что авторы связывают как с усилением водного стресса, так с механическим воздействием (Rosen, 1982).
Колючие растения могут защищать соседние растения, делая их недоступными для крупных фитофагов. В условиях интенсивного выпаса на субальпийских лугах Кавказа значительное число видов растений сохраняется и образует генеративные органы только близь колючих видов растений рода Cirsium, которые поддерживают флористическое богатство растительных сообществ в этих условиях (Callaway et al., 2000).
Растения часто конкурируют за опылителей, однако привлечение опылителей одними видами может повышать вероятность опыления их соседей. Thomson (1978) обнаружил, что Hieracium florentinum посещается опылителями значительно чаще, если произрастает вместе с Hieracium auranticum, чем в индивидуальных зарослях. Аналогичная закономерность показана для безнектарного Podophyllum peltatum, произрастающего с Pedicularis canadensis, который значительно эффективнее привлекает опылителей (Laverty, 1992).
6 • Положительные отношения
между растениями
Многие из рассмотренных выше примеров изменения среды растениями показывают возможность благоприятного воздействия одних растений на другие. Существует два типа подходов к изучению таких положительных отношений:
6. Положительные отношения между растениями
255
1) Пространственная сопряженность, хотя она не отличает благоприятствование от случаев приуроченности нескольких видов к одним и тем же элементам изначально неоднородной среды. Одно из первых наблюдений пространственной структуры отмечено Phillips (1909), который обнаружил, что проростки сосны Pinus monophylia встречаются преимущественно под полынью Artemisia tridentata и редко на открытых участках. Интересные наблюдения проведены в Патагонских Андах (Чили). На двух высотных уровнях (700 и 900 м) исследован микроклимат и видовой состав растений, обитающих внутри и вне крупных подушечных растений Azorella monantha. Показано, что температура почвы под подушками была существенно выше, чем вне их, и различия увеличивались с высотой. На высоте 700 м отмечено 27 видов растений внутри подушек и 29 видов вне их, в то время как на высоте 900 м ситуация изменилась — здесь уже 34 вида растений встречались внутри и только 18 — вне подушек, что связывается с усилением неблагоприятных воздействий среды (Arroyo et al., 2003).
2) Эксперименты по удалениям и имитациям. Только полевые эксперименты часто позволяют отличить эффект благоприятствования от совместной приуроченности растений к отдельным микроместообитаниям. Однако эксперименты с удалением имеют и ряд сложностей для интерпретации. Например, благоприятный эффект от затенения может перекрываться корневой конкуренцией, и необходимы специальные эксперименты для разграничения эффектов конкуренции и благоприятствования. Необходимо иметь в виду и долговременные последствия совместного произрастания. Например, удаление вида не вызывает уменьшения участия другого вида, когда последний может пользоваться условиями среды, уже созданными первым видом (накопление опада, аккумуляция азота и т. п.) и не исчезающими при удалении. Так, полевые и лабораторные эксперименты Callaway et al. (1991) показали, что лучшее развитие однолетних злаков под Quercus macrocarpa в Калифорнии обусловлено увеличением богатства почвы за счет дубового опада. Перспективно использование механических моделей, например технических конструкций, создающих затенение, аналогичное наблюдаемому в природе под покровом растений-нянь, но исключающих корневую конкуренцию.
В целом для природных сообществ характерны сложные комбинации как положительных, так и отрицательных взаимодействий. Конкурентные взаимодействия между растениями опосредуются и могут нивелироваться
256
Глава 5. Типы взаимоотношений растений в фитоценозах
сложными отношениями с другими организмами. Так, например, специальные эксперименты по затенению и исключению фитофагов в Чилийской маторрали показали, что приуроченность ряда видов однолетних и многолетних растений к зарослям кустарников связана как с защитой от фитофагов и распределением семян, так и с уменьшением испарения в этих условиях (Jaksic, Fuentes, 1980; Fuentes et al., 1984).
Положительные отношения носят чаще односторонний характер (один вид положительно воздействует на другой, второй не оказывает такого же воздействия на первый). Взаимно положительные отношения возможны, например, при изменении доступности ресурсов под влиянием отдельных видов растений. В Испании бобовый кустарник Retama sphaerocarpa благоприятно влияет на произрастающую совместно с ним шандру Marrubium vulgare. Эксперименты показали и обратное положительное влияние шандры на кустарник (Pugnaire et al., 1996). Другой пример взаимно положительных отношений — на береговых дюнах в Калифорнии простратная форма кустарника Baccharis pilularis положительно влияет на появление и приживание всходов и рост кустарникового азотфиксирующего бобового — Lupinus arboreus, который в дальнейшем обогащает почву азотом, используемым и соседями (Rudgers, Maron, 2003).
Воздействия одних растений на другие возможно через окружающую микрофлору. Каждый вид растений создает свои комплексы микроорганизмов в ризосфере, которые могут как положительно, так и отрицательно влиять на соседние растения (Wardle, Nicholson, 1996).
Многие эксперименты с удалениями позволили определить, в каких условиях преобладают конкурентные отношения, а в каких — благоприятствование, т. е. выявить взаимодействие положительных и отрицательных отношений. Например, эксперименты по удалениям на засоленных маршах Новой Англии (Bertness, 1991; Bertness, Shumway, 1993) показали, что конкурентные отношения преобладают в более благоприятных условиях, в то время как в условиях сильного абиотического стресса четко выражены эффекты благоприятствования. Во многих случаях мы наблюдаем суммарный эффект наложения положительных и отрицательных воздействий растений друг на друга. Так, в степях Патагонии Aguiar et al. (1992) нашли, что различные кустарники защищают злаки от ветра и высыхания, но этот эффект сильно проявляется лишь при исключении корневой конкуренции.
Баланс положительных и отрицательных воздействий может зависеть от жизненной стадии растений. Например, McAuliffe (1988) показал, что молодые особи Larrea tridentata — типичного пустынного кустарника — ассоциированы с отмершими особями Ambrosia dumosa, ключевого
6. Положительные отношения между растениями
257
вида для приживания всходов Larrea. Возможно, циклические смены растений-нянек с другими видами очень широко распространены в природе.
Один и тот же вид растений может по-разному воздействовать даже на близкородственные растения. Например, для переходной зоны между кустарниковой степью и горными лесами на массиве Great Basin в США экспериментально было показано, что Artemisia tridentata конкурирует за воду с Pinus ponderosa и Pinus Jeffery i, но улучшает приживаемость молодых растений Pinus monophylla, которые были приурочены к зарослям полыни. Здесь удаление кустарника способствовало увеличению выживаемости Pinus ponderosa, но уменьшало выживаемость Р. топо-phylla (Callaway et al., 1996).
Выраженность конкуренции и положительных отношений зависит от суровости условий среды. Bertness и Callaway (1994) предположили, что роль положительных отношений возрастает, а конкуренции уменьшается с возрастанием неблагоприятности среды. Так, Belsky (1994) сообщает о более сильном конкурентном воздействии деревьев в саваннах на растущие под их пологом злаки во влажные годы и большей роли благоприятствования в сухие. Благоприятствование часто наблюдается на отдельных этапах сукцессий, когда, например, происходит смена травяной стадии древесной. Сумах (Rhus copallina) часто выступает пионерным видом деревьев при вторжении на сухие луга, а под его пологом лучше развиваются другие виды деревьев. Однако такое благоприятствование не является облигатным, оно лишь ускоряет сукцессию, которая в противном случае происходит медленнее.
В эволюционном аспекте следствием наличия положительных отношений между растениями могут быть (Brooker, Callaghan, 1998): 1) уменьшение дальности распространения диаспор (выгоднее быть рядом), 2) компактная морфология растений, 3) ограниченный отклик на улучшение условий среды, 4) увеличение интенсивности микоризных связей между растениями, 5) увеличение тенденций к клональному росту.
Глава
Состав и структура фитоценозов
1 • Состав фитоценозов
В этом разделе мы рассмотрим несосудистые компоненты растительных сообществ (цианеи, водоросли, лишайники и мохообразные), а также флористическую насыщенность сообществ и механизмы ее поддержания.
1.1 • Свободноживущие цианеи и водоросли в составе наземных экосистем
Цианеи и водоросли встречаются в составе практически всех наземных фитоценозов. Они заселяют поверхность более крупных растений и других субстратов, а также обитают в почве и на ее поверхности. Насчитывается несколько сотен видов наземных цианей и водорослей (табл. 6.1). Наиболее часто в почвах России и сопредельных стран встречается более 50 видов цианей и водорослей из трех отделов — зеленые (Chlorophyta), желто-зеленые (Xanthophyta) и диатомеи (Bacillariophyta). Именно эти группы образуют ведущий комплекс видов наземных водорослей.
Наземная альгофлора весьма своеобразна. Лишь немногие виды водорослей развиваются одновременно в водоемах и в почве. Так, по данным Э. А. Штины (1997), из 1140 видов, найденных в почвах наиболее изученной Кировской области, только 12 % видов оказались общими для почв и водоемов.
Таблица в.1 • Общее число видов почвенных и эпифитных водорослей и цианей для территории бывшего СССР (по: Штина, 1991, 1998)
Группа Всего отмечено Встречено более трех раз Ведущий комплекс (отмечено более 100 раз)
Цианеи 409 222 28
Chlorophyta 631 303 15
Xanthophyta 185 115 7
Bacillariophyta 256 78 4
Euglenophyta 19 2 -
Chrysophyta 3 - -
Pyrrophyta 4 - -
Rhodophyta 1 - -
Всего 1508 720 54
260
Глава 6. Состав и структура фитоценозов
Таблица 6.2 • Биомасса и продукция водорослей в наземных фитоценозах (с. м. — сухая масса, в остальных случаях — живая, или сырая, масса)
Сообщество Биомасса (сырая, кг/га) Продукция (кг/га) Источник
Тундры 32-227 до 1000 Перминова, 1980
Темнохвойные леса 7-30 - Штина, 1959
Низинные луга 8-20 220-420 Штина, 1959; Панкратова, 1981
Пашня 50-1500 - Broady, 1979
Пустыня (Негев) до 1970 1470 (с. м.) Shachak, Steinberger, 1980
Такыры 500-1400 (с.м.) - Новичкова-Иванова, 1980
Высокогорья (Памир) до 72 (с. м.) 750 (с. м.) Базова, 1978
Высокогорья (Кавказ) 12-16 - Лейнсоо и др., 1987
Цианеи и водоросли входят в состав практически всех изученных ценозов. Их обычная численность составляет 1000-10 000 000 клеток на 1 г почвы. Биомасса этой группы уступает таковой для сосудистых растений, но в пустынях достигает высоких величин (табл. 6.2). Только для водорослей из наземных растений отношение биомассы к годичной продукции значительно меньше 1, составляя 0,12-0,6, в отдельных случаях продукция может превышать биомассу в 35 раз. Это связано с коротким жизненным циклом водорослей и быстрым оборотом поколений. Время оборота биомассы в благоприятных естественных условиях составляет от 1-3 до 6-7 дней. Необходимо отметить, что значительную долю продукции наземных водорослей составляют прижизненные выделения (до 89 % ассимилированного углерода), что подчеркивает их большое значение в почвообразовании. Водоросли играют большую роль в закреплении почв. Длина нитей водорослей составила для почв Гис-сарского хребта 88 м/см3 почвы, на песках — 65 м/см3 (Штина, 1991). Наземные цианеи могут вносить также существенный вклад в фиксацию атмосферного азота (10-25 кгN/ra в год, см. п. 3 гл. 2).
Распределение по почвенному профилю. Цианеи и почвенные водоросли населяют преимущественно поверхность почвы, но многие могут проникать на значительную глубину, переходя на гетеротрофный способ питания. Довольно много водорослей концентрируются в ризосфере, где
1. Состав фитоценозов
261
а) д)
Рис. 6.1 • Схема жизненных форм почвенных водорослей (по: Штина, Голлербах, 1976). Жизненные формы (в скобках — типичные представители): а — Ch-форма (Chlorococcum, Chlorella); б — С-форма (Cylindrospermum); в — Х-форма (Xanthophyta); г — В-форма (Bacillariophyta); д — P-форма (Phormodium, Plectonema); е — М-форма (Microcoleus, Schizothrix, Hydrocoleus); ж — Н-форма (Hormidium, Ulothrix, Tribonema); з — N-форма (Nostoc); и — V-форма (Vaucheria)
их численность в 1,5-4,6 раз выше, чем в окружающей почве вдали от корней. Такой ризосферный эффект на светло-каштановой почве усиливался от весны к осени по мере снижения влажности почвы (Алексахи-на, 1981). По данным Э. А. Штины и М. М. Голлербаха (1976), наибольшая глубина проникновения живых водорослей составила: в тундровой почве — 10 см, в подзолистой — 150 см, в торфяно-болотной — 80 см, в черноземе — 150 см. Для фотосинтеза водорослей характерна низкая компенсационная точка по освещенности (около 0,04-0,1 % от полного света). Такая освещенность наблюдается в разных типах почв в зависимости от их гранулометрического состава на глубине от 1 мм до 2 см (Штина, Голлербах, 1976).
Основная физиологическая особенность наземных водорослей и цианей — способность к высыханию и длительному пребыванию в сухом
262
Глава 6. Состав и структура фитоценозов
Рис. 6.2 • Соотношение жизненных форм водорослей в различных фитоценозах (по: Штина, Голлербах, 1976): а — дерново-осоково-моховая пятнистая тундра, Таймыр; б — ельник на подзолистой почве, Московская область: в — разнотравно-злаковая степь на мощном черноземе, Курская область: г — полынно-типчаковая степь на светло-каштановой почве, Центральный Казахстан; д — горная степь в Иссык-Кульской котловине, Тянь-Шань. Цифры над столбцами указывают число отмеченных видов
состоянии. Например, для Nostoc commune показано восстановление фотосинтетической активности после 107-летнего хранения в гербарии (Cameron, 1962). Наименее засухоустойчивы диатомеи и желто-зеленые
1. Состав фитоценозов
263
водоросли. Для почвенных водорослей отмечается также высокая морозостойкость.
В зависимости от характера колоний, их положения в почве и наличия слизистых выделений различают несколько жизненных форм почвенных водорослей и цианей, названных по первым буквам типичных родов данной жизненной формы (рис. 6.1). Соотношение между жизненными формами специфично для почв разных природных зон (рис. 6.2).
Развитие водорослей обычно отрицательно связано с сомкнутостью покрова сосудистых растений, поскольку водоросли занимают преимущественно открытые участки почвы. Например, в почвах зональных степей при снижении проективного покрытия высших растений с севера на юг биомасса водорослей увеличивается в 8-10 раз, при этом усиливается роль цианей, образующих напочвенные разрастания (Кузяхметов, 1991). С другой стороны, в экспериментах J. Р. Grime (1998) с выращиванием травянистых растений в микрокосме было показано, что водорослевая пленка на поверхности почвы подавляет прорастание мелкосемянных видов, но позволяет прорастать и приживаться всходам крупносемянных злаков.
Водоросли являются пионерами зарастания обнаженных субстратов. При этом наиболее ранние группировки чаще образуют одноклеточные зеленые водоросли, а не цианеи (Штина, 1985). Многие водоросли развиваются как эпифиты, особенно на мохообразных. Обычны азотфиксирую-щие ассоциации цианей со мхами. Местами пышного развития почвенных водорослей служат также копролиты дождевых червей и кротовины.
Отдельные группы водорослей существенно различаются по экологическим свойствам. Так, цианеи лучше представлены на нейтральных и слабощелочных почвах (особенно в аридных условиях), в то время как зеленые водоросли более характерны для кислых почв. Интересно отметить, что желто-зеленые водоросли могут служить индикаторами ненарушенных и незагрязненных почв, так как эта группа водорослей первой исчезает при различных формах антропогенных нарушений почвенного покрова (Штина, 1990). В отдельных случаях водоросли выступают индикаторами микроусловий в почвах. Например, есть виды, специфичные для муравейников в фисташковых сообществах в Бадхызе (Новичкова-Иванова, 1991).
Почвенные водоросли потребляются многими группами беспозвоночных, такими как амебы, инфузории, клещи, коллемболы, нематоды, дождевые черви, личинки насекомых, энхитреиды, моллюски. Искусственное внесение водорослей в почву сопровождается резким увеличением численности беспозвоночных с коротким циклом развития и снижением численности водорослей (Некрасова, 1980).
264
Глава 6. Состав и структура фитоценозов
1.2 • Лишайники
Как симбиотические организмы, лишайники обладают рядом специфических черт, позволяющих им играть особую роль в составе современных растительных сообществ. В связи с большей долей грибного компонента в талломе большинства лишайников эффективность их фотосинтеза на 1-2 порядка ниже, чем у цветковых растений в тех же условиях. Например, в высокогорьях Alectoria ochroleuca фиксирует около 0,5 мг СО2 г/час, в то время как травянистые растения там же фиксируют 10-40 мг СО2 г/час. В целом у лишайников в оптимальных условиях интенсивность фотосинтеза варьирует в пределах 0,2-2 мг СО2 г/час (Лархер, 1978). У многих лишайников имеются механизмы концентрирования СО2, что связано с низкой компенсационной точкой по СО2 у этой группы организмов (Palmqvist, 2000). Фотобионт составляет обычно 5-10% объема таллома, поэтому большие потери на дыхание не дают возможности иметь высокие показатели нетто-фотосинтеза. Отношение максимального фотосинтеза к дыханию в талломе Peltigera в целом составляет около 1,6, а у изолированного фотобионта этого лишайника — около 8,0 (Hale, 1983).
Фотосинтез лишайников зависит от увлажнения их таллома. У видов с цианеями для восстановления фотосинтеза после иссушения необходима жидкая вода, а у видов с зелеными водорослями — достаточно парообразной при высокой относительной влажности воздуха. Оптимум фотосинтеза наблюдается при неполном насыщении водой, так как полное насыщение затрудняет газообмен. Например, для 22 видов лишайников умеренных дождевых лесов Новой Зеландии (гл. об. Pseudo-cyphellaria, Sticta) отмечено максимальное содержание воды (на сухой вес) 357-3360%, оптимальная для фотосинтеза влажность составляла 86-1300% (Lange et al, 1993).
Температурные границы фотосинтеза лишайников обычно сдвинуты в область более низких значений, чем у сосудистых растений. Так, верхняя компенсационная точка (значение фактора, при котором фотосинтез уравновешивает дыхание) у ряда видов лишайников около +35 °C, в то время как у арктических лишайников — около +20 °C (Stereo-caulon alpinum +22 °C, Parmelia encausta +18 °C) (Lange et al, 1986). Оптимальная температура фотосинтеза арктических видов лишайников +5 + +10 °C, а нижняя компенсационная точка у многих видов лежит существенно ниже 0°С (Stereocaulon alpinum —24 °C, Cetraria nivalis —20 °C, Nephroma arcticum, Cladina stellaris —5~,—10 °C). В самых холодных областях Антарктиды со среднегодовой температурой ниже -20 °C встречается лишайник Rinodina endophragmia — один из наиболее холодостойких видов нашей планеты.
1. Состав фитоценозов
265
I.00 0.75 0.50 +0.25 О -0.25
26-27 сентября 1967
интенсивность обмена СО2
Баланс за день: Ассимиляция — дыхание 1,32-0,78=0,54 мг СОг/г сух.м
Рис. 6.3 • Углеродный баланс пустынных лишайников (фотосинтез — 1,32 мг/г в сутки, дыхание — 0,78 мг/г в сутки, остаток (прирост) — 0,54 мг/г в сутки) и содержание воды в их талломах на примере Ramalina maciformis в пустыне Негев (по: Lange, 1970)
Дыхание еще более устойчиво к низким температурам и отмечено у Thamnolia vermicularis, Dactilina arctica при -26 °C. Такая морозостойкость связана с тем, что вода замерзает между гифами грибов и лед не повреждает клетки. Поэтому лишайники могут долго сохранять жизнеспособность при низких температурах: например, Alectoria ochroleuca 3,5 года хранилась при -60 °C и затем полностью восстановила свой фотосинтез (Larson, 1978). Многие виды выносят 6 часов при -196 °C (Lange, Карреп, 1972). С другой стороны, талломы многих видов повреждаются при замерзании во влажном состоянии. В связи с устойчивостью фотосинтеза к низким температурам у лишайников возможен существенный прирост в зимнее время. По данным А. С. Са-лазкина (1937), примерно 20% прироста кормовых лишайников Арктики наблюдается под снегом (по данным Tranquillini, 1957, снег пропускает около 30 % света при мощности 5 см и 5 % света при мощности 20 см).
Мощность снега существенно влияет на рост и выживаемость лишайников. В Арктике и высокогорьях эпилитные лишайники рода Rhizocar-роп поселяются на камнях и выживают лишь при небольшой мощности снежного покрова. Эксперименты по переносу камней в разные режимы
266
Глава 6. Состав и структура фитоценозов
Рис. 6.4 • Пустыня Намиб: лишайниковая приморская пустыня в окрестностях Alexander Вау (ЮАР) (Theloschistes capensis s. I.)
снежности в альпийском поясе Колорадо показали (Benedict, 1990), что виды типового подрода Rhizocarpon отмирали в течение 5-8 лет при длительности залегания снега более 40-43 недель. Это было вызвано исчерпанием у них запасов углеводов в связи с длительным дыханием под снегом при увлажнении талой водой. Для эпифитов снег также имеет большое значение. Показано, например, что нижняя граница распространения Parmelia olivacea по стволу деревьев тесно связана с локальной высотой снежного покрова. В экспериментах талломы этого вида не выживали под снегом более 2 месяцев (Sonesson et al., 1994).
В аридных условиях лишайники могут существовать за счет кратковременного увлажнения росой в предутренние часы (рис. 6.3). Лишайники в приморский пустынях в Южной Африке и Намибии получают большую часть влаги за счет туманов с океана (рис. 6.4).
В связи с низкой эффективностью фотосинтеза годичный прирост лишайников характеризуется низкими величинами (Hale, 1983). В линейном выражении он составляет от долей до нескольких десятков мм в год (Rhizocarpon geographicum — 0,25-0,5 мм/год, Cetraria pinas-
1. Состав фитоценозов
267
tri — 1,15 мм/год, Lobaria pulmonaria — 4,8 мм/год, Peltigera spp. — 10-30 мм/год, максимальная величина у Ramalina menziesii — 90 мм/год). Годичная продукция лишайников обычно составляет первые проценты от их биомассы. Например, в Арктике у Dactylina arctica — до 8 %, в альпийских сообществах Кавказа у Cetraria islandica — 1-2%. Максимальный прирост отмечен у Lobaria oregana, L. pulmonaria и Ramalina menziesii — до 30-50% в год от биомассы (Honegger, 2001). Доминант лесотундровых сообществ Cladina stellaris в оптимальных условиях имеет прирост около 5 мм/год, или 35 г/м (Chernov, 1985), максимальные величины — до 0,17 г/г в год, хотя средний прирост значительно ниже (den Herden et al., 2003). Поэтому на восстановление сбитых оленьих пастбищ требуется от 30 до 50 лет.
Биохимия лишайников также своеобразна. Зеленые фотобионты передают микобионту продукты фотосинтеза в виде многоатомных спиртов, а цианеи — в виде глюкозы. Эти вещества быстро переводятся в маннитол и таким образом становятся недоступными для обратного потребления фотобионтом. Маннитол может служить и долговременным запасным веществом (Palmqvist, 2000). Но среди запасных веществ наиболее распространен лишайниковый крахмал — лихенин. Его особенно много в кустистых лишайниках Cetraria islandica и Alectoria ochroleuca (45-50 % сухой массы). Пищеварительный тракт северных оленей переваривает лихенин примерно на 78 % за счет бактериальных ферментов (Андреев, 1934).
Особенностью многих видов лишайников является синтез так называемых лишайниковых кислот — низкомолекулярных органических кислот, присутствующих практически только у лишайников. Всего их известно около 250, из них 75 свойственны исключительно лишайникам и не образуются грибом в отсутствии водоросли. Их содержание составляет 0,1-2% сухой массы, но в отдельных случаях может быть существенно выше. Так, у Parmelia tinctorum содержание леканоровой кислоты может достигать 36 % сухой массы. Лишайниковые кислоты могут выполнять различные функции. Многие из них являются антибиотиками и используются в медицине, как, например, Cetraria islandica — от простудных заболеваний. Согласно Vartia (1973), более 50% лишайниковых веществ обладает антимикробными свойствами. Видимо, эти вещества обеспечивают долголетие талломов (сходные грибы без фотобионтов живут значительно меньше). Под эпигейными (напочвенными) лишайниками может происходить уменьшение численности бактерий, что ведет к уменьшению скорости разложения органики, накоплению и сохранению гумуса в почве. Лишайниковые кислоты также способствуют защите талломов от поедания животными, что может быть связано
268
Глава 6. Состав и структура фитоценозов
с их горьким вкусом и детеррентными свойствами. Это было показано в опытах с улиткой Helix hortensis (Fraenkel, 1959; Gerson, Seaward, 1977), которая не поедает некоторые виды лишайников с высоким содержанием лишайниковых кислот, но активно поедает их после обработки слабым раствором соды. Для слизня Pallifera varia (Lawrey, 1980, 1983) показано пищевое предпочтение видов лишайников с низкой питательной ценностью, но и низким содержанием лишайниковых кислот. Кроме того, лишайниковые кислоты могут являться агентами выветривания горных пород, образуя комплексные связи с катионами.
Основными источниками ЭМП для лишайников выступают атмосферные осадки, пыль и субстрат. Изучение распространения Usnea longissima по ветвям и деревьям ели в Норвегии показало, что лишайники предпочтительно поселяются на ветвях, которые богаче ЭМП даже в пределах одной кроны (Gauslaa et al., 1998). Связанный азот лишайники могут получать за счет азотфиксации цианей или других азотфиксирующих бактерий. Фиксация азота при низких температурах незначительна, она максимальна при 15-20 °C. В целом вклад лишайников в азотный баланс экосистем не очень велик: 0,1-10 кг N/ra в год в северных экосистемах, эпифиты в лесах Орегона (Lobaria огедапа) фиксируют до 6 кг N/ra, в тропических лесах виды Sticta, Leptogium, Pannaria — 1,5-8 кг N/ra в год (Forman, 1975). Особую группу составляют нитрофильные лишайники, живущие в условиях значительного поступления связанного азота (птичьи базары, верхние части камней — рис. 6.5).
Лишайники обладают высокой адсорбционной способностью, особенно по отношению к катионам. Поэтому они могут накапливать в своих талломах значительное количество тяжелых металлов в загрязненных районах. Например, близ металлургического комбината в Приморье виды рода Cladonia содержали в золе до 8,8 % свинца, 0,02-0,03 % кадмия и 0,64 % цинка. Лишайники аккумулируют также значительное количество радионуклидов, передаваемых в дальнейшем по пищевым цепям. Оленеводы в тундре, регулярно питающиеся мясом северных оленей, получают в год в 100-1000 раз большие дозы радиации, чем другие люди, что в 2-5 раз превышает нормы, установленные на атомных станциях (Криволуцкий, Покаржевский, 1996). С другой стороны, лишайники очень чувствительны к газообразному загрязнению атмосферы, главным образом SO2, HF и окислами азота. На этом свойстве основан метод лихеноиндикации, когда по видовому составу лишайниковых сообществ (преимущественно эпифитов в городских парках) определяют кумулятивный уровень загрязнения атмосферы.
По приуроченности к субстрату выделяют напочвенные (эпигей-ные) лишайники, эпифитные (эпифиты в узком смысле — на стволах
1. Состав фитоценозов
269
Рис. 6.5 • Сообщества нитрофильных лишайников на верхних частях камней, часто используемых птицами как присады, в альпийском поясе Тебердинского заповедника (справа — Lecanora rubina крупным планом). Наибольшее поступление азота с экскрементами характерно для верхушки камня, где лишайники отсутствуют
растений, а также эпифиллы — на листьях и эпиксилы — на обнаженной древесине), а также наскальные (эпилитные), которые могут развиваться на разнообразных минеральных субстратах, включая метеориты. Например, в Гренландии на метеоритах отмечены представители 10 родов (Xanthoria, Lecanora, Lecidea и др.) (Hansen, Graff-Petersen, 1986).
В фитоценозах лишайники часто имеют сложные отношения с мохообразными. Ряд видов может развиваться на мхах и подавлять их рост. Например, Jahns (1982) показал динамику отношений между Baeomyces rufus и мхами Dicranella. Мелкие соредии Baeomyces поселяются между стебельками мхов, а затем образуют сплошное слоевище. Под его покровом мхи отмирают, а потом и все слоевище превращается в соредиозную массу, а на свободном месте вновь поселяются мхи. С другой стороны, эксперименты В. С. Ипатова и Т.Н.Тарховой (1983) по взаимным пересадкам в сосняках куртин Cladonia arbuscula и Pleurozium schreberi показали высокую устойчи-
270
Глава 6. Состав и структура фитоценозов
Рис.в.в • Растительные сообщества с высоким участием лишайников: а — лишайниковая арктическая тундра, окрестности Тикси, Якутия; б — альпийская лишайниковая пустошь, Тебердинский заповедник, Карачаево-Черкесская Республика; в — лиственничная лесотундра в верховьях Колымы, Магаданская область; г — Usnea spp. на пихте (Abies delavayi), горные леса провинции Юннань, Китай
вость исходных пятен мхов и лишайников к внедрению других растений. При подсадке лишайников в моховые куртинки они быстро зарастали подушками мхов. Подсаженные же куртинки мхов в пятна лишайников погибали в них от высокой температуры (там она была на 10-20 °C выше, чем в исходных моховых куртинках, см. п. 5.2 гл. 5).
Роль лишайников наиболее значительна в тундрах, северной тайге, аридных областях (рис. 6.4) и высокогорьях, значительного участия
1. Состав фитоценозов
271
достигают лишайники-эпифиты во влажных лесах (рис. 6.6). Биомасса лишайников в сообществах с их участием варьирует от нескольких грамм до 1,5 кг/м2 Наибольшие величины отмечены для кобрезиевого луга на Тянь-Шане — 1,4 кг/м2 и в лесотундре — 1,3 кг/м2, типичные величины для лишайниковых сообществ составляют 200-800 г/м2 (2-8 т/га). Биомасса эпифитов в лесах оценивается величинами 70-300 г/м2
Для сравнительной оценки видового разнообразия лишайников часто используют лишайниковый коэффициент — отношение числа видов лишайников к числу видов сосудистых растений на той же территории. В тропиках он составляет около 0,1, для Эстонии — 0,54, для Таймыра — 4,0, а для Антарктиды превышает 100.
Лишайники служат пищей для ряда беспозвоночных и позвоночных животных. Гусеницы многих видов бабочек подсемейства лишайниц (Lithosiinae) питаются лишайниками. В рационе северного оленя обычно 20-40 % составляют лишайники (главным образом Cladonia, Cetraria, Alectoria). В сыром состоянии лишайники рода Cladonia издают сильный запах, который северный олень чувствует из-под снега высотой 40-60 см (Формозов, 1990). Другие копытные также используют лишайники, особенно в зимний период, например, для кавказского тура (Capra caucasica) большую роль в зимнем питании играет Cetraria islandica.
Скорость разложения отмерших частей талломов лишайников в большинстве случаев значительно ниже, чем опада сосудистых растений, что может быть обусловлено содержанием защитных лишайниковых кислот. Так, в сосняках Эстонии потери в весе опада кустистых лишайников за 2 года составили 48-63 %, а в альпийских сообществах Северного Кавказа — 14-28% (Паринкина, Пийн, 1989; Воронина и др., 1986). Но относительная скорость разложения больше, чем прироста, поэтому отмершие части составляют незначительный процент от общей фитомассы лишайников. По данным F. Е. Wielgolaski (1978), основанным на определении ассимиляционных способностей разных частей таллома, отмершие части составляют лишь около 2 % таллома кустистых лишайников.
Велика роль лишайников как пионеров заселения скальных субстратов. На карбонатных скалах происходит медленное растворение поверхности скалы за счет кислых продуктов обмена. Ризины эпилитных лишайников могут проникать в известняк на глубину от 0,3 до 16 мм у разных видов. На гранитах ризины проникают по слюдяным кристаллам, они не только оказывают механическое воздействие на породу при высыхании и набухании, но и являются агентами биогеохимического выветривания. Это выветривание происходит под воздействием углекислоты, щавелевой кислоты, лишайниковых кислот и ведет к образованию вто
272
Глава 6. Состав и структура фитоценозов
ричных глинистых минералов. Кроме того, в процессе жизнедеятельности лишайники аккумулируют значительные количества биогенных элементов (N, Р, К, S) из породы, осадков и пыли. При поселении на мелкоземе лишайников может происходить подкисление субстрата и консервация органики в нем. Так, по данным О. М. Паринкиной и Т. Пийна (1978), под пятнами лишайников по сравнению с голым грунтом уменьшается pH (на 0,2-0,9), в 2-3 раза увеличивается содержание С и N.
Широко распространенные лишайниковые сосняки часто являются элементами циклического пироклимакса, так как их поддержание возможно лишь при периодическом воздействии огня или выпаса оленей. Сплошной лишайниковый покров препятствует приживанию всходов сосны. Только при его разрушении наблюдается возобновление этой породы. Поддержание лишайниковых сосняков может быть связано с невысокой интенсивностью выпаса. Так, по экспериментальным данным, полученным в Финляндии (Vare et al., 1995), отсутствие выпаса ведет к полному господству Cladina stellaris, а выпас оленей увеличивает участие мохообразных (особенно Pleurozium schreberi, Dicranum spp.).
Видовое богатство эпифитных лишайников может служить индикатором состояния лесов и степени их нарушенности. Так, сравнение видового состава и разнообразия эпифитных лишайников на елях в Финляндии и Швеции показало существенное снижение их разнообразия в эксплуатируемых лесах по сравнению со старовозрастными (Esseen et al., 1996; Kuusinen, Siitonen, 1998). Состав эпифитных группировок используется в этих странах для выявления наиболее ценных старовозрастных лесов с целью их охраны.
1.3 • Мохообразные
Мохообразные (антоцеротовые, печеночники и мхи) включают около 20 000 видов растений, существенно отличающихся по своим экологофизиологическим свойствам и роли в наземных сообществах от сосудистых растений.
Фотосинтез многих мохообразных может происходить при более низкой освещенности, чем у сосудистых растений; этим обусловлено их высокое участие в нижних ярусах бореальных лесов. У некоторых пещерных видов нижняя компенсационная точка фотосинтеза по освещенности соответствует светимости луны. Водные бриофиты, в отличие от сосудистых растений, эффективно используют СО2, растворенный в воде, а не бикарбонат-ион, что дает им возможность успешно развиваться при затенении и в условиях быстрого течения, с низким содержанием карбонатов. С этим может быть связано пышное развитие водных
1. Состав фитоценозов
273
Рис. 6.7 • Растительные сообщества с высоким участием мохообразных: а — субарктическое сфагновое болото, Абиско, Швеция; б — ельник-зеленомошник, Приокско-Террасный заповедник, Московская область; в — дно холодных ручьев в альпийском поясе, Азгек, Карачаево-Черкесская Республика; г — мохово-дриадовая тундра, окрестности Тикси, Якутия
и околоводных мхов в холодных горных ручьях, текущих по силикатным горным породам (рис. 6.7 в).
Большинство мохообразных способны высыхать и длительное время пребывать в воздушно-сухом состоянии. При намокании они быстро (за 1-2 часа) восстанавливают процессы фотосинтеза и дыхания, в отличие от сосудистых пойкилогидрических растений, которым для восстановления фотосинтеза требуется более суток (Proctor, Tuba, 2003).
274
Глава 6. Состав и структура фитоценозов
Элементы минерального питания мохообразные получают из осад-<ов, стекающих со стволов вод, лишь незначительную часть — из почвы, ще ризоиды служат преимущественно органами закрепления. Листья мохообразных обладают высокой катионообменной способностью, поглодал катионы металлов из растворов и обменивая их на ионы водорода. Эффективность адсорбции катионов листьями мхов примерно в 100 раз выше, чем анионов. С этим связано подкисление вод, прошедших через ихи (болотные воды), а также почвы под куртинками мхов. С другой зтороны, в случае выпадения сильно кислых осадков катионы вымыва-отся из листьев, что приводит к гибели многих видов мохообразных, сказавшихся неустойчивыми к этому виду антропогенных воздействий.
Роль мохообразных особенно высока в тундрах, болотах, бореальных песах, в некоторых пустынях (рис. 6.7). Болота, наряду с некоторыми другими экосистемами, являются источником кислорода в атмосфере [Вставка X]. 3 тропических лесах насчитывается 1500-2000 видов мхов и столько же печеночников, при этом низменные леса бедны видами, а горные (на высотах 1500-3000 м) очень богаты. Биомасса эпифитных мохообразных в горных песах достигает 11 т/га, в низменных — 1,7 т/га. В горных лесах эпифитные мохообразные представляют важный средообразующий элемент, так как, пропитанные дождевой водой, они в значительной степени обуславливают постоянство высокой влажности воздуха (Gradstein, Poes, 1989).
Вставка X. Какие природные экосистемы выделяют в атмосферу больше кислорода, чем поглощают?
Растения в экосистемах образуют органическое вещество и выделяют кислород в соответствии с суммарным уравнением фотосинтеза:
пСО2 + пН2О = (СН2О)П ф +nO21 •
В процессе дыхания самих растений и разложения органики гетеротрофами органика окисляется, и на ее полное окисление затрачивается ровно такое же количество кислорода, какое было выделено в процессе фотосинтеза. Поэтому все сбалансированные экосистемы не выделяют дополнительный кислород в атмосферу. Лишь те сообщества, в которых идет прогрессирующее накопление органики, обогащают атмосферу земли кислородом. К ним относятся системы с отложением органики на дно водоемов (океан, континентальные озера и т. п.) или в торфяной залежи (болота), а также молодые растущие леса. Далеко не все экваториальные леса выделяют кислород в атмосферу. Большая роль амазонских лесов в обогащении атмосферы кислородом связана не столько с их высокой продукцией, сколько с выносом Амазонкой и ее притоками большого количества органических веществ в океан с их последующей аккумуляцией на дне.
2. Флористическая насыщенность и неполночленность фитоценозов
275
Моховой покров оказывает значительное влияние на сосудистые растения, и особенно на прорастание семян и приживание всходов. Сохранение семян и приживание всходов различаются в моховых куртинках и вне их. Более успешное появление всходов в колониях бриофитов отмечено для дюн (Bonnot, 1971), лесов (Nakamura, 1986), пустынь (St. Clair et al., 1985). Менее успешное (по сравнению с открытыми местами) семенное возобновление наблюдалось на лугах, в лесах и пустошах. Интересно отметить, что прорастание семян Rhododendron ponticum в лесах Ирландии происходило только в моховых куртинках определенной структуры, оно не наблюдалось как в отсутствии бриофитов, так и при слишком толстом моховом покрове (Gross, 1981). Напротив, полевые эксперименты по удалению мохового покрова на меловых лугах приводили к увеличению численности всходов большинства видов растений (During, van Tooren, 1990).
С другой стороны, сосудистые растения могут оказывать положительное влияние на рост мхов. Так, в двухлетнем эксперименте с выращиванием мохообразных под пологом травянистых растений было показано, что покров мхов увеличивался с увеличением покрова сосудистых растений за счет создания лучшего микроклимата. Влияние было видоспецифичным. Festuca pratensis благоприятно влияла на Rhytidiadel-phus squarrosus и Brachytecium rutabulum, Trifolium pratense — только на Brachythecium, в то время как Prunella не оказывала влияния на мхи (Ingerpuu et al., 2005).
Мохообразные обычно плохо поедаются фитофагами в связи с низкой питательной ценностью. Однако для норвежского лемминга зеленые мхи часто составляют основу рациона (Oksanen, Oksanen, 1981).
2 • Флористическая насыщенность и флористическая неполночленность фитоценозов
Флористической насыщенностью называют число видов на единицу площади, в то время как под флористическим богатством обычно понимают общее число видов в пределах контура растительного сообщества. Число видов растет с увеличением исследуемой площади, а зависимость между этими показателями обычно описывается одним из двух уравнений:
S = с х А\
S = а 4- b х In (А),
(1)
(2)
276
Глава 6. Состав и структура фитоценозов
Таблица 6.3 • Примеры флористической насыщенности различных растительных сообществ (среднее число видов сосудистых растений на 1 м2 и 100 м2)
Страна, регион Сообщество 1 м2 100 м2
США (Kilburn, 1966) прерии 12,4 39,2
дубовый лес 16,3 76,4
Великобритания (Hopkins, 1955) луг 17,9 44,3
буковый лес 4,2 18,8
сосняк 10,3 31,0
Новая Зеландия (Wilson, Sykes, 1988) смешанный лес - 24,4
выжигаемый луг - 18,0
засоленные марши - 6,2
Китай (Tsuyuzaki et al., 1990) высокогорные болота 3,4-9,1 -
Голландия луг на дюнах 25,4 -
Россия, Кавказ (Onipchenko, Semenova, 1995) пестроовсяницевый луг 24,4 56,8
альпийское болото 9,9 23,0
Швейцария, Альпы (Onipchenko, Semenova, 1995) Seslerietum (альпийский луг) 26,8 50,6
альпийское болото 11,1 23,2
где S — число видов, А — площадь, а, 6, с, z — регрессионные параметры. Уравнение (1) было предложено Аррениусом (Arrhenius, 1921), а уравнение (2) — Глизоном (Gleason, 1922). Параметры с и а показывают флористическую насыщенность (число видов на 1 м2), а параметры z и b — скорость увеличения числа видов с ростом площади (крутизну кривой).
Многие исследователи рассчитывали эти параметры в масштабе от квадратных метров до континентов и изучали их зависимость от свойств сообществ и географического положения. Например, для больших флористических районов Северной Америки изучено изменение параметра z. Было показано уменьшение этого коэффициента при увеличении широты местности от тропиков к тундрам, а также увеличение z с ростом среднегодовой температуры и потенциальной эвапотранспирации (Qian et al., 2007).
Bartha и Ittzes (2001) проанализировали влияние отдельных факторов на форму кривой «виды—площадь». Число видов на большой пло
2. Флористическая насыщенность и неполночленность фитоценозов
277
щади (правая часть кривой) соответствует видовому пулу сообщества (флористическому богатству), число видов в левой части кривой (на маленькой площадке или единичной площади) зависит от плотности особей (или размеров растений), а прогиб средней части — от пространственной структуры (расположения особей — случайного, равномерного или агрегированного) и относительного участия видов растений.
Флористическая насыщенность изменяется от нескольких до многих десятков видов на квадратный метр в разных сообществах (табл. 6.3). Она зависит от многих факторов. На островах число видов на единицу площади ниже, чем на материке (островной эффект, см. ниже). Если размеры растений более крупные, число видов на небольших площадках обычно невелико. Сообщества, занимающие крайние позиции в диапазоне того или иного фактора, обычно отличаются низкой флористической насыщенностью, как, например, высокогорные болота в Китае, на Кавказе и в Альпах, имеющие сходные низкие показатели числа видов на квадратный метр (табл. 6.3). Как отмечалось выше (п. 3.1 гл. 5), наиболее флористически богатые сообщества земного шара — финбош в Южной Африке и кустарниковые пустоши Австралии — развиваются на бедных почвах и характеризуются большим возрастом. Однако их высокая флористическая насыщенность проявляется на больших площадях (от сотен квадратных метров до нескольких гектаров) (рис. 6.8).
В состав естественных растительных сообществ входят далеко не все виды, которые потенциально могут произрастать в тех или иных экологических условиях, из-за ограниченности поступления диаспор и воздействия уже представленных видов. В связи с этим Л. Г. Раменский (1924) ввел представления о разной степени флористической неполночленности фитоценозов. Он выделял:
1) абсолютно полночленные сообщества — сообщества, в состав которых входят все виды, способные в них произрастать (в значительной степени абстрактная категория);
2) туземно полночленные сообщества, включающие все виды местной флоры, способные в них произрастать;
3) практически полночленные сообщества — сообщества, в которые могут внедряться виды туземной флоры, но они не могут достигать там большого участия, т. е. от внедрения структура сообществ существенно не изменяется;
4) явно неполночленные сообщества — сообщества, восприимчивые к внедрению видов, которые могут достигать большого участия (доминировать), таким образом принципиально изменяя структуру исходного сообщества.
278
Глава 6. Состав и структура фитоценозов
Рис. 6.8 • Наиболее
флористически богатые сообщества:
слева — кустарниковые пустоши в Западной Австралии (доминанты Banksia speciosa, Conospermum sp. — фото К. Е. Werger, с любезного разрешения автора), справа — южноафриканский финбош (национальный парк «Западный берег»)
Флористическая насыщенность сообществ зависит от общего «пула» видов данной территории, способных существовать в условиях этого сообщества. Специальные исследования в Эстонии (Partel et al., 1996) показали, что число видов, присутствующих в сообществе, положительно
скоррелировано с потенциальным числом видов флоры региона, способных произрастать в данных условиях (эта способность была определена дологическим шкалам Элленберга). Флористическая насыщенность
2. Флористическая насыщенность и неполночленность фитоценозов
279
50
□0 40
150
б)
—1—
100
150
•1
*2
12 *13
0 100 200
300
Актуальный видовой пул
Региональный видовой пул
Рис. 6.9 • Связь между показателями видового разнообразия для растительных сообществ 14 типов (показаны номерами) в Эстонии: а — связь между флористическим богатством сообществ разных типов и флористической насыщенностью — числом видов на 1 м2; б — связь между региональным пулом видов (виды флоры Эстонии, способные произрастать в экологических условиях рассматриваемых сообществ) и актуальным видовым пулом (флористическим богатством сообщества) (по: Partel et al., 1996)
сообщества (число видов на 1 м2) положительно скоррелирована с общим числом видов, отмеченным в данном сообществе (рис. 6.9).
Согласно взглядам М. Zobel et al. (2000), растительные сообщества можно разделить на три группы с точки зрения лимитирования их состава: 1) абиотически лимитированные (неблагоприятные экологические условия препятствуют вселению других видов), 2) лимитированные поступлением диаспор, 3) лимитированные конкуренцией. Разграничить эти механизмы и соответственно увеличить их видовое разнообразие можно с помощью следующих воздействий: в случае 1 — созданием более благоприятных микроусловий, в случае 2 — добавлением диаспор (подсев семян), в случае 3 — при создании нарушений полога (Partel et al., 2000).
Объяснена одновершинная зависимость разнообразия от продукции растительных сообществ (см. п. 3.1 гл. 5) как следствие семенного лимитирования (М. Partel и М. Zobel, 2007). Анализ среднеевропейской флоры показал, что на бедных почвах виды имеют высокую семенную продуктивность, но у них слабее выражены приспособления к дальнему разносу диаспор (зоохорные, гидрохорные или анемохорные), что ограничивает их колонизационную способность. Напротив, виды богатых почв чаще имеют поиспособленид^^пяпь^р^.хоя^.тродз7 эупнее, что снижает общую семенную продукцию, ограничивающую
280
Глава 6. Состав и структура фитоценозов
55 Г
50- ф
45-
40-
35- •
30- •
25-
20- е
15-
ЮI-----1---•-----1---1---1----1----1---1----1
О 500 1000 1500 2000 2500 3000 3500 4000 4500
Рис.в.10 • Зависимость средней флористической насыщенности (число видов сосудистых растений на 25 м2 ордината) от площади, занимаемой сообществами в высокогорьях Тебердинского заповедника(абсцисса, гектары) (по: Онипченко, Павлов, 2009)
колонизационную способность. Минимальные ограничения колонизационной способности отмечаются на средних уровнях богатства почвы, что и приводит к увеличению разнообразия в таких условиях.
Флористическая насыщенность и флористическое богатство сообществ может прямо зависеть от занимаемой этим сообществом площади. Исследования альпийских сообществ в Тебердинском заповеднике выявили тесную связь между площадью сообществ на территории заповедника и числом видов в них на площадках 5 х 5 м (рис. 6.10): площадь, занимаемая сообществом, объясняет 89 % варьирования флористической насыщенности. Сообщества с самой низкой флористической насыщенностью — альпийские ковры и альпийские болота — занимают и наименьшую площадь в высокогорных ландшафтах заповедника. Напротив, среди альпийских сообществ наибольшей насыщенностью отличаются пестроовсяницевые луга, они же занимают наибольшие площади в альпийском поясе.
Поступление диаспор определяет состав сообществ на разном масштабе флористического богатства. Vandvik и Goldberg (2005) выделили несколько областей на кривой зависимости флористического разнообразия от поступления диаспор (рис. 6.11). На самых низких значениях нижний порог разнообразия не зависит от поступления диаспор, он определяется не семенным поддержанием уже существующих растений (этот порог определяется экспериментально при предотвращении естественного поступления диаспор). Затем идет область, в которой разнообразие поддерживается естественным поступлением диаспор до наблюдаемого в природе уровня. Выше этого — область ограничения разнообразия поступлением диаспор, которая выявляется в экспериментах с подсевом, когда дополнительный подсев увеличивает флористическую насыщенность. На высоком уровне разнообразие уже не зависит от притока дополнительных диаспор (формирование полночленного сообщества), и начинается область лимитирования ниш (биотический контроль). К сожалению,
3. Факторы поддержания флористического состава фитоценозов
281
Рис.в.11 • Связь между поступлением диаспор (абсцисса) и флористическим богатством (ордината) модельного сообщества. Прерывистые стрелки показывают пороговые значения, вне которых локальное разнообразие сообщества не зависит от поступления диаспор (определяются в экспериментах с предотвращением семенного дождя и дополнительным подсевом). Тонкие стрелки указывают на естественный поток диаспор и флористическое богатство. Черные стрелки слева показывают области разного лимитирования флористического богатства (по: Vandvik и Goldberg, 2005)
эксперименты, оценивающие изменения сообществ при предотвращении естественного поступления диаспор, весьма редки.
Разнообразие растений влияет на разнообразие других компонентов экосистемы. Например, в экспериментах (Stephan et al., 2000) было показано, что активность и разнообразие почвенных бактерий пропорциональны логарифму видового богатства растений. Среди исследованных видов растений наибольший эффект на видовое разнообразие бактерий оказал Trifolium repens, который может рассматриваться как ключевой вид в изученной системе.
3 • Факторы поддержания флористического состава фитоценозов
Механизмы формирования состава сообществ и поддержания их флористического разнообразия являются основными проблемами синэкологии растений. Большое влияние на изучение этих вопросов оказал принцип конкурентного исключения Гаузе (Gause, 1934), который в современной формулировке звучит так: «число видов, неограниченно долго сосуще
282
Глава 6. Состав и структура фитоценозов
ствующих в однородной среде с постоянными численностями, не может превышать числа плотностно-зависимых лимитирующих факторов» (Романовский, 19896). Развитие синэкологии растений в последние деся-гилетия в значительной степени связано с попыткой объяснить, каким образом в реальных растительных сообществах сосуществуют десятки и сотни видов растений при сходных потребностях всех зеленых растений в ограниченном числе ресурсов (см. п. 3.1 гл. 5). Большинство исследова-гелей выдвигали гипотезы, которые объясняли несоответствие реальных сообществ различным условиям, при которых должен выполняться принцип Гаузе. Альтернативный подход связан с представлениями об экологической эквивалентности (одинаковости видов), которая также может объяснять видовое разнообразие растительных сообществ (нейтральная геория Hubbell, см. ниже). В этом разделе будут кратко обобщены существующие представления о различных механизмах сосуществования растений с отсылкой к соответствующим разделам настоящей книги или другим источникам, где они описаны более подробно.
Все факторы, влияющие на состав фитоценозов и их флористическое богатство, можно разделить на две большие группы: исторические и (рецентные) экологические. Первые связаны с историей территории, действием экологических факторов в предшествующий период и островным эффектом (MacArthur, Wilson, 1967). Все они определяют ту или иную степень флористической неполночленности территорий или отдельных фитоценозов. В узком смысле эти факторы определяли возможность поступления диаспор тех или иных видов растений в рассматриваемые сообщества в настоящее время или в недалеком прошлом.
3.1 • Исторические факторы
На формирование современных растительных сообществ оказывали влияния различные события в истории Земли, связанные с формированиями флор, миграциями видов из других областей, недавними катастрофами и т. п. (см. п. 4 гл. 9). Большое значение имеет и степень изоляции той или иной территории, вызывающая островной эффект (MacArthur, Wilson, 1967) [Вставка XI].
МакАртур и Уилсон (MacArthur, Wilson, 1967) создали теорию, объясняющую видовое богатство островов на основе равновесия между
вселением новых видов и вымиранием уже существующих. Согласно их теории, число видов в островных биотах увеличивается при увеличении размеров острова
Вставка XI. Основные закономерности островной биогеографии
3. Факторы поддержания флористического состава фитоценозов
283
и его абсолютной высоты (как показателя разнообразия экологических условий) и уменьшается с увеличением расстояния до материка, являющегося источником вселяющихся на остров видов. Эти закономерности могут быть применимы не только к океаническим островам, но и к сообществам, развивающимся на изолированных экотопах (озера в тундре, небольшие болота среди лесных массивов, альпийские сообщества на изолированных вершинах гор и т. п.). Учет этих закономерностей важен также при анализе антропогенно фрагментированных участков фитоценозов. Например, сравнительное изучение состава 143 изолированных участков лугов по берегам Рейна в Нидерландах, проведенное в 1956 и 1988 гг., показало, что общее число видов на участке возрастало с увеличением его размера, вариабельности среды внутри участка и с уменьшением степени изолированности участка (расстояния до соседнего). Вымершие за период наблюдений популяции были в среднем мельче и располагались дальше от соседних, т. е. скорость вымирания вида на данном участке отрицательно скоррелирована с наличием вида на соседних участках (Ouborg, 1993).
Рис- XI.1 • Как островные экосистемы могут рассматриваться не только настоящие острова в океане (а), но, например, и изолированные участки высокогорных сообществ, приуроченные к вершинам гор, склоны и основания которых покрыты лесом (б)
284
Глава 6. Состав и структура фитоценозов
Флористическая насыщенность отдельных участков сообществ зависит также от времени существования сообществ в аналогичных условиях и их общей распространенности в регионе. Так, например, считается, что сухие карбонатные луга Европы значительно богаче видами, чем луга на кислых почвах, в связи с тем, что луговые и степные сообщества на карбонатных породах имели очень широкое распространение на просторах Евразии и длительное время для видообразования и коэволюции, а травяные сообщества на кислых почвах — нет (Grubb, 1986; Bengtsson et al., 1994).
3.2 • Современные факторы
Основное внимание экологов привлекали современные экологические механизмы поддержания видового разнообразия. Обзору таких факторов посвящено значительное число работ (Braakhekke, 1980, 1985; Shmi-da, Ellner, 1984; Абросов, 1984; Боголюбов, 1989; van der Maarel, 1988; Fowler, 1990; Wilson, 1990; Zobel, 1992). Наиболее полный анализ дал Palmer (1994), который приводит около 120 различных (частично пересекающихся) гипотез разных авторов, объясняющих механизмы поддержания видового богатства растительных сообществ.
Экологические механизмы сосуществования видов в фитоценозе можно условно разделить на 5 основных групп:
1. Расхождение по ресурсам и абиотическая гетерогенность среды, как пространственная — вертикальная и горизонтальная, так и временная — сезонная, флуктуационная, постепенные климатические изменения.
2. Влияние «нарушений»: масс-эффект, «gap»-napaflnrMa и регенерационная ниша, альтернативные стратегии жизненного цикла.
3. Влияние фитофагов.
4. Положительные взаимодействия между растениями.
5. Механизмы сбалансированной конкуренции и замедления конкурентного исключения: принципы равного шанса (карусель видов), начальная неоднородность структуры (мозаичность), циклическая конкурентная сеть, стабилизирующая коэволюция.
Естественно, что рассматриваемые механизмы не являются альтернативными, они могут сложным образом переплетаться в каждом сообществе, однако некоторая условная схематизация необходима для удобства изложения.
3. Факторы поддержания флористического состава фитоценозов
285
3.2.1 • Расхождение по ресурсам и абиотическая гетерогенность среды
Первая группа механизмов тесно связана с представлениями об экологической нише у растений (Гиляров, 1978; Bergh, van der Braakhekke, 1978; Hurlbert, 1981; Newman, 1982; Работнов, 19846; Миркин, 1985; Онипченко, 1987 и др.). Сюда можно отнести различия между видами по формам и источникам потребляемых ресурсов и по их пространственной локализации и времени использования.
Очевидно разделение растений по источникам потребляемого азота на две группы: имеющие симбиотические связи с азотфиксирующими прокариотами (см. п. 1 гл. 2) и не имеющие такого симбиоза. Несомненно, эти группы растений занимают в ценозах различные ниши, в пользу чего свидетельствует успешное внедрение бобовых при их искусственном подсеве на луга (Работнов, 1992). Виды растений также существенно различаются по способности поддерживать ассоциативную азотфикса-цию в ризосфере и филлосфере (Умаров, 1984; п. 2 гл. 2).
Основным источником почвенного азота для растений являются минеральные соединения (аммоний и нитраты) и низкомолекулярные органические соединения (аминокислоты), образующиеся при минерализации органического вещества. Примером частичного расхождения видов по источникам азота могут быть альпийские растения высокогорий Колорадо (Miller, Bowman, 2002). Festuca brachyphylla поглощала глицин интенсивнее, чем неорганические соединения азота. Carex rupestris и Ко-bresia myosuroides приблизительно одинаково потребляли аммонийные и нитратные формы азота. Виды разнотравья (Mertensia, Campanulaceae; Bi start a, Polygonaceae) имели наиболее высокую, а осоковые — наиболее низкую нитратредуктазную активность как показатель способности усваивать нитраты. При этом все виды могли использовать все изученные источники азота, хотя и с разной интенсивностью поглощения. Другие эксперименты также подтвердили, что специализация растений по потреблению нитратов или аммония в большинстве случаев не является полной, но часто наблюдается предпочтительное использование той или иной формы азота. Например, Arnica montana и Cirsium dissectum лучше используют нитратный азот, a Calluna vulgaris — аммонийный (de Graaf et al., 1998).
Частичную дифференциацию растений можно наблюдать и по источникам фосфора — лишь некоторые виды могут поглощать фосфор из фосфоритов (Работнов, 19856). Лишь микоризным растениям доступны фосфаты, содержащиеся внутри почвенных агрегатов с микропорами, куда не могут проникнуть корни растений, но свободно проникают гифы
286
Глава 6. Состав и структура фитоценозов
микоризных грибов (Simard et al., 2003). Кроме того, грибы, образующие микоризы разных типов (п. 1 гл. 3), имеют разные источники фосфора в почве.
Соотношение между поступлением различных ресурсов определяет видовой состав и флористическое богатство растительных сообществ (см. теорию D. Tilman в п. 3.1 гл. 5). W. G. Braakhekke и D. А. Р. Hooft-man (1999) выдвинули интересную гипотезу о том, что наибольшее видовое разнообразие на лугах наблюдается в случае оптимального для всего сообщества соотношения между поступающими ЭМП. Гипотеза была проверена на 74 площадках, где определяли параметры разнообразия и концентрации N, Р и К в надземной биомассе. Высокое разнообразие не было свойственно площадкам с экстремальными значениями отношений N/Р, Р/К, К/N, что поддерживает обсуждаемую гипотезу. Сенокошение может приближать баланс ЭМП к оптимальному, так как нелимитирующий ресурс удаляется с биомассой быстрее, чем лимитирующий.
Пространственное разделение ниш растений по источникам почвенных ресурсов — воды и элементов минерального питания — наиболее очевидно в случае разной глубины укоренения. Более ярко такое разделение проявляется в аридных областях (рис. 6.12), где могут сосуществовать виды разных экологических групп — фреатофиты (их корни достигают уровня почвенно-грунтовых вод), трихогигрофиты (поглощают воду капиллярного подъема от зеркала грунтовых вод) и омброфиты (используют воду атмосферных осадков из верхних горизонтов почвы) (Горышина, 1979). Однако, как было сказано в п. 5.3 гл. 5, растения могут активно изменять водный режим почв за счет гидравлического лифта. Изучение структуры подземных органов в большинстве фитоценозов показывает значительные различия по глубине укоренения и расположению основной массы всасывающих корней разных видов растений в различных горизонтах почвы. Роль таких различий для устойчивого сосуществования видов растений показана как экспериментально (опыты O’Brien с соавторами, см. Newman, 1982), так и на математических моделях (Berendse, 1979). Мохообразные и лишайники занимают в сообществах особые ниши, так как получают воду и элементы минерального питания преимущественно с атмосферными осадками, а не из почвы. Особые ниши в фитоценозах занимают также паразитные и полупаразитные растения, получающие воду и элементы минерального питания от растений-хозяев.
Исходная пространственная гетерогенность среды дает возможность сосуществовать большому числу видов растений, отличающихся соотношением потребляемых ресурсов (см. взгляды Д. Тильмана, п. 3.1 гл. 5). Вторичная неоднородность среды, создаваемая самими растениями (см.
3. Факторы поддержания флористического состава фитоценозов
287
многолетние
м
1.0
0.5 поверхность почвы
0.5
1.0
1.5
2.0
2.5
Рис. 6.12 • Средний размер и расположение корневых систем и надземных побегов растений аридных местообитаний (медианные и максимальные величины по большой выборке, главным образом североамериканских пустынь) (по: Schenk, Jackson, 2002; Schenk, 2006)
п. 5 гл. 5), может также повышать видовое разнообразие сообществ. Так, в экспериментальных посевах монокультур и смесей культурных растений Palmer и Maurer (1997) изучили разнообразие сорных растений, которое оказалось существенно выше в случае смешанных посевов.
Большую роль в сосуществовании видов играет изменчивость экологических факторов во времени, при которой каждый вид имеет периоды «конкурентного оптимума условий», в то время как другие периоды недостаточны для его конкурентного исключения. Для сосуществования видов важны разные по масштабу времени формы динамики экологических, в том числе почвенных, факторов — как кратковременная сезонная и флуктуационная динамика, так и постепенные климатические изменения.
Существует много примеров сезонного (фенологического) дополнения видов: сосуществование видов контрастной экологии (околоводного сусака — Butomus umbellatus и лугово-степного подмаренника настоящего — Galium verum) на лугах Волго-Ахтубинской поймы с резко-переменным водным режимом (Раменский и др., 1956); фенологическое дополнение видов растений с СЗ и С4 типом фотосинтеза на лугах Северной Каролины (Fowler, Antonovics, 1981); синузии эфемероидов и растений летней вегетации в широколиственных лесах (Кожевников, 1931; Смирнова, 1987) и др. Развитие растений в разные периоды одного сезона сопряжено с различиями в потреблении почвенных ресурсов. Разногодичные флуктуации условий среды также могут способствовать сосуществованию видов растений с различной экологией, таких как, например, степной вид типчак — Festuca valesiaca и болотный ситняг Heleocharis palustris, чередующие свое доминирование на некоторых типах лугов в сухие и влажные годы соответственно (Работнов, 1992). При редких
288
Глава 6. Состав и структура фитоценозов
неблагоприятных условиях (например, засуха в гумидных районах) наиболее чувствительные виды сильнее уменьшают свое участие, чем более устойчивые. В работе S. М. Buckland et al. (1997) было показано, что устойчивость к засухе луговых растений Великобритании, оцениваемая по содержанию воды в листьях в период сильной засухи 1995 г., была положительно скоррелирована с глубиной укоренения растений.
В работах G. 1. Agren и Т. Fagerstrom (1980, 1984) на математических моделях показано, что в сообществе могут устойчиво сосуществовать виды, конкурирующие за общие ресурсы, но чередующие периоды своей высокой семенной продуктивности по годам в связи со стохастическим характером изменения условий среды. Постепенные климатические изменения, часто носящие циклический характер, с длительностью циклов, измеряемых столетиями и тысячелетиями, также способствуют сосуществованию видов растений, имеющих различные климатические опти-мумы, но высокую толерантность к широкому диапазону условий (Wilson, 1990). Последнее особенно характерно для сообществ высокогорий, где климатические ритмы проявляются наиболее ярко, но большинство видов растений имеет большую продолжительность жизни особей или клонов, а конкурентное вытеснение в условиях низкой продуктивности крайне замедлено.
3.2.2 • Влияние нарушений
Для поддержания видового разнообразия фитоценозов большое значение имеют различные типы нарушений, вызванных как внутренними (вывалы деревьев), так и внешними (роющая деятельность животных, солифлюкция и др.) для фитоценоза причинами (Grime, 1977, 1979; White, 1979; Pickett, 1980; Denslow, 1985; Sousa, 1984; van der Maarel, 1993) (cm. n. 3.7.2 гл. 5). Нарушением считается неизбирательное удаление (гибель) отдельных особей, освобождающее участок пространства с высокой доступностью ресурсов для колонизирующих его растений. Отчуждение фитомассы животными, носящее чаще всего избирательный характер, лучше рассматривать в отдельной группе механизмов, связанных с воздействием фитофагов. В группу механизмов, связанных с нарушениями, целесообразно включать «масс-эффект», концепцию регенерационной ниши и <^ар»-парадигму, а также представления об альтернативных стратегиях жизненного цикла (см. п. 3.7 гл. 5).
Зависимость числа видов от интенсивности нарушений — обычно одновершинная кривая. Например, наибольшее флористическое богатство в прериях наблюдалось на участках с умеренными нарушениями, вызванными различными причинами (Collins, Barber, 1986). Однако при
3. Факторы поддержания флористического состава фитоценозов
289
рассмотрении этой зависимости необходимо учитывать и фактор времени, поскольку стабилизация видового состава при определенном уровне нарушений происходит не очень быстро. По данным эксперимента по гамма-облучению дубово-березового леса в Brookhaven National Laboratory, проводимого с 1961 г., вначале флористическое разнообразие близ источника значительно снизилось. Но по данным Olsvig (1979; Peet et al., 1983), в 1976 г. наибольшее флористическое богатство наблюдалось уже непосредственно вблизи источника радиации во внутреннем кольце радиусом 10-30 м от источника.
Термин «масс-эффект» был использован в работе A. Shmida и S. Е11-пег (1984). Он объясняет присутствие в сообществе или на его отдельных участках видов, чьи особи приживаются лишь в местах с ослабленным конкурентным воздействием соседей (например, в местах нарушений), и их популяции поддерживаются за счет постоянного потока диаспор с соседних сообществ или участков (детальнее рассмотрен в п. 4.2.5 гл. 6). Широкое распространение получила предложенная Р. Grubb (1977) концепция регенерационной ниши. Она предполагает возможность успешного сосуществования видов, имеющих сходные экологические потребности во взрослом состоянии, но существенно различающиеся по условиям приживания их всходов и подроста. В применении к пространственной структуре эта концепция близка к <^ар»-парадигме, рассматриваемой в п. 4.2.6 гл. 6.
3.2.3 • Влияние фитофагов
Высокое видовое богатство фитоценозов может поддерживаться за счет сложной трофической структуры биогеоценоза. Как одно из проявлений принципа Гаузе показано, что устойчиво сосуществовать может неограниченное число видов продуцентов, каждый из которых имеет специализированного фитофага, играющего роль плотностно-зависимого фактора регуляции численности потребляемого растения (Боголюбов, 1989; Tilman, 1982,1986). Кроме того, высокое видовое разнообразие растений может поддерживаться неспециализированным фитофагом, предпочитающим наиболее обильные (доминирующие) виды растений (Paine, 1966; Harper, 1969; Murdoch, 1969; Shmida, Ellner, 1984) (см. п. 6 гл. 4).
Первые три группы гипотез можно рассматривать с точки зрения компромиссов (трейдофф, англ, trade-offs) между различными способностями растений. По мнению D. Tilman (2000), сосуществование видов может быть обусловлено наличием компромиссов (трейдоффов): 1) между конкурентоспособностью и колонизационной способностью (см. 4.2.7 гл. 6), 2) между конкурентоспособностью и восприимчивостью к болезням и фитофагам, 3) между способностью довольствоваться средним уровнем
290
Глава 6. Состав и структура фитоценозов
ресурсов и способностью к быстрому поглощению ресурсов во время их всплесков (пульсов) (см. 3.7 гл. 5), 4) между способностью видов конкурировать за разные ресурсы в гетерогенной среде (см. 3.1 гл. 5).
3.2.4 • Положительные взаимодействия между растениями
При наличии положительных взаимодействий между доминантами и редкими видами растений также возможно поддержание высокого видового разнообразия. В работах по теоретической экологии этот механизм обычно рассматривается как выделение одними видами метаболитов, стимулирующих развитие других (Боголюбов, 1989). В чистом виде этот механизм вряд ли имеет существенное значение для формирования структуры фитоценозов. Однако его аналогом может быть транспорт ЭМП и органических веществ по единой микоризной сети (см. п. 1.9 гл. 3). Несомненно, что изменения в абиотической и биотической среде, описанные в п. 5 гл. 5 и п. 6 гл. 5, также могут способствовать поддержанию видового разнообразия.
Отсутствие конкурентного исключения может быть связано с «гнездовой» структурой положительных отношений в сообществе, когда один или несколько видов оказывают положительное влияние на группу других видов. В результате исследования ряда растительных сообществ в Мексике было выявлено, что число видов, положительно влияющих на другие, колеблется от 13 до 31, а число видов, на которые оказывается положительное влияние, — от 30 до 65 на сообщество. Один из видов (Mimosa luisana) положительно влиял на 48 других видов растений! (Verdu, Valiente-Banuet, 2008)
3.2.5 • Сбалансированная конкуренция
В эту группу объединены разнообразные гипотезы, связанные с представлениями о небольших различиях в конкурентоспособности видов, а также о циклических конкурентных сетях (Goldberg, Werner, 1983; Shmida, Ell-ner, 1984; Wilson, 1990; Aarssen, 1992).
Гипотеза «лотереи» была предложена J. Н. Connell (1979). Согласно его взглядам, сообщества прикрепленных организмов могут поддерживать высокое видовое разнообразие в случае, если все виды имеют равные возможности занять свободные места и затем удерживать их от вторжения других видов. Исследования тропических лесов в Панаме показали, что большинство образующих их видов являются «генерали-стами» и могут занимать любые прорывы полога (Hubbell, Foster, 1986, 1987; см. ниже). К близким выводам пришел Е. van der Maarel (1993), изучавший закономерности смены видов малолетних растений в альварах
3. Факторы поддержания флористического состава фитоценозов
291
Рис. в.13 • Два варианта пространственной структуры сообщества из двух конкурирующих видов (более сильный конкурент показан темным цветом). Слева — одновидовые пятна (небольшая поверхность контактов двух видов, замедленное конкурентное исключение), справа — диффузное распределение более слабого конкурента, большее число межвидовых контактов с возможностью более быстрого конкурентного исключения
Швеции. Им была предложена модель «карусели», описывающая недетерминированную последовательность смен видов на мелких площадках в течение нескольких лет при сохранении общей структуры сообщества (см. п. 4.2.5 гл. 6). Видимо, неконкурентное сосуществование при наличии свободных микрозон может объяснить часто высокое видовое разнообразие несомкнутых фитоценозов, развивающихся, например, на скалах и осыпях (Shmida, Ellner, 1984).
Начальная неоднородность (мозаичность) горизонтальной структуры фитоценозов может резко замедлить конкурентное исключение, так как внутри одновидовых пятен преобладает внутривидовая конкуренция, а область межвидовых контактов резко снижается (Wilson, 1990; рис. 6.13). Высокая стабильность такой мозаики была показана и на математических моделях (А. С. Комаров, личное сообщение).
Гипотеза стабилизирующей коэволюции, предложенная L. W. Ааг-ssen (1983, 1989), предполагает усиление отбора на возрастание конкурентоспособности видов при уменьшении их участия в сообществе («преимущество быть редким»). Чем выше численность популяции какого-либо вида, тем чаще особи этого вида произрастают вблизи друг друга и вступают во внутривидовую конкуренцию, в то время как при низкой численности популяции преобладают межвидовые контакты.
Высокое видовое разнообразие может поддерживаться также при наличии циклической конкурентной сети (вид А вытесняет вид В, В — С, С — А и т.п.; рис.6.14) и циклических микросукцессий (Watt, 1947; Wilson, 1990; Aarssen, 1989,1992). Возможность существования циклических конкурентных сетей в луговых сообществах была экспериментально
292
Глава 6. Состав и структура фитоценозов
Рис.в.14 • Схема циклической конкурентной сети. Вид А конкурентно вытесняет вид В, В вытесняет С, а С вытесняет А с микроучастков их произрастания
подтверждена в работе D. R. Taylor и L W. Aarssen (1990). В ней показано, что такие (нетранзитивные) конкурентные сети с участием различных генотипов могут способствовать поддержанию видового разнообразия.
Ряд экспериментов, проведенных с растениями карбонатных лугов в Англии, также показали важность внутривидовой изменчивости доминирующих видов для поддержания флористического богатства этих лугов (Fridley et al., 2007; Whitlock et al., 2007). Исход конкуренции между Koeleria macrantha и Carex caryophyllea на бедных почвах зависел от того, каким генотипом был представлен тот или иной вид растений. Показано, что внутри-популяционная Генетическая дифференциация может поддерживать сосуществование видов за счет нетранзитивных конкурентных отношений между
ними (Fridley et al., 2007). В эксперименте с совместным выращиванием 6 видов трав (Festuca ovina, Koeleria macrantha, Carex caryophyllea, Succisa pratensis, Leontodon hispidus, Campanula rotundifolia), где каждый вид был представлен 16, 4 или 1 биотипом, выживаемость растений всех видов, кроме Succisa pratensis, была выше в варианте их представленности 16 биотипами по сравнению с 4. Авторы также подтвердили нетранзитивность конкурентных отношений между видами, представленными разными генотипами (Whitlock et al., 2007).
3.2.в • Нейтральная теория S. Р. Hubbell
Нейтральная теория исходит из предпосылки о функциональной эквивалентности видов. Основные положения этой теории были сформулированы S. Р. Hubbell (2005) в последней четверти XX в. под влиянием исследования тропического леса на Barro Colorado Island (ВС1) в Панаме (рис. 6.15). S.P. Hubbell полагает, что виды под действием отбора приобретают сходные черты, дающие им приспособленность к наиболее распространенным условиям среды, независимо от того, сколько видов вырабатывают те же особенности. Согласно этой гипотезе, небольшое число пионерных видов в сомкнутых лесах ВС1 отражает небольшую площадь прорывов полога. Два важных фактора предотвращают конкурентное исключение: ограниченность распространения и приживания, а также локальная гетерогенность абиотической и биотической среды, в которой произрастают отдельные особи каждого вида. Семена большинства деревьев распространяются не далее 20 м от материнского
3. Факторы поддержания флористического состава фитоценозов
293
Рис.в.15 • Barro Colorado Island (BCI) — основной объект исследования тропических лесов с позиций нейтральной теории: а — схема Панамского канала, красным цветом выделен BCI (www.stri.si.edu, с любезного разрешения Carlos Caballero); б — аэрофотоснимок тропического леса на этом острове (фото Christian Ziegler, см. Gewin, 2006; under Creative Commons Attribution License)
294
Глава 6. Состав и структура фитоценозов
растения, а семена пионерных видов сохраняют всхожесть 2-3 года. Большое значение имеет плотностно-зависимая смертность семян и проростков, показанная для большинства видов. Чем выше плотность особей одного вида, тем выше их смертность за счет воздействия фитофагов, патогенных грибов и т. п. Это приводит к большей вероятности приживания новых растений на значительном расстоянии от материнского. Среди 20 ближайших соседей данного дерева только 20% принадлежало к тому же виду (Hubbell, 2005). Таким образом, центральный механизм поддержания высокого флористического богатства тропических лесов — плотностно-зависимая смертность. Этот же механизм может действовать и в лесах умеренного пояса (Hille Ris et al., 2002; ter Steege, Zagt, 2002).
3.2.7 • Отбор на редкость и массовость
Помимо конкурентных могут существовать другие механизмы, регулирующие участие видов в составе ценоза. По представлениям Hoft с соавторами (1993), как редкость, так и массовость (обильность) видов имеют свои издержки. Издержки редкости — это сложность полового размножения в связи с низкой встречаемостью растений, в то время как издержки массовости — большая подверженность хищникам, паразитам, болезням. Участие каждого вида в составе сообщества может рассматриваться как компромисс между этими издержками. При этом организмы с бесполым размножением имеют существенно сниженные издержки редкости (Taper, 1993).
А. Жигон и Р. Марти (Gigon, Marti, 1994) предприняли попытку выяснить, каковы основные причины редкости большинства видов на флористически богатом сухом лугу на севере Швейцарии, в составе которого насчитывалось более 100 видов сосудистых растений. Они выделили 4 возможных причины редкости отдельных видов:
1) Приуроченность к особым микроучасткам — колеям от сенокосилок (Carlino simplex, Aster amellus, Asperula cynanchica), колониям полевок (Dactylis glome rata, Lolium perenne, Galium album), нарушениям в местах пороев кабанов, нор гадюк (Sedum acre, Gentiana cilia-ta, Arabis hirsute, Myosotis arvensis, Campanula rapunculus, Arenaria leptoclados).
2) Поступление диаспор извне и развитие неполночленных популяций, не способных к самоподдержанию в условиях сенокошения (Aster amellus, Buphthalmum salicifolium, Peucedanum cervaria, Seseli liban-otis, Verbascum thapsus, Pinus Silvestris и другие деревья).
3) Регулирующее воздействие других организмов: преимущественное поедание полевками бобовых (9 видов), кабаном — Orchis pollens, поражение фитопатогенным грибом Linum catharticum.
4. Пространственная структура фитоценозов
295
4) Конкурентное воздействие доминантов. Для проверки этого положения проведены эксперименты с обрубкой корней и удалением соседей для 3 редких видов (Dactylis glomerata, Centaurea jacea, Leucanthemum vulgare) и 3 сходных по жизненной форме доминантов (Bromus erectus, Salvia pratensis, Scabiosa columbaria). Численность побегов видов первой группы возросла в 6-17 раз, а второй — лишь в 4-5 раз по сравнению с контролем. Еще более значительная разница получена по численности генеративных побегов и цветков. На основании полученных результатов авторы предлагают удалять соседей редких видов для поддержания популяции последних.
Авторы также подчеркивают, что антропогенная гомогенизация среды (типа городских газонов) ведет к уменьшению возможного числа сосуществующих видов (Gigon, Leutert, 1996). Аналогичный анализ видов с малым участием в склерофилльных и дождевых лесах Австралии (55 и 116 видов соответственно) показал, что большая часть этих видов (91 % и 95 % соответственно) значительно более обильны в каких-либо других местообитаниях (Murray et al., 1999). Таким образом, многие редкие виды являются таковыми на границах своих синэкологических ареалов.
4 • Пространственная структура фитоценозов
В понятие «структура растительных сообществ» разные авторы вкладывают различный смысл. В. В. Мазинг (1973) рассматривал три возможные трактовки слова «структура»:
1) структура = состав (набор элементов и количественные соотношения между ними);
2) структура = строение (пространственное расположение элементов); в этом значении мы и будем рассматривать структуру растительных сообществ в настоящей главе;
3) структура = совокупность связей между компонентами системы (функциональная структура).
4.1 • Вертикальная структура (ярусность)
Во многих сообществах размеры растений разных видов существенно отличаются по высоте, что служит основой для выделения ярусов. Существует два основных подхода к определению ярусов:
296
Глава 6. Состав и структура фитоценозов
Количество азота в экосистемном цикле (условные единицы)
Рис.6.1в • Теоретическая зависимость развития ярусов (масса листвы) лесных фитоценозов от богатства почвы азотом (условные единицы). Растения нижних ярусов более эффективно используют азот, но обитают в условиях перехвата света верхними ярусами (по: Kull et al., 1995)
1) ярус как совокупность растений одной или сходных жизненных форм (так называемые вложенные ярусы; в таком случае, например, древесный ярус образован деревьями от поверхности почвы до верхушек деревьев),
2) ярус как слой концентрации надземных органов растений.
Выделение ярусов целесообразно при резких отличиях в тех или иных характеристиках растительного покрова по высоте (биомассе, листовой поверхности и т. п.). При плавном изменении этих показателей, носящем одновершинный характер, выделение ярусов нецелесообразно (на лугах, в тропических лесах и т. п.). Лучше всего ярусность выражена в одновозрастных бореальных лесах.
К. Kull et al. (1995) исследовали влияние почвенного богатства на вертикальную структуру (ярусность) на примере лесов Эстонии. Степень развития основных ярусов (древесного, травяного, мохового) зависела от богатства почв азотом (рис. 6.16). Рассмотрено два пути увеличения эффективности фотосинтеза: 1) увеличение интенсивности фотосинтеза за счет обогащения листьев белками и другими азотсодержащими компонентами, 2) улучшение эффективности использования азота, т. е. увеличение отношения фотосинтез/содержание азота за счет увеличения длительности жизни листьев и т. п. Первый путь более выгоден в условиях высокой доступности азота, а второй — на бедных почвах. Исходя их этих посылок, построена модель, предсказывающая изменение биомас
4. Пространственная структура фитоценозов
297
сы листьев в разных ярусах в зависимости от богатства почвы азотом для лесных сообществ. Она показывает, что наибольшее участие в листовой поверхности для мохового яруса должно наблюдаться на бедных почвах, среднего яруса — на умеренно богатых почвах, а на богатых почвах древесный ярус будет абсолютно преобладать по массе листвы, что и подтвердилось при анализе лесов в Эстонии (Kull et al., 1995).
Растения, образующие разные ярусы, могут развиваться частично автономно от растений других ярусов. Например, при спуске с гор Северного Урала В. Б. Сочава (1930) наблюдал следующие смены доминирующих видов в сообществах: альпийские лужайки (Festuca ovina — Polytrichum commune), ерники (Betula папа — Festuca ovina — Polytrichum commune), лиственничники (Larix sibirica — Betula папа — Festuca ovina — Polytrichum commune). Он ввел представление об инкумбации (наложении) ярусов, наблюдаемой при спуске с гор, и обратном процессе — декумба-ции (выпадении) ярусов, наблюдаемой при подъеме в горы. Аналогичные закономерности наблюдаются и в горах Кавказа, где у верхней границы леса Rhododendron caucasicum произрастает под пологом березы, а выше образует самостоятельные сообщества рододендроновых стлаников. Сходные ряды отмечаются и для северной границы леса в Арктике.
4.2 • Горизонтальная структура сообществ
Горизонтальная структура большинства растительных сообществ гетеро-генна. При рассмотрении элементов пятнистости растительного покрова часто различают мозаичность (пятна, не связанные с изменением среды) и комплексность (пятна, различающиеся по свойствам среды, прежде всего почвенной). Безусловно, это разделение весьма условно, так как любые изменения в структуре покрова влекут за собой те или иные различные по длительности существования изменения в среде (см. п. 5 гл. 5).
Существует несколько классификаций пространственных мозаик по причинам их возникновения. Так, весьма сходны представления Kershaw (1963) и Whittaker (1970), выделивших: 1) морфологическую (рост, распространение растений), 2) средовую и 3) социологическую (вызванную отношениями между растениями) мозаичность (Anand, 1994).
Т. А. Работнов (1992) выделяет следующие типы мозаичности по причинам, ее вызывающим:
1) эдафотопическая — связана с неоднородностью почвы (эдафотопа), в том числе повышением—понижением в микрорельефе;
2) эпизодическая, обусловленная неоднородностью в поступлении диаспор;
3) ценобиотическая — связана с воздействием одних видов на другие;
298
Глава 6. Состав и структура фитоценозов
4) кленовая — образование клонов в результате вегетативного размножения;
5) зоогенная, обусловленная деятельностью животных;
6) антропическая — связана с деятельностью человека (выборочная рубка, кострища и т. п.).
4.2.1 • Эдафотопическая мозаичность
Связь между распространением отдельных видов растений и свойствами почв внутри растительных сообществ (т. е. в масштабе десятков сантиметров — десятков метров) хорошо известна и иллюстрируется многими примерами. Приведем лишь немногие из них.
Сопряженное исследование распространения растений и изменения свойств почв в сосняке-зеленомошнике на Валдае выявило несколько значимых связей в пределах пробной площади 3600 м2, разбитой на площадки 2 х 2 м. Участие вейника тростниковидного (Calamagrostis arundinacea) резко увеличивалось на участках с повышенным pH горизонта В, где располагается большая часть его корней (Рысин, Рысина, 1987). В то же время участие напочвенного зеленого мха Aulacomnium palustre резко снижалось с увеличением pH подстилки (рис. 6.17).
В горных условиях, где мощность почвы может ограничивать развитие корневых систем растений, обнаруживается довольно четкая приуроченность отдельных видов внутри сообщества (в масштабе десятков сантиметров) к участкам почвы разной мощности. Например, все часто встречающиеся виды альпийских лишайниковых пустошей (АЛП) можно разделить на 3
Рис. 6.17 • Изменение участия (баллы) растений (ордината)
в сосняке-зеленомошнике на Валдае в зависимости от свойств почвы.
Вертикальные линии показывают ошибку средних значений (Онипченко, 2011)
4. Пространственная структура фитоценозов
299
Рис.в.18 • Средняя глубина почвы (ордината) под различными видами растений в альпийских лишайниковых пустошах. Виды, отмеченные сверху столбиков неперекрывающимися буквами, значимо отличаются по средним глубинам почвы (р < 0,05). Сокращенные названия видов: Ас — Alchemilla caucasica, Ad — Antennaria dioica, Al — Arenaria lychnidea, Am — Anthemis marshalliana, As — Anemone speciosa, Av — Agrostis vinealis, BL — лишайники и мохообразные, BS — обнаженная почва, Bv — Bromus variegatus, Cc — Carum caucasicum, Cco — Campanula collina, Ct — Campanula tridentata, Cu — Carex umbrosa, Fo — Festuca ovina, Hv — Helictotrichon versicolor, Me — Minuartia circassica, Ok — Oxytropis kubanensis, Tp — Trifolium polyphyllum, Vv — Vaccinium vitis-idaea (по: Батчаева и др., 2003)
группы (рис. 6.18). К первой группе относятся виды, встречающиеся при мощности почвы (мощность измерялась при помощи металлического щупа до первого камня) менее 17 см. Сюда относятся Helictotrichon versicolor, Bromus variegatus, Carex umbrosa, Minuartia circassica, Oxytropis kubanensis и другие виды. Участки, лишенные растений, а также пятна мхов и лишайников также характеризуются незначительной мощностью почвы. Ко второй группе растений, приуроченных к почвам средней мощностью 17-20 см, можно отнести Antennaria dioica, Campanula tridentata, Festuca ovina, Trifolium polyphyllum, Alchemilla caucasica, Anemone speciosa. Три вида растений, часто встречающихся на участках с мощностью почвы более 20 см, относятся к третьей группе — Campanula collina, Vaccinium vitis-idaea, Agrostis vinealis. В рассмотренные группы входят растения разных жизненных форм, но интересно отметить, что последнюю группу (растения на наиболее глубоких участках почвы) образуют виды, обладающие наиболее значительной вегетативной подвижностью.
