/








































































































































































































































































































































































































































































































































Автор: Яковлев В.Н.
Теги: физиология сравнительная физиология медицина
ISBN: 5-87456-436-5
Год: 2005




Текст
НОРМАЛЬНАЯ
ФИЗИОЛОГИЯ
МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ И СОЦИАЛЬНОГО
РАЗВИТИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
ВОРОНЕЖСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ
АКАДЕМИЯ ИМ. Н.Н. БУРДЕНКО
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
УЧЕБНЫЕ МОДУЛИ
ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ СТУДЕНТОВ
Эе издание, переработанное и дополненное
Рекомендовано Учебно-методическим объединением
по медицинскому и фармацевтическому образованию
вузов России в качестве учебного пособия для студентов
медицинских вузов
Под редакцией
заслуженного работника высшей школы РФ
доктора медицинских наук, профессора
В.Н. Яковлева
Воронеж
2005
УДК 612 (07)
Нормальная физиология: Учебные модули для самостоятельной работы студентов:
Учебное пособие. - 3-е изд., перераб. / Под ред. В.Н. Яковлева.- Воронеж: Изд-во им. Е.А.
Болховитинова, 2005. - 528 с.
ISBN 5-87456-436-5
Пособие предназначено для повышения эффективности самостоятельной работы сту-
дентов при подготовке к практическим и итоговым занятиям во внеаудиторное время. Оно со-
держит теоретический материал занятия, расположенный в логической структуре, раскры-
вающей важнейшие механизмы физиологических процессов, а также содержит профильный
материал для студентов лечебного, педиатрического и стоматологического факультетов. Для
самоконтроля усвоенного материала и выработки мышления в пособие включены нетиповые
задачи и эталоны ответа к ним. Предложенный теоретический материал облегчает использо-
вание лекций и учебника при подготовке студентов к практическим занятиям н к экзамену.
Пособие предназначено для российских и иностранных студентов лечебного, педиатрического
и стоматологического факультетов медицинских вузов. Оно может также быть полазно для
врачей, интересующихся физиологическим базисом своей специальности.
Учебное пособие рекомендовано к изданию Проблемной учебно-методической комиссией по
нормальной физиологии при Всероссийском учебно-научно-методическом центре по непре-
рывному медицинскому и фармацевтическому образованию Минздрава РФ и Учебно-
методическим объединением медицинских и фармацевтичестх вузов.
Авторский коллектив:
проф. В.Н. Яковлев, проф. И.Э. Есауленко, доц. А.В. Сергиенко, доц. Е.В. Дорохов,
доц. А.В. Карпова, доц. Ю.А. Горшенев, асе. НД. Афанасьева, асе. Т.С. Голованова,
асе. Л. А. Киреева, асе. ВЛ. Черкашина.
Рецензенты:
академик РАМН, профессор Н.А. Агаджанян
(Российский университет дружбы народов, Москва),
профессор В.П. Дегтярев
(Московский государственный медико-стоматологический университет),
профессор В.И. Кобрин
(Государственная академия им. Маймонида, Москва)
© Авторский коллектив, 2005
ISBN 5-87456-436-5
ОГЛАВЛЕНИЕ
Предисловие.................................................................5
От редактора................................................................б
Список основных сокращений..................................................7
Часть I. Общая физиология .............................................................................9
Тема 1. Введение в физиологию. Осиовиые понятия
физиологии. Принципы формирования и организации
физиологических функций.................................................9
Тема 2 Биопотенциалы. Механизмы процесса
возбуждения и его распространения......................................31
Тема 3. Основные закономерности раздражения
возбудимых тканей......................................................42
Тема 4. Нейрон как структурно-функциональная
единица ЦНС. Функции глии..............................................47
Тема 5. Рефлекторный принцип деятельности нервной
системы. Нервные центры. Процессы возбуждения и
торможения в ЦНС.......................................................64
Тема 6. Координирующая и интегрирующая деятельность ЦНС.
Методы изучения ЦНС...................................................72
Тема 7. Частная физиология ЦНС: функции отделов головного
мозга, его кровоснабжение. Гематоэнцефалический
барьер Спинномозговая жидкость.........................................84
Тема 8. Вегетативная (автономная) нервная система.....................112
Тема 9. Эндокринная система...........................................124
Тема 10. Физиология мышц..............................................157
Тема 11. Роль различных отделов ЦНС в регуляции
мышечного тонуса и фазных движений....................................171
Часть П. Частная физиология.....................................................................187
Тема 12. Физико-химические свойства крови.............................187
Тема 13. Эритроцитарная и лейкоцитарная системы крови.................194
Тема 14. Физиологическая система регуляции агрегатного
состояния крови. Группы крови.........................................210
Тема 15. Физиологические функции сердца...............................228
Тема 16. Регуляция сердечной деятельности.
Кровообращение в миокарде.............................................236
Тема 17. Нагнетательная функция сердца. Внешние
3
проявления деятельности сердца. Методы исследования сердца.244
Тема 18. Регуляция гемодинамики...........................253
Тема 19. Физиология дыхания. Внешнее дыхание.
Газы крови, их транспорт. Газообмен между кровью и тканями.267
Тема 20. Регуляция дыхания.................................285
Тема 21. Регуляции кислотно-основного состояния............295
Тема 22. Общая характеристика функциональной
системы питания и физиологической системы
пищеварения. Пищеварение в полости рта и желудка..........305
Тема 23. Пищеварение в тонком и толстом кишечнике.........327
Тема 24. Обмен веществ и энергии. Питание..................338
Тема 25. Физиология выделения. Мочеобразование и
мочевыделение. Выделительная функция кожи, легких,
пищеварительного тракта...................................354
Тема 26. Терморегуляция...................................375
Часть III. Интегративная физиология 389
Тема 27. Общая физиология анализаторов. Тактильный,
болевой, вкусовой, обонятельный, интероцептивный,
температурный и двигательный анализаторы..................389
Тема 28. Зрительный анализатор.............................412
Тема 29. Слуховой и вестибулярный анализаторы.............427
Тема 30. Физиология высшей нервной деятельности.
Условные рефлексы. Типы ВИД...............................439
Тема 31. Физиологические основы психических
функций человека..........................................448
Тема 32. Физиологические основы целенаправленного
поведения человека........................................469
Тема 33. Физиология воспроизведения.......................483
Тема 34. Физиология адаптации.............................498
Ответы к нетиповым задачам...................................508
Литература для самоподготовки студентов......................514
Основной учебно-методический комплекс.....................514
Дополнительная литература.................................514
4
ПРЕДИСЛОВИЕ
В вузовской педагогике есть ряд проблем, в решении которых педагогиче-
ские коллективы кафедр находятся в постоянном долгу перед студентами. К
ним относятся, например, отбор учебного материала по данному предмету, ко-
ординация его с другими предметами и индивидуализация преподавания. Дан-
ное учебное пособие, созданное коллективом кафедры нормальной физиологии
Воронежской медицинской академии, является попыткой частичной «выплаты»
этого долга. Пятнадцатилетний опыт учебно-методической работы позволил
сотрудникам кафедры создать вариант педагогической системы, направленный
на активизацию самостоятельной работы студентов.
Эффективность самостоятельной работы студентов и теоретической подго-
товки их к занятию обеспечивается заданием 1, в котором имеется тщательно
разработанная структура теоретического материала, последовательно раскры-
вающая тему данного занятия. Универсальность этого блока достигается диф-
ференцировкой материала при помощи разных шрифтов, указывающих на сте-
пень его объема и сложности, а также включением профильного материала для
студентов лечебного, педиатрического и стоматологического факультетов. Это
дает возможность использовать его студентами разных факультетов, а также
дифференцировать материал для студентов одной группы с разным уровнем
подготовки. В целом этот блок предназначен для облегчения усвоения и ис-
пользования материала лекций и учебников прн подготовке студента к занятию.
Второе издание пособия было апробировано в течение трех лет в учебном
процессе на кафедре нормальной физиологии ВГМА и в других медицинских
вузах России. Третье издание существенно переработано, особенно в отноше-
нии теоретического материала, его иллюстративности. Самоконтролю усвоен-
ного материала служит разработанный авторами комплекс нетиповых задач.
Более совершенная структура переработанного пособия увеличивает возмож-
ность использования его студентами и преподавателями других медицинских
вузов.
Положительно оценивая данное учебно-методическое издание, хочется по-
желать коллективу кафедры дальнейших успехов на пути совершенствования
преподавания нормальной физиологии, действительно являющейся «научным
стержнем, на котором держатся все медицинские науки».
Академик РАМН Н. А. Агаджанян
5
ОТ РЕДАКТОРА
Данное пособие предназначено для повышения эффективности самостоя-
тельной работы студентов при подготовке к практическим, итоговым занятиям
и экзамену.
Стержневым компонентом пособия является Задание 1, в котором представ-
лен теоретический материал данного занятия, расположенный в логической по-
следовательности, раскрывающей структуру учебного материала и главные ме-
ханизмы физиологических процессов. Он составлен и переработан в данном из-
дании иа основе источников отечественной и зарубежной, научной и педагоги-
ческой литературы и по спектру рассматриваемых вопросов соответствует дей-
ствующей программе по нормальной физиологии. Этот материал путем ис-
пользования разных размеров шрифтов дифференцирован на факультативный и
обязательный, что позволяет изменять объем его усвоения как в связи со спе-
цификой факультета, так и с уровнем индивидуальной подготовки студента.
Материал, представленный мелким шрифтом и в скобках, имеет факультатив-
ный характер. В пособии представлены также профильные материалы для сту-
дентов трех факультетов: лечебного, педиатрического н стоматологического. В
целом теоретический материал облегчает работу студентов с учебником и лек-
циями по данной теме. Лектору он позволяет «разгрузить» лекционный курс от
феноменологических и классификационных вопросов и облегчает переход к
чтению проблемных лекций. Та часть материала, которая выносится на само-
стоятельную работу студентов, определяется лектором в конце лекции по дан-
ной теме. На самостоятельную работу студентов может быть вынесена и тема
какого-либо занятия целиком.
Третье издание пособия переработано коллективом авторов, в нем отражен
опыт использования предыдущего издания в учебном процессе на кафедре нор-
мальной физиологии Воронежской медицинской академии при работе со сту-
дентами в течение трех лет. Существенным отличием третьего издания от пре-
дыдущего является полная его ориентация на внеаудиторную самостоятельную
работу, внесенный в него иллюстративный материал и блок нетиповых (ситуа-
ционных) задач с эталонами ответов. Мы надеемся, что студенты смогут найти
крупицы ценного за обложкой этой книги.
Авторы понимают, что любая выполненная крупная работа по совершенст-
вованию учебного процесса не может быть лишена недостатков, н будут благо-
дарны за критические замечания как преподавателей, так и студентов.
Профессор В.Н. Яковлев.
6
Список основных сокращений
(а-в) - артериовенозная разница
АВ - альвеолярная вентиляция
АВ-узел - атриовентрикулярный узел
АД - артериальное давление
АДГ - антндиуретический гормон
АКТГ - адренокортикотропный гормон
АПК - антиген-представляющая клетка
АРД - акцептор результата действия
Ата - атмосфера абсолютная
АТФ (АДФ) - адеиозинтрнфосфат (аде-
нозинднфосфат)
АЦ - аденилатциклаза
ВНС - вегетативная нервная система
ВИП - вазоактивный интестинальный
пептид
ВИД - высшая нервная деятельность
ВОО - величина основного обмена
ВП - вызванные потенциалы
ВПСП ~ возбуждающий постсннаптиче-
ский потенциал
ГАМК - гамма-аминомасляная кислота
ГДФ (ГТФ)- гуанознндифосфат (гуано-
зинтрифосфат)
ГИП (ЖИЛ) - гастроннгибнрующий
(желудочный ингибирующий) пептид
ГКГС - главный комплекс гистосовме-
стимости
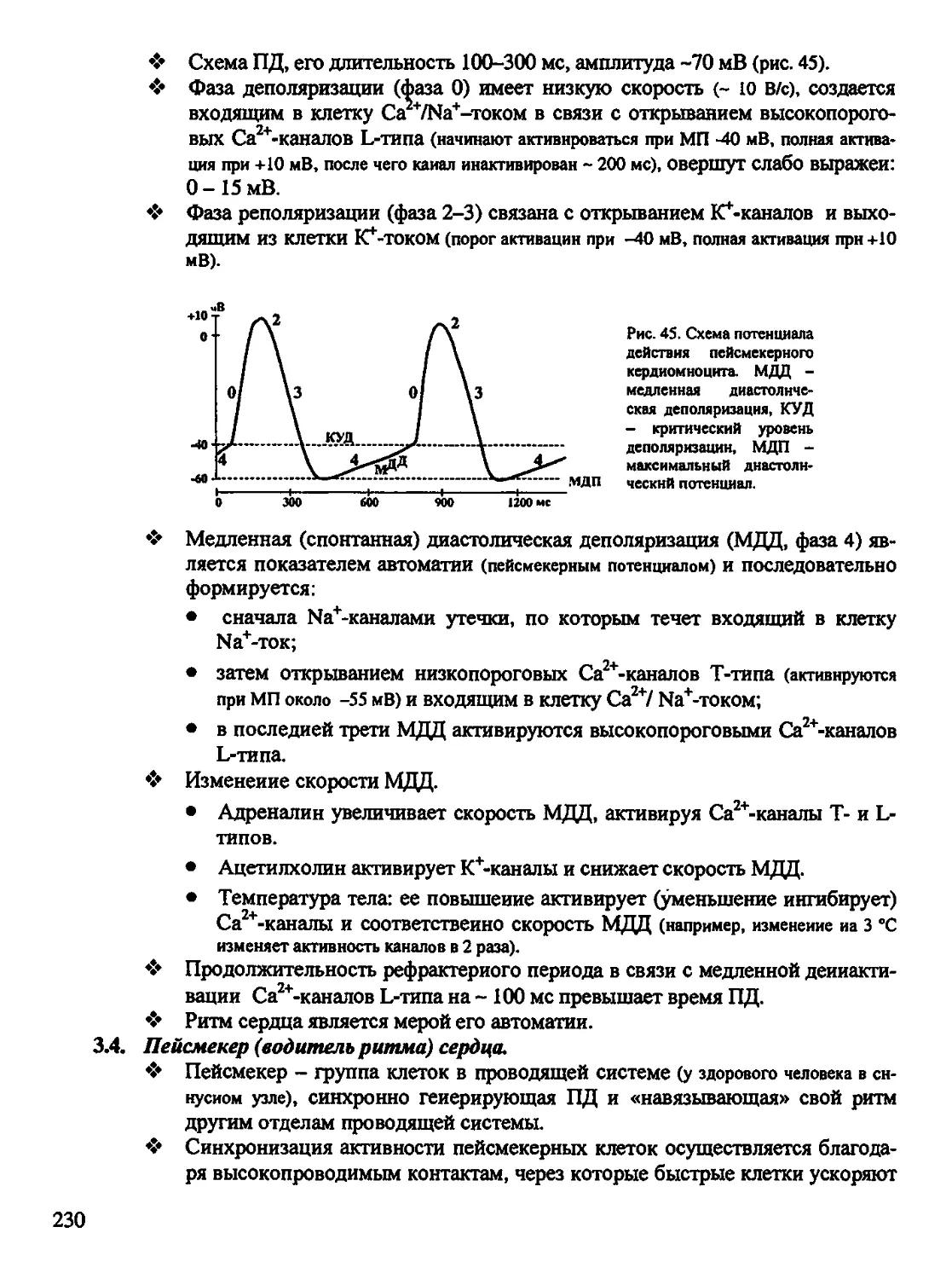
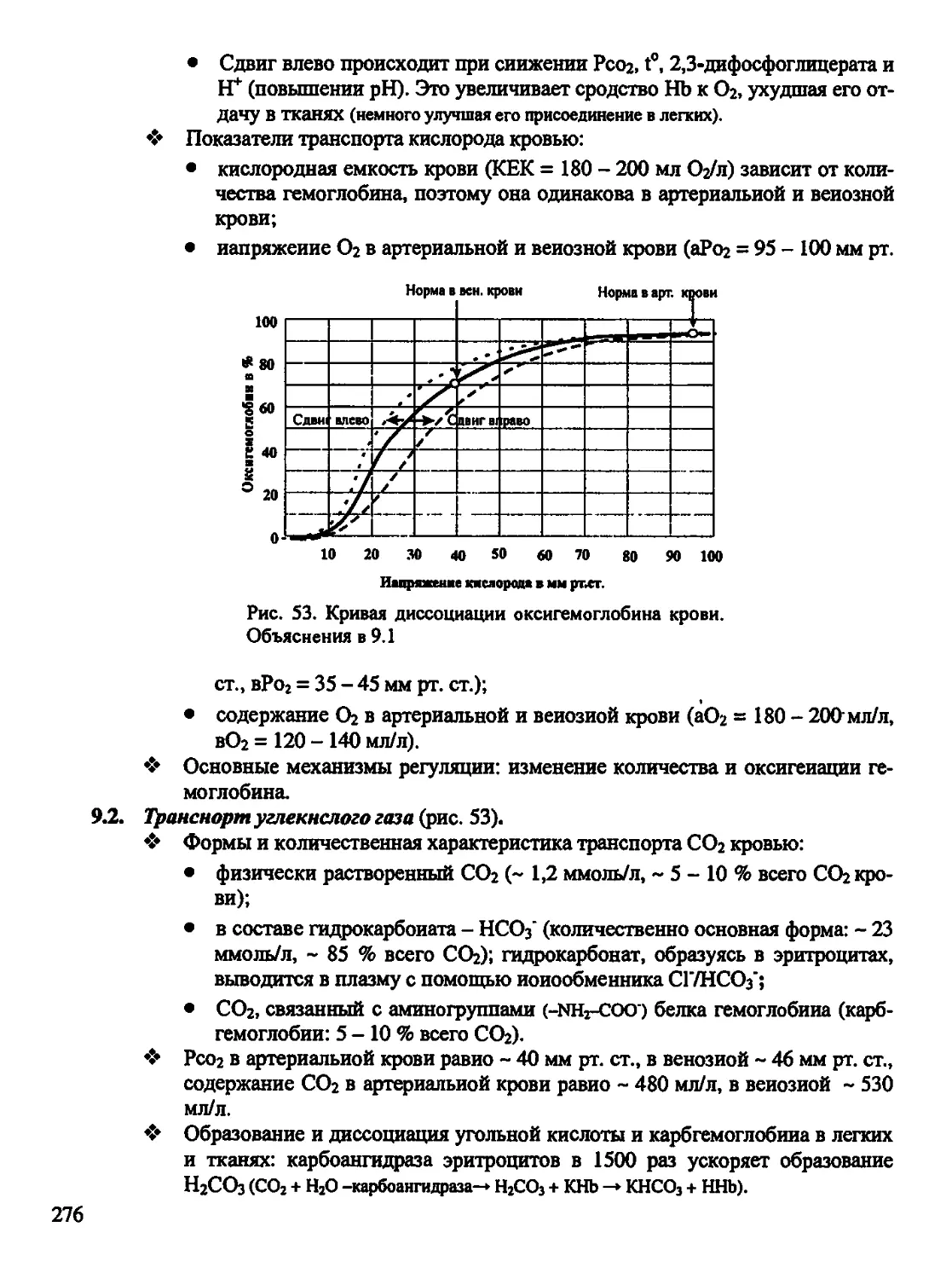
ДАГ - диацилглицерол
ДЕ - двигательная единица мышц
ДК - дыхательный коэффициент
ДНК - дезоксирибонуклеиновая кислоте
ДО - дыхательный объем
ЖЕЛ - жизненная емкость легких
ИЛ - интерлейкин
ИФз - инозитолтрифосфат
КМ - кальмодулин
КОС - кислотно-основное состояние
КУД - критический уровень деполяриза-
ции
ЛГ -лютеинизирующий гормон
ЛПВП (ЛПНП и ЛПОНП)-липопротеи-
ды высокой плотности (низкой и очень
низкой плотности)
ЛЦМ - легкие цепи миозина
МДД - медленная диастолическая депо-
ляризации
МОД - минутный объем дыхания
МОК - минутный объем кровообра-
щения
МП(П) - мембранный потенциал (покоя)
МСГ - меланоцитостимулирующий гор-
мон
НАД(Н) - никотииамиддннуклеотид
(восстановленный)
ОЕЛ - общая емкость легких
ООЛ - остаточный объем легких
ОПС - общее периферическое сопротив-
ление
ОФВ| - объем форсированного выдоха за
1 сек
ОЦК - объем циркулирующей крови
ПД - потенциал действия
ПК - протеинкиназа (фосфопротиенки-
наза)
ПОг - потребление кислорода
РАСК - регуляция агрегатного состоя-
ния крови
РНК - рибонуклеиновая кислота (ин-
формационная - мРНК, транспортная -
тРНК, рибосомная -рРНК)
РОад (выд) - резервный объем вдоха
(выдоха)
РП - рецепторный потенциал
РФ - ретикулярная формация
СА-узел - синоатриальный узел
СМЖ - спинномозговая жидкость
СО - систолический объем
СОЭ - скорость оседания эритроцитов
СКК - стволовая клетка крови
СТГ- соматотропный гормон
Тз СП) - тироксин (трнйодтнронин)
ТПСП - тормозной постсинаптический
потенциал
ТРФр - трансформирующий ростковый
фактор бета
ТТГ - тиреотропный гормон
УФ - ультрафиолет
7
ФБС - фаза быстрого сна
ФМС - фаза медленного сна
ФНО - фактор некроза опухолей
ФОЕ - функциональная остаточная ем-
кость
ФС - функциональная система
ФСГ - фолликулостимулирующий
гормон
ХЦК-ПЗ - холецистокинин-панкреози-
мин
ЦНС - центральная нервная система
цАМФ (цГМФ) - циклический адено-
зинмонофосфат (гуанозннмонофосфат)
ЧД - частота дыхания
ЧСС - частота сердечных сокращений
ЭКГ - электрокардиограмма
ЭП - эритропоэтин
ЭПД - эфферентная программа действия
ЭПС - эндоплазматическая сеть
ЭЭГ - электроэнцефалограмма
ЮГА - юкстагломерулярный аппарат
почки
ВВ - буферные основания крови
BD (BE) - дефицит (избыток) буферных
оснований крови
CD (кластеры дифференцировки) - спе-
цифические антигены (функцнональ-
ныамолекулы) на клеточной мембране
лимфоцитов, макрофагов и др клеток
Ig - иммуноглобулины
НЬ- гемоглобин
Рсог - напряжение углекислого газа
Роз - напряжение кислорода
SB - стандартный бикарбонат
Xs - постоянная длины мембраны (рас-
стояние, на котором амплитуда потен-
циала уменьшается на 63%)
т - постоянная аремени мембраны (со-
противление х емкость) - это время, за
которое амплитуда потенциала снижает-
ся на 67% от начальное величины
ВНИМАНИЕ! Материал пособия, набранный мелким шрифтом
(кроме методов исследования и профильного материала), является
факультативным, его незнание не снижает оценку. Он может быть
использован при подготовке реферативных докладов, в качестве
справочного материала и при изучении клинических дисциплин.
8
Медицине нельзя научить, медицине
можно только научиться.
(М.Я. Мудрев)
Невежество ближе к спасению, чем
полузнание.
(Авиценна)
ЧАСТЬ I
ОБЩАЯ ФИЗИОЛОГИЯ
Тема 1. Введение в физиологию. Основные понятия физиологии. Прин-
ципы формирования и организации физиологических функций
Цель: а) знать методологические принципы в физиологии; физиологическую функцию, ее параметры
и норму, взаимоотношения функции и структуры, методы исследования физиологических
функций; общую характеристику функций клетки, ткани, органа, организма; единство организ-
ма и внешней среды, принцип системогенеза, понятие о гомеостазе и гомеокииезе;
б) уметь использовать эти знания для понимания функций целостного организма, его реакций на
изменение внутренней и внешней среды (реализуется при изучении курса физиологии).
Мотивация темы: «Физиология - это научный стержень, на котором держатся все медицинские науки»
К. Бериар.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала (большой материал данного задания ие
предусматривает детального его усвоения и запоминания при подготовке к этому
занятию, а рассчитан на многократное обращение к нему при изучении курса
нормальной физиологии).
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по дайной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Нормальная физиология - наука о функциях (механизмах жизнедеятельности)
целостного здорового организма и регуляции этих функций в условиях приспособ-
ления организма к изменяющейся среде.
1.1. Главные задачи физиологии, ее связь с другими науками.
❖ Изучение функционирования организма, его частей, их взаимодействия.
❖ Изучение адаптации организма к внешним условиям.
❖ Изучение особенностей функционирования организма в онтогенезе.
❖ Создание теории здоровья и здорового образа жизни.
9
< • Связь физиологии с другими науками (физикой, биологией, биохимией, ана-
томией, гистологией, патофизиологией, клиническими науками и др.).
1.2. Основные исторические этаны развития физиологии.
❖ Древняя физиология (6 в. до н. э. -15 в.) формировалась во врачебных и фило-
софских школах Гиппократа, Галена, Аристотеля и др.; основным методом
исследования было наблюдение.
& Экспериментальная физиология (У. Гарвей, 1628): физиология как самостоя-
тельная наука, период бурного накопления экспериментальных данных,
преобладание аналитического направления. («Гарвей был первым, отворившим
дверцы тесной клетки средневековой схоластики, в которой, подобно птице, томилась в
продолжении многих веков физиологическая мысль, н выпустил ее на простор широкого
экспериментального исследования» П.М. Никифоровский.)
❖ Современная экспериментальная физиология (20-21 в.в.), ее особенности:
эксперименты на человеке, системный подход в физиологических исследо-
ваниях, изучение влияния социальных факторов на функции организма -
физиология труда, экологическая физиология.
1.3. Физиология как научная основа здорового образа жизни, оценки здоровья и
работоспособности человека («Здоровый нищий счастливее больного короля». А. Шо-
пенгауэр).
❖ «Здоровье - это состояние полного физического, духовного и социального
благополучия, а не только отсутствие болезней» (ВОЗ).
❖ Основные показатели здоровья:
• нормальные показатели функций организма в состоянии покоя (стати-
ческое здоровье);
• нормальные физиологические резервы организма (динамическое здоро-
вье);
• достаточный уровень морально-волевых и мотивационных установок
личности (психическое здоровье);
• социальная активность, деятельное отношение к миру (социальное здо-
ровье).
1.4. Методы исследования в физиологии («Знание - дочь опыта». Леонардо да Винчи): 1)
наблюдение; 2) острый и хронический эксперимент; 3) использование функ-
циональных нагрузок; 4) клинико-физиологические и лабораторные методы; 5)
кибернетическое (математическое) моделирование.
2. Диалектико-материалистические основы физиологии («Медицина является родиной н
источником материализма». Л. Фейербах).
2.1. Позиция материалиста в физиологии и медицине связана с двумя главными
положениями: во-первых, с признанием объективного характера изучаемых
функций, т.е. существованием их вне нашего сознания; во-вторых, с признани-
ем познаваемости всех, в том числе психофизиологических функций. Диалек-
тическая позиция предусматривает познание функций организма в движении,
развитии, взаимосвязи друг с другом, их соподчинении.
2.2. Методологические принципы в физиологии.
❖ Принцип целостности: организм во взаимодействии со средой выступает
как единый целостный объект («Как все в единстве сплетено, одно в другом вопло-
10
щено». и. Гете). Организм характеризуется большой активностью целого по
отношению к частям, подчинением частей целому; при этом часть может
существовать только в составе целостного организма («Нельзя познать части, не
познав целого». Б. Паскаль). Целостность организма формируется деятельно-
стью ряда физиологических систем: нервной, эндокринной, иммунной,
кровообращением. Ведущую роль при этом играет нервная система.
• Нервизм - направление в физиологии и медицине, которое признает за
нервной системой главную роль в регуляции процессов жизнедеятель-
ности организма в норме и патологии.
• Единство организма и внешней среды («Организм и внешняя среда - нерас-
членимое единство». И м. Сеченов). Организм является открытой системой,
обменивающейся с внешней средой информацией, веществом и энерги-
ей. Выдающуюся роль в этом обмене играют физиологические системы
- нервная, пищеварения, дыхания, выделения, терморегуляции.
Принципы эволюции функций - это закономерности изменения функций в
эволюционном процессе («... путеводная нить физиологического анализа», по Л.А.
Орбели, 1933). Отметим некоторые из этих принципов (по Ю.В. Наточину, 1984).
• Принцип мультифуикциональности. В процессе эволюции увеличивает-
ся количество функций различных органов. Например, у миксин почки
выполняют функцию регуляции объема жидкости, у миног добавляется
функция осморегуляции, у высших позвоночных - инкреторная и мета-
болическая функции.
• Принцип интенсификации функций. Например, функции фильтрации и
реабсорбции на 1 г почек у млекопитающих в 10 - 100 выше, чем у
низших позвоночных.
• Принцип смены функций. Например, кроветворная функция почек у
рыб сменяется функцией регуляции кроветворения у высших позвоноч-
ных.
• Принцип надстройки функций. Новые функции не заменяют, а наслаи-
ваются на старые, подчиняя их. Например, помимо осморазведения мо-
чи, у млекопитающих и птиц развивается (и становится доминирующей)
функция осмоконцентрации мочи. Это связано с надстройкой новой
структуры - мозгового вещества почек.
Детерминизм - учение о всеобщей связи и обусловленности объективных
явлений.
• Выделены различные связи: жесткие причинно-следственные (цепные,
разветвленные, сетчатые), обратные связи (отрицательные и положи-
тельные), связи-отражения с образованием памяти, связи-
взаимодействия, связи-субординации и др.
• Принцип причинности является ядром детерминизма и лежит в основе
всех форм детерминирования.
• Введено понятие полной причины, состоящей из специализирующей
причины (фактора, который вызывает специфические свойства следствия), внеш-
них и внутренних условий (которые способствуют реализации причинного фак-
11
тора), пускового фактора (повода, т.е. внутреннего или внешнего фактора, кото-
рый определяет время возникновения следствия под влиянием совокупных условий).
* Сложный характер полной причины (например, ее вариабельность в связи с
условиями и пусковыми факторами) создает вероятностный характер причин-
но-следственных отношений (вероятностный детерминизм).
❖ Системный подход в физиологии (И.П. Павлов, П.К. Анохнн). Системность яв-
ляется всеобщим и неотъемлемым свойством материи (Л. Берталанффи, 1962).
Она характеризует преобладание организованности над хаотичностью. Ор-
ганизм человека относится к сложным, целостным, открытым системам,
способным к антиэнтропийности и самоорганизации. («Живой организм пред-
ставляет собой маленький островок антиэнтропни в бескрайнем океане энтропии». Н. Ви-
нер). Система - это комплекс взаимодействующих элементов. Свойства и
функции системы не являются суммой свойств и функций ее элементов.
При деятельности системы всегда имеются системные свойства, которых
нет у элементов системы. С другой стороны, включаясь в новую систему,
элемент (исполнительный орган) приобретает новые свойства, отсутст-
вующие у него в изолированном состоянии или при функционировании в
старой системе (А.А. Ухтомский). Совокупность устойчивых отношений и
связей между элементами называется структурой системы. Качество сис-
темы определяется, во-первых, элементами (их природой, свойствами, количест-
вом), во-вторых, структурой, т.е. связью, взаимодействием элементов. («Все,
что находится во взаимосвязи, должно рассматриваться в этой связи». Ян Каминский).
3. Физиологическая функция,
3.1. Функция - специфическая жизнедеятельность клеток, тканей, органов, орга-
низма в целом, проявляющаяся как физиологический процесс или совокупность
процессов. Функциональные изменения направлены на сохранение живой сис-
темы и её адаптацию к условиям среды.
3.2. Параметры и норма функции.
❖ Норма функции является объективной категорией. Ее механизмы генетиче-
ски детерминнроваиы, и по наследству передается определенная норма фи-
зиологических реакций. Но генотип определяет норму реакций лишь как
возможность ее проявления. Реализация генотипа происходит в определен-
ных условиях внешней среды, так возникает конкретная индивидуальная
(фенотипическая) норма.
❖ Параметры нормы (нормативы) представляют собой её количественную
характеристику. Для целей физиологии и медицины нормативы должны, с
одной стороны, наиболее полно отражать многообразие индивидуальной
нормы (иначе отдельные здоровые индивидуумы попадут в зону патологии), С другой
стороны, при полном охвате индивидуальных различий часть больных по-
падут в зону нормы. Поэтому обычно выбирается интервал нормы (норма-
тив), в который укладываются показатели 95% здоровых людей. Кроме то-
го, нормативы разрабатываются для как можно более однородной совокуп-
ности людей: по полу, возрасту и т.д., что уменьшает разброс индивиду-
альных показателей.
3.3. Взаимоотношение функции и структуры, их единство.
12
❖ Под структурой в медицине обычно понимают строение, пространствен-
ное расположение, последовательность в живых объектах (это соответствует
философской категории «внешняя форма»). Современная биология рассматривает
структуру как совокупность устойчивых связей элементов системы, обес-
печивающая ее целостность при различных внешних и внутренних измене-
ниях (это соответствует категории «внутренняя форма»).
❖ Функциональные изменения при этом можно рассматривать как выражение
внутренних перестроек структуры живого. Структура и функция составля-
ют динамическое единство («Функция без структуры немыслима, структура без
функции бессмысленна». В.Х. Василенко).
4. Клетка и ее функции.
4.1. Клетка - элемент органа (ткани), состоящий из системы органелл, способный к
самовоспроизведению, метаболизму, раздражимости, адаптации, изменчивости.
Клетка является наименьшей самостоятельной единицей живого.
4.2. Основные структурно-функциональные компоненты клетки.
Плазмолемма (внешняя клеточная мембрана, толщина - 10 нм) — барьерная (раз-
граничительная), транспортная, рецепторная функции, формирование био-
потенциалов. Важнейшими функциональными единицами ее являются
ионные каналы и насосы, белки-транспортеры, ферменты, рецепторы.
Наружная поверхность плазмолеммы может быть многократно увеличена
за счет мнкроворсннок. Поверхность плазмолеммы покрыта гликокаликсом
- слоем толщиной около 50 нм, состоящим из олигосахаридов, связанных
ковалентно с гликолипидами и гликопротеинами мембраны. Функции
глнкокаликса: межклеточное узнавание и взаимодействие, пристеночное
пищеварение, барьерная (молекулярное снто, затрудняющее поступление в клетку
крупных молекул, микробов, внрусоа).
❖ Цитоплазма содержит гиалоплазму, мембранные органеллы - эндоплазма-
тическую сеть, митохондрии, пластинчатый комплекс, лизосомы, перокси-
сомы, немембранные органеллы - рибосомы, клеточный центр, цитоскелет.
• Гиалоплазма (цитозоль) обеспечивает химическое взаимодействие кле-
точных структур друг с другом, осуществляет внутриклеточные транс-
портные процессы, обмен углеводов, белков, липидов и др. веществ, ме-
сто депонирования гликогена, жиров, пигментов, кислороднезавнсимого
образования АТФ.
• Митохондрии. В крнстах происходит утилизация кислорода и образо-
вание основного количества АТФ в клетке. В матриксе осуществляется
цикл Кребса (продукция НАДН и СО2), (3-окнсление жирных кислот, депо-
нирование Са2+, с участием митохондриальных ДНК н рибосом синте-
зируются некоторые мембранные белки митохондрий.
• Рибосомы (активны при образовании полисом) осуществляют считывание
кода мРНК (процесс трансляции), синтез пептидов и белков в цитозоле и
гранулярной эндоплазматической сети.
• Гранулярная эндоплазматическая сеть (ЭПС) обеспечивает синтез бел-
ков (мембранных, цитозольных, лизосомных, секретируемых) на ПОЛИСОмах сети,
затем эти белки через поры попадают в просвет ЭПС, где модифициру-
13
ются и транспортируются в комплекс Гольджи. Рибосомы, связанные с
ЭПС, обеспечивают в основном синтез белков, используемых в других
клетках. Свободные рибосомы, расположенные в гиалоплазме, образу-
ют преимущественно белки для собственных нужд клетки.
• Агранулярная ЭПС — синтез липидов (фосфолипидов, жирных кислот, стероид-
ных гормонов), гликогена, депонирование и освобождение Са2+, детокси-
кация экзогенных и эндогенных веществ, модификация синтезируемых
белков (гликозилирование, фосфорилирование, карбоксилирование и др.), об-
разование липопротеинов, образование мембран для других органелл
(например, лизосом и пероксисом).
• Пластинчатый комплекс (комплекс Гольджи) - модификация белков,
их сортировка с помощью рецепторов и упаковка в отщепляющиеся пу-
зырьки, которые или секретируются из клетки, или образуют первичные
лизосомы; синтез полисахаридов, образование мембран для других ор-
ганелл клетки (например, лизосом и пероксисом).
• Лизосомы (первичные, вторичные - фаголизосомы и аутофагосомы, остаточные
тельца) - внутриклеточное «пищеварение» с помощью кислых гидролаз
(pH среды около 5,0): питательная, защитная функции, обновление внутри-
клеточных структур.
• Пероксисомы - окисление мочевой кислоты, D-аминокислот и 2-
гидроксикислот с помощью кислорода и образования пероксида водо-
рода (Н2О2), который обезвреживается под действием каталазы. Основ-
ные функции - защита от образующегося в клетке Н2О2, метаболизм
липидов и вышеназванных кислот.
• Клеточный центр (центросома) состоит из центриолей и связанных с
ними микротрубочек. Центриоли в делящихся клетках формируют вере-
тено деления, в неделящихся клетках определяют полярность клеток
эпителия.
• Цитоскелет - опорно-двигательный аппарат клетки состоит из ните-
видных структур: микрофиламентов, микрофибрилл и микротрубочек.
Микрофиламенты содержат сократительные белки (актин, миозин),
тропомиозин и др. белки, обеспечивают в основном двигательную
функцию: подвижность клетки и внутриклеточные движения везикул,
митохондрий, токи цитоплазмы и др. Микрофибриллы выполняют пре-
имущенственно опорно-каркасную функцию. Микротрубочки образуют
эластический каркас, обеспечивающий лабильную форму клетки, и
обеспечивают передвижение вдоль них везикул и белков (например, ак-
сонный транспорт). В целом цитоскелет обеспечивает форму и подвиж-
ность клетки, распределение и перемещение компонентов клетки,
транспорт веществ в клетку и из нее; участвует в межклеточных соеди-
нениях.
❖ Ядро (оболочка, хроматин, нуклеоплазма, ядрышко) - хранение генетической ин-
формации в ДНК и передача ее при делении клеток, генетический контроль
клеточных процессов на основе контроля синтеза белков, в ядрышке про-
исходит синтез рРНК (с участием РНК-полнмеразы I), образование субъединиц
14
рибосом, синтез тРНК (с участием РНК-полимеразы Ш), синтез кофермента
НАД* (из НАМ, поступающего из цитоплазмы).
• Сохранение наследственной информации обеспечивается деятельно-
стью репарационных ферментов, ликвидирующих спонтанные мутации
генов (например, комплекса ферментов - эксцизионной эндонуклеазы, ДНК-
полимеразы и ДНК-лигазы).
• Передача генетической информации обеспечивается репликацией моле-
кул ДНК при митозе (а периоде S) клетки, позволяющей дочерним клет-
кам получать одинаковый геном.
• Экспрессия генов, приводящая к синтезу белков, осуществляется по
схеме: транскрипция и процессинг (синтез иа матрице ДНК с участием РНК-
полимеразы И первичного транскрипта и превращение последнего в мРНК путем уда-
ления иекодирующих участков - интронов и блокады 5’- и З'-коицов мРНК, делающей
ее устойчивой к эндонуклеазе) —> перенос генетической информации в соста-
ве мРНК из ядра в цитоплазму —> трансляция (считывание информации с
мРНК и сборка пептидной цепи иа рибосомах при взаимодействии мРНК, тРНК и
рРНК) —> посттрансляционные изменения пептидов в агранулярной ЭПС
и аппарате Гольджи.
• Важным регулятором экспрессии генов являются гормоны (стероидные и
тироидиые гормоны, витамины А и D3), которые активируют ядерные рецеп-
торы (факторы транскрипции), соединенные со специальными участками
ДНК.
• Роль ранних генов. Активация ранних генов (протоонкогенов) является
первичной реакцией генома клетки на различные раздражители. Их ак-
тивация наступает при действии на клетку гормонов, медиаторов, ин-
терлейкинов и др. веществ. При этом образуются внутриклеточные фак-
торы экспрессии ранних генов - цАМФ, высокий уровень Са2+, проте-
инкиназа С и др. Активация ранних генов начинается через 5-7 мин по-
сле действия раздражителей и достигает максимума через 60-90 мин,
продолжительность жизни их мРНК невелика - от нескольких минут до
часов. Ранние гены (протоонкогены - c-fos, fos-B, fra-1 и др.) являются уни-
версальными регуляторами транскрипции генома. Кодируемые ими
белки обеспечивают выборочную активацию отдельных генов в геноме.
В связи с этим они являются ключевым звеном в осуществлении при-
способительных реакций организма и перехода кратковременной адап-
тации в долговременную.
4.3. Транспортная функция клеточной мембраны.
❖ Функциональная структура мембраны.
• Функции липидов (фосфолипиды - лецитин, фосфатидилсерии, фосфатидилииозит
и сфингомиелин, гликолипиды, холестерол) — барьерная, транспортная, микро-
окружение белков, сопротивление, емкостные свойства и текучесть
мембраны.
• Функции белков - переносчики, ионные каналы и насосы, ферменты,
циторецепторы, клеточные контакты.
15
• функции углеводов - в составе гликопротеинов участвуют в рецепции,
реакциях иммунитета и распознавания клеток.
• Компоненты мембраны удерживаются нековалеитными связями, по-
этому они обладают подвижностью и могут диффундировать в пределах
липидного бислоя преимущественно в продольном направлении.
❖ Виды и механизмы транспортных процессов.
• Пассивный транспорт (по градиенту концентрации с уменьшением энергии
транспортируемого вещества).
•ф- Простая диффузия (через липидный бнслой н ионные каналы) по градиен-
ту концентрации с уменьшением энергии транспортируемого веще-
ства. Через липидный бислой переносятся малые молекулы: непо-
лярные - О2, N2, этанол, эфир и др., полярные незаряженные моле-
кулы - Н2О, мочевина, глицерин, СО2, аммиак и др. Неорганиче-
ские иоиы (Na+, К+, Са2+, СГ и др.) переносятся с участием ионных
каналов. При этом, кроме градиента концентрации, играет роль
электрический градиент между отрицательной внутриклеточной и
положительной внеклеточной средами - мембранный потенциал,
который усиливает транспорт катионов и препятствует транспорту
анионов в клетку и наоборот.
Транспортируемы» вещества
Простая Облегченная Первично Вторично
диффузия диффузия активный активный
транспорт транспорт
Рнс. 1. Схема различных видов транспорта через клеточную мембрану.
Вторично-активный транспорт показан на примере глюкозы (Гл).
ф Облегченная диффузия используется для переноса более крупных
полярных незаряженных молекул (например, глюкозы, аминокислот) и
осуществляется с использованием белков-переносчнков, которые
избирательно связывают транспортируемые молекулы и за счет
конформационных изменений переносят их через мембрану. Она
характеризуется специфичностью, ббльшей скоростью, явлением
16
насыщения (когда заняты все переносчики), регулируется с помощью
гормонов (например, инсулином при транспорте глюкозы).
Активный транспорт (против градиента концентрации с увеличением энергии
транспортируемого вещества).
Первично-активный, транспорт непосредственно использует энер-
гию АТФ и осуществляется деятельностью ионных насосов: калий-
натриевого, кальциевого, протонного, белок которых обладает
свойствами как переносчика, так и фермента АТФазы.
ф- Вторично-активный транспорт: используется энергия градиента
какого-либо нона (чаще Na4), предварительно созданная деятельно-
стью ионного насоса, для транспорта других ионов (иоиообменники),
глюкозы, аминокислот. Транспорт происходит с участием белков-
переносчиков.
Микровезикулярный транспорт (эндоцнтоз н экзоцитоз) являются вари-
антом активного транспорта, служит для переноса через мембрану
крупномолекулярных веществ (например, белков).
Эндоцитоз выполняет питательную, защитную, регуляторную
функции (последняя связана с изменением количества рецепторов мембраны и
эндоцитозом гормонов, не проникающих через мембрану). При эндоцитозе
локальный участок плазматической мембраны впячивается и от-
шнуровывается, формируя мелкие (пиноцитозные) или крупные
(фагоцитозные) пузырьки. Обычно эндоцитозные пузырьки слива-
ются с первичными лизосомами, образуя вторичные лизосомы, в
которых переваривается макромолекулярное содержимое пузырь-
ков. Выделяют три вида эндоцитоза - пиноцитоз, опосредованный
рецепторами эндоцитоз и фагоцитоз.
При пиноцитозе происходит неспецифический захват внекле-
точной жидкости и растворенных в ней веществ (в том числе и
макромолекул), которые используются внутри клетки или пе-
реносятся сквозь клетку.
При опосредуемом рецепторами эндоцитозе вещества (напри-
мер, липопротеины, незрелый тироглобулин, гормоны, иммуноглобулины,
антигены) сначала связываются со специфическими белками-
рецепторами плазматической мембраны, которые далее кон-
центрируются в окаймленных ямках мембраны (окаймление со-
вершается особым белком - кластрииом, обладающим свойством АТФазы).
После впячивання и отшнуровывания образуются окаймленные
пузырьки (эндосомы), взаимодействующие с лизосомами.
При фагоцитозе (И.И. Мечников, 1883) крупные клеточные час-
тицы (обломки своих клеток, инфекционные агенты и др.) захватывают-
ся «профессиональными» клетками (микро- и макрофагами) с
дальнейшим внутриклеточным перевариванием.
Экзоцитоз используется для выделения из клеток медиаторов, гор-
монов, ферментов, которые используются в организме в регулятор-
ных, питательных и других целях. Их выделение из клетки проис-
17
ходит в составе экзоцитозных пузырьков, образующихся в аппарате
Гольджи. В этих пузырьках упаковываются белки, образовавшиеся
в рибосомах ЭПС. Низкомолекулярные вещества (медиаторы, некото-
рые гормоны и др.) попадают в пузырьки преимущественно путем вто-
рично-активного транспорта. Пузырьки транспортируются с помо-
щью микрофиламентов и микротрубочек к плазмолемме, сливаются
с ней (процесс слияния активируется фосфолипидом лизолецитином), и со-
держимое пузырьков выделяется во внеклеточную среду. Важным
регулятором экзоцитозной активности является уровень внутрикле-
точного Са2+.
4.4. Рецепторная функция клеткп осуществляется специальными белками, кото-
рые находятся преимущественно на плазмолемме и в ядре (для проникающих в
клетку гормонов).
❖ Главные виды рецепторов плазмолеммы.
• Ионотропные рецепторы («рецепторы-каналы») - в своем составе имеют
ионные каналы, расположены преимущественно в области синапсов,
участвуют в передаче возбуждающих н тормозных влияний (Н-
холинорецепторы, ГАМКа- рецепторы, глутаматные НМДА-рецепторы, гли-
циновые рецепторы, 5-НТ- рецепторы).
• Метаботропные рецепторы - связаны с G-белком мембраны. Через
них действуют около 80 % гормонов и медиаторов, которые плохо про-
никают в клетку и формируют внутриклеточные эффекты с помощью
вторых посредников (подробнее в теме 9, задание 1:2.3).
• Рецепторы, регулирующие поступление молекул в клетку (например,
липидов в составе липопротеинов низкой и высокой плотностн).
• Адгезивные рецепторы (семейства интегрииов, кадгеринов, иммуноглобу-
линов, кластеров дифференцировки - CD2, CD54, CD58 и др., селектинов) -
осуществляют связь «клетка - клетка» и «клетка - структуры межкле-
точной среды» (например, с базальной мембраной).
• Регуляция рецепторной функции плазмолеммы. Эта функция на клеточ-
ном уровне может быть снижена (десенситизация рецепторов) или уси-
лена (сенситизация рецепторов).
О" Десенситизация рецепторов возникает при усилении сигнала нлн
увеличении временя его действия на рецептор. Она может быть
связана с действием протеннфосфокиназ, фосфорилирующих ре-
цепторы, что снижает их чувствительность к сигнальным молеку-
лам. Другим механизмом является эндоцнтоз комплексов «сигналь-
ная молекула+рецептор», что уменьшает количество рецепторов на
плазмолемме (интернализация рецепторов). Такне рецепторы могут раз-
рушиться в лизосомах или опять встраиваться в мембрану при ос-
лаблении стимула.
у" Сенситизация рецепторов может быть связана с увеличением син-
теза рецепторов и, следовательно, их количества на плазмолемме, с
увеличения сопряженности рецептора с G-белком и вторыми по-
18
средниками, а также с протеолизом белков, блокирующих рецепто-
ры.
• Рецепторы ядра клетки стимулируются сигнальными молекулами, легко
проникающими в клетку - стероидными и тироидными гормонами, ви-
таминами Dj и А. Они представляют собой белки (факторы транскрип-
ции), прочно связанные с ДНК хроматина. Каждый рецептор содержит
несколько специфических участков (доменов): ДНК-связывающий, гор-
монсвязывающий, активирующий транскрипцию и др. Через эти рецеп-
торы стимул становиться фактором регуляции транскрипции (синтеза
мРНК) в каких-либо генах.
4.5. Энергетическая функция клетки осуществляется в митохондриях и гиало-
плазме путем образования макроэргических связей АТФ, энергия которых (7,3
ккал/моль) используется для различных видов работы клетки: биосинтетической,
сократительной, электрической, осмотической и др.
❖ Митохондрии играют главную роль в энергетическом обмене. Они окис-
ляют пируват и жирные кислоты (через ацетилСоА) в цикле трикарбоновых
кислот, образуя активные формы водорода (в составе НАДИ, частично ФАДНа).
Их электроны поступают в цепь дыхательных ферментов внутренней мем-
браны митохондрий, а протоны - в матрикс. Энергия, освобождающаяся
при транспорте электронов в дыхательной цепи используется для создания
протонного градиента по обе стороны внутренней мембраны. Энергия это-
го градиента во время обратного потока протонов через протонный канал
АТФ-синтетазы используется для фосфорилирования АДФ, в результате
чего образуется АТФ. (Например, при окислении одной молекулы глюкозы с участи-
ем митохондрий реально образуется 27 молекул АТФ.) Часть энергии АТФ резерви-
руются в виде креатиифосфата и ионных градиентов клетки
❖ Энергетическая функция гиалоплазмы связана с гликолизом. При этом
окисление одной молекулы глюкозы приводит в конечном балансе к обра-
зованию двух молекул АТФ и молекулы НАДН, водород которой может
использоваться в митохондриях. Образование АТФ в гиалоплазме проис-
ходит как в аэробных, так и в анаэробных условиях. Таким образом, хотя
гликолиз имеет малую эффективность образования АТФ по сравнению с
митохондриями, важной его особенностью является малая зависимость от
кислородного состояния клетки.
4.6. Биосинтетическая функция клетки осуществляется взаимосвязанным ком-
плексом органелл: рибосом, ЭПС и аппарата Гольджи. Синтез и модификация
белков рассмотрен выше (4.2, роль рибосом, ЭПС и пластинчатого комплекса). Синтез
гликогена и липидов (в том числе фосфолипидов и холестерола) происходит в аграну-
лярной ЭПС и завершается в комплексе Гольджи. При этом гликоген остается
внутриклеточно, а липиды и холестерол используются преимущественно в кле-
точных мембранах или в составе липопротеидов секретируются из клетки.
(Часть стероидных гормонов синтезируются в митохондриях надпочечников.)
4.7. Функции внутриклеточного переваривания (питательная, защитная, обновле-
ние клеточных структур) обеспечивается первичными и вторичными лизосома-
ми. Объектом переваривания служат как продукты межклеточной среды, так и
собственные компоненты клетки.
19
4.8. Детоксикационная функция: 1) детоксикация аммиака путем образования
глутамина и мочевины; 2) в агранулярной ЭПС гидроксилирующая система пу-
тем конъюгации с УДФ-глюкоронатом переводит образовавшиеся в организме
и поступившие в него биоактивные и токсичные вещества в водорастворимые и
малотоксичные, выделяющиеся с желчью и мочой; 3) антиоксидантная система:
ферменты (супероксиддисмутаза, каталаза, глутатиоипероксидаза и др.) и метаболиты (ас-
корбат, токоферол, цистеин, мочевина и др.) обезвреживают активные радикалы ки-
слорода (супероксидный анион, пероксид водорода, гидроксильный анион, синглетный ки-
слород).
4.9. Специализированные функции клетки являются следствием ее дифференци-
ровки (например, сокращение миоцита, генерация нервного импульса в нейроне, синтез имму-
ноглобулинов в плазматических клетках). Они изучаются в различных разделах курса
физионогии.
4.10. Жизненный путь клетки.
❖ ' Клеточный цикл не половых клеток состоит из интерфазы и митоза.
• Интерфаза имеет последовательные периоды: Gi (Go), S и Gj.
ф- Период Gi (от нескольких часов до суток, следует за митозом) — в эту фазу
клетка синтезирует белки, РНК, ферменты синтеза предшественни-
ков ДНК.
ф- Период Go - клетка может выйти из цикла, длительно быть неак-
тивной (например, клетки иммунной памяти), ИЛИ функционировать как
дифференцированная клетка в обычных условиях (например, гепато-
цит), или достигать необратимой дифференцировки (например, нейро-
ны, кардиомиоциты).
"Ф" Период S (синтетический период, обычно 8-12 часов) — происходит син-
тез и копирование ДНК с постепенным удвоением ее количества
(процесс репликации), удваивается и число хромосом, увеличивается
синтез рРНК и белка, разделяются центриоли.
"Ф" Период Ga (премитотический период, 2-4 часа) —увеличивается синтез
мРНК и белка (особенно тубулинов для митотического веретена), накапли-
вается АТФ для обеспечения митоза.
"Ф" В движении клетки по митотическому циклу важную роль играют белки цикли-
ны, которые активируют ферменты киназы (циклинзависимые киназы). Киназы, в
свою очередь, активируют ферменты синтеза ДНК, белков и аккумуляции энер-
гии. Если клетка не готова к переходу в следующую фазу, цикл останавливается
в сверенных точках, главные из которых действуют в зоне перехода фазы G, в S
и G; в митоз. Остановка цикла происходит за счет действия синтезируемых бел-
ков (р1б, p21WAF"', р27 и др.).
• Митоз имеет следующие основные фазы: профазу, метафазу, анафазу,
телофазу, в результате которых образуются две дочерние клетки, иден-
тичные материнской (в половых клетках осуществляется мейоз, в ходе которого
образуются гаплоидные гаметы).
❖ Физиологические механизмы гибели клетки.
• Концепция апоптоза - активного, генетически запрограммированного
процесса гибели клетки.
20
‘Ф’ Пусковые факторы (нарушение рецепторного аппарата клетки, некоторые
цитокины, глюкокортикоиды, активные радикалы кислорода, этанол, низкокало-
рийная диета и др.) приводят к нескольким путям активации апоптоза.
Один из них реализуется через активацию гена р53, продукт кото-
рого белок р53 является фактором транскрипции многих генов. (Ген
р53 запускает апоптоз, если путем остановки клеточного цикла и активации фер-
ментов репарации ДНК, не может восстановить ее нормальную структуру.) В
случае запуска апоптоза происходит транскрипция генов (bax, bak,
bad), приводящая к образованию белков-стимуляторов апоптоза.
Через ряд промежуточных реакций они активируют ферменты (эн-
донуклеазы, сериновые и цистеиновые протеазы), вызывающие поврежде-
ние ядра, плазмолеммы, цитоплазмы и распад клетки на отдельные
фрагменты с их последующим фагоцитозом. Противодействуют
апоптозу «гены-спасители» (bcl-2, bcl-x), кодирующие белки-
ингибиторы апоптоза.
ф" Значение апоптоза заключается в регуляции гисто- и органогенеза
во внутриутробном развитии, инволюции зрелых органов и тканей,
регуляции популяции клеток, уничтожении старых и генетически
измененных клеток В зрелых тканях и др. (Уменьшение способности к
апоптозу ниже физиологического уровня - путь к аутоиммунным заболеваниям и
злокачественной трансформации клеток).
• Концепция антиген-маркера стареющих клеток.
ф' В клеточной мембране стареющих клеток (эритроцитах, лейкоцитах,
тромбоцитах, клетках кожи, легких и др.) экспрессируется скрытый по-
верхностными гликопротеидами белок полосы Ш.
ф- Аутоантитела к этому маркёру старения служат мостиком, через
который старая клетка присоединяется к фагоциту, далее она унич-
тожается путем аутофагоцитоза.
5. Физиологическая характеристика ткаии.
5.1. Ткань - система клеток и внеклеточных структур, объединенных общей функ-
цией, строением и (или) происхождением.
5.2. Основные виды и функции тканей.
❖ Эпителиальные ткани.
• Покровный эпителий - барьерная функция, защита от механических,
химических, инфекционных и других воздействий, обмен веществ меж-
ду организмом и внешней средой (процессы всасывания, секреции, экскреции),
участие в терморегуляции.
• Железистый эпителий - секреция гормонов, пищеварительных соков,
слезной жидкости, спинномозговой жидкости, сального секрета.
❖ Кровь и лимфа - газообмеиная, трофическая, защитная, транспортная
функции, гуморальная связь между органами и тканями.
❖ Соединительные ткани - механическая, опорная, формообразующая, тро-
фическая, защитная (макрофаги, антитела), пластическая (регенерация, заживление
ран) и другие функции.
21
♦ > Мышечные ткани (скелетная, сердечная, гладкомышечная) - поза и движе-
ние тела, тонус и сокращение гладкомышечных органов, сокращение серд-
ца.
❖ Нервная ткань - восприятие раздражения, проведение и передача возбуж-
дения, генерация возбуждения и торможения, анализ и синтез информации,
интеграция функций, формирование целостности организма и др.
5.3. Межклеточные взаимодействия. Осуществляются как в пределах ткани, так и
между тканями и органами. Для них характерна двунаправленность сигнала.
При этом всегда происходит изменение активности всех взаимодействующих
клеток, что может запустить самостоятельные цепи событий. Важнейшую (но
не единственную) роль играет рецепторный аппарат клетки (см. выше 4.4). Меж-
клеточные взаимодействия можно разделить на два главных типа - контактные
и неконтактные.
❖ Контактные межклеточные взаимодействия (формируются при образо-
вании межклеточных соединений - контакте клеток друг с другом).
• Адгезионные соединения: простые контакты, образованные слоями гли-
кокаликса по всей поверхности контактирующих клеток с участием
белков-рецепторов (интегрины, кадгерины и др.); сцепляющие контак-
ты, осуществляемые с участием фибрилл цитоскелета, которые подхо-
дят к месту контакта: адгезивный поясок, десмосомы, соединяющие
клетки; полудесмосомы, соединяющие эпителиальные клетки с базаль-
ной мембраной.
• Запирающие соединения (плотные контакты) осуществляют специальные
интегральные белки, которые образуют ячеистые сети, проходящие в
виде поясков через апикальные поверхности клеток, они практически не
имеют межклеточных щелей и не пропускают молекул (например, в одно-
слойном эпителии н эндотелии).
• Проводящие соединения (в основном обеспечивают передачу информации).
Ф- Щелевые контакты (имеются во всех группах тканей, например, нексус в
мышечной ткани, электрический синапс в нервной ткани) - образованы гек-
сагональными белковыми структурами (коинексонами), образую-
щие поры, которые пропускают ионы и некоторые молекулы (мм < 2
кД: глюкозу, АТФ и др.). Осуществляют метаболическую и электриче-
скую кооперацию клеток.
ф- Химические синапсы - осуществляют передачу с помощью химиче-
ских посредников (медиаторов) возбуждающих, тормозных и трофи-
ческих влияний от нейрона на иннервируемую клетку (нервную,
мышечную или железистую) через ионотропные и метаботропные ре-
цепторы.
Через ионотропные рецепторы межклеточные взаимодействия
осуществляются в химических синапсах возбуждающих и тор-
мозных нейронов по схеме: нервный импульс —» экзоцитоз ме-
диатора из пресинаптического окончания —» действие его на
ионотропные каналы постсинаптической мембраны —»возбуж-
22
дающий или тормозной постсинаптические потенциалы (под-
робнее в теме 4; задание 1:4.3).
Через метаботропные рецепторы межклеточные взаимодей-
ствия осуществляются по схеме действия гормонов: сигнал
(раздражитель) —» рецептор —» второй посредник -» ответ (см.
ниже в 5.3).
❖ Неконтактные межклеточные взаимодействие. Осуществляются через
образование информационных молекул (гормонов, антигенов, аутоантител
и др.), переносимых жидкостями организма - межклеточной, кровью, лик-
вором, лимфой. Действие их на клетки той же ткани, где они образуются
называется паракринией, действие их на клетки других тканей называется
эндокринией. Общая схема неконтактного межклеточного взаимодействия
следующая.
• Сигнал (первый посредник) — информационные молекулы (ннформоны), ко-
торые вырабатываются в одних клетках и специфически действуют на
другие клетки (клетки-мишеии). К ним относятся гормоны, цитокины,
продукты обмена арахидоновой кислоты (простагландины, лейкотриены
и др.), антигены, аутоантитела к клеточным рецепторам гормонов и др.
• Рецепторы - белковые или гликопротеиновые молекулы, восприни-
мающие действие сигнала (информационных молекул) и включающие вто-
рые посредники. Они расположены преимущественно на плазмолемме и
в ядре (см. выше 4.4).
• Вторые посредники - молекулы, передающие сигнал непроникающего
в клетку раздражителя иа эффекторы (исполнительные молекулы), изме-
няющие функции клетки. К иим относятся циклические нуклеотиды
(цАМФ, цГМФ), Са2+, инозитол-3-фосфат, днацилглицерол и др. Образо-
ванию вторых посредников обычно предшествует активация G-белка
мембраны, который, в свою очередь, активирует (Gs) или тормозит (GJ
ферменты образования посредников (например, аденилатциклазу).
• Ответ - изменения функций клеток, связанные с фосфорилированием,
дефосфорилированием и протеолизом имеющихся в клетке белков (кана-
лов, ферментов, рецепторов, структурных белков и др.), а также с изменением
скорости образования белков через регуляцию процессов транскрипции
и трансляции.
5.4. Внеклеточный матрикс (высокомолекулярные межклеточные вещества).
Основные межклеточные вещества: коллагены, ламинины, фибронектин,
нидоген, протеогликаны, тромбосподин и др. - находятся в виде геля, спо-
собны к самоагрегации, в результате которой возникают супрамолекуляр-
ные структуры, взаимодействующие между собой и с окружающими клет-
ками. Конденсированной их формой является базальная мембрана - спе-
циализированный компонент внеклеточного матрикса. Она служит местом
прикрепления клеток, влияет иа их дифференцировку и миграцию, является
барьером для макромолекул.
❖ Функции внеклеточного матрикса: участвует в адгезии клеток, связывает
факторы роста и другие цитокины, регулирует клеточный рост и диффе-
23
ренцировку, приводит к гидратации и создает эластичность внеклеточного
матрикса.
6. Общая физиологическая характеристика органов.
6.1. Орган - часть организма, эволюционно сложившаяся система тканей и интегра-
ция функциональных элементов, объединенных общей функцией, строением и
развитием.
6.2. Функциональный элемент органа - структурно-функциональный комплекс,
состоящий из клеток всех тканей органа, объединенных кровообращением и
иннервацией (А.М. Чернух, 1979).
❖ Рабочая часть, состоящая из специфических (паренхиматозных) клеток.
❖ Соединительная ткань, обеспечивающая условия выполнения функций
паренхиматозным клеткам.
❖ Микроциркуляторная единица: пространственно-повторяющийся комплекс
сосудов микроциркуляции.
❖ Примеры функционального элемента органа: печеночная долька, нефрон,
нейронная колонка, фолликул щитовидной железы.
6.3. Специализация органов (их физиологические особенности) изучаются в курсе
нормальной физиологии.
7. Общая характеристика организма.
7.1. Организм - элемент вида, система органов и интеграция физиологических сис-
тем, способная к самоорганизации, саморегуляции, самовоспроизведению и от-
вечающая иа изменения условий среды как единое целое.
7.2. Физиологические системы и их основные функции.
❖ Нервная - прием, обработка и хранение информации, создание целостности
организма, связь его с внешней средой и адаптация к ней, целенаправлен-
ное поведение, познание внешнего мира и самого человека.
❖ Эндокринная - гуморальная регуляция и координация функций органов,
тканей, целостного организма.
❖ Иммунная - защита организма от чужеродных антигенов внешнего и внут-
реннего происхождения (обеспечение антигенного гомеостаза), участие в эндок-
ринных, метаболических, терморегуляторных, психофизиологических и
других процессах.
Системы кровообращения и лимфообращения - микроциркуляция и обмен
веществ в тканях, осуществление всех функций крови, сенсорная и термо-
регуляторная функции, взаимосвязь всех органов.
❖ Система крови - дыхательная, трофическая, выделительная, иммунная, ре-
гуляторная, транспорт теплоты, регуляция кислотно-основного состояния
(КОС).
❖ Пищеварительная - секреторная, ферментативная переработка пищи, мо-
торная, всасывательная, экскреторная, иммунобиологическая, регуляция
КОС.
< • Мочевыделительная - выделение конечных продуктов обмена, сохранение
гомеостаза, поддержание водно-сопевого баланса, регуляция КОС, инкре-
торная функция.
24
❖ Система воспроизведения - образование половых клеток и гормонов, оп-
лодотворение, развитие эмбриона и плода, вскармливание ребенка.
❖ Покровная (кожа) - барьерная, защитная, терморегуляторная, выделитель-
ная, сенсорная.
7.3. Целостность организма.
❖ Главную роль в формировании целостности организма играют нервная, эн-
докринная, иммунная системы и кровообращение.
❖ Функциональная система (ФС), временно объединяющая функции орга-
низма для достижения какого-либо полезного результата, является важ-
нейшим физиологическим механизмом формирования целостности орга-
низма.
❖ Системогенез - процесс формирования в онтогенезе ФС организма. Ос-
новные принципы системогенеза - гетерохрония (разновременное созревания
различных ФС в ходе онтогенеза), консолидация элементов (процесс объединения
фрагментов конкретной ФС, развивающихся в разных местах организма), минимальное
обеспечение функций (ФС начинает функционировать с минимального набора функ-
ций, обеспечивающих выживание, а затем усложняется за счет включения приобретенных
реакций).
7.4. Нервная и гуморальная регуляция функций организма, их неразрывное един-
ство.
❖ Нервная регуляция осуществляется с участием безусловных и условных
рефлексов, высшая форма нервной регуляции реализуется в рамках дея-
тельности функциональных систем.
❖ Гуморальная регуляция осуществляется преимущественно с помощью тка-
невых и дистантных гормонов.
• К тканевым гормонам относятся различные виды веществ - амины (на-
пример, гистамин), пептиды (например, брадикинин), гликопептиды (напри-
мер, цитокины - интерлейкины, интерфероны, колониестимулирующие фак-
торы и др.), продукты обмена арахидоновой кислоты (простагландины,
тромбоксан Л2, лейкотриены). Тканевые гормоны действуют преимущест-
венно паракринно (действуют на рецепторы рядом лежащих клеток) н ау-
токринно (выделяются клеткой и действуют на рецепторы этой же клетки).
• Дистантные гормоны выделяются специализированными клетками
эндокринных желез и некоторых органов, а действуют преимуществен-
но в других органах и тканях (см. тему 9).
❖ Единство нервной и гуморальной регуляции:
• нейроны некоторых структур ЦНС (например, ядер гипоталамуса) образуют
гормоны - либерины и статины, регулирующие деятельность эндокрин-
ной системы;
• циторецепторы различных структур нейрона (особенно синапсов) являются
объектом действия гормонов, изменяющих их функциональную актив-
ность;
• механизмы действия нейромедиаторов в синапсах с метаботропными
рецепторами аналогичны действию гормонов.
25
7.5. Основные кибернетические принципы регуляции функций организма осуще-
ствляются с помощью прямых и обратных связей (нервных и гуморальных).
❖ Прямые связи передают информацию (команду) от регулирующего центра
к эффекторному (исполнительному) органу, изменяющую его функции в
сторону приспособления организма к среде (например, импульсация по эффе-
рентному отделу нервной системы, связь эндокринной железы через гормон с ор-
ганом-мишенью).
❖ Обратные связи (отрицательные и положительные) передают информацию
в регулирующий центр об эффективности осуществляемого им предшест-
вующего регулирующего воздействия, что позволяет внести коррекцию в
эту регуляцию (например, обратная афферентация в рефлексе и функциональной
системе, действие гормона периферической железы на секрецию тропного гормо-
на аденогипофизом).
• Отрицательная обратная связь («+» «-» взаимодействие) является ме-
ханизмом стабилизации функциональных параметров организма, улуч-
шения его устойчивости (при этом регулятор системы изменяет знак возмущаю-
щего сигнала на противоположный; например, повышение концентрации гормона в
крови приводит к торможению его секреции и наоборот).
• Положительная обратная связь является механизмом самоусилеиия
(саморазвития) физиологического процесса (при этом регулятор системы ие из-
меняет знак возмущающего сигнала, а усиливает его: например, фаза деполяризации
потенциала действия, увеличение концентрации фолликулостимулирующего гормона
перед овуляцией).
7.6. Саморегуляция постоянства внутренней среды организма («Постоянство внут-
ренней среды есть условие свободной жизни». К. Бернар).
❖ Гомеостаз - постоянство внутренней среды и устойчивость основных фи-
зиологических функций организма, а также совокупность физиологических про-
цессов, обеспечивающих это постоянство (К. Бернар, 1878; У. Кеннон, 1929). Термин
гомеокинез подчеркивает относительность постоянства внутренней среды,
существование изменения ее показателей, например, в результате действия
суточных биоритмов.
❖ Основные принципы саморегуляции.
• Регуляция по отклонению параметров гомеостаза осуществляется иа ос-
нове отрицательной обратной связи.
• Регуляция по возмущению позволяет включать (с помощью экстерорецеп-
тров) регуляцию еще до отклонения параметров гомеостаза (например, уси-
ление теплопродукции с холодовых рецепторов кожи возникает еще до снижения тем-
пературы ядра тела).
• Регуляция по прогнозированию включается до действия раздражителя,
когда еще только возникает ситуация, приводящая к его появлению (на-
пример, повышение теплопродукции в теплом помещении, если прогнозируется дея-
тельность организма в холодных условиях).
8. Профильные материалы для студентов лечебного, педиатрического и стомато-
логического факультетов.
8.1. Возрастная периодизация онтогенеза человека.
• > Антенатальный этап.
26
• Начальный период (ранний эмбриогенез): 1-я неделя, происходит в яйцеводе и
матке, от зиготы до начала имплантации бластоцисты.
• Эмбриональный (зародышевый) период: 2 - 8-я недели, завершение импланта-
ции бластоцисты, закладка эмбриональных зачатков тканей и органов, начало
развития плаценты, переход от гистотрофного к гематотрофному типу пита-
ния.
• Плодный (фетальный) период: 9 - 40-я недели, завершение образования пла-
центы (12-я неделя), формирование тканей и органов, системы мать - плод,
кровообращения плода.*
(Некоторые авторы выделяют перинатальный период - от 28-й недели внутриутроб-
ного развития до 7-10-го дия жизни.)
❖ Постнатальный этап.
• Период новорожденности: первые 10 дней жизни (по другим классификациям -
первые 30 дней).
• Грудной возраст: с 11-го дня жизни до 1 года (по другим классификациям - 2 -
12 мес.; младенческий возраст).
* Раннее детство: 1 — 3 года (по другим классификациям - ясельный возраст или
преддошколъны й).
• Первое детство: 4-7 лет (по другим классификациям - дошкольный возраст).
• Второе детство: для мальчиков - 8 - 12 лет, для девочек - 8 - 11 лет (по дру-
гим классификациям - младший школьный возраст).
• Подростковый возраст, или пубертатный период: для мальчиков -13-16 лет,
для девочек - 12-15 лет (по другим классификациям - средний и старший школьный
возраст, отрочество, период полового созравания).
• Юношеский возраст илн ювенильный: для юношей - 17 - 21 год, для девушек
- 16 - 20 лет.
• Зрелый возраст, I период: для мужчин - 22 - 35 лет, для женщин - 21 - 35 лет.
• Зрелый возраст, П период: для мужчин - 36 - 60 лет, для женщин - 36 - 55
лет.
• Пожилой возраст: для мужчин - 61 - 74, для женщин - 56 - 74 года.
• Старческий возраст: 75 - 90 лет.
• Долгожители - старше 90 лет.
8.2. Критические периоды онтогенеза. Развитие организма является нелинейным процес-
сом. Периоды эволюционного морфофункционального созревания чередуются с пе-
риодами переломных скачков в развитии (критических периодов), связанных как с
внутренними (биологическими), так н с внешними (социальными) факторами. Крити-
ческие периоды - этапы развития, характеризующиеся, с одной стороны, качественны-
ми морфофункциональными преобразованиями основных физиологических систем н
целостного организма, с другой стороны, усложняющимся взаимодействием биологи-
ческих и социально-психологических факторов развития. Они сопровождаются напря-
жением адаптационных механизмов, повышенной чувствительностью к различным по-
вреждающим факторам: радиации, алкоголю, никотину, лекарствам, гипоксин, вирусам
и др. В эти периоды легче возникает дезадаптация и нарушение здоровья. Выделяют
следующие критические периоды.
• Оплодотворение (образование зиготы).
27
• Имплантация бластоцисты (7 - 8-е сутки).
• Развитие зачатков органов и формирование плаценты (3 - 8-я недели).
• Интенсивное развитие головного мозга (15 - 20-я недели).
• Формирование основных физиологических систем организма, дифференци-
ровка полового аппарата (20 - 24-я недели).
• Рождение и период новорожденное*™ оказывают сильное стрессовое воздей-
ствие, вызванное родовым актом и резко изменившимися условиями среды.
• Раннее постнатальное развитие, характеризующееся интенсивным морфо-
функциональным созреванием, когда отсутствие воздействия каких-либо фак-
торов среды может привести к иесформированности функций (например, зри-
тельного анализатора, речи).
• Возраст 7—10 лет, когда одновременно изменяются базовые механизмы орга-
низации ВИД, вегетативных функций, обмена веществ. При этом происходят
значительные перестройки организации когнитивных (познавательных) про-
цессов за счет формирования механизмов локальной активации ЦНС, а нисхо-
дящие влияния регулируют все органы и обмен веществ. Важным внешним
фактором является резкая смена социальных условий - начало обучения в
школе.
• Половое созревание (11 - 16 лет) характеризуется изменением эндокринной
функции гипоталамуса, что приводит к изменению взаимодействия корковых
и подкорковых структур. При этом значительно снижается эффективность
центральных регуляторных механизмов^ определяющих произвольную регу-
ляцию и саморегуляцию. С другой стороны повышаются социальные требова-
ния к подросткам, возрастает их самооценка, что приводит к несоответствию
социально-психических факторов и функциональной возможности организма.
• Пожилой и старческий возраст характеризуется снижением функциональных
резервов организма, что увеличивает его чувствительность к экстремальным
факторам.
8.3. Теории старения (для студентов лечебного факультета, а также для всех тех, кто на-
деется дожить до старости). («...Среди тысяч людей только один умирает естественной
смертью, остальные погибают вследствие безрассудной манеры бытия». Маймонид.)
♦ ♦♦ Старение- процесс возникновения в организме возрастных изменений, ведущих
к недостаточности функций организма, ограничивающих его физиологические ре-
зервы и приспособительные возможности. Для развития старения характерна ге-
терохронность (различие по времени старения отдельных органов и тканей) и ге-
теротопность (неодинаковая выраженность старения в разных органах и различ-
ных структурах одного и того же органа).
♦ ♦♦ При всем разнообразии концепций и теорий старения в них прослеживаются три
доминирующих положения:
• Старение - генетически запрограммированный процесс, заключительная ста-
дия генетической программы индивидуального развития. При этом факторы
среды имеют незначительное влияние на его развитие.
• Старение - результат постепенного накопления повреждений, возникающих в
ходе самой жизни («катастрофа ошибок»), что приводит к снижению надежно-
сти организма, т.е. старение является вероятностным (стохастическим) про-
цессом. В этом случае запрограммировано не старение, а биологические свой-
ства организма.
28
• Старение определяется примерно в равной степени двумя группами факторов:
генетическими и экологическими, т.е. имеются два компонента - генетически
запрограммированный и вероятностный, стохастический.
Подробный обзор теорий и концепций старения провел Л.З. Тель (2001). В зави-
симости от того уровня, на котором преимущественно действует пераопричина
старения выделяют три главных группы теорий старения - молекулярные, клеточ-
ные и организменные.
• К молекулярным теориям старения относятся генетические, метаболические и
конформационные.
ф* Примером генетических теорий является теория геномных нарушений и
близкая к ней теория накопления ошибок.
• ф* К метаболическим теориям относятся свободнорадикальная, пигментного
обмена (накопление маркера старения - липофусцина) и кальциевая (на-
копление Са2+ в лизосомах, синапсах, цитозоле). Свободнорадикальная
теория старения - одна из наиболее интенсивно развивающихся в на-
стоящее время. Свободные радикалы кислорода (супероксидный анион,
гидроксильный радикал и др.) при избыточном их образовании (окисли-
тельный стресс) повреждают макромолекулы (ДНК, белки, липиды, кол-
лаген) и клеточные мембраны. В организме имеется мощная антиокси-
дантная система (ферменты - супероксиддисмутаза, каталаза и др., мета-
болиты - витамины А, Е и С, мелатонин и др.), активность которой прямо
коррелирует с продолжительностью жизни, а недостаточность или исто-
щение которой приводит к укорочению жизни. К более старым вариантам
метаболической теории старения относятся теории «изнашивания», рас-
траты жизненной энергии, аутоинтоксикации.
ф* Конформационные теории связывают первопричину старения с конфор-
мационными изменениями вторичной, третичной и четвертичной струк-
туры макромолекул.
• Современные варианты клеточной теории старения связаны с «лимитом
ХеЙфлика» и концепцией апоптоза. ХеЙфлик установил, что культивируемые
штаммы нормальных клеток имеют ограниченное число делений, в процессе
которых клетки стареют и гибнут. Клетки человека способны к 40 - 60 деле-
ниям, что дает возможность продолжительности жизни около 110 лет. При
синдроме преждевременного старения (прогерии) клетки способны к 2 - 10
делениям. Важная роль апоптоза (см. выше 4.10) в старении не вызывает со-
мнения, однако их взаимосвязь неоднозначна. В органах, в которых нет или
почти нет стволовых клеток (например, в головном мозге) имеется возрастное
снижение чувствительности клеток к апоптозу. В результате старые клетки
приобретают способность накапливать молекулярные повреждения, которые
нарушают их функцию и делают потенциально опасными для организма. В ор-
ганах, образуемых из обратимо дифференцированных клеток (например, пе-
чень), напротив, сохраняется высокий уровень ответа на индукторы апоптоза
(например, на окислительный стресс).
• К организменным теориям старения относятся адаптационно-регуляторные
теории. В них старение рассматривается как функция целостного организма в
его непрерывном взаимодействии с внешней средой, придается важное значе-
ние процессам нарушения регуляции, возрастному повышению порога (т.е.
снижению чувствительности) гипоталамуса к регуляторным сигналам, в ре-
29
зультате чего развивается нарушение репродуктивных, адаптационных (гипе-
радаптоз) и метаболических процессов (ожирение, атеросклероз, преддиабет).
По мнению авторов этих теорий (ВЛ. Дильман, 1987; В.В. Фролькис, 1988) старе-
ние ие запрограммировано, является побочным продуктом выполнения гене-
тической программы развития.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3: 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1-2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [12], [13], [14], [16], [17].
3.2. Дополнительная литература (см. с. 514 - 522): [1], [5], [9], [13]. [19], [30], [35],
[46], [47], [70], [130], [138], [146], [176-179], [183], [189] [190], [222] [225], [226].
[251-254].
33. Наглядные средства обучения. Использование страницы кафедры в Интернете -
www.vsma.ac.ru/~nDhvs : слайды для компьютерной демонстрации из «Атласа по
физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты остаточ-
ного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицамв, диа-
проектором и слайдами (по графику кафедры).
33. Консультации преподавателе еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов по данной теме буд ет осуществ-
ляться иа итоговом занятии и экзамене.
30
Тема 2. Биопотенциалы. Механизмы процесса возбуждения и его рас-
пространения
Целы а) знать мембранный потенциал, его происхождение, ионные градиенты клетки, их механизмы;
локальный ответ, критический уровень деполяризации мембраны; потенциал действия, его фазы
н происхождение, соотношение фаз возбудимости с фазами потенциала действия; рефрактер-
ность, ее причины; механизмы распространения возбуждения;
б) уметь использовать эти знания для понимания функциональной роли мембранных потенциа-
лов и фазового изменения возбудимости в тканях организма.
Мотивация темы: изучение электрофизиологических проявлений аозбуждения необходимо для пони-
мания мышечной, в том числе сердечной, деятельности, функций нервной системы и методов их
исследования. Механизмы нарушения возбуждения нервной и мышечной систем яаляются важ-
ным фактором развития сердечно-сосудистых, неврологических заболеваний и терминальных
(пограничных с жизнью) состояний.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором а конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Возбудимость как специализированная форма раздражимости.
1.1. Возбудимость - способность специализированной ткани (нервной, мышечной,
железистой) отвечать процессом возбуждения на действие раздражителей. Воз-
буждение - процесс электрического, сократительного или секреторного ответа
специализированной ткани, в котором обязательным компонентом является бы-
строе колебание электрического потенциала клеточной мембраны.
1.2. Виды электрических ответов возбудимых тканей.
❖ Местные: препотенциал (локальный ответ), возбуждающий и тормозной
постсинаптические потенциалы, рецепторный потенциал.
❖ Импульсный, активно распространяющийся (потенциал действия).
2. Основные исторические этапы изучения электрических явлений в организме.
2.1. Косвенное доказательство наличия животного электричества (Л. Гальвани, 1771 - 1793).
2.2. Прямое доказательстао наличия разности потенциалов в мышце (К. Маттеуччи, 1842).
23. Регистрация потенциала действия (Э. Дюбуа-Реймон, 1870).
2.4. Кабельная теория распространения потенциала действия (Л. Германн, 1885).
2.5. Мембранная теория происхождения биопотенциалов (Ю. Бернштейн, 1902).
2.6. Экспериментальное обоснование ионных механизмов потенциала действия (А. Ходжкин, Э.
Хаксли, 1963).
3. Мембранный потенциал покоя (МПП) - общий признак всех живых клеток
(возбудимых и невозбудимых).
3.1. Мембранный потенциал покоя - это трансмембранный потенциал между дву-
мя прилегающими к плазмолемме водными фазами (внеклеточной и внутрикле-
точной), регистрируемый до начала действия раздражителя (рис. 2). При этом
внутриклеточная прилегающая жидкость заряжена отрицательно по отношению
31
Рис. 2. Схематическое изображение мембранных потенциалов.
1 - мембранный потенциал покоя (трансмембранный потенциал);
2 - поверхностный мембранный потенциал, экранированный слоем
противоионов.
к внеклеточной прилегающей жидкости, потенциал которой принимается рав-
ным нулю. Величина МПП от -10 мВ в эритроцитах до -100 мВ в волокнах
Пуркинье сержа; функциональная роль - создает электрическое поле высокого
напряжения, влияющее на функциональную активность компонентов мембраны
(особенно белков), создает основу для других биопотенциалов клетки.
Рис. 3. Сдвиги мембранного
потенциала покоя (МПП).
1 - деполяризация,
2 - реполяризация,
3 - гиперполяризация.
ЗЛ. Сдвиги мембраниого потенциала (рис. 3).
❖ Деполяризация - сдвиг в позитивную сторону.
32
❖ Гиперполяризация - сдвиг в негативную сторону.
❖ Реполяризация - возврат МП из состояния деполяризации к исходному
уровню.
❖ Овершут - «перескок через нуль» с перезарядкой мембраны.
3.3. Условия возникновения мембранного потенциала.
Ионные градиенты клетки между цитозолем и внеклеточной средой (подробнее
в табл. 1).
* Величины градиентов: К+ в клетке ~ в 33 раза больше, чем во внеклеточной
среде, Na+ в клетке - в 14 раз меньше, СГ в клетке ~ в 20 раз меньше.
• Механизмы формирования градиентов: К*, Na^-насос формирует гради-
енты Na+ н К*, градиент С1‘ создается в результате использования энер-
гии градиента Na+ или К1' в процессах антипорта и симпорта, предпола-
гают также существование хлорного насоса.
Таблица 1. Ионные градиенты между клеткой и внеклеточной средой
Иои Внутри клетки, ммоль/л Вне клетки, Градиент кон- Равновесный потенциал, Бион. мВ Электродвижу- щая сила, мВ, ЭДС= МПП— Енои
ммоль/л центрации Внутри/Вие
Na* 10 145 1/14 +60 -140
к* 150 4,5 33/1 -94 +14
Са" 0,00005 U 1/24000 +140 -210
СГ б 114 1/20 -80 0
НСОз" 8 24 1/3
❖ Различная проницаемость мембраны для ионов определена наличием ион-
ных каналов, их количеством и состоянием.
• Ионные каналы - это интегральные белки мембраны, состоящие из не-
скольких (до пяти) субъединиц, образующих пору и способные с боль-
шей или меньшей избирательностью (селективностью) пропустить в клет-
ку или из клетки неорганические ионы по концентрационному и элек-
трическому градиентам (рис. 4).
• В структуре канала выделяют наружное и внутреннее устья, участок,
выполняющий роль «селективного фильтра» (d = 0,3 - 0,6 нм), через ко-
торый ион может пройти после частичной или полной утраты своей
гидратной оболочки.
• Каналы, кроме каналов утечки, имеют воротный механизм, который
определяет закрытое (потенциально активное), открытое (активированное)
или закрытое (инактивированное) состояние канала. Состояние «ворот»
(проницаемость канала) регулируется изменением электрического поля
(поляризации) мембраны - потенциалуправляемые каналы, или влияни-
ем эндогенных н экзогенных химических веществ (нейромедиаторов,
гормонов, лекарств) - хемоуправляемые каналы.
33
ф- Потенциалуправляемые каналы имеют воротную «частицу» в виде
диполя, на концах которого располагаются разноименные заряды.
По времени срабатывания ворот (от миллисекунд до секунд) каналы
подразделяют на быстрые и медленные. Активность канала может
быть изменена различными веществами (например, ионами кальция,
высокоэнергетическнми фосфатнами группами), которые присоединя-
ются к белкам на внутренней стороне канала.
ф Хемоуправляемые каналы (синонимы: «канал-рецептор», «ионотропный ре-
цептор») находятся в составе рецептора, на который действуют био-
активные вещества: нейромедиаторы - ацетилхолин, ГАМК, глута-
мат и др., гормоны, лекарства (например, М-холннорецептор,
ГАМКА-рецептор и др.)
пи ишууед [рЙДО
3
4
1
2
Рнс. 4. Схематическое изображение ионных каналов мембраны.
А - селективный фильтр, Б - активационные ворота, С - инактнвацион-
ные ворота. 1 - каналы утечки без воротного механизма, 2 - 4 - каналы с
воротным механизмом: 2 - канал закрыт, потенциально активен, 3 - канал
открыт, 4 - канал закрыт, инактивирован, 5 - лнпидный бислой мембраны
• Механочувствительные каналы (МЧК) изменяют проводимость в ответ
на деформацию мембраны при действии механических раздражителей,
гидростатического и осмотического давления. Натяжение мембраны
может передаваться к каналу прямо через липидный бислой и (или) ци-
тоскелет клетки. Выделены различные виды МЧК: каналы, активируе-
мые и ингибируемые растяжением мембраны; катионные (калиевые,
кальциевые, неселективные) и анионные каналы и др. Доказано, что они
могут создавать токи, достаточные для изменения электрического по-
тенциала мембраны и активации потенциалуправляемых каналов.
• Проницаемость каналов можно изменить (увеличить или уменьшить) в
результате действия на их белки различных веществ (Са2+, цАМФ, ино-
зитол-3-фосфата, G-белков плазмолеммы, протеинфосфокиназ и др.),
через них действуют на клетку медиаторы и гормоны (например, механиз-
мом модуляции актианости является фосфорилирование аминокислотных остатков
белковых субъединиц канала).
• Через ионный канал в течение 1 с может проходить 12-20 млн. ионов.
Поэтому ионные токи каналов во много раз превосходят ионные токи,
связанные с работой ионных насосов и ионообменников.
34
❖ В состоянии физиологического покоя проницаемость мембраны (Р) опре-
деляется в основном каналами утечки, оиа очень низкая для Na4, средняя
для СГ и более высокая для К4. Если Рк4 принять за 1, то Рк4: Ра': Р Na+ = 1
: 0,4:0,04.
3.4. Механизмы возникновения МПП.
❖ Диффузия К4 из клетки по каналам утечки до равновесного потенциала
(Ек+= -94 мВ) является главным механизмом формирования МПП (К* как по-
ляризующий ион). Равновесный потенциал (ЕЯОн) - это потенциал, при кото-
ром возникает равенство двух сил - силы перемещения иона по химиче-
скому градиенту и противоположной по направлению электростатической
силы; при этом прекращается диффузия иона. Равновесный потенциал pac-
г. 1
считывается по уравнению Нернста: с к =------In
где: R - газовая
постоянная (мера внутренней энергии вещества), Т - абсолютная температура (у человека
310° К), F - число Фарадея (количество электрических зарядов в I М вещества, Z - ва-
лентность иона с учетом знака. Кв, Кн - концентрация иоиа в ммолях соответственно
внутри и снаружи клетки.
❖ Асимметричная работа К4, Na4-Hacoca (на 2 К* перемещаемого в клетку, из
нее выводится 3 Na4) создает поляризацию мембраны (примерио -10 мВ) и
является вторым механизмом формирования МП.
❖ Небольшая диффузия Na4 по каналам утечки внутрь клетки (Ема. = +60 мВ)
делает реальный МПП несколько ниже, чем Ек* (Na4 как деполяризующий ион).
❖ Ионы СГ пассивно распределены относительно мембраны в соответствии с
МПП и ЕсГ. Распределение иоиов хлора при этом стремится уравнять ЕсГ
с МПП, поэтому СГ не могут существенно влиять на МПП. Однако при
изменении МП (например, при потенциале действия) перемещение ионов хло-
ра существенно влияет на МП (например, вход СГ в кардиомиоцит в фазе началь-
ной реполяризации ПД).
* •* Суммарный вкляд этих ионов а возникновение мембранного потенциала рассчитывается
по уравнению Гольдмана - Ходжкина - Катца:
RT, pjtf4 ]« + ?„„ [мГр + рДсг]?
М1111 =----In---г 1-------г 1--------1 1 где: Р - проницаемость плазмолеммы
zF PK[K^ + P„ANa^ + Pcl[cr^
для ионоа, другие обозначения см. аыше а формуле Нернста.
4. Препотенцнал (локальный ответ).
4.1. Препотенциал - локальный потенциал, который возникает при действии суб-
пороговых раздражителей в тех же участках мембраны, где и потенциал дейст-
вия.
4.2. Схема препотенциала (ИО)'. расположен в субпороговой области, имеет фазу
деполяризации и реполяризации.
4.3. Механизм возникновения препотенциала (ЛО).
❖ Начальная чисто физическая деполяризация мембраны.
❖ Добавочная деполяризация, связанная с открытием потенциалзависимых
№4-каналов и входящим в клетку Ыа4-током.
Открытие потенциалзависимых К4-каналов и увеличение выходящего из
клетки К4-тока вызывает фазу реполяризации.
35
❖ Во время ЛО входящий в клетку Ыа+-ток меньше, чем выходящий из клет-
ки К+-ток плюс ток утечки.
4.4. Свойства препотепциала (ЛО).
• ❖ Амплитуда зависит от силы раздражителя (закон силы).
* ❖ Способен к суммации.
❖ Во время ЛО повышена возбудимость.
❖ Распространение его происходит с затуханием амплитуды на небольшие
расстояния (в пределах 1 мм).
4.5. Основная функциональная роль препотепциала (ЛО): деполяризует МП до
критического уровня деполяризации, что приводит к развитию потенциала
действия.
5. Критический уровень деполяризации мембраны (КУД, Ек).
5.1. КУД - это тот уровень, при котором деполяризация мембраны принимает реге-
неративный (самоусиливающийся) характер, свидетельствующий о развитии
потенциала действия.
S.2. При этом входящий в клетку Ыа+-ток равен выходящему из клетки К+-току, что
характеризует электрическую нестабильность мембраны.
53. Раздражитель, деполяризующий МП до КУД называется пороговым раздра-
жителем.
5.4. Величина потенциала, равная разности между КУД и МП, называется порого-
вым потенциалом (1111 = МП - КУД), ои характеризует возбудимость клетки
(чем меньше ПП, тем больше возбудимость и наоборот).
б.
Потенциал действия (ПД).
6.1.
6.2.
63.
ПД- это электрический компонент возбуждения нервных и мышечных клеток,
представляющий собой быстрое колебание их МП, сопровождающееся, как
правило, перезарядкой мембраны. Имеются единичные данные о ПД железистых клеток.
6.4.
Условие возникновения ПД - дей-
ствие раздражителей пороговой и
сверхпороговой силы в электровоз-
будимых участках мембраны.
Схема ПД (рис. 5): деполяризация,
овершут и пик, реполяризация, сле-
довые потенциалы; амплитуда ПД
от 70 мВ в клетках гладких мышц
до 120 мВ в миокарде и длитель-
ность от 1 мс в нервных волокнах
до 300 мс в миокарде.
Восходящая фаза ПД (передний
фронт ПД: деполяризация плюс пе-
до поляризация
Рис. 5. Схема потенциала действия.
резарядка), ее механизмы.
❖ Деполяризация, достигшая уровень КУД, открывает быстрые m-ворот в по-
тенциалзависимых Na^-каналах и формирует входящий Na+-TOK (до 6 мА/см2),
который резко преобладает иад выходящим К+-током, что приводит к даль-
нейшей деполяризации мембраны.
36
❖ Деполяризации при этом имеет регенеративный (самоусиливаюшийся) ха-
рактер (пример положительной обратной связи), что обеспечивает стандартную
величину ПД при действии пороговых и сверхпороговых раздражителей.
❖ Входящий Na+-TOK не только доводит МП до нуля, но и вызывает переза-
рядку (реверсию заряда) мембраны (около +20 мВ).
❖ Деполяризация мембраны вызывает инактивацию Иа+-каналов (закрытие в
них медленных h-ворот), поэтому пнк ПД не достигает величины равновес-
ного Иа+-потенциала, равной около +55 мВ.
6.5. Нисходящая фаза ПД (реполяризация мембрапы), ее механизмы.
❖ Деполяризация в восходящей фазе открывает потенциалзависимые К+-
каналы н усиливает выходящий из клетки К+-ток.
❖ Преобладание К+-тока над входящим Иа+-током приводит к восстановле-
нию поляризации мембраны (выходит из клетки К* столько, сколько в нее вошло
Na4). Работающая клетка создает вокруг себя зону повышенной концентра-
ции К*.
* Реполяризация ликвидирует инактивацию Иа+-каналов (h-ворота открыва-
ются, ш-ворота закрываются).
6.6. Следовые потенциалы, их иоппые механизмы. После высоковольтной части
(спайка) ПД развиваются в разной последовательности и степени выраженности
следовые потенциалы.
Следовая деполяризация (следовой отрицательный потенциал) связана с резким
снижением повышенного К+-тока по механизму отрицательной обратной
связи.
Следовая гиперполяризация (следовой положительный потенциал) связана с дли-
тельным небольшим избытком К+-тока и элекгрогениой деятельностью К+,
Na+-Hacoca.
6.7. Активация К*,Ыа*-пасоса легко восстанавливает нарушенные во время ПД
ионные градиенты, которые в результате одиночного ПД нарушаются незначи-
тельно (от 0,0001 % в гигантском аксоне кальмара до 1 % в тонких волокнах типа С). Клетка
без «подзарядки» может генерировать около 100 тысяч ПД.
6.8. Функциональная роль ПД: в нервной ткани - основной способ передачи ин-
формации, в мышечной ткани - пусковой механизм сокращения миоцитов.
7. Изменение возбудимости при возбуждении.
7.1. Основные факторы, влияющие на возбудимость: сдвиги КУД н МП, увели-
чивая или уменьшая пороговый потенциал, соответственно снижают или уве-
Рис. 6. Кривая изменения возбудимости
(Б) в течение потенциала действия (А).
МПП - мембранный потенциал покоя,
КУД - критический уровень деполяриза-
ции. 1 - абсолютная рефрактерность; 2 -
относительная рефрактерность; 3 - су-
пернормальная возбудимость; 4 - суб-
нормальная возбудимость.
37
личивают возбудимость клетки; увеличение инактивации Иа^-каналов сдвигает
КУД в позитивную сторону, увеличивая пороговый потенциал и снижая возбу-
димость.
7.2. Фазные изменения возбудимости клетки в течение ПД (рис. б).
❖ Фаза абсолютной рефрактерности характеризует полную невозбудимость
клетки (высокую инактивацию Ма*-каналов); она соответствует восходя-
щей и части нисходящей (до уровня примерно на 20 мВ позитивней КУД) фазам
ПД, обеспечивает прерывистый характер возбуждения, лимитирует ла-
бильность.
• Лабильность есть скорость, с которой клетка успевает закончить пол-
ный период ПД во времени.
• Показателем лабильности является максимальное число ПД, которое
возбуждаемая структура способна генерировать в полном соответствии
с ритмом раздражения (Л = 1 сек/абсолютная рефрактерность в сек).
❖ Фаза относительной рефрактерности соответствует интервалу времени, в
течение которого МП в фазе реполяризации находится в диапазоне КУД ±
10 - 15 мВ; новый ПД в этой фазе можно получить при действии сверхпо-
роговых раздражителей.
❖ Фаза супернормальной возбудимости соответствует препотенциалу н сле-
довой деполяризации; ПД можно вызвать субпороговым раздражителем.
❖ Фаза субнормальной возбудимости соответствует следовой гиперполяриза-
ции; ПД можно вызвать сверхпороговым раздражителем.
8. Непрерывное распространения ПД происходит в мышечных и немиелиновых во-
локнах (рис. 7).
8.1. Этан злектротоннческого распространения возникшего ПД.
Б
Рис. 7. Непрерывное распространение потенциала действия в нерв-
ном волокне. А - возникновение ПД в нервном волокне. Б - рас-
пространение ПД в обе стороны от места возникновения. 1 - об-
ласть деполяризации (преобладает входящий в клетку натриевый
ток), 2 - соседняя область, в которой локальный ток от области ПД
вызывает деполяризацию до критического уровня, 3 - область ре-
поляризации на месте предшествующего ПД.
38
*•* Сначала происходит пассивное (без реакции ионных каналов мембраны) распро-
странение ПД как электротонического (физического) потенциала в соседние
участки мембраны.
❖ Эффективность электротонического распространения зависит от физиче-
ских (кабельных) свойств волокон (большого сопротивления мембраны, низкой ем-
кости мембраны, низкого сопротивления цитоплазмы волокна) и количественно ха-
рактеризуется постоянной длины мембраны (Хщ) - расстоянием, на котором
амплитуда биопотенциала уменьшается до 37 % исходной величины (это
расстояние наибольшее в миелиновых волокнах - до 5 мм).
8.2. Этап возникновения ПД на соседнем участке.
❖ Деполяризация мембраны до КУД приводит к возникновению ПД в сосед-
нем участке.
❖ При этом в результате регенеративной деполяризации сохраняется посто-
янство амплитуды ПД в процессе распространения (распространение без зату-
хания).
♦ 5» Скорость распространения ПД прямо пропорциональна крутизне его Переднего
фронта.
9. Профильные материалы дли стоматологического факультета. Электрохимиче-
ские потенциалы (ЭХП) в ротовой полости.
9.1. Возникновение ЭХП связано с наличием в ротовой полости трех сред, контактирую-
щих друг с другом: ротовая жидкость, насыщенная Ог и СОг, выполняет роль электро-
лита; твердая среда (зубы и зубные протезы) - роль электродов; мягкая среда (десиа,
слизистая оболочка) - роль проводящего материала.
9.2. ЭХП в ротовой полости способствует ее самоочищению. По величине ЭХП можно ус-
тановить состояние зуба, процесс рассасывания цемента и необходимость замены про-
теза.
93. В разных точках ротовой полости ЭХП имеют различные величины.
❖ ЭХП у здоровых (кариесрезистентных) людей равен от 5 до 160 мВ (по отношению
к ртутно-каломельному электроду).
❖ Каждый вид зубов и его поверхности имеют различные величины ЭХП: наиболее
высокую - в режущем крае резцов, наиболее низкую - в пришеечной области зу-
бов.
9.4. Разность ЭХП в разных точках ротовой полости вызывает возникновение между ними
электрического тока.
Разность ЭХП в 1 - 12 мВ считается нормальной.
Разность ЭХП в 60 мВ приводит к патологическим изменениям - гальванозу, при
котором гальванические токи нарушают функциональное состояние полости рта.
9.5. Условия возникновения гальванических микротоков.
❖ Использование металлов при протезировании и пломбировании.
❖ Твердая (зубной протез) и жидкая (слюна, десневая жидкость) среды образуют
электрод-систему, на границе которой возникает разность потенциалов.
♦ ♦♦ В нейтральной среде (pH 7,0) электрохимическая реакция сопровождается избыт-
ком Н*, поэтому у больных с протезами из нержавеющей стали или хромово-
кобальтового сплава поваляется ощущение кислоты в полости рта.
❖ В кислой среде происходит процесс растворения металлического протеза и воз-
никновение анодных участков коррозии.
9.6. Гальванизм в стоматологии как совокупность симптомов (металлический вкус, чув-
39
ство жжения, изменение вкусовой чувствительности, сухость во рту или слюнотечение,
воспаление слизистой и др.).
❖ Зависимость симптомов от силы тока: при 80 мкА явления гальванизма выражены
сильно, при 25 - 80 мкА возникают слабые ощущения, при 5 мкА симптомов нет.
❖ Физико-химические основы гальванизма - увеличение концентрации ионов ме-
таллов в слюне в результате электрохимических коррозионных процессов.
Токсическое действие ионов металлов на функции слизистой оболочки рта приво-
дит к понижению и извращению вкусовой чувствительности на сладкое, соленое,
кислое как следствие изменений в рецепторах, нарушению образования речи;
влияние микроэлементов на слизистую оболочку желудка и кишечника вызывает
обострение хронических желудочно-кишечных заболеваний.
9.7. Гальванизация в стоматологии для лечебных целей; действие постоянного тока низко-
го напряжения (30 - 80 В) и небольшой силы (до 50 мА) в области слизистой оболочки
рта.
Физиологические механизмы гальванизации.
• Местные реакции - продолжительная гиперемия (60 мин и более): расшире-
* ние сосудов, ускорение кровотока, увеличение проницаемости стенки сосуда,
активация обмена веществ, регенерация эпителия и соединительной ткани.
• • Рефлекторные реакции иа раздражение рецепторов ротовой полости, приво-
дящие к изменениям функции внутренних органов и показателей гемодинами-
ки (АД и ЧСС).
♦♦♦ Лекарственный электрофорез ткани зуба (эмали, дентина, пульпы) и периодонта.
♦♦♦ Электрообезболивание твердых тканей и пульпы зуба.
• Величина постоянного тока для обезболивания - 15 - 20 мкА.
• Механизм - анэлектротон (блокада деполяризации мембраны болевых рецеп-
торов под анодом и прекращение болевой импульсации).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3: 3.1 и 3.2). Объем реферата 2-3 стр., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. стр. 514, использовать имеющиеся
у вас учебники и пособия): [1] -[9], [11], [13], [14], [16], [17].
3.2. Дополнительная литература (см. стр. 514-522): [25], [30], [40], [93], [128], [165],
[222], [225], [236], [245], [251-254].
33, Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphvs: слайды для компьютерной демонстрации из «Атласа
по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; учебное по-
собие «Общая физиология возбудимых тканей» /В.Н. Яковлев, И.Э. Есауленко. -
Воронеж, ВГМА, 2003 - 73 с; тесты остаточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
ЗЛ. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться иа практи-
ческом занятии по дайной теме, итоговом занятии н экзамене.
40
Задание 4. Решение ситуационных задач (ответы см. стр. 508-513).
Задача 2-1. В локальном участке гипоксии головного мозга (например, при тромбозе артерии
мозга) в результате дефицита кислорода в нейронах произошло резкое изменение уровня АТФ и
началась деполяризация клеточной мембраны. В плазмолемме мембраны имеется K*,Na+-Hacoc,
потенциалуправляемые №*-каналы, а также К*-каналы, активируемые снижением в гиалоплазме
концентрации АТФ и повышением уровня Nat Напишите механизмы, связанные с реакцией ион-
ных каналов, которые будут препятствовать в этих условиях деполяризации мембраны.
Задача 2-2. При сердечной недостаточности больные принимают сердечные гликозиды, которые
ингибируют K*,Na*-Hacoc. В плазмолемме кардиомиоцита, кроме этого насоса, имеется Na7Ca2*-
ионообменник, с помощью которого цитозольный Са2* выводится из клетки во внеклеточную
жидкость в обмен на внеклеточный Na+, а также имеются активируемые нонами кальция К*-
каналы. Ответьте на следующие вопросы.
1) Как в результате действия сердечных глюкозидов изменится уровень Na* в цитозоле и по-
рог возбудимости (пороговый потенциал) кардиомиоцита?
2) Как изменится концентрация Са2* в цитозоле кардиомиоцита?
3) Как изменится скорость скорость фазы реполяризации потенциала действия кардиомио-
цита?
Задача 2-3. При высокой активности нейронов (генерации в них потенциалов действия с высокой
частотой) концентрация К* в межклеточной жидкости в связи с фазой реполяризации увеличива-
ется в 3-4 раза, что может привести к формированию в них ПД даже на субпороговые раздражи-
тели. После высокой активности нейронов в них возникает гиперполяризации клеточной мембра-
ны. Объясните: 1) Почему во время высокой активности нейронов их возбудимость может повы-
шаться? 2) Почему через некоторое время после высокой активности нейронов может возникать
гиперполяризация их клеточной мембраны?
Задача 2—4. При снижении концентрации Na* в плазме крови и межклеточной жидкости до 90-
100 ммоль/л, возникает тяжелая неврологическая симптоматика вплоть до паралича бульбарных
центров (сердечно-сосудистого, дыхательного и др.). Объясните эти яаления, исходя из меха-
низмов формирования потенциалов действия нейронов в условиах резкой гипонатриемии.
Задача 2-5. В эксперименте при изучении механизмов развития потенциала действия на мембра-
не гигантского аксона кальмара повысили внеклеточную концентрацию Na* и создали более вы-
сокий градиент, чем обычно существует между вне- и внутриклеточной средой (10:1). Объясните,
как это отразится на пороговом потенциале, возбудимости аксона и скорости развития в нем ПД.
41
Тема 3. Основные закономерности раздражения возбудимых тканей
Цель: а) знать основные закономерности раздражения возбудимых тканей: законы силы, «всё или ни-
чего», крутизны (аккомодация); оценку возбудимости - порог раздражения (пороговая сила, по-
роговое время); понятие о хронаксии; лабильность; парабиоз; полярный закон; электроток, ка-
толическую депрессию;
б) уметь использовать знание закономерностей раздражения при изучении физиологии цен-
тральной нервной системы, сердца, скелетных и гладких мышц.
Мотивация темы: знание закономерностей процессов возбуждения в тканях необходимо врачу для по-
нимания нарушений функций нервной и мышечной систем при различных заболеваниях. Эти
закономерности используются для электродиагностики поражений нервов и мышц и физиотера-
певтического лечения заболеваний.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены а «Графике н
содержанки самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1« Роль силы раздражителя.
1.1. Классификация раздражителей по силе', пороговые, субпороговые, сверхпо-
роговые.
1.2. Взаимосвязь между силой раздражителя и возбудимостью ткани (порого-
вая сила как мера возбудимости: чем больше сила порогового раздражителя,
тем меньше возбудимость и наоборот).
13. Закон силы (силовых отношений) - ответная реакция в определенном диапа-
зоне находится в прямой зависимости от силы раздражителя. По этому закону
осуществляется: 1) формирование на мембране клетки препотенциала, рецеп-
торного и постсинаптического потенциалов; 2) сокращение скелетной мышцы,
реакция нервной ткани на раздражители.
1.4. Закон «всё или ничего» - ответная реакция ие зависит от силы сверхпороговых
раздражителей: пороговый и сверхпороговый раздражители дают одинаковую
Пп On.
39 35 29 25 29 35-37 вРеия
Рис. 8. Схема оптимума н песси-
мума силы раздражения (по Н.Е.
Введенскому, 1886). Цифры -
расстояние в см между первичной
и вторичной катушками индукци-
онного аппарата (чем меньше
расстояние, тем больше раздра-
жение), 39 - порог раздражения,
Opi - оптимум в начале тетаниза-
ции мышцы. Орг - оптимум в
процессе тетанизации после пес-
симума раздражения, Ps - песси-
мум раздражения.
42
амплитуду ответа. По этому закону осуществляется: 1) формирование потен-
циала действия; 2) ответ ткани (миокард).
1.5. Оптимум и пессимум силы (частоты') раздражения (Н. Е. Введенский, 1896)
(рис. 8).
❖ Оптимум силы (частоты) - это наименьшая сила (или частота) раздражи-
теля, дающая максимальную реакцию. При этом максимальное сокращение
дает узкий диапазон силы раздражения: более слабые и более сильные раз-
дражители дают меньшую силу ответа Главным его механизмом является
улучшение синаптической передачи (подробно в теме 4; задание 1:4.4).
❖ Пессимум силы (частоты) - это действие сверхсильных раздражителей,
сопровождающееся уменьшением ответной реакции. Главным его меха-
низмом является ухудшение синаптической передачи - синаптическая де-
прессия (подробно в теме 4; задание 1:4.4).
2. Роль скорости нарастания силы раздражители (закон крутизны, э. дюбуа-Реймон, 1848).
2.1. Для возникновения возбуждения (ПД) необходима определенная скорость на-
растания силы раздражителя (например, силы электрического тока), ниже которой
возбуждение вообще не возникает. Это явление приспособления возбудимых
структур к медленно нарастающим по силе раздражителям называется аккомо-
дацией (рнс. 9).
2.2. Механизмы ак-
комодации.
❖ Медленная
деполяриза-
ция, вызы-
вая развитие
инактивации
№+-каналов,
приводит к сдвигу КУД в позитивную сторону.
❖ Активация потенцналуправляемых К+-каналов и выход К+ из клетки пре-
пятствуют деполяризации (сдвигу МП в позитивную сторону).
❖ В результате увеличивается пороговый потенциал и снижается возбуди-
мость.
2.3. Скорость аккомодации: возбудимые структуры с высокой аккомодационной
способностью (например, двигательные волокна) отвечают только на раздражители с
выраженной динамикой их силы; структуры с низкой аккомодационной спо-
собностью (например, чувствительные волокна) отвечают на малодинамичные раз-
дражители.
3. Роль длительности действия раздражителя (рнс. 10).
3.1. Связь между силой и временем действия раздражителя (кривая силы-времени
Гоорвега - Вейса). В сверхпороговом диапазоне, чем больше сила раздражите-
ля, тем меньше время его действия, необходимое для возбуждения. При очень
кратковременном действии раздражителя возбуждение не возникает, как бы нн
была велика его сила. <" '
3.2. На кривой силы-времени имеется точка R (реобаза), соответствующая порого-
вому раздражителю (реобаза - старое название порогового раздражителя), и определяет-
Рис. 9. Схема аккомодации. Раздра-
жители с большой скоростью изме-
нения силы (А и Б) вызывают по-
тенциал действия. Раздражитель с
малой скоростью нарастания силы
не вызывает потенциал действия
(аккомодация).
43
Полезное время
Рис. 10. Кривая силы - длительности.I
- сила раздражения, t - время; R - рео-
база; 2R - удлоенная реобаза.
ся полезное время (минимальное времени, необходимое для действия порого-
вого раздражителя). Поскольку реобаза со-
ответствует пологой части кривой и точное
определение полезного времени затруднено
было предложено определять хронаксию.
ЗЛ Хронаксия (Л. Лалик, 1909) - наименьшее
время, необходимое для действия раздражи-
теля удвоенной пороговой силы (две реоба-
зы); ее нормальные величины для скелетных
мышц конечностей 0,1 - 0,7 мс.
3.4. Хронаксиметрия - метод определения хро-
наксин, используется в клинике (например, для
диагностики периферического паралича) И В фи-
зиологии труда (например, для диагностики утом-
ления).
4. Раздражение возбудимых тканей постоянным током. Катэлектротон и аиэлек-
тротои.
4.1. Закон полярности раздражения (Э. Пфлюгер, 1859).
❖ При действии слабого тока непосредственно на мышцу сокращение возни-
кает при замыкании тока только под катодом и отсутствует при размыка-
нии тока, при средних токах сокращение возникает под катодом при замы-
кании тока и под анодом прн размыкании тока.
Л Б
Катэлектрон Анэлектрон
Рис. 11. Схема изменения мембранного потенциала покоя (МПП) и критического
уровня деполяризации (КУД) под катодом (А) и анодом (Б) при длительном дейст-
вии тока субпороговой сияы.х - пороговый потенциал (по Д.П.Матюшкнну,1984).
❖ Механизм возбуждения под катодом прн замыкании тока связан с деполя-
ризацией мембраны, достигающей КУД.
❖ Механизм возбуждения под анодом при размыкании тока связан с ликвида-
цией гиперполяризации, в результате чего МП достигает уровня КУД, ко-
торый во время действия тока сместился вниз (рис. 11Б).
4.2. Катэлектротон: под катодом сначала увеличивается возбудимость, проводи-
мость, лабильность, затем возникает католическая депрессия (В.Ф. Вериго, 1883)
- снижение этих показателей (рис. НА).
4.3. Анэлектротон: под анодом сначала снижаются возбудимость, проводимость,
лабильность, затем эти показатели повышаются.
5. Парабиоз как обратимые фазные изменения возбудимости, проводимости и ла-
бильности тканей (Н.Е. Введенский, 1901).
44
5.1. Парабиоз возникает при действии на нерв (нервно-мышечного препарата) деполя-
ризующих раздражителей, например, КС1, анестетиков, охлаждения (до парабиоза
ответная реакция осуществляется по закону силы).
30 20 29 26 29 26 22 26
Рис. 12. Парабиотическне фазы в нервно-мышечном препарате (по Н.Е. Введенскому, 1901).
Цифры - расстояние в см между первичной и вторичной катушками индукционного аппарата, чем
меньше цифра, тем бояьше раздражение.
I - уравнительная фаза: величина тетанического сокращения мышцы при различной силе раздра-
жения примерно одинакова.
П - парадоксальная фаза, прн более сильном раздражении амплитуда тетануса меньше, чем при
слабом раздражении.
Ш - тормозная фаза: В - одиночные слабые раздражения (26) парабиотического участка способны
вызывать мышечные сокращения; А - более сильное и длительное раздражение (23) на фоне раз-
дражения В оказывает тормозное действие на парабиотическнй участок.
5.2. Механизмы парабиоза связаны с постепенно нарастающей инактивацией Na+-
каналов, что приводит к увеличению продолжительности ПД (увеличивается время
абсолютной и относительной рефрактерности) и снижению его амплитуды.
5.3. Фазы парабиоза (рнс. 12).
❖ Уравнительная фаза характеризуется примерно одинаковой ответной ре-
акцией на действие раздражителей малой, средней и большой силы (это свя-
зано с уменьшением лабильности парабиотического участка н трансформацией частого
ритма сильных раздражителей в низкочастотный ритм слабых раздражителей).
❖ Парадоксальная фаза характеризуется тем, что амплитуда ответной реак-
ция на слабые раздражители больше, чем на сильные раздражителя (это свя-
зано с дальнейшим снижением лабильности; в этих условиях низкочастотные ритмы сла-
бых раздражителей проходлт участок парабиоза, а высокочастотные ритмы сильных раз-
дражителей почти полностью блокируются).
❖ Тормозная фаза характеризуется отсутствием ответной реакции как на сла-
бые, так и сильные раздражители (это состояние связано с высоким уровнем инак-
тивации Ыа+-каналов, приводящей к полной потере возбудимости).
5.4. Если факторы, вызывающие парабиоз, не приводят к грубым структурным из-
менениям, то фазы парабиоза проходят обратное развитие: возбудимость, про-
водимость и лабильность этого участка восстанавливаются.
45
6. Профильные материалы для студентов стоматологического факультета. Элек-
троодонтометрия, ее значение для диагностики заболеваний и лечения зубов.
6.1. Принцип метода: определяются пороги раздражения зубов при действии постоянного
тока.
❖ Порог раздражения для здоровых зубов 2-6 мкА.
❖ Снижение порога раздражения (менее 2 мкА, т.е. повышение возбудимости) про-
исходит при парадонтозе.
♦ ♦♦ Повышение порога раздражения (более 6 мкА, т.е. снижение возбудимости) воз-
никает при пульпитах, при гибели пульпы порог более 100 - 200 мкА.
6.2. Клиническое значение электроодонтометрии - оценивают степень поражения пуль-
пы и эффективность лечения ее заболеваний.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [14], [17].
3.2. Дополнительная литература (см. с. 514 - 522): [25], [30], [40], [128], [165], [222],
[225], [245], [253].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nDhvs: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; учебное
пособие «Общая физиология возбудимых тканей» /В.Н. Яковлев, И.Э. Есауленко. -
Воронеж, ВГМА, 2003 - 73 с.; тесты остаточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными табяицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508-513).
Задача 3-1. У больного патологический процесс, захвативший участок нервного ствола, вызвал
там инактивацию натриевых каналов, что привело к нарушению проведения нервных импульсов
(ПД) через этот участок. Вы решили восстановить проводимость в нерве, используя действие по-
стоянного тока. Какой электрод должен быть приложен к пораженному участку и почему?
Задача 3-2. Продолжительность спайка ПД (его высоковольтной части без следовых потенциа-
лов) в нервном волокне равна 1 мс, в мышечном волокне - 5 мс. Определите теоретически воз-
можную лабильность этих волокон.
Задача 3-3. В медицинской практике через мышечную ткань человека (и нервные волокна в ней)
пропускают токи высокой частоты (100000 Гц) н мощностью около 200 Вт. Хронаксия мышцы,
которая подвергается действию этих электромагнитных колебаний, равна 0,1 мс. Будет или не бу-
дет в этих условиях сокращение мышцы и почему?
46
Тема 4. Нейрон как структурно-функциональная единица нервной сис-
темы. Функции глии
Цель: а) знать основные функции ЦНС, структурно-функциональные элементы нейрона; рецепторы:
классификация, основные свойства, механизмы возбуждения, кодирование силы раздражителя,
регуляция рецепторов; механизм проведения нервного импульса по нервным волокнам, харак-
теристика волокон А, В, С; синапс, его строение, классификацию синапсов, механизм передачи
возбуждения: медиаторы, их синтез н секреция, взаимодействие с рецепторами постснналтиче-
ской мембраны, постсинаптические потенциалы; механизмы формирования возбуждения в ней-
роне: аксонный холмик и суммация постсинаптических потенциалов; трофическую функцию
нейрона, аксонный транспорт; функции глии;
б) уметь использовать эти знания для анализа деятельности ЦНС при регуляции функций цело-
стного организма.
Мотивация темы: изучение обшей физиологии ЦНС важно для анализа жизнедеятельности здорового
и больного организма с точки зрения принципов целостности, нервизма, взаимоотношения ор-
ганизма и внешней среды, т.е. для формирования правильного мышления врача. Вместе с тем
эти знания необходимы в деятельности врача для диагностики поражения нервной системы, по-
нимания механизмов ее заболеваний и их лечения (иглоукалывание, массаж и др.).
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логическую
структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором а конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов» на стенде. Остальные фрагменты задания
1 могут быть использованы для коррекции материала записанной лекции и подготовки ре-
фератов.
1. Общая характеристика нервной системы («Великий функциональный организатор» К.
Бернар).
1.1. Нервная система - физиологическая система, специализированная на приеме,
переработке и сохранении информации об окружающем мире и внутренней
среде организма и ее использовании для взаимодействия целостного организма
с внешней средой.
❖ Главные функции нервной системы: 1) регуляция функций различных
структур организма; 2) интеграция частей организма в единое целое; 3)
взаимодействие организма с окружающей средой (в том числе социальной)
и адаптация к ней; 4) формирование целенаправленного поведения; 5) по-
знание окружающего мира и самого организма.
❖ Отделы нервной системы.
• Анатомически выделяют: 1) ЦНС, включающую в себя спииной и го-
ловной мозг; 2) периферическую нервную систему, включающую в себя
нервы и ганглии (узлы).
• Физиологически выделяют: 1) соматическую (анимальную) нервную
систему, иннервирующую скелетные мышцы и регулирующую их со-
кращение; 2) автономную (вегетативную) нервную систему, ре-
гулирующую деятельность внутренних органов, сосудов и желез; 3)
сенсорную (афферентную) нервную систему, обеспечивающую поступ-
47
ление в ЦНС информации, используемой для регуляции соматических,
вегетативных функций, процессов адаптации и ВНД; 4) кору больших
полушарий и подкорковые образования, обеспечивающие высшую ре-
гуляцию вегетативных и соматических функций, процессов адаптации, а
также целенаправленное поведение и субъективный мир человека.
1.2. функциональные уровни организации нервной системы.
• i' Клеточный уровень: нейроны и глиоциты.
❖ Нейронные цепи: рефлекторный принцип, нервные центры.
❖ Системная организация: взаимодействие сенсорной, ассоциативной, моду-
лирующей, интегративно-пусковой систем.
2. Общая характеристика клеточного функционального уровни нервной системы.
2.1. Нейрон как функциональная единица нервной системы.
❖ Виды нейронов.
• По характеру отростков: псевдоуниполярные (например, нейроны спи-
нальных узлов), биполярные (например, обонятельные нейроны-рецепторы, бипо-
лярные нейроны сетчатки, нейроны спирального ганглия), мультиполярные (на-
пример, мотонейроны).
• По звеньям рефлекторного пути: афферентные, вставочные (интернейро-
ны), эфферентные.
• Релейные (проекционные) нейроны: нейроны сенсорных путей с длинными
аксонами, переключающиеся, например, в ядрах таламуса.
• Импульсные (генерируют потенциал действия) и неимпульсные нейроны (пе-
редают влияние с помощью локальных потенциалов: рецепторного, постсинаптиче-
ского).
• По виду медиатора: холинергические, адренергические, дофаминерги-
ческие, серотонинергические и другие нейроны.
• Нейросекреторные нейроны - способны трансформировать нервный
импульс в секрецию гормонов (например, часть иейроиов гипоталамуса).
• По модальности - характеру воспринимаемого и передаваемого сигна-
ла: механорецепторные, хеморецепторные, зрительные, обонятельные,
вкусовые и др.
❖ Структурно-функциональные элементы нейрона: рецепторы, отростки,
сома, синапсы.
2.2. Клетки нейроглии (Р. Вирхов, 1856).
❖ Виды глиальных клеток: астроциты, олигодендроциты, леммоциты, микро-
глиоциты, эпендимоциты.
❖ Нейроны и глиоциты действуют как единый функциональный комплекс,
доля глиоцитов в этом комплексе наиболее высокая (10:1) в ЦНС человека.
3. Функции сенсорных рецепторов.
3. 1. Рецепторы - специализированные структуры, воспринимающие раздражители
внешней среды и внутренней среды организма и трансформирующие энергию
раздражения в рецепторный потенциал, который затем преобразуется в нервные
импульсы.
32. Классификация рецепторов.
48
• •• По структуре рецепторов: свободные нервные окончания (болеаые, температурные, не-
которые механорецепторы), инкапсулированные нервные окончания (тельца Пачини,
Мейсснера н Руффини, колбы Краузе, мышечные веретена, сухожильные рецепторы Голь-
джи), спецнализнроаанный нейрон (например, фоторецепторы, центральные хеморецепто-
ры), эпителиальная волосковая клетка (например, слуховые н вестибулярные рецепторы).
❖ По расположению в организме: экстерорецепторы и интерорецепторы, цен-
тральные и периферические рецепторы.
❖ В зависимости от вида раздражителя: механо-, хемо-, термо-, фоторецеп-
торы, ноцицепторы.
❖ По психофизиологическому критерию: зрительные, слуховые, вкусовые,
обонятельные, тактильные.
❖ По степени специфичности: моно- и полимодальиые рецепторы.
❖ По скорости адаптации: быстро- и медленно адаптирующиеся, смешан-
ные.
❖ По электрофизиологическому критерию: первичные рецепторы (рецептор-
ный потенциал и потенциал действия возникают в одной клетке) и вторич-
ные рецепторы - РП возникает в волосковой клетке или фоторецепторе, а
ПД - в нейроне (например, зрительные, слуховые, вестибулярные).
3.3. Рецепторы как сенсорные преобразователи.
❖ Первичное преобразование энергии раздражителя состоит в отборе раздра-
жителей - рецепторы взаимодействуют преимущественно с теми раздражи-
телями, к которым они эволюционно приспособлены; на этом этапе во мно-
гих рецепторах происходит усиление сигнала.
❖ Формирование рецепторного потенциала связано с открытием ионных ка-
налов и появлением ионных токов, среди которых преобладает входящий в
клетку Na+-TOK, приводящий к деполяризации мембраны рецептора. (Ампли-
туда деполяризации находится между мембранным потенциалом покоя н нулевым потен-
циалом.)
❖ Превращение рецепторного потенциала в потенциал действия происходит,
если величина РП достаточна для деполяризации мембраны до КУД в тех
участках мембраны, в которых возможно возникновение ПД.
3.4. Кодирование силы раздражителя в рецепторе и афферентном нейроне: в
рецепторе сила раздражителя кодируется амплитудой рецепторного потенциа-
ла, а на аксонном выходе афферентного нейрона (т.е. в аксонном холмике н аксоне) —
частотой нервных импульсов. (Частотное кодирование осуществляется с участием быст-
рых потенциалзависимых К+-каналов, которые открываются небольшой деполяризацией и при
ее увеличении аыше -50 мВ - инактивируются. В связи с этим слабые раздражители открыва-
ют эти каналы, и выходящий К*-ток, препятствующий формированию ПД. уменьшает их часто-
ту, а сильные раздражители инактивируют эти каналы, поэтому частота ПД возрастает.)
3.5. Свойства рецепторов и регуляция их функции.
*1* Высокая возбудимость рецепторов (например, пороговая сила для слухового ре-
цептора 5-Ю'18 Дж).
❖ Снижение возбудимости (адаптация рецепторов) возникает при действии
сильных и (или) длительных раздражителей. Одним из механизмов адапта-
ции связан с накоплением Са2+ в рецепторе, которые активируют Са2+-
зависимые К+-каналы; выходящий из клетки К+-ток усиливает процесс ре-
поляризации, в результате чего уменьшается амплитуда РП.
49
❖ Повышение возбудимости (сенситизация) рецепторов при длительном дейст-
вии раздражителей возможно, например, в болевых рецепторах.
❖ Спонтанная активность рецепторов связана с нестабильностью их МП;
имеющаяся при этом флюктуирующая деполяризация в случаях достиже-
ния КУД, генерирует потенциалы действия (спонтанную активность ней-
ронов).
❖ Нейрогуморальная регуляция возбудимости рецепторов. Нервная регуля-
ция рецепторов связана с эфферентной иннервацией рецепторов тормоз-
ными и возбуждающими нейронами из вышележащих центров (например,
слуховых рецепторов из нейронов верхней оливы). Действие гормонов также может
изменить чувствительность рецепторов (например, повышение возбудимости зри-
тельных и слуховых рецепторов под действием адреналина).
4. Синапсы («Концепция синапса - сердце нейронной доктрины». С. Палей)
4.1. Синапсы - специализированные межклеточные контакты, обеспечивающие пе-
редачу возбуждающих или тормозных влияний от нейрона на иннервируемую
клетку (нервную, мышечную, железистую).
4.2. Классификация синапсов.
❖ По виду соединяемых клеток: межнейронные, иейроэффекторные, нейро-
рецепторные.
❖ По характеру действия: возбуждающие и тормозные.
❖ По способу передачи сигнала: химические, электрические, смешанные.
❖ По природе медиатора: адренергические, холинергические, глутаматерги-
ческие и др.
4.3. Химические синансы (составляют подавляющее большинство синапсов нервной
системы человека).
❖ Структурные элементы синапса (рис. 13).
• Пресинаптическое окончание имеет везикулы разной формы и величи-
ны (40 - 300 нм) с молекулами медиатора (а также митохондрии, агранулярную
ЭПС, микрофиламенты, цитоскелет). (Везикулы образуются в гладкой ЭПС и аппарате
Гольджи и с помощью аксонного транспорта поступают в пресинапс.)
Количество медиатора (несколько тысяч молекул, концентрация ~ 100
ммоль/л), содержащегося в одной везикуле, называется квантом ме-
диатора. (В везикуле есть также АТФ, ионы, ферменты н др. белки, Н*-насос в
мембране.)
ф- В пресинапсе имеется два пула везикул: 1) небольшой пул (10 - 15)
везикул находится вблизи пресинаптической мембраны, содержит
запас доступного медиатора, который может быстро освободиться;
2) большой пул находится в удалении от пресинаптической мем-
браны, содержит мобилизационный запас медиатора, который мо-
жет быстро (за 5 - 12 с) переместиться в малый пул.
• Синаптическая щель (-50 нм) содержит поперечные гликопротеидные
филаменты (глнкокаликс).
• Постсинаптическая мембрана имеет рецепторы, с которыми соединяют-
ся молекулы медиатора.
50
•> Механизмы синаптической передачи формируют два процесса - превраще-
ние электрического сигнала в химический и химического сигнала обратно в
электрический.
• Превращение электрического сигнала в химический.
Ф- Поступление нервного импульса в пресинаптическое окончание от-
крывает в плазмолемме Са2+-каналы(Ы-типаиР-типа).
Ф’ Вход Са стимулирует экзоцитоз везикул (квантов медиатора). (Для эк-
зоцитоза одной везикулы надо 4 Са2*, для этого концентрация Са2* должна быть
более 100 мкмоль/л. Имеется два варианта экзоцитоза: 1) классический экзоцитоз
с полным слиянием везикулы с пресииаптической мембраной и выходом всего
содержимого медиатора; 2) экзоцитоз без полного слияния с образованием вре-
менного канала и частичным освобождением медиатора.)
ВД-._____/ . \ Рис. 13. Схематическое изображе-
ние химического синапса.
1 - пресинаптическое окончание, 2
- синаптическая щель, 3 - постси-
наптическая мембрана, ПД - по-
тенциал действия, Р - рецептор
постсинаптической мембраны,
ВПСП - возбуждающий постси-
наптический потенциал, ТПСП -
тормозной постсинаптический
потенциал.
ВПСП
ТПСП
Ф- Характеристика медиаторов.
По химическому строению: ацетилхолин, моноамины (норадре-
налин, серотонин, дофамин), аминокислоты (глутамат, аспартат,
глицин, ГАМК), пептиды (например, вещество Р, ВИП и др.).
Один и тот же медиатор в разных синапсах может передавать
как возбуждающее, так и тормозное влияние (например, ацетилхо-
лин, норадреналин).
Есть медиаторы, передающие, как правило, тормозное влияние
(например, ГАМК в постнатальном периоде, глицин) или возбуж-
дающее влияние (например, глутамат).
Доказана возможность выделения в одном синапсе более чем
одного медиатора (наиболее частая совместная локализация медиато-
ров: ацетилхолнн+ГАМК - в нейронах нижней оливы, мотонейроны спин-
ного мозга, амакриновые клетки сетчатки и др.; ГАМК+глицин - в нейронах
спинного мозга и мозжечка; ГАМК + глутамат - в нейронах гиппокампа,
зубчатой фасции, пирамидных нейронах). (Синапс ГАМК + глутамат при
слабых сигналах работает как ГАМКергический, а при сильных - как глу-
таматергический. Кроме того, предварительное выделение ГАМК подавляет
51
спонтанную активность постсинаптического нейрона - «обнуление»,
«сброс», что улучшает восприятие слабых сигналов.)
Кроме медиатора, в синаптическую щель выделяются полипеп-
тиды, которые модулируют действие медиатора (например, эн-
кефалин, холецистокинин, соматостатин и др.).
Ф’ Диффузия медиатора через синаптическую щель (- 50 нм) к пост-
синаптической мембране осуществляется за десятые доли милли-
секунды.
• Превращение химического сигнала обратно в электрический (возбуж-
дающий и тормозной постсинаптические потенциалы).
Ф- Секретируемый медиатор действует на рецепторы постсинаптиче-
ской мембраны.
Ф’ Постсинаптические рецепторы являются сложными белками, со-
стоящими из нескольких (до пяти) субъединиц. Они обычно скон-
центрированы в отдельных местах мембраны, образуя кластеры
(пучки). (Рецепторы к медиатору имеются также вне постсииаптической мем-
бране - на пресннаптической мембране, соме н дендритах, на клетках глии н ка-
пиллярах.)
Ф’ Рецепторы разделяют на две группы - ионотропные н метаботроп-
ные.
Ионотропные рецепторы (рецепторы «быстрого ответа») имеют в
своем составе ионный канал и передают сигнал на постсинап-
тическую клетку с помощью изменения ионной проницаемости
мембраны, формируя ВПСП и ТПСП (например, Н-
холинорецепторы, ГАМК*- и ГАМКс-рецепторы, глициновые, НМДА-
глутаматные н б-НТ3-серотониновые рецепторы).
Метаботропные рецепторы (рецепторы «более медленного ответа»)
передают сигнал на постсинаптическую клеткус-номощью G-
белка мембраны и вторых посредников (аналогично действию гор-
монов). Они также могут изменять активность ионных каналов в
клетке, действуя на них опосредованно - через G-белок или (и)
вторые посредники (см. тему 8). (В данной теме рассматриваются
только ионотропные рецепторы.)
При действии медиатора в ионотропных рецепторах происхо-
дит открывание ионных каналов и возникновение ионных то-
ков, формирующих возбуждающий или тормозной постсинап-
тические потенциалы (ВПСП и ТПСП) (рис. 14).
□ Формирование ВПСП в возбуждающих синапсах.
о Ионные токи, формирующие ВПСП, имеют в качестве
преобладающего компонента входящий в клетку Na+-
ток.
о Схема ВПСП: расположен в субпороговой области,
имеет фазы деполяризации и реполяризации; его па-
раметры - амплитуда несколько мВ, длительность от 5
- 10 мс до сотен мс.
52
о Функциональная роль: ВПСП характеризует собой ло-
кальное возбуждение, суммация ВПСП деполяризует
мембрану до критического уровня и является причи-
ной возникновения нервного импульса (ПД) в постси-
наптической клетке.
о Миниатюрные Постсинаптические потенциалы возникают при
спонтанном выделении 1-2 квантов медиатора в секунду, их ам-
плитуда - 0,15 мВ, они поддерживают возбудимость синапса в
условиях функционального покоя.
Рис. 14. Схема постсинаптических потенциалов.
ВПСП - возбуждающий постсинаптический по-
тенциал, ТПСП - тормозной постсцнашический
потенциал, МПП - мембранный потенциал покоя,
КУД - критический уровень деполяризации.
□ Формирование ТПСП в тормозных синапсах...
о Постсинаптическое торможение.
- Ионные токи, создающие ТПСП: входящий в
клетку СГ-ток или.(и) выходящий из клетки К+-
ток; ТПСП представляет собой обычно гиперпо-/
ляризацию мембраны амплитудой -0,24 мВ (щЖ их
суммации до 10 мВ), продолжительностью от 2 - 10
мс до сотен мс.
- Функциональная роль: ТПСП уменьшают возбу-
димость нейрона' и вероятность возникновения
ПД.
о Пресинаптическое торможение.
- Формируется в аксо-аксональных синапсах, кото-
рые образуют тормозные нейроны на аксонных
окончаниях возбуждающих афферентных нейро-
нов. Обнаружены в ядрах тройничного нерва, зад-
них столбах спинного мозга, некоторых ядрах та-
ламуса.
ГАМК открывает СГ-канал ГАМК-рецептора. Вы-
ход СГ из аксона сопровождается деполяризацией
мембраны. ПД, идущий к возбуждающему синап-
53
су через этот участок, распространяется пассивно
с уменьшением амплитуды (так как вход Na* в первой
фазе ПД будет при открытых СГ-каиалах будет нейтрализо-
ван сопутствующим входом СГ - связь разных зарядов), а,
следовательно, уменьшается выброс медиатора в
возбуждающем синапсе. (Снижение амплитуды ПД
всего иа 5 % приводит к уменьшению ВПСП на 50 %.)
Функциональная роль: позволяет затормозить от-
дельные синаптические входы на нейроне, регу-
лирует приток сенсорной импульсацин в ЦНС.
Рис. 15. Схематическое изо-
бражение постсииаптичес-
кого торможения (по 3. Зи-
бернагал, А. Десполулос, ,
1991). А - без торможения, Б -
пресинаптическое тормо-
жение, а - аксоаксональный
сниапс, б - синапс, функция
которого тормозится.
• Объемное распространение медиатора. Синапсы «закрытого» (глиальными оболочка-
ми) типа имеются в небольшом количестве. Большинство синапсов «открытого» типа,
медиатор которых распространяется в трехмерном диффузионном потоке. Он действу-
ет иа рецепторы пресииаптической мембраны этого же нейрона (аутокрниио), на ре-
петиторы других нейронов и глиоцитов (паракрииио) и даже иа клетки капилляров
(участвует в регуляции локального кровотока).
❖ Саморегуляция синаптической передачи осуществляется с участием рецеп-
торов пресииаптической мембраны. Например, в адренергических синапсах
на пресииаптической мембране имеются аг- и Р-адренорецепторы. Высокие
концентрации норадреналина в синаптической щели через аз-
адренорецепторы тормозят выделение медиатора из пресинаптического
окончания, низкие концентрации норадреналина через Р-адренорецепторы
стимулируют экзоцитоз медиатора.
4.4. Основные свойства химических синапсов.
❖ Одностороннее проведение возбуждения связано с тем, что медиатор вы-
деляется из пресинаптического окончания, а постсинаптические потенциа-
лы формируются только на постсинаптической мембране.
❖ Синаптическая задержка (0,5 -1,0 мс)’ обусловлена здесь самой низкой ско-
ростью проведения возбуждения (- 2 мм/с).
❖ Низкая лабильность, равная 100 - 150 передаваемым импульсам в секунду
(в сравнении с 1000 импульсами в аксоне).
54
Трансформация ритма возбуждения связана с необходимостью суммации
ВПСП (в нервно-мышечном синапсе скелетных мышц отсутствует).
Синаптическая пластичность - свойство синапсов изменять (увеличивать
или уменьшать) эффективность передачи сигнала с нейрона на нейрон в зави-
симости от характера их предшествующей активации.
• Увеличение эффективности синаптической передачи может быть в ви-
де кратковременной и долговременной потенциации.
Ф Кратковременная потенциация (КП) - кратковременное (от 1 с до
2-5 мин) повышение амплитуды ВПСП в течение н после кратко-
временной ритмической высокочастотной стимуляции синапса. Ее
механизмы связывают с накоплением в пресннаптнческом оконча-
нии остаточного Са2+, выделенного во время предшествующих пе-
редач нервного импульса через данный синапс. Предполагают так-
же активацию экзоцитоза медиатора в результате фосфорилирова-
ния Са2+-завнсимой протеннкиназой белков синаптических везикул
(синапсина I). В результате увеличивается экзоцитоз медиатора. КП
является одним из механизмов возникновения феномена проторе-
ния пути (см. тему 6; 1.3).
"Ф’ Долговременная потенциация (ДП) - увеличение эффективности
синаптической передачи в некоторых отделах ЦНС (например, гиппо-
кампе, коре больших полушарий) в ответ на высокочастотную стимуля-
цию (например, 100 - 400 Гц в течение секунд-минут). Ее электрофизиоло-
гической основой является длительное (десятки минут - дни) увеличе-
ние амплитуды ВПСП, ее метаболической основой является вклю-
чение регуляторных систем клетки - кальциевой, циклического
АМФ, инозитолполифосфатной (подробно они рассмотрены как вторые
посредники в теме 9; задание 1:2.2). Долговременная потенциация имеет
важное значение в процессах обучения, механизмах памяти, услов-
ных рефлексах.
• Уменьшение эффективности синаптической передачи - кратковремен-
ная и долговременная синаптическая депрессия.
Ф Кратковременная депрессия развивается параллельно кратковре-
менной потенциации и взаимодействует с ней. Ее механизмы свя-
зывают с истощением в пресннаптнческом окончании резерва
медиатора.
"Ф’ Долговременная депрессия (ДД) развивается при низкочастотной
стимуляции (1 - 5 Гц в сравнении с 100 - 400 Гц для долговременной по-
тенциацин), ее развитию способствует несовпадение по времени ак-
тивации пре- и постсннаптического нейронов, а также невысокие
концентрации Са2+. ДД лежит в основе угасания условных рефлек-
сов и забывания. (ДД может смениться долговременной потенцнацией при
переходе от низко- к высокочастотной стимуляции, например, при восстановле-
нии условных рефлексов).
55
❖ Высокая чувствительность синапсов к фармакологическим веществам
(агонисты и антагонисты синаптических рецепторов, ингибиторы и актива-
торы ферментов синтеза и распада медиаторов).
❖ Трофическая функция синапсов осуществляется в результате неквантовой утечки медиато-
ра в синапсе и выделения наряду с медиатором полипептидов-спутников, оказывающих
трофическое действие на иннервируемую клетку.
4.5. Электрические синапсы являются вариантом щелевых межклеточных контак-
тов с низким омическим сопротивлением, они более характерны для пренаталь-
ного периода.
❖ Особенности структуры: узкая синаптическая щель (2-4 им) с попереч-
ными белковыми мостиками, имеющими каналы, пропускающими элек-
тролиты и низкомолекулярные вещества.
❖ Механизмы передачи возбуждения: токи ПД свободно проникают в пост-
сннаптическую клетку и деполяризуют в ней мембрану до критического
уровня (нет необходимости в медиаторе).
❖ Основные свойства: быстрая и надежная передача возбуждения, высокая
лабильность и низкая утомляемость, однако в них плохо выражена одно-
сторонность передачи и возможность регуляции передачи, среди них прак-
тически нет тормозных синапсов.
5. Возникновение возбуждения (потенциала действия) в нейроне.
5.1. В первых афферентных нейронах причиной нервного импульса является рецеп-
торный потенциал, в других нейронах - ВПСП.
5.2. Для возникновения ПД в нейроне необходима пространственная и временная
суммация ВПСП.
5.3. Нервный импульс (ПД) возникает в нейроне, если суммация возбуждающих
влияний (ВПСП) алгебраически преобладает над суммацией тормозящих влия-
ний (ТПСП), и это преобладание может деполяризовать мембрану до критиче-
ского уровня.
5.4. Наиболее легко ПД возникает в аксонном холмике, где наиболее низкий поро-
говый потенциал (-10 мВ) (триггерная или пусковая роль аксонного холмика).
53. Распространение ПД из холмика происходит по аксону (передача возбуждающего
или тормозного влияния на иннервируемую клетку) и в тело нейрона (сигнал для изменения
метаболизма нейрона в связи с импульсной его активностью).
б. Проведение возбуждения в нервных волокнах.
6.1. Структурная характеристика нервных волокон: имеют осевой цилиндр (нервный
отросток) и глиальную оболочку, образованную леммоцитами в периферической нервной сис-
теме и олигодендроцитами в ЦНС. Если мембрана глиальной клетки одним слоем окружает
осевой цилиндр, то образуются безмиелиновые волокна, если же она многократно спирально
закручивается вокруг осевого цилиндра и образуют слой миелина (миелиновую муфту), форми-
руются миелиновые волокна. Участки осевого цилиндра между миелиновыми муфтами (межуз-
ловыми сегментами) называются перехватами Ранвье.
6.2. Классификация нервных волокон (по Эрлангеру - Гассеру, 1937).
• ❖ Аа - миелиновые афферентные волокна от механорецепторов кожи, мы-
шечных и сухожильных рецепторов, эфферентные волокна к скелетным
мышцам (d= 12-20мкм, V = 70- 120м/с).
56
❖ Ар - миелиновые афферентные волокна от кожных рецепторов прикосно-
вения н давления н от части мышечных рецепторов (d = 5 -12 мкм, V = 30 - 70
м/с).
❖ Ау-миелиновые эфферентные волокна к мышечным рецепторам (d = з - 5
мкм, V = 15 - 30 м/с).
❖ А5-миелиновые афферентные волокна от части тактильных, температур-
ных н болевых рецепторов кожи, а также суставных рецепторов (d = 2 - 5
мкм, V = 12 - 30 м/с).
4 В - миелиновые преганглнонарные волокна вегетативной нервной системы
(d = 1 - 3 мкм, V = 5 - 14 м/с).
❖ С - немиелиновые постганглионарные волокна вегетативной нервной сис-
темы, афферентные волокна от некоторых болевых, тепловых н висцераль-
ных рецепторов (d = 0,3 -1,3 мкм, V = 0,5 - 2,3 м/с).
63. Проведение возбуждения но нервному волокну.
❖ Распространение локальных потенциалов (рецепторного, постсннаптиче-
ского, препотенциала) происходит электротоническн на малые расстояния
• (до 1 - 2 мм). Прн этом используются физические («кабельные») свойства
волокна (сопротивление и емкость его мембраны н цитоплазмы) н практи-
чески отсутствует реакция ионных каналов мембраны.
❖ Проведение потенциала действия происходит с использованием как элек-
тротонического, так н активного механизма (реакции ионных каналов и ге-
нерации ПД).
• Непрерывный тип проведения ПД происходит в немнелиновых волок-
нах С: начинается с этапа электротоннческого проведения возникшего
ПД в соседний участок, где происходит деполяризация мембраны до
КУД н возникает «новый» ПД н т.д. (рнс. 7; с. 38)
Рис. 16. Сальтаториое распространение потенциала действия в миелиновых нерв-
ных волокнах. А - возникновение ПД в перехвате Ранвье среднего участка волок-
на; Б - распространение ПД в обе стороны от места возникновения; стрелками по-
казаны токи на электротоиическом этапе распространения ПД.
