





















































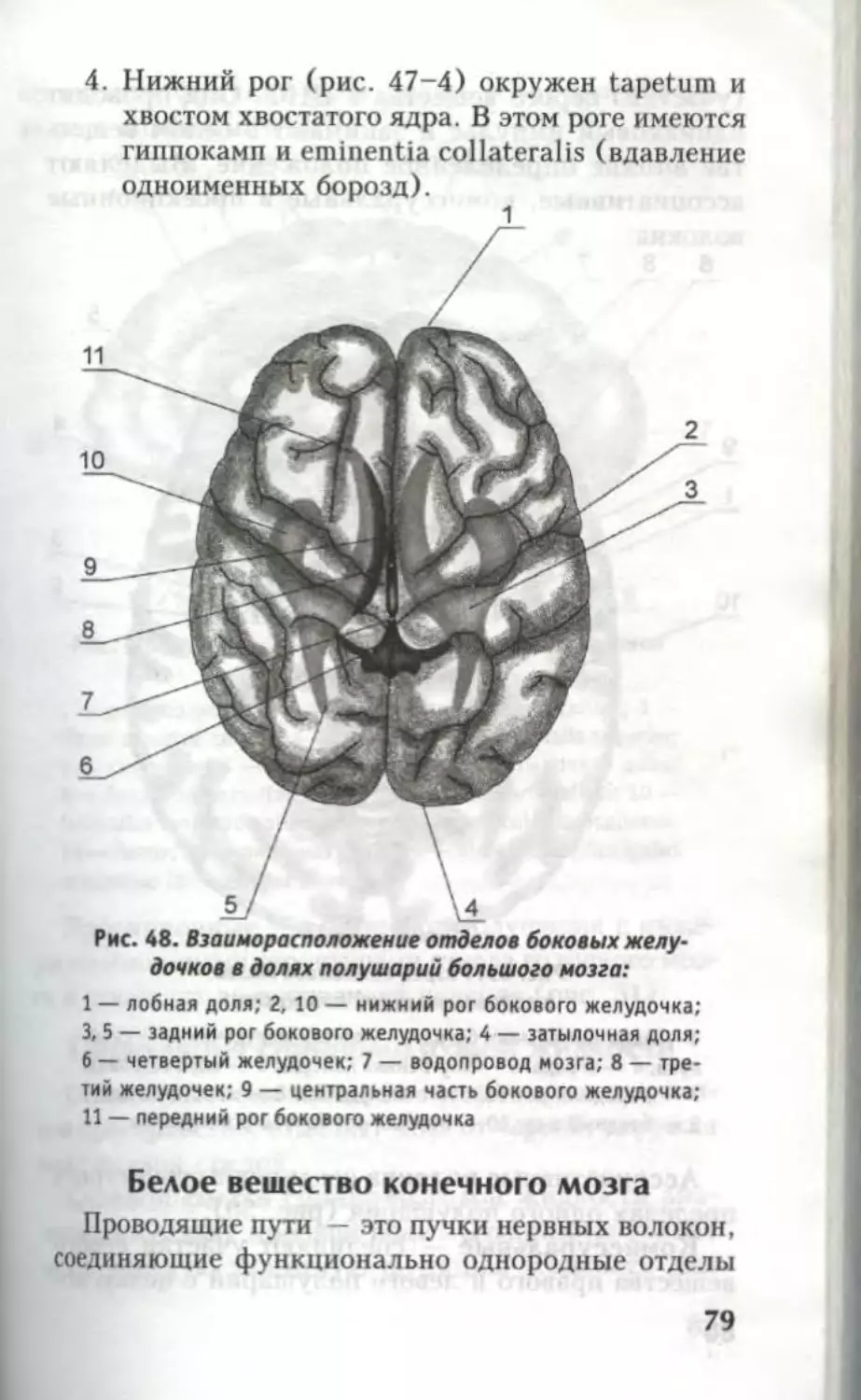
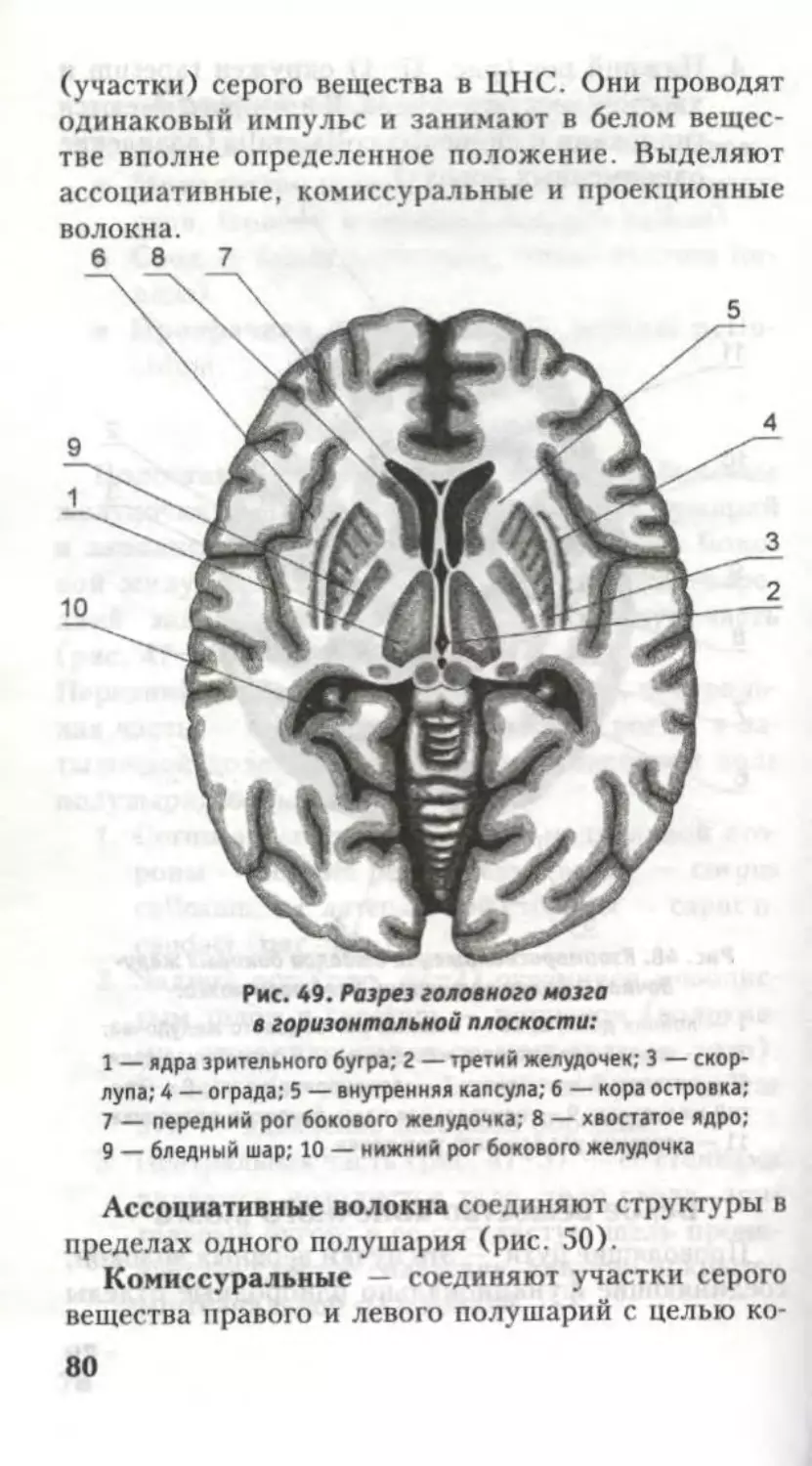
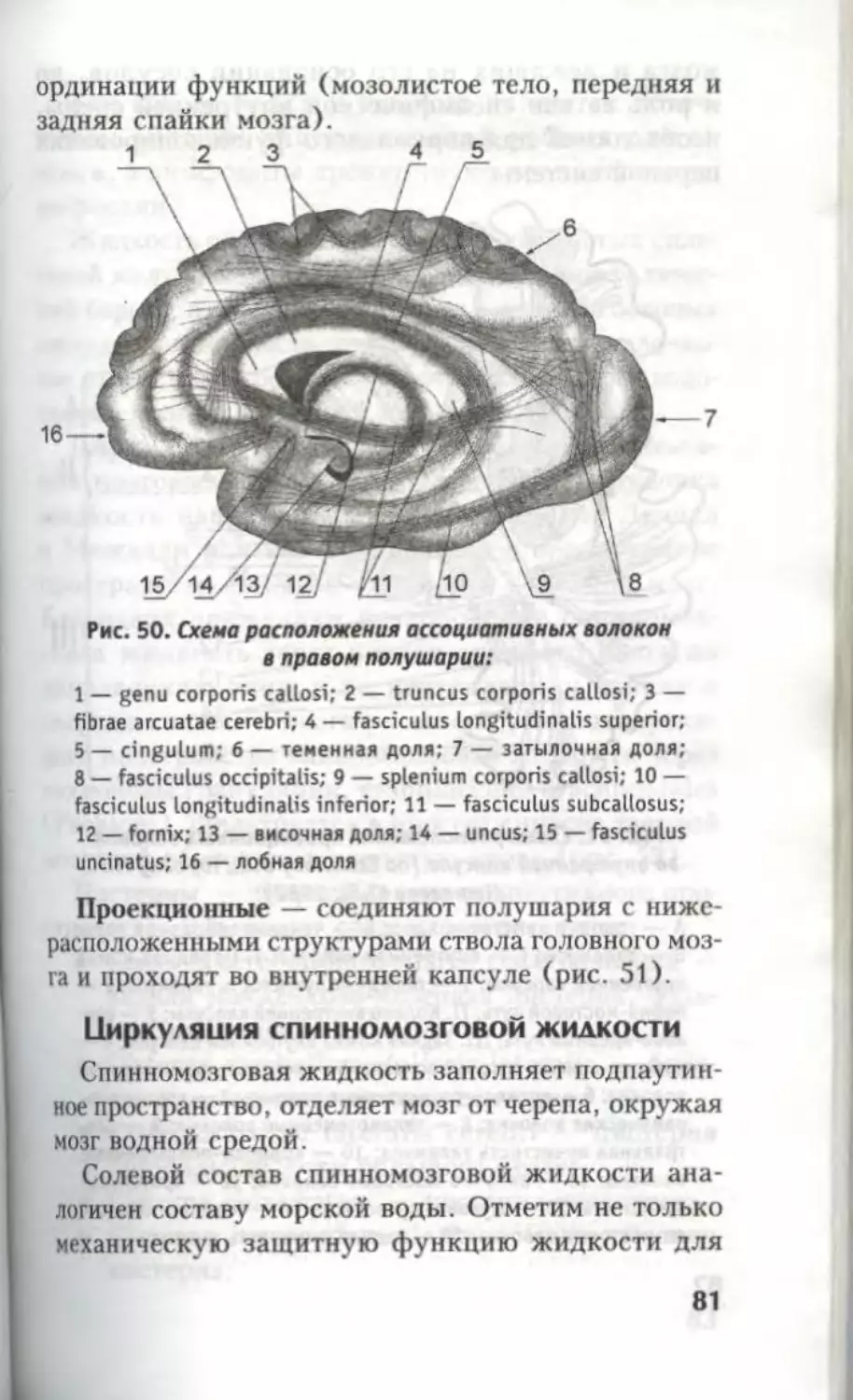


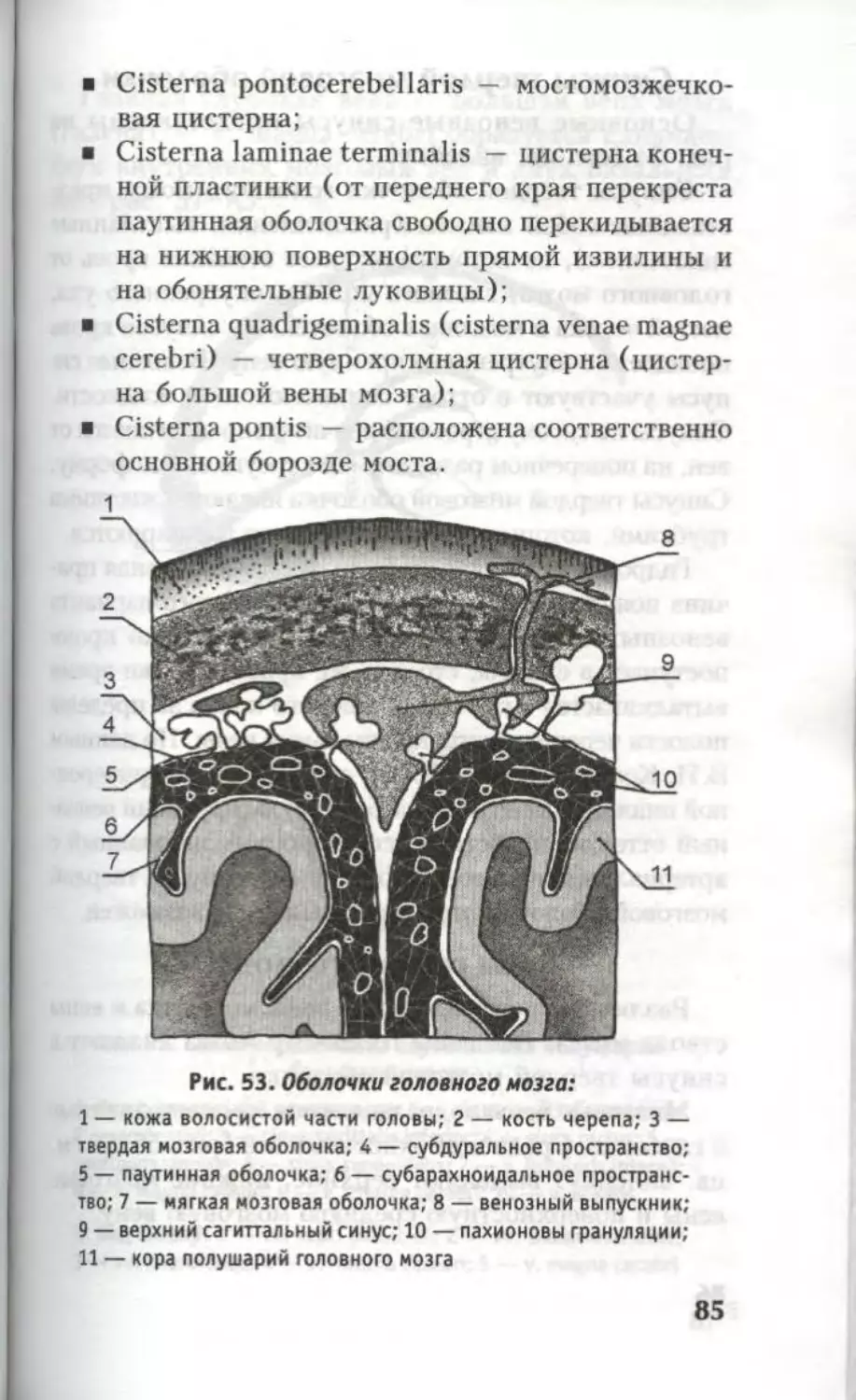

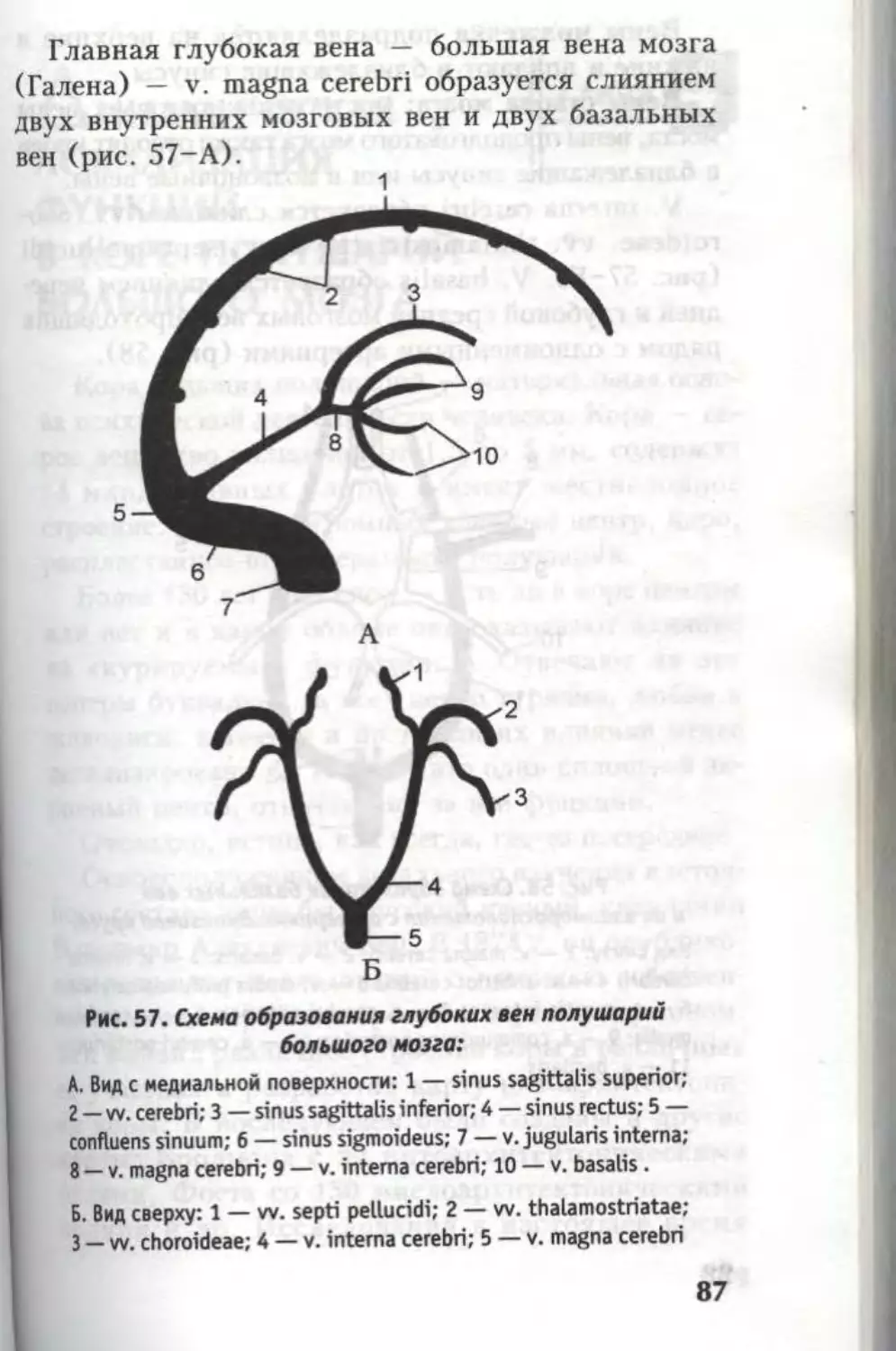
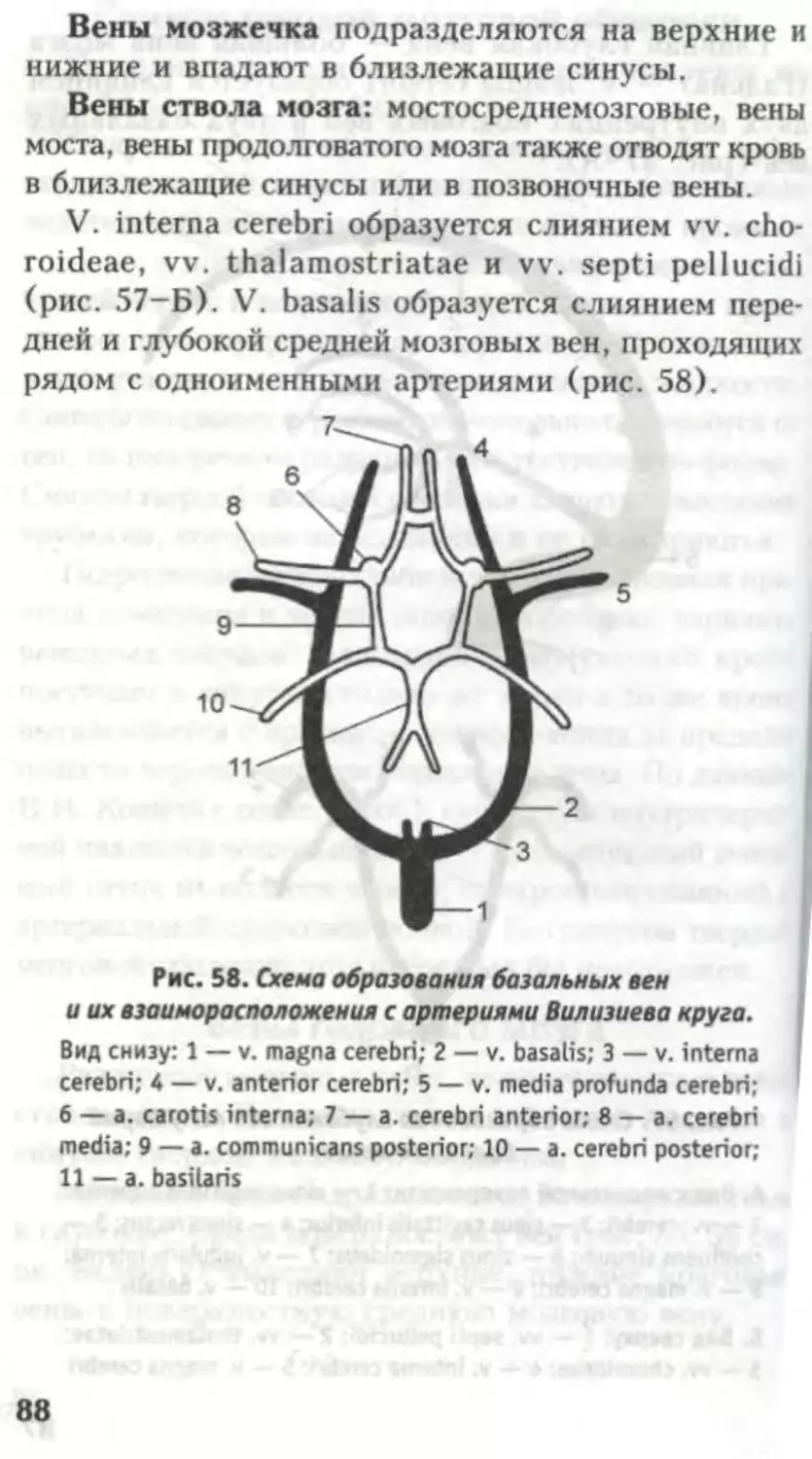















































































































Автор: Кондрашев А.В. Каплунова О.А.
Теги: анатомия анатомия человека сравнительная анатомия антропология медицина физиология физиология человека нервная система
ISBN: 978-5-699-33394-3
Год: 2010




А. В. Кондрашев, О. А. Каплунова
АНАТОМИЯ
НЕРВНОЙ
СИСТЕМЫ
МЕДИЦИНСКИЙ АТЛАС
МЕДИЦИНСКИЙ АТЛ AC
АНАТОМИЯ
НЕРВНОЙ
СИСТЕМЫ
А. В. Кондрашев, 0. А. Каплунова
ЭКСМО
МОСКВА
2009
УДК 611.8(075.8)
ББК 28.706
К 19
Рецензенты:
Афонин А. А., дм.н., проф., зам. директора по науке ГУ НИИ аку-
шерства и педиатрии
Бабаев М. В., проф., зав. кафедрой лучевой диагностики и лучевой
терапии Ростовского государственного медицинского универ-
ситета
Терентьев В. И, проф., зав. кафедрой внутренних болезней № 1 Рос-
товского государственного медицинского университета
Кондрашев А. В.
К 19 Анатомия нервной системы / А. В. Кондрашев, О. А Кап-
лунова. — М.: Эксмо, 2010. — 224 с. : ил. — (Медицинский
атлас).
ISBN 978-5-699-33394-3
Анатомия центральной и периферической нервной системы - один
из самых трудных вопросов всей нормальной анатомии человека. Быва-
ет недостаточно классических учебников и атласов, чтобы хорошо по-
нять весь материал. Настоящий атлас - не совсем обычный. Авторы по-
следовательно проводят читателя по всем разделам анатомии нервной
системы, давая развернутые текстовые пояснения к схемам и цветным
рисункам.
Приведенная в атласе информация в удобной для усвоения форме
дает ответы на все пункты Государственной программы по неврологии.
Книга рекомендована УМО по медицинскому и фармацевтическо-
му образованию в качестве учебного пособия для студентов медицин-
ских вузов. Она будет полезна также студентам ссузов, интернам, орди-
наторам. аспирантам и юным врачам.
УДК 611.8(075.8)
ББК 28.706
© Кондрашев А. В., Каплунова О. А., 2008
© ООО «Наука-Спектр», 2008
© ООО «Издательство «Эксмо», 2009
ISBN 978-5-699-33394-3
ВЕДЕНИЕ
Данное учебное пособие предназначено для
самостоятельной работы студентов при изучении
анатомии центральной и периферической нервной
системы, учитывая дефицит учебной литературы и
известные трудности при освоении этого раздела
анатомии.
Настоящее пособие не может заменить учебник и
атлас по анатомии, но предназначено для повторе-
ния наиболее сложных теоретических вопросов по
неврологии перед зачетами или государственным
экзаменом по нормальной анатомии.
При подготовке данного пособия был использо-
ван лекционный курс кафедры нормальной анато-
мии Ростовского государственного медицинского
университета.
В учебное пособие включены общие и частные
вопросы анатомии мозга, нервов, органов чувств,
вегетативной нервной системы; в нем систематизи-
руются имеющиеся в литературе данные, восполня-
ются некоторые пробелы в материалах учебников.
Информация о наружном и внутреннем стро-
ении различных отделов мозга, органов чувств,
о ветвях и зонах иннервации нервов, развитии
и аномалиях развития обобщена и соответствует
ответам на все пункты экзаменационных вопросов
по неврологии.
В пособии имеются 146 схем и рисунков, большая
часть которых являются оригинальными, в том чис-
3
ле схемы проводящих путей спинного и головного
мозга, а также проводящих путей анализаторов.
В пособии приводятся краткие пояснения, указа-
ны русские и латинские термины, соответствующие
современной анатомической номенклатуре.
Учебное пособие предназначено для студентов
всех факультетов медицинских вузов, ординаторов
и начинающих врачей — неврологов.
Морфология
и ОНТОГЕНЕЗ
ЦЕНТРАЛЬНОЙ
Глава
НЕРВНОЙ СИСТЕМЫ, ЕЕ РОЛЬ
В ОРГАНИЗМЕ. СПИННОЙ МОЗГ
Органам и системам органов свойственны до-
статочно узкие, специфические функции, которые
обеспечивают жизнедеятельность организма. Объ-
единение (интеграция) органов и систем обеспечи-
вается:
1) структурным объединением всех частей орга-
низма в единое анатомическое образование;
2) связью всех частей организма при помощи:
жидкостей, циркулирующих в сосудах, по-
лостях, пространствах (гуморальная связь,
регуляция);
нервной системы (нервная регуляция).
Неврология изучает нервную систему, Systema
nervosum, которая состоит:
Systema nervosum centrale
Meninges
Medulla spinalis
Оболочки головного
и спинного мозга
Encephalon
Truncus encephali
Hemispherium
cerebri
Cerebellum
5
Systema nervosum periphericum
Ganglia Plexus Nervi (spinalis Plexus
(spinalis et cerebrales) et cerebrales)
Функции нервной системы сложны и многооб-
разны:
регулирующая — нервная система обеспе-
чивает связь между организмом и внешней
средой;
интегрирующая — объединяет функции всех
органов;
адаптивная — определяет приспособительное
поведение организма к окружающей среде;
двигательная — обеспечивает целенаправлен-
ные и автоматические движения;
психическая — обусловливает индивидуаль-
ные особенности человека.
В отличие от других интегративных систем нервная
система выполняет свои функции очень быстро (в со-
тые доли секунды), прицельно и кратковременно.
Основным анатомическим элементом нервной
системы является нейрон, нейроцит (рис. 1).
Различают следующие морфологические формы
нейронов: униполярный нейрон, биполярный, псев-
доуниполярный и мультиполярный.
Серое вещество спинного и головного мозга
состоит из групп клеточных тел нейронов. В от-
личие от него белое вещество образовано, глав-
ным образом, миелинизированными аксонами или
нервными тканями, выходящими из клеточных тел
нейронов.
Жировые изоляционные миелиновые оболочки
повышают скорость передачи нервных импульсов.
Передача нервного импульса с одного нейрона на
6
другой, к мышечному волокну, железе осуществля-
ется посредством синапсов (рис. 2).
Через синапс нервный импульс проходит в одном
направлении — от аксона одного нейрона к телу дру-
гого (аксосоматическая связь, филогенетически более
древняя), или от аксона одного нейрона к дендриту
другого (аксодендритическая связь, более молодая).
Передача импульса происходит при помощи медиа-
торов (ацетилхолин, катехоламины — адреналин и
норадреналин, некоторые аминокислоты, пептиды и
другие биологически активные вещества).
Рис. 1. Схема строения
нейроцита:
1 — ядро шванновской
клетки; 2 — дендрит;
3 — миелиновая обо-
лочка; 4 — мышечное
волокно; 5 — шваннов-
ская клетка в области
нервно-мышечного си-
напса
7
Нервная система — комплекс нейронов, связан-
ных бесчисленным количеством синапсов. Нервное
возбуждение распространяется от одного нейрона к
другому, третьему и так далее.
Рис. 2. Межнейрональный синапс:
1 — нервное волокна (аксон); 2 — синаптические пузырьки;
3 — синаптическая щель; 4-хеморецепторы постсинаптической
мембраны; 5 — постсинаптическая мембрана; 6 — синаптичес-
кая бляшка; 7 — митохондрия
Нервная система состоит из трех видов элемен-
тов, различных по функции:
I. Рецептор (восприниматель), связан с аффе-
рентным (центростремительным), чувствительным
нейроном, проводящим возбуждение к центру (это
начало анализа по И.П. Павлову).
II. Кондуктор (проводник), вставочный, ассо-
циативный нейрон, осуществляющий замыкание,
переключение возбуждения с центростремитель-
ного на центробежный нейрон (это уже начало
синтеза).
III. Эфферентный (центробежный) нейрон
обеспечивает ответную реакцию (двигательную или
секреторную). Имеется и постоянная обратная связь,
«обратная афферентация» по П.К. Анохину.
8
В основе деятельности нервной системы лежит
рефлекс (И.М. Сеченов). Рефлекс это ответ-
ная реакция организма на раздражение, которая
происходит при участии ЦНС. Нейроны в нервной
системе, вступая в контакт друг с другом, образуют
рефлекторные дуги, по которым нервный импульс
проходит только в одном направлении. В зависи-
мости от сложности рефлекторного акта различают
простые и сложные рефлекторные дуги. Для осу-
ществления безусловных рефлексов образуются про-
стые рефлекторные дуги. Простейшая рефлекторная
дуга состоит из двух нейронов. Один — связан с
рецептором, другой посылает нейрит к мышце или
железе. Значительно чаще имеется третий, «вста-
вочный* нейрон.
В простой рефлекторной дуге нейроны по фун-
кции различные:
1. Чувствительные (афферентные), тела их лежат
вне ЦНС, в чувствительных узлах нервов (рис. 3).
Их периферический отросток начинается рецеп-
тором.
Рис. 3. Схема простой рефлекторной дуги:
1 — чувствительный нейрон (ложноуниполярная клетка спинно-
мозгового узла); 2 — вставочный нейрон (чувствительное ядро
заднего рога); 3 —двигательный нейрон (клетки двигательного
ядра переднего рога)
9
Рецепторы в зависимости от локализации раз-
личают: экстерорецепторы (в коже), интерорецеп-
торы (во внутренних органах), проприорецепторы
(в аппарате движения).
2. Вставочные нейроны (ассоциативные) нахо-
дятся в ЦНС, передают возбуждение с чувствитель-
ного нейрона на двигательный нейрон.
3. Двигательные (эфферентные) нейроны, их
тела располагаются в ЦНС и в вегетативных уз-
лах. Аксоны этих нейронов продолжаются в виде
нервных волокон к рабочим органам (мышцам).
Для осуществления условных рефлексов обра-
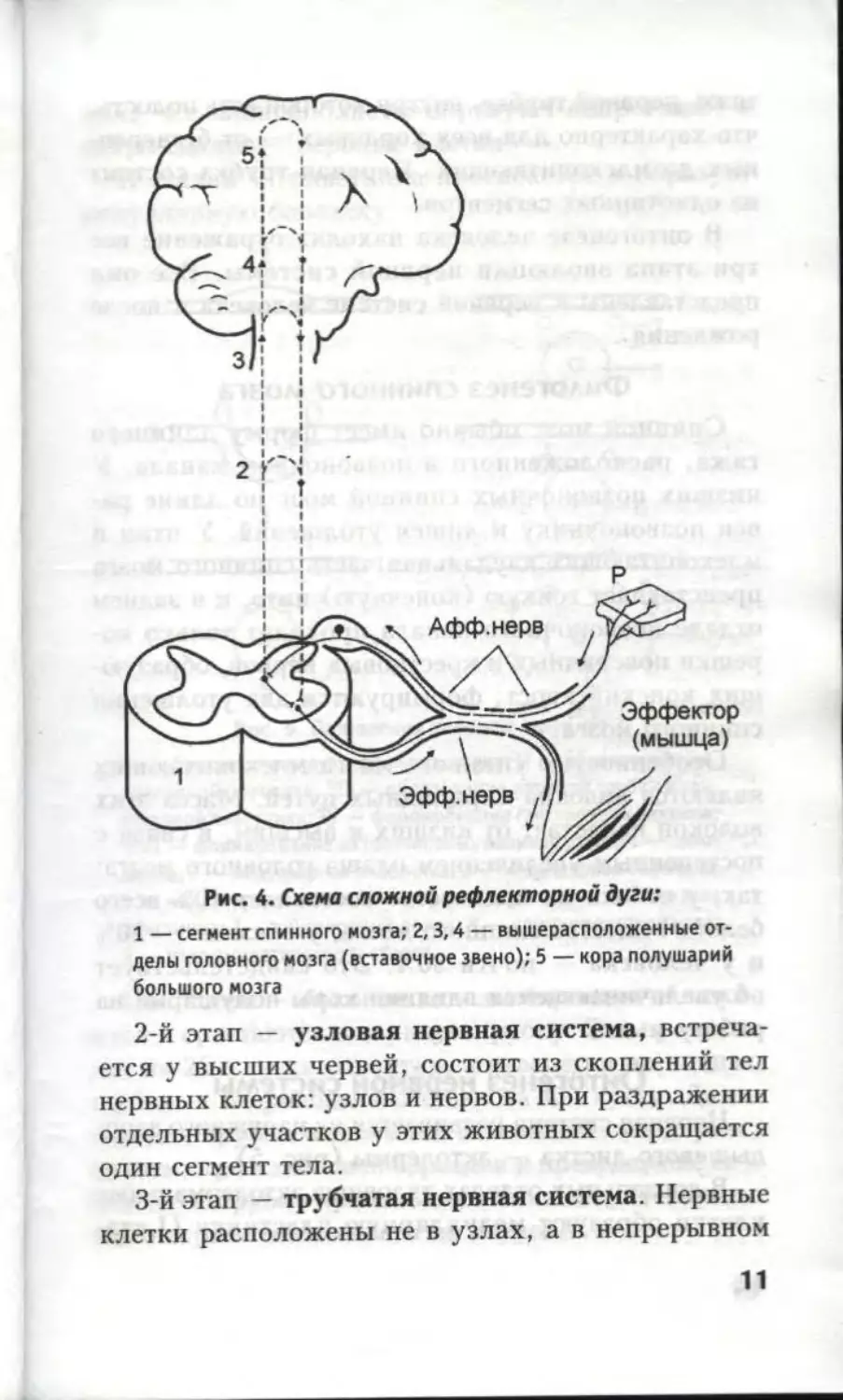
зуются сложные рефлекторные дуги. Усложнение
рефлекторных дуг происходит за счет вставочного
звена (рис. 4). Ассоциативные нейроны образуют
многочисленные ядра в пределах спинного и голо-
вного мозга.
Между нервным центром и рабочим органом при
осуществлении любого рефлекса устанавливается
двусторонняя связь. Достигая эффекторов, распо-
ложенных в мышце или железе, нервные импульсы
вызывают ответную реакцию на раздражение. Име-
ющаяся обратная связь позволяет контролировать
правильность исполнения команд, поступающих из
нервных центров, и вносить коррекцию в выполне-
ние ответных реакций организма.
Филогенез нервной системы
Различают три этапа развития нервной системы
у многоклеточных животных.
1-й этап — сетевидная нервная система, встре-
чается у кишечнополостных (гидр), состоит из при-
митивных сетчатых структур. Раздражение любого
участка тела у этих животных приводит к общей
двигательной сократительной реакции.
10
Рис. 4. Схема сложной рефлекторной дуги:
1 — сегмент спинного мозга; 2,3,4 — вышерасположенные от-
делы головного мозга (вставочное звено); 5 — кора полушарий
большого мозга
2-й этап — узловая нервная система, встреча-
ется у высших червей, состоит из скоплений тел
нервных клеток: узлов и нервов. При раздражении
отдельных участков у этих животных сокращается
один сегмент тела.
3-й этап — трубчатая нервная система. Нервные
клетки расположены не в узлах, а в непрерывном
11
тяже, нервной трубке, внутри которой есть полость,
что характерно для всех хордовых — от бесчереп-
ных до млекопитающих. Нервная трубка состоит
из однотипных сегментов.
В онтогенезе человека находят отражение все
три этапа эволюции нервной системы. Все они
представлены в нервной системе человека и после
рождения.
Филогенез спинного мозга
Спинной мозг обычно имеет форму длинного
тяжа, расположенного в позвоночном канале. У
низших позвоночных спинной мозг по длине ра-
вен позвоночнику и лишен утолщений. У птиц и
млекопитающих каудальная часть спинного мозга
представляет тонкую (конечную) нить, и в заднем
отделе позвоночного канала проходят только ко-
решки поясничных и крестцовых нервов, образую-
щих конский хвост; формируются два утолщения
спинного мозга.
Особенностью спинного мозга млекопитающих
являются волокна пирамидных путей. Масса этих
волокон нарастает от низших к высшим, в связи с
постепенным увеличением плаща головного мозга:
так, у собаки их количество составляет 10% всего
белого вещества спинного мозга, у обезьян 20%
и у человека - почти 30%. Это свидетельствует
об увеличивающемся влиянии коры полушарий на
работу мышц.
Онтогенез нервной системы
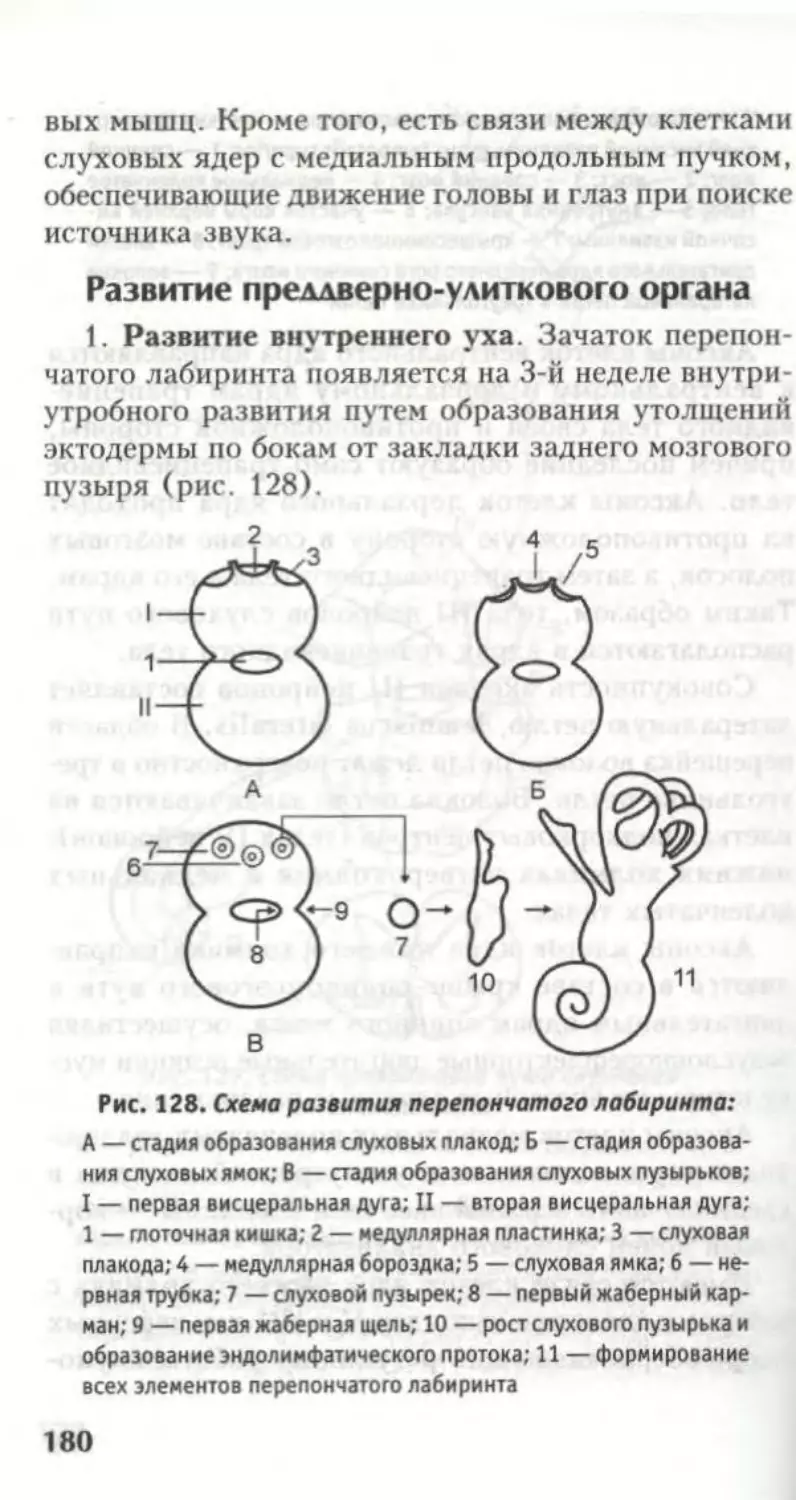
Нервная система развивается из наружного заро-
дышевого листка — эктодермы (рис. 5).
В дорзальных отделах туловища эктодермальные
клетки образуют медуллярную пластинку (I ста-
12
дия). Ее спонгиобласты образуют нейроглию, а
нейробласты — нервные клетки.
II стадия пластинка прогибается и образует
медуллярную бороздку.
Рис. 5. Онтогенез нервной трубки:
I — образование медуллярной пластинки; II — образование ме-
дуллярной бороздки; III — образование нервной трубки и ган-
глиозной пластинки; IV — формирование ганглиозных валиков;
V, VI — формирование из ганглиозных валиков узлов; 1 — экто-
дерма; 2 — медуллярная пластинка; 3 — медуллярная бороздка;
4 — медуллярный валик; 5 — нервная трубка; 6 — ганглиозная
пластинка; 7 — ганглиозный валик; 8 — спинномозговой узел;
9 — узел симпатического ствола.
III стадия боковые отделы бороздки сближа-
ются и срастаются в нервную трубку. Это происхо-
дит на 25-е сутки развития, а отверстия на концах
трубки невропоры зарастают на 30-е сутки. Далее
нервная трубка отделяется от кожной эктодермы,
причем отрыв мозговой бороздки и превращение ее в
мозговую трубку происходит мгновенно. Механизм
этого отрыва загадочен и не изучен.
13
По обе стороны нервной трубки эктодерма об-
разует нервный гребень — медуллярные валики.
Одни клетки из них мигрируют в брюшную полость,
образуя вегетативные узлы и мозговое вещество над-
почечников. Другие клетки формируют ганглиоз-
ную пластинку, делящуюся на ганглиозные валики.
Из последних формируются чувствительные узлы
спинномозговых и черепных нервов соматической
и вегетативной нервной системы.
Итак, нервная система развивается из нервной
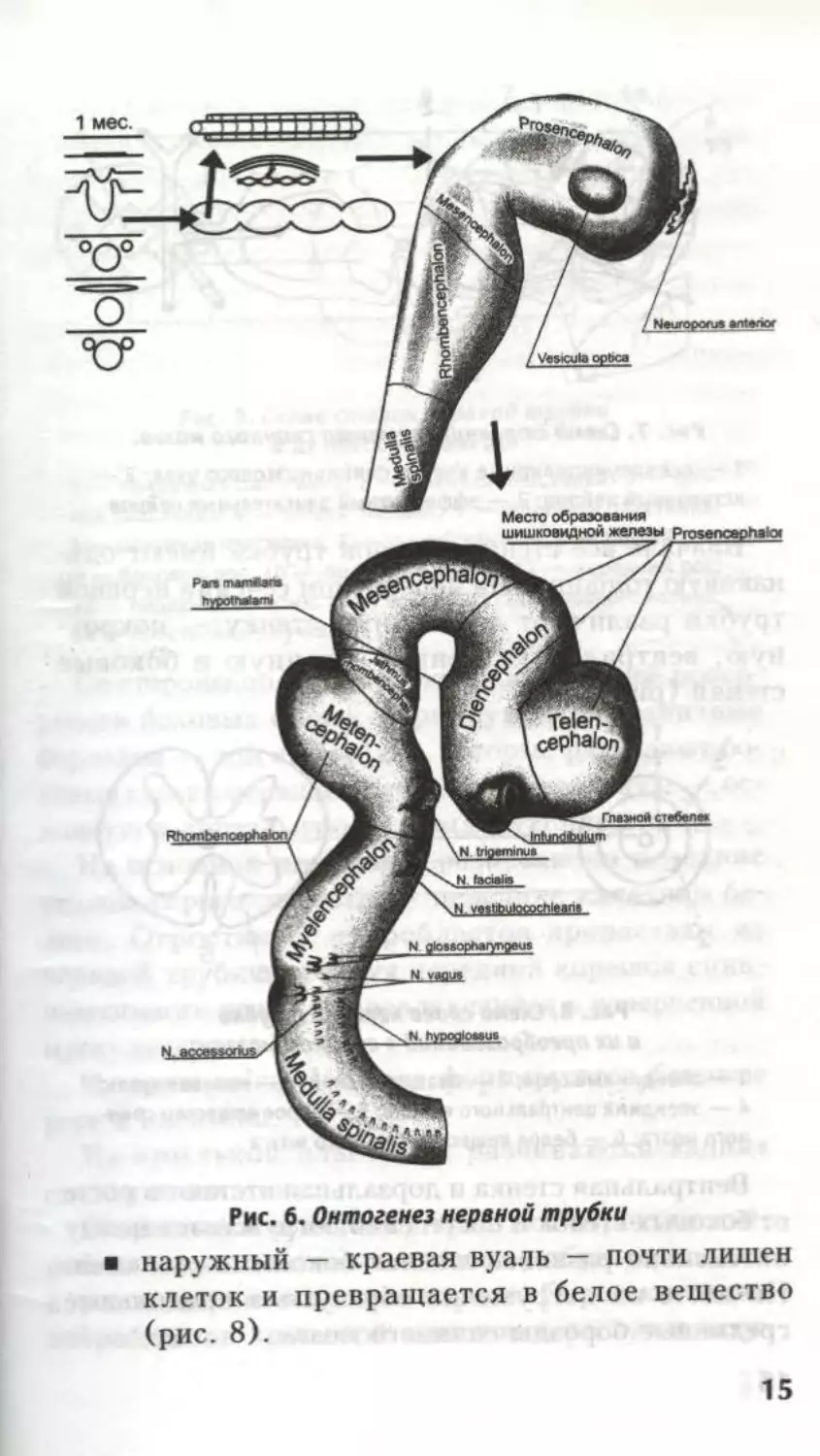
трубки и медуллярных валиков. Из краниальной
части нервной трубки развивается головной мозг:
вначале три мозговых пузыря, затем пять — бу-
дущие отделы головного мозга, сохраняющие
полость — остаток полости первичной мозговой
трубки (рис. 6), и органы чувств, из каудальной —
спинной мозг, из медуллярных валиков — чувс-
твительные и вегетативные узлы, а также хромаф-
финная ткань.
Эмбрион человека имеет метамерное строение.
Метамер — ряд последовательно расположенных
участков тела с повторяющимися системами морфо-
функциональных образований. Нервный сегмент
(рис. 7) — поперечный отрезок спинного мозга и
связанные с ним правый и левый спинномозговые
нервы, развивающиеся из одного невротома (не-
вромера).
Онтогенез спинного мозга
В период образования нервная трубка имеет три
слоя:
внутренний — эпендимный слой (будущая
выстилка мозговых полостей);
средний — плащевой слой, из которого раз-
вивается серое вещество мозга;
14
Место образования
Рис. б. Онтогенез нервной трубки
наружный — краевая вуаль — почти лишен
клеток и превращается в белое вещество
(рис. 8).
15
1
2
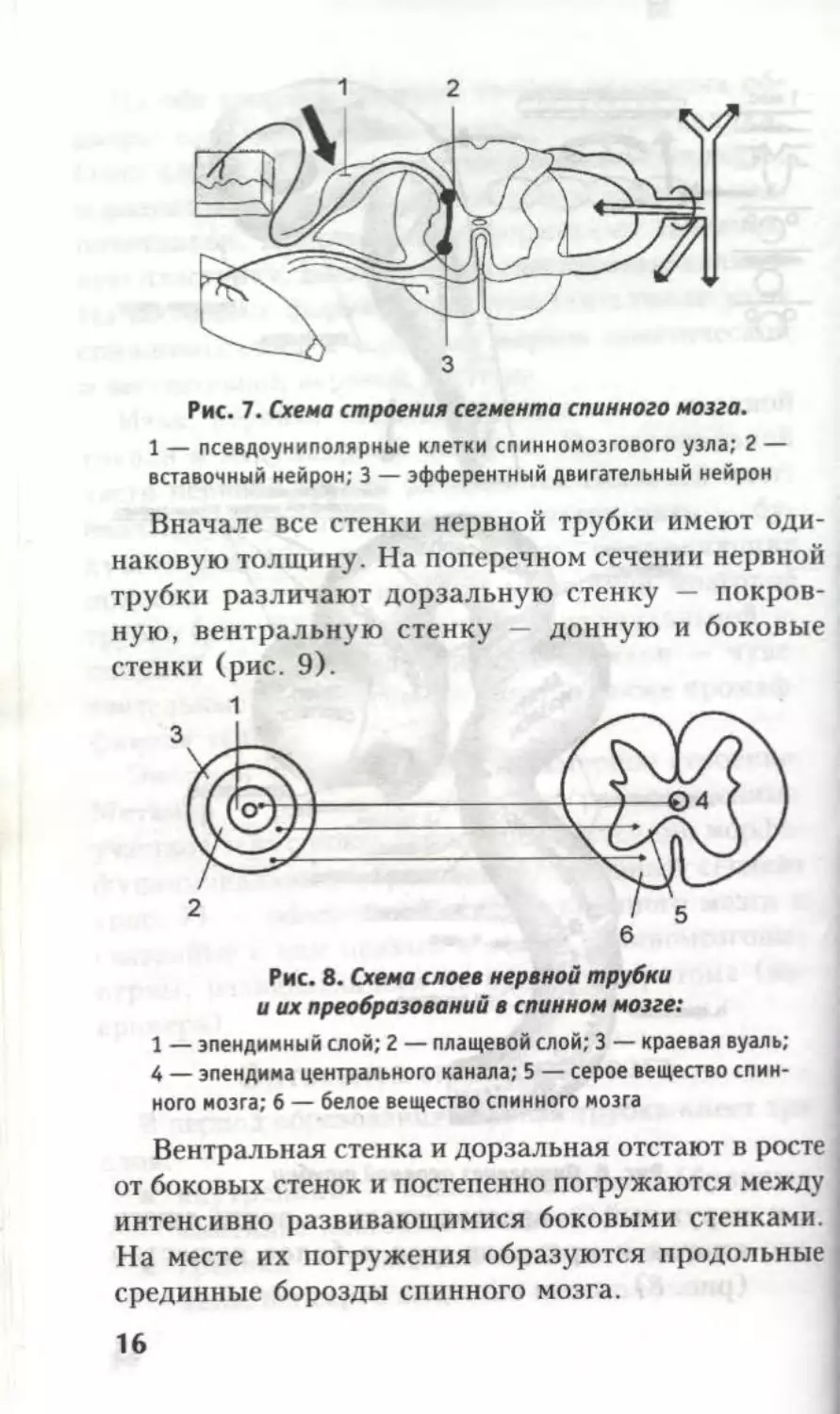
Рис. 7. Схема строения сегмента спинного мозга.
1 — псевдоуниполярные клетки спинномозгового узла; 2 —
вставочный нейрон; 3 — эфферентный двигательный нейрон
Вначале все стенки нервной трубки имеют оди-
наковую толщину. На поперечном сечении нервной
трубки различают дорзальную стенку — покров-
ную, вентральную стенку — донную и боковые
стенки (рис. 9).
Рис. 8. Схема слоев нервной трубки
и их преобразований в спинном мозге:
1 — эпендимный слой; 2 — плащевой слой; 3 — краевая вуаль;
4 — эпендима центрального канала; 5 — серое вещество спин-
ного мозга; 6 — белое вещество спинного мозга
Вентральная стенка и дорзальная отстают в росте
от боковых стенок и постепенно погружаются между
интенсивно развивающимися боковыми стенками.
На месте их погружения образуются продольные
срединные борозды спинного мозга.
16
8
7
14
Рис. 9. Схема стенок нервной трубки
и их преобразований:
1 — покровная пластинка; 2 — боковая пластинка; 3 — дон-
ная пластинка; 4 — sulcus limitans; 5 — крыльная пластинка;
6 — основная пластинка; 7 — задний канатик. 8 — задний рог;
9 — боковой рог; 10 — боковой канатик; 11 — передний рог;
12 — передний канатик; 13 — нейробласт ганглиозного валика;
14 — собственные пучки
Со стороны полости трубки на внутренней повер-
хности боковых стенок формируются пограничные
бороздки — sulcus limitans, которые разделяют бо-
ковые стенки нервной трубки на вентральную — ос-
новную и дорзальную — крыльную пластинки.
Из основной пластинки развиваются передние
столбы серого вещества и передние канатики бе-
лого. Отростки ее нейробластов прорастают из
нервной трубки, образуя передний корешок спин-
номозгового нерва, направляющийся к исчерченной
мускулатуре.
Из зоны sulcus limitans формируются боковые
рога и канатики.
Из крыльной пластинки развиваются задние
столбы и канатики.
В крыльную пластинку врастают отростки ней-
робластов ганглиозных валиков, образуя задние
корешки спинномозговых нервов. Отростки одних
нейробластов крыльной пластинки растут и всту-
17
пают в контакт с двигательными клетками основ-
ной пластинки. Отростки других — перемещаются,
образуя собственные пучки спинного мозга или
проекционные волокна. Так устанавливаются двух-
сторонние связи спинного мозга с головным мозгом
и замыкаются простые рефлекторные дуги.
На ранних стадиях развития эмбриона нервная
трубка тянется по всей длине тела. С образованием
позвоночника спинной мозг заключается в спинно-
мозговой канал. До трех месяцев эмбрионального
периода развития спинномозговой канал и спинной
мозг совпадают в размерах, и нервы выходят через
межпозвоночные отверстия напротив мест своего
образования. После трех месяцев рост позвоночника
происходит более интенсивно, чем спинного мозга.
Спинной мозг перемещается вверх, а его конец вы-
тягивается в тонкий тяж — filum terminale. Спин-
номозговые нервы каудальных отделов удлиняются
и проходят в виде пучка нервов, так называемым
конским хвостом (40 корешков нижних сегментов
и конечная нить).
Верхняя граница спинного мозга — foramen
magnum или перекрест пирамид. Нижний уровень у
новорожденных расположен у III поясничного поз-
вонка, к концу 1 года — II поясничного позвонка, с
4 лет как у взрослых — между I и II поясничными
позвонками. Сегмент спинного мозга — отрезок, соот-
ветствующий паре спинномозговых нервов. Выделяют
8 шейных сегментов, 12 грудных, 5 поясничных,
5 крестцовых и 1 копчиковый сегмент (рис. 10).
Правило Шипо
В верхнем шейном отделе сегменты спинного
мозга располагаются соответственно позвонкам, в
нижнем шейном и верхнем грудном — на 1 позвонок
18
выше, в среднем грудном — на 2, в нижнем груд-
ном — на 3 позвонка выше (рис. 10). Крестцовые
сегменты находятся на уровне XII грудного — II по-
ясничного позвонка.
Рис. 10. Схема расположения сегментов спинного мозга
и спинномозговых нервов по отношению к позвонкам
В спинном мозге различают 2 утолщения. Шейное
утолщение состоит из 4 нижних шейных и 1 груд-
19
ного сегмента и соответствует месту отхождения
нервов, направленных к верхней конечности. По-
яснично-крестцовое утолщение состоит из 4 нижних
поясничных и 2 верхних крестцовых сегментов и
соответствует отхождению нервов для иннервации
нижней конечности.
Аномалии развития спинного мозга
1. Амиелия — отсутствие спинного мозга, часто
сочетается с анэнцефалией.
2. Миелоцеле — выбухание спинного мозга через
расщелину позвоночника.
3. Миелоцистоцеле — спинномозговая грыжа (до
4 случаев на 10 тысяч новорожденных) — свя-
зана с дефектом в дужках позвонков — spina
bifida. Грыжевой мешок содержит истонченное
вещество спинного мозга, его центральный
канал, оболочки и корешки нервов.
4. Разделение спинного мозга по длине на 2 части
костной или фиброзной перемычкой. Как при
спинномозговой грыже, так и при разделении
спинного мозга наблюдается нарушение де-
ятельности тазовых органов и нижних конеч-
ностей.
5. Гидромиелия — водянка спинного мозга.
Центральный канал расширен, переполнен
спинномозговой жидкостью, задние канатики
истончены.
6. Дипломиелия — удвоение спинного мозга в
шейном или поясничном отделе, реже — на
всем протяжении. Аномалия является резуль-
татом раздельного смыкания каждой половины
нервной трубки.
7. Недоразвитие клеток передних или задних
рогов.
20
Внутреннее строение спинного мозга
Спинной мозг имеет сегментарное, билатеральное
строение. Внутреннее его ядро образовано серым
веществом, состоящим из нейронных клеток, не-
миелинизированных аксонов, глиальных клеток и
кровеносных сосудов. Наружное белое вещество
состоит из пучков миелинизированных аксонов, пе-
редающих импульсы к спинному мозгу и от него.
