/



























































































































































Теги: общая экология биоценология гидробиология биогеография общая микробиология биология экология микробиология
ISBN: 5-12-001076-8
Год: 1990




Текст
Ю. А. Горбенко
ЭКОЛОГИЯ
И ПРАКТИЧЕСКОЕ
ЗНАЧЕНИЕ
МОРСКИХ
КИЕВ
НАУКОВА ДУМКА
1990
АКАДЕМИЯ НАУК УКРАИНСКОЙ ССР
ИНСТИТУТ БИОЛОГИИ ЮЖНЫХ МОРЕЙ ИМ. А.О.КОВАЛЕВСКОГО
ЭКОЛОГИЯ И ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ
МОРСКИХ МИКРООРГАНИЗМОВ
Киев Наукова думка 1990
Уда 574:579.2(26)
Экология и практическое значение морских микроорганизмов / Горбенко Ю.А.;
Отв. ред. Миронов О.Г., АН УССР. Ин-т биологии южных морей им. А.О.Ковалев-
ского. - Киев: Наук, думка, 1990. - 160 с. - ISBN 5-12-001076-8.
В монографии обобщены данные о микробиоцеиотическом происхождении
морской взвеси и микробиогеином осаждении карбонатов в море, выдвигается
гипотеза об образовании морского песка на взвеси. Рассматриваются взаимо-
связи био- и а биопараметров изучаемой системы с основными компонентами
загрязнения моря.
Для экологов, гидробиологов, гидрологов, микробиологов.
Ил. 8. Табл. 52. Библиогр.: с. 153-157.
Ответственный редактор О.Г.МИРОНОВ
Утверждено к печати ученым советом
Института биологии южных морей им. А. О. Ко вал веского
Редакция биологической литературы
Редактор Л. Е. Мечева
Г 1903040100-546
М221(04)-90
ISBN 5-12-001076 8
© Ю.А.Горбенко, 1990
ПРЕДИСЛОВИЕ
Микроорганизмы играют ведущую роль в энергетике морских экосис-
тем, динамике их функциональных состояний, в биологической про-
дуктивности моря и процессах биологического самоочищения.
Исследования экосистемы морских микроорганизмов, проведен-
ные на основании многолетних системных наблюдений в море, позво-
ляют выявить структуру и взаимодействие ее элементов, а также рас-
смотреть некоторые вопросы биоценологии. В результате обработки
многочисленных экспериментальных данных с помощью методов ма-
тематической статистики был изучен ряд взаимоотношений в экосис-
теме морских микроорганизмов.
Благодаря регулярным наблюдениям в бухте у Севастополя (3-
6 раз в месяц) установлено, что существуют корреляционные взаимо-
связи между всеми изучаемыми био- и абиотическими параметрами. В
процессе жизнедеятельности микроорганизмов происходит образование
органического вещества, осаждение карбонатов в перифитоне в виде
СаСОэ и отложение ряда элементов, в том числе многих металлов.
Определено, что изучаемый комплекс множества био- н абиоти-
ческих параметров морской среды, сообщества перифнтонных микро-
организмов н его метаболитов, по всем основным критериям может
быть отнесен к экосистеме (Горбенко, 1977). Структура н динамика
указанной экосистемы после включения в нее исследованного нами
сообщества микроорганизмов взвеси, а также фактора Солнца были
изучены с помощью статистического анализа временных рядов на-
блюдений.
Применяя кластерный и корреляционный методы, установили, что
в совокупности всех рассматриваемых параметров могут быть выде-
лены различно связанные между собой группы. Анализ корреляцион-
ных зависимостей в разные годы наблюдений свидетельствует о том,
что, несмотря на большую изменчивость, взаимные корреляции внутри
групп, а также некоторые межгрупповые связи сохраняются в течение
ряда лет и являются стабильными. Периодичность в динамике множест-
ва параметров изучали методом Фурье. Выявлена годовая периодич-
ность, характерная почти для всех параметров. Исходя из фазовых
отношений обнаружены сезонная и многолетняя сукцессии изучаемых
параметров.
3
Определены причины многолетней изменчивости сухой массы
сообщества перифитонных микроорганизмов и числа живых диато-
мовых планктона. В обоих случаях фактором, определяющим их из-
менения, является активность Солнца. При этом многолетняя изменчи-
вость сухой массы стимулируется (г “ 0,922), а число пиков диатомо-
вых в течение года подавляется солнечной активностью (г =» -0,89) .
Установлено также, что на сезонные изменения массы слизи воздей-
ствует температура воды в море, а на годовой цикл динамики диато-
мовых — продолжительность солнечного сияния (Горбенко, Кры-
шев, 1985).
Цель наших исследований - Изучение экологии морской экосис-
темы1 микроорганизмов, включающей в себя микроорганизмы сли-
зистой пленки обрастаний, выявление основных закономерностей ее
развития, динамики и функционирования, а в конечном итоге выяс-
нение причинных связей в системе.
С помощью математических методов были выявлены важные вза-
имоотношения, в том числе и причинные связи, а также структура и
динамика био- и абиопараметров изучаемой экосистемы, для орга-
низмов которой биотопом служат взвешенное вещество, поверхность
твердых и плотных Погруженных предметов, а также воды Мирового
океана.
За оказанную в работе помощь до сбору и обработке материалов
выражаю благодарность сотрудникам Института биологии южных мо-
рей Л.Я.Татаренко, В.П.Горбенко, И.Н.Гутник, Ю.Л.Ковальчук, а так-
же доктору физико-математических Наук И. И.Крыше в у и доктору
технических наук Ю.В.Подвинцеву.
Принято экосистему считать синонимом сухопутного биогеоценоза (Шварц,
1970). Поскольку водные экосистемы нельзя отождествлять с биогеоценозом,
их следует называть биогидроценозами (Мордухай-Болтовской, 1975).
4
ГЛАВА 1. ИСТОРИЯ ВОПРОСА
К перифитону относятся микро- и макроорганизмы, обитающие на пог-
руженных в море твердых, неживых предметах естественного и искус-
ственного происхождения за пределами придонного слоя воды1. Этот
слой вместе с донным, мягким или рыхлым грунтом является биото-
пом другого сообщества - бентоса (Карзинкин, 1926-1927, Дуплаков,
1933, Горбенко, 1977).
В результате изучения гетеротрофных бактерий, обитающих в мор-
ской воде, пришли к выводу, что комплекс микроорганизмов, форми-
рующихся на голых камнях, взятых с суши и погруженных в море,
в основном аналогичен микрообрастанию на стеклянных и металли-
ческих (нержавеющая сталь) пластинах.
При этом было замечено, что в первые часы пребывания стеклян-
ных пластин в море на них встречаются бактерии, передвигающиеся
по поверхности стекла. Через несколько часов некоторые из них прик-
реплялись к стеклу (Морское обрастание о борьба с ним, 1957). О сход-
стве диатомовых водорослей перифитона, обитающих на естественных
субстратах и на поверхности пластин обрастания, известно давно (Прош-
кина-Лавренко, 1963). Однако, микрообрастания на стеклянных и
металлических пластинах несколько отличались. На первых было мень-
ше диатомовых, чем на вторых, очевидно, из-за их лучшей освещен-
ности.
В экспедициях в Средиземное море На нис "Академик Вавилов”
в июне - сентябре 1959 г. ив октябре 1960 г. - январе 1961 г. мы изу-
чали распространение гетеротрофных бактерий планктона во всей тол-
ще моря. Изучая оседание и развитие морских микроорганизмов на
различных Противообрастаемых красках с ядовитыми компонентами,
обнаружили, что поверхность красок заселяли в основном бактерии, чис-
ленность которых чрезвычайно быстро возрастала и была намного
большей, чем на стеклянных пласгинах-контроле, т.е. бактерии из
окружающей морской воды на поверхность красок привлекала орга-
ническая основа (канифоль) их состава. Ядовитые компоненты этих
покрытий, как мы убедились, на бактерии не влияли (Горбенко, 1963).
1 В то же время к обрастанию можно отнести перифитон на погруженных в море
предметах искусственного происхождения.
5
Вместе с тем на красках встречалось очень мало диатомовых во-
дорослей, они, как правило, были мелкими, иногда на длинных тяжах
’’ножках” и явно угнетались ядами красок. Простейшие организмы на
красках встречались также редко из-за действия ядов.
Для наблюдения за динамикой и развитием микрообрастания на
пластинках в море вместо обычного стекла использовали толстую
стеклянную ппастшу с вырезом посередине*, которая обрастала мик-
роорганизмами, а покровное стекло покрывало прорезь. Таким обра-
зом, рыхлая и нежная слизистая пленка микрообрастаний не поврежда-
лась. Поскольку пластина была погружена в море постоянно, а извле-
калась только на 2 ч для наблюдении под микроскопом, то мы получи-
ли возможность ’’погрузиться под воду с микроскопом” и наблюдать
в динамике жизнь микроорганизмов первичной слизистой пленки
обрастаний в естественных условиях их обитания. Воду под покровным
стеклом меняли каждые 30 мин с помощью пипетки со свежей мор-
ской водой из моста погружения пластины.
При изучении микроперифитона в Мексиканском заливе у берегов
Кубы (июнь-сентябрь 1965 г.), установили, что в этой части Океана
динамика мнкрообрастания аналогична таковой в Черном море. Но
поскольку температура морской воды в этом районе колеблется в
пределах IB-29 С, то и биоциклы проходят значительно быстрее.
Так, сплошной слой диатомовых водорослей здесь вырастал за 5-7 сут,
тогда как в Черном море летом в самое жаркое время года (июле-
августе) - только за 8-10 сут.
Исследуя стеклянные и металлические пластины (нержавеющая
сталь) в прикубинском районе Атлантического океана, мы наблюдали
примерна такие же характерные различия в их микрообрастании, как
и в Черном море. На пластинах из металла число диатомовых водорос-
лей приблизительно соответствовало количеству гетеротрофных бакте-
рий, но на стеклянных пластинах диатомей было в 10-50 раз больше,
чем бактерий. Вместе с тем количество бактерий на стекле и металле
В первые 11 сут после погружения в море не отличалось, а колебания
численности были аналогичны (Горбенко, 1977),
Пробные системные исследования с параметрами перифитона и
Планктона были проведены в ноябре — декабре 1965 г. В 1966 г. была
поставлена l-я серия пластин для исследования динамики микрообрао
таний. Число изучаемых параметров 20. Впоследствии количество пара^
метров было увеличено до 50 и более. Соответственно была освоена
серия методов определения их величины. Вели в 1966 г. мы все наблю-
дения проводили ежедневно, то в связи с увеличением числа наблюде-
ний за ряд последующих лет дискретность их стала равна 5 сут. Затем,
начиная с 1973 г., экспериментальным путем был выбран оптимальный
перерыв между наблюдениями - 10 рут. Результаты указанных иссле-
дований используются для изучения ряда процессов, происходящих
в море.
1 Камера Горяева для подсчетов элементов крови (Горбенко, 1977).
6
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
Настоящая работа написана на основании материалов 19-летних наблю-
дений, начатых в 1966 г. у подводного стенда в северной части Севасто-
польской бухты, в 1000 м от открытой части Черного моря, на расстоя-
нии 0,5-1,5 м от поверхности, на глубине 4-5 м. Наблюдения проводи-
лись не реже 1 раза в 10 сут. В данном разделе рассматриваются общие
снедения Но методике и материалам, имеющим отношение ко всем
разделам исследований. Для лучшего понимания результатов работы
. в ряде некоторых разделов текста методика приводится дополни-
тельно.
Объекты исследования (факторы). Сообщество организмов обрас-
таний (Сооб), сообщество перифитонных микроорганизмов (СПМ),
сообщество микроорганизмов планктона (СМП) и взвеси (СМВ)1,
; фактор карбонатного равновесия (ФКР), фактор солнца (ФС), фактор
морской среды (ФМС), фактор загрязнения (ФЗ), фактор кристалли-
ческих веществ (ФКВ).
Предметы исследований (параметры). Сооб - балянусы, ботрнл-
люсы, мидии, мшанки - измеряются в милиграммах на 100 см*2, гид-
роиды - в процентах на 100 см“2 (всего 5 параметров).
I СПМ — число гетеротрофных (по числу колоний на поверхности
• культуральной среды, см" 2 ), палочковидных и кокковидных бактерий,
| живых и мертвых диатомовых водорослей (количество клеток на
; 1 см'2 поверхности пластин). Простейшие организмы - подвижные
(свободноживушие) и прикрепленные (количество организмов на
1 см'2 поверхности пластин), растворенное органическое вещество
(ЮВ) (в единицах экстинции Е на волне 270 нм — Ej-») и pH, транс-
формированные СПМ (РОВ СПМ и pH СПМ), карбонаты и сухая масса
слизистой пленки (после высушивания при 100 v, мг и г). Карбонаты
определяли, используя микрометод газового анализа (Горбенко,
1977),-всего 11 параметров.
СМВ - число гетеротрофных бактерий (по числу колоний в 1 мл
s взвеси), численность живых и мертвых диатомовых, простейшие под-
! вижные и прикрепленные (в 1 мл взвеси), pH и РОВ, трансформиро-
1 О правомочности выведения на уровень ценоза 2 последних сообществ микро-
организмов сказано далее.
7
ванные СМВ (после 2-часового взаимодействия с взвешенным вещест-
вом РОВ СМВ и pH СМВ), РОВ - по EJTO. Карбонаты и сухая масса СМВ
(в мг и г после высушивания при 100 ° С) - всего 9 параметров.
СМП - число гетеротрофных бактерий (количество колоний в
1 мл воды). Численность живых и мертвых диатомовых водорослей
(число в 1 мл воды) - 3 параметра*.
ФС - продолжительность сияния солнца (освещенность) - число
дней сияния солнца, активность солнца (по числу Вольфа — W)> энерго-
баланс, доля баланса в инфракрасной области, альбедо видимого спект-
ре, суммарная радиация - всего 7 параметров.
ФМС - содержание РОВ (по экстинции Е на волне 260 нм,
Емо). Окисляемость в щелочной среде (мг-л"’), pH, нитриты-
азот (мкг-л-1), фосфор - минеральный и органический (мкг-л"1),
температура (°C), соленость (%о), шедочность(мг-экв-л’*) - всего 9
параметров.
ФЗ - хлороформный экстракт из морской воды (ХлЭк), фенол-
содержащие соединения (мг. л-’) - 2 параметра. Пробы воды для
определения брали на глубине 25-30 см.
ФКР - углекислота СОа (мл-л-1), гидрокарбонат-ионы - НСОз,
карбонат-ионы (мг-нон- л~ *) - всего 3 параметра. Параметры ФКР
рассчитывались по pH, температуре, щелочности и солености морской
воды.
ФКВ — кварц и кальцит - % сухой массы слизистой пленки - 2 па-
раметра. Для определения указанны» минералов сухая масса СПМ
подвергалась рентген-дифрактометрическому анализу на приборе
”ДРОН-1,5”.
Таким образом, исследовали 8 факторов, состоящих из 51 парамет-
ра. Это позволило изучить н понять взаимодействие и поведение
элементов относительно большой экосистемы микроорганизмов в бух-
те, имеющей характер олиготрофного водоема с признаками мезо-
трофии (Горбенко, 1977). В этом регионе проявляется большинство
процессов и явлений, происходящих во всем море, а также процессы,
протекающие и в Океане, поскольку Черное море является частью Ми-
рового океана.
Многие интересные и веяные закономерности, взаимосвязи и явле-
ния были обнаружены В результате совместных работ с Д.Ф.Ермако-
вой и ДМ Витюком, а также Э.С Тримонксом.
Наши исследования являются экспериментом в естественных усло-
виях, который заключается в том, что в различные сезоны в динамике
изучаются все возможные параметры морской среды» влияющие на
жизнедеятельность сообщества обрастаний- Поскольку эти параметры
взаимосвязаны и постоянно изменяются, то чем больше мы проводим
* В некоторых главах настоящей книги численности микроорганизмов СПМ
СМВ, СМП приводятся как биомасса (мг.см , мг мл ).
|*№падвний за ними, тем больше узнаем о каждом из них и обо всех
MNBCW, В результате накапливается столько информации об изучаемой
уДОаме, что после соответствующей ее обработки становятся понят-
Вмми многие важные закономерности этой системы. К ним относятся:
ЙДОгура и взаимосвязи элементов системы, процессы, происходящие
I'Wfc зависимость от факторов морской среды и солнца, удается даже
МИВНть причины взаимодействия некоторых параметров. Материалы,
Доученные в течение ряда лет наблюдений, позволяют обнаружить
*вые явления, закономерности ц свойства ряда изучаемых элементов
Жосястемы.
ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
3.1. О БИОЦЕНОЛОГИИ
Как Показали наши исследования, комплекс микроорганизмов обраста-
ний (диатомовые водоросли, бактерии и простейшие) имеет общий
биотоп (поверхность твердых и плотных предметов в море). Микро-
организмы комплекса связаны друг с другом и с параметрами окру-
жающей морской среды не только в отдельности, ио и в целом - их
суммарными метаболитами (FOB и pH, карбонаты микроперифи-
тона и др.).
В процессе формирования микроорганизмов первичной слизистой
пленки образуются четкие биосукцессии. Все выявленные взаимосвязи
перифитонных микроорганизмов подтверждены (как за 1, так и за
много лет наблюдений) корреляционным анализом. Вследствие этого
указанный комплекс микроорганизмов как отвечающий всем крите-
риям ценоза был отнесен нами к сообществу перифитонных микро-
организмов — СПМ (Горбенко, 1977).
Хотя количество корреляционных связей между микроорганиз-
мами планктона меньше, чем в перифитоне из-за большой расплывча-
тости и изменчивости местообитания планктона, планктонные микробы
вместе с тем взаимосвязаны с СПМ, взвесью, с абиотическими факто-
рами моря и даже с параметрами Солнца. Все зто свидетельствует о
том, что эти организмы отвечают основным критериям биоценоза,
поэтому комплекс микроорганизмов планктона мы отнесли к сооб-
ществу, которое назвали СМП (Горбенко, 1977; Горбенко, Крышев,
1985). Прн этом установлено, что оно не имеет такой стабильной
структуры й тесных взаимосвязей, как СПМ.
Если о бактериях и диатомовых водорослях СПМ уже неоднократ-
но сообщалось (Горбенко, 1977; Подвинцев, Горбенко, 1977а, б;
Подвинцев, Горбенко, 1979; Крышев, Горбенко, 1979 и др.), то иифор-
мации о простейших организмах СПМ пока мало. Поэтому в настоя-
щей работе приводятся сведения н об этой группе организмов.
10
3.2. О ПРОСТЕЙШИХ ОРГАНИЗМАХ СПМ И ИХ ВЗАИМОСВЯЗИ X
В настоящем разделе обсуждаются результаты комплексных наблюде-
ний за микроорганизмами в море, в том числе и за простейшими. Мы
определяли образ жизни этих организмов (прикрепленные или подвиж-
ные формы), их численность и родовую принадлежность. В число прос-
тейших входили и мелкие жгутиковые формы водорослей, внешне
неотличимые от первых. Идентификацию организмов до рода проводи-
ли по существующим определителям (Kent, 1980-1982; Kahl, 1933;
Lepsi, 1960).
Средн прикрепленных форм простейших обнаружены организмы
следующих родов: Actinomonas, Monosiga, Salpingaeca, Folliculina, мно-
гочисленные Vorticellae, а также Zoothamnion, Carchesium, Platicola,
Podophria, Acineta, Doxophyllum, Ephelota, Stentor, Dendrosoma. Боль-
шинство из них наблюдались в обрастании (Брайко, 1985). Были учте-
ны также подвижные формы простейших, которые, как правило, нахо-
дились в движении, останавливаясь на миг только изредка. Встречав-
днеся нам организмы были очень похожи на представителей следую-
щих родов: Actinophrys, Bod о, Monas. Oikomonas, Pleuromonas, Salpin-
gaeca, Spongomonas, Phisomonas, Heteromita, Anthopysa, Petalomonas,
Chrysomonas, Anisonema, Peridinium, Gymnodinium, Prorocentrum,
Trachelocerca, Urotricha, Pyrsonympha, Geliozoa, Epiclintes, Euplotes,
Rhynchomonas, Flabelula, Unda.
В первые дни экспонирования пластин в море на них, как правило,
появлялись мелкие формы прикрепленных простейших - представители
родов Monas, Actinomonas, Monosiga, Salpingaeca. В то же время среди
прикрепленных форм обитали и подвижные - представители родов Mo-
nas, Bodo, Oicotnonas, Phisomonas, Heteromyta, Authophysa, Rhynchomonas
и подвижные Salpingaeca. Это были в основном жгутиковые формы.
Через 10-19 сут появлялись более крупные прикрепленные орга-
низмы - представители родов Vorticellae, Acineta, Zoothamnion, Sten-
tor и подвижные, в основном ресничные инфузории - Anthophysa,
Urotricha, Trachelocerca, Pyrsonympha, Geliozoa, Epiclintes.
После 20-30 сут на поверхности погруженного предмета обнаружи-
вались колониальные прикрепленные организмы Zoothamnion, Vorti-
cella, Folliculina, Carchesium, а также Acineta, Loxophyllum и подвижные
Euplotes, EpicHnthes, амебы Flabelula, Unda и др.
С помощью расчета взаимных корреляционных функций мы уста-
новили наличие корреляции простейших организмов со всеми осталь-
ными элементами СПМ, а также величину временных задержек этих
корреляций (табл. 1). При этом подвижные формы простейших корре-
лировали 19 связями с компонентами СПМ. Из них одновременными
были 11 корреляций - с диатомовыми живыми и мертвыми, гетеро-
трофными и палочковидными бактериями и pH СПМ. Остальные связи
осуществлялись с задержкой на 10-30 сут.
11
Таблица 1. Корреляционные взаимосвязи простейших организмов СПМ
Параметр Простейшие
Подвижные Неподвижные (прикрепленные)
1977 г. 1978 г. 1979 г. 1980 г. 1977 г. 1978 г. 1979 г. 1980 г.
Диатомовые IL.4JL 0.60 0.44 - -0,30 0,35 -0.32 -0.49 живые 0 0 0 20 0 -10 -10 Диатомовые 0,39 - -0,49 - - 0,30 0,32 0.33 мертвые 0 10 0 30 -15" Гетеротрофные 0,34 0,35 -0,44 0,40 - 0/18 0,30 -0.31 бактерии 0 0 10 0 -36" -20 -10 Палочковидные 0,46 0.31 0.52 0.59 -0.30 -0,35 -0.44 - бактерии 0 0 0 0 —10 —10 20 Кокковидные 0,44 - - -0,39 0,59 0,49 0,31 0.36 бактерии -30 10 - 20 20 10 0 Сухая 0,38 _ _ _ Q.48 - - 0.40 масса 30 10 -20 V « 0.30 - 0,46 0,34 - 0.44 Карбонаты 30 10 20 - 20 “ -0.45 -0.38 - -0,54 -0,30 0,38 -0,37 рн 0 10 10 10 10 30 - 0,59 0,39 0,33 0,62 ГОВ 1(Г -20~ “0“ Ю
Примечание. В числителе - коэффициент корреляции; в знаменателе - времен-
ные задержки корреляции, сут; 0 - корреляция обоих параметров наступает
одновременно; знак перед числом - один параметр в корреляции (первый)
запаздывает по отношению ко второму; если величина в знаменателе положитель-
ная - первый параметр в корреляции опережает второй.
Прикрепленные формы, в свою очередь, коррелировали 30 связями
с остальными биокомпонентами СПМ. При этом одновременными были
4 связи простейших - с диатомовыми живыми и мертвыми, кокковид-
ными бактериями и РОВ СПМ. Остальные 26 связей осуществлялись
с задержкой от 10 до 30 сут (Горбенко, Крышев, 1985).
Достоверные корреляционные связи простейших почти во все годы
исследований наблюдались для живых диатомовых водорослей гетеро-
трофных и палочковидных бактерий. Численность кокковидных бакте-
рий с pH СПМ коррелировала 6 лет из 8, с диатомовыми мертвыми —
5 лет из 8, карбонаты с РОВ - 4 года из 8, а с сухой массой СПМ - 3
года из 8.
Подвижные простейшие коррелировали с гетеротрофными и палоч-
ковидными бактериями 4 года из 4, с диатомовыми мертвыми, кок-
ковидными бактериями и pH СПМ — 2 года из 4, с сухой массой и кар-
бонатами - 1 год из 4, а с РОВ СПМ ни одной корреляции ие наб-
людалось.
12
. Прикрепленные формы простейших все 4 года из 4 коррелировали
с диатомовыми живыми, кокковидными бактериями, pH и РОВ СПМ,
3 года из 4 — с диатомовыми мертвыми СПМ, гетеротрофными, палоч-
ковидными бактериями и карбонатами СПМ, 2 года из 4 - с сухой
массой СПМ. Большее количество корреляций неподвижных форм
; Простейших организмов свидетельствует об их более важном значе-
ним в СПМ.
к’ .
t
( '
I КОМПЛЕКС МИКРООРГАНИЗМОВ ВЗВЕСИ
| В КАЧЕСТВЕ ЦЕНОЗА
Исследования изучаемой экосистемы свидетельствовали о Том, что
микроорганизмы, подобные перифитонным, встречаются не только в
перифитоне или обрастании. Еще в 1967 г., изучая микроорганизмы
Г взвеси, мы заметили, что структура и цвет слизистой пленки СПМ на
пластинах, высушенных на воздухе, аналогичны морской взвеси, соб-
ранной в море, нанесенной на пластины и высушенной. Поэтому мор-
ская взвесь была включена в общий комплекс наших системных ис-
следований. Было установлено, что на взвеси, так же как в перифито-
! не, обитают диатомовые водоросли (живые и мертвые), гетеротрофные
. бактерии и простейшие (подвижные и прикрепленные формы). Они
взаимосвязаны между собой и с окружающей морской средой, обра-
зуют метаболиты — РОВ и pH взвеси, а на поверхности взвешенного
вещества осаждают карбонаты. Однако для того, чтобы убедиться,
Что комплекс микроорганизмов взвеси является (или нет) сообщест-
вом микроорганизмов, необходимо было установить, отвечает ли он
всем признакам сообщества. Для этого мы провели корреляционный
анализ всех имеющихся числовых данных, собранных в течение 6 лет
наблюдений. Оказалось, что коррелируют между собой почти все па-
раметры микроорганизмов взвеси.
Обитают эти микробы на парящих в толще воды частицах взвеси
, в очень изменяющихся условиях морской среды, в отличие от значи-
тельно более комфортных условий обитания микроорганизмов пери-
фитона (СПМ). Поэтому и количество парных корреляций между
микроорганизмами взвеси гораздо меньше, чем в перифитоне, и вели-
чина их в целом несколько меньше (табл. 2). Подобная закономер-
ность проявлялась и у микроорганизмов планктона (Горбенко, 1977).
Этого, однако, достаточно, чтобы констатировать факт наличия взаи-
мосвязей внутри сообществ микроорганизмов планктона и взвеси.
Обнаружены корреляции микроорганизмов взвешенного вещест-
ва также с их метаболитами, ГОВ и pH взвеси (табл. 3). Все 9 изу-
чаемых параметров взвеси в различные годы коррелируют с 74 био-
и абиотическими параметрами изучаемой . морской экосистемы
(табл. 4).
13
Таблица 2. Корреляционные связи микроорганизмов взвеси за период наблмде-
иий с 1970 г. «>1982 г.
Параметр Коэффициент корреляции (г)
1970 г. 1971г. 1979-1980 гг. 1981г. 1982 г.
Диатомовые живые — jy»- томовые мертвые 0,76 0,46 0,95 0,82 0,84 Диатомовые живые - гете- ротрофные бактерии -0,34 0,35 Диатомовые мертвые— ге- теротрофные бактерии 0,30 Д иатомовые живые - под- вижные формы простейших 0,3 2 Диатомовые мертвые - прикрепленные формы простейших 0,46 Гетеротрофные бактерии - подвижные формы прос- тейших -0,30 Гетеротрофные бактерии - прикрепленные формы простейших -0,33 -0,31 даатомовые живые - при- крепленные формы прос- тейших 0,50 Подвижные формы простей- ших- неподвижные 0,58 Д иатомовые мертвые - не- подвижные формы прос- тейших 0,43 0,45 039 л 63 65 44 43 45 rjnjn «0,30 при л «42
Таблица 3. Корреляционные связи микрооргзнмзмов взвеси с их метаболитами
(pH и РОВ взвеси)
Параметр Коэффициент корреляции (г)
1971г. 1972 г. 1974 г. 1979-1980 гг. 1981г
Диатомовые живые—
РОВ взвеси 0,39-0,55 -
Диатомовые мертвые -
РОВ взвеси -0,36-0,53 -
ГН взвеси - РОВ взвеси — -0,40
етеротрофные бакте-
рии - pH взвеси - 0,49
Под вижные формы
простейших - карбона-
Йа взвеси - -
рмкреплеиные формы
Простейших- карбонаты - -
Подвижные формы прос-
тейших- pH - -
0,38
0,40
-0,31
-0,31 -0,33
44 43
’’min *0,30 при л «42
14
Таблица 4. Корреляция параметров взвеси с параметрами морской воды, 1968-1974 гг.
Параметр Коэффициент корреляции (П
1968 г. 1969т. 1970 г. 1971 г. 1972 г. 1973 г. 1974 г.
Диатомовые живые освещенность -0,38 - - 0,36 — - - фосфор минеральный - - 0,37 0,54 - - 0,44 фосфорорганический - - - 030 0,51 — pH морской воды - - - 0,32 - нсо; и со?- - - - дэв - - - диатомовые живые планктона - - 0,53 0,60 0,64 . - 0,47 кислород вод ы - - 0,39 - 0,43 ' - 033 фенол - - 0,51 - - -033 зоопланктон _ _ _ о,36 — - ' - температура воды ______ -038 диатомовые мертвые планктона _ _ _ 034 - - - РОВ морской воды -, - - _ _ _ -0,32 взвесь в воде ______ 032 масса 10 мл взвеси - - 0,48 - _ _ _ хлороформный экстракт (ХлЭк) ______ 0,59 Диатомовые мертвые СО, НСО;, СО?” 035 фосфор минеральный - - 0,30 - _ _ _ фосфорорганический _ _ _ о, 41 - - - кислород - - 0,31 0,45 - освещенность _ _ _ о,48 - - - прозрачность _ _ _ 0,53 - - - диатомовые живые планктона - - 0,43 _ _ _ _ масса 10 мл взвеси - - -034 _ _ _ _ Гетеротрофные бактерии хлор - -032 - - - - - соленость _____ —039 — К
04 Охоммпмее неба. 4 .
паи*
кислород
дрыивжЖ
00.
PQBCMB
' крмашй
февря
doj-
рн- нсо;
РОВ - прозре’вость
pH
прозрачное»
температура морской вода
pH-СО’,-
массе 10 мл взвеси
pH морской вода
температура вода
диатомовые левые планктона
диатомовые мертвые планктона
температура морской воды
СО в вода
соленость
0г
зоопланктон
СО,
шелон,
ХлЭк
,30 при я « 0,42
Кмффяцмят коррвляци (г)
ms г. 1989 г. 1970 г. 1971г. 1972 г. 1973 г.| 1974 г.
__ — -038
-оз* XV — —
— ••V озз —- -Ml -035 М5
834 — — — -озз 0,35 —
— — озо — —
-038 — — — — — 033
W — 0Д5 — озо — —
032 озо — —• — — М5
«А» «V — озо — — —
— — — 0.63 — — 0,34
031 0,35 — «V —— —- 037
— -032 — — — — -031
-мз -озо «V — V — 0,42
-оз* V. — — — 0,31-0,67
— 039 М9 — — — 035
035 V. •V — 037 «V
— 0,32 — — -036 —
— -03* — — — 0,46 —
— озо — — — —
-034 — — — — 0,36 0,41
0,3* — 0,43 мз —
— — -032 — • 034 -0,33 —
— * —0,35 — —
— -X» -036 —- — —
(у* 4^1 >" i t. 5
' «— — V,
— — —
— —• —
032 озо
0,31 035
— 0,32 —
V V.
— 0,30 —
— —
- 0,32
— —0,34 XXX»
— 0,30 V
71 67 63
мо 0,77 035
0,55 хм»
0,65
— ОДО 0,44 -
— — 0,39 —
•— — —
-0,35 V.
—. —. -0,30 м—
V, V 0,46 V.
0,53 0Д8 •* —
— — —
— —
—
65 62 45 46
О I I I I I I I I I I 111$
I
I
ОСО I I I I I I I о I I I <0
iiii i i i i i i i i i i3
SIRS! 2 ЗЭ
cncnco tn иклсл-
IIII I ooo I о I cJcfcj’O
iiii 1351।। । । । । p
Карбонаты взвеси коррели-
руют с микроорганизмами взве-
си и их метаболитами (табл. 5).
Кроме того, они коррелируют
и с параметрами морской сре-
ды обычными корреляционными
связями (табл, б), а на 10-45-е
сутки наблюдений корреляции
функций (Крышев, Горбенко,
1971) на этих высоких частотах
в основном оказались тесными и
взаимными (табл. 7).
Как установила А.Н.Бобко-
ва, органические компоненты
взвеси, белки, углеводы и орга-
ническое вещество взаимоскор-
релированы (табл. 8).
На ранних стадиях разви-
тия взвеси в море (10-45 сут)
взаимные корреляции имеют
большую величину, чем впослед-
ствии, поэтому в начале наблю-
дений за свежеосевшей взвесью
связи между компонентами взве-
си сильнее, чем в более длитель-
ные промежутки времени (нес-
колько месяцев, год).
Изучение взаимосвязей па-
раметров морской экосистемы
(Подвинцев, Горбенко, 1979) с
помощью факторного метода и
частных корреляций позволило
установить, что изучаемые 42 па-
раметра могут быть (по стати-
стическим данным) расположе-
ны в виде 9 группировок - фак-
торов с сильными, средними н
слабыми статистическими корре-
ляционными связями, в кото-
рых сильные связи образуют яд-
ра структур. При этом парамет-
ры взвоеп входят в состав яд-
(из 9). А эле-
мент вдарен Ь гетеротрофные
ТйкТепШ!. жишПе и мертые диа-
17
Таблица 5. Корреляция карбонатов, отлагающихся на взвеси, с микроорганизма-
ми взвеси и их метаболитами
Параметр Коэффициент корреляция (г)
1969 г. 1970 г. 1971 г. 1972 г. 1973 г. 1974 г.
Гетеротрофные бактерии — — -0,33 -0,31 — —
РОВ взвеси — — 0,37 -0,32 — -0,43
pH взвеси — -0,36 -0,49 — -0,52
Диатомовые мертвые — — 0,40 — — —
Диатомовые живые 0,36 — -0,51 . — • —
п 67 63 65 62 45 46
rmjn = 0,300 прИЛ х 42
Таблица 6. Корреляция карбонатов взвеси с параметрами взвеси н морской среды
Параметр Коэффициент корреляции (г)
1968г. 1969г. 1970г. 1971г. | 1972г. 1973г. 1974г.
Диатомовые жи-
вые взвеси — — — — -0,32 —
Гетеротрофные
бактерии планк-
тона 0,41 — — -0,49 — —
pH морской воды — — -0,44 0,70 — -0,37 -0,38
Масса 10 мл взвеси 0,40 — — — —
Температура воды 0,36 — —* — 0,59 -0,53
Щелочность воды 0,40 — — —- 0,34
о, 0,36 — — — — 0,52
Зоопланктон — — — 0,46 — — 0,5
Кислород — — — — 0,64 — —
НСО.’иСОГ — — — — 0,33-0,38 —- —
Фосфор органичес- •
кий воды — — — — -0,55 — —
Прозрачность — — — — -0,34 —
Минеральная
часть взвеси — —- — — •— -0,69 —
СО, и НСО," — — — . — 0,32-0,39 —•
ХлЭк — — — — — 0,31 —
Кремний в воде — — — — — 0,35 -0,65
Соленость — — — — — — 0,54
Фенол — — — — — 0,37
п 71 67 63 65 62 45 46
rmin ~ 0>30 при п - 0,42
томовые водоросли, pH и карбонаты взвеси — связаны не только меж-
ду собой, но также с остальными параметрами морской экосистемы
и с несколькими параметрами солнца (Подвинцев, Горбенко, 1979).
Многочисленные корреляционные связи элементов взвеси, уста-
новленные различными методами и в разные годы, свидетельствуют о
том. что эти элементы взаимосвязаны с окружающей морской средой,
18
Таблица 7. Взаимные корреляционные функции параметров
взвеси, 10-45 сут наблюдений (1971 г.)
Параметры Коэффициент корреляции (г)
Диатомовые живые - диатомовые мертвые 0,54
Диатомовые живые - РОВ взвеси 0,55
Диатомовые мертвые взвеси- РОВ взвеси 0,53
Дптомовые живые - освещенность 0,72
Диатомовые живые - фосфор органический
и минеральный 0,64-0,65
Дптомовые живые - НСО, и COJ в море 0,64-0,70
Диатомовые живые - pH морской воды 0,73
Диатомовые мертвые - фенол 0,75
Гетеротрофные бактерии — зоопланктон 0,75
Гетеротрофные бактерии - ХлЭк 0,75
Гетеротрофные бактерии - кислород 0,60
Гетеротрофные бактерии - СО, 0,54
РОВ взвеси - кислород 0,63
РОВ взвеси- фенол 0,70
РОВ взвеси - pH морской воды 0,55
РОВ взвеси-НСО7 0,65
/’min = 0,24 при я = 70
Таблица 8. Корреляционные связи органических компонентов
взвеси (но данным АН.Бобковой), 1979-1981 гг.
Параметр Коэффициент корреляции (г)
1979- 1980 гг. 1981 г.
Белки - органические вещества 0,44 0,89
Белки - углеводы 0,51 0,91
Органическое вещество - углеводы 0,47 0,83
я “43
г min = 0,30 при я “ 42
т.е. взвесь — один из типичных представителей морской среды, сущест-
венно зависящий от нее.
Поскольку диатомовые, бактерии и простейшие, а также их мета-
болиты — pH, РОВ взвеси, карбонаты, отлагающиеся там, взаимосвя-
заны друг с другом и со средой, имеют общий биотоп, а судя по мно-
гим корреляциям их метаболитов, отвечают на воздействия среды как
единое целое, то есть все основания считать, что комплекс микроорга-
низмов взвеси (МВ) является сообществом, которое было назва-
но СМВ.
19
34. О ПОДОБИИ МИКРОЦЕНОЗОВ ПЕРИФИТОНА И ВЗВЕСИ
Многолетние исследования динамики экосистемы микроорганизмов в
регионе Севастопольской бухты, включающей в себе сообщества мик-
роорганизмов СПМ и СМВ, позволили сделать сравнительный анализ
основных характеристик этих сообществ. Определили видовой состав
диатомовых водорослей во взвеси, представленный 37 видами и 14
родами.
Диатомовые водоросли, встречающиеся
Севастопольской бухты
Вид
Asterionella japonica
Achnauthes bravipes Ag.
A. longipes Ag.
Amphora arcus Greg.
A. bigibba Grun.
A. inflexa (Bieb) H.LSmith
A. insecta
A. hyalina Kiits
Amphora sp. (1,2, 3)
Amphiprora sp. (1, 2, 3) Ehr.
Bacilbria peradoxa
Campylodiscus Thuretii Bred. GmeL
CaJoncissp. CL
Cocconeis scutellum Ehr.
Cymbelbsp.
Chaetoceros sp. Ehr.
CeratauBna Bergonii Perag
Coscinodiscus sp. Ehr.
Cyctotelh caspia. Griin
Dtploneis sp. Ehr.
Diatoma sp. D.C.
Detonub confervaceae (Cl) Gran.
Fraginaria diHcatissima Pr - Lavr.
Gyro sig ma sp. Hass.
Grammatophora marina Kilts.
Licmophora flabellate Ag.
Licmophora sp. Ag. (1,2)
LeptocyHndrus minimus Gran.
Melosira moniliformis таг. Subghbosa Gran.
Melosira sulcata (Ehr.) Kiitz.
M. moniUfornis (O. Mufl) Ag.
M. juergensii Ag.
Navicula sp. Воту (1-6)
Nitzchia closterium (Ehr) W. SM.
N. longissima (Bred) Ralfs.
N. sigma W. Sm.
N. seriata CL
N. delicathsima CL
N. reverse W. Sm.
Nitzchia sp. (1 -3)
N. temiirostris Mcrs. 1
в морской взвеси
Количество
Звида
Звида
2 вида .
6 ВИДОВ
3 вида
3 вида
2П
Pleurorigma sp. W. Sm. (1-3)
Stauroneb sp. Eta-.
Surirelb gemma Eta.
Synedra sp. Eta. (1-3) .
StrtateHa unipunctata (Lyngb.) Ag.
Sceletonema oostatum (Grov) Cl.
Synedra bacutas Greg.
RMzosolenia fragilisslma Bergen
Rhizosolenia calcar avic Scnultze
Rhabdonema adriaticwn Ktttz.
Звида
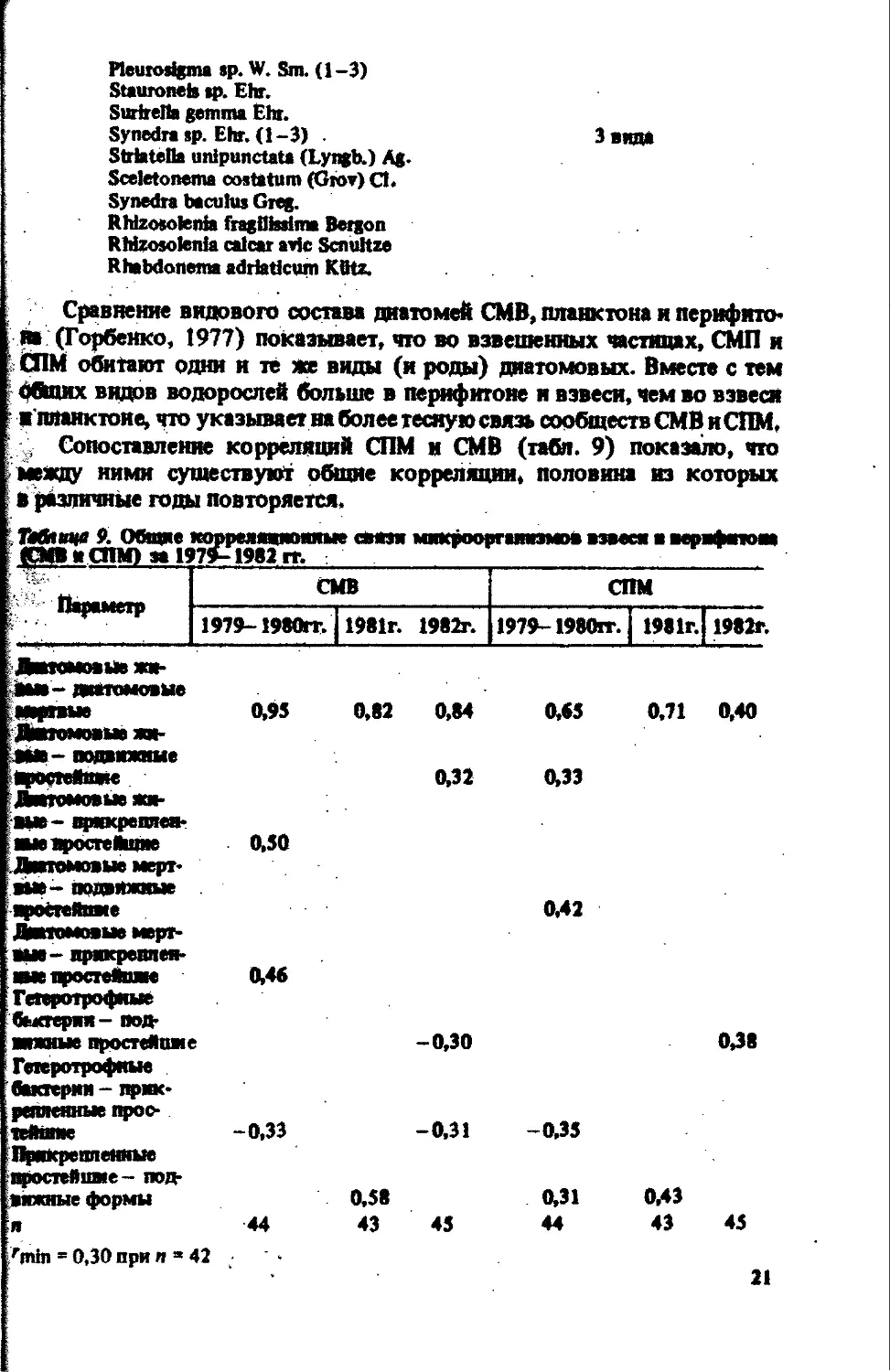
Сравнение видового состава диатомей СМВ, планктона н перифито* на (Горбенко, 1977) показывает, что во взвешенных частицах, СМП н СПМ обитают одни и те же виды (и роды) диатомовых. Вместе с тем общих видов водорослей больше в перифитоне и взвеси, чем во взвеси ипланктоие, что указывает на более тесную связь сообществ СМВ и СПМ. , Сопоставление корреляций СПМ и СМВ (табл. 9) показало, что между ними существуют общие корреляции, половина из которых в различные годы повторяется. . Мгмм 9. Общие тюмеяаоаяоииые омзи микромгаинзмов взвеси в перифитона (СМВ И СПМ) за 197»-1982 гг.
Параметр СМВ СПМ
1979-1980гг. 1981г. 1982г. 1979-1980гг. 1981г. 1982г.
Диатомовые жи- аы*-диатомовые аортные 0,95 0,82 0,84 0,65 0,71 0,40 Дитмовыв ям- ина- подвижные щюстейпже 0,32 0,33 Дмтомовые жи- вые- прикреплен- ные вроете Hume 0,50 Диатомовые мерт- вые- подвижные простейше 0,42 Диатомовые мерт- вые- прикреплен- ные простейшие 0,46 Гетеротрофные бактерии- под- вижные простеИиме -0,30 0,38 Гетеротрофные бактерии - прик- репленные прос- тейшие -0,33 -0,31 -0,35 Прикреплемше простейшие - под- вижные формы 0,58 0,31 0,43 я 44 43 45 44 43 45 fmin = 0,30 при и » 42 •
21
Таблица Id ОрпктюаА состав сухой массы СПМ к СМВ за 1980-1981 гг.
(по манным А.Н.Бобковой)
Состав СПМ СМВ
Органическое вещество (% сухой массы СПМ и СМВ) 30-80 24-46
Углеводы (% органического вещества) 10-28 6-13
Белок (% органического вещества) 18-30 6—15
Содержание свободных аминокислот (% органического вещества) 0,15-0,27 0,02-0,05
Пигменты (% органического вещества) 0,6 -3,0 0,2 -0,3
Сумма хлорофиллов а, в, с (% органического вещества)* 0,3 -2,0 0,07-0,2
Сумма каротиноидов (% органического ве- щества) 0,3 -1,5 0,07—0,2
* Как в СПМ, так и в СМВ доминирует хлорофилл а.
Таблица 11. Корреляционные связи биохимических характеристик взвеси и пери-
фитона СМВ и СПМ за' 1979-1981 гг. (по данным АМБобковой)
Параметры Коэффициенты корреляции (г)
1979—1980 гг. 1981г.
Белки СПМ - белки СМВ 0,71 0,75
Белки СПМ - органическое вещество СМВ 0,37 0,73
Углеводы СПМ - углеводы СМВ 0,31 0,85
Углеводы СПМ - белки взвеси СМВ 0,51 0,78
Органическое вещество СПМ - органическое вещесг-
во СМВ 0,53 0,75
Углеводы СПМ - органическое вещество СМВ 0,35 0,72
Органическое вещество СПМ - белки СМВ 0,60 0,77
Белки СПМ - углеводы СМВ — 0,64
Органическое вещество СПМ - углеводы СМВ — 0,71
я 43 45
“0.30 при л "42
На связь органических компонентов СПМ и СМВ указывает биохи-
мический характер взвеси и перифитона, проведенный А.Б .Бобковой,
Состав отдельных компонентов в обоих сообществах идентичен, только
содержание их различно (табл. 10). А.Н.Бобкова установила, что бел-
ки, углеводы и органическое вещество СПМ и СМ В тесно коррелируют
Друг с другом (табл. 11).
Это не характерно дли экологических связей изучаемой систе-
мы, так как корреляции в основном были слабыми или средними
(0,3 < г < 0,69) из-за множественности взаимосвязей и зависимостей
в море (Горбенко, 1977; Горбенко, Крышев, 1985).
Микроорганизмы СМВ, перифитона и метаболиты, образуемые
ими, коррелируют с одними и теми же параметрами указанных сооб-
Ществ и окружающей морской среды (табл. 12). Карбонаты, отлагаю-
22
^fyuifu 1Z Общие корремаюпые связи микроорганизмов сообщества взвеси
;? и перифитона СПМ в СМВ с параметрами морской среды за 1971 г.
Параметр Коэффициент корреляции (г)
pH СМВ и СПМ - pH морской среды 1 pH СМВ и СПМ-СО! . pHСМВиСПМ-нсо~ , pH СМВ и СПМ - фенол Дштомовые живые СПМ и СМВ - сухи масса СПМ и СМВ i, Автомовые живые СПМ и Cttt - гетеротрофные бактерии ;.С!Щи(МВ : В- 0,39 и 0,44 0,36 и 0,40 —0,37 и-0,49 —0,32 и-0,40 -0,36 и —0,43 0,33 и 0,39 63
[ " 0«30 ПР" я ° 42
’ ПЯяйца 13. Общие корреляционные связи карбонатов взвеси с другими парамет-
IftoM СПМ и СМВ ЗВ 1968-1971 гт.
Параметр Коэффициент корреляции (г)
1968 г. 1969 г. 1970 г. 1971г.
' Диатомовые живые - СПМ 1 Диатомовые живые- СМВ -0,38 0,39 -0,31 0,36 —
' Гетеротрофные бактерии — СИМ U,41 ОрЗЗ
Гетеротрофные бактерии - СМВ 0,64 — 0,34
Охи масса взвеси 0,44 — —
; pH воды — — -0,44 —
pHСПМ — 0,35 —
Диатомовые мертвые - СПМ — — 0,35
' Автомевые мертвые - СМВ — -и- 0,40
; pH емв 031 0,46 —• —
Г*',' 71 67 63 65
rmin ” 0,30 при я = 42
г
: внеся микроорганизмами взвеси, в различные годы коррелируют с
I одними и теми же параметрами СМВ и СПМ (табл. 13). Существуют
; корреляции карбонатов СПМ и СМВ с сухой массой и РОВ трансфор-
F Мнрованным, а также с гетеротрофными бактериями и диатомовыми
| водорослями и pH метаболитов всех изучаемых сообществ (табл. 14).