Помимо растений, от почвенной неоднородности зависит распределение многих групп организмов. Ettema и Wardle (2002) приводят многочисленные примеры таких зависимостей. В агроэкосистеме почвенная микробная биомасса и биомасса коллембол показали пространственную зависимость в масштабе > 200 м, отражающую зависимое от положения
300
Глава 6. Состав и структура фитоценозов
в ландшафте содержание органического вещества в почве (Fromm et al., 1993). Аналогично Crist (1998) показал для низкотравной степи, что пространственная мозаика термитов в масштабе 330 м была связана с топографическим градиентом и мозаикой растительности. Пространственная агрегированность в масштабах сантиметров и метров часто структурируется отдельными растениями и варьируется в зависимости от их жизненной формы, размеров и структуры. Mitchell et al. (2000) показали, что на участке сфагнового покрова 40 х 60 см раковинные амебы были агрегированы в пятна 1-10 см. Grundmann, Debouzie (2000) с использованием иглы-бура с диаметром 1 мм показали, что бактерии Nitrobacter были пространственно агрегированы в пятна 2-4 мм в диаметре, что авторы связывают с размерами почвенных агрегатов и распределением тонких корней растений.
4.2.2 • Влияние растений на горизонтальную неоднородность почв Растения, в свою очередь, могут оказывать различное влияние на горизонтальную структуру почвенного покрова, повышая или понижая его неоднородность. Повышение неоднородности обычно связано с индивидуальным воздействием особей и групп растений на свойства почвы (эти воздействия подробно рассмотрены в п. 5 гл. 5). С другой стороны, конкуренция за почвенные ресурсы может приводить к снижению гетерогенности почв за счет более эффективного поглощения ЭМП из участков почв, обогащенных ими.
Гетерогенность среды обитания в сообществе с позиций «самого растения» можно изучать экспериментально с использованием двух подходов: 1) подсадка тестовых растений в сообщество и наблюдение за их развитием; 2) взятие почвенных образцов из разных мест и выращивание на них тестовых растений в лабораторных условиях (Bell, Lechowicz, 1991).
Пространственная пятнистость свойств почвы характеризуется двумя основными параметрами — контрастностью пятен и их размерами. Под каждым типом растительности формируются почвы со своими показателями пространственного варьирования.
По наблюдениям Е. А. Дмитриева и В. П. Самсоновой (1975, 1978, 1979), гетерогенность лесных почв проявляется преимущественно на расстояниях менее 1 м (0,3-0,8 м). Аналогичные закономерности отмечены и в лесах Великобритании (Farley, Fitter, 1999). Гетерогенность на лугах и прериях выражена в меньшей степени (Wilson, Kleb, 1996; Kleb, Wilson, 1997), поэтому при внедрении древесных растений на участки прерии пространственная гетерогенность почвенных ресурсов обычно повышается. Так, в исследовании М. Partel и S. Wilson (2002) гетерогенность почв, взаимно пересаженных между прерией и лесом, изменяется уже
4. Пространственная структура фитоценозов
301
в течение вегетационного периода, т. е. находится под сильным влиянием растительности. Большое значение в этих процессах имеют особенности структуры корней травянистых и древесных растений. По наблюдениям с помощью миниризотронов в прерии и лесу (из Populus tremuloides), общая длина тонких корней в лесу была существенно ниже, чем в прерии (0,7 и 1,4 см/см2 соответственно), а коэффициент вариации длины корней в лесу был существенно выше, чем в прерии (92 % и 65 % соответственно). Размер характерных скоплений корней был выше в лесу (8-12 см), чем в прерии (3-4 см). Скорость оборота тонких корней была выше в лесу (49 %), чем в прерии (20 %) (Partel, Wilson, 2002), что хорошо согласуется с данными других исследователей.
В пустынных сообществах США характерные размеры почвенных неоднородностей составляли 1-3 м, что было связано с размерами одиночных кустарников (Schlesinger et al., 1996). В то же время для полынной степи основная вариабельность почвенных свойств была отмечена на расстоянии до 1 м, а при увеличении расстояния до 7 м дисперсия свойств оставалась практически постоянной (Jackson, Caldwell, 1993).
Проведенные нами сравнительные исследования варьирования почвенных свойств на участках различных сообществ, отличающихся по богатству почв (Онипченко и др., 1998), показали, что более высокое содержание того или иного ЭМП не связано однозначно с его большей пространственной вариабельностью. Таким образом, обсуждаемое в экологии правило Тейлора (см. Soberon, Loevinsohn, 1987), постулирующее увеличение вариабельности экологических параметров при увеличении их средних значений, не находит очевидных подтверждений для почв малонарушенных сообществ. В нашем случае пестроовсяницевые луга, образованные плотнодерновинными злаками (Festuca varia, Nardus stricta и др.), отличались как наибольшим содержанием большинства ЭМП в почве, так и наименьшей их вариабельностью в пространстве среди 4 изученных сообществ. Эти луга могут служить хорошим примером «гомогенизирующего» влияния растений на свойства почвы, обусловленного сплошной пронизанностью верхних горизонтов тонкими корнями злаков, быстро потребляющими ЭМП из любых обогащенных участков. Преимущественный рост корней на участках, богатых ЭМП, был показан в специальных экспериментах в степных сообществах (Jackson, Caldwell, 1996).
В работе V. R. Kelly и С. D. Canham (1992) исследована пространственная неоднородность поступления света (интервал измерений 1 см) и почвенных ресурсов (содержание воды и азота, интервал измерений 10 см) в сообществах залежей. Было показано, что для всех трех характеристик коэффициент вариации уменьшается с увеличением их средних значений, т. е. на богатых
302
Глава 6. Состав и структура фитоценозов
Рис. 6.19 • Примеры горизонтальной гетерогенности почвенных свойств в кустарниковом сообществе Ulex-Erica в Испании, карты содержания: а — органического вещества в почве; б — азота аммония; в — азота нитратов (по: Gallardo et al., 2005)
участках ресурсы распределяются более равномерно. Ряд других исследований (Gross et al., 1995; Cain et al., 1999) также показали наименьшую вариабельность содержания азота в почве при больших величинах его содержания. Все эти данные свидетельствуют о том, что правило Тейлора не работает в наземных экосистемах, где избирательная активность корней растений способствует более равномерному распределению почвенных ресурсов в условиях их большей доступности.
Другой пример изучения пространственной неоднородности почв в Испании приводят Gallardo et al. (2005). Методами пространственной статистики исследована пространственная структура содержания аммония, нитратов и органического вещества в почвах 6 различных растительных сообществ. Размеры неоднородностей сильно варьировали между сообществами и были выше для аммония, чем для нитратов. Получена положительная корреляция содержания аммония и органики в почве, но не найдена корреляция между содержанием аммония и нитратов. В этом исследовании средние значения и дисперсия почвенных свойств были положительно скоррелированы между 6 сообществами. Пример пространственной структуры для кустарникового сообщества Ulex-Erica приведен на рис. 6.19.
В природных растительных сообществах нечасто встречаются регулярные пространственные структуры. Для их формирования необходимо сочетание положительных обратных связей на небольшом расстоянии и отрицательных — на более значительном. Для растительных сообществ это может наблюдаться в случае благоприятных отношений между ближайшими соседями и конкуренции с более удаленными. Примерами могут служить полоски лесов, образуемые на верхней границе
4. Пространственная структура фитоценозов
303
леса хвойными породами в Скалистых горах, а также регулярное размещение кустарников в аридных регионах (Rietkerk, van de Koppel, 2008).
4.2.3 • Ценобиотическая мозаичность
Как положительные, так и отрицательные отношения между видами растений могут создавать характерные формы мозаичности. Примером может служить мелкая мозаичность кустистых лишайников и сосудистых растений на альпийских пустошах Кавказа.
ст
Рис. в.20 • Внешний вид (слева) и вертикальный профиль (справа) альпийской лишайниковой пустоши. На профиле показано значительное развитие корней растений под пятнами лишайников. Растения: 1 — Helictotrichon versicolor, 2 — Cetraria islandica, 3 — Carex umbrosa, 4 — Festuca ovina, 5 — Plantago atrata (по: Онипченко, 1985)
304
Глава 6. Состав и структура фитоценозов
Альпийские лишайниковые пустоши имеют весьма своеобразную горизонтальную структуру: мелкие пятна до 10 см в диаметре и отдельные куртинки лишайников чередуются с дерновинами и отдельными побегами сосудистых растений (рис. 6.20). Нами (Онипченко, 1985; Павлов и др., 1998) было предложено следующее объяснение механизмов формирования такой структуры. В условиях малой мощности почв и их бедности элементами минерального питания растения развивают мощную корневую систему в горизонтальном направлении, занимающую большую площадь, чем могут занять их надземные побеги. Таким образом, в надземной сфере образуются как бы пустоты, которые заселяются лишайниками, получающими элементы минерального питания из атмосферы. Лишайники доминируют по биомассе, но практически не конкурируют с сосудистыми растениями за почвенные ресурсы. Бедность условий среды лимитирует надземную биомассу сосудистых растений, и они «оставляют свободные места», занимаемые кустистыми лишайниками. Сосудистые растения формируют надземный «каркас» сообщества, без которого эпи-гейные (напочвенные) лишайники не могут существовать, так как легко сдуваются с наветренных склонов сильными ветрами. Из этих предположений вытекает несколько экспериментально проверяемых следствий (рис. 6.21): 1) удаление лишайников не должно вызвать существенных изменений в структуре покрова сосудистых растений; 2) увеличение богатства почв должно приводить к увеличению надземной биомассы сосудистых растений, смыканию их полога и уменьшению участия лишайников в составе фитоценоза; 3) при изоляции участков дернины под пятнами лишайников путем обрубки корней эти участки должны быстрее заселяться сосудистыми растениями по сравнению с участками лишайников, не изолированными обрубкой (контролем). Для проверки этих положений и были проведены три цикла экспериментов (Павлов и др., 1998; Onipchenko, 1994; Onipchenko et al., 2004).
В эксперименте с удалением лишайников кустистые эпигейные лишайники были аккуратно извлечены пинцетом с небольших площадок. Проективного покрытия сосудистых растений оставалось около 40 % в течение всего времени наблюдений (10 лет). Голые участки почвы, бывшие под пятнами лишайников, практически не уменьшились в размерах и не зарастали сосудистыми растениями, а местами поверхность почвы разрыхлялась под действием «морозного рыхления». При внесении элементов минерального питания существенные изменения биомассы растений и структуры сообщества произошли лишь в вариантах с внесением N. Флористический состав сообщества не изменился в течение 5 лет эксперимента, но увеличение надземной биомассы было весьма существенным
4. Пространственная структура фитоценозов
305
Рис. 6.21 • Схема эксперимента по изучению механизмов формирования горизонтальной структуры альпийских пустошей: 1 — удаление лишайников, 2 — внесение в почву элементов минерального питания, 3 — корневая обрубка под пятнами лишайников (по: Onipchenko, 1994)
(трехкратным), особенно для злаков и осок. Биомасса лишайников уменьшилась на вариантах NP и N. При обрубке участков дернины под пятнами лишайников эти участки дернины были изолированы от корней соседних растений. Численность побегов и всходов на экспериментальных площадках медленно возрастала с небольшими флуктуациями в течение 17 лет наблюдений. Значимое увеличение суммарной численности побегов сосудистых растений по сравнению с контролем отмечалось с 10-го года наблюдений. В целом результаты проведенных экспериментов свидетельствуют в пользу обсуждаемой гипотезы о ведущей роли бедности почв и корневой конкуренции в формировании мелкомозаичной горизонтальной структуры альпийских пустошей.
Горизонтальная структура и динамика сообществ зависит от характера взаимоотношений между видами. Yodzis (1978) ввел представление о двух типах контроля — контроле основателем и контроле доминированием. В первом случае изначально занятое особью какого-либо вида место не может быть занято другими растениями до естественной гибели первопоселенца («основателя»). Если возможно конкурентное вытеснение (замещение) на данном месте, то это контроль доминированием. В реальных сообществах оба типа контроля имеют место. Чем меньше время
306
Глава 6. Состав и структура фитоценозов
жизни модулей более слабого вида и скорость горизонтального разрастания потенциального доминанта, тем менее выражен контроль доминированием в сообществе. Контроль доминированием был показан Rydin (1985) для сообщества сфагновых мхов. Контроль основателем может поддерживать высокое видовое разнообразие сообщества в широком диапазоне условий (Herben, 1995), он создает более гомогенную тонкодисперсную горизонтальную структуру сообщества (Herben, Нага, 1997).
Важность контроля основателем экспериментально проверена на смешанном посеве растений разных функциональных групп (Korner et al., 2008). Авторы исследовали влияние последовательности заселения видов на формирование видового состава. Задержка в посеве на 3 недели приводила к существенным различиям в составе сообщества, сохраняющимся в течение 4 укосов (двух вегетационных сезонов). Успех в раннем развитии корневых систем обеспечивал доминирование видов в дальнейшем, он перевешивал эффект взаимодействий в надземной сфере. Первые поселяющиеся функциональные группы видов обеспечивали доминирование в образовавшемся сообществе на 70 % (бобовые), 98 % (разнотравье) и 94 % (злаки) через 2 месяца после посева. Наибольшая итоговая биомасса, близкая к таковой на синхронном посеве, отмечена при начальной посадке бобовых, существенно меньшая — при начальной посадке видов злаков или разнотравья (Korner et al., 2008).
Другой пример ценобиотической мозаичности связан с пространственным распределением подроста и взрослых деревьев Quercus тасго-carpa (Dickie et al., 2005). Было показало, что лучше всего подрост развивается на расстоянии 12-20 м (в среднем 15 м) от взрослых деревьев, где его развитию благоприятствуют эктомикоризные связи со взрослыми деревьями. На более близком расстоянии на первый план выступает конкуренция.
4.2.4 • Клональная мозаичность и разделение труда
Вегетативное разрастание растений происходит с разной скоростью, а захват нового пространства происходит на разное время. Lovett Doust (1981; Silvertown, Wilson, 2000) предложил различать две стратегии вегетативного разрастания растений: фаланга (phalanx) (например, у белоуса) и партизаны (guerilla) (например, у полевицы). В первом случае медленное разрастание сопряжено с длительным удержанием территории, а во втором происходит быстрый захват освободившихся удаленных участков, но в то же время быстрое замещение более конкурентоспособными видами.
Вегетативно-подвижные (клональные) растения часто имеют преимущества при использовании пятнистой (по ресурсам) среды. Пятна разных ресурсов (например, влага в почве и «солнечные окна») могут не совпадать и даже
4. Пространственная структура фитоценозов
307
Рис. 6.22 • Схема «разделения труда» между раметами. Изолированные раметы, или неклональные растения (верхний ряд), развивают относительно большую биомассу корней при недостатке почвенных ресурсов и большую биомассу листьев при недостатке света. Напротив, связанные столонами раметы (нижний ряд) развивают большую массу листьев на лучше освещенных участках и большую биомассу корней на участках, обогащенных почвенными ресурсами, поскольку между этими раметами происходит компенсация дефицитных ресурсов путем передачи продуктов фотосинтеза, воды и элементов минерального питания (по: Stuefer, 1997)
чередоваться наподобие шахматной доски. В этом случае преимущества клональных растений зависят от возможности транспорта продуктов фотосинтеза и ЭМП между раметами (особями вегетативного происхождения). Этот транспорт зависит от строения проводящей системы растений и не всегда носит реципрокный (взаимный) характер. Возможность реципрокного транспорта воды и ассимилятов между раметами показана для Trifolium repens и Potentilla anserine(Stuefer, 1996; Stuefer et al., 1996). На рис. 6.22 схематично
308
Глава 6. Состав и структура фитоценозов
показано, как вегетативно-подвижные растения могут эффективно использовать гетерогенную среду за счет разделения труда между раме-тами для получения ресурсов. Неклональные (неподвижные) растения в случае недостатка почвенных ресурсов вынуждены развивать большую биомассу подземных органов для получения этих ресурсов, а в случае недостатка света — большую листовую поверхность. Напротив, в условиях соединения побегов растений корневищами или столонами специализация рамет, например у Trifolium repens, вызывает лучшее развитие поглощающих органов в условиях обилия ресурсов (листьев при освещении, корней на богатых участках почвы). Побеги, находящиеся при недостаточном освещении, получают продукты фотосинтеза от хорошо освещенных побегов, снабжая их, в свою очередь, водой и ЭМП (Stuefer et al., 1996).
Но такое «разделение труда» наблюдается лишь в условиях пространственной гетерогенности почв. В экспериментах с рядом видов корневищных осок (de Kroon et al., 1996) с использованием дейтерия было показано, что при равномерном поступлении воды к раметам транспорт воды между ними практически отсутствовал. При пятнистом поступлении воды наблюдался интенсивный перенос воды от лучше обеспеченных рамет к хуже обеспеченным, причем как в акропетальном (от основания к верхушке), так и в базипетальном (от верхушки к основанию) направлениях.
Растения, образующие плотные клоны, могут существенно изменять условия произрастания как для побегов своего вида, так и для других растений. Примером может служить клональный вид Scirpus holoschoenus в Италии, который образует кольца с периферическим разрастанием. Большая масса подземных органов внутри кольца состоит почти целиком из отмерших корней. Появление всходов Scirpus и их рост были резко подавлены во внутренней части кольца. Но рост Pulicaria dysenterica и Mentha aquatica, их семенная продукция и биомасса отдельных растений были существенно выше во внутренней (дегенеративной) зоне кольца Scirpus, что, вероятно, связано со специфическими изменениями почвенных свойств в результате произрастания этого вида. Поглощение литиевой метки корнями Scirpus из почвы снаружи кольца было примерно на два порядка выше, чем корнями из внутренней части кольца. Это исследование подтверждает представления об отрицательной обратной связи при развитии Scirpus — этот вид ухудшает условия произрастания для особей своего вида, но улучшает для других видов (Bonanomi et al., 2005).
Как отмечалось в п. 3.2.5 гл. 6, горизонтальная структура сообществ влияет на конкурентные отношения между растениями: наличие одновидовых мозаик уменьшает межвидовую конкуренцию. Это было четко
4. Пространственная структура фитоценозов
309
показано в экспериментах J. van Andel, Т. Dueck (1982), которые выращивали 3 вида однолетних крестовников (Senecio vulgaris, S. sylvaticus, S. viscosus) в монокультурах, гомогенной смеси (особь каждого вида была в соседстве только с особями двух других) и в гетерогенной смеси видов, образующих одновидовые группы по 8 особей. Было показано, что в монокультуре и группах урожай семян для каждого вида не менялся, а в гомогенном посеве более быстро развивающийся S. vulgaris имел на 250 % больше семян, в то время как другие виды резко снижали урожай семян в этом варианте: S. sylvaticus — на 300 % и S. viscosus — на 215 % по сравнению с их монокультурами и одновидовыми пятнами.
Аналогичные эксперименты с 4 видами однолетников провели Р. Stoll и D. Prati (2001). Было показано, что в условиях конкуренции у менее конкурентоспособных видов агрегированное распределение особей увеличивало надземную продукцию по сравнению со случайным распределением, а у более конкурентоспособных — уменьшало.
4.2.5 • Теория «масс-эффекта» и модель «карусели»
Как отмечено выше, термин «масс-эффект» (синонимы: пространственная автокорреляция, висинизм и др.) был предложен A. Shmida и S. ЕПпег (1984) для объяснения присутствия в сообществе или на его отдельных участках видов, популяции которых поддерживаются за счет постоянного потока диаспор с соседних сообществ или участков (см. п. 3.2.2 гл. 6). Примером могут служить всходы и ювенильные особи деревьев на сенокосном лугу у опушки леса. Семена деревьев поступают в большом количестве из леса, но при сенокосном использовании луга не могут развиться там во взрослые растения, хотя и постоянно встречаются в составе лугового сообщества. В пространственной структуре сообщества «масс-эффект» проявляется в большей вероятности встречи того или иного вида растений на площадке, если на соседней площадке присутствуют особи этого вида. Следствием этого является большее флористическое сходство близко расположенных площадок по сравнению с удаленными. Поэтому количественно роль «масс-эффекта» в формировании структуры сообществ можно оценить по степени уменьшения флористического сходства между площадками при увеличении расстояния между ними на местности. Примером может служить сравнение кривых «флористическое сходство — расстояние» для альпийских сообществ четырех типов (рис. 6.23) на площадках 50 х 50 см. В этом случае альпийские пустоши представляют собой наиболее «гомогенное» сообщество, а пест-роовсяницевые луга — наиболее «гетерогенное». Аналогичные эффекты уменьшения флористического сходства с расстоянием могут быть вызваны градиентами изменения почвенных свойств, однако в исследованных
310
Глава 6. Состав и структура фитоценозов
Рис. 6.23 • Изменение флористического сходства (коэффициент Съеренсена, %, ордината) между площадками 50 х 50 см при увеличении расстояния между ними (метры, абсцисса). Сообщества: АЛП — альпийская лишайниковая пустошь, АК — альпийский ковер, ПЛ — пестроовсяницевый луг, ГКЛ — гераниево-копеечниковый луг (по: Онипченко, Покаржевская, 1994)
Таблица 6.4 • Классификация видов по их мобильности внутри сообщества на основании частоты встречаемости на мелких площадках в пространстве и времени (по: van der Maarel, 1996)
Встречаемость во времени
низкая (<33%) средняя (33-66%) высокая (>66%)
Встречаемость в про-странстве низкая (<25%) случайные локальные
средняя (25-75%) флуктуирующая (большая разница по годам) пульсирующие
средняя (25-75%) аккумулирующая (небольшая разница по годам) циркулирующие
высокая (>75%) константные
4. Пространственная структура фитоценозов
311
Arenaria serpyllifoUa «случайный» F срцд = 1.4
F F (уыул
1986 0 0
X987 4 4
X988 .. . 0 4
X989 0 4
X990 £ 5 9
X99X ♦ 2 XX
X992 .. 0 XX
X993 X 12
X994 ±. • X X3
Veronica spicata
«локальный»
Fw = 6.8
F F (уыул
X 9 8 8 •••••••++•+•••••••••••••••• ••+++••••••• 7 7
X 9 8 9 •••••••++•+••••••••••••••• +++•••• 7 7
X990 •••••••++•+•••••••••••••••••••++••••••• 5 7
X99X ...........♦ ..............±. ♦♦♦.............. 9 9
X992 •••••• • + • •••••••••••••••••••• • • • •. 5 XO
X993 ... .±.±. <*>.<*>*...........+ ..++++........ XO X3
X994 .......................................... 8 14
Trifolium striatum « пульсирующий » F w = 6.1
F F«ywyn
1986 9 9
X987 X XO
X988 2 XX
X989 О XX
X990 ..♦♦♦♦♦♦♦ . +♦♦♦♦♦♦♦+♦......... 21 25
X99X 0 24
X992 0 24
X993 ♦.♦.♦♦♦♦♦.. 22 29
X994 0 29
Anthyllis vuliieraria « циркулирующий » F = 20.0
F F (уыул
X986 ......21 21
X987 ♦♦♦♦.♦±♦♦.±1....±...♦±*.♦♦♦♦.♦ 23 29
X988 ♦♦♦♦♦♦♦♦♦.♦♦.!..++++++++.+ 29 32
X989 ♦♦♦♦♦♦♦♦♦♦.♦.♦..♦♦♦♦.....!♦♦♦♦♦♦.♦.♦ 25 33
X990 .....22 35
X99X *±...♦♦♦♦♦♦♦♦ 33 38
X992 ♦...♦....♦.................. ♦........ 8 38
X993 .....♦........................... 6 38
X994 24 38
Рис.в.24 • Примеры изменения встречаемости видов растений разных типов мобильности на 40 площадках 10 х 10 см в альварных сообществах о. Оланд (Швеция). Присутствие вида отмечено «+», присутствие на ранее не занятой этим видом площадке — «±», точкой отмечено отсутствие надземных частей растений. Показана текущая, средняя и кумулятивная пространственная встречаемость (по: van der Maarel, 1996)
312
Глава 6. Состав и структура фитоценозов
альпийских сообществах Северо-Западного Кавказа почвенные свойства не показали аналогичных изменений (Онипченко и др., 1998). Это позволяет заключить, что пространственная гетерогенность внутри альпийских фитоценозов (в масштабе 1-5 м) обусловлена не почвенной неоднородностью, а другими причинами, среди которых важную роль играет «масс-эффект», связанный с особенностями вегетативного размножения растений и распространением их диаспор.
Модель «карусели», предложенная Е. van der Maarel (1996), говорит о том, что виды травянистых растений могут сменять во времени друг друга на мелких площадках случайным образом. Он приводит классификацию видов в зависимости от их пространственной и временной встречаемости на мелких площадках (табл. 6.4).
«Случайные» виды имеют низкую пространственную и временную встречаемость, к ним относятся малолетние растения, появляющиеся на разных площадках в разное время, как, например, Arenaria serpyllifo-Па (рис. 6.24). «Локальные» виды имеют низкую пространственную встречаемость, но длительное время произрастают на отдельных площадках, как, например, Veronica spicata. «Пульсирующие» виды резко увеличивают свою встречаемость в отдельные годы, снижая ее в последующие, а «циркулирующие», напротив, имея высокую пространственную встречаемость, могут резко снижать ее в отдельные годы.
4.2.в • Возрастное состояние и «gap^-парадигма
Особое значение для организации фитоценозов имеет мозаичность, связанная с изменением возрастного состояния доминантов. Для лесов этот подход в первой половине XX в. разрабатывал шведский ученый Rutger Sernander (1866-1944), однако его работы были малоизвестны, так как публиковались на шведском языке. Впервые детально описал этот тип мозаичности для разных типов монодоминантных сообществ A. Watt (1947), который показал, что эти сообщества состоят из разновозрастных пятен, образуемых доминантами, и что устойчивость сообщества в целом обусловлена постоянно идущими «микросукцессиями» в каждой его точке. Классическим объектом у A. Watt (1947) были заросли орляка (Pteridi-um aquilinum). Разрастаясь на новую территорию с помощью корневищ, растения проходили несколько стадий: пионерную, стадию построения, зрелую и стадию разрушения (рис. 6.25). По краю заросли эти стадии образовывали четкую систему колец, в то время как внутри заросли отмечалась мозаика участков, находящихся на разных стадиях развития орляка. Аналогичные закономерности A. Watt (1947) описал для вересковой пустоши, овсяницевого луга, букового леса и других сообществ.
4. Пространственная структура фитоценозов
313
Край заросли орляка
Высота вам
Внутренняя часть заросли
пионерная стадия зрелая стадия
построения разрушения
Рис. 6.25 • Схема, иллюстрирующая связь между стадиями развития краевого фронта орляковой заросли (Pteridium aquilinum) и пространственной структурой внутренней части заросли в Великобритании (Lakenheath Warren): а — изменение высоты вай и глубины залегания корневища от края заросли в сторону центра; б — относительный размер и направление роста корневищ орляка, основные ветви корневища параллельны с краю заросли и образуют сеть внутри нее; в — смена стадий развития участков корневища с краю заросли и мозаика стадий внутри заросли (по: Watt, 1947)
В дальнейшем были изучены сходные смены с участием двух или нескольких видов растений. Пример такой циклической смены двух группировок приводит Н. Godinez-Alvarez et al. (2002; Moore, 2002). В пустынях Мексики наблюдаются смены участков с доминированием гигантского кактуса Neobuxbaumia tetetzo и участков с кустарником Mimosa luisana. Всходы этого кактуса выживают только под пологом кустарника М. luisana, который погибает, когда кактус достигает значительных размеров. На месте
314
Глава 6. Состав и структура фитоценозов
Рис. 6.26 • Схема элементов мозаично-ярусной структуры естественных лесов: синхронно развивающиеся популяционные локусы древесных видов на местах бывших прорывов полога (по: Смирнова, 2004)
отмерших кактусов снова поселяется М. luisana, таким образом происходит смена пятен кактусов и кустарниковой мимозы. Питающиеся плодами кактуса летучие мыши с вероятностью 72% откладывают экскременты под покровом мимозы, тем самым способствуя смене участков кустарников на заросли кактуса. Циклическая динамика двух доминантов вересковых болот с участием огня описана в п. 3.9.1.2 гл. 9.
Этот подход значительно позднее был подхвачен многими исследователями лесов, особенно тропических (Ричардс, 1952), и в дальнейшем получил название «новой лесной парадигмы». Основные положения этой парадигмы следующие (по: В. Н. Короткову, 1991):
1. Вне зависимости от географического положения и флористического состава естественные леса имеют сходные принципы мозаичноярусной организации.
2. Естественные леса представляют собой сукцессивную (последовательно сменяемую) мозаику разновозрастных элементов.
4. Пространственная структура фитоценозов
315
Таблица в.5 • Масштабы нарушений, вызывающих мозаичность в лесах (по: Clark, 1990)
Размеры мозаик, м2 Причины возникновения
100000-10000000 Ураганы, паводки, землетрясения, пожары
1000-100000 Штормовые ветры, групповые вывалы в результате болезней
100-1000 Вывалы отдельных деревьев
10-100 Опадение отдельных ветвей
3. Элементы мозаично-ярусной структуры выделяются по скоплениям синхронно развивающихся популяционных локусов древесных видов. Скопления формируются в естественных лесах после нарушений, приводящих к образованию прорывов в сплошном пологе леса (рис. 6.26).
4. Размеры прорывов в пологе леса определяют видовой состав успешно возобновляющихся древесных видов и их количественные сочетания.
5. Онтогенез деревьев в лесных ценозах, включающий стадии молодости, зрелости, старения и смерти, определяет популяционную жизнь многих «подчиненных» видов автотрофов и ряда гетеротрофов.
6. Устойчивое существование лесного массива возможно лишь при условии закономерного сочетания элементов мозаично-ярусной структуры, находящихся на разных стадиях развития.
Прорывы полога («gaps», «окна», нарушения) в лесах могут иметь различные размеры в зависимости от факторов, их вызывающих (табл. 6.5). Экологические условия в местах выпадения деревьев резко меняются: усиливается минерализация органических соединений в почве, увеличивается освещенность и т. п. Увеличение поступления света зависит от соотношения размеров «окна» и высоты полога окружающего древесного яруса (рис. 6.27).
Выпадение деревьев из древостоя может происходить двумя путями: со сломом ствола (бурелом) или с выворачиванием части корневой системы и почвы (ветровал) (рис. 6.28). Во втором случае происходит существенное нарушение почвенного покрова и формируется ветровальнопочвенный комплекс (Скворцова и др., 1983), в котором можно выделить следующие элементы (Смирнова, 2004):
316
Глава 6. Состав и структура фитоценозов
D
D:H Освещенность, % от полной
0:1 10
0,5:1 20-45
1:1 45-70
2:1 65-90
Рис. 6.27 • Изменение условий освещенности в «окнах» в зависимости
от соотношения диаметра (D) и высоты полога (Н) (по: Minckler, Woerheide, 1965)
а) область вне вывала без нарушений почвы и напочвенного покрова;
б) вывальный ком, формирующий в дальнейшем локальное повышение микрорельефа;
в) вывальная яма с удаленными верхними горизонтами почвы, характеризующаяся повышенным увлажнением;
г) валежина (упавший ствол), который в процессе разложения увеличивает локальную гумусированность и часто служит субстратом для лучшего приживания всходов древесных пород (см. п. 3.2 гл. 7).
В зависимости от размеров прорывов полога их зарастание происходит по-разному. Мелкие прорывы могут заполняться боковым разрастанием крон соседних деревьев, которое усиливается в направлении лучшего поступления света в связи с чувствительностью фитохромов в побегах к отношению красный дальний красный свет. В небольших «окнах» с большей вероятностью развиваются теневыносливые деревья, а в более крупных способны произрастать светолюбивые породы.
Для возобновления каждого вида деревьев требуется своя площадь «окон». Она больше для светолюбивых пород, меньше для теневыносливых, но зависит от изменения светолюбия в ходе онтогенеза (см. п. 3.4 гл. 7). В тропических лесах Мексики эта площадь составила для светолюбивой Cecropia obtusifolia 5 га, а для теневыносливого Astrocaryum тех-
4. Пространственная структура фитоценозов
317
Рис. 6.28 • Выпадение старых деревьев из древостоя: а — со сломом ствола, без нарушения почвенного покрова (Betula pendula); б — с выворачиванием части корневой системы и значительным нарушением почвенного покрова (Pinus silvestris), Приокско-Террасный заповедник, Московская область
icanum — 0,06 га (Martinez-Ramos et al., 1988, 1989; Коротков, 1991). Для основных лесообразующих пород широколиственных лесов Восточной Европы О. В. Смирнова и др. (1988, 1989) рассчитали минимальную площадь, на которой может осуществляться полный оборот поколений.
Таблица 6.6 • Минимальные размеры популяционных локусов видов деревьев широколиственных лесов европейской части России, обеспечивающих нормальный оборот поколений (по: Чистякова, 1994)
Популяционная характеристика Бук Дуб Ясень Липа Клен остролистный Клен полевой Ильм Граб
Площадь кроны взрослого дерева, м2 150-200 100-300 150-300 100-300 100-300 50-150 120-300 70-100
Минимальный размер локусов, м2: имматурных виргинильных генеративных 20-50 150-200 200-400 300-500 1000 1500-2500 50-100 250-300 1000-1500 200 300 400-500 20 70-100 400-500 10 20 100-200 50 250 400-500 30 100 400-500
Площадь устойчивого существования популяций, га 1,0-5,4 12,8- 42,5 5,0-13,3 1,1-2,7 1,1-1,8 0,4-1,1 1,0-1,8 0,2-1,2
318 Глава 6. Состав и структура фитоценозов
4. Пространственная структура фитоценозов
319
Эта площадь минимальна для наиболее теневыносливых пород (Acer platanoides, Acer campestre, Carpinus betulus, Ulmus glabra) — до 2 га, в то время как для дуба она составляет несколько десятков гектаров (табл. 6.6).
Наличие прорывов в пологе характерно не только для лесных фитоценозов. Аналогичные структуры могут наблюдаться в травяных сообществах, где они связаны главным образом с деятельностью землероев. Произрастание отдельных видов травянистых растений может быть связано с различными по размерам и интенсивности нарушений участками (см. п. 7 гл. 4).
4-2-7 • Модель Тильмана: пространственная структура и колонизационная способность
Tilman (1994) предложил модель, учитывающую пространственные ограничения в возможностях поступления семян в сообщество. Он показал, что в однородной пространственной среде может сосуществовать большое число видов растений (в модели 40, а теоретически не ограничено), отличающихся по двум параметрам: колонизационной способности (вероятности попадания диаспоры на освободившийся участок) и длительности жизни особей. Эти свойства растений должны быть отрицательно скоррелированы, т. е. чем выше колонизационная способность, тем меньше длительность жизни. В этом случае вид с наибольшей длительностью жизни не способен быстро освоить все освободившиеся участки, на которые с большей вероятностью вселяются короткоживущие виды. Модель проверена на примере участка прерии с доминированием Schizachyrium scoparium, который имеет низкую пороговую концентрацию по нитратному азоту в почве и может рассматриваться в качестве суперконкурента (см. п. 3.1 гл. 5). В плотный посев этого вида практически не внедряются другие виды, в то время как естественные сообщества с доминированием Schizachyrium scoparium флористически очень богаты, что автор объясняет ограниченными возможностями семенной репродукции. Tilman (1994) делает вывод о возможности увеличения участия редких видов за счет подсева их семян. От <^ар»-парадигмы эти взгляды отличаются представлением о континуальном изменении свойств колонизационной способности, а не наличии двух-трех групп видов, различающихся по этим параметрам. Кроме того, Tilman et al. (1997) показали, что доминирующие виды устойчивых сообществ, обладающие большой конкурентоспособностью при поглощении ресурсов, но малой колонизационной способностью, весьма уязвимы к антропогенной фрагментации ландшафтов. Они исчезают одними из первых из состава сообществ, если таковая имеет место.
320 Глава 6. Состав и структура фитоценозов
На основании этих представлений была создана модель конкуренции в пространственной среде (Lehman, Tilman, 1998). Ее поведение сильно зависело от соотношения смертности и колонизационной способности входящих в нее видов. При анализе поведения модели было сделано два важных вывода:
1) коэффициент вариации (по времени) участия вида в сообществе уменьшался с увеличением степени доминирования вида, т. е. более редкие виды сильнее флуктуировали по годам;
2) с уменьшением конкурентоспособности (доминирования) увеличивается степень агрегированности пространственного размещения особей вида. Таким образом, виды с более агрегированным пространственным размещением сильнее изменяют свою численность по годам.
Глава _____
/ Онтогенез
растении в фитоценозах
Латентный период (жизнеспособные диаспоры) Виргинильный период
• Генеративный период
• Вегетативное размножение растений. Клональные растения
• Длительность жизни растений
1 • Основные понятия и термины периодизации онтогенеза
Популяционный подход в экологии растений предполагает рассмотрение различных аспектов структуры популяций, образующих растительные сообщества. В настоящем разделе будут рассмотрены лишь онтогенетические (возрастные) состояния растений. Т. А. Работнов (1950) и А. А. Ура-нов (1967) предложили следующую периодизацию онтогенетических состояний, широко используемую в нашей стране:
1. Первичный покой (латентный период).
2. Виргинильный (прегенеративный) период — от прорастания семени до образования генеративных побегов — подразделяется на проростки, или всходы (р), ювенильные особи (j), имматурные (im) и взрослые вегетативные (v) особи.
3. Генеративный период, часто подразделяемый на три подпериода (gl, g2, g3).
4. Постгенеративный, или сенильный, период часто подразделяется на субсенильный (ss) и собственно сенильный (s).
В связи со значительной ролью вегетативного размножения в поддержании популяций многих видов растений важно бывает различать особи семенного и вегетативного происхождения. Генетой называют особь, происходящую из семени, и совокупность всех ее вегетативных потомков, или рамет (Harper, 1977). Генеты, как правило, различаются генетически, в то время как раметы генетически идентичны (если не учитывать соматические мутации).
Доли особей вегетативного и генеративного происхождения сильно различаются в клонах разных растений. Например, по наблюдениям над зарослями арктической осоки Carex bigelowii, в популяциях которой практически отсутствуют проростки, ее генетическая изменчивость была очень высока, и на 85-88 изученных рамет приходилось 41-55 различных генет. Это свидетельствует о большой роли генеративного размножения, по крайней мере в недалеком прошлом (Jonsson et al., 1996).
Кроме широкого представления о популяции как о любой совместно обитающей группе организмов одного вида в отечественной литературе часто используют термин ценопопуляция, под которой понимают совокупность особей одного вида в пределах фитоценоза (термин В. В. Петровского, 1961). Каждая (цено)популяция может быть охарактеризована своим онтогенетическим (возрастным) составом, часто называемым также спектром.
2. Латентный период (жизнеспособные диаспоры)
323
2 • Латентный период
(жизнеспособные диаспоры)
2.1 • Размеры семян
Размеры семян у покрытосеменных растений варьируют больше чем на 10 порядков. Одними из наиболее мелких семян характеризуются орхидные (например, Goodyera repens — 2 х 10-6 г, рекорд у орхидных — 0,31 х 10"6 г, а число семян на коробочку достигает 4 млн (Arditti, Ghani, 2000)). С другой стороны, наиболее крупные семена Lodoicea maldivica (сейшельская пальма) достигают 2 х 104 г. Размеры семян у растений одного вида обычно меньше варьируют, чем другие органы растений, хотя у ряда видов встречается гетерокарпия и гетероспермия.
Виды растений с мелкими семенами встречаются чаще, чем с крупными. Изучение большого числа видов показало наличие следующей (фрактальной) зависимости: частота видов с данным размером семени обратно пропорциональна квадратному корню из размеров семени (Hegde et al., 1991).
Мелкие семена — часто результат отбора на непоедаемость позвоночными: например, показано, что семена весом менее 3 мг практически не поедаются. В целом размер семян является компромиссом между: а) потребностью всходов в питании (чем больше, тем лучше), б) защитой от фитофагов (чем больше, тем привлекательнее для фитофагов), в) дальностью распространения (чем легче, тем дальше), г) общим числом семян, образуемых растением. Наблюдается тенденция к увеличению размеров семян с увеличением размеров самого растения. Эта тенденция была подтверждена при сравнительном анализе умеренных флор пяти регионов (Leishman et al., 1995). Очень крупные семена растений тропических островов (сейшельская пальма) могут быть связаны с давлением отбора на преимущество семян с большим запасом веществ при отсутствии обратного давления со стороны фитофагов (Janzen, 1983). Крупные семена пальм (кокосы) часто содержат жидкий эндосперм, помогающий решать проблемы недостатка воды на ранних стадиях развития проростка.
Крупносемянные виды вкладывают большую часть ассимилятов в защитные структуры семени. Так, у Erigeron canadense при массе семени 0,072 мг семенная кожура составляла 15% его массы, а у Тгадородоп pratense (масса семени 10,3 мг) — уже 61 % (Fenner, 1983; Kitajima, 2007).
Масса семени во многом зависит от систематической принадлежности таксона. Анализ большого числа наблюдений показал, что 55 % дисперсии логарифма массы семени приходится на уровень порядка и выше, 12 % — на уровень семейств внутри порядков, 26 % — между родами внутри семейств, 8% — между видами внутри родов (Westoby et al., 1997).
324
Глава 7. Онтогенез растений в фитоценозах
2.2 • Происхождение семенных банков в почве
Почва служит важным хранилищем покоящихся диаспор растений — семян, плодов, вегетативных зачатков, спор. Устойчивое существование и поддержание разнообразия многих растительных сообществ было бы невозможно без такого хранилища генетической информации.
По источникам поступления почвенные семенные банки отдельных видов растений могут быть отнесены к одному из трех типов (Работнов, 1992):
1. Автохтонные — семена поступают от растений, плодоносящих в непосредственной близости от мест их погребения, эти растения входят в состав современных растительных сообществ данной территории.
2. Инвазионные — семена приносятся к месту их погребения различными агентами (ветер, животные и пр.) из других растительных сообществ; растения, их образующие, не встречаются поблизости.
3. Реликтовые — принадлежат растениям, в настоящее время отсутствующим или не плодоносящим в непосредственной близости от мест погребения, но эти растения обсеменялись на данном месте раньше, в предшествующие сукцессионные стадии развития растительного сообщества. Для этих растений нехарактерен дальний перенос семян, поэтому они практически не могли попасть на данную территорию в результате заноса.
Интересным примером сохранения реликтового семенного банка в почве может служить присутствие в почвах некоторых прибрежных лугов в Великобритании семян практически исчезнувшего здесь вида — фиалки персиколистной (Viola persicifolia). Семена этого вида адаптированы к прорастанию в условиях резкопеременного водного режима пойменных сообществ, который, однако, был кардинально изменен человеком при строительстве плотин, что и привело к исчезновению этого вида. Несмотря на это, Viola persicifolia несколько раз была обнаружена в семенном банке луговых сообществ (Pullin, Woodell, 1987; Thompson, 1992).
2.3 • Покой семян
Как показали теоретические исследования Rees (1997), покой семян (отложенное прорастание) может быть эволюционно выгоден растениям в следующих случаях:
1) изменения среды (например, выпадение осадков) непредсказуемы;
2) семена способны определять наступление этих изменений в среде;
3) между близкородственными потомками наблюдается конкуренция, действие которой смягчается при разновременном прорастании семян.
2. Латентный период (жизнеспособные диаспоры)
325
Таблица 7.1 • Классификация типов покоя семян (по: Nikolaeva, 1977; Baskin, Baskin, 1998)
Тип покоя Причина Выход из покоя
Эндогенный покой
Физиологический Физиологический механизм ингибирования (ФМИ) прорастания Тепловая и/или холодная стратификация
Морфологический Недоразвитый зародыш Благоприятные условия для роста зародыша и прорастания
Морфофизиологический ФМИ прорастания и недоразвитый зародыш Тепловая и/или холодная стратификация
Экзогенный покой
Физический Непроницаемость для воды покровов семени или плода Открытие специализированных структур
Химический Ингибиторы прорастания, синтезирующиеся в структурах плода и проникающие в семена Вымывание ингибиторов
Механический (обычно сочетается с физиологическим) Механические структуры, ограничивающие рост зародыша Тепловая и/или холодная стратификация
Возможность сохранения жизнеспособности семян в почве обусловлена различными механизмами их покоя (табл. 7.1). Эндогенный покой связан с процессами, происходящими в зародыше семян и/или эндосперме, в то время как экзогенный — с покровными структурами семени и плода.
Покой семян может быть врожденным, а может формироваться в процессе их хранения или пребывания в почве (индуцированный покой). Такой покой вызывается, например, затенением семян листьями растений и снижением при этом физиологически важного соотношения красный : дальний красный свет в поступающем к семенам свете (Wesson, Wareing, 1969; Silvertown, 1982). Индуцированный покой связан с синтезом в семенах ингибиторов прорастания, он изменяется в зависимости
326
Глава 7. Онтогенез растений в фитоценозах
от сезона. Например, у свежесобранных (осенью) семян пустынного однолетника Eriogonum abertianum покой отсутствовал, он индуцировался зимой и почти исчезал к следующей осени. Таким образом, всхожесть семян сильно колебалась в зависимости от сезона (Baskin et al., 1993).
Выход семян из покоя и последующее их прорастание может стимулироваться следующими факторами:
1. Колебание температуры, которое более значительно у поверхности почвы. Наиболее важны колебания температуры для прорастания семян растений влажных, часто околоводных местообитаний, таких, например, как жерушник исландский (Rorippa islandica), горец перечный (Polygonum persicaria), которые не прорастают при постоянных температурах. У многих других растений колебания температур увеличивают всхожесть семян (Thompson, Grime, 1983).
2. Воздействие света.
3. Образование в почве нитратов. Эти факторы образуют так называемую систему, обеспечивающую семенам возможность определить их нахождение на нарушенном участке почвы, в прорыве полога растительности («gap-detection»). Прорастание в таких участках делает более вероятным успешное приживание всходов и развитие взрослого растения. Наличие микроучастков с обнаженной почвой способствует приживанию всходов многих видов растений. Например, на залежах в Мичигане было показано резкое увеличение приживания всходов травянистых монокарпиков на участках с искусственным удалением подстилки и надземных частей растений по сравнению с контрольными (ненарушенными) участками (см. Luken, 1990). В других случаях, наоборот, не полностью разложившиеся остатки растений могут быть хорошими субстратами для приживания всходов. В елово-тсуговых лесах на северо-западе Америки 94-98 % всходов деревьев отмечается по старым стволам деревьев, покрывающим только 6-11 % почвы в этом лесу (Harmon, Franklin, 1989). Таким образом, разлагающиеся бревна играют важную роль для приживания всходов деревьев в этих лесах как свободные от конкуренции субстраты, хорошо сохраняющие влагу.
4. Воздействие огня и дыма — комплексный фактор, вызывающий прорастание семян многих растений аридных областей, где пожары являются регулярными. Во время интенсивных пожаров почва выступает практически единственным убежищем, где сохраняются жизнеспособные семена. Активное прорастание семян после пожаров может быть связано со следующими факторами: 1) растрескиванием от огня твердых оболочек, 2) тепловой стимуляцией зародыша,
2. Латентный период (жизнеспособные диаспоры) 327
3) высокотемпературным иссушением семенных покровов, делающим их более проницаемыми для воды и газов, 4) воздействием этилена, аммония, двуокиси азота и других компонентов дыма или дымовых экстрактов. Так, у 12 из 28 изученных видов растений южноафриканского финбоша обнаружено стимулирование прорастания семян после обработки дымом. Дым сам по себе вызывал усиление прорастания семян растений из семейств Asteraceae, Ericaceae, Restionaceae, а дымовые экстракты — у видов Proteaceae (Brown, 1993). Аналогичные закономерности отмечены для многих растений западной Австралии, где из 94 видов растений с плохо прорастающими семенами 45 видов увеличили всхожесть после обработки холодным дымом. Усиление прорастания отмечено также при проращивании на фильтровальной бумаге, обработанной дымом, или поливе водой, содержащей компоненты дыма (Dixon et al., 1995; Keeley, Fotheringham, 1998).
5. Промывной водный режим почвы. Прорастание семян ряда пустынных однолетников происходит только после сильных осадков, пропитавших почву на значительную глубину и дающих возможность однолетникам завершить цикл своего развития до полного истощения запасов воды в почве. Механизмом «узнавания» такой ситуации у семян, находящихся в самых верхних почвенных слоях (смачиваемых и небольшими осадками), является вымывание ингибиторов прорастания фильтрующимися водами (Работнов, 1992). При искусственном полоскании семян этих растений в воде их покой быстро прерывается.
2.4 • Пространственная структура семенных банков
По расположению в почве (глубине погребения) семянной банк можно условно разделить на два типа: 1) активный — семена находятся в приповерхностном слое почвы, где возможно успешное прорастание семени и приживание всхода, и 2) потенциальный — семена располагаются в более глубоких слоях почвы, их успешное прорастание возможно лишь при выносе этих слоев ближе к поверхности при вспашке, нарушениях, в результате роющей деятельности землероев и т.п. (Работнов, 1992).
Жизнеспособные семена в большинстве случаев находятся преимущественно в поверхностных горизонтах почвы до глубины 5-10 см, на лугах отдельные всхожие семена найдены до глубины 40-50 см, существуют единичные находки в нижних горизонтах почвы до глубины 100-150 см. Более глубокому погребению семян в естественных условиях способствуют деятельность землероев, дождевых червей, отложение наилка
328
Глава 7. Онтогенез растений в фитоценозах
Рис.7.1 • Вертикальное распространение всхожих семян в почвах альпийских сообществ Северо-Западного Кавказа, Тебердинский заповедник (по слоям 2 см в процентах от общей численности всхожих семян).
Растительные сообщества: ALH — альпийская лишайниковая пустошь, FVG — пестроовсяницевый луг с доминированием плотнодерновинных злаков, GHM — гераниево-копеечниковый луг, SBC — альпийский ковер в условиях значительного снегонакопления, RC — рододендроновый стланик с большой мощностью опадогенного горизонта, FEN — альпийское приручьевое болото с хорошо выраженной аккумуляцией торфа (по: Semenova, Onipchenko, 1994)
в поймах, вывалы деревьев в лесах, отложение органических материалов в гидроморфных условиях, растрескивание почвы при иссушении в сухой период в сезонно-влажном климате (Работнов, 1992). Экспериментально показано, что искусственное погребение семян в почву существенно увеличивает длительность сохранения их жизнеспособности (Thompson et al., 1993).
Пример вертикального распределения всхожих семян в высокогорных почвах различных растительных сообществ Северо-Западного Кавказа по нашей работе (Semenova, Onipchenko, 1994) приведен на рис. 7.1. Наибольшее число всхожих семян в более глубоких слоях почвы отмечено на высокопродуктивном гераниево-копеечниковом лугу, где наиболее высока численность роющих грызунов — кустарниковых полевок, в рододендроновых стланиках с мощным органогенным горизонтом почвы, образовавшимся из опада Rhododendron caucasicum, и на альпийских болотах в гидроморфных условиях со значительным торфонакоплением.
Помимо почвы некоторые виды растений могут сохранять семена длительное время на надземных частях: например, в Северной Америке ряд видов сосен накапливают семена в шишках до пожара, во время которого они раскрываются. Некоторые травянистые растения умеренной зоны также могут иметь кратковременные надземные семенные банки. Например, Lithospermum officinale на меловых лугах в Голландии, а так-
2. Латентный период (жизнеспособные диаспоры)
329
Рис. 7.2 • Среднее число видов цветковых растений (флористическая насыщенность) на площадках 10 х 10 см для почвенного семенного банка (1) и современного растительного сообщества (2). Сокращения: FVG — пестроовсяницевый луг, GHM — гераниево-копеечниковый луг, SBC — альпийский ковер, RC — рододендроновый стланик, FEN — альпийское приручьевое болото, MSC — влажная альпийская осыпь, DSC — сухая альпийская осыпь (по: Semenova, Onipchenko, 1994)
же Carlina vulgaris. Семена поступают в почву весной при перегнивании стебля. При небольшом снежном покрове семена таких растений могут активно потребляться птицами (Willems, 1995).
Горизонтальное распределение семян в почвах обычно носит агрегированный характер. Наибольшее поступление семян отмечается вблизи материнских растений, хотя здесь и наиболее велико их потребление семеноядны-ми фитофагами. При переносе ветром разные субстраты, в зависимости от характера их поверхности, в разной степени улавливают семена, сохранность семян также сильно варьирует в разных условиях. В работе В. L. Gartner et al. (1983) приводится следующий пример неоднородности распределения всхожих семян Eriophorum vaginatum под разными пятнами растений в тундре (среднее число семян на квадратный метр):
пятна голого грунта 0
моховой покров 1200
пятна лишайников 0
кочки пушицы 63
Флористическая насыщенность (число видов растений на едини-
цу площади) для современных растительных сообществ и их семенных
330
Глава 7. Онтогенез растений в фитоценозах
банков обычно существенно различаются, прежде всего в связи с разными размерами взрослых растений и семян. Однако при рассмотрении на небольших по размерам площадках эти величины могут быть весьма сходны. Например, в сомкнутых альпийских сообществах эти показатели оказались весьма близки для площадок 10 х 10 см, однако на болоте и осыпях видовое разнообразие семян в почве было примерно вдвое выше, чем в современных сообществах (рис. 7.2). В целом же численность видов растений, сохраняющих в почве жизнеспособные диаспоры, сопоставима с флористическим богатством современных растительных сообществ, определенным по растениям в активных фазах онтогенеза.
2.5 • Длительность сохранения жизнеспособных семян
Семена разных видов растений сохраняют свою жизнеспособность в почве различное время. В работе К. Thompson и J. Р. Grime (1979) было предложено различать 4 типа семенных банков: I — временный банк с осенним прорастанием семян, II — временный банк с весенним прорастанием се-
Рис. 7,3 • Четыре типа семенных банков растений сезонного умеренного климата. Семена, готовые к прорастанию и прорастающие, показаны светлой областью, жизнеспособные, но покоящиеся семена — черной (по: Thompson, Grime, 1979). I — временный банк с осенним прорастанием семян, II — временный банк с весенним прорастанием семян, III — кратковременный устойчивый семенной банк (семена сохраняют всхожесть до 5 лет), IV — долговременный устойчивый семенной банк (семена сохраняют всхожесть более 5 лет)
2. Латентный период (жизнеспособные диаспоры)
331
О 0-0.5 0-1 0-1.5 0-2 0-2.5 0-3 0-3.5
Рис. 7.4 • Связь между массой диаспор и дисперсией их трех измерений для: а — семян 44 видов и б — плодов 53 видов растений. Черные кружки (•) — виды с устойчивым семенным банком в почве (длительность сохранения жизнеспособности не менее 5 лет); крестики (х) — виды с устойчивым семенным банком, сохраняющие жизнеспособные диаспоры менее 5 лет; ромбики (ф) — виды, длительность сохранения семян которых не определена. Пунктирная линия ограничивает область видов с длительно сохраняющимися диаспорами (по: Thompson et al., 1993)
мян, III — кратковременный устойчивый семенной банк (семена сохраняют всхожесть до 5 лет), IV — долговременный устойчивый семенной банк (семена сохраняют всхожесть более 5 лет) (рисЛЗХ С большой вероятностью оценить возможность растений Европы образовывать устойчивые семенные банки в почве можно по форме их семян (рис 7 Л; Thompson
332
Глава 7. Онтогенез растений в фитоценозах
et al., 1993). Семена, характерные для устойчивых семенных банков, часто имеют небольшие размеры, плотную и гладкую оболочку. Напротив, для видов, не образующих устойчивые семенные банки, более типичны крупные, часто удлиненные или уплощенные семена. На основании изучения 97 видов британской флоры было показано, что хорошее предсказание возможности сохранения семян в устойчивом семенном банке может быть сделано на основании веса семян и их формы, выраженной как дисперсия из трех размеров (у сферических = 0). В целом семена сохраняются в почве тем лучше, чем они мельче и чем их форма ближе к сферической, хотя есть и отдельные исключения (рис. 7.4). Однако эти закономерности не являются универсальными для всех регионов мира. Детальные исследования семян 101 вида австралийской флоры (Leish-man, Westoby, 1998) не подтвердили эту закономерность для растений Австралии.
Обычно длительно сохраняют жизнеспособность семена травянистых растений, хотя значительные постоянные семенные банки отмечены и для многих тропических деревьев. Из кустарников умеренных широт таким свойством обладает североамериканский вид Prunus pensylvan-ica, занесенный в Европу, семена которого могут сохраняться в почве 60-100 лет (Marks, 1974; Thompson, 1992).
Для оценки длительности сохранения всхожих семян в почве используют непрямые и прямые методы. Первые связаны с нахождением семян в почвах известного возраста, хотя и не исключают более позднее попадание семян в эти субстраты. Вторые основаны на прямом слежении за судьбой специально погребенных семян. Обширная база данных по численности и длительности сохранения семян отдельных видов растений северо-западной Европы была составлена К. Thompson et al. (1996); приводимые ниже данные частично взяты из этой базы.
По непрямым данным, семена полевых сорняков могут сохраняться в почве под лесом, имеющим возраст не менее 60-70 лет. В тундре, в слоях, близких к мерзлотным, найдены живые семена Luzula parviflo-ra, Carex bigelowii; радиоуглеродный возраст семенной кожуры составил около 200 лет. При археологических раскопках фундаментов зданий обнаружены семена ряда обычных видов растений, предположительно сохранявшихся там более 660 лет (Lamium album, Lamium purpureum, Taraxacum officinale) и более 460 лет (Glechoma hederacea и Trifolium repens).
Среди прямых данных наиболее известны эксперименты американских исследователей W. J. Beal и J. W. Т. Duvel (Toole, Brown, 1946; Kivilaan, Bandurski, 1981). Доктор Beal поместил семена 20 видов в песок в ши-рокогорлых стеклянных бутылках, которые были закопаны в землю гор
2. Латентный период (жизнеспособные диаспоры)
333
лышком вниз. Опыт был заложен в 1879 г., его результаты прослежены за 100 лет. Через 80 лет три вида сохранили всхожие семена (Oenotera sp., Rumex crispus, Verbascum blattaria), а через 120 лет проросли семена Verbascum blattaria, отмечен единственный проросток Malva rotundifolia (Telewski, Zeevaart, 2002).
Методически более корректные опыты доктора Дювеля (Duvel) (семена были помещены в почву в цветочные горшки и закопаны на разную глубину), к сожалению, продолжались лишь 39 лет (1902-1941). Из 107 исследуемых видов через 20 лет всхожие семена имел 51 вид, а через 39 лет — 39 видов (Toole, Brown, 1946).
Иногда оценить длительность сохранения всхожести семян можно после природных катастроф. Так, после погребения пеплом слоем около метра пахотной почвы в результате извержения вулкана в Японии через 10 лет в этой почве оставалось около 17 % всхожих семян от исходного семенного банка, относящихся к 25 видам (Tsuyuzaki, 1991).
Длительность сохранения жизнеспособных семян в почве положительно скоррелирована с содержанием в семенах орто-дигидроксифе-нолов, которые могут служить как защитными веществами против фитофагов, так и антиоксидантами. Наиболее высокая концентрация этих веществ отмечена в порядке Myrtales (Onagraceae и близкие семейства) (Hendry et al., 1994). С другой стороны, семена многих анемохорных (распространяемых ветром) видов обычно недолго сохраняют свою жизнеспособность. Это может быть связано (Lokesha et al., 1992) с накоплением в них липидов как запасных веществ. Липиды обладают высокой энергетической ценностью, что минимизирует вес семян при фиксированном содержании энергии, но липиды относительно быстро окисляются, что сокращает длительность жизни семян (Thompson, 2000).
Значительная гибель семян в почве может быть обусловлена воздействием фитопатогенных грибов. В экспериментах (Leishman et al., 2000) жизнеспособность семян двух (Medicago lupulina, Rubus fruticosus) из четырех исследованных видов через год хранения в почве значимо возрастала при обработке ее фунгицидами по сравнению с контролем.
2.6 • Почвенные банки в различных сообществах и природных зонах
Почвенные семенные банки характерны практически для всех наземных растительных сообществ различных природных зон. Ниже приводятся обобщенные данные по содержанию всхожих семян на квадратный метр (по сводкам: Работнов, 1982, 1986; Leek et al., 1989) в почвах различных природных зон:
334
Глава 7. Онтогенез растений в фитоценозах
арктические пустыни (Северная Земля) 300-700
тундры (Таймыр) 1500-4000
леса умеренного пояса (зрелые) 200-3000
северные степи до 10000
южные степи 2000-5000
прерии США 300-800
дождевые тропические леса 170-900
равнинные луга 10000
высокогорные луга, пустоши и болота 300-10000
агроценозы до 100000
Содержание семян в почвах пустынь сильно варьирует в разных регионах, в отдельных случаях оно превышает таковое даже для агроценозов. Так, в пустыне Мохаве (Mojave), Калифорния, семенной банк составил около 106000 с/м2 (в весовом отношении 38 г/м2), что значительно выше, чем в других североамериканских пустынях (Price, Joyner, 1997).
В холодных антарктических пустынях оба обитающих там вида сосудистых растений (Coiobanthus quitensis, Deschampsia antarctica) образуют значительные семенные банки в почве (100-1600 семян на м2) (McGraw, Day, 1997).
В пределах геохимических катен наиболее благоприятные условия для сохранения семян (высокая влажность, низкое содержание кислорода) наблюдаются в гидроморфных условиях, поэтому численность всхожих семян в почвах болот и заболоченных лугов обычно существенно выше, чем в автономных сообществах.
2.7 • Стратегии растений, сукцессии и смены семенных банков
Состав и соответствие семенных банков современной растительности существенно меняются в ходе сукцессий. В лесных сообществах, развивающихся после луговой стадии, семенной банк обычно обедняется. Например, в посадках ситхинской ели (Picea sitchensis) разного возраста найдено следующее количество всхожих семян (Hill, Stevens, 1981):
Возраст ели
5-10 лет
17-18 лет
29-30 лет
41-43 года
Число семян на 1 м2
2300
680
600
380
2. Латентный период (жизнеспособные диаспоры)
335
Таблица 7.2 • Изменение численности всхожих семян вереска и полевицы (семян/м2) в почве верещатника и березняка на разных стадиях развития березового древостоя (по: Hester et al., 1991)
Вид Возраст березы, лет
0 17 28 37 63
Calluna vulgaris 52900 43 800 34 900 17 900 9420
Agrostis capillaris 0 + 397 962 4750
В ходе вторичных (восстановительных) сукцессий происходит не только снижение численности семян в почвенном банке, но и изменение его состава. Так, по наблюдениям в Шотландии (Hester et al., 1991), при зарастании вересковых пустошей березой в напочвенном покрове сменяются следующие доминанты: вереск (Calluna vulgaris) —> черника (Vaccinium myrtillus) -> луговик извилистый (Deschampsia flexuosa) —> полевица (Agrostis capil-laris). При этом в почвенном семенном банке уменьшается численность семян вереска и возрастает — полевицы (табл. 7.2).
При изучении состава семенных банков фитоценозов, находящихся на разных стадиях сукцессии или при разных режимах нарушений, было выявлено, что в зрелых (климаксовых) растительных сообществах доминирующие виды растений не образуют семенного банка. Быстро теряют всхожесть и обычно летом не обнаруживаются в почве соответствующих лесов семена основных лесообразующих пород (сосна, пихта, ель, дуб), в почвах степей не сохраняются семена ковылей, а в почве альпийских лугов — семена основных доминирующих видов (Festuca varia, Geranium gymnocaulon, Hedysarum caucasicum), несмотря на большой ежегодный урожай семян этих видов (рис. 7.5; Semenova, 2004).
В работе Hopfensperger (2007) на основании анализа 108 публикаций по сходству состава семенных банков и современных сообществ трех типов (луга, леса, болота) (использован коэффициент Съеренсе-на) показано, что наименьшим сходством характеризуются леса (31 %), наибольшим — луга (54 %), средним — болота (47 %). Сходство уменьшалось с увеличением времени от последнего нарушения для лесов и болот, но увеличивалось — для лугов.
Напротив, наибольшее участие в составе семенных банков имеют растения начальных стадий сукцессий. Поэтому соответствие между семенным банком и составом фитоценоза может быть очень различным. Оно минимально для малонарушенных климаксовых лесов и максимально для часто нарушаемых сообществ, образуемых однолетниками (приморские марши, агроценозы, заросли однолетних солянок и т. п.). Таким
336
Глава 7. Онтогенез растений в фитоценозах
Продукция семян + ALH AFVG -GHM • SBC
Рис. 7-5 • Соответствие между запасом жизнеспособных семян в почве (семенной банк, семян/м2, ордината) и среднегодовым урожаем семян (продукция семян, семян/м2, абсцисса) в различных растительных сообществах альпийского пояса Тебердинского заповедника (по: Semenova, 2004). Сообщества: ALH — альпийская лишайниковая пустошь, FVG — пестроовсяницевый луг, GHM — гераниево-копеечниковый луг, SBC — альпийский ковер. Виды растений: Ас — Alchemilla caucasica, As — Anemone speciosa, Ao — Anthoxanthum odoratum, Ct — Campanula tridentata, Ca — Carex atrata, Cp — Carex pyrenaica, Cs — Carex sempervirens, Cc — Carum caucasicum, Cm — Carum meifolium, Cpu — Cerastium purpurascens, Coc — Corydalis conorhiza, Cu — Carex umbrosa, Fb — Festuca brunnescens, Fo — Festuca ovina, Fv — Festuca varia, Gg — Geranium gymnocaulon, Gsu — Gnaphalium supinum, He — Hedysarum caucasicum, Lh — Leontodon hispidus, Ls — Luzula spicata, Mca — Matricaria caucasica, Ns — Nardus stricta, Ok — Oxytropis kubanensis, Pco — Pedicularis comosa, Pn — Pedicularis nordmanniana, Pa — Phleum alpinum, Ro — Ranunculus oreophilus, Sc — Scorzonera сапа, Sa — Senecio aurantiacus, Sp — Sibbaldia procumbens, Ts — Taraxacum stevenii, Vg — Veronica gentianoides
2. Латентный период (жизнеспособные диаспоры)
337
образом, возможность образования семенных банков тесно связана с эко-лого-ценотическими стратегиями растений (Работнов, 1992; Onipchenko et al., 1998). Растения-виоленты (по: Раменский, 1935), или конкуренты (по классификации Grime, 1979), формирующие большую биомассу в благоприятных условиях (низкая интенсивность нарушений и хорошее поступление ресурсов), образуют значительное число крупных семян, не сохраняющих свою жизнеспособность длительное время. Напротив, эксплеренты (рудералы) характеризуются быстрым ростом и коротким жизненным циклом. Они накапливают жизнеспособные семена длительное время, сохраняя покой до нарушений растительного покрова, которые дают им возможность успешно пройти свой жизненный цикл. Пати-енты (стресс-толеранты) обычно имеют небольшой запас семян в почве.
2.8 • Споры мохообразных и папоротников в почвах
Содержание спор и вегетативных зачатков споровых растений исследовано значительно меньше, чем почвенные семенные банки (Milberg, 1991). В табл. 7.3 приведены некоторые примеры оценки численности жизнеспособных спор папоротникообразных в почвах нескольких растительных сообществ, основанные на подсчете числа образовавшихся заростков в лабораторных условиях (поэтому приводимые величины могут рассматриваться как минимальные).
Имеющиеся данные показывают значительно большее содержание спор в почвах по сравнению с семенами. Эти споры, видимо, могут сохраняться весьма продолжительное время. Так, по наблюдениям на старом (возраст 35 лет) лугу в Швеции число спор папоротников было существенно выше на глинистой почве, чем на песчаной, споры лучше сохранялись на неудобренных участках, чем на удобряемых, а наиболее
Таблица 7-3 • Содержание жизнеспособных спор папоротников в почвах (данные разных исследователей, оценки по числу образовавшихся заростков в лабораторных условиях; см. Milberg, 1991)
Места взятия проб Число видов Число заростков на 1 м2
Посадка ананаса в Малайзии 9 12000
Лес в Швейцарии > 2 530000
Листопадный лес, северо-восток США 5 5000000
Луг в Швеции 3 56000-170000
338
Глава 7. Онтогенез растений в фитоценозах
высокое содержание живых спор отмечено на заповедном (без выпаса и кошения) варианте луга. Очень важно, что споровый банк был представлен 3 видами папоротников (Thelypteris phegopteris, Cystopteris frag-ilis, Gymnocarpium dryopteris), не встречающихся на самом лугу или его лесных окрестностях. По данным разных авторов, споры папоротников сохраняют жизнеспособность в почве не менее 20-50 лет (Milberg, 1991).
В почвах присутствует и значительный запас жизнеспособных диаспор мохообразных (как в виде спор, так и в виде вегетативных зачатков). Исследования в хвойных лесах Швеции показали, что среди видов мохообразных даже ненарушенных участков леса в почве преобладали диаспоры пионерных видов, быстро осваивающих нарушенные участки (Polytrichum juniperinum, Bartramia ithyphylla и др.). Таким образом, для споровых банков отмечаются те же закономерности, что и для семенных (Jonsson, 1993). Одним из механизмов покоя, препятствующих осеннему прорастанию спор мохообразных, может быть необходимость для прорастания довольно узкого интервала температур, не наблюдающихся осенью (например, у Physcomitrium sphaericum) (During, 1997).
3 • Виргинильный период
3.1 • Прорастание семян
Виргинильный период начинается с прорастания семян и приживания всходов. Размеры семян связаны с экологическими условиями прорастания. Предполагалось, что крупные семена увеличивают вероятность приживания всходов в затененных местообитаниях, что может быть связано с несколькими особенностями: 1) всходы крупносемянных видов дольше выживают в условиях затенения, 2) всходы крупносемянных видов больше биомассы вкладывают в надземные органы, 3) всходы крупносемянных видов имеют большую высоту, 4) всходы крупносемянных видов более способны к этиоляции в условиях затенения. Эти предположения были проверены, например, на 23 видах растений Австралии с различным размером семян. Было показано, что всходы растений крупносемянных видов выживали дольше лишь в условиях 99 %-го затенения, а предположение 2 не подтвердилось. Предположения 3 и 4 подтвердились: всходы крупносемянных видов были крупнее и выше в условиях 95 %-го затенения, способность к этиоляции у них также была значительно выше. Большие семена адаптивны в условиях резкого градиента интенсивности света (прорыв слоя опада, травяные сообщества) или в случае частого возникновения прорывов в пологе (Leishman, Westoby, 1994а, 1994b).
3. Виргинильный период
339
6
а)
• Betula lenta
ф Rhus glabra
Betula populifolia
Liriodendron tulipifera
Ф Pinus strobus
Acer rubrum
• Gleditsia triacanthos
1 _ Quercus rubra •
Castanea mollissima
0’------------’-----------1------------’----------’ ф I
-1,0 0 1,0 2,0 3,0 4,0
средняя масса запасов веществ в семени (мг, логариф. шкала)
6)
Castanea mollissima
Quercus rubra
• Gleditsia triacanthos
• Pinus strobus
Acer rubrum
Liriodendron tulipifera
Rhus glabra
Betula populi foli
• Betula lenta
1,0*----------«-----------1-----------1-----------'-----------'
-1,0 0 1,0 2,0 3,0 4,0
средняя масса запасов веществ в семени (мг, логариф. шкала)
Рис. 7.в • Связь между массой запасных веществ в семенах деревьев Северной Америки (абсцисса, логарифмический масштаб) и смертностью всходов (а), максимальной высотой всходов через 12 недель после выращивания в условиях сильного затенения (б) (по: Grime, Jeffrey, 1965)
Аналогичные эксперименты с проростками 9 видов деревьев Северной Америки показали, что интенсивность отмирания всходов была отрицательно скоррелирована с их массой и высотой проростков через 12 недель в условиях затенения (рис. 7.6) (Grime, Jeffrey, 1965; Grime, 1979).
340
Глава 7. Онтогенез растений в фитоценозах
Размеры семян имеют большое значение при интенсивной внутривидовой конкуренции, особенно у гетерокарпичных видов растений. Например, Black (1958; Miles, 1979) провел эксперименты с одновидовыми посевами Trifolium subterraneum. Он посадил семена со средним весом 4 мг и 10 мг в чистые и смешанные посевы. В чистых посевах масса растений, выращенных из разных семян, не отличалась, а в смешанных средняя масса растений из крупных семян составила 0,32 г, а растений из мелких семян — 0,06 г, последние растения получали лишь 2 % поступающего света. Сходный эксперимент был проведен в Южном Уэльсе со смешанными посевами однолетников с разной массой семян при разной плотности. Значение массы семени для доминирования в смешанном посеве возрастало при увеличении плотности. Численность трех видов, имевших наиболее крупные семена, составила 49 % от общей при низкой плотности посева и 83% — при высокой плотности (Turnbull et al., 1999).
Reader (1993) провел эксперименты по выявлению факторов, влияющих на появление всходов 12 видов растений залежей с различной массой семян. Удаление покрова трав резко увеличивало частоту появления всходов у мелкосемянных видов и видов с твердой кожурой, в то время как изоляция от семеноядов (главным образом муравьев) резко увеличивала число появляющихся всходов крупносемянных видов. Автор делает вывод, что фитофагия лимитирует размножение крупносемянных видов, а затенение — мелкосемянных.
Изучение фенологии прорастания семян различных видов в высокотравных лугах Японии показало, что более крупные семена прорастают позже, чем мелкие (Masuda, Washitani, 1990).
В ходе прорастания происходит изменение массы семени и всходов: в первые 1-4 дня наблюдается снижение массы (сухой) по сравнению с семенем, затем — экспоненциальный рост (до 40-50-го дня), затем некоторое замедление. Относительную скорость роста (RGR) часто определяют в период экспоненциального роста (2-4 недели) (Swanborough, Westoby, 1996). Относительная скорость роста (RGR) в целом отрицательно скоррелирована с размерами семян. Интересная закономерность обнаружена при сравнении размеров меристем крупно- и мелкосемянных видов. Показано, что у большинства крупносемянных видов объем корневых меристем был больше, чем побеговых, а у мелкосемянных видов — наоборот, причем разница в объеме меристем часто была десятикратной (Swanborough, Westoby, 1996). Крупносемянные виды обычно имеют запасающие семядоли, в то время как мелкосемянные — тонкие фотосинтезирующие семядоли (Ibarra-Manriquez et al., 2001; Zanne et al., 2005; Kitajima, 2007).
Прорастание семян зависит при прочих равных условиях и от их ориентации в почве. Специальные эксперименты показали, что всхожесть
3. Виргинильный период
341
семян Cichorium intybus, Daucus carota, Potentilla recta и других видов была ниже при положении зародышевого корешка вертикально вниз. У других видов (Erysimum cheiranthoides и Lycopus uniflorus) положение семени не влияло на всхожесть (Bosy, Aarssen, 1995).
3.2 • Приживание всходов
Приживанию всходов многих видов растений благоприятствует наличие микроучастков с обнаженной почвой. Например, на залежах в Мичигане было показано резкое увеличение приживания всходов травянистых монокарпиков на участках с искусственным удалением подстилки и надземных частей растений по сравнению с контрольными (ненарушенными) участками (см. Luken, 1990).
В других случаях, наоборот, не полностью разложившиеся остатки растений могут быть хорошими субстратами для приживания всходов (рис. 7.7). В елово-тсуговых лесах на северо-западе Америки 94-98 % всходов деревьев отмечается по старым стволам деревьев, покрываю-
Рис. 7.7 • Возобновление ели (Picea abies) по разлагающемуся стволу старого дерева этого же вида; ельник-зеленомошник, Приокско-Террасный заповедник, Московская область
342
Глава 7. Онтогенез растений в фитоценозах
щим только 6-11 % почвы в этом лесу (Harmon, 1989 и др.). Harmon и Franklin (1989) показали, что разлагающиеся бревна играют важную роль как свободные от конкуренции субстраты для приживания всходов деревьев в этих лесах. Но в других случаях всходы разных видов деревьев лучше приживаются на разных элементах ветровально-почвенного комплекса (см. п. 4.2.6 гл. 6). Так, в хвойно-широколиственных лесах Миннесоты (Webb, 1988) было показано, что возобновление хвойных чаще происходило на разлагающейся древесине пней и стволов деревьев, кленов (Acer saccharum, A. rubrum) — только на ненарушенных вывалами участках, березы (Betula papyrifera) и осины (Populus tremuloides) — на пнях и буграх.
Большое значение для приживания всходов имеет и пространственное расположение взрослых особей своего и других видов. Например, в экспериментах Bergelson (1990) семена Capsella bursa-pastoris и Senecio vulgaris подсевали в созданный покров из Роа annua с одинаковой плотностью побегов мятлика, но различным их расположением: случайным или агрегированным. В случае агрегированного распределения мятлика приживаемость всходов и плотность побегов подсеваемых трав были на второй год (вторая генерация) в 4-8 раз выше, чем при более равномерном распределении.
3.3 • Длительность виргинильного периода
Возраст образования первых генеративных побегов зависит от биологических особенностей вида, условий среды и взаимоотношений между растениями, особенно конкурентных. Растения отличаются большим варьированием этого возраста даже в пределах одного вида в сходных условиях обитания (временная поливариантность развития, по Л. А. Жуковой, 1986). Классическим примером может служить возраст зацветания травянистых монокарпиков (часто неправильно именуемых двулетниками). По наблюдениям Т. А. Работнова (1960а) за Libanotis montana на лугах в пойме Оки, из отмеченных 39 особей этого вида ни одно растение не зацвело до 4-летнего возраста, в возрасте 5 лет зацвело 2 растения, в возрасте 6 лет — 6, 7 лет —1,8 лет —1,9 лет — 3, 11 лет — 4 растения; оставшиеся 19 особей не зацвели до 12-летнего возраста.
Травянистые поликарпики также зацветают в разном возрасте. Например, по наблюдениям в лесах Швеции, возраст первого цветения составил для Hepatica nobilis 4-13 лет, для Sanicula еигораеа — 8-16 лет (Inghe, Tamm, 1985). Здесь также наблюдается поливариантность онтогенеза, способствующая высокой устойчивости популяций к неблагоприятным изменениям среды.
3. Виргинильный период
343
Рис. 7.8 • Идеальные типы кривых выживания (а) и реальные кривые выживания (б, в, г) одновозрастных групп растений (когорт) Erucastrum gallicum на склонах известковых карьеров в Нью-Йорке. Метеорологические условия отдельных лет: б — обильные осадки весь сезон, в — весенняя засуха, г — периодические дожди (по: Klemow, Raynal, 1983)
Смертность растений на ранних этапах развития зависит от локальных условий среды и также может сильно варьировать у одного вида. В большинстве случаев у многолетников наиболее высока смертность на ранних стадиях развития. Но даже у однолетников все 3 основных типа кривых выживания (I — низкая ранняя смертность и высокая поздняя, II — постоянная смертность в разных возрастных состояниях, III — высокая ранняя смертность и низкая у особей более поздних возрастных состояний) могут наблюдаться у одного вида в зависимости от метеорологических условий отдельных лет (рис. 7.8). При линейном снижении логарифма численности смертность постоянна, она не зависит от возрастного состояния. Так, по наблюдениям над однолетником Erucastrum
344
Глава 7. Онтогенез растений в фитоценозах
gallicum в США, кривая I типа была отмечена в год с обильными осадками, II типа — при сменах дождливых и сухих периодов, а близкая к III типу — при сильной весенней засухе (Klemow, Raynal, 1983; Luken, 1990).
Смертность всходов может быть выше, чем в других стадиях жизненного цикла. Согласно обзору Ehrlen и Lehtila (2002), средняя длительность жизни древесных растений — 99 лет, а травянистых многолетников — 22 года. У этих растений для поддержания популяций может быть достаточно редких лет, в которые происходит успешное приживание всходов. Например, у Hepatica nobilis приживание всходов может происходить раз в десятилетие (Inghe, Tamm, 1985). По обзорным данным Moles и Westoby (2004), причинами смертности всходов являются: фитофаги (38 %), засуха (35 %), фитопатогенные грибы (20 %), физическое повреждение (4,6%), конкуренция с другими проростками (1,6%) и конкуренция с окружающими взрослыми растениями (1,3%) (Eriksson, Ehrlen, 2008).
3.4 • Изменение свойств растений в онтогенезе
В онтогенезе растений изменяются многие эколого-физиологические характеристики особей. Наряду с часто большей уязвимостью молодых особей они чаще более теневыносливы (хотя и не всегда). Примером могут служить изменения теневыносливости деревьев в онтогенезе, полученные О. И. Евстигнеевым (1991) (табл. 7.4). В случае деревьев умеренного пояса происходит увеличение минимального количества света, необходимого для роста особей при переходе к более старшим возрастным состояниям. При этом у светолюбивых пород (береза, осина) взрослые вегетативные особи требуют в 10 раз больше света, чем ювенильные, в то время как у теневыносливых (липа, клен) — всего в 2-3 раза.
В ходе онтогенеза изменяются также особенности водного режима. Так, для кустарника Chrysothamnus nauseosus из полуаридных сообществ штата Юта в США эффективность использования воды убывала в ряду: проростки > ювенильные = мелкие генеративные > крупные генеративные растения (Donovan, Ehleringer, 1992). Растения разных возрастных состояний могут существенно различаться по устойчивости к засолению. Например, всходы Avicennia marima (обычное дерево мангровых зарослей) имели более низкий осмотический потенциал, чем морская вода, что позволяло им поглощать воду из моря (Downton, 1982). С другой стороны, у Typha domingensis прорастание семян и приживание всходов происходят только в воде с низкой соленостью, в то время как взрослые растения этого вида могут выносить высокую соленость воды (Beare, Zedler, 1987). У Prosopis flexuosa (бобовое) устойчивость к за-
4. Генеративный период
345
Таблица 7.4 • Световые минимумы (% от полного света) деревьев в разном онтогенетическом состоянии (по: Евстигнеев, 1991). Возрастные состояния: j — ювенильные, iml, im2 — имматурные, v2 — взрослые вегетативные растения
Вид Возрастные состояния
j iml im2 v2
Betula pendula 3,1 4,5 10,7 30,0
Populus tremula 2,7 4,3 9,9 27,7
Quercus robur 1,2 2,6 5,0 15,1
Tilia cordata 0,6 0,7 0,8 1,3
Acer platanoides 0,3 0,4 0,5 0,8
солению также возрастает от ювенильных растений к взрослым (Catalan et al., 1994). Холодостойкость ювенильных и взрослых растений также может существенно различаться. Например, гибель от холода проростков Digitalis purpurea ограничивает распространение этого вида на север в Германии, в то время как взрослые растения более морозостойки (Bru-elheide, Heinemeyer, 2002; Facelli, 2008).
В онтогенезе может происходить изменение жизненной формы растений, например переход от гемикриптофитов к геофитам, наблюдаемый у ряда злаков, таких как Bromus inermis (Работнов, 1992). Одни и те же внешние воздействия могут по-разному сказываться на особях разных возрастных состояний в условиях конкуренции. Например, по данным Т. А. Работнова (1950), взрослые особи Anemone fasciculata на субальпийских лугах Кавказа отрицательно реагировали на раннее скашивание, в то время как молодые особи — положительно. Напротив, на внесение удобрений взрослые особи реагировали положительно, а молодые — отрицательно.
4 • Генеративный период
4.1 • Семенная продуктивность, урожай семян и репродуктивное усилие
Согласно Т. А. Работнову (19606), различают семенную продуктивность (число семян, формирующихся особью или побегом растения) и урожай семян (число или масса семян, образующихся на единицу площади).
346
Глава 7. Онтогенез растений в фитоценозах
Основные факторы, лимитирующие семенную продуктивность растений, можно ранжировать по мере их важности:
1. Наличие ресурсов, особенно почвенных (внесение ЭМП часто резко увеличивает семенную продуктивность).
2. Доступность пыльцы (для перекрестноопыляющихся видов).
3. Влияние фитофагов и неблагоприятных условий в процессе созревания семян.
Часто наблюдается линейная зависимость весовой продукции семян от вегетативной биомассы растения: существует минимальная необходимая пороговая вегетативная биомасса для начала репродукции, выше которой масса семян составляет постоянную долю от вегетативной биомассы (Weiner, 1990).
В экспериментах искусственное опыление часто увеличивает завя-зываемость семян растений, но не всегда ведет к увеличению семенной продуктивности или урожая семян. При недостатке ресурсов часть завязавшихся семян абортируется. Кроме того, увеличение семенной продуктивности в один сезон часто ведет к истощению растения и снижению образования генеративных структур в следующий сезон (см. ниже). Отношение числа созревающих семян к числу семязачатков (S/О), по наблюдениям в США, для однолетников составило около 85 %, а для многолетников — 50 % независимо от системы скрещивания (само- и пере-крестноопыляющиеся виды). Это отношение, видимо, определяется генетически, в то время как среда влияет на семенную продуктивность через численность цветков и побегов (Wiens, 1984). Дальний разнос пыльцы у ветроопыляемых растений не всегда связан с эффективным опылением растений на больших расстояниях. Так, Sork et al. (2002) показали на основании анализа всходов 21 взрослого дерева Quercus lobata, что эффективное число доноров пыльцы для одного дерева было меньше 4 (деревьев), а среднее эффективное расстояние переноса пыльцы — 65 м (Koenig, Ashley, 2003).
Опыление цветков сокращает образование новых цветков на растении, а повреждение зеленых плодов — наоборот, увеличивает. В экспериментах с Silene latifolia средняя продукция составила 12,2 цветка на неопыленные растения и 4,3 цветка — на опыленные. Новая генерация цветков возникала на неповрежденных растениях через 29,5 дней, а на растениях с удаленными на 100 % зелеными плодами — через 20,1 дней (Wright, Meagher, 2003).
Для оценки вклада ассимилятов (продуктов фотосинтеза) в размножение используют показатель «репродуктивное усилие» (РУ). Его часто рассчитывают как долю генеративных органов (включая поддерживающие их
4. Генеративный период
347
Таблица 7-5 • Зависимость репродуктивного усилия Chamaesyce hirta (%) от уровня поступления ресурсов (богатства почвы) и плотности посевов в горшках (по: Snell, Burch, 1975)
Уровень ресурсов Число растений на горшок
2 8 32 64
1 14 8 7 7
2 27 18 И 10
4 32 29 25 14
8 26 25 22 16
структуры — цветоножки и т.п.) от массы надземных побегов. Наибольшие величины РУ отмечены у следующих видов (см. Willson, 1983):
Astrocaryum mexicanum — 37% (Sarukhan, 1980),
Lupinus arboreus — 26% (Pitelka, 1977),
Solidago speciosa — 35% (Abrahamson, Gadgil, 1973),
Arisaema atrorubrus — 39% (Abrahamson, 1979).
Репродуктивное усилие может значительно различаться у одного вида в различных местообитаниях. Как показали эксперименты с Chamaesyce hirta (Snell, Burch, 1975), оно увеличивается при увеличении уровня ресурсов и уменьшении плотности растений в посевах (табл. 7.5).
Для травянистого многолетника Heloniopsis orientaiis, имеющего большую амплитуду вертикального распространения в горах Японии, показана зависимость РУ от высоты над уровнем моря и типа сообщества, в котором он произрастает (Kawana, Masuda, 1980; Wilson, 1983). РУ возрастает с 10% на абсолютной высоте 100 м до 26% на высоте 2600 м, но при этом семенная продукция даже несколько уменьшается с высотой в связи с неблагоприятностью условий высокогорий.