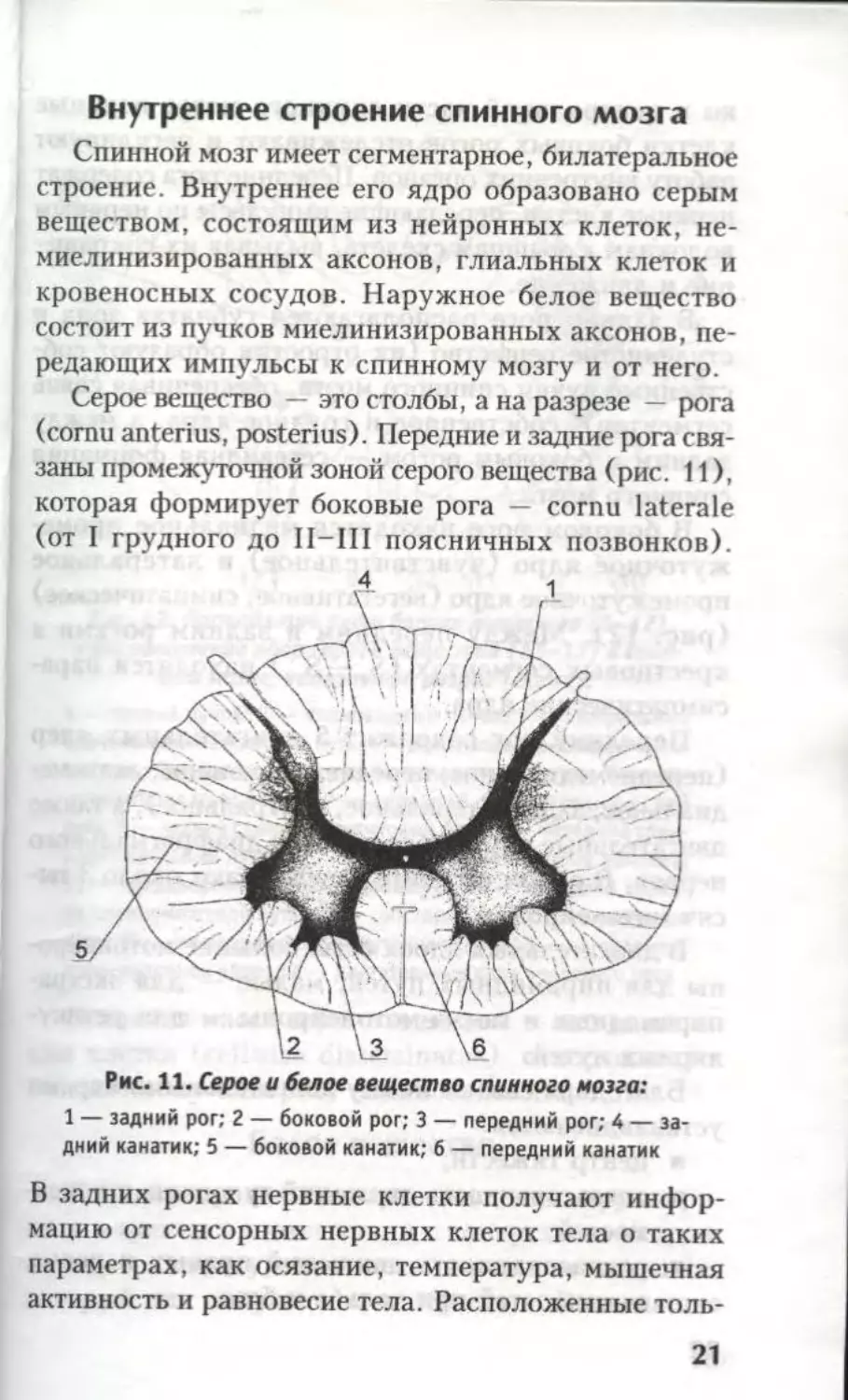
Серое вещество — это столбы, а на разрезе — рога
(cornu anterius, posterius). Передние и задние рога свя-
заны промежуточной зоной серого вещества (рис. 11),
которая формирует боковые рога — cornu laterale
(от I грудного до П-Ш поясничных позвонков).
Рис. 11. Серое и белое вещество спинного мозга:
1 — задний рог; 2 — боковой рог; 3 — передний рог; 4 — за-
дний канатик; 5 — боковой канатик; 6 — передний канатик
В задних рогах нервные клетки получают инфор-
мацию от сенсорных нервных клеток тела о таких
параметрах, как осязание, температура, мышечная
активность и равновесие тела. Расположенные толь-
21
ко в центральной части спинного мозга нервные
клетки боковых рогов отслеживают и регулируют
работу внутренних органов. Передние рога содержат
нервные клетки, передающие импульсы по нервным
волокнам к мышцам скелета, вызывая их сокраще-
ние и движение.
В заднем роге располагаются губчатая зона и
студенистое вещество (их отростки образуют соб-
ственные пучки спинного мозга, обеспечивая связь
сегментов), собственное и грудное ядра, а между
задним и боковым рогом — сетевидная формация
спинного мозга.
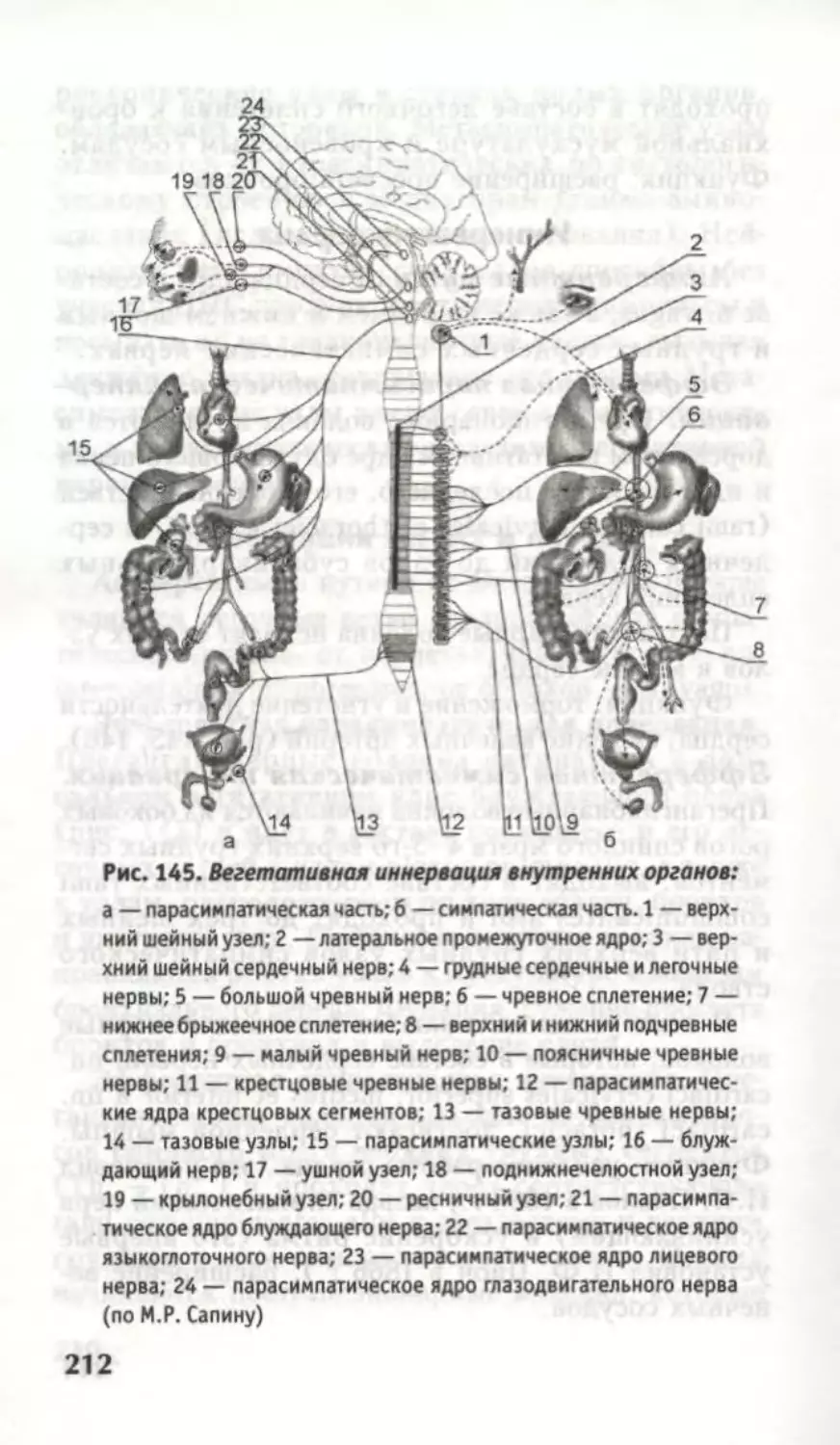
В боковом роге находятся медиальное проме-
жуточное ядро (чувствительное) и латеральное
промежуточное ядро (вегетативное, симпатическое)
(рис. 12). Между передним и задним рогами в
крестцовых сегментах (SH —S1V) находятся пара-
симпатические ядра.
Передний рог содержит 5 двигательных ядер
(переднемедиальное, переднелатеральное, заднеме-
диальное, заднелатеральное, центральное), а также
двигательные ядра добавочного и диафрагмального
нервов. В одном сегменте насчитывают около 3 ты-
сяч мотонейронов.
В двигательных ядрах есть: большие мотонейро-
ны для пирамидных путей, малые — для экстра-
пирамидных и гамма-мотонейроны — для ретику-
лярных путей.
Благодаря связям между двигательными ядрами
устанавливаются:
центр тяжести;
согласованность движений туловища и конеч-
ностей;
согласованность движений правых и левых
конечностей при ходьбе и беге.
22
Рис. 12. Проводящие пути белого вещества (1-12)
и расположение ядер серого вещества (13-17) в спин-
ном мозге; поперечный разрез (схема):
1 — тонкий пучок; 2 — клиновидный пучок; 3 — латеральный
корково-спинномозговой путь; 4 — красноядерно-спинномозго-
вой путь; 5 — латеральный спинно-таламический путь; 6 — кры-
шеспинномозговой путь; 7 — передний спинноталамический
путь; 8 — задний спинномозжечковый путь; 9 — передний спин-
номозжечковый путь; 10 — ретикулоспинномозговые волокна;
11 — преддверно-спинномозговой путь; 12 — передний корко-
во-спинномозговой путь; 13 — собственное ядро; 14 — грудное
ядро; 15 — медиальное промежуточное ядро; 16 — латеральное
промежуточное ядро; 17 — двигательные ядра переднего рога
Основная масса серого вещества — это рассеян-
ные клетки (cellulae disseminatae), относящиеся к
собственному аппарату спинного мозга.
Белое вещество
Миелинизированные нервные волокна группиру-
ются в тракты в соответствии с конкретным направ-
лением — к мозгу или от него — и типом импульса,
который они получают или передают. Восходящие
23
тракты передают нервные импульсы обо всех ощуще
ниях, возникающих в теле, вверх по спинному мозг)
к головному мозгу. Нисходящие тракты передают
импульсы от мозга к скелетным мышцам, вызывал
произвольные и непроизвольные движения.
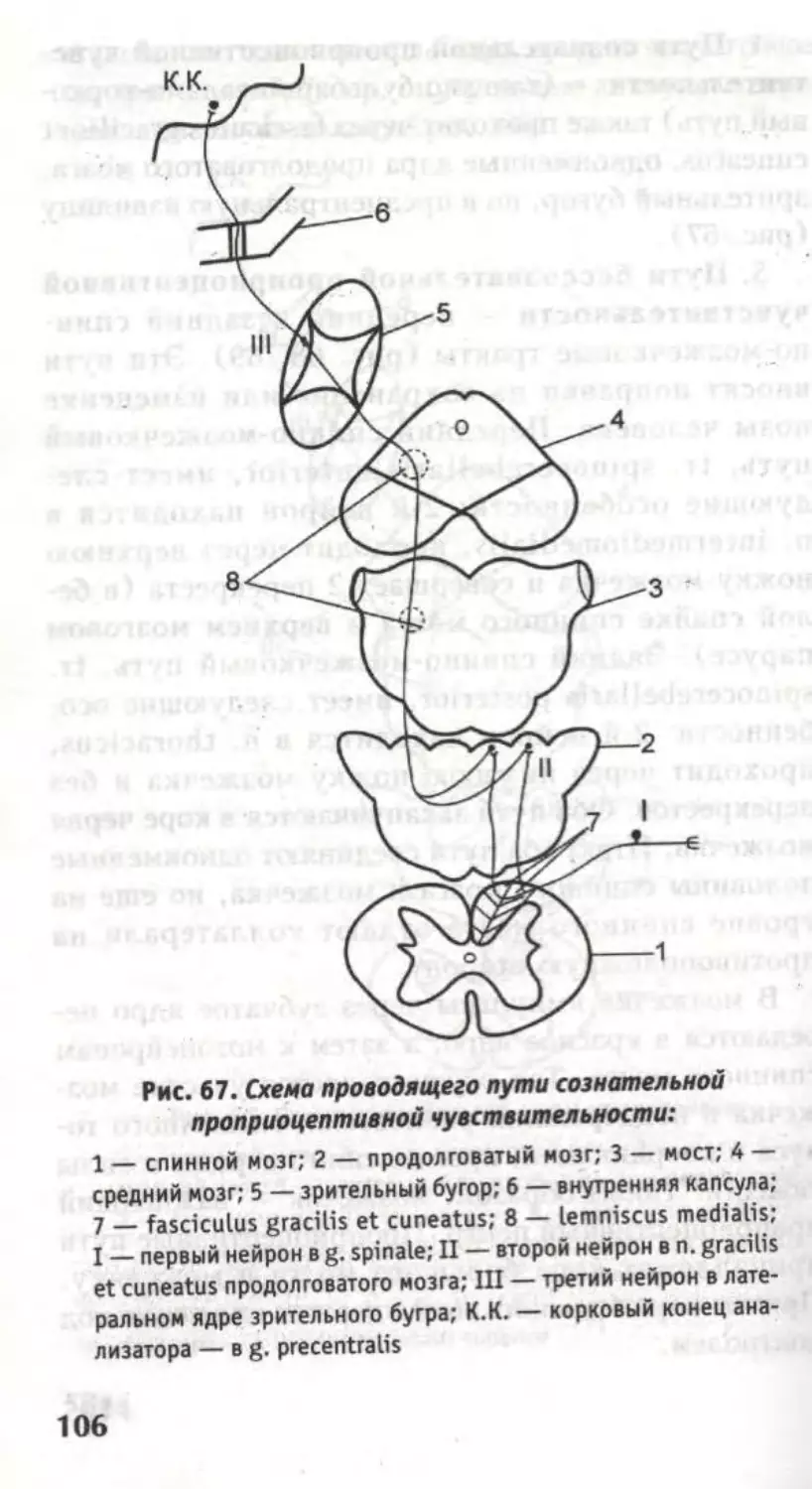
Пути заднего канатика
1. Тонкий пучок (fasciculus gracilis) расположен
медиально, в нем проходят волокна, идущие
от нижней половины тела, нижних конечнос-
тей через 19 нижних спинномозговых узлов и
далее к продолговатому мозгу.
2. Клиновидный пучок (fasciculus cuneatus) рас-
положен латерально, в нем проходят волокна
от верхней части тела через верхние 12 спин-
номозговых узлов к продолговатому мозгу.
Оба пучка проводят сознательную тактильную,
проприоцептивную чувствительность и чувство
стереогноза.
3. Задний собственный пучок (fasciculus proprius
posterior).
Пути бокового канатика
4. Боковой собственный пучок (fasciculus proprius
lateralis).
5. Передний спинномозжечковый путь (tr. spino-
cerebellaris anterior).
6. Задний спинномозжечковый путь (tr. spino-
cerebellaris posterior).
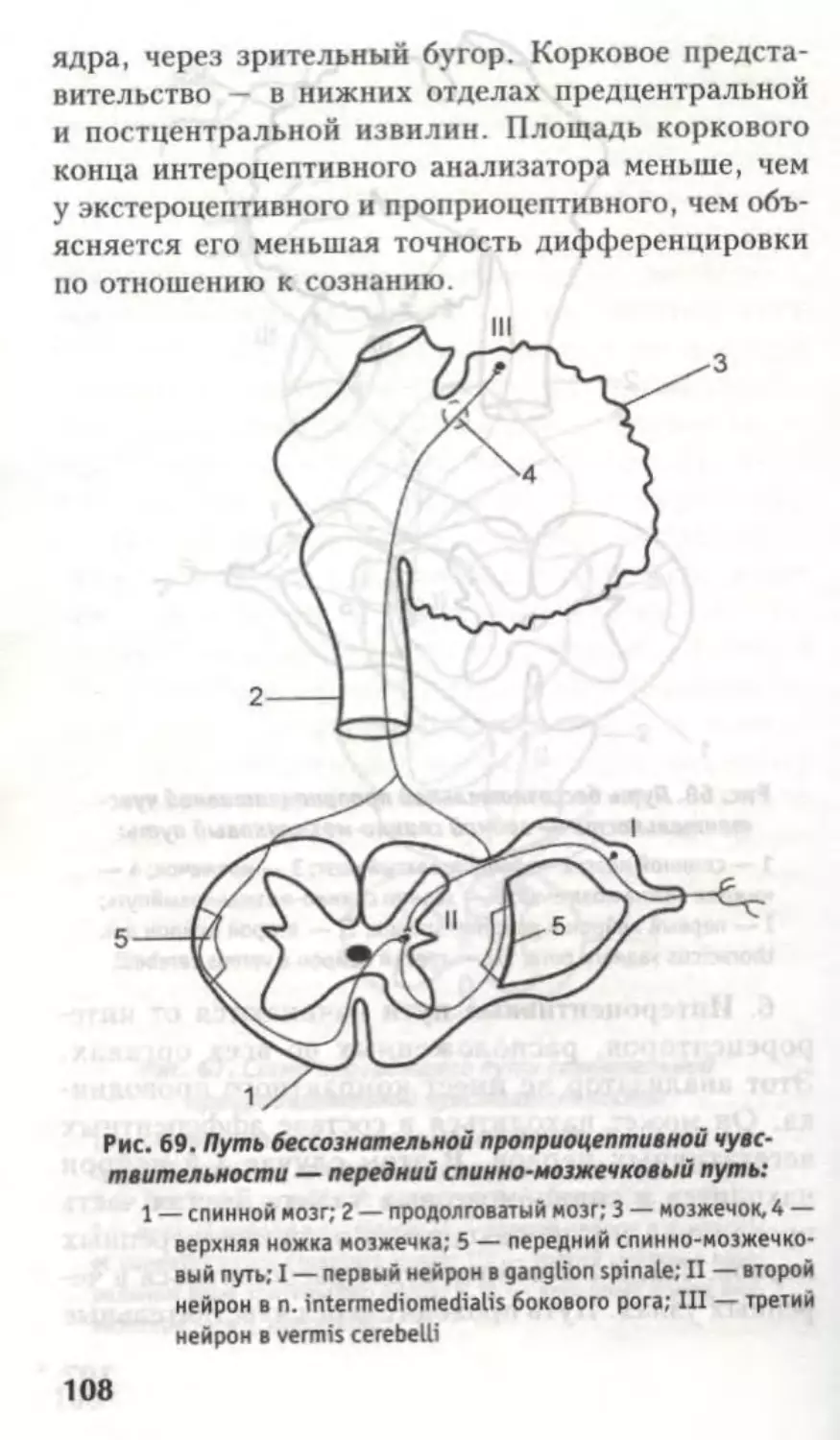
Оба проводят бессознательную проприоцептив-
ную чувствительность.
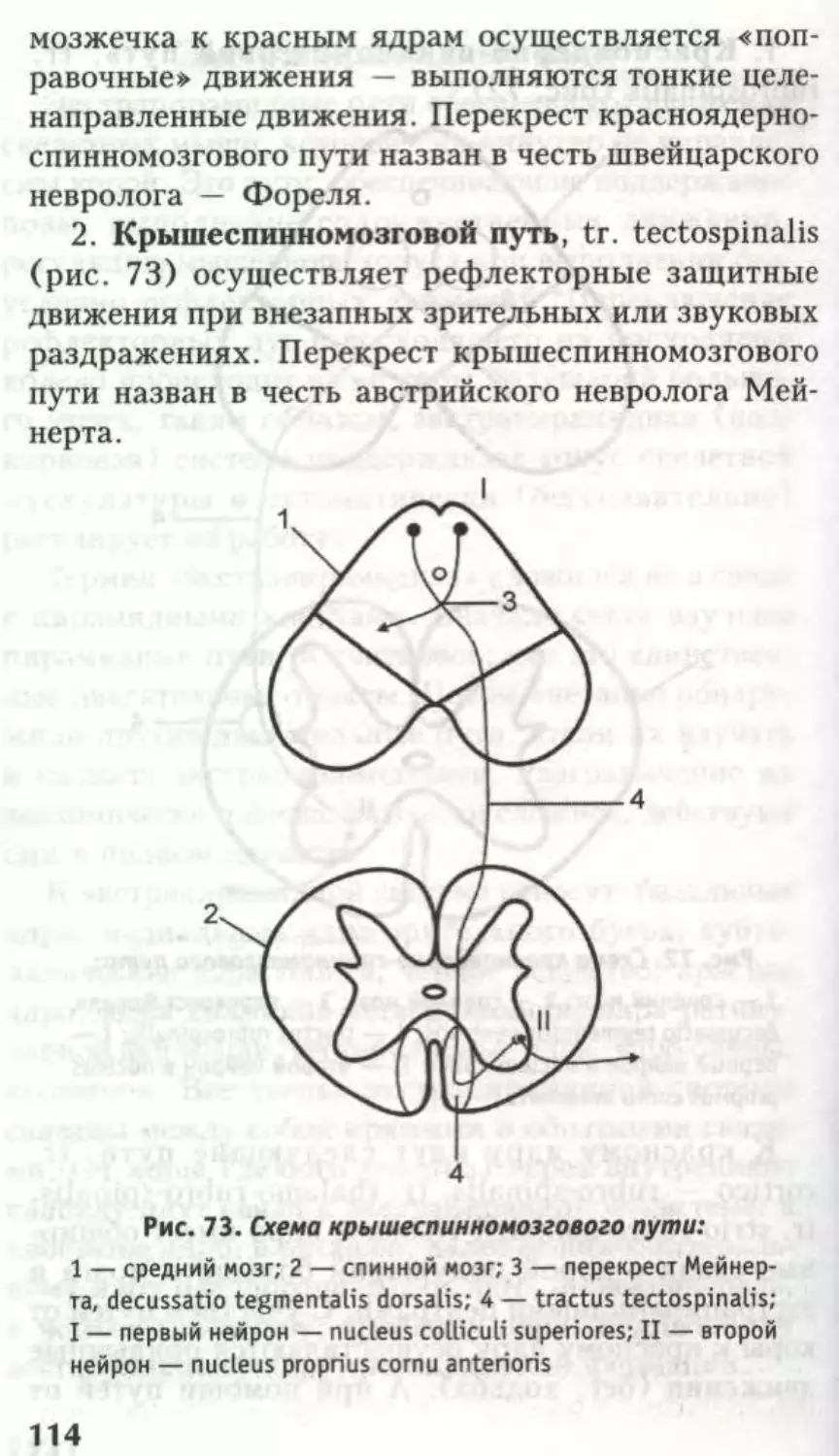
7. Спиннокрышечный путь (tr. spinotectalis).
8. Латеральный спинноталамический путь (tr.
spinothalamicus lateralis) — проводит созна-
24
тельную температурную и болевую чувстви-
тельность.
9. Латеральный корково-спинномозговой путь
(tr. corticospinalis lateralis) — сознательный
двигательный, пирамидный путь.
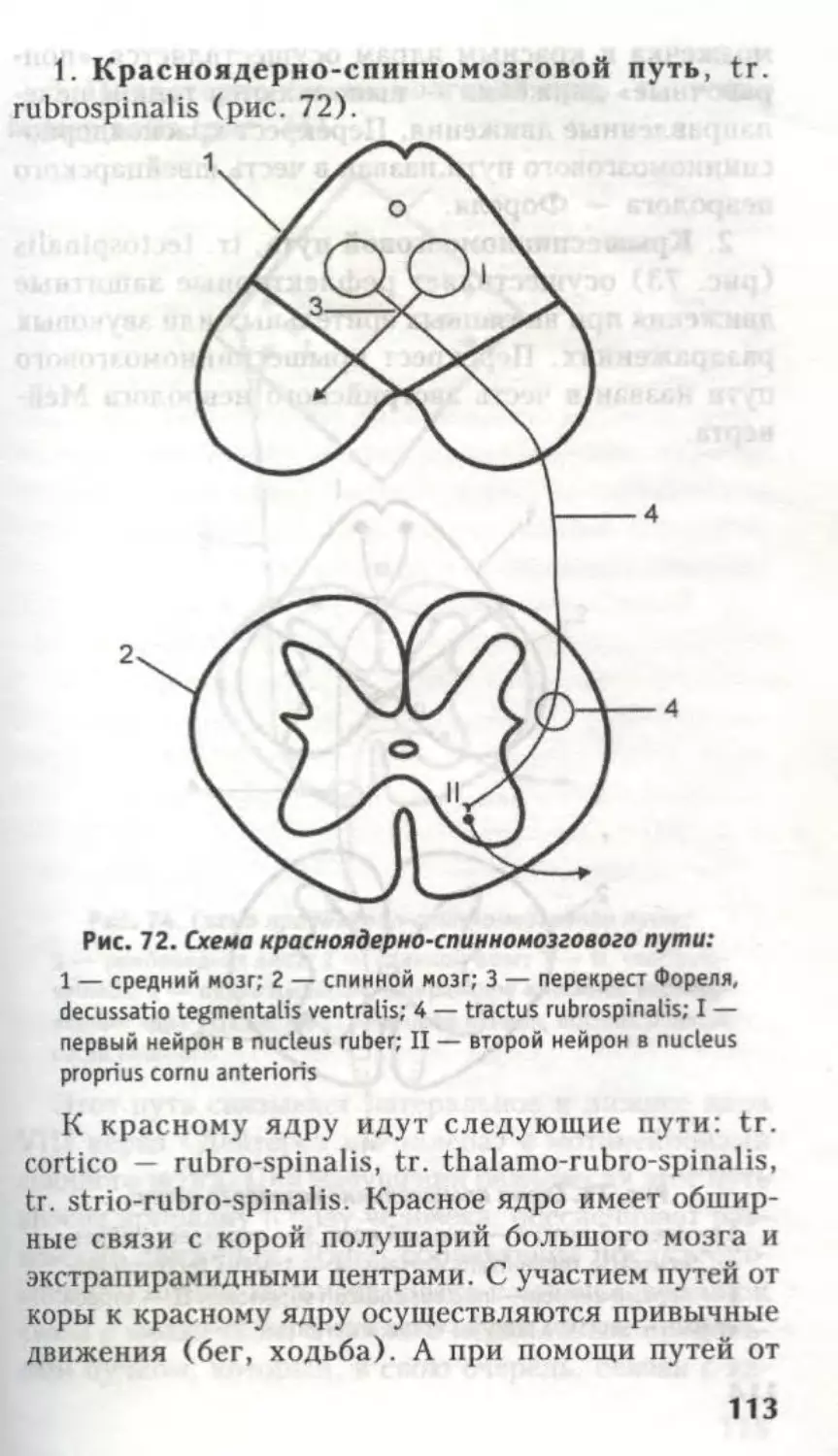
10. Красноядерно-спинномозговой путь (tr.
rubrospinalis).
И. Оливо-спинномозговые волокна (fibrae
olivospinales).
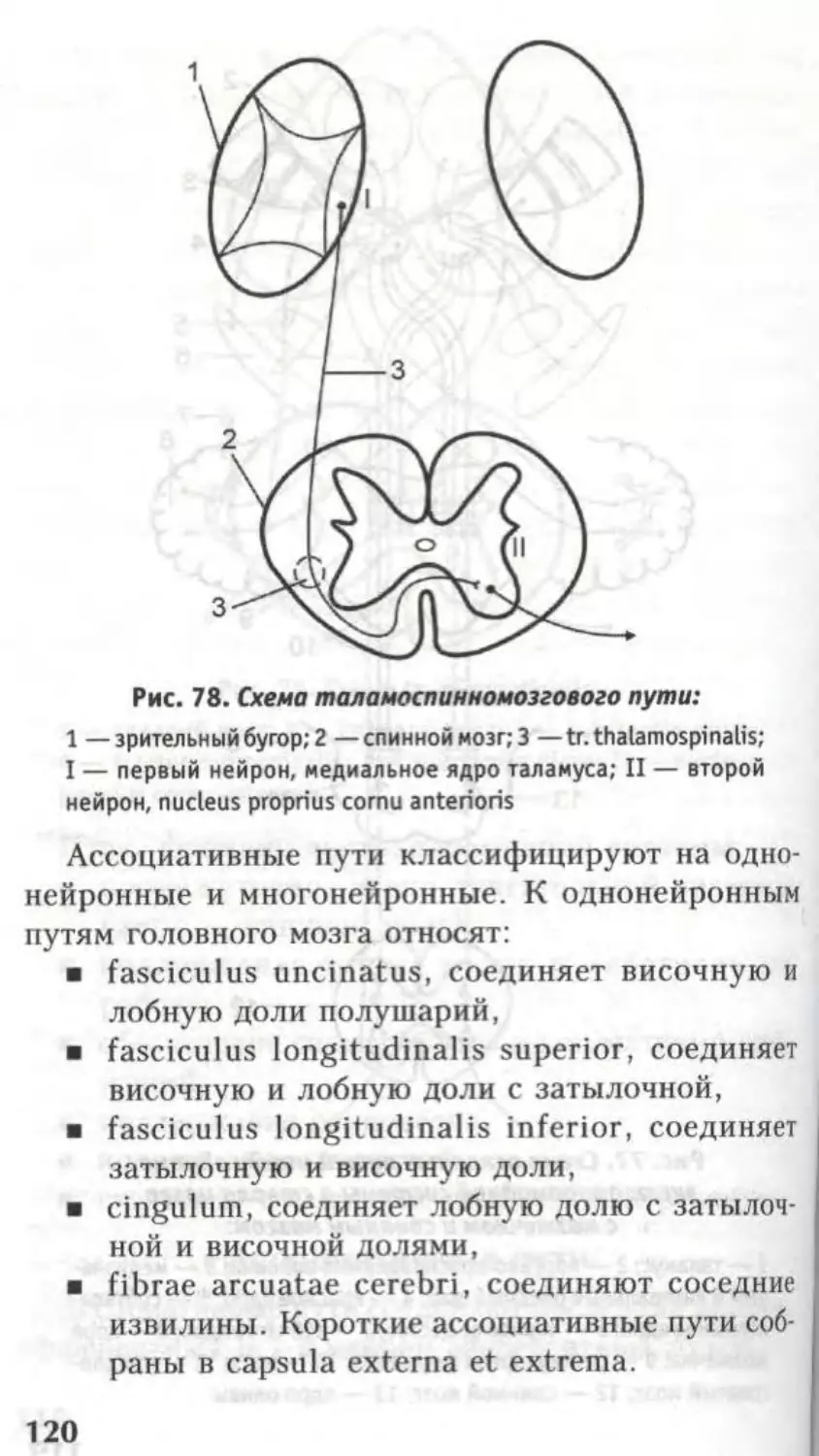
12. Таламо-спинномозговой (tr. thalamospi-
nalis).
Пути 10-12 являются бессознательными, двига-
тельными, экстрапирамидными.
Пути переднего канатика:
14. Передний собственный пучок (fasciculus
proprius anterior).
15. Передний корково-спинномозговой путь (tr.
corticospinalis anterior) — сознательный, дви-
гательный пирамидный путь.
16. Крыше-спинномозговой путь (tr. tecto-
spinalis).
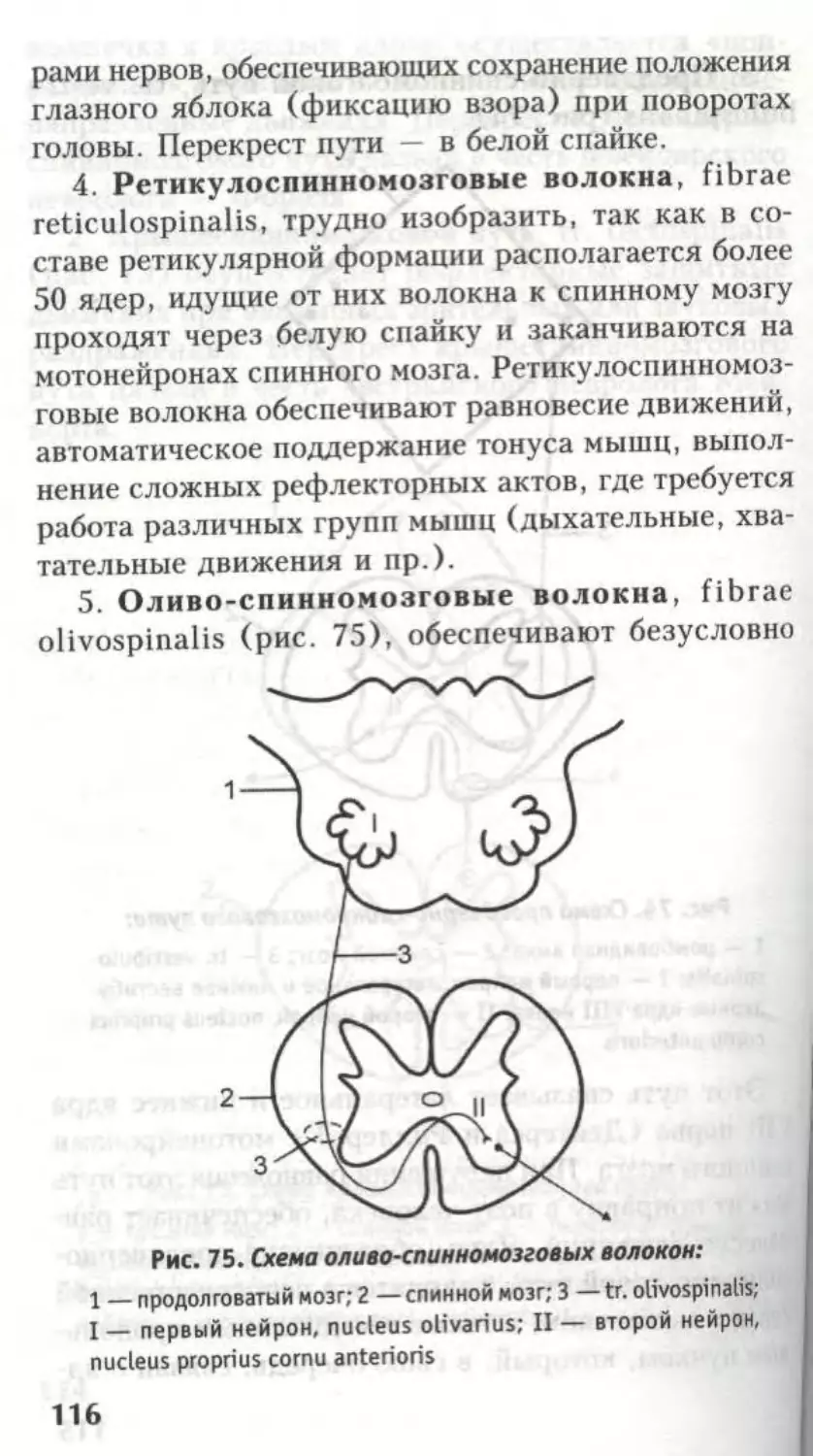
17. Ретикулоспинномозговые волокна (fibrae
reticulospinalis).
18. Преддверно-спинномозговой путь (tr. ves-
tibulospinalis).
Многие авторы считают, что лишь 15 и 16 пути
расположены собственно в переднем канатике,
в то время как остальные занимают близкое к
нему положение в боковом канатике.
Пути 16-18 являются бессознательными, двига-
тельными, экстрапирамидными.
19. Передний спинноталамический путь (tr. spino-
thalamicus anterior) — проводит сознательную
тактильную чувствительность.
25
20. Медиальный продольный пучок (fasciculus
longitudinalis medial is) имеется лишь в шей-
ных сегментах.
Сегментарный аппарат спинного мозга — это со-
вокупность нервных структур, обеспечивающих вы-
полнение врожденных рефлексов, к нему относятся:
заднекорешковые волокна, собственные пучки, ядра
передних рогов, рассеянные клетки, клетки студе-
нистого вещества, губчатой и терминальной зон.
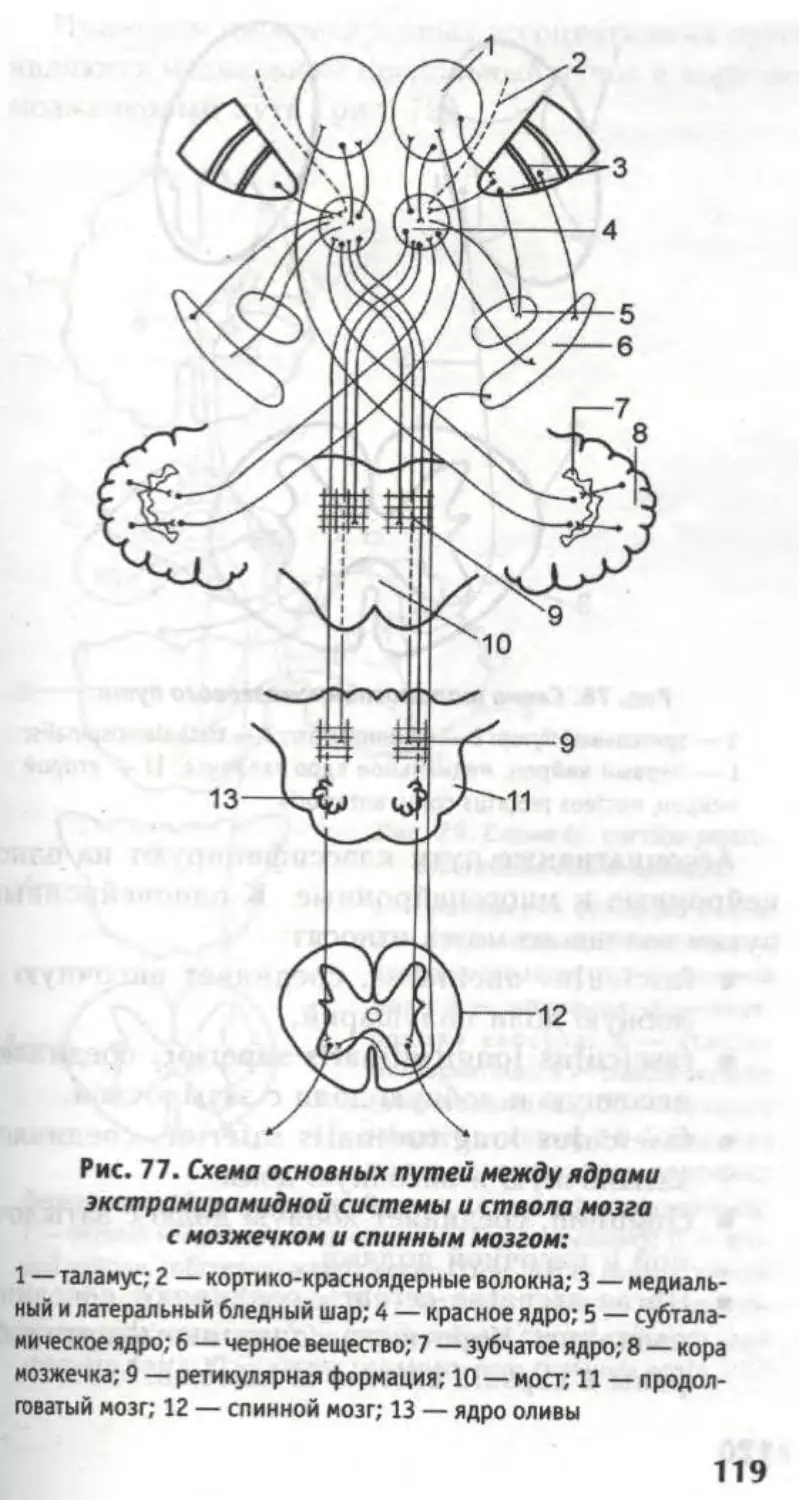
Проводниковый аппарат спинного мозга обеспечи-
вает двухстороннюю связь спинного мозга с интег-
рационными центрами головного мозга (корой моз-
жечка, корой полушарий большого мозга, верхними
холмиками четверохолмия). Этот аппарат представ-
лен чувствительными и двигательными путями.
Интеграционный (надсегментарный) аппарат
спинного мозга включает восходящие и нисходящие
пути, а также ядра: собственное, грудное и меди-
альное промежуточное.
Возрастные особенности
спинного мозга
1. У детей раннего возраста передние рога серого
вещества преобладают над задними рогами.
2. Центральный канал у новорожденных относитель-
но шире, чем у взрослых, внизу заканчивается
конечным желудочком. Этот желудочек макси-
мально расширяется к 2 годам, потом редуциру-
ется вплоть до полной облитерации у взрослых.
3. Проводящие пути у новорожденных находятся
на разной степени миелинизации, например,
боковой корково-спинномозговой путь полно-
стью миелинизируется к 4 годам.
4. У новорожденных хорошо выражены суб- и
эпидуральные пространства, которые увели-
чиваются с увеличением возраста.
26
Морфология
ромбовидного,
СРЕДНЕГО
Глава
И ПРОМЕЖУТОЧНОГО МОЗГА
Онтогенез головного мозга
Головной мозг развивается из головного отде-
ла нервной трубки. У 3-4-недельного эмбриона
головной мозг состоит из 3 мозговых пузырей, от-
деленных друг от друга небольшими сужениями
(рис. 13).
£7)
/j / Рис. 13. Схема
( \ развития мозговых
пузырей:
J k I — Rhombencephalon;
J 1дЗ) II— Mesencephalon;
( II J 'J Г III — Prosencephalon;
] [ Г 4 ) 1— Myelencephalon;
iV] \ ~ Metencephalon;
v 3 — Mesencephalon;
\5/ 4 — Diencephalon; 5 —
Telencephalon
К концу 4-й недели стадия 3 мозговых пузы-
рей при следующей дифференцировке переходит
в стадию 5 мозговых пузырей, дающих начало 5
главным отделам головного мозга. Одновременно
с этим нервная трубка изгибается в сагиттальном
направлении, образуя теменной, затылочный и мос-
27
товой изгибы, способствуя обособлению 5 мозговых
пузырей (рис. 14).
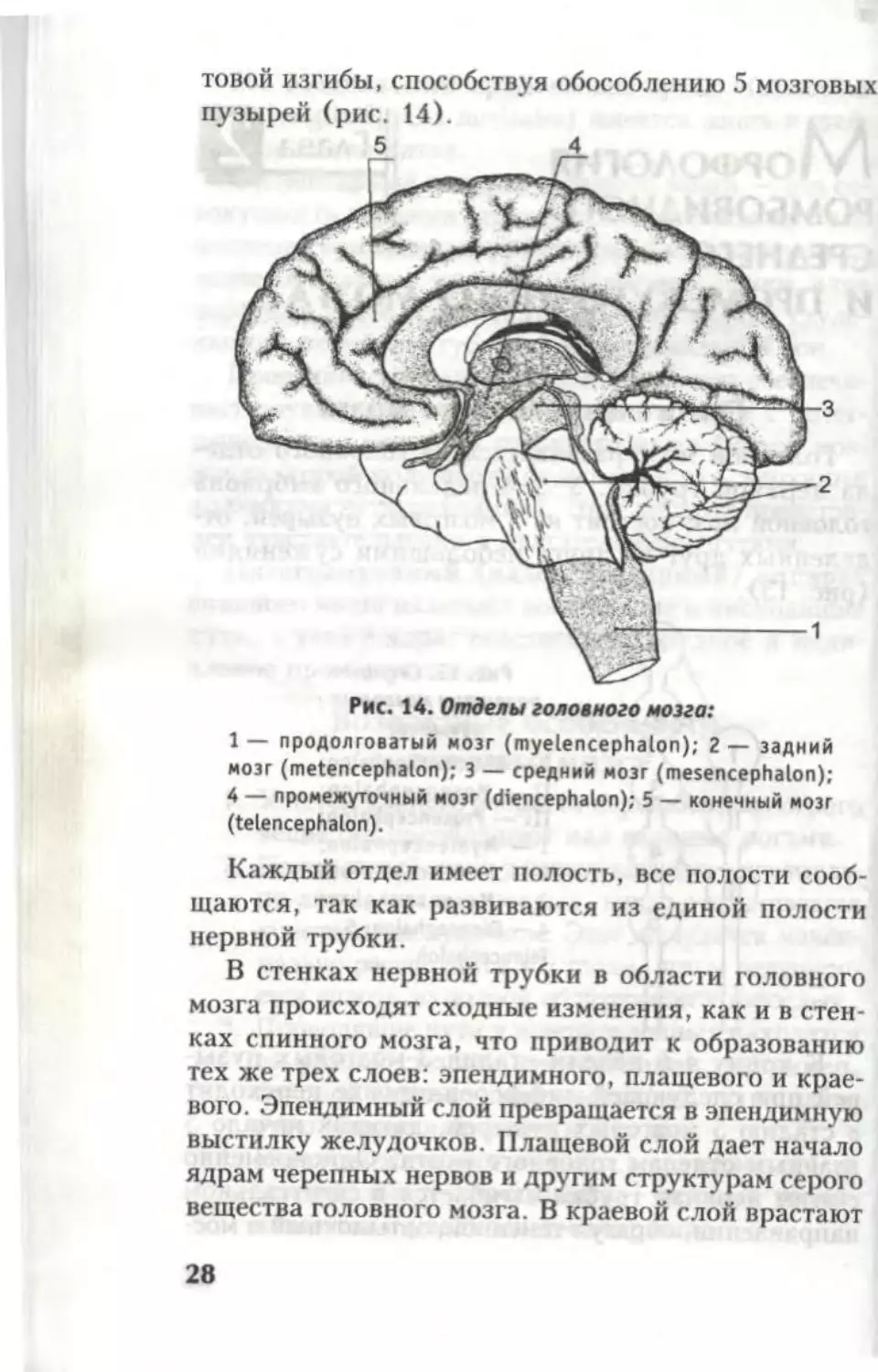
Рис. 14. Отделы головного мозга:
1 — продолговатый мозг (myelencephalon); 2 — задний
мозг (metencephalon); 3 — средний мозг (mesencephalon);
4 — промежуточный мозг (diencephalon); 5 — конечный мозг
(telencephalon).
Каждый отдел имеет полость, все полости сооб-
щаются, так как развиваются из единой полости
нервной трубки.
В стенках нервной трубки в области головного
мозга происходят сходные изменения, как и в стен-
ках спинного мозга, что приводит к образованию
тех же трех слоев: эпендимного, плащевого и крае-
вого. Эпендимный слой превращается в эпендимную
выстилку желудочков. Плащевой слой дает начало
ядрам черепных нервов и другим структурам серого
вещества головного мозга. В краевой слой врастают
28
продольно расположенные миелинизированные во-
локна, соединяя спинной мозг с отделами головного
мозга. Таким образом, из этого слоя развивается
белое вещество головного мозга.
Отделы головного ллозга
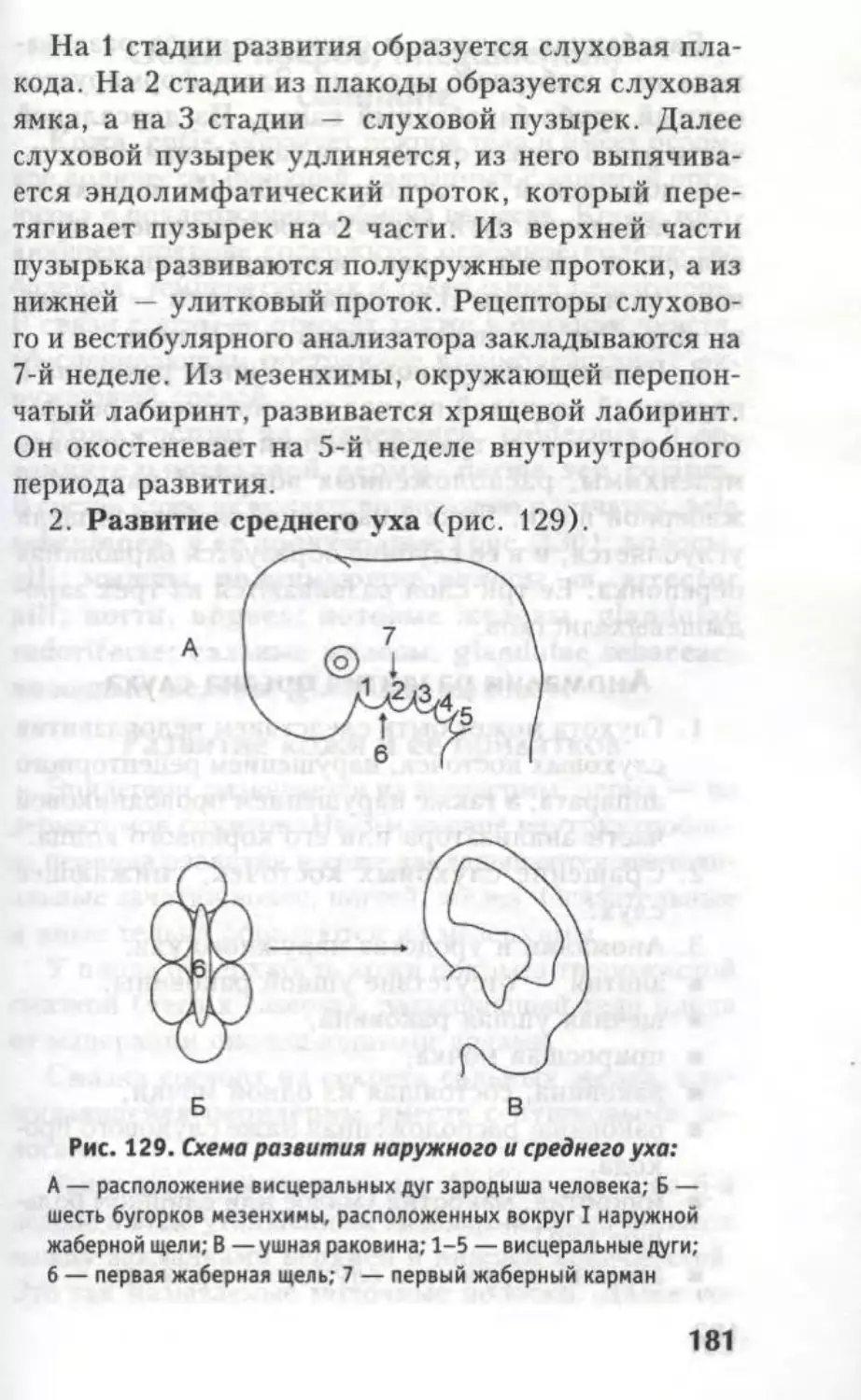
Стадии 3 мозго- вых пузырей Стадии 5 мозговых пузырей Полость моз- гового пузыря
1. Ромбовидный мозг — Rhomben- cephalon 1. Продолговатый мозг Myelencephalon II. Задний мозг — Metencephalon IV желудочек — Ventriculus quar- tus
II. Средний мозг — Mesence- phalon III. Средний мозг — Mesencephalon) Водопровод мозга — Aque- ductus cerebri
III. Передний мозг — Prosen- cephalon IV. Промежуточный мозг — Diencepha- lon) V. Конечный мозг — Telencephalon Третий желудо- чек — Ventricu- lus tertius
Так же, как при образовании спинного мозга, эле-
менты головного мозга развиваются из различных
отделов нервной трубки. Так, из покровной плас-
тинки развиваются крыша III желудочка, нижние
ножки мозжечка, эпиталамус. Донная пластинка
редуцируется. Из крыльной пластинки развиваются
дорзальные отделы ствола, а из основной пластин-
ки — вентральные отделы ствола мозга.
Вес головного мозга
Расположенный в черепе головной мозг содержит
свыше 12 миллиардов нейронов и 50 миллиардов
опорных глиальных клеток, но весит при этом не-
много.
У новорожденного вес головного мозга составляет
360-370 г.
В 9 месяцев — 720-740 г (удваивается).
29
В 3 года — 1100-1200 г (утраивается).
У взрослых — 1400-2200 г.
Не надо думать, что вес мозга и умственные спо-
собности находятся в пропорциональной зависимос-
ти. Так, самый большой мозг (3350 г) принадлежал
умственно неполноценному человеку.
Вместе со спинным мозгом головной мозг обес-
печивает и регулирует множество бессознательных
процессов, происходящих в теле человека, а также
координирует большинство произвольных движе-
ний. Более существенным является то, что мозг
управляет сознанием и разнообразными интеллек-
туальными функциями, начиная с мышления, обу-
чения и творчества.
Ствол мозга (truncus encephali)
Ствол мозга — это головной мозг без полушарий
мозжечка и полушарий большого мозга.
Принято считать, что структуры ствола мозга
сохраняют сегментарность и содержат элементы
ретикулярной формации. Этим ствол и отличается
от других отделов мозга.
В состав ствола мозга входят: добавочный, за-
дний, средний мозг. Существует также мнение,
что в состав ствола мозга входит промежуточный
мозг.
Ствол мозга — наиболее старая часть головного (
мозга в фило- и онтогенезе. Это сегментированная
часть мозга, причем сегменты отделяются выходами ,
черепных нервов. j
Молодой, надсегментарной частью мозга яв- <
ляются полушария большого мозга. Все команды
полушария получают от ствола, и, наоборот, все ,
ответные импульсы от полушарий к периферии t
проходят через ствол. г
30
Продолговатый мозг, бульбус —
myelencephalon, medulla oblongata,
bulbus cerebri
Поражение этой части ствола, называемой луко-
вицей, бульбусом, приводит чаще к смерти, реже
к бульбарным параличам. Еще в III веке до н.э.
Герофил учил, что жизнь человека находится на
острие пера (calanus scriptorius), а расположено
оно в нижней части ромбовидной ямки.
В IX веке в области продолговатого мозга были
найдены центры кровообращения, дыхания и тер-
морегуляции. Кровоизлияние в этом отделе мозга
кроме прочего дает стойкое повышение температу-
ры, приводящее к летальному исходу.
В состав myelencephalon входят:
1. Продолговатый мозг, medulla oblongata:
а) пирамиды, pyramis medullae oblongatae;
б) оливы, oliva;
в) тонкий пучок, fasciculus gracilis;
г) клиновидный пучок, fasciculus cuneatus.
2. Нижние ножки мозжечка, pedunculus сеге-
bellaris inferior.
3. Нижний мозговой парус, velum medullare
inferius.
4. Нижняя часть четвертого желудочка, ventri-
culus quartus (рис. 15, 16).
Наружные дугообразные волокна связывают тон-
кие и клиновидные ядра продолговатого мозга с
мозжечком и формируют нижнюю ножку мозжечка
(рис. 17).
Внутренние дугообразные волокна после пере-
креста идут внутри ствола к таламусу (бульботала-
мический путь). Он является основой медиальной
петли, lemniscus medialis, к которой присоединяют-
31
ся другие волокна (все экстероцептивные, проприо-
цептивные и интероцептивные восходящие пути).
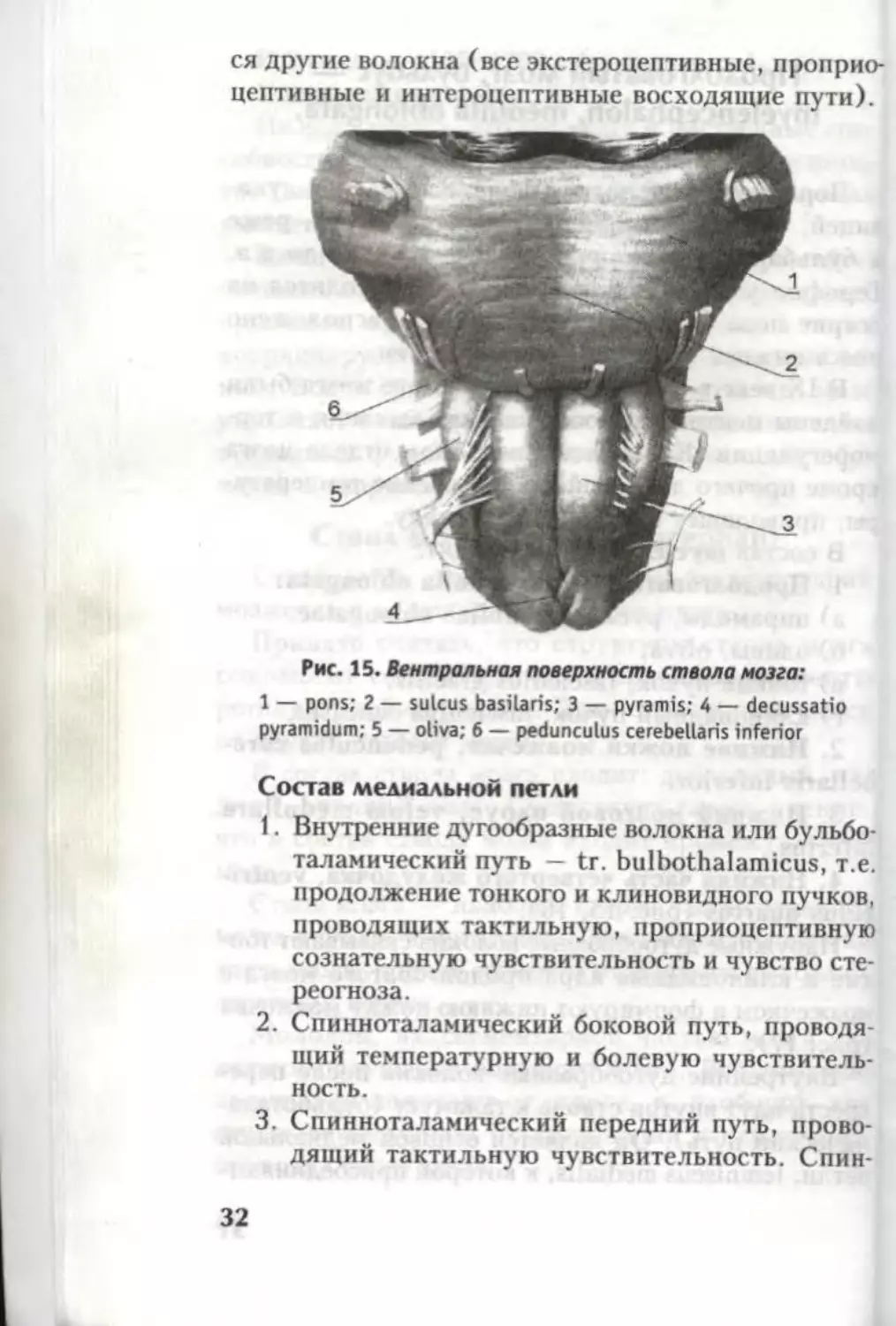
Рис. 15. Вентральная поверхность ствола мозга:
1 — pons; 2 — sulcus basilaris; 3 — pyramis; 4 — decussatio
pyramidum; 5 — oliva; 6 — pedunculus cerebellaris inferior
Состав медиальной петли
1. Внутренние дугообразные волокна или бульбо-
таламический путь — tr. bulbothalamicus, т.е.
продолжение тонкого и клиновидного пучков,
проводящих тактильную, проприоцептивную
сознательную чувствительность и чувство сте-
реогноза.
2. Спинноталамический боковой путь, проводя
щий температурную и болевую чувствитель-
ность.
3. Спинноталамический передний путь, прово-
дящий тактильную чувствительность. Спин-
32
ноталамические пути (1 и 2) объединяются в
lemniscus spinalis.