57
• Сальтаторный (скачкообразный) тип проведения ПД происходит в миели-
новых волокнах (рис. 16): возникший в перехвате Ранвье ПД проходит
электротонически межперехватный сегмент (миелиновую муфту), деполя-
ризует мембрану в соседнем перехвате, где генерируется «новый» ПД.
Если соседние перехваты поражены, то ПД, имеющий большую ампли-
туду (~ 90 мВ), может вызвать ПД в 4 - 5-м перехвате (гарантийный
фактор нейрона).
❖ Преимущества сальтаторного типа проведения ПД: наибольшая экономич-
ность, так как возбуждается только мембрана перехватов (составляет 1 % площади
волокна); большая скорость проведения в связи с электротоническим распро-
странением потенциала в области миелиновых муфт.
6.4. Особенности проведения возбуждения по нервным волокнам.
❖ Двустороннее проведение ПД от места возникновения.
* ** Изолированное проведение возбуждения (обусловлено тем, что петли тока в меж-
клеточной жидкости нерва почти не проникают в невозбуждеиные волокна нерва из-за
большого сопротивления их оболочек).
❖ Возможность функционального блока проведения возбуждения (Н.Е. Введен-
ский, 1901). При действии различных факторов на нерв (анестетиков, гипоксии,
воспаления, охлаждения, электротока), вызывающих длительную деполяризацию
клеточной мембраны, возникает полный блок проведения нервных импуль-
сов (состояние парабиоза), связанный с инактивацией более 50% натриевых
каналов. При этом протяженность парабиозического участка должна пре-
высить постоянную длины мембраны (Хт), иначе ПД может распростра-
ниться через этот участок электротонически. После действия этих факторов
проведение возбуждения по волокнам нерва восстанавливается, если не
произошли грубые структурные изменения. (Это свойство нервных волокон
широко используется в медицинской практике, например, при местной анестезии с помо-
щью новокаина).
❖ Большая скорость проведения возбуждения (от 0,5 до 120 м/с).
❖ «Изумительно долгая неутомляемость нерва» (Н.Е. Введенский, 1883) обуслов-
лена большой экономичностью ПД, при котором используется только одна
миллионная часть запасов ионных градиентов клетки.
6.5. Проведение возбуждения в нервных стволах В естественных условиях каждое волокно нерва воз-
буждается от своего источника (например, эфферентное - от аксонного холмика, афферентное - от
рецептора), и ПД в них проводятся асинхронно. Кроме того, чувствительные и двигательные волокна
проводят импульсы в противоположном направлении. Суммарная электрическая активность нерва (ней-
рограмма) создается электрической активностью составляющих его волокон и зависит от числа возбу-
жденных волокон, степени шунтирования местных токов невозбужденными волокнами, син-
хронности проведения ПД в волокнах, методики отведения электрической активности нерва. В связи с
этим анализ нейротраммы представляет трудную задачу.
7. Трофическая функция нейрона.
7.1. Аксонный транспорт (осуществляет передвижение вдоль аксона нейроплазмы, органелл,
макромолекул и низкомолекулярных веществ).
❖ Быстрый аксонный транспорт происходит в двух направлениях.
• Антероградный транспорт (от тела клетки к аксонным окончаниям, V =
250 - 400 мм/сутки) доставляет везикулы, митохондрии, гликопротеины,
ферменты, липиды, медиаторы и другие вещества. С промежуточной
58
скоростью (4 - 50 мм/сутки) транспортируются митохондриальные и
миозиноподобные белки, актии, клатрин, кальмодулин.
• Ретроградный транспорт (от аксонных окончаний к телу нейрона, V = 200
- 300 мм/сутки) доставляет везикулы, содержащие остатки разрушенных
структур, фрагменты мембран, нейроростовые факторы, лизосомальные
ферменты, ацетилхолинэстеразу, вирусы, столбнячный экзотоксин.
• Механизм: транспорт осуществляется вдоль микротрубочек с помощью
белка кинозина, связывающего органеллу с микротрубочкой и преобра-
зующего энергию АТФ в механическую работу.
❖ Медленный аксонный транспорт происходит в антероградном направлении
и состоит из передвижения всего столба аксоплазмы (V = 1 — 2 мм/сутки);
этим транспортом перемещаются образованные в ЭПС белки микротрубо-
чек и микрофиламентов, каналов, насосов, ферментов цитозоля, т-белки
(сборка микротрубочек) и др.
❖ Функциональная роль аксонного транспорта.
• Поддержание структуры и функции аксона и его окончаний, аксонный
рост и образование новых синапсов.
• Трофическое влияние на иннервируемую клетку: регуляция ее обмена
веществ, размножения, дифференцировки.
7.2. Импульсное нейротрофическое влияние. Генерация, распространение и передача нервного
импульса (ПД) изменяет не только электрические свойства клеточной мембраны, но и функ-
ционально-структурные свойства клетки в целом. Это обусловлено тем, что в реализации био-
электрических процессов участвуют биоэнергетические, синтетические, пластические, ультра-
структурные, генетические процессы. Связывающим звеном биоэлектрических и обеспечиваю-
щих их процессов являются вторые посредники (цАМФ, ИФ3, ДАГ, NO, Са2+), образующиеся
при передаче в синапсах как с ионотропными, так и особенно, с метаботропными рецепторами.
Примером нарушения импульсного трофического влияния, возникающего при гиподинамии ор-
ганизма (аксонный транспорт сохранен!), является гипотрофия скелетных мышц (ионотропные
рецепторы в синапсе) и сердца (метаботропные рецепторы). В здоровом организме импульсное
трофическое влияние и аксонный транспорт взаимодействуют друг с другом.
8. Функциональная роль нейроглии.
8.1. Барьерная (разграничительная) функция.
❖ Образование гемато-эицефалического барьера: уплощенные отростки аст-
роцитов образуют периваскулярную мембрану.
❖ Образование нейроликворного барьера, отделяющего нейроны от ликвора,
осуществляют эпендимная глия и отростки астроцитов.
❖ Образование перинейрональных оболочек вокруг тела нейрона и области
синапсов обеспечивают астроциты.
❖ Образование гематоликворного барьера в сосудистых сплетениях мозга
осуществляют хориоидные эпендимоциты.
8.2. Метаболическая и регуляторная функции осуществляются астроцитами и
олигодендроцитами.
❖ Регулируют метаболизм нейронов.
❖ Обеспечивают связь между нейронами и локальным кровотоком.
❖ Поддерживают внеклеточную концентрацию К+, поглощая его избыток,
связанный с высокой импульсной активностью нейронов.
59
* ** Участвуют в метаболизме медиаторов (ГАМК, глутамата, катехоламинов и др.),
удаляя их из синаптической щели и транспортируя в нейроны.
❖ Участвует в образовании памяти, синтезируя некоторые белки памяти.
❖ Астроциты образуют нейроростовые факторы и оказывают сильное влия-
ние на развивающиеся нейроны.
8.3. Образование миелиновых и немиелиновых оболочек нервных волокон осуще-
ствляют леммоциты в периферической нервной системе и олигодендроциты в
ЦНС.
8.4. Защитная (иммунная) функция осуществляется астроцитами и микро-
глиоцитами: они имеют свойства антигенпредставляющих клеток, фагоцитар-
ную активность, вырабатывают цитокины.
8.5. Опорная функция: астроциты формируют опорный каркас ЦНС, внутри кото-
рого располагаются нейроны, другие глиоциты и волокна, в эмбриогенезе на-
правляют миграцию развивающихся нейронов по особым каналам, формируе-
мым отростками астроцитов.
9. Профильные материалы для студентов лечебного факультета. Структурно-
функциональные особенности нервных клеток н глнн прн старении организма.
9.1. Структурные изменения нейронов.
❖ Снижение общего числа нейронов (до 40 - 70 %).
• > Дистрофические процессы в нейронах и их отростках: вакуолизация, накопление
липидов и пигмента липофусцина в цитоплазме, сегментарная демиелинизация
аксонов.
❖ Уменьшение числа аксодендритических синапсов и увеличение числа дендроден-
дритических синапсов.
❖ Уменьшение количества активных синапсов и резервных синапсов (шипикового
аппарата).
9.2. Функциональные изменения нейронов.
❖ Снижение синтеза АТФ и активности К4, Na+-Hacoca.
❖ Снижение лабильности нейрона.
❖ Замедление проведения возбуждения через синапс.
Ослабление синтеза медиатора в синапсах (например, ацетилхолина), компенса-
торно снижается и активность ферментов их распада (например, ацетилхолинэ-
стераза).
❖ Снижение синтеза белка, диффузионных итранспортных процессов, в том числе
аксонного транспорта в нейронах с накоплением «пигмента старости» липофус-
цина.
❖ Снижение функционального резерва нейронов и их трофического алияния.
9.3. Структурно-функциональные изменения глии при старении организма.
Ф Относительное увеличение количества глиальных клеток по отношению к нейро-
нам.
❖ Умеренное снижение функций микроглии, компенсаторная гиперфункция астро-
цитов.
❖ Усиление интенсификации белкового и энергетического обменов в клетках глии,
их трофической функции: передачи пластических веществ от глии к нейрону.
❖ Участие глии в удалении пигмента липофусцина из нейронов.
❖ Увеличение обратного захвата глиоцитами медиаторов нейронов.
❖ Усиление роли глии в образовании и закреплении временных связей.
60
10. Профильные материалы для студентов педиатрического факультета.
10.1. Структурно~функционалъные особенности ЦНС в антенатальном периоде.
❖ У плода количество нейронов ЦНС достигает максимума к 20 - 24-й неделе и ос-
тается в постнатальном периоде без резкого снижения до пожилого возраста. Ней-
роны имеют малые размеры и площадь синаптической мембраны.
❖ Аксоны развиваются раньше дендритов, отростки нейронов интенсивно растут и
ветвятся. Наблюдается увеличение длины, диаметра и миелинизации аксонов к
концу антенатального периода.
❖ Филогенетически старые пути миелинизируются раньше, чем филогенетически
новые; например, вестибуло-спинальные пути с 4 мес. внутриутробного развития,
руброспииальиые пути с 5 - 8 мес., пирамидные пути после рождения.
Имеется равномерное распределение Na+ и К+-каналов в мембране миелиновых и
немиелиновых волокон.
❖ Возбудимость, проводимость, лабильность нервных волокон значительно ниже,
чем у взрослых.
❖ Синтез большинства медиаторов начинается в период внутриутробного развития.
ГАМК в антенатальном периоде яаляется возбуждающим медиатором и через
Са2+-механизм оказывает морфогенные эффекты - ускоряет рост аксонов и денд-
ритов, синаптогенез, экспрессию циторецепторов.
К моменту рождения заканчивается процесс дифференциации нейронов ядер про-
долговатого и среднего мозга, моста.
❖ Имеется структурно-функциональная незрелость глиальных клеток.
10.2. Особенности ЦНС в периоде новорожденности.
❖ Возрастает степень миелинизации нервных волокон, их количество составляет 1/3
уровня взрослого организма (например, миелинизирован руброспинальиый путь).
❖ Уменьшается проницаемость клеточных мембран для ионов. Нейроны имеют бо-
лее низкую амплитуду мембранного потенциала - около 50 мВ (у взрослых - 70
мВ).
❖ На нейронах синапсов меньше, чем у взрослых, мембрана нейрона имеет рецепто-
ры к синтезируемым медиаторам (ацетилхолин, ГАМК, серотонин, норадреналин
и дофамин). Содержание медиаторов в нейронах мозга новорожденных низкое и
составляет 10 - 50 % от такового у взрослых.
Отмечается развитие шипикового аппарата нейронов. ВПСП и ТПСП имеют
ббльшую длительность и меньшую амплитуду, чем у взрослых. Количество тор-
мозных синапсов на нейронах меньше, чем у взрослых.
Повышается возбудимость корковых нейронов.
Исчезает (точнее, резко уменьшается) митотическая активность и возможность ре-
генерации нейронов. Продолжается пролиферация и функциональное созревание
глиоцитов.
10.3. Особенности ЦНС в грудном возрасте.
❖ Созревание ЦНС быстро прогрессирует. Наиболее интенсивная миелинизация
нейронов ЦНС происходит в конце первого года после рождения (например, к 6
мес. завершается миелинизация нервных волокон полушарий мозжечка).
Возрастает скорость проведения возбуждения по аксонам.
Наблюдается уменьшение продолжительности ПД, происходит укорочение абсо-
лютной и относительной рефрактерных фаз нервных волокон (длительность абсо-
лютной рефрактерности 5 - 8 мс, относительной - 40 - 60 мс в раннем постна-
тальном онтогенезе, у взрослых соответственно 0,5 - 2 и 2 - 10 мс).
Кровоснабжение мозга у детей относительно больше, чем у взрослых.
61
10.4. Особенности развития ЦНС в другие возрастные периоды.
Структурно-функциональные изменения в нервных волокнах.
• Увеличение диаметров осевых цилиндров (к 4 - 9 годам).
• Миелинизация во всех периферических нервных волокнах близка к заверше-
нию к 9 годам, а пирамидных путей заканчивается к 4 годам.
• Происходит концентрация ионных каналов в области перехватов Ранвье, рас-
стояние между перехватами увеличивается. Непрерывное проведение возбуж-
дения сменяется сальтаторным, скорость его проведения после 5-9 лет почти
не отличается от скорости у взрослых (50 -70 м/с).
• Отмечается низкая лабильность нервных волокон у детей первых лет жизни, с
возрастом она увеличивается (у детей 5-9 лет приближается к норме взрос-
лых - 300 - 1000 импульсов).
♦♦♦ Структурно-функциональные изменения в синапсах.
• Значительное созревание нервных окончаний (нервно-мышечных синапсов)
происходит к 7 - 8 годам.
• Увеличиваются терминальные разветвления аксоня и суммарная площадь его
окончаний.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). Прн
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3: 3.1 и 3.2). Объем реферата 2-3 стр., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. стр. 514, использовать имеющиеся
у вас учебники и пособия): [1-9], [11], [12], [13], [14], [15], [16].
3.2. Дополнительная литература (см. стр. 514 - 522): [6], [28], [30], [47], [48], [56],
[67], [69], [97], [126], [134], [152], [169], [187], [222], [225], [227], [236], [242], [245],
[251-254].
3.3. Наглядные средства обучения. Использование страницы кафедры в Интернете -
www.vsma.ac.ru/~nDhvs: Слайды для компьютерной демонстрации из «Атласа по
физиологии человека» / В.Б. Брии. - Владикавказ: СОГМА. - 1999; тесты остаточ-
ного уровня знаний по дайной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. стр. 508 - 513).
Задача 4-1. В течение 5 часового нарастающего дефицита кислорода (гипоксии) в нейронах уча-
стка головного мозга резко нарушилось образование АТФ в митохондриях. Как в этих условиях
изменится аозбудимость и импульсная активность нейронов? Обоснуйте свои ответы.
Задача 4—2. Из раствора, окружающего нервное волокно, удален ион натрия. Для сохранения
электронейтральности в раствор введен катион холина в эквимолярном количестве. Как отразится
62
эта замена на величину мембранного потенциала покоя и способность волокна генерировать по-
тенциалы действия? Обоснуйте свои ответы. При ответе учтите, что мембрана нервного волокна
полностью не проницаема для холина.
63
Тема 5. Рефлекторный принцип деятельности нервной системы. Нерв-
ные центры. Процессы возбуждения и торможения в ЦНС
Цель: а) знать рефлекторный принцип деятельности и основные функции ЦНС; понятие о рефлексах и
их классификация, рефлекторный путь; понятие о нервном центре; закономерности и особенно-
сти возбуждения в ЦНС; основные принципы распространения возбуждения в ЦНС; процесс
торможения в ЦНС, современные представления о механизмах центрального торможения, ос-
новные виды торможений;
б) уметь использовать эти знания для понимания функциональной деятельности ЦНС.
Мотивация темы: рефлекторные реакции являются важнейшим механизмом регуляции функции здо-
рового организме, а также процессов патогенеза и выздоровления в больном организме. Возбу-
ждение и торможение являются главными процессами в ЦНС, лежащими в основе функцио-
нальной активности и свойств нервных центров. Нарушение соотношения этих процессов имеет
важное значение в развитии многих заболеваний. Некоторые физиологические свойства нерв-
ных центров (например, их пластичность) используется в практической медицине для разработ-
ки методов восстановления пораженных функций.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Рефлекторный принцип деятельности нервной системы.
1.1. Общая характеристика.
❖ Рефлекс - реакция организма, осуществляемая с участием нервной систе-
мы в ответ на раздражение рецепторов. Рефлекс проявляется активацией
или торможением деятельности органов и является основным механизмом
приспособления организма к условиям среды.
❖ Основные этапы изучения проблемы рефлекса: Р. Декарт (17 в.) - механи-
стическая концепция рефлекса (под действием раздражителя на органы чувств натя-
гиваются «нервные нити», идущих к мозгу нервов, и открываются клапаны, через которые
из полостей мозга выходят в нервы «животные духи» - потоки мелких частиц, устрем-
ляющиеся к мышцам и раздувающие их); И. Прохазка (18 в.) - биологическая
концепция рефлекса; И.М. Сеченов (19 в.) - психофизиологическая кон-
цепция рефлекса; И.П. Павлов (20 в.) - условный рефлекс как механизм по-
ведения организма; П.К. Анохин (20 в.) - теория функциональных систем.
1.2. Рефлекторный путь (рис. 17).
❖ Рецепторное звено (восприятие и кодирование раздражителей); совокупность ре-
цепторов, с которых формируется рефлекс, называется рецептивным полем
рефлекса или рефлексогенной зоной.
* *• Афферентное звено (частотно-спектральное перекодирование н проведение возбуж-
дения, аксонный транспорт).
Центральное звено (анализ н синтез афферентной нмпульсацин, перекодирование ин-
формации и выработка команды).
❖ Эфферентное звено (проведение возбуждения, аксонный транспорт).
64
❖ Эффекторное звено (приспособительные изменения функции исполнительных орга-
нов).
❖ Обратная афферентация (Ч. Белл, 1826) и ее значение (информация о ре-
зультате рефлекса, коррекция рефлекса и формирование самонастраиваю-
щегося нервного контура регуляции - рефлекторного кольца).
13. Классификация рефлексов.
❖ По звеньям рефлекторного пути.
• По виду рецепторов:
•ф- Экстероцептивные рефлексы.
Рис. 17. Рефлекторный путь соматического (А) и вегетативного (Б) рефлексов.
1 - рецептор, 2 - афферентные волокна, 3 - центр, 4 - эфферентные волокна,
5 - эффектор, Г - ганглионарный нейрон. Пунктиром показана обратная аф-
ферентация.
ф Интероцептивные рефлексы (висцероцептивные, проприоцептив-
ные, вестибулоцептивные).
• По центральному звену.
ф По уровню его замыкания:
истинные рефлексы (замыкание через ЦНС);
периферические рефлексы (замыкание через ганглии);
аксон-рефлексы (осуществляются в пределах одного нейрона по разветвле-
ниям аксона, их существование дискутируется).
ф По уровню основного замыкания истинных рефлексов: спинальные,
бульбарные, мезенцефальные, мозжечковые, диэнцефальные и кор-
ковые рефлексы.
• По эфферентному звену: соматические и вегетативные рефлексы.
• По эффекторному звену: легочные, сердечные, мигательные и др. реф-
лексы.
❖ По биологическому значению.
• Пищевые, питьевые рефлексы.
• Половые рефлексы.
• Защитные рефлексы.
• Ориентировочно-исследовательские рефлексы.
65
• Гомеостатические рефлексы.
• Локомоторные и позно-тонические рефлексы.
❖ По характеру связи между нейронами.
* Безусловные рефлексы (жесткие, генетически-детерминированные связи).
• Условные рефлексы (гибкие, вероягностио-детерминированные связи).
•Z* По отношению к физиологическим системам.
• Собственные (внутрисистемные) рефлексы - рецепторное и эффекторное
звенья находятся в пределах одной физиологической системы.
• Сопряженные (межсистемные) рефлексы - рецепторное и эффекторное
звенья находятся в разных физиологических системах.
1.4. Особенности проведения нервного импульса в рефлекторной дуге.
❖ Одностороннее проведение от рецептора к эффектору обусловлено одно-
сторонним проведением возбуждения в синапсах рефлекторного пути.
❖ Латентное время рефлекса составляет время, необходимое для формирова-
ния рецепторного потенциала, проведения возбуждения в афферентном и
эфферентном звеньях, передачи возбуждения в центральных и нервно-
эффекторных синапсах.
2. Физиологическая характеристика иервиых центров.
2.1. Нервный центр - совокупность нейронов, расположенных в разных структурах
ЦНС, необходимая для осуществления какого-либо рефлекса (или функции) орга-
низма.
❖ Главная часть (ядро) центра - это «группа нейронов, деятельность которой
является необходимым н достаточным условием для обеспечения данной
функции организма» (А.А. Ухтомский). Ее нейроны наиболее специализиро-
ваны в функциональном отношении, их разрушение полностью выключает
данцый рефлекс (функцию).
❖ Вспомогательные части центра расположены в разных отделах ЦНС, они
увеличивают диапазон приспособления данной функции к различным ус-
ловиям среды. Их нейроны имеют более широкие рецептивные поля, высо-
кую степень многофункциональности (полимодальности).
❖ Итак, в физиологическом смысле под нервным центром понимают «созвез-
дие» нервных центров, расположенных на разных этажах ЦНС и регули-
рующих ту или иную функцию (А.А. Ухтомский).
2.2. Пластичность нервных центров.
❖ Пластичность нервных центров - это способность их изменять свое функ-
циональное значение н восстанавливать утраченную функцию; формирует-
ся преимущественно на основе вспомогательных частей центра, их много-
функциональности. (Важную роль в пластичности играют холинергические проекции
переднебазальиого мозга. Их разрушение ингибирует пластические реакции.)
❖ Пластичность нервных центров увеличивается от спинного мозга до коры
(наиболее пластичные структуры мозга - кора, гиппокамп, зубчатая фасция).
❖ Пластичность нервных центров наибольшая у детей и уменьшается с воз-
растом.
3. Процесс возбуждении в ЦНС.
66
3.1. Закономерности формирования возбуждения в ЦНС.
Ф Суммация возбуждения - нервные центры могут «суммировать чувстви-
тельные по одиночке недействительные раздражения до импульса, дающе-
го движение, если эти раздражения достаточно часто следуют друг за дру-
гом» (И.М. Сеченов). В основе суммации возбуждения в центрах лежит про-
странственная и временная суммация возбуждения в нейроне (рис. 18).
• Временная суммация ВПСП осуществляется в одном и том же синапсе
при частой передаче через него нервных импульсов (интервалы между им-
пульсами должны быть короче продолжительности одного ВПСП).
• Пространственная суммация ВПСП осуществляется в области не-
скольких близлежащих синапсов, если в них в одно и то же время (в пре-
делах продолжительности ВПСП) происходит передача импульса.
Рис. 18. Схематическое изображение временной и пространственной сум-
мации ВПСП при возбуждении нейрона. ВПСП - возбуждающий постси-
наптический потенциал. ПД - потенциал действия.
❖ Облегчение возбуждения - это способность нервного центра при одновре-
менной стимуляции с даух рецепторных зон данать возбуждение больше,
чем сумма двух его возбуждений при раздельной стимуляции этого центра
(В1+2 > В,+ В2). Формируется на основе суммации возбуждения в центрах,
возникает при действии слабых и средних по силе раздражителей.
❖ Окклюзия возбуждения - нервный центр при одновременной стимуляции с
даух рецепторных зои дает возбуждение меньшее, чем сумма даух его воз-
буждений прн раздельной стимуляции этого центра (Вн2 < В| + В2). Обуслов-
лена неспособностью нейронов, генерирующих импульсы (фаза абсолютной
рефрактерное™!), отвечать иа стимуляцию с других афферентных входов.
Окклюзия возникает при действии сильных раздражителей.
❖ Посттетаническая потенциация - феномен усиления рефлекторного отве-
та нервного центра после предшествующего небольшого его раздражения;
67
механизмы потенциации связаны с кратковременным повышением эффек-
тивности синаптической передачи (подробнее см. тему 4; задание 1:4.3).
3.2. Особенности возбуждения в ЦНС.
❖ Трансформация ритма возбуждения.
• Понижающая трансформация, в ее основе лежит необходимость про-
странственной н временной суммации ВПСП для возбуждения нейрона
(формирования в нем ПД).
• Повышающая трансформация основана на мультипликации возбужде-
ния (рнс. 19А). Возбуждение (ПД), распространяющееся по линейной
цепи нейронов, за счет разветвления аксонов н включения других ин-
тернейронов конвергирует на эфферентный нейрон этой цепн, что при-
водит к формированию высокоамплитудного и длительного ВПСП, на
«гребне» которого возникает пачка потенциалов действия.
❖ Низкая лабильность нервных центров, их утомляемость, чувствительность
к гипоксии и сннаптоактивным веществам.
Рис. 19. Нейронная схема мультипликации (А) и реверберации (Б) возбуждения в ЦНС.
❖ Тонус покоя нервных центров создается в результате афферентной импуль-
сацин (фоновая активность рецепторов) н влияния гуморальных факторов.
4. Основные принципы распространения возбуждения в ЦНС.
4. 1. Одностороннее проведение возбуждения (от афферентного входа к эфферентному вы-
ходу, обусловлено односторонним проведением в синапсах).
4. 2. Центральная задержка проведения возбуждения (создается синаптической задерж-
кой в многочисленных синапсах центра).
43. Дивергенция возбуждения - распространение возбуждения от одного нейрона
(центра) к нескольким нейронам (центрам); участвует в мультипликации и ирра-
диации возбуждения (см. рнс. 20Б).
44. Конвергенция возбуждения - распространение возбуждения от нескольких ней-
ронов (центров) к одному нейрону (центру); участвует в интегративной функции
нервной системы (в ее основе лежит мультисннаптичность нейронов) (см. рнс. 20А).
43. Реверберация возбуждения - распространение возбуждения по кольцевым це-
пям нейронов; является механизмом его пролонгирования (рнс. 19Б).
5. Процесс торможения в ЦНС (и.М. Сеченов, 1862).
5.1. Торможение - активный процесс, возникающий в нервной системе и приводя-
щий к ослаблению или предотвращению возбуждения.
5.2. Основные виды торможения, их механизмы («Возбуждение - это дикий камень,
ожидающий скульптора» А.А. Ухтомский).
68
Рис. 20. Нейронная схема конвергенции (А)
и дивергенции (Б) возбуждения в ЦНС.
❖ Первичное торможение как деятельность тормозных нейронов (Дж. Экклс).
• Постсинаптическое торможение (см. тему 4; задание 1:4.3).
• Пресинаптическое тор-
можение (см. тему 4; задание
1: 4.3).
• Примечание: недавно в коре и
гиппокампе были обнаружены
специальные тормозные ней-
роны, которые способны тор-
мозить другие тормозные ней-
роны (дают растормаживаю-
щий эффект).
❖ Вторичное (пессимальное)
торможение, по Введенско-
му, возникает при высоко-
частотной и длительной активности возбуждающих нейронов, при которой
развивается кратковременная синаптическая депрессия (ее механизм связыва-
ют с истощением резерва доступного медиатора).
5.3. Торможение в нервных центрах (рис. 21).
Ф Возвратное торможение - сильное возбуждение нервного центра сопро-
вождается самоторможением его через тормозные интернейроны Реишоу.
Это механизм ограничения перевозбуждения центров (отрицательная обратная
связь).
❖ Латеральное торможение - возбужденный центр через тормозные интер-
нейроны окружает себя зоной торможения. Это механизм концентрации
возбуждения (одновременная отрицательная индукция).
♦> Реципрокное торможение - возбуждение одного центра сопровождается
торможением другого центра, осуществляющего антагонистический реф-
лекс; это важнейший механизм координации деятельности центров.
Рис. 21. Нейронная схема возвратного (А), латерального (Б) и реци-
прокного (В) торможения. Черным цветом обозначены тормозные
нейроны.
5.4. Взаимоотношения между процессами возбуждения и торможения.
❖ Иррадиация и концентрация возбуждения.
❖ Одновременная положительная и отрицательная индукция.
• > Последовательная положительная и отрицательная индукция.
69
б. Профильные материалы для студентов лечебного факультета. Изменение реф-
лекторной деятельности при старении организма.
6.1. Уменьшение силы и увеличение латентного времени рефлекторной реакции (например,
ахиллового, брюшных, конъкжтивального рефлексов).
6.2. Снижение коркового контроля рефлексов (положительный хоботковый губной реф-
лекс, снижение или отсутствие подошвенного рефлекса и появление рефлекса Бабин-
ского).
63. Снижение концентрации нервных процессов и преобладание процессов торможения в
нервных центрах.
6.4. Ослабление восстанавливающей роли тормозного процесса, но облегчение развития
охранительного торможения.
6.5. Компенсаторное повышение чувствительности нервных центров к гуморальным влия-
ниям (за счет усиления проницаемости гематоэнцефалического барьера и повышения
чувствительности рецепторного аппарата к некоторым фармакологическим и биоак-
тивным веществам).
7. Профильные материалы для студентов педиатрического факультета. Особенно-
сти рефлекторных реакций плода, новорожденных и детей в грудном возрасте (см. тему
И; задание 1: И).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [12-16].
3.2. Дополнительная литература (см. с. 514 - 522): [6], [28], [30], [47], [97], [126],
[134], [152], [169], [222], [225], [236], [242], [245], [251-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphvs: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
33. Консультации преподавателей еженедельно по индивидуальному графику.
3.6 . Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 5-1. Выдающийся отечественный физиолог А.А. Ухтомский писал в одной из работ:
«Возбуждение - это дикий камень, ожидающий скульптора». Как называется «скульптор», спо-
собный отшлифовать процесс возбуждения?
Задача 5-2. Что произойдет, если в синаптических окончаниях клеток Реншоу спинального мо-
торного центра вместо глицина стал бы выделяться глутамат?
70
Задача 5-3. JSpa одинаковых по силе раздражителя вызывают два двигательных рефлекса. Реф-
лекторный путь первого рефлекса 30 см, преобладающий тип волокон АД. Рефлекторный путь
второго рефлекса 20 см, преобладающий тип волокон Аа. Скорость ответа рецепторов н эффек-
торных мышц в обоих рефлексах одинакова. Измерение латентного времени обоих рефлексов по-
казало, что у первого рефлекса оно меньше, чем у второго. С чем это может быть связано?
71
Тема 6. Координационная и интегративная деятельность НДС. Мето-
ды изучения ЦНС
Цель: а) знать принципы координационной деятельности ЦНС: переключения, реципрокности,
обратной связи, проторения пути, общего «конечного пути», доминанты; многоуровневую
регуляцию ЦНС соматических н вегетативных функций и взаимодействие этих уровней в
процессе регуляции функций; современное представление об интегративной деятельности
ЦНС; системную организацию функций мозга по принципу взаимодействия проекционных,
ассоциативных, интегративно-пусковых систем; функциональный элемент мозга, знаковую
функцию мозга: гнозис, праксис; методы исследования функций ЦНС;
б) уметь использовать эти знания для понимания функционирования мозга в процессах са-
морегуляции функций целостного организма, его поведения;
в) иметь представления о методах исследования : стереотаксический метод, электроэнцефа-
лография, вызванные потенциалы.
Мотивация темы: «Мозг оказался местом страшных раздоров, борьбы н насилия, господства одно-
го центра н угнетения других... Побеждает и правит тот, кто нужнее в данный момент, чья
поддержка необходима всему организму» И.П. Павлов.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую
логическую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения,
определяются лектором в конце читаемой по данной теме лекции, и обозначены в
«Графике н содержании самостоятельной работы студентов». Остальные фрагменты
задания 1 могут быть использованы для коррекции материала записанной лекции и
подготовки рефератов.
1. Основные прввципы координационной деятельности ЦНС.
1.1. Координационная деятельность ЦНС - это согласование и соподчинение
деятельности различных ее центров. (Иерархия центров формируется как нв одном
уровне, так и на разных уровнях ЦНС, что обеспечивает устойчивость и динамичность
нервной регуляции.)
1.2. Принцип общего «конечного пути» (Ч. Шеррингтон, 1906) - это использова-
ние одного нервного центра (например, непосредственно выходащего на эффектор)
другими центрами, реализующими через него свое функциональное значе-
ние. При этом на основе принципа доминанты происходит «борьба» этих
центров за общий конечный путь.
U. Принции проторения пути - если возбуждение в ЦНС несколько раз про-
шло по одним и тем же цепям нейронов, то в дальнейшем оно легче всего
пойдет по этому (проторенному) пути. В основе проторения пути лежат крат-
ковременная и долговременная потенциация (см. тему 4; задание 1:4.4).
1.4. Принции переключения - способность ЦНС переключать одну и ту же аф-
ферентную импульсацию на центры различных рефлексов, как безусловных
(И. М. Сеченов), так и условных (И. П. Павлов).
1.5. Принцип рециирокности (Ч. Шеррингтон, 1905) - возбуждение одного центра
сочетается с торможением другого, осуществляющего функционально про-
тивоположный рефлекс.
72
1.6. Принцип обратной связи - результат любой рефлекторной деятельности с
помощью обратной афферентации оценивается нервным центром. Прн этом
обратная связь может как усилить, так и уменьшить текущую рефлекторную
деятельность.
1.7. Принцип доминанты (А.А. Ухтомский, 1923).
❖ Доминанта - временно господствующий в ЦНС возбужденный нерв-
ный центр, который направляет (соподчиняет, объединяет) работу других
центров. Это совокупность возбужденных центров, которые временно
кооперируются для выполнения биологически или социально важной
функции организма. При этом доминанта направляет поведение орга-
низма в сторону удовлетворения главной потребности.
❖ Свойства доминанты.
• Доминантный центр имеет повышенную возбудимость н может от-
вечать на субпороговые раздражители.
• Доминантный центр конвергирует возбуждение с других центров,
что резко увеличивает рецептивное поле доминанты.
• Доминантный центр способен суммировать возбуждение, конверги-
руемое с других центров, н усиливать свой функциональный ответ.
• Доминантный центр способен тормозить другие центры, действие
которых препятствует удовлетворению доминирующей потребности.
• Доминантный центр чувствителен к гуморальным раздражителям.
1.8. Принцип кортикализации функций - в процессе эволюционного развития
к коре больших полушарий перешли в усовершенствованном виде функции,
которые в более примитивном виде выполняли ниже расположенные цен-
тры. (Например, регуляция таких двигательных функций, как локомоция - прыжки, ходь-
ба, бег - и выпрямительные рефлексы, у низших позвоночных полностью обеспечивается
стволом мозга, и удаление больших полушарий практически их не изменяет. У кошек пере-
резка ствола между средним и промежуточным мозгом существенно не влияет на выпрями-
тельные рефлексы, но нарушает в остром периоде локомоцию, которая в дальнейшем час-
тично восстанавливается. Выключение коры больших полушарий у обезьян и у человека
приводит к потере не только локомоции, но и выпрямительных рефлексов.)
1.9. Многоуровневая регуляция ЦНС соматических и вегетативных функций
осуществляется на спинальном, бульбарном, среднемозговом, ретнкулярно-
стволовом, диэнцефальном, мозжечковом, корковом уровнях. В процессе
регуляции функции происходит взаимодействие между различными уров-
нями ЦНС. При этом вышележащий уровень включает регулируемую функ-
цию в более сложные проявления жизнедеятельности организма и увеличи-
вает диапазон ее адаптивной регуляции.
2. Современное представление об интегративной деятельности ЦНС (П.К. Ано-
хин, 1975; О.С. Андрианов, 1995; К.В. Судаков, 2000).
2.1. Интегративная деятельность ЦНС - это функциональное объединение
деятельности ее центров; при этом происходит и координация их деятельно-
сти.
2.2. Организация интегративной деятельности.
73
❖ Уровень нейрона (интегративная функция нейрона базируется на его мультисинап-
тичности и процессе конвергенции).
❖ Уровень микросистем нейронов. Микросистема - минимальная сово-
купность нейронов, их отростков, глии и сосудов, которая обеспечивает
элементарные процессы интеграции. Примером являются нейронные
колонки. Исследования с погружением микроэлектродов перпендику-
лярно поверхности коры показали, что все встречаемые на пути нейро-
ны отвечают на раздражитель только одного вида (например, тактильный).
Напротив, при погружении электрода под углом на его пути попадались
нейроны разной модальности. Был сделан вывод, что в коре мозга име-
ются функциональные объединения нейронов, расположенные в цилин-
дрике диаметром 0,5 - 1,0 мм, включающем все слои коры и содержа-
щем несколько сотеи нейронов. Такие структуры были названы нейрон-
ными колонками. Они обнаружены в моторной и сенсорной коре. Со-
седние нейронные колонки могут частично перекрываться, а также
взаимодействовать друг с другом по механизму латерального торможе-
ния и осуществлять саморегуляцию по типу возвратного торможения.
❖ Интеграция на уровне нервных центров осуществляется по принципу
доминанты.
❖ Уровень больших интегративных систем мозга. Их взаимодействие ле-
жит в основе системной организации функций мозга.
• Сенсорная (проекционная) система (кора затылочной, частично теменной и
височной долей) обрабатывает информацию от рецепторов, раздражае-
мых стимулами внешней и внутренней среды, формирует ощущение.
Афферентные пути в сенсорную кору поступают преимущественно
от специфических сенсорных ядер таламуса.
• Ассоциативные (синоним - межсенсорные) системы интегрируют весь
мозг в целостно работающую систему. Они включают участки но-
вой коры большого мозга, которые расположены рядом с сенсорны-
ми и двигательными зонами, но не выполняют непосредственно чув-
ствительных или двигательных функций. Формируют функции вос-
приятия, гнозиса, праксиса, мышления, речи И др. (Ассоциативная кора
является филогенетически наиболее молодой частью новой коры, получившей
наибольшее развитие у человека, составляя у него 70-75% неокортекса). Выде-
ляют три ассоциативные системы мозга: таламолобную, таламоте-
менную и таламовисочную.
ф" Таламолобная система (ассоциативная кора лобной доли + медиодор-
сальное ядро таламуса). Главной ее функцией является формирова-
ние программ целенаправленного поведения особенно в новой
для человека обстановке. Реализация этой общей функции осно-
вывается на следующих функциях таламолобной системы.
Формирование доминирующей мотивации, обеспечиваю-
щей направление поведения человека. Эта функция основа-
на на двусторонних связях лобной коры с лимбической сис-
74
темой и ролью последней в регуляции высших эмоций че-
ловека, связанных с его социальной деятельностью и твор-
чеством.
Обеспечение вероятностного прогнозирования, что выра-
жается изменением поведения в ответ иа изменения обста-
новки окружающей среды и доминирующей мотивации.
Функция запоминания временной последовательности со-
бытий.
Самоконтроль действий путем постоянного сравнения ре-
зультата действия с исходными намерениями, что связано с
созданием аппарата предвидения (акцептор результата действия,
по П.К. Анохину).
Таламотеменная система (ассоциативная кора теменной доли + задние
ассоциативные ядра таламуса).
Функция гнозиса: различные виды узнавания - формы, ве-
личины, значения предмета, понимание речи, познания за-
кономерностей и тл,
Функция формирования в сознании трехмерной модели те-
ла («схема тела») и внешнего мира является вариантом гно-
стической функции. При этом используется информация от
различных сенсорных систем - соматосенсорной, тактиль-
ной, зрительной, вестибулярной и др.
Функция праксиса осуществляется преимущественно ней-
ронными сетями, находящимися в надкраевой извилине (по-
ля 39 и 40), они обеспечивают хранение и реализацию про-
граммы двигательных автоматизированных и целенаправ-
ленных актов (например, рукопожатие, причесывание, зажигание
спички и др.).
ф- Таламовисочная система (ассоциативная височная кора и задняя
группа ассоциативных ядер таламуса) включает слуховой центр ре-
чи Вернике, находящийся в верхней височной извилине (поле 22
левого полушария), обеспечивающий речевой гнозис - распознава-
ние и хранение устной речи как собственной, так и чужой. Об-
работка несловесной информации (особенности голоса, мело-
дия) осуществляется преимущественно аналогичным полем
правого полушария. На границе височной, теменной и затылоч-
ной долей (поле 39) находится центр чтения письменной речи,
обеспечивающий распознавание и хранение образов письменной
речи.
Лимбикоретикулярная (активирующая) система, аккумулируя возбу-
ждение с различных сенсорных систем, оказывает активирующее
влияние на все большие системы мозга. В этом процессе использует-
ся восходящее активирующее влияние ретикулярной формации
ствола мозга на различные области коры, базальные ядра, задний ги-
поталамус. Лимбическая система (структуры древней н старой коры, мнн-
75
далины височной доли, гипоталамуса и др.) оказывают активирующее влия-
ние через формирующиеся эмоции, одной из функций которых явля-
ется мобилизация психофизиологических и физиологических резер-
вов организма.
• Интеграционнопусковая система сют на выходе интегрирующей
деятельности мозга (общий конечный путь) и осуществляет запуск дви-
гательных программ, реализующих поведение, в нее входят пре-
имущественно моторные зоны коры. Премоторная кора (поле 6) полу-
чает информации нз базальных ядер и мозжечка и создает програм-
мы сложных движений. В премоторной коре находятся также двига-
тельные центры, связанные с социальными функциями человека:
центр письменной речи в заднем отделе средней лобной извилины
(поле б), моторный центр устной речи Брока в заднем отделе нижней
лобной извилины (поле 44). Эти двигательные программы реализуют-
ся как произвольные движения через пирамидные пути коры прецен-
тральной извилины.
2.3. Основные механизмы интегративной деятельности.
❖ Генетически обусловленные межнейронные связи как механизм инте-
грации (например, безусловные рефлексы и инстинкты).
Вероятностные, возникающие при жизни, временные связи (например,
условный рефлекс).
❖ Сочетания жестких, генетически детерминированных блоков (модулей) и
гибких, вероятностно детерминированных звеньев мозга.
Функциональная система как «единица интегративной деятельности це-
лого организма».
3. Функциональная система как «единица интегративной деятельности целого ор-
ганизма» (П.К. Анохин).
3.1. Функциональная система - динамическое объединение различных отделов
нервной системы, физиологических систем и их компонентов, взаимодейст-
вие которых обеспечивает достижение полезных для организма результатов.
При этом полезный результат является системообразующим фактором, «ви-
зитной карточкой» функциональной системы.
3.2. Виды функциональных систем (ФС).
❖ ФС с внутренним звеном саморегуляции обеспечивают поддержание
гомеостаза внутренними, как правило, генетически детерминированны-
ми механизмами саморегуляции (например, ФС регуляции артериального дав-
ления).
❖ ФС с относительно пассивным внешним звеном саморегуляции (напри-
мер, ФС регуляции газового состава крови, имеющая внешнее звено, обеспечивающее
газообмен организма с виешией средой).
❖ ФС с активным поведенческим звеном саморегуляции (например, ФС пита-
ния, размножения, достижения социально полезных результатов).
3.3. Компоненты функциональной системы (рис. 22).
❖ Афферентный синтез.
76
• Пусковая афферентация определяется несоответствием какого-либо
показателя гомеостаза данным условиям организма.
• Доминирующая мотивация обеспечивает выбор в качестве объекта
регуляции наиболее важного из измененных в данный момент пара-
метров.
• Мобилизация памяти - включение в регуляцию генетически детер-
минированных и приобретенных приспособительных реакций.
• Обстановочная афферентация - оценка состояния физиологиче-
ских систем организма и условий внешней среды.
Рис. 22. Схема функциональной системы (по П.К. Анохину).
ОА - обстановочная афферентация, ПА - пусковая афферентация.
❖ Принятие решения - выбор из нескольких возможных вариантов дейст-
вия единственный вариант (ограничение степеней свободы).
❖ Выработка эфферентной программы действия - совокупность возбу-
жденных вегетативных, соматических и психофизиологических цен-
тров, способных включить эффекторные органы, необходимые для дос-
тижения полезного результата.
❖ Выработка акцептора результата действия: нейронной модели по-
лезного результата, который должен быть достигнут.
❖ Эфферентное возбуждение: вегетативные, соматические и эндокрин-
ные каналы регуляции.
❖ Действие, его результаты и их параметры.
❖ Обратная афферентация и роль акцептора результата действия в пре-
кращении или коррекции деятельности функциональной системы.
77
4. Методы исследования функций ЦНС.
4.1. Перерезка, раздражение и разрушение.
4.2. Стереотаксическая методика Метод точного введения электродов, микропипе-
ток, термопар в глубокорасположенные структуры мозга с помощью стереотакси-
ческого прибора. Координаты структур мозга определены в специальных стерео-
таксических атласах и выражены в трёхмерной системе координат. Согласно этим
координатам с помощью микрометрических винтов вводят электрод в нужную точ-
ку структуры мозга.
Стереотаксическая методика используется для изучения деятельности различных
глубинных структур мозга. Через введённые электроды можно регистрировать био-
электрическую активность мозга (например, ЭЭГ, вызванные потенциалы), раздра-
жать или разрушать структуры мозга. С помощью введённых канюль можно подво-
дить химические вещества к различным структурам мозга и в его желудочки. Эта
методика помогла определить локализацию многих нервных центров и изучить их
функции, понять принцип деятельности мозга как единого целого.
43. Нейронография - регистрация импульсной активности отдельных нейронов с по-'
мощью микроэлектродов (d - 1 мкм), подведенных вплотную к клетке, в экспери-
менте и в некоторых случаях при нейрохирургических операциях у человека (при
этом используется стереотаксический аппарат).
4.4. Электроэнцефалография (ЭЭГ) - метод регистрации суммарной электрической ак-
тивности головного мозга. Отражает алгебраическую сумму ВПСП, ТПСП и следо-
вых потенциалов множества нейронов, деятельность которых синхронизируется
или десинхронизируется структурами ствола мозга. На ЭЭГ выделяют четыре ос-
новных ритма (рис. 23).
Рис. 23 . Схематическое изображение основных ритмов энцефало-
граммы и реакции десиихронизации ЭЭГ.
❖ Альфа-ритм (частота 8 - 13 Гц, амплитуда до 70 мкВ) является регулярным
ритмом, механизм его синхронизации связан с деятельностью таламуса. Он
доминирует у здоровых людей старше 9 лет в состоянии физического и эмо-
ционального покоя.
❖ Бета-ритм (14 - 30 Гц, амплитуда до 30 мкВ) характеризуется нерегулярными
низкоамплитудными волнами, которые сменяют альфа-ритм при сенсорной
стимуляции, интеллектуальном и эмоциональном напряжении. Эта смеиа на-
78
зывается десинхронизацией ЭЭГ. Ее механизм связан с влиянием активирую-
щей ретикулярной формации мозга и лимбической системы.
❖ Тета-ритм (3 - 7 Гц, до 200 мкВ) при бодрствовании занимает незначитель-
ную часть ЭЭГ. Он хорошо выражен при длительном эмоциональном напря-
жении и неглубоком сне, а также у ребенка до 7 лет. Происхождение тета-
ритма связывают с активностью синхронизирующей системы моста головного
мозга.
❖ Делыпа-ритм (1 - 3 Гц, до 300 мкВ) при бодрствовании встречается на ЭЭГ в
виде кратковременных эпизодов. Постоянно регистрируется во время глубоко-
го сна, занимая до 80 % ЭЭГ, а также у плода и грудного ребенка. Его проис-
хождение связывают с активностью бульбарной синхронизирующей системы.
❖ Гамма-ритм (свыше 30 Гц, амплитуда до 15 мкВ) наблюдается при решении
задач, требующих максимальной концентрации внимания.
4.5. Вызванные потенциалы (ВП) мозга - это электрический ответ различных макро-
структур нервной системы в ответ на сенсорную стимуляцию. В медицинской
практике обычно регистрируют ВП коры головного мозга в ответ на раздражение
зрительных, слуховых, тактильных рецепторов. Для выделения низкоамплитудных
(около 15 мкВ) компонентов ВП на фоне ЭЭГ используют прием компьютерного
усреднения - многократного суммирования участков ЭЭГ, следующих за подачей
стимула.
❖ ВП представляет собой комплекс чередующихся позитивных (направленных
вниз) и негативных (направленных вверх) волн общей продолжительностью
около 300 мс (рис. 24). Обычно фиксируют 4 негативных и 4 позитивных ком-
понента.
.•«Ле
Рис. 24. Схематическое изо-
бражение вызванного потен-
циала на вспышку света. П -
позитивные, Н - негативные
компоненты, цифровые ин-
дексы означают порядок сле-
дования компонентов. Начало
записи совпадает с моментом
подачи раздражителя.
I , I________I________I________I
0 100 200 300 400 мс
Компоненты ВП в интервале 20 - 100 мс обусловлены преимущественно аф-
ферентным возбуждением, поступающим по лемнисковой, быстропроводящей
системе через специфические ядра таламуса, частично через его ассоциатив-
ные ядра и базальные ядра. Эту часть ВП называют первичным ответом (ран-
ним комплексом). Он легче всего фиксируется (с меньшим латентным перио-
дом, большей амплитуды) в сенсорной коре, соответствующей данному рецеп-
торному входу.
Компоненты ВП в интервале 100 - 300 мс обусловлены преимущественно не-
специфическим афферентным притоком через РФ ствола и неспецифические
79
ядра таламуса. Эту часть ВП называют вторичным ответом (поздним ком-
плексом).
4.6. Хронорефлексометрия. При раздражении рецепторных зон раздражителями (свето-
вым, звуковым и др.) возникают соматические и вегетативные реакции, латентное
время которых регистрируют с помощью хронорефлексометров. Среднее время ре-
акции на звук - 0,14 с, свет - 0,18 с, боль - 0,8 с. Метод применяется в физиологии
труда и спорта, в диагностике профзаболеваний, поражений ЦНС и др.
4.7. Функциональная компьютерная томография (получение «срезов» мозга с помощью ком-
пьютера).
❖ Позитронно-эмиссионная томография - это метод функционального изотоп-
ного картирования мозга. Основана на введение в кровоток изотопов (О|!, N11.
F" и др.) в соединении с дезоксиглюкозой. Чем активнее участок мозга, тем
больше поглощает он меченой глюкозы. Ее радиоактивное излучение регист-
рируется детекторами, расположенными вокруг головы. Информация от де-
текторов поступает на компьютер, который создает «срезы» мозга, отражаю-
щие неравномерность распределения изотопа в связи с метаболической актив-
ностью мозговых структур.
•** Функциональная магнитно-резонансная томография основана на том, что при
потере О2 гемоглобин приобретает парамагнитные свойства. Чем выше мета-
болическая активность мозга, тем больше объемный и линейный кровоток в
данном участке мозга и тем меньше соотношение парамагнитного дезоксиге-
моглобина к оксигемоглобину. В мозге существует много очагов активации,
что отражается в неоднородности магнитного поля. Этот метод позволяет вы-
явить активно работающие участки мозга.
5. Профильные материалы для студентов лечебного факультета. Особенности
координационной н интегративной деятельности ЦНСнрн старении.
5.1. Особенности интегративных нейрофизиологических механизмов.
♦ > Ослабление субординационных влияний в ЦНС за счет ослабления коркового
контроля.
❖ Ограничение потока информации, направленного к объектам регуляции.
♦ > Изменение характера обратной информации, поступающей в центры («дезин-
формация» центров о состоянии на периферии в связи с изменением их возбу-
димости).
Увеличение несоответствия центральных программ и результатов их реализа-
ции, числа некорректируемых «ошибок» в деятельности организма (теряется
тонкое приспособление программы при ее реализации в конкретных условиях
среды).
5.2. Изменение ЭЭГ при старении организма.
❖ Замедление частоты a-ритма после 60 лет до 6 - 8 Гц, снижение его амплиту-
ды меньше 30 мкВ.
• •• Замедление частоты p-ритма и снижение его амплитуды соответственно
меньше 14 Гц и 10 мкВ.
• 1* Снижение амплитуды 0- и 6- ритмов до 30 мкВ.
Ф Причины изменения ЭЭГ у пожилых людей связаны с морфофункциональной
перестройкой в ЦНС (больше в кортико-ретикулярной цепи), изменением ме-
таболизма, гипоксическими явлениями, обусловленными снижением кровото-
ка мозга.
80
♦ ** Изменения ЭЭГ отражают понижение лабильности нейронов и сдвиг
нейродинамических процессов в сторону торможения.
6. Профильные материалы для студентов педиатрического факультета.
6.1. Особенности координационной и интегративной деятельности ЦНСу ребенка
после рождения.
♦ ♦♦ Поддержание вертикальной позы требует сложной нервно-мышечной коорди-
нации, она несовершенна до 1-го года.
♦ ♦♦ Совершенствование процесса ходьбы, бега, прыжков (смена быстрого движе-
ния на медленное) развивается со 2-го по 12-й год.
Усиление концентрации и координации процессов возбуждения и торможения,
увеличение точности и меткости движений развивается до 13 - 14 лет, при-
ближаясь к уровню взрослых только к 20 годам.
♦ ♦♦ Формирование навыков трудовой деятельности (начало с 3-летнего возраста).
❖ Повышение уровня произвольных движений, дальнейшее развитие высших
отделов ЦНС (памяти).
6.2. Особенности электроэнцефалограммы в онтогенезе.
♦♦♦ Антенатальный период.
• У плода спонтанная ЭЭГ появляется с 3 - 5 мес. На ЭЭГ регистрируются
медленные колебания, разделенные длительными (10 с - 3 мин) паузами.
Они обусловлены активностью подкорковых структур (кора головного
мозга развивается с 6-го мес.).
• К 8 мес. колебания на ЭЭГ становятся непрерывными с частотой 1 - 2 и 3 -
4 Гц, их амплитуда периодически уменьшается (проявление фазы «быстро-
го» сна). Эти колебания обусловлены уже участием нейронов коры боль-
ших полушарий.
Неонатальный период.
• У бодрствующих новорожденных ЭЭГ носит непрерывный характер, реги-
стрируются волны низкой амплитуды (30 - 50 мкВ) с частотой 1 - 2 и 4 - 6
Гц (5- и 0-ритмы).
• Отмечается наличие реакции активации на сильный раздражитель в виде
уплощения ЭЭГ (показатель функционирования ретикулокортикальных н
таламо-кортикальных связей).
♦♦♦ Особенности ЭЭГ в грудном возрасте.
• Устойчивая активность на ЭЭГ впервые появляется у детей около 3 мес.
Регистрируются волны с частотами 3 - 5 Гц и около 1 Гц.
• Свойственный человеку a-ритм ЭЭГ отмечается в 3 - 4 мес.
❖ Особенности ЭЭГу детей других возрастных групп.
• В возрасте от 1 года до 16 лет на ЭЭГ происходит постепенное становле-
ние а-рнтма, и доминирование его сначала в затылочных, а затем и в пе-
редних отделах мозга к 9 - 15-летнему возрасту. На этом фоне происходит
постепенная редукция 0- и 5-ритмов, формируется реакция десинхрониза-
ции ЭЭГ (смена a-ритма на р-ритм).
• К 16 - 18 годам характер ЭЭГ становится как у взрослых.
7. Профильные материалы для студентов стоматологического факультета.
7.1. Афферентные влияния с рецепторов полости рта (см. тему 26; задание 1: 11).
81
7.2. Метод вызванных потенциалов (ВП) и его использование в определении
локализации проекционных зон зубов н языка в ЦНС.
❖ С помощью метода регистрации ВП (первичных ответов) установлена топо-
графия первичных сенсорных зон анализаторов, связанных с областью рта
(вкусового, температурного, болевого, тактильного, проприоцептивного). Эти
зоны находятся в постцентральной и частично прецентральной извилине в об-
ласти проекции лица.
❖ При раздражении соответствующих рецепторов ВП раньше регистрируются в
ядрах таламуса; например, установлено, что на механические и термические
стимулы языка ВП регистрируются в латеральных, а на вкусовые - в медиаль-
ных, вентральных задних ядрах таламуса.
73. Электроэнцефалография и ее использование в стоматологии.
❖ В стоматологии ЭЭГ используется для объективной оценки функционального
состояния головного мозга и выяснения роли центральных механизмов в про-
исхождении глоссалгнй (проявляется ощущениями покалывания, онемения,
жгучих болей в полости рта, чаще языка).
Изменения ЭЭГ в зависимости от формы глоссалгии имеют признаки избы-
точной синхронизации a-ритмов, отсутствие зональных различий на ЭЭГ и
частые высокоамплитудные заостренные волны.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема
реферата выбирается с использованием учебных элементов (задание 1).
При подготовке реферата использовать список основной н дополни-
тельной литературы (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., рег-
ламент 5-7 мин.
Задание 3. Для выполнения заданий 1 н 2 использовать следующие средства для
самоподготовки студентов.
3.1. Основной учебно-методический комплекс (см с 514, использовать имеющие-
ся у вас учебники и пособия): [1-9], [11 - 14], [16].
3.2. Дополнительная литература (см. с. 514 - 522): [4], [12], [16], [30], [86], [97],
[120], [146], [152], [169], [173], [176], [177], [188], [196], [222], [225], [226],
[245], [253].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Ин-
тернете - www.vsma.ac.ru/~nDhvs: Сяайды для компьютерной демонстрации из
«Атласа по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999;
Учебное пособие В.Н. Яковлева, И.Э. Есауленко «Физиология головного моз-
га». - Воронеж, ВГМА, 2001. - 75 с.; тесты остаточного уровня зиаинй по дан-
ной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами,
диапроектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться иа
практическом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 6-1. Физиолог в опытах на собаках изучал двигательные рефлексы. По правилам экс-
периментальной работы животным за сутки до опыта пища не дается, вода не ограничивается.
82
Однажды по халатности лаборанта в опыт попали накормленные животные. К удивлению фи-
зиолога в ответ на раздражение афферентного нерва задней конечности у собак возникло не
сокращение мышц конечности, а акт дефекации. Какой новый принцип работы головного моз-
га был открыт в этих опытах и как объяснить результаты приведенных опытов.
83
Тема 7. Частная физиология ЦНС: функции отделов головного мозга,
его кровоснабжение. Гематоэнцефалический барьер. Спинномоз-
говая жидкость
Цель: а) знать функции различных отделов головного мозга: ствола (продолговатый мозг, мост, сред-
ннй мозг), промежуточного мозга, мозжечка, лимбической системы, базальных ядер, новой ко-
ры больших полушарий.
б) уметь использовать эти знания для понимания функциональной деятельности ЦНС.
Мотивация темы: см. тему 6.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, опре-
деляются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
N.B.: в данной теме не изложена роль головного мозга в регуляции движений, она
подробно рассмотрена в теме 11. При нодготовке к занятию вы встретите многочис-
ленные ссылки на материал других тем, этот материал на данном занятии имеет фа-
культативный характер.
1. Общая характеристика головного мозга. Головной мозг является главным отделом
ЦНС, выполняющим высшую регуляцию двигательных, висцеральных, эндокринных функ-
ций и психофизиологических процессов. Он состоит из конечного мозга (кора больших по-
лушарий, белое вещество, базальные ядра), промежуточного, среднего, заднего (мост и моз-
жечок) и продолговатого мозга (рис. 25). Часть этих структур (от трех до пяти) называют
стволом мозга. В физиологической и клинической литературе ствол мозга обычно ограничи-
вают тремя отделами: продолговатым мозгом, мостом и средним мозгом, совместная дея-
тельность которых формирует основные стволовые функции - рефлекторную, интегратив-
ную, нейрорегуляторную и проводниковую.
2. Функции ствола головного мозга.
2.1. Функции ствола, реализуемые ядрами черепных нервов. В стволе мозга нахо-
дятся ядра Ш - ХП пар черепных нервов, через которые осуществляются чувст-
вительные (сенсорные), двигательные (соматические) и вегетативные (парасим-
патические) функции.
❖ Ядра глазодвигательного нерва (п. oculomotorius, Ш) расположены в среднем
мозге. Двигательное ядро сокращает верхнюю, нижнюю, внутреннюю пря-
мые, нижнюю косую мышцы глаза и мышцу, поднимающую верхнее веко,
участвуя в глазодвигательных рефлексах. Добавочное (парасимпатическое)
ядро, иннервируя сфинктер зрачка и ресничную мышцу, осуществляет
рефлексы сужения зрачка н аккомодации глаза.
❖ Ядро блокового нерва (n. trochlearis, IV) находится в среднем мозге. Иннер-
вируя верхнюю косую мышцу, оно осуществляет поворот глазного яблока
вниз и кнаружи.
❖ Тройничный нерв (n. trigeminus, V) имеет двигательное и чувствительные
ядра. Двигательное ядро расположено в мосту, иннервирует жевательную
мускулатуру н вызывает движение нижней челюсти вверх, вниз, в стороны
84
и вперед, а также напрягает мягкое небо и барабанную перепонку. Чувст-
вительные ядра (среднемозговое, мостовое, спинальное) получают от ко-
жи, слизистых оболочек, органов лица и головы тактильную, температур-
ную, висцеральную, проприоцептивную, болевую импульсацию, входят в
проводниковый отдел соответствующих анализаторов и участвуют в раз-
личных рефлексах, например, жевательном, глотательном, чихательном.
Ядро отводящего нерва (n. abducens, VI) расположено в мосту. Сокращая
наружную прямую мышцу глаза, вызывает поворот его наружу.
Ядра яйцевого нерва (n. facialis, VII) находятся в мосту. Двигательное ядро
вызывает сокращении мимической и вспомогательной жевательной муску-
латуры, регулирует передачу звуковых колебаний в среднем ухе в резуль-
тате сокращения стременной мышцы. Чувствительное ядро одиночного
пути, иннервируя вкусовые луковицы передней 2/3 языка, анализирует
вкусовую чувствительность, участвует в моторных и секреторных пищева-
рительных рефлексах. Верхнее слюноотделительное (парасимпатическое)
ядро стимулирует выделение секретов подъязычной, подчелюстной слюн-
ных и слезной желез.
Большие
полушария ------
Промежуточный
мозг (таламус
и гипоталамус)
Средний мозг
Мост
Рис. 25. Основные отделы головного мозга.
Чувствительные ядра преддверноулиткового нерва (n. vestibulocochlearis,
VIII) расположены в продолговатом мозге. Вестибулярные ядра, иннерви-
руя рецепторы вестибулярного аппарата, участвуют в регуляции позы и
равновесия тела (статические и статокинетические рефлексы), в вестибуло-
глазных и вестибуловегетативных рефлексах, входят в проводниковый от-
дел вестибулярного анализатора. Улитковые ядра, иннервирующие слухо-
вые рецепторы, участвуют в слуховом ориентировочном рефлексе, входят в
проводниковый отдел слухового анализатора.
85
❖ Ядра языкоглоточного нерва (n. glossopharyngeus, IX) расположены в про-
долговатом мозге. Двойное (двигательное) ядро вызывает поднимание
глотки и гортани, опускание мягкого неба и надгортанника в глотательном
рефлексе. Чувствительное ядро одиночного пути получает вкусовую, так-
тильную, температурную, болевую и интероцептивную чувствительность
от слизистой оболочки глотки, задней трети языка, барабанной полости и
каротидного тельца, входит в состав соответствующих анализаторов, уча-
ствует в рефлексах жевания, глотания, в секреторных и моторных пищева-
рительных рефлексах, в сосудистых и сердечных рефлексах (из каротидно-
го тельца). Нижнее слюноотделительное (парасимпатическое) ядро сти-
мулирует секрецию околоушной слюнной железы.
❖ Ядра блуждающего нерва (n. vagus, X) расположены в продолговатом мозге.
Двойное (двигательное) ядро, сокращая мышцы неба, глотки, верхней час-
ти пищевода и гортани, участвует в рефлексах глотания, рвоты, чихания,
кашля, в формировании голоса. Чувствительное ядро одиночного пути, ин-
нервируя слизистую оболочку неба, корня языка, дыхательных путей, аор-
тальное тельце, органы шеи, грудной и брюшной полости, участвует в ка-
честве афферентного звена в глотательном, жевательном, дыхательных,
висцеральных рефлексах. Оно входит в проводниковый отдел интероцеп-
тивного, вкусового, тактильного, температурного и болевого анализаторов.
Заднее (парасимпатическое) ядро, иннервируя сердце, гладкую мускулату-
ру и железы органов шеи, грудной и брюшной полостей, участвует в сер-
дечных, легочных, бронхиальных, пищеварительных рефлексах.
• > Двигательное ядро добавочного нерва (n. accessorius, XI) расположено в про-
долговатом и спинном мозге. Сокращая грудино-ключично-сосцевидную и
трапециевидную мышцы, оно вызывает наклон головы набок с поворотом
лица в противоположную сторону, поднимание плечевого пояса вверх,
приведение лопаток к позвоночнику.
❖ Двигательное ядро подъязычного нерва (n. hypoglossus, XII) расположено в
продолговатом мозге; иннервируя мышцы языка, вызывает его движение в
рефлексах жевания, сосания, глотания и осуществлении речи.
Таким образом, с участием ядер черепных нервов реализуется сенсор-
ная и рефлекторная (соматическая и вегетативная) функции ствола мозга.
2.2. Сложные цепные ритмические рефлексы ствола мозга (рефлексы жевания,
глотания, рвоты, кашля, чихания), в каждом из которых задействованы ядра не-
скольких черепных нервов, осуществляются с интегрирующим участием рети-
кулярной формации, они рассмотрены в физиологических системах пищеваре-
ния и дыхания (см. тему 19; задание 1: 12.1 и тему 22; задание 1:8.2 и 9.1).
2.3. Роль ствола мозга в регуляции мышечного тонуса и установочных рефлек-
сов, (см. тему 11; задание 1:4.3 и 4.4).
2.4. Ретикулярная формация (РФ) ствола мозга
❖ РФ образована нейронами, расположенными в центральных отделах ствола
мозга как диффузно, так и в виде ядер. Нейроны РФ имеют длинные мало-
ветвящиеся дендриты и хорошо ветвящийся аксон, который позволяет ней-
рону установить контакты примерно с 25000 других нейронов. Нейроны
РФ принимают импульсы от сенсорных путей, идущих от разных рецепто-
86
ров. Это полимодальные нейроны, имеющие большие рецепторные поля. У
нейронов РФ длительный латентный период ответа на периферическую
стимуляцию, связанный с проведением возбуждения до них через много-
численные синапсы. Они имеют тоническую активность, равную в покое 5
- 10 имп/с. Нейроны РФ обладают высокой чувствительностью к некото-
рым веществам кровн (например, адреналину, СОт) и лекарствам (барбиту-
ратам, аминазину и др.).
Ретикулярная формация имеет связи с многочисленными структурами
ЦНС (рис. 26).
• Афферентные входы поступают в РФ преимущественно от трех источ-
ников: 1) от температурных н болевых рецепторов по волокнам спино-
ретикулярного пути и тройничного нерва; 2) от сенсорной и, частично,
других зон коры головного мозга по кортикоретикулярным путям им-
пульсация поступает в ядра, дающие начало ретикулоспинальным пу-
тям, а также в ядра, которые проецируются на мозжечок; 3) от ядер
мозжечка по мозжечковоретикулярному пути.
РФ ствола мозга
Кора большого
мозга
Тройничный
нерв
Мозжечок
Спиноретику-
лярный тракт
Кора большого
мозга
t - . I
Таламус
* Гипоталамус
Базальные ядра
» Мозжечок
Ретикулос нина-
* льные тракта
Афферентные входы
Эфферентные выходы
Рис. 26. Схема основных афферентных и эфферентных связей
ретикулярной формации ствола мозга.
• Эфферентные выходы из РФ проецируются: 1) в спинной мозг по лате-
ральному и медиальному ретикулоспинальным путям; 2) к верхним от-
делам головного мозга (неспецифическим ядрам таламуса, заднему ги-
поталамусу, полосатому телу) идут восходящие пути, начинающиеся в
ядрах продолговатого мозга и моста; 3) к мозжечку идут пути, которые
начинаются в латеральном н парамедианном ретикулярных ядрах и в
ядре покрышки моста. Многообразие связей и структур РФ определяет
ее многочисленные функции, которые можно объединить в трн главные
87
группы: соматические (двигательные), сенсорные (восходящее влияние
на большой мозг) и вегетативные.
<• Соматические функции РФ проявляются в ее координирующем влиянии
на двигательные ядра черепных нервов, нисходящем влиянии на моторные
спинальные центры и активность мышечных рецепторов (см. тему 11, задание
1:4.3 и 4.4).
❖ Восходящее влияние РФ на большой мозг может быть как активирующим,
так и тормозным. Импульсы ретикулярных нейронов продолговатого моз-
га, моста и среднего мозга поступают к неспецифическим ядрам таламуса,
и после переключения в них проецируется в различные области коры.
• Активирующее влияние восходящей РФ показано в опыте по перерезке
ствола мозга между верхними и нижними буграми четверохолмия (Ф.
Бремер, 1935). Такое животное вело себя как спящее: у него был нарушен
контакт с внешним миром, оно не реагировало на световые и обоня-
тельные раздражители (спящий мозг, по Бремеру). Такое же состояние го-
ловного мозга остается, если при перерезке ствола сохранить основные
афферентные пути (например, от лица по тройничному нерву), и повре-
дить только восходящие пути РФ. Прямое доказательство влияния РФ
по восходящим путям на бодрствующее состояние мозга было получено
в хронических опытах с раздражением РФ через погружные электроды
у сонных животных (X. Мегун, Дж. Моруцци, 1949). Стимуляция РФ вызы-
вала пробуждение животного. У него возникала ориентировочная реак-
ция, a-ритм и более медленные ритмы сменялись высокочастотным р-
ритмом, что называется реакцией десинхронизации ЭЭГ и свидетельст-
вует об активном состоянии коры головного мозга. Она определялась в
различных зонах коры, что свидетельствует о генерализованном влия-
нии РФ. Активирующее влияние восходящей РФ участвует в регуляции
цикла «сон-бодрствование» и уровня сознания.
• Тормозное влияние РФ на большой мозг изучено значительно хуже. По-
казано, что раздражением некоторых точек РФ ствола мозга можно пе-
ревести животное из бодрствующего состояния в сонное (В. Гесс, 1929,
Дж. Моруцци, 1941). При этом на ЭЭГ возникает реакция синхронизации
биоритмов.
❖ Вегетативные функции ретикулярной формации. РФ поддерживает то-
нус вегетативных центров, интегрирует симпатические и парасимпатиче-
ские влияния для реализации потребностей целостного организма, передает
модулирующее влияние от гипоталамуса и мозжечка к внутренним орга-
нам. РФ является важнейшим компонентом жизненно важных центров
продолговатого мозга - сердечно-сосудистого и дыхательного.
2.5. Нейрорегуляторная функция ствола головного мозга. Кроме нейронов, пе-
редающих свое влияние с помощью медиаторов, имеются группы нейронов, ко-
торые выделяют нейрорегуляторы (амины и пептиды), изменяющие метаболи-
ческую активность и функциональное состояние иннервируемых нейронов, их
способность отвечать на действие медиатора, модулируя таким образом синап-
тическую передачу. Изучение нейронов с идентифицированными нейромодуля-
88
торами позволило выделить в стволе мозга нейрорегуляторные системы, кото-
рые действуют как «регуляторы регуляторов».
❖ Норадренергическая система ствола мозга. Норадренергические нейроны
(их медиатор - норадреналин) имеются преимущественно в двух ядрах
ствола - голубом пятне (ГП) среднего мозга и вентролатеральном ядре ре-
тикулярной формации продолговатого мозга. Главной структурой является
ГП (locus coeruleus), содержащее более 50% всех норадренергических нейро-
нов ствола мозга. ГП имеет обширные афферентные и эфферентные связи
со всеми отделами ЦНС.
• Нейроны ГП имеют фоновую импульсную активность (10-20 Гц), зави-
сящую от уровня бодрствования. На мембране нейронов имеются ре-
цепторы, вызывающие их торможение (рецепторы к норадреналину, ад-
реналину, серотонину, опиоидным пептидам, ГАМК, глицину) или воз-
буждение (рецепторы к ацетилхолину, пептиду Р, глутамату).
• Стимуляция ГП, как правило, угнетает спонтанную активность иннер-
вируемых нейронов (действие через Р-адреиорецепторы), ио у части
нейронов (например, ядра VII черепного нерва) норадреналин через а-
адреиорецепторы вызывает возбуждение. При этом интернейроны более
чувствительны к норадреналину, чем мотонейроны. В соответствующих
стимуляции участках мозга происходит сужение артерий и снижение
локального кровотока.
• Норадренергическая система контролирует деятельность сенсорных
систем по проведению болевой и неболевой импульсации, а также
моторных систем, угнетая фоновую активность нейронов их ядер. Она
участвует в регуляции деятельности висцеральных систем, эмоциональ-
ных состояний тоски и страха, механизмов памяти и внимания, фаз цик-
ла «сои - бодрствование», индуцируя фазу быстрого сна. Иннервируя
ядра гипоталамуса, нейроны ГП включаются в центральное звено
стресс-формирующей системы, участвуя в развитии общего адаптаци-
онного синдрома.
❖ Серотонинергическая система ствола мозга образована скоплением серо-
тонинергических нейронов (СТ-нейроиов), образующих ядра шва, цен-
тральное серое вещество и ретикулярные магиоцеллюляриые ядра. Она
имеет хорошо выраженные афферентные и эфферентные связи со всеми
отделами ЦНС. Фоновая активность СТ-нейроиов наиболее высока во вре-
мя бодрствования, снижается во время «медленного» сна и прекращается в
фазе «быстрого» сна.
Стимуляция СТ-системы ствола мозга тормозит нейроны сенсорных
путей спинного мозга, ствола и таламуса. Среди них особенно сильно угне-
таются нейроны, передающие болевую информацию. СТ-система тормозит
также симпатические и парасимпатические нейроны спинного мозга, ио
оказывает возбуждающее действие на его мотонейроны. Полагают, что СТ-
система участвует в контроле проведения болевой чувствительности, в ре-
гуляции цикла «сои - бодрствование», потребления пищи и воды, тормозит
сексуальное и агрессивное поведение.
89
❖ Дофаминергическая система ствола мозга представлена черным вещест-
вом среднего мозга, нейроны которого передают свое влияние с помощью
медиатора дофамина. В функциональном плане черное вещество включа-
ют в стрнопаллцдарную систему (см. тему 11; задание 1: 6).
2.6. Проводниковая функция ствола головного мозга выполняется восходящими
и нисходящими путями, часть из которых идет без переключения, другая часть
переключается в стволовых центрах.
❖ Восходящие (афферентные) пути являются частью проводникового отдела
анализаторов, передающих информацию от рецепторов в сенсорную кору.
В стволе мозга выделяют две восходящие системы: специфическую и не-
специфическую.
• Специфическую афферентную (лемнисковую) систему составляют вос-
ходящие пути, переключающиеся в вентральных задних (латеральных и
медиальных) ядрах, латеральном и медиальном коленчатых телах тала-
муса.
ф Медиальная петля образуется аксонами нейронов тонкого (Голля) и
клиновидного (Бурдаха) ядер, которые проводят от рецепторов ко-
нечностей, туловища и шеи проприоцептивную, тактильную и вис-
церальную чувствительность. Волокна медиальной петли переклю-
чаются в заднелатеральных вентральных ядрах таламуса.
• ф Тройничная петля (тригеминально-таламический путь от ядер V
нерва) проводит от головы проприоцептивную и тактильную чувст-
вительность и переключается заднемедиальном вентральном ядре.
• ф Латеральная петля (или слуховая петля) образована волокнами слухо-
вого пути, которые переключаются в медиальном коленчатом теле
таламуса.
• ф Спинномозговая петля (волокна спиноталамических путей) прово-
дит тактильную и локализованную болевую чувствительность, пе-
реключается в заднелатеральных вентральных ядрах таламуса.
• ф К специфической проводящей системе в функциональном отноше-
нии относятся проводящие пути зрительного анализатора, волокна
которого не входят в состав лемнисков, но переключаются в сен-
сорном релейном ядре таламуса - латеральном коленчатом теле.
• ф Функциональной особенностью специфической проводящей систе-
мы является высокая скорость проведения возбуждения, ее нейро-
ны имеют небольшие рецептивные поля, они преимущественно мо-
номодальны. В центрах переключения афферентных нейронов (в
стволе, таламусе) имеется выраженная топографическая проекция
периферии. В результате обеспечивается быстрая передача инфор-
мации с тонким различением свойств раздражителя. При этом в
корковом отделе соответствующего анализатора с коротким ла-
тентным периодом возникают вызванные потенциалы, называемые
первичным ответом (см. тему 6; задание 1:4.5).
• Неспецифическую (экстралемиисковую) восходящую систему состав-
ляют преимущественно волокна бокового спиноталамического и спино-
90
ретикулярного путей и часть тригемиио-таламических волокон, прово-
дящих температурную и болевую чувствительность. Они прямо или че-
рез нейроны активирующей РФ ствола переключаются в неспецифиче-
ских ядрах таламуса и далее проецируются в различные области коры,
особенно лобную орбитальную кору.
Функциональной особенностью неспецифической системы явля-
ется медленное проведение информации с плохой локализацией раз-
дражителя и анализом его свойств. Рецептивные поля нейронов боль-
шие, нейроны полимодальны, связанны с несколькими видами чувстви-
тельности, топография проекции периферии в центрах не выражена.
При проведении возбуждения через эту систему на ЭЭГ фиксируются с
большим латентным периодом вызванные потенциалы, называемые
вторичным ответом (см. тему 6; задание 1: 4.5). Неспецифическая сис-
тема получает коллатеральные волокна от специфической системы, что
обеспечивает связь этих двух афферентных систем.
• Кроме проведения возбуждения по специфической и неспецифической
афферентным системам часть сенсорной импульсации поступает в кору
мозжечка по дорсальному спииомозжечковому пути от рецепторов
мышц и связок, и вентральному спиномозжечковому пути от рецепто-
ров сухожилий, кожи и внутренних органов, а также по вестибуломоз-
жечковому пути от вестибулярных рецепторов. Из коры мозжечка через
его зубчатые и промежуточные ядра обработанная информация переда-
ется по церебеллоталамическому пути в двигательные вентральные ядра
таламуса, из которых после переключения проецируется в соматосен-
сорную, моторную и премоторные зоны корю.
Нисходящие проводниковые пути ствола мозга в функциональном отно-
шении можно объединить в несколько групп.
• Двигательные пирамидные пути, начинающиеся от клеток Беца коры
прецентральной извилины, иннервируют мотонейроны передних рогов
спинного мозга (кортикоспинальный путь) или мотонейроны двига-
тельных ядер черепных нервов (кортикобульбарный путь), обеспечивая
произвольные сокращения мышц конечностей, туловища, шеи и головы
(подробнее см. тему 11; задание 1: 8).
• Моторные центры ствола мозга и их пути являются важнейшим компо-
нентом экстрапирамидной системы, основной функцией которой явля-
ется произвольная и рефлекторная регуляция мышечного тонуса, позы и
равновесия. К этой системе на уровне ствола мозга относятся кортико-
рубральиый и кортикоретикуляриый пути, оканчивающиеся на мотор-
ных центрах ствола, от которых идут отмеченные выше рубро- и рети-
кулоспинальный пути.
• В стволе мозга проходят нисходящие пути, обеспечивающие двигатель-
ные функции мозжечка. К ним относятся мощный кортикомостомоз-
жечковый путь, по которому в мозжечок поступает импульсация от дви-
гательной и других областей коры. Обработанная в коре мозжечка и его
ядрах информация поступает на моторные ядра ствола (красное, вести-
91
булярные, ретикулярные), что включает мозжечок в функции экстрапи-
рамидной системы.
• Через ствол мозга проходит начинающийся в четверохолмии тектоспи-
нальиый путь, который обеспечивает двигательные реакции организма в
ориентировочных зрительном и слуховом рефлексах.
3. Функции промежуточного мозга. Промежуточный мозг состоит из таламической облас-
ти и гипоталамуса. Таламическая область включает в себя таламус и коленчатые тела, кото-
рые в нейрофизиологии рассматриваются совместно, и эпиталамус (эпифиз).
3.1. Таламус (thalamus - зрительный бугор) представляет собой парный ядерный ком-
плекс, включающий до 60 ядер, которые можно разделить иа следующие три
главные группы ядер: релейные, ассоциативные и неспецифические. Все ядра
таламуса в разной степени обладают тремя общими функциями: переключаю-
щей, интегративной и модулирующей.
Рис. 27. Основные афферентные и эфферентные связи
ядер таламуса
♦♦♦ Релейные ядра таламуса (синонимы: специфические, переключательные) разделя-
ют на сенсорные и несенсорные.
• Сенсорные ядра переключают потоки афферентной импульсации в
сенсорные зоны коры (рис. 27). В них также происходит перекодирова-
ние и обработка информации. Выделяют три основных структуры.
ф- Вентральные задние ядра (заднелатеральное и заднемедиальное)
являются главным реле для переключения соматосенсорной аффе-
92
рентной системы, импульсы которой поступают по волокнам меди-
альной петли и по волокнам других афферентных путей (см. выше
2.6). В них переключаются тактильная, проприоцептивная, вкусо-
вая, висцеральная, частично температурная и болевая чувствитель-
ность. В этих ядрах имеется топографическая проекция периферии;
при этом функционально более тонко организованные части тела
(например, язык, лицо) имеют бблыпую зону представительства.
Импульсация из вентральных задних ядрах проецируется в сомато-
сенсорную кору постцентральной извилины, где формируются со-
ответствующие ощущения. Электростимуляция вентральных зад-
них ядер вызывает парестезии (ложные ощущения) в разных частях
тела, иногда нарушение «схемы тела» (искаженное восприятие частей
тела).
ф- Латеральное коленчатое тело является реле для переключения зри-
тельной нмпульсацин в затылочную кору, где она используется для
формирования зрительных ощущений.
ф- Медиальное коленчатое тело является реле для переключения слу-
ховой и, возможно, вестибулярной нмпульсацин в височную кору.
• ф' Переключательную функцию в сенсорных ядрах обеспечивают та-
ламокортикальные (релейные) нейроны, длинный аксон которых
непосредственно иннервируют нейроны сенсорной коры. Особен-
ностью этой передачи является ее высокая точность с минималь-
ным искажением входного сигнала. Регуляция этой передачи осу-
ществляется с помощью тормозных н возбуждающих интернейро-
нов сенсорных ядер. (Например, через них осуществляется корковый кон-
троль проведения возбуждения через сенсорные ядра таламуса.)
Несенсорные релейные ядра таламуса (передние и вентральные ядра)
переключают в кору несенсорную импульсацию, поступающую в тала-
мус из разных отделов головного мозга.
• ф' В передние ядра импульсация поступает в основном из мамилляр-
ных тел гипоталамуса. Нейроны передних ядер проецируются в
лимбическую кору. От нее аксонные связи идут к гиппокампу и
опять к гипоталамусу, в результате чего образуется нейронный
круг, движение возбуждения по которому обеспечивает формиро-
вание эмоций (см. ниже 5.1). В связи с этим, передние ядра таламуса
рассматриваются как часть лимбической системы и обозначаются
как «лимбические» ядра таламуса (рис. 28).
• ф' Вентральные ядра участвуют в регуляции движения, выполняя та-
ким образом моторную функцию. В них переключается импульса-
ция от базальных ядер, зубчатого ядра мозжечка, красного ядра
среднего мозга, которая затем проецируется в моторную и премо-
торную кору (поля 4 и 6). Через эти ядра таламуса происходит пере-
дача в моторную кору сложных двигательных программ, образо-
ванных в мозжечке и базальных ядрах.
93
• Специфические ядра не только образуют корковые проекции, но и по-
лучают нисходящие корковые волокна из своей проекционной зоны, что
создает структурную основу для кольцевых взаиморегулирующих от-
ношений между корой и таламусом.
❖ Ассоциативные ядра таламуса принимают импульсацию ие от проводни-
ковых путей анализаторов, а от других ядер таламуса. Эфферентные выхо-
ды от этих ядер направляются преимущественно в ассоциативные поля ко-
ры (рис. 27). В свою очередь, кора мозга посылает волокна к ассоциатив-
ным ядрам, регулируя их функции. Главной из них является интегративная
функция, которая выражается в объединении деятельности как таламиче-
ских ядер, так и различных зон ассоциативной коры полушарий мозга.
• Подушка получает главные входы от коленчатых тел и неспецифиче-
ских ядер таламуса. Эфферентные пути от нее идут в височио-темеиио-
затылочные зоны коры, участвующие в гностических (узнавание пред-
метов, явлений), речевых и зрительных функциях (например, в интегра-
ции слова со зрительным образом), а также в восприятии «схемы тела».
Электростимуляция подушки доминантного полушария приводит к ре-
чевым нарушениям типа аномии (нарушение называния предметов), разруше-
ние подушки вызывает нарушение «схемы тела».
• Медиодорсальное ядро получает входы от гипоталамуса, миндале-
видного тела, гиппокампа, таламических ядер, центрального серого ве-
щества ствола. Проекция этого ядра распространяется на ассоциатив-
ную лобную и лимбическую кору. Оно участвует в формировании эмо-
циональной и поведенческой двигательной активности, а также в запо-
минании. Разрушение этих ядер устраняет у больных страх, тревогу, на-
пряженность, страдание от боли, но возникает лобный синдром: сниже-
ние инициативы, безразличие, гипокинезия.
• Латеральные ядра получают зрительную и слуховую импульсацию
от коленчатых тел и соматосенсорную импульсацию от вентрального
ядра. Интегрированная сенсорная информация от этих источников далее
проецируется в ассоциативную теменную кору и используется в функ-
ции гнозиса, праксиса, формировании «схемы тела».
❖ Неспецифические ядра составляют эволюционно более древнюю часть та-
ламуса, включающую парные интраламинарную ядерную группу и ретику-
лярные ядра. Эти ядра содержат нейроны «ретикулярного типа» и функ-
ционально рассматриваются как продолжение ретикулярной формации на
уровне промежуточного мозга.
• Неспецифические ядра имеют многочисленные входы как от других
ядер таламуса, так и внеталамнческие: по латеральному спиноталамиче-
скому, спиноретикулоталамическому путям, текто- и тегментоталами-
ческим путям, проводящим преимущественно болевую и температур-
ную чувствительность. В неспецифические ядра поступает непосредст-
венно или через ретикулярную формацию также часть импульсации по
коллатералям от всех специфических сенсорных систем. Кроме того, в
неспецифические ядра поступает импульсация из моторных центров
94
ствола (красное ядро, черное вещество), ядер мозжечка, от базальных
ядер и гиппокампа, а также от коры мозга, особенно лобных долей. Не-
специфические ядра имеют эфферентные выходы на другие таламиче-
ские ядра, кору больших полушарий, а также нисходящие пути к другим
структурам ствола мозга.
• Благодаря этим связям неспецифические ядра таламуса выступают в
роли интегрирующего посредника между стволом мозга и мозжечком с
одной стороны, и новой корой, лимбической системой и базальными
ядрами с другой стороны, объединяя их в единый функциональный
комплекс. На кору мозга неспецифический таламус оказывает преиму-
щественно модулирующее (изменяющее состояние) влияние. Разруше-
ние неспецифических ядер не вызывает грубых расстройств эмоций,
восприятия, сна и бодрствования, образования условных рефлексов, но
нарушает тонкую регулировку поведения. В связи с этим их главной
функцией считается модулирующее влияние, обеспечивающее «плавную
настройку» высшей нервной деятельности.
3.2. Гипоталамус включает в себя преоптическую область и область перекреста
зрительных нервов, серый бугор и воронку, сосцевидные (мамиллярные) тела.
❖ В гипоталамусе выделяют, по данным разных авторов, от 15 до 48 парных ядер,
которые подразделяются на 3 - 5 групп. Обычно выделяют 3 основные группы
ядер: 1) передняя группа содержит медиальное преоптическое, супрахиазматиче-
ское, супраоптическое, паравентрнкулярное и переднее гипоталамическое ядра; 2)
средняя группа включает дорсомедиальное, вентромедиальное, аркуатное (дуго-
образное) и латеральное гипоталамические ядра; 3) задняя группа включает суп-
рамамилляриое, премамилляриое, мамиллярные ядра, заднее гипоталамическое н
перифорниатное ядра, субталамическое ядро Луиса.
❖ Важной физиологической особенностью гипоталамуса является высокая
проницаемость его гематоэнцефалического барьера для различных ве-
ществ, в том числе и для полипептидов. Это обусловливает большую чув-
ствительность гипоталамуса к сдвигам во внутренней среде организма и
способность реагировать на колебания концентрации гуморальных факто-
ров. В гипоталамусе, по сравнению с другими структурами головного моз-
га, имеется самая мощная сеть капилляров (1100 - 2600 на мм2) и самый боль-
шой уровень локального кровотока.
❖ Ядра гипоталамуса образуют многочисленные связи друг с другом, а так-
же с выше- и нижележащими структурами ЦНС. Главные афферентные
пути в гипоталамус идут от лимбической системы, коры больших полуша-
рий, базальных ядер и ретикулярной формации ствола. Основные эффе-
рентные пути из гипоталамуса идут в ствол мозга - его ретикулярную
формацию, моторные и вегетативные центры, в вегетативные центры спин-
ного мозга, от мамиллярных тел к передним ядрам таламуса и далее, в лим-
бическую систему, от супраоптического и паравентрикулярного ядер к
нейрогипофизу, от вентромедиального и аркуатного ядер к аденогипофизу,
а также эфферентные выходы к лобной коре н полосатому телу.
❖ Гипоталамус является многофункциональной системой, обладающей ши-
рокими регулирующими и интегрирующими влияниями.Однако важней-
95
шие функции гипоталамуса трудно соотнести с его отдельными ядрами.
Как правило, отдельно взятое ядро имеет несколько функций, а отдельно
взятая функция локализуется в нескольких ядрах. В связи с этим физиоло-
гия гипоталамуса рассматривается обычно в аспекте функциональной спе-
цифики его различных областей и зон.
• Гипоталамус как центр интеграции вегетативных функций (см. тему 8;
задание 1: 8.2).
• Гипоталамус как центр регуляции эндокринной системы (см. тему 9;
задание 1: 2.5).
• Роль гипоталамуса в терморегуляции (см. тему 26; задание 1:4.3).
• Роль гипоталамуса в регуляции поведения. Гипоталамус объединяет от-
дельные жизненно важные функций в сложные комплексы, обеспечи-
вающие различные формы биологически целесообразного поведения:
пищевого, полового, питьевого, агрессивно-оборонительного и др., на-
правленные на выживание индивидуума и вида. Однако в осуществле-
нии даже биологических форм поведения гипоталамус обеспечивает
только базовые механизмы. Эмоциональный компонент поведения осу-
ществляется с обязательным участием лимбической системы, корковых
структур, а социализация биологического поведения - с участием новой
коры, особенно лобной доли.
-$ Пищевое поведение (см. тему 22; задание 1: 4).
-$ Половое поведение (см. тему 33; задание 1: 6).
-$ Поведение «сон - бодрствование» (см. тему 31; задание 1: 2.8).
ф- Питьевое поведение. Электрическое раздражение точек гипотала-
муса, расположенных сзади н наружно от супраоптических ядер
вызывает резко выраженную активацию питьевого поведения и по-
требления воды (полидипсию). Разрушение этой зоны, напротив,
приводит к отказу от приема воды (адипсии). Эта зона гипоталаму-
са была названа «центром жажды». В дальнейшем было показано,
что часть нейронов «центра жажды» обладают осморецептивными
свойствами и стимулируются повышением осмотического давления
крови. На активность «центра жажды» влияют также импульсы от
периферических (сосудистых и тканевых) осморецепторов и кон-
центрация некоторых веществ крови (например, АДГ).
-$ Агрессивно-оборонительное поведение. Агрессивные и оборони-
тельные реакции были получены при раздражении различных зон
гипоталамуса; передней и задней, вентромедиальной и латеральной
(в. Гесс, 1928). Вероятно, это связано с тем, что имеются различные
виды агрессивности, направленные на борьбу за самосохранение, за
лидерство в группе, за территорию и др. Типичный симптомоком-
плекс этих реакций состоит в ярких проявлениях отрицательных
эмоций (гнев, ярость, страх), резкими вегетативными эрготропными
сдвигами, попытками к нападению или бегству. Вместе с тем агрес-
сивное поведение может быть без эмоционального возбуждения
96
(«холодная атака») или сопровождаться положительной эмоцией.
Перерезка ствола мозга ниже гипоталамуса ликвидирует агрессив-
ное поведение. В осуществлении агрессивно-оборонительных реак-
ций гипоталамус взаимодействует с центральным серым веществом
среднего мозга. В этой структуре были обнаружены «нейроны аг-
рессии» (Д. Адамс, 1968), которые через гипоталамус запускают ре-
акции агрессии и не возбуждаются при других реакциях. При по-
вреждении этих нейронов резко увеличивается порог электрическо-
го раздражения гипоталамуса для вызова реакции ярости. На агрес-
сивное поведение выраженное стимулирующее влияние оказывают
андрогены, особенно тестостерон (пороговая величина 1-2 мкг/л).
ф- Перерезка ствола мозга сразу выше гипоталамуса (У. Кеннон) пока-
зала, что у животных легко возникает агрессивное поведение с ре-
акциями ярости и вегетативными проявлениями симпатического
характера. Однако они не направлены иа конкретный объект
(«ложная агрессия и ярость»). Эти данные свидетельствуют о важ-
ной роли вышележащих отделов мозга в формировании «осмыс-
ленного» агрессивного поведения. Большое значение в этом аспекте
имеет миндалевидный комплекс височной доли, раздражение ко-
торых резко активирует (а разрушение - тормозит) агрессию. Огра-
ничивающее действие на агрессивное поведение и соотношение его
с морально-правовыми нормами оказывает лобная кора (в частно-
сти, орбитальные зоны).
3.3. Эпифиз - эндокринная железа, расположенная в области промежуточного моз-
га (см. тему 9; задание 1: 11).
4. Функции мозжечка. Мозжечок - это отдел головного мозга, образующий вместе с мостом
задний мозг. Составляя 10% массы головного мозга, мозжечок включает в себя более поло-
вины всех нейронов ЦНС. Это свидетельствует о больших возможностях обработки инфор-
мации н соответствует главной функции мозжечка как органа координации и контроля слож-
ных автоматизированных движений. В осуществлении этой функции важную роль играют
обширные связи мозжечка с другими отделами ЦНС и рецепторным аппаратом.
4.1. Роль мозжечка в регуляции движения (см. тему 11; задание 1:5).
4.2. Роль мозжечка в регуляции вегетативных функций (см. тему 8; задание 1:8.3).
5. Лимбическая система. Под лимбической системой понимают функциональное объ-
единение различных структур конечного, промежуточного и среднего мозга, обеспе-
чивающее эмоционально-мотивационные компоненты поведения и интеграцию вис-
церальных функций организма.
5.1. Структурно-функциональная организация. В лимбическую систему включа-
ют образования древней и старой коры (обонятельный бугорок, гиппокамп,
крючок, парагиппокампальная, зубчатая и поясная извилины), подкорковые яд-
ра (миндалевидное тело, ядра перегородки), мамиллярные тела гипоталамуса,
передние ядра таламуса, а также орбитальную лобную кору.
❖ Афферентные входы в лимбическую систему осуществляются от различ-
ных областей головного мозга, а также через гипоталамус от РФ ствола, ко-
торая считается главным источником ее возбуждения. Важным стимули-
97
рующим афферентным входом являются также импульсы от обонятельных
рецепторов по волокнам обонятельного нерва (n. olfactorii, I), поступающие
в различные структуры лимбической системы.
❖ Эфферентные выходы из лимбической системы осуществляются через ги-
поталамус (особенно его мамиллярные тела) на нижележащие вегетатив-
ные и соматические центры ствола и спинного мозга. Другой эфферентный
выход проводит возбуждение из лимбической системы в новую кору (пре-
имущественно ассоциативную). Через нее лимбическая система включается
в регуляцию высших психических функций.
❖ Кольцевые нейронные связи объединяют различные ее структуры
(рис. 28) и дают возможность длительной циркуляции (реверберации) воз-
буждения, которая является механизмом его пролонгирования, повышения
проводимости синапсов и запоминания информации. Реверберация возбу-
ждения создает условия для сохранения единого функционального состоя-
ния структур замкнутого круга и навязывания этого состояния другим
структурам мозга. Важнейшим циклическим образованием лимбической
системы является лимбический круг Пейпеца, идущий от гиппокампа через
свод к мамиллярным телам, от них к передним ядрам таламуса, от него в
поясную извилину и от нее через парагиппокампову извилину обратно к
гиппокампу. Этому кругу придается большое значение в формировании
эмоций, обучении и памяти. Другой лимбический круг (от миндалевидного
тела к мамиллярным телам гипоталамуса, от них к лимбической области
среднего мозга и обратно к миндалине) имеет важное значение в форми-
ровании агрессивно-оборонительных, пищевых и сексуальных реакций.
5.2. Функции лимбической системы. Получая информацию о внешней среде и о
внутренней среде организма, лимбическая система после сравнения и обработ-
ки этой информации запускает вегетативные, соматические и поведенческие
реакции, обеспечивающие адекватное приспособление организма к внешней
среде и сохранение гомеостаза. В этом приспособлении задействованы сле-
дующие функции лимбической системы.
Неясная извилина
Парагиппокампова
извилина
Тала
даере)
Гиппокамп
Рис. 28. Схема основ-
ных внутренних свя-
зей лимбической сис-
темы. А - круг Пейпе-
ца, Б - круг через
миндалину (амигдалу),
ГТ/МТ - мамиллярные
тела гипоталамуса,
СМ - средний мозг
(лимбическая об-
ласть).
Регуляция висцеральных функций. В связи с этой функцией лимбическую
систему иногда называют «висцеральным мозгом». Оиа осуществляется
98
преимущественно через гипоталамус, который является диэнцефалическим
звеном лимбической системы. О тесных эфферентных связях лимбической
системы с внутренними органами свидетельствуют разнообразные измене-
ния их функций при раздражении лимбических структур, особенно минда-
лин. При этом эффекты имеют различный знак в виде активации или угне-
тения висцеральных функций: происходит повышение или понижение сек-
реции различных гормонов аденогипофизом (особенно АКТГ и гонадотро-
пинов), частоты сердечных сокращений, моторики и секреции желудка и
кишечника.
> Формирование эмоций (переживаний, в которых отражается субъективное отношение
человека к предметам внешнего мира и результатам собственной деятельности) - важ-
нейшая функция лимбической системы (см. также тему 31; задание 1:1).
• Важную роль в лимбической системе при формировании эмоций игра-
ют поясная извилина и миндалевидное тело. Электрическая стимуляция
миндалевидного тела у человека вызывает преимущественно отрица-
тельные эмоции - страх, гнев, ярость. Напротив, его двустороннее уда-
ление у обезьян резко снижает их агрессивность, повышает тревож-
ность, неуверенность в себе. У таких животных нарушается способность
оценивать информацию (особенно зрительную и слуховую), поступающую
из окружающей среды, и связывать эту информацию со своим эмоцио-
нальным состоянием. В результате нарушается нормальное взаимодей-
ствие организма с окружающей средой, в том числе зоосоциальные от-
ношения с другими особями в группе. Наряду с этим миндалевидное те-
ло участвует в процессе сравнения конкурирующих эмоций, выделения
доминирующей эмоции (и мотивации) и, следовательно, влияет на вы-
бор поведения.
• Поясная извилина, имеющая многочисленные связи, как с новой корой,
так и со стволовыми центрами, выполняет, вероятно, роль главного ин-
тегратора различных систем мозга, участвующих в формировании эмо-
ций.
Роль лимбической системы в обучении преимущественно связана с основ-
ным лимбическим кругом Пейпеца (рис. 28). Среди структур лимбической
системы, ответственных за память и обучение, главную роль играют гиппо-
камп и связанные с ним задние зоны лобной коры. Их деятельность совер-
шенно необходима для консолидации памяти - перехода кратковременной
памяти в долговременную. Повреждение гиппокампа у человека вызывает
резкое нарушение усвоения новой информации, образования промежуточ-
ной и долговременной памяти.
• Электрофизиологической особенностью гиппокампа является то, что в
ответ на сенсорное раздражение, стимуляцию ретикулярной формации и
заднего гипоталамуса в гиппокампе развивается синхронизация элек-
трической активности в виде низкочастотного тета-ритма (4 — 7 Гц). При
этом в новой коре, напротив, возникает десинхронизация в виде высо-
кочастотного бета-ритма (14 - 30 Гц). Пейсмекером тета-ритма считают
перегородку (ее медиальное ядро). По мнению большинства исследовате-
99
лей, он связан с обработкой мозгом информации и организацией памя-
ти.
• Другой электрофизиологической особенностью гиппокампа является
его уникальная способность в ответ на стимуляцию отвечать длитель-
ным (часы, дни н даже недели) увеличением амплитуды возбуждающих
постсинаптических потенциалов, что приводит к облегчению синапти-
ческой передачи и запоминанию информации. Ультраструктурным про-
явлением участия гиппокампа в механизмах памяти является увеличе-
ние числа шипиков на дендритах его пирамидных нейронов, что свиде-
тельствует об усилении синаптической передачи возбуждающих и тор-
мозных влияний.
• Вместе с тем, в одной из форм обучения (однократном обучении) важ-
ную роль играет миндалевидное тело благодаря свойству индуцировать
сильные отрицательные эмопии, что способствует быстрому и прочно-
му формированию временной связи.
❖ Сенсорная функция лимбической системы. В лимбической системе нахо-
дится корковый отдел обонятельного анализатора (наиболее вероятные
структуры - парагиппокампальная извилина, ее крючок, гиппокамп). Его глав-
ный эфферентный выход через свод, мамиллярные тела, передние ядра ги-
поталамуса на другие структуры лимбической системы объясняет наличие
резко выраженного эмоционального компонента в обонятельном воспри-
ятии. На этом базируется метод лечебного воздействия пахучих веществ -
ароматерапия.
6. Функции базальных ядер. Это совокупность трех парных образований, расположенных в
конечном мозге в основании больших полушарий: филогенетически древней части - бледно-
го шара (pallidum), более позднего образования - полосатого тела (striatum: хвостатое ядро +
скорлупа) и наиболее молодой части - ограды (claustrum), расположеной между скорлупой и
островковой корой. В функциональном отношении к базальным ядрам подключают структу-
ры промежуточного н среднего мозга: субталамические ядра и черное вещество (substantia
nigra), в совокупности все они составляют стриопаллидарную систему.
6.1. Роль базальных ядер в регуляции движений (см. тему 11; задание 1:6).
6.2. Влияние базальных ядер на высшую нервную деятельность.
❖ Влияние полосатого тела. Раздражение хвостатого ядра полосатого тела
(влияние передается через таламус) вызывает появление в коре больших
полушарий высокоамплитудных медленных ритмов, характерных для фазы
медленного сна - так называемую «каудатную депрессию». При этом на-
блюдается задержка поведенческих реакций (например, ориентировочной,
пищедобывательной): животное как бы «застывает» в одной позе. Разру-
шение полосатого тела уменьшает время сна в цикле «сон - бодрствова-
ние».
❖ Роль полосатого тела в организации поведения. Кошки с удаленной корой,
но сохраненными хвостатыми ядрами в упрощенном виде могут осуществ-
лять некоторые формы поведения: передвижение в пространстве, поиск
пищи и ранее выработанные простые условные рефлексы. Дополнительное
разрушение хвостатых ядер прекращает эти формы поведения.
100
❖ Стимуляция бледного шара вызывала активацию центра голода и пищевого
поведения, увеличение объема кратковременной памяти. Разрушение блед-
ного шара сопровождалось развитием сонливости, эмоциональной тупости,
затруднялось осуществление имеющихся и выработка новых условных
рефлексов.
❖ Таким образом, базальные ядра, кроме регуляции двигательных функций,
включаются в регуляцию цикла «сон - бодрствование», в формирование
условных рефлексов, в сложные формы восприятия (например, осмысление
текста), организацию эмоционально-мотивационной сферы и адекватного
приспособительного поведения.
7. Функции новой коры больших полушарий. Новая кора (неокортекс) представляет со-
бой слой серого вещества общей площадью 1100 - 1900 см2, покрывающий большие полу-
шария. Она составляет около 72% всей площади коры. В коре имеется около 14 млрд, нейро-
нов, количество глиоцитов примерно в 10 раз больше. Кора головного мозга является в фи-
логенетическом плане наиболее молодой нервной структурой, у человека она осуществляет
высшую регуляцию функций организма и психофизиологические процессы, обеспечиваю-
щие различные формы поведения.
7.1. Структурно - функциональная характеристика, В направлении с поверхности
вглубь коры различают б горизонтальных слоев.
❖ I. Молекулярный слой имеет очень мало клеток, но большое количество ветвящих-
ся дендритов пирамидных клеток, формирующих сплетение, расположенное па-
раллельно поверхности. На этих дендритах образуют синапсы афферентные во-
локна, приходящие от ассоциативных и неспецифических ядер таламуса.
* ** II. Наружный зернистый слой составлен в основном звездчатыми клетками и час-
тично малыми пирамидными клетками. Волокна клеток второго слоя расположе-
ны преимущественно вдоль поверхности коры, образуя кортикокортикальные свя-
зи.
♦ ♦♦ Ш. Наружный пирамидный слой состоит в основном из пирамидных клеток сред-
ней величины. Аксоны этих клеток, как и зернистые клетки П слоя, образуют кор-
тикокортикальные ассоциативные связи.
❖ IV. Внутренний зернистый слой по характеру клеток (звездчатые клетки) и распо-
ложению их волокон аналогичен наружному зернистому слою. В этом слое имеют
синаптические окончания афферентные волокна, идущие от нейронов специфиче-
ских ядер таламуса и, следовательно, от рецепторов сенсорных систем.
♦ ♦♦ V. Внутренний пирамидный слой образован средними и крупными пирамидными
нейронами, причем гигантские пирамидные клетки Беца расположены в двига-
тельной коре. Аксоны этих клеток образуют эфферентные кортикоспинальные и
кортикобульбарный двигательные пути.
♦ t* VI. Слой полиморфных клеток образован преимущественно веретенообразными
клетками, аксоны которых образуют кортикоталамические пути.
Оценивая в целом афферентные и эфферентные связи коры, можно отметить,
что в слоях I и IV происходит восприятие и обработка поступающих в кору сигна-
лов. Нейроны II и Ш слоев осуществляют кортикокортикальные ассоциативные
связи. Покидающие кору эфферентные пути формируются преимущественно в V -
VI слоях. Более детально деление коры на различные поля проведено на основе
цитоархитектонических признаков (формы и расположения нейронов) К. Бродма-
ном (1909), который выделил 52 поля, многие из которых характеризуются функ-
циональными и нейрохимическими особенностями.
101
7.2. Кортикализация функций (см. тему 6; задание 1:1.8).
7.3. Локализация функций в коре головного мозга. При разработке этой проблемы
были сформулированы две противоположные по смыслу концепции: 1) концеп-
ция узкого локализационизма, стремящаяся поместить функцию в одну отдель-
но взятую структуру; 2) концепция функциональной равноценности (эквипо-
тенциальности) различных корковых структур. Современная концепция локали-
зации функций базируется на принципе многофункциональности (но не равно-
ценности!) корковых полей. Свойство мультифункциональности позволяет дан-
ной корковой структуре включаться в обеспечение различных форм деятельно-
сти, реализуя при этом основную, генетически присущую ей функцию (О.С.
Андрианов, 1976). Степень мультифункциональности различных корковых
структур не одинакова; например, в полях ассоциативной коры она выше, чем в
первичных сенсорных полях, а в корковых структурах выше, чем в стволовых.
В основе мультифункциональности лежит многоканальность поступления в ко-
ру мозга афферентного возбуждения, перекрытия афферентных возбуждений
особенно на таламическом и корковом уровнях, модулирующее влияние раз-
личных структур (например, неспецифического таламуса, базальных ядер) на
корковые функции, взаимодействие корково-подкорковых и межкорковых пу-
тей проведения возбуждения. Результаты изучения локализации функций по-
зволили условно разделить кору головного мозга на различные функциональ-
ные зоны - картирование коры. Детальные функциональные карты (например. К.
Клейста, 1959) используются в неврологической практике, однако надо иметь в
виду упрощенный характер этих схем. Одним из наиболее крупных вариантов
функционального разделения коры головного мозга является выделение в ией
сенсорной, ассоциативной и двигательной областей.
7.4. Сенсорные области коры - зоны, в которые проецируются сенсорные раздра-
жители (синонимы: проекционная кора, корковые отделы анализаторов). Онн расположе-
ны преимущественно в теменной, височной и затылочной долях. Афферентные
пути в сенсорную кору поступают преимущественно от специфических сенсор-
ных ядер таламуса (вентральных задних и коленчатых тел).
4* Зоны сенсорной коры, раздражение илн разрушение которых вызывает
четкие и постоянные изменения чувствительности организма, называются
первичными сенсорными областями (ядерными частями анализаторов, по И.П.
Павлову). Они состоят преимущественно из мономодальных нейронов и
формируют ощущения одного качества. В первичных сенсорных зонах
обычно имеется четкое пространственное (топографическое) представи-
тельство частей тела, их рецепторных полей. Вокруг первичных сенсорных
зон находятся менее локализованные вторичные сенсорные зоны, полимо-
дальные нейроны которых отвечают на действие нескольких раздражите-
лей.
❖ Важнейшей сенсорной областью является теменная кора постцентраль-
ной извилины и соответствующая ей часть парацентральной дольки на ме-
диальной поверхности полушарий (поля 1 - 3), которую обозначают как
первичная соматосенсорная область (SI). Здесь имеется проекция кожной
чувствительности противоположной стороны тела от тактильных, болевых,
температурных рецепторов, интероцептивной чувствительности н чувстви-
102
тельности опорно-двигательного аппарата от мышечных, суставных, сухо-
жильных рецепторов. Проекция участков тела в этой области характеризу-
ется тем, что проекция головы и верхних отделов туловища расположена в
нижнелатеральных участках постцентральной извилины, проекция нижней
половины туловища и ног - в верхнемедиальных зонах извилины, проекция
ннжней части голени и стоп расположена в коре парацентральной дольки
на медиальной поверхности полушарий (рис. 29). При этом проекция наи-
более чувствительных участков (язык, губы, гортань, пальцы рук) имеют
относительно большие зоны по сравнению с другими частями тела. Пред-
полагается, что в зоне тактильной чувствительности языка расположена
проекция и вкусовой чувствительности.
Рис. 29. Схема чувствительного н двигательного гомункулусов (по
У. Пенфильду, Т. Расмуссену, 1950). Разрез полушарий во фрон-
тальной плоскости. А - проекция общей чувствительности в коре
постцентральной извилины. Б - проекция двигательной системы в
коре прецентральной извилины.
Кроме SI выделяют меньших размеров вторичную соматосенсорную
область (Sn), расположенную на верхней стенке боковой борозды, на гра-
нице ее пересечения с центральной бороздой. Функции филогенетически
более древней sn изучены плохо. Известно, что локализация поверхности
тела в ней менее четкая, импульсация сюда поступает как от противопо-
ложной, так и от «своей» стороны, предполагают ее участие в сенсорной и
моторной координации двух сторон тела.
Другой первичной проекционной зоной является слуховая кора (поля 41,42),
которая расположена в глубине латеральной борозды (кора поперечных ви-
сочных извилин Гешля). В этой зоне в ответ на раздражение слуховых ре-
цепторов кортиева органа формируются звуковые ощущения, изменяю-
щиеся по громкости, тону и другим качествам. Здесь имеется четкая топи-
ческая проекция: в разных участках коры представлены различные участки
кортиева органа. К проекционной коре височной доли относится предпо-
ложительно также центр вестибулярного анализатора в верхней и средней
височных извилинах (поля 20 и 21). Обработанная сенсорная информация
103
используется для формирования «схемы тела» и регуляции функций моз-
жечка (височномостомозжечковый путь).
❖ Еще одна первичная проекционная область новой коры расположена в за-
тылочной коре - первичная зрительная область (кора части клиновидной
извилины и язычковой дольки, поле 17). Около поля 17 расположена кора
вторичной зрительной области - поля 18 и 19 (подробно в теме 28; задание 1:5).
Основная часть информации об окружающей среде и внутренней среде
организма, поступившая в сенсорную кору, передается для дальнейшей ее
обработки в ассоциативную кору.
7.5. Ассоциативные области коры (синонимы: межсенсорная, межаналнзаторная кора)
включают участки новой коры большого мозга, которые расположены рядом с
сенсорными и двигательными зонами, но не выполняют непосредственно чув-
ствительных или двигательных функций. Границы этих областей обозначены
недостаточно четко, неопределенность преимущественно связана с вторичными
(высшими) проекционными зонами, функциональные свойства которых явля-
ются переходными между свойствами первичных проекционных и ассоциатив-
ных зон. Ассоциативная кора является филогенетически наиболее молодой ча-
стью новой коры, получившей наибольшее развитие у приматов и человека. У
последнего она составляет около 50% всей коры или 70% неокортекса.
Основной физиологической особенностью нейронов ассоциативной коры,
отличающей их от нейронов первичных зон, является полнсенсорность (поли-
модальность): они отвечают с почти одинаковым порогом не на один, а на не-
сколько раздражителей - зрительные, слуховые, кожные н др. Полнсенсорность
нейронов ассоциативной коры создается как ее кортико-кортикальными связями
с разными проекционными зонами, так и главным ее афферентным входом от
ассоциативных ядер таламуса, в которых уже произошла сложная обработка
информации от различных чувствительных путей. В результате этого ассоциа-
тивная кора представляет собой мощный аппарат конвергенции различных сен-
сорных возбуждений, позволяющий произвести сложную обработку информа-
ции о внешней и внутренней среде организма и использовать ее для осуществ-
ления высшнх психофизиологических функций (восприятия, внимания, мышле-
ния и др.).
По таламокортикальным проекциям выделяют три ассоциативные системы
мозга: таламотеменную, таламолобную и таламовисочную (см. тему 6; зада-
ние 1: 2.2).
Психофизиологические функции, осуществляемые ассоциативной корой,
инициируют поведение организма, обязательным компонентом которого явля-
ются произвольные целенаправленные движения, осуществляемые при обяза-
тельном участии двигательной коры.
7.6. Двигательные области коры (см. тему 11; задание 1:8).
7.7. Взаимоотношения правого и левого полушарий у человека проявляются в
двух главных формах - функциональной асимметрии больших полушарий и со-
вместной их деятельности.
❖ Функциональная асимметрия полушарий является важнейшим психофи-
зиологическим свойством головного мозга человека. Современные иссле-
104
дования показали, что в формировании одних психофизиологических
функций доминирует левое полушарие, в формирование других - правое
(концепция частичного доминирования полушарий Р. Сперри, 1981). Выделяют
психическую, сенсорную и моторную межполушарную функциональную
асимметрии мозга.
• Психическая асимметрия. В качестве примера психической асимметрии
приведем формирования устной речи (центр Брока), которое контроли-
руется левым полушарием как у правшей (в 95%), так и у левшей (в
70%). Другие виды психической асимметрии изложены в теме 31; зада-
ние 1.
• Сенсорная асимметрия. При зрительном восприятии правое полуша-
рие воспринимает зрительный образ целостно, сразу во всех подробно-
стях, легче решает задачу различения предметов и опознания бессмыс-
ленных предметов, которые трудно описать словами. Левое полушарие
оценивает зрительный образ расчленено, аналитически, при этом каж-
дый признак (форма, величина и др.) анализируются раздельно. Легче
опознаются знакомые предметы и решаются задачи сходства предметов,
зрительные образы лишены конкретных подробностей и имеют высо-
кую степень абстракции.
• Моторная асимметрия выражается в предпочтительном использовании
одной руки (доминирует праворукость). Остальные формы моторной асим-
метрии (например, предпочтительное использование одной ноги в отдельных двига-
тельных актах) имеет индивидуальный, а не видовой характер.
Функциональная асимметрия больших полушарий, обеспечивая но-
вый, более высокий уровень регуляции сложных функций мозга, одно-
временно повышает требования к совмещению деятельности двух по-
лушарий.
Парность в деятельности больших полушарий обеспечивается наличием
комиссуральной системы (мозолистого тела, передней, задней, гиппокампальной и
хабенулярной комиссур, межталамического сращения), которые анатомически со-
единяют два полушария головного мозга. Показано, что возбуждение из
участка раздражения одного полушария передается через комиссуральную
систему не только в симметричный участок другого полушария, но также и
в несимметричные участки коры (например, из зрительной коры одного полушария
в моторную кору противоположного полушария). Использование метода условных
рефлексов показало, что если рефлекс вырабатывается, например, на так-
тильное раздражение кожи одной стороны тела, то он легко воспроизво-
дится при таком же раздражении симметричных участков кожи другой сто-
роны тела, т.е. в процессе выработки рефлекса происходит «перенос» вре-
менной связи в другое полушарие. Предварительная перерезка мозолистого
тела прекращала (или сильно затрудняла) «перенос» условного рефлекса.
Вместе с тем, перерезка всех комиссур больших полушарий полностью не
исключает возможность межполушарного взаимодействия при выработке
временной связи. Эти результаты свидетельствуют о том, что элементар-
105
ные формы взаимодействия двух полушарий могут осуществляться через
четверохолмие и ретикулярную формацию ствола.
8. Гематоэнцефалический барьер.
8.1. Структура гематоэнцефалического барьера (ГЭБ). ГЭБ, отделяющий нейро-
ны головного мозга от крови, имеет отличительные признаки сравнительно с
другими гистогематическими барьерами. Эндотелиальные клетки капилляров
имеют очень маленькое количество пор, плотные контакты между клетками,
малую способность к пнноцитозу, почти не содержат окошек. Базальная мем-
брана с её клетками (перицитами) хорошо выражена, имеет большую толщину (до
500 нм). Главная отличительная особенность ГЭБ связана с наличием в нем гли-
ального компонента. Главным видом глиальных клеток, участвующих в обра-
зовании ГЭБ, являются астроциты, которые своими отростками образуют свое-
образные футляры вокруг капилляров, покрывая около 85% их площади.
Структура ГЭБ показана на рис. 30. Выделяют две главных функции ГЭБ - за-
щитную и регуляторную.
Эндотелий
Базальная
мембрана
Рис. 30. Структурная организация гематоэнцефалического барьера в сенсомотор-
ной коре мозга крыс. Электронограмма. Ув. 15000. АЦ - астроцит, ПЦ - перицит,
ГЭБ - гематоэнцефалический барьер. (Препарат предоставлен профессором В.П.
Федоровым).
8.2. Защитная функция ГЭБ заключается в предупреждении или ограничении по-
ступления в головной мозг различных веществ крови как эндогенного, так и эк-
зогенного происхождения. Эта функция проявляется постоянно в физиологиче-
ских условиях, а также при развитии патологии. Примером защиты мозга от
эндогенных веществ крови является билирубин, который практически отсутст-
вует в ткани мозга не только у здорового человека, но и у больного желтухой
(кроме новорожденных). ГЭБ непроницаем и для многих других физиологически
активных веществ крови: серотонина, адреналина, ацетилхолина, ГАМК, до-
фамина, тироксина, инсулина и др. Особый интерес для медиков представляет
малая проницаемость ГЭБ для многих антибиотиков (например, пенициллина,
106
стрептомицина, левомицетина, тетрациклина). Для повышения эффекта их действия иа
ЦНС врачи используют введение лекарств в обход ГЭБ непосредственно в лик-
вор, учитывая, что барьер «ликвор - мозг» имеет бблыпую проницаемость.
8.3. Регулирующая функция ГЭБ заключается в контроле поступления в головной
мозг веществ, необходимых для его метаболизма и функции. Она осуществля-
ется через механизмы избирательной проницаемости барьера, в результате чего
достигается гомеостазирование внутренней среды мозга, что создает опти-
мальные условия для его функционирования. Не случайно в онтогенезе «за-
крытие» ГЭБ (его почти полная функциональная зрелость) совпадает с критиче-
ским периодом развития коры головного мозга плода (12-16 недели).
В качестве примера отметим регулирующую роль ГЭБ в отношении ме-
диаторного и электролитного гомеостаза мозга. ГЭБ непроницаем для боль-
шинства медиаторов, выделяющихся в синапсах головного мозга. Эти медиа-
торы поглощаются астроцитами н используются для повторного применения.
С другой стороны ГЭБ непроницаем для медиаторов, циркулирующих в крови,
например, серотонина, ГАМК, адреналина, дофамина, ацетилхолина, но обыч-
но проницаем для их метаболических предшественников. Если же биологиче-
ски активные вещества попадают в головной мозг, минуя ГЭБ, например, при
введении в ликвор, они часто оказывают противоположный эффект: адреналин
вызывает парасимпатические, а ацетилхолин - симпатические эффекты, инсу-
лин вызывает гипергликемию, гистамин - повышение АД и частоты сердеч-
ных сокращений. Регулирующая функция ГЭБ создает возможность образова-
ния и поддержания собственного пула медиаторов независимо от концентра-
ции в крови этих физиологически активных веществ. Головной мозг способен
обеспечить высокий уровень гомеостаза в своей внутренней среде ионов ка-
лия, магния, несколько меньше кальция. При выраженном изменении их кон-
центрации в крови амплитуда изменения концентрации в ЦСЖ значительно
меньше. В связи с этим, головной мозг более устойчив к повышению или сни-
жению концентрации этих электролитов в крови, чем другие органы, напри-
мер, скелетные мышцы и сердце. Напротив, ГЭБ достаточно проницаем для
воды, и поэтому сдвиги осмолярности, прежде всего в связи с изменением
концентрации натрия в кровн, сильно влияют на функции головного мозга,
вызывая его отек или обезвоживание.
8.4. «Безбарьерные» области мозга. В некоторых областях головного мозга ГЭБ
слабо выражен н имеет ультраструктурные особенности. Одна из них связана
со строением капилляров, которые в этих зонах относятся к фенестрированно-
му типу, т.е. имеют большие поры диаметром 70 - 80 нм. Другой особенностью
является слабовыраженный «астроцитарный чехол» капилляров, в результате
чего нейроны непосредственно контактируют с базальной мембраной капилля-
ра. «Безбарьерные» зоны расположены в основном в гипоталамусе (ядра серого
бугра, субфорникальный орган и супрахиазматическая область), а также иа границе
продолговатого и спинного мозга (area postrema), в нейрогипофизе н эпифизе.
Высокая проницаемость барьера в этих зонах имеет два главных физиологиче-
ских последствия. Первое из них связано центральной хеморецепцией, позво-
ляющей ЦНС получить информацию о составе крови и СМЖ н участвовать в
регуляции потребления воды (субфорникальный орган, area postrema), осуще-
107
ствлении биоритмов (супрахиазматическое ядро). Второе связано с тем, что
«безбарьерные» области мозга обеспечивают попадание в кровь секретируе-
мых в ЦНС гормонов, например в срединном возвышении ножки гипофиза,
нейрогипофизе, эпифизе.
9. Кровоснабжение головного мозга.
9.1. Приток крови к мозгу осуществляется по внутренним сонным и позвоночным
артериям (соотношение кровотока в них 4:1), они образуют на основании мозга вил-
лизиев круг, функциями которого являются: 1) гашение пульсовых колебаний
(равномерность притока); 2) создание первой линии анастомозов между сосудами
притока (реализуется при нарушении притока крови в патологических условиях). Отток
крови из мозга осуществляется по венам, впадающим в венозные синусы. В
них имеется большое количество анастомозов и связей с венозными сплете-
ниями головы и спинномозгового канала, что препятствует возникновению
венозного застоя.
9.2. Регуляция мозгового кровотока.
❖ Нейрогенные механизмы регуляции.
• Симпатические влияния из верхних шейных ганглиев преимущественно
на магистральные сосуды мозга; они поддерживают сосудистый тонус
покоя, при максимальной импульсации повышают его иа 30 %. Норад-
ренергическая иннервация внутримозговых сосудов имеет не симпати-
ческое происхождение, а, вероятно, осуществляется нейронами голубо-
го пятна среднего мозга. (Разрушение этой структуры ведет к увеличению крово-
тока в некоторых отделах мозга на фоне снижения концентрации норадреналина.)
• Парасимпатические влияния (из ядра VII нерва) на магистральные и пи-
альные артерии приводят к снижению тонуса сосудов.
❖ Метаболические механизмы регуляции: повышение концентрации К+, НГ,
СОг, аденозина, оксида азота (NO) и снижение напряжения Oi приводит к
расширению мозговых сосудов, противоположная динамика вызывает су-
жение сосудов.
❖ Гормональные влияния: вазопрессин, ангиотензин-2, простагландины Fzo
повышают тонус (суживают сосуды), простагландины Ег снижают тонус сосу-
дов.
9.3. Ауторегуляция мозгового кровотока - способность мозга сохранять постоян-
ный кровоток, несмотря на колебания системного АД в интервале от 70 до 170
мм рт. ст. Исследование ауторегуляции мозгового кровотока в динамике показа-
ло наличие двух фаз; пассивной и активной. Во время непродолжительной (в
течение 5 с) пассивной фазы кровоток мозга изменяется в том же направлении,
что и САД. Если САД изменяется медленно (более 20 с), то эта фаза не выра-
жена. В активной фазе ауторегуляции мозговой кровоток, несмотря на про-
должающееся изменение САД, возвращается к нормальной величине. Актив-
ная фаза ауторегуляции имеет два компонента: быстрый, для развития которо-
го достаточно 2 - 3 с, и медленный, для развития которого требуется 30 - 40 с.
Быстрый компонент имеет нейрогенный механизм, его можно выключить бло-
кадой а-адренорецепторов. Медленный компонент активной фазы ауторегуля-
108
ции обеспечивается преимущественно миогенными и метаболическими меха-
низмами.
10. Спинномозговая жидкость (СМЖ).
10.1. Внеклеточная жидкость головного мозга имеет двойственное происхождение
(рис. 31). Часть жидкости образуется путем фильтрации в капиллярах ткани го-
ловного мозга. Этот процесс ограничен низкой проницаемостью эндотелия для
воды и выражен меньше, чем'в других органах. Другая часть внеклеточной
жидкости образуется в сосудистых сплетениях желудочков мозга. Она называ-
ется спинномозговой жидкостью или ликворам. Общее количество СМЖ у
взрослого человека 110 - 160 мл. Она расположена преимущественно в желу-
дочках мозга (- 30 мл) и в подпаутинном пространстве, представляющем систему
из ликвороносных каналов н субарахноидальных ячей (- 100 мл). Продолжением
подпаутинного пространства вглубь мозга являются околососудистые про-
Рис. 31. Схема циркуляции межклеточной и спинномозговой жидкости.
странства Вирхова - Робина, через которые СМЖ и ее вещества размером до 20
нм сообщаются с жидкостью межклеточного пространства мозга, объем которо-
го равен 15 - 20% объема мозга. Давление СМЖ в горизонтальном положении
человека равно 8 - 13 мм рт. ст. (100 - 180 мм вод. ст.). Состав СМЖ отличается
от состава плазмы более низкой концентрацией питательных веществ (глюкозы,
аминокислот, электролитов) н очень низким содержанием белков (0,1 г/л). Ко-
личество лейкоцитов (лимфоцитов н моноцитов) в ликворе в тысячи раз мень-
109
ше, чем в крови (0 - 5-106/л). Состав СМЖ в различных участках ликворного
пространства неодинаков (например, для К*. СГ, белка).
10.2. Образование, отток и резорбция СМЖ.
❖ Главным местом образования СМЖ (70% и более) являются сосудистые спле-
тения желудочков мозга, имеющие площадь до 300 см2 и высокий уровень
кровотока (- 300 мл/100 гмин.). Гематоликворный барьер состоит из капилля-
ров, имеющих фенестрированный эндотелий и слой эпителиальных клеток,
представляющих собой модифицированные эпендимоциты. Первым этапом
образования СМЖ является фильтрация плазмы через фенестрированные
капилляры. В результате ультрафильтрат поступает в соединительную
ткань под эпителий ворсинок. Однако состав СМЖ является почти посто-
янным, несмотря на колебания различных веществ (например. К*, Са2*, амино-
кислот, катехоламинов и др.) в фильтрируемой плазме. Стабилизация состава
СМЖ связана с процессами секреции и реабсорбции, осуществляемыми в
эпителии ворсинок сосудистого сплетения. Объем образуемого СМЖ в су-
тки равен около 500 мл, в результате чего его обновление составляет 3-7
раз в сутки. Стимуляция адренергических волокон из верхнего шейного
симпатического узла снижает образование СМЖ на 30%, а холинэргиче-
ская стимуляция повышает образование СМЖ, не изменяя кровоток.
❖ Основной отток СМЖ происходит в церебральные и спинальные арах-
ноидальные ворсинки (пахионовы грануляции). Особенно много их в области
верхней сагиттальной пазухи. Кроме этого пути, отток СМЖ осуществля-
ется путем реабсорбции в сосудистых сплетениях желудочков (- 10% СМЖ),
через арахноидальную мембрану и, возможно, капилляры мозга (преимуще-
ственно паравентрикуляриой области). При давлении СМЖ = 8,3 мм рт. ст. име-
ется равновесие между продукцией и резорбцией СМЖ. При повышении
давления до 15 мм рт. ст. резорбция СМЖ линейно возрастает. Резорбция
СМЖ осуществляется в результате процессов осмоса, простой и облегчен-
ной диффузии, активного транспорта и везикулярного транспорта.
10.3. Функции СМЖ.
'Z' Механическая функция защиты мозга создается субарахноидальным лик-
ворным пространством - своеобразной «гидравлической подушкой» мозга.
Масса головного мозга (- 1500 г) уменьшается в ликворе до 50 г.
❖ Функция центральной гуморальной регуляции связана с тем, что в ликвор
поступают биоактивные вещества (медиаторы, гормоны и др.) как из крови
сосудистых сплетений, так и из нейронов и глиальных клеток. С током
СМЖ они поступают к нейронам различных структур ЦНС. Особенно вы-
раженное влияние они могут оказать на околожелудочковые органы (сре-
динное возвышение, нейрогипофиз, эпифиз, субфорникальный и субкомис-
суральный органы стенок Ш желудочка, area postrema продолговатого моз-
га и др.), нейроны которых контактируют с ликвором (его активными ве-
ществами) непосредственно или через тонкий слой эпендимных клеток.
Посредством этих механизмов ликвор участвует в регуляции эндокринной
гипоталамо-адеиогипофизарной системы, водно-солевого обмена и чувства
НО
жажды, регуляции сердечно-сосудистой системы и дыхания, возникнове-
ния сиа и поддержания бодрствования и др.
❖ Защитная и иммунологическая функции СМЖ связаны с его бактерицид-
ными свойствами - иммуноглобулинами G и А, системой комплемента,
лимфоцитами и моноцитами.
❖ Транспортная функция - характеризуется участием в перемещении раз-
личных органических и неорганических веществ в пределах ЦНС и выве-
дением в венозную систему высокомолекулярных продуктов метаболизма.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основиой и дополнительной ли-
тературы (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методнческий комплекс (см. с. 514, использовать имеющейся у
вас учебники и пособия): [1-9], [11], [12], [14-16], [18].
3.2. Дополнительная литература (см. с. 514 - 522): [14], [16], [30], [33], [94], [97],
[103], [104], [111], [133], [134], [161], [169], [173], [222], [225], [232], [233], [242],
[245], [251-253].
33. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/-nphvs: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брнн. - Владикавказ: СОГМА. - 1999; Учебное
пособие В.Н. Яковлев, И.Э. Есауленко «Физиология головного мозга». - Воронеж,
ВГМА, 2001. - 75 с.; тесты остаточного уровня знаний.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
111
Тема 8. Вегетативная (автономная) нервная система
Цель: а) знать структурно-функциональные особенности вегетативной нервной системы (ВНС), влия-
ния симпатического, парасимпатического и метаснмпатнческого отделов на иннервируемые ор-
ганы, относительный антагонизм и синергизм их влияний, принципы организации афферентно-
го, эфферентного звена вегетативных рефлексов; организацию вегетативных ганглиев, механиз-
мы передачи в ннх возбуждения, функциональные различия преганглионарных и постганглио-
нарных волокон, медиаторы ВНС и основные виды их рецепторов; вегетативные центры и их
роль в регуляции вегетативных функций, вегетативные компоненты поведения при формирова-
нии целостных поведенческих актов;
б) уметь использовать эти знания для объяснения вегетативных компонентов поведения орга-
низма в разных условиях его существования, оценки возрастных особенностей вегетативной ре-
гуляции функций, понимания роли вегетативной системы в обеспечении гомеостаза.
Мотивация темы: знание механизмов регуляции вегетативных функций организма позволяет с помо-
щью физиологических и фармакологических воздействий управлять гомеостатическими про-
цессами в организме. Знание адаптационно-трофических влияний симпатического отдела ВНС
важно для понимания механизмов обеспечения адаптаций организма. Понимание роли ВНС в
обеспечении эмоциональных реакций и стресса позволяет внести необходимую коррекцию в ве-
гетативные процессы и избежать нежелательных сдвигов гомеостаза, нарушения функций внут-
ренних органов.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекцин н подготовки рефератов.
1» Общая характеристика вегетативной нервной системы.
1.1. Вегетативные и соматические функции организма. Ф. Биша (1801) предложил
разделять функции на вегетативные, общие с растениями - рост, обмен веществ, раз-
множение, и анимальные (соматические) - функции скелетной мускулатуры. И. Рейл
(1807) ввел понятие вегетативная нервная система. К. Бернар рассматривал ее как сис-
тему непроизвольной иннервации. Дж. Ленглн (1898) выделил симпатический, пара-
симпатический и энтериновый отделы, предложил название «автономная нервная сис-
тема».
1.2. Вегетативная нервная система (ВНС) - это часть нервной системы, которая
регулирует деятельность внутренних органов, просвет сосудов, обмен веществ,
обеспечивая гомеостаз и адаптацию организма.
1.3. Функциональные особенности ВНС
❖ Относительная автономность ВНС: возможность регуляции (в меиее со-
вершенном виде) при полном нарушении связи с ЦНС, низкая возможность
произвольной регуляции.
❖ Генерализованный характер возбуждения в эффереитно-эффекториом от-
деле.
♦ ♦♦ Низкая лабильность и скорость проведения возбуждения, большой латент-
ный период вегетативных рефлексов.
❖ Основные функции вегетативной нервной системы: Обеспечение и восста-
новление гомеостаза, участие в процессах адаптации.
112
1.4. Основные объекты управления ВИС.
* Гладкие миоциты различных физиологических систем.
❖ Клетки желез внешней и внутренней секреции.
❖ Сердечная мышца.
1.5. Отделы ВИС: симпатический, парасимпатический, метасимпатический.
Рис. 32. Механизмы передачи влияний в нервно-эффекторных синапсах вегета-
тивной нервной системы. Пунктирные стрелки отражают ауторегуляцию в си-
напсах и их реципрокные отношения. ПК - протеинкиназа, ФЛ-С - фосфопипа-
за-С, АЦаза - аденилатциклаза, ГЦаза - гуанилагциклаза, другие сокращения
см. в списке сокращений.
2. Симпатический отдел вегетативной нервной системы (область иннервации - все ор-
ганы и ткаии).
2.1. Спинальные центры - преганглионарные нейроны боковых рогов спинного
мозга Св - L3, их фоновая активность -3 Гц. (Спиноцилиарный центр - Cg-Thi,
слюнной центр - Th^, сердечный центр - Th|_3, сосуды и потовые железы - Cu-Lj и
др.) Они получают влияния от афферентных входов в спинной мозг и вышеле-
жащих структур головного мозга (особенно нейронов ретикулярной формации).
2.2. Преганглионарные волокна - аксоны преганглионарных нейронов относятся к
типу В, скорость проведения возбуждения - 10 м/с. ПД с длительной следовой
деполяризацией.
2.3. Ганглии симпатической нервной системы (паравертебральные и превертеб-
ральные: верхний и нижний брызжеечные, чревный узлы) содержат ганглио-
нарные нейроны и интернейроны.
113
❖ Механизм передачи возбуждения в симпатических ганглиях с преганглио-
нариого иа ганглионарный нейрон: медиатор ацетилхолин действует на Н-
холинорецептор, содержащий в своем составе Na+-, К+-канал, открывание
которого формирует входящий Na+-TOK и ВПСП. (Ганглиоблокаторы - бензогек-
соний, гексаметоний и др.)
❖ В ганлиях имеется также тормозные адренергические интернейроны (МИФ-
клетки), которые формируют иа ганлионарных нейронах ТПСП. В регуляции вы-
деления медиатора участвуют также М-холинорецепторы и опиатные рецепторы
на ганглионарных нейронах.
2.4. Постганглионарные волокна - аксоны ганглионарных нейронов, относятся к
типу С, скорость проведения возбуждения - 1 м/с. ПД с длительной следовой
гиперполяризацией. Соотношение числа пре- и постганглиоиарных волокон
1:100 обеспечивает дивергенцию и иррадиацию возбуждения.'Эта посттанглио-
иарная иррадиация и мультипликация возбуждения способствует мобилизации
функциональных резервов организма.
23. Нейроэффекторные синапсы (разветвления аксона ганлионарных нейронов имеют мно-
гочисленные варикозные утолщения, каждый из которых образует синапс на исполнительных
клетках).
• 1* Симпатические адренергические синапсы (медиатор: норадреналин - 90%, адре-
налин - 7%, дофамин - 3%, выделяется также комедиатор - иейропептид Y) (см. рис.
32).
• Медиатор норадреналин действует на а- и 3-адренорецепторы постси-
наптической мембраны. Эффект действия норадреналина зависит от ти-
па адренорецепторов (например, медиатор через а-рецепторы вызывает сужение
кровеносных сосудов, а через pi-рецепторы - их расширение), количества рецеп-
торов и их соотношения иа клетке.
• Действие норадреналина через а-адренорецепторы («1 и аг, каждый из
который имеет несколько подтипов) (блокаторы: фентоламин, дроперидол н др.).
Ф Локализация и реализуемые через них эффекты: сосуды (at, аг) -
сужение, миоциты кишечника и желудка (at) - снижение тонуса,
гладкомышечные сфинктеры (ai) - сокращение, мочеточник (at, аг)
- сокращение, Р-клетки поджелудочной железы (аг) - торможение
секреции инсулина и др.
ф Механизмы действия.
Эффект норадреналина через а,-рецепторы реализуется пре-
имущественно через Gq-белки мембраны —» |ФЛ-С —» )ИФз и
ДАТ —»|Са2+ +КМ —» |киназа ЛЦМ —» сокращение ГМК сосу-
дов, сфинктеров, радиальной мышцы радужки и др.
Эффект норадреналина через аг-рецепторы реализуется пре-
имущественно через Gt-белки мембраны —» 4-АЦ —» |цАМФ —»
|ПК-А:
□ в ГМК происходит ^концентрации Са2+ и активация кина-
зы ЛЦМ, что сопровождается сокращением;
□ в эндокринных клетках происходит снижение активность
ферментов синтеза гормонов (например, инсулина, ангиотен-
114
зина П и др.), в жировой ткани торможение ферментов ли-
полиза (липазы), в печени - гликогенолиза (фосфорилазы);
□ в пресннаптическом окончании торможение синтеза и сек-
реции медиатора норадреналина.
• Действие норадреналина через Р-адренорецепторы (через ₽| сильное,
через $2— слабое действие) (блокаторы: пропраиол, атенолол и др.).
Ф Локализация н реализуемые через них эффекты: сердце (Pi)— увели-
чение силы и частоты сокращений, сосуды и бронхи (32>3i) - рас-
ширение, миоциты кишечника (₽2>₽0, мочевого пузыря, матки (Ра)
- расслабление, жировая ткань (pi) - активируется липолиз, печень
(р 2) - гликогенолиз н глюконеогенез и др.
Ф Механизмы действия: норадреналин через Р-адренорецепторы и за-
тем мембранный Gj-белок активирует АЦ и повышает уровень
цАМФ, который стимулирует ПК-A, фосфорилирующую различ-
ные белки:
□ в сердце фосфорилирование Са2+-насоса ЭПС приводит к
его ингибированию, что увеличивает уровень Са2+ в цито-
золе, его соединение с тропонином и затем силу сокраще-
ния;
□ в ГМК фосфорилирование киназы ЛЦМ делает ее (киназу)
менее чувствительной к комплексу Са2++КМ. В результате
происходит расслабление ГМК
□ фосфорилирование ферментов повышает их активность
(например, ферментов синтеза гормонов - либеринов и
статинов гипоталамуса, тропных гормонов аденогипофиза,
инсулина и др., фосфорилазы в печени и липазы в жировой
ткани);
□ в пресннаптическом окончании адренергических синапсов
стимулируется синтез н секреция медиатора.
• Инактивация медиатора: около 80% норадреналина поступает путем эндоцитоза об-
ратно в пресинаптическое окончание, небольшая часть разрушается моноаминооксн-
дазой пре- и постсннаптической мембран и катехол-О-метилтрансферазой постсинап-
тической мембраны, часть диффундирует в межклеточное пространство и кровь.
Симпатические холинергические синапсы (имеются в небольшом количестве).
• Медиатор ацетилхолин действует на М- и Н-холино-рецепторы постси-
наптической мембраны (активируются также мускарином и никотином соответст-
венно).
• М-холинорецепторы: локализация н реализуемые через них эффекты -
клетки потовых желез (усиление секреции), гладкие миоциты сосудов
скелетных мышц (расширение сосудов). Механизмы действия см. ниже
3.5.
• Н-холинорецепторы расположены в синапсах на хромаффинных клет-
ках надпочечников (секреция катехоламинов). Особенность этой области в
том, что нейросекреторные синапсы образуют преганглионарные во-
локна, а хромаффинные клетки являются аналогом ганглионарного ней-
115
рона. Механизм действия: см. выше 2.3 (скопления хромаффинных клеток
расположены также на поверхности аорты, в каротидном тельце, в симпатических
ганглиях).
❖ Ауторегуляция секреции норадреналина в синапсе осуществляется через
рецепторы на пресинаптической мембране (см. рис. 29).
• Высокая концентрация норадреналина в синаптической щели и адрена-
лина в крови вызывает через аг-адренорецепторы торможение освобо-
ждения норадреналина из пресинаптического окончания.
• Низкая концентрация норадреналина в синаптической щели и адрена-
лина в крови вызывает через (^-адренорецепторы усиление секреции
норадреналина из пресинаптического окончания.
• Через М-холинорецепторы происходит реципрокное торможение аце-
тилхолином секреции норадреналина при возбуждении парасимпатиче-
ского отдела.
2.6. Основное значение симпатического отдела заключается в обеспечении сроч-
ной адаптации путем мобилизации физиологических и психических резервов
организма.
2.7. Понятие о симпатоадреналовой системе (Л.А. Орбели, 1926;): возбуждение
симпатического отдела, который иннервирует мозговой слой надпочечников,
стимулирует секрецию адреналина в кровь. Адреналин возбуждает через рети-
кулярную формацию симпатические центры, что еще больше увеличивает его
секрецию. Формируется положительная обратная связь, увеличивающая влия-
ние симпатической нервной системы на организм.
3. Парасимпатический отдел вегетативной нервной системы (область иннервации бо-
лее узкая, например, ее нет в сосудах кожи).
3.1. Парасимпатические центры в стволе головного мозга - ядра Ш (цилиарная
мышца и сфинктер зрачка), VH (слезные, подчелюстные и подъязычные слюнные же-
лезы), IX (околоушные слюнные железы, сосуды слизистой рта), X (органы грудной и
брюшной полостей, кроме органов малого таза) пар черепных нервов, в спинном
мозге - сегменты Si - S4 (органы малого тазе и наружные половые органы) содер-
жат преганлионарные нейроны..
3.2. Преганглионарные волокна - аксоны преганлионарных нейронов, относятся к
типу В, скорость проведения - 10 м/с. ПД с длительной следовой деполяриза-
цией.
3.3. Ганглии парасимпатической нервной системы.
❖ Расположены вблизи иннервируемого органа либо в стенке органа (интра-
мурально).
<• Механизм передачи возбуждения в ганглиях: медиатор ацетилхолин дейст-
вует на Н-холинорецептор (блокатор - гексометоний и др.), содержащий в своем
составе Na+, К+-канал, открытие которого формирует входящий №+-ток и
ВПСП.
3.4. Постганглионарные волокна - аксоны ганглионарных нейронов, относятся к
типу С, скорость проведения возбуждения 1 м/с. ПД с длительной следовой ги-
перполяризацией. Соотношение пре- и постганглионарных волокон 1:1.
116
3.5. Нейроэффекторные синапсы (окончания постганглионарного нейрона на исполнитель-
ных клетках) - медиатор ацетилхолин действует на М-холинорецепторы постси-
наптической мембраны (блокатор - атропин).
❖ Локализация М-холинорецепторов н реализуемые через них эффекторы:
сердце (Мз) - снижение частоты и силы сокращений, сосуды (расширение),
бронхи (сужение), мочевой пузырь (сокращение), слюнные, слезные, желу-
дочные железы (Mi) - стимуляция секреции н др.
❖ Механизмы действия ацетилхолина через М-холинорецепторы (см. рис.
32).
• В ГМК желудка, кишечника, мочевого пузыря, бронхов и др. эффект
ацетилхолина реализуется через Gq-белок —» f ФЛ-С —< |ИФз и ДАТ —<
|Са2+ +КМ —» Ткиназа ЛЦМ —» сокращение ГМК. (В сердце кальциевый
механизм с включением тропомиозина выражен слабо, но может привести к «парадок-
сальному» увеличению ритма и силы сокращений.)
• В кровеносных сосудах эффект ацетилхолина реализуется через цито-
зольную гуаннлатцикпазу, которая активируется образующимся в эндо-
телии NO и приводит к увеличению уровня цГМФ н активности ПК-G.
Последняя ингибирует киназу легких цепей миозина, что приводит к
расслаблению ГМК сосудов.
• В проводящей системе сердца эффект ацетилхолина также реализуется
через гуанилатциклазиую систему. При этом протеинкиназа G активи-
рует К+-канал. Выходящий из клетки К+-ток тормозит медленную диа-
столическую деполяризацию (изложена в теме 15: 3.3), что приводит к
уменьшению ритма и проводимости сердца.
❖ Инактивация медиатора: большая часть разрушается ацетилхолинэстеразой
(холин и ацетат захватывается пресинапсом), часть диффундирует в меж-
клеточное пространство и кровь.
3.6. Значение парасимпатического отдела вегетативной нервной системы.
❖ Обеспечивает вместе с метасимпатическим отделом сохранение и восста-
новление гомеостаза.
❖ Регулирует процессы анаболизма и запасания резервов.
4. Метасимпатический отдел автономной нервной системы (Дж. Ленгли, 1898; А.Д.
Ноздрачев, 1980).
4.1. Метасимпатическая нервная система - это комплекс микроганглиев, распо-
ложенных в стенках полых органов, обладающих собственной моторной актив-
ностью (сердце, желудочно-кишечный тракт, мочеполовая система). Имеет са-
мую узкую область иннервации - только органы, обладающие собственной мо-
торной активностью.
4.2. Организация рефлекторного пути в метасимпатическом отделе (все нейроны в
интрамуральных ганглиях).
❖ Рецепторы и волокна афферентных нейроны (афферентная часть дуги).
❖ В ганглиях афферентные, вставочные н эфферентные нейроны («центр»),
❖ Аксоны эфферентных нейронов, нервно-эффекторный синапс (эфферент-
ная часть дуги).
117
4.3. Нейроэффекторные синапсы имеют различные медиаторы (АТФ, серотонин,
пептиды, ацетилхолин, норадреналин).
4.4. Влияние метасимпатического отдела (активирующее и тормозное) зависит
от вида медиаторов, типа рецепторов, включаемых внутриклеточных посредни-
ков, исходного состояния органа.
❖ На моторику иннервируемых органов.
❖ На секрецию и всасывание.
❖ На регуляцию местного кровотока.
❖ На секрецию местных эндокринных клеток.
4.5. Физиологическое значение метасимпатического отдела.