При исследовании репродуктивного усилия выделяют два аспекта: 1) аллокация (вклад биомассы) в генеративные структуры и 2) снижение роста вегетативных структур (соматическая стоимость репродукции). Существуют разные мнения о границах репродуктивных структур. Одно из разумных определений дали Thompson и Stewart (1981): репродуктивные структуры — это все структуры, не представленные на вегетативном растении, но имеющиеся у генеративных особей. Высокое репродуктивное усилие показывают виды с высокой смертностью взрослых особей. Теория жизненного цикла предсказывает, что монокарпики должны иметь более высокое репродуктивное усилие, чем поликарпики. С возрастом
348
Глава 7. Онтогенез растений в фитоценозах
репродуктивное усилие (РУ) обычно увеличивается, что было показано для Astrocaryum mexicanum (Pinero et al., 1982), Solidago pauciflosculosa (Pritts, Hancock, 1983), Drosera closterostigma (Karlsson, Pate, 1992b), Fa-gus sylvatica (Comps et al., 1994). Соматическая стоимость репродукции может выражаться в снижении ростовых характеристик, запасов питательных веществ, размеров вегетативных частей генеративных растений, что в дальнейшем ведет к снижению репродукции и увеличению смертности (Karlsson, Mendez, 2005; Obeso, 2002).
Фотосинтез генеративных органов растений, особенно зеленых плодов, может иметь большое значение для обеспечения плодов и семян ас-симилятами. Так, Bazzaz et al. (1979) показали, что у внетропических деревьев доля собственного фотосинтеза зеленых плодов составляет от 2,3 % (Quercus macrocarpa) до 64,5 % (Acer platanoides). Особенно велика эта доля у плоских анемохорных плодов, расширенные части которых не только служат для их распространения, но и активно фотосинтезируют. Зародыши растений ряда тропических видов бобовых (Caesalpinia spp., Enterolobium cyclocarpum и др.) имеют зеленые семядоли, фотосинтезирующие сквозь покровы и теряющие свою зеленую окраску при созревании (Janzen, 1983).
4.2 • Перерывы в цветении и вторичный покой растений
Образование генеративных органов и семян требует от растений больших затрат продуктов фотосинтеза и элементов минерального питания, что тормозит образование новых генеративных структур в следующий сезон. Поэтому, переходя в генеративный период, многолетние поликарпиче-ские растения обычно цветут не ежегодно, а с перерывами, в некоторые годы оставаясь в вегетативном состоянии. Например, особи Sanicula еи-гораеа, по наблюдениям на постоянных площадках в Швеции, обычно не образуют генеративных побегов в год, следующий после цветения (Inghe, Tamm, 1985). У вегетативно-неподвижных альпийских растений Северо-Западного Кавказа большинство особей цвело ежегодно, небольшая часть с перерывами в несколько лет, при этом чем длительнее был перерыв, тем реже он наблюдался (рис. 7.9). У исследованных видов вероятность повторного цветения на следующий год составляла: у Anemone speciosa — 77 %, у Campanula tridentata — 67 %, у Carum caucasicum — 80%. У A. speciosa из 71 генеративной особи 14 лет без перерыва цвело 5, у С. caucasicum из 109 растений — 2 особи, в то время как у С. tridentata ни одна розетка не образовывала генеративные побеги без перерыва 10 и более лет. В целом для генеративных особей изученных видов ти-
4. Генеративный период
349
Рис. 7.9 • Распределение длительностей перерывов в цветении генеративных особей трех видов растений Северо-Западного Кавказа по материалам 14-летних наблюдений (по: Онипченко, Комаров, 1997). По оси абсцисс — длительность перерывов в цветении (годы), по оси ординат — число случаев (логарифмический масштаб)
пичным является чередование нескольких лет беспрерывного цветения с кратковременными (чаще 1-2 года) перерывами (Онипченко, Комаров, 1997). Интенсивное плодоношение ослабляет также вегетативное размножение и рост растений. В семенные годы уменьшается толщина годичных колец и прирост корневищ.
В ходе онтогенеза у ряда видов сосудистых растений возможен переход в состояние вторичного покоя, когда растения сохраняют жизнеспособные подземные органы, но не образуют надземных побегов. Классический пример приводит Т. А. Работнов (1964) для Chaerophyllum prescottii, растения которого после нескольких лет развития в вегетативном состоянии образуют клубень и могут переходить во вторичный покой. При нарушениях почвы (порой) возможен быстрый выход из покоя и образование генеративного побега. В этом случае адаптивное значение вторичного покоя связано с использованием периодов с ослабленной конкуренцией. Переход в покоящееся состояние отмечен при травмировании корней у женьшеня (Куренцова, 1944). Наиболее изучен переход во вторичный покой у ряда видов орхидных. В частности, по наблюдениям на постоянных квадратах в Швеции, было показано, что Orchis mascula может образовывать генеративные побеги, пребывая до этого несколько
350
Глава 7. Онтогенез растений в фитоценозах
лет в состоянии вторичного покоя (Tamm, 1972). Экспериментальные исследования европейских орхидей Cypripedium calceolus и Cephalanthera longifolia показали, что физиологические стрессовые воздействия (дефолиация, затенение) увеличивают вероятность перехода взрослых особей в состояние вторичного покоя без увеличения итоговой смертности. Таким образом, переход во вторичный покой может рассматриваться как адаптивная реакция растений (Shefferson et al., 2005). 10-летние наблюдения за орхидеей Cleistes bifaria (Кегу et al., 2005) показали, что длительность стадии вторичного покоя составляет 1-4 года (в среднем 1,4 года), выживаемость особей в этом состоянии — 0,86 (0,77-0,94), средняя доля особей в состоянии вторичного покоя — 30% (16-47%). Выживаемость особей этого вида в покоящемся состоянии была положительно скоррелирована с количеством осадков в текущем году. Morrow, Olfelt (2003) показали, что Solidago missoutiensis может возобновить свой рост из покоящегося корневища, не образовывавшего надземные побеги после их повреждения фитофагами 10 лет назад (Morrow, Olfelt, 2003).
4.3 • Типы регенерационных стратегий
Ф. Грайм (Grime, 1985) предложил различать 5 основных типов регенерационных стратегий, связанных с адаптацией к различным типам местообитаний.
1. Вегетативное распространение (V) — характерно для растений как продуктивных, так и бедных местообитаний, подверженных нарушениям низкой интенсивности.
2. Сезонное возобновление в прорывах полога (S) — часто встречается в местообитаниях, подверженных сезонно предсказуемым нарушениям под действием климата или биотических факторов.
3. Регенерация, включающая постоянный семенной или споровый банк (Bs), характерна для местообитаний, подверженных пространственно предсказуемым, но по времени непредсказуемым нарушениям (образование прорывов в пологе).
4. Регенерация, включающая дальний разнос многочисленных диаспор (W), типична для труднодоступных (утесы, пни и т. п.) или подверженных пространственно непредсказуемым нарушениям местообитаний.
5. Регенерация, включающая устойчивую виргинильную стадию (Bj), характерна для низкопродуктивных местообитаний, подверженных низкой интенсивности нарушений.
5. Вегетативное размножение растений. Клональные растения
351
5 • Вегетативное размножение растений.
Клональные растения
Цветковые растения могут сочетать различные формы бесполого (веге-тативное размножение, вивипария) и семенного размножения. Каждый из этих типов имеет характерные размеры пропагул, влияющих на выживание потомков. Т. Callaghan и U. Emanuelsson (1985; Callaghan, 1987) оценили эти величины для травянистых растений тундр и бореальных лесов (табл. 7.6). В результате вегетативного размножения образуется небольшое число генетически практически идентичных потомков, вероятность выживания которых существенно выше, чем потомков, образовавшихся из семян.
Размеры и масса семени растений обычно отрицательно скоррелированы с их способностью к вегетативному размножению (рис. 7.10). Крупносемянные виды реже размножаются вегетативным путем. С другой стороны, многие клональные растения сочетают стратегию дальней колонизации легкими семенами и ближней колонизации вегетативными потомками, что позволяет им успешно произрастать в антропогенно-нарушенных ландшафтах. Так, анализ связи особенностей размножения растений Великобритании с использованием базы данных экологической флоры показал, что виды, сочетающие вегетативное и семенное размножение, имеют наиболее обширные ареалы в пределах Британских островов и континентальной Европы по сравнению с видами, размножающимися только семенами или только (преимущественно) вегетативно (Fitter, Peat, 1994).
Относительный вклад в семенное и вегетативное размножение может изменяться в разных популяциях одного и того же вида в зависимости от их плотности. При высокой плотности растений и большой роли конкуренции растения больше вкладывают в семенное размножение (дальше
Таблица 7-6 • Сравнение трех типов размножения травянистых растений тундр и бореальных лесов Северной Европы (по: Callaghan, Emanuelsson, 1985)
Показатель Половое размножение Бесполое размножение
вивипария вегетативное
Сухая масса пропагул, мг 0,02-7,98 0,9-3,9 1,1-39
Число пропагул на особь или побег 11-40000000 11,5-40 0,1-3,97
Вероятность выживания 0-0,07 0,1-0,6 0,43-0,97
352
Глава 7. Онтогенез растений в фитоценозах
размножением размножения
Рис. 7.10 • Средняя масса семян травянистых поликарпиков во флоре окрестностей Шеффильда (Великобритания) (по: Hodgson, Mackey, 1986)
разнос в пространстве/времени), а при низкой плотности более интенсивно вегетативное размножение, служащее для освоения ближайшего пространства. Это было показано, например, Е. Л. Любарским и В. И. Полуяновой (1984) для Ranunculus repens и Rautiainen et al. (2004) для Potentilla anserina.
Анализ встречаемости вегетативно-подвижных (клональных) растений в различных сообществах (van Groenendael et al., 1997) показал, что клональные растения чаще встречаются: а) в арктических сообществах, чем в умеренных, б) в водных биотопах, чем в наземных, в) в затененных местах, чем в открытых, г) на бедных почвах, чем на богатых. Как было отмечено выше (см. п. 4.2.4 гл. 6), вегетативно-подвижные растения могут эффективнее использовать гетерогенную среду за счет «разделения труда» между раметами для получения ресурсов.
Среди древесных растений многие виды способны к вегетативному размножению при помощи корневых отпрысков или новых побегов, образующихся из спящих почек при основании стебля. Сравнение высоты растений, способных и не способных к вегетативному размножению, для видов Южной Африки (Midgley, 1996) показало, что растения тех видов, которые размножаются только семенами, значительно выше, чем виды, способные к вегетативному размножению.
6. Длительность жизни растений
353
6 • Длительность жизни растений
Растения обычно продолжают расти в течение всей жизни. Если нараста-ние и отмирание происходят одновременно (как у многих корневищных растений), то установить общую длительность жизни таких особей бывает сложно. Лишь для деревьев с четко выраженными годичными кольцами хорошо установлен максимальный возраст, который может достигать нескольких тысячелетий. Например, для Pinus longaeva (Р. aristata Engelmann var. longaeva) известны особи возрастом более 4800 лет (Brown, 1996) (рис. 7.11). Гигантские секвойи в Калифорнии живут несколько меньше, их возраст обычно не превышает 2000 лет (рис. 7.11 в).
Мнения о длительности жизни вегетативно-подвижных растений противоречивы. Ряд авторов рассматривают накопление соматических нарушений как основную причину их старения, другие рассматривают эти растения как потенциально бессмертные (Watkinson, 1992). Для дер-новинной Carex curvula с очень незначительным годичным приростом (доли мм) расчеты, проведенные в Швейцарии с использованием анализа ДНК для установления границ особей, показали минимальный возраст одного клона 2000 лет (Steinger et al., 1996). По данным Vasek (1980), возраст клонов кустарника Larrea thdentata превышает 10000 лет, т. е. длительность последнего межледниковья. Возраст клона североамериканской осины (Populus tremuloides), занимающего площадь 81 га и состоящего из 47 000 стеблей, также оценивается в 10 000 лет (Kemperman, Barnes, 1976; Steinger et al., 1996).
Один из методов оценки длительности жизни растений — сравнение старых и современных фотографий с хорошо различимыми особями растений. Такая работа была проведена для участков Большого Каньона в Колорадо (Bowers et al., 1995). Сравнивая 355 пар снимков, сделанных с 1872 г., авторы показали, что особи многих видов живут более 100 лет (пустынные кустарники и суккуленты: Ambrosia dumosa, Atriplex canescens, Opuntia acanthocarpa, Yucca angustissima и др.).
Длительность жизни многолетних травянистых растений в природных условиях оценить весьма трудно. Для их оценки используют подходы математической демографии, которые позволяют рассчитывать длительность жизни по данным краткосрочных наблюдений за возрастным состоянием и смертностью большого числа особей. Например, для 29 видов двудольных и 11 видов злаков прерии в Канзасе было показано, что средняя длительность жизни растений на уровне всходов составляла от 0,6 до 6,5 лет, а максимальная длительность жизни особей двудольных растений варьировала от 3 до 25 лет, злаков — от 5 до 39 лет (Lauenroth, Adler, 2008). Высокогорные растения живут значительно дольше. Ана-
354
Глава 7. Онтогенез растений в фитоценозах
Рис. 7.11 • Древесные растения с большой длительностью жизни отдельных особей и клонов: а, б — Pinus longaeva на верхней границе леса в Южной Неваде, абсолютная высота 3300 м (фото Stan Shebs, Wikimedia Commons, public domain); в, г — Sequoia sempervirens (в — фото Miguel Vieira, Wikimedia Commons, public domain); д — роща Populus tremuloides в Канаде (фото Tewy, Wikimedia Commons, under GNU Free Documentation License)
логичный демографический анализ показал, что средняя длительность жизни особей, достигших генеративного состояния, составила 70 лет для Carum caucasicum, 65 лет для Campanula tridentata и 390 лет для Anemone speciosa. Эта величина имеет большой разброс внутри вида; расчеты показывают, что около 1 % генеративных особей могут достигать значительно большей длительности жизни, составляющей около 87,
6. Длительность жизни растений 355
123 и 1400 лет для Carum caucasicum, Campanula tridentata и Anemone speciosa соответственно (Sizov et al., 2004).
В разных группах растений старение (наступление сенильного периода) происходит по-разному. У монокарпиков он наступает очень быстро. Старение и отмирание деревьев происходит часто в силу изменения биомеханических характеристик ствола, уменьшения доли фотосинтезирующих тканей, уменьшения прочности ствола при повреждении его центральной части и т. п. Вопрос о наличии сенильного периода у травянистых поликарпиков остается открытым. Во многих случаях для растений отмечено квазисенильное состояние, морфологически сходное с сенильным, но обусловленное не возрастом, а сильным угнетением (Работнов, 1975). При переносе таких особей в более благоприятные условия они вновь переходят в генеративное состояние.
Глава
Биомасса
и продукция фитоценозов
Запасы и структура фитомассы сообществ различных типов. Аллокация
Продукция фитоценозов и факторы, ее лимитирующие
Крупнотравье как система с максимальной продукцией
Энергосодержание фитомассы и энергетическая продукция фитоценозов
1 • Запасы и структура фитомассы сообществ различных типов. Аллокация
|При изучении запасов фито массы, Вставка XII. Основные термины продукции и биологического круга-и понятия, связанные ворота используется ряд терминов,
с продуктивностью наземных экосистем значения которых несколько разнят------------------------------------ся у разных авторов. Здесь мы приведем их определения, которые были использованы при выполнении международной биологической программы в 1960-1980 гг. (см. Гришина, 1974; Базилевич и др., 1978; Работнов, 1992; Базилевич, 1993; Богатырев, Телес-нина, 2010).
Биомасса — масса живых организмов в единице объема или площади среды. Биомасса растений включает в себя также неживые ткани (например, сердцевину стволов деревьев, гиалиновые клетки сфагновых мхов), имеющие важное функциональное значение и окруженные живыми тканями. Как синоним биомассы растений используется также понятие «урожай на корню» (standing crop).
Мортмасса (некромасса) — масса отмерших, но частично сохранивших анатомическую структуру частей организмов, главным образом растений, в наземных экосистемах. В лесах выделяют опад (листья и другие короткоживущие части растений), отпад (отмершие многолетние части растений — крупные ветви, стволы и т.п.), в травяных сообществах — ветошь (отмершие надземные части растений, обычно сохраняющие связь с живыми растениями). В почве выделяют также отмершие подземные органы растений (корни, корневища и др.).
Фитомасса — суммарная масса живых и отмерших частей растений на единицу площади (фитомасса = биомасса + мортмасса).
Продукция — количество органического вещества, образуемое на единицу площади за единицу времени. Различают первичную продукцию — продукцию автотрофов и вторичную — продукцию животных-фитофагов. Продукцию растений чаще оценивают как нетто-продукцию (без учета дыхания растений, синоним — чистая продукция) в отличие от брутто-продукции, в которую включено дыхание растений.
Основные термины, используемые при изучении фитомассы, даны во Вставке XII. Запасы биомассы растений в разных типах сообществ различаются на несколько порядков (табл. 8.1). По соотношению надземной и подземной биомассы можно выделить сообщества, где преобладает надземная масса (полярные пустыни, леса), и те, где ббльшая часть биомассы сосредоточена в почве (тундра, луга, степи, пустыни). Соотношение подземной и надземной биомассы зависит также от воздействия
358
Глава 8. Биомасса и продукция фитоценозов
Таблица 8.1 • Запасы биомассы в сообществах основных природных зон европейской части России и сопредельных стран (усредненные данные, г/м2; по Базилевич, 1993; Базилевич, Титлянова, 2008 и др.)
Типы сообществ Всего Надземная Подземная Отношение надземной биомассы к подземной
Полярные пустыни (Земля Франца-Иосифа) 160 130 30 4,4
Тундры 1700 620 1080 0,57
Ельники 21000 16100 4900 3,3
Сосняки 13000 10600 2400 4,4
Широколиственные леса 33000 26 200 6800 3,9
Степи 1300 220 1080 0,20
Пустыни 1500 300 1200 0,25
Рис. 8.1 • Аллокация (распределение) биомассы (%) между органами травянистых растений низкогорий (слева) и высокогорий (справа) в Альпах; средние данные для 49 видов (по: Korner, Renhardt, 1987)
2. Продукция фитоценозов и факторы, ее лимитирующие
359
фитофагов. Так, в большинстве типов травяных сообществ преобладает подземная фракция биомассы, а на высокогорных лугах Новой Зеландии, сформировавшихся без воздействия травоядных млекопитающих, — надземная (Meurk, 1978).
Наполненность надземной среды органами растений невелика, она обычно не превышает 0,5-1 % (на лугах часто 3,5-4,5 дм3 на 1 м2) (Работнов, 1992), подземными органами — 10%.
Аллокацией биомассы называют распределение биомассы или продуктов фотосинтеза между отдельными органами растений. Отдельные типы аллокации свойственны различным жизненным формам растений, аллокация меняется также в онтогенезе и зависит от условий произрастания. Например, изучение распределения биомассы между различными органами для высокогорных и низкогорных растений показало, что относительная доля листьев и запасающих подземных органов остается постоянной, но доля стеблей и генеративных органов уменьшается, а тонких корней — возрастает у высокогорных растений (см. рис. 8.1) (Komer, Renhardt, 1987).
2 • Продукция фитоценозов и факторы, ее лимитирующие
Первичная нетто-продукция различных сообществ варьирует от долей грамма до 2-4 кг сухой массы на квадратный метр в год (табл. 8.2,
Таблица 8.2 • Чистая первичная продукция, биомасса и время оборота биомассы для разных типов экосистем (по: Риклефс, 1979; Базилевич, Титлянова, 2008 и др.; усредненные данные)
Типы экосистем Продукция, г/м2 в год Биомасса, г/м2 Время оборота биомассы, годы
Тропический дождевой лес 2000 45000 22,5
Листопадный лес умеренной зоны 1200 30000 25
Бореальный лес 1000 17000 17
Степи умеренной зоны 800 1300 1,6
Кустарниковые пустыни 70 700 10
Болота и марши 2500 15000 6
Озера и реки 500 20 0,04 (15 дней)
Открытый океан 125 3 0,024 (9 дней)
360
Глава 8. Биомасса и продукция фитоценозов
Рис. 8.2 • Продукция сухого органического вещества зональных и интразональных (болота) типов растительных сообществ различных термических поясов (по: Базилевич, 1993). Пояса и типы растительных сообществ: П — полярный (I — тундры); Б — бореальный (II — темнохвойные леса, III — широколиственно-хвойные леса); СБ — суббореальный (IV — широколиственные леса, V — луговые и настоящие степи, VI — сухие степи, VII — пустыни); СТ — субтропический (VIII — широколиственные леса, IX — ксерофитные леса и редколесья, X — пустыни); 1 — зональные сообщества, 2 — интразональные сообщества (болота)
см. также п. 3 гл. 8). Для наземных сообществ наибольшая скорость оборота биомассы свойственна травяным экосистемам, а также сообществам наземных и эпифитных водорослей (см. п. 1.1 гл. 6).
Продукция растительных сообществ определяется и лимитируется многими факторами. Среди важнейших можно отметить водный режим, поступление ЭМП, длительность вегетационного периода и интенсивность нарушений. В масштабе природных зон увлажнение играет очень большую роль. Это наглядно проявляется при сравнении продукции зональных типов сообществ с продукцией сообществ, приуроченных к тем же природным зонам, но развивающихся в гидроморфных условиях (понижениях рельефа) с достаточным увлажнением. Такое сравнение про-
2. Продукция фитоценозов и факторы, ее лимитирующие
361
первичная нетто-продукция за месяц (кг/м2) длительность вегетационного периода, месяцы годичная первичная нетто-продукция (кг/м2 в год)
Влажный тропический лес др 12 2.5(1.&-3.0)
Умеренный листопадный лес 0.24 5 1.2 (1.0-1.5)
Бореальный лес ИННВН °-21 5 1.1 (0.3-2.0)
Тропические травяные сообщества HHHIHH0-25 10 2.5 (0.2-4.0)
Луга и стели умеренного пояса °-17 6 1.0 (0.2-1.5)
Альпийские сообщества (северной умеренной зоны) °-2° 2 0.4 (0.2-0.6)
Рис. 8.3 • Сравнение месячной и годичной первичной нетто-продукции (сухая масса) различных биомов. С учетом длительности вегетационного периода месячная продукция основных биомов сходна от влажных тропических лесов до альпийских сообществ (по: Korner, 1999)
ведено Н. И. Базилевич (1993) (рис. 8.2) для природных зон территории бывшего СССР, которая показала, что при продвижении на юг и увеличении поступления солнечной радиации продукция зональных сообществ лимитируется преимущественно осадками, а продукция болот непрерывно возрастает. Наибольшие величины продукции (около 5 кг/м2 в год) отмечены для околоводных тростниковых (Phragmites australis) зарослей в пойме Амударьи.
С другой стороны, С. Korner (1999) показал, что различия в продукции растительных сообществ различных регионов связаны главным образом с различиями в длительности вегетационного периода. Перерасчет продукции на месяц вегетации приводит к сходным показателям (170-250 г/м2 сухого вещества в месяц) для широкого спектра сообществ, не испытывающих недостатка влаги, — от тропических лесов до альпийских фитоценозов (рис. 8.3).
Низкое содержание в почве элементов минерального питания растений может существенно сдерживать продукцию растительных сообществ. Elser et al. (2007) провели анализ большого числа работ по лимитированию первичной продукции пресноводных, морских и наземных сообществ N и Р. Лимитирование Р одинаково широко распространено среди этих трех типов, а совместное лимитирование N и Р — среди наземных и пресноводных сообществ. Таким образом, пресноводные, морские
362
Глава 8. Биомасса и продукция фитоценозов
Рис. 8.4 • Миниризотроны для изучения роста корней: а — схема устройства мин и ризотрона (http://www.archiroot.org.uk, с любезного разрешения Lionel Dupuy); б — установленная в поле трубка мин и ризотрона (фото Daniel LeCain, U.S. Department of Agriculture; Agriculture Research Service, Fort Collins, Colorado USA, с любезного разрешения автора); в — проведение наблюдений на мин и ризотроне CI-600 (фото предоставлено CID Bio-Science, Camas, WA USA, www.cid-inc.com)
и наземные сообщества не отличаются принципиально по лимитированию ЭМП. Продукция луговых сообществ может быть колимитирована N и Р, а тундровых — больше N, чем Р (Elser et al., 2007).
При изучении продукции наземных экосистем особые слож
ности вызывает оценка продукции корней, особенно тонких, и изучение изъятия первичной продукции фитофагами (см. также п. 4 гл. 4). Для изучения роста и развития тонких корней обычно применяют метод миниризотронов (рис. 8.4) — прозрачных трубок, вкопанных в почву под разным углом, в сочетании с видеокамерой, снимающей изображение с их поверхности, на которой видны корни растений. Скорость оборота корневых систем в разных типах сообществ существенно различается, она всегда выше для тонких корней по сравнению с толстыми (рис. 8.5). Исследования ряда ученых показали, что в случае увеличения доступности азота в почве
2. Продукция фитоценозов и факторы, ее лимитирующие
363
Рис. 8.5 • Скорость оборота (продукция, деленная на биомассу, год-1) биомассы корней растений разных типов сообществ в трех широтных поясах (по: Gill, Jackson, 2000)
обычно происходит уменьшение биомассы тонких корней, но увеличивается скорость их обновления, т. е. уменьшается длительность жизни (Hendricks et al., 1993). Использование миниризотронов позволило оценить длительность жизни и продукцию тонких корней Acer saccharum в лесу, где этот вид являлся основным доминантом. Средняя длительность жизни оказалась около 240 дней, а годичная продукция тонких корней оценивается в 600 г/м2 (Atkinson, 1992).
Сезонные периоды образования биомассы побегов и корней могут существенно различаться. Так, при сравнении времени продукции корней и надземных органов в прерии и осиновом лесу в Канаде было показано, что пик продукции корней в лесу наблюдался на 45 дней позже, чем пик продукции листьев, а в прерии — на 15 дней позже. Продолжительность продукции корней была на 40 % дольше, чем побегов, а в целом продукция корней составляла 80-90 % общей продукции в этих сообществах (Steinaker, Wilson, 2008).
364
Глава 8. Биомасса и продукция фитоценозов
Значительная доля продукции растений поступает через корневые системы к микоризным грибам и другим почвенным консортам (см. п. 1.7 гл. 3). Hogberg и Read (2006) показали, что примерно половина выделяемого из почвы углекислого газа образуется при дыхании корней, микоризных грибов и других связанных с корнями микроорганизмов, а не при разложении отмерших частей растений и почвенной органики. На лугах при затенении растений дыхание почвы снижалось на 40 % (Craine et al., 1999). При кольцевании (прекращении нисходящего тока) в сосняке (Hogberg et al., 2001) почвенное дыхание снизилось на 37 % в течение 5 дней и на 56% в течение 14 дней. При этом образование плодовых тел эктомикоризных грибов снизилось на 98 %. Показано, что у ряда видов деревьев тонкие корни живут дольше, чем раньше считалось. Например, для Pinus taeda время оборота тонких (менее 1 мм в диаметре) корней оценивается в 4 года (Matamala et al., 2003; Hogberg, Read, 2006).
Погодные условия отдельных лет и воздействие фитофагов оказывают существенное влияние на продукцию растительных сообществ. Например, в работе Frank (2007) изучено влияние трехлетней засухи на надземную и подземную продукцию выпасаемых сообществ Йеллоустонского национального парка. Подземная продукция (по наблюдениям в миниризотронах) в слое 0-20 см снизилась на невыпасаемых участках на 39 %, на выпасаемых — на 49 %, надземная же значимо не изменилась. Надземная продукция была максимальна в первый месяц после снеготаяния, снижалась во второй месяц и была крайне незначительна в оставшееся время. Подземная продукция была высока в первый месяц после схода снега, но поддерживалась на высоком уровне (без засухи) — около 150 г на 1 м2 в месяц — в следующие 3 месяца и существенно снижалась в конце вегетационного периода (Frank, 2007).
Для пастбищных экосистем разработано несколько методик оценки надземной продукции: 1) оценка по необходимому потреблению животными (базовый бюджет, рассчитанный на основе физиологических свойств животных), 2) огораживание участков и учет на коротких временных интервалах, 3) срезание биомассы, имитирующее стравливание, и оценка отавности (скорости восстановления биомассы), 4) использование нормированной плотности животных. Большинство методов дает заниженные данные (причин много: непоступление экскрементов и т.п.), более адекватным является метод передвижных изгородей, огораживающих участки на короткое время, — метод 2 (McNaughton et al., 1996).
При увеличении возраста лесных насаждений снижается их надземная нетто-продукция (в зрелых лесах — до 76 % от молодых). Выдвигаются 3 причины такого снижения: 1) при увеличении возраста деревьев
3. Крупнотравье как система с максимальной продукцией
365
изменяется баланс между фотосинтезирующими и дышащими органами растений, 2) уменьшается доступность ЭМП в почвах, 3) возрастают устьичные ограничения, снижающие интенсивность фотосинтеза. Анализ различных исследований и моделирование процессов накопления биомассы показали, что причины 2 и 3 являются основными для лесов умеренного и холодного климатов, а в условиях жаркого климата причины 1 и 3 выходят на первый план (Gower et al., 1996).
3 • Крупнотравье как система с максимальной продукцией
Во внетропических районах Евразии наиболее высокой продукцией среди наземных экосистем отличается своеобразный тип растительных сообществ — крупнотравье. Эти сообщества встречаются на Камчатке, Сахалине, в Японии и на близлежащих территориях. По данным В. Л. Морозова (1994), средняя высота этих травяных сообществ превышает 2 м, среди основных доминантов — Angelica ursina (до 5 м высоты!), Filipendu-la camtschatica, Reynoutria sachalinensis, Petasites amplus (рис. 8.6). Суммарный индекс листовой поверхности (ИЛП) достигает 10-12, все листья крупнотравных растений сосредоточены на значительной (обычно более 1 м) высоте над поверхностью почвы (рис. 8.7). Годичная продукция этих сообществ может достигать почти 4 кг сухого вещества на квадратный метр (табл. 8.3). Эта высокая продукция обусловлена комплексом факторов: благоприятные условия увлажнения, высокая доступность ЭМП
Таблица 8.3 • Запасы биомассы (г/м2) и годичная продукция двух крупнотравных сообществ (по: Морозов, 1994)
Общая биомасса Надземная биомасса Подземная биомасса Отношение надземной биомассы к подземной Годичная продукция
Камчатка, Fiiipenduia camtschatica
3100 920 2180 0,42 1810
Сахалин, Reynoutria sachalinensis
6220 2720 3500 0,78 3820
366
Глава 8. Биомасса и продукция фитоценозов
Рис. 8.6 • Дальневосточное крупнотравье: а — Reynoutria sachalinensis, б — Angelica ursina, в — Petasites amplus (б, в — фото П. В. Крестова, с любезного разрешения автора)
3. Крупнотравье как система с максимальной продукцией
367
Рис. 8.7 • Распределение листовой поверхности по высоте у сахалинского крупнотравья на пойме (1) и террасах (2). Сообщества с доминированием: а — Angelica ursina, б — Anthriscus sylvestris, в — Filipendula camtschatica, г — Petasites ampius, d — Aconogonon weyrichii, e — Reynoutria sachaiinensis (по: Морозов, 1994)
из вулканических отложений, благоприятный световой режим (большая длительность солнечного сияния при высокой облачности) и умеренные температуры, не вызывающие больших затрат на дыхание. В зимнее время мощность снега составляет 70-200 см, что предотвращает глубокое промерзание почвы (Морозов, Белая, 1988).
368
Глава 8. Биомасса и продукция фитоценозов
4 • Энергосодержание фитомассы и энергетическая продукция фитоценозов
Помимо оценок запасов биомассы и продукции, выраженных в единицах массы, возможно оценить запасы и продукцию экосистем в энергетиче-
Таблица 8.4 • Содержание энергии в сухом органическом веществе растений (по: Лархер, 1978)
Растительный материал, вещества Содержание энергии
ккал/г кДж/г
Мхи и лишайники 3,4-4,6 14,2-19,3
Травянистые растения: побеги 3,8-4,3 15,9-18,0
корни 3,2-4,7 13,4-19,7
семена 4,4-5,0 18,4-21,0
Листопадные деревья: листья 3,9-4,8 16,3-20,1
древесина ствола 3,9-4,2 16,3-17,6
корни 4,0-4,7 16,3-18,6
Средиземноморские склерофиллы: листья 4,8-5,2 20,1-21,8
древесина ствола 4,5-4,7 18,9-19,7
корни 4,2-4,7 17,6-19,7
Вечнозеленые хвойные деревья: хвоя 4,9-5,0 20,5-21,0
древесина ствола 4,7-4,8 19,7-20,1
Вересковые кустарнички: листья 5,0-5,6 21,0-23,5
оси побегов 5,1-5,8 21,4-24,3
Вещества растений: глюкоза 3,7 15,5
крахмал, целлюлоза 4,2 17,6
белок 5,5 23,0
липиды 9,3 38,9
лигнин 6,3 26,4
изопрен 11,2 46,9
4. Энергетическая продукция фитоценозов
369
ских единицах. Энергосодержание тканей растений изменяется в незначительных пределах (Лархер, 1978; см. табл. 8.4), составляя в среднем около 4-4,5 ккал/г сухой массы. Наибольшие величины отмечены для побегов вересковых кустарничков (5,0-5,8 ккал/г), а наименьшие — для листьев и побегов травянистых растений (3,8-4,3 ккал/г).
В энергетических величинах первичная годичная продукция различных сообществ варьирует от 30 ккал/м2 в пустынях до 70000 ккал/м2 в тропических лесах (усредненные данные по типам сообществ). Энергетическая эффективность перевода первичной продукции во вторичную составляет для беспозвоночных фитофагов в среднем 9,6 %, а для травоядных млекопитающих — 0,4% (McNaughton et al., 1991) (см. п. 4 гл. 4).
На основании анализа энергосодержания в годичной продукции можно рассчитать энергетическую эффективность функционирования растительного сообщества, или его КПД (коэффициент полезного действия). Этот показатель рассчитывается как доля (процент) энергии, запасенной в продуцированной биомассе, по отношению к поступившей солнечной энергии. Возможны несколько вариантов расчетов, связанных с учетом: 1) нетто- или брутто-продукции, 2) общей поступающей солнечной радиации или только ее фотосинтетически активной части (ФАР), 3) поступления энергии в течение всего года или только вегетационного периода. При учете нетто-продукции и ФАР в течение всего года максимальные показатели из наземных сообществ характерны для тропических лесов (около 1,5%), для большинства других сообществ они не превышают 1 %. При учете нетто-продукции и ФАР за вегетационный период величина энергетической эффективности обычно составляет 1-2%, для интенсивных сельскохозяйственных культур — до 6 %, для крупнотравья на Камчатке — до 4,9 %, на Сахалине — до 8,7 %.
v
Сезонная динамика
Флуктуации (разногодичная изменчивость фитоценозов)
Сукцессии
Эволюция сообществ и историческая динамика
„„весь
| Динамика фитоценозов
д
инамика фитоценозов — это их изменение во времени. В зависимости от масштаба времени и характера изменений выделяют суточную, сезонную, флуктуационную (разногодичную), сукцессионную, историческую и эволюционную динамику растительных сообществ. Суточная динамика, связанная с солнечной ритмикой и суточными изменениями физиологических процессов, здесь специально не рассматривается.
1 • Сезонная динамика
Сезонная динамика связана с сезонным ходом температуры, освещенности, осадков. Она выражена везде, кроме некоторых экваториальных областей. Так, Reich et al. (2004) на обширном фактическом материале (более 40 000 листьев с 1445 деревьев 23 видов в течение 9 лет в Амазонии) исследовали сезонную динамику развертывания и отмирания листьев с целью проверки гипотезы об отсутствии цикличности этих процессов в условиях стабильного климата экваториальных лесов. Авторы подтвердили отсутствие сезонности в ритмике развития листьев и показали, что максимальная длительность жизни листьев составляла более 8,4 лет.
1.1 • Температура как фактор сезонной динамики
Низкие температуры влияют на растения двояко: путем прямого сдерживания или нарушения физиологических процессов и косвенно через уменьшение микробиологической активности почв. Так, в опытах, проведенных в Англии с райграсом многолетним (Lolium регеппе), при температуре менее 5,4 °C наблюдалось прямое торможение ростовых процессов, а в интервале температур 5,4-8,3 °C — косвенное сдерживание за счет недостатка минерализующегося азота. При внесении азотных удобрений рост райграса происходил и в этом интервале температур (Blackman, 1936). Сдерживание микробиологических процессов за счет низких температур имеет большое значение в замедлении биологического круговорота в тундрах и высокогорьях.
372
Глава 9. Динамика фитоценозов
Рис. 9.1 • Corydalis conorhiza и ее снежные корни: а — внешний вид цветущего растения; б — вытаивающие из снега корни; в — корни в талой воде, стекающей со снежника; г — отмершие снежные корни образуют корочки вокруг развивающихся побегов после схода снега
В то же время высокогорные растения долгоснежных местообитаний (например, Ranunculus adoneus в Колорадо) могут поглощать основное количество аммонийного азота из почвы в процессе снеготаяния при температурах почвы от 0 до 8 °C. Напротив, наибольшее поглощение
фосфатов у этого вида происходило в конце периода вегетации и было
связано с максимумом развития арбускулярной микоризы в этот период
(Mullen et al, 1998).
Еще более специализированы к поглощению соединений азота при
низких температурах корни ряда высокогорных растений Кавказа, раз-
вивающиеся в снегу (Onipchenko et al, 2009; рис. 9.1). С помощью ста-
бильного изотопа В * * * * * * 15 N для Corydalis conorhiza было показано активное
1. Сезонная динамика
373
потребление минеральных форм азота из снежника, в то время как поверхность почвы была промерзшая, и другие растения не могли использовать этот азот.
Расхождение видов по периодам вегетации и потреблению ресурсов возможно и у однолетних растений. Так, на лугах с доминированием однолетников в Калифорнии (средиземноморский климат) выделяют группу раннесезонных и позднесезонных однолетников. В экспериментах по изменению свойств почв видами этих функциональных групп (Hooper, Vitousek, 1998) было показано, что первая группа видов сильнее всего снижала концентрацию минеральных форм азота в почве в феврале, а вторая — в мае, что может служить примером фенологического расхождения видов по использованию почвенных ресурсов.
Низкие температуры могут сдерживать семенное размножение растений и их распространение в более холодные области. Например, липа (Tilia cordata) естественно распространена на юге Британских островов, но в посадках она прекрасно развивается и в Центральной Шотландии. Однако в этом случае у липы резко подавлено семенное размножение, так как при температуре ниже 15 °C замедляется прорастание пыльцы и развитие пыльцевой трубки, что приводит к ингибированию развития зародыша (Pigott, Huntley, 1981).
Размер генома у ряда групп растений связан с их холодостойкостью и возможностью развиваться ранней весной. Многими авторами отмечалось, что тропические растения, как правило, имеют меньшие размеры генома, чем растения умеренного пояса. По данным Grime et al. (1985), скорость весеннего развертывания листьев у луговых растений Англии и размер генома положительно скоррелированы. Это связано с тем, что низкие температуры сильнее тормозят деление, чем рост клеток, поэтому в условиях низких весенних температур более адаптивно иметь крупные клетки, рост которых происходит быстро. Кроме того, было показано, что размер генома растений Великобритании положительно скоррелирован с морозоустойчивостью надземных побегов. Эта закономерность выявлена в специальном эксперименте с раннелетним промораживанием участков дернины пяти фитоценозов в течение двух дней (MacGillivray, Grime, 1995).
Заложение генеративных почек растений в холодных областях происходит в разное время. В зависимости от температурных условий местообитания и биологических особенностей отдельных видов закладка генеративных почек у высокогорных растений может происходить либо в год цветения, либо в год, предшествующий цветению, либо за два года до него (рис. 9.2). Поэтому численность генеративных побегов может
374
Глава 9. Динамика фитоценозов
Рис. 9.2 • Заложение генеративных почек у Ranunculus glacialis происходит за два года до цветения, у Primula glutinosa и Saxifraga oppositifolia — в предшествующий цветению сезон, у Cerastium uniflorum — в сезон цветения. Голубым цветом показан снежный покров (по: Reisigl, Keller, 1987)
быть связана не с экотопическими условиями текущего сезона, а с условиями предшествующих лет.
1.2 • Световой режим и сезонная динамика
Длительность светового дня имеет значение не только для индукции цветения, но для продукции растений. Так, в экспериментах на дикорастущих и культурных злаках умеренного пояса было показано, что при удлинении светового периода (но сохранении суммарного количества ФАР) продукция может возрастать почти вдвое, еще сильнее увеличивается листовая поверхность (Нау, 1990).
В осенне-зимний период при низком уровне поступления света многие процессы в фитоценозах умеренных широт не прекращаются. В это время наиболее активно происходит фотосинтез эпифитных мхов и лишайников. Свет может проникать и под снег (см. п. 1.2 гл. 6).
Длительность вегетационного периода совместно с обеспеченностью растений элементами минерального питания влияют на длительность жизни листьев (компромисс: долгоживущие листья имеют меньшую эффективность фотосинтеза, но более экономны с точки зрения
1. Сезонная динамика
375
затрат ЭМП). Это приводит к различной реакции листьев листопадных и вечнозеленых растений на увеличение длительности вегетационного периода: листопадные виды увеличивают, а вечнозеленые — уменьшают длительность жизни листьев при увеличении вегетационного периода. Это было показано, например, для высокогорных видов долгоснежных местообитаний в Японии (листопадного Sieversia pentapetala и вечнозеленых Phyllodoce aleutica и Rhododendron aureum) (Kikuzawa, Kudo, 1995).
1.3 • Водный режим и сезонная динамика
Для ритмики фитоценозов большое значение имеет как режим поступления осадков, так и режим поемности (на лугах). Особенно важен режим поемности в аридных регионах. Например, для низовий Волги, расположенных в зоне пустынь и полупустынь, до урегулирования стока можно было четко различить три периода сезонного развития: 1) до паводка, 2) паводок, 3) после паводка. В связи с высокой толерантностью взрослых особей и снижением их физиологической активности в неблагоприятных условиях здесь на одном квадратном метре могут сосуществовать растения весьма контрастной экологии, например типчак (Festuca valesiaca) и ситняг болотный (Eleocharis palustris) (Раменский и др., 1956) (см. также п. 3.2.1 гл. 6).
1.4 • Снеговой режим
Снеговой режим часто определяет длительность вегетационного сезона не только в отдельных регионах, но и на близкорасположенных участках, приуроченных к разным формам мезорельефа. При неравномерном пространственном распределении снега (например, в гумидных высокогорьях) сход снежного покрова во многом определяет начало сезонного развития растений. Для горно-тундровых сообществ Арктики U. Molau (1993) выявил, что время цветения растений связано с рядом важных характеристик их репродуктивной сферы. Он выделил две принципиально различающиеся группы видов — ранне- и позднецветущие (табл. 9.1). Для раннецветущих растений основная проблема — эффективное опыление в условиях неустойчивых температур, поэтому они производят относительно много пыльцы. Позднецветущие, напротив, рискуют не успеть завершить созревание плодов до окончания вегетационного периода, с чем и связаны их репродуктивные особенности.
В холодных областях (тундры, высокогорья) почвы под снегом (более 50-60 см глубины) имеют зимнюю температуру не ниже -5 °C. При близких к 0 °C температурах в этих почвах наблюдается микробиоло-
376
Глава 9. Динамика фитоценозов
Рис. 9.3 • Зависимость зимней эмиссии СО2 от среднесуточной температуры поверхности почвы (То) в типичной тундре за период 1 октября 2001 г. — 31 мая 2002 г. (поселок Лаврентия; данные Д. Карелина)
Таблица 9.1 • Сравнение особенностей размножения
ранне- и позднецветущих видов горно-тундровых растений (по: Molau, 1993)
Раннецветущие Позднецветущие
Часто вечнозеленые Часто летнезеленые
Бесполое размножение редко Часто вивипария, апомиксис, луковички
Самоопыление редко Самоопыление часто
Отношение пыльца: семяпочки
Высокое Низкое
Отношение семена: семяпочки
Низкое Высокое
«Рискуют пыльцой» «Рискуют семенами»
гическая активность — дыхание, минерализация азота и т. п. (рис. 9.3). Обычно почвы сильно промерзают поздней осенью (до глубокого снега), а затем зимой оттаивают под снегом. Поэтому продукция сообществ, приуроченных к участкам со значительным снежным покровом, существенно выше, чем малоснежных участков (Brooks et al., 1997). Таким образом,
1. Сезонная динамика 377
характер снежного покрова четко отражается на пространственном варьировании богатства почвы ЭМП и продукции растительных сообществ.
1.5 • Выделение фенологических периодов
В отличие от фиксированных (календарных) сезонов сроки наступления фенологических периодов колеблются год от года в зависимости от погодных условий. Существует несколько подходов к выделению фенологических периодов:
1) по метеорологическим условиям — например, наступление весны может быть определено по переходу среднесуточной температуры через О °C;
2) по индикаторным видам. Этот подход широко распространен в географической фенологии, наступление сезонов определяется по фенологическим фазам широко распространенных видов (цветение орешника, яблони и др.). На основании широкой сети данных фенологи строят изофены — линии на карте, соединяющие точки с одновременным наступлением тех или иных фенофаз индикаторных видов;
3) по фенологическим кривым. Фенологические кривые показывают изменение по сезонам числа видов в сообществе, находящихся в той или иной фенологической фазе (рис. 9.4, 9.5). Чаще фенологические кривые имеют одновершинный характер, но встречаются и другие варианты. Например, для широколиственных лесов Т. К. Горыши-на (1969) приводит двувершинную кривую цветения (первый, более
Рис. 9.4 • Кривая цветения (1, А — число цветущих видов) и относительная освещенность (%) на уровне травяного покрова (2, Б) для травянистых растений в липо-дубовом лесу (по: Горышина, 1969)
378
Глава 9. Динамика фитоценозов
май
июнь июль август сентябрь
—1 —«—2 —а—з - -х -4 —ж- 5
Рис. 9.5 • Фенологические кривые альпийских пустошей Северо-Западного Кавказа и фенологические периоды, выделенные на их основе (по: Онипченко, 1983). По оси абсцисс — число видов, находящихся в разных фенологических состояниях. Обозначения кривых: 1 — вегетация до цветения, 2 — бутонизация, 3 — цветение, 4 — созревание плодов и семян, 5 — рассеивание созревших плодов и семян
высокий пик в мае, второй — в начале июля; рис. 9.4). В случае одновершинных кривых фенологической весной можно считать период, когда большинство видов растений еще не зацвели, ранним летом — период, когда большинство видов находятся в цветущем состоянии, поздним летом — период, когда у большинства видов созревают семена, а осенью — период, когда большинство видов рассеивают созревшие семена (рис. 9.5).
Наступление отдельных фенологических фаз, их частота и длительность определяются компромиссом многих факторов — климатических условий, доступности ресурсов, развитием опылителей, фитофагов и распространителей плодов и семян. Например, задержка в развертывании листьев у отдельных деревьев в муссонных лесах Индии ведет к резкому увеличению повреждения их фитофагами (Murali, Sukumar, 1993). Для двух видов деревьев (Metrosideros collina и Sophora chrysophylla) на Гавайях, опыляющихся одной группой насекомых, Carpenter (1976) показал расхождение по периодам цветения в случае совместного произрастания этих видов и перекрывание — при раздельном произрастании (Fenner, 1998).
1. Сезонная динамика
379
В связи с массовым цветением отдельных видов растений, так же как и с другими фенологическими явлениями (например, осенним отмиранием листьев), существенно могут меняться цветовые свойства сообществ, или их аспекты. Изучение смен аспектов не только позволяет понять механизмы фенологического расхождения видов растений внутри сообщества, но и имеет очень большое значение для дешифрирования аэро-и космических снимков. Ведь сообщества одного и того же типа на одном снимке могут иметь совершенно разные спектральные характеристики, если их сезонное развитие различается, например, при разном времени схода снежного покрова.
1.6 • Феноритмотипы и проблема вечнозелености
И. В. Борисова (1972) предложила разделить растения внутри сообщества на несколько феноритмотипов — групп растений со сходной длительностью начала и конца вегетации, а также с одинаковой направленностью смен основных фенологических состояний. По длительности вегетации различают: 1) длительновегетирующие (более 2 месяцев) растения, включая летнезеленые, летнезимнезеленые (длительность жизни листьев менее 1 года, часть листьев позднелетней генерации перезимовывает в зеленом состоянии) и вечнозеленые (длительность жизни листа более 1 года) и 2) коротковегетирующие (менее 2 месяцев), включая эфемеры (однолетники) и эфемероиды (многолетники). Соотношение видов, относящихся к разным феноритмотипам, в составе сообществ может быть индикатором их сезонной ритмики.
Ритмика развития и длительность жизни листьев на деревьях и кустарниках сильно варьируют даже в умеренном климате. К. Kikuzawa (1989) для лесов Японии выделил 6 типов листьев по ритмике их развития (рис. 9.6). Затенение и недостаток ЭМП обычно увеличивают длительность жизни листьев.
В условиях умеренного пояса значительное число видов являются зимнезелеными. Растения многих видов злаков и осок зимуют с зелеными базальными частями листовой пластинки. По данным Е. И. Лапшиной (1928), на лугах северо-запада европейской части России из 80 видов 44 вида перезимовывали частично в зеленом состоянии. Основное функциональное значение большой длительности жизни листьев зимнезеленых растений заключается в экономии ЭМП в условиях их недостатка. Вечнозеленые листья могут служить хранилищами ЭМП, запасенными в благоприятный период. Так, в старых листьях кустарника чапарра-ли Adenostoma fasciculatum накапливаются ЭМП, полученные в течение дождливого зимнего периода и используемые в период весеннего раз-
380
Глава 9. Динамика фитоценозов
летнезеленые
а) быстрое развертывание б) промежуточное в) сукцессивное
со (До-'—•—>———Ь- —•—«—•——♦ 00—1—-—•—•—•—
2 М И И А С О М И И А С О МИИАСО
о с зимующими листьями
q г) зимнезеленые д) гетероптозис е) вечнозеленые
Рис. 9.6 • Основные типы сезонной динамики листьев древесных растений широколиственных лесов острова Хоккайдо (Япония) (по: Kikuzawa, 1989).
Темные кружки показывают число листьев, прикрепленных к побегу в данный момент времени. Светлые кружки показывают кумулятивное число листьев, развернувшихся на побеге. Примеры видов: а — Quercus mongolica;
б — Magnolia obovata (М. hypoleuca); в — Alnus hirsute; г — Daphne kamtchatica; d — Euonymus alatus; e — Daphniphyllum macropodum
вертывания листьев (Mooney, Rundel, 1979; Koide, 1991). Аналогичные данные были получены для брусники, где листья накапливают значительное количество азота, который весной перемещается в растущие побеги. В целом можно отметить следующие особенности:
1) вечнозеленые листья экономят ЭМП,
2) вечнозеленые листья увеличивают длительность вегетационного периода,
3) склерофилльность (жестколистность) вечнозеленых листьев препятствует водному дефициту,
4) у вечнозеленых листьев ниже потери на дыхание.
Но листья вечнозеленых растений, в отличие от зимнезеленых листьев, не всегда способны к фотосинтезу в зимнее (холодное) время. Например, у растений субальпийского пояса Кавказа зимой у летнезимнезеленых видов (например, Festuca varia) наблюдался фотосинтез с низкой интенсивностью, а у вечнозеленого Rhododendron caucasicum — нет (Нахуцришвили,
2. Флуктуации (разногодичная изменчивость фитоценозов)
381
1974, 1975). Этот вид рододендрона зимует в условиях глубокого снежного покрова, искусственное удаление которого в зимнее время ведет к быстрому высыханию и отмиранию листьев. С другой стороны, вечно- и зимнезеленые растения подлеска широколиственных лесов более эффективно фотосинтезируют при низких температурах и в условиях низкой освещенности, чем летнезеленые виды (Landhausser et al., 1997).
Исследования склерофилльных растений в восточной Австралии (Wright, Cannon, 2001) показали, что жесткость листьев (сила их разрыва) была положительно скоррелирована с длительностью их жизни.
Экологически противоположную группу образуют коротковегети-рующие растения. Роль эфемеров наиболее велика в аридных регионах с коротким периодом достаточного увлажнения, а роль эфемероидов — в лесах, но не всегда листопадных. Так, Anemone nemorosa обильно представлена в богатых ельниках на западе европейской части России, хотя эфемероиды могут отсутствовать в ряде березовых лесов. Здесь основным фактором развития этой группы растений является высокое богатство почвы и отсутствие ее промерзания зимой. Развивающиеся вскоре после схода снега эфемероиды активно поглощают ЭМП, прежде всего нитраты, из насыщенной водой почвы, что имеет очень важное значение в удержании ЭМП от вымывания их из почвы талыми водами в весенний период, когда корни других растений еще не очень активны. Эта роль показана для эфемероидов широколиственных лесов (Peterson, Rolfe, 1982а, 1982b; Anderson, Eickmeier, 1998), причем возможности «депонирования» ЭМП снижались при уменьшении доступности света. Таким образом, эфемероиды могут быть отнесены к растениям эксплерентной (рудеральной) стратегии, в то время как вечнозеленые растения — к па-тиентам (стресс-толерантам).
По мнению Т. К. Горышиной (1971; Горышина, Митина, 1970), некоторые виды лесных растений могут рассматриваться как скрытые эфемероиды. Например, сныть Aegopodium podagraria около 50 % продуктов фотосинтеза накапливает в течение 2 недель весной при высокой доступности света (до развертывания листьев деревьев) и имеет низкий уровень фотосинтеза теневых листьев в течение лета.
2 • Флуктуации (разногодичная изменчивость фитоценозов)
Флуктуациями называются кратковременные (в масштабе нескольких лет) изменения структуры растительных сообществ. Эти изменения, в отличие от сукцессий, характеризуются: 1) ненаправленностью (т. е. отсутствием
382
Глава 9. Динамика фитоценозов
многолетних трендов изменения структуры сообществ в том или ином направлении), 2) обратимостью (возможностью возвращения к состоянию, близкому к исходному), 3) отсутствием существенных изменений флористического состава. Чаще всего флуктуациями считают изменения, происходящие в пределах одного солнечного цикла (около 11 лет).
Флуктуации могут вызываться различными причинами. Т. А. Работнов (1983) предложил различать несколько типов флуктуаций по причинам, их вызывающим: экотопические, антропические, зоогенные, фито-циклические, фитопаразитарные.
2.1 • Экотопические флуктуации
Экотопические флуктуации вызваны изменчивостью метеорологических и гидрологических условий. Изменения экотопа могут вызывать цепочки различных механизмов воздействия на растительный покров. Например, засуха обычно сопряжена с повышением температуры и усилением засоления в аридных регионах, а также может вызывать массовое развитие саранчовых, потребляющих значительную долю надземной продукции.
Август 1947 Август 1948
• верховые злаки, преимущественно Bromus erectus, реже Arrhenatherum elatius и Dactylis glomerata
Lotus corniculatus
Centaurea jacea
Medicago sativa
Salvia pratensis
Daucus carota
Pieris hieracioides
Рис. 9.7 • Изменение структуры сухого карбонатного луга
на постоянном квадрате (площадь — 1 м2) в Швейцарии в засушливый 1947 г. и «нормальный» 1948 г. (по: Ludi, Zoller, 1949)
2. Флуктуации (разногодичная изменчивость фитоценозов)
383
Примером воздействия засухи может служить резкое изменение структуры лугов на карботантных почвах в Швейцарии (рис. 9.7 — по LQdi, Zoller, 1949): снижение участия отдельных доминирующих видов (Bromus erectus), практическое исчезновение других (Lotus corniculatus) при лучшем развитии глубокоукореняющихся растений (Salvia pratensis, Daucus carota) (LQdi, Zoller, 1949; Gigon, 1997).
Наибольшее влияние оказывает засуха на растительные сообщества, развивающиеся на маломощных почвах. Например, на сухих приморских лугах (альварах) в Эстонии в течение 5 лет была исследована динамика участия сосудистых растений, мохообразных и лишайников на постоянных площадках в связи с динамикой метеорологических факторов (Ingerpuu, Kupper, 2007). Флуктуации общего покрытия мохообразных, как и доминирующего вида Ctenidium molluscum, были связаны с общим количеством осадков. Покрытие сосудистых растений было значимо связано с индексом увлажнения (осадки х число дней с осадками / средняя температура лета) (рис. 9.8). Наиболее консервативным поведением отличались кустистые лишайники, их покрытие по годам изменялось незначительно.
Участки сообществ на разных элементах мезорельефа показывают разную интенсивность экотопических флуктуаций. Более резко выражены флуктуации в западинах и гидроморфных элементах рельефа, где уровень увлажнения значительно сильнее колеблется по годам, чем на возвышенных элементах мезорельефа, откуда поступающая с осадками вода стекает или фильтруется в нижерасположенные участки.
Южные осцилляции — Эль-Ниньо, связанные с изменением температуры и системы ветров над южной частью Тихого океана, оказывают существенное влияние на метеорологические условия, структуру и продукцию растительности многих регионов мира. Эль-Ниньо происходит нерегулярно раз в 3-6 лет и вызывает резкое увеличение осадков в некоторых частях мира и резкие засухи — в других. В противоположные (Ла-Нинья) годы погодные условия сменяются на контрастные по отношению к Эль-Ниньо годам. На сухих островах Калифорнийского залива покрытие растений в нормальные годы колеблется между 0 и 4 %, в то время как в годы Эль-Ниньо оно составляет 54-89% (Polis et al., 1997). Возобновление ряда кактусов и кустарников (Larrea) в пустыне Сонора происходит почти исключительно в годы Эль-Ниньо. С другой стороны, Эль-Ниньо вызывает сильные засухи и пожары в амазонских тропических лесах (Holmgren et al., 2001).
Для прерий в Миннесоте (США) показано интересное влияние засухи на годичную продукцию растений: сильная засуха вызывает резкие
384
Глава 9. Динамика фитоценозов
Рис. 9.8 • Флуктуации ежегодных осадков (темные кружки), летнего индекса увлажнения (осадки х число дней с осадками / средняя температура лета; треугольники) и проективное покрытие мхов (a), Ctenidium molluscum (б), лишайников (в), сосудистых растений (г) на контрольных участках. Годы отмечают период между двумя измерениями (с сентября предыдущего года до октября текущего). Различными буквами отмечены значимые различия (по: Ingerpuu, Kupper, 2007)
периодические колебания продукции в последующие годы с периодом 2 года (наблюдались в течение 9 лет после засухи) (Haddad et al., 2002; рис. 9.9). При этом аналогичной периодичности в выпадении осадков не наблюдалось, как и в динамике численности основных групп фитофагов. Рассмотрена гипотеза депонирования азота в один год в ветоши и соответствующее снижение его доступности в следующий год. Она
2. Флуктуации (разногодичная изменчивость фитоценозов)
385
Рис. 9.9 • Динамика продукции растений в Cedar Creek (Миннесота, США) в течение 1981-2000 гг. на контрольных участках прерий и на сукцессионных площадках. Показаны средние значения и ошибки среднего. Периодические колебания с циклом 2 года хорошо выражены на обоих участках после засухи (по: Haddad et al., 2002)
подтверждается экспериментами с выжиганием и с внесением азота, которые снимают наблюдающийся на контроле осцилляционный режим. По другой гипотезе засуха синхронизирует продукционную двухлетнюю циклику основных доминирующих видов (чередование годов с низкой и высокой семенной продукцией) (Haddad et aL, 2002).
Согласно рассмотренной в п. 4.2.7 гл. 6 модели конкуренции в пространственной среде, коэффициент вариации (по времени) участия вида в сообществе уменьшается с увеличением степени доминирования вида, т. е. участие более редких видов должно сильнее изменяться по годам. В работе Hobbs et al. (2007) получены подтверждения этой модели. Исследователи провели 20-летние наблюдения за динамикой однолетнего злаковника в Северной Калифорнии на контрольных участках, участках исключения гоферов и огороженных от наземных фитофагов участках. Нарушения почвы гоферами существенно увеличивались при исключении крупных травоядных. Динамика сообщества определялась преимущественно режимом осадков (за период наблюдений было 2 пика). Видовое богатство снижалось во время засух и увеличивалось после обильных осадков, при этом виды реагировали на изменение условий индивидуально. Встречаемость 18 видов из 36 значимо коррелировала с количеством осадков. Коэффициент вариации встречаемости групп видов был самым низким для однолетних трав (наиболее обильных) и самым высоким — для многолетнего разнотравья, группы с наименьшим участием. Таким образом, коэффициент вариации был положительно связан с рангом видов, т. е. отрицательно — с их участием (см. рис. 9.10) (Hobbs et aL, 2007).
386
Глава 9. Динамика фитоценозов
600 -г
500-
0 5 10 15 20
25
ранг видов
Рис.9.10 • Связь ранга видов (виды ранжированы по участию в составе сообщества: чем выше ранг, тем ниже участие) с коэффициентом вариации их проективного покрытия по годам (по: Hobbs et al., 2007)
Продукция сообществ в целом и отдельных видов растений значительно колеблется по годам, особенно в близких к экстремальным условиях. Пример исследования таких колебаний приводится в работе F. S. Chapin и С. Korner (1996) для тундр на Аляске (табл. 9.2). Необходимо подчеркнуть, что колебания продукции отдельных видов по годам были значительно сильнее, чем колебания общей продукции сообщества, что свидетельствует о лучшем развитии отдельных видов в разные по метеорологическим условиям годы.
В ряде работ предпринималась попытка связать размах флуктуационных колебаний продукции и надземной биомассы с видовым разнообразием растительных сообществ. На основании 13-летних наблюдений на постоянных площадках старовозрастных залежей в Миннесоте D. Tilman (1996) показал, что степень варьирования биомассы по годам отрицательно скоррелирована с видовым богатством участков. На флористически богатых участках варьирование отдельных видов по годам было весьма значительным, в то время как общая надземная биомасса колебалась менее значительно (компенсационный эффект). Однако видовое разнообразие в этом случае не являлось причиной меньших колебаний биомассы, поскольку продукция таких участков была лимитирована доступностью элементов минерального питания в почве, которая
2. Флуктуации (разногодичная изменчивость фитоценозов)
387
Таблица 9.2 • Разногодичная динамика общей продукции арктической тундры (Аляска) и отдельных видов этого сообщества (CV — коэффициент вариации по годам, %) (по: Chapin, Korner, 1996)
Виды Продукция по годам (% от средней) CV,%
1968 1969 1970 1978 1981
Eriophorum vaginatum 77 58 148 101 116 31
Betula папа 30 52 55 248 121 79
Ledum palustre 106 138 62 103 91 25
Vaccinium vitis-idaea 135 172 96 28 71 50
Общая продукция 93 110 106 84 107 10
не очень резко изменялась по годам. С другой стороны, на флористически бедных, но высокопродуктивных участках продукция в большей степени лимитирована условиями увлажнения, которые очень значительно изменялись по годам в связи с колебанием осадков.
2.2 • Антропические (антропогенные) и зоогенные флуктуации
Антропогенные флуктуации обусловлены различиями в форме и интенсивности воздействия человека, например в сроках и частоте сенокошения. Существенные изменения в составе растительных сообществ могут происходить и при изменении численности фитофагов и/или роющих животных, которые вызывают зоогенные флуктуации. В годы пика численности грызунов резко увеличивается изъятие ими биомассы растений и степень нарушенности почвенного покрова. Примером может служить периодически наблюдаемый «залежный режим» в степях Монголии при высокой численности полевки Брандта, когда происходит отмирание дер-новинных злаков (типчак, ковыли) и разрастание корневищных злаков, связанное с разрыхлением почвы землероями и улучшением поступления ЭМП. Восстановление степной структуры сообществ происходит в течение нескольких лет до новой вспышки численности полевки.
2.3 • Фитоциклические флуктуации
Фитоциклические флуктуации связаны с особенностями жизненного цикла компонентов фитоценозов и/или неравномерным по годам их семенным или вегетативным размножением. Классический пример — цикли-
388
Глава 9. Динамика фитоценозов
Рис. 9.11 • Массовое цветение Tri folium pratense на субарктических лугах в северной Швеции
ческое участие клеверов (особенно Trifolium pratense) в составе фитоценозов. На ряде типов лугов раз в несколько лет возникают «клеверные годы», когда участие клеверов резко возрастает (рис.9.И). Это явление может быть связано с балансом элементов минерального питания в почве, уменьшением соотношения N : Р, поскольку искусственное внесение РК (фосфорно-калийных) удобрений вызывает аналогичное увеличение участия клеверов (Работнов, 1983).
2. Флуктуации (разногодичная изменчивость фитоценозов)
389
Рис. 9.12 • Anthriscus sylvestris и флуктуации его биомассы по годам на постоянных площадках Bibery в Англии (по: Grime et al., 1994)
Другим примером фитоциклических флуктуаций может быть массовое цветение монокарпичных зонтичных на лугах. Так, участие Anthriscus sylvestris по биомассе менялось по годам более чем в десять раз на постоянных площадках в Bibury (Англия) (рис. 9.12; Grime et al., 1994).
2.4 • Типы флуктуаций по степени их выраженности
Т. А. Работнов (1972) предложил различать следующие типы флуктуаций по степени выраженности: скрытые, осцилляционные, дигрессионно-де-мутационные.
При скрытых флуктуациях происходят лишь количественные изменения в участии отдельных видов растений, выявляемые при специальных учетах на постоянных площадках. Примером таких флуктуаций может служить изменение численности генеративных побегов растений по годам, зависящее от многих внутренних и внешних факторов. На рис. 9.13 показана динамика численности генеративных побегов Campanula tri-dentata и Festuca ovina на постоянных площадках альпийских пустошей в Тебердинском заповеднике за 25 лет наблюдений. У обоих видов численность побегов на участке 62 м2 колебалась примерно в диапазоне 500-3000, однако у Festuca ovina в 1999 г. (после засушливого 1998 г.) генеративные побеги совсем не образовывались, a Campanula tridentata ежегодно образовывал не менее 500 побегов.
390
Глава 9. Динамика фитоценозов
годы
Рис. 9.13 • Динамика численности генеративных побегов (ордината) двух видов растений на альпийских лишайниковых пустошах в Тебердинском заповеднике в течение 25 лет (абсцисса). Вслед за засушливыми сезонами 1998 и 2000 гг. следует спад численности побегов обоих видов, a Festuca ovina в 1999 г. совсем не образует генеративных побегов
Изменения численности побегов по годам у отдельных видов могут быть сходными (в значительной мере синхронными), если эти виды имеют сходные экологические требования для закладки и развития побегов. В таком случае коэффициенты корреляции между численностями побегов этих видов по годам будут положительными и виды образуют корреляционные плеяды, примеры которых для гераниево-копеечниковых лугов приведены на рис. 9.14. В случае контрастных экологических требований (например, один вид лучше развивается в сухие, другой — во влажные годы) участие этих видов в составе сообществ скоррелировано отрицательно, они входят в различные корреляционные плеяды.
2. Флуктуации (разногодичная изменчивость фитоценозов)
391
Рис. 9.14 • Корреляционные плеяды видов растений альпийских гераниево-копеечниковых лугов Тебердинского заповедника (по: Чередниченко, 2001). Выделено три плеяды с разным числом видов (20, 3 и 1). Сплошной линией соединены виды, численности побегов которых по годам значимо положительно скоррелированы (р < 0,05), т. е. изменения численности побегов видов, образующих одну плеяду, в значительной степени синхронизированы по годам. Длительность наблюдений — 16 лет. Виды растений: 1 — Carex atrata, 2 — Anthemis cretica, 3 — Agrostis vinealis, 4 — Leontodon hispidus, 5 — Deschampsia flexuosa, 6 — Minuartia circassica, 1 — Ranunculus oreophilus, 8 — Veronica gentianoides, 9 — Carum meifolium, 10 — Hedysarum caucasicum, 11 — Geranium gymnocaulon, 12 — Gentiana pyrenaica, 13 — Campanula tridentata, 14 — Minuartia aizoides, 15 — Luzula multiflora, 16 — Sibbaldia procumbens, 17 — Nardus stricta, 18 — Anthoxanthum odoratum, 19 — Potentilla verna, 20 — Scorzonera сапа, 21 — Festuca brunnescens, 22 — Phleum alpinum, 23 — Matricaria caucasica, 24 — Festuca ovina
При осцилляциях в сообществе сосуществует несколько видов растений, способных доминировать в различных условиях. Они часто встречаются в луговых фитоценозах. Например, А. П. Шенников (1930) описал в пойме Волги близ Ульяновска следующие осцилляции, связанные со сменой доминантов во влажные и сухие годы. Во влажные годы на этих лугах доминирует Alopecurus pratensis, а в сухие — Bromus in-ermis и Agropyron repens, хотя все эти виды присутствуют в любой год
392
Глава 9. Динамика фитоценозов
Рис. 9.15 • Флуктуации покрытия вереска Calluna vulgaris на двух площадках (показаны разной штриховкой) на альварах острова Оланд (южная Швеция). Вереск практически полностью отмирал после засух в 1975, 1982 и 1992 гг. (по: Rosen, 1995)
наблюдений, но с разным участием. Такого рода осцилляции делают бессмысленной классификацию луговой растительности по доминирующим видам.
Дигрессионно-демутационные флуктуации являются переходными к сукцессиям. Они включают фазу нарушения и последующее восстановление близкого к исходному состояния. Часто наблюдаются на лугах после засухи, вымокания и т. п. Примером может служить изменение участия вереска (Calluna vulgaris) в составе альварных сообществ на острове Оланд (Rosen, 1995; рис. 9.15). Вереск практически исчезал после сильных засух, а затем постепенно восстанавливал свое высокое участие.
Другим примером дигрессионно-демутационных флуктуаций могут служить изменения в составе луговых фитоценозов в пойме Оки (Работнов, 1984а). Луга с доминированием Alopecurus pratensis или Digraphis arundinacea довольно устойчивы по годам, однако в годы длительного заливания полыми водами они могут угнетаться и резко снижать свое участие на 2-3 года, когда их место занимают демутационные эксплерен-
2. Флуктуации (разногодичная изменчивость фитоценозов)
393
ты — Ranunculus repens и Agrostis stolonifera, более устойчивые к затоплению и быстро увеличивающие свое участие при угнетении основных доминантов.
К дигрессионно-демутационным флуктуациям можно отнести и различные случаи пирогенных циклов, которые для удобства мы рассмотрим в разделе сукцессий.
2.5 • Сочетание различных механизмов флуктуационной динамики в естественных фитоценозах
В естественных растительных сообществах разногодичная динамика каждого вида может определяться комплексом различных факторов, при этом ведущие факторы могут значительно различаться у разных видов. В этом разделе мы рассмотрим примеры конкретных исследований флуктуационной динамики, в которых изучены комплексные механизмы
изменения участия отдельных видов по годам.
Многие виды рода Chiono-chloa — крупные плотнодерно-винные злаки в горах Новой Зеландии — образуют генеративные побеги через год, что ведет к большой вариабельности численности этих побегов по годам (рис. 9.16). На основании длительных (И лет) наблюдений и математических моделей исследованы механизмы нерегулярного (хаотического) чередования лет с высокой численностью генеративных побегов и семян на примере Chionochloa pallens (Rees et al., 2002). Показано, что численность побегов, с одной стороны, имеет «зубчатый» характер (год подъем — год спад),
год наблюдений
Рис. 9.16 • Динамика численности генеративных побегов Chionochloa pallens на горе Hutt в Новой Зеландии; каждая линия соответствует отдельным растениям (по: Rees et al., 2002)
что связано с внутренней ди-
намикой ресурсов растений. С другой стороны, число побегов в годы подъема также сильно варьирует стохастическим образом и зависит
от температуры в середине лета предшествующего года, когда закла-
394
Глава 9. Динамика фитоценозов
Рис. 9.17 • Схема основных факторов, формирующих асинхронную флуктуационную изменчивость покрытия различных видов растений на сухом лугу в северо-западной Швейцарии. Виды, на покрытие которых данный фактор действует положительно, помещены в столбцах под знаком «+», отрицательно — под знаком «-» (по: Gigon, 1997)
дываются цветочные почки. Высокие температуры в этот период благоприятно влияют на их заложение. После цветения генеративный побег отмирает, но из боковой почки развивается вегетативный побег, который может зацвести через 3-5 лет (в зависимости от метеорологических условий). Коэффициент вариации численности генеративных побегов по годам у видов рода Chionochloa достигает 151-308 %, что считается одним из самых высоких показателей в мире. Высокая изменчивость цветения по годам, видимо, имеет эволюционное происхождение, так как во время массового цветения уменьшается потеря семян из-за семеноядов. На видах этого рода отмечено три вида насекомых, которые совместно могут потреблять более 90 % урожая семян. У Chionochloa pollens в годы низкой численности потребляется около 80% семян, в годы высокой — менее 10% (Rees et al., 2002).
На Ротамстедской станции в Англии была исследована динамика участия отдельных видов в составе экспериментальных лугов в течение
3. Сукцессии
395
60 лет (1920-1979). Выделено 3 группы видов по их динамике: 1) стабильные, 2) имеющие постоянный тренд увеличения участия и 3) «взрывные», т. е. показавшие увеличение, а затем снижение участия (Silvertown et al., 2002). Проверялись две гипотезы: 1) увеличение участия «взрывных» видов связано с большей скоростью роста популяции в периоды ослабления конкурентных отношений; 2) снижение участия этих видов в дальнейшем связано с недостаточной генетической изменчивостью, ведущей к меньшей пластичности видов при увеличении интенсивности конкуренции. В целом обе гипотезы подтвердились. Семенная продукция «взрывных» видов была значимо выше, чем других. С другой стороны, в 7 случаях из 7 «взрывные» виды имели ббльшую долю самоопыления, чем другие, что снижало их генетическое разнообразие. Таким образом, динамика отдельных видов растений в сообществах определяется как их экологическими, так и генетическими признаками (Silvertown et al., 2002).
Сочетание флуктуаций разных типов приводит к возможности сосуществования многих видов растений в пределах одного сообщества. Для сухих лугов на северо-западе Швейцарии A. Gigon (1997) приводит обобщенную схему сочетания флуктуаций разных типов и продолжительности, позволяющую представить их разнонаправленные последствия (рис. 9.17).
3 • Сукцессии
3.1 • Определение сукцессий
Существуют различные определения сукцессий как смен растительных сообществ. Т. А. Работнов (1983: 204) приводит следующее определение, подчеркивающее отличие сукцессий от флуктуаций: «Сукцессии — необратимые, направленные, т. е. происходящие в определенном направлении, изменения растительного покрова, проявляющиеся в смене одних фитоценозов другими».
С. J. Burrows (1990) определяет растительную сукцессию как последовательные изменения в видовом составе на каком-либо участке. Г. Вальтер (1982) рассматривает сукцессию как последовательный ряд (во времени или пространстве) фитоценозов, а сами фитоценозы — как ее стадии.
Учение о сукцессиях особенно активно развивалось в Северной Америке. В его развитии большую роль сыграл Н. С. Cowles (1869-1939), который говорил о рядах развития сообществ — сукцессионных сериях, которые приводят к возникновению климатических формаций. Термин
396
Глава 9. Динамика фитоценозов
климакс первым ввел в употребление американский эколог Н. N. Whitford в 1901 г. Но, безусловно, наибольшую роль в развитии теории сукцессий и представлений о климаксе сыграл Ф. Клементс (1874-1945).
3.2 • Взгляды Ф. Клементса
Ф. Клементс (Clements, 1916,1928) различал первичные и вторичные сукцессии. Первичные протекают на новых субстратах (вулканические отложения, морены, новые дюны и т. п.), вторичные — там, где растительный покров ранее существовал, но был нарушен или уничтожен, однако сохранилась почва. Это подразделение несколько условно, и R. Peet (1992) предложил свои определения: первичные сукцессии — это те, где главный механизм включает долговременные изменения среды, а во вторичных главные механизмы — популяционные процессы.
По субстратам, на которых начинается первичная сукцессия, Ф. Клементс различал гидросерии (зарастание водоемов, включая галосерии — соленых водоемов и оксисерии — пресных водоемов) и ксеросерии (зарастание твердых минеральных субстратов, включая литосерии — прочных каменистых субстратов (скал и крупнокаменных осыпей) и псам-мосерии — рыхлых субстратов (песка, глинистых отложений и т. п.)). По мнению Ф. Клементса, сукцессии растительного покрова проходят через дискретные и детерминированные (строго определенные) фазы и завершаются климаксом, одним для различных субстратов в пределах обширных географических регионов (моноклимакс). Реальная пестрая мозаика растительного покрова объяснялась Ф. Клементсом несколькими причинами: незавершенностью сукцессий из-за специфических эда-
Рис. 9.18 • Сообщества разного возраста, развивающиеся на сходных субстратах (снижение биомассы и продукции при прогрессирующем обеднении почвы): дюнные ландшафты Cooloola в Квинсленде (Австралия). Рост девушки (на фото а и б) для масштаба — 1,65 м. Сравнение высокоствольных и низкоствольных лесов, развитых на дюнах разного возраста: а, в — высокоствольный лес на относительно молодых дюнах (возраст около 10 000-20 000 лет); б, г — низкоствольный лес на древних дюнах (возраст около 200000-400000 лет); д, е — лесные и кустарниковые сообщества на террасах в Новой Зеландии (Waitutu): д — высокоствольный лес (Weinmannia racemosa, Nothofagus solandri, Metrosideros umbellatum и Dacrydium cupressinum; возраст 80000-100 000 лет), e — разреженный лес с зарослями кустарников (Halocarpus biformis, Halocarpus bidwillii, Nothofagus solandri Leptospermum scoparium; возраст около 300 000 лет) (no: Walker et al., 1981; Wardle et al., 2004)
3. Сукцессии
397
фических условий (субклимакс), «нарушениями» климаксовых сообществ за счет постоянного воздействия внешних нарушающих факторов, в том числе антропогенных (дисклимакс). Кроме того, рассматривался «панклимакс» — объединение физиономически сходных климаксов — два или более родственных климаксов или формаций с одинаковыми общими климатическими условиями, одинаковой жизненной формой и общими
398
Глава 9. Динамика фитоценозов
родами доминантов, что приблизительно соответствует зональной растительности.
При рассмотрении климаксовых сообществ необходимо различать два подхода к их устойчивости. Фитоценотический подход рассматривает способность всех основных компонентов фитоценоза к самовозобновлению в климаксовом сообществе, что обеспечивает его долговременную устойчивость. С этой точки зрения многие естественные сообщества могут рассматриваться как климаксовые. Геохимический подход постулирует стабильность условий среды, базирующуюся на долговременной сбалансированности геохимических потоков. Даже при небольшой несбалансированности происходит кумулятивное накопление и/или потери тех или иных биогенов, ведущие к смене экологических условий и дальнейшей смене сообществ. Однако такая сбалансированность точно никем не показана. Это связано с небольшим (тысячелетия) временем существования многих сообществ умеренной зоны. Интересные наблюдения проведены в Австралии, где на сходных (климатически и геологически) участках дюн в зависимости от их возраста формируются различные сообщества. Долговременное существование лесов вело к постепенному обеднению почвы ЭМП, прежде всего невозобновимым фосфором (так называемая ретрогрессивная динамика). Леса, развитые на дюнах возрастом 10000-20000 лет, имели значительно большую высоту и биомассу, чем на дюнах возрастом 200000-400000 лет (Walker et al., 1981; Wardle et al., 2004) (рис. 9.18). Высокая продукция амазонских лесов поддерживается значительным поступлением фосфора с пылью, приносимой из Африки (Swap et al., 1992; Polis, 1999).
Сравнение «ретрогрессивной» динамики длительно не нарушаемых лесных сообществ в Австралии, Новой Зеландии, на Аляске, Гавайских островах и в Швеции выявило, что видовое разнообразие деревьев достигало максимума вместе с биомассой леса и в дальнейшем снижалось по мере снижения содержания фосфора в почве. В то же время общее видовое разнообразие сосудистых растений в ходе ретрогрессии продолжало увеличиваться (Wardle et al., 2004, 2008).
Ф. Клементс (Clements, 1916, 1928) выделял следующие этапы первичных сукцессий:
1) процесс, инициализирующий сукцессию (образование незаселенных растениями участков);
2) миграция растений на эти участки за счет поступления диаспор. С увеличением возраста доступного субстрата увеличивается число видов, его заселяющих (см. рис. 9.19);
3) приживание и размножение растений;
3. Сукцессии
399
Рис. 9.19 • Связь между временем и числом видов, присутствующих в ходе первичных сукцессий: а — рукотворные пруды в Англии (по: Godwin, 1923); б — песчаные дюны в Онтарио (по: Morrison, Yarranton, 1973); в — вулканический остров Кракатау — (по: Docters van Leeuwen, 1936; см. Miles, 1979)
4) конкуренция;
5) изменение условий среды;
6) стабилизация.
3.3 • Классические примеры первичных сукцессий и современные взгляды на них
Сукцессии на моренах ледника на Аляске, по данным Crocker, Major (1955), включают следующие стадии (рис. 9.20):
1) пионерная — образована мхами (Rhacomitrium canescens, R. lanugi-nosum) с разреженным покровом сосудистых растений разных жизненных форм (Chamaenerion latifolium, Equisetum variegatum, Dryas drummordi, Salix arctica);
2) заросли арктических ив (Salix barclayi\ S. sitchensis, S. alexensis);
3) заросли кустарниковой ольхи (Alnus viridis);
4) лес из ситхинской ели (Picea sitchensis) (примерно через 80 лет после отступления ледника);
5) климаксовый лес (Picea sitchensis, Tsuga heterophylla, T. mertensiana).
Длительность сукцессии — около 200 лет. В ходе сукцессии происходит подкисление почвы (исходный материал — карбонатные породы), pH снижается с 8 до 5, происходит накопление органики и общего азота
400
Глава 9. Динамика фитоценозов
Рис. 9.20 • Первичные сукцессии после отступления ледника на Аляске: а — ледник Mendenhall Glacier (фото Greg Wiles, с любезного разрешения автора);
б — ледник Гулькана (Gulkana Glacier) (фото М.С. Блинникова, с любезного разрешения автора);
3. Сукцессии
401
в — Chamaenerion latifolium (фото М.С. Блинникова, с любезного разрешения автора); г, д — Alnus viridis; е — лес из Picea sitchensis (фото Bureau of Lard Management, USA, Wikimedia); ж — побеги Picea sitchensis (фото Dr. Phil Gates, с любезного разрешения авторе)
402
Глава 9. Динамика фитоценозов
в почве. Наиболее богатые почвы по основным ЭМП — на стадии ольховых зарослей, в дальнейшем под елью идет обеднение почвы за счет выноса ЭМП из почвенного профиля и образование подзолистого горизонта. Однако в более поздней работе С. L. Fastie (1995) содержится пересмотр этой классической смены. Автор показывает, что в разные периоды времени пионерные стадии сукцессий после отступления ледников на Аляске могли значительно различаться, в том числе и по доминирующим видам; это зависело от текущей климатической обстановки и наличия диаспор растений. По его детальным наблюдениям, уже пионерные сообщества в местах отступления ледника до 1840 г. были образованы главным образом Picea sitchensis с примесью Tsuga heterophylla, в период 1840-1900 гг. — Alnus viridis, а после 1900 г. — Populus trichocarpa. Автор предостерегает от толкования пространственных рядов как временных даже в случае отступления ледников.
Зарастание дюн на озере Мичиган описано по Olson (1958). Побережье и прилегающие территории озера Мичиган представляют собой систему разновозрастных дюн в связи с понижением уровня озера. Возраст старых дюн сопоставим с голоценом — 10000-12000 лет. По пространственному градиенту от побережья озера внутрь дюнной системы (увеличению возраста дюн) наблюдаются следующие стадии:
1) пионерная — образована злаком Ammophila breviligulata (% 100 лет);
2) вечнозеленые кустарники и дерновинные злаки (~ 300 лет);
3) сосновый лес из Pinus banksiana и P.strobus (через ~350 лет);
4) климаксовый дубовый лес из Quercus velutina.
Максимум видового разнообразия наблюдался при возрасте дюн около 285 лет, а далее число видов убывало в связи с выпадением пионерных видов (Lichter, 1998).
Более поздние исследования не подтвердили сукцессионный характер этих смен, связанный с прогрессирующими изменениями среды самими растениями (Johnson, Miyanishi, 2008). Olson (1958) детально изучил возраст разных дюн и показал, что дюны одного возраста имеют очень разный растительный покров, зависящий от мезорельефа и истории нарушений. Представления о благоприятном изменении свойств почв видами ранних стадий сукцессий для видов более поздних стадий не подтвердились экспериментально. Lichter (2000) провел пересадки и показал, что такие «позднесукцессионные» виды, как Juniperus communis, Pinus strobus и Quercus rubra, могут хорошо расти на молодых дюнах со слабо развитыми почвами. Физическая стабилизация дюн имеет большое
3. Сукцессии
403
Рис. 9.21 • Поднимающиеся из воды острова в южной Швеции
значение для приживания растений, а затем различные другие факторы нарушений определяют структуру растительного покрова (Johnson, Miyanishi, 2008).
Освобождающиеся из озер острова в Швеции (описаны по Rydin, Borgegard, 1991). В связи с тектоническим поднятием Скандинавского полуострова в течение относительно небольшого исторического периода в озерах, соединенных с морем, образуются острова, которые постепенно заселяются растениями (рис. 9.21). Время их подъема можно определить по старым картам. На изученных островах за 100 лет (1886-1985) выяв-
404
Глава 9. Динамика фитоценозов
Рис. 9.22 • Вулканический остров Суртсей (Surtsey) близ Исландии: а — вид острова с пляжа, на переднем плане — Нопскепуа peploides, фото Erling Olafsson, с любезного разрешения автора; б, в — первое семенное растение, найденное через 2 года после извержения, — Cakile edentula; б — фото Paul Slichter, в — фото Dave Ingram, http://lslandNature.ca, с любезного разрешения авторов
лено 112 видов. Среди пионерных видов преобладали автогамные однолетники. Семена заносятся на острова по воде, воздуху, а также птицами.
Вулканический остров Суртсей (Surtsey) расположен близ Исландии и имеет диаметр около 2 км (рис. 9.22). Образование острова связано с мощным извержением, произошедшим в 1963 г. Первое семенное растение отмечено на острове в 1965 г. (Cakile edentula). Через 23 го-
3. Сукцессии
405
Рис. 9.23 • Схема архипелага Кракатау до и после извержения (а) (с любезного разрешения Vic Camp, from the How Volcanoes Work website http://www.geology.sdsu.edu/how_volcanoes_work/index.html); Casuarina equisetifolia (б) и пионерные виды деревьев Pongamia pinnata (в) (фото J. М. Garg, Wikimedia Commons, under GNU Free Documentation License)
да (1986 г.) всего было выявлено 23 вида сосудистых растений (Burrows, 1990). Значительная суровость климата и удаленность источников диаспор объясняют медленные процессы заселения острова. В целом первичные сукцессии на лавовых полях в Исландии включают стадии: 1) Racomitrium lanuginosum (начинается с отдельных пятен на лавах возрастом менее 20 лет), сплошной моховой покров образуется на участках возрастом 20-100 лет; 2) колонизация сосудистыми растениями (возраст 100-600 лет), на смену которой приходит стадия 3) дифференциации растительного покрова согласно вариациям условий среды по склонам разной экспозиции (возраст — более 600 лет) (Cutler et al., 2008).
406
Глава 9. Динамика фитоценозов
В тропиках заселение идет значительно быстрее. Классическим примером может быть вулканический архипелаг Кракатау, расположенный между Явой и Суматрой (Индонезия). Сильное извержение 1883 г. на месте более крупного острова образовало небольшой остров Rakata, где после извержения высшие растения отсутствовали (рис. 9.23 а). Всего за 100 лет после извержения на этом острове отмечено около 400 видов, при этом уже через 15 лет отмечена богатая флора на побережье. Особенно хорошо приживались бобовые родов Canavalia и Pongamia (рис. 9.23 б), а также Casuarina equisetifolia (рис. 9.23 в), имеющие симбиоз с азотфиксаторами. Эти деревья быстро сформировали сомкнутый лес. Текущее число видов (около 200), отмеченное на острове, стабилизировалось (Burrows, 1990).