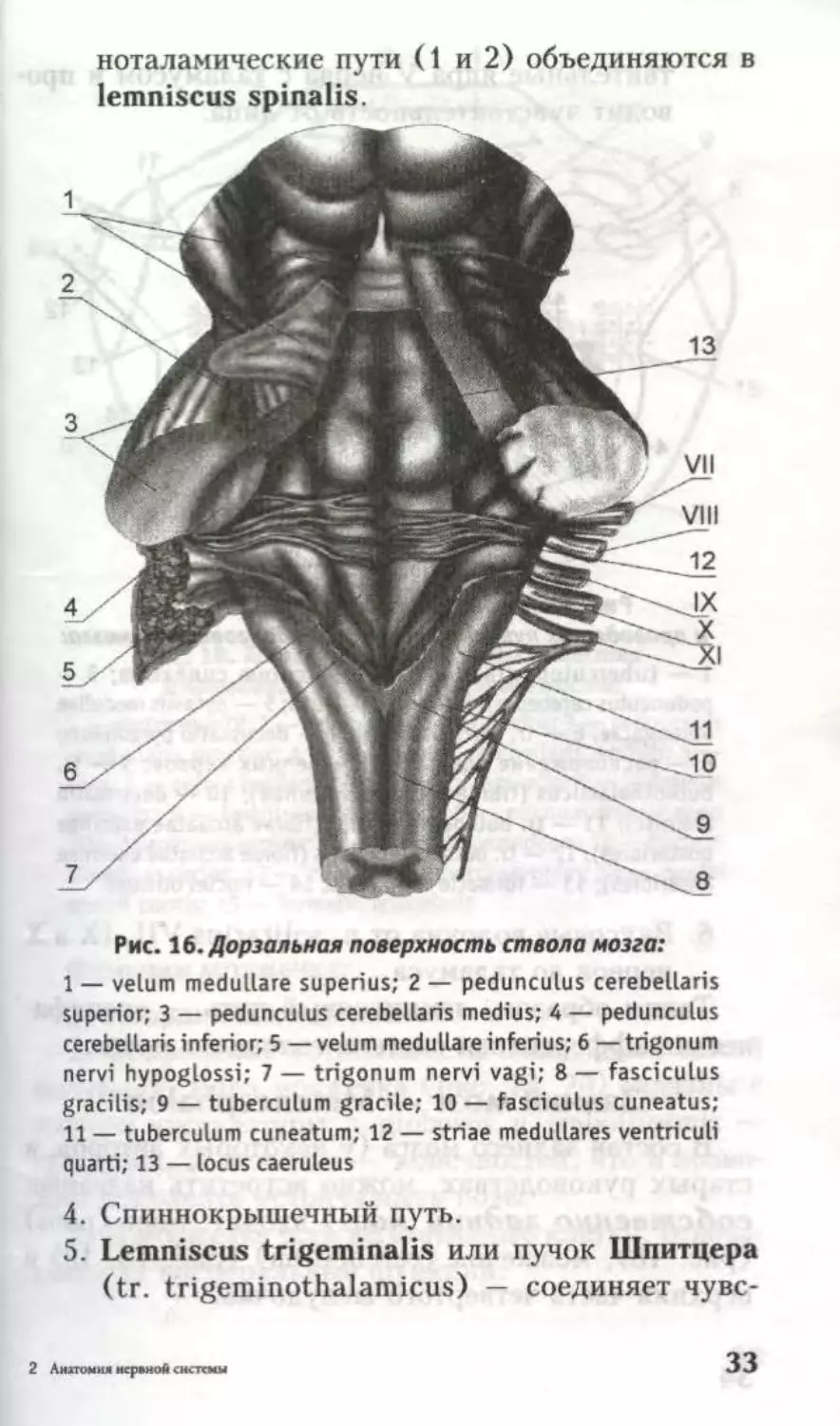
Рис. 16. Дорзальная поверхность ствола мозга:
1 — velum medullare superius; 2 — pedunculus cerebellaris
superior; 3 — pedunculus cerebellaris medius; 4 — pedunculus
cerebellaris inferior; 5 — velum medullare inferius; 6 — trigonum
nervi hypoglossi; 7 — trigonum nervi vagi; 8 — fasciculus
gracilis; 9 — tuberculum gracile; 10 — fasciculus cuneatus;
11 — tuberculum cuneatum; 12 — striae medullares ventriculi
quarti; 13 — locus caeruleus
4. Спиннокрышечный путь.
5. Lemniscus trigeminalis или пучок Шпитцера
(tr. trigeminothalamicus) — соединяет чувс-
2 Анатомия нервной системы
33
твительные ядра V нерва с таламусом и про
водит чувствительность от лица.
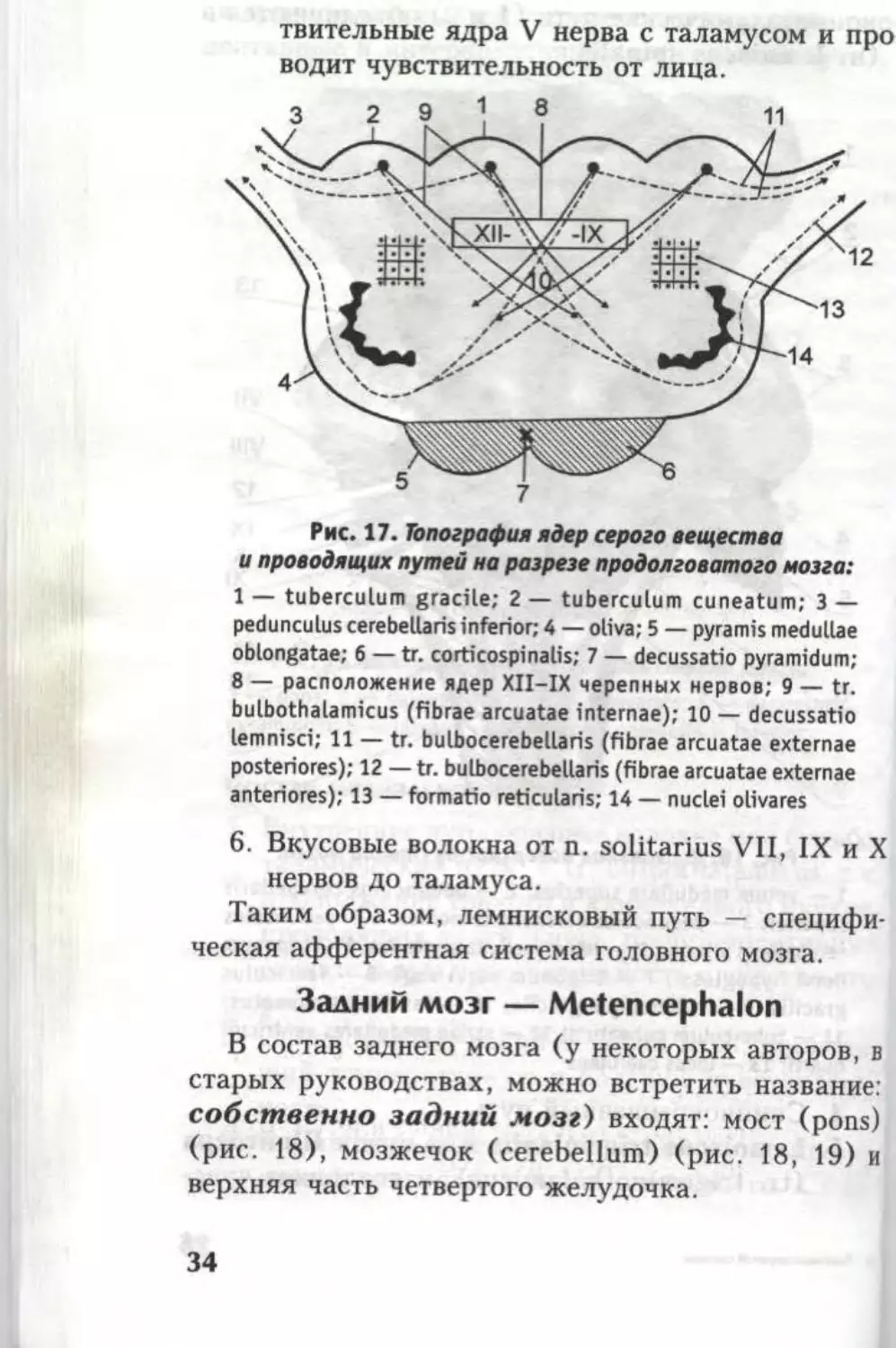
Рис. 17. Топография ядер серого вещества
и проводящих путей на разрезе продолговатого мозга:
1 — tuberculum gracile; 2 — tuberculum cuneatum; 3 —
pedunculus cerebellaris inferior; 4 — oliva; 5 — pyramis medullae
oblongatae; 6 — tr. corticospinalis; 7 — decussatio pyramidum;
8 — расположение ядер XII—IX черепных нервов; 9 — tr.
bulbothalamicus (fibrae arcuatae internae); 10 — decussatio
lemnisci; 11 — tr. bulbocerebellaris (fibrae arcuatae externae
posteriores); 12 — tr. bulbocerebellaris (fibrae arcuatae externae
anteriores); 13 — formatio reticularis; 14 — nuclei olivares
6. Вкусовые волокна от n. solitarius VII, IX и X
нервов до таламуса.
Таким образом, лемнисковый путь — специфи-
ческая афферентная система головного мозга.
Задний мозг — Metencephalon
В состав заднего мозга (у некоторых авторов, в
старых руководствах, можно встретить название:
собственно задний мозг) входят: мост (pons)
(рис. 18), мозжечок (cerebellum) (рис. 18, 19) и
верхняя часть четвертого желудочка.
34
Рис. 18. Топография ядер серого вещества
и проводящих путей на разрезе моста:
1 — ventriculus IV; 2 — velum medullare superius; 3 — pedunculus
cerebellaris superior; 4 — pedunculus cerebellaris medius; 5 —
pons; 6 — sulcus basilaris; 7 — tr. corticospinalis; 8 — lemniscus
medialis; 9 — lemniscus lateralis; 10 — расположение ядер
VIII-V черепных нервов; 11 — corpus trapezoideum; 12 — nucleus
olivaris superior; 13 — nuclei corporis trapezoidei; 14 — nuclei
proprii pontis; 15 — formatio reticularis
Функции мозжечка:
1. Координация движений.
2. Сохранение устойчивости позы, даже самой
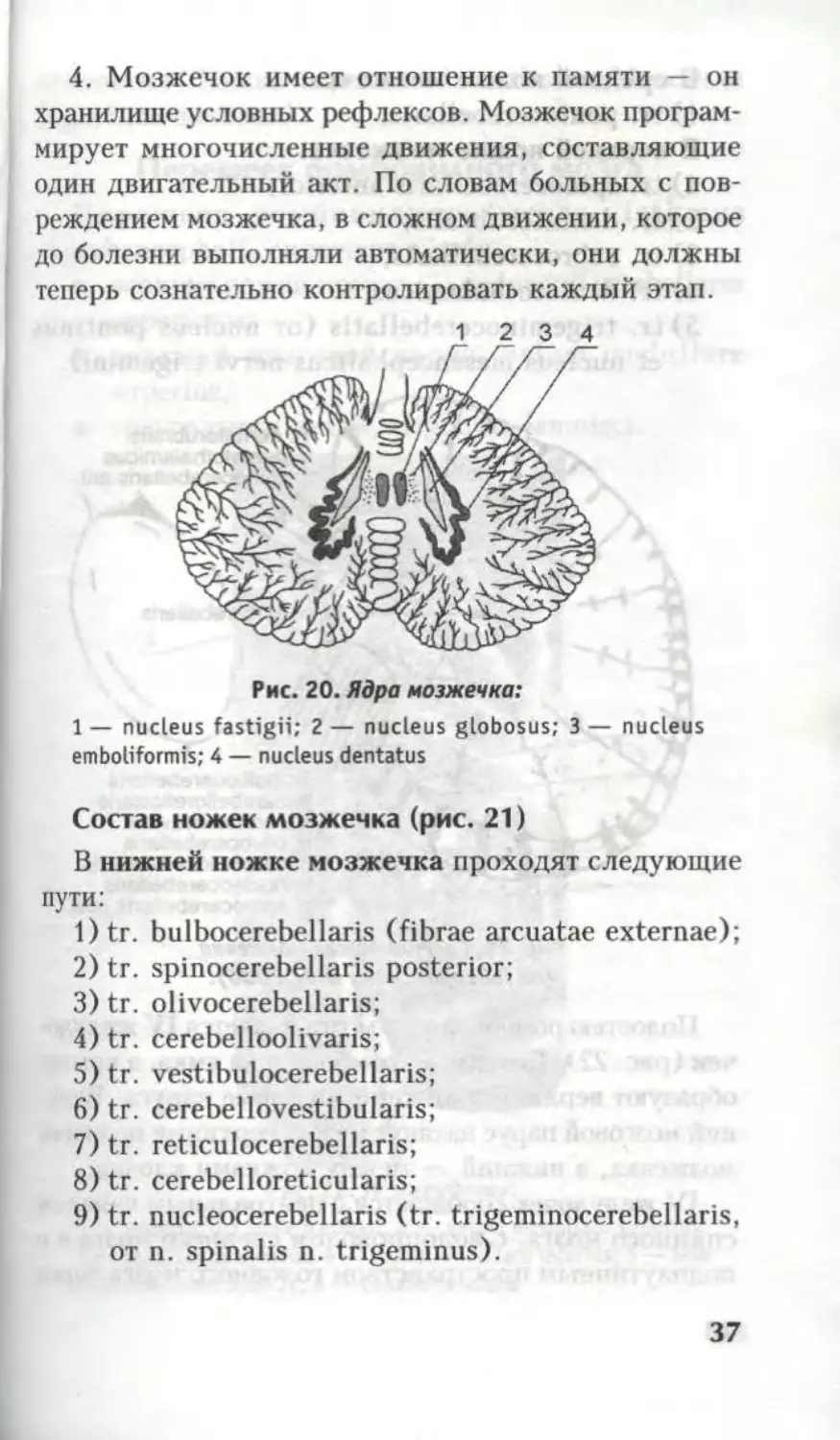
вычурной. Ядра мозжечка (рис. 19, 20) связаны с
тонусом мускулатуры: n. globosus, n. emboliformis —
туловища, n. dentatus — конечностей, что и позво-
ляет сохранять устойчивость позы.
3. В мозжечке есть вегетативные клетки, отвеча-
ющие за висцеральные функции.
35
9
8 4 1
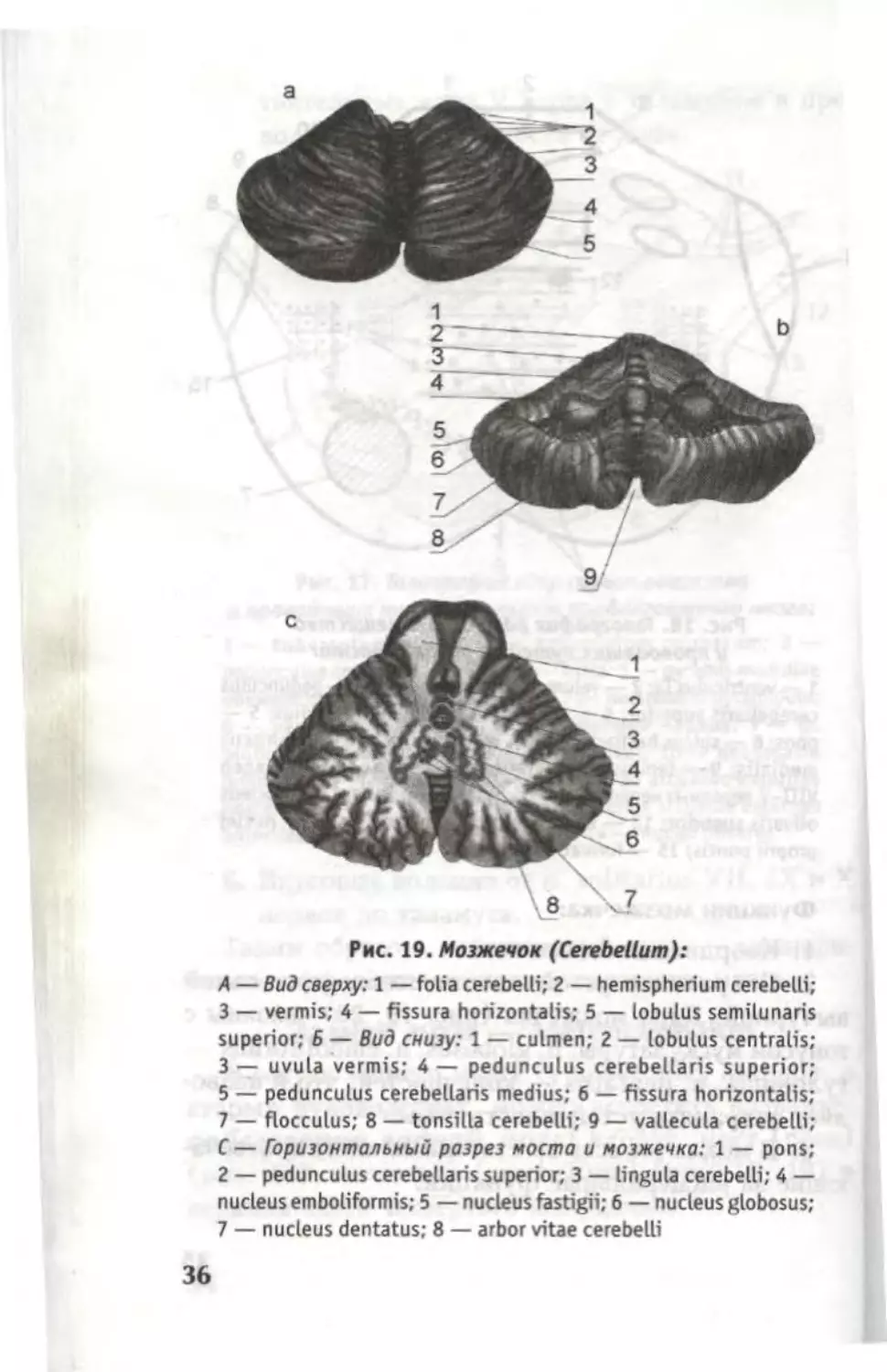
Рис. 19. Мозжечок (Cerebellum):
А — Вид сверху: 1 — folia cerebelli; 2 — hemispherium cerebelli;
3 — vermis; 4 — fissura horizontalis; 5 — lobulus semilunaris
superior; Б — Вид снизу: 1 — culmen; 2 — lobulus centralis;
3 —- uvula vermis; 4 — pedunculus cerebellaris superior;
5 — pedunculus cerebellaris medius; 6 — fissura horizontalis;
7 — flocculus; 8 — tonsilla cerebelli; 9 — vallecula cerebelli;
C — Горизонтальный разрез моста и мозжечка: 1 — pons;
2 — pedunculus cerebellaris superior; 3 — lingula cerebelli; 4 —
nucleus emboliformis; 5 — nucleus fastigii; 6 — nucleus globosus;
7 — nucleus dentatus; 8 — arbor vitae cerebelli
36
4. Мозжечок имеет отношение к памяти — он
хранилище условных рефлексов. Мозжечок програм-
мирует многочисленные движения, составляющие
один двигательный акт. По словам больных с пов-
реждением мозжечка, в сложном движении, которое
до болезни выполняли автоматически, они должны
теперь сознательно контролировать каждый этап.
Рис. 20. Ядра мозжечка:
1 — nucleus fastigii; 2 — nucleus globosus; 3 — nucleus
emboliformis; 4 — nucleus dentatus
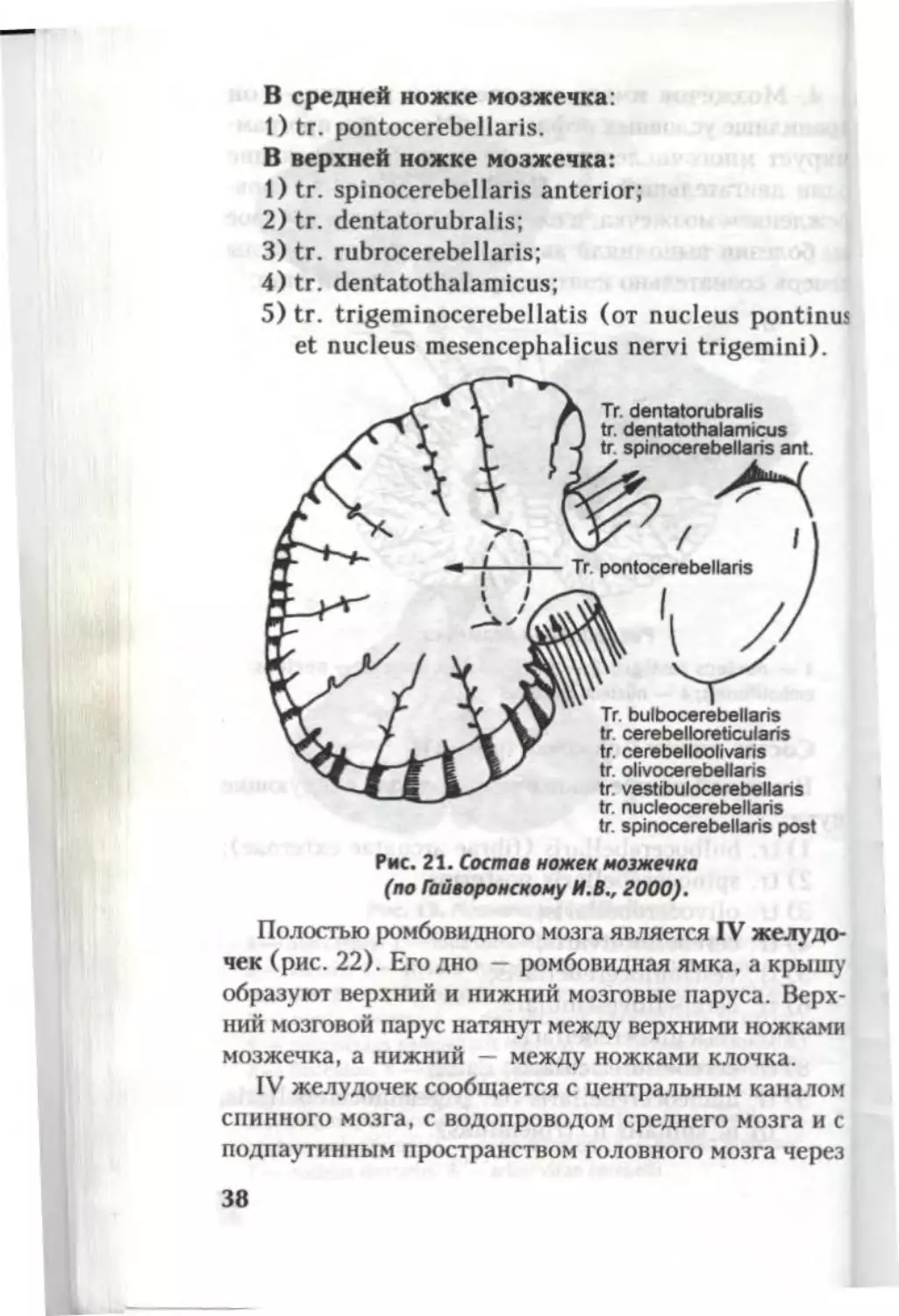
Состав ножек мозжечка (рис. 21)
В нижней ножке мозжечка проходят следующие
пути:
1) tr. bulbocerebellaris (fibrae arcuatae externae);
2) tr. spinocerebellaris posterior;
3) tr. olivocerebellaris;
4) tr. cerebelloolivaris;
5) tr. vestibulocerebellaris;
6) tr. cerebellovestibularis;
7) tr. reticulocerebellaris;
8) tr. cerebelloreticularis;
9) tr. nucleocerebellaris (tr. trigeminocerebellaris,
от n. spinalis n. trigeminus).
37
В средней ножке мозжечка:
1) tr. pontocerebellaris.
В верхней ножке мозжечка:
1) tr. spinocerebellaris anterior;
2) tr. dentatorubralis;
3) tr. rubrocerebel laris;
4) tr. dentatothalamicus;
5) tr. trigeminocerebellatis (от nucleus pontinus
et nucleus mesencephalicus nervi trigemini).
Tr. dentatorubralis
tr. dentatothalamicus
tr. spinocerebellaris ant.
Tr. pontocerebellaris
Tr. bulbocerebellaris
tr. cerebelloreticularis
tr. cerebelloolivaris
tr. olivocerebellaris
tr. vestibulocerebellaris
tr. nucleocerebellaris
tr. spinocerebellaris post
Рис. 21. Состав ножек мозжечка
(по Гайворонскому И.В., 2000).
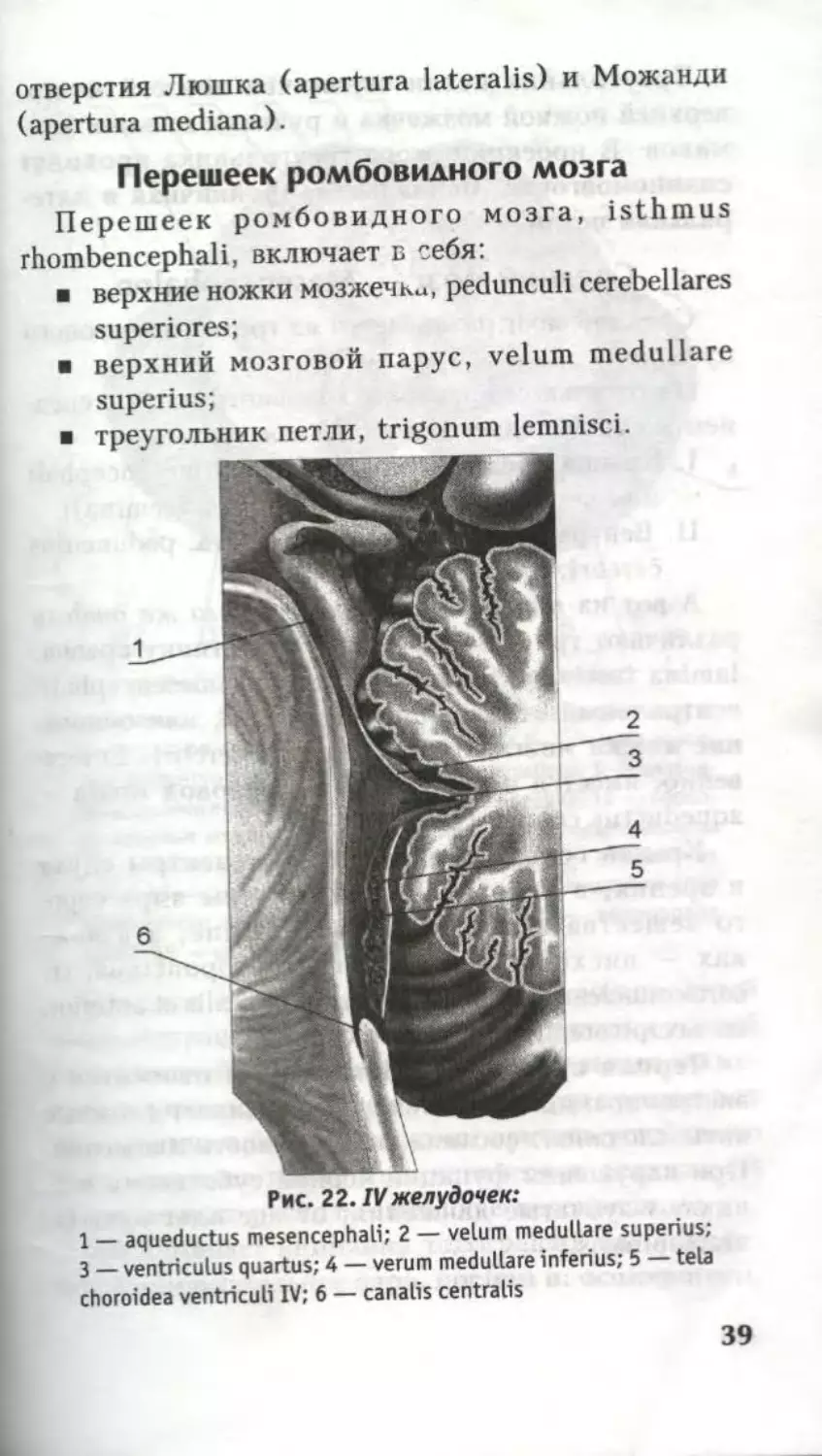
Полостью ромбовидного мозга является IV желудо-
чек (рис. 22). Его дно — ромбовидная ямка, а крышу
образуют верхний и нижний мозговые паруса. Верх-
ний мозговой парус натянут между верхними ножками
мозжечка, а нижний — между ножками клочка.
IV желудочек сообщается с центральным каналом
спинного мозга, с водопроводом среднего мозга и с
подпаутинным пространством головного мозга через
38
отверстия Люшка (apertura lateralis) и Можанди
(apertura mediana).
Перешеек ромбовидного мозга
Перешеек ромбовидного мозга, isthmus
rhombencephali, включает в себя:
верхние ножки мозжечки, pedunculi cerebellares
super lores;
верхний мозговой парус, velum medullare
superius;
треугольник петли, trigonum lemnisci.
Рис. 22. IVжелудочек:
1 — aqueductus mesencephali; 2 — velum medullare superius;
3 — ventriculus quartus; 4 — verum medullare inferius; 5 — tela
choroidea ventriculi IV; 6 — canalis centralis
39
Треугольник петли ограничен ножкой мозга,
верхней ножкой мозжечка и ручками нижних хол-
миков. В проекции этого треугольника проходят
спинномозговая, медиальная, тройничная и лате-
ральная петли.
Средний мозг — Mesencephalon
Средний мозг развивается из третьего мозгового
пузыря.
На тотальном препарате головного мозга в сред-
нем мозге выделяется дорсальная часть:
1. Крыша среднего мозга: tectum mesencephali
или четверохолмие (lamina quadrigemina);
II. Вентральная часть: ножка мозга: pedunculus
cerebri.
А вот на поперечном разрезе этого же отдела
различают три основные части: пластинку крыши,
lamina tecti; покрышку, tegmentum mesencephali;
вентральный отдел pedunculi cerebri, или основа-
ние ножки мозга, basis pedunculi cerebri. Естест-
венно, имеется и полость водопровод мозга
aqueductus cerebri (рис. 23).
Крыша содержит подкорковые центры слуха
и зрения, в покрышке расположены ядра серо-
го вещества и проходят восходящие, а в нож-
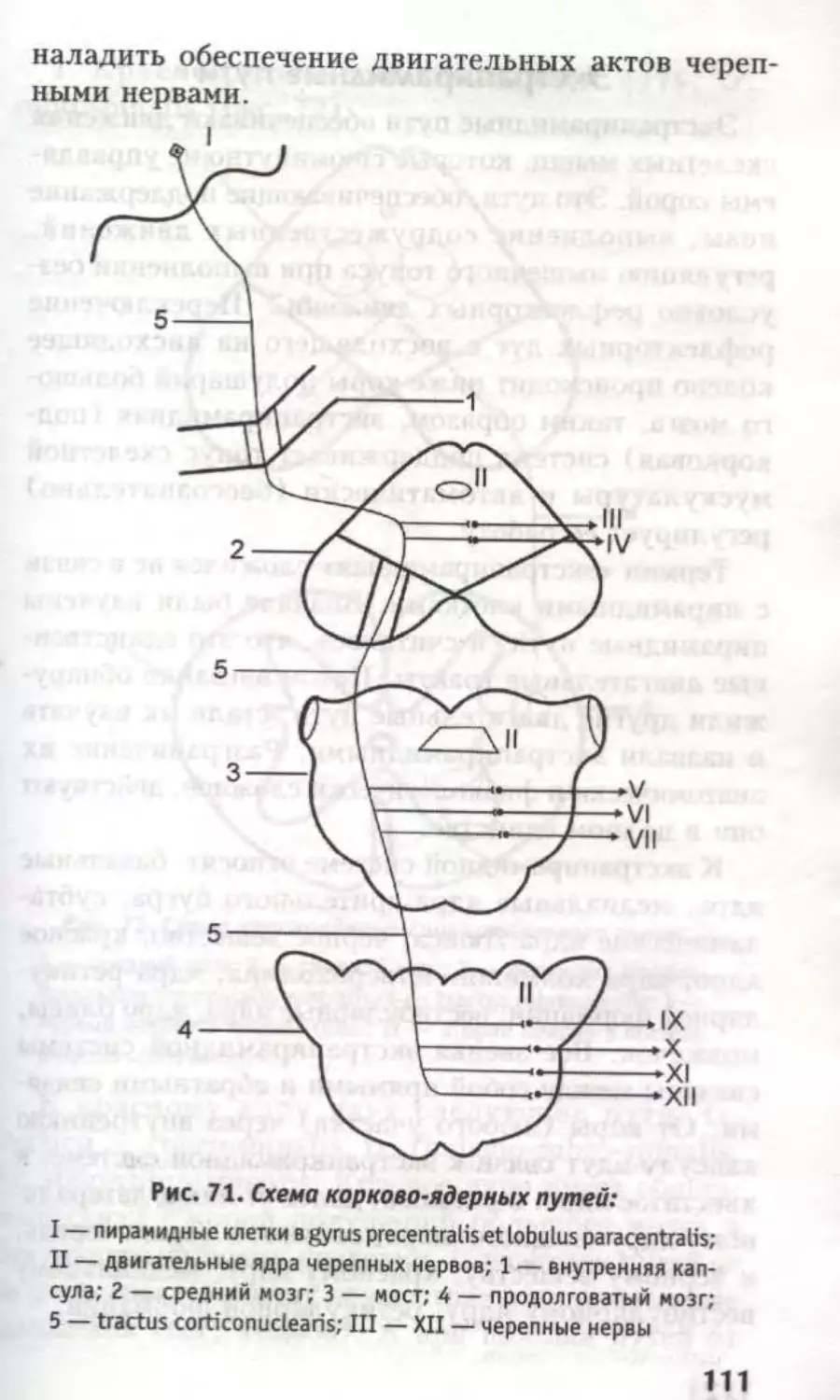
ках — нисходящие пути: tr. frontopontinus, tr.
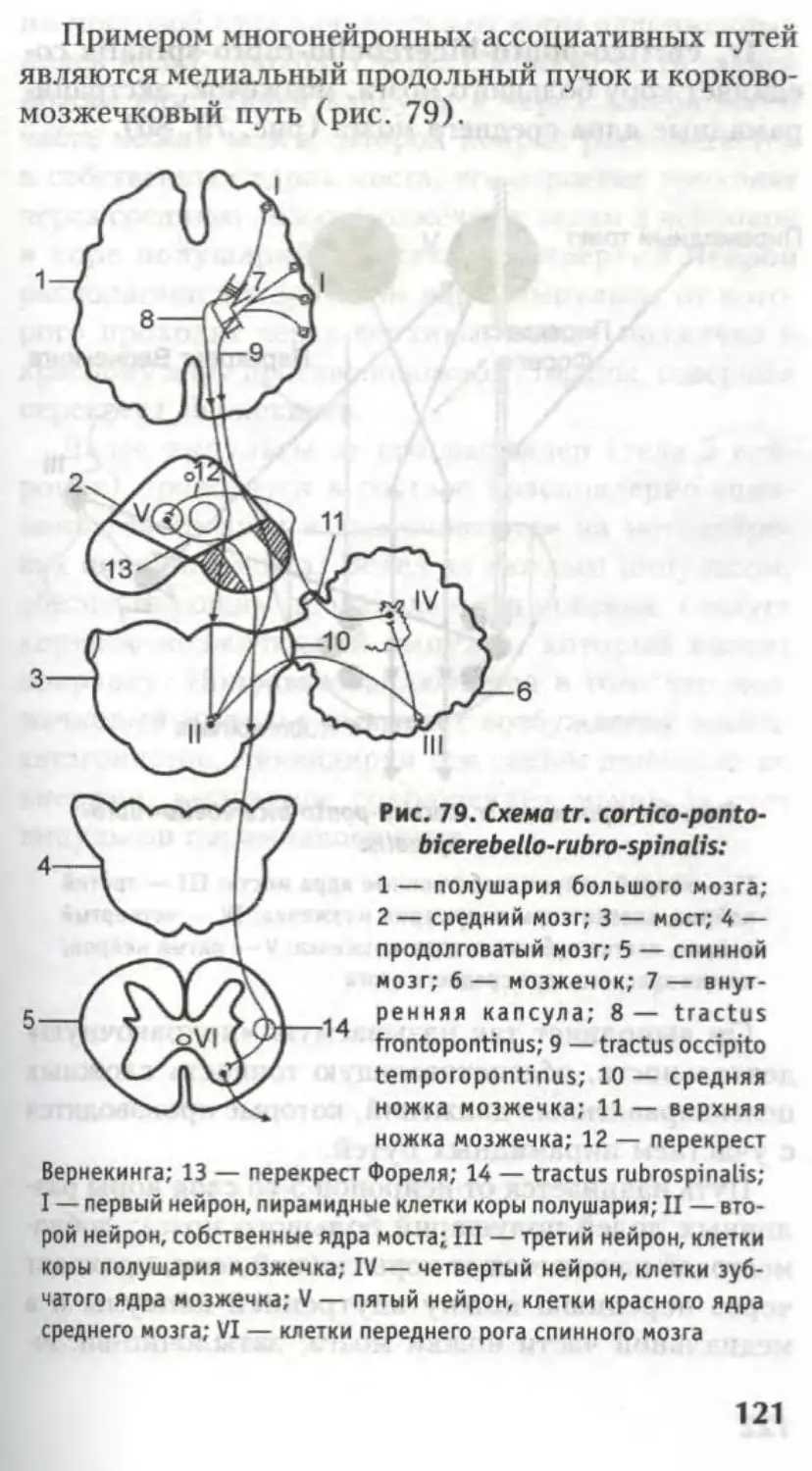
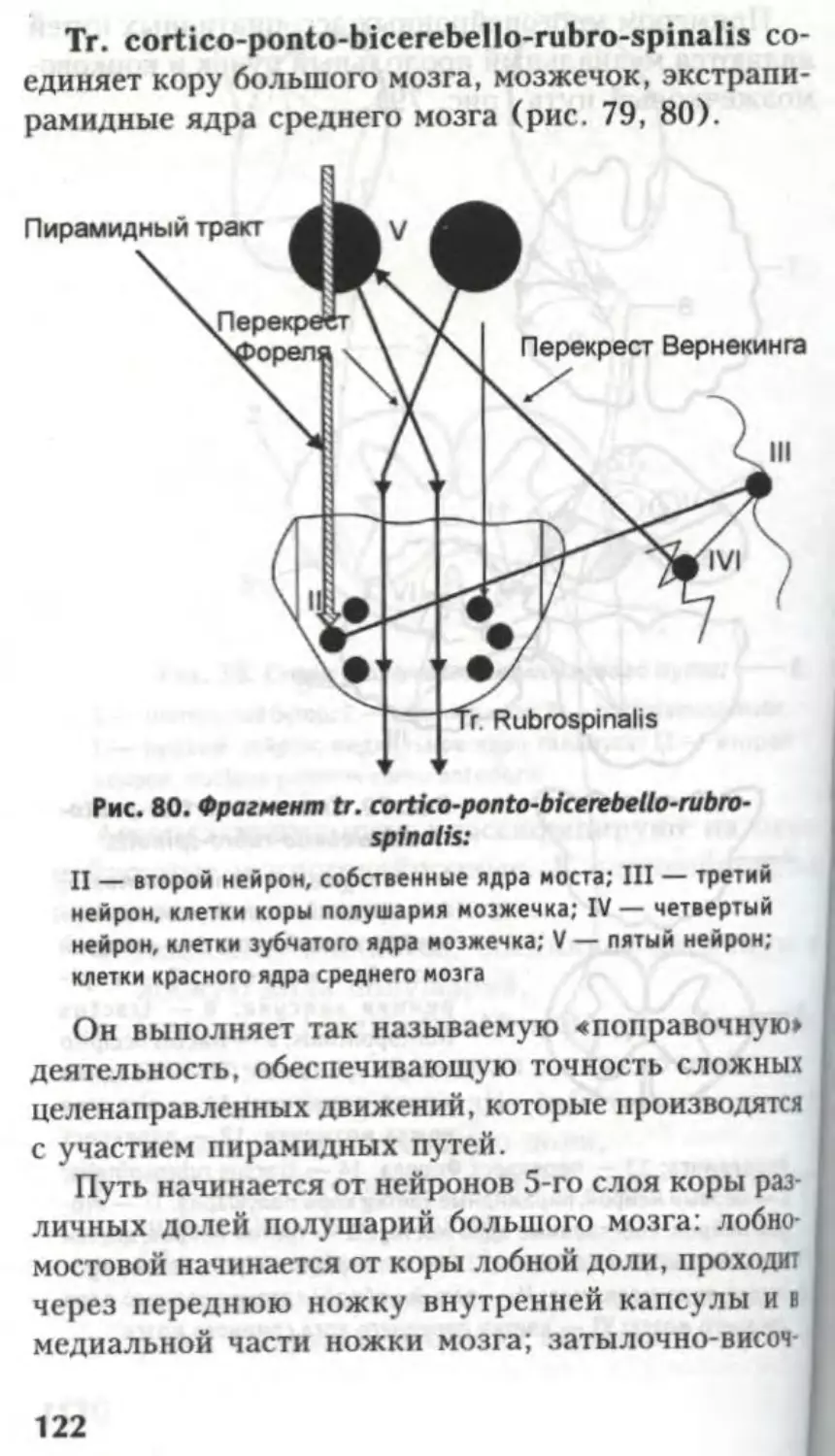
corticonuclearis, tr. corticospinal is lateralis et anterior,
tr. occipitotemporopontinus (рис. 24).
Черная субстанция Зёммеринга относится к
экстрапирамидной системе, обеспечивает сложные
акты глотания, сосания, пластичность движений.
При нарушении функций черной субстанции воз-
никают зубчатые движения; от нее идет путь tr.
nigrospinalis.
40
13 14 15 16 17 18 19
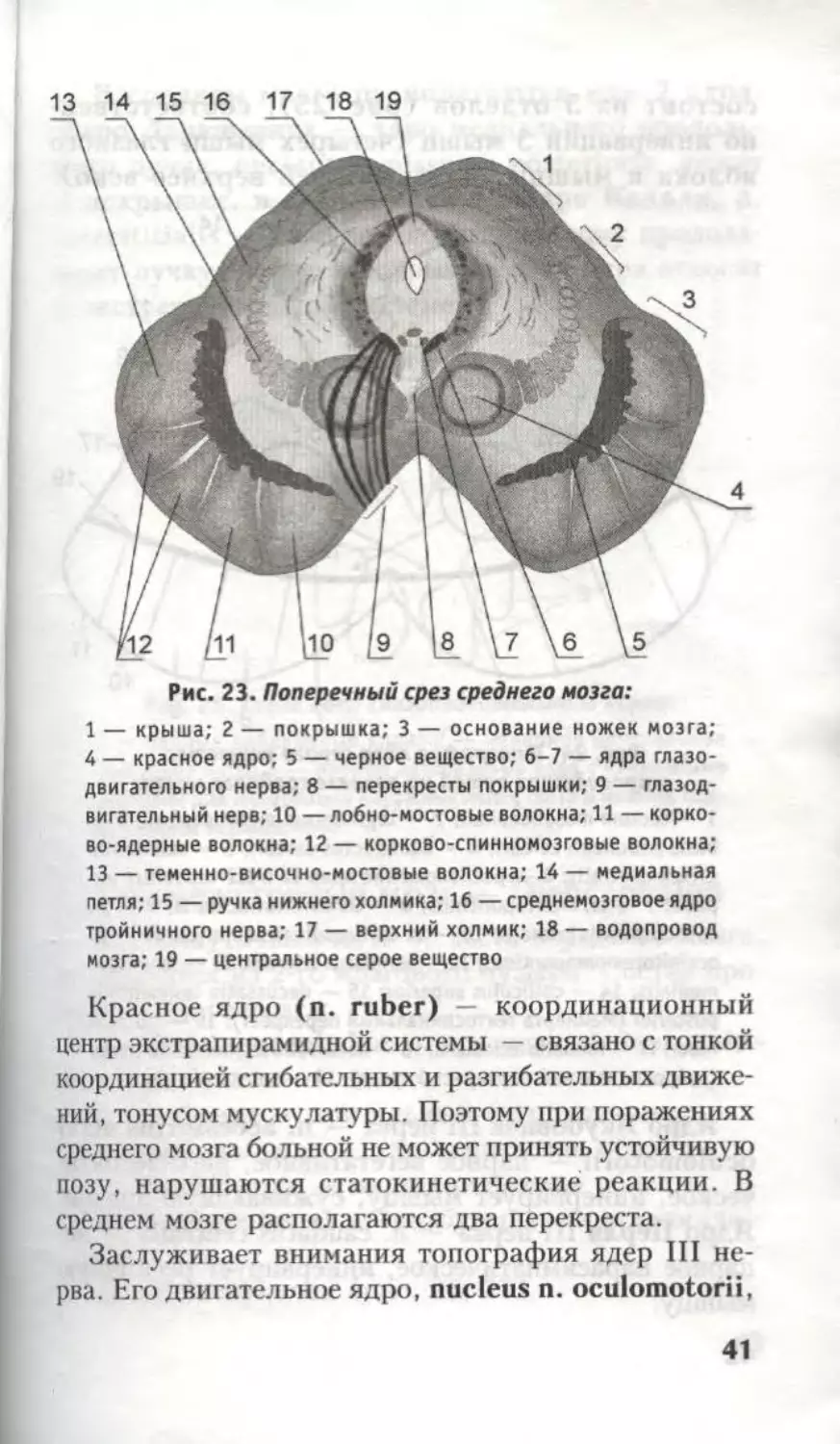
Рис. 23. Поперечный срез среднего мозга:
1 — крыша; 2 — покрышка; 3 — основание ножек мозга;
4 — красное ядро; 5 — черное вещество; 6-7 — ядра глазо-
двигательного нерва; 8 — перекресты покрышки; 9 — глазод-
вигательный нерв; 10 —лобно-мостовые волокна; 11 — корко-
во-ядерные волокна; 12 — корково-спинномозговые волокна;
13 — теменно-височно-мостовые волокна; 14 — медиальная
петля; 15 — ручка нижнего холмика; 16 — среднемозговое ядро
тройничного нерва; 17 — верхний холмик; 18— водопровод
мозга; 19 — центральное серое вещество
Красное ядро (n. ruber) координационный
центр экстрапирамидной системы — связано с тонкой
координацией сгибательных и разгибательных движе-
ний, тонусом мускулатуры. Поэтому при поражениях
среднего мозга больной не может принять устойчивую
позу, нарушаются статокинетические реакции. В
среднем мозге располагаются два перекреста.
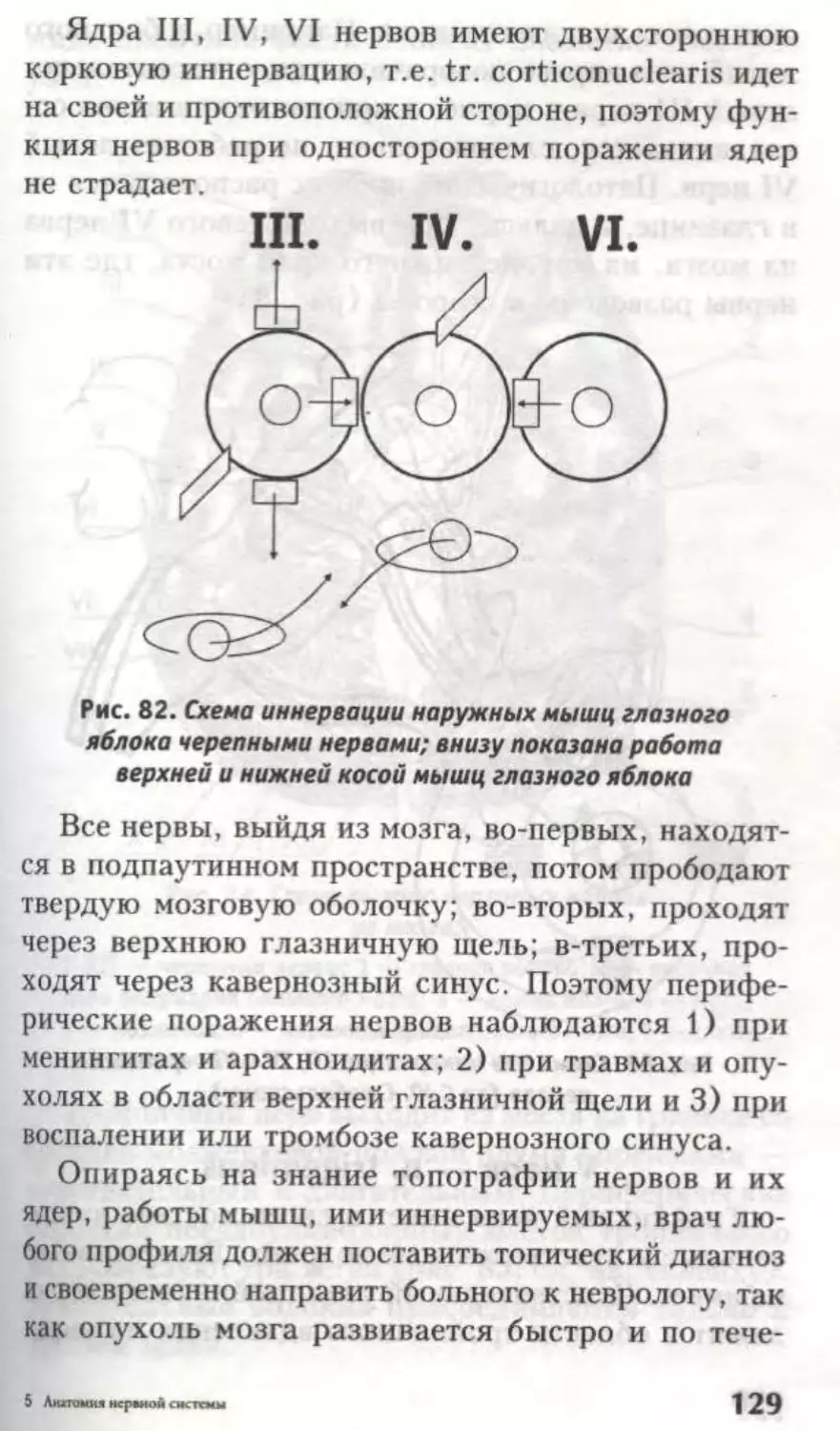
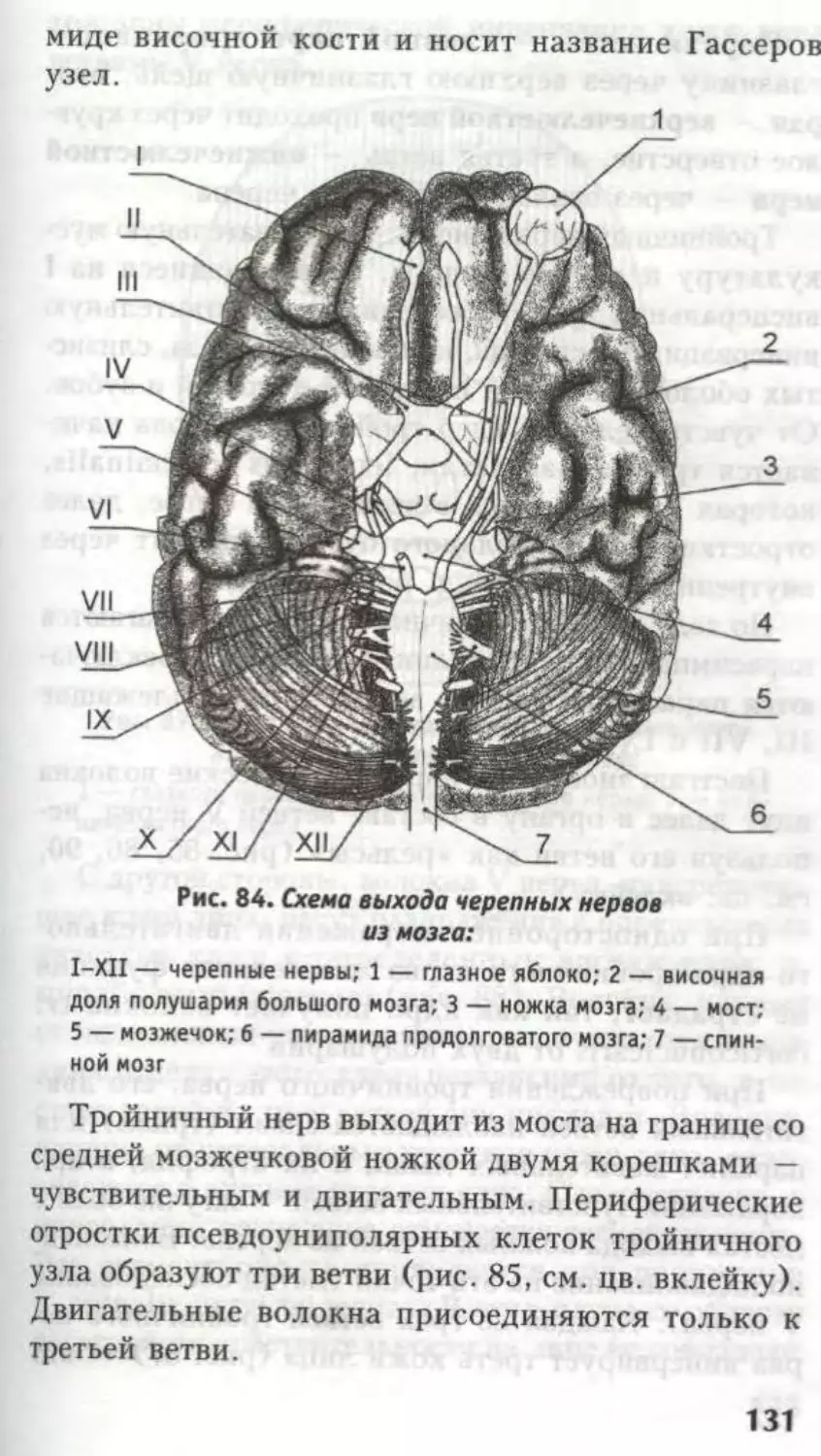
Заслуживает внимания топография ядер III не-
рва. Его двигательное ядро, nucleus n. oculomotor!i,
41
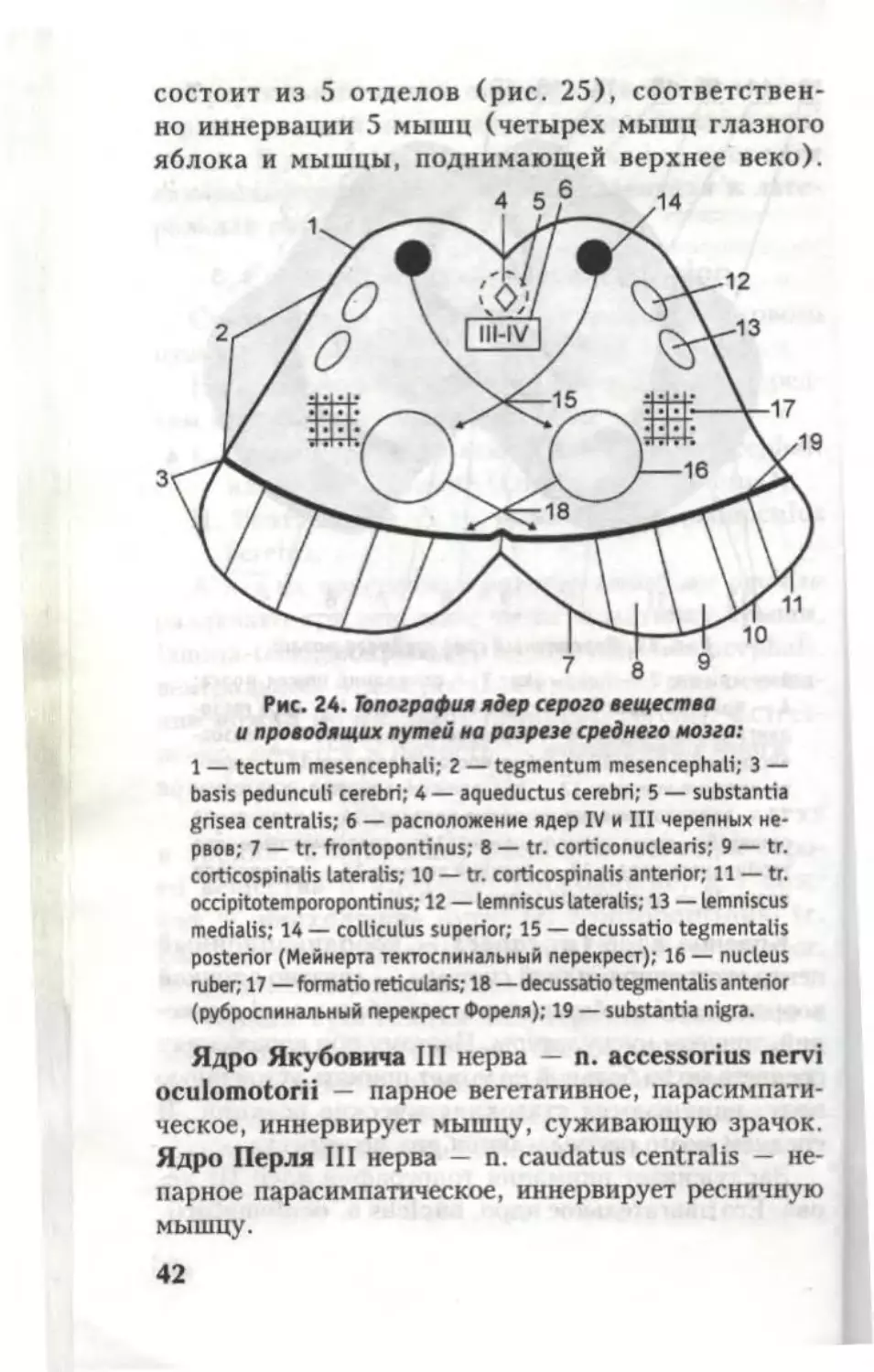
состоит из 5 отделов (рис. 25), соответствен-
но иннервации 5 мышц (четырех мышц глазного
яблока и мышцы, поднимающей верхнее веко).
7 8 9
Рис. 24. Топография ядер серого вещества
и проводящих путей на разрезе среднего мозга:
1 — tectum mesencephali; 2 — tegmentum mesencephali; 3 —
basis pedunculi cerebri; 4— aqueductus cerebri; 5 — substantia
grisea centralis; 6 — расположение ядер IV и III черепных не-
рвов; 7 — tr. frontopontinus; 8 — tr. corticonuclearis; 9 — tr.
corticospinalis lateralis; 10 — tr. corticospinalis anterior; 11 — tr.
occipitotemporopontinus; 12 — lemniscus lateralis; 13 — lemniscus
medialis; 14 — colliculus superior; 15 — decussatio tegmentalis
posterior (Мейнерта тектоспинальный перекрест); 16 — nucleus
ruber; 17 — formatio reticularis; 18 — decussatio tegmentalis anterior
(руброспинальный перекрест Фореля); 19 — substantia nigra.
Ядро Якубовича III нерва — n. accessorius nervi
oculomotorii — парное вегетативное, парасимпати-
ческое, иннервирует мышцу, суживающую зрачок.
Ядро Перля III нерва — n. caudatus centralis — не-
парное парасимпатическое, иннервирует ресничную
мышцу.