S В последующих исследованиях мы определяли кальций и кварц,
' Ожигающиеся в перифитоне и во взвеси, методом рентгендифракто-
f Метрического анализа на приборе ’’ДРОН-1,5” (Горбенко, Тримонис,
: 1980). Результаты определений показали, что зти кристаллические
; вещества коррелируют не только между собой во взвеси и погружен-
ь вых в море предметах, но также с параметрами окружающей морской
. Среды (табл. 15).
Совместно с А.Ю.Митропольским (Горбенко, Митропольский,
1976) были изучены качественный состав и количество микроэлемен-
тов в слизистой пленке СПМ и в морской взвеси. Результаты этой
23
Таблица 14. Общие корреляте* содержим карбонатов, отлагающихся микроор-
Ганизмамк СИМ СМВ, с параметрами планктона, перифитона и взвеем за 1968-
1981 гг.
Параметр Коэффициент корреляции (г)
СМВ сим
Сухая масса -0,30-0,71 0,68-0,88
Гетеротрофные бактерии
СПМ —> 0,30-0,64
СМВ 0,30-0,63
СМП -0,57—0,98 0,31-0,41
Диатомовые живые
СПМ -0,52—0,59 —
СМВ — 0,39-0,50
СМП -0,53—0,87 0,36-0,38
РОВ трансформированное 0,25—0,75
СПМ mU — 0,33-0,45
pH СМВ -0,36—0,57 —
сям — -0,30—0,54
Число наблюдений л 43-71 43-71
'нИц п₽и Уровне достоверности 95 % и ДЙИНОМЛ 0,30 0,30
Таблица 13. Корреляции рдоиси гающихец микроортавкамвый сП средн м кальция и кварца, отла- Ц И СМВ, с параметрами
Параметр I Коэффициент J корреляции (г)
КварцСМВ- кварцСПМ
Кварц СМВ - кальцит СМВ
Кварц СМВ - кальцит СПМ
Кальцит СМВ - температура морской воды
Кварц СМВ - температура морской воды
Калыагт СМВ - pH морской воды
Кварц СМВ-pH морской воды
Кварц СМВ - ЮВ СИМ
Кварц СПМ - РОВ СМВ
'min “0,330 Пр* я “3?
0,377
0,336
0,675
-0,650
-0,656
-0,412
-0,356
-0,538
-0,421
Еаботы показали, что в обоих указанных сообществах имеются едина-
овые металлы и микроэлементы- При этом большинство микро-
элементов осаждались микроорганизмами СМВ и СПМ примерно в
одинаковых количествах.
В совместных исследованиях с И-И.Крышевым (Крышев, Горбен-
ко, 1982) определяли периодические составляющие параметров СМВ
и СПМ путем анализа временных рядов с помощью разложения в ряд
Фурье. Результаты данного исследования показали, что ход динамики
Таблица 16. Периодические компоненты динамики микросообществ перифитона
• взвеси, по многолетним данным (Севастопольская бухта, 1967-1976 гг.)>
(Крышев, Горбенко, 1982)
Параметр Доминирующие гармоники, сут Критические величины амплитуды Среднее значение параметра
Период Фаза' Ч> Ампли- туда Я 95% 99%
-,ч. СМВ
Диатомовые 360 -85 121000 46000 сут 57000 сут 217000 экзмл-1
живые 180 -80 78000
120 -70 49000
Гетеротроф- 360 -20 540000 346000 сут 445000 сут 164-10“' экз-мл-*
тае бактерии 120 -20 470000
РОВ взвеси, 1100 50 0,033 0,015 0,018 0,322 (по Е17.)
330 -260 0,024
120 30 0,016
pH взвеси 1460 -760 0,04 0,017 0,020 8,05
360 -240 0,03
: •! 1 180 -45 0,03
Карбонаты, % 1000 750 1.8 3.6 0,5 17,9 мг п"‘
630 -26 1,8 3,6
« * 360 -26 3,8 16,7
180 26 1,2 1,8
120 -19 1,0 1,2
! СПМ
Диатомовые 1100 -160 22600 15100 сут 18700 сут 10’ экз-см-’
живые 360 -185 16500
• 100 -20 15500
30 -20 36300
Гетеротроф 1100 -310 4900 3300 сут 4100 сут 23600 экзем-’
иые бактерии 360 -260 5000
30 -20 6300
К» пернфи- 360 -200 0,036 0,015 0,018 0,124 (Ело)
. WW, Е»» pH перифи- 850 -190 0,03 0,017 0,020 8,23
тона 360 -230 0,03
180 25 0,02
30 -20 0,04
Карбонаты, 36QO 900 0,12 6,8 0,04 0,17 мг-см"’
.МММ"* 365 240 0,15 10,1
160 150 0,05 1,4
30 28 0,17 15,0
численности основных параметров СМВ и СПМ - диатомовые водорос-
ли живые, гетеротрофные бактерии РОВ СПМ и РОВ СМВ, pH СПМ и
pH СМВ - имеет период, фазу и амплитуду. При этом каждый из пара-
метров отдельно имеет общий параметр с аналогичным параметром в
другом сообществе и равен 360 сут (табл. 16).
На основании сказанного (а также табл. 9—16) установлено, что
существует тесная взаимосвязь микросообществ перифитона и взвеси.
25
Это позволяет говорить, что они аналогичны, а также о том, что СПМ и
СМВ по большинству признаков подобны. Выявленное подобие свиде-
тельствует о том, чю все закономерности развития микроорганизмов
в СМВ н СПМ примерно одинаковы.
Вместе с тем имеются различия между указанными сообществами:
1. За 5 лет наблюдений (1977-1981 гг.) отношение численности
гетеротрофных бактерий и живых диатомовых водорослей в СМВ было
равно 1,12, а в СПМ - 0,88, т.е. в СМВ численность гетеротрофных
бактерий несколько превышала численность диатомей, а в СПМ, наобо-
рот, было больше диатомовых.
2. Существуют различия в органическом составе (табл; 10). Орга-
нического вещества и углеводов в перифитоне было в 2 раза больше,
чем во взвеси. Белка в СПМ соответственно оказалось в 2,5 раза боль-
ше. А количество пигментов, каротиноидов и аминокислот во взвеси
было меньше, чем в перифитоне, примерно в 10 раз. Сумма хлорофил-
лов (а, б, с) в СПМ была также на порядок больше, чем в СМВ, но и
в том и другом сообществе доминировал хлорофилл в.
3. Сырая взвесь по консистенции несколько отличается от сырого
соскоба слизистой пленки СПМ. После центрифугирования (как отме-
чает А.Н.Бобкова) зта пленка на стекле держится комочком, а взвесь
хорошо размазывается. Цвет взвеси также немного темнее.
4. Огромно различие в величине субстрата. Размер частицы взвеси
примерно 0,2 мкм - 1 мм, а субстратом для СПМ обычно являются
крупные предметы от нескольких сантиметров до сотен и тысяч мет-
ров, как, например, набережная у моря.
5. Кроме того (что очень важно), частичка взвеси с микроорганиз-
мами (пока она взвесь) все время носится ”по воле волн” иа любые )
расстояния и глубину, где условия окружающей среды иные, т.е. она j
существует в изменяющихся условиях моря. А микроорганизмы пери-
фитона обитают на постоянном, строго определенном участке погру-
женного предмета, на одной и той же глубине, в несравненно лучших
условиях, чем морская взвесь.
Таким образом, хотя оба эти микросообщества подобны, они
имеют четкие отличия, что позволяет считать их различными сооб-
ществами.
Многие вопросы биоценологии уже достаточно хорошо изучены
(Дуплаков, 1933, Карзинкин, 1926-1927; Очерки по истории эко-
логии, 1970; Проблемы методологии, системных исследований, 1970;
Методика изучения биогеоценозов..., 1975, и др.), однако сущест-
вует еще много неизученных. Ниже мы остановимся на некоторых
из них.
26
ха ВСЕГДА ЛИ КОМПЛЕКС МИКРООРГАНИЗМОВ
РЧ16ЛЯЕТСЯ СООБЩЕСТВОМ?
^ Впервые определение сообщества дал К.Мёбиус (Mobius, 1877). Биоце-
&Йгз — это сообщество живых организмов, занимающих определенную
*Ирриторию, приспособленных к внешней среде и друг другу, связан-
Вых в единое целое, изменяющееся при изменении условий среды или
. численности отдельных видов. На примере многолетнего изучения
^Сообществ микроорганизмов перифитона и взвеси критерии биоцено-
зы, рассмотренные в литературе, можно несколько дополнить.
F " 1. В окружающей морской воде должны обитать микроорганизмы,
£ которые при формировании ценоза на биотопе, если не способствова-
то хотя бы не препятствовали развитию друг друга в процессе жиз-
недеятельности.
Г"" 2. Чтобы сообщество нормально развивалось, функционировало
было устойчивым, необходима его стабильность, т.е. определенный
^сйстав организмов, их внутренние и внешние связи, ход сукцессий,
^метаболизм и т.п. Причем все это изменяется и повторяется в опреде-
яенных пределах, поскольку элементы ценоза находятся в динамичес-
равновесии.
!₽т 3. Стабильность состава и качества элементов сообщества может
^' Ойхраняться, если обеспечивается определенное постоянство биотопа.
В? Имболее благоприятным во всех отношениях биотопом можно наз-
например, поверхность твердых и плотных (неживых) предметов,
|4 Погруженных в море, — она относительно долго не изменяется.
4. Окружающая среда должна быть также в меру постоянной. Наи-
бояее благоприятной средой с относительным постоянством компо-
Жйтов (pH, температура, солевой состав, регулируемый карбонатным
равновесием, содержание РОВ, состав и количество элементов, ионов,
ЧККроэлементов и т.п.) является природная вода морей и океанов, ко-
^Торая представляет собой биотоп и среду обитания для морских орга-
НЕЗмов.
’ Можно ли считать, учитывая все сказанное, что естественные сооб-
«Щвства IM, СМВ и СМП й комплексы микроорганизмов, обитающие
разлагающемся или разрушающемся трупе животного или растения,
^ЙИбо развивающиеся при культивировании организмов в лабораторных
8«осудах однотипны?
Пусть какой-нибудь природный объект, например труп морского
^Животного или рыбы, подвергается естественным превращениям, свя-
занным с его смертью. Вначале его будут разрывать на части хищные
|»Ж|бы, прежде всего крупные (акулы, барракуды), потом мелкие (лоц-
^'мИы, скумбрии и др.). Затем начнут действовать нематоды и простей-
МйИе организмы. Только после них в деструкцию остатков активно
ЧНслючаются бактерии и другие микроорганизмы, после которых от
организмов останутся только костные части скелета, а также частицы
27
трудноусваиваемых органических веществ. Эти частицы являются не
чем иным, как детритом. Несомненно, что они будут заселены бакте-
риями из окружающей воды и, если ие разложатся совсем, то будут
служить микросубстратом для образования взвеси. Примерно так же,
как разложение и распад трупов животных и рыб, происходит деструк-
ция водорослей - микрофитов и других морских организмов. В итоге
также образуются трудноусвояемые частицы детрита.
Таким образом, при указанных превращениях органического ма-
териала в море происходят его необратимые изменения, и в результате
он почти полностью исчезает. Во время этих превращений одни организ-
мы сменяют другие, им незачем устанавливать связи друг с другом,
поскольку привлекает их подходящее для них состояние разрушающе-
гося органического вещества или посмертные метаболиты. Нет у этих
гидробионтов также общего биотопа, поскольку он не постоянен, а все
время необратимо изменяется.
Второстепенное значение имеют для них факторы среды, потому
что все функции указанных организмов-консументов и редуцентов
направлены в основном на использование трупа животного или рыбы.
Поэтому виды и популяции, например микроорганизмов, не являются
единым целым, реагирующим на изменение среды, а напротив, они ма-
ло или совсем не зависят друг от друга.
Из-за необратимых изменений на трупе (биотоп) происходит заме-
на групп и популяций организмов, а не периодически повторяющиеся
сукцессии, повторение которых исключается, так как от этого биото-
па при благоприятных условиях вскоре ничего ие останется.
В итоге из-за того, что во время деструкции остатков органическо-
го вещества макро- и микроорганизмами не соблюдаются необходи-
мые критерии, характерные для биоценоза, комплекс их нельзя отнести
к сообществу.
В связи со сказанным, очевидно, недостаточно только увидеть груп-
пу совместно обитающих в природе микро- или макроорганизмов,
чтобы считать их сообществом. Необходимо вначале убедиться в том,
что данный комплекс отвечает основным критериям, обязательным для
биоценоза. Случается, однако, поступают совсем иначе - так, Т.Фен-
шель (Fenschel, 1970) получал растительный детрит путем разложения
’’черепашьей травы”. Бактерии и микроводоросли, обнаруженные при
этом, он отнес к сообществу.
Ю.И.Сорокии (Sorokin, 1978) отмечал, что в ходе метаболической
деструкции водных сообществ организмов и их фекалий микрофлора
и простейшие формируют агрегаты органических частиц, которые явля-
ются субстратом (убежищем) для микросообществ, основную роль в
которых играют бактерии, инфузории, жгутиковые, а также.диатомо-
вые водоросли и мелкие рачки. Оба автора к микросообществу отнесли
комплекс микроорганизмов, обнаруженных на детрите, без проверки
того, отвечает ли он условиям, предъявляемым к биоценозу.
28
г
На основании всего сказанного выше можно утверждать, что мик-
роорганизмы ’’черепашьей травы” и осуществляющие метаболическую
деструкцию водных сообществ к биоценозу не относятся. Так как бы-
ли рассмотрены процессы разложения — в одном случае растений, а в
"другом - по-видимому, животных (’’водных сообществ”), следова-
тельно, эти микроорганизмы представляют собой только комплекс
сменяющихся микроорганизмов, осуществляющих деструкцию.
Возникает другой вопрос, не являются ли микросообществом мик-
роорганизмы, культивируемые с какой-то целью в лабораторных сосу-
дах? Для культивирования гетеротрофных бактерий, как известно, в
сосуды обычно добавляется какой-нибудь источник углерода, авто-
трофным бактериям требуется определенный набор минеральных со-
лей, а для олигокарбофилов берут природную воду без всяких доба-
вок. Среды засевают каким-нибудь природным материалом либо куль-
турой бактерий. При этом маловероятно, что среда будет благоприят-
ЙЙк для всех бактерий, помещенных в колбы. Обычно там развиваются
Только некоторые из них, часто не взаимосвязанные с остальными орга-
низмами и совсем не обязательно совместимые с ними.
Ik- В результате жизнедеятельности микроорганизмов, для которых
культуральная среда благоприятна, она изменяется и поэтому уже
Отличается от природной. Кроме того, при культивировании бактерий
образуется много метаболитов, обычно ядовитых для них самих. Из
ВОДЫ потребляется кислород, а вода насыщается углекислотой и часто
Яодкисляется от этого, т.е. любая из культуральных сред, постоянно
Изменяясь, в конце концов становится не только неблагоприятной для
Микроорганизмов, но ядовитой и даже убивающей их.
Ц О каких связях между бактериями тут может идти речь? Какое
Постоянство среды будет соблюдаться? Какая может быть взаимосвязь
бактерий и убивающей их среды? Совершенно ясно, что в лабораторных
колбах с культуральной средой образуется не сообщество, а только
комшекс микроорганизмов.
Конечно, бывают случаи, когда в аквариумах создаются маленькие
экосистемы. Это происходит в том случае, если там есть водоросли-про-
дуценты, моллюски, рыбы я ракообразиые-консументы, бактерии-ми-'
яерализаторы, редуценты, песок, ил и т.п. В таких уравновешенных
экосистемах, однако, культивировать бактерии для эксперимента или
Промышленных целей невозможно, так как при изъятии оттуда или до-
бавке туда еще каких-нибудь микро- или макроорганизмов экосистема
может разрушиться.
Обитая в морской среде, микросообщества взвеси, перифитона и
планктона взаимосвязаны между собой и с абиофакторами среды. Поэ-
тому мы изучали также структуру экологических связей, их тесноту и
как взаимодействуют параметры посредством этих связей в эко-
29
ГЛАВА 4. ВЗАИМОСВЯЗИ БИО-
И АБИОТИЧЕСКИХ ФАКТОРОВ ЭКОСИСТЕМЫ
41. ИЗУЧЕНИК ВЗАИМОСВЯЗИ pH МОРСКОЙ воды,
МЕТАБОЛИТОВ МИКРООРГАНИЗМОВ ПЕРИФИТОНА И ВЗВЕСИ
С ПОМОЩЬЮ КОРРЕЛЯЦИОННОГО АНАЛИЗА
Концентрация водородных ионов в морской воде тесно связана с про-
цессами создания и распада органического вещества, определяющими
содержание угольной кислоты, которая непосредственно влияет на pH
(Алекин, 1966). Как отмечал С.Зо Белл (Zo Bell, 1946), морские бак-
терии, интенсивно используя органические вещества морской воды,
выделяют огромное количество СО2 в воду.
Расположение Севастопольской бухты, глубоко вдающейся в сушу,
множество гидротехнических сооружений и судов благоприятствуют
развитию микроорганизмов перифитона, численность которых здесь
велика. Большое количество органических и минеральных веществ,
поступающих в воду бухты с суши, способствует образованию значи-
тельной массы взвеси, обильно населенной микроорганизмами (Гор-
бенко, 1977).
Задача настоящей работы заключается в определении роли мета-
болитов микроорганизмов в изменении pH морской воды. Для этого
было сделано следующее дополнение к методике. Измерение pH мор-
ской воды, pH метаболитов сообщества перифитонных микроорганиз-
мов (pH СПМ) и pH метаболитов (pH взвеси) проводили регулярно
3—6 раз в месяц в течение 1968-1973 гг. За величину pH СПМ принима-
ли отрицательный логарифм концентрации водородных ионов в свежей
морской Воде после 2-часового взаимодействия в лабораторных сосу-
дах этой воды с пленкой СПМ, наросшей на стеклянных пластинах в
море; pH взвеси соответственно определяли после такого же взаимо-
действия воды с определенным количеством взвеси, собранной в море.
Концентрацию водородных ионов определяли на потенциометре|
ЛПУ-01 со стеклянным электродом сразу После отбора проб и спустя)
2 ч (для контроля возможного изменения pH со временем). Как пока-
зали контрольные измерения, после 2-часовой выдержки значения pH
изменялись не более чем на 0,04, т.е. изменения не выходили за предел
30
ТМяши 17. Обычйне Оху) и частные Oxyz) корреляционные связи pH с мор-
ем»* иодо* (1), метаболитами СПМ (2) и нзнесью (3) (Севастопольская бухта)
ж Год Коэффициент корреляции Чисяо наблюде- ний,»
ги Гц.» 'ц '1М ГМ-1
, 1968 0,694 0,503 0,574 0,215 0,660 0,444 71
1969 0 0 0,406 0 0 0 69
1970 0 0 0,494 0 0 0 70
* 1971 0,500 0,322 0,529 0,378 0,485 0,300 68
«’ 1972 0,393 0,412 0 0 0,303 0,329 68
. 1973 0,546 0,378 0,691 0,602 0,433 0,092 36
гЦрИ л ' 36 rmin 0,330
ры ошибки наблюдений. Данные наблюдений были проанализированы с
Ьрмошью корреляционного анализа, при котором вначале вычислили
^*рные, а затем частные корреляции (Горбенко, 1979). Годовые коле-
£|ния pH морской воды достигали 8,23-8,39, pH СПМ - 8,20-8,42, pH
Установлено, что в результате жизнедеятельности микроорганиз-
мов СПМ морская вода, содержащая следы органики, обычно под-
кисляется. Только в нескольких случаях в течение года морская вода
JBHRia более щелочная, чем в контроле. Микроорганизмы взвеси, кон-
тактируя с водой, подкисляют ее. Последнее объясняется тем, что
'морские микроорганизмы (в основном бактерии и диатомовые водо-
лосли), в изобилии развиваясь на взвеси и погруженных предметах,
*1 процессе метаболизма образуют в основном углекислоту, кислород,
Маличные органические кислоты, снижающие pH окружающей среды.
^Подщелачивание воды может происходить при использовании диатомо-
: йыми водорослями СПМ углекислоты из морской воды при интенсив-
ном фотосинтезе весной, летом и осенью (Горбенко, 1977).
Установив факт влияния на морскую воду микроорганизмов СПМ
и взвеси, мы попытались определить их воздействие на pH воды. При
сопоставлении кривых изменения pH в 1971 г. и другие годы наблюде-
ний связь между pH воды, взвеси и СПМ не обнаружена, ио с помощью
корреляционного анализа удалось выяснить, что эти величины коррели-
руют между собой (табл. 17).
За 6 лет наблюдений корреляция pH морской воды и метаболитов
СПМ (ги) обнаруживалась 4 раза, pH воды и взвеси (rjj) —5 раз, а
pH СПМ и взвеси (г и) — 4 раза. Меньше всего корреляционных связей
было в 1969 н 1970 гг. (только г 1Э). За это время каждая из указан-
вдх корреляций однажды имела значение, близкое к тесной корреля-
ционной связи (г = 0,7).
Полученная информация свидетельствует о том, что изучаемые па-
раметры - pH воды, перифитона и взвеси - по-видимому, взаимосвя-
заны. Для более детального анализа этого предварительного вывода
были рассчитаны частные корреляции, с помошью которых можно бо-
31
лее точно наметить причинные связи по известным формулам (Рокиц-
кий, 1967) (табл. 17).
В 1968 г. на корреляцию pH воды и СПМ (г13) почти не влиял pH
взвеси (3). На корреляцию pH СПМ и взвеси (г и) pH воды (1) влиял
также слабо. В то же время корреляция pH воды и взвеси (г]3) в зна-
чительной мере зависела от величины pH СПМ, которая во взаимосвя-
зи указанных параметров была, по-видимому, определяющим пара-
метром (табл. 17). В 1969 и 1970 гг. наблюдалась умеренная корреля-
ция только pH воды и взвеси (г13). В 1971 г. элиминация одного из
изучаемых параметров уменьшала корреляцию двух оставшихся, т.е.
все параметры были взаимосвязаны. В 1972 г., напротив, исключение
1 параметра немного увеличивало корреляцию 2 остальных, т.е. изучае-
мые параметры почти не зависели друг от друга. В 1973 г. на корреля-
цию pH воды и СПМ (г») оказывал слабое влияние pH взвеси (3),
Корреляция pH воды и взвеси (г13) также почти ие зависела от pH
СПМ (Горбенко, 1977). Корреляцию pH СПМ и взвеси (ги) осуществ-
ляла pH морской воды (1), а при ее исключении связь г исчезала.
Следовательно, можно предположить, что pH морской воды в данный
период наблюдений была определяющим параметром во взаимосвязи
трех рассматриваемых параметров.
Таким образом, в итоге 6-летннх наблюдений в большинстве случа-
ев установлено влияние величины pH. Есть все основания считать,
что от pH метаболитов СПМ и, может быть, немного больше от pH взве-
си в значительной мере зависит pH морской воды Севастопольской
бухты. Это влияние определяется наличием значительного количества
твердого субстрата и благоприятных условий для образования взвеси,
так как на взвеси и на поверхности предметов обильно развиваются
морские микроорганизмы, влияющие посредством своих метаболитов
на изменение pH воды в бухте. Намного больше информации можно
получить, изучая систему со многими параметрами.
ФАКТОРНОЕ ПРЕДСТАВЛЕНИЕ МОРСКИХ
МИКРОБИОЛОГИЧЕСКИХ СИСТЕМ
В гидробиологических и других работах при изучении зависимости
и связей организмов друг с другом и со средой обитания нередко упот-
ребляют термин ’’фактор” среды, организма, био- или абиотический.
Применяют для этого также понятия: ’’показатели”, ’’характеристики”,
’’элементы”, ”параметры”. Четкого разделения при этом понятий
’’фактор”, "показатель” или ’’характеристика” нет. Нередко пишут,
например, температурным фактор, имея в виду, однако, один показа-
тель абиосреды - температуру. Под биогеиом подразумевают азот или
фосфор, или оба элемента вместе, в то время как в математике во
избежание путаницы существует строгое определение фактора и пара-
32
метра. Имеется также факторный метод, с помощью которого можно
получить весьма интересные результаты, используя многомерный ста-
тистический анализ сложных биосистем.
Факторный метод предполагает, что поведение таких систем опре-
деляется действием небольшого числа реально существующих или не-
которых гипотетических факторов (например, рассмотрение морских
экосистем связано с действием биотического, абиотического, гидро-
химического и других факторов). При этом воздействие фактора на
сложную систему может проявляться в изменении группы параметров.
Поэтому фактор является более обобщенным термином по отношению
К наблюдаемым параметрам. В связи со сказанным здесь и во всех
наших работах четко разделяются понятия ’’фактор” и ’’параметр” (под
параметром мы подразумеваем какое-то одно наблюдение, показатель,
характеристику или элемент).
Параметры наблюдаются и изменяются (кислород, pH, температу-
ре). Факторы же непосредственно не наблюдаются и не изменяются -
они проявляются в процессе решения задач, а затем интерпретируют-
ся (Подвинцев, Горбенко, 19776). Конечно, можно в начале исследо-
Вания выделить какие-то отдельные факторы и к ним отнести подходя-
щие параметры, но факторы, адекватно отражающие состояние окру-
жающей морской воды, можно получить только после специального
исследования. При этом далеко не всегда их количество и состав будут
-соответствовать первоначальному вычленению.
Дополнение к методике состояло в следующем. В качестве исход-
ных данных были использованы 20 наблюдений одною года за 17 па-
рямет рами в море. Проводились они почти каждые 5 и 10 сут ежеме-
сячно в течение года одновременно на погруженных в море пластинах и
В окружающей морской среде.
Параметры: Х^ — палочковидные формы бактерий (ФБ); Х2 -
кокковидные ФБ; Х3 - диатомовые водоросли (живые); Х4 - диато-
мовые мертвые; Xs - гетеротрофные ФБ; Х6 - растворенное органи-
ческое вещество (РОВ) СПМ; Х2 - pH метаболитов СПМ; Х8 - сухая
масса СПМ; Х9 - карбонаты СПМ; - концентрация РОВ морской
Воды; - температура воды; Х12 - величина pH морской воды;
Х13 - СО’"; Х14 - НСО3; Х13 - СО2; Х16 - соленость; Хи - содер-
жание 02 в воде.
Предварительно разбили на факторы (F) указанные параметры
(фактор - функция от параметров): Fj (Хп Х2, Х3, Х4, Xs) - биоти-
ческий фактор СПМ; F3 (Хб> Х-j, Х$, Х9) - биокоспый фактор мета-
болизма СПМ; F3(yj0,Xii, Х12) - фактор состояния морской воды;
F<(X13, Х14, X15, Х16, Х17) - фактор карбонатного равновесия, соле-
ности и кислорода. Первые два фактора можно отнести к биотическим,
последние два - к абиотическим.
Результаты решения статистических критериев Андерсона и Томпсо-
на (Подвинцев, Горбенко, 19776) представлены в факторной модели №1.
33
Факторная модель W1
Xi Fi Fi F3 Fi
xt 0,98 -0,25 0,06 -0,1
*2 0,96 -0,35 0,28 0,22
Хз 0,52 0,21 -0,34 -0,23
Х4 0,63 0,22 0,11 0,15
XS 0,64 0,05 -0,12 -0,04
Xi -0,45 0,99 -0,29 -0,24
Хз -0,55 0,26 0,32 0,38
*в 0,54 0,36 0,07 -0,12
Х9 0,48 0,59 -0,1 -0,03
Хю 0,19 -0,34 0,72 -0,28
Хц -0,03 0,01 0,82 -0,12
Хю -0,16 0,32 0,93 0,42
Хц -0,09 0,1 1,17 0,54
Хц 0,06 -0,39 0,08 0,8
Хи 0,09 -0,13 -0,92 -0,07
Хю 0,2 0,03 0,43 0,53
*17 -0,25 0,4 -0,03 0,46
Как следует из модели № 1, наибольший вкладе факторF, вносят
не только параметры Х>, Хз, Хз, Х4, Хз , но также параметры Х7 и
Х9 (pH СПМ и сухая масса СПМ). Имеется вклад параметров Х1Э и
Х„ (СОГ и СО2) в фактор F3 Ойо. Хп, Х13). Это свидетельствует
о том, что необходимо перенести указанные параметры в соответствую-
щие факторы и свести задачу к проверке следующей гипотезы о фак-
торном представлении: Fj (Xj, Xj, X3, X4, Xs, X«, X7, Xg). F3 (X«,
X,); F3 (XI0, XH, Xn, XJ3, X,s); F4 (X„, X„, X17). Решение дан-
ной задачи приведено в факторной модели N* 2.
Факторная модель N* 2
X Fi Fi F3 Fi
Xt 0,91 -0,01 -0,27 -0,26
Xi 0,74 -0,01 -0,08 -0,02
Хз 0,84 -0,17 -0,23 -0,16
х9 0,67 0,19 0,09 0,12
Хз 0,64 0,06 -0,23 -0,08
Хз -0,07 -0,4 0,72 0,56
хя 0,51 0,39 0,02 0,15
х6 -0,47 0,36 0,05 0,02
Хо 0,54 0,45 -0,04 -0,02
Хю -0,03 0,04 0,37 -0,55
*1, 0,07 -0,09 0,74 -0,26
0,06 0,02 0,99 0,45
*В 0,02 0,05 1,12 0,43
*15 -0,12 0,07 0.91 -0,06
*14 -0.04 -0,27 0.07 0,74
*t6 0.23 -0,08 -0,24 0,59
*17 -0.25 0,35 0,17 0,69
34
Из решения следует, что параметры Х6 и Х9 (РОВ СПМ и карбона-
ТЫ СПМ) необходимо перевести в фактор F,, параметр Хч (pH СПМ) -
* фактор F3, а Хю (концентрация РОВ морской воды) - в фактор
F4. После этого следует перейти к проверке гипотезы о существовании
В адекватности остальным параметрам изучаемой биосистемы следую-
1ШХ факторов: F4 (Хь Х2, Х3, Х4, Xs, Х„ Х9> Х6); F2 (Х7, Хп,
Ха, Х13, XiSJ; F3 (Xw, X14, Хц,Хп). Окончательное решение задачи
Представлено в факторной модели № 3.
Факторная модель № 3
Xi F, 1'3
Xi 0,82 -0,23 -0,04
Xi 0,75 -0,14 0,08
Х3 0,55 0,08 -0,15
Х4 0,83 0,11 0,09
Xs 0,67 -0,22 0
Xt 0,89 0,03 -0,1
х9 0,91 0,05 0,04
Xt 0,36 0,29 0,06
Хч -0,42 0,74 0,19
Xu 0,18 0,59 -0,49
Ха 0,26 0,86 0,07
Хи . 0,21 0,92 -0,76 0
Ха -0,25 0,23
хю 0,22 0,12 -0,42
х» -0,32 0,08 -0,05 0,58
хи -0,23 0,63
Ха 0 0,15 0,57
Это решение в задаче заключительное, так как вклады факторов в
параметры своих групп максимальны для всех параметров. Сущест-
венно повышалась адекватность решения в факторной модели № 3
во сравнению с исходным решением в модели № 1.
Последнее решение находится на критическом уровне адекват-
ности по критерию Томпсона с вероятностью Р = 0,98. Из этого сле-
дует, что достаточно 3 факторов, чтобы представить все 17 параметров.
Анализ решения приводит к следующим выводам:
1. Не подтверждается исходное предположение о существовании 4
факторов в механизме взаимодействия изучаемых параметров в море.
2. Установлено, что механизм взаимодействия этих параметров -
функция 3 факторов: F3 (Х3, Х2, Х3, X4t Xs, Х6, Хл, Х9) - биотичес-
кого фактора СПМ; Р2(Хч, Хп, Х12, Х33, Х«) - фактора карбонат-
Ного равновесия, pH и температуры, и F3 (Xi0, Xl4, Х16, Xn) - факто-
ра Состояния воды. Первый фактор биотический, второй ~ биоабио-
риеский, а третий - абиотический.
3. В состав биафактора F3 (факторная модель № 3) вошли пара-
метры Х3 (сухая масса СПМ) и Х9 (карбонаты СПМ), которые ранее
35
предварительно были отнесены к фактору метаболитов СПМ — F2 (Хб,
Х2, Хя, Х9). В действительности же оба указанных параметра тесно
связаны с жизнедеятельностью микроорганизмов СПМ, что было уста-
новлено ранее (Горбенко. 1977). Поэтому такой перенос закономерен.
4. Параметр Х6 (РОВ СПМ) тоже вошел в F, - биотический фак-
тор. Однако большая величина погрешности d6 = 0,89 (Подвинцев,
Горбенко, 19776) объясняется тем, что фотометрический метод, с по-
мощью которого получен этот параметр (Горбенко, 1977), по-видимо-
му, имеет ошибку измерения больше, чем другие параметры. Такая же
ошибка характерна и для другого параметра Хю (РОВ морской воды)
в факторе /'3 (Хю, Х^, Ху6, Хю), определяемом тем же методом.
5. Параметр Х7 (pH метаболитов СПМ) хотя и обусловливается
метаболическими процессами в’ СПМ, но вместе с тем ои вошел в абио-
фактор карбонатного раэвовесия с pH и температурой воды — F2 (Х9,
Хц, Хп, Хгз, Xt!t), поскольку, очевидно, сильнее взаимодействует с
биоабиофактором F2, чем с биофактором Fj.
6. Факторный метод с математической точностью позволяет осу-
ществить корректный перенос наугад выбранных факторов в факторы,
адекватные изучаемой биосистеме и окружающей морской среде.
С помощью факторного метода и корреляционного анализа можно
изучить также значительно более сложную систему. При этом в системе
определяются не только действующие факторы, но выявляется также
ее структура, выражающаяся в группировках, взаимосвязанных между
собой.
<3. ОПРЕДЕЛЕНИЕ ГРУППИРОВОК И ФАКТОРОВ
МЕТОДАМИ КОРРЕЛЯЦИОННОГО И ФАКТОРНОГО АНАЛИЗОВ
Кроме уже известных из предыдущих исследований параметров, в дан-
ной работе мы изучали ряд солнечных и атмосферных параметров, а
также магнитные характеристики. Поскольку А Л.Чижевский (1976)
доказал большое влияние солнечной активности на микроорганизмы
суши, то вполне логичным являлось предположение о зависимости мор-
ских микроорганизмов, а также абиофактора моря от этого параметра.
Необходимая информация о данных параметрах была собрана в Севас-
топольской бухте и в прилегающих к ней районах суши (Подвинцев,
Горбенко,1979).
Особенности методики заключались в следующем. В течение 1971 г.
провели 60 наблюдений (л = 60). Изучали 54 биопараметра морской
среды, а также абиотические параметры моря, солнца и атмосферы,;
которые регистрировали с дискретностью 5—7 сут. Затем составили:
матрицу наблюдений. После проверки гипотез по критерию Кульбака
12 параметров (Х43 - Хм), оказавшихся ниже критического уровня
(Гкр = 0.95), не несли существенной информации об изучаемой систе-
36
Таблица 18. Статистические характеристики параметров экосистемы
Обозна чаше j Наименование Единица измерения г
Хх Гетеротрофные бактерии планктона (СПМ) мг.мл^* 12,74
х. Гетеротрофные бактерии перифитона (СПМ) мг-см^? 11,24
Xi Гетеротрофные бактерии взвеси (СМВ) мгчмл^; 6,66
Ха Диатомовые водоросли взвеси (живые) МГ'МЛ - 2,17
(СМВ)
xs Палочковидные бактерии перифитона (СПМ) МГ-емЗ? 8,OS
xf Кокковидные бактерии перифитона (СПМ) МГ-СМ * 7,7
Xi Диатомовые водоросли плактона (живые) мгмл^1 7,81
(СМП)
Xi Диатомовые водоросли перифитона (живые) мгсм-1 2,71
(СПМ)
Xi Диатомовые водоросли взвеси СМВ мгмл-1 2,18
(мертвые)
X* Диатомовые водоросли планктона (мертвые) мгмл 9,6
(СМП)
*11 Диатомовые водоросли перифитона (мертвые) мгсм-1 6,01
(СПМ)
xn РОВ, трансформированное перифитоном Емо 0,068
(СПМ)
*13 РОВ морской воды Емч, 0,361
XM Сухая масса пленки перифитона (СПМ) 15 см 2 1,77
x« Карбонаты взвеси (СМВ) % 2,61
Xk pH морской воды ед. 8,34
Xn pH взвеси (СМВ) ед. 8,101
XW pH метаболитов перифитона (СПМ) ед. 8,3
xn Нитриты-азот (N - NOj) в морской воде мгл* х»
X» Окисляемое» морской воды мг-л* 1,48
xn Углекислота в морской воде (COj) мл-л 1 1,707
Xn Гидро карбонат-ионы (НСОЭ) мгэквл^: 2,73
Xn Карбонат-ионы (СО3 ) мгэкв-л 1 2,76
XM Кислород в морской воде (О2) мл-л 1 6,3
Xn Температура морской воды Т’С 14,86
Xn Щелочность морской воды мгэквд 1 3,283
Xv Содержание фенола в воде мг-л * Хэт
Xn Содержание ХлЭк в воде мг-л 1 1,27
Xv Энергетический баланс Безразмерная 0,474
Xn Влажность воздуха % 72,4
Xn Альбедо видимого спектра Безразмерная 0,173
Xn Доля баланса в инфракрасной области Безразмерная -0,129
Xn Число Вольфа Безразмерная 66,2
Xn Температура воздуха ТС 13,0
Xn Атмосферное давление мм рт. ст. 765,2
Xn Количество часов сияния солнца ч 6,45
Xn Облачность Баллы 4,99
X» Суммарная радиация Безразмерная 0,742
xn РОВ, трансформированное взвесыо (СМВ) Е»о 0,292
xn Соленость морской воды %о 17,97
Xn Прозрачность морской воды м Х<и
x* Суммарная среднесуточная радиация к Джем 2х 1323,0
хсут 1
37
мв и были отброшены как второстепенные. Это следующие параметры:
относительная масса (%) карбонатов СПМ, фосфор минеральный,
направление и скорость ветра, индекс магнитный планетарный, продол-
жительность светового и лунного дня, рассеянная радиация у поверх-
ности моря, относительная и отраженная радиация на ту же плоскость
(видимый спектр), международная геомагнитная характеристика -
средняя магнитная активность для каждого дня, средняя солнечная
амплитуда. В табл. 18 приведены средние значения (Xt) и их раз-
мерность.
С помощью корреляционных матриц обработали 42 достоверных
параметра (табл. 18) с целью выявления частных корреляций между
каждой парой параметров, исключая влияние всех остальных. Факто-
ры, действующие в изучаемой системе, определяли методом косо-
угольного факторного анализа.
Биопараметры (Xi - Хц) и абиопараметры (Х)3 - Хи\ Хэ« -
Xi3, Хи) измеряли и регистрировали по известным методам
(Горбенко, 1977). Значения солнечных параметров Хм,Хзэ,Хм, Х»,
X» - Хп были зарегистрированы в Карадагской актинометри-
ческой лаборатории. Параметр X» представляет собой относительную
часть суммарной радиации, падающей на горизонтальную плоскость у
поверхности моря. Параметр солнечной активности X» (число Воль-
фа - И) б*1111 взят из литературных источников (Geophysical Research,
1971). Метеопараметры атмосферы (X», Хм - Хэт, X4S, Хщ, Хм,
Хм) взяты из архива Севастопольского отделения Государственного
океанографического института гидрометслужбыСССР, геомагнитные па-
раметры состояния ионосферы (Хп, XS3, Хм) - из журнала ’’Геофи-
зические исследования” (1971), (Подвинцев, Горбенко, 1979).
После вычислений частные корреляции были разбиты на 3 полу-
интервала: 1) (гц) >0,561; 2)0,561 > (гд) >0,515; 3)0,515 > (г//) >
> 0,379. Вероятности связи параметров в каждой из этих групп оказа-
лись следующими: Pt >0,995 ; 0,995 >Р2 >0,99; 0,99 >Р3 >0,95.
Все 42 изучаемых параметра обозначаем числами в кружках, свя-
зи между ними - прямыми линиями, сильно взаимосвязанные пара-
метры (по величине частных корреляций) - жирными линиями. В ре-
зультате эти параметры оказались разбитыми на 9 группировок (Л,
В, С, D, Е, F, G, Н, /), каждая из которых состоит из нескольких сильно
взаимосвязанных параметров (рис. 1). Уровень вероятности парамет-
ров, взаимосвязь которых менее сильная, немного ниже - они обозна
Чены двойными линиями. Тонкие линии и стрелы показывают, что па-
раметры со слабыми связями также входят в состав 9 группировок,
Но еще на более низком уровне вероятности, чем остальные. Таким
образом, каждая группировка представляет собой основу из 2-6 пара-
метров с сильными и средними связями между ними, окруженную
оболочкой из многочисленных параметров со слабыми связями. При
этом все группировки непосредственно или через параметры со сред
38
Fnc. 1. Структура группировок параметров биосистемы (обозначение в тексте)
Ней и слабой связями взаимосвязаны (рис. 1). В итоге выявлена струк-
тура изучаемой морской системы со слабыми, средними и сильными
связями, имеющими величину коэффициентов частной корреляции
др = 0,379-0,910 с вероятностью Р= 0,95 ± 0,999.
!? При более подробном рассмотрении структуры изучаемой системы
Выясняется, что центральной группировкой ее является группировка
Л В ней сосредоточены З из 7 рассматриваемых параметра солнца
.(энергобаланс - доля баланса в инфракрасной области -
39
суммарная радиация - Х»), вошедших в морскую систему, а также со-
леность воды (Х<). Значимы и группировки А и В с параметрами
солнца - альбедо видимого спектра (Х31) и активность солнца по чис-
лу W (Х33) (рис. 1).
Все группировки рассматриваемой системы можно разбить на 2 час-
ти — группы параметров А, В, С, D, Е, которые взаимосвязаны связями
среднего уровня, и группировки параметров (F, G, Н, Т), связанные
друг с другом и с названными выше параметрами слабыми связями.
Следует отметить, что такие известные биогенные параметры, как нит-
риты-азот (Х1«) и фосфор минеральный (Х44), в изучаемой системе
имеют второстепенное значение. Параметр X*» не вошел в изучаемую
систему как малозначимый, а параметр Х]9 хотя и относится к системе,
но связей с другими параметрами и группировками не имеет. Это сви-
детельствует о том, что в Севастопольской бухте оба эти параметра не
являются лимитирующими.
Между солнечными параметрами группы D - суммарная радиация
(Хзз), энергобаланс (Х29) и доля баланса в инфракрасной области
(Х33) - имеются самые тесные по величине корреляционные связи
(г/у = 0,72 ± 0,91).
Параметры Х39 и X3J непосредственно влияют на соленость воды
(Хдо) в море. При этом чем выше энергобаланс (Х19), тем больше со-
леность воды (г// = 0,76). В то же время увеличение доли баланса в
инфракрасной области (Х33) приводит к уменьшению солености (гу =
= —0,75). Связь эта определяется испарением морской воды при ее
нагреве - тем самым осуществляется один из важнейших круговоро-
тов веществ на нашей планете. Это влечет за собой увеличение соленос-
ти. При этом возрастание температуры воды летом приводит к увеличе-
нию испарения. Зимой испарение значительно меньше. То же происходит
и с соленостью воды. На основании сказанного можно считать, что испа-
рение воды в море, влекущее за собой увеличение солености, происхо-
дит в результате воздействия солнечных параметров - (XJ9) и (Х^).
Параметр (Хзз) влияет на 6 других параметров, связанных группи-
ровками А, В, Е, F, И, I. Параметры солнца - энергобаланс (XJ9) и сум-
марная радиация (Хзз) - влияют на группу Е. Имеющийся в группи-
ровке А солнечный параметр - число Вольфа W - fX33), отражающий
солнечную активность, действует на группировки В, Е, F.
В ядрах группировок А И В параметры солнца влияют на ряд дру-
гих параметров: число Вольфа (ИО - на гетеротрофные бактерии СПМ
и СМВ (Xj и Х3) с коэффициентами корреляции rit33 « 0,53 и гхзз =
“ 0,54. Альбедо видимого спектра (X3i) влияет на pH морской воды
(Х1в) и на содержание карбонатов взвеси (Хц). В то же время облач-
ность в баллах (Х37) влияет на альбедо видимого спектра (Х31), задер-
живая попадание этих лучей в море.
Испытывая на себе влияние солнца, соленость морской воды (Хад)
в свою очередь влияет на ее прозрачность (Х« ). Между этими величи-
40
дами существует отрицательная зависимость (г 40.41 = —0,540). Но проз-
рачность (Хц) зависит еще от атмосферного давления (Х35), а корре-
ляция между этими параметрами г 35.41 = —0,64. Однако механизм влия-
ния солености (Х«о) и атмосферного давления (X3S) на прозрачность
(Х41) пока неясен.
Давление атмосферы (Х3$) также влияет на биомассу кокковид-
ных бактерий (Х«) с корреляцией гЛ6 «— 0,56. Предполагаемая причи-
на этой связи следующая. Увеличение интенсивности фотосинтеза фито-
планктона в море при повышении атмосферного давления, т.е. уста-
новлении антициклона и безоблачной погоды, может задерживать раз-
витие кокковидных форм бактерий СПМ. Численность их, а значит, и
биомасса от этого будут уменьшаться. Понижение давления атмосфе-
ры при циклоне, наоборот, влечет за собой увеличение облачности и
снижение интенсивности фотосинтеза, что может привести к вспышке
развития кокковидных форм бактерий-
Концентрация РОВ морской воды в группе A (Хц) влияет на
биомассу кокков (Х6) и гетеротрофных бактерий (Х3) с корреляцией
Пз* “ 0,60 и rJ3_3 • -0,51. Это свидетельствует о том, что биомасса
указанных микроорганизмов в существенной мере зависит от содержа-
ния РОВ в море. На увеличение биомассы кокков (Хв) влияет также
доля баланса в инфракрасной области спектра (Х33) так как, по-внди-
Мому, указанные бактерии реагируют на эти невидимые тепловые
лучи солнца.
Важно отметить, что большую роль в изучении морской системы иг-
мет именно температура воздуха (Хм), а не воды (X3J), что, казалось
бы, естественнее для морских процессов. Воздух атмосферы в основном,
ПР-видимому, нагревается в результате действия солнечного параметра -
альбедо видимого спектра (Х31), в меньшей степени зависит от темпера-
туры (Хм), следовательно, на температуру воздуха (Хм) влияет облач-
ность (Х37), которая, скрывая солнце, уменьшает в конечном счете (Хм).
Температура воздуха (Хм)> как выяснилось, обусловливает в
определенной мере осаждение карбонатов на взвеси (Хи). Она влияет
также на РОВ СПМ (Хи). окисляемость морской воды (Х30), на со-
держание углекислоты (Хц) и кислорода (Хц) (АлекИН. 1966).
Р ТО же время температура роды (Хц) в изучаемой системе играет
Явно второстепенную роль. Она связана только с единственным пара-
метром - содержанием нефтепродуктов в воде (Хм)- Гетеротроф-
ные бактерии СПМ (Хз) испытывают влияние активности солнца
Йм) > pH метаболитов взвеси (Хи) И щелочности воды (Хц).
Па количество осаждающихся карбонатов взвеси (Хц) в группи-
ровке В влияет не только температура воздуха (Хм) И альбедо вц-
димого спектра (Хц), к»К отмечалось выше, но и облачность в бал-
лах (Х37), что связано, возможно, с интенсивностью фотосинтеза фи-
топланктона в воде,- pH воды (Хц), живыми и мертвыми диатомовы-
ми водорослями СПМ (Х8 и Хц) из группировки "С”, а кроме того, с
41
биомассой кокков (Х6), РОВ СПМ (Хи), pH СПМ (Xi8),cCO2 (Х21)
и кислородом (Х24).
От pH морской воды (Х1в) зависит концентрация фенола (Х27),
что пока трудно объяснить, а также биомасса гетеротрофных бактерий
взвеси СМВ (Х3), мертвые диатомовые СМП (Уш), РОВ морской
воды (Хп) и СО|~ (Х23).
В группе параметров С существует множество связей биомассы
живых диатомовых СПМ (Х8) с 7 различными параметрами, в число
которых входят: гетеротрофные бактерии СПМ (Xi)^ кокковидные
бактерии СПМ (Х6), pH метаболитов СПМ (Xie), альбедо видимого
спектра (Х31), температура воздуха (Хм), влажность воздуха (Хэо)
и прозрачность воды (Х41). Мертвые диатомовые СПМ (Хц) связаны
только с живыми диатомовыми СПМ (Х8) и кислородом (Хм).
Важно подчеркнуть, что параметры карбонатного равновесия в
группе ”Г” - НСО3 (Х22), СО?" (Х23) и щелочность воды (Х2в) свя-
заны со всеми группами параметров, кроме ”С” и ”6”, а также с актив-
ностью солнца W (Х33), суммарной радиацией солнца (Х42), бакте-
риями гетеротрофами СПМ (Х2), РОВ морской воды (Х13), pH воды и
pH СМВ (Х16, Хп), т.е. на карбонатное равновесие влияют не только
био- и абиотические параметры, их изменение определяют также пара-
метры солнца, что до сих пор не было известно.
Живые и мертвые диатомовые СМВ (Х4, Х4) в группе G связа-
ны с 4 группами параметров (А, В, С, также с гетеротрофными
бактериями СМП (Xi), карбонатами СМВ (Xi$), pH метаболитов СПМ
(Х18) и с кислородом (Хм). Сухая масса СПМ (Хм) и pH СМВ (Х17),
от которого, по-видимому, зависит изменение этой массы в группе Н,
связаны 7 группировками (Л, В, D, Е, F, G, Н) с 8 параметрами, среди
которых гетеротрофные бактерии СПМ и СМВ (Х2, Х3), диатомовые
живые СМВ (Х4),а также НСО3 (Х22), щелочность воды (Х26), давле-
ние атмосферы (Хэ$) и суммарная среднесуточная радиация солн-
ца (Х42).
В группе параметров I имеются связи с другими группами (А, С,
D, Е, F), а диатомовые СМП (Х7) связаны с РОВ СМВ (Х^), который
в свою очередь связан с содержанием ХлЭк (Х28). С ХлЭк связаны
также соленость (Х^), температура воды (Х25) и доля баланса в ин-
фракрасной области (Хя).