Извержение вулкана Святой Елены в США произошло в 1980 г. и оставило огромные лавовые и пепловые поля, где существовавшая ранее растительность (высокогорные сообщества и хвойный лес) была полностью уничтожена. После извержения на этих территориях были заложены постоянные площадки для детального изучения первичных сукцессий. В связи с неблагоприятными условиями вулканического субстрата (сухость, подвижность, значительные перепады температур, бедность азотом) зарастание происходит весьма медленно. Наиболее обильным пионером зарастания таких субстратов выступает азотфиксирующий Lupinus lepidus — небольшой зимнезеленый малолетник. Показано, что видовое разнообразие увеличивалось с возрастом субстрата и колоний Lupinus lepidus, при этом особенно положительно на присутствие люпина реагировали мхи (del Moral, Rozzell, 2005). Фитофаги, активно потребляющие семена люпина, снижали скорость первичной сукцессии (Fagan, Bishop, 2000).
За исключением последних примеров, связанных с регулярными наблюдениями за ходом сукцессий после извержений, большинство классических работ, посвященных первичным сукцессиям, базируется на анализе пространственных рядов (расстояния от края ледника, берега водоема и т. п.), рассматриваемых как временные, что может приводить к ошибочным выводам. Это связано с возможной неоднородностью субстратов, разными экологическими условиями в разное время и неравномерностью поступления диаспор во времени. Детальные палеоботанические исследования позволяют уточнить реальный ход сукцессионных смен.
Последовательным сторонником теории Ф. Клементса был С. М. Разумовский (1981), который считал гумусонакопление главным фактором сукцессионной динамики, ведущей к мезофитизации и мезотрофизации растительных сообществ. По его мнению, и сухие пески, и мелкие озера
3. Сукцессии
407
в процессе сукцессий могут преобразовываться в мезотрофные климак-совые леса. Однако эти представления никак не подтверждаются прямыми наблюдениями и результатами палеоэкологических исследований. Даже первичные сукцессии не всегда ведут к увеличению плодородия почв. Например, в Якутии, лёссовидные почвообразующие породы более плодородны, чем зональные торфянистые тундровые почвы. Эти породы оказываются лучшим субстратом для развития продуктивных растений. Снятие верхнего горизонта почвы (распространенный агротехнический прием) способствует образованию луговых сообществ, которые затем (при накоплении органики в почве) переходят в тундровые. Естественная динамика болотных массивов, связанная с накоплением торфа, ведет к уменьшению их продуктивности и переходу из евтрофной в мезотроф-ную и олиготрофную стадии. Неизвестны случаи произрастания продуктивных лесных массивов на более или менее мощных торфяных залежах. Да и классические гидросерии на небольших лесных водоемах значительно пересмотрены при детальных палеоботанических исследованиях. Например, Klinger (1996) показал, что, хотя зарастание водоемов в таежной зоне Канады начинается с их берегов, дальнейшее наступление болотной растительности идет в двух направлениях — зарастание водоема и заболачивание леса (рис. 9.24). При этом жизненность деревьев резко снижается, а развитие подроста подавляется на заболоченной территории, что никак не согласуется с представлениями о «наступлении леса на болото».
Длительность первичных сукцессий оценивается столетиями и первыми тысячелетиями. Так, хвойный лес после отступания ледника в Альпах может появиться уже через 100 лет, на Аляске — через 200 лет, а для развития леса из Machilus thunbergi на лавовых отложениях в Японии требуется около 700 лет; дубовый лес на дюнах Мичигана появляется примерно через 1000 лет после образования дюн.
Возможны различные варианты изменения биомассы сообществ в ходе первичных сукцессий. Peet (1992) рассматривает три основных варианта:
1) асимптотический рост (в случае лимитирования продукции ЭМП);
2) достижение пика, а затем небольшое снижение (в случае лимитирования продукции светом);
3) достижение пика, а затем резкое снижение, происходящее вследствие изменения структуры популяций или обеднения почвы.
Разнообразие растений в ходе сукцессий обычно возрастает, достигает максимума на средних стадиях и снижается при конкурентном воздействии позднесукцессионных видов. Guo (2003) исследовал, насколь-
408
Глава 9. Динамика фитоценозов
Рис. 9.24 • Схема развития болотного массива через зарастание озера и заболачивание окружающего леса. Основные стадии включают: а) поселение торфообразующих мхов (главным образом Sphagnum) по берегам водоема; б) аккумуляцию торфа как в процессе зарастания озера, так и в ходе заболачивания леса, приводящую к подъему уровня грунтовых вод; в) продолжающееся заболачивание в двух направлениях, ведущее к формированию олиготрофного болотного массива (по: Klinger, 1996)
ко разнообразие и биомасса изменяются синхронно в ходе сукцессии на одном месте на примере послепожарной сукцессии в горах Южной Калифорнии. На ранних стадиях сукцессии и разнообразие, и биомасса возрастали и были положительно связаны. На поздних стадиях сукцессии биомасса продолжала возрастать, а разнообразие снижалось, поэтому наблюдалась отрицательная связь между разнообразием и биомассой. Автор делает вывод, что зависимость между биомассой и разнообразием связана с сукцессионной стадией сообществ.
3.4 • Взгляды В. Н. Сукачева
на классификацию сукцессий
В. Н. Сукачев (1934, 1938) выделял три основных типа сукцессионных процессов.
Сингенез — «процесс заселения территории растениями, процесс борьбы между ними за территорию и средства жизни и процесс ежи-
3. Сукцессии
409
вания растений и формирования взаимоотношений между ними» (Сукачев, 1948: 105). Взгляды на сингенез несколько различаются у разных авторов, использующих это понятие. По Б. М. Миркину (1984), сингенез — это сукцессии, протекающие вследствие размножения растений без существенного изменения ими условий местообитаний, т. е. 2-4-й этапы первичных сукцессий по Клементсу (см. п. 3.2 гл. 9). Т. А. Работнов (1992) считает целесообразным включать в это понятие процессы, происходящие при внедрении в сложившиеся фитоценозы новых для них видов растений. Но поскольку при этом формируются новые сообщества (новые комбинации видов), в природе до того не существовавшие, эти процессы лучше рассматривать как эволюцию растительных сообществ (см. п. 4 гл. 9).
Эндоэкогенез (эндоэкогенетические сукцессии) — это сукцессии, идущие под влиянием внутренних механизмов с кумулятивным изменением среды. Эти сукцессии соответствуют Клементсовскому механизму смен — изменение среды каждой предшествующей стадией сукцессии в неблагоприятном для себя направлении, но благоприятном для видов последующей стадии.
Экзогенез — сукцессии, идущие под влиянием изменения (долговременного) внешних условий, например подтопления, засоления и т. п.
3.5 • Дальнейшее развитие представлений о климаксе в США
Учение Ф. Клементса стало господствующим на американском континенте в середине XX в. Однако постепенно представления о климаксе претерпели существенные изменения с позиций признания континуальной природы растительного покрова. Вначале было предложено различать более дробные климаксовые экосистемы (концепция поликлимакса) и разделять климаксы по факторам их поддержания, включая такие внешние факторы, как огонь (пирогенный климакс), выпас и др. Соединил представления о непрерывности растительного покрова с теорией климакса R. Whittaker (см. Уиттекер, 1980), предложивший концепцию климакс-мозаики (climax pattern). Согласно R. Whittaker, непрерывным (континуальным) является как ход сукцессий, так и пространственная структура растительного покрова, сложенного климаксовыми сообществами. В этом случае в каждой точке земной поверхности может сформироваться свое наиболее устойчивое и самоподдерживающееся сочетание видов растений (климакс), которые наиболее оптимально используют условия среды, включая имеющиеся ресурсы, на рассматриваемом локальном
410
Глава 9. Динамика фитоценозов
участке. Поэтому климаксовые сообщества являются локальными и уникальными.
Дальнейшее развитие представлений о сукцессиях было связано с выявлением механизмов сукцессионных смен.
3.6 • Взгляды Коннела и Слатиера.
Механизмы сукцессий
Коннел и Слатиер (Connell, Slatyer, 1977) предложили 3 модели сукцессионных смен, отличающиеся по взаимодействиям видов предшествующей и последующей стадий сукцессий (модели стимуляции, толерантности и ингибирования).
Модель стимуляции. Каждая предшествующая стадия сукцессии улучшает условия среды для наступления следующей, что соответствует классическим представлениям Ф. Клементса. В этом случае удаление видов на какой-либо стадии сукцессии должно замедлить наступление следующей стадии.
Модель толерантности предполагает, что виды, первыми заселившие пространство, существенно не улучшают и не ухудшают среду, но в условиях сомкнутых сообществ более теневыносливые и конкурентоспособные виды следующих стадий вытесняют первых поселенцев. Удаление видов какой-либо стадии не влияет на скорость развития видов следующих стадий.
Модель ингибирования. Пионерные виды конкурентно сдерживают наступление видов более поздних стадий, но они более подвержены влиянию вредителей и болезней, поэтому рано или поздно сменяются видами более поздних стадий. Доказательства этой модели содержатся в ряде работ. Так, например, Niering, Egler (1955), Niering, Goodwin (1974) показали, что покров кустарников задерживает инвазию деревьев до 45 лет в ходе вторичных сукцессий. В полевых опытах McCormik (1968) было показано, что удаление пионерных однолетников приводит к более быстрому росту и раннему цветению многолетников следующих стадий сукцессий при восстановлении прерий. Эта теоретическая модель сукцессии имеет и практическое применение. Например, в США широко используются некоторые быстрорастущие кустарники и травы средних стадий сукцессий в лесах для подавления развития деревьев на полосах отчуждения ЛЭП. Для стимуляции развития этих кустарников используют селективные гербициды (Luken, 1990).
Одним из важнейших факторов, определяющих ход и скорость сукцессий, является поступление диаспор, особенно для позднесукцессионных видов. Естественные сукцессии могут быть многократно ускоре-
3. Сукцессии
411
Рис. 9.25 • Изменение
синтаксономического
спектра «агростепи» в Башкирии в ходе 5-летней сукцессии после посева. Синтаксоны (классы и порядки): Sec-Ch — Secalietea+Chenopodietea (виды сегетальных и рудеральных сообществ), Ar — ArtemisietaНа vulgaris, On — Onopordetalia acanthi, MA — Molinio-Arrhenatheretea (луговые), FB — Festuco-Brometea (степные) (по: Абдуллин, Миркин, 1995)
ны при искусственном посеве семян растений климаксовых сообществ на участки, где предварительно были уничтожены раннесукцессионные виды-рудералы. Этот подход широко используется при восстановлении степной растительности на залежах и эродированных участках по методике, детально разработанной Д. С. Дзыбовым в 70-х гг. XX в. [Вставка XIII] (обзор см.: Дзыбов, 2010). Сложную смесь семян степных растений заготавливают (убирают) комбайнами в несколько сроков на существующих степных участках и высевают с использованием сельскохозяйственной техники. Уже через 2-3 года формируется степное сообщество (точнее, «агростепь»), видовой состав, структура и флористическая насыщенность которого близки к естественным степям региона. Доля видов классов сорно-рудеральной растительности уменьшается, а класса Festuco-Brometea — увеличивается, как это было показано в аналогичных экспериментах в Башкирии (рис. 9.25; Абдуллин, Миркин, 1995).
Вставка XIII. Разработка теории и технологии восстановительной сукцессии в России (метод агростепей Д. С. Дзыбова, текст автора)
Экспериментальное изучение теории восстановительной сукцессии (демутации) травяных экосистем проводится в России с 1974 г. (обзор публикаций см.: Дзыбов, 2010). Одновременно разраба
тывалась и технология верификации (воплощения) теоретических положений в практику. Такой подход способствовал как объективизации новых научных положений о сукцессии, так и решению актуальных задач ускоренного восстановления биоразнообразия и борьбы с опустыниванием на всех континентах Земли ускоренными темпами. В результате долголетних исследований сформулированы следующие положения и принципы вторичной восстановительной сукцессии и ее ускорения в десятки раз по сравнению с самовосстановлением:
412
Глава 9. Динамика фитоценозов
____ ___________________________________________1) главным тормозящим факто-Продолжение Вставки XIII____________Р°м вторичной сукцессии явля-
ется конкуренция со стороны быст-ро развивающихся рудеральных и сегетальных видов, обусловленная продвинутостью в росте и развитии особей этих видов — первопоселенцев на месте уничтожения климаксовой (зональной, поясной) растительности;
Рис- XIII.1 • Ускоренное восстановление степной растительности методом агростепи Д. С. Дзыбова (2010): а — уборка посевной смеси комбайном в зоне сухой степи; б — посев смеси; в — массовое цветение растений агростепи (Stipa pennata, S. pulcherrima, Echium russicum, Dracocephalum austriacum и др.); г — геоботаническое описание агростепи (аспектирует Dracocephalum austriacum, Filipendula vulgaris, Jurinea arachnoidea, Pedicularis sibthorpii) (фото из Дзыбов, 2010)
2) многочисленные промежуточные серийные стадии сукцессии, обусловленные межвозрастной борьбой особей предыдущих стадий, находящихся в возрастном диапазоне «имматурные — виргинильные — генеративные”, существенно снижают шансы внедрения особей последующих мигрантов в возрастном состоянии «всходы — ювенильные», отличающихся низкой конкурентоспособностью в плотных ценозах;
3. Сукцессии
413
3) в ходе сукцессий конкурентное преимущество всегда оказывается на стороне особей более взрослого возрастного состояния, независимо от стратегии вида;
4) искусственное ускорение вторичной сукцессии, в основе которой лежит ослабление межвозрастной конкурентной борьбы, приводит к блокированию (исключению) промежуточных «бурьянистых» или субклимаксовых стадий с формированием фитоценозов, флористически и ценотически идентичных зональным;
5) новые теоретические положения об особенностях сукцессионного процесса, проверенные экспериментально и в производственных условиях в методе агростепей, всегда дают положительные результаты, независимо от почвенно-растительной зоны. При этом необходимы два вполне доступных условия: а) экотоп (восстанавливаемая территория) должен быть свободен от диаспор видов сорно-полевой флоры (достигается обычной вспашкой и культивацией); б) сложный банк семян в естественной смеси внесен одноразово на специально подготовленную территорию, что обеспечивает равные возможности роста и развития растениям эталонной степи — источника диаспор (рис. XIII.1).
Изложенная теория и технология метода агростепей обеспечивает получение в течении двух-трех лет поликомпонентного фитоценоза, близкого по составу и структуре к исходному сообществу — донору диаспор, быстрое восстановление биоразнообразия и борьбу с опустыниванием земель в геометрической прогрессии.
Поступление диаспор растений, в свою очередь, зависит от близости участков позднесукцессионных или климаксовых сообществ к сериальному. Glenn-Lewin (1980) изучал скорость восстановления участка прерии, нарушенного перевыпасом, который непосредственно примыкал к ненарушенному участку. За 30 лет сукцессии виды прерии проникли полосой 10-20 м на бывшее пастбище за счет вегетативного размножения, что соответствует скорости распространения 30-60 см/год (van der Valk, 1992). Таким образом, скорость сукцессии зависит от размеров участков сообществ, на которых она происходит, и от их окружения.
В экспериментальных луговых микрокосмах исследовали влияние богатства почвы, нарушений при дефолиации, микроклимата и порядка заселения видами на итоговую структуру сообществ. Показано сильное влияние условий среды на видовое разнообразие и подверженность внедрению новых видов. Видовой состав, в свою очередь, на 96 % определялся порядком заселения растений, что подчеркивает важную роль исторической составляющей в формировании структуры сообществ (Ejr-naes et al., 2006).
Характер взаимоотношений между видами претерпевает значительные изменения в ходе сукцессии. Так, по наблюдениям на вырубках
414
Глава 9. Динамика фитоценозов
Рис. 9.26 • Ammophila arenaria на песчаных прибрежных дюнах Голландии
в Сьерра-Неваде (Калифорния), ассоциированность между двумя обычными видами (Arctostaphylos patula и Cirsium vulgare) менялась от нейтральной (2-й год вырубки) через отрицательную (3-й год) к нейтральной (4-й год) и снова отрицательной на 5-6-й год (Rejmanek, LepS, 1996). Авторы объясняют это сменой характера отношений между видами: на 3-й год всходы Arctostaphylos погибают близ взрослых особей Cirsium, а на 5-6-й год уже Cirsium погибает близ развившихся кустов Arctostaphylos.
Степень расхождения экологических ниш по потребляемым ресурсам увеличивается в ходе сукцессии. Parrish и Bazzaz (1982) провели эксперименты с совместным выращиванием пар видов, характерных для различных стадий сукцессий по схеме de Wit (см. п. 3.4 гл. 5). При выращивании совместно видов начальных стадий сукцессий их суммарный урожай равнялся среднему урожаю в монокультуре, в случае совместного выращивания видов средних стадий сукцессий суммарный урожай был на 20 % выше, чем в монокультурах, а видов поздних стадий прерии — на 70% выше.
3. Сукцессии
415
Большую роль в сукцессионных сменах могут играть гетеротрофные организмы — фитопатогенные грибы, наземные и почвенные фитофаги и др. В ходе сукцессий на дюнах в почве накапливаются патогенные микроорганизмы, сильно влияющие на рост (подавляют его) растений более ранних (но не поздних!) стадий сукцессий (Dobson, Crawley, 1994). Например, для пионера зарастания песчаных дюн — Ammophila arenaria — в Нидерландах показана самая высокая жизненность на свежих песчаных наносах чистого песка (рис. 9.26). При стабилизации дюн в почве увеличивается численность патогенных грибов и нематод. Эксперименты по выращиванию этого вида на субстратах разного сукцессионного возраста показали, что его жизненность снижалась при увеличении сукцессионного возраста субстрата (Van der Goes, van der Putten, 1992). В то же время эти патогенные организмы не угнетали рост Festuca rubra — вида, более типичного для следующих стадий сукцессии (Van der Putten, Peters, 1997). В экспериментах (De Deyn et al., 2003) было показано, что почвенная мезофауна (главным образом насекомые-корнееды) избирательно подавляет развитие раннесукцессионных доминантов, чем способствует ускорению сукцессии и повышению видового разнообразия на площадках. При внесении почвенных инсектицидов раннесукцессионные виды разнотравья резко усиливали свои позиции. Наиболее сильное воздействие на ход сукцессий оказывают почвенные беспозвоночные средних стадий сукцессий.
Микоризные грибы могут ускорять ход сукцессий, не только улучшая рост растений поздних стадий, но и подавляя развитие раннесукцессионных безмикоризных растений специфическими метаболитами, выделяемыми в почву (см. п. 1.5 гл. 3).
3.7 • Сукцессионная теория Тильмана
D. Tilman (1988) в своей книге «Plant strategies and the dynamics and structure of plant communities» разработал теоретическую модель сукцессий с учетом смен видов с различной аллокацией (распределением биомассы между основными органами растений; см. п. 1 гл. 8). Основные положения этой теории представлены в настоящем разделе.
Пороговая концентрация ресурса (7?*, см. п. 3.1 гл. 5) возрастает с увеличением уровня потерь биомассы (при отчуждении внешними агентами или нарушениях), поэтому в местах с высоким уровнем нарушений выживают лишь виды с высокой R*, т. е. требующие большего количества ресурсов для своего развития.
Относительная скорость роста (прирост массы на единицу имеющейся массы за единицу времени, RGR) значительно различается у видов
416
Глава 9. Динамика фитоценозов
Рис. 9.27 • а — ресурсные изоклины по свету и элементам минерального питания (условные единицы) для 6 модельных видов растений, точки показывают двувидовые равновесия, каждое из которых не было стабильным; б — долговременный исход конкуренции между этими видами на градиенте богатства почвы; в — изменение доступности света (на уровне почвы) и элементов минерального питания в конкурентной системе из 6 видов растений на градиенте увеличения поступления ЭМП (по: Tilman, 1988)
разных жизненных форм. Максимальная RGR среди исследованных видов (Grime, Hunt, 1975) отмечена для однолетника Роа annua — 0,38 день-1, наименьшая — для Picea sitchensis — 0,03 день-1. В целом всходы однолетних растений растут быстрее, чем многолетних, а травянистые растения — быстрее, чем деревья. Этот ряд: однолетники — многолетники — древесные растения есть ряд увеличения вклада (аллокации) в нефотосинтезирующие структуры. Аллокация ассимилятов в нефотосинтезирующие органы уменьшает максимальную скорость роста растений, поскольку рост связан с интенсивностью фотосинтеза листьев. Аллока-
3. Сукцессии
417
Рис. 9.28 • Все возможные варианты аллокации биомассы в листья (AL), стебли (AS) и корни (AR) изображены в виде схематического прямоугольного треугольника. В любой точке внутри этого треугольника сумма аллокаций AL + AS + AR = 1. Каждая диагональная линия — это линия постоянной аллокации в корни. Точки около прямого угла соответствуют растениям с высокой аллокацией в корни, у верхнего угла — в стебли, у правого угла — в листья (по: Tilman, 1988)
ция в листья различается у разных групп растений в десятки раз. Так, у деревьев она может составлять 2-3 %, а у водных плавающих трав — до 80-98 % (Korner, 1993). В то же время эти величины, как и отношение биомассы корней к биомассе побегов, меняется в различных экологических условиях и в пределах одного вида. По наблюдениям за 8 видами растений на залежах в Миннесоте при 15-кратном увеличении доступности азота соотношение корни побеги увеличивалось как максимум в 2,5 раза. Chapin (1980) приводит данные, что при 100-кратном увеличении доступности азота это соотношение увеличивается для разных видов в 1,5-12 раз.
Модель ALLOCATE создана Д. Тильманом для изучения межвидовой конкуренции в случае, когда виды растений различаются между собой только распределением ресурсов (аллокацией) между корнями, стеблями и листьями, но идентичны по остальным параметрам (например, интенсивности фотосинтеза на единицу листовой биомассы и т.п.).
Вначале рассмотрим картину с конкуренцией между 6 видами, различающимися по ресурсным изоклинам (см. п. 3.1 гл. 5), при разном уровне богатства почвы (рис. 9.27). Вид А (наибольшая аллокация в корни) побеждает в конкуренции на самых бедных почвах, а вид D (большая доля стеблей и листьев) — на самых богатых. Виды F и Е вытесняются из любых местообитаний. Вид с аллокацией, адаптивной в одной части градиента почвенных ресурсов, становится неконкурентоспособным в другой.
Все возможные картины аллокации представлены в виде треугольной схемы на рис. 9.28. Для изучения потенциальной возможности существования и роста видов в монокультуре просчитаны соответствующие
418
Глава 9. Динамика фитоценозов
О .1 .2 .3 .4 .5 .6 .7 .8 .9 1.0
0 .1 .2 .3 .4 .5 .6 .7 .8 .9 1.0
аллокация в листья
аллокация в листья
Рис. 9.29. Возможные жизнеспособные аллокации при одновидовом произрастании растений (без межвидовой конкуренции) при разном уровне поступления элементов минерального питания (ЭМП, условные единицы). Область жизнеспособных аллокаций не заштрихована, а нежизнеспособных — заштрихована. При более высоком уровне богатства почвы растения с более разнообразными аллокациями могут существовать (незаштрихованная область больше). Все расчеты сделаны при уровне смертности 0,2 за неделю (по: Tilman, 1988)
модели при разном уровне поступления почвенных ресурсов (рис. 9.29). Этот анализ показал, что не все теоретически возможные варианты аллокации жизнеспособны. При этом на богатых почвах разнообразие вариантов аллокаций выше, чем на бедных.
3. Сукцессии
419
Рис. 9.30 • Изоклина (черная кривая) лучших конкурентных аллокаций при разном уровне поступления почвенных ресурсов (показаны цифрами, условные единицы от 25 — очень бедные почвы до 1000 — очень богатые почвы) и среднем уровне потерь (смертность 0,2 за неделю) — результат вычислительного эксперимента с 200 «видами» растений, различающимися только по аллокации биомассы в стебли, листья и корни (по: Tilman, 1988)
Рис. 9.31 • Максимальная относительная скорость роста растений (RGR, неделя-1) с различной аллокацией (результаты расчетов в модели ALLOCATE) (по: Tilman, 1988)
аллокация в листья
аллокация в листья (AL)
Далее для 200 вариантов аллокаций («видов») изучены последствия конкуренции при разном почвенном плодородии. Показано (рис. 9.30), что при данном уровне поступления ЭМП конкурентное преимущество может иметь вид со строго определенной «морфологией» (аллокацией). С ростом доступности почвенных ресурсов лучшим конкурентом будет вид с более высокой аллокацией в стебли и меньшей — в корни. Аллокация в листья имеет максимум на среднем уровне богатства почвы ЭМП.
Д. Тильман считает, что относительная скорость роста (RGR) прямо пропорциональна аллокации в листья (рис. 9.31), поэтому максимальный прирост наблюдается при среднем уровне плодородия и низком уровне нарушений.
420
Глава 9. Динамика фитоценозов
О .1 Я Л .4 .5 .6 .7 Л .9 1.0 аллокация в листья
О .1 .2 .3 .4 .5 .6 .7 .8 .9 1.0 аллокация в листья
Рис. 9.32 • Лучшие конкурентные аллокации растений в условиях разного богатства почвы (числа у изоклин) и разной интенсивности нарушений (фигуры а-г по интенсивности потерь). Высокая интенсивность нарушений ведет к доминированию растений с высокой аллокацией в листья, но низкой — в стебли и корни (по: Tilman, 1988)
Анализ влияния нарушений (усиление смертности) на адаптивную аллокацию проведен для разных уровней потерь (доля погибших особей за 1 неделю) (рис. 9.32). Он показал, что увеличение интенсивности нарушений, связанных с отчуждением биомассы (большей смертности особей), ведет к конкурентному успеху видов с большей аллокацией в листья, но с низкой — в стебли и корни. Нарушение ведет к одновременному воз-
3. Сукцессии
421
Изменения вдоль градиента свет : почвенные ресурсы
Низкое* Поступление ЭМП в почву «Высокое
Низкое* Содержание ЭМП в почве »Высокое
Низкая* Общая биомасса растений «Высокая
Высокое*Проникновение света к поверхности почвы-йНизкое Низкая * Высота взрослых растений «Большая
Низкая*-«Высокая*-Относительная скорость роста—►Низкая Мелкие « Размер семян * Крупные
Ранний< Возраст первой репродукции---►Поздний
Высокая-*----------Аллокация в корни * Низкая
Низкая * Аллокация в стебли —Высокая
Рис. 9.33 • Предсказанная моделью ALLOCATE зависимость морфологических свойств и параметров жизненного цикла растений от продуктивности сообщества (богатства почвы) при невысоком и постоянном уровне нарушений. Условия меняются от бедных местообитаний с высокой доступностью света к продуктивным сообществам на богатых почвах с низким проникновением света на уровень почвы (по: Tilman, 1988)
растанию поступления света и ЭМП, а в условиях высокой доступности всех ресурсов конкурентное преимущество имеют виды с наибольшей скоростью роста, а этот показатель определяется аллокацией в листья.
Анализ полученных результатов показывает, что каждая из жизнеспособных аллокаций является лучшей (в смысле конкурентного преимущества) в отдельном типе местообитаний, определяемом по интенсивности поступления ресурсов и интенсивности отчуждения биомассы (нарушений).
Модель ALLOCATE предсказывает следующие изменения в морфологических и популяционных признаках растений на градиентах увеличения богатства почвы и интенсивности нарушений (см. рис. 9.33,9.34). Высота растений увеличивается при увеличении богатства почв (доступности ЭМП). На богатых почвах более адаптивны более крупные семена, в то время как в бедных местообитаниях размер семян согласно модели не имел важного значения для конкурентоспособности. Вклад в многолетние органы у растений богатых почв должен возрастать. Большинство из этих закономерностей хорошо согласуется с данными из географии растений.
422
Глава 9. Динамика фитоценозов
Изменения вдоль градиента нарушений на богатых почвах
Низкий*—Уровень нарушений (потери биомассы)—^Высокий Низкое *------Содержание ЭМП в почве-------► Высокое
Высокая *-----Общая биомасса растений------► Низкая
Низкое «-Проникновение света к поверхности почвы-^Высокое Большая*------Высота взрослых растений-----► Низкая
Низкая < Относительная скорость роста »Высокая
Поздний------► Возраст первой репродукции----► Ранний
Высокая« Аллокация в стебли • Низкая
Низкая «—--------Аллокация в листья---------►Высокая
Рис. 9.34 • Предсказанная моделью ALLOCATE зависимость морфологических свойств и параметров жизненного цикла растений от уровня нарушений (интенсивности отчуждения биомассы) при постоянном уровне поступления ЭМП в почву
Рассмотренные выше закономерности касаются итога конкуренции между видами с различной аллокацией в условиях разного богатства почв и интенсивности нарушений. D. Tilman (1988) показывает также, что этот окончательный итог конкуренции устанавливается после довольно длительного процесса перехода доминирования от одного вида к другому. Этот процесс, происходящий без кумулятивных изменений в почвенном плодородии, автор называет переходной динамикой (transient dynamics). Автор делает очень важный вывод, что немедленный отклик системы на экспериментальное воздействие может существенно отличаться от долговременного эффекта, поэтому краткосрочные наблюдения могут привести к неверным выводам. Эксперимент, длящийся недостаточно долго, чтобы выявить переходную динамику, покажет преимущество не видов, наиболее конкурентоспособных в данной среде, а видов, обладающих здесь максимальной скоростью роста.
Вычислительный эксперимент (рис. 9.35, 9.36) показал смену видов с различными аллокациями для условий разного богатства почв. Общая тенденция переходной динамики: доминирование переходит от видов, типичных для мест с высоким уровнем нарушений, к видам с оптимальной для данных условий аллокацией. При этом флуктуационные колебания
3. Сукцессии
423
Рис. 9.35 • Популяционная динамика смешанного (100 видов с различными аллокациями) посева на бедных почвах при уровне нарушений 0,2 неделя-1 Показаны только 8 видов со значительным участием. Виды обозначены латинскими буквами и имеют следующую структуру аллокаций (доля листьев, стеблей и корней соответственно): А: 0,55, 0,05, 0,40; В: 0,50, 0,025, 0,475; С: 0,55, 0,025, 0,425; D: 0,60, 0,05, 0,35; Е: 0,60, 0,025, 0,375; F: 0,65, 0,025, 0,325; G: 0,70, 0,025, 0,275; Н: 0,75, 0,025, 0,225. Каждый из 8 видов доминировал в какой-либо период времени в течение моделируемых 40 лет (показаны только 10). К 40-му году вид В вытеснил все остальные виды (по: Tilman, 1988)
424
Глава 9. Динамика фитоценозов
Рис. 9.36 • Траектории смены доминантов с различными аллокациями в ходе конкурентного вытеснения на почвах разного богатства (а-г в порядке увеличения почвенного богатства ЭМП). Каждая точка показывает «вид», доминирующий на отдельном этапе смен, виды обозначены буквами. Стрелки показывают направления смен. В некоторых случаях вид может доминировать больше чем 1 раз. Во всех случаях исходная смесь состояла из одних и тех же 100 видов (результаты анализа модели ALLOCATE; по Tilman, 1988)
и общей биомассы, и доминирования отдельных видов значительно выше на богатых почвах.
Под сукцессией Tilman (1988) понимает динамику растительности, связанную с преобразованием среды (рост почвенного плодородия). По его представлениям, первичные сукцессии — это сукцессии в более или менее чистом виде, а для вторичных характерно наложение сук-
3. Сукцессии
425
Рис. 9.37 • Роль переходной динамики и долговременного накопления ЭМП в почве в сменах видов с различной аллокацией на почвах разного исходного богатства. Уровень ЭМП = 1000 в модели достигается через 100 лет. На бедных почвах (а) переходная динамика длится около 10 лет, сменяясь на 90 лет сукцессионной динамикой, связанной с накоплением ЭМП в почве. На умеренно богатых почвах (б) период переходной динамики — 7 лет. На богатых почвах (в) все смены видов представляют собой переходную динамику, смена видов заканчивается через 6 лет (расчеты на модели ALLOCATE; по Tilman, 1988)
сукцессия (изменение свойств почв)
цессионной и переходной динамики (рис. 9.37). Автор отмечает также, что на сукцессионную динамику накладывает значительный отпечаток различная скорость поступления диаспор отдельных видов (см. выше). В целом Tilman (1988) полагает, что не стоит искать универсальный ключ ко всем сукцессиям. Существует общая тенденция большей роли трансформации почв при первичных сукцессиях и большей роли переходной
426
Глава 9. Динамика фитоценозов
динамики при вторичных. Можно предсказать, что все сукцессии идут от низких растений к высоким, а вторичные сукцессии — от быстрорастущих видов к более медленнорастущим.
Проверка описанной модели на вторичных сукцессиях на залежах Миннесоты (США) показала большую роль переходной динамики и скорости колонизации для объяснения смен видов. Получено хорошее согласование смен доминирующих видов со структурой их биомассы: в ходе смен (при переходной динамике) доминирование переходит к видам с большей аллокацией в корни (отношение массы корней к массе побегов, главным образом листьев): Agrostis scabra 0,42 -> Agropyron repens 1,0 Poa pratensis 1,2 Schizachyrium scoparium 1,5 —> Andropogon gerardi 2,3. Сравнение аллокации и относительной скорости роста (RGR) 28 видов разных стадий сукцессий на залежах (Gleeson, Tilman, 1994) также показало, что на поздних стадиях сукцессий преобладают виды с высокой аллокацией в корни и низкой RGR. В то же время всходы могут иметь существенно отличающуюся картину аллокации от взрослых особей.
Даже при вторичных сукцессиях идет обогащение почвы гумусом и ЭМП. Распашка прерий в Миннесоте (США) и возделывание сельскохозяйственных культур привели к потерям 75% азота и 89% почвенного углерода по сравнению с естественными почвами. Сукцессии на заброшенных полях (залежах) приводят к медленному восстановлению почвенного запаса, но для 95 %-го восстановления запасов азота требуется 180 лет, а углерода (органического вещества) — 230 лет (Knops, Tilman, 2000).
3.8 • Сукцессии и стратегии растений Ф. Грайма
Ф. Грайм (Grime, 2001) рассматривает различные варианты сукцессий с позиции разработанной им теории стратегий растений (см. п. 3.7.2 гл. 5). Во многих случаях, по мнению автора, сукцессии идут со сменой видов рудеральной стратегии на конкурентов, а затем усиливается роль стресс-толерантов. Однако в суровых условиях (например, в тундре или высокогорьях) из состава сукцессионных сообществ могут быть полностью исключены рудералы и конкуренты, а сукцессионную динамику целиком определяют виды стресс-толерантной стратегии. Пример графического представления первичной сукцессии на моренах после отступления ледников на Аляске в ординационном треугольнике стратегий приведен на рис. 9.38.
Роль видов растений, относящихся к различных стратегиям, может сильно меняться в ходе первичных и вторичных сукцессий (рис. 9.39 по Grime, 1987). Растения разных жизненных форм занимают различные области в ординационном треугольнике (рис. 9.39 а-д). В лесных сообществах
3. Сукцессии
427
Рис. 9.38 • Ход первичной сукцессии на ледниковых моренах на Аляске (Crocker, Major, 1955) в ординационном треугольнике CSR стратегий Ф. Грайма. Величина общей биомассы сообществ отражена размерами кружков. Вовлеченность в сукцессию растений разных жизненных форм может быть оценена с помощью схем, приведенных на рис. 9.39 (по: Grime, 2001)
Рис. 9.39 • Сукцессионные смены с точки зрения изменения стратегий видов (по: Grime, 1987). Верхний ряд — области растений разных жизненных форм в треугольнике стратегий, средний ряд — сукцессии разных типов с позиций смены основных стратегий растений (стрелки показывают направление смен, цифры — длительность смен в годах, размеры черных кружков пропорциональны надземной биомассе), нижний ряд — роль различных регенерационных стратегий растений в ходе смен растительных сообществ, представленных в среднем ряду (1У — многочисленные мелкие далеко распространяющиеся диаспоры, V — вегетативное размножение, Bs — устойчивый семенной банк в почве, Bsd — устойчивый банк проростков): е, л — вторичная сукцессия после вырубки леса на умеренно богатых почвах; ж, м — вторичная сукцессия после вырубки леса на более бедных почвах; з, н — первичная сукцессия на выходах скал; и, о — изменение сообщества при ежегодной вспашке и внесении удобрений в посевах однолетних культур; к, п — циклические смены при периодической вырубке подроста Fraxinus excelsior (верх) и выжигании вересковых пустошей (низ)
428
Глава 9. Динамика фитоценозов
(рис. 9.39 е-ж) сукцессии идут по типичному варианту R —> С —> S, итоговая биомасса зависит от богатства почвы. Напротив, в ходе первичных сукцессий на скалах (рис. 9.39 з) стресс-толеранты сменяются более конкурентоспособными видами. При наличии периодических нарушений смены состава сообществ происходят с частичным возвратом в исходное состояние, в этом случае лучше говорить о флуктуационной, а при ежегодных нарушениях (посевы сельскохозяйственных культур, сенокосные луга) — о сезонной динамике сообществ (см. выше). Эти типы динамики также могут быть отражены в ординационном треугольнике стратегий Ф. Грайма (рис. 9.39 и, к, о, п).
3.9 • Факторы сукцессионнной динамики
3.9.1 • Пожары
3.9.1.1 • Распространение пожаров
в растительных сообществах земного шара
Влияние пожаров вызывает циклические изменения в составе и структуре сообществ. В зависимости от интенсивности пожаров, длительности и характера послепожарных смен их можно отнести как к флуктуационной, так и к сукцессионной динамике. Для удобства изложения все формы пожарной динамики рассмотрены в этом разделе.
Многие растительные сообщества земного шара сформировались и поддерживают свое существование за счет периодического воздействия огня (рис. 9.40). К таким сообществам можно отнести североаме-
Рис. 9.40 • Распространение пожаров по земному шару в 1998 г., по данным Европейского космического агентства. Большинство пожаров приходится на травяные сообщества с доминированием С4 злаков (по: Bond et al., 2005: ATSR World Fire Atlas)
3. Сукцессии
429
риканскую чапарраль, южноафриканский финбош, вересковые пустоши в Шотландии, многие хвойные леса Северной Америки и Евразии (лишайниковые сосняки и т.п., а по мнению Komarek (1983) — почти все леса с доминированием видов Pinus и Quercus обязаны своим развитием пожарам), многие луга и степи, тропические и субтропические саванны, большинство типов растительности Австралии.
Пожары во многом изменили облик природных зон Земли. В средиземноморском климате в отсутствие пожаров могут произрастать широколиственные леса, а их современная замена кустарниками обязана деятельности огня. Внедрение злаков на Гавайи вызвало увеличение частоты пожаров и уничтожение естественных лесов (Bond, van Wilgen, 1996).
W. J. Bond (2005) предлагает различать растительность, находящуюся в равновесии с локальным климатом, и растительность, контролируемую потребителями (главным образом крупными млекопитающими и огнем). Последняя занимает значительную часть суши Земли. Степень этого контроля оценивается по разности между расчетной биомассой сообществ (по климатическим данным) и реальной биомассой. Климат может поддерживать леса на вдвое большей площади, чем они занимают сейчас. С другой стороны, по мнению Flannery (1995), современное широкое распространение пожаров является артефактом вымирания травоядной мегафауны. С использованием верифицированной модели глобальной динамики растительности земного шара составлена карта растительности в условиях отсутствия пожаров. Огромные территории, особенно в Африке и Южной Америке, могли бы быть покрыты лесами в отсутствие огня, площадь лесов могла увеличиться с 27 % сейчас до 56 %, т. е. удвоиться (рис. 9.41). Широкое распространение пожаров связано с экспансией С4 злаков 6-8 млн лет назад, задолго до интенсификации пожаров человеком. Большой возраст таксонов, адаптированных к воздействию огня, свидетельствует о широком распространении пожаров до появления человека (Bond et al., 2005).
Пожары поставляют в атмосферу огромное количество продуктов горения органических веществ, только в тропиках 2700-6800 миллионов тонн органических веществ сгорает ежегодно (главным образом в саваннах) (Crutzen, Andreae, 1990). Огонь является одним из наиболее эффективных потребителей первичной надземной продукции растительных сообществ, своеобразным «фитофагом». При этом многие защитные приспособления растений против реальных фитофагов (жесткость побегов, большая длительность жизни листьев, колючки и опушение) улучшают «потребление» растений и их опада огнем. Воздействие огня и выпаса имеет многие противоположные влияния на цикл азота. Например,
430
Глава 9. Динамика фитоценозов
б)
Сомкнутость деревьев при отсутствии пожаров
20 40 60 80 100
20 40 60 80 100
Наблюдаемая сомкнутость деревьев в 2000 г.
i
5 10 40 100
Рис. 9.41 • Сомкнутость древесного яруса (%) в различных районах земного шара, смоделированная для условий реального климата при отсутствии пожаров (а), с учетом пожаров (б) и наблюдаемая по данным спутниковой съемки в 2000 г. (в) (The Food and Aqriculture Organization of the United Nations (FAO), 2001). Градации наблюдаемой сомкнутости: 40-100% — сомкнутые леса, 10-40% — сомкнутые саванны и разреженные леса, 5-10% — разрозненные деревья. Карта совмещает естественные леса и плантации (по: Bond et al., 2005)
3. Сукцессии
431
как показано наблюдениями в высокотравной прерии (Johnson, Matched, 2001), ежегодное выжигание увеличивает прирост корней на 25%, а выпас уменьшает прирост корней (примерно на 30 %). Выжигание увеличивает отношение С N в корнях, а выпас — уменьшает (около 60 и 40 соответственно). В этих условиях минерализация азота при выпасе ускоряется, а при выжигании — замедляется.
Естественными причинами возгорания растительности чаще всего являются грозы, реже — искры от падающих по скалам камней и извержения вулканов. Однако в современных условиях большинство пожаров возникает по вине человека. Чем выше отношение площади побегов к их объему, сильнее рассечение листьев, тем лучше горят растения, поскольку эти особенности обеспечивают наиболее быстрое высыхание растительных материалов. Основные свойства растительных сообществ, способствующие возникновению пожара: большое накопление горючих материалов, их низкая влажность, оптимальное расположение растений в пространстве, низкая скорость разложения, высокий уровень накопления вторичных метаболитов. Чем меньше влажность ветоши, тем выше температура ее горения.
Для описания режимов пожаров используется 3 основных характеристики: а) частота горения, б) сезон горения и в) интенсивность огня (температура или количество выделяемой энергии при горении). При рассмотрении влияния пожаров на растительность выдвинуто две основные гипотезы. Одна из них считает основным фактором характер горения (интенсивность, пространственная структура огня), а другая основное значение придает частоте горения, т. е. интервалам времени между пожарами. Для травяных сообществ очень важным фактором выступает сезон горения. Например, в экспериментах по выжиганию высоких злаковников в Южной Африке один из доминантов Themeda triandra относительно уменьшала свое участие после осенних пожаров, а другой доминант Tris-tachya leucothrix увеличивала при осенних и уменьшала — при зимних палах.
Интенсивность выделения тепла при горении в различных сообществах составляет (кВт/м2) (по: Bond, van Wilgen, 1996):
злаковники 1000-3000
саванны 500-10 000
кустарники (чапарраль и др.) 500-20000
хвойные леса 500-100 000
432
Глава 9. Динамика фитоценозов
Рис. 9.42 • Схематизированный план кустов вереска (Calluna vulgaris) и живых и отмерших кочек пушицы (Eriophorum vaginatum) на площадке 2 х 4 м влажной пустоши в северо-восточной Шотландии (по: Keatinge, 1975)
3.9.1.2 • Примеры сообществ, подверженных периодическому воздействию огня
Вересковые пустоши в Шотландии
Среди сосудистых растений этих растительных сообществ, развивающихся на сфагновом покрове в условиях значительного увлажнения, доминируют вереск Calluna vulgaris и пушица Eriophorum vaginatum. Сосуществование этих видов во многом обусловлено пожарами, происходящими с частотой раз в 12-20 лет (по: Keatinge, 1975). Вереск укореняется в пределах дерновин пушицы, а его дальнейшее разрастание приводит к отмиранию значительного числа этих дерновин (рис. 9.42). Дальнейшее развитие вереска приводит к накоплению его отмерших побегов, а также к накоплению горючих материалов, возгорающихся от внешних (часто антропогенных) причин. В ходе пожара вереск выгорает практически полностью, а подземные органы пушицы остаются жизнеспособными. В первые годы после пожара пышно разрастается пушица, в вереск медленно возобновляется из семенного банка. Его дальнейшее развитие ведет к угнетению пушицы и новому циклу возгорания. Структура сообществ зависит от частоты выжигания пустошей. Если периодичность пожаров менее 10 лет — доминирует пушица, а при периодичности 20 лет — вереск (Keatinge, 1975).
Чапарраль в Калифорнии
Кустарниковые сообщества (чапарраль) встречаются на западе США в условиях средиземноморского климата с жарким летом и прохладной зимой
3. Сукцессии
433
Рис. 9.43 • Чапарраль: а — общий вид горной чапаррали в Южной Калифорнии (фото Antandrus, Wikimedia Commons, under GNU Free Documentation License); б — один из основных доминантов чапаррали — Adenostoma fasciculatum (Rosaceae) (фото Stan Shebs, Wikimedia Commons, under GNU Free Documentation License); в — пожар в чапаррали (фото Richard W. Halsey, California Chaparral Institute, с любезного разрешения автора)
(среднезимний минимум температур 1,7-7, 2 °C), количество осадков варьирует от менее 500 до более 1000 мм в год. Доминанты — вечнозеленые кустарники, 1-1,5 м высотой, основным доминантом часто выступает Adenostoma fasciculatum (рис. 9.43). Пожары обычны с интервалом 10-40 лет, они интенсивные с высокой (до 700 °C) температу
434
Глава 9. Динамика фитоценозов
рой. Надземные части кустарников почти нацело сгорают (рис. 9.43 б, в). В первые после пожара годы наблюдается пышное развитие травяного покрова, начало отрастания кустарников из оснований побегов и возобновление из семян. Прорастание семян «пожарных однолетников» стимулируется образующимися в почве нитратами (Thanos, Rundel, 1995). Всходы Adenostoma fasciculatum появляются на 2-3-й год после пожара, главным образом на местах, испытавших менее интенсивный прогрев почвы (Odion, Davis, 2000). Затем происходит постепенное увеличение покрытия кустарников, отложение их трудно разлагаемого опада, усиление конкурентного воздействия на травы и увеличение численности грызунов. Все это приводит к формированию маловидовых зарослей кустарников с постепенным накоплением горючего материала — отмерших стеблей и мощного слоя опада — до нового пожара.
Какие факторы, связанные с пожаром, вызывают такие смены? Специальными экспериментами, проведенными в приморской чапаррали в Калифорнии, было показано:
1. Прямое воздействие огня увеличивает появление всходов кустарников и полукустарников, но уменьшает число всходов однолетников.
2. Только удаление кустарников не влияет на появление всходов, но увеличивает выживаемость однолетников.
3. Исключение млекопитающих-фитофагов (также наблюдаемое при пожарах) улучшает выживаемость всходов однолетников и увеличивает появление всходов всех видов (Tyler, 1996).
При защите чапаррали от огня в течение длительного времени происходит деградация кустарникового покрова, появление всходов деревьев и формирование открытых дубовых или сосновых редколесий с травяным покровом, похожим на таковой в чапаррали. При более частых пожарах (с интервалом менее 10 лет) наблюдается некоторое ослабление роли кустарников, постоянное присутствие травяного яруса, усиление роли злаков.
Южноафриканский финбош
Флористически богатые кустарниковые сообщества (финбош) на юге Африки существуют в условиях регулярных пожаров (рис. 9.44). В состав этих сообществ входят представители различных семейств цветковых растений, но наиболее типичны виды вересковых (Ericaceae), протейных (Proteaceae) и рестиевых (Restionaceae). Максимальное видовое разнообразие здесь наблюдается при редких (в экспериментах через 40 лет) пожарах, в то время как при более частых (15—26 лет) флористическая
3. Сукцессии
435
Рис. 9.44 • Южноафриканский финбош: а — Berzelia abrotanoides (Bruniaceae) в заповеднике Hottentots Holland; б — приморский финбош на атлантическом побережье, на переднем плане — Didelta carnosa (Asteraceae); в — Mimetes fimbriifolius (Proteaceae) в столовых горах; г — Orothamnus zeyheri (Proteaceae) — редкий вид, увеличивающий численность после пожара (г — фото Colin Paterson-Jones, с любезного разрешения автора)
насыщенность снижается (Schwilk et al., 1997). Огонь имеет большое значение в сохранении редких и исчезающих видов растений, адаптированных к его воздействию. Так, например, охрана популяции Orothamnus zeyheri (Proteaceae) (рис. 9.44 г) в Южной Африке от огня привела по
436
Глава 9. Динамика фитоценозов
чти к полному вымиранию этого вида и только применение палов способствовало восстановлению численности популяции за счет массового появления и приживания всходов (Boucher, 1981). Численность фитофа-гов-семеноядов после пожара в финбоше обычно резко снижается, что дает лучшую возможность для прорастания семян и приживания всходов. Массовое цветение однодольных геофитов после пожара — одно из частых явлений на юге Африки. С другой стороны, выгоревшие участки часто привлекают большое количество травоядных млекопитающих в связи с улучшенным качеством биомассы отрастающих растений.
3.9.1.3 • Семенное и вегетативное возобновление растений после пожаров
По восстановлению после пожара растения подразделяются на две большие группы:
1) способные к вегетативному отрастанию из сохранившихся побегов, спящих почек и т. п.;
2) восстанавливающиеся из семян; вегетирующие растения погибают после пожара.
Первая группа по сравнению со второй характеризуется более медленным ростом побегов, развитием подземных органов запасания, задержкой в цветении. У второй группы выше репродуктивное усилие. Эти группы имеют конкурентное преимущество при разной частоте пожаров и размерах выгорае-мых территорий. Clark (1991) обратил внимание на разные стратегии адаптации растений этих групп. Для первой группы это максимизация эпизодов размножения за время жизни растения, а для вто-
Рис. 9.45. Одревесневший клубень (lignotuber) у Eucalyptus rossii (фото Brian Walters, с любезного разрешения автора)
3. Сукцессии
437
Рис. 9.46 • Примеры шишек и плодов, накапливающих надземный семенной банк на древесных растениях и раскрывающихся при воздействии пожаров (слева — до пожара, справа — открытые после пожара): а — Widdringtonia (Cupressaceae); б — Накеа (Proteaceae); в — соцветие Protea (Proteaceae) (по: Bond, van Wilgen, 1996)
рой — максимизация вероятности достижения репродуктивной зрелости к моменту следующего горения (Bond, van Wilgen, 1996).
Одним из приспособлений к переживанию пожаров у древесных растений может служить образование одревесневающих клубней (lignotubers), типичных для ряда южноафриканских и многих австралийских растений, особенно эвкалиптов (рис. 9.45). Эти органы (полу)погружены в почву и переносят воздействие огня, а затем образуют надземные побеги (Trabaud, 1987).
Плоды и стробилы многих деревьев и кустарников раскрываются лишь после воздействия огня, а до этого накапливаются на взрослых растениях в течение многих лет (надземный семенной банк). Особенно это свойственно соснам, кипарисам и многим растениям семейства Proteaceae (рис. 9.46).
438
Глава 9. Динамика фитоценозов
Основным фактором приживания всходов и их развития после пожара являются погодные условия, прежде всего интенсивность осадков. Прорастание семян многих растений после пожаров задерживается до наступления влажного сезона (Bond, van Wilgen, 1996). Семена многих растений активно прорастают после пожара, что может быть связано с:
1) растрескиванием от огня твердых оболочек;
2) тепловой стимуляцией зародыша;
3) высокотемпературным иссушением семенных покровов;
4) воздействием этилена или аммония как компонентов дыма;
5) воздействием других компонентов дыма или дымовых экстрактов.
Например, при изучении прорастания семян 28 видов растений финбоша у 12 видов обнаружено стимулирование прорастания семян после обработки дымом. Дым сам по себе вызывал усиление прорастания семян представителей Asteraceae, Ericaceae, Restionaceae, а дымовые экстракты — у видов Proteaceae. Наиболее резкое усиление прорастания семян отмечено для Syncarpha vestita (Helichrysum vestitum) (Asteraceae) (Brown, 1993).
Аналогичные закономерности отмечены для многих растений западной Австралии, где из 94 видов растений с плохо прорастающими семенами 45 видов увеличили всхожесть после обработки холодным дымом. Усиление прорастания отмечено также при проращивании на фильтровальной бумаге, обработанной дымом, или поливе водой, содержащей компоненты дыма (Dixon et al., 1995). Двуокись азота как важный компонент дыма может быть индуктором прорастания семян растений из областей, подвергающихся периодически пожарам (Keeley, Fotheringham, 1998).
Развитие растений после пожаров во многом связано с обогащением почвы ЭМП, поступающими с золой. При пожарах в австралийских сообществах на каждые 100 г сухого органического вещества может откладываться 15,9 г золы, общее поступление золы может составить 239 г/м2 (Pate, Dell, 1984). Из золы поступает небольшое количество азота и фосфора, умеренное количество кальция и магния, существенное количество калия по отношению к содержанию этих элементов в верхнем слое почвы. В ходе возобновления растений в течение 18 месяцев после пожара они аккумулируют значительно больше N и Mg, примерно такое же количество Р, Са и К, которое содержалось в поступившей золе. Развивающиеся после пожара эфемеры содержат существенно больше Р, чем другие растения (1,05 мг/г против 0,32-0,49 мг/г). Практически все растения песчаных равнин с высокой эффективностью реутилизиру-ют ЭМП (особенно Р) из отмирающих листьев и побегов, чего, однако, не происходит в случае пожаров. Некоторые геофиты эффективно накапливают ЭМП в клубнях или других запасающих органах в течение
3. Сукцессии
439
многих лет, реутилизируя 80-99 % Р, N, К из отмирающих листьев. Значительное количество ЭМП аккумулируется в семенах. Kuo et al. (1982) показали, что семена составляют 1-16% массы плодов, аккумулируя при этом 83-98% Р, 43-76% N, более 50% Zn и Си от общего содержания в плодах. Содержание Р в семенах составляет 9-21 мг/г сухой массы, что существенно выше, чем у многих растений других регионов (Pate, Dell, 1984).
3.9.1.4 • Пожары в травяных сообществах и тундрах
Пожары на лугах и в степях обычно происходят при накоплении горючего материала более 500 кг/га. Оптимальная влажность ветоши при этом 7-20%, при влажности 30-40% загорания обычно не происходит. Дер-новинные злаки могут страдать от отсутствия пожаров в связи с сильным накоплением ветоши и затенением новых растущих побегов. Многие злаки интенсивно цветут после пожара (Bond, van Wilgen, 1996).
Ежегодное выжигание (весеннее) сухих лугов в Европе и некоторых типов прерий в США приводит к увеличению их продукции, но снижению флористической насыщенности. Например, после 15-летнего ежегодного выжигания карбонатных лугов в Швейцарии при увеличении продукции произошло резкое уменьшение участия одного из доминантов — Bromus erectus при увеличении участия другого доминанта — Brachypodium pinnatum (Ryser et al., 1995). Частое выжигание может вести к прогрессирующему обеднению почв в связи с потерями азота и серы при горении и вымыванию биогенов из почвы после пожара. В то же время ежегодное выжигание сухих и влажных прерий с Pinus palustris в Северной Кароли-
Таблица 9.3 • Среднее число видов сосудистых растений в двух типах прерий при различных режимах выжигания (по: Wolf, Peet, 1980)
Размер площадок (м2) Влажная прерия Сухая прерия
ежегодное выжигание выжигание через 3-4 года ежегодное выжигание выжигание через 3-4 года
0,25 23 10 22 7
1 39 17 31 9
25 54 48 48 33
625 84 69 70 60
440
Глава 9. Динамика фитоценозов
не (США) поддерживало большую флористическую насыщенность, чем выжигание раз в 3-4 года (Wolf, Peet, 1980; Peet et al., 1983) (табл. 9.3).
С другой стороны, выжигание растительных сообществ, ранее не испытывавших воздействие огня (например, высокогорных лугов в Новой Зеландии), ведет к быстрой деградации этих сообществ, потере большого количества биомассы и биогенов, смене растениями, адаптированными к нарушениям. Последствия таких пожаров четко проявляются в растительном покрове даже через 20 лет после горения (Calder et al., 1992; Gitay et al., 1992).
В сухие годы пожарам подвержены также тундровые сообщества. Например, в 2007 г. (самый теплый вегетационный сезон за последние 129 лет!) на северных территориях Аляски наблюдались частые пожары, один из наиболее сильных произошел в долине реки Анактувук. Он продолжался около трех месяцев, при этом выгорело более 100000 га тундры (Jones et al., 2009). Восстановление покрова сосудистых растений наблюдается в тундре после пожара примерно за 10 лет (Wein, Bliss, 1973; Racine et al., 2004), восстановление уровня мерзлоты — за 10-25 лет (MacKay, 1995; Racine et al., 2004). Анализ причин пожаров в тундрах Аляски с 1959 по 2007 гг. показал, что в подавляющем большинстве случаев они возникали от молний Qones et al., 2009).
3.9.1.5 • Пожары в лесах
Лесные пожары обычно разделяют на верховые (огонь захватывает кроны деревьев) и низовые (огонь распространяется по подстилке, валежнику и нижним ярусам леса). Древесные породы значительно отличаются по устойчивости к воздействию огня. Толщина коры — основной фактор устойчивости деревьев к огню, поскольку перегрев камбия вызывает его гибель. Поэтому взрослые деревья более устойчивы, чем подрост (Bond, van Wilgen, 1996). Развитие ряда видов деревьев связано с периодическими пожарами. Например, Pinus resinosa (одна из основных лесообразующих пород в ряде районов США) требует для своего возобновления пожара. В лесах из этой породы довольно часты низовые пожары, но только редкие мощные верховые пожары ведут к существенному омоложению популяции этого вида. До поселения европейцев в Северной Америке сухие участки с доминированием Populus tremu-loides или Pinus banksiana горели в естественных циклах примерно через 50 лет, а леса из Pinus resinosa, Р. strobus — через 150-350 лет (Peet, 1992).
Пожары могут изменять структуру и состав древостоев. Так, при частых пожарах ель (более чувствительная к пожарам порода) выпада
3. Сукцессии
441
ет и уступает место сосне. Эвкалиптовые леса Тасмании накапливают значительное количество горючего материала и естественно горят с периодичностью 100-300 лет. Если они не горят более 350 лет, эвкалипты не могут возобновляться семенным путем, и леса сменяются малогорючими многовидовыми дождевыми лесами, под пологом которых эвкалипты не могут возобновляться. С другой стороны, если эвкалиптовые леса горят часто до достижения деревьями репродуктивного возраста, то эти леса сменяются зарослями хорошо горючих склерофилльных деревьев и кустарников (Peet, 1992). Исторические исследования показали, что периодическое выжигание австралийскими аборигенами участков лесов до прихода европейцев было широко распространено и явилось одним из факторов поддержания участия некоторых видов растений в составе лесов (Bowman, 1998).
В хвойных лесах северной Швеции изучено содержание и свойства угля в сукцессионной послепожарной серии площадок, возрастом от 1 года до 350 лет, с момента последнего пожара. Масса угля в почве варьировала в пределах 984-2074 кг/га. Фенольные соединения, выделяемые Empetrum hermaphroditum, эффективно адсорбировались углем, имеющим возраст до 100 лет, но не больше. Микробиальная биомасса вблизи кусочков угля была существенно выше, чем вне их. Подчеркивается важная роль угля как сорбента аллелопатически активных веществ, выделяемых шикшей и отрицательно воздействующих на развитие подроста сосны (Zackrisson et al., 1996). Эксперименты, проведенные с выращиванием сеянцев Pinus sylvestris и Betula pendula с добавлением и без добавления угля и опада Vaccinium myrtillus (богатого фенолами) на разных лесных почвах, показали, что рост березы был существенно выше с углем, а поглощение ею азота при добавлении угля увеличивалось в 6,2 раза. На рост сосны добавление угля имело меньшее воздействие. Заростки папоротников появлялись на почве из-под вересковых кустарничков только при добавлении угля (Wardle et al., 1998).
Ход послепожарных (пирогенных) сукцессий связан с доминированием нитрофильных видов на начальных стадиях. Типичным представителем является Chamaenerion angustifolium, анемохорные семена которого распространяются на большие расстояния. Развитие травянистых растений имеет большое значение в сохранении и накоплении ЭМП в почве после пожара, так как при отсутствии быстро сформированного покрова значительная часть биогенов вымывается из почв и безвозвратно теряется для экосистемы (см. п. 3.9.2 гл. 9). На смену травянистым растениям в таежной зоне приходят мелколиственные породы, лишь затем сменяющиеся медленнорастущими хвойными (рис. 9.47).
442
Глава 9. Динамика фитоценозов
Рис. 9.47 • Пирогенные сукцессии в сосняках: а — иван-чай (Chamaenerion angustifolium) — пионер послепожарных сукцессий; б — горелый лес с иван-чаем на пятый год после пожара, Тебердинский заповедник, Северо-Западный Кавказ; в — сосняк-зеленомошник до пожара, Приокско-Террасный заповедник, Московская область;
3. Сукцессии
443
г — сосняк-зеленомошник через 2 года после низового пожара, там же; д — этот же участок через 8 лет после пожара, подрост березы и осины достигает высоты 1,5-2 м, ослабленные огнем сосны погибли и выпали через 3-5 лет после пожара
444
Глава 9. Динамика фитоценозов
3.9.2 • Вырубки лесов
После вырубки лесов происходят разнообразные изменения в растительном покрове, зависящие от исходного типа леса, характера вырубки и дальнейшего использования территории. Удаление древесного яруса приводит к резкому изменению климатических условий — увеличивается поступления света к нижним ярусам, уменьшается эвапотранспира-ция, что в таежных лесах часто ведет к заболачиванию, усиливающемуся на просеках и лесосеках. На местах локальных вырубок часто наблюдается развитие мхов (сообщества с доминированием Polytrichum commune, а также Sphagnum). В почве резко увеличивается интенсивность минерализации органики и нитрификация, связанные с прекращением поглощения ЭМП деревьями, разложением корней деревьев и увеличением температуры почвы за счет лучшего прогрева. Обычно пышно развивающиеся травы поглощают значительную часть доступных форм ЭМП, образовавшихся в почве, однако при отсутствии травяного покрова происходит усиление вымывания и потеря многих биогенов из экосистемы. В специальном эксперименте с применением гербицидов на вырубках на стационаре Hubbard Brook в США (Borman, Likens, 1979) было показано, что вынос с фильтрующимися водами NO3 составил 114 кг/га в год, Са — 78 кг/га, К — 30 кг/га. Концентрации нитратов в сточных водах увеличились в 40 раз, калия — в 11, кальция — в 5,2 раза по сравнению с исходным лесом. Нитрификация приводит к подкислению почвы (pH изменился с 5,1 до 4,3), что в свою очередь способствует вымыванию катионов. В целом вынос азота на вырубке смешанного леса в Hubbard Brook увеличился в 160 раз по сравнению с исходным лесом.
3.9.3 • Выпас и сенокошение
Влияние выпаса на растительные сообщества и их компоненты рассмотрено в п. 10 гл. 4. Выпас связан с воздействием крупных фитофагов, изменения в интенсивности этого воздействия также приводят к сукцессионным сменам растительных сообществ. Наблюдаемый во многих регионах перевыпас ведет к развитию процессов пастбищной дигрессии, которая имеет общие и частные закономерности для сообществ разных природных зон. К общим закономерностям можно отнести развитие процессов эрозии почв, разрастание непоедаемых видов растений, резкое падение продуктивности, переход к доминированию растений с коротким жизненным циклом, высокой относительной скоростью роста, высокой аллокацией биомассы в листья (см. п. 3.7 гл. 9). При увеличении интенсивности выпаса выше определенного порога происходит смена видов и доминирующих стратегий защиты от фитофагов — от видов, устойчивых к поеданию (способных отрастать и т. п.), к видам, избегающим
3. Сукцессии
445
поедания (имеющим эффективные средства механической или химической защиты) (Briske, 1996).
В то же время воздействие выпаса на различные сообщества может приводить к сукцессиям разного типа. Выпас в лесах (и искусственное увеличение численности диких копытных, например серн, в Альпах) приводят к уничтожению подроста лиственных деревьев, часто — к снижению верхней границы леса и т. п. В пустынях происходит разбивание «пустынной корки» из мхов и водорослей, что ведет к ослаблению азотфиксации, усилению поступления осадков в почву и созданию микроусловий для приживания всходов растений. Наиболее катастрофические смены растительности происходят при завозе копытных на острова или в регионы, где их раньше не было и виды местной флоры не были адаптированы к существенному отчуждению биомассы (Новая Зеландия, острова Тихого океана и т. п.).
Можно различать два основных аспекта сукцессионной динамики растительности при выпасе: изменение фитоценозов под влиянием выпаса и при его прекращении (демутация). Примером такой демутации может служить вторичная сукцессия, происходящая при прекращении выпаса на местах стоянок скота в альпийском поясе Центральных Гималаев (Rikhari et al., 1993). Путем изучения разновозрастных (от 2 до 40 лет) мест бывших стоянок скота авторами выявлены виды растений ранних стадий сукцессий (2-3 года) — Rumex nepalensis, Cirsium wallichii, Poa pratensis, средних стадий (12-20 лет) — Plantago major, Polygonum filicaule и климаксовых лугов (30-40 лет и окружающая растительность) — Danthonia cachemyriana, Trachydium roylei (рис. 9.48). В ходе сукцессии надземная биомасса постепенно уменьшалась с 350-370 г/м2 до 146-200, при этом общая численность побегов растений увеличилась в 2,5-3 раза. Флористическая насыщенность увеличивалась в ходе сукцессии, а флористическое разнообразие (индекс Шеннона) достигало максимума на средних стадиях (Rikhari et al., 1993).
Сенокошение — один из древних видов хозяйственной деятельности человека; например, на Британских островах оно активно производилось уже в железном веке. Сенокошение часто имеет положительный эффект, увеличивая флористическое богатство травяных сообществ. Коса или сенокосилка могут выступать своеобразными «фитофагами с трофической адаптацией» (см. п. 6 гл. 4), поскольку, срезая растения на одной высоте (обычно 3-5 см), они удаляют большую долю надземной фитомассы у более высоких, обычно доминирующих видов растений. Более низкие растения повреждаются меньше, что позволяет им устойчиво существовать в сообществах, подвергаемых периодическому сенокошению, и обусловливает высокое флористическое богатство косимых участков.
446
Глава 9. Динамика фитоценозов
Относительное проективное покрытие (%)
3. Сукцессии
447
С другой стороны, сенокошение на материковых лугах, где отсутствует принос веществ с наилком, может приводить к прогрессирующему обеднению почвы и снижению продукции лугов в связи с постоянным выносом биогенов со скошенными растениями за пределы экосистемы. В этих сообществах потери биогенов должны компенсироваться внесением удобрений. В экспериментах на Ротамстедской станции в варианте без внесения удобрений за 90 лет при ежегодном скашивании средний урожай снизился почти втрое (с 28,7 до 10,9 ц/га), в то время как при внесении компенсирующего количества удобрений существенных изменений в урожайности не происходило (Brenchley, Warington, 1958; Работнов, 19856).
В целом скашивание изменяет конкурентоспособность видов, сезонную ритмику их развития, влияет на семенную продуктивность растений (обычно отрицательно) и на приживание всходов (чаще положительно), что приводит к омоложению популяций на косимых участках по сравнению с некосимыми. Изменение травостоев под влиянием скашивания зависит от сроков и частоты кошения. На лугах, длительно использующихся как сенокосы, большинство видов растений обсеменяется либо до, либо после скашивания. Для улучшения состава сенокосных лугов обычно рекомендуют внедрение сенокосооборотов, поскольку при чередовании сроков кошения создаются возможности для обсеменения большого числа видов растений. Возможно и чередование сенокошения с выпасом, которое традиционно применялось на среднегорных лугах Кавказа.
Сенокошение на лугах часто приводит к увеличению эвапотранс-пирации, усилению роли более ксерофильных видов растений. Травяные болота при регулярном скашивании могут изменяться в луговые сообщества, при этом разные режимы кошения приводят к формированию разных по составу травостоев на исходно однородной территории. Примером могут служить изменения, происшедшие на крупноосоковом болоте в Нидерландах (данные Gomes; см. Luken, 1990) (рис. 9.49). В ис-
Рис. 9.48 • Изменение относительного проективного покрытия доминирующих видов растений в ходе сукцессии на заброшенных стоянках скота в альпийском поясе Центральных Гималаев (по: Rikhari et aL, 1993) в зависимости от их возраста (времени последнего использования).
Для примера приведены 2 серии площадок. Виды растений: RN — Rumex nepalensis, РР — Роа pratensis, RH — Ranunculus hirtellus, PM — Plantago major, CA — Carex alpina, TR — Trachydium royeli, C\N — Cirsium wallichii, PF — Polygonum filicaule, PN — Potentilla nepalensis, GE — Geum datum, DC — Danthonia cachemyriana
448
Глава 9. Динамика фитоценозов
Рис.9.49 • Изменения осокового болота в Нидерландах под влиянием разных режимов сенокошения и внесения удобрений: а — исходное сообщество (без кошения), доминант Carex riparia', б — трехкратное кошение в мае, июле и сентябре (Agrostis stolonifera, Ranunculus repens); в — кошение в мае и сентябре — полидоминантное сообщество (6-10 содоминантов); г — однократное кошение в августе — Alopecurus pratensis; д — кошение в июле и сентябре — полидоминантное сообщество; е — удобрение N50P20K20 — Alopecurus pratensis (данные Oomes; см. Luken, 1990)
ходном сообществе и варианте без скашивания абсолютно доминировала Carex riparia, однократное скашивание в августе привело к доминированию Alopecurus pratensis, на вариантах с двукратным скашиванием в мае
3. Сукцессии
449
и сентябре, а также в июле и сентябре образовывалось полидоминант-ное сообщество с высоким участием 6-10 видов растений, трехкратное сенокошение в мае, июле и сентябре привело к доминированию экспле-рентов — Agrostis stolonifera и Ranunculus repens. Интересно отметить, что внесение невысоких доз комплексных удобрений (N50P20K20) имело сходный эффект с однократным скашиванием, на этом варианте также сформировалось сообщество с доминированием лисохвоста (Alopecurus pratensis).
Реакция отдельных видов на скашивание может быть связана с ритмикой развития и длительностью жизни их листьев. Так, на лугах в Швейцарии, развитых на карбонатных породах, была исследована связь фенологии листьев с доминированием отдельных видов при разных режимах скашивания и выжигания (Kahlert et al., 2005). На контрольном варианте без удаления биомассы три вида (Brachypodium pinnatum, Bromus erectus и Salvia pratensis) содоминировали. Bromus с относительно короткоживущими листьями и большей листовой продукцией в конце сезона доминировал на участках ежегодного кошения в июне, он практически исчез из участков с зимним выжиганием. Brachypodium продуцировала долгоживущие листья в начале сезона и не переносила кошения в июне, но доминировала на участках зимнего выжигания (в это время у нее оставалась только ветошь). Salvia имел долгоживущие листья, которые быстро отмирали осенью, он доминировал на участках с кошением в октябре (Kahlert et al., 2005).
Другой пример изменения структуры естественных лугов под влиянием скашивания приводит И. Г. Вершинин (1961). В пойме Енисея до кошения преобладали луга с доминированием Calamagrostis langsdorffii, на 6-10-й год после кошения стали преобладать Anthriscus sylvestris и Bromus inermis, а на 13-16-й год доминирование перешло к Alopecurus pratensis, при этом вейник практически выпал из состава травостоя (рис. 9.50). Таким образом, во многих сообществах при внедрении сенокошения увеличивается участие злаков, виды которых обычно хорошо переносят частичное отчуждение биомассы. Эту закономерность подтверждают и наблюдения на разновозрастных залежах в Чехии, где сенокошение также увеличивало участие злаков в травостое (Koblihova-Baumova, 1990).
Но для некоторых травяных сообществ, виды которых неадаптированы к регулярному отчуждению биомассы, сенокошение может приводить к их быстрому разрушению. Например, при ежегодном сенокошении крупнотравные сообщества на Камчатке быстро деградируют и сменяются лугами. Сохранение крупнотравных сообществ без их существенной
450
Глава 9. Динамика фитоценозов
Рис. 9.50 • Смена доминирующих видов луговых сообществ в пойме Енисея после начала сенокошения (по: Вершинин, 1961): а — Calamagrostis langsdorffii (исходный доминант); б — Anthriscus sylvestris (доминирует через 6-10 лет после начала кошения); в — Alopecurus pratensis (доминант после 13-16 лет кошения)
деградации возможно лишь при кошении не чаще 1 раза в 3 года (Морозов, 1994; Белая, Морозов, 2000).
Значительные изменения в составе фитоценозов происходят и при прекращении их использования в качестве сенокосов. В этом случае обычно наблюдается накопление большого количества ветоши, что увеличивает подверженность пожарам, увеличивается теплоизоляция почвы, в северных районах может происходить подъем уровня вечной мерзлоты, усиливается заболачивание в связи со снижением эвапотранспирации. В степях уменьшается роль плотнодерновых злаков при увеличении участия корневищных злаков, а также кустарников. В подзоне настоящих степей наблюдается доминирование кустарника Caragana frutex и снижение флористической насыщенности сообществ. В некоторых случаях при прекращении сенокошения заболачивания не происходит в связи с развитием высокотравья, виды которого отличаются высокой транспирацией. Например, низинные болота с Carex acutiformis в Швейцарии перешли в заросли высокотравья (Filipendula ulmaria, Urtica dioica и др.) при прекращении кошения (Gigon, Bocherens, 1985).
3.9.4 • Внесение удобрений
Внесение минеральных и органических удобрений осуществляется человеком в широких масштабах на землях сельскохозяйственного использования, включая сенокосные и пастбищные луга. С площадей, где производится внесение удобрения, содержащиеся в них ЭМП попадают
3. Сукцессии
451
во многие растительные сообщества, вызывая их сукцессионные изменения. Для изучения влияния удобрений на состав и динамику сообществ проведены многочисленные эксперименты в различных природных зонах, однако наиболее многочисленные исследования осуществлены на лугах (см., например, Работнов, 1973; Silvertown et al., 2002). В результате этих исследований было показано, что реакция отдельных видов растений на внесение ЭМП зависит от многих условий, в том числе:
1) от степени лимитирования продукции данным ЭМП;
2) от эффективности использования данным видом растений ЭМП (например, на лугах при внесении азотных удобрений обычно усиливается роль злаков, имеющих более низкую концентрацию азота в надземных органах, т. е. тратящих меньше азота на построение единицы биомассы);
3) от наличия в сообществе более конкурентоспособных видов. Например, Anthoxanthum odoratum положительно реагировал на внесение комплексных (NPK) удобрений в отсутствие сильных конкурентов и отрицательно — в их присутствии (Работнов, 1992).
Внесение азотных удобрений оказывает наибольшее влияние на структуру сообществ, часто ведет к резкому увеличению их продукции и снижению флористического разнообразия. Обратная сукцессия при прекращении внесения удобрений сильно замедлена: например, бывшие участки стоянок скота на Кавказе сохраняют высокопродуктивную рудеральную растительность более 70 лет. Для низкотравной прерии в США последействие внесенных азотных удобрений отмечается более 20 лет (Vinton, Burke, 1995). Внесение удобрений, особенно азотных, в тундровые и некоторые высокогорные фитоценозы способствует усилению в них роли злаков, т. е. олуговению этих сообществ.
В работе Pennings et al. (2005) была предпринята попытка оценить предсказуемость (постоянство) реакции отдельных видов на внесение азотных удобрений в разных условиях (в 9 различных районах США). Было выбрано 20 видов травянистых растений, каждый из которых был представлен не менее чем в 6 различных экспериментах с внесением азота. Из 20 видов один (Spartina alternifolia) во всех случаях увеличивал, а 5 видов (Erigeron strigosus, Juncus gerardii, J. roemerianus, Lespedesa capitata, Solidago rigida) — во всех случаях уменьшали свое участие при обогащении почвы азотом. Восемь видов показали предсказуемую реакцию в зависимости от географического положения, окружающих видов, продукции сообщества (наиболее часто) и других характеристик. Авторы делают вывод, что реакция вида на внесение азота часто предсказуема, но она зависит от многих абиотических и биотических факторов.
452
Глава 9. Динамика фитоценозов
в)
Рис. 9.51 • Экспериментальное внесение удобрений (Park Grass Experiment) на Ротамстедской опытной станции: а — внешний вид экспериментального поля — аэрофотоснимок; б — некоторые экспериментальные площадки показывают большое разнообразие травянистых растений, свойственное лугам до начала эксперимента; в — изменение флористической насыщенности площадок на разных вариантах внесения удобрений (с любезного разрешения Stephen James: Copyright Rothamsted Research Limited; copying in any form is prohibited)
Suding (неопубл.) на основании анализа большого числа экспериментов выделила 4 особенности растений, связанные с их положительным откликом на азотные удобрения: 1) отсутствие связей с азотфиксирую-щими прокариотами («не бобовость»), 2) однодольность, 3) клональность, 4) значительная высота (верхняя треть полога). Для Spartina al tern ifolia характерны все четыре признака. Spartina patens по-разному реагировала на внесение азота в зависимости от соседей: она уменьшала участие в соседстве с Spartina alternifolia или Distichlis spicata, но увеличивала в паре с Juncus gerardii (Pennings et al., 2005).
3. Сукцессии
453
Изменение общей продукции при внесении удобрений зависит от многих факторов, в том числе от сукцессионной стадии и характерной интенсивности нарушений. Часто большее увеличение продукции наблюдается не для климаксовых сообществ, а для сериальных. Для высокотравной прерии в США показано, что увеличение листовой продукции при внесении азотных удобрений зависело от частоты выжигания прерий: на участках, не выжигаемых 15 лет, продукция увеличилась на 9%; на нечасто выжигаемых участках — на 45 %, а на ежегодно выжигаемых участках — на 68 %. Причины этих различий связаны с разным уровнем потерь азота при выжигании (Seastedt et al., 1991).
Самым длительным экологическим экспериментом с внесением удобрений в луговые сообщества является эксперимент на Ротамстедской опытной станции в южной Англии, где на площади 4 гектара в 1856 г. был заложен Park Grass Experiment (рис. 9.51). До этого территория в течение многих столетий служила сенокосом и пастбищем. При отсутствии экспериментальных воздействий состав растительного сообщества, разнообразие и продукция находились в квазистабильном состоянии, но были подвержены колебаниям в связи с климатическими факторами. Азот, фосфор и калий — основные лимитирующие продукцию почвенные ресурсы в Ротамстеде, в то же время кислотность почвы, изменяющаяся при известковании и внесении удобрений, оказывает большое влияние на состав и разнообразие растительных сообществ. Leontodon hispidus — предпочитаемое скотом кормовое растение пастбищ — увеличило свое участие при снятии пастбищной нагрузки в течение первых 50-70 лет эксперимента. Участие злаков снижалось, а разнотравья — увеличивалось на контрольных участках за этот период времени. На площадках, получавших аммонийный азот, абсолютным доминантом стал Hoicus lanatus, а при нитратном азоте доминировали Alopecurus pratensis и Arrhen-atherum eiatius. При внесении неазотных удобрений усиливалась роль бобовых и уменьшалась — злаков. Для популяций Anthoxanthum odoratum была показана генетическая дифференциация по отношению к кислотности (Snaydon, 1970), особи с известковавшихся площадок лучше росли на менее кислой почве, и наоборот. Эксперименты с взаимными пересадками Anthoxanthum odoratum также показали экотопическую дифференциацию особей по вариантам эксперимента. Мучнистая роса (Erisyphe graminis) лучше развивалась на площадках с внесением азота, а ржавчина Puccinia poae-nemoralis — на площадках с большей высотой растительного покрова (Tilman et al., 1994). В целом долговременные эксперименты на Ротамстедской станции показали: 1) в каждом варианте внесения удобрений образуется луг с характерным набором видов
454
Глава 9. Динамика фитоценозов
растений, 2) при внесении удобрений не происходит уменьшения продукции в условиях ежегодного сенокошения, в отличие от контрольных участков (без внесения удобрений), 3) наиболее резкое обеднение видового состава произошло при внесении NPK, когда азот вносили в виде сульфата аммония, что вызывало резкое подкисление почвы (рис. 9.51). В разных вариантах эксперимента число видов на 200 м2 варьировало от 3 до 44. Самое высокое число видов оставалось на неудобряемых участках, флористическая насыщенность уменьшалась монотонно с увеличением биомассы на площадках, не получавших азот или получавших его в форме нитрата натрия. Помимо азота, внесение фосфора также снижало видовое разнообразие, а внесение РК снижало разнообразие еще сильнее. (Crawley et al., 2005).
При внесении удобрений увеличивается также эффективность использования воды растениями, уменьшается транспирационный коэффициент — количество транспирированной воды на единицу продукции. Мезофиты обычно более отзывчивы на внесение азотных удобрений, чем ксерофиты. В связи с этим может наблюдаться мезофитизация сухих лугов за счет усиления конкурентоспособности мезофитов. Например, в экспериментах X. Элленберга (Ellenberg, 1978) при внесении на сухой луг азотных удобрений биомасса ксерофита Bromus erectus возросла с 87 до 114 г/м2, а мезофита Arrhenaterum elatius — со 167 до 961 г/м2, что привело к абсолютному доминированию последнего.
Несколько иное воздействие на луговые сообщества имеют фосфорные удобрения. Их внесение также может приводить к снижению флористической насыщенности сообществ. Анализ результатов многолетнего внесения фосфорных удобрений в луговые ценозы показал, что в результате существенно увеличивается выравненность сообществ, т. е. равномерность распределения видов по участию (Wilson et al., 1996). Бобовые растения обычно отрицательно реагируют на внесение азотных удобрений и положительно — на фосфорные и калийные.
3-9-5 • Кислотные дожди и SO2
Во второй половине XX в. промышленность (прежде всего тепловые электростанции) существенно увеличила выброс в атмосферу сернистого газа (SO2) и окислов азота при сокращении выбросов пыли (часто карбонатной). Это привело к увеличению кислотности осадков, которое вызвало гибель больших лесных массивов близ индустриальных зон в Европе и Северной Америке. Кислые осадки вызвали гибель мохового покрова на болотах (обратный катионитный эффект, см. п. 1.3 гл. 6), подкислили почву, усиливая вынос катионов и увеличивая подвижность токсичных
4. Эволюция сообществ и историческая динамика
455
анионов алюминия. На месте больших площадей еловых лесов в горах Чехии образовались злаковые луга с очень бедным видовым составом растений (Nardus stricta, Anthoxanthum odoratum, Deschampsia flexuosa, Festuca rubra) (Herben et al., 1993).
Ряд исследований были посвящены изучению влияния повышенной концентрации SO2 на рост отдельных видов растений. Например, при выращивании в течение 83 дней на повышенном фоне SO2 реакция была различной у видов разной стратегии (по: Грайму). Стресс-толеранты оказались наиболее устойчивыми, в то время как конкуренты и рудералы — менее устойчивы. Наиболее сильное снижение общей биомассы отмечено у Urtica dioica (на 81 %), Cerastium fontanum (на 77 %), Bromus steriiis (на 53%) (Ashenden et al., 1996).
В неблагоприятных условиях среды действие двуокиси серы усиливается. Так, Abies nordmanniana повреждалась двуокисью серы на бедных почвах, но не повреждалась на богатых (Jung, Winter, 1992). При низких температурах снижение фотосинтеза у Vicia faba под действием SO2 было существенно больше, чем при высоких (Kropff et al., 1990). Повышение содержания SO2 в атмосфере снижает морозостойкость травянистых растений (Davison, Bailey, 1982), увеличивает поражаемость растений фитофагами. В почве под влиянием увеличения концентрации SO2 уменьшается численность бактерий и возрастает — грибов (Steubing et al, 1989; Mansfield, 1999).
4 • Эволюция сообществ
и историческая динамика
4.1 • Эволюция сообществ
Эволюция растительных сообществ, по определению Б. М. Миркина (1984), — это изменения растительности, которые ведут к формированию сообществ новых типов (новых комбинаций совместно обитающих растений), не существовавших ранее.
Природная эволюция сообществ — очень длительный процесс. Он связан с появлением (видообразование, естественная миграция) и исчезновением видов на отдельных территориях. Но человек в результате своей деятельности запустил довольно быстрые процессы антропогенной эволюции.
Б. М. Миркин (1984) предложил следующую классификацию форм антропогенной эволюции растительности:
456
Глава 9. Динамика фитоценозов
Рис. 9.52 • Заросли Imperata cylindrica (Роасеае) в Японии (фото Keisotyo, Wikimedia Commons, under GNU Free Documentation License)
А. Целенаправленная эволюция:
а) замена естественной растительности на искусственные сообщества;
б) интродукция видов из других районов и включение их в состав естественных или искусственных сообществ;
в) конструирование новых типов искусственных сообществ (лесных, полевых, парковых, рекультивационных на техногенных субстратах).
Б. Стихийная эволюция:
а) непреднамеренный занос видов растений, грибов и животных из других районов; например, среди широкого спектра антропогенных воздействий на лесные экосистемы Европы как наиболее важное рассматривается широкое распространение фитопатогенных грибов (Rackham, 2008);
б) уничтожение видов или снижение их генетического разнообразия за счет уничтожения части форм;
4. Эволюция сообществ и историческая динамика
457
в) смещение границ природных зон вследствие хозяйственной деятельности человека;
г) возникновение новых устойчивых вторичных типов сообществ: например, сведение лесов в тропической Азии привело к разрастанию агрессивного местного злака Imperata cylindrica, образующего монодоминантные сообщества, покрывающие около 35 млн га (среди них 4 % от площади Индонезии) (рис. 9.52) (Garrity et al., 1996); затраты на борьбу с этим злаком могут составить многие миллиарды долларов (Chapin et al., 2000);
д) формирование новых сообществ на антропогенных субстратах вследствие самозарастания: например, техногенные отвалы в Англии, содержащие высокие концентрации свинца, меди и цинка, медленно зарастали устойчивыми расами Agrostis tenuis, Anthoxanthum odoratum, Festuca ovina, Plantago lance-olata, формируя сообщества, ранее не отмеченные в природе.
Занос человеком растений в новые для них местообитания приобрел особенный размах в последние столетия. Для многих регионов отмечаются процессы синантропизации флоры и растительности — увеличение роли видов, которые не присущи естественным сообществам. Так, во второй половине XX в. доля синантропных видов составляла (Jager, 1977) во флоре: Средней Европы — 16%, Японии — 14%, Новой Зеландии — 25 %, острова Крит — 33 %, острова Гран-Канария — 40 %, Азорских островов — 60 %. Таким образом, наиболее значимы заносные виды на островах, где их число примерно втрое выше, чем на аналогичной территории материков (Lonsdale, 1999).