42
В среднем мозге располагаются еще 2 ядра.
Ядро Даркшевича — ядро медиального продоль-
ного пучка, nucleus comissurae posterioris, лежит
в покрышке, в задней спайке. Ядро Кахаля, п.
interstitialis — дает начало медиальному продоль-
ному пучку, лежит в покрышке. Эти ядра относят
к экстрапирамидной системе.
Рис. 25. Схема ядер глазодвигательного нерва:
1 — парные добавочные ядра; 2 — непарное центральное
хвостовое ядро; 3 — парные латеральные ядра с клеточными
группами для иннервации наружных мышц глаза и мышцы, под-
нимающей верхнее веко
Промежуточный мозг — Diencephalon
Промежуточный мозг — часть переднего мозга,
развивается из 2-го мозгового пузыря. Состав про-
межуточного мозга:
A. Thalamencephalon — зрительный мозг, тала-
мический мозг:
Thalamus — зрительный бугор, таламус;
Metathalamus — заталамическая область;
Epithalamus — надталамическая область.
Б. Hypothalamus — подбугорье, подталамичес-
кая область.
В. III желудочек (рис. 26, 27).
43
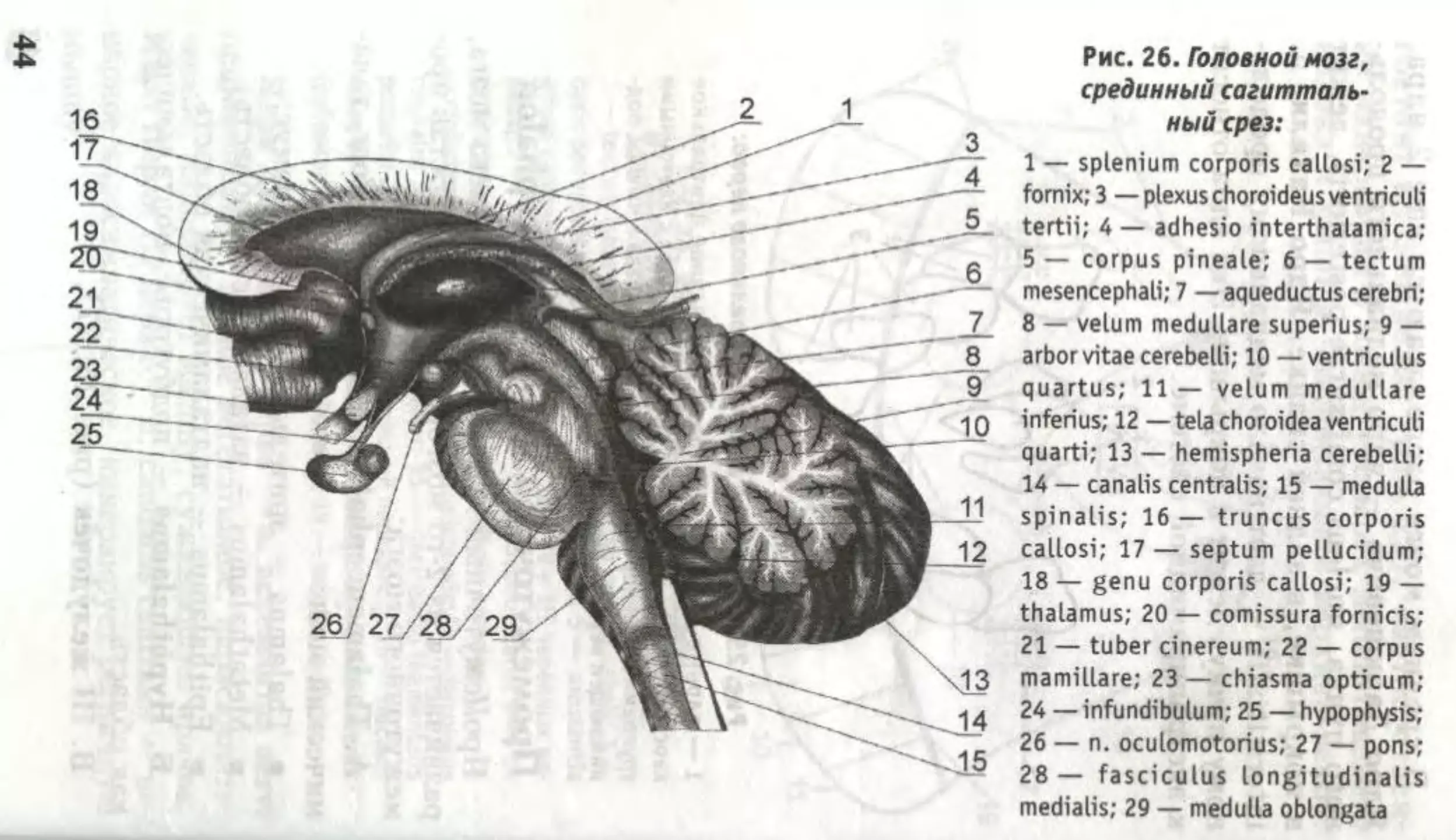
£
Рис. 26. Головной мозг,
срединный сагитталь-
ный срез:
1 — splenium corporis callosi; 2 —
fornix; 3 — plexus choroideus ventriculi
tertii; 4 — adhesio interthalamica;
5 — corpus pineale; 6 — tectum
mesencephali; 7 —aqueductus cerebri;
8 — velum medullare superius; 9 —
arborvitae cerebelli; 10 — ventriculus
quartus; 11 — velum medullare
inferius; 12 — tela choroidea ventriculi
quarti; 13 — hemispheria cerebelli;
14 — canalis centralis; 15 — medulla
spinalis; 16— truncus corporis
callosi; 17 — septum pellucidum;
18 — genu corporis callosi; 19 —
thalamus; 20 — comissura fornicis;
21 — tuber cinereum; 22 — corpus
mamillare; 23 — chiasma opticum;
24 — infundibulum; 25 — hypophysis;
26 — n. oculomotorius; 27 — pons;
28— fasciculus longitudinalis
medialis; 29 — medulla oblongata
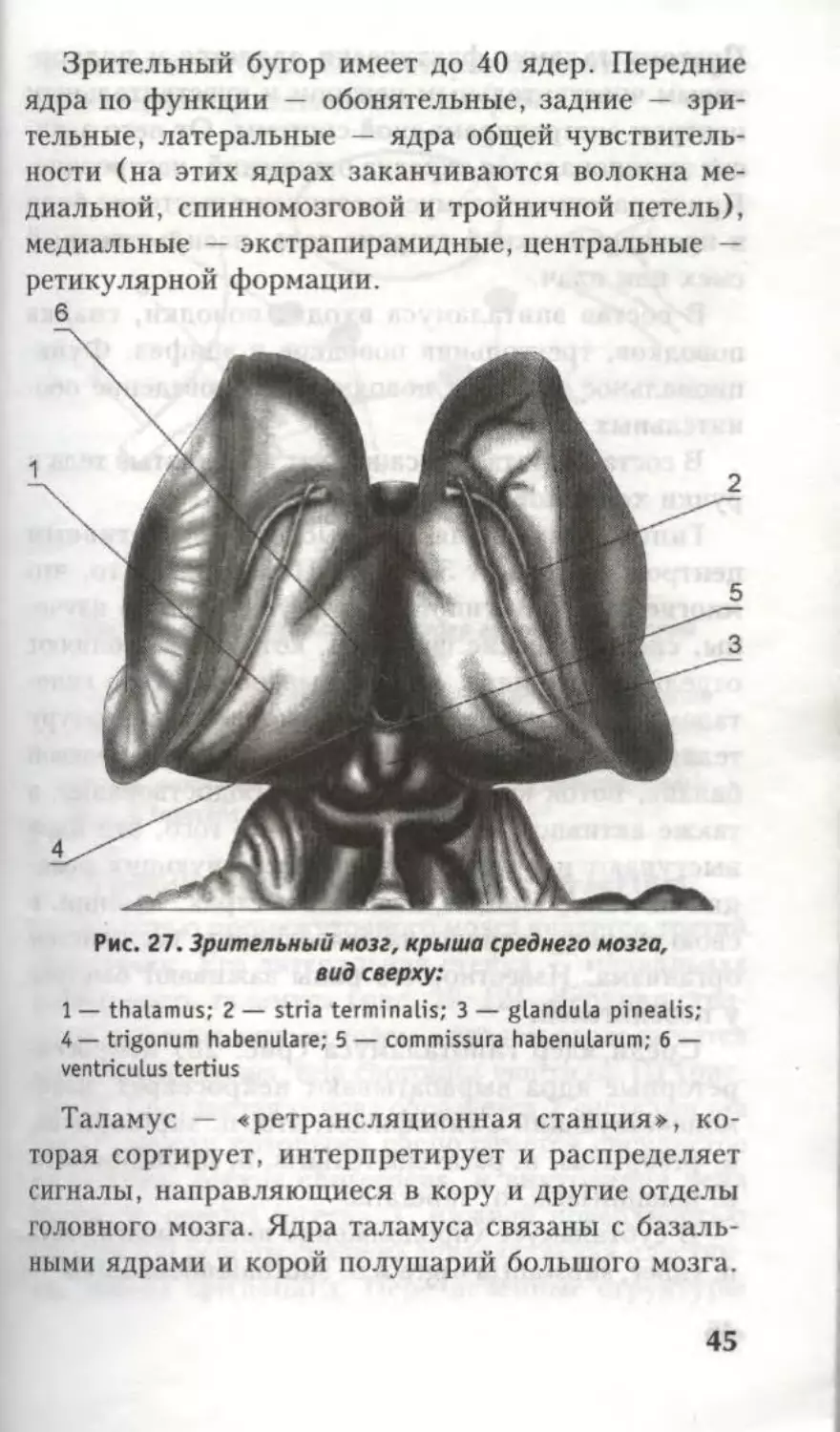
Зрительный бугор имеет до 40 ядер. Передние
ядра по функции — обонятельные, задние — зри-
тельные, латеральные — ядра общей чувствитель-
ности (на этих ядрах заканчиваются волокна ме-
диальной, спинномозговой и тройничной петель),
медиальные — экстрапирамидные, центральные —
ретикулярной формации.
Рис. 27. Зрительный мозг, крыша среднего мозга,
вид сверху:
1 — thalamus; 2 — stria terminalis; 3 — glandula pinealis;
4 — trigonum habenulare; 5 — commissura habenularum; 6 —
ventriculus tertius
Таламус «ретрансляционная станция», ко-
торая сортирует, интерпретирует и распределяет
сигналы, направляющиеся в кору и другие отделы
головного мозга. Ядра таламуса связаны с базаль-
ными ядрами и корой полушарий большого мозга.
45
Поэтому таламус фактически является и подкор-
ковым чувствительным центром и чувствительным
центром экстрапирамидной системы. От него зави-
сит эмоциональная окраска ощущений, настроение.
При поражении таламуса возникают жестокие боли
в противоположной стороне тела, насильственный
смех или плач.
В состав эпиталамуса входят поводки, спайка
поводков, треугольник поводков и эпифиз. Функ-
циональное значение поводков — проведение обо-
нятельных импульсов.
В состав метаталамуса входят коленчатые тела и
ручки холмиков четверохолмия.
Гипоталамус является высшим вегетативным
центром, содержит 32 ядра. Несмотря на то, что
многие функции гипоталамуса уже хорошо изуче-
ны, специфические функции, которые выполняют
отдельные его ядра, еще не выяснены. Ядра гипо-
таламуса отслеживают и регулируют температуру
тела, количество потребляемой пищи, водно-солевой
баланс, поток крови, цикл сна и бодрствования, а
также активность гормонов. Кроме того, эти ядра
выступают посредниками соответствующих реак-
ций на такие эмоции, как гнев и страх. Эмоции, в
свою очередь, влияют на деятельность всех систем
организма. Известно, что раны заживают быстрее
у победителей.
Среди ядер гипоталамуса (рис. 28) нейросек-
реторные ядра вырабатывают нейросекрет, взаи-
модействующий с гипофизом, это: n. supraopticus,
n. preopticus, n. paraventricularis, n. infundibularis,
n. mammillaris, n. tuberalis.
В субталамусе (продолжение ножек мозга) есть
n. ruber, substantia nigra и n. subthalamicus Luyis —
46
Люисово тело (рис. 28), которое относят к экстра-
пирамидным двигательным центрам.
Рис. 28. Схема расположения ядер гипоталамической
области:
1 — thalamus; 2 — chiasma opticum; 3 — hypophysis; 4 — tectum
mesencephali; 5 — pedunculus cerebri; 6 — n. supraopticus;
7 — n. preopticus; 8 — n. infundibularis; 9 — n. paraventricularis;
10 — n. mammillaris; 11 — n. subthalamicus (Люисово, Luyis);
12 — n. tuberalis
Третий желудочек, ventriculus tertius
Полостью промежуточного мозга является третий
желудочек. Его латеральная стенка — медиальная
поверхность таламуса (рис. 29-12). Верхняя стен-
ка — под мозолистым телом и сводом располагается
сосудистая основа, tela choroidea ventriculi III (рис.
29-2). Сосудистая основа образуется 2 листками pia
mater, между которыми располагается сосудистое
сплетение, plexus choroideus, и внутренняя вена
мозга, v. cerebri interna. С нижней поверхностью
сосудистой основы сращена эпителиальная пластин-
ка, lamina epithelialis. Перечисленные структуры
47
входят в состав гематоэнцефалического барьера при
образовании спинномозговой жидкости.
Нижняя стенка третьего желудочка: верхняя
поверхность ножки мозга, сосцевидные тела, серый
бугор с воронкой и гипофизом, углубление воронки
(рис. 29-10), зрительный перекрест и углубление
зрительное (рис. 29-9), задняя продырявленная
субстанция.
Передняя стенка: передняя спайка мозга (рис. 29-5),
межжелудочковое отверстие Монроя, столбы свода,
терминальная пластинка.
12 3 4
Рис. 29. Головной мозг, срединный сагиттальный срез:
1 — corpus fornicis; 2 — tela choroidea ventriculi III; 3 — recessus
pinealis; 4 — glandula pinealis; 5 — comissura cerebri anterior;
6 — adhesio interthalamica; 7 — n. opticus; 8 — chiasma opticum;
9 — recessus opticus; 10 — recessus infundibuli; 11 — hypophysis
cerebri; 12 — thalamus
Задняя стенка: задняя спайка мозга, спайка по-
водков, эпифиз, поводок, апертура водопровода
мозга и шишковидное углубление (рис. 29-3).
48
Сетевидная формация, formatio
reticularis
Сетеввдная формация известна с 1845 г., описана
Дейтерсом (O.F.C. Deiters) в 1885 г. В настоящее
время продолжается ее изучение.
Сетевидная формация располагается между за-
дними и боковыми рогами шейных сегментов спин-
ного мозга, в покрышке мозгового ствола, в цен-
тральном ядре зрительного бугра. Представляет
собой комплекс анатомически и функционально
взаимосвязанных нейронов, окруженных множес-
твом волокон, идущих в различных направлени-
ях к ядерным структурам и проводящим путям
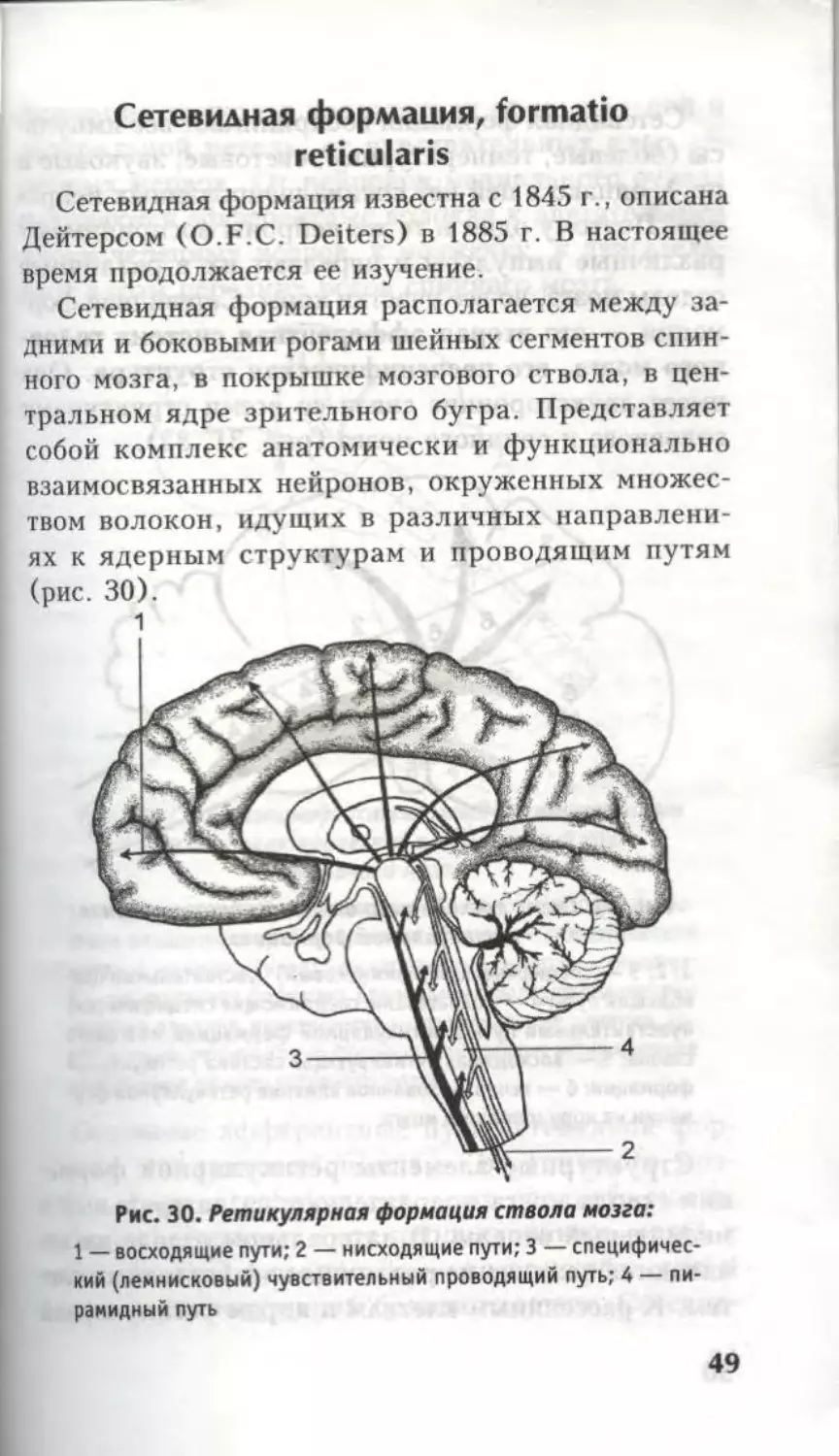
(рис. 30).
1
Рис. 30. Ретикулярная формация ствола мозга:
1 — восходящие пути; 2 — нисходящие пути; 3 — специфичес-
кий (лемнисковый) чувствительный проводящий путь; 4 — пи-
рамидный путь
49
Сетевидная формация воспринимает все импуль-
сы (болевые, температурные, световые, звуковые и
пр.), однако в ней нет специализированных нейро-
нов. Поэтому одни и те же нейроны воспринимают
различные импульсы и передают их в различные
отделы мозга, во все участки коры. Сетевидная фор-
мация — это вторая афферентная система голов-
ного мозга, его неспецифическая структура. Она
имеет двухсторонние связи со всеми структурами
головного и спинного мозга (рис. 31, 32).
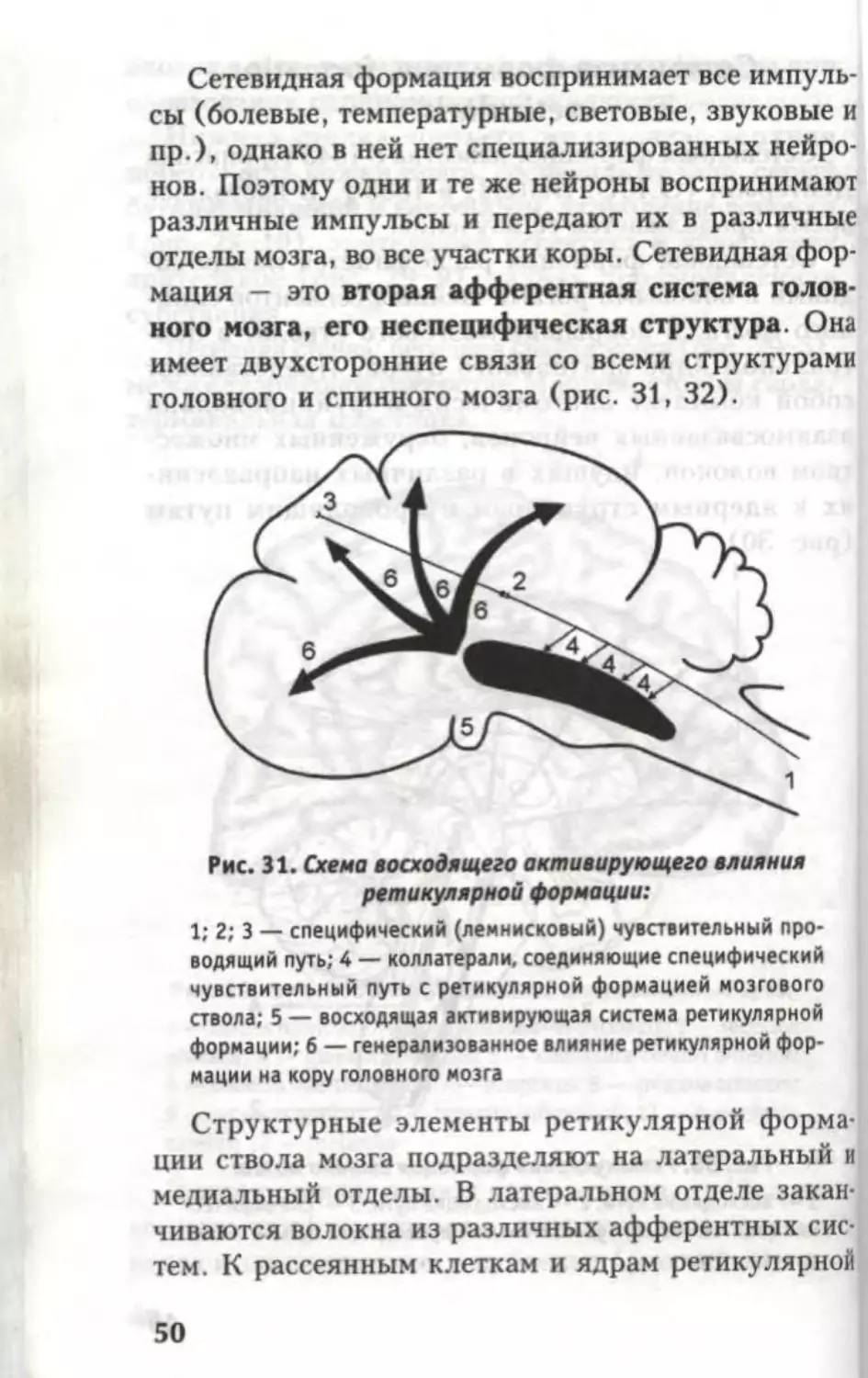
Рис. 31. Схема восходящего активирующего влияния
ретикулярной формации:
1; 2; 3 — специфический (лемнисковый) чувствительный про-
водящий путь; 4 — коллатерали, соединяющие специфический
чувствительный путь с ретикулярной формацией мозгового
ствола; 5 — восходящая активирующая система ретикулярной
формации; б — генерализованное влияние ретикулярной фор-
мации на кору головного мозга
Структурные элементы ретикулярной форма-
ции ствола мозга подразделяют на латеральный и
медиальный отделы. В латеральном отделе закан-
чиваются волокна из различных афферентных сис-
тем. К рассеянным клеткам и ядрам ретикулярной
50
формации подходят коллатерали от медиальной и
латеральной петель, от чувствительных ядер че-
репных нервов. От нейронов медиального отдела
начинаются эфферентные волокна к двигательным
ядрам черепных нервов, к мозжечку, к двигатель-
ным ядрам передних рогов спинного мозга.
Рис. 32. Схема взаимодействия между симпатическим
тонусом и электрической активностью коры
головного мозга:
1 — чувствительный нерв, на который наносится стимул (бо-
левое раздражение); 2 — спинной мозг; 3 — симпатические
нервы; 4 — надпочечник; 5 — каротидный синус; 6 — гипофиз;
7 — ретикулярная формация. Сплошными стрелками обозначены
нервные влияния, пунктирными — гормональные влияния, ко-
торые через ретикулярную формацию оказывают активирующее
воздействие на кору головного мозга
Основные афферентные пути сетевидной фор-
мации: tr. spinoreticularis — от спинного моз-
га, tr. tegmentothalamicus — от среднего мозга,
reticulothalamicus — от продолговатого мозга и
моста, tr. thalamocorticalis — ко всем областям и
слоям коры полушарий большого мозга. Сетевид-
51
ная формация активизирует кору большого мозга
и мозжечок.
Кора полушарий большого мозга, в свою оче-
редь, посылает по tr. corticoreticularis импульсы
в ретикулярную формацию в составе пирамидных
путей. Основным эфферентным трактом является
tr. reticulospinalis. Этот путь проводит тонические
импульсы к гамма-мотонейронам спинного мозга.
Сетевидная формация регулирует моторное звено,
обеспечивая координацию движений, синхронность
мышечных сокращений, обеспечивает нестандарт-
ные движения, рефлекс равновесия, устанавливает
антигравитационный тонус мускулатуры, удержи-
вающий тело над землей. Сетевидная формация
перераспределяет мышечный тонус, что в кризис-
ных ситуациях приводит к мобилизации скрытых
резервов организма.
Установлена роль голубоватого пятна и ядер
шва в регуляции сна и бодрствования. Голубоватое
пятно, locus caeruleus, находится в верхнелате-
ральной части ромбовидной ямки. Нейроны этого
ядра продуцируют норадреналин, активирующий
вышележащие отделы мозга. Особенно высока ак-
тивность нейронов голубоватого пятна во время
бодрствования, во время глубокого сна она угасает
почти полностью.
Ядра шва, (nuclei raphes), располагаются по
срединной линии продолговатого мозга. Нейроци-
ты этих ядер вырабатывают серотонин, который
вызывает процессы разлитого торможения и со-
стояние сна.
Ядра ретикулярной формации продолговатого
мозга имеют связи с вегетативными ядрами IX,
X нервов и симпатическими ядрами спинного моз-
га. Поэтому они участвуют в регуляции сердечной
52
деятельности, дыхания, тонуса сосудов, секреции
желез и так далее.
Ядра Кахаля и Даркшевича, относящиеся к ре-
тикулярной формации среднего мозга, к медиаль-
ному продольному пучку (fasciculus longitudinalis
medialis), имеют связи с ядрами III, IV, VI, VIII,
IX, X и XI пар черепных нервов. Они координи-
руют работу этого пучка, обеспечивая сочетанные
повороты головы и глаз при изменении позы или
при поиске источника звука, фиксацию взгляда.
(Данные движения совершенно необходимы при
трудовых и игровых актах).
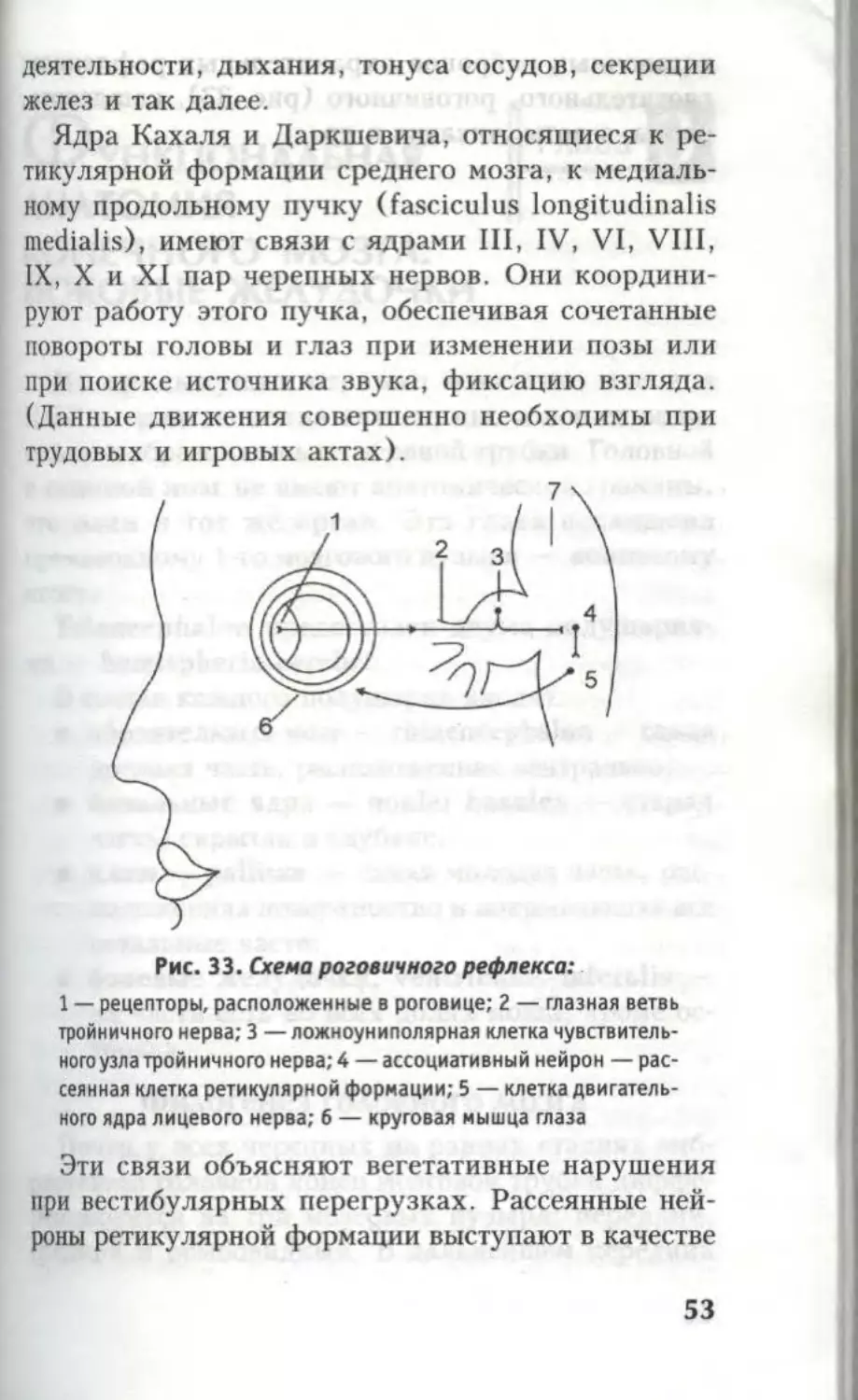
Рис. 33. Схема роговичного рефлекса:
1 — рецепторы, расположенные в роговице; 2 — глазная ветвь
тройничного нерва; 3 —ложноуниполярная клетка чувствитель-
ного узла тройничного нерва; 4 — ассоциативный нейрон — рас-
сеянная клетка ретикулярной формации; 5 — клетка двигатель-
ного ядра лицевого нерва; б — круговая мышца глаза
Эти связи объясняют вегетативные нарушения
при вестибулярных перегрузках. Рассеянные ней-
роны ретикулярной формации выступают в качестве
53
вставочных нейронов охранительных рефлексов
глотательного, роговичного (рис. 33), кашлевого
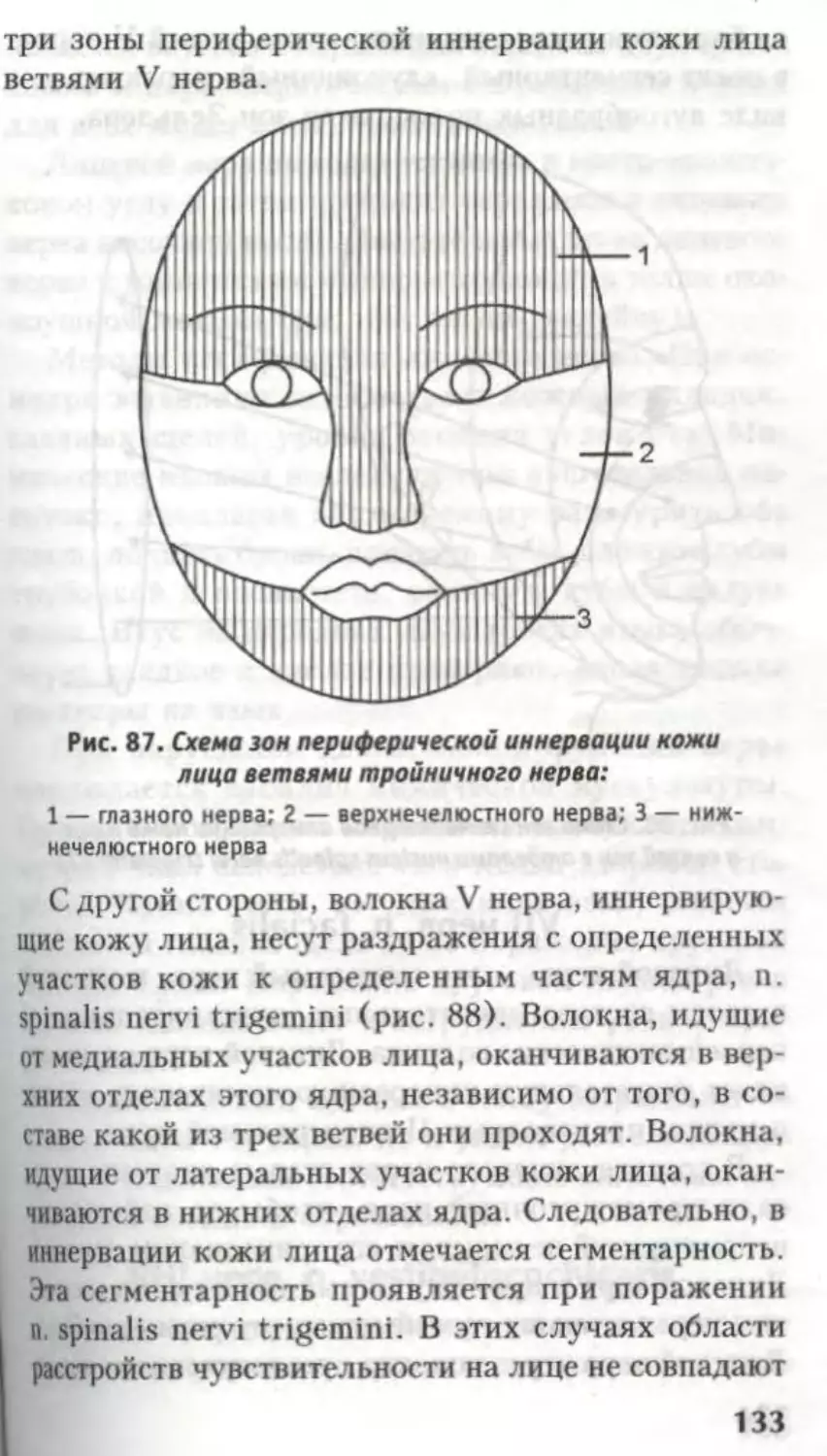
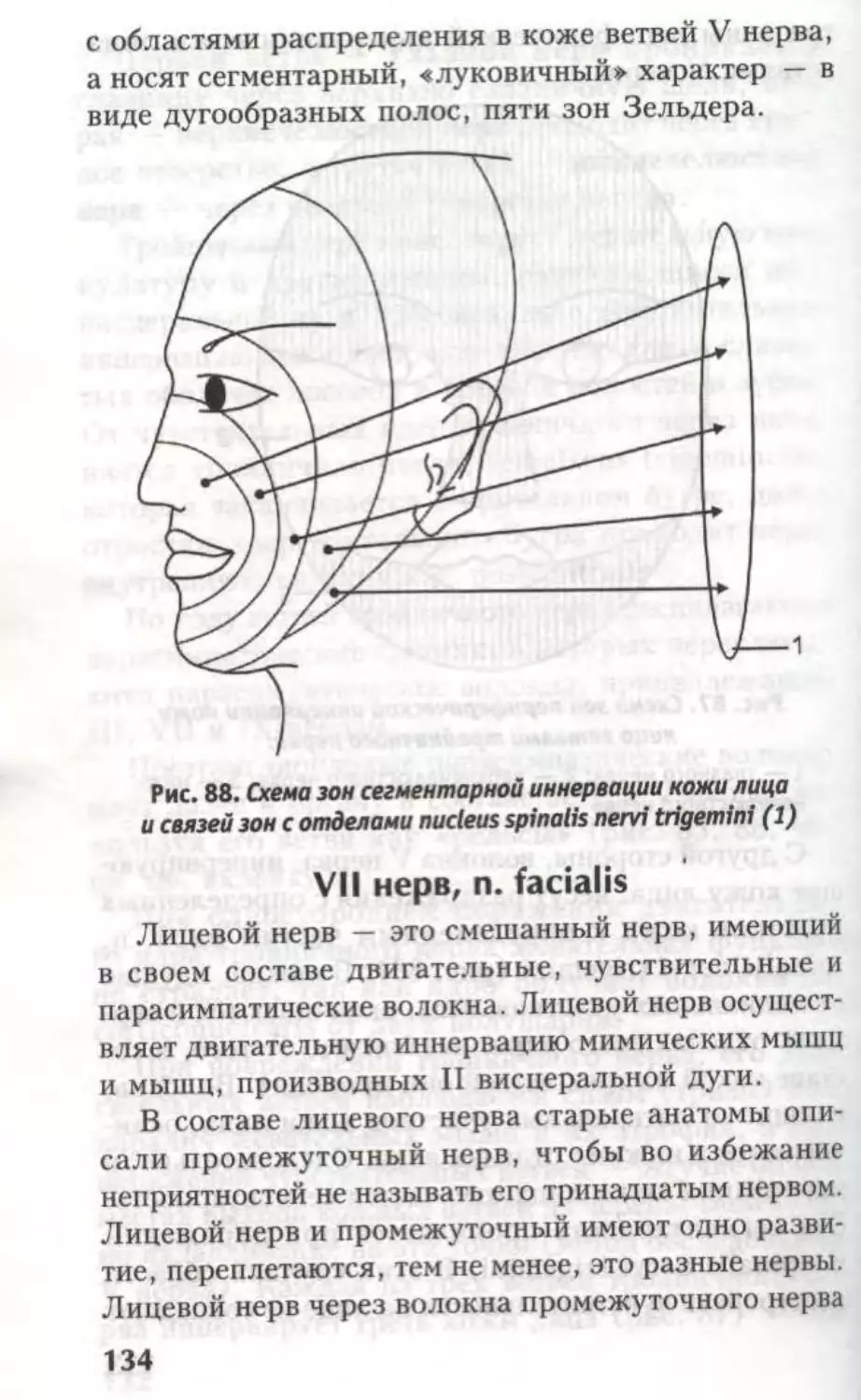
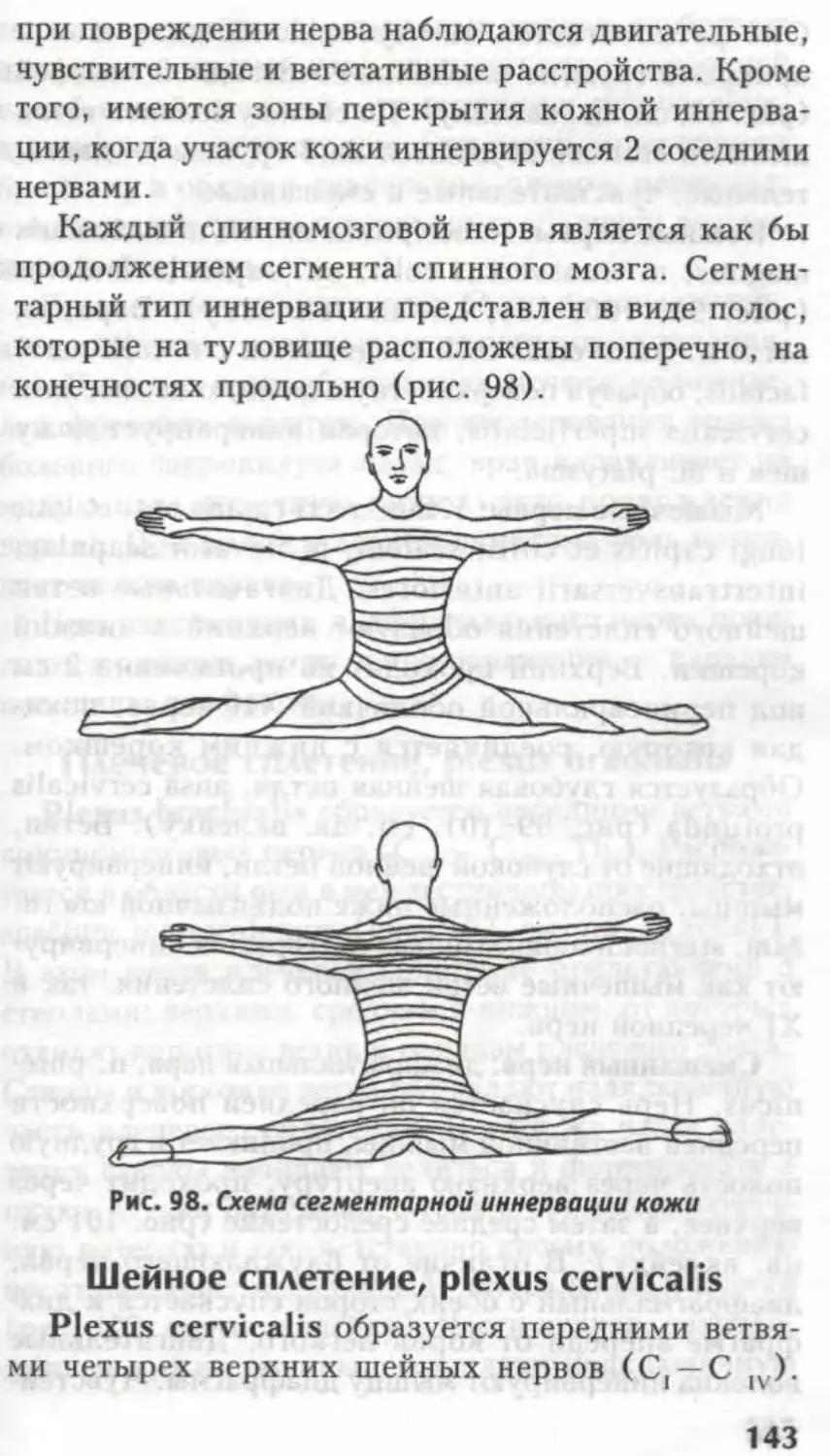
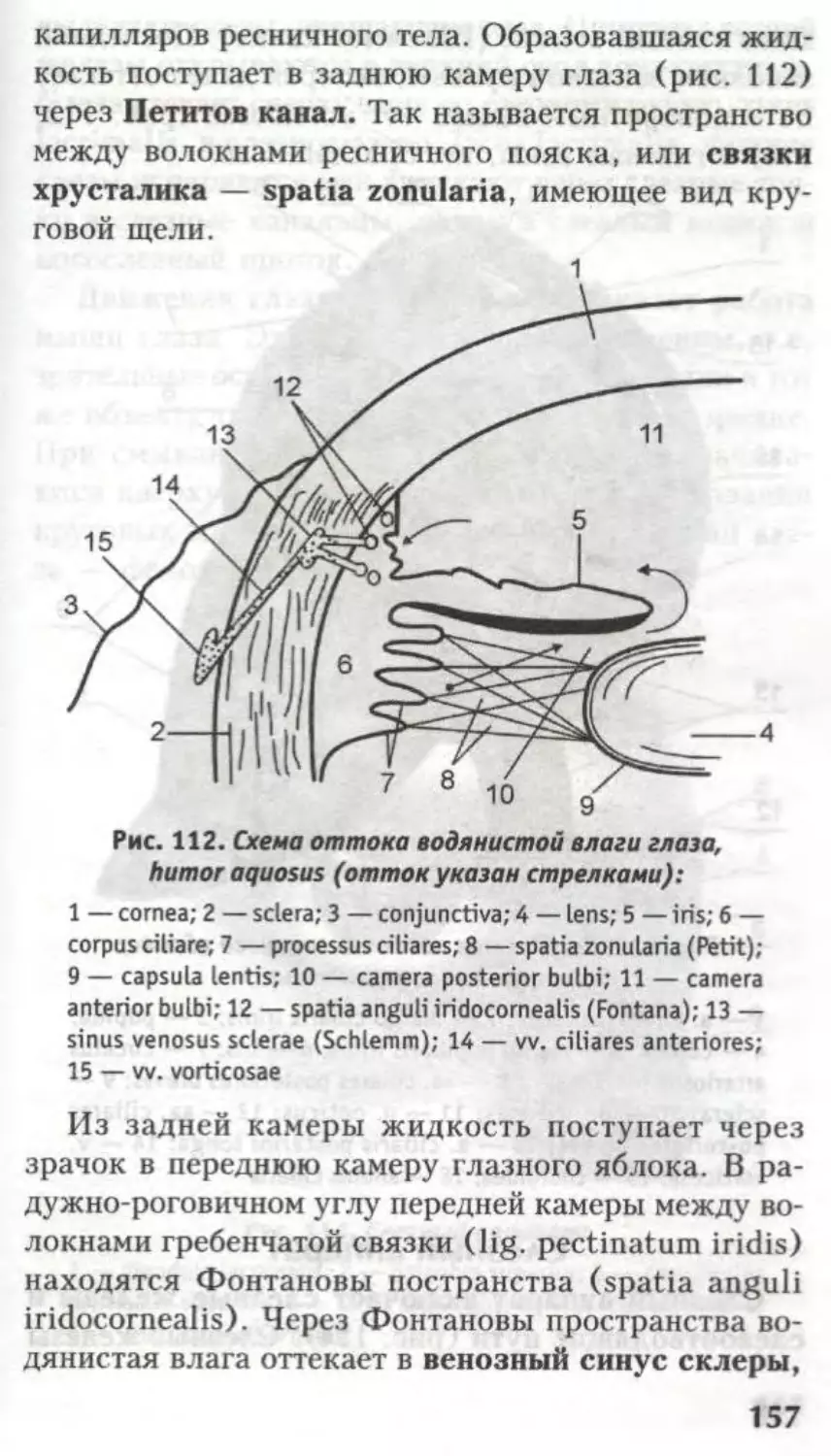
рвоты, зевоты, чихания и др.
ФуНКиОНААЬНАЯ
АНАТОМИЯ
КОНЕЧНОГО МОЗГА.
Боковые жеаудочки
а
Глава
Из предыдущих страниц известно, что вся
ЦНС — родственница эктодермы, а все ее вели-
колепие образовалось из нервной трубки. Головной
и спинной мозг не имеют анатомической границы,
это один и тот же орган. Эта глава посвящена
производному 1-го мозгового пузыря — конечному
мозгу.
Telencephalon представлен двумя полушария-
ми — hemispheria cerebri.
В состав каждого полушария входят:
обонятельный мозг — rhinencephalon — самая
древняя часть, расположенная вентрально;
базальные ядра — nuclei basales — старая
часть, скрытая в глубине;
плащ — pallium — самая молодая часть, рас-
положенная поверхностно и покрывающая все
остальные части;
боковые желудочки, ventriculus lateralis —
их части есть во всех долях мозга, кроме ос-
тровка.
Филогенез головного мозга
Почти у всех черепных на ранних стадиях эмб-
риогенеза головной конец мозговой трубки диффе-
ренцируется на три мозговых пузыря: передний,
средний и ромбовидный. В дальнейшем передний
55
и ромбовидный снова разделяются, каждый на две
части. В результате образуется 5 отделов: конечный
мозг, промежуточный, средний, задний и продол-
говатый. Распределение серого и белого вещества
в головном мозге у низших черепных такое же, как
и в спинном мозге: желудочки выстланы серым ве-
ществом, белое вещество лежит снаружи, являясь
продолжением слоев спинного мозга. Кроме того,
в мозжечке, а у высших позвоночных и в конечном
мозге, снаружи белого вещества появляется серое
вещество — кора.
У круглоротых головной мозг весьма примити-
вен. Продолговатый мозг составляет самый обшир-
ный отдел. Конечный мозг развит слабо, состоит
из двух половин, каждая имеет передний больший
отдел — обонятельную долю и меньший задний -
собственно полушария.
У хрящевых рыб мозжечок по величине равен
переднему мозгу. В промежуточном мозге развит
только эпифиз. Конечный мозг не разделен на по-
лушария, впереди переходит в обонятельные доли.
У костистых рыб разделение конечного мозга едва
намечено.
У амфибий продолговатый мозг укорочен. Выра-
жены зрительные бугры, гипофиз. Конечный мозг
преобладает над остальными отделами и состоит из
двух удлиненных полушарий.
У рептилий мозжечок развит сильнее, чем у ам-
фибий, особенно у крокодилов. Конечный мозг
состоит из обонятельных долей и полушарий.
В полушариях выражены полосатые тела и плащ с
настоящей корой (пирамидными клетками).
У птиц общая масса мозга увеличивается. Боль-
ших размеров достигает мозжечок. Кора полушарий
развита слабо. Обонятельные доли незначительны.
56
Головной мозг млекопитающих характеризуется
высоким развитием плаща и дифференцировкой коры
полушарий. Хорошо развит мозжечок, появляется
мост, четверохолмие. Полушария разрастаются во
всех направлениях, образуются отдельные доли.
Главное в филогенезе головного мозга позвоноч-
ных — развитие конечного мозга и его плаща. У
амфибий появляется закладка плаща, который у
рыб только намечается; у рептилий уже образуется
настоящая кора, у млекопитающих конечный мозг
значительно превосходит все остальные отделы
головного мозга.
Изучение абсолютного и относительного веса
головного мозга представителей различных классов
позвоночных показывает:
вес мозга по сравнению с размерами тела на-
растает в очень незначительной степени;
только у немногих (самых крупных) животных
абсолютный вес мозга больше, чем у челове-
ка;
по весу мозга человек значительно превосхо-
дит наиболее близко к нему стоящих челове-
кообразных обезьян;
относительный вес мозга велик у приматов и
затем у животных с незначительными разме-
рами тела.
Онтогенез конечного мозга
Конечный мозг или первый мозговой пузырь
(telencephalon), первоначально единый, быстро
начинает преобразовываться, поскольку его боковые
отделы растут быстрее, чем средняя часть. Поэтому
между растущими полушариями образуется глубо-
кая щель. Базальные ядра возникают путем впячи-
вания основания полушарий в боковые желудочки
57
мозга. Мозолистое тело развивается из покровной
пластинки. Кора полушарий возникает в результа-
те миграции клеток плащевого слоя через краевую
вуаль на поверхность.
В первой половине антенатального периода раз-
вития кора не имеет борозд и извилин — гладкий
мозг. Первичные борозды на поверхности мозга
появляются в начале второй половины внутриутроб-
ного периода развития, а вторичные и третичные
борозды и извилины возникают в последние его
месяцы.
Аномалии развития головного мозга
Аномалии развития головного мозга чаще всего
бывают связаны с аномалиями развития черепа и
проявляются параличами, судорогами, олигофре-
нией.
1. Экзэнцефалия — открытые полушария, от-
сутствуют покровные кости черепа.
2. Краниосхизис — расщепление черепа, сопро-
вождаемое недоразвитием или отсутствием
мозга.
3. Анэнцефалия — отсутствие головного мозга
и отсутствие костей свода черепа; сочетание
анэнцефалии с отсутствием спинного мозга
(амиелией) наблюдается у нежизнеспособных
плодов.
4. Энцефалоцеле — черепно-мозговая грыжа,
выпадение наружу части мозга при дефектах
в черепе.
5. При маленьком черепе (микроцефалии) раз-
вивается малый мозг (микроэнцефалия).
6. При макроцефалии (большом черепе), как
правило, наблюдается накопление жидкости
в желудочках мозга — гидроцефалия.
58
7. Микрогирия, полигирия — мелкие, множествен-
ные, бессистемно расположенные извилины.
8. Макрогия — утолщение основных извилин,
уменьшение их количества, нарушение струк-
туры коры.
9. Порэнцефалия — полости, кисты в ткани моз-
га, очаговая аплазия в конечном мозге, обра-
зующиеся в результате нарушения смыкания
нервной трубки.
10. Прозэнцефалия — мозг имеет 1 полушарие и
1 полость, или мозг разделен сзади, но лобные
доли слиты.
И. Гипоплазия лобных долей.
12. Аплазия мозолистого тела.
13. Диффузная аплазия участков мозга.
14. Аринэнцефалия — аплазия обонятельных
структур.
15. Ателэнцефалия — отсутствие больших по-
лушарий и базальных ядер при сохранности
черепа; развитие мозга остановилось на стадии
трех мозговых пузырей.
Плат, pallium
Плащ — филогенетически молодой отдел го-
ловного мозга. Он покрывает ствол и составляет
надсегментарный отдел головного мозга. Непосред-
ственных связей с периферией не имеет, а осущест-
вляет их посредством ствола мозга или спинного
мозга.
К плащу относятся:
кора большого мозга — cortex cerebri;
белое вещество полушарий — substantia
alba.
Кора — серое вещество, это огромное экранное
ядро, распластанное по поверхности полушарий.
59
Эта поверхность покрыта бороздами и складками
(извилинами). Причем две трети коры находится
в бороздах, а одна треть — на поверхности между
бороздами в извилинах.
Различают шесть слоев (пластинок) коры
(рис. 34).
Рис. 34. Схема слоев коры:
1 — молекулярная пластинка (концевые сетеобразные разветв-
ления отростков нервных клеток, прилежат к pia mater); 2 — на-
ружная зернистая пластинка (маленькие нейроциты); 3 — на-
ружная пирамидная пластинка (малые и средние пирамидные
нейроциты); 4 — внутренняя зернистая пирамидная пластинка
(маленькие нейроциты); 5 — внутренняя пирамидная пластинка
(большие пирамидные клетки); 6 — мультиформная пластинка
(граничит с белым веществом); 7 — белое вещество
Складчатость — это универсальное приспособле-
ние природы для увеличения поверхности. В эво-
60
люции развиваются органы, постоянно работающие.
В этом отношении мозг и череп несравнимы. Мозг
бурно развивается в течение тысячелетий, увеличи-
вается и приобретает складчатость.
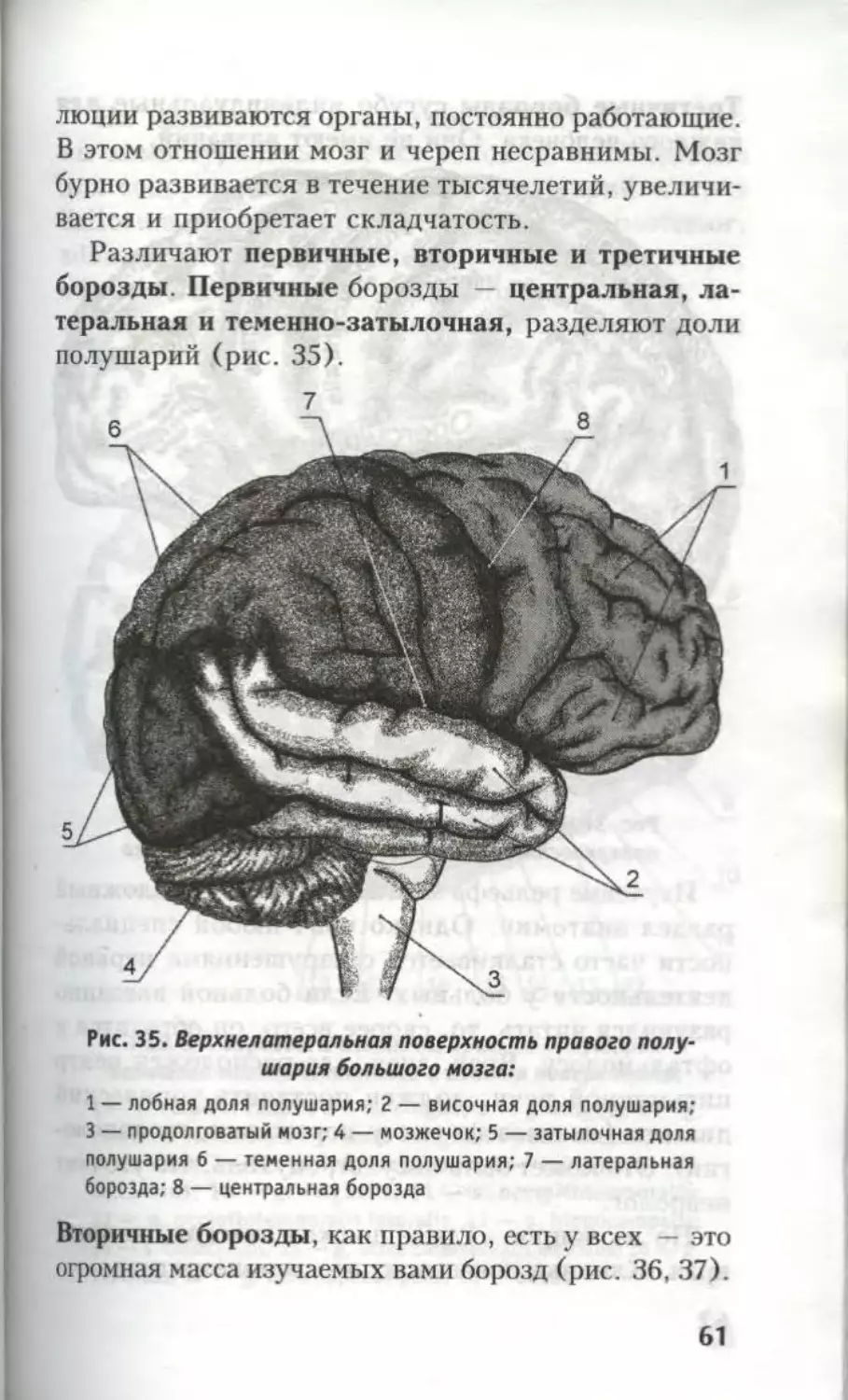
Различают первичные, вторичные и третичные
борозды. Первичные борозды — центральная, ла-
теральная и теменно-затылочная, разделяют доли
полушарий (рис. 35).
Рис. 35. Верхнелатеральная поверхность правого полу-
шария большого мозга:
1 — лобная доля полушария; 2 — височная доля полушария;
3 — продолговатый мозг; 4 — мозжечок; 5 — затылочная доля
полушария 6 — теменная доля полушария; 7 — латеральная
борозда; 8 — центральная борозда
Вторичные борозды, как правило, есть у всех - это
огромная масса изучаемых вами борозд (рис. 36, 37).
61
Третичные борозды сугубо индивидуальные для
каждого человека. Они не имеют названий.
Рис. 36. Борозды и извилины верхнелатеральной
поверхности правого полушария большого мозга
Изучение рельефа мозга — достаточно сложный
раздел анатомии. Однако врач любой специаль-
ности часто сталкивается с нарушениями нервной
деятельности у больных. Если больной внезапно
разучился читать, то, скорее всего, он обратится к
офтальмологу. Врач, зная, где расположен центр
письменной речи, должен поставить топический
диагноз (диагноз по месту поражения, по тополо-
гии). Это может быть инсульт, опухоль, что уточнит
невролог.
Но на вопрос «Где?» должен ответить каждый
врач. Клиника показывает это демонстративно.