❖ Повышает надежность регуляции висцеральных функций, делает ее авто-
номной, относительно независимой от ЦНС.
❖ Разгружает высшие отделы ЦНС от обработки излишней информации (ко-
личество нейронов в ганглиях метасимпатического отдела превышает количество нейро-
нов в спинном мозге).
5. Рефлексы вегетативной нервной системы.
5.1. Рецепторное звено: вегетативные рефлексы возникают при стимуляции раз-
личных рецепторов.
❖ Интерорецепторы.
• Механорецепторы (баро-, волюморецепторы, репепторы растяжения
легких и др.) информируют о величине давления в просвете органа, его
объеме.
• Хеморецепторы сосудов и тканей обеспечивают контроль за постоянст-
вом состава внутренней среды.
• Терморецепторы сосудов кожи, спинного мозга и гипоталамуса обес-
печивают сохранение температурного гомеостаза.
• Осморецепторы осуществляют контроль постоянства осмотического
давления.
• Болевые рецепторы сигнализируют о действии повреждающих ткань
раздражителей.
❖ Проприорецепторы - один из наиболее мощных физиологических регуля-
торов вегетативных функций, информируют о физической нагрузке.
Экстерорецепторы (температурные, болевые, органы чувств) передают влияния
внешних факторов иа вегетативные функции.
5.2. Афферентные пути.
❖ От интероцепторов по волокнам X, IX пары черепных нервов, чревных и
тазовых нервов, в спинном мозгу пути Голля и Бурдаха.
❖ От экстерорецепторов (тактильных, температурных и болевых) - спи-
но-таламические и спино-ретикулярные тракты.
❖ От проприорецепторов - пути Голля и Бурдаха, спинномозжечковые пути.
5.3. Уровни замыкиния вегетативных рефлексов.
❖ Ганглионарный (вне ЦНС, периферические рефлексы).
❖ Спинально-стволовой (истинные рефлексы).
5.4. Особенности организации эфферентного звена по сравнению с соматическими
рефлексами.
118
❖ Эфферентный ганглионарный нейрон вне ЦНС (в превертебральном, пара-
вертебральном или внутриорганном ганглиях).
❖ Разные последствия перерезки передних корешков:
• автономное функционирование внутренних органов за счет ганглио-
нарных механизмов управления;
• атрофия в скелетных мышцах.
❖ Разные типы нервных волокон: в вегетативном - преганглионарные типа В,
постганглионарные - типа С; в соматическом - преимущественно Аа и Ау.
5.5. Виды вегетативных рефлексов, их функциональное значение.
❖ Висцеро-висцералъные рефлексы.
• Собственные - реализуются в пределах одной физиологической систе-
мы, обеспечивают саморегуляцию функций (например, с барорецепто-
ров сосудов на деятельность сердца).
• Сопряженные - возникают с ннтерорецепторов одной системы, а реали-
зуются в другой, обеспечивая их функциональные взаимосвязи (напри-
мер, рефлекс Гольца, возникающий с рецепторов органов брюшной полости и тормо-
зящий деятельность сердца).
❖ Висцеро-сенсорные рефлексы - изменяют активность рецепторов (например,
гастролингвальный рефлекс, зоны Захарьина-Геда).
❖ Висцеро-моторные рефлексы - ограничивают активность мышц и физиче-
скую работу, формируют мышечную защиту прн патологии.
❖ Сенсо-висцеральные рефлексы- обеспечивают адаптацию при действии
факторов внешней среды (например, глазосердечный рефлекс Даннни - Ашне-
ра, рефлексы с терморецепторов кожи на сердце и дыхание).
❖ Моторно-висцеральные рефлексы - обеспечивают синхронное изменение
функций внутренних органов при физической работе.
5.6. Виды рефлекторных влияний.
♦ ♦♦ Пусковые (например, запуск секреции потовых желез, пищеварительных желез).
♦ ♦♦ Корригирующие (например, изменение ритма сердца).
♦ ♦♦ Адаптационно-трофические (Л.А. Орбели, А.Г. Гииецинский).
• Прямые влияния путем аксонного транспорта веществ к управляемому органу.
• Гуморальные влияния реализуются через катехоламины в крови и ликворе, влияют на
выделение гормонов и иейропептидов, чувствительность к ним тканей и иа метабо-
лизм органов.
6. Тонус вегетативных нервных центров - это непрерывная импульсация их нейронов в состоянии фи-
зиологического покоя (в симпатических центрах выражен меньше, чем в парасимпатических).
Формирование тонуса обеспечивают: афферентация от рецепторов кожи, скелетных
мышц, внутренних органов, вестибулярного аппарата, действие гуморальных факторов на
центральные и периферические хеморецепторы (изменения содержания Оз СОз, pH, мета-
болитов, гормонов).
♦♦♦ Функциональная роль тонуса: создает определенный уровень активности в эффекторах
(например, в миоцитах сосудов), позволяет плавно изменять функцию эффектора и обес-
печивает повышение чувствительности к управляющим сигналам.
Признаками симпатикотонии являются: тахикардия, тенденция к повышению АД, повы-
шенная физическая работоспособность, беспокойный сон, склонность к запорам.
*♦* Признаками парасимпатикотонии являются: брадикардия, склонность к обморокам, хо-
лодная кожа, гиперсаливация, малая выносливость, астения.
119
7. Взаимодействие между отделами вегетативной иервиой системы при регуляции
физиологических функций.
7.1. Концепция функционального антагонизма симпатического и парасимпатиче-
ского отделов: противоположные влияния в органах, имеющих двойную иннер-
вацию, например, в сердце, кишечнике, бронхах.
7.2. Концепция разных стратегий.
Ф Парасимпатическая обеспечивает запас энергетических резервов, усилива-
ет анаболические процессы, поддерживает гомеостаз.
❖ Симпатическая мобилизует резервы, усиливает катаболизм, нарушает го-
меостаз.
73. Концепция синергизма и относительного антагонизма симпатического н па-
расимпатического отделов.
❖ Действуют на разные объекты (круговая н радиальная мышцы радужной
оболочки), обеспечивая адекватную освещенность сетчатки в разных усло-
виях.
❖ Обеспечивают оптимальную секрецию слюнных желез на пищевые или от-
вергаемые раздражители (изменяется состав и количество выделяемой
слюны).
8. Высшая центральная регуляции вегетативных функций.
8.1. Ретикулярная формация ствола мозга как модулятор активности вегетативных
центров. РФ:
❖ поддерживает тонус вегетативных центров;
❖ интегрирует симпатические и парасимпатические влияния для обеспечения
потребностей целостного организма (например, сердечно-сосудистый центр
обеспечивает стабилизацию гемодинамики, согласованно меняя сердечный вы-
брос н сопротивление сосудов при разных состояниях);
❖ передает модулирующие влияния от мозжечка и гипоталамуса к органам.
8.2. Гипоталамус как интегратор и модулятор вегетативной н эндокринной актив-
ности.
❖ Показано наличие в гипоталамусе двух функционально зон (В. Гесс, 1928),
регулирующих н интегрирующих деятельность симпатических н пара-
симпатических центров.
• Стимуляция задней области гипоталамуса вызывало комплекс вегета-
тивных реакций, характерный для симпатической нервной системы:
увеличение частоты н силы сердечных сокращений, подъем АД, повы-
шение температуры тела, расширение зрачков, гипергликемию, тормо-
жение перистальтики кишечника и др., что свидетельствует о роли зад-
него гипоталамуса в интеграции различных симпатических центров. Эта
область была обозначена Гессом как эрготропная система мозга,
обеспечивающая мобилизацию и расходование энергетических ресурсов
организма при активной его деятельности.
• Раздражение преоптической н передней областей гипоталамуса сопро-
вождалось признаками активации парасимпатической нервной систолы:
урежением ритма сердца, снижением АД, сужением зрачков, увеличе-
нием перистальтики и секреции желудка, кишечника н др. Эта область
120
гипоталамуса была обозначена Гессом как трофотропная система,
обеспечивающая процессы отдыха, восстановления и накопления энер-
гетических ресурсов организма. Однако в дальнейшем было показано,
что эрготропная и трофотропная области перекрывают друг друга, и
можно только говорить о преобладании их в заднем и переднем гипота-
ламусе соответственно. Вместе с тем, на уровне гипоталамуса происхо-
дит не только интеграция деятельности различных вегетативных цен-
тров, но и включение их как компонент в более сложные физиологиче-
ские системы различных форм биологического поведения, направлен-
ных на выживание организма, поддержание гомеостаза и сохранение
вида. Например, вазомоторные вегетативные реакции, обеспечивающие
саморегуляцию АД, осуществляются сосудодвигательным центром про-
долговатого мозга, тогда как вазомоторные реакции, связанные с термо-
регуляцией, эмоциями, агрессивно-оборонительным поведением, реали-
зуются на уровне гипоталамуса и более высоких центров.
❖ На уровне гипоталамуса осуществляется сомато-висцеральная интеграция
при поведенческих актах.
8.3. Мозжечок выступает как интегратор соматических и вегетативных процессов
(Л.А. Орбели, 1935). Эксперимент и клиническая практика показали, что пораже-
ния мозжечка, кроме двигательных расстройств, сопровождаются различными
нарушениями вегетативных функций: сердечно-сосудистой и пищеварительной
систем, дыхания, гемопоэза и др. Было выявлено, что на вегетативные функции
в большей степени влияет старый и древний мозжечок (преимущественно
червь), куда поступает часть импульсации от интерорецепторов. Эфферентные
влияния на вегетативную сферу мозжечок оказывает, в основном, через ядра
шатра. Они реализуются через ядра РФ ствола и могут быть возбуждающими,
тормозящими и смешанными. (Конкретные механизмы переработки интероцептивной
импульсации мозжечком неизвестны.) Можно предположить, что регуляция вегета-
тивных функций мозжечком направлена преимущественно на обеспечение дви-
гательной функции.
8.4. Лимбическая система осуществляет связь вегетативных процессов с эмоцио-
нальными состояниями: страх, ярость, гнев, радость и т.д. (вегетативные прояв-
ления эмоций).
8.5. Новая кора больших полушарий (особенно лобные отделы).
❖ Имеется корковое представительство внутренних органов (В.Н. Чернигов-
ский).
❖ Из одних и тех же областей коры осуществляется эфферентное влияние на
двигательные функции и вегетативное обеспечение движением, что обес-
печивает интеграцию соматических и вегетативных функций при формиро-
вании целенаправленного поведения.
❖ Доказана условнорефлекторная регуляция деятельности внутренних орга-
нов (К.М. Быков), которая позволяет осуществлять опережающую регуляцию
(по прогнозированию).
❖ Корковое управление вегетативными процессами происходит также при
гипнотическом внушении, самовнушении, биоуправлении.
121
9. Профильные материалы для студентов лечебного факультета. Возрастные изме-
нения вегетативной нервной системы при старении организма.
9.1. Изменения в рецепторном звене.
❖ Происходит снижение чувствительности механорецепторов, например, бароре-
цепторов дуги аорты и каротидного синуса, которые обеспечивают депрессорный
контроль за уровнем АД (может развиваться гипертензия), механорецепторов же-
лудочно-кишечного тракта (уменьшение моторики).
❖ Повышается чувствительности хеморецепторов - это приводит к росту влияния
гуморальных факторов (например, вазопрессина, ангиотензина II) по сравнению с
нервными влияниями.
9.2. Изменения в центральном звене.
❖ Происходит инволюция нейронов: снижение числа нейронов в ЦНС и в ганглиях
(до 50 % от исходного), числа циторецепторов на нейронах и их чувствительности
(в том числе к нейромедиаторам и гормонам).
❖ Затрудняется выработка вегетативных условных рефлексов и увеличивается их
латентное время, замедляется восстановление возбудимости.
❖ Снижается лабильность нейронов как в ЦНС, так и в вегетативных ганглиях.
❖ Возрастает чувствительность к гуморальным факторам (регресс функции).
9.3. Изменения в эфферентном звене.
❖ Ослабляются нервные рефлекторные влияния иа органы, адренергические влия-
ния приобретают затяжной характер.
❖ Усиливаются гуморальные (в том числе виесинаптические) влияния.
Снижается диапазон адаптационных возможностей организма.
Ослабляются трофические влияния и возникают трофические нарушения в тканях.
10. Профильные материалы для студентов педиатрического факультета.
10.1. Незрелость вегетативной нервной системы у новорожденного.
& Нейроны ЦНС.
• Имеют низкий уровень МПП (около -20 мВ).
• Нейроны поливалентны: чувствительны и к ацетилхолину, и к адреналину.
• Не завершена миелинизация преганглионарных волокон, поэтому скорость
проведения ПД низкая.
♦ ♦♦ Созревание холинергических механизмов передачи иа уровне ганглиев начинается
со 2-й недели.
• В первые дни после рождения для передачи в ганглиях используются симпа-
топодобные вещества.
• Позднее формируются холинорецепторы, которые концентрируются в постси-
иаптической мембране (вместо диффузного распределения по всей поверхно-
сти мембраны эфферентного нейрона).
* ** У нейронов симпатических ганглиев имеется автоматия - следствие высокой про-
ницаемости их плазмолеммы для Na*.
10.2. Гетерохронность созревания отделов вегетативной нервной системы.
♦ ♦♦ В первые годы жизни ведущую роль в регуляции играет симпатический отдел, ак-
тивность парасимпатического отдела начинает проявляться с 3 мес. и формирует-
ся к 7 годам.
♦ ♦♦ Симпатическое управление сердца появляется раньше, чем парасимпатическое
(после 6 мес. формируется парасимпатическое влияние).
122
♦ > Парасимпатическая регуляция желудочно-кишечного тракта созревает раньше,
чем симпатическая (последняя появляется после отнятия от груди).
10.3. Тонус центров блуждающих нервов после рождения практически отсутствует, фор-
мируется вплоть до пубертатного периода под влиянием:
•> интероцептивной импульсации (прежде всего от хемо- и барорецепторов сосудов);
проприорецепторов (увеличение двигательной активности способствует формиро-
ванию тонуса).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-11], [12], [14-16], [18].
3.2. Дополнительная литература (см. с. 514 - 522): [3], [5], [27] ,[28], [30], [35], [43],
[47], [69], [122-124], [146], [152], [169], [178], [179], [183], [187], [193], [196], [197],
[198], [222], [226], [245], [251-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ni/~nDhvs: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; учебное
пособие «Физиология головного мозга» /В.Н. Яковлев, И.Э. Есаулеико. - Воронеж,
ВГМА, 2001 - 75 с.; тесты остаточного уровня знаний по данной теме
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором н слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по нндивидуаяьиому графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться иа практи-
ческом занятии по дайной теме, итоговом занятии и экзамене.
123
Тема 9. Эндокринная система
Цель: а) знать структурно-функциональную организацию эндокринной системы, образование, выделе-
ние, перенос н распад гормонов, их основные механизмы действия, саморегуляцию эндокринной
системы, функциональные связи желез внутренней секреции и нервной системы, нейросекреты
гипоталамуса; основные виды влияний гормонов на органы и системы организма; методы иссле-
дования эндокринной функции;
б) уметь оценивать роль эндокринных механизмов в регуляции гомеостаза, адаптации организма
к условиям внешней среды, в процессах высшей нервной деятельности н поведения.
Мотивация темы: гормональная регуляция является обязательным элементом поддержания гомеостаза,
участвует в реализации высших психических функций: мотивации, эмоции, сиа, памяти. Адап-
тивные реакции организма, в том числе стресс, протекают при обязательном участии гормонов.
Нарушения функции эндокринных желез являются одной из наиболее распространенных форм
патологии, с которой сталкивается врач любой специальности. Знания в области эндокринологии
необходимы как для своевременной и точной диагностики нарушений, так и для корректного
применения гормонов в лечебной практике.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Общая характеристика эндокринной системы.
1.1. Эндокринная система - совокупность взаимосвязанных и различным образом
организованных эндокринных клеток, осуществляющая централизованное
управление функций организма с помощью сигнальных молекул - гормонов.
Гормоны - внеклеточные сигнальные регуляторы, образующиеся специализи-
рованными эндокринными клетками и обладающие дистантным действием.
1.2. Виды объединения эндокринных клеток.
❖ Формируют эндокринные железы.
♦ ♦♦ Образуют часть железистого (например, островки поджелудочной железы, гонады)
или нежелезистого (например, почки, сердце) органа.
< ♦ Эндокринные клетки диффузно расположены в органах (гастроинтестинальная
система).
❖ Образование эндокринной системы как результат взаимодействия эндок-
ринных структур.
1.3. Виды действия гормонов.
❖ Эндокринное (дистантное) действие - гормоны эндокринной железы дей-
ствуют на клетки-мишени других органов и тканей.
❖ Паракринное действие - гормоны эндокринной клетки действуют на сосед-
ние клетки в пределах одного органа (например, гастроинтестинальные гормоны,
простагландины).
♦ ♦♦ Аутокринное действие - гормоны эндокринной клетки действуют на эту же
клетку (например, простагландины).
1.4. Химическая характеристика гормонов.
124
❖ Основные классы гормонов.
• Белково-пептидные гормоны (например, гормоны гипофиза).
• Производные аминокислот (из тирозина образуются адреналин, норадреналин,
дофамин, тироидные гормоны, из триптофана - мелатонин, серотонин).
• Стероидные гормоны (кортикостероиды, половые гормоны, 1,25-(ОН)2-витамни
Вз).
❖ Основные фрагменты молекулы гормонов.
• Адресный фрагмент обеспечивает связывание гормона с циторецепто-
ром (например, а-субъединнца лютеинизирующего гормона). (Снижение этого срод-
ства приводит к гормонорезистентности.)
• Актон обеспечивает специфичность действия гормона (например, р-
субъединица лютеинизирующего гормона).
• Вспомогательные фрагменты служат для изменения его действия, опре-
деляют его иммунологические свойства.
2. Физиологическая организация эндокринной функции.
2.1. Продукция гормонов.
❖ Биосинтез гормонов.
• По схеме синтеза белка (прямой генетический контроль) на полисомах обра-
зуется прогормои - предшественник одного или нескольких гормонов,
который с помощью сигнального гидрофобного пептида поступает
внутрь ЭПС, где происходит его обработка, завершаемая в аппарате
Гольджи упаковкой молекул гормона в секреторные гранулы (например, в
клетках аденогипофиза образуется пептид - проопиомеланокортии, из которого обра-
зуется несколько гормонов: АКТГ, МСГ, {3-липотропин, {3-зидорфин).
• Опосредованный контроль через образование ферментов, синтезирую-
щих небелковые гормоны. Стероидные гормоны прн этом не накапли-
ваются в клетке, катехоламины накапливаются в секреторных гранулах,
тироидные гормоны в коллоиде фолликула накапливаются внеклеточно.
❖ Секреция гормонов.
• Экзоцитоз секреторных гранул (пептидные гормоны, катехоламины) стимули-
руется Са2+, поступившим в цитозоль через Са2+-каналы при деполяри-
зации мембраны клетки или (и) из гладкой ЭПС при действии инозитол-
трифосфата.
• Освобождение гормона из белковосвязанной формы (тироксин из иодиро-
ванного тироглобулина).
• Свободная диффузия гормонов, растворимых в липидах мембраны (на-
пример, кортикостероиды, половые гормоны).
2.2. Циркуляторный транспорт гормонов (основные формы их транспорта в
крови).
❖ Свободная, несвязанная форма составляет от сотых долей процента (на-
пример, Т4 и Т3) до 100 % (например, инсулин). Только эта форма гормона дей-
ствует на клетки-мишени.
❖ Специфическое связывание с глобулинами (70 - 80 %) обеспечивает соз-
дание запаса и связывание избытка гормона в крови, защиту его от инак-
125
тивации и от потери через почки. (Виды глобулинов - транскортнн, тироксин-
связывающий глобулин, прогестин, тестостерои-эстрогенсвязывающий глобулин.)
❖ Неспецифическое связывание с альбуминами крови (5 - 10 %). Благода-
ря низкой прочности связи оии являются основным источником свобод-
ных гормонов В крови. (Небольшая часть гормона связана с форменными элемен-
тами крови.)
♦ ♦♦ Периферическая конверсия гормонов - превращение гормонов в органах и тканях, от-
даленных от эндокринной железы, в результате которого изменяется активность гормо-
на (например, превращение тироксина в трииодтиронин в печени, тестостерона в ди-
гидротестостерон - в простате).
Рнс. 33. Общая схема действия гормонов через вторые посредники.
Г - гормон, Р - рецептор, Ф - фермент
23. Физиологические эффекты гормонов.
Клетки-(ткани-, органы-)мишеии, избирательно реагирующие иа гормо-
ны, имеют циторецепторы - белковые приемники и преобразователи
гормонального сигнала. (Есть данные, что на один рецептор может действовать
более, чем один гормон.)
• Циторецепторы расположены на клеточной мембране или в ядре (500-
30000 рецепторов в клетке).
• Максимальный эффект гормона наблюдается при его концентрации,
обеспечивающей занятость всего 5 - 10 % общего числа рецепторов (фе-
номен «резервных» рецепторов).
• Изменение числа рецепторов является способом регуляции гор-
монального ответа (активация и ингибирование синтеза рецепторов, интернализа-
ция рецепторов путем эндоцитоза).
♦♦♦ Гормоны, плохо проникающие в клетку (белково-пептидные, катехоламины), со-
единяются с рецепторами иа клеточной мембране и вызывают внутрикле-
точные эффекты через вторые посредники (см. рис. 33). Большинство гор-
монов после присоединения к рецепторам активируют регулируемый гуа-
нилнуклеотидами G-белок мембраны. (Активация состоит в обмене ГДФ на ГТФ в
а-субъедииице G-белка. При этом происходит его распад на а-субъеднинцу н Ру-димер,
имеющие самостоятельные регуляторные эффекты. Более изучено действие а-
субъединнцы, являющейся ферментом ГТФазой. Гидролиз ГТФ до ГДФ освобождает
энергию, используемую для передачи регуляторного сигнала. ГДФ-а-субъединица соеди-
126
няется с Ру-димером, вновь образуя ГДФ-О-белок.) Имеется несколько видов G-
белков (G„ G„ Gq, Go), которые влияют на ферменты (например, адеиилатцикла-
зу, фосфолипазу С), образующие различные вторые посредники (например,
цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки ьГогут как акти-
вировать (GJ, так и ингибировать (Gi) образование вторых посредников.
Вторые посредники, в свою очередь, активируют соответствующие им про-
теинкиназы, фосфорилирующие различные клеточные белки по остаткам
серина и треонина, что изменяет их функциональную активность (чаще уве-
личивает). Наряду с этим активированные G-белки оказывают выраженное
влияние на проницаемость кальциевых и калиевых каналов мембраны.
В действии гормонов, плохо проникающих в клетки, можно выде-
лить 4 системы: аденилатциклазную, фосфоинозитидную, гуанилатцик-
лазную и тирозинкиназную.
Рис. 34 Схема действия гормоиов через аденилатцнклазную систему.
Г - гормон, Р - рецептор, Ф - фермент, ФДЭ - фосфодиэстераза.
• Аденилатциклазная система. Эту систему используют гормоны, осуще-
ствляющие свой эффект через аденилатциклазу и синтез второго по-
средника - циклического АМФ (3,5-цАМФ). Действие гормона может как
активировать, так и тормозить его образование (см. рис. 34).
• ф- Комплекс «гормон + рецептор» активирует Gq-белок, который акти-
вирует аденилатциклазу и образование цАМФ (например, так действу-
ют адреналин через |3|/|)грецепторы, АДГ через У2-рецепторы, глюкагон, АКТГ,
ТТГ, ФСГ, ЛГ, МСГ, коргиколиберин, соматолиберии, кальцитонин, хорногона-
дотропин человека, паратгормон и др.).
• ф- Комплекс «гормон + рецептор» активирует Gi-белок, который тор-
мозит аденилатциклазу и образование цАМФ (например, так действуют
адреналин через Огрецепторы, ангиотензин П через АТ-рецепторы, аденозин че-
рез А|-рецепторы, соматостатин, опиоиды через Д-, 8- и к-рецепторы, ацетилхо-
лин через Мг-холииореиепторы гладкомышечных клеток).
• ф цАМФ влияет на функции клетки через изменение активности про-
теинфосфокиназы-А, фосфорилирующей различные клеточные
127
белки: мембранные (изменяется ее проницаемость), ферменты (изменяется
метаболизм клетки), ядерные (изменение транскрипции), рибосом (изменение
трансляции), микротрубочек (двигательные эффекты, изменение формы клет-
ки) и Др.
• Фосфоинозитидная система. Эту систему используют гормоны, кото-
рые, действуя через Gq-белок, активируют в мембране фосфолипазу С
(см. рис. 35). Она образует из фосфолипида мембраны (фосфатидилинозн-
толдифосфата) два посредника - инозитол-1,4,5-трифосфат (ИФз) и 1,2-
диацилглицерол (ДАТ).
Рнс. 135. Схема действия гормонов через фосфоинозитидную систему.
Г - гормон, Р - рецептор, ФЛ-С - фосфолипаза С, ФИФз - фосфатиди-
линозитолдифосфат, ИФз - ннозитолтрифосфат, ДАГ - диацилглнцерол
•ф ИФз, переходя в цитоплазму, действует на рецепторы гладкой ЭПС,
что приводит к открыванию Са2+-каналов и выходу Са2+ в цитозоль.
Са2* при этом выступает в качестве третьего посредника, который
связывается с белком кальмодулином и активирует Са2+-кальмоду-
линзависимую протеинкиназу, фосфорилирующую эффекторные
белки, влияющие на функции клеток, их пролиферацию и диффе-
ренцировку (регулирует активность аденилатциклазы и синтез цАМФ, фосфо-
липазы-Аг н синтез простагландинов, тромбоксана, простациклина,
гуанилатциклазы и синтез цГМФ, фосфодиэстеразы и разрушение цАМФ и
цГМФ; участвует в регуляции секреции инсулина, гормонов щитовидной желазы,
гипофиза и надпочечников, кишечной секреции и секреции нейромедиаторов).
(Роль Са2* как посредника действия гормонов обусловлена его сильным влиянием
при низких концентрациях 10"8 - 10 М на метаболизм и функции клетки.
Гормоны могут увеличивать концентрацию Са2* в цитозоле как в результате
открывания Са^-каналов клеточной мембраны, так выхода Са2*из гладкой ЭПС.
В клетке есть специальные белки, связывающие Са2* прн низкой концентрации -
кальмодулин, гуанилатциклаза, тропонин С и др. Присоединение Са2* к этим
белкам изменяет их конфигурацию и свойства. Это приводит к изменению
многочисленных функций клеток - метаболической, секреторной,
сократительной, биоэлектрической и др.)
128
ДАГ, оставаясь на внутренней поверхности мембраны, активирует
протеинкиназу С, фосфорилирующую эффекторные белки (например,
активирует киназу легкой цепи миознна), а также синтез простагландинов
из арахидоновой кислоты, что изменяет функции клетки.
•ф- Через ИФз и ДАГ действуют адреналин (ai-рецепторы), вазопрес-
син (Vi), ацетилхолин (Mi), ангиотензин П (АТ,), гистамин (Hi), се-
ротонин (5-НТ2), холецистокинин, окситоцин, ВИП и др.
• Гуанилатциклазная система. Некоторые гормоны (например, ацетилхо-
лин, цитокины) действуют через гуанилатциклазу, образующую второй
посредник - циклический ГМФ (цГМФ). Гуанилатциклаза существует в
двух формах - мембраносвязаниой и растворимой в цитозоле.
Мембраносвязанная гуанилатциклаза является частью рецептора
плазмолеммы, ее активация гормоном (например, Ыа*-уретическим) уве-
личивает образования цГМФ, который стимулирует протеинкиназу G, фосфори-
лирующую белки).
Растворимая в цитозоле гуанилатциклаза содержит в своем составе
гем, с которым связывается сигнальная молекула - NO (оксид азота
образуется из аргинина в ряде клеток - эндотелии, нейронах и др. -
под действием ацетилхолина, механических раздражителей и др.).
Активированная оксидом азота гуанилатциклаза через протеинки-
иазу G фосфорилирует белки (например, так действует ацетилхо-
лин, вызывая расширение сосудов).
• Тирозинкиназная система характеризуется особыми рецепторами, ко-
торые путем фосфорилирования и дефосфорилирования сигнальных
белков по тирозиновым остаткам, регулируют процессы роста, диффе-
ренцировки, выживания, метаболизма и смерти клеток. Эту систему ис-
пользуют некоторые гормоны - инсулин, инсулиноподобные факторы
роста (или соматомедины), факторы роста эпидермиса, тромбоцитов и
др. Многие рецепторы к таким гормонам содержит в своем составе ти-
розинкиназу, которая при действии гормона аутофосфорилирует внут-
реннюю часть рецептора по остаткам тирозина Активированный таким
образом рецептор передает через сложный и недостаточно изученный
комплекс посредников фосфорилирующее и дефосфорилирующее влия-
ние по остаткам тирозина иа многие сигнальные молекулы цитоплазмы
и ядра. Фосфорилирование и дефосфорилирование белков может при-
водить как к их активации, так и инактивации.
• Выше представлены преимущественные пути действия гормонов через отдельные
системы посредников. В действительности гормоны оказывают преимущественно раз-
ветвленные эффекты. Это связано с тем, что комплекс Г+Р действует с разной степе-
нью на несколько G-белков. В свою очередь активированные G-белки действуют иа
несколько эффекторов (ферментов, каналов).
Клеточные эффекты липорастворимых гормонов, легко проникающих че-
рез клеточные мембраны (стероидные и тироидные гормоны, 1,25-(ОН)г-
витамин D3).
129
• Гормон соединяется с ядерным рецептором. Рецепторы представляют
собой белки (факторы транскрипции), прочно связанные с ДНК хрома-
тина и дополнительными ядерными белками. Каждый рецептор содер-
жит несколько специфических участков (доменов): ДНК-связывающий
домен, гормонсвязывающий домен (гидрофобный «карман»), домен, акти-
вирующий транскрипцию и др.
• Активируются процессы транскрипции (образование мРНК с помощью фер-
мента РНК-полимеразы П) И трансляции (взаимодействие рРНК, тРНК и мРНК,
индуцирующее синтез белков и дифференцировку клеток). Например, глюкокор-
тикоиды стимулируют гены, обеспечивающие синтез Рг-адренорецеп-
торов, тироидные гормоны - гены Pi- и Рг-адренорецепторов. (Изучаются
ядерные белки, способные как активировать, так и ингибировать действие на геном
комплекса «гормон+рецептор».)
❖ Основные эффекты гормонов в целостном организме.
• Метаболический: гормоны - основные регуляторы активности фермен-
тов.
• Нейрологический: гормоны через циторецепторы нейронов и глии дей-
ствуют на формирование процессов возбуждения и торможения, памя-
ти, проницаемость гематоэнцефалического барьера.
• Морфогенетический: Половые гормоны, СТГ, тироксин влияют на про-
цессы роста тела и формообразования, созревания гонад и половой
дифференцировки, высвобождения гамет.
• Кинетический: сокращение гладких мышц, секреция в эндо- и экзок-
ринных железах, изменение пигментации.
2.4. Катаболизм и экскреция гормонов.
❖ Катаболизм гормонов - их необратимая ферментативная инактивация и
превращение в растворимую форму - происходит в разных органах, пре-
имущественно в печени, почках, легких.
• Белково-пептидные гормоны разрушаются пептидазами печени, почек и других орга-
нов.
• Стероидные гормоны инактивируются преимущественно в микросомальной фракции
(путем восстановления двойной связи кольца А н кетогрупп и процессов декарбокси-
лирования). Продукты распада переходят в растворимую форму путем образования
эфиров с глюкуроновой и серной кислотами.
• Аминокислотные гормоны: 1) катехоламины (адреналин, норадреналин) инактивиру-
ются в разных тканях путем дезаминирования (моноамннооксидаза) и метилирования
(катехол-О-метилтрансфераза); 2) тироидные гормоны - путем реакции декодирова-
ния, дезаминирования остатков аланина, образования эфиров с глюкуроновой и серной
кислотами.
❖ Экскреции гормонов из организма: выделяются преимущественно (~ 90 %)
инактивированные гормоны и небольшое количество активных гормонов.
По почечно-мочевому пути выделяются - 80 % гормонов, по печеночно-
кишечному пути - 20 % гормонов.
❖ Показателем скорости катаболизма и экскреции гормонов является период
их полураспада (Т«): для катехоламинов - 1 - 3 минуты; для белково-
130
пептидных гормонов - 10 - 20 минут; для стероидных - 0,5 - 1,5 часа; для
тироидных - около 4 суток.
2.5. Регуляция эндокринной функции. Можно выделить три главных пути регуля-
ции: трансгипофизарный через аденогипофиз, трансгипофизарный через нейро-
гипофиз и парагипофизариый (минуя гипофиз).
❖ Трансгипофизарная регуляция через аденогипофиз осуществляется с помо-
щью пептидергических нейронов гипоталамуса, гормоны которых регули-
руют образование тропных к периферическим железам гормонов аденоги-
пофиза (рис. 36).
• Трансформация нервного импульса в нейросекреторный процесс в ней-
ронах передней и средней групп ядер гипоталамуса (медиальном преопти-
ческом, интерстициальном, аркуатном, перивентрикулярном и паравентрикулярном и
др.) приводит к секреции тропных к гипофизу гормонов (как правило, пеп-
тидов) двух видов - либеринов и статинов. На этот процесс оказывают
сильное влияние норадренергические (голубое пятно), серотонинергиче-
ские (ядра шва) и дофаминергические нейроны ствола мозга.
-ф- Либерины стимулируют выработку гормонов аденогипофиза.
Кортиколиберин (пептид из 41 аминокислот, образуется в паравентри-
куларных, аркуатных, дорсомедиальных, вентромедиальных ядрах) стиму-
лирует секрецию АКТГ (в небольшой степени и других гормонов, об-
разующихся из общего предшественника - проопиомелаиокортина: МСГ,
р-эидорфина, р-липотропнна).
Его секрецию стимулируют
адреналин, норадреналин и
серотонин (а также ацетилхолин,
ангиотензин П, нейропептид Y,
глутамин), тормозят - глюкор-
ТИКОИДЫ (а также АДГ, пептид Р,
опиоиды).
Тиролиберин (трипептид, обра-
зуется в параветприкулярных, суп-
рахиазматических, дорсомедиаль-
ных и серобугорных ядрах) сти-
мулирует секрецию ТТГ и
пролактина гипофизом. Сти-
мулируют секрецию тироли-
берина эстрогены, стресс, ох-
лаждение организма. Тормо-
зят секрецию - соматостатин
и дофамин гипоталамуса, со-
матотропный гормон, высо-
кий уровень в крови глюко-
кортикоидов, перегревание
организма (а также СТГ, норадреналин через а-адренорецепторы, серо-
тонин, эндорфин).
Либерины
ГЛ,ТЛ,КЛ,
СЛ,ПЛ(?),МЛ
МСГ
Пролактин
Статины
СС,ПС(?),
МС
Неэядоафишпяе 1
пвшмпшгеаж ]
Функционалъио-
мстабадяческие
отпеты
Рис. 36. Схема трансгипо-
физарной регуляции через
аденогипофиз.
131
Соматолиберии (пептид из 40-44 аминокислот, образуется в аркуатных
и вентромедиальных ядрах) стимулирует секрецию соматотропного
гормона гипофиза. Секрецию соматолиберина стимулируют
низкий уровень СТГ в крови, серотонин, норадреналин, эндор-
фины, тормозит - высокий уровень СТГ.
ГонадоЛиберии (трипептид, образуется в мелкоклеточных нейронах ар-
куатных ядрах тонического полового центра у мужчин и женщин и цикли-
ческом центре преоигической области у женщин) стимулирует секре-
цию ФСГ и ЛГ гипофиза. Секреция гонадолиберина имеет им-
пульсный характер (с периодом у мужчин 3-4 часа, у женщин в фолли-
кулярную фазу цикла - 1,5 часа, в лютеиновую - 3-4 часа), и только она
обладает стимулирующим эффектом (постоянное введение гормона
с лечебной целью дает тормозной эффект). Стимулируют секрецию
гонадолиберина норадренергические нейроны среднего мозга,
адреналин (а также ангиотензин П, серотонин, нейропептид Y). Тормо-
зят секрецию - серотонин- и дофаминергические нейроны,
опиатные пептиды, ГАМК, вещество Р, ВИП, холецистокинин.
Пролакголиберин точно не доказан, его функцию выполняет,
вероятно,тиролиберин.
Меланолиберин стимулирует секрецию МСГ гипофиза.
ф Статины тормозят выработку тропных гормонов аденогипофиза.
Соматостатин (пептид из 14 аминокислот образуется в паравентрикуляр-
ных ядрах) тормозит секрецию СТГ, ПТ, АКТГ гипофиза.
Пролактостатин (или/и дофамин) тормозит секрецию пролак-
тина гипофиза.
Меланостатии тормозит секрецию МСГ гипофиза.
• Транспорт либеринов и статинов.
•ф Из нейронов они секретируются в кровь фенестрированных капил-
ляров срединного возвышения (воронки) гипоталамуса.
•ф Переносятся с кровью к аденогипофизу и поступают через сеть ка-
пилляров к эндокринным клеткам аденогипофиза. (Двойная сеть ка-
пилляров препятствует поступлению этих гормонов в системный кровоток.)
• В аденогипофизе либерины и статины регулируют образование троп-
ных гормонов (АКТГ, ПТ, ФСГ, ЛГ, СТГ, пролактина, МСГ), изменяя
порог их реакции на гормоны периферических желез.
• Тропные гормоны аденогипофиза действуют на зависимые железы и
ткани.
• Саморегуляция активности гипоталамо-гипофизарной системы осуще-
ствляется с помощью отрицательных обратных связей.
ф Главное значение имеет длинная связь: влияние гормона перифери-
ческой железы на продукцию тропных гормонов аденогипофиза.
При этом порог реакции клеток аденогипофиза регулируется либе-
ринами и статинами
•Ф Допускается наличие коротких связей: влияние тропных гормонов
аденогипофиза на продукцию либеринов и статинов гипоталамуса.
132
(Вопрос о влиянии гормонов периферических желез на секрецию либеринов и
статинов также еще ждет своего разрешения.)
❖ Трансгипофизарная регуляция через нейрогипофиз.
• В супраоптических и паравентрикулярных ядрах гипоталамуса образу-
ются АДГ (вазопрессин) и окситоцин. (В гликопепгиде-предшественнике они
находятся вместе.)
• В результате аксонного транспорта эти гормоны поступают в нейроги-
пофиз и депонируются в нем (окончания аксонов расположены на базальной
мембране капилляров).
• Из нейрогипофиза АДГ и окситоцин секретируются в околокапилляр-
ное пространство и затем поступают в кровь, а также через Ш желудо-
чек мозга в цереброспинальную жидкость (влияние на поведение и память).
❖ Парагипофизарный путь.
• Прямой нервный контроль секреции гормонов осуществляется в мозго-
вом слое надпочечников, эпифизе, гипоталамусе (его нейросекреторных
ядрах); в этих железах происходит преобразование нервного импульса в
нейросекреторный процесс.
• Эндокринная (негипофизарная) регуляция осуществляется в результате
влияния гормонов периферических эндокринных желез друг на друга
(например, торможение кортизолом секреции тироксина, соматостатином секреции
инсулина и глюкагона).
• Гуморальная регуляция секреции инсулина, альдостерона, паратгормона
осуществляется соответственно уровнем глюкозы, Na+ и Са2+ в крови.
3. Методы исследования желез внутренней секреции.
3.1. Удаление железы или подавление ее активности.
3.2. Пересадка железы или введение гормонов.
3.3. Определение количества гормонов и их метаболитов в биологических жидко-
стях (крови, моче, ликворе) биохимическими, иммунорадиологическими и имму-
нофлюоресцентными методами анализа.
3.4. Функциональные нагрузочные пробы (например, тест на толерантность к глюкозе).
4. Физиология гипофиза.
4.1. Гормоны аденогипофиза (белково-пептидные).
*** Соматотропный гормон (СТГ) (образуется сомаготропоцитами аденогипофиза,
концентрация в плазме 1-10 мкг/л).
• Влияние СТГ на опорно-двигательный аппарат.
ф’ Стимулирует транспорт аминокислот и глюкозы через мембрану
клеток, синтез белков (в том числе антител), липолиз (совместно с глюко-
кортикоидами).
ф’ Вызывает гипертрофию тканей.
ф’ Стимулирует синтез в клетках печени семейства полипептидов, по-
лучивших название соматомедины (наиболее изучен иисулиноподобиый
фактор роста I или соматомедин С), через которые осуществляет свои
влияния на мишени. Эффекты соматомединов:
133
стимулируют зону роста эпифизарного хряща за счет увеличения
поглощения сульфата (образование хондроитиисульфата), аминокислот
(увеличение синтеза белков) и увеличения количества митозов;
стимулируют регенерацию тканей, заживление ран, компенса-
торную гипертрофию после удаления одного из парных орга-
нов;
стимулируют секрецию соматостатина в гипоталамусе.
• Метаболические эффекты СТГ.
- ф- Метаболизм белков. СТГ имеет метаболический эффект на скелет-
ные и сердечную мышцы, в клетки которых стимулирует транспорт
аминокислот и глюкозы, в них активируется синтез ДНК, РНК и
белков.
• ф- Метаболизм жиров. СТГ обладает катаболическим эффектом на
жировую ткань. Он стимулирует мобилизацию из нее жирных ки-
слот, уменьшая содержание в ней триглицеридов, и повышая со-
держания жирных кислот и глицерола в крови. СТГ увеличивает
окисление жирных кислот в печени до кетоновых тел.
• ф- Метаболизм углеводов. СТГ оказывает антиинсулиновый эффект и
способен вызывать гипергликемию. Она связана преимущественно
со способностью СТГ вызывать повышение в плазме крови жирных
кислот, что активирует глюконеогенез в печени. В результате анти-
инсулинового эффекта СТГ подавляет транспорт глюкозы в жиро-
вую ткань.
• ф- Метаболизм электролитов. СТГ вызывает почечную реабсорбцию
ионов кальция, фосфата и натрия, что участвует в его влиянии на
костную и хрящевую ткани.
• Регуляция продукции СТГ.
• ф- Основным стимулятором продукции СТГ является соматолиберии
гипоталамуса, а основным ингибитором - соматостатин (см. выше
2.5, регуляция через аденогипофиз), через которые преимущественно
и действуют различные физиологические и патологические факто-
ры.
ф" Активирует секрецию СТГ: сон (пик секреции во время фазы медленного
сив), умеренная мышечная нагрузка, стресс, голодание, гипоглике-
мия, аминокислоты аргинин и лизин, эстрогены, опиоидные пепти-
ды, АДГ.
• ф- Ингибируют секрецию СТГ соматомедин С, мелатонин; секреция
СТГ снижена в конце беременности, несмотря на высокий уровень
эстрогенов, и у тучных людей.
*** Адренокортикотропный гормон (АКТГ) (образуется кортикотропоцитами аде-
ногипофиза, суточный ритм секреции: максимум перед пробуждением, минимум перед
сном; содержание в плазме 25 -100 нг/л).
• Эффекты АКТГ (мишень - пучковая и сетчатая зоны коры надпочечни-
ков).
134
•Ф" Гипертрофия и гиперплазия коры надпочечников. Усиление про-
дукции глюкокортикоидов (в результате усиления синтеза мРНК и фермен-
тов стероидогенеза), в меньшей степени увеличивается продукция минералкор-
тикоидов.
ф" Вненадпочечниковое действие АКТГ: активация липолиза, усиле-
ние секреции инсулина, гиперпигментация кожи (АКТГ как аналог
МСГ по действию на меланоциты), ускоряет запоминание (фрагмент
АКТГ^о).
• Регуляция продукции АКТГ
Ф" Активатор секреции кортиколиберин, выделение которого усили-
вают стрессоры: боль, травма, кровопотеря, переохлаждение, пере-
гревание, инфекции, психические переживания, физическая нагруз-
ка и др.
•Ф" Ингибиторы секреции АКТГ: глюкокортикоиды, мелатонин.
•Ф" Роль отрицательных обратных связей.
Короткая: низкий уровень АКТГ стимулирует секрецию кор-
тиколиберииа в гипоталамусе, высокий - тормозит.
Длинная: низкий уровень глюкокортикоидов в крови стимули-
рует секрецию кортиколиберина гипоталамуса и АКТГ адено-
гипофиза, высокий - тормозит.
Тиротроппый гормоп (ТТГ) (образуется тиреотропоцитами аденогипофиза; пик
секреции ночью; концентрация в плазме 0,2-4 мЕД/л).
• Эффекты ТТГ (клетка-мишень - тироциты щитовидной железы). ТТГ дей-
ствует на рецепторы тироцитов и через вторые посредники (цАМФ, ино-
зитол-3-фосфат, Са2+) стимулирует все этапы синтеза и секреции тироид-
ных гормонов в щитовидной железе.
• Регуляция продукции ТТГ.
Активаторы секреции ТТГ: тиролиберии, переохлаждение, низкие
концентрации тиреоидных гормонов в крови, норадреналин (через а-
адренорецепторы), серотонин, эстрогены.
•ф" Ингибиторы секреции ТТГ: соматостатин, СТГ, мелатонин, дофа-
мин, глюкокортикоиды.
•ф- Роль отрицательных обратных связей в саморегуляции.
Короткие: низкий уровень ТТГ стимулирует секрецию тиро-
либерииа, высокий - тормозит.
Длинные: низкий уровень тироидных гормонов в крови стиму-
лирует секрецию тиролиберина и ТТГ, высокий - тормозит.
Пролактин (ПРЛ) (образуют лактотропоцигы аденогипофиза, импульсный тип секре-
ции, пик - в ночные часы; концентрация в плизме женщин 8-20 мкг/л, мужчин 5-8 мкг/л).
• Эффекты ПРЛ (органы-мишени - молочные железы и гонады).
• ф" Стимулирует пролиферацию протоков, развитие долек, гиперпла-
зию молочных желез, секрецию молока за счет активации синтеза
казеина и а-лактоглобулина, жиров, углеводов (эффект реализуется в
присутствии кортизола, инсулина, тироидных гормонов, СТГ, эстрогенов и про-
гестерона).
135
В физиологических концентрациях способствует нормальному раз-
витию и функционированию овариальных фолликулов. При корм-
лении грудью высокий уровень ПР Л в плазме вызывает ановуля-
ториое действие (лактогенная аменорея).
Ф Влияет на поведение («гормон материнской любви»),
•Ф В мужском организме ПРЛ усиливает действие лютеинизирующего
гормона на образование андрогенов в клетках Лейдига, стимулиру-
ет выработку подвижных сперматозоидов и функции простаты.
• Регуляция продукции ПРЛ.
Ф Стимулирует секрецию тиролиберин гипоталамуса (выполняет функ-
цию пролактолнберина), серотонин, ангиотензин П, ТТГ.
Ф Физиологические активаторы секреции: беременность и грудное
вскармливание (нейроэндокринный рефлекс: при сосании груди афферентные
импульсы, поступающие в гипоталамус, вызывают выброс ПРЛ в кровь, концен-
трация гормона возрастает примерно в 40 раз), сои - максимальное количе-
ство в первые часы глубокого сна, стресс, ТТГ.
Ф Ингибиторы секреции ПРЛ: дофамин (выполняет функцию пролактоста-
тина), ГАМК, прекращение вскармливания ребенка грудью.
❖ Фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон
(ЛГ) (образуют гонадотропоциты аденогипофиза; концентрация в плазме ФСГ 4-40
МЕ/л; ЛГ 5 - 30 МЕ/л).
• Эффекты ФСГ.
Ф У женщин стимулирует рост и созревание фолликулов и повышает
их чувствительность к ЛГ, увеличивая синтез рецепторов ЛГ и их
чувствительность; увеличивает вызванную ЛГ секрецию эстроге-
нов.
Ф У мужчин в период полового созревания обеспечивает рост и раз-
витие интерстициальных клеток Лейдига, которые продуцируют
андрогены. В половозрелом возрасте стимулирует сперматогенез и
синтез циторецепторов, связывающих андрогены.
• Эффекты ЛГ.
Ф У женщин обеспечивает созревание яйцеклетки и овуляцию, разви-
тие желтого тела, продукцию эстрогенов и прогестерона (совместно с
пролактином).
Ф У мужчин стимулирует образование андрогенов в клетках Лейдига,
через андрогены поддерживает гаметогенез.
• Гипоталамический контроль регуляции продукции ФСГ и ЛГ (имеется
импульсное выделение гормонов с периодом 90 -120 мни).
Ф Гонадолиберин гипоталамуса стимулирует секрецию ФСГ и ЛГ в
аденогипофизе. Центры секреции гоиадолиберина в гипоталамусе:
тонический (активен у мужчин н женщин), расположен в аркуатных
ядра, обеспечивает базовую секрецию ФСГ и ЛГ, имеющую
импульсной характер.
136
циклический (функционирует только у женщин после полового созрева-
ния), расположен в преоптической области, обеспечивает пре-
довуляторный выброс ЛГ и ФСГ,
•ф- Отрицательные обратные связи обеспечивают стабилизацию уровня
гормонов.
Короткая: низкие концентрации ФСГ и ЛГ стимулируют сек-
рецию гипоталамического гонадолиберина, высокие - тормо-
зят.
Длинная: низкие концентрации гормонов половых желез (у муж-
чин тестостерона, у женщин эстрогенов) стимулируют секрецию го-
надолиберина, ЛГ н ФСГ гипоталамуса, высокие - тормозят.
• ф- Положительная обратная связь в женском организме обеспечивает
предовуляторный пик секреции гонадолиберина, выброс ЛГ и ову-
ляцию в ответ на повышение уровня эстрогенов.
• ф- Ингибиторы секреции ФСГ и ЛГ: мелатонин, стресс, пролактин,
прогестерон, ингибин (пептид, образующийся в яичниках и семенниках).
❖ Меланоцитостимулирующий гормон (а-МСГ, синоним.: интермедин) обра-
зуют меланотрофы аденогипофиза; он имеет общий предшественник (проопиомеланокор-
тин) с АКТГ, p-эндорфнном и ^-липотропным гормонами.
• Эффекты а-МСГ (мишень - пигментные клетки).
•ф- Стимулирует в меланоцитах эпидермисе кожи синтез меланина
(ферменты тирозиназа и ДОФА-оксидаза), образование его гранул (мелано-
сом), которые накапливаются в кератоцитах в виде «защитного»
зонтика над ядром. Обеспечивая пигментацию кожи и волос, регу-
лирует степень УФ-облучения эпидермиса и сопряженные с этим
процессы синтеза провитамина D3 н распада фолиевой кислоты.
’С’ Участвует в обеспечении фоторецепции сетчатки (пигментный
слой).
•ф- Участвует в механизмах памяти (фрагмент МСГ,.,»).
• Регуляция секреции МСГ.
•ф- Стимулируют секрецию МСГ: меланолиберин гипоталамуса, сен-
сорные стимулы - звук, прикосновение, грудное вскармливание,
травма.
ф' Ингибиторы секреции МСГ: меланостатин, кортизол по механизму
отрицательной обратной связи.
4.2. Гормоны нейрогипофиза (поступают в него из гипоталамуса, см. выше 2.4).
❖ Антидиуретический гормон (АДГ) или вазопрессин (нонапептнд супраопти-
ческих и паравентрикуларных ядер гипоталамуса, концентрация в плазме 1-13 нг/л).
• Эффекты АДГ (вазопрессина).
"Ф* Увеличивает реабсорбцию воды в дистальных канальцах и собира-
тельных трубочках почек, что приводит к уменьшению диуреза (эф-
фект реализуется через Ут-рецепторы, повышение уровня цАМФ и увеличение
числа водных каналов в клеточной мембране).
137
"ф- В более высоких концентрациях усиливает сокращения гладких
мышц и вызывает сужение сосудов (эффект реализуется через V,-
рецепторы —> ИФ3/ДАГ —»Са2*).
ф- Активирует центр жажды и питьевое поведение (одновременно тормозит
секрецию ренина).
•ф- Участвует в механизмах формирования памяти (стимулирует обу-
чение).
• Регуляция секреции АДГ.
• ф- Главным регулятором секреции АДГ является осмоляльность плаз-
мы, которая контролируется осморецепторами гипоталамуса, сосу-
дов, тканей. Гиперосмия (>290 мосм/кг) стимулирует секрецию АДГ,
гипоосмия (<280 мосм/кг) тормозит секрецию АДГ.
- ф- Второй регулятор секреции АДГ - колебания объема циркулирую-
щей крови (ОЦК) и АД, которые регистрируются барорецепторами
аорты, сонной артерии, легочной артерии, волюморецепторами
предсердий и вен. Гиповолемия (снижение ОЦК более чем на 5 %) и
снижение АД стимулируют секрецию, гиперволемия (увеличение
ОЦК) и повышение АД тормозят секрецию АДГ.
ф" Другие факторы, влияющие на секрецию АДГ.
Стимулируют секрецию АДГ симпатоадреналовая система (через Р-адрено-
рецепторы), ангиотензин И, гистамин, ацетилхолин.
Тормозят секрецию АДГ натрийуретический гормон предсердий, этанол,
переохлаждение.
Окситоцин (образуется в паравентрнкулярных и супраоптических ядрах гипоталамуса,
концентрация в плазме 3,2 мМЕ/л).
• Эффекты окситоцина реализуются через поступление Са2+.
• ф- Усиливает сократительную активность матки во время родов (эстро-
гены увеличивают чувствительность матки к окситоцину, а прогестерон снижа-
ет).
• ф- Усиливает сокращение гладкой мускулатуры протоков молочных
желез и выделение молока.
- ф- Стимулирует продукцию простагландинов в эндометрии, которые
совместно с окситоцином вызывают начало родов (мобилизуют Са2*).
• ф- Участвует в механизмах формирования памяти (нарушает запомина-
ние).
• Регуляция секреции окситоцина: рефлекторная стимуляция секреции с
рецепторов растяжения шейки матки и влагалища во время родов, сос-
ков молочных желез при кормлении грудью; условнорефлекторная - на
крик ребенка и кормление.
5. Физиология щитовидной железы.
5.1. Функции тироцитов и образуемых ими фолликулов.
❖ Образуют тироидные гормоны (производные тирозина): тироксин (тетрайод-
тиронин или Т4) - около 100 нмоль/сут. и трийодтиронин (Тз) - около 5
нмоль/сут.
138
Синтез тироидных гормонов осуществляется с участием двух парал-
лельно протекающих процессов - образованием тироглобулина и по-
ступлением аниона йода в тироцит.
Ф- Синтез тироглобулина осуществляется в гранулярном ЭПС тироци-
тов, затем он переностя в аппарат Гольджи для процесса иодирова-
ния.
ф' Извлечение йодида из крови тироцитами осуществляется в резуль-
тате симпорта 2Na+ + Г, затем происходит его окисление при уча-
стии тиропероксидазы, после чего он может иодировать тироглобу-
лин. (Часть Г поступает в коллоид полости фолликула через ионный канал на
апикальной мембране и участвуют там в иодировании тироглобулина.)
ф- В аппарате Гольджи происходит иодирование части тирозольных
остатков в недрах молекулы тироглобулина с образованием моно-
и дииодтирозинов. Далее они конденсируются с образованием тет-
раиодтиронииа (Та) и трииодтиронина (Тз). Иодированный тирог-
лобулин, упакованный в пузырьках, путем экзоцитоза секретирует-
ся в коллоид полости фолликула. Итак, тиреоидные гормоиы со-
держатся в составе тироглобулина, который является формой их
внеклеточного хранения.
'Ф' Рециклизауия тироглобулина. В коллоиде фолликула имеется тироглобулин раз-
личной степени йодирования. Незрелый тироглобулин захватывается тироцитом
путем рецепторзависимого эндоцитоза (рецептором незрелого тироглобулина яв-
ляется N-ацетилглюкозамнн), где и подвергается дополнительному йодированию
и созреванию (конденсации). Образовавшийся зрелый тироглобулин секретиру-
ется в полость фолликула.
Секреция тироидных гормонов.
ф- Зрелый тироглобулин путем пиноцитоза перемещается в тироцит,
где эидоцитозиые пузырьки соединяются с лизосомами (образуют-
ся фаголизосомы).
ф' В них йодированный тироглобулин расщепляется пептилазами с
образованием аминокислот, моно- и дийодтирозинов и молекул
тетра- и трийодтиронина (Та и Тз), последние секретируются в
кровь. Все вышеизложенные процессы в тироците стимулируются
ТТГ.
ф’ Вместе с гормонами из железы в кровь выделяется небольшое количество тирог-
лобулнна и тиропероксидазы, которое увеличивается при воспалительных н зло-
качественных поражениях железы н сопровождается появление аутоантител к
ним.
Периферическая конверсия тироидных гормонов. Основное количество
трийодгиронниа (- 80%) образуются в клетках-мишенях (особенно в
печени и почках) из поступившего туда Та путем отщепления одного Г
под действием тироксиндейодиназ. (Отщепление Г от наружного фенольного
кольца Та приводит к образованию высокоактивного Тз, а от внутреннего тирозильно-
го кольца - к малоактивному реверсионному Т3 _ рТ3.) Таким образом, клетки-
мишени, увеличивая или уменьшая превращение Та в Тз могут сами ре-
гулировать эффективность действия этих гормонов.
139
❖ Циркуляторный транспорт гормонов
• Концентрация в сыворотке крови общего количества Т4 = 60-155
нмоль/л, Тз = 1,2-3,5 нмоль/л. Концентрация свободных (физически рас-
творенных) тироидных гормонов очень низка (Т<= 9-28 пмоль/л, т3 = 3,8-7,4
пмоль/л), ио именно свободные формы гормона действует на ткани-
мишени
• Основное количество тироидных гормонов (более 99,5%) связана с бел-
ками крови, образуемые печенью: тироксинсвязывающим глобулином,
преальбумииом и альбумином. Эти белки создают резерв ТГ в организ-
ме, связывают избыточное их количество, поступающее из железы, или
принятое в лечебных целях, предупреждают их потерю через почки.
ф Тироксинсвязывающий глобулин прочно связывает и транспорти-
рует - 75% Т4 и - 85% Тз, образуя относительно инертный резерв
тироидных гормонов в крови.
ф Преальбумин связывает 10-15% Т4 и менее 5% Тз, прочность связи
значительно меньше, чем с ТСГ.
ф Альбумин связывает ~ 10% Т4 и несколько меньше Тз. Благодаря
очень низкой прочности связи, ои (и преальбумин) является основ-
ным источником свободных ТГ, образуя лабильный резерв, кото-
рый может быть использован при функциональных нагрузках.
❖ Физиологические эффекты тироидных гормонов первично реализуются
преимущественно через геном клетки, однако у них имеются и эффекты,
реализуемые непосредственио иа уровне клеточной мембраны и цитоплаз-
мы.
• Геномные эффекты ТГ осуществляются через ядерные рецепторы, при
этом ТГ играют роль регуляторов геииой экспрессии.
ф ТГ проникают в клетку путем простой диффузии и проникают в яд-
ро, где с высоким сродством и специфичностью связываются с
ядерными рецепторами. Они представляют собой белки (факторы
транскрипции), прочно связанные с ДНК хроматина и дополни-
тельными ядериыми белками.
Каждый рецептор содержит несколько специфических участков
(домеиов): ДНК-связывающий домен, гормонсвязывающий до-
мен, активирующий транскрипцию домен и др.
• Влияние рецепторов ТГ на ДНК-чувствительные участки усили-
вают дополнительные ядерные белки. В качестве них выступают,
например, рецепторы витаминов А и D. Фосфорилирование ре-
цептора (по остаткам серина, треонина) увеличивает его связывание с
ДНК.
Рецепторы ТГ кодируются двумя протоонкогенами с-егЬА, расположеными в
локусах в двух хромосом: 3 и 17. Н связи с этим имеется несколько изоформ
рецепторов. Экспрессия генов-рецепторов ТГ, а значит, и соотношение изо-
форм зависит от вида клеток, их функционального состояния. С этим в опре-
деленной степени н связаны особенности действия ТГ в клетках различных
тканей и в клетках одной ткани при различных функциональных состояниях.
140
Мутация, изменяющая только один из доменов белка-рецептора ТГ может
резко изменить эффективность действия гормона (например, мутация домена,
активирующего транскрипцию, приведет к тому, что эффект гормона будет за-
блокирован, хотя рецептор и будет связывать гормон и присоединяться вместе
с ним к ДНК хроматина).
’Ф’ Сигнал с комплекса «ТГ + тироидный рецептор + чувствительный к
нему участок ДНК» передается на транскрипционный ген-промотор
(ТАТА-бокс), связанный с РНК-полимеразоЙ П, обеспечивающей
синтез мРНК на соответствующем структурном гене (процесс
Транскрипции). (Активация транскрипции тироидными гормонами может
быть усилена набором ядерных белков под общим названием коакгиваторы; к
ним относятся, например, коактиватор стероидных гормонов-1, коактиватор сис-
темы цАМФ).
"Ф’ Последующие эффекты действия ТГ связаны с набором активиро-
ванных структурных генов н функциональным значением обра-
зующихся белков.
Негеномные эффекты тироидных гормонов осуществляются без уча-
стия ядерных рецепторов, они реализуются через рецепторы плазмо-
леммы, цитозоля и митохондрий. Они возникают быстро (через секунды
- минуты) и могут быть вызваны в условиях блокады транскрипции ге-
нов н синтеза белка. Рецепторы для ТГ (обычно ферменты) при этом яв-
ляются и эффекторными молекулами. Негеномные эффекты ТГ осуще-
ствляют с участием систем вторых посредников, используемых другими
гормонами: аденилатциклазной, фосфоинозитидной и др. Во многих
случаях негеномные эффекты ТГ пролонгируются и усиливаются ге-
номными эффектами.
'Ф* При действии ТГ на плазматическую мембрану показано повышение транспорта
глюкозы и аминокислот, активности Са2+-насоса, адренорецепторов, Ыа*-каналов,
Са2+-каналов L-типа и др.
* Ф* В цитозоле ТГ активируют протеинкиназу С, протеинкиназу А, но тормозят об-
разование активного фермента гликолиза - пируваткиназы
* Ф* В митохондриях ТГ в физиологических концентрациях активирует дыхание, в
высоких патологических концентрациях разобщают окисление и фосфорилиро-
вание, что лежит в основе калорогеиного эффекта ТГ.
* Ф* В гладкой ЭПС активируется Са2+-иасос, что уменьшает концентрацию Са2+ в
цитозоле. В шероховатой ЭПС стабилизируется специфические мРНК и пролон-
гируется синтез белка.
'Ф* В гладкомышечных клетках сосудов увеличивается расслабление (уменьшает
сосудистый тонус).
’Ф' В цитоскелете стимулируется полимеризация растворимого G-актина с образова-
нием F-актина (т.е. актиновых филаментов).
Действие ТГ на физиологические системы.
Ф Влияния на нервную систему (для деятельности нейронов особенно важен
Тз, который образуется в глиоцитах из Т4, извлекаемого из крови).
ТГ стимулируют в нейронах и глиоцитах экспрессию генов, ко-
дирующих наиболее важные для деятельности нервной ткани
141
белки: тубулины, основной белок миелина, синаптосины, фактор
роста нервов и др.
ТГ стимулируют развитие мозга. Их действие особенно важно в
критический период развития головного мозга (последний три-
местр беременности - первые недели после рождения), когда в
значительной мере завершаются основные процессы морфогенеза
мозга: дифференцировка клеток, миелинизация волокон, форми-
рование синапсов н др.
Стимулируют психическое развитие (формирование синапсов,
миелинизацию аксонов, транспорт аминокислот через ГЭБ), при
недостатке - ослабление памяти и замедление умственных про-
цессов.
ТГ повышают возбудимость ЦНС н ее симпатического отдела
(усиливают активность катехоламинов).
ТГ активируют гипоталамические центры и повышают аппетит
и потребление пищи.
• Ф Влияния на сердечно-сосудистую систему: увеличивают сердечный
выброс, систолическое и пульсовое давление, частоту сердечных
сокращений н кровоток в большинстве тканей, уменьшают сосуди-
стый тонус.
• Ф Влияние на гемопоэз: ТГ усиливают эритропоэз (Т4 > Т3), увеличивая
образование Рг-адренорецепторов на плазмолемме эритроидных
клеток н стимулируя секрецию эритропоэтина.
• Ф Влияние на систему дыхания: стимулируют развитие легких, уве-
личивают частоту, глубину дыхания н минутный объем дыхания.
• Ф Пищеварение: ТГ усиливают моторику и секрецию желудка н ки-
шечника, ускоряют всасывание глюкозы в кишечнике.
• Ф ТГ усиливают теплопродукцию и повышают температуру тела.
ф Обмен веществ: ТГ в физиологических концентрациях стимулиру-
ют синтез белка н катаболизм холестерола, гликогенолиз, липолиз и
окисление жирных кислот, увеличивают концентрацию глюкозы в
кровн, усиливают диурез, выведение Na+ и К* с мочой.
• ф Репродуктивная система: ТГ необходимы для ее созревания и нор-
мальной функции. Они стимулируют выработку гоиадолиберина,
лютеинизирующего гормона, активируют клетки Сертоли и выра-
ботку тестостерона в яичках, увеличивают чувствительность яич-
ников к гонадотропным гормонам, а эндометрия - к эстрогенам.
Инактивация тироидных гормонов происходит в различных тканях. Период полураспада
Т< составляет 4-7 суток, Т3 -1-3 суток.
• В тканях -мишенях путем последовательного дейодировання с образованием дийодти-
ронинов, монойодтироиинов и тиронина, практически не обладающих гормональной
активностью.
• Около 20% ТГ инактивируются в печени путем конъюгации с глюкуроновой и серной
кислотами и в виде глюкуроиатов и сульфонатов экскретируются с желчью.
142
• Небольшое количество ТГ инактивируется почками путем дейодироваиия и сульфати-
рования, с последующей экскрецией метаболитов с мочой.
❖ Регуляция продукции тироидных гормонов. Тироциты щитовидной железы
входят в систему трансгипофизарной регуляции через аденогипофиз: гипо-
таламус -» аденогипофиз -» тироциты фолликулов.
• Основным регулятором секреции ТГ является тиреотропный гормон
(Tn), образуемый тиротропоцитами аденогипофиза (пик секреции ночью)
(см. выше 2.5).
ф- ТТГ действует на рецепторы тироцитов и через аденилатциклазную
и фосфоинозитидную системы стимулируют все этапы образования
н секреции ТГ.
Основным стимулятором секреции ТТГ гипофиза является ти-
ролнберин (ТЛ) гипоталамуса (трипептид паравентрикуляриых, супра-
хиазматических, дорсомедиальных и серобугориых ядер). Эффект ТЛ осу-
ществляется через рецепторы на тиротропоцитах гипофиза с
включением вторых посредников: инозитол-3-фосфата, Са2+,
цАМФ. Стимулируют секрецию ТТГ и ТЛ низкий уровень Т3/Т4 в
крови, эстрогены, норадреналин, стресс, охлаждение организма и
др.
» Тормозит секрецию ТТГ гипофиза и ТЛ гипоталамуса сомато-
статин н дофамин гипоталамуса, соматотропный гормон, серото-
нин, высокий уровень Т3/Т4 в крови, высокие (патологические и фар-
макологические) уровни глюкокортикоидов, перегревание организ-
ма, , эндорфины.
• Некоторое влияние на секрецию ТГ тироцитами оказывает вегетативная
иннервация: симпатические (адренергические и ВИП-пептидергичес-
кие) влияния через аденилатциклазную систему увеличивают секрецию
ТГ; парасимпатические влияния оказывают тормозящий эффект.
5.2. Функции парафолликулярных клеток щитовидной железы (образуют поли-
пептид кальцитонин, концентрация в плазме до 100 hi/л, Т1/2 около 5 суток, катаболизм в
печени, почках, костной ткани).
❖ Кальцитонин по влиянию на фосфорно-кальциевый обмен является анта-
гонистом паратгормона и холекальциферола (bht.Dj).
• Механизм действия кальцитонина обусловлен преимущественно акти-
вацией аденилатциклазы н накоплением цАМФ, что приводит к тор-
можению активного транспорта Са2* из клеток.
• Кальцитонин усиливает образование и минерализацию костной ткани,
стимулируя функцию остебластов (поглощение Са2* н фосфатов нз кро-
ви) и подавляя активность остекластов
• Кальцитонин тормозит всасывание Са2* и фосфатов в тонком кишечни-
ке и реабсорбцию их в почках, увеличивая выделение Са2*, фосфатов с
мочой, что способствует снижению уровня Са2* в крови.
143
• Кальцитонин увеличивает силу и уменьшает частоту сердечных сокра-
щений, тормозит моторную функцию желудка, кишечника и секрецию
желудка.
• Кальцитонин является стресс-лимитирующим фактором, ограничивая
активацию гипоталамо-гипофизарио-надпочечниковой системы при
действии стрессоров.
❖ Активируют секрецию кальцитонина гиперкальциемия (а также глюкагон,
гастрин, катехоламины, эстрогены), тормозят — гипокальциемия, соматостатин.
6. Околощитовидные железы.
6.1. Физиологические эффекты паратгормона - повышает уровень Са2+ в крови в
результате действия следующих механизмов.
❖ Повышение активности остеоцитов и остеокластов (усиление резорбции кост-
ной ткани).
❖ Увеличение реабсорбции Са2+ в дистальных почечных канальцах и сниже-
ния реабсорбции фосфатов в проксимальных почечных канальцах (фосфа-
турия).
❖ Усиление всасывания Са2+ в тонком кишечнике (действует совместно с витами-
ном D3).
❖ Эффект паратгормона связан со стимуляцией адеиилатциклазы, увеличени-
ем уровня цАМФ и активности протеинфосфокиназы А в клетках-мишенях.
6.2. Регуляция продукции паратгормона (суточный ритм секреции: в ночные часы в 2 - 3
раза больше, чем в дневное время).
❖ Основной стимулятор секреции - гипокальциемия (ускоряет трансформацию
пропаратгормоиа в активный гормон).
❖ Ингибитор секреции - снижение уровня Mg2* в крови.
7. Эндокринные функции островков Лангерганса поджелудочной железы: а-клеткн
(25 %) продуцируют глюкагон; р-клеткн (60 %) - инсулин; 5-клетки (10 %) - соматостатин (все поли-
пептиды).
7.1. Эндокринные функции /1-клеток, вырабатывающих инсулин (в крови находится в
свободной форме, концентрация в плазме 6-24 мМе/л).
❖ Продукция инсулина (40 - 50 елУсут.).
• Биосинтез инсулина (ген в хромосоме 11).
ф На полирибосомах гранулярной ЭПС образуется препрогормон (мм
= 11500 Да).
Ф Затем он направляется в цистерны ЭПС, где от него отщепляется
префрагмент и образуется одноцепочечный проинсулин (мм = 9000
Да), который переходит в аппарат Гольджи.
ф В аппарате Гольджи от проинсулииа отщепляется С-пептид и обра-
зуется инсулин (мм = 5734 Да), который упаковывается в гранулы.
• Секреция инсулина стимулируется глюкозой крови, которая проникает
в (3-клетки преимущественно с помощью транспортера ГЛЮТ 2, что
приводит к повышению уровня АТФ и закрытию АТФ-зависимых КФ
каналов. Возникает деполяризация мембраны до КУД и возникникает
ПД. Вход Са2+ в клетку во время ПД запускает экзоцитоз гранул инсу-
144
лина н открытие lC-каналов. Секреция инсулина осуществляется в
кровь панкреатической вены.
Циркуляторный транспорт. Инсулин, поступая в кровь воротной вены,
доставляется в печень и далее в системный кровоток. Инсулин не имеет
белка-носителя в плазме (в отличие от иисулиноподобных факторов роста), и по-
этому Ti/2 его не превышает 10 мин.
Физиологические эффекты инсулина (основные мишени - мышечная и жиро-
вая ткань, печень).
• Клеточные механизмы действия инсулина. Инсулин соединяется с ин-
сулиновым рецептором (IR) плазмолеммы н активирует его тирозннки-
назный участок, что приводит к аутофосфорилированию Р-субъединицы
рецептора. Активация рецептора приводит к образованию в клетке
мультиферментиого комплекса, способного осуществить фосфорилиро-
вание и дефосфорилирование тирозиновых остатков в молекулах бел-
ков, что и определяет влияние инсулина на .метаболические процессы.
ф В формировании фосфорилирующего эффекта участвует белок-посредник IRS-1,
который обеспечивает проведение сигнала на различные молекулы, среди кото-
рых наиболее изучен фермент фосфатидилинозитол-3-кииаза, которая стимули-
рует перемещение белка-переносчика глюкозы ГЛЮТ-4 из цитозольных везикул
в плазматическую мембрану.
Ф’ Дефосфорилирующий эффект осуществляется путем активирования инсулино-
вым рецептором тирозиновых протеинфосфатаз, одной из мишенью которых яв-
ляется IRS-белок, что приводит к его инактивации и, следовательно, ингибиро-
ванию эффектов инсулина.
• Влияние инсулина на обмен углеводов.
Ф" Стимулирует транспорт глюкозы через мембраны мышечных и жи-
ровых клеток (на них увеличивается количество белков-переносчиков ГЛЮТ
4, образующих гидрофильные трансмембранные каналы для глюкозы, которые в
40 раз уаеличивают транспорт глюкозы в клетки).
Ф’ Снижает уровень глюкозы в крови (гипогликемия).
ф- Усиливает синтез гликогена (активация глнкогенсннтетазы).
Ф" Активирует глюкокиназу (усиление использования глюкозы).
ф- Активирует глюкозо-6-фосфатдегидрогеназу (использование глю-
козы в пентозофосфатном пути).
ф- Ингибирует глюкозо-6-фосфатазу в печени и почках (уменьшает
выход глюкозы в кровь).
Ф" Ингибирует глюконеогенез, снижая образование через геном клю-
чевого фермента глюконеогенеза (фосфоэнолпируваткарбоксикиназу).
• Влияние инсулина на липидный обмен.
ф- Тормозит липолиз и стимулирует липогенез (усиливает синтез
жирных кислот и триглицеридов).
ф- Снижает кетогенез за счет стимуляции окисления кетоновых тел
(ацетоацетата, f-оксибутирата) в цикле трикарбоновых кислот.
• На обмен белков инсулин оказывает анаболическое влияние.
145
Усиливает транспорт аминокислот и синтез белков в мышечной и
жировой ткани, в печени.
Тормозит протеолиз и оказывает антикатаболическое действие.
❖ Катаболизм инсулина происходит во многих тканях, ио преимущественно
в печени, почках и плаценте, в результате восстановления дисульфидных
мостиков (фермент глютатион-иисулин-трансгидрогеиаза) и гидролиза цепей (фер-
мент инсулиназа).
❖ Регуляция продукции инсулина.
• Гуморальная регуляция.
•ф' Повышение глюкозы в крови - основной стимулятор секреции ин-
сулина (пороговая величина глюкозы - около 5 ммоль/л, максимальный ответ -
при концентрации около 25 ммолъ/л). На р-клетках имеются белок-транспортер
глюкозы - ГЛЮТ 2.
ф- Стимулируют секрецию инсулина аминокислоты лейцин, аргинии,
лизин.
• Эндокринная регуляция.
Глюкагон, глюкокортикоиды, эстрогены, прогестерон и гастроинте-
стинальные гормоны (особенно секретин, холецистокннин-панкреозимии)
стимулируют секрецию инсулина. Последние подготавливают ин-
сулиновую систему к предстоящему при пищеварении всасыванию
глюкозы в кровь.
•ф' Соматостатин и катехоламины ингибирует секрецию инсулина.
• Нервная регуляция (вегетативная нервная система).
Парасимпатические влияния стимулируют секрецию инсулина
(ацетилхолин через М-холинорецепторы) и катехоламины.
•ф' Симпатические влияния тормозят секрецию инсулина через
аг-адренорецепторы и стимулируют - через рг-адреиорецепторы
(суммарный эффект - тормозящий).
7.2. Эндокринная функция а-клеток, вырабатывающих глюкагон (30 - 120 иг/л
плазмы, период полураспада 5-10 мин).
Физиологические эффекты глюкагона.
• Влияние иа обмен углеводов и белков.
ф- Стимулирует продукцию глюкозы в печени в результате гликоге-
нолиза (активирует фосфорилазу).
• ф' Тормозит гликогеисинтетазу и синтез гликогена.
ф- Стимулирует катаболизм белков и глюконеогенез - образование
глюкозы из аминокислот и лактата.
• ф' Усиливает вход в печень аланина и образование мочевины.
• Влияние иа обмен липидов.
•ф' Усиливает липолиз в жировой ткани (через аденилатциклазу и цАМФ).
ф' Увеличивает поставку в печень свободных жирных кислот.
• Усиливает сокращение сердца.
• Влияние иа секрецию других гормонов.
146
Ф' Стимулирует Р-клетки и секрецию инсулина.
Ф’ Стимулирует мозговой слой надпочечников и секрецию катехола-
минов.
•Ф Стимулирует секрецию кальцитонина в щитовидной железе.
❖ Регуляция секреции глюкагона.
• Секреция стимулируется гипогликемией, аминокислотой аргинином,
свободными жирными кислотами, холецистокинин-панкреозимином,
симпатической нервной системой (через 0-адренорецепторы).
• Секреция тормозится гипергликемией, соматостатином, секретином.
73. Эндокринная функция 3-клеток, вырабатывающих соматостатин.
❖ Эффекты соматостатина.
• Тормозит секрецию инсулина и глюкагона.
• Снижает всасывание глюкозы в кишечнике.
• Тормозит секрецию соляной кислоты, пепсинов и моторику в желудке,
секрецию ферментов поджелудочной железы.
❖ Регуляция секреции: стимуляторами секреции служат глюкагон и все акти-
ваторы секреции инсулина.
8. Физиология надпочечников (корковый и мозговой слои).
8.1. Корковый СЛОЙ (клубочковая зона вырабатывает минералкортикоиды, пучковая зона -
глюкокортикоиды, сетчатая зона - преимущественно андрогены).
Альдостерон (минералокортикоид, в плазме 0,14 - 0,23 нмоль/л).
• Физиологические эффекты альдостерона.
ф- Основные органы-мишени: почки, потовые и слюнные железы, же-
лудок и толстая кишка.
"Ф Механизм действия альдостерона связан с увеличением синтеза и
активацией уже существующих переносчиков Na+ (Na7H*-
ионообменннка, К*, Na*-Hacoca) и Н*-насоса.
"Ф В канальцах почек альдостерон увеличивает реабсорбцию Na+ и
усиливает секрецию К+ и Н*.
"Ф В потовых и слюнных железах увеличивает реабсорбцию Na+.
"Ф Влияет на поддержание водного и электролитного баланса.
• Регулирует объем жидкости (например, ОЦК) вместе с АДГ.
Регулирует осмотическое давление за счет изменения реабсорб-
ции Na+ в почках.
• Ф Регулирует АД: задержка Na+ в организме увеличивает ОЦК и уве-
личивает чувствительность гладких миоцитов артериол к сосудо-
суживающим веществам (например, к адреналину).
• Регуляция продукции альдостерона.
ф Стимулируют секрецию увеличение уровня ангиотензина П, высо-
кий уровень К+ и низкий уровень Na+ в крови (а также простациклин и
АКТГ).
"Ф Тормозят секрецию высокое содержание Na+, дофамин, Na+-
уретический гормон предсердий.
147
•ф Влияние позы на величину секреции: в положении стоя секреция в
2-4 раза больше, чем лежа, это обеспечивает поддержание ОЦК и
АД на уровне, оптимальном для мозгового кровоснабжения.
*•* Глюкокортикоиды (основной гормон - кортизол, продукция 20 - 25 мг/сут, 95 %
гормона транспортируется в кровн в связи с транскортином).
• Эффекты глюкокортикоидов.
ф- Влияние на углеводный обмен: повышение уровня глюкозы в крови
как результат усиления образования в печени глюкозы из амино-
кислот и жирных кислот (глюконеогенез) и уменьшение использова-
ния глюкозы и аминокислот в мышцах.
• ф Влияние на жировой обмен: повышение уровня жирных кислот в
крови как результат активации липолиза и одновременного усиле-
ния отложения жира в некоторых частях тела (лицо, туловище).
• ф Влияние на белковый обмен: торможение синтеза белков в мышцах,.
отрицательный азотистый баланс, преобладание катаболического
эффекта иад анаболическим.
- ф- Разрешающее (пермиссивное) действие для ряда гормонов (напри-
мер, реализация эффектов катехоламинов связана с активацией кортизоном ге-
нов, регулирующих синтез Рг и Рг-адренорецепторов).
* Ф Участие в стабилизации АД (увеличивает силу сокращений сердца, сосуди-
стый тонус, а также ОЦК за счет слабого минералкортикондного эффекта).
• ф Участие в организации стресс-синдрома (повышают резистентность к
стрессорам).
• Ф Оказывают влияние на ЦНС и ВНД (увеличение возбудимости
нервной системы, изменение эмоционального состояния).
* Ф В фармакологических дозах глюкокортикоиды оказывают противовоспалитель-
ное н противоаллергическое действие (связано с освобождением лнпокортина,
обеспечивающего стабилизацию мембран лизосом и снижение проницаемости
капилляров); иммунодепрессантное действие (снижение продукции В- н Т-лим-
фоцитов, антител, лимфоцитолиз).
• Регуляция продукции глюкокортикоидов (суточный ритм: минимум - перед
сном, максимум - в 6 - 8 часов утра).
• Ф Кортиколиберин и АКТГ стимулируют синтез и секрецию.
• Ф Саморегуляцию обеспечивают отрицательные обратные связи.
Длинные: повышение уровня глюкокортикоидов в крови тор-
мозит синтез и секрецию кортиколиберина и АКТГ, снижение
уровня глюкокортикоидов увеличивает их секрецию.
Короткие: снижение уровня АКТГ стимулирует продукцию
кортиколиберина, повышение - тормозит.
Андрогены (дегидроэпиандростерон н его сульфат - основные андрогены коры надпо-
чечников, нз андрогенов в надпочечниках образуется небольшое количество эстрогенов).
• Половые различия: у взрослого мужчины здесь образуется только 5 %
всех андрогенов; у женщин в фолликулярную фазу цикла - более поло-
вины, после овуляции - менее половины общего количества андрогенов.
• Преобразования надпочечниковых андрогенов в тестостерон, прогесте-
рон и эстрогены происходит в периферических тканях-мишенях: под-
148
кожной жировой клетчатке, волосяных фолликулах, в молочных желе-
зах, во время беременности в плаценте.
• Эффекты андрогенов надпочечников.
ф- В раннем антенатальном периоде (12 - 20 недель) обеспечивают
вместе с семенниками развитие наружных гениталий по мужскому
типу.
ф- Обусловливают опускание яичек в мошонку (стимулируют рост семя-
выносящих протоков, семенных канальцев, связок).
ф- Являются основным источником андрогенов в препубертатном пе-
риоде (участвуют в запуске полового созревания).
• Регуляцию секреции андрогенов надпочечников стимулируют АКТГ и
пролактин.
8.2. Мозговой слой надпочечников («специализированный симпатический ганглий», хро-
маффинные клетки которого выделяют катехоламины: адреналин ~ 80 %, норадреналин - 18 %,
дофамин - 2 %).
Рецепция катехоламинов (циторецепторы клеточной мембраны).
• Рецепторы: адреналин н норадреналин действуют на <х-
адренорецепторы (а, и aj н Р-адренорецепторы (0, и |У мембран различ-
ных клеток возбудимых и невозбудимых тканей. (Рецепторы дофамина рас-
положены преимущественно в мозге: гипоталамусе, гипофизе, черном веществе.)
• Сродство катехоламинов к адренорецепторам.
Ф" К a-адренорецепторам имеют высокое сродство как адреналин, так
н норадреналин. Через эти рецепторы в гладких миоцитах развива-
ются преимущественно стимулирующие эффекты, а в секреторных
клетках - преимущественно тормозные.
"Ф* К 0-адренорецепторам имеет высокое сродство адреналин и невы-
сокое - норадреналин. Через эти рецепторы осуществляются пре-
имущественно стимулирующие эффекты на клеточный метаболизм
и секрецию гормонов, а на гладкие миоциты - тормозные эффекты
(на сердце - стимулирующий эффект).
• Влияние гормонов на количество адренорецепторов.
Ф" Эстрогены увеличивают, а прогестерон уменьшает число а-
адренорецепторов в матке и соответственно усиливают или умень-
шают ее сократительную активность.
Ф" Тироидные гормоны и глюкокортикоиды увеличивают число 0-
адренорецепторов в тканях (особенно в сердце) и повышают их
чувствительность к катехоламинам.
• Влияние двигательной активности на соотношение а- и 0-адрено-
рецепторов: прн гипокинезии увеличивается число а-адренорецепторов,
прн физической нагрузке - 0-адренорецепторов (например, в коронарных со-
судах).
❖ Физиологические эффекты катехоламинов (срочная мобилизация резер-
вов организма).
149
• Мобилизация через ^-адренорецепторы энергетических резервов орга-
низма (усиление гликогенолиза в печени и в мышцах, липолиза в жировой ткани,
окислительных процессов и образования энергии, повышение секреции инсулина и
глюкагона, ренина, тироидных и половых гормонов, АДГ и мелатонина).
• Повышение активности сердечно-сосудистой системы (рост минутного
объема, ЧСС, АД, ОЦК).
ф Через 0|-адренорецепторы увеличивается частота и сила сердечных сокращений.
"Ф" Через -адренорецепторы обеспечивается сужение сосудов и перераспределение
крови между органами, выброс крови из депо и увеличение доставки О; к тканям.
"Ф" Через Рт рецепторы происходит расширение сосудов скелетных мышц.
• Торможение тонуса и перистальтики кишечника (через а- и
Р-адренорецепторы), желудка (через Р-адреиорецепторы), сокращение их
сфинктеров (а-адренорецепторы).
• Дыхательная система: рост вентиляции легких, снижение сопротивле-
ния в бронхах (через Рт-адреиорецепторы расслабляются гладкие миоциты брон-
хов).
• Увеличивается возбудимость нервной н мышечной систем, чувстви-
тельность анализаторов.
• Формируются адаптационно-трофические влияния на ткани.
• Совместно с симпатической нервной системой (симпатоадреналовая
система) обеспечивает быстрое реагирование при стрессе.
❖ Катаболизм и экскреция катехоламинов.
• Катаболизм происходит преимущественно в печени, почках, легких, а
также в синапсах в результате реакций окисления (моноамниооксидаза) и
метилирования (катехол-О-метилтрансфераза).
• Продукты катаболизма (ванилилмнндальная кислота, метадренални и иорметад-
реиалнн) выводятся главным образом с мочой (их определение в моче исполь-
зуется с диагностическими целями).
Нервная регуляция секреции катехоламинов (мозговой слой надпочечника яв-
ляется гуморальным усилителем симпатической нервной системы - частью симпатоадре-
наловой системы. М.Н. Чебоксаров, 1910; Л.А. Орбели; У. Кенион).
• Активация симпатических центров в грудных сегментах спинного моз-
га.
• Передача сигнала с аксона преганглионарного нейрона на хромаффин-
ные клетки мозгового слоя приводит к их активации н выбросу в кровь
путем экзоцитоза катехоламинов н пептидов-спутников (модуляторов).
9. Половые железы (см. тему 33; задание 1: 5).
10. Гормональная функции плаценты (см. тему 33; задание 1: 6).
11. Эпифиз (гормоны: мелатонин н серотонин, образующиеся из триптофана, полипеп-
тиды).
11.1. Физиологические эффекты гормонов эпифиза («Эпифиз - тесные башмаки для ги-
пофиза»).
150
❖ Тормозят секрецию гонадолиберина и гонадотропинов (в онтогенезе тор-
мозят половое созревание).
❖ Тормозят секрецию ГН' и СТГ, является антагонистом МСГ (обесцвечивают
меланофоры).
❖ Обеспечивают вместе с супрахиазматическим ядром гипоталамуса форми-
рование суточных ритмов («биологические часы»),
❖ При стрессе тормозит освобождение гонадолиберинов, ФСГ и ЛГ.
❖ Нейрофизиологическое влияние: мелатонин через мелатониновые рецепто-
ры (А, В, С) и ГАМК-рецепторы тормозных нейронов оказывает «транкви-
лизирующее» влияние. Наиболее высокая плотность мелатониновых ре-
цепторов на нейронах супрахиазматического ядра, гиппокампа, стриатум,
коры больших полушарий, фоторецепторах.
❖ Мелатонин и серотонин являются эндогенным антиоксидантами (ликвиди-
руют свободные радикалы кислорода).
11.2. Регуляция продукции мелатонина осуществляется от фоторецепторов —> суп-
рахиазматические ядра —> симпатические центры верхних грудных сегментов
—> верхний шейный ганглий —> пианелоциты (аденилатциклазная система).
❖ Торможение образования мелатонина из серотонина происходит при дей-
ствии света (норадреналин + (3-адренорецепторы).
❖ Активация синтеза и секреции мелатонина происходит в темноте (70 % всей
продукции гормона с 23 до 7 часов).
Функция эпифиза изменяется в соответствии с фазами менструального
цикла: максимальный уровень мелатонина у женщин во время менструа-
ции, минимальный - во время овуляцнн.
12. Тимус (центральный орган иммунной системы, максимальная функция - до 12 лет, после чего по-
степенно развивается атрофия).
12.1. Эпителиальные клетки образуют ряд полнпептидных гормонов, главными из
которых являются тимопоэтин и тимозины, действующие не только в тимусе,
но и в других тканях.
❖ Тимозины стимулируют образование на Т-лимфоцитах специфических ре-
цепторов (CD4*, CD8* н др.),.
❖ Тимопоэтин влияет на ранние этапы дифференцировки Т-
предшественников в Т-лимфоциты.
❖ Активируют образование лимфоцитами цитокинов (интерлейкинов, интер-
феронов, колониестимулирующих и туморнекрозирующих факторов), ко-
торые регулируют функции иммунокомпетентных клеток.
12.2. Физиологические эффекты гормонов тимуса.
❖ Стимулируют в тимусе дифференцировку предшественников в Т-
лимфоциты, созревание и пролиферацию Т-лимфоцитов (образуются Т-
хелперы, Т-киллеры, Т-супрессоры), стимулируют селекцию Т-лимфоцитов
(уничтожаются путем апоптоза агрессивные лимфоциты, которые могут реагировать на
аутоантигены организма). Подробнее см. тему 13 А: задание 1; 3.
❖ Ускоряют рост организма и увеличивают мышечную силу (после 12 лет эта
функция переходит к половым гормонам).
❖ Ускоряют рост костей, задерживают в них Са2+ (синергисты кальцитонина и
СТГ).
151
❖ Тормозят половое развитие.
❖ Влияют на разрушение ацетилхолина в нервно-мышечных синапсах (инги-
бируют передачу нервного импульса на мышечные клетки).
12.3. Регуляция функций тимуса.
❖ Глюкокортикоиды и половые гормоны оказывают тормозящее влияние на
тимус и вызывают его возрастную инволюцию.
❖ Тироидные гормоны стимулируют функцию тимуса.
13. Эндокринные функции почек, сердца и желудочно-кишечного тракта изложены
в соответствующих темах занятий (темы 15,22 и 25).
14. Профильный материал дли студентов лечебного факультета.
13.1. Общие изменений эндокринной функции при старении.
Ъ Снижается продукция гормонов большинства желез: щитовидной, половых, под-
желудочной, эпифиза, коры надпочечников (компенсаторно нарастает продукция
гормонов аденогипофиза, например, гонадотропинов при климаксе).
❖ Нарушается связывание гормонов в крови как следствие изменения белко-
во-синтетической функции печени.
♦ > Снижается число рецепторов и их способность связывать гормоны.
❖ Нарушается регуляция в системе гипоталамус-гипофиззависимая железа в резуль-
тате снижения чувствительности гипоталамуса к сигналам обратной связи.
13.2. Гипоталамус.
Ухудшается кроваток в гипоталамусе и гипофизе. Снижается активность нейро-
секреторных процессов.
Ослабляется секреторный ответ на рефлекторные воздействия, но усиливается от-
вет на гуморальные раздражения.
& Старение гипоталамуса происходит неравномерно: максимальные изменения ре-
гистрируются в аркуатных ядрах и передней группы ядер гипоталамуса.
Нарастает уровень вазопрессина в крови и увеличивается чувствительность сосу-
дов к нему (особенно коронарных).
13.3. Аденогинофиз.
Секреция гонадотропинов (ФСГ и ЛГ) при старении нарушается раньше всего.
.• Исчезают ритмические колебания выброса гонадолиберинов и гонадотропи-
нов у женщин во время климакса. Уровень ФСГ в крови возрастает в 10 - 15
раз, ЛГ - в 3 раза, что является результатом снижения уровня эстрогенов в
крови. Через несколько лет продукция гипофизарных гонадотропинов и про-
лактина снижается.
• У мужчин уровень ФСГ и ЛГ при старении остается неизменным или повыша-
ется из-за снижения уровня андрогенов в крови.
Ъ Снижается секреция СТГ, отсутствует ночной пик секреции.
❖ Снижается секреция АКТГ, в том числе и при стрессе.
13.4. Половые железы. Возрастные изменения у мужчин и женщин (см. тему 34; задание 1:
П)
13.5. Тимус - после начала полового созревания подвергается возрастной инволюции, еже-
годно около 3 % его ткани замещается жировой.
15. Профильные материалы для студентов педиатрического факультета.
152
14.1. Развитие эндокринной системы в антенатальном периоде: продукция гормонов
опережают по времени формирование циторецепторов, гипоталамический контроль
устанавливается после 4 мес. и в разное время для разных желез.
14.2. Развитие гиноталамуса.
4* Закладка ядер происходит на 2 - 3-м мес., активность ядер проявляется с 20 - 26
недель, окончательное созревание происходит в постнатальном периоде.
4» Портальная сеть формируется с 14 недель и к рождению полностью сформирова-
на.
4* Связи гипоталамуса с гипофизом в первые три месяца не функционируют, чувст-
вительность появляется с 20 недель.
14.3. Развитие гипофиза.
4> Продукция СТГ.
• Секреция в антенатальном периоде начинается с 9-й недели, достигает макси-
мума к 30 - 34 неделям. В это время концентрация СТГ в - 20 раз больше, чем
у взрослого; рецепторы для СТГ в тканях плода не активны, основная роль
СТГ у плода - мобилизация жиров при недостаточном снабжении глюкозой.
• В постнатальном периоде с 3 мес. появляется ночной пик секреции, снижается
концентрация СТГ в крови. Секреция зависит от уровня глюкозы в крови: при
гипогликемии повышается, при гипергликемии снижается. Если возникает не-
достаток секреции СТГ, то он проявится только после 3 лет задержкой роста
(карликовость без нарушения интеллекта).
❖ Продукция АКТГ.
• Впервые появляется с 8 недели, его концентрация растет с 20-й недели, уро-
вень в 4 - 5 раз больше, чем у взрослого.
• Контроль развития коры надпочечника устанавливается со второй половины
антенатального периода.
• АКТГ матери через плаценту не проникает, тогда как глюкокортикоиды мате-
ри проходят к плоду и могут влиять на гипоталамус и гипофиз плода.
❖ Продукция ТТГ.
• Секреция у плода появляется с 13 недель, концентрация в 20 раз больше, чем у
взрослых, усиливает активность щитовидной железы, отрицательная обратная
связь формируется с 4 мес.
• Неонатальный пик секреции ТТГ через 30 мин после родов, концентрация
гормона повышена в 10 - 15 раз, возвращается к норме к 2 - 3-му дню.
4» Продукция гипофизарных гонадотропинов (ФСГ, ЛГ и пролактина) (см. тему 34;
задание 1: 2 и 4).
4» Продукция АДГ.
• В антенатальном периоде его концентрация и активность низкие, осмо- и во-
люморегуляцию осуществляет в основном организм матери.
• В постиатальном периоде секреция появляется после 4 месяцев и к концу пер-
вого года становится такой же, как у взрослых.
14.4. Развитие надночечника.
4* Антенатальный период.
• С 6-й недели развивается кора надпочечника под влиянием плацентарных
гормонов - АКТГ, гонадотропинов. Мозговой слой формируется позже, пре-
153
обладает синтез норадреналина, тогда как у взрослых - адреналина. Уровень
катехоламинов у плода в конце беременности в 6 - 30 больше, чем у взрослых.
• Скорость созревания коры надпочечников у плода мужского пола выше. Син-
тез гормонов осуществляется в результате совместной активности фетопла-
центарной единицы. Соотношение кортизол/кортизон у плода 1:4.
♦♦♦ Глюкокортикоиды стимулируют:
• развитие легких и синтез сурфактанта (предупреждает у новорожденных ате-
лектазы легких);
• развитие молочных желез в пубертатном периоде;
• синтез рецепторов катехоламинов и обмен катехоламинов;
• синтез белков в печени.
143. Развитие щитовидной железы,
*♦* Продукция и транспорт гормонов у плода.
• Раньше начинается синтез тироглобулина, позднее появляется его способность
связывать йод.
• Основная форма гормона до 32-й недели - Т4. Основная транспортная форма
до 12-й недели - с альбумином, затем с тироглобулином.
• Регуляция секреции у плода до 20 недель автономна, так как ТТГ матери через
плаценту не проникает, а собственные гипоталамус и гипофиз еще не созрели.
После 20-й недели секреция переходит под контроль ТТГ.
• Неонатальный криз: через 30 мин после родов резко возрастает секреция гор-
монов, к норме возврат через 2-3 дня.
•t* Роль тироидных гормонов.
• У плода действие гормонов (например, на термогенез и метаболизм) выражено
меньше, чем в постнатальном периоде, это связано с недостаточным и неоди-
наковым по времени созреванием циторецепторов в разных тканях.
• Влияние на развитие мозга: в критические периоды развития (последние 3 ме-
сяца развития плода и первые недели после родов): при дефиците гормонов
снижен транспорт аминокислот в мозг, скорость синтеза белков мозговой тка-
ни, нарушена дифференцировка коры и других отделов мозга, сииаптогенез. В
хроническом варианте это приводит к умственной отсталости - кретинизму.
• При дефиците гормонов щитовидной железы тормозится образование корти-
колнберина и снижается резистентность организма к стрессовым факторам.
14.6. Развитие паращитовидных желез и секреция кальцитонина (регуляция уровня
кальция).
♦ ♦♦ У плода активность паратгормона появляется после 10-й, кальцитонина - после
14-й недели. Материнские паратгормон и кальцитонин через плаценту не прохо-
дят, усиливает секрецию кальцитонина эстрогены плаценты.
♦ ♦♦ Функциональная незрелость паращитовидных желез и почек (реабсорбция Са2+) в
первые два дня после рождения может вызвать гипокальциемию и тетанию ново-
рожденных.
❖ Развитие скелета в постнатальном периоде сопровождается активацией всасыва-
ния Са2+ в кишечнике и реабсорбцией его в почках (паратгормон и витамин D3),
усилением отложения Са2+ в костях и торможением резорбции костной ткани
(кальцитонин).
14.7. Развитие половых желез в антенатальном периоде (см. тему 34; задание 1: 2).
154
14.8. Островковый аппарат поджелудочной железы.
•*• р-Клетки.
• Появляются у плода с 12 недель. До 16 недель продукция инсулина происхо-
дит на низком уровне, и гомеостаз глюкозы обеспечивает мать. Секреция ин-
сулина стимулируется глюкозой и аргинином.
• Созревание рецепторов к инсулину происходит в последние недели антена-
тального и первые недели постнатального периода. Раньше оии образуются в
печени, позже - в мышцах.
• Через плаценту инсулин проникает в минимальных количествах.
♦> а-Клетки.
• Глюкагон определяется у плода с 12 недель, через плаценту не проходит.
• В первые часы после рождения секреция глюкагона снижается, спустя 4-10
часов - усиливается.
• Активируя распад гликогена в печени и распад жиров, глюкагон обеспечивает
энергетический гомеостаз, противодействует постнатальной гипогликемии.
14.9. Гормональный канал связи матери и ребенка через молоко матери (см. тема 33, за-
дание 1: 10.3).
16. Профильные материалы для студентов стоматологического факультета.
15.1. Гормональные факторы, участвующие в регуляции биосинтеза и секреции слюн-
ных желез.
•Z' Их роль минимальна в антенатальном и раннем постнатальном периоде, но отчета
ливо проявляется с периода полового созревания.
Тестостерон увеличивает синтез ферментов в слюнных железах, поэтому у муж-
чин их активность в слюны выше, чем у женщин. При кастрации снижается ак-
тивность а-амилезы, кислой и щелочной фосфатаз в слюне.
Тироидные гормоны щитовидной железы увеличивают активность в слюне моно-
аминооксидазы, пероксидазы, амилезы.
* Гипофизарно-надпочечниковая система стимулирует активность ферментов
слюнных желез. Секретин повышает активность а-амилазы.
15.2. Паротин (белковый гормон слюнных желез, вырабатывается в протоках околоушных
и подчелюстных желез).
❖ Активирует рост и обызвествление зубов и скелета, стимулирует отложение Са2* в
дентине (влияние паротина на процессы окостенения осуществляется как в ре-
зультате прямого действия на ткани, так и опосредованно через гормоны гипофи-
за). Препятствует развитию кариеса.
♦♦♦ Стимулирует развитие мезенхимальных тканей и прорезывание зубов.
15.3. Другие биологически активные вещества слюнных желез.
❖ Калликреин-кининовая система (действующее вещество - брадикинин) снижает
тонус сосудов и АД, повышает проницаемость капилляров (в том числе в слюн-
ных железах) и усиливает секрецию слюны.
Л* Факторы роста нервной ткани, эпителиальной ткани, фактор гранулоцитоза
(влияют на кроветворение).
Фактор, тормозящий активность тимуса.
15.4. Взаимосвязь слюнных желез с эндокринными железами.
❖ Снижают включение йода в щитовидной железе.
❖ Участвуют в регуляции обмена Са2* вместе с околощитовидными и щитовидной
железами.
155
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 н 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [14-16].
3.2. Дополнительная литература (см. с. 514 - 522): [5], [13], [15], [17], [20], [30], [35],
[43], [47], [54], [63], [65], [89], [ПО], [100], [107], [147], [148], [150], [152], [159],
[178], [180], [183], [190], [196], [222-225], [229], [236], [245], [248], [251] -[254].
33. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphvs: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором н слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 9-1. Больной, длительно принимавший глюкокортикоиды в дозировке, значительно пре-
вышающей их продукцию в организме в норме, прекратил их прием без согласования с врачом.
На третьи сутки (период полураспада глюкокортикоидов 1,5 часа) он поступил с аппендицитом в
хирургическое отделение. Во время операции, проводимой под местным обезболиванием с обыч-
ной степенью травматизации, внезапно развился шок с летальным исходом. Используя свои зна-
ния по физиологии эндокринной системы, объясните причины необычной реакции больного на
операционную травму.
Задача 9-2. У больного обнаружена феохромоцитома - гормонально активная опухоль из хро-
маффинных клеток мозгового слоя надпочечников. У него имеются следующие симптомы: гипер-
тония (АД=200/110 мм рт.ст.), тахикардия (105 уд./мин), гипергликемия (глюкоза 12 ммоль/л),
небольшая глюкозурия. Известно, что феохромоцитома может быть с преимущественной секре-
цией норадреналина или адреналина. Используя свои знания по физиологии эндокринной систе-
мы, объясните, какой тип секреции у данного больного - норадреналиновый или адреналиновый?
Задача 9-3. У мужчины двадцатилетиего возраста рост тела 120 см, нормальные пропорции тела,
сохранен интеллект. Используя свои знания по физиологии эндокринной системы, объясните
возможные причины и механизмы низкого роста у данного человека.
156
Тема 10. Физиология мышц
Цель: а) знать физиологические свойства мышц, типы мышечных сокращений, фазы одиночного со-
кращения, тетанус, зависимость амплитуды сокращения от частоты раздражения, силу и работу
мышц, закон средних нагрузок, особенности двигательных единиц различных мышц, теорию
мышечного сокращения н расслабления, биоэлектрические, химические и тепловые процессы а
мышцах; физиологические особенности гладких мышц;
б) уметь использовать эти знания для понимания функций поперечно-полосатых и гладких
мышц в целостном организме н его физиологических системах;
в) иметь представление о методе нсследоваиня - электромнографнн; приобрести практический
навык по применению динамометрии.
Мотивация темы: нет настолько здорового человека, который мог бы позволить себе мышечную без-
деятельность.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по дайной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1, Общая характеристика мышц (составляют - 50 % массы тела).
1.1. Виды мышц: поперечно-полосатые мышцы (скелетные и сердечная), гладкие
мышцы.
1.2. Физические свойства мышц: эластичность и упругость, напряжение (тонус),
пластичность (неупругая эластичность).
1.3. Физиологические свойства мышц: возбудимость, автоматия (гладкие и сер-
дечная мышца), проводимость, сократимость.
1.4. Основные функции мышц: формирование позы и перемещение тела в про-
странстве, насосная функция сердца и сосудистый тонус, дыхание и бронхиаль-
ный тонус; передвижение содержимого полых органов (перистальтика, тонус сфинк-
теров), выработка тепла; депо гликогена, резерв белков и воды, механическая
защита внутренних органов.
2. Физиология скелетных мышц.
2.1. Структурно-функциональная характеристика мышечных волокон.
❖ Мышечное волокно - сократительная единица скелетной мышцы (d = 1 о — 100
мкм, 1 = 5 - 18 см, IO46 волокон в мышце), состоит из миосимпласта и стволовых
клеток-саттелитов (камбиальный резерв), окруженных базальной мембра-
ной. Благодаря стволовым клеткам, мышцы способны к физиологической
регенерации (обновлению мышечных волокон) и репаративной регенера-
ции (восстановлению мышечных волокон после гибели симпласта).
❖ Миофибриллы - сократительный аппарат мышечных волокон, расположены
в МИОСИМПЛасте (1-2 тыс.миофнбрилл, составляют - 50 % объема волокна, d - 1 мкм,
1 = длине волокна).
❖ Саркомер - сократительная единица миофибрнллы (d - 1 мкм, 1 = 2 - 3 мкм).
• Протофибрилла или миофиламенты (их - 3000 в саркомере).
157
Актиновые нити или F-актии (- 2000 в саркомере, d - 6 нм, 1 - 1,0 мкм,
содержат - 400 молекул G-актина), имеют миозинсвязывающие участки,
расположенные друг от друга на расстоянии 10 нм.
ф Миозиновые нити (~ 1000 в саркомере, d - 10 нм, 1 = 1,0 - 1,5 мкм, содержат
- 250 молекул миозина). Фрагменты миозина: легкий меромиозин (d - 2 нм, 1 -
100 нм); тяжелый меромиозин (1 - 80 нм); шейка (d — 2 нм, 1-60 нм); двойная
головка (1 - 29 нм, содержит неактиную в покое АТФазу, две легких цепи мио-
зина); имеются шарнирные участки между легким и тяжелым меромиозинами и в
области шейки тяжелого меромиозина.
• Регуляторные белки - тропонин и тропомиозин (лежат вдоль актиновой спи-
рали).
ф Тропонин имеет три субъединицы: С - обеспечивает высокое срод-
ство к Са2*, Т - связь с тропомиозином, I - ингибирование взаимо-
действия актина и миозина. Тропонин присоединен к актину и тро-
помиозину так, что тропомиозин блокирует миозинсвязывающие
участки актиновой нити.
ф Присоединение Са2* к С-субъединице тропонина прекращает бло-
каду тропомиозином образования актомиозиновых мостиков.
• Т-система, состоящая из Т-трубочки и прилегающих к ней двух цис-
терн гладкой ЭПС.
ф Т-трубочки - впячивание плазмолеммы вглубь мыщечного волокна
на границе А- и 1-дисков. Их мембрана имеет потенциалзависимые
рецепторы дигидропиридина.
ф Цистерны ЭПС содержат депонированные ионы Са2* (- ю ммоль/л),
связанные с белком кальсеквестрином. В мембране цистерн имеют-
ся Са2*-иасос и рианодиновые рецепторы, содержащие Са2*-канал.
ф Т-система передает сигнал потенциала действия плазмолеммы на
цистерны ЭПС.
2.2. Теория мышечного а»фащения и расслабления (теория скольжения нитей,
X. Хаксли и А. Хаксли, 1954; актиновый тип регуляции сокращения).
❖ Электромеханическое сопряжение.
• Распространение ПД по сарколемме в Т-трубочке приводит к активации
ее потенциалзависимых рецепторов дигидропиридина.
• Сдвиг этих рецепторов открывает Са2*-канал рецепторов рианодина
цистерн саркоплазматической сети.
• Выход Са2* из цистерн приводит к увеличению его концентрации в ци-
тозоле с 10'7<’8) до 10*4 М (пороговая концентрация сокращения 10'7
М).
• Связывание Са2* с С-субъединицей тропонина'увеличивает степень
спирализации тропомиозина, что открывает миозинсвязывающие участ-
ки актиновых нитей.
❖ Скольжение нитей (сокращение саркомера).
• АТФаза миозиновой головки вызывает гидролиз АТФ до АДФ и неор-
ганического фосфата (Ф„), но продолжает удерживать оба продукта. В
158
таком состоянии головка связывается с актиновой нитью, образуя с ее
нитью угол около 90°.
• Отсоединение АДФ и Ф„ от головки миозина сопровождается основным
выделением свободной энергии (силовой удар). В результате головка по-
ворачивается в шарнирной области до угла 45° (наименьшая энергия ак-
томиозиновой связи), осуществляя гребковое движение, что вызывает пе-
ремещение актиновой нити вдоль миозиновой на 1 % длины саркомера
(на - 10 нм). (В каждом «гребке» участвует - 500 головок миозиновой нити. Во
время сильного сокращения частота «гребков» составляет около 5 раз в секун-
ду.)
• Присоединение АТФ к головке миозина уменьшает ее сродство к акти-
новой нити, что вызывает разъединение актомиозиновых мостиков. Да-
лее головка присоединяется в новом месте - ближе к Z-линии, и цикл
повторяется.
• При максимальном сокращении (до 50 % длины саркомера) необходимо
около 50 циклов образования и разъединения актомиозиновых мости-
ков.
2.3. Расслабление миофибрилл.
❖ Для расслабления миоцита необходимы два главных условия: наличие дос-
таточного уровня АТФ и низкая концентрация Са2+ (10'7 М и ниже).
❖ Присоединение АТФ к головкам миозина приводит к разрушению акто-
миозиновых мостиков.
❖ Низкий уровень Са2+в саркоплазме создается активацией Са2+-насоса и пе-
ремещением Са2+ в цистерны гладкой ЭПС, где он связывается с белком
кальсеквестрином. Снижение концентрации Са2+ в саркоплазме до 1О’”’8) М
восстанавливает (с участием тропонина) блокаду тропомиозином миозинсвязы-
вающих участков актиновых нитей. Наступает расслабление миоцита.
2.4. Химические и тепловые процессы при сокращении мышцы.
❖ Расход энергии: на процесс сокращения используется - 70 % энергии, на
процесс расслабления -15 %, на работу К+, Иа+-натриевого насоса - 5 %, на
синтезы -10 %.
❖ Образование энергии (АТФ - прямой источник энергии для функции
мышц: 1 моль АТФ освобождает 30 кДж или 7,3 ккал).
• Фосфагенный путь (резерв креатинфосфата + АТФ) и конверсия 2АДФ
в АТФ и АМФ с участием фермента аденилаткиназы может обеспечить
- 5 с максимальной двигательной активности.
• Гликолиз может обеспечить 1-2 мин максимальной двигательной ак-
тивности.
• Аэробное окисление глюкозы может обеспечить длительную активность
средней мощности.
❖ Сокращение мышц сопровождается выделением тепла (КПД мышечного со-
кращения - 50%).
• Тонус покоя составляет - 18 % энергии основного обмена.
159
• Терморегуляторный тонус повышает теплопродукцию до 50%, холодо-
вая мышечная дрожь - на короткое время в 5 раз.
• Физическая работа (рабочая добавка) освобождает до 4000 ккал/сут.
3. Физиологические особенности поперечно-полосатых мышц.
3.1. Нервно-мышечная передача.
•Ь Структура нервно-мышечного синапса. Важной особенностью этого синапса, от-
личающего его от синапсов ЦНС, является наличие в синаптической щели базаль-
ной мембраны мышечного волокна. К ней прикреплены молекулы ацетилхолинэ-
стеразы, разрушающие медиатор, она влияет на коицентрацю рецепторов в зоне
постсинаптической мембраны.
❖ Механизм проведения возбуждения через синапс.
• В фазическом мышечном волокне быстрых и медленных двигательных
единиц имеется один синапс с длинным пресинаптическим окончанием
(1-2 мм).
• ф Везикулы расположены преимущественно в активных зонах пресн-
наптической мембраны, в одной везикуле содержится около 4 ты-
сяч молекул ацетилхолина (квант медиатора). Ацетилхолин образу-
ется в пресннапсе из ацетил-КоА н холина под действием фермента
холинацетилтрансферазы.
ф Потенциал действия, достигая пресинаптического окончания, от-
крывает в его плазмолемме потенциалзависимые Са2+-каналы. Вход
Са2+ в пресинапс индуцирует процесс экзоцитоза медиатора. При
передаче возбуждения выделяется в синаптическую щель 100 - 300
квантов ацетилхолина.
• ф Ацетилхолин действует на а-субъединицы Н-холинорецептора (его
субъединицы: 2а,6,8,у) постсинаптической мембраны (синоним: концевая
пластинка), открывая в нем ионный канал Na+- н К*-проводимости.
(Постсииаптическая мембрана имеет складки, что увеличивает количество ре-
цепторов. На одной концевой пластинке 10-20 млн рецепторов.)
ф Входящий Na+-TOK формирует ВПСП (потенциал концевой пла-
стинки) на постсннаптической мембране.
ф Характерна высокая амплитуда (30 - 40 мВ) одиночного ВПСП, спо-
собного генерировать ПД в миоците.
ф Взаимодействие ацетилхолина и рецептора кратковременно (десятые
доли мс), так как ацетилхолинэстераза синапса гидролизирует ми-
диатор.
ф В покое наблюдается спонтанное выделение 1-2 квантов медиато-
ра в секунду и формирование миниатюрных ВПСП.
• Тонические мышечные волокна.
ф Тонические мышечные волокна, создающие у амфибий тонус мышц, заменены в
мышцах млекопитающих деятельностью медленных фазических мышечных во-
локон, и тонические волокна остались только в некоторых мышцах (например,
глазодвигательных мышцах и мышцах среднего уха).
ф Одно Волокно имеет 3-10 синапсов и иннервируется обычно несколькими мо-
тонейронами. Высокое входное сопротивление сарколеммы обеспечивает боль-
160
шую величину ВПСП и большую эффективность элекгротоиического распро-
странения возбуждения.
ф' ВПСП без генерации ПД непосредственно запускает мышечное сокращение по
типу функциональной ацетилхолиновой контрактуры, создающей длительное со-
кращение с минимальными энергетическими затратами.
3.2. Двигательные единицы (ДЕ).
❖ ДЕ - совокупность мышечных волокон, иннервируемых разветвлениями
одного мотонейрона. Виды двигательных единиц: медленные, малоутом-
ляемые; быстрые, легко утомляемые (а также быстрые, устойчивые к утомлению).
В мышце имеются, как правило, все виды ДЕ, но в разных соотношениях.
❖ Функциональные особенности медленных ДЕ (красные волокна).
• Иннервируются небольшими н высоко возбудимыми а-мотонейронамн
с низкой скоростью проведения возбуждения по аксону и частотой им-
пульсацин 6-10 Гц.
• Количество мышечных волокон в ДЕ сравнительно небольшое, они
имеют меньшее количество миофибрилл и развивают меньшую силу
сокращения.
• Волокна имеют низкую активность миозиновой АТФазы и низкую ско-
рость сокращения.
• Имеют хорошее кровоснабжение, много митохондрий, миоглобина, вы-
сокий аэробный обмен, поэтому обладают низкой утомляемостью. Спо-
собны выполнять длительную маломощную работу.
• Дают гладкий тетанус при небольшой частоте разряда мотонейрона
(-16 Гц).
• В регуляции движения обеспечивают мышечный тонус и позу, а также
способность к длительной циклической работе - бег, плавание и др. (на-
пример у марафонцев их количество в мышцах достигает 85%).
❖ Функциональные особенности быстрых ДЕ (белые волокна).
• Иннервируются крупными, менее возбудимыми а-мотонейронами с вы-
сокой скоростью проведения ПД по аксону н частой импульсацией - 50
Гц.
• Количество мышечных волокон в ДЕ сравнительно большое.
• Миоциты содержат большое число миофибрилл и развивают большую
силу сокращения.
• Имеют высокую активность миозиновой АТФазы и развивают высокую
скорость сокращения.
• Имеют слаборазвитую капиллярную сеть, мало митохондрий, миогло-
бина, но содержат много гликолитических ферментов, большой запас
креатинфосфата и гликогена, анаэробный тип энергообеспечения.
• Способны развивать большую мощность, но быстро утомляются.
• Дают гладкий тетанус при большей частоте разряда мотонейрона (~ 30
Гц).
161
• В регуляции движения обеспечивают преимущественно фазический
компонент - перемещение организма и его частей в пространстве с
большой скоростью и мощностью (например, у спринтеров и прыгунов
количество быстрых ДЕ в мышцах достигает 90%).
••• Функциональные особенности быстрых, устойчивых к утомлению ДЕ.
• По структурно-функциональным свойствам занимают среднее положение между мед-
ленными и быстрыми ДЕ.
• Вероятно, используются в быстрых ритмических движениях (ходьба, бег).
33. Типы сокращений.
Изометрическое (изменяется тонус), изотоническое (изменяется двина) и ауксо-
тоиическое (изменяется длина н тонус) сокращение.
❖ Одиночное сокращение, его фазы: латентная, сокращения и расслабления.
❖ Суммация сокращений: зубчатый и гладкий тетанус.
• Механизм возникновения тетануса в естественных условиях связан с
высокой частотой генерации ПД а-мотонейронами двигательных цен-
тров: десятки Гц в состоянии покоя, - 100 Гц во время двигательной ак-
тивности. В связи с этим каждый последующий ПД попадает или в на-
чало фазы расслабления мышцы (зубчатый тетанус), или в фазу сокра-
щения мышцы (гладкий тетанус).
• Зависимость амплитуды от частоты раздражения. Наибольшая амплиту-
да сокращения мышцы имеется во время гладкого тетануса в узком диа-
пазоне частоты раздражения, при котором в волокне имеется оптималь-
ное соотношение Са2+ и молекул тропонина - 4:1 (оптимум раздраже-
ния); при увеличение частоты раздражения амплитуда снижается (пес-
симум раздражения).
4. Сила и работа мышц.
4.1. Сила мышцы.
❖ Абсолютная и максимальная произвольная сила скелетных мышцы 6-17
кг/см2.
❖ Факторы, определяющие силу сокращения мышцы.
• Соотношение медленных и быстрых ДЕ (композиция мышцы).
• Число активных ДЕ, включение ДЕ в соответствии с возбудимостью их
мотонейронов («правило размера»),
• Режим работы активных ДЕ (одиночное сокращение, зубчатый тетанус
и гладкий тетанус).
• Величина физиологического сечения мышцы (сумма поперечного сече-
ния ее всех волокон; в перистых мышцах оиа больше геометрического
сечения).
а Длина мышцы и сила сокращения (кривая зависимости «длина-сила»).
‘Ф’ Сила сокращения мышцы (точнее, саркомера) пропорциональна зоне
перекрытия актиновых и миозиновых нитей, т. е. количеству обра-
зующихся актомиозиновых мостиков. Наибольшая сила развивает-
ся при длине мышцы в состоянии покоя.
162
ф- При укорочении или удлинении мышцы на 50 % зона перекрытия
нитей и сила сокращения резко снижаются.
4.2. Работа мышц.
❖ Виды работы.
• Динамическая (в изотоническом режиме).
• Статическая (в изометрическом режиме).
• Уступающая (в условиях удлинения мышцы).
❖ Закон средних нагрузок: мощность мышцы (сила х скорость сокращения)
максимальна при умеренной нагрузке.
❖ Функционально-структурные изменения мышцы в зависимости от нагруз-
ки: гипертрофия и атрофия.
4.3. Утомление мышцы (периферические механизмы утомления).
❖ Накопление продуктов обмена (кислоты, ионы калия).
❖ Энергетический дефицит.
• > Снижение эффективности нервно-мышечной передачи.
4.4. Методы исследования мышц.
❖ Динамометрия. Для измерения силы мышц используют динамометры, Принцип
метода: регистрирующие максимальное напряжения мышцы, которое деформиру-
ет стальную скобу. Степень деформации отмечается поворотом стрелки, которая
на шкале указывает развиваемое усилие в кг. Кистевым динамометром определя-
ется сила мышц кисти, становым - сила мышц разгибателей спины. Для изучения
мышечной силы в рабочих профессиональных движениях и спортивной медицине
в рукоятке инструментов помещают пружинные и тензометрические датчики, ко-
торые воспринимают мышечные усилия и передают соответствующую информа-
цию к записывающей аппаратуре.
❖ Электромиография (ЭМГ) - метод исследования скелетных мышц, основанный
на регистрации их биопотенциалов. Для отведения биопотенциалов мышц челове-
ка используют накожные и погружные (игольчатые) электроды, запись биопотен-
циалов проводится на электромиографе. Электромиограмма при записи с игольча-
тых электродов представляет собой алгебраическую сумму потенциалов действия
отдельных волокон двигательной единицы (локальная ЭМГ), при записи с накож-
ных электродов - сумму потенциалов действия разных двигательных единиц (ин-
терференционная ЭМГ). Амплитуда волн ЭМГ колеблется от 1-2 до 3000 мкВ
(обычно 100 -200 мкВ), частота - 15-20 имп/с. ЭМГ анализируется по амплитуде
и частоте биопотенциалов. Между амплитудой зубцов ЭМГ и развиваемой мыш-
цей силой существует примерно линейная зависимость, а частота биопотенциалов
ЭМГ в определенной степени пропорциональна напряжению мышцы.
5. Физиологическая характеристика гладких мышц (составляют около 10% массы те-
ла).
5.1. Структурные особенности гладкомышечных клеток (ГМК). Веретенообразные одноядерные
клетки длиной от 20 до 500 мкм, толщиной 5-20 мкм, образующие мышечные пучки, форми-
рующие слои гладкой мускулатуры. Клетки соединены между собой высокопроводимыми кон-
тактами (нексусами), в одних мышцах их много (например, сфинктер зрачка, матка во время
беременности), в других мало или почти иет (продольная мышца кишки, семявыносящий про-
ток). Саркомеры отсутствуют. Актиновые нити образуют трехмерную сеть, нити которой со-
единены с сарколеммой и друг с другом с помощью плотных телец (а-актинин, аналог Z-
линий). Тяжелые цепи миозина находится в деполимеризованном состоянии (миозиновые нити
образуются в процессе сокращения), их головки содержат двойные легкие цепи, которые пре-
163
дотвращают связывание миозиновых головок с актиновой нитью. ГМК способны к гипертро-
фии и регенерации. Са2+ депонирован в цистернах гладкой ЭПС, которая слабо выражена, и в
мелких пузырьках (кавеолах), находящихся под плазмолеммой.
5.2. Физиологические особенности ГМК (миозиновый тип регуляции сокращения).
❖ Величина мембранного потенциала равна -50 - (-60) мВ. В некоторых мио-
цитах (например, матки, кишечника, некоторых сосудов) ОН нестабилен, его ВОЛНЫ
деполяризации могут достигать КУД с генерацией ПД. Миоциты имеют
большую внутриклеточную концентрацию Na+ и СГ, хорошие кабельные
свойства (Х„ = 1 - 3 мм).
❖ Нервно-мышечная передача образуется разветвлением аксонов пост-
ганглионарных нейронов вегетативной нервной системы.
• В концевых частях аксона имеются варикозные расширения, содержа-
щие медиатор. Расстояние между ними и миоцитами («синаптическая
щель») равно 10 - 2000 нм.
• Типичные постсинаптические мембраны (концевые пластинки) не вы-
ражены, рецепторы к медиаторам более равномерно распределены по
миоциту. Медиатор, выделенный из варикозных расширений, путем
диффузии поступает к нескольким ГМК, и эффектором является не от-
дельный миоцит, а пучок миоцитов, соединенных нексусами.
• Основными медиаторами синаптической передачи в ГМК является аце-
тилхолин (действует на М-холинорецепторы) И норадреналин (действует на а- и
Р-адреиорецепторы). В некоторых синапсах имеются другие, плохо изучен-
ные медиаторы (ВИП, атф, пептиды). Эффектом передачи может быть
возбуждение или торможение.
ф- Возбуждение обусловлено развитием возбуждающего синаптиче-
ского потенциала - деполяризацией длительностью 500 - 1000 мс,
вызванной преимущественно входящим в клетку На+-током (напри-
мер, при действии ацетилхолина на М-холинорецепторы в миоцитах кишечника
или норадреналина на а-адренорецепторы в миоцитах сосудов). Деполяриза-
ция ГМК может быть вызвана также ПД, поступившем в иее через
нексус. Деполяризация открывает потенциалзависимые Са -
каналы плазмолеммы.
• ф- Торможение обусловлено тормозным синаптическим потенциалом
- гиперполяризацией продолжительностью 800 - 1000 мс, вызван-
ной повышением К+-проницаемости и выходящим из клетки Ис-
током (например, действие ацетилхолина на М-холинорецепторы в миоцитах
некоторых сосудов, норадреналина на а-адренорецепторы миоцитов кишечника).
❖ Механизмы сокращения ГМК.
• Сокращение запускается Са2+, их пороговая концентрация в цитозоле
равна 10~7 М, максимальное сокращение происходит при концентрации
10 s М Са2+.
• Имеется два основных источника Са2+.
Ф Важную роль играет внеклеточный Са2+ (отличие от скелетных мышеч-
ных волокон). При действии медиаторов и гормонов Са2+ поступает в
ГМК через различные Са2+-каналы мембраны: хемоуправляемые,
164
потенциалзависимые и механочувствительиые. Вход Са2+ через эти
каналы увеличивает его концентрацию в цитозоле.
ф" Другим источником Са2+ являются цистерны гладкой ЭПС (и кавео-
лы). Мобилизация Са2+ из нее осуществляется в результате актива-
ции Са2+-каналов инозитолтрифосфатом, образуемым фосфолипа-
зой С, активируемой гормонами и медиаторами (например, при дейст-
вии норадреналина на а-адреиорецепторы ГМК сосудов или при действии аце-
тилхолина на М-холинорецепторы ГМК бронхов).
• Са2+ в цитозоле соединяется с белком кальмодулином, и этот комплекс
активирует специальную протеинкиназу, фосфорилирующую легкие
цепи миозииа (киназу легких цепей миозина - ЛЦМ). Фосфорилирован-
ная цепь перестает ингибировать взаимодействие актина с миозином.
Активация при этом миозиновой АТФазы приводит к скольжению ак-
тина и миозина относительно друг друга, что приводит к укорочению
ГМК и развитию силы. АТФазная активность миозина ГМК в - 10 раз
меньше, чем в скелетных миоцитах. Поэтому скорость и сила сокраще-
ния В ГМК меньшие. (Кроме фосфорилирования легкой цепи миозииа происходит
и фосфорилирование тяжелой цепи миозина, что приводит к сборке из миозина тол-
стых филаментов.)
• Некоторые медиаторы и гормоны (например, адреналин через
аг-рецепторы, ацетилхолин через Мг-рецепторы, серотонин через
5-НТг-рецепторы, ангиотензин П через ATi-рецепторы) вызывают со-
кращение ГМК в результате ингибирования аденилатциклазы —> сниже-
ния уровня цАМФ —» торможения протеинкиназы А. В результате сни-
жается фосфорилирование киназы ЛЦМ, что делает ее более чувстви-
тельной к комплексу Са2++кальмодулин и способствует сокращению
ГМК. (Предполагают также, что протеинкиназа А активирует выход Са2+ из гладкой
ЭПС и ингибирует Са2+-насосы цистерн гладкой ЭПС и клеточной мембраны, в ре-
зультате чего накапливается Са2* в цитозоле.)
❖ Механизмы расслабления ГМК.
• Расслабление ГМК возникает при уменьшении концентрации внутри-
клеточного Са2+ до 10'7 М и ниже, что снижает активность киназы ЛЦМ.
При этом постоянно активная протеиифосфатаза дефосфорилирует ее.
Основным путем снижения уровня Са2+ является удаление его из клетки
во внеклеточную среду через ионообменники (например, Ca27Na*-
ионообмеиник) и Са2+-насос клеточной мембраны. Часть Са2+ депонирует-
ся в цистерны гладкой ЭПС с помощью Са -иасоса.
• Расслабление ГМК при действии ряда медиаторов и гормонов (например,
адреналина через 0-адренорецепторы, АДГ через У2-рецепторы, гистамина через
Н2-рецепторы) осуществляются в результате стимуляции аденилатциклазы
—<• повышения уровня цАМФ —<• активации протеинкиназы А, которая
фосфорилирует киназу ЛЦМ, что делает ее (киназу) менее чувствитель-
ной к комплексу Са2+ + кальмодулин. В результате происходит расслаб-
ление ГМК.
5.3. Адекватные раздражители гладких мышц: нейромедиаторы, гормоны (дис-
тантные и тканевые), растяжение.
165
6. Профильные материалы для студентов лечебного факультета. Возрастные изме-
нения возбудимости, проводимости и сократимости мытц.
6.1. Уменьшается возбудимость мышц (снижается величина мембранного потенциала, воз-
растает инактивация Ыа+-каналов).
6.2. Снижается лабильность мышц в результате удлинения ПД и абсолютной рефрактерно-
сти (~ в 3 раза). Замедляется скорость проведения ПД по миоциту.
63. Масса мышц к 70 - 90 годам снижается почти вдвое преимущественно за счет умень-
шения диаметра мышечных волокон, снижается сила сокращения мышц.
7. Профильные материалы для студентов педиатрического факультета.
7.1. Онтогенез нервно-мытечного синапса,
♦♦♦ На 4-й неделе эмбрионального развития миотомы, дающие начало поперечнопо-
лосатой мускулатуре, состоят из миобластов, которые размножаются и мигрируют
в прилежащие области. На 5 - 10-й неделе миобласты теряют способность к деле-
нию и сливаются в миосимпласты - миотрубки. В них начинается синтез сократи-
тельных белков и образование миофиламентов, холинорецепторов и нексусов. С
20-й недели нексусы исчезают, и миотрубки превращаются в мышечные волокна.
♦X* Структурно-функциональное созревание нервно-мышечиого синапса.
• Пресинаптическая часть.
Ф Двигательные нервные окончания появляются в мышцах на 13 - 14-й не-
деле развития плода, их формирование продолжается и после рождения.
ф Созревание пресинапса у новорожденного проявляется увеличением тер-
минального разветвления аксона и площади концевых бляшек.
Ф Темп созревания нервных окончаний к 7 - 8 годам увеличивается в связи
с появление более быстрых и точных движений. Усиливается синтез аце-
тилхолина в мотонейронах и количество квантов медиатора, выделяюще-
гося в синаптическую щель
• Постсинаптическая часть.
Ф При контакте окончания аксона с миотрубкой в этом месте появляются
скопления митохондрий, рибосом, микротрубочек. На поверхности миот-
рубки образуются выемки, в которых помещается окончание аксона; об-
разуется примитивный нервно-мышечный синапс.
*$ В процессе развития моторной иннервации устраняются первичная мно-
жественная иннервация мышечного волокна и разлитая чувствительность
мембраны к ацетилхолину, она остается.только в концевой пластинке. На
ней образуются складки и увеличивается количество Н-холинорецепто-
ров.
*$ Появляется синтез ацетилхолинэстеразы в постсинапсе и повышается
скорость гидролиза ацетилхолина. Возрастает амплитуда потенциала кон-
цевой пластинки н увеличивается надежность передачи возбуждения че-
рез синапс.
♦♦♦ Синаптическая передача.
• Длительность синаптической задержки у новорожденного - 4,5 мс (у взросло-
го ~ 0,5 мс).
• Лабильность синапсов новорожденных - 20 имп/с (у взрослых - 120 нмп/с).
Скорость передачи через синапс достигает значений взрослого к 7 - 8 годам.
7.2. Резвитие мышц и их иннервация в ностнатальном периоде.
166
♦ ♦♦ Развитие мышц.
• Рост мышечной массы происходит, в основном, за счет увеличения продоль-
ного и поперечного размеров миоцитов, количества миофибрилл, общее число
волокон увеличивается незначительно (~ на 10 %).
• Масса мышц: у новорожденного - 23 % массы тела, в конце 1-го года - 16 % ,
к 8 годам - 27 %, к 15 годам - 33 %, к 17 - 18 годам - 44 % (как у взрослого).
• Рост мышечных волокон в длину осуществляется благодаря почкам роста на
их концах, примыкающих к сухожилиям. К миофибриллам на концах при-
страиваются новые саркомеры.
• Интенсивное развитие внутримышечной соединительной ткани и образование
миофибрилл происходит в первые 3 года. Созревание миоцитов завершается к
6 годам. Развитие мышц, их соединительной ткани, иннервации и сосудистой
системы происходит до 25 - 30 лет.
♦♦♦ Иннервация мышц.
• Рецепторный аппарат развивается раньше, чем моторный (с 10 - 12 недель у
плода), и его развитие в основном завершается к рождению.
• Формирование афферентной иннервации завершается к 7 - 8 годам жизни.
• Становление эфферентной иннервации приближается к уровню взрослых к 11
- 13 годам, а полностью завершается в 18 - 20 лет.
7.3. Функции мышцу детей в разные возрастные периоды,
♦ > Участие в терморегуляции - главная функция скелетной мускулатуры новорож-
денных. В связи с этим имеется постоянная активность скелетной мускулатуры,
даже во время сна.
♦ ♦♦ Постоянная активность мышц новорожденных является стимулом бурного роста
мышечной массы, конечностей, суставов.
♦ ♦♦ У грудного ребенка терморегуляционная функция скелетной мускулатуры умень-
шается, а двигательная - возрастает.
♦ ♦♦ Интенсивное развитие и увеличение тонуса разгибателей, свойственное взрослому
организму, происходит к 5 годам.
7.4. Физиологические свойства мышц в онтогенезе,
♦ 5* Возбудимость.
• Низкая возбудимость мышц плода (большой порог, хронаксия в 1,5-10 раз
выше хронаксии мышц взрослых).
• МПП мышечных волокон новорожденных равен -20 - (-40) мВ, что связано с
низким уровнем содержания К* в клетке и большим, чем у взрослых, содержа-
нием Na+.
• Имеется низкая амплитуда и большая продолжительность ПД (длительные фа-
зы абсолютной и относительной рефрактерности).
• В процессе роста происходит изменение биофизических свойств мембраны
мышечного волокна (уменьшение ионной проницаемости, увеличение актив-
ности ионных насосов), что приводит к увеличению МПП и ПД.
Ъ Низкая скорость проведения возбуждения мышечных волокон плода и новорож-
денных с возрастом постепенно увеличивается в связи с утолщением миоцитов и
увеличением амплитуды ПД
♦ ♦♦ Чрезвычайно низкая лабильность мышц плода (3 - 4 Гц) определяет их ярко вы-
раженные тонические свойства. Низкая лабильность скелетных мышц и иннерви-
167
рующих их волокон в раннем детском возрасте постепенно увеличивается с воз-
растом.
7.5. Сократимость, сила, работоспособность, выносливость, утомление мышц.
❖ Сократимость мышц.
• В мышцах эмбрионов слабо выражена способность взаимодействия между ак-
тином и миозином.
• У плода и новорожденных имеются медленные одиночные сокращения мышц
(более длительные фазы укорочения н расслабления).
• У новорожденных одинаковая скорость сокращения быстрых и медленных ДЕ,
начало их дифференцировки происходит в первые дни жизни.
❖ Сила мышц.
• С возрастом сила сокращения увеличивается вследствие нарастания общего
поперечного сечения миофибрилл и повышения плотности «упаковки» мио-
фибрилл, а позднее и увеличения плотности расположения мышечных волокон
в мышцах.
• До 10 - 12 лет мышечная сила преобладает у девочек, в период полового со-
зревания - у мальчиков. Наибольший прирост силы у мальчиков в 15 - 18 лет
(приближение к нижней границе азрослых).
❖ Работоспособность мышц увеличивается в процессе онтогенеза в результате
улучшения координации и точности движений, уменьшения энергетических за-
трат на единицу работы двигательного аппарата.
♦♦♦ Утомление мышц.
• Чем меньше ребенок, тем быстрее он утомляется, что связано с особенностями
ЦНС, так как сама мышца способна длительное время работать без утомления.
• «Активный отдых» имеет большое значение для профилактики утомления у
детей разного возраста.
♦♦♦ Выносливость.
• С возрастом увеличивается способность работы с неснижающейся мощно-
стью на фоне нарастающего утомления.
• Наибольший прирост выносливости в 7 - 10 лет, в 16 - 19 лет она составляет
85 % от показателя взрослых, а в 20 - 29 лет наивысший уровень выносливо-
сти. Половые различия выносливости: у мальчиков всех возрастов выносли-
вость выше, чем у девочек.
8. Профильные материалы для студентов стоматологического факультета.
8.1. Функциональное назначение отдельных жевательных мышц (см. тему 22; задание
1: 13.14).
8.2. Контрактура жевательной мускулатуры и ее последействие.
♦♦♦ При развитии утомления жевательных мышц может наступить их замедленное
расслабление, что носит название контрактуры жевательных мышц.
♦♦♦ При этом частично или полностью ограничивается движение нижней челюсти, а
следовательно, акт приема пищи, ее механическая обработка, а также дыхательная
и речевая функция полости рта.
83. Физические свойства жевательных мышц. Сила и работа жевательной мускула*
туры. Гнатодинамометрия.
♦♦♦ Жевательная мускулатура принадлежит к силовым мышцам, т.е. развивает при
сокращении преимущественно силу. Абсолютная сила всех жевательных мышц ~
168
390 кг. При усиленном смыкании челюстей в пародонте возникает ощущения боли
и происходит рефлекторное прекращение дальнейшего увеличения давления» хотя
сила мышц еще не исчерпана.
❖ Гнатодинамометрия - метод, позволяющий определить выносливость опорных
тканей зуба с помощью специальных приборов - гнатодинамометров. При закры-
вании рта через специальные пластинки для зубов передается на пружину давле-
ние, которое регистрируется по шкале. Результаты гнатодинамометрии зависят от
функционального состояния пародонта, а также жевательных мышц и зубов. Мак-
симальная сила мышц в области жевательных зубов 90 - 150 кг, фронтальных зу-
бов 60-75 кг. В зависимости от вида зубов: у резцов 10 - 25 кг, у моляров 30 - 90
кг.
8.4. Особенности электромиографии жевательных мышц.
❖ Электромиографня (ЭМГ) позволяет зарегистрировать электрическую активность
мышц. В стоматологии ЭМГ записывают при физиологическом покое, произволь-
ном сжатии челюстей, жевании, глотании и других состояниях. При анализе ЭМГ
определяют время биоэлектрической активности и покоя, их соотношение, сред-
нюю амплитуду биопотенциалов и другие показатели. Метод позволяет выявить
ранние проявления дисфункции жевательных мышц и косвенно - функциональное
состояние пародонта (например, при пародонтозе регистрируется низкоамплитуд-
ная ЭМГ).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 н 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [13-16].
3.2. Дополнительная литература (см. с. 514 - 522): [24], [26], [27], [30], [35], [47],
[57], [165], [190], [203], [222], [225], [245], [251-254].
3.3. Наглядные средства обучения. Использование страницы кафедры в Интернете -
www.vsma.ac.ru/~nphvs : Слайды для компьютерной демонстрации из «Атласа по
физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты конечно-
го уровня знаний.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться иа практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 10 - 1. Каким должен быть интервал между раздражающими импульсами тока, чтобы в
скелетной мышце произошла: 1) частичная суммация сокращения (зубчатый тетанус); 2) полная
суммация сокращения (гладкий тетанус)? Длительность фазы укорочения мышцы - 50 мс, фазы
расслабления - 75 мс, фаза абсолютной рефрактерности в потенциале действия миоцита - 5 мс,
относительной рефрактерности - 2 мс.
169
Задача 10-2. При миастении, которая характеризуется патологической слабостью и утомляемо-
стью скелетных мышц, уменьшается вероятность взаимодействия медиатора ацетилхолина с ре-
цепторами постсинаптической мембраны нервно-мышечных синапсов. Причина заключается в
том, что у больных появляются антитела к этим рецепторам, которые блокируют их и способст-
вуют их разрушению. Основываясь на ваших знаниях механизмов передачи в нервно-мышечном
синапсе, предложите виды лекарств (по характеру действия), которые в этих условиях могли бы
улучшить синаптическую передачу. При ответе пусть вас не лимитируют возможности современ-
ной фармакологии.
170
Тема 11. Роль различных отделов ЦНС в регуляции мышечного тонуса и
фазных движений
Цель: а) знать роль двигательного анализатора в оценке положения тела и движений; значение отде-
лов мозга в осуществлении моторных функций: роль спинного, продолговатого и среднего
мозга в регуляции мышечного тонуса н фазных движений, механизмы поддержания равнове-
сия тела, участие ретикулярной формации ствола в поддержании и перераспределении мы-
шечного тонуса; мозжечок, его корригирующее и стабилизирующее действие на моторную
функцию, участие в организации двигательных программ; роль базальных ядер в формирова-
нии мышечного тонуса, сложных двигательных актов и двигательных программ; роль коры
больших полушарий в регуляции движений;
б) уметь использовать эти знания для понимания роли движения в регуляции гомеостаза,
адаптации и целенаправленного поведения человека;
в) приобрести практические навыки исследования клинически важных проприоцептивных и
кожно-мышечных рефлексов.
Мотивация темы: см. тему 10.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Общая характеристика движения.
1.1. Физиологическая роль движения (от элементарных безусловных рефлексов до
двигательных компонентов социальных форм поведения).
1.2. Виды движений: 1) активные: непроизвольные, произвольные, автоматизиро-
ванные; 2) пассивные (без сокращения мышц); 3) локомоция как совокупность
согласованных движений, с помощью которых человек перемещается в про-
странстве.
13. Компоненты двигательного акта.
❖ Моторный: тонический (медленные двигательные единицы) и фазический
(быстрые двигательные единицы).
❖ Вегетативный: безусловно- и условнорефлекторный.
❖ Сенсорный (двигательный анализатор).
2. Двигательный анализатор.
2.1. Рецепторный отдел.
❖ Мышечные веретена прикреплены параллельно экстрафузульным (рабо-
чим) мышечным волокнам, возбуждаются при увеличении длины мышцы
(являются датчиками длины мышцы). (Они состоят из нескольких инкапсулиро-
ванных интрафузальных мышечных волокон, вокруг которых спиралевидно обвивается
окончание сенсорного нейрона. Имеется два типа мышечных веретен: ядерносумчатые,
которые реагируют иа скорость растяжения мышцы, и ядерноцепочечные, являющиеся
датчиками степени растяжения мышцы.)
❖ Сухожильные рецепторы Гольджи являются датчиками силы сокращения
(напряжения) мышцы.
171
❖ Суставные рецепторы дают информацию о движении на основе изменения
положения суставов.
2.2. Проводниковый отдел.
❖ Первые афферентные нейроны (волокна Аа и Ар) от суставных рецепторов (в
меньшей степени от мышечных и сухожильных) входят в состав тонкого и
клиновидного путей спинного мозга и переключаются в продолговатом
мозге на вторые нейроны. (Для мышц лица - волокна V черепного нерва, трой-
ничный ганглий.)
❖ Вторые нейроны в ядрах Бурдаха и Голля продолговатого мозга, их аксоны
входят в состав медиальной петли. (Для мышц лица вторые нейроны в средне-
мозговом ядре V нерва.)
❖ Третьи афферентные нейроны в заднелатеральных вентральных ядрах та-
ламуса.
* ♦• Афферентные нейроны от мышечных и сухожильных рецепторов преимущественно пере-
ключаются в задних рогах спинного мозга на нейронах, образующих спинно-мозжечковые
и спинно-оливный пути. Их импульсация используется в мозжечке для регуляции тонуса
мышц, позы, равновесия и произвольных движений.
2.3. Корковый отдел.
❖ Преимущественно в коре, образующей центральную борозду и прилегаю-
щую часть прецентральной ИЗВИЛИНЫ. (Сенсорный контроль движения осуществ-
ляется также в соматосенсорной области II, расположенной на границе пересечения цен-
тральной борозды с верхним краем височной доли)
❖ Корковое представительство мышц тела (рис. 29, с. 103): в верхнем участке
прецентральной извилины проецируются мышцы нижних конечностей и
туловища, в среднем и нижнем участках - мышцы верхних конечностей и
головы. Величина корковой проекции различных мышц пропорциональна
количеству в них рецепторов, что совпадает со сложностью н точностью
осуществляемых движений. Особенно велики размеры проекции мышц
языка, кисти руки н мимических мышц (У. Пенфилд, 1937).
❖ Результатом деятельности двигательного анализатора является кинестезия
- ощущение положения и перемещения тела в пространстве, которое ис-
пользуется для контроля и коррекции движений.
2.4. В сенсорном обеспечении движения участвуют также вестибулярный, так-
тильный, слуховой, зрительный анализаторы.
3. Спинальные механизмы регулиции мышечного тонуса и фазных движений.
3.1. Моторные центры содержат тонические и фазические а-мотонейроны, у-
мотонейроны, возбуждающие и тормозящие интернейроны. Только а-
мотонейроны непосредственно вызывают сокращение мышц, поэтому цен-
тральный вопрос регуляции движений - регуляция активности а-мотонейронов.
3.2. Регуляция активности а-мотонейронов.
❖ Стимулирующее влияние с мышечных веретен, активирующая роль у-
мотонейронов.
❖ Тормозящее влияние с сухожильных рецепторов Гольджи.
❖ Влияние с экстеро- и ннтерорецепторов.
❖ Саморегуляция через тормозные клетки Реншоу.
172
Латеральное торможение осуществляет локализацию возбуждения в мо-
торном центре.
❖ Реципрокное торможение осуществляет координацию деятельности функ-
ционально противоположных моторных центров (например, сгибателей и
разгибателей).
❖ Влияние с супраспннальных двигательных центров (см. ниже пп. 4 - 7).
33. Спинальные соматические рефлексы.
Л* Проприоцептивные (сухожильные, миотатические) рефлексы.
• Сгибательно-локтевой рефлекс (с двуглавой мышцы, центр в С4.5).
• Разгибательно-локтевой рефлекс (с трехглавой мышцы, С6.7).
• Коленный рефлекс (с четырехглавой мышцы бедра, Lm).
• Ахиллов рефлекс (сгибание стопы, с икроножной мышцы, S,.2).
❖ Кожно-мышечные рефлексы.
• Брюшные рефлексы: верхний (ThM), средний (ТЬмо), нижний (ТЬ|И2).
• Кремастерный рефлекс (L,.;).
• Подошвенный рефлекс (S,.2).
• Анальный рефлекс (84.5).
❖ Шейные позно-тонические рефлексы.
• Наклон головы вперед увеличивает тонус сгибателей рук и разгибате-
лей ног.
• Наклон головы назад увеличивает тонус разгибателей рук и сгибателей
ног.
• При повороте и наклоне головы в сторону увеличивается тонус разгиба-
телей на стороне поворота.
❖ Ритмические рефлексы (чесательный, потирания, шагания) сопровождают-
ся многократным повторным сгибанием и разгибанием конечностей.
3.4. Двигательные функции спинального организма (синдром полного поперечного раз-
рушения спинного мозга).