Многие виды одичали из культуры (в Средней Европе — Acorus calamus, Aristolochia clematitis, Impatiens parviflora и др.; рис. 9.53). Ряд видов расширяет свой ареал, внедряясь в естественные ценозы. Такие растения вслед за Н. С. Камышевым (1959) называют агриофитами. Некоторые виды интенсивно расширяют свой ареал. Например, свербига Bunias ori-entalis в настоящее время входит в состав многих луговых и степных сообществ в Средней России. Ее распространение идет с юго-востока европейской части России.
Лишь немногие из заносных видов становятся агриофитами (табл. 9.4). Lohmeyer, Sukopp (1992) оценивают долю успешно внедряющихся видов от всех занесенных в европейские страны в 1-3%.
По времени заноса растения делятся на: археофиты занесены в доисторическое время и в галло-римский период (до великих географических открытий XVI в.), а неофиты занесены относительно недавно (с XVI в.). Lohmeyer и Sukopp (1992) приводят следующую динамику появления
458
Глава 9. Динамика фитоценозов
Рис. 9.53 • Агриофиты в Средней России: а — Acorus calamus (родина — Восточная Азия); б — Impatiens glandulifera (Восточная Азия); в — Galinsoga ciliata (Северная Америка); г — Heracleum sosnowskyi (Западный Кавказ); д — Bunias orientalis (Юго-Восточная Европа); е — Chamomilla suaveolens (Северо-Восточная Азия и Северная Америка)
агриофитов в Средней Европе (см. табл. 9.5). Наибольшее число агрио-фитов в этом регионе появилось в XIX в.
Изучение биотопической приуроченности заносных сорняков в Центральной Европе показало, что археофиты более обычны в посевах более древних сельскохозяйственных культур (зерновые злаки), в то время как неофиты наиболее многочисленны в посевах недавно интродуцированных культур (кукуруза, рапс) (Pysek et al., 2005). Среди наиболее древних агриофитов в Средней Европе — такие виды, как Echinochloa crus-galli, Sisymbrium officinale, Artemisia absintium, Poa bulbosa. По происхождению среднеевропейские агриофиты делятся на выходцев из других рай-
4. Эволюция сообществ и историческая динамика
459
Таблица 9.4 • Число занесенных, прижившихся и внедряющихся в естественные ценозы видов (по: Lohmeyer, Sukopp, 1992)
Категории видов Британские острова Германия Нидерланды
Всего завезено видов 32000 12000 7000
Прижилось видов 322 385 220
Внедрилось в естественные ценозы ? « 200 75
Таблица 9.5 • Число видов агриофитов, появившихся
в Средней Европе в разное время (по: Lohmeyer, Sukopp, 1992)
Годы Число видов %
до 1500 г. 50 29,7
1500-1599 13 7,7
1600-1699 13 7,7
1700-1799 21 12,4
1800-1899 50 29,7
1900-1991 22 13,0
онов Европы и из Западной Азии (54%), из Северной Америки (30%), из других районов Азии (9 %).
В связи с близостью экологических условий и очень активными транспортными связями значительная часть неофитов в Европе происходит из Северной Америки, а неофиты в Северной Америке — из Европы. По данным W. М. Lonsdale (1999), значительно больше видов было занесено из Старого Света в Новый, чем наоборот. В США огромные площади заняты несколькими европейскими видами (рис. 9.54). Так, под влиянием выпаса и выжигания сформировались сообщества «однолетних злаков-ников» из однолетних видов рода Bromus (В. tectorum и др.). Bromus tectorum на западе Северной Америки покрывает около 40 млн га там, где частота пожаров увеличилась более чем в 10 раз (Whisenant, 1990). Внедрение Татапх на берега рек в США увеличило эвапотранспирацию и принесло ущерб 65-180 млн долларов ежегодно за счет снижения поступления воды для муниципальных и сельскохозяйственных нужд. Кроме того, усиление отложения наилка в кустах Tamarix привело к сужению русла каналов и возрастанию ущерба от наводнений на 50 млн долларов ежегодно (Zavaleta, 2000; Chapin et al., 2000). В целом экономический
460
Глава 9. Динамика фитоценозов
Рис. 9.54 • Европейские агриофиты в Северной Америке: а — Lythrum salicaria; б — Bromus tectorum; в — Tamarix sp.; г — Hypericum perforatum; d — Acer platanoides; e — Alliaria petiolata
ущерб от заносных видов в США оценивается в 34 млрд долларов в год (Pimentel et al., 2005; Theoharides, Dukes, 2007).
Выделяют четыре основных стадии инвазии растений: 1) занос диаспор, 2) колонизация (преодоление абиотического фильтра), 3) прижива
4. Эволюция сообществ и историческая динамика
461
ние (преодоление биотического фильтра) и 4) широкое распространение по ландшафту. Занос диаспор может быть многократным, что повышает генетическое разнообразие интродуцентов. Но возможно и широкое распространение при точечном заносе, например, для Salix babylonica в Новой Зеландии (Маск, 1995). В планетарном масштабе значительно больше видов вселилось из Евразии в Африку и Америку, чем наоборот (59 % вселенцев в другие регионы происходят из Евразии) (Theoharides, Dukes, 2007).
Многие исследователи пытались выяснить, какие особенности определяют успех приживания заносных видов на новом месте и внедрение их в естественные или нарушенные растительные сообщества. Инвазионные виды очень разнообразны по своим эколого-биологических характеристикам, но некоторые признаки, способствующие вселению в новые местообитания, оказались широко представленными в этой группе растений.
Большинство заносных видов характеризуются рудеральной (экс-плерентной) стратегией (см. п. 3.7 гл. 5) — коротким жизненным циклом, большой семенной продукцией. Многим свойственна способность к вегетативному размножению или апомиксис. В последнем случае отпадает необходимость в специализированных опылителях, а в случае раздельнополых видов — в особях другого пола (например, занесенная в 1840 г. в Англию Elodea canadensis представлена только женскими особями). Важным может являться наличие покоя у семян. Например, завезенный водный гиацинт Eichhornia crassipes с востока Южной Америки имеет покоящиеся семена в иле, сохраняющие жизнеспособность не менее 15 лет. Освоению бедных субстратов способствует наличие связей с азотфикса-торами, как в случае с завозом на Гавайи Myrica faya (см. п. 1.2 гл. 2).
Специальный анализ свойств видов, позволяющих им активно внедряться в новые сообщества, не выявил каких-либо специфических черт у активных «колонизаторов». Например, среди наиболее массовых видов нарушенных участков в районе Шеффильда (Англия) преобладали виды, широко распространенные в регионе в целом. Поступление значительного количества диаспор на новые участки во многом определяло успех внедрения этих видов (Hodkinson, Thompson, 1997). В работе Blumenthal et al. (2009) было показано, что большинство агрессивных интродуцентов из Европы в США обладают быстрым ростом (конкурентная стратегия по Грайму) и в то же время лишены большинства своих вирусных и грибных патогенов, которые встречаются в их исходных местообитаниях в Европе. Именно сочетание этих двух признаков делает возможным успешную колонизацию в новых условиях. Интересно отметить, что в «домашних» условиях виды конкурентной стратегии имеют
462
Глава 9. Динамика фитоценозов
примерно в 4 раза больше грибных патогенов и в 7 раз больше вирусных, чем виды, относимые к стресс-толерантам или стресс-толерантам-руде-ралам (Seastedt, 2009).
Особое внимание было уделено рассмотрению свойств сообществ, наиболее подверженных внедрению новых видов. Основной вывод из многочисленных экспериментов, посвященных этой проблеме, довольно прост — «растительное сообщество становится более подверженным внедрению новых видов, когда в нем возрастает количество неиспользуемых ресурсов» (Davis et al., 2000: 229). Особенно интересен эксперимент, в котором на участках лугов в Бакстоне создавался градиент двух факторов (богатство почв и уровень нарушений) и подсевались семена 54 видов растений, исходно отсутствовавших в составе сообщества (Burke, Grime, 1996). Наибольшее покрытие подсеянных видов наблюдалось при сочетании самого высокого уровня богатства и самого высокого уровня нарушений, т. е. в условиях наибольшего количества ресурсов, не(до)использованных исходным сообществом. В этом случае покрытие внедрившихся видов достигало максимума (30 %) (рис. 9.55). Было показано, что приживаемость растений прямо связана с общей площадью нарушений, особенно для мелкосемянных видов (крупносе-мянные — Anthriscus sylvestris, Heracleum sphondylium, Vicia cracca — приживались вне зависимости от нарушений). Авторы делают вывод, что именно динамика ресурсов, а не видовое разнообразие или продукция, определяет чувствительность сообществ к внедрению новых видов. Из этих исследований вытекает несколько следствий, позволяющих оценивать возможный успех вселения новых видов в сообщество:
1) сообщества с периодическим обогащением ресурсами более восприимчивы к инвазиям;
2) наиболее восприимчивы сообщества в период сразу после резкого поступления ресурса;
3) восприимчивость сообществ к инвазиям возрастает при нарушениях, воздействии фитофагов, паразитных грибов и т. п.;
4) нет прямой связи между флористическим богатством сообществ и их восприимчивостью к инвазиям, а также между продукцией сообществ и восприимчивостью к инвазиям (Davis et al., 2000; Grime, 2001) — во флористически богатых сообществах число заносных видов оказалось даже выше, чем во флористически бедных (Lonsdale, 1999). Последнее следствие подтверждается также детальным анализом вселения заносных видов в США (Stohlgren et al., 2008). Здесь число успешно занесенных видов превосходит число вымерших за то же время примерно в 24 раза (всего в США естественных видов 15 233, занос-
4. Эволюция сообществ и историческая динамика
463
Рис. 9.55 • Изменение покрытия внедряющихся в сообщество видов в эксперименте с градиентами богатства и нарушенное™ почв. В эксперименте использовано 5 уровней богатства почвы (с F1 по F5) и 5 уровней интенсивности нарушений (с DI по D5). Покрытие внедрившихся после подсева видов наибольшее (около 30%) в правом верхнем углу, т.е. при сочетании высокого почвенного плодородия с высокой интенсивностью нарушений (по: Grime, 2001)
ных — 3363, вымерло за период наблюдений — 140). В более флористически богатые районы внедряется больше видов, чем в менее богатые.
Среди агриофитов в Средней Европе преобладают виды следующих семейств: Asteraceae — 19%, Роасеае — 6,9 %, Brassicaceae — 6,9 %, Rosaceae — 6,1 % (Lohmeyer, Sukopp, 1992). Всего в Европе отмечено 580 видов натурализовавшихся экзотов, относящихся к 113 семействам. Их больше в Южной, чем в Северной Европе (Weber, 1997).
Особенности ценотической обстановки и близость экологических условий к исходным во многом определяют успех видов на новых территориях. Наибольшее число интродуцентов отмечается в несомкнутых, часто нарушаемых сообществах. Так, в Средней Европе наиболее обильны агриофиты в прибрежных сообществах вдоль рек — 142 вида, на скалистых обнажениях — 67 видов, в приморской полосе — 36 видов (Lohmey-er, Sukopp, 1992).
Устойчивость лесов к внедрению заносных видов часто связывают с высоким светолюбием интродуцентов. Однако Martin et al. (2009) показали, что в тропических и умеренных лесах мира встречается по крайней мере 139 видов теневыносливых интродуцентов (рис. 9.56). Важными свойствами лесных интродуцентов можно считать короткое время оборота поколений, длительный период рассеивания семян, небольшие размеры семян и длительное сохранение их жизнеспособности (Rejmanek, Richardson, 1996; Mehrhoff, 1998). В Северной Америке 99% древесных интродуцентов были интродуцированы преднамеренно для целей озеленения (Martin et al., 2009).
Анализ числа заносных видов на разных территориях показал, что заповедники и национальные парки имеют в среднем вдвое меньше интродуцентов, чем прилегающие территории (Lonsdale, 1999).
464
Глава 9. Динамика фитоценозов
Рис- 9,56 • Теневыносливые агриофиты: а — Veronica fiiiformis (происходит с Кавказа, заселяет затененные кладбища в Средней Европе); б — Vinca minor (родина — Средиземноморье, колонизирует темные леса в Средней Европе и на Северном Кавказе); в — Rhododendron ponticum («возвращается» на Британские острова и Среднюю Европу, где обитал до оледенения); г — Taxus baccata (родина — Европа, Северная Африка, Юго-Западная Азия; занесен в Новую Зеландию, где внедряется в естественные тенистые леса)
4. Эволюция сообществ и историческая динамика
465
Взаимодействие заносных видов с естественным почвенным населением может быть разнообразным. Интродуценты могут получать преимущество от отсутствия специализированных корневых фитофагов, но могут использовать новые микоризные связи для получения веществ от других растений, что имеет место у Centaurea maculosa (С. stoebe ssp. micranthos). Эксперименты показывают, что почвенная биота чаще оказывает негативное влияние на аборигенные, чем на заносные виды растений. Но бывают и обратные ситуации: например, американские почвы сильнее ингибируют рост Ammophila, чем европейские (Reinhart, Callaway, 2006). Внедряющаяся в леса северо-востока США Alliaria peti-olata содержит вещества, подавляющие развитие грибов арбускулярной микоризы у лиственных деревьев, что ведет к гибели молодых деревьев и способствует распространению этого вида (Stinson et al., 2006; Theo-harides, Dukes, 2007).
Виды, расширяющие свои ареалы в Европе на север в связи с потеплением климата, меньше подвержены фитопаразитарным инфекциям и воздействию почвенных фитофагов, поскольку расселение последних отстает от расселения хозяев. Van Grunsven et al. (2007) изучили воздействие голландских почв на 3 пары видов растений, в каждой паре один вид был давно местным, а другой появился недавно из Южной Европы. Нестерилизованные почвы больше подавляли рост местных видов, чем недавних пришельцев. По сравнению со стерильными почвами снижение биомассы местных видов составило 35%, а пришельцев — 13%.
Вселение новых видов (как растений, так и животных) может существенно изменять режим нарушений в сообществах. Так, внедрение некоторых злаков в травяные ценозы увеличивает вероятность пожаров. Завезенные козы на многие острова усиливали роль интродуцированных злаков, в то время как удаление коз вызывало восстановление аборигенной древесной растительности и уменьшение роли заносных видов (Spatz, Mueller-Dombois, 1975; Mueller-Dombois, Spatz, 1975; Mack, D’Antonio, 1998).
Многие виды успешно «возвращаются» в места их доледникового произрастания: например, Rhododendron ponticum — в Англию и Среднюю Европу. Незначительное число видов проникает в сомкнутые леса (Jmpatiens parviflora, Vinca minor) (Lohmeyer, Sukopp, 1992). Среди массовых агриофитов в Северной Америке 52 % видов обладают высокой теневыносливостью (Martin et al., 2009). Не обнаружено агриофитов в ряде болотных и высокогорных сообществ (Lohmeyer, Sukopp, 1992).
Среди заносных видов в Европе во второй половине XX в. появилось значительное число галофитов. Их современное расселение с морских побережий вглубь Британских островов и континентальной Европы (особенно Германии) связано с засолением обочин дорог при их зимней
466
Глава 9. Динамика фитоценозов
обработке солью. К таким видам относятся Suaeda maritima, Spergularia maritime, Plantago maritima, Aster tripolium и др.
Во многих случаях агриофиты не только внедряются в существующие сообщества, но и доминируют в новых фитоценозах. Например, в Средней Европе обычны заросли Helianthus tuberoses по берегам рек, растительные сообщества на скалах с доминированием Isatis tinctoria и Iris germanica, густые заросли Reynoutria japonica, монодоминантные сообщества с доминированием Impatiens glandulifera на участках влажных и богатых почв. Последний вид был занесен из Гималаев, он часто вытесняет околоводные виды за счет эффективного семенного размножения, поскольку шмели предпочитают посещать цветки этого вида, так как они выделяют примерно в 10 раз больше нектара, чем местные виды (Chittka, Schiirkens, 2001; Миркин, Наумова, 2002). Внедряющиеся виды могут значительно уменьшать флористическую насыщенность сообществ за счет конкурентного подавления аборигенов. Такое снижение отмечено, например, при внедрении Reynoutria japonica и Helianthus tuberosus. В то же время при внедрении Solidago gigantea снижения флористической насыщенности не наблюдалось (Lohmeyer, Sukopp, 1992).
Одним из основных факторов успешного внедрения новых видов является «избегание» естественных фитофагов и фитопатогенов, сдерживающих численность этих видов в условиях естественного обитания. Фауна фитофагов на агриофитах существенно обеднена по сравнению с аборигенными видами и зависит от давности вселения агриофитов. Так, например, для агриофитных родов деревьев отмечено небольшое число видов насекомых-фитофагов в Средней Европе: Castanea — И, Aesculus — 9, Juglans — 7, Robinia — 2. Для сравнения на аборигенных видах родов Quercus и Betula известно более чем по 300 видов насекомых-фитофагов. Такие же различия наблюдаются в пределах одного рода: на аборигенной Impatiens noli-tangere известно 19 видов фитофагов, а на заносных /. parviflora и /. glandulifera — 8 и 6 видов соответственно (Lohmeyer, Sukopp, 1992). Количественные оценки повреждаемости фитофагами также различаются для аборигенных и заносных видов. В работе Han et al. (2008) авторы сравнили 12 пар видов (в каждой паре один аборигенный, другой — заносный) в высокотравной прерии Северной Америки по степени повреждения их листьев фитофагами. В целом повреждение фитофагами листьев аборигенных видов было на 83,6 % выше, чем листьев инвазионных видов. Кроме того, выжигание и внесение азотных удобрений резко увеличивают поражаемость аборигенных видов ржавчинными грибами, редко встречающимися на заносных видах.
За исключением нескольких случаев катастрофических заносов грибов, эти организмы слабо исследованы как интродуценты. Видимо, очень
4. Эволюция сообществ и историческая динамика
467
многие виды непаразитных грибов были занесены с почвой и другими материалами в новые области. Иногда фитопатогенные грибы специально завозят для борьбы с заносными растениями, как, например, Phragmid-ium violaceum для борьбы с Rubus fruticosus в Австралии. Считается, что 65-85 % всех фитопатогенных грибов являются интродуцентами (Pimentel et al., 2001). Заносы сапротрофов не менее обычны, но их значение мало исследовано. Примером может служить занос в Европу и Северную Америку красивого макромицета Clathrus archeri. Часто грибы заносятся вместе с растениями-хозяевами, как, например, микоризные грибы эвкалиптов в Испании. Причины быстрого распространения заносных грибов также мало исследованы, они могут быть сходны с таковыми для растений. Например, на Cryphonectria parasitica паразитирует вирус, подавляющий рост гриба, но этот вирус не был занесен в Северную Америку вместе с грибом, что и вызвало катастрофическую гибель каштановых лесов. С другой стороны, наличие этого вируса в Европе не остановило инвазию гриба, но уменьшило его патогенное действие, не вызывающее гибель каштанов (Anagnostakis, 2001; Desprez-Loustau et al., 2007).
4.2 • Историческая динамика в кайнозое
Растительный покров земного шара претерпевал значительные изменения в различные периоды истории Земли. Большое разнообразие цветковых растений, существовавшее к началу кайнозоя, способствовало формированию богатой и разнообразной растительности. Начало кайнозоя (65,5 млн лет назад) связано с падением Юкатанского астероида, имевшего диаметр около 10 км, которое уничтожило около 75 % видов организмов, населявших Землю (Polis, 1999). Кайнозой подразделяется на 2 (3) периода и 7 эпох (табл. 9.6).
Таблица 9.6 • Подразделения кайнозоя
Период Эпоха Время начала, лет назад
четвертичный голоцен 10000
плейстоцен 2 600000
третичный неоген плиоцен 5300000
миоцен 23000000
палеоген олигоцен 33900000
эоцен 55 800000
палеоцен 65 500000
468
Глава 9. Динамика фитоценозов
Рас. 9.57 • Глобусы Земли на границе мелового и третичного периода (а) и в эоцене (50 млн лет назад) (б) (рис. Ron Blakey, Colorado Plateau Geosystems, с любезного разрешения автора)
Палеоцен (рис. 9.57, 9.58)начался с момента падения Юкатанского астероида, а его нижняя граница не столь четко определена. В этот период начинается широкое распространение древних млекопитающих, однако их современные отряды сформируются в эоцене. Климат раннего
4. Эволюция сообществ и историческая динамика
469
Рис. 9.58 • Ландшафты палеоцена: а — тропический дождевой лес на отрогах Скалистых гор в Колорадо через 1,4 млн лет после падения Юкатанского астероида, в подлеске — саговники, а высокие деревья с досковидными корнями имеют крупные листья с острыми верхушками, что свидетельствует о влажном и теплом климате (©Bailey Archives, Denver Museums of Nature & Science, картина Jan Vriesen, с любезного разрешения музея); б — тропический лес в Северной Америке, на переднем плане хищное млекопитающее Chriacus, на дереве — Ptilodus и Peradectes (рис. John Barber из Gould, 2001)
470
Глава 9. Динамика фитоценозов
ID-
S'
6-
о 4'
< 2-
0-
-2-
Ра! | Ео | 01 | Mio
Плиоцен | Плейстоцен
Голоцен
60 50 40 30 20 10 5 4 3 2 1 500 250
10 5 2 1 0,15 0
Миллионы лет назад
Тысячи лет назад
Рис. 9-59 • Изменение температуры на земном шаре в кайнозое (по рис.: Robert A. Rohde, Wikimedia Commons, under GNU Free Documentation License: http://en.wikipedia.org/wiki/Geologic_temperature_tecord)
палеоцена был несколько суше и холоднее, чем в предшествующем меловом периоде, однако в течение всего периода температура повышалась и к его концу достигла палеоцен-эоценового термического максимума (рис. 9.59).
Большую часть третичного периода в целом на территории Европы, как и большей части суши, сохранялся тропический климат. Зональность была выражена в слабой степени, а комплекс арктотретичной флоры в нынешних высоких широтах включал многие рода, современный ареал которых охватывает теплые умеренные и субтропические регионы (Taxodium, Sequoia, Magnolia). Для территории современной Гренландии указывается 282 вида третичной флоры, включая виды родов Ginkgo, Castanea, Platanus (Вальтер, 1982). В начале палеоцена широкое распространение получили разнообразные папоротники, первыми колонизировавшие обширные пространства после падения астероида.
Начало эоцена маркируется появлением первых современных млекопитающих, а его конец — массовым вымиранием многих видов, что многие исследователи связывают с падением крупных метеоритов. Эта эпоха была наиболее теплой в кайнозое, в начале периода наблюдался термический максимум эоцена (рис. 9.59), когда средняя температура на Земле была примерно на 6 °C выше современной. Этот максимум мог быть обусловлен временной замкнутостью Арктического океана (см. рис. 9.61). После этого максимума температура в эоцене медленно снижалась к олигоцену. Кроме экстремально сухих участков суши, ее
4. Эволюция сообществ и историческая динамика
471
Рис- 9-60 • Эоцен Средней Европы: заболоченный лес (Taxodium, Chamaecyparus) с древним тапиром на переднем плане (картина 3. Буриан, см. Аугуста, Буриан, 1961)
Рис. 9.61 • Схематичная карта мира в начале эоцена. Точками показаны области сухого климата, косой штриховкой — муссонные области. Стрелки отмечают направление океанических течений, а цифры — температуру поверхности воды (°C). А — Лабрадорское море, В — Гренландское море, С — Тасманово море, D — перешеек Дрейка (по: Tallis, 1991)
472
Глава 9. Динамика фитоценозов
Рис. 9.62 • Глобусы Земли в олигоцене (а) и миоцене (б) (рис. Ron Blakey, Colorado Plateau Geosystems, с любезного разрешения автора)
бблыиая часть была покрыта лесами. В этот период зональность была выражена в меньшей степени, тропические леса на севере доходили до 45° с. ш., а участки суши около полюсов могли быть покрыты леса
4. Эволюция сообществ и историческая динамика
473
ми умеренного пояса. Полюса были лишены льда, пальмы, крокодилы и другие тропические организмы обитали на территории современной Европы и Северной Америки (Greenwood, Wing, 1995). В конце периода Индия вплотную приблизилась к Азии, Антарктида отсоединилась от Австралии, в Европе началось образование Альп. Для оценки температуры в прошлом широко используется метод анализа краев листьев у древесных двудольных. Процент листьев с ровным (не зубчатым) краем положительно скоррелирован со среднегодовой температурой (Greenwood, 2005).
Олигоцен характеризовался большим снижением температуры и образованием ледников в Антарктиде. В начале периода Европа соединилась с Азией, что вызвало миграцию многих азиатских видов в Европу и вымирание части европейских видов. Похолодание и образование шапок льда вызвало снижение уровня океана. В олигоцене продолжалось интенсивное горообразование в системе Альпийской складчатости (Пиренеи, Альпы, Кавказ, Гиндукуш, Гималаи) (рис. 9.62). Южная Америка отсоединяется от Антарктиды, вокруг которой формируются циркумполярные течения, охлаждающие континент. В связи с похолоданием многие субтропические леса сменяются на листопадные, резко увеличивается площадь открытых сообществ (саванн и степей), что ведет к развитию многих групп крупных травоядных животных (рис. 9.63). Одной из причин снижения продукции растительного покрова и смены лесов травяными сообществами могло быть снижение концентрации СО2 в атмосфере. В эоцене и олигоцене содержание СО 2 постепенно снижалось, но было существенно выше современного (2000-500 ppm). Затем в миоцене оно было постоянно низким, на уровне около 250 ppm. Это могло быть связано со значительным поглощением СО 2 при выветривании алюмосиликатов, выход которых на дневную поверхность в больших масштабах был связан с интенсивным горообразованием на различных континентах в эти эпохи. Снижение концентрации СО 2 до нижнего предела лимитировало продукцию растений, вело к смене лесов с глубоко укореняющимися видами деревьев открытыми травяными сообществами, что в свою очередь снижало интенсивность выветривания минералов (Pagani et al., 2009).
В миоцене в целом продолжилось общее похолодание, хотя в его первой половине наблюдалось некоторое потепление, вызвавшее таяние ледников в Антарктиде. Южная и Северная Америка еще остаются изолированными, продолжаются горообразовательные процессы в Европе, Азии, Северной и Южной Америке (рис. 9.62). Возвышающиеся Гималаи закрывают доступ влажным воздушным массам с Индийского океана, что приводит к усилению аридности климата Центральной Азии. Травяные биомы получают широкое распространение, злаки становятся одним
474
Глава 9. Динамика фитоценозов
4. Эволюция сообществ и историческая динамика
475
Рис. 9.63 • Ландшафты олигоцена (по рис.: Peter Barrett из Palmer, Barrett, 2009): а — прибрежный тропический лес на территории современного Египта (1 — цапля Ardea, 2 — пастушок Sarothrura, 3 — узконосая обезьяна Apidium, 4 — хоботное Phiomia, 5 — обезьяна Aegyptopithecus); б — горный субтропический лес на берегу реки в Колорадо (6 — карликовый опоссум Herpetotherium, 7 — белянка Oligodonta, 8 — сетчатокрылое Marquettia, 9 — дневная бабочка Prodryas, 10 — Mahonia (Berberidaceae), 11 — лошадь Mesohippus, 12 — жук-стафилин Bledius, 13 — Rosa)
Таблица 9.7 • Роды древесных растений, хорошо представленные в плиоценовой флоре севера Центральной Европы, и их современное распространение (по: Sauer, 1988)
Рода, присутствующие в настоящее время во флоре региона
Голосеменные Abies, Larix, Picea, Pinus, Taxus
Покрытосеменные Acer, Alnus, Betula, Carpinus, Cornus, Corylus, Fagus, Fraxinus, Ilex, Populus, Prunus, Quercus, Tilia, Ulmus
Рода, не представленные в настоящее время в регионе, но сохранившиеся в других регионах
Средиземноморье
Голосеменные Cedrus, Tetraclinis
Покрытосеменные Castanea, Celtis, Juglans, Liquidambar, Ostrya, Pla-tanus, Pterocarya, Styrax, Zelkova
Восточная Азия
Голосеменные Cephalotaxus, Chamaecyparis, Cunninghamia, Ginkgo, Glyptostrobus, Keteleeria, Pseudolarix, Sciadopitys, Thuja, Torreya, Tsuga
Покрытосеменные Actinidia, Ailanthus, Castanea, Cinnamomum, Coccu-lus, Corylopsis, Diospyros, Engelhardtia, Eucommia, Juglans, Koelreuteria, Lindera, Liquidambar, Lirioden-dron, Magnolia, Morus, Ostrya, Paulownia, Pterocarya, Sapindus, Styrax, Zelkova
Восточная Северная Америка
Голосеменные Chamaecyparis, Taxodium, Tsuga
Покрытосеменные Asimina, Berchemia, Carya, Castanea, Celtis, Diospyros, Juglans, Lindera, Liquidambar, Liriodendron, Magnolia, Morus, Nyssa, Ostrya, Persea, Platanus, Robinia, Sabal, Sapindus, Sassafras, Styrax
Западная Северная Америка
Голосеменные Chamaecyparis, Sequoia, Torreya, Tsuga
Покрытосеменные Celtis, Juglans, Platanus, Sapindus, Styrax
476
Глава 9. Динамика фитоценозов
4. Эволюция сообществ и историческая динамика 477
Рис. 9.64 • Ландшафты миоцена: а — разреженный лес в Индии (млекопитающие: 1 — Hippopotamodon (семейство свиньи), 2 — хоботное Platybelodon, 3 — древние гиены Percrocuta, 4 — антилопа Protragocerus (по рис.: Peter Barrett из Palmer, Barrett, 2009)); б — субтропический миоценовый лес в Средней Европе (деревья — дубы, буки, клены и другие широколиственные, а также секвойи и пальмы; животные — мастодонты (Trilophodon), олени (Paleomeryx, Dicrocerus), древние фламинго (Paleolodus)) (картина 3. Буриан, см. Аугуста, Буриан, 1961)
из доминирующих семейств во флоре земного шара. Продолжается развитие многих групп фитофагов, особенно копытных (рис. 9.64).
Климат плиоцена приближался к современному (см. рис. 9.59), окончание этого периода связано с началом оледенений на континентальных просторах Северной Америки и Евразии. Важнейшим событием тектоники плиоцена является соединение Северной и Южной Америки, которое привело к взаимной миграции многих видов растений и животных. Флора Европы конца плиоцена близка к современной флоре востока США или Восточной Азии, т. е. значительно богаче видами (табл. 9.7; Sauer, 1988). Тропические леса уменьшают свои площади, широкое распространение получают хвойные бореальные и листопадные леса и травяные биомы (рис. 9.65).
Плейстоцен — эпоха оледенений, которая началась около 2,6 млн лет назад (рис. 9.66). Континентальные оледенения достигали в отдельных местах 40° широты, а перигляциальная мерзлотная зона простиралась еще на сотни километров. Толщина покровных ледников составляла до 3,5 км, они аккумулировали значительное количество воды, и понижение уровня океана доходило до 100 м. Снижение температуры и площади океанов вызывало уменьшение количества осадков, что приводило к увеличению площадей аридных территорий. В плейстоцене выделяется несколько ледниковых эпох, чередующихся с межледниковьями, когда климат приближался к современному. В начале плейстоцена оледенения были более частыми (менее продолжительными), чем в его конце (см. рис. 9.59). Оледенения носят локальные названия для разных местностей, для Восточной Европы последнее оледенение — валдайское (вюрмское — в Альпах, висконсинское — в Северной Америке), а максимальное — днепровское (рисское). В каждое межледниковье в Европе происходила новая миграция видов с юга и востока, но каждый раз флора обеднялась по сравнению с плиоценовой, поскольку между ледником и горами Альпийской складчатости (Альпы, Крым, Кавказ) преобладали безлесные пространства. На юге Северной Америки существовало несколько рефугиумов, где сохранялась флористически богатая лесная растительность. Из этих рефугиумов виды возвращались на более север-
478
Глава 9. Динамика фитоценозов
Рис. 9.65 • Открытый ландшафт плиоцена (Танзания) (животные — индикаторы отрытых пространств: 1 — антилопа дик-дик Madoqua, 2 — цесарка Numida, 3 — африканский слон Loxodonta, 4 — черный носорог Diceros, 5 — жираф Giraffa (рис. Peter Barrett из Palmer, Barrett, 2009))
ные территории в межледниковья. В связи с этим современная флора умеренного пояса Северной Америки значительно богаче видами таковой в Европе (см. табл. 9.7).
Обширные пространства Евразии и Северной Америки были заняты тундростепной растительностью, которая поддерживала своеобразную мамонтовую фауну (крупных травоядных млекопитающих). Наряду с мамонтом, шерстистым носорогом, большерогим оленем и другими вымершими ныне животными эти сообщества населяли такие разобщенные в настоящее время виды, как сайгаки и северные олени. Особенно большие площади тундростепей были представлены на северо-востоке Азии севернее современного полярного круга, значительная часть этих территорий в настоящее время затоплена водами Северного Ледовитого океана. На остатках этой суши (остров Врангеля) небольшая популяция мамонтов дожила до середины голоцена.
4. Эволюция сообществ и историческая динамика
479
Рис. 9.66 • Ландшафты плейстоцена: а — разреженный широколиственный лес на юге Европы (1 — бурый медведь, 2 — олень Eucladoceros giulii, 3 — кабан St/s scrofa, 4 — лисица Vulpes praeglacialis, 5 — лань Dama) (рис. Peter Barrett из Palmer, Barrett, 2009); б — перигляциальный ландшафт в северной Испании (животные: мамонты, шерстистый носорог (Coelodonta antiquitatis), европейский пещерный лев (Panthera leo spelaea)) (по: Sedwick, 2008; картина Mauricio Anton, under Creative Commons Attribution License)
Голоцен представляет собой последнее межледниковье. Найдены свидетельства очень быстрого потепления в раннем дриасе (примерно 11 000 лет назад), которое вызвало таяние ледника в течение нескольких десятилетий. Это, в свою очередь, обусловило очень быструю перестройку состава и структуры растительных сообществ. Возможно, что быстрая
480
Глава 9. Динамика фитоценозов
Рис. 9.67 • Схема миграции видов: 1 — из основного ареала, варианты: а — в форме широкого сплошного фронта, б — в форме узких прорывов, в — путем создания форпостов при дальнем заносе диаспор; 2 — из локальных рефугиумов (по: Pearson, 2006; Зитте и др., 2007)
замена открытых перигляциальных территорий лесами и вызвала массовое вымирание крупных травоядных млекопитающих плейстоцена. Анализируя озерные отложения в различных точках североатлантического региона, Williams et al. (2002) установили быструю (менее 100 лет) смену растительности в раннем дриасе. В другой работе Yu (2003) показал быструю смену (за столетие) еловых лесов сосновыми в этот период, вызванную голоценовым потеплением (Post, 2003). С другой стороны, деятельность первобытных охотников также во многом обусловила вымирание представителей мамонтовой фауны.
Миграция растений на освободившиеся от ледника территории вызывает много дискуссий, так как часто расчеты, проведенные на основании наблюдаемой дальности разноса семян и возраста генеративного состояния, дают значительно заниженные величины по сравнению с историческими данными. В работе McLachlan et al. (2005) на основании молекулярных данных было показано, что скорость миграции двух ви-
4. Эволюция сообществ и историческая динамика
481
дов деревьев (Fagus grandifolia, Acer rubrum) в постледниковый период (менее 100 м в год) была существенно ниже, чем раньше предполагалось на основании пыльцевых данных (100-1000 м в год). Молекулярные данные свидетельствуют о наличии нескольких более близких к леднику рефугиумов, из которых происходила постледниковая миграция (Pearson, 2006) (рис. 9.67). С другой стороны, Clark (1998; Bennett, 1998) объяснял более быстрое распространение деревьев на север важностью редких случаев очень дальнего заноса семян. В настоящее время на моделях, учитывающих турбулентность и восходящие токи воздуха, показана возможность дальнего переноса анемохорных семян в условиях небольшой скорости ветра и нагрева земной поверхности в ясные дни (Tackenberg, 2003). Дальность разноса с учетом этого процесса на 2 порядка больше, чем без его учета. Вероятность захвата семян такими потоками оценивается как 1-5%, что существенно выше, чем предполагалось ранее для дальнего транспорта (Nathan et al., 2002). Значительно медленнее могут расселяться многие лесные мирмекохорные травы. Например, для Asarum canadensis показана дальность разноса семян муравьями до 35 м, что в соответствии с диффузной моделью дает величину 10-11 км за 16000 лет, в то время как реальный последниковый ареал этого вида простирается на многие сотни километров, что свидетельствует о значительно большей роли событий случайного заноса для расселения таких видов в межледниковые периоды (Cain et al., 1998).
Безлесье тундростепей, в свою очередь, было обусловлено влиянием крупных фитофагов мамонтовой фауны — как в настоящее время популяции слонов в Африке уменьшают участие деревьев в саваннах (см. п. 10.1 гл. 4). Современная таежная и тундровая растительность сформировались при значительно более низком прессе крупных фитофагов, чем тундростепи. Zimov et al. (1995) считают, что исчезновение мамонтовой фауны привело к смене степной растительности влажными моховыми тундрами в Сибири и на Аляске (Bardgett, 2005). Реликтовые остатки тундростепной растительности встречаются в Якутии (берега Лены; рис. 9.68), на Чукотке, острове Врангеля и в других районах Северо-Восточной Азии (Юрцев, 1981). В условиях интенсивного выпаса в холодных областях (тундровая зона, высокогорья) формируются сообщества с большим участием злаков и/или осоковых (кобрезии). С этим может быть связано развитие горно-луговых растительных сообществ в гумид-ных высокогорьях Альп и Кавказа, в то время как на Урале, в горах Сибири и Дальнего Востока преобладают горно-тундровые сообщества с высоким участием кустарничков и кустарников, мхов и лишайников. Прекращение выпаса на высокогорных лугах Альп ведет к развитию
482
Глава 9. Динамика фитоценозов
Рис.9.в8 • Степи с доминированием Stipa krylovii в Центральной Якутии (окрестности г. Якутска) (а, б), растения якутских степей: Koeleria cristata (в), Orostachys malocophylla (г), Veronica incana (д)
более мощного кустарникового покрова, усилению роли кустарничков, мхов и лишайников (Spatz et al., 1978).
Наиболее теплым был атлантический период голоцена (5-8 тыс. лет назад), так называемый климатический оптимум голоцена (см. рис. 9.59). В этот период границы распространения многих видов деревьев шли
4. Эволюция сообществ и историческая динамика
483
Рис. 9.69 • Вид на язык ледника Morteratsch в Швейцарских Альпах с позиции его границы в 1970 г. Верхний снимок сделан в 1985 г., нижний — в 2002 г. Видно значительное отступание его в течение последнего столетия и процессы зарастания морены (фото Jurg Alean: SwissEduc, http://www.swisseduc.ch, с любезного разрешения автора)
значительно дальше к северу, чем сейчас. Последние тысячелетия также характеризуются значительными климатическими изменениями. Первые века новой эры были относительно теплыми (например, эпоха великого переселения народов IV-V вв.), а в XIII-XIX вв. наблюдалось похолодание, известное под названием малого ледникового периода. Ледники в Альпах и на Кавказе отступают с середины XIX в., особенно ускорилось их таяние в последние десятилетия (рис. 9.69).
484
Глава 9. Динамика фитоценозов
Рис. 9.70 • Карта распространения памятников ымыяхтахской культуры в Северо-Восточной Азии (по: Федосеева, 1980)
Начиная с плейстоцена человек является важнейшим фактором преобразования природы, в том числе растительного покрова. Большинство современных растительных сообществ испытывали в прошлом и испытывают в настоящее время различные формы прямого или косвенного антропогенного воздействия. Большая часть территории суши была населена людьми в конце плейстоцена и в голоцене. Например, даже наиболее холодные области Северо-Восточной Азии, долины Лены, Яны, Индигирки и Колымы были плотно населены 2-4 тыс. лет назад охотниками и рыболовами ымыяхтахской культуры (рис. 9.70), существенно изменявшими популяции диких животных-фитофагов. Другим примером древнего воздействия человека на существующие ныне сообщества могут служить карбонатные луга (альвары) в Эстонии. Их современное видовое разнообразие положительно скоррелировано с плотностью поселений позднего железного века (800-1000 лет назад) как по общему видовому богатству, так и по флористической насыщенности на 1 м2 (Partel et al., 2007). Использование отдельных видов (например, вяз на веточный корм скоту, липу на лапти) человеком приводило к изменению их участия в составе сообществ. Эти процессы происходили и в недалеком
4. Эволюция сообществ и историческая динамика
485
прошлом: например, даже заповедники в лесной зоне Советского Союза имели большие планы на заготовку бересклета (для нужд шинной промышленности) в конце 40-х гг. прошлого столетия, что резко снизило участие этого кустарника во многих лесных фитоценозах.
Переложное земледелие, длительное время существовавшее на значительной части лесных территорий, вызывало периодическую смену леса и пашни, относительная площадь которых во многом зависела от плотности населения. Например, лесистость Московской области была в несколько раз ниже современной во времена Ивана Грозного, но существенно выше современной после «смутного времени» начала XVII в., когда население центральных областей России резко уменьшилось.
Леса Средней Европы также формировались человеком в зависимости от хозяйственных нужд в ту или иную эпоху (Вальтер, 1982). Их массовая раскорчевка происходит с VIII в. н. э., в XIV-XV вв. человек сажал преимущественно дубовые леса, поскольку желуди являлись основным кормом для широко разводимых свиней. После завоза картофеля с XVI в. роль желудей для откорма свиней падает, наиболее массово разводят бук, из древесины которого получают уголь, в больших количествах необходимый для металлургии. Но с переходом металлургии на каменный уголь преимущество при посадках получают ель и сосна, используемые на деловую древесину и для производства бумаги. Таким образом, структура лесного покрова Европы, как и ряда других регионов, формировалась и формируется преимущественно под воздействием целенаправленной деятельности человека.
4.3 • Современное изменение климата и его влияние на растительный покров
4.3.1 • Баланс углерода и увеличение содержания СО? в атмосфере
В современный период климат Земли претерпевает значительные изменения, которые большинство исследователей связывают с деятельностью человека (рис. 9.71). Увеличение концентрации ряда газов (СО2, СН4 и др.) вызывает «парниковый эффект», при котором более длинноволновая радиация, отраженная от земной поверхности, задерживается атмосферой, содержащей эти газы. Однако повышение концентрации СО2 в атмосфере ведет и ко многим другим последствиям, различным образом отражающимся на растительном покрове.
Концентрация СО2 в атмосфере в 1991 г. составляла 355 /гл/л, или ppm — частей на миллион, в 2012 г. она достигла 395 ppm (данные обсерватории Mauna Loa Observatory, Hawaii), что примерно на 41 % выше,
486 Глава 9. Динамика фитоценозов
СО2
370 г I
Е ./
§ 330 - Jfflr
!320
О 310 1 ‘ * 1
° 1960 1970 1980 1990
МШИ
И Температура, Т
1900 1930 1960 1990
Озон
со _ И Пыль s 201-----------
1926 1940 1960 1980 1997
5 Q L---1--1---1--1--1--L
° 1965 1975 1985 1995
Нитраты
Рис. 9.71 • Атмосферные компоненты глобального изменения климата в конце XX в.: а — концентрация С02 (по: Keeling, 1993); б — изменение температуры с сильными региональными отклонениями, показанными на примере Швейцарских Альп (по: Beniston et al., 1997); в — содержание озона на высоте более 1900 м в Центральной Европе (единицы Добсона, 1 еД = 100 hPa cm) (по: Staehelin et al., 1998); г — содержание пыли в воздухе у поверхности Земли (по: Moulin et al., 1997); д — поступление нитратов с осадками на поверхность ледников (по: Doscher et al., 1995); е — выбросы N20 (по: Mosier, Kroeze, 1998; Korner, 2000)
1960 1970 1980 1990
4. Эволюция сообществ и историческая динамика
487
чем в доиндустриальный период (280 ppm — 1750-1800 гг.). Она возрастает ежегодно на 1,8 /хл/л, или на 0,5%. К 2050 г. предполагаемая концентрация достигнет 530-600 /хл/л (Campbell, Grime, 1993). Содержание углерода в атмосфере сопоставимо с таковым в растительном мире и соответствует для поверхности суши слою 1,3 мм чистого углерода. Надземный растительный углерод обновляется примерно за 14-19 лет, а углерод биомассы океана — за 2-6 дней. Ежегодное антропогенное сжигание ископаемого и древесного топлива составляет около 7 млрд т (около 1 % атмосферного запаса С) и превышает 10% ежегодного естественного обмена между атмосферой и биосферой (Кбгпег, 2000). Большая часть углерода биомассы растений (600 млрд т) сосредоточена в лесах и другой древесной растительности, примерно половина которой находится в тропиках и субтропиках. В сельскохозяйственных культурах сосредоточено около 2 %, в океанических организмах — 0,2 %. Океанические сообщества малочувствительны к увеличению СОг, но изменение температуры может вызвать огромные изменения в циркуляции вод мирового океана (Кбгпег, 2000).
В истории Земли концентрация СО2 существенно менялась от 6000 //л/л в палеозое (500 млн лет назад) до 200 /хл/л в позднем плейстоцене (15 тыс. лет назад) (Вегпег, 1997; Poorter, Navas, 2003).
Проведено значительное число лабораторных и природных экспериментов по изучению влияния повышения уровня СОг на растения и фитоценозы. На уровне отдельных растений показаны следующие эффекты (Campbell, Grime, 1993; Poorter, Navas, 2003):
1) возрастание интенсивности фотосинтеза и уменьшение фотодыхания; быстрорастущие травянистые СЗ виды реагируют сильнее, чем медленнорастущие СЗ травы или С4 растения, САМ и древесные растения показали умеренную реакцию;
2) уменьшение проводимости устьиц;
3) возрастание эффективности использования азота;
4) возрастание листовой поверхности и скорости развертывания листьев, при этом удельная листовая поверхность может снижаться (листья становятся более толстыми);
5) задержка отмирания листьев;
6) усиление симбиотической азотфиксации;
7) уменьшение доли листьев и увеличение доли корней в общей биомассе;
8) увеличение концентрации неструктурных углеводов в листьях, но уменьшение — азота и белков.
488
Глава 9. Динамика фитоценозов
Микоризные и безмикоризные растения могут по-разному реагировать на увеличение СО2. В опытах (Diaz et al., 1993) было показано, что при выращивании участков дернины богатого луга в условиях удвоения концентрации СО2 безмикоризные Rumex obtusifolius и Cardamine flexuosa уменьшали рост побегов и демонстрировали симптомы азотного голодания. Их фотосинтез был более интенсивным, но большая часть ассимилятов выделялась через корни и поступала к несимбиотическим почвенным микроорганизмам, которые при этом сильно иммобилизировали ЭМП из почвы, вызывая их недостаток у растений. В то же время у микоризного растения бедных почв — Calluna vulgaris — таких процессов не отмечено, он несколько увеличил свою биомассу в сравнении с контролем.
К. Thompson (1994) показал, что в условиях усиления различных форм воздействия человека на ландшафты (на примере Англии и Нидерландов) в последние десятилетия виды S-стратегов (по Грайму) сокращают свои ареалы, уменьшается площадь сообществ с их участием, в то время как виды, расширяющие свои ареалы, в том числе большинство заносных, по системе Грайма относятся к С- и R-стратегам. Усиление роли видов С- и R-стратегий связано как с эвтрофикацией и увеличением доли нарушенных земель, так и с их положительной реакцией на потепление и увеличение СО2 в атмосфере, что было показано в работе R. Hunt et al. (1993). Виды конкурентной стратегии в наибольшей степени увеличивали свою биомассу при повышении концентрации СО2 (рис. 9.72).
На уровне отдельных сообществ повышение содержания СО2 часто ведет к увеличению общей продукции. Например, луговые сообщества в ответ на повышение СО2 в среднем увеличат продукцию на 16% (разброс от -20 % до +90 %); если исключить удобряемые луга, то разброс между 0 и 10% (Когпег, 2000).
Между повышением содержания СО2 и повышением температуры наблюдается также непрямая положительная связь. Повышение СО2 ведет к большей эффективности использования воды, меньшей открытости устьиц, что в свою очередь уменьшает транспирацию и транспирационное охлаждение поверхности листьев и, как следствие, вызывает еще большее повышение температуры (Press, 1999). Например, увеличение биомассы луговых растений при увеличении СО2 было обусловлено увеличением влажности почвы, связанным с более экономным использованием воды растениями (уменьшением транспирационных потерь). В экспериментах с Carex flacca и Bromus erectus чем ниже была влажность почвы, тем сильнее увеличивалась биомасса растений при увеличении концентрации СО2. Во всех вариантах при повышенном СО2 влажность почвы была выше, чем в контроле (Volk et al., 2000).
4. Эволюция сообществ и историческая динамика
489
Рис. 9.72 • Треугольник стратегий Ф. Грайма (см. п. 3.7.2 гл. 5), показывающий ожидаемое потенциальное увеличение в биомассе изолированных, хорошо развивающихся растений в возрасте около 50 дней при повышенной концентрации С02 (540 /хл/л) (по: Hunt et al., 1993)
Таким образом, увеличение СО2, с одной стороны, вызывает увеличение продукции (отрицательная обратная связь с СО2 в атмосфере), а с другой — увеличение температуры и усиление микробиологической деструкции органики (положительная обратная связь с СО2 в атмосфере) (Pendall et al., 2004).
Изменения площади природных зон и отдельных элементов растительного покрова приводят к различным изменениям в атмосферных процессах. Потепление в Арктике ведет к цепочке процессов с положительной обратной связью: меньше льда в океане и больше кустарниковой и древесной растительности на суше — ниже альбедо (доля отраженной радиации) и больше нагрев. Бореальные леса увеличивали площади до 1990 г., а затем снизили в связи с рядом факторов (усиление засухи, вспышки численности насекомых-фитофагов, большая частота пожаров и др.) (Goetz et al., 2005). Напротив, в Сахели (область к югу от Сахары) опустынивание ведет к увеличению альбедо и уменьшению эвапотранс-пирации, что снижает поступление конвекционного тепла в атмосферу,
490
Глава 9. Динамика фитоценозов
уменьшает интенсивность восходящих атмосферных процессов и снижает муссонное поступление осадков (Chapin et al., 2008).
Увеличение содержания СО2 в атмосфере может приводить и к различным изменениям в животном населении, непрямо воздействующим на фитоценозы. Так, в экспериментах С. Korner (см. Thebaud, Johnston, 1997) в ответ на увеличение СО2 в атмосфере было показано уменьшение повреждаемости листьев насекомыми-фитофагами и резкое увеличение активности дождевых червей. При увеличении СО2 вдвое на карбонатных лугах продукция копролитов дождевых червей увеличилась с 16 до 22 кг/га в год (Zaller, Arnone, 1997).
4.3.2 • Увеличение температуры
В течение большей части последнего тысячелетия температура на поверхности Земли медленно понижалась, однако с середины XIX в. наблюдается ее резкое увеличение (рис. 9.73; Hughes, 2000)
Увеличение температуры оказывает прямое и косвенное влияние на продукцию растительных сообществ и изменяет запас углерода в различных компонентах экосистем. Усиление фотосинтеза сопряжено с усилением дыхания, итоговое влияние этих процессов на продукцию может быть различным. Фотосинтез растений при повышении температуры может увеличиваться, уменьшаться или не изменяться. Непрямое действие повышения температуры (рис. 9.74) может быть более важным, чем прямое. Механизмы отклика сообществ на потепление клима-
1.0
1000 1200 1400 1600 1800 2000
Годы
Рис. 9.73 • Изменение температуры воздуха в Северном полушарии за последние 1000 лет. Белой линией отмечен тренд температуры за период 1000-1850 гг. (по: Hughes, 2000)
4. Эволюция сообществ и историческая динамика
491
Рис. 9.74 • Основные факторы, влияющие на изменение первичной нетто-продукции (NPP) и запасы углерода в связи с глобальным потеплением в наземных экосистемах (Tjoelker, Zhou, 2007)
та весьма разнообразны, включая увеличение вегетационного периода, увеличение доступности ЭМП, изменение видового состава и водного режима сообществ. Рост травянистых растений в тундре более отзывчив на увеличение температуры, чем древесных. В высокотравной прерии продукция увеличилась на 25 % при увеличении температуры (Luo et al., 2009). Усредненная величина для 20 полевых экспериментов — увеличение надземной продукции на 19% (Rustad et al., 2001). В других случаях возможно снижение продукции при повышении температуры. В кратковременных экспериментах повышение температуры обычно сильнее стимулирует дыхание, чем фотосинтез, увеличивая отношение дыхание фотосинтез. Однако растения способны настраивать интенсивность дыхания на температуру, поэтому долговременные эксперименты показывают относительную стабильность этого отношения (Gifford, 1994, 1995). Дыхание почвы в большинстве случаев увеличивается при увеличении температуры. В эксперименте на болотах северного Висконсина увеличение температуры увеличивало роль кустарников по отношению к граминои-дам (осоки, злаки и ситниковые) и граминоидов по отношению к видам
492
Глава 9. Динамика фитоценозов
разнотравья (Weltzin et al., 2000). В высокогорьях увеличение роли кустарников и уменьшение роли трав при повышении температуры ведет к снижению поступления опада в почву и снижению запаса почвенной органики на альпийских лугах (Saleska et al., 2002). Метаанализ экспериментов в 12 экосистемах показал среднее увеличение скорости минерализации азота в почвах на 46 % при повышении температуры (Rustad et al., 2001; Luo, 2007).
Виды с ограниченным распространением, особенно полярные и горные виды, сокращают свой ареал в связи с глобальным потеплением, риск их вымирания наиболее высок. На основании анализа большого числа долговременных рядов наблюдений Parmesan (2006) показал, что 59 % видов из исследованных 1598 изменили свою фенологию и/или ареалы за последние 20-140 лет. Расширение ареалов на север идет со средней скоростью 6,1 км за десятилетие, в горах подъем составляет 6,1 м за десятилетие (Parmesan, Yohe, 2003). Отмечается удлинение вегетационного периода в высоких широтах Северного полушария на 1,1-4,9 дней за каждые 10 лет с 1951 г. (Menzel et al., 2003). Однако реакции различных видов на потепление не синхронны, что ведет к десинхронизации многих процессов в сообществе (Visser, Both, 2005). Пингвины исчезают из своих наиболее северных местообитаний, в то время как антарктические растения Deschampsia antarctica и Colobanthus quitensis расширяют свой ареал и увеличивают покрытие (Smith, 1994). С 1979 по 2001 г. в Нидерландах отмечено 77 новых видов эпифитных лишайников, имеющих более южное распространение (van Herk et al., 2002; Parmesan, 2006).
4.3.3 • Поступление окислов азота и эвтрофикация
Увеличение СО2 в атмосфере часто сопряжено с увеличением выбросов окислов серы и азота, вызывающих эвтрофикацию. Например, поступление минеральных форм азота с осадками на Ротамстедскую станцию в Южной Англии за период 1888-1913 гг. составило 4,1 -5,0 кг/га, а за период 1969-1978 гг. — 15,1-18,2 кг/га азота в год (Tamm, 1991).
Усилившееся поступление связанного азота в экосистемы вызывает эффекты, сходные с внесением азотных удобрений (см. п. 3.9.4 гл. 9). Например, в составе пустошей в Нидерландах уменьшается участие вереска в связи с его большей подверженностью фитофагам и меньшей морозостойкостью при обогащении почвы азотом, и увеличивается участие злаков (Deschampsia flexuosa, Molinia caerulea). Совместное воздействие эвтрофикации и повышения концентрации СО2 обычно увеличивает первичную продукцию растительных сообществ, а в болотных сообществах эти процессы могут приводить к увеличению эмиссии метана в атмосферу. Например, в экспериментах Hutchin et al. (1995; Lee, 1998), увеличе
4. Эволюция сообществ и историческая динамика
493
ние содержания СОг до 550 /хл/л ведет к удвоению выделения метана, видимо, в связи с увеличением поступления ассимилятов в ризосферу Eriophorum vaginatum и усилением активности метаногенных микроорганизмов в ней.
4.3.4 • Влияние увеличения поступления УФ-радиации
Наибольшее влияние на организмы имеет УФ-В-радиация (длина волны 280-320 нм), которая составляет примерно 0,5 % солнечной энергии, поступающей на поверхность Земли. В связи с разрушением озонового слоя наибольшее относительное увеличение поступления УФ-В-радиации наблюдается в высоких широтах, а по времени года — зимой и ранней весной (Caldwell et al., 1999).
Воздействие УФ на наземные растения — это не только стресс, но и фактор, влияющий на морфогенез, продукцию вторичных метаболитов, структуру растительности. Многие исследователи полагают, что развитие метаболизма полифенольных соединений тесно связано с воздействием УФ. У отдельных растений увеличение поступления УФ вызывает следующие изменения (Rozema et al., 1991):
1) уменьшение высоты, размеров, длины листьев;
2) увеличение толщины листа, интенсивность ветвления;
3) изменение угла отхождения листа;
4) изменение ритмики развития растений и ускорение процессов старения.
Растения различаются по устойчивости к воздействию УФ-В. Брио-фиты затененных мест значительно чувствительнее деревьев (в связи с высокой концентрацией полифенольных соединений у последних). Ксерофиты более устойчивы, чем мезофиты, поскольку имеют более толстые покровные ткани. Так, у кактуса Cereus giganteus толщина эпидермиса и кутикулы достигает 1 мм и пропускает лишь 0,1 % поступающего УФ-В-излучения, в то время как пропускание ФАР составляет 60-80 % (Gwynn-Jones et al., 1999). Тропические растения более устойчивы, чем растения умеренных областей, в связи с исходно более высоким поступлением УФ-В-радиации в тропиках (Sullivan, Rozema, 1999).
Увеличение поступления УФ-В вызывает изменение химического состава растений (синтез защитных соединений) и приводит к изменению отношений растений со своими симбионтами. У бобовых существенно снижается нитрогеназная активность, количество и диаметр клубеньков, а у растений с арбускулярной микоризой уменьшается число арбускул, что снижает эффективность симбиоза (van de Staaij et al., 1999). УФ-В-радиация резко снижает азотфиксацию эпифитных цианей на поверхности
494
Глава 9. Динамика фитоценозов
мохообразных в Арктике, но не в Субарктике. Она подавляет азотфикса-цию цианей в цефалодиях Peltigera aphthosa, но не внутренних циано-бионтов у Peltigera didactyla (Solheim et al., 2006). Увеличение синтеза защитных фенольных соединений в большинстве случаев снижает поражаемость растений фитофагами (Caldwell et al., 1999).
УФ-излучение может увеличивать скорость разложения опада за счет его фотодеградации. С другой стороны, УФ-излучение подавляет развитие поверхностной микрофлоры и увеличивает концентрацию фенольных соединений в листьях до их отмирания, что может снижать скорость разложения опада (Paul et al., 1999). При разложении опада под действием УФ-В-радиации усиливается эмиссия СО, особенно заметная в индустриально не загрязненных областях (саванны Африки) (Caldwell et al., 1999).
Увеличение содержания в атмосфере СО2 и усиление УФ-излучения часто оказывают противоположное действие на растения. Однако в целом увеличение УФ-излучения вряд ли окажет существенное влияние на продукцию и структуру наземных фитоценозов (Rozema et al., 1991).
4.3.5 • Повышение концентрации озона в приземных слоях атмосферы
При антропогенном загрязнении атмосферы, с одной стороны, происходит разрушение озонового слоя, а с другой — концентрации озона в нижних частях атмосферы и высокогорьях могут возрастать.
Виды растений сильно отличаются друг от друга по устойчивости к озону. Под действием озона обычно сильнее уменьшается продукция семян и репродуктивное усилие, чем страдает вегетативная сфера. Толерантность к озону связана с устьичной проводимостью обратной связью. Растения богатых почв более устойчивы, чем растения бедных. Повышенное содержание озона может изменять конкурентные отношения между растениями, например, сильнее угнетая клевер, чем злаки в смешанных посевах. В Калифорнии было показано, что участие клевера отрицательно скоррелировано со средней концентрацией оксидантов в воздухе (Westman, 1979, 1985; Davison, Barnes, 1998).
Список литературы
Абатуров Б. Д. Млекопитающие как компонент экосистем (на примере растительноядных млекопитающих в полупустыне). М.: Наука, 1984. 286 с.
Абдуллин М. Р., Миркин Б. М. Опыт создания «агростепей» в башкирском степном Зауралье // Бюлл. МОИП. Отд. биол. 1995. Т. 100. №5. С. 77-84.
Абросов Н. С. Факторы поддержания видового разнообразия экосистем // Биофизические методы исследования экосистем. Новосибирск: Наука, 1984. С. 6-24.
Абросов Н.С., Ковров Б. Г., Черепанов О. А. Экологические механизмы сосуществования и видовой регуляции. Новосибирск: Наука, 1982. 302 с.
Алексахина Т. И. Водоросли в светло-каштановой почве под лесными полосами // Повышение устойчивости защитных насаждений в полупустыне. М.: Наука, 1981, с. 172-178. Алехин В. В. Основные понятия и основные единицы в фитоценологии // Советская ботаника, 1935. №5. С. 21-34.
Андреев В.Н. Кормовая база ямальского оленеводства // Советское оленеводство, 1934. Вып. 1, с. 99-164.
Аугуста Й., Буриан 3. По путям развития жизни. Прага: Артия, 1961. 175 с.
Ахметжанова А. А., Онипченко В. Г. Реакция растений альпийского гераниево-копеечни-кового луга на увеличение доступности почвенных ресурсов: оценка изменения биомассы // Бюлл. Московск. о-ва испытателей природы, отд. биол. 2005. Т. 110. Вып. 1. С. 52-59.
Базилевич Н. И. Биологическая продуктивность экосистем Северной Евразии. М.: Наука, 1993. 293 с.
Базилевич Н. И., Титлянова А. А. Биотический круговорот на пяти континентах: Азот и зольные элементы в природных наземных экосистемах. Новосибирск: Издательство СО РАН, 2008. 382 с.
Базилевич Н. И., Титлянова А. А., Смирнов В. В., Родин Л. Е., Нечаева Н. Г, Левин Ф. И. Методы изучения биологического круговорота в различных природных зонах. М.: Мысль, 1978. 184 с.
Базова Г А. Почвенные водоросли высокогорий Памира. Душанбе: Дониш, 1978. 171 с.
Байкалова А. С., Онипченко В. Г. Микосимбиотрофизм альпийских растений Тебердинского заповедника // Опыт исследования растительных сообществ в заповедниках (Тр. ЦНИЛ Главохоты РСФСР). М., 1988. С. 93-107.
Батчаева О. М., Захаров А. А., Онипченко ВГ Зависимость распространения растений от глубины почвы в альпийских фитоценозах Тебердинского заповедника // Бюлл. Моск, о-ва испыт. природы, отд. биол. 2003. Т. 108. №4. С. 53-57.
Белая Г. А., Морозов В. Л. Дальневосточное крупнотравье как особый тип растительности // Ботанический журнал. 2000. Т. 85. № 10. С. 75-81.
Биокомплексные исследования в Казахстане. Ч. 3. Комплексная характеристика основных растительных сообществ пустынных степей Центрального Казахстана. Л.: Наука, 1976. 292 с.
496
Список литературы
Богатырев Л. Г., Телеснина В. М. Словарь терминов и показателей, используемых при изучении биологического круговорота. М.: МАКС Пресс, 2010. 184 с.
Боголюбов А. Г. О принципе конкурентного исключения и механизмах поддержания видового разнообразия сообществ // Биологические науки. 1989. №11. С. 5-18.
Борисова И. В. Сезонная динамика растительного сообщества // Полевая геоботаника. Л.: Наука, 1972. Т. 4. С. 5-94.
Брызгалова В. А. Влияние ржавчинного грибка Puccinia suaveolens (Pers.) Rostr. на развитие сорняка Cirsium arvense 11 Болезни растений, 1928. T. 17. №3-4. С. 101-118.
Вальтер Г. Общая геоботаника. М.: Мир, 1982. 263 с.
Вальтер Г. Растительность земного шара. Эколого-физиологическая характеристика. Т. 1. Тропические и субтропические зоны. М.: Прогресс, 1968. 552 с.
Вершинин И. Г. Заливные луга р. Енисея в Игарском районе и приемы их использования // Тр. НИИСХ Крайнего Севера. Норильск, 1961. Т. 10. С. 251-298.
Воронина И. Н., Онипченко В. Г., Игнатьева О. В. Компоненты биологического круговорота на альпийских лишайниковых пустошах северо-западного Кавказа // Почвоведение, 1986. №1. С. 29-37.
Гиляров А. М. Современное состояние концепции экологической ниши // Успехи современной биологии, 1978. V. 85. №3. Р. 431-446.
Голубев В. Н. Рецензия: Т. А. Работнов «Фитоценология» // Биологические науки. 1980. №6. С. 108-112.
Горы шина Т. К. Ранневесенние эфемероиды лесостепных дубрав. Л.: Изд-во ЛГУ, 1969. 232 с.
Горышина Т.К. Сезонная динамика фотосинтеза и продуктивности некоторых летневегети-рующих травянистых растений лесостепной дубравы // Ботанический журнал. 1971. Т. 56. №1. С. 62-75.
Горышина Т.К. Экология растений. М.: Высшая школа, 1979. 368 с.
Горышина Т.К., Митина М.Б. Эколого-физиологические особенности весенних и летних листьев сныти Aegopodium podagraria L. и их роль в продуктивности травостоя лесостепной дубравы // Вести. Ленингр. ун-та, сер. Биология, 1970. № 2, С. 71-77.
Гришина Л. А. Биологический круговорот и его роль в почвообразовании. М.: Изд-во Моск, ун-та, 1974-128 с.
Гузев В. С., Куличевская И. С., Звягинцев Д. С. Сканирующая электронная микроскопия при изучении взаимодействия микроорганизмов с растениями // Микроорганизмы как компонент биогеоценоза. Е.Н. Мишустин (отв. ред.). М.: Наука, 1984. С. 92-107.
Дзыбов Д. С. Агростепи. Ставрополь: Аргус, 2010. 256 с.
Дмитриев Е.А., Самсонова В. П. О пространственном варьировании содержания обменного кальция в дерново-подзолистой почве под ельником // Биологические науки. 1975; No И. С.112-118.
Дмитриев Е.А., Самсонова В.П. Квазипериодичность в изменении некоторых свойств дерново-подзолистой почвы под ельником // Биологические науки. 1979, No 4. С. 92-97.
Дмитриев Е.А., Самсонова В. П. Пространственная изменчивость некоторых свойств в профиле дерново-подзолистой почвы под лесом // Структура почвенного покрова и использование почвенных ресурсов (ред. Фридланд В. М.). М.: Наука, 1978. С. 61-65.
Добринский Л. Н., Давыдов В. А., Кряжимский Ф. В., Малафеев Ю. М. Функциональные связи мелких млекопитающих с растительностью в луговых биогеоценозах. М.: Наука, 1983. 161 с.
Душков В. Ю. О связи усыхания вязовых насаждений с азотным голоданием // Почвоведение, 1980. №3, С. 74-82.
Евстигнеев О. И. Отношение лиственных деревьев к свету // Биологические науки. 1991. Т. 8. С. 20-29.
Жук А. В. Морфогенез и происхождение гаусторий у видов рода Cuscuta (Cuscutaceae)// Ботанический журнал. 1997. Т. 86. №5. С. 1 — 15.
Список литературы
497
Жукова Л. А. Поливариантность онтогенеза луговых растений // Жизненные формы в экологии и систематике растений. М.: МГПИ им. В. И. Ленина, 1986. С. 104-114.
Журавлева Н.А. Физиология травянистого сообщества: принципы конкуренции. Новосибирск: Наука, 1994. 168 с.
Звягинцев Д. С., Добровольская Т Г, Лысак Л. В. Растения как центры формирования бактериальных сообществ // Журнал общей биологии. 1993. Т. 54. №2. С. 183-200.
Зитте П., Вайлер Э. В., Кадерайт Й. В., Брезински А., Кернер К. Ботаника. Т. 4. Экология. (На основе учебника Э. Страсбургера, 35 издание). М.: Академия, 2007. 249 с.
Ипатов В. С., Тархова Т. Н. Взаимовлияние моховых и лишайниковых синузий в зеленомошно-лишайниковых сосняках // Экология. 1983. № 1, С. 20-26.
Калакуцкий Л. В., Добрица С. В., Парийская А. Н. Актинориза и ее роль в биологической азотфиксации // Тез. докл. 5 Всес. биохим. съезда. М., 1985. Т. 1. С. 99-100.
Камышев Н. С. К классификации антропохоров // Ботанический журнал. 1959. Т. 44. №11. С. 1613-1616.
Каратыгин И. В. Грибные организмы и их роль в эволюции экосистем // Ботанический журнал. 1994. Т. 79. № 2. С. 13-20.
Каратыгин И. В. Коэволюция грибов и растений. Санкт-Петербург: Гидрометеоиздат, 1993. 118 с. (Труды БИН РАН. вып.9).
Карпов В. Г. Экспериментальная фитоценология темнохвойной тайги. Ленинград: Наука, 1969. 336 с.
Кожевников А. В. О перезимовке и ритме развития травянистых растений широколиственных лесов // Бюлл. Моск, о-ва испытателей природы. Отд. биол. 1931. Т. 40. № 1-2. С. 73-108.
Колчинский Э.И. Эволюция биосферы. Л.: Наука, 1990. 235 с.
Коротков В. Н. Новая парадигма в лесной экологии // Биологические науки. 1991. № 8. С. 7-20.
Котт С. А. Сорные растения и борьба с ними М.: Сельхозгиз, 1948. 261 с.
Криволуцкий Д.А., Покаржевский А.Д. Роль наземных животных в биогенной миграции элементов. Животные в биогеоценозах // Чтения памяти академика В. Н. Сукачева, в. 14. Шилов И. А. (ред.). М.: б. и., 1996. С. 34-100.
Кузнецов А. Н. Тропический диптерокарповый лес. М.: ГЕОС, 2003. 140 с.
Кузяхметов Г. Г. Водоросли зональных почв степи и лесостепи // Почвоведение. 1991. №9, С. 63-71.
Куренцова Г. Е. Некоторые данные к биологии и культуре жень-шеня (Panax ginseng С. А. Меу.) // Советская ботаника, 1944. № 1. С. 47-50.
Кучерук В. В. Типы убежищ млекопитающих и их распространение по природным зонам внетропической Евразии // Вопросы географии. М., 1960. Вып. 48. С. 121-134.
Лапшина Е. И. О перезимовывании высших растений по наблюдениям в окрестностях Петергофа //Тр. Петергоф, естественно-научного ин-та, 1928. Вып. II 5. С. 36-98.
Ларин И. В., Агабабян Ш. М., Работнов Т А. и др. Кормовые растения сенокосов и пастбищ СССР. М.-Л.: Сельхозгиз, 1950-1956. Т. 1-3, 687,947,879 с.
Лархер В. Экология растений. М.: Мир, 1978. 384 с.
Лейнсоо Т.А., Онипченко В. Г., Штина Э.А. Почвенные и эпифитные водоросли // Биогеоценозы альпийских пустошей (на примере северо-западного Кавказа). Работнов Т.А. (ред.). М.: Наука, 1987. С. 31-35.
Любарский Е. Л., Полуянова В. И. Структура ценопопуляций вегетативно-подвижных растений. Казань: Изд-во Казан, университета, 1984. 139 с.
Мазинг В. В. Консорции как элементы функциональной структуры биоценозов // Труды МОИП. 1966. Т. 27. С. 117-127.
Мазинг В. В. Что такое структура биогеоценоза // Проблемы биогеоценологии. Т. А. Работнов, Е. М.Лавренко (ред.). М.: Наука, 1973. С. 148-156.
498
Список литературы
Макаров М. И., Онипченко В. Г, Малышева Т. И., Булатникова И. В., Вертелина О. С. Биологический круговорот и роль растений в формировании свойств почв альпийских экосистем Северо-Западного Кавказа // Вестник Моск, ун-та, Сер 17 Почвоведение, 1999. №3. С. 29-37.
Манаков К.Н. Элементы биологического круговорота на Полярном Севере. Л.: Наука, 1970. 160 с.
Марков М. В. Популяционная биология растений. Казань: Изд-во Казанского университета, 1986. 111 с.
Мина В. Н. Выщелачивание некоторых веществ атмосферными осадками из древесных растений и его значение в биологическом круговороте // Почвоведение, 1970. №6. С. 7-17.
Миркин Б. М. Антропогенная динамика растительности // Итоги науки и техники (ВИНИТИ). Ботаника, 1984. Т. 5. С. 139-236.
Миркин Б.М. Теоретические основы современной фитоценологии. М.: Наука, 1985. 137 с.
Миркин Б. М., Наумова Л. Г. Адвентивизация растительности в призме идей современной экологии // Журнал общей биологии. 2002. Т. 63. №6. С. 500-508.
Миркин Б.М., Наумова Л. Г. Наука о растительности. Уфа: Гилем, 1998. 413 с.
Михневский В. К. Роль азота и азотфиксирующих культур в повышении плодородия дерново-подзолистых почв // В кн.: Окультуривание почв в нечерноземной зоне. М.: Московский рабочий, 1970. С. 132-140.
Молчанов А. А. Лес и окружающая среда. М.: Наука, 1968. 247 с.
Морозов В. Л. Феномен природы крупнотравье. М.: Наука, 1994. 229 с.
Морозов В. Л., Белая Г. А. Экология дальневосточного крупнотравья. М.: Наука, 1988. 255 с.
Нахуцришвили Г. Ш. Об экологии высокогорных растений в зимнее время // Известия АН ГССР, сер.биол., 1975. Т. 1. №3. С. 260-264.
Нахуцришвили Г. Ш. Экология высокогорных растений и фитоценозов Центрального Кавказа. Ритмика развития, фотосинтез, экобиоморфы. Тбилиси: Мецниереба, 1974. 194 с. Некрасова К. А. Трофические связи почвенных животных и водорослей (на примере коллем-бол) // Почвенная фауна и биологическая активность осушенных и рекультивируемых торфяников. М.: Наука, 1980. С. 160-166.
Новиков ТА. Теоретические основы и методы изучения питания и трофических связей млекопитающих и птиц // Труды Петергофского биологического института, 1976. Вып. 24. С. 45-65.
Новичкова-Иванова Л. М. Альгосинузии муравейников как компонент зоогенной мозаичности травяного покрова фисташников // Ботанический журнал. 1991. Т. 76. № И. С. 1504-1512.
Новичкова-Иванова Л. Н. Пустынные водоросли фитоценозов Сахаро-Гобийской пустынной области. Л.: Наука, 1980. 255 с.
Онипченко В. Г. Механизмы обособления экологических ниш у наземных растений // Журнал общей биологии. 1987. Т. 48. №5. С. 687-695.
Онипченко В. Г. Микосимбиотрофизм и вегетативная подвижность растений альпийских сообществ северо-западного Кавказа // Вестник Моск, ун-та, сер. 16 Биология, 1996. №1. С. 43-46.
Онипченко В. Г. Роль почвы в формировании и сохранении разнообразия растений // Роль почвы в формировании и сохранении биологического разнообразия / Отв. ред. Г. В. Добровольский, И. Ю. Чернов. М.: Товарищество научных изданий КМК, 2011. С. 86-155.
Онипченко В. Г. Сезонная динамика фитоценоза альпийской пустоши на Северном Кавказе // Бюлл. Моск, о-ва испытателей природы, отд. биол. 1983. Т. 88. №5. С. 106-114.
Онипченко В. Г. Структура, фитомасса и продуктивность альпийских лишайниковых пустошей // Бюлл. Моск, о-ва испытателей природы, отд. биол. 1985. Т. 90. № 1. С. 59-66.
Список литературы
499
Онипченко В. Г., Вертелина О. С., Макаров М. И. Пространственная гетерогенность высокогорных фитоценозов и свойств почвы // Почвоведение, 1998. №6. С. 689-695.
Онипченко В. Г., Комаров А. С. Длительность жизни и динамика популяций растений в высокогорьях: опыт оценки на примере трех альпийских видов северо-западного Кавказа // Журнал общей биологии. 1997. Т. 58. №6. С. 64-75.
Онипченко В. Г., Павлов В. Н. Флористическая насыщенность альпийских сообществ зависит от занимаемой ими площади //Доклады Академии наук, 2009. Т. 427, п 5. С. 710-712.
Онипченко В. Г., Покаржевская Г. А. Изучение пространственной структуры альпийских сообществ с позиций теории «масс-эффекта» // Бюлл. Моск.о-ва испытателей природы, отд. биол. 1994. Т. 99. №3. С. 58-64.
Павлов В. Н. История кафедры геоботаники // Кафедра геоботаники Московского университета: 75 лет со дня основания / Под ред. В. Н. Павлова, И. А. Губанова и С. А. Баландина. М., 2004. С. 5-32.
Павлов В.Н., Онипченко В. Г., Аксенова А. А., Волкова Е.В., Зуева О. И., Макаров М.И. Экспериментальное изучение организации альпийских фитоценозов северо-западного Кавказа // Журнал общей биологии. 1998. Т. 59, No 5, С. 453-476.
Панкратова Е.М. Роль азотфиксирующих синезеленых водорослей (Цианобактерий) в накоплении азота и повышении плодородия почвы: Автореф. дис. д-ра биол. наук. М., 1981. 39 с.
Панфилов Д. В., Шамурин В. Ф., Юрцев Б. А. О сопряженном распространении шмелей и бобовых в Арктике // Бюлл. Московск. о-ва испытателей природы, отд. биол. 1960. Т. 65. Вып. 3. С. 53-62.
Паринкина О.М., Пийн Т. Почвенный микробоценоз как индикатор условий среды под покровом напочвенных лишайников //Лихеноиндикация состояния окружающей среды. Матер. Всес. Конф., 3-5 октября 1978 г. Таллин, 1978. С. 107-113.
Паринкина О. М., Пийн Т. X. Разложение напочвенных кустистых лишайников в условиях бореальной зоны // Экология. 1989. №3. С. 31-39.
Перминова Г. Н. Биомасса и продукция водорослей в тундровых почвах // Ботанический журнал. 1980. Т. 65. №6. С. 859-863.
Петровский В. В. Синузии как формы совместного существования растений // Ботанический журнал. 1961. Т. 46. № 11. С. 1615-1620.
Пианка Э. Эволюционная экология. М.: Мир, 1981. 400 с.
Работнов ТА. Влияние минеральных удобрений на луговые растения и луговые фитоценозы. М.: Наука, 1973. 178 с.
Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. отан. ин-т АН СССР. Сер. 3 Геоботаника. М.-Л., 1950. Вып. 6. С. 7-204.
Работнов Т.А. Жизнеспособные семена в почвах природных биогеоценозов СССР // Теоретические и прикладные аспекты биогеографии: Сб. ст. / Отв. ред. И. А. Соколов. М.: Наука, 1982. С. 35-59.
Работнов Т.А. Жизнеспособные семена в почвах природных биогеоценозов // Бюлл. МОИП. Отд. биол. 1986. Т. 91. Вып. 3. С. 3-18.
Работнов Т.А. Изучение флюктуаций (разногодичной изменчивости) фитоценозов // Полевая геоботаника. Л.: Наука, 1972. Т. 4. С. 95-136.
Работнов Т.А. Изучение ценотических популяций в целях выяснения «стратегии жизни» видов растений // Бюллетень Моск. О-ва Испыт. Природы, отд. биол., 1975. Т. 80. №2. С. 5-17.
Работнов Т.А. История фитоценологии. М.: Аргус, 1995. 158 с.
Работнов Т.А. К биологии монокарпичных многолетних луговых растений // Бюл. МОИП. Отд. биол., 1964. Т. 69. Вып. 6. С. 47-55.
500
Список литературы
Работнов ТА. К биологии порезника промежуточного (Libanotis intermedia Rupr.) // Сб. работ по геоботанике, ботанической географии, систематике растений и палеогеографии. (Тр. МОИП. Отд. биол. Т. 3: Секция ботаники). М., 1960а. С. 221-228.
Работнов Т.А. Луговедение: Учебник для университетов. 2-е изд., перераб. М.: Изд-во Моск, ун-та, 1984а. 319 с.
Работнов Т.А. Методы изучения семенного размножения травянистых растений в сообществах // Полевая геоботаника. М.; Л.: Изд-во АН СССР, 19606. Т. 2, С. 20-40.
Работнов Т.А. О конкуренции между растениями в растительных сообществах // Бюлл. Моск, о-ва испытателей природы. Отд. биол. 19846. Т. 89. №5, С. 82-94.
Работнов Т.А. О типах стратегии растений // Экология. 1985а. №3. С. 3-12.
Работнов Т.А. Фитоценология. 3-е изд., перераб. и доп. М.: Изд-во Моск, ун-та, 1992. 351 с.
Работнов Т.А. Фитоценология: Учебное пособие для биологических специальностей вузов. 2-е изд., перераб. М.: Изд-во Моск, ун-та, 1983. 292 с.
Работнов Т.А. Экология луговых трав. М.: Изд-во Моск, ун-та, 19856. 176 с.
Работнов Т.А. Экспериментальная фитоценология: Учебно-методическое пособие. М.: Изд-во Моск, ун-та, 1998. 240 с.
Разумовский С.М. Закономерности динамики биоценозов. М.: Наука, 1981. 231 с.
Райс Э.Л. Аллелопатия. М.: Мир, 1978. 392 с.
Раменский Л. Г. Введение в комплексное почвенно-геоботаническое изучение земель. М.: Сельхозгиз, 1938. 620 с.
Раменский Л. Г. О принципиальных установках, основных понятиях и терминах производственной типологии земель, геоботаники и экологии // Советская ботаника, 1935. №4. С. 25-40.
Раменский Л. Г. О сравнительном методе экологического изучения растительных сообществ // Дневник 12 съезда русских естествоиспытателей и врачей, отдел 2. Вып. 9. М., 1910. С. 389-390.
Раменский Л. Г. Основные закономерности растительного покрова и методы их изучения // Вестник опытного дела Средне-Черноземной области. Воронеж, 1924. С. 37-73.
Раменский Л. Г.у Цаценкин И. А.у Чижиков О. Н.у Антипин Н. А. Экологическая оценка кормовых угодий по растительному покрову. М.: Гос. Изд-во с/х лит-ры, 1956. 472 с.
Риклефс Р. Основы общей экологии. М.: Мир, 1979. 424 с.
Романовский Ю. Э. Современное состояние концепции стратегии жизненного цикла // Биологические науки. 1989а. № 11, С. 18-31.
Романовский Ю. Э. Конкуренция за флуктуирующий ресурс: эволюционные и экологические последствия //Журнал общей биологии. 19896. Т. 50. №3. С.304-315.
Романовский Ю. Э. Стратегии жизненного цикла: синтез эмпирического и теоретического подходов // Журнал общей биологии. 1998. Т. 59. №6. С. 565-585.
Рысин Л. П.у Рысина Г. П. Морфоструктура подземных органов лесных травянистых растений. М.: Академия наук СССР, 1987. 207 с.
Сабинин Д.А. О внутривидовой борьбе в искусственных и естественных насаждениях растений // Внутривидовая борьба у животных и растений. М.: Изд-во Моск, ун-та, 1948. С. 33-46.
Салазкин А. С. Быстрота роста кормовых лишайников // Советский оленевод. 1937. Вып. 11. С. 43-54.
Селиванов И. А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 232 с.
Симонян С.А., Берсегян А. М. К познанию фитоценологической роли микромицетов в различных типах растительности Армении // Микология и фитопатология, 1974. Т. 8. №5. С. 315-322.
Скворцова Е. Б.у Уланова Н. Г, Басевич В. Ф. Экологическая роль ветровалов. М.: Лесная промышленность, 1983. 192 с.
Список литературы
501
Смирнова О. В. (ред.) Восточноевропейские леса: история в голоцене и современность. В 2 кн. М.: Наука, 2004. Кн. 1-479 с., Кн. 2-575 с.
Смирнова О. В. Структура травяного покрова широколиственных лесов. М.: Наука, 1987. 207 с.
Смирнова О. В., Попадюк Р.В., Чистякова А. А. Популяционные методы определения минимальной площади лесного ценоза // Ботанический журнал. 1988. Т. 73. № 10. С. 1423-1433.
Смирнова О. В., Чистякова А. Л., Попадюк Р. В. Популяционные механизмы динамики лесных ценозов // Биологические науки. 1989. №11. С. 48-58.
Сочава В. Б. Пределы лесов в горах Ляпинского Урала // Труды Ботан. музея АН СССР. 1930. Вып. 22. С. 1-48.
Сукачев В. Н. Главнейшие понятия из учения о растительном покрове // Растительность СССР. Т. 1. М.-Л.: Изд-во АН СССР, 1938. С. 15-37.
Сукачев В. Н. Некоторые общие теоретические вопросы фитоценологии // Вопросы ботаники. Вып. 1, М.-Л., 1954. С. 291-309.
Сукачев В. Н. О некоторых современных проблемах изучения растительного покрова // Ботанический журнал. 1956. Т. 41. №4. С. 176-186.
Сукачев В.Н. Советское направление в фитоценологии // Вестник АН СССР, 1948. Т. 2. С. 101-108.
Сукачев В.Н. Что такое фитоценоз? // Советская ботаника, 1934. №5. С.4-18, 47-50.
Тюрюканов А. Н., Абатуров Ю.Д., Летова А. Н. Экспериментальное изучение роли корневых систем растений в миграции радиоизотопов // Методы изучения продуктивности корневых систем и организмов ризосферы. Междунар. симп. 28.08-12.09.1968. Л.: Наука, 1968. С. 222-225.
Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
Умаров М. М. Ассоциативная азотфиксация в биогеоценозах // Почвенные организмы как компоненты биогеоценоза. М.: Наука, 1984. С. 185-199.
Уранов А. А. Онтогенез и возрастной состав популяций // Онтогенез и возрастной состав популяций цветковых растений. М.: Наука, 1967. С. 3-8.
Уранов А. А. Фитогенное поле // Проблемы современной ботаники. М.: Наука, 1965. Т. 1., С. 251-254.
Федосеева С. А. Ымыяхтахская культура Северо-Восточной Азии. Новосибирск: Наука, 1980. 224 с.
Филин В. Р., Филина Н. И. О цианеях в коробочке анреэи скальной // Бюлл. Московск. о-ва испытателей природы, отд. биол. 1989. Т. 94. Вып. 1. С. 93-101.
Филина Н. И., Филин В. Р. Спорогенез и строение спородермы Tetraphis pellucida Hedw. (Tetraphidaceae, Musci) // Биологические науки. 1989. №5. С. 64-71.
Формозов А. Н. Снежный покров как фактор среды, его значение в жизни млекопитающих и птиц СССР. 2-е изд. М.: Изд-во МГУ, 1990. 287 с.
Харборн Дж. Введение в экологическую биохимию. М.: Мир, 1985. 312 с.
Холодный Н.Г.К познанию снижения урожая зерновых культур, обусловленного дождевой погодой // Избранные труды. Т. 3, Киев: АН УССР, 1957. С. 421-428.
Чередниченко О. В. Естественная разногодичная динамика геранево-копеечниковых лугов Тебердинского заповедника // Заповедное дело. 2001. Вып. 9. С. 53-70.
Чистякова А. А. Реконструкция возрастной пространственной структуры популяций деревьев // Восточно-европейские широколиственные леса. О. В. Смирнова (ред.). М.: Наука, 1994. С. 132-143.
Шенников А.П. Волжские луга Средневолжской области. Л., Ульяновск: Окрземупр. и окр-план, 1930. 386 с.