62
На медиальной поверхности полушария вид-
ны комиссуральные пучки волокон, необходимые
для содружественной работы полушарий: corpus
callosum, comissura cerebri anterior et posterior,
adhaesio interthalamica. Sulcus cinguli уходит вверх,
образуя ветвь — ramus marginalis sulci cinguli, ко-
торая отделяет предклинье от околоцентральной
дольки.
Рис. 37. Борозды и извилины правого полушария
большого мозга; медиальная и нижняя поверхности:
1 — s. corporis callosi; 2 — s. cinguli; 3 — g. cinguli; 4 — g.
frontalis superior; 5 — ramus marginalis sulci cinguli; 6 —
precuneus; 7 — s. parietooccipitalis; 8 — cuneus; 9 — s.
calcarinus; 10— g. lingualis; 11— s. occipitotemporalis;
12 — g. occipitotemporalis lateralis; 13 — s. hippocampalis;
14 — s. collaterals; 15 — g. occipitotemporalis medialis; 16 — g.
dentatus; 17 — g. parahippocampalis; 18 — uncus
63
G. cinguli продолжается в перешеек, isthmus
gyri cinguli, на нижней поверхности полушарий
перешеек продолжается в g. parahippocampalis
и далее в uncus. Все вместе образуют сводчатую
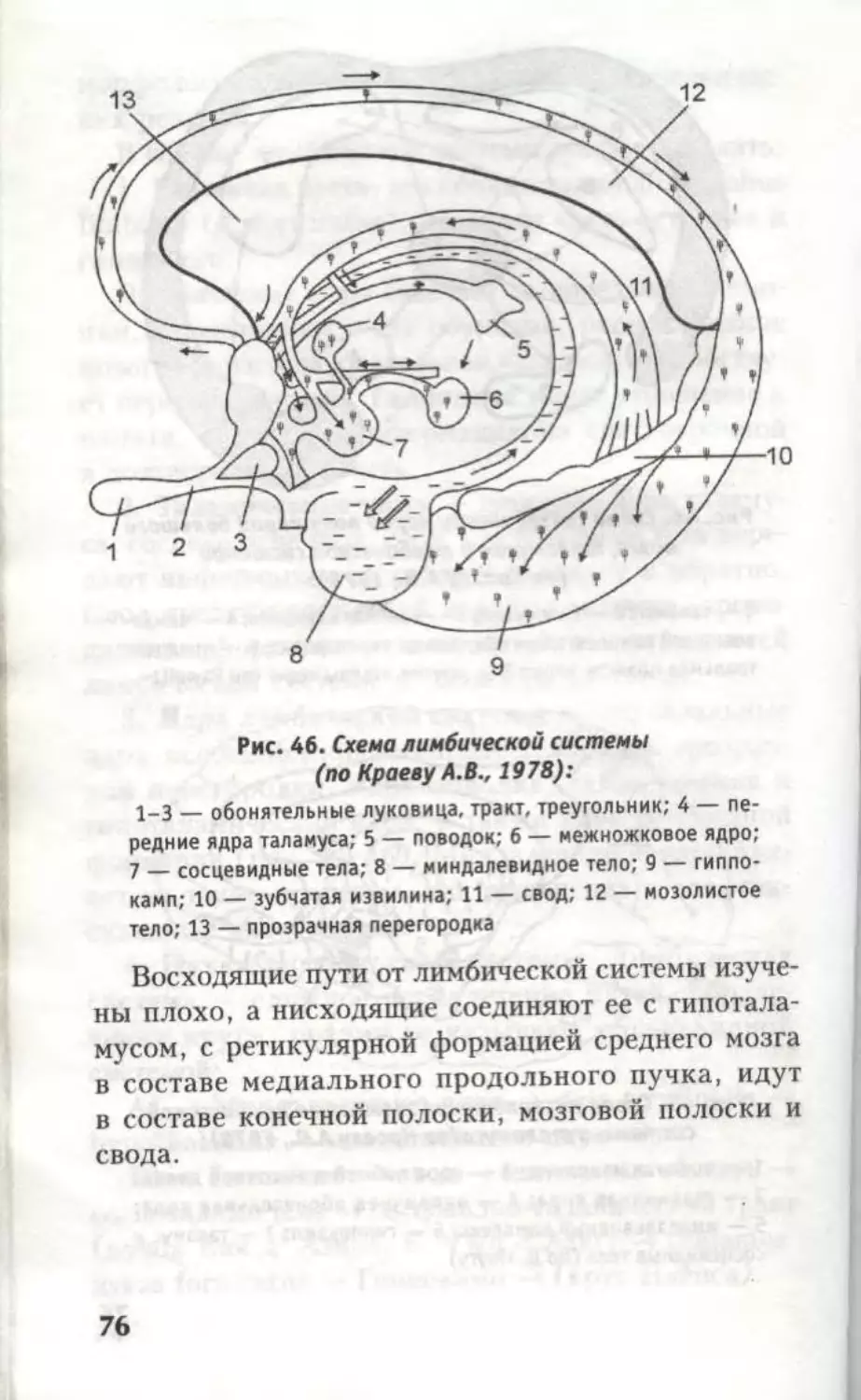
извилину, g. fornicatus или лимбическую долю,
lobus limbicus (Брока), относящуюся к обоня-
тельному мозгу и лимбической системе. Парал-
лельно g. occipitotemporalis medialis располагает-
ся g. occipitotemporalis lateralis. Граница между
g. occipitotemporalis lateralis и g. temporalis inferior
проходит не по борозде, а по нижнему краю лате-
ральной поверхности полушария (рис. 38).
Обонятельный мозг
Rhinencephalon — это древняя часть конечного
мозга. Можно сказать и по другому — конечный
мозг в филогенезе развился под влиянием обонятель-
ного мозга, подобно тому, как промежуточный мозг
развивался, испытывая влияние органа зрения.
Обоняние — дистантный орган чувств. Потеря
обоняния не делает человека инвалидом (если, ко-
нечно, он не работает парфюмером), но без обоня-
ния информация от внешнего мира обеднена эмоци-
ями. Обоняние — главный орган чувств у водных
и наземных животных.
При выходе животного мира на сушу преиму-
щество в развитии получили те виды, поведение
которых сформировалось на основе органа обо-
няния. На его структурах у них сформировались
важнейшие поведенческие рефлексы: половой,
пищевой, ориентировочный и оборонительный.
У некоторых животных они и сейчас в основном
определяют поведение.
Когда у древних людей стало появляться слож-
ное социальное поведение, оно, так же, как и при-
64
1
Рис.38. Нижняя поверхность большого мозга:
1 — s. olfactorius; 2 — g. rectus; 3 — gg. orbitales; 4 — s.
temporalis inferior; 5 — s. occipitotemporalis; 6 — s. rhinalis;
7 — s. hippocampalis; 8 — s.collaterals; 9 — g. temporalis
medius; 10— uncus; 11 — g. parahippocampalis; 12 — g.
occipitotemporalis medialis; 13 — g. occipitotemporalis lateralis,
она же на латеральной поверхности полушария — g. temporalis
interior; 14 — g. linqualis
обретаемые человеком высшие функции нервной
системы, стало базироваться на уже имеющихся
структурах обонятельного мозга. Именно поэтому
у современного человека тесно связаны обоняние и
высшая нервная деятельность. Становление их шло
параллельно. Конечно, эти рефлексы у человека во
3 Аютомма нервной системы
65
многом подавлены воспитанием, культурой, тради-
циями, обычаями, мировоззрением и так далее. Тем
не менее, они очень важны и для поведенческих
реакций современного человека — Homo sapiens.
Итак, обонятельный мозг мы изучаем в пределах
конечного мозга.
Топографически в обонятельном мозге различают
два отдела: периферический и центральный.
Периферический отдел или обонятельная доля
включает в себя:
обонятельную луковицу, bulbus olfactorius;
обонятельный путь, tractus olfactorius;
обонятельный треугольник, trigonum olfacto-
rium;
переднее продырявленное вещество, substantia
perforate anterior.
Центральный отдел включает в себя:
гиппокамп (древняя обонятельная кора),
hippocampus;
кору вокруг обонятельной луковицы (старая
обонятельная кора);
сводчатую и зубчатую извилины, g. fornicatus,
g. dentatus (новая обонятельная кора);
свод и прозрачную перегородку (проводящие
пути обонятельного мозга).
Таким образом, обонятельный мозг развивался
и, развиваясь, получал новые участки коры.
Подкорковые центры — базальные
ядра (nuclei basales)
Различают следующие базальные ядра
(рис. 39-42):
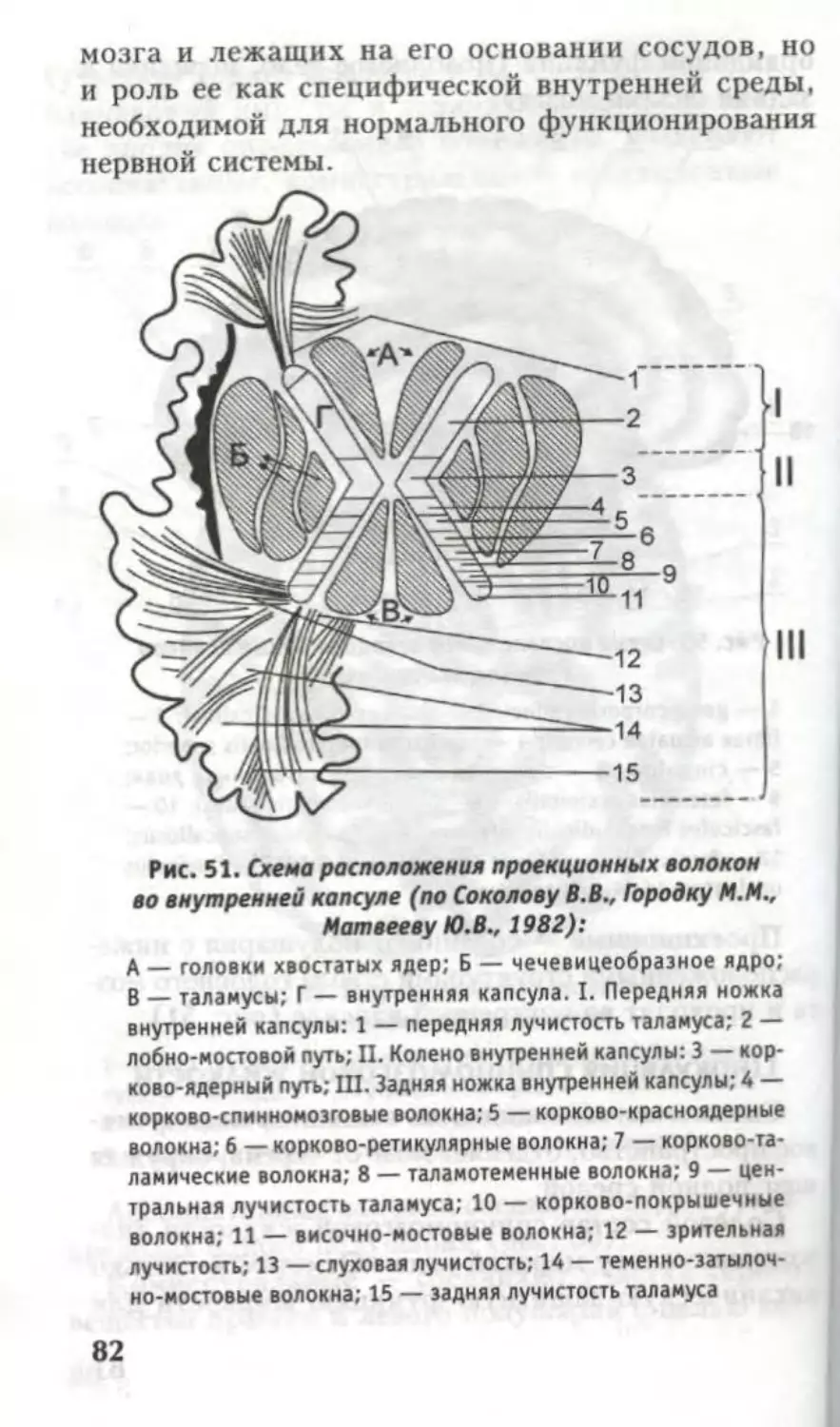
1. Полосатое тело, corpus striatum:
хвостатое ядро, n. caudatus (имеет caput,
corpus, cauda);
66
чечевицеобразное ядро, n. lentiformis (состо-
ит из скорлупы, putamen, и медиального и
латерального бледных шаров, globus pallidus
medialis et lateralis).
Неврологи давно полагали, что хвостатое ядро
и скорлупа — одно ядро. Действительно, по совре-
менному представлению, так оно и есть. Изогнутое
ядро дважды попадает в срез (рис. 39). N. caudatus
и п. putamen (филогенетически более новые обра-
зования), объединяются под названием стриатум,
a globus pallidus medialis et lateralis (филогене-
тически более старое образование) — называются
паллидум.
хвостатое ядро (1) впереди сливается со скорлупой (2)
2. Claustrum — ограда.
3. Corpus amygdaloideum — миндалевидное тело,
представляет комплекс вторичных ядер.
67
О4'
00
1
19
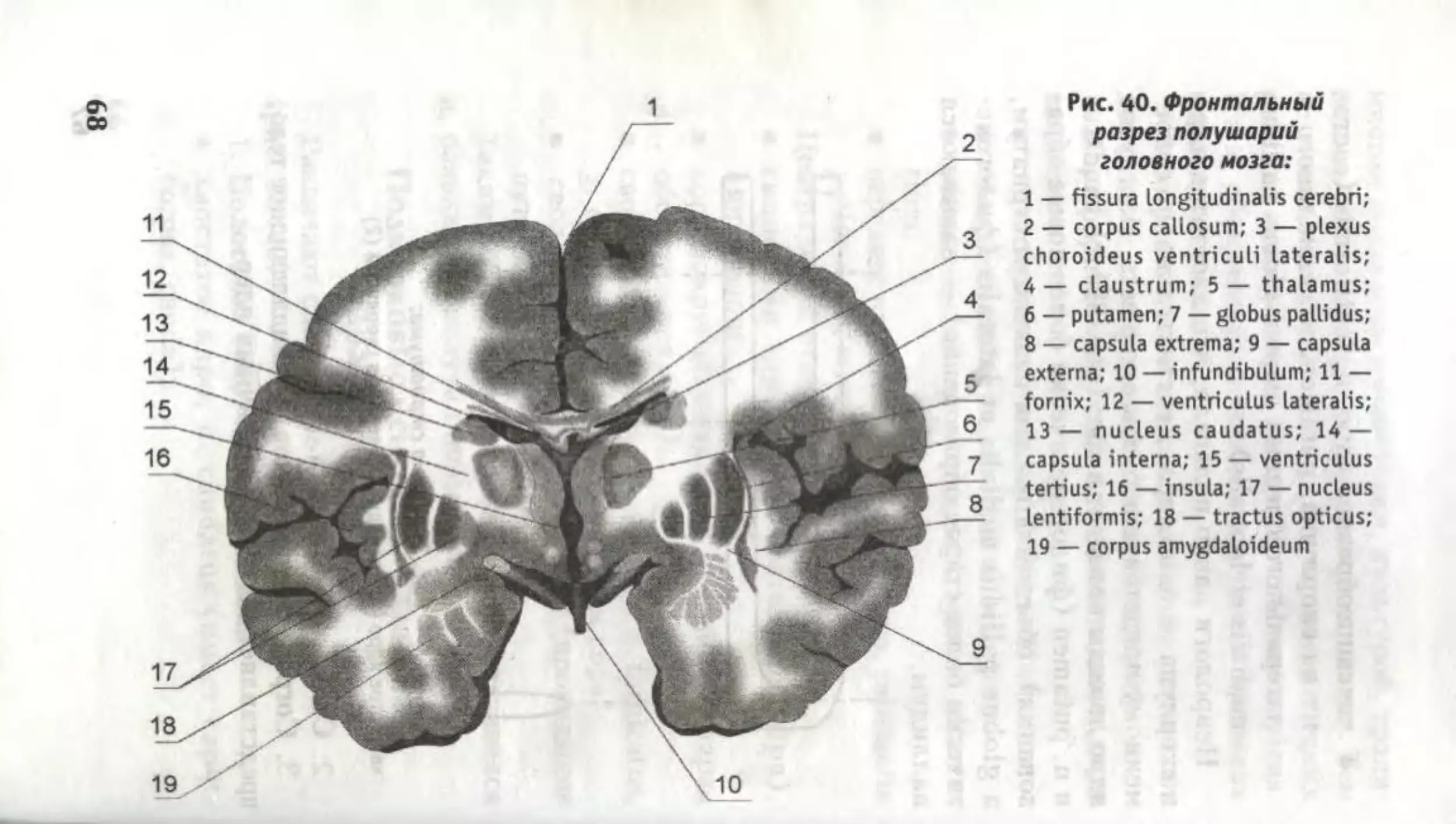
Рис. 40. Фронтальный
разрез полушарий
головного мозга:
1 — fissura Longitudinalis cerebri;
2 — corpus callosum; 3 — plexus
choroideus ventriculi lateralis;
4 — claustrum; 5 — thalamus;
6 — putamen; 7 — globus pallidus;
8 — capsula extrema; 9 — capsula
externa; 10 — infundibulum; 11 —
fornix; 12 — ventriculus lateralis;
13— nucleus caudatus; 14 —
capsula interna; 15 — ventriculus
tertius; 16 — insula; 17 — nucleus
lentiformis; 18 — tractus opticus;
19 — corpus amygdaloideum
В настоящее время происходит переход на но-
вый термин: <Стприопаллидарная система*:
По филогенетической давности и функциональ-
ному значению стриопаллидарную систему подраз-
деляют:
Archistriatum — с. amygdaloideum;
Paleostriatum — globus pallidus;
Neostriatum — n. caudatus et putamen.
Следовательно, как старый отдел — паллидарная
система тормозится и регулируется молодой — стри-
арной системой.
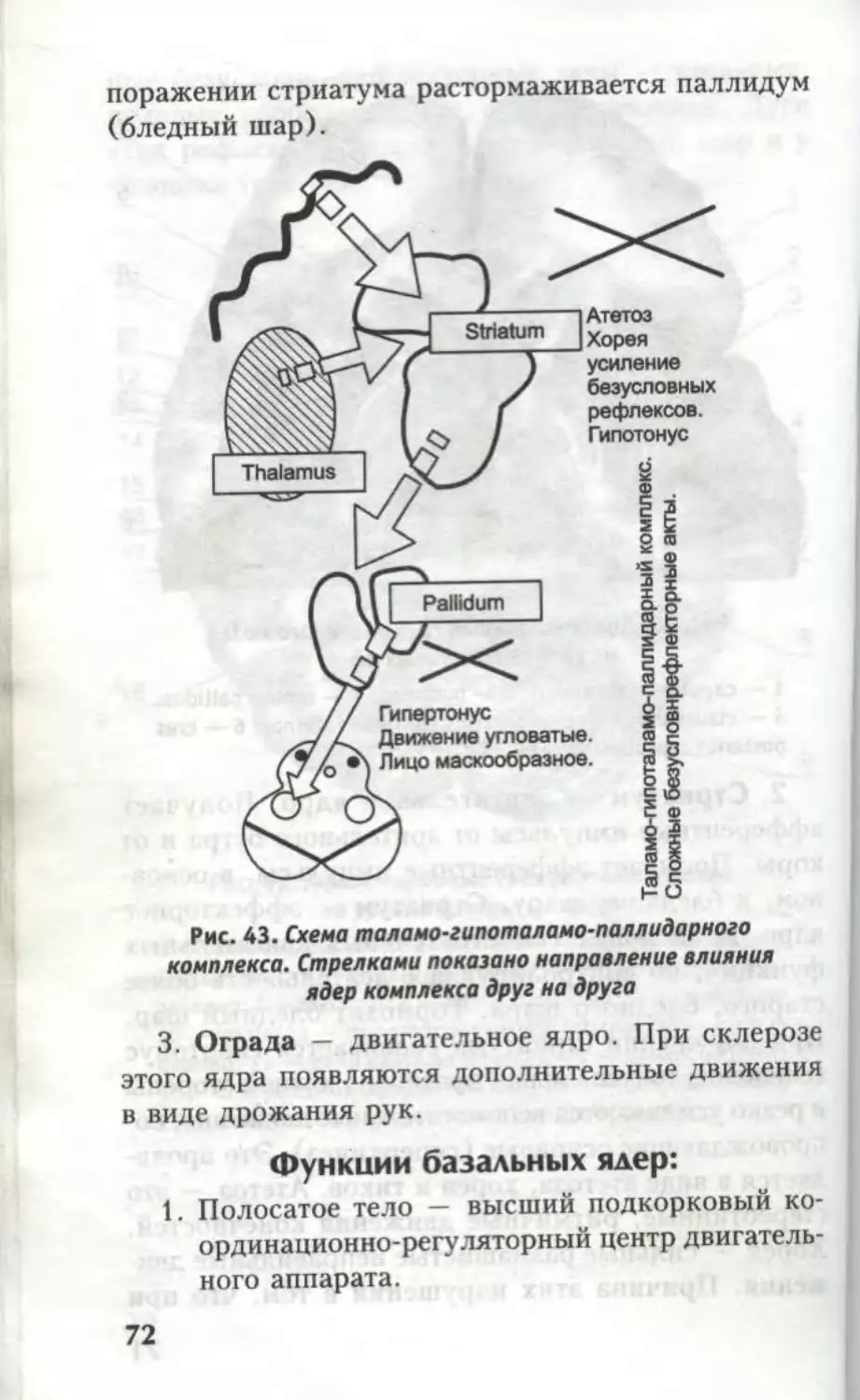
1. Бледный шар — двигательное ядро. Раздра-
жение этого ядра вызывает сокращение мышц шеи,
конечностей, туловища (преимущественно на про-
тивоположной стороне). Бледный шар эффекторно
связан с центрами среднего и заднего мозга, тормо-
зит красное ядро. При поражении бледного шара
красное ядро освобождается от его влияния. Разви-
вается гипертонус (повышение тонуса) мышц проти-
воположной стороны и гипокинез; лицо маскообраз-
ное, движения угловатые, однообразные (симптомы
паркинсонизма). Таламо-гипоталамо-паллидарный
комплекс у высших животных обеспечивает слож-
69
ные безусловно-рефлекторные акты — пищевые,
половые, оборонительные, ориентировочные. Дуги
этих рефлексов проходят через бледный шар и у
человека (рис. 43).
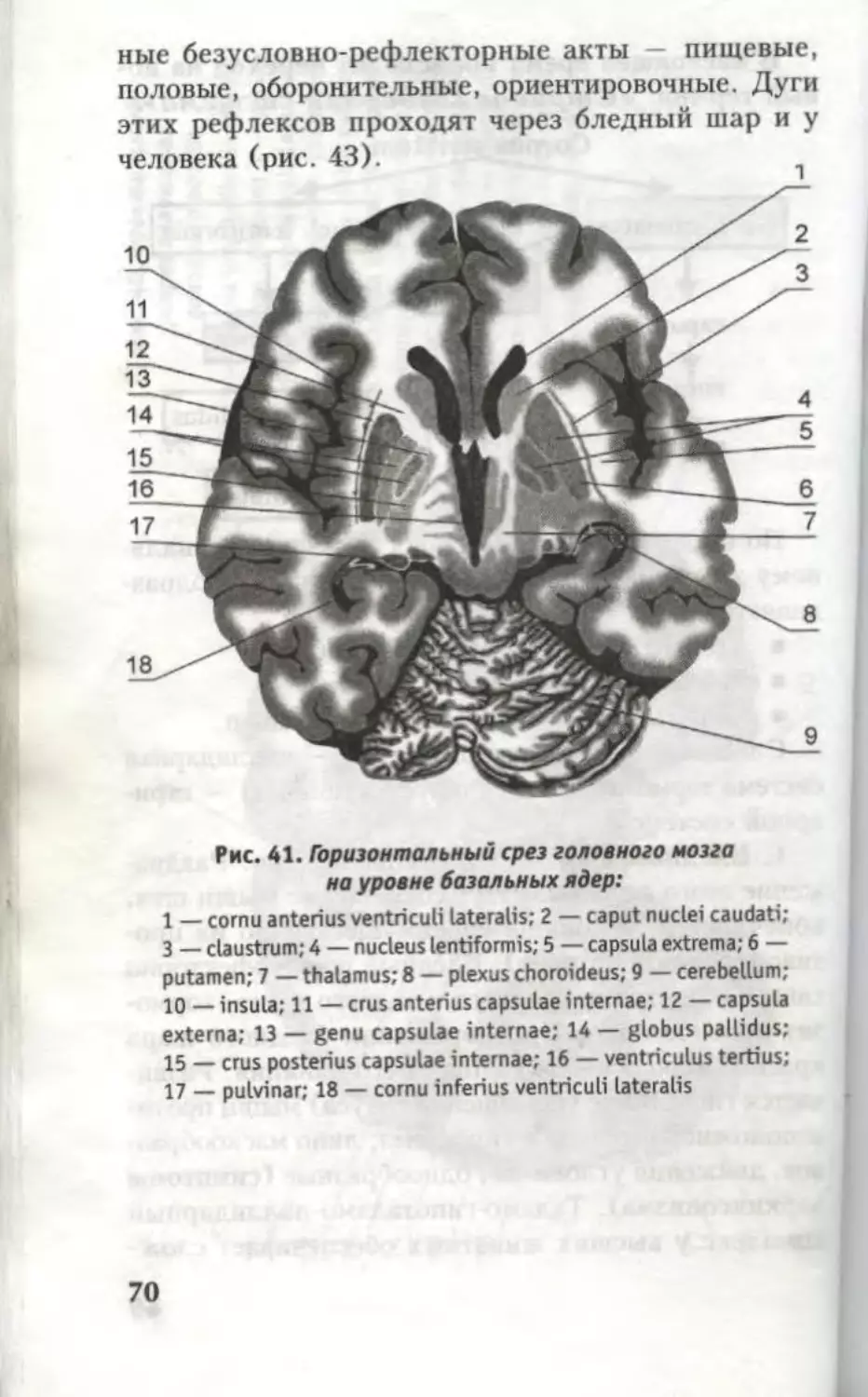
Рис. 41. Горизонтальный срез головного мозга
на уровне базальных ядер:
1 — cornu anterius ventriculi lateralis; 2 — caput nuclei caudati;
3 — claustrum; 4 — nucleus lentiformis; 5 — capsula extrema; 6 —
putamen; 7 — thalamus; 8 — plexus choroideus; 9 — cerebellum;
10 — insula; 11 — crus anterius capsulae internae; 12 — capsula
externa; 13 — genu capsulae internae; 14 — globus pallidus;
15 — crus posterius capsulae internae; 16 — ventriculus tertius;
17 — pulvinar; 18 — cornu inferius ventriculi lateralis
70
Рис. 42. Горизонтальный срез головного мозга
на уровне базальных ядер:
1 — caput nuclei caudati; 2 — putamen; 3 — globus pallidus;
4 — claustrum; 5 — crus anterius capsulae internae; 6 — crus
posterius capsulae internae
2. Стриатум — двигательное ядро. Получает
афферентные импульсы от зрительного бугра и от
коры. Посылает эфферентные импульсы, в основ-
ном, к бледному шару. Стриатум — эффекторное
ядро, не имеющее самостоятельных двигательных
функций, но контролирующее деятельность более
старого, бледного шара. Тормозит бледный шар.
При поражении стриатума развивается гипотонус
(снижение тонуса) мышц противоположной стороны
и резко усиливаются вспомогательные движения, со-
провождающие основные (гиперкинез). Это прояв-
ляется в виде атетоза, хореи и тиков. Атетоз — это
стереотипные, ритмичные движения конечностей.
Хорея - сильные размашистые неправильные дви-
жения. Причина этих нарушений в том, что при
71
поражении стриатума растормаживается паллидум
(бледный шар).
Рис. 43. Схема таламо-гипоталамо-паллидарного
комплекса. Стрелками показано направление влияния
ядер комплекса друг на друга
3. Ограда — двигательное ядро. При склерозе
этого ядра появляются дополнительные движения
в виде дрожания рук.
Функции базальных ядер:
1. Полосатое тело — высший подкорковый ко-
ординационно-регуляторный центр двигатель-
ного аппарата.
72
2. Базальные ядра — высшие вегетативные ко-
ординационные центры, регулирующие обмен
веществ, теплообразование, теплоотдачу и
сосудистые реакции.
3. Базальные ядра осуществляют сложнейшие
безусловные рефлексы (оборонительный, ори-
ентировочный, пищевой, половой).
4. Базальные ядра — это центры, интегрирую-
щие безусловно-рефлекторные двигательные
и вегетативные реакции в единый целостный
акт поведения.
Действие полосатого тела на органы, иннервируе-
мые вегетативной нервной системой, осуществляется
посредством связей с гипоталамусом.
Лимбическая система
Лимбическая система — это комплекс струк-
тур среднего, промежуточного и конечного мозга,
расположенных преимущественно на медиальной
поверхности полушария и составляющих субстрат
для проявления наиболее общих реакций организма
(сна, бодрствования, эмоций, памяти, мотиваций
и так далее). Термин «лимбическая система* ввел
Мак-Лейн (Me Lean) в 1952 г., подчеркивая связь с
большой лимбической долей Брока — lobus limbicus
(g. fornicatus).
Лимбическая система, развившаяся с древних
времен, влияет на подсознательное, инстинктивное
поведение человека, похожее на поведение живот-
ных, связанное с выживанием и размножением. Но
у людей многие из этих врожденных, примитивных
форм поведения регулируются корой полушарий
большого мозга. Лимбическая система базируется на
обонятельных структурах мозга, так как на ранних
этапах эволюции именно обонятельный мозг был
73
морфологической основой важнейших поведенчес-
ких реакций.
В составе лимбической системы можно выделить:
1. Корковая часть, это обонятельная доля, lobus
limbicus (g. fornicatus), передняя часть островка и
гиппокамп.
Лимбическая кора отвечает за поведение и эмо-
ции, а гиппокамп — за обучение, распознавание
нового. Парагиппокампальная извилина способству-
ет перемене эмоций. Гиппокамп имеет отношение к
памяти, переводит информацию из краткосрочной
в долгосрочную память.
2. Таламическая часть — передние ядра таламу-
са, сосцевидные тела, свод. Сосцевидные тела пере-
дают информацию от свода к таламусу и обратно.
Свод представляет собой нервные волокна, прово-
дящие информацию от гиппокампа и других частей
лимбической системы к сосцевидным телам.
3. Ядра лимбической системы — это базальные
ядра, особенно миндалевидное тело, ядра прозрач-
ной перегородки, ядра поводка, таламические и
гипоталамические ядра, а также ядра сетевидной
формации (рис. 44-46). Миндалевидное ядро вли-
яет на такие процессы, как отношение к еде, сек-
суальный интерес, гнев.
4. Пучки лимбической системы. Лимбическая
система — сложное переплетение путей, образу-
ющих круги, потому ее называют кольцевидной
системой:
А) -> Миндалевидное ядро —> stria terminalis —>
hypothalamus —» миндалевидное ядро ->
Б) —> Гиппокамп —>свод —> септальная область -»
сосцевидные тела -» сосцевидно-таламический тракт
(пучок Вик д’ Азира, F. Vicq d’Azyr) —> thalamus
gyrus fornicatus —» Гиппокамп —> (круг Пэйпса).
74
Рис. 44. Схема связей между корой полушарий большого
мозга, таламусом и лимбической системой
(по КраевуА.В., 1978):
1 — таламус; 2 — гиппокамп; 3 — поясная извилина; 4 — минда-
левидный комплекс; 5 — прозрачная перегородка; 6 — предцен-
тральная область коры; 7 — другие отделы коры (по Pawell)
Рис. 45. Схема расположения элементов лимбической
системы и таламуса(по Краеву А.В., 1978):
1 — поясная извилина; 2 — кора лобной и височной долей;
3— глазничная кора; 4 — первичная обонятельная кора;
5 — миндалевидный комплекс; 6 — гиппокамп; 7 — таламус и
сосцевидные тела (по Д. Плугу)
75
Рис. 46. Схема лимбической системы
(по КраевуА.В., 1978):
1-3 — обонятельные луковица, тракт, треугольник; 4 — пе-
редние ядра таламуса; 5 — поводок; 6 — межножковое ядро;
7 — сосцевидные тела; 8 — миндалевидное тело; 9 — гиппо-
камп; 10 — зубчатая извилина; 11 — свод; 12 — мозолистое
тело; 13 — прозрачная перегородка
Восходящие пути от лимбической системы изуче-
ны плохо, а нисходящие соединяют ее с гипотала-
мусом, с ретикулярной формацией среднего мозга
в составе медиального продольного пучка, идут
в составе конечной полоски, мозговой полоски и
свода.
76
Рис. 47. Схема строения отделов боковых желудочное:
1 — передний рог; 2 — задний рог; 3 — центральная часть;
4 — нижний рог, С.с. — corpus callosum, Cap.n.c. — caput nuclei
caudati, S.p. — septum pellucidum, Ca. av. — calcar avis, Thai —
thalamus, Pl.ch.v.I — plexus chorioideus ventriculi I, Cau.n.c. —
cauda nuclei caudati, S.hip. — hippocampus, E.c. — eminentia
collaterals. Cor. — corpus nuclei caudati
Функции лимбической системы
Л. С. — центр интеграции вегетативных и сома-
тических компонентов реакций высокого ранга:
мотивационных и эмоциональных состояний,
сна, ориентировочно-исследовательской актив-
ности, и в конечном счете — поведения.
Л. С. — центральный орган памяти.
Л. С. обеспечивает сохранение человеком ин-
дивидуальных и видовых особенностей, чувс-
тва «Я», личности.
77
Средний отдел конечного мозга
К среднему отделу конечного мозга относятся:
Мозолистое тело — corpus callosum (имеет:
genu, truncus, и splenium corporis callosi).
Свод — fornix (columnae, corpus et crura for-
nicis).
Прозрачная перегородка — septum pellu-
cidum.
Боковые желудочки
Полостью конечного мозга являются боковые
желудочки. Они находятся в глубине полушарий
и заполнены спинномозговой жидкостью. Боко-
вой желудочек имеет несколько отделов: пере-
дний, задний, нижний рога и центральную часть
(рис. 47-49).
Передний рог находится в лобной доле, централь-
ная часть — в теменной доле, задний рог — в за-
тылочной доле и нижний рог — в височной доле
полушария большого мозга.
1. Cornu anterius ограничен с медиальной сто-
роны — septum pellucidum, сверху — corpus
callosum, и с латеральной стороны - caput п.
caudati (рис. 47-1).
2. Задний рог (рис. 47-2) ограничен мозолис-
тым телом и tapetum — покровом (волокна-
ми, отщепившимися от мозолистого тела).
В этом роге имеется птичья шпора — calcar
avis — вдавление шпорной борозды.
3. Центральная часть (рис. 47-3) — ее стенками
являются мозолистое тело, тело свода, зри-
тельный бугор; в ее сосудистую щель прони-
кает сосудистое сплетение, изнутри покрытое
эпителиальной пластинкой.
78
4. Нижний рог (рис. 47-4) окружен tapetum и
хвостом хвостатого ядра. В этом роге имеются
гиппокамп и eminentia collateral is (вдавление
одноименных борозд).
1
Рис. 48. Взаиморасположение отделов боковых желу-
дочков в долях полушарий большого мозга:
1 — лобная доля; 2, 10 — нижний рог бокового желудочка;
3, 5 — задний рог бокового желудочка; 4 — затылочная доля;
6 — четвертый желудочек; 7 — водопровод мозга; 8 — тре-
тий желудочек; 9 — центральная часть бокового желудочка;
11 — передний рог бокового желудочка
Белое вещество конечного мозга
Проводящие пути — это пучки нервных волокон,
соединяющие функционально однородные отделы
79
(участки) серого вещества в ЦНС. Они проводят
одинаковый импульс и занимают в белом вещес-
тве вполне определенное положение. Выделяют
ассоциативные, комиссуральные и проекционные
волокна.
6 8 7
Рис. 49. Разрез головного мозга
в горизонтальной плоскости:
1 — ядра зрительного бугра; 2 — третий желудочек; 3 — скор-
лупа; 4 — ограда; 5 — внутренняя капсула; 6 — кора островка;
7 — передний рог бокового желудочка; 8 — хвостатое ядро;
9 — бледный шар; 10 — нижний рог бокового желудочка
Ассоциативные волокна соединяют структуры в
пределах одного полушария (рис. 50).
Комиссуральные — соединяют участки серого
вещества правого и левого полушарий с целью ко-
80
ординации функций (мозолистое тело, передняя и
задняя спайки мозга).
Рис. 50. Схема расположения ассоциативных волокон
в правом полушарии:
1 — genu corporis callosi; 2 — truncus corporis callosi; 3 —
fibrae arcuatae cerebri; 4 — fasciculus longitudinals superior;
5 — cingulum; 6 — теменная доля; 7 — затылочная доля;
8 — fasciculus occipitalis; 9 — splenium corporis callosi; 10 —
fasciculus longitudinalis inferior; 11 — fasciculus subcallosus;
12 — fornix; 13 — височная доля; 14 — uncus; 15 — fasciculus
uncinatus; 16 — лобная доля
Проекционные — соединяют полушария с ниже-
расположенными структурами ствола головного моз-
га и проходят во внутренней капсуле (рис. 51).
Циркуляция спинномозговой жидкости
Спинномозговая жидкость заполняет подпаутин-
ное пространство, отделяет мозг от черепа, окружая
мозг водной средой.
Солевой состав спинномозговой жидкости ана-
логичен составу морской воды. Отметим не только
механическую защитную функцию жидкости для
81
мозга и лежащих на его основании сосудов, но
и роль ее как специфической внутренней среды,
необходимой для нормального функционирования
нервной системы.
Рис. 51. Схем расположения проекционных волокон
во внутренней капсуле (по Соколову В. В., Городку М.М.,
Матвееву Ю.В», 1982):
А — головки хвостатых ядер; Б — чечевицеобразное ядро;
В — таламусы; Г — внутренняя капсула. I. Передняя ножка
внутренней капсулы: 1 — передняя лучистость таламуса; 2 —
лобно-мостовой путь; II. Колено внутренней капсулы: 3 — кор-
ково-ядерный путь; Ш. Задняя ножка внутренней капсулы; 4 —
корково-спинномозговые волокна; 5 — корково-красноядерные
волокна; 6 — корково-ретикулярные волокна; 7 — корково-та-
ламические волокна; 8 — таламотеменные волокна; 9 — цен-
тральная лучистость таламуса; 10 — корково-покрышечные
волокна; 11 — височно-мостовые волокна; 12 — зрительная
лучистость; 13 — слуховая лучистость; 14 — теменно-затылоч-
но-мостовые волокна; 15 — задняя лучистость таламуса
82
Так как ее белки и глюкоза являются источ-
ником энергии для нормальной работы клеток
мозга, а лимфоциты препятствуют проникновению
инфекции.
Жидкость образуется из сосудов сосудистых спле-
тений желудочков, проходя через гематоэнцефаличес-
кий барьер, и обновляется 4-5 раз в день. Из боковых
желудочков жидкость оттекает через межжелудочко-
вое отверстие в третий желудочек, затем через водо-
провод мозга в четвертый желудочек (рис. 52).
Циркуляции жидкости способствует пульса-
ция мозговых артерий. Из четвертого желудочка
жидкость направляется через отверстия Люшка
и Можанди (Lushka и Magendii) в подпаутинное
пространство, омывая спинной и головной мозг.
Благодаря движениям позвоночника спинномоз-
говая жидкость течет позади спинного мозга по
направлению вниз, а по центральному каналу и
спереди спинного мозга — наверх. Из подпаутин-
ного пространства спинномозговая жидкость через
пахионовы грануляции, granulationes arachnoidales
(Pachioni), фильтруется в просвет синусов твердой
мозговой оболочки, в венозную кровь (рис. 53).
Цистерны — это расширения подпаутинного про-
странства. Различают следующие цистерны:
Cisterna cerebellomedullaris, cisterna magna —
задняя мозжечково-мозговая цистерна, боль-
шая цистерна;
Cisterna cerebellomedullaris lateralis — боко-
вая мозжечково-мозговая цистерна;
Cisterna fossae lateralis cerebri — цистерна
латеральной ямки большого мозга;
Cisterna chiasmatica — цистерна перекреста.
Cisterna interpeduncularis — межножковая
цистерна;
83
1 13
Рис. 52. Схема путей циркуляции спинномозговой
жидкости:
1 — пахионовы грануляции; 2 — боковой желудочек; 3 — полу-
шарие большого мозга; 4 — мозжечок; 5 — четвертый желудо-
чек; 6 — спинной мозг; 7 — субарахноидальное пространство;
8 — корешки спинномозговых нервов; 9 — сосудистое сплете-
ние; 10 — намет мозжечка; 13 — верхний сагиттальный синус
Cisterna ambiens — охватывающая цистерна
(на дне щели между затылочными долями по-
лушарий и верхней поверхностью мозжечка);
Cisterna pericallosa — околомозолистая цис-
терна (вдоль верхней поверхности и колена
мозолистого тела);
84
Cisterna pontocerebellaris — мостомозжечко-
вая цистерна;
Cisterna laminae terminal is — цистерна конеч-
ной пластинки (от переднего края перекреста
паутинная оболочка свободно перекидывается
на нижнюю поверхность прямой извилины и
на обонятельные луковицы);
Cisterna quadrigeminalis (cisterna venae magnae
cerebri) — четверохолмная цистерна (цистер-
на большой вены мозга);
Cisterna pontis — расположена соответственно
основной борозде моста.
Рис. 53. Оболочки головного мозга:
1 — кожа волосистой части головы; 2 — кость черепа; 3 —
твердая мозговая оболочка; 4 — субдуральное пространство;
5 — паутинная оболочка; 6 — субарахноидальное пространс-
тво; 7 — мягкая мозговая оболочка; 8 — венозный выпускник;
9 — верхний сагиттальный синус; 10 — пахионовы грануляции;
И — кора полушарий головного мозга
85
Синусы твердой мозговой оболочки
Основные венозные синусы представлены на
рис. 54 (см. цв. вклейку).
Синусы твердой оболочки головного мозга пред-
ставляют собой каналы (расщепления, выстланные
эндотелием), по которым оттекает венозная кровь от
головного мозга, глазного яблока, внутреннего уха,
костей черепа и мозговых оболочек. Из синусов кровь
попадает во внутреннюю яремную вену. Венозные си-
нусы участвуют в оттоке спинномозговой жидкости.
Синусы по своему строению значительно отличаются от
вен, на поперечном разрезе имеют треугольную форму.
Синусы твердой мозговой оболочки являются жесткими
трубками, которые не спадаются и не расширяются.
Гидродинамическая стабилизация — основная при-
чина появления в черепе такого необычного варианта
венозных сосудов. Благодаря этому сколько крови
поступает в синусы, столько же крови в то же время
выталкивается с противоположного конца за пределы
полости черепа в экстракраниальные вены. По данным
В.И. Кошева с соавт. (2006), существует внутричереп-
ной пиальный венозный насос — пульсирующий веноз-
ный отток из полости черепа, синхронизированный с
артериальной пульсовой волной. Без синусов твердой
мозговой оболочки этот насос был бы невозможен.
Вены головного мозга
Различают мозговые вены, вены мозжечка и вены
ствола мозга. Все вены головного мозга впадают в
синусы твердой мозговой оболочки.
Мозговые вены подразделяются на поверхностные
и глубокие. Среди поверхностных вен (рис. 55, 56 см.
цв. вклейку) выделяют верхние, нижние мозговые
вены и поверхностную среднюю мозговую вену.
86
Главная глубокая вена — большая вена мозга
(Галена) — v. magna cerebri образуется слиянием
двух внутренних мозговых вен и двух базальных
вен (рис. 57-А).
1
Рис. 57. Схема образования глубоких вен полушарий
большого мозга:
К. Вид с медиальной поверхности: 1 — sinus sagittalis superior;
2 — w. cerebri; 3 — sinus sagittalis inferior; 4 — sinus rectus; 5 —
confluens sinuum; 6 — sinus sigmoideus; 7 — v. jugularis interna;
8 — v. magna cerebri; 9 — v. interna cerebri; 10 — v. basalis.
Б. Вид сверху: 1 — w. septi pellucidi; 2 — w. thalamostriatae;
3 — w. choroideae; 4 — v. interna cerebri; 5 — v. magna cerebri
87
Вены мозжечка подразделяются на верхние
нижние и впадают в близлежащие синусы.
Вены ствола мозга: мостосреднемозговые, вены
моста, вены продолговатого мозга также отводят кровь
в близлежащие синусы или в позвоночные вены.
V. interna cerebri образуется слиянием vv. cho-
roideae, vv. thalamostriatae и vv. septi pellucidi
(рис. 57-Б). V. basalis образуется слиянием пере-
дней и глубокой средней мозговых вен, проходящих
рядом с одноименными артериями (рис. 58).
Рис. 58. Схема образования базальных вен
и их взаиморасположения с артериями Вилизиева круга.
Вид снизу: 1 — V. magna cerebri; 2 — v. basalis; 3 — v. interna
cerebri; 4 — v. anterior cerebri; 5 — v. media profunda cerebri;
6 — a. carotis interna; 7 — a. cerebri anterior; 8 — a. cerebri
media; 9 — a. communicans posterior; 10 — a. cerebri posterior;
11 — a. basilaris
88
Динамическая
локализация
ФУНКЦИЙ
В КОРЕ ПОЛУШАРИЙ
БОЛЬШОГО МОЗГА
Q
Глава
Кора больших полушарий — материальная осно-
ва психической деятельности человека. Кора - се-
рое вещество толщиной от 1,5 до 5 мм, содержит
14 млрд нервных клеток и имеет шестислойное
строение. Кора — огромный ядерный центр, ядро,
распластанное по поверхности полушарий.
Более 130 лет идет спор — есть ли в коре центры
или нет и в каком объеме они оказывают влияние
на «курируемые» функции: 1. Отвечают ли эти
центры буквально за все (центр туризма, любви к
живописи, к театру и пр.), или их влияние менее
детализировано. 2. Кора — это один сплошной эк-
ранный центр, отвечающий за все функции.
Очевидно, истина, как всегда, где-то посередине.
Основоположником детального изучения клеточ-
ного состава коры был русский ученый, киевлянин
Владимир Алексеевич Бец. В 1874 г. он опублико-
вал результат исследований с помощью собствен-
ного метода серийных срезов и окраски кармином.
Бец выявил различное строение коры в различных
ее участках и разработал карту цитоархитектони-
ки коры. В последующем были созданы и другие
карты: Бродмана с 52 цитоархитектоническими
полями, Фогта со 150 миелоархитектоническими
полями и др. Исследования в настоящее время
89
продолжаются в Институте мозга в Москве и в
других странах.
Представления о локализации функций в коре го-
ловного мозга имеют большое практическое значение
для решения задач топики поражений в полушариях
мозга. Повседневный клинический опыт показыва-
ет, что существуют определенные закономерности
зависимости расстройств функций от расположения
патологического очага. Исходя из этого, клиницист и
решает задачи топической диагностики. Однако так
дело обстоит с простыми функциями: движением и
чувствительностью. Функции более сложные, филоге-
нетически молодые, не могут быть узколокализован-
ными; в осуществлении сложных функций участвуют
весьма обширные области коры, даже вся кора.
Работы В.А. Беца были внимательно изучены
И.П. Павловым. С учетом этих данных, Иваном
Петровичем Павловым были созданы основы нового
и прогрессивного учения о локализации функций
в головном мозге. Павлов рассматривал кору по-
лушарий большого мозга как совокупность корко-
вых концов анализаторов. Павлов создал учение об
анализаторах. По Павлову, анализатор — нервный
механизм, анализирующий явления внешнего и внут-
реннего мира путем разложения сложного комплекса
раздражений на отдельные элементы. Он начинается
воспринимающим аппаратом и кончается в мозгу,
т.е. анализатор включает рецепторный аппарат,
проводник нервных импульсов и корковый центр.
Павлов доказал, что корковый конец анализа-
тора — это не строго очерченная зона. В нем есть
ядро и рассеянные элементы. Ядро — место кон-
центрации нервных клеток, где происходит высший
анализ, синтез и интеграция. На его периферии, в
рассеянных элементах, совершаются простые анализ
90
и синтез. Площади рассеянных элементов соседних
анализаторов перекрывают друг друга (рис. 59).
Рис. 59. Схема зон перекрытия рассеянных элементов
соседних анализаторов
По Павлову — работа II сигнальной системы
неразрывно связана с функциями всех анализато-
ров, поэтому невозможно представить локализацию
сложных функций II сигнальной системы в ограни-
ченных корковых полях. Павловым заложены осно-
вы учения о динамической локализации функций в
коре. Представления о динамической локализации
функций в коре предполагают возможность исполь-
зования одних и тех же корковых структур в разно-
образных сочетаниях для обслуживания различных
сложных корковых функций. Так, ассоциативные
пути объединяют анализаторы, способствуя высшей
синтетической деятельности коры мозга. Сегодня
ученые знают, что раздражение трансформируется в
возбуждение, передающееся в корковый конец ана-
лизатора. Не ясно другое — где и как возбуждение
трансформируется в ощущение? Какие структуры
отвечают за это? Так, при раздражении зрительного
поля в области шпорной борозды возникают «про-
стые» галлюцинации в виде световых или цветовых
91
пятен, искр, теней. Раздражение наружной поверх-
ности затылочной доли дает «сложные» галлюцина-
ции в виде фигур, движущихся предметов.
В двигательной зоне коры обнаружены клетки,
дающие разряд импульсов на зрительные, слуховые,
кожные раздражения, а в зрительной зоне коры вы-
явлены нейроны, отвечающие электрическими раз-
рядами на осязательные, звуковые, вестибулярные
и обонятельные раздражители. Кроме того, были
найдены нейроны, которые отвечают не только на
“свой» раздражитель, как теперь говорят, раздра-
житель своей модальности, своего качества, но и
на один-два чужих. Их назвали полисенсорными
нейронами.
Положение некоторых корковых
анализаторов
I. Анализаторы, воспринимающие раздра-
жения из внутренней среды.
1. Двигательный анализатор. До И.П. Павлова в
коре различали двигательную зону. Павлов доказал,
что двигательная зона есть корковый конец двига-
тельного анализатора. Как же анализатор может быть
двигательным, если анализаторы воспринимают чувс-
тва? Павлов объяснил это следующим образом. ЦНС
получает информацию о первоначальной позе челове-
ка. В зависимости от исходной позы требуются раз-
личные движения и мышечная сила для реализации
желаемого действия. Поэтому сигналы от проприо-
рецепторов поступают в предцентральную извилину
(рис. 60-2, см. цв. вклейку) — это афферентный
путь. Вставочным нейроном двигательного анализато-
ра являются пирамидные клетки Беца в 5-м слое коры
этой извилины. Исполнителями являются мотоней-
роны спинного мозга, отростки которых доходят до
92
исчерченной мускулатуры: по tr. corticonuclearis до
мышц головы, шеи, некоторых органов (языка, глаза,
глотки, неба, гортани); по tr. corticospinalis — до
мышц шеи, туловища, конечностей. Соматотопичес-
кая проекция на предцентральную извилину мышц
различных областей тела носит название «моторный
гомункулюс Пенфилда». Тело человека проецируется
на извилине «вверх ногами» и на противоположном
полушарии (рис. 59, см. цв. вклейку).
2. Ядро целенаправленных движений, праксии,
находится в g. supramarginalis, обеспечивает вы-
полнение сложных, целенаправленных движений
в определенной последовательности, выученной в
течение жизни. У правшей этот центр слева. При
его поражении возникает апраксия, нарушение целе-
направленных действий.
3. Ядро сочетанного поворота головы и глаз (в
другую сторону) располагается в средней лобной
извилине (рис. 63-5, см. цв. вклейку). При пораже-
нии этого центра возникает паралич взора, голова
и глаза повернуты в сторону очага поражения.
4. Ядро вестибулярного анализатора находится
в височной и теменной долях. При поражении воз-
никает атаксия, расстройство равновесия.
5. Ядро анализатора, получающего импульсы от
внутренних органов, расположено в нижних отделах
предцентральной и постцентральной извилин.
Корковые концы анализаторов
внешнего мира
1. Ядро слухового анализатора — в средней
части верхней височной извилины, извилины Гешля.
При его раздражении возникают слуховые галлюци-
нации. Поражение центра ведет к снижению слуха
на противоположной стороне.
93
2. Ядро зрительного анализатора находится
по сторонам от шпорной борозды. Причем ядро
правого полушария связано путями с латеральной
половиной сетчатки правого глаза и медиальной
половиной левого глаза. При раздражении возни-
кают простейшие зрительные галлюцинации. На
латеральной поверхности затылочной доли нахо-
дится центр зрительной памяти. При поражении
центра зрительная память утрачивается, так же
как и способность ориентироваться в незнакомой
местности.
3. Ядро обонятельного и вкусового анализатора
находится в пределах обонятельного мозга — uncus
и парагиппокампальная извилина (рис. 61, см. цв.
вклейку). Вкусовое восприятие нарушается также
при поражении нижних отделов g. postcentralis.
Центры двусторонние. Раздражение ведет к появ-
лению обонятельных и вкусовых галлюцинаций.
При поражении их снижаются обоняние и вкус
с двух сторон или возникает нарушение узнавания
запахов.
4. Ядро анализатора общей чувствительности
(температурной, болевой, тактильной) находится
в g. postcentralis; связано с противоположной по-
ловиной тела. В верхний отдел извилины поступа-
ет информация от рецепторов кожи ноги, в сред-
ний — от руки, в нижний — от головы. Размеры
проекционных зон соматосенсорной коры прямо
пропорциональны количеству рецепторов, нахо-
дящихся в кожных покровах. Этим объясняется
наличие наиболее крупных соматосенсорных зон,
соответствующих лицу и кисти (сенсорный гомунку-
люс Пенфилда — рис. 62, см. цв. вклейку). Раздра-
жение этого центра вызывает парестезии — чувство
ползания мурашек, ощущение онемения, покалыва-
94
ния. При раздражении — утрата чувствительности
на противоположной стороне тела.
5. Ядро стереогнозии — одного из видов кожной
чувствительности — находится в верхней темен-
ной дольке (у правшей — слева). Его поражение
сопровождается утратой узнавания предмета на
ощупь — астереогнозия.
Перечисленные корковые анализаторы I сигналь-
ной системы имеются и у человека, и у животных.
Они необходимы для восприятия раздражений из
внешнего мира и внутренней среды. Речь является
одной из поздних (филогенетически новых) функ-
ций больших полушарий.
Корковые концы анализаторов
II сигнальной системы
Речь является только человеческой функцией.
Цитоархитектонические поля, имеющие наибольшее
отношение к речевой функции, существуют только у
человека. Человек мыслит всегда словами на родном
языке. С помощью ускоренной киносъемки были
обнаружены колебания голосовых связок, когда че-
ловек не говорил, даже когда спал. Словесное мыш-
ление и речь относятся ко 11 сигнальной системе.
Слова, по выражению Павлова, являются «сиг-
налами сигналов», они отвлечены от действитель-
ности, обобщены и составляют личное, человеческое
мышление. Например, человек видит определенный,
объективно существующий предмет, в области шпор-
ной борозды возникает субъективный образ предме-
та — это световой сигнал, полученный от внешнего
мира, от предмета, на сетчатке глаза. В дальнейшем
достаточно назвать предмет, т.е. произнести слово
(«сигнал сигнала»), как в сознании возникает его
образ — человек получил сигнал сигнала.
95
Так как речь, сознание — филогенетически но-
вые, молодые функции мозга, корковые концы рече-
вых анализаторов наименее локализованы. Однако
в коре выделяют определенные участки, которым
присущи речевые функции. Слово надо изобразить с
помощью работы исчерченной мускулатуры — гор-
тани, губ, щек, языка, неба при разговоре; мышц
рук — при письме; мышц глазного яблока — при
чтении. Поэтому двигательные анализаторы устной
и письменной речи располагаются рядом с двига-
тельной областью (рис. 63, см. цв. вклейку).
1. Ядро двигательного анализатора артикуляции
речи — центр Брока расположен в заднем отде-
ле нижней лобной извилины. При поражении его
возникает моторная афазия, больной не способен
говорить, но понимает обращенную к нему речь.
При частичной моторной афазии больной говорит
с трудом, допускает ошибки — аграмматизмы, ко-
торые сам замечает.
2. Ядро слухового анализатора устной речи
тесно связано с корковым центром слухового анали-
затора и расположено в верхней височной извилине,
но в ее задних отделах — центр Вернике. Пора-
жение этого отдела ведет к появлению сенсорной
афазии — нарушению понимания устной речи, в
том числе и своей речи. Вместо одних слов боль-
ной произносит другие. Такие больные не осознают
своего дефекта, обижаются на окружающих, что те
их не понимают.
О функциях, которыми данная область коры
обладает в норме, часто можно судить по расстрой-
ствам или нарушениям поведения, наступающим
при повреждении области. Самая распространенная
причина повреждений — это закупорка артерии,
питающей кору, и гибель тканей, снабжаемых ею.
96
Рис. 54. Ве-
нозные пазухи
твердой моз-
говой оболочки
и их связь пос-
редством вен
выпускников
(vv, emissariae)
с поверхност-
ными венами
головы:
А. Вид сбоку:
1 — V. emissaria
parietalis, 2 — si-
nus sagittalis su-
perior, 3 — sinus
sagittalis inferior,
4 — v. tempora-
lis superficialis,
5 — sinus rectus,
6 — v. ophthal-
mica superior,
7 — sinus caver-
nosus, 8 — plex-
us pterygoideus,
9 — v. angularis,
10 — v. jugularis
interna, 11 — v.
facialis, 12 — v.
emissaria occipi-
talis, 13 — con-
fluens sinuum,
14 — sinus trans-
versus, 15 — v.
auricularis pos-
terior, 16 — si-
nus petrosus superior, 17 — sinus sigmoideus, 18 — v. emissaria
mastoidea, 19 — v. retromandibular's;
Б. Вид сверху: 1 — sinus cavernosus, 2 — plexux basilaris, 3 — sinus
petrosus inferior, 4 — sinus petrosus superior, 5 — sinus sigmoideus,
6 —sinus occipitalis, 7 — sinus transversus, 8 — confluens sinuum,
9 — sinus sagittalis superior
Рис. 55. Взаимоотношение верхних и нижних поверхнос-
тных вен на медиальной и нижней поверхностях полу-
шария с ветвями передней и задней мозговых артерий
(по Д. Б. Бекову)
Рис. 56. Взаимоотношение поверхностных вен на ла-
теральной поверхности полушария с ветвями средней
мозговой артерии, выходящими из боковой щели мозга.
Калибр крупных вен увеличивается по мере уменьшения
калибра крупных соседних артерий (по Д. Б. Бекову)
Рис. 59. Дви-
гательный го-
мункулус. Схема
проекции частей
тела человека на
область корко-
вого конца дви-
гательного ана-
лизатора (кора
предцентраль-
ной извилины).
Разрез полуша-
рия — фрон-
тальный.