Сначала в первые 3-5 недель имеется спинальный шок, характеризующийся атонией
мыши и отсутствием сухожильных и кожно-мышечиых рефлексов.
❖ После шока при высоких (шейный, верхнегрудной) уровнях разрушения повы-
шается тонус мышц ног и туловища, происходят судорожные подергивания
мышц, усиливаются спинальные рефлексы, наблюдаются и сложные реф-
лекторные движения парализованных конечностей (например, перекрест-
ный разгибательный и шагательный рефлексы).
❖ При низких уровнях поражения (нижнегрудной н верхиепоясничный) атония и
арефлексия в нижних конечностях сохраняется и после шока (не восстанавли-
ваются также функции дефекации и мочеиспускания), что свидетельствует о зави-
симости функций нижних спинальных сегментов от верхнегрудиых н шей-
ных сегментов.
❖ Невозможно поддержание вертикальной позы.
❖ Отсутствуют произвольные движения.
4. Роль ствола мозга (продолговатый мозг + мост + средний мозг) в регуляции дви-
жений.
173
4.1. Двигательные функции ядер череиных нервов (эфферентные звенья безусловных,
условных двигательных рефлексов н произвольных движений).
❖ ХП (подъязычный) - движение языка: жевание, артикуляция.
❖ XI (добавочный) - сокращение грудино-ключично-сосцевидной и трапе-
циевидной мышц: наклон головы в сторону, поворот головы, подъем пле-
чевого пояса вверх.
❖ IX, X (языкоглоточный, блуждающий) - сокращение мышцы мягкого нёба,
глотки, надгортанника, гортани, голосовых связок (рефлексы глотания,
рвоты, чихания, кашля, фонация).
❖ VII (лицевой) - сокращение мимических, жевательных мышц, стремечко-
вой мышцы среднего уха.
❖ VI (отводящий) - сокращение латеральной мышцы глаза (поворот глазного
яблока кнаружи).
❖ V (тройничный) - сокращение жевательной, височной, крылонёбной мышц,
мышцы, натягивающей барабанную перепонку и мышцы нёба.
❖ IV (блоковый) - сокращение верхней косой мышцы глаза, поворот глазного
яблока вниз н кнаружи.
❖ Ш (глазодвигательный) - сокращение прямой верхней мышцы глаза (пово-
рот глазного яблока вверх), прямой внутренней мышцы (поворот глазного яблока
внутрь), прямой нижней мышцы (поворот глазного яблока книзу), мышцы, под-
нимающей веко.
4.2. Сложнокоординированные двигательные рефлексы ствола мозга. В них уча-
ствуют ядра нескольких черепных нервов, иногда н моторные спинальные цен-
тры, объединяющиеся в единый функциональный комплекс с помощью ретику-
лярной формации мозга. К ним относятся рефлекторные акты жевания, сосания,
глотания, рвоты, дакания, кашля, чихания (см. темы 19 н 22).
4.3. Роль ствола мозга в регуляции мышечного тонуса (рис. 37).
❖ Разгибательная (экстензорная) система.
• Вестибулоспинальный путь.
Ф Начинается от вестибулярных ядер (латерального, медиального, верхнего)
продолговатого мозга.
ф- Прямо и через интернейроны возбуждает а- н у-мотонейроны раз-
гибателей (реципрокно тормозит мотонейроны сгибателей).
ф- Повышает тонус разгибателей конечностей, туловища, шеи.
• Медиальный ретикулоспинальный путь.
Начинается от медиальных ядер ретикулярной формации моста.
Через интернейроны спинного мозга возбуждает а- и у-магоней-
роны разгибателей (реципрокно тормозит мотонейроны сгибателей).
ф’ Повышает тонус разгибателей (преимущественно туловища н про-
ксимальных отделов конечностей).
❖ Сгибательная (флексорная) система.
• Латеральный ретикулоспинальный путь.
ф’ Начинается от ретикулярного гигантоклеточного ядра продолгова-
того мозга.
174
v* Через интернейроны возбуждает а- и у-мотонейроны сгибателей
(реципрокно тормозит мотонейроны разгибателей).
Ф Повышает тонус сгибателей (преимущественно конечностей).
Средний
мозг
Мост
Продол-
говатый
мозг
Повышение
тонуса раз-
бателейи
снижение -
сгибателей
Повышение
тонуса сги-
бателей и
снижение -
разгибателей
Спинной мозг
Рис. 37. Схема основных
моторных центров ствола
и их путей, регулирующих
тонус мышц туловища и
конечностей. КЯ - красное
ядро и рубро-спинальный
путь, РЯМ - ретикулярные
ядра моста и медиальный
ретикулоспинальный путь,
ВЯ - вестибулярные ядра
и вестибулоспинальный
путь, РГЯ - ретикулярное
гигантоклеточное ядро и
латеральный ретикуло-
спинальный путь, МН -
спинальные мотонейроны.
Объяснения в 5.1.
• Руброспинальный путь (Монакова).
Начинается от красных ядер покрышки среднего мозга.
•ф* Через интернейроны возбуждает а- н у-мотонейроны сгибателей
(реципрокно тормозит разгибатели).
Ф" Повышает тонус сгибателей.
❖ Децеребрационная ригидность (Ч. Шеррингтон, 1896).
• Возникает при перерезке между продолговатым и средним мозгом (или
их функциональном разобщении).
• Проявляется у человека резким повышением тонуса разгибателей ниж-
них конечностей, туловища, шеи.
• Механизмы ригидности.
Выключено активирующее влияние красных ядер на сгибатели, что
относительно увеличивает влияние вестибулярных ядер на разгиба-
тели.
Выключено тормозное влияние коры мозга и мозжечка на вестибу-
лярные ядра.
В развитии децеребрационной ригидности основную роль играет
возбуждение у-мотонейронов (перерезка у животных задних ко-
решков спинного мозга снимает ригидность).
4.4. Ретикулярная формация среднего мозга осуществляет координацию функ-
ций ядер глазодвигательных (III, IV, VI) нервов, обеспечивая содружественные
движения глаз в горизонтальном н вертикальном направлениях. В парамедиан-
ной РФ продолговатого мозга имеются нейроны, управляющие горизонтальны-
ми движениями глаза. Эти же нейроны, но опосредованно, через нейроны РФ
среднего мозга, управляют вертикальными движениями глаз. При двусторонних
175
поражениях этих отделов РФ становится невозможным произвольное выполне-
ние горизонтальных и вертикальных движений глаз.
4.5. Роль ствола мозга в регуляции установочных рефлексов: сохранение нор-
мального положения тела и равновесия (Р. Магнус, 1924).
❖ Статические (позно-тонические) рефлексы.
• Рефлексы позы (возникают на изменение положения головы, центра
тяжести и равновесия тела).
ф- Шейные рефлексы (см. выше 3.3).
ф Вестибулярные рефлексы (с рецепторов вестибулярного аппарата).
Нейтральное положение головы - горизонтальное.
Височная ось головы поднята на 45° вверх от горизонтали -
максимальный тонус разгибателей.
Височная ось головы опущена на 45° от горизонтали - тонус
разгибателей минимальный.
ф Вертикальная поза человека (голова вверх, шея несколько вен-
трально): шейные и вестибулярные рефлексы усиливают друг дру-
1 га; повышен тонус разгибателей ног и сгибателей рук.
• Выпрямительные рефлексы возникают на изменение нормальной позы
(с рецепторов шеи, лабиринта, кожи, сетчатки).
ф Сначала шейный тонический рефлекс и восстановление
нормального положения головы.
- ф Затем цепь рефлексов с перераспределением тонуса мышц конеч-
ностей и туловища с восстановлением нормальной позы тела.
• Рефлексы компенсаторного движения глаз удерживают поле зрения при измене-
нии положения головы.
❖ Статокинетические рефлексы (при линейных и угловых ускорениях, с ре-
цепторов вестибулярного аппарата).
• Рефлексы прямолинейного ускорения (с рецепторов отолитового аппа-
рата).
ф Лифтный рефлекс: при подъеме сначала повышается тонус сгиба-
телей ног, затем - разгибателей ног и восстанавливается исходное
соотношение тонуса этих мышц, при прекращении подъема или в
начале опускания лифта - противоположные изменения тонуса
мышц.
ф Рефлекс приземления: преобладание тонуса сгибателей при контак-
те ног с опорой.
• Рефлексы вращения (с рецепторов ампул полукружных каналов) включают всю
мускулатуру тела и глазных мышц.
Ф Глазной нистагм (способствует сохранению зрительной ориента-
ции) при вращении.
При ускорении вращения: 1) фаза медленного отклонения
глазных яблок в сторону, противоположную вращению (с ре-
цепторов полукружных каналов); 2) фаза быстрого возвраще-
176
ния глазных яблок в исходное состояние по ходу вращения
(связана с корковыми центрами).
При замедлении вращения - обратная последовательность
движения глаз.
4.6. Роль ствола мозга в осуществлении локомоции (ходьба, бег, прыжки).
❖ Локомоторная область среднего мозга - клиновидное ядро.
❖ Локомоторная область гипоталамуса (его субмамиллярная зона).
❖ Сильная их зависимость от корковых моторных центров у человека.
4.7. Клинически важные рефлексы ствола мозга.
4г Глоточный и нёбный рефлексы: рецепторы мягкого неба и глотки —* афферентные волок-
на IX и X нервов —* двигательные ядра IX и X нервов (продолговатый мозг) —* эфферент-
ные волокна IX н X нервов —> глотание.
♦♦♦ Роговичный рефлекс: рецепторы роговицы —* афферентные волокна 1-й ветви V нерва —•
двигательное ядро VII нерва (мост) —* сокращение круговой мышцы глаза.
4г Зрачковый рефлекс: рецепторы сетчатки —* афферентные волокна П нерва, верхнее дву-
холмие —» парасимпатические ядра Ш нерва (средний мозг) —»парасимпатические волокна
Ш нерва, ресничный ганглий —► круговая мышца радужной оболочки, сужение зрачка.
4г Глазной нистагм (см. выше 4.5).
5. Роль мозжечка в регуляции движений.
5.1. Функциональная структура мозжечка.
❖ Древний мозжечок состоит из клочка, узелка и ннжней части червя, имеет афферентный
вход от вестибулярных ядер.
4г Старый мозжечок состоит из парафлоккулярного отдела и верхней части червя, имеет
афферентные входы от спинного мозга а составе переднего и заднего спиноцеребеллярных
путей (от мышечных рецепторов).
4г Новый мозжечок состоит из полушарий, получает афферентные входы от разных областей
коры (в том числе и от двигательной) через мостоцеребеллярный путь и от мышечных ре-
цепторов через нижнюю оливу по оливоцеребеллярному пути.
Ф Белое вещество мозжечка содержит три парных ядра (шатра, промежуточные и зубча-
тые), это главный эфферентный выход мозжечка на моторные центры головного мозга.
4г Нейронная характеристика коры мозжечка. Тормозные нейроны Пуркинье
(средний слой) создают основное функциональное влияние мозжечка (рис.
38).
• Афферентные входы на клетки Пуркинье.
ф- Афферентный вход по лиановидным волокнам от мышечных, кож-
ных рецепторов и двигательной коры оказывает сильное прямое
возбуждающее действие на клетки Пуркинье и более слабое тор-
мозное действие через корзинчатые и звездчатые клетки.
ф- Афферентный вход по моховидным волокнам от мышечных, кож-
ных, вестибулярных рецепторов, коры больших полушарий и рети-
кулярной формации оказывает на нейроны Пуркинье через клетки-
зерна возбуждающее действие и через звездчатые клетки более сла-
бое тормозное действие. Итак, клетки Пуркинье, будучи тормозны-
ми нейронами, получая преимущественно возбуждающее аффе-
рентное влияние, превращают его в более или менее выраженное
тормозное эфферентное влияние.
177
• Эфферентные выходы клеток Пуркинье (выделяют три симметричные
вертикальные зоны).
• ф- Клетки Пуркинье коры медиальной (червячной) зоны и флоккуло-
нодулярной доли проецируются преимущественно иа ядра шатра,
далее иа вестибулярные ядра Дейтерса и ретикулярную формацию
продолговатого мозга и моста, далее по вестибуло- и ретикулоспи-
нальным путям на моторные спинальные центры (тонус мышц).
ф’ Клетки Пуркинье промежуточной (околочервячной) зоны проеци-
руются преимущественно иа промежуточные ядра (шаровидное и
пробковидное), далее иа красные ядра среднего мозга и через руброс-
пинальный тракт на спинальные моторные центры. Менее выражен
путь от промежуточных ядер к вентролатеральным ядрам таламуса
и далее к двигательной коре.
ф Клетки Пуркинье коры латеральной зоны полушарий проецируют-
ся на зубчатые ядра и далее к вентролатеральным ядрам таламуса, и
затем в двигательную кору.
52. Главные двигательные функции мозжечка (рис. 39).
от нижней олн- на моторные от ядер моста
вы (лнановид- центры голов* (моховидные
ные волокна) кого мозга волокна)
Рис. 38. Схема основных меж*
нейронных связей мозжечка.
КП - клетки Пуркинье, кз -
клетки-зерна, кк - корзинчатые
клетки, зк - звездчатые клетки,
темным цветом выделены тор-
мозные иейроиы, «+», «-» -
возбуждающие и тормозные
влияния. Объяснения в 5.1.
❖ Регуляция мышечного тонуса, позы и равновесия.
• Осуществляется преимущественно древним, частично старым и новым
мозжечком, входящими в медиальную червячную зону. Она получает
информацию от вестибулярных, мышечных, кожных, зрительных и слу-
ховых рецепторов о состоянии мышц и положенин тела в пространстве.
• Через эфферентные выходы по вестибуло-, рубро-, ретикулоспиналь-
ным путям перераспределяется тонус мышц и изменяется поза, что по-
зволяет сохранить равновесие тела.
• Функциональный тест: сохранение равновесия в позе Ромберга.
4г Координация позы и целенаправленных движении.
178
• Выполняется старым и новым мозжечком, входящими в промежуточ-
ную (околочервячную) зону.
• Сюда поступает информация от моторной коры (программа движения)
и от рецепторов мышц и кожи (выполнение движения).
• Сравнивая эту информацию, промежуточная зона через эфферентные
выходы (на двигательную кору и руброспинальный путь) проводит ко-
ординацию позы и движения и корректирует направление движения.
• Функциональный тест: пальценосовая и пяточно-коленная пробы.
А Б В
Рис.39. Схема функций мозжечка (по Р. Шмидту с измене-
ниями). А - регуляция тонуса, позы и равновесия; Б - ко-
ординация позы и целенаправленных движений; В - про-
гоаммипование пооизвольных движений.
Участие в программировании целенаправленных движений.
• Выполняет новый мозжечок, входящий в латеральную зону его полу-
шарий.
• Сюда поступает импульсация из ассоциативной коры больших полуша-
рий (замысел движения).
• В коре мозжечка она перерабатывается в программу движения.
• Эта программа передается через зубчатое ядро в таламус и в моторную
кору.
• Через пирамидную и экстрапирамидную систему она реализуется в
сложное целенаправленное движение.
• Контроль и коррекция медленных программированных движений осу-
ществляется на основе обратной афферентации, а быстрых (баллистиче-
ских) - путем изменения программы.
179
• Функциональный тест: проба на адиадохокинез.
6. Роль базальных ядер в регуляции движений.
6.1. Структура базальных ядер: полосатое тело (неостриатум: хаостатое ядро + скорлупа), блед-
ный шар (паллидум), ограда (клауструм), в функциональном плане в стриопаллидариую систе-
му аключают субталамическое ядро и черное вещество.
6.2. Связи базальных ядер (рис. 40).
❖ Афферентная информация поступает в полосатое тело из: 1) всех зои ко-
ры; 2) неспецифических ядер таламуса; 3) черного вещества.
❖ Главные эфферентные выходы: 1) от полосатого тела к бледному шару; 2)
от бледного шара к таламусу и далее к двигательной коре; 3) от бледного
шара к центрам ствола (красное ядро, ретикулярная формация) и через
нижнюю оливу к мозжечку; 4) от полосатого тела к черному веществу.
63. Функциональная рояь полосатого тела.
❖ Влияние иа бледный шар: преимущественно тормозное (медиатор - ГАМК),
частично возбуждающее.
Спинной мозг Мозжечок
Рис. 40. Схема основных
афферентных и эффе-
рентных связей базаль-
ных ядер. ЧВ - черное
вещество, СТЯ - субта-
ламическое ядро, ВЯ -
вентральные ядра, ИЛЯ -
интраламинарные ядра,
«+»,«-» - возбуждающие
и тормозные влияния.
Объяснения в 6.2 - 6.4.
Взаимосвязь с черным веществом.
• Полосатое тело оказывает тормозное влияние иа черное вещество (ме-
диатор — Г АМК).
• Черное вещество оказывает модулирующее влияние на полосатое тело
(медиатор - дофамии).
❖ Влияние иа таламус: полосатое тело и бледный шар передают через ядра
таламуса информацию (например, двигательные программы) в двигательную КО-
РУ-
❖ Влияние на моторную кору: улучшает координацию сложных движений,
осуществляемых через двигательную кору, за счет «вытормаживания» не-
нужных в данных условиях степеней свободы движения.
180
• I* Стимуляция полосатого тела вызывает: 1) двигательные реакции: поворот
головы, туловища, сгибание конечностей; 2) задержку поведенческой реак-
ции («застывание» в одной позе).
❖ Поражение полосатого тела вызывает:!) насильственные избыточные
движения (гиперкинезы); 2) нарушение двигательных компонентов поведе-
ния, выработки условных рефлексов.
6.4, Функциональная роль бледного шара («...Когда-то в прошлом единственная столица
двигательных актов». И.А. Бернштейн).
Бледный шар оказывает модулирующее (изменяющее активность) влияние иа
двигательную кору, красное ядро, ретикулярную формацию, мозжечок.
❖ Стимуляция бледного шара вызывает: 1) элементарные двигательные реак-
ции в виде сокращения мышц конечностей, шеи и лица; 2) стимуляцию
центра голода в латеральном гипоталамусе (пищевое поведение).
❖ Выключение бледного шара приводит к снижению двигательной активно-
сти: адинамия, сонливость, эмоциональная тупость, нарушение выработки
условных рефлексов.
6.5. В целом двигательные функции базальных ганглиев выражаются:
❖ в выработке сложных двигательных программ, которые реализуются через
моторную кору и обеспечивают двигательные компоненты условных реф-
лексов и более сложных форм поведения;
❖ в контроле параметров движения: силы, амплитуды, скорости и направле-
ния.
7. Роль таламуса в регуляции движения.
В несенсорных релейных ядрах (вентральные переднее н латеральное ядра) переключаются
импульсы из мозжечка и стриопаллидарной системы, которые затем проецируются в
моторную кору (поля 4, 6, 8, 44). Через эти ядра осуществляется передача в двига-
тельную кору сложных двигательных программ, образованных в мозжечке и базаль-
ных ядрах. (Стимуляция моторной коры с вентролатерального ядра таламуса способствует разви-
тию долговременной потенциации в моторной коре - формирование двигательной памяти.)
8. Корковый уровень регуляции движения.
8.1. Моторная кора.
❖ Первичная моторная область (М-I): прецентральная извилина и парацен-
тральная долька (поле 4) создает способность к тонким координированным
движениям, особенно пальцев рук.
❖ Премоторная зона - задние участки лобных извилин, осуществляет выс-
шие двигательные функции, связанные с планированием и координацией
произвольных движений: навыки письма (поле б), поворот глаз и головы в
противоположную сторону (поле 6), речедвигательный центр Брокб (поле
44).
• •• Добавочная моторная область (М-П) (часть верхней лобной нзвилнны на медиальной по-
верхности около парацентральной дольки) участвует в формировании позы, ритмических
движений и некоординированных движений.
❖ Корковое представительство мышц человека (см. выше 2.3).
❖ Функциональная клеточная организация коры: двигательные колонки - ко-
ординируют мотонейроны, иннервирующие отдельные мышцы или мото-
нейроны мышц, действующих на один сустав.
181
❖ Двигательная кора осуществляет свои функции через пирамидную и экст-
рапирамидную системы, оканчивающихся на одних и тех же мотонейронах
двигательных центров черепных нервов и спинного мозга
8.2. Пирамидная система (произвольные движения, целенаправленные двигатель-
ные программы, тонкие координированные движения конечностей, контроль
силы движения).
❖ Кортикоспинальный путь (около 1 млн. волокон).
• От пирамидных клеток V слоя через внутреннюю капсулу, ножки мозга
и мост в пирамиды продолговатого мозга, где происходит перекрест 75
- 90 % волокон.
• Спннной мозг: латеральный кортикоспинальный путь состоит из пере-
крещенных волокон; медиальный кортикоспинальный путь состоит из
неперекрещенных в пирамидах волокон, онн частично перекрещивают-
ся в спинном мозге. Этот путь оканчивается в шейном и грудном отде-
лах.
• В спинальных моторных центрах волокна оканчиваются преимущест-
венно на интернейронах, частично непосредственно на а-мото-
нейронах.
❖ Кортикобульбарный путь (около 20 млн. волокон).
• Повторяет ход кортикоспинального тракта до нижнего края верхней
оливы.
• В области ствола мозга волокна образуют перекрест и направляются к
мотонейронам ядер черепных нервов (Ш - VII, IX - ХП).
83. Экстрапирамидную систему составляют кортикорубральные и кортикорети-
кулярные пути, начинающиеся приблизительно от тех же зон, которые дают на-
чало пирамидным путям. Они влияют на моторные центры спинного мозга не
непосредственно, а через соответственно руброспннальные и ретикулоспиналь-
ные пути. В экстрапирамидную систему входят базальные ядра и мозжечок, ко-
торые также влияют на а- и у-мотонейроны не прямо, а через моторные центры
ствола мозга. Через эту систему осуществляется рефлекторная и произвольная
регуляция тонуса и позы, скорости, ритма и пластичности движений, выполне-
ние заученных двигательных актов: ходьба, бег, речь, письмо, моторные прояв-
ления эмоций
8.4. Имеется выраженное взаимодействие пирамидной и экстрапирамидной
систем и способность их к компенсации двигательных функций. Например,
при перерезке Пирамидных путей у приматов полностью сохраняются все дви-
жения, кроме тонких координированных и независимых движений отдельных
пальцев рук. (Поскольку обе системы работают в функциональном единстве некоторые спе-
циалисты призывают к пересмотру понятий «пирамидная»» и «экстрапнрамидная» системы, од-
нако альтернативная концепция еще не предложена.)
9. Общая схема организации целенаправленных движений.
9.1. План движения.
❖ Побуждение к движению создается в лимбической системе (роль эмоций).
❖ Замысел движения создается в ассоциативной коре.
182
9.2. Программа движения формируется в базальных ядрах, мозжечке, премоторной
коре.
9.3. Выполнение движения.
❖ Двигательная кора.
❖ Стволовые моторные центры, базальные ядра, мозжечок.
❖ а-Мотонейроны спинальных моторных центров н моторных ядер черепных
нервов.
❖ Двигательные единицы мышц.
9.4. Типы целенаправленных движений.
❖ Медленные (следящие): коррекция осуществляется во время движения на
основе обратной афферентации.
❖ Баллистические: коррекция осуществляется по оценке результата движе-
ния путем изменения программы.
10. Профильные материалы для студентов лечебного факультета. Особенности дви-
гательной активности в пожилом и старческом возрасте.
10.1. Уменьшение амплитуды и плавности движений, нарастание ригидности. Неловкость
при ходьбе (шаркающая, мелкими шагами походка).
10.2. Снижение мышечного тонуса, сухожильных рефлексов, точности и автоматичности
движений.
10.3. Снижение скорости двигательных реакций в произвольных и непроизвольных двига-
тельных актах, особенно при выполнении сложных движений.
10.4. Бедность мимики, тихая, недостаточно модулированная речь.
11. Профильные материалы для студентов педиатрического факультета.
11.1. Двигательная активность плода.
4 « Сгибание шеи, ведущее к отстранению поверхности головы от раздражителя в от-
вет на раздражение периораяьной области лица (на 8-й неделе).
4 * Генерализованные движения рук и ног при раздражении кожи (одновременное со-
кращение сгибателей и разгибателей).
* 1* Появление двигательных реакций при раздражении проприорецепторов (с 9-й не-
дели), хватательной реакции (с 12-й недели), мимических движений на вкусовые и
обонятельные раздражители (в более поздние сроки).
❖ Тоническое сокращение мышц-сгибателей, формирующее позу плода, обеспечи-
вающую минимальный его объем в матке, поддерживается за счет раздражения
кожных рецепторов и проприорецепторов.
❖ Периодические генерализованные сокращения мышц-разгибателей ощущаются
матерью как шевеление плода (4 - 8 в час) с 4 - 4,5 мес.
♦ ♦♦ Дыхательные движения начинаются с 14-й недели; начиная с 6,5 - 7 мес. плод
может дышать длительное время.
11.2. Характеристика движений новорожденного.
♦♦♦ Спонтанные периодические движения.
• Некоординированность, нецелеиаправленность, генерализованность движе-
ний.
• Появление координированных ритмических движений, отведения и приведе-
ния с участием всех конечностей, затем изолированных движений.
• Преобладание периодов двигательной активности над периодами покоя.
183
• У бодрствующего новорожденного имеются сосательные движения и движе-
ния головы, направленные на поиск матери.
Тонус мышц новорожденного поддерживается:
• импульсами от проприорецепторов и кожных терморецепторов;
• импульсами от рецепторов легких, активирующихся при вдохе;
• для новорожденного характерна ортотоническая поза, которая является след-
ствием преобладания тонуса сгибателей.
♦♦♦ Генерализированность рефлексов, обширность рефлексогенных зон.
• Не выражен контроль со стороны моторной коры больших полушарий.
• Иррадиация возбуждения в ЦНС связана со слабостью процессов торможения,
недостаточной миелинизацией аксонов.
• Быстрое совершенствование уже сформированных к рождению рефлексов под
действием внешней среды, появление новых рефлексов, преимущественно за-
щитного характера.
• Тесная связь развития локомоторной функции с формированием установоч-
ных рефлексов.
11.3. Двигательные рефлексы новорожденного.
❖ Коленный рефлекс характеризуется сгибанием коленного сустава в связи с преоб-
ладанием тонуса сгибателей, преобразуется в разгибательный на 2 месяце.
♦ ♦♦ Хватательный рефлекс (рефлекс Робинсона) - схватывание и удержание предме-
та при прикосновении к ладони, исчезает иа 2 - 4-м мес.
♦ ♦♦ Обхватывание (рефлекс Моро) - отведение рук в стороны и разгибание пальцев с
последующим возвращением рук в исходное положение, исчезает после 4-го мес.
♦ ♦♦ Ползание (рефлекс Бауэра) - в положении лежа на животе ребенок поднимает го-
лову и совершает ползающие движения; если подставить к подошвам ладонь, то
ребенок начнет активно отталкиваться ногами от препятствия; исчезает к 4-му
мес.
♦ ♦♦ Рефлекс Кернига - у лежащего на спине ребенка сгибают ногу в тазобедренном и
коленном суставах, а затем пытаются разогнуть ногу в коленном суставе. Рефлекс
положителен, если это удается сделать; исчезает после 4-го мес.
♦ ♦♦ Лабиринтный рефлекс - вызывается изменением положения головы в пространст-
ве: у ребенка, лежащего на спине, повышен тонус разгибателей шеи, спины, ног;
при переворачивании на жнвот увеличивается тонус сгибателей шеи, спины, ко-
нечностей.
♦ ♦♦ Хоботковый рефлекс - выпячивание губ хоботком в результате сокращения кру-
говой мышцы рта при легком ударе пальцем по губам ребенка, исчезает к концу
1-го года жизни.
♦ ♦♦ Поисковый рефлекс (поиск груди матери) - поглаживание кожи в области угла рта
вызывает опускание губ, отклонение языка и поворот головы в сторону раздражи-
теля, исчезает к концу 1-го года.
♦ ♦♦ Подошвенный рефлекс (рефлекс Бабинского) - изолированное тыльное разгибание
большого пвльца и подошвенное сгибание остальных, которые иногда веерооб-
разно расходятся, при раздражении подошвы по наружному краю стопы в направ-
лении от пятки к пвльцам, исчезает после 12 мес.
11.4. Двигательная активность в грудном возрасте.
♦> Созревание моторных центров ЦНС.
• Интенсивное созревание ЦНС в первые месяцы жизни.
184
• К 6-му мес. заканчивается морфологическое становление полосатого тела,
миелинизации волокон мозжечка.
• К концу 1-го года происходит интенсивная миелинизация волокон ЦНС и раз-
витие пирамидных слоев в моторной коре (например, гигантских пирамидных
клеток Беца).
• Быстрое созревание нервной системы и скелетной мускулатуры ведет к увели-
чению двигательной активности, что, в свою очередь, стимулирует рост и раз-
витие мышечной и нервной системы (взаимодействие структуры и функции).
❖ Тонус мышц.
• Имеется гипертонус мышц в первые месяцы жизни, обусловленный незрело-
стью ЦНС и преобладанием возбуждения над торможением.
• Уменьшение общего тонуса мышц на 1-м году происходит в результате по-
вышения роли моторной коры, мозжечка, полосатого тела в регуляции движе-
ний.
• Формируется тонус разгибателей на 2-м мес. жизни, и к 6 мес. имеется равный
тонус мышц-антагоннстов.
Рефлексы грудного ребенка включают рефлексы новорожденного и вновь форми-
рующиеся.
• Туловищно-выпрямительный - при соприкосновении стоп ребенка с опорой
наблюдается выпрямление головы, формируется с 1 мес.
• Рефлексы Ландау: верхний - ребенок в положении на животе поднимает голо-
ву и верхнюю часть туловища, опираясь на плоскость руками; иижннй - в по-
ложении на животе ребенок разгибает и поднимает ноги, формируется к 5 -
6-му мес.
11.5. Двигательные навыки в грудном возрасте,
♦ ♦♦ Двигательные навыки ребенка 2-5 мес.
• Возникают движения руками в направлении видимому предмету и захватыва-
ние предмета (с 2 мес.).
• Появляется способность к тоническому сокращению шейной мускулатуры н
поддержанию головки в положении лежа на животе (к концу 2-го мес.), окон-
чательное формирование способности держать головку (на 3-м мес.).
• Формируется навык ползания (в 3 мес.) способность переворачиваться со спи-
ны на живот, затем обратно (в 4 - 5 мес.).
❖ Двигательные навыки ребенка 5-9 мес.: переступание при поддержке; способ-
ность садиться, вставать на четвереньки, вставать, стоять и опускаться, поддержи-
ваясь за опору.
♦♦♦ Двигательные навыки ребенка 9-12 мес.
• Двигательная активность верхних конечностей становится координированной,
движения - более точными (к 10-му мес.).
• Предварительное приспособление пальцев рук к форме предмета, который ре-
бенок намерен схватить. Появление различий в действиях правой и левой рук.
• Нестабильность вертикальной позы. Появление попыток ходить к концу 1-го
года.
• Образование новых двигательных навыков методом «проб и ошибок», путем
подражания.
185
• Используется взаимодействие зрительного и двигательного анализаторов для
образования новых двигательных навыков.
11.6. Двигательные навыки и их развитие в возрасте 1 -14 лет.
Движения второго года жизни: совершенствование ходьбы, появление способно-
сти к бегу, перешагиванию предметов, подъему по лестнице, прыжки в длину.
❖ Движения третьего года жизни.
• Подпрыгивания на месте, отрывая ноги от поверхности.
• Совершенствование прыжков, перешагивание.
• Действие рук как стабилизаторов и усилителей скорости движения.
• Ведущим уровнем организации произвольных движений становится теменно-
премоторный.
❖ Движения в возрасте3-6лет.
• Развитие игровой деятельности: бег, прыжки, гимнастические упражнения.
• Обучение рисованию, игре на музыкальных инструментах.
• Освоеиие точных движений, связанных с развитием мелких мышц кисти; со-
вершенствование координации всех форм двигательной активности.
Движения в последующие возрастные периоды.
• Заметное увеличение точности и меткости движений после 7 лет, что связано с
интенсивным созреванием двигательного анализатора, корково-подкорковых и
внутрикортикальных связей между двигательными и ассоциативными облас-
тями.
• Значительное увеличение скорости однократного движения и приближение к
величинам взрослых в 13 - 14 лет, некоторое замедление увеличения скорости
в 16 - 17 лет, максимальная скорость движения к 20 - 30 годам.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3, 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [14-16].
3.2. Дополнительная литература (см. с. 514 - 522): [24], [26], [27], [29], [30], [57],
[133], [145], [165], [169], [203], [222], [225], [236], [245], [251-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/-nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; Учебное
пособие В.Н. Яковлева, И.Э. Бсаулеико «Физиология головного мозга». - Воронеж,
ВГМА, 2001. - 75 с.; тесты остаточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором н слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
186
ЧАСТЬ!!
ЧАСТНАЯ ФИЗИОЛОГИЯ
Тема 12. Физико-химические свойства крови («Кровь - сок совсем особенного свой-
ства». И. Гвте).
Цель: а) знать понятие о системе крови, ее основные функции; количество и состав крови и плазмы,
функциональную роль электролитов и белков плазмы; осмотическое н онкотическое давление,
кислотно-основное состояние крови, основные механизмы их регуляции; гематокрит; скорость
оседания эритроцитов и факторы, влияющие на нее; гемолиз и его виды; основные константы
крови и их регуляцию; возрастные изменения физико-химических свойств крови;
б) уметь использовать эти знания для объяснения функций системы крови в целостном орга-
низме в разных условиях внешней и внутренней среды;
в) иметь представление о методах исследования: технике взятия крови, определении осмоти-
ческой резистентности эритроцитов и скорости оседания эритроцитов.
Мотивация темы: кровь является важнейшим компонентом внутренней среды организма, относитель-
ное постоянство которой, в том числе физико-химических показателей крови (осмотического
давления, pH и др.), является необходимым условием жизнедеятельности организма. Измене-
ния физико-химических свойств крови являются важным механизмом в патогенезе многих за-
болеваний, нспрльзуются для их диагностики и контроля лечения.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Понятие о системе крови (Г.Ф. Ланг, 1939), ее основные функции.
1.1. Компоненты системы крови: 1) органы кроветворения, 2) циркулирующая
кровь, 3) органы кроверазрушения, 4) аппарат иейрогуморальиой регуляции
(создает устойчивые связи между элементами системы).
1.2. Основные функции крови.
❖ Транспортная: дыхательная, трофическая, экскреторная, терморегуляция
(транспорт тепла).
❖ Защитная: иммунитет, гемостаз, реакция буферов.
❖ Регуляторная: гуморальная регуляция.
2. Количество и состав кровн.
2.1. Абсолютное и относительное количество крови: у здорового человека ~ 5,0
л; 70 мл/кг; - 7 % массы тела, что называется нормоволемией; понятие о гипер-
и гиповолемии.
2.2. Состав крови.
❖ Форменные элементы (98% их объема составляют эритроциты);
• клетки - лейкоциты;
• постклеточные структуры - эритроциты, тромбоциты;
187
• гематокрит - отношение объема эритроцитов к объему крови (муж. 40 -
48 %, жеи. 36 - 42 %), изменение его величины характеризует степень
сгущения или разведения крови.
❖ Плазма крови (межклеточная жидкость крови: 2,8 - 3,0 л, воды - 90 %).
• Минеральный состав плазмы (в ммоль/л).
•ф- Основные катионы плазмы: Na* - 140; К* - 4,5; Са2* - 2,3; их функцио-
нальная роль: влияние на осмотическое давление, перераспределе-
ние воды между внеклеточным и клеточным отсеками, возбуди-
мость и сократимость клеток, свертывание крови.
•ф- Основные анионы плазмы: CI* - 102; бикарбонаты - 22; фосфаты - 2; их
функциональная роль: осмотическое давление, перераспределение
воды, возбудимость клеток, буферы крови.
• Белки плазмы (65 - 85 г/л).
• ф- Виды белков: альбумины - 55 %, глобулины (сц, а2, ₽, у) - 41 %,
фибриноген - 4 %, а также белковые гормоны и ферменты; основ-
ные места их образования: печень, лимфоциты; плазма без фибри-
ногена называется сывороткой.
• ф- Функции белков: питательная, транспортная, создание онкотиче-
ского давления, гормональная, ферментная, буферная и иммунная
функции, участие в свертывании крови, агрегации эритроцитов.
• Низкомолекулярные органические вещества: промежуточные и конеч-
ные продукты обмена, витамины, микроэлементы, олигопептиды и др.
3. Основные физико-химические показатели крови и механизмы их регуляции.
3.1. Осмотическое и онкотическое давление крови.
❖ Величина осмотического (- 285 мосм/кг, 7,3 а™, 5600 мм рт. ст.) и онкотического,
обусловленного белками, давления (- 28 мм рт. ст.) называется нормоосмия;
повышение и снижение его - соответственно гиперосмия и гипоосмии
❖ Осмотическое давление крови, учитывая легкий транспорт воды и электро-
литов между сосудистым и интерстициальным отсеками, влияет на обмен
воды между этими отсеками и клеточным отсеком. Онкотическое давление
первично и преимущественно влияет иа обмен жидкости между сосуди-
стым и интерстициальным отсеками и, следовательно, на регуляцию объе-
ма крови. Это обусловлено тем, что онкотическое давление и концентрация
белков в крови в 7-10 раз больше, чем в интерстициальной жидкости, а
транспорт белков через стенку капилляра затруднен.
❖ Регуляция постоянства осмотического давления.
• Осморегулирующий рефлекс: периферические (сосудистые и тканевые)
и центральные (гипоталамические) осморецепторы, основной нервный
центр - гипоталамус, эфферентное гормональное звено - ангидиурети-
ческий гормон, альдостерон, Ма*-уретический гормон сердца; главный
эффекторный орган - почки.
• Функциональная система, обеспечивающая постоянство осмотического
давления крови (подробно в теме 25; задание 1:13).
3.2. Реакция крови (кислотно-основное состояние крови).
188
❖ Величина pH крови (7,4 ± 0,04; ~ 40 нмоль/л Н4).
❖ Регуляция постоянства pH крови.
• Гемические механизмы: деятельность гемоглобинового, бикарбонатно-
го, белкового, фосфатного буферов плазмы и эритроцитов. Их общая
буферная мощность 48 ммоль/л крови.
• Главные висцеральные механизмы регуляции: выведение через легкие -
17 000 ммоль СОг (потенциального донора Н*) в сутки и через почки - 70
ммоль Н*, что эквивалентно суточной продукции «нелетучих» кислот
(серной, фосфорной и др.).
• Функциональная система, обеспечивающая постоянство КОС крови (из-
ложена в теме 21).
33. Вязкость (внутреннее трение) крови.
❖ Показатели вязкости: кровь - 5,5, плазма - 1,8 сП (сантипуаз).
❖ Факторы, влияющие на вязкость.
• Клетки крови: вязкость прямо пропорциональна количеству клеток. Од-
нако в сосудах микроциркуляции вязкость крови равна вязкости плаз-
мы, этот эффект связан с образованием пристеночного слоя плазмы и
легкой деформируемостью эритроцитов. Увеличение количества лейко-
цитов, объем каждого из которых в 2,5 раза превышает объем эритроци-
та, резко повышает сопротивление кровотоку в капиллярах.
• Белки крови: вязкость крови прямо пропорциональна концентрации
белков.
• Количество воды: сгущение крови повышает вязкость, гидремия - сни-
жает.
• Артериальное давление: при повышении АД вязкость снижается (в ре-
зультате разрушения вязкости, связанной со структурой кровн), при снижении АД
- повышается.
❖ Значение вязкости для циркуляции крови: гидродинамическое сопротивле-
ние прямо пропорционально вязкости.
4. Гемолиз как нарушение осмотического равновесия клетки и окружающей среды.
4.1. Предгемолитические формы эритроцитов: куполообразная, сферическая.
4.2. Виды гемолиза.
❖ По локализации: внутриклеточный и внутрисосудистый.
По причинному фактору: 1) осмотический (например, снижение активности Na4,
К+-насоса при старении эритроцитов); 2) химический (например, действие окислите-
лей, восстановленный глутатион-SH при этом является «тиоловым щитом» эритроцитов);
3) биологический (например, образование цитолитического комплекса Сзь-С, системы
комплимента при иммунных процессах); 4) механический (например, при применении
аппарата искусственного кровообращения).
4.3. Границы осмотической резистентности эритроцитов', начало гемолиза -
0,50-0,45% раствор NaCl, полный гемолиз - 0,40-0,35% раствор NaCl; увеличе-
ние числа молодых эритроцитов (ретикулоцитов) увеличивает осмотическую
резистентность, старых эритроцитов - снижает.
5. Скорость оседания эритроцитов (СОЭ).
189
5.1. Для объяснения СОЭ обычно используется концепция агрегации эритроцитов и
их оседание в соответствии с законом Стокса (осаждение частиц в вязкой сре-
де). Согласно закону Стокса скорость оседания прямо пропорциональна квадра-
ту радиуса эритроцита и разницы между плотностью эритроцитов и плазмы и
обратно пропорциональна вязкости плазмы.
5.2. Агрегация эритроцитов («монетные столбики», их сети и конгломераты неправильной
формы) вызывает уменьшение площади клеточной поверхности, контактирую-
щей с плазмой, что приводит к снижению трения и способствует оседанию
эритроцитов. Агрегации эритроцитов (увеличению СОЭ) способствует нейтра-
лизация отрицательного поверхностного заряда -СОО‘ групп их мембраны, на-
пример, при увеличении грубодисперсных белков (у-глобулинов и фибриноге-
на) в плазме.
Примечание: агрегационная концепция СОЭ и предлагаемые механизмы агрегации (нейтрали-
зация заряда эритроцитов, образование между ними макромолекулярных мостиков) не соответ-
ствуют ряду современных экспериментальных данных, и суть процесса СОЭ пока не находит
однозначного объяснения.
6. Методы исследования физико-химических свойств крови.
6.1. Определение скорости оседания эритроцитов. Прн определении СОЭ используется
свойство крови, смешанной с цитратом натрия, не свертываться при стоянии, а разде-
ляться на два слоя: иижний - эритроциты, верхний - плазма. Оседание эритроцитов
происходит с различной скоростью в зависимости от изменения физико-химических
свойств крови. Норма: у мужчин 1-10 мм/ч, у жеищии - 2 - 15 мм/ч.
6.2. Определение границ осмотической стойкости эритроцитов (унифицированная
методика). Осмотическую стойкость эритроцитов можно количественно определить с
помощью гипотонических растворов хлорида натрия разной концентрации. Для этого
применяют растворы, различающиеся по концентрации на 0,05%, в последовательном
ряду от 0,85% до 0,10%. Концентрация гипотонического раствора, при которой в нор-
ме начинается гемолиз у человека равна 0,50-0,45%, а полный гемолиз происходит при
0,40-0,35%. Молодые формы эритроцитов отличаются более высокой осмотической
устойчивостью, старые эритроциты и некоторые патологические формы эритроцитов
менее устойчивы к осмотическому повреждению.
6.3. Определение гематокрита. Гематокрит показывает процентное отношение общего
объема эритроцитов к объему крови. В промытые гепарином и высушенные гематок-
ритные капилляры до метки «100» набирают кровь, затем помещают в специальную
иасадку, центрифугируют 30 мии при 3000 об/мин, и отмечают, какую часть градуиро-
ванного капилляра занимают эритроциты. В норме гематокрит у мужчин равен 40-48
%, у женщин - 36-42%, у новорожденных - 50-55%.
7. Профильный материал для студентов лечебного факультета.
7.1. Этапы внутриутробного кроветворения.
❖ 1-й этап: желточный (мезобластический) - с 3-й до 9-й недели, гемопоэз проис-
ходит вне эмбриона в сосудах желточного мешка, здесь из стволовых клеток обра-
зуются примитивные первичные эритробласты (мегалобласты), содержащих НЬР.
♦ ♦♦ 2-й этап: печеночный (гепатолиенальный) начинается с 6-й недели и длится поч-
ти до рождения, достигает максимума на 3 - 5-м мес. В печени сначала осуществ-
ляется как мегалобластический, так н нормобластический эритропоэз, а с 7-го мес.
- нормобластический эритропоэз. Кроме эритропоэза в печеин осуществляется
гранулоцитопоэз, мегакарноцитопоэз, моноцитопоэз и лимфопоэз. На этом этапе
190
(11-я неделя - 7-й мес.) в селезенке осуществляется эритропоэз, гранулоцитопоэз,
моноцитопоэз, лимфопоэз.
❖ 3-й этап: костномозговой (медуллярный) начинается с конца 3-го мес. и продол-
жается в постнатальном периоде.-В костном мозге костей (начиная с ключицы) из
стволовых клеток осуществляется эритропоэз по нормобластическому типу, гра-
нулоцитопоэз, моноцитопоэз, мегакариоцитопоэз и лимфопоэз. Лимфопоэз осу-
ществляется также в селезенке, тимусе, лимфоузлах, нёбных миндалинах, пейеро-
вых бляшках кишечника.
7.2. При старении организма происходят следующие изменения.
❖ Снижение относительной массы крови (до 67 мл/кг); буферных оснований (бикар-
бонатов крови); концентрации белков крови и коэффициента альбуми-
ны/глобулины за счет гипоальбуминемии.
❖ Возникают гиперглобулинемия и гиперфибриногенемия, увеличение остаточного
азота и холестерина крови.
♦ ♦♦ При функциональных нагрузках происходят более резкие сдвиги физико-
химических показателей крови и более длительный период восстановления.
8. Профильный материал для студентов педиатрического факультета.
8.1. Этапы внутриутробного кроветворения (см. выше 7.1).
8.2. Особенности кроветворения у детей разного возраста.
♦♦♦ Постепенное уменьшение миелоидной ткани в костном мозге.
❖ Имеется функциональная лабильность кроветворного аппарата, возможен частич-
ный возврат к мегалобластическому типу кроветворения у грудных детей.
8.3. Физико-химические свойства крови новорожденных детей.
♦ ♦♦ Снижены pH и буферные основания крови (декомпенсированный ацидоз в 1-й
день и затем компенсированный ацидоз).
♦ ♦♦ Низкая величина СОЭ (1-2 мм/ч).
❖ Снижено онкотическое давление крови.
♦ ♦♦ Увеличены вязкость и плотность в течение 1-й недели (вязкость 10 - 15; плот-
ность 1,060 - 1,080), величина вязкости к концу 1-го мес. достигает уровня взрос-
лых (4 - 5).
8.4. Изменение количества крови, гематокрита, состава плазмы в онтогенезе.
♦ ♦♦ Количество крови: более высокая относительная масса крови у новорожденных и
грудных детей (до 15 %) и постепенное снижение ее до уровня взрослых (7 %) к 6
- 9 годам; некоторое увеличение количества крови в период полового созревания.
♦ ♦♦ Величина гематокрита: 1) более высокая величина гематокрита (54 %) у новоро-
жденных; 2) более низкая величина гематокрита в грудном возрасте и в детстве до
15 лет (35 - 39 %); 3) «нормализация» величины гематокрита в конце пубертатно-
го периода (40 - 45 %).
❖ Состав плазмы: 1) снижение количества белков в период новорожденное™ (48 -
56 г/л) и постепенное увеличение его до уровня взрослых (70 - 80 г/л) к 3 - 4 го-
дам; 2) относительно уменьшенное содержание глобулинов и увеличенное содер-
жание альбуминов в течение 1-го года жизни; 3) аминокислот у детей первых лет
жизни на 35 % меньше, чем у взрослых; 4) общее количество липидов в крови но-
ворожденных снижено, но на 4 - 7-й день их количество возрастает примерно
вдвое; 5) содержание глюкозы в крови грудных детей ниже, чем у взрослых (4,0 -
4,6 ммоль/л) и достигает уровня взрослых (3,5 - 5,7 ммоль/л) у детей 12 - 14 лет.
♦ ♦♦ В крови плода отсутствует карбоангидраза, у новорожденных ее активность низка;
уровень, характерный для взрослых, устанавливается к 5 годам.
191
83. Динамика показателей физико-химических свойств крови детей различного воз-
раста и последовательность их изменений в онтогенезе,
❖ Показатели, быстро «нормализующиеся» в периоде новорожденное*™: повышение
до уровня взрослых величин гематокрита, вязкости, плотности крови, СОЭ и сни-
жение pH.
❖ Показатели, «нормализующиеся» в периодах детства: повышение до уровня
взрослых содержания буферных оснований крови, белка в плазме и соотношения
белковых фракций, онкотического давления.
♦ ♦♦ Показатели, «нормализующиеся» в подростковом возрасте: повышение до уровня
взрослых гематокрита и снижение относительной массы крови.
9. Профильный материал для студентов стоматологического факультета.
9.1. Взаимосвязь слизистой оболочки рта с органами кроветворения во внутриутробном
развитии: слизистая оболочка рта формируется у эмбриона к 12-му дню и выполняет
функцию кроветворения; по мере развития плода эта функция переходит к печени, се-
лезенке и костному мозгу.
9.2. Сравнительная характеристика физико-химических констант крови и слюны: по
сравнению с кровью слюна имеет более низкое осмотическое давление (50 - 100
мосм/кг), вязкость (1,1 - 1,32), буферную емкость (~ 8 ммоль/л); близкую к нейтраль-
ной pH (5,6 - 7,6); более высокое содержание К* (~ 20 ммоль/л); более низкое содержа-
ние Na+ (~ 30 ммоль/л), СГ (~ 30 ммоль/л).
93. Взаимосвязь физико-химических свойств крови и функций органов ротовой полос-
ти.
♦ ♦♦ Существует прямая зависимость концентрации солей в слюне (NaCl, КС1 и др.) от
концентрации их в циркулирующей крови. Однако при изменении осмотического
давления и ионного состава крови эта зависимость компенсаторно изменяется в
результате изменения степени реабсорбции ионов Na* н СГ в слюнных протоках.
♦ ♦♦ Вода, поступающая из крови в слюну через межклеточные пространства ацинар-
ных клеток, пассивно сопровождает движение солей.
♦ ♦♦ Пассивным путем переносятся из крови в слюну липиды, мочевина, глюкоза; из-
менение концентрации их в слюне повторяет динамику концентрации их в крови.
♦ ♦♦ Белки проникают в слюну из крови путем пиноцитоза.
При дефиците Fe происходят изменения слизистой оболочки ротовой полости,
развивается атрофия слизистой языка, десен, щек.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3, 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [12], [13-16].
3.2. Дополнительная литература (см. с. 514 - 522): [3], [27], [30], [35], [42], [43],
[47], [107], [220], [222], [225], [238], [252-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/-nphys: Слайды дяя компьютерной демонстрации из «Атла-
192
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. -1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
ЭЛ. Консультант преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студ ентов будет осуществляться на практи-
ческом занятии но данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 507 - 513).
Задача 12-1. Пациент с длительным неполноценным питанием (частичным голоданием) пришел
на прием к врачу. При осмотре н пальпации у него были обнаружены отеки (задержка воды в ин-
терстициальном пространстве подкожной клетчатки) в области ног. Какие физико-химические
изменения крови могут быть при этом причиной отека, и почему он локализуется в области ног?
Задача 12-2. В организме человека артериальная кровь достоверно отличается на 0,04-0,06 еди-
ниц pH от венозной крови. Какая кровь имеет более низкую величину pH н почему?
193
Тема 13. Эритроцитарная и лейкоцитарная системы крови
Цель: а) знать эритроцитарную систему крови, строение, функции, количество эритроцитов и гемо-
глобина; физиологический эритроцитоз; нервную и гуморальную регуляцию эритропоэза; ви-
ды и количество лейкоцитов, их функции, лейкоцитарную формулу, физиологический лейко-
цитоз, причины и механизмы его развития; нервную и гуморальную регуляцию лейкопоэза;
иммунитет, иммунный ответ, его регуляцию, иммунитет как регуляторную систему; возрас-
тные изменения эритроцитарной и лейкоцитарной систем;
б) уметь применять эти знания для анализа процессов жизнедеятельности организма и оцени-
ки лабораторных показателей эритроцитарной и лейкоцитарной систем здорового человека;
в) иметь представление о методах исследования количества эритроцитов, содержания гемо-
глобина, цветового показателя крови, количества лейкоцитов и их процентного соотношения.
Мотивация темы: эритроцитарная система обеспечивает дыхательную функцию крови, участвует в регу-
ляции КОС и осуществлении деятельности всех физиологических систем организма. Лейкоци-
тарная система обеспечивает защитную функцию крови - иммунный ответ в здоровом и боль-
ном организме. Реакции эритроцитарной и лейкоцитарной систем являются важным механиз-
мом адаптации организма к различным условиям среды. Нарушение функций эритроцитарной
и лейкоцитарной систем приводит к патологическим процессам и заболеваниям системы кро-
ви (анемии, лейкоцитозы и лейкопении, лейкозы и др.).
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции.
1. Эритроцитарная система.
1.1. Эритроцитарная система - физиологическая система, состоящая из эритро-
цитов циркулирующей крови, органов их образования и разрушения, объеди-
ненных в систему аппаратом нейрогуморальиой регуляции. (Более узким понятием
является эритрон, предложенный У. Каслом (19290, под которым понимается совокупность
образующихся, циркулирующих и депонированных эритроцитов.)
1.2. Образование эритроцитов (~ 2 *10п в сутки, резерв увеличения до 8 раз).
❖ Место эритропоэза: миелоидная ткань (~ 1,8 кг) костного мозга.
♦♦♦ Схема эритропоэза: стволовая клетка крови (СКК) —» полипотентный предшественник
миелопоэза (КОК-ГЭММ) —> взрывообразующая единица эритропоэза (БОЕ-Э) —> унипо-
тентный предшественник эритропоэза (КОК-Э) —> проэритробласт —»базофильный, поли-
хромвтофильиый, оксифильный эритробласты —> ретикулоцит —> эритроцит. Из проэрит-
робласта образуется 16-32 эритроцитов. Время образования эритроцитов ~ 6 суток.
1.3. Эритроциты циркулирующей крови.
❖ Особенности структуры эритроцитов.
• Размеры: d ~ 8 мкм, толщина - 2,2 мкм, S - 145 мкм2, V - 85 мкм3
• Форма двояковогнутого диска дает иаилучшее отношение S/V (в 1,7 раза
больше, чем в шаре), оиа поддерживается субмембранными белками (спек-
трином, актином и др.), обеспечивает высокую эластичность эритроцитов.
❖ Основные функции эритроцитов.
• Транспортная.
194
Ф Транспорт Ог осуществляется в результате присоединения 4-х мо-
лекул Ог к молекуле гемоглобина. (Главной особенностью гемоглоби-
на является способность присоединять и отдавать Ог в узком диапазоне
его парциального давления 0 - 100 мм рт.ст.)
Транспорт СОг: поступивший в эритроцит СОг с помощью карбо-
ангидразы превращается в угольную кислоту, Н+ которой акцепти-
руется гемоглобином, а гидрокарбонат с помощью иоиообменника
СГ/НСОз" переносится в плазму в обмен на СГ.
Транспорт биологически активных веществ, например, гормонов
(суммарная S эритроцитов крови - 3500 м2).
• Регуляция кислотно-осиовиого состояния: гемоглобиновый буфер (со-
ставляет 35 % буферной емкости крови), продукция НСО'з при связывании
гемоглобином протона угольной кислоты.
• Защитная: перенос иа поверхности эритроцитов иммуноглобулинов,
компонентов системы комплемента, иммунных комплексов; участие в
гемостазе.
Функциональная характеристика гемоглобина («Теперь вся сила в гемоглобине.
Сказав это, «Пьер и Константин» умолк. Замолчали и горожане, каждый по-своему раз-
мышляя о таинственных силах гемоглобина». И. Ильф, Е. Петров).
• Внутриэритроцитариая локализация гемоглобина обеспечивает умень-
шение онкотического давления и в капиллярах - вязкости крови, отсут-
ствие фильтрации гемоглобина в почках (при полном гемолизе вязкость
плазмы увеличивается в 2 раза - до 3,1 сП).
• Состав молекулы гемоглобина: 4 гема (4Fe+2 + 4 протопорфирииа IX) +
глобин (4 пептидных цепи).
• Виды гемоглобина в зависимости от состояния гема и глобина.
Ф Оксигемоглобин (НЬОг) - присоединивший 4 молекулы О2 (в арте-
риальной крови - 98 %, в венозной - 65 %). Присоединение первой
молекулы Ог, увеличивает сродство ко второй Ог и т.д.
•ф Дезоксигемоглобин (ННЬ) - гемоглобин, отдавший кислород (в ар-
териальной крови - 2 %, в венозной - 35 %).
Ф Метгемоглобин (MetHb) - имеет окисленные атомы железа (Fe3+),
не присоединяющие Оз (в норме < 3 %). При этом в оставшемся
«нормальном» НЬА кривая диссоциации смещена влево, что за-
трудняет отдачу Оз.. (Метгемоглобииредуктаза эритроцитов восстанавливает
MetHb в НЬ с использованием АТФ.)
Ф Карбгемоглобин (НЬСОг) - присоединивший СОг к глобину (нахо-
дится в венозной крови).
Ф Карбоксигемоглобин (НЬСО) - СО имеет сродство к НЬ в 240 раз
выше, чем О2, и блокирует присоединение Ог. (Например, если в кро-
ви Рсо=0,5 мм рт.ст., то 50% гемоглобина будет связано с СО.)
Ф В зависимости от полипептцдного состава глобина: HbAi (2а , 20)
- 97 % всего НЬ; НЬА2 (2<х, 28) - 2 %; HbF (2а, 2у) -1 %, имеет наи-
большее сродство к Ог.
195
• Функции гемоглобина.
ф- Транспорт Ог: 1 молекула НЬ присоединяет 4 молекулы О2; 1 г НЬ
связывает 1,34 МЛ Ог. (Присоединение кислорода к НЬ сопровождается коо-
перативным эффектом: по мере присоединения молекул Ог увеличивается срод-
ство гема к кислороду - при присоединении последней молекулы в 100 раз боль-
ше, чем первой.)
ф’ Транспорт СО2: в тканях СО2 присоединяется карбаминовой связью
к глобину, так переносится -10 % СОг к легким.
• Виутриэритроцитарная регуляция сродства НЬ к Ог..
•Ф’ Накопление в эритроците промежуточного продукта гликолиза
2,3-дифосфоглицерата уменьшает сродство НЬ к кислороду, стаби-
лизируя дезоксигенированное состояние гемоглобина.
*Ф* Величина сродства НЬ к кислороду определяется тирозином Р-цепн (в 41-м по-
ложении). (Если он заменен на фенилаланин, то сродство уменьшается в 2 раза).
❖ Количественная характеристика эритроцитов крови (см. 1.6).
1.4. Разрушение эритроцитов.
❖ Продолжительность жизни ~ 90 дней.
❖ Места разрушения эритроцитов в организме.
• Неэффективный эритропоэз в миелоидной ткани (разрушается макро-
фагами до 10 % образующихся эритроцитов).
• Внутриклеточный гемолиз осуществляется мононуклеарной системой
фагоцитов селезенки, печени, костного мозга (~ 85 % циркулирующих
эритроцитов).
• Внутрисосудистый гемолиз (- 15 % циркулирующих эритроцитов). (Не-
большое количество плазменного НЬ = 3 - 10 мг/л связывается белком плазмы гаптог-
лобином и затем поглощается печенью.)
❖ Механизмы разрушения эритроцитов:
• Осмотический механизм: снижение активности К+, Na+-nacoca —> на-
копление Na+ и Н2О в клетке —> образование сферической формы и за-
стревание эритроцитов в синусоидных капиллярах селезенки —> фагоци-
тоз.
• Иммунный механизм: в мембране старых эритроцитов экспрессируется
белок полосы Ш («метка» для их уничтожения), аутоантитела к нему
служат мостиком, присоединяющим старый эритроцит к фагоциту. За-
тем старый эритроцит уничтожается путем фагоцитоза.
• Небольшая часть старых эритроцитов (до 15 %) гемолизируется в кро-
вотоке в результате осмотического шока и действия цитолитического
комплекса (С!ь.,Сб-С9) системы комплемента, образование которого акти-
вируется присоединением аутоантитела к эритроциту.
Ф Превращение гемоглобина в организме.
• Отщепление гема и образование свободного билирубина в клетках мо-
нонуклеарной фагоцитарной системы и выделение его в кровь, превра-
щение в печени свободного билирубина в связанный и выделение его в
желчный капилляр.
196
• Гидролиз глобина лизосомнымн протеазами до аминокислот.
• Освобождающиеся прн распаде гемоглобина ионы железа вновь (на
95 %) используются прн образовании эритроцитов.
1.5. Нейрогуморальная регуляция эритроцитарной системы.
Эритробластический островок костного мозга образован макрофагом и прилегающими к
его отросткам дифференцирующимися эритроидными клетками. Макрофаг обеспечивает
эритроидные клетки ферритином, эритропоэтином, пластическими веществами, фагоцити-
рует вытолкнутые ядра. По мере созревания эритробласты передвигаются по длине отро-
стка в направлении венозного синуса. В кровь выходят ретикулоциты и эритроциты.
Роль цитокинов (вырабатываются клетками стромы, макрофагами, лимфоцита-
ми, действуют как факторы роста и дифференцировки клеток). На ранних этапах
(СКК —» ...—» КОК-Э) стимулируют эритропоэз фактор СКК, ИЛ-1, ИЛ-3,
ИЛ-6, ИЛ-9, КСФ-ГМ н др. На следующих этапах (от КОК-Э до эритроци-
та) основным регулятором является эритропоэтин. Тормозит эритропоэз
ИЛ-8, эритроцитарный кейлон, ингибитор эритропоэза.
❖ Роль эритропоэтина (ЭП, гликопротеин - основной гормон, контроли-
рующий интенсивность эритропоэза).
• Основные места синтеза: почки - 85 - 90 % , макрофаги печени и кост-
ного мозга.
• Клеткн-«мишени» для ЭП: все клетки красного ростка от КОК-Э до
эритроцитов.
• Основной пусковой фактор продукции ЭП: гипоксия (рассогласование
доставки и потребления Оз).
• Механизм действия: вторым посредником является цАМФ, изменение
концентрации которой в ЭП-чувствительных клетках стимулирует син-
тез НЬ (созревание) и пролиферацию эритроидных клеток.
❖ Железо является лимитирующим фактором эритропоэза (необходимо для
синтеза гема).
• Суточная потребность в железе для эритропоэза 20 - 25 мг, 95 % этого
количества организм получает из разрушенных эритроцитов, а 5 % (су-
точная потребность в экзогенном железе у мужчин - 1 мг, у женщин в
связи с менструальным циклом - 2,8 мг) всасывается в тонком кишеч-
нике (резерв повышения всасывания небольшой - а 1;5 - 2 раза, поэтому лаже не-
большая хроническая кровопотеря приводит к железодефицитной анемии).
• Всосавшееся железо в крови связывается с белком трансферрином,
окисляется до Fe3+ церрулоплазмином и в такой форме транспортирует-
ся в макрофаги костного мозга и печени, где депонируется в виде фер-
ритина и гемосидерина (большие нерастворимые конгломераты ферритина,
образованные в лизосомах). (В организме человека 3-5 г Fe, в эритроцитах
содержится 65% этого количества.)
❖ Роль витаминов: Вн и фолиевая кислота (В<>) необходимы для синтеза нук-
. леиновых кислот в эритроидных клетках; С и Bj влияют на синтез гема; Вг
и пантотеновая кислота (Вз) участвуют в образовании липидного бислоя
плазмолеммы эритроцита. Витамины Е, С и РР защищают эритроциты от
перекисного окисления.
197
❖ Роль гормонов: активируют эритропоэз адреналин, глюкокортикоиды, ти-
роксин, тестостерон; ингибируют - эстрогены.
•> Роль нервной системы в регуляции динамического равновесия образования
и разрушения эритроцитов.
• Рецепторы каротидного синуса и аортальной зоны как кислородные
сенсоры.
• Центральное звено - гипоталамус, вегетативные центры (при юаимодей-
ствии с высшими центрами).
• Эфферентное звено - симпатический отдел нервной системы оказыва-
ет: 1) непосредственное влияние на красный росток костного мозга (на
плазмолемме его клеток имеются ^-адренорецепторы); 2) на почки (выработка эри-
тропоэтина); 3) иа выработку гормонов, регулирующих эритропоэз: адре-
налин мозгового слоя надпочечников, тироидные гормоны щитовид-
ной железы.
1.6. Методы исследования эритроцитарной системы.
❖ Методика подсчета эритроцитов в крови. Принцип методики: эритроциты кровн,
разведенные в слабом гипертоническом илн изотоническом растворе NaCl, счи-
тают в пяти больших (80-тн малых) квадратах счетной камеры и пересчитывают
на 1 л кровн, исходя из объема и разведения крови. Автоматический подсчет эрит-
роцитов с помощью гематологических анализаторов и счетчиков: большинство
приборов работает на кондуктометрическом методе - разведенная кровь пропус-
кается через микроотверстие прибора, проходящая через него клетка увеличивает
сопротивление между электродами, и возникающий импульс передается на счет-
ное устройство. Норма: муж.-4-5,Ь1012/л,жен.-3,7-4,710|2/л.
♦ t* Определение концентрации гемоглобина в крови (унифицированный метод). Прин-
цип метода: гемоглобин окисляется в метгемоглобин, который вступает в реакцию
с ацетонциангидрином н образует цианметтемоглобнн, интенсивность окраски ко-
торого, определяемого спектрофотометрическн, пропорциональна концентрации
гемоглобина. Норма: муж. - 140 - 160 г/л, жен. - 120 - 140 г/л. Концентрация ни-
же 110 г/л для любого возраста и пола считается анемией.
♦ ♦♦ Определение цветового показателя (ЦП). Принцип расчета: ЦП позволяет оце-
нить степень насыщения эритроцитов гемоглобином. При идеальном содержании
эритроцитов и гемоглобина в одном эритроците содержится 33 пг гемоглобина.
Величина 33 пг представляет собой нормальное содержание гемоглобина в эрит-
роците и условно принимается за единицу, что обозначается как ЦП. Практически
ЦП вычисляют делением показателя концентрации гемоглобина, выраженной в
г/л, на число из первых трех цифр количества эритроцитов в 1 л крови с после-
дующим умножением полученного результата на 3. В норме ЦП у взрослых равен
0,85 - 1,05; у новорожденных-0,9- 1,3.
2. Лейкоцитарная система.
2.1. Образование лейкоцитов.
♦♦♦ Места лейкопоэза в организме: миелоидная ткань костного мозга (образование грану-
лоцитов, моноцитов, В-лнмфоцитов и предшественников Т-лимфоцитов) и лимфоидная
ткань (дифференцировка и размножение В- и Т-лимфоцитов, плазмоцитов). 4
♦♦♦ Схема лейкопоэза.
• Гранулоцитопоэз: СКК —» КОК-ГММЭ —» бипотентные предшественники (нейтро-
фильно-эозниофильиый, гранулоцитарно-моноцитарный, гранулоцитарио-эритроци-
198
тарный) —> унипотентные предшественники нейтрофилов, базофилов и эозинофилов;
соответствующие миелобласты —> промиелоциты —>миелоциты —> метамиелоциты —>
палочкоядерные гранулоциты —> сегментоядерные гранулоциты (нейтрофилы, эози-
нофилы, базофилы).
• Моноцитопоэз: СКК —> полипотентный предшественник миелопоэза —> бипотентный
предшественник гранулоцитов и моноцитов —> унипотентный предшественник моно-
цитов —> монобласт —> промоноцит —>моноцит.
• Лимфопоэз: СКК —> полипотентный предшественник лимфопоэза (КОК лимф.) —>
предшественники Т- и В-лимфоцитов (пре-КОК-Т, пре-КОК-В) —> Т- и
В-лимфобласты —> Т- н В-лимфоциты.
2.2. Лейкоциты циркулирующей крови.
❖ Виды и функции лейкоцитов (их процентное соотношение составляет лей-
коцитарную формулу).
• Нейтрофилы (48 - 78 % от общего числа лейкоцитов): их зернистость содер-
жит вещества, обладающие высокой бактерицидной активностью (лизо-
цим, катионные белки, дефензины, лактоферрин, коллагеназу, миелопероксидазу, об-
разующую хлорноватистую кислоту, и др.); иа плазмолемме имеются рецепто-
ры к 1g, белкам комплемента, молекулам адгезии, цитокинам. Основные
функции нейтрофилов - фагоцитоз и уничтожение микробов и клеточ-
ных обломков; продукция факторов хемотаксиса; гранулоцитарного и
моноцитарного КСФ, ИЛ-6 (интерферона р2) и др. Фагоцитоз сопровожда-
ется резким повышением нейтрофилами потребления Оз («респираторный
взрыв»), до 90% которого идет на продукцию свободных радикалов ки-
слорода, повреждающих объекты фагоцитоза.
• Базофилы (0 - 1,0 %): «фармакологические бомбы» - синтезируют и де-
понируют в гранулах биоактивные вещества (гепарин, гистамин, лейкотрие-
ны, фактор хемотаксиса эозинофилов и др.); на плазмолемме имеют Рз—
адренорецепторы, рецепторы к IgE, стимуляция которых вызывает де-
грануляцию базофилов; участвуют в регуляции агрегатного состояния
крови, тонуса и проницаемости сосудов, тонуса бронхов; стимулируют
синтез IgE, участвуют в аллергических реакциях (например, анафилактиче-
ский шок, бронхиальная астма).
• Эозинофилы (0,5 - 5,0 %): гранулы содержат антипаразитарный щелочной
белок, простагландины, лейкотриены, гистаминазу, ингибитор деграну-
ляции тучных клеток и базофилов; на мембране имеются рз~
адреиорецепторы, рецепторы к IgE, IgG, IgM и белкам комплемента (на-
пример, С3, и С31); эозинофилы участвуют в уничтожении гельминтов (ас-
карид и др.), тормозят функцию базофилов.
• Лимфоциты (20 - 35 %): обеспечивают гуморальный и клеточный имму-
нитет; регулируют деятельность клеток других типов в иммунных реак-
циях, процессах пролиферации и регенерации тканей, дифференцировки
клеток, секретируют цитокины (подробнее см. ниже п.З).
• Моноциты (3-11 %): фагоцитоз в кислой среде (обеспечение иеспеци-
фической защиты организма против микробов, опухолевых клеток и за-
раженных вирусами клеток); участие в иммунных реакциях (в качестве
антигенпредставляющих и эффекторных клеток); синтез цитокинов, не-
199
которых компонентов системы комплемента, факторов, принимающих
участие в гемостазе.
❖ Циркулирующий и пристеночный пулы лейкоцитов: миграция лейкоцитов
из сосудистого русла в периферические ткани особенно активно происхо-
дит на уровне посткапиллярных венул; оиа невелика у гранулоцитов, силь-
но выражена у моноцитов и лимфоцитов. Миграция происходит в несколь-
ко стадий: лейкоциты адгезируются на поверхности эндотелия, мигрируют
за пределы сосуда за счет сократительной активности цитоскелета; направ-
ленность миграции обусловлена факторами хемотаксиса.
❖ Количественная характеристика лейкоцитов (см. ниже 2.5).
❖ Физиологические лейкоцитозы (увеличение количества лейкоцитов крови
выше 9*10’/л).
• Виды: пищевой, миогенный, эмоциональный, при беременности.
• Механизмы: 1) ведущим является перераспределительный механизм -
мобилизация лейкоцитов краевого сосудистого пула и зрелого пула ко-
стного мозга; 2) может присоединяться продукционный механизм (на-
пример, при миогеином лейкоцитозе).
• Отличие от реактивных (продукционных) лейкоцитозов: небольшое
увеличение лейкоцитов в крови (до 1,5 раз), отсутствие сдвига влево в
нейтрофильном ряду.
Лейкопения (снижение содержания лейкоцитов ниже 4*10%; наблюдается, как правило,
при патологических состояниях).
23. Разрушение лейкоцитов.
♦> Продолжительность жизни лейкоцитов: гранулоциты (погибают после един-
ственной вспышки активности), моноциты н короткоживущие лимфоциты - от
нескольких часов до 5 - 10 суток; долгоживущие лимфоциты - от несколь-
ких месяцев до 5 лет.
Разрушаются лейкоциты в тканях, куда они мигрировали.
2.4. Нейрогуморальная регуляция лейкоцитарной системы.
& Влияние цитокинов.
• Трансформацию СКК в полипотентный (КОК-ГММЭ) н бипотентные
предшественники миелопоэза стимулируют фактор СКК, ИЛ-1, ИЛ-3
(или мульти-КСФ), ИЛ-6, ИЛ-11, КСФ-ГМ, тормозят - ИЛ-8, ИЛ-10, лей-
коцитарные кейлоны.
• Образование монопотентных предшественников и более дифференци-
рованных клеток активируют соответствующие колониестимулирую-
щие факторы (КСФ) и другие факторы:
Ф образование нейтрофилов активирует КСФ-Г, ИЛ-8, продукты рас-
пада нейтрофилов; тормозит - нзоферритин, выделяющийся из зре-
лых нейтрофилов;
ф* образование эозинофилов активирует КСФэоз. (или ИЛ-5);
Ф- образование базофилов (и тучных клеток) активирует КСФбаз.;
ф" образование моноцитов активирует КСФмон., тормозит выделяю-
щийся из моноцитов простагландин Е.
200
• Трансформацию СКК в предшественник Т-лимфоцитов и в дифферен-
цированные Т-лимфоциты стимулируют ИЛ-1, ИЛ-2, ИЛ-4, ИЛ-5, ИЛ-9,
ИЛ-12, гормоны тимуса и др; тормозит - ТРФр.
• Трансформацию СКК в предшественник В-лимфоцитов и в дифферен-
цированные В-лимфоциты стимулируют ИЛ-1, ИЛ-2, ИЛ-4, ИЛ-6, ИЛ-7,
ИЛ-11 и др.; тормозит - трансформирующий ростковый фактор ТРФр.
❖ Влияние гормонов на лейкопоэз.
• Глюкокортикоиды стимулируют пролиферацию нейтрофилов, тормозят
образование эозинофилов и лимфоцитов, выход моноцитов из костного
мозга.
• Адреналин стимулирует пролиферацию моноцитов и выход нейтрофи-
лов из костного мозга в кровь.
• Тимозин, СТГ, окситоцин стимулируют пролиферацию Т-лимфоцитов.
❖ Нервная регуляция лейкопоэза.
• Рецепторное звено - сосудистые и тканевые хеморецепторы.
• Центральное звено - гипоталамус и вегетативные центры при взаимо-
действии с высшими центрами.
• Эфферентное звено - вегетативная нервная система:
"Ф- симпатические влияния и адреналин через Рг-адренорецепторы
(нейтрофилез, перераспределительный лейкоцитоз);
-ф- парасимпатические влияния (лимфоцитоз, эозинопения, перерас-
пределительная лейкопения).
2.5. Методики исследования лейкоцитов.
••• Методика подсчета лейкоцитов. Кровь для разведения и лизиса эритроцитов
смешивают с раствором уксусной кислоты. Лейкоциты считают по всей сетке в
100 больших (не разделенных на малые) квадратах счетной камеры и пересчиты-
вают на 1 л крови, исходя из объема и разведения крови. Автоматический подсчет
лейкоцитов проводят с помощью гематологических анализаторов и счетчиков:
разведенная кровь пропускается через микроотверстие прибора, проходящая через
него клетка увеличивает сопротивление между электродами, и возникающий им-
пульс передается иа счетное устройство. Норма: количество лейкоцитов крови
равно4-9-10’/л.
❖ Лейкоцитарная формула определяется путем последовательного подсчета всех
видов лейкоцитов (считают 200 лейкоцитов в мазке крови) с выявлением их про-
центного соотношения (норма см. выше 2.2). В лейкоцитарной формуле можно
определить ядерный индекс (ЯИ) в нейтрофильном ряду: Метамиелоциты + Па-
лочки)/Сегменты, который равен 0,05 - 0,08; сдвиг влево (ЯИ > 0,08) свидетельст-
вует об увеличении молодых форм нейтрофилов, вправо (< 0,05) - старых форм.
3. Физиологическая система иммунного ответа обеспечивает сохранения антигенно-
го гомеостаза организма и участвует в регуляции других, неиммунных функций ор-
ганизма.
3.1. Физиологические функции иммунной системы: 1) генетический контроль, т.е.
уничтожение генетически измененных (мутантных) клеток; 2) защита от сапро-
фитов (в крови всегда имеются антитела к ним); 3) регуляция функций орга-
низма (подробно ниже в 3.6).
201
3.2. Органы иммунного ответа.
* t* Центральные органы (в них происходит преимущественно антигеннезависимая фаза
развития лимфоцитов).
• Костный мозг - из стволовых клеток образуются В-лимфоциты, моно-
циты (макрофаги) и предшественники Т-лимфоцитов.
• Тимус - предшественники Т-лимфоцитов превращаются в Т-
лимфоциты. Гематотимнческнй барьер в корковой части тимуса защи-
щает развивающиеся Т-лимфоциты от антигенов нз внутренней среды
организма. В дальнейшем при переходе в мозговую часть тимуса
Т-лимфоциты, способные распознавать чужеродный антиген в комплек-
се с собственными белками главного комплекса гистосовместимости
(ГКГС), остаются в организме. Т-лимфоциты, не способные реагировать
с собственными белками ГКГС («бесполезные» лимфоциты) н способные
взаимодействовать с собственными антигенами в комплексе со своими
белками ГКГС («агрессивные» лимфоциты) уничтожаются с помощью
апоптоза и макрофагов в мозговой части тимуса.
Периферические органы (в них происходит аитигензависимая фаза развития лимфо-
цитов).
• Селезенка (фильтр в системе кровообращения) — здесь происходит распозна-
вание попавшего через кровеносное русло антигена рецепторами Т-, В-
лимфоцитов, их активация, пролиферация и дифференцировка, секреция
цитокинов, продукция 1g (антител).
• Лимфатические узлы (фильтр в системе лимфообращения) - здесь происхо-
дит распознавание поступившего по лимфатическим путям антигена ре-
цепторами Т- и В-лимфоцнтов; их активация, пролиферация, дифферен-
цировка, секреция цитокинов, продукция 1g.
• Ассоциированная со слизистыми оболочками и кожей лимфоидная
ткань (миндалины, пейеровы бляшки, аппендикс и др.) - содержит
В-лнмфоциты, осуществляющие образование IgA, секретируемых на
поверхность слизистой и кожи, и Т-лимфоцнты, осуществляющие им-
мунный надзор за зараженными и мутантными клетками.
3.3. Иммунокомпетентные клетки.
❖ Антиген-представляющие клетки (АПК) представляют антиген иммунной
системе.
• «Профессиональные» АПК: макрофаги, дендритные АПК кожи и сли-
зистых (преобразованные моноциты), В-лимфоциты (содержат главный ком-
плекс гистосовместимости П класса - ГКГС-П).
• «Непрофессиональные» АПК - это все клетки организма (содержат глав-
ный комплекс гистосовместимости I класса - ГКГС-1).
• Роль ГКГС в иммунном ответе заключается в том, что иммунокомпетентные
клетки могут распознавать большинство чужеродных антигенов только в том
случае, если они представлены в комплексе с ГКГС.
202
• Обработка (процессинг) и представление антигена являются сложными
процессами, протекающими с участием многих ферментов и регулятор-
ных белков.
❖ Лимфоциты.
• В-лимфоциты (составляют 10 - 20 %) участвуют в гуморальном имму-
нитете, дифференцируются в плазматические клетки, секретирующие
иммуноглобулины; часть В-лимфоцитов при иммунных реакциях пре-
вращаются в В-клетки памяти.
• Т-лимфоциты (70 - 80 %, поверхностный маркер CD3).
ф’ Т-хелперы (Тх, поверхностный маркер CD4):
• Тх; образуют цитокины ИЛ-2, у-интерферон, стимулируя клеточ-
ный иммунитет;
» Тхз образуют цитокины ИЛ-4, ИЛ-5, ИЛ-10, ИЛ-13, стимулируя
гуморальный иммунитет.
• ф Т-супрессоры (Тс) предположительно угнетают активность
В-лимфоцитов, Т-киллеров и Тх.
ф- Т-киллеры (Тк, поверхностный маркер CD8) - цитотоксические лимфо-
циты, являются эффекторными клетками в клеточном иммунитете
(распознают антиген в комплексе с ГКГСI).
ф- Т-клегки памяти (Тп) содержат информацию о встрече иммунной
системы с чужеродным антигеном.
• Нулевые лимфоциты (5 - 10 %): к ним относятся NK-клетки (на-
туральные киллеры), участвующие в противоопухолевом иммунитете, а
также предшественники Т- и В-лимфоцитов.
3.4. Иммунный ответ (формируется с преобладанием клеточного или гуморально-
го иммунитера).
❖ Клеточный иммунитет.
• Образуется в ответ на появление в организме внутриклеточных антиге-
нов в любой клетке (опухолевые белки и белки внутриклеточных паразитов - ви-
русов, микобактерий туберкулеза, грибов, простейших).
• Антиген расщепляется в цитоплазме до пептидов (8-13 аминокислот) и
транспортируется в гладкую ЭПС, где связывается с молекулами ГКГС-
I и поступает на поверхиость плазмолеммы для представлеиия Т-
лимфоцитам.
• Антигенный комплекс на плазмолемме распознается цитотоксическими
лимфоцитами - Т-киллерами и натуральными киллерами (этому предшест-
вует образование и активация Txi).
• Активированные Т-киллеры размножаются и поражают антигенизме-
ненную клетку:
ф- контактным путем, присоединяясь к клетке-«мишени» с помощью
молекул адгезии, и секретируют белок перфорин, который в мем-
бране «мишени» образует поры (d -10 им); через эти поры входят
Na+ и вода, что приводит к осмотическому шоку и гибели клетки
203
(через поры входят также сериновые протеазы - гранзимы, запус-
кающие программу гибели клетки);
ф" гуморальным путем, секретируя токсические медиаторы (фактор некроза опухо-
лей, интерферон у, интерлейкин-1, радикалы О2 н др.), повреждающие клетки-
-«мишени».
❖ Гуморальный иммунитет.
• Образуется в ответ на появление в организме внеклеточных (экзоген-
ных) антигенов (например, внеклеточно паразитирующих бактерий - стафилокок-
ков, стрептококков, кишечной палочки и др., инъекции белковых препаратов).
• Антигены поступают в профессиональные АПК (макрофаги, В-лимфоциты,
дендритные АПК), проходят обработку в их эндосомах и (в комплексе с бел-
ками ГКГС-П) поступают на поверхность плазмолеммы.
• Антигенный комплекс распознается Тхз, которые стимулируют проли-
ферацию В-лимфоцитов.
• Активированные В-лимфоциты поступают в лимфоидную ткань (особен-
но тех зон, которые послужили входными воротами для антигена), где превраща-
ются в плазматические клетки, секретирующие 1g. В молекуле 1g име-
ются Fab-фрагменты, связывающие их с антигеном, и Fc-фрагмент, с
помощью которого 1g может присоединиться к нейтрофилу, макрофагу,
лимфоциту, фактору комплемента.
• Имеются 1g следующих пяти типов:
Ф" IgM (8 - 10 % всех 1g) - полиреактивные естественные антитела, первая линия
быстрой защиты, главный класс 1g иа ранних стадиях первичного иммунного от-
вета, а также у плода, активируют систему комплемента.
Ф" IgG (~ 75 %) - вырабатываются в основном при вторичном иммунном ответе
(защищают от бактерий, вирусоа, токсинов), главный опсонизирующий 1g при
фагоцитозе, активируют систему комплемента, единственные 1g, которые прохо-
дят плацентарный барьер.
Ф" IgA (15-20 %) имеются в двух формах: в плазме и секретах экзокринных желез
(основное количество в слезной жидкости); блокируют прикрепление бактерий к
слизистой оболочке, опсонируют микробы.
Ф’ IgE (- 0,004 %) - вырабатываются в слизистых пищеварительного и дыхательно-
го путей, вызывают дегрануляцию тучных клеток, защищают от гельминтов, ак-
тивируют систему комплемента.
Ф’ IgD (<!%)- активируют систему комплемента.
❖ Формирование иммунологической памяти - это образование долгоживущих
лимфоцитов (Т- и В-клеток памяти) после контакта организма с чужерод-
ными антигенами, что позволяет иммунной системе более быстро и сильно
отвечать иа повторный контакт с этим же антигеном.
❖ Иммунологическая толерантность характеризует способность иммунной
системы не реагировать на антигены собственного организма. Лимфоциты,
которые реагируют иа эти антигены в комплексе с ГКГС, т.е. потенциально
аутореактивные Т-лимфоциты, гибнут во время внутриутробного развития
плода.
3.5. Нейрогуморальная регуляция иммунного ответа.
❖ Нервная регуляция.
204
• Афферентное звено: допускается возможность стимуляции рецепторов
продуктами, выделяющимися из клеток иммунной системы - цитоки-
нами, цАМФ, Са2+.
• Центральное звено: гипоталамус и вегетативные центры (при взаимодей-
ствии с высшими отделами ЦНС).
• Эфферентное звено:
• Ф’ на лимфоцитах и макрофагах имеются а- и р-адреиорецепторы, М-
холинорецепторы, серотониновые и опиатные рецепторы и др.;
• ф- симпатическая нервная система действует через а-
адреиорецепторы (преобладают на Тс) и p-адренорецепторы (пре-
обладают на Тх), оказывая как стимулирующий, так и ингибирую-
щий эффекты иа иммунитет (при симпатэктомии снижается образование
ig);
• ф- парасимпатическая нервная система, действуя через М-
холинорецепторы, стимулирует синтез Ig (особенно IgG), пролифе-
рацию лимфоцитов (однако тормозит исходно активированную пролифера-
цию).
❖ Эндокринная регуляция.
• Гормоны тимуса (тимозин и тимопоэтин) вызывают дифференцировку
предшественников Т-лимфоцитов в иммунокомпетентные клетки.
• Глюкокортикоиды в физиологическом диапазоне концентраций пре-
имущественно ингибируют Т-супрессоры и таким образом стимулиру-
ют иммунные процессы (например, синтез антител); высокие концентрации
тормозят пролиферацию и функции лимфоцитов (особенно Тх), макро-
фагов-моноцитов (образование в них ИЛ-1 и ФНОа) и натуральных киллеров.
• Гормоны щитовидной железы (Т4, Тз, кальцитонин) и мелатонин эпифиза
стимулируют В-лимфоциты и образование Ig.
• Половые гормоны в высоких дозах тормозят (а в низких - стимулируют)
функциональную активность лимфоцитов.
• СТГ стимулирует продукцию антител и ИЛ-2, активирует макрофаги и
натуральные киллеры, через тимус стимулирует образование Т-
лимфоцитов.
• Окситоцин увеличивает пролиферацию Т-лимфоцитов.
• Пролактин активирует макрофаги, повышает образование ИЛ-2.
3.6. Иммунитет как регулирующая система.
❖ Регуляция деятельности нервной системы.
• Влияние на синаптическую передачу. На нейронах коры, мозжечка, гип-
покампа обнаружены рецепторы к цитокинам. Интерлейкины 1р, 2, 6
могут непосредственно регулировать выход медиаторов и эффектив-
ность их рецепторов. Например, ИЛ-1р подавляет синтез и выход глу-
тамата, но повышает активность ГАМКд-рецепторов.
• Влияние на формирование памяти: ИЛ-1р и ИЛ-6 подавляют долго-
временную потенциацию в гиппокампе и консолидацию памяти.
205
❖ Участие в эндокринной регуляции организма.
• Иммунокомпетентные клетки способны образовывать различные гор-
моны: АКТГ, ТТГ, СТГ, вазопрессин, окситоцин, пролактин, вазоактив-
ный интестинальный полипептид.
• Цитокины иммунной системы влияют на образование гормонов в орга-
низме (например, а- и Р-интерфероны стимулируют синтез глюкокор-
тикоидов, тироксина, меланина); ИЛ-1 стимулирует гипоталамо-
гипофизарную ось (синтез кортиколиберина и АКТГ) и участвует в процессах
адаптации организма.
❖ Влияние на систему крови.
• Участие в балансе образования и разрушения клеток крови.
"О’ Цитокины, выделяемые иммунокомпетентными клетками, участ-
вуют в активации гемопоэза. Например, ИЛ-3 стимулирует образо-
вание гранулоцитов, моноцитов и эритроцитов, ИЛ-5 - эозинофи-
лов, ИЛ-8 - нейтрофилов.
"О’ Иммунные механизмы участвуют в разрушении клеток крови в здо-
ровом организме. Например, экспрессированный белок полосы Ш
мембраны старых эритроцитов является антигеном и «меткой» для
их аутоиммунного разрушения.
• Фактор некроза опухолей усиливает образование тромбина и снижает
фибринолиз, что способствует образованию фибринового тромба (на-
пример, в очаге воспаления).
❖ Влияние на терморегуляцию организма: цитокины (ИЛ-1, ИЛ-6, ФНОа),
выделяемые иммунокомпетентными клетками, стимулируют «холодовые»
нейроны гипоталамуса и увеличивают теплопродукцию в организме.
Влияние на обмен веществ: ФНОа тормозит образование ферментов синте-
за жира и препятствует образованию липидов в адипоцитах жировой ткани,
стимулирует распад гликогена и продукцию лактата в мышечной ткани,
лейкоцитарной системы.
4. Профильный материал для студентов лечебного факультета. Изменения гемопо-
эза при старении организма.
4.1. Этапы антенатального кроветворения (см. тему 12; задание 1: 8.1).
4.2. Эритроцитарная и лейкоцитарная системы.
♦ ♦♦ Уменьшение общей массы красного костного мозга и его пролиферативной ак-
тивности.
Тенденция к уменьшению количества эритроцитов и гемоглобина крови.
❖ Физиологическая лейкопения старости, эозннопения.
❖ Уменьшение функционального резерва эритро- и лейкопоэза в экстремальных ус-
ловиях.
4.3. Система иммунитета.
❖ Гуморальный иммунный ответ у стариков на первичное введение антигена резко
снижен преимущественно за счет уменьшения образования IgG.
't* Клеточный иммунитет снижается меньше, чем гуморальный; отмечено уменьше-
ние цитолитической активности киллеров.
206
Ф Изменение неспецифических факторов иммунитета: снижается активность лизо-
цима крови, фагоцитирующая и переваривающая активность нейтрофилов.
5. Профильный материал для студентов педиатрического факультета.
5.1. Этапы антенатального кроветворения (см. тему 12; задание 1: 8.1).
5.2. Роль селезенки и лимфоидной ткани в антенатальном кроветворении.
Ф Роль селезенки. Гемопоэз в селезенке осуществляется с 11-й недели и продолжает-
ся после рождения. В ней осуществляются эритропоэз, гранулоцитопоэз, моноци-
топоэз и лимфопоэз, которые к концу 7-го мес. заметно ослабевают, кроме лим-
фопоэза. Лимфопоэз становится преобладающим, более 90 % лимфоцитов имеют
признаки Т- или В-лимфоцитов. Источником В-лимфоцитов и предшественников
Т-лимфоцитов являются печень и костный мозг.
Ф Роль тимуса. Лимфопоэз начинается с 10-й недели из стволовых гемопоэтических
клеток, поступающих в тимус из эмбриональной печени, а затем и из костного
мозга; образуются различные субпопуляцнн Т-лимфоцитов.
Ф Роль лимфатических узлов (нёбных миндалин, пейеровых бляшек). Лимфопоэз
начинается с 12-й недели и продолжается в постнатальном периоде. Здесь раз-
множаются В-лимфоциты, поступившие из печени и костного мозга, а также Т-
лимфоциты, поступившие из тимуса.
53. Особенности кроветворения у детей разного возраста.
Ф У новорожденных эритропоэз относительно в 5 раз выше, чем у взрослых.
Ф У детей раннего возраста кроветворение сначала происходит в костном мозге по-
всеместно, начиная с 4 лет происходит постепенное уменьшение количества мие-
лоидной ткаии относительно массы тела.
❖ В период полового созревания кроветворение сохраняется только в красном кост-
ном мозге губчатого вещества тел позвонков, ребер, грудины, костей голени и
бедренных костей.
Ф У детей раннего возрастаимеется функциональная лабильность кроветворного ап-
парата и возможность активации печеночного типа кроветворения.
5.4. Изменение количества и качества эритроцитов у плода, новорожденных и детей
грудного возраста.
Ф У плода происходит постепенное увеличение количества эритроцитов, уменьше-
ние их диаметра н объема, количества ядросодержащих клеток.
Ф У новорожденных наблюдается увеличение количества эритроцитов в 1-й день
(6, Ь 10,2/л), усиленное их разрушение на 2 - 3-й день (физиологическая желтуха);
резкое снижение количества эритроцитов в течение 1-го месяца до 4,74012/л, уко-
роченный срок жизни (12 дней), вариабельность размеров н формы (анизо- и пой-
килоцитоз).
Ф У детей грудного возраста происходит дальнейшее снижение количества эритро-
цитов на протяжении 1-го полугодия, затем постепенное увеличение до 4,24012/л.
5.5. Особенности гемоглобина плода и новорожденных и динамика его перехода в гемо-
глобин взрослых.
Ф Эмбриональный гемоглобин (НЪР) преобладает до 9 - 12 недель, замещается фе-
тальным гемоглобином (HbF) к 3-му мес. внутриутробного развития.
Ф Гемоглобин взрослых (НЬА) появляется в крови плода иа 4-м мес.; количество его
до 8 мес. не превышает 10 %.
Ф У новорожденных HbF составляет ~ 70 %, НЬА - 30 %.
5.6. Изменение лейкоцитов и лейкоцитарной формулы в онтогенезе.
207
Лейкоциты у плода появляются к концу 3-го месяца, происходит постепенное
уменьшение содержания молодых форм лейкоцитов и увеличение общего количе-
ства до 6 - 13»109/л к рождению.
❖ У новорожденных имеется физиологический лейкоцитоз - 11х109/л, много незре-
лых форм лейкоцитов, сдвиг ядерного индекса влево.
♦ ♦♦ У детей грудного возраста количество лейкоцитов - 9-109 /л, низкая двигательная
и фагоцитарная активность.
♦ ♦♦ Изменение лейкоцитарной формулы: одинаковое процентное содержанке нейтро-
филов и лимфоцитов имеется в 4 - 6-й дни после рождения (1-й перекрест кривых
их содержания) н в 4 - 6 лет (2-й перекрест); между перекрестами: на 2 - 3-мес.
лимфоциты достигают максимума (60 - 65 %), нейтрофилы - минимума (25 - 27
%), затем количество лимфоцитов уменьшается, а нейтрофилов увеличивается до
43 - 44 %; уменьшается сдвиг ядерного индекса влево.
❖ Развитие системы Т- и В-лимфоцитов: дифференцировка Т- и В-лнмфоцитов про-
исходит в конце внутриутробного развития и сразу после рождения; у новорож-
денных доля Т-лнмфоцитов от всех лимфоцитов больше, чем у взрослых; у детей
после 2 лет - такая же, как у взрослых
6. Профильный материал для студентов стоматологического факультета.
6.1. Взаимосвязь слизистой оболочки рта с органами кроветворения. Слизистая обо-
лочка рта формируется у эмбриона к 12-му дню н выполняет функцию кроветворного
органа. По мере развития плода эта функция переходит к печени, селезенке и костному
мозгу. Единство источника развития соединительной тканн н кроветворных органов
(мезодерма) объясняет скопление лейкоцитов в слизистой оболочке рта. Патологиче-
ские процессы в различных участках слизистой оболочки рта нередко являются пер-
вичными признаками поражения кроветворной системы.
6.2. Часть лимфоидной ткани локализуется в ротовой полости (лимфоидное кольцо) и
участвует в защитной функции слизистой оболочки полости рта и целостного организ-
ма.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3, 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [12], [14-16].
3.2. Дополнительная литература (см. с. 514- 522): [3], [5], [27], [30], [35], [43], [47],
[49], [54], [58], [71], [72], [77], [78], [81], [89], [95], [116], [135], [139], [151], [152],
[157], [174], [184], [220], [222], [225], [238], [250], [251], [253], [254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nDhvs: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором н слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
208
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 13-1. Перед вами анализы крови трех здоровых мужчин. Известно, что один из них при-
надлежит спортсмену, сдавшему кровь после интенсивной физической нагрузки, второй - взят у
человека через 1,5 часа после приема пищи, третий - у человека находящегося в состоянии фи-
зиологического покоя. Определите, какой анализ принадлежит каждому из испытуемых. Обос-
нуйте свой ответ.
Эритроциты (1012/л) Гемоглобин (г/л) Лейкоциты (Ю’/л)
1-й анализ: 4,7 140 11
2-й анализ: 5,7 175 12
3-й анализ 4,8 145 4,8
Задача 13-2. В анализе крови у людей высокогорных районов обнаруживают увеличение количе-
ства эритроцитов и гемоглобина как у тех, кто только что прибыл в этот район, так и тех, кто по-
стоянно живет в высокогорных условиях. У последних обычно увеличено и количество ретикуло-
цитов в мвзке крови. Объясните главный механизм увеличения эритроцитов и гемоглобина в том
и другом случае.
Задача 13-3. У человека с кровоточащим полипом толстого кишечника в течение года потеря же-
леза с гемоглобином превышала его всасывание в кишечнике, в результате чего произошло исто-
щение запасов депонированного в организме железа. Как изменится по сравнению с нормой
(больше - меньше) в его крови количество гемоглобина, эритроцитов, цветовой показатель и кон-
центрация эритропоэтина? Какой из двух показателей (эритроциты, гемоглобин) будет изменен в
меньшей степени?
Задача 13-4. При некоторых заболеваниях в тканях органи -ма вследствие дегрануляции тучных
клеток освобождается большое количество биологически активного вещества - гистамина. Лей-
коцитарная система включается в ликвидацию высокого уровня гистамина. За счет какого вида
лейкойкоцитов и как будет изменена лейкоцитарная формула при этом? Какой физиологический
смысл этой реакции?
209
Тема 14. Физиологическая система регуляции агрегатного состояния
крови. Группы крови
Цель: а) знать свертывающую и противосвертывающую системы крови как компоненты физиологи-
ческой системы поддержания ее жидкого состояния и способности к образованию тромба;
факторы, ускоряющие и замедляющие свертывание крови; регуляцию системы гемостаза, ее
возрастные изменения и клинико-физиологические показатели; группы крови (система АВО и
Rh), правила переливания крови, кровезамещающие растворы;
б) применять эти знания для понимания состояния и регуляции систем свертывания и проти-
восвертывания крови в организме, для анализа результатов исследования системы гемостаза и
групп крови;
в) приобрести умения в оценке состояния нормы: времени свертывания крови, кровотечения,
рекальцификации, протромбинового времени и индекса, а также в определении группы крови
по системе АВО и Rh.
Мотивация темы: жидкое состояние крови и способность к свертыванию при нарушении целостности
кровеносных сосудов являются необходимыми условиями жизни организма. В настоящее вре-
мя более 50 % людей умирают от болезней, в развитии которых важным звеном является на-
рушение свертывания крови. Исследования в области гемокоагуляции помогают ответить иа
вопросы, связанные с профилактикой, патогенезом и лечением атеросклероза, инфаркта мио-
карда, гипертонической болезни, инсульта, тромбоза, различных форм кровоточивости и дру-
гих заболеваний. Кровезамещающая терапия является важным компонентом лечения тяжелых
заболеваний; поэтому знания групп крови, правил переливания крови необходимы врачу лю-
бой специальности.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов» на стенде. Остальные фрагменты задания
1 могут быть использованы для коррекции материала записанной лекции.
1. Общая характеристика системы регуляции агрегатного состояния крови
(РАСК).
1.1. Система РАСК - физиологическая система, обеспечивающая жидкое состоя-
ние крови и способность ее образовывать тромб при нарушении сосудистой
стенки.
1.2. Система РАСК состоит из:
♦♦♦ свертывающей системы крови, включающей в себя: 1) сосудисто-
тромбоцитарный гемостаз; 2) коагуляционный гемостаз;
❖ противосвертывающей системы крови, включающей в себя: 1) антикоагу-
лянты; 2) фибринолиз.
1.3. Гемостатический потенциал (как общий, так и локальный) является показателем
баланса свертывающей и противосвертывающей систем; его виды: нейтраль-
ный - свертывающая система функционально уравновешена с противосверты-
вающей, положительный - преобладание свертывающей системы (риск тромбо-
за), отрицательный - преобладание противосвертывающей системы (риск крово-
течения).
210
Свертывание крови
Противосвертывение крови
Сосудисто-
тромбоцитар-
ный гемостаз
Коагуляци-
онный
гемостаз
Антикоагу-
лянты
Фибринолиз
3*
Гемостатический потенциал
Нейтральный, положительный, отрицательный.
Обший и локальный
Рис. 41. Схема системы регуляции агрегатного состояния крови.
2. Главными взаимосвязанными элементами свертывающей н противосверты-
вающей систем крови являются сосудистая стенка, тромбоциты и плазменные
факторы (материал носит справочный характер).
2.1. Роль сосудистой стенки. Эндотелий сосудистой стенки синтезирует и секретирует как тром-
богенные, так и антитромбогенные факторы.
Тромбогенные факторы: фактор Виллебрандта (транспортная субъединица фактора УШ
плазмы), тромбоцитарный ростовой фактор, ингибитор активатора плазминогена (ингиби-
рует фибринолиз), ангиотензин IV, эндотелии-1.
♦♦♦ Антитромбогенные факторы: оксид азота (NO) и простациклин (простагландин Ij)
уменьшают адгезию и агрегацию тромбоцитов; антитромбин Ш (плазменный кофактор ге-
парина) ингибирует тромбин (Па), IXa, Ха, Х1а; тканевой активатор плазминогена активи-
рует фибринолиз, тромбомодулин блокирует рецепторы дня тромбина на эндотелиоцитах.
2.2. Роль тромбоцитов. Тромбоциты выделяют факторы, обзначаемые арабскими цифрами и буквой
Р (от platelet - пластинка).
❖ Pj - акцелератор-глобулин (идентичен фактору V плазмы) связывается с мембранной
тромбоцита, служит рецептором для Ха, предотвращая его инактивацию антитромбином
III и поэтому ускоряет образование тромбина.
4» Pj - акцелератор тромбина, ускоряет превращение фибриногена в фибрин.
♦ ♦♦ Р3 - тромбоцитарный тромбопластин (фосфолипиды мембраны) активирует факторы свер-
тывания и защищает их от действия антитромбина Ш, как матрица обспечивает взаимо-
действие факторов свертывания.
♦ ♦♦ Р4 - антигепариновый фактор, способный быстро связываться с гепарином и инактивро-
вать его.
♦ ♦♦ Р5 - тромбоцитарный фибриноген соединяется с рецепторами мембраны тромбоцитов, вы-
зывая их агрегацию.
♦ ♦♦ Р« - тромбостении, комплекс сократительных белков, участвующих в активации тромбо-
цита (превращение из дисковидной в сферическую форму с длинными отростками), реак-
ции освобождения биоактивных веществ (экзоцитоз гранул тромбоцитов), ретракции
тромба.
Р? - тромбоцитарный антиплазмин тормозит фибринолиз.
♦ ♦♦ Р« - активатор фибринолиза.
♦ ♦♦ р9 - фибринстабилизирующий фактор, переводящий фибрин тромба в нерастворимую
форму (аналогичен плазменному ХШ).
• 5» Рм - серотонин (5-гидрокситриптамин) действует на рецепторы тромбоцита, стимулируя
реакцию освобождения, оказывает сосудосуживающее действие.
❖ Рц - АДФ выделяется в реакции освобождения и, действуя на рецепторы к АДФ, стиму-
лирует ее, вызывает необратимую агрегацию тромбоцитов.
211
Фактор Виллебраидта - присоединяется к рецепторам тромбоцитов, участвуя в их адге-
зии и агрегации, служит белком-переносчиком для фактора VIII плазмы.
❖ Тромбоксан Аг - продукт обмена арахидоновой кислоты, вызывает сильный спазм сосу-
дов и резко стимулирует агрегацию тромбоцитов.
♦♦♦ Фибронектин и тромбоспондин - белки, модулирующую адгезию и агрегацию тромбо-
цитов.
0-Тромбо глобулин - препятствует синтезу простациклина сосудистой стенкой.
23. Роль плазменных факторов. В плазме имеются как факторы свертывания, так и антитромбоген-
ные факторы - антикоагулянты и факторы фибринолиза.
♦ ♦♦ Факторы свертывания обозначены римскими цифрами по мере их открытия, активация
фактора обозначается добавлением буквы «а».
• I - фибриноген (димер из 6-ти полипептидов, образуется в печени). Под действием
тромбина (Па), расщепляющего пептидные связи Арг-Гли, от фибриногена отщепля-
ются четыре низкомолекулярных пептида и он превращается в фибрик-моиомер, спо-
собный к аутополимеризации за счет свободных связей.
• II - протромбин (сериновая протеаза, образуется в печени при участие вит.К). Под
действием протромбииазы от него отщепляются а-, 0- и у-тромбины (Па): а-тромбии
очень активен, ио быстро нейтрализуется антитромбином Ш; 0-тромбии менее акти-
вен, но устойчив к действию гепарина и антитромбина Ш; у-тромбин не имеет сверты-
вающей активности, ио обладает фибринолитическим действием.
• III - тканевой тромбопластин (апопротеин 1П в комплексе с фосфолипидами, вхо-
дит в состав мембран клеток многих тканей; эритроциты, лейкоциты и тромбоциты
содержат только липидный комплекс). При контакте с Vila и Са2* способен активиро-
вать X (по быстрому пути)._Является матрицей для развертывания реакций образова-
ния протромбииазы, что обеспечивает высокую локальную концентрацию факторов
свертывания.
• IV - ионы Са (поступает с пищей, концентрация в плазме 2,00-2,75 ммоль/л). Они
обеспечивают связывание по карбоксильным группам факторов свертывания (П, VII,
IX, X) с фосфолипидами мембран тромбоцитов (Р3) и клеток поврежденных тканей
(Ш). Участвуют в активации протромбииазы, переходе фибриногена в фибрин, агрега-
ции тромбоцитов и ретракции тромба. Связывают гепарин и ингибируют фибринолиз.
• V - проакцелерин (образуется в печени). Необходим для образования протромбииазы,
входит в ее состав. Активированный и связанный с рецепторами тромбоцитов фактор
Va (акцелерин) в свою очередь является рецептором для Ха, что, создает оптимальные
условия для взаимодействия Ха и протромбина. Сам проакцелерин активируется
тромбином (Па).
* VI - активированный Va, исключен из классификации.
• VH - нроконвертин (сериновая протеаза, образуется в печени с участием вит. К).
Принимает участие в образовании протромбииазы по быстрому пути, активирует X и
- действие его на протромбин. Сам активируется факторами Ш, Па.
• VIII - антигемофильный глобулин А (образуется в печени, селезенке, лейкоцитах,
эндотелии, почках; в кровн циркулирует в виде комплекса из 3-х субъединиц: VIIIk -
коагулирующая субъединица, Vniar - основной антигенный маркер, VUI-фВ - фактор
Виллебранда образуется эндотелием, тромбоцитами, является транспортной субъеди-
ницей и участвует в агрегации тромбоцитов). Фактор УШ является рецептором для
1Ха и в 500 раз увеличивает его действие на переход X —► Ха. Сам фактор УШ акти-
вируется тромбином (Па). (При недостатке возникает гемофилия А.)
9 IX - антигемофильный глобулин В или Кристмас-фактор (сериновая протеаза, об-
разуется в печени при участие вит.К; концентрация в плазме 0,004 г/л; Т[/г= 13-30 час).
Активирует переход X в Ха и УП и Vila. Сам активируется Ха, Vila. Па.
212
• X - фактор Стюарта - Праузра (сериновая протеаза, образуется в печени при уча-
стие вит.К). Является центральным компонентом протромбиназы, переводит протром-
бин в тромбин (Па), расщепляя в его молекуле связи Арг-Изолиз. Сам активируется
IXa, Villa, Р3 по медленному пути и Vila, Ш по быстрому пути (активируется также
трипсином и ферментом из яда гадюки).
• XI - плазменный предшественник тромбопластина или антигемофильный фак-
тор С (предположительно образуется в печени). Необходим для активации IX в IXa.
Сам активируется факторами ХПа, XIV, XV.
• XII - фактор контакта (Хагемана) (сериновая протеаза, предположительно образу-
ется эндотелиоцитами, лейкоцитами, макрофагами). Активируется отрицательно заря-
женными поверхностями, адреналином, прекалликреином (XIV), контактом с волок-
нами коллагена. Запускает образование протромбиназы по медленному пути, активи-
рует XI, VII, XIV. Активирует фибринолиз и систему комплемента. (При недостатке
XII резко удлиняется время свертывания, но нет кровоточивости. Вероятно, есть дру-
гие пути включения медленной системы коагуляции.)
• XIII - фибринстабилизирующий фактор (глутаминтрансфераза) (образуется фиб-
робластами, мегакариоцитами и др. клетками). Катализирует образование специфиче-
ских изопептидных связей между Е-аминогруппой лизила и амидной группой глутами-
нила, соединяя полимеризованные мономеры фибрина тромба в полимер, устойчивый
к фибринолизу. Сам ХШ активируется тромбином (Па) (а также адреналином, глюко-
кортикоидами, АДГ).
• XIV - фактор Флетчера (прекалликреин) (образуется гранулоцитами, клетками ки-
шечника, почек, слюнных, слезных и потовых желез) участвует в активации факторов
XII, IX, Vn и плазминогена Сам активируется ХПа, при активации превращается в
фермент калликреин (XlVa), образующий из кининогена (XV) кинины (брадикииии,
каллидин).
• XV - фактор Фитцжеральда (высокомолекулярный кининоген) образуется в тка-
нях. При его активации калликреином (XlVa) превращается в кинины (брадикинин,
каллидин). Участвует в активации ХП, XI и плазминогена (фибрииолиз).Таким обра-
зом, факторы XIV, XV и их активированные формы образуют кинин-калликреиновую
систему, образующую кииины - брадикинин и каллидин. Ее главные функции: акти-
вация свертывания крови и фибринолиза, вазодилятация.
♦♦♦ Антитромбогенные плазменные факторы образуют антикоагулянтную и фибриноли-
тическую системы
• Антикоагулянтная система состоит из первичных и вторичных антикоагулянтов (они
изложены в 4.1).
• Фибринолитическая система имеет два звена: плазминовое (главное) и неплазминовое
(подробно изложена в 4.2).
3. Свертывающая система крови (осуществляет остановку кровотечения - гемо-
стаз).
3.1. Сосудисто-тромбоцитарный гемостаз (осуществляет остановку кровотечения в со-
судах микроциркуляции d < 200 мкм).
❖ Функциональная роль тромбоцитов.
• Тромбоциты - фрагменты цитоплазмы мегакариоцитов, развивающихся
в миелоидной ткани (СКК -» КОК-ГЭММ -> КОКмег. -» Мегакариобласт ->
Промегакариоцит —> Мегакариоцит Тромбоциты). Образование тромбоцитов
стимулируют ИЛ-1, ИЛ-3, КСФмег., тормозит - ТРФр из тромбоцитов.
Количество тромбоцитов в крови 180 - 320 109/л , 2/3 тромбоцитов цир-
кулирует в крови, 1/3 - депонирована в селезенке.
213
• Тромбоциты содержат в гранулах и выделяют при активации факторы,
участвующие в процессах свертывания и противосвертывания крови (их
список выше в 2.2): 1) вызывающие спазм сосудов - серотонин (Рю), адре-
налин, тромбоксан Аг; 2) вызывающие освобождение из тромбоцитов
активных веществ - АДФ (Рп), серотонин (Рю); 3) вызывающие адгезию
и агрегацию тромбоцитов - АДФ (Рп), фибриноген (Р5), фибронектин,
фактор Виллебранда, тромбоксан Ат; 4) способствующие свертыванию
крови - тромбоцитарный тромбопластин (Рз), фибриноген (Рз), акселе-
ратор-глобулин (Pi), тромбостенин (Ре), фибринстабилизирующий фак-
тор (Р?); 5) тормозящие процессы противосвертыванне крови - антиге-
париновый фактор (Ра), антиплазмин (Р?).
• Тромбоциты имеют на плазматической мембране рецепторы к факто-
рам свертывания и противосвертывания крови (фибриногену, тромбину,
Va, Ха, фактору Виллебранда, фибронектину, АДФ, серотонину и др.), в
том числе и к выделяющимся из тромбоцитов.
• Тромбоциты оказывают амиотрофическое влияние: эндотелий посто-
янно поглощает тромбоциты (-15 % циркулирующих тромбоцитов в сутки); их
вещества (напр., фактор роста эндотелия) используются эндотелием для вос-
становления структурных и функциональных свойств сосудистой стен-
ки.
❖ Фазы сосудисто-тромбоцитарного гемостаза.
• Спазм поврежденных сосудов:
• ф- запускается рефлекторным механизмом - медиатор норадреналин,
а-адренорецепторы (первичный спазм);
• ф- поддерживается адреналином, серотонином, тромбоксаном А2, вы-
деляющимися из тромбоцитов, эндотелином-I сосуда (вторичный
спазм).
• Адгезия (прилипание) тромбоцитов: они имеют рецепторы, с помощью
которых способны прикрепляться к фактору Виллебранда, коллагену,
фибронектину в зоне повреждения сосуда.
• ' Агрегация (окучивание, образование конгломерата) тромбоцитов.
• ф Активация тромбоцитов запускается действием на их рецепторы
АДФ, серотонина, тромбина, адреналина (через аг-адренорецепторы).
Через кальциевый механизм (выделение Са2* из плотных трубочек в цито-
плазму) происходит изменение формы (тромбоциты приобретают сфериче-
скую форму с длинными отростками) и запускается секреторный процесс
(реакция освобождения).
• ф- Реакция освобождения из тромбоцитов активных веществ (адрена-
лина, серотонина, АДФ, тромбопластина, антигепарнна, фибрино-
гена, тромбоксана А2 и др.) происходит как самоусиливающийся
процесс (положительная обратная связь; аутокриниая и паракринная регуля-
ция).
ф- Эти вещества запускают процесс агрегации, активируя рецепторы
тромбоцитов к фибриногену, фактору Виллебранда и коллагену.
214
Онн связывают тромбоциты друг с другом и с субэндотелием. Фи-
нальным этапом агрегации является связывание тромбоцитов путем
присоединение фибриногена к его активированным рецепторам.
• ф Агрегация тромбоцитов сначала носит обратимый характер, и ее
ингибиторы (простациклин, NO, простагландины Е| и Дг. тканевой
активатор плазминогена) могут перевести тромбоциты в неактивное
состояние; в дальнейшем агрегация становится необратимой и за-
канчивается распадом тромбоцитов.
• Ретракция (сокращение) тромбоцитарного тромба осуществляется сокра-
тительным белком тромбоцитов - тромбостенином (Рб), стабилизация -
фнбринстабилизирующим фактором тромбоцитов (Ре).
• Показателем активности сосудисто-тромбоцитарного гемостаза являет-
ся время кровотечения (норма до 4 мин).
3.2. Коагуляционный гемостаз (проферментно-ферментный каскад, А.А. Шмидт, 1861; П. Мо-
равитц, 1904) осуществляется с использованием преимущественно факторов свер-
тывания плазмы, тромбоцитов, эндотелия сосудов и поврежденных тканей (оии
приведены как справочный материал выше в 2.2). Коагуляция крови осуществляется в 3
фазы: 1-я фаза может протекать по внутреннему и внешнему путям, 2-я и 3-я
фазы являются общими.
*♦* Общая характеристика плазменных факторов крови. Восемь из пятнадцати факторов
свертывания крови являются ферментами (П, VII, IX - XIV). Из иих все, кроме XIII, пред-
ставляют собой сериновые протеазы, активирующие другие факторы путем расщепления в
молекуле белка связей Арг-Лиз. Четыре из этих факторов (II, VII, IX, X) имеют в своем
составе участки с большим количеством остатков у-карбоксилглутаминовой кислоты, для
ее синтеза необходим вит.К. Эти участки связывают ионы Са2* с помощью которых эти
факторы прикрепляются к фосфолипидам тромбоцитов и клеток поврежденных тканей,
имеющим отрицательный заряд. (Для остановки кровотечения достаточной 10-15% от
нормальной концентрации большинства факторов, например, II, V- XI).
❖ Фазы коагуляционного гемостаза.
• Фаза I - образование протромбииазы (комплекс, состоящий из активи-
рованного фактора X, акцелерина, тромбоцитарного тромбопластина и
Са2+: Ха + V + Р3 + Са2*).
•ф’ Внутренний (медленный) путь (5-8 мин):
повреждение эндотелия вызывает активацию фактора контакта
(ХП —» ХПа) (она может быть усилена при активации прекалликреииа -
XIV и перехода его в калликреин - XlVa);
ХПа вызывает активацию плазменного предшественника тром-
бопластина (XI —» Х1а);
Х1а активирует антигемофильный глобулин В (IX —» 1Ха), ко-
торый образует комплекс с антигемофильным глобулином А
(УШ), Р3 тромбоцитов и Са2* (при этом Vin является рецептором для
1Ха, увеличивая егр активность в 500 раз);
1Ха в составе вышеназванного комплекса активирует X, обра-
зуется протромбнназа (Ха + va+ Рз+Са2*).
• ф Внешний (быстрый) путь (5 - 10 с):
215
повреждение тканей приводит к освобождению тканевых фос-
фолипидов (Ш);
они активируют проконвертин (VH —> Vila), образуется ком-
плекс его с Ш и Са ,
конвертин (VHa) в составе этого комплекса активирует переход
X —> Ха, образуется тканевая протромбиназа.
• Фаза II - под действием протромбиназы (с участием Са2+, акцелерина
Va и Р3) из протромбина (П) образуется тромбин (Па) (2-5 с). (Р3 тром-
боцитов повышает активность протромбиназы в 1000 раз!)
• Фаза III - образование фибринового тромба (2-5 с).
• ф- Тромбин приводит к отщеплению от фибриногена (I) четырех низ-
комолекулярных пептидов, и фибриноген превращается в фибрин-
мономер (Im), имеющий свободные связи.
Рис. 42. Схема свертывания крови по коагуляционному гемостазу
(объяснения в 3.2).
ф- Благодаря этим связям, молекулы (Ьп) образуют сеть волокон фиб-
рина - фибрин-полимер S (растворимый фибрин, чувствителен к плазмину).
• ф- Из фибрина S под действием фибринстабилизирующего фактора
(ХШа), образующего специфические изопептидные связи между
полимеризованными молекулами фибрина-мономера, образуется
нерастворимый фибрин i (нечувствителен к плазмину), составляющий
фибриновый тромб. (Его трехмерная сеть захватывает из кровотока формен-
ные элементы - преимущественно эритроциты - и не пропускает их во внесосу-
дистое пространство.)
• Посткоагуляционная фаза (~ 70 мин). Ретракция тромба (уменьшение
его объема - на 50 %) осуществляется с участием сократительных белков
216
тромбоцитов, находящихся в тромбе, и приводит к окончательному ге-
мостатически полноценному тромбу.
❖ Показателями активности коагуляционного гемостаза (по внутреннему пу-
ти) являются время свертывания крови (норма - 5 - 10 мин) и показатели
тромбоэластограммы.
4. Противосвертывающей система крови.
4,1. Антикоагулянты (блокируют различные фазы свертывания крови).
❖ Первичные антикоагулянты имеются в крови до начала свертывания.
• Антитромбин III (в основном образуется в эндотелии н в печени). Плазменный
кофактор гепарина, при отсутствие гепарина его активность уменьшает-
ся в 700 раз. В активной форме составляет 90% всей антитромбиновой и
70% антикоагулятной активности крови. Ингибирует тромбин (Па), а
также Ха, IXa, Х1а.
• Гепарин (сульфатированные полисахариды, образуются тучными клетками особенно
печени и легких). Образует комплекс с антитромбином Ш, превращая его в
активный антикоагулянт. В малых концентрациях ингибирует образова-
ние протромбиназы (I фазу), в высоких концентрациях тормозит все три
фазы коагуляции, активирует фибринолиз. Комплекс гепарин+антитро-
мбин Ш может фиксироваться на мембране эндотелиальных клеток,
обеспечивая тромборезнстентность сосудистой стенки.
• ai-Антитрипсин (содержание в плазме 3 г/л) ингибирует тромбин (Па),
ХПа, IXa, Х1а.
• Протеин С (сериновая протеаза, образуется в печение с участием вит.К).
Инактивирует Va, VHI, ускоряет фибринолиз. Сам активируется тром-
бином, протеином S, тромбомодулином.
• Тромбомодулин (образуется в эндотелии) блокирует рецепторы для тром-
бина на эндотелиоцитах, активирует протеин С.
• Антитромбопластины ингибируют фактор Ш и VHa.
❖ Вторичные антикоагулянты образуются в процессе свертывания крови и
фибринолиза.
• Атитромбин I - это фибрин, который адсорбирует и инактивирует
тромбин, Va, Ха.
• Антитробин VI - это продукты фибринолиза, которые блокируют фи-
бириноген и фнбрин-мономер, тромбин и Х1а.
4.2. Фибринолиз (препятствует образованию и осуществляет лизис фибрина тром-
ба, образующегося в процессе постоянного локального гемостаза, может осу-
ществляться по двум вариантам - с участием плазмина и без участия плазми-
на).
❖ Плазминовый вариант фибринолиза осуществляется в двух фазах.
• В 1-й фазе плазменный профермент плазминоген превращается в ак-
тивную сериновую протеазу - плазмин. Плазминоген (одноцепочечный
гликопротеин, образуемый в печени, костном мозге, почках и др. органах; концентра-
ция в плазме 0,2 г/л), имеет лизинсвязывающие участки, через которые
присоединяется к фибрину (высокое сродство) и фибриногену (низкое
217
сродство), и поэтому в иорме всегда входит в состав образующего фиб-
ринового тромба. В его активации участвуют:
"Ф* тканевые активаторы плазминогена (ТАП): сериновые протеазы из
клеток различных тканей (например, урокиназа почек, Р8 тромбо-
цитов, протеазы нейтрофилов) и эндотелия их сосудов;
-ф- плазменные активаторы плазминогена (ПАП): ХПа, калликреин
(XTVa), факторы комплемента С8 и Сз, протеин С и др.
•ф- экзогенные вещества: стрептокиназа стрептококков, лекарства.
• Во 2-й фазе образовавшийся плазмин расщепляет фибрин тромба (по
Лиз-Арг и Лиз-Лиз пептидным связям) до пептидов и аминокислот (продукты
деградации фибрина).
• Ингибиторы фибринолиза:
аз-антиплазмин (гликопротеин, синтез в печени) - самый активный ин-
Плазминоген
Тканевые активаторы
плазминогена
Плазменные активаторы ——►
плазминогена
Экзогенные вещества f
Плазмин
а,- антиплазмин
макротобулин
а,-антитрипсин
Фибрин
Продукты лизиса
фибрина
Неплазминовый фибринолиз
Рис. 43. Схема фибринолиза (объяснения в 4.2)
> активация, I торможение
гибитор фибринолиза, образует прочные комплексы с плазмином,
лишая его ферментативной активности, и с фибрином, делая его
нечувствительным к плазмину;
•ф’ аз-макроглобулин (гликопротеин) оказывает более медленное, ио дли-
тельное ингибирующее действие иа плазмин, образуя с ним кова-
лентный комплекс;
•ф’ <Х|-антитрипсин (синтезируется в печени) — оказывает медленное ин-
гибирующее действие иа плазмин (быстрое на трипсин).
Неплазминовый вариант осуществляется фибринолитическими протеазами лейкоцитов,
тромбоцитов, эритроцитов и антитромбином Ш в комплексе с гепарином, которые могут
непосредственно расщеплять фибрин.
5. Нейрогуморальная регуляция агрегатного состояния кровн.
5.1. Нервная регуляция.
❖ Основное рецепторно-афферентное звено: хеморецепторы каротидной и
аортальной зон, реагирующие иа тромбин, плазмин и другие факторы свер-
218
тывающей и противосвертывающей систем; афферентные волокна IX и X
черепных иервов.
❖ Главные центральные структуры.
• Гипоталамус как интегратор вегетативных и эндокринных влияний иа
систему РАСК.
• Парасимпатические центры ствола и спинного мозга, симпатические
центры спинного мозга.
❖ Эфферентное звено.
• Симпатическая нервная система преимущественно через а-
адреиорецепторы ускоряет свертывание крови (активация ХП, высво-
бождение из сосудистой стенки и тканей тромбопластина, повышение
тромбопластической активности жирных кислот) и усиливает фибрино-
лиз за счет выделения из эндотелия тканевого активатора плазминогена
и урокиназы.
• Парасимпатическая нервная система ускоряет свертывание крови за
счет выброса тромбопластина и усиливает фибринолиз за счет выброса
активаторов плазминогена из сердца н сосудов, а также гепарина н ан-
титромбина-Ш.
5.2. Эндокринные влияния: адреналин, глюкокортикоиды, СТГ, АДГ, кальцитонин,
тестостерон, прогестерон первично вызывают гиперкоагуляцию и вторично ак-
тивируют фибринолиз.
S3. Гуморальные факторы: ионы Са2+ связывают гепарин и тормозят фибрино-
лиз; жирные кислоты снижают активность гепарина и выброс активаторов
плазминогена нз эндотелия; протеаза адипсии, вырабатываемая в жировой тка-
ни, участвует в тромбообразовании.
6.
Основные методики исследования системы
гемостаза.
6.1. Определение временп кровотечения (метод
Дьюке). Принцип: определение времени спон-
танной остановки кровотечения проводят по-
сле повреждения сосудов микроциркуляции
(прокол кожи мякоти ногтевой фаланги паль-
ца с помощью скарификатора). Норма: 2-5
мин. Оценка результатов: повышение времени
кровотечения свидетельствует об уменьшении
«Ограничение исследования кро-
ви только вне организма заводит
в тупик. В конце концов, перед
нами всегда встает вопрос: что
это собственно все значит, кото-
рый заставляет нас отворачивать-
ся от пробирки и возвращаться к
живому организму». А.А. Шмидт,
1882.
количества тромбоцитов н (или) нарушении их функций, уменьшение времени крово-
течения - о сильном спазме сосудов микроциркуляции и большом освобождении тка-
невого тромбопластина (Ш).
6.2. Определение времени свертывания кровп (унифицированный двупробирочный метод
Ли-Уайта). Принцип: определение времени спонтанного свертывания нестабилизиро-
ванной крови прн 37° С в несилнконированной и силиконированной пробирках. Нор-
ма: 5 - 7 мин в несилнконированной и 15-25 мин в силиконированной пробирках.
Оценка результатов: уменьшение времени свертывания наблюдается при дефиците
факторов свертывания (I, П, УШ) или (и) увеличении в крови антикоагулянтов. Уко-
рочение времени указывает на тенденцию к гиперкоагуляции.
219
Рис. 44. Схема тромбоэластограммы.(ТЭГ)
R - время реакции, К - время от конца R до
расхождения плеч ТЭГ до 20 мм, МА - мак-
симальная амплитуда расхождения плеч.
6.3. Определение времени рекальцификации плазмы (унифицированный метод). Прин*
цип: определяют время свертывания цитратной плазмы после добавления хлорида
кальция (рекальцификация). Тест характеризует преимущественно 1-ю фазу свертыва-
ния. Норма: 60 - 120 с. Оценка результатов: увеличение времени говорит о снижении
свертывания крови, снижение времени
- об увеличении свертывания крови.
6.4. Определение толерантности плаз-
мы к гепарину (унифицированный ме-
тод Gorstmen). Принцип: определяют
время рекальцификации цитратной
плазмы в присутствии гепарина. Нор-
ма: 11 - 16 мин. Оценка результатов:
если после добавления гепарина время
свертывания больше, чем в контроле,
это указывает на понижение толерант-
ности к гепарину и на снижение свертывающих свойств крови. Уменьшение времени
свертывания в этих условиях говорит о повышении толерантности к гепарину и повы-
шении свертывающей активности крови.
63. Определение протромбинового времени (унифицированный метод Квика). Принцип:
определяют время свертывания цитратной плазмы в присутствии тканевого тромбо-
пластина и хлорида кальция. Тест позволяет судить об образовании протромбиназы по
внешнему пути. Норма: 12 - 20 с. Оценка результатов: увеличение времени свертыва-
ния говорит о снижении плазменных факторов П, V, УП, X более чем на 60% от нор-
мы, или концентрации фибриногена менее 1 г/л, т.е. о тенденции к гипокоагуляции
крови. Уменьшение времени свертывания говорит об увеличении концентрации этих
факторов или фибириногена и тенденции к гиперкоагуляции крови.
6.6 . Тромбоэластограмма (ТЭГ) как показатель свертывающей и противосвертывающей
систем.
❖ Время реакции R (8-10 мин) - интервал от начала записи до расхождения плеч
ТЭГ на 1 мм, что соответствует образованию первых волокон фибрина.
❖ Время свертывания К (6 - 8 мии) - интервал от расхождения плеч ТЭГ от 1 до 20
мм, характеризует образование фибринового сгустка определенной прочности.
Максимальная амплитуда та (45 - 60 мм), наибольшее расхождение между пле-
чами ТЭГ, характеризует максимальную эластичность сгустка.
7. Группы крови человека.
7.1. Групповые антигены и антитела,
❖ Групповые антигены (агглютиногены): известно несколько десятков анти-
генов в эритроцитах, которые лают -10 млн групповых сочетаний.
❖ Групповые антитела (агглютинины и гемолизины плазмы).
• Нормальные антитела (синонимы: врожденные, полные) имеются в норме в
постнатальном периоде, как правило, в системе AB0, относятся к IgM.
(Реагируют с антигенами при комнатной температуре в солевой среде, инактивируют-
ся при нагревании.)
• Изоиммунные антитела (синонимы: приобретенные, неполные) образуются в
ответ на поступление соответствующего антигена, относятся IgG. (Они
термостабильны, ие реагируют с антигенами в солевой среде, определяются непрямой
пробой Кумбса).
220
• Если при переливании крови нормальные или изоиммуниые антитела
соединяются с групповыми антигенами на мембране эритроцитов, про-
исходит их агглютинация и активация системы комплемента с образо-
ванием цитолитического комплекса (С,ь, Cs-C<>), вызывающего гемолиз
эритроцитов (может возникнуть гемотрансфузионный шок).
7.2. Системы групп крови (по убывающей антигенной силе).
❖ Система АВО (К. Ландштейнер, 1901).
• В эритроцитах имеются агглютиногены (антигены) А, В. Это олигоса-
хариды, связанные с сфинголипидами и белками мембраны эритроци-
тов. В плазме имеются нормальные антитела - агглютинины а (анти-А) и
Р (анти-В), относящиеся к IgM (выработанные в процессе иммунизации аглюти-
нины относятся к igG). В системе АВО имеется 4 (I -IV) группы крови.
• ф- (I) 0 а|3 (- 33 % людей).
(П) А р (~ 38 %). Кроме людей с Ai - сильным антигеном (1,3 млн
молекул А в эритроците), есть люди с Аг - слабым антигеном (0,25
мли молекул А в эритроците), имеется риск ложного определения
первой группы крови (!).
• ф- (Ш) В а (- 21 % людей).
• ф- (TV) АВ 0 (- 8 % людей).
• Образование агглютиногенов А и В в эритроидных клетках кодируется тремя генами»
расположенными в хромосоме 9. Они контролируют образование ферментов синтеза
олигосахаридов в конечном счете формирующих систему АВО (материал носит спра-
вочный характер).
Н-ген (аллели НН, Hh) кодирует образование фермента фукозилтрансферазы, об-
разующей фукозо-а1,2-галактозу-р, которая является предшественником как А,
так и В агглютиногенов. При генотипе hh образуется неактивная трансфераза н
предшественник не синтезируется.
Ген А (аллели АА, Аа) кодирует А-спецнфическую трасферазу, продукт реакции
которой - N-ацетил-галактозамнн - присоединяется к предшественнику.
Геи В (аллели ВВ, ВЬ) кодирует В-специфическую трансферазу, присоединяю-
щую к предшественнику галактозу.
I (0) группа крови возникает, когда в локусе АВО отсутствуют гены А и В (в этом
случае локус Н-гена может иметь любую аллель: НН, Hh, hh), няи при наличии в
локусе Н-гена аллели hh, в этом случае не образуется предшественник агглюти-
ногенов А и В, и тогда может быть любая аллель в локусе АВО.
Ф П (А) группа крови возникает, когда в локусе Н-гена имеются аллели НН или Hh
(предшественник образуется), а в локусе АВО имеются аллели АА или Аа (обра-
зуется агглютиноген А).
ф Ш (В) группа крови возникает, когда в локусе Н-гена имеются аллели НН или h
(предшественник образуется), а в локусе АВО имеются аллели ВВ или ВЬ (обра-
зуется агглютиноген В).
IV (АВ) группа кровя возникает, когда в локусе Н-гена имеются аллели НН или
Hh (предшественник образуется), а в локусе АВО имеются аллели АА или Аа и
ВВ или ВЬ (образуются аггяютииогены А'и В).
❖ Система Резус (Rh) (К. Ландштейнер, А. Винер, 1940) состоит из - 40 антигенов,
наиболее сильным из которых является D (способен иммунизировать человека).
Антиген имеется в эритроцитах 86 % людей (Rh*) и отсутствует у 14 % лю-
221
дей (Rh'); врожденных антител нет, могут быть антитела изоиммунного
происхождения (IgG-анти D). Rh-конфликт наступает: 1) при переливании
Rh+ крови Rh" реципиенту; 2) при развитии Rh+ плода у Rh" матери (Rh-
фактор появляется у плода с 3-го мес., достигает иммунологической активности к концу
беременности и через плаценту поступает к матери. Образующиеся у матери Rh-антитела
проникают в плод. Если отец Rh* н мать Rh“, то плод в 50-100% Rh*); 3) Rh” кровь ма-
тери после Rh-конфликта с плодом переливается Rh+реципиенту.
Система Келл-, имеется активный фактор Келл у 8 % людей и мало активный фактор Чел-
лано у 99 % людей; антитела изоиммунного происхождения.
♦ ♦♦ Система Даффи-, имеются два антигена - более активный у 66 % людей н менее активные
у 83 % людей, антитела изоиммунного происхождения.
❖ Другие системы: MNS, Кидд, Лютеран, I, Левис и др.
73. Методы исследования групп крови.
❖ Определение группы крови в системе АВО.
Принцип метода: для определения групп крови используется явление агглю-
тинации исследуемой крови со стандартными сыворотками. При встрече одно-
именных агглютиногенов н агглютининов (А+а; В+р) происходит агглютинация
эритроцитов.
В углубления планшеты под соответствующими обозначениями наносят раз-
ными пипетками по капле (0,1 мл) сынороткн Оар (I), Ар (II), Ва(Ш) групп кровн
двух серий. Из проколотого пальца стеклянной палочкой переносят на пластинку 6
маленьких капель (- 0,01 мл) крови каждую рядом со стандартной сывороткой
(количество кровн должно быть в - 10 раз меньше сыворотки). Разными палочками
размешивают каплю крови с сывороткой до равномерного розового цвета. План-
шету периодически покачивают н реакция оценивают через 5 мин. Если происхо-
дит реакция агглютинации капля становится прозрачной, а эритроциты склеивают-
ся в виде комочков. Чтобы реакцию агглютинациии не маскировали образующиеся
иногда «монетные столбики» эритроцитов, через 3 мин от начала реакции в каж-
дую пробу надо добавить 1 каплю изотонического раствора NaCl. Группа кровн
устанавливается по наличию илн отсутствию агглютинации. Результат реакций в
каплях с сыворотками одной и той же группы двух серий должны совпадать. (Тем-
пература в помещении должна быть в пределах 15 - 25°С.) Трактовка результатов:
• агглютинация не наступила ии в одной капле сывороток - группа крови 0 сф
(0;
• агглютинация произошла с сыворотками I и Ш групп - группа крови Ар (II);
• агглютинация наступила с сыворотками I и П групп - группа крови Bet (Ш);
• агглютинация наступила с сыворотками X П и Ш групп. Для исключения не-
спецнфической агглютинации необходимо провести реакцию с сывороткой IV
группы, если агглютинация не наступит, исследуемая кровь ABO (IV) группы.
В настоящее время рекомендована методика определения групп кровн с помо-
щью цоликлонов анти-А и анти-В, которая имеет более высокую специфичность и
выраженность реакции агглютинации, поэтому можно пользоваться одной серией
реагентов.
В углубления планшеты двумя пипетками наносят по капле цолнклона анти-А и
аити-В, рядом с каплями антител наносят по маленькой капле крови (в 10 раз
меньше капли цоликлонов), и капли смешивают. Результат реакции оценивают в
течение 2,5 мнн. Трактовка результатов:
222
• агглютинации нет ни с цоликлоном анти-А, ни с цоликлоном анти-В - группа
крови 0аР(1),
• агглютинация только с цоликлоном анти-А - группа крови Ар(П),
• агглютинация только с цоликлоном анти-В - группа крови Ва(Ш),
• агглютинация имеется как с цоликлоном анти-А, так и с цоликлоном анти-В -
группа крови AB0(IV). В этом случае для исключения спонтанной агглютина-
ции необходимо провести контрольное исследование крови путем добавления
капли изотоническим раствором 'хлорида натрия, в котором агглютинация
должна отсутствовать.
❖ Определение резус-принадлежности.
Для определения резус-принадлежности используют явление агглютинации,
возникающее при контакте резус-агглютиногена эритроцитов с антирезус-
агглютинином.
На планшету под соответствующими обозначениями наносят по одной капле
контрольной сыворотки (стандартная сыворотка группы крови испытуемого) и
стандартной антирезусиой сыворотки. Рядом помещают по капле исследуемой
крови (количество крови должно быть примерно вдвое меньше капли сыворотки).
После размешивания наблюдают в течение 5 минут за реакцией.
Трактовка результатов: если в пробе со стандартной антирезус-сывороткой
произошла агглютинация - иследуемая кровь резус-положительная; если агглюти-
нация не произошла - кровь резус-отрицательная. В контрольной сыворотке агг-
лютинации не должно быть.
❖ При неблагоприятном трансфузионном и акушерском анамнезе определение
изоиммунных антител непрямой пробой Кумбса.
7.4. Правила переливания крови.
По системе АВО и Rh:
• переливать можно только одпогрупппую (!) по системе АВО кровь;
• нельзя переливать Rh+ кровь Rh’ реципиенту;
• при неблагоприятном анамнезе провести пробу Кумбса на изоиммун-
иые антитела.
❖ Перед переливанием крови обязательно проводится:
• определение группы крови по системе АВО и Rh-принадлежности крови
как донора, так и реципиента;
• проба иа совместимость: в 2-3 капли сыворотки реципиента, взятой по-
сле ретракции сгустка крови, добавляют маленькую каплю донорская
кровь, агглютинация должна отсутствовать;
• биологическая проба: струйно переливают 5 - 10 мл крови и в течение
3-х мин убеждаются в отсутствии реакций несовместимости, такую
процедуру проводят 3 раза, после можно продолжать трансфузию;
7.5. Физиологические механизмы действия переливаемой крови.
Заместительное действие (А.М. Филомафитский, 1848).
• Заполнение сосудистой системы.
• Стимуляция редепонирования крови.
223
• Срок жизни компонентов перелитой крови: эритроцитов - 30 - 120 су-
ток, белков -18 - 36 суток.
❖ Стимулирующее действие.
• ЦНС: 1-е сутки торможение, потом усиление выработки условных реф-
лексов, повышение работоспособности нейронов.
• Гипоталамус: стимуляция пищевого центра.
• Кровообращение: повышение АД, сосудистого тонуса, мобилизация
крови из депо.
• Обмен веществ: повышение основного обмена, дыхательного коэффи-
циента, секреции инсулина; положительный азотистый баланс.
❖ Гемостатическое действие (свежей крови).
• Поступает большое количество факторов свертывания крови.
• Активируется синтез прокоагулянтов в печени.
• Количество тромбоцитов сначала уменьшается, потом увеличивается.
• Время свертывания крови уменьшается.
❖ Иммунологическое действие.
• Активируется фагоцитоз, синтез антител.
• С кровью поступают 1g (пассивный иммунитет).
❖ Питательное действие: преимущественно за счет поступления белков и
липидов.
❖ Эндокринный эффект: стимуляция секреции гормонов адено- и нейроги-
пофиза, коры и мозгового слоя надпочечников, щитовидной железы и ин-
сулярного аппарата.
7.6. Принципы приготовления и классификация кровезаменителей.
Принципы приготовления кровезамещающих растворов: 1) изоосмия, 2) изоио-
иия, 3) реокоррекция, 4) гемокоррекция (обеспечение транспорта ОД
❖ По функциональным признакам выделяют следующие виды кровезамени-
телей.
• Гемодинамические: среднемолекулярные (например, полиглкжии), низко-
молекулярные (например, реополиглюкин), препараты желатины (желатиноль).
• Дезинтоксикациоииые: гемодез, полидез (поливинил).
• Для парентерального питания: белковый гидролизии (иапр., амииопеп-
тид), растворы аминокислот (например, полиамии), жировая имульсия.
• Регуляция водно-солевого обмена и кислотно-основного состояния (фи-
зиологический раствор, раствор Рингера, лактосол и др.).
• Препараты с функцией переноса Ох растворы модифицированного ге-
моглобина (например, геленпол), эмульсии фторуглеродов (например, перфто-
раи).
8. Профильный материал для студентов лечебного факультета.
8.1. Возрастные изменения компонентов системы гемостаза и «разбалансирование»
их в старости.
224
Повышение активности свертывающей системы: увеличение содержания антиге-
мофильного глобулина А (VIII), фибриногена (I), фибринстабилизирующего фак-
тора (ХШ), тромбофилический тип тромбоэластограммы.
* Компенсаторное (менее выраженное) повышение активности противосвертываю-
щей системы: увеличение содержания гепарина, но снижение его активности, по-
вышение фибринолитической активности.
❖ Снижение «надежности» системы гемостаза в экстремальных условиях.
8.2. Изменения в групповых системах крови при старении: увеличивается вероятность
пояаления изоиммунных антител к различным эритроцитарным антигенам в связи с
переливаниями крови и беременностями в течение жизни.
9. Профильный материал для студентов педиатрического факультета. Функцио-
нальные особенности системы гемостаза у детей в онтогенезе.
9.1. Изменение тромбоцитов в онтогенезе: в первые часы после рождения содержание
тромбоцитов примерно равно содержанию у взрослых, к 7 - 9-му дню происходит его
снижение - до 17010%, а к концу 2-й недели их количество вновь возрастает до пер-
воначальной величины и в дальнейшем практически не меняется. У новорожденных
менее выражены, чем у взрослых, агрегация тромбоцитов и выделение ими фосфоли-
пидов (Р3) и серотонина.
9.2. Становление системы свертывания крови в онтогенезе, ее особенности в анте- и
постнатальном периоде.
❖ Особенности системы свертывания в антенатальном периоде.
• До 16 - 20 недель кровь плода не способна свертываться из-за отсутствия в
плазме крови фибриногена, протромбина, акцелерина.
• Фибриноген появляется на 4 - 5-м мес. внутриутробной жизни (0,6 г/л), низкая
активность фибринстабилизирующего фактора.
• Отмечена высокая концентрация гепарина к 7-му мес. (почти в 2 раза выше,
чем у взрослых), снижение ее до уровня взрослых к рождению.
• Время свертывания с 6 мес. такое же, как у взрослых.
• Факторы свертывающей и противосвертывающей систем крови синтезируют-
ся печенью плода, их низкий уровень обусловлен незрелостью клеточных
структур, участвующих в биосинтезе этих факторов.
❖ Особенности системы свертывания в постнатальном периоде.
• Для новорожденных характерна низкая концентрация многих факторов свер-
тывающей системы крови (П, VII, IX - ХШ), антикоагулянтов и плазминогена,
но соотношение их концентраций и время свертывания крови такие же, как у
взрослых.
• Происходит постепенное повышение концентрации тромбогенных и анти-
тромбогенных факторов, которая достигает нормы взрослых к 14-15 лет.
9.3. Формирование групповых признаков крови в онтогенезе.
❖ Формирование групповых признаков: агглютиногены А и В формируются у плода
к 2 - 3-му мес., агглютинины (а, р) - к моменту или сразу после рождения.
❖ Наблюдается низкая способность к агглютинации эритроцитов у новорожденных
и «нормализация» ее к 10-летнему возрасту.
❖ Rh-антиген определяются у плода на 3-м мес., и достигает иммунологической ак-
тивности к концу беременности; активность Rh-антигена во внутриутробном пе-
риоде выше, чем у взрослых.
225
9.4. Резус-конфликт между организмом матери и плода возникает при повторной
беременности Rh* женщины Rh* плодом.
10. Профильный материал для студентов стоматологического факультета.
10.1. В слюне ~ 70 % людей могут содержаться агглютиногены системы АВО. (Олигосахари-
ды, образующие эти антигены, находятся в составе гликопротеинов на поверхности
секреторных клеток и связаны с муцином слюны. В клетках слюнных желез код фер-
мента (фукозилтрансферазы), образующего предшественник антигенов А и В, имеется
в специальном гене Se, находящемся в секреторном локусе (в эритроидных клетках
этот код в гене Н). Клетки слюнных желез, имеющие в секреторном локусе аллели SeSe
или Sese, образуют предшественник антигенов, а имеющие аллели sese - не образуют
этот предшественник.)
10.2. Физиологические обоснования мероприятий при длительном кровотечении при опера-
циях в ротовой полости и способы его предотвращения.
❖ В качестве местных средств остановки кровотечения применяются тампонирова-
ние, холод, гемостатическая губка, адреналин в растворах для местной анестезии.
❖ Повышение активности системы свертывания: введение раствора CaCh, отдель-
ных факторов свертывания при их дефиците, внкасола и др.
❖ Снижение активности противосвертывающей системы (введение
аминокапроновой кислоты и др.).
❖ При большой кровопотере восполнение объема циркулирующей крови путем пе-
реливания коллоидных кровезамещающих жидкостей или крови (последнее по
жизненным показаниям).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3.1 и 3.2). Объем реферата 2-3 стр., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. стр. 514, использовать имеющиеся
у вас учебники и пособия): [1-9], [11], [13-16].
3.2. Дополнительная литература (см. стр. 514 - 522): [3], [27], [30], [39], [43], [47],
[55], [73], [77], [88], [95], [99], [142], [195], [222], [225], [252-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999:; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
ЗЛ. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. стр. 508 - 513).
Задача 14-1. У больного гемофилией А (недостаток в крови фактора УШ) определили количество
тромбоцитов, время свертывания крови и время кровотечения. Количество тромбоцитов и их
226
функциональная активность оказались нормальными. Как будет изменено время свертывания
крови и время кровотечения? Обоснуйте свой ответ.
Задача 14-2. Когда в эксперименте животному впервые внутривенно медленно ввели тромбин,
исследователи с удивлением обнаружили, что время свертывания крови у него не только не
уменьшилось, а резко увеличилось. Используя свои знания по системе регуляции агрегатного со-
стояния крови, объясните результаты эксперимента.
Задача 14-3. На стекло нанесены 4 капли стандартной сыворотки IV группы крови, в которые
последовательно внесено небольшое количество крови (в соотношении 1:10). В первую каплю -
крови I группы, во вторую - П-й группы, в третью - Ш-й группы, в четвертую - IV-й группы.
Произойдет ли агглютинация в этих каплях? Объясните, почему.
Задача 14-4. Пациенту с потерей 25% циркулирующей крови (около 1,5 л) врач решил применить
переливание крови. У пациента определена группа крови П, Rh+. Взяв пять ампул (1 л) крови I
группы, врач убедился, что ее срок годности не превышен, а группа ампульной крови соответст-
вует обозначенной на этикетке. В начале переливания крови пациенту бьшн проведены пробы на
индивидуальную совместимость и биологическую совместимость. Однако после переливания
крови у пациента развился тяжелый гемотрансфузиониый шок в результате гемолиза эритроци-
тов. Какую ошибку допустил врач? Обоснуйте свой ответ. Прежде чем проверить свой ответ, ре-
шите задачу 14-5, поскольку ответ к этим задачам дан один.