Шенников А.П. Природные факторы распределения растений в экспериментальном освещении // Журнал общей биологии. 1942. Т. 3. №5-6. С. 331-361.
502
Список литературы
Шилов И. А. Экология. М.: Высшая школа, 1997. 512 с.
Шилов И. А. Функциональная роль животных в биогеоценозах // Животные в биогеоценозах. Чтения памяти академика В. Н. Сукачева, в. 14. Шилов И. А. (ред.). М.: б. и., 1996. Р.3-12.
Штина Э.А. Почвенные водоросли как пионеры зарастания техногенных субстратов и индикаторы состояния нарушенных земель // Журнал общей биологии. 1985. Т. 46. № 4. С. 435-443.
Штина Э.А. Почвенные водросли как экологические индикаторы // Ботанический журнал. 1990. Т. 75. №4. С. 441-453.
Штина Э.А. Роль водорослей в биогеоценозах суши // Альгология (Киев). 1991. Т. 1. № 1. С. 23-35.
Штина Э. А. Сообщества водорослей основных типов почв СССР и их диагностическое значение // Ботанический журнал. 1959. Т. 44. №8. С. 1062-1074.
Штина Э.А. Флора водорослей бассейна реки Вятки. Киров: Вят. гос. с.-х. акад., 1997. 96 с.
Штина Э.А., Голлербах М.М. Экология почвенных водорослей. М.: Наука, 1976. 143 с.
Штина Э.А. Состав почвенных водорослей на территории бывшего СССР // Проблемы ботаники на рубеже ХХ-ХХ1 вв.: Тез. Докл. 11(Х) съезда РБО (26-29 мая, Санкт-Петербург. Т. 2. СПб., 1998. С. 125.
Юрцев Б. А. Продукционные стратегии и жизненные формы растений // Жизненные формы в экологии и систематике растений. М.: б. и., 1986. С. 9-23.
Юрцев Б. А. Реликтовые степные комплексы северо-восточной Азии (проблемы реконструкции криоксеротических ландшафтов Беринги). Новосибирск: Наука, 1981. 168 с.
Aarssen L. W. Causes and consequences of variarion in competitive ability in plant communities // Journal of Vegetation Science, 1992. V.3. №2. P. 165-174.
Aarssen L. W. Competitive ability and species coexistence: a «plant’s eye» view // Oikos, 1989. V. 56. №3. P. 386-401.
Aarssen L. W. Ecological combining ability and competitive combining ability in plants: towards a general evolutionary theory of coexistence in systems of competition // American Naturalist, 1983. V. 122. №6. P. 704-731.
Aarssen L. IV., Epp G.A. Neighbour manipulations in natural vegetation: a review // Journal of Vegetation Science, 1990. V. 1. № 1. P. 13-30.
Abrahamson IV. G. Patterns of resource allocation in wildflower populations of fields and woods // American Journal of Botany, 1979. V.66. № 1. P. 71-79.
Abrahamson IV. G., Gadgil M. Growth form and reproductive effort in goldenrods // American Naturalist, 1973. V. 107. №957. P.651-661.
Adlassnig IV., Peroutka M., Lambers H., Lichtscheidl /. K. The roots of carnivorous plants // Plant and Soil, 2005. V.274. № 1-2. P. 127-140.
Adler L. S. Host effects on herbivory and pollination in a hemiparasitic plant // Ecology, 2002. V. 83. №10. P. 2700-2710.
Adler L. S., Irwin R. E. Ecological costs and benefits of defenses in nectar // Ecology, 2005. V.86. №11. P. 2968-2978.
Aerts R. Nutrient resorption from senescing leaves of perennials: are there general patterns? // Journal of Ecology, 1996. V. 84. №4. P. 597-608.
Aerts R. The role of various types of mycorrhizal fungi in nutrient cycling and plant competition // Mycorrhizal Ecology (Ecological Studies. V. 157), van der Heijden M. G. A., Sanders 1. R. (eds.). Berlin e. a.: Springer, 2003. P. 117-133.
Agrawal A. A. Communication between plants: this time it’s real // Trends in Ecology and Evolution, 2000. V. 15. № 11. P. 446.
Agrawal A. A. Herbivory and maternal effects: mechanisms and consequences of transgenera-tional induced plant resistance // Ecology, 2002b. V. 83. № 12. P. 3408-3415.
Список литературы
503
Agrawal A. A. Optimal foraging and phenotypic plasticity in plants // Trends in Ecology and Evolution, 2002a. V. 17. №7. P. 305.
Agren G. /., Fagerstrom T. Coexistence of plant species: a theoretical investigation // Ecol. Bull., 1980. V.32. P. 547-551.
Agren G. /., Fagerstrom T. Limiting dissimilarity in plants: randomness prevents exclusion of species with similar competitive abilities // Oikos, 1984. V. 43. №3. P. 369-375.
Agrios G.N. Plant pathology. 3rd Edition. San Diego e. a., California: Academic Press, 1988. 803 p.
Aguiar M.R., Soriano A., Sala O.E. Competitiion and facilitation in the recruitment of seedlings in Patagonian steppe // Functional Ecology, 1992. V. 6. № 1. P. 66-70.
Aksenova A. A., Onipchenko V.G., Blinnikov M.S. Plant interaction in alpine tundra: 13 years of experimental removal of dominant species // Ecoscience, 1998. V. 5. №2. P. 258-270.
Alexander /., Watling R. Macrofungi of Sitka spruce in Scotland // Proceedings of the Royal Society of Edinburgh. Section B. Biological Sciences, 1987. V.93. № 1-2. P. 107-115.
Alexander T, Toth R., Meier R., Weber H. C. Dynamics of arbuscule development and degeneration in onions, bean, and tomato with reference to vesicular-arbuscular mycorrhizas in grasses // Canadian Journal of Botany, 1989. V. 67. №8. P. 2505-2513.
Allen E. B., Allen M. F. Competition between plants of different successional stages: mycorrhizae as regulators // Canadian Journal of Botany, 1984. V. 62. № 12. P. 2625-2629.
Allen M.E The ecology of mycorrhizae. Cambridge: Cambridge Univ. Press, 1991. 184 p.
Ameloot E., Verheyen К., Hermy M. Meta-analysis of standing crop reduction by Rhinanthus spp. and its effect on vegetation structure // Folia Geobotanica, 2005. V. 40. № 2-3. P. 289-310.
Anagnostakis S. The effect of multiple importations of pests and pathogens on a native tree // Biological Invasions, 2001. V. 3. №3. P. 245-254.
Anand M. Pattern, process and mechanism the fundamentals of scientific inquiry applied to vegetation science // Coenoses, 1994. V.9. №2. P. 81-92.
Andersen D. C., MacMahon J. A. Population dynamics and bioenergetics of a fossorial herbivore, Thomomys talpoides (Rodentia: Geomyidae) in a spruce-fir sere // Ecological Monographs, 1981. V. 51. №2. P. 179-202.
Anderson I. C., Chambers S. M., Cairney J. W. G. Distribution and persistence of Australian Pisolithus species genets at native sclerophyll forest field sites // Mycological Research, 2001. V. 105. №8. P. 971-976.
Anderson V.J., Briske D.D. Herbivore-induced species replacement in grasslands: Is it driven by herbivory tolerance or avoidance? // Ecological Applications, 1995. V. 5. № 4. P. 1014-1024.
Anderson W. B., Eickmeier W. G. Physiological and morphological responses to shade and nutrient additions of Ciaytonia virginica (Portulacaceae): implications for the «vernal dam» hypothesis // Canadian Journal of Botany, 1998. V. 76. №8. P. 1340-1349.
Andersson T. Influence of stemflow and throughfall from common oak Quercus robur on soil chemistry and vegetation patterns // Canadian Journal of Forest Research, 1991. V. 21. №6. P. 917-924.
Anthony P.A., Holtum J. A. M.Jackes B. R. Shade acclimation of rainforest leaves to colonization by lichens // Functional Ecology, 2002. V. 16. №6. P. 808-816.
Antonovics J. Plant veneral diseases: insights from a messy metaphor // New Phytologist, 2005. V. 165. №1. P. 71-80.
Aphalo P. J., Ballare C. L. On the importance of information-acquiring systems in plant-plant interactions // Functional Ecology, 1995. V. 9. № 1. P. 5-14.
Appanah S., Gentry A. H., and LaFrankie J. V. Liana diversity and species richness of Malaysian rain forests // Journal of Tropical Forest Science, 1992. V. 6. № 2. P. 116-123.
504
Список литературы
Applegate R. D., Rogers L. L., Casteel D.A., Novak J. M. Germination of cow parsnip seeds from grizzly bear faces // Journal of Mammalogy, 1979. V. 60. № 3. P. 655.
Arditti J., Ghani A. K. A. Numerical and physical properties of orchid seeds and their biological implications // New Phytologist, 2000. V. 145. №3. P. 367-421.
Arnold A. E., Lutzoni F. Diversity and host range of foliar fungal endophytes: are tropical leaves biodiversity hotspots? // Ecology, 2007. V.88. №3. P. 541-549.
Arrhenius O. Species and area // Journal of Ecology, 1921. V. 9. № 1. P. 95-99.
Arrogo M. T.K., Cavieres L.A., Penaloza A., Arrogo-Kalin M.A. Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes // Plant Ecology, 2003. V. 169. № 1. P. 121-129.
Arslanian R.L., Harris G.H., Stermitz F.R. New quinolizidine alkaloids from Lupinus argenteus and its hosted root parasite Castilleja sulphurea: stereochemistry and conformation of some naturally occurring cyclic carbinolamides // Journal of Organic Chemistry, 1990. V.55. №4. P. 1204-1210.
Ashenden T. IV., Hunt R., Bell S.A., Williams T. G., Mann A., Booth R. E., Poorter L. Responses to SO2 pollution in 41 British herbaceous species // Functional Ecology, 1996. V. 10. №4. P. 483-490.
Atkinson D. How long is the life span of a root? // Trends in Ecology and Evolution, 1992. V. 7. №6. P. 173-174.
Atsatt P R. Host-parasite interactions in higher plants // Physiological Plant Ecology. 111. Responses to the chemical and biological environment (Encyclopedia of Plant Physiology. New Series, 12C). Lange O. L. et al. (eds.). Berlin e. a.: Springer, 1983. P. 519-535.
Agres E., Dromph К. M., Cook R., Ostle N., Bardgett R. D. The influence of below-ground herbivory and defoliation of a legume on nitrogen transfer to neighbouring plants // Functional Ecology, 2007. V. 21. № 2. P. 256-263.
Baker H. G., Baker I. Amino acids in nectar and their evolutionary significance // Nature, 1973. V.241. P. 543-545.
Baker К. F. Evolving concepts of biological control of plant pathogens // Annu. Rev. Phytopathol., 1987. V.25. P. 67-85.
Bakker E. S., Ritchie M. E., OlffH., Milchunas D. G., Knops J. M. H. Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size // Trends in Ecology and Evolution, 2006. V.9. №7. P. 780-788.
Ball M.C., Egerton J.J.G., Lutze J.L., Gutschick V.P., Cunningham R.B. Mechanisms of competition: thermal inhibition of tree seedling by grass // Oecologia, 2002. V. 133. №2. P. 120-130.
Ballare C.L., Scopel A. L. Phytochrome signalling in plant canopies-testing its population-level implications with photoreceptor mutants of Arabidopsis 11 Functional Ecology, 1997. V. 11. №4. P. 441-450.
Ballare C.L., Scopel A. L, Jordan E. T, Vierstra R.D. Signalling among neighboring plants and the development of size inequalities in plant populations // Proceedings of the National Academy of Sciences, USA, 1994. V.91. P. 10094-10098.
Bardgett R.D. The biology of soil. Oxford e.a.: Oxford University Press, 2005. 242 p.
Bardgett R.D., Wardle D.A. Herbivore-mediated linkages between aboveground and belowground communities // Ecology, 2003. V.84. №9. P. 2258-2268.
Barea J. M., Azcon R., Azcon-Aguilar C. Interactions between mycorrhizal fungi and bacteria to improve plant nutrient cycling and soil structure // Microorganisms in soils: roles in genesis and functions. Buscot F., Varma A. (eds.). Berlin e.a.: Springer, 2005. P. 195-212. Borneo A., Yom-Tov Y., Friedman j. Does ingestion by birds affect seed germination? // Functional Ecology, 1991. V.5.M3. P.394-402.
Список литературы
505
Borneo, Yom-Tov and Friedman, 1992 Borneo, A., Yom-Tov, Y., and Friedman, J. (1992). Effect of frugivo-rous birds on seed dispersal and germination of multi-seeded fruits. Acta Oecol. 13, 209-219.
Barrios E., Herrera R. Ecology of nitrogen fixation in Campsiandra laurifolia Benth // New horizons in nitrogen fixation. Palacios R. et al. (eds.). Dordrecht: Kluwer, 1993. P. 595.
Bartha S., Ittzes P. Local richness-species pool ratio: a consequence of the species-area // relationship // Folia Geobotanica, 2001. V. 36. № 1. P. 9-23.
Baskin С. C., Baskin J.M. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination. London: Academic Press, 1998-700 p.
Baskin С. C., Chesson P. L, Baskin J. M. Annual seed dormancy cycles in two desert winter annuals // Journal of Ecology, 1993. V. 81, No 3. P. 551-556.
Bausenwein U., Millard P, Thornton B., Raven J. A. Seasonal nitrogen storage and remobilization in the forb Rumex acetosa 11 Functional Ecology, 2001. V. 15. №3. P. 370-377.
Baylis G. T S. Fungi, phosphorus and the evolution of plant roots // Search, 1972. V. 3, p 257-258.
Baylis G. T S. The magnolioid mycorrhiza and mycotrophy in root systems derived from it // Endomycorrhizas. Sanders F. E., Mosse B., Tinker P. B. (eds.). London: Academic Press, 1975. P. 373-389.
Bazzaz F.A., Carlson R. IV., Harper J.L Contribution to reproductive effort by photosynthesis of flowers and fruits // Nature, 1979. V. 279. P. 554-555.
Beare P.A., Zedler J.B. Cattail invasion and persistence in a coastal salt marsh: the role of salinity // Estuaries, 1987. V. 10. № 1. P. 165-170.
Behmer S. T, Lloyd С. M., Raubenheimer D., Stewart-Clark J., Knight J., Leighton R. S., Harper F A., Smith J.A.C. Metal hyperaccumulation in plants: mechanisms of defence against insect herbivores // Functional Ecology, 2005. V. 19. № 1. P. 55-66.
Bell G., Lechowicz MJ. The ecology and genetics of fitness in forest plants. 1. Environmental heterogeneity measured by explant trials // Journal of Ecology, 1991. V. 79, No 3. P. 663-685.
Belsky A J. Does herbivory benefit plants? A review of the evidence // American Naturalist, 1986. V. 127. №6. P. 870-892.
Belsky A J. Influences of trees on savanna productivity: test of shade, nutrients, and tree-grass competition // Ecology, 1994. V. 75. №4. P. 922-932.
Belyea L. R., Lancaster J. Assembly rules within a contingent ecology // Oikos, 1999. V. 86. № 3. P. 402-416.
Benedict J. B. Lichen mortality due to late-lying snow: results of a transplant study // Arctic and Alpine Research, 1990. V. 22. № 1. P. 81-89.
Bengtsson J., Fagerstrom T, Rydin H. Competition and coexistence in plant communities // Trends in Ecology and Evolution, 1994. V.9. №7. P. 246-250.
Beniston M., Diaz H. F., and Bradley R. S. Climatic change at high elevation sites; A review // Climatic Change, 1997. V.36. №3-4. P. 233-251.
Benner J. IV, Vitousek P M. Development of diverse epiphyte community in response to phosphorus fertilization // Ecology Letters, 2007. V. 10. № 1. P. 628-636.
Bennett K. D. The power of movement in plants // Trends in Ecology and Evolution, 1998. V. 13. №9. P. 339-340.
Bennett R. N., Wallsgrove R. M. Secondary metabolites in plant defense mechanisms // New Phytologist, 1994. V. 127. №4. P. 617-633.
Bentivenga S.P., Hetrick B.A.D. Relationship between mycorrhizal activity, burning, and plant productivity in tailgrass prairie // Canadian Journal of Botany, 1991. V. 69. № 12. P. 2597-2602.
Benzing D. H. Vascular epiphytes. General biology and related biota. Cambridge: Cambridge University Press, 1990. 354 p.
506
Список литературы
Berendse F. Competition between plant populations with different rooting depth // Oecoiogia (Berlin), 1979. V.43, No 1. P. 19-26.
Berendse E Competition between plant populations with different rooting depths. II. Pot experiments // Oecoiogia, 1981. V. 48. № 3. P. 334-341.
Berendse E interspecific competition and niche differentiation between Plantago lanceolata andAnthoxanthum odoratum in a natural hayfieid //Journal of Ecology, 1983. V. 71. № 2. P. 379-390.
Berendse E, Aerts R. Competition between Erica tetralix L. and Molinia caerulea (L.) Moench as affected by the availability of nutrients // Acta Oecoiogica, 1984. V. 5. № 1. P. 3-14.
Berendse E, Elberse IV. Th., Geerts R. H. E. M. Competition and nitrogen loss from plants in grassland ecosystems // Ecology, 1992. V. 73. № 1. P. 46-53.
Bergelson J. Life after death: site pre-emption by the remains of Poa annua 11 Ecology, 1990. V. 71. №6. P. 2157-2165.
Bergh J. P, van der Braakhekke IV. G. Coexistence of plant species by niche differentiation // Verh. Kon. ned. akad. wetensch. Afd. natuurk., 1978; R. 2. №70. P. 125-136.
Bergman B., Johansson C., Soderback E. The Nostoc-Gunnera symbiosis // New Phytologist, 1992. V. 122. №3. P. 379-400.
Bernays E. A., Chapman R. E Plant chemistry and acridoid feeding behavior // Biochemical aspects of plant and animal coevolution. J. Harborne (ed.) London, New York: Academic Press, 1978. P. 99-141.
Bernays E. A., Cooper Driver G., Bilgener M. Herbivores and plant tannins // Advances in Ecological Research, 1989. V. 19. P. 263-302.
Berner R.A. The rise of plants and their effect on weathering and atmospheric CO2 // Science, 1997. V.276. №5312. P. 544-546.
Bertness M. D. interspecific interactions among high marsh perennials in a New England salt marsh // Ecology, 1991. V. 72. № 1. P. 125-137.
Bertness M. D., Callaway R. Positive interactions in communities // Trends in Ecology and Evolution, 1994. V.9. №5. P. 191-193.
Bertness M. D., Shumway S. IV. Competition and facilitation in marsh plants // American Naturalist, 1993. V. 142. №4. P. 718-724.
Bezemer T. M., van Dam N. M. Linking aboveground and belowground interactions via induced plant defenses // Trends in Ecology and Evolution, 2005. V. 20. № 11. P. 617-624.
Bidartondo M. I. The evolutionary ecology of myco-heterotrophy // New Phytologist, 2005. V. 167. №2. P. 335-352.
Bidartondo M. /., Burghardt B., Gebauer G., Bruns T. D., Read D.J. Changing partners in the dark: isotopic and molecular evidence of ectomycorrhizal liaisons between forest orchids and trees // Proceedings of the Royal Society London, Biological Science (B), 2004. V. 271. №1550. P. 1799-1806.
Biondini M., Klein D.A., and Redente E. E Carbon and nitrogen losses through root exudation by Agropyron cristatum, A. smithii, and Bouteloua gracilis 11 Soil Biology and Biochemistry, 1988. V. 20. №4. P. 477-482.
Black J. N. Competition between plants of different initial seed sizes in swards of subterranean clover (T subterraneum L.) with particular reference to leaf area and the light microclimate // Australian Journal of Agricultural Research, 1958. V.9. №3. P. 299-318.
Blackburn IV. H. impacts of grazing intensity and specialized grazing systems on watershed characteristics and responses // Developing strategies for rangeland management. National Research Councii/National Academy of Sciences. Boulder, CO: Westview Press, 1984. P. 927-983.
Blackman G. E. The influence of temperature and available nitrogen supply on the growth of pasture in the spring // Journal of Agricultural Science, 1936. V. 26. №4. P. 620-647.
Список литературы
507
Blum J. D., Klaue A., Nezat C.A., Driscoll С. T Johnson С. E., Siccama T G., Eagar C, Fahey T./., Likens G. E. Mycorrhizal weathering of apatite as an important calcium source in basepoor forest ecosystems // Nature, 2002. V. 417. №6890. P. 729-731.
Blumenthal D., Mitchell С. E., PySek P, and JaroSfk V. Synergy between resource availability and pathogen release in plant invasion // Proceedings of the National Academy of Sciences, 2009. V. 106. № 19. P. 7899-7904.
Boddey R. M., Urquiaga S., Reis V, Dobereiner J. Biological nitrogen fixation associatrd with sugarcane // Plant and Soil, 1991. V. 137. № 1. P. 111 -117.
Bode H. R. Beitr2ge zur Kenntnis allelopathischer Erscheinungen bei einigen Juglandaceen // Planta, 1958; B. 51, S. 440-480.
Bonanomi G., Rietkerk M., Dekker S. C., Mazzoleni S. Negative plant-soil feedback and positive species interactions in a herbaceous plant community // Plant Ecology, 2005. V. 181. № 2. P. 269-278.
Bond IV./. Keystone species // Biodiversity and ecosystem function. Schulze E.-D., Mooney H. A. (eds.). Berlin: Springer, 1993. P. 237-253.
Bond IV./. Large parts of the world are brown or black: a different view on the «Green World» hypothesis // Journal of Vegetation Science, 2005. V. 16. №3. P. 261-266.
Bond IV. /., Slingsby P Seed dispersal by ants in shrublands of the Cape province and its evolutionary implications // South African Journal of Science, 1983. V. 79. P. 231-/233.
Bond WJ., van Wilgen В. IV. Fire and plants. London e.a.: Chapman and Hall, 1996. 272 p.
Bond IV. /., Woodward E L, Midgley G. E The global distribution of ecosystems in a world without fire // New Phytologist, 2005. V. 165. №2. P. 525-538.
Bonfante P, Perotto S. Strategies of arbuscular mycorrhizal fungi when infecting host plants // New Phytologist, 1995. V. 130. № 1. P. 3-21.
Bonilla D., Roda E Nitrogen cycling responses to disturbance: trenching experiments in an evergreen oak forest // Nutrient cycling in terrestrial ecosystems. Harrison A. E, Ineson P, Heal O.W. (eds.). London e.a.: Chapman and Hall, 1990. P. 179-189.
Bonkowski M. Protozoa and plant growth: the microbial loop in soil revisited // New Phytologist, 2004. V. 162. №3. P. 617-631.
Bonkowski M., Griffiths B. S., Scrimgeour C. Substrate heterogeneity and microfauna in soil organic ‘hotspots’ as determinants of nitrogen capture and growth of rye-grass // Applied Soil Ecology, 2000. V. 14. № 1. P. 37-53.
Bonnot EJ. Sur la place et le rdle des Bryophytes dans la v6g6tation des dunes // Colloques phytosociologiques 1: dunes, 1971. P. 149-158.
Bormann E FL, Likens G. E. Pattern and process in a forest ecosystem. New York: Springer, 1979. 253 p.
Bosy /., Aarssen L. IV. The effect of seed orientation on germination in a uniform environment: differential success without genetic or environmental variation // Journal of Ecology, 1995. V. 83. №5. P. 769-773.
Bosy J.L., Reader RJ. Mechanisms underlying the suppression of forb seedlings by grass (Poa pratensis) litter // Functional Ecology, 1995. V.9. №4. P. 635-639.
Boucher C. Autecological and population studies of Orobanthus zeyheri in the Cape of South Africa // Biological aspects of rare plant conservation. Synge H. (ed.). New York: Wiley, 1981. P. 343-353.
Bousquet /., Girouard E., Strobeck C., Dancik В. P, Lalonde M. Restriction fragment polymorphisms in the rDNA region among seven species of Alnus and Betula papyrifera 11 Plant and Soil, 1989. V. 118. №1-2. P. 231-240.
Bowen G. D. Tree roots and the use of soil nutrients // Nutrition of Plantation Forests. Bowen G. D., Nambiar E. K. S. (eds.). London: Academic Press, 1984. P. 146-179.
Bowers J.E., Webb R.H., Rondeau RJ. Longevity, recruitment and mortality of desert plants in Grand Canyon, Arizona, USA // Journal of Vegetation Science, 1995. V. 6. № 4. P. 551-564.
508
Список литературы
Bowers М.А. Influence of herbivorous mammals on an old-field plant community: years 1 -4 after disturbance // Oikos. 1993. V.67. №1.P. 129-141.
Bowman D.M.J.S. The impact of Aboriginal landscape burning on the Australian biota // New Phytologist, 1998. V. 140. №3. P. 385-410.
Bowman W.D., Schardt J. C., Schmidt S.K. Symbiotic N 2-fixation in alpine tundra: ecosystem input and variation in fixation rates among communities // Oecologia, 1996. V. 108. № 2. P. 345-350.
Braakhekke W. G. On coexistence: a causal approach to diversity and stability in grassland vegetation // Agricultural Research Reports. Centre for Agrobiological Research, Wageningen, 1980. V.902. 164 p.
Braakhekke W. G. On coexistence: a causal approach to diversity and stability in grassland vegetation // Agricultural Research Reports. Centre for Agrobiological Research, Wageningen, 1980. V.902. 164 p.
Braakhekke W. G. The significance of competition for plant diversity // Z. zool. Syst. Evolut.-forsch., 1985. V.23. №4. P. 315-327.
Braakhekke W. G., Hooftman D.A.P. The resource balance hypothesis of plant species diversity in grassland // Journal of Vegetation Science, 1999. V. 10, No 2. P. 187-200.
Bradshaw A., Chadwick M.y Jowett D.y Lodge R., Snaydon R. Experimental investigations into the mineral nutrition of several grass species III. Phosphate level // Journal of Ecology, 1960a. V. 48. №3. P. 631-637.
Bradshaw A., Lodge R., Jowett D., Chadwick M. Experimental investigations into the mineral nutrition of several grass species II. pH and calcium level // Journal of Ecology, 1960b. V.48.M1. P. 143-150.
Bradshaw A., Chadwick M. Jowett D., Snaydon R. Experimental investigations into the mineral nutrition of several grass species IV. Nitrogen level // Journal of Ecology, 1964. V. 52. №3. P. 665-676.
Brasier С. M. The unexpected element: Mycovirus involvement in the outcome of two recent pandemics, Dutch elm disease and chestnut blight // Pests, Pathogens and Plant Communities. Burdon J. J., Leather S. R. (eds.). Oxford: Blackwell Scientific, 1990. P. 289-307.
Braun-Blanquet J. Pflanzensoziologie. 3 Auf. Wien, New York: Springer, 1964. 865 S.
Brenchiey W. E., and Warrington K. The Park Grass plots at Rothamsted 1856-1949. Harpenden (United Kingdom): Rothamsted Experimental Station, 1958. 144 p.
Briske D. D. Strategies of plant survival in grazed systems: a functional interpretation // Foraging strategies of grazing animals. Hodgson J., Illius A. W. (eds.). Wallingford: CAB International, 1996. P. 37-67.
Brisson /., Reynolds J. F. The effects of neighbors on root distribution in a creosotebush (Larrea tridentata) population // Ecology, 1994. V. 75. №6. P. 1693-1702.
Broady P.A. Qualitative and qualitative observations on green and yellow-green algae in some English soils // British Phycological Journal, 1979. V. 14. №2. P. 151-160.
Bronstein J. L.y Alarcon R., GeberM. The evolution of plant-insect mutualisms // New Phytologist, 2006. V. 172. №3. P. 412-428.
Brooker R. W, Callaghan T. V. The balance between positive and negative plant interactions and its relationship to environmental gradients: a model // Oikos, 1998. V. 81. №1. P. 196-207.
Brooks P.D., Schmidt S.K., Williams M. W. Winter production of CO2 and N2O from alpine tundra: environmental controls and relationship to inter-system C and N fluxes // Oecologia, 1997. V. 110. №3. P. 403-413.
Brown J.H., Heske EJ. Control of a desertgrassland transition by a keystone rodent guild // Science, 1990. V.250. P. 1705-1707.
Brown N. A. C. Promotion of germination of finbos seeds by plant-derived smoke // New Phytologist, 1993. V. 123. №3. P. 575-584.
Список литературы
509
Brown Р. М. OLDLIST: A database of maximum tree ages // Conference on Tree Rings, Environment, and Humanity: Relationships and Processes, Tucson, Arizona. Dean J.S., Meko D. M. and Swetnam T.W. (eds.). Radiocarbon (Special issue), 1996. P. 727-731.
Brown V.K., GangeA. C. Differential effects of above- and below-ground insect herbivory during early plant succession // Oikos, 1989. V. 54. № 1. P. 67-76.
Brown V.K., GangeA. C. Effects of root herbivory on vegetation dynamics // Plant Root Growth: An Ecological Perspective. Atkinson D. (ed.). Oxford e. a.: Blackwell SP, 1991. P. 453-470.
Brown V. K., GangeA. C. Secondary plant succession — how is It modified by insect herbivory? // Vegetatio, 1992. V. 101. № 1. P. 3-13.
Bruelheide H., HeinemeyerA. Climatic factors controlling the eastern and altitudinal distribution boundary of Digitalis purpurea L. in Germany // Flora, 2002. V. 197. №6. P. 475-490.
Brundrett M. C. Coevolution of roots and mycorrhizas of land plants // New Phytologist, 2002. V. 154. №2. P. 275-304.
Brundrett M. C. Coevolution of roots and mycorrhizas of land plants // New Phytologist, 2002. V. 154. №2. P. 275-304.
Buchmann S.L., Nabhan G.P. The forgotten pollinators. Washington, DC: Island Press, 1996. 292 p.
Buckland S. M., Grime J.P., Hodgson J. G., Thompson K. A comparison of plant responses to the extreme drought of 1995 in northern England // Journal of Ecology, 1997. V. 85. №6. P. 875-882.
Bultman T. L, Borowicz K. L, Schneble R. M., Coudron T. A., Bush L. P. Effect of a fungal endophyte on the growth and survival of two Euplectrus parasitoids // Oikos, 1997. V. 78. №1. P. 170-176.
Bultman T. L, McNeill M. R., Goldson S. L. Isolate-dependent impacts of fungal endophytes in a multitrophic interaction // Oikos, 2003. V. 102. №3. P. 491-496.
Burdon J. J. The role of parasites in plant populations and communities // Biodiversity and ecosystem function. Schulze E.-D., Mooney H. A. (eds.). Berlin: Springer, 1993. P. 165-179.
Burke M.J. IV., Grime J.P. An experimental study of plant community invasibility // Ecology, 1996. V. 77. №3. P. 776-790.
Burrows C.J. Processes of vegetation change. London e.a.: Unwin Hyman, 1990. 551 p.
Cain M. L., Damman H., Muir A. Seed dispersal and the holocene migration of woodland herbs // Ecological Monographs, 1998. V. 68. №3. P. 325-347.
Cain M.L., Subler S., Evans J.P., Fortin M.-J. Sampling spatial and temporal variation in soil nitrogen availability // Oecoiogia, 1999. V. 118, No 4. P. 397-404.
Calder J. A., Wilson J.B., Mark A. E, Ward G. Fire, succession and reserve management in a New Zealand snow tussock grassland // Biological Conservation, 1992. V. 62. № 1. P. 35-45.
Caldwell M. M. Water parasitism stemming from hydraulic lift: A quantitative test in the field // Israel Journal of Botany, 1990. V. 39. P. 395-402.
Caldwell M. M., Searles P. S., Flint S. D., Barnes P. IV. Terrestrial ecosystem responses to solar UV-B radiation mediated by vegetation, microbes and abiotic photochemistry // Physiological plant ecology. Press M. C., Scholes J. D., Barker M. G. (eds.). Oxford: Blackwell Science, 1999. P. 241-262.
Callaghan T. V. Plant population processes in arctic and boreal regions // Ecological Bulletins // Copenhagen, 1987. V. 38. P. 58-68.
Callaghan T. V, Emanuelsson U. Population structure and processes of tundra plants and vegetation // The population structure of vegetation. White J. (ed.) Handbook of Vegetation Science. V. 3. Dordrecht: Junk, 1985. P. 399-439.
Callaway R.M. Positive interactions among plants // Botanical Review, 1995. V. 61. №4. P. 306-349.
510
Список литературы
Callaway R. М., DeLucia Е. Н., Moore D., Nowak R., Schlesinger W. H. Competition and facilitation: contrasting effects of Artemisia tridentata on desert vs. montane pines // Ecology, 1996. V. 77. №7. P. 2130-2141.
Callaway R. M., Kikvidze Z., Kikodze D. Facilitation by unpalatable weeds may conserve plant diversity in overgrazed meadows in the Caucasus Mountains // Oikos, 2000. V. 89. № 2. P. 275-282.
Callaway R.M., King L. Temperature-driven variation in substrate oxygenation and the balance of competition and facilitation // Ecology, 1996. V. 77, No 4. P. 1189-1195.
Callaway R. M., Nadkarni N. M., Mahall В. E. Facilitation and interference of Quercus dou-glasii on understory productivity in central California // Ecology, 1991. V. 72. №4. P. 1484-1499.
Callaway R.M., Reinhart К. O., Moore G. W., Moore D.J., Pennings S. C. Epiphyte host preferences and host traits: mechanisms for species-specific interactions // Oecologia, 2002. V. 132. №2. P. 221-230.
Callaway R. M., Ridenour W. M., Laboski T, Weir T, Vivanco J. M. Natural selection for resistance to the allelopathic effects of invasive plants // Journal of Ecology, 2005. V. 93. № 3. P. 576-583.
Cameron R. E. Species of Nostoc Vaucher occurring in the Sonoran desert in Arizona // Transactions of the American Microscopical Society., 1962. V. 81. №4. P. 379-384.
Campbell B. D., Grime J. P. An experimental test of plant strategy theory // Ecology, 1992. V. 73. №1. P. 15-29.
Campbell B. D., Grime J. P A comparative study of plant responsiveness to the duration of episodes of mineral nutrient enrichment // New Phytologist, 1989. V. 112. №2. P. 261-267.
Campbell B. D., Grime J. P. Prediction of grassland plant responses to global change // Proceedings of the 17 Inter. Grassland Congress, 1993. P. 1109-1118.
Canadell /., Jackson R.B., Ehleringer J.R., Mooney H.A., Sala O.E., Schulze E.-D. Maximum rooting depth of vegetation types at the global scale // Oecologia, 1996. V. 108, No 4. P. 583-595.
Cantrill D.J., Douglas J. G. Mycorrhizal conifer roots from the Lower Cretaceous of the Otway Basin, Victoria // Australian Journal of Botany, 1988. V. 36. №3. P. 257-272.
Carey E. V, Marler MJ., Callaway R.M. Mycorrhizae transfer carbon from a native grass to an invasive weed: evidence from stable isotopes and physiology // Plant Ecology, 2004. V. 172. №1. P. 133-141.
Carlisle A., Brown A.H.F., White EJ. The organic matter and nutrient elements in the precipitation beneath a sessile oak (Quercus petraea) canopy // Journal of Ecology, 1966. V. 54. №1. P. 87-98.
Carlsson U., Elmqvist T., Wennstrom A., Ericson L. Infection by pathogens and population age of host plants // Journal of Ecology, 1990. V. 78. №4. P. 1094-1105.
Carow Th., Furst R. Fleischfressende Pflanzen. Nuedlingen: Verlag Thomas Carow, 1990. 72 S.
Carpenter F. L. Plant-pollinator interactions in Hawaii: pollination energetics of Metrosideros collina (Myrtaceae) // Ecology, 1976. V. 57. № 6. P. 1125-1144.
Carroll G. C. The biology of the endophytism in plants with particular reference to woody perennials // The Microbiology of the Phyllosphere. N.J. Fokkema and J. van den Heuvel (eds.). Cambridge: Cambridge University Press, 1986. P. 205-222.
Carson W. P, Peterson C.J. The role of litter in an old-field community: impact of litter quantity in different seasons on plant species richness and abundance // Oecologia, 1990. V. 85. №1. P. 8-13.
Carthew S. M., Goldingay R. L. Non-flying mammals as pollinators // Trends in Ecology and Evolution, 1997. V. 12. №3. P. 104-108.
Список литературы
511
Catalan L, Balzarini M, Taleisnik E, Serena R, Karlin U. Effects of salinity on germination and seedling growth of Prosopis flexuosa (D. C.) // Forest Ecology and Management, 1994. V. 63. №2-3. P. 347 — 357.
Chaneton E.J., Facelli J.M. Disturbance effects on plant community diversity: spatial scales and dominance hierarchies // Vegetatio, 1991. V. 93. №2. P. 143-155.
Chapin F. S. Ill The mineral nutrition of wild plants // Annual Review of Ecology and Systematics, 1980. V. 11. P. 233-260.
Chapin F S. Ill, Korner C. Arctic and alpine biodiversity: its patterns, causes and ecosystem consequences // Functional roles of biodiversity: a global perspective. Mooney H. A. et al. (eds.). Chichester e. a.: John Wiley and Sons, 1996. P. 7-32.
Chapin F S. Ill, Randerson J. T, McGuire A. D., Foley J. A., Field С. B. // Changing feedbacks in the climate-biosphere system // Frotiers in Ecology and Environment, 2008. V. 6. № 6. P. 313-320.
Chapin F. S. Ill, Zavaleta E. S., Eviner V. T, Naylor R. L, Vitousek P. M., Reynolds H. L, Hooper D. U., Lavorel S., Sala О. E., Hobbie S. E., MackM. C., Diaz S. Consequences of changing biodiversity // Nature, 2000. V.405. №6783. P. 234-242.
Chernov Y.l. (ed.) The living tundra. Cambridge: Cambridge University Press, 1985. 215 p. Chittka L, Schurkens S. Successful invasion of a floral market // Nature, 2001. V. 411. P. 653. Choudhury D. Herbivore induced changes in leaf-litter resource quality: A neglected aspect of herbivory in ecosystem nutrient dynamics // Oikos, 1988. V. 51. №3. P. 389-393.
Clark D. B. The role of disturbance in the regeneration of neotropical moist forests // Reproductive Ecology of Tropical Forest Plants. Bawa K. S., Hadley M. (eds.). Paris: UNESCO, 1990. P. 291-315.
Clark D. B., Clark D. A. Distribution and effects of tree growth of lianas and woody hemiepiphytes in a Costa Rican tropical wet forest // Journal of Tropical Ecology, 1990. V. 6. № 3. P. 321-331.
Clark J.S. Disturbance and tree life history on the shifting mosaic landscape // Ecology, 1991. V. 72. №3. P. 1102-1118.
Clark J. S. Why trees migrate so fast: Confronting theory with dispersal biology and the paleo record // American Naturalist, 1998. V. 152. №2. P. 204-224.
Clarkson D. T Factors affecting mineral nutrient acquisition by plants // Ann. Rev. Plant Physiol., 1985. V.36. P. 77-115.
Clay K. Comparative demography of three graminoids infected by systemic, clavicipitaceous fungi // Ecology, 1990. V. 71. № 2. P. 558-570.
Clay K. Fungal endophyte infection and the population dynamics of grasses // Population biology of grasses. Cheplick G. P. (ed.). Cambridge: Cambridge Univ. Press, 1998. P. 255-285.
Clay K., Schardl C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses // American Naturalist, 2002. V. 160, Suppl.. P. 99-127.
Clements F E. Competition in plant societies. News Service Bulletin, Carnegie Institution of Washington, Washington, 1933.
Clements EE. Plant succession and indicators. New York: Wilson, 1928. 463 p.
Clements F E. Plant succession: an analysis of the development of vegetation. Washington: Carnegie Inst., 1916. №242. 512 p.
Cocks P S. The influence of density and nitrogen on the outcome of competition between two annual pasture grasses (Hordeum leporinum Link and Lolium rigidum Gaud.) // Australian Journal of Agricultural Research, 1974. V. 25. № 2. P. 247-258.
Coley P.D. Gap size and plant defenses // Trends in Ecology and Evolution, 1993. V. 8. № 1. P. 1-2.
Coley P.D., Bryant J.P., Chapin FS. Ill Resource availability and plant antiherbivore defense // Science, 1985. V.230. №4728. P. 895-899.
512
Список литературы
Collins S. L, Barber S. C. Effects of disturbance on diversity in mixed-grass prairie // Vegetatio, 1985. V. 64. №2-3. P. 87-94.
Comps B., ThiebautB., Barriere G., Letouzey J. Repartition de la biomasse entre organs vegetatifs et reproducteurs chez le hetre europeen (Fagus sylvatica L), selon le secteur de la couronne et 1’age des arbres // Annals of Forest Science, 1994. V. 51. № 1, s. 11-26.
Connell J. H. Apparent versus «real» competition in plants // Perspectives on plant competition. Grace J.B., Tilman D. (eds.). San Diego e. a.: Academic Press, 1990. P. 9-26.
Connell J. H. Tropical rain forests and coral reefs as open non equilibrium systems // Population dynamics /Symp. Brit. Ecol. Soc. Anderson R. M., Turner B. D. and Taylor L. R. (eds.). Oxford e.a.: Blackwell, 1979. V.20. P. 141-163.
Connell J. H., Slaty er R. 0. Mechanisms of succession in natural communities and their role in community stability and organization // American Naturalist, 1977. V. 111. №982. P. 1119-1144.
Cooper E.J., Wookey P.A. Floral herbivory of Dryas octopetala by Svalbard reindeer // Arctic, Antarctic, and Alpine Research, 2003. V. 35. № 3. P. 369-376.
Coppock D. L. Vegetation and pastoral dynamics in the southern Ethiopian rangelands: implications for theory and management // Range Ecology at Disequilibrium. Behnke R. H., Scoones 1. and Kerven C. (eds.). London: Oversees Development Institute, 1993. P. 42-62.
Craine J. M., Berin D. M., Reich P. B., Tilman D. G., Knops /. M. H. Measurement of leaf longevity of 14 species of grasses and forbs using a novel approach // New Phytologist, 1999. V. 142. №3. P. 475-481.
Crawford J. IV., Harris J. A., Ritz K., Young 1. M. Towards an evolutionary ecology of life in soil // Trends in Ecology and Evolution, 2004. V.20. №2. P.81-87.
Crawley MJ. Herbivory // Physiological plant ecology. Press M. C., Scholes J.D., Barker M. G. (eds.). Oxford: Blackwell Science, 1999. P. 199-217.
Crawley M.J., Johnston A.E., Silvertown /., Dodd M., de Mazancourt C., Heard M. S., Hen-mann D. E, Edwards G. R. Determinants of species richness in the Park Grass Experiment // American Naturalist, 2005. V. 165. №2. P. 179-192.
Cresswell J. E. Capture rates and composition of insect prey of the pitcher plant Sarracenia purpurea 11 American Midland Naturalist, 1991. V. 125. № 1. P. 1-9.
Crews T. E. The presence of nitrogen fixing legumes in terrestrial communities: Evolutionary vs ecological considerations // Biogeochemistry, 1999. V. 46. № 1-3. P. 233-246.
Crist T. O. The spatial distribution of termites in shortgrass steppe: a geostatistical approach // Oecoiogia, 1998. V 114. №3. P. 410-416.
Crocker R. L., Major J. Soil development in relation to vegetation and surface age at Glacier Bay, Alaska // Journal of Ecology, 1955. V. 43. № 2. P. 427-448.
Crutzen P./., Andreae M. O. Biomass burning in the tropics: impact on atmospheric chemistry and biogeochemical cycles // Science, 1990. V. 250. P. 1669-1678.
Curtis T. P, Sloan IV. T, Scannell J. IV Estimating prokaryotic diversity and its limits // PNAS USA, 2002. V.99. № 16. P. 10494-10499.
Cutler N. A., Belyea L. R., Dugmore A J. The spatiotemporal dynamics of a primary succession // Journal of Ecology, 2008. V.96. №2. P. 231-246.
Cyr H., Pace M. L. Magnitude and patterns of herbivory in aquatic and terrestrial ecosystems // Nature, 1993. V.361. P. 148-150.
Dahlgren J., Oksanen L., Sjodin M., Olofsson J. Interactions between gray-sided voles (Clethri-onomys rufucanus) and bilberry (Vaccinium myrtillus), their main winter food plant // Oecoiogia, 2007. V. 152. №3. P. 525-532.
Dai X. Impact of cattle dung deposition on the distribution pattern of plant species in an alvar limestone grassland // Journal of Vegetation Science, 2000. V. 11. №5. P. 715-724.
Список литературы
513
Dale Н., Press М. С. Elevated atmospheric СО2 Influences the interaction between the parasitic angiosperm Orobanche minor and its host Tri folium repens 11 New Phytologist, 1998. V. 140. №1. P. 65-73.
Dale J. E., Egley G. H. Stimulation of witchweed germination by run-off water and plant tissues // Weed Science, 1971. V. 19. №6. P. 678-681.
Damle M., Giri A. P, Sainani M.N., Gupra V.S. Higher accumulation of proteinase inhibitors in flowers than leaves as a possible basis for differential feeding preference of Helicoverpa armigera on tomato (Lycopersion esculentum Mill, Cv. Dhanashree) // Phytochemistry, 2005. V.66. №21. P. 2659-2667.
D'Antonio С. M., Mahall В. E. Root profiles and competition betwwen the invasive, exotic perennial, Carpobrotus edulis, and two native shrub species in California coastal scrub // American Journal of Botany, 1991. V. 78. №7. P. 885-894.
Davidson D. W. The effects of herbivory and granivory on terrestrial plant succession // Oikos, 1993. V. 68. №1. P. 23-35.
Davidson D. W. The role of resource imbalances in the evolutionary ecology of tropical arboreal ants // Biological Journal of the Linnean Society, 1997. V. 61. №2. P. 153-181.
Davies D. M., Graves J. D. Interaction between arbuscular mycorrhizal fungi and the hemiparasitic angiosperm Rhinanthus minor during co-infection of a host // New Phytologist, 1998. V. 139. №3. P. 555-563.
Davies D. M., Graves J. D., Elias С. O., Williams P. J. The impact of Rhinanthus spp. on sward productivity and composition: implications for the restoration of species-rich grasslands // Biological Conservation, 1997. V. 82. № 1. P. 87-93.
Davis M.A., Grime J.P., Thompson K. Fluctuating resources in plant communities: a general theory of invasibility // Journal of Ecology, 2000. V. 88. № 3. P. 528-534.
Davison A. W, Bailey l.F. SO2 pollution reduces the freezing resistance of ryegrass // Nature, 1982. V.297. P. 400-402.
Davison A. W, Barnes J.D. Effects of ozone on wild plants // New Phytologist, 1998. V. 139. №1. P. 135-151.
Dawson E. Y. The giant of Galapagos // Natural History, 1962; November. P. 52-57.
De Bruyn £., Scheirs/., Verhagen R. Nutrient stress, host plant quality and herbivore performance of a leaf-mining fly on grass // Oecologia, 2002. V. 130. №4. P. 594-599.
De Deyn G. B., Raaijmakers С. E., Zoomer H. R., Berg M. P, de Ruiter P C., Verhoef H. A., Bezemer T M., van derPutten W. H. Soil invertebrate fauna enhances grassland succession and diversity // Nature, 2003. V. 422. №6933. P. 711-716.
de GraafM. С. C., Bobbink R., Roelofs J. G. M., Verbeek P. J. M. Differential effects of ammonium and nitrate on three heathland species // Plant Ecology, 1998. V. 135, No 2. P. 185-196.
de Jong T.J., Klinkhamer P. G.L., van Staalduinen MJ. The consequences of pollination biology for selection for mass or extended blooming // Functional Ecology, 1992. V. 6. № 5. P. 606-615.
de Kroon H., Fransen B., van Rheenen J. W. A., van Dijk A., Kreulen R. High levels of inter-ramet water translocation in two rhizomatous Carex species, as quantified by deuterium labelling // Oecologia, 1996. V. 106. № 1. P. 73-84.
de Kroon H., MommerL., NishiwakiA. Root competition: towards a mechanistic understanding // Root ecology (Ecological Studies. V. 168). de Kroon H., Visser E. J. W. (eds.). Berlin, Heidelberg: Springer, 2003. P. 215-234.
De Nooij M. P The role of weevils in the infection process of the fungus Phomopsis subordinaria in Plantago lanceolata // Oikos, 1988. V. 52. № 1. P. 51-58.
De Wit С. T On competition // Verslagen van Landbouwkundige Onderzoekingen, 1960. V. 66.8. P. 1-82.
Dean W. RJ., Milton SJ. Patch disturbances in arid grassy dunes: antelope, rodents and annual plants // Journal of Arid Environments, 1991. V. 20. №3. P. 231-237.
514
Список литературы
Dearing М. D.y Foley IV. /., McLean S. The influence of plant secondary metabolites on the nutritional ecology of herbivorous terrestrial vertebrates // Annu. Rev. Ecol. Syst., 2005. V.36. P. 169-189.
Dearnaley J.D. IV. Further advances in orchid mycorrhizal research // Mycorrhiza, 2007. V. 17. №6. P. 465-486.
del Arco J. M.y Escudero A.y Garrido M. V. Effects of site characteristics on nitrogen retranslocation from senescing leaves // Ecology, 1991. V. 72. №2. P. 701-708.
del Moral R. Competition as a control mechanism in subalpine meadows // American Journal of Botany, 1983. V. 70. № 2. P. 232-245.
del Moral R., Muller С. H. The allelopathic effects of Eucalyptus camaldulensis 11 Amer. Midi. Naturalist, 1970. V.83. P. 254-282.
del Moral R.y Rozzell L. R. Long-term effects of Lupinus lepidus on vegetation dynamics at Mount St. Helens // Plant Ecology, 2005. V. 181. №2. P. 203-215.
DeLuca T. H.y Zackrisson O., Nilsson M. C., Sellstedt A. Quantifying nitrogeg fixation in feather moss carpets of boreal forest // Nature, 2002. V. 419. P. 917-920.
Den Dubbelden К. C., Oosterbeek B. The availability of external support affects allocation patterns and morphology of herbaceous climbing plants // Functional Ecology, 1995. V. 9. №4. P. 628-434.
den Herden M.y Kytbviita M-M.y Niemela P. Growth of reindeer lichens and effects of reindeer grazing on ground cover vegetation in a Scots pine forest and a subarctic heathland in Finnish Lapland // Ecography, 2003. V. 26. №1. P. 3-12.
Denslow J. S. Disturbance-mediated coexistence of species // The ecology of natural disturbance and path dynamics. Pickett S.T. A., White P.S. (eds.). Orlando e.a.: Academic Press, 1985. P. 307-323.
Desprez-Loustau M.-L.y Robin C.y Buee M.y Courtecuisse R.y Garbaye /., Suffert F.y Sache /., Rizzo D. M. The fungal dimension of biological invasions // Trends in Ecology and Evolution, 2007. V.22. №9. P. 472-480.
DeWalt S.J.y Schnitzer S.A.y and DenslowJ. S. Density and diversity of lianas along a chronosequence in a central Panamanian tropical forest //Journal of Tropical Ecology, 2000. V. 16. №1. P.1-19.
Diaz S.y Grime J. P.y Harris J.y McPherson E. Evidence of a feedback mechanism limiting plant response to elevated carbon dioxide // Nature, 1993. V. 364. P. 616-617.
Dickie LA.y Koide R. T.y Steiner К. C. Influences of established trees on mycorrhizal, nutrition, and growth of Quercus rubra seedlings // Ecological Monographs, 2002. V. 72. № 4. P. 505-521.
Dickie l.A.y Schnitzer S. A.y Reich P B.y Hobbie S.E. Spatially disjunct effects of co-occuring competition and facilitation // Ecology Letters, 2005. V. 8. № 11. P. 1191-1200.
Dighton J. Fungi in ecosystem processes. New York, Basel: Marcel Dekker, 2003. 420 p.
Dighton /. Summary chapter: nutrient supply to plants // Nutrient cycling in terrestrial ecosystems. Harrison A.F., Ineson P., Heal O.W. (eds.). London e.a.: Chapman & Hall, 1990. P. 441-445.
Dijk E.y Willems J. H.y van Andel J. Nutrient responses as a key factor to the ecology of orchid species // Acta Botanica Neerlandica, 1997. V. 46. №4. P. 339-363.
Dillenburg L.R.y Teramura A.H.y Forseth LN.y and Whigham D.F. Photosynthetic and biomass allocation responses of Liquidambar styraciflua (Hamamelidaceae) to vine competition // American Journal of Botany, 1995. V.82. №4. P. 454-461.
Dillenburg L. R.y Whigham D. F.y Teramura A. H.y and Forseth I. N. Effects of vine competition on availability of light, water, and nitrogen to a tree host (Liquidambar styraciflua) 11 American Journal of Botany, 1993. V.80. №3. P. 244-252.
Список литературы
515
Dirzo R., Harper J. L. Experimental studies on plant-slug interactions. II. Effect of grazing by slugs on hight density monocultures of Capsella bursa-pastoris and Poa annua 11 Journal of Ecology, 1980. V.68. №3. P.999-1011.
Dixon K. W., Roche S., Pate J. S. The promotive effect of smoke derived from burn native vegetation on seed germination of Western Australian plants // Oecoiogia, 1995. V. 101. №2. P. 185-192.
Dobson Л., Crawley M. Pathogens and the structure of plant communities // Trends in Ecology and Evolution, 1994; V.9. № 10. P. 393-398.
Dobson A. P., Hudson P. J. Parasites, disease and the structure of ecological communities // Trends in Ecology and Evolution, 1986. V. 1. № 1. P. 11-15.
Docters van Leeuwen W.M. Krakatau: 1883-1933. A Botany // Annales du Jardin Botanique de Buitenzorg, 1936. V. 46-47. P. 1-506.
Donovan L.A., Ehleringer J. R. Contrasting water-use patterns among size and life history classes of a semiarid shrub // Functional Ecology, 1992. V.6. №4. P. 482-488.
Dorr I. How Striga parasitizes its host: а ТЕМ and SEM study // Annals of Botany, 1997. V. 79. №5. P. 463-472.
Doscher A., Gaggeler H. IV, Schotterer U., Schwikowski M. A 130 years deposition record of sulfate, nitrate and chloride from a high-alpine glacier // Water, Air, and Soil Pollution, 1995. V.85. №2. P. 603-609.
Downton IV./. S. Growth and osmotic relations of the mangrove Avicennia marina, as influenced by salinity // Australian Journal of Plant Physiology, 1982. V.9. №5. P. 519-528.
Downum K. R. Light-activated plant defense // New Phytologist, 1992. V. 122. №3. P. 401-420.
During HJ. Bryophyte diaspore banks // Advances in Bryology, 1997. V. 6. P. 103-134.
During HJ., van Tooren B.E Bryophyte interactions with other plants // Botanical Journal of the Linnean Society, 1990. V. 104. № 1-3. P. 79-98.
Duthoit C.M. G. Slugs and food preferences // Plant Pathology, 1964. V. 13. № 2. P. 73-77.
Eckstein R. L., Karlsson P. S., Weih M. Leaf life span and nutrient resorption as determinants of plant nutrient conservation in temperate-arctic regions // New Phytologist, 1999. V. 143. №1. P. 177-189.
Edwards C.A., Lofty J.R. Biology of earthworms. 2nd Ed. London: Chapman and Hall, 1977. 333 p.
Edwards P. J. Insect herbivory and plant defense theory // Toward a more exact ecology. The 30th Symposium of the British Ecological Society London 1988. Grubb P. J. and Whittaker J. B. (eds). Cambridge: University Press, 1989. P. 275-297.
Ehrlen J., Lehtila K. How perennial are perennial plants? // Oikos, 2002. V. 98. № 2. P. 308-322.
Eissenstat D. M. A comparison of phosphorus and nitrogen transfer between plants of different phosphorus status // Oecoiogia, 1990. V.82. №3. P. 342-347.
Eissenstat D. M., Caldwell M. M. Competitive ability is linked to rates of water extraction: a field study of two aridland tussock grasses // Oecoiogia, 1988b. V. 75. № 1. P. 1-7.
Eissenstat D. M., Caldwell M. M. Seasonal timing of root growth in favorable microsites // Ecology, 1988a. V.69. №3. P. 870-873.
Eissenstat D. M., Wells С. E., Yanai R. D., Whitebeck J. L. Building roots in a changing environment: implication for root longevity // New Phytologist, 2000. V. 147. № 1. P. 33-42.
Ejrnaes R., Bruun H.H., Graae В J. Community assembly in experimental grasslands: suitable environment or timely arrival? // Ecology, 2006. V.87. №5. P. 1225-1233.
Elias T S. Extrafloral nectaries: their structure and distribution // The biology of nectaries. Bentley B., Elias T.S. (eds.). New York: Columbia University Press, 1983. P. 174-203.
Elias T. S. Morphology and anatomy of foliar nectaries of Pithecellobium macradenium (Legu-minosae) // Botanical Gazette, 1972. V. 133. № 1. P. 38-42.
Ellenberg H. Physiologisches und okologisches Verhalten derselben Pflanzenarten // Berichte der Deutschen botanischen Gesellschaft, 1953; B. 65, S. 351-362.
516
Список литературы
Ellenberg Н. Uber einige Fortschritte der kausalen Vegetationskunde // Vegetatio, 1954. V. 5/6. №1, S. 199-211.
Ellenberg H. Vegetation Mitteleuropas mit den Alpen in kausaler, dynamischer und historischer Sicht. 2 Aufl. Stuttgart: Ulmer, 1978. 982 S.
Elliott G.N., Chen W.M., Chou J.H., Wang H. C., Sheu S. Y, Perin L, Reis V.M., Moulin L, Simon M. F. et al. (2007). Burkholderia phymatum is a highly effective nitrogen-fixing symbiont of Mimosa spp. and fixes nitrogen ex planta // New Phytologist. V. 173. № 1. P. 168-180.
Ellison A.M., Gotelli N.J. Evolutionary ecology of carnivorous plants // Trends in Ecology and Evolution, 2001. V. 16. № 11. P. 623-629.
Ellison A. M., Gotelli N.J. Nitrogen availability alters the expression of carnivory in the northern pitcher plant, Sarracenia purpurea // Proceedings of the National Academy of Sciences, U.S.A., 2002. V.99. №7. P.4409-4412.
Ellison A.M., Gotelli N.J., Brewer J.S., Cochran-Stafira D.L., Kneitel J.M., Miller T. E., Worley A. C., Zamora R. The evolutionary ecology of carnivorous plants // Advances in Ecological Research, 2003. V. 33. P. 1-74.
Elser J. J., Bracken M. E. S., Cleland E. E., Gruner D. S., Harpole W. S., Hillebrand H., Ngai J. T, Seabloom E. W., Shurin J. B. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems // Ecology Letters, 2007. V. 10. №12. P. 1135-1142.
Emerman S. H., Dawson T. E. Hydraulic lift and its influence on the water content of the rhizosphere: An example from sugar maple, Acer saccharum 11 Oecologia, 1996. V. 108. №2. P. 273-278.
Engelbrecht L. Cytokinin activity in larval infected leaves // Biochemie und Physiologic der Pflanzen, 1971; B. 162. № 1. P. 9-27.
Eriksson O., Ehrlen J. Seedling recruitment and population ecology // Seedling Ecology and Evolution. Leek M. A., Parker V. T., Simpson R. L. (eds.). Cambridge e. a.: Cambridge Univ. Press, 2008. P. 239-254.
Espinosa-Garcia F. J., Langenheim J. H. The endophytic fungal community in leaves of a coastal redwood population-diversity and spatial patterns // New Phytologist, 1990. V. 116. № 1. P. 89-97.
Esseen P. A., Renhorn K.-E., Pettersson R. B. Epiphytic lichen biomass in managed and old-growth boreal forests: effect of branch quality // Ecological Applications, 1996. V. 6. № 1. P. 228-238.
Ettema C.H., Wardle D.A. Spatial soil ecology // Trends in Ecology and Evolution, 2002. V. 17, No 4. P. 177-183.
Evans R. The erosional impacts of grazing animals // Progress in Physical Geography, 1998. V.22. №2. P. 251-268.
Evans R.D., Ehleringer J.R. A break in the nitrogen cycle in aridlands? Evidence from <5I5N of soil // Oecologia, 1993. V.94. №3. P. 314-317.
Facelli J. M. Specialized strategies I: seedlings in stressfull environments // Seedling Ecology and Evolution. Leek M. A., Parker V. T., Simpson R. L. (eds.). Cambridge e. a.: Cambridge Univ. Press, 2008. P. 56-78.
Facelli J. M., Pickett S. T.A. Plant litter: its dynamics and effects on plant community structure // Botanical Review, 1991. V. 57. № 1. P. 1-32.
Fagan W. F., Bishop J. G. Trophic interactions during primary succession: herbivores slow a plant reinvasion at Mount St. Helens // American Naturalist, 2000. V. 155. № 2. P. 238-251.
Fahey B. D., Jackson R. Hydrological impacts of converting native forests and grasslands to pine plantations, South Island, New Zealand // Agricultural and Forest Meteorology, 1997. V.84. №1. P. 69-82.
Список литературы
517
Fahnestock J. Т, Knapp А. К. Plant responses to selective grazing by bison: Interactions between light, herbivory and water stress // Vegetatio, 1994. V. 115. № 2. P. 123-131.
Farley R.A., Fitter A. H. Temporal and spatial variation in soil resources in a deciduous woodland //Journal of Ecology, 1999. V.87. №4. P. 688-696.
Fastie C. L. Causes and ecosystem consequences of multiple pathways of primary succession at Glacier Bay, Alaska // Ecology, 1995. V. 76. №6. P. 1899-1916.
Feeny P. Plant apparency and chemical defense // Recent Advances in Phytochemistry, 1976. V. 10. P. 1-40.
Feeny P. Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter moth caterpillars // Ecology, 1970. V. 51. №4. P. 565-581.
Fenner M. Relationships between seed weight, ash content and seedling growth in twenty-four species of Compositae // New Phytologist, 1983. V.95. №4. P. 697-706.
FennerM. The phenology of growth and reproduction in plants // Perspectives in Plant Ecology, Evolution and Systematics, 1998. V. 1. № 1. P. 78-91.
Fenner M., Hanley M.E., Lawrence R. Comparison of seedling and adult payability in annual and perennial plants // Functional Ecology, 1999. V. 13. №4. P. 546-551.
Finlay R.D., Read D.J. The structure and function of the vegetative mycelium of ectomycor-rhizal plants. 1. Translocation of l4C-labelled carbon between plants interconnected by a common mycelium // New Phytologist, 1986. V. 103, No 1. P. 143-156.
Fischer D. G., Hart S. C., Rehill B. J., Lindroth R. L., Keim P, Whitham T. G. Do high-tannin leaves require more roots? // Oecologia, 2006. V. 149. №4. P.668-675.
Fisher B. L., Sternberg L. S. L., Price D. Variation in use of orchid extrafloral nectar by ants // Oecologia, 1990. V. 83. № 2. P. 263-266.
Fisher P.I., Graf F, Petrini L.E., Sulton В. C., and Wookey P.A. Fungal endophytes of Dryas octopetata from a high arctic polar semi desert and from the Swiss Alps // Mycologia, 1995. V.87. №3. P.319-323.
Fitter A. H. Darkness visible: reflections on underground ecology // Journal of Ecology, 2005. V.93. №2. P. 231-243.
Fitter A. H., Peat H.J. The Ecological Flora Database // Journal of Ecology, 1994. V. 82. №2. P. 415-425.
Flannery T. F. The future eaters: An ecological history of the Australian lands and people. Sydney: Reed Books, 1995. 423 p.
FolgaraitP. J., Johnson H.L., Davidson D. W. Responses of Cecropia to experimental removal of Mullerian bodies // Functional Ecology, 1994. V. 8. № 1. P. 22-28.
Fonseca C.R.S. Herbivory and the long-lived leaves of an Amazonian ant-tree // Journal of Ecology, 1994. V.82. №4. P. 833-842.
Forman R. T. Canopy lichens with blue-green algae: a nitrogen source in a Colombian rain forest // Ecology, 1975. V. 56. № 5. P. 1176-1184.
Fowler N.L. Competition and coexistence in a North Carolina grassland. 11. The effects of the experimental removal of species //Journal of Ecology, 1981. V. 69. №3. P. 843-854.
Fowler N. L. The effects of competition and environmental heterogeneity on three coexisting grasses // Journal of Ecology, 1990. V. 78. № 2. P. 389-402.
Fowler N. L.,Antonovics J. Competition and coexistence in a North Carolina grassland. 1. Patterns in undisturbed vegetation //Journal of Ecology, 1981. V. 69. №3. P. 825-841.
Fraenkel G. S. The raison d’etre of secondary plant substances // Science, 1959. V. 129. № 3361. P. 1466-1470.
Francis R., Read D.J. The contributions of mycorrhizal fungi to the determination of plant community structure // Plant and Soil, 1994. V. 159. № 1. P. 11-25.
Frank D.A. Drought effects on above- and belowground production of a grazed temperate grassland ecosystem // Oecologia, 2007. V. 152. № 1. P. 131-139.
518
Список литературы
Frank D. A., McNaughton S.J. Evidence for the promotion of aboveground grassland production by native large herbivores in Yellowstone National Park // Oecoiogia, 1993. V. 96. №2. P. 157-161.
FranklandJ. C. Autecology and the mycelium of a woodland litter decomposer // The Ecology and Physiology of the Fungal Mycelium. D. H. Jennings and A. D. M. Rayner (eds.). Cambridge: Cambridge University Press, 1984. P. 241-260.
Fransen B., de Kroon H., Berendse F Soil nutrient heterogeneity alters competition between two perennial grass species // Ecology, 2001. V. 82. №9. P. 2534-2546.
Fraser L. H., Grime J. P. Primary productivity and trophic dynamics investigated in a North Derbyshire dale // Oikos, 1997. V.80. №3. P. 499-508.
Fretwell S. D. The regulation of plant communities by food chains exploiting them // Perspectives in Biology and Medicine, 1977. V.20. №2. P. 169-185.
Frey-Klett P, Garbage J., Tarkka M. The mycorrhiza helper bacteria revisited // New Phytologist, 2007. V. 176. №1. P. 22-36.
Fricke K. Licht- und Schattenholzarten, ein wissenschaftlich nicht begriindes Dogma // Central-blatt fiir das gesamte Forstwesen, 1904; B. 30, S. 315-325.
Fridley J. A., Grime J. R, Bilton M. Genetic identity of interspecific neighbours mediates plant responses to competition and environmental variation in a species-rich grassland // Journal of Ecology, 2007. V.95. №5. P. 908-915.
Friedman/., Hutchins A., Li С. K, and Perry D. A. Actinomycetes inducing phytotoxic or fungistatic activity in a Douglas-fir forest and in an adjacentareaof repeated regeneration failure in southwestern Oregon // Biologia Plantarum, 1989. V. 31. №6. P. 487-495.
Friese C. F, Allen M. F The spread of VA mycorrhizal fungal hyphae in the soil: inoculum types and external hyphal architecture // Mycologia, 1991. V. 83. №4. P. 409-419.
Fromm H., Winter K, Fisser/., Hantschel R., Beese F The influence of soil type and cultivation system on the spatial distributions of the soil fauna and microorganisms and their interactions // Geoderma, 1993. V. 60. № 1-4. P. 109-118.
Fuentes E.R., Otaiza R.D., Alliende M. C., Hoffman A., Poiani A. Shrub clumps of the Chilean matorral vegetation: Structure and possible maintenance mechanisms // Oecoiogia, 1984. V.62. №3. P. 405-411.
Fuerst E.R, Putnam A.R. Separating the competitive and allelopathic components of interference: theoretical principles // Journal of Chemical Ecology, 1983. V.9. P. 937-944.
Gali-Muhtasib H., Smith С. C., Higgins J. J. The effect of silica in grasses on the feeding behavior of the prairie vole Microtus ochrogaster 11 Ecology, 1992. V. 73. № 5. P. 1724-1729.
Gallardo A., Parama R., Covelo F Soil ammonium vs. nitrate spatial pattern in six plant communities: simulated effect on plant populations // Plant and Soil, 2005. V. 277. №1-2. P. 207-219.
Gange A. C. Positive effects of endophyte infection on sycamore aphids // Oikos, 1996. V. 75, N. 3. P. 500-510.
Gange A. C., Brown V. K., Farmer L. M. A test of mycorrhizal benef it in an early successional plant community // New Phytologist, 1990. V. 115. № 1. P. 85-91.
GangeA. C., Brown V. K. Actions and interactions of soil invertebrates and arbuscular mycorrhizal fungi in affecting the structure of plant communities // Mycorrhizal Ecology (Ecological Studies. V. 157). van der HeiJden M. G. A. and Sanders 1. R. Berlin e. a. Springer, 2003. P. 321-344.
Gange A. C., Stagg P G., Ward L. K. Arbuscular mycorrhizal fungi affect phytophagous insect specialism // Ecology Letters, 2002. V. 5. № 1. P. 11-15.
Gardener C.J., Mclvor J. G., and Jansen A. Passage of legume and grass seeds through the digestive tract of cattle and their survival in feces // Journal of Applied Ecology, 1992. V.30. №1. P. 63-74.
Список литературы
519
Gardener М. С., Gillman М. Р. The taste of nectar — a neglected area of pollination ecology // Oikos, 2002. V.98. №3. P. 552-557.
Gardner G. The reproductive capacity of Fraxinus excelsior on the Derbyshire limestone // Journal of Ecology, 1977. V. 65. № 1. P. 107-118.
Garrity D. P, Soekadi M., Van Noordwijk M., De La Cruz R., Pathak P S., Gunasena H. P. M., Van So N., Huijun G., Majid N. M. The Imperata grasslands of tropical Asia: area, distribution and typology // Agroforestry Systems, 1996. V. 36. № 1-3. P. 3-29.
Gartner B.L. Is the climbing habit of poison oak ecotypic? // Functional Ecology, 1991a. V. 5. №5. P. 696-704.
Gartner B. L. Structural stability and architecture of vines vs. shrubs of poison oak, Toxicodendron diversilobum 11 Ecology, 1991b. V. 72. №6. P. 2005-2015.
Gartner B. L, Chapin F. S., Shaver G. R. Demographic patterns of seedling establishment and growth of native graminoids in an Alaskan tundra disturbance // Journal of Applied Ecology, 1983. V.20. №3. P. 965-980.
Gause G.F. The struggle for existence. Baltimore, MD: Williams and Wilkins, 1934. 163 p.
Gauslaa K, Ohlson M., RolstadJ. Fine-scale distribution of the epiphytic lichen Usnea longissima on two even-aged neighbouring Picea abies trees // Journal of Vegetation Science, 1998. V.9. Nel. P.95-102.
Gebauer G., Meyer M. 15 N and 13 C natural abundance of autotrophic and myco-heterotrophic orchids provides insight into nitrogen and carbon gain from fungal association // New Phytologist, 2003. V. 160. Ne 1. P. 209-223.
Gehring C.A., Whitham T. G. Interactions between aboveground herbivores and the mycorrhizal mutualists of plants // Trends in Ecology and Evolution, 1994. V. 9. Ne 7. P. 251-255.
Gehring C.A., Whitham T. G. Reduced mycorrhizae on Juniperus monosperma with mistletoe: the influence of environmental stress and tree gender on a plant parasite and a plant-fungal mutualism // Oecologia, 1992. V. 89. Ne 2. P. 298-303.
Gemma J. N., Koske R. E. Mycorrhizae in recent volcanic substrates in Hawaii // American Journal of Botany, 1990. V. 77. P. 1193-1200.
Gentry A. H. The distribution and evolution of climbing plants // The biology of vines. Putz F. E. and Mooney H. A. (eds.). Cambridge: Cambridge University Press, 1991. P. 3-49.
Gernns H., von Alten H., Poehling H.-M. Arbuscular mycorrhiza increased the activity of a biotrophic leaf pathogen — is a compensation possible? // Mycorrhiza, 2001. V. 11. Ne5. P. 237-243.
Gerson U., Seaward M. R. D. Lichen-invertebrate associations // Lichen ecology. Seaward M. R. D. (ed.). London: Academic Press, 1977. P. 69-118.
Gewin V Beyond neutrality — ecology finds its niche // PLoS Biology, 2006. V. 4. №8, p. e278. doi: 10.137 l/journal.pbio.0040278.
Giamoustaris A., Mithen R.F. The effect of modifying glucosinolate content of leaves of oilseed rape (Brassica napus ssp. oleifera) on its interaction with specialist and generalist pests // Annals of Applied Biology, 1995. V. 126. №2. P. 347-363.
Gibson С. C., Watkinson A. R. Host selectivity and the mediation of competition by the root hemiparasite Rhinanthus minor // Oecologia, 1991. V. 86. № 1. P. 81-87.
Gibson С. C., Watkinson A. R. The role of the hemiparasitical annual Rhinanthus minor in determining grassland community structure // Oecologia, 1992. V. 89. № 1. P. 62-68.
Gifford R.M. The global carbon cycle: a viewpoint on the missing sink // Australian Journal of Plant Physiology, 1994. V. 21. № 1. P. 1-5.
Gifford R. M. Whole plant respiration and photosynthesis of wheat under increased CO2 concentration and temperature: long-term vs short-term distinctions for modeling // Global Change Biology, 1995. V. 1. №4. P. 249-263.
520
Список литературы
Gigon А. К hierarchic approach in causal ecosystem analysis. The calcifuge-calcicole problem in alpine grasslands // Potentials and limitat. ecosys. anal. (Ecological Studies). Schulze E.-D., Zwolfer H. (eds.). Berlin: Springer, 1987; Bd. 61. P. 228-244.
Gigon A. Fluktuationen des Deckungsgrades und die Koexistenz von Pflanzenarten in Trespen-Halbtrockenrasen (Mesobromion) // Phytocoenologia, 1997; Bd. 27. №2, S. 275-287.
Gigon A., Bocherens J. Wie rasch verandert sich ein nicht mehr gemahtes Ried im Schweizer Mittelland // Berichte Geobot. Inst. Rubel, 1985; Bd. 52, S. 53-65.
Gigon A., Leutert A. The Dynamic keyhole-key model of coexistence to explain diversity of plants in limestone and other grasslands // Journal of Vegetation Science, 1996. V. 7. №1. P. 29-40.
Gigon A., MartiR. Seltenheit, Konkurrenz und Naturschutz von Pflanzen in Trespen-Halbtrocken-rasen bei Schaffhausen // Verhandlungen der Gesellschaft fur Okologie, 1994; Bd. 23, S. 231-236.
Giladi 1. Choosing benefits or partners: a rewiev of the evidence for the evolution of myrme-cochory // Oikos, 2006. V. 112. № 3. P. 481-492.
Gill R.A., Jackson R.B. Global patterns of root turnover for terrestrial ecosystems // New Phytologist, 2000. V. 147. № 1. P. 13-31.
Giller К. E., Ormesher Awah E M. Nitrogen transfer from Phaesolus bean to intercropped maize measured using 15N-enrichment and 15N-isotope dilution methods // Soil Biology and Biochemistry, 1991. V. 23. №4. P. 339-346.
Gitag H., Lee W. G., Allen R.B., Wilson J. B. Recovery of Chionochloa rigida tussocks from fires in South Island, New Zealand // Journal of Environmental Management, 1992. V.35. №4. P. 249-259.
Gleason H.A. On the relation between species and area // Ecology, 1922. V. 3. № 2. P. 158-162.
Gleason H.A. The individualistic concept of the plant association // Bulletin of the Torrey Botanical Club, 1926. V.53. № 1. P. 7-26.
Gleason H.A. The structure and development of the plant association // Bulletin of the Torrey Botanical Club, 1917. V.44. № 10. P. 463-481.
Gleeson S. K., Tilman D. Plant allocation, growth rate and successional status // Functional Ecology, 1994. V. 8. №4. P. 543-550.
Glenn-Lewin D. C. The individualistic nature of plant community development // Vegetatio, 1980. V. 43. №1-2. P. 141-146.
Godinez-Alvarez H., Valiente-Banuet A., and Rojas-Martinez A. The role of seed dispersers in the population dynamics of the columnar cactus Neobuxbaumia tetetzo 11 Ecology, 2002. V. 83. №9. P. 2617-2629.
Godwin H. Dispersal of pond floras // Journal of Ecology, 1923. V. 11. № 2. P. 160-164.
Goetz S.J., Prince S. D., Small J. Advances in Satellite Remote Sensing of‘Environmental Variables for Epidemiological Applications // Remote sensing and G1S in‘epidemiology. S. 1. Hay, S.E.Randolph, D.J.Rogers (eds). London: Academic Press, 2000. P. 283-311.
Goldberg D. E. Influence of competition at the community level: an experimental version of the null models approach // Ecology, 1994. V. 75. №5. P. 1503-1506.
Goldberg D. E., Novoplanskg A. On the relative importance of competition in unproductive environments //Journal of Ecology, 1997. V.85. №4. P.409-418.
Goldberg D. E., Werner P.A. Equivalence of competitors in plant communities: a null hypothesis and a field experimental approach // American Journal of Botany, 1983. V. 70. №7. P. 1098-1104.
Gomez J. M. Importance of direct and indirect effects in the interaction between a parasitic angiosperm (Cuscuta epithgmum) and its host plant (Hormathophglla spinosa) 11 Oikos, 1994. V.71.№1. P.97-106.
Список литературы
521
Goss MJ. Consequences of the activity of roots on soil // Plant Root Growth: An Ecological Perspective. Atkinson D. (ed.). Oxford e.a.: Blackwell SP, 1991. P. 171-186.
Gottsberger G. Seed dispersal by fish in the inundation regions of Humaita, Amazonia // Biotropica, 1978. V. 10. №3. P. 170-183.
Gottsberger G.t Morawetz IV. Development and distribution of the epiphytic flora in an Amazonian savanna in Brazil // Flora, 1993, 188. №2. P. 145-151.
Gould SJ. (ed.) The book of life: an illustrated history of the evolution of life on‘Earth, second edition. New York: W.W. Norton and Company. 2001. 256 p.
Goulson D. Bumblebees: their behaviour and ecology. Oxford e. a.: Oxford Univ. Press, 2003. 235 p.
Gower S. T.t McMurtrie R. E., Murty D. Aboveground net primary production decline with stand age: potential causes // Trends in Ecology and Evolution, 1996. V. 11. №9. P. 378-382.
Grace J. A clarification of the debate between Grime and Tilman // Functional Ecology, 1991. V.5.№5. P.583-587.
Gradstein S. R., Poes T. Bryophytes // Tropical rainforest ecosystems. Lieth H. and Werger M. J. A. (eds.). Amsterdam: Elsevier Sci. Publ., 1989. P. 311-325.
Graves J. D. Host-plant responses to parasitism // Parasitic plants. Press M. C., Graves J. D. (eds.). London e.a.: Chapman and Hall, 1995. P.206-225.
Graves J. D., Watkins N. K., Fitter A. H., Robinson D., and Scrimgeour C. Intraspecific transfer of carbon between plants linked by a common mycorrhizal network // Plant and Soil, 1997. V. 192. №2. P. 153-159.
Greenwood D. R. Leaf form and the reconstruction of past climates // New Phytologist, 2005. V. 166. №2. P. 355-357.
GreenwoodD. R.t Wing S. L. Eocene continental climates and latitudinal temperature-gradients // Geology, 1995. V. 23. № 11. P. 1044-1048.
Gregory P. J., McGowan M., and Biscoe P V. Water relations of winter wheat. 2. Soil water relations // Journal of Agricultural Science, 1978. V. 91. № 1. P. 103-116.
Grime J. P Benefits of plant diversity to ecosystems: immediate, filter and founder effects // Journal of Ecology, 1998. V.86. №6. P. 902-910.
Grime J. P Dominant and subordinate components of plant communities: implication for succession, stability and diversity // Colonization, succession and stability. Gray A. J., Crawley M. J. and Edwards P.J. (eds.). Oxford: Blackwell Publ., 1987. P. 413-428.
Grime J. P Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory // American Naturalist, 1977. V. 111. №982. P. 1169-1194.
Grime J. P Factors limiting the contribution of pteridophytes to a local flora // Proceedings of the Royal Society of Edinburgh, 1985. V. 86B. P. 403-421.
GrimeJ.P. Plant strategies and vegetation processes. Chichester e.a.: J. Wiley and Sons, 1979. 371 p.
Grime J. P. Plant strategies, vegetation processes, and ecosystem properties. 2nd Edition. Chichester: John Wiley and Sons, 2001. 417 p.
GrimeJ.P. Vegetation classification by reference to strategies // Nature, 1974. V. 250. P. 26-31.
Grime J. P, Cornelissen J. H. C., Thompson K., Hodgson J. G. Evidence of a causal connection between anti-herbivore defence and the decomposition rate of leaves // Oikos, 1996. V.77. №3. P. 489-494.
Grime J.P, Hodgson J. G., HuntR. Comparative plant ecology: a functional approach to common British species. London e.a.: Unwin Hyman, 1988. 742 p.
Grime J. P, Hunt R. Relative growth rate: its range and adaptive significance in a local flora // Journal of Ecology, 1975. V.63. №2. P. 393-422.
Grime J.P., Jeffrey D. IV. Seedling establishment in vertical gradients of sunlight // Journal of Ecology, 1965. V.53. №3. P.621-642.
522
Список литературы
Grime J.P., Mackey J.M.L., Hillier S.H., Read DJ. Floristic diversity in a model system using experimental microcosms // Nature, 1987. V. 328. №6129. P. 420-422.
Grime J.P., Shacklock J.M.L., Band S.R. Nuclear DNA contents, shoot phenology and species co-existence in a limestone grassland community // New Phytologist, 1985. V. 100. №3. P. 435-445.
Grime J.P., Thompson K., Hunt R. et al. Integrated screening validates primary axes of specialisation in plants // Oikos, 1997. V. 79. №2. P. 259-281.
Grime J.P., Willis A. J., Hunt R., Dunnett N.P. Climate-vegetation relationships in the Bibury Road Verge experiments // Insight from foresight: long-term experiments in agricultural and ecological sciences. Leigh R.A., Johnston A. E. (eds.). Wallingford: CAB International, 1994. P. 271-285.
Gross J. R. The establishment of Rhododendron ponticum in the Killarny oalwoods, S. W. Ireland // Journal od Ecology, 1981. V. 69. №3. P. 807-824.
Gross K. L, Pregitzer K. S., Burton A J. Spatial variation in nitrogen availability in three succes-sional plant communities //Journal of Ecology, 1995. V.83, No 3. P.357-367.
Groves R. H., Williams J. D. Growth of skeleton weed (Chondrilla juncea L.) as affected by growth of subterranean clover (Trifolium subterraneum L.) and infection by Puccinia chondrillina Bubak & Syd. // Australian Journal of Agricultural Research, 1975. V. 26. №6. P. 975-983.
Grubb P The maintenance of species-richness in plant communities: the importance of the regeneration niche // Biological Review, 1977. V. 52. № 1. P. 107-145.
Grubb P J. A positive distrust in simplicity — lessons from plant defenses and from competition among plants and among animals // Journal of Ecology, 1992. V. 80. №4. P. 585-610.
Grubb P J. Problems posed by sparse and patchily distributed species in species-rich plant communities // Community Ecology. Diamond J., Case T. (eds.). New York: Harper and Row, 1986. P. 207-226.
Grubb P J. Root competition in soils of different fertility: a paradox resolved? // Phytocoenologia, 1994. V.24. P. 495-505.
Grubb P.J., Green H.E., Merrifield R.C.J. The ecology of chalk heath: its relevance to the calcicole-calcifuge and soil acidification problems // Journal of Ecology, 1969. V. 57. № 1. P. 175-212.
Grummer G. Die gegenseitige Beeinflussung Hoherer Pflanzen-Allelopathie. Jena: Fischer, 1955. 162 S.
Grundmann G. £., Debouzie D. Geostatistical analysis of the distribution of NH4 and NO2- oxidizing bacteria and serotypes at the millimeter scale along a soil transect // FEMS Microbiology Ecology, 2000. V. 34. № 1. P. 57-62.
Guo D., Mitchell RJ., Withington J.M., Fan P.-Р., Hendricks J J. Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: root branch order predominates // Journal of Ecology, 2008. V.96. №4. P. 737-745.
Guo Q. Temporal species richness-biomass relationships along successional gradients // Journal of Vegetation Science, 2003. V. 14. № 1. P. 121-128.
Guo Q., Brown J. H. Temporal fluctuations and experimental effects in desert plant communities // Oecologia, 1996. V. 107. №4. P. 568-577.
Guo Q., Thompson D. B., Valone T /., and Brown J. H. The effects of vertebrate granivores and folivores on plant community structure in the Chihuahuan Desert // Oikos, 1995. V. 73. №2. P. 251-259.
Gurevitch J., Collins S. L. Experimental manipulation of natural plant communities // Trends in Ecology and Evolution, 1994. V.9. №3. P. 94-98.
Gurevitch /., Morrow L. £., Wallace A., Walsh 1. S. A meta-analysis of field experiments on competition // American Naturalist, 1992. V. 140. №4. P. 539-572.
Список литературы
523
Gurevitch /., Unnasch R. S. Experimental removal of a dominant species at two levels of soil fertility // Canadian Journal of Botany, 1989. V. 67. № 12. P. 3470-3477.
Gwynn-Jones D., Lee /., Johanson U., Phoenix G., Callaghan T, Sonesson M. The responses of plant functional types to enhanced UV-B radiation // Stratospheric ozone depletion: the effects of UV-B radiation on terrestrial ecosystems. Rozema J. (ed.). Leiden: Backhuys Publishers, 1999. P. 173-185.
Hacker S. D., Bertness M. D. Morphological and physiological consequences of a positive plant interaction // Ecology, 1995. V. 76, No 7. P. 2165-2175.
Haddad N. M., Tilman D., Knops M. H. Long-term oscillations in grassland productivity induced by drought // Ecology Letters, 2002. V. 5. № 1. P. 110-120.
Hale M.E.Jr. The biology of lichens. 3rd Edit. Baltimore: Edward Arnold, 1983. 190 p.
Hamilton R. G., Lloyd R. M. An experimental study on the effects of earthworms on the ecological success of fern gametophytes // American Fern Journal, 1991. V.81. № 1. P. 95-99.
Han X., Dendy S.P., Garrett K.A., Fang L., Smith M.D. Comparison of damage to native and exotic prairie plants by natural enemies // Plant Ecology, 2008. V. 198. №2. P. 197-210.
Handel S. N., Fisch S. B., Schatz G. E. Ants disperse a majority of herbs in a mesic forest community in New York State // Bulletin of the Torrey Botanical Club, 1981. V. 108. № 4. P. 430-437.
Hanley M. E., Lamont В. B., Fairbanks M. M., Rafferty С. M. Plant structural traits and their role in anti-herbivore defence // Perspectives in Plant Ecology, Evolution and Systematics, 2007. V. 8. №4. P. 157-178.
Hansen E. S., Graff-Petersen P Lichens growing on the Ella Island Meteorite, Central East Greenland // Lichenologist, 1986. V. 18. № 1. P. 71-78.
Hanslin H. M., Karlsson P S. Nitrogen uptake from prey and substrate as affected by prey capture level and plant reproductive status in four carnivorous plant species // Oecologia, 1996. V. 106. №3. P. 370-375.
Harley C. D. G., Bertness M. D. Structural interdependence: an ecological consequence of morphological responses to crowding in marsh plants // Functional Ecology, 1996. V. 10. №5. P. 654-661.