Интересно, что указанные связи с ХлЭк были бы понятны, если бы
это соединение было образовано морскими огранизмами, но, как
известно, это постороннее вещество, принесенное в море извне. Для
объяснения данного парадокса необходимо точно разделять в морской
воде ХлЭк и нефтепродукты от автохтонных углеводородов. А пока
вместе с аллохтонной частью ХлЭк определяют жиры и липиды мор-
ских организмов, а в особенности планктонных.
Bceio одна связь биомассы живых диатомовых СМП (Х7) с мерт-
выми /тиатомовыми СМП (Х,,,) не объясняет полностью функциониро-
4?
вание продуцентов в море. Об этом же свидетельствует и тот факт,
что кислород (Х24), на изменение содержания которого в море влияет
доля баланса в инфракрасной области (Х33) > связан только с мертвыми
диатомовыми СМВ (Х9) и СПМ (Хц); Хотя известно, что процесс
фотосинтеза в море осуществляют только живые диатомовые и потому
кислород должен быть связан именно с ними. Причиной этого в данном
случае может быть и ошибка наблюдений, поскольку при переводе
численности диатомовых в биомассу мы принимаем во внимание толь-
ко доминантные виды диатомей, не учитывая биомассу всех обнару-
женных видов этих микроводорослей. Очевидно, при изучении больших
морских экосистем следует основываться на численности живых диато-
мовых, а не на их биомассе.
Получив указанные выше группировки, отражающие структуру
их в море, мы провели факторный анализ изучаемых параметров. Для
этого, использовав в качестве исходных факторов полученные группи-
ровки, мы выбрали нулевое приближение структуры по всем этим
9 факторам:
FA (Х«, Хп, Хм, Х2, Хз, Хзз)
FB (Хн, Х1б, Хзь Х37, Х„, Х5, Х>0)
Fc (Х8, Хп, Хзо)
FD (Хз9,Хэз(Хза,Хад;
FB (Хза,Х4,,Хм, Х12)
FF (Хзз, Хзз. Хза, Хм, Х42)
Fg (Х4, Х„ Х„ Хза)
FH (Х14, Х17)
Ft (Х7, X», Хзз, Хза).
В результате проведения нескольких необходимых процедур
последовательного улучшения решений (что было подробно показано
р Предыдущем разделе этой книги) была найдена наилучшая фактор-
ная модель изучаемой морской системы, о которой мы подробно рас-
скажем ниже. Полученное факторное решение оказалось близким к
структуре изучаемой морской системы, определенной частными корре-
ляциями. После расчленения на отдельные факторы сохранились основ-
ные ядра группировок с сильными и средними связями образующих
их параметров (рис. 1), которые не изменились или дополнились па-
раметрами, окружающими ядра других группировок. Все параметры,
Образующие факторы, стали равнозначны. При этом число факто-
ров оказалось равным количеству группировок, которых насчиты-
(Ищось 9. Таким образом, можно констатировать, что в изучаемой
Системе понятие ’’группировка” в общих чертах аналогично поня-
тию "фактор”.
4.4. ВЗАИМОСВЯЗИ ПАРАМЕТРОВ В «АКТОРАХ
Фактор 1 (группа Я). Основное ядро группы А, которое как и в других
группах, состоит из параметров со средними и сильными связями, до-
полнилось несколькими параметрами из окружения ядра: углекисло-
той, растворенной в морской воде (JV2i) и окисляемостью морской
воды (Х20), но Из группы исчезла температура воздуха (^эд)- Взаи-
мовлияние параметров этого важного фактора заключается в следую-
щем. Активность солнца (Х33) влияет на гетеротрофные бактерии СПВ и
СМВ (Х2, Х3), а также на кокковидные формы бактерий СПМ (Хв).
Все эти микроорганизмы используют РОВ (Xi3) из воды, с количест-
вом которого связана окисляемость морской воды (Хм). В результа-
те жизнедеятельности бактерий образуется углекислота (Х21) - один
из важнейших параметров карбонатного равновесия, которая в океане
в основном продуцируется бактериями (Zo Bell, 1946).
Фактор 2 (группировка В). Ядро группировки В дополнилось па-
раметрами из окружения ядра: биомассой палочковидных бактерий
СПМ (Х3) и мертвыми диатомовыми СМП (Хю). Взаимодействие па-
раметров в факторе 2 может быть следующим. Альбедо видимого
спектра солнца (X3i) влияет на биомассу палочковидных бактерий
(Х3). Они используют останки мертвых диатомей СМП (Хю) или их
посмертные метаболиты. Отмирание этих диатомовых (Х10) может
быть связано с содержанием фенола (Х27) в воде. Указанные парамет-
ры, очевидно, влияют на pH морской воды (Х1в) и карбонаты СМВ
(Хи). В свою очередь, альбедо видимого спектра солнца (Х37) зави-
сит от облачности в баллах (Х37), которая может задерживать лучи
солнца при освещении моря.
Фактор 3 (группа С). Основное ядро группы С дополнилось из
окружения ядра лишь влажностью воздуха (Х»). Кроме того, биомас-
са живых диатомовых СПМ (Хв) обусловливает величину биомассы
мертвых диатомей СПМ (Хп) Роль влажности воздуха в развитии диа-
томовых неясна.
Фактор 4 (группировка D). Ядро этой основной группировки, сос-
тоящее из 3 параметров солнца, дополнилось параметрами из других
групп: суммарной среднесуточной радиацией (Х«2), являющейся, воз-
можно, производной от суммарной радиации (Хда). Этот фактор,
по-видимому, является основной движущей силой и источником энер-
гии в изучаемой морской системе, вследствие того, что он включает
в себя 4 параметра солнца из имеющихся 7. Все эти параметры: доля
баланса в инфракрасной области спектра (X»), энергобаланс (Х29),
суммарная радиация (Хда) и суммарная среднесуточная радиация
(Х<2), нагревая воду в море и обеспечивая энергией его жизнь, вызы-
вают изменение солености (Хдо). Группировка D, влияя в основном на
все остальные группы (факторы) (кроме G) системы, является гла-
венствующей.
44
г
Фактор 5 (группа Е). Ядро этой группы дополнилось параметрами
РОВ СПМ (Хи) и pH СПМ (Хи). Взаимодействие параметров следую-
щее. Давление атмосферы, изменяя облачность, может изменять осве-
щенность морской воды, влияя на ее прозрачность (X<i), что в свою
очередь влияет на pH метаболитов СПМ (Хц) и РОВ СПМ (Х«).
Фактор 6 (группировка F). К этой группировке добавились пара-
метры из других групп: количество часов солнечного сияния (Хз»),
температура воды (X2S) и воздуха (Х34). Взаимосвязь параметров в
труппе прослеживается отчетливо. Освещенность (Хз») и связанная с
Ней суммарная среднесуточная радиация солнца (Х«) в группах F и
И (рис. 1), no-видимому, обусловливают изменение температуры воз-
; духа и воды (Хм, Хм). Последние и такие параметры, как знергоба-
; лайс (X»), доля баланса в инфракрасной области (X»), а также сум-
• мерная среднесуточная радиация (Х«) из фактора 4, нагревая воду,
являются косвенной причиной изменения основных компонентов кар-
f боиатного равновесия (НСО3 - Хи и СО’" - Х23),а также щелочное-
\ Ти воды (Х2в), связанной с ними.
' Карбонатное равновесие изменяет соотношение катионов и ионов
i море, вызывая осаждение или растворение солей. Значит, Подтверж-
дается известный факт, что карбонатное равновесие - основной регуля-
' тор гидрохимического режима в море, а кроме того, влияющий и на
биопараметры.
Фактор 7 (группа F). Основное ядро этой группы дополнилось из
; окружения ядра следующими параметрами: биомассой гетеротрофных
' бактерий СМП (Xt) и кислородом (Х24). Взаимосвязь параметров это-
го фактора, не выявившаяся в группе, где кислород воды (Хм) был
связан с мертвыми диатомовыми взвеси СМВ (Х«) и СПМ (Хц), в
данном случае проявилась отчетливо. Образование кислорода в этой
Когстеме в основном зависит от живых диатомовых СМВ (Х4), а они
определяют биомассу мертвых диатомей (Х9). Гетеротрофные бак-
; терии планктона, очевидно, зависят от массы отмерших диатомо-
вых (Х9).
Фактор 8 (группировка Н). Ядро этой группировки, являющее-
ся также фактором, осталось неизменным. О взаимосвязи пара-
> метров ядра группы можно сказать лишь, что pH взвеси СМВ (Х|2),
' по-видимому, влияет на изменение сухой массы слизистой пленки
СПМ (Хм).
Фактор 9 (группа /) Основное ядро этой группы, как и предыду-
щей (Я), осталось неизменным. Взаимосвязь параметров здесь, очевид-
но, состоит в том, что содержание ХлЭк в воде (Хм) и РОВ (Хз»)
каким-то образом влияет на биомассу диатомовых живых СМП (X-j).
Так, по характеру и назначению комплекса параметров, входящих
в состав факторов, последние можно расположить по значимости и
роли, которую они играют в изучаемой системе и соответственно
назвать их.
45
Fi - биоабнотический. Содержит 1 параметр солнца. Возможно,
выполняет функции внешнего регулятора в системе.
F2 - биоабнотический, также содержит 1 солнечный параметр.
F3 - биоабнотический.
- абиотический, в котором содержится 4 параметра солнца.
Является основным источником тепла и энергии в морской системе.
Fs — биокосноабиотический (биокосный параметр, т.е. связан-
ный с метаболитами биопараметров).
F6 - абиотический. Содержит 1 параметр солнца, параметры темпе-
ратуры, карбонатного равновесия и гидрохимического режима мор-
ской воды. Поэтому наряду со внешним регулятором системы факто-
ром Fx фактор Ft можно считать внутренним регулятором в изучаемой
системе.
Fy - биокосный.
Ft - биокосный.
F9 — биоабиокосный.
В результате проведенных исследований структура морской систе-
мы, выявленная частными корреляциями, подтвердилась и факторным
анализом. С помощью обоих методов удалось определить ие только
взаимосвязь, но и взаимодействие параметров и факторов системы
(Подвинцев, Горбенко, 1979). Большой интерес также представляют
взаимоотношения ’’внутри” и ’’вне” сообщества обрастаний (Сообр.)
с компонентами внешней по отношению к Сообр. морской среде. I
Одним из основных путей изучения взаимодействия био- и абиоти-
ческих параметров элементов в биосистеме является применение мате-]
магического эксперимента. Суть его в том, что математическим путем
поочередно увеличивали (и уменьшали) величину одного из парамет-j
ров изучаемой системы, констатируя, как повлияет это воздействие]
на остальные элементы. | |
Такой эксперимент сэ множеством параметров и их взаимоотно-1
шениями можно провести только с помощью математических мето-|
дов. В природных условиях моря или в лаборатории проведение его]
невозможно. I
46
: ГЛАВА & ИЗУЧЕНИЕ ВЗАИМООТНОШЕНИЙ В ЭКОСИСТЕМЕ
С ПОМОЩЬЮ МАТЕМАТИЧЕСКОГО ЭКСПЕРИМЕНТА
&1. ИЗМЕНЕНИЕ КОЛИЧЕСТВА МИКРООРГАНИЗМОВ
НА РАННИХ СТАДИЯХ РАЗВИТИЯ БИОЦЕНОЗА ОБРАСТАНИЙ
>' Изучив Детально все данные о динамике численности отдельных групп
микроорганизмов СПМ за 1 типичный год на первых стадиях биоти-
’ ческой сукцессии, равной 30 сут (табл. 19), мы подробно рассмотрели
: изменение численности каждой группы микроорганизмов СПМ, чтобы
‘ выбрать среди них самые информативные.
j Результаты этой работы показали, что порядок смены сукцессий
j микроорганизмов в рассматриваемый период оставался прежним. Из-
f менение численности отдельных групп микроорганизмов в ходе биоти-
< ческой сукцессии в целом имело общую тенденцию к увеличению от
: 1-х до 30-х суток, но при этом варьировало по-разному.
' От 10-х к 30-м суткам численность диатомовых водорослей обычно
^неуклонно повышалась (в 10 из 12 месяцев) и развитию их, очевидно,
! Шчего не мешало. В это же время гетеротрофные бактерйи, по-види-
мому, испытывали влияние диатомовых, особенно после 20-х суток,
когда количество микроводорослей интенсивно возрастало и они свои-
ми телами начинали покрывать всю поверхность погруженного предме-
та и бактерий на его поверхности. Поэтому численность бактерий-ге-
теротрофов обычно возрастала с 10-х на 20-е сутки месяца, а затем сни-
жалась с 20-хна 30-е сутки (в 8 из 12 месяцев).
Количество палочковидных форм бактерий, в отличие от предыду-
щих микроорганизмов, изменялось по-разному. Одну половину года
(преимущественно в холодный период) оно возрастало с 10-х на 20-е
сутки, а потом снижалось на 30-е сутки (5 месиз 12). Другую полови-
ну года (преимущественно в теплый период) оно неуклонно новы-
шалось с 10-х до 30-х суток (в 5 мес из 12). В данном случае, по-ви-
димому, как-то влияла сезонность, поскольку в один период года
на палочковидные формы воздействовали диатомовые водоросли,
в другой - нет. Число кокковидных бактерий изменялось еще ме-
нее определенно - чаще возрастало с 10-х на 30-е сутки (5 мес из
12) преимущественно в теплое время года, также как и коли-
47
Таблица 19. №меиеине численности (ig) микроорганизмов
перифитона (обрастаний) в различные сроки экспозиции в
м»- Микроорганизмы Время экспонирования
сяц пластин в море, сут
10 f . 20 30
1 Гетеротрофные бактерии (1) 4,27 4,47 4,74
Диатомовые водоросли (2) 4,51 5,83 5,85
Палочковидные бактерии (3) 5,64 5,77 5,77
Кокковидные бактерии (4) 5,63 5,87 5,87
II 1 4,04 4,45 4,37
2 4,67 5,48 5,5
3 5,66 5,86 5,8
4 5,53 5,80 5,72
III 1 4,13 4,46 4,28
2 4,87 5,42 5,56
3 5,54 5,65 5,96
4 5,66 5,77 6,2
IV 1 4,10 4,00 4,46
2 4,08 4,88 5,2
3 5,60 5,65 5,58
4 5,65 5,90 5,88
V 1 3,90 3,99 3,75
2 4,27 4,78 5,51
3 5,12 5,14 5,99
4 5,28 5,78 5,78
VI 1 4,00 4,50 4,39
2 4,30 4,86 5,00
3 4,90 5,46 5,47
4 5,56 5,90 6,00
VII 1 3,75 4,52 3,99
2 5,06 4,91 5,51
3 5,13 5,09 5,47
4 5,28 5,64 5,67
vin 1 3,48 4,55 4,63
2 4,38 5,30 5,41
3 5,26 5,90 5,97
4 5,59 6,00 6,10
IX 1 3,39 3,9 3,37
2 5,34 4,97 5,68
3 5,57 5,70 5,20
4 5,65 5,66 5,59
X 1 4,И 4,37 4,03
2 4,89 5,32 5,30
3 5,40 3,89 5,73
4 5,45 5,31 5,80
XI 4,13 4,72 4,39
3,96 5,08 5,12
3 5,76 6,03 5,25
4 5,52 5,94 6,02
X|I 1 3,98 4,26 4,30
2 3,24 4,79 4,67
3 5,02 5,62 6,64
4 5,71 5,87 5,59
48
честно палочковидных бактерий, а в другое время изменялось по-
разному.
Таким образом, судя по колебанию численности микроорганизмов
СПМ в период биотической сукцессии, наиболее определенной она
была у диатомовых водорослей и гетеротрофных бактерий СПМ, по-
видимому, связанных между собой. Учитывая это, в математическом
эксперименте в качестве основных представителей микроорганизмов
СПМ мы использовали диатомовые водоросли и гетеротрофные бакте-
рии СПМ. Затем из 40 параметров изучаемой системы 1 типичного года
наблюдений с помощью математических методов (по критерию Кульба-
ка) выбрали 25 наиболее взаимосвязанных. В число их в качестве
объектов исследований входили сообщества обрастаний (Сообр.), мик-
роорганизмы перифитона (СПМ), планктона (СМП) и взвеси (СМВ).
В указанных сообществах в качестве предметов исследований
были живые (а также мертвые) диатомовые водоросли и гетеротроф-
ные бактерии. Кроме того, одновременно изучали абиотические пара-
метры окружающей среды1 (табл. 20, 21).
Изучаемая биосистема может быть условно разбита на 7 факторов
со следующими параметрами.
1. Фактор макроорганизмов обрастаний: балянусы, мидии, гидрои-
ды, мшанки, ботриллюсы (5 параметров).
2. Фактор гетеротрофных бактерий СПМ, СМВ и СМП (3 пара-
метра) .
3. Фактор живых диатомовых водорослей СПМ, СМВ и СМП (3 па-
раметра) .
4. Фактор мертвых диатомовых водорослей СПМ, СМВ и СМП
(3 параметра).
Как отмечалось выше, СПМ и СМВ отвечали всем признакам био-
ценоза. Что касается СМП, также сформированного в основном бакте-
риями И диатомовыми водорослями, то количество и теснота корре-
ляционных связей, по нашим определениям, как ’’внутри”, так и ’’вне”
его были меньшими, чем в СПМ и СМВ (Горбенко, 1977). Значит,
СМП, образованное из микроорганизмов, обитающих в толще воды
и свободно парящих или плавающих, менее четко организовано, чем
СМВ и СПМ, обитающих на границе раздела вода — твердая поверх-
ность предмета (или относительно плотная поверхность взвеси). Тем не
менее СМП соответствует всем основным критериям ценоза, и его
уверенно можно отнести к сообществу.
5. Фактор морской воды: концентрация РОВ, содержание кислоро-
да, щелочность воды, фосфор минеральный, нитриты - азот и pH воды
(6 параметров).
Данные наблюдений за параметрами солнца и атмосферы были предоставлены
нам сотрудниками астрономических и гидрометеорологических учреждений
Крыма и г.Севастополя, за что мы искрение благодарны им.
49
Таблица 20. Изменение величины абиотических параметров в
иоде Севастопольской бухты в течение одного года наблюдений
Параметр I Пределы । колебаний Размерность
Концентрация РОВ 0,314-0,534 По экстинции Е иа
волне 260 нм
Кислород 4,55-7,44 млл
Фосфор минеральный 0-32,0 мкг л *
Нитриты-азот 0-10,4 мкг л *
Щелочность 3,15-3,37 мгэквл '
₽н 8,12-8,50
COj (1(Г5), 0,92-3,12 моль-л”1
нср5 сю"’) 2,46-2,99 мг нон-л" *
со; по *) 1,67-4,19 мг-иоил
ХлЭк 0-6,67 мг.л *
Фенол — мгл
Таблице 21. Изменение величины биотических параметров иа поверхности погру-
женных в море предметом в поде Севастопольской бухты в течение одного года
наблюдений
Параметр Пределы колебаний Размерность
Балянусы 0-567,69 —1 мг-100 см пластины
Мидии 0-13,49 То же
Мшанки 0-0,36 33
Ботрии люсы 0-3,20 ♦» 33
Гидроиды 0-9,60 Площадь 100 см .занимав-
мая гидроидами (%)
Диатомовые водоросли живые
СМП 16,8-10”*—6,80-10"’ мг-мл“*
СПМ 0,16 10"*- 3,78 мг.см”’
СМВ 0,13-10”* - 8,06 Диатомовые водоросли мертвые мг. мл"1
СМП 6,3 10”в - 56,0 10"5 мг-мл”1
СПМ 0-4,65 10”’ . мгсм”’
СМВ 0,66 10”’- 2,65-10”’ мг-мл”1
Гетеротрофные бактерии
СМП 0,892 10”в - 0.18540”4 мгмл"*
СПМ 0,556 10-5 - 0,525-10”4 мг-см^’
СМВ 0.28240"4 - 0,899 10~3 мг-мл"*
6. Фактор загрязнения: концентрация нефтепродуктов и фенолов
в море (2 параметра).
7. Фактор карбонатного равновесия: углекислота (СО2), гидро-
карбонат-ионы (HCOj), карбонат-ионы (СО*-) (3 параметра).
Предварительная математическая обработка комплекса из 7 фак-
торов с 25 параметрами показала, что они состоят в тесной статисти-
ческой связи с вероятностью Р = 0,99, т.е. все указанные факторы с па-
раметрами объединяются в единую биосистему.
50
6.2. ИНТЕРПРЕТАЦИЯ РЕЗУЛЬТАТОВ
МАТЕМАТИЧЕСКОГО ЭКСПЕРИМЕНТА
После проведения соответствующих расчетов была составлена табл. 22 -
25x25 параметров. По диагонали в ней располагаются параметры-воз-
будители, которые мысленно можно поднять вверх таблицы и кото-
рым мы давали приращение в 1 % от их средней Величины. По вертика-
ли размещены градиенты возбуждаемых параметров-возбудителей,
которые изменяют (увеличивают или уменьшают) свою величину. При
значениях градиента до 0,3 ответная реакция данного параметра на дей-
ствие параметра-возбудителя считается слабой, от 0,3 до 0,89 - средней,
от 0,9 до 1,9 - сильной, а от 2,0 и более - очень сильной.
В ответ на внешнее воздействие одного из параметров-возбудите-
лей величиной 1 % на остальные параметры оии либо изменялись (до
5,4 %), либо изменений не обнаруживалось (табл. 22). При этом биоти-
ческие и биокосные (мертвые диатомовые водоросли) параметры от
Внешнего воздействия в целом изменялись мало. Вместе с тем влияние
некоторых из них на биопараметры было сильным, в ряде случаев пре-
вышая 1 %.
Путем суммирования градиентов по вертикали удалось найти са-
мые сильные (и слабые) параметры-возбудители и знак, с которым они
Преимущественно влияют на указанную биосистему (табл. 23). По дан-
ным табл. 23 установили, что действие изучаемых параметров на био-
массу в основном положительное (15 случаев), но бывает отрицатель-
ное (10 случаев).
Основными параметрами (по сумме градиентов), регулирующи-
ьв связи в системе, являются гетеротрофные бактерии СМП и баля-
нусы. Затем следуют мшанки и гетеротрофные бактерии СМВ, а потом
мертвые диатомеи СМП, СПМ и СМВ, за которыми следуют гидрокар-
бонат-ионы и гидроиды. Отметим, что из 13 сильнейших параметров-
возбудителей 6 относятся к СПМ И СМВ (табл. 23), т.е. микроорганиз-
мы Перифитона и близкие к ним взвеси играют важную роль в изучае-
мой экосистеме.
Если механизм влияния HCOj на другие параметры еще не совсем
дрен, то микро- и макроорганизмы воздействуют на гидрокарбонаты
сдоими метаболитами, т е. связями экологического метаболизма.
Важно отметить, ЧТО В целом наиболее регулирующее влияние в систе-
ма имеют гетеротрофные бактерии, живые и мертвые диатомовые и
балянусы с мшанками. В то время как абиопараметры морской среды,
В том числе и компоненты загрязнения, играют второстепенную роль.
Суммируя градиенты по горизонтали, мы нашли самые возбуди-
мые параметры в системе и знак, с которым они претерпевают внешнее
Воздействие параметров возбудителей (табл. 24).
Для возбудимых параметров, так же как н для возбудителей, ха-
рактерно доминирование градиентов с положительным знаком (18),
51
Таблица 22. Ответ возбуждаемых параметров на воздействие параметрои-воэбуди fl теней (пояснения в тексте)
№ п/п Г' i 2 3 4 5 6 7 8 11 10 И 12 13 14 15 16 17
1 Баля- 0,8 0,5 -0,02 -0,07 -0,42 0,16 -0,1 II -0,25 -0,2 -0,18 -0,17 -0,13 -0,06 0,23 -0,18 -0,1
2 нусы 0,77 Мидии 0,11 0,15 0,03 0,19 0,18 -0,09 Ц -0,22 0,05 -0,11 -0,16 0,06 0,19 0,1 -0,13 -0,03
3 0,26 0,22 Мшанки 0,21 -0,14 0,14 0,22 0,23 Ц -0,18 -0,03 -0,17 -0,11 -0,06 -0,02 -0,03 0,01 0,06
4 0,07 0,1 0,25 Ботрии- 0,04 0,05 -0,17 -0,24 ! 0,07 0,01 -0,05 0,26 -0,09 0,17 -0,09 0,2 -0,05
5 - 0,12 0,05 -0,2 ЛЮСЫ 0,06 Гид- 0,16 0,12 -0,08 И -0,08 -0,00 0,02 -0,09 0,56 0,34 0,06 0,14 0,03
6 0,06 0,01 0,12 0,12 роиды 0,04 Гетеро- 0,08 -0,03 || 0,23 -0,13 0,01 0,03 -0,12 0,04 -0,23 -0,01 0,03
7 0,25 0,14 0,25 -0,18 0,1 трофы СМП 0,01 Гетеро- 0,42 1 0,1 0,02 0,5 0,08 0,03 -0,07 -0,04 -0,16 -0,13
8 -0,17 -0,07 0,18 -0,23 -0,07 0,07 трофы СПМ 0,43 Мертвые 0,22 0,4 0,24 0,13 0,21 0,01 -0,23 -0,17 0,1
9 -0,25 -0,26 -0,41 0,17 -0,14 0,63 0,21 диатомо- вые СМП ] 0,14 | Гетеро- 0,36 0,24 0,34 0,23 -0,02 -038 -0,05 0,12
10 -0,2 0,09 0,21 -0,1 0,00 -0,56 -0,09 0,48 Ц трофы СМВ 0,36 Живые 0,08 0,22 0,39 0,64 0,1 0,11 -0,03
11 -0,17 -0,1 -0,001 -0,13 0,06 -0,28 0,42 0,29 Ц 0,23 диато- мовые СМП 0,1 Живые 0,62 -0,09 -0,04 -0,11 0,04 -0,08
12 -0,17 -0,19 -0,32 0,35 -0,09 0,4 0,17 0,06 fl 0,34 0,22 диато- мовые СМВ 0,62 Мертвые 0,1 -0,06 -0,21 0,1 0,13
13 -0,12 0,09 0,13 -0,17 0,61 -0,45 -0,05 0,27 1 0,23 0,4 -0,09 диато- мовые СМВ 0,09 Живые 03 0,16 0,23 -0,05
14 -0,06 0,16 -0,22 0,25 0,29 0,4 0,02 -0,06 1 -0,02 0,06 -0,03 -0,06 диато- мовые СПМ 0,51 Мертвые 0,18 0»13 0,04
15 0,02 0,1 -0,12 -0,05 0,05 -0,06 -0,00 -0,25 fl -0,33 0,06 -0,14 0,18 диатомо- вые СПМ 0,13 0,22 РОВ -0,13 0,09
16 0,31 0,11 1,25 -0,36 0,5 -2,1 -0,73 0,28 0,52 0,00 -0,1 -0,04 0,23 0,1 0,25 -0,05 pH -0,00
17 - 0,05 0,05 0,14 -0,08 0,04 -0,1 -0,15 0,1 fl 0,07 0,03 0,08 -0,00 -0,00 0,08 0,01 Щело*
18 -0.1 -0,18 0,32 0.1 3 а 12 -0,07 -0,02 -0,07 0,12 0,1 0,28 0,08 fl 0,16 -0,03 0,00 0,03 0,00 0,00 0,12 -0,26 ность -0,27
19 -0,2 -0,39 -0,57 0,27 0,12 0,78 0,16 -0,18 fl 0,39 0,33 0,25 0,16 0,29 0,24 -0,48 0,41 -0,00
20 -0,37 0,03 -0,2 1,02 0,54 0,22 fl 0,31 -0,00 0,3 0,15 -0,05 -0,27 -0,25 -0,26 -0,19
21 22 -0,15 0,1 - 0,3 0.04 0,14 0,3 0,00 -0,09 0,34 -0,21 -0,04 -0,11 0,09 fl -0,2 fl 0,13 0,05 -0,04 0,23 -0,00 -0,08 -0,07 0,08 -0,00 -0,11 0,05 0,3 -0,05 0,36 -0,07 -0,29 -0,02 -0,03
24 (М9 9 14 0,49 3,15 -1,75 0,98 -5,4 -1,27 1,45 fl j 0,72 -0,24 -0,21 0,42 -0,42 0,26 0,53 -0,82 -0.9
94 0 39 -1.19 0,17 -0,27 2,08 0,75 -0,05 fl j 0,4 0,21 0,39 0,05 0,11 -0,17 -0,51 -0,59 0,3
25 0^04 0,28 1.13 0,24 0,23 -2,00 -0.76 0,08 fl [ -0,32 -0,16 -0,34 -0,00 -0,11 0,13 0,43 0,67 -0,18
53
52
Окончание табл. 22.
Жп/п 18 19 20 21 22 23 24 j 25
1 —0,26 -0,21 -0,34 -0,06 0,09 -0,17 - 0,24 0,13 2 0,01 -0,13 -0,33 -0,18 0,09 -0,12 -0,19 0,1 3 -0,03 -0,17 -0,03 -0,25 -0,06 -0,12 -0,12 0,12 4 - 0,09 0,09 — 0,19 0,01 0,33 - 0,25 -0,31 0,22 5 0,17 0,23 -0,1 -0,01 -0,1 0,02 0,05 -0,01 6 0,14 0,1 0,44 0,24 - 0,04 0,24 0,23 - 0,24 7 0,26 -0,02 0,19 -0,05 -0,07 0,22 0,27 -0,3 8 0,12 0,04 0,34 0,09 -0,23 0,29 0,33 0,28 9 0,34 0,41 0,32 0,06 0,08 0,43 0,47 - 0,4 10 - 0,22 0,31 -0,02 0,04 0,19 0,03 0,12 -0,08 11 -0,13 0,21 0,29 0,05 -0,1 0,18 0,26 - 0,23 12 0,2 0,19 0,17 -0,14 0,11 0,11 0,13 -0,1 13 - 0,14 0,26 - 0,08 0,06 - 0,13 - 0,07 0,00 0,00 14 0,17 0,27 -0,25 -0,01 0,33 -0,06 -0,07 0,03 15 0,2 —0,4 -0,18 -0,06 0,35 -0,18 -0,27 0,21 16 - 0,26 - 0,42 - 0,26 0,0 -0,28 -0,681 0,65 0,66 17 -0,32 -0,1 0,1 -0,01 -0,02 -0,04 0,02 0,09 18 Фосфор -0,08 0,47 0,02 0,05 0,36 0,31 -0,34 мине- ральный 19 0,24 01 -0,1 -0,1 0,11 0,03 0,09 -0,05 20 0,74 -0,1 NOj-N -0,07 - 0,04 0,54 0,52 - 0,47 21 0,05 0,04 0,18 Нефть -0,06 0,14 0,05 -0,19 22 - 0,02 - 0,06 - 0,11 -0,04 Фенол Q.07 - 0,02 - 0,05 23 -2,28 0,19 0,74 1,28 -0,57 СО, 1 -1 24 1,17 0,05 0,46 - 0,32 0,19 0,94 НСОЗ -0,09 25 - 0,12 -0,00 - 0,42 0,25 -0,24 - 0,95 -0,9 СО1,"
а отрицательных - значительно меньше (7). Наиболее возбудимыми
оказались компоненты карбонатного равновесия (СО2, HCOj и СО’-)-
Из них 2-й и 3-й параметры были и среди параметров-возбудителей, т.е.
они, по-видимому, не только зависят от ряда параметров среды, но
также являются регуляторами в ней. Интересна группа гетеротроф-
ные бактерии СМВ и мертвые диатомеи СМП, СМВ, СПМ, которые на
4-7-м местах среди сильнейших параметров-возбудителей и на 4-6-м
и 9-м - среди самых возбудимых. Среди сильных возбудителей и са-
мых возбудимых параметров находятся также живые диатомовые
СМП, СПМ и кислород, т.е. эти параметры играют в системе двоякую
роль — регуляторов и исполнителей.
Среди самых сильных параметров-возбудителей менее всего возбу-
димы балянусы, гетеротрофные бактерии СМП, мшанки и гидроиды,
в то время как самые слабые возбудители - фенол, pH воды, мидии,
ХлЭк, РО0 и щелочность - менее всего возбудимы.
При детальном просмотре вертикальных строк табл- 22 можно ви-
деть, как изменяются параметры системы при воздействии параметров-
возбудителей. В большинстве случаев параметры-возбудители влияют
на возбуждаемые параметры и положительно, и отрицательно. Липп
54
Таблица 23. Действие параметров-возбудителей на биосистему*
Самые сильные Самые слабые
Параметр Градиент Параметр Градиент
Гетеротрофные бактерии СМП -5,21 Фенол -0,20
Баляну сы -5,10 Фосфор минеральный -0,06
Мшанки 3,74 РОВ морской воды -0,31
Гетеротрофные бактерии СМВ 3,13 Гетеротрофные бактерии СПМ 0,37
Мертвые диатомовые СМП 2,91 Мидии 0,87
Мертвые д иатомовые СПМ 2,63 ХлЭк 0,94
Мертвые диатомовые СМВ 2,49 Углекислота (COj) 0,96
Гидрокарбонат-иоиы (НСОз) 2,38 Щелочность -0,78
Гидроиды 2,14 pH -1,07
Карбонат-ионы (СО3) -1,79 Живые диатомовые СМВ 1.11
Живые диатомовые СМП -1,65 Нитриты-азот 1,28
Кислород (От) 1,54 Ботрнплюсы -1,22
Живые диатомовые СПМ 1,49
* По сумме градиентов.
Таблица 24. Степень возбуждения некоторых параметров*
Более возбудимые Менее возбудимые
Параметр Градиент Параметр Градиент
COj _ -4,14 Мшанки -0,05 HCpJ 3,55 Щелочность -0,10 СО? -3,50 pH воды -0,16 Гетеротрофные бактерии СМВ 2,64 Фенол 0,23 Мертвые диатомовые СМП 2,31 Мидии 0,34 Мертвые диатомовые СМВ 2,12 Ботрнплюсы 0,34 Живые диатомовые СМП 2,07 ХлЭк 0,36 Кислород 2,01 РОВ воды -0,65 Мертвые диатомовые СПМ 2,00 Фосфор минеральный 0,86 Нитриты-азот 1,85 Гидроиды 1,22 Гетеротрофные бактерии СПМ 1,82 Гетеротрофные бактерии СМП 1,24 Живые диатомовые СПМ 1,68 Балянусы -1,25 Живые диатомовые СМВ 1,29
* По сумме градиентов.
мсса мертвых диатомовых планктона, перифитона и взвеси только
положительно влияет на систему. Также положительным влиянием на
другие параметры характеризуется биомасса гидроидов.
По горизонтальным строкам табл. 22 можно подробно проследить,
как реагирует каждый элемент системы на внешнее воздействие. Как
отмечалось выше, чаще всего изучаемые элементы системы испытыва-
ют на себе положительное и отрицательное влияние параметров-возбу-
дителей. Биомасса мшанок не подвержена влиянию остальных пара-
метров-возбудителей, если градиент превышает или равен 0,3, в то вре-
55
Таблица 25. Количественное влияние параметров-возбудителей на возбуждаемые
параметры (по вертикальным строкам пбл. 22)
Параметр Количество возбуждае- мых пара- метров Степень возбуждения
Балянусы 5 Средняя
Мидии 4
Мшанки 10 Средняя, сильная н очень силь-
Ботриллюсы 4 ная Средняя и сильная
Гидроиды 3 То же
Гетеротрофные бактерии планктона 13 Средняя, сильная и самая силь-
Гетеротрофные бактерии перифитона 7 ная Средняя и сильная
Гетеротрофные бактерии взвеси 9 Средняя
Живые диатомовые водоросли 4
планктона Живые диатомовые водоросли взвеси 5 -•
Живые диатомовые водоросли пери- фитона Мертвые диатомовые водоросли 4
3 Средняя и сильная
планктона Мертвые диатомовые водоросли 3 Средняя
взвеси Мертвые диатомовые водоросли па- 4 *»
рифитона РОВ 6 »
pH воды 4 **
щелочность воды 2 Средняя и сильная
Фосфор минеральный в воде 5 Средняя, сильная и очень силь-
Кислород воды 4 ная Средняя
Нигриты-азот воды 9
Содержание ХлЭк в воде 2 Средняя и сильная
Содержание фенола в воде 4 Средняя
Углекислота воды 6 Средняя и сильная
Гндрокарбонат-ионы в воде 8 То же
Карбонат-ионы в воде 6
мя как сама она является сильным параметром-возбудителем (табл. 23).
Только положительный эффект оказывает внешнее воздействие на со-
держание гидроидов и биомассу гетеротрофных бактерий планктона,
мертвые диатомовые планктона, перифитон, а также концентрацию
фенола. Отрицательное влияние претерпевает лишь щелочность. Таким
образом, элементы биомассы обрастаний при внешнем воздействии
могут испытывать положительное и отрицательное влияния, только
положительное или только отрицательное либо почти совсем не реаги-
ровать на него (табл. 24). О степени влияния макроорганизмов обрас-
таний, а также бактерий, диатомовых, гидрохимических показателей
36
Таблица 26, Ответная количественная реакция возбуждаемых параметров на дей
стене параметров-возбудителей (по горизонтальным строкам табл. 22)
Параметр
Количество
возбуждений
Степень возбуждения
Балянусы
Мидии
Мшанки
Ботрнплюсы
Гидроиды
Гетеротрофные бакт ерии планктона
Гетеротрофные бактерии перифи-
тона
Гетеротрофные бактерии взвеси
Живые диатомовые водоросли
планктона
Живые диатомовые водоросли взвеси
Живые диатомовые водоросли пери-
фитона
Мертвые диатомовые водоросли
планктона
Мертвые диатомовые водоросли
взвеси
Мертвые диатомовые водоросли пе-
рифитона
FOB
pH воды
Щелочность воды
Фосфор минеральный в воде
Кислород воды
Нитриты-азот воды
Содержание ХлЭк в воде
Содержание фенола в воде
Углекислота воды
Гидрокарбонат-ионы в воде
Карбокатионы в воде
4 Средняя
2 Слабая
О Возбуждения нет
2 Слабая
з ”
1
3 Средняя
11 Средне-сильная
6
2 Слабая
4
4
5
3
3
11 Средняя, сильная и очень силь-
ная
1 Слабая
4
6
11 Средняя, сильная
2 (лабая
3
20 Средняя, сильная, очень силь-
ная и самая сильная
13 Средняя, сильная и очень силь-
ная
10 То же
И карбонатного равновесия на элементы изучаемой биосистемы,в каж-
дом отдельном случае можно судить по табл. 25-28.
На содержание углекислоты в системе влияют все макроорганиз-
мы. Большая часть макроорганизмов действует также на pH и нитриты-
азот. Оказывают они влияние также на гидрокарбонат-ионы, мертвые
диатомовые взвеси и другие элементы (табл. 27).
Диатомеи влияют на гидрохимические характеристики морской
воды (в том числе на кислород и углекислоту). От них зависит масса
мертвых клеток, соответствующих ио биотону диатомовым, они
влияют на бактерии, друг на друга и на некоторые микроорганизмы
(табл. 28). Гетеротрофные бактерии играют весьма важную роль в
изучаемой системе. Все они действую! в основном на i идрохимитескне
57
Таблица 27. Влияние микро- и макрооргаиязмов обрастания и гидрохимических
показателей на другие параметры*
Параметр Баля- нусы Мидии Мшанки Ботрил- люсы Гидрои- ды
СО2 4- 4- 4- 4- 4-
рн + + 4- +
N-NO2 4- + 4- — —
о2 — — + — —
НСОэ — 4- 4- — ——
соГ — 4- — —
Мертвые диатомовые водоросли — — 4- 4- —
взвеси Живые диатомовые водоросли — — — 4-
перифитона Гетеротрофные бактерии взвеси — 4- — —
Мидии + — — — *—
Балянусы — 4- 4- — —•
ХлЭк — — 4- — —
Фенол — — — 4- —-
* Здесь и в табл. 28-30 знак (+) показывает на какие возбуждаемые парамет-
ры влияют параметры-возбудители; знак (-) - отсутствие связи.
Таблица 28. Влияние живых диатомовых на другие параметры
Параметр Живые диатомовые водоросли
планк- тона 1 взвеси 1 г ерифи- тона
Мертвые диатомовые водоросли планктона 4- — —
Гетеротрофные бактерии взвеси 4- — —
Живые диатомовые водоросли перифитона 4- — —
Оз 4- — —
Гетеротрофные бактерии перифитона — 4- —•
Мертвые диатомовые водоросли взвеси — 4- —
N-NOj — 4- —
HCOj — 4- —•
COj" — 4- —
Гидроиды — — 4-
Живые диатомовые водоросли планктона — — 4-
Мертвые диатомовые водоросли планктона — — 4*
СО2 — — 4-
характеристики и компоненты карбонатного равновесия, а также на
мертвые и живые диатомеи, на параметры загрязнения и на некоторые
макроорганизмы (табл. 29). Большое значение в системе также имеют
гидрохимические характеристики с параметрами карбонатного равно-
весия (табл. 39). Они влияют одна на другую, на гетеротрофные бакте-
рии, особенно на бактерии взвеси, на диатомовые водоросли и на мак-
i юргашгмы.
58
Таблица 29. Влияние гетеротрофных бактерий на другие параметры
Параметр Гетеротрофные бактерии
планк- тона взвеси перифи тона
N-NO, ч- 4-
НСОз + + +
pH + + 4
СО2 + 4
сог + 4 +
Мертвые диатомовые водоросли взвеси 4 —
0, + 4* —
Живые диатомовые водоросли планктона + +
Гетеротрофные бактерии взвеси — —
Мертвые диатомовые водоросли планктона — — +
Живые диатомовые водоросли взвеси — — +
Живые диатомовые водоросли перифитона — —
Балянусы + — —
Мертвые диатомовые водоросли перифитона + — —
ХлЭк + — —
РОВ — + —
Таблица 30. Влияние гидрохимических характеристик на другие параметры
Параметр СО2 НСОз cot- 02 N-NO2 pH Фосфор мине- ральный
Гетеротрофные бак- 4 4 4 4- — 4
терии взвеси С02 — 4- — 4 4 4
рн 4 4 4 4 —- — —-
Фосфор минеральный 4- 4 4 — 4 —
НСОз 4 — —• — + 4 4
СО?" + 4 + 4
нЛю; 4 4 + — — — 4-
Мертвые диатомовые — 4 — — + — —
водоросли планктона 02 — — — + —
Живые диатомовые — 4 - — —
водоросли планктона РОВ 4
Гетеротрофные бак- - - — - 4 - —
терии планктона Ботрнплюсы 4 — — — —. ...
Гетеротрофные бак- — — + — — —
терии перифитона Щелочность __ .... — — — 4
Балянусы — -• — — 4 — —
Мидии — — — 4 — —
С помощью математического эксперимента выявлено взаимовлия-
ние ряда параметров друг на друга, когда поочередно один из них
является параметром-возбудителем, а другой возбуждаемым пара-
59
метром. Положительно в этом случае влияют друг на друга: 1)баляну-
сы и мидии, 2) гидроиды и живые диатомовые перифитона, 3) живые
диатомовые планктона и гетеротрофные бактерии взвеси, 4) живые
диатомовые водоросли планктона и перифитона, 5) живые диатомовые
взвеси и гетеротрофы перифитона, 6)живые диатомовые перифитона
и гидроиды, 7) гетеротрофные бактерии планктона и нитриты-азот,
8)гетеротрофные бактерии взвеси и нитриты-азот, 9) гетеротрофные
бактерии взвеси и гидрокарбонат-ионы, 10)гетеротрофные бактерии
взвеси и углекислота, 11) гетеротрофные бактерии взвеси и кислород,
12) гетеротрофные бактерии перифитона и живые диатомовые взвеси,
13)углекислота и фосфор минеральный, 14)углекислота и гидрокар-
бонат-ионы, 15)углекислота и нитриты-азот, 16)гидрокарбонат-ионы
и фосфор минеральный, 17) гидрокарбонат-ионы и нитриты-азот,
18)карбонат-ионы и pH, 19) кислород и pH, 20) кислород и живые
диатомовые планктона, 21)нитриты-азот и фосфор минеральный.
В связи с этим мы имеем все основания считать, что 21 пара элемен-
тов системы (из 65 пар параметров с градиентом более или равным
0,3, табл. 22) не только состоит друг с другом в прямой положитель-
ной связи, когда А влияет на В (или В действует на А), но и усиливает
его. Здесь может иметь место также обратная положительная связь,
когда элемент А действует на элемент В, усиливая его, а элемент В в
ответ на это влияет на элемент А, в свою очередь вызывая его усиление.
Оказывают положительное или отрицательное влияние друг на дру-
га: 22) pH и гидрокарбонат-ионы. Указанные элементы состоят друг
с другом в прямой отрицательной и в прямой положительной связях,
но вполне возможно, что здесь замаскирована и обратная положитель-
но-отрицательная связь, когда элемент А влияет на элемент В, усили-
вая его, а элемент В в ответ на это действует на А, уменьшая его.
В целом отрицательно влияют друг на друга: 23)балянусы и нитри-
ты-азот. 24) мидии и нитриты-азот, 25)углекислота и карбонат-ионы,
26) гетеротрофные бактерии взвеси и карбонат-ионы.
В данном случае мы имели дело с прямой отрицательной связью,
когда элемент А действует на В (или В -* А), ослабляя или уменьшая
его. Вполне возможно также, что в данном случае имеет место и обрат-
ная отрицательная связь, когда А влияет на В, уменьшая его, а В в ответ
действует на А, тоже заставляя его уменьшаться.
Б.З. ОСНОВНЫЕ ИТОГИ МАТЕМАТИЧЕСКОГО ЭКСПЕРИМЕНТА
Следует отметить, что взаимосвязь элементов биосистемы с сообщест-
вом обрастания (Горбенко, 1977) подтвердилась и в настоящем экспери-
менте. Это выражается в том, что в большинстве случаев параметры сис-
темы отвечали на воздействие параметров-возбудителей, которым искус-
ственно было дано приращение 1 %. Только в 21 случае из 600 градиенты
60
оказались равными 0, т.е. на воздействие не реагировали. Что касается
остальных градиентов, то они отличались от 0, чаще всего были слабы-
ми (меньше 0,3), но тем не менее взаимодействие элементов изучаемой
системы удалось проследить достаточно отчетливо.
В системе выявлены основные параметры-возбудители, действую-
щие на остальные элементы системы положительно или отрицательно
(табл, 23). К ним в первую очередь относятся гетеротрофные бакте-
рии планктона, влияющие на 13 параметров системы из 24, — вызывая
возбуждение последних, они были первыми и по сумме градиентов
(табл. 25). Эти бактерии влияли на представителей всех 7 изучаемых
био- и абиотических факторов системы (гетеротрофные бактерии
взвеси, живые диатомовые планктона и перифитона, балянусы и мерт-
вые диатомовые взвеси и перифитона, а также на нитриты-аэот, кисло-
род, углекислоту, гидрокарбонат- и карбонат-ионы, нефть). При этом
влияние их на остальные элементы системы было наиболее сильным,
в 5 случаях превышая 1 % и достигая даже наибольшего значения
градиента, равного 5,4 % (табл. 22).
Следовательно, гетеротрофные бактерии планктона, биомасса
которых в морской воде в 1971 г. колебалась от 0,892-10“* до
0,185-Ю4 мг-мл-1 и была намного меньше биомассы диатомовых водо-
рослей, обитающих там же (табл. 21), являлись главными регулятора-
ми изучаемой в эксперименте системы. Их действие на живые элементы
системы осуществляется не только путем минерализации бактериями
отмерших останков гидробионтов, но также, очевидно, посредством
большого количества органических и неорганических метаболитов.
Среди них обнаружены витамины, а также, по-видимому, сильно дей-
ствующие в малых дозах эктокрины, на которые указывал СЛукас
(1965). Гетеротрофные бактерии могут влиять на параметры абиоти-
ческого фактора в изучаемой системе через углекислоту, в больших
количествах образуемую ими, и изменение pH среды.
Следующий по силе параметр-возбудитель в системе - это
мшанки, биомасса которых, однако, относительно невелика (от 0 до
0,36 мг-100 см"’, табл. 21). Они действовали на 10 параметров из
24 и были вторыми по сумме градиентов. При этом действие мшанок
на остальные параметры также было сильным и в 4 случаях превышало
1 %, достигая 3,15 % (табл. 23). Мшанки влияли на представителей
б (из 7) факторов системы, кроме фактора живых диатомовых, а
именно - на балянусы, гетеротрофные бактерии и мертвые диатомо-
вые взвеси, pH, углекислоту, кислород, нитриты-аэот, гидрокарбонат-
и карбонат-ионы, ХлЭк. Их влияние иа балянусов на ранних стадиях
развития сукцессии отражает отдельные моменты биоценотических
взаимодействий. Проявляются они прежде всего через трофическую
связь. Известно, что увеличение биомассы мшанок на эксперименталь-
ных пластинах свидетельствует о наличии большого количества их
личинок в планктоне, которые на ползающе-плавающей стадии могут
61
служить пищей баляяусам. Это, очевидно, и обусловливает положитель-
ное влияние мшанок на поселение усоногих раков. Высокая плотность
мшанок, наряду с чрезвычайно быстрым темпом роста, ведет к изме-
нению экологического климата сообщества обрастаний, что в конеч-
ном счете влияет не только иа биотические, но и на абиотические пара-
метры среды (Брайко,1983, 1985).
Сильным параметром-возбудителем являются также гетеротроф-
ные бактерии взвеси. Биомасса их за год колебалась от 0,282-10"4 до
0,9-Ю*3 мг-мл"1 (табл. 21). Они влияли на 9 параметров из 24 или
на 6 факторов из 7, кроме фактора макроорганизмов. Это — мертвые
диатомовые взвеси, нитриты-азот, гидрокарбонат- и карбоиат-ионы, pH,
СОа, кислород и РОВ. Гетеротрофы взвеси развивались иа ее поверх-
ности в больших количествах и каким-то образом действовали на гид-
рохимические характеристики, биогены и компоненты карбонатного
равновесия в море.
К сильному параметру-возбудителю можно отнести также нитри-
ты-азот. Они влияли на 9 параметров из 24: балянусы, мидии, гетеро-
трофные бактерии планктона И взвеси, а также на мертвые диатомовые
пла>жтона, фосфор минеральный, углекислоту, гидрокарбонат- и кар-
бонат-ионы, т.е. иа представителей 6 из 7 изучаемых факторов, кроме
фактора загрязнения (фенол и ХлЭк). Как параметры-возбудители на
гидрохимические характеристики и компоненты карбонатного равно-
весия влияют гетеротрофные бактерии планктона, взвеси и перифито-
на. Все они действуют на СОа, НСОа, СОа3", pH и нитриты-азот, по-ви-
димому, в определенной мере регулируя их Величину в системе. Вместе
с тем компоненты карбонатного равновесия, биогены и pH как пара-
метры-возбудители в изучаемой системе влияют друг на Друга, т.е.
взаимодействуют между собой.