1 — gyrus pre-
centralis; 2 —
височная доля
мозга; 3 — sulcus
lateralis; 4 — ven-
triculus Lateralis; 5 — fissura Longitudinalis cerebri (no Penfield;
Rasmussen; 1950)
Рис. 60. Схема локализации корковых концов анали-
заторов в коре большого мозга (латеральная поверх-
ность правого полушария):
1 — постцентральная извилина — локализация коркового
конца кожного анализатора (осязательная; болевая и
температурная чувствительность); 2 — предцентральная
извилина — локализация коркового конца двигательного
анализатора (проприоцептивных раздражений); 4 — верхняя
теменная долька — локализация коркового конца анализатора
частного вида кожной чувствительности (узнавание
предметов на ощупь — стереогнозия); 5 — средняя часть
верхней височной извилины — локализация коркового конца
слухового анализатора; 9 — нижние отделы предцентральной
и постцентральной извилин — локализация коркового конца
анализатора импульсов, идущих от внутренностей и сосудов
Рис. 61. Схема локализации корковых концов анали-
заторов в коре большого мозга (медиальная и нижняя
поверхность правого полушария):
2 — парацентральная долька — локализация коркового конца
двигательного анализатора (проприоцептивных раздражений);
6 — края шпорной борозды — локализация коркового конца
зрительного анализатора; 7 — парагиппокампальная извилина —
локализация коркового конца обонятельного анализатора;
8 — крючок — локализация корковых концов обонятельного
и вкусового анализаторов
Рис. 62. Чувствительный гомункулус. Схема проекции
частей тела человека на область коркового конца ана-
лизатора общей чувствительности (кора постцент-
ральной извилины). Разрез полушария — фронтальный:
1 — gyrus postcentralis; 2 — височная доля мозга; 3 — sulcus
lateralis; 4 — ventriculus lateralis; 5 — fissura longitudinals
cerebri (no Penfield; Rasmussen; 1950)
Рис. 63. Схема локализации корковых концов анали-
заторов в коре большого мозга (латеральная поверх-
ность правого полушария):
1 — задняя часть верхней височной извилины — локализация
коркового конца слухового анализатора устной речи; 2 — задняя
часть нижней лобной извилины — локализация коркового конца
двигательного анализатора артикуляции речи; 3 — угловая
извилина нижней теменной дольки — локализация коркового
конца зрительного анализатора письменной речи; 4 — задний
отдел средней лобной извилины — локализация коркового
конца анализатора письменной речи; 5 — средняя лобная
извилина — локализация коркового конца двигательного
анализатора, относящегося к сочетанному повороту головы и
глаз в противоположную сторону
6
Рис. 85. Схема связей парасимпатических волокон III,
VII и IX черепных нервов с парасимпатическими узлами
и ветвями V черепного нерва:
1 — n. ophthalmicus; 2 — n. maxillaris; 3 — п. mandibuLaris;
4 — n. frontalis; 5 — n. lacrimalis; 6 — n. supraorbitalis; 7 — n.
nasociliaris; 8 — gangl. ciliare; 9 — n. zygomaticus; 10; 11 — n.
infraorbitalis; 12 — nn. alveolares superiores; 13 — n. buccalis;
14 — gangl. pterygopalatinum; 15 — n. lingualis; 16 — n. al-
veolaris inferior; 17 — n. mentalis; 18 — gangl. oticum; 19 — n
auriculotemporalis; 20 — n. petrosus major; 21 — gangl. sub-
mandibulare; 22 — n. petrosus minor; 23 — n. chorda tympani;
24 — radix motoria
Рис. 86. Тройничный нерв:
1 — тройничный узел; 2 — нижнечелюстной нерв; 3 — большой
каменистый нерв; 4 —лицевой нерв; 5 — ушно-височный нерв;
6 — язычный нерв; 7 — щечный нерв; 8 — нижний альвеолярный
нерв; 9 — подбородочный нерв; 10 — нерв крыловидного
канала; И — крылонебный узел; 12 — узловые ветви; 13 —
верхние альвеолярные нервы; 14 — подглазничный нерв; 15 —
скуловой нерв; 16 — соединительная ветвь; 17 — слезный нерв;
18 — надглазничные нерв; 19 — лобный нерв; 20 — короткие
ресничные нервы; 21 — ресничный узел; 22 — носоресничный
нерв; 23 — глазной нерв; 24 — верхнечелюстной нерв
1
Рис. 89. Поверхностные нервы головы и шеи:
1 — rami temporalis VII n.; 2 — n. supraorbitalis V нерва; 3 — rr.
zygomatici VII n.; 4 — n infraorbitalis V нерва; 5 — rr. buccales VII
n.; 6 — n. facialis; 7 — n. mentalis; 8 — r. marginalis mandibularis
VII n.; 9 — r. colli VII n.; 10 — n. transversus colli; 11 — nn. su-
praclaviculares; 12 — n. accessorius; 13 — n. auricularis magnus;
14— n. occipitalis minor; 15 — n. occipitalis major; 16 — n.
auriculotemporalis
Рис. 90. Нервы головы и шеи:
1 — п. opticus; 2 — п. oculomotorius; 3 — n. ophthalmicus;
4 — n. maxillaries; 5 — n. mandibulahs; 6 — n. occipitalis ma-
jor; 7 — radix inferior ansa cervicalis; 8 — ramus superior ansa
cervicalis XII n.; 9 — n. auricularis magnus; 10 — n. accessorius;
11 — n. vagus; 12 — rami muscularis; 13 — nn. supraclavicu-
lares; 14 — n. phrenicus; 15 — n. frontalis; 16 — n. lacrimalis;
17 — n. infraorbitalis; 18 — rami alveolares superiores posteriors;
19 — ramus alveolaris superior medius; 20 — n. lingualis; 21 — n.
alveolaris inferior; 22 — n. hypoglossus
Рис. 91. Нервы шеи:
1 — m. styloglossus; 2 — truncus sympathicus; 3 — п. vagus;
4 — n. cardiacus cervicalis superior; 5 — r. cardiacus cervicalis
superior; 6 — n. laryngeus recurrens; 7 — n. cardiacus cervicalis
inferior; 8 — m. scalenus anterior; 9 — n. hypoglossus; 10 — gan-
gl. cervicale superior; 11 — n. vagus
Рис. 92. Топография
языкоглоточного, блужда-
ющего
и добавочного нервов:
1 — добавочный нерв; 2 —
нижний узел блуждающего
нерва; 3 — внутренняя сонная
артерия; 4 — языкоглоточный
нерв; 5 — глоточная ветвь
блуждающего нерва; 6 —
общая сонная артерия; 7 —
правый блуждающий нерв; 8 —
возвратный гортанный нерв;
9 — плечеголовной ствол;
10 — правый главный бронх;
И — левая правая легочная
вена; 12 — легочной ствол;
13 — пищеводное сплетение;
14 — левый блуждающий
нерв; 15 — левая доля печени;
16 — чревный ствол; 17 —
брюшная часть аорты; 18 —
двенадцатиперстная кишка
▼А
Рис. 94. Схема ветвей спинномозгового нерва:
1 — ложноуниполярная клетка спинномозгового узла; 2 —
чувствительное ядро заднего рога; 3 — двигательное ядро
переднего рога; 4 — симпатическое ядро бокового рога; 5 —
спинномозговой нерв; 6 — задняя ветвь; 7 — менингеальная
ветвь; 8 — передняя ветвь; 9 — белая соединительная ветвь;
10 —серая соединительная ветвь; синяя линия —чувствительные
волокна; красная линия — двигательные волокна; черная
сплошная линия — симпатические преганглионарные волокна;
черная пунктирная линия — симпатические постганглионарные
волокна
Рис. 95. Об-
ласти рас-
простране-
ния кожных
нервов туло-
вища:
1 — ПП. SU-
praclavicu lares
(ветви plexus
cervicalis);
2 — п. cutaneus
brachii lateralis
(ветвь n. axil-
laris); 3 — n.
cutaneus brachii
medialis (ветвь
plexus cervica-
lis); 4 — n. cu-
taneus brachii
posterior (ветвь
n. radialis); 5 —
rr. cutanei later-
als (от задних
ветвей грудных
нервов); 6 —
nn. clunium su-
periors (задние ветви поясничных нервов); 7 — г. cutaneus lateralis
(ветвь n. iliohypogastricus); 8 — n. cutaneus femoris lateralis (ветвь
plexus lumbalis); 9 — n. cutaneus femoris posterior (ветвь plexus sacra-
lis); 10 — nn. clunium inferiors (ветви n. cutaneus femoris posterior);
11 — nn. clunium medii (задние ветви крестцовых нервов); 12 — rr.
cutanei dorsales mediates (от задних ветвей грудных нервов)
Рис. 96. Задние ветви
спинномозговых не-
рвов; слева — кожные
ветви, справа — мы-
шечные
Рис. 97. Схема иннервации мышц волокнами, идущими
от различных сегментов, в составе одного нерва (1)
или двух нервов (2).
Рис. 99. Кожные нервы шеи:
1 — п. occipitalis major; 2 — ramus colli nervi facialis; 3 — ansa
cervicalis superficialis; 4 — n. occipctalis minor; 5 — n. auricu-
laris magnus; 6 — n. transversus colli; 7 — nn. supraclaviculares;
8 — n. accessorius
Рис. 100. Поверхностные нервы головы и шеи:
1 — rami temporalis; 2 — plexus parotideus; 3 — rami zygomatici;
4 — n. occipitalis major; 5 — n. auricularis magnus; 6 — n. oc-
cipitalis minor; 7 — ramus marginalis mandibulae; 8 — ramus
colli; 9 — rami inferiores nervi transverus colli; 10 — n. trans-
versus colli; 11 — nn. supraclaviculares; 12 — n. supraorbitalis;
13 — n. frontalis; 14 — rami palpebrales; 15 — n. infraorbitalis;
16 — rami labiates superiores; 17 — rami buccales; 18 — n. fa-
cialis; 19 — rami mentales
Рис. 101. Диафрагмальный нерв:
1 — n. accessorius; 2 — n. hypoglossus; 3 — plexus cervicalis;
4 — ansa cervicalis profunda; 5 — n. phrenicus; 6 — plexus bra-
chialis; 7 — n. vagus
8
7
Рис. 102. Нервы
1 плечевого спле-
2 тения:
1 — plexus brachialis;
д 2 — clavicula; 3 — v.
$ axillaris; 4 — a. axilla-
r is; 5 — nn. pectorales
6 medialis et lateralis;
6 — n intercostobrachi-
alis; 7 — n. thoracicus
longus; 8 — n. thora-
codorsalis; 9 — n. axil-
laris; 10 — n. cutaneus
brachii medialis; 11 — n.
radialis; 12 — n. ulnaris;
13 — n. cutaneus ante-
brachii medialis; 14 — n.
medianus; 15 — n. mus-
culocutaneus; 16 —fasc.
lateralis; 17 — fasc. me-
dialis; 18 — fasc. poste-
rior (по M. P. Сапину)
Рис. 103. Нервы передней поверхности правой
верхней конечности:
а — поверхностные нервы: 1 — nn. supraclaviculares; 2 — n.
cutaneus brachii medialis; 3 — v. basilica; 4 — n. cutaneus ante-
brachii medialis; 5 — v. intermedia cubiti; 6 — n. cutaneus brachii
lateralis superior; 7 — v. cephalica; 8 — n. cutaneus antebrachii
lateralis; 9 — ramus superficialis n. radialis ;
6 — глубокие нервы: 1 — fasciculus lateralis; 2 — fasciculus
medialis; 3 — n. cutaneus brachii medialis; 4 — n. ulnaris; 5 —* n.
musculocutaneus; 6 — n. medianus; 7 — vv. brachiales; 8 — n.
radialis; 9 — rami musculares n. medianus; 10 — ramus superfi-
cialis n. radialis; 11 — nn. digitales palmares proprii; 12 — nn.
digitales palmares communes
Рис. 104. Нервы задней поверхности правой верхней
конечности:
а — поверхностные нервы: 1 — rami cutanei n. supraclavicuta-
ris; 2 — n. cutaneus beachii lateralis superior; 3 — n. cutaneus
brachii posterior; 4 — n. cutaneus antebrachii medialis; 5 — n.
cutaneus antebrachii lateralis; 6 — ramus superficiatis n. radialis;
7 — nn. digitales dorsales; 8 — ramus dorsalis n. ulnaris; 9 — nn.
digitales dorsales;
b — глубокие нервы: 1 — n. suprascapularis; 2 — rami muscu-
lares; 3 — n. axillaris, 4 — n. radialis; 5 — rami musculares; 6 — n.
cutaneus antebrachii posterior; 7 — ramus profundus n. radialis;
8 — n. interosseus antebrachii posterior; 9 — ramus superficialis
n. radialis; 10 — n. ulnaris, 11 — ramus dorsalis n. ulnaris
Рис. 105. Плечевое сплетение и передние ветви груд-
ных нервов; сбоку (большая грудная и косые мышцы
живота удалены):
1 — n. phrenicus; 2 — plexus brachialis; 3 — nn. pectorales me-
dialis et lateralis; 4 — n. thoracicus longus; 5 — nn. intercostales;
6 — n. subcostalis; 7 — n. iliohypogastricus; 8 — n. ilioinguinalis;
9 — n. medianus; 10 — n. ulnaris; 11 — n. cutaneus antebrachii
medialis; 12 — fasciculus lateralis; 13 — n. musculocutaneus;
14 — fasciculus posterior; 15 — fasciculus medialis; 16 — n.
dorsalis scapulae
Рис. 106. Межреберные
артерии, вены и нервы:
1 — лп. intercostales.
1
Рис. 107. Поясничное и крестцовое сплетения:
1 — п. subcostalis; 2 — n. iliohypogastricus; 3 — n. ilioinguinalis;
4 — n. cutaneus femoris lateralis; 5 — n. genitofemoralis; 6 — n.
femoralis; 7 — n. obturatorius.
Рис. 108. Нервы
нижних конечное-
шей, спереди:
1 — plexus lumbalis;
2 — n. cutaneus femo-
ris Lateralis; 3 — plex-
us sacralis; 4 — rami
cutanei anteriores;
5 — n. saphenus; 6 —
n. peroneus superficia-
Lis; 7 — nn. digitales
dorsales pedis; 8 — n.
peroneus profundus;
9 — n. fem oralis ;
10 — n. obturatorius;
11 — n. genitofemo-
ralis; 12 — ramus cu-
taneus n. obturatorius;
13 — rami musculares
n. femoralis; 14 — n.
saphenus; 15 — n.
peroneus communis;
16 — rami musculares
n. peroneus profun-
dus; 17 — n. peroneus
superficialis; 18 — n.
peroneus profundus;
19 — n. cutaneus dor-
salis medialis; 20 — n.
cutaneus dorsalis in-
termedius; 21 — n. cu-
taneus dorsalis latera-
lis; 22 — nn. digitales
dorsales pedis
22
Рис. 109. Не-
рвы нижних
конечностей,
сзади:
1 — п. gluteus
superior; 2 — n.
gluteus inferior;
3 — n. puden-
dus;4 — n.ischi-
adicus; 5 — lig.
sacrotuberale;
6 — n. cutaneus
femoris pos-
terior; 7 — rami
musculares n.
ischiadicus; 8 —
n. peroneus com-
munis; 9 — n.
tibialis; 10 — n.
cutaneus surae
lateralis; 11;
21 — n. surah’s;
12 — n. tibia-
lis; 13 — nn.
clunium superi-
ores; 14 — nn.
clunium medii;
15 — nn. clu-
nium inferiors;
16 — n. cuta-
neusfemoris post-
erior; 17 — n.
cutaneus surae
medialis; 18 —
n. saphenus;
19 —n. cutaneus
surae lateralis;
20 — rami cu-
tanei cruris me-
diates; 22 — n.
cutaneus dorsalis
lateralis
1
2
7
6
5
3
Рис. 124. Разрез через улитко-
вый канал:
1 — ductus сосhlean’s; 2 — scala ves-
tibuli; 3 — modiolus; 4 — ganglion
spirale cochleae; 5 — периферические
отростки клеток ganglion spirale coch-
leae; 6 — scala tympani; 7 — костная
стенка улиткового канала; 8 — lamina
spiralis ossea; 9 — membrana vestibu-
laris; 10 — organum spirale seu orga-
num Cortii; 11 — membrana basilaris
24
Рис. 137. Вегетативная иннервация внутренних органов:
а — парасимпатическая часть, б — симпатическая часть
1 — верхний шейный узел; 2 — латеральное промежуточное ядро;
3 — верхний шейный сердечный нерв; 4 — грудные сердечные
и легочные нервы, 5 — большой чревный нерв; 6 — чревное
сплетение; 7 — нижнее брыжеечное сплетение; 8 — верхний
и нижний подчревные сплетения; 9 — малый чревный нерв;
10 — поясничные чревные нервы; 11 — крестцовые чревные
нервы; 12 — парасимпатические ядра крестцовых сегментов;
13 — тазовые чревные нервы; 14 — тазовые узлы; 15 —
парасимпатические узлы; 16 — блуждающий нерв; 17 — ушной
узел, 18 — поднижнечелюстной узел; 19 — крылонебный узел;
20— ресничный узел, 21 — парасимпатическое ядро блуждающего
нерва; 22 —парасимпатическое ядро языкоглоточного нерва, 23 —
парасимпатическое ядро лицевого нерва; 24 — парасимпатическое
ядро глазодвигательного нерва (по М. Р. Сапину)
Рис. 138. Ход волокон спинномозговых нервов и связь
их с симпатическим стволом:
1 — г. posterior nervi spinalis; 2 — чувствительные волокна;
3 — ganglion spinale; 4 — radix dorsalis; 5 — radix ventralis;
6 — funiculus lateralis; 7 — cornu posterius; 8 — cornu lateralis;
9 — funiculus posterior; 10 — sulcus medianus posterior; 11 —
canalis centralis; 12 — comissura anterior grisea; 13 — fissura
mediana anterior; 14 — funiculus anterior; 15 — cornu anterius;
16 — орган (кишка); 17 — ganglion visceralis; 18 — fibrae post-
ganglionares OTg. visceralis; 19 — fibrae preganglionares Kg. vis-
ceralis; 20 — fibrae postganglion a res OTg. tr. sympathici; 21 — r.
communicans albus (fibrae preganglionares к g. tr. sympathici);
22 — r. communicans albus (fibrae preganglionares к g. visceralis);
23 — nervus spinalis; 24 — fibrae postganglionares; 25 — gangli-
on tr. sympathici; 26 — r. interganglionaris; 27 — чувствительные
волокна; 28 — r. communicans griseus (fibrae postganglionares к
n. spinalis); 29 — r. anterior nervi spinalis; 30 — двигательные
волокна клеток переднего рога спинного мозга
Рис. 139. Шейный и грудной отделы
симпатического ствола:
1 — gangl. cervicale superius; 2 — gangl. cervicale medium;
3 — gangl. cervicothoracicum; 4 — plexus subclavius; 5 — gangl.
thoracica; 6 — r. communicansgriseus; 7 — n. splanchnicus major;
8 — n. splanchnicus minor (по M. P. Сапину)
Рис. 140. Поясничный и крестцовый отделы
симпатического ствола:
1 — gangl. lumbalia; 2 — rr. communicantes (поперечные);
3 — gangl. sacralia; 4 — gangl. impar; 5 — r. communicans (gri-
seus); 6 — nn. splanchnici Lumbales (по М.Р. Сапину)
Рис.141. Шейный,
грудной, брюшной
и тазовый отде-
лы вегетативной
нервной системы:
1 — plexus pul-
monalis; 2 — plexus
coeliacus; 3 — plexus
gastrici; 4 — plexus
entericus; 5 — plex-
us mesentericus su-
perior; 6 — plexus
mesentericus infe-
rior; 7 — plexus
d e f e r e n t i a I i s ;
8 — pl. cavernosus
penis; 9 — gan-
glion cervicale su-
per us; 10 — plexus
caroticus commu-
nis; 11 — n. vagus;
12— ganglion
cervicale medium;
13 —ganglion cervi-
cothoracicum (stel-
latum); 14 — gan-
glia thoracica trunci
sympathici; 15 —
plexus esophageus;
16 — plexus aor-
ticus abdominalis;
17 — n. splanchni-
cus major; 18 — n.
splanchnicus minor;
19 — plexus rena-
lis; 20 — ganglia
lumbalia trunci sym-
pathici; 21 — plexus
recta Lis medius
11
Рис. 142.
Чревное
сплетение:
1 — gangl. со-
eliacum (левый);
2 — plexus
suprarenalis;
3 — gangl.
aorticorenale
(левый);
4 — gangl.
mesentericum
superius; 5 —
plexus inter-
mesentericus;
6 — gangl.
aorticorenale
(правый); 7 — plexus renalis; 8 — gangl. coeliacum (правый); 9 —
truncus vagalis posterior; 10 — n. splanchnicus major; 11 — truncus
vagalis anterior (по М.Р. Сапину)
Рис. 144. Вегетативные
сплетения таза:
1 — plexus hypogastricus supe-
rior; 2 — n. hypogastricus; 3 —
plexus hypogastricus inferior;
4 — plexus vesicalis; 5 — plexus
prostaticus; 6 — pl. cavernosus
penis; 7 — nn. splanchnici pelvini
(по М.Р. Сапину)
Зоны Брока и Вернике были идентифицированы
таким образом более 100 лет назад — вскрытия
умерших с характерными афазиями и дефектами
речи при жизни показывали повреждения именно
этих областей левого полушария.
3. Ядро двигательного анализатора письменной
речи находится в заднем отделе средней лобной
извилины рядом с центром сочетанного поворота
глаз и головы и центром движения руки. Понятно,
что процесс письма сопряжен с движениями глаз,
головы и рук. При его поражении расстраивается
письмо — возникает аграфия.
4. Ядро зрительного анализатора письменной
речи расположено рядом с ядром зрительного ана-
лизатора и в угловой извилине. При его поражении
развивается алексия — расстройство понимания
письменных и печатных знаков. В тяжелых случаях
больной не может читать.
На первый взгляд кажется, что, подобно боль-
шинству других органов тела, мозг имеет совершен-
ную билатеральную симметрию. В действительности
многие из более специализированных функций при-
сущи только либо одному полушарию, либо другому.
Нет даже анатомической симметрии, а асимметрия
зависит от доминирующей руки. Ученые полагают,
что уменьшение функциональной асимметрии мозга
ведет к снижению интеллектуальных способностей.
Правое и левое полушария нашего мозга постоянно
обмениваются информацией. Перерезка мозолистого
тела не вызывает заметного снижения интеллекта,
но является причиной таких казусов, когда больной
одной рукой подносит зажигалку и открывает кран,
чтобы зажечь газ, а другой рукой закрывает кран.
Вообще же, клиника, характерная для больных с
функционально разобщенными полушариями, чрез-
4 Анатомия нсрпной системы
97
вычайно сложна. Ее проявления тяжелы и полно-
стью инвалидизируют человека.
Итак, следует запомнить, что все речевые ана-
лизаторы у правшей находятся в левом полушарии
и наоборот. Причем все речевые анализаторы за-
кладываются в обоих полушариях, а развивают-
ся с одной стороны и функционально являются
асимметричными. Правое полушарие у правшей
является «немым». Ученые говорят: «Наши мысли,
облеченные в слова, шагают под звуки левого мар-
ша». Но в последнее время стало ясно, что и правое
полушарие отнюдь не глухо к речи и не является
немым. От него зависит умение ориентироваться в
пространстве, целостность зрительного восприятия,
образная память, музыкальное письмо, музыкаль-
ные способности. Известно, что французский ком-
позитор Равель вследствие паралича потерял дар
речи и способность писать, но продолжал успешно
сочинять музыку и сам записывал на бумагу свои
произведения нотными знаками.
В соответствии с этим основная деятельность
левого полушария получила название логико-вер-
бального мышления, оно также отвечает за все дви-
гательные функции, а правого — пространственно-
образного.
Творческое вдохновение — это напряженная ра-
бота всего мозга. Мало того, ученые все чаще при-
ходят к выводу, что та или иная структура мозга,
тесно связанная с какой-либо функцией, может в
то же время участвовать и в реализации других
функций. Функции отдельных областей головного
мозга хорошо изучены, а взаимосвязь между ними
еще до конца не ясна.
В височной доле происходит распознавание зву-
ков, их тональности и громкости. Височная доля
98
играет важную роль в запоминании. В затылочной
доле происходит распознание и интерпретация зри-
тельных образов. Теменная доля отвечает за вос-
приятие ощущений (осязание, температура, боль).
Новая, филогенетически молодая кора лобной доли
мозга имеет самое непосредственное отношение к
интеллектуальной деятельности человека, она уп-
равляет речью, мыслительными операциями и эмо-
циями, выученными движениями. Из врачебной
практики известно, что повреждение лобной доли,
наиболее совершенного отдела коры мозга, лишает
больного возможности предвидеть ход событий и
соответствующим образом строить свое поведение,
больные теряют волю, целеустремленность, дели-
катность. Академик Анохин считал, что лобная
кора не имеет отношения к отдельным функциям
мозга: памяти, восприятию, мотивации, эмоциям, но
осуществляет их интеграцию в целенаправленные,
пластичные поведенческие реакции.
Проводящие пути
Глава
Вышепреведенные данные о проводящих путях
спинного мозга, ствола мозга и анализаторах в коре
полушарий большого мозга являются основой для
целостного представления о топографии восходящих
и нисходящих путей.
В нервной системе нервные клетки образуют
рефлекторные дуги, включающие рецептор, кон-
дуктор и двигательный нейрон. Различают простые
(2-3-нейронные) дуги и сложные (многонейрон-
ные), в которых увеличено количество вставочных
нейронов.
Проводящие пути — части сложных дуг, это
пучки нервных волокон, соединяющие функцио-
нально неоднородные отделы серого вещества в
ЦНС, проводящие одинаковые импульсы и зани-
мающие в ЦНС определенное положение. Иссле-
дуют проводящие пути методом расщипывания или
перерождения.
Проводящие пути классифицируют на ассоциа-
тивные, комиссуральные, проекционные.
Ассоциативные пути соединяют ядра одной сто-
роны и одного уровня ЦНС. В полушариях про-
ходят в capsula externa et extrema. В стволе мозга
переключаются на неспецифических ядрах formatio
reticularis. А в спинном мозге располагаются в
fasciculus proprius и вовлекают большое количество
мышц в ответ на рефлекторное раздражение.
100
Комиссуральные пути соединяют ядра разных
сторон одного уровня ЦНС.
Проекционные пути соединяют различные ядра
различных сторон и уровней ЦНС, выходят за пре-
делы ЦНС. С их помощью картина внешнего мира
как бы проецируется на кору, как на экран, и там
оценивается.
В свою очередь, проекционные проводящие пути
подразделяют на восходящие и нисходящие. Чувс-
твительные восходящие (афферентные) пути могут
быть экстероцептивными, проприоцептивными и
интероцептивными. Двигательные нисходящие (эф-
ферентные) пути подразделяют на пирамидные и
экстрапирамидные.
Чувствительные (восходящие,
афферентные) пути
1. Путь температурной и болевой чувствитель-
ности, tr. gangliospinothalamocorticalis lateralis
(рис. 64). 1-й нейрон этого пути — в спинномозго-
вом узле, 2-й нейрон — в n. proprius задних рогов
спинного мозга. Отростки 2 нейронов переходят на
противоположную сторону в области белой спай-
ки. 3-й нейрон располагается в латеральном ядре
зрительного бугра. Отростки 3 нейронов проходят
через середину задней ножки внутренней капсулы и
заканчиваются в корковом конце анализатора — в
постцентральной извилине.
2. Тактильная чувствительность — ее прово-
дят fasciculus gracilis et cuneatus, а также tr. spino-
thalamicus anterior (рис. 65).
1-й нейрон этого пути находится в g. spinale, 2-й
нейрон — в двух местах — в продолговатом мозге и
в спинном — substantia gelatinosa. Перекрест пути
101
совершают также в двух местах. Природа разнесла
локализацию вторых нейронов и перекресты этих
Рис. 64. Схема проводящего пути температурной
и болевой чувствительности:
1 — спинной мозг; 2 — продолговатый мозг; 3 — мост; 4 — сред-
ний мозг; 5 — зрительный бугор; б — внутренняя капсула; 7 — tr.
spinothalamicus lateralis; 8 — lemniscus medialis; I — первый
нейрон в g. spinale; II — второй нейрон в n. proprius заднего
рога; III — третий нейрон в латеральном ядре зрительного бугра;
К.К. — корковый конец анализатора — eg. postcentralis
путей (феномен бивариантности). 3-й нейрон распо-
лагается в зрительном бугре. Корковый конец — в
g. postcentralis.
102
3. Путь стереогноза — (ганглио-бульбарно-тала-
мо-корковый путь) проходит через fasciculus gracilis
Рис. 65. Схема проводящего пути тактильной
чувствительности:
1 — спинной мозг; 2 — продолговатый мозг; 3 — мост; 4 —
средний мозг; 5 — зрительный бугор; 6 — внутренняя кап-
сула; 7 — tr. spinothalamicus anterior; 8 — fasciculus gracilis
et cuneatus; 9 — lemniscus medialis; I — первый нейрон в
g. spinale; II — второй нейрон в substantia gelatinosa заднего
рога и в n. gracilis et cuneatus продолговатого мозга; III — тре-
тий нейрон в латеральном ядре зрительного бугра; К.К. — кор-
ковый конец анализатора — eg. postcentralis
103
et cuneatus, одноименные ядра продолговатого моз-
га, зрительный бугор, внутреннюю капсулу в верх-
нюю теменную дольку (рис. 66).
Рис. 66. Схема проводящего пути стереогноза:
1 — спинной мозг; 2 —- продолговатый мозг; 3 — мост; 4 —
средний мозг; 5 — зрительный бугор; 6 — внутренняя капсула;
7 — fasciculus gracilis et cuneatus; 8 — lemniscus medialis;
I — первый нейрон в g. spinale; II — второй нейрон в n. gracilis
et cuneatus продолговатого мозга; III — третий нейрон в лате-
ральном ядре зрительного бугра; К.К. — корковый конец ана-
лизатора — в lobulus parietalis superior
104
4. Путь сознательной проприоцептивной чувс-
твительности — (ганглио-бульбарно-таламо-корко-
вый путь) также проходит через fasciculus gracilis et
cuneatus, одноименные ядра продолговатого мозга,
зрительный бугор, но в предцентральную извилину
(рис. 67).
5. Пути бессознательной проприоцептивной
чувствительности — передний и задний спин-
но-мозжечковые тракты (рис. 68, 69). Эти пути
вносят поправки на сохранение или изменение
позы человека. Передний спинно-мозжечковый
путь, tr. spinocerebellaris anterior, имеет сле-
дующие особенности: 2-й нейрон находится в
п. intermediomedialis, проходит через верхнюю
ножку мозжечка и совершает 2 перекреста (в бе-
лой спайке спинного мозга и верхнем мозговом
парусе). Задний спинно-мозжечковый путь, tr.
spinocerebellaris posterior, имеет следующие осо-
бенности: 2-й нейрон находится в n. thoracicus,
проходит через нижнюю ножку мозжечка и без
перекрестов. Оба пути заканчиваются в коре червя
мозжечка. Итак, оба пути соединяют одноименные
половины спинного мозга и мозжечка, но еще на
уровне спинного мозга отдают коллатерали на
противоположную сторону.
В мозжечке импульсы через зубчатое ядро пе-
редаются в красное ядро, а затем к мотонейронам
спинного мозга. Так осуществляется участие моз-
жечка в поддержании равновесия, мышечного то-
нуса и координации, преодолении инерции и силы
тяжести. Таким образом, мозжечок — важнейший
проприоцептивный центр. Проприоцептивные пути
принадлежат коре большого мозга и мозжечку.
Причем кора большого мозга держит мозжечок под
контролем.
105
Рис. 67. Схема проводящего пути сознательной
проприоцептивной чувствительности:
1 — спинной мозг; 2 — продолговатый мозг; 3 — мост; 4 —
средний мозг; 5 — зрительный бугор; 6 — внутренняя капсула;
7 — fasciculus gracilis et cuneatus; 8 — lemniscus medialis;
I — первый нейрон в g. spinale; II — второй нейрон в n. gracilis
et cuneatus продолговатого мозга; III — третий нейрон в лате-
ральном ядре зрительного бугра; К.К. — корковый конец ана-
лизатора — в g. precentralis
106
3
Рис. 68. Путь бессознательной проприоцептивной чувс-
твительности — задний спинно-мозжечковый путь:
1 — спинной мозг; 2 — продолговатый мозг; 3 — мозжечок; 4 —
нижняя ножка мозжечка; 5 — задний спинно-мозжечковыйпуть;
I — первый нейрон в ganglion spinale; II — второй нейрон в п.
thoracicus заднего рога; III — третий нейрон в vermis cerebelli
6. Интероцептивные пути начинаются от инте-
рорецепторов, расположенных во всех органах.
Этот анализатор не имеет компактного проводни-
ка. Он может находиться в составе афферентных
вегетативных нервов. В этом случае 1-й нейрон
находится в спинномозговых узлах. Другая часть
проводника может находиться в составе черепных
нервов. Тогда 1-й нейрон будет располагаться в че-
репных узлах. Путь проходит через чувствительные
107
ядра, через зрительный бугор. Корковое предста-
вительство — в нижних отделах предцентральной
и постцентральной извилин. Площадь коркового
конца интероцептивного анализатора меньше, чем
у экстероцептивного и проприоцептивного, чем объ-
ясняется его меньшая точность дифференцировки
по отношению к сознанию.
Рис. 69. Путь бессознательной проприоцептивной чувс-
твительности — передний спинно-мозжечковый путь:
1 — спинной мозг; 2 — продолговатый мозг; 3 — мозжечок, 4 —
верхняя ножка мозжечка; 5 — передний спинно-мозжечко-
вый путь; I — первый нейрон в ganglion spinale; II — второй
нейрон в п. intermediomedialis бокового рога; III — третий
нейрон в vermis cerebelli
108
Двигательные (нисходящие,
эфферентные) пути
Пирамидные пути
Любой поведенческий акт обеспечивается испол-
нительными органами — мышцами. Пирамидные
пути проводят импульсы от пирамидных клеток
коры мозга через двигательные ядра спинного моз-
га или черепных нервов к скелетным мышцам.
1. Корково-спинномозговые пути, tr. corti-
cospinalis lateralis et anterior, начинаются от боль-
ших пирамидных клеток (клеток Беца) 5-го слоя
коры предцентральной извилины и околоцентраль-
ной дольки (рис. 70). Перекрест происходит в про-
долговатом мозге для бокового пути и посегментно в
белой спайке спинного мозга для переднего. Причем
в продолговатом мозге совершают перекрест 80%
волокон, а в спинном — 20%. Конец пути — мо-
тонейроны спинного мозга, отростки которых в
составе передних корешков спинномозговых нервов
достигают скелетной мускулатуры туловища и ко-
нечностей.
2. Корково-ядерный путь, tr. corticonuclearis,
начинается также от больших пирамидных клеток
предцентральной извилины и околоцентральной
дольки (рис. 71). 2-й нейрон пути — тела клеток
двигательных ядер черепных нервов, отростки ко-
торых достигают исчерченной мускулатуры головы
и некоторых органов. Этот путь дает ответвления
на свою и противоположную сторону, кроме XII
и VII нервов, к ядрам которых подходят волокна
только с противоположной стороны. По сути дела,
этот тракт представляет собой небольшой руче-
ек, отделяющийся от Ниагары пирамидного пути,
109
чтобы, направившись к ядрам черепных нервов,
проинформировать их об идущих вниз командах и
Рис. 70. Схема корково-спинномозговых путей:
I — пирамидные клетки в gyrus precentralis et lobulus para-
centralis; II — nucleus proprius cornu anterioris; 1 — внутренняя
капсула; 2 — средний мозг; 3 — мост; 4 — продолговатый мозг;
5 — спинной мозг; 6 — tractus corticospinalis; 7 — decussatio
pyramidum; 8 — tractus corticospinalis lateralis; 9 — tractus
corticospinalis anterior.
110
наладить обеспечение двигательных актов череп-
ными нервами.
Рис. 71. Схема корково-ядерных путей:
I — пирамидные клетки в gyrus precentralis et lobulus paracentralis;
II — двигательные ядра черепных нервов; 1 — внутренняя кап-
сула; 2 — средний мозг; 3 — мост; 4 — продолговатый мозг;
5 — tractus corticonuclearis; III — XII — черепные нервы
111
Экстрапирамидные пути
Экстрапирамидные пути обеспечивают движения
скелетных мышц, которые сиюминутно не управля-
емы корой. Это пути, обеспечивающие поддержание
позы, выполнение содружественных движений,
регуляцию мышечного тонуса при выполнении без-
условно рефлекторных движений. Переключение
рефлекторных дуг с восходящего на нисходящее
колено происходит ниже коры полушарий большо-
го мозга, таким образом, экстрапирамидная (под-
корковая) система поддерживает тонус скелетной
мускулатуры и автоматически (бессознательно)
регулирует ее работу.
Термин «экстрапирамидная» сложился не в связи
с пирамидными клетками. Вначале были изучены
пирамидные пути, и считалось, что это единствен-
ные двигательные тракты. Потом внезапно обнару-
жили другие двигательные пути, стали их изучать
и назвали экстрапирамидными. Разграничение их
анатомически и физиологически сложное, действуют
они в полном единстве.
К экстрапирамидной системе относят: базальные
ядра, медиальные ядра зрительного бугра, субта-
ламические ядра Люиса, черное вещество, красное
ядро, ядра холмиков четверохолмия, ядра ретику-
лярной формации, вестибулярные ядра, ядро оливы,
мозжечок. Все звенья экстрапирамидной системы
связаны между собой прямыми и обратными связя-
ми. От коры (любого участка) через внутреннюю
капсулу идут связи к экстрапирамидной системе: в
хвостатое ядро, в putamen, далее от них к латераль-
ному ядру бледного шара, от него, в свою очередь,
к черному веществу, красному ядру, медиальному
вестибулярному ядру, ретикулярной формации.
112
1. Красноядерно-спинномозговой путь, tr.
rubrospinalis (рис. 72).
Рис. 72. Схема красноядерно-спинномозгового пути:
1 — средний мозг; 2 — спинной мозг; 3 — перекрест Фореля,
decussatio tegmentalis ventralis; 4 — tractus rubrospinalis; I —
первый нейрон в nucleus ruber; II — второй нейрон в nucleus
proprius cornu anterioris
К красному ядру идут следующие пути: tr.
cortico — rubro-spinalis, tr. thalamo-rubro-spinalis,
tr. strio-rubro-spinalis. Красное ядро имеет обшир-
ные связи с корой полушарий большого мозга и
экстрапирамидными центрами. С участием путей от
коры к красному ядру осуществляются привычные
движения (бег, ходьба). А при помощи путей от
113
мозжечка к красным ядрам осуществляется «поп-
равочные» движения — выполняются тонкие целе-
направленные движения. Перекрест красноядерно-
спинномозгового пути назван в честь швейцарского
невролога — Фореля.
2. Крышеспинномозговой путь, tr. tectospinal is
(рис. 73) осуществляет рефлекторные защитные
движения при внезапных зрительных или звуковых
раздражениях. Перекрест крышеспинномозгового
пути назван в честь австрийского невролога Мей-
нерта.
Рис. 73. Схема крышеспинномозгового пути:
1 — средний мозг; 2 — спинной мозг; 3 — перекрест Мейнер-
та, decussatio tegmentalis dorsalis; 4 — tractus tectospinalis;
I — первый нейрон — nucleus colliculi superiores; II — второй
нейрон — nucleus proprius cornu anterioris
114
3. Преддверно-спинномозговой путь, tr. vesti-
bulospinalis (рис. 74).
Рис. 74. Схема преддверно-спинномозгового пути:
1 — ромбовидная ямка; 2 — спинной мозг; 3 — tr. vestibulo-
spinalis; I — первый нейрон, латеральное и нижнее вестибу-
лярные ядра VIII нерва; II — второй нейрон, nucleus proprius
cornu anterioris
Этот путь связывает латеральное и нижнее ядра
VIII нерва (Дейтерса и Роллера) с мотонейронами
спинного мозга. При нарушении равновесия этот путь
вносит поправку в позу человека, обеспечивает рав-
новесие движений. Ядра, образующие преддверно-
спинномозговой путь, находятся в непосредственной
связи с мозжечком, а также с медиальным продоль-
ным пучком, который, в свою очередь, связан с яд-
115
рами нервов, обеспечивающих сохранение пол ожени?
глазного яблока (фиксацию взора) при поворота?
головы. Перекрест пути — в белой спайке.
4. Ретикулоспинномозговые волокна, fibrat
reticulospinalis, трудно изобразить, так как в со
ставе ретикулярной формации располагается более
50 ядер, идущие от них волокна к спинному мозг}
проходят через белую спайку и заканчиваются нг
мотонейронах спинного мозга. Ретикулоспинномоз
говые волокна обеспечивают равновесие движений,
автоматическое поддержание тонуса мышц, выпол-
нение сложных рефлекторных актов, где требуется
работа различных групп мышц (дыхательные, хва-
тательные движения и пр.).
5. Оливо-спинномозговые волокна, fibrae
olivospinalis (рис. 75), обеспечивают безусловно
Рис. 75. Схема оливо-спинномозговых волокон:
1 — продолговатый мозг; 2 — спинной мозг; 3 — tr. olivospinalis;
I — первый нейрон, nucleus olivarius; II — второй нейрон,
nucleus proprius cornu anterioris
116
рефлекторное поддержание тонуса мышц и двига-
тельные акты, направленные на сохранение равно-
весия. Волокна начинаются от ядра оливы продолго-
ватого мозга, совершают перекрест в белой спайке и
заканчиваются на мотонейронах спинного мозга.
6. Корково-среднемозговые волокна, fibrae
corticomesencephalici, соединяют кору полушарий
большого мозга с такими структурами среднего
мозга, как substantia nigra, tegmentum mesencephali
et tectum mesencephali. От черного вещества (Зем-
меринга) идет путь tr. nigrospinalis (рис. 76, 77)
между боковым и передним канатиком спинного
мозга, совершает перекрест через белую спайку и
заканчивается в мотонейронах спинного мозга. Путь
отвечает за пластичность движений.
Волокна от globus pallidus направляются в thalamus,
n. ruber, substantia nigra, corpus subthalamicus. Эф-
ферентные волокна от globus pallidus доходят до
передних рогов спинного мозга через n. ruber по дви-
гательной цепи, состоящей из двух звеньев: tr. strio-
rubralis и tr. rubrospinalis, а также через substantia
nigra, по цепи, состоящей из трех звеньев: tr. strio-
nigralis, tr. nigro-reticularis и tr. reticulo-spinalis
(рис. 76, 77).
Студенты должны понимать, что tr. cortico —
strio-thalamo — rubro-spinalis это и есть tr. rubro-
spinalis, но с вышестоящими отделами, как и tr.
cortico-ponto-bicerebello-rubro-spinalis, сведения о
котором будут даны несколько ниже.
7. Таламоспинномозговой (рис. 78), tr. thala-
mospinalis начинается от медиальных ядер зритель-
ного бугра, проходит в боковом канатике, совершает
перекрест в белой спайке и заканчивается на мото-
нейронах передних рогов спинного мозга.
117
1
Рис. 76. Схема tr. nigrospinalis:
1 — средний мозг; 2 — спинной мозг; 3 — substantia nigra;
4 — tractus nigrospinalis; I — substantia nigra; II — nucleus
proprius cornu anterioris.
Итак, функции экстрапирамидной системы:
промежуточное звено двигательной системы
(кора — спинной мозг);
поддержание тонуса мышц и «настроя» на
работу;
обеспечение синхронности и автоматизма дви-
жений;
поддержание равновесия;
координация движений;
стартовые и защитные двигательные реакции.
Ассоциативные пути
Ассоциативные пути соединяют последний нейрон
афферентных и 1-й нейрон эфферентных путей.
118
Рис. 77. Схема основных путей между ядрами
зкстрамирамидной системы и ствола мозга
с мозжечком и спинным мозгом:
1 — таламус; 2 — кортико-красноядерные волокна; 3 — медиаль-
ный и латеральный бледный шар; 4 — красное ядро; 5 — субтала-
мическое ядро; 6 — черное вещество; 7 — зубчатое ядро; 8 — кора
мозжечка; 9 — ретикулярная формация; 10 — мост; 11 — продол-
говатый мозг; 12 — спинной мозг; 13 — ядро оливы
119
1
Рис. 78. Схема таламоспинномозгового пути:
1 —зрительный бугор; 2 —спинной мозг; 3 —tr. thalamospinalis;
I — первый нейрон, медиальное ядро таламуса; II — второй
нейрон, nucleus proprius cornu anterioris
Ассоциативные пути классифицируют на одно-
нейронные и многонейронные. К однонейронным
путям головного мозга относят:
fasciculus uncinatus, соединяет височную и
лобную доли полушарий,
fasciculus longitudinalis superior, соединяет
височную и лобную доли с затылочной,
fasciculus longitudinalis inferior, соединяет
затылочную и височную доли,
cingulum, соединяет лобную долю с затылоч-
ной и височной долями,
fibrae arcuatae cerebri, соединяют соседние
извилины. Короткие ассоциативные пути соб-
раны в capsula externa et extrema.
120
Примером многонейронных ассоциативных путей
являются медиальный продольный пучок и корково-
мозжечковый путь (рис. 79).
IV
III
6
Рис. 79. Схема tr. cortico-ponto-
bicerebello-rubro-spinatis:
1 — полушария большого мозга;
2 — средний мозг; 3 — мост; 4 —
продолговатый мозг; 5 — спинной
мозг; 6 — мозжечок; 7 — внут-
ренняя капсула; 8 — tractus
frontopontinus; 9 — tractus occipito
temporopontinus; 10— средняя
ножка мозжечка; 11— верхняя
ножка мозжечка; 12 — перекрест
Вернекинга; 13 — перекрест Фореля; 14 — tractus rubrospinalis;
I — первый нейрон, пирамидные клетки коры полушария; II — вто-
рой нейрон, собственные ядра моста; III — третий нейрон, клетки
коры полушария мозжечка; IV — четвертый нейрон, клетки зуб-
чатого ядра мозжечка; V — пятый нейрон, клетки красного ядра
среднего мозга; VI — клетки переднего рога спинного мозга
121
Tr. cortico-ponto-bicerebello-rubro-spinalis со-
единяет кору большого мозга, мозжечок, экстрапи-
рамидные ядра среднего мозга (рис. 79, 80).
Рис. 80. Фрагмент tr. cortico-ponto-bicerebello-rubro-
spinalis:
II — второй нейрон, собственные ядра моста; III — третий
нейрон, клетки коры полушария мозжечка; IV — четвертый
нейрон, клетки зубчатого ядра мозжечка; V — пятый нейрон;
клетки красного ядра среднего мозга
Он выполняет так называемую «поправочную»
деятельность, обеспечивающую точность сложных
целенаправленных движений, которые производятся
с участием пирамидных путей.
Путь начинается от нейронов 5-го слоя коры раз-
личных долей полушарий большого мозга: лобно-
мостовой начинается от коры лобной доли, проходит
через переднюю ножку внутренней капсулы и в
медиальной части ножки мозга; затылочно-височ-
122
но-мостовои путь начинается от коры одноименных
долей мозга, проходит через среднюю часть задней
ножки внутренней капсулы и через латеральную
часть ножки мозга. Второй нейрон располагается
в собственных ядрах моста, его отростки проходят
через среднюю ножку мозжечка к телам 3 нейронов
в коре полушарий мозжечка. Четвертый нейрон
располагается в зубчатом ядре, импульсы от кото-
рого проходят через верхнюю ножку мозжечка к
красному ядру противоположной стороны, совершая
перекрест Вернекинга.
Далее импульсы от красных ядер (тела 5 ней-
ронов) проводятся в составе красноядерно-спин-
номозгового пути и заканчиваются на мотонейро-
нах спинного мозга. Вслед за каждым импульсом,
обеспечивающим произвольные движения, следует
корково-мозжечковый импульс, который вносит
поправку. Поправка заключается в том, что моз-
жечковый импульс вызывает возбуждение мышц-
антагонистов, ликвидируя тем самым движение по
инерции, вызванное сокращением мышц за счет
импульсов пирамидного пути.
ч
ЕРЕПНЫЕ НЕРВЫ
(NERVI CRANIALES)
Глава
Развитие черепных нервов
Развитие черепных нервов тесным образом связа-
но: 1) с развитием ЦНС, ее дифференцировкой из
первичной нервной трубки, 2) с развитием мускула-
туры и кожных покровов (производных сомитов),
3) с первичными закладками внутренних органов
и сердечно-сосудистой системы.
Факторами, определяющими сложность разви-
тия черепных нервов, являются: развитие органов
чувств и висцеральных дуг и редукция головных
сомитов.
Двигательные нервы возникают путем прораста-
ния в закладки мышц нервных волокон из двига-
тельных ядер в развивающемся головном мозге.
Чувствительные нервы возникают путем прорас-
тания отростков нервных клеток, находящихся в
нервных узлах. Одни отростки этих клеток врастают
в мозг, другие — в кожные покровы или слизистые
оболочки. Чувствительные узлы черепных нервов
развиваются так же, как и спинномозговые узлы,
посредством миграции нервных клеток из гангли-
озных валиков.
I и II пары черепных нервов представляют собой
выросты мозга, являются самим мозгом, вынесен-
ным на периферию: I пара вырост обонятельного
(конечного) мозга, II пара — промежуточного. По
своему строению и происхождению они занимают
особое положение среди черепных нервов, так как
124
не имеют ядер в мозге и, являясь чувствитель-
ными нервами, не имеют чувствительных узлов.
Итак, эти нервы - сам мозг, они не нуждаются
в ядрах.
Ill, IV, VI пары в своем развитии связаны со
средним мозгом (ядро VI пары позже смещается
в мост) и тремя головными (предушными) миото-
мами, из которых развиваются мышцы глазного
яблока (рис. 81). Первому предушному миотому
соответствует III нерв, второму — IV нерв и тре-
тьему — VI нерв.
V, VII, IX, X, XI пары черепных нервов в своем
развитии связаны с ромбовидным мозгом и висце-
ральными дугами (рис. 81). Это нервы жаберного
происхождения.
Рис. 81. Схема развития черепных нервов:
Ш-ХП — черепные нервы; 1-5 — висцеральные дуги; б — пре-
душные миотомы; 7 — заушные миотомы
I висцеральная дуга — мандибулярная. С нею
связано развитие V пары черепных нервов. На ее
125
основе развивается жевательный аппарат: жеватель-
ные мышцы, мышцы дна рта.
II висцеральная дуга — гиоидная. С нею связано
развитие VII пары, от которой далее отделяется VIII
нерв. На основе этой дуги формируются подъязыч-
ная кость, мимическая мускулатура.
III висцеральная дуга — ей соответствует IX
нерв, из дуги развивается шилоглоточная мышца.
IV висцеральная дуга — ей соответствует X
нерв, развиваются мышцы гортани, мышцы глот-
ки, неба.
V висцеральная дуга — ей соответствует XI нерв,
развиваются грудино-ключично-сосцевидная и тра-
пециевидная мышцы.
XII пара черепных нервов развивается путем
слияния верхних шейных спинномозговых нервов
и связана с заушными миотомами, из которых об-
разуются мышцы языка.
Строение черепных нервов в принципе не отли-
чается от строения спинномозговых, хотя имеется
ряд особенностей: 1) ни один из черепных нервов
не соответствует полному спинномозговому нерву,
не имеет двух корешков, которые выходят из мозга
раздельно, а потом соединяются; 2) черепные нер-
вы, как и спинномозговые, имеют в своем составе
двигательные, чувствительные и вегетативные во-
локна, но не все нервы смешанные.
Рассмотрим план строения черепного нерва, со-
держащего только чувствительные волокна, соот-
ветствующие заднему корешку спинномозгового
нерва. Такой нерв обязательно имеет узел с чувс-
твительными псевдоуниполярными клетками вне
мозга и чувствительные ядра в головном мозге,
соответствующие чувствительным ядрам задних
рогов спинного мозга.
126
Черепной нерв может содержать только двига-
тельные или двигательные и вегетативные волокна,
соответствуя переднему корешку спинномозгового
нерва. В этом случае нерв имеет двигательные и
вегетативные ядра в мозге, как и у спинномозгово-
го нерва. Однако в составе спинномозгового нерва
проходят симпатические вегетативные волокна, а
в составе черепных нервов — парасимпатические.
И, наконец, есть черепные нервы, которые в своем
составе имеют чувствительные, двигательные и па-
расимпатические волокна.
I пара — nervi olfactorii
Обонятельные нервы (их около 20) — чувстви-
тельные нервы, не имеющие анатомически оформ-
ленных чувствительных ядер и узлов. Состоят из
отростков чувствительных клеток обонятельного
эпителия — filia olfactoria. Малая прочность тонких
обонятельных нервов и фиксация их в отверстиях
lamina cribrosa твердой мозговой оболочкой обус-
ловливает разрывы или прижатия при травмах,
опухолях и отеке мозга и т.п., что приводит к сни-
жению или потере обоняния.
Обоняние проверяют с помощью набора пахучих
эссенций отдельно для каждой стороны.
II пара — nervus opticus
Зрительный нерв представляет собой часть само-
го мозга, поэтому он не нуждается в ядре. Являясь
нервом специальной чувствительности — не имеет
анатомически оформленного узла. Формируется из
отростков мультиполярных клеток сетчатки. Каж-
дый зрительный нерв состоит приблизительно из
миллиона волокон, которые отправляют сигналы
от сетчатки в головной мозг. По ходу нерва разли-
127
чают 4 части: 1) внутриглазная, pars intraocularis,
2) глазничная, pars orbitalis, 3) канальная, pars
canalis и 4) внутричерепная, pars intracranialis. 2-я,
3-я и 4-я части нерва окружены оболочками голо-
вного мозга и спинномозговой жидкостью.
У офтальмологов есть таблицы для проверки
остроты зрения и приборы для определения полей
зрения.
Полное повреждение зрительного нерва ведет
к слепоте, частичное — к выпадению отдельных
участков поля зрения — появлению скатом.
Ill, IV, VI пары — n. oculomotorius,
n. trochlearis, n. abducens
Глазодвигательный нерв имеет в своем составе
двигательные и вегетативные волокна. Известно,
что двигательное ядро состоит из 5 групп клеток.
Волокна из отдельных групп ядра иннервируют
определенные мышцы глазного яблока: верхнюю
прямую, мышцу, поднимающую верхнее веко, ниж-
нюю косую, медиальную и нижнюю прямую мыш-
цы. Парасимпатическое ядро, n. accessorius nervi
oculomotorii, или ядро Якубовича, иннервирует
мышцу, суживающую зрачок, m. sphincter pupil 1ае,
а ядро Перля — n. caudatus centralis, иннервиру-
ет ресничную мышцу, гл. ciliaris, участвующую в
аккомодации.
Блоковый и отводящий нервы — чисто двига-
тельные. Блоковый иннервирует верхнюю косую
мышцу, а отводящий — наружную прямую мышцу
глазного яблока (рис. 82, 83). Итак, три нерва
регулируют произвольные движения глазного яб-
лока, а глазодвигательный нерв к тому же конт-
ролирует степень кривизны хрусталика в процессе
фокусировки и сужение зрачка при ярком свете.
128
Ядра III, IV, VI нервов имеют двухстороннюю
корковую иннервацию, т.е. tr. corticonuclearis идет
на своей и противоположной стороне, поэтому фун-
кция нервов при одностороннем поражении ядер
не страдает.
Рис. 82. Схема иннервации наружных мышц глазного
яблока черепными нервами; внизу показана работа
верхней и нижней косой мышц глазного яблока
Все нервы, выйдя из мозга, во-первых, находят-
ся в подпаутинном пространстве, потом прободают
твердую мозговую оболочку; во-вторых, проходят
через верхнюю глазничную щель; в-третьих, про-
ходят через кавернозный синус. Поэтому перифе-
рические поражения нервов наблюдаются 1) при
менингитах и арахноидитах; 2) при травмах и опу-
холях в области верхней глазничной щели и 3) при
воспалении или тромбозе кавернозного синуса.
Опираясь на знание топографии нервов и их
ядер, работы мышц, ими иннервируемых, врач лю-
бого профиля должен поставить топический диагноз
и своевременно направить больного к неврологу, так
как опухоль мозга развивается быстро и по тече-
5 Анатомия нервной СНСТЮО4
129
нию всегда злокачественная. Например, у больного
левый глаз вправо поворачивается, следовательно,
левый III нерв в норме, а при движении влево ос-
танавливается, следовательно, не работает левый
VI нерв. Патологический процесс располагается не
в глазнице, а дальше, при выходе левого VI нерва
из мозга, на уровне нижнего края моста, где эти
нервы разведены в стороны (рис. 84).
Рис. 83. Схема зон иннервации III, IV и VI черепных
нервов (по С.Ю. Стебельскому)
V нерв — п. trigeminus
Тройничный нерв имеет в своем составе двига-
тельные и чувствительные волокна. Вегетативных
не имеет. Чувствительный узел тройничного нерва
лежит в области тройничного вдавления на пира-
130
миде височной кости и носит название Гассеров
узел.
Рис. 84. Схема выхода черепных нервов
из мозга:
I-XII — черепные нервы; 1 — глазное яблоко; 2 — височная
доля полушария большого мозга; 3 — ножка мозга; 4 — мост;
5 — мозжечок; б — пирамида продолговатого мозга; 7 — спин-
ной мозг
Тройничный нерв выходит из моста на границе со
средней мозжечковой ножкой двумя корешками —
чувствительным и двигательным. Периферические
отростки псевдоуниполярных клеток тройничного
узла образуют три ветви (рис. 85, см. цв. вклейку).
Двигательные волокна присоединяются только к
третьей ветви.
131
Первая ветвь — глазной нерв проникает
глазницу через верхнюю глазничную щель,
рая — верхнечелюстной нерв проходит через
лое отверстие, а третья ветвь — нижнечелкм
нерв — через овальное отверстие черепа.
Тройничный нерв иннервирует жевательную мус
кулатуру и другие мышцы, развивающиеся из
висцеральной дуги. Обеспечивает чувствительну
иннервацию кожи лица, конъюнктиву глаза, слизис
тых оболочек носовой и ротовой полостей и зубов
От чувствительных ядер тройничного нерва начи
нается тройничная петля, lemniscus trigeminalis
которая заканчивается в зрительном бугре, дале<
отростки ядер зрительного бугра проходят чере;
внутреннюю капсулу к g. postcentralis.
По ходу ветвей тройничного нерва располагаются
парасимпатические ганглии, в которых переключа-
ются парасимпатические волокна, принадлежащие
III, VII и IX нервам.
Постганглионарные парасимпатические волокна
идут далее к органу в составе ветвей V нерва, ис-
пользуя его ветви как «рельсы» (рис. 85 , 86, 90,
см. цв. вклейку).
При одностороннем поражении двигательно-
го ядра тройничного нерва жевательная функция
не страдает, так как ядро получает волокна tr.
corticonuclearis от двух полушарий.
При повреждении тройничного нерва, его дви-
гательных ветвей наблюдается спазм (тризм) или
паралич жевательных мышц и их атрофия, а при
поражении чувствительных ветвей — жгучие боли в
местах выхода кожных ветвей из черепа. Болезнен-
но надавливание на эти точки (метод обследования
V нерва). Каждая из трех ветвей тройничного не-
рва иннервирует треть кожи лица (рис. 87) - это
132
зоны периферической иннервации кожи лица
ями V нерва.