Harley J.L. The mycorrhizal associations // Cell Interactions. Berlin e.a., 1984. P. 148-186.
Harley J. L., Smith S. E. Mycorrhizal Symbiosis. London, New York: Academic Press, 1983. 483 p.
Harmon M. E., Franklin J. F. Tree seedlings on logs in Picea-Tsuga forests of Oregon and Washington // Ecology, 1989. V. 70. № 1. P. 48-59.
Harmon M.E. (1989) Retention of needles and seeds on logs in Picea sitchensis — Tsuga heterophylla forests of coastal Oregon and Washington // Canadian Journal of Botany, 1989. V.67. №6. P. 1833-1837.
Harper J. L. The role of predation in vegetational diversity // Brookhaven Symposium Biology, 1969. №22. P. 48-62.
Harper J.L. The population biology of plants. London: Academic Press, 1977. 892 p.
Hartnett D. C., Samanus R.J., Fischer E., Hetrick B.A.D. Plant demographic responses to mycorrhizal symbiosis in tailgrass prairie // Oecologia, 1994. V.99. № 1. P. 21-26.
Hartnett D. C., Wilson G. W. T. Mycorrhizae influence plant community structure and diversity in tailgrass prairie // Ecology, 1999. V.80. №4. P. 1187-1195.
Hatcher P. E., Paul N. D., Ayres P. G., Whittaker J. B. The effect of an insect herbivore and a rust fungus individually, and combined in sequence, on the growth of two Rumex species // / New Phytologist, 1994. V. 128. № 1. P. 71-/78.
Haukioja E. On the role of plant defenses in the fluctuation of herbivore populations // Oikos, 1980. V.35. №2. P. 202-213.
Hawkins H.J., Johansen A., George E. Uptake and transport of inorganic and organic nitrogen by arbuscular mycorrhizal fungi // Plant and Soil, 2000. V. 226. №2. P. 275-285.
524
Список литературы
Hay R. К. М. The influence of photoperiod on dry-matter production of grasses and cereals // New Phytologist, 1990. V. 116. № 2. P. 233-254.
Heath M. C. Evolution of resistance to fungal parasitism in natural ecosystems // New Phytologist, 1991. V. 119. №3. P. 331-343.
Hedin P. A., Miles L. R., Thompson A. C., Minyard J. P. Constituents of the cotton bud. Formulation of a boll weevil feeding stimulant mixture // Journal of Agricultural and Food Chemistry, 1968. V.16. №3. P. 505-513.
Hegarty E. E. The climbers — lianes and vines // Ecosystems of the world 14B. Tropical rain forest ecosystems. Biogeographical and ecological studies. H. Lieth and M. J. A. Werger (eds.). Amsterdam: Elsevier, 1989. P. 339-353.
Hegde S. G., Lokesha R., Ganeshaiah K. N., Shaanker R. U. Seed size distribution in plants: an explanation based on fractal geometry // Oikos, 1991. V. 62. №1. P. 100-101.
Heide-Jorgensen H. S. Parasitic flowering plants. Leiden, Boston: Brill, 2008. 438 p.
Heil M. Indirect defence via tritrophic interactions // New Phytologist, 2008. V. 178. № 1. P.41-61.
Hendricks J J., Nadelhoffer KJ., Aber J. D. Assessing the role of fine roots in carbon and nutrient cycling // Trends in Ecology and Evolution, 1993. V. 8. №5. P. 174-178.
Hendry G.A. E, Thompson K., Moss C.J., Edwards E., Thorpe P. C. Seed persistence: a correlation between seed longevity in the soil and ortho-dihydroxyphenol concentration // Functional Ecology, 1994. V. 8, No 5. P. 658-664.
Heppell К. B., Shumway D. L., Koide R. T The effect of mycorrhizal infection of Abutilon theophrasti on competitiveness of offspring // Functional Ecology, 1998. V. 12. №2. P. 171-175.
Herben T Founder and dominance control: neglected concepts in the community dynamics of clonal plants // Abstracta Botanica. Budapest, 1995. V. 19. P. 3-10.
Herben T, Brezina S., Skalova H., Hadincova V., Krahulec E Variation in plant performance in a grassland: Species-specific and neighbouring root mass effects // Journal of Vegetation Science, 2007. V. 18. № 1. P. 55-62.
Herben T, Hara T Competition and spatial dynamics of clonal plants // The ecology and evolution of clonal plants, de Kroon H., van Groenendael (eds.). Leiden: Backhuys Publishers, 1997. P. 331-357.
Herben T, Krahulec E, Hadincova V, Kovarova M. Small scale spatial dynamics of plant species in a grassland community during six years // Journal of Vegetation Science, 1993. V. 4. №2, p 171-178.
Herms D.A., Mattson W. J. The dilemma of plants: to grow or defend // Quarterly Review of Biology, 1992. V.67. №3. P. 283-335.
Herrera С. M. Habitat-shaping, host plant use by a hemiparasitic shrub, and the importance of gut fellows // Oikos, 1988. V. 51. № 3. P. 383-386.
Hessing M. B. Geitonogamous pollination and its consequences in Geranium caespitosum // American Journal of Botany, 1988. V. 75. №9. P. 1324-1333.
Hester A. J., Gimingham C.H., Miles J. Succession from heather moorland to birch woodland 111. Seed availability, germination and early growth // Journal of Ecology, 1991. V. 79, No 2. P. 329-344.
Hill M.O., Stevens P.A. The density of viable seeds in soils of forest plantations in upland Britain //Journal of Ecology, 1981. V.69. №2. P. 693-709.
Hill R.D. What are hemoglobins doing in plants? // Canadian Journal of Botany, 1998. V. 76. №5. P. 707-712.
Hille Ris. Lambert J., Clark J. S., and Beckage B. Density-dependent mortality and the latitudinal gradient in species diversity // Nature, 2002. V. 417. №6890. P. 732-735.
Список литературы
525
Нinsinger Р, Plassard С., Tang С., Jail lard В. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: a review // Plant and Soil, 2003. V. 248. №1. P. 43-59.
Hiol EH., Dixon R.K., Curl E.A. The feeding preference of a mycophagous collembolan varies with the ectomycorrhiza symbiont // Mycorrhiza, 1994. V. 5. №2. P. 99-103.
Hirsch G., Braun U. Communities of parasitic microfungi // Fungi in vegetation science. W. Win-terhoff (ed.). Dordrecht e.a.: Kluwer Academic Publishers, 1992. P.225-250.
Hjalten J., Danell K., and Ericson L. Effects of simulated herbivory and intraspecific competition on the compensatory ability of birches // Ecology, 1993. V. 74. №4, P. 1136-1142.
Hoagland В. IV., Collins S. L. Gradient models, gradient analysis, and hierarchial structure in plant communities, Oikos, 1997. V. 78. №1. P. 23-30.
HobbieJ.E., Hobbie E.A. N-15 in symbiotic fungi and plants estimates nitrogen and carbon flux rates in Arctic tundra // Ecology, 2006. V. 87. №4. P. 816-822.
Hobbie S. E. Effects of plant species on nutrient cycling // Trends in Ecology and Evolution, 1992. V. 7. №10. P. 336-339.
Hobbs R.J., Yates S., Mooney H.A. Long-term data reveal complex dynamics in grassland in relation to climate and disturbance // Ecological Monographs, 2007. V. 77. № 4. P. 545-568.
Hochuli D. F. Does silica defend grasses against invertebrate herbivores? // Trends in Ecology and Evolution, 1993. V. 8. № 11. P. 418-419.
Hochuli D. F. The ecology of plant/insect interactions: implications of digestive strategy for feeding by phytophagous insects // Oikos, 1996. V. 75. № 1. P. 133-141.
Hodge A., Campbell C. D., Fitter A. H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material // Nature, 2001. V. 413. P. 297-299.
Hodgson J. G., Mackey J. M. The ecological specialization of dycotyledonous families within a local flora: some factors constraining optimization of seed size and their possible evolutionary significance // New Phytologist, 1986. V. 104. №3. P. 497-515.
Hodkinson D.J., Thompson K. Plant dispersal: the role of man // Journal of Applied Ecology, 1997. V.34. №6. P. 1484-1496.
Hoft F.A., Valone T.J., Brown J.H. Competition theory and the structure of ecological communities // Evolutionary Ecology, 1993. V. 7. №2. P. 142-154.
Hdgberg P, Nordgren A., Buchmann N., Taylor A. F. S., Ekblad A., Hdgberg M. N., Nyberg G., Read D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration // Nature, 2001. V. 411. №6839. P. 789-792.
Hdgberg P, Read D.J. Towards a more plant physiological perspective on soil ecology // Trends in Ecology and Evolution, 2006. V. 21. № 10. P. 548-554.
Hdgberg R. A., Jackson R.B. Global patterns of root turnover for terrestrial ecosystems // New Phytologist, 2000. V. 147. № 1. P. 13-31.
Holdworth D. K., Mark A. F. Water and nutrient inputoutput budgets: effects of plant cover at seven sites in upland snow tussock grasslands of Eastern and Central Otago, New Zealand // Journal of the Royal Society of New Zealand, 1990. V. 20. № 1. P. 1-24.
Holmgren M., Scheffer M., Ezcurra E., Gutierrez J.R., Mohren G.M.J. El Nino effects on the dynamics of terrestrial ecosystems // Trends in Ecology and Evolution, 2001. V. 16. №2. P. 89-94.
Honegger R. The symbiotic phenotype of lichen-forming ascomicetes // Fungal Associations. Hock B. (ed.): The Mycota IX. Berlin, Heidelberg: Springer-Verlag, 2001. P. 165-188.
Hood M. E., Antonovics J. Species descriptions show signs of disease // Proceedings of the Royal Society London Series B, 2003. V. 270, Suppl. 2, p. S156-S158.
Hook P. B., Lauenroth IV. K. Root system response of a perennial bunchgrass to neighborhoodscale soil water heterogeneity // Functional Ecology, 1994. V.8. №6. P. 738745.
526
Список литературы
Hooper D. U., Vitousek Р. М. Effects of plant composition and diversity on nutrient cycling // Ecological Monographs, 1998. V. 68. № 1. P. 121-149.
Hopfensperger K. N. A review of similarity between seed bank and standing vegetation across ecosystems // Oikos, 2007. V. 116. №9. P. 1438-1448.
Hopkins B. The specie-sarea relations of plant communities // Journal of Ecology, 1955. V. 43. №2. P. 409-426.
Horner-Devine M.C., Leibold M.A., Smith V.H., Bohannon B.J.M. Bacterial diversity patterns along a gradient of primary productivity // Ecology Letters, 2003. V. 6. №6. P. 613-622.
Horton J. L., Hart S. C. Hydraulic lift: a potentially important ecosystem process // Trends in Ecology and Evolution, 1998. V. 13, No 6. P. 232-235.
Howe H. E, Westley L. C. Ecological relationships of plants and animals. New York, Oxford: Oxford University Press, 1988. 273 p.
Hubbell S. P. Neutral theory in community ecology and the hypothesis of functional equivalence // Functional Ecology, 2005. V. 19. № 1. P. 166-172.
Hubbell S. P, Foster R. B. Canopy gaps and the dynamics of a neotropical forest // Plant ecology. Crawley M.J. (ed.). Oxford: Blackwell SP, 1986. P. 77-96.
Hubbell S. P, Foster R. B. The spatial context of regeneration in a neotropical forest // Colonization, succession and stability. Gray A. J., Crawley M.J., Edwards P.J. (eds.). Oxford e.a.: Blackwell SP, 1987. P.395-412.
Hughes L. Biological consequences of global warming: is the signal already apparent? // Trends in Ecology and Evolution, 2000. V. 15. № 2. P. 56-61.
Hughes L., Westoby M., Johnson A. D. Nutrient costs of vertebrate- and ant-dispersed fruits // Functional Ecology, 1993. V. 7. № 1. P. 54-62.
Hull R.J., Leonard O.A. Physiological aspects of parasitism in mistletoes (Arceuthobium and Phoradendron). 1. The carbohydrate nutrition of mistletoe. Plant Physiology, 1964a. V. 39. №6. P. 996-1007.
Hull R.J., Leonard O.A. Physiological aspects of parasitism in mistletoes (Arceuthobium and Phoradendron). 11. The photosynthetic capacity of mistletoe // Plant Physiology, 1964b. V. 39. №6. P. 1008-1017.
Hunt R., Hand D. W., Hannah M.A., Neal A. M. Further responses to CO2 enrichment in British herbaceous species // Functional Ecology, 1993. V. 7. №6. P. 661-668.
Huntly N. Inouye R. Pocket gophers in ecosystems: patterns and mechanisms // BioScience, 1988. V. 38. №11. P. 786-793.
Hurlbert S. H. A gentle depilation of the niche: Dicean resource sets in resource hyperspace // Evolutionary Theory, 1981. V. 5. P. 177-184.
Huss-Danell K. Actinorhizal symbioses and their N2 fixation // New Phytologist, 1997. V. 136. №3. P. 375-405.
Hutchin P. R., Press M. C., Lee J. A., Ashenden T. W. Elevated concentrations of CO2 may double methane emissions from mires // Global Change Biology, 1995. V. 1. №2. P. 125-128.
Hutchings M.J.John E.A. Distribution of roots in soil, and root foraging activity // Root ecology (Ecological Studies. V. 168). de Kroon H., Visser E.J. W. (eds.). Berlin, Heidelberg: Springer, 2003. V. 33-60.
Hutchinson G.E. Concluding remarks, Cold Spring Harbor Symp. // Quantitative Biology, 1957. V.22. P. 415-422.
lason G.R., Hester A. J. The response of heather (Calluna vulgaris) to shade and nutrients — predictions of the carbon-nutrient balance hypothesis //Journal of Ecology, 1993. V. 81. №1. P. 75-80.
Ibarra-Manriquez G., Ramos M. M., Oyama K. Seedling functional types in lowland rain forest in Mexico // American Journal of Botany, 2001. V. 88. № 10. P. 1801-1812.
Ibisch P.L. Neotropische Epiphytendiversitat — das Beispiel Bolivien.------Wiehl: Martina-
Galunder-Verlag, 1996. 357 S.
Список литературы
527
Inderjit, Weiner J. Plant allolochemical interference or soil chemical ecology? // Perspectives in Plant Ecology, Evolution and Systematics, 2001. V. 4. № 1. P.3-12.
Ingerpuu N., Kupper T. Response of calcareous grassland vegetation to mowing and fluctuating weather conditions // Journal of Vegetation Science, 2007. V. 18. № 1. P. 141-146.
Ingerpuu N., Liira/., Partel M. Vascular plants facilitated bryophytes in a grassland experiment // Plant Ecology, 2005. V 180. № 1. P. 69-75.
Ingham E. R., Trofymow J. A., Ames R. N., Hunt H. W, Morley C. R., Moore J. C., and Coleman D. C. Trophic interactions and nitrogen cycling in a semiarid grassland soil. Part 11. System responses to removal of different groups of soil microbes or fauna // Journal of Applied Ecology, 1986. V 23. №2. P. 615-630.
Ingham J. L Phytoalexins and other natural products as factors in plant disease resistance // Botanical Review, 1972. V. 38. №3. P. 343-424.
Inghe О., Tamm С. O. Survival and flowering of perennial herbs. IV The behavior of Hepatica nobilis and Sanicula еигораеа on permanent plots during 1943-1981 // Oikos, 1985. V.45. №3. P. 400-420.
Jackson R. B., Caldwell M. M. Integrating resource heterogeneity and plant plasticity: modeling nitrate and phosphorus uptake in a patchy soil environment // Journal of Ecology, 1996. V. 84. №6. P. 891-903.
Jackson R. B., Caldwell M. M. Shading and the capture of localized soil nutrients: nutrient contents, carbohydrates, and root uptake kinetics of a perennial tussock grass // Oecoiogia, 1992. V.91.№4. P. 457-462.
Jackson R. B., Caldwell M. M. The scale of nutrient heterogeneity around individual plants and its quantification with geostatistics // Ecology, 1993. V. 74, No 2. P. 612-614.
Jager E.J. Veranderung des Artenbestandes von Floren unter dem Einfluss des Menschen // Biol. Rdsch., 1977; Bd. 15, S. 287-300.
Jahns H. M. The cyclic development of mosses and the lichen Baeomyces rufus in an ecosystem // Lichenologist, 1982. V. 14. №3. P. 261-265.
Jakobsen I. Transport of phosphorus and carbon in arbuscular mycorrhizas // Mycorrhiza: structure, function, molecular biology, and biotechnology. 2nd ed. Varma A. and Hock B. (eds.). Berlin: Springer, 1999. P. 305-332.
Jakobsen I. Transport of phosphorus and carbon in arbuscular mycorrhizas // Mycorrhiza: structure, function, molecular biology, and biotechnology. 2nd ed. Varma A. and Hock B. (eds.). Berlin: Springer, 1999. P. 305-332.
Jaksic F. M., Fuentes E. R. Why are native herbs in the Chilean matorral more abundant beneath bushes: microscale or grazing? // Journal of Ecology, 1980. V. 68. № 2. P. 665-669.
Janzen D. H. Physiological ecology of fruits and their seeds // Functional Responses to Chemical and Biological Environments. O. L. Lange, P. S. Nobel, С, B. Osmond, H. Ziegler (eds.). Encyclopedy of Plant Physiology (New Series), 1983. V. 12C. P. 625-655.
Jarzen D.M. The occurrence of Gunnera pollen in the fossil record // Biotropica, 1980. V. 12. №2. P. 117-123.
Jasper D. A., Abbott L K., Robson A. D. Soil disturbance reduces the infectivity of external hyphae of vesiculararbuscular mycorrhizal fungi // New Phytologist, 1989. V. 112, No 1. P. 93-99.
Jeschke D. IV., Pate J. S. Mineral nutrition and transport in xylem and phloem of Banksia prionotes (Proteaceae), a tree with dimorphic root morphology // Journal of Experimental Botany, 1995. V.46. №8. P. 895-905.
Jobbagy E. G., Jackson R.B. The uplift of soil nutrients by plants: biogeochemical consequences across scales // Ecology, 2004. V. 85. №9. P. 2380-2389.
Joffre R., Rambal S. Soil water improvement by trees in the rangelands of southern Spain // Oecoiogia Plantarum, 1988. V. 9. P. 405-422.
528
Список литературы
Johansen A., Jakobsen I. Jensen E.S. Hyphal transport of 15N-labelled nitrogen by a vesicular-arbuscular mycorrhizal fungus and its effect on depletion of inorganic soil N // New Phytologist, 1992. V. 122. № 2. P. 281-288.
Johansen A., Jakobsen /., Jensen E. S. Hyphal N transport by a vesicular-arbuscular mycorrhizal fungus associated with cucumber growth at three nitrogen levels // Plant and Soil, 1994. V. 160. №1. P.1-9.
Johnson C. N. Interactions between mammals and ectomycorrhizal fungi // Trends in Ecology and Evolution, 1996. V. 11. № 12. P. 503-507.
Johnson E.A., Miyanishi K. Testing the assumptions of chronosequences in succession // Ecology Letters, 2008. V. 11. №5. P. 419-431.
Johnson L. C., Matchett J. R. Fire and grazing regulate belowground processes in tallgrass prairie // Ecology, 2001. V.82. № 12. P. 3377-3389.
Johnson M. G., Tingey D. T, Phillips D. L, and Storm MJ. Advancing fine root research with minirhizotrons // Environmental and Experimental Botany, 2001. V. 45. №3. P. 263-289.
Johnson N.C., Gehring C.A. Mycorrhizas: symbiotic mediators of rhizosphere and ecosystem processes // The Rhizosphere. An ecological Perspective. Cardon Z. G., Whitbeck J. L. (eds.). Amsterdam e.a.: Elsevier AP, 2007. P. 73-100.
Johnston A., Dormaar J. E, Smoliak S. Long-term grazing effects on fescue grassland soils // Journal of Range Management, 1971. V. 24. №3. P. 185-188.
Jones В. M., Kolden C.A., Jandt R., Abatzoglou J. I, Urban E, Arp C. D. Fire behavior, weather, and burn severity of the 2007 Anaktuvuk River Tundra Fire, North Slope, Alaska // Arctic, Antarctic, and Alpine Research, 2009. V. 41. №3. P. 309-316.
Jones DA. Cyanogenic glycosides and their function // Phytochemical Ecology. J.B. Harborne (ed.). London: Academic Press, 1972. P. 103-124.
Jonsson B. G. The bryophyte diaspore bank and its role after small-scale disturbance in a boreal forest //Journal of Vegetation Science, 1993. V. 4, No 6. P. 819-826.
Jonsson O., Jonsdottir /., Cronsberg N. Clonal diversity and allozyme variation in populations of the arctic sedge Carex bigelowii (Cyperaceae) // Journal of Ecology, 1996. V. 84. № 3. P. 449-459.
Joshi G. С. P. С. P, Kothyari В. P. New host of Dendrophthoe falcata (Linn, f.) Etting // Indian Journal of Forestry, 1985. V.8. P. 235.
Jung /., Winter K. Mineral nutrient deficiency increases the sensitivity of photosynthesis to sulphur dioxide in needles of a coniferous tree, Abies nordmanniana // Oecologia, 1992. V.90. №1. P. 70-73.
JCirgens A., El-Sayed A.M., Suckling D.M. Do carnivorous plants use volatiles for attracting prey insects? // Functional Ecology, 2009. V. 23. №5. P. 875-887.
Kahlert B. R., Ryser P, Edwards P. J. Leaf phenology of three dominant limestone grassland plants matching the disturbance regime // Journal of Vegetation Science, 2005. V. 16. №4. P. 433-442.
Kannadan S., Rudgers J. A. Endophyte symbiosis benefits a rare grass under low water availability // Functional Ecology, 2008. V. 22. №4. P. 706-713.
Karanika E. D., MamolosA. P, Alifragis D.A., Kalburtji K. L. Veresoglou D. S. Arbuscular mycorrhizas contribution to nutrition, productivity, structure and diversity of plant community in mountainous herbaceous grassland of northern Greece // Plant Ecology, 2008. V. 199. №2. P. 225-234.
Karlsson P. S., Mendez M. The resource economy of plant reproduction // Reproductive Allocation in Plants. Reekie E. G., Bazzaz F. A. (eds.). Amsterdam e. a.: Elsevier AP, 2005. P.1-49.
Karlsson P. S., Pate J. S. Contrasting effects of supplementary feeding of insects or mineral nutrients on the growth and nitrogen and phosphorus economy of pygmy species of Drosera // Oecologia, 1992a. V. 92. № 1. P. 8-13.
Список литературы
529
Karlsson Р. S., Pate J. S. Resource allocation to asexual gemma production and sexual reproduction in south-western Australia pygmy and micro stil-form species of sundew (Drosera spp., Droseraceae)// Australian Journal of Botany, 1992b. V. 40. №3. P. 353-364.
Kawano S., Masada J. The productive and reproductive biology of flowering plants. VII. Resourse allocation and reproductive capacity in wild populations of Heloniopsis orientalis (Thunb.) C.Tanaka (Liliaceae) // Oecologia, 1980. V.45. №3. P. 307-317.
Kays S., Harper J.L. The regulation of plant and tiller density in a grass sward // Journal of Ecology, 1974. V.62. № 1. P. 97-105.
Keatinge T. Plant community dynamics in wet heathland // Journal of Ecology, 1975. V. 63. № 1. P. 163-172.
Keddy P, Fraser L. H.y Wisheu 1. C. A comparative approach to examine competitive response of 48 wetland plant species // Journal of Vegetation Science, 1998. V. 9. № 6. P. 777-786.
Keddy P.A. Competition. London: Chapman and Hall, 1989. 198 p.
Keddy P.A. Competitive hierarchies and centrifugal organisation in plant communities // Perspectives on plant competition. Grace J. B., Tilman D. (eds.). San Diego e. a.: Academic Press, 1990. P. 265-290.
Keddy P.A.y MacLellan P. Centrifugal organization in forests // Oikos, 1990. V. 59. № 1. P. 75-84.
Keeley J.E., Fotheringham C.J. Smoke-induced seed germination in California chaparral // Ecology, 1998. V. 79, No 7. P. 2320-2336.
Keeling C. D. NATO lecture 2: Surface ocean CO2 // The Global Carbon Cycle. M. Heimann (ed.). New York: Springer-Verlag, 1993. P. 413-430.
Kelly V.R., Canham C.D. Resource heterogeneity in old fields // Journal of Vegetation Science, 1992. V.3. P. 542-552.
Kemperman J. A., Barnes В. V. Clone size in American aspens // Canadian Journal of Botany, 1976. V. 54. № 22. P. 2603-2607.
Kennedy C.E.J., Southwood T.R.E. The number of species of insect associated with British trees. A reanalysis // Journal of Animal Ecology, 1984. V. 53. № 2. P. 455-478.
Kershaw K.A. Pattern in vegetation and its causality // Ecology, 1963. V. 44. № 2. P. 377-388.
Kery M., Gregg К. B., Schaub M. Demographic estimation methods for plants with unobservable life-states // Oikos, 2005. V. 108. № 2. P. 307-320.
Khalid A. N., Iqbal S.H. Mycotrophy in a vascular stem parasite Cuscuta reflexa 11 Mycorrhiza, 1996. V. 6. №1. P. 69-71.
Kielland K. Amino acid absorption by arctic plants: implications for plant nutrition and nitrogen cycling // Ecology, 1994. V. 75. №8. P. 2373-2383.
Kikuzawa K. Ecology and evolution of phenological pattern, leaf longevity and leaf habit // Evolutionary trends in plants, 1989. V. 3. P. 105-110.
Kikuzawa K., Kudo G. Effects of the length of the snow-free period on leaf longevity in alpine shrubs: a cost-benefit model // Oikos, 1995. V. 73. № 2. P. 214-220.
Kikvidze Z. Neighbour interaction and stability in subalpine meadow communities // Journal of Vegetation Science, 1996. V. 7. № 1. P. 41-44.
Kilburn P.D. Analysis of species-area relation // Ecology, 1966. V. 47. №5. P. 831-843.
Kile G. A. Armillaria root rot in Eucalypt forests: Aggravated endemic disease! // Pacific Science, 1983. V.37. №4. p. 459-464.
Killingbeck К. T. Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency // Ecology, 1996. V. 77. №6. P. 1716-1727.
Kitajima K. Seed and seedling ecology // Functional Plant Ecology. 2nd edit. Pugnaire F. I., Valladares F. (eds.). Boca Raton e.a.: CRC Press, 2007. P.549-579.
Kivilaan Л., Bandurski R. S. The one hundred-year period for the Dr. Beal’s seed viability experiment // American Journal of Botany, 1981. V. 68. №9. P. 1290-1292.
Klapp E. Wiesen und Weiden. 4th Auf. Berlin und Hamburg: Parey, 1971. 620 S.
530
Список литературы
Kleb Н. R., Wilson S. D. Vegetation effects on soil resource heterogeneity in prairie and forest // American Naturalist, 1997. V. 150, No 3. P. 283-298.
Kleier C., Lambrinos J. G. The importance of nurse associations for three tropical alpine life forms // Arctic, Antarctic, and Alpine Research, 2005. V. 37. №3. P. 331-336.
Klemow К. M., Raynal D. J. Population biology of an annual plant in a temporally variable habitat //Journal of Ecology, 1983. V.71. №3. P.691-703.
Klinger L. F. The myth of the classic hydrosere model of bog succession // Arctic and Alpine Research, 1996. V. 28. № 1. P. 1-9.
Klinkhamer P. G.L., de Jong T.J. Attractiveness to pollinators: a plant’s dilemma // Oikos, 1993. V. 66. №1. P. 180-184.
Klironomos J. N., Kendrick B. Relationships among microarthropods, fungi, and their environment // Plant and Soil. 1995. V. 170. №1. P. 183-197.
Knight S. E. Cost of carnivory in the common bladderwort, Utricularia macrorhiza // Oecoiogia, 1992. V. 89. №3. P. 348-355.
Knoch T R., Faeth S. H., Arnott D. L. Endophytic fungi alter foraging and dispersal by desert seed-harvesting ants // Oecoiogia, 1993. V.95. №4. P. 470-473.
Knops J.M.H., Tilman D. Dynamics of soil nitrogen and carbon accumulation for 61 years after agricultural abandonment // Ecology, 2000. V. 81. № 1. P. 88-98.
Koblihova-Baumova H. Mowing // Succession in abandoned fields studies in Central Bohemia, Czechoslovakia. J. Osbornov^, M. Kov£rov£, J. Leps, arid K. Prach (eds.). Dordrecht: Kluwer Academic Publishers, 1990. P. 94-97.
Koenig W. D., Ashley M. V. Is pollen limited? The answer is blowin’ in the wind // Trends in Ecology and Evolution, 2003. V. 18. №4. P. 157-159.
Koh S., Hik D. S. Herbivory mediates grass-endophyte relationships // Ecology, 2007. V. 88. №11. P. 2752-275.
Koide R. T. Nutrient supply, nutrient demand and plant response to mycorrhizal infection // New Phytologist, 1991. V. 117. №3. P. 365-386.
Komarek E. V. Fire as an anthropogenic factor in vegetation ecology // Man’s impact on vegetation. W. Holzner, M.J.A.Werger and l.lkusima (eds.). The Hague e.a.~Dr. W.Junk Publishers, 1983. P. 77-82.
Koptur S. Extrafloral nectary-mediated interactions between insects and plants // Insect-plant interactions. V.IV. Bernays E.A. (ed.). Boca Raton: CRC Press, 1992. P.81-129.
Korner C. Alpine plants: stressed or adapted? // Physiological plant ecology. Press M. C., Scholes J.D., Barker M. G. (eds.). Oxford: Blackwell Science, 1999. P. 297-311.
Kbrner C. Biosphere responses to CO 2 enrichment // Ecological Applications, 2000. V. 10. №6. P. 1590-1619.
Kbrner C. Scaling from species to vegetation: the usefulness of functional groups // Biodiversity and ecosystem function. Schulze E.-D., Mooney H. A. (eds.). Berlin: Springer, 1993. P. 117-140.
Kbrner C., Renhardt U. Dry matter partitioning and root/leaf area ratios in herbaceous perennial plants//Oecoiogia (Berlin), 1987. V.74. №3. P. 411-418.
Kbrner C., Stocklin /., Reuther-Thiebaud L., Pelaez-Riedl S. Small differences in arrival time influence composition and productivity of plant communities // New Phytologist, 2008. V. 177. №3. P. 698-705.
Kourtev P. S., Ehrenfeld J. G., Haggblom M. Exotic plant species alter the microbial community structure and function in the soil // Ecology, 2002. V83, No 11. P. 3152-3166.
Kranz J. Fungal diseases in multispecies plant communities // New Phytologist, 1990. V. 116. №3. P. 383-405.
Kropff MJ., Smeets W.L.M., Meijer E.M.J., ban der Zalm A J. A., Bakx EJ. Effects of sulphur dioxade on leaf photosynthesis: the role of temperature and humidity // Physiologia Plantarum, 1990. V.80. №4. P. 655-661.
Список литературы
531
Krupa S., Fries N. Studies on ectomycorrhizae of pine. 1. Production of volatile organic compounds // Canadian Journal of Botany, 1971. V. 49. №8. P. 1425-1431.
Kull 0., Aan A., Soelsepp T. Light interception, nitrogen and leaf mass distribution in a multilayer plant community // Functional Ecology, 1995. V.9, No 4. P. 589-595.
Kuo /., Hocking P./., Pate /. S. Nutrient reserves in seeds of selected Proteaceous species from south-western Australia // Australian Journal of Botany, 1982. V. 30. №2. P. 231-249.
Kuusinen M., Siitonen J. Epiphytic lichen diversity in old-growth and managed Picea abies stands in southern Finland //Journal of Vegetation Science, 1998. V.9. №2. P. 283-292.
Lambers H., Cramer M.D., Shane M. W., Wouterlood M., Poot P, Veneklaas E.J. Introduction: structure and functioning of cluster roots and plant responses to phosphate deficiency // Plant and Soil, 2003. V. 248. № 1. P. ix-xix.
Landeweert R., Hoffland E., Finlay R. D., Kuyper T. W, van Breemen N. Linking plants to rocks: ectomycorrhizal fungi mobilize nutrients from minerals // Trends in Ecology and Evolution, 2001. V. 16. №5. P. 248-254.
Landhausser S.M., Stadt KJ., Lieffers V.J. Photosynthetic strategies of summergreen and evergreen understory herbs of the boreal mixedwood forest // Oecologia, 1997. V. 112. №2. P. 173-178.
Lange O.L., Budel B., Heber U., Meyer A., Zellner H., Green T. G.A. Temperate rainforest lichens in New Zealand: high thallus water content can severely limit photosynthetic CO 2 exchange // Oecologia, 1993. V.95. №3. P. 303-313.
Lange O. L., Kappen L. Photosynthesis of lichens from Antarctica // Antarctic Research Series, 1972. V.20. P. 83-95.
Lange O. L., Kilian E., Ziegler H. Water vapor uptake and photosynthesis of lichens: performance differences in species with green and blue-green algae as phycobionts // Oecologia, 1986. V. 71. №1.P. 104-110.
Lange O.L., Schulze E.-D., Koch W. Experimentell-Okologische Untersuchungen an Flechten der Negev-Wiiste. 11. CO2 -Gaswechsel und Wasserhaushalt von Ramalina maciformis (Del.) Bory, am natiirlichen Standort wShrend der sommerlichen Trockenperiode // Flora, 1970; B. 159. № 1, S. 38-62.
Lanta V., Leps J. Effects of species and functional group richness on production in two fertility environments: an experiment with communities of perennial plants // Acta Oecologica, 2007. V.32. №1. P. 93-103.
Lapeyrie E, Picatto C., Gerard J., Dexheimer J. T. E. M. study of intracellular and extracellular calcium oxalate accumulation by ecto-mycorrhizal fungi in pure culture or in association with Eucalyptus seedlings // Symbiosis, 1990. V.9. №2. P. 163-166.
Larson D. W. Patterns of lichen photosynthesis and respiration following prolonged frozen storage // Canadian Journal of Botany, 1978. V. 56. № 17. P. 2119-2123.
Laube S., Zotz G. Which abiotic factors limit vegetative growth in a vascular epiphyte? // Functional Ecology, 2003. V. 17. № 5. P. 598-604.
Lauenroth W. K, Adler P B. Demography of perennial grassland plants: survival, life expectancy and life span // Journal of Ecology, 2008. V.96. №5. P. 1023-1032.
Laverty T M. Plant intercations for pollinator visits: a test of the magnet species effect // Oecologia, 1992. V.89. №4. P. 502-508.
Law R., Lewis D. H. Biotic environments and the maintenance of sex: some evidence from mutualistic symbioses // Biological Journal of the Linnean Society, 1983. V.20. №3. P. 249-276.
Lawrey J. D. Correlations between lichen secondary chemistry and grazing activity by Pallifera varia// Bryologist, 1980. V.83. №3. P. 328-334.
Lawrey J. D. Lichen herbivore preference: a test of two hypotheses // American Journal of Botany, 1983. V. 70. №8. P. 1188-1194.
532
Список литературы
Lawton J. Н, Schroder D. Effect of plant type, size of geographic range and taxonomic isolation on number of insect species associated with British plants // Nature, 1977. V. 265. P. 137-140.
Leake J. Johnson D., Donnelly D., Muckle G., Boddy L., ReadD. Networks of power and influence: the role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning // Canadian Journal of Botany, 2004. V.82. №8. P. 1016-1045.
Leake J.R. The biology of myco-heterotrophic (’saprophytic’) plants // New Phytologist, 1994. V. 127, No 2. P. 171-216.
Leek M.A., Parker V. T, Simpson R.L. (ed.) Ecology of soil seed banks. London e.a.: Academic Press, 1989. 462 p.
Lee D. W.. O’Keefe J., Holbrook N. M., Field T. S. Pigment dynamics and autumn leaf senescence in a New England deciduous forest, eastern USA // Ecological Research, 2003. V. 18. №6. P 677-694.
Lee J. A. Unintentional experiments with terrestrial ecosystems: ecological effects of sulfur and nitrogen pollutants //Journal of Ecology, 1998. V.86. №1. P. 1-12.
Leffler A J., Peek M.S., Ryel R.J., Ivans C. Y, Caldwell M.M. Hydraulic redistribution through the root systems of senesced plants // Ecology, 2005. V. 86. № 3. P. 633-642.
Lehman C. L., Tilman D. Competition in spatial habitats // Spatial ecology: The role of space in population dynamics and interspecific interactions. Tilman D., Kareiva P. (eds.). Princeton: Princeton Univ. Press, 1998. P. 185-203.
Lehtila K.. Syrjanen K. Compensatory responses of two Melampyrum species after damage // Functional Ecology, 1995. V.9. №3. P. 511-517.
Lehtonen P, Helander M., Wink M., Sporer E, Saikkonen K. Transfer of endophyte-origin defensive alkaloids from a grass to a hemiparasitic plant // Ecology Letters, 2005. V. 8. №12. P. 1256-1263.
Leigh Hodge A., Fitter A. H. Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material // New Phytologist, 2009. V. 181. №1. P. 199-207.
Leishman M.R., Masters GJ., Clarke I.P., Brown V.K. Seed bank dynamics: the role of fungal pathogens and climate change // Functional Ecology, 2000. V. 14. №3. P. 293-299.
Leishman M. R., Westoby M. Seed size and shape are not related to persistence in soil in Australia in the same way as in Britain // Functional Ecology, 1998. V. 12, No 3. P. 480-485.
Leishman M. R., Westoby M. The role of large seed size in shaded conditions — experimental evidence // Functional Ecology, 1994a. V. 8. № 2. P. 205-214.
Leishman M. R., Westoby M. The role of seed size in seedling establishment in dry soil conditions — experimental evidence from semi-arid species //Journal of Ecology, 1994b. V.82. №2. P.249-258.
Leishman M. R., Westoby M., and Jurado E. Correlates of seed size variation — a comparison among five temperate floras // Journal of Ecology, 1995. V. 83. № 3. P. 517-529.
Lekberg V, Koide R. T. Is plant performance limited by abundance of arbuscular mycorrhizal fungi? A meta-analysis of studies published between 1988 and 2003 // New Phytologist, 2005. V. 168. №1. P. 189-204.
Lerat S., Gauci R., Catford J. G., Vierheilig H., Piche Y., and Lapointe L. 14 C transfer between the spring ephemeral Erythronium americanum and sugar maple saplings via arbuscular mycorrhizal fungi in natural stands // Oecologia, 2002. V. 132. №2. P. 181-187.
Li A. R., Guan K.-Y. Arbuscular mycorrhizal fungi may serve as another nutrient strategy for some hemiparasitic species of Pedicularis (Orobanchaceae) // Mycorrhiza, 2008. V. 18. №8. P 429-436.
Li X.-L., George E., Marschner H. Extension of the phosphorus depletion zone in VA-mycorrhizal white clover in a calcareous soil // Plant and Soil, 1991. V. 136. № 1. P. 41-48.
Список литературы
533
Lichter J. Colonization constraints during primary succession on coastal Lake Michigan sand dunes // Journal of Ecology, 2000. V. 88. № 5. P. 825-839.
Lichter J. Primary succession and forest development on coastal Lake Michigan sand dunes // Ecological Monographs, 1998. V. 68. №4. P. 487-510.
Lindblad P, Bergman B. The cycad-cyanobacterial symbiosis // Handbook of symbiotic cyanobacteria. Rai A. N. (ed.) Boca Raton, Florida: CRC Press, 1990. P. 137-159.
Linhart Y. B. Disease, parasitism and herbivory: multidimentional challenges in plant evolution // Trends in Ecology and Evolution, 1991. V. 6. № 12. P. 392-396.
Liste H.-H., White J. C. Plant hydraulic lift of soil water — implications for crop production and land restoration // Plant and Soil, 2008. V.313, N |. P. 1-17.
Loehle C. Jones R. H. Adaptive significance of root grafting in trees // Functional Ecology, 1990. V. 4. №2. P. 268-271.
Lohmeyer IV., Sukopp H. Agriophyten in der Vegetation Mitteleuropas // Schriftenreihe fiir Vegetationskunde, Bonn-Bad, Godesberg, 1992; H. 25. 186 S.
Lokesha R., Hegde S. G., Shaanker R. U., Ganeshaiah K. N. Dispersal mode as a selective force in shaping the chemical composition of seeds // American Naturalist, 1992. V. 140. №3. P. 520-525.
Lonsdale IV. M. Global patterns of plant invasions and the concept of invasibility // Ecology, 1999. V.80. №5. P. 1522-1536.
Looijen R. C., van Andel J. Ecological communities: conceptual problems and definitions // Perspectives in Plant Ecology, Evolution and Systematics, 1999. V. 2. №2. P. 210-222.
Loope L. L., Medeiros A. C. Biotic interactions in Hawaiian high elevation ecosystems // Tropical alpine environments: plant form and function. Rundel P. W., Smith A. P., Meinzer F. C. (eds.). Cambridge: Cambridge Univ. Press, 1994. P. 337-354.
Louda S. M. Limitation of the recruitment of the shrub Haplopappus squarrosus (Asteraceae) by flower- and seed-feeding insects // Journal of Ecology, 1982. V. 70. № 1. P. 43-53.
Lovett Doust L. Population dynamics and local specialization in a clonal perennial (Ranunculus repens) 1. The dynamics of ramets in contrasting habitats // Journal of Ecology, 1981. V.69. №3. P. 743-755.
Ludi IV, Zoller H. Einige Beobachtungen iiber die Diirreschaden des Sommers 1947 in der Nordschweiz und am schweizerischen Jurarand // Berichte Geobot. Inst. Riibel, Ziirich 1948. Zurich, 1949; S. 69-85.
Ludwig F, Dawson T. E., Kroon H., Berendse F, Prins H. H. Hydraulic lift in Acacia tortilis trees on an East African savanna // Oecoiogia, 2003. V. 134. №3. P. 293-300.
Luken J. O. Directing ecological succession. London, New York: Chapman and Hall, 1990. 251 p.
Lundquist P-О. Carbon cost of nitrogenase activity in Frankia-Alnus incana root nodules // Plant and Soil, 2005. V. 273. № 1-2. P. 235-244.
Luo Y. Terrestrial carbon-cycle feedback to climate warming // Annual Review of Ecology, Evolution, and Systematics, 2007. V. 38. P. 683-712.
Luo Y. Q., Sherry R., Zhou X. H., and Wan S. Q. Terrestrial carbon-cycle feedback to climate warming: experimental evidence on plant regulation and impacts of biofuel feedstock harvest // Global Change Biology Bioenergy, 2009. V.l.Nsl. P. 62-74.
Luttge U. (Ed.) Vascular plants as epiphytes. Evolution and ecophysiology. Berlin ea.: Springer, 1989 (Ecological studies. V. 76). 270 p.
Lutz HJ. Vegetation on trenched plot twenty-one years after establishment // Ecology, 1945. V. 26. № 2. P. 200-202.
MacArthur R.H. Geographical ecology: patterns in the distribution of species. N.Y. et al.: Harper and Row, 1972. 269 p.
MacArthur R. H., Wilson E. O. The theory of island biogeography. Princeton: Princeton Univ. Press, 1967. 203 p.
534
Список литературы
MacGillivray С. W, Grime J.P. et ISP Team. Testing predictions of the resistance and resilience of vegetation subjected to extreme events // Functional Ecology, 1995a. V. 9. №4. P. 640-649.
MacGillivray C. W, Grime J. P. Genome size predicts frost resistance in British herbaceous plants: implications for rates of vegetation response to global warming // Functional Ecology, 19956. V. 9. №2. P. 320-325.
Mack M. C., D'Antonio С. M. Impacts of biological invasions on disturbance regimes // Trends in Ecology and Evolution, 1998. V. 13. №5. P. 195-198.
Mack R. N. Understanding the processes of weed invasions: the influence of environmental stochasticity // Weeds in a changing world. Proceedings No. 64. Stirton С. H. (ed.). Brighton: British Crop Protection Council, 1995. P. 65-74.
Mackay J.R. Active layer changes (1968 to 1993) following the forest-tundra fire near Inuvik, N. W.T., Canada // Arctic and Alpine Research, 1995. V. 27. №4. P. 323-336.
Malo J. Suarez F. Herbivorous mammals as seed dispersers in a Mediterranean dehesa // Oecologia, 1995. V. 104. №2. P. 246-255.
Mandahar C. L., Gill P. S. The epidemiological role of pollen transmission of viruses // Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz, 1984. V. 91. P. 246-249.
Mansfield T.A. SO2 pollution: a bygone problem or a continuing hazard? // Physiological plant ecology. Press M. C., Scholes J.D., Barker M. G. (eds:). Oxford: Blackwell Science, 1999. P. 219-240.
Mark A. F, Dickinson K.J.M. Maximizing water yield with indigenous non-forest vegetation: a New Zealand perspective // Frotiers in Ecology and Environment, 2008. V. 6. № 1. P. 25-34.
Marks P L The role of pin cherry (Prunus pensylvanica L.) in the maintenance of stability in northern hardwood ecosystems // Ecological Monographs, 1974. V. 44. №1. P. 73-88.
Marks S., Clay K. Physiological responses of Festuca arundinacea to fungal endophyte infection // New Phytologist, 1996. V. 133. №4. P. 727-733.
Marquis R.J., Whelan C. Plant morphology and recruitment of the third trophic level: subtle and and little-recognized defenses? // Oikos, 1996. V. 75. № 2. P. 330-334.
Marschner H. Mineral nutrition of higher plants. 2nd ed. London: Academic Press, 1995. 889 p.
Marschner H., Romheld V. In vivo measurement of root-induced pH changes at the soil-root interface: effect of plant species and nitrogen source // Zeitschrift fur Pflanzenphysiologie, 1983; Bd. 111. P. 241-251.
Martin P H., Canham C. D., Marks P. L. Why forests appear resistant to exotic plant invasions: intentional introductions, stand dynamics, and the role of shade tolerance // Frotiers in Ecology and Environment, 2009. V. 7. №3. P. 142-149.
Martinez-Ramos M., Alvares-Buylla E., Sarukhan J. Tree demography and gap dynamics in a tropical rain forest // Ecology, 1989. V. 70. № 3. P. 555-558.
Martinez-Ramos M., Sarukhan Pinero D. The demography of tropical trees in the context of forest gap dynamics: the case of Astrocaryurn mexicanum at Los Tuxtlas tropical rain forest // Plant Population Ecology. Oxford: Blackwell Sci. Publ., 1988. P. 293-313.
Martorell C., Ezcurra E. The narrow-leaf syndrome: a functional and evolutionary approach to the form of fog-harvesting rosette plants // Oecologia, 2007. V. 151. №4. P. 561-573.
Marvier M.A. A mixed diet improves performance and herbivore resistance of a parasitic plant // Ecology, 1998. V.79. №4. P. 1272-1280.
Massey F. P, Ennos A. R., Hartley S. E. Herbivore specific induction of silica-based plant defences // Oecologia, 2007. V. 152. №4. P. 677-683.
Masuda M., Washitani 1. A comparative ecology of the seasonal schedules for reproduction by seeds in a moist tall grassland community // Functional Ecology, 1990. V. 4, N. P. 169-182.
Список литературы
535
Matamala R., Gonzalez-Meier M.A., JastrowJ. D., Norby RJ., Schlesinger IV H. Impacts of fine root turnover on forest NPP and soil C sequestration potential // Science, 2003. V. 302. P. 1385-1387.
Matthies D. Interactions between the root hemiparasite Melampyrum arvense and mixtures of host plants: heterotrophic benefit and parasite-mediated competition // Oikos, 1996. V. 75. №1. P. 118-124.
Mattson IV./. Herbivory in relation to plant nitrogen content // Annual Review of Ecology and Systematics, 1980. V. 11. P. 119-161.
McAuliffe J. R. Markovian dynamics of simple and complex desert plant communities // American Naturalist, 1988. V. 131. №4. P. 459-490.
McCall A. C., Irwin R. E. Florivory: the intersection of pollination and herbivory // Trends in Ecology and Evolution, 2006. V.9. № 12. P. 1351-1365.
McConnaughay K. D. M., BazzazF.A. The occupation and fragmentation of space: consequences of neighbouring roots // Functional Ecology, 1992. V. 6. №6. P. 704-710.
McCormik J. Succession // Student Publication of the Graduate School of Fine Arts, University of Pennsylvania, 1968; VIA 1. P. 22-35, 131-132.
McFarlane K.J., Yanai R.D. Measuring nitrogen and phosphorus uptake by intact roots of mature Acer saccharum Marsh., Pinus resinosa Ait., and Picea abies (L.) Karst // Plant and Soil, 2006. V. 279. № 1-2. P. 163-172.
McGraw J.B., Day T.A. Size and characteristics of a natural seed bank in Antarctica // Arctic and Alpine Research, 1997. V.29. №2. P. 213-216.
McKendrich S. L., Leake J. R., Taylor D. L., Read D.J. Symbiotic germination and development of myco-heterotrophic plants in nature: ontogeny of Corallorhiza trifida Chatel and characterization of its mycorrhizal fungi // New Phytologist, 2000. V. 145. №3. P. 523-537.
McKendrick S. L., Leake J. R., Read D. J. Symbiotic germination and development of myco-heterotrophic plants in nature: transfer of carbon from ectomycorrhizal Salix repens and Betula pendula to the orchid Corallorhiza trifida through shared hyphal connections // New Phytologist, 2000. V. 145. №3. P. 539-548.
McKey D. Legumes and nitrogen: The evolutionary ecology of a nitrogen-demanding lifestyle // In: Sprent J. I. and McKey D. (eds) Advances in Legume Systematics. V. 5: The Nitrogen Factor. Royal Botanic Gardens, Kew, 1994. P. 211-228.
McLachlan J. S., Clark J. S., and Manos P. S. Molecular indicators of tree migration capacity under rapid climate change // Ecology, 2005. V. 86. №8. P. 2088-2098.
McLellan A. J., Fitter A. H., Law R. On decaying roots, mycorrhizal colonization and the design of removal experiments // Journal of Ecology, 1995. V. 83. № 2. P. 225-230.
McNaughton S.J. Grasses and grazers, science and management // Ecological Applications, 1993. V. 3. №1. P. 17-20.
McNaughton S.J. Physiological and ecological implications of herbivory // Physiological and ecological implications of herbivory. Functional Responses to Chemical and Biological Environments. 0. L. Lange, P. S. Nobel, С. B. Osmond, H. Ziegler (eds.). Encyclopedy of Plant Physiology (New Series), 1983. V. 12C. P. 657-678.
McNaughton S.J., Milchunas D. G., Frank D.A. How can net primary productivity be measured in grazing ecosystems? // Ecology, 1996. V. 77. №3. P. 974-977.
McNaughton S.J., Oesterheld M., Frank D. A., Williams K.J. Primary and secondary production in terrestrial ecosystems // Comparative analyses of ecosystems. Cole J., Lovett G., Findlay S. (eds.). New York e.a.: Springer, 1991. P. 120-139.
McNaughton S.J., Tarrants J.L., McNaughton M.M., Davis R.H. Silica as a defense against herbivory and a growth promotor in African grasses // Ecology, 1985. V. 66. № 2. P. 528-535.
McRill M., Sagar G. R. Earthworms and seeds // Nature, 1973. V. 243. P. 482.
536
Список литературы
Meeks J. С. Cyanobacterial-bryophyte associations // Handbook of symbiotic cyanobacteria. Rai A. N. (ed.). Boca Raton, Florida: CRC Press, 1990. P. 43-63.
Mehrhoff L. J. The Biology of Plant Invasiveness // New England Wild Flower Society, 1998; V 2. №3. P. 8-10.
Menzel A. Jakobi G., Ahas R., Scheifinger H., Estrella N. Variations of the climatological growing season (1951-2000) in Germany compared with other countries // International Journal of Climatology, 2003. V. 23. № 7. P. 793-812.
Meunier J D., Colin F., and Alarcon C. Biogenic silica storage in soils // Geology, 1999. V. 27. №9. P. 835-838.
Meurk C. D. Alpine phytomass and primary production in Central Otago, New Zealand // New Zealand Journal of Ecology, 1978. V. 1. № 1. P. 27-50.
Mevi-Schutz /., Erhardt A. Amino acids in nectar enhance butterfly fecundity: a long-awaited link // American Naturalist, 2005. V. 165. №4. P. 411-419.
Michelsen A, Schmidt I. K., Jonasson S., Dighton /., Jones H. E., Callaghan T. V. Inhibition of growth, and effects on nutrient uptake of arctic graminoids by leaf extracts — allelopathy or resource competition between plants and microbes? // Oecoiogia, 1995. V. 103, No 4. P. 407-418.
Michelsen A., Schmidt I. K. Jonasson S., Quarmby C., Sleep D. Leaf 15 N abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non- and arbuscular mycorrhizal species access different sources of soil nitrogen // Oecoiogia, 1996. V. 105. №1. P. 53-63.
Midgley J J. Why the world’s vegetation is not totally dominated by resprouting plants; because resprouters are shorter than reseeders // Ecography, 1996. *V. 19. № 1. P. 92-95.
Milberg P Fern spores in a grassland soil // Canadian Journal of Botany, 1991. V. 69, No 4. P. 831-834.
Milchunas D. G., Lauenroth W. K. Quantitative effects of grazing on vegetation and soils over a global range of environments // Ecological Monographs, 1993. V. 63. №4. P. 327-366.
Miles J. Vegetation dynamics. London: Chapman and Hall, 1979. 80 p.
Milewski A. V, Young T. P., Madden D. Thorns as induced defenses: experimental evidence // Oecoiogia, 1991. V. 86. №1. P. 70-75.
Miller A. E., Bowman W. D. Variation in nitrogen-15 natural abundance and nitrogen uptake traits among co-occurring alpine species: do species partition by nitrogen form? // Oecoiogia, 2002. V. 130, No 4. P. 609-616.
Miller R. M., Reinhardt D. R., Jastrow J. D. External hyphal production of vesicular-arbuscular mycorrhizal fungi in pasture and tallgrass prairie communities // Oecoiogia, 1995. V. 103. №1. P. 17-23.
Minckier L.S., Woerheide D. Reproduction of hardwoods 10 years after cutting as affected by site and opening size // Journal of Forestry, 1965. V. 63. № 2. P. 103-107.
Mirkin B.M. Which plant communities do exist? // Journal of Vegetation Science, 1994. №5. P. 283-284.
Mitchell C.E., Tilman D., Groth J. V. Effects of grassland plant species diversity, abundance, and composition on foliar fungal disease // Ecology, 2002. V. 83. №6. P. 1713-1726.
Mitchell D.J. The use of vegetation and land use parameters in modeling catchment sediment yields // Vegetation and Erosion. Thornes J. B. (ed.). Chichester: Wiley, 1990. P. 289-316.
Mitchell E.A. D., Borcard D., Buttler A. J., Grosvernier Ph., Gilbert D., Gobat J.-M. Horizontal distribution patterns of testate amoebae (Protozoa) in a Sphagnum magellanicum carpet // Microbial Ecology, 2000. V.39. №4. P. 290-300.
Moen J. Diffuse competition — diffuse concept // Oikos, 1989. V. 54. № 2. P. 260-263.
Molau U. Relationships between flowering phenology and life history strategies in tundra plants // Arctic and Alpine Research, 1993. V. 25. №4. P. 391-402.
Список литературы
537
Moles А. Т, Westoby М. What do seedlings die from and what are the implications for evolution of seed size? // Oikos, 2004. V. 106. № 1. P. 193-199.
Molish H. Der Einfluss einer Pflanze auf die andere — Allelopathie. Jena: Fisher, 1937. 106 S.
Monreal M., Berch S. M., Berbee M. Molecular diversity of ericoid mycorrhizal fungi // Canadian Journal of Botany, 1999. V. 77. № 11. P. 1580-1594.
Mooney H.A., Rundel P. W. Nutrient relations of the evergreen shrub, Adenostoma fasciculatum in the California chaparral // Botanical Gazette, 1979. V. 140. P. 109-113.
Moora M., Zobel M. Effect of arbuscular mycorrhiza on inter- and intraspecific competition of two grassland species // Oecologia, 1996. V. 108. № 1. P. 79-84.
Moore A. W. Azolla: biology and agronomic significance // Botanical Review, 1969. V. 35, N. 1. P. 17-35.
Moore P. D. Express delivery by bat // Nature, 2002. V. 420. № 6911. P. 34-35.
Moore P.D. Measuring competition in plant communities // Nature, 1982. V. 298. №5874. P. 514-515.
Morrison R.G., Yarranton G.A. Diversity, richness and evenness during a primary sand-dune succession at Grand Bend, Ontario // Canadian Journal of Botany, 1973. V. 51. № 12. P. 2401-2411.
MorrowP.A., OlfeltJ.P. Phoenix clones: recovery after long-term defoliation-induced dormancy // Ecology Letters, 2003. V. 6. № 2. P. 119-125.
Mosier A. R., Kroeze C. A new approach to estimate emissions of nitrous oxide from agriculture and its implications for the global N2O budget // Global Change Newsletter, The international Geosphere-Biosphere Programme, 1998. V. 34. № 1. P. 8-14.
Mosse B., Stribley D. P., LeTacon F. Ecology of mycorrhizae and mycorrhizal fungi // Advances in Microbial Ecology, 1981. V. 5. P. 137-210.
Mouissie A. M., van der Veen C.E.J., Veen (Ciska) G.E, van Diggelen R. Ecological correlates of seed survival after ingestion by Fallow Deer // Functional Ecology, 2005. V. 19. №2. P. 284-290.
Moulin C., Guillard E, Dulac F, and Lambert С. E. Long-term daily monitoring of Saharan dust load over ocean using Meteosat 1SCCP-B2 data 1. Methodology and preliminary results for 1983-1994 in the Mediterranean // Journal of Geophysical Research, 1997. V. 102, N D14. P. 16947-16958.
Moyer-Henry K. A., Burton J. W., Israel D. W, Rufty T. W. Nitrogen transfer between plants: A 15 N natural abundance study with crop and weed species // Plant and Soil, 2006. V. 282. №1-2. P. 7-20.
Mueller-Dombois D., Spatz G. The influence of feral goats on the lowland vegetation in Hawaii Volcanoes National Park // Phytocoenologia, 1975. V. 3. № 1. P. 1-29.
Mullen R. B., Schmidt S. K., Jaeger С. H. Ill Nitrogen uptake during snowmelt by the snow buttercup, Ranunculus adoneus 11 Arctic and Alpine Research, 1998. V. 30. № 2. P. 121-125.
Muller С. H. The role of chemical inhibition (allelopathy) in vegetational composition // Bull. Torrey Botanical Club, 1966. V.93. №5. P. 332-351.
Muller C.B., God fray H. C.J. Predators and mutualists influence the exclusion of aphid species from natural communities // Oecologia, 1999. V. 119. № 1. P. 120-125.
MOller J., Dulieu H. Enhanced growth of non-photosynthesizing tobacco mutants in the presence of a mycorrhizal inoculum // Journal of Experimental Botany, 1998. V. 49. №321. P. 707-711.
МйИег-Schwarze D., Brashear H., KinnelR., Hintz K.A., LioubomirovA., Ski bo C. Food processing by animals: do beavers leach tree bark to improve palatability? // Journal of Chemical Ecology, 2001. V.27. №5. P. 1011-1028.
Murali K. S., Sukumar R. Leaf flushing and herbivory in a tropical deciduous forest, southern India // Oecologia, 1993. V.94. № 1. P. 114-119.
538
Список литературы
Murdoch IV. IV. Switching in general predators: experiments on predator specificity and stability of prey populations // Ecological Monographs, 1969. V.39. №4. P. 335-354.
Murphy S. D. Biochemical and physiological aspects of pollen allelopathy // Chemical Ecology of Plants: Allelopathy in Aquatic and Terrestrial Ecosystems, inderjit, A. U. Maiiik (eds.). Basel e.a.: Birkhauser Veriag, 2002. P.245-260.
Murray B. R.y Rice B. L.y Keith D.A., Myerscough P. J.y Howell J.y Floyd A. G.y Mills K.y Westoby M. Species in the tail of rank-abundance curves // Ecology, 1999. V. 80. №6. P. 1806-1816.
Musselman L.J.y Press M.C. introduction to parasitic plants // Parasitic plants. Press M. C., Graves J.D. (eds.). London e.a.: Chapman and Hail, 1995. P. 1-13.
Musser R. O.y Hum-Musser S. M.y Eichenseer H.y Peiffer M.y Ervin G.y Murphy J. B.y Felton G. IV. Caterpillar saliva beats plant defenses // Nature, 2002. V. 416. №6881. P. 599-600.
Mutikainen P.y Delph L. F Effects of herbivory on male reproductive success in plants // Oikos, 1996. V.75. №3. P. 353-358.
Mwendere E.J., Mohamed Saleem M.A. infiltration rates, surface runoff, and soil loss as influenced by grazing pressure in the Ethiopian Highlands // Soil Use and Management, 1997. V. 13. №1. P. 29-35.
Myers J.A., Veilend M.y Gardescu S.y Marks P L. Seed dispersal by white-tailed deer: implications for long-distance dispersal, invasion, and migration of plants in eastern North America // Oecologia, 2004. V. 139. № 1. P. 35-44.
Nagashima H.y Terashima /., and Katoh S. Effects of plant density on frequency distributions of plant height in Chenopodium album stands: analysis based on continuous monitoring of height-growth of individual plants // Annals of Botany, 1995. V. 75. №2. P. 173-180.
Nakamura T. Bryophyte and lichen succession on fallen logs and seedling establishment in Tsuga-Abies forests of central Japan // Symposia Bioiogica Hungarica, 1986. V. 35. P. 485-495.
Nathan R.y Katul G. G.y Horn H. S.y Thomas S. M.y Oren R.y Avissar R.y Pacala S. IV., Levin S.A. Mechanisms of long-distance dispersal of seeds by wind // Nature, 2002. V. 418. № 6896. P. 409-413.
Nathan R.y Muller-Landau C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment//Trends in Ecology and Evolution, 2000. V. 15. № 7. P. 278-285.
Negi G. C. S.y Rikhari H. C.y Ram /., Singh S. P Foraging niche characteristics of horses, sheep and goats in an alpine meadow of the Indian Central Himalaya // Journal of Applied Ecology, 1993. V.30. №3. P. 383-394.
Neveu N.y Grandgirard /., Nenon J.P.y Cortesero A.M. Systemic release of herbivore-induced plant volatiles by turnips infected by concealed root-feeding larvae Delia radicum L. // Journal of Chemical Ecology, 2002. V.28. №9. P. 1717-1732.
Newingham B. A.y Callaway R. M.y BassiriRad H Allocating nitrogen away from a herbivore: a novel compensatory response to root herbivory // Oecologia, 2007. V. 153. №4. P. 913-920.
Newman E. I. Niche separation and species diversity in terrestrial vegetation // The plant community as a working mechanism. Newman E.I. (ed.). Oxford e.a.: Blackwell SP, 1982. P.61-77.
Newsham K. K.y Fitter A. H.y Watkinson A. R. Multi-functionality and biodiversity in arbuscuiar mycorrhizas // Trends in Ecology and Evolution, 1995. V. 10. № 10. P. 407-411.
Newsham K. K.y Watkinson A. R.y West H. M.y Fitter A. H. Symbiotic fungi determine plant community structure: changes in a lichen-rich community induced by fungicide application // Functional Ecology, 1995. V.9. №3. P.442-447.
Nicolson S. IV. Direct demonstration of nectar reabsorption in the flowers of Grevillea robusta (Proteaceae)// Functional Ecology, 1995. V.9. №4. P.584-588.
Niering IV. A., EglerF.E. A shrub community of Viburnum ientago, stable for twenty-five years // Ecology, 1955. V.36. №2. P. 356-360.
Список литературы
539
Niering IV. A., Goodwin R.H. Creation of relatively stable shrublands with herbicides: arresting succession on rights-of-way and pastureland // Ecology, 1974. V. 55. №4. P. 784-795.
Nikolaeva M. G. Factors controlling the seed dormancy pattern // The physiology and biochemistry of seed dormancy and germination. Khan A. A. (Ed.). Amsterdam: North-Holland, 1977. P. 51-74.
Nilsson M.-C. Separation of allelopathy and resource competition by the boreal dwarf shrub Empetrum hermaphroditum Hagerup // Oecoiogia, 1994. V.98, No 1. P. 1-7.
Nilsson М.-C., Hogberg P, Zackrisson 0., Fengyou W. Allelopathic effects of Empetrum hermaphroditum on development and nitrogen uptake by roots and mycorrhizas of Pinus silvestris 11 Canadian Journal of Botany, 1993. V. 71. P. 620-628.
Nobel P. S. Extreme temperatures and thermal tolerances for seedlings of desert succulents // Oecoiogia, 1984. V.62. №3. P. 310-317.
Northup R.R., Yu Z., Dahlgren R.A., Vogt K. A. Polyphenol control of nitrogen release from pine litter // Nature, 1995. V.377. P. 227-229.
Novotny V, Basset V, Miller S. E., Drozd P, Cizek L. Host specialization of leaf chewing insects in a New Guinea rainforest // Journal of Animal Ecology, 2002a. V. 71. №3. P. 400-412.
Novotny V, Basset V, Miller S. E., Weiblen G. D., Bremer B., Cizek L, Drozd P. Low host specificity of herbivorous insects in a tropical forest // Nature, 2002b. V. 416. P. 841-844.
Noy-Meir I., Oron T. Effects of grazing on geophytes in Mediterranean vegetation // Journal of Vegetation Science, 2001. V. 12. №6. P. 749-760.
Nunez-Farfan J., Fornoni J., Valverde P. L. The evolution of resistance and tolerance to herbivores // Annual Review of Ecology, Evolution, and Systematics, 2007. V.38. P. 541-566.
O'Dowd D.J. Pearl bodies of a neotropical tree, Ochroma pyramidale: ecological implications // American Journal of Botany, 1980. V.67. №4. P. 543-549.
O'Keefe D. M., Sylvia D. M. Chronology and mechanisms of P uptake by mycorrhizal sweet potato plants // New phytologist, 1992. V. 122. №4. P. 651-659.
Obeso J. R. The cost of reproduction in plants // New Phytologist, 2002. V. 155. № 3. P. 321 -348.
Odion D. C., Davis F. W. Fire, soil heating, and the formation of vegetation patterns in chaparral // Ecological Monographs, 2000. V. 70. №1. P. 149-169.
Ohnmeiss T. E., Baldwin I. T. The allometry of nitrogen to growth and an inducible defense under nitrogen-limited growth //Ecology. V. 75. №4. P. 995-1002.
Oksanen L. Predation, herbivory, and plant strategies along gradients of primary productivity // Perspectives on plant competition. Grace J. B. and Tilman D. (eds.). San Diego e. a.: Academic Press, 1990. P. 445-474.
Oksanen L, Fretwell S. D., Arruda J., and Niemela P. Exploitation ecosystems in gradients of primary productivity // American Naturalist, 1981. V. 118. №2. P. 240-261.
Oksanen L, Oksanen T. Lemmings (Lemmus lemmus) and grey-sided voles (Clethrionomys rufocanus) in interaction with their resources and predators on Finnmarksvidda, northern Norway // Reports of the Kevo Subarctic Research Station, 1981. V. 17. P. 7-31.
Olff H., Ritchie M. E. Effects of herbivores on grassland plant diversity // Trends in Ecology and Evolution, 1998. V. 13. № 7. P. 261-265.
Oliveira P. S. The ecological function of extrafloral nectaries: herbivore deterrence by visiting ants and reproductive output in Caryocar brasiliense (Caryocaraceae) // Functional Ecology, 1997. V. 11. №3. P. 323-330.
Olson J. S. Rates of succession and soil changes on southern Lake Michigan sand dunes // Botanical Gazette, 1958. V. 119. №3. P. 125-170.
Olsson P.A., Tyler G. Occurence of non-mycorrhizal plant species in south Swedish rocky habitats is related to exchangeable soil phosphate // Journal of Ecology, 2004. V. 92. № 5. P. 808-815.
Olsvig L. S. Pattern and diversity analysis of the irradiated oak-pine forest, Brookhaven, New York // Vegetatio, 1979. V.40. №2. P.65-78.
540
Список литературы
OmarM., Heather W. A. Effect of saprophytic phylloplane fungi on germination and development of Melampsora larici-populina // Transactions of the British Mycological Society, 1979. V. 72. №2. P. 225-231.
Onipchenko V. G. The spatial structure of the alpine lichen heaths (ALH): hypothesis and experiments // In: Onipchenko V. G., Blinnikov M. S. (eds.). Experimental investigation of alpine plant communities in the Northwestern Caucasus / VerOffentlichungen des Geobotanischen Institutes der ETH, Stiftung Riibel, Ziirich, 1994; H. 115. P. 100-111.
Onipchenko V. G., Blinnikov M. S., Gerasimova M.A., Volkova E. V, Cornelissen J. H. C. Experimental comparison of competition and facilitation in alpine communities varying in productivity // Journal of Vegetation Science, 2009. V. 20. № 4. P. 718-727.
Onipchenko V. G., Cherednichenko О. V, Zakharov A. A. Structure and dynamics of closed alpine communities. Spatial structure // Alpine ecosystems in the Northwest Caucasus (ed. Onipchenko V.G.). Dordrecht e.a.: Kluwer Academic Publishers, 2004. P.99-125.
Onipchenko V. G., Makarov ML, van der Maarel E. Influence of alpine plants on soil nutrient concentrations in a monoculture experiment // Folia Geobotanica, 2001. V. 36, n 3. P. 225-241.
Onipchenko V. G., Makarov M. L, van Logtestijn R. S. P, Ivanov V B., Akhmetzhanova A. A., Tekeev D. K., Ermak A. A., Salpagarova F. S., Kozhevnikova A. D., Cornelissen J. H. C. New nitrogen uptake strategy: specialized snow roots // Ecology Letters, 2009. V. 12. № 8. P. 758-764.
Onipchenko V. G., Semenova G. V. Comparative analysis of the floristic richness of alpine communitiesin the Caucasus and the Central Alps // Journal of Vegetation Science, 1995. V. 6. № 2. P. 299-304.
Onipchenko V G., Semenova G. V, van der Maarel E. Population strategies in severe environments: alpine plants in the northwestern Caucasus // Journal of Vegetation Science, 1998. V. 9. №1. P. 27-40.
Onipchenko V G., Zobel M. Mycorrhiza, vegetative mobility and responses to disturbance of alpine plants in the Northwestern Caucasus // Folia Geobotanica, 2000. V. 35. № 1. P. 1-11.
Oomes M.J.M., Mooi H. The effect of cutting and fertilizing on the floristic composition and production of an Arrhenatherion elatioris grassland // Vegetatio, 1981. V. 46-47. № 1. P. 233-239.
Osborne B.A., Sprent J. 1. Ecology of the Nostoc-Gunnera symbiosis // Cyanobacteria in Symbiosis. Rai A. N., Bergman B., Rasmussen U. (eds.). Dordrecht e. a.: Kluwer, 2002. P. 233-251.
Osonubi О., Окоп I. E. and Bamiduro T. A. Effect of different fungal inoculation periods on performance of Gmelina seedlings under dry soil conditions // Forest Ecology and Management, 1990. V.37. №4. P. 223-232.
Ouborg N.J. Isolation, population size and extinction: the classical and metapopulation approaches applied to vascular plants along the Dutch Rhine-system // Oikos, 1993; V 66. № 2. P. 298-308.
Ozinga W. A., van Andel /., McDonnell-Alexander M. P. Nutritional soil heterogeneity and mycorrhyza as determinants of plant species diversity // Acta Botanica Neerlandica, 1997. V. 46, No 3.P. 237-254.
Paavolamen L., Kitunen V, and SmolanderA. Inhibition of nitrification in forest soil by monoterpenes // Plant and Soil, 1998. V. 205. № 2. P. 147-154.
Pagani M., Caldeira K., Berner R., Beerling D.J. The role of terrestrial plants in limiting atmospheric CO2 decline over the past 24 million years // Nature, 2009. V. 460. P. 85-88. Paine R. T. Food web complexity and species diversity // American Naturalist, 1966. V. 100. №1. P. 65-75.
Palmer D., Barrett P. Evolution: the story of life. L.: Mitchell Beazley, 2009. 368 p.
Список литературы
541
Palmer М. W. Variation in species richness: towards a unification of hypotheses // Folia Geobotanica et Phytotaxonomica, 1994. V. 29. №4. P. 511-530.
Palmer M. IV., Maurer T.A. Does diversity beget diversity? A case study of crops and weeds // Journal of Vegetation Science, 1997. V.8. №2. P. 235-240.
Palmer S. C. E, Hester A. J., Elston D.A., Gordon I. J., Hartley S. E. The perils of having tasty neighbors: grazing impacts of large herbivores at vegetation boundaries // Ecology, 2003. V. 84. №11. P. 2877-2890.
Palmqvist K. Carbon economy in lichens // New Phytologist, 2000. V. 148. № 1. P. 11-36.
Parker C. The influence of Striga spp.on sorghum under varying nitrogen // Proceedings of the 3rd International Symposium on Parasitic Weeds. 1CARDA/ Internationa Parasitic Weeds Research Group, 7-8 May, 1984. Parker C. et al. (eds.). Allepo, Syria. P. 90-98.
Parker V. T. Conceptual problems and scale limitations of defining ecological communities: a critique of the Cl concept (Community if Individuals) // Perspectives in Plant Ecology, Evolution and Systematics, 2001. V.4. №2. P. 80-96.
Parmesan C. Ecological and evolutionary responses to recent climate change // Annual Review of Ecology, Evolution, and Systematics, 2006. V. 37. P. 637-669.
Parmesan C., Yohe G. A globally coherent fingerprint of climate change impacts across natural systems // Nature, 2003. V 421. P. 37-42.
Parniske M. Cue for the branching connection // Natura, 2005. V. 435. P. 750-751.
Parrish J.A.D., Bazzaz F.A. Competitive interactions in plant communities of different succes-sional ages // Ecology, 1982. V 63. №2. P. 314-320.
Partel M., Helm A., Reitalu T, Liira J., Zobel M. Grassland diversity related to the Late Iron Age human population density //Journal of Ecology, 2007. V.95. №3. P. 574-582.
Partel M., Wilson S. D. Root dynamics and spatial pattern in prairie and forest // Ecology, 2002. V.83, No 5. P. 1199-1203.
Partel M., Zobel M. Dispersal limitation may result in the unimodal productivity-diversity relationship: a new explanation for a general pattern // Journal of Ecology, 2007. V 95. №1. P. 90-94.
Partel M., Zobel M., Liira J., Zobel K. Species richness limitations in productive and oligotrophic plant communities // Oikos, 2000. V.90. № 1. P. 191-193.
Partel M., Zobel M., Zobel K., van derMaarelE. The species pool and its relation to species richness: evidence from Estonian plant communities // Oikos, 1996. V. 75. № 1. P. 111-117.
Paschke M. W., Dawson J. O. Avian dispersal of Frankia 11 Canadian Journal of Botany, 1993. V71.№8. P.1128-1131.
Paschke M. W., Dawson J. O. Frankia abundance in soils beneath Betula nigra and other non-actinorhizal woody plants // Acta Oecologica, 1992. V. 13. №3, 407-415.
Pate J. S. Mineral relationships of parasites and their hosts // Parasitic plants. Press M. C., Graves J.D. (eds.). London e.a.: Chapman and Hall, 1995. P.80-102.
Pate J. S., Dell B. Economy of mineral nutrients in Sandplain species // Kwongan: Plant life of the Sandplain. J. S. Pate, J. S. Beard (eds.). Nedlands: University of Western Australia, 1984. P. 227-252.
Paul L. R., Chapman В. K., Chanway С. P Nitrogen fixation associated with Suillus tomentosus tuberculate ectomycorrhizae on Pinus contorta var. latifolia // Annals of Botany, 2007. V. 99. №6. P. 1101-1109.
Paul N. D., Ayres P. G. The impact of pathogen (Puccinia lagenophora) on populations of groundsel (Senecio vulgaris) overwintering in the field. 1. Mortality, vegetative growth and the development of size hierarchies //Journal of Ecology, 1986a. V. 74. № 4. P. 1069-1084.
Paul N. D., Ayres P. G. The impact of pathogen (Puccinia lagenophora) on populations of groundsel (Senecio vulgaris) overwintering in the field. 11. Reproduction // Journal of Ecology, 1986b. V. 74. №4. P. 1085-1094.
542
Список литературы
Paul N., Callaghan T, Moody S., Gwynn-Jones D., Johanson U., Gehrke C. UV-B impacts on decomposition and biogeochemical cycling // Stratospheric ozone depletion: the effects of UV-B radiation on terrestrial ecosystems. Rozema J. (ed.). Leiden: Backhuys Publishers, 1999. P. 117-133.
Pearson /., Stewart G. R. The deposition of atmospheric ammonia and its effects on plants // New Phytologist, 1993. V. 125. № 2. P. 283-305.
Pearson R. G. Climate change and the migration capacity of species // Trends in Ecology and Evolution, 2006. V. 21. № 3. P. 111 -113.
Peet R. K. Community structure and ecosystem function // Plant succession: theory and prediction. Glenn-Lewin D. C., Peet R. K., Veblen T.T. (eds.). London: Chapman and Hall, 1992. P. 103-151.
PeetR.K., Glenn-Lewin D. C., Wolf J. W Prediction of man’s impact on plant species diversity // Man’s impact on vegetation. W. Holzner W., Werger M.J.A., Ikusima 1. (eds.). The Hague: Junk, 1983. P. 41-54.
Pendall E., Bridgham S., Hanson P/., Hungate B., Kicklighter D. IV, Johnson D. IV, Law В. E., Luo E, Megonigal J.P, Olsrud M., Ryan M. G., Wan S. Below-ground process responses to elevated CO2 and temperature: a discussion of observations, measurement methods, and models // New Phytologist, 2004. V. 162. № 2. P. 311-322.
Pennings S. C., Callaway R. M. Impact of parasitic plant on the structure and dynamics of salt marsh vegetation // Ecology, 1996. V. 77. №5. P. 14*10-1419.
Pennings S. C., Callaway R. M. Parasitic plants: parallels and contrasts with herbivores // Oecoiogia, 2002. V. 131. №4. P. 479-489.
Pennings S. C., Clark С. M., Cleland E. E., Collins S. L., Gough L., Gross K. L., Miichunas D. G., Suding K. N. Do individual plant species show predictable responses to nitrogen addition across multiple experiments? // Oikos, 2005. V. 110. № 3. P. 547-555.
Peroutka M., Adlassnig IV, Volgger M., Lendl T, Url IV. G., and Lichtscheidl I. K. Utricularia: a vegetarian carnivorous plant? Algae as prey of bladderwort in oligotrophic bogs // Plant Ecology, 2008. V. 199. № 2. P. 153-162.
Perry L. G., Thelen G. C, Ridenour IV. M., Weir T L., Callaway R. M., Paschke M. IV, VivancoJ. M. Dual role for an allelochemical: (+)-catechin from Centaurea maculosa root exudates regulates conspecific seedling establishment // Journal of Ecology, 2005. V. 93. № 6. P. 1126-1135.
Petermann J.S., Fergus A. J. E, Turnbull L.A., Schmid B. Janzen-Connell effects are widespread and strong enough to maintain diversity in grasslands // Ecology, 2008. V. 89. № 9. P. 2399-2406.
Peterson D. L., Rolfe G. L. Nutrient dynamics of herbaceous vegetation in upland and floodplain forest communities // American Midland Naturalist, 1982a. V. 107. № 2. P. 325-339.
Peterson D. L., Rolfe G. L. Seasonal variation in soil nutrients in floodplain and upland forests of central Illinois // Soil Science Society of America Journal, 1982b. V. 46. № 6. P. 1310-1315.
Peterson R. L. Adaptation of root structure in relation to biotic and abiotic factors // Canadian Journal of Botany, 1992. V. 70. №4. P. 661-675.
Petrini O. Ecological and physiological aspects of host-specificity in endophytic fungi // Endophytic fungi in grasses and woody plants: systematics, ecology and evolution. Redlin S. C. and Cams L. M. (eds.). St. Paul: APS Press, 1996. P. 87-100.
Petrini O., Fisher P. J. Fungal endophyte in Salicornia perennis 11 Transactions of the British Mycological Society, 1986. V. 87. №4. P. 647-651.
Petrini O., Hake U., and Dreyfuss M. M. An analysis of fungal communities isolated from fruticose lichens // Mycologia, 1990. V. 82. №4. P. 444-451.
Phillips F.J. A study of pinyon pine // Botanical Gazette, 1990. V. 48. P. 216-223.
Phillips O. L., Martinez R. V, Mendoza A. M., Baker T R., Vargas P. N. Large lianas as hyperdynamic elements of the tropical forest canopy // Ecology, 2005. V. 86. № 5. P. 1250-1258.
Список литературы
543
Phoenix G. К., Press М. С. Linking physiological traits to impacts on community structure and function: the role of root hemiparasitic Orobanchaceae (ex-Scrophulariaceae) // Journal of Ecology, 2005. V. 93. № 1. P. 67-78.
Pickett S. TA. Non-equilibrium coexistence of plants // Bull. Torr. Bot. Club, 1980. V. 107. №2, 238-248.
Pigott C. D., Huntley /. P. Factors controlling the distribution of Tilia cordata at the northern limits of its geographical range. III. Nature and causes of seed sterility // New Phytologist, 1981. V.87. №4. P.817-839.
Pimentel D., McNair S., Janecka J., Wightman J., Simmonds C., O'Connell C., Wong E., Russel L., Zern J., Aquino I, Tsomondo T. Economic and environmental threats of alien plant, animal and microbe invasions // Agriculture, Ecosystems and Environment, 2001. V. 84. № 1. P. 1-20.
Pimentel D., Zuniga R., Morrison D. Update on the environmental and economic costs associated with alien-invasive species in the United States // Ecological Economics, 2005. V. 52. № 3. P. 273-288.
Pinero D., Sarukhan J., Alberdi P The costs of reproduction in a tropical palm, Astrocaryum mexicanum 11 Journal of Ecology, 1982. V. 70. №2. P. 473-481.
Pirozynski K.A., Malloch D. W. Seeds, spores and stomachs: coevolution in seed dispersal mutualisms // Coevolution of fungi with plants and animals. Pirozynski K. A., Hawksworth D. L. (eds.). London: Academic Press, 1988. P. 227-246.
Pitelka L. F. Energy allocation in annual and perennial lupines (Lupinus: Leguminosae) // Ecology, 1977. V. 58. №5. P. 1055-1065.
Pleasants J. M. Evidence for short-distance dispersal of pollinia in Asclepius syriaca L. // Functional Ecology, 1991. V. 5. № 1. P. 75-82.
Polis G.A. Why are parts of the world green? Multiple factors control productivity and the distribution of biomass // Oikos, 1999. V.86. № 1. P. 3-15.
Polis G. A., Hurd S. D., Jackson S. I, Pinero F. S. El Nino effects on the dynamics and control of an island ecosystem in the Gulf of California // Ecology, 1997. V. 78. №6. P. 1884-1897.
Poorter H., Navas M.-L. Plant growth and competition at elevated CO2: on winners, losers and functional groups // New Phytologist, 2003. V. 157. №2. P. 175-198.
Post E. Climate-vegetation dynamics in the fast lane // Trends in Ecology and Evolution, 2003. V. 18. №11. P. 551-553.
PreisserE. L., Dugaw C.J., Dennis B., Strong D. R. Plant facilitation of a belowground predator // Ecology, 2006. V. 87. № 5. P. 1116-1123.
Press M., Barker M., Scholes J. (eds.). Physiological plant ecology. British Ecological Society Symposium. Oxford: Blackwell Science, 1999. №39. 480 p.
Press M. C. Carbon and nitrogen relations // Parasitic plants. Press M. C., Graves J. D. (eds.). London e.a.: Chapman and Hall, 1995. P. 103-124.
Press M. C. Dracula or Robin Hood? A functional role for root hemiparasites in nutrient poor ecosystems // Oikos, 1998. V. 82. №3. P. 609-611.
Press M. C. The functional significance of leaf structure: a search for generalizations // New Phytologist, 1999. V. 143. № 1. P. 213-219.
Press M. C., Graves J. D., Stewart G. R. Physiology of the interaction of angiosperm parasites and their higher plant hosts // Plant, Cell and Environment, 1990. V. 13. № 1. P. 91-104.
Press M.C., Nour J. J., Bebawi EE, Stewart G.R. Antitranspirant-induced heat stress in the parasitic plant Striga hermonthica — a novel method of control // Journal of Experimental Botany, 1989. V.40. №214. P.585-559.
Press M. C., Phoenix G. K. Impacts of parasitic plants on natural communities // New Phytologist, 2005. V. 166. №3. P.737-751.
Press M. C., Seel W. E. Interactions between hemiparasitic angiosperms and their hosts in the subarctic // Ecological Bulletins, 1996. V. 45. P. 151-158.
544
Список литературы
Price М. V., Joyner J. W. What resources are available to desert granivores: seed rain or soil seed bank? // Ecology, 1997. V. 78, No 3. P. 764-773.
Price P. W., Bouton С. E., Gross P, McPheron B.A., Thompson J. N., Weis A. E. Interaction among three trophic levels: Influence of plants on interactions between insect herbivores and natural enemies // Annual Review of Ecology and Systematics, 1980. V. 11. P. 41-65.
Pritts M. P, Hancock J. F. Seasonal and lifetime allocation patterns of the woody goldenrod, Solidago pauciflosculosa Michaux. (Compositae) // American Journal of Botany, 1983. V. 70. №2. P. 216-221.
Proctor M. С. E, Tuba Z. Poikilohydry and homoihydry: antithesis or spectrum of possibilities? // New Phytologist, 2003. V. 156. №3. P. 327-349.
Proulx M., Mazumder A. Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems // Ecology, 1998. V. 79. №8. P.2581-2592.
Pugnaire F. I., Haase P, Puigdefabregas J. Facilitation between higher plant species in a semiarid environment // Ecology, 1996. V. 77. №5. P. 1420-1426.
Pullin A. S., Woodell S.R.J. Response of the fen violet, Viola persicifolia Schreber, to different management regimes at Woodwalton Fen National Nature Reserve, Cambridgeshire, England // Biological Conservation, 1987. V. 41. №3. P. 203-217.
Putz EE. The natural history of lianas on Barro Colorado Island, Panama // Ecology, 1984. V.65. №6. P. 1713-1724.
Puustinen S., Jarvinen O., Tiilikkala K. Asymmetric competition between a hemiparasitic plant and a cyst nematode on a shared host plant // Ecoscience, 2001. V. 8. № 1. P. 51-57.
Pysek P, Jarosik V, Chytry M., Kropac Z., Tichy L., Wild J. Alien plants in temperate weed communities: prehistoric and recent invaders occupy different habitats // Ecology, 2005. V.86. №3. P. 772-785.
Pywell R.F., Bullock J.M., Walker K.J., Coulson S.J., Gregory S.J., Stevenson MJ. Facilitating grassland diversification using the hemiparasitic plant Rhinanthus minor 11 Journal of Applied Ecology, 2004. V.41. №5. P. 880-887.
Qian H., Fridley J. D., Palmer M. W. The latitudinal gradient of species-area relationship for vascular plants of North America // American Naturalist, 2007. V. 170. №5. P. 690-701. Quested H. Parasitic plants — impact on nutrient cycling // Plant and Soil, 2008. V. 311. № 1-2. P. 269-272.