Следует отметить также в качестве параметров-возбудителей ба-
лянусы, биомасса которых в 40 и более раз Превышает биомассу дру-
гих биопараметров (от 0 до 567,69 мг-100 см"1 пластины, табл. 21).
Поэтому действуют они на другие организмы посредством своих ме-
таболитов. Все указанные выше элементы системы, играющие роль
сильных параметров-возбудителей, очевидно, в основном являются
ее регуляторами. На углекислоту, pH И на нитриты-азот оказывают
влияние также балянусы, мидин, ботриллюсы и гидроиды (табл. 27).
Обнаружены и самые возбудимые параметры в системе (табл. 24).
К ним в Первую очередь относится углекислота, на которую влияют
20 параметров системы из 24. Данный параметр, очевидно, является про-
изводной от остальных элементов системы, которые при приращении 1 %
наиболее сильно действуют на углекислоту, вызывая увеличение (или
уменьшение) ее на 1-5 %. На углекислоту оказывают влияние мшанки
(градиент 3,15 %), ботриллюсы (1,75 %), гетеротрофные бактерии
планктона (5,4 %), гетеротрофные бактерии перифитона (1,27 %),
мертвые диатомовые планктона (1,45 %), фосфор минеральный
62
(2,28 %), ХлЭк (1,28 %), гидрокарбонат-ионы (1 %), карбонат-ионы
(1%) (табл. 23).
Затем следуют менее возбудимые гидрокарбонат-ионы, на которые
Влияют 13 параметров из 24, в том числе мшанки (градиент 1,19 %),
гетеротрофные бактерии планктона (2,08 %), фосфор минеральный
(1,17 %). Сильно возбуждаются и карбонат-ионы, на которые действу-
ют 10 параметров из 24. Из иих оказывают сильное влияние мшанки
(градиент 1,13 %) и гетеротрофные бактерии планктона (2,00 %)
(табл. 25). Таким образом, от компонентов карбонатного равновесия
(С02, НСОэ, СОз”) зависят равновесие солевого состава в море, вы-
падение в осадок и другие процессы. С этими компонентами связаны
Круговороты веществ в море и ряд живых и неживых элементов
юучаемой системы обрастания. Сами они являются производными
от других элементов системы.
Сильно возбудимыми параметрами являются: гетеротрофные бак-
терии взвеси (на них действуют 11 параметров из 24), нитриты-азот
(11 параметров из 24) и pH воды (11 параметров из 24) (табл. 24,
26). Значит, один из самых важных параметров в море - pH - в опре-
деленной мере величина производная от других элементов системы, а
именно от мшанок (градиент 1,25), гетеротрофных бактерий планкто-
яа (градиент 2,1), перифитона (градиент 0,73) и компонентов карбо-
натного равновесия (градиент 0,68-0,66). Что же касается параметра
нитриты-аэот — важного биогенного элемента, то его величина также
регулируется рядом параметров системы, из которых главные - гете-
ротрофные бактерии планктона (градиент 1,02), фосфор минеральный
(градиент 0,74) и компоненты карбонатного равновесия (градиент
0,47—0,54). Таким образом, в результате математического экспери-
мента выявлены параметры морской системы, которые, имея важное
значение в системе, в значительной степени являются производными
других параметров.
Нам удалось обнаружить также относительно независимые пара-
метры, которые почти или совсем не возбуждаются другими парамет-
рами системы. К ним в первую очередь относятся мшанки. Также
почти не возбудимы гетеротрофные бактерии планктона, на которые
влияет всего I параметр из 24 (табл. 26). Вместе с тем оба указанных
элемента системы являются самыми сильными параметрами-возбуди-
телями, т.е., не испытывая на себе влияние остальных параметров, они
тем не менее действуют на большинство из них. Следовательно, биомас-
сы гетеротрофных бактерий и мшанок, очевидно, наиболее важные
элементы в системе, от которых существенно зависят остальные пара-
метры системы. Большую роль в системе играют также гетеротрофные
бактерии перифитона и взвеси, нитриты-азот и компоненты карбонат-
ного равновесия. К слабо возбудимым параметрам относятся ботрил-
люсы, гидроиды, мидии, живые диатомовые взвеси и содержание
ХлЭк в воде.
63
Одним из важнейших результатов математического эксперимента
является возможность выявления прямых положительных и отрица-
тельных связей в системе. Обнаружение взаимовлияния ряда элементов
дает основание предполагать, что в изучаемой системе действуют также
обратные положительные и реже обратные отрицательные связи.
На основании экспериментальных данных можно представить ха-
рактер связей (или структуру) изучаемой системы, принимая во вни-
мание только градиент > 0,3 (табл. 22). В.В.Меншуткин и Ю.И.Соро-
кин (1975) называли подобные градиенты коэффициентами связи.
Поскольку каждый из них, информируя нас о взаимодействии двух
элементов системы, одновременно отражает величину и знак соответ-
ствующих связей, то эти градиенты в определенной мере являются
также связями. Из 300 возможных связей с градиентом > 0,3 имеет-
ся всего 80 прямых (42 положительных и 38 отрицательных) и 27,
по-видимому, обратных (21 положительная и 6 отрицательных)1
(табл. 24).
В связи со сказанным мы считаем, что результаты математическо-
го эксперимента достоверно можно отнести к природным условиям
моря. Это значит, что при данных условиях, т.е. увеличении (или умень-
шении) изучаемых параметров примерно на 1 % эта система в общих
чертах ’’работает” и в море. Основные данные проведенного экспери-
мента могут быть использованы при создании управляемых морских
систем (Горбенко, Подвинцев, 1979). Для определения взаимосвязей
и действия одних параметров на другие нами применялись и иные мето-
ды математической статистики.
6.4. СТРУКТУРА ВЗАИМОСВЯЗЕЙ ПАРАМЕТРОВ ЭКОСИСТЕМЫ.
ИЗУЧЕННАЯ С ПОМОЩЬЮ АНАЛИЗА КОРРЕЛЯЦИОННЫХ ФУНКЦИЙ
Однолетние наблюдения. Определение структуры взаимосвязей боль-
шой морской экосистемы, состоящей кроме СПМ, СМВ и СМП также
из гидрохимических характеристик и параметров Солнца, требует
расчета взаимных корреляционных функций не менее чем за период
однолетних наблюдений (Горбенко, Крышев, 1985).
После расчета 1500 функций за все сезоны одного года наблюдений
в море (число наблюдений за год 72, количество био- и абиотических
параметров 35) с помощью кластерного анализа определили, что пара-
метры изучаемой экосистемы объединяются в отдельные группировки,
Выделенные по тесноте корреляционных связей внутри их по отноше-
нию к межгрупповым связям, и выстраиваются следующим образом
(табл. 31): 1-я группировка в 1-м ряду ограничена параметрами 1-7;
1 Каждая обратная связь состоит из двух прямых связей - положительной и
отрицательной.
64
2-я - во 2-м ряду параметрами 8-14; 3-я - в 3-м ряду параметрами
15-21; 4-я — в 4-м ряду параметрами 22-28; 5-я - в 5-м ряду пара-
метрами 29-35.
Каждая выделенная группа разделена диагональю. Ниже ее распо
лагаются величины корреляционных функций (корреляций) с разным
знаком, выше - соответственно величины: 1 (опережения (со знаком
(+), который обычно не ставится); 2)запаздывания (со знаком (-))
или одновременности в наступлении корреляций (с цифрой 0). Опере-
жение или запаздывание считается по отношению к первой величине
в корреляции (левая часть табл. 31), ко второй - номера этих пара-
метров (вверху табл. 31). Числа в таблице вне группировок обозна-
чают величину корреляции и временных задержек в наступлении меж-
групповых корреляций.
1 группа - компоненты карбонатного равновесия, фенол и pH мета-
болитов. Характеризуется самым большим количеством и тесной вза-
имосвязью корреляционных функций внутри группы, которых было
21 (максимально возможное число), а между группами - соответст-
венно 65. Запишем их в виде дроби: связь внутри группы - в числи-
теле, вне ее - в знаменателе, получим 21/65 (так будем записывать
аналогичные связи и для других групп). Все связи этой группы насту-
пают одновременно. Теснота связей по модулю г = I 10,32 + 0,951.
П группа - параметры СИМ. В группе уже появились пропуски
связей между группами, которых также стало меньше и уменьшилась
теснота связей. Число связей внутри и вне группы 17/32, недостовер-
ных - 4. Все связи наступают почти одновременно, только 2 из них с
опережением 10 и 20 сут. Теснота связей г = 10,31 4 0,751.
III группа - продуценты, консументы и параметры, обусловливаю-
щие развитие продуцентов. Теснота связей в группе меньше, чем в пре-
дыдущей, но вне группы их намного больше. Число связей 18/75, недо-
стоверных - 3. Из них одновременно наступают 2; с опережением 10-
70 сут - 11, с запаздыванием - 5-46 сут. Теснота связей г = 10,31 4
4 0,641.
IV группа - микроорганизмы самоочищения моря и их метаболи-
ты. Группа характеризуется еще меньшей теснотой связей и меныним
их количеством между группами, Число связей 16/61, из них недосто-
верны 5. Одновременно наступают 6, с опережением 10-35 сут - 5, с
запаздыванием 5-20 сут — тоже 5. Теснота связей г =10,31 4 0,541.
V группа - биогены и абио дара метры. В группе еще меньше теснота
связей н их количество как вне, так и внутри группы. Число связей
J1/55, из них одновременных 2, наступают с опережением 10-40 сут -
5, с запаздыванием 20-70 сут - 4. Теснота связей г = 10,31 4 0,451.
Фактически активность солнца - независимая величина, которая
действует в основном на биопараметры почти всех групп, определяя
процессы их жизнедеятельности. Что касается прозрачности, то она,
безусловно, функция многих био- и абиопараметров в море. Поэтому
65
Таблица 31. Группировки параметров изучаемой экосистемы (объяснения в текс
ьг п.п. 1 Элементы системы Груп па Среднее Размерность 1
1 pH, 1 8,33
2 со23” 17,53 мгл'1 0,95
3 НСОз 166,3 мгл*’ -0,84
4 со2 0,39 мл-л~* -0,91
5 Фенолбподобиые (фенол) вещества мг-л”1 -0,42
6 pH СПМ 8,22 0,44
7 pH СМВ 8,05 0,39
8 Диатомовые живые СПМ I! 113000 кл-см”2 —
9 Диатомовые мертвые СПМ 4960 кл-см”2 —
10 Гетеротрофные бактерии СПМ 2230 колоний-см-2 —
И Палочковидные бактерии СПМ 553000 кл-см 2 —
12 Кокковидные бактерии СПМ 486000 кл-см” —
13 Сухая масса СПМ 1,0 МГ'СМ~* —
14 Карбонаты СПМ 0,04 мг см 2 —
15 Диатомовые живые СМП Ш 690 кл-мл”* 0,40
16 Диатомовые живые СМВ 230000 кл-мл" 0,32
17 Зоопланктон — — —
18 Продолжительность сияния Солнца 6,5 ч-сут”1 -0,36
19 Температура воды 14,5 °C —
20 Диатомовые мертвые СМП 80 кл-мл 1 —
21 Диатомовые мертвые СМВ 110000 кл-мл 1 -0,36
22 РОВ морской воды IV 0,360 EJ® —
23 Гетеротрофные бактерии СМП 5070 колониймл -0,35
24 Гетеротрофные бактерии СМВ 1490000 колоний, мл 1 —
25 ХлЭк 1,27 мг-л 1 —
26 РОВ СПМ 0,492 Ejd —
27 РОВ СМВ 0,757 Его —
28 Карбонаты СМВ 17,9 % —
29 Нитриты-азот V 3,9 мкг-л”1 —
30 Фосфор минеральный 10,8 мкг-л * —
31 Фосфор органический 48,7 мкг-л * —
32 Кислород в воде 6,36 мл-л 1 —
33 Соленость воды 17,91 %О —
34 Прозрачность воды — М —
35 Солнечная активность 9,8 ЧИСЛО —
эта характеристика, как и солнечная активность связана со всеми
выявленными группами, кроме перифитона (II группа) в данном ис-
следовании (табл. 31). Таким образом, уже выделение только одно*
петних взаимных корреляционных функций в отдельные группиров-
ки в море (Севастопольская бухта) отражает их большое значение В
экологии изучаемой экосистемы и иерархичный характер по ряду
признаков.
Результаты данной работы также доказывают, что параметрь
группы I регулируют биохимические, биологические, гидрохимические
геологические и другие процессы в море вследствие изменения карб<
натного равновесия, а также влияния на биопроцессы фенолоподобны
6fi
те)
2 3 4 5 6 7 8 J 10 и 12
0 0 0 0 0 0 — — — —
0 0 0 0 0 — 5 0 — __
-0,84 0 0 0 0 — 5 - — —
-0,87 0,79 0 0 0 — — — —
-0,44 0,50 0,43 0 0 — 5 — 0 —
0,40 -0,49 -0,32 -0,40 0 - -10 0 -
0,36 -0,37 -0,33 -0,32 0,38 0 — — —
— — — — — -0,32 0 10 0 0
0,36 -0,48 — -0,45 — — 0,75 10 0 0
— — — — 0,35 — 0,51 0,39 20 20
-0,31 — 0,45 -0,43 0,50 0,39 0,42 0
— — — — — — 0,63 0,34 0,37 0,62
— — — — — — 0,43 0,38 0,40 0,52
— — — — — - —. 0,31 0,39
0,37 — -0,36 -0,32 -0,36 — — — -0,34 — —
0,37 0,36 — 0,51 0,53 — -0,41 - -0,31 -0,42 -0,37
— — *** 0,42 — -0,40 — — — - 0,32
-0,36 — -0,32 — 0,32 -0,37 - - — - -
— — — -0,32 — — — — — .—
— -0,34 — 0,33 -0,40 — — — —
-0,35 0,40 *— 0,47 — -0,36 — 0,34 — -
— -0,31 0,32 — — — -0,43 0,33 — — - 0,41
— — 0,44 -0,33 — 0,32 — — — 0,32 -
— 0,33 0,42 -0,33 — 0,38 — — — — —
— — 0,36 — - -0,33 -0,37 — - — -0,35
— —. — — — — — — — — 0,36
0,31 -0,35 — -0,37 — -0,32 — — — 0,32 -
— — — — - 0,37 -0,49 — — — — —
— -0,31 -0,37 — — — — — 0,33 — —
— — - — - 0,32 — — - -0,37 -
— — — — — — —
-0,32 — -0,35 — 0,38 — — — -0,32 .
-0,33 — 0,35 — — — — — — — —
— 0,34 — -0,40 0,32 — — — — — -
- — — 0,32 — -0,35 — — — — —
веществ и pH метаболитов СМВ и СПМ, играющих, очевидно, роль экто-
кринов (Лукас, 1965).
Наиболее малозависимая от других 11 группа, так как она сла-
бее всех связана с остальными группами. Очевидно, ее экологическое
значение в море в том, что она является важной составной частью био-
ценоза (перифитона) на всех погруженных в море предметах и опреде-
ляет осаждение на них карбонатов, металлов и других веществ. Эта
группа, безусловно, участвует в образовании органического вещества,
а также в процессах самоочищения воды.
Группа 111 обеспечивает первичные процессы продуктивности в
море, имея больше всего связей с остальными группами.
67
Продолжение табл. 31
№ п.п. Элементы системы ! Груп- па Среднее Размерность 13
1 2 РИ2_ С'Оз I 8,33 17,53 1 МГ-Л -
3 НСОз 166,3 мг-л* —
4 со2 0,39 млл~; —
5 Фенолоподобные (фенол) вещества мг-п —
6 pH СПМ 8,22 —
7 pH СМВ 8,05 —
8 Диатомовые живые СПМ П 113000 кл.см^ 0
9 Диатомовые мертвые СПМ 4960 кием"2 0
10 Г етеротрофные бактерии СПМ 2230 колоннй<м” —
11 Палочковидные бактерии СПМ 553000 кл.см 2 0
12 Кокковидные бактерии СПМ 486000 клсм 0
13 Сухая масса СПМ 1,0 мт см
14 Карбонаты СПМ 0,04 МГ'СМ 0,91
15 Диатомовые живые СМП III 690 kh.mjTJ —
16 Диатомовые живые СМВ 230000 кл. мл —.
17 Зоопланктон — — —
18 Продолжительность сияния Солнца 6,5 ч-сут-1 -
19 Температура воды 14,5 °С_ -
20 Диатомовые мертвые СМП 80 кл-мл^1 —
21 Диатомовые мертвые СМВ 110000 кл-мл 1 —
22 РОВ морской воды IV 0,360 Е260 .
23 Гетеротрофные бактерии СМП 5070 КОЛОНИЙ-МП -
24 Гетеротрофные бактерии СМВ 1490000 колоний-ми-1 —
25 ХлЭк 1,27 мг-л-1 —
26 РОВСПМ 0,492 Езто 0,34
27 РОВ СМВ 0,757 Е270 -
28 Карбонаты СМВ 17,9 % 0,31
29 Нитриты-азот V 3,9 мкг-л"* —
30 Фосфор минеральный 10,8 Мкг. л-* —
31 Фосфор органический 48,7 Мкг л-1 — 4
32 Кислород в воде 6,36 мл-л~’ —
33 Соленость воды 17,91 %0 —
34 Прозрачность воды М 1
35 Солнечная активность 9,8 Число — *
Параметры IV группы выполняют важнейшие функции в процесса
самоочищения в море, по крайней мере от ХлЭк, который разлагав
гетеротрофные бактерии планктона и взвеси, а продукты разложен!1
пополняют содержание РОВ в море. Элементы группы имеют множес
во связей с другими группами.
Группа V включает в себя важнейшие биогены - азот и фосфо]
необходимые для развития многих микроводороспеЙ и бактери:
В группе находится основной для жизни большинства организмов М
таболит водорослей - кислород и другие абиопараметры. Поэтому пр
однолетнем анализе параметры группы имели много межгрупповы
связей.
64
14 15 16 17 18 19 20 21 22 23 24
— 0 0 — -35 — — 65 — -15
— 0 15 — -35 — — 25 — -.
— — -10 — — — -15 -5 5 — 15
— 0 — — -30 — — — 20 -15 20
— 45 -10 45 — 55 — 0.47 40 15
— -10 -10 — 0 — -5 — —
- — — 30 -10 — 35 -0,36 — 45 0
— — -15 — — — — — 20 — 10
— — —• — — — 0 0 —
— — 15 — — — — —
0 -25 -15 — — — — — -20 —
0 — -15 -10 — — — — 20 10
0 — — — — — — •— — —
— 20 — — — — — — — —.
0,46 0 55 10 65 60 — 10 __ —
0,50 0,64 55 10 65 -20 0 40 -5 —
— 0,40 0,58 -45 60 5 70 — -35
— -0,50 -0,47 -0,43 60 -5 — —
— -0,40 -0,44 0,34 -0,31 -5 — —
—. 0,31 0,41 0,45 0,31 -0,32 -40 — 15
— — 0,34 0,32 — — 0,31 — — —
— 0,33 -0,38 — — — — — 5 0
— — 0,33 0,32 — -0,39 — 0,54 35
— — — — — — — — 0,44 0,31
— -0,33 — — — — -0,31 — 0,41 0,52 0,45
— — -0,32 0,31 — — — — -0,31 -0,39 -0,36
-0,34 — -0,39 0,42 — — 0,34 — — 0,36 —
0,31 0,34 0,32 — — — 0,37 — — -0,37
- — — -0,32 -0,33 -0,31 -0,31 -0,33 —
— — • — — -0,31 — 0,42 — — 0,32
— 0,38 0,43 0,35 — — — — — —
— — -0,43 -0,40 — — 0,36 -0,11 — — —
— -0,32 0,36 *•— — — — — 0,39
— 0,34 0,40 0,40 — — 0,34 0,37 -0,32 0,43 0,35
- -0,34 0,34 — — 0,35 0,31 — — — 0,33
Что касается активности Солнца, то больше всего этот параметр
коррелирует (4 из 10) с параметрами Ш группы (продуценты и консу-
менты) - живыми диатомовыми водорослями планктона и взвеси.
Это значит, что от изменения солнечной активности в определенной
мере зависит численность этих микроводороспей, а следовательно, и
их продуктивность. С остальными (I, IV и V) группами активность
имеет по 2 корреляции. Теснота всех этих связей г = 10,32 Т 0,441.
Задержка во времени 5-55 сут.
Важно отметить, что диатомовые живые и мертвые СМП, живые
Диатомовые СМВ, гетеротрофные бактерии и карбонаты СМВ, а также
PH СМВ в корреляции опережают величину повышения активности
69
Окончание табл. 31
N*n.n. Элементы системы Груп- па Среднее Размерность 25
1 Рн, I 8,33 —
2 СО2" 17,53 мг.л * —
3 HCOj 166,3 мгл 1 —
4 COj 0,39 мл-л 1 10
5 Фенолоподобные (фенол) вещества мгл 1 —
6 pH СПМ 8,22 —
7 pH СМВ 8,05 15
8 Диатомовые живые СПМ П 113000 КЛ.СМ 2 —
9 Диатомовые мертвые СПМ 4960 кл.см 2 —
10 Гетеротрофные бактерии СПМ 2230 КОЛОНИЙ-СМ —
11 Палочковидные бактерии СПМ 553000 кл.см 2 —
12 Кокковидные бактерии СПМ 486000 кл-см 2 —
13 Сухая масса СПМ 1.0 мг.см 2 —•
14 Карбонаты СПМ 0,04 мг.см 2 —
15 Диатомовые живые СМП III 690 кл.мл~1 55
16 Диатомовые живые СМВ 230000 кл-мл 1
17 Зоопланктон — — —
18 Продолжительность сияния Солнца 6,5 чхут * —
19 Температура воды 14,5 °C —
20 Диатомовые мертвые СМП 80 кл.мл *
21 Диатомовые мертвые СМВ 110000 кл.мл * —
22 РОВ морской воды IV 0,360 Езео -10
23 Гетеротрофные бактерии СМП 5070 КОЛОНИЙ.МЛ
24 Гетеротрофные бактерии СМВ 1490000 колоний.мл 1
25 ХлЭк 1,27 мг.л 1
26 РОВ СПМ 0,492 Ело -0,1
27 РОВ СМВ 0,757 Егто — 1
28 Карбонаты СМВ 17,9 % -0,1
29 Нитриты-азот V 3,9 мкг-л~* —
30 Фосфор минеральный 10,8 мкг-л ' — 1
31 Фосфор органический 48,7 МКГ’Л 1 0,1
32 Кислород в воде 6,36 _ ———* 1 мл «л —• ч
33 Соленость воды 17,91 %о — Ч
34 Прозрачность воды — М 0,1
35 Солнечная активность 9,8 число “•1
Солнца на 5-40 сут. Поэтому данные параметры могут служить инд
торами наступления времени вспышек солнечной активности и, сл<
вателъно, начала времени действия ее на земные организмы (Горбе*
Крышев, 1985).
Многолетние наблюдения. Поскольку логично было рассмотр
как изменяются корреляционные взаимосвязи не только в течс
1 года, но и в другие годы, были рассчитаны взаимные коррелят
ные функции за 1969- 1973 гг. Из числа параметров исключили л
зоопланктон, так как наблюдения за ним в указанные годы (кр
1971 г.) отсутствовали, но ввели параметр - содержание креМ1
Всего было рассчитано около 2500 взаимных корреляционных ф]
70
26 27 28 29 30 31 32 33 34
— — — — —
— 10 — — — — — -55 . —
— 10 — 5 — — 50 60
— — — 5 — — — -55
— -25 — — — — 50 — -20
.— —• 15 — — — — — 20
- 0 0 - -5 — 50 —
— — — — — — — — —
— — — — — — —• — —
— — — -5 — — —► —
— 0 *— — 10 — 20 — —
0 — — — — —
0 — 0 — — —
— -10 0 — — — — — —
- — 35 — — -50 — -55 25
-15 -30 -10 — — -60 60 -70 70
-5 45 — — — -5 5 15 —
— — 5 5 *—• —- —
— — 50 — —
— -35 -35 -25 30 — 0 35 —
— — — -10 — — 55 -25 -25
0 —. — — — — — — 55
5 -5 — -5 — — — — -20
0 —- 0 — -5 — — 0 -35
10 — -20 — — -55 — — -45
-10 0 -10 — — 20 — —
-0,34 0 — 10 — — — 60
0,31 0,35 — — — — 15
0,32 — — 0 — 20 —
— -0,31 — 0,45 25 -65 -70 -50
— 0,34 -0,31 — 30
-0,44 — — — 0,31 — 0 10
— — — — -0,39 0,33 —
— 0,38 0,35 —. 0,31 С,31 3,34
— — 0,44 — — -0,40 — 0,35 —
Ний. С помощью кластерного анализа мы убедились, что во все годы
наблюдений также выделяются I-V группы с теми же параметрами,
Л и в течение 1 года наблюдений (Горбенко, Крышев, 1985). Вмес-
те с гем, как и следовало ожидать, количество и соотношение внутрен-
них связей и их численность по отношению к межгрупповым связям
оказались иными. В среднем за год отношение числа внутренних свя-
зей к внешним составляло: в I группе - 19/12, во II - 15/7, в III -
10,9, в IV- 13/1, в V - 10/0.
Затем был проведен также многомерный спектральный анализ
элементов изучаемой системы, подобный вычислению частных кор-
реляций только за 1 год, для чего с помощью ЭВМ пришлось рас-
71
Таблица 3Z Отношение внутригруппо-
вых связей параметров экосистемы (чис-
литель) к межгрупповым связям (зна-
менатель) в различные сроки наблюде-
ний при однолетнем (А), многолетнем
(Б) и многомерном анализе (В) взаим-
ных корреляционных функций
Вид анализа Группа
1 II III IV V
А 21 17 18 16 Ц 65 ЗТ 75 61 55 Б 19 15 10 11 10 12 7 9 1 0 В 21 17 12 11 2 26 8 10 5 0 ь jo 16 и Д -1 34 15 31 22 18
считать 595 взаимных корреля-
ционных функций. С помощью
этого метода удалось определить
влияние на каждую пару корре-
лирующих параметров экосисте-
мы всех остальных ее парамет-
ров. Связи коррелирующей пары
элементов, на которые не влия-
ют остальные 33 параметра изу-
чаемой системы, были названы
нами прямыми связями.
Спектральный многомерный
анализ также подтвердил уста-
новленный выше факт, что струк-
тура параметров изучаемой эко-
системы характеризуется 5 взаи-
мосвязанными группировками с
такими же параметрами, как в однолетних и многолетних структурах
группировок (Горбенко, Крышев, 1985).
Соотношение в группировках внутренних и внешних связей, полу-
ченное в результате многомерного анализа, следующее: I группа -
21/26, II- 17/8, III- 12/10, IV- 11/5,V- 2/0, Каждый из 3 математи-
ческих методов, примененный для выявления структуры экосистемы,
на основе ее общей стабильности выявлял некоторые особенности,
присущие системе с различным характером временных рядов. Поэтому
были разными количество связей и их отношение друг к другу.
Если свести вместе соотношение внутренних и внешних связей
I-V групп, то можно заметить некоторые тенденции в их изменении
(табл. 32). Наиболее отчетливо проявляется общая тенденция - умень-
шение величины внутригрупповых связей при увеличении номера груп-
пы, т.е. чем меньше величина взаимных корреляционных функций в
группе, тем меньше число внутренних корреляционных связей. Ска-
занное подтверждается и при усреднении связей (табл. 32). Что касает-
ся межгрупповых связей, то никаких четких изменений их в связи с
уменьшением номера группы не наблюдается. Р целом же на основании
данных всех видов анализа можно отметить, что по количеству связей
между группами (от большего числа к меньшему) они располагаются
следующим образом: J, III, IV, V и II.
Поскольку II группа имеет наименьшее число таких связей, то
подтверждается сказанное выше, что она самая независимая, т.е. разви-
тие ее определяется в основном внутренними биоценотическнми связя-
ми. Что касается группы V, то она, по сути, объединена формально
и состоит из а биопара метров, у которых связей между группами мог-
ло быть и больше, что и подтвердилось при расчете однолетних кор-
реляций.
72
Так, по результатам однолетних и многолетних наблюдений уста-
новлено, что 35 изучаемых параметров экосистемы Севастопольской
бухты могут быть объединены в 5 группировок-факторов, каждая из
Которых состоит из определенного числа элементов-параметров, тесно
связанных внутри этих групп, находящихся, в свою очередь, во взаи-
мосвязи.
Одним из интересных и важных результатов проведенной работы
является тот факт, что элементы изучаемой экосистемы оказались
Объединены математическими метода ми в группировки не просто ме-
ханически, а на основании выполняемой ими функции (или специаль-
ности), другими словами, с учетом их экологической ниши в море.
При этом группы I, III и IV в основном, по-видимому, ответственны за
такие важные процессы, как регуляция (I группа), первичная продук-
тивность (III группа) и самоочищение (IV группа).
Таким образом, сделав расчет взаимных корреляционных функций
множества основных параметров какой-нибудь экосистемы, а затем
разбив их с помощью кластерного анализа на отдельные группы-факто-
ры, связанные между собой, мы имеем возможность узнать, какую
роль играют указанные группы в море. Это значит, что данный метод
Может быть рекомендован для выделения экологических ниш биоти-
ческих параметров и роли в морской экосистеме абиотических.
Следует отметить, что в факторном анализе, описанном в разделе
4.2., использован совершенно иной математический метод, а количество
Параметров равнялось 17. Тем не менее с помощью указанного анализа
Удалось выделить фактор /д, в котором были все элементы группиров-
ки II и РОВ СПМ. Был обособлен также фактор F2, в котором с по-
мощью факторного анализа были объединены 4 параметра, общих с
теми же параметрами, что и в группе I. Из 4 параметров фактора
F3 2 были общими с таковыми в группе V.
При определении эко ниши групп 1-V мы обратили внимание на то,
Что во взаимосвязи группировок существует соподчинение. Оно не
Проявилось в однолетних взаимных корреляционных функциях, где
Все группы просто связаны между собой (рис. 2, А), но четко вырази-
лось В многолетних и многомерных корреляционных функциях (рис. 2,
Б, В), почти не различаясь. Поэтому рассмотрим их вместе.
Соподчинение заключалось в следующем-1 группа, находящаяся на
самом высоком уровне изучаемой экосистемы, посредством парамет-
ров карбонатного равновесия либо величины pH, непосредственно
вязанной с ним, либо другими параметрами, влияет на группы III и
IV (продуценты, консументы, микроорганизмы самоочищения моря
от ХлЭк и их параметры). I группа взаимосвязана с группой П (пара-
метры СПМ). Группа III (продуценты и консументы) не влияет на
группу II и взаимосвязана с группой IV (параметры самоочищения).
II и IV группы взаимосвязаны. Группа V (биогены, соленость и кисло-
род) влияет на IV группу (очевидно,посредством соединений азота и
73
Рис. 2. Соподчинение группировок - параметров морской экосистемы
микроорганизмов: А — при однолетнем, Б — многолетнем, Л — многомер-
ном анализах (объяснение в тексте)
фосфора, обусловливающих жизнедеятельность бактерий - гетеротро-
фов СМП и СМВ, от которых зависит использование нефтепродуктов) и
взаимосвязана с группой Ш (рис. 2).
Что касается связи СПМ с группой IV (самоочищения), то осу-
ществляется она через РОВ, компоненты которого могут использо-
ваться бактериями СПМ. Существующая также связь гетеротрофных
бактерий II и IV групп, очевидно, метаболическая. С группой I компо-
нентами карбонатного равновесия и pH метаболитов СПМ связана
П группа — СПМ, т.е. в изучаемой экосистеме СПМ играет какую-то
еще промежуточную роль кроме собственного развития, выявить ко-
торую, используя однолетний и многолетний анализы корреляционных
функций, пока не удается. В итоге, в настоящей экосистеме регулятором
является группа I. Группа III определяет процесс первичной продукции,
IV группа — самая зависимая от других групп — обусловливает процесс
самоочищения моря от ХлЭк, группа V обеспечивает биогенами группы
III и IV. Роль группы II пока не совсем определена.
5.5. ПРОГНОЗИРОВАНИЕ БИО- И АБИОТИЧЕСКИХ ПАРАМЕТРОВ
В настоящее время становится все более актуальной проблема изучения
и прогноза морских экосистем на основе матриц экспериментальных
панных, накопленных в результате регулярных наблюдений за состоя-
и
нием Солнца, атмосферы и морской среды с учетом антропогенных
загрязнений (Подвинцев, Горбенко, 1983).
Для того, чтобы стало возможным применение математического
аппарата, связанного с привлечением информационных мер при поста-
новке и решении задач, необходимо следующее: 1)формирование под-
множества параметров, описывающих изучаемую систему; 2)решение
и оценка множественных временных рядов. В качестве материала ис-
следований были использованы наблюдения за 1971 г. с 42 параметра-
ми моря, атмосферы, земли и Солнца (табл. 18). В настоящем разделе
биопараметры Xt — Хц названы г- Уп.
В ряде литературных источников приходится встречаться с тем, что
некоторые авторы берут один или несколько абиотических параметров
морской (или пресноводной) среды, которыми обычно бывают биоге-
ны (азотистые и фосфорные соединения), и для обнаружения каких-
нибудь зависимостей сопоставляют их чаще всего с биотическими па-
раметрами той же среды. Поэтому, имея многолетние временные ряды
с многочисленными био- и абиотическими параметрами, мы задались
целью проверить, насколько правомочны такие сопоставления. Для
оценки квадратов множественных корреляций, как и раньше, выбрали
Ркр s 0,95. При этом последовательно для каждого параметра Yx - Уп
создавались различные варианты вектора и биопараметров, чтобы
выяснить их взаимосвязь с биопараметрами (табл. 33). Если в строке
вычисленные квадраты множественных корреляций превышали кри-
тический уровень (Ркр = 0,95), то они признавались значимыми (в
табл. 33 эти значения подчеркнуты).
Анализ табл. 33 позволяет сделать следующие выводы:
1)еслй в системе использовать все исходные 42 параметра, в том
числе и малоинформативные, то молодо будет описать лишь 4 из il
биопараметров (У3 — гетеротрофные бактерии взвеси, У$ — палочко-
видные бактерии перифитона, У7 и У8 - живые диатомовые планктона
И перифитона);
2) сформированная из 31 параметра система, в которой исключены
малоинформативные параметры, имеет наибольшую информацию о
биопроцессах, и каждыми 30 из них можно описать каждый 31-й;
3)отказ от учета фенола и ХлЭк приводит к тому, что параметры
У» — гетеротрофные микроорганизмы планктона, У7 и У4 - живые
диатомовые планктона и взвеси - становятся незначительными;
4) учет только 1—2 любых абиопараметров не позволит провести
Никаких Исследований, поскольку эти параметры, как правило, не ока-
жутся связанными с другими параметрами;
5)изучение указанных биопараметров только по фенолу и ХлЭк
или ГС воды, нитритам и азоту Недопустимо, так как оно не дает
достаточной для этого информации;
6)выявлена возможность описания изучаемых 11 биопараметров
в исследуемой системе по pH, РОВ, фосфору минеральному, нигритам-
75
Таблица 33. Множественные корреляции био- н абнопараметров окружающей
среды
Система абио- параметров у> y2 Уз Ул Уз У6 у-1 Уз Уз УЮ Ун
Полная систе- ма (42 пара- метра) 0,86 0,81 0,90 0,85 0,89 0,82 0,93 0,91 0,87 0,83 0,85
Сформиро- ванная систе- ма (31 пара- метр) 0,84 0,79 0,88 0,83 0,88 0,80 0,92 0,89 0,84 0,81 0,83
pH морской воды 0,06 0,14 0,09 0,52 0,39 0,19 0,57 0,01 0,36 0,28 0,11
pH, РОВ, фос- фор минераль- ный, нитриты, азот, Т °C морской воды 0,52 0,72 0,65 0,61 0,60 0,47 0,69 0,25 0,54 0,41 0,62
Без учета гид- рохимических характерис- тик (14 пара- метров) 0,71 0,54 0,74 0,61 0,37 0,35 0,56 0,42 0,73 0,54 0,52
Без фенола и ХлЭк (29 па- раметров) 0,73 0,78 0,82 0,72 0,86 0,79 0,74 0,88 0,83 0,80 0,81
Фенол н ХлЭк (2 параметра) 0,16 0,15 0,21 0,09 0,13 0,10 0,17 0,26 0,08 0,25 0,35
ХлЭк (1 па- раметр) 0,14 0 0,15 0,06 0,12 0 0,10 0,02 0,07 0,02 0,03
Температура воды (1 пара- метр) 0,48 0,27 0,37 0,11 0,30 0 0,10 0,12 0,37 0,16 0,24
Нитриты-азот (1 параметр) 0,23 0,16 0,21 0,08 0,08 0 0,27 0,02 0,01 0,07 0,02
азоту и Т 0 С морской воды. Установлено, что этими абиотическими па-
раметрами вместе можно описать только численность гетеротрофных
бактерий перифитона (У2).
Кратко остановимся на постановке задачи прогноза биопараметров
на 1990-2000 гг. Рассматривается сформированная система из 42 пара-
метров. Кроме рассмотренных выше биопараметров Kj - Гц в нее
вошли следующие абиопараметры: и Х41 (табл. 18). Отметим, что
Yit ¥3, У4, У7, У9, yi0 в табл. 33 и 34 измерялись в десятичных лога-
рифмах численности бактерий и микроводорослей, находившихся в
1 мл морской воды или взвеси. Параметры У2, К5, У6, У8, Уп приве-
дены также в логарифмах численности, но на 1 см поверхности погру-
женного предмета.
Даны матрицы экспериментальных наблюдений в 1971 г. за био-
(l'l97t) и абиотическими (Лущ) параметрами окружающей среды
76
Таблица 34. Результаты прогноза биопараметров на 1990-2000 гг.
Год Число, месяц Г1 У’ УЭ Уд у5 у6 у7 У8 У9 Ую У и
1990 15.01 4,22 4,73 6,92 4,85 6,03 5,0 1,74 3,52 4,12 1,89 2,86
16.04 3,52 4,6 6,47 4,9 5,72 5,42 2,23 4,59 4,55 1,85 3,28
16.07 5,74 3,19 6,26 6,57 4,53 3,8 1,99 3,65 6,62 2,5 3,41
15.09 4,18 3,28 5,34 6,61 6,18 6,25 138 4,04 6,42 0,95 3,47
2000 15.01 4,18 4,48 6,57 4,68 6,63 5,73 2,21 4,56 4,15 1,57 3,53
15.04 3,38 4,6 6,61 4,38 5,86 5,98 2,31 4,92 4,07 1,84 3,11
16.07 3,54 3,18 6,22 6,94 4,39 3,48 2,75 5,07 4,17 2,76 4,26
15.09 4,41 3,73 6,06 5,51 6,52 6,55 0,7 2,48 4,97 0,82 3,16
Проверка прогноза
1969 15-01 4,09 4,08 6,87 4,25 6,33 5,19 1,91 4,05 4,67 1,95 3,37
16.04 4,07 5,46 6,95 5,18 5,74 4,7 2,1 4,46 5,25 2,27 3,42
11.07 2,57 4,13 5,69 5,84 4,74 5,72 2,66 5,62 4,9 2,56 3,66
12.09 3,95 5,19 5,63 2,71 6,15 6,94 0,88 4,96 3,73 1,87 3,23
1973 19.01 4,29 3,68 6,71 4,99 6,12 5,13 2,81 4,41 5,2 1,95 3,51
19.04 2,86 4,14 6,28 3,48 5,31 6,54 3,31 4,95 3,53 2,18 2,4
19.07 4,37 3,11 6,21 6,17 5,35 4,46 3,17 4,07 6,32 2,28 3,56
11.10 3,52 3,3 5,67 4,87 5,39 5,76 2,69 4,47 5,18 1,75 3,07
1978 20.01 4,4 4,38 6,08 4,88 5,32 5,35 2,53 4,91 3,36 1,51 2,75
20.04 3,45 5,05 6,85 4,12 5,9 6,02 2,89 4,59 4,1 2,45 2,51
20.07 3,64 3,98 6,3 4,52 5,75 5,07 2,67 4,23 5,5 2,55 3,23
11.09 3,87 4,22 5,23 6,27 5,85 4,99 2,19 3,97 6,71 1,77 3,86
Вероят- 0,7 0,78 0,73 0,97 0,7 0,85 0,74 0,8 0,72 0,73 0,9
гость (Р)
размерами 60 х 11 и 6 х 31 соответственно. Даны матрицы контроль-
ных проверок по наблюдениям в каждом году К/, Xt (f = 1969, 1973,
1978), даты которых приведены в табл. 34. Даны матрицы экспертных
оценок основных абиопроцессов в 1990 и 2000 rr. - Xt990 и Хыоо-
Требуется на основе вышеприведенных матриц составить наилучший
прогноз матриц биопараметров У199о и К2000.
При составлении матриц экспертных оценок состояния окружаю-
щей среды X199Q и Хюю учитывали прогноз активности Солнца (W),
загрязнения нефтью и засоления (5%о) Черноморского бассейна на рас-
сматриваемые годы. Анализ этих данных показывает увеличение со-
лености и уровня загрязнения среды. В 1990 г. ожидается максимум
солнечной активности, а в 2000
Результаты прогноза по модели
и его контрольной проверки при-
ведены в табл. 34. Проверка
прогноза показала высокий уро-
вень адекватности с вероят-
ностью Р = 0,7 4 0,9. При провер-
ке прогноза (табл. 34) наиболее
адекватными оказались биопара-
г. — ее средний уровень (табл. 35).
Таблица 35. Результаты прогноза соле-
ности, активности солнца и содержания
нефти в море до конца XX в. (данные
Севастопольского отделения ГОИН)
Параметр 1990 г. 2000 г.
S, %о 18,062 18,187
W (число Вольфа) 134 70
Нефть (ХлЭк) — —
77
метры У2 (гетеротрофные бактерии перифитона), У4 (живые диато-
мовые водоросли взвеси), Y6 (кокковндные бактерии перифитона),
Yt (диатомовые живые перифитона). Затем следуют менее адекват-
ные параметры — Y3 (гетеротрофные бактерии взвеси), Y1 (живые
диатомовые водоросли планктона), У9 (диатомовые мертвые взвеси),
У10 (диатомовые мертвые планктона). Самая низкая точность прог-
ноза у параметров У1 (гетеротрофные бактерии планктона) и У5 (па-
лочковидные бактерии перифитона). Не во всех случаях критерий
Фишера был более 1, что указывает на общую адекватность модели.
Из табл. 34 следует, что в 1990 и 2000 гг. 15.01 У1 мало будут отли-
чаться от соответствующих значений в январе 1969, 1973 и 1978 гг.
В итоге установлено, что сформированная система (из 31 абиоти-
ческого параметра) имеет наибольшую величину информации о биопа-
раметрах и по 31 абиопараметру можно описать любой из 11 биопара-
метров. Обнаружено, что абиопараметры pH, РОВ, фосфор минераль-;
ный, нитриты-азот и Т °C морской воды дают возможность описать
только 1 параметр из 11 - численность гетеротрофных бактерий пери-
фитона (У2). Остальные 10 биопараметров ими не описываются. Это
имеет значение потому, что некоторые авторы в исследованиях с био-
параметрами используют в основном указанную группу абиопарамет-
ров или отдельные из них, что оказывается не совсем корректным.
Наибольшую величину адекватности имеют биопараметры перифи-
тона — гетеротрофные и кокковидные бактерии, живые и мертвые
диатомовые водоросли (У2, Уб, У8, Уп) Самый надежный прогноз
дают микроорганизмы, развивающиеся на твердых предметах в море,
где наиболее стабильные и благоприятные условия для их жизнедея-
тельности по сравнению с обитанием в толще воды или на частицах
взвеси, поэтому наблюдаются здесь и более устойчивые закономернос-
ти развития микроорганизмов перифитона (Горбенко, 1977).
При изучении данных табл. 34 оказалось, что увеличение солености,
загрязнения нефтью и постоянные колебания солнечной активности в
1990-2000 гг. в целом еще не достигают такой величины, чтобы су-
щественно повлиять иа численность прогнозируемых групп микроор-
ганизмов планктона, перифитона и взвеси в морской среде. Поэтому
число одних (гетеротрофные бактерии планктона и взвеси, живые
диатомовые взвеси и мертвые диатомовые перифитона) возрастает,
количество других (гетеротрофные и кокковидные бактерии перифи‘>
тона, живые диатомовые планктона и Перифитона, мертвые диатомо-
вые планктона) уменьшается, численность третьих (палочковидные
бактерии перифитона и мертвые диатомовые взвеси) в 1990 г. увели-
чивается (или уменьшается), а в 2000 г. уменьшается (или увеличива-
ется). Эго значит, что колебания числа всех указанных групп микро-
организмов и в 1990-2000 гг., по-видимому, будут происходить еще
примерно так же^ как и в настоящее время (Подвинцев, Горбенко,
1983).
78
Способ прогнозирования с помощью анализа Фурье. На основе
указанного способа установили (Горбенко, Крышев, 1985),что в дина-
мике параметров морской экосистемы имеют место гармоники с раз-
личными значениями периодов, изменяющихся от 1 мес до 10 и более
лет. При этом для многих элементов системы характерно сразу нес-
колько периодических компонент.
При выявлении гармонических составляющих множества био- и
абиотических параметров экосистемы (период, фаза, амплитуда) мы
рассчитывали также безразмерную величину вклада каждой домини-
рующей гармоники в дисперсию ряда (<?):
4 (1)
где р| — вклад в дисперсию; Rk - амплитуда; к — номер соответствую-
щей гармоники.
Оказалось, что для биопараметров вклады значимых гармоник в
дисперсию, как правило, не превышают 10 %, что свидетельствует о
заметной случайной составляющей в динамике указанных парамет-
ров. Так, для мертвых диатомей СПМ вклад годовой гармоники по вы-
ражению (1) составляет 7,8 %, а для диатомовых живых и мертвых
СПМ — соответственно 1,4 и 6,6, %. Вклад бактерий СПМ гетеротроф-
ных и кокковидных составляет 3,6 и 4,9 %, а гетеротрофов СМВ -
6,5 %. Для диатомей живых СМП и СМВ, а также для мертвых СМВ
эти величины больше - 22,2 ; 16,5 и 15,6 % соответственно. В то же
время вклад абиопараметров значительнее. Так, температура и осве-
щенность (годовая гармоника) дают в дисперсию *» 79 %, кислород —
54, СО2з- — 54, СО2 — 43, НСО3 - 36, нитриты азота — 30 и pH морской
воды - 20 %.
Если принять во внимание, что для изучаемых элементов системы
существуй» гармоники с периодом Тк, которые дают основной вклад
в дисперсию, то в таких случаях, по-видимому, данные наблюдения
можно представить в виде следующего выражения:
x(t) = xe+ S Rk cos {t-4>k), (2)
k=t lk
t ie t ~ время; - фаза; m - число доминирующих гармоник. Его
можно использовать для прогноза, а также для приближенной оценки
динамики ряда наблюдений, например при восстановлении недостаю-
щих данных.
Поэтому по расчетным данным периодических составляющих пара-
метров динамика кислорода, растворенного в воде, в соответствии с
выражением (2) приближенно может быть записана следующим ра-
венством:
x(t) = 6,36 + 0,88- cos-^- (t - 90е). (3)
79
Рис. 3. Расчетная кривая динамики кислорода в море: а — сред-
немесячные значения О, в 1979 г., б - то же в 1980 г.
Расчетная кривая удовлетворительно описывает ход реальных
параметров О2 за ряд лет (рис. 3), если были соблюдены необходимые
условия для расчета (Горбенко, Крышев, 1985). Таким образом, с
помощью указанного метода можно успешно прогнозировать большин-
ство параметров экосистемы.
80
ГЛАВА 6. ИЗМЕНЕНИЯ СРЕДЫ,
ВЫЗЫВАЕМЫЕ МИКРООРГАНИЗМАМИ
&1. ВЛИЯНИЕ ФАКТОРОВ СРЕДЫ
НА ИЗМЕНЕНИЕ ВЕЛИЧИНЫ pH И КОНЦЕНТРАЦИИ РОВ
Для составления требующейся системы уравнений регрессии (Подвин
цев, Горбенко, 1981) использовали матрицу из 45 наблюдений за
27 био- и абиотическими параметрами морской среды. Наблюдения
проводили каждые 5; 10; 20 и 30 сут в бухте у Севастополя в течение
1 года. Объектами и предметами наблюдений были следующие факторы
и входящие в них параметры.
1) Биофактор с параметрами Xi, А'6, У, (численность гетеро-
трофных бактерий СПМ, живых и мертвых диатомовых водорослей
СПМ) — 3 параметра.
2)Фактор метаболитов микроорганизмов спараметрамиХ2.Х3, А'4)
XJ( Х9, Ало (РОВ СПМ, pH СПМ, сухая масса слизистой пленки СПМ,
количество карбонатов СПМ, РОВ СМВ, pH СМВ) - 6 параметров.
3)Физико-химический фактор с параметрами Xg, Хц, Хц, X1S,
Xl4, Xu, Xu, Xn, Xl8, Xis (температура воды, концентрация компо-
нентов карбонатного равновесия — СО2, НСО2Э-, кислород; окисляв-
мость воды - Перманганатная и щелочная; щелочность; содержание
фосфора минерального и нитритов азота) - 10 параметров.
Остальные 6 параметров - содержание карбонатов в СПМ и СМВ;
число гетеротрофных бактерий СМП и СМВ; масса 10 мл взвеси; соле-
ность воды по информационной мере Кульбака (1967) - были отбро-
шены как малозначимые в данной системе.
Затем была построена и решена система регрессионных уравнений
для РОВ (переменная Уров) и pH (переменная УрН):
ГроВ = 0,647 - 0,0148X1 - 0,05Х3 + 0,0бО47Х4 - 0,0032Xs +
+ 0,0183Х6 + 0,00485Xg + 0,56Х9 - 0,00245Хп - 0,001 ЗХ12 +
+ 0,0108X14 + 0,0077X1$ -0,00634Х16 + 0,0639Х17 +O,O27Xig + 0,03Х18;
ГРН = 6,15 + 0,109X1 + 0,016Х2 - 0,00054Х4 + 0,155Х6 - 0,182Х7 +
+ 0,015Х8 + 0,15X10 - 0,0019Xu + 0,288Х13 + 0,027Xi4 + O,O225X1S +
0,104Х17 -0,151Х19.
81
Оба уравнения оказались адекватными с вероятностью F = 0,993.
Коэффициент множественной корреляции Кров со всеми остальными
параметрами в уравнении равен 0,836, для Урц его величина соответ-
ственно равна 0,776, т.е. матрица исходных наблюдений достаточно пол-
но учитывает информацию о взаимосвязи параметров в море.