Задача 14-5. Условия задачи такие же, как в 14-4. Какие эритроциты будут гемолизироваться -
перелитой крови или пациента? Обоснуйте свой ответ. Прежде чем проверить свой ответ, решите
задачу 14-4, поскольку ответ к этим задачам дан один.
227
Тема 15. Физиологические функции сердца
Цель: а) знать общую морфофункциональную характеристику системы кровообращения и ее роль в
поддержании жизнедеятельности организма; физиологические свойства сердца и их механиз-
мы (автоматия, возбуждение, проведение, сокращение); проводящую систему сердца, ее функ-
циональные особенности; эндокринную функцию сердца;
б) уметь использовать полученные знания для понимания механизмов функционироаания
сердца в целостном организме при его различных состояниях.
Мотивация темы: «Когда я присутствую на вскрытии умершего от заболевания сердца, то в половине
случаев я ие могу понять, от чего он умер, а в другой половине случаев - как он еще мог
жить» (С.П. Боткин).
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции.
1. Общая характеристика системы кровообращения.
1.1. Кровообращение (circulatio sanguinis) - физиологическая система, осу-
ществляющая перемещение крови в кровеносной системе, что обеспечивает
обмен веществ в органах и тканях организма.
1.2. Основные исторические этаны:
• •• Ациркуляторная теория К. Галена (2 в): кровь образуется в печени, поступает в сердце, из
него в органы, где потребляется.
♦ ♦♦ Циркуляторная теория У. Гарвея (1628).
• 2* Открытие капиллярного кровообращения (Ж. Мальпиги, 1661).
♦ ♦♦ Открытие законов гидродинамики (Д. Бернулли, 1738; Ж. Пуазейль, 1840).
* 2* Открытие сосудодвигательного центра (Ф.В. Овсянников, 1871).
Ф Выяснение газотранспортной функции крови (И.М. Сеченов, 1891).
♦ ♦♦ Изобретение аппарата искусственного кровообращения (С.С. Брюхонеико, 1923).
1.3. Основные компоненты системы кровообращения: 1) сердце; 2) сосуды боль-
шого и малого кругов; 3) кровь (изложена в темах 12-14); 4) аппарат нейрогумо-
ральной регуляции.
1.4. Роль и место системы кровообращения в поддержании жизнедеятельно-
сти организма,
& Основные функции системы кровообращения: 1) транспортная: трофиче-
ская и выделительная; 2) регуляторная; 3) иммунная; 4) афферентная; 5)
терморегуляторная.
❖ Кровообращение как интегрирующая система участвует в формировании
целостности организма.
2. Сердце как компонент системы кровообращения.
2.1. Структура сердца (масса - 300 г).
*2* Камеры сердца: предсердия и желудочки - осуществляют поток крови.
*♦* Клапаны сердца: створчатые (двухстворчатый, трехстворчатый); полулунные (аортальный
и пульмональный) - определяют направление потока крови.
228
*•* Сократительные (рабочие) кардиомиоциты (d=10-25 мкм, С=100—150 мкм) составляют 99
% массы миокарда (обладают возбудимостью, проводимостью и сократимостью; автома-
тия in vivo не выражена).
♦t* Проводящие (атипичные) кардиомиоциты формируют проводящую систему сердца (обла-
дают автоматией, проводимостью, сократимость отсутствует или слабо выражена; Т-
трубочки не выражены, миофибрилл мало):
• Клетки узлов проводящей системы:
ф’ Р-клетки (водители ритма или пейсмекерные клетки) расположены в центральной
части узлов, обладают автоматией и проводиостью, сократимость отсутствует;
ф’ Переходные клетки расположены в периферической части узлов, проводят ПД от
Р-клеток к клеткам проводящих путей и рабочим кардиомиоцитам, автоматия у
них плохо выражена.
• Клетки проводящих путей (пучка Гиса его ножек, волокон Пуркинье, межузловых пу-
тей) передают возбуждение на рабочие кардиомиоциты; имеют хорошую проводи-
мость, слабую автоматик) и очень слабую сократимость.
♦♦♦ Секреторные кардиомиоцнты расположены в предсердиях и выполняют эндокринную
функцию.
Межклеточные контакты хорошо выражены в области вставочных дисков, соединяющих
кардиомиоциты преимущественно «конец в конец», имеют пальцевые выросты и вдавле-
ния:
• Функциональную связь в вставочных дисках создают щелевые контакты (синонимы:
высокопроводимые контакты или нексусы, их сопротивление - 5 Ом/см2), которые со-
ставляют 10 - 20 % площади вставочных дисков.
• Механическую связь в области вставочных дисков создают десмосомы и промежуточ-
ные контакты (прикрепления актиновых нитей к плазмолемме).
• С помощью вставочных дисков кардиомициты образуют мышечные волокна.
2.2. Основные физиологические функции сердца: автоматия, возбудимость,
проводимость, сократимость образуют интегральную функцию - нагне-
тательную функцию.
3. Автоматия сердца.
3.1. Автоматия - способность сердца самостоятельно генерировать потенциалы
действия и переходить в состояние возбуждения в результате процессов, разви-
вающихся в самом миокарде. Создается преимущественно Р-клетками узлов
проводящей системы.
3.2. Компоненты проводящей системы:
❖ Синоатриальный узел (СА-узел) расположен в правом предсердии около
устья полых вен.
❖ Межузловые (передний, средний и задний) и межпредсердный пути соединяют
СА-узел с АВ-узлом.
❖ Атриовентрикулярный узел (АВ-узел) расположен в правом предсердии
около предсердно-желудочковой перегородки, имеет три зоны: предсерд-
ноузловую (А-N), собственно узел (N), переход от узла к пучку Гиса (N-H).
❖ Пучок Гиса, его ножки, ветви левой ножки, волокна Пуркинье состоят из
клеток Пуркинье, проводящих ПД к рабочим кардиомиоцитам.
3.3. Потенциал действия Р-клеток СА-и АВ-узлов.
♦ ♦♦ Максимальный диастолический потенциал (МДП) в клетках СА-узла -50.. .-60 мВ, в клет-
ках АВ-узла -60.. .-70 мВ.
ООП
❖ Схема ПД, его длительность 100-300 мс, амплитуда -70 мВ (рис. 45).
❖ Фаза деполяризации (фаза 0) имеет низкую скорость (~ ю в/с), создается
входящим в клетку Ca2+/Na+-TOKOM в связи с открыванием высокопорого-
вых Са2+-каналов L-типа (начинают активироваться при МП -40 мВ, полная актива-
ция при +10 мВ, после чего канал инактивирован ~ 200 мс), овершут слабо выражен:
0-15 мВ.
❖ Фаза реполяризации (фаза 2-3) связана с открыванием К*-каналов и выхо-
дящим из клетки К*-ТОКОМ (порог активации при -40 мВ, полная активация прн +10
мВ).
Рис. 45. Схема потенциала
действия пейсмекерного
кердиомноцита. МДД -
медленная диастоличе-
ская деполяризация, КУД
- критический уровень
деполяризации, МДП -
максимальный диастоли-
ческий потенциал.
❖ Медленная (спонтанная) диастолическая деполяризация (МДД, фаза 4) яв-
ляется показателем автоматик (пейсмекерным потенциалом) и последовательно
формируется:
• сначала №+-каналами утечки, по которым течет входящий в клетку
Иа+-ток;
• затем открыванием низкопороговых Са2+-каналов Т-типа (активируются
при мп около -55 мВ) и входящим в клетку Са2+/ Иа+-током;
• в последней трети МДД активируются высокопороговыми Са2+-каналов
L-типа.
❖ Изменение скорости МДД.
• Адреналин увеличивает скорость МДД, активируя Са2+-каналы Т- и L-
типов.
• Ацетилхолин активирует К+-каналы и снижает скорость МДД.
• Температура тела: ее повышение активирует (уменьшение ингибирует)
Са2+-каналы и соответственно скорость МДД (например, изменение иа 3 °C
изменяет активность каналов в 2 раза).
❖ Продолжительность рефрактерного периода в связи с медленной деииакти-
вации Са2+-каналов L-типа на - 100 мс превышает время ПД.
❖ Ритм сердца является мерой его автоматик.
3.4. Пейсмекер (водитель ритма) сердца.
♦ ♦♦ Пейсмекер - группа клеток в проводящей системе (у здорового человека в си-
нусном узле), синхронно генерирующая ПД и «навязывающая» свой ритм
другим отделам проводящей системы.
❖ Синхронизация активности пейсмекерных клеток осуществляется благода-
ря высокопроводимым контактам, через которые быстрые клетки ускоряют
230
возбуждение медленных, а медленные за счет сопротивления и емкости их
мембран «притормаживают» быстрые. Парасимпатические влияния на
сердце увеличивают синхронизацию пейсмекерных клеток.
❖ Градиент автоматии (Г. Станниус, 1880; В. Гаскел, 1887).
• Синоатриальный узел (60 - 80 им/мин) состоит из Р-клеток, промежу-
точных клеток и является пейсмекером сердца у здорового человека.
• Атриовентрикулярный узел (40 - 60 им/мин), Р-клеток значительно
меньше, они находятся в нижней части узла (N-H).
• Пучок Гиса (30 - 40 им/мин).
• Волокна Пуркинье (около 20 им/мин).
4. Проводимость и проведение возбуждения в сердце.
4.1. Свойство проведения ПД имеют как атипичные, так и типичные кар-
диомиоциты, переход ПД из одной клетки в другую обеспечивают нексусы (они
занимают -15 % плошали вставочных дисков). Скорость проведения ПД прямо про-
порциональна скорости фазы деполяризации (крутизне переднего фронта ПД) и ве-
личине диаметра кардиомиоцита.
4.2. Проведение ПД в различных отделах проводящей системы.
❖ СА-узел (около 15 см/с).
❖ Межузловые тракты (70 см/с), рабочие кардиомиопиты предсердий (60
см/с).
❖ АВ-узел имеет самую низкую скорость проведения (3 см/с), что формирует
АВ-задержку - 0,04 с, связанную с небольшим количеством нексусов и
большим продольным сопротивлением миоцитов; эта задержка создает по-
следовательность сокращений желудочков после предсердий и блокирует
проведение через узел импульсов с частотой более 180 - 200 в мин.
❖ Пучок Гиса (100 - 150 см/с), волокна Пуркинье (200 - 400 см/с) имеют са-
мую высокую скорость проведения возбуждения, что обеспечивает высо-
кую степень синхронности сокращения всех рабочих миоцитов желудоч-
ков.
Возможны врожденные добавочные путн проведения возбуждения из предсердий в желу-
дочки, мннуя предсердно-желудочковый узел: например, пучки Кента, соединяющие
предсердия н желудочки; пучок Джеймса, соединяющий предсердия с нижней частью
предсердно-желудочкового узла (они создают риск аритмии).
< • Проведение возбуждения из проводящей системы в рабочие кардиомиоци-
ты непосредственно осуществляют клетки Пуркинье: их ПД по форме и
механизмам сходен с ПД рабочих кардиомиоцитов (кроме фазы мдд>, но име-
ет большую длительность и рефрактерный период до 500 мс.
4.3. Функциональное значение проводящей системы.
❖ Создает ритмическую генерацию потенциалов действия.
❖ Обеспечивает последовательные сокращения предсердий и желудочков.
❖ Создает синхронизацию возбуждения и сокращения кардиомиоцитов же-
лудочков.
S. Возбудимость и возбуждение сердца.
5.1. Потенциал действия рабочих кардиомиоцитво желудочков и предсердий
(рис. 46).
731
❖ Схема ПД, его длительность 200 - 300 мс, амплитуда 100 - 120 мВ, овер-
шут 20 -30 мВ (МПП равен -80...-90 мВ).
❖ Фаза деполяризации (фаза 0) обусловлена открыванием быстрых ^-кана-
лов и входящим Na+-TOKOM (скорость фазы 100 - 300 В/с, блокатор каналов лидока-
ин).
❖ Фаза начальной быстрой реполяризации (фаза 1) связана с активацией К+-
Кана-ЛОВ И ВЫХОДЯЩИМ ИЗ клетки К+—ТОКОМ. (В реполяризации определенную роль
могут играть ионы СГ, потенциалзависимые каналы которых активируются при МП более
позитивным, чем -65 мВ.)
❖ Фаза медленной реполяризации (фаза 2, «плато») связана с открыванием
высокопороговых Са2+-каналов L-типа и входом в клетку Са2+ (блокатор ка-
налов верапамил). Входящий Са2*-ток примерно равен выходящему из клетки
К+-току.
❖ Фаза конечной быстрой реполяризации (фаза 3) формируется резким пре-
обладанием выходящего из клетки К+-тока, т.к. через - 200 мс происходит
инактивация Са2+-каналов
❖ Фаза покоя (фаза 4) имеет
стабильный МП (отсутствует
спонтанная деполяризация).
5.2. Изменение возбудимости
кардиомиоцита в течение
потенциала действия.
❖ Фаза абсолютной
рефрактерности (- 270 мс),
новый ПД невозможен.
❖ Фаза относительной
рефрактерности (- 30 мс),
новый ПД возможен при
действии сильных
раздражителей, его амплитуда и скорость проведения снижены.
Фаза супернормалъной возбудимости (- 10 мс), новый ПД возможен при
действии субпороговых раздражителей.
❖ Экстрасистола - внеочередное возбуждение и сокращение сердца на до-
полнительные раздражения.
Уязвимый период во время перехода абсолютной в относительную рефрактерность: кар-
диомиоциты неоднородны по рефрактерности и миокард электрически нестабилен (риск
фибрилляции предсердий или желудочков).
53. Общее время возбуждения предсердий (0,06 - 0,10 с), желудочков (0,06 - 0,09
с): сначала возбуждаются межжелудочковая перегородка и папиллярные мыш-
цы, затем верхушка сердца, стенки желудочка и последним миокард основания
желудочков.
6. Сократимость и сокращение сердца.
6.1. Функциональная структура кардиомиоцита.
••• Миофибриллы занимают - 50 % объема миоцита.
Саркомеры н протофибриллы (актиновые и миозиновые нити).
Т-система (Т-трубочки и цистерны саркоплазматической сети).
ив
Рис. 46. Схема ПД рабочего кардиомици-
та. КУД - критический уровень деполя-
ризации, МПП - мембраный потенциал
покоя. Объяснения в 5.1.
232
6.2. Электромеханическое сопряжение.
❖ Потенциала действия, распространяющийся по сарколемме, открывает в
фазе «плато» Са2*-каналы (L-типа). Поступление Са2* из межклеточной сре-
ды в миоциты приводит к активации Са2*-каналов рианодиновых рецепто-
ров гладкой ЭПС и выходу Са2* из ее цистерн (Са2*-индуцируемая мобилизация
Са2* из цистерн; блокатор каналов веропамил, активатор кофеин). (Без внеклеточного Са2*
сердце останавливается в течение 1 мии.)
❖ Кроме этого, ионы Са2* выходят в гиалоплазму (цитозоль) также через
ИФз-зависимые Са2*-каналы мембраны цистерн, активируемые через Р-
адренорецепторы кардиомиоцитов норадреналином и адреналином.
❖ Повышение концентрации Са2* в гиалоплазме с 10'7 до 10"5 М (пороговая
концентрация для сокращения) запускает сокращение.
6.3. Механизмы сокращения кардиомиоцитов (актиновый тип регуляции сокращения).
❖ Связывание Са2* с тропонином увеличивает степень спирализации тропо-
миозина, что открывает миозинсвязывающие участки актиновых нитей.
❖ АТФаза миозиновой головки вызывает гидролиз АТФ до АДФ и неорга-
нического фосфата (Ф„), но продолжает удерживать оба продукта. В таком
состоянии головка связывается с актиновой нитью, образуя с ее нитью угол
около 90°.
❖ Отсоединение АДФ и Фн от головки миозина сопровождается основным
выделением свободной энергии (силовой удар). В результате головка пово-
рачивается в шарнирной области до угла 45° (наименьшая энергия актомиози-
новой связи), осуществляя гребковое движение, что вызывает перемещение
актиновой нити вдоль миозиновой на 1 % длины саркомера (на - 10 нм).
❖ Присоединение АТФ к головке миозина вызывает разъединение актомио-
зиновых мостиков. Далее головка присоединяется в новом месте - ближе к
Z-линии, и цикла повторяется.
❖ При максимальном сокращении (до 50 % длины саркомера) необходимо
около 50 циклов образования и разъединения актомиозиновых мостиков.
6.4. Механизмы расслабления кардиомиоцитов.
❖ Возврат концентрации Са2* к исходному уровню (10"7 -10’8 М).
• Депонирование Са2* (~ 80%) в цистерны ЭПС в результате активацш^
Са2*-насоса играет главную роль в удалении Са2* из гиалоплазмы.
• Са2*, поступивший из внеклеточный жидкости (~ 20%), удаляется из
гиалоплазмы с помощью ЗНа*/Са2*-ионообмениика и Са2*-насоса плаз-
молеммы. (Работа иоиообмеииика требует высокого трансмембранного градиента
Na*, создаваемого К,№-насосом, если деятельность насоса,тормозится, например гли-
козидами, концентрация Са2* в гиалоплазме будет повышаться.)
❖ При снижении уровня Са2* до порогового уровня (10'7 -10'8 М) происходит
рассоединение актомиозиновых мостиков.
6.5. Некоторые особенности сокращения сердца.
❖ Миокард сокращается в соответствии с законом «всё или ничего», что обу-
словлено синхронной активностью пейсмекерных клеток и свободным рас-
пространением ПД из одного миоцита в другой через высокопроводящие
контакты.
233
❖ Сердце действует в режиме одиночных сокращений, что связано с длитель-
ной фазой абсолютной рефракгерности потенциала действия, которая «пе-
рекрывает» фазу сокращения миокарда (новое сокращение возможно толь-
ко во время диастолы).
Л* При одной н той же частоте нервной стимуляции сердца оно может развивать разную силу
сокращения, это связано со способность кардиомноцитов модулировать количество посту-
пающего в гиалоплазму Са2* на один ПД.
7. Гормональная функция сердца - образование натрийуретического гормона (сек-
реторными кардиомноцитамн преимущественно правого предсердия, а также нейронами ЦНС, клет-
ками эндотелия сосудов).
7.1. Регуляция продукции.
❖ Рецепторы к На+-уретическому гормону расположены на клетках нефрона,
сосудов, коре надпочечников, плаценте.
❖ Стимуляция его продукции происходит прн:
• растяжения кровью предсердий (увеличение пред- или постнагрузки
сердца);
• повышении концентрации ангиотензина П, вазопрессина, катехолами-
нов (повышение АД н частоты сердечных сокращений);
• гипернатрнемнн.
❖ Ингибируют продукцию противоположные явления.
7.2. Физиологические эффекты.
❖ Механизм действия: Ыа+-уретический пептид действует через ANP-
рецептор (тнрозннкиназа, интегрированная с гуанилатциклазой) —► повышение
уровня цГМФ —► увеличение активности протеинкиназы G.
❖ В почках:
• увеличивается выведение с мочой Na+ в результате снижения его реаб-
сорбции (угнетает синтез альдостерона, ренина и АДГ);
• усиливается клубочковая фильтрация (в результате расширения афферентных
н сужения эфферентных артериол).
♦♦♦ Расширение крупных сосудов и коронарных артерий, снижение АД в результате торможе-
ния секреции ренина и образования ангиотензина П; подавление секреции из эндотелия
сосудосуживающих эндотелннов.
*♦* В ЦНС: торможение центров жажды и солевого аппетита в результате снижения секреции
вазопрессина.
8. Профильный материал (см. тему 18).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). Прн
подготовке реферата использовать список основной н дополнительной ли-
тературы (задание 3, 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мнн.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11-16].
234
3.2. Дополнительная литература (см. с. 514 - 522): [7], [8], [27], [30], [35], [43], [47],
[52], [87]. [113], [140], [143], [152], [153], [207], [208], [210], [219]. [222], [225], [245],
[251-254].
33. Дистанционные средстеа обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. -1999; тесты ос-
таточного уровня знаний по д анной теме.
3.4. Учебная каината для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
33. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться иа практи-
ческом занятии по д анной теме, итоговом занятии н экзамене.
235
Тема 16. Регуляции сердечной деятельности. Кровообращение в мио-
карде
Цель: а) знать регуляцию сердечной деятельности (многенную, гуморальную, нервную), особен-
ности и механизмы регуляции кровоснабжения миокарда;
б) уметь использовать эти знания для объяснения изменений функций миокарда н показа-
телей его работы прн различных физиологических состояниях организма;
в) ознакомиться с методиками рефлекторного (сенсокардиальные и внсцерокарднальные
J рефлексы) влияния на сердечную деятельность.
Мотивация темы: см. тему 15.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логи-
ческую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения,
определяются лектором в конце читаемой по данной теме лекции, и обозначены в
«Графике и содержании самостоятельной работы студентов». Остальные фрагменты
задания 1 могут быть использованы для коррекции материала записанной лекции и
подготовки рефератов.
1. Общая характеристика регуляции сердечной деятельности.
1.1. Регуляция деятельности сердца - это приспособление минутного сердеч-
ного выброса к метаболической потребности организма.
1.2. Основные виды регуляторных влияний на сердце.
♦ > Хронотропные - влияние на ритм сердца («+» и «-»).
❖ Инотропные - влияние на силу сердечных сокращений («+» и «-»).
Батмотропные - влияние на возбудимость сердца («+» и «-»).
❖ Дромотропные - влияние на проводимость сердца («+» и «-»).
1.3. Гетерометрическая регуляция осуществляется в результате изменения
длины волокон миокарда в ответ на изменение притока крови к сердцу; го-
меометрическая регуляция осуществляется при исходно одинаковой длине
волокон миокарда (постоянном кровенаполнении камер сердца).
2. Миогенные механизмы саморегуляции сердца.
2.1. Закон сердца (О. Франк, 1895; Э. Старлинг, 1912).
❖ Формулировка: сила сокращения желудочков сердца прямо пропорцио-
нальна длине их мышечных волокон перед сокращением (положительный
инотропный эффект, гетерометрическая регуляция).
❖ Механизмы: число актомиозиновых мостиков наибольшее при растяже-
нии саркомера на - 10 % длины покоя; растяжение через механочувст-
вительные Са2+-каналы стимулирует выход Са2+ из гладкой ЭПС.
❖ Физиологический смысл закона сердца - приспособление сердца к пред-
нагрузке: увеличение притока крови при физической работе, мобилиза-
ции крови из депо, горизонтальном положении тела и др.
* •* Предел действия закона сердца (до 20 % растяжения миоцитов, что соответствует дав-
лению в левом желудочке 30 - 40 мм рт. ст.).
2.2. Феномен Анрепа (1912).
236
❖ Формулировка: сила сокращения левого желудочка прямо пропорцио-
нальна повышению давления в аорте (положительный инотропный эффект, го-
меомегрическая регуляция).
❖ Механизмы: 1) повышение АД в аорте увеличивает коронарный крово-
ток, улучшает метаболизм сержа и силу его сокращений; 2) повышение
АД в аорте приводит к повышению конечнодиастолического давления,
и серже реагирует в соответствии с законом Старлинга.
❖ Физиологическое значение эффекта Анрепа (приспособление сердца к
постнагрузке - нагрузке «давлением»),
2.3. Эффект «лестницы» (Г. Боуднч, 1871).
❖ Формулировка: при повышении частоты сокращения сердца увеличива-
ется и сила его сокращения (положительный хроноинотропный эффект, разно-
видность гомеометрической регуляции). Механизм связывают с накоплением
Са2+ в гиалоплазме.
❖ Физиологический смысл эффекта «лестницы»: связь ЧСС и их силы наи-
более эффективно увеличивает МОК.
3. Внутрисердечные периферические рефлексы (метасимпатическая нервная сис-
тема).
3.1. Рефлекторная дуга периферических рефлексов в серже:
❖ механо-, хеморецепторы миокарда, особенно в предсердиях и левом же-
лудочке;
❖ афферентные, вставочные, эфферентные (адренергические, холинэрги-
ческие и др.) нейроны ганглиев сержа;
❖ эффектор - кардиомиоциты различных отделов сердца.
3.2. Кардиостимулирующий внутрисердечный рефлекс возникает:
❖ при раздражении рецепторов предсердий (увеличение притока крови);
❖ при снижении кровенаполнения желудочков и давления в аорте;
при сочетанном действии двух предыдущих факторов.
3.3. ’ Кардиоингибирующий внутрисердечный рефлекс возникает при раздраже-
нии рецепторов предсердий и желудочков на фоне переполнения желудоч-
ков кровью и повышения давления в аорте.
3.4. Физиологическое значение внутрисердечных рефлексов - осуществляют
коррекцию механизмов саморегуляции сержа, стабилизацию сердечного
выброса (их роль повышается в трансплантированном сердце).
4. Нервная регуляция (по типу истинных рефлексов).
4.1. Дуга рефлексов.
❖ Рецепторы сержа и сосудов (собственные рефлексы), другие рецепторы
организма (сопряженные рефлексы).
❖ Сердечно-сосудистый центр продолговатого мозга (В.Ф.Овсянников,1871).
• Афферентная импульсация поступает в центр от:
"Ф- рецепторов сосудов и сержа в ядро ожночного пути (IX и X
нервов);
у- хеморецепторной зоны дыхательного центра при изменении хи-
мического состава крови (ацидоз, гипоксемия, гиперкапния);
237
-ф- от других рецепторов организма через коллатерали в ретикуляр-
ную формацию.
• Кардиостимулирующая (и прессорная) зона центра расположена в
заднебоковых отделах продолговатого мозга на уровне нижнего угла
ромбовидной ямки.
ф- В нее входят структуры продолговатого мозга (ядро одиночного пу-
ти, латеральное и парамедианное ретикулярные ядра, хеморецепторная зона
дыхательного центра), раздражение которых вызывает повышение
ритма сердца н АД.
-ф- Нейроны этой зоны имеют эфферентный выход на симпатиче-
ские нейроны Thu - для сердца (н Th, -1< - для сосудов).
• Кардиоингибирующая (и депрессорная) зона находятся в передних от-
делах продолговатого мозга у нижнего угла ромбовидной ямки.
ф- В нее входят структуры ствола (гигантоклеточное ретикулярное ядро,
ретикулярное вентральное ядро, каудальное н оральное ядра моста, заднее
ядро X нерва), раздражение которых вызывает торможение функ-
ции сердца и снижение АД.
ф- Нейроны этой зоны имеют эфферентный выход на па-
расимпатические нейроны заднего ядра X нерва - для сердца (IX
н VII нерва - для некоторых сосудов головы) и оказывают тормозной
эффект на спинальные симпатические нейроны, иннервирующие
сердце (н сосуды).
ф- Имеется выраженный тонус покоя кардноингибирующей зоны,
снижение ее тонуса стимулирует работу сердца.
4.2. Влияние блуждающих нервов на сердце (К. Фолькман, 1838; Э. и Г. Вебер, 1845).
Правый X нерв иннервирует преимущественно правое предсердие н синусный узел
(преобладает влияние на ритм).
Левый X нерв иннервирует преимущественно левое предсердие и предсердно-
желудочковый узел (преобладает влияние на атриовентрикулярную проводимость).
Преганглионарные нейроны находятся в парасимпатическом ядре X нерва; постганг-
лионарные нейроны - в нервном сплетении сердца.
❖ Медиатор нервно-мышечного синапса - ацетилхолин действует на Мг-
холинорецепторы кардиомиоцитов. Происходит уменьшение автоматии,
возбудимости, проводимости желудочков н сократимости. Изучаются
несколько механизмов этого действия.
• Ацетилхолин может прямо активировать К^-каналы сарколеммы,
минуя вторые посредники, что объясняет его короткий латентный
период и краткое последействие. Более длительную акгнвапию К*-
каналов ацетилхолин оказывает через G-белок, стимулируя гуанн-
латцнклазу, увеличивая образование цГМФ и активность протеин-
киназы G. Повышение выхода К* из клетки приводит к:
ф- увеличению поляризации мембраны, что снижает возбудимость;
ф- замедлению скорости МДД (замедление ритма);
238
замедлению проведения в атриовентрикулярном узле (в результа-
те уменьшения скорости деполяризации);
ф укорочению фазы «плато» (что уменьшает входящий в клетку Са2+-ток)
и снижению силы сокращения (преимущественно предсердий).
Ф Вместе с тем укорочение фазы «плато» в кардиомиоцитах предсердий приво-
дит к уменьшению периода рефрактерности, т.е. повышению возбудимости
(риск предсердных экстрасистол, например, во время сна).
• Ацетилхолин оказывает через Gj-белок тормозящее действие на аде-
нилатциклазу, снижая уровень цАМФ и активность протеинкиназы
А. В результате уменьшается проводимость Са2+-каналов и входя-
щий в клетку Са2+-ток, что приводит к снижению силы сокращения.
• Ацетилхолин через М-холинорецепторы на пресинаптических окон-
чаниях симпатических волокон уменьшает в них освобождение но-
радреналина и, следовательно, симпатических влияний на миокард.
❖ Тонус покоя кардиоингибирующей зоны сердечно-сосудистого центра
обусловлен: импульсацией с рецепторных зон дуги аорты, каротидного
синуса н сердца; импульсацией в ретикулярную формацию с различных
сенсорных систем; гуморальными факторами (гормоны, СОг); влиянием
из дыхательного центра.
4.3. Влияние на сердце симпатических нервов (И.Ф. Циои, 1867).
♦ ♦♦ Преганглионарные нейроны в боковых рогах Th, - Th5 спинного мозга. Ганглионарные
нейроны преимущественно в звездчатом (нижний шейный + верхний грудной) ганг-
лии. Оии иннервируют практически равномерно сократительный миокард и проводя-
щую систему сердца.
❖ Медиатор нервно-мышечного синапса - норадреналин действует пре-
имущественно через Pi-адренорецепторы (в меньшей степени через а,- н 02-
адренорецепторы, имеющиеся в небольшом количестве). Происходит увеличение
автоматии, возбудимости, проводимости и сократимости миокарда.
Эффект осуществляется с более длительным латентным периодом и по-
следействием:
❖ Механизмы: стимуляция норадреналином [^-адренорецепторов приво-
дит через Gs-белок к активации аденилатциклазы, повышению образо-
вания цАМФ и активности ПК А, фосфорилирующей различные белки:
• в результате активации Са2+-каналов Т-типа и Iz-типа (через G,-белок и
протеинкиназу А) в атипичных кардиомиоцитах увеличивается ско-
рость МДД («+» хронотропный эффект);
• повышение активности Са2+-каналов и торможение Са2+-насоса цис-
терн ЭПС, а также увеличение входа Са2+ в фазе «плато» (через откры-
тые Са2*-каналы L-типа) в рабочих миоцитах приводит к повышению
уровня Са2+ в гиалоплазме (кроме того, происходит торможение ингибирую-
щей субъединицы тропонина) («+» инотропный эффект);
• активация Са2+- и №+-каналов приводит к уменьшению порогового
потенциала, уменьшению времени развития ПД и рефрактерного пе-
риод («+» батмотропный и дромотропный эффекты).
239
❖ Тонус центров симпатической иннервации сердца в покое не выражен.
4.4. Интеро- и экстероцептивные рефлекторные влияния на сердце.
❖ Внутрисердечные рефлексогенные поля (кардио-кардиальный рефлекс).
• Рефлекс Бейнбриджа (1915): вливание в вену крови или физиологиче-
ского раствора в объеме, достаточном для растяжения правого пред-
сердия, вызывает «+» хронотропный эффект (увеличение ЧСС и
МОК) и почти не влияет на систолический объем. Наиболее важна
при этом стимуляция рецепторов правого предсердия в области
устья полых вен (при двусторонней перерезке блуждающих нервов эффект ис-
чезает).
• При раздражении механорецепторов левого и правого желудочков
бопее типичной реакцией является «-» инотропный эффект.
• При внутривенном вливании химических веществ (спирта, алкалоидов и др.) с эн-
докардиальных рецепторов желудочков сердца и коронарных сосудов возникает
рефлекс Бецольда - Яриша с триадой: брадикардия, гипотензия, апноэ.
❖ Внутрисосудистые рефлекторные поля (вазо-кардиальные рефлексы).
• С барорецепторов дуги аорты и синокаротидных зон (например, при
повышении АД) возникает брадикардия и снижение сердечного вы-
броса (при снижении АД - противоположные эффекты).
• С хеморецепторов дуги аорты и синокаротидных зон типичной реф-
лекторной реакцией на гипоксию являются тахикардия и вазоконст-
рикция.
❖ Висцеральные рефлекторные поля (висцеро-кардиальные рефлексы):
прн раздражении рецепторов брюшины (рефлекс Гольца) или эпигастраль-
ной области (рефлекс Тома - Ру) возникает брадикардия вплоть до оста-
новки сердца. (Ускользание сердца от вагуса обусловлена Ка4-каналами атипичных
кардиомиоцитов, открываемыми при гиперполяризации.)
❖ Проприоцептивные рефлекторные поля (моторно-кардиальные рефлек-
сы): при мышечной нагрузке возникает тахикардия и увеличение сер-
дечного выброса.
❖ Экстероцептивные рефлекторные поля (сенсо-кардиальные рефлексы):
например, рефлекторная брадикардия при надавливании на глазные яб-
локи (рефлекс Данини - Ашиера), при раздражении рецепторов носовой по-
лости, хоподовых рецепторов кожи живота.
5. Гуморальная экстракардиальная регуляция.
5.1. Влияние гормонов на деятельность сердца.
❖ Катехоламины: (адреналин и норадреналин) через Pi-адренорецепторы
и активацию аденилатциклазной системы оказывают «+» ино-, хроно-,
батмо- и дромотропный эффекты: механизмы действия аналогичны
симпатическому влиянию (см. выше 4.3).
❖ Тироксин и глюкокортикостероиды через экспрессию и репрессию со-
ответствующих генов увеличивают на кардиомиоцитах количество Рг
адренорецепторов и уменьшают количество Мг-холииорецепторов, что
сопровождается повышением силы и частоты сердечных сокращений.
240
(Кроме того, тироксин активирует Ыа+-каналы и замедляет их инактивацию, что спо-
собствует увеличению возбудимости, а также активирует Са2*-насос цистерн ЭПС,
что, снижая уровень Са2* в цитозоле, способствует расслаблению сердца.)
❖ Глюкагон усиливает сокращение сердца через активацию аденилат-
циклазной системы в кардиомиоцитах.
5.2. Влияние электролитов на деятельность сердца.
❖ Влияние К*.
• Увеличение уровня внеклеточного К+ ведет к повышению калиевой
проницаемости мембраны, что может приводить как к ее деполяри-
зации, так и гиперполяризации. Умеренная гиперкалиемия (до 6
ммоль/л) чаще вызывает деполяризацию и повышает возбудимость
сердца. Высокая гиперкалиемия (до 13 ммоль/л) чаще вызывает гипер-
поляризацию, что угнетает возбудимость, проводимость и автома-
тию вплоть до остановки сердца в диастоле.
• Гипокалиемия (меньше 4 ммоль/л) снижает К+-проиицаемость мембра-
ны и активность К+,№+-насоса, поэтому возникает деполяризация,
вызывающая повышение возбудимости и автоматии, активацию ге-
теротропных очагов возбуждения (аритмию).
❖ Влияние Со2*.
• Гиперкальциемия ускоряет диастолическую деполяризацию и ритм
сердца, повышает возбудимость и сократимость, очень высокая кон-
центрация может привести к остановке сердца в систоле.
• Гипокальциемия снижает диастолическую деполяризацию и ритм.
6. Интеграции механизмов, регулирующих работу сердца.
6.1. Гипоталамус осуществляет:
❖ прямые нервные связи с парасимпатическим и симпатическим центрами
(передний и задний гипоталамус соответственно) и интеграцию этих вегетатив-
ных центров в регуляции сердца;
❖ интеграцию вегетативной и эндокринной регуляции сердечной деятель-
ности (гипоталамо-гипофизарная регуляция, симпатоадреналовая сис-
тема);
❖ включение сердца как вегетативного компонента пищевой, половой, аг-
рессивно-оборонительной и других поведенческих актов.
6.2. Кора больших полушарий головного мозга - высшая ступень в иерархии
механизмов регуляции сердца.
Кардиоактивные зоны коры мозга (моторная и премоторная зоны, поясная из-
вилина, орбитальная поверхность лобиых долей, передняя часть височной доли) час-
тично совпадают с зонами корковой проекции сердечной афферентации.
❖ Через кору мозга регуляция сердца включается для обеспечения высших
форм целенаправленного поведения человека.
❖ Кора головного мозга включает реакцию сердца в периферические ком-
поненты психофизиологических процессов (эмоций, внимания и др.).
❖ Кора мозга создает возможность условиорефлекториого влияния иа
сердечную деятельность (например, предстартовое состояние).
241
7. Кровоснабжение миокарда.
7.1. Структурные особенности.
♦> Левая коронарная артерия снабжает левый желудочек, межжелудочковую перегородку
и большую часть левого н правого предсердий.
Правая коронарная артерия снабжает правый желудочек н часть левого н правого
предсердий.
& Плохо выражены межартернаяыше анастомозы (пропускают 3 - 5 % кровотока).
7.2. Количественная характеристика коронарного кровотока.
❖ В покое через коронарные артерии сердца протекает 60 - 80 мл кро-
ви/100 г/мин, т.е. - 5 % МОК (250 мл/мин), потребляется 6 - 10 мл
Qz/100 г/мин. При физической нагрузке эти показатели увеличиваются в
4-7 раз.
❖ Сердце извлекает из артериальной крови ~ 50% От (в организме в сред-
нем 25 - 30 %).Поэтому увеличение потребления От в сердце обеспечи-
вается в основном увеличением коронарного кровотока и в меньшей
степени увеличением экстракции От из крови.
Базальный уровень потребления О2, сохраняющий структуру сердца, около 1,5
мл/100г/мин.
7.3. Резкие колебания коронарного кровотока в течение кардиоцикла.
'Ъ В период систолы левого желудочка его сосуды пережимаются, крово-
ток составляет - 15 % общей величины в кардиоцнкле, в период диасто-
лы кровоток составляет - 85 %; в предсердиях и правом желудочке эти
колебания слабо выражены.
Роль диастолического давления в аорте для коронарного кровотока оп-
ределена тем, что оно является входным давлением для коронарных
сосудов.
7.4. Регуляция коронарного кровотока.
❖ Миогенная регуляция обеспечивает ауторегуляцию коронарного крово-
тока в диапазоне АД = 70 -145 мм рт. ст.
❖ Миоциты коронарных артерий имеют а- и Р-адренорецепторы и М-
холинорецепторы, количество которых больше в крупных артериях.
❖ Нейрогенный тонус составляет в покое около 20 % тонуса.
• Симпатическая стимуляция через а-адренорецепторы вызывает су-
жение коронарных сосудов н снижение кровотока (первая кратко-
временная фаза); увеличивая работу сердца и образование вазоак-
тивных метаболитов, вызывает расширение коронарных сосудов и
увеличение коронарного кровотока (вторая более длительная фаза).
(Расширению сосудов способствует и слабый эффект норадреналина на рг
адренорецепторы.)
• Парасимпатическая стимуляция вызывает расширение сосудов че-
рез М-холинорецепторы (первая кратковременная фаза); уменьшая
работу сердца, приводит к сужению коронарных артерий (вторая бо-
лее длительная фаза) (роль блуждающего нерва в регуляции коронарного кро-
вотока признается ие всеми исследователями).
❖ Гуморальная регуляция.
242
• Вазоактивные метаболиты:
Ф- повышение концентрации аденозина, оксида азота (NO), К*, Н+,
СОг, снижение Ог приводит к расширению коронарных артерий;
ф- противоположная динамика метаболитов приводит к сужению
коронарных артерий.
• Гистамин, брадикинин, простагландины Е, Ыа+-уретический пептид
приводят к расширению коронарных сосудов.
• Адреналин через ед-адренорецепторы суживает (слабый эффект),
через Р-адренорецепторы расширяет коронарные сосуды (преобла-
дающий эффект).
• Ангиотензин П н вазопрессин суживают коронарные сосуды.
8. Профильный материал (см. тему 18).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема
реферата выбирается с использованием учебных элементов (задание 1).
Прн подготовке реферата использовать список основной и дополни-
тельной литературы (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с.,
регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для
самоподготовки студентов.
3.1. Основной учебно-методнческий комплекс (см. с. 514, использовать имеющие-
ся у вес учебники и пособия): [1-9] [11-16].
3.2. Дополнительная литература (см.с. 514 - 522): [7], [8], [27], [30], [43], [52],
[113], [140], [143], [152], [153], [207], [208], [210], [219], [222], [225] [245] [251-
254].
3.3. Дистантные средства обучения. Использования страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из
«Атласа по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999;
тесты остаточного уровня знания по данной теме.
3.4. Учебная комната для самонодготовки, оснащенная учебными таблицами,
диапроектором и слайдами (по графику кафедры).
33. Консультации преподавателей еженедельно по индивидуальному графику.
3.6 . Контроль самостоятельной работы студентов будет осуществляться на
практическом занятии по данной теме, итоговом занятии й экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 16—1. После длительного лечения fj-адреноблокаторами пациент резко прекратил т
прием. Используя свои знания по физиологии ретуляции сердца, ответьте, как изменится дея-
тельность сердца после отмены препарата. Объясните свой ответ.
243
Тема 17. Нагнетательная функция сердца. Внешние проявления дея-
тельности сердца. Методы исследовании сердца
Цель: а) знать сердечный цикл н его фазовую структуру; систолический н минутный объемы крови,
сердечный индекс, работу сердил; внешние проявления сердечной деятельности (электриче-
ские, звуковые, механические), их происхождение и методики исследования (электрокардио-
графия, фонокардиография, ультразвуковая кардиография н др.);
б) уметь объяснить происхождение тонов сердца, артериального давления, пульса, дать функ-
циональную оценку основным показателям нагнетательной функции сердца, выбрать адекват-
ные методики исследования сердца;
в) ознакомиться с практическими навыками: на уровне знаний: электрокардиографией н фазо-
вым анализом сердечного цикла, исследованием сердечного выброса; иа уровне умения: с ана-
лизом ЭКГ здорового человека, аускультацией тонов сердца, пальпацией пульса.
Мотивация темы: см. тему 1S.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике н
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Сердечный цикл (см. рис. 47).
1.1. Систола предсердий (0.1 с; зубец Р на ЭКГ).
❖ Сокращение миокарда начинается с зоны СА-узла (устья полых вен), что
блокирует поступление крови в полые вены.
❖ Сокращение стенок предсердий создает IV тон сердца (регистрируется на фо-
нокардиограмме).
❖ Давление в предсердиях увеличивается с 0 до 5 в правом, с 5 до 12 мм рт.
ст. в левом.
❖ Состояние клапанов: створчатые открыты, полулунные закрыты.
❖ Гемодинамика: в желудочки переходит последняя порция (~ 30 мл) крови, а
основное количество (80 - 90 мл) уже имеется в желудочках к моменту
систолы предсердий.
❖ Остальное время цикла (0,7 с) предсердия находятся в диастоле и заполня-
ются кровью.
1.2. Систола желудочков (0,33 с, зубцы QRST на ЭКГ).
❖ Период напряжения (0,08 с).
• Фаза асинхронного сокращения (0,05 с):
ф давление в желудочках не меняется и равно диастолическому
(- 0 в правом, - 5 мм рт. ст. в левом);
ф состояние клапанов: створчатые открыты, полулунные закрыты;
ф внутрижелудочковой гемодинамики нет.
• Фаза изометрического сокращения (0,03 с):
244
Ф давление в желудочках быстро растет: в левом желудочке до 70 -
80, в правом до 10 - 15 мм рт. ст.;
ф состояние клапанов: створчатые закрываются, полулунные закры-
ты;
Ф вибрация створчатых клапанов и напряжение миокарда желудочков
формируют I (систолический) тон;
Ф внутрижелудочковой гемодинамики нет.
Период изгнания крови.
Фаза быстрого изгнания
(0,12 с):
ф давление в левом же-
лудочке 120 - 130, в
правом до 20 - 25 мм
рт. ст. (пик АД достига-
ется в конце фазы);
Ф состояние клапанов:
створчатые закрыты,
полулунные отбыты;
гемодинамика: про-
исходит выброс кро-
ви (около 2/3 систо-
лического объема) в
аорту и легочный
СТВОЛ (максимальный
систолический поток на
аортальном клапане около
1,5 м/с), кровь посту-
ПРЕДСЕВДИЯ ЖЕЛУДОЧКИ
диастола
Направление потоп кровя
-систола
Рнс. 47. Схема сердечного цикла.
Объяснения в задании 1. 1.
пает в предсердия.
• Фаза медленного изгнания (0,13 с):
•ф давление в левом желудочке 100 - 110, в правом - до 20 - 25 мм рт.
ст.;
Ф состояние клапанов: створчатые закрыты, полулунные открыты;
Ф гемодинамика: происходит выброс крови (около 1/3 систолического
объема) в аорту и легочный ствол, кровь поступает в предсердия;
ф конечносистолический объем желудочков равен - 60 мл.
••• Систолическую функцию желудочков (по данным УЗИ-исследования сердца) характери-
зуют фракция выброса, которая в норме составляет 55-70% коиечиодиастолическбго объ-
ема крови, и фракция укорочения - разница между конечнодиастолическим н конечно-
систолическим размерами желудочка, выраженная в процентах к конечиодиастолическому
размеру (норма > 30%).
13. Диастола желудочков (0,47 с, сегмент Т - Р на ЭКГ)
❖ Период расслабления (0,12 с).
• Протодиастолическая фаза (0,04 с):
Ф интервал времени между прекращением изгнания крови и захлопы-
ванием полулунных клапанов;
245
• ф- давление в желудочках несколько меньше давления в аорте и ле-
гочном стволе;
ф- состояние клапанов: створчатые закрыты, полулунные закрывают-
ся;
ф- вибрация полулунных клапанов формирует П (диастолический) тон.
• Фаза изометрического расслабления (0,08 с):
•ф- давление в обоих желудочках быстро падает почти до 0 мм рт. ст.;
•ф- состояние клапанов: створчатые закрыты, полулунные закрыты.
❖ Период наполнения желудочков кровью с).
• Фаза быстрого наполнения (0,08 с) связана с повышением объема и
давления крови в предсердиях во время предшествующей систолы же-
лудочков :
ф- давление в правом желудочке - 0, левом - 5 мм рт. ст.;
• ф' состояние клапанов: створчатые открыты, полулунные закрыты;
ф- гемодинамика: происходит быстрое наполнение желудочков кро-
вью (до 70 % объема), (максимальный диастолический поток на митральном
клапане около 100 см/с);
• ф- вибрация стеиок желудочковЧформирует Ш той (регистрируется на фо-
нокардиограмме).
• Фаза медленного наполнения (0,17 с):
• ф- давление в правом желудочке - 0, левом - 5 мм рт. ст.;
ф- состояние клапанов: створчатые открыты, полулунные закрыты;
ф- гемодинамика, происходит уменьшение скорости наполнения же-
лудочков кровью.
• Фаза наполнения желудочков в связи с систолой предсердий: поступает
- 30 мл крови, конечнодиастолический объем желудочков 120 -130 мл.
Диастолическую функцию желудочков (по данным УЗИ сердца) характеризует отношение
Е/А (норма > 1), где Е - скорость потока крови на митральном клапане при пассивном на-
полнении желудочка (см/с), А - при систоле предсердий.
1.4. Методы исследования сердечного цикла.
❖ Поликардиография - метод исследования сердечного цикла, основанный на
синхронной регистрации ЭКГ, ФКГ и сфигмограммы сонной артерии. По-
зволяет рассчитать длительность периодов и фаз сердечного цикла, индекс
напряжения миокарда, скорость опорожнения желудочков и другие
показатели.
❖ Зондирование сердца (Форсманн, 1929) - введение катетера в полости правого
или левого сердца с целью изучения внутрисердечной гемодинамики, дав-
ления и газового состава крови в полостях сердца и других показателей.
❖ Ультразвуковое исследование сердца (см. ниже 6.4).
2. Энергетическое обеспечение нагнетательной функции сердца.
2.1. Основные метаболиты, в равных долях используемые сердцем для синтеза
АТФ: жирные кислоты, глюкоза, лактат.
246
2.2. Основные пути образования АТФ: окислительное фосфорилирование (самый
мощный); гликолиз (устойчив к гипоксии); использование креатинфосфата (са-
мый быстрый).
2.3. Основные пути использования АТФ: процесс сокращения (- 70 %); транспорт
Са2+ (-15 %); работа К+, Na+-Hacoca (- 5 %); процессы синтеза (- 10 %).
2.4. Потребление Ог миокардом: в покое 6 - 8 мл Ог/ЮО г в 1 мин, при физиче-
ской нагрузке до 30 мл Oj/lOO г в 1 мин.
3. Работа сердца (1,1 Дж за цикл).
3.1. Работа по перемещению объема крови против давления:
❖ левый желудочек (0,93 Дж);
❖ правый желудочек (0,14 Дж).
3.2. Работа по сообщению крови ускорения (около 0,03 Дж).
3.3. Коэффициент полезного действия сердца (15-40 %).
4. Методы исследовании и количественная оценка нагнетательной функции серд-
ца.
4.1. Систолический объем крови - объем крови, который сердце выбрасывает в
аорту и легочный ствол за один сердечный цикл (- 70 мл):
❖ Зависимость от ЧСС: при увеличении ее до 145 в мин заметного снижения
наполнения сердца и систолического объема не происходит, при более вы-
соком ритме систолический объем снижается;
• > зависимость от конечного систолического и диастолического объемов: уве-
личение конечнодиастолического и уменьшение конечносистолического
объемов крови приводит к увеличению систолического объема;
❖ влияние на систолический объем пола (у мужчин он больше на -10%, чем у
женщин) и положения тела в пространстве: переход в горизонтальное по-
ложение увеличивает систолический объем за счет увеличения притока
крови к сердцу;
❖ функциональный резерв сердца по этому показателю двукратный.
4.2. Минутный объем крови (МОК) - количество крови, которое выбрасывает
сердце в аорту (или в легочный ствол) за минуту (- 5 л/мин):
❖ прямо пропорциональная зависимость от систолического объема крови и
частоты сердечных сокращений;
❖ функциональный резерв по МОК шестикратный;
❖ влияние возраста и пола на функциональные резервы по МОК: у мужчин
выше, чем у женщин; снижается у пожилых и старых людей;
❖ систолический и минутный объем крови правого и левого желудочков
сердца строго согласован и одинаков (возникающие рассогласования ликвидиру-
ются в течение нескольких циклов).
4.3. Методы определения минутного объема крови:
❖ Метод Фика - определяют поступивший за минуту в организм Оз и его ар-
териовенозную разницу: МОК = Поступивший в 1 мин Ог / (a-в) Ог л. Ко-
личество вдыхаемого Ог определяется, например, с помощью метаболи-
метра. Для анализа артериовенозной разницы по Ог кровь берут из артерии
и путем катетеризации из правого предсердия или желудочка, где венозная
кровь равномерно смешана.
247
❖ Метод разведения основан на записи кривой разведения различных инди-
каторов (красителей, радиоактивных изотопов и др.), введенных в вену или
полость сердца.
❖ Интегральная реография основана иа регистрации изменения сопротивле-
ния тканей грудной клетки току сверхвысокой частоты в связи с выбросом
систолического объема крови. При этом уменьшение сопротивления про-
порционально величине систолического объема. По формуле, которая учи-
тывает амплитуду систолической волны реограммы, время изгнания, рас-
стояние между электродами и ряд других факторов, определяют величину
СО, а умножив ее на число сердечных сокращений, получают МОК.
❖ Ультразвуковое исследование сердца (см. 6.4).
5. Звуковые проявления сердечной деятельности.
5.1. Тоны сердца (звуковые колебания частотой 15 - 400 Гц).
❖ I (систолический) тон сердца (0,10 - 0,17 с) возникает в периоде напряжения
(фаза изометрического сокращения) систолы желудочков в результате виб-
рации створок створчатых клапанов (а также напряжения миокарда желудочков,
открытия полулунных клапанов и колебания стеиок аорты и легочной артерии).
❖ II (диастолический) тон сердца (0,06 - 0,08 с) возникает в протодиастоличе-
ской фазе диастолы желудочков в результате вибрации полулунных клапа-
нов (аортальный клапан захлопывается быстрее пульмонального, поэтому имеется нор-
мальное расщепление И тона).
III тон сердца (0,03 - 0,06 с) возникает в фазе быстрого наполнения диастолы
желудочков в результате вибрации их стенок.
❖ IV тон сердца возникает в систолу предсердий в результате вибрации их
стеиок.
5.2. Методы исследования тонов сердца.
❖ Аускультация.
• Акустическая проекция створчатых и полулунных клапанов:
Ф’ проекция верхушки сердца-митральный клапан;
• ф второе межреберье справа от грудины - аортальный клапан;
• ф второе межреберье слева от грудины - клапан легочной артерии;
• ф конец грудины, область прикрепления мечевидного отростка -
трехстворчатый клапан.
• Звуковая характеристика I и П тонов сердца: первый более продолжи-
тельный и глухой, второй более короткий и высокий.
❖ Фонокардиография - метод графической регистрации тонов и шумов серд-
ца; ФКГ здорового человека состоит из колебаний I, П, а также нередко Ш
и IV тонов сердца.
6. Механические пронвлення сердечной деятельности.
6.1. Сердечный толчок возникает в результате движения верхушки сердца вверх и
вперед при систоле желудочков, пальпируется в левом пятом межреберье по
срединно-ключичной линии.
6.2. Артериальный пульс: механизм образования, функциональная характеристика,
методы исследования: пальпация и сфигмография.
248
6.3. Венный пульс: механизм образования; метод исследования - флебография.
6.4. Методы регистрации механической деятельности сердца.
❖ Баллистокардиография - метод графической регистрации слабых смеще-
ний тела, вызванных сокращением сердца и движением крови в крупных
сосудах.
❖ Эхокардиография (УЗИ сердца) - метод визуализации полостей и структур
сердца при помощи ультразвуковых волн, отраженных от его различных
структур. Эхокардиографы имеют ультразвуковые датчики, одновременно
посылающие импульсный сигнал (2,25-бмГц) и воспринимающие в течение
пауз отраженные от структур сердца волны. Полученный эхосигнал посту-
пает на электронный усилитель, который подает информацию на регистра-
тор и на экран осциллографа. Прибор снабжен маркером для определения
временных интервалов и расстояния между структурами сердца. Возможна
синхронизация изображения сердца с зубцами ЭКГ, а также использование
эффекта Доплера, что позволяет изучить особенности кровотока в иссле-
дуемом участке. Позволяет определить толщину миокарда, размеры и объ-
емы камер сердца, кинетику клапанов, характеризовать систолическую и
диастолическую функции желудочков (см. выше 1.1 и 1.2) и другие показате-
ли.
7. Электрокардиография (ЭКГ) - метод графической регистрации во времени изме-
нений разности потенциалов электрического поля сердца в ее проекции на ось отве-
дения (А.Уоллер, 1887; В. Эйнтховен, 1903, Нобел, премия 1924). Метод позволяет исследовать
процессы автоматии, возбуждения, его проведения в сердце и не отражает прямо со-
кращение миокарда.
7.1. Для регистрации ЭКГ используются классические (двухполюсные от конечно-
стей: I правая рука - левая рука, П правая рука - левая нога, Ш левая рука - ле-
вая нога), усиленные (однополюсные от конечностей: aVR, aVL, aVF) и груд-
ные (однополюсные от грудной клетки: Vi - V6) отведения.
7.2. Дипольпо-векторная теория электрокардиографии. Диполь формирует раз-
ность потенциалов между возбужденными (-) и невозбуждеиными (+) миоци-
тами, вектор направлен от «-» к «+». В процессе возбуждения сердца возникает
три суммарных вектора: вектор деполяризации предсердий, вектор деполяриза-
ции желудочков, вектор реполяризации желудочков. Во время кардиоцикла
вектор меняется по величине и направлению. Максимальная величина вектора
достигает в период зубца R и достигает около 2 мВ/см. Если вектор проециру-
ется на положительную полуось отведения записывается положительный зубец,
если на отрицательную полуось - отрицательный зубец, если перпендикулярно,
то зубец не формируется.
7.3. Компоненты ЭКГ. В течение сердечного цикла записываются (рис. 48):
❖ пять постоянных зубцов (Р, Q, R, S, Т) н непостоянный зубец U;
❖ сегменты (участки ЭКГ между двумя соседними зубцами, расположенные в
норме иа изолинии): P-Q и S-T;
❖ интервалы (участки ЭКГ, включающие сегмент и один или несколько зуб-
цов): P-Q, Q-T.
249
7.4. Происхождение компонентов ЭКГ:
❖ зубец Р- возбуждение предсердий (0,05-0,25 мВ; 0,06-0,10 с);
Ф сегмент P-Q - предсердия возбуждены и нет разности потенциалов, в это
время происходит проведение возбуждения через предсердно-
желудочковый узел, по пучку Гиса и волокнам Пуркинье (0,06 - 0,10 с), ко-
торое ие сопровождается возникновением зубцов из-за малой массы прово-
дящей системы (~ 1%); (формирование диполя и вектора между возбужденными
предсердиями и невозбуждеиными желудочками не происходит из-за изолирующих
свойств фиброзной предсердно-желудочковой перегородки);
❖ зубец Q - возбуждение межжелудочковой перегородки и верхушки сердца
(до V» зубца R; до 0,03 с, всегда отрицательный);
Рис. 48 Электрокардиограмма и ее компоненты.
❖ зубец R - возбуждение основной массы мускулатуры желудочков, кроме
основания и субэпикардиального слоя (0,3 - 2,0 мВ; до 0,03 с, всегда положитель-
ный);
з убец S отражает состояние, когда возбуждены все отделы желудочков,
кроме их основания (у предсердно-желудочковой перегородки) (0 - 0,6 мВ; до
0,03 с, всегда отрицательный);
❖ сегмент S-Т- возбуждены все отделы желудочков, отсутствует разность
потенциалов (0,02 - 0,12 с);
❖ зубец Т - процесс реполяризации желудочков, начинающийся с верхушки
сердца и субэпикардиального слоя, миоциты которого имеют более корот-
кую фазу «плато» и рефрактерность (0,2 - 0,6 мВ; 0,10 - 0,25 с).
7.5. Анализ ЭКГ здорового человека включает в себя анализ сердечного ритма (регуляр-
ность, локализация пейсмекера сердца - синусный ритм, ЧСС); анализ проведения воз-
буждения по предсердиям, атриовентрикулярной системе, рабочему миокарду желу-
дочков: анализ желудочкового комплекса
❖ Анализ сердечного ритма включает в себя оценку регулярности сердечных со-
кращений, подсчет их частоты и определение источника возбуждения (пейсмеке-
ра) сердца.
250
• Для оценки регулярности сокращений сердца (правильный или неправильный
ритм сердца) измеряют продолжительность интервалов R-R в пяти последова-
тельно записанных сердечных циклах и вычисляют его среднюю величину.
Определяют в процентах наибольшее отклонение интервала R-R от средней
величины. Ритм считается правильным, если разброс измеренных интервалов
R-R не превышает 10% от средней величины.
• При правильном ритме, зная величину одного интервала, можно рассчитать
частоту сердечных сокращений. В норме у взрослого человека она равна 60-75
в минуту.
• У здорового человека пейсмекером сердца является синоатриальный узел, и
регистрируется синусовый ритм сердца. Признаком его являются: наличие во
П стандартном отведении положительных, одинаковой формы зубцов Р, каж-
дый из которых предшествует комплексу QRS.
Анализ функции проводимости в сердце включает в себя оценку времени прове-
дения возбуждения по предсердиям (длительность зубца Р), проведение по атрио-
вентрикулярной системе (длительность сегмента P-Q), проведение по рабочему
миокарду желудочков (длительность комплекса QRS).
Анализ зубца Р. Продолжительность зубца Р свидетельствует о проведении воз-
буждения в предсердиях. Его полярность является важным признаком локализа-
ции пейсмекера сердца (напр., положительный зубец Р в 1 и П отведениях харак-
теризует синусовый ритм). Зубец Р в норме всегда положителен в отведениях 1,П,
aVL, Vj-Vc; всегда отрицателен в aVR. Увеличение амплитуды зубца Р может сви-
детельствовать об увеличении массы предсердий (гипертрофия).
Анализ желудочкового комплекса QRST:
• Зубец Q всегда отрицателен, непостоянен. Оценка его амплитуды и длитель-
ности имеет большое клиническое значение (их увеличение является ЭКГ-
признаком инфаркта миокарда).
• Зубец R всегда положителен. Изменение его амплитуды позволяет оценить
амплитуду зубца Q, а его сопоставление с амплитудой зубцов R в других отве-
дениях - положение электрической оси сердца.
• Зубец S всегда отрицателен, непостоянен. В клиническом плане имеет значе-
ние для определения электрической оси сердца, гипертрофии миокарда
желудочков.
• Анализ сегмента S-Т имеет важное клиническое значение. Смещение его ниже
(более 0,5 мм) или выше (более 1 мм) изолинии является грозным патологиче-
ским признаком (ишемия миокарда).
• Анализ зубца Т включает определение направления (полярности) формы и ам-
плитуды. Их изменения могут свидетельствовать о нарушении коронарного
кровотока и обменных процессов в сердце. Зубец Т (как и зубец Р) всегда по-
ложителен в отведениях I, П, aVL, Va-Ve; всегда отрицателен - в aVR.
По ЭКГ можно определить электрическую ось сердца, которая является результи-
рующим вектором ЭДС сердца во фронтальной плоскости и в норме, как правило,
соответствует анатомической оси сердца и, следовательно, дает представление о
положении сердца в грудной полости. Приблизительно электрическую ось сердца
определяют по величине зубцов R в трех стандартных отведениях. При нормаль-
ном положении оси зубец R наибольший во П стандартном отведении. При откло-
нении электрической оси сердца влево (левограмма) зубец R наибольший в I отае-
251
дении; при отклонении вправо (правограмма) - в Ш отведении. Более точно элек-
трическую ось сердца характеризует угол альфа, образованный электрической
осью и осью I отведения, проведенной через условный электрический центр серд-
ца. Этот угол можно определить путем геометрического построения в треугольни-
ке Эйнтховена, зная величину зубцов QRS в двух стандартных отведениях, или по
специальным таблицам.
8. Профильный материал (см. тему 18).
Задание 2; Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7
мин.
Здддние З, Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас лекцию, учебник, практикумы и пособия): (1-9], [12-16].
3.2. Дополнительная литература (см.с. 514 - 522): [7], [8], [25], [27], [30], [43], [52],
[113], [140], [152], [153], [207], [208], [209], [210], [222], [245], [251-254].
3.3. Дистантные средства обучения. Использования страницы кафедры в Интернете -
www vsma.ac ru/-nphys : Слайды для компьютерной демонстрации из «Атласа по
физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты остаточ-
ного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуедьиому графику.
3.6. Контроль самостоятельной работы студентов по данной теме будет осуществ-
ляться на итоговом занятии и экзамене.
252
Тема 18. Регуляция гемодинамики
Цель: а) знать основные законы гидродинамики, функциональную классификацию кровеносных со*
судов, факторы, обеспечивающие движение крови по сосудам высокого и низкого давления;
виды кровяного давления; факторы, определяющие его величину, артериальный и венозный
пульс, их происхождение; характеристику микроциркуляторного русла, капиллярный крово-
ток н его особенности; роль микроциркуляции в обмене веществ между кровью н тканями;
механизмы регуляции тонуса сосудов, рефлекторную регуляцию системного артериального
давления, значение сосудистых рефлексогенных зон, сосудодвигательиый центр; функцио-
нальную систему поддержания кровяного давления и органного кровотока; лимфатическую
систему, ее функции, лимфообразование и лимфоток, их механизмы и регуляцию;
б) применять этн знания двя понимания функционального состояния сердечно-сосудистой
системы н кровоснабжения различных органов прн физиологических нагрузках и покое, выбо-
ра адекватных методик исследования различных параметров гемодинамики;
приобрести умения измерения артериального давления аускультативным методом; а также
знания принципов методик определение венозного давления, плетизмографии, реографии, ка-
пилляроскопии.
Мотивация темы: от уровня гемодинамики зависят трофика, метаболизм тканей, их энергообеспече-
ние, удаление продуктов обмена. Состояние кровообращения различных тканей в пределах
одного органа и различных органов в пределах организма при разных видах жизнедеятельно-
сти не одинаково. Знание механизмов управления и взаимосвязи локальной и системной гемо-
динамики позволяет разрабатывать рациональные режимы труда и отдыха, принципы профи-
лактики и лечения сердечно-сосудистых н других заболеваний.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике н
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции.
1. Основные законы гемодинамики.
1.1. Объемный кровоток (Q = dP/R) прямо пропорционален градиенту давлению
крови (для большого круга он практически равен давлению в аорте) н обратно пропор-
ционален сопротивлению сосудов (закон Пуазейля).
1.2. Линейная скорость кровотока (V - Q/tcj^) прямо пропорциональна объемному
кровотоку и обратно пропорциональна радиусу сосудов во 2-й степени, т.е. пло-
щади сечения сосудов.
1.3. Сопротивление кровотоку в сосуде (R = 81т|/7ег*) прямо пропорционально длине
сосуда (1), вязкости крови (я) и обратно пропорционально радиусу сосуда в 4-й
степени.
2. Функциональная характеристика сосудистого русла.
2.1. Сосуды высокого давления (артерии).
❖ Характеристика артерий:
• Эластические артерии (аорта, сонные, подмышечные, подвздошные)
обеспечивают принятие сердечного выброса без резкого повышения АД
и за счет своего растяжения создают энергию, которая поддерживает
кровоток во время диастолы сердца.
253
• Мышечные артерии (чем дальше от сердца, тем относительно больше количество
гладкомышечных клеток в стенки сосуда) создают активный сосудистый тонус
в результате сокращения гладких миоцитов.
❖ Артериальное давление (см. ниже п. 4).
❖ Суммарная площадь поперечного сечения: артерии - 20, аорта - 4 см2.
❖ Содержат небольшой объем крови (10 - 15 %).
❖ Доля в сосудистом сопротивлении - 25 %.
❖ Линейная скорость кровотока от 50 до 20 см/с.
❖ Основные функци - создание градиента давления крови в большом и малом
кругах кровообращения, сглаживание пульсации.
2.2. Сосуды сопротивления (артериолы, прекапилляры).
❖ Создают наибольшее сосудистое сопротивление (- 40 %).
❖ Давление кровн в них от 70 до 30 мм рт. ст.
❖ Суммарная площадь поперечного сечения - 40 см2.
❖ Линейная скорость кровотока от 20 до 5 см/с.
❖ Содержат наименьший объем крови (~ 2 %).
❖ Основная функция: стабилизация системного АД, перераспределение кро-
вотока, сглаживание пульсации.
2.3. Обменные сосуды - капилляры (входят в состав сосудов микроциркуляции: артериолы,
прекапилляры, капилляры, посткапилляры, венулы).
❖ Самая низкая линейная скорость кровотока (0,5 - 1 мм/с).
❖ Самая большая площадь суммарного сечения (- 4000 см2) и суммарная
площадь сосудистой стенки (S s 700 м2, однако в физиологических условиях
функционирует до 10% капилляров).
❖ Давление крови от 30 до 12 мм рт. ст.
❖ Содержат объем кровн 5 - 10 %.
❖ Доля в сосудистом сопротивлении - 25 %.
Основная функция - транскапиллярный обмен веществ.
• Диффузионно-осмотический механизм обмена метаболитами и водой
между внутрисосудистым и межклеточным отсеками (практически не влия-
ет на объем циркулирующей крови).
• Пиноцитоз и экзоцитоз с образованием временных транскапиллярных
каналов для транспорта белков.
• Фильтрационно-реабсорбционный механизм (рис. 49).
ф’ Силы фильтрации: капиллярное давление (~ 30 мм рт. ст.) + онкоти-
ческое давление интерстиции (- 7 мм рт.ст.) на артериальном конце
капилляра превышают силу реабсорбции (онкотическое давление
кровн ~ 25 мм рт.ст.) примерно на 12 мм рт. ст.; происходит фильт-
рация (~ 20 л/сут).
Сила реабсорбции иа венозном конце капилляра (онкотическое дав-
ление крови ~ 25 мм рт.ст.) превышает силы фильтрации: капил-
лярное давление (~ 10 мм рт.ст.) + онкотическое давлениие интер-
стнции (~ 7 мм рт.ст.) примерно на 9 мм рт. ст., здесь происходит
реабсорбция (~ 18 л/сут).
254
Ф" Гидростатическое давление межклеточной жидкости может быть нулевым (не
влияет на фильтрацию и реабсорбцию), иметь положительную величину (спо-
собствует реабсорбции) или отрицательную величину (способствует филь-
трации).
Рис. 49. Схематическое изображение процессов фильтрации и
реабсорбции в кровеносном капилляре: а - артериальный ко-
нец, в - венозный конец капилляра.
Ф’ Уравнение Стерлинга: Q = [(P„, + Р„) - (Р„ ± Рщжре,)] х ККФ, где Q - количест-
во фильтруемой или реабсорбируемой жидкости, Рш - давление крови в капил-
лярах, Рот- онкотичесое давление в межклеточной жидкости ткани, Рм - онкоти-
ческое давление в крови, Рннпрст - гидростатическое давление в интерстнции (по-
ложительное или отрицательное), ККФ - коэффициент капиллярной фильтрации
(мл/мин в 100 г ткани на 1 мм рт.ст.). Сдаиги уравнения могут быть в сторону
увеличения фильтрации и повышения реабсорбции.
2.4. Сосуды большого объема (венулы, вены).
❖ Содержат самый большой объем крови (70 - 80 %).
❖ Суммарная площадь поперечного сечения - 250 см2, полые вены - 7 см2.
❖ Самое низкое давление кровж (от 10 до 0 мм рт. ст.).
❖ Доля в сосудистом сопротивлении - 10 %.
❖ Линейная скорость кровотока от 3 до 18 см/с.
❖ Основные функции:
• Возврат к сердцу кровж осуществляется в результате:
Ф’ наличия небольшого градиента давления (- 10 мм рт. ст.);
ф- венозной констрикции и действия венозных клапанов;
ф- мышечного насоса (сокращения скелетной мускулатуры и действия
венозных клапанов);
Ф* дыхательного насоса (присасывающее действие грудной клетки,
обусловленное отрицательным плевральным давлением);
Ф’ предсердного насоса (сдвиг предсердно-желудочковой перегородки к верхушке
сердца при систоле желудочков).
• Депонирование и редепонирование крови (- 1 л крови):
Ф* большая растяжимость венозного резервуара (в 20 раз больше, чем ар-
терий: давление 1 мм рт. ст. увеличивает объем резервуара на 100 мл);
ф- венозные синусы селезенки вмещают - 0,5 л крови и - 20 % эрит-
роцитов;
255
в депонировании крови играет роль также венозная система печени,
КОЖИ, легких И других органов (например, при вертикальной позе в венах
ног депонируется около 0,5 л крови).
2.5. Шунтирующие сосуды (артериоло-венулярные анастомозы) осуществляют регуля-
цию капиллярного кровотока, возврат крови к сердцу.
2.6. Сосуды резорбции (лимфатическая система, см. ниже п. 8).
3. Сосудистый тонус и его регуляция.
3.1. Сосудистый тонус, его виды.
❖ Пассивный тонус обусловлен физическими свойствами сосуда (его упругой
эластичностью). Он прямо пропорционален давлению крови и радиусу со-
суда (уравнение Лапласа: Т = аРг).
❖ Активный тонус обусловлен сокращением мышечного слоя сосуда, он иг-
рает главную роль в регуляции сосудистого тонуса. Активный тонус сосу-
дов образован миогенным (базальным) тонусом, который модулируется ме-
стными вазоактивными метаболитами, гуморальными и нейрогенными ме-
ханизмами, способными его изменить (увеличить или уменьшить) в не-
сколько раз.
3.2. Миогенный (базальный) тонус составляет в покое до 80 % активного тонуса.
Он обусловлен преимущественно сокращением пейсмекерных миоцитов сосу-
да, обладающих автоматией, и сохраняется при исключении нервных и гумо-
ральных алияний.
3.3. Вазоактивные метаболиты и тканевые гормоны в регуляции сосудистого
тонуса (образуются в эндотелии сосудов и прилегающих клетках тканей).
❖ Снижают тонус и расширяют сосуды следующие факторы: повышение
уровня К+, СО2, Н+, аденозина, NO, простагландины Е2, D2, Iz (простацик-
лин), лейкотриены С4, D4 и Е4, гистамина, брадикинина и снижение Poj. эн-
дотелиновый фактор гиперполяризации и др.
❖ Повышают тонус и суживают сосуды следующие факторы: повышение
концентрации серотонина (через 5-НТ-рецепторы) и эндотелии I, тромбок-
саны А2 и В2, простагландины F2 и Н2 и др.
3.4. Вазоактивные дистантные гормоны (гуморальная регуляция сосудистого то-
нуса):
❖ Адреналин через а-адренорецепторы (ТФЛ-С -»"ГИФз/ДАГ -♦ ТСа2‘+ КМ -> Тки-
иаза ЛЦМС и 4.АЦ -» J-цАМФ -> 4.ПК-А -> Ткиназа ЛЦМС) повышает тонус сосу-
дов, через р2-адренорецепторы (ТАЦ — "ГцАМФ — Тпк-А -» 4-кииаза ЛЦМС)
снижает тонус сосудов.
❖ Ангиотензин II действует через ATi-рецепторы (ТФЛ-С -» "ГИФз/ДАГ ->
ТСа2*+КМ -» Ткиназа ЛЦМС, другое влияние - иигибироввиие аденилатциклазной сите-
мы), повышает тонус в 50 раз больше, чем адреналин, образуется с помо-
щью почек и эндотелия.
❖ Вазопрессин через V i -рецепторы (ФЛ-С -» ИФ3/ДАГ -» Са2* -» ТСа^-ь КМ -» Тки-
наза ЛЦМС) повышает тонус.
• > Альдостерон вызывает задержку Na* в крови и увеличивает чувствитель-
ность сосудистой стенки к адреналину, повышает тонус сосудов.
❖ Тиреоидные гормоны, Na*-уретический гормон и ВИП снижают тонус.
256
3.5. Рефлекторные механизмы регуляции сосудистого тонуса (нейрогенный то-
нус обусловлен сокращением под действием нервного импульса преимущест-
венно миоцитов, не имеющих автоматию; составляя в покое - 20 %, он может
увеличивать в физиологических условиях общий сосудистый тонус в 3 раза.
❖ Рецепторно-афферентное звено в регуляции.
• Механорецепторы и хеморецепторы сосудистого русла:
•ф рецепторы каротидной зоны, нерв Геринга (ветвь IX нерва) (см. тему 16;
задание 1:4.4);
Ф рецепторы аортальной зоны, нерв Циона - Людвига (ветвь X нерва);
Ф рецепторы других кровеносных сосудов.
• Механорецепторы сердца, афферентные волокна X нерва:
ф левого желудочка (рефлекс Бецольда - Яриша) (см. тему 16; задание 1:
4.4);
Ф левого предсердия (рефлекс Китаева): при возбуждении рецепторов
левого предсердия происходит рефлекторное сужении артерий лег-
ких.
• Другие рецепторы и сенсорные системы организма (сопряженные реф-
лексы).
❖ Сердечно-сосудистый центр продолговатого мозга (В.Ф. Овсянников. 1871)
(см. тему 16; задание 1:4.1.).
❖ Эфферентное звено в регуляции.
• Симпатические адренергические (норадреналин) влияния через а,-
рецепторы повышают тонус сосудов, через Рг-рецепторы - снижают (см.
тему 8; задание 1:2.4).
• Симпатические холинергические влияния (через М-холинорецепторы)
снижают тонус в некоторых сосудах скелетных мышц (см. тему 8; задание
1:2.4).
• Парасимпатические влияния через М-холинорецепторы снижают тонус
(например, сосуды мозга И легких). Механизмы сосудорасширяющего действия
ацетилхолина связаны с высвобождением из эндотелия NO и торможением через пре-
сиивптические М-холинорецепторы секреции норадреналина в адренергических си-
напсах.
❖ Саморегуляция системного АД осуществляется на основе отрицательной
обратной связи (регуляция по отклонению). Главными рецепторами явля-
ются барорецепторы каротидного тельца (импульсация от них начинается
при давлении около 60 мм рт.ст. и увеличивается при повышении давления
до 160 мм рт.ст.) и рецепторы аортального тельца (импульсация от них при
давлении около 100 мм рт.ст. и увеличивается при повышении давления до
200 мм рт.ст.). Если АД крови превышает нормальный уровень, то импуль-
сация от этих рецепторных зон, поступая в сердечно-сосудистый центр, по-
вышает активность кардиоиигибирующей и депрессорной зон. В результате
этого на сердце и сосуды увеличиваются парасимпатические влияния и
уменьшаются симпатические влияния: уменьшается сердечный выброс и
сосудистый тонус, повышенное АД снижается до нормального уровня. При
257
снижении АД крови происходят противоположные регуляторные эффекты,
которые также приводят к нормализации АД.
4. Артериальное давление (АД) как клинико-физиологический показатель системной
гемодинамики.
4.1. Систолическое давление (АДс) - наибольшее АД во время систолы желудоч-
ков; имеет прямую зависимость от величины систолического выброса и обрат-
ную зависимость от эластичности артерий. Нормальный диапазон АДс в боль-
шом круге от 100 до 140 мм рт. ст., в малом круге 20 - 25 мм рт. ст.
4.2. Диастолическое давление (АДд) - наименьшее АД во время диастолы желу-
дочков; имеет прямую зависимость от периферического сопротивления сосу-
дов. Нормальный диапазон АДд в большом круге 65 - 85 мм рт. ст., в малом
круге 10—15 мм рт. ст.
4.3. Пульсовое давление (АДп) - разница между систолическим и диастолическим
АД; увеличивается в связи со снижением АДд и (или) повышением АДс. Нор-
мальный диапазон АДп в большом круге кровообращения 30 - 50 мм рт. ст.
4.4. Среднее артериальное давление (АДср) - уровень АД который, будучи посто-
янным в течение кардиоцикла, даст тот же гемодинамический эффект, как и при
реальных колебаниях АД; рассчитывается по формуле: АДср = АДд + 1/ЗаДп
(норма 90 - 100 мм рт. ст.).
5. Регулвцнв системной гемодинамики.
5.1. Системная гемодинамика - это кровообращение в крупных сосудах большого
круга.
5.2. Основные регулируемые параметры: 1) минутный объем крови; 2) перифери-
ческое сопротивление сосудов; 3) объем циркулирующей крови; 4) системное
АД.
5.3. Регуляция минутного объема крови.
Регуляция систолического объема (инотропные влияния - см. тему 16; задание 1:
4.2 и 4.3).
❖ Регуляция частоты сердечных сокращений (хронотропные влияния - см. тему 16;
задание 1:4.2 и 4.3).
♦> Роль венозного возврата к сердцу (см. выше 2.4).
5.4. Регуляция периферического сопротивления сосудов (см. выше 2.2 и 3).
53. Регуляция объема циркулирующей крови
❖ Фильтрационио-реабсорбционный механизм (см. выше 2.3).
❖ Почечный механизм:
• прессорный диурез: увеличение (снижение) диуреза в результате повы-
шения (снижения) фильтрационного давления в клубочках при артери-
альной гипертензии (гипотензии), превышающей верхнюю (нижнюю)
границу ауторегуляции кровотока в почках;
• волюмо-осморегулирующий рефлекс (регуляция реабсорбции воды и
Na+ в почках при изменении ОЦК и осмотического давления крови)
(подробнее см. в теме 25; задание 1:13.2 и 13.3).
❖ Депонирование и редепонирование крови (см. выше 2.4).
5.6. Регуляция системного артериального давления осуществляется в результате
деятельности функциональной системы, полезным результатом деятельности
258
которой является необходимая в данных условиях величина АД (обычно это
нормальный уровень АД). При изменении величины АД пусковая афферента-
ция от барорецепторов сосудов (преимущественно от каротидного и аортально-
го тельца) поступает в сердечно-сосудистый центр продолговатого мозга. При
взаимодействии с высшими отделами ЦНС он формирует эфферентную про-
грамму действия, которая через нервный и гуморальный каналы изменяет на-
гнетательную функцию сердца, сосудистый тонус и объем циркулирующей
крови, что создает тот уровень АД, нейронная модель которого сформирована в
акцепторе результата действия.
6. Регуляции региональной гемодинамики.
6.1. Региональная гемодинамика - это кровообращение в отдельных органах.
6.2. Ауторегуляция кровотока в органах - способность органов сохранять посто-
янный кровоток, несмотря на колебания системного АД. В ее осуществлении
участвуют как нейрогенные механизмы (быстрая фаза ауторегуляции), так и
миогенные (эффект Бейлисса) и метаболические механизмы (медленная фаза
ауторегуляции) (подробнее механизмы см. тему 7. задание 1:9).
6.3. Функциональная (рабочая, артериальная) гиперемия осуществляется за счет
изменения концентрации вазоактивных веществ (повышение уровня К*, СОг,
Н*, аденозина, NO и снижение напряжения Ог), вызывающих расширение сосу-
дов в работающем органе
6.4. Функциональная венозная гиперемия, ее роль в депонировании кровн.
6.5. Функциональная «ишемия» возникает при перераспределении кровотока (не
приводит к структурно-функциональным нарушениям).
6.6. Особенности кровообращения в некоторых органах: мозг (см. тему 7; задание 1:
9), легкие (см. тему 19; задание 16 8.3), кожа (см. тему 26; задание 1: 3.3), почки (см. тему
25; задание 1: 2.3).
7. Методы исследования гемодинамики.
7.1. Косвенный метод определения артериального давления. Аускультативным методом
Короткова можно измерять систолическое и диастолическое давление. На обнаженное
плечо испытуемого, иа 1-2 см проксимальнее локтевого сгиба накладывают манжетку
сфигмоманометра. В локтевой ямке находят пульсирующую лучевую артерию и уста-
навливают над ней фонендоскоп. Нагнетанием воздуха в манжетку в ней создается
давление выше артериального, пульс исчезает. Открывают клапан и, выпуская воздух
из манжетки, выслушивают сосудистые тоны в лучевой артерии, которые возникают в
результате турбулентного движения прорвавшейся кровн. Момент пояаяения тонов
соответствует систолическому давлению. Когда при снижении давления в манжетке
ток крови становится ламинарным, тоны исчезают. Этот момент соответствует диасто-
лическому давлению
7.2. Артериальная осциллография - косвенный метод определения уровня артериального
давления. Она позволяет, кроме максимального и минимального давления, определить
среднее АД и судить об эластичности сосудистых стеиок. Принцип заключается в ре-
гистрации пульсаций крупной артерии в условиях ее компрессии или декомпрессии.
Первые пульсации возникают в те мгновения, когда максимальное АД начинает пре-
вышать давление воздуха в манжетке. По мере снижения давления в манжете осцилля-
ции все больше увеличиваются и достигают максимальной амплитуды. При дальней-
шем уменьшении компрессии сосудов величина пульсаций постепенно снижается до
полного исчезновения.
259
7.3. Метод определения времени полного кругооборота крови, т.е. времени прохождение
кровью большого и малого кругов. Принцип метода: в локтевую вену вводят индика-
тор (например, флюоресцеин или радиоактивный изотоп) и определяют, через какой про-
межуток времени он появляется в одноименной вене другой стороны. Время полного
кругооборота крови (рука-рука) в среднем равно 20 сек, на кровоток в малом круге
приходится 4-7 сек.
7.4. Методы изучения сосудистых реакций:
❖ Плетизмография - метод регистрации изменения объёма органа или части тела,
вызванного изменениями его кровенаполнения. Применяется в основном для
оценки сосудистого тонуса. Наиболее часто используется механическая плетизмо-
графия и электроплетнзмографня (см. ниже реография). При механической пле-
тизмографии исследуемую конечность или ее часть герметизируют в специальном
сосуде (рецепторе), заполненном водой или воздухом. Колебания объёма при этом
с помощью датчика превращаются в электрические колебания н записываются ре-
гистратором (например, электрокардиографом).
❖ Реография - метод исследования пульсового кровенаполнения органов н частей
тела, основанный на регистрации колебаний сопротивления тканей переменному
току высокой частоты. Изменения сопротивления возникают в результате различ-
ного кровенаполнения тканей в систолу и диастолу. В систолу кровенаполнение
увеличивается, а сопротивление уменьшается, в диастолу - наоборот. Реография
проводится с помощью реографа с регистрацией реограммы на электрокардиогра-
фе синхронно с ЭКГ. В зависимости от места наложения электродов различают
реоэнцефалографию, реокардиографию, реографию печени, конечностей и т.д.
8. Лимфатическая система.
8.1. Общая характеристика лимфатической системы-, лимфатические капилляры, лимфатиче-
ские сосуды (большой диаметр, ие выражена базальная мембрана), лимфоузлы, грудной и пра-
вый лимфатические протоки, впадающие в левый и правый венозные углы (слияние подклю-
чичных н внутренних яремных вен), клапаны, симпатическая и парасимпатическая иннервация.
8.2. Механизмы образования лимфы («Лимфа - зеркало интерстициальной жидкости». К.
Бернар).
❖ Фильтрационно-реабсорбционный механизм (Э. Старлинг, 1909): преоблада-
ние фильтрации над реабсорбцией, т.е. действие остаточной силы фильтра-
ционного давления.
❖ Диффузия белка по градиенту концентрации из межклеточного простран-
ства в лимфатический капилляр через межклеточные щели вместе с водой и
путем пиноцитоза через эндотелиальную клетку (онкотическое давление!), объ-
ясняет образование лимфы при нулевом и отрицательном интерстици-
альном давлении.
❖ Количество образующейся лимфы (2-4 л/сут), ее электролитный состав,
pH, глюкоза, мочевина близки к плазме крови, белков в 1,5 раза меньше,
количество лейкоцитов около 12 • 109/л, среди них 85% лимфоцитов, эрит-
роцитов - 2 • 109/л .
8.3. Механизмы передвижения лимфы.
❖ Первоначальное гидростатическое давление (2 - 3 мм рт. ст.) осуществляет
перемещение лимфы в лимфатических капиллярах и посткапиллярах и за-
пускает сокращение лимфангионов.
❖ Лимфатический насос: ритмические сокращения (10 - 15 в 1 мин) лимфо-
сосудов, роль клапанов, симпатические влияния, гистамин.
260
• > Передвижению лимфы способствуют дыхательный иасос и массажное дей-
ствие тканей: сокращение скелетных и гладких мышц, пульсация артерий.
8.4. Основные функции лимфатической системы.
❖ Дренажная функция: возврат белков (- 200 г/сутки), воды и электролитов из
межклеточного пространства в кровь.
❖ Детоксикационная функция- переход в лимфу из межклеточного простран-
ства патологически измененных белков, их конгломератов, токсинов и кле-
ток с обезвреживанием их в лимфоузлах.
❖ Участие в иммунологических реакциях (циркуляция лимфоцитов, лимфоидная
ткань узлов, фильтрационная система синусов лимфоузлов)
❖ Всасывание и перенос питательных веществ (например, липидов).
❖ Участие в осуществлении гуморальной связи между клетками через ткане-
вую жидкость.
9. Профильный материал для студентов лечебного факультета.
9.1. Особенности кровообращения в антенатальном периоде.
❖ Эмбриональный период (2 - 8-я неделя, желточное кровообращение, неполное об-
разование плаценты).
• Двухкамерное сердце формируется на 4-й неделе, 3-камерное сердце (два
предсердия, желудочек) - к концу 5-й недели, 4-камерное сердце - к 7-й неде-
ле. Параллельно происходит закладка и формирование сосудов.
• Частота сердечных сокращений- 6-я неделя - 110 уд/мин, 8 - 12-я недели - 170
уд/мин
• Кровообращение обеспечивает движение крови в теле эмбриона и желточном
мешке.
♦♦♦ Фетальный период (с 3-го месяца до рождения кровообращение плода и матери
разделено, контактируя через плаценту).
• Роль плацентарного кровообращения
"Ф Плацента выполняет функции легких, кишечника, почек. Плацентарная
мембрана имеет площадь - 7 м2, -толщину 2-6 мкм. Объем протекающей
материнской крови через лакуны плаценты - 800 мл/мин, крови плода че-
рез капилляры ворсинок - 130 мл/мин на 1 кг массы тела плода (около 50
% крови плода).
• ф От плаценты оттекает по пупочной вене наиболее оксигенированная
кровь (НЬО2 - 80 %);
• ф От пупочной вены отходит веточка к печени, через которую она получает
наиболее оксигенированную кровь; после чего пупочная вена прев-
ращается в венозный (аранциев) проток, впадающий в ннжнюю полую
вену, в которую поступает также кровь от нижней части тела и печени
(НЬОз - 25%). Смешанная венозная кровь полой вены имеет насыщение
Оз около 60 %.
ф Кровь по нижней полой вене попадает в правое предсердие, и основная ее
часть при помощи заслонки нижней полой вены поступает через овальное
отверстие в левое предсердие, желудочек и аорту
- ф По верхней полой вене венозная кровь (НЬОз - 25 %) попадает в правое
предсердие (частично смешивается с кровью нижней полой вены), пре-
имущественно поступает в правый желудочек и легочный ствол, далее 1/3
261
крови поступает в легкие, а 2/3 крови легочного ствола через артериаль-
ный (боталлов) проток поступает в аорту дистальнее отхождения сосудов
сердца, головы и верхних конечностей. Поэтому головной мозг, сердце и
верхние конечности снабжаются более оксигенированной кровью и быст-
рее развиваются, чем нижняя часть тела
Ф От подвздошных артерий отходят две пупочные артерии, по которым (в
составе пупочного канатика) ббльшая часть крови возвращается в плацен-
ту (НЬО2~ 60 %).
Ч* Благодаря овальному отверстию и артериальному протоку правое и левое
сердце у плода соединены параллельно, а не последовательно (как после
рождения), и 4-камерное сердце функционально работает как 2-камерное.
• Нагнетательная функция.
*0* Правые предсердие и желудочек непосредственно участвуют в гемодина-
мике большого круга за счет овального отаерстия между предсердиями и
артериального протока между аортой и легочным стволом.
Ч* Имеется высокое давление крови в правом желудочке.
Ф Величина систолического объема в последней трети внутриутробного пе-
риода - 1 - 3 мл; минутного объема - 150 - 450 мл; ЧСС - 140 - 130
уд/мин
• Регуляция деятельности сердца плода.
Ф Внутрисердечные механизмы регуляции.
автоматах водителя ритма является главным фактором регуляции сер-
дечной деятельности;
малое значение имеет гетерометрическая и гомеометрическая саморе-
гуляция.
Ф Внесердечные механизмы (рефлексы с баро- и хеморецепторов синокаро-
тидной зоны и дуги аорты) выражены слабо.
• Особенности ЭКГ плода, низкая амплитуда зубцов, отсутствие зубцов Q и Т.
• Особенности тонов сердца и фонокардиограммы плода:
Ч* Систолический тон сердца появляется с 18 - 20-й недели, диастолический
тон - несколько позже;
Ф Имеется короткий первый тон на ФКГ.
• Регуляция сосудистого тонуса плода: тонус сосудов слабый и обусловлен в
основном тонусом их гладкой мускулатуры; преобладает его гуморальная ре-
гуляция (например, низкий уровень pH и РО2 снижает тонус сосудов)
9.2. Изменения функции сердца при старении организма.
❖ Снижение регенерации митохондрий, синтеза АТФ и креатинфосфата.
♦ ♦♦ Уменьшение числа пейсмекерных клеток.
Уменьшение эластичности миофибрилл н роли гетерометрического механизма ре-
гуляции (закона Старлинга).
’?• Уменьшение числа ^-адренорецепторов и положительного инотропного эффекта
катехоламинов на сердце.
Снижение рефлексов на сердце с барорецепторов и повышение - с хеморецепто-
ров.
Снижение максимальной частоты сердечных сокращений (ЧСС = 220 - возраст).
262
❖ Уменьшение ударного и минутного объемов на 25 % и функционального резерва
сердца по МОК.
♦ ♦♦ При нагрузках увеличивается период врабатывания сердца (снижение лабильно-
сти сердечного центра) и затягивается восстановительный период.
9.3. Возрастные изменения нервной и гуморальной регуляции тонуса сосудов, артери-
ального и еенозного давления.
♦ ♦♦ Увеличение рефлекторных влияний на сосудистый тонус с хеморецепторов н
уменьшение - с барорецепторов (возможна ортостатическая гипотензия - потеря
равновесия, обмороки вследстаие ишемии головного мозга).
❖ Увеличение в связи с повышением концентрации вазопрессина и ангиотензина П
общего периферического сопротивления сосудов (в 1,3 - 1,8 раза).
❖ Увеличение скорости распространения пульсовой волны в связи с уменьшением
эластичности сосудов.
Л * Увеличение систолического и диастолического АД до 60 лет, затем преимущест-
венно увеличивается систолическое АД в связи с уменьшением эластичности аор-
ты.
♦ ♦♦ Снижение венозного давления в связи с расширением венозного русла, снижение
тонуса н эластичности вен (венозный стаз, уменьшение венозного возврата).
♦ ♦♦ Увеличение емкости сосудистого русла, снижение сердечного выброса приводит к
увеличению времени кругооборота крови, что частично компенсируется увеличе-
нием (а~в)О1
9.4. Особенности адаптации кровообращения к деятельности в различных условиях
при старении.
❖ Уменьшены физиологические резервы кровообращения по МОК, СО, ЧСС
♦ ♦♦ Удлинен период врабатывания и восстановительный период при нагрузках.
♦ ♦♦ Более выражена гипертензивная реакция при нагрузках.
♦ ♦♦ В регуляции кровообращения снижена доля высоколабильных рефлекторных ме-
ханизмов и повышена роль гуморальных механизмов В результате повышается
чувствительность к стрессовым ситуациям.
9.5. Особенности коронарного кровообращения в старости.
❖ Повышен тонус коронарных сосудов в связи с увеличением концентрации вазо-
прессина и ангиотензина II
❖ Уменьшено кровоснабжение миокарда вследствие атеросклеротического измене-
ния сосудов, изменения их реакции на нервные и гуморальные влияния в сторону
преобладания прессорных эффектов.
9.6. Особенности мозгового кровообращения.
♦♦♦ Мозговой кровоток с возрастом снижается незначительно.
♦♦♦ Снижаются функциональные возможности ауторегуляции мозгового кровотока
вследствие атеросклеротического поражения сосудов.
10. Профильный материал для студентов педиатрического факультета.
10.1. Особенности кровообращения плода (см. выше 91.).
10.2. Система кровообращения в неонатальном онтогенезе и периоде новорожденно-
сти.
♦5» Перестройка кровообращения у новорожденных.
• Перевязка пупочных сосудов прекращает плацентарное кровообращение, за-
пустевает венозный проток, резко падает давление в правом предсердии.
263
• Включается дыхание, полностью включается малый круг кровообращения (его
сопротивление снижается в 5 раз), снижается давление в правом желудочке,
запустевает артериальный проток.
• Резко увеличивается приток крови из легких в левое предсердие, повышается
давление в нем, что вызывает захлопывание клапана овального отверстия.
• Кровообращение полностью разделяется иа два круга, сердце функционирует
как 4-камерное.
❖ Величина АД и формула его расчета у детей разного возраста.
• Тотчас после рождения систолическое АД повышается с 60 до 90 мм рт. ст.,
затем через 2-3 часа снижается до 60 - 70 мм рт ст. и увеличивается в первые
две недели Диастолическое АД в первые сутки около 40 мм рт. ст.
• Формула расчета АД у детей до года- 76 + 2 м (м - количество месяцев после
рождения), после года 100 + 0,5 г (г - годы жизни).
❖ Увеличивается скорость распространения пульсовой волны в артериях вследствие
увеличения их упругости (от 510 до 780 см/с)
❖ Особенности регуляции деятельности сердца новорожденного.
• Гомеометрическая саморегуляция преобладает над гетерометрической.
• Парасимпатические влияния усиливаются и преобладают над симпатически-
ми Усиливаются рефлекторные влияния с баро- и хеморецепторов синокаро-
тидной зоны и дуги аорты
• Имеется низкая чувствительность сердца к гуморальным факторам регуляции
❖ Особенности ЭКГ новорожденного ребенка: электрическая ось сердца смещена
вправо за счет большой массы правого желудочка; укорочен интервал PQ и время
комплекса QRS.
❖ Особенности ФКГ новорожденного ребенка- Появляется Ш и IV тоны на ФКГ
Имеется более короткий, чем у взрослых, I тон и более длительный II тон
103. Последующие возрастные периоды.
♦♦♦ Функциональные особенности сердца
• Уменьшение ЧСС и увеличение длительности сердечного цикла за счет удли-
нения диастолы с возрастом
• Увеличение систолического и минутного объемов
• Увеличение давления крови в левом желудочке и снижение в правом.
❖ Варианты форм подросткового сердца, особенности функции сердечно-
сосудистой системы, характерные для каждого из этих вариантов.
• Нормальное сердце у гармонично развитых подростков, наилучшее соответст-
вие между ростом тела и сердца.
• «Малое» или «капельное» сердце - рост сердца отстает от роста тела- имеется
низкий систолический объем, тахикардия, понижение АД, функциональный
систолический шум.
• «Гипертрофированное» сердце, рост сердца опережает рост тела, имеется уве-
личение минутного объема, низкая частота сокращений, повышение АД,
функциональный систолический шум, возможны функциональные шумы в
возрасте 1 - 17 лет
♦♦♦ Функциональные особенности сосудов в грудном возрасте.
264
• Увеличение просвета легочных сосудов, уменьшение ОПСС в малом круге, но
увеличение в большом круге.
• Систолическое АД = 90 - 100, диастолическое АД - 42 мм рт ст.
• Увеличение скорости пульсовой волны за счет повышения упругости сосудов
• Включение клапанов вен в регуляцию гемодинамики (предупреждение обрат-
ного тока венозной крови), увеличение депонирующей возможности венозной
системы (печени, селезенки)
• В последующие возрастные периоды увеличение ОПСС и АД. Увеличение
объемного кровотока через скелетные мышцы и почки и снижение кровотока
через головной мозг. Повышение АД в период полового созревания (юноше-
ская гипертензия).
♦♦♦ Регуляция кровообращения.
• В течение 1-го года жизни усиливается влияние парасимпатического отдела,
роль рефлексов с баро- и хеморецепторов дуги аорты и каротидного синуса.
• Совершенствуется механизм перераспределения кровообращения различных
органов в зависимости от их функционального состояния (рабочая гиперемия
и «ишемия»).
• Увеличивается роль гетерометрического механизма регуляции.
• Увеличиваются парасимпатические влияния, уменьшается ЧСС
• Сокращается латентный период сосудистых реакций, возникает отчетливая
депрессорная направленность
• Повышаются адаптационные возможности сердца к физическим нагрузкам
♦♦♦ Типы ЭКГ в возрасте 1-7 лет и 12 - 18 лет.
• В возрасте от 1 - 7 лет увеличение амплитуды зубцов R и Т, отрицательный
или двухфазный зубец Т в Ш отведении, уменьшение амплитуды зубца Q, уве-
личение интервала PQ и комплекса QRS, возможность синусовой аритмии
• В возрасте 8-12 лет снижение амплитуды зубцов Р и R от I к Ш отведению,
возможны отрицательные зубцы Р и Т в Ш отведении, постепенное смещение
электрической оси сердце влево, возможно расщепление комплекса ORS в Ш
отведении.
11. Профильный материал для студентов стоматологического факультета.
11.1. Нервная и гуморальная регуляция тонуса сосудов ротовой полости,
❖ Нервные механизмы (преобладающая регуляция).
• Симпатическая иннервация: сосудодвигательный центр (продолговатый мозг)
—> верхний шейный симпатический гаиглий —» в нервно-мышечном синапсе
норадреналин: а-адреиорецепторы - сужение сосудов, Pi-адреиорецепторы
(слабый эффект) - расширение сосудов.
• Парасимпатическая иннервация: сосудодвигательиый центр —» волокна VII,
IX, X нервов, медиатор ацетилхолин —» М-холинорецепторы —» расширение
сосудов
Гуморальные механизмы.
• Вазоактивные метаболиты: повышение концентрации аденозина, К4*, COz» Я*,
снижение Рог приводит к расширению сосудов, противоположная динамика
этих метаболитов суживает сосуды.
265
• Гистамин, брадикинин, простагландины Е вызывают расширение сосудов.
• Адреналин через а-адренорецепторы вызывает сужение сосудов, через 3-
адреиорецепторы - расширение сосудов.
• Ангиотензин П и вазопрессин вызывают сужение сосудов.
❖ Миогенный тонус сосудов предупреждает отек пульпы, его снижение при функ-
циональной нагрузке (жевание) улучшает трофику парадонта (рабочая гипе-
ремия).
11.2. Рефлекторные изменения деятельности сердца и тонуса сосудов, обусловленные
раздражением слизистой оболочки полости рта и зубов.
❖ Болевое раздражение- у симпатикотоников - тахикардия и повышение АД, а у па-
раснмпатикотоииков - брадикардия и понижение АД
❖ Раздражение рецепторов тройничного нерва может формировать кардиоингнби-
рующий и депрессорный эффекты
Сильно выражено психоэмоциональное влияние на сосудистый тонус
113. Причины изменения кровяного давления при различных манипуляциях в полости
рта.
❖ Психоэмоциональное напряжение (активация симпатоадреналовой системы)
❖ Болевое раздражение может вызывать как симпатические эффекты (кардиостиму-
лирующий и прессорный), так и парасимпатические эффекты (кардиоингибнрую-
щий и депрессорный)
11.4. Методы исследования гемодинамики в стоматологии.
❖ Исследование функционального состояния сосудов слизистой оболочки полости
рта: капилляроскопия, капиллярография.
❖ Реография и ее использование в стоматологии.
♦ ♦♦ Плетизмография: изменение плетизмограммы под влиянием раздражений органов
полости рта
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3.1 н 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 н 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см с 514, использовать имеющиеся у
вас учебники и пособия). [1-16].
3.2. Дополнительная литература (см с. 514 - 522)- [3], [27], [30], [43], [45], [62], [64],
[113], [124], [125], [146], [152], [153], [178], [179], [182], [183], [193], [197], (206].
[209], [210], [222], [225], [226], [234], [240], [244], [[245], 251-254].
33. Дистантные средства обучения. Использования страницы кафедры в Интернете -
www vsma ас ni/~nphys Слайды для компьютерной демонстрации из «Атласа по
физиологии человека» / В.Б Брин - Владикавказ СОГМА - 1999, тесты остаточ-
ного уровня знаний по данной теме
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры)
3.5. Консультации преподавателей еженедельно по индивидуальному графику
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
266
Тема 19. Физиология дыхания. Внешнее дыхание. Газы крови, их транс-
порт. Газообмен между кровью и тканями
Цель' а) знать значение дыхания для организма, его этапы; дыхательный цикл, механизм вдоха и вы-
доха, вентиляцию легких, легочные объемы и емкости, методы их определения; функцию ды-
хательных путей, регуляцию их просвета, газообмен в легких, транспорт О2 и СО2 кровью, га-
зообмен между кровью и тканями, тканевое дыхание, роль миоглобина, недыхательные функ-
ции легких,
б) уметь использовать эти знания для оценки функциональных показателей системы внешнего
дыхания и транспорта газов кровью здорового человека,
в) ознакомиться с практическими навыками на уровне знания исследования внешнего дыха-
ния и транспорта кислорода кровью (спирография, пневмотахометрия, оксигемометрия, опре-
деление петли «поток - объем»); на уровне умения спирометрия
Мотивация темы: «Пока дышу - надеюсь» (латинская поговорка)
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов» Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов
1. Общая характеристика системы дыхания.
1.1. Система дыхания - физиологическая система, обеспечивающая процесс по-
ступление Ог в организм, доставку и использование его в тканях, а также выве-
дение конечного продукта дыхания - СОг из организма.
1.2. Значение для организма.
❖ Прямое метаболическое действие Ог:
• Через цитохром С и цитохромоксидазу митохондрий кислород, являясь
акцептором электронов, обеспечивает окислительное фосфорилирова-
ние - синтез АТФ, через цитохром Р450 гладкой ЭПС - биосинтетиче-
ский и детоксикационный эффекты.
• Образование продуктов неполного восстановления Ог - свободных ра-
дикалов (супероксидный анион, пероксид водорода, гидроксильный радикал и др.).
Их физиологические эффекты' влияние на вязкость и текучесть мем-
бран, вазоконстрикция, пролиферация лимфоцитов, участие в механиз-
мах фагоцитоза и др
• > Напряжение Ог и СОг является важным фактором регуляции сосудистого
тонуса, кислотно-осиовного состояния, эритропоэза, а также самой систе-
мы дыхания
❖ Легкие участвуют в регуляции кислотно-основного состояния, терморегу-
ляции и выполняют другие негазообменные функции (см ниже п 12).
1.3. Основные этапы дыхания: 1) легочная вентиляция; 2) газообмен в легких; 3)
транспорт газов кровью; 4) газообмен между кровью и тканями; 5) тканевое
дыхание.
267
2. Легочная вентиляция рассматривается в п.п. 3-7: дыхательный цикл, легочная и
альвеолярная вентиляция, методы исследования внешнего дыхания, работа, совер-
шаемая при дыхании, функции дыхательных путей.
3. Дыхательный цикл.
3.1. Биомеханическая характеристика легких.
Растяжимость легких (мера их податливости) - способность легких увеличивать свои раз-
меры и объем при действии приложенной силы. Легкие более растяжимы при небольших
и средних размерах, чем при больших Растяжимость легкого и грудной клетки в норме
равна 200 мл/см вод ст (В патологии растяжимость легких может как увеличиваться, иапример
при эмфиземе, так и уменьшаться, например при фиброзе, пневмонии )
❖ Эластичность - есть мера упругости легких, сила отдачи, которую создает растя-
жение легких. Чем больше эластичность, тем больше требуется приложить давле-
ние для достижение заданного объема легких.
❖ Растяжимость и эластичность легких связаны обратно пропорциональной зависи-
мостью' чем больше растяжимость, тем меньше эластичность и наоборот.
3.2. Механизмы вдоха (inspiratio).
♦** Перед вдохом после спокойного выдоха эластическая тяга грудной клетки (направлена на-
ружу) и легких (направлена внутрь) имеют противоположное направление, их сумма равна
нулю
❖ Сокращение инспираторных мышц.
• Сокращение диафрагмы увеличивает вертикальный объем грудной
клетки и обусловливает - 70 % дыхательного объема.
• Сокращение наружных межреберных мышц увеличивает фронтальный
и сагиттальный размеры грудной клетки.
• Сокращение вспомогательной мускулатуры (лестничных, грудино-
ключично-сосцевидных, грудных мышц) происходит при легочной венти-
ляции > 50 л/мин
Эластическая тяга грудной клетки при вдохе, ие превышающем 70 % жизненной емкости
легких, направлена на увеличение объема грудной клетки и легких (способствует вдоху)
❖ В результате действия этих факторов происходит увеличение объема груд-
ной клетки.
❖ Связь изменений объема грудной клетки с объемом легких, приводящая к
увеличению объема легких, обеспечивается:
• отрицательным давлением в плевральной щели, равным -6 мм рт ст.
при спокойном вдохе (плевральное давление равно пищеводному давлению
и может быть измерено с помощью баллонного катетера);
• силой «сцепления» молекул НзО в слое жидкости между грудной и ле-
гочной плеврой.
❖ При увеличении объема легких в них, по закону Бойля, давление воздуха
понижается и становится отрицательным (-2 мм рт. ст при спокойном вдохе,
до -80 мм рт. ст. во время попытки максимального вдоха при закрытых дыхатель-
ных путях - проба Мюллера), поэтому атмосферный воздух поступает в лег-
кие
3.3. Механизмы выдоха (expiratio).
268
❖ Эластическая тяга легких всегда и комплекса «грудная клетка + легкие»
при вдохе направлена во внутрь, поэтому выдох в покое (до уровня функцио-
нальной остаточной емкости) происходит пассивно.
❖ Активный выдох требует сокращения экспираторных мышц (внутренних
межреберных, передней брюшной стенки).
❖ При спокойном выдохе уменьшается отрицательное плевральное давление
(-2 мм рт. ст.), а внутрилегочное давление становится положительным (+2
мм рт. ст. при спокойном выдохе, до +100 мм рт. ст. во время попытки макси-
мального выдоха при закрытых дыхательных путях - проба Вальсальвы), что
приводит к выведению воздуха из легких.
♦ ♦♦ После спокойного выдоха алгебраическая сумма эластической тяги грудной клетки + эла-
стической тяги легких = 0, дыхательная система находится в покое.
4. Легочная и альвеолириая вентиляции.
4.1. Легочные объемы и емкости, их функциональная и количественная характери-
стика (рис. 50).
❖ Частота дыхания (ЧД =12 -18 в 1 мнн) характеризует возбудимость дыхатель-
ного центра.
❖ Дыхательный объем [в покое до (VT) - 0,5 л] - это объем вдыхаецрго (или вы-
дыхаемого) воздуха, зависит от возбудимости дыхательногацентра, растя-
жимости и эластичности легких, состояния грудной клетки и бронхиальной
проходимости. (ДО в наибольшей мере связан с физической нагрузкой, но обыч-
но не превышает 50% от ЖЕЛ.)
❖ Резервный объем вдоха [РОвд (IRV) - 2,5 л] - это количество воздуха, кото-
рый максимально можно вдохнуть после спокойного вдоха; характеризует
силу инспираторных мышц, растяжимость и эластичность легких, состоя-
ние легочной ткани.
❖ Резервный объем выдоха [РОвыд (ERV) - 1,2 л] - это количество воздуха, ко-
торый максимально можно выдохнуть после спокойного выдоха; в ос-
новном зависит от бронхиальной проходимости.
❖ Жизненная емкость легких (ЖЕЛ (VC) - 4,2 л] - это количество воздуха, ко-
торое можно максимально выдохнуть после максимального вдоха; отра-
жает силу дыхательной мускулатуры, растяжимость и эластичность легких,
площадь дыхательной мембраны, бронхиальную проходимость. Получен-
ный показатель в норме не отличается иа ±15 % от должной ЖЕЛ; ДЖЕЛ
(в мл) = 25 (20 у жен.) х Рост (см). '
❖ Остаточный объем легких [ООЛ (RV) - 1,2 л] - это воздух, оставшийся в лег-
ких после максимального выдоха; зависит от эластичности легких, силы
выдоха, бронхиальной проходимости (спирографическн ие определяется).
❖ Функциональная остаточная емкость [ФОЕ (FRC) = ООЛ + РОвыд - 2,4 л] - это
воздух, имеющийся в легких после спокойного выдоха, составляет - 30%
ЖЕЛ. (Представляет собой «буфер» между атмосферным и альвеолярным возду-
хом, предотвращающий выраженные колебания Рог и Рсог в альвеолярном воздухе
в течение дыхательного цикла.)
❖ Общая емкость легких [ОЕЛ (TLC) = ЖЕЛ + ООЛ - 5,5 л] - это воздух, имею-
щийся в легких после максимального вдоха; зависят в основном от меха-
нических свойств легких и грудной клетки, силы инспираторных мышц.
269
❖ Минутный объем дыхания (МОД = ДО х ЧД - 7 л/мин) является количествен-
ной характеристикой легочной вентиляции.
4.2. Физиологическое (или функциональное) мертвое пространство состоит из:
❖ анатомического мертвого пространства, равного - 140 мл или 2 мл/кг мас-
сы тела (вентиляция мертвого пространства ~ 2,2 л/мин);
❖ альвеолярного мертвого пространства (вентилируемые, но не перфузируе-
мые альвеолы), равного 10-15 мл.
43. Альвеолярная вентиляция (АВ, V,i).
❖ АВ - объем вдыхаемого (или выдыхаемого) воздуха, участвующего в газооб-
мене в легких в единицу времени; она является показателем эффективности
легочной вентиляции и должна быть не менее 70 % МОД (4,5 - 5 л/мин).
❖ Состав альвеолярного воздуха относительно постоянен, так как в течение
дыхательного цикла обменивается только - 1/7 его часть.
5. Методы исследования внешнего дыхаиив.
5.1. Спирография и спирометрия.
❖ Спирография позволяет определить следующие объемы и емкости: ДО,
РОвд, РОвыд, ЖЕЛ, МОД, ФЖЕЛ, ОФВ| (см. 4.1 и рис. 50).
Скоростные показатели сопрограммы:
• форсированная жизненная емкость легких (ФЖЕЛ) - объем воздуха, выдыхаемый че-
ловеком с максимальной силой и скоростью после максимального вдоха, объем фор-
сированного выдоха рассчитывается за 1-ю, 2-ю и 3-ю секунды, ОФВ] - 3,5 л;
• индекс Тиффно (отношение объема форсированного выдоха за 1-ю секунду к ЖЕЛ,
выраженное в процентах) = ОФВ|/ЖЕЛ • 100 % > 70 %, отражает в основном проходи-
мость крупных бронхов. .
Рис. 50. Спирограмма легких. Объяснения в 4.1.
❖ Петля «поток-объем» - это изменение скорости потока (л/с) воздуха при
максимальном вдохе и выдохе, которое регистрируется при изменении
объема легких в диапазоне ЖЕЛ (рис. 51). Показатели экспираторной части
кривой:
07П
• П-пик - максимальная величина скорости потока выдыхаемого воздуха
(~ 9 л/с), она зависит от силы сокращения экспираторных мышц и со-
стояния бронхов.
• П75 % - величина скорости потока выдыхаемого воздуха на уровне 75 %
ЖЕЛ (~ 8 л/с), является показателем проходимости крупных (сегмен-
тарных) бронхов.
л/с
Рис 51. Петля «поток-объем»
Объяснения в 5.1.
• П50 % — величина скорости потока выдыхаемого воздуха на уровне 50 %
ЖЕЛ (- 5 л/с), является показателем проходимости средних бронхов.
• П25 % - величина скорости потока выдыхаемого воздуха на уровне 25 %
ЖЕЛ (~ 2 л/с), является показателем проходимости мелких бронхов.
5.2. Методы плетизмографии и дилюции газов позволяют определять ООЛ, а
вместе со спирографией - ФОБ, ОЕЛ.
5.3. Пневмотахометрия и инемотахография позволяют определить скорость по-
тока (в л/с) вдыхаемого и выдыхаемого воздуха, оценить состояние тонуса'
бронхов, определить работу дыхания, эластическое и иеэластическое сопротив-
ление дыханию.
5.4. Определение состава вдыхаемого, выдыхаемого и альвеолярного воздуха про-
изводят методами газового анализа воздуха (СОз - капнографами, Оз - оксйге-
мографами). - . •
б. Работа, совершаемая при дыхании (- 0,25 кгм/мин).
6.1. Преодоление сил, препятствующих изменению объема легких (направлена на
преодоление эластического сопротивления легких н грудной клетки, составляет - 70%
величины общей работы). ' ’ ’
❖ Эластическое сопротивление легких.
• Около 65% эластической тяги легких формирует поверхностное натя-
жение водной пленки альвеол (сила, возникающая на границе двух фаз
271
«альвеолярный газ - водная пленка»), направленное на уменьшение по-
верхности и объема альвеол;
• ф главным регулятором эластической тяги легких являются сурфак-
танты (липопротеиды, вырабатываемые альвеолоцитами П типа), кото-
рые в ~б раз снижают поверхностное натяжение водной пленки
альвеол, увеличивая растяжимость легких (в состав сурфактантов входит
витамин Е, который защищает их от агрессивных форм кислорода);
ф пленка сурфактанта (- 50 нм толщиной) при выдохе утолщается и пре-
дотвращает спадение альвеол и респираторных бронхиол (поверхно-
стное натяжение при выдохе равно 0,005 Н/м);
ф «недостаточность» истонченной пленки сурфактанта при вдохе
уменьшает растяжимость альвеол и ограничивает их увеличение
(поверхностное натяжение при вдохе в 10 раз выше - 0,05 Н/м).
• Соотношение эластических и коллагеновых волокон в легких также
влияет на их растяжимость: коллагеновые волокна растягиваются до
10% длины, эластические волокна - в 20-30 раз больше.
❖ Эластическая тяга грудной клетки при вдохе менее 70 % ЖЕЛ направлена
наружу и облегчает вдох, при ббльшей глубине вдоха направлена внутрь и
препятствует вдоху.
6.2. Преодоление сил сопротивления движению воздуха (иеэластическое сопротив-
ление дыханию, составляет 30 % общей величины работы, увеличивается при повыше-
нии частоты дыхания).
❖ Неэластическое сопротивление образуется:
• аэродинамическим сопротивлением потока воздуха в различных участ-
ках дыхательных путей;
• трением тканей дыхательного аппарата.
❖ Бронхиальный тонус оказывает выраженное влияние на неэластическое со-
противление: сужение бронхов повышает сопротивление, расширение
бронхов - снижает.
❖ Увеличение частоты и глубины дыхания повышает неэластическое сопро-
тивление.
6.3. Кислородная цена дыхания: в покое на вентиляцию легких затрачивается ~ 3
% потребленного организмом От, при максимальной физической работе - до 20
%.
7. Воздухопроводящая функция дыхательных путей (основной вид транспорта - кон-
векция).
7.1. К воздухопроводящим путям относят: внелегочные пути (нос, глотка, гортань, тра-
хея) и внутрилегочиые пути (17 генераций бронхов, в которых имеются потоки воздуха, и
18 - 22 генерации респираторных бронхов, где осуществляется диффузия газов).
7.2. Роль дыхательных путей в сопротивлении дыханию: при вдохе наибольшее
сопротивление оказывает полость носа (- 54 %), при выдохе - бронхи (особен-
но сегментарные) - 50 %.
7.3. Регуляция просвета дыхательных путей.
❖ Нейрогенная регуляция (основной регулятор в норме).
• Парасимпатические влияния (заднее ядро X нерва):
272
Ф- через ацетилхолин и М-холинорецепторы (Тфосфолипаза С -»
ТИФэ/ДАГ -»(Са2*—*Ткиназа ЛЦМ) вызывают сужение бронхов, напри-
мер, при выдохе;
Ф" через пептид Р вызывают сужение бронхов;
ф- через медиатор ВИП вызывают расширение бронхов.
• Симпатические влияния через норадреналин и р2-рецепторы вызывают
расширение бронхов, например, при вдохе ((цАМФ -»(протеинкиназа А -»
фосфорилирование киназы ЛЦМ уменьшает ее чувствительность к Са2*, слабый эф-
фект).
❖ Гуморальная регуляция.
• Бронходилатацию вызывают:
Ф- адреналин через р2-адренорецепторы ((цАМФ -»(протеинкиназа А -»
фосфорилирование киназы ЛЦМ уменьшает ее чувствительность к Са24); плот-
ность Рг-адренорецепторов увеличивается по мере уменьшения диа-
метра бронхов;
Ф" простагландин Е2;
Ф- глюкокортикоиды увеличивают синтез р2-адренорецепторов в мио-
цитах бронхов, тормозят дегрануляцию тучных клеток.
Ф" гистамин через Н2-рецепторы эпителиальных клеток увеличивает секрецию NO,
' простагландина Е?, эпителиального расслабляющего фактора;
• Бронхоконстрикцию вызывают вещества из тучных клеток:
ф- гистамин через Н|-рецепторы (фосфолипаза С -» (ИФз / ДАГ -> (Са2* -»
(киназа ЛЦМ);
ф- брадикинин;
Ф" простагландины F2a, D2;
Ф" лейкотриены Da и С4 (старое название: медлеинореагируюшая субстанция)
действуют в 1000 раз сильнее, чем гистамин (фосфолипаза С -» (ИФ3/
ДАГ — (Са2*—« (киназа ЛЦМ).
Примечание: другие функции дыхательных путей см. п. 12.
8. Г азообмен в легких (вид транспорта - диффузия).
8.1. Газовый состав вдыхаемого (Ог- 21 %, 159 мм рт. ст.; СО2- 0,03 %, 0,22 мм рт. ст.), вы-
дыхаемого (Ог- 16 %, 115 мм рт. ст.; СО2-4,5 %, 33 мм рт. ст.) и альвеолярного (О2- 14
%, 100 мм рт. ст.; СО2- 5,5 %, 40 мм рт. ст.) воздуха, последний имеет относительно
постоянный состав.
8.2. Газообмен осуществляется в ацинусах (- 300 тысяч) состоящих из респираторных
бронхиол (18-22 генерация), альвеолярных ходов и альвеол (- 300 мли).
83. Особенности легочного кровообращения.
❖ Легкие являются единственным органом, через который проходит весь ми-
нутный объем крови. Легочные сосуды обладают большой растяжимостью
и могут вместить минутный объем крови в 5 раз больше, чем в покое. В го-
ризонтальном положении объем крови (-600 мл) в сосудах легких выше,
чем стоя (это способствует развитию отека легких в патологии). (При активном вдохе
кровенаполнение легких повышается до 1000 мл, при активном выдохе - снижается до 200
мл.)
273
❖ Легочные сосуды являются сосудами малого давления: систолическое АД
равно 20 - 25, диастолическое - 10 - 15, среднее - 14 -18 мм рт.ст. Поэтому
на кровоток легких в вертикальном положении сильно влияет гидростати-
ческое давление столба крови (в легких нулевое гидростатическое давление крови
находится на уровне правого предсердия, т.е. корня легкого; на каждые 1,3 см выше корня
легких артериальное н венозное давления снижаются на 1 мм рт.ст., ниже кория легкого
повышаются).
• Ь На кровоток в легких влияет также альвеолярное давление (Рал), которое в зависимости от
зоны легкого может быть выше, равно или ниже артериального (Рар) и венозного (Рв) дав-
лений. В зависимости от соотношений Рал, Рар, и Рв в легких выделяют в положении стоя
три функциональные зоны (сверху вниз). В 1-й зоне (верхушки легких, - 5 см) Рал > Рар >
Рв. В результате компрессии сосудов микроциркуляции кровоток в этой зоне минимален н
имеется только во время систолы правого желудочка. Во 2-й зоне (- 10 см) Рар > Рал >
Рв, кровоток осуществляется в результате разности давления между артериальным и аль-
веолярным давлением н существенно зависит от последнего. В 3-й зоне (- 20 см) Рар > Рв
> Рал, кровоток осуществляется в результате разницы между артериальным и венозным
давлением н существенно не зависит от альвеолярного давления.
❖ Нейро-гуморальная регуляция тонуса легочных сосудов.
• Симпатоадреналовые влияния: норадреналин и адреналин действуют
через а-адренорецепторы (а>> а2), вызывая сокращение сосудов, и через
Рз-адренорецепторы, вызывая расширение сосудов. (При невысоком уровне
сосудистого тонуса симпатоадреналовые влияния оказывают сосудосуживающий эф-
фект, при высоком - сосудорасширяющий эффект.)
• Парасимпатические влияния: ацетилхолин через М-холинорецепторы
вызывает снижение сосудистого тонуса при исходно повышенном его
уровне (при низком тонусе отмечен и сосуднстосужнвающнй эффект).
• Сосуды легких, в отличие от сосудов большого круга, сокращаются при
снижении Ро2, накоплении Н+ и действии гистамина. Гипоксическое
сужение легочных сосудов способствует балансиройке кровотока и вен-
тиляции: невентилируемые альвеолы выключаются из кровотока. (Гипок-
сия легких в патологии вызывает легочную артериальную гипертензию, повышает на-
грузку на правый желудочек и приводит к его гипертрофии - легочное сердце.)
• Эндотелий, контактируя с кровью всего объема сердечного выброса,
играет исключительную роль в регуляции сосудистого тонуса. Он выде-
ляет пептиды эндотелины как сосудорасширяющего, так и сосудосужи-
вающего действия, активные вазодилятаторы - простациклин и эндоте-
лиальный расслабляющий фактор, с помощью ангитензинпревращаю-
щего фермента образует сильный вазоконстриктор - ангиотензин П.
❖ В легочно-сердечном комплексе происходит небольшой сброс крови спра-
во-налево через мелкие веиы бронхов и сердца, поэтому Ро2 в артериальной
крови на 10-15 мм рт.ст. меньше, чем в альвеолах и их капиллярах (альвео-
лярно-артериальный градиент).
8.4. Диффузионный барьер в легких (площадь - 80 м2, толщина - 1 мкм).
❖ Газообмен осуществляется путем простой диффузии по закону Фика: диф-
фузия газа прямо пропорциональна градиенту его парциального давления и
площади барьера, обратно пропорциональна толщине барьера V = К8(Р,-
Pi)/L, где S - площадь барьера, L - его толщина, Р - парциальное давяенне Ch или СО2 в
альвеолах и крови легочных капилляров, К - коэффициент диффузии.
274
❖ При спокойном дыхании максимальный диффузионный градиент между
альвеолярным воздухом и капиллярной кровью для Ро2 равен 60 мм рт. ст.,
для Рсо2- 6 мм рт. ст. В норме диффузия газов в ацинусах осуществляется
уже в первой трети легочных капилляров. Величина диффузионной спо-
собности легких - 25 МЛ О2/мин • 1 мм рт. ст.
8.5. Для идеального обмена Ог и СОг надо, чтобы соотношение между вентиляцией
и кровотоком в легких было равно 1. Однако в норме имеется неодинаковое
отношение вентиляции и кровотока в различных отделах легких в верти-
кальном положении: в верхних отделах вентиляция превышает кровоток
(В/К-З), в средних отделах они примерно равны (В/К - 0,9), в нижних отделах
кровоток превышает вентиляцию (В/К - 0,7).
КРОВЬ
ЛЕГКИЕ'
О2—
СО2-<-
Артериальная
Эритроцит
Ка
fyCOj«-H2O+C
НСРз ,CI-
Венозная
ТКАНИ
CO 2 1
СГ НСОз
Ог
СО
O/HHb-HHbO^HbOg ->ннь
Н2О
СГ НСО^Н-НСОз’ CI-
Рис. 52. Транспорт кислорода н углекислого газа кровью. Темный
кружочек - НСО37СГ -иоиообмениик, Ка - карбоангидраза.
Объяснения в 9.1-9.2.
9. Транспорт газов кровью (главный вид транспорта - конвекция).
9.1. Транспорт кислорода от легких к тканям (рис. 52).
❖ Физически растворенного Ог в крови 0,3 об % или 3 мл Ог/л крови (при Рог
= 100 мм рт.ст.). Основное количество Ог (~ 190 мл/л крови) транспортиру-
ется в связи с гемоглобином в форме оксигемоглобина.
❖ Кривая диссоциации оксигемоглобина (рис. 53). Оксигемоглобина в артери-
альной крови 96-98 %, в венозной крови - 70%. На диссоциацию оксиге-
моглобина влияют следующие факторы крови: pH, Рсо2, температура, про-
межуточный продукт гликолиза в эритроцитах 2,3-дифосфоглицерат.
• Сдвиг вправо происходит при повышении Рсог, t°, 2,3-дифосфоглице-
рата и Н+ (снижении pH). Это уменьшает сродство НЬ к О2, улучшая его
отдачу в тканях (немного затрудняя его присоединение в легких);
275
• Сдвиг влево происходит при снижении Рсог, t°, 2,3-дифосфоглицерата и
Н+ (повышении pH). Это увеличивает сродство НЬ к Ог, ухудшая его от-
дачу в тканях (немного улучшая его присоединение в легких).
❖ Показатели транспорта кислорода кровью:
• кислородная емкость крови (КЕК = 180 - 200 мл Ог/л) зависит от коли-
чества гемоглобина, поэтому она одинакова в артериальной и венозной
крови;
• напряжение Ог в артериальной и венозной крови (аРог = 95 - 100 мм рт.
Напряженке кяслорада в мм ртхг.
Рис. 53. Кривая диссоциации оксигемоглобина крови.
Объяснения в 9.1
ст., вРо2 = 35 - 45 мм рт. ст.);
• содержание Ог в артериальной и венозной крови (аОг = 180 — 200 мл/л,
вОг = 120 -140 мл/л).
❖ Основные механизмы регуляции: изменение количества и оксигенации ге-
моглобина.
9.2. Транспорт углекислого газа (рис. 53).
❖ Формы и количественная характеристика транспорта СОг кровью:
• физически растворенный СОг (~ 1,2 ммоль/л, ~ 5 - 10 % всего СОг кро-
ви);
• в составе гидрокарбоиата - HCOj’ (количественно основная форма: - 23
ммоль/л, - 85 % всего СОг); гидрокарбонат, образуясь в эритроцитах,
выводится в плазму с помощью иоиообменника СГ/НСОз';
• СО2, связанный с аминогруппами (-NHy-COO ) белка гемоглобина (карб-
гемоглобии: 5 -10 % всего СОг).
❖ Рсог в артериальной крови равно - 40 мм рт. ст., в венозной - 46 мм рт. ст.,
содержание СОг в артериальной крови равно - 480 мл/л, в венозной ~ 530
мл/л.
❖ Образование и диссоциация угольной кислоты и карбгемоглобииа в легких
и тканях: карбоангидраза эритроцитов в 1500 раз ускоряет образование
НгСОз (СОг + Н2о -карбоангидраза— Н2СО3 + КНЬ — КНСО3 + ННЬ).
276
9.3. Методы исследования газового состава крови.
❖ Полярографическая методика (микроАструп) (см. тему 20, задание 1; 8).
•I' Оксигемометрия и оксигемография позволяет оценить кислородтраиспортную
функцию крови. Они основаны на том, что в красной части спектра (620-680 нм)
коэффициент поглощения света для восстановленного гемоглобина в несколько
раз больше, чем для оксигемоглобина. При этом получают величины насыщения
гемоглобина кислородом в процентах. Для того, чтобы вычислить содержание Ог
в пробе крови, нужно знать количество в ней гемоглобина. Используя кислород-
ную емкость 1 г гемоглобина (1,34 мл О2), можно вычислить содержание О2 в кро-
ви. Комбинированные оксигемометры, кроме кюветного определения оксигемог-
лобина в пробах крови, снабжены ушным датчиком для проведения непрерывной
бескровной оксигемометрии. При этом прибор регистрирует относительную вели-
чину насыщения гемоглобина крови кислородом по отношению к исходной вели-
чине, установленной на 96% при дыхании испытуемого воздухом, или 100% - при
дыхании кислородом. В некоторых оксигемометрах вносится поправка на величи-
ну оптической плотности ткани «без крови» и регистрируют абсолютные величи-
ны оксигемоглобина циркулирующей крови». Оксигемометры, снабженные само-
писцем, позволяют проводить оксигемометрию - записывать динамику изменения
оксигемоглобина в крови.
10. Г азообмен между кровью и тканями (главный вид транспорта - диффузия).
10.1. Диффузионные градиенты Ро2 и Рсо2 между кровью и тканью:
❖ Ро2 в притекающей к тканям крови - 95 мм рт. ст., в межклеточной жидко-
сти - 45 мм рт. ст., на поверхности клеток - 20 мм рт. ст., в митохондриях -
1 мм рт. ст.; эти градиенты обеспечивают поступление кислорода из крови
в клетки тканей;
❖ напряжение СО2: в притекающей к тканям крови - 40 мм рт. ст., в клетках
- 60 мм рт. ст., что обеспечивает поступление СО2 из клеток тканей в
кровь.
10.2. Количественно обмен О2 между кровью и тканями характеризует артериове-
нозная (a-в) разница по О2, равная 50 мл О2/л крови, и коэффициент использо-
вания О2 (КИО2), характеризующий долю О2, поступившего из крови в клетки
ткани, он равен 25 - 40 %.
103. Основной механизм регуляции - сдвиги кривой диссоциации оксигемоглобина,
изменение объемного кровотока в тканях и органах.
11. Тканевое дыхание (основной вид транспорта-диффузия).
11.1. Основные пути внутриклеточного потребления О}.
❖ Митохондриальный путь (40 - 85 % всего О2), 4-электроиное восстановле-
ние О2 до воды под действием цитохромоксидазы, основная функция - ак-
кумуляция энергии в виде АТФ.
❖ Микросомальный путь в гладкой ЭПС (10 - 40 % всего потребляемого Оз),
монооксигеназная реакция (с участием цитохрома Р450) внедряет атом кислоро-
да в молекулу окисляемого вещества, что приводит к образованию поляр-
ных (т.е. водорастворимых) веществ. Основные функции этого пути - синтез и
инактивация стероидных гормонов, детоксикация ксенобиотиков.
❖ Образование активных продуктов неполного восстановления О2 (суперок-
сидный анион, перекись водорода, гидроксильный радикая и др.) — 5 - 15 % (в нейтро-
филах - до 90%) всего потребляемого О2. Функциональная роль: фагоцитар-
277
ная активность лейкоцитов, вазомоторное действие, лизис клеточных мем-
бран (например, при овуляции) и др.
11.2. Миоглобин (много в красных мышцах и миокарде) депонирует и транспортирует Ог в
клетке: обладает высоким сродством к О2 (Pso= 8 мм рт. ст.) и отдает его только
при низком Рог в клетке (меньше 10 мм рт. ст., например, при сокращении
мышц).
11.3. Потребление Ог является показателем тканевого дыхания в организме: ПО2 =
(a-в) Ог х МОК л/мин; представляет собой наиболее адекватный показатель фи-
зической нагрузки, в целом организме минимальное ПО2 равно 0,2 л/мин, в по-
кое - 0,3 л/мин, максимальное ПОг - 3,0 л/мин характеризует состояние сер-
дечно-сосудистой и дыхательной систем.
11.4. Основной механизм регуляции потребления кислорода: изменение активно-
сти дыхательных ферментов и перераспределение использования Ог в клетке
между основными путями (митохондриальным и микросомальным).
12. Негазообменные функции легких.
12.1. Очищение воздуха и дыхательных путей.
❖ Фильтрация воздуха в полости носа.
♦ 1* Осаждение корпускулярных примесей (инерционное и гравитационное осаждение,
броуновское движение).
❖ Экзокринная функция дыхательных путей.
• Слизистая оболочка полости носа образует 100 - 500 мл секрета в су-
тки, передвигаемого к глотке;
• Слизистая трахеи и бронхов образует трахеобронхиальный секрет (10 -
50 мл/сутки), его вязкость до 10 000 раз больше воды:
" Ф увеличивает секрецию гистамин, ацетилхолин, лейкотриен С4;
ф выведение секрета по направлению к глотке осуществляется с по-
мощью «реснитчатого эскалатора» со скоростью 15 - 30 мм/мин в
трахее, 3-10 мм/мин в бронхах;
Ф коллатеральная вентиляция через межальвеолярные поры, бронхо-
альвеолярные и межбронхиальные каналы способствует удалению
секрета.
❖ Защитные дыхательные рефлексы:
• Рефлекс кашля («сторожевой пес» легких).
Ф Запускается с рецепторов глотки, гортани, трахеи и бронхов.
Ф Центр кашля расположен в продолговатом мозге (функционально объ-
единяет чувствительные, двигательные ядра IX и X нервов и парасимпатическое
ядро X нерва, инспираторные и экспираторные отделы дыхательного центра).
Ф Через эфферентные пути (медиальные и латеральные ретикулоспинальные
пути к мотонейронам дыхательных мышц и двигательные волокна n.vagus к
мышцам гортани и IX нерва к мягкому небу) запускается трехфазная ис-
полнительная реакция: 1) глубокий вдох; 2) форсированный выдох
на фоне закрытой голосовой щели (внутриплевральное давление -
100 мм рт. ст.); 3) внезапное открытие голосовой щели, напряжение
мягкого нёба, поток воздуха выходит через рот (до 12 л/с или 120 м/с).
• Рефлекс чихания.
278
ф- Запускается с рецепторов слизистой оболочки полости иоса.
ф Центр чихания продолговатого мозга (функционально объединяющий
спинальное ядро V нерва, двигательные ядра IX и X нервов и парасимпатическое
ядро X нерва, инспираторные и экспираторные отделы дыхательного центра);
Через эфферентные пути (медиальные и латеральные ретикулоспинальные
пути к мотонейронам дыхательных мышц и двигательные волокна n.vagus к
мышцам гортани и IX нерва к мягкому небу) запускается трехфазная эф-
фекторная реакция: 1) глубокий вдох; 2) форсированный выдох иа
фоне закрытой голосовой щели; 3) внезапное открытие голосовой
щели, опускание мягкого нёба, поток воздуха направляется в носо-
вую и частично в ротовую полость.
• Ринобронхиальный рефлекс: с рецепторов преддверия носа (например, при вдыхании
холодного воздуха, нырянии в холодной воде) происходит сужение бронхов.
12.2 . Кондиционирование вдыхаемого воздуха.
❖ Согревание воздуха (на это затрачивается - 10 % общей теплоотдачи).
❖ Увлажнение воздуха (выделяется в сутки около 440 мл воды).
❖ Независимо от внешних условий в бронхах 10-й и ббльшей генерации тем-
пература воздуха равна 37°С, влажность - 100 %.
12.3 . Иммунологические защитные механизмы.
❖ Неспецифические гуморальные факторы (лизоцим, интерферон, пирогены
и др.).
❖ Специфические гуморальные факторы (В-лимфоциты, иммуноглобулины,
например, 1g А).
Клеточные неспецифические механизмы (микрофаги и макрофаги).
❖ Клеточные специфические механизмы (Т-лимфоциты).
12.4 . Контроль активности БАВ крови (серотонина, брадикинина, норадреналина,
простагландинов, гистамина и др.) путем инактивации их эндотелием капилля-
ров легких.
12.5 . Эндокринная функция легких.
❖ Превращение ангиотензина I в ангиотензин П превращающим ферментом
эндотелиальных клеток капилляров.
Тучные клетки (составляют около 2 % массы легкого и поверхности альвеол) проду-
цируют гистамин, серотонин, простагландины, гепарин, лейкотриены Da,
С5 и др.
❖ «Нейроэндокринные» эпителиальные клетки выделяют пептидные гормо-
ны: соматостатин, кальцитонин, холецитокинин, энкефалины, вещество Р и
др., включающиеся в регуляцию функций легких и целостного организма.
12.6 . Регуляция системы гемостаза.
❖ Увеличение потенциала коагуляции: продукция тромбопластина, тромбок-
сана Аз (вызывает агрегацию тромбоцитов).
❖ Снижение потенциала коагуляции: продукция гепарина, простациклина,
активация фибринолиза, поглощение из крови фибрина.
12.7 . Участие в депонировании крови: может депонироваться до 15% крови.
13. Профильный материал для студентов лечебного факультета.
13.1. Структурно-функциональная характеристика системы дыхания плода.
Особенности дыхательной системы плода.
279
• Легкие не являются органом внешнего дыхания, альвеолы и бронхи запоянены
жидкостью, которая секретируется альвеолоцитами 2-го типа. Отсутствует
смешивание легочной и амниотической жидкостей за счет закрытой голосовой
щели.
• Плацента является основным органом дыхания во внутриутробном развитии:
Ог поступает из крови матери в кровь плода, а СОг - наоборот.
• Большая толщина плацентарной мембраны (в 10 - 15 раз превышает толщину
легочной мембраны) не позволяет выравнивать Рог и Рсог по обе ее стороны.
♦ Особенности транспорта газов кровыо.
• Транспорт кислорода.
Ф Низкое Рог оксигенированной крови пупочной вены (30-50 мм рт. ст.), со-
держание оксигемоглобина (65 - 80%) и кислорода (100 - 150 мл/л кро-
ви), еще меньшее содержание его в сосудах сердца и мозга и других орга-
нах
ф- Высокое сродство гемоглобина плода (HbF) с Ог создает сдвиг кривой
диссоциации оксигемоглобина влево.
Ф К концу беременности происходит снижение содержания оксигемоглоби-
на до 40 %.
• Транспорт углекислого газа.
Ф Низкое Рсог в артериальной крови плода (35 - 45 мм рт. ст.) за счет ги-
первентиляции беременных.
Ф Отсутствие карбоангидразы в крови плода, приводящее к исключению из
газообмена 38 - 42 об % СОг, содержащегося в гидрокарбонате.
Ф Диффузия СО2 через плацентарную мембрану осуществляется за счет фи-
зически растворенного в плазме СОг.
Ф К концу беременности происходит увеличение содержания СОг в крови
плода до 600 мл/л.
• Адекватному обеспечению тканей плода Ог способствуют следующие факто-
ры:
Ф интенсивный тканевой кровоток (в 2 раза больше, чем у взрослого);
Ф преобладание анаэробных окислительных процессов над аэробными:
Ф небольшие энергетические затраты плода.
• Внутриутробные дыхательные движения способствуют развитию легких, ды-
хательной мускулатуры, венозному притоку крови к сердцу.
13.2. Изменение легочных объемов, вентиляции легких и резерва дыхания к старости.
Ф Легочные объемы и емкости-
• снижаются величины ДО, ЖЕЛ, РО вдоха и РО выдоха;
• увеличивается ООЛ и физиологическое мертвое пространство за счет частич-
ного залустевания и облитерации легочных капилляров
Ф Нарушение равномерности вентиляции легких приводит к уменьшению поступле-
ния воздуха преимущественно в иижние отделы легких, где кровообращение мак-
симально.
Ф Уменьшение альвеолярной вентиляции приводит к снижению количества Ог, уча-
ствующего в газообмене.
Ф Компенсаторно увеличиваются МОД и частота дыхания в покое.
280
♦> Уменьшается эффективность дыхания за счет увеличения его кислородной цены.
13.3. Газообмен в легких и транспорт газов кровью характеризуется наличием физиологи-
ческой гипоксемии старости (снижение а-Ро2 и НЬОг) в результате:
❖ уменьшения величины диффузионной способности легких;
❖ увеличения числа артериовенозных шунтирующих сосудов в малом круге;
♦♦♦ снижения перфузии легких за счет уменьшения легочного кровотока.
14. Профильный материал для студентов педиатрического факультета.
14.1. Структурно-функциональная характеристика системы дыхания плода (см. вы-
ше^ 1).
14.2. Система дыхания у новорожденных
❖ Структурные особенности органов дыхания новорожденных
* Узость верхних дыхательных путей (носовых входов, отсутствие придаточных
пазух носа и нижнего носового прохода, небольшая глотка)
• Недостаточное развитие слизистых желез трахеи и бронхов.
• Низкая дифференцировка ацинусов, меньшие размеры и количество альвеол.
• Низкая растяжимость легких (мало эластических волокон).
• Горизонтальное расположение ребер ограничивает экскурсию грудной клетки,
что обусловливает диафрагмальный тип дыхания.
• Высокая растяжимость грудной клетки.
Внешнее дыхание новорожденного.
• Вентиляция легких новорожденного.
Ф Относительно большая величина МОД (500 - 1300 мл/мин), альвеолярной
вентиляции (400 - 500 мл/мин) в покое, низкий резерв их увеличения.
Ф Высокая частота дыхания (40 - 60 в 1 мин), нерегулярный ритм (частое
дыхание чередуется с редким), небольшой дыхательный объем (10 - 25
мл).
* Ф* Образование функциональной остаточной емкости легких (до 75 - 80 мл)
и удаление жидкости из альвеол Жизненная емкость легких равна 120 -
150 мл.
* Ф* Высокое аэродинамическое сопротивление дыхательных путей.
* Ф* Глубокие периодические вдохи улучшают аэрацию легких, равномерное
распределение в них воздуха, препятствуют образованию ателектазов
• Газообмен в легких между кровью и альвеолярным воздухом.
Ф Увеличение диффузионной поверхности за счет газообмена не только в
альвеолах, ио и в альвеолярных ходах.
Ф Начинает функционировать малый круг кровообращения, легочный кро-
воток новорожденного относительно больше, чем у взрослого человека
(более широкая сеть капилляров).
Ф Низкое отношение вентиляции к перфузии, неравномерность вентиляции
альвеол.
• Транспорт газов кровью у новорожденного
Ф Транспорт кислорода.
Высокое Р02 (~ 120 мм рт ст) и низкое Рсог (~ 23 мм рт. ст.) в арте-
риальной крови
281
Невысокое содержание оксигемоглобина (- 90 %) в момент рожде-
ния с последующим увеличением до 98 % в первые сутки, этому спо-
собствует большое сродство к Ог фетального гемоглобина (HbF - 70
%).
Большая кислородная емкость крови (210 - 260 мл О,/л) с после-
дующим снижением до 140 - 150 мл О,/л крови во второй половине
1-го мес. за счет разрушения эритроцитов (физиологическая желтуха
новорожденных) и снижения содержания гемоглобина (происходит
замена HbF на НЬА).
Большее потребление организмом Ог обеспечивается более высоким
уровнем вентиляции легких, кровообращения в легких и в организме.
Более высокая устойчивость тканей (в том числе н нервной) к гипок-
сии.
Ф Транспорт СОг осуществляется преимущественно в виде физически рас-
творенного и связанного с гемоглобином СОг, так как карбоангидраза, ак-
тивирующая образование и распад НСОз', у новорожденных появляется в
конце 1-й недели, содержится в малом количестве и активность ее неве-
лика.
14.3. Система дыхания у детей грудного и более старших возрастов.
❖ Развитие легких в онтогенезе, типы дыхания у детей.
• Формирование ацинусов и увеличение количества альвеол до 7 - 8 лет (осо-
бенно в первые 3 года), увеличение объема альвеол с 7 - 8 лет.
• Увеличение плотности капилляров в легких и их диффузионной способности.
• Постепенная смена тахипнотнческого типа дыхания на брадипнотический.
• Переход с брюшного на смешанный тип дыхания происходит к 3 годам, в воз-
расте 14 - 17 лет у девушек формируется грудной тип дыхания, у юношей -
брюшной.
❖ Вентиляция легких детей грудного и более старших возрастов.
• Увеличение МОД в 10 раз (особенно на 1-м году и в периоде полового созре-
вания).
• Уменьшение частоты и стабилизация ритма дыхания, отношение ЧСС к часто-
те дыхания у новорожденного - 1, в 1 год 2 - 3, у взрослого 3-4.
• Увеличение ДО с возрастом (1 год - 70 мл, 6 лет - 160 мл, 14 лет - 300 мл, у
взрослых - 300 - 800 мл).
• Увеличение ЖЕЛ (1 месяц - 130 мл, 6 лет - 1200 мл, 14 лет - 2700 мл, взрос-
лые - 3500 - 5000 мл).
• Снижение аэродинамического сопротивления дыханию за счет увеличения
просвета дыхательных путей.
• Снижение эластического сопротивления дыханию с возрастом за счет увели-
чения в легких доли эластических волокон относительно коллагеновых.
Уменьшение энергетической цены дыхания.
❖ Газообмен в легких.
• Имеется высокий диффузионный градиент Рог между альвеолярным воздухом
и кровью за счет постепенного увеличения интенсивности альвеолярной вен-
тиляции.
282
• Увеличивается вентиляционно-перфузионное отношение, что соответствует
высокому уровню обмена веществ.
• Имеется относительно большая площадь диффузионной поверхности за счет
более плотной капиллярной сети легких у детей.
• Высокое Рог (105 - 108 мм рт. ст.) и низкое Рсог (- 35 мм рт. ст.) в артериаль-
ной кровн.
& Особенности транспорта Ог и СОг кровью.
• Замена HbF на НЬА к 2 - 3 мес.
• Высокий уровень НЬО2 (- 98 %), меньшая кислородная емкость крови связана
с пониженным содержанием гемоглобина в крови.
• Невысокая (а-в)Ог из-за более быстрого движения крови по капиллярам.
• Снижение с возрастом устойчивости к кислородному голоданию связано с
увеличением интенсивности аэробных окислительных процессов.
15. Профильный материал для студентов стоматологического факультета.