Рис. 87. Схема зон периферической иннервации кожи
лица ветвями тройничного нерва:
1 — глазного нерва; 2 — верхнечелюстного нерва; 3 — ниж-
нечелюстного нерва
С другой стороны, волокна V нерва, иннервирую-
щие кожу лица, несут раздражения с определенных
участков кожи к определенным частям ядра, п.
spinalis nervi trigemini (рис. 88). Волокна, идущие
от медиальных участков лица, оканчиваются в вер-
хних отделах этого ядра, независимо от того, в со-
ставе какой из трех ветвей они проходят. Волокна,
идущие от латеральных участков кожи лица, окан-
чиваются в нижних отделах ядра. Следовательно, в
иннервации кожи лица отмечается сегментарность.
Эта сегментарность проявляется при поражении
и. spinalis nervi trigemini. В этих случаях области
расстройств чувствительности на лице не совпадают
133
с областями распределения в коже ветвей V нерва,
а носят сегментарный, «луковичный» характер — в
виде дугообразных полос, пяти зон Зельдера.
Рис. 88. Схема зон сегментарной иннервации кожи лица
и связей зон с отделами nucleus spinalis nervi trigemini (1)
VII нерв, n. facialis
Лицевой нерв — это смешанный нерв, имеющий
в своем составе двигательные, чувствительные и
парасимпатические волокна. Лицевой нерв осущест-
вляет двигательную иннервацию мимических мышц
и мышц, производных II висцеральной дуги.
В составе лицевого нерва старые анатомы опи-
сали промежуточный нерв, чтобы во избежание
неприятностей не называть его тринадцатым нервом.
Лицевой нерв и промежуточный имеют одно разви-
тие, переплетаются, тем не менее, это разные нервы.
Лицевой нерв через волокна промежуточного нерва
134
является вкусовым нервом для передних двух третей
языка и парасимпатическим секреторным нервом
для всех желез лица, кроме околоушной.
Лицевой нерв выходит из мозга в мосто-мозжеч-
ковом углу и затем проходит через канал лицевого
нерва височной кости. Двигательные ветви лицевого
нерва к мимическим мышцам проходят в толще око-
лоушной железы (рис. 89, см. цв. вклейку).
Методы исследования лицевого нерва. При ос-
мотре выявляют асимметрию кожных складок,
глазных щелей, уровня стояния углов рта. Ми-
мические мышцы исследуют при двигательной на-
грузке, предлагая обследуемому зажмурить оба
глаза, поднять брови, показать зубы, сложить губы
трубочкой и посвистеть, сомкнуть губы и надуть
щеки. Вкус на передних двух третях языка обыч-
но на сладкое и кислое проверяют, капая жидкие
растворы на язык.
При нарушении двигательной функции нерва
наблюдается паралич мимической мускулатуры.
При одностороннем поражении наступает асим-
метрия лица вследствие тяги мышц здоровой сто-
роны. Кроме этого, не смыкаются веки, остается
открытой глазная щель из-за поражения круговой
мышцы глаза. Поражение круговой мышца рта и
щечной мышцы ведет к затруднению речи и при-
ема пищи.
Патологический процесс в мосту затрагивает не
только ядра VII нерва, но и рядом расположенное
ядро VI, добавляя соответствующие симптомы по-
ражения соседних образований.
VIII нерв, n. vestibulocochlearis
Преддверно-улитковый нерв является чувстви-
тельным, проводит слуховые импульсы от спи-
135
рального органа и информацию о положении тела
согласно ориентации головы и движении тела в
пространстве.
Улитковый (спиральный) узел находится в спи-
ральном канале улитки, преддверный узел — во
внутреннем слуховом проходе. Выйдя из внутрен-
него слухового прохода, VIII нерв входит в мост в
области мосто-мозжечкового угла.
Поражение вестибулярной части VII нерва прояв-
ляется головокружением, нарушением устойчивости
тела в покое, нарушением координации движений,
а поражение слуховой части проявляется сни-
жением, искажением слуха или глухотой. В норме
человек должен услышать одним ухом шепот на
расстоянии 4-6 метров.
Вестибулярный аппарат проверяют на специаль-
ном кресле. После 10 оборотов на кресле в одну сто-
рону и 10 — в другую испытуемый должен пройти
прямо 10 метров. •
IX нерв, п. glossopharyngeus
Языкоглоточный нерв по функции смешанный,
имеет двигательные, чувствительные и парасимпа-
тические волокна. Языкоглоточный нерв выходит из
продолговатого мозга из дорсо-латеральной бороз-
ды, а из полости черепа через яремное отверстие,
в пределах которого находятся его чувствительные
узлы. Затем нерв располагается между внутренней
сонной артерией и внутренней яремной веной, про-
ходит к шилоглоточной мышце и распадается на
концевые язычные ветви.
IX нерв является чувствительным нервом для
задней трети языка, мягкого неба, среднего уха и
глотки, двигательным нервом для шилоглоточной
мышцы и секреторным для околоушной железы.
136
X нерв, n. vagus
Блуждающий нерв, также смешанный нерв.
X нерв иннервирует мышцы глотки, мягкого неба,
гортани, их слизистую оболочку, все органы груд-
ной и брюшной полостей до сигмовидной кишки.
Блуждающий нерв выходит из дорсолатераль-
ной борозды продолговатого мозга, через яремное
отверстие покидает череп. В области шеи нерв идет
в составе сосудисто-нервного пучка вместе с об-
щей сонной артерией и внутренней яремной веной
в пределах сонного треугольника. Через верхнюю
апертуру попадает в грудную полость, располагается
сначала в верхнем средостении, а потом в заднем
(рис. 90-92, см. цв. вклейку). Проходит в брюшную
полость через пищеводное отверстие, причем левый
блуждающий нерв образует переднее сплетение пи-
щевода и желудка, а правый — заднее. Зона иннер-
вации нерва простирается до сигмовидной кишки.
Изолированное поражение IX или X нерва встре-
чается редко. При вовлечении в процесс двух нервов
наблюдаются нарушения глотания (пища попадает
в гортань или выливается в носовую полость), го-
лос приобретает гнусавый оттенок. Полное выпа-
дение функций блуждающего нерва несовместимо
с жизнью.
XI нерв, n. accessorius
Добавочный нерв является двигательным нер-
вом. Соответственно двум двигательным ядрам у
нерва имеются черепной и спинномозговой кореш-
ки. Спинномозговой корешок поднимается через
большое затылочное отверстие, соединяется с че-
репным корешком, и они вместе покидают череп
через яремное отверстие.
137
Добавочный нерв иннервирует мышцы: груди-
но-ключично-сосцевидную и трапециевидную. При
поражении нерва развивается паралич и атрофия
этих мышц.
При двухстороннем поражении голова свисает
на грудь.
XII нерв, n. hypoglossus
Подъязычный нерв также является двигатель-
ным нервом. Из продолговатого мозга выходит
корешками через вентро-латеральную борозду,
из полости черепа — через canalis hypoglossalis.
Спускается между внутренней сонной артерией и
внутренней яремной веной в поднижнечелюстной
треугольник (рис. 90, 91, см. цв. вклейку).
Здесь нерв принимает верхний корешок от пе-
редних ветвей 1-2-го шейных спинномозговых
нервов, идущий на образование глубокой шейной
петли. На протяжении 2 см этот корешок следует
вместе с подъязычным нервом, используя его обо-
лочку в качестве кабеля. После выхода верхнего
корешка глубокой шейной петли подъязычный нерв
образует дугу и вступает в толщу языка, иннерви-
руя его мышцы.
При поражении нерва наступает паралич мышц
языка противоположной стороны. При высовыва-
нии языка заметно его отклонение в сторону пора-
жения, так как здоровые мышцы выдвигают язык
сильнее.
Спинномозговые
НЕРВЫ (NERVI SPINALES)
Глава
Развитие спинномозговых нервов
При образовании нервной трубки отростки нейро-
бластов основной пластинки прорастают в исчер-
ченную мускулатуру (рис. 93), образуя передние
двигательные корешки. Отростки нейробластов ган-
глиозных валиков врастают в крыльную пластинку
нервной трубки, образуя задние чувствительные ко-
решки. Слияние корешков с образованием спинномоз-
гового нерва происходит на 5-6-й неделе развития.
Рис. 93. Схема расположения невротомов, миотомов и
дерматомов после образования конечностей
Зародыш имеет метамерное строение. Метаме-
ры — это ряд последовательно расположенных
участков тела, в которых в той или иной степени
повторяются системы морфофункциональных об-
разований. Сегментами нервной трубки являются
невротомы. Напротив 1-го невротома располагается
миотом и дерматом. До 4-5-й недели внутриутроб-
ного развития сохраняется четкая система: невро-
том — миотом — дерматом.
В конце 4-5-й недели появляются почки конеч-
ностей. При этом происходит перемещение того,
139
что лежало друг напротив друга, а нервные ве-
точки вытягиваются за перемещающимися мыш-
цами (рис. 93). Так как почки верхних конечнос-
тей закладываются на уровне 4-го шейного - 1-го
грудного сегментов, а почки нижних — на уровне
поясничных и крестцовых сегментов, то из нервных
отростков именно этих сегментов образуются пле-
чевое, поясничное и крестцовое сплетения.
Исчерченная мускулатура способна сокращаться
в 8 недель, а в 2-3 месяца эти сокращения носят
рефлекторный характер. В это же время диафрагма
начинает совершать тренировочные дыхательные
движения.
Спинномозговые нервы - это парные образова-
ния периферической нервной системы, формирую-
щиеся соединением передних и задних корешков,
выходящие из позвоночного канала через межпозво-
ночные отверстия и иннервирующие определенный
участок тела (метамер). Спинномозговые нервы
образуют сплетения и нервные стволы. У человека
31 пара спинномозговых нервов: 8 пар шейных
(С( — Сн), 12 — грудных (Tht ThI2), 5 пояснич-
ных (L( — L5), 5 — крестцовых (S, S5) и 1 пара
копчиковых (Cot).
Спинномозговые нервы в своем составе имеют
различное количество нервных волокон, которое
определяется размерами иннервируемой области,
насыщенностью рецепторными аппаратами и диф-
ференцировкой скелетных мышц. Самыми толстыми
являются нижние шейные, поясничные и крестцовые
спинномозговые нервы, иннервирующие верхние и
нижние конечности. Задние корешки спинномоз-
говых нервов, за исключением I шейного нерва,
гораздо толще передних, что свидетельствует о пре-
обладании в составе нерва чувствительных волокон
140
над двигательными волокнами. Корешки спинно-
мозговых нервов вблизи спинного мозга проходят
в подпаутинном пространстве и окружены мягкой
мозговой оболочкой. В области межпозвоночных
отверстий они вместе со спинномозговым узлом
плотно облачаются твердой мозговой оболочкой,
переходящей в пределах ствола спинномозгового
нерва в периневральное влагалище.
Каждый спинномозговой нерв, выйдя из меж-
позвоночного отверстия, делится на 4 ветви: ме-
нингеальную, г. meningeus, заднюю, г. dorsalis,
переднюю, г. ventralis и белую соединительную
ветвь, г. communicans albus. Менингеальная ветвь
спинномозгового нерва содержит чувствительные и
симпатические волокна. Она иннервирует оболоч-
ки спинного мозга и их сосуды (рис. 94, см. цв.
вклейку).
Задняя и передняя ветви являются смешанными
и иннервируют кожу, мышцы и скелет в области
туловища и конечностей. Они в своем составе име-
ют чувствительные, двигательные и симпатичес-
кие волокна. Чувствительные волокна начинаются
от рецепторов кожи, мышц, сухожилий, связок,
надкостницы и костей. Двигательные волокна за-
канчиваются в скелетных мышцах. Симпатические
волокна иннервируют потовые железы, мышцы, под-
нимающие волосы, гладкую мускулатуру сосудов.
Задние ветви сохраняют сегментарное строение.
Они иннервируют глубокие мышцы и кожу задней
поверхности шеи и спины и делятся на медиаль-
ную и латеральную ветви (рис. 95, 96, см. цв.
вклейку).
Передние ветви спинномозговых нервов, так же
как и задние, смешанные по функции, в основном
утрачивают свойственное им вначале метамерное
141
строение. Сегментарный ход передних ветвей спин-
номозговых нервов сохраняется только на туловище,
где не происходило смещение метамеров. Здесь
развиваются межреберные нервы. В шейном отделе,
поясничном и крестцовом отделах передние ветви
утратили метамерное строение, соединяются друг с
другом петлями и образуют сплетения.
Сплетение, plexus — это переплетающиеся пере-
дние ветви спинномозговых нервов, которые обра-
зуются за счет смещения дерматомов и миотомов и
иннервируют шею, конечности и переднюю повер-
хность туловища.
Различают 4 сплетения: шейное, плечевое, по-
ясничное и крестцовое. Нервы, отходящие от этих
сплетений, могут быть чувствительными, двигатель-
ными или смешанными. В них есть симпатические
волокна. Поэтому клиническая картина поражения
складывается из двигательных, чувствительных и
вегетативных расстройств.
Аксоны, выходящие из соседних сегментов, мо-
гут идти к мышцам в составе 1-го или 2-го нервов
(рис. 97, см. цв. вклейку). Кроме того, в составе
1-го нерва могут находиться волокна, идущие от
1-го, 2-го, или 3-го сегментов.
Следует также уточнить понятие периферической
и сегментарной иннервации. Каждый спинномозго-
вой нерв распределяется в пределах определенного
участка кожи или в определенных мышцах, т.е. в
своей зоне. Такая иннервация называется перифе-
рической или зональной (рис. 98, см. цв. вклейку).
Неврологи с помощью иглоукалывания определяют
места повреждения нервов; отсутствие чувстви-
тельности в той или иной области может выявить
нарушения в нервных отделах, удаленных от ис-
следуемой зоны. Так как все нервы смешанные, то
142
при повреждении нерва наблюдаются двигательные,
чувствительные и вегетативные расстройства. Кроме
того, имеются зоны перекрытия кожной иннерва-
ции, когда участок кожи иннервируется 2 соседними
нервами.
Каждый спинномозговой нерв является как бы
продолжением сегмента спинного мозга. Сегмен-
тарный тип иннервации представлен в виде полос,
которые на туловище расположены поперечно, на
конечностях продольно (рис. 98).
Рис. 98. Схема сегментарной иннервации кожи
Шейное сплетение, plexus cervicalis
Plexus cervicalis образуется передними ветвя-
ми четырех верхних шейных нервов (С, —С 1V).
143
Оно располагается на глубоких мышцах шеи и
прикрыто грудино-ключично-сосцевидной мышцей
(рис. 99, см. цв. вклейку). По составу волокон ветви
шейного сплетения делятся на 3 группы — двига-
тельные, чувствительные и смешанные.
Кожные нервы: N. occipitalis minor; n. auricularis
magnus; n. transversus colli; nn. supraclaviculares
(рис. 90, 100, 101, см. цв. вклейку). Верхняя
ветвь n. transversus colli соединяется c r. colli nervi
facialis, образуя поверхностную шейную петлю, ansa
cervicalis superficialis, которая иннервирует кожу
шеи и m. platysma.
Мышечные нервы: К mm. recti capitis ant. et lat.;
longi capitis et colli; scaleni; m. levator scapulae;
intertransversarii anteriores. Двигательные ветви
шейного сплетения образуют верхний и нижний
корешки. Верхний проходит на протяжении 2 см
под периневральной оболочкой XII нерва, поки-
дая которую, соединяется с нижним корешком.
Образуется глубокая шейная петля, ansa cervicalis
profunda (рис. 89-101, см. цв. вклейку). Ветви,
отходящие от глубокой шейной петли, иннервируют
мышцы, расположенные ниже подъязычной кости.
Mm. sternocleidomastoideus et trapezius иннервиру-
ют как мышечные ветви шейного сплетения, так и
XI черепной нерв.
Смешанный нерв: диафрагмальный нерв, n. phre-
nicus. Нерв спускается по передней поверхности
передней лестничной мышцы, проникает в грудную
полость через верхнюю апертуру, проходит через
верхнее, а затем среднее средостение (рис. 101 см.
цв. вклейку). В отличие от блуждающего нерва,
диафрагмальный с обеих сторон спускается к диа-
фрагме впереди от корня легкого. Двигательные
волокна иннервируют мышцу диафрагмы. Чувстви-
144
тельные ветви диафрагмальных нервов прободают
диафрагму: правый нерв проходит рядом с верхней
полой веной, а левый — у верхушки сердца, меж-
ду плеврой и перикардом. Эти ветви иннервируют
брюшину в области диафрагмы, плевру, перикард,
пищевод, соединительнотканную оболочку печени,
желчный пузырь.
При печеночной патологии болит не сама печень,
а ее оболочка, снабженная нервными окончания-
ми. Поэтому при заболеваниях печени положите-
лен френикус-симптом. При обследовании голова
больного запрокинута назад, врач надавливает на
малую надключичную ямку (место прохождения
нерва). При положительном симптоме боль возни-
кает только справа.
При раздражении диафрагмального нерва появ-
ляются одышка, икота, а при поражении — паралич
половины диафрагмы.
Плечевое сплетение, plexus brachialis
Plexus brachialis образуется передними ветвями
спинномозговых нервов (Cv — CVI11, Th,). Распола-
гается в области шеи в межлестничном пространстве,
spatium interscalenum (рис. 102, см. цв. вклейку).
В этом месте плечевое сплетение представлено 3
стволами: верхним, средним и нижним, от которых
отходят короткие ветви к мышцам плечевого пояса.
Стволы и короткие ветви составляют надключичную
часть плечевого сплетения. В этой же части спле-
тения стволы начинают делиться и формировать 3
пучка. Пучки окружают с трех сторон подключич-
ную артерию и соответственно своему положению
носят название: медиальный, латеральный и задний
(рис. 102, см. цв. вклейку). Части пучков, располо-
женные ниже ключицы, составляют подключичную
145
часть плечевого сплетения, которая разделяется на
его длинные ветви.
Короткие ветви и их зоны иннервации:
N. dorsalis scapulae иннервирует m. levator
scapulae, mm. rhomboidei.
N. thoracicus longus — m. serratus anterior.
N. suprascapularis — mm. supraspinatus и
infraspinatus; капсулу плечевого сустава.
Nn. pectorales medialis et lateralis — m.
pectoralis major et minor.
N. subclavius иннервирует m. subclavius.
N. subscapularis — m. subscapularis, teres
major.
N. thoracodorsalis — m. latissimus dorsi.
N. axillaris — mm. deltoideus, teres minor,
плечевой сустав; его ветвь — n. cutaneus brachii
lateralis superior — иннервирует кожу над дельто-
видной мышцей.
Длинные ветви и их зоны иннервации (рис. 103,
104, см. цв. вклейку):
N. musculocutaneus иннервирует все передние
мышцы плеча; его ветвь — n. cutaneus antebrachii
lateralis — кожу предплечья с латеральной сто-
роны.
N. medianus — иннервирует передние мышцы
предплечья (за исключением m. flexor carpi ulnaris и
половины т. flexor digitorum profundus), thenar (за
исключением т. adductor pollicis, глубокой головки
т. flexor pollicis brevis), первую и вторую тт.
lumbricales, кожу I, II, III и половины IV пальцев
на ладонной поверхности кисти.
N. ulnaris иннервирует m. flexor carpi ulnaris
и половину т. flexor digitorum profundus, т.
adductor pollicis, глубокую головку т. flexor pollicis
brevis, все mm. interossei, третью и четвертую mm.
146
lumbricales, hypothenar, кожу V, IV и половины III
пальцев на тыле кисти, а также V и половины IV
пальцев на ладонной поверхности кисти.
Nn. cutaneus brachii et antebrachii mediales —
кожу плеча и предплечья с медиальной стороны
N. radialis — задние мышцы плеча и предпле-
чья, кожу задней и заднелатеральной поверхности
плеча, задней поверхности предплечья, I, II и по-
ловины III пальцев на тыле кисти.
При ручном методе извлечения плода у новорож-
денного может произойти разрыв ветвей, отходящих
от 5-6-го шейных сегментов. Эти ветви образуют п.
suprascapularis и n. axillaris, которые иннервируют га.
supraspinatus, m. infraspinatus и m. deltoideus. Плечо
при этом свисает книзу, приведено и повернуто вов-
нутрь, так называемая «рука, просящая взятку».
При повреждении n. dorsalis scapulae развивает-
ся «крыловидная лопатка». При этом не работают
ромбовидные мышцы, а лопатку перетягивает пе-
редняя зубчатая мышца. «Крыловидная лопатка»
наблюдается также при повреждении n. thoracicus
longus при удалении молочной железы.
При повреждении n. musculocutaneus невозможно
сгибание в локтевом суставе, развивается атрофия
бицепса.
При повреждении лучевого нерва возникает «сви-
сающая кисть», так как не работают разгибатели
кисти.
Поражение локтевого нерва вызывает образова-
ние «когтистой лапы», так как не функционируют
и атрофируются межкостные мышцы и западают
межкостные промежутки; не сгибаются 4-й и 5-й
пальцы, а 1-й — не приводится.
При повреждении срединного нерва развивается
«обезьянья кисть» вследствие атрофии мышц тена-
147
pa. He сгибаются 1-й, 2-й и 3-й пальцы. Такую кисть
называют еще рукой моления или рукой акушера.
Межреберные нервы,
nn. intercostales
Межреберные нервы — это передние ветви 11
верхних грудных нервов (рис. 105, 106, см. цв.
вклейку); передняя ветвь 12-го грудного нерва назы-
вается подреберным нервом, n. subcostalis. Верхние
6 межреберных нервов иннервируют кожу и мышцы
груди, плевру, а также молочные железы, а ниж-
ние кожу и мышцы живота, а также брюшину.
Правый подреберный нерв в верхних отделах
иннервирует плевру, а ниже — брюшину в правой
паховой области. В связи с этим иногда право-
стороннюю плевропневмонию принимают за ап-
пендицит, поскольку боли иррадиируют по ходу
правого n. subcostalis и полностью симулируют все
аппендикулярные симптомы. Картина крови при
этом, естественно, также воспалительная. Поэтому
хирургу необходимо прослушивать легкие, чтобы
больному с плевропневмонией не сделать ненужную
операцию.
Поясничное сплетение, plexus lumbalis
Plexus lumbalis образуется передними ветвями
L, — L|V и веточкой от XII грудного нерва. Пояс-
ничное сплетение располагается в толще большой
поясничной мышцы. Нервы, начинающиеся от по-
ясничного сплетения, выходят из-под латерального
или медиального края большой поясничной мышцы
или прободают ее спереди (рис. 107, 108, см. цв.
вклейку). Они направляются к передней брюшной
стенке, к наружным половым органам и к нижней
конечности.
148
Rami muscuiares — к квадратной мышце по-
ясницы, поясничным мышцам.
N. iliohypogastricus — иннервирует внутрен-
нюю косую и поперечную мышцы живота, кожу
верхней части ягодицы и кожу передней брюшной
стенки выше лобковой области.
N. ilioinguinalis проходит в паховом канале, ин-
нервирует содержимое пахового канала, мышцы живота
и кожу лобка, мошонки или большой половой губы.
N. genitofemoralis появляется на передней по-
верхности большой поясничной мышцы, его г. femo-
ral is иннервирует кожу бедра под паховой связкой,
а г. genitalis — половые органы.
N. cutaneus femoris lateralis иннервирует кожу
латеральной поверхности бедра.
N. femoralis (рис. 107, 108, см. цв. вклейку)
проходит через мышечную лакуну на бедро, в бедрен-
ном треугольнике распадается на мышечные ветви к
передним мышцам бедра и кожные ветви к передней
поверхности бедра. Его ветвь — подкожный нерв,
n. saphenus, проходит в приводящем канале, выходит
через его переднее отверстие, на голени располагает-
ся рядом с большой подкожной веной; иннервирует
кожу голени и стопы с медиальной стороны.
N. obturatorius (рис. 107, 108, см. цв. вклей-
ку) выходит из-под медиального края большой
поясничной мышцы, направляется в малый таз и
покидает его через запирательный канал; иннерви-
рует все приводящие мышцы, тазобедренный сустав,
ш. obturatorius и кожу над ними.
Повреждение запирательного нерва вызывает
затруднение приведения бедра.
Повреждение бедренного нерва вызывает атро-
фию четырехглавой мышцы бедра, больной не мо-
жет разогнуть голень и согнуть бедро.
149
Крестцовое сплетение, plexus sacralis
Plexus sacralis образуется передними ветвями
I I S — S
bIV 1-V O, oIV
Располагается на передней поверхности груше-
видной мышцы; его ветви покидают малый таз че-
рез надгрушевидное и подгрушевидное отверстия
(рис. 107, 109, см. цв. вклейку).
Короткие ветви:
Rami musculares к внутренней запирательной,
грушевидной мышцам и квадратной мышце бедра.
N. gluteus superior иннервирует m. gluteus
medius, gluteus minimus, tensor fasciae latae.
N. gluteus inferior иннервирует m. gluteus
maximus и капсулу тазобедренного сустава.
N. pudentus выходит из полости малого таза
через подгрушевидное отверстие и через малое се-
далищное отверстие попадает в fossa ischiorectalis.
Иннервирует мышцы и кожу промежности, наруж-
ные половые органы.
Длинные ветви:
N. ischiadicus (рис. 109, см. цв. вклейку) вы-
ходит из полости малого таза через подгрушевидное
отверстие, в ягодичной области располагается под
нижней частью большой ягодичной мышцы. В ниж-
ней трети бедра или в подколенной ямке делится
на свои конечные ветви: большеберцовый и общий
малоберцовые нервы. Его rr. musculares иннерви-
руют заднюю группу мышц бедра.
N. tibialis (рис. 109, см. цв. вклейку) проходит
в голено-подколенном канале, позади медиальной
лодыжки делится на конечные ветви — nn. plantares
lateralis et medialis. Большеберцовый нерв иннерви-
рует задние мышцы голени.
N. plantaris medialis иннервирует мышцы меди-
альной группы подошвы кроме m. adductor hallucis и
150
латеральной головки m. flexor hallucis brevis, flexor
digitorum brevis, первую и вторую mm. lumbricales.
Nn digitales plantares proprii иннервируют кожу
обращенных друг к другу I-IV пальцев.
N. plantaris lateralis иннервирует третью и чет-
вертую mm. lumbricales, m. quadratus plantae, m.
flexor digiti minimi, m. abductor digiti minimi, все
mm. interossei, m. adductor hallucis и латераль-
ную головку m. flexor hallucis brevis. Nn. digitales
plantares proprii иннервируют кожу обращенных
друг к другу сторон IV-V пальцев.
N. peroneus (fibularis) communis отдает кож-
ную ветвь — n. cutaneus surae lateralis, которая
вместе с такой же медиальной ветвью от большебер-
цового нерва образует n. suralis и далее n. cutaneus
pedis dorsalis lateralis.
N. peroneus (fibularis) superficialis (рис. 108, см.
цв. вклейку) проходит через canalis musculoperoneus
superior, иннервирует латеральные мышцы голени; его
кожные ветви: n. cutaneus dorsalis medialis иннерви-
рует медиальную сторону стопы, I пальца и края II и
III пальцев, a N. cutaneus dorsalis intermedius — кожу
обращенных друг к другу сторон III—V пальцев.
N. peroneus (fibularis) profundus (рис. 108, см.
цв. вклейку) прободает межмышечную перегородку
голени. Иннервирует переднюю группу мышц го-
лени, голеностопный сустав, короткий разгибатель
пальцев; его ветви — nn. digitales dorsales иннерви-
руют кожу I межпальцевого промежутка.
Повреждение общего малоберцового нерва, ветви
которого иннервируют передние и задние мышцы
голени, приводит к их атрофии, к возникновению
у больного свисающей стопы (конской стопы) и
петушиной походки (чтобы не задевать носком,
больной высоко поднимает ногу).
151
Повреждение большеберцового нерва приводит
к атрофии задних мышц голени. При этом развива-
ется когтистая или пяточная стопа. Больной ходит
на пятках, стопа, пальцы находятся в состоянии
разгибания, своды стоны углублены.
Копчиковое сплетение, plexus coccygeus, обра-
зуется передними ветвями Sv, СО] Его ветви, пп.
anococcygei, иннервируют кожу у верхушки копчика
и анального отверстия.
Органы чувств
(ORGANA SENSUUM)
Глава
Учение об органах чувств —
эстезиология
Органами чувств называют морфофункциональ-
ные образования, воспринимающие энергию внешне-
го воздействия, трансформирующие ее в нервный
импульс и передающие этот импульс в мозг. Павлов
назвал органы чувств анализаторами (в широком
понимании). Каждый орган чувств имеет:
периферический прибор, воспринимающий
внешнее воздействие и трансформирующий
его в нервный импульс (рецептор);
проводящий путь, передающий нервный им-
пульс в нервный центр (кондуктор);
нервный центр в коре мозга (корковый конец
анализатора).
По восприятию разноудаленных объектов разли-
чают: 1) контактную чувствительность (это осяза-
ние, давление, боль, вкус) и 2) дистантную чувстви-
тельность (зрение, слух, равновесие, обоняние).
Различают три основных типа органов чувств:
К первому относят орган зрения и орган обоняния.
В их составе есть первично-чувствующие клетки,
воспринимающие и передающие раздражение.
Ко второму типу относят орган вкуса, орган
равновесия и слуха. У них есть специализированные
эпителиальные клетки, передающие раздражение к
вторично-чувствующим клеткам.
153
К третьему типу с невыраженной органной ор-
ганизацией относят нервные окончания, восприни-
мающие чувство давления и прочие.
Орган зрения — organum visas
К зрительному анализатору относятся:
I. Oculus (ophthalmos) — глаз, состоящий из
глазного яблока (bulbus oculi) и зрительного нерва
(nervus opticus). Глазное яблоко состоит из трех
оболочек (фиброзной, сосудистой и сетчатки) и
структур, образующих ядро (рис. 110).
Рис. 110. Глазное яблоко; разрез в горизонтальной
плоскости (схема):
1 — cornea; 2 — camera anterior bulbi; 3 — lens; 4 — iris; 5 —
camera posterior bulbi; 6 — conjunctiva; 7 — m. rectus lateralis;
8 — sclera; 9 — choroidea; 10 — retina; 11 — fovea centralis;
12 — n. opticus; 13 — excavatio disci; 14 — axis bulbi externus;
15 — m. rectus medialis; 16 — поперечная ось глазного яблока;
17 — corpus ciliare; 18 — zonula ciliaris; 19 — axis opticus
154
II. Вспомогательный аппарат, к которому отно-
сятся мышцы глазного яблока, слезный и защитный
аппарат.
III. Проводящий зрительный путь и центры
зрения.
Оболочки глазного яблока:
1. Фиброзная оболочка — tunica fibrosa bulbi —
обусловливает форму глазного яблока, выполняет
защитную функцию. В ней различают прозрачную
часть — роговицу (cornea) и склеру (sclera).
2. Сосудистая оболочка - tunica vasculosa
bulbi — имеет большое количество сосудов. В ней
имеются три части:
радужка — iris,
ресничное тело — corpus ciliare,
собственно сосудистая оболочка — choroidea
(рис. 111).
3. Внутренняя (чувствительная) оболочка глаз-
ного яблока — tunica interna bulbi, состоящая из
сетчатки — retina и сосудов сетчатки. Сетчатка имеет
2 части: наружный пигментный слой, pars pigmentosa,
и слой фоторецепторов, pars nervosa — 130 млн.
палочек — рецепторов светового зрения и 6-7 млн.
колбочек — рецепторов цветового зрения.
Содержимым глазного яблока, составляющим
его ядро, являются: водянистая влага, хрусталик
и стекловидное тело. Все образования, входящие
в состав ядра глазного яблока, в норме прозрачны
и выполняют светопроводящую и светопреломляю-
щую функции.
Водянистая влага (humor aquosus) находится
в передней и задней камерах глазного яблока.
Задняя полость глазного яблока заполнена свелым
гелем, называемым стекловидным телом. Продуци-
155
руемые ресничным телом водянистая влага и стек-
ловидное тело поддерживают постоянное внутреннее
давление, благодаря чему глазное яблоко сохраняет
свою форму.
Водянистая влага осуществляет трофику всех
бессосудистых образований глаза (роговицы, хрус-
талика, стекловидного тела), поставляя им кисло-
род, глюкозу и протеины, а по составу аналогична
спинномозговой жидкости.
Рис. 111. Сосудистая оболочка глаза, передний отдел;
внутренняя поверхность:
1 — corona ciliaris; 2 — choroidea; 3 — zonula ciliaris; 4 — lens;
5 — sclera; 6 — plicae ciliares; 7 — processus ciliares; 8 — facies
posterior iridis; 9 — pars ciliaris retinae; 10 — ora serrata; 11 —
retina; 12 — orbiculus ciliaris
Циркуляция водянистой влаги
Водянистая влага образуется путем ультрафиль-
трации крови через стенку ресничных отростков и
156
капилляров ресничного тела. Образовавшаяся жид-
кость поступает в заднюю камеру глаза (рис. 112)
через Петитов канал. Так называется пространство
между волокнами ресничного пояска, или связки
хрусталика — spatia zonularia, имеющее вид кру-
говой щели.
1
Рис. 112. Схема оттока водянистой влаги глаза,
humor aquosus (оттокуказан стрелками):
1 — cornea; 2 — sclera; 3 — conjunctiva; 4 — lens; 5 — iris; 6 —
corpus ciliare; 7 — processus ciliares; 8 — spatia zonularia (Petit);
9 — capsula lentis; 10 — camera posterior bulbi; 11 — camera
anterior bulbi; 12 — spatia anguli iridocornealis (Fontana); 13 —
sinus venosus sclerae (Schlemm); 14 — vv. ciliares anteriores;
15 — w. vorticosae
Из задней камеры жидкость поступает через
зрачок в переднюю камеру глазного яблока. В ра-
дужно-роговичном углу передней камеры между во-
локнами гребенчатой связки (lig. pectinatum iridis)
находятся Фонтановы постранства (spatia anguli
iridocornealis). Через Фонтановы пространства во-
дянистая влага оттекает в венозный синус склеры,
157
sinus venosus sclerae (Шлеммов канал). Последний
заполнен венозной кровью, которая далее оттекает
по водоворотным венам, vv. vorticosae (рис. 112,
113) в глазные вены, vv. ophthalmicae.
Рис. 113. Сосудистая оболочка глазного яблока;
наружная поверхность:
1 — a. ciliaris anterior; 2 — margo ciliaris iridis; 3 — pupilla;
4 — cornea; 5 — margo pupillaris iridis; 6 — iris; 7 — circulus
arteriosus iridis major; 8 — aa. ciliares posteriores breves; 9 —
sclera; 10 — nn. ciliares; 11 — n. opticus; 12 — aa. ciliares
posteriores breves; 13 — a. ciliaris posterior longa; 14 — v.
vorticosa; 15 — choroidea; 16 — anulus ciliaris
Слезный аппарат
Слезный аппарат включает слезные железы и
слезоотводящие пути (рис. 114). Слезные железы
158
выделяют слезы, очищающие глаз. Протоки слезной
железы открываются в верхний свод конъюнктивы.
Слеза стекает сверху вниз по слезному ручью, rivus
lacrimalis, в слезное озеро, lacus lacrimalis, лишние
слезы испаряются или вытекают через слезные точ-
ки в слезные канальцы, потом в слезный мешок и
носослезный проток.
Движения глазных яблок обеспечивает работа
мышц глаза. Эти движения содружественны, т.е.
зрительные оси обоих глаз направлены на один и тот
же объект, что обеспечивает бинокулярное зрение.
При смыкании век обоих глаз глаза поворачива-
ются кверху, что объясняется связями иннервации
круговых мышц глаза и нижних косых мышц гла-
за — феномен Белла (Bell).
Рис. 114. Слезный аппарат:
1 — glandula lacrimalis; 2 — palpebra superior; 3 — canaliculus
lacrimalis superior; 4 — lacus lacrimalis; 5 — saccus lacrimalis;
6 — ductus nasolacrimalis
159
Проводя ши й путь зрительного
анализатора
Рецепторами органа зрения и одновременно те-
лами I нейронов являются палочки и колбочки
сетчатки. Биполярные клетки — тела II нейронов,
а ганглиозные клетки сетчатки — тела III нейро-
нов. Аксоны ганглиозных клеток образуют зритель-
ный нерв, зрительный перекрест (перекрещиваются
только волокна, расположенные медиально) и зри-
тельный тракт (рис. 115).
Зрительный тракт делится на три пучка, подхо-
дящих к подкорковым центрам зрения — к верхним
холмикам четверохолмия, к латеральным колен-
чатым телам и задним ядрам таламуса (телам IV
нейронов).
От клеток ядра верхнего холмика начинается
крышеспинномозговой путь, осуществляющий без-
условнорефлекторные двигательные реакции мус-
кулатуры на внезапные сильные световые раздра-
жения.
Аксоны нейронов латеральных коленчатых тел
проходят через заднюю часть задней ножки внут-
ренней капсулы и заканчиваются в корковом конце
зрительного анализатора — на медиальной повер-
хности затылочной доли по сторонам от шпорной
борозды.
Аксоны клеток задних ядер таламуса передают
зрительную информацию в интеграционный центр
промежуточного мозга — медиальные ядра таламу-
са, а затем в двигательные центры экстрапирамид-
ной системы, в лимбическую систему и гипотала-
мус, обеспечивая эмоциональные реакции, работу
мышц и внутренних органов в ответ на зрительные
раздражения.
160
Рис. 115. Схема проводящего пути зрительного
анализатора:
R — рецепторы и I — первый нейрон — палочки и колбочки сет-
чатки; II — второй нейрон — биполярные клетки сетчатки; III —
третий нейрон — ганглиозные клетки сетчатки; IV — четвертый
нейрон — клетки ядер верхних холмиков среднего мозга, лате-
рального коленчатого тела, подушки таламуса (colliculus superior,
corpus geniculatum laterale, pulvinarthalami); V — корковый конец
зрительного анализатора — клетки коры по сторонам шпорной
борозды (sulcus calcarinus); 1 — спинной мозг; 2 — средний
мозг; 3 — таламус; 4 — внутренняя капсула; 5 — латеральное
коленчатое тело; 6 — кора затылочной доли; 7 — крышеспин-
номозговой тракт (tractus tectospinalis); 8 — клетки двигатель-
ного ядра переднего рога спинного мозга; 9 — зрительный нерв;
10 — зрительный перекрест; 11 — зрительный тракт
6 Аил ом ия нервной системы
161
Филогенез органа зрения
Глаза приматов отличаются от глаз других мле-
копитающих.
Они расположены ближе к носу, что дает сте-
реоскопическое зрение.
В глазнице у них появляется задняя стенка,
что улучшает фиксацию глазного яблока.
Формируется желтое пятно сетчатки, дающее
большую остроту зрения.
Формируется цветовое зрение, позволяющее
животным отличать плоды и листья разной
спелости.
Глаза приматов научились определять мимику
членов стада.
У человека глаза отличаются от глаз обезьян
и других млекопитающих белым цветом склеры,
поэтому люди определяют, куда смотрит другой
человек.
Онтогенез органа зрения
Глаз развивается из разных источников. Сетчатка
и зрительный нерв формируются из зачатка нервной
трубки, сначала переднего, а затем промежуточного
мозга. Из мозговой эктодермы на 3-й неделе внут-
риутробного развития образуются глазные пузырьки
(рис. 116). Затем пузырьки впячиваются вовнутрь
и образуются двустенные глазные бокалы (4-я не-
деля), внутренняя стенка которых преобразуется
в pars nervosa, а наружная — в pars pigmentosa
сетчатки. Глазной стебелек становится зрительным
нервом. Часть наружной эктодермы напротив глаз-
ного бокала под индукционным влиянием глазного
бокала утолщается, инвагинирует и отшнуровыва-
ется, образуя хрусталик (5-я неделя).
162
3
2
2
Рис. 116. Схема развития глаза:
I — образование глазного пузырька: 1 — мозговая эктодерма;
2 — наружная эктодерма; 3 — глазной пузырек; 4 — глазной
стебелек; II — образование глазного бокала; III — формиро-
вание хрусталика из наружной эктодермы; IV — инвагинация
хрусталика в глазной бокал: 2 — наружная эктодерма; 3 — хрус-
талик; 4 — глазной стебелек; 5 — полость глазного бокала
Вокруг закладки глазного бокала мезенхима сгу-
щается в два слоя, из которых образуются фиброзная
и сосудистая оболочки глаза. Передняя часть фиброз-
ной оболочки становится прозрачной роговицей.
Мезенхима, проникающая внутрь глазного бока-
ла, образует стекловидное тело, его сосуды и сосуды
хрусталика. Эпителий конъюнктивы развивается
из наружной эктодермы. На 3-м месяце из свода
конъюнктивы образуются 6 отростков, из которых
163
развивается слезная железа. Веки формируются из
складок кожи на 7-й неделе; на 9-й неделе их края
срастаются эпителиальным швом, который исчезает
к 7 месяцам.
Развитие сосудов глазного яблока
Первые кровеносные сосуды появляются при
длине зародыша в 4,5 мм в мезенхиме, окружающей
глазной бокал (рис. 117—а). Сосуды проникают в
мезенхимную капсулу хрусталика из заднего отдела
глазного яблока через формирующееся стекловидное
тело — артерия стекловидного тела, a. hyaloidea
(рис. 117-6).
Рис. 117. Развитие сосудов глазного яблока.
(Объяснение в тексте)
В стекловидном теле от нее отходят боковые вет-
ви, которые оплетают заднюю поверхность хруста-
лика и продолжаются в сосуды, расположенные на
передней поверхности развивающегося хрусталика,
164
куда они также проникают из развивающейся сосу-
дистой оболочки, образуя на передней поверхности
хрусталика зрачковую мебрану, membrana pupil laris
(рис. 117-г). На 8-й неделе хрусталик уже сфор-
мировался и его капсула обильно кровоснабжается,
особенно в заднем отделе. Хорошо кровоснабжается
зрачковая мембрана, расположенная на передней
поверхности хрусталика.
На 4-м мес. внутриутробного развития централь-
ная ветвь артерии стекловидного тела на задней
поверхности хрусталика делится по магистраль-
ному типу вплоть до капилляров. Капилляры на-
правляются радиально к экватору хрусталика и
переходят в посткапилляры, расположенные строго
параллельно в месте прикрепления ресничного по-
яска (рис. 117-в). Часть артериол развивающейся
радужки вступают в зрачковую мембрану. Сосу-
дистый рисунок зрачковой мембраны образован
дистальными петлями этих артериол с широким
основанием и узкой вершиной, которые лепес-
ткообразно сходятся к центру будущего зрачка
(рис. 117-г).
Артерия стекловидного тела, arteria hyaloidea,
к 6-8 месяцам внутриутробного периода развития
полностью редуцируется. В дальнейшем обнаружена
редукция сосудистой оболочки хрусталика и сосу-
дов зрачковой мебраны (рис. 118). На 8-м месяце
внутриутробного развития эти сосуды исчезают.
Аномалии развития органа зрения
1. Анофтальм — отсутствие одного или двух
глаз.
2. Циклопия — наличие одного или двух глаз
в одной орбите по средней линии в области
лба. Над глазом находится хобот — видоиз-
165
мененный нос. Циклопия сочетается с другими
тяжелыми пороками развития, поэтому циклоп
нежизнеспособен.
3. Микрофтальм, макрофтальм — слишком ма-
ленькие или большие глазные яблоки.
4. Астигматизм — искажение изображения на
сетчатке при неправильной кривизне роговицы
и (или) хрусталика.
в г
Рис. 118. Редукция сосудистой оболочки хрусталика:
а — 4 мес.; 6 — 5 мес.; в — 7 нес.; г — 8 мес. развития; инъ-
екция сосудов водной взвесью черной туши
166
5. Врожденная катаракта — врожденное помут-
нение роговицы.
6. Кератоконус — роговица имеет форму конуса.
7. Врожденная глаукома — недоразвитие Шлем-
мова канала, Фонтановых пространств или
Петитова канала.
8. Колобома — щель, недостающий сегмент в ра-
дужке или других оболочках глазного яблока.
9. Нарушение сроков обратного развития сосу-
дов сосудистой облочки хрусталика (их пере-
систенция) приводят к тяжелым аномалиям:
Клокетов канал (J. G. Cloquet), колобома
сосудистой оболочки, передний и задний лен-
тиконусы хрусталика, микрофакия. Все они,
как правило, сопровождаются врожденной
слепотой. Она же может возникать при нару-
шениях в проводящих путях или в корковом
конце анализатора.
10. Дальнозоркость, близорукость.
И. Сохранение ветвей a. hyaloidea, мешающих
прохождению света через стекловидное тело.
12. Врожденный вывих хрусталика, афакия (его
отсутствие), бифакия (удвоение хрусталика).
13. Сохранение мигательной перепонки (третьего
века птиц). Эпикант (вертикальная полулун-
ная складка у внутреннего угла глаза).
14. Врожденный птоз (опущение века); врожден-
ный энтропион (край века завернут к глазу,
ресницы повреждают глаз); эктропион (веко
завернуто конъюнктивой наружу).
15. Кошачий глаз — зрачок в виде щели. Мно-
жественные зрачки.
16. Альбинизм (1 случай на 20 тысяч новорож-
денных) — при этом цвет радужки серо-крас-
ный, красный зрачок.
167
17. Врожденный меланоз — при этом серая скле-
ра, темная окраска глазного дна.
18. Гетерохрония радужек.
19. Пятнистая гиперпигментация радужки.
20. Врожденное косоглазие при недостаточности
мышц глаз.
21. Дальтонизм — отсутствие какого-либо вида
колбочек в сетчатке (красно-, сине- и зелено-
чувствительных). Дальтон не видел красного
цвета — протаноп. Врубель — дейтараноп (не
видел зеленого цвета). Если нет всех трех видов
колбочек — мир предстает черно-белым.
Преддверно-улитковый орган, organum
vestibulocochleare
Орган слуха и равновесия является перифери-
ческой частью анализатора гравитации, равновесия
и анализатора слуха. Располагается в пределах
одного анатомического образования — лабиринта
и состоит из наружного, среднего и внутреннего
уха (рис. 119).
I. Наружное ухо (auris externa) состоит из
ушной раковины, auricula, наружного слухового
прохода, meatus acusticus externus, и барабанной
перепонки, membrana tympanica. Наружное ухо
играет роль слуховой воронки для улавливания и
проведения звука.
Между наружным слуховым проходом и барабан-
ной полостью располагается барабанная перепонка,
membrana tympanica. Барабанная перепонка упру-
га, малоэластична, тонка (0,1-0,15 мм толщиной),
в центре вогнута вовнутрь. Мембрана имеет три
слоя: кожный, фиброзный и слизистый. В ней есть
ненатянутая часть, pars flaccida — Шрапнелева мем-
брана, не имеющая фиброзного слоя, и натянутая
168
часть, pars tensa. В практических целях мембрану
делят на квадранты.
Рис. 119. Преддверно-улитковый орган (схема):
1 — наружный слуховой проход; 2 — слуховая труба; 3 — ба-
рабанная перепонка; 4 — молоточек; 5 — наковальня; б —
улитка
II. Среднее ухо (auris media) состоит из барабан-
ной полости, cavitas tympani, слуховой трубы, tuba
auditiva, и сосцевидных ячеек, cellulae mastoideae.
Среднее ухо — это система воздухоносных полостей
в толще каменистой части височной кости.
Барабанная полость имеет вертикальный размер
10 мм и поперечный размер — 5 мм. У барабан-
ной полости имеется 6 стенок (рис. 120): лате-
ральная — перепончатая (paries membranaceus),
медиальная — лабиринтная (paries labyrinthicus),
передняя — сонная (paries caroticus), задняя — со-
сцевидная (paries mastoideus), верхняя — пок-
рышечная (paries tegmentalis) и нижняя — ярем-
169
ная (paries jugularis). Нередко в верхней стенке
встречаются щели, в которых слизистая оболочка
барабанной полости прилежит к твердой мозговой
оболочке.
Рис. 120. Схема строения стенок барабанной полости:
1 — paries tegmentalis; 2 — paries mastoideus; 3 — paries
jugularis; 4 — paries caroticus; 5 — paries labyrinthicus; 6 — a.
carotis interna; 7 — ostium tympanicum tubae auditivae; 8 —
canalis facialis; 9 — aditus ad antrum mastoideum; 10 — fenestra
vestibuli; 11 — fenestra cochleae; 12 — n. tympanicus; 13 — v.
jugularis interna
Барабанная полость делится на три этажа: над-
барабанный карман, recessus epitympanicus, сред-
ний — mesotympanicus и нижний — подбарабанный
карман, recessus hypotympanicus. В барабанной
полости находятся три слуховые косточки: мо-
лоточек, наковальня и стремечко (рис. 121), два
170
сустава между ними: наковальне-молоточковыи,
art. incudomallearis, и наковальне-стременной, art.
incudostapedialis, и две мышцы: напрягающая бара-
банную перепонку, m. tensor tympani, и стременная,
m. stapedius.
2
Рис. 121. Слуховые косточки:
1 — malleus; 2 — incus; 3 — stapes
Слуховая труба — канал длиной 40 мм; имеет
костную часть, pars ossea и хрящевую часть, pars
cartilaginea; соединяет носоглотку и барабанную
полость двумя отверстиями: ostium tympanicum
tubae auditivae и ostium pharyngeum tubae auditivae.
При глотательных движениях щелевидный просвет
трубы расширяется и свободно пропускает воздух
в барабанную полость.
III. Внутреннее ухо (auris interna) имеет кост-
ный и перепончатый лабиринт. В состав костного
лабиринта, labyrinthus osseus, входят полукружные
каналы, преддверие и канал улитки (рис. 122).
171
9
Рис. 122. Схема строения костного лабиринта:
1 — cochlea; 2 — cupula cochleae; 3 — vestibulum; 4 — fenestra
vestibuli; 5 — fenestra cochleae; 6 — crus osseum simplex; 7 —
crura ossea ampullares; 8 — crus osseum commune; 9 — canalis
semicircularis anterior; 10 — canalis semicircularis posterior;
11 — canalis semicircularis lateralis
Перепончатый лабиринт, labyrinthus membra-
naceus, имеет полукружные протоки, маточ-
ку, мешочек и проток улитки (рис. 123). Внут-
ри перепончатого лабиринта находится эндолимфа,
а снаружи — перилимфа.
Эндолимфатический проток, расположенный в
водопроводе преддверия, и эндолимфатический
мешочек, находящийся в расщеплении твердой моз-
говой оболочки, защищают лабиринт от чрезмерных
колебаний.
На поперечном срезе костной улитки видны три
пространства: одно — эндолимфатическое и два —
перилимфатических (рис. 124, см. цв. вклейку).
Поскольку они поднимаются по завиткам улитки,
их называют лестницами. Срединная лестница, scala
media, заполненная эндолимфой, имеет на срезе треу-
172
гольное очертание и называется улитковым протоком,
ductus cochlearis. Пространство, расположенное выше
улиткового протока, называется лестницей преддве-
рия, scala vestibuli; пространство, расположенное
ниже — барабанной лестницей, scala tympani.
Рис. 123. Схема строения перепончатого лабиринта:
1 — ductus cochlearis; 2 — sacculus; 3 — utriculus; 4 — ductus
semicircularis anterior; 5 — ductus semicircularis posterior;
6 — ductus semicircularis lateralis; 7 — ductus endolymphaticus
в aquaeductus vestibuli; 8 — saccus endolymphaticus; 9 —
ductus utriculosaccularis; 10 — ductus reunions; 11 — ductus
perilymphaticus в aquaeductus cochleae
Путь проведения звука
Звуковые волны улавливаются ушной раковиной,
направляются в наружный слуховой проход, вызы-
вают колебания барабанной перепонки. Колебания
перепонки передаются системой слуховых косточек
окну преддверия, далее перилимфе по лестнице пред-
дверия до вершины улитки, затем через просветлен-
ное окно, геликотрему, — перилимфе барабанной
лестницы и затухают, ударяясь о вторичную бара-
банную перепонку в улитковом окне (рис. 125).
Через преддверную мембрану улиткового прото-
ка колебания перилимфы передаются эндолимфе и
173
основной мембране улиткового протока, на которой
располагается рецептор слухового анализатора —
Кортиев орган.
Рис. 125. Схема распространения звука
в органе слуха:
1 — membrana tympanica; 2 — malleus; 3 — incus; 4 — stapes;
5 — membrana tympanica secundaria; 6 — scala tympani; 7 —
ductus cochlearis; 8 — scala vestibuli
Проводящий путь вестибулярного
анализатора
Рецепторы вестибулярного анализатора: 1) ам-
пулярные гребешки, crista ampullaris — восприни-
мают направление и ускорение движения; 2) пятно
маточки, macula utriculi — земное притяжение, по-
ложение головы в покое; 3) пятно мешочка, macula
sacculi — рецептор вибрации.
174
Тела I нейронов располагаются в преддверном
узле, g. vestibulare, который находится на дне внут-
реннего слухового прохода (рис. 126). Центральные
отростки клеток этого узла образуют преддверный
корешок VIII нерва, n. vestibularis, и заканчиваются
на клетках вестибулярных ядер VIII нерва — телах
II нейронов: верхнее ядро — ядро В.М. Бехтерева
(есть мнение, что только это ядро имеет непосредс-
твенную связь с корой), медиальное (главное) —
G.A Schwalbe, латеральное — O.F.C. Deiters и
нижнее — Ch.W, Roller. Аксоны клеток вестибу-
лярных ядер формируют несколько пучков, которые
направляются в спинной мозг, в мозжечок, в состав
медиального и заднего продольных пучков, а также
в таламус.
В спинной мозг идут аксоны клеток ядер Дейтер-
са и Роллера, образуя преддверно-спинномозговой
путь. Он заканчивается на клетках двигательных
ядер передних рогов спинного мозга (тела III ней-
ронов).
В мозжечок направляются аксоны клеток ядер
Дейтерса, Швальбе и Бехтерева, образуя преддвер-
но-мозжечковый путь. Этот путь проходит через
нижние ножки мозжечка и заканчивается на клетках
коры червя мозжечка (тело III нейрона).
Аксоны клеток ядра Дейтерса направляются в
состав медиального продольного пучка, который
связывает вестибулярные ядра с ядрами III, IV, VI
и XI черепных нервов и обеспечивает сохранение
направления взгляда при изменении положения
головы.
От ядра Дейтерса аксоны также направляются в
состав заднего продольного пучка, который связы-
вает вестибулярные ядра с вегетативными ядрами
III, VII, IX и X пар черепных нервов, что объяс-
175
няет вегетативные реакции в ответ на чрезмерные
раздражения вестибулярного аппарата.
Рис. 126. Схема проводящего пути вестибулярного
анализатора:
R — рецепторы — чувствительные клетки ампулярных гребеш-
ков и клетки пятен маточки и мешочка, crista ampullaris, macula
utriculi et sacculi; I — первый нейрон — клетки преддверного
узла, ganglion vestibulare; II — второй нейрон — клетки верх-
него, нижнего, медиального и латерального вестибулярных ядер,
n. vestibularis superior, inferior, medialis et lateralis; III — тре-
176
тий нейрон — латеральные ядра таламуса; IV — корковый конец
анализатора — клетки коры нижней теменной дольки, средней
и нижней височных извилин, lobulus parietalis inferior, gyrus
temporalis medius et inferior; 1 — спинной мозг; 2 — мост;
3 — мозжечок; 4 — средний мозг; 5 — таламус; 6 — внутренняя
капсула; 7 — участок коры нижней теменной дольки и средней
и нижней височной извилин; 8 — преддверно-спинномозго-
вой тракт, tractus vestibulospinalis; 9 — клетка двигательного
ядра переднего рога спинного мозга; 10 — ядро шатра моз-
жечка, п. fastigii; И — преддверно-мозжечковый тракт, tractus
vestibulocerebellaris; 12 — к медиальному продольному пучку,
ретикулярной формации и вегетативному центру продолговатого
мозга, fasciculus longitudinalis medialis; formatio reticularis,
n. dorsalis nervi vagi
Нервные импульсы к корковому концу вестибу-
лярного анализатора проходят следующим образом.
Аксоны клеток ядер Дейтерса и Швальбе переходят
на противоположную сторону в составе преддверно-
таламического тракта к телам III нейронов — клет-
кам латеральных ядер таламуса. Отростки этих
клеток проходят через внутреннюю капсулу в кору
височной и теменной долей полушария.
Проводящий путь слухового
анализатора
Рецепторы, воспринимающие звуковые раздра-
жения, располагаются в Кортиевом органе. Он на-
ходится в улитковом протоке и представлен волос-
ковыми сенсорными клетками, расположенными на
базальной мембране.
Тела I нейронов находятся в спиральном узле
(рис. 127), расположенном в спиральном кана-
ле улитки. Центральные отростки клеток этого
узла образуют улитковый корешок VIII нерва,
п. cochlearis, и заканчиваются на клетках вент-
рального и дорзального улитковых ядер VIII нерва
(тела II нейронов).
177
V
6
IV
Рис. 127. Схема проводящего пути слухового
анализатора:
R — рецепторы — чувствительные клетки спирального орга-
на; I — первый нейрон — клетки спирального узла, ganglion
spirale; II — второй нейрон — переднее и заднее улитковые
ядра, n. cochlearis dorsalis et ventralis; III — третий нейрон —
передние и задние ядра трапециевидного тела, n. dorsalis et
ventralis corporis trapezoidei; IV — четвертый нейрон — клетки
ядер нижних холмиков среднего мозга и медиального коленча-
того тела, n. colliculus inferior et corpus geniculatum mediate;
178
V — корковый конец слухового анализатора — клетки коры вер-
хней височной извилины, gyrus temporalis superior; 1 — спинной
мозг; 2 — мост; 3 — средний мозг; 4 — медиальное коленчатое
тело; 5 — внутренняя капсула; б — участок коры верхней ви-
сочной извилины; 7 — крышеспинномозговой тракт; 8 — клетки
двигательного ядра переднего рога спинного мозга; 9 — волокна
латеральной петли в треугольнике петли
Аксоны клеток вентрального ядра направляются
к вентральному и дорзальному ядрам трапецие-
видного тела своей и противоположной стороны,
причем последние образуют само трапециевидное
тело. Аксоны клеток дорзального ядра проходят
на противоположную сторону в составе мозговых
полосок, а затем трапециевидного тела к его ядрам.
Таким образом, тела III нейронов слухового пути
располагаются в ядрах трапециевидного тела.
Совокупность аксонов III нейронов составляет
латеральную петлю, lemniscus lateralis. В области
перешейка волокна петли лежат поверхностно в тре-
угольнике петли. Волокна петли заканчиваются на
клетках подкорковых центров (телах IV нейронов):
нижних холмиках четверохолмия и медиальных
коленчатых телах.
Аксоны клеток ядра нижнего холмика направ-
ляются в составе крыше-спинномозгового пути к
двигательным ядрам спинного мозга, осуществляя
безусловнорефлекторные двигательные реакции мус-
кулатуры на внезапные слуховые раздражения.
Аксоны клеток медиальных коленчатых тел про-
ходят через заднюю ножку внутренней капсулы в
среднюю часть верхней височной извилины — кор-
ковый конец слухового анализатора.
Имеются связи клеток ядра нижнего холмика с
клетками двигательных ядер V и VII пар черепных
ядер, обеспечивающие регуляцию работы слухо-
179
вых мышц. Кроме того, есть связи между клетками
слуховых ядер с медиальным продольным пучком,
обеспечивающие движение головы и глаз при поиске
источника звука.
Развитие преддверно-улиткового органа
1. Развитие внутреннего уха. Зачаток перепон-
чатого лабиринта появляется на 3-й неделе внутри-
утробного развития путем образования утолщений
эктодермы по бокам от закладки заднего мозгового
пузыря (рис. 128).