Quested H. M., Press M. C., Callaghan T. V. Litter of hemiparasite Bartsia alpina enhances plant growth: evidence for a functional role in nutrient cycling // Oecologia, 2003. V. 135. №4. P. 606-614.
Quested H. M., Press M. C., Callaghan T. И, Cornelissen J. H. C. The hemiparasitic angiosperm Bartsia alpina has the potential to accelerate decomposition in sub-arctic communities // Oecologia, 2002. V. 130. № 1. P. 88-95.
Racine С. H, Jandt R.R., Meyers C.R., and Dennis J. Tundra fire and vegetation change along a hillslope on the Seward Peninsula, Alaska, U. S. A. // Arctic, Antarctic, and Alpine Research, 2004. V. 36. № 1. P. 1-10.
Rackham O. Ancient woodlands: modern threats // New Phytologist, 2008. V. 180. №3. P. 571-586.
Raffa K.F., Berryman A. A. The role of host plant resistance in the colonization behavior and ecology of bark beetles // Ecological Monographs, 1983. V. 53. № 1. P. 27-49.
Rai A.N. Cyanobacteria in symbiosis // Handbook of symbiotic cyanobacteria. Rai A. N. (ed.). Boca Raton, Florida: CRC Press, 1990. P. 1-7.
Rai A. N., Soderback E., Bergman B. Cyanobacterium — plant symbioses // New Phytologist, 2000. V. 147. №3. P. 449-481.
Ramsell J., Paul N. D. Preferential grazing by molluscs of plant infected by rust fungi // Oikos, 1990. V.58. №2. P. 145-150.
Список литературы
545
Rautiainen Р, Koivula К., Hyvarinen М. The effect of within-genet and between-genet competition on sexual reproduction and vegetative spread in Potentilla anserina ssp. egedii 11 Journal of Ecology, 2004. V. 92. № 3. P. 505-511.
Read DJ. The structure and function of the ericoid mycorrhizal root // Annals of Botany, 1996. V. 77. P. 365-374.
Read DJ. Towards ecological relevance — progress and pitfalls in the path towards an understanding of mycorrhizal functions in nature // Mycorrhizal Ecology (Ecological Studies. V. 157), van der Heijden M. G. A. and Sanders 1.R. (eds.). Berlin e.a.: Springer, 2003. P. 3-29.
Read DJ., Duckett J. G., Francis R., Ligrone R., Russell A. Symbiotic fungal associations in «lower» land plants // Philosophical Transactions of the Royal Society B: Biological Sciences, 2000. V.355. № 1398. P. 815-831.
Read D.J., Francis R., Finlay R.D. Mycorrhizal mycelia and nutrient cycling in plant communities // Ecological interactions in soil. Fitter A. H. et al. (eds.). Oxford: Blackwell SP, 1985. P. 193-217.
Read DJ., Stribley D.P. Diffusion and translocation in some fungal culture systems // Trans. Br.Mycol. Soc., 1975. V.64. P. 381-388.
Reader RJ. Control of seedling emergence by ground cover and seed predation in relation to seed size for some old-field species // Journal of Ecology, 1993. V. 81. № 1. P. 169-175.
Reader RJ. Relationship between species relative abundance and plant traits for an infertile habitat // Plant Ecology, 1998. V. 134, No 1. P. 43-51.
Redecker D., Morton J. B., Bruns T. D. Ancestral lineages of arbuscular mycorrhizal fungi (Glo-males) // Molec. Phylogenet. Evol., 2000. V. 14. № 2. P. 276-284.
Rees M. Evolutionary ecology of seed dormancy and seed size // Plant life histories, Silvertown J., Franco M., Harper J. L. (eds.). Cambridge: Cambridge University Press, 1997. P. 121-142.
Rees M., Kelly D., Bjornstad O. N. Snow tussocks, chaos, and the evolution of mast seeding // American Naturalist, 2002. V. 160. № 1. P. 44-59.
Reich P. B., Uhl C., Walters M. B., Prugh L., Ellsworth D. S. Leaf demography and phenology in amazonian rain forest: a census of 40 000 leaves of 23 tree species // Ecological Monographs, 2004. V. 74. № 1. P. 3-23.
Reichman O. J., Benedix J. H., Jr., Seastedt T. R. Distinct animal-generated edge effects in a tailgrass prairie community // Ecology, 1993. V. 74. №4. P. 1281-1285.
Reichman O.J., Seabloom E. W. The role of pocket gophers as subterranean ecosystem engineers // Trends in Ecology and Evolution, 2002. V. 17. № 1. P. 44-49.
Reichman O.J., Smith S. C. Responses to simulated leaf and root herbivory by a biennial, Tragopogon dubius 11 Ecology, 1991. V. 72. № 1. P. 116-124.
Reinhart К. O., Callaway R.M. Soil biota and invasive plants // New Phytologist, 2006. V. 170. №3. P. 445-457.
Reisigl H., Keller R. Alpenpflanzen im Lebensraum. Stuttgart, New York: Gustav Fischer Verlag, 1987. 149 S.
Rejmanek M., LepS J. Negative associations can reveal interspecific competition and reversal of competitive hierarchies during succession // Oikos, 1996. V. 76. № 1. P. 161-168.
Rejmanek M., Richardson D. M. What attributes make some plant species more invasive? // Ecology, 1996. V.77. №6. P. 1655-1661.
Reynolds H.L., Hungate B.A., Chapin F.S.lll, D'Antonio C.M. Soil heterogeneity and plant competition in an annual grassland // Ecology, 1997. V. 78. №7. P. 2076-2090.
Richards J. H., Caldwell M.M. Hydraulic lift: substantial nocturnal water transport between soil layers by Artemisia tridentata roots // Oecoiogia, 1987. V. 73. №4. P. 486-489.
Richards P. W. The tropical rain forest. Cambridge: Cambridge University Press, 1952. 450 p.
546
Список литературы
Ridenour W. М., Callaway R. M. The relative importance of allelopathy in interference: the effects of an invasive weed on a native bunchgrass // Oecologia, 2001. V. 126, No 3. P. 444-450.
Ridley H.N. The dispersal of plants throughout the world. Kent: Reeve and Co, 1930. 744 p.
Rietkerk M., van de Koppel J. Regular pattern formation in real ecosystems // Trends in Ecology and Evolution, 2008. V.23. №3. P. 169-175.
Rikhari H. C., Negi G. S. C., Ram /., Singh S. P. Human-induced secondary succession in an alpine meadow of Central Himalaya, India // Arctic and Alpine Research, 1993. V. 25. №1. P. 8-14.
Rilling M. C., Mummey D.L. Mycorrhizas and soil structures // New Phytologist, 2006. V. 171. №1. P. 41-53.
Roberts R. G., Robertson J. A., Hanlin R. T. Fungi occurring in the achenes of sunflower (Helianthus annuus) 11 Canadian Journal of Botany, 1986. V. 64. №9. P. 1964-1971.
Robertson A. I. Plant-animal interactions and the structure and function of mangrove forest ecosystems // Australian Journal of Ecology, 1991. V. 16. №4. P. 433-443.
Rodrigues K. F., Samuels G. Preliminary study of endophytic fungi in a tropical palm // Myco-logical Research, 1990. V. 94. №6. P. 827-830.
Rodriguez A., Clapp J. P., Dodd J. C. Ribosomal RNA gene sequence diversity in arbuscular mycorrhizal fungi (Glomeromycota) // Journal of Ecology, 2004. V.92. №6. P. 986-989.
Rogers W.E., Nelson R.R. Penetration and nutrition of Striga asiatica 11 Phytopathology, 1962. V. 52. №10. P. 1064-1070.
Romero G. Q., Mazzafera P., Vasconcellos-Neto /., Trivelin P. С. O. Bromeliad-living spiders improve host plant nutrition and growth // Ecology, 2006. V. 87. № 4. P. 803-808.
Rosen E. Periodic droughts and long-term dynamics of alvar grassland vegetation on Gland, Sweden // Folia Geobotanica et Phytotaxonomica, 1995. V. 30. №2. P. 131-140.
Rosen E. Vegetation development and sheep grazing in limestone grasslands of south Gland, Sweden // Acta phytogeographica Suecica. Uppsala: Almqvist and Wiksell International, 1982. V.72. -108 c.
Rosendahl S. Communities, populations and individuals of arbuscular mycorrhizal fungi // New Phytologist, 2008. V. 178. №2. P. 253-266.
Roslin T. Herbivory passing the limits // Trends in Ecology and Evolution, 2002. V. 17. №8. P. 355.
RossowL.J., Bryant J.R, Kielland K. Effects of above-ground browsing by mammals on mycorrhizal infection in an early successional taiga ecosystem // Oecologia, 1997; V 110. № 1. P. 94-98.
Rovira A. D., Campbell R. Scanning electron microscopy of microorganisms on the roots of wheat // Microbial Ecology, 1974. V. 1. № 1. P. 15-23.
Rowe N., Speck T Plant growth forms: an ecological and evolutionary perspective // New Phytologist, 2005. V. 166. № 1. P. 61-72.
Rozema J., van de Staaij J., Bjorn L. O., Caldwell M. UV-B as an environmental factor in plant life: stress and regulation // Trends in Ecology and Evolution, 1991. V. 12. № 1. P. 22-28.
Rudgers J.A., Maron J.L. Facilitation between coastal dune shrubs: a non-nitrogen fixing shrub facilitates establishment of a nitrogen-fixer // Oikos, 2003. V. 102. № 1. P. 75-84.
Ruess R. IV. The role of large herbivores in nutrient cycling of tropical savannas // Determinants of Tropical Savannas. Walker В. H. (ed.). Oxford: 1RL Press, 1987. P. 67-91.
Ruhren S., Handel S. N. Jumping Spiders (Salticidae) enhance the seed production of a plant with extrafloral nectarines // Oecologia, 1999. V. 119. №2. P. 227-230.
Rundel R. W. Ecological success in relation to plant form and function in the woody legumes // In: Stirton С. H. and Zarucchi J. L. (eds.) Advances in Legume Biology Monogr. Syst. Bot. Missouri Bot. Gard., 1989. V.29. P. 377-398.
Список литературы
547
Russell Е. IV. Soil conditions and plant growth. London: Longman, 1973. 849 p.
Russell G., Grace J. The effect of windspeed on the growth of grasses // Journal of Applied Ecology, 1979. V. 16. №2. P. 507-514.
Rustad L. E., Campbell J. L, Marion G. M., Norby RJ., Mitchell MJ., Hartley A., Cornelissen J., Gurevitch J. and GCTE-NEWS. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming // Oecologia, 2001. V. 126. №4. P. 543-562.
Rusterholz H. P, ErhardtA. Can nectar properties explain sex-specific flower preferences in the Adonis Blue butterfly Lysandra bellargus? 11 Ecological Entomology, 2000. V. 25. № 1. P. 81-90.
Rydin H. Effect of water level on desiccation of Sphagnum in relation to surrounding Sphagna // Oikos, 1985. V.45. №3. P. 374-379.
Rydin H., Borgegard S. Plant characteristics over a century of primary succession on islands: Lake Hjalmaren // Ecology, 1991. V. 72. №3. P. 1089-1101.
Ryel RJ. Hydraulic redistribution // Progress in Botany. Esser K. et al. (eds.). Berlin e.a.: Springer, 2004. V.65. P. 413-435.
Ryser P, Langenauer R., Gigon A. Species richness and vegetation structure in a limestone grassland after 15 years management with six biomass removal regimes // Folia Geob-otanica et Phytotaxonomica, 1995. V. 30. №2. P. 157-167.
Sachs J. L., Simms E. L. The origin of uncooperative rhizobia // Oikos, 2008. V. 117. № 7. P. 961-966.
Saleska S.R., Shaw M.R., Fisher M.L., Dunne J. A., Still CJ., Holman M.L., and Harte J. Plant community composition mediates both large transient decline and predicted long-term recovery of soil carbon under climate warming // Global Biogeochemical Cycles, 2002. V. 16. №4. P. 1055 (18 p.) doi: 10.1029/2001GB001573.
Salibi D. O., Norton В. E. Survival of perennial grass seedlings under intensive grazing in semi-arid rangelands // Journal of Applied Ecology, 1987. V. 24. №1. P. 145-151.
Salonen V., Lammi A. Effects of root hemiparasitic infection on host performance: reduced flower size and increased flower asymmetry // Ecoscience, 2001. V. 8. №2. P. 185-190.
Salonen V., Setala H., and Puustinen S. The interplay between Pinus sylvestris, its root hemiparasite, Melampyrum pratense, and ectomycorrhizal fungi: Influences on plant growth and reproduction // Ecoscience, 2000. V. 7. №2. P. 195-200.
Salonen V., Vestberg M., Vauhkonen M. The effect of host mycorrhizal status on host plant-parasitic plant interactions // Mycorrhiza, 2001. V. 11. №2. P. 95-100.
Sanders I. R., Koide R. T., and Shumway D. L. Mycorrhizal stimulation of plant parasitism // Canaian Journal of Botany, 1993. V. 71. №9. P. 1143-1146.
Sarukhdn J. Demography problems in tropical systems // Demography and evolution in plant populations. Solbrig O.T. (ed.). Oxford: Blackwell, 1980. P. 161-188.
Sauer J. D. Plant Migration: The Dynamics of Geographic Patterning in seed Plant Species. Berkeley: University of California Press, 1988. 282 p.
Sawyer N.A., Chambers S.M., Cairney J. IV. G. Distribution of Amanita spp. genotypes under eastern Australian sclerophyll vegetation // Mycological Research, 2003. V. 107. № 10. P. 1157-1162.
Schadler M., Roeder M., Brandl R., Matthies D. Is palatability of root-hemiparasitic plant influenced by its host species? // Oecologia, 2005. V. 146. №2. P. 227-233.
Schaefer H. M., Wilkinson D. M. Red leaves, insects and coevolution: a red herring? // Trends in Ecology and Evolution, 2004. V. 19. № 12. P. 616-618.
Schat H., van Beckhoven K. Water as a stress factor in the coastal dune system // Ecological responses to environmental stresses. Rozema J., Verkleij J. A. C. (eds.). Dordrecht e. a.: Kluwer Academic Publichers, 1991. P. 76-89.
548
Список литературы
Schenk HJ. Root competition: beyond resource depletion // Journal of Ecology, 2006. V. 94. №4. P. 725-739.
Schenk HJ., Jackson R. B. Rooting depths, lateral root spreads and below-ground / aboveground allometries of plants in water-limited ecosystems // Journal of Ecology, 2002. V.90. №3. P. 480-494.
Schlesinger W.H., Raikes J. A., Hartley A.E., Cross A. F. On the spatial pattern of soil nutrients in desert ecosystems // Ecology, 1996. V. 77. № 2. P. 364-374.
Schnitzer S.A., Bongers F. The ecology of lianas and their role in forests // Trends in Ecology and Evolution, 2002. V. 17. №5. P. 223-230.
Schoonhoven L. M., van Loon J J. A., Dicke M. Insect-plant biology. Oxford e. a.: Oxford University Press, 2005-421 p.
Schowalter T. D. Insect ecology. An ecosystem approach. 2nd edit. Amsterdam e. a.: Elsevier AP, 2006-572 p.
Schreiner R. P., Koide R. T Stimulation of vesicular-arbuscular mycorrhizal fungi by mycotrophic and nonmycotrophic plant root systems // Applied and Environmental Microbiology, 1993. V. 59. №8. P. 2750-2752.
Schultz J.C. How plants fight dirty // Nature, 2002. V. 416. №6878. P. 267.
Schultz J.C. Plant responses induced by herbivores // Trends in Ecology and Evolution, 1988. V. 3. №2. P. 45-49.
Schupp E. IV. Quantity, quality and the effectiveness of seed dispersal by animals // Vegetatio, 1993. V. 107. №8. P. 15-29.
Schussler A., Schwartzott D., Walker C. A new fungal phylum, the Glomeromycota: phylogeny and evolution // Mycological Research, 2001. V. 105. № 12. P. 1413-1421.
Schuster L. Uber den Sammeltrieb des Eichelhahers (Garrulus) 11 Vogelwelt, 1950; Bd. 71, S.9-17.
Schwab S.M., Menge J. A., and Tinker P.B. Regulation of nutrient exchange between host and fungus in VA mycorrhizae // New Phytologist, 1991. V. 117. №3. P. 387-398.
Schwilk D. IV., Keeley J. E., Bond IV. J. The intermediate disturbance hypothesis does not explain fire and diversity pattern in fynbos // Plant Ecology, 1997. V. 132. № 1. P. 77-84.
Schwinning S., Weiner J. Mechanisms determing the degree of size asymmetry in competition among plants // Oecoiogia, 1998. V. 113. № 1. P. 447-455.
Seastedt T. Traits of plant invaders // Nature, 2009. V. 459. P. 783-784.
Seastedt T.R., Briggs J.M., and Gibson DJ. Controls of nitrogen limitation in tailgrass prairie // Oecoiogia, 1991. V. 87. № 1. P. 72-79.
Sedwick C. What killed the woolly mammoth? // PLoS Biology, 2008. V. 6. № 4, p. e99. DOI: 10.1371 /journal.pbio.0060099.
Seel IV. £., Jeschke IV. D. Simultaneous collection of xylem sap from Rhinanthus minor and the hosts Hordeum and Trifolium: hydraulic properties, xylem sap composition and effects of attachment // New Phytologist, 1999. V. 143. № 2. P. 281-298.
Seel IV. £., Parsons A. N., and Press M. C. Do inorganic solutes limit growth of the facultative hemiparasite Rhinanthus minor L. in the absence of a host? // New Phytologist, 1993. V.124. №2. P. 283-289.
Seel IV. £., Press M. C. Influence of the host on three sub-arctic annual facultative root hemiparasites. 1. Growth, mineral accumulation and above-ground dry-matter partitioning // New Phytologist, 1993. V. 125. № 1. P. 131-138.
Seel IV. E., Press M. C. Influence of the host on three sub-Arctic annual facultative root hemiparasites: 11 Gas exchange characteristics and resource use-efficiency // New Phytologist, 1994. V. 127. №1. P. 37-44.
Selosse M.-A., FaccioA., Scappaticci G., Bonfante P. Chlorophyllous and achlorophyllous specimens of Epipactis microphylla (Neottieae, Orchidaceae) are associated with ectomycor-rhizal septomycetes, including truffles // Microbial Ecology, 2004. V. 47. № 4. P. 416-426.
Список литературы
549
Selosse М.-А., Richard F., Не X., Simard S. W. Mycorrhizal networks: des liaisons dangereuses? // Trends in Ecology and Evolution, 2006. V. 21. № 11. P. 621-628.
Semenova G. V. Soil seed banks // Alpine ecosystems in the Northwest Caucasus (ed. Onipchenko V. G.). Dordrecht e.a.: Kluwer Academic Publishers, 2004. P. 194-212.
Semenova G. V, Onipchenko V. G. Soil seed banks // In: Onipchenko V. G., Blinnikov M. S. (eds.). Experimental investigation of alpine plant communities in the Northwestern Caucasus / Verdffentlichungen des Geobotanischen Institutes der ETH, Stiftung RObel, Ziirich, 1994; H. 115. P. 69-82.
Serjeant K., Tang R., Anfang N., Beggs J. R., Goddard M. R. Yeasts associated with the New Zealand Nothofagus honeydew system // New Zealand Journal of Ecology, 2008. V. 32. №2. P. 209-213.
Shachak M., Steinberger Y. An algae-desert snail food chain: energy flow and soil turnover // Oecologia, 1980. V.46.№3. P. 402-411.
Shahjahan M. Erigeron flowers as a food and attractive odor source for Peristenus pseudopai-lipes, a braconid parasitoid of the tarmshed plant bug // Environmental Entomology, 1974. V. 3. №1. P. 69-72.
Shane M. W., Lambers H. Cluster roots: A curiosity in context // Plant and Soil, 2005. V. 274. №1-2. P. 101-125.
Sharkey T. D., Lerdau M. T Atmospheric Chemistry and hydrocarbon emissions from plants // Ecological Applications, 1999. V.9. №4. P. 1107-1108.
Shefferson R. P., Kull T, Tali K. Adult whole-plant dormancy induced by stress in long-lived orchids // Ecology, 2005. V. 86. № 11. P. 3099-3104.
Shmida A, Ellner S. Coexistence of plant species with similar niches // Vegetatio, 1984. V. 58. №1. P. 29-55.
Silvertown ]., McConway KJ., Hughes Z., Biss P., MacnairM., Lutman P. Ecological and genetic correlates of long-term population trends in the Park Grass Experiment // American Naturalist, 2002. V. 160. №4. P. 409-420.
Silvertown J., Wilson B. Spatial interactions among grassland plant populations // Geometry of ecological interactions. Dieckmann U., Law R., Metz J. A. J. (eds.). Cambridge: Cambridge Univ. Press, 2000. P. 28-47.
Silvertown J. W. Introduction to plant population ecology. London, New York: Longman, 1982. 209 p.
Simard S. W., Durall D. M. Mycorrhizal networks: a review of their extent, function, and importance // Canadian Journal of Botany, 2004. V.82. №8. P. 1140-1165.
Simard S. W., Jones M. D., Durall D. M. Carbon and nutrient fluxes within and between mycorrhizal plants // Mycorrhizal Ecology (Ecological Studies. V. 157). van der Heijden M. G.A. and Sanders l.R. (eds.). Berlin e.a.: Springer, 2003. P.33-74.
Simard S. W., Molina R., Smith J. E., Perry D. A., and Jones M. D. Shared compatibility of ectomycorrhizae on Pseudotsuga menziesii and Betula papyrifera seedlings grown in mixture in soils from southern British Columbia // Canadian Journal of Forest Research, 1997b. V.27. №3. P. 331-342.
Simard S. W., Perry D.A., Jones M. D., Myrold D. D., Durall D. M., and Molina R. Net transfer of carbon between ectomycorrhizal tree species in the field // Nature, 1997a. V. 388. P. 579-582.
Sizov I. £., Mikhailova N. V, Mikhailov A. V, Onipchenko V. G., Komarov A. S. Age structure and adaptation of the populations of three non-clonal alpine perennial plants // Alpine ecosystems in the Northwest Caucasus (ed. Onipchenko V. G.). Dordrecht e. a.: Kluwer Academic Publishers, 2004. P. 181-194.
Skene K.R. Cluster roots: some ecological considerations // Journal of Ecology, 1998. V. 86. №6. P. 1060-1064.
550
Список литературы
Skogsmyr /. Male reproductive investment and veneral diseases in plants: the case of the sticky catchfly, Lychnis viscaria // Oikos, 1993. V.66. №2. P. 209-215.
SIdma K. Insect juvenile hormone analogues // Annual Review of Biochemistry, 1971. V. 40. P. 1079-1102.
Smallwood K. S., Morrison M. L. Estimating burrow volume and excavation rate of pocket gophers (Geomyidae) // Southwestern Naturalist, 1999. V.44. №2. P. 173-183.
Smith F.A., Smith S. E., Timonen S. Mycorrhizas // Root ecology (Ecological Studies. V. 168), de Kroon H., Visser E.J.W. (eds.). Berlin, Heidelberg: Springer, 2003. P. 257-295.
Smith R. I. L. Vascular plants as bioindicators of regional warming in Antarctica // Oecologia, 1994. V.99. №3-4. P. 322-328.
Smith S.E., Read D.J. Mycorrhizal Symbiosis // 2nd edition. San Diego e.a.: Academic Press, 1997. 605 p.
Snaydon RW. Rapid population differentiation in a mosaic environment. 1. The response of Anthoxanthum odoratum populations to soils // Evolution, 1970. V. 24. № 2. P. 257-269.
Snell T. IV, Burch D. G. The effects of density of resource partitioning in Chamaesyce hirta (Euphorbiaceae) // Ecology, 1975. V. 56. №3. P. 742-746.
Soberon MJ., Loevinsohn M. Patterns of variations in the numbers of animal populations and the biological foundations of Taylor’s law of the mean // Oikos, 1987. V. 48, No 3. P. 249-252.
Soderstrom B., Finlay R.D., Read D.J. The structure and function of the vegetative mycelium of ectomycorrhizal plants. IV. Qualitative analysis of carbohydrate contents of mycelium interconnecting host plants // New Phytologist, 1988. V. 109. №2. P. 163-166.
Solheim B., Zielke M., Bjerke J. IV., Rozema J. Effects of enhanced UV-B radiation on nitrogen fixation in arctic ecosystems // Plant Ecology, 2006. V. 182. № 1-2. P. 109-118.
Sonesson M., Osborne C., Sandberg G. Epiphytic lichens as indicators of snow depth // Arctic and Alpine Research, 1994. V. 26. №2. P. 159-165.
Sork V. L., Dyer R.J., Davis F. IV., Smouse P. E. Mating patterns in a savanna population of valley oak (Quercus lobata Nee) // Fifth Symposium on Oak Woodlands: Oaks in California’s Changing Landscape, 22-25 October 2001, USDA Forest Service General Technical Report. R. Standiford, D. McCreary, and K. L. Purcell (eds.). San Diego, California, 2002. P. 427-439.
Soudzilovskaia N.A., Onipchenko V.G. Experimental investigation of fertilization and irrigation effects on an alpine heath, northwest Caucasus, Russia // Arctic, Antarctic, and Alpine Research, 2005. V.37. №4. P. 602-610.
Sousa IV. P. The role of disturbance in natural communities // Annual Review of Ecology and Systematics, 1984. V. 15. P. 353-391.
Spatz G., Mueller-Dombois D. Succession patterns after pig digging in the grassland communities on Mauna Loa, Hawaii // Phytocoenologia, 1975. V.3. №2/3. P. 346-373.
Spatz G., Weis S., Dolar D. M. Der Eifluss von Bewirtschafts2nderungen auf die Vegetation von Almen im Gasteiner Tai // Veroffentlichungen des Osterreichischen MaB-Hochgebirgsprogramms Hohe Tauern. Bd. 2. Innsbruck: Wagner, 1978; S. 163-180.
Spehn E.M., Scherer-Lorenzen M., Schmid B., Hector A., Caldeira M. C., Dimitrakopoulos P. G., Finn J. A., Jumpponen A., O'Donnovan G., Pereira J. S., Schulze E.-D., Troumbis A. K, Korner C. The role of legumes as a component of biodiversity in a cross-European study of grassland biomass nitrogen // Oikos, 2002. V.98. №2. P. 205-218.
Spencer-Barreto M.M., Gagnaire J., Piebin R., Beaudoing G., Duhoux E. Stem mamillae of Sesbania rostrata brem development into adventitious roots and role in mineral absorption // Acta Oecologica Oecologia Plantarum, 1989. V. 10. №4. P. 411-422.
Sperry J. S., Holbrook N. M., Zimmermann M. H., and Tyree M. T. Spring filling of xylem vessels in wild grapevine // Plant Physiology, 1987. V. 83. №2. P. 414-417.
Список литературы
551
Spiller D.A., Agrawal A. A. Intense disturbance enhances plant susceptibility to herbivory: natural and experimental evidence // Ecology, 2003. V. 84. № 4. P. 890-897.
Sprent J. I. Nitrogen fixation and growth of non-crop legume species in diverse environments // Perspectives in Plant Ecology, Evolution and Systematics, 1999. V. 2. №2. P. 149-162.
St. Clair L.L., Webb B.L, Johansen J.R., Bebecker G. T Cryptogamic soil crusts enhancement of seedling establishment in disturbed and undisturbed areas // Reclamation and Revegetation Research, 1985. V. 3. №2. P. 129-136.
Staehelin J., Renaud A, Bader J., McPeters R., Viatte P, Hoegger B., Bugnion V., Giroud M., and Schill H. Total ozone series at Arosa (CH): Homogenization and data comparison // Journal of Geophysical Research, 1998. V. 103. P. 5827-5841.
Stanton M. L, Rejmanek M., Galen C. Changes in vegetation and soil fertility along a predictable snowmelt gradient in the Mosquito Range, Colorado, U. S. A. // Arctic and Alpine Research, 1994. V.26. V.4. P. 364-374.
Steinaker D. E, Wilson S. D. Phenology of fine roots and leaves in forest and grassland // Journal of Ecology, 2008. V.96. №6. P. 1222-1229.
Steinauer E. M., Collins S. L Feedback loops in ecological hierarchies following urine deposition in tailgrass prairie // Ecology, 2001. V.82. №5. P. 1319-1329.
Steinger T, Kbrner Ch., Schmid B. Long-term persistance in a changing climate: DNA analysis suggests very old ages of clones of alpine Carex curvula 11 Oecoiogia, 1996. V. 105. № 1. P. 94-99.
Stensmyr M. C, Urru /., Coll и /., Celander M., Hansson B. S., Angioy A.-M. Rooting smell of dead-horse arum florets // Nature, 2002. V. 420. №6916. P. 625-626.
Stephan A., Meyer A. H., Schmid B. Plant diversity affects culturable soil bacteria in experimental grassland communities // Journal of Ecology, 2000. V. 88, No 6. P. 988-998.
Steubing L, Fangmeier A., Both R., Frankelfeld M. Effects of SO2, NO2, and O3 on population development and morphological and physiological parameters of native herb layer species in a beech forest // Environmental Pollution, 1989. V. 58. №4. P. 281-302.
Stevens M. H. H., Carson W. P. Plant density determines species richness along an experimental fertility gradient // Ecology, 1999. V. 80, No 2. P. 455-465.
Stewart C. N.Jr., Nilsen E. T. Drosera rotundifolia growth and nutrition in a natural population with special reference to the significance of insectivory // Canadian Journal of Botany, 1992. V. 70. №7. P. 1409-1416.
Stewart G. R., Schmidt S. Evolution and ecology of plant mineral nutrition // Physiological plant ecology (eds. Press M. C., Scholes J.D., Barker M. G.). Oxford: Blackwell Science, 1999. P.91-114.
Stinson K.A., Campbell S.A., Powell J. R., Wolfe В. E., Callaway R. M., Thelen G. C, Hallett S. G., Prati D., Klironomos J. N. Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms // PLoS Biology, 2006. V. 4. № 5. P. 140-145. doi: 10.1371 /journal.pbio.0040140.
Stohlgren T. J., Barnett D. T, Jarnevich C. S., Flather C., Kartesz J. The myth of plant species saturation // Ecology Letters, 2008. V. 11. №4. P. 313-326.
Stoll P, Prati D. Intraspecific aggregation alters competitive interactions in experimental plant communities // Ecology, 2001. V.82. №2. P.319-327.
Stone J. E. Water conduction in roots and root grafts of Pinus resinosa. Ithaca: Cornell Univ., 1974. 180 p.
Stone J. K., Bacon C. W., White J. E, Jr. An overview of endophytic microbes: endophytism defined // Microbial Endophytes. Bacon C. W. and White J. E, Jr. (eds.). New York, Basel: Marcel Dekker, 2000. P. 3-29.
Stoutjesdijk P. High surface temperatures of trees and pine litter in the winter and their biological importance // International Journal of Meteorology, 1977. V. 2. №21. P. 325-331.
552
Список литературы
Stoutjesdijk Р. On the measurement of the radiant temperature of vegetation surfaces and leaves // Wentia, 1966. V. 15. P. 191-202.
Stoutjesdijk P, Barkman JJ. Microclimate, vegetation, and fauna. Uppsala: Opulus Press, 1992. 216 p.
Strauss S. Y Floral characters link herbivores, pollinators, and plant fitness // Ecology, 1997. V.78. №6. P. 1640-1645.
Strauss S. Y., Agrawal A. A. The ecology and evolution of plant tolerance to herbivory // Trends in Ecology and Evolution, 1999. V. 14. №5. P. 179-185.
Streams FA., Shahjahan M., LeMasurier H. G. Influence of plants on the parasitization of the tarnished plant bug by Leiophron pallipes //Journal of Economic Entomology, 1968. V. 61, 996-99.
Strong D.R., Levin D.A. Species richness of plant parasites and growth form of their host // American Naturalist, 1979. V. 114. № 1. P. 1-22.
Stuefer J. F. Division of labour in clonal plants? On the response of stoloniferous herbs to environmental heterogeneity. Ph. Dissertation, Utrecht Universityy (the Netherlands), 1997. 119 p.
Stuefer J. F. Potential and limitations of current concepts regarding the response of clonal plants to environmental heterogeneity // Vegetatio, 1996. V. 127, No 1. P. 55-70.
Stuefer J.F., De Kroon H., During H.J. Exploitation of environmental heterogeneity by spatial division of labour in a clonal plant // Functional Ecology, 1996. V. 10, No 3. P. 328-334.
Sullivan j., Rozema J. UV-B effects on terrestrial plant growth and photosynthesis // Stratospheric ozone depletion: the effects of UV-B radiation on terrestrial ecosystems. Rozema J. (ed.). Leiden: Backhuys Publishers, 1999. P. 39-57.
Sundstrbm K.-R., Huss-Danell K. Effects of water stress on nitrogenase activity in Alnus incana 11 Physiologia Plantarum, 1987. V. 70. №2. P. 342-348.
Suttle К. B. Pollinators as mediators of top-down effects on plants // Ecology Letters, 2003. V. 6. №8. P. 688-694.
Swanborough P, Westoby M. Seedling relative growth rate and its components in relation to size: phylogenetically independent contrasts // Functional Ecology, 1996. V. 10. №2. P. 176-184.
Swap R., Garstang M., Greco S., Talbot R., Kallberg P. Saharan dust in the Amazon Basin // Tellus B, 1992. V.44. №2. P. 133-149.
Swift M. J., Boddy L. Animal-microbial interactions during wood decomposition // Invertebratemicrobe Interactions. J. M. Anderson, A. D. M. Rayner (Eds.). Cambridge: Cambridge University Press, 1984. P. 89-131.
Sydes C., Grime J. P Effects of tree leaf litter on herbaceous vegetation in deciduous woodland. I. Field investigations // Journal of Ecology. 1981a. V. 69. №1. P. 237-248.
Sydes C., GrimeJ.P. Effects of tree leaf litter on herbaceous vegetation in deciduous woodland II. An experimental investigation // Journal of Ecology. 1981b. V. 69. №1. P. 249-262.
Tackenberg O. Modeling long-distance dispersal of plant diaspores by wind // Ecological Monographs, 2003. V. 73. №2. P. 173-189.
Takemoto T, Ogawa S., Nishimoto N., Arihara S., Bue K. [Insect moulting activity of crude drugs and plants] // Yakugaku Zasshi, 1967. V. 87. № 11. P. 1414-1418.
Tallis J. H. Plant Community History: Long-Term Changes in Plant Distribution and Diversity. London: Chapman and Hall, 1991. 398 p.
Tamm С. O. Growth and plant nutrient concentration in Hylocomium proliferum in relation to tree canopy // Oikos, 1950. V. 2. № 1. P. 60-64.
Tamm С. O. Nitrogen in terrestrial ecosystems // Ecological Studies. Berlin e.a.: Springer Verlag, 1991. V.81. 115 p.
Tamm С. O. Survival and flowering of some perennial herbs. II. The behaviour of some orchids on permanent plots // Oikos, 1972. V. 23. №1. P. 23-28.
Список литературы
553
Taper М. L. A new look at competition theory and the structure of ecological communities // Trends in Ecology and Evolution, 1993. V.8. №9. P. 308-309.
Tateno M. Benefit to N 2-fixing alder of extending growth period at the cost of leaf nitrogen loss without resorption // Oecologia, 2003. V. 137. №3. P. 338-343.
Taylor B. R. Air-drying depresses rate of leaf litter decomposition // Soil Biology and Biochemistry, 1998. V.30. №3. P. 403-412.
Taylor D. L.y Bruns T D. Independent, specialized invasions of ecto-mycorrhizal mutualism by two non-photosyntetic orchids // PNAS, 1997. V.94. №9. P. 4510-4515.
Taylor D. L.y Bruns T D. Population, habitat and genetic correlates of mycorrhizal specialization in the «cheating» orchids Corallorhiza maculata and C. mertensiana 11 Molecular Ecology, 1999. V.8. №10. P. 1719-1732.
Taylor D.L., Bruns T.D., Leake J.R., Read D.J. Mycorrhizal specificity and function in myco-heterotrophic plants // Mycorrhizal Ecology (Ecological Studies. V. 157). van der Heijden M.G.A. and Sanders 1.R. (eds.). Berlin e.a.: Springer, 2003. P.375-413.
Taylor D. R.y Aarssen L. W. Complex competitive relationships amongs genotypes of three perennial grasses: Implications for species coexistence // American Naturalist, 1990. V. 136. №3. P. 305-327.
Tedersoo L.y Pellet P.y Koljalg U.y Selosse M.-A. Parallel evolutionary paths to mycoheterotro-phy in understorey Ericaceae and Orchidaceae: ecological evidence for mixotrophy in Pyroleae // Oecologia, 2007. V. 151. №2. P. 206-217.
Telewski F. W.y Zeevaart J.A.D. The 120-yr period for Dr. Beal’s seed viability experiment // American Journal of Botany, 2002. V.89. №8. P. 1285-1288.
Tepedino V. J. The importance of bees and other insect pollinators in maintaining floral species composition // Great Basin Naturalist Memoirs, 1979. №3. P. 139-150.
ter Steege H.y Zagt R. Density and diversity // Nature, 2002. V. 417. №6890. P. 698-699.
Tewksbury J.J., Nabhan G. P. Directed deterrence by capsaicin in chillies // Nature, 2001. V. 412. P. 403-404.
Thanos C. A.y Rundel P. W. Fire-followers in chaparral: nitrogenous compounds trigger seed germination // Journal of Ecology, 1995. V. 83. №2. P. 207-216.
Thebaud C.y Johnston A. Plant responses to global changes in CO2: unfinished business // Trends in Ecology and Evolution, 1997. V. 12. № 11. P. 425-426.
Theodose T.A., Jaeger C.H. Ill, Bowman W.D., Schardt J.C. Uptake and allocation of 15 N in alpine plants: implications for the importance of competitive ability in predicting community structure in a stressful environment // Oikos, 1996. V. 75, No 1. P. 59-66.
Theoharides K.A., Dukes J.S. Plant invasion across space and time: factors affecting nonindige-nous species success during four stages of invasion // New Phytologist, 2007. V. 176. №2. P. 256-273.
Thomas R.J., Logan K.A.B., Ironside A. D.y Bolton G.R. Measurement of the accumulation of N by grass in a continuously grazed upland pastures // Nutrient cycling in terrestrial ecosystems. Harrison A. F., Ineson P., Heal O. W. (eds.). London e. a.: Chapman & Hall, 1990. P. 428-440.
Thompson K. Predicting the fate of temperate species in response to human disturbance and global change // NATO Advanced Research Workshop om Biodiversity, Temperate Ecosystems and Global Change. Boyle T. J. B., Boyle С. E. B. (eds.). Berlin, Heidelberg: Springer, 1994; N 1 20. P. 61-76.
Thompson K. The functional ecology of seed banks // Seeds. The ecology of regeneration in plant communities. Fenner M. (eds.). Wallingford: CAB International, 1992. P. 231-258.
Thompson K. The functional ecology of soil seed banks // Seeds: the ecology of regeneration in plant communities. 2nd ed., Fenner M. (ed.): CAB International. 2000. P. 215-235.
554
Список литературы
Thompson К., Bakker J., Bekker R. The soil seed banks of North West Europe. Cambridge: Cambridge Univ. Press, 1996. 288 p.
Thompson K., Band S. R., Hodgson J. G. Seed size and shape predict persistence in soil // Functional Ecology, 1993. V. 7. P. 236-241.
Thompson K., Grime J. P. A comparative study of germination responses to diurnally-fluctuating temperatures // Journal of Applied Ecology, 1983. V. 20. № 1. P. 141-156.
Thompson K., Grime J. P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats // Journal of Ecology, 1979. V. 67. №3. P. 893-921.
Thompson K., Stewart A. J. A. The measurement and meaning of reproductive effort in plants // American Naturalist, 1981. V. 117. №2. P. 205-211.
Thomson J. D. Effects of stand composition on insect visitation in two-species mixtures of Hieracium 11 American Midland Naturalist, 1978. V. 100. №2. P. 431-440.
Thrall P. H., Biere A., Antonovics J. Plant life-history and disease susceptibility the occurrence of Ustilago violacea on different species within the Caryophyllaceae // Journal of Ecology, 1993. V. 81. №3. P. 489-498.
Tilman D. Biodiversity: population versus ecosystem stability // Ecology, 1996. V. 77. №2. P. 350-363.
Tilman D. Causes, consequences and ethics of biodiversity // Nature, 2000. V. 405. № 6783. P. 208-211.
Tilman D. Competition and biodiversity in spatially structured habitats // Ecology, 1994. V. 75. №1. P. 2-16.
Tilman D. Evolution and differentiation in terrestrial plant communities: the importance of the soil resource:light gradient // Community Ecology. Diamond J. and Case T.J. (eds.). New York e.a.: Harper and Row, 1986. P. 359-380.
Tilman D. Plant strategies and the dynamics and structure of plant communities // Monogra-graphs in population biology (v. 26). Princeton: Princeton Univ. Press, 1988. 362 p.
Tilman D. Resource competition and community structure. Princeton: Princeton Univ. Press, 1982. 297 p.
Tilman D., Dodd M. E., SHvertown /., Poulton P. R., Johnston A. E., Crawley MJ. The Park Grass Experiment: insights from the most long-term ecological study // Long-term experiments in agricultural and ecological sciences. R. A. Leigh, A. E. Johnston (eds.). Wallingford: CAB International, 1994. P. 287-303.
Tilman D., Lehman C. L., Yin C. Habitat destruction, dispersal, and deterministic extinction in competitive communities // American Naturalist, 1997. V. 149. №3. P. 407-435.
Tjoelker M. G., Zhou X. The many faces of climate warming // New Phytologist, 2007. V. 176. №4. P. 739-742.
Toole E. H., Brown E. Final results of the Duvel seed experiment // Journal of Agricultural Research, 1946. V.72. P. 201-210.
Towne E. G., Hartnett D. C., Cochran R. C. Vegetation trends in tailgrass prairie from bison and cattle grazing // Ecological Applications, 2005. V. 15. № 5. P. 1550-1559.
Trabaud L. Fire and survival traits of plants // The role of fire in ecological systems. Trabaud L. (ed.). The Hague: SPB Academic Publishing, 1987. P. 65-89.
Tranquillini W. Standortsklima, Wasserbilanz und CO2-Gaswechsel junger Zirben (Pinus cembra) an der alpinen Waldgrenze // Planta, 1957. V. 49. №6. P.612-661.
Trappe J. M. Phylogenetic and ecological aspects of mycotrophy in the angiosperms from an evolutionary standpoint // Ecophysiology of va mycorrhizal plants. Safir G. R. (ed.). Boca Raton: CRC Press, 1987. P. 5-25.
Trappe J. M., Luoma D. L. The ties that bind: fungi in ecosystems // The fungal community: its organization and role in the ecosystems. Carroll G. C., Wicklow D. T. (eds.). New York e. a.: Marcel Dekker, Inc., 1992. P. 17-27.
Список литературы
555
Traveset A., Riera N., Mas R. E. Passage through bird guts causes interspecific differences in seed germination characteristics // Functional Ecology, 2001. V. 15. №5. P. 669-675.
Traveset A., Willson M.F. Effect of birds and bears on seed germination of fleshy-fruited plants in temperate rainforest of southeast Alaska // Oikos, 1997. V.80. № 1. P. 89-95.
Trenbath B. R. Biomass productivity of mixtures // Advances in Agronomy, 1974. V. 26. P. 177-210.
Trudgill S. T. Soil and vegetation systems. 2nd ed. Oxford: Clarendon Press, 1988. 211 p.
Tsang A., Maun M.A. Mycorrhizal fungi increase salt tolerance of Strophostyles helvola in coastal foredunes // Plant Ecology, 1999. V. 144. № 2. P. 159-166.
Tsuyuzaki S. Survival characteristics of buried seeds 10 years after the eruption of the Usu volcano in Northern Japan // Canadian Journal of Botany, 1991. V. 69. № 10. P. 2251 -2256.
Tsuyuzaki S., Urano S., Tsujii T Vegetation of alpine marshland and its neighboring areas, northern part of Sichuan Province, China // Vegetatio, 1990. V.88. № 1. P. 79-86.
Turnbull L.A., Rees M., and Crawley MJ. Seed mass and the competition/colonization trade-off: a sowing experiment // Journal of Ecology, 1999. V.87. №5. P.899-912.
Tyler С. M. Relative importance of factors contributing to postfire seedling establishment in maritime chaparral // Ecology, 1996. V. 77. № 7. P. 2182-2195.
Uren N. C. Types, amounts, and possible functions of compounds released into the rhizosphere by soil-grown plants // The rhizosphere: biochemistry and organic substances at the soil-plant interface. Pinton R. et al. (eds.). New York: Marcel Dekker, 2001. P. 19-40.
Valiente-Banuet A., Bolongaro-Crevenna A., Briones O., Ezcurra £., Rosas M., Nuez H., Barnard G., Vazquez E. Spatial relationships between cacti and nurse shrubs in a semi-arid environment in central Mexico // Journal of Vegetation Science, 1991. V. 2. № 1. P. 15-20.
van Andel J., Bakker J. P, Grootjans A. P. Mechanisms of vegetation succession: a review of concepts and perspectives // Acta Botanica Neerlandica, 1993. V. 42. №4. P. 413-433.
van Andel /., Dueck T The importance of the physical pattern of plant species in replacement series // Oikos, 1982. V. 39. № 1. P. 59-62.
Van de Merendonk S., van Lenteren J. V Determination of mortality of greenhouse whitefly Tiraleurodes vaporarium (Westwood) (Homoptera: Aleyrodidae) eggs, larvae and pupae on four host-plant species: eggplant (Solanum melongena L), cucumber (Cucumis sativus L), tomato (Lycopersicum esculentum L.) and paprika (Capsicum annuum L.) // Med. Fac. Landbouww. Rijksuniv., 1978; Gent 43. P. 421-429.
van de Staaij /., Rozema /., Aerts R. The impact of solar UV-B radiation on mutualistic plant/micro-organism interactions at the soil-root interface // Stratospheric ozone depletion: the effects of UV-B radiation on terrestrial ecosystems. Rozema J. (ed.). Leiden: Backhuys Publishers, 1999. P. 159-171.
Van der Goes P. С. E. M., van der Putten W. H. Nematode problems in Ammophila arenaria (marram grass) // Meded. Facult. Landbouww., 1992; RUGent 57/3a. P. 847-856.
van der Huge J. С. H. Why are there no insects and so few higher plants, in the see? New thoughts on an old problem // Functional Ecology, 1996. V. 10. №4. P. 546-547.
van der Heijden G.M., Healey J. R., and Phillips O.L. Infestation of trees by lianas in a tropical forest in Amazonian Peru // Journal of Vegetation Science, 2008. V. 19. №6. P. 747-756.
van der Heijden M.G.A., Bardgett R. D., van Straalen N. M. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems // Ecology Letters, 2008. V. ll.№3. P. 296-310.
van der Heijden M.G.A., Klironomos J. N., Ursic M., Moutoglis P, Streitwolf-Engel R., Boiler T, Wiemken A., Sanders 1. R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity // Nature, 1998. V. 396. №6706. P. 69-72.
van der Krift TA J., Berendse F. The effect of plant species on soil nitrogen mineralization // Journal of Ecology, 2001. V.89, No 4. P. 555-561.
556
Список литературы
van der Krift T.A.J., Kuikman P./., Berendse F. The effect of living plants on root decomposition of four grass species // Oikos, 2002. V. 96, No 1. P. 36-45.
van der Maarel E. Pattern and process in the plant community: Fifty years after A. S. Watt // Journal of Vegetation Science, 1996. V. 7. № 1. P. 19-28.
van der Maarel E. Plant species turnover and minimal area in a limestone grassland // Abstracta Botanica, 1993. V. 17. № 1-2. P. 173-178.
van der Maarel E. Species diversity in plant communities in relation to structure and dynamics // Diversity and pattern in plant communities. During H. J., Werger M. J. A., Willems J. H. (eds.). The Hague: SPB Academic Publishing, 1988. P. 1-14.
Van der Maarel E., Leertouwer J. Variation in vegetation and species diversity along a local environmental gradient // Acta Bot. Neerl., 1967. V. 16. №6. P. 1-11.
van derMeijden E. Plant defense, an evolutionary dilemma: contrasting effects of (specialist and generalist) herbivores and natural enemies // Entomologia Experimentalis et Applicata, 1996. V. 80. №1. P. 307-310.
van der Meljden E., Klinkhamer P. G. L. Conflicting interests of plants and the natural enemies of herbivores // Oikos, 2000. V. 89. № 1. P. 202-208.
Van der Pijl L. Principles of dispersal in higher plants. 3rd Edition. Berlin e.a.: Springer, 1982.
216 p.
van der Patten IV. H. Plant-soil feedback as a selective force // Trends in Ecology and Evolution, 1997. VI2, No 5. P. 169-170.
van der Putten IV. H., Peters В. A. M. How soil-borne pathogens may affect plant competition // Ecology, 1997. V. 78, No 6. P. 1785-1795.
van der Putten W.H., Vet L.E.M., Harvey J. A., Wackers F.L. Linking above- and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists // Trends in Ecology and Evolution, 2001. V. 16. № 10. P. 547-554.
van der Valk A. G. Establishment, colonization and persistence // Plant succession: theory and prediction. Glenn-Lewin D. C., Peet R. K., Veblen T. T. (eds.). London, New York: Chapman and Hall, 1992. P. 60-102.
van Groenendael J.M., Klimes L, Klimesova /., Hendriks R.J.J. Comparative ecology of clonal plants // Plant life histories. Silvertown J., Franco M., Harper J. L. (eds.). Cambridge: Cambridge University Press, 1997. P. 191-209.
van Grunsven R.H.A., van der Putten W.H., Bezemer T.M., Tamis W.L.M., Berendse F., Veenendaal E. M. Reduced plant-soil feedback of plant species expanding their range as compared to natives //Journal of Ecology, 2007. V.95. №5. P. 1050-1057.
van Herk С. M., AptrootA., van Dobben H. F. Long-term monitoring in the Netherlands suggests that lichens respond to global warming // Lichenologist, 2002. V. 34. № 2. P. 141-54.
van Omnieren RJ., Whitham T G. Changes in interactions between juniper and mistletoe mediated by shared avian frugivores: parasitism to potential mutualism // Oecologia, 2002. V. 130. №2. P. 281-288.
Van Tichelen К. K, Vanstraelen Г, Colparet J. V Nutrient uptake by intact mycorrhizal Pinus sylvestris seedlings: a diagnostic tool to detect copper toxicity // Tree Physiology, 1999. V. 19. №3. P. 189-196.
Van Tol R. IV. H. M., Van Der Sommen A.T.C., Boff M. I. C., Van Bezooijen J., Sabelis M. IV, Smits P. H. Plants protect their roots by alerting the enemies of grubs // Ecology Letters, 2001. V. 4. №4. P. 292-294.
Vander Wall S. B., Kuhn К. M., Gworek J. R. Two-phase seed dispersal: linking the effects of frugivorous birds and seed-caching rodents // Oecologia, 2005. V. 145. № 2. P. 282-287.
Vandermeer J. The ecology of intercropping. Cambridge: Cambridge University Press, 1989. 237 p.
Vandvik V, Goldberg D. E. Distinguishing the roles of dispersal in diversity maintenance and in diversity limitation // Folia Geobotanica, 2005. V. 40. № 1. P. 45-52.
Список литературы
557
Vare Н., Ohtonen R., Oksanen J. Effects of reindeer grazing on understorey vegetation in dry Pinus sylvestris forests // Journal of Vegetation Science, 1995. V.6. №4. P. 523-530.
Vartia K. 0. Antibiotics in lichens // The Lichens. Ahmadjian V. and Hale M. E. (eds.). New York: Academic Press, 1973. P. 547561.
Vasek E C. Creosote bush: Long-lived clones in the Mohave Desert // American Journal of Botany, 1980. V.67. №2. P. 246-255.
Vasek F. C., Lund L. J. Soil characteristics associated with a primary plant succession on a Mojave Desert dry lake // Ecology, 1980. V. 61. № 5. P. 1013-1028.
Vazquez de Aldana B. R., Geerts R. H. E. M., Berendze F. Nitrogen losses from perennial grass species // Oecoiogia, 1996. V. 106. №2. P. 137-143.
VellendM., Myers J. A., Gardescu S., Marks P. L. Dispersal of Trillium seeds by deer: implications for long distance migration of forest herbs // Ecology, 2003. V. 84. №4. P. 1067-1072.
Venkataraman G.S. Nonsymbiotic nitrogen fixation and organic matter in the tropics // 12th International Congress of Soil Science, New Delhi, India, 1982; Rev. Soil Res., Part 1. P. 205-235.
Verdu M., Valiente-Banuet A. The nested assembly of plant facilitation networks prevents species extinctions // American Naturalist, 2008. V. 172. №6. P. 751-760.
Vessey J.K., Pawlowski K, Bergman B. Root-based N 2-fixing symbiosis: Legumes, actinorhizal plants, Parasponia sp. and cycads // Plant and Soil, 2004. V. 266. № 1. P. 205-230.
Vicari M., Bazely D. R. Do grasses fight back? The case for antiherbivore defences // Trends in Ecology and Evolution, 1993. V. 8. №4. P. 137-141.
Vinton M.A., Burke /. C. Interactions between individual plant species and soil nutrient status in shortgrass steppe // Ecology, 1995. V. 76. № 4. P. 1116-1133.
Visser M. E., Both C. Shifts in phenology due to global climate change: the need for a yardstick // Proceedings of the Royal Society B: Biological Sciences, 2005. V. 272. № 1581. P. 2561-2569.
Vitousek P. M. Biological invasions and ecosystem processes: towards an integration of population biology and ecosystem studies // Oikos, 1990. V. 57. № 1. P. 7-13.
Volk M., Niklaus P.A., Korner C. Soil moisture effects determine CO2 responses of grassland species // Oecoiogia, 2000. V. 125. № 3. P. 380-388.
Von Liebig J. Chemistry in its application to agriculture and physiology. London: Taylor and Walton, 1840. 195 p.
Vrieling K, de Vos H., van Wijk C.A.M. Genetic analysis of the concentration of pyrrolizidine alkaloids of Senecio jacobaea 11 Phytochemistry, 1993. V. 32. №5. P. 1141-1144.
Walker J., Thompson С. H., Fergus J. F, Tunstall B. R. Plant succession and soil development in coastal sand dunes of subtropical eastern Australia // Forest succession: concepts and applications. West D. C., Shugart H. H., Botkin D. B. (eds.). New York: Springer, 1981. P. 107-131.
Wallander H., Wickman T, Jacks G. Apatite as a P source in mycorrhizal and non-mycorrhizal Pinus sylvestris seedlings // Plant and Soil, 1997. V. 196. № 1. P. 123-131.
Walter H. Allgemeine Geobotanik. 3 neubear. Auf. Stuttgart: Ulmer, 1986. 279 S.
Wan C., Sosebee R. E., McMichael B. L. Does hydraulic lift exist in shallow-rooted species? A quantitative examination with a half-shrub Gutierrezia sarothrae 11 Plant and Soil, 1993. V. 153. №1. P. 11-17.
Wang B., Qi и Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants // Mycorrhiza, 2006. V. 16. № 5. P. 299-363.
Wardle D.A., Bardgett R.D., Walker L.R., Peltzer D.A., Lagerstrom A. The response of plant diversity to ecosystem retrogression: evidence from contrasting long-term chronosequences // Oikos, 2008. V. 117. № 1. P. 93-103.
558
Список литературы
Wardle D.A., Nicholson K.S. Synergistic effects of grassland plant species on soil microbial biomass and activity: implications for ecosystem-level effects of enriched plant diversity // Functional Ecology, 1996. V. 10. №3. P. 410-416.
Wardle D.A., van der Putten W. H. Biodiversity, ecosystem functioning and above-ground-belowground linkages // Biodiversity and ecosystem functioning: synthesis and perspectives. Loreau M., Naeem S., Inchausti P. (eds.). Oxford e.a.: Oxford University Press, 2002. P. 155-168.
Wardle D. A., Walker L R., Bardgett R. D. Ecosystem properties and forest decline in contrasting long-term chronosequences // Science, 2004. V. 305. №5683. P. 509-513.
Wardle D.A., Zackrisson O., Nilsson M.-C. The charcoal effect in boreal forests: mechanisms and ecological consequences // Oecologia, 1998. V. 115. №3. P. 419-426.
Warming E. Plantesamfund — Grundtraek af den okologiske Plantegeografi. P. G. Philipsens Forlag, Kjobenhavn, 1895. 335 pp.
Waser N. M., Price M. V. What plant ecologists can learn from zoology // Perspectives in Plant Ecology, Evolution and Systematics, 1998. V. 1. №2. P. 137-150.
Watkins N.K., Fitter A. H., Graves J.D., Robinson D. Carbon transfer between C3 and C4 plants linked by a common mycorrhizal network, quantified using stable carbon isotopes // Soil Biol. Biochem., 1996. V.28. №4-5. P.471-477.
Watkinson A. Plant senescens // Trends in Ecology and Evolution, 1992. V. 7. № 12. P. 417-420. Watt A. S. Contribution to the ecology of bracken (Pteridium aquilinum). Vll. Bracken and litter.
1. The origin of rings // New Phytologist, 1956. V. 55. P. 369-388.
Watt A. S. Contribution to the ecology of bracken (Pteridium aquilinum). Vll. Bracken and litter. 3. The cycle of change // New Phytologist, 1970. V. 69. P. 431-449.
Watt A. S. Pattern and process in the plant community //Journal of Ecology, 1947. V. 35. № 1-2. P. 1-22.
Watteau E, Berthelin J. Microbial dissolution of iron and aluminium from soil minerals: efficiency and specificity of hydroxamate siderophores compared to aliphatic acids // European Journal of Soil Biology, 1994. V. 30. № 1. P. 1-9.
Webb S. L. Windstorm damage and microsite colonization in two Minnesota forests // Canadian Journal of Forest Research, 1988. V. 18. №9. P. 1186-1195.
Weber E. F. The alien flora of Europe: a taxonomic and biogeographic review // Journal of Vegetation Science, 1997. V. 8. №4. P. 565-572.
Wein R. W., Bliss L. C. Changes in arctic Eriophorum tussock communities following fire // Ecology, 1973. V.54. №4. P. 845-852.
Weiner J. Following the growth of individuals in crowded plant populations // Trends in Ecology and Evolution, 1995. V. 10. № 10. P. 389-390.
Weiner J. The influence of competition on plant reproduction // Plant reproductive ecology: patterns and strategies. Doust J. L., Doust L. L. (eds.). New York, Oxford: Oxford Univ. Press, 1990. P. 228-245.
Welden C. W., Slauson W. L The intensity of competition versus its importance: an overlooked distinction and some implications // Quarterly Review of Biology, 1986. V. 61. № 1. P. 23-44.
Wells С. E., Glenn D. M., Eissenstat D. M. Soil insects alter fine root demography in peach (Prunus persica) 11 Plant, Cell and Environment, 2002. V. 25. №3. P. 431-439.
Weltzin J. F, Pastor J., Harth C., Bridgham S. D., Updegraff K., Chapin С. T Response of bog and fen plant communities to warming and water-table manipulations // Ecology, 2000. V. 81.№12. P. 3464-3478.
Wennstrom A. Systemic diseases on hosts with different growth patterns // Oikos, 1994. V. 69. №3. P. 535-538.
Wennstrom A., Ericson L Environmental heterogeneity and disease transmission within clones of Lactuca sibirica // Journal of Ecology, 1992. V.80. № 1. P. 71-77.
Список литературы
559
Wenny D. G., Levey D. J. Directed seed dispersal by bellbirds in a tropical cloud forest // Proceedings of the National Academy of Sciences USA, 1998. V. 95. № 11. P. 6204-6207.
Wesson G., Wareing P. F. The role of light in the germination of naturally occurring population of buried weed seeds // Journal of Experimental Botany, 1969. V. 20. № 2. P. 414-425.
West N. E., Rea К. H., Harniss R. 0. Plant demographic studies in sagebrush-grass communities of southeastern Idaho // Ecology, 1979. V. 60. № 2. P. 376-388.
Weste G, Cooke D., and Taylor P. The invasion of native forest by Phytophthora cinnamomi. 11. Postinfection vegetation patterns, regeneration, decline in inoculum and attempted control // Australian Journal of Botany, 1973. V. 21. № 1. P. 13-29.
Westman W. E. Air pollution injury to coastal sage scrub in the Santa Monica Mountains, Southern California // Water, Air and Soil Pollution, 1985. V. 26. №1. P. 19-41.
Westman W.E. Measuring the inertia and resilience of ecosystems // BioScience, 1978. V. 28. №11. P. 705-710.
Westman W. E. Oxidant effects on California coastal sage scrub // Science, 1979. V. 205. № 4410. P. 1001-1003.
Westoby M. К leaf-height-seed (LHS) plant ecology strategy scheme // Plant and Soil, 1998. V. 199. №2. P. 213-227.
Westoby M. Comparing Australian ecosystems to those elsewhere // Bioscience, 1988. V. 38. №8. P. 549-556.
Westoby M., Leishman M., Lord J. Comparative ecology of seed size and dispersal // Plant life histories. Silvertown J., Franco M., Harper J. L. (eds.). Cambridge: Cambridge University Press, 1997. P. 143-162.
Westover К. M., Kennedy A. C., Kelley S. E. Patterns of rhizosphere microbial community structure associated with co-occuring plant species // Journal of Ecology, 1997. V. 85. №6. P. 863-873.
Whigham D. E, McCormick M. K., O'Neill J. P. Specialized seedling strategies 11: orchids, bromeli-ads, carnivorous plants, and parasites // Seedling Ecology and Evolution. Leek M. A., Parker V.T., Simpson R. L. (eds.). Cambridge e.a.: Cambridge Univ. Press, 2008. P. 79-100.
Whisenant S. G. Changes fire frequences on Idaho’s Snake River Plains: ecological and management implications // Proceedings symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management, Las Vegas, NV, April 5-7, 1989. McArthur E. D., Romney E. M., Smith S. D., Tueller P. T. (eds.). Ogden Utah: U. S. Forest Service, Intermountain Research Station, General Technical Report 1NT-276, 1990. P.4-10.
White /. Demographic factors in populations of plants // Demography and evolution in plant populations. O.T.Solbrig (ed.). Oxford e.a.: Blackwell, 1980. P. 21-48.
White J. E, Jr., Reddy P V, Bacon C. W. Biotrophic endophytes of grasses: a systematic appraisal // Microbial Endophytes. Bacon C. W. and White J. F., Jr. (eds.). New York, Basel: Marcel Dekker, 2000. P. 49-62.
White P Pattern, process, and natural disturbance in vegetation // Bot. Review, 1979. V. 45. P. 229-299.
Whitlock R., Grime J. P, Booth R., Burke T. The role of genotypic diversity in determining grassland community structure under constant environmental conditions // Journal of Ecology, 2007. V.95. №5. P. 895-907.
Whitmore T. C. (ed.) Tree flora of Malaya: a manual for foresters. Том 2. Kuala Lumpur: Longman, 1973. 444 p.
Whittaker R. H. Communities and Ecosystems. — New York: Macmill, 1970. 162 p.
Wielgolaski EE. Primary production of alpine communities in Norway estimated by CO2-exchange and harvesting techniques // Proc. 4th Int. Congr. Photosynthesis. London, 1978. P. 245-257.
560
Список литературы
Wiens D. Ovule survivorship, brood size, life history, breeding systems, and reproductive success in plants // Oecoiogia, 1984. V.64. № 1. P. 47-53.
Wilkinson D.M. Mycorrhizal evolution // Trends in Ecology and Evolution, 2001. V. 16. №2. P. 64-65.
Willems A. The taxonomy of rhizobia: an overview // Plant and Soil, 2006. V. 287. № 1-2. P.3-14.
Willems J. H. Soil seed bank, seedling recruitment and actual species composition in an old and isolated chalk grassland site // Folia Geobotanica et Phytotaxonomica, 1995. V. 30. №2. P. 141-156.
Willems J. H.y Huijsmans K. G. A. Vertical seed dispersal by earthworms: a quantitative approach // Ecography, 1994. V. 17. №2. P. 124-130.
Williams J. W., Post D. M., Cwynar L. C., Lotter A. E, and Levesque A. J. Rapid and widespread vegetation responses to past climate change in the North Atlantic region // Geology, 2002. V.30. №11. P. 971-974.
Willis A. Braunton Burrows: the effects on vegetation of the addition of minetai nutrients to the dune soils // Journal of Ecology, 1963. V. 51. № 2. P. 353-374.
Willis Л., Yemm E. Braunton Burrows: mineral nutrient status of the dune soils // Journal of Ecology, 1961. V.49. №2. P. 377-390.
Wilson A. D.y Harrington G. N. Grazing ecology and animal production // Management of Australia’s rangelands. Harrington G. N., Wilson A. D., Young M. D. (eds.). East Melbourne: CS1RO, 1984. P. 63-77.
Willson M.E Plant reproductive ecology. New York: Wiley, 1983-282 p.
Wilson D. Fungal endophytes which invade galls: insect pathogens, benign saprophytes, or fungal inquilines? // Oecoiogia, 1995. V. 103. №2. P. 255-260.
Wilson J.B. Mechanisms of species coexistence: twelve explanations for Hutchinson’s ’Paradox of the plankton’: evidence from New Zealand plant communities // New Zealand Journal of Ecology, 1990. V. 13. № 1. P. 17-42.
Wilson J.B. Shoot competition and root competition // Journal of Applied Ecology, 1988. V. 25. №1. P. 279-290.
Wilson J. B., Sykes M. T Some tests for niche limitation by examination of species diversity in the Dunedin area, New Zealand // New Zealand Journal of Botany, 1988. V. 26. №2. P. 237-244.
Wilson J. B., Wells T.C.E., Trueman I. C., Jones G., Atkinson M. D., Crawley M.J., Dodd M. E.y Silvertown J. Are there assembly rules for plant species abundance? An investigation in relation to soil resources and successional trends // Journal of Ecology, 1996. V. 84. №4. P. 527-538.
Wilson S. D.y Kleb H. R. The influence of prairie and forest vegetation on soil moisture and available nitrogen // The American Midland Naturalist, 1996. V. 136. №2. P. 222-231.
Witmer M. C., Cheke A. S. The dodo and the tambalacoque tree: an obligate mutualism reconsidered // Oikos, 1991. V. 61. № 1. P. 133-137.
Wolf J. W.y Peet R. K. Production and diversity in southeastern coastal plain savannas // Bull. Ecol. Soc. Amer., 1980. V. 61. №2. P. 77.
Wolfe L. M. Why alien invaders succeed: support for the escape-from-enemy hypothesis // American Naturalist, 2002. V. 160. №6. P. 705-711.
Woodman G. W.y Fernandes G. W. Differential mechanical defence: herbivory, evapotranspiration and leaf-hairs // Oikos, 1991. V. 60. № 1. P. 11-19.
Wright I. J., Cannon K. Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora // Functional Ecology, 2001. V. 15. №3. P. 351-359.
Wright J. W.y Meagher T. R. Pollination and seed predation drive flowering phenology in Silene latifolia (Caryophyllaceae) // Ecology, 2003. V. 84. №8. P. 2062-2073.
Список литературы
561
Хи J. Т, Ми С. The relation between growth of Gastrodia data protocorms and fungi // Acta Botanica Sinica, 1990. V. 32. № 1. P. 26-31.
Yanai R. D., Eissenstat D. M. Coping with herbivores and pathogens: a model of optimal root turnover // Functional Ecology, 2002. V. 16. № 6. P. 858-869.
Yemm E. IV., Willis A. J. The effects of maleic hydrazide and 2,4-dichlorophenoxyacetic acid on roadside vegetation // Weed Research, 1962. V. 2, No 1. P. 24-40.
Yoda K., Kira T., Ogawa H., Hozumi K. Self-thinning in overcrowded pure stands under cultivated and natural conditions //J.Biol. Osaka City Univ., 1963. V. 14. №4. P. 107-129.
Yodzis P. Competition for space and the structure of ecological communities // Lecture Notes in Biomathematics (Berlin: Springer), 1978. V. 25. P. 1-191.
Yu S.-Y. Centennial-scale cycles in mid-Holocene sea level along the southeastern Swedish Baltic coast // Geological Society of America Bulletin, 2003. V. 115. № 11. P. 1404-1409.
Zackrisson O.t Nilsson М.-C., and Wardle D.A. Key ecological function of charcoal from wildfire in the Boreal forest // Oikos, 1996. V. 77. № 1. P. 10-19.
Zaller J.G., Arnone III J. A. Activity of surface-casting earthworms in a calcareous grassland under elevated atmospheric CO2 // Oecologia, 1997. V. 111. № 5. P. 249-254.
Zamora R., Hodar J. A., Gomes J. M. Plant-herbivore interaction: beyond a binary vision // Handbook of Functional Plant Ecology. Pugnaire F. 1. and Valladares F. (eds.). New York, Basel: Marcel Dekker, 1999. P. 677-718.
Zanne A., Chapman C.A., Kitajima K. Evolutionary and ecological correlates of early seedling morphology in East African trees and shrubs // American Journal of Botany, 2005. V. 92. №6. P. 972-978.
Zavaleta E. S. The emergence of waterfowl conservation among Yup’ik hunters in the Yukon-Kuskokwim Delta, Alaska // Human Ecology, 2000. V. 27. №2. P. 231-266.
Zeevalking H. J., Fresco L E M. Rabbit grazing and diversity in a dune area // Vegetatio, 1977,V. 35. №3. P. 193-196.
ZimovS.A., Chuprynin V. I., OreshkoA. R, Chapin E S. Ill, Chapin M. C., and Reynolds J. F. Effects of mammals on ecosystem change at the Pleistocene-Holocene boundary // Arctic and Alpine Biodiversity: Patterns, Causes and Ecosystem Consequences. Chapin F. S. Ill, Korner Ch. (eds.). Berlin: Springer-Verlag, 1995. P. 127-135.
ZobelM. Plant species coexistence — the role of historical, evolutionary and ecological factors // Oikos, 1992. V.65. №2. P. 314-320.
Zobel M., Otsus M., Liira J., Moora M., Mols T Is small-scale species richness limited by seed availability or microsite availability // Ecology, 2000. V. 81. № 12. P. 3274-3282.
Zotz G. Gefassepiphyten in temperaten Waldern // Bauhinia, 2002; B. 16, S. 13-22.
Zotz G. Vascular epiphytes in the temperate zones — a review // Plant Ecology, 2005. V. 176. №2. P. 173-183.
Zotz G., List C. Zufallsepiphyten — Pflanzen auf dem Weg nach oben // Bauhinia, 2003; В. 17, S. 25-37.
Предметный указатель
К-стратеги 212
R-стратеги 212, 213
RYT (суммарный относительный урожай) 189-191
Абрин 114
агеотропный рост 40
агриофиты 457-460, 463-466 азотфиксация 24, 25, 27, 29, 30, 32,
34, 38, 40-44, 102, 173, 246, 250, 252, 268, 285, 445, 487, 493, 494
— ассоциативная 173, 250
— симбиотическая 24
азотфиксирующие прокариоты 23-25, 27, 119, 285, 452
аккумуляция азота 255
алкалоиды 85, 86, 89, 112, 113, 119, 120, 142, 164
аллелопатия 158, 223, 224, 227-232 аллокация 124, 175, 347, 357-359, 415-423, 425, 426, 444
альвары 156, 241, 242, 290, 311, 383, 392, 484
альгофлора 259
анаморфы 58
анастомозы 73
антоцианы 118
антропогенная эволюция 455
апомиксис 461
арабитол 74
арбускулы 49, 50, 62, 493
ареал 15, 17, 132, 137, 214-216, 295, 351, 457, 465, 470, 480, 481, 488, 492
— синэкологический 214-216
— физиологический 215
— фитоценотический 215
— экологический 215
аридйость 473
аридные регионы 155, 236, 243, 303, 375, 381, 382
археофиты 457, 458
аспект 206, 214, 232, 249, 257, 445
аэренхима 247, 248
Бактериальные помощники микориз 69, 71
биогеоценоз 21, 289
биомасса 357
биотрофы 22
биоценоз 18
болота 19, 65, 98, 100,215, 220, 246, 274, 276, 277, 280, 283, 328, 334, 335, 359, 360, 447, 454, 491
— низинные 182, 450
— олиготрофные 182
бруцин 115
Вегетативное размножение 427, 461
везикулы 32, 49, 71
веламен 172
ветошь 128, 431, 439, 449, 450
виоленты 198, 208, 212-214,
216, 337
водопроницаемость 134, 155, 240
Предметный указатель
563
вторичные метаболиты 111, 118, 431, 493
выветривание 55, 56, 70, 248, 268, 271, 473
выделение 18, 19, 22, 28, 33 — активное 224 — пассивное 224
вытаптывание 151, 154, 200
Галактитол 166 галлообразователи 85, 127 галофит 465 гаустории 160, 162, 164, 168 гейтеногамия 128, 142 гелсемин 142 генеты 56, 189, 322 геоботаника 15, 16, 20, 252 геофиты 153, 205, 345, 436, 438 гетероцисты 39-41 гиалиновые клетки 38, 357 гибберлиновая кислота 164 гидравлический лифт 243, 244, 286 гидроюглон 229 гильдия видов 218 гипотеза «недостатка ресурсов» 119 — оптимальной защиты 120 — уязвимости 120, 129 — «экспонированности» 119 гифосфера 56 гликопротеины 114, 116 глицин 225 гломалин 62 годичный прирост 75, 266, 353 голопаразиты 159, 162 гормоны 115, 123, 164 горчичные масла 68 госсипол 114, 115 гранулометрический состав почвы 245, 261
гумидные районы 155, 288
Двуокись азота 438 демография растений 16 демутация 445 денитрификация 156
детерминант 22
дефолиация 28, 105, 122, 128, 165, 201, 350, 413
диаспоры 20, 22, 32, 143, 257, 277, 279-282, 289, 294, 297, 309, 312,319, 323, 324, 330, 331, 338, 350, 398, 402, 405, 406, 410,413, 425, 427, 460, 461, 480
Жасмонат 122
жизненность растений 65, 87, 93, 100, 128, 162
Закон Либиха 179
залежный режим 135, 387
занос диаспор 460, 461
засоление 60, 65, 68, 134, 155, 162, 245, 246, 248, 344, 382, 409
засуха 200, 201, 206, 209, 211, 241, 489
заурохория 147
защитный индекс 122
зоохория 143, 144, 149-151, 279 синзоохория 143, 148-150 эндозоохория 143, 148-151 эпизоохория 143, 147, 150
Избегание 103, 104, 116, 128, 194, 466
избыточное потребление 196 изопрен 224, 368
ингибиторы прорастания 162,
325, 327
индекс листовой поверхности 175, 365
— полезности микориз 64
инсектициды 75, 126, 127, 129-131, 137,415
интродуцент 230, 461, 463, 465-467 информационные сигналы 223
Кайнозой 467, 470
капиллярный подъем влаги 155, 245
564
Предметный указатель
капсаицин 112, 113 каротин 114 катехин 230
кластерные корни 33, 34, 66, 68, 246 клональные растения 232 колонизация 460, 461 компенсационная точка 261, 264, 272
кониин 112
конкурентная иерархия 208, 220 конкурентный отклик 211 — эффект 211
конкуренты 184, 191, 200, 203, 205, 208, 222, 291
конкуренция 179 — побеговая 193, 194 консорт 22, 193, 194, 364 консорция 21, 22 континуум 17, 20 копролиты 102, 137, 144, 263, 490 копрофаги 145, 156 коралловидные корни 40 корневищные злаки 450 корневое избегание 194 — перемешивание 194 корневые выделения 28, 43, 162, 166, 194, 224-227, 230, 248
коэффициент усвоения энергии 124 ксерофит 454, 493
Лианы 107, 158, 164, 173-178 лихенин 267 лихеноиндикация 268 лишайники 38, 39, 44, 81, 169-171, 173, 234, 240, 259, 264-272, 286, 299, 303-305, 329, 368, 374, 383, 384, 481,482, 492 лишайниковые кислоты 267, 268, 271 лишайниковый коэффициент 271 лолитремы 85
Макромицеты 96, 467 маннитол 74, 166, 267
мезофит 454, 493
мезофитизация 454
ментол 114
метан 492, 493
метод истощения 216
механические аналоги корней 195
миграция растений 480
микогетеротрофы 54, 62, 76, 77,
79, 80
микориза
везикулярно-арбускулярная (арбускулярная микориза) 465, 493
— эрикоидная 180
микоризная сеть в почве 72-75, 290
микоризосфера 56
микосимбиотрофизм 48, 81
микофаги 68, 76, 82
микромицеты 96
миксотрофия 79
минеры 123, 127, 129, 198
миоцен 467, 472, 473, 477
мирмекохория 145, 147, 481
мирмикофильные деревья 110
модель 180, 185, 194, 206, 220, 221
— Тильмана 178, 185, 186, 319
монокарпик 205
монокультура 166, 189-191, 193,
194, 209-211, 228, 250-252,
287, 309, 414, 417
монотерпены 224
морозостойкость 263, 265, 455, 492
морфологический индекс 200, 202
муцигель 225
мюллеровы тела 172
Небелковые аминокислоты 114
небулофиты 240
нематодоциты 75
неофиты 457-459
несимметричность конкуренции 186
неупругая деформация 155
никотин 116, 122
нитратредукция 196
нитрификация 224, 230, 250, 444
Предметный указатель
565
нитрогеназа 27, 32, 42 нитрофильные виды 441
Оборот биомассы 260, 359, 360 озон 224, 486, 494 окислительно-восстановительный потенциал почв 247, 248 оксалат кальция 105 олигоцен 467, 470, 472, 473, 475 опад 24, 36, 38, 56, 57, 60, 118, 125, 129, 169, 170, 172, 196, 200-202, 224, 229, 231, 237, 245, 246, 248, 255, 271, 328, 338, 357, 429, 434, 441, 492, 494
опыление 17, 89, 137, 139-143, 254, 346, 375
оскулы 164
отложение экскрементов 151, 155, 156
относительная скорость роста 175, 195, 196, 200-203, 340, 415, 419, 426, 444
отношение корни:побеги 426 — «корни:побеги» 162, 196, 198 — корни побеги 417
— «красный : дальний красный свет» 186
отношения контактные 158 — непрямые трансбиотические 158, 178
Палеоцен 467-470 паразитоиды 86, 105, 110, 111, 123 парниковый эффект 485 пастбищная дигрессия 444 патиенты 199, 208, 212, 214, 216, 337, 381 пелотоны 59 пептиды 114 перамин 85 пинен 114 пирогенные сукцессии 441 пищевые детерренты 105, 111, 117, 118
пластичность 186, 195, 203, 205, 395 плейстоцен 467, 477, 479, 480, 484, 487
плиоцен 467, 477, 478 плотнодерновые злаки 450 подкисление 33, 239, 246, 272, 274, 399, 444, 454
подстилка (опадогенный горизонт) 55, 56, 95, 96, 102, 231,237, 247, 298, 326, 341, 440 пожары 428-443, 450, 459, 465, 489 — верховые 440 — низовые 440, 443 покой 461 полупаразиты 65, 66, 68, 81, 145, 158-160, 162-169, 218, 286 пороговая концентрация ресурса 179, 180, 186, 195, 212-214, 415 порой 75, 135, 136, 150, 294, 349 похитители нектара 142 правило Law—Lewis 84 — Сукачева 187 превышение урожая нетрансгрессивное 191 ----трансгрессивное 191 продукция
нетто-продукция 227, 491 — первичная 218, 429, 491, 492 пространственная сопряженность 255 протокорм 59
Реабсорбция нектара 142 репродуктивное усилие 213, 214, 436, 494 ресурс 178 ресурсы 48 реутилизация 36, 38, 166 ризины 271 ризоплан 24, 42 ризосфера 24, 27, 28, 42, 43, 60, 110, 230, 240, 246-248, 256, 260, 261, 285, 493 рицин 114
566
Предметный указатель
рудералы 200, 203, 205, 208, 212, 213, 337, 411, 426, 455, 462
Салицилат 122 сапонины 114 сапротрофы 22, 46, 76, 79, 88, 95, 96, 224, 467 сверлильщики 127 связи 18, 22 — топические 169 — трофические 22 семенная продуктивность 447 — продукция 64, 104, 142, 279, 308, 347, 385, 395, 461 семенные банки 427, 432, 437 семенояды 85, 127, 130, 144, 340, 436 сенокос 447, 450, 453 сенокошение 192, 444, 445, 447-450, 454 сердечные глюкозиды 114 сеть Гартига 54, 55 симбиоз актинориза 24 — ризобиальный 24, 25, 28 — цианобактериальный 24, 25 симбиотрофы (микосимбиотрофы, эндофиты) 46 синэкология 16, 17, 281, 282 склерофилльность 105, 380 смертность растений 187 срастание корней 158, 159 — стволов 158 стравливание 151, 153, 364 стресс-толеранты 200, 201, 203, 205, 208, 209, 212, 216, 337, 381, 426, 428, 455, 462 стригол 162 стрихнин 115 сукцессия 35, 65, 66, 69, 72, 94, 129, 130, 154, 174, 228, 246, 257, 334, 335, 381,392, 393, 395, 396, 398-400, 402, 405-415, 424-428, 442, 445, 447, 451 — вторичная 174, 426, 427, 445 — первичная 426-428
супервид 186
сфера средообразующего воздействия 233
схема De Wit 189, 220, 414
Танины 116-120, 151
телеоморфы 58
терпеноиды 114, 120, 123
терпены 60, 114, 117, 122
типы стратегий растений
функциональные типы 17, 199 токсины 111, 114-117, 119, 120, 129, 153
толерантность 103, 288, 375, 410, 494
транспирационный коэффициент 454
транспирация 165, 175, 241, 243, 244, 450, 488
траншейная обрубка 218, 220, 222 трегалоза 69, 74, 80
триада Герли—Коха 230
Удельная листовая поверхность 199, 487
удобрения 27, 42, 63, 92, 98, 100, 173, 182, 184, 185, 187, 192, 198, 215, 224, 228, 252, 345, 371, 388, 427, 447-454, 466, 492
устьичная проводимость 494
Фар 493
фенилуксусная кислота 233
фенология 449, 492
филлоплан 24, 42, 96
финбош 182, 429, 434-436, 438
фитоалексин 89, 115
фитогенное поле 233
фитолиты 105, 107, 122, 249
фитомасса 175, 445
фитометр 216
фитопатогенные грибы 46, 60, 65, 76, 88, 89,91-96, 193, 194, 294, 333, 344, 415, 456, 467
Предметный указатель
567
фитосоциология 16
фитофаги 19, 21, 22, 28, 41, 60, 61, 65, 85-87, 93, 102-105, 107, 109-119, 122-134, 142, 147, 152, 162, 164, 165, 167, 171, 172, 175, 178, 196, 198, 200, 201, 209, 222, 228, 253, 254, 256, 275, 284, 288, 289, 294, 323, 329, 333, 340, 344, 346, 350, 357, 359, 362, 364, 369, 378, 384, 385, 387, 406, 415, 429, 434, 436, 444, 445, 455, 462, 465, 466, 477, 481, 484, 489, 490, 492, 494
фитоценология 12, 13, 15-17 фитоценотипы 198 флавоноиды 118, 119, 226 флористическая насыщенность 72, 130, 131, 133, 152, 182, 259, 275-280, 284, 329, 411, 434, 439, 440, 445, 450, 452, 454, 466, 484
флуктуации 211
— ресурсов 212
форофит 169, 170, 172, 178 фосфорные удобрения 63, 65,
173, 454
фотобионт 38, 39, 264, 267 фотосинтез 59, 71, 73, 75, 87, 100, 107, 109, 119, 123, 159, 160, 162, 165, 167, 170-172, 196, 201, 234, 235, 241, 243, 261, 264-267, 272-274, 287, 296, 307, 308, 346, 348, 359, 365, 374, 380, 381, 416, 417, 455, 487, 488, 490, 491
- СЗ 172
- С4 172
- САМ 172, 235, 243
фототоксины 115
фреатофит 240, 286
фунгистатические вещества 66, 68 фунгициды 63, 72, 89, 129, 228, 333
Целлюлозные оболочки 117
цефалодии 494
цианогенные глюкозиды 111,
112, 119
циклический пироклимакс 272
Чапарраль 429, 431-434
Шоровые солончаки 65
Эвапотранспирация 243, 444, 447, 450, 459, 489 эволюция сообществ 455 эвтрофикация 488, 492 эдификатор 213 экдизоны 115 эккрисотрофы 22, 46, 47 экологическая ниша 285, 414 ---- реализованная 215 ----фундаментальная 215 экологические факторы 64, 65, 178, 282, 284, 287
----нересурсные 191
----ресурсные 178, 233
— фильтры 19, 20
экология растений 15-17, 287, 322 экосистема 12, 18, 21, 22, 24, 25, 46, 73, 100, 101, 153, 259, 268, 274, 281, 283, 302, 357, 359, 360, 362, 364, 365, 368, 409, 411, 441, 444, 447, 456, 490-492
эксплеренты 199, 208, 212-214, 337, 381,392, 449, 461
экссудат 71
экстрафлоральные нектарники 109, 110, 172
элайосомы 144, 145, 147
эндофиты 46, 81-87, 93, 165
эоцен 173, 467, 468, 470, 471, 473
эпипаразиты 80
эпифиллы 170, 171, 269
эпифиты 38, 39, 59, 158, 169-173, 259, 263, 266, 268, 271, 272, 274, 360, 374, 492, 493
568 Предметный указатель
эргот-алкалоиды 85
эрозия 135, 155, 200, 240, 444
этилен 122, 223, 327, 438
эфемероиды 74, 104, 120, 205, 287, 379, 381
эфемеры 104, 205, 379, 381, 438
эффект Janzen-Connell 228 эффекта Сукачева 186, 187 эффективность улавливания
пыльцы 142
Юглон 229
Владимир Гертрудович ОНИПЧЕНКО
___________________________________________________Об автору Доктор биологических наук, профессор, заведующий кафедрой геоботаники биологического факультета Московского государственного университета имени М. В. Ломоносова. Окончил МГУ в 1980 г. Научная и преподавательская деятельность В. Г. Онипченко, помимо МГУ, связана с Карачаево-Черкесским государственным университетом имени У. Д. Алиева и Тебердинским заповедником (Карачаево-Черкесия). Основная сфера научных интересов — исследование структурнофункциональной организации альпийских экосистем Кавказа. В. Г. Онипченко — автор более 300 опубликованных работ, включая статьи в международных журналах экологического профиля, а также ряд монографий на русском и английском языках; среди них: «Alpine vegetation of the Teberda Reserve, the Northwest Caucasus» и «Alpine ecosystems in the Northwest Caucasus». В разные годы являлся членом редколлегии журналов «Биологические науки», «Oecoiogia Montana», «Бюллетень МОИП. Отдел биологический» и др., а также членом редакционного совета журнала «Растительность России». Исполнитель и руководитель проектов РФФИ, совместных международных проектов с университетами Уппсалы (Швеция; 1994-1998), Шеффилда (Великобритания; 2000-2001), Отаго (г. Данидин, Новая Зеландия; 2002), Утрехта и Амстердама (Нидерланды; 1998-2001,2005-2012), а также с Институтом биологии Китайской академии наук (г. Чэнду; 2004-2012). Под его руководством защищено 20 кандидатских диссертаций.