Анализ уравнения системы Кров показывает, что содержание РОВ
определяется численностью микроорганизмов СПМ и СМП, некоторы-
ми показателями жизнедеятельности - метаболитами этих микроорга-
низмов и СМВ, а также биогенными веществами и компонентами кар-
бонатного равновесия в море. Карбонатное равновесие обусловливает
изменение солевого состава в море, а кроме того, как нами обнаруже-
но, и РОВ, т.е. РОВ — функция не только биотических факторов (Хай-
лов, 1971), но и ряда абиотических.
Анализ уравнения системы Урц показывает, что pH зависит от тех
же факторов, что и концентрация РОВ в море. Поэтому, по нашему
мнению, недостаточно считать, что колебания величины pH в море свя-
заны только с наличием дериватов угольной кислоты (Алекин, J966).
Эта величина является также функцией биопараметров. Уравнения
} РОВ и Крн позволяют целенаправленно изменять среду путем регули-
рования определенных параметров (Подвинцев, Горбенко, 1981).
Представляет немалый интерес также изучение осаждения каль-
цита и кварца микроорганизмами СПМ и СМВ на погруженных в море
предметах.
&Z О МИКРОБИОГЕННОМ ОТЛОЖЕНИИ КВАРЦА
И КАЛЬЦИТА МИКРООРГАНИЗМАМИ
В течение 1974-1976 гг. в море (Севастопольская бухта) выставляли
стеклянные пластины, с которых в конце каждого месяца соскаблива-
ли наросшую слизистую пленку СПМ. Высушенная до постоянного ве-
са сухая масса СПМ подвергалась рентген-дифрактометрическому ана-
лизу на приборе”ДРОН-1,5”для качественного и количественного опре-
деления ее минерального кристаллического состава (Горбенко, Три-
монис, 1980). Одновременно определяли численность гетеротрофных
бактерий и живых диатомовых водорослей морской воды (СМП) и
СПМ, содержание карбонатов, осаждаемых СПМ, сухую массу СПМ,
концентрацию РОВ морской воды и РОВ СПМ. Измеряли также темпе-
ратуру и pH морской воды (общее число параметров - 13).
Кроме кристаллических компонентов пленки СПМ определяли и
аморфные, например кремнезем. Известно, что панцирь диатомовых
водорослей состоит из кремнезема (SiO2 + хН2О), близкого к опалу-
(Диатомовый анализ, 1949). Поскольку на сухую массу диатомей
приходится 50 % SiO2 (Виноградов, 1944), с помощью таблиц удель-
ных весов этих микроводорослей (составленных в лаборатории фито-
82
Таблица 36. Содержание соединений кремния и кварца в слизистой пленке микро
организмов, % сухой массы пленки
Дата Карбонаты | Кальцит Кварц Кремнезем
29.03.74 10,88 17,0 11,0 5,2
29.04.74 17,49 21,0 10,0 2,2
30.05.74 11,27 7.0 0 4,2
28.06.74 12,27 5,0 0 3,8
30.07.74 18,84 30,0 6,0 1,5
30.08.74 15,48 16,0 0 1,6
30.09.74 24,95 25,0 11,0 4,6
30.10.74 21,80 27,0 8,0 1,2
29.11.74 14,0 8,0 3,0 1,1
30.12.74 20,95 15,0 11,0 6,0
30.01.75 16,90 14,0 9,0 5,7
28.02.75 10,60 10,0 8,0 8,2
31.03.75 18,50 22,0 8,0 1,3
30.06.75 11,56 8,0 2,0 5,6
30.07.75 11,79 13,6 4,0 2,8
29.08.75 16,84 12,0 5,0 0,5
29.09.75 17,30 13,0 6,0 0,3
30.10.75 17,81 15,0 5,0 10,0
29.11.75 23,20 26,0 6,0 1,2
29.01.76 13,03 22,0 6,0 2.5
30.03.76 13,92 8,0 3,0 4,8
30.04.76 14,46 12,0 3,0 1,8
30.07.76 18,98 20,0 7,0 2,0
X * 1р 16,3816,65 18,3319,0 6,84±4,0 4,6214,0
О 15,9 7,9 7,4 7,2
Примечание Среднее значение (х) и доверительный интервал (7р) рассчитаны без
"выскакивающих” значений, которыми оказались наименьшие варианты (5 -
для кальцита, 4 - для кварца и 8 - для кремнезема).
планктона ИнБЮМ АН УССР), а также зная состав доминантных форм
диатомовых, мы теоретически рассчитали количество кремнезема,
находившегося в составе их створок. Этот минерал в отличие от кварца
кристаллического, определяемого аналитически, является аморфным.
Данные статистически обработаны. Вычислили 78 возможных пар-
ных корреляций, из которых достоверными оказались 16. Для вычисле-
ния 24 частных корреляций использовали несколько умеренных пар-
ных корреляций с величиной меньше rmjn при Р = 95 %.
Определили, что из состава минеральных веществ в сухой массе
слизистой пленки СПМ в основном отлагались кальций и кварц в фор-
ме кристаллов и аморфный кремнезем. При этом количество кальцита
было в 2,7 раза больше чем кварца, и в 4 раза больше, чем кремнезема
(табл. 36). Be'--, карбонат в пленке, как отмечалось ранее, микробио-
генного происхождения, по кристаллической форме - это кальцит, о
чем свидетельствуют средние значения (тзбл.36) и тесная корреляция
(г = 0,75) между карбонатом и кальцитом (табл. 37). Судя но доволь-
на
Таблица 37. Парные корреляции кальцита, кварца и других парамет-
ров в морской среде (число наблюдений, п = 23)
Параметр Кальцит - кварц Кальцит- карбонаты Кварц - РОВ СПМ Кварц - pH морской воды Кварц- температур3 воды Кварц- карбонаты Кремнезем - диатомовые живые СПМ Кремнезем - сухая масса СПМ Кремнезем - гетеротрофные бактерии CJ Кремнезем - РОВ СПМ Сухая масса СПМ - РОВ СПМ Гетеротрофные бактерии СПМ - темпера Планктон - температура воды Температура воды - pH воды Сухая масса СПМ — карбонаты СПМ РОВ морской воды - РОВ СПМ РОВ морской воды - температура воды 'min при уровне достоверности 95 % и дат Коэффициент корреляции (г) 0,63 0,75 0,44 -0,41 -0,43 0,48 0,41 -0,69 1М -0,47 -0,41 0,57 тура воды 0,74 -0,44 0,43 0,47 0,54 0,30 том п — 0,41
Таблица 38. Частные и парные корреляции кальцита, кварца и других парамет-
ров морской среды
Параметр Номер параметра, п Корреляция
парная частная
Кальцит 1 г 12 =0,63 г 123=0,45
Кварц 2 '13= 0,75 ^112= 0,35
Карбонаты СПМ 3 я 0,48 '23.1=0,019
Кварц 1 ец =-0,41 '123= “0,27
pH воды 2 г13=-0,43 г 112 = —0,32
Температура воды 3 '23= 0,43 'гзл = 0,32
Кварц 1 г п=-0,30 '123=-0,19
РОВ морской воды 2 ^13=-0,43 г112= — 0,34
Температура воды 3 '23= 0,30 'д|в0,19
Кремнезем 1 '12=-0,34 г из = — 0,025
Карбонаты СПМ 2 rJ3=-0,69 'ш =-0,64
Сухая масса СПМ 3 '23= 0,47 '23.1= 0,30
Кремнезем 1 '12 = -0,47 '123=-0,37
Гетеротрофные бактерии СПМ 2 г 13= -0,69 '1X2=-0,65
Сухая масса СПМ 3 ^23” 0,30 '23.1=-0,04
Кремнезем 1 '12= - 0/7 ' 12.3= -0,39
Гетеротрофные бактерии СПМ 2 'J3=-0,41 г>3£= ~ 0,28
РОВ HIM 3 '23= 0,33 'гзд = 0,18
Кремнезем 1 '12 = -0,69 '123= -0,61
Сухая масса СПМ 2 '13=~-0/1 г112= ~ 0,03
РОВ СПМ 3 '23=0,57 '232 = 0,44
Сухая масс? СПМ 1 г12 ~ 0,30 '123=0,14
Гетеротрофны- бактерии СПМ 2 '13=0,57 '1М= 0,52
РОВ СПМ 3 ^23“ 0,33 '23.1 = 0,20
84
но высокой величине коэффициента корреляции кальцита и кварца
(г = 0,63), отложение одного из них зависит от другого (табл. 37).
Осаждение кальцита зависит еще только от карбонатов СПМ.
Отложение кремнезема обусловлено живыми диатомовыми, об-
щей сухой массой, гетеротрофными бактериями и РОВ СПМ (табл. 37),
т.е. происхождение его биотическое. В то же время осаждение кварца
определяется главным образом абиотическими параметрами (карбо-
натами, кальцитом, pH и температурой воды) и только одним биоти-
ческим РОВ СПМ (общим с кремнеземом). С помощью частных корре-
ляций эти данные удалось уточнить. Установлено, что отложение квар-
ца зависит от кальцита, но совершенно не зависит от карбонатов
(табл. 38). Вместе с тем температура и pH морской воды в равной
мере влияют на осаждение кварца. Хотя отложение кварца обусловли-
вают РОВ и температура воды в море, этот процесс больше зависит от
последнего параметра (табл. 38). Осаждение кремнезема определяет-
ся в основном сухой массой СПМ и не зависит от карбонатов СПМ.
На отложение этого минерала влияют гетеротрофные бактерии и жи-
вые диатомовые СПМ (табл. 38). Остальные био- и абиотические па-
раметры изучаемой биомассы также коррелируют между собой
(табл. 37, 38).
ОБ ОБРАЗОВАНИИ КАЛЬЦИТА, КВАРЦА И КРЕМНЕЗЕМА
И ГИПОТЕЗЕ ПРОИСХОЖДЕНИЯ МЕЛКОГО ПЕСКА
В МОРЯХ И ОКЕАНЕ
Генезис кальцита может происходить следующим образом: аморфные
частицы карбоната, образованные микроорганизмами в результате ряда
химических реакций, сопровождающих метаболизм СПМ (Горбенко,
1977), под влиянием кварца и каких-то параметров окружающей мор-
ской среды, не попавших в число изучаемых нами, кристаллизуются
в кальцит. Этот минерал, как установлено нами, может составлять до
1/5 части сухой Массы слизистой пленки СПМ.
Кремнезем образуется из створок диатомовых, масса которых
составляет часть сухой массы СПМ, а изменение численности этих
микроводорослей зависит и от бактерий-гетеротрофов. Количество
кремнезема составляет 1/20 часть сухой массы пленки СПМ.
Генезис кварца в СПМ не зависит от наличия кремнезема. Однако
возможно, что кварц кристаллизуется из кремнезема под действием
кальцита, pH и температуры воды, а также РОВ СПМ либо выделяется
из раствора морской воды, превращаясь на поверхности пластин со сли-
зистой пленкой СПМ в кристаллическую форму. Доля его в массе
пленки СПМ составляет примерно 1/14 часть (Горбенко, Тримонис,
1980). Судя по имеющимся данным (в нашей совместной работе с
Э.С.Тримонисом), примерно в таких же количествах и в таких же
85
соотношениях, как в перифитоне погруженных в море предметов
(табл. 36), кальцит и кварц отлагаются на морской взвеси.
Если кальцит на взвеси имеет вид порошка, то кварц напоминает
мелкие песчинки разных оттенков. Эти частички, похожие на песок,
не принесены извне на взвесь, а автохтонного происхождения. Об их
автохтонности свидетельствуют корреляционные связи с био- и абио-
тическими параметрами среды, которые ие образуются со случайно
попадающим на взвесь песком. Когда удавалось исследовать взвесь
через несколько месяцев экспонирования ловушки (т.е. ’’постарев-
шую” взвесь), то обнаруживали, что эти песчинки - кристаллы кварца
имеют тенденцию к увеличению размеров, т.е. к росту.
Так как из растворенного состояния в морской воде кварц в мик-
росообществе взвеси (а также в перифитоне) отлагается в кристалли-
ческой форме, мы выдвигаем гиоптезу об одном из возможных путей
образования морского песка. Смысл гипотезы в следующем. Вследст-
вие того что частицы кварца, обнаруженные нами во всех пробах
взвешенного вещества в продолжение ряда лет наблюдений, вырастают
на частицах взвеси от 0 до каких-то размеров, то они вместе с кальци-
том в определенной мере управляют плавучестью и возрастом взвешен-
ных в воде частиц взвеси. Например, образовавшиеся в море микро-
биоценотическим путем взвешенные частицы при парении в морской
воде постепенно тяжелеют от увеличения в их составе количества
кальцита и кварца. При достижении какой-то критической массы час-
тицы взвеси теряют плавучесть и начинают опускаться на дно. На дне
эти частицы прекращают свое существование как взвеси и становятся
частью ила или проглатываются бентосными животными. При избытке
углекислоты в иле (и по другим причинам) кальцит и карбонаты на
бывших частицах взвеси растворяются, а крупинки кварца в виде мел-
ких песчинок освобождаются, образуя морской песок.
Есть возможность рассчитать примерное количество образующего-
ся таким образом биогенного песка. Как мы отмечали выше, доля
кварца во взвеси составляет примерно 1/14 часть сухой массы слизис-
той пленки СПМ. Поскольку СМВ и СПМ подобны, можно предполо-
жить, что такая же примерно часть кварца отлагается и на взвеси. Как
подсчитал Д.М.Витюк (1983), в океане каждый год оседает 5,4 млрдт
взвеси (в пересчете на сухую массу), поэтому 1/14 часть ее составит
около 386 млн т. Это и будет примерное количество мелкого песка,
ежегодно образующегося в Мировом океане на оседающей взвеси. Раз-
мер песчинок настоящего биогенного песка примерно 0,1-0,25 мм и
меньше. Он относится к мелкозернистому песку. Различные оттенки
этого песка зависят от включений в кварц различных минераль-
ных веществ, также отлагающихся на взвеси (Горбенко, 1977).
Известны еще среднезернистые пески (песчинки 0,25--0,5 мм) и круп-
нозернистые (0,5—1 мм) (Курс общей геологии, 1976). На основании
сказанного мы считаем, что часть (если не большее количество) мелко-
86
зернистого песка в море имеет биогенное происхождение, дополняя
песок, образовавшийся путем выветривания и разлома горных пород.
6.4. ОСАЖДЕНИЕ КАРБОНАТОВ
МИКРООРГАНИЗМАМИ ВЗВЕСИ
И ПЕРИФИТОНА
В результате жизнедеятельности сообществ микроорганизмов СМВ и
СПМ в толще воды на частицах взвеси и на предметах, погруженных в
море, постоянно осуществляется один из важнейших процессов - осаж-
дение карбонатов. На ход этого процесса влияют также факторы среды,
в частности компоненты карбонатного равновесия морской среды
(Горбенко,1977).
В 1-е сутки после погружения в море, например стеклянных плас-
тин, количество отложившихся на них карбонатов составляет только
тысячные, сотые и десятые доли миллиграмма на 15 см2 пластины. На
ЗО-е сутки оно повышается уже до 20 мг и более на ту же площадь,
что составляет около 28 %, в то время как на взвеси количество карбо-
натов может достигать до 30 % ее сухой массы и даже более. Карбонаты
(в основном в виде СаСО3), образованные при участии микроорганиз-
мов, могут отлагаться в результате протекания, по крайней мере, 25
химических реакций (Горбенко, 1977). Основная из всех этих реак-
ций - известная реакция связывания углекислоты, постоянно и непре-
рывно образующейся в процессе жизнедеятельности микроорганиз-
мов, в том числе СМВ и СПМ:
Н2О + СО2 -> Н2СО3
Н2СО3 ** Н+ + НСО3 (I стадия диссоциации)
НСО3 Н+ + CO2- (II стадия диссоциации).
Карбонат-ионы (СО2-) реагируют с катионами кальция, а также
других тяжелых металлов,всегда имеющимися в море и образующими
на поверхности взвеси и перифитона карбонаты. В их осаждении по-
верхность указанных морских материалов, по-випимому, играет роль
катализатора процесса карбонатосаждения, происходящего на границе
раздела фаз морская вода - плотная поверхность, являющейся место-
обитанием СПМ и СМВ.
Для микроорганизмов перифитона и взвеси биологическое значе-
ние процесса отложения карбонатов заключается в снижении вредной
Кислотности, вызываемой избыточным количеством СО2 вблизи их
клеток. Способствуют осаждению карбонатов также диатомовые водо-
росли, используя в процессе фотосинтеза углекислоту из окружаю-
щей воды.
При изучении данных об осаждении карбонатов на взвеси и пласти-
нах обрастания, например за 1968 и 1969 гг. (табл. 39, 40), четко выра-
женной сезонности не установлено. Вместе с тем в течение ряда лет
87
Таблица 39. Количество карбонатов, отлагающихся на пластинах обрастания в море (мг С*СО, на 15 см2)
Месяц Время экспонирования в море, сут
5 10 15 20 25 30 10 20 30
1968 г. 1969 г.
Январь 0,270 0,151 0,207 1,634 2,150 8,250 0,031 0,455 0,747
Февраль 0,079 0,167 0,542 1,760 3,025 3,500 0,109 0,448 0,662
Март 0,126 0,627 0,972 1,530 6,015 6,550 0,037 0,318 5,491
Апрель 0,050 0,246 0,950 2,050 2,800 1,380 0,114 0,403 6,475
Май 0,017 0,088 0,084 0,020 0,901 0,733 2,175 0,325 4,553 4,635 2,598
Июнь 0,042 0,460 1,050 3,200 4,680 0,011 7,557
Июль 0,113 1,520 3,750 10,530 20,950 18,000 0,291 7,332 9,683
Август 0,266 0,720 5,250 9,675 5,825 22,550 0,244 2,329 6,148
Сентябрь 0,419 2,220 6,750 9,300 12,137 8,680 0,062 8,566 9,035
Октябрь 0,067 0,208 0,815 2,780 8,450 12,375 0,415 12,157 26,319
Ноябрь 0,088 0,543 1,650 4,500 3,325 16,050 0,078 3,773 6,979
Декабрь 0,016 0,021 0,052 3,526 7,000 5,200 0,245 2,722 6,787
Таблица 40. Количество карбонатов, осаждающихся со взвесью на дне сосуда-ло-
вушки, г на 314 см2 дна__________________________________________
Месяц Время экспонирования в море, сут
10 20 30
А Б А Б А 1 Б
Январь 0,51 0,14 1968 г. 0,92 0,23 2,47 0,47
Февраль 0,68 0,11 2,32 0,37 2,20 0,41
Март 2,83 0,54 2,80 0,52 2,35 0,51
Апрель 2,14 0,98 2,19 0,18 2,50 0,18 0,13
Май 1,73 0,11 1,73 0,08 2,46
Июнь 2,63 0,18 2,76 0,12 0,99 3,62 0,51
Июль 2,61 0,27 6,82 10,70 1,74
Август 0,74 0,07 6,00 0,52 11,71 2,37
Сентябрь 1,68 0,11 4,27 0,53 7,12 1,07 0,58
Октябрь 0,75 0,13 3,02 0,32 2,90
Ноябрь 1,84 0,16 2,89 0,36 3,61 0,30 2,51
Декабрь 2,85 0,40 6,72 0,64 12,00
Январь 0,81 0,05 1969 г. 0,93 0,05 2,52 0,55 1,29
Февраль 0,90 0,13 4,13 1,06 5,24
Март 2,07 0,26 4,14 0,51 3,80 0,52
Апрель 2,53 0,18 0,16 6,76 0,70 6,43 0,83
Май 1,47 4,47 0,50 5,37 0,69
Июнь 5,05 0,49 0,57 5,80 0,64 9,76 1,62
Июль 4,00 8,45 1,62 8,42 2,03
Август 1,38 0,32 3,10 0,56 2,65 0,77
Сентябрь 3,17 0,53 4,59 1,38 5,10 1,18
Октябрь 4,88 0,86 5,34 1,86 0,16 10,53 4,00
Ноябрь 3,65 0,91 0,51 1,86 0,52
Декабрь 2,29 0,06 9,45 2,77 5,10 1,10
Примечание. А - сухая масса взвеси, Б - содержание карбонатов на ней.
88
наблюдений иногда выделяются 2 периода наиболее интенсивного кар-
бонатосаждения - январь - июнь и июль - декабрь. В 1-й половине
года, как правило, отлагается меньше карбонатов, чем во 2-й. На
взвеси в январе - июне 1968 и 1969 гг. обнаруживалось немного мень-
ше карбонатов, чем во 2-й половине года. В перифитоне погруженных
предметов эти различия были выражены сильнее, а в июле - декабре
карбонатов отлагалось в 2,13-3,11 раза больше, чем в 1-й полови-
не года.
Что касается причин указанного явления, то поскольку вода в
Севастопольской бухте в январе - июне холодней, чем в последующее
время, то косвенно на процесс карбонатосаждения можег влиять тем-
пература морской воды. Однако за многие годы наблюдений нам не
удалось обнаружить достоверных корреляций температуры воды с
карбонатами, поэтому влияние температуры — причина косвенная и,
может быть, второстепенная. Основную же роль в этом процессе играл
биотический фактор и, следовательно, биотические параметры морской
среды, а именно микроорганизмы СМП, СМВ и СПМ и метаболиты
двух последних ценозов (табл. 41, 42).
Это, несомненно, является прямым подтверждением вывода о том,
что в бухте у Севастополя, в Черном море, а возможно, и в других мо-
рях и Мировом океане карбонаты имеют микробиогенное происхож-
дение (Горбенко,1977).
Осаждение карбонатов в перифитоне. Ранее (Горбенко, 1977) мы
нашли корреляции количества карбонатов СПМ с живыми диатомовы-
ми водорослями, бактериями, с метаболитами и сухой массой пленки
СПМ (которая по массе обычно составляет 19,5 ± 3,28 % сырой массы)
и с карбонатами СМВ (табл. 41). Затем все временные ряды парамет-
ров многолетних наблюдений мы, используя корреляционный анализ
(Горбенко, Крышев, 1985), исследовали с помощью различных матема-
тических методов (табл. 41).
В результате анализа однолетних взаимных корреляционных
функций были выявлены 4 корреляции карбонатов перифитона (СПМ),
3 из которых (с кокковидными, палочковидными бактериями и сухой
массой СПМ) уже были обнаружены ранее (Горбенко, 1977), свиде-
тельствуя о стабильности этих связей. Связь карбонатов СПМ и СМВ
подтверждает подобие указанных микросообществ, что отмечалось
выше.
На следующем, более высоком уровне корреляционного анализа -
вычисление корреляций за много лет наблюдений - совершенно четко
установлено следующее. Осаждение карбонатов на предметах, погру-
женных в море, обусловливается всеми микроорганизмами СПМ,
образующими массу СПМ, которая является интегральным компонен-
том этого микросообщества, в основном определяющим указанный
процесс. На осаждение карбонатов также влияет концентрация водо-
родных ионов в воде, выраженная величиной pH (табл. 41).
89
Таблица 41. Корреляции количества карбонатов в перифитоне
с параметрами экосистемы
Параметр Коэффициент
корреляции (г)
Палочковидные бактерии * 0,380
Кокковидные бактерии 0,375
Гетеротрофные бактерии 0,491
Живые диатомовые водоросли 0,452
РОВ СПМ 0,359
Сухая масса 0,784
Однолетние корреляции **
Палочковидные бактерии 0,310
Кокковидные бактерии 0,390
Сухая масса СПМ 0,910
Карбонаты взвеси (СМВ) 0,310
Многолетние корреляции (по усредненным данным)
Простейшие (неприкрепленные) организмы 0,370
Диатомовые живые 0,596
Диатомовые мертвые 0,665
Гетеротрофные бактерии 0,740
Палочковидные бактерии 0,587
Кокковидные бактерии 0,570
Сухая масса СПМ 0,922
pH морской воды - 0,603
Многомерные корреляции (гм)
Гетеротрофные бактерии СПМ 0,370
Кокковилные бактерии СПМ 0,580
Сухая масса СПМ 0,920
Карбонаты СМВ 0,560
Влияние других параметров
Без сухой массы СПМ 0,29
Без сухой массы СПМ 0,08
'’min = 0.300 при п - 42
* Горбенко, 1977; ** Горбенко, Крышев, 1985.
На одном из самых высоких уровней корреляционного анализа -
многомерном анализе корреляционных функций — выявляются пря-
мые корреляции, на которые не влияют все остальные параметры эко-
системы. Либо напротив — обнаруживаются косвенные связи, которые
не устанавливаются без других, дополнительных параметров. В резуль-
тате этого анализа были выявлены только 4 связи. Оказалось, что ге-
теротрофные бактерии СПМ коррелируют с карбонатами СПМ посред-
ством сухой массы СПМ, а при ее элиминации эта корреляция стано-
вится только чуть ниже уровня достоверности (табл. 41). Поэтому
можно считать, что связь карбонатов СПМ с бактериями-гетеротрофами
СПМ все же существует. Результаты исследований, изложенные выше,
сви.<егсльггпуюг о том. что карбонаты СПМ — чисто биотический
параметр, на который из внешних параметров морской среды влияет
только pH воды. В то же время не связаны с карбонатами СПМ кокко-
видные бактерии этого микросообщества, так как при элиминации
сухой массы СПМ указанная связь становится практически равной О
(г = 0,08).
Две независимые прямые связи, по-видимому, самые важные -
связь карбонатов СПМ с карбонатами СМВ может свидетельствовать
о том, что процесс осаждения карбонатов в перифитоне и на взвеси
регулируется одними и теми же параметрами окружающей среды либо
осуществляется аналогичными микроорганизмами ценоза. Прямая
связь содержания карбонатов СПМ с сухой массой СПМ - не только
одна из самых важных в процессе карбонатосаждения, но и одна из са-
мых интересных.
На графиках кривых динамики карбонатов и сухой массы СПМ за
15 и более лет их ход точно повторяет друг друга. Причем, отклонения
от параллельности очень редки, т.е. даже грубым методом сравнения
в микроценозе двух биокосных параметров в продолжение многих
лет между ними обнаруживалась тесная корреляционная взаимосвязь,
впоследствии подтвержденная точным расчетом. В наших многолет-
них исследованиях множества параметров тесные корреляции осталь-
ных параметров морской экосистемы (г > 0,7) встречаются намно-
го реже.
Так как масса СПМ в основном определяет количество осаждае-
мых карбонатов в перифитоне, возникает вопрос, а какие факторы
влияют на изменение сухой массы СПМ? Сопоставив все имеющиеся
параметры за 1968—1977 гг. наблюдений и рассчитав скорость роста
слизистой пленки СПМ (е) (Горбенко, Крышев, 1985), мы обнаружи-
ли, что данный параметр, солнечная активность W (число Вольфа) и
сухая масса СПМ (Хм) взаимосвязаны. При этом коэффициенты кор-
реляции активности Солнца с сухой массой СПМ и скоростью роста
пленки составляли соответственно г = 0,92 и г = 0,94, указывая на их
тесную взаимосвязь. В то же время коэффициенты корреляции In АЛМ
и In f с температурой морской воды составляли соответственно г -
» 0,80 и г = 0,94. Затем была построена соответствующая математичес-
кая модель.
После расчета всех коэффициентов в соответствующих уравнениях
модели за 10-летний период наблюдений, а также по сезонам нами бы-
ли получены общие математические формулы, определяющие измене-
ния сухой массы Хм и скорости роста пленки СПМ (£) в зависимости
от изменений активности Солнца и температуры воды в море:
¥м = (8,7‘10-3 IV + 0,14)еаГ,
$ = (3,510-4 W + 0,07)eaf,
где И' - число Вольфа; а =0,065 (°C) соответствует увеличению ско-
рости процесса примерно в 2 раза при увеличении температуры на
91
10 °C (правило Вант-Гоффа); t — температура морской воды; е-
транснендентное’ число: -J'—— (1 + -*)" =2,718281828...
Указанные уравнения отражают тесную взаимосвязь многолетних
изменений сухой массы СПМ и среднегодовых величин солнечной ак-
тивности за период с 1969 по 1977 г. Соответствующая корреляцион-
ная взаимосвязь за 10-летний период времени положительна, т.е. в годы
с повышенными значениями чисел Вольфа (солнечной активности)
наблюдается стимуляция роста слизистой пленки СПМ. В то же время
сезонный ход скорости роста пленки СПМ определяется величинами
среднемесячных температур воды в море. В свою очередь, как отмеча-
лось выше, сухая масса СПМ тесно коррелирует с карбонатами СПМ
(г = 0,92), определяя процесс карбонатотложения. Таким образом,
солнечная активность является фактором, определяющим многолет-
ние изменения сухой массы СПМ на погруженных в море предметах.
Л температура воды — фактор, влияющий па сезонные колебания ско-
рости роста слизистой пленки СПМ (Горбенко, Крышев, 1985). В итоге
есть основание считать, что выявленные в течение продолжительного
времени причинные зависимости изменений массы слизистой пленки
СПМ, ее скорости роста и отложений карбонатов в перифитоне, их
четко повторяющийся ход и тесная взаимосвязь закономерны.
Какое же место занимает процесс осаждения карбонатов СПМ на
погруженных в море предметах - перифитоне? Если суммировать
количество осаждаемых карбонатов на 30-е сутки за все месяцы 1968 и
1969 гг. (табл. 39), то получится соответственно 109,97 мг и 88,480 мг
на 15 см2 пластины. За 2 года на 1 см2 это составит 13,198 мг/см2.
За 1000 лет соответственно отложится 6 599 мг/см2, или 6,6 г/см2.
За 2500 лет (возраст современного Черного моря (Страхов и др.,
1954)) соответственно осело бы 16,497 г~ 16,5 г/см2.
Для определения массы 1 см3 карбоната усредним сухую массу
различных карбонатов (известняк, кальцинированная сода, мел и мра-
мор). Средний удельный вес их составляет примерно 1,8 г/см3. Следо-
вательно, на поверхности погруженных в море предметов, в той его
части, где в СПМ входили все его живые компоненты и необходимые
условия морской среды (в шельфовой части кислородной зоны Черно-
го моря), за 1000 лет на площади 1 см2 успело бы нарасти карбоната
СПМ 6,6 г: 1,8, т.е. 3,67 см, а за 2500 лет —соответственно 16,5 г/см2 :1,8,
т.е. 9,17 см.
Осаждение карбонатов на морской взвеси. Как установлено ранее
(Горбенко, 1977), черты осаждения карбонатов на взвеси и на
погруженных в море предметах сходны. 1. В СПМ и СМВ примерно
одинаковы колебания количества осаждающихся карбонатов. 2. В тече-
ние 1 дня наблюдений в обоих ценозах отлагается примерно одинаковое
1 Т.е. число, выраженное непериодической бесконечной десятичной дробью.
*?2
количество карбонатов. 3. С увеличением возраста взвеси и микро-
перифитона количество карбонатов на них возрастает. 4. Кривые ди-
намики осаждения карбонатов СПМ и СМВ в основном повторяют
друг друга. 5. В 1968-1969 гг. обнаружены корреляции содержания
карбонатов на взвеси и в перифитоне (г = 0,506 и г = 0,355), т.е., оче-
видно, механизмы их осаждения на различных морских субстратах
подобны (табл. 42).
При изучении структуры указанных выше параметров (раздел 4.2),
состоящей из 9 группировок, карбонаты СМВ оказались в одной из
основных — ”В” (рис. 1) (Подвинцев, Горбенко, 1979). Центральное
ядро группы ”В” составляют: альбедо видимого спектра Солнца, являю-
щееся, возможно, регулятором; облачность, которая, по-видимому,
как-то влияет на видимый спектр, пропуская или задерживая его;
карбонаты СМВ; pH морской воды и содержание фенола в море. Все
они связаны самыми сильными связями (г > 0,561) с вероятностью
Pi > 0,995. Кроме сильных связей между отдельными группами были
обнаружены и связи средней силы (0,561 >г >0,515) с вероятностью
0,995 > Р2 > 0,99. Умеренными связями (0,515 > г > 0,379) с вероят-
ностью 0,99 > Р3 > 0,95 карбонаты СМВ связаны также с диатомовыми
водорослями СМВ, кокковидными бактериями СПМ, РОВ СПМ, pH
СПМ, долей баланса в инфракрасной области спектра Солнца, проз-
рачностью, атмосферным давлением СО2 и О2 (табл. 42). Таким обра-
зом, карбонаты СМВ взаимодействуют с 4 группами изучаемой экосис-
темы (А, Е, С и D), что указывает на большую разветвленность связей,
а следовательно, и на зависимость этого параметра от многих пара-
метров системы. По результатам проведенных исследований на карбо-
наты СМВ в основном влияют абиопараметры (8 единиц) и в меньшей
степени - биопараметры (6 единиц) (Подвинцев, Горбенко, 1979).
В наших последующих исследованиях (Горбенко, Крышев, 1985)
с помощью вычисления однолетних корреляций удалось обнаружить
11 корреляционных связей карбонатов СМВ с другими элементами
экосистемы, которые в основном были биотическими параметрами
СМВ, СПМ и СМП и их метаболитами. На карбонаты СМВ влияли диа-
томовые СМП живые и мертвые, диатомовые живые и гетеротрофные
бактерии СМВ, сухая масса и карбонаты СПМ, а также метаболиты —
pH и РОВ СПМ, pH и РОВ СМВ и ХлЭк (вернее, его автохтонная часть).
Корреляционные связи более высокого порядка - многолетние -
позволили выявить более стабильные и постоянные зависимости — ко-
личество их 7. Из них только 2 были с параметрами абиотической мор-
ской среды, а остальные биотические - с параметрами СМВ, СМП и
СПМ и их метаболитами. Связи с диатомовыми СМП, гетеротрофными
бактериями СМВ, РОВ СПМ и СМВ, а также с ХлЭк уже были выявлены
в однолетних корреляциях, т.е. они оказались постоянными и стабиль-
ными (табл. 42). Обнаружили также связи с гетеротрофными бактерия-
ми СМП и РОВ морской воды. В итоге карбонаты взвеси определялись,
93
Таблица 42. Корреляции количества карбонатов на взвеси с параметрами эко-
системы
Параметр* *
Коэффициент корреляции (г)
Карбонаты СПМ*
pH морской воды**
Альбедо видимого спектра
Облачность
Температура воздуха
Диатомовые водоросли СПМ (живые)
Диатомовые водоросли СПМ (мертвые)
Кокковидные бактерии СПМ
Диатомовые водоросли СМВ (мертвые)
РОВ СПМ
£НСПМ
Прозрачность
Доля баланса в инфракрасной области
света Солнца
Атмосферное давление
0,355-0,506
rmin = 0,300
0,561-0,910
0,561-0,910
0,561-0,910
0,515-0,561
0,515-0,561
0,515-0,561
0,379-0,515
0,379-0,515
То же
''min “ 0,500 при п = 16
Однолетние корреляции***
pH СПМ -0,37
pH СМВ -0,49
Сухая масса СПМ 0,31
Карбонаты СПМ 0,31
Диатомовые живые СМП 0,34
Диатомовые мертвые СМП 0,32
Диатомовые живые СМВ 0,32
Гетеротрофные бактерии СМВ -0,37
ХлЭк -0,31
РОВ СПМ 0,31
РОВ СМВ 0,35
Многолетние корреляции ****
Диатомовые живые СМП 0,485
РОВ морской воды 0,390
Гетеротрофные бактерии СМП -0,330
Гетеротрофные бактерии СМВ -0,445
ХлЭк -0,383
РОВ СПМ 0,340
РОВ СМВ 0,338
Многомерные корреляции (гу)****
pH СМВ 0,70
Карбонаты СПМ 0,56
Диатомовые живые СМВ 0,35
Диатомовые мертвые СМВ 0,34
ХлЭк 0,43
РОВ СМВ 0,60
Влияние других параметров
Без pH СМВ 0,150
Без диатомовых живых СМВ 0,160
Без зоопланктона 0,280
»» »*
''min = 0,300 при п “42
• Горбенко, 1977; ** Подвинцев, Горбенко, 1979; **♦ Горбенко, Крышев, 1985;
**** По усредненным данным (Горбенко, Крышев, 1985).
94
а вернее обусловливались,биопараметрами — диатомеями, бактериями
и их метаболитами, а также содержанием РОВ морской воды. Послед-
ние, по-видимому, используются в качестве источника углерода бакте-
риями и определялись хлороформным экстрактом, играющим какую-
то роль в карбонатосаждении на взвеси.
Многомерный анализ однолетних наблюдений на еще более высо-
ком уровне позволил выявить основные корреляции элементов ’’ске-
лета” — группировок параметров экосистемы, а также косвенные кор-
реляционные связи. Установили, что корреляции карбонатов СМВ с
живыми диатомовыми СМВ не осуществляются без pH СМВ, а корреля-
ции карбонатов с мертвыми диатомовыми СМВ — без диатомовых жи-
вых СМВ и зоопланктона.
Основными прямыми связями, обусловливающими отложение кар-
бонатов СМВ, являются их связи с pH и РОВ СМВ и ХлЭк, которые
повторялись в одно- и многолетних корреляциях, а также с карбоната-
ми СПМ. Можно предположить, что микроорганизмы СМВ используют
автохтонную часть ХлЭк, а образующиеся при этом метаболиты СМВ и
карбонаты СПМ вместе с микроорганизмами СМВ способствуют осаж-
дению карбонатов СМВ. Таким образом, осаждение карбонатов на
взвеси, так же как и карбонатов в перифитоне, определяется главным
образом биотическими компонентами системы и только в незначитель-
ной мере — абиотическими.
Вследствие того что с карбонатами СМВ коррелировало намного
больше параметров экосистемы, чем с карбонатами СПМ, и общее ко-
личество корреляций первых было почти в 2 раза больше, чем вторых,
поэтому, очевидно, главным по значимости процессом в море надо счи-
тать осаждение карбонатов на взвеси, а затем — карбонатов в пери-
фитоне.
Как отмечалось выше, карбонаты в зависимости от времени экспо-
нирования взвеси в сосуде-ловушке составляют от 0 до 30 % и более
сухой массы взвешенного вещества, динамику осаждения которого
можно видеть в табл. 43. По данным этой таблицы, экая процентное
содержание карбонатов, мы рассчитали, сколько их осаждается со
взвесью (табл. 40).
Какое же место занимают карбонаты СМВ при осаждении в море?
После суммирования количества карбонатов, осевших в сосуд-ловуш-
ку за 30 сут в каждый месяц года, количество их, осажденных со
взвесью за 1968 г., составило 10,78 г., а за 1969 г. — 15,10 г. За 2 года
это составляет 25,88 г, или « 26,000 г. За 1000 лет соответственно на
314 см2 (площадь дна сосуда) осело бы 13000 г, т.е. это равно 41,400 г
на 1 см2, а за 25000 лет - соответственно 103,500 г. На каждом 1 см2,
где оседала бы взвесь, например в районе шельфа, высота оседания чис-
того карбоната за 2500 лет составила бы 103,500 : 1,8 = 57,5 см. За
1000 лет - соответственно 41,400 : 1,8 = 23,0 см2. Следовательно,при
наличии во взвеси всех компонентов СМВ (т.е. цельного микроценоза)
95
Таблица 43. Количество осаждающейся взвеси на дне сосуда-ловушки в обычных условиях мор- ской среды в шельфовор
Время экспонирования в море, сут части кислородной зоны Черного моря, где самые благоприятные условия для
Месяц 10 20 30
А Б А Б А Б развития микроорганизмов потому что с суши смыва- ется достаточное количест- во органических и мине- ральных веществ (Слевич, 1977), на дне отложились бы указанные количества карбонатов. Как известно (Стра- хов и др., 1954), в мидие- вом и фазеолиновом чер- номорских илах макси- мальное накопление СаСО3 за 2500 лег составляет 20- 50 г/см1. Поэтому, если с морской взвесью за этот же срок на каждом 1 см* отложилось бы не 103,500 г карбоната кальция, что по- казали наши данные, а толь- ко половина (из-за небла гоприятных условий сре ды), то и в этом случае ко- личество карбонатов СМВ превышало бы величину карбонатосаждения ми- диями и фазеолинамн, ука-
1968 г. Январь 3,7 0,51 5,0 0,92 10,0 2,47 Февраль 4,0 0,68 10,0 2,32 10,0 2,20 Март 10,0 2,83 10,0 2,80 10,0 3,35 Апрель 10,0 2,14 10,0 2,19 10,0 2,50 Май 10,0 1,73 10,0 1,73 10,0 2,46 Июнь 14,0 2,63 12,0 2,76 16,0 3,62 Июль 10,0 2,61 30,0 6,82 25,0 10,70 Август 5,0 0,74 17,5 6,00 27,0 11,71 Сентябрь 7,5 1,68 14,5 4,27 26,0 7,12 Октябрь 2,0 0,75 8,0 3,02 7,0 2,90 Ноябрь 7,0 1,84 10,0 2,89 13,0 3,61 Декабрь 10,5 2,85 29,0 6,72 32,0 12,00 1969 г. Январь 3,5 0,81 4,0 0,93 8,5 2,52 Февраль 4,5 0,90 13,0 4,13 17,0 5,24 Март 9,5 2,07 19,0 4,14 18,0 3,80 Апрель 14,0 2,53 26,0 6,76 24,0 6,43 Май 7,0 1,47 21,0 4,47 22,0 5,37 Июнь 23,5 5,05 27,0 5,80 34,0 9,76 Июль 18,0 4,00 50,0 8,45 36,0 8,42 Август 7,0 1,38 9,0 3,10 13,0 2,65 Сентябрь 13,0 3,17 18,0 4,59 20,0 5,10 Октябрь 20,0 4,88 14,5 5,34 37,0 10,53 Ноябрь 11,0 3,65 7,0 1,86 8,3 1,86 Декабрь 9,5 2,29 34,0 9,45 17,0 5,10 Примечание. К — объем свежей взвеси, мл; Б - сухая масса этой же взвеси, г на 314 см2 дна.
зывая на важнейшую роль взвеси в этом процессе на шельфе Черного
моря. Учитывая, что взвесь отлагается по всему морскому дну, а глу-
бина этого моря (немногим более 2 тыс. м) не достигает критического
уровня (4—5 тыс. м), где карбонаты растворяются в воде, то величина
карбонатосаждения с морской взвесью в открытых частях Черного
моря будет намного больше 7,2 г/см1, что отложилось за 2500 лет
(Страхов и др., 1954). Учитывая также, что в перифитоне твердых и
плотных неживых предметов за 2500 лет могло отложиться 16,5 г/ см1
карбонатов (Горбенко, 1977), есть основание считать, что микроорга-
низмы СМВ играют весьма важную роль в осаждении карбонатов в
Черном море.
Если предположить,что в Мировом океане условия для осаждения
карбонатов в перифитоне и взвеси не хуже, чем в Черном море, в кото-
96
ром большую часть занимает сероводородная зона, то приведенные
Нами количественные данные можно сравнивать также с карбонаг-
осаждением в Океане. По данным А.П.Лисицына (1974), в Беринговом
море моллюски накапливают 57,7 г СаСО3 на 1 м2 поверхности-субст-
рата в год, что считают высоким показателем.
Как было отмечено, за 2 года наших ежедекадных наблюдений на
дне сосуда-ловушки на 30-е сутки каждого месяца оседало 26,0 г на
314 см2. На 1 см2 в среднем за год это составляет 26,0 г : 2 : 314 =
= 41,400 мг, на 1 м2 - соответственно 41,400 мг-10000 см2 = 414 г/м2,
т.е. количество карбонатов во взвеси в Черном море при сходных с
Беринговым морем условиях больше в 7 раз количества карбонатов,
осаждаемых моллюсками, которых принято считать основными орга-
низмами, накапливающими и осаждающими карбонаты в морских во
доемах (Страхов и др., 1954; Лисицын, 1974; и др.).
Масса карбонатов, отлагающихся во взвеси в Черном море за
1000 лет, достигает 41,400 г/см2. Это составляет 23,00 см в высоту и
в 6 раз превышает количество карбонатов, осаждающихся хемогенным
путем на Багамской банке за тот же срок (Лисицын, 1974). По данным
этого же автора, суммарная скорость осаждения карбонатов на Багам-
ской банке 80 см/1000 лет. Такое возможно потому, что Багамская
банка, вероятно, единственное место в Мировом океане, в котором
существуют аномально благоприятные условия для осаждения карбо-
натов биогенным и хемогенным путями. Возможно также, что эти дан-
ные несколько преувеличены.
Если по данным наших исследований суммировать величины осаж-
дения карбонатов взвеси и в перифитоне также за 1000 лет, то полу-
чим следующее: карбонаты перифитона, осаждающиеся за 1000 лет
(6,6 г/см2), плюс карбонаты взвеси за этот же срок (41,4 г/см2) в сум-
ме составляют 48,10 г/см2. Высота слоя карбонатов соответственно
23,00 + 3,67 = 26,67 см. Эти величины примерно в 3 раза меньше та-
ковых, полученных на Багамской банке, однако при сравнении резуль-
татов наших исследований с данными других ученых оказывается, что
по масштабам осаждения карбонатов в морях и Океане (Страхов и др.,
1954; Лисицын, 1974; и Др.) отложения в Черном море превосходят
все автохтонные источники отложения карбонатов в морских водоемах
по количеству, объему и распространению.
Следовательно, можно предположить, что путем осаждения со
взвесью и в перифитоне в морях и Океане микроорганизмами СМВ
и СПМ отлагается наибольшее количество карбонатов. При этом карбо-
натосаждение происходит постоянно.
97
ГЛАВА 7. МИКРОБИОЦЕНОТИЧЕСКОЕ ОБРАЗОВАНИЕ
МОРСКОЙ ВЗВЕСИ
Известно, что взвесь — это мельчайшие частицы, взвешенные в морской
воде автохтонного (А) и аллохтонного (Б) происхождения. (А) -
это взвесь, образованная в основном из остатков животных и расте-
ний, в состав которой входят минеральные и органические вещества;
обычно называется детритом (Лисицын, 1964; Родина, 1966; Соро-
кин, 1970; Хайлов, 1971; Хорн, 1972; Емельянов, Шимкус, 1973;
Романкевич, 1977).
Взвешенное вещество образуется из продукции фитопланктона
(Винберг, 1969; Лисицын, 1974; Романкевич, 1977; Вигюк, 1983;
и др.), а также из хлопьевидных колоний — агрегатов морских микро-
организмов. Бактерии при этом, развиваясь на частицах взвеси, исполь-
зуют адсорбированные на них органические вещества из состава РОВ в
морской воде. Затем органика снова оседает и снова используется
бактериями и т.д. В результате действует поток: РОВ -* поверхность
взвеси. Без бактерий агрегаты не образуются (Сущеня, 1968; Сорокин,
1970; Хайлов, Финенко, 1971; Романкевич, 1977).
Наибольшая часть взвеси является продуктом распада отмерших
животных и растительных организмов, их частей тела, фекалий и мета-
болитов, т.е. взвесь в основном представляет собой детрит (Финенко,
1965; Родина, 1966; Fenshel, 1970; Riley, 1970; Хайлов, 1971; Пети-
па, 1981).
Путь аллохтонного образования взвеси включает в себя ряд нап-
равлений:
терригенное - продукты размыва горных пород суши и сточные
воды, стекающие в море;
вулканическое — материал извержений;
золовое - занос пылевых частиц и песка ветром;
космическое — метеоритная пыль (Лисицын, 1974; Романкевич,
1977; Вигюк, 1983; и др.).
Есть сведения и о других возможных путях образования взвеси в
море:
в результате микроциркуляции поверхностных вод Ленгмюра;
посредством пузырьков воздуха;
непоср.лстпенно из РОВ на границе раздела вода - воздух (Riley,
%
1970; Скопинцев, 1971, Хайлов, Финенко, К/1; Хайлов; 19'’]; Soro-
kin, 1978; и Др.). К сожалению, каких-либо количественных данных об
аллохтонном образовании взвеси в море нам не встречалось.
7.1. ОБЩИЕ ПРИЗНАКИ МОРСКОЙ ВЗВЕСИ
Как Показали наши многолетние исследования, неськнря иа большие
различия условий окружающей морской среды в различных районах
Мирового океана, можно выделить ряд наиболее важных признаков,
общих для всей взвеси, независимо от места ее сбора в океане.
1. Размеры взвеси составляют 0,2 мкм и более (поскольку вели-
чины 0,1 мкм уже могут достигнуть молекулы РОВ)1 до 1 мм (Суше-
на, 1968; Тихий океан, 1970; Петина, 1981; Витюк, 1983; Лисицын,
1984; Мельников, 1986). Изредка встречаю!ся частицы величиной до
50 мм и больше (Кондратьева, 1975).
2. По морфологическим признакам взвесь в основном состоит из
очень мелких частиц, которые могут формироваться в ветвистые це-
почки. Взвесь крупнее 1 мкм (ее в океане больше всего) представля-
ет собой рыхлые и плотные аморфные агрегаты, фрагменты встречают-
ся реже (Мельников, 1976; Хайлов, 1971, Витюк, 1983,и др.).
3. Состав взвеси - это почти всегда смесь органического и неорга-
нического вешесгва. По мнению большинства исследователей, мине-
ральная часть ее обычно количественно преобладает над органической
(Тихий океан, 1970; Романкевич, 1977; и др.).
4. Органическое вещество взвеси, как правило, это сумма живого
и неживого вещества в различных соотношениях (Сущеня, 1968; Ви-
тюк, 1983).
5. Содержание взвеси в океане в среднем составляет 1 мг/л (Ли-
сицын, 1974; Витюк, 1970, 1983).
6. Цвет взвеси в основном 2 оттенков - желтовато-бурый и серый
(Тихий океан, 1970; Лисицын, 1974; Витюк, 1983).
7. Во взвеси всегда обнаруживаются карбонаты (в основном
СаСО3). Количество их составляет 0,5-33 % сухой массы взвеси
(Емельянов, Шимкус, 1974; Витюк, 1975; Горбенко, 1977; и др.).
8. Существует сезонность изменения количества взвеси (Лисицын,
1974; Тихий океан, 1970; Витюк, 1983).
9. Во взвеси, как правило, содержатся соединения кремния (Емелья-
нов, Шимку с, 1974; Лисицын, 1974; Романкевич, 1977; и др.).
10. Практически на всех глубинах океана на частицах взвеси
встречаются бактерии, количество которых составляет 0,5—10 % (Ти-
хий океан, 1970; Романкевич, 1977; Сорокин, 1977; Витюк, 1983;
и др.).
1 Например, полисахаридные клубки (Хайлов 1971)
99
Л. Диатомовые и другие представители микроводорослей также
встречаются иа взвеси, но в меньших количествах, чем бактерии. Они
обнаруживаются в основном до глубины 2-3 тыс. м. Ниже их коли-
чество незначительно, а на глубине 5 тыс. м диатомей не обнаружива-
ется вовсе (Тихий океан, 1970; Вигюк, 1983; и др.).
12. В состав органического вещества взвеси входят белки, жиры
(липиды) и углеводы, а также аминокислоты и пигменты (Тихий
океан, 1970; Романкевич, 1977: Витюк, 1983; и др.).