Рис. 128. Схема развития перепончатого лабиринта:
А — стадия образования слуховых плакод; Б — стадия образова-
ния слуховых ямок; В — стадия образования слуховых пузырьков;
I — первая висцеральная дуга; II — вторая висцеральная дуга;
1 — глоточная кишка; 2 — медуллярная пластинка; 3 — слуховая
плакода; 4 — медуллярная бороздка; 5 — слуховая ямка; 6 — не-
рвная трубка; 7 — слуховой пузырек; 8 — первый жаберный кар-
ман; 9 — первая жаберная щель; 10 — рост слухового пузырька и
образование эндолимфатического протока; 11 — формирование
всех элементов перепончатого лабиринта
180
На 1 стадии развития образуется слуховая пла-
кода. На 2 стадии из плакоды образуется слуховая
ямка, а на 3 стадии — слуховой пузырек. Далее
слуховой пузырек удлиняется, из него выпячива-
ется эндолимфатический проток, который пере-
тягивает пузырек на 2 части. Из верхней части
пузырька развиваются полукружные протоки, а из
нижней — улитковый проток. Рецепторы слухово-
го и вестибулярного анализатора закладываются на
7-й неделе. Из мезенхимы, окружающей перепон-
чатый лабиринт, развивается хрящевой лабиринт.
Он окостеневает на 5-й неделе внутриутробного
периода развития.
2. Развитие среднего уха (рис. 129).
Рис. 129. Схема развития наружного и среднего уха:
А — расположение висцеральных дуг зародыша человека; Б —
шесть бугорков мезенхимы, расположенных вокруг I наружной
жаберной щели; В —ушная раковина; 1-5 — висцеральные дуги;
б — первая жаберная щель; 7 — первый жаберный карман
181
Барабанная полость и слуховая труба развива-
ются из I жаберного кармана. Здесь формируется
единый трубо-барабанный канал. Из дорсальной
части этого канала образуется барабанная полость,
а из дорсальной — слуховая труба. Из мезенхимы
I висцеральной дуги образуются молоточек, нако-
вальня, m. tensor tympani, и иннервирующий ее V
нерв, из мезенхимы II висцеральной дуги — стремя,
m. stapedius и иннервирующий ее VII нерв.
3. Развитие наружного уха. Ушная раковина и
наружный слуховой проход развиваются в резуль-
тате сращения и трансформации шести бугорков
мезенхимы, расположенных вокруг I наружной
жаберной щели. Ямка I наружной жаберной щели
углубляется, и в ее глубине образуется барабанная
перепонка. Ее три слоя развиваются из трех заро-
дышевых листков.
Аномалии развития органа слуха
1. Глухота может быть следствием недоразвития
слуховых косточек, нарушением рецепторного
аппарата, а также нарушением проводниковой
части анализатора или его коркового конца.
2. Сращение слуховых косточек, снижающее
слух.
3. Аномалии и уродства наружного уха:
анотия — отсутствие ушной раковины,
щечная ушная раковина,
приросшая мочка,
раковина, состоящая из одной мочки,
раковина, расположенная ниже слухового про-
хода,
микротия, макротия (малое или слишком боль-
шое ухо),
атрезия наружного слухового прохода.
182
Обший покров, integumentum
commune
Кожа, cutis, образует покров тела и имеет огром-
ное количество функций, связанных с защитой орга-
низма и поддержанием обмена веществ. Кроме того,
в общем покрове содержится огромное количество
болевых, температурных и тактильных рецепторов.
В связи с этим ее относят также к органам чувств,
обеспечивающим постоянное взаимодействие с ок-
ружающей средой.
Кожа состоит из эпидермиса, epidermis, и со-
единительнотканной дермы, derma seu corium.
В состав кожи включают подкожную клетчатку, tela
subcutanea, и ее производные (рис. 130): волосы,
pili; мышцы, поднимающие волосы, m. arrector
pili; ногти, ungues; потовые железы, glandulae
sudoriferae; сальные железы, glandulae sebaceae;
молочные железы, glandulae mammae.
Развитие кожи и ее придатков
Эпидермис развивается из эктодермы, дерма — из
дерматомов сомитов. На 3-м месяце внутриутробно-
го периода развития в коже закладываются эпители-
альные зачатки волос, ногтей, желез. Осязательные
и иные тельца образуются из мезенхимы.
У плода поверхность кожи покрыта творожистой
смазкой (vernix caseosa), защищающей тело плода
от мацерации околоплодными водами.
Смазка состоит из секрета сальных желез, слу-
щивающейся перидермы вместе с пушковыми во-
лосами.
Закладки молочных желез формируются на 6-й
неделе в виде утолщенных эктодермальных полосок
между закладками верхней и нижней конечностей.
Это так называемые молочные полоски. Далее со-
183
храняется только верхняя часть полоски. В виде
аномалии развития у человека образуются добавоч-
ные молочные железы или грудные соски — поли-
мастия.
Рис. 130. Разрез кожи:
1 — выводной проток glandula sudorifera; 2 — m. arrector pili;
3 — radix pili; 4 — bulbus pili; 5 — papilla pili; 6 — fundus
folliculi pili; 7 — retinaculum cutis; 8 — glandula sebacea; 9 —
glandula sudorifera; 10 — жировые дольки
Если у мужчины молочная железа (чаще с одной
стороны) развивается по женскому типу, то это
называется гинекомастией.
Аномалии развития кожи
Ихтиоз — чрезмерное ороговение кожи, между
участками ороговения находятся кровоточа-
щие трещины.
184
Очаговая аплазия кожи.
Дистрофия ногтей, волос.
Альбинизм. Частичный альбинизм — вити-
лиго.
Кисты, невусы, белая прядь.
Гипертрихоз — избыток волос.
Алопеция — отсутствие волос.
Орган вкуса — organum gustus
Вкусовой анализатор обеспечивает восприятие
вкусовых раздражений, проведение нервных им-
пульсов до вкусовых нервных центров, анализ и
интеграцию поступившей в них информации. Вкусо-
вые рецепторы представлены вкусовыми луковица-
ми (почками), расположенными в ротовой полости,
преимущественно в слизистой языка, и составляю-
щими орган вкуса.
Вкусовые клетки функционально специализиро-
ваны: сладкое воспринимается кончиком языка, кис-
лое — боковой поверхностью языка, горькое — кор-
нем языка, соленое — всей поверхностью языка.
Проводящий путь вкусового
анализатора
I нейроны вкусового пути представлены псевдо-
униполярными клетками, расположенными в трех
различных узлах (афферентных нервов языка): в
коленчатом узле, ganglion geniculi лицевого нерва,
в нижнем узле, ganglion inferius языкоглоточного
нерва и в нижнем узле, ganglion inferius блужда-
ющего нерва (рис. 131).
Периферические отростки клеток коленчатого
узла направляются к вкусовым луковицам передних
двух третей языка в составе барабанной струны
лицевого нерва.
185
к-----------------7
IV
Рис. 131. Схема проводящего пути вкусового
анализатора:
R — рецепторы — чувствительные клетки сосочков язы-
ка (papillae vallatae, foliatae, fungiformes); I — первый ней-
рон — клетки коленчатого узла VII нерва, нижнего узла IX
и X HepeoB(ganglion geniculi, ganglion inferius IX, ganglion
inferius X); II — второй нейрон — ядра одиночного пути (п.
tractus solitarii); III — третий нейрон — клетки латеральных
ядер зрительного ядра (thalamus); IV — корковый конец ана-
лизатора — клетки коры крючка и парагиппокампальной изви-
лины (uncus, gyrus parahippocampalis); 1 — барабанная стру-
186
на; chorda tympani VII нерва — иннервирует передние 2/3 язы-
ка; 2 — язычные ветви; rr. linguales IX нерва — иннервируют
заднюю треть языка; 3 — глоточная ветвь, г. pharyngealis X
нерва — иннервирует надгортанник и дужки; 4 — ромбовидная
ямка; 5 — таламус; 6 — внутренняя капсула; 7 — участок коры
крючка и парагиппокампальной извилины
Периферические отростки клеток нижнего узла
языкоглоточного нерва направляются к вкусовым
луковицам задней трети языка в составе язычной
ветви языкоглоточного нерва. Периферические от-
ростки нижнего узла блуждающего нерва направля-
ются к вкусовым луковицам глотки, надгортанника,
неба и корня языка в составе глоточных ветвей
этого нерва.
Центральные отростки клеток всех трех узлов
подходят к клеткам ядер одиночного пути, n. tractus
solitarii (тела II нейрона т.е. к ядрам афферентных
нервов языка). Аксоны клеток этих ядер совершают
перекрест и направляются в составе ядерно-талами-
ческого тракта к таламусу (тела III нейронов). Ак-
соны III нейронов в составе таламо-коркового пути
подходят к корковому концу вкусового анализато-
ра — крючку и парагиппокампальной извилине.
Имеется связь III нейронов вкусового анализатора
через медиальные ядра таламуса с двигательными
центрами экстрапирамидной системы, с лимбической
системой, в частности, с миндалевидным ядром.
При поражении барабанной струны, лицевого
нерва, язычной ветви языкоглоточного нерва воз-
никают расстройства вкусовой чувствительности на
одноименной стороне языка.
При поражении латеральных ядер таламуса, та-
ламо-коркового тракта, коркового конца вкусового
анализатора отмечается частичное снижение вку-
са на противоположной стороне. При поражении
187
миндалевидного тела больной не может обозначить
вкус, хотя его ощущает (вкусовая агнозия).
Полная утрата вкуса носит название агевзия,
снижение вкуса — гипогевзия, извращение вку-
са — дисгевзия.
Развитие органа вкуса связано с развитием сли-
зистой языка из материала I, II, III и IV висцераль-
ных дуг, а мышц языка — из заушных миотомов,
чем и объясняется иннервация языка V, VII, IX, X
и XII черепными нервами.
Орган обоняния, organum olfactorium
Обонятельный анализатор обеспечивает воспри-
ятие обонятельных раздражений, проведение нерв-
ных импульсов до обонятельных центров, анализ и
интеграцию поступившей в них информации.
Рецепторы обонятельного анализатора распо-
лагаются в обонятельной области слизистой обо-
лочки носа и представляют собой периферические
отростки обонятельных клеток (рис. 132). Сами
обонятельные клетки являются телами I нейрона
обонятельного анализатора (рис. 133, 134).
Центральные отростки обонятельных клеток со-
ставляют обонятельные нервы, nn. olfactorii, кото-
рые проникают в полость черепа через отверстия
lamina cribrosa решетчатой кости. Обонятельные
нервы идут к обонятельной луковице и вступают
в контакт с митральными клетками обонятельной
луковицы (тела II нейрона).
Аксоны вторых нейронов идут в составе обоня-
тельного тракта, разделяются на медиальный пу-
чок — к обонятельной луковице противоположной
стороны, латеральный пучок — к корковому кон-
цу анализатора и промежуточный пучок, который
подходит к телам III нейронов. Тела III нейронов
188
располагаются в обонятельном треугольнике, яд-
рах прозрачной перегородки и переднем проды-
рявленном веществе.
Рис. 132. Обонятельная область (окрашенныйучасток
слизистой латеральной стенки полости носа
и перегородки носа):
1 — bulbus olfactorius; 2 — nn. olfactorii (lateralis); 3 —tractus
olfactorius; 4 — concha nasalis superior; 5 — nn. olfactorii
(medialis); 6 — septum nasi; 7 — concha nasalis inferior; 8 —
concha nasalis media
Аксоны III нейронов направляются в корковый
конец обонятельного анализатора тремя путями:
от клеток в обонятельном треугольнике длинным
путем над мозолистым телом, от ядер прозрачной
перегородки идет средний путь через свод и от пе-
реднего продырявленного вещества короткий путь
ведет сразу в крючок.
189
z
Рис. 133. Проводящие пути обонятельного анализатора:
R — рецепторы — периферические отростки чувствительных
клеток слизистой оболочки обонятельной области носовой
полости; I — первый нейрон — чувствительные клетки сли-
зистой оболочки обонятельной области носовой полости;
II — второй нейрон — митральные клетки обонятельной лу-
ковицы (bulbus olfactorius); III — третий нейрон — клетки
обонятельного треугольника, переднего продырявленного ве-
щества и ядер прозрачной перегородки (trigonum olfactorium,
septum pellucidum, substantia perforata anterior); IV —- кор-
ковый конец обонятельного анализатора — клетки коры
крючка и парагиппокампальной извилины (uncus et gyrus
parahippocampalis); 1 — обонятельная область носовой по-
лости (pars olfactoria tunicae mucosae nasi); 2 — обонятельные
нервы (nn. olfactorii); 3 — обонятельная луковица; 4 — обо-
нятельный тракт и его три пучка: медиальный, промежуточный
и латеральный (tractus olfactorius, stria olfactoria lateraris,
intermedia et medialis); 5 — короткий путь — к корковому
концу анализатора; 6 — средний путь — через пластинку
прозрачной перегородки, свод и бахромку морского конька к
коре; 7 — длинный путь — над мозолистым телом в составе
поясного пучка; 8 — сосцевидные тела и путь от них к таламусу
(fasciculus mamillothalamicus); 9 — ядра таламуса; 10 — вер-
хние холмики среднего мозга и путь к ним от сосцевидных тел
(fasciculus mamillotegmentalis)
190
Длинный путь обеспечивает обонятельные ас-
социации, средний — поиск источника запаха, а
короткий — двигательную защитную реакцию на
резкий запах. Корковый конец обонятельного ана-
лизатора находится в крючке и парагиппокампаль-
ной извилине.
Рис. 134. Схема проводящего пути обонятельного
анализатора
191
Особенностью обонятельного анализатора яв-
ляется то, что нервные импульсы первоначально
поступают в кору, а потом из коры в подкорковые
центры: сосочковые тела и передние ядра таламуса,
связанные между собой сосочково-таламическим
пучком.
Подкорковые центры в свою очередь связаны
с корой лобных долей, двигательными центрами
экстрапирамидной системы, лимбической системой
и ретикулярной формацией, обеспечивая эмоцио-
нальные реакции, защитные двигательные реакции,
изменение тонуса мускулатуры и т.п. в ответ на
обонятельные раздражения.
Развитие органа обоняния
Закладка органа обоняния занимает самый пе-
редний край нервной пластинки. Затем закладка
периферической части обонятельного анализатора
отделяется от зачатка ЦНС и перемещается в обо-
нятельную часть развивающейся носовой полости.
На 4-м месяце внутриутробного периода развития
в обонятельной части клетки дифференцируются
на опорные и обонятельные. Отростки обоня-
тельных клеток прорастают через еще хрящевую
lamina cribrosa в обонятельную луковицу. Так
происходит вторичное соединение органа обоня-
ния с ЦНС.
Аномалии развития органа
обоняния
1. Аринэнцефалия — отсутствие центральной и
периферической частей обонятельного мозга.
2. Дефекты обонятельных нервов.
3. Ослабление, отсутствие обонятельного воспри-
ятия.
192
При заболеваниях слизистой оболочки полости
носа, опухолях основания мозга и лобной доли
отмечается патологическое снижение обоняния (ги-
посмия) или полная его потеря (аносмия). При
аллергических состояниях возможно обострение
обоняния (гиперосмия).
7 Анатомия нервной системы
Автономная часть Глава |
ПЕРИФЕРИЧЕСКОЙ
НЕРВНОЙ СИСТЕМЫ
(ВЕГЕТАТИВНАЯ НЕРВНАЯ
СИСТЕМА), PARS AUTONOMICA
SYSTEMATIS NERVOSI PERIPHERIC!
Pars autonomica systematis nervosi peripherici —
часть нервной системы, иннервирующая сердце,
сосуды, внутренние органы, железы.
В 1880 г. французский анатом Мари Биша (Marie
Francois Xavier Bichat) создал стройное учение
о том, что есть органы животной и растительной
жизни, и иннервируются они анимальными и веге-
тативными нервами.
Позже английский гистолог и физиолог Джон
Ленгли (John Newport Langley) дал вегетативной
системе термин «автономная нервная система»,
подчеркивая, что она управляет непроизвольными
функциями.
Ленинградский ученый Леон Абгарович Орбели
(1882-1958) создал теорию об адаптационно-тро-
фической функции вегетативной нервной системы
(ВНС).
Функция ВНС не подконтрольна нашему созна-
нию, но находится в подчинении высших вегетатив-
ных центров, расположенных во всех отделах ЦНС,
в том числе в коре полушарий большого мозга. Од-
нако локализация ядер, отвечающих за вегетативные
функции, до сих пор не уточнена.
194
Вегетативная нервная система, так же как и ани-
мальная, выполняет свои функции по принципу
рефлексов. Вегетативные реакции в виде сокраще-
ний гладких мышц, сердечной мышцы происходят
в ответ на раздражение рецепторов внутренних
органов или тканей.
I нейроном вегетативной рефлекторной дуги
(рис. 135) является чувствительная клетка, ко-
торая располагается в спинномозговом узле или
краниальных чувствительных узлах V, VII, IX, X
черепных нервов.
II нейроном такой дуги служит ассоциативная
клетка одного из вегетативных ядер спинного мозга
(латеральное промежуточное ядро торако-люмбаль-
ного отдела или крестцовые парасимпатические
ядра) или вегетативных ядер черепных нервов.
III нейрон вегетативной рефлекторной дуги —
эффекторный, располагается за пределами ЦНС, в
вегетативных узлах 1-го, 2-го и 3-го порядка. Ак-
сон этого нейрона заканчивается в рабочем органе
(гладкой мышце, железе, сердечной мышце).
Морфологические отличия анимальной нервной
системы и вегетативной показаны на рис. 136.
На основании физиологических, фармакологичес-
ких и морфологических особенностей ВНС подразде-
ляется на 2 части: симпатическую (pars sympathica)
и парасимпатическую (pars parasympathica).
Функциональные различия
Симпатическая часть затормаживает органы
пищеварения, выделения, остальные возбуждает;
вызывает расширение зрачков, усиливает окисли-
тельные процессы, потребление питательных ве-
ществ, усиливает дыхание, учащает сердцебиение,
увеличивает поступление кислорода к мышцам. Ее
функция — трофическая.
195
196
1
Рис. 135. Схема веге-
тативной рефлектор-
ной дуги:
1 — radix dorsalis; 2 —
j ganglion spinale; 3 — n.
intermediolateralis; 4 — пре-
ганглионарные волокна в
составе переднего кореш-
ка; 5 — n. spinalis; 6 — г.
communicans albus; 7 —
ganglion trunci sympathici;
I 8 — r. communicans griseus;
) 9 — постганглионарные во-
локна в составе спинномоз-
гового нерва; 10 — постганг-
лионарные волокна в составе
внутренностного нерва; 11 —
ganglion plexus autonomicum;
12 — постганглионарные во-
локна в составе висцеральных
и сосудистых сплетений; 13 —
постганглионарные волокна к
кровеносным сосудам (по М.Р.
Сапину)
Рис. 136. Схема рефлекторных дуг: анимальной
(слева) и вегетативной (справа):
1 — nucleus intermediolateralis; 2 — ganglion spinale; 3 — ramus
communicans albus; 4 — ganglion trunci sympathici; 5 — ramus
communicans griseus; 6 — преганглионарное симпатическое во-
локно; 7 — постганглионарное симпатическое волокно; 8 — аф-
ферентное волокно; 9 — n. splanchnicus major (minor); 10 — n.
vagus; 11 —узел брюшного аортального сплетения; 12 — рецеп-
тор в стенке кишки; 13 — интрамуральный парасимпатический
узел; 14 — эффектор в скелетной мышце; 15 — рецепторы
в коже и подкожной жировой клетчатке; 16 — эфферентное
волокно анимальной рефлекторной дуги; 17 — афферентное
волокно анимальной рефлекторной дуги; 18 — ассоциативный
нейрон анимальной рефлекторной дуги
197
Анимальная Вегетативная
I нейрон заканчивается в за- дних рогах спинного мозга в боковых рогах
II нейрон не выходит за пре- делы спинного мозга выходит
III нейрон— в передних рогах спинного мозга в узлах 1-го, 2-го или 3-го порядка
От центра к рабочему ор- гану нервное волокно не прерывается Волокно обязательно пре- рывается
Территория иннервации ог- раничена (мышцы, кожа) Обширная территория ин- нервации (внутренние орга- ны, сосуды, мышцы, кожа, так как в них есть сосуды, обеспечивающие обменные процессы).
Степень сохранения сег- ментарное™ отчетливая Полностью отсутствует
— Феномен мультипликации (Лэнгли) — количество пост- ганглионарных волокон боль- ше, чем преганглионарных
Диаметр нервов— 13-14 мкм; волокна миелиновые (мякотные) Диаметр нервов — 5-7 мкм; предузловые волокна — ми- елиновые, постганглионар- ные — безмякотные
Скорость проведения воз- буждения —100-120 м/с 0,3-10 м/с
— Нервы образуют сплетения, обвивающие стволы арте- рий.
Парасимпатическая часть активизирует пище-
варительную и выделительную системы, тормозит
окислительные процессы, вызывает сужение зрачка,
торможение сердечной деятельности, опорожнение
198
полостных органов — усиление перистальтики; ее
функция — охраняющая органы.
В целом, ВНС выполняет адаптационно-трофи-
ческую функцию, изменяет тонус органов. Антаго-
низма между симпатической и парасимпатической
частями ВНС нет. Во-первых, объекты приложе-
ния импульсов от этих двух частей вегетативной
нервной системы разные; во-вторых, соотношение
симпатических и парасимпатических импульсов ди-
намически меняется на различных фазах функции
органа. Они могут действовать и как антагонисты,
и как синергисты. Поэтому правильнее говорить о
синергизме частей ВНС.
Симпатическую иннервацию имеют все (без ис-
ключения) органы и ткани. Парасимпатическую
иннервацию не получают (есть такое мнение) кро-
веносные сосуды (кроме коронарных), потовые
железы, мышцы, поднимающие волосы, скелетные
мышцы и мозговое вещество надпочечников.
Фармакологические различия
Межнейронная передача импульса в ВНС совер-
шается с участием ряда медиаторов. Медиатор в
преганглионарных волокнах для симпатической и
парасимпатической нервной системы один — аце-
тилхолин. Медиаторы в постганглионарных волок-
нах для симпатической системы — адреналин или
норадреналин, а для парасимпатической — ацетил-
холин. Различия в медиаторах позволяют клиницис-
там избирательно воздействовать на симпатическую
и парасимпатическую части ВНС.
Морфологические различия
Вегетативная нервная система имеет центральный
и периферический отделы.
199
К центральному отделу относятся сегментарные
аппараты и надсегментарные аппараты. Симпа-
тические и парасимпатические центры относят к
сегментарным аппаратам. Надсегментарные центры
находятся во всех пяти отделах мозга.
В симпатической части центральные нейроны
располагаются только в спинном мозге, в латераль-
ных промежуточных ядрах боковых рогов, начиная
от VIII шейного сегмента и до II поясничного сег-
мента (рис. 137, см. цв. вклейку).
В парасимпатической части центральные ней-
роны расположены в двух местах: в краниальном
отделе и в сакральном.
В краниальном отделе парасимпатические цен-
тральные нейроны располагаются в среднем мозге
(мезенцефалическая часть), продолговатом мозге и
мосту (бульбарная часть) и представлены парасим-
патическими ядрами ill, VII, IX и X пар черепных
нервов.
Сакральный отдел представлен парасимпатичес-
кими ядрами, расположенными во 2-4-м крестцовых
сегментах Su — S|V
Высшие вегетативные (надсегментарные) центры
объединяют симпатическую и парасимпатическую
части ВНС и располагаются в стволе и полушариях
большого мозга:
кора полушарий большого мозга,
базальные ядра,
ядра гипоталамуса,
серое вещество вокруг водопровода,
ретикулярная формация,
лимбическая система,
клетки мозжечка.
Гипоталамус, являясь посредником между нерв-
ной и эндокринной системой, обеспечивает интег-
200
рацию симпатических и парасимпатических реф-
лексов, контролирует работу вегетативных центров
ствола мозга.
Проекционные центры в различных отделах
коры полушарий большого мозга составляют вы-
сший уровень регуляции, который предназначен
для вегетативного обеспечения целенаправленной
деятельности человека. Установлены связи лобной
и височной долей полушарий большого мозга с яд-
рами гипоталамуса.
Периферический отдел ВНС состоит из преган-
глионарных волокон, вегетативных узлов и пост-
ганглионарных волокон. К нему относятся узлы,
ветви, нервы и сплетения.
I. Узлы
К симпатической системе относятся узлы 1-го и
2-го порядка (рис. 135-137, см. цв. вклейку). Узлы
1-го порядка образуют truncus sympathicus. Их 20-25
пар: 3 шейных, 10-12 грудных, 3-5 поясничных,
4-5 поясничных, 1 — копчиковый.
Узлы 2-го порядка висцеральные, расположены
в сплетениях. Так, в брюшном аортальном сплетении
находятся: ganglia phrenica, ganglia coeliaca, ganglia
aortorenalia, ganglia renalia, ganglion mesentericum
superius, ganglion mesentericum inferius.
К парасимпатическим узлам относятся узлы 3-го
порядка. Это краниальные парасимпатические
узлы (g. pterygopaiatinum, g. submandibulare,
g. sublinguale, g. oticum, g. ciliare) и интрамураль-
ные узлы.
II. Ветви
1. Межузловые ветви (rami interganglionares) со-
единяют узлы симпатического ствола между собой.
201
2. Белые соединительные ветви (rami communi-
can tes al bi) соединяют спинномозговые нервы (вось-
мой шейный, все грудные и два поясничных) с
узлами симпатического ствола.
Эти ветви присутствуют у тех узлов, которые со-
ответствуют сегментам спинного мозга, содержащим
симпатические ядра. Будучи преганглионарными,
по гистологическому строению они миелиновые
(рис. 138, см. цв. вклейку).
3. Серые соединительные ветви (rami communi-
cantes grisei) соединяют узел симпатического ствола
со спинномозговым или черепным нервом. Будучи
постганглионарными, по гистологическому строе-
нию они безмиелиновые.
Серые соединительные ветви в составе спинно-
мозговых или черепных нервов направляются к
сосудам мышц, сосудам кожи, обеспечивая трофику
этих образований.
III. Нервы
П арасимпатическая система своих нервов
не имеет, использует анимальные нервы. Исключе-
ние — nn. splanchnic! pelvici, nn. ciliares breves.
Симпатические нервы подразделяются на не-
рвы крупных сосудов, органные и внутренностные
симпатические нервы.
Шейный отдел симпатического ствола представ-
лен 3 узлами (верхним, средним и нижним), их
межузловыми ветвями и располагается на глубоких
мышцах шеи (рис. 139, см. цв. вклейку). ,
От верхнего шейного узла отходят:
серые соединительные ветви к спинномозговым
нервам шейного сплетения;
внутренний сонный нерв (n. caroticus internus),
образующий одноименное сплетение;
202
1
наружные сонные нервы (nn. carotici externi),
образующие сплетение по ходу наружной сон-
ной артерии. Внутреннее и наружное сонные
сплетения образуют общее сонное сплетение;
яремный нерв (n. jugularis) обеспечивает связи
с IX, X, XI и XII черепными нервами;
гортанно-глоточные ветви (rr. laryngopharyngei)
соединяются с IX и X черепными нервами и
образуют глоточное и пищеводное сплетения;
верхний шейный сердечный нерв (n. cardiacus
cervicalis superior)
От среднего шейного узла отходят:
серые соединительные ветви к спинномозговым
нервам плечевого сплетения;
средний шейный сердечный нерв (n. cardiacus
cervicalis inferior).
От нижнего шейного узла (шейно-грудного
или звездчатого) отходят:
серые соединительные ветви к спинномозговым
нервам плечевого сплетения;
подключичная петля, ansa subclavia, соединя-
ющая нижний и средний шейные узлы симпа-
тического ствола; ветви от петли и нижнего
шейного узла образуют pl. subclavius;
позвоночный нерв (n. vertebralis), образую-
щий одноименное сплетение, иннервирующее
сосуды головного мозга;
нижний шейный сердечный нерв (n. cardiacus
cervicalis inferior).
От грудных узлов симпатического ствола
отходят:
серые соединительные ветви к межреберным
нервам (рис. 138, 139, см. цв. вклейку);
ветви, образующие plexus aorticus thoracicus,
plexus pulmonalis, plexus oesophagealis;
203
грудные сердечные нервы (nn. cardiaci
thoracici), отходящие от верхних 5-6-го груд-
ных узлов симпатического ствола;
большой внутренностный нерв (n. splanchnicus
major), начинается от 5-9-го грудных узлов
симпатического ствола (рис. 139, см. цв. вклей-
ку), проходит между медиальной и промежу-
точной ножками диафрагмы, состоит преиму-
щественно из постганглионарных волокон и
заканчивается в узлах чревного сплетения;
малый внутренностный нерв (n. splanchnicus
minor), начинается от 10-И-го грудных уз-
лов симпатического ствола, проходит через
медиальную ножку диафрагмы, состоит пре-
имущественно из постганглионарных волокон
и заканчивается в узлах чревного сплетения.
От малого чревного нерва берет начало почеч-
ная ветвь, г. renalis, которая заканчивается в
аортопочечном узле чревного сплетения;
низший внутренностный нерв (n. splanchnicus
imus), идет рядом с малым внутренностным
нервом, начинается от 12-го грудного узла
симпатического ствола и заканчивается в по-
чечном сплетении.
От поясничных узлов симпатического
ствола отходят:
серые соединительные ветви к нервам пояснич-
ного сплетения (рис. 140, см. цв. вклейку);
поясничные внутренностные нервы (nn.
splanchnic! lumbales), которые идут к чрев-
ному сплетению, а также к почечному, надпо-
чечниковому и брюшному аортальному спле-
тениям.
От тазовых узлов поясничного ствола
отходят:
204
серые соединительные ветви к нервам крест-
цового сплетения;
крестцовые внутренностные нервы (пп.
splanchnic! sacrales), которые идут к подчрев-
ным сплетениям для симпатической иннерва-
ции органов малого таза
IV. Сплетения
Вегетативные сплетения оплетают артериальные
стволы, артерии, доставляя вегетативные волокна
к органам (рис. 141, см. цв. вклейку). В своем
составе они имеют не только симпатические, па-
расимпатические и чувствительные волокна, но и
узлы. Например, pl. caroticus internus, externus et
communis, pl. pulmonalis, pl. aorticus thoracicus,
pl. subclavius.
В брюшной полости самое крупное сплетение — pl.
aorticus abdominalis (рис. 141-142, см. цв. вклейку).
От узлов брюшного аортального сплетения отходят
многочисленные ветви во все стороны, как лучи
солнца. Отсюда старое название этого сплетения —
солнечное, pl. solans. К узлам сплетения подходят
большие и малые внутренностные нервы и внутрен-
ностные поясничные нервы. Ветви брюшного аор-
тального сплетения продолжаются на артерии, ответ-
вляющиеся от брюшной аорты, формируя вторичные
сплетения: pl. phrenici, pl. gastrici, pl. coeliaccus, pl.
lienalis, pl. pancreaticus, pl. hepaticus, pl. suprarenelis,
pl. renalis, pl. testicularis (ovaricus), pl. mesentericus
superior et inferior, pl. intermesentericus.
Брюшное аортальное сплетение продолжается на
общие подвздошные артерии (рис. 143), формируя
подвздошные сплетения (pl. iliaci). От него отходят
4-5 нервов, образующих непарное верхнее подчрев-
ное сплетение, pl. hypogastricus superior. Верхнее
205
подчревное сплетение образуют поясничные и крест-
цовые внутренностные нервы.
Рис. 143. Схема брюшного и тазового отделов
вегетативной нервной системы:
1 — plexus aorticus abdominalis; 2 — plexus iliacus; 3 — plexus
hypogastricus superior; 4 — plexus hypogastricus inferior; 5 —
plexus coeliacus; 6 — gangl. phrenica; 7 — gangl. coeliaca; 8 —
plexus lienalis; 9 — plexus hepaticus; 10 — plexus suprarenalis;
11 — gangl. aortorenalia; 12 — gangl. renalia; 13 — plexus
renalis; 14— gangl. mesentericum superius; 15 — plexus
mesentericus superior; 16 — plexus intermesentericus; 17 —
gangl. mesentericum inferius; 18 — plexus mesentericus inferior;
19 — нервы, отходящие от верхнего подчревного сплетения;
20 — nn. hypogastrici
206
От верхнего подчревного сплетения отходят 2
подчревных нерва, nn. hypogastrici, которые по бо-
кам прямой кишки и на m. levator ani образуют пар-
ное нижнее подчревное сплетение, pl. hypogastricus
inferior или pl. pelvicus (рис. 143; 145, см. цв. вклей-
ку). К этому сплетению также подходят крестцовые
внутренностные нервы (симпатические) и тазовые
внутренностные нервы (парасимпатические). От
нижнего подчревного сплетения отходят ветви, ко-
торые образуют сосудистые и органные сплетения:
pl. rectalis medius et inferior, pl. prostaticus. pl.
deferentialis, pl. uterovaginalis, pl. vesicalis, pl.
cavernosus penis (clitoridis).
Развитие вегетативной
нервной системы
Одновременно с образованием спинного мозга
из эктодермы развиваются спинномозговые и ве-
гетативные узлы. Исходным материалом для них
служат клеточные элементы ганглиозной пластинки,
дифференцирующиеся в нейробласты, глиобласты
и хромаффинобласты.
Большая часть последних смещается на перифе-
рию, образуя вегетативные узлы и хромаффинную
ткань: мозговое вещество надпочечников и пара-
ганглии.
Общность развития анимальной и вегетативной
нервных систем, а также сегментарность нервной
системы позволили обосновать явление, названное
именами Захарьина и Геда (зоны Захарьина-Теда).
А именно: заболевание органа сопровождается изме-
нением чувствительности и болями в соответствую-
щей области. Например, при стенокардии отмечается
иррадиация болей в левую руку, при заболевании
почек — в поясницу, печени — в правую лопатку,
207
при язве желудка - между лопатками. Суть явления
в следующем: один и тот же спинномозговой узел
получает чувствительное (болевое) раздражение из
кожи и органа, развивающихся из одного сегмента
или расположенных на уровне одного сегмента. Пере-
ключение патологического раздражения с висцераль-
ных рецепторов на клетки болевой чувствительности
происходит в задних рогах спинного мозга или в зри-
тельном бугре. Нередко возникающая генерализация
висцеральных раздражений, особо у детей, может
привести к ощущению болей в зонах, соответствую-
щих другим органам. Поэтому зоны Захарьина-Геда
не всегда имеют диагностическое значение.
Проводяшие пути вегетативной
нервной системы
Афферентные пути
1. Афферентными путями вегетативного рефлек-
са могут служить афферентные пути температурной,
болевой, тактильной чувствительности или специ-
фической чувствительности — зрения, вкуса.
2. Афферентные пути от интерорецепторов про-
ходят следующим образом:
от рецепторов органов головы, шеи, грудной
полости по чувствительным ветвям черепных
нервов (X, IX, VII, V). Тогда: 1-й нейрон нахо-
дится в чувствительных узлах черепных нервов,
2-й нейрон — в чувствительных ядрах черепных
нервов, от них начинается tr. nucleothalamicus,
3-й нейрон — в зрительном бугре, корковый
конец анализатора — нижний отдел предцент-
ральной и постцентральной извилин;
от рецепторов органов брюшной полости
и таза по внутренностным нервам. Тогда:
208
1-й нейрон находится в спинномозговом узле,
2-й нейрон — в задних рогах спинного мозга,
3-й нейрон — в зрительном бугре, корковый
конец анализатора — нижний отдел пред- и
постцентральных извилин.
Эфферентные пути вегетативной
нервной системы
Морфология их сложна и мало изучена. Можно
составить следующую схему.
1-й нейрон расположен в коре полушарий боль-
шого мозга (лобная и височная доли, по мнению
других авторов — вся кора). Далее через базальные
ядра или непосредственно ко вторым нейронам.
2-й нейрон расположен в высших вегетативных
центрах, в частности, в ядрах гипоталамуса. От-
ростки этих нейронов объединяются в дорзальный
продольный пучок, fasciculus longitudinalis dorsalis
(Шютца). Волокна пучка заканчиваются в стволе
мозга на парасимпатических ядрах черепных не-
рвов, в спинном мозге — на симпатических ядрах
грудных и поясничных сегментов и на парасимпа-
тических ядрах крестцового отдела.
3-й нейрон расположен в симпатических и пара-
симпатических ядрах.
4-й нейрон находится в вегетативных узлах, пос-
тганглионарные волокна которых достигают испол-
нительного органа.
Таким образом, автономность вегетативной нерв-
ной системы относительна, так как имеется связь
коры полушарий большого мозга с ядрами гипота-
ламуса.
В последнее время в вегетативной нервной
системе выделяют третью — метасимпатическую
часть. Это обширные нервные сплетения и мик-
209
роскопические узлы в стенках полых органов,
обладающих моторикой. Метасимпатические узлы
отличаются от парасимпатических по гистологи-
ческому строению и медиаторам (гамма-амино-
масляная кислота и пуриновые основания). Ней-
родиты метасимпатических узлов способны без
участия ЦНС продуцировать нервные импульсы и
посылать их на гладкомышечные клетки, вызывая
движение органа, сокращение его стенок. Мета-
симпатические узлы имеют связь с симпатически-
ми и парасимпатическими частями вегетативной
нервной системы.
Иннерваиия легких и плевры
Афферентными путями от висцеральной плевры
являются легочные ветви грудного отдела симпа-
тического ствола, от париетальной плевры — пп.
intercostales и n. phrenicus, от бронхов — n. vagus.
Эфферентная парасимпатическая иннервация.
Преганглионарные волокна начинаются в дор-
сальном вегетативном ядре блуждающего нерва
(рис. 145) и идут в составе последнего и его ле-
гочных ветвей к узлам plexus pulmonalis, а также
к узлам, расположенным по ходу трахеи, бронхов
и внутри легких. Постганглионарные волокна на-
правляются от этих узлов к мускулатуре и железам
бронхиального дерева. Функция: сужение просвета
бронхов и бронхиол и выделение слизи.
Эфферентная симпатическая иннервация. Пре-
ганглионарные волокна выходят из боковых ро-
гов спинного мозга верхних грудных сегментов
(Thn — ThVI) и проходят через соответствующие
rami communicantes albi к звездчатому и верхним
грудным узлам симпатического ствола. От них
начинаются постганглионарные волокна, которые
210
проходят в составе легочного сплетения к брон-
хиальной мускулатуре и кровеносным сосудам.
Функция: расширение просвета бронхов.
Иннервация сердца
Афферентные пути от сердца идут в соста-
ве п. vagus, а также в среднем и нижнем шейных
и грудных сердечных симпатических нервах.
Эфферентная парасимпатическая иннер-
вация. Преганглионарные волокна начинаются в
дорсальном вегетативном ядре блуждающего нерва
и идут в составе последнего, его сердечных ветвей
(rami cardiaci cervicales et thoracici n. vagi) и сер-
дечных сплетений до узлов субэпикардиальных
сплетений сердца.
Постганглионарные волокна исходят от этих уз-
лов к мышце сердца.
Функция: торможение и угнетение деятельности
сердца; сужение венечных артерий (рис. 145, 146).
Эфферентная симпатическая иннервация.
Преганглионарные волокна начинаются из боковых
рогов спинного мозга 4-5-го верхних грудных сег-
ментов, выходят в составе соответственных rami
communicantes albi и проходят до трех шейных
и пяти верхних грудных узлов симпатического
ствола.
В этих узлах начинаются постганглионарные
волокна, которые в составе сердечных нервов, пп.
cardiaci cervicales superior, medius et inferior и nn.
cardiaci thoracici, достигают сердечной мышцы.
Функция: усиление работы сердца (это установил
И.П. Павлов в 1888 г., назвав симпатический нерв
усиливающим) и ускорение ритма (это впервые
установил И.Ф. Цион в 1866 г.), расширение ве-
нечных сосудов.
211
24
Рис. 145. Вегетативная иннервация внутренних органов:
а — парасимпатическая часть; 6 — симпатическая часть. 1 — верх-
ний шейный узел; 2 — латеральное промежуточное ядро; 3 — вер-
хний шейный сердечный нерв; 4 — грудные сердечные и легочные
нервы; 5 — большой чревный нерв; 6 — чревное сплетение; 7 —
нижнее брыжеечное сплетение; 8 — верхний и нижний подчревные
сплетения; 9 — малый чревный нерв; 10 — поясничные чревные
нервы; 11 — крестцовые чревные нервы; 12 — парасимпатичес-
кие ядра крестцовых сегментов; 13 — тазовые чревные нервы;
14 — тазовые узлы; 15 — парасимпатические узлы; 16 — блуж-
дающий нерв; 17 — ушной узел; 18 — поднижнечелюстной узел;
19 — крылонебный узел; 20 — ресничный узел; 21 — парасимпа-
тическое ядро блуждающего нерва; 22 — парасимпатическое ядро
языкоглоточного нерва; 23 — парасимпатическое ядро лицевого
нерва; 24 — парасимпатическое ядро глазодвигательного нерва
(по М.Р. Сапину)
212
Рис. 146. Схема иннервации сердца (А) и расположение
субэпикардиальных сплетений спереди (Б) и сзади (В):
1 — truncus sympathies; 2 — n. vagus; 3 — шейные узлы сим-
патического ствола; 4 — грудные узлы симпатического ствола;
5 — n. cardiacus cervicalis superior; 6 — n. cardiacus cervicalis
medius; 7 — n. cardiacus cervicalis inferior; 8 — r. cardiacus
cervicalis superior; 9 — r. cardiacus cervicalis inferior; 10 — rr.
cardiaci thoracici; 11 — aorta; 12 — truncus pulmonalis; 13 — по-
верхностное экстракардиальное сплетение; 14 — глубокое экс-
тра кард и аль ное сплетение; 15-18 — шесть субэпикардиальных
сплетений (Воробьева)
213
Иннервация слезной железы
и слюнных желез
Афферентным путем для слезной железы
является п. lacrimalis (ветвь n. ophthalmicus от
п. trigeminus), для поднижнечелюстной и подъ-
язычной — n. lingualis (ветвь n. mandibularis от п.
trigeminus) и chorda tympani (ветвь n. intermedins),
для околоушной — n. auriculotemporalis и п.
glossopharyngeus.
Эфферентная парасимпатическая иннер-
вация слезной железы (рис. 145). Центр лежит
в верхнем отделе продолговатого мозга и связан с
ядром промежуточного нерва (nucleus salivatorius
superior). Преганглионарные волокна идут в составе
n. intermedius, далее n. petrosus major до ganglion
pterygopalatinum.
Отсюда начинаются постганглионарные волокна,
которые в составе n. maxillaris и далее его ветви, п.
zygomaticus, через связи с n. lacrimalis достигают
слезной железы.
Эфферентная парасимпатическая ин-
нервация поднижнечелюстной и подъязыч-
ной желез. Преганглионарные волокна идут от
nucleus salivatorius superior в составе n. intermedius,
далее chorda tympani и n. lingualis до ganglion
submandibulare, откуда начинаются постганглио-
нарные волокна, достигающие желез.
Эфферентная парасимпатическая иннер-
вация околоушной железы. Преганглионарные
волокна идут от nucleus salivatorius inferior в со-
ставе n. glossopharyngeus, далее n. tympanicus, п.
petrosus minor до ganglion oticum. Отсюда начина-
ются постганглионарные волокна, идущие к железе
в составе n. auriculotemporalis V нерва. Функция:
214
усиление секреции слезной и названных слюнных
желез; расширение сосудов желез.
Эфферентная симпатическая иннерва-
ция всех названных желез. Преганглионарные
волокна начинаются в боковых рогах верхних груд-
ных сегментов спинного мозга и заканчиваются в
верхнем шейном узле симпатического ствола. Пост-
ганглионарные волокна начинаются в этом узле и
доходят до слезной железы в составе plexus caroticus
internus, до околоушной — в составе plexus caroticus
externus и до поднижнечелюстной и подъязычной
желез — через plexus caroticus externus и затем через
plexus facialis. Функция: задержка отделения слюны
(сухость во рту).
Иннервация внутренних мыши
глазного яблока
Афферентным путем от внутренних, глад-
ких мышц глазного яблока (m. sphincter pupillae,
т. ciliaris и т. dilatator pupillae) являются п.
oculomotorius и n. ophthalmicus.
Эфферентная парасимпатическая иннер-
вация т. shincter pupillae и т. ciliaris. Преганг-
лионарные волокна идут от nucleus accessorius и
n. caudatus centralis nervi oculomotorius в составе
III нерва до ganglion ciliare. От узла начинаются
постганглионарные волокна, идущие в составе пп.
ciliares breves до ресничной мышцы и сфинктера
зрачка. Функция: сужение зрачка и аккомодация
глаза к дальнему и близкому видению.
Эфферентная симпатическая иннервация
m. dilatator pupillae. Преганглионарные волокна
начинаются в боковых рогах верхних грудных сег-
ментов спинного мозга и заканчиваются в верхнем
шейном узле симпатического ствола. Постганглио-
215
парные волокна начинаются в этом узле и доходят до
m. dilatator papillae в составе n. caroticus interims,
plexus caroticus internus и plexus ophthalmicus.
Функция: расширение зрачка, а также сужение
сосудов глаза.
Иннервация органов желудочно-кишеч-
ного тракта (до сигмовидной кишки)
Афферентные пути от этих органов идут в
составе n. vagus, n. splanchnicus major et minor,
plexus hepaticus, plexus coeliacus, грудных и пояс-
ничных спинномозговых нервов и n. phrenicus.
Эфферентная парасимпатическая иннер-
вация. Преганглионарные волокна начинаются в
дорсальном вегетативном ядре блуждающего нерва и
идут в составе последнего, его ветвей (rami coeliaci)
до терминальных узлов в толще органов. Функция:
усиление перистальтики полых органов, усиление
секреции желез.
Эфферентная симпатическая иннервация.
Преганглионарные волокна выходят из боковых
рогов спинного мозга верхних грудных сегментов и
проходят через соответствующие rami communicantes
albi и симпатический ствол без перерыва в составе п.
splanchnicus major et minor до промежуточных узлов
(ganglia coeliaca et ganglion mesentericum superius
et inferius). От них начинаются постганглионарные
волокна, которые проходят в составе plexus aorticus
abdominalis, plexus coeliacus, plexus hepaticus — к
печени; plexus aorticus abdominalis, plexus coeliacus,
plexus gastricus — к желудку; plexus aorticus
abdominalis, plexus mesentericus superior — к ки-
шечнику (до середины colon transversum); plexus
aorticus abdominalis, plexus mesentericus inferior —
к кишечнику до сигмовидной кишки.
216
Функция: замедление перистальтики, угнетение
секреции желез.
Иннервация органов малого таза
Афферентные пути от этих органов идут
в составе plexus mesentericus inferior, plexus
hypogastricus superior et inferior, nn. splanchnici
pelvini.
Эфферентная парасимпатическая иннер-
вация. Преганглионарные волокна начинаются в
парасимпатических ядрах крестцовых сегментов
(Su — SIV), идут в составе nn. splanchnici pelvini
до внутриорганных узлов. Функция: возбуждение
перистальтики сигмовидной и прямой кишки, рас-
слабление ш. sphincter ani internus; сокращение m.
detrusor urinae, расслабление m. sphincter vesicae.
Эфферентная симпатическая иннервация.
Преганглионарные волокна выходят из боковых
рогов поясничных сегментов спинного мозга и про-
ходят через соответствующие rami communicantes
albi, rr. interganglionares до крестцовых узлов сим-
патического ствола, далее в составе n. splanchnicus
lumbalis et sacralis, plexus hypogastricus superior
et inferior и сплетений органов малого таза до их
гладкой мускулатуры.
АКЛЮЧЕНИЕ
Чтобы понять, как функционирует организм че-
ловека на различных этапах развития и в различных
условиях, недостаточно знать его строение, необхо-
димо знать те структуры и функции, которые дела-
ют его единым целым, способным самовосстанавли-
ваться, саморегулироваться, приспосабливаться к
постоянно меняющейся внутренней и внешней среде.
Система, позволяющая объединиться различным
структурам организма в единое целое, — нервная
система. В настоящем учебном пособии изложено
современное представление о структуре и функциях
этой системы. Приведенный в пособии лекционный
материал существенно дополнит знания о нервной
регуляции организма.
Авторы надеются, что пособие поможет разо-
браться в сложных взаимосвязях нервной систе-
мы и пробудит интерес к углубленному изучению
этого интереснейшего и все еще малоизученного
раздела анатомии. В нервной системе много белых
пятен, которые необходимо ликвидировать, изучив
строение и функции структур и клеток, входящих
в эту интегрирующую систему. Для молодых спе-
циалистов, пытливых и интересующихся тайнами
жизни, нервная система — благодатная почва для
изучения и пробы своих сил. Успехов и удач вам,
читатель!
218
Литература
1. Анатомия человека. В 2 томах. Т. 1. Под ред.
акад. РАМН, проф. М. Р. Сапина. Изд. 3-е. М.: Меди-
цина, 1996. — 560 с.
2. Беков Д. Б. Атлас венозной системы головного
мозга человека. М. : Медицина. — 1965. — 358 с.
3. Г(шворонский И. В. Нормальная анатомия чело-
века. Т. 1: Учебник для мед. вузов. — СПб: Спец, лит.,
2000. - 560 с.
4. Гайворонский И. В. Нормальная анатомия чело-
века. Т. 2: Учебник для мед. вузов. — СПб: Спец, лит.,
2000. - 560 с.
5. Гайворонский И. В., Косоуров А. К. Аномалии и
пороки развития человека. СПб, 2001. — 96 с.
6. Зернов Д. Руководство по описательной анатомии
человека. Т. II. — М.—Л. — 146 с.
7. Лазюк Г. И. Тератология человека. М.: Медици-
на. - 1991. - 480 с.
8. Колесников Л. Л., ЧукбарА. В, Развитие, возрас-
тные изменения и аномалии развития органов человека.
М.: Медицина XXI, 2004. — 144 с.
9. Кошев В, И., Петров Е. С., Волобуев А. Н. Це-
ребральное сердце: миф или реальность? Клиническая
анатомия и экспериментальная хирургия. Ежегодник
Российской ассоциации клинических анатомов. — Вып.
6. — Оренбург, 2006. — С. 35-40.
10. Крылова Н. В., Искренно И. А. Анатомия в схе-
мах и рисунках. Спинной, головной мозг и черепные
нервы. Уч. пособие. — М.: изд. Университета дружбы
народов, 1986. — 166 с.
Крылова Н. В., Наумец Л. В. Анатомия орга-
нов чувств (в схемах и рисунках). Атлас —пособие. М.:
МИА, 2003. - 96 с.
219
12. Привес М.Г., Лысенков Н.К., Бушкович В.И.
Анатомия человека. — 10-е изд., перераб. и доп. — СПб.:
Гиппократ, 1997. — 704 с.
13. Соколов В. В., Городок М. М., Матвеев Ю. В,
Методические указания к практическим занятиям по
ЦНС для педиатрического факультета медицинских ин-
ститутов. Ростов-на-Дону, 1982. — 58 с.
14. Соколов В. В,, Городок М. М., Матвеев Ю.В.
Схематические рисунки топографии ядерных образова-
ний и проводящих путей спинного и головного мозга
человека. Ростов-на-Дону, 1982. — 43 с.
15. Сомов Е. Е. Клиническая анатомия органа зрения
человека. Уч. пособие. — Санкт-Петербург, 1992. — 65 с.
16. Стебельский С. Ю. Анатомия сосудистой и пе-
риферической нервной систем человека: Атлас схем. —
Киев: Вища шк., 1984. — 167 с.
17. Турыгин В. В. Ассоциативные, комиссуральные
и проекционные проводящие пути головного и спинного
мозга. — Челябинск, 1983. — 132 с.
18. Швырев А. А. Малый анатомический атлас. —
Ростов-на-Дону: Мини Тайп, Феникс, 2005. — 255 с.
Оглавление
Введение.................................3
Глава 1. Морфология и онтогенез центральной
нервной системы, ее роль в организме.
Спинной мозг ............................5
Филогенез нервной системы ...........10
Филогенез спинного мозга.............12
Онтогенез нервной системы ...........12
Онтогенез спинного мозга.............14
Аномалии развития спинного мозга.....20
Внутреннее строение спинного мозга ....21
Возрастные особенности спинного мозга .26
Глава 2. Морфология ромбовидного, среднего
и промежуточного мозга..................27
Онтогенез головного мозга ...........27
Ствол мозга .........................30
Продолговатый мозг ..................31
Состав медиальной петли: ............32
Задний мозг..........................34
Состав ножек мозжечка................37
Перешеек ромбовидного мозга .........39
Средний мозг.........................40
Промежуточный мозг...................43
Третий желудочек ....................47
Сетевидная формация..................49
Глава 3. Функиональная анатомия конечного
мозга. Боковые желудочки................55
Филогенез головного мозга............55
Онтогенез конечного мозга ...........57
Аномалии развития головного мозга: ..58
Плащ ................................59
Обонятельный мозг ...................64
Подкорковые центры — базальные ядра .66
221
Лимбическая система ....................73
Средний отдел конечного мозга...........78
Боковые желудочки ......................78
Белое вещество конечного мозга..........81
Циркуляция спинномозговой жидкости .....81
Синусы твердой мозговой оболочки .......86
Вены головного мозга....................86
Глава 4. Динамическая локализация функций в коре
полушарий большого мозга ..................89
Положение некоторых корковых
анализаторов............................92
Корковые концы анализаторов
внешнего мира: .........................93
Корковые концы анализаторов II сигнальной
системы.................................95
Глава 5. Проводящие пути .................100
Чувствительные пути ....................101
Двигательные пути ......................109
Ассоциативные пути .....................118
Глава 6. Черепные нервы (nervi craniales).124
Развитие черепных нервов ...............124
I пара.................................127
II пара................................127
III, IV, VI пары.......................128
V нерв.................................130
VII нерв...............................134
VIII нерв..............................135
IX нерв................................136
X нерв.................................137
XI нерв................................137
XII нерв...............................138
Глава 7. Спинномозговые нервы.............139
Развитие спинномозговых нервов .........139
Шейное сплетение.......................143
Плечевое сплетение.....................145
Межреберные нервы ......................148
222
Поясничное сплетение..................148
Крестцовое сплетение..................150
Глава 8. Органы чувств ..................153
Орган зрения ..........................154
Преддверно-улитковый орган............168
Общий покров .........................183
Развитие кожи и ее придатков..........183
Аномалии развития кожи ................184
Орган вкуса...........................185
Орган обоняния, organum olfactorium ...188
Глава 9. Автономная часть периферической
нервной системы (вегетативная
нервная система).........................194
Морфологические различия .............199
I. Узлы ..............................201
II. Ветви.............................201
III. Нервы ...........................202
IV. Сплетения .......................205
Развитие вегетативной нервной системы.207
Проводящие пути вегетативной нервной
системы ..............................208
Иннервация легких и плевры ...........210
Иннервация сердца ....................211
Иннервация слезной железы
и слюнных желез ......................214
Иннервация внутренних мышц
глазного яблока.......................215
Иннервация органов желудочно-кишечного
тракта (до сигмовидной кишки) ........216
Иннервация органов малого таза .......217
Заключение...............................218
Литература...............................219
223
Учебное издание
МЕДИЦИНСКИЙ АТЛАС
Кондрашев Александр Васильевич
Каплунова Ольга Антониновна
АНАТОМИЯ НЕРВНОЙ СИСТЕМЫ
Учебное пособие
Директор редакции И. Федосова
Ответственный редактор О. Шестова
Художественный редактор М. Левыкин
Редакционно-издательские работы
выполнены ИП С. Шаповаловым
ООО - Издательство -Эксмо-
127299, Москва, ул. Клары Цеткин, д. 18/5. Тел. 411-68-86, 956-39-21.
Home раде: www.eksmo.ru E-mail: infoQeksmo.ru
Оггтош* торговля юшгаын «Эксмо-.*
ООО «ТД -Эксмо- 142702. Московская обл , Ленинский р-н. г. Видное.
Белокаменное ш., д. 1, многоканальный тел 411 -50-74.
E-mail: receptionBeksmo-Mte.ru
По вопросам прлобретотшя княг -Эксмо» зарубежными оптовыми
покупателями обращаться а отдел зарубежных продаж ТД •Эксмо»
E-mail intemationelBeksmo-Mle.ru
International Sates: International wholesale customers should contact
Foreign Sates Department ot TraOng House - Eksmo- ter their orders.
intemetionalBeksmo-Mle.ru
По вопросам заказа книг корпоративным клиентам, в том числе в специальном
оформлении, обращаться по тел 411-68-59доб 2115,2117,2118.
Е-тай: vtpxskaxBeksmo.ru
Оптовая торговля бумажно-б aww» я канцелярскими товарами для школы
и офиса •Канц-Эксмо»: Компания -Камц-Эксмо- 142700. Московская обл., Ленин-
ский р-н. г Видное-2. Белокаменное ш.. д. 1, а/я 5. Тел./факс *7 (495) 745-28-87
(многоканальный), e-mail kancBeksmo-Mte.ru. сайт www.kanc-eksmo.ru
Полны» асоорпавент книг издательства «Эгсмо» для оптовых покупателе»:
В Санкт-Петербурга: ООО СЗКО. пр-т Обуховской Обороны, д 84Е.
Тел. (812) 365-46-03/04. В Нижнем Новгорода: ОООТД -Эксмо НН», ул. Маршала
Воронова, д. 3. Тел. (8312) 72-36-70 В Казани: Филиал ООО «РДЦ Самара»,
ул. Фрезерная, д 5. Тел (843) 570-40-45/46. В Самаре: ООО -РДЦ-Самара»,
пр-т Кирова, д 75/1. литера «Е». Тел (846) 269-66-70
В Ростова-на-Дону: ООО -РДЦ-Ростов», пр. Стачки, 243А. Тел. (863) 220-19-34.
В Екатеринбурге: ООО -РДЦ-Екатеринбург-, ул. Прибалтийская, д. 24а.
Тел. (343) 378-49-45 В Киеве: ООО -РДЦ Эксмо Украина-. Московский пр-т, д. 9
Тел./факс (044) 495-79-80/81 Во Львове: ТП ООО «Эксмо-Запад», ул. Буэкова, д 2.
Тел./факс: (032) 245-00-19. В Симферополе: ООО «Эксмо Крым-, ул. Киевская,
д. 153. Тел./факс (0652) 22-90 03. 54-32-99. В Казахстане; ТОО «РДЦ Алматы»,
уп. Домбровского, д. За Тел./факс (727) 251 -59-90/91. rdc-almatyBmail.ru
Подписано в печать 23.09.2009. Формат 84x108 ’/32.
Печать офсетная Усл. печ. л. 11.76 + вкл.
Тираж 2000 экз Заказ № 7492
Отпечатано с готовых файлов заказчика в ОАО «ИПК
Ульяновский Дом печати*. 432980, г. Ульяновск, ул. Гончарова, 14
редлагаемый атлас — удобное, наглядное,
компактное пособие по анатомии нервной
системы, сочетающее в себе преимущества класси-
ческого атласа, учебного пособия и справочника.
В атласе вы найдете:
90 цветных детальных иллюстраций с поясне-
ниями, прекрасно исполненных на плотной
глянцевой бумаге;
русские и латинские термины, данные в соответ-
ствии с современной анатомической номенкла-
турой;
необходимое разъяснение сложных вопросов
анатомии мозга, нервов, органов чувств и вегета-
тивной нервной системы.
Атлас будет полезен и необходим студентам вузов,
интернам, ординаторам, аспирантам, а также
молодым врачам.