15.1. Ротовое дыхание, его особенности: Ротовое дыхание, по сравнению с носовым, созда-
ет меньшее сопротивление (особенно при вдохе), но и меиьшее согревание и очищение
вдыхаемого воздуха.
15.2. Роль полости рта и органов дыхания в формировании речи.
Ф Работа аппарата дыхания во время речи называется речевым дыханием. Органы,
непосредственно участвующие в образовании речи:
• органы дыхания (легкие, трахея н бронхи);
• активные - подвижные, способные менять объем и форму речевого тракта и
создавать в нем препятствия для выдыхаемого воздуха (гортань, глотка, мяг-
кое нёбо, язык, губы);
• пассивные - неподвижные, лишенные этой возможности (зубы, твердое иёбо,
полость носа и придаточные пазухи).
Периферические механизмы образования речи:
• энергетический (дыхательные мышцы, гладкие мышцы трахеи и бронхов)
обеспечивает движение воздуха в дыхательных путях;
• генераторный (тоновый - гортань, шумовой - создание щелей в полости рта)
обеспечивает фонацию (голос);
• резонаторный (рот и глотка, носоглотка с придаточными полостями) обеспе-
чивает артикуляцию (звуки).
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 н 3.2). Объем реферата 2-3 стр., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебник и пособия): [1-9], [11], [12], [14], [15], [16].
283
3.2. Дополнительная литература (см. стр. 514 - 522): [27], [30], [34], [43], [53], [62],
[66], [71], [84], [85], [101], [152], [154], [174], [179], [192], [194], [196], [204], [206],
[222], [225], [244], [245], [246], [251 - 254].
3.3. Дистанционные средства обучения. Использование страницы кафедры а Интер-
нете - www.vsma.ac.ru/-nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брни. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебпая комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 19-1. При хроническом отравлении угарным газом (СО) человек почувствовал слабость,
быструю утомляемость. Ои обратился в больницу. При обследовании у него определили показа-
тели внешнего дыхания (они оказались в пределах нормы) и показатели кислородного состояния
артериальной крови — содержание Ог, физически растворенного в плизме От и Poj. Что произой-
дет с этими показателями (увеличатся, уменьшатся или существенно ие изменятся) и почему?
Задача 19-2. Объясните, почему выдыхаемый воздух по сравнению с альвеолярным воздухом имеет более
высокое содержание О2 и более низкое содержание СО;.
284
Тема 20. Регуляция дыхания
Цель: а) знать структуры ЦНС, обеспечивающие дыхательную периодику; рецепторы легких, их
роль в саморегуляции дыхания, рефлексы Герннга н Брейера, значение рецепторов дыхатель-
ных мышц в компенсацнн дыхательных нагрузок; роль гипоталамуса, лимбической системы и
коры больших полушарий в регуляции дыхания; влияние на частоту и глубину дыхания газо-
вого состава и pH артериальной крови; значение центральных и периферических хеморецеп-
торов в обеспечении газового гомеостаза, изменение вентиляции легких при гиперкапнии и
гипоксии; дыхание при физической работе, повышенном и пониженном барометрическом
давлении, резервные возможности системы дыхания; первый вдох новорожденного ребенка,
возрастные особенности регуляции дыхания;
б) уметь использовать эти знания для анализа деятельности системы дыхания в аспекте регу-
ляции функций целостного организма в различных условиях жизнедеятельности;
в) ознакомиться с практическими навыками на уровне знаний: определение возбудимости ды-
хательного центра.
Мотивация темы: важнейшая функция регуляции дыхания - поддержание постоянства показателей
внутренней среды организма: Рог и Pcoj, а также pH крови. Нарушение регуляции дыхания
часто приводит к необратимым изменениям в организме и служит причиной его гибели. Зна-
ние механизмов, обеспечивающих регуляцию акта дыхания в физиологических условиях, по-
зволяет дать правильную оценку эффективности дыхания на разных его этапах в условиях
нормы н патологии.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задааие 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержанки самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Общая характеристика регуляции внешнего дыхания.
1.1. Регуляция дыхания - это согласование внешнего дыхания с метаболической
потребностью организма в различных условиях.
1.2. Основные регулируемые показатели:
❖ Напряжение Оз (Рог) в артериальной крови (90 - 100 мм рт. ст.).
❖ Напряжение СОг (Рсог) в артериальной крови (38 - 42 мм рт. ст.).
❖ Регуляция минутного объема дыхания (МОД), т.е. глубины и частоты ды-
хания.
1.3. Общая схема регуляции внешнего дыхания (рис. 54).
❖ Основные звенья регуляции.
• Хеморецепторное звено обеспечивает дыхательный центр информацией
о необходимом уровне легочной вентиляции в данное время.
• Механорецепторное звено обеспечивает дыхательный центр информа-
цией о реальной вентиляции легких в данное время (по активности дыха-
тельной мускулатуры).
• Центральное звено (дыхательный центр продолговатого мозга и вышележащие
центры, регулирующие легочную вентиляцию) обеспечивает коррекцию венти-
ляции легких в зависимости от метаболических потребностей организ-
ма.
285
• Исполнительное звено (мотонейроны спинного мозга: Сы - диафрагма, ТЪц -
инспираторные мышцы, Thg-io - экспираторные мышцы) обеспечивает дыха-
тельный цикл, не обладает автоматизмом и, будучи изолированным, не
может поддерживать дыхание.
Ф Основные кибернетические типы регуляции дыхания.
1.4.
• Регуляция по отклонению
(отрицательная обратная
связь с хеморецепторов при
отклонении Pcoj, Р02, pH ар-
териальной крови).
• Регуляция по возмущению
(например, влияния на ды-
хательный центр с про-
приорецепторов и мотор-
ной коры при физической
работе).
• Регуляция по прогнозирова-
нию (условно-рефлек-
торная регуляция дыхания,
функциональная система
регуляции газового состава
кровн).
Типы легочного дыхания (у здорово-
Рис. 54. Схема регуляции дыхания
го человека).
❖ Эупноэ - нормальное дыхание
в покое, сопровождающееся чувством комфорта.
❖ Тахипноэ - увеличение частоты дыхания выше 20 в 1 мин.
♦ > Брадипноэ - снижение частоты дыхания ниже 12 в 1 мин.
❖ Гиперпноэ - увеличение МОД прн повышенной продукции COj.
❖ Гипопноэ - снижение МОД при сниженной продукции СОа.
❖ Гипервентиляция - увеличение МОД, не соответствующее продукции СОг,
ведет к гипокапнии и гипероксии.
❖ Гиповентиляция - снижение МОД, не соответствующее продукции СОг,
ведет к гиперкапнии и гипоксемии.
❖ Апноэ - временная остановка дыхания, обусловленная главным образом
снижением физиологической стимуляции дыхательного центра (например,
после произвольной гипервентиляцни).
❖ Диспноэ (одышка) - нарушение частоты, глубины и ритма дыхания, сопро-
вождающееся субъективным ощущением недостатка воздуха или затруд-
ненным дыханием (например, при тяжелой физической нагрузке).
2. Дыхательный центр (центральный дыхательный механизм).
2.1. Бульбарный дыхательный центр (в ретикулярной формации в области дна IV желу-
дочка у нижнего угла ромбовидной ямкн) (Флураис, 1822; Н.А. Миславскнй, 1885; Р. Баум-
гартен, 1956).
286
❖ Инспираторная область включает в себя дорсомедиальную группу (ДМГ)
нейронов н ростральную часть вентролатеральной группы (ВЛГ) нейронов.
Она содержит:
• Ранние инспираторные нейроны - интернейроны, которые активны в
начале вдоха (расположены в ВЛГ).
• Полные инспираторные нейроны - эфферентные нейроны центра, ин-
нервирующие мотонейроны мышц вдоха, активны в течение всего вдоха
(расположены в ДМГ и ВЛГ).
• Поздние инспираторные нейроны - эфферентные нейроны центра, ин-
нервирующие мотонейроны мышц вдоха, активны в конце вдоха (распо-
ложены в ДМГ и ВЛГ).
• Инспираторно-тормозные нейроны - предполагаемые тормозные ин-
тернейроны инспираторной зоны, способные затормозить активность
. полных и поздних инспираторных нейронов.
❖ Экспираторная область включает в себя ВЛГ нейронов, преимущественно
ее каудальную часть. Она содержит:
• Постинспираторные нейроны - интернейроны, активные в первой по-
ловине выдоха, тормозят как инспираторные, так и экспираторные ней-
роны (расположены в ростральной ВЛГ).
• Экспираторные нейроны - эфферентные нейроны центра, иннерви-
рующие мотонейроны мышц выдоха, активны во второй половине вы-
доха, преимущественно при усиленном выдохе (расположены в каудальной ВЛГ).
• Преинспираторные нейроны - интернейроны, блокирующие возбужде-
ние экспираторных нейронов, способствуют смене выдоха на вдох.
2.2. Пневмотаксический центр моста (Лэмсден, 1923).
❖ Расположен в верхняя часть моста (медиальное парабрахнальное ядро).
❖ Эффект: тормозит инспираторные нейроны, ограничивает длительность
вдоха и повышает частоту дыхания.
❖ Перерезка в нижней части моста приводит к удлинению вдоха, при допол-
нительной перерезке блуждающих нервов происходит апнейзис (глубокие
длительные вдохи и короткие выдохи).
2.3. Лпнейстический центр расположен в нижней части моста, оказывает постоянную стимуля-
цию инспираторных нейронов, способствует вдоху и пролонгирует его.
2.4. Периодическая деятельность (автоматия) дыхательного центра.
❖ Автоматия дыхательного центра - способность его обеспечить смену
вдоха и выдоха за счет своих внутренних механизмов при постоянной им-
пульсации с хеморецепторов. Автоматия дыхательного центра находится
под выраженным произвольным корковым контролем.
❖ Генерация дыхательного ритма. Выделяют трн фазы активности дыха-
тельных нейронов в течение дыхательного цикла: инспираторная, постин-
спираторная и экспираторная фазы.
• Инспираторная фаза (соответствует вдоху) обусловлена последовательной
активацией ранних, полных и поздиих инспираторных нейронов, что
сопровождается линейным нарастанием их суммарной активности. Рез-
кое уменьшение их активности (смена вдоха на выдох), как полагают, свя-
287
зано с активацией особых тормозных нейронов, возбуждение которых
осуществляется от нейронов пневмотаксического центра моста н от ре-
цепторов растяжения легких (см. ниже 3.2).
• Постинспираторная фаза соответствует первой половине выдоха (пас-
сивная экспирация) н обусловлена особыми постинспираторными нейро-
нами, которые тормозят как инспираторные, так и экспираторные ней-
роны. Они, вероятно, обеспечивают интервал времени, необходимый
для выведения воздуха из легких в связи с их эластической тягой.
• Экспираторная фаза соответствует второй половине выдоха (активная
экспирация) и обусловлена активацией экспираторных нейронов, иннер-
вирующих мотонейроны мышц выдоха. В конце выдоха происходит
возбуждение преннспираторных нейронов, которые тормозят импульса-
цию экспираторных нейронов (прекращают выдох). Возможным источ-
ником возбуждения преннспираторных нейронов являются ирритант-
ные рецепторы легких, возбуждающиеся при уменьшении объема лег-
ких во время выдоха (инспираторно-облегчающнй рефлекс Геринга - Брейера).
(При частом дыхании экспираторная фаза может быть ие аыражеиа, и постннспира-
торная фаза может непосредственно переходить в следующую фазу инспирации.)
3. Рефлекторная регуляция дыхании.
3.1. Влияние с хеморецепторов на дыхательный центр.
❖ Гуморальные факторы, участвующие в регуляции дыхания: Ро2, Рсо2 и pH
крови:
• стимулируют легочную вентиляцию гиперкапния (МОД увеличивается до
80 л/мнн), гипоксемия и ацидоз (МОД увеличивается до 30 л/мнн);
• уменьшают легочную вентиляцию гипокапния, гипероксия и алкалоз.
❖ Периферические хеморецепторы:
• локализуются в сосудах (особенно в артериях), тканях внутренних ор-
ганов, их концентрация максимальна в синокаротидной и аортальной
зонах;
• афферентная импульсация от них проводится по нерву Циона - Людви-
га (аетвь X иераа) и нерву Геринга (ветвь IX нерва);
• имеют высокую чувствительность к изменению Роз в артериальной
крови (особенно к его снижению), в меньшей степени реагируют на измене-
ния Рсоз н pH;
• имеют короткое латентное время действия на дыхательный центр (- 5 с).
❖ Центральные хеморецепторы (хеморецепторная зона дыхательного цен-
тра):
• локализуются на переднебоковой поверхности продолговатого мозга и
моста в виде трех пар скопления нейронов;
• отличаются высокой чувствительностью к изменению pH (пороговые ко-
лебания pH -0,01) и Рсог в ликворе;
• имеют длительное латентное время действия на дыхательный центр (~
25 с).
288
3.2. Влияние с механорецепторов на дыхательный центр.
❖ С механорецепторов легких регулируется частота и глубина дыхания.
• Рецепторы растяжения легких (медленно адаптируются) расположены в
гладких мышцах трахеи и бронхах, реагируют на увеличение объема
легких при вдохе; с них возникает инспираторно-тормозящий рефлекс
Геринга — Брейера (если дыхательный объем > 1 л).
• Ирритантные рецепторы расположены в эпителии бронхов, реагируют
на быстрое изменение объема легких, на механические воздействия
(пыль) и на пары химических веществ:
-ф- это быстро адаптирующиеся рецепторы, порог раздражения у ннх
выше, чем у рецепторов растяжения легких;
ф" с них формируются рефлексы кашля, сужения бронхов при выдохе,
инспираторно-облегчающий рефлекс Геринга - Брейера при спаде-
нии легких, который укорачивает выдох и способствует смене его
на вдох.
• Юкстаальвеолярные рецепторы (J-рецепторы) локализуются в интерсти-
ции альвеол у капилляров, реагируют на давление жидкости в межкле-
точном пространстве легких, с ннх формируется одышка и торможение
у-мотонейронов (ограничение физической нагрузки при угрозе левожелудочковой
недостаточности и отека легких).
❖ Проприорецепторы дыхательных мышц участвуют в компенсации дыха-
тельных нагрузок:
• локализуются в дыхательной мускулатуре, преимущественно в межре-
берных мышцах;
• усиливают сокращение дыхательной мускулатуры при увеличении со-
противления дыханию, ослабляют - при уменьшении сопротивления
дыханию.
4. Влиииие на дыхательный центр высших отделов ЦНС.
4.1. Гипоталамус осуществляет:
♦> связь дыхания с обменом веществ и терморегуляцией в организме;
❖ регуляцию дыхания для обеспечения поведенческих актов, направленных
на удовлетворение биологических потребностей (агрессивно-
оборонительной, пищевой, половой и др.).
4.2. Лимбическая система осуществляет связь дыхания с вегетативной регуляцией
органов и с эмоциями.
4.3. Кора больших полушарий осуществляет:
❖ связь дыхания с изменениями внешней и внутренней среды (через сенсорную
кору);
❖ условно-рефлекторную и произвольную регуляцию дыхания;
❖ корковое дублирование автоматии дыхательного центра;
❖ регуляцию дыхания для обеспечения социальных форм поведения;
❖ особенности дыхания при образовании речи (участие в механизмах фона-
ции и артикуляции: см. тему 31; задание 1: б.З).
289
4.4. Функциональная система поддержания газового состава крови, ее централь-
ные и периферические компоненты.
❖ Полезный результат имеет не один, а несколько параметров - pH, Рсог,
РО2.
❖ Рецепторное звено: 1) пусковая афферентация с кислородных и гиперкап-
нических сенсоров; 2) обстановочная афферентация с интро- и экстероре-
цепторов.
Центральное звено (центры регуляции систем дыхания, кровообращения и
крови).
❖ Эфферентное звено (вегетативная и соматическая нервная система, эндок-
ринная система).
❖ Эффекторное звено: 1) взаимодействие физиологических систем дыхания,
кровообращения и крови, обеспечивающее газовый гомеостаз крови; 2) по-
веденческие механизмы, направленные иа поддержание газового состава
крови.
5. Особенности дыхания в различных условиях.
5.1. Дыхание при физической работе.
❖ Увеличение легочной вентиляции в результате:
• условно-рефлекторной активации дыхательного центра (предстартовое
состояние);
• стимуляции дыхательного центра с проприорецепторов работающих
мышц и из двигательной коры (пусковые факторы);
• возбуждения центральных и периферических хеморецепторов при из-
менении газового состава и pH крови.
Транспорт газов крови увеличивается в результате:
• ускорения кровотока (увеличение ЧСС, СО, МОК);
• использования резервных и депонированных эритроцитов костного
мозга и селезенки;
• рабочей гиперемии мышц (они получают до 80% МОК).
❖ Увеличение потребления Ог мышцами в результате:
• увеличения артерио-венозной разницы по Ог (сдвиг кривой диссоциации ок-
сигемоглобина вправо), увеличения КИОг;
• активации тканевого дыхания мышц.
4* Потребление кислорода организмом (л/мин): в покое - 0,25; при ходьбе - 2,0; тяжелой фи-
зической работе - до 4,0; кислородный долг может достигать 20 л.
52. Дыхание при пониженном барометрическом давлении.
❖ Снижение парциального давления Ог во вдыхаемом воздухе приводит к
формированию гипоксемии.
❖ Раздражение дыхательного центра с периферических кислородных хеморе-
цепторов приводит к увеличению легочной и альвеолярной вентиляции.
❖ Происходит активация эритропоэза, увеличение концентрации гемоглобина
и кислородной емкости крови.
290
❖ Кривая диссоциации оксигемоглобина сдвигается в легких влево (улучше-
ние оксигенации гемоглобина), а в тканях - вправо (увеличение отдачи Ог
тканям).
❖ Повышаются МОК и число функционирующих капилляров.
❖ Устойчивость клеток к гипоксии увеличивается за счет перестройки мета-
болических путей использования Ог в клетке.
5.3. Дыхание нри повышенном барометрическом давлении.
❖ Увеличение сопротивления дыхательных путей потоку воздуха приводит к
увеличению работы дыхательных мышц.
❖ Увеличение растворимости газов в крови вызывает:
• опасность кислородного и азотного отравления;
• опасность газовой эмболии при ускоренной декомпрессии.
❖ Профилактика кессонной болезни (медленная декомпрессия).
5.4. Дыхание нри изменении газовой среды.
❖ Физиологические основы действия карбогена', смеси 96 % Ог и 4 % СОг
(увеличивается поступление Ог в организм и стимулируется дыхательный
центр с хеморецепторов иа гиперкапнический стимул).
• Применяется для возбуждения дыхательного центра при отравлении токсинами, ле-
карствами, СО, при асфиксии новорожденных.
• Улучшает газотранспортную функцию крови, увеличивая содержание физически рас-
творенного Ог (-2 об %) и оксигемоглобина за счет повышения Рог в альвеолярном
воздухе.
• Оказывает через центральные хеморецепторы возбуждающее действие на дыхатель-
ный центр, вызывая учащенне и углубление дыхания, несмотря на уменьшение или
ликвидацию гипоксического стимула.
❖ Физиологические основы гипербарической оксигенации (ГБО).
• Изменение кислородного режима крови и тканей в условиях ГБО: каж-
дая 1 ата кислорода вызывает увеличение физически растворенного Ог
иа - 2 об % или - 700 мм рт. ст.; при 3 ата количество Ог в плазме крови
(6 об %) становится равным артерио-венозной разнице по Ог.
• Адаптационно-метаболическая теория ГБО (А.Н. Леонов, 2003).
•ф Метаболический эффект:
прямой - через цитохромоксидазу и цитохром Р450: энергетиче-
ский, синтетический и детоксикационный;
опосредованный - через продукцию свободных радикалов и их
действия на структуру ферментов и других белков («конформаци-
онная сила» ГБО).
• ф Функциональный эффект: изменение функции физиологических
систем.
• ф Морфогенетический эффект: процессы дифференцировки, проли-
ферации, регенерации и др.
Ф Варианты адаптационных эффектов: стимулирующий, ингиби-
рующий, корригирующий.
Ф Адаптационное действие ГБО является основой положительного
клинического применения ГБО.
291
• Токсическое действие ГБО:
Ф общетоксическое (формирование гистотоксической гипоксии);
Ф нейротоксическое (возникает при высоких давлениях в виде эпилептиформ-
ных судорог);
Ф пульмотоксическое (возникает при невысоких давлениях, ио длительном или
частом применении ГБО);
ф допустимая продолжительность дыхания здорового человека при повышенном
давлении О} (А.Г. Жиронкии, 1973): при 10 кПа (воздух) - 20 кПа - ие ограниче-
но; 50 кПа - до 133 часов; 100 кПа (1 ата) - до 21 часа; 150 кПа (1,5 ата) - до 2-х
часов; 250 кПа (2,5 ата) - до 0,5 часа; 300 кПа (3 ата) - до 0,3 часа.
6. Профильный материал для студентов лечебного факультета.
6.1. Механизмы нарушения дыхания при пневмотораксе.
♦♦♦ Пневмоторакс - наличие воздуха в плевральной полости при нарушении ее гер-
метичности. Приводит к частичному или полному спадению легкого и выключе-
нию его из газообмена.
Нарушение дыхания при этом связаио с отсутствием нли резким уменьшением от-
рицательного плеврального давления и сил «сцепления» в слое плевральной жид-
кости. Поэтому увеличение объема грудной клетки при вдохе почти не сопровож-
дается увеличением объема легких и формированием отрицательного внутриле-
гочного давления, вызывающего перемещение атмосферного воздуха в легкие.
6.2. Физиологические основы действия карбогена (см. выше 5.4).
63. Основные физиологические механизмы изменения дыхания при подъеме на высоту
(см. выше 5.2).
6.4. Физиологические механизмы водолазной ияи кессонной болезни (см. выше 5.3).
6.5. Изменение снабжения тканей О2 у пожилых и старых людей при физической на-
грузке и кислородном голодании
4» Более выражена кислородная задолженность и более длителен восстановительный
период. Это связано:
• с исходной постоянной гипоксемией старости: снижением в артериальной
крови содержания Ог до 180 мл/л и Рог до 75 мм рт. ст.;
• со снижением резервов увеличения МОД, альвеолярной вентиляции и сердеч-
ного выброса.
♦ ♦♦ Компенсация недостаточности системы дыхания происходит за счет увеличения
экстракции Ог из крови, внутриклеточного гликолиза, лучшего сопряжения дыха-
ния и фосфорилирования.
6.6. Регуляция дыхания при старении»
♦ ♦♦ Повышается чувствительность периферических хеморецепторов к Рог и Рсог.
Снижается чувствительность механорецепторов легких и мышц (ослабление реф-
лексов Геринга - Брейера).
♦ > Уменьшается резерв дыхания (снижена максимальная вентиляция легких).
♦ ♦♦ Увеличивается время восстановления исходной величины потребления Ог после
физической нагрузки.
* t* Ослабляются условнорефлекторная регуляция дыхания, безусловные рефлексы с
проприорецепторов, защитные рефлексы с верхних дыхательных путей.
♦ ♦♦ Происходят деструктивные изменения в рецепторах и эфферентных нервных
окончаниях, ослабевают рефлексы, реализуемые с участием блуждающих нервов.
7. Профильный материал для студентов педиатрического факультета.
292
7.1. Механизм первого вдоха и выдоха новорожденного.
♦ ♦♦ Пусковые факторы, возбуждающие дыхательный центр:
• повышение Рсог, Н*, снижение Рог стимулируют центральные хеморецепторы,
периферические (артериальные) хеморецепторы еще не функционируют;
• резко усиливается поток афферентных импульсов от тактильных рецепторов,
Херморецепторов кожи;
• возрастает импульсацня с проприо**, вестибулорецепторов, возникающая во
время родов и сразу после рождения;
♦ ♦♦ Прекращается торможение дыхательного центра раздражением жидкостью рецеп-
торов, расположенных в области ноздрей («рефлекс ныряльщика»).
♦ ♦♦ Происходит инспираторное возбуждение мышц вдоха (диафрагма), снижается
внутриплевральное давление (на 20 - 80 см вод. ст.).
Высокие энергетические затраты (при первом вдохе затрачивается энергии в 10 -
15 раз больше, чем при последующих).
♦ ♦♦ Формируется функциональная остаточная емкость легких (объем вдоха больше,
чем выдоха).
♦ ♦♦ Выдох осуществляется с активным участием экспираторных мышц.
7.2. Регуляция дыхания новорожденного.
♦ ♦♦ Нерегулярность дыхательного ритма.
Относительно низкая реакция дыхания на стимуляцию центральных хеморецепто-
ров и .особенно периферических хеморецепторов (начинают функционировать с 1-
го мес. жизни).
❖ Высокая реактивность дыхательного центра на импульсы рецепторов растяжения
легких и ирритантных рецепторов (усиление рефлексов Геринга-Брейера).
Нейроны дыхательного центра в антенатальном периоде и в первые дни после ро-
ждения обладают пейсмекерными свойствами, что способствует поддержанию
вентиляции легких у новорожденного.
♦ ♦♦ Функциональная незрелость пиевмотаксического центра.
❖ Высокая устойчивость бульбарных центров к гипоксии.
♦ ♦♦ Высокая координация деятельности дыхательного центра с центрами сосания и
глотания: во время кормления частота дыхания соответствует частоте сосатель-
ных движений (центр сосания обычно навязывает дыхательному центру свою, бо-
лее высокую частоту возбуждения); во время глотания дыхательные пути пере-
крываются и отделяются от глотки мягким нёбом и надгортанником, голосовые
связки смыкаются.
7.3. Регуляция дыхания у детей разных возрастов. Дыхание при физической работе.
♦♦♦ Особенности регуляции дыхания:
• высокая возбудимость дыхательного центра и высокая вентиляция легких
поддерживается при сравнительно низком Рсог и высоком Рог в артериальной
крови;
• низкая реактивность периферических хеморецепторов;
• формирование произвольной регуляции дыхания (увеличение коркового кон-
троля);
• участие дыхания в речевой функции.
❖ Дыхание при физической работе.
293
• Повышение легочной вентиляции происходит преимущественно за счет уве-
личения частоты, а ие глубины дыхания; эффективность его ниже, чем у взрос-
лых.
• Меньшее увеличение (а-в)О2 и КИО2 тканями по сравнению со взрослыми
приводит к большему усилению работы сердца для достаточного обеспечения
тканей кислородом.
• Повышение эффективности и экономичности кислородного обеспечения орга-
низма с возрастом к 20 годам (в период полового созревания эти показатели
снижаются, ухудшается качество их регулирования).
8. Профильный материал для студентов стоматологического факультета.
8.1. Функциональная связь процессов дыхания, жевания и глотания,
❖ Охлаждение горячей пищи за счет дыхания через рот (продувание воздуха над
пищей).
❖ Защитный дыхательный рефлекс (изменение дыхания) в процессе жевания и оста-
новка дыхания при глотании пищевого комка. Во время глотания челюсти смыка-
ются, мягкое нёбо поднимается, сокращающиеся нёбно-глоточные мышцы обра-
зуют перегородку между ртом и носовой полостью, вход в гортань закрывается
надгортанником, голосовые связки закрывают голосовую щель; пищевой комок
при сокращении мышц глотки попадает в пищевод.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре-
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли-
тературы (задание 3; 3.1 и 3.2). Объем реферата 2-3 стр., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11], [12], [14], [15], [16].
3.2. Дополнительная литература (см. стр. 514 - 522): [27], [30], [34], [43], [44], [53],
[66], [84], [101], [118], [152], [154], [179], [192], [194], [196], [204], [222], [225], [237],
[244], [245], [246], [251-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508-513).
Задача 20-1. В результате локальной травмы позвоночника у человека, нырявшего в реку, про-
изошло выключение грудного дыхания при сохранении диафрагмального дыхания и функций
мышц рук. Какие сегменты спинного мозга при этом поражены?
294
Тема 21. Регуляции кислотно-основного состояния (КОС)
Цель: а) знать общую схему организации функциональной системы регуляции КОС, ее центральные
и исполнительные (физико-химические и физиологические) механизмы деятельности; роль
легких, почек и органов пищеварения в поддержании pH крови; показатели КОС и принципы
методов их определения;
б) уметь использовать эти знания для понимания процессов регуляции кислотно-основного
гомеостаза организма в различных условиях жизнедеятельности, оценивать лабораторные по-
казатели системы КОС здорового человека;
в) ознакомиться с практическими навыками на уровне знаний: методикой определения показа-
телей газового состава и кислотно-основного состояния крови.
Мотивация темы: одним из важнейших условий гомеостаза является постоянство pH внутренней сре-
ды организма. От величины pH зависит активность ферментов, направление и интенсивность
окислительно-восстановительных реакций, процессы катаболизма и синтеза белков, жиров и
углеводов, проницаемость мембран, чувствительность рецепторов, способность гемоглобина
связывать и отдавать Oj. Организм имеет мощные механизмы поддержания КОС, знание кото-
рых позволяет понять формирование защитно-приспособительных реакций при патологии
КОС, определить методы профилактики и коррекции его нарушений.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции.
1. Общая характеристика кислотно-основного состояния.
1.1. КОС - соотношение кислых и основных веществ в организме (кислотами считают-
ся вещества, освобождающие в процессе диссоциации Н*, а основаниями - вещества, связы-
вающие эти ионы).
1.2. Функциональное значение КОС.
❖ Метаболический аспект: от КОС зависит нормальное функционирование
ферментов и структурных белков, активность витаминов, микроэлементов,
липидов.
❖ Физиологический аспект: КОС влияет на скорость транспорта веществ че-
рез мембрану клеток, возбудимость клеточной мембраны, функции физио-
логических систем (тонус сосудов, стимуляция дыхательного центра, диссоциация ок-
сигемоглобина, эффекты гормонов и др.).
13. Основные сдвиги КОС.
❖ Ацидоз - нарушение КОС, характеризующееся появлением в крови абсо-
лютного или относительного избытка кислот. Может быть без существен-
ного изменения pH крови (компенсированный ацидоз), а также со снижением
pH крови (суб- и декомпенсированный ацидозы).
❖ Алкалоз - нарушение КОС с абсолютным или относительным увеличением
количества оснований в крови. Может быть без существенного изменения
pH кровн (компенсированный алкалоз), а также с повышением pH крови (суб- и
декомпенсированный алкалозы).
1.4. Общая схема функциональной системы регуляции КОС (рис. 55).
295
2. Концентрация Н* (pH) крови - результат деятельности функциональной систе-
мы КОС, ее системообразующий фактор.
2.1. Характеристика pH крови как физиологической константы.
❖ pH - отрицательный логарифм концентрации Н* (рН= -lg[H*J).
❖ Нормальные величины pH крови: артериальной - 7,40 ± 0,04, веиозиой -
7,36 ± 0,05, чему соответствует 40 ± 1,4 нмоль Н7л.
❖ Совместимые с жизнью величины pH и концентрации Я* крови (pH = 7,4 ± 0,4,16 - 100
нмоль Н+/л). При 6,8<рН>8,0 наступает гибель организма.
Рис. 5S. Упрощенная схема функциональной системы регуляции ки-
слотно-основного состояния.
2.2. Основные источники образования Н* н оснований в организме.
❖ Источники ИГ.
• Образование СОг равно 13 000 - 20 000 ммоль/сут, СОг является потен-
циальным донором Н* в реакциях СОг+н2о -> н2со3 -> н*+ нсо3‘.
• Образование нелетучих (серной, соляной, фосфорной, молочной и др.)
кислот при метаболизме серосодержащих аминокислот (Нг5О4), катион-
ных аминокислот - лизина, аргинина, гистидина (HCI), фосфобелков и
фосфолипидов (Н2РО4) (-235 ммоль/сут).
❖ Источники оснований в крови: эндогенное образование бикарбоната (НСО3)
при метаболизме анионных аминокислот (аспартата, глутамата) и органи-
ческих кислот (-160 ммоль/сут).
❖ «Чистая» продукция нелетучих кислот 235 - 160 = 75 ммоль/сут (1 ммоль
Н*7 кг массы тела в сутки).
23. Уравнение Гендерсона-Гасселбалха используется при расчетах показателей
КОС.
296
pff=pK+lg-p^-
№
где pK = 6,1 (pK - это значение pH, при котором
концентрация диссоциированной и недиссо-
циированной форм кислоты одинакова, для
Н2СО3 равна 6,1); [Н2СО3] = Рсо2 х 0,03 (0,03
ммоль/л/мм рт.ст. - константа растворимости
СОг в воде).
3. Рецепция колебаний концентраций водородных ионов крови.
3.1. Периферические хеморецепторы:
❖ расположены в сосудах - наибольшая плотность в синокаротидном н аор-
тальном тельцах, возможно их наличие в тканях;
❖ имеют меньшую чувствительность к сдвигам pH (пороговые колебания pH
~ 0,1) и напряжению СО2;
❖ латентное время рефлекса через дыхательный центр -4 с.
3.2. Центральные хеморецепторы:
❖ расположены в области вентролатеральной поверхности продолговатого
мозга и моста, содержащей три парных скопления рецепторов;
❖ имеют высокую чувствительность к изменению pH (пороговые колебания
pH ~ 0,01) и напряжению СО2 в ликворе;
❖ латентное время рефлекса - 25 с.
4. Центральное Звено функциональной системы КОС.
4.1. Афферентный синтез.
❖ Пусковая и обстановочная афферентация с периферических и центральных
хеморецепторов (Н+, Ро2, Рсо2).
❖ Обстановочная афферентная импульсация с интерорецепторов, сигнализи-
рующая о состоянии органов, наиболее важных для регуляции КОС (лег-
кие, почки, органы желудочно-кишечного тракта, кости).
♦ > Обстановочная афферентная импульсация с экстерорецепторов о состоянии
внешней среды.
4.2. Акцептор результата действия - нейронная модель величины pH крови и
других показателей КОС, необходимых в данных условиях.
4.3. Эфферентная программа действия - совокупность возбужденных нервных
центров, способных включить исполнительные механизмы регуляции КОС:
❖ Дыхательный центр ствола головного мозга.
❖ Гипоталамус н нейроэндокринные эфферентные влияния на метаболизм и
функции внутренних органов.
❖ Симпатические и парасимпатические центры регуляции желудочно-
кишечного тракта, почек, костной ткани, гемопоэза.
4.4. Исполнительные механизмы регуляции КОС - физико-химические и физио-
логические (см. ниже).
5. Физико-химические исполнительные механизмы регуляции КОС.
5.1. Эффект разведения образующихся кислот и оснований в водных пространст-
вах организма: во внутриклеточной жидкости (- 43 % массы тела, - 33 л), меж-
клеточной жидкости (- 12 %, - 10 л), плазме крови (- 5 ~ 3,5 л); это первая
линия защиты, ее мощность небольшая.
5.2. Буферные механизмы.
297
❖ Клеточные буферы составляют около 88 % буферной емкости организма:
• Белковый буфер: его общая емкость в 20 раз превышает емкость белко-
вого буфера крови.
• Фосфатный буфер: его емкость в 400 раз превышает емкость фосфат-
ного буфера крови.
• Бикарбонатный буфер: его общая емкость в 2,5 раза превышает емкость
бикарбоиатного буфера крови (в клетках -8 ммоль НСО37кг).
• Растворение сильных кислот и оснований в липидах, препятствующих
их диссоциации.
❖ Буферы крови.
• Бикарбонатный буфер (мощность в крови - 53 %, в эритроцитах -
30%).
ф- Смесь слабой угольной кислоты (1,2 ммоль/л) и сопряженного
сильного основания - бикарбоната (24 ммоль/л) в соотношении
1/20.
- ф- Реакция буфера с иекарбоновыми кислотами и их основаниями: Н* +
НСОз' -> Н2СО2 -» Н2О + СОг; ОН + Н2СО2 -» НСО< + Н2О.
• ф- В связи с тем, что гемоглобиновый, белковый и фосфатный буферы
опосредуют свое влияние через бикарбонатный буфер, именно он в
итоге определяет pH крови.
• ф" Кроме того, бикарбонатный буфер тесно связан с функциями лег-
ких и почек: концентрация угольной кислоты сильно зависит от
функции легких, бикарбоната крови - от деятельности почек.
• Гемоглобиновый буфер (мощность в крови - 35 %, в эритроцитах - 55 %; его дей-
ствие тесио связано с осуществлением транспорта О2 и СОг).
- ф- Гемоглобиновая буферная система состоит из нескольких подсис-
тем (ННЬ/ННЬО2; КНЬ/ННЬ; КНЬОг/ННЬО2), главной из которых является
ННЬ/ННЬО2.
ф’ Присоединение О2 к молекуле ННЬ настолько изменяет ее конфи-
гурацию, что она в 80 раз больше по сравнению с ННЬ отщепляет
Н+ (т.е. ННЬОгпри этом является кислотой, а ННЬ - основанием).
ф’ В его буферных свойствах большую роль играет аминокислота гис-
тидин (8% аминокислотного состава), имидазольное КОЛЬЦО которого
обладает выраженными амфотерными свойствами.
• Белковый буфер (мощность в крови - 7 %).
• ф" Роль белков обусловлена их амфотерными свойствами.
ф- Глобулины играют бблыпую роль в нейтрализации оснований, об-
ладая более выраженной кислой диссоциацией.
ф’ Альбумины, содержащие много диаминокислот, диссоциируют как
основания, в ббльшей мере нейтрализуют кислоты.
• Фосфатный буфер (мощность в крови - 5 %, в эритроцитах - 10 %).
298
ф* Смесь слабой кислоты - однозамещенного (кислого) фосфата и
сильного основания - двузамещениого (основного) фосфата в соот-
ношении 1/4.
ф- Реакция буфера с кислотами и основаниями: IT+HPCU2’-» Н2РО4‘;
НСОз’ + Н2РО4-> н2о + со2 + нроЛ
ф- Поддерживает концентрацию бикарбонатов в плазме крови при ре-
акциях буфера с избытком Н2СОз.
6. Физиологические исполнительные механизмы регуляции КОС.
6.1. Роль легких в регуляции КОС.
❖ Выводят 13 000 - 20 000 ммоль СО2 (потенциального донора Н4) в сутки.
❖ Изменение вентиляции легких является наиболее срочным и мощным из
физиологических механизмов регуляции КОС.
• Гипервентиляция является механизмом компенсации ацидоза (иапри-
мер, объем вентиляции увеличивается в 2 раза при снижении pH иа 0,1).
• Гиповентиляция является механизмом компенсации алкалоза (напри-
мер,объем вентиляции уменьшается в 2 раза при повышении pH иа 0,1).
6.2. Роль почек в регуляции КОС Почки обеспечивают выведение Н4 в количестве,
равном их образованию в организме из нелетучих кислот (серной, фосфориой и
др.): 60 —100 ммоль/сут.
❖ В клетках канальцев почек происходит реакция гидратации СО2 и диссо-
циации угольной кислоты на Н+ и бикарбонат (СО2 + Н2О -> Н2СОз -»
НСОз’ + Н4). Протоны секретируются из клеток канальцев в мочу в обмен
на реабсорбцию Na4; сопряженио с этим происходит переход бикарбоната
из клеток канальцев в плазму крови (1 Н4 -> в мочу, 1 НСОз’ -» в кровь).
❖ Имеются 3 фазы секреции Н4 в мочу (в проксимальных канальцах секреция
происходит преимущественно в результате ионообмеиа с Na4, в дисталь-
ных канальцах и собирательных трубках - с помощью Н4-насоса) (рис. 56).
• Фаза «реабсорбции» бикарбонатов (в проксимальных канальцах).
ф Секретированные Н4 в первичной моче взаимодействуют с про-
фильтрованными из плазмы крови НСОз’.
ф’ В результате образуется Н2СОз, которая карбангидразой разлагает-
ся на СО2 и Н2О (они могут остаться в моче или реабсорбироваться).
ф- При этом иа каждый секретируемый Н4 и разрушенный НСОз’ мочи
в кровь из клеток канальцев поступает такое же количество НСОз’.
ф- Таким образом почки возвращают в кровь то количество НСОз’ (~
3500 ммоль/сут), которое фильтруется из плазмы.
• Фаза титруемых кислот (выделяется 20 - 30 ммоль НУсут в проксимальных ка-
нальцах и собирательных трубочках).
ф- Секретированные Н4 в канальциевой моче взаимодействуют с про-
фильтрованным щелочным фосфатом (НРО42’) с образованием ки-
слого фосфата (Н2РО4’) и закислением мочи (максимально до
рН=4,5).
299
V Кроме выделения кислого фосфата почки могут выделять профильтрованные ор-
ганические кислоты (например, лактат, ацетоацетат).
Интенсивность фазы титруемых кислот при ацидозе может увели-
чиваться в 1,5 раза.
F фильтрация S секреция R реабсорбция
Рис. 56. Роль почек в регуляции кислотно-
основного состояния: I фаза «реабсорбции»
бикарбоната, П фаза титруемых кислот, Ш
фаза аммониогенеза.
• Фаза аммониогенеза (выделяется 30 - 50 ммоль Н*/сут).
В клетках проксимальных и дистальных канальцев образуется ам-
миак преимущественно в результате дезамидирования глутамина н
дезаминирования глутамата.
Ф Аммиак секретируется в просвет канальцев, присоединяет секрети-
рованный Н+, и образуется ион аммония (NH3 + Н+ —> NlV), при
этом закисления мочи практически не происходит.
ф" Интенсивность аммониогенеза может быть увеличена прн ацидозе
до 10 раз.
6.3. Роль желудочно-кишечного тракта в регуляции КОС.
❖ Роль желудка.
300
• Секреция Н* в желудочный сок сопряжена с поступлением бикарбона-
тов в плазму крови и формированием щелочного резерва крови (рис. 58,
с. 314).
• Секреция желудочного сока компенсаторно увеличивается при ацидозах
и тормозится при алкалозах.
• Потеря желудочного сока (например, прн многократной рвоте) вызывает алкалоз.
❖ Роль кишечника.
• Секреция бикарбоната в панкреатический и кишечный соки сопряжена с
поступлением Н* в плазму крови.
• Выработка панкреатического и кишечного сока компенсаторно усили-
вается при алкалозах и тормозится при ацидозах.
• Потеря панкреатического и кишечного соков (поносы) вызывает ацидоз.
❖ В печени происходит:
• утилизация молочной кислоты с образованием гликогена, разрушение
кетоновых тел;
• связывание жирных кислот при образовании триглицеридов и фосфор-
ной кислоты при образовании фосфолипидов;
• продукция Н* при синтезе мочевины: 2NH/ + СО2 -> мочевина + Н2О + 2Н+.
* Характер питания человека.
• Потребление растительной пищи (окисление органических кислот) спо-
собствует защелачиванию крови (например, цитрат + 4,5О2 5СО2 +
ЗН2О + НСОзЭ-
• Потребление белковой животной пищи (увеличение синтеза мочевины)
способствует закислению внутренней среды организма.
6.4. Роль костной ткани (нонообмен между кровью н костной тканью).
❖ При хронических ацидозах Н+ поступают в костную ткань в обмен на Са2+,
К , Na (может возникать остеопороз, гиперкалиемия; например, при снижении pH крови
иа 0,1 концентрация К* в плазме увеличивается иа 0,6 ммоль/л).
❖ При алкалозах Н* костной ткани замещаются на Са , К+, Na+ крови (напри-
мер, при повышение pH крови иа 0,1 концентрация К* в плазме снижается иа 0,6 ммоль/л).
7. Клинико-физиологические показатели КОС.
7.1. pH крови (см. выше п. 2.1) - актуальная величина pH.
7.2. Напряжение углекислого газа (PcoJ - актуальная величина Рсоз крови, опре-
деленная без доступа воздуха при температуре 37 °C В артериальной крови = 40
± 2 мм рт. ст., в венозной крови = 46 ± 3 мм рт. ст. (совместимые с жизнью отклоне-
ния 15 - 100 мм рт. ст.).
7.3. Бикарбоиаты крови (SB и АВ).
❖ Стандартный бикарбонат (SB = 24 ± 2,5 ммоль/л) - рассчитанное содержа-
ние НСОз" в плазме крови при стандартных условиях (полное насыщение гемо-
глобина кислородом, Рсог = 40 мм рт. ст. и 37 °C).
❖ Актуальный бикарбонат (АВ = 21 ± 2,5 ммоль/л): истинная концентрация
НСОз' в плазме крови, рассчитанная при фактическом состоянии Рсоз и Роз
крови.
301
7.4. Буферные основания крови (ВВ = 48 + 5 ммоль/л): сумма всех анионов плазмы,
обладающих буферными свойствами в стандартных условиях (Рсог = 40 мм рт.
ст., t=37 °C).
7.5. Избыток и дефицит буферных оснований крови (BE и BD) - разница между
фактической величиной буферных оснований крови пациента и значением бу-
ферных оснований, рассчитанных в стандартных условиях; нормальный диапа-
зон ± 2,5 ммоль/л. (Показывает то количество сильной кислоты или основания, которое надо
добавить, чтобы pH крови стал равен 7,4)
7.6. Титрационная кислотность и содержание аммония в моче: титрацнонная
кислотность мочи - 20 - 30 ТЕ/сут; содержание аммония в моче - 30 - 50
ммоль/сут.
8. Метод определения показателей КОС в кровп с помощью газоанализатора кровп (метод
микроАструпа) основан на потенциометрическом принципе. В камере, куда в микродозах по-
мещают исследуемую кровь, находятся электроды, имеющие избирательную чувствитель-
ность к Н+ (электрод pH), молекулам Ог (электрод Р02) и СОг (электрод Рсог). Поляризацион-
ные напряжения, которые возникают иа электродах, пропорциональны величинам концентра-
ции исследуемых веществ. На цифровом индикаторе непосредственно отсчитывается величи-
на pH в единицах, а величины напряжений газов - в мм рт. ст. Непосредственно определяе-
мые показатели - pH, рОг, рСОг, НЬ. Расчетные показатели - SB, АВ, ВВ, BE и BD, оксиге-
моглобин.
9. Профильный материал для студентов лечебного факультета.
Изменение КОС при старении организма,
9.1. Уменьшается буферная емкость крови преимущественно за счет снижения содержания
бикарбонатов и концентрации белков плазмы крови.
9.2. Развивается компенсированный метаболический ацидоз в результате:
♦♦♦ снижения потребления Ог. участвующего в газообмене, в результате снижения
альвеолярной вентиляции и уменьшения поступления воздуха в нижние отделы
легких, где кровоток максимален;
❖ формирования физиологической гипоксемии старости при снижении перфузии
легких в результате уменьшения легочного кровотока;
более высокой кислородной задолженности и длительного восстановительного
периода.
9.3. Прогрессирующее снижение почечного кровотока, клубочковой фильтрации и реаб-
сорбции приводит к уменьшению способности почек к экскреции Н* мочи.
10. Профильный материал для студентов педиатрического факультета.
10.1. Особенности КОС крови плода и новорожденного ребенка,
❖ pH крови сдвинут в кислую сторону (7,13 - 7,23), что обусловлено образованием
иедоокнсленных продуктов обмена (метаболический ацидоз).
❖ Близкие к взрослым цифры pH устанавливаются в течение 3-5 суток после рож-
дения.
❖ На протяжении всего детства сохраняется небольшой компенсированный ацидоз,
постепенно убывающий с возрастом.
10.2. Роль почек в регуляции КОС
❖ В основе лежат 3 фазы секреции Н* почками, но общая эффективность их меньше,
чем у взрослого человека.
4 * Удельный вес каждого процесса иной по сравнению со взрослым организмом: ос-
новную роль играет фаза аммониогенеза; роль экскреции кислых фосфатов неве-
302
лика, потому что ребенок при кормлении молоком матери обладает весьма огра-
ниченными ресурсами фосфатов.
❖ Почки удовлетворительно поддерживают КОС при грудном вскармливании, пере-
ход на искусственное вскармливание вызывает избыточное образование кислых
продуктов.
Компенсация сдвигов КОС у ребенка осуществляется в менее широких пределах,
поэтому у детей наблюдается склонность к ацидозам.
10.3. Роль легких в регуляции КОС (см. Профильный материал для педиатрического фа-
культета по темам 18 и 19).
11. Профильный материал для студентов стоматологического факультета.
11.1. Реакция слюны как физиологическая константа.
♦ ♦♦ pH слюны относится к менее строгим константам, чем pH крови. Ее величина мо-
жет на десятые доли отличаться от стабильного значения pH крови (7,4) за счет
постоянной реабсорбции избытка ионов Н+ и Na+ из просвета слюнных желез в
кровь (снижение кислотности слюны) и поступления НСОз’ из сыворотки крови в
слюну путем активного транспорта (повышение щелочности слюны).
❖ Смешанная слюна является основным регулятором КОС в полости рта (см. табл.
2). Величина pH ротовой жидкости без какой-либо стимуляции отличается посто-
янством. В течение суток имеются колебания pH слюны: утром он ниже, чем в се-
редине дня и имеет тенденцию к повышению вечером. Ночью pH смешанной
слюны ниже, чем днем. Наряду с суточным ритмом изменений pH ротовой жидко-
сти, отмечено снижение его значений с возрастом. В разных участках полости рта
pH различен: на слизистой оболочке твердого неба он на 0,7 - 1,2 ед. щелочнее,
чем в других областях. В области нижней губы он на 0,3 - 0,8 ед. щелочнее, чем в
области верхней.
❖ pH поддерживается буферными системами слюны (бикарбонатным, фосфатным,
белковым), но их буферная мощность меньше, чем в крови. От pH слюны зависит
перераспределение минеральных компонентов между слюной и эмалью зубов.
При сдвиге pH слюны в кислую сторону уменьшается ее насыщенность фосфором
и кальцием, и слюна приобретает деминерализующие свойства, что может привес-
ти к появлению очага деминерализации в эмали зуба (кариеса).
❖ Кислотосодержащие продукты и напитки (сахар, шоколад, фрукты, соки и др.) вы-
зывают сдвиг pH .слюны в кислую сторону, что при длительном их присутствии в
полости рта может вызывать эрозию твердых тканей зубов. Продукты (орехи, сы-
ры и др.), в которых присутствуют аммонийсодержащие вещества, способны из-
менять КОС ротовой жидкости в щелочную сторону на 0,5 - 0,7. Тенденция к за-
щелачиванию ротовой жидкости и камнеобразованию яаляется приспособитель-
ной реакцией, направленной на снижение патогенного действия кислот и фермен-
тов, выделяемых микроорганизмами.
❖ Профилактика и коррекция нарушений КОС в полости рта заключается в приме-
нении рациональных методов личной гигиены с использованием методов оценки
КОС (pH-метрия), употреблении продуктов, изменяющих pH ротовой жидкости,
обладающих бактерицидным действием и др.
❖ Методы определения реакции слюны в стоматологии.
Определение pH слюны можно провести с помощью аппарата микроАструпа (см.
выше п.8) и pH-метра. Используются pH-метры, снабженные чувствительным к Н*
измерительным электродом и вспомогательным электродом сравнения. При этом
исследуемую жидкость помещают в маленькую кювету. Для определения pH непо-
средственно во рту используют металлооксидные измерительные электроды из сурьмы
303
или специальные оливы, в которых запаяны измерительный и сравнительный электро*
ды. Существует радиометрический метод определения pH во рту (на расстоянии).
Таблица 2. pH н некоторые показатели кровн, слюны и жидкости зубного нелета у лю-
дей (по А.И. Воложину и др., 1998, с изменениями)___________________
Вещества, показате- ли Артери- альная кровь Слюна Жид- кость зубного налета
Паротидная Подчелюст- ная Смешанная
Н С Н с Н с
pH 7,4±0,04 6,8 7,7 6,5 7,4 7,1 6,8-7,5 6,5
Бикарбонаты (ммоль/л) 24±2,5 1,0 22-30 2-4 14-16 5 15-20 -
Кальций (ммоль/л) 2,5 1.0 1,0 1,3 1,8 2,7 2,3 6,5
Фосфор (неорг., ммоль/л) 1,0 10 3 5,4 2 6 4 14
Аммоний (мг%) 0,08-0,11 0,9 0,06 0.7 0,04 12 4-8 18
Мочевина (мг%) 14-40 30 25 10 5 20 17 -
Общий белок (г/л) 65-85 2,5 2,9 1.1 1.5 2,85 2.9 1,49
Примечание: Н - нестимулнрованнва слюна; С - стимулированная слюна.
Задание 2. Подготовить реферативные сообщения на тему данного занятия. Тема ре*
ферата выбирается с использованием учебных элементов (задание 1). При
подготовке реферата использовать список основной и дополнительной ли*
тературы (задание 3; 3.1 и 3.2). Объем реферата 2-3 стр., регламент 5-7
мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [14-16].
32. Дополнительная литература (см. с. 514 - 522): [27], [30], [36], [53], [66], [84],
[141], [154], [155], [196], [204], [215], [222], [225], [235], [245], [251-254].
33. Дистанционные средства обучения. Использование страницы кафедры в Интер*
иете - www.vsma.ac.ni/-nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
33. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
304
Тема 22. Общая характеристика функциональной системы питания и
физиологической системы пищеварении. Пищеварение в по-
лости рта н желудка
Цель: а) знать функциональную систему питания, основные функции пищеварительной системы,
иейрогуморальные принципы ее регуляции, методы исследования, пищевую мотивацию (пи-
щевой центр, голод, насыщение, аппетит), типы и основные этапы пищеварения, пищевари-
тельный конвейер и его функции (секреция, моторика, всасывание); эндокринную функцию
пищеварительного тракта; инкрецию ферментов; периодическую деятельность органов пище-
варения; пищеварение в полости рта и желудка; глотание, его фазы; иепищеварктельиые
функции полости рта и желудка; методики изучения пищеварительного тракта;
б) уметь использовать эти знания для понимания роли пищеварения в деятельности целостно-
го организма;
в) иметь представления о методах исследования пищеварения у человека.
Мотивация темы: «... Над всеми явлениями человеческой жизни господствует забота о насущном хле-
бе. Он представляет собой ту древнейшую связь, которая соединяет все живые существа, в том
числе и человека, со всей остальной, окружающей нх природой. ...Точное знание судьбы пи-
щи в организме должно составить предмет идеальной физиологии будущего» Из Нобелевской
речи И.П. Павлова.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Функциональней система питании обеспечивает постоннство питательных ве-
ществ в крови, имеет следующие звеньн (рис. 57).
1.1. «Внешнее» звено - пищевой центр, формирующий пищевую мотивацию и по-
ведение (сопровождается активацией моторной и теменной коры, миндалины,
медиальных ядер гипоталамуса).
1.2. «Внутреннее» звено - питательные вещества крови и тканевой обмен веществ.
1.3. Связывающее звено - физиологическая система пищеварения.
2. Общаи характеристика пищеварении.
2.1. Пищеварение - физиологическая система, обеспечивающая поступление пищи в
организм, ее механическую и химическую обработку до мономеров и всасывание их
во внутреннюю среду организма. (Система пищеварения выполняет функцию преобразования
пищевых вещесте в питательные вещества.)
2.2. Типы пищеварения.
❖ По месту действия ферментов: 1) полостное (внеклеточное); 2) мембран-
ное (пристеночное); 3) внутриклеточное (лизосомальное).
❖ По источникам ферментов: 1) собственное (ферменты организма); 2) сим-
биотное (ферменты микрофлоры); 3) аутолитическое (ферменты пищи).
2.3. Пищеварительный конвейер.
❖ Этапы: ротовой, пищеводный, желудочный, тонкокишечный и толстоки-
шечный.
❖ Функции: секреция, моторика, химический гидролиз, всасывание.
305
❖ Процессы в пищеварительном конвейере: физические - размельчение, на-
бухание, растворение; химические - денатурация, гидролиз; физиологиче-
ские - секреция, моторика, всасывание.
3. Основные принципы и механизмы регуляции пищеварения.
Рис. 57. Упрощенная схема функциональной системы питания.
1 - внешнее звено; 2 - внутреннее звено; 3 - связывающее звено
3.1. Нервная регуляция.
❖ Соматическая нервная система (регулирует жевание, 1-ю и частично 2-ю
фазы глотания, дефекацию).
❖ Вегетативная нервная система (регулирует все этапы).
• Симпатический отдел (норадреналин) тормозит моторику (p./fe-
адренорецепторы) и тонус (а-адренорецепторы) желудка' и кишечника, со-
кращает их сфинктеры (а-адренорецепторы), стимулирует секрецию фер-
ментов в пищеварительных железах, тормозит всасывание. (Постганглио-
иарные симпатические волокна преимущественно иннервируют нейроны интраму-
ральных ганлнев и только иа миоциты сфинктеров выходят непосредственно, коме-
диатором норадреналина является соматостатин.)
• Парасимпатический отдел (ацетилхолин, М-холииорецептоы) - стимулирует
моторику и тонус, расслабляет сфинктеры, стимулирует образование
водно-электролитного компонента секретов, процесс всасывания, сек-
рецию гастроэнтеропанкреатических гормонов.
• Метасимпатический отдел обеспечивает местную саморегуляцию пи-
щеварительных органов (см. тему 8, задание 1; 4).
3.2. Гуморальная регуляция.
* Гастроэнтеропанкреатическая эндокринная система.
• Диффузно расположенные в желудочно-кишечном тракте эндокринные
клетки (более 20 видов). Образуют около 30 полипептидных гормонов
и аминов местного (например соматостатин, гистамин), внутрисистемного и
организменного действия (гастрин, секретин, холецистокинин и др.); некото-
306
рые из иих выполняют роль нейромедиаторов (бомбезин, вещество Р, ВИП,
энкефалин).
• Эндокринные клетки имеют рецептивный полюс, обращенный в по-
лость пищеварительного канала, на который влияют вещества химуса и
пищеварительные ферменты, изменяя секрецию гормонов.
• Главные клетки-мишени для интестинальных гормонов: 1) гладкие
миоциты пищеварительной трубки; 2) гландулоциты; 3) эндокринные
клетки; 4) интрамуральные и центральные нейроны.
• > Влияние гормонов эндокринных желез - адреналина, глюкокортикоидов и
др. (Например, глюкокортикоиды в фармакологических дозах повышают секрецию НС1 и
снижают секрецию слизи в желудке.)
3.3. Видьгрегуляторных влияний.
❖ Для состояния покоя (натощак) характерна периодическая деятельность ор-
ганов пищеварения (подробнее в теме 23; задание 1: 5).
❖ Пусковое влияние (преимущественно нервно-рефлекторное).
❖ Корригирующие нервные и гуморальные влияние текущей деятельности по
типу отрицательной обратной связи.
4. Пищевой центр.
4.1. Структура пищевого центра.
❖ Гипоталамус - «ядро» пищевого центра.
❖ Лобная кора и лимбическая система - социализация пищевого поведения и
связь с эмоциями.
❖ Базальные ядра - формирование двигательных программ пищевого пове-
дения.
❖ Ретикулярная формация ствола - координация более простых рефлексов в
сложные рефлекторные акты (например, глотание, жевание), формирование
их ритмической активности.
• > Вегетативные и соматические центры ствола и спинного мозга - испол-
нительные нервные центры.
4.2. Главные функции пищевого центра.
❖ Формирование пищевой мотивации и связанных с ней эмоций (голод, аппе-
тит, насыщение).
❖ Удовлетворение пищевой мотивации в результате пищевого поведения.
❖ Регуляция системы пищеварения.
43. Голод («Голод - лучшая приправа к еде». Сократ).
❖ Центр голода локализован в латеральных ядрах гипоталамуса. Разрушение
этих ядер вызывает афагию; стимуляция - гиперфаппо.
❖ Стимулируют центр голода снижение питательных веществ в крови - глю-
козы, жирных кислот, аминокислот - теория голодной крови И.П. Павлова,
тирокси, гастрин и нейропептид Y гипоталамуса, стимуляция перифериче-
ских и центральных холодовых рецепторов, а также механорецепторов же-
лудка - теория пустого желудка У. Кеннона.
4.4. Аппетит - активатор пищевого поведения.
❖ Избирательный аппетит.
Расстройства аппетита: анорексия, булемия, извращение аппетита.
307
4.5. Насыщение.
❖ Центр насыщения локализован в вентромедиальных ядрах гипоталамуса.
• Разрушение этих ядер вызывает гиперфагию, стимуляция - афагию.
• Стимулируют центр насыщения некоторые вещества крови (глюкоза,
аминокислоты, жирные кислоты, кислоты цикла Кребса), повышение
температуры крови, пептидный гормон жировой ткани - лептин.
❖ Виды (фазы) насыщения.
• Сенсорное насыщение (предрезорбционное, первичное).
ф Формируется во время приема пищи с рецепторов полости рта, пи-
щевода, желудка.
ф Центр насыщения стимулируется как рефлекторным путем, так и
гуморально в результате мобилизации депонированных питатель-
ных веществ (гликогена, липидов).
ф Позволяет окончить прием пнщи еще до всасывания питательных
веществ из тонкого кишечника.
• Обменное насыщение (пострезорбцнонное, вторичное).
Ф Формируется в результате всасывания питательных веществ в
кровь.
ф Стабилизируется уровень питательных веществ в крови и пролон-
гируются промежутки между приемами пищи.
ф Восстанавливаются депо питательных веществ в организме.
4.6. Голод, насыщение, аппетит являются средствами оценки пищевой мотивации
и приема необходимого количества питательных веществ.
5. Пищеварительные функции системы пищеварения («Пнща, которая не переваривается,
съедвет того, кто ее съел». Фврадж).
5.1. Моторная (двигательная) функция - осуществляется на воех этапах.
❖ Мышечный слой пищеварительного канала.
• Поперечно-полосатая мускулатура: главные и вспомогательные жева-
тельные мышцы, мышцы глотки, верхней и частично средней трети пи-
щевода, наружный сфинктер прямой кишки.
• Гладкая мускулатура (от пищевода до наружного сфинктера прямой
кишки), межмышечное и подслизистое нервные сплетения.
❖ Типы двигательной активности пищеварительного тракта, их функцио-
нальная роль.
• Жевание (размельчение пищи с помощью зубов и жевательных мышц).
• Перистальтика - волнообразные сокращения, впереди пищевого комка
волна расслабления, сзади - сокращения (функции передвижения и пе-
ремешивания пищи, удаления непереваренных продуктов, ритм 3 - 15 в
1 мин). В ее основе лежит базальный ритм спонтанной децоляризации
мембранного потенциала гладких миоцитов, при периодическом дости-
жении уровня КУД возникают пачки ПД и сокращения.
ф Пищевой комок через рецепторы возбуждает сенсорные нейроны в межмышеч-
ном сплетении.
308
* у* Сенсорные нейроны через интернейроны возбуждают холннэргическне мотоней-
роны, которые сокращает циркулярную мышцу сзади (орально) пищевого комка
и продольную мускулатуру впереди комка.
* у* Сенсорные нейроны через интернейроны возбуждают мотонейроны, которые че-
рез медиаторы АТФ н ВИП расслабляют циркулярную мышцу впереди комка;
продольная мускулатура сзади комка при этом расслаблена.
* у* Поэтому впереди комка идет волна расслабления (продольная мышца сокращена,
циркулярная расслаблена), сзади комка - волна сокращения (продольная мышца
расслаблена, циркулярная сокращена).
• Ритмическая сегментация - фрагментарное сокращение циркулярных
мышц (перемешивание пищи с соками, ритм 12 - 18 в 1 мин).
• Маятникообразные движения (перемешивание пищи, ритм - 10 в 1
мин).
• Тонические сокращения сфинктеров (имеется около 35 сфинктеров,
обеспечивающих функциональное разобщение отделов, остановку хи-
муса, его однонаправленное движение).
5.2. Секреторная функция (-Юл пищеварительных соков а сутки, общая площадь секреции -
около 30 м2) - процесс образования гландулоцитами секрета и выделение его в
пищеварительный канал. Железы имеют фенестрированный (хорошо проницае-
мый) тип капилляров и большой кровоток (например, в поджелудочной железе - 100
мл/100 г в мин.), который при пищеварении увеличивается в 3-4 раза
❖ Фазы секреторного процесса.
• Поступление исходных веществ из кровотока через базальную мембра-
ну в гландулоциты путем диффузии, активного транспорта, пииоцитоза.
• Синтез секрета: белков в гранулярной ЭПС, полисахаридов в аппарате
Гольджи.
• Оформление белковых продуктов в гранулы в аппарате Гольджи.
• Накопление гранул в апикальном полюсе клетки.
• Выведение секрета из клетки происходит, как правило, по мерокриио-
вому типу (без нарушения структуры клетки) с использованием нескольких
механизмов - экзоцитоза (запускается увеличением уровня Са2* в цитозоле),
диффузии, ионообмена.
❖ Инкреция пищеварительных ферментов (см. ниже 6.3).
5.3, Химический гидролиз пищи.
❖ Полостное пищеварение (Р. Реомюр, 1753) осуществляется ферментами пище-
варительных соков, оно обеспечивает преимущественно начальную стадию
гидролиза (разрушает - 50 % связей).
❖ Мембранное пищеварение (А М. Уголев, 1957) осуществляется мембранными
ферментами микроворсинок и ферментами гликокаликса до мономеров (бо-
лее 22 ферментов осуществляют гидролиз - 50 % связей).
❖ Внутриклеточное пищеварение (И.И. Мечников, 1883) осуществляет гидролиз
всосавшихся олигопептидов в лизосомах энтероцитов и лейкоцитов, им-
мигрированных в межклеточное пространство кишечной стенки.
5.4. Всасывательная функция.
309
❖ Всасывание - процесс транспорта компонентов пищи из полости пищева-
рительного тракта во внутреннюю среду: межклеточную жидкость, кровь и
лимфу.
❖ Топография всасывания.
• Через клетки (их мембраны) и по межклеточным щелям.
• Основное место - тонкий кишечник, где площадь всасывания - 200 м2,
небольшое всасывание в полости рта, желудке, толстом кишечнике.
Механизмы всасывания (при всасывании одного вещества может быть использовано
несколько механизмов).
• Простая и облегченная диффузия по концентрационному (химическо-
му), осмотическому и электрическому градиентам: вода, короткие жир-
ные кислоты, небольшое количество витаминов С, Вт, Ва, моносахари-
дов, аминокислот, Na+ и др.
• Вторично-активный транспорт (использование энергии градиента
Na*): основное количество моносахаридов и аминокислот, СГ, НСОз’,
витамины (Bi, Вз, С, Н).
• Первично-активный транспорт - насосы.
• Эндоцитоз - всасывание небольшого количества пептидов.
• Экзоцитоз - выход из энтероцитов хиломикронов при всасывании ли-
пидов.
б. Непищеварительиые функции системы пищеварения.
6.1. Экскреторная функция.
❖ Выделение конечных продуктов обмена: билирубина, желчных кислот, не-
большого количества мочевины, креатинина.
❖ Выделение экзогенных веществ: соли тяжелых металлов, лекарственные
препараты.
6.2. Эндокринная функция (см. выше 3.2).
6.3. Инкреция (эндосекреция) пищеварительных ферментов из гландулоцитов в
, кровь и лимфу.
❖ Включаются в метаболизм всего организма.
❖ Могут модулировать секрецию ферментов (например, трипсиноген крови
тормозит секрецию трипсиногена поджелудочной железы н стимулирует секрецию пепси-
ногена желудка).
❖ Частично разрушаются эндотелием сосудов и частично выводятся с мочой,
потом, молоком.
♦ ♦♦ В патологии могут иметь патогенетическое и диагностическое значение.
6.4. Регуляция кислотно-основного состояния (см. тему 21; задание 1:6.3).
6.5. Регуляция гемопоэза.
❖ Всасывание витамина Ви (протекторное действие R-белка слюны и гаст-
ромукопротеииа париетальных клеток).
& Всасывание ионов железа в тонком кишечнике.
6.6. Защитная функция.
❖ Барьерные механизмы обусловлены способностью слизистой оболочки
пищеварительной трубки препятствовать проникновению во внутреннюю
среду бактерий, токсинов и полимеров пищи.
310
❖ Бактерицидное действие пищеварительных соков создается соляной и
желчными кислотами, лизоцимом, пектиновыми веществами и др.
❖ Иммунологические механизмы.
• Пищеварительный тракт имеет клетки системы мононуклеарных фаго-
цитов, которые представляют антигеи иммунокомпетентным клеткам.
• Пищеварительный тракт имеет организованную лимфоидную ткань
(лимфоузлы, миндалины) и лимфоидную ткань, ассоциированную со
слизистой оболочкой кишечника, лимфоциты которой узнают антиген и
формируют гуморальный и клеточный типы иммунного ответа.
7. Методы исследования пищеварения.
7.1. Хронический эксперимент.
❖ Р. де Грааф (1662) предложил метод наложения фистулы протока поджелу-
дочной железы.
❖ В.А. Басов (1842) предложил фистулу желудка.
❖ Р. Гейденгайн (1878) разработал операцию изолированного желудочка иа
большой кривизне без сохранения его иннервации.
❖ И.П. Павлов (1894) предложил изолированный желудочек с сохраненной
иннервацией, опыт «мнимого» кормления, фистулы протоков слюнной и
поджелудочной желез.
❖ Е.С. Лондон (1919) создал метод ангиостомии - наложения постоянных
фистул на крупные вены внутренних органов, позволяющий исследовать
химический состав оттекающей от органа крови.
7.2. Клинико-физиологические методики-, мастикациография, жевательные пробы,
исследование секреции слюны (капсула Лешли - Красногорского), желудочно-
го, поджелудочного, кишечного соков и желчи, электрогастрография, использо-
вание функциональных нагрузок (например, введение гистамина при изучении
секреции желудка).
7.3. Клинико-инструментальные методики-, рентгеновские методики, эндоско-
пия, ультразвуковое исследование и др.
7.4. Клинико-лабораторные методики-, анализ слюны, желудочного сока и желчи,
кала.
8. Пищеварение в полости рта.
8.1. Анализ вкусовых свойств пищи и разделение их на пищевые и отвергаемые
(участвуют зрительный, обонятельный, тактильный, температурный анализато-
ры и кортико-лимбические структуры).
8.2. Механическая обработка пищи - жевание («Хорошо пережевано - наполовину пе-
реварено». Пословица).
❖ Роль жевательных мышц:
• Главные мышцы осуществляют основные жевательные движения: опус-
кание нижней челюсти (двубрюшная, подбородочно-подъязычная, че-
люстно-подъязычная), ее поднимание и смыкание зубов (жевательная,
височная, крыловидная мышцы).
• Вспомогательные мышцы (языка, щек, губ) удерживают пищу между
жевательными поверхностями.
311
❖ Зубы осуществляют откусывание, разламывание, раздавливание и растира-
ние пищи.
❖ Рефлекторный акт жевания является сложным ритмическим безусловным
рефлексом с хорошо выраженным произвольным контролем.
• Афферентное звено рефлекса осуществляется с тактильных, вкусовых,
температурных рецепторов слизистой оболочки рта и проприорецепто-
ров аппарата жевания. Оно обеспечивается в основном сенсорными во-
локнами и ядрами тройничного нерва (V).
• Центр жевания (центральный генератор ритма жевания) находится В ретику-
лярной формации продолговатого мозга и моста. Он вызывает ритмиче-
ское возбуждение мотонейронов мышц, поднимающих и опускающих
нижнюю челюсть. Генератор ритма жевания может быть запущен не
только с рецепторов, ио и от двигательной области лобной коры, что
обеспечивает произвольный контроль жевания.
• Основное эфферентное влияние центра жевания осуществляется через
двигательное ядро V нерва.
❖ Мастикациография - метод исследования биомеханики жевательного акта,
позволяет зарегистрировать 5 фаз: 1) покой; 2) введение пищи в рот; 3)
ориентировочная; 4) основная; 5) формирование пищевого комка (подробнее
ниже в 13.7).
83. Секреторная функция полости рта.
❖ Количество и состав слюны.
• В сутки секретируется 1,5 - 2,0 л слюны: при низкой скорости секреции
0,25 - 0,5 мл/мин, при высокой скорости секреции - 4 мл/мин; pH = 5,8
-7,4.
• Слюна по сравнению с плазмой имеет в 2 - 3 раза меньшие плотность,
вязкость и осмотическое давление, концентрацию электролитов (кроме,
К+, Са2+, НСОз') и мочевины, в ~ 20 раз меньше белков и глюкозы.
• В слюие имеются высокоактивная а-амилаза и низкая активность дру-
гих ферментов (липазы, протеазы, нуклеазы, ингибитора трипсина и др.); глико-
протеин муцин; хорошая бактерицидная активность за счет лизоцима,
Ig А, лейкоцитов.
❖ Физиологическое значение слюны: участвует в увлажнении и ослизнении
пищевой массы, формировании пищевого комка, переваривании углеводов,
возникновении вкусовых ощущений, необходима для устной речи.
❖ Регуляция слюноотделения осуществляется преимущественно нервными
механизмами по типу безусловных и условных рефлексов.
• Важнейшим рецепторным полем являются вкусовые, тактильные, боле-
вые, температурные рецепторы полости рта.
• Афферентная импульсация по волокнам V, VII, IX и X черепных нервов
поступает в продолговатый мозг, где переключается на бульбоспиналь-
ный центр слюноотделения. Он включает в себя верхнее и нижнее слю-
ноотделительные ядра продолговатого мозга, иннервирующие слюнные
312
железы парасимпатическими волокнами VII и IX нервов, и нейроны
сегментов Th> -Thj, осуществляющих симпатическую иннервацию.
• Парасимпатические влияния (ацетилхолин, М-холинорецепторы) приводят к
выделению большого количество жидкой слюны с высокой концентра-
цией электролитов и низкой - муцина.
• Симпатические влияния через p-адреиорецепторы стимулируют секре-
цию ферментов слюиы (слабый эффект через а-адренорецепторы на секрецию во-
ды, Na*, СГ).
8.4. Химическая обработка пищи в ротовой полости', начальный гидролиз крах-
мала и гликогена под действием а-амилазы ограничен временем акта жевания
(~ 18 с) и продолжается в желудке.
8.5. Всасывательная функция слизистой ротовой полости-, всасывание неболь-
шого количества воды, глюкозы и лекарственных веществ (ограничено време-
нем акта жевания).
9. Глотание - переход пищевого комка из полости рта в желудок.
9.1. Рефлекс глотания является сложным цепным безусловным рефлексом с про-
извольным контролем 1-й фазы.
❖ В процессе передвижения пищевого комка из полости рта в пищевод про-
исходит последовательное возбуждение рецепторов корня языка, мягкого
нёба, глотки и пищевода. Импульсация по чувствительным волокнам IX и
X черепных нервов поступает в центр глотания.
❖ Центр глотания, расположенный в продолговатом мозгу и мосту, включает
в себя чувствительное ядро одиночного пути и двойное (двигательное) ядро
IX, X нервов, прилегающие зоны ретикулярной формации. Этот центр
функционально объединяет нейроны примерно двух десятков ядер ствола,
шейных и грудных сегментов спинного мозга.
❖ В результате этого обеспечивается строго координированная последова-
тельность сокращения мышц, участвующих в акте глотания: челюстно-
подъязычных, языка, мягкого нёба, глотки, гортани, надгортанника и пи-
щевода.
❖ Центр глотания функционально связан с центрами жевания и дыхания:
рефлекс глотания останавливает акт жевания и дыхание (обычно в фазе вдоха).
9.2. Фазы акта глотания (Ф. Мажанди, 1817).
❖ Ротовая фаза (произвольная) приводит к формированию пищевого комка
объемом 5-15 см3, который выносится на корень языка.
❖ Глоточная фаза (быстрая, непроизвольная) сопровождается напряжением
мягкого нёба, подъемом гортани и опусканием надгортанника, в результате
комок пищи перемещается к началу пищевода (давление в глотке - 45 мм
рт. ст., в пищеводе - 30 мм рт. ст.).
❖ Пищеводная фаза (медленная, непроизвольная).
• Передвижение пищи в пищеводе осуществляется с помощью пери-
стальтики (скорость - 3 см/с).
• Парасимпатические влияния стимулируют перистальтику пищевода,
сокращают верхний (поперечнополосатый) пищеводный сфинктер и рас-
слабляют инжний (гладкомышечный) пищеводный сфинктер (НПС).
313
Рис. 58. Схема секреции соляной кислоты па-
риетальными экзокриноцитами желез желудка
1- внутриклеточный каналец; черный кружочек
- Н+,К+-насос; светлый кружочек - НСО37СГ-
ионообменник; КА - карбоангидраза
• Симпатические влияния тормозят перистальтику пищевода (Р-
адренорецепторы) и повышают тонус НПС (а-адренорецепторы).
• Время прохождения плотной пищи 3-9 с, жидкой 1-2 с.
• НПС имеет тонус покоя (10-30 мм рт. ст.; максимальный - ночью) выше, чем в же-
лудке (S мм рт. ст.), что препятствует поступлению (рефлюксу) желудочного сока в
пищеаод. Повышают тонус НПС гастрин, мотилии, гистамин, понижают тонус - сек-
ретин, холецистокинин, ГИП.
10, Пищеварение в желудке («Здоровье всего тела куется в кузнице желудка». М. Сервантес).
10.1. Секреторная функция желудка (осуществляется собственными, пилорическими и кар-
диальными железами, суммарная поверхность секреции около 5 м2).
❖ Количество (1,5 - 2,5 л/сут) и состав желудочного сока.
• Соляная кислота (па-
риетальные экзокрииоциты
собственных желез):
ф концентрация:
-120 ммоль/л; 40 -
60 Т.Е.; pH ~ 1;
продукция - 2,8
ммоль/ч;
ф основные зоны
секреции: дно и
тело желудка
(особенно малая
кривизна);
ф механизм секре-
ции (рис. 58): ис-
точником Н+ яв-
ляется диссоциа-
ция Н2СОз; Н+ пе-
реносится в просвет внутриклеточного канальца и затем желудка с
помощью К*, НЛ-насоса, СГ - в результате ионообмена на НСОз’.
• Пепсиногены и липаза (главные экзокрииоциты собственных и пилорических же-
лез):
ф пепсиногены 1-й группы образуются в области дна желудка, пре-
вращаются в пепсин, имеют более кислый оптимум pH (1,5 - 2,0);
ф пепсиногены 2-й группы образуются в пилорическом отделе, пре-
вращаются в гастриксин, оптимум pH менее кислый (2 - 3,5);
ф образование ферментов происходит в гранулярной ЭПС, доработка
и упаковка в гранулы в аппарате Гольджи, выделение из клетки пу-
тем экзоцитоза;
ф небольшая часть пепсиногена ннкретнруется в кровь (~ 100 мкг/л) к выделяется с
мочой в виде уропепснногена.
• Муцин (поверхностные эпителиоциты и мукоциты желез):
314
гликопротеиды (содержат до 80 % углеводов) образуются в области дна,
тела и пилорического отдела, образуют оболочку толщиной 1 мм,
прилегающую к слизистой;
Ф’ защищают слизистую, активируют пепсиногены.
• Гидрокарбонат (НСОз-) секретируется поверхностными эпителиоцита-
ми, оказывает нейтрализующее НС1 действие под слоем муцина.
❖ Регуляция желудочной секреции.
• Регуляция секреции НС1:
"Ф- стимулируют три главных фактора: гистамин (через Н2-рецепторы и
цАМФ), парасимпатические влияния (ацетилхолин через Mi-
холннорецепторы, (цАМФ и Са24), гастрин и гастриносвобождающий
пептид (через (цАМФ и Са24), , а также механическое и химическое
раздражение химусом, продукты переваривания белков;
'Ф' тормозят: симпатические влияния (через а-адреиорецепторы), сомато-
статин, ГИП, кислый химус pH < 3 (а также ВИП, глюкагон, простаглан-
дин Ег, альдостерон).
• Регуляция секреции пепсиногенов:
'Ф' стимулируют: парасимпатические (через М-холииорецепторы) и симпа-
. тические (через Р-адренорецепторы) влияния, гастрин и гастрииосвобо-
ждающий пептид, панкреатический пептид;
"Ф- ингибируют: соматостатин, ГИП.
• Регуляция секреции слизи и бикарбоната:
"Ф- стимулируют: парасимпатические влияния, гастрии и гастриносво-
бождающий пептид, гистамин (через Н2-рецепторы);
ф' тормозят: соматостатин, АКТГ и глюкокортикоиды.
❖ Фазы желудочной секреции.
• «Мозговая» фаза: условно- и безусловиорефлекторный компоненты,
пусковой механизм («запальный» сок).
• Желудочная фаза вызывается действием пищи на слизистую оболочку
желудка, нужна для обеспечения соответствия количества и качества
сока пищевому содержимому; корригирующий механизм.
• Кишечная фаза вызывается с рецепторов 12-перстной и тощей кишки
(нервный путь) и после всасывания продуктов гидролиза веществ (гу-
моральный путь); корригирующий механизм.
❖ Приспособление желудочной секреции к различным видам пищи и пище-
вым режимам.
10.2. Химическая обработка иищи в желудке.
❖ Роль на.
• Активирует пепсиногены, отщепляя С-концевой ингибирующий пеп-
тид.
• Устанавливает оптимум pH для действия пепсина.
• Вызывает денатурацию и набухание белков.
315
• Оказывает бактерицидный эффект (в норме желудочный сок стерилен, при
ахлоргидрии содержит - 10’ микробов/мл).
❖ Роль пепсина и гастриксина: расщепляют белки до крупных полипептидов,
частично до аминокислот. Разрушается - 10 % связей (преимущественно обра-
зованных карбоксильными группами ароматических незаменимых аминокислот - фенила-
ланина, триптофана).
❖ Роль а-амилазы слюны: расщепляет крахмал и гликоген с образованием а-
декстрина, мальтозы н небольшого количества глюкозы в участках желу-
дочного химуса, не пропитавшихся соляной кислотой. Разрушается - 15 %
связей.
❖ Роль желудочной липазы и липазы языка.
• Имеют оптимум pH 4 - 5, устойчивы к НС1 и пепсину.
• Активность невелика, они переваривают в основном эмульгированные
жиры молока, яичного желтка с короткоцепочечными (< 10 атомов углеро-
да) жирными кислотами.
10.3. Моторная функция желудка (время основной эвакуации пищи из желудка 1-3 часа,
полной эвакуации - до 10 часов).
❖ Функция резервуара (дно и часть тела желудка - «проксимальный» желу-
док).
• Натощак объем желудка -50 мл, максимальный физиологический объ-
ем 1500 мл.
• Релаксацию «проксимального» желудка во время приема пищи вызы-
вают симпатические влияния (через 0-адренорецепторы), вагусные нехоли-
нергическне влияния (медиатор ВИП), гастрин, ГИЛ.
• Сокращение «проксимального» желудка вызывают вагусные холинер-
гические влияния.
❖ Перемешивание и передвижение пищи осуществляют тело и антрально-
пилорнческий отдел - «дистальный» желудок).
• Виды движений желудка:
• ф- тонические волны желудка продолжительностью до 2 мин форми-
руются пейсмекером на малой кривизне вблизи кардан;
ф- перистальтика (ритм - 3 нмп/мнн, скорость 1 см/с) запускается
пейсмекером на границе верхней и средней трети тела желудка (те-
ло желудка является ключевой структурой, определяющей моторную ак-
тивность гастродуоденального комплекса);
• ф- систолические сокращения антрально-пнлорнческого отдела запус-
каются пейсмекером пилорического отдела (ритм - 3 имп/мин),
приводят к эвакуации химуса в 12-перстную кишку, а также к из-
мельчению химуса, отбрасыванию назад не растертых частиц.
• Регуляция моторики желудка:
ф- стимулируют моторику желудка: парасимпатические влияния (аце-
тилхолин), гастрин, мотилнн, серотонин;
316
-ф- тормозят: симпатические влияния и адреналин, ГИП, соматостатин,
секретин, ХЦК, серотонин на фоне повышенной двигательной ак-
тивности.
❖ Рвотный рефлекс - непроизвольный выброс содержимого пищеварительно-
го тракта преимущественно через рот. Возникает при раздражении рецеп-
торов корня языка, глотки, желудка, кишечника, брюшины, вестибулярного
аппарата, непосредственно рвотного центра.
• Афферентная импульсация поступает в центр рвоты преимущественно
по чувствительным волокнам X, IX, а также VIII (вестибулярная часть)
нервов.
• Центр рвоты расположен в дорсальном отделе ретикулярной форма-
ции продолговатого мозга, его нейроны имеют М- и Н-холинорецеп-
торы. Рвотный центр регулируется хеморецепторной пусковой зоной
дна IV желудочка, которая находится вне гемато-энцефалического барь-
ера, ее нейроны имеют ДгСдофаминовые)-, 5-НТ(серотониновые)-,
Н(гистаминовые)-рецепторы, стимуляция которых веществами крови (на-
пример, апоморфииом) вызывает рвоту (блокада вышеназванных рецепторов ле-
карствами подавляет рвотный рефлекс).
• Эфферентные импульсы из рвотного центра поступают по блуждаю-
щему и чревному нервам к желудку (сокращение привратника, расслабление
дна), пищеводу (расслабление сфинктеров), тонкому кишечнику (повыше-
ние тонуса, антиперистальтика), а через моторные спинальные центры по со-
матическим нервам - к диафрагме и мышцам брюшной стенки, сокра-
щение которых приводит к изгнанию содержимого желудка (при этом
мягкое небо поднимается, голосовая щель закрывается).
• Рвота сопровождается урежением и углублением дыхания, повышени-
ем слюноотделения, тахикардией.
10.4. Всасывательная функция: в условиях рецептивной релаксации в проксималь-
ном отделе желудка происходит небольшое всасывание воды, минеральных со-
лей, глюкозы, аминокислот, хорошо всасывается алкоголь.
11. Профильный материал для студентов лечебного факультета.
11.1. Изменение аппетита и насыщения при старении.
Происходит повышение порогов, т.е. снижение вкусовой чувствительности к ки-
слому, горькому, соленому и особенно к сладкому (избыточное потребление слад-
кого).
❖ Имеется тенденция к избыточному питанию, ожирению (уменьшение энергетиче-
ских затрат на фоне нормальной или повышенной возбудимости пищевого цен-
тра).
11.2. Изменение секреторной и моторной функций пищеварительной системы при
старении организма.
♦> Секреторная функция.
• В полости рта:
ф" уменьшение количества секреторных клеток в слюнных железах, сниже-
ние объема секреции слюны (сухость во рту);
317
снижение активности амилазы слюны, что уменьшает переваривание уг-
леводов в желудке.
• В желудке:
ф уменьшение количества желез, их париетальных и главных клеток;
снижение образования количества секрета и выделения свободной НС1 иа
40 - 50%;
"Ф снижение продукции пепсина и его снижение протеолитической активно-
сти.
Ф Моторная функция.
• Ротовая полость: снижение эффективности жевания в связи с уменьшением
количества зубов.
• Пищевод:
ф удлинение и искривление пищевода; атрофия его гладкой и поперечнопо-
лосатой мускулатуры;
ф снижение тонуса пищевода и величины внутрипищеводиого давления,
ослабление силы и скорости перистальтических движений;
ф «вялость» глотания (замедление прохождения пищи).
• Желудок:
ф жировое перерождение мышечных волокон, увеличение количества со-
единительной ткани;
ф ослабление двигательной активности желудка во время пищеварения.
12. Профильный материал для студентов педиатрического факультета.
12.1. Питание в антенатальном периоде.
Ф Гистотрофное питание: за счет запасов цитоплазмы яйцеклетки, слизистой обо-
лочки матки, желточного мешка (до 2 - 3 мес.).
Ф Гемотрофное или трансплацентарное питание: питательные вещества поступа-
ют из крови матери через плаценту, ие требуется расщепления питательных ве-
ществ, возможен их гидролиз ферментами плаценты (с 2 - 3 мес. и до рождения).
Ф Амниотрофное питание (с 4 - 5 мес. и до рождения).
• Амниотическая жидкость поступает в пищеварительный тракт плода за счет
дыхательных, сосательных и глотательных движений; пищеварение сначала
происходит аутолитически, позже появляется собственный тип пищеварения
(полостное, пристеночное, внутриклеточное).
• Происходит подготовка пищеварительного аппарата к постиатальиому лакто-
трофиому питанию: формируются эндокринные клетки, закладываются пери-
ферические и центральные механизмы регуляции пищеварительных функций.
12.2. Типы питании в постнатальпом периоде.
Ф Лактотрофное питание: молоко матери, донора.
• Питательные вещества молока и молозива матери гидролизуются по типу ау-
толитического пищеварения.
• Внутриклеточное и мембранное пищеварение сформировано в большей мере,
чем полостное.
ф Искусственное питание (смеси, коровье молоко).
• Резко усиливает секреторную деятельность пищеварительных желез, раньше
включается собственный тип пищеварения.
318
• Не обеспечивает иммунную защиту (не содержит нативных 1g А).
❖ Смешанное питание (лактотрофное + другие питательные вещества), введение
прикорма (рекомендуется с 4 мес.).
••• Дефинитивное питание (замещает лактотрофное и смешанное питание в 1 год),
для него характерен собственный тип пищеварения.
12.3. Пищеварение в полости рта. Сосание. Глотание. Дыхание. Становление жевания.
Особенности слюноотделения.
❖ Сосание - безусловный рефлекс, обеспечивающий в грудном возрасте поступле-
ние в организм питательных веществ, формируется у плода 4 мес.
Глотание.
• У грудного ребенка, в отличие от взрослых, сосание, глотание и дыхание мо-
гут происходить одновременно.
• Пища не попадает в гортань из-за более высокого, чем у взрослых, расположе-
ние ее входа.
• Глотательный рефлекс формируется раньше, чем сосательный.
❖ Дыхание во время кормления со смешанного типа меняется на грудной, при опу-
щенной диафрагме молоко легче проходит по пищеводу в желудок.
❖ Жевание.
• Становится эффективным к 1,5 - 2 годам.
• Прорезывание молочных зубов с б до 30 мес.; они заменяются постоянными с
5-6 до 12-18 лет;«зубымудрости» появляются в 18-25 лет.
Особенности слюноотделения.
• У новорожденных слюнные железы выделяют мало слюны, она необходима
для герметизации губ с грудью матери при лактотрофном питании.
• С 4 - б мес. слюноотделение усиливается из-за введения прикорма, неумение
глотать слюну приводит к слюнотечению, которое прекращается в 1 - 1,5 года.
• Ведущее значение в регуляции слюноотделения имеет безусловный рефлекс с
рецепторов корня языка и слизистой, на его основе формируются натуральные
условные слюноотделительные рефлексы.
• Особенности качественного состава слюны: pH слюны 6,0 - 7,8; низкая актив-
ность амилазы (1/3 уровня взрослого), достигает уровня взрослого в 2 года;
высокая липолитическая активность (реализуется в желудке); активность ли-
зоцима меньше, чем у взрослого.
12.4. Пищеварение в желудке, его особенности у детей первого года жизни.
❖ Физиологическая незрелость желудочных желез у новорожденных и меньшее их
количество; особенности состава желудочного сока.
• Более раннее появление в желудке пепсиногенов, чем НС1, связано с тем, что
главные клетки начинают функционировать раньше, чем париетальные.
• Выделяется фермент реннин (сычужный фермент), который створаживает мо-
локо, предотвращая быстрый его выход из желудка и подготовляя казеин к
действию пепсина.
• Снижение pH желудочного сока в первые часы до 1 - 2 ед., затем нарастание
до 4 - б к концу 1-й недели и снижение до 3 - 4 к концу первого года.
319
• Общая титрационная кислотность желудочного сока у новорожденных 3-10
ТЕ, к году - 12 - 21 ТЕ, свободная НС1 соответственно 0,8 - 4,5 и 6 - 10 (ни-
же, чем у взрослых).
• Интенсивность секреции соляной кислоты ниже, чем у взрослых, и зависит от
типа питания: минимальная при грудном вскармливании и увеличивается в 2 -
4 раза при смешанном и раннем искусственном вскармливании.
Активность ферментов желудочного сока у новорожденных и грудных детей.
• Активность ферментов при грудном вскармливании ниже, чем при вскармли-
вании коровьим молоком.
• Фитолитическая активность появляется к 2 - 3 мес.; зоолитическая активность
- в 5 - 6 мес.
• Протеолитическая активность к концу 1-го года жизни возрастает в 3 раза, но
остается в 2 раза ниже, чем у взрослого. Переход от фетального пепсина к пеп-
сину и гастриксину происходит в возрасте 2 мес.
• Высокая активность липазы желудочного сока осуществляет гидролиз эмуль-
гированного жира молока.
♦♦♦ Моторная деятельность желудка.
• Имеется слабая перистальтика. Порция молока задерживается в желудке на 2 -
3 ч после кормления, смесь с коровьим молоком - на 3 - 4 ч.
• Увеличение белков и жиров в пище замедляет эвакуацию до 5 - 6 ч.
13. Профильный материал для студентов стоматологического факультета.
13.1. Роль ротовой полости в формировании функциональной системы питания.
❖ Блок афферентного синтеза.
• В ЦНС идет информация от вкусовых, обонятельных, тактильных, темпера-
турных, болевых рецепторов. Первыми возбуждаются тактильные рецепторы,
затем температурные и еще позже вкусовые.
• Изменение функции вкусового анализатора является признаком нарушений не
только в полости рта, но и в других отделах организма.
• Расстройства вкуса: агевзия - потеря вкусовой чувствительности; гипогевзия -
снижение вкусовой чувствительности; гипергевзия - повышение вкусовой чув-
ствительности; парагевзия - извращение вкуса; дисгевзия - расстройства тон-
кого анализа вкусовых веществ.
❖ Блок эфферентной программы действия: нервные центры, регулирующие секре-
цию и моторную ротовой полости, являются обязательным компонентом ЭПД
функциональной системы питания.
13.2. Роль рецепторов полости рта в процессах сепсорпого насыщения.
❖ В формировании сенсорного насыщения участвуют проприорецепторы жеватель-
ных мышц, вкусовые и тактильные рецепторы языка, а также рецепторы глотки,
пищевода, желудка и обонятельные рецепторы.
❖ Количество активных вкусовых рецепторов меняется в зависимости от состояния
системы пищеварения, т.е. имеется их функциональная мобильность. Например,
после приема пищи порог вкусовых ощущений повышается (чувствительность
снижается). Одним из механизмов рефлекторных влияний от желудка при раздра-
жении его пищей является «гастролингвальный рефлекс», вкусовые рецепторы
выступают в роли эффекторов.
320
13.3. Роль рецепторов ротовой полости в регуляции секреторной п моторной функций
желудочно-кишечного тракта.
❖ С рецепторов полости рта начинаются рефлекторные реакции, влияющие на сек-
рецию и моторику желудочно-кишечного тракта. Стимулируется образование же-
лудочного и панкреатического соков («мозговая» фаза секреции).
С рецепторов полости рта запускается рефлекторный акт глотания. На некоторое
время снижается тонус и моторика желудка (желудок как резервуар). Двлее сти-
мулируется моторика желудка и выход желчи из пузыря.
13.4. Состав и свойства слюны. Физиологическая роль ее компонентов. Реакция слюны
как физиологическая константа, методы ее определения.
♦ ♦♦ Слюна - это смесь секретов трех пар слюнных желез (околоушных, подъязычных,
поднижнечелюстных) и мелких желез полости рта.
Физиологическая роль слюны: см. выше 8.3.
♦ ♦♦ Реакция слюны (pH) как физиологическая константа, методы ее определения (см.
тему 21; задание 1: 11.1).
❖ Состав слюны.
• Вода - 99,4 % (0,58 % сухой остаток), pH - 5,8 - 7,8, относительная плотность
- 1,001 - 1,017, вязкость - 1,1 - 1,32 сантиПуаз, осмоляльность - 50 - 250
мосм/кг, белок 1,5 -4,0 г/л.
• Электролиты (создают осмотическое давление, при секреции увлекают за со-
бой воду):
Калий - 20 - 30 ммоль/л
Натрий - 7 - 90 ммоль/л
Кальций - 1 - 4 ммоль/л
Хлор - 10 - 45 ммоль/л
Фосфор — 6,2 ммоль/л
Бикарбонаты - 7 - 60 ммоль/л
Мочевина----1,7 ммоль/л
Глюкоза----0,2 ммоль/л
• К ферментам слюны собственно железистого происхождения относятся ами-
лаза, пероксидаза, лактатдегидрогеназа, лизоцим, аминотрансферазы, инги-
битор трипсина, кислая и щелочная фосфатаза и др.
133. Качественные особенности химического состава секретов, выделяемых различ-
ными слюнными железами.
♦♦♦ Околоушная железа выделяет серозный (белковый) секрет, содержащий много
хлорида калия и натрия, содержит каталазу, не содержит щелочной фосфатазы,
очень мало тромбопластина, имеет низкую фибринолитическую активность. Сек-
рет содержит витамины, антибиотики, что указывает на экскреторную функцию
железы.
*!* Поднижнечелюстная железа выделяет серозно-слизистый секрет, в котором мно-
го органических веществ (муцин, амилаза). Обладает наибольшей фибринолити-
ческой активностью.
❖ Подъязычная железа выделяет серозно-слизистый секрет, имеющий аысокую ак-
тивность щелочной и кислой фосфатаз, много муцина и тромбопластина.
13.6. Ротовая жидкость, ее отличие от слюны и физиологическое значение.
*♦* Ротовая жидкость - это смешанная слюна, кроме секретов желез и десневой
жидкости содержит микроорганизмы и продукты их жизнедеятельности, десква-
мированный эпителий, слизь, лимфоциты и нейтрофилы, остатки пищи. Содержит
белки (около 17 видов), углеводы, свободные аминокислоты, компоненты фибри-
нолиза (плазминоген, проактиватор и активатор плазминогена), прокоагулянты.
321
❖ Кроме ферментов железистого происхождения содержит лейкоцитарные фермен-
ты (лактатдегидрогеназа, мальтаза, лизоцим, хондроитин-сульфатаза, липаза, про-
теиназы, пептидазы, альдолаза, пероксидаза) н микробные ферменты (каталаза,
гексокиназа, аминотрансферазы, сахараза, гиалуронидаза и др.). Физиологическое
значение ротовой жидкости такое же, как и слюны.
13.7. Механизм слюнообразования, влияние кровоснабжения на секрецию слюнных же~
лез.
❖ Кровоснабжение слюнных желез осуществляется ветвями наружной сонной арте-
рии. Симпатические волокна, идущие вдоль артерий из верхнего шейного узла,
вызывают сужение сосудов, а парасимпатические волокна в составе барабанной
струны н нижнего каменистого нерва вызывают расширение сосудов.
Секреция слюны (саливация) - сложный процесс, в котором участвуют клетки как
ацинусов, так и протоков. В клетках ацинусов секреторный цикл включает 5 фаз:
1) поступление исходных веществ в железистую клетку; 2) синтез первичного
продукта; 3) транспорт и созревание секрета; 4) накопление секрета в клетке; 5)
выделение секрета из клетки. Так образуется первичная слюна, состав которой
близок к плазме крови. Вторичная слюна образуется при движении первичной
слюиы в протоках (вставочных, стриарных, междольковых). В них происходит ре-
абсорбция Na+, К*, СГ, секреция бикарбоната, мочевины.
♦ ♦♦ Механизм выделения секрета: слюные железы являются мерокринными - секре-
ция происходит без разрушения клетки или отрыва части цитоплазмы.
❖ Кровоснабжение слюнных желез обильное, развиты артериоловенулярные ана-
стомозы, венулы снабжены сфинктерами, закрытие которых увеличивает давление
в капиллярах. Это увеличивает использование веществ крови для синтеза секрета.
Кровообращение в железах оказывает влияние на количество и электролитный со-
став слюны. Однако связь между кровотоком и секрецией не прямо пропорцио-
нальная, так как секреция является активным процессом. (Еще Людвиг показал, что
давление в секреторных капиллярах может быть выше, чем в кровеносных.)
13.8. Иннервация слюнных желез, влияние симпатических и парасимпатических нервов
на секрецию слюнных желез.
♦ ♦♦ Парасимпатическая иннервация осуществляется от верхнего и нижнего слюноот-
делительных ядер.
• Ог верхнего слюноотделительного ядра преганглионарные волокна в составе
барабанной струны идут до поднижнечелюстного и подъязычного узлов, где
переключаются на постганглионарные секреторные волокна, которые в соста-
ве язычного нерва подходят к поднижнечелюстной и подъязычной слюнным
железам.
• Ог нижнего слюноотделительного ядра возбужение передается по преганг-
лионарным волокнам, идущим в составе нижнего каменистого нерва (ветвь
языкоглоточного) к ушному узлу, где переключается на постганглионарные
волокна, которые в составе ушно-височного нерва иннервируют околоушную
слюнную железу.
• Стимуляция парасимпатических волокон (ацетилхолин, М-холи-норецепторы)
вызывает обильную секрецию жидкой слюны, содержащей много солей и мало
органических вещеста.
♦ 5» Симпатические центры расположены в боковых рогах Thi - Thj.
322
• Возбуждение от них поступает по преганглионарным волокнам в верхний
шейный ганглий, а затем по постганглионарным волокнам по ходу ветвей на-
ружной сонной артерии достигает слюнных желез.
• Стимуляция симпатических волокон (норадреналин, p/a-адренорецеторы) вы-
зывает выделение небольшого количества густой слюны, богатой органиче-
скими веществами с малым содержанием солей.
❖ Денервация слюнных желез приводит к непрерывной паралитической сек-
реции.
13.9. Приспособительный характер слюноотделения на различные пищевые и отвер-
гаемые вещества.
❖ У человека имеется непрерывный тип секреции слюны (-0,3 мл/мии). Она связана
с речевой функцией, обеспечивая во время речи увлажнение полости рта. Одним
из сильных раздражителей слюноотделения является сухость пищи.
❖ Слюноотделение приспосабливается к тем потребностям организма, которые для
него в данный момент наиболее существенны. При раздражении вкусовых рецеп-
торов выделяется слюна, богатая ферментами; при раздражении терморецепторов
- жидкая, бедная органическими веществами. При значительной потере жидкости
слюиообразование снижено.
13.10. Регуляция деятельности слюнных желез.
Регуляция слюноотделения (см. выше 8.3 и 13.8).
❖ В регуляции слюноотделения играют роль и вышележащие центры. Раздражение
переднего отдела гипоталамуса и коры около боковой борозды усиливает слюно-
отделение. Показано также тормозящее влияние коры на слюноотделительный
центр.
❖ Опережающая регуляция слюноотделения осуществляется по типу условных реф-
лексов на раздражители, связанные с приемом пищи, - зрительные, слуховые,
обонятельные (натуральные условные рефлексы) и иа другие раздражители, непо-
средственно не связанные с приемом пищи, например, свет лампочки, звук метро-
нома (искусственные условные рефлексы). Показано влияние на слюноотделение
гормонов гипофиза, адреналина, мииералкортикоидов, инсулина.
13.11. Методы получения секрета слюнных желез.
Получение смешанной слюны у человека осуществляется с помощью губок, ватных
тампонов, помещенных в ротовую полость, а также вакуумных сифонов. Используется
также свободное вытекание слюны в градуированный сосуд.
Для сбора слюны из отдельных желез проводят канюлирование их протоков поли-
этиленовыми трубками. Чистый секрет околоушной железы можно получить также с
помощью двухкамерной капсулы Лешли-Красногорского. Наружная камера служит для
присасывания капсулы к слизистой оболочке щеки, а через внутреннюю камеру, распо-
ложенную в зоне протока, собирают отделяемый секрет.
Полученные секреты отдельных желез или смешанной слюны исследуют в количе-
ственном и качественном отношении.
13.12. Методы обследования слюнных железу человека.
❖ Эхосиалография основана иа разной степени поглощения ультразвука различными
тканями. При этом каждый слой ткани создает отраженный импульс, который иа
экране осциллографа создает изображение исследуемого органа. Эхосиалография
позволяет получить послойное изображение слюнной железы и дает представле-
ние о ее макроструктуре (величине, форме, плотности).
❖ Термовизиография основана на разной степени инфракрасного излучения тканя-
ми. На кинескопе тепловизора создается картограмма температуры лица и шеи. В
323
области лица температура колеблется от 23 до 35°С и выше, в области слюнных
желез 31-35°С. Метод помогает диагностировать воспалительные процессы в об-
ласти слюнных желез.
* 1* Радиосиалография позволяет исследовать секреторную функцию слюнных желез.
Вводят внутривенно 7-11 МБк 49Тс0д (пертехнетат). В течение 20 мии он концен-
трируется в железе. При этом записывается кривая радиоактивности. На 30 мину-
те дают 5 г сахара и наблюдают секреторный отрезок сиалограммы. Падение ра-
диоактивности на околоушной железе состааляет 32-36% в течение 4±1 мин.
13.13. Методы исследования протоков слюнных желез у человека.
❖ Зондирование. Исследование проходимости главных слюнных протоков проводят
с помощью введения в них специальных зондов. Этот метод позволяет установить
направление хода протоков, наличие сужения, присутствие слюнного камня.
❖ Сиалография. Метод рентгенологического исследования слюнных протоков путем
заполнения их контрастным веществом (например, йодлиполом, урографином),
который в количестве 1,0-1,5 мл вводится через иглу или канюлю с помощью
шприца. Контраст проникает в мельчайшие разветвления протоков (до V поряд-
ка). Метод позволяет выявлять поражение протоков и паренхимы железы.
13.14. Функциональная характеристика жевательного аппарата. Его роль в механиче-
ской обработке пищи в полости рта.
Л* Собственно жевательные мышцы.
• Основные: жевательная (поднимает нижнюю челюсть, выдвигает вперед и
смещает в свою сторону); височная (обеспечнвет подъем и возвращение на ме-
сто выдвинутой вперед нижней челюсти); латеральная крыловидная (выдвига-
ет нижнюю челюсть вперед при двустороннем сокращении); медиальная кры-
ловидная (при одностороннем сокращении смещает челюсть в противополож-
ную сторону, при двустороннем - поднимает ее).
• Дополнительные (опускают нижнюю челюсть): подбородочно-подъязычная;
челюстно-подъязычная; переднее брюшко двубрюшной мышцы.
♦ ♦♦ Вспомогательные жевательные мышцы (губ, щек, языка) обеспечивают удержа-
ние пищи в ротовой полости, расположение ее между жевательными поверхно-
стями зубов и формирование пищевого комка.
♦ ♦♦ Роль зубов.
• Участие в акте жевания: резцы (8 зубов) осуществляют откусывание; клыки (4
зуба) - разрывание, разламывание; малые коренные зубы (8 зубов) - размина-
ние и растирание пищи; большие коренные зубы (12 зубов) - разминание и
растирание пищн.
• В стоматологии имеется понятие «жевательный центр зубного ряда», под ко-
торым понимается область зубочелюстной системы, где наиболее эффективно
осуществляется механическая переработка принятой пищи. Таким центром яв-
ляются малые и большие коренные зубы той стороны, на которой происходит
пережевывание пнщи. При жевании правый и левый жевательные центры мо-
гут действовать попеременно. Функция жевания может полноценно осуществ-
ляться при использовании жевательного центра только одной стороны.
• Участие в механизмах артикуляции, создают форму звуковой полости, спо-
собствуя внятности речи.
13.15. Формирование пищевого комка (5 - 15 мл) осуществляется в результате акта жева-
ния и секреции слюны. Пищевой комок, консистенция которого подготоалена для гло-
324
тания, выносится на корень языка, что приводит к смене рефлекса жевания на рефлекс
глотания.
13.16. Акт жевания и его саморегуляция. Роль проприорецепторов жевательных мышц,
механорецепторов слизистой оболочки и периодонта в регуляции жевательного
акта.
♦ ♦♦ Акт жевания - совокупность процессов, осуществляемых жевательной мускула-
турой и зубами, приводящих к механическому измельчению пищи, смешиванию
ее со слюной и формированию пищевого комка, пригодного для глотания.
♦ ♦♦ Акт жевания является сложным ритмическим рефлексом с выраженным произ-
вольным контролем. В покое нижняя челюсть поднята, но не до конца, так как ос-
тается щель 1-6 мм. Тонус мышц, поднимающих и опускающих нижнюю че-
люсть, вызванный растяжением их проприорецепторов, уравновешен с учетом
массы челюсти. При попадании пищи в рот афферентное возбуждение с вкусовых,
тактильных, температурных рецепторов ротовой полости рефлекторно тормозит
сокращение мышц, поднимающих нижнюю челюсть. Начинает преобладать дей-
ствие мышц, опускающих челюсть, в результате чего рот открывается.
♦ ♦♦ Растяжение при этом рецепторов мышц, поднимающих челюсть, вызывает реф-
лекторную реакцию «отдачи», т.е. сокращение этих мышц и закрытие рта. Смыка-
ние (окклюзия) зубных рядов раздражает рецепторы парадонта, что рефлекторно
ограничивает сокращение мышц, поднимающих челюсть. Если пища находится в
ротовой полости, то рот опять открывается. Центральный генератор ритма жева-
ния (центр жевания) находится в ретикулярной формации продолговатого мозга и
моста. Он вызывает ритмическое возбуждение мотонейронов мышц, поднимаю-
щих и опускающих нижнюю челюсть. Запуск генератора ритма осуществляется не
только с рецепторов полости рта, но и из двигательного поля лобной коры. По-
этому человек может произвольно замедлить или участить жевательные движе-
ния, изменить их характер.
13.17. Мастикациография. Мастикациограмма и ее анализ.
Мастикациографня является методом изучения биомеханики жевательного акта.
Возникающие при жевании пищи изменения давления воздуха в резиновой манжетке,
фиксированной под нижней челюстью, регистрируется на кимографе в виде мастика-
циограммы.
На мастикациограмме выделяют 5 фаз: 1 фаза - фаза покоя, фиксируется изоли-
ния; 2 фаза - введение пищи в рот, ей соответствует первый подъём кимограммы, вы-
сота которой зависит от степени открытия рта, а крутизна - от продолжительности вве-
дения пищи в полость рта; 3 фаза - ориентировочная (фаза адаптации) характеризуется
нисходящей кривой; 4 фаза - основная, наблюдаются равномерные подъёмы и сниже-
ния кривой, амплитуда н частота которых зависят от консистенции пищи и полноцен-
ности жевательного аппарата; 5 фаза - формирование пищевого комка и проглатывание
пнщи. Графически это выражается волнообразной кривой с некоторым уменьшением
размаха волн.
В норме время одного жевательного акта равно 16,0±1 с., количество жевательных
движений в нём - 18±8. По характеру мастнкацнограммы можно судить об изменениях
в жевательной системе, об эффективности ортопедических и других лечебных меро-
приятий.
13.18. Жевательная проба по Рубинову: 0,8 г лесного ореха жуют до появления рефлекса
глотания. Выплевывают, высушивают и просеивают через снто с ячейками 2,4 мм. В
норме жевание до 14 с, остатка иа сите нет. Жевательная проба позволяет судить об
эффективности жевания до и после протезирования, а также определить функциональ-
ную ценность протеза и степень адаптации к нему.
325
13.19. Основные функции зубов и нарадонта.
❖ Функции зубов (см. выше 13.14).
❖ Функции пародонта.
• Тактильная чувствительность воспринимает консистенцию пищи, предотвра-
щает попадание твердых частиц в желудок.
• Опорно-удерживающая (фиксирует зубы в челюсти).
• Распределяет н регулирует жевательное давление.
• Пластическая (восстановление тканей - кость, вторичный цемент).
• Трофическая - питает цемент и стенки альвеолы.
13.20. Всасывательная функция слизистой оболочки полости рта.
♦ ♦♦ Всасывательная функция слизистой ротовой полости ие имеет важного значения в пище-
варении. Это обусловлено кратковременностью пребывания пищи во рту н малым количе-
ством мономеров.
❖ Всасывание обусловлено наличием постоянно увлажненного эпителия, близко
расположенными к поверхности слизистой оболочки кровеносными сосудами.
Слизистая проницаема для йода, Na+, К*, некоторых аминокислот, гидрокарбона-
та, алкоголя, антибиотиков, валидола, нитроглицерина. Нормальная слизистая
всасывает лекарственные вещества быстрее, чем патологически измененная.
♦ ♦♦ Механизмы всасывания: простая и облегченная диффузия по градиенту концен-
трации, осмос (вода), активный транспорт с использованием ионных градиентов и
насосов.
Задание 2. Подготовить реферативные доклады иа тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мии.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-16].
3.2. Дополнительная литература (см. с. 514 - 522): [13], [30], [31], (35], [47], [83],
[91]. [98], [107], [108], [129], [137], [170], [179], [185], [196], [199], [205], [211], [212],
[222], [225], [226], [228], [251-254].
33. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/-nphys: слайды для компьютерной демонстрации из «Атласа
по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты оста-
точного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии н экзамене.
326
Тема 23. Пищеварение в тонком н толстом кишечнике
Цель: а) знать механизмы секреторной, ферментативно-перевариваюшей, всасывательной и мотор-
ной функции тонкого и толстого кишечника; роль поджелудочной железы и печени в пищева-
рении; состав и свойства поджелудочного сока, желчи и кишечного секрета, механизмы их
секреции; регуляцию панкреатической, кишечной секреции, образования желчи и ее выделе-
ния в двенадцатиперстную кишку; полостной и мембранный гидролиз питательных веществ;
роль моторной деятельности тонкого и толстого кишечника в пищеварении и механизмы ее
регуляции; значение микрофлоры в кишечнике; механизмы всасывания различных веществ в
тонком и толстом кишечнике, регуляцию всасывания, методики изучения всасывания; возрас-
тные особенности пищеварения в кишечнике; периодическую деятельность органов пищева-
рения;
б) уметь использовать эти знания для объяснения роли функции кишечника в функциональной
системе питания в целостном организме.
Мотивация темы: см. тему 22.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
I. Пищеварение в тонком кишечнике.
1. Секреторные процессы, участвующие в тонкокишечном пищеварении (t - 5 м).
1.1. Внешнесекреторная функция поджелудочной железы (клетки ацинусов образуют
нейтральный секрет с большим содержаигием ферментов и СГ, клетки протоков - щелочной
секрет с большим содержанием НСО3“).
❖ Количество и свойства поджелудочного сока (~ 1»5 л/сут, рН=8 - 8,5;
НСОз' * 120 ммоль/л, изотоничен плазме).
❖ Основные ферменты панкреатического сока.
• Протеазы (переваривают белки):
“у- трипсин образуется из трипсиногена под действием энтерокиназы и
трипсина (разрывает только пептидные связи, содержащие карбоксильные
группы лизина и аргинина), трипсин-ингибитор защищает поджелудоч-
ную железу от самопереваривания;
• ф химотрипсин образуется из химотрипсиногена под действием трип-
сина (наиболее активен при гидролизе связей, образованных карбоксильными
группами фенилаланина, тирозина и триптофана);
Ф карбоксипептидазы А и В (из прокарбоксипептидаз под действием трипси-
на).
• Липазы: панкреатическая липаза расщепляет триглицериды на жирные
кислоты и моиоглицериды, холестеролэстераза - холестеролэстеры на
холестерол и жирные кислоты, фосфолипаза А расщепляет фосфолипи-
ды.
327
• а-Амилаза разрушает 1,4-связи в крахмале и гликогене с образованием
преимущественно мальтозы.
• Нуклеазы расщепляют РНК и ДНК на нуклеотиды.
❖ Регуляция панкреатической секреции.
• Фазы отделения поджелудочного сока: 1) мозговая (через X нерв; менее
выражена, чем в желудке); 2) желудочная (ваго-вагальные рефлексы из тела
и антральной части желудка); 3) кишечная (основная).
• Гуморальные влияния иа панкреатическую секрецию:
•Ф- стимулируют: секретин (секреция HjO, НСОз'), ХЦК-ПЗ (секреция
ферментов), гастрин и гастриносвобождающий пептид (а также серо-
тонин, субстанция Р, инсулин);
•ф- тормозят: соматостатин, глюкагон, ГИП, панкреатический пептид.
• Нервные влияния (менее выражены, чем гуморальные):
•ф- парасимпатические (X нерв) стимулируют секрецию ферментов и
НСОз' (ацетилхолнн+М-холинорецепторы —»ИФУДАГ —• )Са2+);
"Ф* симпатические (чревные нервы) тормозят секрецию (норадрена-
лии+Р-адреиорецепторы -»|АЦ -> (цАМФ -»|Са24);
•Ф- пептидергические (стимулирующие и тормозные).
• Приспособительная секреция поджелудочного сока иа различные виды
пиши н пищевые рационы.
1.2. Желчесекреторная функция печени.
❖ Количество и состав печеночной желчи (~ 1 л/сутки, рН=8,2; НСОз' - 30
ммоль/л, изотонична плазме).
• Желчные кислоты (35 ммоль/л): в печени из холестерина образуются
холевая и хенодезоксихолевая кислоты (2:1); оии конъюгируют с обра-
зованием гликохолевой и таурохолевой кислот (4:1), которые секрети-
руются в желчь.
• Холестерин (3 ммоль/л). (Его растворимость в желчи создается тем, что холесте-
рин входит в состав мнцелл, образованных желчными кислотами и фосфолипидами.
Холестериновые камни составляют 85% всех желчных камней).
• Билирубин (1,5 ммоль/л).
• Фосфолипиды (особенно лецитин).
❖ Образование желчи (холерез).
• Секреция из гепатоцита в желчный капилляр желчных кислот, холесте-
рина, билирубина, электролитов (Na+, СГ, НСОз') - активный транспорт,
воды - по осмотическому механизму.
• Печеночно-кишечная циркуляция желчных кислот.
•Ф- В сутки по кругу «печень -► желчь -> тонкий кишечник -> реаб-
сорбция в конце подвздошной кишки -» портальная кровь -» пе-
чень» циркулирует - 24 г желчных кислот (10-15 циклов/сутки).
•ф- Вновь образуется в печени 0,5 г желчных кислот в сутки и столько
же покидает организм через кишечник.
• Обмен билирубина.
328
ф- В сутки из гемоглобина образуется - 230 мг свободного билируби-
на, который транспортируется к печени в связи с альбумином (в поч-
ках не фильтруется).
ф В печени свободный билирубин переходит в связанный путем
конъюгации с УДФ-глюкуроновой кислотой (под действием глюкоро-
нилтрансферазы).
ф Связанный билирубин (моно- и диглюкоронид) поступает с желчью в
кишечник, где гидролизуется до билирубина и восстанавливается
микрофлорой в уробилиноген и стеркобилиноген, в толстом ки-
шечнике они окисляются в уробилин и стеркобилин и покидают ор-
ганизм (- 200 мг/сут).
ф- Часть билирубина, уробилиногена и стеркобилиногена (- 30 мг/сутки)
опять поступает в печень по портальной веие, образуя печеночно-
кишечную циркуляцию.
ф- Часть стеркобилиногена, уробилина и стеркобилина поступает че-
рез геморроидальные вены в полую вену и, минуя печень, вьщеля-
ется почками (под названием уробилиновые тела мочи, - 2 мг/сутки).
❖ Желчевыделение (холекинез).
• Желчевыводящие капилляры, ходы и протоки - общий печеночный, пу-
зырный, общий желчный имеют сфинктеры.
• Функции желчного пузыря.
ф Резервуарная функция 60 - 80 мл желчн (в патологических условиях до
1000 мл).
"Ф Концентрирующая: за счет реабсорбции Na+, СГ и воды концен-
трация желчных кислот и холестерина увеличивается в 8 - 10 раз.
•ф- Выделение желчи (сокращение протоков, пузыря, расслабление
сфинктеров).
Стимулируют: парасимпатические влияния, ХЦК-ПЗ, гастрин,
липиды пищи, сульфат магния;
Тормозят: симпатические влияния, панкреатический пептид,
глюкагон.
❖ Функции желчи.
• Эмульгирует жиры с образованием мицелл.
• Повышает панкреатическую секрецию и активность липазы.
• Способствует всасыванию липидов и жирорастворимых витаминов.
• Стимулирует образование и выделение желчи, моторику кишечника.
• Инактивирует пепсины.
• Обладает бактерицидным действием.
1.3. Тонкокишечная секреция (кишечные железы Бруннера и Либеркюна, общая площадь
секреции -14 м2).
❖ Количество и свойства кишечного сока (2 л/сут, рН=8; НСО3' - 100
ммоль/л, изотоиичеи плазме).
❖ Ферментный состав (более 20 ферментов, большая часть остается в зоне
щеточной каемки и гликокаликса, часть выходит в полость кишечника):
329
• протеазы, дипептидазы, аминопептидаза;
• энтерокиназа (активирует трипсиноген);
• карбогидразы: мальтаза, лактаза, сахараза, 1,6-глюкозидаза;
• фосфатаза, нуклеаза, лизоцим н др.
❖ Секреция щелочной слизи, содержащей гликопротеины.
❖ Регуляция кишечной секреции.
• Особенность: практически отсутствует стимуляция во время приема
пищи (химус поступает через 1 - 6 ч).
• Стимуляция секреции местными механическим и химическим (НС1, про-
дукты белкового и липидного гидролиза) раздражителями химуса, а также
ВИП.
• Тормозит секрецию соматостатин.
• Рефлексы метасимпатической нервной системы (медиаторы: ВИП, ацетил-
холин и др.) и центральные вегетативные рефлексы.
2. Химическая обработка пищи в тонком кишечнике (почти полностью происходит в 12-
ти перегной и тощей кишках; одни энтероцит снабжает питательными веществами около 100 тысяч
клеток организма).
2.1. Переваривание белков.
❖ В тонкий кишечник поступают белки, полипептиды пищи и пищеваритель-
ных соков (всего около 120 г/сутки).
❖ В полостном пищеварении трипсин и химотрипсин гидролизуют белкн до
пептидов.
❖ Карбокси- и аминопептидазы гидролизуют пептиды до дипептидов, отщеп-
ляя от них по одной аминокислоте.
Гликокёшикс покрывает энтероцнты пленкой около 1 мкм и заполняет про-
странство между микроворсинками. Он является молекулярным ситом, не
пропускающим белки н полипептиды в зону пристеночного пищеварения.
❖ В пристеночном пищеварении дипептиды гидролизуются дипептидазой до
аминокислот.
Некоторое количество дипептидов всасывается и гидролизуется в эитероцитах лизосо-
мальными пептидазами.
2.2. Переваривание углеводов.
❖ В тонкий кишечник поступают полисахариды (крахмал, гликоген), продук-
ты их частичного гидролиза (а-декстрины, мальтоза, глюкоза), а также сахароза,
лактоза моносахариды.
В полостном пищеварении крахмал н гликоген под действием панкреати-
ческой амилазы и кишечной 1,6-глюкозидазы гидролизуются до а-
декстрина, мальтозы, глюкозы.
❖ В пристеночном пищеварении дисахариды (сахароза, мальтоза, лактоза)
расщепляются под действием соответствующих ферментов (сахаразы,
мальтазы, лактазы) до моносахаридов (глюкозы, фруктозы, галактозы).
23. Переваривание лииидов.
330
❖ В кишечник из желудка поступают крупноэмульгированные липиды (d = 1 -
2 мкм), которые с помощью желчных кислот и лецитина превращаются в
мицеллы (d = 3 - 6 нм), способные пройти через гликокаликс.
❖ В полости кишечника триглицериды под действием панкреатической липа-
зы гидролизуются до жирных кислот, глицерола и 2-моноацилглицерола.
❖ Фосфолипиды гидролизуются под действием фосфолипазы А, эфиры холе-
стерина - под действием холестеролэстеразы.
3. Всасывательная функция тонкого кишечника.
3.1. Происходит на громадной площади (S - 200 м1), имеется сопряжение процессов
переваривания и всасывания, обусловленное тем, что молекулы ферментов и
переносчиков на мембране пространственно сопряжены.
3.2. Всасывание продуктов гидролиза белков (аминокислоты и часть дипептидов).
❖ В апикальной мембране (микроворсинках щеточной каемки) имеется 5 ви-
дов переносчиков аминокислот (для кислых, основных, нейтральных кислот, нмн-
нокислот, ₽ и у-аминокнслот).
❖ На апикальной мембране происходит активный транспорт с использовани-
ем градиента Na+, на базальной мембране - облегченная диффузия.
❖ Часть дипептидов всасывается (с использованием переносчиков и энергии
градиента Н+) и гидролизуется в лизосомах.
3.3. Всасывание продуктов гидролиза углеводов.
❖ Всасываются только моносахариды с использованием активного транспор-
та (энергия градиента Na+), облегченной и простой диффузии.
• Глюкоза и галактоза через апикальную мембрану переносятся путем ак-
тивного транспорта, через базальную мембрану переходят путем про-
стой и облегченной диффузии.
• Фруктоза транспортируется путем облегченной диффузии, манноза и
пентоза - простой диффузии.
<• Регуляция всасывания углеводов:
• усиливают парасимпатические влияния, глюкокортикоиды, тироксин,
инсулин;
• тормозят симпатические влияния, соматостатин.
3.4. Всасывание продуктов гидролиза жиров.
❖ Мицеллы (d = 3 - 6 им) свободно проникают в просветы (50 - 100 нм) между
микроворсинками и достигают поверхности энтероцита.
❖ Липидное содержимое мицеллы (жирные кислоты, глицерол, 2-моноацил-
глицерол, продукты переваривания фосфолипидов и эфиров холестерина)
диффундируют с помощью переносчиков (возможно, активным транспор-
том) внутрь энтероцита (освободившиеся из мицеллы желчные кислоты всасываются
в конце подвздошной кишкн с помощью Na’-завнснмого активного транспорта).
❖ В агранулярной ЭПС энтероцнтов происходит ресинтез триглицеридов,
фосфолипидов и холестеролэфиров, которые в аппарате Гольджи упаковы-
ваются в липопротеиновую оболочку, образуя преимущественно хиломик-
роны (d до 1000 нм) и липопротеины высокой плотности, которые транспор-
тируются из энтероцита путем экзоцитоза в лимфатические сосуды кишеч-
331
ника. (В энтероцитах происходит синтез холестерола de novo, поэтому тонкий кишечник
играет важную роль в его обмене.)
* Короткоцепочечные жирные кислоты (< 10 атомов углерода) и глицерол могут
всасываться вне состава мицелл и поступать в кровь.
❖ Регуляция всасывания липидов:
• усиливают парасимпатические влияния, секретин, ХЦК-ПЗ, глюкокор-
тикоиды;
• тормозят симпатические влияния, адреналин.
❖ В норме всасывается около 98 % липидов пищи.
3.5. Всасывание воды (около 8 л/сут).
❖ Вода может диффундировать в обе стороны, поэтому кишечный химус изо-
тоничен плазме крови.
❖ При всасывании моносахаридов, аминокислот, Na'*' вода следует за ними в
соответствии с осмотическим градиентом.
3.6. Всасывание ионов.
Поступление Na+ в энтероцит через апикальную мембрану происходит по
электрохимическому градиенту с участием переносчика, из энтероцита че-
рез базальную и латеральную мембраны с помощью Na+, К+-насоса (всасыва-
ется в сутки - 25 г Na*).
❖ Всасывание СГ совпадает (через потенциал) с транспортом Na+, но СГ вса-
сывается больше, так как есть еще обмен СГ на НСОз".
❖ Всасывание Са2+ происходит с помощью Са2+-связывающего белка и 1,25-
(ОН)г-витамина Dj, паратгормон активирует всасывание. Выход Са2+ из эн-
тероцита происходит с участием Са2*Жа+-ионообменника и Са2+-насоса.
3.7. Всасывание витаминов.
❖ Некоторые водорастворимые витамины (С, Вь Вг, Н) переносятся через
апикальную мембрану вторичноактивным транспортом с использованием
энергии градиента Na+, через базальную мембрану - диффузией.
❖ Жирорастворимые витамины (А, Е, D, К), содержащиеся в мицеллах, вса-
сываются вместе с липидами.
4. Моторная функция тонкого кишечника (основные пейсмекеры в пилороантраль-
ном отделе желудка и около дуоденального сосочка).
4.1. Перемешивание химуса осуществляется с помощью мелкой перистальтики,
ритмической сегментации, маятникообразного движения и сокращения ворси-
нок.
4.2. Передвижение химуса осуществляется с помощью пропульсивной (глубокой)
перистальтики. Химус находится в тонкой кишке 4 - б ч.
4.3. Эвакуация химуса в толстый кишечник происходит при рефлекторном открыва-
нии илеоцекального сфинктера, градиент ритмов перистальтики в илеоцекаль-
ной зоне 14/7 в 1 мин.
4.4. Регуляция моторики тонкого кишечника:
❖ стимулируют: парасимпатические влияния, гастрин, мотилин, грубая и
жирная пища;
❖ тормозят: симпатические влияния, адреналин.
П. Пищеварение в толстом кишечнике (С = 1,5 -1,8 м).
332
1. Секреторная функция толстого кишечника.
1.1. Количество и состав кишечного сока (0,05 л/сут, рН=8,7).
❖ Малая активность ферментов (пептидазы, липазы, амилазы, нуклеазы, ще-
лочной фосфатазы).
❖ Большое количество слизи (бокаловидные клетки).
1.2. Регуляция секреции.
❖ Основной стимул - механическое раздражение.
❖ Преобладают местные механизмы регуляции: рефлексы метасимпатиче-
ской нервной системы, секреция ВИП, энтерогастрона, серотонина.
2. Химическая обработка питательных веществ в толстом кишечнике (преимущест-
венно в проксимальных отделах).
2.1. Роль ферментов кишечного сока: компенсаторно включаются при нарушении
тонкокишечного пищеварения.
• Роль микрофлоры кишечника (бифидум бактерии, кишечная палочка, молочнокислые
бактерии и др., - 2-10 ’/1 г содержимого):
❖ расщепление ферментов, поступивших из тонкого кишечника (трипсина,
амилазы и др.);
❖ ферментативное расщепление клетчатки (до 40 %), крахмала (~ 40 r/сут), угле-
воде? слизи с образованием летучих жирных кислот - уксусной, пропионо-
вой и масляной кислот, используемых эитероцитами в качестве основного
энергетического материала; при этом образуется водород, который связы-
вается метанообразующими бактериями с образованием метана;
❖ ферментативное образование NHj из мочевины (- 260 ммоль/сутки);
❖ образование глутамата и ГАМК, которые используются как медиаторы в
нейрогуморальиой регуляции;
❖ ферментативное превращение желчных кислот в дезоксихолевую и литохо-
левую, а также билирубина в уробилиноген и стеркобилиноген;
❖ образование небольшого количества витаминов К, группы В, биотина, Ви
при наличии в пище кобальта;
❖ расщепление углеводов до кислых продуктов (уксусной, пропионовой,
масляной кислот, СОг), гнилостное разложение белков (индол, скатол, гис-
тамин, тиронин и др.).
3. Всасывательная функция толстого кишечника (преимущественно в проксимальных от-
делах, механизмы см. выше I; 3).
3.1. Хорошая способность к всасыванию воды (-1400 мл/сут, но не более б л в су-
тки; формирование кала).
3.2. Всасывание электролитов (Na* - 200 ммоль/сут, СГ - 100 ммоль/ сут; НСО3‘ - 70
ммоль/сут) и мочевины (- 100 ммоль/сут), секреция К*.
33. Всасывание образованных микрофлорой аммиака (- 260 ммоль/сут), уксусной,
пропионовой и масляной кислот (- 300 ммоль/сут).
3.4. В небольших количествах всасываются глюкоза, аминокислоты (возможность пи-
тательных клизм).
4. Моторная функция толстого кишечника.
333
4.1. Питательные вещества принятой пищи начинают поступать в толстый кишеч-
ник через 6 - 7 ч, заполняют его через 24 ч (~ 1500 мл химуса в сутки), полное
опорожнение происходит за 2 - 3 дня.
4.2. Резервуарная функция обеспечивается гаустрацией, тонусом сфинктеров, пла-
стическим тонусом.
4.3. Функция перемешивания содержимого обеспечивается гаустрацией, маятнико-
образными движениями, мелкой перистальтикой и антиперистальтикой.
4.4. Функция передвижения химуса осуществляется в результате глубокой пропуль-
сивной перистальтики, возникающей 3-4 раза в сутки. Снижение тонуса сиг-
мы способствует переходу кала в прямую кишку, повышение тонуса ведет к за-
порам.
4.5. Регуляция моторики толстого кишечника.
❖ Стимулируют моторику механические и химические раздражители химуса
(например, пищевые волокна) через парасимпатические влияния.
❖ Тормозят моторику симпатические влияния, серотонин, адреналин.
4.6. Акт дефекации (норма - от 2 раз в сутки до 1 раза в двое суток).
❖ Раздражение механорецепторов прямой кишки (давление 30 - 40 мм рт.
ст.), афферентная импульсации по волокнам тазового и срамного нервов.
❖ Центральное звено - аноспинальный центр (S24), а также моторные центры
диафрагмы и брюшных мышц.
❖ Эфферентное звено - парасимпатические волокна тазового нерва, сомати-
ческие волокна диафрагмальных, межреберных, подвздошно-подчревных и
подвздошно-паховых нервов.
❖ Эффекторное звено:
• расслабление внутреннего анального сфинктера (гладкая мышца, не-
произвольный контроль);
• расслабление наружного анального сфинктера (поперечнополосатая
мышца, произвольный контроль с 2 лет);
• перистальтика толстой кишки;
• сокращение мышц брюшного пресса и диафрагмы.
5. Периодическая деятельность органов пищеварения (В.Н. Болдырев, 1904).
5.1. 90-минутный биоритм (20 мин активности и 70 мин покоя), возникающий между
приемами пищи после того, как произошло полное опорожнение желудка. Име-
ет два компонента.
❖ Моторный компонент («голодные сокращения желудка», миграционный
моторный комплекс) стартует в 12-перстной кишке в области фатерова со-
сочка. Возникают сначала нерегулярные, затем регулярные сокращения,
которые охватывают желудок, нижний отдел пищевода и тонкий кишеч-
ник. Эти сокращения активируются мотилииом (тормозятся гастрином и гнета-
мином).
❖ Секреторный компонент (35 - 40 мл) включает в себя выход в кишечник
желчи и панкреатического сока с большим количеством белка. Он регули-
руется вегетативной нервной системой и ХЦК-ПЗ.
5.2. Физиологическое значение периодической деятельности.
❖ Возбуждение центра голода и пищевого поведения.
334
❖ Переход на эндогенное питание веществами пищеварительных соков.
❖ Экскреция конечных продуктов обмена.
6. Профильный материал дли студентов лечебного факультета.
6.1. Изменение секреторной и моторной функций пищеварительной системы при
старении организма.
♦ > Секреторная функция.
• В поджелудочной железе.
ф- Снижение массы железы на 50 %, числа ацинусов, островков и Р-клеток;
"Ф- Снижение внешнесекреторной функции железы:
снижение объема секреции и активности панкреатических фер-
ментов (липазы, трипсина после 60 лет, амилазы после 70 лет на
50-80%);
более выраженное снижение протеолитической активности пан-
креатического сока, чем липолитической.
" Ф- Нарушение гуморального механизма регуляции деятельности железы
(снижение концентрации секретина и ХЦК-ПЗ).
• В тонком кишечнике.
• ф- Уменьшение количества функционирующих желез, замена секреторных
' клеток соединительнотканными.
• ф- Уменьшение числа ворсинок на слизистой оболочке.
"Ф* Снижение функции секретирующих клеток (например, снижение выра-
ботки ХЦК-ПЗ).
• В толстом кишечнике: изменение кишечной микрофлоры - уменьшение ко-
личества молочнокислых микробов, снижение ферментативной активности
кишечной палочки, развитие гнилостной микрофлоры.
Моторная функция тонкого и толстого кишечника: увеличение общей длины из-за
атрофии мышечного слоя, снижение мышечного тонуса кишечной стенки, замед-
ление перистальтики и продвижения химуса, склонность к запорам.
6.2. Функции печени при старении.
Снижение регенерационной способности и массы органа.
❖ Уменьшение функциональной активности клеток печени:
• снижение белковосинтетической функции (преимущественно синтеза альбу-
минов и протеолитических ферментов);
• снижение гликогенсинтезирующей и гликогендепонирующей функции;
• снижение синтеза фосфолипидов, увеличение синтеза холестерина;
• снижение выделительной функции печени;
• снижение фагоцитирующей способности макрофагов печени.
•> Желчный пузырь:
• снижение эвакуаторно-моторной функции желчного пузыря;
• увеличение объема остаточной желчи, что способствует образованию желчных
камней и желчекаменной болезни.
7. Профильный материал дли студентов педиатрического факультета.
7.1. Пищеварение в двенадцатиперстной кишке.
••• Поджелудочная железа.
335
• Масса поджелудочной железы увеличивается в течение 1-го года в 3 раза, пан-
креатическая секреция - в 10 раз.
• У новорожденных низкая реактивность гландулоцитов ацинусов к стимулято-
рам секреции (секретину и ХЦК-ПЗ); через месяц возрастает секреция трипси-
ногена, карбоксипептидаз.
• Смешанное и искусственное вскармливание повышает объем секреции, выде-
ление ферментов и их активность.
❖ Печень.
• Печень новорожденных относительно велика и составляет 4 % от массы тела
(у взрослых - 2,5 %), к концу 1-го года абсолютная масса печени удваивается.
• Желчеобразование у новорожденных достаточно интенсивное (на 1 кг массы
тела в 4 раза больше, чем у взрослых).
• Желчь богата муцином и пигментами, но бедна желчными кислотами и холе-
стерином, что вызывает недостаточное усвоение жиров прн прикорме моло-
ком.
• В составе желчи преобладает таурохолевая кислота, которая усиливает пан-
креатическую секрецию, обладает антисептическим действием.
7.2. Пищеварение в тонкой кишке,
❖ Кишечник у детей относительно длиннее, чем у взрослых. Слизистая оболочка
более тонкая, ворсинок меньше, кишечник имеет хорошо развитую кровеносную и
лимфатическую систему.
* ?* Секреторная и всасывательная функции тонкого кишечника.
• Интенсивное мембранное пищеварение компенсирует низкую интенсивность
полостного пищеварения. В гидролизе питательных веществ участвует отно-
сительно большая часть тонкой кишки.
• Лактаза тонкой кишки более легко гидролизует лактозу коровьего молока, чем
женского. (При недостатке лактазы, нерасщепленная лактоза повышает осмо-
тическое давление химуса, что приводит к диарее и обезвоживанию.)
• Белки женского молока перевариваются и всасываются в кишечнике полнее
(90 - 95 %), чем коровьего (60 - 70 %);
• Слизистая тонкой кишки обладает в грудном возрасте высокой проницаемо-
стью (высокомолекулярные вещества молока - иммуноглобулины, гормоны -
поступают в организм ребенка).
• Повышение активности энтерокиназы и щелочной фосфатазы и задержка в ор-
ганизме Са2+ н Mg2+ при раннем прикорме овощами.
• Переход на дефинитивное питание (крахмал, гликоген) усиливает синтез маль-
тазы, сахаразы и снижает синтез лактазы.
❖ Моторная функция тонкого кишечника: при грудном вскармливании химус эва-
куируется за 12 - 13 ч; при искусственном - более медленно.
7.3. Пищеварение в толстой кишке. Микрофлора желудочно-кишечного тракта.
❖ Слизистая оболочка толстой кишки ие вырабатывает полостных ферментов. Пи-
щеварение осуществляется ферментами, поступившими из тонкого кишечника.
❖ Кишечник плода стерилен, заселение микроорганизмами происходит в первые 24
- 48 ч жизни: 1-я фаза - асептическая (длится 10 - 20 ч); 2-я фаза - развитие мик-
роорганизмов (2-4 сут); 3-я фаза - стабилизация микрофлоры (2 недели).
336
❖ Расщепление остатков пищи идет за счет деятельности микробной флоры кишеч-
ника. Микрофлора кишечника обладает защитными, антитоксическими функция-
ми, влияет на скорость обновления энтероцитов, участвует в инактивации физио-
логически активных веществ и ферментов.
❖ Дефекация.
• Кишечник освобождается от первородного квла (мекония) в первые часы по-
сле рождения; меконий состоит из остатков плодных вод, слизи, спущенного
эпителия, через 3-4 дня замещается типичным калом.
• Количество дефекаций у новорожденного 5 - 7, у грудных детей 2-3 раза в
сутки, произвольный контроль устанавливается к 2 годам.
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-16].
3.2. . Дополнительная литература (см. с. 514 - 522): [13], [27], [30], [35], [43], [47],
[83], [91], [98], [107], [108], [129], [137], [179], [185], [196], [199], [205], [211], [212],
[222], [225], [226], [228], [245], [251-254].
3-3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ni/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 23-1. У человека произошло полное травматическое повреждение спинного мозга в об-
ласти L|. Какие и почему возникнут у него изменения в акте дефекации? Анализ вести на основе
его физиологических механизмов.
Задача 23-2. У больного с воспалительным процессом одного из органов произошло резкое уве-
личение в крови активности трипсина. Какой из органов у него воспален и какой физиологиче-
ских процесс лежит в основе увеличения этого фермента в крови?
337
Тема 24. Обмен веществ н энергии. Питание
Цель: а) знать обмен веществ между организмом и внешней средой как основное условие жизни и
сохранения гомеостаза, пластическую и энергетическую роль питательных веществ, баланс
их прихода и расхода, азотистое равновесие, механизмы его поддержания, регуляцию пита-
тельных веществ в организме; значение воды, минеральных веществ и микроэлементов для
организма, регуляцию водного и минерального обмена, физиологическую роль витаминов;
энергетический баланс организма, приход и расход энергии, основной обмен и факторы, его
определяющие, специфически-линамическое действие пищи, энергетические затраты при
разных видах труда в различном возрасте; физиологические нормы питания, потребность
организма в белках, жирах, углеводах и минеральных солях; основы рационального пита-
ния; теории сбалансированного и адекватного питания, альтернативные концепции питания.
6) уметь использовать эти знания для анализа энергетических затрат организма, потребности
в питательных веществах при деятельности организма в разных условиях;
в) иметь представления о методах исследования энергетических затрат человека (прямая и
непрямая калориметрия), принципах составления пищевого рациона; вычисления должных
величин основного обмена.
Мотивация темы: «Все, что природа накопить сумела, незримо входит и в природу тела». Авиценна.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Обмен веществ - характерный признак жизни. Он обеспечивает непрерывное обра-
зование, обновление, разрушение клеточных структур, синтез и разрушение различ-
ных химических соединений. Включает в себя два процесса.
1.1. Анаболизм (ассимиляция) - усвоение организмом питательных веществ, в результате
которого они становятся частью биологических структур или откладываются в виде
депо (сопровождается ростом и увеличением массы тела).
1.2. Катаболизм (диссимиляция) - образование в организме из сложных веществ более
простых веществ, с использованием их преимущественно для энергетических целей
(сопровождается снижением массы тела).
2. Обмен белков (составляют 15 - 20 % массы тела).
2.1. Функции белков: 1) пластическая - белкн мембран, коллаген, эластнн, кератин
и др.; 2) энергетическая - на долю белков приходится 12 - 18 % энергообеспе-
чения (окисление 1 г белков освобождает 4 ккал); 3) каталитическая — ферменты по
химической природе являются белками; 4) транспортная - перенос с помощью
белков билирубина, лнпндов, кислорода, железа н др.; 5) гормональная - белко-
во-пептидные гормоны составляют - 80 % всех гормонов; 6) сократительная -
актин и мнознн; 7) защитная - иммуноглобулины, интерферон, фибриноген,
плазмин; 8) регуляция работы генов - факторы транскрипции.
2.2. Биологическая ценность пищевых белков.
❖ Полноценные белки содержат весь необходимый набор незаменимых ами-
нокислот в соотношениях, обеспечивающих нормальные процессы синтеза.
К ним относятся преимущественно белки животного происхождения - мя-
338
са, яиц, молока, рыбы, которые должны составлять не менее 30 % суточно-
го рациона. (Наиболее полноценные, эталонные белки - белки яиц и молока.)
❖ Неполноценные белки не содержат полного набора незаменимых аминокис-
лот (валин, изолейцин, лейцин, лизин, метионин, триптофан, треонин, фенилаланин).
Например, белки пшеницы (глнаднн), ячменя (гордеин) и кукурузы (зеин)
содержат мало лизина, а желатина - мало цистеина н триптофана.
♦ ♦♦ Два или несколько неполноценных белков, в которых отсутствуют разные незаменимые
аминокислоты, могут суммарно обеспечить потребность организма во асех незаменимых
аминокислотах.
2.3. Азотистый баланс - соотношение количества азота, поступившего с пищей
(белки) и выделенного из организма (мочевина, мочевая кислота, креатинин и др.). Так
как 1 г азота содержится в 6,25 г белка, то, зная количество выделенного азота с
мочой, можно определить количество усвоенного белка.
❖ Азотистое равновесие (нейтральный баланс азота): у взрослого человека
при адекватном питании количество введенного азота с пищей равно выве-
денному; прн увеличении потребления белка устанавливается на новом,
более высоком уровне.
❖ Положительный азотистый баланс: синтез белка преобладает над распа-
дом (прн увеличении массы тела, во время беременности, в период роста
организма, прн усиленных тренировках в связи с ростом мышечной массы).
❖ Отрицательный азотистый баланс: количество выведенного азота боль-
ше, чем поступающего с пищей (белковое голодание, питание неполноцен-
ными белками, различные заболевания).
2.4. Промежуточный обмен белков.
❖ Синтез белков.
• Аминокислоты, всосавшиеся из кишечника в кровь, используются в раз-
ных органах на пластические и энергетические нужды, не депонируют-
ся. Заменимые аминокислоты образуются в реакциях трансаминирова-
ния в различных органах.
• Центральным органом синтеза белков, используемых в других органах и
тканях, является печень. Высокий уровень синтеза белка имеется в лим-
фоидной ткани в связи с образованием иммуноглобулинов.
❖ Распад белков.
• В сутки у человека распадается - 400 г белков (средний период полу-
распада белков организма равен 80 дней). При этом 2/3 образующихся
аминокислот используются для синтеза нового белка, а 1/3 безвозвратно
теряется в окислительном метаболизме и должна поступать с пищей.
• Наиболее медленно распадаются белки мышц (Тщ - 180 суток), наибо-
лее быстро - белково-пептидные гормоны (Тщ несколько минут).
❖ Потребность в белках.
• Коэффициент изнашивания Рубнера - наименьшие потерн белка для ор-
ганизма в состоянии покоя - 0,33 г на 1 кг массы тела (т.е. - 23 г бел-
ка/сутки).
• Физиологический минимум белка превышает коэффициент изнашива-
ния и равен 30 - 45 г белка в сутки.
339
• Физиологический оптимум белка равен 80 - 100 г белка в сутки (1 г/кг
массы тела).
2.5. Нейрогуморальная регуляция обмена белков.
•> Синтез белка стимулируют СТГ, тироксин и трийодтиронин, глюкокорти-
коиды в печени.
❖ Распад белка увеличивают глюкокортикоиды (в мышцах и лимфоидной ткани) и
глюкагон.
3. Обмен липидов (составляют 10 - 20 % массы тела).
3.1. Функции липидов: 1) пластическая - важный компонент клеточных мембран;
2) энергетическая - при окислении липиды дают максимальное образование
энергии (1 г жиров - 9 ккал); 3) механическая - фиксация положения органов,
уменьшение травмирования органов; 4) терморегуляторная - теплоизолирую-
щие свойства подкожной клетчатки, теплопродукция в бурой жировой ткани; 5)
используются для синтеза биологически активных веществ (простагландинов, про-
стациклинов, тромбоксанов, лейкотриенов, стероидных гормонов).
3.2. Промежуточный обмен липидов. (Клетки тканей человека в связи с функцией тепло-
кровна и интенсивной локомоцией имеют высокую потребность в окислении ненасыщенных и
насыщенных жирных кислот. Особенно важны для энергетики насыщенные жирные кислоты с
С16.1а, которые легко вступают в митохондриях в р-окисление).
❖ Синтез липидов.