Выделенные признаки являются наиболее характерными для
морской взвеси, следовательно, и для большей чаши взвеси в океане.
А взвеси, имеющие много сходных признаков, очевидно, имеют и об-
щие механизмы образования. Рассмотрим, возможно ли образование
большей части взвешенного вещества морской воды из живого ве-
щества.
7.2. О ВОЗМОЖНОСТИ ПОПОЛНЕНИЯ ВЗВЕСИ
ИЗ ЖИВОГО ОРГАНИЧЕСКОГО ВЕЩЕСТВА
На основе более чем 20 тыс. определений концентрации взвеси в толще
вод Мирового океана А.П.Лисицын (1974) показал, что она составляет
примерно 1 мг/л. Исходя из этой усредненной величины и объема Ми-
рового океана, было рассчитано, что в его водах находится приблизи-
тельно 1370 млрд т (в пересчете на сухую массу) взвеси. Более полови-
ны взвешенного вещества приходится на Тихий океан. Какое же коли-
чество живых организмов в океане и какова их продукция?
Хотя приведенные в табл. 44 данные являются приближенными,
как и все подобные, рассчитанные для Мирового океана, тем не менее
нельзя не обратить внимание на колоссальную способность бактерий
к воспроизводству, выраженную достаточно большой величиной коэф-
фициента П/Б. Безусловно, роль бактерий в био- и абиотических про-
цессах в море еще до конца не выяснена.
Продуктивность фитопланктонных организмов примерно в 3 раза
меньше, чем бактерий. Утилизируя энергию Солнца, они трансформи-
руют ее в органическое вещество своих тел. Затем эта энергия много-
кратно используется и трансформируется на различных трофических
уровнях — консументами и редуцентами — во всей толще воды и дна,
а также в различных биохимических процессах жизнедеятельности орга-
низмов. При этом в толще воды используется 95 % аккумулированной
энергии, а оставшаяся часть, примерно 5 % — бентосными животными.
Только 0,2 % оставшейся после рассеяния в биопроцессах энергии ини-
циирует процессы преобразования донных отложений и формирования
осадочных пород (Богоров, 1974).
Для того чтобы рассмотреть возможность пополнения морской
взвеси (масса которой относительно постоянна) посредством живого
10П
органического вещества мор-
ской воды, мы предположили,
что биомасса последнего - ве-
ТаЗлица 44. Биомасса и продукция живых
организмов Мирового океана, млрд т по
сырой массе (Богоров, 1974)
личина более или менее пос- организм тоянная. Продукция - нао- борот, величина переменная, Биомас- са Продук- ция Коэффи- циент П/Б
вследствие того что она еже- Продуценты годно почти вся расходуется, Фитопланктон 1,5 550 366 а потом снова воспроизво- Фитобентос 0,2 0,2 1 датся. Как отмечалось выше, Консументы сырая масса живого орга- Зоопланктон 21,5 53 2,5 нического вещества равна Зообентос 10,0 3 1/3 34,27 • 10’ т. Сухая мае- Нектон 1,0 0,2 1/5 са соответственно составляет Редуценты 5,63 -10’ т, а масса золы ЖИ- Бактерии 0,07 70 1000 вого - 2,89-10’ т (Богоров, Суммарное со- 34,27 676,4 - 1974) держание
При сравнении массы взвешенного вещества с живой органикой в
Океане нами было принято, что сухая масса живого вещества равна
5,6 млрд т (Горбенко, 1986). Эту же величину использовали Н.И.Ба-
зилевич с соавт. (1972), а также Е.А.Романкевич (1977). Наши расче-
ты, однако, показали, что баланс пополнения взвеси из всех известных
ее источников с общим содержанием взвешенного вещества в Океане
не сходится (Горбенко, 1986).
Если учесть, что морская взвесь — это органоминеральное вещест-
во, можно в расчетах использовать также минеральную часть, коли-
чество золистых веществ которой составляет немного более поло-
вины биомассы живого вещества (Богоров, 1974). Таким образом,
сухая биомасса живого вещества составит 5,63-10’ + 2,89-10’ =
= 8,53 млрдт.
Допустим, что все живое ежегодно переходило бы во взвесь,
тогда для ее восстановления понадобилось бы 1370 • 10’ млрд г:
8,53-10’ млрдт = 160,61 лет, т.е. примерно 161 год. Иначе говоря, все
живые организмы Океана не могли бы стать основным источником
пополнения взвеси, поскольку для воспроизводства органической
части взвеси требуется 10-20 лет (Романкевич, 1977).
В действительности значительное количество живых организмов —
рыбы, водоросли, моллюски и другие — изымается человеком для пи-
щевых и технических целей. Часть одних организмов используется для
пиши других. Происходит, как известно, и естественная смерть мор-
ских организмов, составляющих живое вещество Океана.
101
7.3. РОЛЬ ОТМИРАНИЯ МОРСКИХ ГИДРОБИОНТОВ
Вопрос о естественном отмирании мирских организмов исследовате-
ли ИнБЮМ изучали главным образом в Черном море.
Смертность фитопланктона в Черном море колеблется примерно
от 5 до 48 % (в среднем 27 %) (Кондратьева, 1967). Выше слоя темпе-
ратурного скачка (термоклина) количество отмирающих клеток в
2,4 раза меньше, чем живых. Под термоклином это соотношение увели-
чивается до 12 % (слой 12-25 м) и 60 % (75-100м). В среднем оно
составляет 36 % (Павлова, Миронов, 1970). В северо-западной части
моря смертность фитопланктона иногда достигает 100 % (Нестеро-
ва, 1969).
Нами установлено, что в бухте у Севастополя за 12—14 лет наблю-
дений в планктоне ежегодно отмирает до 71 % диатомовых водорослей,
в перифитоне - 75 и во взвеси - до 90 %*. По данным нескольких лет
наблюдений, в этом районе отмирает до 60 % клеток фитопланктона,
а к концу цветения некоторых их популяций смертность достигает
100 % (Сеничева, 1960). Если усреднить все приведенные выше данные,
то смертность фитопланктона, фитоперифитона и фитовзвеси в Черном
море составляет примерно 61 %. По данным А.А.Калугиной-Гутник
(1975), среди макрофитов ежегодно отмирает: филлофлоры — до
50 %, цистозиры — 80 и ульвы — 100 %. При условном усреднении эта
величина составляет 77 %.
Смертность зоопланктона ниже, чем фитопланктона. Выше термо-
клина количество мертвых зоопланктонных животных составляет
1,5 %, под термоклином в слое 10-25 м - до 2,5 %. В слое 100-150 м
содержится до 10 % мертвых организмов (Павлова, Миронов, 1970).
Определили, что в пелагиали морей смертность ряда зоопланкто-
неров составляет 22-36 %, в среднем 29 % (Залезинская, 1968). При
условном усреднении данных о смертности зоопланктона получаем
среднюю величину его отмирания, равную 10,0 %. Условное усреднение
всех приведенных выше данных об отмирании организмов, дающих
основную биомассу в море, показывает, что отмирание живого органи-
ческого вещества в Черном море составляет приблизительно 60 %.
Можно считать, что условия существования морских организмов
в Океане лучше, чем в Черном море (например, сероводородная зона
в Океане встречается редко), поэтому ежегодную естественную смерт-
ность их следует принять равной примерно 50 %.
Таким образом, детрит может образоваться только из половины
биомассы живого вещества (8,53 млрд т : 2 «4,27 млрд т), являющей-
ся реальным источником взвеси в Океане. Однако детрит ие может
быть основным источником взвеси, так как составляет лишь 1/321 <
1 Эти числа могут быть немного преувеличены вследствие попадания в ловушку >
для взвеси из толщи воды створок мертвых диатомей. ’
102
f
часть взвешенного вещества в Океане (1370 млрд т : 4,27 млрд т =
= 321). Ряд исследователей (Лисицын, 1974; Романкевич, 1977; Ви-
тюк, 1983) существенную роль в образовании взвеси отводят фито-
планктону.
7.4. ВОЗМОЖНОСТИ ПОПОЛНЕНИЯ МОРСКОЙ ВЗВЕСИ
ПРОДУКЦИЕЙ ФИТОПЛАНКТОНА
Данные о первичной продукции, полученные некоторыми исследо-
вателями (Базилевич, Родин, Розов, 1971; Кобленц-Мишке, Ведер-
ников, 1977; Marine Organic Chemistry, 1981), представлены ниже.
Средняя годовая (валовая) продукция фитопланктона (по сухой массе), млрд т-год”’ Литературный источник
105 Г.Г. Винберг (1960)
60 Н.И.Баэилевич и др. (1971)
95 Е.А.Ромаикевич (1977)
80 П.А.Моисеев (1969)
100 Е.Стимаи-Нильсен (1957)
НО В.Г.Богоров (1967)
88 О. И. Кобленц- Мишке, В.В.Ведерников
(1977)
77 Б.А. Скопинцев (1971)
50 Marine organic Chemistry (1981)
Средняя величина первичной продукции, полученная по данным этой
таблицы, составляет 85 ± 18 млрд т • год-1 (по сухой массе). Как
известно (Винберг, 1960; Богоров, 1967; Романкевич, 1977), собст-
венные траты фитопланктона на обмен составляют 30-40 % продукции,
в среднем около 35 %.
Наибольшая часть продукции (до 90 % и более) в открытых частях
Океана и морях используется зооорганизмами-фитофагами. Вместе с
тем в прибрежных районах потребление состаиляет около 10 %. При
усреднении величина выедания продукции фитопланктона, изменя-
ясь в пределах 10-90 %, в среднем составит около 50 %. За вычетом
трат на обмен (35 %) из общего количества продукции (100 %) ос-
тается 15 %.
По данным А.П.Лисицына (1974), Е.А.Романкевич (1977), на дно
Океана ежедневно оседает от 3-8 до 25 %, или в среднем около 15 %
взвешенного вещества от суммы первичной продукции. Таким об-
разом, судя по литературным данным, первичная продукция фи-
топланктона в Океане практически ничем не пополняется, а если
какая-то часть взвеси и образуется из фитопланктона, то она ничтож-
но мала.
103
7.6. АЛЛОХТОННЫЕ ИСТОЧНИКИ ВЗВЕСИ
И ДИСБАЛАНС ЕЕ СОДЕРЖАНИЯ С ПОПОЛНЕНИЕМ В ОКЕАНЕ
По расчетам А.ПЛисицына (1974), большое количество взвешенного
материала (до 18,5 млрд т-год"1) поставляют в Океан реки. Леднико-
вый и эоловый материал, а также абразия берегов и дна увеличивают
указанную величину до 22,13 млрд т • год-1 (без учета растворивших-
ся веществ — 3,2 млрд т-год"1). Вместе с вулканогенным и космо-
генным материалом пополнение составит около 24,7 млрд т-год"1.
Следует учесть, однако, что 80-90 % (в среднем 85 %) терригенного
стока осаждается в периферической части Океана, особенно вблизи
устьев рек (Лисицын, 1974), что составляет около 16 млрд т. Следова-
тельно, потенциальным источником взвеси в Океане являются терри-
генные, эоловые, космические и вулканические стоки, и это состав-
ляет 24,7-16,0 = 8,7, т.е. применно 9 млрд т-год"1.
В итоге, 1370 млрд т взвеси, поступающей в Океан за год, по-
полняется 4,27 млрд т детрита и 9 млрд т эолового, терригенного,
космического и вулканического материала, что примерно равно
13 млрд т-год"1. Это составляет только 1/105 всего взвешенного ве-
щества в Океане,и для сведения баланса этого недостаточно.
7.6. ОБРАЗОВАНИЕ ВЗВЕСИ В МОРЕ
В 1968-1982 гг. в Севастопольской бухте Черного моря у подводного
стенда на глубине 2 м от поверхности воды и в 3-4 м от дна в начале
каждого месяца подвешивали сосуд объемом 10 л для сбора взвеси.
Регулярно через 5-10 сут из сосуда брали часть осевшего взвешенного
вещества и исследовали его. Определяли количество осевшей взвеси,
ее состав, цвет и структуру. С помощью микроскопа в сообществе
микроорганизмов взвеси (СМВ), а также в сообществах перифитона
(СПМ) и планктона (СМП) подсчитывали состав и количество диатомо-
вых водорослей,простейших организмов, а на культуральной агарово-
белковой среде - число колоний гетеротрофных бактерий. В окружаю-
щей морской вода анализировали содержание РОВ, нитритов азота и
фосфора (органического и минерального). Для вычисления концентра-
ции компонентов карбонатного равновесия (СОа, HCOj, СО’э") опре-
деляли температуру воды, соленость, pH и щелочность. После 2-часо-
во го контакта взвеси и слизистой пленки СПМ с морской водой, окру-
жающей их, в воде определяли метаболиты РОВ СПМ и СМВ, pH СПМ
И СМВ. В высушенной до постоянной массы пленке СПМ определяли
количество карбонатов, микроэлементов и других веществ. Все чис-
ловые данные обрабатывали с помощью корреляционного анализа.
Для этого вычислили более 500 корреляций, из которых достоверных
с величиной Р = 0,30 ± 0,95 оказалось около 200.
104
Природные наблюдения показали, что морская взвесь представляет
собой слизистые частицы в аморфных ветвистых агрегатах разного раз-
мера (5 - 10 - 60 мкм). Эти агрегаты, в свою очередь, состоят из более
мелких частиц размером 0,2-3,0 мкм. Реже обнаруживали хлопья и
уплотненные частицы, еще реже фекалии планктонных рачков и совсем
редко - фрагменты мертвых планктонных организмов, которые не
всегда попадаются в изучаемой порции взвеси, подтверждая наш вывод
о том, что детрит составляет весьма малую часть взвеси.
Морфологически взвешенное вещество Черного моря по большин-
ству из 12 признаков (отмечавшихся выше) соответствует взвеси из
Океана. Только в заливах и бухтах морских водоемов из-за большой
их продуктивности частички взвеси более крупных размеров, чем в
открытых акваториях морей и Океана.
Во взвеси обычно встречаются также различные по цвету (от
прозрачных до мутноватых или желтых, красных, черных и т.д.) крис-
таллы органических и минеральных веществ. Кроме того, наблюдают-
ся капельки нефтепродуктов, ’’чистые” и обросшие микробной слизью.
Свежие капли нефти прозрачны и покрыты только следами микроб-
ной слизн, а старые окутаны слизью, иногда целиком, и цвет их корич-
невый или черный. Встречаются обросшие слизью пузырьки газа н про-
ростки водорослей-макрофитов, чаще всего красных.
В процессе наблюдений обнаружили, что в различных пробах взвесь
отличается в основном только величиной и формой частиц либо разме-
ром и цветом. Преобладали обычно частицы размером 10—15 мкм, но
иногда взвесь формировалась в тяжи длиною до 0,5 мм. Совсем редко
летом наблюдали тяжи взвеси до 0,5 см и немного более. При этом
фекальные комки н уплотненная часть взвеси по цвету темнее осталь-
ной взвеси. Цвет взвеси изменяется Не резко от светло-песочного до
желто- и буро-песочного, иногда с зеленоватым оттенком, реже взвесь
темная н еще более редко — черная нз-за сульфидов металлов, образо-
вавшихся в результате сульфатредукции взвешенного вещества. Сле-
дует отметить, что почерневшая в результате сульфатредукции взвесь,
кратковременно выдержанная на воздухе, снова приобретает свой
Первоначальный светло- или желто-песочный цвет с прозеленью, т.е.
процесс сульфатредукции, по-видимому, обратим. В теплое время
года заметно, как от 10-х к 30-м суткам наблюдений (т.е. с увели-
чением возраста взвеси) цвет взвеси изменяется от более светлого
к темному.
Частицы взвеси почти сплошь усеяны бактериями (А.ГРодина,
1966). На прозрачных или полупрозрачных частицах видно, что бакте-
рии располагаются в основном одним слоем. Часть из них всегда пере-
мещается между частицами взвеси. Бактерии могут быть различной
формы — палочковидные, нити, кокки и реже сардины. Обычны пред-
ставители Подвижных бактерий - вибрионы различных размеров и
форм - от запятой до клетки с одним витком.
105
Встречаются малоподвижные спириллы и изредка очень подвиж-
ные и гибкие спирохеты. Значительно меньше (по количеству) диа-
томовых водорослей и еше меньше простейших организмов (прикреп-
ленные и подвижные формы).
Среди простейших чаще всего наблюдались жгутиковые формы
родов: Bodo, Rhinchomonas и другие (Fenschel, 1970). Реже встреча-
лись инфузории и еще реже амебы. Иногда во взвеси мы наблюдали
синезелепые водоросли нитевидной формы и извитые клетки Spiruli-
па tenuissma. Иногда в жаркое время года в собранной взвеси проис-
ходит процесс сульфатредукции и к обычному ее запаху (рыбьего жи-
ра или нефти) примешивается запах сероводорода, а на взвеси появ-
ляются различные виды серных бактерий рода Beggiatoa. На крупных
частицах взвешенного вещества встречались науплиусы, раки Награс-
ticidae и нематоды. Диатомовые водоросли, главным образом беито-
планктонные формы, представлены в основном 51 видом и родом, при-
веденных ранее (раздел 1.4).
На основе материалов и результатов наблюдений, проведенных
нами за природной взвесью, удалось подойти к расшифровке механиз-
ма образования ее в море.
7.7. МЕХАНИЗМ ОБРАЗОВАНИЯ МОРСКОЙ ВЗВЕСИ
Взвешенное вещество в морской воде образуется в результате смены
4 стадий биотической сукцессии при развитии СМВ на поверхности
частиц - микросубстрата (пыль, вулканический пепел, терригенные
крупицы, трудноразлагаемые частицы детрита), непрерывно попа-
дающих в море или образующихся там.
I стадия. Попавшие в воду мельчайшие частицы - мик ро субстрат -
адсорбируют на своей поверхности молекулы органических веществ
и уже в первые часы заселяются бактериями. На частицы их привле-
кает не только адсорбированное органическое вещество, но и преиму-
щества, предоставляемые им твердой поверхностью частиц, действую-
щей на границе раздела с водой, как катализатор. Кроме того, в этом
месте обеспечиваются более благоприятные условия для использова-
ния бактериями органического вещества, на которое они могут целе-
направленно ’ действовать ферментами, не вымывающимися водой.
На плотном субстрате, кроме того, подвижная клетка бактерий или
диатомовых водорослей может выбрать более подходящее место для
осуществления процессов жизнедеятельности, т.е. проявляются преиму-
щества жизни микроорганизмов на твердой (или плотной) поверх-
ности (Zo Bell, 1943).
Примерно к концу I стадии, продолжающейся всего несколько
дней, на частицах нарастает слизистая пленка светло-песочного цвета,
реже буроватого. Иногда в это время на взвеси появляются мелкие
106
жгутиковые простейшие организмы родов Bodo, Rhinchomonas и др.,
питающиеся бактериями и пленкой.
II стадия. Наступает приблизительно через 3-4 дня летом и 5-7 -
зимой.На частицах взвеси появлялись диатомовые водоросли, после че-
го светло-песочная взвесь становилась бурой, частицы росли, увеличи-
ваясь в размерах. В начале стадии или немного позже на взвеси обнару-
живались синезеленые водоросли, раки Harpacticidae и нематоды. В этот
Период микроорганизмы начинают осаждать из морской воды карбона-
ты, кремний, микроэлементы и другие неорганические компоненты.
Ill стадия. Характеризуется тем, что мелкие частицы взвеси, кото-
рые образовались в I и II стадиях, достигнув размера примерно 3,0-
3,5 мкм, сливаются обычно в более плотные структуры величиной
10 мкм и более. Цвет структур, как правило, серо-бурый, потому что
на частицах развивалось типичное СМВ. Стадия долговременная.
IV стадия - заключительная. Она наступает, когда отдельные час-
тицы взвеси достигают размеров 50-100 мкм и становятся плотнее и
тяжелее. От этого плавучесть их теряется, и они начинают оседать на дно,
становясь осадком. В сосуде для сбора взвеси, погруженном в море,
количество частиц таких размеров обычно больше, чем мелких. Разме-
ры частиц, склонных к оседанию, 30-60 мкм (Riley, 1970). Однако в
Севастопольской бухте размеры таких частиц в 2 раза больше. Причина
этого-в более высокой продуктивности бухты, являющейся зоной
шельфа, в которой условия для образования взвешенного вещества
достаточно благоприятны. Из-за мощных течений и больших глубин осе-
дание его на дно продолжается несколько лет (Riley, 1970). Поэтому’
возникает вопрос, как доказать, что она образуется именно так, а не
вдаче? Для этого необходимо было получить взвесь опытным путем.
Точно повторить процесс образования взвеси в эксперименте со
Всеми сукцессиями и другими изменениями во времени - дело чрезвы-
чайно трудное и почти невозможное. Большое количество связей, мно-
жество био- и абиотических факторов морской среды, экологическая
обстановка в море, течение, компоненты загрязнения в лабораторных
условиях воспроизвести нельзя. Однако,если природный процесс возни-
кновения взвеси воспроизвести только в основных чертах, то можно
получить взвесь, подобную морской.
7.& ОБРАЗОВАНИЕ ВЗВЕСИ ЭКСПЕРИМЕНТАЛЬНЫМ ПУТЕМ
В подготовительной методической работе путем постановки вспомога-
тельных опытов были выявлены: 1)наиболее подходящий субстрат для
образования взвеси; 2) размер субстрата и оптимальное количество
опытных сосудов и воды в них; 3)вид РОВ для культуральной среды и
его концентрация; 4)время экспозиции опыта; 5) характер и количест-
во микрофлоры для засева колб.
107
В Hioie в качестве РОВ были использованы пептон, агар-агар (ис-
пользуемый для приготовления твердых культуральных сред) и раст-
воримый крахмал. Лучшим микросубстратом - основой для образо-
вания взвеси — оказались толченое стекло и графит, которые исти-
рали до пылевидного состояния. Были выбраны сосуды для экспери-
мента и количество морской воды в них. Поскольку в предваритель-
ных опытах не удалось выявить, какой из микросубстратов лучше —
графит или стекло, провели 2 серии экспериментов. 1-я серия опытов
повторялась 5 раз, число повторностей в каждом опыте — 3. Экспери-
менты проводили в летне-осенний период на среде с пептоном,
концентрация которого для быстроты ответа была максимальной
(100 мг.л-1). Микросубстратом в данном случае служили графит и
стекло, а колбы экспонировали при температуре морской воды 18-
20 °C. 2-ю серию опытов проводили в осенне-зимний период на средах
не только с пептоном, но и с безазотистыми компонентами РОВ -
агар-агаром и растворимым крахмалом, концентрации которых были
ниже, чем в 1-й серии опытов. Микросубстратом служил только гра-
фит, который способствовал получению более четких результатов, чем
стекло. Температура морской воды была 12-14 °C. Число опытов
такое же, как и в 1-й серии экспериментов.
В опытах использовали колбы объемом 5 л, в контроле - 3 л.
В каждую колбу наливали по 2 л свежей морской воды, пропущенной
через бумажные фильтры. Туда же вносили по 150 мг-л-1 стеклянной
или графитовой пыли. РОВ вносили в количестве 30,50 и 100 мг-л-1,
поскольку в прибрежных водах морей и Океана его содержится от
5- 10 до 50-100 мг-л-1 (Хайлов, 1971). Микрофлору для засева сред
смывали со свежей морской взвеси и фильтровали ее через бумажный
фильтр. Всего в каждую колбу вносили по 15 мл этой суспензии.
Срок экспонирования колб составлял 2—7 сут. По истечении это-
го времени вода в колбах просветлялась,pH среды понижалась,а взвесьи
микроорганизмы выпадали в осадок. На 8-10-е сутки и позже насту-
пало отравление микроорганизмов собственными продуктами обмена
веществ (самоотравление), и условия их существования в изолирован-
ных сосудах резко ухудшались. После окончания опыта взвесь микро-
скопировали, а затем пропускали через мембранные фильтры ”Сын-
пор № 4”. Высушенные до постоянной массы фильтры (при t = 100 °C)
взвешивали на аналитических весах, а осадок анализировали.
Наблюдения за развитием микроорганизмов в экспериментальных
сосудах показали, что в 1-й серии опытов уже на 2-е сутки вода во всех
сосудах помутнела от количества развившихся там бактерий. На 3-4-е
сутки экспонирования черные до опыта крупицы графита посветлели
из-за образовавшейся пленки микроор»анизмов. Вместе с тем 1/4 часть
этих крупиц оставалась взвешенной в толще воды у дна, не оседая обычно
до 6-7-х суток опыта. В то же время частицы стеклянной пыли были в
основном только в осадке, и пленка на них была намного тоньше.
108
§
2S
'g
ро
§ о со
н
£
g 54 о 5 о
X п
5
X
&
X
к X
сх
ООО
ООО
Г1 N N
A A A
00 00 00
[исходная
Количество карб<> иатов на взвеси Y (% прироста)
1 Прирост вещест- । ва взвеси X । (мг сухой массы) i
Содержание компонента в среде, мл-л~’
oooo о
m xt-У rn <*>
A_ A A_ A A
сч гпст'сч ГЦ
о <э о о о
»П m *Г> П V>
Обнаружили, что на части-
цах искусственной взвеси раз-
вился почти сплошной слой
бактерий, так же как и на при-
родных взвешенных частицах.
Бактерии тоже активно пере-
мещались между частицами
взвеси. На всех образцах обна-
руживались одни и те же мик-
роорганизмы; палочковидные
бактерии, вибрионы. Встреча-
лись спириллы с 4-5 вит-
ками, похожие на Rhodospiril-
lum rubrum, нитевидные клет-
ки бактерий — подвижные и
неподвижные, обычные мел-
кие жгутиковые формы прос-
тейших организмов, питаю-
щиеся компонентами слизис-
той пленки.
В опытных сосудах с гра-
фитом и стеклом, растертых
до пылевидного состояния,
микрофлоры было значитель-
но больше, чем в контроле.
Этот факт свидетельствует, что
поверхность твердых предме-
тов или плотных частиц спо-
собствует развитию микро-
организмов (Zo Bel!, 1943).
В то же время микроорга-
шгзмов было заметно боль-
ше в сосудах с графитом,
чем со стеклом. В послед-
них, в частности, наблюда-
лось намного меньше вибрио-
нов, реже встречались спирил-
лы и простейшие организмы.
Через 7-10 сут взвесь осаж-
далась, а жидкость в колбах
светлела и подкислялась до
pH 7,5 и ниже, т.е. для микро-
организмов условия развития
становились неблагоприятны-
ми (табл. 45).
109
Результаты эксперимента свидетельствуют, что во всех образцах
количество осадка и взвеси на частицах неорганической пыли увеличи-
валось. Это сопровождалось снижением pH, а в опытных колбах еще и
накоплением карбонатов, отлагаемых микроорганизмами, и более
сильным подкислением среды, чем в контрольных образцах. Увеличе-
ние органического вещества и карбонатов в колбах с графитом было
наибольшим.
Итак, опыт показал, что в морской воде с бактериями, графито-
вой и стеклянной пылью, незначительно (на 0,01 %) обогащенной пеп-
тоном, за относительно короткое время образуется типичная взвесь,
по ряду основных признаков соответствующая взвешенному веществу
морской воды. Эту взвесь, которая образовалась при развитии СМВ на
минеральных частицах в течение 7 сут, мы назвали микробиоцено-
тической взвесью.
В контрольных колбах после помутнения среды обнаруживали
тонкую муть, напоминающую мельчайшую взвесь, но без карбонатов.
Поскольку одним из постоянных компонентов взвеси являются карбо-
наты, отлагающиеся на ней, то взвешенное вещество из контрольных
сосудов можно считать нетипичным. Частички такой взвеси в море,
по-видимому, могут возникать из бактериальных агрегатов (Сорокин,
1970) и также служить микросубстратом для образования микробио-
ценотиЧеской взвеси (через некоторое время на этих частицах в резуль-
тате развития СМВ отложатся карбонаты и станут обычной взвесью).
В проведенном эксперименте установили, что более благоприятной
основой для образования взвеси в лабораторных условиях является
графит и менее благоприятной - стекло.
Накопление в контроле большего количества ’’нетипичной” взвеси,
чем в опытном сосуде со стеклянной Пылью, можно объяснить отноше-
ние поверхности сосуда к его объему, величина которого в малых со-
судах выше, чем в больших. Как известно, это один из факторов, спо-
собствующих развитию бактерий в лабораторных сосудах (Whippl,
1901).
7.9. СКОРОСТЬ ОБРАЗОВАНИЯ
МИКРОБИОЦЕНОТИЧЕСКОЙ ВЗВЕСИ
И БАЛАНС ее в океане
На основании полученных данных (1-й эксперимент, табл. 45) можно
рассчитать скорость роста частиц взвешенного вещества в Океане, если
предположить, что СМВ развивалось примерно так же, как на графито-
вой и стеклянной пыли. Скорость нарастания взвеси
t '
110
где Го — начальная скорость; Гк — конечная скорость; t — время, сут.
Для частиц микросубстрата ь море, которые качественно подобны
пыли из графита, Ио = 0; Гк = 95,8 мг, или 0,0958 г за 7 сут (табл. 45);
за 1 сут нарастало V! = 0,0958 : 7 = 0,014 г-сут"1.
Для упрощения расчета предположим, что на 1/365 части всего
взвешенного вещества Мирового океана развитие происходит так же,
как на крупицах графитовой и стеклянной пыли в эксперименте: 1/365
от 1370-Ю’ т - 3,75-10’ т = 3,75-10” г-сут”1. Если на 0,3 г графитовой
пыли за сутки наросло 0,014 г сухой массы взвеси, то на 3,75-10” г
нарастало бы 0,173-10” г, или 173-10* т-сут”1. За год ее нарастет соот-
ветственно 63,14-10’ т. Это значит, что если бы с такой же скоростью
СМВ развивалось бы не на 1/365 части взвеси в Океане, а иа 22/365, то
вся взвесь (1370 млрд т) могла бы обновиться за 1 год.
Для частиц микросубстрата взвеси, качественно подобных стек-
лянной пыли, Ио =0; =0,0033; V2 =0,0005 г-сут”1.
Учитывая, что на 0,3 г пыли из стекла за сутки нарастало 0,0005 г
взвешенного вещества, то на 3,75 • 10” г взвеси это составило бы
0,006-10” г = 6-10* т-сут”1. За год это составит 2,19-10’ т. Таким
образом, на 1/365 части взвеси в Океане может нарасти 2,19-10’ -
63,14-10’ млрд т сухой массы взвеси. В среднем за год это составит
32,66-10’ т * 33-10’ т-год”1. Если предположить, что с такой же ско-
ростью масса взвеси будет нарастать не на 1 /365 ее части, а на 42/365,
то вся взвесь в Океане микробиоценотическим путем могла бы восста-
новиться уже в течение 1 года.
Вследствие того что в 100 мг пептона содержится намного больше
органического азота, чем в морской воде, мы повторили указанный
эксперимент с меньшим его количеством и с безазотистыми органи-
ческими веществами (агар-агаром и растворимым крахмалом). Кроме
того, вследствие неблагоприятных качеств из опыта было решено
исключить стеклянную пыль.
II серию опытов ставили в осенне-зимнее время (по той же методи-
ке, что и в 1-м эксперименте). Температура воды в море и экспери-
менте была намного ниже, чем в I серии опытов, составляя 12-14 С,
и условия эксперимента были более жесткими, что не могло не отра-
зиться на его результатах. Через 3-4 сут вода в опытных колбах заму-
тилась, а крупицы графита посветлели из-за того, что они обросли сли-
зистой пленкой СМВ.
Наблюдения под микроскопом показали, что взвесь в первые сут-
ки после ее образования состояла из мельчайших частиц, которые поз-
же склеивались в аморфные комочки, такие же, как в море. Часть
образовавшейся взвеси, как и в предыдущем эксперименте с графи-
том, находилась на дне колб, а часть - у дна.
В слизистой пленке обнаруживались малоподвижные кокковидные
и изогнутые формы морских бактерий. Если в период минимальной
численности диатомовых водорослей в море, когда проводили I зкспе-
111
римент, диатомовые на взвеси в колбах не встречались, то время нас-
тоящих экспериментов совпало с осенним максимумом числа диато-
мей в море. Поэтому на частицах образовавшейся взвеси мы обнару-
жили несколько видов диатомей родов Navicula, Nitzschia и др. Из
простейших наблюдались инфузории и округлые формы, похожие на
роды Bodo, Rhinchomonas. Встречались нитевидные бактерии. Спирил-
лы во время данных наблюдений выявлены не были.
Из 3 применявшихся в опытах источников углерода наиболее бла-
гоприятным для микроорганизмов оказался агар-агар. Во взвеси из
среды с агар-агаром прирост органического вещества наибольший. При
этом отложилось и наибольшее количество карбонатов. Из-за незначи-
тельных изменений в период II серии опытов влияние температуры
не наблюдалось. Концентрация РОВ в колбах также мало влияла на
образование взвеси. В эксперименте во всех колбах среда подкисля-
лась микроорганизмами: наиболее интенсивно - в среде с раствори-
мым крахмалом, наименее - на агар-агаре.
В контроле легкое помутнение среды и следы прироста органичес-
кого вещества были обнаружены только на пептоне. Причем карбона-
тов в этом веществе, как и в контроле I серии опытов, обнаружено не
было. В контроле с агар-агаром и крахмалом среда не помутнела и
прироста органического вещества в ней не обнаружили. Слабое подкис-
ление среды отметили только в контроле с крахмалом. На остальных
средах в контроле pH почти не менялась.
Таким образом, на различных источниках углерода, с содержанием
азота и без него, в концентрации, равной содержанию РОВ в прибреж
ных частях моря, на пыли графита, являющейся микросубстратом для
формирования взвеси, в результате жизнедеятельности СМВ образу-
ется взвесь, подобная природной взвеси, как и в 1 -м эксперименте.
Какова же скорость образования взвешенного вещества в данном
опыте, достаточна ли она для воспроизведения всей массы взвеси в
морях и Океане? Эту скорость рассчитывали по данным табл. 45, как и
В предыдущем эксперименте:
50 мг пептона-л-1; « 0,001 г.сут” 1
30 мг агар-агара-л-1; 0,0046 г-сут- *
50 мг агар-агарал-1; Ке * 0,0049 г-сут-1
30 мг крахмала л”1; » 0,00035 г-сут- *
50 мг крахмала-л-1; = 0,0005 г-сут-1
Соответственно рассчитываем, сколько могло бы нарасти вещества
в Океане, которое стало бы микросубстратом для формирования взве-
си (1/365 от 1370 млрд т =а 3,75-10” г-сут-1). В опыте с пептоном
На 0,3 г графитовой пыли скорость нарастания взвеси составляв!
0,001 г-сут-1, а на 3,75-10** г-сут-1 с такой же скоростью наросло бН
12,5-1012 г-сут-1, или 12,5-10* т-сут-1. В опыте с 30 мг агар-агара эта
Величина составила бы 57,5-10* т-сут-?. В опыте с 50 мг агар-агара она
была бы равна соответственно 61,25-10* т-сут-1 .вопытесЗОмгкрахма-
112
ла - 4,37-Ю6 т-сут-*, в опыте с 50 мг крахмала - 6,25-Ю® т-сут-1.
Средняя величина прироста массы взвеси в проведенных опытах состав-
ляет 28,37-10® т-сут-*. За год с такой же скоростью на 1/365 части
взвеси нарастало бы 10,35-10’ т-год-1, что составляет 1/132 части об-
щего количества взвешенного вещества в Океане. Таким образом,
если бы на 132/365 части всей взвеси Мирового океана микробиоце-
нотическим путем и со скоростью такой же, как в эксперименте, обра-
зовалась взвесь, то уже за 1 год вся ее масса в Океане возобнови-
лась бы.
При сравнении скорости образования взвеси на среде с пептоном
в I серии опытов (Kj = 0,014 г-сут-*) и во II (К4 s 0,001 г-сут-*)
(табл. 45) отметили, что примерно на порядок выше, чем К4.
Ясно, что это следствие более жестких условий 2-го эксперимента, что
отмечалось выше, а также и того, что длительность 1-го эксперимента
составляла 7 сут, а 2-го - всего 2 сут.
В целом же скорости образования взвеси в обоих экспериментах,
проведенных в различные сезоны года, различались мало. На среде с
агар-агаром в концентрации 30—50 мг-л-* этот показатель всего в 3 ра-
за меньше, чем на среде со 100 мг-л“ ’ пептона. На среде с растворимым
крахмалом в концентрации 30-50 мг-л-* и с графитом скорость была
Почти такой же, как на среде с пептоном, но со стеклянной пылью
(I серия опытов).
Зная о почти неограниченных возможностях микроорганизмов
растворять любые органические и неорганические вещества, мы не сом
деваемся в том, что взвесь в эксперименте будет образовываться на
среде почти с любым органическим веществом. А микросубстратом
может быть любое неорганическое или трудно используемое микро-
организмами органическое вещество. Изменяться будут только сроки
экспонирования в опыте, скорость образования взвешенного вещества,
а может быть, еще температура среды и концентрация РОВ в ней. Та-
ким образом, теоретически обоснована и экспериментально доказана
возможность образования взвеси в море и Океане автохтонно - микро-
Ьиоценотическим путем.
Вследствие того что пока не известен какой-либо иной путь обра-
зования и пополнения взвеси в морских водоемах, мы имеем все осно-
вания считать, что наибольшая часть взвеси в Океане образуется ука-
занным путем. Меньшая часть взвеси поступает с суши, из рек, возду-
ха, вулканов и космоса, а также пополняется за счет детрита, кото-
рый, безусловно, является частью взвеси. Вследствие того что проис-
хождение взвеси и детрита различное, аналогию между ними проводить
нельзя. Как показали результаты настоящей работы, взвесь и детрит,
по-видимому, следует рассматривать как различные морские материа-
лы, подобные микросообществам.
По результатам 1-го эксперимента для воспроизводства всей
взвеси в Океане за 1 год достаточно, чтобы только 44/365 части были
113
использованы в качестве микросубстрата для развития СМВ, которое
проходило бы с такой же скоростью, как в эксперименте. По данным
2-го эксперимента достаточно было бы, чтобы 132/365 части взвешен-
ного вещества были использованы. В среднем (по данным обоих экспе-
риментов) это составит 88/365, или «» 0,25, т.е. микробиоценотическое
образование взвеси, происходящее с такой же скоростью, как в экспе-
рименте, только на 1/4 части взвешенного вещества Океана, уже за год
по своим масштабам могло бы возобновить всю массу взвеси в Миро-
вом океане.
В действительности в морях и Океане условия для образования
взвеси хуже, чем в эксперименте. Если концентрация РОВ - главного
источника углерода для бактерий СМВ - в прибрежных районах мор-
ских водоемов колеблется примерно от 10 до 25 мг л" ’, а у поверхнос-
ти раздела фаз - выше 25 мг-л” 1 (в поверхностной пленке воды она
может составлять даже 100 мг-л“ *), то в среднем в Океане и морях она
составляет 1,6—3,0 мг-л-1 (Хайлов, 1971; Романкевич, 1977). Темпе-
ратура воды в Океане также примерно в 2-5 раз ниже, чем в экспе-
-рименте (18-20 °C). Следовательно, скорость нарастания массы
взвеси на частицах — микро субстрате — будет в целом меньше экспе-
риментальной. Как было отмечено выше, органическая часть взвеси в
Океане обновляется за 10-25 лет (Романкевич, 1977). Учитывая боль-
шие возможности образования микробиоценотической взвеси и отно-
сительно высокую скорость этого процесса, время обновления взве-
шенного вещества в Океане, по нашему мнению, составляет не более
5-10 лет.
ГЛАВА 8. ИСПОЛЬЗОВАНИЕ НЕКОТОРЫХ РЕЗУЛЬТАТОВ
ЭКОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
КАК ИНДИКАТОРОВ ЯВЛЕНИЙ И ПРОЦЕССОВ
В МОРЯХ И ОКЕАНЕ
&1. ВЛИЯНИЕ КОНВЕКТИВНОГО ПЕРЕМЕШИВАНИЯ ВОД
В СРЕДИЗЕМНОМ МОРЕ
НА ИЗМЕНЕНИЕ ЧИСЛЕННОСТИ ГЕТЕРОТРОФНЫХ БАКТЕРИЙ
Из-за особенностей гидрометеорологического режима, в котором от-
четливо выражены лишь два времени года - зима и лето, Средиземное
море является примером крупного морского бассейна, где зимнее
конвективное перемешивание распространяется на большие глубины,
а в ряде случаев - до дна. Скорость этого процесса, однако, мала
(Schott, 1915! Валло, 1948; Pollak, 1951; LaCombe, Tchernia, 1962;
1972, 1974; Овчинников с соавт., 1976). Наша задача заключалась в
том, чтобы в зимний и летний сезоны в море изучить распределение
гетеротрофных бактерий по глубинам и сопоставить полученные дан-
ные с гидрологическими характеристиками морской среды.
Особенности методики. В летней (июль—октябрь 1959 г.) и зим-
ней (ноябрь-декабрь I960 г. - январь 1961 г) экспедициях нис’’Ака-
демик Вавилов” в центральной и восточной частях Средиземного моря
мы выполнили соответственно 40 и 53 микробиологические станции,
на которых по всем стандартным горизонтам, где брали батометричес-
кие пробы (0, 10, 25, 50, 75, 100, 150, 200, 250, 300, 400, 500, 750,
1000, 1250, 2000, 2500, 3000, 3500 м), учитывали колонии гетеротроф-
ных бактерий из морской воды, вырастающих на культуральной сре-
да - сухом питательном агаре (СПА).
В качестве дополнительного материала использовали данные о бак-
териях-гетеротрофах, полученные аналогичным образом Э.Я.Анищен-
ко на 92 станциях нис "Академик Ковалевский”, выполненных в сен-
тябре - октябре 1958 г-> июле-сентябре 1959 г., июне-июле I960 г.,
феврале 1962 г. в основном в летнее время (карта микробиологичес-
ких станций приведена на рис. 4).
Гетеротрофные бактерии учитывали методом проращивания мем-
бранных фильтров в чашках Петри с культуральной агарово-белковой
11$
Таблица 46. Численность колоний гетеротроф-
ных бактерий в 40 мл воды летом и зимой
на различных станциях
Глуби- на, м Станция, N*
5 566 368 569 372 71
Лето Зима Лето Зима Лето Зима
0 87 400 25 282 160 410
10 16 344 6 254 31 310
25 12 330 166 272 25 540
50 20 185 6 140 41 274
75 1 320 40 182 95 200
100 76 370 233 — 175 260
150 120 290 44 147 150 255
200 10 230 94 226 248 332
250 4 164 6 И 45 284
300 290 192 175 72 6 320
400 2 252 4 35 15 242
500 3 272 75 265 401 410
750 — 186 — 210 191 120
1000 7 272 — 196 40 272
1250 0 — — 9 — 330
1500 3 30 — — 38 —
2000 2 110 — — 16 —
2500 205 264 — — 205 —
3000 — 394 — — 2 —
средой, через которые пред-
варительно фильтровали во-
ду из гидрологических ба-
тометров (30-40 мл). Этот
метод широко применял
А.Е.Крисс (1959). В нас-
тоящей работе использова-
ли числовые данные рас-
пределения бактерий-гете-
ротрофов в водной толще
моря.
Все имеющиеся данные
о численности гетеротроф-
ных бактерий в море В раз-
личные сезоны и годы мож-
но в основном разделить
на ’’летнее” и ’’зимнее”
распределение. Это связано
с тем, что в гидрометео-
рологическом смысле в мо-
ре отчетливо выделяются
лишь лето и зима (что от-
мечалось выше).
Основная особенность
летнего распределения гетеротрофов в том, что перепад величин макси-
мума и минимума их численности на разных горизонтах большой (от
нулей и однозначных до дву- и трехзначных чисел), но величины дос-
таточно малы (обычно одно- и двузначные, редко трехзначные). Зимнее
распределение, наоборот, характеризуется малыми перепадами (только
от дву- до трехзначных чисел), но значительно большей (на 1—2 поряд-
ка) величиной численности колоний бактерий (обычно только трех-
значные числа), т.е. перепады численности гетеротрофов зимой ’’размы-
ваются”. При этом, как правило, стратификация слоев остается неиз-
менной, т.е. не зависит от сезона (табл. 46).
Вследствие того что зимой характеристики водных масс выравни-
ваются в результате смешивания вод по горизонтам, мы считаем, что
изменение числа колоний гетеротрофов по глубинам отражает этот
процесс.
8.2. ЛЕТНЕЕ РАСПРЕДЕЛЕНИЕ ЧИСЛЕННОСТИ БАКТЕРИЙ
Численность бактерий по станциям летне-осеннего рейса нис ’’Акаде-
мик Вавилов” в 1959 г. располагалась следующим образом. На 22 стан-
циях из 40, разбросанных по акваториям Тирренского моря, залива
116
Сирт, Ионического моря у берегов Греции, между островами Родос и
Крит, а также южнее о-ва Крит, в морях Леванта и Эгейском (ст. IP 324,
328 , 329, 353, 358, 359, 361, 363, 365, 366, 368, 370, 372, 379, 381,
383, 394, 396, 400, 414, 417, 418), обнаружили отчетливое летнее рас-
пределение колоний гетеротрофных бактерий (табл. 47).
В летне-осеннее время 1958, 1959, 1960 и 1962 гг. в рейсах на
нис ’’Академик Ковалевский” Э.Я.Анищенко выполнила 66 станций
(ст. № 1-48, поскольку в разных морях некоторые номера станций
повторяются). Эти станции располагались в Тирренском, Ионическом,
Критском, Андриатическом и Леваита морях, в проливе Отранто и
центральной части Средиземного моря.
На 11 станциях, выполненных в рейсах на нис ’’Академик Вавилов”
летом-осенью 1959 г. в восточной части Ионического моря, в Тир-
ренском море, Тунисском проливе и на юге моря Леванта (ст. № 304,
306, 320, 321, 329, 332, 335, 340, 342, 390, 391), слои воды были пере-
мешаны до 25-381 м. В наибольшей части водной толщи наблюдалось
летнее распределение бактерий-гетеротрофов. Подобное явление наблю-
далось и на 2 летних станциях (август 1959 г.), выполненных в рей-
сах на нис ’’Академик Ковалевский” в Ионическом море (ст. № 18, 19).
Иной тип распределения гетеротрофных бактерий обнаружили на
7 станциях в Тирренском, Ионическом морях и море Леванта, выпол-
ненных в рейсах на нис ’’Академик Ковалевский” летом 1959 г.
(ст. If 9, 13, 14, 18, 19, 29). До глубины 50—400 м наблюдалось летнее
распределение, а ниже иа всю глубину станций (628-1727 м) слои
воды были перемешаны, как зимой. Это объясняется, по-видимому,
мощными глубинными течениями, протекающими под указанными
станциями (Овчинников и др., 1976).
Интересно отметить, что на станциях (ст. № 344, 348, 350, 352,
355, 388, 392), выполненных летом экспедицией нис ’’Академик Вави-
лов” в юго-западной, северо-западной и центральной частях залива
Сирт, а также в северо-западной части моря Леванта, обнаружили ти-
пично зимнее распределение гетеротрофов, а следовательно, и переме-
шивание всей толщи воды - от поверхности до дна (рис. 4, табл. 46).
Одной из причин такого явления может быть то, что 5 первых из ука-
занных станций относительно мелководные (их глубина 75-196 м,
лишь глубина ст. № 350-500 м) и находятся в струе наибольшей ско-
рости движения Северо-Африканского течения (Овчинников и др.,
1976); это, по-видимому, способствует сильному смешиванию вод.
Станции № 388, 392 (глубина 772-3000 м) располагаются примерно
в центре моря Леванта, рядом с конвергенцией № 7, в месте столкнове-
ния поверхностных течений и прохождения глубинных вод, отчего,
по-видимому, перемешивание вод может распространяться от поверх-
ности до больших глубин даже в летнее время.
Зимнее распределение бактерий наблюдалось также на ст. № 22 на
юге Адриатического моря (глубина 300 м).выполненной летом 1959 г.
117
море
119
в рейсе на нис ’’Академик Ковалевский", в том месте, где проходят
глубинные воды (Овчинников и др., 1976). В целом на 80 станциях
из 106, выполненных в летне-осеннее время 1958—1960, 1962 гг.,
наблюдалось типично летнее распределение гетеротрофных бактерий.
Эго свидетельствует о летней вертикальной структуре морских вод,
почти не перемешанных в это время года.
&3. ЗИМНЕЕ РАСПРЕДЕЛЕНИЕ ЧИСЛЕННОСТИ БАКТЕРИЙ
Третий осенне-зимний рейс нис ’’Академик Вавилов" начался 5.11.1960 г.
В это время еще стояла ясная, почти без ветров погода. На станциях,
выполненных в Мраморном (ст. №514, 515) и Эгейском морях
(ст. № 516, 517, 518, 519), наблюдалось типично летнее распределение
бактерий (рис. 4, табл. 46). Погода начала ухудшаться 10 ноября,
усилился ветер и началось волнение моря. До начала января 1961 г.
уже почти все время штормило.
Анализируя последствия такой погоды в изучаемой части Среди-
земного моря, подвергшегося зимней конвекции вод, мы обнаружи-
ли ряд общих черт в изменившемся распределении гетеротрофных
бактерий.
1. На 41 станции из 53 (исключая 6 станций, указанных выше,и 6
станций в Северной части моря Леванта), выявилось зимнее распреде-
ление бактерий, т.е. в большей части водной толщи происходило пере-
мешивание вод.
2. По микробиологическим данным смешивание вод в море распро-
страняется неравномерно. В открытых частях моря и некоторых проли-
вах скорость его больше, а в северной части моря Леванта конвекция
мала или ее совсем нет.
3. Отчетливо распространяется перемешивание вод на глубину со
временем 10.11.1960 г., т.е. подтверждаются литературные данные о нача-
ле зимнего конвективного перемешивания средиземноморских вод.
Более детально перемешивание вод происходило так. Утром
10.11.1960 г. в 9 ч 00 мин была выполнена ст. № 519, которая находи-
лась севернее о-ва Крит. День был прохладный, но ясный, и распреде-
ление численности бактерий по всем горизонтам было летним. Пример-
но в12 ч 00 мин - 13 ч 00 мин подул холодный ветер и погода начала
ухудшаться. В 23 ч 00 мин была выполнена ст. № 521 на юго-восток
от о-ва Крит. Погода по-прежнему ухудшалась, началось небольшое
волнение. Изменившаяся за указанные сутки погода, ветер и понижен-
ная температура воздуха, по-видимому, привели к конвективному пе-
ремешиванию поверхностных слоев воды, и зимнее распределение ге-
теротрофов на этой станции наблюдалось до глубины 150 м.
Предположим, что перемешивание вод произошло примерно с
12 ч 00 мин до 23 ч 00 мин, т.е. в течение 12 ч. После деления глубины
120
перемешивания (150 м) на время продолжения конвекции (12 ч) ока-
залось, что примерная скорость конвекции (Икои) вод в данном слу-
чае ГКоН ~ 0,35 см. с-1. Поскольку ранее считали, что Ккон = 0,01-
0,001 см с"1 (Овчинников и др. 1976), величина, полученная нами,
была в 35-350 раз больше.
Заметим, что перемешивание вод на указанной станции произошло
только из-за конвекции водных масс, поскольку в этом районе летом
не обнаруживали станций с зимним распределением (етеротрофных
бактерий и на расположенных наиболее близко ст. № 27, 32 (нис ’’Ака-
демик Ковалевский”) летом было летнее распределение гетеротрофов.
Ст. № 536 была выполнена 13.11 - через 3 сут от начала зимнего се-
зона в море. Располагалась она и центре моря, севернее о-ва Крит.
Перемешивание вод на станции наблюдалось до глубины 2060 м. Ско-
рость конвективного перемешивания V - 0,794 см.с”1 - это в 79-
794 раза больше ранее принятой. В этом районе летом также не обнару-
живали станций с зимним распределением гетеротрофных бактерий.
На ст. № 368, расположенной рядом, летом было типично летнее распре-
деление бактерий. Таким образом была рассчитана скорость конвек-
ции и на других станциях, где (ст. № 550) уже через 5 сут конвектив-
ное Перемешивание достигало глубины 2974 м.
В целом на ст. № 521,536, 539, 550 скорость конвективного пере-
мешивания составляла V - 0,35-0,79 см-с’1, т.е. во много сотен раз
больше, чем считалось ранее.
Глубокое Перемешивание наблюдалось на 24 станциях, выполнен-
ных в основном в зимнее время (ст. № 22, 3888, 392 - летние; 111,
112, 115-117, 521, 536, 539, 550, 564, 566, 572, 575, 577, 585, 586,
590-594 - зимние) (рис. 4).
В море станции, где происходило сильное и слабое перемешивание,
чередовались, т.е. конвективное перемешивание в море зимой было
локальное. Кроме того, на некоторых станциях, выполненных зимой
(ст. № 595, 596, 598, 599, 601, 602) и расположенных в северо-западной
и северной частях моря Леванта, наблюдалось типично летнее распре-
деление бактерий. В этом районе, очевидно, конвективного перемеши-
вания не происходило.
Из 26 станций, выполненных в рейсе на нис ’’Академик Ковалев-
ский” 3-22.02.1962 г„ только на 5 (ст. № 111, 112, 115, 116, 117)
обнаружили зимнее перемешивание. На 14 (ст. № 25, 102, 104, 105,
107-109, 113, 114, 118-121) наблюдали типично летнее распределение
бактерий, на остальных (ст. № 103, 110, 122- 126) - смешанное летне-
зимнее (рис. 4). Таким образом, можно сделать следующие выводы.
1. Февраль характеризуется смешанным распределением гетеро-
трофных бактерий, и, очевидно, его следует считать переходным ме-
сяцем.
2. Зимний сезон в море продолжается примерно с 10 ноября по
январь, т.е. всего около 3 мес.
121
Основные результаты о зимнем перемешивании средиземномор-
ских вод, полученные по данным распределения и изменения числен-
ности гетеротрофных бактерий, были изложены на итоговой конферен-
ции по изученности Средиземного моря (1-7.10.1962 г.). Поскольку
результаты наших исследований были получены на основе микро-
биологического метода (не гидробиологического!) и в то время зна-
чительно опережали общее мнение по данному вопросу, то они были
отвергнуты.
В 1968 г. была организована международная экспедиция, названная
МЕДОС (1969, 1970), в состав которой вошли известные океанологи
Англии, Франции, Италии и США. Используя существующие материалы
по изучению Средиземного моря, а также результаты собственных де-
тальных исследований гидро метео режима этого моря зимой, было
установлено, что сильное зимнее охлаждение воздуха способствует
быстрому охлаждению поверхностных слоев моря. В результате интен-
сивной конвекции уже через неделю после наступления похолодания
и возникновения сильных ветров в холодный период года перемеши-
вание вод достигало глубины 1500-2400 м. Известно, что скорость
конвекции составляет не более ЕКГ’-МО73 см.с~1,ав придонном
слое воды и того меньше - 1 •10’6-0,18,10-‘* см.с-’ (Овчинников с
ооавт., 1976), но по результатам экспедиции МЕДОС она оказалась
равной 0,25-0,40 см-с"1. Это в 25—400 раз больше принятой раньше.
Таким образом, результаты микробиологических исследований
1958-1961 гг., сообщенные нами на конференции в 1962 г., подтвер-.
дались через 7—8 лет иностранными гидрологами, но приоритет остал-
ся не за нашей страной.
8.4. СВЯЗЬ ЧИСЛЕННОСТИ ГЕТЕРОТРОФНЫХ БАКТЕРИЙ
С ВЕРТИКАЛЬНОЙ СТРУКТУРОЙ ВОД В ТИХОМ ОКЕАНЕ
В разделе использованы данные А.Е.Крисса (1959) о численнос-
ти гетеротрофных бактерий во всех глубинах водной толщи цент-
ральной части Тихого океана, полученные в 26-м рейсе нис ’’Витязь”
(рис. 5). Рассматривая распределение гетеротрофных бактерий в свя-
зи с гидрологическими явлениями, А.Е.Крисс считал, что воды с пони-
женной и повышенной численностью бактерий — разнородные воды.
Поэтому слои, для которых характерно большое количество бактерий,
он относил к богатым органикой, например Коралловое море. Воды,
бедные бактериальной жизнью, считал водами, обедненными органи-
ческим веществом, например, антарктические.
Интерпретируя данные о содержании бактерий-гетеротрофов в
море или океане, мы исходили из других предпосылок, например,
что при смешивании разнокачественные воды уплотняются, а на грани-
це раздела наблюдаются максимальные градиенты плотности (Зубов,
122
1Ю J?O !ЗО М НО 160 170 180 ПО 160 150 140 130 120 ПО 100 90 80 70 60
ПО 120 730 140 750 160 170 180 ПО 160 150 140 130 120 110 100 90 80 70 60 50
Рис. 5. Карта микробиологических станций в Тихом океане по разрезам 172° в.д. и 174° з.д. (Крисс, 1959)
Рис. 6. Кривые бактериальных максимумов в Тихом океане по разрезу 172° в.д.
1957). Это обстоятельство замедляет скорость движения взвеси, детри-
та и других частиц, концентрация которых здесь становится выше,
чем вне границ раздела (Богоявленский, 1955). Все это привлекает
бактерии, которые также скапливаются в зонах уплотнения, создавая
максимумы численности по сравнению с выше- и нижележащими слоя-
ми воды (рис. 6, 7).
Соединив на графиках (рис. 6, 7) относительные максимумы чис-
ла бактерий на станциях по разрезу 172° в.д. и 174° з.д., начиная с са-
мого верхнего от поверхности воды, который, может находиться на глу-
бинах 0-50 м, мы получили картину вертикального распределения
слоев морской воды с повышенным числом гетеротрофных бактерий
(Горбенко, 1964).
Как показано на рис. 6, 7, ход линий, соединяющих относительные
бактериальные максимумы на разрезах по глубинам, имеет одну и ту
же тенденцию - к подъему или опусканию. При этом выявляется сло-
истая структура толщи воды, что отмечалось многими исследова-
телями (Муромцев, 1958, Галеркин с соавт., 1982; Степанов, 1983; и
др.). Для того чтобы понять ход бактериальных линий в толще воды,
мы сравнили налги данные с существующими литературными данными
(Степанов, 1760, 1983; Бурков, 1980; Галеркин с соавг., 1982; и др.),
я на их ое’ч зе Ръша составлена габл. 47. В поверхностных слоях воды
524
Мирового океана гидрологические параметры определяли инструмен-
тальным методом и по сносу судов, глубинную циркуляцию и структу-
ру вод - расчетным путем.
Если ранее считали, что дивергенция и конвергенция характерны
только для поверхностных слоев воды, то в насюяшее время указан-
ные явления обнаружнва.ггт на глубинах 1000 и 2000 м (Степанов,
1983). Этот факт соответствует высказыванию Н.Н.Зубова (1957) о
том, что при смешивании вод возникает их уплотнение, которое ведет
. опусканию более плотных слоев воды, чем окружающие, а следо-
вательно, к распространению конвергенции на большую глубину.
Установлено расположение ряда фронтов в Океане и вергенций,
имеющихся не только на поверхности, но и на глубинах 1000 и 2000 м
(Степанов, 1960, 1983; Галеркин с соавт., 1982; Федоров, 1983; и др.).
Вместе с тем, в результате корректировки данных (Степанов, 1983)
уже не учитывают Экваториальную и Южную субтропические конвер-
генции. А список вергенций Мирового океана, составленный В.И.Сте-
пановым (1960), к 80-м годам сократился. К сожалению, так же мало
расширилось описание гидрологии глубоководных впадин Тонга и Кер-
мадек, которые изучала В.В.Леонтьева (1960).
125
Таблица 47. Конвергенции, дивергенции и фронты Мирового океана
Коорди- наты Мировой океан* Коорди- наты Тихий океан** Коорда наты Индийский океан***
°с.ш.
“сдп.
50-38 Субполярная дивергенция
46-33 Субполярная конвергенция
(Субполярный фронт)
33-24 Субтропическая конвергенция
9-6 Субэкваториальный фронт
8 Субэкваториальная дивергенция
4 Экваториальная конвергенция
2-1 Экваториальный фронт
О
° юли.
0-4 Экваториальная дивергенция
(Экваториальный фронт)
5-10 Субэкваториальная дивергенция
2-Ю Субэкваториальный фронт
18-27 Субтропическая конвергенция
(22-31)
36-39 Субтропическая дивергенция
38 Субполярная конвергенция
41—50 Субантарктический фронт
46 Субполярная дивергенция
42-60 Антарктический фронт
°с.ш.
56-45 Субполярная дивергенция
31—19 Субтропическая конвергенция
22—8 Тропическая дивергенция
9-1 Тропическая конвергенция
О
°ю.ш.
1-5 Тропическая дивергенция
12-28 Субтропическая конвергенция
65- 70 Антарктическая дивергенция
7,5 Тропический фронт
0
°ю.ш
2,0-7,5 Тропическая дивергенция
24- 37 Субтропическая конвергенция
32,5 Субантарктическая конверген-
ция (Субантарктический фронт)
45 Антарктическая конвергенция
(Южный полярный фронт)
62,5 Антарктическая дивергенция
Степанов, I960,1983;
** Тихий океан, 1982;
*** Индийский океан, 1982; Бурков, 1980.
Районы опускания бактериальных кривых на разрезах 172 °в.д. и
174° з.д. (на 1-м разрезе наибольшее количество станций, а следова-
тельно, он наиболее информативный) мы условно обозначили буква-
ми ”А, Б, В, Г, Д, Ж, 3, И, К”. Районы подъема кривых - соответствен-
но ”Л, М, Н, П, Р, Т, У, Ф”. На рис. 6, 7 показано, что в районах опуска-
ния вод ”А, Б, Г, Д, Ж, 3, И, К” кривые опускаются примерно от
10- 100 до 1600-2800 м. В районе бактериальных кривых ”В” опуска-
ние начинается с 400 м и прослеживается примерно до 2000 м (рис. 6).
В районах подъема ”П, Р, Т, У, Ф” кривые поднимаются из глубин
больше 2000 м - до 10—25 м от поверхности. Подъем бактериальных
кривых”Л” и ”М”отмечается только до 400—500 м от поверхности.
Выявленную структуру бактериальных кривых мы сравнили с литера-
турными данными (табл. 48) для той же широты Тихого океана. Срав-
нение показало, что району опускания кривых ”А” соответствуют
Южная субполярная конвергенция и Субантарктический фронт; подъе-
му кривых ”Л” — район Субтропической дивергенции, а району опуска-
ния бактериальных максимумов ”Б” и ”В” - Субтропическая кон-
вергенция. Южной тропической и субэкваториальной дивергенциям
соответствует подъем бактериальных кривых ”П”. Здесь же распола-
гается Южный тропический фронт. Подъем иа экваторе ”Р” соответ-
ствует Экваториальной дивергенции, а опускание линий бактериальных
максимумов ”Д” - Экваториальному фронту. Северной Субэквато-
риальной (или тропической) дивергенции соответствуют подъемы
кривых ”Р” и ”Т”, а район подъема кривых ”П” - Субэкваториально-
му фронту. Опускание линий бактериальных максимумов ”И” и ”К”
соответствует Северной субтропической конвергенции.
Сказанное выше свидетельствует, что опускание бактериальных
кривых соответствует районам конвергенций, а подъемы их — дивер-
генций.'На основании этого мы считаем, что районы ”Г, И, 3”, где ли-
нии бактериальных максимумов опускаются, являются конвергенция-
ми. Соответственно места подъема их - ”М, Н, У, Ф” - это районы дивер-
генций, которые также еще не определены. Итак, бактериальные кривые
являются индикатором вер» жальной структуры вод в Тихом океане.
а х СВЯЗЬ РАСПРЕДЕЛЕНИЯ ЧИСЛЕННОСТИ
ГЕТЕРОТРОФНЫХ БАКТЕРИЙ
С ВЕРТИКАЛЬНОЙ СТРУКТУРОЙ ВОД В ИНДИЙСКОМ ОКЕАНЕ
По нашему методу с помощью бактериальных кривых М.Н.Лебедева
(1962) построила разрез по глубинам Индийского океана. Материалом
для работы послужили микробиологические данные, собранные во
2-м рейсе нис ”Обь” на 61 станции по разрезу от Индии до Антарктиды
примерно по 89-100° в.д. Мы использовали эти данные, предваритель-
но откорректировав их (рис. 8).
127
Рис. 8. Кривые бактериальных максимумов в Индийском океане по разрезу
89-100“ вл.
Районы опускания бактериальных кривых отмечены буквами Л, С,
Е, G, I, L, N, Р и S, районы подъема - В, D, F, Н, К, М, О, R (имеются
также нечетко выраженные опускание и подъем между Н и Z, которые
мы не учитывали). Как видно из рис. 6,7, структура бактериальных кри-
вых в Индийском океане отличается от таковой в Тихом океане, где
подъемы и опускания линий бактериальных максимумов широко,
равномерно и симметрично распространяются в северное и южное
полушария.
Поскольку Индийский океан примерно с 23° с.ш. с севера ограни-
чен сушей, то, возможно, поэтому бактериальные кривые в Северном
полушарии до 10° ю.ш. несколько сжаты. В результате на расстоянии
от 23° с.ш. до 7° ю.ш. (30°) располагаются 4 района опускания кривых
И 4 района подъема. Южнее 7° ю.ш. и до Антарктиды, наоборот, эти
характеристики растянуты. От 7° ю.ш. и до 66ь ю.ш. (60°) располо-
жены только 5 районов опускания вод и 4 района подъема. Интересно,
что от 15° ю.ш. до 66° ю.ш. на глубине от 0 до 75 м ход бактериальных
линий прямолинейный.
Установлено, что водная толша океана состоит из 6-9 слоев, распо-
ложенных между максимумами бактериальных кривых. Эти слои, ве-
роятно, соответствуют поверхностным, промежуточным и глубинным
водам. В 5 районах Л, I, N, Р и 5 опускание бактериальных линий прос-
леживается от самой поверхности. В 4 районах - С, Е, G, L - опускание
кривых с поверхности не наблюдается. Кривые бактериальных макси-
мумов поднимаются к поверхности воды только в точках О и R и лишь
частично - в точках К и М. В районах D, F, Н кривые поднимаются на
расстояние 75-250 м от поверхности (рис. 8).
При сопоставлении данных наших исследований с литературными
(табл. 48) можно отметить следующее. Район опускания бактериаль-
128
ных линий у берегов Индии (S) по местоположению близок к Суб-
тропической конвергенции в Мировом океане. Экваториальная кон-
вергенция и тропический фронт почти точно совпадают с районом опус-
кания бактериальных кривых N. Подъем линий на экваторе ”М” точно
соответствует экваториальной дивергенции и близок к экваториально-
му фронту. Южная тропическая дивергенция и Южный тропический
фронт совпадают с районом подъема бактериальных кривых К. Опус-
кание линий бактериальных максимумов (G) соответствует Южной
субтропической конвергенции. Субантарктический и Антарктический
фронты почти совпадают с опусканием бактериальных линий в районе
”Е”. К этому же району относится, по-видимому, и Южная субполярная
конвергенция. Подъем кривых в районе D (Мировой океан) соответ-
ствует Субполярной дивергенции. Расположение Антарктической ди-
вергенции соответствует району подъема бактериальных линий ”В”.
Мы считаем, что районы опускания бактериальных кривых в Ин-
дийском океане А, С, I, L, Р соответствуют еще не описанным гидро-
логическим районам конвергенций. А районы подъема линий бакте-
риальных максимумов D, F, Н, О, R указывают на места дивергенций.
Все это является следствием того, что бактерии скапливаются в местах
уплотнения вод на границе между слоями водной толщи, где накапли-
ваются органические вещества.
Рельеф дна, по-видимому, влияет на вертикальное движение вод-
ных масс. Отмечено, что на разрезе по 172° в.д. в Тихом океане (рис. 6)
1-й подъем дна за 35° ю.ш. влияет на подъем кривых Л. Район опуска-
ния и подъема донного рельефа на 34-17° ю.ш. соответствует опуска-
нию кривых ”Б” и последующему подъему ”М”. Далее очередной
подъем на 15-18° ю.ш. тесно связан с подъемом бактериальных линий
”Н”. Сильному опусканию дна (от 15° до 5° ю.ш.) соответствует район
опускания кривых ”Г”, подъема ”П” и опускания ”Д”. Далее подъему
донного рельефа на 2° ю.ш. отвечает подъем бактериальных линий ”П”.
На неполном разрезе по 174° зд. (рис. 7) бактериальные линии
также соответствуют неровностям дна. Кроме того, над глубоковод-
ными впадинами Кармадек и Тонга наблюдается опускание кривых
бактерий, отчетливо заметное в районе ”В”. Между впадинами, где дно
имеет выпуклость на 22—25° ю.ш., отмечается четкий подъем бакте-
риальных линий ”М”,
Таким образом, микробный метод подтверждает мнение ряда
исследователей (Галеркин и соавт., 1982) о том, что профиль дна влия-
ет на циркуляцию вод океана и, очевидно, на вертикальные перемеще-
ния вод.
Известно, что вертикальные структуры вод, течений, водных масс
в океане расположены симметрично (Krummel, 1911; Зубов, 1956,
1957; Степанов, 1960, 1983; и др.). Подобная симметрия характерна
и для вертикальной структуры бактериальных линий на разрезах через
Индийский и Тихий океаны (Лебедева, 1962; Горбенко, 1964). Так,
129
если на разрезе через центральную часть Тихого океана по 172° з.д.
ось симметрии провести по 5° с.ш., то район хода кривых между подъе-
мами Р и Т оказывается центром симметрии (рис. 6, 7). При этом
район подъема кривых Р становится симметричен району ”Т”, опуска-
ния ”Д” - опусканию ”3”; подъема ”П” - подъему ”У”, опускания
”Г” — опусканию ”И”; район подъема ”Н” - подъему ”Ф”, а опуска-
ния ”В” — опусканию ”К”. Это значит, что по всему разрезу располо-
жение кривых бактериальных максимумов симметрично оси симмет-
рии, смещенной к северу до 5° с.ш.
В Индийском океане также прослеживается симметрия структу-
ры бактериальных линий.Ось симметрии пройдет по опусканию ”L”,
т.е. примерно по 4° ю.ш. (рис. 8). Тогда район подъема линий ”К”
симметричен району ”М”, опускания I - опусканию N, подъема Н -
подъему О, опускания G - опусканию Р, район подъема F - подъему
R, опускания Е — опусканию S, т.е. распределение бактериальных мак-
симумов в Индийском океане, так же как и в Тихом, симметрично рас-
положению кривых, центр симметри которых смещен на 4° ю.ш. Что
касается явлений, называемых фронтами, то В.Н.Степанов (1960)
считал, что фронтами можно назвать конвергенции, и был против того,
чтобы к ним относили дивергенции.
Сопоставив вертикальную структуру линий бактериальных макси-
мумов с местоположением фронтов, описанных в литературе (Галер-
кин с соавт., 1982; Индийский океан, 1982; Степанов, 1983), мы уста-
новили, что Северному субэкваториальному (или Тропическому)
фронту в Мировом океане соответствуют район опускания кривых ”Ж”
и нечеткий подъем между ”Р” и ”Т” (рис. 6). Северному экваториаль-
ному фронту отвечает подъем бактериальных линий в точке ”Р”, Южно-
му экваториальному фронту - опускание бактериальных кривых D
(рис. 6), Субантарктическому фронту (или Южному полярному) -
подъемы кривых ”Л”, F, D (рис. 6, 8) и опускание линий ”А”, ”Е”
(рис. 6, 8), антарктическому (или Полярному) фронту - опускание
”Е” и подъемы кривых D и В, а также опускание ”С” (рИс. 6, 8).
Таким образом, если кривые бактериальных максимумов фикси-
руют районы подъема и опускания вод, то фронт может соответство-
вать как конвергенции, так и дивергенции либо тому и другому вместе.
В определенной степени сказанное подтверждается данными гидроло-
гов. К.Н.Федоров (1983) считал, что в зоне прибрежного апвеллинга
происходит интенсивное опускание вод со стороны фронтального раз-
дела, обращенного к берегу. А в сторону открытого моря слой скачка
приближен к самой поверхности, чему способствует наблюдаемая здесь
дивергенция.
Как отмечал В.Н.Степанов (1960), дивергенции течений на картах
показывают редко, так как их труднее обнаружить, чем конвергенции.
В то же иремь с помощью бактериального метода, применяемого нами,
районы подъема вод, а следовательно, и дивергенции определяются
130
так же просто, как и районы конвергенции. Что же касается их распро-
странения в водной толще, то, по мнению Н.П.Зубова (1957), посколь-
ку при смешивании воды уплотняются, а значит, становятся тяжелее,
то поэтому они могут распространяться и на глубины. Бактериальный
метод, как можно видеть на рис. 6-8, подтверждая сказанное, отража-
ет вергенции, по-видимому, на всем их протяжении. Кроме того, пос-
кольку вся вертикальная структура вод по микробиологическим дан-
ным представляет собой конвергенции, чередующиеся с дивергенция-
ми, то вергенции, судя по стандартным горизонтам, имеют слоистое
строение, указывающее либо только иа подъем вод, либо только на
опускание.
Судя по бактериальному методу, смещения поверхностной части
конвергенции и дивергенции относительно глубинной почти не су-
ществует, т.е. мнение В.Н.Степанова (1983) о гом, что в некоторых
случаях субтропическая конвергенция (Южная и Северная) и тропи-
ческая (Северная и Южная) дивергенция на глубине намного уходят
от ее поверхностной части, нашими данными не подтверждается.
Если вергенции в поверхностных слоях океана и морей гидрологи
определяют по сносу судов и с помощью приборов (Галеркин с соавт.,
1982), то сведения о циркуляции вод в глубинных слоях этих природ-
ных бассейнов они получают пока только расчетным путем, с помощью
математических моделей. В то же время микробиологическим методом
может быть определена вертикальная циркуляция вод, а построение
пересекающихся разрезов позволяет представить вергенции и циркуля-
цию вод в них в трехмерном пространстве.
&& ОПРЕДЕЛЕНИЕ С ПОМОЩЬЮ МИКРОБИОЛОГИЧЕСКОГО МЕТОДА
РАЙОНОВ ЦИРКУЛЯЦИИ ВОД В СРЕДИЗЕМНОМ МОРЕ
Как отмечалось в начале настоящего раздела об индикации вертикаль-
ной структуры вод с помощью микроорганизмов, было сделано около
200 микробиологических станций, на которых учитывались гетеротроф-
ные бактерии водной толщи Средиземного моря (рис. 4).
Для того чтобы иайти и изучить купола дивергенции и вогнутые
линзы конвергенций, кривые бактериальных максимумов рассматри-
вались в 3 районах центральной и восточной части моря, где можно
было сделать перекрещивающиеся разрезы (рис. 4). Обнаружив таким
образом купол или воронку, мы предположили, что это районы дивер-
генции или конвергенции (Лебедева, Горбенко, 1964).
Дивергенции соответствуют ложбинам динамического рельефа в
центральных частях циклонических круговоротов, те отдельные пото-
ки расходятся, воды в них вращаются против часовой стрелки и подни-
маются из глубины, Конвергенции соответствуют гребням динами-
ческого рельефа в центральных частях антициклонических круговоро-
131
тов, где отдельные потоки сходятся, а воды вращаются по часовой
стрелке и опускаются на глубину (Степанов, I960; Океанология;
Физика океана, 1978).
В изучаемой части Средиземного моря нами обнаружены следую-
щие районы вращения вод. Бактериальные линии по разрезу от Тунис-
ского пролива к Неаполю, а затем от Мессинского пролива к о-ву
Сардиния указывают на подъем вод в центре Тирренского моря на
ст.№ 10, 12, где воды вращаются против часовой стрелки (рис. 4).
В северной части Ионического моря по пересекающимся разрезам
от 5 до 19 и от 15 до 17 станции наблюдается подъем вод с циклони-
ческим вращением. Юго-восточнее также обнаруживается циклоничес-
кое вращение вод с центром по линии ст. № 24 и 36. Южнее о-ва Крит,
судя по куполообразному изгибу бактериальных кривых, вероятно,
проходит область дивергенции с циклональным вращением вод.
Еще восточнее — в море Леванта, учитывая пересекающиеся разрезы
в районе ст. № 388 и 28, бактериальные линии уходят воронкой вниз,
т.е. в этом месте намечается, по-видимому, конвергенция с антицикли-
ческим вращением вод (Лебедева, Горбенко, 1964). В результате нам
удалось выявить 5 основных циклонических круговоротов (рис. 4),
из которых 4 соответствуют таковым на схеме среднемесячной цирку-
ляции вод (Овчинников с соавт., 1976).
Подводя итог сказанному, отметим, что способность морских
гетеротрофных бактерий скапливаться в местах повышения содержа-
ния органического вещества (что происходит на границе раздела, напри-
мер 2 слоев разнокачественных вод) позволяет (по-разному интер-
претируя эту закономерность) обнаруживать различные океанологи-
ческие явления. Таким образом, например, удалось определить реаль-
ную скорость конвекции в Средиземном море, а по перекрещиваю-
щимся разрезам в трехмерном пространстве — от поверхности вод и
до дна — найти основные районы горизонтальной циркуляции воды в
центральной и восточной частях этого моря. На основе опубликованных
данных о численности гетеротрофных бактерий в Индийском и Тихом
океанах (Крисс, 1976), была выявлена вертикальная структура распо-
ложения бактериальных максимумов в указанных водоемах. По всем
признакам эта структура указывает районы подъема и опускания вод
(т.е. конвергенций и дивергенций), а также, возможно, океанические
фронты и профиль дна океанов.
Ш
ГЛАВА 9. ХАРАКТЕРИСТИКА ХЛЭК И ФЕНОЛОВ,
НАХОДЯЩИХСЯ В МОРЕ
9.1. О ПРИРОДЕ ХЛЭК И ЕГО ВЗАИМОСВЯЗЯХ
С БИО- И АБИОФАКТОРАМИ ИЗУЧАЕМОЙ СИСТЕМЫ
Как отмечал О.Г.Миронов (1973), в общей проблеме антропогенного
загрязнения морей и океанов одно из ведущих мест занимают углево-
дороды, и в первую очередь нефть и нефтепродукты. Поэтому нельзя
рассматривать какую-либо морскую экосистему, не учитывая влияния
на нее нефти и фенола, тоже являющегося загрязнителем. Вместе с тем
липиды и углеводороды в значительных количествах имеются в мор-
ских микро- и макроорганизмах. После гибели указанных гидробион-
тов (либо другим путем) эти вещества оказываются в морской воде,
а из липидов могут образовываться углеводороды (Миронов, 1973).
Поскольку углеводороды биогенного происхождения по строению
и составу близки к нефтяным фракциям, отделить их от нефти чрезвы-
чайно трудно (Миронов, 1973), особенно когда для этого применяют
наиболее употребительные растворители — хлороформ или 4-хлористый
углеводород. Конечно, вместе с углеводородами и нефтью указанным
способом экстрагируются и липиды биогенного происхождения. В на-
ших исследованиях изучали хлороформный экст£ »кт (или вытяжку)
из морской воды. В ней содержатся нефть и ее продукты, углеводоро-
ды и липиды. Как оказалось, вытяжка четко взаимосвязана с био- и
абиофакторами изучаемой экосистемы.
Для вычисления частных корреляций между 42 параметрами
и обществ микроорганизмов и окружающей среды применяли методы
информационной статистики, а также факторный анализ (Подвинцев,
Горбенко, 1979). Этим же методом обнаружили тесные корреляции
хлороформенного экстракта (ХлЭк) с параметрами Солнца и морской
воды (табл. 48). С помощью анализа Фурье (Горбенко, Крышев,
1985) выявили следующие периодические компоненты ХлЭк: периоды
Т = 1800, 365, 200, 130, 35 и 20 сут; амплитуда 0,96-0,31. Соответ-
ственно вклад каждой гармоники в дисперсию ряда составлял 12,6%;
7,7; 5,4; 3,3; 1,5; 1,3 %. Такие же периоды примерно: 1)у диатомо-
вых живых СПМ (Т = 1800, 365, 180, 120,90 и 30 сут); 2)в содержании
Таблица 48. Корреляция ХлЭк с параметрами экосистемы
Параметр Коэффициент корреляции (И Параметр экосистемы Коэффи- циент кор- реляции (г)
А* РОВ морской воды 0,379-0,515 В ♦** Однолетние корреляции
Доля баланса в инфра- 0,379-0,515 Диатомовые живые СПМ -0,37
красной области спектра Кокки СПМ -0,35
Солнца Диатомовые живые СМП 0,33
РОВ СМВ 0,561-0,515 Диатомовые мертвые СМП -0,31
Температура морской воды 0,379-0,515 Углекислота 0,36
'min = 0.35 pH СМВ -0,33
Б** РОВ морской воды 0,41
РОВ морской воды 0,532 Бактерии-гетеротрофы СМП -0,52
pH СПМ 0,561 Бактерии-гетеротрофы СМВ 0,45
Кислород 0,537 Многолетние
корреляции****
Диатомовые живые СМП 0,631 РОВ морской воды 0,50
Диатомовые живые СПМ -0,363 Бактерии-гетеротрофы СМП 0,42
Диатомовые мертвые СПМ -0,383 Бактерии-гетеротрофы СМВ 0,44
Фосфор минеральный -0,339 РОВ СПМ -0,34
Температура воды -0,354 Карбонаты СМВ 0,38
Нефтеокисляющие (н.о.) 0,581 РОВ СМВ 0,46
бактерии СМП и СПМ .
Н.о и гетеротрофные бак- 0,626 rmin “ 0,30
терии СПМ
Н.о бактерии и РОВ СМВ 0,630
Нои гетеротрофные бак- 0,678
терии СМВ —
'min =0,35
А*- Подвинцев, Горбенко (1979); Б**- Горбенко,Татаренко (1983); В***-
Горбенко, Крышев (1985); ****- по усредненным данным (Горбенко, Крышев,
1985).
кислорода в море (Т = 1800, 365, 120 и 20 сут); 3)СОз~ (Т = 1800,
365, 200 и 40 сут); 4)фенола (Т = 1800, 365, 140 и 24 сут) ; 5)гетеро-
трофных бактерий СПМ (Т = ... 1800,900, 365,30 сут); ^палочковид-
ных бактерий СПМ (Т = ... 1800, 340, 180, НО, 30 сут); 7)РОВ СПМ
(Т = ... 1800, 365, 170 сут).
Этот факт свидетельствует, что изменение содержания ХлЭк в море
имеет определенные периоды и амплитуду колебаний, которые в це-
лом сходны с другими морскими параметрами изучаемой экосистемы.
В течение 1979-1980 гг. (14 мес) мы изучали экологическое зна-
чение ХлЭк в море и влияние морской биосистемы микробов СПМ,
СМВ и СМП на него (Горбенко, Татаренко, 1983). Корреляционный
анализ данных, для которого было рассчитано 80 корреляций, пока-
зал, что между изучаемыми параметрами существуют достоверные кор-
реляции (табл. 48). Корреляционные связи ХлЭк с диатомовыми жи-
выми планктона (СМП) и микроперифитона (СПМ) могут свидетель-
134
Таблица 49. Многомерные корреляции ХлЭк с параметрами экосистемы (Горбен-
Параметр Коэффициент корреляции (г) Параметры, влияющие на корреляцию Величина кор- реляции без исключаемых параметров
Диатомовые живые СПМ 0,34 Без гетеротрофных бакте- рий СПМ 0,25
Без кокков СПМ 0,28
Гетеротрофные бактерии СМП 0,56 Без кислорода 0,33
Гетеротрофные бактерии 0,70 Нет
СМВ Кокки СПМ 0,46 Без РОВ СПМ 0,23
Без диатомовых живых СПМ 0,27
Карбонаты СМВ 0,43 Нет
Углекислота -0,35 Без гетеротрофных бакте- рий СМП 0,24
РОВ морской воды 0,37 Без гетеротрофных бакте- рий СМП 0,21
Без гетеротрофных бакте- рий СМВ 0,23
Кислород 0,60 Нет
РОВ СПМ 0,56 Без гетеротрофных бакте- рий СМП 0,19
rmin = 0,30 rmin = ОЗ®
ствовать о непосредственной связи определенной (или наибольшей)
части данного экстракта с численностью диатомей.
Нами обнаружены (Горбенко, Татаренко, 1983; Подвинцев, Гор-
бенко, 1979) корреляции ХлЭк с РОВ, pH, с температурой воды и кис-
лородом, а также корреляционные связи нефтеокисляюших бактерий
с гетеротрофными бактериями и другие корреляции (табл. 48).
В наших исследованиях (Горбенко, Крышев, 1985) вначале были
выявлены достоверные корреляции корреляционных функций по одно-
летним наблюдениям, которых оказалось 9. Провели также анализ
корреляций по многолетним наблюдениям (1969—1975 гг.), подтвер-
дивший, что большинство корреляций, проявляющихся в течение года
наблюдений (их оказалось 6), выявляется и в последующие годы. За-
вершал работу многомерный анализ данных того же года наблюдений.
С его помощью обнаружили 9 корреляционных связей, большинство из
них однотипны с корреляциями однолетнего анализа (табл. 48,49).
Чтобы выяснить, как на любую пару взаимных корреляций влияют
остальные 33 функции при многомерном анализе, было рассчитано око-
ло 600 корреляций, из которых достоверными (г > 0,3) оказались
только 230 (Горбенко, Крышев, 1985).
Из табл. 48. следует, что в однолетних корреляциях ХлЭк взаимо-
связан с диатомовыми СМП и СПМ, углекислотой, РОВ морской воды,
135
а также гетеротрофными бактериями СМП н СМВ, кокками СПМ и
pH СМВ. Судя по этим корреляционным связям, продуцентами авто-
хтонной части ХлЭк могли быть диатомовые водоросли СМП и СПМ, из
которых со вторыми обычно коррелируют кокковидные формы бак-
терий, взаимосвязанные с диатомовыми.
Бактерии-гетеротрофы СМП и СМВ могут также участвовать в
образовании автохтонной части ХлЭк (липидов, жироподобных ве-
ществ) в ходе различных реакций, например конденсации, проходя-
щих одновременно с деструкцией этого экстракта. При этом в соот-
носительных количествах образуются углекислота и РОВ СМВ, из кото-
рых РОВ СМВ может также пополнять концентрацию РОВ морской во-
ды. Из многолетних корреляций исчезли корреляции ХлЭк с диатомо-
выми водорослями. Это, очевидно, результат переключения знака кор-
реляций (Горбенко,1977).
Судя по многолетним корреляциям, гетеротрофные бактерии СМП,
СМВ, а также, возможно, и СПМ (корреляции которых с ХлЭк
оказались слабыми) используют ХлЭк, в результате образуются
метаболиты РОВ СМВ и РОВ СПМ, которые пополняют концентрацию
РОВ морской воды. В процессе использования гетеротрофами СМВ
ХлЭк на взвеси отлагаются карбонаты, коррелирующие с ним
(табл. 49).
Поскольку частные корреляции параметров изучаемой экосистемы
вычисляли по результатам наблюдений того же года, что и однолетние,
многомерный анализ позволил уточнить все обнаруженные ранее кор-
реляционные связи и выявить прямые, на которые корреляции всех
остальных параметров не влияли. Вместо исчезнувших корреляцион-
ных связей ХлЭк с диатомовыми СМП и СПМ, обнаруживаемых при од-
нолетнем анализе, появились корреляции с карбонатами СМВ, кисло-
родом и РОВ СПМ (табл. 49).
Установлено, что умеренная корреляция ХлЭк с диатомовыми жи-
выми СПМ без гетеротрофных и кокковидных бактерий почти не изме-
нялась при их исключении. А это указывает на то, что диатомеи явля-
ются продуцентами части ХлЭк. В то же время корреляционная связь
ХлЭк с кокками СПМ без РОВ СПМ и диатомовых живых СПМ стано-
вилась недостоверной и, следовательно, исчезала. Корреляционных свя-
зей ХлЭк с углекислотой, а также с РОВ СПМ без гетеротрофных бакте-
рий также не обнаружено. В то же время величина корреляционной
связи ХлЭк с гетеротрофными бактериями СМП без кислорода умень-
шается, но остается достоверной. Определили также прямые связи
ХлЭк с кислородом (г • 0,60), гетеротрофными бактериями СМВ
(г ® 0,43) и карбонатами СМВ, на которые не влияют остальные Па-
раметры.
В результате проведенной работы можно сделать следующие вы
воды: на ХлЭк влияют гетеротрофные, кокковидные бактерии и диа-
томовые водоросли СПМ. Однако в процессе образования и деструкции
136
автохтонной части на ХлЭк оказывают влияние главным образом гете-
ротрофные бактерии СМВ и СМП, а также кислород морской воды.
В процессе использования ХлЭк бактернями-гетеротрофами СМВ на
взвеси отлагаются карбонаты СМВ. Судя но корреляционным связям,
гармоническим составляющим, сезонности и другим признакам, прису-
щим только автохтонному биотическому (или биокостному) компо-
ненту, в Севастопольской бухте содержание автохтонных биогенных
углеводородов, жиров н липидов выше, чем в сбрасываемой в море
нефти. Объяснить это явление можно тем, что в бухте находится боль-
шое количество взвеси, большая поверхность твердых предметов и
донных грунтов, а также органических и минеральных веществ, по-
падающих в море с суши и обеспечивающих интенсивное развитие
микроорганизмов СМП, СМВ, СПМ, которые участвуют в продуциро-
вании ХлЭк.
а.2. МИКРООРГАНИЗМЫ, ИСПОЛЬЗУЮЩИЕ ХЛЭК
И УЧАСТВУЮЩИЕ В САМООЧИЩЕНИИ МОРЯ В ПРИРОДНЫХ УСЛОВИЯХ
Из морской воды и донных грунтов юго-западной части Черного моря
у Крыма было выделено 138 культур бактерий, окисляющих нефть,
из которых 55 росли на минеральной среде с нефтепродуктами или
нефтью, постепенно разрушая их. Эти бактерии являются деструктора-
ми нефтепродуктов в морских условиях. На основании полученных
данных был сделан вывод о том, что нефтеокисляющне (или углеводо-
родокнсляющие) бактерии в Черном и других морях разлагают нефть
и ее продукты. Установлено, что в морской воде преобладают бактерии
родов Pseudobacterium, Micrococcus, Sarcina, Pseudomonas, Achromo-
bacter, Bacterium, Vibrio, Spirillum, Bacilium, а в меньшем количестве
встречались роды Vibrio, Chromobacterium (Миронов, 1971, 1973).
Бактерии не только окисляют нефтепродукты и нефть, но и играют
главную роль в деструкции тяжелых нефтяных фракций (нефтяных
комков) (Миронов, Георга-Копулос, 1981). Эги микроорганизмы
также используют биогенные липиды и углеводороды. Бактериальные
организмы, окисляющие нефть, нефтепродукты и другие вещества,
входят в одну из самых важных экологических групп морских микро-
организмов - группу гетеротрофных бактерий, которые являются
главными минерализаторами-редуцентами органического вещества в
морях и океане. Обитают они, как известно, во всей толще воды - от
поверхности до дна, на поверхностной пленке - нейстоне, на частицах
взвеси и на поверхности погруженных в море предметов естественного
и искусственного происхождения. Очень подходит бактериям-гетеро-
трофам и донный грунт морских водоемов. Вместе с бактериями но
всех указанных биотопах живут простейшие организмы, а на i дубине,
куда еще проникает свет, и диатомовые водоросли.
137
Таблица 50. Результаты расчета частных корреляций
Параметр Коэффициент корреляции
ХлЭк (1), РОВ (2), pH г 12 0,532 г 16 = 0,561 г 26= 0,429
СПМ (6)* г 12.6 = 0,394 Г142 = 0,438 г261 ~ 0,187
ХлЭк (1), РОВ (2), кис- г 12 =0,532 г 13 =0,537 Гзз ~ 0,369
пород (3) rlZ3 =0,428 г = 0,437 г23,1 =0,116
ХлЭк (1), кислород (3), г 16 = 0,544 г13 = 0,537 Г6Э = 0,700
рн г 16,3 ~ 0,310 г)а6= 0,250 Г611 = 0,578
СПМ (6), ХлЭк (1),рН Пт = -0,428 г 16 =0,544 г76 = -0,440
СПМ (6) Нефтеокисляющие бак- терии СПМ (7), 07.6 = -0,243 г 16,7 = 0 459 г Тед = 0,266
ХлЭк (1), диатомовые г и = -0,363 г 19 = -0,383 гк = 0,953
СПМ: живые (8) и мерт- вые (9) г 189 =0,003 г19.8 = 0,135 гем = 0,942
* Для каждой группы, состоящей из 3 параметров, в верхней строке приведены
парные корреляции, в иижией - частные.
В данном разделе мы рассмотрим роль бактерий-гетеротрофов в
процессе самоочищения моря от нефтепродуктов. В состав этих бакте-
рий входят и нефтеокнсляющие бактерии. Для изучения влияния СМП,
СПМ и СМВ на нефтепродукты исследовали несколько факторов и
параметров. Биотический фактор — живые и мертвые диатомовые во-
доросли, гетеротрофные бактерии, а также относящиеся к гетеротроф-
ным нефтеокисляющим бактерии, которые развиваются на среде с
нефтепродуктами. Кроме того, изучали метаболиты микроорганиз-
мов — РОВ и pH СПМ (6 параметров). Абиотический фактор - содер-
жание ХлЭк, pH, концентрации РОВ, нитриты и нитраты, фосфор общий
и минеральный, кислород (8 параметров). Наблюдения проводили в
1979- 1980 гг.1 (14 мес).
Видовой состав нефтеокисляющих бактерий СМП, СПМ и СМВ,
а также донных осадков определяли по руководству Д.Берджи (Ber-
gey, 1948, 1957). Числовые данные были обработаны с помощью мето-
дов корреляционного анализа парных и частных корреляций (Горбен-
ко, Татаренко, 1983). Соответствующие расчеты показали, что числен-
ность нефтеокисляющих бактерий СПМ и СМВ находится в корреля-
ционной связи с гетеротрофными бактериями этих же сообществ.
Данных настоящих исследований, полученных в течение 14 мес,
оказалось недостаточно, чтобы выявить достоверные корреляции неф-
теокисляющих и гетеротрофных бактерий с содержанием нефти в море.
Такая корреляция четко проявилась для бактерий-гетеротрофов (Гор-
бенко, Крышев, 1985) (табл. 48, 49). Установленный факт свидетель-
ствует, что при изучении микробного процесса самоочищения моря от
1 Учет не<Ьтеокисляюших бактерий и определение ХлЭк проведены Р.В.Соснов-
ской, содержа ше нитритов, нитратов, фосфора и кислорода в море - М.В.Ки-
рнковоГ.
:.<я
нефти, определяемого в хлороформенном экстракте, необходимо учи-
тывать не только нефтеокисляющие, но и гетеротрофные бактерии
СМП, СПМ и СМВ. Поскольку ХлЭк коррелирует с РОВ СМВ, pH СПМ,
Кислородом и РОВ морской воды, то эти корреляции, очевидно, указы-
вают на использование нефти бактериями-гетеротрофами СМВ и СПМ
с привлечением кислорода. В результате изменяется pH воды, образует-
ся РОВ СМВ, в свою очередь, пополняющее концентрацию РОВ мор-
ской воды.
Вычисление некоторых частных корреляций позволило уточнить
полученные данные (табл. 50). Было установлено, что РОВ морской
воды, pH СПМ и кислород примерно в одинаковой мере связаны с
ХлЭк в море.
Ниже приведен список видов бактерий, окисляющих ХлЭк
Из воды (СМП) Штамм
Vibrio comma (Schroeter) Bergey et al. 2
Micrococcus galophilus Bergey et al.
Pseudomonas periphyta Zo Bell and Upham
M.varians Migula
Ps. membranula Zo Bell and Upham
Achromobacter sinuosum (Wright) Bergey et al.
Из взвеси (СМВ)
Vibrio comma (Schroeter) Bergey et al, 1
V .comma (Schroeter) Bergey et al. 3
V .comma (Schroeter) Bergey et al. 4
Из перифитона (СПМ)
Vibrio liquefaciens (Migula) Bergey et al.
V .comma (Schroeter) Bergey et al. 6
Из донных осадков
Vibrio aquatilis Giinther
V .comma (Schroeter) Bergey et ai. 5
Achromobacter geniculatum (Wright) Bergey ci ci.
Как свидетельствует список, во всех изучаемых биотопах встреча-
ются бактерии V.comma различных штаммов. В ценозах СМВ и СИМ
вибрионы были единственными представителями бактерий. В донных
осадках доминировали бактерии рода Vibrio. Совсем иной состав
бактериальных форм был в СМИ — кроме одного вила г.ибрионов там
обнаружены бактерии родов Micrococcus, Pseudomonas. Achromobacter.
Полученные данные подтверждают, что ХлЭк, в состав которого
входят нефть и ее продукты, под влиянием гетеротрофных бактерий и
кислорода воды в данном случае подвергается деструкции в аэробных
условиях. Причем деструкция и минерализация происходят в основном
на границе раздела фаз вода - поверхность частицы взвеси, вода - по-
верхность твердого, погруженного в море предмета, вода — донный
грунт, а также, по-видимому, только в меньшем количестве, на Гранине
раздела вода — поверхность капли нефтепродукта. Этот факт был дока
зан в экспериментах (Zo Bell с соавт., 1943), и которых было чстко
139
показано, что бактерии потребляют углеводороды нефти нз вод-
ной среды только при добавлении в нее песка или измельченного
асбеста.
Выделенные нами нз воды н донного грунта нефтеокисляющие
гетеротрофные бактерии, по родовому составу сходны с бактериями,
обнаруженными в тех же биотопах О.Г.Мироновым (1971). Судя по
видовому составу этих бактерий, минерализацию нефти и ее продуктов
на взвеси и в перифитоне осуществляют в основном бактерии рода
Vibrio. Поскольку они доминируют и в донных осадках, то микроб-
ная деструкция нефтепродуктов происходит и там.
Вследствие того что в Севастопольской бухте вибрионы встреча-
ются во всех биотопах от поверхности до дна, то они, по-вндимому,
являются одним из основных деструкторов ХлЭк (Горбенко, Тата-
ренко, 1983).
9.3. ПРИРОДА ФЕНОЛА И ЕГО ВЗАИМОСВЯЗИ
С БИО- И АБИОФАКТОРАМИ ЭКОСИСТЕМЫ
Фенолы, попадающие в море, могут иметь аллохтонное н автохтонное
происхождение (Подвинцев, Горбенко, 1977). Они образуются при
коксовании угля и других видах его переработки. Фенолы использу-
ются практически во всех отраслях промышленности и народного хо-
зяйства, мировое производство их приближается к 2 млн т в год, и как
следствие, большую часть промышленных стоков составляют фенол-
содержащие воды.
Другим значительным источником поступления фенолов в море
являются потери нефти при ее транспортировке и переработке, а также
при разведке и добыче в морских водоемах (мировая добыча нефти
более 2 млрд т в год). Поскольку в состав нефти входит фенол (ру-
мынская нефть - до 10 % фенола, а бориславская - до 24 %), то нефтя-
ное загрязнение сопровождается также фенольным.
Фенолы автохтонного происхождения образуются в море в процес-
се метаболизма различных водных организмов. При расщеплении бел-
ковых аминокислот, содержащих бензольное кольцо, накапливается
монофенолкарбоновая кислота, которая выводится животными во
внешнюю среду с мочой. Фенолподобные соединения образуются
также при отмирании в воде наиболее распространенных видов водо-
рослей и прн массовом развитии синезеленых водорослей. Многие
микроорганизмы при расщеплении различных субстратов в экспери-
менте синтезируют фенолы. Таким путем они, очевидно, образуются
и в природных условиях.
140
9.4. ОСНОВНЫЕ ПАРАМЕТРЫ,
ОПРЕДЕЛЯЮЩИЕ СОДЕРЖАНИЕ ФЕНОЛА В МОРЕ
(РЕГРЕССИОННЫЙ АНАЛИЗ)
Методика работы следующая. В течение года (каждые 10, 20, 30 сут)
было проведено 36 наблюдений за 19 различными параметрами одно-
временно, среди которых был фенол. Эти параметры и явились исход-
ными данными, где:
Y- содержание фенола в морской воде (мг-л~*);
Х{ - концентрация РОВ СПМ (Е270 на волне 270 нм);
Х2 - pH метаболитов СПМ;
Х3 - численность диатомовых водорослей живых СПМ (1g чис-
ленности) ;
Х4 - число мертвых диатомовых СПМ (1g);
Xs - число гетеротрофных бактерий СМП (1g числа колоний);
Х6 — концентрация РОВ в морской воде (по Е2да);
Хт— температура морской воды (°C);
Х8 - pH воды;
Х9 - РОВ СМВ (по Е27О);
Xt0 - pH СМВ;
Хц - Количество гетеротрофных бактерий СПМ (1g числа ко-
лоний) ;
Хп - содержание карбонатов СМВ (% сухой массы);
Х13 - содержание карбонатов СПМ (% сухой массы);
Х)4 — содержание карбонатов СПМ (мг« 15 см"2 пластины об-
растания) ;
XiS — сухая масса слизистой пленки СПМ (мп 15 см"2 пластины);
Xi6 - сухая масса 10 мл взвеси (г);
Х!7 - содержание нитритов азота в воде (мкг-л" *);
Х18 - содержание нефти в воде (мг-л" ’);
Xi9 — содержание фосфора минерального в воде (мкг-л" *).