• В процессе всасывания в кишечнике происходит ресинтез липидов, ко-
торые преимущественно в составе хиломикронов и ЛПВП поступают в
лимфу и далее в кровоток (см. рис. 59). Липиды ЛПВП (преимущественно
ненасыщеныые жирные кислоты) используются в тканях, а липиды хиломик-
ронов (преимущественно триглицериды) поступают в печень, имеющую
рецепторы к белкам хиломикрон. Печень оптимизирует жирные кисло-
ты, поступающие из хиломикрон, превращая их преимущественно в на-
сыщенные жирные кислоты с Cie-ia, которые легко вступают в митохон-
дриях в р-окисление.
• Печень является ключевым органом в синтезе липидов (триглицеридов,
фосфолипидов, жирных н желчных кислот), которые в составе Л1ЮНП посту-
пают в кровь и вместе с ЛПВП частично превращаются в ЛППП и
ЛПНП (желчные кислоты поступают в желчь). ЛПВП н ЛПНП используются в
различных органах с помощью рецепторзависимого эндоцитоза.
• В жировой ткани происходит синтез жира из жирных кислот и глице-
рол-3-фосфата.
• Во многих тканях образуются биологически активные метаболиты жир-
ных кислот: простагландины, тромбоксаны, простациклин, лейкотриены.
• В надпочечниках н половых железах синтезируются стероидные гормо-
ны.
❖ Распад липидов.
• В крови распад липидов, находящихся в хиломикронах и ЛПОНП (в ос-
новном триглицеридов с образованием жирных кислот), происходит ПОД действи-
ем липопротеиновой липазы эндотелием сосудов (активируется гепари-
340
ном). Ненасыщенные жирные кислоты соединяются с альбуминами и
Л ЛОНП - ЛИПОПРОТЕИНЫ очой» НИЗКОЙ ПЛОТНОСТИ, ЛППЛ • липопротеины промежуточной ПЛОТНОСТИ. ЛПНП •
ЛИПОПРОТЕИНЫ НИЗКОЙ ПЛОТНОСТИ (АТЕРОСЕННЫ), ЛПВЛ • ЛИПОПРОТЕИНЫ ВЫСОКОЙ ПЛОТНОСТИ (МПИАТЕРОгаННЫ)
Al. АП. AIV.B48, В100, С1, СП.СШ.Е - АПОБЕЛКИ ЛИПОПРОТЕИНОВЫХ КОШЦНМСОв, ОБПАДАЮЩЖ ТРАНСПОРТ
НЫМИ ФУНКЦИЯМИ, ДЛЯ НИХ ИМЕЮТСЯ СООТВЕСТВУКХЦИЕ РЕЦЕПТОРЫ НА МЕМБРАНЕ КЛЕТОК
-РЕЦЕПТОР К АПОБЕЛКАМ
Рис. 59. Схема обмена липидов в организме
• В жировой ткани распад триглицеридов происходит с образованием
жирных кислот и глицерола, большая часть которых поступает в кровь.
• В бурой жировой ткани распад жира сопровождается окислением жир-
ных кислот в митохондриях до СОз и Н2О. При этом имеется разобще-
ние окисления и фосфорилирования и образуется большое количество
тепла.
• Жирные кислоты, поступившие в кровь, окисляются в разных тканях с
образованием большого количества энергии, половина которой аккуму-
лируется в АТФ.
❖ Потребность в липидах.
• Суточная потребность в липидах составляет 50 -100 г.
• В организме человека не синтезируются полиненасыщенные жирные
кислоты (иапример, линолевая, а-линолеиовая и арахидоновая). Они
имеются в больших количествах в растительных маслах. Поэтому 1/3
поступающих с пищей липидов должна быть за счет растительных жи-
ров.
3.3. Регуляция обмена липидов.
Нервная регуляция.
• Симпатические влияния усиливают распад (норадреналин + р-адренорецеп-
торы, )цАМФ) и тормозят синтез (а-адреиорецепторы, 1цАМФ) триглицеридов
в жировой ткани.
341
• Парасимпатические влияния способствуют отложению жира (ацетилхолин
+ М-холинорецепторы, (цАМФ).
❖ Гуморальная регуляция.
• Активируют мобилизацию жира и его окисление адреналин (₽-
адренорецепторы, (цАМФ) и гипогликемия (через торможение секреции инсу-
лина); другие гормоны (СТГ, ТТГ, АКТГ, глюкагон, вазопрессин) у человека
оказывают очень слабое действие.
• Тормозят мобилизацию жира и активируют его синтез инсулин (осо-
бенно в жировой тканн), глюкокортикоиды (депонирование жира в об-
ласти лица и шеи), адреналин (аг-адренорецепторы, (цАМФ)
4. Обмен углеводов (составляют 0,6 % массы тела, суточная потребность 300-350 г).
4.1. Функции углеводов: 1) энергетическая - основной источник энергии в орга-
низме, углеводы обеспечивают не менее 50 % суточного энергообеспечения
(окисление 1 г углеводов освобождает 4 ккал); 2) образуют депо (в виде гликогена)
легко мобилизируемого энергетического материала; 3) пластическая - продук-
ты промежуточного обмена углеводов используются для синтеза аминокислот,
липидов, полисахаридов; 4) углеводы, входящие в гликопротеиды, определяют
их видовую и тканевую специфичность.
4.2. Промежуточный обмен углеводов.
❖ Глюкоза поступает в кровь нз кишечника в процессе пищеварения и нз пе-
чени в результате гликогенолиза и глюконеогенеза. Использование глюко-
зы происходит во всех органах: например, в печени (синтез гликогена и
липидов), в мышцах (окисление глюкозы и синтез гликогена), ткани мозга
(окисление), жировой ткани (окисление и синтез липидов).
❖ Глюкоза поступает в клетки различных тканей с помощью белков-
транспортёров глюкозы — ГЛЮТ|-5 (в жировую и мышечную под контролем инсу-
лина) и после перехода в глюкозо-б-фосфат используется по одному из 4
главных путей.
• Количественно основной путь - аэробное окисление до СОг н Н2О. При
этом нз 1 моля глюкозы реально образуется 25 молей АТФ (теоретически
38 молей АТФ), в которых аккумулируется - 44 % всей энергии распада
глюкозы (последняя равна 2880 кДж/моль). Основная функция этого пути -
освобождение и запасание энергии.
• Синтез гликогена, который особенно характерен для печени, мышц н
почек.
• Пентозофосфатный путь, в котором образуются: 1) НАДФ-Нг, служащий
источником водорода для восстановительных синтезов (например, синтез
липидов в жировой ткани и печени); 2) рнбозо-5-фосфат, который использует-
ся для синтеза ДНК, РНК, АТФ и др.
• Глюкозо-б-фосфат превращается под действием его фосфатазы в глюко-
зу, которая выделяется в кровь (характерно для печени).
❖ Прн интенсивном использовании глюкозы или (и) недостатке углеводов в
питании глюкоза синтезируется из продуктов метаболизма (пирувата, оксало-
342
ацетата, фумарата, глицерола), которые образуются при окислении аминокислот
и жирных кислот.
4.3. Нейрогуморальная регуляция обмена углеводов.
❖ Активирует использование глюкозы (снижает ее уровень в крови) иисулии,
он способствует использованию глюкозы в тканях (особенно мышечной и жиро-
вой) и депонированию ее в виде гликогена (особенно в печени и мышцах) и син-
тезу аминокислот (белков). Парасимпатические влияния (М-холинорецеп-
торы) и симпатические влияния (0-адренорецепторы) стимулируют секре-
цию инсулина, симпатические влияния через а-адренорецеп-торы тормозят
секрецию инсулина.
❖ Стимулируют продукцию глюкозы (повышают ее уровень в крови) симпа-
тические влияния и ряд контринсулярных гормонов: глюкагон в печени,
адреналин через 02-адренорецепторы в скелетных миоцитах и сердце в ре-
зультате гликогенолиза, глюкокортикоиды (в результате гликогенолиза), глюко-
кортикоиды, глюкагон (в результате глюконеогенеза), СТГ (активирует секре-
цию глюкагона), тироксин (стимулирует глюкозо-6-фосфатазу).
5. Обмен воды и минеральных веществ.
5.1. Вода (60 % от массы тела у взрослого, 40 - 50 л).
❖ Функции воды: 1) универсальный растворитель, обеспечивающий протека-
ние всех метаболических реакций организма; 2) конвекционный транспорт
веществ (гемодинамика); 3) участие в терморегуляции (транспорт тепла,
испарение как механизм теплоотдачи); 4) эффект разведения эндогенных и
экзогенных токсинов, образующихся кислот и оснований.
❖ Водные отсеки организма: внутриклеточная вода (71 % всей воды, - 33 л),
внеклеточная вода (~ 21 %, - 10 л), внутрисосудистая вода (~ 8 %, - 3,5 л).
❖ Баланс воды: потребление (экзогенная вода 2,1 л/сут и эндогенная - 0,4 л/
сут); выделение воды (с кожи и поверхности альвеол 0,9 л/сут, через почки
- 1,5 л/сут, с калом - 0,1 л/сут). Минимальная суточная потребность в воде
- 1,7 л.
❖ Регуляция обмена воды тесно связана с обменом натрия и осуществляется в
результате волюмрегулирующего и осморегулирующего рефлексов с уча-
стием гормонов - АДГ, альдостерона, натрийуретического пептида сердца
(подробнее см. тему 24; задание 1: 13).
5.2. Макроэлементы (натрий, кальций, калий, хлор, кислород, углерод, водород, азот, фосфор,
магний). Для их всасывания, транспорта в крови н хранения обычно требуются специфические
белки-носители. Экскреция происходит с мочой, потом и калом.
<• Калий.
• Функции К*: 1) главный внутриклеточный катион, определяющий осмо-
тическое давление в клетках; 2) его диффузия из клетки является глав-
ным механизмом формирования мембранного потенциала и фазы репо-
ляризации в потенциалах возбудимых клеток; 3) влияет на сосудистый
тонус.
• Содержание К* (в ммолях) во всем организме 3700 - 4600 (- 175 г), в ске-
летных мышцах - 2800, коже - 360 - 500, костях - 218, печени - 135 -
200, в внеклеточной жидкости - 60. Основным депо К+ являются скелет-
ные мышцы.
343
• Потребность К* в сутки 90-100 ммоль (~ 4,5 г), всасывается в кишеч-
нике - 90 % К+ пищи.
• Концентрация К* в межклеточной жидкости в основном определяется
двумя факторами: 1) pH внеклеточной жидкости: уменьшение (увеличе-
ние) pH на 0,1 приводит к увеличению (уменьшению) К* в крови на 0,6
ммоль/л; 2) реабсорбцией и секрецией К* в почечных канальцах.
• Все пищеварительные соки, молоко н пот содержат К* в бблыпей кон-
центрации, чем плазма крови.
• Выделение К* из организма осуществляется главным образом почками с
мочой (80 - 90 ммоль/сут), калом (~ 10 ммоль/сут).
• Для баланса К* в организме необходимо, чтобы его количество, выде-
ленное с мочой, было равно количеству К+, всосавшемуся в кишечнике.
• Основным регулятором обмена К* в организме является альдостерон.
❖ Натрий.
• Функции Na*: 1) главный катион внеклеточных жидкостей, это опреде-
ляет его роль в формировании осмотического равновесия этих жидко-
стей и обмена воды между клеточным и внеклеточным пространствами;
2) участвует в формировании фазы деполяризации ПД и других биопо-
тенциалов; 3) участвует в регуляции сосудистого тонуса, повышает его.
• Баланс Na*. Организм человека содержит - 4200 ммоль (~ 95 г) Na+: во
внеклеточных жидкостях - 2100 (50 %), в костях находится - 1800 (42
%), внутри клеток 260 (8 %) Na+.
• Поступает Na* в организм в количестве 3 - 6 г в сутки; поступление
Na+ меньше 1 г в сутки приводит к его отрицательному балансу с разви-
тием гипонатриемии.
• Наибольшая часть поступившего в организм Na+ находится в коже и
подкожной клетчатке, мышцах, хрящах; эти ткани рассматриваются как
депо Na+.
• Почки являются главным органом, определяющим натриевое равновесие
в организме, прн этом главным регулятором является альдостерон.
• ' Na+ выделяется нз организма с мочой (~ 90 %), пбтом н калом.
❖ Кальций.
• Функции Ct/*: 1) пластическая - входит в состав костей (10 % массы орга-
на), зубов (эмаль - 34 %, дентин - 24 %); 2) влияет на возбудимость нервной и
мышечной тканей; 3) необходим для свертывания крови; 4) регулирует
проницаемость клеточной мембраны; 5) запускает мышечное сокраще-
ние, секрецию ферментов, гормонов и медиаторов; 6) является посред-
ником при реализации эффектов многих гормонов; 7) влияет на репли-
кацию ДНК и клеточное деление.
• Содержание С</* в организме значительно превышает количество всех
других минеральных веществ и равно - 25 молей (1,1 кг), 99 % этого ко-
личества приходится на долю костей н зубов. В крови содержится - 2,5
344
ммоль/л Са2+, половина которого находится в ионизированном виде,
другая половина - в комплексе с альбуминами.
• Потребность в Сс?+ взрослого человека равна 0,5 - 0,8 г/сут (- 8 мг/кг
массы), у детей, беременных и кормящих матерей она в 2 - 4 раза боль-
ше. Лучшими источниками Са2+ являются молоко, сыры, творог, фасоль,
зеленый лук. Ухудшает всасывание Са2+ из кишечника большое содер-
жание в пищи жиров, фосфатов и щавелевой кислоты, улучшают всасы-
вание белки пищи, лактоза.
• Регулируют уровень Сс?+ в крови паратгормон и 1,25-(OH)2-D3 (они по-
вышают уровень Са2+ в результате мобилизации его из костей, увеличе-
ния реабсорбции в почках и всасывания в кишечнике), а также кальци-
тонин (снижает уровень Са2+ в результате перехода его в кости, выделе-
ния с мочой и торможения всасывания в кишечнике).
• Выделение Сс?+ из организма происходит как с мочой (- 30 %, 0,25 г/сут),
так и через кишечник.
❖ Хлор.
• Функции СГ: 1) участие в регуляции водно-солевого обмена и осмотиче-
ского давления во внеклеточной и клеточной жидкостях; 2) участие в
формировании биопотенциалов клеток; 3) входит в состав соляной ки-
слоты желудочного сока.
• Содержание СГ в организме около 3 молей (ПО г), суточная потреб-
ность - 5 г, он поступает в организм в основном за счет NaCl пищи, наи-
большее количество СГ (до 60 % всего СГ) находится в коже и подкожной
клетчатке.
• Выделение СГ из организма происходит главным образом с мочой (около
140 ммоль/сут), а также с потом.
❖ Магний.
• Содержание в организме около 1 моля (-140 г). Распределение по орга-
нам (%): в костях - 53, мышцах - 27, в других органах и тканях - 18, во
внеклеточной жидкости - 1 (в сыворотке крови 0,75 - 1,0 ммоль/л, 1/3
связана с белками).
• Функции магния: является кофактором многих ферментов, участвует в
обмене энергии, белков, углеводов и жиров, уменьшает возбудимость
клеток и замедляет синаптическую передачу, расслабляет гладкую мус-
кулатуру, снижает АД; является физиологическим антагонистом каль-
ция.
• В кишечнике всасывается (преимущественно пассивно) - 37% магния
пищи. Суточная потребность - 15 ммоль.
53. Микроэлементы (железо, йод, медь, цинк, кобальт, молибден, селен, хром, фтор, кремний и
др.). Их функции связаны с тем, что они входят в состав многих ферментов: (например, в окси-
дазах - медь, молибден, в глутатионпероксидазе - селей, в лактатдегидрогеиазе и карбоангид-
разе - цинк); гормонов (например, в Т3, Т<- йод), гемоглобина и цитохромов - железо (см. тему
13; задание 1:1.5); витаминов (например, B,2 - кобальт).
6. Обмен витаминов.
345
6.1. Жирорастворимые витамины. Для их всасывания необходимы пищевые жиры и желчь,
в транспорте участвуют липопротеины крови или специфические белки. Эти витамины депони-
руются в печени и частично в жировой ткани, выделяются через желчь и при кишечно-
печеночной рециркуляции частично экскретируются с калом, некоторые их метаболиты выде-
ляются с мочой. Первичные механизмы действия жирорастворимых витаминов осуществляются
на уровне генома, в хроматине которого имеются их белковые рецепторы. Через них витамины
действую иа транскрипцию, доказано также их влияние на рецепторы проникающих в клетку
гормонов (например, тироксина).
• > А (ретинол, образуется в организме при расщеплении p-каротина) влияет на функ-
цию зрения, обеспечивает адаптацию глаза к свету (входит в состав родопсина),
необходим для роста и дифференцировки эпителиальной, иервиой и кост-
ной ткани (при его недостатке возникает ночная слепота, ксерофтальмия, нарушение
роста).
❖ D3 (кальциферол) - основной гормональный регулятор обмена Са2+ и фос-
фора. Обеспечивает нормальное развитие скелета и хорошее состояние им-
мунной системы. Его образование прямо пропорционально уровню УФ-
облучеиию эпидермиса кожи, что, в свою очередь, регулируется содержа-
нию в ней меланина (при недостатке D3 у детей развивается рахит, у взрослых - ос-
теомаляция).
❖ Е (токоферол) - главный антиоксидант организма, влияет иа функцию раз-
множения, повышает продукцию гонадотропинов гипофиза и половых
гормонов (при его недостатке у детей развивается анемия).
❖ К (филлохиион) активирует в печени синтез факторов свертывания - про-
тромбина (II), а также VII, IX и X (при его недостатке возникает кровото-
чивость).
6.2. Водорастворимые витамины. После всасывания в кишечнике запасаются в связанном с
ферментами или транспортными белками виде (депо витамина В12 печень) и выделяются с мо-
чой, когда их уровень в плазме превышает почечный порог.
❖ Bj (тиамии) - кофермент пируват- и а-кетоглутаратдегидрогеназ, участвует
во всех видах обмена, нормализует работу сердца, повышает секрецию и
моторику желудка, обспечивает нормальную функцию аксона, в частности
аксонный транспорт (при его недостатке возникают полиневриты).
❖ В2 (рибофлавин) - кофермент дегидрогеназ (например, ацетил-СоА-
дегидрогеназы), влияет иа рост и развитие плода и ребенка (при его недос-
татке возникает хейлит - воспаление красной каймы губ).
❖ Вз (пантотеновая кислота) - кофермент реакции переноса ацильной группы,
необходим для синтеза жирных кислот, стероидных гормонов, ацетилхоли-
на.
❖ В« (пиридоксин) - как кофермент участвует в реакциях переамииирования
и декарбоксилирования, обезвреживания амииов, образования фосфолипи-
дов (миелинизация нервных вожжой), аксоном транспорте, распаде гликогена,
является липотропным фактором.
❖ В$ (фолат) - коферментные формы витамина включают метильные группы
в реакции синтеза нуклеиновых кислот (пуринов и пиримидинов), поэтому ои
необходит для всех процесса, связанных с быстрой клеточной пролифера-
цией, например, для сперматогенеза, гемопоэза по иормобластическому
346
типу (при недостатке возникает мегалоОластическая анемия), изоыточное уч<-
облучение кожи разрушает фолат.
❖ Ви (цианкобаламин) участвует в образовании коферментных форм Вв (син-
тез нуклеиновых кислот и метионина), обеспечивает сгорание в цикле Кребса
продуктов окисления жирных кислот, регулирует гемопоэз по нормобла-
стическому типу (при недостатке возникают мегалобластическая анемия, фуникуляр-
ный миелоз).
❖ С (аскорбиновая кислота) участвует в окислительно-восстановительных
процессах, образовании коллагена, кортикостероидов, некоторых медиато-
ров, всасывании железа, является антиоксидантом (при его недостатке развива-
ется цинга).
❖ Н (биотин) - кофермент карбоксилаз, участвует в реакциях переноса СОг,
способствуя его включению в оксалоацетат цикла Кребса и жирные кисло-
ты; синтезируется кишечной микрофлорой (белок сырых яиц авидни блокирует
его всасывание).
❖ РР (никотиновая кислота) - кофермент дегидрогеназ, участвует в реакциях
клеточного окисления (переноса водорода) (при его недостатке возникает пелла-
гра).
7. Энергетический баланс организма (1 ккал - 4,19 кДж).
7.1. Энергетический баланс оранизма - это состояние равновесия между его энер-
гетическими затратами и энергетической ценностью потребляемой пищи. Энер-
гетические затраты осуществляются на основной обмен и более высокий уро-
вень физиологической активности организма (физическую и психоэмоциональную ак-
тивность, факультативный термогенез, рост, беременность, лактацию и др.)
7.2. Основной обмен (ОО) - это минимальный (совместимый с жизнью) уровень
' энергозатрат организма, который определяется при стандартных условиях: ут-
ром, в покое, лежа, натощак, при температуре комфорта (-20 °C). (При неполном
соблюдении этих условий говорят об энергозатратах покоя.)
❖ Физиологическое значение - поддержание минимально необходимого для
жизни клеток уровня окислительных процессов н деятельности постоянно
работающих органов и систем (сердце, почки, дыхательные мышцы, тонус
сосудов и др.).
❖ Величина основного обмена (ВОО) равна 1 ккал/кг массы тела в час, -7100
кДж/сутки или -1700 ккал/сутки (почти половина ВОО затрачивается иа функции
печенн и головного мозга).
❖ Факторы, определяющие ВОО.
• Размеры тела: ВОО прямо пропорциональная массе - 1 ккал/кг массы
тела в час и поверхности тела - 960 ккал/м2.
• Состав тела. В массе тела выделяют два компонента: нежировую (метабо-
лически активную) массу и жировую (метаболические пассивную) массу, поэто-
му увеличение жировых депо - снижает, а увеличение мышечной массы
- увеличивает ВОО.
• Пол. У женщин ВОО на 5-10 % ниже, чем у мужчин, т.к. масса депони-
рованного жира (-15%) у женщин выше, чем у мужчин (-12%).
347
• Возраст. BOO у детей относительно выше, чем у взрослых, до двухлет-
него возраста (особенно у новорожденных) и в периоде полового созре-
вания; в пожилом возрасте ВОО снижается в связи с увеличением жиро-
вой массы тела.
• Гормоны: тиреоидные гормоны и адреналин оказывают стимулирующее
влияние на ВОО (например, при тиреотоксикозе ВОО повышается до 75%, при ги-
потиреозе снижается на 30%).
• Климатические условия: у жителей тропиков ВОО равен - 1500
ккал/сутки, у жителей Арктики - 2800 ккал/сутки.
• Специфически динамическое действие пищи (максимально при приеме бел-
ковой пищи) повышает ВОО на 15-20%; обусловлено преимущественно
действием тиреоидных гормонов, секреция которых при приеме пищи
увеличивается в 2 раза.
7.3. Рабочий обмен. Энергетические затраты организма в наибольшей мере зависят
от интенсивности мышечной работы: учитывая особенности профессии, выде-
ляют по энергозатратам пять видов труда.
❖ Умственный труд сопровождается энергозатратами -1,5 ккал/кг массы/час
(2400 - 2700 ккал/сутки).
❖ Легкий физический труд -1,7 ккал/кг/час (2800 - 3000 ккал/сутки).
❖ Средний физический труд -1,9 ккал/кг/час (3100 - 3300 ккал/сутки).
❖ Тяжелый физический труд -2,2 ккал/кг/час (3400 - 3800 ккал/сутки).
❖ Очень тяжелый физический труд -2,5 ккал/кг/час (3900 - 4300 ккал/сутки).
7.4. Важнейшим показателем баланса энергии является масса тела.
❖ Баланс энергии у здорового человека должен обеспечивать нормальную ве-
личину массы тела в соответствии с его возрастом, ростом и функциональ-
ным состоянием (например, беременностью). Сдвиг баланса в сторону превы-
шения потребления энергии над ее расходом ведет к повышении массы те-
ла, сдвиг в противоположную сторону - к снижению массы тела.
❖ Массу тела у взрослых с 20 до 60 лет можно оценить, вычислив индекс
массы тела (ИМТ или индекс Кетле): ИМТ = масса тела (кг)/рост (м2). В
норме ИМТ равен 18,5 - 25 н прямо коррелирует с количеством жира в ор-
ганизме. Более высокие показатели ИМТ свидетельствуют об ожирении,
более низкие - об энергетической недостаточности, что свидетельствует о
риске для здоровья. (ИМТ не дает верную информация при сильно развитой мускула-
туре, беременности, развитии отеков.)
7.5. Методы определения энергетических затрат в организме.
❖ Прямая калориметрия (камерный биокалориметр) - непосредственно учитывается
тепло, выделенное организмом.
*!• Непрямая калориметрия - расчет теплопродукции по газообмену (количество по-
требленного Оз и выделенного СОг).
• Камерный (закрытый) метод Шатерникова.
• Некамерные (открытый, полуоткрытый, закрытый) методы.
• Полный газовый анализ:
Ф определить количество поглощенного Ог и выделенного СОг;
348
"Ф- вычислить дыхательный коэффициент (ДК) - отношение выделенного
СО2 к объему поглощенного О2 (при окислении углеводов -1,0; при окис-
лении жиров - 0,7; при окислении белков -0,8);
‘Ф’ определить по таблице калорический эквивалент кислорода (количество теп-
ла, освобождающееся после потребления организмом I л Oj) с учетом ДК и, умно-
жив на количество потребленного в сутки кислорода, вычислить энерге-
тические затраты в организме. (Потребление организмом 1 л О2 сопровождается
выделением 5.1 ккал тепла.)
• Неполный газовый анализ:
‘Ф’ определить количество поглощенного кислорода;
"Ф- использовать для расчета калорический эквивалент кислорода при усред-
ненном дыхательном коэффициенте - 0,85 - 0,90.
8. Питание - процесс поступления, переваривания, всасывания и усвоения в организме
пищевых веществ, необходимых для покрытия энергетических и пластических по-
требностей организма, образования его физиологически активных веществ.
8.1. Значение питания для жизнедеятельности организма.
• > Пластическое обеспечение функций.
❖ Энергетическое обеспечение функций.
❖ Регуляция функций организма.
8.2. Питательные вещества - те органические и неорганические вещества пищи,
которые усваиваются организмом в процессе обмена веществ.
❖ Белки, жиры, углеводы и их производные.
❖ Вода, соли, микроэлементы.
❖ Витамины.
• > Пищевые волокна (частично усваиваются благодаря микрофлоре кишечника).
8.3. Взаимозаменяемость питательных веществ.
❖ Правило изодинамии Рубнера: питательные вещества в энергетическом ас-
пекте могут заменять друг друга в соответствии с их калорическими коэф-
фициентами.
❖ Критическая оценка этого правила: не учитываются пластические потреб-
ности организма.
8.4. Нормы питания.
❖ Суточная потребность организма в белках, углеводах, жирах, минеральных
солях, витаминах.
❖ Потребность организма в питательных веществах зависит от географиче-
ских и климатических условий, от возраста, профессии, состояния орга-
низма (усиленный рост, беременность, период лактации).
8.5. Основные принципы рационального питания.
❖ Энергетическая ценность питания должна соответствовать энергетическим
затратам организма.
❖ Состав питательных веществ должен соответствовать физиологическим по-
требностям в них организма.
❖ Питание должно быть разнообразно, содержать свежие натуральные про-
дукты растительного и животного происхождения.
❖ Суточное количество пищи должно поступать в организм порционно (регу-
лярность, кратность и чередование приемов пищи).
349
❖ Концепция здорового питания учитывает роль различных питательных ве-
ществ (их количество и соотношение) как факторов профилактики заболеваний.
9. Теории и концепции питания (достоинства и недостатки).
9.1. Теория сбалансированного нитания.
❖ Приток питательных веществ должен точно соответствовать их расходу.
❖ Научно обоснованное соотношение питательных веществ в зависимости от
условий деятельности организма.
❖ Пища состоит из нескольких компонентов, различных по физиологическо-
му значению (пищевых, балластных н токсических веществ).
❖ Концепция об идеальной (рафинированной) пищи, мономерных диетах и па-
рентеральном питании могут быть использованы в определенных условиях
(например, при лечении больных, в космонавтике), но не могут быть использованы
для регулярного питания здоровых н больных людей.
9.2. Теория адекватного питания (А.М. Уголев, 1985).
❖ Для нормального питания организма должен быть не только поток первич-
ных питательных веществ, но и потоки вторичных нутриентов, экзогенных
и эндогенных регуляторных веществ, токсинов и балластных веществ.
❖ В питании организма важное значение имеет эндоэкология, создающаяся
взаимодействием организма н его микрофлоры.
❖ Баланс питательных веществ достигается за счет освобождения их нз
структур пнщи в результате полостного, мембранного н внутриклеточного
пищеварения, а также синтеза новых веществ, в том числе незаменимых.
❖ Необходимым компонентом пнщи являются не только питательные, но н
балластные вещества пнщн - клетчатка, гемицеллюлоза, пектин, легнин
(суточная потребность 20 - 30 г).
9.3. Альтернативные концепции питания (вегетарианство, раздельное питание, сыроеде-
ние, их критическая оценка).
10. Профильный материал для студентов лечебного факультета.
10.1. Физиологические основы диетотерапии.
Диета - рацион питания, который предусматривает определенное количественное
и качественное соотношение пищевых веществ, способы кулинарной обработки,
интервалы в приеме пищи. Она должна:
• обеспечивать физиологические потребности больного человека в пластиче-
ских, энергетических и регуляторных веществах;
• индивидуализировать питание, учитывая особенности усвоения пищи у здоро-
вого и больного человека;
• обеспечивать пищеварение при нарушении образования пищеварительных
ферментов;
• учитывать взаимодействие пищевых веществ в желудочно-кишечном тракте;
• стимулировать восстановительные процессы в органах и тканях;
• компенсировать пищевые вещества, теряемые организмом больного;
• направленно изменять режим питания;
• учитывать местное и общее воздействие пищи на организм;
350
• использовать в питании методы щажения, тренировки, разгрузки и контраст-
ных дней.
10.2. Энергетические затраты организма в старческом возрасте.
❖ Уменьшение интенсивности основного обмена после 40 лет в связи со снижением
функции щитовидной, поджелудочной, половых желез.
❖ Уменьшение рабочей прибавки в связи со снижением двигательной активности,
атрофией мышц.
❖ Снижение величины максимального потребления Ог.
❖ Сохранение возбудимости пищевого цента при снижении энергетических затрат
(тенденция к дисбалансу между приходом и использованием питательных ве-
ществ).
11. Профильные материалы для студентов педиатрического факультета.
11.1. Особенности обмена веществ и энергии у детей. Виды синтеза.
❖ Преобладание анаболических процессов над катаболическими (накопление массы
тела, рост и дифференцировка тканей).
♦ ♦♦ Высокая интенсивность обменных процессов.
❖ Энергетическая стоимость построения 1 г тканей: до 4 мес. - 4 - 6 ккал, у более
старших - 6,3 ккал.
❖ Увеличение массы тела (наибольшая скорость в первые 3 месяца - 30 г/сутки, до 1
года ~ 10 г/сутки, затем снижение).
❖ Энергетическая стоимость роста: наибольшая в первые 3 мес. ~ 140 ккал/сутки, до
года ~ 60 ккал/сутки, низкая от 3 лет до периода полового созревания - 30
ккал/сутки, в период полового созревания увеличивается до 110 ккая/сутки).
❖ Виды синтеза: синтез роста - увеличение массы организма в период усиленной
пролиферации, роста и дифференцировки клеток; синтез самообновления - синтез
структур, разрушающихся в процессе диссимиляции; функциональный синтез -
синтез ферментов, гормонов, пептидов, образование иммунных тел; синтез реге-
нерации - синтез в тканях после травмы или истощения. Наибольшие изменения
претерпевает синтез роста, меньшие - синтез самообновления и функциональный.
11.2. Обмен белков. Потребность в белках и незаменимых аминокислотах
❖ Положительный азотистый баланс.
❖ Потребность в белках: до 3 мес. - 2,5 г/кг в сутки, с 4 мес. - увеличивается, после
3 лет уменьшается.
❖ Высока потребность в незаменимых аминокислотах (особенно в лейцине, фенила-
ланине, лизине, валине, треонине).
❖ Снижение содержания общего белка и его фракций в плазме крови.
❖ Обмен белков особенно важен для создания интеллектуального индекса, синтеза
антител, адаптации к стрессорам.
11.3. Обмен жиров. Потребность в жирах Особенности жировой ткани новорожден-
ного.
* ♦* Особенностью жировой ткани новорожденного является наличие бурого жира (8
% от массы тела), являющегося важнейшим местом теплопродукции.
❖ Состав жира у новорожденного отличается от материнского, так как плод образу-
ет свой жир из неэстерифицированных жирных кислот н глюкозы, поступающих
через плаценту.
Доля жира в теле новорожденного зависит от массы тела при рождении: при 1500
г - 3 %; при 2500 г - 8 %; при 3500 г - 16 %.
351
❖ Потребность в жирах: 5,5 - 6,5 г/кг в 1-й год жизни, 3,5 - 4,0 г/кг - в 1 - 6 лет, 1,6
- 1,8 г/кг в подростковом возрасте.
Жиры - основной источник энергии новорожденных, они имеют высокую концен-
трацию жирных кислот в крови.
♦ ♦♦ Динамика содержания жира в организме: максимально в 6 месяцев (26 % от массы
тела), минимально в возрасте 6-9 лет, у девочек увеличивается в период полово-
го созревания.
11.4. Обмен углеводов. Потребность в углеводах. Усвояемость углеводов.
❖ Потребность в углеводах - 13 г/кг в 1-й год, 16 г/кг - в 1 - 3 года, далее снижение
до 6,5 г/кг, у подростков 7,3 г/кг.
❖ Высокая интенсивность образования глюкозы из гликогена (увеличена концен-
трации глюкагона в плазме крови).
♦ ♦♦ При использовании глюкозы в организме увеличена доля анаэробного гликолиза у
новорожденных и детей 1 года жизни.
♦ 5» Концентрация глюкозы крови (~ 4,1 ммоль/л при рождении, снижение до 2,9
ммоль/л - физиологическая гипогликемия, повышение к концу 1-й недели до 4,1
ммоль/л).
♦ ♦♦ Высокая толерантность к углеводной нагрузке.
11.5. Особенности водно-солевого обмена у детей.
❖ Относительно большое содержание воды у новорожденного.
Увеличение объема внутриклеточной воды, снижение объема межклеточной воды
с возрастом.
❖ Интенсивное выведение воды через кожу и легкие.
❖ Низкое чувство жажды (склонность к обезвоживанию).
Высокая потребность в Na+, К+, Са2+, фосфатах.
Невысокая эффективность осмо- и волюмрегулирующих рефлексов из-за низкой
чувствительности центральных и периферических осморецепторов.
11.6. Особенности энергетического обмена у детей.
♦♦♦ Основной обмен.
• Относительная величина основного обмена у новорожденного в 2 раза выше,
чем у взрослого; в период полового созревания в 1,5 раза выше; у мальчиков на
10 % больше, чем у девочек.
• Большое значение имеют эндогенные источники энергии: гликогеи - при рож-
дении, использование жиров через несколько часов после рождения (ДК падает
до 0,73), через сутки увеличивается до 0,9 (вновь используются углеводы).
❖ Рабочий обмен.
• Основной обмен составляет - 60 %; рабочая добавка ~ 40 %.
• Затраты энергии на рост и увеличение массы депонированных веществ - 15 %
(у взрослого 0 %), на работу мышц - 15 % (у взрослого ~ 25 %).
• Специфически динамическое действие пищи - 0 - 5 % (у взрослого 10 - 20
%), эиергопотери с экскрементами - 5-10%(-5%у взрослого).
• Снижен диапазон изменений энергетического обмена при физических нагруз-
ках.
• Энергозатраты иа достижение полезного результата у ребенка выше, чем у
взрослого, в связи с несовершенством регуляции.
352
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной н дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 н 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной у чебно-методи пески й комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11-16].
ЗЛ. Дополнительная литература (см. с. 514 - 522): [13], [30], [83], [107], [108], [170],
[179], [180], [196], [222], [225], [226], [228], [245], [248], [252-254]. .
3.3. Дистанционные средства офктетшя. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/-nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брни. - Владикавказ: СОГМА. -1999; тесты ос-
таточного уровне знаний по данной теме.
3.4. Учебная комната Для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
353
Тема 25. Физиология выделения. Мочеобразование и мочевыделение.
Выделительная функция кожи, легких, пищеварительного
тракта
Цель: а) знать ’функции органов выделения (почек, кожи, легких, пищеварительного тракта), их уча-
стие в поддержании гомеостаза; нефрон как морфофункцноиальную единицу почкн; кровооб-
ращение в почке и особенности его регуляции; процессы мочеобразовання (фильтрация, реаб-
сорбция и секреция, осмокоицентрированне и разведение), их механизмы и нейрогуморальную
регуляцию; состав первичной и конечной мочи; функции чашечно-лохаиочиой системы, моче-
точников, мочевого пузыря, мочеиспускательного канала; их рефлекторную регуляцию; роль
почек в поддержании азотистого баланса, осмотического давления, pH крови, объема крови;
невыделительные функции почек; адаптивные изменения функций почек при различных усло-
виях внешней среды; возрастные изменения мочеобразовання н мочеотделения;
б) ознакомиться с практическими навыками иа уровне знаний: методики количественной
оценки механизмов мочеобразовання (клиренсов различных веществ).
Мотивация темы: главным органом выделения являются почкн. Основная их функция - экскреторно-
гомеостатическая, выражающаяся в регуляции азотистого, водно-солевого, кислотно-
основного гомеостаза. Кроме того, деятельность почек имеет большое значение для регуляции
сосудистого тонуса, эритропоэза и метаболизма в организме. Выключение функции почек
(острая и хроническая почечная недостаточность) приводит к смертельному исходу в резуль-
тате нарастания содержания потенциально токсичных азотистых соединений, Na*, К*, сульфа-
тов и фосфатов, увеличения объема внеклеточной жидкости, ацндоза и др. нарушений.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логическую
структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Физиологическая система выделения - совокупность органов, взаимосвязанная
деятельность которых обеспечивает постоянство ионного состава, осмоляльности,
pH, объема жидкости сосудистого, интерстициального и внутриклеточного отсеков,
концентрации конечных продуктов обмена во внутренней среде организма.
1.1. Выделительные функции системы дыхания.
❖ С выдыхаемым воздухом из организма выводятся различные вещества:
СОг, аммиак, ацетон, этанол и др. и испаряется около 400 мл воды в сутки.
❖ В составе трахеобронхиального секрета из организма выводятся продукты
деградации сурфактанта, мочевина, 1g А и др.
1.2. Выделительная функция системы пищеварения.
❖ Слюнные железы: выделение солей тяжелых металлов, лекарств, родани-
стого калия и др.
♦ ♦♦ Печень: экскреция билирубина и продуктов его превращения в кишечни-
ке, холестерина, желчных кислот, продуктов распада гормонов, лекарств,
ядохимикатов и др.
❖ Желудок: в составе желудочного сока выводятся конечные продукты ме-
таболизма (мочевина, мочевая кислота), лекарственные и ядовитые вещества
(ртуть, йод, салициловая кислота, хинин).
354
Кишечник: экскреция из крови солей тяжелых металлов, Mg", Са" (50 %
выделяемого организмом), воды; выделение продуктов распада пищевых ве-
ществ, которые ие подверглись всасыванию в кровь, и веществ, посту-
пивших в просвет кишечника со слюной, желудочным, поджелудочным
соками и желчью.
1.3. Выделительная функция кожи.
❖ В составе пота из организма выделяются вода (в обычных условиях 0,3 - 1,0
л/сутки, при гиперсекреции до 10 л/сутки), мочевина (5 - 10 % выделяемого количест-
ва), мочевая кислота, креатинин, электролиты.
❖ Сальные железы за сутки выделяют - 20 г секрета, который на 2/3 состоит
из воды, а 1/3 составляют холестерин, сквален, аналоги казеина, продукты
обмена половых гормонов, кортикостероидов, витаминов и ферментов.
1.4. Система мочеобразования и мочевыделения как главный компонент ФСВ
(см. ниже).
2. Общая характеристика системы мочеобразования и мочевыделения.
2.1. Основные компоненты: почки - корковое и мозговое (наружное н внутреннее) вещество, экст-
раренальные мочевыводящие пути (чашечки, лоханки, мочеточники, мочевой пузырь, мочеис-
пускательный канал).
2.2. Нефрон как структурно-функциональная единица почки (- 1 млн. в почке).
❖ Части нефрона и их функция.
• Почечное тельце: сосудистый клубочек (50 - 100 капилляров), капсула и моче-
вое пространство - осуществляет функцию фильтрации.
• Почечные канальцы (длина 35 - 50 мм, общая длина - 100 км) осуществляют
функции реабсорбции, секреции, осмоконцентрирования и разведения.
ф Проксимальный извитой каналец (25 % длины), у эпителиоцитов
хорошо выражена щеточная кайма.
ф- Петля Генле (50 % длины): нисходящая часть (проксимальный пря-
мой каналец и тонкий нисходящий каналец) и восходящая часть
(тонкий и толстый канальцы, плотное пятно).
ф- Дистальный извитой каналец (15 % длины) через связывающий ка-
налец открывается в собирательную трубку.
ф- Собирательная трубка (в функциональном плане относят к нефро-
ну, 10 % длины). Имеет светлые клетки, обеспечивающие реаб-
сорбцию воды и темные клетки, которые секретируют Н1', закисляя
мочу.
• Юкстагломерулярный (околоклубочковый) аппарат (ЮГА) имеет инкретор-
ную функцию, включает в себя три главных совокупности клеток.
ф- Юкстагломерулярные (зернистые) клетки представляют собой мо-
дифицированные гладкомышечные клетки приносящей и частично
выносящей артериол, образуют ренина, секреция которого повыша-
ется при снижениидавления крови в приносящей артериоле и по-
вышении симпатических влияний на ЮГА почки.
ф- Клетки плотного пятна дистального извитого канальца - натриевые
рецепторы, реагирующие на осмотическое давление канальцевой
мочи.
355
ф- Юкставаскулярные клетки (Гормагтига) образуют скопление между
плотном пятном и клубочком в углублении между приносящей и
выносящей артериолами, предположительно содержат ангиотензи-
иазу (разрушение ангиотензина П) и являются резервом по секре-
ции ренина.
❖ Основные виды нефронов.
• По расположению почечного тельца (клубочка).
ф- Поверхностные нефроны (~ 15%) берут начало преимущественно
от поверхностных, но также интракортикальных и юкстамедулляр-
ных почечных телец и соприкасаются с поверхностью почки.
ф- Внутрикорковые нефроны (~ 70 %) начинаются от почечных телец
средней части коркового слоя, имеют короткие канальцы, в них
преобладают функции фильтрации, реабсорбции, секреции, менее
выражена осмокоицеитрация мочи.
ф- Юкстамедуллярные нефроны (~ 15 %) начинаются от почечныъх
телец в корковой зоне, прилегающей к мозговому веществу, в них
резко выражена функция концентрирования и разведения мочи.
• По длине петли:
Ф’ короткопетлевые - сгиб петли расположен в корковом или наруж-
ном мозговом веществе;
Ф- длиннопетлевые - сгиб петли расположен во внутреннем мозговом
веществе;
Ф- все три вида нефронов могут быть короткопетливые и длиннопет-
левые, но поверхностные и внутрикорковые нефроны преимущест-
венно короткопетлевые, а юкстамедуллярные - преимущественно
длиииопетлевые.
3. Кровообращение в почке.
3.1. Особенности сосудистой сети почек.
❖ Двойная сеть капилляров (позволяет пространственно разобщить функцию
фильтрации и реабсорбции):
• приносящие артериолы дают первую капиллярную клубочковую сеть с высо-
ким давлением крови 60 мм рт. ст., диаметр выносящей артериолы на 1/3 меньше
приносящей), что обеспечивает фильтрацию;
• выносящая нз клубочка артериола распадается на вторую капиллярную сеть
вокруг канальцев с низким давлением крови (~ 10 мм рт. ст.), что обеспечивает
реабсорбцию.
❖ От выносящих артериол отходят также прямые сосуды, которые повторяют
ход петель Гейле в мозговом веществе; кровоток по прямым сосудам со-
ставляет - 2 % почечного кровотока; эти сосуды играют важную роль в
противоточном осмоконцентрирующем механизме.
3.2. Характеристика кровотока в почках.
❖ Высокий уровень кровотока: 1000 - 1200 мл/мии иа 1,73 м2 поверхности
тела или 450 мл/100 г ткани почки в 1 мин, что составляет 20 - 25 % сер-
дечного выброса.
356
❖ Высокий уровень потребляемого кислорода (6 - 10 %, - 18 мл/мин) при
низкой величине артериовенозной разницы - 14 мл Ог/л крови.
❖ Ауторегуляция почечного кровотока в диапазоне 80 - 180 мм рт. ст. сис-
темного артериального давления.
❖ Два круга кровообращения в почках.
• Кровоток в корковом веществе (составляет 90 %, осуществляется приносящи-
ми артериолами корковых нефронов, отходящими от дистальных участков
междольковых вртерий).
• Кровоток в мозговом веществе (составляет 10 %, осуществляется по принося-
щим артериолам юкстамедуллярных нефронов, отходящих от проксимальных
участков междольковых артерий, в также по прямым артериолам, отходящими
непосредственно от дугоаых артерий).
• Создается возможность (она реализуется при централизации кровотока) путем
сужения междольковой артерии между приносящими артериолами корковых и
юкстамедуллярных нефронов выключить кровоток в корковом веществе и зна-
чительно уменьшить фильтрацию.
3.3. Определение почечного кровотока непрямым методом (клиренс по параамино-
гиппуровой кислоте - ПАТ).
❖ Создается постоянная низкая (3 - 4 мг %) концентрация в плазме тест-
вещества (ПАГ, диодраст и др.), от которого кровь освобождается в почках
путем секреции.
❖ Определяют концентрацию ПАГ в плазме (Рпаг), моче (Лпаг) и минутный
диурез(Vмин)*
❖ Вычисляют коэффициент очищения по формуле Спдг = (Плат / Рпаг) ' VMKH,
который отражает почечный плазмоток (~ 600 мл/мин).
❖ Расчет кровотока делают исходя из плазмотока и величины гематокрита и
пересчитывают на стандартную поверхность тела (1,73 м2).
4. Классификация функций почек.
4.1. Функция мочеобразования: процессы клубочковой ультрафильтрации, каналь-
цевой реабсорбции и секреции; нейрогуморальная регуляция мочеобразования;
осмоконцентрирование мочи (противоточно-множительный механизм); осмотическое
разведение мочи.
4.2. Выделительные (экскреторно-гомеостатические) функции: 1) азотвыдели-
тельная; 2) осмо- и волюмрегулирующая; 3) регуляция ионного состава крови;
4) регуляция кислотно-основного состояния.
4.3. Невыделительные функции: 1) инкреторная; 2) метаболическая.
5. Клубочковая ультрафильтрация (переход бесклеточной и безбелковой части
крови из капилляров клубочка в полость капсулы).
5.1. Основные факторы, определяющие фильтрацию.
❖ Площадь клубочкового фильтра (- 1,5 м2/100 г почки).
❖ Гидравлическая проницаемость фильтра (зависит от количества и диамет-
ра пор).
❖ Эффективное фильтрационное давление (~ 15 мм рт. ст.).
5.2. Характеристика клубочкового фильтрационного барьера.
357
❖ Компоненты: клетки эндотелия капилляров (поры около 80 нм), базальная
мембрана и подоциты (поры до 6 нм).
❖ Общая поверхность пор, через которые идет фильтрация, составляет до 30
% общей поверхности капилляров.
❖ Через фильтр свободно проходят молекулы d < 3,6 нм (ММ < 10 кД), напри-
мер, электролиты, мономеры, пептиды; не проходят молекулы d > 8,8 нм
(ММ > 80 кД), например, глобулины, фибриноген, а также клетки крови.
❖ Частично проходят молекулы 3,6 < d < 8,8; например, незначительно
фильтруется альбумин (0,01 %) и внеэритроцитарный гемоглобин, если он
не связан с гаптоглобином (3 %).
S3. Механизм фильтрации.
❖ Фильтрационное давление (10 - 20 мм рт. ст.) равно разнице между дав-
лением крови в капиллярах клубочка (Р, - 50 мм рт. ст.) и силами, препятст-
вующими фильтрации (онкотическое давление крови Р2 = 20 - 25 мм рт. ст. и гидро-
статическое давление фильтрата в капсуле Р3 = 10 - 18 мм рт. ст.): ФД = Р| — Р2 - Рз.
❖ Состав ультрафильтрата: пептиды, глюкоза, аминокислоты, гормоны,
витамины, органические кислоты. Концентрация одновалентных ионов
практически равна плазменным, двухвалентных ионов на 20 - 40 % мень-
ше за счет связи их с белками плазмы; отсутствуют клетки крови и белки
(незначительно фильтруются альбумины - 10 r/суткн), вещества, связанные с бел-
ками (иапрнмер, свободный билирубин).
❖ Количество фильтрата (первичной мочи): 150 л/сут или 100 мл/мин, что
составляет 20 % проходящей через клубочек плазмы (фильтрационная
фракция).
5.4. Определение скорости клубочковой фильтрации (СКФ).
♦ ♦♦ В качестве тест-вещества (фильтруется, но не реабсорбируется и не секретируется)
используется эндогенный креатинин или вводится полисахарид инулии.
❖ Определяют концентрацию тест-вещества в плазме (Р), в моче (U) и ми-
нутный диурез (V).
❖ По формуле Стест в-во — (U-гест в-во^Ртсст в-во) VMHH определяют СКФ; напри-
мер, Скреат = 100 - 130 мл/мин в пересчете на поверхность тела 1,73 м2.
6. Канальцевая реабсорбция (процесс перехода веществ из канальцевой мочи во вне-
клеточную жидкость и далее в кровь).
6.1. Общая характеристика,
❖ Основной энергопотребляемый процесс в почках, большая площадь реаб-
сорбции (до 60 м2) за счет щеточной каёмки.
• 5* Топография реабсорбции:
Ф в проксимальных извитых канальцах: глюкоза, аминокислоты, полипептиды, вн*
тамииы, вода, мочевина, мочевая кислота, Na*, К*. Са2*, Mg2*, SO42’.
НРОЛ СГ, НСОз’;
Ф в петле Генле реабсорбция сопряжена с осмоконцентрированием мочи:
в нисходящей части: вода, мочевая кислота;
в толстом канальце восходящей части: Na*, К*, Са2*, Mg2*, СГ;
Ф в дистальных извитых канальцах: вода, Na*, Са2*, СГ;
Ф в собирательных трубках: Na*, К*. СГ, вода, мочевина.
6.2. Реабсорбция натрия (99,4 % профильтрованного).
358
♦♦♦ Топография: проксимальные канальцы ~ 65 %, толстый восходящий отдел петли - 25 %;
дистальные канальцы, собирательные трубки - 9 %; контролируется альдостероном 5 -
10 % реабсорбции Na+.
❖ Механизм: на апикальной мембране по электрохимическому градиенту с
использованием переносчика (антипорт Na*-H*; симпорт Na*-K*-2Cr; Na*-
глюкоза; Ыа*-аминокислоты) и натриевых каналов; на базальной мембране -
К , Na—насос. (Стимулируют реабсорбцию На+аигиотензин-П в малых концентраци-
ях, норадреналин, эндотелии, инсулин; тормозят реабсорбцию Na* - Ыа*-уретический
гормон, паратгормон, дофамин, ангиотеизии-П а больших концентрациях.)
6.3. Реабсорбция калия (- 85 % профильтрованного).
♦♦♦ Топография: проксимальные канальцы - 75 %, восходящая часть петли и дистальный ка-
налец- 10 %.
❖ Механизм: на апикальной мембране симпорт Na*-K+-2CF; на базальной
мембране диффузия через каналы, часть К* (а также Na+ и СГ) проходит че-
рез межклеточные щелн.
6.4. Реабсорбция белка (100 % профильтрованного, - 10 г/сутки).
Топография-, в проксимальных канальцах практически полностью (выделяется с мочой -
50 мг/сутки).
Механизм: пнноцитоз у основания микроворсннок апикальной мембраны,
гидролиз белка до аминокислот протеазами лизосом, на базальной мем-
бране облегченная диффузия аминокислот.
6.5. Реабсорбция пептидов (100 % профильтрованных).
♦♦♦ Топография: проксимальные канальцы.
❖ Механизм: разрушаются пептидазами апикальной мембраны и далее реаб-
сорбируются как аминокислоты (или реабсорбируются с помощью переносчиков и
разрушаются внутри клетки пептидазами).
6.6. Реабсорбция аминокислот (95 - 99 % профильтрованных).
♦♦♦ Топография: проксимальные канальцы.
❖ Механизм: на апикальной мембране происходит вторичный активный
транспорт с использованием градиента Na+ и пяти типов переносчиков
(для кислых, основных, нейтральных аминокислот, нмннокнслот, всех остальных амино-
кислот); на базальной мембране - облегченная диффузия.
6.7. Реабсорбция глюкозы (100 % профильтрованной, - 1000 ммоль).
♦ ♦♦ Топография:* проксимальных канальцах практически полностью.
❖ Механизм: на апикальной мембране происходит вторичный активный
транспорт с использованием переносчика и энергии градиента Na+, на ба-
зальной мембране - облегченная диффузия.
❖ Порог реабсорбции глюкозы - 10 ммоль/л крови, выше которого она по-
является в конечной моче.
6.8. Реабсорбция анионов хлора (> 99 % профильтрованного).
Топография: проксимальные канальцы, восходящий отдел петли Гейле, дистальные ка-
нальцы и собирательные трубкн.
*♦* Механизм: на апикальной мембране происходит с помощью переносчика, используя
симпорт с Na*, в собирательных трубках диффузия осуществляется по концентрацион-
ному градиенту, иа базальной мембране - по электрическому градиенту; часть СГ (а так-
же Na* и К*) диффундируют через межклеточные щели.
6.9. Реабсорбция ионов кальция (99 % профильтрованного).
359
♦♦♦ Топография: проксимальные канальцы, толстый сегмент восходящей части петли Генле,
дистальные канальцы.
Механизм: на апикальной мембране и через межклеточные шунты происходит пассивно
по электрическому градиенту, на базальной мембране - с помощью кальциевого насоса
(Са^-АТФаза) и 3Na+/ Са2+-обменника.
6.10. Реабсорбция ионов магния (94 % профильтрованного).
♦♦♦ Топография: проксимальные канальцы мало проницаемы, толстый сегмент восходящей
части петли Генле играет основную роль.
*♦♦ Механизм: на апикальной мембране и через межклеточные шунты происходит пассивно
по электрическому градиенту, на базальной мембране - в результате активного транс-
порта.
6.11. Реабсорбция мочевины (50 - 60 % профильтрованной).
♦♦♦ Топография: проксимальные канальцы, собирательные трубки.
Механизм: реабсорбция происходит пассивно с током реабсорбируемой воды (моча в
сравнении с плазмой концентрируется по мочевине в 70 раз).
6.12. Реабсорбция воды (99,2 % профильтрованной).
Топография: в проксимальных канальцах - 70 %, нисходящем отделе петли ~ 18 %, дис-
тальных канальцах - 5 %, собирательных трубках ~ 7 % (контроль АДГ).
♦♦♦ Механизмы: через водные каналы в проксимальных и дистальных канальцах вода следу-
ет за Na+, в нисходящей части петли и собирательных трубках вода следует против высо-
кой осмоляльности интерстиция.
6.13. Реабсорбция гидрокарбоната (см. тема 21; задание 1; 6.2).
6.14. Определение скорости реабсорбции, В практической медицине определяют
максимальную реабсорбцию глюкозы (при уровне ее в крови - 25 ммоль/л) как раз-
ницу между количеством профильтрованной глюкозы (концентрация ее в плаз-
ме, умноженная на величину фильтрации по Скреат) и величиной выделенной
глюкозы с мочой (концентрация в моче, умноженная на минутный диурез);
нормальная величина 375 ± 65 мг/мин.
7. Канальцевая секреция (процесс перехода веществ из крови через канальцы в мочу,
а также из клеток канальцев в мочу).
7.1. Топография секреции:
♦♦♦ в проксимальных канальцах: Н+, NH3, мочевая кислота, холин, серотонин, пенициллин,
парааминогиппуровая кислота, фенолрот и др.;
в тонкой части петли: мочевина;
♦♦♦ в дистальных канальцах и собирательных трубках: Н*, NH3 и К* при избыточном поступ-
лении с пищей.
7.2. Секреция К* (при избыточном поступлении с пищей).
Топография: дистальные канальцы и собирательные трубки.
❖ Механизм: К+, Na+-Hacoc на базальной мембране, диффузия через каналы
на апикальной мембране, контролируется альдостероном; часть К+ секре-
тируется по межклеточным щелям через плотные контакты.
73. Секреция Д* из клетки канальца в мочу (~ 75 ммоль/сутки).
Топография: проксимальные, дистальные канальцы, собирательные трубки.
❖ Механизм: Н* освобождается при диссоциации образующейся Н2СО3; на
апикальной мембране клеток проксимальных канальцев выделяется в про-
свет канальцев путем ионообмена (антипорта) с Na+, в собирательных
трубках - с участием ГТ-насоса.
7.4. Секреция NH3 из клеток канальцев в мочу (* 50 ммоль/сутки).
360
*•* Топография: проксимальные и дистальные канальцы, собирательные трубки.
*'* Механизм: NHj образуется в клетке в реакциях дезамидирования глутами-
на и дезаминирования глутамата, иа апикальной мембране происходит
диффузия Ь1Нз в просвет канальца.
7.5. Определение скорости секреции. В практической медицине определяется мак-
симальная канальцевая секреция с помощью параамииогиппуровой кислоты
или диодраста (на фоне их высокой концентрации в крови) как величина, равная разно-
сти между количеством выделенного с мочой вещества (концентрация в моче, ум-
ноженная нв минутный диурез) и количеством профильтрованного вещества (концен-
трация в плазме с учетом доли свободно растворимого в плазме вещества, умноженная на вели-
чину фильтрации). Нормальная величина для диодраста 52 ± 9 мг/мин.
8. Осмоконцентрирование мочи (противоточио-миожительный механизм).
8.1. Нарастающий градиент осмотического давления в межклеточной жидкости
от границы коры и мозгового слоя (~ 300 мосм/кг) до вершины пирамид (~ 1200
мосм/кг) является основой концентрирующего механизма и создается:
❖ активной реабсорбцией в межклеточную жидкость интерстиции Na*/2C1"
/К* в толстом сегменте восходящей части петли (симпорт с помощью перенос-
чика, стимулируется АДГ);
❖ реабсорбцией мочевины из нижних отделов собирательных трубок (место
высокой проницаемости для мочевины);
❖ задержкой этих веществ в межклеточной жидкости с помощью циркуля-
ции между сосудами (прямые сосуды) и канальцевой (восходящая часть петли -
собирательная трубка), образующими противоточную систему.
8.2. В нисходящий тонкий отдел петли Генле, имеющий высокую проницае-
мость для воды и низкую для электролитов:
❖ поступает моча из проксимального канальца, изотоничная плазме (~ 300
мосм/кг);
❖ при продвижении фильтрата вода выходит в интерстиций против осмоти-
ческого давления, а электролиты и мочевина остаются в канальце;
❖ в результате увеличивается осмотическое давление фильтрата в нисходя-
щей части петли от 300 до 1200 мосм/кг в области сгиба петли;
< • при этом объем фильтрата уменьшается иа 18 % (на - 35 л).
8.3. В восходящую часть петли Генле, имеющую высокую проницаемость для ио-
нов и низкую для воды:
❖ поступает гиперосмотичный фильтрат (до 1200 мосм/кг);
❖ в тонком сегменте происходит пассивная реабсорбция NaCI, в толстом
сегменте - активная реабсорбция Na*, К*, СГ из просвета канальца в ин-
терстиций (см. выше 7.6);
❖ в результате в дистальный каналец поступает гипотоничная жидкость (~
200 мосм/кг), богатая мочевиной; здесь продолжается реабсорбция Na*,
СГ и воды, поэтому осмотическое давление существенно не меняется, а
объем жидкости уменьшается.
8.4. В собирательные трубки поступает гипо- или изотоничная моча, богатая мо-
чевиной:
❖ здесь происходит осмотическое концентрирование мочи (максимально до
1200 мосм/кг), так как вода реабсорбируется ие только следуя за Na*, СГ и
361
мочевиной, но и против высокого осмотического давления интерстиция;
часть мочевины также реабсорбируется в интерстиций и поступает в вос-
ходящую часть петли (рециклизация мочевины);
❖ реабсорбция воды в собирательных трубках контролируется АДГ.
8.5. Осмоконцентрирующую способность почек можно оценить: 1) пробой с су-
хоедением (Фольгарда) - максимальная плотность мочи не ниже 1,027, осмо-
ляльность не ниже 900 мосм/кг; 2) определением осмотического индекса (ОИ =
UM0CU/ Риоси), который в норме - 2, при осмоконцентрации - 4.
9. Осмотическое разведение мочи (водный диурез). Возникает при водной нагрузке (а
также нарушении секреции или действия АДГ).
9.1. Снижение секреции АДГ при водной нагрузке приводит к блокаде реабсорбции
воды в собирательных трубках при сохраняющейся реабсорбции Na+ и СГ в
нефроне.
9.2. Максимальная осмоляльность в интерстиции снижается при этом до 500
мосм/кг.
9.3. Максимальный водный диурез равен от 12 л/сут (контроль АДГ при нормальном объ-
еме фильтрата) до 30 л/сутки при увеличении фильтрации в результате повышен-
ного приема жидкости.
9.4. Минимальное осмотическое давление мочи снижается до 50 мосм/кг.
9.5. Методы исследования: 1) проба с разведением (водная нагрузка - 1 л) - снижается относитель-
ная плотность мочи до 1,002, осмотическое давление до 40-50 мосм/кг; 2) определение осмоти-
ческого индекса (UUOCM / Рмоем) показывает его снижение до 0,2 (норма - 2).
10. Количество и состав конечной мочи.
10.1. Количество: 1,0 - 1,5 л/сутки; облигатный диурез 0,7 - 0,8 л/сутки, дневной
диурез равен 2/3 - 3/4 суточного.
10.2. Физико-химические свойства: удельный вес 1,010 - 1,025; осмоляльность 600 -
800 мосм/кг (крайние пределы 50 - 1400 мосм/кг); реакция мочи: при смешан-
ной пище - кислая или нейтральная (рН=5,0 - 7,0); при растительной пище -
щелочная.
10.3. Состав мочи:
❖ форменные элементы: эритроциты до 1000 в мл, лейкоциты до 4000 в мл,
единичные эпителиальные клетки; глюкоза и белок практически отсутст-
вуют (глюкоза 0,06-0,8 ммоль/л; белок до 50 мг/сутки);
♦ ♦♦ азотистые вещества: мочевина 330 - 580 ммоль/сутки, аммиак 10 - 107 ммоль/суткн,
креатинин 5,3 - 17,7 ммоль/сутки, мочевая кислота 1,5 - 4,5 ммоль/сутки, аминокислоты
3,6 - 14,3 ммоль/сутки, некоторые ферменты (урокиназа н др.);
♦ ♦♦ соли: хлориды 110 - 250 ммоль/суткн, натрий 40 - 220 ммоль/сутки, калий 25 - 125
ммоль/суткн, фосфаты 13-42 ммоль/суткн, оксалаты 0,114 - 0,456 ммоль/сутки.
11. Нейрогуморальная регуляция мочеобразования.
11.1. Нервная регуляция мочеобразования.
*** Иннервация почки: почки иннервируются симпатическими волокнами (Тю - L2) и пара-
симпатическими волокнами (преимущественно X нерва); в почках иннервируются эндо-
телий и миоциты сосудов, клетки капсулы н канальцев (базальные мембраны).
❖ Влияние на сосуды.
362
• Симпатические влияния осуществляются через а-адренорецепторы афферент-
ных и эфферентных артериол (сужение сосудов) и Р-адренорецепторы аффе-
рентных (расширение сосудов, секреция ренина).
• Парасимпатические влияния осуществляются через М-холинорецепторы (рас-
ширение сосудов). Основной эффект ацетилхолина реализуется через М-хо-
линорецепторы эндотелиальных клеток и усиление продукции NO, который
через гуанилатциклазу сосудистых миоцитов и увеличение цГМФ приводит к
расслаблению сосудов.
•> Влияние на фильтрацию.
• Симпатические и парасимпатические влияния оказывают неоднозначные эф-
фекты на фильтрацию в почках. Отмечено как увеличение, так и снижение
фильтрации, что связывают с различными ответами приносящих и выносящих
артериол в каждом конкретном случае.
• Симпатические влияния могут увеличивать фильтрацию через повышение
системного АД, однако в обычных условиях это влияние «выключено» ауторе-
гуляцией почечного кровотока.
❖ Влияние на канальцы.
• Симпатические влияния увеличивает реабсорбцию Na* и воды, уменьшают
диурез.
• Парасимпатические влияния снижают реабсорбцию Na* и воды, повышают
экскрецию воды, Na*, СГ, К*, Са2* и фосфатов.
11.2. Гуморальная регуляция мочеобразования.
❖ Антидиуретический гормон гипоталамуса.
• Стимулируют секрецию АДГ гиперосмия, вертикальная поза, симпатические
влияния и адреналин (в патологии - гиповолемия, гипотензия и др.).
• Ингибируют секрецию АДГ гипоосмия, горизонтальное положение (в патоло-
гии - гиперволемия, гипертензия, этанол).
• Клетки-мишени для АДГ: клетки дистальных канальцев и собирательных тру-
бок почек (преимущественно через У2-рецепторы и |цАМФ); миоциты сосу-
дов (через Vi-рецепторы, ИФз/ДАГ сужение сосуда).
• Почечные эффекты АДГ: увеличение реабсорбции воды и уменьшение диуре-
за; под контролем АДГ реабсорбируется - 8 % профильтрованной воды (- 12
л/сутки).
• Механизм действия АДГ: через У2-рецепторы (ТцАМФ и активности протеинки-
назы-А) происходит фосфорилирование аквопоринов (белков водных каналов
апикальной мембраны) и резкое повышение проницаемости для воды (через
Vi-рецепторы - снижение реабсорбции воды, что модулирует эффект через Vr-
рецепторы).
• Физиологический ингибитор АДГ - простагландин Ег.
❖ Альдостерон коры надпочечников.
• Стимулируют его секрецию гиперкалиемия и гипонатриемия, ангиотензин П
(например, при гиповолемии).
• Ингибируют секрецию альдостерона гипернатриемия и гипокалиемия, на-
трийуретический пептид, гиперволемия.
• Мишени: светлые и темные клетки собирательных трубок почек.
363
• Почечные эффекты альдостерона и их механизмы.
ф’ Увеличивает реабсорбция Na+ в кровь (альдостерон контролирует реаб-
сорбцию - 2 % профильтрованного Na*, -20 г/сутки), активируя образование
Na^-каналов на апикальной мембране н K*,Na*-Hacoca базальной
мембраны (светлых) клеток собирательных трубок.
ф Увеличивает секрецию К* в мочу, активируя образование молекул
К*,№*-насоса базальной мембраны и К*-каналов апикальной мем-
браны (темных) клеток собирательных трубок.
ф Увеличивает секрецию Н* в мочу, активируя образование молекул
Н*-насоса апикальной мембраны (темных) клеток собирательных
трубок.
❖ Ангиотензин II (см. ниже 16.1).
❖ Натрийуретический пептид предсердий.
• Стимулируют его секрецию гиперволемия, повышение АД, адреналин, АДГ,
ацетилхолин.
• Мишени: клубочки и собирательные трубки.
• Почечные эффекты натрийуретического пептида и их механизмы.
•ф Увеличение клубочковой фильтрации (на 30 - 50 %) в результате
сужения выносящей артериолы и расширения приносящей артерио-
лы.
Повышение экскреции Na* с мочой в результате увеличения его
фильтрации и снижения реабсорбции, связанной с торможением
секреции альдостерона (физиологический антагонист альдостеро-
на).
❖ Адреналин и норадреналин надпочечников (действуют аналогично симпа-
тическим влияниям).
12. Процессы мочевыведения н мочеиспускания.
12.1. Движение мочи в канальцах нефрона осуществляется по градиенту гидроста-
тического давления от 18 мм рт. ст. в капсуле до 0 мм рт. ст. в конце собира-
тельной трубки.
12.2. Основные функции экстраренальных мочевыводящих путей-, порционный
транспорт мочн, резервуарная функция, рудиментарная реабсорбционная спо-
собность.
12.3. Основные силы, транспортирующие мочу, сокращение гладких мышц данно-
го участка, расслабление мышц нижележащего участка, увеличение забрюшин-
ного давления (до 70 мм рт. ст.).
12.4. Роль чашечек.
* В фазе наполнения входной сфинктер чашечки открыт, выходной закрыт,
фаза наполнения длится 2-3 дыхательных цикла.
❖ Фаза опорожнения совпадает с вдохом; входной сфинктер закрыт, выход-
ной открыт.
❖ Из экстраренальных путей чашечки обладают наибольшей способностью
к реабсорбции, которая реализуется при повышении давления мочи до 50
- 55 мм рт. ст.
364
12.5. Роль лоханки.
❖ Наполнение лоханки приводит к возбуждению механорецепторов, и реф-
лекторно расслабляется лоханочно-мочеточниковый сегмент.
Опорожнение лоханки совпадает с непроизвольным глубоким вдохом, ко-
торый повышает забрюшинное давление.
❖ Повышение давления в верхней трети мочеточника рефлекторно сокраща-
ет лоханочно-мочеточниковый сегмент, и опорожнение лоханки прекра-
щается.
12.6. Роль мочеточников.
❖ Транспорт мочи происходит с помощью перистальтики.
❖ Ритмически взаимодействуют три функциональные секции (цистовды): на-
пример, если верхняя секция находится в фазе сокращения, то средняя - в
фазе расслабления, а нижняя - в фазе сокращения.
❖ Нижняя треть мочеточника имеет более низкую возбудимость и способна
накапливать ббльшую порцию мочи.
❖ Устье мочеточника работает как клапан, обеспечивая свободное поступ-
ление порции мочи в пузырь и препятствуя ее обратному движению.
12.7. Роль мочевого пузыря.
❖ Резервуарная функция пузыря обеспечивается:
• высоким пластическим тонусом m. detrusor;
• симпатическими влияниями (центр в Lu), которые расслабляют мышцу (через
^-адренорецепторы) и сокращают сфинктер пузыря (через а>-
адреиорецепторы).
❖ Функцию опорожнения пузыря обеспечивает центр мочеиспускания в 82-4.
• Раздражение механорецепторов пузыря вызывается растяжением его стенок (а
не давлением мочи как таковым).
• Первые позывы возникают прн объеме пузыря - 150 мл и резко увеличивают-
ся при объеме 200 - 300 мл. Высокая скорость наполнения пузыря стимулиру-
ет афферентную импульсацию.
• Парасимпатические влияния (nn. splanchnic! pelvici) через ацетилхолин и М-
холинорецепторы сокращают мышцу пузыря (при этом расслабляется его
сфинктер).
• Произвольный контроль мочеиспускания осуществляется двигательной корой
через мотонейроны поперечно-полосатой мышцы сфинктера мочеиспуска-
тельного канала (иннервируемой ветвью n. pudendus).
4* Показателями эффективности сократительной функции пузыря являются:
• максимальная скорость потока мочи не менее 15 мл/с в первой трети времени
' мочеиспускания (метод флуометрнн);
• остаточное количество мочи не более 10 % объема пузыря перед мочеиспус-
канием (но не более 30 мл) (ультразвуковое исследование).
13. Азотовыделительная функция почек.
13.1. Почки выводят из организма основное количество конечных продуктов азоти-
стого обмена: мочевины, креатинина, мочевой кислоты, индикана (например, 90 %
мочевины и весь креатинин).
365
13.2. Показатели нормальной азотвыделительной функции почек: азотсодержащие
вещества в крови не должны быть выше нормы (в ммоль/л: мочевина - 2,5 - 8,3, креа-
тинин - 0,04 - 0,09, остаточный азот крови - 14,3 - 28,5), а в суточной моче не ниже
нормы (мочевина - 330 - 580, креатинин - 5,3 -17,7 в сутки).
14. Осмо- и волюмрегулирующая функция почек.
14.1. Эта функция осуществляет поддержание осмотического давления внеклеточной
жидкости в очень узких пределах (285 + 2 мосм/кг) и (в менее узких пределах) объ-
ема циркулирующей жидкости, что достигается изменением выделения Na4 и
воды с мочой.
14.2. Осморегулирующая функция.
❖ Коррекция гиперосмии.
• Стимуляция центральных осморецепторов в супрахиазменной зоне гипотала-
муса и периферических рецепторов в сосудах, печени, почках и других орга-
нах приводит к:
ф- повышению секреции АДГ в супраоптических и паравентрикуляр-
ных ядрах гипоталамуса (при 295 мосм/кг в крови уже максимальная секре-
ция);
Ф’ раздражению центра жажды в гипоталамусе (область III желудочка) и
приему воды.
• Влияние гиперосмии на клетки клубочковой зоны в надпочечниках приводит к
понижению секреции альдостерона.
• Стимуляция клеток предсердия приводит к увеличению секреции натрийуре-
тического пептида.
• В результате происходит повышение реабсорбции воды в почках, увеличение
экскреции Na4 с мочой н прием жидкости, что способствует коррекции гипе-
росмии.
❖ Коррекция гипоосмии.
• Снижение активности центральных и периферических осморецепторов приво-
дит к:
ф уменьшению образования АДГ в ядрах гипоталамуса (при 280 мосм/кг
в крови секреция АДГ прекращается);
ф- активации центра солевого аппетита в гипоталамусе;
ф- снижению секреции натрийуретического пептида в предсердиях;
ф- стимуляции образования альдостерона в надпочечниках.
• В результате происходит увеличение реабсорбции и уменьшение экскреции
Na4 в почках, повышение приема соли, увеличение выделения воды через поч-
ки, что способствует коррекции гипоосмии.
143. Волюмрегулирующая функция.
❖ Коррекция гиповолемии.
• Влияние с механорецепторов объема и давления (предсердий, правого
желудочка, легочных артерий и вен, полой вены, каротидных и аортальных те-
лец, афферентных артериол почки) приводит:
ф- в гипоталамусе к повышению секреции АДГ, возбуждению центра
жажды н солевого аппетита;
366
Ф в почках к гиперсекреции ренина и затем повышенному образова-
нию ангиотензина П;
ф’ в надпочечниках к гиперсекреции альдостерона;
"Ф в предсердиях к снижению секреции натрийуретического пептида.
• В результате происходит повышение реабсорбции воды и Na*, возможно сни-
жение фильтрации в почках, увеличение приема жидкости, что способствует
коррекции гиповолемии.
❖ Коррекция гиперволемии.
• Влияние с механорецепторов объема и давления приводит:
ф в гипоталамусе к снижению секреции АДГ;
ф в почках к снижению секреции ренина и образования ангиотензина
П;
ф в надпочечниках к снижению секреции альдостерона;
ф в предсердиях к увеличению секреции натрийуретического пептида.
• В результате увеличивается экскреция воды и Na* в почках, что способствует
коррекции гиперволемии.
14.4. Роль почек в регуляции ионного состава крови (почки регулируют не только общий
показатель - осмотическое давление кровн, но н концентрацию отдельных иоиов, т.е. ионный
состав крови).
❖ Регуляция концентрации Na+ осуществляется с участием:
• альдостерона, который повышает реабсорбцию Na* из собирательных
трубок в кровь;
• натрийуретического пептида предсердий, который увеличивает экскре-
цию Na+ с мочой;
• паратгормона и тирокальцитонина, которые повышают экскрецию Na* с мочой.
❖ Регуляция концентрации К+ осуществляется с участием:
• альдостерона, который способствует секреции К* в мочу и экскреции
его из организма;
• пврвтгормона, который увеличивает экскрецию К* с мочой.
❖ Регуляция концентрации Са2* и фосфата (НРО42‘) осуществляется с уча-
стием:
• холекальциферола (витамина D3), который способствует реабсорбции
Са2* и НРО4 в дистальных канальцах;
• паратгормона, который повышает реабсорбцию Са2* в дистальном неф-
роне, а также ингибирует реабсорбцию Са2* и НРОд2' в проксимальных
канальцах;
• тирокальцитонина, который способствует экскреции с мочой Са2* и НРО/".
❖ Регуляция концентрации Mg2* осуществляется с участием:
• паратгормона, который способствует реабсорбции Mg2* в толстом вос-
ходящем колене петли Генле;
• минералкортикоидов, больших доз тирокальцитонина, АДГ, которые
способствуют экскреции Mg2* с мочой.
15. Регуляции кислотно-осиовного состоянии (см. тему 21; задание 1:6.2).
367
16. Инкреторные функции почек.
16.1. Регуляция сосудистого тонуса.
❖ Прессорная ренин-ангиотензиновая система.
• Ренин (протеаза) секретируется миоэпителиоидными клетками аффе-
рентных артериол ЮГА н поступает в кровоток.
• Секрецию ренина стимулируют снижение почечного и системного АД,
объема крови, концентрации Na+ мочи в области плотного пятна, увели-
чение симпатических влияний и действие адреналина на почки (через ₽,/
рг -адренорецепторы).
• Ренин отщепляет от ангиотензиногена (а2-глобулина, образуемого пече-
нью) неактивный декапептид - ангиотензин I, который под действием
ангиотеизинпревращающего фермента (АПФ) в легких и других органах
переходит в активный октапептид - ангиотензин П.
• Эффекты ангиотензина П.
Ф’ Сосуды: прямое сосудосуживающее действие, повышение АД.
• ф" Почки: в невысокой концентрации локально суживает артериолы
почек, уменьшает в них кровоток и клубочковую фильтрацию, ин-
гибирует секрецию ренина.
• ф" ЦНС: стимулирует в продолговатом мозге прессорную зону сосу-
додвигательного центра, в гипоталамусе - центр жажды и солевой
аппетит.
• ф" Надпочечники: оказывает прямое стимулирующее действие иа вы-
работку альдостерона.
❖ Депрессорная система.
• Калликреин-кининовая система (ренальный антигипертензивный меха-
низм).
- ф- Почки (как и другие органы) образуют пептидазу калликреин, который
действует на пептид кининоген (образуемый клетками дистального ка-
нальца), отщепляя от него активные кинины - нонапептид брадикинин
и декапептид каллидин (физиологические антиподы ангиотензина П, разруша-
ются киназой).
ф- Выработку кининов стимулируют увеличение АД и объема крови,
ангиотензин П, мннералкортикоиды.
- ф- Эффекты кининов: сильное сосудорасширяющее действие особенно
на афферентные артериолы, натрийуретический и диуретический
эффекты, усиление продукции простагландинов.
• Почечные простагландины (в обычных условиях их действие минимально и
резко увеличивается при уменьшении объема циркулирующей крови, кровото-
ка почек, фильтрационного давления).
"Ф" Синтез простагландинов (из арахидоновой кислоты) происходит пре-
имущественно в интерстициальных клетках стромы мозгового ве-
щества (стимулируют синтез ангиотензин II, кинины, катехоламины, АДГ).
•ф" Образуются простагландины как сосудорасширяющие (ПрЕ2 н про-
стациклин), так и сосудосуживающие (тромбоксаны, ПрЕго).
368
•ф- Основные эффекты простагландинов: увеличение почечного крово-
тока (расширение приносящей артериолы), экскреции Na*, продукции ки-
нинов; увеличение (LipEj, простациклин) или снижение (ПрРзо) выра-
ботки ренина.
16.1 . Регуляция эритропоэза.
Эритропоэтин (гликопротеин) вырабатывается преимущественно клетками
интерстиция мозгового вещества почек (85 - 90 % всего ЭП в здоровом организ-
ме), резервным органом образования является печень.
❖ Стимулируют продукцию ЭП: снижение напряжения Ог в ткани почек на
венозном конце капилляра, усиление симпатических влияний.
❖ Почечный кислородный сенсор локализован в эндотелии капилляров про-
ксимальных канальцев коркового и мозгового вещества.
❖ ЭП, синтезированный в почках, поступает в кровь н действует на клетки-
мишенн красного ростка миелоидной ткани, активируя их пролиферацию
и синтез в них гемоглобина (подробнее см. 12, задание 1.1.5).
16.2 . Регуляция кальциевого обмена (через активацию витамина Dj).
В проксимальных извитых канальцах (под действием la-гидроксилазы) из 25-
гидроксн-Из образуется активная форма витамина D3 - 1а,25-
дигидроксихолекальциферол. Стимулируют его секрецию гипокальцие-
мия и гипофосфатемия.
Действие холекальциферола (через протеинкиназу С активирует белки Са^-иасоса
и транспортеров Саг+ в клеточной мембране, через экспрессию генов увеличивает синтез
этих белков).
• В почках: способствует реабсорбции Са2* в дистальных канальцах и
НРО42' в проксимальных канальцах.
• В тонком кишечнике: усиливает всасывание Са2* и ПРО/'.
• В костях: мобилизует Са2* и НРО42' в результате пролиферации остео-
кластов н резорбции костной ткани.
• Суммарный эффект: способствует повышению концентрации Са2* и
фосфатов в кровн.
17. Метаболическая функция почек.
17.1. Роль почек в обмене углеводов.
❖ При гипергликемии глюкоза извлекается почечными клетками из крови,
при гипогликемии - освобождается в кровоток.
❖ Глюконеогенез в почках более интенсивный, чем в печени; в условиях го-
лодания почки синтезируют половину общего количества глюкозы, обра-
зующейся из аминокислот.
17.2. Роль почек в обмене белков.
❖ Сохраняется фонд аминокислот в крови за счет гидролиза реабсорбиро-
вавшихся белков и пептидов.
❖ Разрушая профильтровавшиеся пептидные гормоны, почки участвуют в
регуляции их уровня в крови.
17.3. Роль почек в обмене липидов.
369
❖ Почки потребляют из крови свободные жирные кислоты и используют их
для образования энергии, триглицеридов и фосфолипидов, которые ис-
пользуются не только в почках, но и в других органах.
❖ Почки поглощают из крови предшественник синтеза холестерина (мевало-
нат), регулируя уровень холестерина в крови.
18. Профильный материал для студентов лечебного факультета.
18.1. «Регресс» основных почечных функций при старении.
❖ Снижение кровотока в почках (на 47 - 73 %).
❖ Снижение уровня клубочковой фильтрации (на 35 - 45 %).
Ъ Уменьшение реабсорбции воды (на - 30 %).
❖ Снижение способности к осмотической концентрации мочи.
18.2. Особенности тока мочи, мочеиспускания и его регуляция у пожилых и старых лю-
дей.
❖ Снижение эластических свойств, атрофия мышечных, разрастание соединитель-
ной ткани приводит к понижению эвакуаторной функции мочевыводящих путей.
❖ Уменьшение количества барорецепторов снижает активность нейрорефлектор-
ных реакций.
❖ Большая частота днскинезнн, анти перистальтических движений, рефлюксов.
❖ Снижение растяжимости стенки мочевого пузыря вызывает учащение позывов
на мочеиспускание.
❖ Ослабление функций сфинктеров мочевого пузыря н мочеиспускательного кана-
ла приводит к недержанию мочи.
18.3. «Искусственная почка» и ее применение в клинике.
❖ «Искусственная почка» является одним из методов очищения крови от веществ,
накапливающихся при почечной недостаточности, основанным на диффузии ве-
ществ через полупроницаемую мембрану.
Гидратцеллюлозная пленка, не пропускающая белки, разделяет текущую кровь и
диализирующий раствор; кровь поступает из артерии и после прохождения через
аппарат возвращается в вену.
❖ Диализирующий растаор подобен плазме по ионному составу и осмотической
концентрации, но не содержит низкомолекулярные вещества и ионы, которые
накапливаются в крови при почечной недостаточности (мочевину, креатинин,
мочевую кислоту, соли К4, Mg2+, Са2*).
❖ В результате из крови в солевой раствор происходит диффузия мочевины, креа-
тинина, мочевой кислоты, Mg2+, К*, Са2* и осуществляется коррекция азотистого
и ионного состава внеклеточной жидкости н ацидоза.
19. Профильный материал для студентов педиатрического факультета.
19.1. Структурно-функциональные особенности почек плода.
❖ Метанефрос (постоянная почка) появляется на 5-й неделе, нефроны незрелые
(клубочки и фильтрующая поверхность малы, канальцы короткие, петля Генле не
развита), образование нефронов продолжается весь внутриутробный период.
❖ Поступление безбелковой жидкости в чашечки лоханки регистрируется на 11 -
12-й неделе, моча гипотонична; количество ее мало: в 5 мес. - 2,2 мл/ч, к рожде-
нию - 26,7 мл/ч.
❖ Выделительная функция почки плода незначительна, эту роль выполняет пла-
цента.
19.2. Структурно-функциональные особенности почек новорожденных.
370
Нефроны функционально незрелы (созревание - к 5 - 7-му году жизни).
❖ Величина почечного кровотока составляет 5 % минутного объема крови (у
взрослых - 25 %). Основная часть крови течет через мозговое вещество (у взрос-
лых - через корковое).
❖ Функциональная незрелость фильтрационного аппарата: мвлая проницаемость и
площадь фильтрующего барьера, низкий уровень кровотока (клиренс по инулину
- 60 мл/мин на 1,73 м2).
Незрелость аппарата реабсорбции (длина проксимальных канвльцев в 10 рвз
меньше, чем у взрослых), поэтому менее интенсивно реабсорбируются амино-
кислоты, ионы и вода, однако белки и глюкоза реабсорбируются полностью.
4 * Низкий уровень секреции веществ из крови в просвет канальцев.
• •• Низкое концентрирование мочи; функция осмоконцентрации формируется в те-
чение 1-го года жизни.
• •• Почки менее чувствительны к гормонам (например, к АДГ) и более подвержены
нервным алияниям по сравнению с почками взрослых.
• •• В меньших пределах происходит компенсация сдвигов кислотно-основного рав-
новесия.
♦ ♦♦ Функциональная незрелость почек новорожденных проявляется при нагрузках:
введение воды может вызвать отек, избыток пищи - гиперазотемию.
19.3. Состав и количество мочи. Причины зкстраренальной иотври воды у новорож-
денного.
❖ У новорожденных детей ночной диурез превышает дневной (у взрослых наобо-
рот).
Моча новорожденных гипотонична (450 мосм/кг) и приближается к уровню
взрослых к концу 1-го года; осмоконцентрация мочи создается в основном соля-
ми, доля мочевины не превышает 15 %.
♦> Экстраренальная потеря воды у детей раннего возраста связана с относительно
большим испарением воды через кожу, легкие, выведением через кишечник.
У новорожденных и грудных детей не развито чувство жажды и имеется сююн-
ность к дегидратации.
19.4. Образование и выделение мочи у детей в другие возрастные периоды.
•> Особенности мочеобразования.
• Скорость клубочковой фильтрации быстро нарастает в течение 1-го года жиз-
ни и медленно увеличивается в дальнейшем, достигая уровня взрослых в 4 го-
да.
• Процессы канальцевой реабсорбции и секреции.
Ф С возрастом происходит увеличение длины канальцев, их избирательной
проницаемости и активности ферментов.
•ф Реабсорбция Na* в раннем постнатальном периоде повышена примерно в
5 рвз по сравнению с взрослым, в результате возможна задержка введен-
ного Na* и развитие отеков.
•ф Уровень реабсорбции и секреции достигает нормы взрослого к концу 1-го
полугодия жизни.
• Осмотическое концентрирование мочи.
•ф Быстрое нарастание концентрационной способности и достижение уровня
взрослых к концу 1-го года.
371
•ф* Создание осмотической концентрации в основном за счет солей (доля мо-
чевины менее 15 % в связи с положительным азотистым балансом).
‘Ф’ Чувствительность собирательных трубок к АДГ приближается к уровню
взрослых к концу 1-го года.
"Ф* Суточный диурез (в мл) у детей старше 1 года можно рассчитать по фор-
муле: 600 + 100 (В-1), где В - возраст в годах.
♦♦♦ Мочеобразование и питьевой режим при искусственном вскармливании детей:
более быстрое развитие концентрирующей способности почек и выделение ги-
пертонической мочи, так как в коровьем молоке больше солей и белков, чем
женском.
♦♦♦ Особенности мочеиспускательного рефлекса у грудных детей.
• Большая частота мочеиспусканий у грудных детей (20 - 25 раз в сутки;
уменьшается до уровня взрослых к 10 - 15 годам).
• Формирование условнорефлекторной регуляции мочеиспускания происходит
к концу первого года жизни, закрепление условного рефлекса к - 2-м годам.
• Увеличение (у детей старше 1 года) порогового объема мочи в мочевом пузы-
ре.
• Преобладание ночного диуреза над дневным диурезом.
193. Особенности почечной регуляции основных физико-химических констант крови у де-
тей.
♦♦♦ Роль почек в регуляции КОС (см. тему 21; задание 1: 10.2).
♦♦♦ Осмо- и волюмрегуляция.
• Низкая способность концентрировать мочу обусловлена незрелостью струк-
тур, обеспечивающих эту функцию почки (см. выше 18.4).
• Организм ребенка затрачивает примерно вдвое больше воды на выведение од-
ного и того же количества осмотически активных веществ по сравнению с
взрослым, что создает риск дегидратации. Дополнительную нагрузку на осмо-
регуляцию дает вскармливание коровьим молоком, в результате чего возраста-
ет потребность в воде для выведения осмотически активных веществ.
• В случае поступления избыточного количества жидкости в организм возника-
ет опасность гидремии, так как у незрелой почки ограничены возможности
выведения воды из-за малой клубочковой фильтрации.
• Имеется незрелость всех звеньев осмо- и волюмрегулирующих рефлексов, ко-
торые формируются до уровня взрослых к началу 2-го года жизни.
• Преобладание процессов ассимиляции над диссимиляцией разгружает экскре-
торную деятельность почек: характерен положительный баланс азота (меньше
образуется и выделяется мочевины), Na+, К+, Са2*, СГ, фосфатов и др.
19.6. Участие вненочечных систем выделения в поддержании гомеостаза. Влияние по-
тоотделения,
♦ ♦♦ Значение потоотделения в выведении продуктов обмена увеличивается с возрас-
том, так как полного развития потовые железы достигают к 7 годам жизни ре-
бенка.
Общее количество потовых желез у новорожденного такое же, как у взрослого (2
- 3,5 млн), плотность в 10 раз выше.
♦ ♦♦ В течение жизни увеличиваются размеры потовых желез и их секреторная актив-
ность.
372
♦ t* У детей функционируют не все потовые железы, так как часть их недоразвита и
не все зрелые функционируют одновременно.
20. Профильный материал для студентов стоматологического факультета.
20.1. Выделительная (экскреторная) функция слюнных желез и слизистых оболочек
полости рта,
❖ Слюнные железы экскретируют из крови йод, бром, соли тяжелых металлов:
ртути, висмута, свинца, золота; лекарства, например, сульфаниламиды, пеницил-
лин и др.
♦♦♦ При недостаточной экскреторной функции почек компенсаторно включаются
слюнные железы, через которые экскретируется мочевина, переходящая в амми-
ак под влиянием уреазы микроорганизмов зубного и язычного налета.
20.2. Роль слюнных желез в экскреции и поддержании гомеостаза организма.
Для слюнных желез характерна экскреторная (неспецифическая) функция, за
счет которой они обеспечивают поддержание гомеостаза организма.
♦5» При недостаточности экскреции мочевины почками слюнные железы выводят
избыток мочевины, помогая поддерживаете этот гомеостатический показатель в
крови.
♦♦♦ При различных заболеваниях слюнные железы могут экскретировать накапли-
вающиеся метаболиты: при подагре в слюну выделяется мочевая кислота, при
желтухе - билирубин, при уремии - мочевина.
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий I и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): (1-9], [11-16].
3.2. Дополнительная литература (см. с. 514 - 522): [30], [36], [79], [119], [141], [144],
[152], [197], [215], [222], [224], [225], [235], [244-246], [247], [249], [251-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 25-1. При отравлении некоторыми веществами (например, флоридзином), несмотря на
нормальный уровень глюкозы в крови она появляется в моче (возникает глюкозурия). Объясните,
какой отдел нефрона и какой молекулярный механизм являются при этом мишенями для токсиче-
ского вещества?
373
Задача 25-2. В почках в области собирательных трубок полностью заблокирован отток мочи.
Давление мочи в канальцах нефрона н капсуле увеличилось на величину фильтрационного давле-
ния. Прекратится ли прн этом фильтрация? Объясните свой ответ.
Задта 25-3. Р/птпык» употребление мочегонных лекарств уменьшает запасы Na+ в организме
н концентрацию его в крови, что приводит к увеличению секреции К* в почках н выведения его из
организма. Объясните механизмы этого явления.
374
Тема 26. Терморегуляция
Цель: а) знать основные физиологические процессы, обеспечивающие поддержание постоянства
температуры тела: теплопродукцию и теплоотдачу, их баланс при сохранении изотермии в
меняющихся температурных условиях среды; становление механизмов терморегуляции в он-
тогенезе, способы измерения температурных показателей и температуру разных участков по-
верхности тела и внутренних органов;
б) уметь применять эти знания при анализе эффектов воздействия иа организм термических
нагрузок (тепловых и холодовых), использовать эти знания для профилактики нарушений
температурного гомеостаза;
в) ознакомиться с практическими навыками иа уровне знания: методы исследования потоот-
деления; на уровне умения: исследование температурной чувствительности.
Мотивация темы: теплокровие является важнейшим условием для обеспечения нормального протека-
ния метаболических процессов. Нарушения температурного гомеостаза (гипо- и гипертермия)
отражаются иа деятельности всех систем организма и прежде всего на интегративных функ-
циях нервной системы. Знание механизмов терморегуляции позволяет давать рациональные
рекомендации по режимам труда и отдыха в разных климатических зонах и разрабатывать на-
учно обоснованные рекомендации по закаливанию, использованию тепловых процедур как
средств реабилитации после заболеваний.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Общая характеристика системы терморегуляции.
1.1. Классификация организмов по способу поддержания температуры тела.
❖ Пойкияотермные (холоднокровные) - это животные и насекомые, темпера-
тура тела которых колеблется в такт с температурой окружающей среды.
(Для повышения температуры тела они используют внешние источники тепла, имеют
обычно высокую теплопроводность тела.)
❖ Гомойотермные (теплокровные) - человек, высшие позвоночные живот-
ные и птицы способны к изотермии: поддержанию постоянной температу-
ры «ядра» тела при значительных колебаниях температуры среды. (Они
имеют мощные внутренние источники тепла и более хорошую теплоизоляцию тела.)
❖ Гетеротермия (суточные или сезонные отклонения температуры превы-
шают ± 0,5 °C) характерна для животных, впадающих в спячку, и некото-
рых птиц. У человека гетеротермия имеется у новорожденных, у взрослых
- во время фазы быстрого сна.
1.2. Роль гомойотермии в эволюционном развитии организмов.
♦ > Значение поддержания постоянства температуры внутренней среды.
• Температура тела влияет на скорость ферментативных реакций и мета-
болизм.
• Температура тела влияет на скорость образования и стабильность свя-
зей в комплексах: гормон - рецептор, медиатор - рецептор, антиген -
антитело, активность ионных каналов и др.
375
• Изменения температуры тела влияют на функции органов: возбуди-
мость ЦНС, сократимость мышц и миокарда.
❖ Преимущества теплокровия.
• Высокая интенсивность обмена веществ делает организм более свобод-
ным в выборе места обитания.
• У гомойотермных организмов в 2 - 3 раза выше скорость деятельности
ЦНС, мышечной и других систем.
Ф Цена теплокровия.
• При тех же размерах тела уровень обмена веществ в 10 н более раз вы-
ше, чем у холоднокровных.
• Теплокровным организмам требуется больше пищи.
• Теплокровные виды имеют меньшую численность н распространен-
ность (особенно в водной среде).
13. Температурные характеристики разных участков тела человека.
❖ «Оболочка» тела в обычных температурных условиях - это слой тканей от
поверхности тела до глубины - 2 см, занимает до 1/2 массы тела.
& Температура оболочки изменяется в зависимости от температуры внешней
среды.
• Для оболочки тела характерен вертикальный градиент температур (лоб
-стопы), который в норме равен 6-8 °C, горизонтальный градиент (тем-
пературная асимметрия кожи конечностей) не превышает 0,6 °C.
• Средневзвешенная температура кожи - 33 °C (среднеарифметическая вели-
чина по результатам измерения на 7 разных участках кожи по специальной формуле,
от нее зависит направление переноса тепла между организмом и средой).
• Проксимально - дистальный градиент для кожи рук составляет 3,5-3,8 °
С, для ног - 4,9-5,7 °C.
❖ Температура «ядра» тела имеет узкие пределы колебаний температуры (на-
пример, в прямой кишке 37,3 ± 0,5 °C). Размеры «ядра» зависят от темпера-
туры окружающей среды: при высокой температуре среды объем «ядра»
увеличивается, а «оболочки» уменьшается, при низкой - увеличивается
толщина «оболочки» и уменьшается «ядро».
❖ Диапазон температуры «ядра» тела, при котором сохраняется жизнеспо-
собность организма равен 20 - 42 °C. Прн температуре - 30 °C происходит
потеря сознания (холодовой наркоз, потребление О2 снижено в 2 раза), 20 - 25 °C -
остановка сердца н дыхания (потребление О2 10-15% от нормы), - 42 °C - теп-
ловой удар). (Повреждающее действие гипертермии связано с тем, что тепловая энергия
молекул становится достаточной для разрушения межмолекулярных связей, обеспечи-
вающих нативные свойства белков -1-7 ккал/моль)
• > Температура комфорта - прн этой температуре минимальна нагрузка на
физиологические системы терморегуляции.
• Для легко одетого человека - это 25 °C прн относительной влажности
воздуха 50 % н температуре стен, равной температуре воздуха; для об-
наженного человека - 28 °C (операционные, бассейны); в воде 31 -
376
35°С (чем больше слой подкожной жировой клетчатки, тем ниже температура ком-
форта: слой жира в 1 мм позволяет снизить температуру комфорта в воде на 1,5 °C).
• Для ощущения комфорта особенно важна температура кожи в области груди (31,5 -
33,5 °C ), а также ее различия с открытами участками кожи, которые не должны пре-
вышать 3 -5°С (разница менее 1,8 создает ощущение жары, более 6 °C - холода).
❖ Зависимость температуры тела от внешних и внутренних факторов.
• Время суток: минимальная температура в 2 - 6 часов утра (во время фа-
зы быстрого фаза сна тонус скелетных мышц и теплопродукция самые
низкие) максимальная между 16 и 18 часами.
• Эмоциональное возбуждение может вызвать повышение температуры
тепа на 2 °C.
• Фаза менструального цикла - перед овуляцией наблюдается повышение
температуры иа 0,5 - 0,7 °C.
• При интенсивной мышечной работе происходит повышение температу-
ры тела до 40 °C.
❖ Методы измерения температуры тела у человека: термометрия и электро-
термометрия, термовизиография.
2. Процессы теплообразования (химическая терморегуляция, обеспечивающая защиту ор-
ганизма от холода).
2.1. Обмен веществ как источник образования тепла.
❖ Несократительный термогенез (может увеличивать теплопродукция до 70%).
• Первичная теплота обусловлена рассеиванием энергии при окисли-
тельном фосфорилировании в митохондриях. КПД при синтезе АТФ не
превышает 45%, остальная энергия субстратов превращается в тепло.
• Вторичная теплота образуется в результате освобождения тепловой
энергии (более 50 %) при использовании АТФ для деятельности органов
(биосинтез, невидимое сокращение мышц, работа насосов, секреция).
• Важную роль в несократительном термогеиезе играет бурая жировая
ткань (особенно у детей). Клетки ее имеют много митохондрий и высо-
кий уровень окисления жирных кислот (в 20 раз выше, чем в белой жировой
ткани). Продукция тепла в ней на единицу массы может быть в 3 раза
выше, чем в работающей мышце. (У взрослых она расположена в области шеи,
между лопатками, в средостении около аорты, полых вен н симпатической цепочки.
Предполагают, что она обогревает крупные сосуды, направляющие кровь в мозг.)
❖ Сократительный термогенез (связан с сокращениями скелетных мышц).
• Непроизвольные мышечные сокращения (преимущественно осуществляются
медленными дангательнымн единицами). Теплопродукция при этом может по-
вышаться в 3 раза.
ф Тонус мышц, формирующий позу (например, сидячая поза увеличивает те-
плопродукция на 40%, а стоячая - на 70% по сравнению с лежачей позой).
ф Терморегуляторный тонус обеспечивают мышцы головы, шеи,
сгибатели конечностей. Теплопродукция повышается до 50% за
счет медленных ДЕ, работающих в режиме зубчатого тетануса.
377
ф' Холодовая мышечная дрожь (обеспечивается нерегулярными периодиче-
скими сериями сокращений мышц антагонистов, начинается раньше всего с же-
вательных мышц, она максимальна при ректальной температуре 34 - 35 °C и пре-
кращается при температуре 30 °C; за несколько минут теплопродукция может
увеличиться в 5 раз, продолжается ие более нескольких часов).
• Произвольные мышечные сокращения (быстрые двигательные единицы, около
2/3 энергии АТФ при этом превращается в тепло). При физической работе ДО 90
% образующегося тепла приходится иа долю скелетных мышц. Тепло-
продукция организма при тяжелом физическом труде может повышать-
ся в 8 раз, кратковременно - в 15 раз.
❖ Отношение величины поверхности тела к массе тела (М. Рубнер): чем
больше поверхность и меньше масса, приходящаяся на единицу поверхно-
сти, тем больше должны быть интенсивность обмена и теплопродукция для
поддержания гомойотермии.
2.2. Роль разных органов в теплопродукции. В условиях покоя тепло выделяют:
органы брюшной полости (особенно печень) - 34 %, мозг - 16 %, мышцы - 16
%, сердце - 11 %, почки - 8 %.
2.3. Регуляция теплопродукции.
❖ Изменение уровня основного обмена: в условиях Крайнего Севера (например,
у эскимосов) основной обмен увеличивается до 2700 ккал/сут, в тропических
условиях снижается до 1500 ккал/сут.
❖ Симпатическая регуляция несократительного термогенеза осуществляет-
ся преимущественно через (J-адренорецепторы.
• Холодовая активация мозгового слоя надпочечников и мобилизация
энергетических резервов: усиление гликогенолиза и липолиза, усиление
окисления глюкозы и жирных кислот.
• Холодовая активация окислительных реакций в буром жире (разобще-
ние окисления и образования АТФ).
•> Эндокринная регуляция несократительного термогенеза.
• Усиление секреции адреналина и тироидных гормонов приводит к уве-
личению интенсивности окислительных процессов, разобщению окис-
ления и фосфорилирования в дыхательной цепи митохондрий, увеличе-
ние образования первичной теплоты.
• Увеличение секреции глюкокортикоидов на холоде усиливает действие
адреналина в результате повышения синтеза p-адренорецепторов.
❖ Соматическая нервная система.
• Экстрапирамидная (непроизвольная) регуляция за счет терморегулятор-
ного тонуса и холодовой мышечной дрожи может увеличивать тепло-
продукцию по сравнению с величиной основного обмена до 3-х раз.
• Пирамидная (произвольная) поведенческая регуляция за счет произволь-
ной двигательной активности (физической нагрузки) может увеличивать
теплопродукцию до 15 раз.
❖ Изменение аппетита и потребления пищи: на холоде - усиление, в жару -
снижение.
❖ Структурные перестройки при долговременной адаптации.
378
• Увеличивается количество бурой жировой ткани с 1-2% до 5%.
• Повышается доля медленных двигательных единиц в мышцах, что уве-
личивает терморегуляторный тонус.
• Увеличивается содержание ненасыщенных жирных кислот в биомем-
бранах.
3. Процессы теплоотдачи (физическая терморегуляция обеспечивают защиту как от холода,
так и от действия жары, в онто- и филогенезе формируется позднее химической терморегу-
ляции).
3.1. Перенос тепла от внутренних органов («ядра») к поверхности тела (кожа,
слизистые оболочки) - внутренние потоки тепла.
* «* Пассивное теплопроведение (эффективность передачи низкая).
❖ Перенос тепла кровью от внутренних органов к коже и слизистым (умень-
шение этого процесса ведет к повышению температуры «ядра» и охлажде-
нию «оболочки»), эффективность передачи высокая.
♦ > Теплоизолирующие свойства оболочки тела зависят:
• от толщины слоя подкожной жировой клетчатки;
• от интенсивности кровотока в сосудах кожи и состояния артериоло-
венулярных шунтов.
3.2. Способы теплоотдачи с поверхности тела (наружные потоки тепла).
Ъ Излучение - осуществляется без непосредственного контакта между тела-
ми.
• В покое обеспечивает до 60 % теплоотдачи (все физические тела при темпе-
ратуре выше абсолютного нуля излучают электромагнитные волны, длина которых за-
висит от температуры поверхности. При физиологической температуре тело человека
излучает инфракрасные волны длиной от 5 до 20 мкм).
• Величина и направление переноса тепла зависит от разницы абсолют-
ных температур кожи и среды СГ*к - Т'с) и от площади излучения.
• Основные участки, с которых осуществляется теплоотдача, - лицо, го-
лова, кисти рук.
• Роль инфракрасной солнечной радиации в поддержании теплового ба-
ланса организма (нагрев организма).
❖ Проведение - прямая передача кинетической энергии молекул от более на-
гретого тела к менее нагретому.
• Величина теплоотдачи по этому пути в покое около 15 %.
• Величина теплопотери зависит:
• ф- от разницы абсолютных температур кожи и среды (Т'к - Т'с) и от
величины поверхности, с которой осуществляется теплоотдача;
• ф- от плотности среды (например, различия в теплопроводности воз-
духа и воды равны 1:25; в воде при 10 °C человек теряет тепло в 15 раз
больше, чем ив воздухе, выживаемость в этих условиях 30 - 90 мин);
• ф от насыщенности воздуха водяными парами (прямая зависимость);
• ф от толщины слоя подкожной жировой клетчатки (обратная зависи-
мость).
379
❖ Конвекция - перенос тепла движущимися частицами среды (воздухом, во-
дой наружу). Величина теплоотдачи в покое около 16 %.
• Естественная конвекция: удаление теплого слоя воздуха или воды с
поверхности теплоотдачи.
• Принудительная конвекция: увеличение скорости смены нагретой около
кожи среды вентилятором, веером н др. (например, теплоотдача при езде на
велосипеде больше, чем при беге).
❖ Испарение. Величина теплоотдачи в покое составляет около 19 %.
• Неощутимое испарение:
ф- с кожи вне потовых желез в сутки испаряется до 700 мл воды;
со слизистых оболочек воздухоносных путей в сутки испаряется до
400 мл воды.
• Ощутимое испарение (потоотделение) обеспечивается мерокриновыми
потовыми железами (для испарения 1 л воды требуется 580 ккал):
ф' начинается в условиях покоя при температуре внешней среды, пре-
вышающей 25 °C;
ф' прн температуре среды, превышающей 33 °C, является единствен-
ным путем отдачи тепла из организма.
• Эффективность испарения зависит от температуры среды и насыщенно-
сти воздуха водяными парами (при напряжении водяных паров более 40 мм рт.
ст. испарение прекращается).
• Ограничение теплоотдачи путем испарения связана с недостаточным
поступлением воды, так как некомпенсированная потеря воды в 10 - 20
% от массы тела смертельна для всех млекопитающих (потеря воды у чело-
века на жвре и при работе в горячих цехах ие должив превышать 2 - 3 % массы тела).
• Методы исследования потоотделения:
ф' по изменению электропроводности кожи;
колориметрический способ (йод-крахмальная проба, визуальный
способ);
Ф’ гравиметрический (весовой) метод.
3.3. Физиологические механизмы регуляции теплоотдачи.
❖ Изменение свойств «оболочки» тела.
• Быстрый путь: изменение интенсивности кровотока в сосудах кожи
(резко изменяет перенос тепла к обояочке тела).
Кровоток может меняться в 100 раз: в условиях температурного
комфорта кровоток в коже равен -10 мл/100 г в мни, прн тяжелой
физической работе и перегревании он может возрастать в 10 - 15
раз и достигать 20% от МОК. При охлаждении кровоток в коже па-
дает до 1мл/100 г в 1 мин (полная вазодилятация увеличивает теплоотдачу с
участка кожи по сравнению с полной вазоконстрикцией в 8 раз).
ф' Противоток в артериальных и венозных сосудах кистей, стоп, лица
действует как поворотно-противоточный теплообменник, сбере-
380
тающий тепло и обеспечивающий разогрев крови, поступающей к
сердцу и мозгу и нормальное функционирование этих органов.
ф Анастомозы шунтируют кровоток и регулируют теплоотдачу путем
изменения толщины изолирующей оболочки и температуры кожи.
ф Симпатическая регуляция кровотока в сосудах кожи осуществляет-
ся через а-адренорецепторы (сосудосуживающие эффекты). Изме-
нения сосудистого тонуса в кожных артериолах при действии низ-
ких и высоких температур являются одним из механизмов обеспе-
чения акклиматизации и закаливания.
• Медленный путь: изменение толщины подкожной жировой клетчатки
как результат изменений аппетита и потребления пищи (изменяет тепло-
изолирующие свойства «оболочки» тела).
Потоотделение (может увеличиваться до 70 мл/мин).
• Регуляция потовых желез осуществляется через холинергические сим-
патические окончания. Ацетилхолин через М-холинорецепторы усили-
вает потоотделение, атропин блокирует эффект (вторым медиатором в
синапсе является ВИП, который увеличивает кровоток в зоне потовой
железы).
• Адреналин, циркулирующий в крови, действует через центральные а-
адренорецепторы и активацию симпатического отдела (например, потоот-
деление при стрессе, чувстве страха).
• Брадикинин стимулирует потоотделение в результате усиления крово-
тока в сосудах потовых желез и повышения проницаемости капилляров.
• Состав и количество пота при термических нагрузках у неадаптиро-
ванных к высоким температурам людей отличается от адаптированных:
у последних пот более гипотоничен, интенсивность отделения выше,
кроме того, у них усилена жажда и прием воды.
Поведенческая терморегуляция позволяет снизить напряжение на физиоло-
гические регуляторные системы.
• Более чем 4-кратное увеличение задержки или отдачи тепла по сравне-
нию с покоем не может быть обеспечено только вышерассмотренными
физиологическими механизмами. В этих случаях поддержание темпера-
турного гомеостаза возможно лишь путем поведенческой регуляции.
• Кора больших полушарий обеспечивает в терморегуляторном поведе-
нии:
"Ф осознание температурного раздражителя и поиск зоны температур-
ного комфорта;
"Ф условнорефлекторную регуляцию теплопродукции и теплоотдачи;
ф произвольное изменение позы и физической активности, изменение
теплоизоляции (в том числе одежды, ее «арктический вариант»
снижает теплоотдачу в 6 раз);
• Поведение организма, осуществляемое на основе высшей нервной дея-
тельности, направлено на поиск и преобразование условий окружающей
381
среды, ограничивающих действия термических факторов по времени
(режим труда в холодных и жарких климатических условиях и др.), по
интенсивности (одежда, жилища, укрытия, создание искусственного
микроклимата и т.д.).
4. Функциональная система терморегуляции.
4.1. Системообразующий фактор - температура крови («ядра» тела).
4.2. Температурный анализатор как звено афферентного синтеза.
❖ Рецепторный отдел.
• Периферические терморецепторы кожи, кожных и подкожных сосудов.
Функциональная роль: ответ на изменение температуры среды и
температуры «оболочки» до изменения температуры «ядра», что
позволяет осуществлять регуляцию по возмущению.
Максимальная плотность и чувствительность терморецепторов на
главных участках теплоотдачи: лицо, голова, шея, минимальная -
иа коже нижних конечностей.
Специализация рецепторов: холодовые, тепловые, полимодальные.
Соотношение холодовых и тепловых рецепторов в коже равно 8:1.
Глубина расположения холодовых рецепторов меньше (0,17 мм),
чем у тепловых (0,3 мм), латентный период ответа у холодовых ко-
роче по сравнению с тепловыми.
ф" В зоне температурного комфорта импульсация от холодовых ре-
цепторов 10-12 Гц, от тепловых - 2-4 Гц. Максимальная чувстви-
тельность и импульсация холодовых рецепторов находится в диапа-
зоне температуры кожи 15 - 34 °C, у тепловых - 34 - 43 °C (при тем-
пературе ниже 20 и выше 44 °C происходит возбуждение болевых рецепторов).
Функциональная мобильность терморецепторов является механиз-
мом настройки температурного анализатора: чем ниже температура
кожи, тем больше активных холодовых точек, прн повышении тем-
пературы растет количество тепловых точек. У адаптированных к
холоду количество активных холодовых точек меньше, чем у не-
адаптированных людей.
Сигналы от периферических терморецепторов интегрируются пре-
имущественно в переднем гипоталамусе.
• Терморецепторы внутренних органов.
Функциональная роль: обеспечивают ответ на изменение теплопро-
дукции и устраняют избыток тепла - регуляция по отклонению.
Терморецепторы ЦНС: максимальное количество содержится в
преоптической области гипоталамуса (соотношение тепловых н холодо-
вых 5:1, дифференциальный порог их чувствительности - 0,011 °C), В СПИННОМ
мозге (чувствительность в 4 раза ниже), ретикулярной формации, коре.
•ф- Терморецепторы сосудов (глубоких вен), желудка, верхних дыха-
тельных путей (преобладают холодовые рецепторы).
❖ Проводниковый отдел: от холодовых рецепторов идет передача импульсов
по волокнам А8 и С, от тепловых - по волокнам С.
382
• От терморецепторов кожи туловища, шеи, конечностей и внутренних
органов идут волокиа 1-х нейронов, находящихся в спинных ганглиях.
Их аксоны переключаются иа 2-е нейроны в задних столбах спинного
мозга, и после перекреста образуют иа противоположной стороне два
главных пути проведения температурной чувствительности.
ф Боковой спииио-таламический путь с преимущественным переклю-
чением прямо или через иейроиы ретикулярной формации в неспе-
цифических ядрах таламуса; часть волокон переключается в специ-
фических ядрах таламуса и обеспечивает осознание в коре темпера-
турного комфорта или дискомфорта.
ф Спинно-ретикулярный путь дает диффузные окончания в ретику-
лярную формацию, иеспецифические ядра таламуса, гипоталамус
(преимущественно передний отдел, где обеспечивается интеграция
сигналов от терморецепторов кожи, самого гипоталамуса и других
структур).
• От терморецепторов кожи головы и лица по афферентным волокнам
тройничного нерва —> тройничный узел (1-е иейроиы) —> спинальное
ядро V нерва (2-е нейроны) —> заднемедиальное вентральное ядро тала-
муса (3-и нейроны). Часть нмпульсацин идет в ретикулярную формацию
и йеспецифические ядра таламуса.
❖ Корковый отдел (постцентральная извилина).
• Обеспечивает восприятие температурных сигналов и осознание дейст-
вия температурного раздражителя (комфорт или дискомфорт).
• Участвует в формировании условиорефлекториых реакций на термиче-
ские раздражения, осуществляет регуляцию по прогнозированию или
упреждению.
4.3. Гипоталамус - важнейшая нервная структура, в которой расположено «ядро»
центра терморегуляции. В него поступает значительная часть импульсации из
проводникового и коркового отделов температурного анализатора, В нем фор-
мируется «установочная точка» и эфферентная программа, обеспечивающая
терморегуляцию. Вместе с тем в центральных механизмах терморегуляции уча-
ствуют и другие отделы ЦНС - кора больших полушарий, продолговатый и
спинной мозг, в которых также обнаружены термочувствительные нейроны.
❖ В гипоталамусе предполагается наличие нескольких видов терморегуля-
торных нейронов.
• Афферентные нейроны, принимающие информацию от перифериче-
ских и центральных терморецепторов. Оии расположены преимущест-
венно в переднем гипоталамусе и участвуют в сопоставлении реальной
температуры тела (и самого гипоталамуса) с установочной точкой (в
норме 37,1 °C), при которой теплопродукция и теплоотдача в организме
уравновешены.
• Термочувствительные нейроны (холодовые и тепловые). Они располо-
жены преимущественно в преоптической области и переднем гипоталя-
мусе (хотя они обнаружены н в других ядрах гипоталамуса, а также в спинном и про-
383
долговатом мозге, коре больших полушарий). Соотношение термочувствитель-
ных и нетермочувствительных нейронов в терморегуляторном центре
гипоталамуса равно 40 : 60, тепловых нейронов в 4-6 раз больше, чем
холодовых. Для термочувствительных нейронов характерна высокая за-
висимость от температуры их импульсации (более 0,6 ими,- с-' • ’С"1) и про-
водимости ионных каналов (Ql0 = 2-6), высокий дифференциальный по-
рог (0,011°С ).
- ф- Повышение температуры активирует Ма+-каналы плазмолеммы те-
пловых нейронов, входящий №+-ток вызывает деполяризацию,
достигающую КУД (нейрон приобретает пейсмекерные свойства).
• ф- Понижение температуры инактивирует К+-каналы плазмолеммы
холодовых нейронов, уменьшение выходящего из клетки К+-тока
приводит к деполяризации мембраны, достигающей КУД.
• Эфферентные нейроны, реализующие эффекторные механизмы термо-
регуляции (теплопродукцию и теплоотдачу) расположены соответст-
венно в заднем и переднем гипоталамусе.
❖ Раздражители гипоталамических нейронов.
• Афферентация от всех терморецепторов.
• Гуморальные сигналы - цитокины, простагландины, гормоны щитовид-
ной железы (Та, Тз), прогестерон, адреналин и др.
• Пирогены: экзогенные (бактериальные, вирусные) и эндогенные (цито-
кины - интерлейкины 1 и 6, ФНОа и интерфероны Р и у, простагланди-
ны Ei) действуют на «холодовые» нейроны гипоталамуса и стимулиру-
ют теплопродукцию, одновременно угнетая теплоотдачу.
Центр теплопродукции (задний гипоталамус: вентролатеральные, промежуточные и
вентромедиальные ядра гипоталамуса).
• Его раздражение вызывает повышение температуры тела в результате
усиления окисления глюкозы и жирных кислот, важную роль при этом
играют ядра гипоталамуса, секретирующие тиролиберин (тиролиберин ->
ТТГ —> тироидные гормоны).
• В дорсомедиальной части заднего гипоталамуса у стенки Ш желудочка
обнаружен моторный центр дрожи, связанный с моторными центрами
продолговатого и спинного мозга и вызывающий сначала повышение
тонуса мышц, а затем - дрожь. Он возбуждается при снижении темпера-
туры гипоталамуса даже на доли градуса.
• Разрушение центра теплопродукции приводит к потере способности
поддерживать температуру тела на холоде (пойкилотермия).
Ф Центр теплоотдачи (передний гипоталамус: паравентрикулярные, супрахиазматиче-
ские и супраоптические ядра, медиальная преоптическая область).
• Раздражение этих структур вызывает увеличение теплоотдачи в резуль-
тате расширения сосудов кожи и повышения температуры кожи, пото-
отделения и тепловой одышки.
• Разрушение приводит к неспособности выдерживать тепловую нагрузку
(перегрев).
384
❖ Эфферентные нейроны гипоталамуса управляют процессами теплопро-
дукции и теплоотдачи:
• от тепловых рецепторов возбуждаются эфферентные нейроны передне-
го гипоталамуса и активируются механизмы, обеспечивающие рост теп-
лоотдачи;
• от холодовых рецепторов возбуждаются эфферентные нейроны заднего •
гипоталамуса и усиливается теплопродукция;
• между этими нейронами имеются перекрестные тормозные связи (ре-
ципрокное торможение).
4.4. Эфферентные (исполнительные) механизмы терморегуляции изложены
выше (см. 2 и 3).
5. Профильный материал для студентов лечебного факультета.
5.1. Особенности терморегуляции у пожилых людей.
❖ Снижение активности желез (щитовидной, половых) приводит к снижению интен-
сивности обменных процессов, интенсивность основного обмена уменьшается со
скоростью 1,5 % в год после 35 - 40 лет, соответственно снижается теплопродук-
ция.
♦ ♦♦ Понижается температура кожи в результате параллельно протекающих процессов
снижения теплопродукции и замедления переноса тепла от «ядра» к коже в ре-
зультате гемодинамических нарушений и декапилляризации тканей. Температур-
ные градиенты и асимметрия более резко выражены.
❖ Снижение активности и количества потовых желез после 60 лет, что вместе с пе-
речисленными выше факторами ухудшает теплоотдачу.
♦ ♦♦ Нарушаются терморегуляторные рефлексы: снижается скорость и сила ответа на
термические раздражения, удлиняется латентный период, затягивается восстанов-
ление, реакции становятся неадекватными.
5.2. Физиологические основы искусственной гипотермии.
❖ Понижение температуры тела на 1 °C приводит к снижению скорости обменных
процессов и потребления Ог на 5 - 6 %, при снижении температуры тела до 30 °C
потребление Ог падает на 50 %, а снижение температуры мозга до 20 °C ведет к
падению потребления Ог в 5-8 раза по сравнению с нормотермией. Это позволяет
замедлить повреждения тканей при прекращении в них кровообращения на более
длительный срок по сравнению с обычной температурой, например, в нейронах
коры срок удлиняется с 5 до 10 мин.
♦♦♦ Гипотермия - это снижение температуры «ядра».
• Различают умеренную гипотермию - температура тела снижается до 32 - 28
°C и глубокую - до 20 °C.
• Гипотермия достигается путем увеличения теплоотдачи либо в охлаждающей
водной среде (ванне), либо перфузией охлажденной крови через аппарат ис-
кусственного кровообращения и сосуды тела.
•ф Для ускорения охлаждения используют лекарства, которые обеспечивают
выключение терморегуляторного тонуса и мышечной дрожи (миорелак-
санты); снижают несократительный термогенез (гвнглиоблокаторы пре-
пятствуют активации симпатоадреналовой системы).
385
•ф* Этим предупреждается перенапряжение механизмов терморегуляции и
сохраняются метаболические резервы организма, необходимые для вос-
становления температуры тела (разогрева).
6. Профильный материал для студентов педиатрического факультета.
6.1. Терморегуляция у плода: изотерм ия поддерживается за счет матери, теплоотдача идет
через плаценту, величина теплопродукции плода составляет до 10 - 15% от теплопро-
дукции матери.
6.2. Особенности терморегуляции новорожденного.
❖ Температурные показатели:
• в первые часы после рождения температура снижена, восстанавливается до 36
- 37 °C через 12 - 24 часа;
• имеется высокая зависимость температуры тела от температуры окружающей
среды: ниже 20 °C наступает быстрое охлаждение, выше 35 °C - перегревание
новорожденного;
• отсутствует четкий суточный ритм колебаний температуры;
• резче выражена температурная асимметрия разных участков.
Теплоотдача относительно большая (в 2,5 раза) по сравнению со взрослыми за
счет.
• большей поверхности теплоотдачи относительно массы тела;
• низкой теплоизоляционной способности кожи из-за слабого развития подкож-
ной жировой клетчатки и интенсивного кровотока в сосудах кожи;
• сильного развития васкуляризации кожи, температурный градиент «ядро -
оболочка» менее выражен, а градиент между кожей и средой больше, чем у
взрослых.
❖ Риск переохлаждения обусловлен:
• недостаточным увеличением теплопродукции - максимум в 2 раза, основной
механизм теплопродукции - несократительный термогенез; особая роль при-
надлежит бурому жиру, он обеспечивает разогрев крови на пути к сердцу и
мозгу;
• отсутствием механизм холодовой дрожи, который включается только в экс-
тремальных условиях и на короткое время;
• несовершенством функционирования холодовых рецепторов.
Легкость перегрева обусловлена:
• высоким уровнем теплопродукции (на 1 кг массы образуется тепла в 1,4 раза
больше, чем у взрослых);
• недостаточностью функции потовых желез (меньшей интенсивности потоот-
деления и более высоким порогом его включения);
• близостью термонейтральной зоны к температуре тела ребенка;
• малой массой тела: для ее нагрева требуется меньше времени и энергии.
6.3. Особенности терморегуляции недоношенных детей.
Более низкий уровень теплопродукции по сравнению с доношенным детьми (в 2
раза).
386
❖ Более высокий уровень теплопотерь из-за плохо выраженного слоя подкожной
жировой клетчатки и высокого соотношения поверхности и массы тела (в 1,4 раза
по сравнению с доношенными детьми).
❖ Выше границы термонейтральной зоны.
❖ Не функционируют или плохо функционируют потовые железы.
♦ ♦♦ Поэтому у недоношенных легко возникает как перегрев, так и переохлаждения, и
необходимо термостатирования среды для снижения нагрузки на терморегулятор-
иые системы (специальные боксы, кювезы).
6.4. Развитие системы терморегуляции.
Увеличивается массы тела, участвующей в теплопродукции, и снижается относи-
тельная поверхность теплоотдачи, что делает нагрузку на систему терморегуляции
менее напряженной.
♦ ♦♦ Улучшаются теплоизолирующих свойств кожи за счет толщины слоя подкожной
жировой клетчатки, совершенствуется регуляция сосудистых реакций на охлаж-
дение и согревание.
❖ Уменьшается нижняя граница термонейтральной зоны, что снижает зависимость
от изменений температуры среды.
❖ К полугоду формируется суточный ритм изменений температуры тела.
❖ Развиваются потовые железы и совершенствуются механизмы теплоотдачи.
7. Профильный материал для студентов стоматологического факультета.
7.1. Топографические особенности показателей температуры органов полости рта. В
полости рта имеется градиент температурной чувствительности:
••• тепловая чувствительность нарастает к задним отделам полости рта, также из-
меняется и температура слизистой оболочки ротовой полости;
♦♦♦ холодовая чувствительность - в обратном направлении, максимальная чувстви-
тельность в начальном отделе важна для быстрого ответа организма на угрозу ох-
лаждения поступающей пищей или водой.
7.2. Пороги чувствительности к температурным раздражителям.
♦ ♦♦ Малая чувствительность слизистой оболочки щек к холоду, еще меньшая к теплу.
Задняя поверхность языка (центральная часть) не чувствительна к температурным
раздражителям.
❖ Центр твердого нёба не чувствителен к теплу.
♦ ♦♦ Максимальная температурная чувствительность на входе в ротовую полость: кон-
чик языка и красная кайма губ. Это обеспечивает анализ температуры поступаю-
щей пищи и запуск защитных рефлексов в случае необходимости.
❖ Температурная чувствительность зубов: резцы: холодовой порог 20 °C, тепловой
порог 52 °C; остальные зубы: холодовой порог 11-13 °C, тепловой порог 60 -
70°С.
7.3. Роль слизистой ротовой полости в обеспечении температурного гомеостаза.
❖ Обеспечивает анализ температуры поступающей пищи.
♦ ♦♦ Выравнивает температуру пищи (действует совместно со слюной).
❖ Запускает терморегуляторные реакции, изменяющие теплоотдачу в ответ на по-
ступление горячей или холодной пищи.
♦ ♦♦ Испарение жидкости со слизистой ротовой полости у человека относительно не-
велико (в отличие от животных, например, собак).
Существует связь между секрецией потовых желез и секрецией слюны: при де-
гидратации организма под действием высоких температур обычно снижается сек-
387
реция слюны, а при потере воды, равной 8 % от массы тела, секреция слюны пол-
ностью прекращается.
7.4. Температурные показатели ротовой полости.
❖ Температура нижней губы 33,1 °C, верхней губы 33,9 °C, под языком близка к
температуре «ядра» тела, температура зубов 30 - 31 °C.
Термический ожог эмали и дентина возникает при превышении температуры этих
тканей до 45 °C, что необходимо учитывать при препаровке полостей зуба с ис-
пользованием высокоскоростных бормашин.
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): [1-9], [11-16].
3.2. Дополнительная литература (см. с. 514 - 522): [1], [3], [12], [21], [30], [38], [43],
[63], [74-76], [150], [152], [159], [180], [196], [197], [221], [222], [225], [226], [230],
[245-247], [251-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508-513).
Задача 26-1. Человека с тяжелым перегреванием организма (t = 40,5 °C) для быстрого охлажде-
ния поместили в ванну с холодной водой (t = 14 °C). Однако сразу после этого, его состояние
ухудшилось, увеличились мозговые симптомы перегревания организма. Используя свои знания
по физиологии терморегуляции, объясните ухудшение состояние этого человека.
Задача 26-2. Два человека с равной поверхностью тела и скоростью плывут в холодной воде (t =
14 °C), затрачивая одинаковое количество энергии. Первый из них имеет нормальное содержание
жира в организме, второй - избыточное. У кого в этих условиях раньше наступит охлаждение ор-
ганизма и почему?
Задача 26-3. Жители Средней Азии при температуре окружающей среды выше 35 °C носят теп-
лые шапки, ватные халаты свободного покроя и пьют горячий чай. Почему это позволяет им легче
переносить жару?
Задача 26-4. Для эффективного охлаждения больного при искусственной гипотермии к клинике
применяют ганглиоблокаторы (блокада передачи нервных импульсов в вегетативных ганглиях) и
миорелаксанты (блокада передачи нервно-мышечном синапсе скелетных мышц). Какие процессы
в терморегуляции при это ингибируются и какой в этом физиологический смысл?
388
ЧАСТЬ Ш
ИНТЕГРАТИВНАЯ ФИЗИОЛОГИЯ
Тема 27. Сенсорные системы (анализаторы). Тактильный, болевой, вку-
совой, обонятельный, интероцептивный, температурный и дви-
гательный анализаторы
Цель: а) знать функциональную организацию сенсорных систем (анализаторов), их значение; пери-
ферический (рецепторный) и проводниковый отделы анализаторов, специфические и неспе-
цифические пути проведения афферентных нмпульсов, их переработка в подкорковых цен-
трах; корковый отдел анализаторов - процессы высшего анализа и синтеза сенсорной инфор-
мации. ее кодирование в различных отделах анализаторов; регуляцию деятельности и адапта-
цию анализаторов; методы их исследования и возрастные особенности; физиологическую ха-
рактеристику вкусового, тактильного, болевого, обонятельного, двигательного, температур-
ного и интероцептивного анализаторов, их рецепторные, проводниковые и корковые отделы;
б) использовать эти знания для понимания процессов жизнедеятельности целостного орга-
низма в его взаимодействии с внешней средой;
Мотивация темы: анализаторы осуществляют восприятие и анализ информации о явлениях, происхо-
дящих как в окружающей организм среде, так и внутри самого организма. Учение об анализа-
торах заложило основы материалистического понимания ощущений, процессов познания,
обучения н поведения. Болевая (ноцицептивная) чувствительность имеет большое значение
для выживания организма, входит в симптомокомплекс многих заболеваний. Знание физиоло-
гических процессов, лежащих в основе деятельности изучаемых на данном занятии анализа-
торов, необходимо для врача любой специальности.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по дайной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Общая физиологии сенсорных систем (анализаторов).
1.1. Общая характеристика анализаторов.
❖ Анализаторы - часть нервной системы, обеспечивающая восприятие и ана-
лиз информации о внешней и внутренней среде организма и формирующая
специфические (для данного анализатора) ощущения.
• Органы чувств (фрагмент анализаторов) - это совокупность рецептор-
ных и специализированных вспомогательных структур, обеспечиваю-
щих восприятие воздействий внешней среды. К ним относят органы
зрения, обоняния, осязания, вкуса, слуха.
• Ощущения н восприятия как конечный результат деятельности анализа-
торов.
Ощущение - это отражение свойств предметов объективного мира,
т.е. сенсорный образ, возникающий прн их действии на органы
389
чувств (рецепторы). Ощущение - субъективный образ объективно
существующего мира.
•ф- Восприятие - истолкование ощущений в соответствии со своим
опытом, т.е. опознание сенсорного образа.
❖ Физиологическое значение анализаторов.
• Анализаторы вводят информацию о внешней и внутренней среде орга-
низма в мозг, что является необходимым условием развития и функ-
ционирования ЦНС.
• Эта информация используется для регуляции гомеостаза, адаптации,
поведения и процессов познания.
❖ Учение об анализаторах как основа материалистической теории отражения
и познания; физиологический идеализм Й. Мюллера (закон специфических
энергий органов чувств) и Г. Гельмгольца (теория символов).
❖ Учение И.П. Павлова об анализаторах; отделы анализаторов: перифериче-
ский, проводниковый, корковый. По Павлову, анализ раздражения, начи-
нающийся в рецепторах и заканчивающийся в коре больших полушарий,
является единым процессом.
1.2. Периферический отдел анализаторов (рецепторы + вспомогательные струк-
туры).
❖ Понятие о рецепторах, их классификация (см. тему 4; задание 1:3.1 и 3.2).
❖ Основные функции рецепторного отдела.
• Обнаружение сигналов.
•ф- Минимальный порог возбуждения - сила раздражителя, вызываю-
щая вероятность восприятия в 50 - 75 % случаев его действия.
"Ф Максимальный порог возбуждения - сила раздражителя, вызываю-
щая болевую реакцию.
• Различение сигнала (дифференциальные пороги).
•Ф Порог интенсивности - минимальное изменение силы раздражите-
ля, которое позволяет ощущать его как новый раздражитель. На-
пример, для обонятельного анализатора дифференциальный порог
интенсивности равен 30 %, для вкусового - 10 %, тактильного - 3
%, зрительного - 1,5 %, слухового - 1 %.
•Ф Пространственные пороги - минимальное расстояние между двумя
раздражителями, при котором они ощущаются раздельно. Напри-
мер, на кончике языка - 1,1 мм, пальца - 2,5 мм, на плече - 68 мм.
"Ф Временные пороги - минимальное время между двумя раздражите-
лями, в течение которого они воспринимаются раздельно (например,
в зрительном анализаторе - 0,04 с для палочек).
• Кодирование информации в рецепторном отделе.
•Ф Кодирование силы раздражителя амплитудой рецепторного потен-
циала.
•ф Кодирование продолжительности действия раздражителя продол-
жительностью рецепторного потенциала (в медленно адаптирую-
щихся рецепторах).
390
ф- Кодирование «начало-конец» раздражения (характерно для быстро
адаптирующихся рецепторов).
ф- Кодирование направления движения стимула (например, в вестибуляр-
ных рецепторах сдвиг киноцилии от стерецилий - возбуждение рецептора, в сто-
рону стереоцилий - торможение рецептора).
1.3. Проводниковый отдел анализаторов.
❖ Проводниковый отдел - цепь афферентных нейронов (от 3 до б), соединяю-
щих рецепторный отдел с сенсорной корой. Его основные функции - про-
ведение сенсорной информации и переработка ее в местах переключения.
❖ Особенности проведения афферентных возбуждений.
• Многоканальность проведения одной и той же информации (принцип
избыточности и надежности).
• Многоуровневость проведения возбуждения: ганглионарный, спиналь-
ный, стволовой и таламический уровни (перекодирование, анализ и син-
тез информации на каждом уровне).
• Наличие «сенсорных воронок»: дивергирующие воронки обеспечивают
детальный и сложный анализ различных свойств раздражителя; конвер-
гирующие воронки уменьшают поступление избыточной информации.
• В каждом анализаторе афферентная импульсация проводится как по
специфическому, так и по неспецифическому путям, но в разных соот-
ношениях.
❖ Специфический (лемнисковый) путь проведения возбуждения переключа-
ется в специфических сенсорных ядрах таламуса (см. тему 7; задание 1:2.6).
❖ Неспецифический (экстралемнисковый) путь проведения возбуждения пе-
реключается в иеспецифических ядрах таламуса (см. тему 7; задание Г. 2.6).
❖ Кодирование информации в проводниковом отделе:
• частотно-спектральное кодирование частотой импульсов и характером
межимпульсных промежутков (структурой импульсного ответа) явля-
ется главной формой кодировки (ее возможности безграничны, так как безгра-
ничны разнообразные вариации пачек импульсов);
• кодирование перекрытием рецептивных полей и полями с on- и off-
центрами;
• кодирование посредством специализированных нейронов-детекторов.
1.4. Центральный (корковый) отдел анализаторов.
❖ Структурно-функциональная характеристика сенсорной коры.
• Первичная проекционная зона: высокодифференцированиые, мономо-
дальные нейроны, расположенные в IV слое коры, организованные в
нейронные колонки; характерна топографическая проекция периферии.
Происходит формирование ощущений (сенсорных образов), осознанное
и подсознательное восприятие действия раздражителей.
• Вторичная проекционная зона: полимодальные нейроны, осуществ-
ляющие взаимодействие анализаторов и более сложную переработку
сенсорной информации.
391
• Дальнейшая обработка сенсорной информации и использование ее для
формирования психофизиологических процессов (восприятия, эмоций,
мышления и др.) происходит в ассоциативной коре (третичные зоны).
• Сенсорная кора оказывает выраженное эфферентное влияние на прове-
дение сенсорного потока через ядра таламуса. Их пропускная актив-
ность организуется с учетом функционального состояния организма.
При этом в одних условиях приоритетно пропускается тактильная
имульсация, в других - проприоцептивная, в третьих - болевая и т.д.
❖ Первичный и вторичный ответы в вызванных потенциалах отражают по-
ступление в кору афферентного возбуждения по специфической и неспе-
цифической системам (подробнее в теме 6; задание 1: 4.5).
❖ В процессах высшего анализа и синтеза афферентных возбуждений про-
странственное кодирование преобладает над временным кодированием.
• Позиционное кодирование: признак раздражителя вызывает возбужде-
ние ограниченной группы нейронов, расположенных в определенном
месте нейронного СЛОЯ (например, глазодоминантные и ориентационные нейрон-
ные колонки).
• Детектирование раздражителя: нейроны-детекторы простых признаков
избирательно отвечают только на определенные параметры раздражи-
теля; детекторы сложных признаков опознают целые образы (например,
детекторы лица).
❖ Ощущения как психологический феномен формируются в результате коль-
цевого пути возбуждения от сенсорной коры —► в ассоциативные зоны ко-
ры —» в лимбико-гиппокампальный комплекс —» в мотивационные структу-
ры гипоталамуса —> возврат возбуждения в кору, включая сенсорную. (Ана-
лизаторы формируют инвариантный сенсорный образ, т.е. независимый от изменения ряда
признаков раздражителя Например, мы узнаем предмет при разной освещенности, разной
локализации в поле зрения и т.д.).
♦ > Закон Вебера - Фехнера: ощущение увеличивается пропорционально лога-
рифму интенсивности раздражителя (Е = a logl+b).
1.5. Адаптация анализаторов - это процесс их приспособления к оптимальному
восприятию раздражителей среды в связи с потребностями организма. Адапта-
ция может выражаться как снижением, так и повышением чувствительности
сенсорной системы.
❖ Адаптационные процессы иа дорецепторном уровне (например, рефлек-
торная реакция зрачка и хрусталика, влияние капсулы тельца Пачини, со-
кращение мышц среднего уха, поворот глаз и головы в сторону раздражи-
теля).
❖ Адаптационные процессы на рецепторном уровне (функциональная мо-
бильность рецепторов).
• Снижение возбудимости рецепторов.
•ф- Кальциевый механизм: повышение концентрации Са2+ в рецепторе
активирует Са2+-зависимые К*-каналы, что препятствует фазе депо-
ляризации и усиливает фазу реполяризации рецепторного потен-
циала.
497
-ф- Фосфорилирование рецепторных молекул, которые утрачивают
свойство активировать G-белки и запускать каскад реакций, приво-
дящих к формированию РП (например, родопсина при действии света, обо-
нятельных молекул при действии цАМФ-зависимой протеинкиназы А).
Инактивация Na’-каналов в зоне перехода рецепторного потенциала
в потенциал действия.
• Повышение возбудимости рецепторов.
Увеличение количества функционирующих рецепторных молекул:
ИХ активация (например, молекул родопсина в темноте) И увеличение ИХ
синтеза.
Увеличение температуры в области рецепторов (активация ионных ка-
налов).
❖ Адаптационные процессы на уровне проводникового отдела.
• Эфферентные тормозящие и возбуждающие влияния на рецепторы от
центров переключения проводникового отдела (например, от нейронов верх-
ней оливы на слуховые рецепторы, формирование полей колбочек с on- и off-
центрами ганглиозными и таламическими нейронами).
• «Воротный» контроль проведения болевой импульсации в задних рогах
спинного мозга.
❖ Адаптационные процессы на корковом уровне.
• Изменение возбудимости корковых сенсорных зон (роль ретикулярной
формации, лимбической системы и охранительного торможения).
• Возбуждающие и тормозные влияния сенсорной коры на центры пере-
ключения афферентной импульсации в таламусе, среднем и продолго-
ватом мозге.
1.6. Взаимодействие анализаторов (основные уровни межанализаторной интегра-
ции):
Спинальный уровень: взаимосвязь тактильной, болевой, температурной,
висцеральной чувствительности в результате конвергенции на нейронах
«Широкого афферентного входа» (зоны Захарьина - Геда и «воротный» контроль
спиииого мозга как пример взаимодействия анализаторов см. ниже З.б и 3.7).
❖ Стволовой уровень: ретикулярная формация ствола, неспецифические и ас-
социативные ядра таламуса объединяют деятельность анализаторов, транс-
формируют их возбуждения в активирующее и модулирующее влияния на
процессы.
❖ Таламокортикальный уровень:
• связь анализаторов через вторичные сенсорные зоны коры и ассоциа-
тивную кору;
• таламотеменная ассоциативная система создает целостные образы ок-
ружающей среды и самого организма («схему тела»);
• таламолобная ассоциативная система оценивает сенсорную информа-
цию и использует ее для построения программ поведения.
2. Тактильный анализатор.
393
2.1. Тактильный анализатор обеспечивает восприятие и анализ информации с ре-
цепторов кожи, видимых слизистых оболочек с последующим формированием
ощущений прикосновения, давления, вибрации.
2.2. Рецепторный отдел (мехаиорецепторы кожи - инкапсулированные нервные
окончания).
• > Рецепторы постоянного по интенсивности прикосновения и давления: дис-
ки Меркеля в голой коже и клетки Меркеля в волосатой коже являются
датчиками силы, реагируют на амплитуду деформации кожи (пороговая де-
формация - 10 мкм, пороговая сила - 45 мН, медленно адаптируются).
❖ Рецепторы прикосновения и вибрации в пределах 40 Гц: тельца Мейсснера
в голой коже и рецепторы волосяных фолликул являются датчиками скоро-
сти деформации кожи, умеренно быстро адаптируются (50 - 500 мс), это
наиболее частый вид рецепторов (43 %), их плотность ~ 140/см2 иа кончике
пальца руки.
❖ Рецепторы быстрых изменений прикосновения, давления, вибрации в пре-
делах 40 — 1000 Гц (максимальная чувствительность при частоте ~ 200-300 Гц): тель-
ца Пачини - в глубоких слоях кожи, сухожилиях, связках, надкостнице,
брыжейке, их пластинки действуют как фильтры медленных колебаний,
эти рецепторы являются датчиками ускорения, очень быстро адаптируют-
ся, их пороговая деформация - 1 мкм.
2.3. Проводниковый отдел тактильного анализатора.
❖ От рецепторов кожи конечностей и туловища идут миелиновые волокна
АР; первые афферентные нейроны расположены в спинальных ганглиях, их
аксоны через задние корешки входят в спиииой мозг и имеют две главные
проекции:
• образуют тонкий и клиновидный пути, идущие в задних канатиках и
переключающиеся (после перекреста) в ядрах Голля и Бурдаха в продолго-
ватом мозге (высокая чувствительность и различение свойств раздражителя),
аксоны нейронов этих ядер образуют медиальную петлю;
• переключаются (перекрест в спиином мозщ) на нейроны III-IV пластин зад-
них столбов, от которых идет передний спинно-таламический путь (низ-
кая чувствительность и различение свойств раздражителя);
• а также участвуют в спинальных сгибательных рефлексах;
• часть тактильной чувствительности (особенно от кожи стоп) идет по заднему спннно-
мозжечковому пути и используется в мозжечке для регуляции тонуса мышц, равнове-
сия и движения.
❖ От рецепторов кожи лица, головы и слизистой оболочки рта идут миелино-
вые волокна АР; первые афферентные нейроны расположены в тройнич-
ном ганглии V нерва. Вторые нейроны расположены в спинальном и сред-
немозговом ядрах V нерва, их аксоны (тройничная петля) направляются в
ядра таламуса.
❖ Третьи афферентные нейроны расположены в специфических заднемеди-
альных вентральных ядрах таламуса, их аксоны идут в корковый отдел.
Имеется топографическая проекция кожи тела в ядрах таламуса. (Небольшая
часть волокон тактильной чувствительности поступает в ретикулярную формацию ствола
и затем в иеспецифические ядра таламуса.)
394
2.4. Корковый отдел тактильного анализатора.
❖ Первичная соматосенсорная область коры (SI) располагается в задней цен-
тральной извилине, сюда поступает афферентная импульсация с противо-
положной стороны тела (лицо представлено билатерально).
• Обладает высокой степенью топографической организации («сенсорный
человечек» Пенфилда; см, рис.29, с. 103).
• Переработанная здесь информация направляется: в моторную кору (ре-
гуляция движения по обратной связи), в теменную ассоциативную кору
(восприятие «схемы тела»), в SI и SII другой стороны (билатеральная
координация), а также к таламусу, ядрам Голля и Бурдаха (эфферент-
ный контроль анализатора).
* •* Вторичная соматосенсорная зона коры (SII), расположенная на верхней стенке сильвиевой
борозды (теменная покрышка), возле слуховой зоны, филогенетически более древняя, чем
зона SI:
• сюда поступает афферентная импульсация как от противоположной стороны, так и от
«своей» стороны;
• имеет менее четкую соматотопическую проекцию поверхности тела;
• вероятно, играет роль в сенсорной и моторной координации активностей двух сторон
тела.
❖ Сенсорная кора имеет колончатый тип строения (В. Маунткасл, 1957), в в ней-
ронных колонках происходит начальная переработка сенсорной информации.
❖ Различение свойств тактильных раздражителей: пространственное разли-
чение - от 2 мм на кончиках пальцев до 70 мм на коже спины; временное
различение лежит в основе ощущения вибрации, движения по шероховатой
конечности; различение интенсивности позволяет различать по силе дав-
ление разные раздражители.
2.5. Методы исследования тактильного анализатора.
❖ Эстезиометрия, циркуль Вебера (пространственные пороги);
❖ Набор волосков Фрея (абсолютные пороги).
3. Болевой (ноцицептивный) анализатор («Боль - сторожевой пес здоровья». Гиппократ).
3.1. Общая характеристика боли.
❖ Болевой (ноцицептивный) анализатор формирует ощущение боли при физиче-
ских и химических воздействиях, оказывающих повреждающее действие на
организм. Отличия боли от других ощущений в том, что она не информи-
рует мозг о качестве раздражителя, а указывает на то, что раздражитель
является повреждающим. Другой особенностью болевой сенсорной систе-
мы является наиболее сложный и мощный ее нисходящий (эфферентный)
контроль (см. ниже 3.7).
♦ ♦♦ Классификация боли' 1) соматическая боль: а) поверхностная (кожная); ранняя и поздняя;
б) глубокая (мышцы, кости, суставы); 2) висцеральная боль (воспаление, дискинезия, на-
рушение кровообращения) во внутренних органах.
• > Компоненты боли:
• сенсорный (боль как ощущение);
• аффективный (эмоциональные, вегетативные и моторные проявления
боли);
395
• вегетативный (нарушение функции внутренних органов);
• потребностно-мотивационный (боль как отрицательная биологическая
потребность, формирующая мотивационное противоболевое поведе-
ние);
• когнитивный (оценка боли - боль как страдание - формируется в лоб-
ной коре).
• > Причины боли: 1) нарушение целостности защитных покровных оболочек тела (кожа, сли-
зистые оболочки) и внутренних отсеков организма (мозговые оболочки, плевра, брюшина
и др.); 2) нарушение кислородного режима органов и тканей до уровня, вызывающего
структурно-функциональные повреждения.