Уравнение регрессии, вычисленное по этим параметрам, имеет
следующий вид:
У = -0,185 + 0,12X1 - 0,18Х2 -О,О136Х5 + 0,424Х6 - 0,0063Х7 +
+ 0,136Х8 + 0,069Хю - 0,001 ЗХ„ + 0,026Xu - О,ОО535Х17 + 0,0027Xig.
Полученное уравнение адекватно с вероятностью Р = 0,97. Все
коэффициенты уравнения значимы по критерию Стьюдента с вероят-
ностью более 0,9. В уравнение не вошли переменные Х3, Х4 — диатомо-
вые СПМ, Х9 — РОВ СМВ, Хц — гетеротрофные бактерии СПМ, Х12 -
карбонаты СМВ, Х13, Х4 — карбонаты СПМ, Х19 — фосфор минераль-
ный. Это значит, что данные параметры не оказывали существенного
влияния на фенол во время наблюдений, а поэтому были элимини-
рованы.
Наибольший статистический вклад в уравнение сделали следующие
параметры:
141
Xi 0,01
X7 1,48
Xs 0,04
X6 0,13
X7 0,10
Xg 1,13
Хю 0,69
X15 0,03
X16 0,08
X17 0,02
Xi» 0,01
Это свидетельствует о том, что указанные параметры в изучаемой
акватории моря обусловливают содержание фенола в воде Севастополь-
ской бухты. Основным из них (по вкладу) является pH СПМ - Х2,
затем pH морской воды -- Xg, pH метаболитов взвеси — Хю- Согласно
результатам наших многолетних исследований, эти параметры в море
обычно взаимосвязаны. К общим показателям СПМ относятся также
РОВ СПМ (Х9) и масса СПМ (Х15). Вместе с тем отдельные группы
микроорганизмов СПМ (живые н мертвые диатомеи СПМ Х3, Х4, гете-
ротрофные бактерии СПМ Хц) в уравнение не вошли, так как не
влияют на содержание фенола.
Следовательно, автохтонное образование фенола в изучаемой аква-
тории моря зависит от продуктов жизнедеятельности микроорганизмов
СПМ и СМВ, являющихся суммарным метаболитом указанных сооб-
ществ, в то время как отдельно микроорганизмы СПМ на образование
фенола не влияют. Автохтонное происхождение этого ароматического
и токсичного вещества зависит также от температуры, концентрации
РОВ морской воды (параметры Х8, Х7, Х6), от числа гетеротрофных
бактерий планктона СМП (Х5) и нитритов азота (Х17) .
Для анализа взаимосвязей 42 параметров экосистемы, в том
числе и фенола, мы применили факторный анализ и методы инфор-
мационной статистики (см. с. 39-42). Обнаружили, что фенол
входит в ядро одной из центральных группировок В вместе с альбедо
видимого спектра Солнца, облачностью, карбонатами СМВ и pH мор-
ской воды, с которыми он связан прямой связью (на нее не влияют
остальные параметры изучаемой экосистемы) (рис. 1). Фенол связан
также с гетеротрофными бактериями СМВ. Таким образом, возможно,
что в указанной группировке альбедо видимого спектра Солнца оказьь
вает регулирующее влияние ьа параметры группы В, в том числе и на
фенол.
С помощью анализа по Фурье (Горбенко, Крышев, 1985) мы опре-
делили, что для динамики содержания фенола в море в 1969-1973 гг.
характерны следующие периодические компоненты: периоды Т = 1800,
365, 140, 24 сут; фазы у = 420, 200, 130, 20 сут; амплитуды А = 0,4-
0,26. Соответственно, вклад каждой гармоники в дисперсию ряда сос-
тавил 9,4; 3,5; 3,4; 3,4. Такие же периоды примерно были не только у
ХлЭк, но также у диатомовых живых СПМ, О2, СО2;-, гетеротрофных
142
и палочковидных бактерий СПМ и РОВ СПМ. Таким образом, выяв-
ленные связи свидетельствуют, что фенол в Севастопольской бухте
в основном является автохтонным компонентом.
Прн изучении структуры взаимосвязей экосистемы с помощью
анализа корреляционных функций (см. с. 65-71) (табл. 31), пос-
ле того как все полученные корреляции были разбиты на отдель-
ные группы, фенол попал в 1 i руппу, которая характеризуется наи-
большим количеством и теснотой корреляционных связей, наступающих
одновременно. По всем признакам эта группа в изучаемой системе
играет роль регулятора. Следовательно, фенол вместе с компонентами
карбонатного равновесия, pH морской воды, СПМ и СМВ также явля-
ется параметром-регулятором. Все параметры в группе I между собой
взаимосвязаны, и структура их взаимосвязей стабильно определяется
в однолетнем и многолетнем анализах, а также в многомерном анализе
тех же однолетних наблюдений, указывая на устойчивость структуры
корреляционных связей в море. Все корреляции фенола с параметрами
экосистемы были сведены нами в табл. 51. Сравнение их с такими же
параметрами ХлЭк (табл. 48) показало, что однолетних и многолетних
корреляций с фенолом 35, а с ХлЭк - 31, многолетних с ХлЭк - 9, а
с фенолом - 12.
Сравнивая корреляционные таблицы ХлЭк и фенола, можно уста-
новить, что у них есть ряд общих корреляций, например с СО2, pH
СПМ, pH СМВ, диатомовыми живыми СМП, температурой воды, гетеро-
трофными бактериями СМП и СМВ, с РОВ СМВ и О2. Этот факт указы-
вает на то, что фенол (которого в море примерно на порядок меньше,
чем ХлЭк, судя по предельно допустимым концентрациям — ПДК)
либо образуется из ХлЭк, либо является его частью (Подвинцев, Гор-
бенко, 1977); при попадании нефтепродуктов в морскую воду фенол
освобождается (табл. 49-51).
Судя по однолетним корреляциям, содержание фенола зависит от
компонентов карбонатного равновесия, pH метаболитов к морской
воды, а также от диатомовых СПМ, СМВ и СМП, разных бактерий
СМВ СПМ и СМП, от РОВ СМВ, кислорода, прозрачности воды и солнеч-
ной активности. Одним из продуцентов фенола, по-видимому, являет-
ся микроценоз взвеси СМВ, а также микроценозы СМП и СПМ. Кисло-
род - агент, способствующий окислению фенола, а активность Солнца-
его регулятор (табл. 51).
В многолетних корреляциях, которые существенно уточняют одно-
летние данные, остались неизменными все первые 6 параметров I груп-
пировки, прочно взаимосвязанные с фенолом, а также диатомовые
СМВ и СМП. Из диатомей основными продуцентами фенола являют-
ся, очевидно, диатомеи взвеси. Роль гетеротрофных бактерий, как и в
однолетних корреляциях, пока не ясна.
Поскольку лучше всего причинные связи можно объяснить с по-
мощью многомерного анализа, результаты его, по-видимому, самые
143
Таблица 51. Корреляции содержания фенола с парамет-
рами экосистемы
Параметр Коэффициент корреляции (г)
А* Палочковидные бактерии СПМ 0,317
Кокковидные бактерии СПМ 0,328
Гетеротрофные бактерии СПМ -0,364
rmin = 0,250
Б * pH морской воды 0,561
Гетеротрофные бактерии СМВ 0,379-0,515
Альбедо видимого спектра солнца 0,379-0,515
rmin = 0,350
Однолетние корреляции
В* pH морской воды -0,42
со|~ -0,44
НСОз 0,50
СО2 0,43
pH СПМ -0,40
pH СМВ -0,32
Диатомовые мертвые СПМ -0,45
Палочковидные бактерии СПМ 0,45
Диатомовые живые СМП -0,32
Диатомовые живые СМВ 0,51
Диатомовые мертвые СМВ 0,47
Гетеротрофные бактерии СМВ -0,33
Гетеротрофные бактерии СМП -0,32
Зоопланктон 0,42
Температура воды -0,32
FOB СМВ -0,37
О2 -0,35
Прозрачность -0,40
Солнечная активность 0,32
** Многолетние корреляции
сог 0,462
НСОз 0,398
СО2 0,390
pH морской воды 0,400
pH СМВ 0,410
pH СПМ 0,427
Диатомовые живые СМП 0,355
Диатомовые живые СМВ 0,442
Диатомовые мертвые СМВ 0,443
Гетеротрофные бактерии СМП 0,515
— . fmin “ 0,300
А* (Горбенко, 1977); В* (Подвинцев, Горбенко, 1979);
В* (Горбенко, Крышев, 1985) ; ** усредненные данные
за 1 год наблюдений (Горбенко, Крышев, 1985).
144
Таблица 52. Многомерные (частные) корреляции содержания фенола с параметра-
ми экосистемы (Горбенко, Крышев, 1985)
Параметр Коэффици- ент корре- ляции (г) Параметры, влияющие на корреляцию Величина корреляции без исклю- чаемых па- раметров
pH морской воды 0,54 Без НСОз 0,16
Без COj 0,19
coi~ 0,60 Без pH воды 0,16
Без НСОз 0,15
НСО3 0,68 Без СОз" 0,39
COj 0,53 Без COV 0,18
Без НСО; 0,22
pH СПМ 0,44 Без pH СМВ 0,17
pH СМВ 0,63 — —
Диатомовые живые СМП 0,49 Без диатомовых живых СМВ 0,12
Без pH морской воды 0,21
Диатомовые живые СМВ 0,65 — —
Диатомовые мертвые СМВ 0,64 Без НСО3 0,24
Гетеротрофные бактерии 0,43 Без диатомовых живых СМВ 0,30
СМВ
Зоопланктон 0,49 — —
Прозрачность 0,42 Без НСО; 0,18
точные и наиболее близки к реальным взаимосвязям в море. Как было
показано выше, параметры 1--6, компоненты карбонатного равнове-
сия, pH морской воды, СПМ и СМВ, а также фенол настолько тесно
взаимосвязаны и так сильно зависят друг от друга, что элиминация
любого из них сильно уменьшает корреляцию почти любой пары осталь-
ных параметров. Исключением являются только связь фенола с гидро-
карбонат-ионами (НСО3), без которой большинство остальных связей
становятся недостоверными (табл. 52). Она зависит от карбонат-ионов
(COj") и при их элиминации остается достоверной. На уровне досто-
верности осталась также связь фенола с гетеротрофными бактериями
СМВ при исключении живых диатомовых СМВ.
Обобщая изложенное выше, можно сказать, что основную роль в
продуцировании фенола играют живые диатомовые взвеси (СМВ),
а также, вероятно, и зоопланктон, так как на прямые связи с ними
фенола другие параметры не влияют. Какое-то значение в этом процес-
се имеют гетеротрофные бактерии СМВ, и особенно pH СМВ, также
прямо связанный с фенолом. Таким образом,фенол имеет следующие
характерные особенности.
1. Концентрация фенола в какой-то мере регулируется альбе-
до видимого спектра Солнца, а возможно, еще и солнечной актив-
ностью.
145
2. С фенолом постоянно связаны компоненты карбонатного равно-
весия, pH СПМ, СМВ и воды. Эти параметры формируются в группу
регуляторов изучаемой экосистемы, следовательно, фенол-параметр
эктокринного характера, относящийся к регуляторам.
3. Для фенола характерны все признаки автохтонного параметра,
имеющего обширные связи с био- и абиотическими параметрами эко-
системы.
4. Фенол попадает в море не только аллохтонным путем, но и об-
разуется из ХлЭк, в частности из нефти. Он, по-видимо му, может об-
разоваться также в результате жизнедеятельности морских микро- и
макроорганизмов.
ЗАКЛЮЧЕНИЕ
На основании материалов, собранных почти за 20 лет наблюдений,
установлено, что элементы сообществ микроорганизмов планкто-
на, перифитона и взвеси — бактерии, диатомовые водоросли и прос-
тейшие организмы - связаны между собой и с окружающей морс-
кой средой.
В результате сравнительного анализа определили, что ценозы СПМ
и СМВ подобны. При этом выделено несколько новых критериев со-
общества микроорганизмов. Все использованные в этой работе крите-
рии ценоза свидетельствуют, что комплекс микроорганизмов, разви-
вающихся на разлагающихся остатках морских животных и растений
при образовании детрита (до тех пор пока не образуется частичка,
трудно используемая микроорганизмами), не является микроценозом.
В процессе деструкции объект ее все время остается комплексом сме-
няющихся форм микроорганизмов, мало или совсем не связанных друг
с другом и внешней средой. Этот комплекс в основном связан только с
субстратом, на который направлено его действие. В результате суб-
страт постепенно уменьшается и в конце концов может совершенно
исчезнуть.
Также не являются сообществом комплексы микроорганизмов,
культивируемые для эксперимента или других целей в изолированных
сосудах, так как среда их обитания все время изменяется в худшую
сторону в результате жизнедеятельности микробов, развивающихся на
ней, и в конце концов их собственные метаболиты могут стать причи-
ной уничтожения всех микроорганизмов в этих сосудах.
Установлено, что хотя детрит и входит в состав морской взвеси,
но отождествлять его со взвесью некорректно. Соответствующий рас-
чет Показывает, что, находясь даже в максимально возможном коли-
честве в воде морей и Океана, детрит составляет только 1/161 часть
массы органо минеральной взвеси, которой в Океане примерно
1370 млрд т.
Вследствие того что время обновления органической части взвеси
(а значит, и общей ее массы) составляет 10—20 лет, детрит нельзя счи-
тать основным источником пополнения взвешенного вещества в мор-
ских бассейнах, так как восполнить всю массу взвеси можно было бы
только за 161 год.
147
Расчет баланса взвешенного вещества в Океане показал, что в сум-
ме все известные автохтонные и аллохтонные источники пополнения
взвеси в нем составляют только 1/100 часть всей ее массы, т.е. имеет
место явный дисбаланс.
Эксперименты с различными органическими веществами подтверж-
дают микробиоценотическое образование морской взвеси. При этом
доказано, что если бы в 1/4 части взвешенного вещества морской воды
(мельчайшие минеральные и органические частицы) новообразование
взвеси прошло аналогичным путем, то тогда для восстановления ее во
всем Мировом океане (1370 млрд т) потребовался бы только 1 год,
а между общим количеством взвеси в Океане и ее воспроизводством
установился бы баланс. Учитывая, однако, что в Океане и морях усло-
вия для образования взвешенного вещества намного хуже, чем в
эксперименте, мы считаем, что время полного обновления взвеси в
Океане составляет не более 5-10 лет.
Исследования показали, что кроме взаимоотношений друг с дру-
гом в море, параметры изучаемой экосистемы испытывают влияние
7 параметров фактора Солнца (энергетический баланс, альбедо види-
мого спектра, доля баланса в инфракрасной области, активность (по
числу Вольфа), количество часов сияния, суммарная радиация, суммар-
ная среднесуточная радиация).
В процессе жизнедеятельности микроорганизмы СПМ и СМВ отла-
гают кварц, а из створок мертвых диатомовых на взвеси и в перифито-
не отлагается кремнезем. Доказано, что на осаждение кварца в основ-
ном влияет кальцит, также всегда отлагающийся вместе с кварцем, а
кроме того — pH, температура воды и РОВ СПМ.
Наблюдая в течение многих лет за морской взвесью, заметили, что
частицы кварца, напоминающие мельчайшие песчинки, с увеличением
возраста взвеси имеют тенденцию к увеличению размеров, т.е. к росту.
Основными факторами отложения кварца на взвеси являются СМВ и
СПМ — в них происходит этот процесс, и они подобны. С одной сторо-
ны, микроорганизмы СМВ, по-видимому, способствуют образованию
щетин кварца, осаждая его в виде ионов из раствора морской воды, а
возникшие частицы потом могут служить центром последующей крис-
таллизации, с другой - они могут растворять кремнеземовые створки
мертвых диатомовых, переводя кварц в ионное состояние.
В связи со сказанным возникла гипотеза об одном из возможных
путей происхождения мелкого кварцевого морского песка. Смысл ее в
том, что поскольку частицы взвеси увеличиваются в размерах, то вмес-
те с ними растут и кристаллы кварца иа них. В результате отложения
микробами СМВ карбонатов и кварцевых частиц частицы взвеси тяже-
леют настолько, что начинают оседать на дно. Так, они накапливаются
или их поедают бентосные животные, попадая в ил с низкой величиной
pH (или по другим причинам), карбонаты взвеси растворяются, частич-
ки распадзюгся, становясь донным осадком, а крупицы кварца осво-
”•18
бождаются, образовывая обычный мелкозернистый песок с размерами
песчинок 0,1—0,25 мм и мельче.
Вследствие того что количество оседающей ежегодно взвеси в океа-
не составляет около 5,4 млрд т (по сухой массе), а кварца в перифито-
не и, следовательно, во взвеси, содержится примерно 1/14 части ее су-
хой массы, то можно рассчитать количество образующегося таким об-
разом в Мировом океане песка. Это составляет приблизительно
386 млн т в год. Установлено, что микробиогенное осаждение карбо-
натов в перифитоне и на взвеси имеет важное, если не первостепенное,
значение в общем процессе карбонатосаждения в Мировом океане.
На основании исследований микробиогенного карбонатосаждения
на предметах, погруженных в море, установили следующее. С такой
же скоростью, как в перифитоне за 1000 лет, в кислородной зоне Чер-
ного моря могло бы отложиться 6,6 г-см" 2 карбонатов, что в высоту
составило бы 3,67 см. За 2500 лет (возраст этого моря) соответственно
отложилось бы 16,5 г-см-2 карбонатов высотой 9,2 см. С морской
взвесью могло бы осесть за 1000 лет экспонирования 41,40 г-см~* кар-
боната высотой 23,00 см, а за 2500 лет - соответственно 103,50 г.см-2
высотой 57,50 см.
При сопоставлении полученных результатов с данными о карбо-
натосаждении в других морях и в Мировом океане оказалось, что
масштабы этого процесса в Черном море (Севастопольская бухта)
достаточно велики. Только на Багамской банке отлагается больше кар-
бонатов, чем в Черном море. Так, за 1000 лет там отложилось до 80 см
карбонатов, очевидно, по причине аномально благоприятных условий
по сравнению с другими районами Мирового океана. Изученный нами
процесс микробиогенного отложения карбонатов мы считаем домини-
рующим по сравнению с другими описанными в литературе аналогич-
ными процессами, которые происходят в морских водоемах.
Интересна особенность морских гетеротрофных бактерий ’’отра-
жать” некоторые важные океанологические характеристики. Предпо-
лагая, что бактерии являются индикаторами структуры и динамики
вод, мы исходили из иных предпосылок, чем А.Е.Крисс (1976), и поэ-
тому пришли к совершенно иным выводам.
На основании имеющихся данных о распределении гетеротрофных
бактерий в морях и океане нами обнаружено, что воды в этих водных
объектах имеют четкую слоистую структуру (5-9 слоев). Гетеро-
трофные бактерии, скапливаясь на границе разделов слоев вод, четко
указывают на вертикальную структуру толщи воды (горизонтальный
ход, районы подъема и опускания слоев).
В результате исследований построена структура вод Тихого и
Индийского океанов от поверхности до дна по разрезам, пересекаю-
щим океаны с севера на юг. При этом на севере океанов обнаружи-
ваются те же вергенции, что и на юге. Были определены также элементы
структуры вод Средиземного моря. Используя данные о численности
149
бактерий-гетеротрофов в Средиземном море, путем сопоставления
пересекающихся разрезов в трехмерном пространстве, установлены
купола подъема и опускания вод, а значит, и направление их вращения,
т.е. горизонтальная циркуляция.
Определили, что в Средиземном море летнее распределение гетеро-
трофных бактерий четко отличается от зимнего. Это происходит от пе-
ремешивания вод зимой в результате процесса конвекции. Подробно
изучив это явление, нами обнаружено, что распределение гетеротроф-
ных морских бактерий в толще воды отражает ход конвективного пе-
ремешивания. В итоге появилась возможность рассчитать фактическую
скорость конвекции. Мы это сделали раньше (1962 г.), чем междуна- ;
родная экспедиция МЕДОК (1969-1970 гг.). По нашим данным, ско-
рость конвекции в море не 0,01-0,001 см-с”1, а в 35-800 раз больше.
Результаты настоящих системных исследований морской экосис-
темы микро- и макроорганизмов в прибрежной зоне моря, продолжаю-
щихся уже 20 лет, а также существующие литературные данные приво-
дят нас к следующим выводам. Параметры Солнца, а также микро-
организмы и факторы среды в прибрежных районах морей и океанов
способствуют интенсивному развитию сообществ планктона (СМП),
перифитона (СПМ) и взвеси (СМВ), которые, как было установ-
лено нами, обусловливают процесс микробиогенного осаждения кар-
бонатов (в основном в виде СаСО3), микробиоценотическое образо-
вание взвеси и другие процессы.
Непрерывно идущий процесс карбонатосаждения, являющийся
следствием нормального метаболизма СПМ и СМВ, проходит в морях
и океане, в основном в прибрежных частях морских бассейнов в облас-
ти шельфа. Отложения известняка на нашей планете, горы и хребты
из СаСО3 с вершинами на месте бывших бухт и заливов древних морей,
где продуктивность наибольшая, залежи мрамора из более ранних от-
ложений карбонатов, кристаллизировавшиеся под большим давлением
поздних отложений за мноше миллионы лет, - все они образовались
на шельфе у берегов в основном в результате жизнедеятельности мик-
роорганизмов СПМ и СМВ.
Как известно, диатомовые водоросли в процессе фотосинтеза обра-
зуют масло, служащее запасным питательным веществом, которое
обеспечивает этим организмам при планктонном образе жизни лучшую
плавучесть. Диатомеи образуют в 3—13 раз больше масла, чем другие
водоросли (Прошкина — Лавренко, 1955). При отмирании диатомей
масло попадает в воду, где во взвешенном состоянии с течением време-
ни может скапливаться, по-видимому, в больших количествах. Возмож-
но, что оно является наибольшей частью автохтонного жира, который
определяется вместе с хлороформным экстрактом в море. При этом
часть ХлЭк разлита по поверхности воды, а диатомовый жир находится
еще и под ее поверхностью (где мы и берем пробы для элюирования
органическими растворителями). Можно предположить, что именно из
1S0
этого жира (и других компонентов ХлЭк), когда они попадают под
землю при геологических катастрофах и фсссилизируются в продол-
жение миллионов лет, образуется нефть.
В связи с постоянно усиливающимся антропогенным загрязнением
морей и океанов нельзя не учитывать влияние, оказываемое на изучае-
мую экосистему основных загрязнителей, к которым относятся угле-
водороды, нефть и ее продукты.
Известно, что обитатели морских водоемов (животные и растения)
образуют в больших количествах липиды и углеводороды, которые
определяются путем элюирования из воды хлороформом или 4-хлорис-
тым углеродом. Поскольку они близки по строению и составу к неф-
тепродуктам и нефти, разделить их при определении чрезвычайно труд-
но. Поэтому мы вынуждены пока изучать влияние на био- и абиопара-
метры изучаемой системы всей указанной смеси (нефть, ее продукты,
углеводороды и липиды), которые выделялись с ХлЭк.
Обнаружили, что ХлЭк связан не только с параметрами морской
воды, но и с долей баланса в инфракрасной области спектра Солн-
ца. Гармонические составляющие (период, фаза, амплитуда) ХлЭк
примерно такие же, как и у диатомовых живых СПМ, кислорода,
фенола, палочковидных бактерий СПМ и РОВ СПМ. Путем применения
различных математических методов удалось установить множество
связей ХлЭк с био- и абиопараметрами экосистемы. Рассчитав частные
корреляции с помощью многомерного анализа, установили, что сущест-
вует взаимовлияние ХлЭк и гетеротрофных, кокковидных бактерий и
диатомовых водорослей СПМ. В основном в образовании автохтонной
части и в деструкции ХлЭк участвуют бактерии-гетеротрофы СМВ и
СМП, а также кислород в морской воде. В процессе образования или
использования ими компонентов ХлЭк на взвеси отлагаются карбо-
наты СМВ.
Результаты проведенных исследований свидетельствуют, что в Се-
вастопольской бухте, по-видимому, содержание автохтонных углеводо-
родов, жиров и липидов выше, чем аллохтонных - нефти, ее продуктов
и углеводородов. Причина - благоприятные условия прибрежной части
шельфа изучаемой акватории Черного моря.
Установлено, что гетеротрофные бактерии (куда входят и нефте-
окисляющие) используют ХлЭк в основном на границе раздела фаз в
мс вода - поверхность частицы взвеси, вода - поверхность твердо-
ю погруженного предмета, вода - донный грунт. В меньшем коли-
честве, по-видимому, ХлЭк утилизируется у раздела фаз вода - кап-
ля ХлЭк.
Вследствие того что одним из важнейших загрязнителей моря
также считают фенол (точнее, фенолоподобные вещества), исследовали
его влияние на компоненты изучаемой системы. Определили, что фе-
нол имеет такие же периоды и фазы, как ХлЭк. В результате исследо-
ваний определили, что фенол и ХлЭк имеют примерно одинаковое ко-
151
личество корреляционных связей. Разбив большинство основных пара-
метров экосистемы на группировки-факторы по величине взаимных
корреляционных функций и тесноте взаимосвязей внутри групп, обна-
ружили, что они выстраиваются в определенном порядке - сверху
вниз. Вверху располагаются группировки, наиболее тесно взаимосвя-
занные и с наибольшей величиной корреляции, ниже - группировки, в
которых число корреляций между группами и внутри их, а также тесно-
та корреляций закономерно уменьшаются. При этом фенол находится
в самой первой и верхней группировке вместе с компонентами карбо-
натного равновесия (СО2, НСО3, COj”), pH морской воды, СМВ и
СПМ. Эта группировка является одним из самых важных регуляторов
процессов, проходящих в изучаемой системе. Фенол представляет со-
бой один из параметров-регуляторов в этой группировке.
Мы предполагаем, что в какой-то мере концентрация фенолоподоб-
ных веществ в море регулируется параметрами Солнца - альбедо ви-
димого спектра и его активностью, в то время как ХлЭк - долей ба-
ланса в инфракрасной области спектра. Судя по многочисленным свя-
зям с другими параметрами и наличию гармонических составляющих,
оба эти параметра ’’загрязнения” в Севастопольской бухте в основном
автохтонного характера. Фенолоподобные вещества, а также ХлЭк,
очевидно, никакого вредного действия на экосистему микроорганиз-
мов не оказывают. Первый из них является параметром-регулятором,
второй - обычным параметром.
Биотопом микроорганизмов, описанных в этой работе, и их биоло-
гической нишей в основном является область шельфа. Это пологий
склон берега, ограниченный изобатой 200 м, воды которого обогаща-
ются минеральными и органическими веществами из моря и с берега.
В результате на шельфе создаются наиболее благоприятные условия
для высокой продуктивности гидробионтов, образования взвесей,
отложения осадков и многих других процессов (Моисеев, 1969; Бего-
ров, 1974; Слевич, 1977). Шельф - место богатых уловов рыбы, добы-
чи полезных ископаемых, нефти и газа.
СПИСОК ЛИТЕРАТУРЫ
Алекин О.А. Химия океана. - Л. : Гидрометеоиздат, 1966. - 246 с.
Алексеев В.Н. Курс качественного полумикроанализа. - М. : Госхимиздат,
1958.- 321 с.
Базилевич Н.И., Родин Л.Е., Розов Н.Н. Сколько весит живое вещество планеты //
Природа. - 1971.- Вып. 1.- С. 27-35.
Богоров В.Г. Биологическая трансформация и обмен энергии и веществ в Океа-
не // Океанология. - 1967. - 7, вып. 5. - С. 11-18.
Богоров В.Г. Планктон Мирового океана. - М. : Наука, 1974.- 320 с.
Богоявленский А.Н. Химическая характеристика вод Курило-Камчатской впади-
ны // Тр. Ин-та океанологии АН СССР. - 1955. - 12. - С. 11-15.
Брайко В.Д Фауна Укра'1ни. Мохуватки. - Киш : Наук, думка, 1983. - Т. 24,
вил. 1. - 119 с.
Брайко В.Д Обрастание в Черном море. - Киев : Наук, думка, 1985. - 124 с.
Бурков ВЛ. Общая циркуляция Мирового океана. - Л. : Наука, 1980. - 254 с.
Валло К. Общая география морей. - М.; Л. : Учпедгиз, 1948. - 492 с.
Винберг Г.Г. Первичная продукция водоемов. - Минск : Изд-во АН БССР, 1960.-
251 с.
Виноградов А.П. Химический элементарный состав организмов моря // Тр. био-
геохим. лаборатории АН СССР. - 1944. - 6. - С. 21- 30.
Витюк ДМ. Взвешенное вещество и его биогенные компоненты. - Киев : Наук,
думка, 1983. - 210 с.
Галеркин ЛИ., Бараш М.Б., Сапожников В.В., Пастернак Ф.А. Тихий океан. -
М. : Мысль, 1982.- 316 с.
Горбенко Ю.А. Образование бактериальной пленки на погруженных в морскую
воду пластинах, покрытых необрастающимн красками // Тр. Севастоп. биол.
станции АН СССР. - 1963. - 16. - С. 447-453.
Горбенко Ю.А. Потребление морскими перифнтонными бактериями органичес-
ких компонентов противообрастаемых красок и метаболиты, образующиеся при
этом /I Биологическое исследование Черного моря и его промысловых ресур-
сов.- Киев : Наук, думка, 1968.- С. 136-143.
Горбенко Ю.А. Экология морских микроорганизмов перифитона. - Киев :
Наук, думка, 1977. - 250 с.
Горбенко Ю.А. О взаимосвязи pH морской воды с метаболитами микроорганиз-
мов перифитона и взвеси в море // Биология моря. - Киев, 1979. - Вып. 48. -
С. 3-7.
Горбенко Ю.А. О степени биологического повреждения канифоли, парафина и
противообрастаемых красок // Обрастание и биокоррозия в водной среде. - М.:
Наука, 1981,- С. 25-30.
Горбенко Ю.А. Микробиоценотическое образование морской взвеси // Изв. АН
СССР.- Сер. биол.- 1986.- Вьш.4.- С. 505-516.
Горбенко Ю.А. О возможности выявления микробиологическим методом райо-
нов подъема и опускания вод в центральной части Тихого океана Ц Океаноло-
гия. - 1964. - 4, вьш. 1. - С. 167- 174.
153
Горбенко Ю.А. Связь колебаний численности гетеротрофных бактерий с конвек- j
тинным перемешиванием вод в Средиземном море и с другими гидрологически-
ми Явлениями // Изв. биол. наук. Сер. биол. - 1988. - № 5. — С. 752- 760.
Горбенко Ю.А., Митропольский А.Ю. Динамика накопления и распределения мик-
роэлементов в морской взвеси //Материалы по минералогии, петрографии и гео-
химии осадочных пород и руд. - Киев : Наук, думка, 1976. - Вып. 4. - С. 49- 55.
Горбенко Ю.А., Подвинцев Ю.В. Анализ взаимодействия элементов в системе
микро- и макроорганизмы обрастаний - био- и абиотические факторы среды
(математический эксперимент) // Биология моря. - Киев, 1979. - Вып. 48. -
С. 28-41.
Горбенко Ю.А., Тримонис Э.С. О биогенном осаждении кальцита и кварца на
пластинах обрастания в море // Экология моря. — 1980. - Вып. 4. — С. 72-77.
Горбенко Ю.А., Татаренко Л.Я. Влияние биосистемы морских микроорганизмов
на содержание нефтепродуктов в море // Там же. - 1983. - Вып. 13. - С. 35-41.
Горбенко Ю.А., Крышев И.И. Статистический анализ динамики экосистемы мор-
ских микроорганизмов. — Киев : Наук, думка, 1985.- 143 с.
Диатомовый анализ. — Л,: Изд-во геол, лит-ры, 1950. — 215 с.
Дуплаков С.Н. Материалы к изучению перифитона // Тр. лимнол. станции в Коси-
но. - 1933.- 16. -С. 51—63.
Емельянов Е.М., Шимкус КМ. Взвешенное вещество в Средиземном море и его
минеральный состав // Океанология.- 1973.- 13, вып.4.— С. 23-31.
Залезинская Л.М. К изучению естественной смертности некоторых организмов
пелагиали Черного моря // Экологическая биогеография контактных зои моря. -
Киев : Наук, думка, 1968.-С. 125-130.
Зубов Н.Н. Уплотнение при смешивании морских вод разной температуры и соле-
ности. - М.: Гидрометеоиздат, 1957.- 112 с.
Зубов Н.Н., Сабинин КД. Вычисление уплотнения при смешении морскихвод.-
М.: Гидрометеоиздат, 1958.- 69 с. .
Индийский океан. — Л.: Наука, 1982. — 388 с.
Карт ин к ин Г.С. Попытка практического разрешения понятия биогеоценоз //
Рус. зоол. журн. - 1926,— 1927.- 7, вып. 4. — С. 211-232.
Калугина - Гутник А.А. Фитобентос Черного моря. - Киев : Наук, думка, 1975.-
248 с.
Кондратьева Т.М. Продукция и суточные изменения фитопланктона в южных мо-
рях: Автореф. дис.... канд. биол. наук. - Одесса, 1967. - 10 с.
Кондратьева Т.М. О нахождении крупных частиц взвешенного органического ве-
щества (’’Морского снега”) в водах Черного моря // Экспедиционные исследо-
вания в Средиземном и Черном морях в ноябре-декабре 1971 г. - Киев: Наук,
думка, 1975. - С. 26-30.
Крисс А.Е. Морская микробиология. - М.: Изд-во АН СССР, 1959.- 455 с.
Крисс А.Е. Микробиологическая океанография. - М.: Наука, 1976. - 289 с.
Крышев И.И., Горбенко Ю.А. О причинах сезонных колебаний численности диато-
мовых водорослей планктона в море // Биология моря. - Киев, 1979.— Вып. 51.—
С. 36-42.
Крышев И.И., Горбенко Ю.А. Периодические процессы в многолетней динамике
элементов морской бносистемы // Экология моря,— 1980.— Вып. 10. — С. 19-27.
Курс общей геологии. - Л.: Недра, 1976,- 535 с.
Лебедева М.Н. Микробиологическая индикация зон конвергенции и дивергенции
в Индийском океане // Океанология. — 1962. — 2, вып. 6. - С 41- 48.
Лебедева М.Н., Горбенко Ю.А. Некоторые черты гидрологической структуры
Средиземного моря по микробиологическим данным // Тр. Севастоп. биол.
станции.- 1964. - 16. - С. 512-527.
Леонтьева В.В. Некоторые данные по гидрологии впадин Тонга и Кармадек //
Гидрометеорологические исследования в первых рейсах ’’Витязя” по программе
МГГ. - Г.-. : Изд-во АН СССР, I960. - С. 72-80.
15*^
Лисицын А.П. Распределение и химический состав взвеси в водах Индийского
океана // Результаты исспед. по программе МГГ. Океанология.— 1964.— № 10.—
С. 31-53.
Лисицын А.П. Осадкообразование в океане. - М.: Наука, 1974.- 438 с.
Лукас С Взаимосвязь между водными организмами через посредство внешних
метаболитов // Океанография. — 1965.— 1, - С. 97- 116.
Мельников И.А. Морфологические особенности частиц органического детрита //
Океанология. - 1976. — 76, вып. 4. - С. 27—35.
Методологические указания № 30. Методы химического анализа загрязненной
морской воды. - М.: Гидрометеоиздат, 1966. — 158 с.
Методика изучения биогеоценозов внутренних водоемов. - М. : Наука, 1975. -
240 с.
Меншуткин В,В., Сорокин Ю.И. Модельное исследование экосистемы Рыбинского
водохранилища II Биология внутренних вод. - Л.: Наука, 1975. - С. 63-75.
Миронов О.Г. Нефгеокисляющие микроорганизмы в море. - Киев : Наук, думка,
1971.- 233 с.
Миронов О.Г. Нефтяное загрязнение и жизнь моря. - Киев : Наук, думка, 1973. -
86 с.
Миронов О.Г., Георга-Ко пуло с Л. А. О самоочищении моря от тяжелых нефтяных
фракций// Гидробиол. жури. - 1981.- 71, № 1.- С. 45-52.
Мордухай-Болтовской Ф.Д Особенности программы и методика биогеоценоти-
ческих исследований внутренних водоемов // Программа н методика биоценоги-
ческих исследований. - М.: Наука, 1975.- С. 109-125.
Моисеев ПА. Биологические ресурсы Мирового океана. - М. : Пищ. пром-сть,
1969.- 338 с.
Морское обрастание н борьба с ним. - М.: Воениздат, 1957. - 501 с.
Муромцев А.М. Основные черты гидрологии Тихого океана. - Л. : Изд-во АН
СССР, 1958.- 386 с.
Нестерова ДА. Об отмирании фитопланктона и возможном значении этого явле-
ния в жизни моря // Вопросы морской биологии. — Киев : Наук, думка, 1969. —
С. 157.
Овчинников ИМ., Плахин. Е А., Москаленко Л.В. и др. Гидрология Средиземного
моря. - Л. : Гидрометеоиздат, 1976. - 375 с.
ОдумЮ. Основы экологии. - М.: Мир, 1975.- 740 с.
Океанология. Физика океана. Гидрофизика океана. - М. : Наука, 1978. - 455 с.
Очерки по истории экологии. - М.: Наука, 1970. - 292 с.
Павлова Е.В., Миронов ГН. Структура пищевых связей, передача и использова-
ние вещества и энергии в планктонных сообществах Черного моря // Биология
моря, - Киев, 1970.- Вып. 19.- С. 41-49.
Петипа Т.С. Трофодинамика копепод в морских планктонных сообществах. -
Киев : Наук, думка, 1981,- 242 с.
Подвинцев Ю.В., Горбенко Ю.А. Регрессионный анализ в задачах исследования мор-
ских экосистем // Биология моря. — 1977. - Вып. 40. - С. 47- 53.
Подвинцев Ю.В., Горбенко Ю.А. Факторное представление морских микробиоло-
гических систем // Там же. - С. 86- 92,
Подвинцев Ю.В., Горбенко Ю.А. Структура взаимосвязи параметров солнца,
атмосферы и морской среды // Комплексные исследования Черного моря :
Мор. гидрофиз. ин-т. - Севастополь, 1979.- С. 162-171.
Подвинцев Ю.В., Горбенко Ю.А. Методы теории информации и статистики при
анализе н прогнозе биопроцессов обрастания в морской среде // Обрастание н
коррозия в морской среде : Биол. повреждения, - М.: Наука, 1981.- С. 227-235.
Подвинцев Ю.В., Горбенко ЮЛ. Информационно-статистическое моделирование
сложных систем в приложении к прогнозу численности микроорганизмов Черно-
го моря на 1990-2000 гт. // Математические методы в биологии. - Киев : Наук,
думка, 1983,- С. 253-260.
155
Проблемы методологии системного исследования. - М. : Мысль, 1970,- 455 с.
Прошкина-Лавренко А.И. Диатомовые водоросли планктона и бентоса Черного
моря,- М.; Л. : Изд-во АН СССР. - 1955. - 222 с.; 1963.- 243 с.
Родино А.Г. Кормовое значение и строение детрита // Биологические ресурсы во-
доемов, пути их реконструкции и использования, - М. : Наука, 1966.- С. 35-42.
Рзкиикий П.Ф. Биологическая статистика. - Минск : Вышейш. шк., 1967. -
327 с.
Романенко В. И., Кузнецов СИ. Экология микроорганизмов пресных водоемов.—
Л. : Наука, 1974.- 195 с.
Романкевич Е.А. Геохимия органического вещества в Океане. — М. : Наука,
1977.- 256 с.
Руководство по морским гидрохимическим исследованиям. - М., 1959. - 278 с.
Сеничева М.И. Сезонная динамика численности, биомассы и продукции фито-
планктона Севастопольской бухты // Экология моря. - I960. - Вьш. 1.- С. 52-59.
Скопиниев Б.А. Современные достижения в изучении органического вещества вод
океанов//Океанология. - 1971.- 11, вып. 6. - С. 48-57.
Слевич СБ. Шельф, - Л. : Гидрометеоиздат, 1977.- 237 с.
Сорокин Ю.И. Об агрегированиости морского бактериопланктона // Докл. АН
СССР. - 1970.- И, вьш. 6.- С. 85-89.
Степанов В.Н. Основные конвергенции и дивергенции вод Мирового океана //
Бюп. океаиогр. комиссии. - I960. - №6,- С. 15-20.
Степанов В.Н. Океаносфера. - М.: Мысль, 1983. — 269 с.
Страхов НМ. Об истинной роли бактерий в образовании карбонатных пород //
Иэв. АН СССР. Сер. геол.- 1948.- №3.- С. 35-41.
Сущеня Л.М. Детрит и его роль в продукционном процессе в водоемах // Гидро-
биол. жури.- 1968. - 4, вып. 2. - С. 41-47.
Тихий океан (Осадкообразование). - М.: Наука, 1970.- Т. 6. Ки. 1. - 427 с.
Федоров КН. Физическая природа и структура океанических фронтов. - Л. :
Гидрометеоиздат, 1983.- 296 с.
Федянин А.С. Бактерии водной толщи Черного моря - продуценты витаминов
группы В // Биология моря. - Киев, 1973. - Вып. 30. - С. 90- 95.
Финенко 3.3. Содержание органического вещества в сестоне Черного и Азовского
морей. Исследование планктона Черного и Азовского морей. - Киев : Наук,
думка, 1965. - 31 с.
Хайлов КМ. Экологический метаболизм-в море. - Киев : Наук, думка, 1971. -
252 с.
Хайлов К.М., Финенко 3.3. Взаимодействие детрита с высокомолекулярными
компонентами РОВ морской воды // Океанология. - 1968. - 8, вып. 6. -
С. 53-61.
Хорн Р. Морская химия. - М. : Мир, 1972. - 399 с.
Чижевский А.Л. Земное эхо солнечных бурь. - М. : Мысль, 1973.- 349 с.
Шварц С.С К истории основных понятий современной экологии II Очерки по
истории экологии.- М.: Наука, 1970.- С. 89-92.
Bergey’s Manual determinative Bacteriology.' - USA, 1948. - 1529 p.; 1957. - 1094 p.
Biddanda B.A. Microbial synthesis of macroparticulate matter // Marine ecology-progress
series. - 1985. - 20. - P. 241 -247.
Fenschel T. Studies on decomposition of organic detritus derived from the turtle grass //
LimnoL and Oceanogr. - 1970. - 15. - P. 45-53.
Journal of the Geophysical Research // Amer. Geophys. Union. U.S. - 1971. - P. 1-36.
Kahl F. Die Tierwelt der Nord und Ostse.- Hamburg, 1933. - 150 p.
Kent W.A. Manual of the Infusoria. - London, 1800-1882. - Vol. 3. - 451 p.
KrilmmelO. Handbuch der Oceanographie. - Stuttgart, 1911. - 320 p.
Lacombe H, Tehernia P. Evolution hydrologique dans la zone de formation d’eau pro-
fonde au large de la cote mediterraneenne grancaise par hiver tres froid// Collog. Intern.
CNRS - Paris, 1962-1963. - P. 51; 1972. - P. 37- 1974. - P. 132 -140.
156
Lepsi J. Fauna Republicii Populate Romine // Acad. Rep. PopuL - I960. - 1, N 2. -
382 p.
Marine Organic Chemistry / Ed E.K.Duursma, R.Dauson. - Amsterdam; Oxford; New
York, 1981. - 521 p.
MEDOC Group (Lacombe H., Tchernia P„ Ribet J., Frasseto R., Smallow J.C., Mil-
ler A.R., Stommel H.). Observation of formation of Deep Water in the Mediterranean
Sea // Nature. - 1969, 1970. - 227. - P. 1037-1045.
Mobius K. Die Auster und die Austem // Wirtschaft mit Karte. - Berlin, 1877. -
S. 20-25.
Pollak M.J.The sources of the seep water of the Eastern Mediterranean Sea // J.Mar.
Res. - 1951. - 10, N 1. - P. 128-135.
Riley G.A. Particulate organic matter in Sea Water // Adv. Mar. Biol - 1970. - 8. -
P. 55-61.
Schott G. Die GewKsser der Mittelmeeres 11 Ann. Hydr. Marit Meteor. - 1915. - 43,
N 1/2.-S. 1-18,49-79.
Sorokin YaJ. Decomposition of Organic Matter and Nutrient Regeneration // Mar.
Ecol - 1978. - 1.- P. 121-130.
Steemann-Nielsen E., Jensen E.A. Primary oceanic production. The autotrophic produc-
tion of organic matter in the oceans // Galatea Repts. - 1957. - 1. - 355 p.
Whippl G. C Changes that plase in the bacterial contents of waters during transporta-
tion // Techn. Quart - 1901. - 14. - P. 61-71.
Zo Bell CE. The effect of solid surfaces upon bacterial activity // J. Bacterio I - 1943. -
46.-P. 23-31.
Zo Bell CE., Harvey CU. A list of Marine Bacteria including Description of sixty New
Species // Bull. Scripps. Inst. Oceanography. - 1944. - 5. - P. 239-249.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ................................................. 3
ГЛАВА 1. ИСТОРИЯ ВОПРОСА .................................. 3
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ.................... 7
ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ........................... Ю
3.1. О биоценологии................................... Ю
3.2. О простейших организмах СПМ и их взаимосвязях.... И
3.3. Комплекс микроорганизмов взвеси в качестве ценоза. 13
3.4. О подобии микроценозов перифитона и взвеси...... 20
3.5. Всегда ли комплекс микроорганизмов является сооб-
ществом ?.......................................... 27
ГЛАВА 4. ВЗАИМОСВЯЗИ БИО- И АБИОТИЧЕСКИХ ФАКТОРОВ
ЭКОСИСТЕМЫ.................................................. 30
4.1. Изучение взаимосвязи pH морской воды, метаболитов мик-
роорганизмов перифитона и взвеси с помощью корреляцион-
ного анализа ........................................ 30
4.2. Факторное представление морских микробиологических
систем.............................................. 32
4.3. Определение групптфовок н факторов методами корреля-
ционного и факторного анализов....................... 36
4.4. Взаимосвязи параметров в факторах............... 44
ГЛАВА 5. ИЗУЧЕНИЕ ВЗАИМООТНОШЕНИЙ В ЭКОСИСТЕМЕ С ПО-
МОЩЬЮ МАТЕМАТИЧЕСКОГО ЭКСПЕРИМЕНТА ......................... 47
5.1. Изменение количества микроорганизмов на ранних стадиях
развития биоценоза обрастаний........................ 47
5.2. Интерпретация результатов математического эксперимента 51
5.3. Основные итоги математического эксперимента..... 60
5.4. Структура взаимосвязей параметров экосистемы, изученная
с помощью анализа корреляционных функций............. 64
5.5. Прогнозирование био-и абиотических параметров..... 74
ГЛАВА 6. ИЗМЕНЕНИЯ СРЕДЫ, ВЫЗЫВАЕМЫЕ МИКРООРГАНИЗ-
МАМИ ...................................................... 81
6.1. Влияние факторов среды иа изменение величины pH и кон-
центрации РОВ ....................................... 81
6.2. О микробиогенном отложении кварца и кальцита микро-
организмами......................................... 82
6.3. Об образовании кальцита, кварца, кремнезема и гипотезе
происхождения мелкого песка в морях и Океане......... 85
6.4. Осаждение карбонатов микроорганизмами взвеси н пери-
___________ фмтона.......................................... 87
158
ГЛАВА 7. МИКРОБ ИОЦЕНОТИЧЕС КОЕ ОБРАЗОВАНИЕ МОРСКОЙ
ВЗВЕСИ................................................... 98
7.1. Общие признаки морской взвеси...................... 99
7.2. О возможности пополнения взвеси из живого органического
вещества.............................................. 100
7.3. Роль отмирания морских гидробионтов............... 102
7.4. Возможности пополнения морской взвеси продукцией
фитопланктона......................................... 103
7.5. Аллохтонные источники взвеси н дисбаланс ее содержа-
ния с пополнением в Океане ........................... 104
7.6. Образование взвеси в море......................... 104
7.7. Механизм образования морской взвеси................106
7.8. Образование взвеси экспериментальным путем........ 107
7.9. Скорость образования микробноценотической взвеси и ба-
ланс ее в Океане....................................... ПО
ГЛАВА 8. ИСПОЛЬЗОВАНИЕ НЕКОТОРЫХ РЕЗУЛЬТАТОВ ЭКОЛО-'
ГИЧЕСКИХ ИССЛЕДОВАНИЙ КАК ИНДИКАТОРОВ ЯВ-
ЛЕНИЙ И ПРОЦЕССОВ В МОРЯХ И ОКЕАНЕ............................ 115
8.1. Влияние конвективного перемешивания вод в Средиземном
море на изменение численности гетеротрофных бактерий ... 115
8.2. Летнее распределение численности бактерий......... 116
8.3. Зимнее распределение численности бактерий......... 120
8.4. Связь численности гетеротрофных бактерий с вертикальной
структурой вод в Тихом океане......................... 122
8.5. Связь распределения численности гетеротрофных бактерий с
вертикальной структурой вод в Индийском океане........ 127
8.6. Определение с помощью микробиологического метода
районов циркуляции вод в Средиземном море............. 131
ГЛАВА 9. ХАРАКТЕРИСТИКА ХЛЭК И ФЕНОЛОВ, НАХОДЯЩИХСЯ
В МОРЕ...................................................... 133
9.1. О природе ХлЭк и его взаимосвязях с био- и абиофакторами
изучаемой системы ................................. I33
9.2. Микроорганизмы, использующие ХлЭк и участвующие в са-
моочищении моря в природных условиях.............. 137 •
9.3. Природа фенола и его взаимосвязи с био- и абиофакторами
экосистемы ....................................... 140
9.4. Основные параметры, определяющие содержание фенола в
море (регрессионный анализ) ...................... Ш
ЗАКЛЮЧЕНИЕ..................................................... I47
СПИСОК ЛИТЕРАТУРЫ............................................. 153
159
Научное издание
ГОРБЕНКО Юрий Александрович
ЭКОЛОГИЯ И ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ
МОРСКИХ МИКРООРГАНИЗМОВ
Художник обложки Н.В.Синеутская
Художественный редактор И.Е.Писарева
Технический редактор Т.К.Валицкая
Оператор В.Н.Курган
Корректоры Л.В.Белокопытова, Л.Н.Валвнева
ИБN* 10144
Сдано в набор 14.06.90. Подл в печ. 26.10.90. Формат 60x84/16. Бум. офс. Н» 1.
Гарн. Пресс Роман. Офс. печ. Усл. печ. л. 9,30. Усл. кр.-отт. 9,53. Уч.-изд. п. 10,70.
Тираж 500 зкз. Заказр-З£эНена 2 р. Ю к.
Оригинал-макет подготовлен в издательстве "Паукова думка”. 252601 Киев 4,
ул. Репина, 3.
Киевская книжная типография научной книги. 252004 Киев 4, ул. Репина, 4.