3.2. Теории боли.
❖ Теория интенсивности (Э. Дарвин, 1794; А. Гольдшейдер, 1886) - нет специфиче-
ских болевых рецепторов и нейронов, боль возникает прн действии сверхсильных
раздражителей на рецепторы различных органов чувств.
❖ Теория специфичности (М. Фрей, 1894) постулирует наличие специфических бо-
левых рецепторов и волокон проведения болевой импульсации.
❖ Современная теория боли базируется преимущественно на теории специ-
фичности с использованием элементов теории интенсивности (центральной
суммации н конвергенции бояевой импульсации) и концепции противоболевой сис-
темы.
3.3. Рецепторный отдел болевого анализатора.
❖ Болевые рецепторы (ноцицепторы) являются свободными окончаниями
чувствительных миелиновых нервных волокон А8 н немиелиновых воло-
кон С. Они найдены в коже, слизистых оболочках, надкостнице, зубах,
мышцах, органах грудной и брюшной полостей и др. (Плотность болевых ре-
цепторов в коже - 200/см2, на границе дентина н эмали зуба - 75000/см2.)
❖ Основные типы болевых рецепторов.
• Механоноцицепторы и механотермические ноцицепторы А8-волокон
реагируют на сильные механические и термические раздражители, про-
водят быструю (б-ЗО м/с) механическую и термическую боль, быстро
адаптируются, их афферентные нейроны имеют малые рецептивные по-
ля.
• Полимодальные ноцицепторы С-волокон реагируют иа механические,
термические (тепловой и холодовой) и химические раздражители, про-
водят (0,5-2 м/с) позднюю, плохо локализованную боль, медленно адап-
тируются, их рецептивные нейроны имеют большие рецептивные поля.
❖ Раздражители болевых рецепторов.
• Механические: сдавливание, растяжение, сгибание, скручивание и др.
(давление более 40 г/мм2).
• Термические: тепловые (при действии t свыше 45 °C), холодовые (при охлаж-
дении ниже 15 °C).
• Химические (возникают из поврежденных клеток, тромбоцитов, плазмы
и окончаний ноцицептивных нейронов).
"Ф- Возбуждают ноцицепторы К+, Н+, серотонин, гистамин, брадики-
нин, АДФ и др.
396
v" Сенсибилизируют ноцицепторы простагланидины и лейкотриены,
вещество Р.
❖ Свойства болевых рецепторов.
• Имеют высокий порог возбуждения (низкую возбудимость).
• Ноцицепторы С-афферентов имеют плохую адаптацию к длительно
действующим раздражителям.
• Возможна сенситизация (сенсибилизация) болевых рецепторов - сниже-
ние порога раздражения при многократной или длительной стимуляции;
сенситизация проявляется в способности ноцицептора отвечать на сти-
мулы субпороговой величины, а также возбуждаться раздражителями
других модальностей.
3.4. Проводниковый отдел болевого анализатора.
♦ > Болевая чувствительность туловища и конечностей (поверхностная и глу-
бокая), а также внутренних органов.
• А5- и С-волокна, первые нейроны находятся в спинальных узлах, их ак-
соны (в области заднего корешка группируются в пучок Лиссауэра) ВХОДЯТ В
спинной мозг и переключаются прямо или через интернейроны в задних
столбах на вторые нейроны (релейные нейроны), аксоны которых вхо-
дят в состав спиноталамических путей. Среди этих нейронов выделяют
специфические болевые нейроны и нейроны «широкого афферентного
входа», на которые конвергируется как болевая, так и неболевая им-
пульсация. Наиболее вероятным медиатором в синапсах медленных
С-афферентов является вещество Р, быстрых А5-аффереитов - глутамат.
Передача возбуждения осуществляется под контролем энкефалических
интернейронов.
• Часть болевой импульсации первых нейронов переключается (через интернейроны) иа
мотонейроны сгибателей и участвует в формировании защитных болевых рефлексов.
• Основная часть болевой импульсации (после переключения в задних столбах)
поступает в восходящие пути, среди которых боковой спинно-
таламический путь проводит большую часть болевых сигналов (его во-
локна переходят на противоположную сторону спинного мозга). В нем выделяют
эволюционно более молодой неоспинно-таламический путь (отсутствует и
низших животных) и древний палеоспинно-таламическнй путь (имеется у
низших животных).
•Ф" Неоспиноталамический путь на уровне ствола мозга проводит бо-
левые сигналы по специфическому пути (спинно-мозговая петля) в
специфические сенсорные (заднелатеральные вентральные) ядра
таламуса, имеющие хорошую топографическую проекцию перифе-
рии тела. (Небольшая часть импульсов поступает в ретикулярную формацию
ствола и далее в иеспецифические ядра таламуса.) Передача возбуждения в
синапсах этого пути осуществляется с помощью быстродействую-
щего медиатора - глутамата. Из специфических ядер таламуса бо-
левые сигналы передаются преимущественно в соматосенсорную
кору SI и SH. Эти особенности неоспинно-таламического пути
формируют его основную функцию - проведение «быстрой» боли и
397
восприятие ее с хорошей локализацией и характеристикой болевых
стимулов.
Палеоспинно-таламический путь проводит белевые сигналы по С-
волокнам по неспецифическому сенсорному пути преимущественно
в неспецифические (интарлиминарные и ретикулярные) ядра тала-
муса прямо (не более 25% волокон) или после переключения в ней-
ронах ретикулярной формации ствола мозга. Передача возбужде-
ния в синапсах этого пути происходит более медленно. Из неспе-
цифических ядер таламуса импульсация поступает не только в со-
матосенсорную, но и другие отделы коры больших полушарий.
Кроме ядер таламуса и ретикулярной формации, волокна этого пути
оканчиваются на нейронах центрального серого вещества и голубо-
го пятна ствола мозга и гипоталамуса. (Небольшая часть импульсации
поступает в специфические сенсорные ядра таламуса.) Через палеоспннно-
таламнческий путь проводится «поздняя», плохо локализуемая
боль, формируются аффективно-мотивационные проявления боле-
вой чувствительности.
Примечание: 1) вышеназванные пути проводят и другие виды чувствительности
(температурную, небольшую часть тактильной); 2) болевая чувствительность час-
тично проводится н по другим восходящим путям (спннно-ретикулярному, тон-
кому и клиновидному, переднему спнино-таламнческому, проприоцептивным пу-
тям спинного мозга и др.).
❖ Поверхностная и глубокая болевая чувствительность лица и полости рта
(тригеминальная зона).
• АЗ- н С-волокна, первые нейроны - в тройничном ганглии V нерва;
вторые нейроны - преимущественно в спинальном ядре V нерва (от ре-
цепторов кожи) н его мостовом ядре (от рецепторов мышц, суставов).
• От этих ядер болевая импульсация (аналогично спиноталамическим путям)
проводится по бульбо-таламическим путям в специфические ядра тала-
муса (от А8-афферентов) н неспецнфические ядра таламуса (от С-
афферентов). (По этим путям проводится и часть болевой импульсации от внутрен-
них органов, поступающих по сенсорным волокнам IX и X нервов в ядро одиночного
пути.)
❖ Роль таламуса в формировании боли.
• Передача и переработка болевой информации в специфических ядрах
таламуса обеспечивает анализ локализации болевого раздражения, его
силы н длительности.
• Передача и переработка болевой информации в неспецнфических ядрах
таламуса обеспечивает мотивационно-аффективный аспект болн.
3.5. Корковый отдел болевого анализатора.
❖ Расположен в соматосенсорной коре - проекционные поля SI и SII. Пер-
вичное поле SI обеспечивает восприятие «быстрой» болн, идентификацию
места ее возникновения на теле. Одиако даже для «быстрой» болн точная
идентификация кожного участка действия болевого раздражителя, требует
параллельного вовлечения в процесс кожных тактильных рецепторов. Поле
SI, которое расположено рядом с моторной корой передней центральной
398
извилины, играет ведущую роль в экстренном включении моторной защит-
ной реакции организма на действие болевого раздражителя.
Невозможность четко локализовать «медленную» боль объясняется
тем, что пути ее проведения через ретикулярную формацию и неспецифи-
ческие ядра таламуса передают возбуждение не только в поля SI и SII боле-
вого анализатора, но и в другие корковые поля.
Соматосенсорное поле SII, расположенное на границе пересечения
центральной борозды с верхним краем височной доли, имеет менее четкую
топографическую проекцию тела, импульсы поступают сюда как от проти-
воположной, так и своей стороны тела. Возможность локализации действия
болевого источника здесь меньше, чем в SI. Однако нейроны данного поля
имеют более мощные двусторонние связи с ядрами таламуса. Это позволя-
ет полю SII избирательно фильтровать проходящее через таламус возбуж-
дение, прежде всего болевого происхождения, причем возможно как уси-
ление, так и ослабление болевого потока.
❖ Лобная кора играет важную роль в восприятии боли, обеспечивая само-
оценку боли (её когнитивный компонент), и формировании целенаправлен-
ного болевого поведения. Перерезка связей между лобной корой и таламу-
сом (операция лоботомия) сохраняет ощущение боли у больных, но она их не
беспокоит (боль не становится страданием).
Лимбическая система (поясная извилина, гиппокамп, зубчатая извилина, миндале-
видный комплекс височной доли) получает болевую информацию от передних
ядер таламуса и формирует эмоциональный компонент боли, запускает ве-
гетативные, соматические н поведенческие реакции, обеспечивающие при-
способительные реакции к болевому раздражителю.
❖ Двигательные компоненты болевого поведения формируются в совмест-
ной деятельности моторной и премоторной коры (поля 4 и б), базальных
ядер и мозжечка.
3.6. Некоторые виды болевых ощущений.
❖ Проекционные (фантомные) боли в области ампутированных конечностей
основаны на законе проекции боли: какая бы часть афферентного пути не
раздражалась, боль ощущается в области рецепторов данного сенсорного
пути. (По современным данным в их пронсхождеиин участвуют и центральные структу-
ры болевой сенсорной системы).
❖ Отраженные боли (зоны Захарьина - Геда) - боль, возникающая при пато-
логии внутренних органов, ощущается на поверхности тела соответствую-
щего дерматома. Их возникновение связано с тем, что нейроны, проводя-
щие болевую импульсацию от рецепторов пораженного органа и рецепто-
ров кожи соответствующего дерматома, сходятся на одном и том же ней-
роне спиноталамического пути (конвергентные нейроны или нейроны
«широкого афферентного диапазона»). Раздражение этого нейрона с рецеп-
торов пораженного органа в соответствие с законом проекции боли приво-
дит к тому, что боль ощущается и в области кожных рецепторов. Зоны За-
харьина - Геда используются в медицине с диагностической и лечебной
целями.
399
3.7. Антиноцицептивная (противоболевая) система - это совокупность взаимо-
связанных структур, относящихся к болевому анализатору, которая обладает
способностью снижать восприятие болевой импульсации вплоть до состояния
аналгезии. В этой системе выделяют несколько компонентов.
❖ Опиоидные пептиды и их рецепторы.
• В последней трети 20 века было выяснено, что опий и его препараты
(например, морфин) оказывают обезболивающий эффект, действуя на спе-
циальные белковые рецепторы нейронов, и что для этих рецепторов
есть свои эндогенные стимуляторы. Ими оказались небольшие пептиды
- продукты протеолиза крупных неактивных пептидов-предшествен-
ников, образующихся в головном мозге, аденогипофизе, мозговом слое
надпочечников, желудочно-кишечном тракте, плаценте. Сейчас нх из-
вестно три. Из проопиомеланокортина образуются а-, [} и у-эндорфины
(состоят из 9-35 аминокислот). Из проэнкефалина А образуется метионин- и
лейцин-энкефалины (состоят из 5-6 аминокислот). Из проэнкефалина-В обра-
зуется динорфины (состоят нз 10-13 аминокислот).
• Опиоидные пептиды действуют на три основных вида опнатиых рецеп-
торов: ц (мю), 8 (дельта) и к (каппа). Наиболее селективным стимулято-
ром ц-рецепторов являются эндорфины (а также морфин), 8-рецепторов
- энкефалины, к-рецепторов - динорфины. Плотность ц-рецепторов и к-
рецепторов высокая в коре больших полушарий и спинном мозгу, сред-
няя - в стволе головного мозга; плотность 8-рецепторов средняя в коре
больших полушарий и спинном мозгу, малая - в стволе мозга.
• Опиоидные пептиды угнетают действие веществ, вызывающих боль, на
уровне ноцицепторов, и нейронов, находящихся в составе цепей, пере-
дающих и воспринимающих болевую импульсацию. Механизмы их
действия связывают с изменением активности аденилатциклазы, фосфо-
липазы С н действием соответствующих вторых посредников (цАМФ,
ИФУ ДАГ. Са24), что уменьшает освобождение медиаторов в синапсе. Эти
пептиды поступают к нейронам болевой сенсорной системы с кровью
(например эндорфины, секретируемые аденогипофизом), с цереброспинальной
жидкостью и выделяются в синаптических окончаниях нейронов проти-
воболевой системы в качестве медиаторов (например, энкефалин). Анал-
гезирующий эффект эндорфинов высокий в головном и спинном мозге,
эффект энкефалинов в этих структурах средний, эффект динорфинов в
головном мозгу низкий, в спинном мозгу - высокий.
❖ Спинальный уровень антиноцицептивной системы. Важнейшим ее меха-
низмом является «воротный контроль» спинного мозга (Р. Мелзак, 1973). Он
осуществляется в задних столбах тормозными нейронами желатинозного
вещества, которые активируются с периферии при стимуляции толстых
миелиновых волокон (например, с тактильных рецепторов, проприорецепторов) и
путем пресинаптического торможения ограничивают передачу болевой
импульсации по спиноталамическому пути (рис. 60). Спинальный воротный
механизм находится также под постоянным влиянием нервных импульсов
структур ствола мозга, которые передаются по нисходящим путям как на
400
нейроны желатинозного вещества, так н релейные нейроны спиноталами-
ческих путей.
НЖВ - тормозные нейроны желатинозного вещества
+ - возбуждение " - торможение
Рис. 60. Спинальный воротный контроль проведения болевой
импульсации: торможение проведения болевых сигналов по во-
локнам С и Л8 при повышение афферентного патока по небо-
левым волокнам Ар и Аа. .
Стволовой уровень антиноцицептивной системы.
• Центральное (околоводопроводное) серое вещество среднего мозга через
ядра шва н гигантоклеточное ядро продолговатого мозга (по рафеспиналь-
иым и ретикулоспинальным путям) активирует тормозные энкефалинергиче-
ские нейроны желатинозного вещества н ингибируют передачу болевой
импульсации в нейронах задних столбов по типу пресинаптического
торможения.
• Г о дубоватое пятно среднего мозга оказывает ингибирующее влияние на
болевую передачу в задних столбах спинного мозга (через норадреналин и
аг-адренорецепторы).
• Ретикулярное ядро таламуса вызывает торможение передачи болевых
импульсов через другие ядра таламуса.
Гипоталамический уровень антиноцицептивной системы.
• Гипоталамус является структурой, контролирующей и регулирующей
антиноцицептивные механизмы стволового уровня за счет влия-
ния норадренергических, дофаминергических, эндорфннергических и
энкефалинергических нейронов различных ядер гипоталамуса.
• Гормон кортиколиберин, образующийся в ядрах гипоталамуса, усили-
вает образование p-эндорфнна в аденогипофизе и секрецию его в крово-
ток.
401
• Стимуляция задней области гипоталамуса вызывает возбуждение сим-
патических центров, в том числе иннервирующих мозговой слой надпо-
чечников. Увеличение секреции катехоламинов при этом сопровождает-
ся повышением секреции и опиоидных пептидов (эндорфинов).
❖ Корковый уровень антиноцицептивной системы.
• Соматосенсорная область коры объединяет и контролирует деятель-
. ность антииоцицептивных систем различного уровня (нарушение этой ин-
тегрирующей функции может вызвать нестерпимую боль.)
• При этом наиболее важную роль в активации спинальных и стволовых
структур аитиноцицептивной системы играет вторая сенсорная область
(SH). Ее нейроны образуют наибольшее количество волокон нисходя-
щего контроля болевой чувствительности, направляющиеся к задним
рогам спинного мозга и ядрам ствола головного мозга. В экспериментах
на животных было показано, что двустороннее удаление областей SB
приводит к более быстрому и интенсивному болевому синдрому, чем у
животных с удаленными SI.
• Соматосенсорная кора (особенно SII) имеет выраженные двухсторонние
связи с ядрами таламуса, через которые она способна ограничить про-
хождение болевого потока через таламус
3.8. Физиологические основы обезболивания и наркоза.
Блокада образования «медиаторов» боли (например, блокаторы синтеза простаг-
ландинов, антигистаминные препараты).
❖ Проводниковая анестезия.
• Местные анестетики (например, новокаин) вызывают инактивацию Na+-
каналов и блокируют проведение ПД.
• Операция хордотомия (перерезка переднебокового канатика спинного
мозга на стороне, противоположной боли) прекращает или резко снижа-
ет проведение основного потока болевой импульсации (как правило, эф-
фект временный).
❖ Применение агонистов опиатных рецепторов (морфина, опиоидные пепти-
ды).
❖ Акупунктурная аналгезия осуществляется в результате механического,
электрического, термического воздействия на биологически активные точ-
ки (скопление сенсорных рецепторов). Основной механизм действия связан
с рефлекторным выбросом из гипофиза эндорфинов в кровоток (после ги-
пофизэкгомии аналгезия ие наступает).
❖ Общее обезболивание (наркоз).
• Наркотические средства подавляют формирование ВПСП в большей
степени, чем ТПСП. Это происходит преимущественно на уровне рети-
кулярной формации и таламуса (например, действие барбитуратов) или на
корковом уровне, особенно в сенсомоторной коре (например, действие эфи-
ра).
402
• Электроаналгезия: используются импульсные токи (100 - 200 Гц, 1 -16 мА,
катод - в лобной области, анод - под сосцевидным отростком), вызывающие като-
лическую депрессию.
3.9. Исследование болевой чувствительности. Алгезиметрия - определение болевого по-
рога с помощью прибора (алгезиметра). который устанавливают в исследуемой точке,
усиливая на нее давление до возникновения отчетливого болевого ощущения. Найден-
ный порог болевой чувствительности выражают в единицах шкалы алгезиметра (в мм).
4. Вкусовой анализатор.
4.1. Вкусовой анализатор обеспечивает восприятие и анализ химических раздра-
жителей при действии их на рецепторы языка и формирование вкусовых ощу-
щений. Его роль - оценка съедобности пищи, участие в рефлекторной регуля-
ции секреции пищеварительных соков. (В формировании вкуса участвуют также так-
тильная и температурная чувствительность ротовой полости и обонятельная чувствительность.)
4.2. Рецепторный отдел вкусового анализатора.
❖ Вкусовые клетки - вторичные контактные хеморецепторы, модифициро-
ванные эпителиальные клетки, имеющие 30 - 40 микроворсииок.
❖ Вкусовые почки (2000 - 10000), каждая содержит 30 - 80 вкусовых клеток.
• Каждая включает в себя рецепторные (вкусовые), опорные и базальные
клетки.
• Дистальные части рецепторных клеток с микроворсинками выходят в
общую камеру, которая через пору на вершине почки сообщается с
внешней средой (вкусовым раздражителем).
• Главное место расположения вкусовых почек - эпителий языка (неболь-
шое количество их имеется в эпителии мягкого неба, надгортанника).
❖ Вкусовые сосочки включают в себя вкусовые почки:
• грибовидные сосочки расположены в теле и на кончике языка, иннер-
вируются лицевым нервом (VII), каждый включает в себя 8-10 вкусо-
вых почек;
• листовидные и желобоватые сосочки расположены в задней трети язы-
ка, иннервируются IX и X черепными нервами, каждый включает в себя
до 200 вкусовых почек.
❖ Топография наибольшей чувствительности языка, формирующая «первич-
ные» вкусовые ощущения: 1) к сладкому - вкусовые рецепторы кончика
языка (наиболее высокие пороги); 2) к соленому - кончик и края языка; 3) к ки-
слому - края языка; 4) к горькому - корень языка (наиболее низкие пороги).
❖ Рецепторный потенциал (РП) вкусового рецептора представлен деполяри-
зацией (амплитуда - до 30 мВ, продолжительность - десятки секунд).
• Возбуждение вкусового рецептора происходит различными путями.
• Ф Сладкие вещества (сахара) через активацию аденилатциклазы уве-
личивают уровень цАМФ, который открывает цАМФ-зависимый
ионный канал, пропускающий внутрь клетки Na* и Са2*, а также ак-
тивирует протинкиназу-А, закрывающую некоторые К*-каналы. В
результате происходит деполяризация мембраны рецептора.
403
• ф Соленые вещества деполяризуют вкусовые клетки в результате
прямой активации Na^-каналов.
• ф Кислые вещества за счет своих ЕГ-ионов блокируют К*-каналы н
увеличивают поток НГ-ионов внутрь клетки через На+-каналы и Не-
зависимые катионные каналы, что приводит к деполяризации ре-
цептора.
• ф Горькие вещества возбуждают рецепторы через: 1) активацию
фосфолипизу-С и образование ИФз, повышение уровня Са2* в клет-
ке; 2) непосредственную блокаду К+-каналов; 3) модуляцию актив-
ности фосфодиэстеразы, вызывающей гидролиз цАМФ и цГМФ.
• РП через синаптический контакт (предполагаемый медиатор серотонин)
генерирует ПД в первом афферентном нейроне.
• Отдельный вкусовой рецептор на действие одних веществ (например,
NaCl) отвечает деполяризацией, а на действие других веществ (например,
сладких н горьких) — гиперполяризацией.
4.3. Проводниковый отдел вкусового анализатора.
❖ От вкусовых клеток отходят волокна VII нерва (барабанная струна в составе
язычного нерва), язычные ветви IX, частично волокна X черепных нервов, од-
но волокно иннервирует один нлн несколько рецепторов, олин рецептор
иннервируется несколькими волокнами.
❖ Вкусовой профиль волокон: одиночное вкусовое волокно отвечает выра-
женной импульсацией (100 - 130 Гц) на действие только одного вкусового
вещества (сладкого, горького н т.д.), на другие вещества импульсация менее
выражена (до 50 Гц).
❖ Первые нейроны находятся в коленчатом узле VII нерва или в нижних уз-
лах (IX и X нервов), их аксоны в продолговатом мозге образуют одиноч-
ный путь, оканчивающийся на вторых нейронах.
Вторые нейроны расположены в ядре одиночного пути, их аксоны:
• достигают специфических вентральных заднемедиальных ядер таламуса
(участвуют в формнрованнн вкусовых ощущений);
• поступают в гипоталамус, миндалину, концевую полоску (формирова-
ние эмоционального компонента вкусового ощущения).
❖ Третьи нейроны находятся в вентральных заднемедиальных ядрах таламуса
(n. VPM), их аксоны проецируются в корковый отдел.
4.4. Корковый отдел вкусового анализатора.
❖ По мнению одних исследователей, он расположен в нижней части постцен-
тральной извилины (поле 43), в зоне проекции языка, по мнению других - в
крючке (поле 34) парагиппокампальной извилины (вкусовая чувствительность
проецируется билатерально).
❖ Здесь формируются вкусовые ощущения 4 качеств: сладкое, кислое, соле-
ное, горькое; считается, что кодирование 4 вкусовых ощущений осуществ-
ляется импульсацией не одного волокна, а нескольких волокон.
404
❖ Одни корковые нейроны отвечают только на вещества с одним вкусовым
качеством, другие реагируют на вкусовые, тактильные и температурные
раздражители языка.
❖ Вкусовой анализатор имеет среднюю чувствительность (она более высокая для
горького - 3 мкмоль/л хинина, для кислого - 300 мкмоль/л НС1, меньшая для сладкого -
40 ммоль/л н соленого - 100 ммоль/л), дифференциальные пороги в диапазоне
средних концентраций минимальны и равны 10 - 20 %.
• Воспринимаемое вкусовое качество вещества зависит от его концентра-
ции (например, поваренная соль низкой концентрации воспринимается сладкой).
• Адаптация вкусового анализатора:
•Ф- имеется высокая способность к адаптации, причем к сладкому и со-
леному она выше, чем к горькому и кислому;
"О’ перекрестная адаптация: изменение чувствительности к одному ве-
ществу при действии другого; имеет место при использовании со-
лей, близких по вкусовому качеству, например, между ВеС12, ВеВг2
и BeS04;
"О’ смещение вкуса происходит при одновременном или последова-
тельном применении нескольких вкусовых раздражителей (напри-
. мер, адаптация к горькому повышает чувствительность к кислому и
соленому, адаптация к сладкому обостряет вкусовое ощущение
всех других вкусовых стимулов);
"О’ восприятие вкуса формируется не только иа основе ощущения че-
тырех вкусовых качеств, но и на основе ощущения прикосновения,
температуры, запаха пищи, т.е. в результате взаимодействия не-
скольких анализаторов.
4.5. Методы исследования вкусового анализатора.
Определение порога вкусовой чувствительности (наименьшей концентрации раствора
вещества, которое при нанесеннн на язык вызывает соответствующее вкусовое ощуще-
ние).
♦> Определение функциональной мобильности вкусового анализатора (фиксируют
возникновение или отсутствие вкусового ощущения после нанесения на сосочки вкусово-
го раздражителя).
5. Обонятельный анализатор.
5.1. Обонятельный анализатор обеспечивает оценку качества внешней среды (на-
пример, воздуха, пиши), участвует в регуляции системы пищеварения, коммуника-
ционных функций, эмоциональных состояний.
5.2. Рецепторный отдел.
❖ Обонятельная область (~ 5 см2) - обонятельный эпителий верхних рако-
вин и соответствующий им участок перегородки носа, имеет 3 вида клеток:
рецепторные, опорные, базальные (предшественники рецепторных клеток).
Рецептор относится к типу первичных (- 10 млн биполярных нейронов, жизнен-
ный цикл - 30 суток, МПП = -55 - 60 мВ), дендрит биполярного нейрона оканчи-
вается булавой с 7 - 12 ресничками, погруженными в слой слизи (секрет бо-
уменовых желез и опорных клеток); немиелиновые аксоны этих нейронов со-
ставляют обонятельный нерв. (В слизи обнаружены запахсвязывающие белки, кото-
405
рые повышают концентрацию пахучих веществ в области рецепторных белков, а также
удаляют пахучие вещества с рецепторов после прекращения стимуляции.)
❖ Адекватный раздражитель - летучие органические молекулы, обладаю-
щие водо- и липидорастворимостью, которые соединяются с белковыми
рецепторами мембраны ресничек рецепторных клеток.
❖ Механизмы возбуждения рецепторов.
• Соединение молекул пахучего вещества с белковым рецептором мем-
браны реснички активирует (через G,-белок) аденилатциклазу и повышает
уровень цАМФ в клетке, что приводит к открыванию цАМФ-зависимых
ионных каналов и входу Na (в меньшей мере Са24) в клетку, что вызывает
деполяризацию - РП, имеющий амплитуду до 25 мВ. (Некоторые пахучие
вещества действуют через фосфолипазу-С—> инозитолтрифосфат/ДАГ).
• РП вызывает генерацию ПД в аксонном холмнке рецепторной клетки.
• Внеклеточным электродом с поверхности обонятельного эпителия
можно зарегистрировать электроольфактограмму - монофазную нега-
тивную волну амплитудой до 10 мВ, суммирующей потенциалы многих
рецепторов.
• Каждый рецептор может возбуждаться многими (20 - 25) пахучими ве-
ществами, отдавая предпочтение некоторым нз иих (обонятельный про-
филь рецепторов); разные рецепторы имеют перекрывающие профили
ответов.
• Пахучие вещества, кроме обонятельных рецепторов, раздражают небольшую часть
нервных окончаний V (полость носа), IX и X (глотка) нервов, и обонятельная инфор-
мация поступает в мозг, минуя обонятельные нервы и тракты.
❖ Адаптация обонятельного рецептора возникает за 2 - 5 с: Са2+, который
вместе с Na* входит в клетку, образует комплекс Са2+ + кальмодулин, пре-
пятствующий открыванию цАМФ-зависимых ионных каналов и формиро-
ванию РП; другой механизм адаптации связан с активацией посредством
цАМФ протеинкиназы А, которая фосфорилирует обонятельные рецептор-
ные молекулы, делая их неспособными активировать С5-белки.
5.3. Проводниковый отдел обонятельного анализатора.
❖ Аксоны (С-волокна) рецепторных нейронов (1-е нейроны) объединяются в
пучки обонятельного нерва (I пара) и направляются в обонятельную луко-
вицу, где оканчиваются (предполагаемый медиатор - днпептид карнозин) с резко
выраженной конвергенцией (1000:1) на дендритах митральных клеток (2-е
нейроны).
❖ Возбудимость митральных клеток регулируется двумя видами вставочных
тормозных нейронов (перигломерулярные и зернистые клетки, которые образуют
тормозные синапсы - медиатор ГАМК - на дендритах митральных клеток); в СВОЮ оче-
редь тормозные нейроны находятся под возбуждающим влиянием эффе-
рентных путей из различных отделов головного мозга.
❖ Аксоны митральных клеток 50 тыс.) образуют обонятельный путь, кото-
рый направляется в обонятельный мозг (и через переднюю комиссуру в противо-
положную нуковицу).
5.4. Корковый отдел обонятельного анализатора.
406
❖ Обонятельный тракт проецируется в следующие отделы обонятельного
мозга: парагиппокампальную извилину, ее крючок, гиппокамп (корковый от-
дел анализатора), переднее продырявленное вещество, подмозолистую об-
ласть. Здесь формируются обонятельные ощущения.
❖ Из обонятельного мозга импульсация направляется:
• в передние ядра таламуса -> в лобноорбитальную кору (оценка обоня-
тельной информации, формирование поведения);
• в мамиллярные тела гипоталамуса в лимбическую кору и ретикулярную
формацию (эмоциональный и вегетативные компоненты обонятельного
восприятия, активация коры мозга);
• в свод —> мамиллярные тела гипоталамуса —> ядра покрышки среднего
мозга —> покрышечно-спинномозговой путь —> двигательные реакции иа
сильные запахи.
❖ Обонятельный анализатор имеет низкий абсолютный порог (одна молекула
пахучего вещества может возбудить рецептор) и высокий дифференциальный по-
рог (- 30 %).
❖ Классификация запахов. Имеется ~ 10 тыс. пахучих веществ, которые груп-
пируются в 7 классов первичных запахов: цветочный, мятный, мускусный,
камфорный, эфирный, едкий, гнилостный; смесь запахов может восприни-
маться как новый запах.
❖ Теория восприятия запахов: наиболее распространена стереохимическая
теория обоняния, согласно которой на мембране рецепторов имеются ре-
цептивные участки (6-7 типов), различающиеся по своей геометрии и
электрофильности, способные связать молекулы пахучих веществ опреде-
ленной формы и размеров.
5.5. Метод исследования обонятельной чувствительности. Ольфактометрия - опреде-
ление порога обонятельной чувствительности к определенному пахучему веществу с
помощью ольфактометра.
6. Интероцептивный анализатор.
6.1. Интероцептивный анализатор воспринимает изменения внутренней среды
организма и поставляет в ЦНС информацию, необходимую для рефлекторной
регуляции работы внутренних органов и гомеостаза.
6.2. Периферический отдел.
❖ Структура рецепторов: свободные нервные окончания нейронов спиналь-
ных и интрамуральных ганглиев, инкапсулированные рецепторы каротид-
ных и аортальных телец.
❖ Виды интерорецепторов: механорецепторы (барорецепторы) в полых ор-
ганах и сосудах, центральные и периферические хеморецепторы, осморе-
цепторы, терморецепторы.
❖ Основные зоны скопления интерорецепторов: каротидное и аортальное
тельца (механо- и хеморецепторы), хеморецепторная зона продолговатого
мозга, гипоталамус (хемо-, осмо- и терморецепторы); в интерстиции (осмо-
и барорецепторы).
6.3. Проводниковый отдел имеет два основных пути проведения висцеральной
чувствительности.
407
❖ От рецепторов внутренних органов периферические волокна (А(3, А8, С)
идут в составе симпатических нервов, их сплетений и стволов до спиналь-
ных ганглиев, где расположены тела первых нейронов, аксоны которых
входят в задние столбы спинного мозга. Там они переключаются на вторые
нейроны, образующие спиноталамические пути. Часть аксонов первых
нейронов идут в задних канатиках по тонкому и клиновидному путям и пе-
реключаются на вторые нейроны в ядрах Голля и Бурдаха продолговатого
мозга. Их аксоны идут в таламус в составе медиальной петли.
❖ Часть рецепторов внутренних органов является окончаниями чувствитель-
ных волокон IX и X нервов. Аксоны первых нейронов, тела которых распо-
ложены в чувствительных ганглиях этих нервов, переключаются на вторые
нейроны в ядре одиночного пути. Аксоны вторых нейронов идут в составе
бульботаламических путей. Значительная часть этой импульсации исполь-
зуется для осуществления висцеро-висцеральных рефлексов.
♦ > Третьи нейроны расположены в ядрах таламуса: специфических (вентраль-
ные задние) и неспецифических (интраламинарные и ретикулярные) ядрах;
в ядрах гипоталамуса: в передней группе (супраоптические и паравентри-
кулярные ядра) и в задней группе (медиальные и латеральные ядра сосцевидных
тел).
6.4. Корковый отдел формирует ощущение внутреннего комфорта или дискомфор-
та.
❖ Сенсорная кора.
• Область SI находится в постцентральной извилине и осуществляет эф-
ферентную регуляцию вегетативных реакций в ответ на поток аффе-
рентной, четко локализованной информации от внутренних органов.
• Область SII - на верхней стенке боковой (сильвиевой) борозды; поступающая сюда
импульсация от внутренних органов не имеет четкой локализации; активирует услов-
ные рефлексы, выработанные на механические раздражения желудка, кишечника, мо-
чевого пузыря, матки.
❖ Лимбическая система: ее структуры - миндалевидное тело и гипоталамус
- имеют двусторонние связи с внутренними органами и лобной корой; по-
сле сравнения и обработки полученной информации лимбическая система
через эфферентные выходы запускает вегетативные, соматические и пове-
денческие реакции, обеспечивающие приспособление организма к сохра-
нению внутренней среды в результате активации или угнетения висцераль-
ных функций.
7. Температурный анализатор (см. тему 26; задание 1:4.2.).
8. Двигательный анализатор (см. тему 11; задание 1:2).
9. Профильный материал для студентов лечебного факультета.
9.1. Тактильный анализатор.
В старческом возрасте тактильная чувствительность снижена в связи с уменьше-
нием количества телец Мейсснера. Женщины всех возрастных групп обладают
лучшей чувствительностью, чем мужчины.
Тактильная чувствительность рецепторов кожи живота в старости выше, чем в
зрелом возрасте, а в области груди и бедра практически одинакова.
408
❖ С возрастом происходит большее снижение вибрационной чувствительности
нижних конечностей, чем верхних, а также наиболее дистальных отделах конеч-
ностей, нежели проксимальных.
9.2. Болевой анализатор.
❖ Данные об изменении болевого порога противоречивы: одни свидетельствуют о
его увеличении после 60 лет, другие - о снижении, а третьи не выявили каких-
либо возрастных изменений.
9.3. Вкусовой анализатор.
❖ У 80 % людей старше 60 лет снижена вкусовая чувствительность, особенно к
сладкому и соленому, что способствует их повышенному потреблению.
Причины снижения восприятия вкуса к старости: частичное разрушение вкусовых
рецепторов на языке, уменьшение слюноотделения, изменение слизистой оболоч-
ки ротовой полости, ухудшение переработки поступающей информации в ЦНС за
счет снижения числа нейронов.
9.4. Обонятельный анализатор
Обонятельная чувствительность снижается после 45-ти лег и резко - после 70-ти в
результате атрофических процессов в слизистой обонятельной области полости
носа; отмечено снижение количества нейронов, силы и подвижности нервных про-
цессов.
10. Профильный материал для студентов педиатрического факультета.
10.1. Функциональные особенности вкусового анализатора у плода и детей.
❖ Формирование вкусовых рецепторов (вкусовых луковиц) начинается у плода на
3-м мес.
У новорожденных вкусовые рецепторы реагируют на все 4 вида вкусовых раздра-
жений (сладкое, кислое, горькое и соленое), имеют высокий порог раздражения;
сладкие вещества вызывают сосательное движение.
❖ Условные рефлексы на вкусовые раздражители вырабатываются на 2-м мес. после
рождения.
❖ Четкое различение вкусовых веществ пищи формируется к концу 1-го года; в до-
школьном возрасте происходит снижение порога вкусовых рецепторов; у подро-
стков вкусовая чувствительность близка к уровню взрослых.
10.2. Особенности тактильного анализатора в раннем онтогенезе.
❖ Инкапсулированные рецепторы (тельца Пачини) формируются у плода с 3 - 4-го
мес.
Ф В первые дни жизни тактильные раздражения всех участков кожи вызывают об-
щую двигательную реакцию; на 2-м мес. появляются локальные реакции.
❖ Условные рефлексы можно выработать на кожно-тактильные раздражители на 3-м
мес. жизни.
❖ Минимальные пороги чувствительности тактильного анализатора имеются в 17 -
20 лег.
10.3. Особенности болевого анализатора в раннем онтогенезе.
♦ > Свободные нервные окончания и реакция на боль формируются у плода на 8-й не-
деле.
4 * Порог болевой чувствительности у новорожденных детей выше; латентный пери-
од больше, ответная реакция выражается общей двигательной реакцией, измене-
нием частоты сердцебиения и дыхания.
Ъ Болевая чувствительность в области лица выше, чем в других участках тела.
409
♦ ♦♦ Через неделю после рождения на болевые раздражители уменьшается общая дви-
гательная реакция, появляются более локальные ответы.
❖ К концу 1-го года жизни ребенок может дифференцировать место болевого раз-
дражения кожи.
♦ ♦♦ После 3 лет становится возможной локализация болевых ощущений, вызванных
раздражением рецепторов внутренних органов.
10.4. Особенности обонятельного анализатора в раннем онтогенезе.
Периферический отдел обонятельного анализатора формируется к 7 - 8 мес. внут-
риутробного развития.
♦ 3» Сразу после рождения обонятельные раздражители воспринимаются двумя вида-
ми рецепторов (обонятельными и рецепторами тройничного нерва). Вещества,
действующие только на обонятельный нерв, называются олъфактивными (анис,
лаванда и др.). Вещества, действующие и на тройничный нерв, называются три-
геминальными (эфир, хлороформ и др.; пороги этих веществ значительно выше).
❖ У новорожденных имеется более быстрая адаптация обонятельных рецепторов,
чем у взрослых, а острота обоняния в 20 - 100 раз ниже.
♦ ♦♦ Условный рефлекс на запаховые раздражители можно выработать с 2 мес. постна-
тального периода.
♦ ♦♦ Ребенок начинает отличать приятные запахи от неприятных на 4-м мес. жизни.
4 » Обонятельный анализатор полностью сформирован к 6 годам жнзнн.
4 * Острота обоняния достигает своего максимума в период полового созревания.
11. Профильный материал для студентов стоматологического факультета. Вкусовой
(оральный) анализатор,
♦♦♦ Рецепторный отдел включает в себя следующие виды рецепторов:
• вкусовые рецепторы (имеют пузырьки, содержащие медиатор серотонин);
• проприорецепторы (мышечные, сухожильные, суставные);
• тактильные рецепторы прикосновения и давления - максимальное их количе-
ство на кончике языка и красной кайме губ, наименьшее - в слизистой оболоч-
ке вестибулярной поверхности десен;
• температурные (высокая чувствительность на кончике языка н красной кайме
губ):
Ф холодовых больше в передних отделах полости рта;
Ф тепловых больше в задних отделах; в области твердого нёба полностью
отсутствует восприятие тепла, а в центральной части задней поверхности
языка отсутствуют температурные рецепторы;
• ноцицепторы - максимальное их количество в тканях зуба, оральная поверх-
ность слизистой оболочки десен обладает наименьшей болевой чувствитель-
ностью;
• временная очередность возбуждения рецепторов при приеме пищи: тактиль-
ные, температурные и вкусовые.
♦♦♦ Проводниковый отдел. От вкусовых рецепторов (см. выше 4.3), тактильных (см.
выше 2.3), температурных (см. тему 25; задание 1: 4.2), болевых рецепторов (см.
выше 3.4) ротовой полости н проприорецепторов мышц лица (см. тему 11; задание
1:2.2).
♦♦♦ Корковый отдел. Первичные проекционные зоны вкусового, болевого, темпера-
турного, тактильного и проприоцептивного анализаторов, связанные с ротовой
областью (ротовой анализатор), находятся преимущественно в коре постцен-
410
тральной извилины (SI, поля 3,1,2) в зоне проекции лица, а также в коре верхней
стенки сильвиевой борозды (SIX) - вторичной соматосенсорной зоне коры.
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). Прн под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 н 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники н пособия): [1-16].
3.2. Дополнительная литература (см. с. 514 - 522): [4], [6], [27], [28], [29], [30-32],
[37], [41], [43], [47], [82], [86], [94], [97], [103], [111], [152], [160], 169], [217], [218],
[222], [225], [239], [244-246], [251-254].
33. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6i. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 522).
Задача 27-1. Операция перерезки задних корешков спинного мозга (ризотомия) на стороне боле-
вого раздражения вызывает резкое снижение ощущения боли, но не ликвидирует его полностью.
Напротив, разрушение соответствующих спинальных ганглиев (ганглиолизис) вызывает полную
аналгезию. Объясните разницу в аиалгезирующнх эффектах этих двух операций.
Задача 27-2. Почему при ощупывании предмета, который может уместиться иа ладони руки, мы
лучше его воспринимаем по сравнению с неподвижным прикладыванием к нему руки?
411
Тема 28. Зрительный анализатор
Цель:
а) знать оптическую характеристику н регуляцию диоптрического аппарата глаза, его рецеп-
торный аппарат - фотохимические и электрические
процессы в нейронах сетчатки; структурно-
функциональную организацию проводникового от-
дела, обработку информации в подкорковых зри-
тельных центрах; корковый отдел анализатора, об-
работку в нем информации и его функции, роль ле-
вого и правого полушария в зрительном воспри-
ятии; восприятие цвета, глубины пространства, по-
ле и остроту зрения; виды н регуляцию движений
«Хочу, когда сверкнет коса,
Когда во прах сойду увечный.
Оставить на земле глаза,
Чтобы на мир смотрели вечно».
NN
глаз; механизмы адаптации зрительного
анализатора;
б) уметь использовать эти знания для анализа жизнедеятельности организма (регуляции дви-
жения, биоритмов, процессов ВНД и др.);
в) приобрести практический навык при исследовании зрачкового рефлекса: иметь представле-
ния об определении остроты зрения, полей зрения и цветового зрения.
Мотивация темы: зрительный анализатор у человека обеспечивает поступление в головной мозг 90%
информации о внешней среде и имеет важнейшее значение для адаптации организма к услови-
ям внешней среды, в том числе и к социальным. Велика также его роль в деятельности функ-
циональных систем, лежащих в основе социальных н биологических форм поведения челове-
ка, в которых зрительный анализатор участвует в пусковой, обстановочной и обратной аффе-
рентации. Нврушение функций зрительного анализатора и его вспомогательных структур воз-
никает не только при глазных заболеваниях, поэтому знание этих функций необходимо для
деятельности врачей разного профиля.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике н
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекция материала записанной лекция н подготовки рефератов.
1. Общая характеристика зрительного анализатора.
1.1. Зрительный анализатор - это сенсорная система, воспринимающая электро-
магнитные излучения с длинами волн видимого диапазона (400 - 760 нм) и фор-
мирующая световые ощущения (обеспечивает поступление в мозг окояо 90% информа-
ции о внешней среде).
1.2. Основные зрительные функции: 1) светочувствительность - способность
различать разные интенсивности диффузного освещения; 2) цветовое зрение -
различение длин воли в пределах видимого спектра; 3) распознавание формы -
требует фокусировки предмета на сетчатке; 4) восприятие движения - требует
воспроизведение движения объекта на рецепторном поле как на экране; 5) вос-
приятие глубины - основано на объединении информации от двух глаз.
13. Отделы зрительного анализатора: 1) периферический отдел - диоптрический
аппарат глаза и сетчатка; 2) проводниковый отдел; 3) корковый отдел.
2. Диоптрический аппарат глаза - система лииз, формирующая на сетчатке перевер-
нутое и уменьшенное изображение внешнего мира (рис. 61).
412
2.1. Компоненты: роговица, камеры с жидкостью, радужная оболочка и зрачок,
хрусталик и его сумка, стекловидное тело, секрет слезных желез.
2.2. Основные оптические показатели (математическое описание преломление лучей в
глазу дал А. Гульстранд, 1911).- преломляющая сила роговицы + передней камеры ~
43 D, уплощенного хрусталика ~ 19,1 D, всего глаза - 58,6 D (фокусное расстояние
глаза - 22,8 мм, расстояние от роговицы до центральной ямки - 24,4 мм, угловому размеру объ-
екта 1° соответствует расстояние - 0,29 мм на сетчатке). Роговица и хрусталик не пропус-
кают ультрафиолетовые лучи.
23. Регуляция диоптрического аппарата глаза.
❖ Рефлекс хрусталика (аккомодация глаза) - это приспособление глаз к чет-
кому видению различно удаленных предметов. Основным компонентом его
является изменение кривизны хрусталика.
• Рефлекторная реакция хрусталика.
Рис. 61. Схема строения глаза и фокусировки зрительного объекта
Кривизна хрусталика зависит от: 1) его эластичности; 2) сил, разви-
ваемых цилиарными мышцами, сосудистой оболочкой и склерой,
воздействующих на сумку хрусталика; 3) внутриглазного давления.
Рефлекторный путь: рецепторы сетчатки (стимул - нечеткое изображе-
ние предмета) —> зрительные нервы и тракты —> непарное парасимпа-
тическое ядро Ш нерва —> цилиарный ганглий, сокращение цилиар-
ных МЫШЦ (ацетилхолин + М-холинорецепторы) —> ослабление натяже-
ния связок сумки хрусталика —> увеличение кривизны хрусталика.
Диапазон аккомодации - 10-14 D, что позволяет фокусировать
предметы, находящиеся на 7-10 см от глаза. Дальняя точка, когда
объект виден уже без аккомодации, равна - 6 м.
• Рефлекс аккомодации глаза включает в себя рефлекторное сужение зрачка, что
уменьшает сферическую аберрацию, и конвергенцию глаз, обеспечивающую
фокусировку близкого объекта.
❖ Рефлекс зрачка - сужение или расширение зрачка, регулирующие световой
поток на сетчатку в зависимости от степени освещенности.
413
• Рецепторы сетчатки (увеличение освещенности) —> зрительные нервы и
тракты -> парные парасимпатические ядра Ш нерва -> цилиарный ганг-
лий —> сокращение сфинктера радужки (ацетилхолин + М-холинорецепторы)
-> сужение зрачка.
• Рецепторы сетчатки (уменьшение освещенности) —> цилиоспинальный центр
(Ся—Thi) -> верхний шейный ганглий -> сокращение радиальной мышцы
радужки (норадреналин + а-адренорецепторы) —> расширение зрачка.
• Диапазон изменения освещенности сетчатки в результате зрачкового
рефлекса - 30 раз, (диаметр зрачка 1,5 - 8,0 мм).
2.4. Варианты рефракции глаза.
❖ Эмметропия - соразмерная рефракция, при которой главный фокус глаза
совпадает с сетчаткой.
❖ Миопия (близорукость) - фокусировка предметов перед сетчаткой (оптическую
силу глаза уменьшают сферическими, рассеивающими, вогнутыми линзами).
❖ Гиперметропия (дальнозоркость) - фокусировка предметов за сетчаткой (опти-
ческую силу глаза увеличивают сферическими, собирательными, выпуклыми линзами).
❖ Астигматизм - неодинаковая диоптрическая сила роговицы в разных
ПЛОСКОСТЯХ (в норме не должен превышать 0,5 D, коррекция осуществляется цилиндри-
ческими рассеивающими или собирательными линзами).
2.5. Внутриглазное давление.
❖ Зависит от соотношения количества вырабатываемой и отводимой из глаза
жидкости (в норме 18 - 26 мм рт. ст.), влияет на постоянство формы глаза и
расположение компонентов диоптрического аппарата.
❖ Жидкость образуется путем ультрафильтрации из капилляров цилиарного
тела и поступает в заднюю, а затем в переднюю камеры глаза.
❖ Отток жидкости осуществляется в трабекулярную часть сосудов в месте
соединения радужной оболочки и роговицы; далее через шлеммов канал - в
венозную систему.
❖ Упругие силы радужки при сужении зрачка облегчают отток жидкости, при
расширении зрачка - затрудняют отток.
❖ К повышению давления наиболее чувствительна механически слабая часть
стенки глаза - место выхода зрительного нерва (решетчатая пластинка).
2.6. Слезная жидкость.
❖ Постоянно вырабатывается слезными железами, близка по составу к ульт-
рафильтрату плазмы кровн, содержит в большой концентрации бактери-
цидные вещества (лизоцим, IgA и др.).
❖ Функции: предохраняет роговицу и конъюнктиву от высыхания, улучшает
оптические свойства роговицы, уменьшает треиие между веками и глазны-
ми яблоками, обладает бактерицидными свойствами, способствует удале-
нию инородных тел.
3. Рецепторный аппарат зрительного анализатора (рис. 62).
3.1. Структура сетчатки (часть промежуточного мозга, вынесенная на периферию): 1) пигмент-
ный слой меланннсодержащих эпителиальных клеток: поглощает свет, участвует в трофике ре-
цепторов (депо витамина А в сетчатке), в их антиоксидантной защите, фагоцитирует продукты
распада фоторецепторов, механически наиболее слабое место (отслойка сетчатки!); 2) слой фо-
торецепторов: рецепция светового раздражителя; 3) слой горизонтальных клеток: тормозные
414
нейроны; 4) слой биполярных клеток: нейроны проведения и конвергенции возбуждения; 5)
слой амакриновых клеток: тормозные нейроны; 6) слой ганглиозных клеток: возникновение по-
тенциала действия, формирование зрительного нерва; слои пронизывают и связывают глиаль-
ные клетки (Мюллеровы клетки).
3.2. Фоторецепторы глаза (палочки н три типа колбочек - вторичные рецепторы).
❖ Расположение рецепторов в сетчатке.
• Колбочки (~ б млн.) расположены в желтом пятне и его центральной ям-
ке, в которой их плотность максимальная (~ 150000/мм2); слой рецепторов
здесь не прикрыт другими нейронами н волокнами зрительного нерва; в
этой области острота зрения максимальная, так как одна колбочка с по-
мощью биполярного нейрона соединяется с одной ганглиозной клеткой
(в других участках сетчатки плотность колбочек низкая ~ 5ООО/мм2).
• Палочки (~ 120 млн) расположены в сетчатке, кроме желтого н слепого
пятен, плотность их возрастает от периферии до желтого пятна (50 —»
150 тыс/мм2);
• В слепом пятне (место выхода зрительного нерва) рецепторов нет.
Колбочка
Горизонтальный
нейрон
Амакриновый
нейрон
Ганглиозный
нейрон
Рнс. 62. Упрошенная схема нейронного строения сетчатки
♦♦♦ Структура фоторецепторов:
• наружный сегмент у палочек содержит около 1000 мембранных дисков, а у колбочек -
складки плазматической мембраны; в состав липопротеидного слоя мембраны этих
дисков и складок включены зрительные пигменты;
• внутренний сегмент содержит митохондрии н К+, Ыа+-насос, восстанавливающий гра-
диенты К+ н Na+;
• тело клетки и аксонный отросток, который синаптически оканчивается на биполярном
и/илн горизонтальном нейронах;
❖ Функции фоторецепторов.
• Функция палочек:
415
• ф имеют высокую чувствительность к свету (в 500 раз выше колбочек) и
приспособлены для ночного зрения;
• ф- обеспечивают периферическое зрение;
• ф воспринимают подвижные объекты.
• Функции колбочек:
•ф- осуществляют центральное зрение и остроту зрения;
•ф- осуществляют цветовосприятие.
33. Фотохимические и электрические процессы в фоторецепторах.
❖ Зрительный пигмент (родопсин) состоит из белка опсина и 11-цис-
ретиналя (альдегид витамина А); длина волны поглощенного пигментом
света зависит от аминокислотного состава его опсина.
• В палочках имеется один вид опсина (у человека имеет максимум поглощения
прн - 500 нм).
• В колбочках имеется три вида опсина: синий, зеленый и красный (мак-
симумы поглощения соответственно ~ 420 нм, - 530 нм и - 560 нм).
В темноте палочки и колбочки
деполяризованы и имеют не-
большой мембранный потенциал
покоя (от -25 до -40 мВ).
• Малая величина МПП обу-
словлена высокой На+-прони-
цаемостью мембраны внеш-
него сегмента и входящим в
клетку На+-током (рнс. 63, А).
• Открытое состояние Na+-Ka-
налов непосредственно удер-
живается цГМФ (образуется из
ГТФ); если цГМФ связан с
Na^-каналом, то канал открыт
(рнс. 63, В).
• Na+, поступивший внутрь
клетки в зоне наружного сег-
мента, выводится из клетки в зоне внутреннего сегмента с помощью К+,
Na+-Hacoca (рис. 63, А).
• Этот входящий и выходящий из фоторецептора Na^-ток называется
темновым током.
❖ На свету палочки и колбочки гиперполяризованы.
• Поглощение света зрительным пигментом (палочку может возбудить один
квант света) запускает цепь реакций (переход 11-цис-ретиналя в транс-ретнналь и
родопсина в метародопсин II. активация в мембране G белка, активация фосфодиэсте-
разы), в результате которых гидролизуется цГМФ н закрываются Na*-
каналы, что приводит к гиперполяризации фоторецепторов (прн макси-
мальном освещение МП сдвигается до -90 мВ) (рнс. 63, С).
416
• При возбуждении (гиперполяризации) фоторецепторов секреция их ме-
диатора (глутамата) уменьшается. При этом биполярные клетки с оп-
центром возбуждаются, т.к. глутамат для них является тормозным ме-
диатором, а биполярные клетки с off-центром тормозятся, т.к. глутамат
для них является возбуждающим медиатором.
❖ Электроретинограмма - суммарный потенциал сетчатки глаза иа действие
света; состоит из нескольких волн (а, b, с, d), отражающих изменения мем-
бранного потенциала разных клеток - фоторецепторов, биполярных, гори-
зонтальных, амакриновых и глиальных клеток.
❖ Роговично-сетчатый потенциал.
• Глаз является электрическим диполем («+» на роговице, «-» иа проти-
воположном полюсе), что используется для записи движения глаз (элек-
троокулография).
• Разность потенциалов связана с тем, что наружные участки пигментных клеток более
отрицательны, чем внутренние сегменты фоторецепторов.
3.4. Функция биполярных клеток сетчатки: 1) проводят возбуждение от рецеп-
тора к ганглиозной клетке безимпульсиым путем (медиатор ацетилхолин); 2)
конвергируют возбуждение на ганглиозных клетках; 3) реагируют на контраст-
ность изображения с помощью полей с on- и off-центрами.
3.5. Функция ганглиозных клеток (выходные нейроны сетчатки, связаны возвратными
ГАМК-путями с фоторецепторами): 1) имеют рецептивные поля с антагонизмом ме-
жду центром и периферией (on-цеитр и off-периферия или наоборот) и участ-
вуют в восприятии освещенности, контраста и цвета; 2) участвуют в темновой и
световой адаптации путем изменения размеров центра и периферии своих круг-
лых рецептивных полей; 3) генерируют ПД.
3.6. Функция горизонтальных и амакриновых клеток: 1) формируют on- и off-
центры путем латерального торможения (медиатор ГАМК, на горизонтальных клетках
есть ГАМКс-рецепторы, сопряженные с СГ-каналами); 2) реализуют эфферентное тор-
мозное влияние на сетчатку.
3.7. Функция клеток глии (Мюллеровы клетки): обладая высокой проницаемостью для
К*, захватывают эти ионы в межклеточном пространстве сетчатки и выводят их
в жидкость стекловидного тела.
4. Проводниковый отдел зрительного анализатора.
4.1. Структурно-топографическая характеристика.
Аксоны ганглиозных клеток образуют зрительный нерв (И пара, - 1,3 млн. волокон, ана-
лог проводящих путей ЦНС).
Зрительные нервы впереди серого бугра гипоталамуса образуют частичный перекрест
(хиазму), где нервные волокна, идущие от носовых половин обеих сетчаток, пересекаются
и переходят на противоположную сторону, а волокна, идущие от височных половин каж-
дой сетчатки, не пересекаются.
После перекреста волокна образуют зрительный тракт, основные проекции которого:
• в латеральные коленчатые тела преимущественно от рецепторов желтого пятна, далее
преимущественно а первичную зрительную кору; имеют сходные с ганглиозными
клетками стимулы н поля с off - н оп-центрамн;
• в верхнее двухолмие и далее в глазодвигательные центры ствола - ядра III, IV, VI нер-
вов (обслуживают рефлексы среднего мозга: зрительный ориентировочный, реакции
417
зрачка и хрусталика); используется импульсация преимущественно от ганглиозных
клеток, связанных с периферией сетчатки и настроенных на детекцию движения;
• в верхнее двухолмие и далее через подушку таламуса в теменную ассоциативную кору
(поле 7), регулирующие произвольные движения глаз контрлатеральиой стороны;
• в супрахиазматическое ядро гипоталамуса (регуляция суточных ритмов).
4.2. Обработка сигналов в верхнем двухолмии.
<• Проекция формируется преимущественно аксонами ганглиозных клеток
периферии сетчатки, настроенных на детекцию движения. Нейроны верх-
него двухолмия отвечают на движения зрительного стимула в определен-
ном направлении.
❖ Здесь имеются также нейроны, центрирующие изображение движущегося
предмета в зоне центральной ямки и управляющие рефлекторным поворо-
том головы н глаз при внезапном появлении на периферии поля зрения
движущегося объекта (зрительный ориентировочный рефлекс).
4.3. Обработка сигналов в латеральных коленчатых телах (ЛКТ).
❖ Входные сигналы в ЛКТ от двух глаз изолированы в разных слоях, отдель-
ные нейроны строго мононуклеарны, имеют рецепторные поля с on- и off-
центрами.
❖ Имеется топографическая проекция сетчатки: проекция центральной ямки
занимает наибольшую часть.
❖ В ЛКТ осуществляется: 1) оценка степени освещенности; 2) оценка контра-
стности; 3) оценка цветовой характеристики стимула.
5. Корковый отдел зрительного анализатора.
5.1. Зрительная кора расположена в затылочных долях больших полушарий и за-
нимает 17 поле (первичная зрительная кора) и 18-19 поля (вторичная зритель-
ная кора). {Зрительная кора имеет более сложное строение: в ией IV и V слой разделяются на
ряд (до трех) подслоев, афферентная импульсация преимущественно поступает к нейронам IV и
III слоев. В зрительную кору левого (правого) полушария проецируют рецепторы левых (пра-
вых) половин сетчатки обоих глаз и, следовательно, изображение с правых (левых) половин по-
лей зрения; интернейроны соединяют через мозолистое тело сенсорные нейроны коры обоих
полушарий.)
Функциональная организация зрительной коры осуществляется как в виде
клеточных слоев, параллельных поверхности мозга, так н в виде нейронных ко-
лонок, перпендикулярных поверхности мозга. Нейроны зрительной коры имеют
прямоугольные рецептивные поля.
5.2. Первичная зрительная кора получает импульсацню преимущественно от ней-
ронов латерального коленчатого тела, в ней хорошо выражена топографическая
проекция сетчатки. Центральная ямка, обеспечивающая остроту зрения, зани-
мает бблыпую часть коркового представительства (в 35 раз больше равного участка
периферии сетчатки) и малые рецептивные поля корковых нейронов. Обработка
зрительной информации осуществляется с участием нейронов-детекторов про-
стых и сложных признаков. В первичной зрительной коре преобладают простые
нейроны. Они реагируют на неподвижные, определенным образом ориентиро-
ванные в рецепторном поле линии (например, детекторы конца линии, угла,
кривизны, разрыва контура и др.). При проекции в зрительной системе какого
либо предмета корковые нейроны реагируют только на его контуры, и не реаги-
руют на внутреннюю однородную часть предмета. Корковые зрительные ней-
418
роны имеют рецептивные поля прямоугольной формы с on- и off-центрами,
прн этом on- и off-зоны расположены параллельно друг другу. (Адекватным раз-
дражителем является узкая полоса, которая полностью совпадает с возбуждающей или тормоз-
ной зонами.) В первичной зрительной коре формируется ощущения яркости, кон-
трастности, цвета, формы, элементарный анализ движения. (Для возникновения
светового ощущения достаточно 5-7 квантов света.)
5.3. Во вторичной зрительной коре преобладают сложные нейроны, получающие
входные сигналы от множества простых нейронов. Они реагируют главным об-
разом на перемещение ориентированных линий в рецепторном поле, среди них
есть детекторы движения в определенном направлении. Другие сложные ней-
роны осуществляет тонкий анализ образов, цвета, зрительное внимание, пере-
мещение взгляда, узнавание знакомой обстановки, знакомого лица, выражения
глаз, движение мимических мышц, формирование чувства пространства и др.
Простые и сложные нейроны зрительной коры группированы в два вида ко-
лонок: глазодомннантные и ориентационные. Глазодоминантная колонка пред-
ставляет собой скопление нейронов в виде прямоугольного столбика (0,5x0,5
мм), пронизывающее все слои коры и дающее возбуждение только при раздра-
жении сетчатки одного глаза (правого или левого). Нейроны переходной зоны
между колонками бинокулярны. Ориентационная колонка реагируют не только
на ориентацию светового раздражителя (например, линии), но и на сдвиг его по
часовой или против часовой стрелки на 180°.
5.4. В ассоциативной коре информация, поступившая из зрительной коры и по-
душки таламуса, используется для формирования сложных зрительных образов
(например, узнавание письменной речи, интеграция слова со зрительным образом,
произвольные движения глаз и др.).
5.5. Восприятие глубины пространства.
❖ Монокулярное восприятие пространства осуществляется путем:
• сравнения величины известных объектов;
• оценки частичного наслоения одних предметов на другие;
• оценки удаленности предметов на основе изменения аккомодации глаза
прн их восприятии;
• использования перспективы (схождение параллельных линий, удаляю-
щихся вдаль);
• параллакса движения (кажущееся относительное смещение близких и более далеких
предметов, если наблюдатель будет двигать головой вправо-влево или вверх-вниз).
❖ Бинокулярное восприятие пространства осуществляется путем:
• оценки удаленности предметов на основе дивергенции и конвергенции
глаз прн их восприятии;
• стереопсиса (стереовосприятия) - наличия небольших различий в изо-
бражениях предмета, проецируемых на сетчатки обоих глаз, что приво-
дит к впечатлению объемности (полное совпадение дает восприятие плоского
изображения; более сильное несовпадение дает даоеиие зрительного образа);
• «физиологического даоення» как механизма оценки глубины: когда изображения фик-
сируемого объекта совпадают на сетчатках обоих глаз, остальные объекты дают не-
совпадающие проекции, но даоеиие их изображения не воспринимается как таковое, а
служит сигналом удаленности этих объектов относительно предмета наблюдения.
419
5.6. Роль правого и левого полушария в зрительном восприятии.
❖ Правое полушарие:
• зрительный образ воспринимает нерасчлененно (целостно), сразу во
всех подробностях;
• легче решает задачу: «В чем различие предъявляемых предметов?»;
• легче опознает неконкретные предметы, которые трудно описать сло-
вами;
• создает предпосылки наглядного мышления.
❖ Левое полушарие:
• зрительный образ оценивает расчлененно, аналитически, при этом каж-
дый признак (форма, величина н др.) анализируются отдельно;
• легче опознает знакомые предметы и решает задачу: «В чем сходство
предъявляемых предметов?»;
• зрительные образы лишены конкретных подробностей, имеют высокую
степень абстракции;
• создает предпосылки логического мышления.
6. Восприятие цвета (трихромазия - нормальное цветовосприятие).
6.1. Трехкомпонентная теория цветного зрения (Т. Юнг, 1802; Г. Гельмгольц, 1859)
предполагает наличие 3 типов колбочек со зрительными пигментами, погло-
щающими различные длины волн света: «синие» (420 нм), «зеленые» (530 нм),
«красные» (560 нм) колбочки. (Цвет - это результат неодинаковой стимуляции колбочек
разного типа. Стимуляция всех типов колбочек дает ощущение, лишенное цвета - «белое».
Одинаковая стимуляция красных и зеленых колбочек, но не синих, даст ощущение желтого.)
6.2. Теория оппонентных цветов (Э. Геринг, 1886). Геринг обратил внимание на вза-
имное уничтожение (ощущение белого) при смешении синего н желтого или
красного и зеленого цветов и предположил существование прн цветовом зре-
нии два оппонентных процесса: один для ощущения синего и желтого, другой
- для зеленого н красного. Таким образом, в теории Геринга постулировано 4
первичных цвета - синий, желтый, зеленый и красный.
Наличие антагонистических нейронов прн цветовом восприятии было пока-
зано при освещении монохроматическим светом с изменяющейся длиной вол-
ны: среди биполярных, горизонтальных, ганглиозных и таламических нейронов
были обнаружены клетки, которые тормозились прн попадании синего света и
возбуждались желтым светом, другие нейроны тормозились зеленым светом и
возбуждались красным светом. Такие нейроны были названы простыми оппо-
нентными клетками.
Часть ганглиозных нейронов, нейронов латерального коленчатого тела н
первичной зрительной коры имеют рецептивные поля, организованные по
принципу антагонизма центра и периферии: в нейронах желтой-синей системы
желтый свет возбуждал нейрон, а синий - тормозил (оп-центр, off-периферия)
или наоборот, в нейронах красно-зеленой системы красный свет возбуждал
нейрон, а зеленый - тормозил (оп-центр, off-периферия) или наоборот. Такие
нейроны были названы дважды оппонентными клетками.
420
В первичной зрительной коре цветоспецифические нейроны особенно чув-
ствительны к границам между разноокрашенными областями. Во вторичной
зрительной коре цветочувствительные нейроны отвечают на раздражители с уз-
ким диапазоном световых тонов. Адекватным раздражителем для других ней-
ронов является цветовой контраст. Во вторичной зрительной коре формируется
относительное постоянство цветового восприятия.
6.3. Нарушения восприятия цвета: врожденные формы цветовой слепоты (старое название -
дальтонизм) связаны с отсутствием генов, кодирующих разные виды опсина в колбочках (гены
красного и зеленого опсинов расположены в Х-хромосоме, ген синего опсина ~ в 7-й хромосо-
ме).
♦ ** Дихромазия (отсутствие восприятия одного цвета).
• Дейтеранопия (частота - 6 %) - «зеленослепые».
• Протанопия (частота - 1,1 %) - «краснослепые».
• Тританопия (частота - 0,01 %) - «синеслепые».
Ахромазия (частота < 0,01 %) - полная цветовая слепота (черно-белое восприятие).
7. Механизмы адаптации зрительного анализатора.
7.1. Зрачковый рефлекс и рефлекс аккомодации хрусталика (см. выше 2.3).
7.2. Фотохимический механизм адаптации: световая адаптация возникает в ре-
зультате фосфорилирования родопсина ферментом родопсинкиназой, фосфоро-
допсин не способен активировать Gj-белок (и каскад реакций, приводящий к возбуж-
дению рецепторов); темновая адаптация возникает в результате ингибирования
родопсинкиназы белком орестином в присутствии Са2+, что приводит к восста-
новлению активности молекул родопсина.
7.3. Функциональная мобильность рецепторов (изменение размеров центра и пери-
ферии рецепторных полей ганглиозных клеток).
7.4. Эфферентное влияние ЦНС на нейроны сетчатки: от зрительной коры и верхне-
го двухолмия идут эфферентные волокна, оканчивающиеся на амакриновых
нейронах сетчатки, оказывая модулирующее влияние на проведение возбужде-
ния от биполярных клеток к ганглиозным.
8. Защитные рефлексы: мигательный, роговичный и слезоотделительный.
8.1. Запускаются с рецепторов конъюнктивы и роговицы (при сильных раздражителях -
также со зрительных и слуховых рецепторов).
8.2. Афферентная импульсация идет преимущественно по глазной ветви тройнич-
ного (V) нерва в его чувствительные ядра (мостовое и спинальное).
8.3. Афферентная импульсация переключается с ядер V нерва на двигательное и
слюноотделительное ядра лицевого (VII) нерва.
8.4. Эффекторное звено: двигательное ядро VII нерва вызывает сокращение круго-
вой мышцы глаза и периодическое смыкание глазной щели; верхнее слюноот-
делительное парасимпатическое ядро (через переключение в крылонебном ганглии)
вызывает стимуляцию секреции слезной жидкости.
9. Движение глаз (глазодвигательный аппарат).
9.1. Функции наружных глазных мышц, их иннервация.
❖ Прямые мышцы:
• нижняя прямая вращает глазное яблоко, направляя зрачок вниз (Ш
нерв);
421
• верхняя прямая вращает глазное яблоко, направляя зрачок вверх (Ш
нерв);
• внутренняя прямая вращает глазное яблоко, направляя зрачок внутрь
(Ш нерв);
• наружная прямая вращает глазное яблоко, направляя зрачок наружу (IV
нерв).
❖ Косые мышцы:
• нижняя косая вращает глазное яблоко, направляя зрачок вбок и кверху
(Ш нерв);
• верхняя косая вращает глазное яблоко, направляя зрачок вниз и вбок
(VI нерв).
9.2. Виды движения глаз.
* *• Зрительный ориентировочный рефлекс (верхнее двухолмие).
❖ Плавные (следящие) движения глаз.
❖ Скачкообразные движения глаз (саккады длительностью 10 - 80 мс), сохраняют
зрительное восприятие (вызывают дезадаптацию рецепторов сетчатки).
❖ Движения глаз при рассматривании сложных изображений имеют периоды
фиксации длительностью 130 — 300 мс (выделение наиболее значимых деталей
изображения).
❖ Бинокулярная координация движений глаз: содружественные движения
глаз вверх-вниз, вправо-влево; конвергентные и дивергентные движения
глаз; вращательные движения глаз.
93. Основные реценторные зоны для регуляции движения глаз: 1) рецепторы сет-
чатки; 2) вестибулярные рецепторы.
9.4 . Центры регуляции движения глаз.
& Вестибулярные ядра осуществляют вестибулоокулярный рефлекс фикса-
ции взора при изменении положения головы.
❖ Ретикулярная формация ствола мозга управляет содружественными гори-
зонтальными и вертикальными движениями глаз, конвергентными и дивер-
гентными движениями глаз.
❖ Мозжечок осуществляет: 1) обработку зрительной информации от зритель-
ного тракта, верхнего двухолмия, а также вестибулорецепторов и проприо-
рецепторов; 2) регуляцию точности выполнения следящих движений, сак-
кад и продолжительности периодов фиксации.
❖ Зрительная, теменная, премоторная кора осуществляет произвольную регу-
ляцию движения глаз: посредством непрямых связей с верхними бугорками
и претектальной областью через мозжечок и вестибулярные ядра обеспечи-
ваются следящие, фиксирующие и саккадические движения глаз.
10. Методы исследования зрительного анализатора.
10.1. Исследование зрачковой реакции и содружественной реакции зрачков на свет.
Исследование зрачковой реакции на свет. Лицо обследуемого обращено к источ-
нику света. Врач прикрывает его оба глаза своими руками, затем через минуту быстро
убирает руку от одного глаза, в результате чего его зрачок быстро суживается. Затем
эту реакцию исследуют на другом глазу. Параметры реакции: латентный период со-
422
кращения зрачка 0,2 - 0,3 с; время сокращения зрачка 0,5 - 1,5 с; ширина зрачка 8 -> 3
мм; затем наступает постепенное расширение зрачка с меньшей скоростью.
Исследование содружественной реакции зрачков на свет. Закрывают рукой один
глаз обследуемого, лицо которого обращено к источнику света. Когда врач отнимает
руку от глаза, в другом глазу также происходит сужение зрачка. При повторном закры-
тии глаза зрачок другого глаза расширяется.
10.2. Определение остроты зрения.
Острота зрения - это способность глаза воспринимать раздельно точки, располо-
женные друг от друга на минимальном расстоянии. В норме глаз способен различать
две точки раздельно под углом 1 мин (международный эталон остроты зрения).
Для определения остроты зрения используют стандартные таблицы с буквенными
знаками или незамкнутыми окружностями, которые расположены в 12 строк. Они рас-
считаны на определение остроты зрения с расстояния 5 м. При этом детали знаков де-
сятого ряда видны под углом 1 мин (острота зрения будет равная 1,0). Если острота
зрения иная, то определяют, в каком ряду таблицы обследуемый различает знаки.
Справа в каждом ряду проставлена острота зрения (V), соответствующая чтению дан-
ного ряда с расстояния 5 м.
10.3. Определение поля зрения,
Поле зрения - это пространство, видимое глазом человека при фиксации азгляда в
одной точке. Различают цветовое (хроматическое) и бесцветное (ахроматическое) поле
зрения. Границы поля зрения каждого глаза исследуют обычно отдельно (монокуляр-
ное поле зрения) с помощью периметра, имеющего дугу, концентричную сетчатой обо-
лочки. В середине её расположен белый неподвижный объект, служащий точкой фик-
сации взгляда. На наружной поверхности нанесены деления от 0 до 90°. Подбородок
испытуемого располагается в выемке штатива периметра. Испытуемый фиксирует од-
ним глазом белый кружок в центре дуги периметра, а другой глаз закрывает рукой. Из-
мерения проводят, перемещая марку (белую или цветную) по внутренней поверхности
дуги периметра от 90° к 0, и просят испытуемого указать тот момент, когда опознавае-
мая марка станет видна фиксированному глазу. Вращая дугу периметра вокруг оси, по-
следовательно измеряют поле зрения в 8-12 меридианах с интервалами 45° или 30°. Ве-
личины поля зрения в градусах для различных цветовых марок заносят по направлени-
ям меридианов измерения в бланки. Ахроматическое поле больше хроматического, что
связано с расположением палочек на периферии сетчатки.
10.4. Определение цветового зрения проводится с помощью Полихроматических таблиц
для исследования цветоощущения / Е.Б. Рабкин. - М.: Медицина, 1971. - 72 с.
11. Профильный материал для студентов лечебного факультета.
11.1. Аккомодация глаза после 40 лет уменьшается и после 60-ти утрачивается в связи с по-
терей эластичности хрусталика, ослаблением цилиарной мышцы и ригидностью цин-
новых связок, что называется пресбиопией (старческой дальнозоркостью).
11.2. Световоспринимающая функция: повышается порог абсолютной световой чувстви-
тельности, уменьшается скорость реакции зрачка под влиянием светового раздражите-
ля, снижается скорость темновой адаптации в результате изменения прозрачности хру-
сталика н увеличения сужения зрачка.
11.3. Снижение остроты зрения после 60 лет происходит в результате:
♦♦♦ уменьшения прозрачности преломляющих сред глаза;
❖ склерозирование сосудов в сосудистой оболочке, приводящего к нарушению тро-
фической функции пигментного слоя, обеспечивающей ресинтез зрительного
пигмента в фоторецепторах; в результате наступает дистрофическое изменение
сетчатки.
423
11.4. Ослабление рефракции происходит в связи с помутнением и уплотнением хрусталика и
роговицы.
11.5. Повышаются пороги цветоощущения, контрастности и цветоразличения, особенно в
голубой части спектра, что объясняется отчасти пожелтением с возрастом хрусталика.
11.6. Поле зрения у пожилых может быть неизменным или слегка концентрически сужен-
ным (для красного и синего цвета).
11.7. Снижение восприятия критической частоты мельканий в результате уменьшения под-
вижности нервных процессов и более медленной обработки зрительной информации в
корковом отделе.
12. Профильный материал для студентов педиатрического факультета.
12.1. Антенатальный период.
Ъ Развития зрительного анализатора начинается на 3-й неделе эмбрионального пе-
риода.
♦♦♦ Кб мес. возникает зрачковый рефлекс на свет.
123. Новорожденные и грудные дети.
♦ ♦♦ Движения глаз в первые дни жизни некоординированы, толчкообразны, замедлен-
ны; периодически наблюдаются сходящееся косоглазие и нистагм, спонтанные
мигания редки (1 - 3 в 1 мин). Фиксация взора появляется не ранее 2-недельного
возраста и составляет 1-2 мин.
На 4-й неделе жизни наблюдается содружестаеннал реакция зрачков, но глаза
почти постоянно закрыты независимо от дейстаия цветового раздражителя; при
слабой освещенности - слабое расширение зрачков из-за недостаточно развитых
мышц радужной оболочки; сужение зрачков при фиксации азора на предметы.
❖ Защитный мигательный рефлекс иа внезапное световое раздражение имеется с
первых дней жизни, защитный рефлекс при приближении предметов к глазам
(смыкание век) появляется с 6-й недели жизни.
♦ ♦♦ Секреция слезной жидкости у новорожденного низкал в результате недостаточной
сформированности слезных желез, слезообразование прн плаче происходит иа 2-м
мес. жизни.
* ?* Цветовое зрение: реакция зрительного анализатора на различные цвета имеется
сразу после рождения: фотостимуляция красным цветом приводит к замедлению
дыхания и ЧСС, к синхронизации биопотенциалов в зрительной коре, воздействие
зеленым цветом - к противоположной реакции. Дифференцировка цветовых раз-
дражителей происходит с 3 - 4 мес. жизни. В 6 мес. дети различают все цвета.
Объемное зрение начинает формироваться с 5 мес. возраста, этому способствует
совершенствование координации движения глаз, фиксация азора на предмете,
улучшение остроты зрения, взаимодействие зрительного анализатора с другими
(особенно с тактильным); к 6 - 9 мес. возникает представление о глубине и отда-
ленности расположения предметов (стереоскопическое восприятие пространства).
♦ ♦♦ Острота зрения у новорожденного низка: в 6 мес. составляет 0,1, в возрасте 1 года
- 0,2. Это связано с незаконченностью развития желтого пятна и коркового отде-
ла. На 1-м году жизни слабо развита темновая и световая адаптация, а также ла-
бильность зрительного анализатора - снижено критическая частота слияния мель-
каний.
123. Дети от 1 года до 5 лет.
♦♦♦ Цветоощущение окончательно формируется к 2 - 3 годам в результате развития
колбочек сетчатки н центральных структур цветоощущения. Поле зрения до 6 лет
значительно сужено.
424
Амплитудные и временные показатели электроретинограммы на световой раздра-
житель достигают максимальных значений к 5 годам.
12.4. Диоптрическая система глаз.
❖ На 1-м году жизни характерна дальнозоркость (гиперметропия 3-4 D), обуслов-
ленная относительно небольшим размером (18,5 мм) переднезадней оси глаза (фо-
кус оптической системы находится за сетчаткой); к 9-12 годам она нормализуется
в результате увеличения переднезаднего размера глазных яблок.
❖ Хрусталик до 10 лет очень эластичен, поэтому имеется максимальная аккомода-
ция; после 10-летнего возраста снижается его эластичность и способность к акко-
модации: с 10 до 15 лет она равна 14 D, ближайшая точка ясного видения в 10 лет
на расстоянии 7 см, в 15 лет - 8 см.
❖ Причины развития близорукости (миопии) у детей дошкольного возраста:
• чрезмерное увеличение переднезаднего размера глазных яблок;
• недостаточная жесткость склеры как один из наследственных факторов;
• длительное рассматривание близких предметов приводит к сокращению на-
ружных мышц глаза;
• профилактика близорукости: 1) соблюдение дистанции рассматриваемых
предметов не менее 35 - 40 см от глаз; 2) при обучении чтению и письму де-
лать перерывы; 3) достаточная освещенность.
Задание 2. Подготовить реферативное доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514) использовать имеющиеся у
вас учебники и пособия): [1-9], [11-15].
3.2. Дополнительная литература (см. с. 514 - 522): [27], [29], [30], [37], [43], [47],
[111], [152]. [171], [173], [175]. [216], [218], [222], [225], [231], [244], [245], [251-
254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиояогии человека» /В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 28-1. Для исследования глазного дна врач капает на конъюнктиву глаза растаор атропина
(блокатора М-холинорецепторов). Используя свои знания по физиологии диоптрического аппара-
та глаза объясните, зачем он применяет атропин и какое нарушение зрительного восприятия будет
у пациента в течение времени действия атропина?
425
Задача 28-2. Объясните, как и почему измениться диаметр зрачка у человека при сильном боле-
вом воздействии?
Задача 28-3. Для более лучшего рассматривания предмета мы приближаем его к глазам. Объсни-
те, почему при этом его можно рассмотреть более детально и почему его фокусировка происходит
не за сетчаткой?
Задача 28-4. Левши в некоторых видах спорта, где необходимо быстрое синтетическое воспри-
ятие ситуации и использование новой руки (например, бокс, фехтование), имеют более быструю
двигательную реакцию, чем правши. Используя свои знания по зрительной сенсорной системе и
регуляции движения, объясните более короткий латентный период двигательной реакции у лево-
руких.'
426
Тема 29. Слуховой и вестибулярный анализаторы
Цель: а) знать структурно-функциональную характеристику слухового анализатора: звукоулавли-
вающие, звукопроводящие и звуковоспринимающие аппараты, проводниковый и корковый
отделы, центральные механизмы анализа звуков, теорию восприятия звуков, бинауральный
слух; роль вестибулярного анализатора в оценке положения и перемещения тела в пространст-
ве, его рецепторный, проводниковый и корковый отделы;
б) уметь использовать этн знания для анализа жизнедеятельности организма, функциональной
оценки слухового и вестибулярного анализаторов по результатам их исследования;
в) ознакомиться с практическими навыками на уровне умения: с методиками исследования
воздушной и костной проводимости (опыты Ринне и Вебера), исследования бинаурального
слуха; на уровне знания: исследование остроты слуха (тональная аудиометрия).
Мотивация темы: Роль слухового анализатора у человека особенно важна в связи с восприятием речи.
Знание процессов проведения и восприятия звука позволяет дать функциональную оценку
деятельности слухового анализатора. Вестибулярный анализатор играет ведущую роль в про-
странственной ориентировке человека. Методы исследования слухового и вестибунярного
анализаторов используются при профотборе (летчики, моряки, космонавты и др.) и в клинике
ЛОР-болезней.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, опре-
деляются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
I. Физиология слухового анализатора.
Слуховой анализатор - сенсорная система, воспринимающая звуковые колебания среды
(частотой от16 - 20 Гц до 16 - 20 кГц) и формирующая звуковые ощущения.
Звук - колебания упругой среды в виде чередующихся волн компрессии и разряжения, образующиеся
вибрирующим объектом. В частотной характеристике звука выделяют тоны и шумы. Простые тоны яв-
ляются гармоническими колебаниями, имеющими одну частоту звучания (например, звук камертона).
Сложные тоны., кроме основной наименьшей частоты (vo), имеют кратные основной (2v0. 3 Vo и др.) допол-
нительные частоты - обертоны (например, гласные звуки, тоны музыкальных инструментов). Тембр (окра-
ска) звука зависит от состава обертонов. Шум - звук, состоящий из несвязанных между собой, неповто-
ряющихся частот (например, согласные звуки, внбрацня машин). Энергетической характеристикой звука
является интенсивность, измеряемая в ваттах (от 1042 - минимальный порог, до 10 вт/м2 - болевой порог)
или звуковое давление, измеряемое в Паскалях (от 2-Ю'5 до 60 Па). Отношение этих интенсивностей равно
1013. Минимальный порог принимают за 0, все остальные интенсивности выражают через десятичный лога-
рифм её отношения к пороговой. При использовании децибел весь диапазон интенсивности воспринимае-
мых звуков до болевого порога укладывается в 130 децибел (дБ).
1. Звукоулавливающий и звукопроводящий аппарат слухового анализатора (обра-
зован наружным и средним ухом, костями черепа и жидкостными каналами улитки).
1.1. Воздушная часть звукопроводящего аппарата.
❖ Ушная раковина и наружный слуховой проход.
• Выполняют функцию коллектора и обеспечивают направленное прове-
дение звука. (При этом ушная раковина за счет концентрации звуковой энергии со-
гласовывает низкое сопротивление в проведении звука окружающей среды и высокое
- в слуховом проходе.)
427
• Выполняют функцию резонатора (слуховой проход имеет резонансные частоты
в области 3,9 и 15 кГц, главная резонансная частота ушной раковины ~ 5 кГц), они
усиливают давление звука на ~ 10 дБ в области высоких частот (> 3
кГц).
• Определяют локализацию источника звука.
Ф Смещение источника звука в вертикальной плоскости определяется в результате
изменения спектра резонансных частот (в связи с формой ушиой раковины).
ф Локализация источника звука спереди-сзади головы связана с тем, что ушная ра-
ковина ослабляет интенсивность звуков, исходящих от источника, расположен-
ного сзади человека.
• Имеют механическую защитную функцию.
❖ Среднее ухо (состоит из барабанной полости и слуховой трубы, пропускает без затуха-
ния низкочастотные звуки до 1 кГц).
• Барабанная полость (находится в основании пирамиды височной кости, её объем
- 2 см3).
ф Барабанная перепонка (малоэластнчная мембрана, отделяет наружный слу-
ховой проход от барабанной полости, S - 70 мм2, толщина ~ 0,1 мм), в зависи-
мости от характера звука колеблется большей нли меньшей площа-
дью (эффективная площадь колебания до 55 мм2) и различными участками,
избирательно реагирует на звуки только слуховой зоны (16 Гц - 20
кГц). Её главная резоиаиасная частота - 1 кГц.
Ф Косточки - молоточек, наковальня, стремя, основание которого ук-
реплено кольцевой связкой в овальном окне (S ~ 3,3 мм2) осуществ-
ляют передачу н усиление звукового давления (в 22 раза, на 25 - 30 дБ).
Вместе с наружным ухом общее усиление звуковой энергии состав-
ляет 35 - 40 дБ, что компенсирует снижение звуковой энергии при
переходе ее нз воздуха в перилимфу улитки. В этом состоит главная
функция среднего уха.
• Мышцы среднего уха (m. tensor timpani и т. stapedius).
"ф- При сильных звуках (> 80 дБ) сокращаются обе мышцы, что умень-
шает проведение как низких, так н высоких звуков (латентное время
рефлекса 10-30 мс).
Ф Сокращение m. tensor timpani, прикрепленного к рукоятки молоточка, напрягает
барабанную перепонку, при этом вся система смещается внутрь и стремя вдавли-
вается в овальное окно и затрудняет восприятие низких и слабых звуков.
ф" Сокращение m. stapedius, прикрепленного к головке стремени, высвобождает
стремя из окна и облегчает восприятие низких и слабых звуков, ио затрудняет
восприятие высоких и сильных звуков; поэтому при восприятии слабых звуков
m. tensor timpani расслаблена, а т. stapedius сокращена.
ф Сокращение т. stapedius возникает также при собственной речи, что снижает ее
маскирующий эффект на воспринимаемую речь.
• Слуховая (евстахиева) труба открывается во время глотания, обеспечи-
вая выравнивание давления по обе стороны барабанной перепонки, а
также имеет дренажную функцию.
1.2. Жидкостная часть звукопроводящего аппарата (улитка в пирамиде височной кос-
ти - костный канал 2,5 завитка, £ - 35 мм).
428
❖ При переходе звука из воздуха в жидкость происходит резкое уменьшение
энергии и амплитуды звука, увеличивается звуковое давление (в 2 раза) и
скорость звука (в 4 раза), частота звука ие изменяется.
❖ Мембраны улитки: тонкая вестибулярная (Рейснерова) и более плотная - ба-
зилярная пластиика, расширяется к верхушке улитки в 5 - 10 раз (в физиоло-
гической литературе термину «пластинка» предпочитают термин «мембрана»).
❖ Каналы улитки:
• верхний (вестибулярная лестница) — от овального окна (точнее, от преддве-
рия);
• нижний (барабанная лестница) - от круглого окна, закрытого вторичной ба-
рабанной перепонкой;
• геликотрема - место соединения верхнего и иижнего каналов (S = 0,4
мм2);
• средний (средняя лестница или улитковый канал) — между вестибулярной мем-
браной и базилярной пластинкой.
* •* Жидкости улитки (перилимфа заполняет пространство между костным и перепонча-
тым лабиринтами, эндолимфа заполняет перепончатый лабиринт).
• Перилимфа заполняет верхний и нижний каналы (Na+ - 140, К+~ 12
ммоль/л; сообщается с перилимфой вестибулярного аппарата).
• Эндолимфа образуется сосудистой полоской улиткового канала и за-
полняет его, имеет более высокую вязкость, по отношению к перилимфе
заряжена положительно +80 мВ (К+~ 145, Na+ - 15 ммоль/л; сообщается с
эндолимфой вестибулярной системы). Высокую концентрацию К+ создает
К+-иасос на апикальной мембране иерецепториых клеток, который вы-
качивает К+ во внеклеточную среду.
• Передача звуковых колебаний по каналам улитки: колебания стремечка
передаются перилимфе преддверия, затем верхнего канала. Колебания
перилимфы верхнего канала через тонкую вестибулярную мембрану пе-
редаются эндолимфе, затем на покровную мембрану и базилярную мем-
брану с органом Корти. Частично колебания перилимфы верхнего ка-
нала передаются в нижний канал через геликотрему. (Колебания мембран
овального и круглого окон находятся в противофазе. Если онн колеблются синфазно -
например, прн разрушении слуховых косточек - возбуждение спирального органа рез-
ко уменьшается.)
1.3. Костная проводимость звуков (предполагаемые механизмы).
❖ Имеет более высокий порог по сравнению с воздушной проводимостью,
при восприятии собственной речи является главным путем проводимости
звука. Предложено два вероятных ее механизма.
❖ Инерционный механизм: при действии низких звуков череп колеблется как
единое целое и, благодаря инерции слуховых косточек (суммарная масса - 53
мг!?), происходит колебание стремечка в овальном окне. (Кость имеет собст-
• венный колебательный резонанс при 600-800 Гц.)
❖ Компрессионный механизм: при действии высоких звуков череп колеблется
отдельными сегментами, возникает периодическое сжатие перепончатого
лабиринта и усиливается давление перилимфы на лабиринтные окна и
429
мембраны улитки. (Интересно отметить, что через череп может передаваться звук час-
тотой до 250 кГц).
2. Рецепторный отдел слухового анализатора - спиральный (кортиев) орган (состо-
ит из эпителиоцитов двух видов: рецепторных - внутренних и наружных и поддерживающих
- фаланговых, столбовых, пограничных и др.).
2.1. Расположен на базилярной пластинке (lamina basilaris, натянута между яимбом спи-
ральной костной пластинки и спиральной связкой), состоящей из поперечно располо-
женных коллагеновых волокон (- 24 тысяч) и гомогенного основного вещества.
Имеет наименьшую ширину (- 0,04 мм), но высокую жесткость у основания
улитки и наибольшую ширину (- 0,5 мм), но низкую жесткость - у верхушки. Ба-
зилярная пластинка является эластической структурой с податливостью, увели-
чивающейся к геликотреме в десятки раз, она находится в ненатянутом (!) со-
стоянии.
2.2. Рецепторные волосковые клетки (вторичные рецепторы, имеют на апикальной по-
верхности 30-60 стереоцилнй разной длины, книоцнлия редуцирована).
❖ Внутренние волосковые эпителиоциты (~ 3500, один ряд клеток) воспринима-
ют звуки разной частоты, преимущественно средней и высокой интенсив-
ности, от них отходят 95% волокон слухового нерва с высокой степени ди-
вергенции - от одного рецептора отходит 10-20 афферентных волокон
(предполагаемый медиатор в нейрорецепторных синапсах - глутамат). Эфферентная ин-
нервация этих рецепторов выражена слабее. (Поражение всех внутренних волосковых кле-
ток ведет к глухоте.)
❖ Наружные волосковые эпителиоциты (- 20 000, 3-5 рядов клеток) имеют со-
кратительные структуры (актин и миозин) и способны сокращаться (их дли-
на в зависимости от частоты генерируемых в ннх токов может изменяться до 2 мкм). Эти
рецепторы имеют афферентную иннервация - 1-2 афферентных волокна на
рецептор и более выраженную возбуждающую и тормозящую эфферент-
ную иннервацию от нейронов верхней оливы ствола мозга (медиаторы в си-
напсах ацетилхолин н ГАМК). Наружные волосковые клетки базальным полю-
сом связаны с базилярной пластинкой, а стереоцилиями, имеющими жест-
кую W- или V-образную конструкцию, - с массивной покровной мембра-
ной. Поэтому любые изменения длины этих фонорецепторов будут вызы-
вать смещение базилярной пластинки. Предполагают, что, «раскачивая»
базилярную пластинку, они обеспечивают высокую чувствительность и из-
бирательность спирального органа. (Этот механизм подтверждается данными ото-
акустической эмнсснн - метода регистрации излучения звука из внутреннего уха в наруж-
ный слуховой проход В его основе лежит обратная связь в системе «наружные волоско-
вые клетки —> барабанная перепонка». При поражении всех наружных волосковых клеток
отоакустическая эмиссия исчезает, а чувствительность слуха снижается на ~ 45 дБ,)
2.3. Тенториальная (покровная) мембрана - коллагеновые волокна и желатиноз-
ная масса, с нижней поверхностью которой связаны волоски наружных и час-
тично внутренних рецепторов (поэтому еб смещения больше воздействуют на наружные
фонорецепторы, чем внутренние). Ее коллагеновые волокна по длине н числу, вероятно, явля-
ются зеркальным отражением таких же волокон базилярной пластинки.
2.4. Электрические явления в улитке.
❖ Улитка - сильно поляризованная структура. Эндолимфа по отношению к
перилимфе имеет заряд, равный +80 мВ, который называется эндокохлеар-
430
ным потенциалом (ЭП). Он создается за счет деятельности сосудистой по-
лоски, образующей эндолимфу. С учетом отрицательного мембранного по-
тенциала клетки ЭП увеличивает вдвое электрический градиент (- 140 мВ)
между вне- и внутриклеточной средой, что делает возможным движение К+
по электрическому градиенту нз эндолимфы внутрь рецепторной клетки
против химического градиента (Величина ЭП зависит от смещения базилярной
мембраны: смещение её вниз увеличивает ЭП на 10%, смещение вверх - уменьшает на
50%.)
❖ Возникновение рецепторного потенциала (РП) связано со сгибанием сте-
реоцилий. (Стереоцилни одной клетки связаны в пучок поперечными нитями. Поэтому
сгибание одной стереоцилни ведет к сгибанию и остальных, что увеличивает количество
открываемых каналов.)
• Если кортиев орган смещается вверх, стереоцилни сгибаются в сторону,
противоположную от стержня улитки. К+-каналы открываются, увели-
чивается входящий в клетку К+-ток и мембрана рецептора деполяризу-
ется. Деполяризация открывает Са2+-каналы, входящие ноны Са2+ за-
пускают секрецию медиатора в синаптическую щель.
• Если кортиев орган смещается вниз, стереоцилни сгибаются в сторону
стержня улитки, lC-каналы закрываются и рецептор гнперполярнзуется.
•> Рецепторный потенциал имеет два варианта:
• переменный РП формируется при действии звука частотой до 1200 Гц и
полностью повторяет его частоту (его амплитуда 5-22 мВ);
• постоянный РП начинает формироваться при частотах звука выше 1000
Гц (его амплитуда во внутренних волосковых клетках 3-7 мВ);
• РП через синаптический контакт (наиболее вероятный медиатор - глутамат)
генерирует импульс в нейроне спирального ганглия.
❖ Кохлеарный микрофонный потенциал генерируется главным образом на-
ружными волосковыми клетками. Если электрод поместить на круглое ок-
но (или в нижний канал) и соединить с усилителем и громкоговорителем,
то последний воспроизведет тот звук, который действует на ухо.
23. Кодирование частоты и интенсивности звука.
❖ Кодирование частоты звука
• Резонансная теория Г. Гельмгольца (1863): учитывая, что базилярная
пластинка имеет поперечные коллагеновые волокна, было предположе-
но, что короткие волокна расположенные ближе к овальному окну, ре-
зонируют в ответ на высокочастотные тоны, а длинные, расположенные
ближе к геликотреме, резонируют в ответ на низкочастотные тоны. (Ос-
новным возражением против этой теории является то, что базилярная мембрана не на-
тянута н резонанс её волокон невозможен.)
• Гидродинамическая теория «бегущей волны» Г Бекеши (1947). Колеба-
ние стремени вызывает в каналах улитки бегущую волну давления, ко-
торая направляется к геликотреме. В результате податливости рейсне-
ровой мембраны и базилярной пластинки скорость распространения
волны невелика и уменьшается по мере её распространения вплоть до
нуля, Поскольку податливость базилярной пластинки нарастает по на-
431
правлению к геликотреме место пластинки, где волна полностью зату-
хает (а перед этим имеет максимальную амплитуду) зависит от частоты
звука: высокие частоты затухают ближе к овальному окну, низкие - к
геликотреме. Частоты менее 800 Гц проходят вдоль всей улитки и зату-
хают около геликотремы.
• В дальнейшем было показано наличие частотной избирательности ре-
цепторов: каждая волосковая клетка имеет высокую чувствительность
(низкий порог) к звукам одной частоты и более низкую к другим частотам.
❖ Кодирование интенсивности звука. При увеличении силы звукового давле-
ния (громкости) происходит повышения амплитуды РП, а также увеличи-
вается количество возбужденных рецепторов за счет вовлечения клеток,
имеющие более высокие пороги (пространственная суммация). При интен-
сивности звука, превышающем 130 дБ, возникает ощущение боли.
3. Проводниковый отдел слухового анализатора.
3.1. Содержит до 6 уровней переключений (кохлеарные ядра, верхняя олива, трапециевид-
ное тело, ядро латерального лемниска, нижнее двухолмие, медиальное коленчатое тело) н не
менее трех перекрестов (верхняя олива, нижнее двухолмие, корковый уровень). Характе-
ризуется выраженной дивергенцией возбуждения, наличием эфферентных воз-
буждающих и тормозных путей от верхней оливы на слуховые рецепторы.
3.2. Функции проводникового отдела.
• > Проведение звуковой информации.
❖ Кодирование продолжительности звука по длительности разряда нейронов
преобладает в нижележащих центрах (кохлеарные ядра, трапециевидные тела,
верхняя олива) и резко уменьшается в вышележащих центрах.
❖ Участие в пространственном восприятие звука происходит в местах кон-
вергенции (верхняя олива, нижнее двухолмие) по различению времени по-
ступления импульсов из правого и левого кортиевого органа.
❖ Эфферентные влияния из верхних олив на наружные слуховые рецепторы
регулируют их активность.
4. Корковый отдел слухового анализатора.
4.1. Первичная слуховая кора: 41-е поле, извилины Гешля височной доли в глубине
сильвиевой борозды и 42-е поле верхней височной извилины - формирует ощу-
щение тонов, шумов, звуков.
4.2. Вторичная слуховая кора: в 22-м поле верхней височной извилины левого по-
лушария (речеслуховой центр Вернике) формируется понимание последовательно-
сти звуков (слов), последовательности слов (фраз), в 22-м поле правого полу-
шария - понимание последовательности тонов (мелодии), интонации, половых
особенностей голоса н др.
4.3. В кодировании звуковой информации важную роль играют нейроны-
детекторы начала звука, окончания звука, чистых тонов, шумов, звуков с из-
меняющейся частотой или амплитудой. Корковые нейроны отвечают на слож-
ные акустические стимулы и их смысловые качества, что создает возможность
восприятия речи.
4.4. Психофизиология слуха.
432
❖ Наибольшая чувствительность слуха в диапазоне 1 - 4 кГц, в котором аб-
солютный порог интенсивности звука минимален (10'*2 вт/м2 или 2 10*5 60 Па).
Резко снижена чувствительность при низких частотах (например, при 100 Гц на
60 дБ). Звук, давлением свыше 130 -140 дБ (107 выше порогового), вызывает
боль и разрушение стереоцилий рецепторов.
❖ Дифференциальные пороги слуховой чувствительности: (при частоте 0,5 - 2
кГц и интенсивности звука 40 - 50 дБ) порог интенсивности звука равен 0,5 - 1,0
дБ, порог частоты 1%, длительности < 10%, угла расположения источника
звука 1 - 3е.
❖ Пространственное восприятие звуковых раздражителей (бинауральный слух)
реализуется на всех уровнях конвергенции (верхняя олива, нижнее двухолмие,
слуховая кора) и создается:
• при действии низких частот (до 1,3 кГц) различением времени поступ-
ления звука в правое и левое ухо (улавливается разница в 30 мкс!);
• при действии более высоких частот - различением интенсивности звука,
поступающего в правое и левое ухо (улавливается разница в 0,5-1 дБ).
• Положение источника звука определяется с точностью до 1 углового
градуса.
5. Методы исследования слухового анализатора.
5.1. Опыт Ринне: измеряют в секундах восприятие звука камертона при воздушной и ко-
стной проводимости (в норме воздушная > костной).
5.2. Опыт Вебера (костная латерализация): ножку звучащего камертона (128 Гц) ставят на
среднюю линию головы в области темени. В норме нет латерализации - звук слышен
посредине головы. При нарушении звукопроводящих путей, например, при закрытии
наружного слухового прохода ватным тампоном, звук отдает в больное ухо, при пора-
жении звуковоспринимающих аппарата - в здоровое ухо.
5.3. Тональная (воздушная и костная) и речевая аудиометрия проводится с помощью
аудиометра. При тональной аудиометрии подают относительно чистые звук (тоны) как
через воздух, так и через кость, и определяют минимальную интенсивность звука в де-
цибелах, которая воспринимается в виде ощущений. Нормальные пороги слуха для
звуков всех частот (от 125 до 10000 Гц) как по воздушной, так и костной проводимости
приведены к нулевой шкале, что упрощает анализ аудиограмм. Находят пороги слы-
шимости для каждой фиксированной частоты звуковых колебаний, постепенно увели-
чивая громкость тонов, и вычерчивают аудиограмму отдельно для правого и левого
уха. В норме она расположена в коридоре 0 ± 10 дБ.
Речевая аудиометрия особенно важна для диагностики центральных поражений
слуха. Речь, заранее записанная на магнитофоне, передаётся через телефон отдельно на
каждое ухо с одинаковой громкостью. Порогом разборчивости речевого слуха является
повторение испытуемым 50% переданных ему сигналов, интенсивность которых в ре-
чевом диапазоне выше порога слуха примерно на 35 дБ. Результаты исследования в ви-
де кривых разборчивости речи записываются на специальных бланках, по осн абсцисс
отмечают интенсивность речи, а по оси ординат - разборчивость речи в процентах.
6. Профильный материал для студентов лечебного факультета.
6.1. Максимальная чувствительность слухового анализатора в 19 - 30 лет. В основе возрас-
тных изменений слуха лежит расстройство звуковоспринимающего аппарата. После 35
лет происходит снижение слуха, особенно в высокочастотном диапазоне (верхняя гра-
433
ница частоты слышимых звуков каждый год снижается на 160 Гц), и к 65 годам верх-
няя граница воспринимающей частоты с нормальным порогом снижается до 1000 Гц.
Ухудшается различение тонов (повышается дифференциальный порог частоты).
6.2. Снижается речевое различение: по данным речевой аудиометрии уровень разборчиво-
сти речи у людей в 70 лет по сравнению с 16-летними падает в 10 раз.
7. Профильный материал для студентов педиатрического факультета.
7.1. Развитие слухового анализатора начинается на 4-й неделе антенатального периода.
Формирование рецепторного отдела происходит к 6 мес., и звуковые раздражители мо-
гут вызывать шевеление плода. Среднее ухо до рождения заполнено слизистой жидко-
стью.
7.2. Восприятие высоты и громкости звуков у новорожденного снижено, улучшается к
концу 2-го мес. жизни; различение звуков различных тональностей возможно на 3-м
мес. жизни. Пороги иа низких частотах меньше, чем на высоких; пространственный
слух (поворот головы на источник звука) возникает на 1-м году жизни.
73. Миелинизация проводникового отдела заканчивается к 4-му году; наибольшая острота
слуха имеется в 14-19 лет; возможно восприятие частот до 32 000 Гц.
II. Физиология вестибулярного анализатора.
1. Вестибулярный анализатор - сенсорная система, оценивающая положение головы,
её направление н скорость перемещения в пространстве.
2. Периферический отдел вестибулярного анализатора. (Расположен в пирамиде ви-
сочной кости, состоит из костного и перепончатого лабиринтов, образующих полукружные
каналы и полости преддверие.)
2.1. Отолитовый аппарат.
❖ Расположен в преддверии (части костного лабиринта), имеет два полостных об-
разования перепончатого лабиринта: маточку (горизонтальное направление) и
мешочек (вертикальное направление), заполненные эндолимфой (сообщается с эн-
долимфой улитки) - концентрация К+ - 145 ммоль/л, эндокохлеарный потен-
циал по отношению к перилимфе +75 мВ, общий поляризационный потен-
циал (МПП) в клетках - 130 мВ. Между костным преддверием и перепон-
чатым лабиринтом расположена перилимфа.
❖ В нем имеется два рецепторных поля - макула мешочка н маточки, содер-
жащие вторичные рецепторы: волосковые клетки, имеющие 40-110 сте-
реоцилий и киноцилию. Они погружены в отолитовую мембрану (желеоб-
разная масса), плотность которой за счет кристаллов кальциевых солей в
2,2 раза больше, чем эндолимфы.
• В каждой макуле имеются группы рецепторов с противоположно ориен-
тированными волосками. (Их фоновая активность равна около 80 Гц).
• Через рецептор постоянно протекает ионный ток, образованный входом
К+ из эндолимфы в клетку через механочувствнтельные каналы мем-
браны волосков (по удвоенному электрическому градиенту) н выходом
К* из клетки в базальном ее полюсе в межклеточную жидкость (по кон-
центрационному градиенту).
• Возбуждение рецепторов вызывается сдвигом отолитовой мембраны,
деформирующей стереоцилни в направлении к кнноцилии, что увели-
чивает величину входящего К+-тока и вызывает деполяризацию мем-
434
браны рецептора. Деполяризация открывает Са2+-каналы иа базальном
полюсе клетки, ионы Са2+ входят в рецептор н стимулируют экзоцитоз
медиатора (сдвиг в противоположном направлении тормозит рецептор).
❖ Адекватными раздражителями отолитового аппарата являются: 1) сила
земного притяжения («ускорение силы тяжести») - главный стимул; 2) ли-
нейное ускорение, особенно вертикальные колебания (порог различения уско-
рения 2-20 см/с); 3) центробежная сила.
• > Функции отолитового аппарата: 1) информация о положении головы в про-
странстве относительно силы земного притяжения; 2) информация о нерав-
номерном движении тела; 3) рудиментарная функция восприятия звуковых сигналов:
при разрушении улитки рецепторы макул воспринимают звуковые колебания с порогом 30
-40 дБ.
2.2. Ампулярный аппарат (Р. Баранн, 1914).
❖ Имеется три рецепторных поля в кристах ампул полукружных каналов: го-
ризонтальном (латеральном), фронтальном (переднем), сагиттальном (заднем).
• Рецепторные поля состоят нз вторичных рецепторов: волосковых кле-
ток I и П типов со стереоцнлиями и одной киноцилией.
• Реснички рецепторов погружены в желатинозную массу - купулу, плот-
ность которой равна плотности эндолимфы. Поэтому при линейном ус-
корении купула, в отличие от отолитовой мембраны, не сдвигается.
• Возбуждение рецепторов вызывается деформацией ресничек при сдвиге
купулы в направлении от стереоцилий к киноцилии, тогда как сдвиг в
противоположном направлении тормозит рецептор (механизмы возбужде-
ния аналогичны рецепторам отолитового аппарата).
❖ Адекватным раздражителем ампулярного аппарата является угловое ус-
корение (порог - 0,005°), которое воспринимается в результате инерции
эндолимфы.
❖ Основная функция ампулярного аппарата - информация о вращении головы
и тела.
2.3. Иннервация рецепторов отолитового и ампулярного аппаратов.
•> Афферентную иннервацию осуществляют волокна (~ 10 ООО) вестибулярной
ветви УШ нерва (проводят сенсорную информацию);
❖ Эфферентная иннервация (~ 400 волокон) осуществляется из различных
участках ствола - вестибулярного ядра Дейтерса, мозжечка, ретикулярной
формации и др. (оказывает тормозное и, в меньшей, степени активирующее
влияние на рецепторы).
3. Проводниковый отдел вестибулярного анализатора.
3.1. Вестибулярный ганглий (первые нейроны) и вестибулярная ветвь преддверно-
слухового нерва (предполагаемые медиаторы в рецепторио-нейроииом синапсе - ацетилхо-
лин, глутамат, ГАМК).
3.2. Вторые нейроны образуют вестибулярные ядра: латеральное (Дейтерса), верхнее
(Бехтерева), медиальное (Швальбе), нижнее (Роллера).
3.3. Третьи нейроны находятся в вентромедиальных и вентральных задних ядер та-
ламуса, медиальных коленчатых телах.
4. Корковый отдел вестибулярного анализатора.
435
4.1. Исследования последнего десятилетия, проведенные иа людях и обезьянах, по-
казали, что вестибулярный аппарат имеет широкое корковое представительст-
во: височная кора, кора островка, передняя часть поясной извилины, нижняя
часть постцентральной извилины.
4.2. «Вестибулярные» корковые нейроны полимодальны, они отвечают также на
проприоцептивные и кожные раздражения. Эти данные указывают на важную
координирующую роль вестибулярной системы в сложной функции простран-
ственной ориентации.
4.3. Корковый отдел формирует ощущение и восприятие движения, осознание по-
ложения тела в пространстве (при закрытых глазах), создает чувство равновесия и
возможность произвольной ориентации в пространстве.
5. Основные рефлексы с участием вестибулярного анализатора.
5.1. Вестибуло-спинальные и вестибуломозжечковые рефлексы участвуют в пере-
распределении тонуса мышц с целью сохранения равновесия (предполагаемый ме-
диатор в центральных синапсах - серотонин).
5.2. Вестибуло-глазиые рефлексы участвуют в движении глаз с целью сохранения
изображения иа сетчатке при изменении положения головы.
5.3. Вестибуло-вегетативные рефлексы (болезнь движения) осуществляют актива-
цию дыхания, моторики желудка, потоотделения, диуреза и др. Они возникают
при действии переменных ускорений частотой около 0,2 Гц, особенно в верти-
кальном направлении (эволюционно значимые частоты при ходьбе, беге, прыжках лежат в
другом диапазоне - 0,5-10 Гц).
6. Методы всследоваввя вестибулярного анализатора.
6.1. Определение порога ощущения противовращения. Эта проба характеризует возбуди-
мость вестибулярного анализатора и проводится на специальном вращательном кресле
Барани при медленном вращении (угловая скорость 0,5-5° в сек). Испытуемый с закры-
тыми глазами при угловой скорости ощущает вращение в определённую сторону. Если
равномерное вращение продолжается, то он скоро перестаёт его ощущать. При оста-
новке возникает ощущение противовращения. Сенсорным порогом считается та наи-
меньшая угловая скорость, после действия которой при остановке возникает ощущение
противовращения. В норме он равен 1,5-4° в секунду.
6.2. Вращательная проба (поствращательный нистагм). Проба проводится на кресле
Барани со скоростью 0,5 оборота в секунду с закрытыми глазами пациента. При этом
угловая скорость равна 180° в 1 сек, что примерно превышает пороговую в 100 раз.
После 10 оборотов кресло внезапно останавливают, просят испытуемого открыть глаза
и регистрируют реакции, вызванные раздражением вестибулярного аппарата: глазной
иистагм, защитные двигательные реакции мускулатуры шеи, конечностей и туловища,
вегетативные реакции (изменение пульса, давления, ЭКГ, дыхания, потоотделения и
др.). В норме продолжительность поствращательного нистагма равна 20-40 с.
7. Профильный материал для студентов лечебного факультета.
7.1. Особенности вестибулярного анализатора в пожилом возрасте. Уменьшение воз-
будимости вестибулярного анализатора наблюдается после 60 лет. Отмечается более
продолжительный нистагм и ощущения противовращения.
7.2. Деятельность вестибулярного анализатора в невесомости.
4* Сначала нарушается анализ положения тела в пространстве из-за невозможности
ориентации на гравитационную вертикаль в результате изменения импульсации с
рецепторов отолитового (и ампулярного) аппарата. Нарушается взаимодействие
436
между зрительным и вестибулярным анализаторами, развивается комплекс вести-
було-вегетативиых реакций (болезнь движения).
❖ Адаптация вестибулярного аппарата к невесомости развивается в течение не-
скольких дней в результате взаимодействия анализаторов на стволовом и корко-
вом уровнях.
8. Профильный материал для студентов педиатрического факультета. Особенности
вестибулярного анализатора в онтогенезе.
8.1. Структурная характеристика. Закладка структур вестибулярного анализатора про-
исходит одновременно со слуховым в виде единого слухового пузырька на 4-Й неделе.
Формирование рецепторного отдела, миелинизация вестибулярного иерва и появление
вестибулярных тонических рефлексов происходит на 4-5м мес. К 22 неделе устанавли-
ваются контакты вестибулярных ядер со спинальными центрами и ядрами глазодвига-
тельных нервов.
8.2. Функциональная характеристика,
♦ ♦♦ Нистагм глаз, статические рефлексы положения (рефлекс Мора), рефлексы иа по-
качивание возникают иа 1 мес. жизни. Покачивание новорожденных детей (осо-
бенно продольные с частотой дыхания матери) благоприятно влияют на их сер-
дечный и дыхательный ритмы.
♦ ♦♦ Выпрямительные рефлексы (обеспечивают удержание головы, сидение и стояние)
и лифтные рефлексы формируются иа 1-м году жизни.
♦ ♦♦ В периоды детства наблюдается увеличение возбудимости рецепторов вестибу-
лярного анализатора и продолжительности нистагма.
♦ ♦♦ Стимуляция вестибулярного анализатора в детстве способствует психическому и
соматическому развитию организма (его нарушения приводят к отставанию ин-
теллектуального развития и ухудшению регуляции движения).
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мии.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся
учебники и пособия): [1-9], [11], [14-15].
3.2. Дополнительная литература (см. с. 514- 522): [6], [16]. [27], [29], [30], [37], [43],
(44]. [47], [111], [131], [149], [152], [167], [171], [173], [217], [218], [222], [225], [236],
[244], [245], [251-254].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/-nphys
3.4. : Слайды для компьютерной демонстрации из «Атласа по физиологии человека»
/В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты остаточного уровня знаний по
данной теме.
3.5. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.6. Консультации преподавателей еженедельно по индивидуальному графику.
3.7. Контроль самостоятельной работы студентов будет осуществляться иа практи-
ческом занятии по данной теме, итоговом занятии по разделу: «Физиология анали-
заторов» и экзамене.
437
Задание 4. Решение ситуационных задач (ответы см. с. 508 - 513).
Задача 29—1. У пациента с воспалительным процессом в носоглотке произошло закрытие евста-
хиевой (слуховой) трубы. Через двое суток он обратился к врачу с жалобой па снижение слуха.
При обследовании наружного уха врач увидел втяженпе барабанной перепонки в сторону бара-
банной полости. Атмосферное давление в течение этих суток пе изменялось. Используя свои зна-
ния по физиологии, объясните причину смещения барабанной перепонки.
Задача 29-2. Прн проведении пробы Ринне у человека в правом ухе костная проводимость равна
18 с, воздушная проводимость 35 с; в левом ухе костная проводимость 20 с, воздушная проводи-
мость 14 с. Соответствует ли порме состояние костной п воздушной проводимости в правом п ле-
вом ухе?
Задача 29—3. Прн проведении пробы Вебера (ножка звучащего камертона расположена на сред-
ней линии головы в области темени) у первого пациента лучшее восприятие звука смещено в сто-
рону здорового уха, а второго испытуемого - в сторону больного уха. Используя свои знания по
физиологии, определите состояние звукопроводящей п звуковоспрппимающей частей слуховой
системы в больном ухе п первого п второго пациентов.
438
Тема 30. Физиология высшей нервной деятельности. Условные рефлек-
сы. Типы ВИД
Цель: а) знать понятие о ВИД, роль условного рефлекса как формы приспособления человека к ме-
няющимся условиям существования, закономерности образования и проявления условных
рефлексов, структурно-функционвльные основы образования условных рефлексов, основы
аналитико-синтетической деятельности коры больших полушарий, роль динамического сте-
реотипа в обучении и приобретении трудовых навыков, типы ВНД, их классификацию, мето-
ды изучения ВНД у человека. Закон силовых отношений и фазовые явления в коре больших
полушарий;
б) уметь использовать эти знания при анализе психофизиологических явлений у человека, оце-
нивать типологические особенности ВНД у человека.
Мотивация темы: условные рефлексы обеспечивают опережающее отражение событий, заблаговре-
менную подготовку к ним. Условные рефлексы на базе второй сигнальной системы позволяют
использовать слово как способ программирования психических н физиологических функций
. при психотерапии, гипнозе. По этой же причине слово может стать патологическим фактором
и причиной ятрогенных заболеваний. Изучение типологических свойств ВНД позволяет эф-
фективно осуществлять профотбор, прогнозировать надежность человека в системах «человек
- машина», оптимизировать условия труда н отдыха, снизить риск развития психосоматиче-
ских заболеваний.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике н
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Общая характеристика высшей нервной деятельности.
1.1. ВНД - это деятельность высших корковых и подкорковых отделов мозга, фор-
мирующая поведение, направленное на наиболее тонкое и точное приспособле-
ние организма к среде.
1.2. И. И. Павлов - создатель учения о ВНД и условных рефлексах (19034936);
главный научный труд «Двадцатилетний опыт объективного изучения высшей
нервной деятельности (поведения) животных, условные рефлексы», 1932.
1.3. Основные методологические принципы учения о ВНД (УШ. Павлов).
❖ Вероятностный детерминизм: условиорефлекторная деятельность ини-
циируется не только внешними раздражителями, но и внутренними про-
цессами - мотивациями и потребностями организма.
❖ Единство анализа и синтеза: «первичного разложения целого на части,
единицы, а затем постепенного сложения целого из единиц, элементов».
❖ Структурный подход - «расположения действия силы в пространстве,
приурочение динамики к структуре», тенденция связать определенные
компоненты условного рефлекса с конкретными структурами мозга.
1.4. Методы исследования ВНД.
❖ Метод условных рефлексов.
♦♦♦ Нейрофизиологические методы:
• нейронография (см. тему 6; задание 1:4.3).
439
• фоновая электроэнцефалография (см. тему 6; задание 1:4.4);
• регистрация вызванных потенциалов (см. тему 6; задание 1:4.5);
• регистрация сверхмедленной активности (например, условнонегатив-
ная волна нли волна ожидания).
❖ Регистрация соматических (электромиография, окулография) н вегетатив-
ных (ЧСС, АД, частота дыхания, кожно-гальванической реакции и др.)
сдвигов при условных рефлексах.
❖ Методы перерезки, разрушения и раздражения мозговых структур.
❖ Функциональная компьютерная томография (см. тему 6; задание 1:4.7).
❖ Тестирование психофизиологических свойств человека.
• Временное выключение одного полушария (используется введение амитала
натрия в одну из сонных артерий либо односторонний электрошок). Эти процеду-
ры, выключая на 30 - 40 мин одно полушарие, позволяют исследовать
функциональную асимметрию полушарий прн реализации разных пси-
хических функций: внимания, восприятия, речи, сознания, эмоций).
• Аппаратурные и бланковые психофизиологические исследования памя-
ти, внимания, эмоций н др.
• Нейрофармакологическне исследования (например, классические опыты И.П.
Павлова и П.М. Никифоровского по влиянию брома и кофеина на процессы возбужде-
ния и торможения, скорость выработки условных рефлексов).
♦♦♦ Термоэнцефалоскопия (термокарты метаболической активности мозговых структур).
2. Условные рефлексы как форма приспособления к изменяющимся условиям су-
ществования.
2.1. Условный рефлекс - это приобретенная реакция организма, формирующаяся на
базе безусловного рефлекса прн сочетании ранее индифферентного (условного)
раздражителя с раздражителем, вызывающим безусловный рефлекс. Условные
рефлексы обеспечивают прогностическую функцию мозга (предвидение собы-
тий) н позволяют заблаговременно подготовиться к наступлению важного для
организма события, отражают временную организацию мира.
2.2. Основные правила образования условных рефлексов.
❖ Нормальное состояние ЦНС: поддержание оптимального уровня актива-
ции, состояния бодрствования.
❖ Предшествование условного сигнала безусловному раздражению.
♦ > Неоднократное сочетание условного и безусловного раздражителей. (Услов-
ный рефлекс может образоваться и при однократном сочетании условного и безусловного
раздражителей, если прн этом имеется сильное эмоциональное подкрепление, например,
при действии болевого раздражителя)
❖ Физиологическая значимость (сила) условного раздражителя должна быть
слабее безусловного раздражителя. (Однако сила условного раздражителя должна
быть достаточной, чтобы вызвать ориентировочный рефлекс. Условный рефлекс может
вырабатываться и на слабые неосознаваемые раздражители, но при этом они должны дей-
ствовать на фоне сильного мотивационного возбуждения, связанного с этими раздражите-
лями.)
❖ Исключение других посторонних раздражений, так как они вызывают ори-
ентировочный рефлекс н торможеине условного рефлекса.
2.3. Свойства условных рефлексов.
440
❖ Сигнальность - предупреждают о безусловном рефлексе, важном для орга-
низма событии.
❖ Обязательное участие высших (корковых и подкорковых) отделов головно-
го мозга.
Расширяют диапазон приспособительных реакций организма.
❖ Имеют индивидуальный характер, не наследуются.
2.4. Проявления условных рефлексов: соматические и вегетативные.
3. Классификация условных рефлексов.
3.1. По характеру безусловного подкрепления.
❖ По потребностям организма.
• Витальные условные рефлексы (пищевые, оборонительные, регуляции
сна).
• Зоосоциальные условные рефлексы (половые, родительские, территори-
альные).
• Условные рефлексы саморазвития или идеальные (игровые, имитаци-
онные, исследовательские).
❖ По наличию или отсутствию подкрепления.
• Положительные или подкрепляемые условные рефлексы.
• Отрицательные или не подкрепляемые условные рефлексы (условный
сигнал приобретает свойства торможения).
❖ По виду подкрепления.
• Рефлексы первого порядка (подкрепление условного сигнала безуслов-
ным раздражением).
• Рефлексы второго и более высоких (у человека 6-7) порядков выраба-
тываются на базе подкрепления другими условными рефлексами перво-
го или последующих порядков.
• Инструментальные рефлексы - для получения подкрепления необходи-
мо совершить активные, произвольные движения (например, нажатие
педали).
3.2. По характеру условного сигнала.
❖ По виду рецепторов, воспринимающих условный сигнал (экстероцептив-
ные и интероцептивные условные рефлексы).
❖ По природе условного сигнала:
• натуральные условные рефлексы, образующиеся на естественные при-
знаки безусловного раздражения (например, запах пищи);
• искусственные условные рефлексы, образующиеся на признаки, кото-
рые не являются свойством безусловного раздражения (например, пищевой
рефлекс иа включение лампочки).
❖ По структуре условного сигнала:
• условные рефлексы на простые раздражители (звук, свет);
• рефлексы на комплексные раздражители (иа сочетание сигналов: комплекс
одновременных или частично перекрывающихся раздражителей);
441
• рефлексы на цепные раздражители (в отличие от комплексных раздражителей
здесь раздражители отделены друг от друга некоторой паузой; вырабатывается ком-
плекс последовательных условных рефлексов на следующие друг за другом сигналы -
динамический стереотип).
33. По соотношению времени действия условного и безусловного раздражите-
ля.
♦♦♦ Наличные рефлексы (в этом случае условный сигнал и безусловное раздражение хотя
бы частично совпадают по времени действия).
•Z* Следовые рефлексы (подкрепление применяется после прекращения действия услов-
ного сигнала, между условным и безусловным сигналами разрыв по времени составляет от
нескольких секунд до 5 мни, что соответствует длительности кратковременной памяти).
4. Механизмы образовании условных рефлексов.
4.1. Стадии образования условных рефлексов.
❖ Стадия генерализации, когда широкий спектр раздражителей, а не только
подкрепляемый, вызывает реакцию. Она осуществляется по механизмам
доминанты. Ее физиологическое значение в том, что она обеспечивает от-
вет без предварительного обучения на все раздражители, сходные с под-
крепляемым.
❖ Стадия специализации. Ее физиологическое значение состоит в том, что
она обеспечивает точный, дифференцированный ответ только на подкреп-
ляемый сигнал, при многократном повторении этот ответ автоматизирует-
ся.
4.2. Структурно-функциональные основы образования условных рефлексов.
❖ Схема замыкания временной связи по И.П. Павлову: афферентная импуль-
сация при действии условного раздражителя поступает в сенсорную кору,
затем через ассоциативную кору выходит на корковое представительство
безусловного рефлекса, а затем поступает в эфферентные пути через сома-
тические и вегетативные центры.
♦> Современные представления о структуре условного' рефлекса дополни-
тельно включают в нее ретикулярную формацию, лимбическую систему,
базальные ядра и другие структуры головного мозга.
4.3. Механизмы образования временной связи.
❖ Нейрофизиологические механизмы.
• Образуются два очага повышенной возбудимости: более слабого - ус-
ловного сигнала, более сильного - безусловного подкрепления. Доми-
нантность последнего создается мотивационным возбуждением (напри-
мер, условный пищевой рефлекс у сытого животного не образуется). Формирование
доминанты приводит к функциональной конвергенции, расширению
рецептивного поля рефлекса, его генерализации.
• Условный раздражитель встраивается в рефлекторную цепь безуслов-
ного рефлекса по принципу «общего конечного пути».
• Между очагами возбуждения происходит иррадиация и межклеточная
реверберация возбуждения.
• Неоднократное сочетание условного раздражителя и подкрепления, а
также реверберация возбуждения приводят к суммации возбуждения.
442
• Формируется феномен проторения пути и долговременной потенциации
с участием гиппокампа, медиаторов и модуляторов синаптической пе-
редачи (см. ниже в 4.3).
• ЭЭГ-признаки образования временной связи:
•ф реакция десинхронизации, т.е. смена а-ритма p-ритмом, является
показателем активации мозговых структур и способствует образо-
ванию долговременной потенции в коре больших полушарий;
6-ритм отражает развитие долговременной потенциации в гиппо-
кампе;
Ф синхронизация электрической активности разных отделов мозга в
диапазоне у-частот отражает установление связи между удаленны-
ми отделами ЦНС при образовании условного рефлекса.
• Дифференцировочное торможение приводит к специализации условно-
го рефлекса.
❖ Нейрохимические механизмы увеличения проводимости через синапсы
(формирование долговременной потенциации).
• Глутамат через быстрые НМДА-рецепторы постсинаптической мембра-
ны (имеющие высокопроницаемый кальциевый канал) вызывает ВХОД Са2+ в
постсинаптический нейрон и активацию Са2+-зависимых протеаз, что
является пусковым механизмом повышения эффективности синаптиче-
ской передачи.
• Длительное (часами) поддержание повышенной синаптической прово-
димости происходит в результате активации глутаматом медленных
квисквалатных рецепторов, которые (через ФЛ-С -> ИФ3 и ДАГ) вызывают
выход Са2+ из агранулярной ЭПС и через геном клетки (активацию ранних
геиов - универсальных регуляторов генома) синтез нейромодуляторных пепти-
дов и белков памяти.
• Секрецию глутамата из пресинаптического окончания усиливают по-
средники (NO, арахидоновая кислота и др.), выделяющиеся постсинаптиче-
ским нейроном, а также глутамат из синаптической щели через рецеп-
торы пресинаптической мембраны (положительная обратная связь).
• В действительности механизмы потенциации намного сложнее. В повышении эффек-
тивности синаптической передачи глутаматные рецепторы взаимодействуют (через
вторые посредники и изменения мембранного потенциала) с адренорецепторами, хо-
лннорецепторами и ГАМК-рецепторами мембраны нейрона. Модулируют синаптиче-
скую передачу нейропептиды и нейрогормоны (эндорфины, энкефалины, ангиотензин
П, вазопрессин, окситоцин).
❖ Улътраструктурные механизмы образования временной связи.
• При образовании условных рефлексов происходит увеличичение синап-
тической поверхности дендритов нейронов, числа и площади аксоши-
пиковых синапсов, что повышает эффективность передачи сигналов
между нейронами.
443
• Происходит увеличение количества концевых разветвлений аксона и их
миелинизация олигодендроцитами, что увеличивает межнейронные свя-
зи и эффективность передачи возбуждения.
5. Торможение условных рефлексов. Виды торможения открыты И.П. Павловым,
который разделил их иа два типа.
5.1. Безусловное (внешнее) торможение.
❖ Гаснущий тормоз возникает при действии нового раздражителя, вызы-
вающего ориентировочный рефлекс, тормозящий условиорефлекторную
деятельность в результате отрицательной индукции. При неоднократном
применении этого раздражителя тормозная реакция угасает.
❖ Постоянный тормоз возникает на повреждающие воздействия (например,
болевые) и вызывает индукционное торможение условнорефлекторной дея-
тельности.
❖ Запредельное торможение возникает под действием сверхсильиых раз-
дражений (пессимум силы) и имеет охранительный характер. Зависит от типа
ВНД (у представителей слабого типа возникает раньше и при более слабых воздействи-
ях), возраста (у детей и стариков возникает легче), функционального состояния
нервной системы (его развитию способствуют переутомление, недосыпание, токсиче-
ские вещества, алкоголь и др. факторы).
5.2. Условное (внутреннее) торможение.
❖ Угасательное торможение возникает при отмене подкрепления условного
сигнала.
❖ Дифференцированное торможение обеспечивает различение близких по
природе сигналов, один из которых подкрепляется, а другой не подкрепля-
ется (например, собаки различают метроном 100 и 104 уд/мин, тоны - 995 и 1000 Гц).
❖ Условный тормоз - добавление к условному сигналу другого раздражителя
и иеподкрепление этой комбинации (например, «свет» - подкрепляется, а «свет +
тон» - не подкрепляется) служит тормозом для основного условного сигнала и
отменяет условный рефлекс иа основной сигнал («тон» стал условным тормозом
для этого и других рефлексов).
❖ Запаздывающее торможение или рефлекс иа время (позволяет включать ответ-
ную реакцию не сразу после действия сигнала, а с некоторой задержкой, например, у со-
бак до трех мни).
❖ Значение всех видов внутреннего торможения заключается в устранении
ненужной в данный момент условнорефлекторной деятельности. Торможе-
ние условных рефлексов связано с возникновением долговременной де-
прессии (см. тему 4; задание 1:4.4).
6. Аналитико-синтетическая и системная деятельность коры больших полушарий
(«Под свойством анализа и синтеза понимается способность нервной системы анализировать, т.е. вы-
делять из внешней среды большее или меньшее количество раздражителей, действующих на орга-
низм, и синтезировать, т.е. объединять те из них, которые в данный момент совпадают с какой-либо
его деятельностью» И.П. Павлов).
6.1. Анализ - это дробление, выделение информации, синтез - объединение, инте-
грация информационных сигналов в нейронных сетях.
6.2. Анализ на уровне деятельности коры больших полушарий осуществляется с
участием дивергентных процессов в ее нейронных сетях, деятельностью иейро-
444
нов-детекторов простых признаков, способностью левого полушария обрабаты-
вать информацию поэтапно, аналитически. Синтез осуществляется, напротив, с
помощью конвергентных процессов, нейроиов-детекторов сложных признаков,
способностью правого полушария обрабатывать информацию одновременно,
синтетически.
6.3. Некоторые проявления аналитико-синтетической и системной деятельности.
❖ Условнорефлекторное переключение - один и тот же условный раздражи-
тель в зависимости от конкретных условий его действия (время дня, разные
помещения и др.) может вызывать разные условные рефлексы. (В переключе-
нии обязательно участие миндалевидные тела височных долей, осуществляющей баланс
доминирующих н конкурирующих мотиваций.)
• > Динамический стереотип (ДС) (И. П. Павлов) - это зафиксированная в памя-
ти единая система из условных и безусловных рефлексов, образующихся
под влиянием многократно повторяющихся в определенной последова-
тельности раздражителей. ДС возникает, если деятельность человека неод-
нократно протекает в однотипных условиях. В ДС конец одного рефлекса
запускает следующий рефлекс и т.д., поэтому не обязательно действие все-
го комплекса раздражителей. При ДС условнорефлекторная деятельность
осуществляется легче и автоматичнее, преимущественно иа подсознатель-
ном уровне с участием подкорковых механизмов, улучшается синхрониза-
ция соматических и вегетативных функций. С этим связана важная роль
ДС в адаптации, обучении и приобретении трудовых навыков.
❖ Функциональная система (см. тему 32; задание 1:4).
7. Типы ВНД (темперамента), их классификация («Темперамент- хороший конь, но плохой
наездник». Гофмиллер).
7.1. Общая классификация для человека и животных.
❖ Используются особенности силы, уравновешенности и подвижности нерв-
ных процессов (И.П. Павлов).
• По силе (оценивается по пределу работоспособности клеток коры н развитию песси-
мального торможения) выделяют сильный и слабый типы.
• По уравновешенности (балансу силы) процессов возбуждения и торможе-
ния - уравновешенный и неуравновешенный.
* По подвижности (скорости смены возбуждения на торможение и наоборот) -
подвижный и инертный.
❖ Используется выраженность информационно-мотивационных систем го-
ловного мозга (П.В. Симонов, 1987); они определяют на какие сигналы и как
отвечать.
• Информационные системы мозга.
•Ф- Лобиая кора - организует поведение на сигналы высоковероятност-
ных событий, в определенных ситуациях.
•ф- Гиппокамп - организует поведение на сигналы с низкой вероятно-
стью событий, в неопределенных ситуациях.
• Мотивационные системы мозга.
•ф- Гипоталамус - выделяет доминирующую потребность и мотива-
цию.
445
ф Миндалевидное тело височной доли - обеспечивает баланс доми-
нирующих и конкурирующих потребностей и мотиваций
❖ Соотношение типов темперамента по классификациям Гиппократа и
Павлова с учетом информационных и мотивационных систем мозга.
• Холерик - «безудержный» тип: сильный, неуравновешенный с преобла-
данием возбуждения, подвижный; четко выделяет доминирующую по-
требность (гипоталамус), игнорирует отвлекающие сигналы (лобная ко-
ра).
• Сангвиник - «живой» тип: сильный, уравновешенный, подвижный тип;
четко выделяет доминирующую мотивацию (гипоталамус), способен
реагировать на сигналы с неясным значением (гиппокамп).
• Флегматик - «спокойный» тип: сильный, уравновешенный, инертный;
имеет сбалансированные потребности без акцента на какую-либо (мин-
далина), реагирует только на высокозиачимые сигналы (лобная кора).
• Меланхолик - «оранжерейный» тип: слабый тип, нерешительный, не-
уравновешенный или уравновешенный, инертный или подвижный; за-
труднен выбор доминирующей мотивации (миндалина), готов реагиро-
вать на широкий круг незначительных сигналов (гиппокамп).
7.2. Частная классификация для человека основана на соотношении первой и
второй сигнальных систем (по И. П. Павлову).
❖ Художественный тип.
• Преобладает активность первой сигнальной системы, базой которой яв-
ляется правое полушарие. Люди этого типа воспринимают явления и
предметы целиком, в виде чувственных образов («захватывают действитель-
ность целиком», по И.П. Павлову).
• У лиц данного типа более высокий уровень невербального интеллекта и
лучше развиты коммуникативные способности.
• Чаще встречаются сильный тип нервной системы и экстраверсия (дея-
тельность направлена на других людей).
Ф Мыслительный тип.
• Более активна вторая сигнальная система и левое речевое полушарие.
• Чаще встречается у меланхоликов, одновременно имеется более высо-
кий уровень тревожности.
• Для данного типа характерен высокий уровень вербального интеллекта,
развитые познавательные способности.
• Чаще встречается интроверсия (деятельность направлена на самого себя).
❖ Средний тип имеет примерно равную активность сигнальных систем.
8. Закон силовых отношений и его изменения прн разных функциональных со-
стояниях организма (пробуждении, засыпании, неврозах, под действием лекарств).
8.1. Фазовые явления в деятельности коры больших полушарий.
❖ Уравнительная фаза - на сильные и слабые, высоко- и малозначимые сиг-
налы возникает одинаковая реакция.
446
❖ Парадоксальная фаза - реакция на малозначимый сигнал более сильная,
чем на высокозначимый.
❖ Ультрапарадоксалъная фаза - неадекватные реакции (например, перед сном
прн засыпании ребенку дают еду - он отворачивается, убирают еду - начинает плакать).
❖ Тормозная фаза - отсутствие реакции на раздражители.
8.2. Экспериментальные неврозы (И. П. Павлов).
❖ Экспериментальные неврозы - это длительные нарушения ВНД вследствие
перенапряжения возбудительного или тормозного нервных процессов, а
также их подвижности.
❖ Для получения неврозов в эксперименте применяют сверхсильные или не-
обычные чрезвычайные раздражители, выработку тонких дифференциро-
вок, нарушения динамических стереотипов.
❖ Экспериментальные неврозы характеризуются нарушением адаптивного
поведения, нарушениями условных рефлексов, возникновением фазовых
состояний, вегетативными расстройствами и нарушениями сна.
9. Профильный материал (см. тему 32; задание 1: 7-9).
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 н 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 н 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющиеся у
вас учебники и пособия): (1-12), [14-16].
3.2. Дополнительная литература (см. с. 514 - 522): [1], [3], [10], [11-14], [16], [18].
[23], [28], [29], [30], [35 ], [43], [47 ], [ 48], [56], [59-61], [67], [86], [90], [97], [Ш].
[112], [120], [121], [132], [134], [136], [145], [152], [156], [161-164], [166], [168].
[173], [175], [176], [177], [179], [188], [196], [200], [201], [213], [214], [226], [236],
[242].
3.3. Дистанциоииые средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации нз «Атла-
са по физиологии человека» /В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
ЗЛ. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
447
Тема 31. Физиологические основы психических функций человека
Цель: а) знать физиологические основы н особенности психических функций человека (эмоции, воспри-
ятие, внимание, сои, память, сознание, мышление, речь), их адаптивную роль и возрастные изме-
нения, личностные особенности психических функций, физиологические методики исследования
психических функций, значение учения о ВНД для теории и практики медицины, педагогики;
б) уметь применять эти знания для рациональной организации умственного труда и отдыха, ис-
пользовать методы физиологического исследования психических функций в практической дея-
тельности.
Мотивация темы: Если со времен Гиппократа и Аристотеля медицина утверждалась во мнении, что
душевные заболевания возникают, как последствия телесных, то с 18 века стали появлять-
ся противоположные концепции. "Если бы органы брюшной полости могли рассказать историю
своих страданий, то мы с удивлением узнали бы, с какой силой душа может разрушать принадле-
жащее ей тело. В истории окончательно расшатанного пищеварения, пораженной в своих тканях
печени или селезенки, в истории заболевания воротной вены или больной матки с ее яичниками
мы могли бы найти свидетельства долгой порочной жизни, врезавшей все свои преступления как
бы неизгладимыми буквами в строение важнейших органов, необходимых для человека" (нем.
психиатр И. Хейнрот, 1818).
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике н
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Физиология эмоций.
1.1. Эмоции - это субъективные реакции на внешние и внутренние раздражители, в
том числе иа результаты собственной деятельности, сопровождаемые выражен-
ными субъективными переживаниями. Проявляются в виде радости, страха,
гнева, стыда, презрения и др.
1.2. Функции эмоций.
❖ Оценочная функция выражается в обобщенной оценке событий: их полез-
ности или вредности. Оценочная функция более очевидна для таких эмо-
ций, как стыд, ненависть, гнев.
♦ ♦♦ Побуждающая функция запускает поведение для удовлетворения домини-
рующей потребности И мотивации. (Побуждающая сила эмоций связана с тем, что
они представляют мотивы в сознании, т.е. делают мотивы осознанными.)
❖ Подкрепляющая функция - влияние эмоций на образование и угасание ус-
ловных рефлексов, обучение И формирование памяти. (Возникновение положи-
тельной эмоции при обучении ияи выработке условного рефлекса служит «наградой», по-
ощряющей дальнейшую деятельность. Появление отрицательной эмоции ведет к прекра-
щению деятельности, к избеганию этого состояния в результате поведения.)
❖ Компенсаторная функция мобилизует физиологические и психические ре-
зервы организма при недостатке информации, необходимой для удовлетво-
рения потребностей.
❖ Переключающая функция изменяет направленность поведения. Она осо-
бенно ярко проявляется при конкуренции мотивов, в результате которой
формируется доминирующая мотивация.
448
❖ Коммуникативная функция обеспечивает неречевые формы коммуникации:
мимика, жест, интонация, поза («язык человеческих чувств»), (До 90% эмоциональ-
ного межличностного общения при устной речи происходит иа невербальном уровне.)
1.3. Классификация эмоциональных ироцессов.
❖ По знаку: положительные и отрицательные.
❖ По интенсивности и длительности: настроение, эмоции, аффект (эмоция,
не контролируемая волей).
❖ По качеству потребностей, запускающих эмоции - биологические и выс-
шие (чувства): социальные и идеальные (например, «умственное наслаждение»
при интеллектуальной деятельности, по Платону).
1.4. Теории эмоций.
❖ Периферическая теория (В. Джеймс, К. Ланге): эмоции обусловлены осозна-
нием изменений в деятельности висцеральных и соматических систем («Мы
плачем не потому, что нам грустно, а нам грустно потому, что мы плачем»),
❖ Центральные теории рассматривают эмоции как результат активации моз-
говых структур.
• Локально-структурные теории: таламическая - возбуждение, поступая
в таламус, делится на два потока: один идет в кору и вызывает субъек-
тивные переживания, а второй идет в гипоталамус и вызывает физиоло-
гические реакции (У. Кеннон, В.М. Бехтерев); гипоталамическая - центры
удовольствия и наказания (В. Гесс, Д. Олдз); лимбическая - см. ниже в 1.5
(Дж. Пейпец, 1937).
• Биологическая теория (П.К. Анохин, 1948): эмоция является результатом
рассогласования или совпадения реального результата деятельности с
прогнозируемым в акцепторе результата действия.
• Потребностно-информационная теория - «отражение мозгом человека
какой-либо актуальной потребности (ее качества и величины) и вероятности
ее удовлетворения, которую субъект непроизвольно оценивает на осно-
ве врожденного и ранее приобретенного опыта» (П.В. Симонов, 1984).
(Оценка вероятности удовлетворения потребности осуществляется как на сознаваемом,
так и неосознаваемом уровнях ВНД.)
1.5. Физиологические механизмы эмоций.
♦ > Запуск эмоций (по П.В. Симонову, 1996): Э = - П (Ин — Ис), где Э - эмоция, ее си-
ла, качество и знак; П - сила и качество потребности; И„ - информация, необхо-
димая для удовлетворения потребности, - информация, существующая у орга-
низма (включает в себя знания, умения, энергетические ресурсы, время, которым
организм располагает для удовлетворения потребности). Положительные эмо-
ции возникают, если возрастает вероятность достижения цели в результате
поступления новой информации (Ис > Ин). Отрицательные эмоции возни-
кают при снижении вероятности достижения цели (Ин > Ис).
❖ Роль различных структур головного мозга в возникновении эмоций.
• Гипоталамус (является критической структурой для возникновения эмоций: пере-
резка ствола ниже гипоталамуса выключает эмоции).
• ф- Гипоталамус является главной структурой, формирующей виталь-
ные (биологические) потребности и эмоции. Стимуляция латераль-
449
ного гипоталамуса формирует положительные эмоции, медиально-
го гипоталамуса - отрицательные эмоции.
ф- Гипоталамус реализует подкрепляющую функцию эмоций. Непо-
средственным подкреплением при обучении является получение
эмоционально значимых стимулов. Это достигается гипоталамусом
на основе принципа максимизации положительных эмоций и ми-
нимизации отрицательных.
• Миндалевидное тело височной доли.
ф- Миндалина обеспечивает выделение доминирующей мотивации и
играет решающую роль в реализации переключающей функции
эмоций, т.е. выборе поведения, соответствующего не только той
или иной мотивации, но и условиям ее удовлетворения (влияние осу-
ществляется через хвостатое ядро).
•ф- Электрическая стимуляция миндалины сопровождается возникно-
вением эмоций страха, гнева, ярости.
"Ф* Удаление миндалины подавляет агрессивность и связанные с ней
эмоции, приводит к нарушению однократного обучения, требующе-
го участия сильной отрицательной эмоции.
• Гиппокамп реагирует на сигналы с низкой вероятностью подкрепления,
расширяет круг извлекаемых энграмм (следов) памяти, компенсирует
недостаток информации в ситуации неопределенности. В гиппокампе
формируется память о пережитых эмоциях.
• Лобнал и височная кора.
-ф- Лобная кора имеет важное значение для формирования высших
эмоций, связанных с социальными отношениями и творчеством, а
также обеспечивает социализацию биологических эмоций.
ф- Височная кора участвует в распознавании эмоциональных реакций
других людей, а также участвует в выражении эмоций.
• Поясная извилина имеет наиболее обширные связи с другими отделами
мозга. Предположительно выполняет функцию высшего координатора
других систем мозга, участвующих в формировании эмоциий.
• Лимбическая система головного мозга (рис. 28. с.98). Большинство выше-
названных структур входят в лимбический круг Пейпеца, идущий от
гиппокампа через свод к мамиллярным телам, от них к передним ядрам
таламуса, от него в поясную извилину и от нее через парагиппокампову
извилину обратно к гиппокампу. Этому кругу придается большое зна-
чение в формировании эмоций, обучения и памяти. Другой лимбический
круг (от миндалевидного тела к мамиллярным телам гипоталамуса, от
них к лимбической области среднего мозга и обратно к миндалине)
имеет важное значение в формировании эмоций, сопровождающих аг-
рессивно-оборонительные, пищевые и сексуальные реакции.
❖ Роль нейромедиаторов в организации эмоций.
• Имеется жесткая зависимость эмоций от биохимического состава внут-
ренней среды мозга.
450
• Стимуляция адренергических (голубое пятно), дофамииергеческих (черное
вещество) и серотонинергических (центральное серое вещество) систем мозга
(увеличение содержания норадреналина, серотонина и дофамина) сопровождается
формированием положительных эмоций.
• Снижение уровня норадреналина и дофамина приводит к возникнове-
нию депрессии с преобладанием чувства тоски, тревоги и страха.
• Ацетилхолин является пусковым медиатором агрессивного поведения,
норадреналин обеспечивает эффекторные проявления, а серотонин ока-
зывает тормозное влияние иа агрессию.
• Влияние эндорфинов и энкефалинов на эмоции реализуется через опи-
атные рецепторы, плотность которых на нейронах лимбической систе-
мы большая (выброс эндорфинов прн ориентировочно-исследовательских реакциях
тормозит отрнцатеяьные эмоции и стимулирует положительные - этим обеспечивает-
ся самоподкрепление исследовательской деятельности).
❖ Функциональная асимметрия полушарий и организация эмоций.
• Левое полушарие контролирует преимущественно положительные эмо-
ции, быстрее реагирует на слайды с выражением радости, уменьшает
степень тревожности.
• Правое полушарие вызывает сдвиг в эмоциональной сфере в сторону
отрицательных эмоций, быстрее реагирует иа слайды с выражением пе-
чали, распознает эмоциональную интонацию речи и окраску голоса.
1.6. В структуре эмоций выделяют собственно эмоциональные переживания и
периферические проявления.
❖ Эти компоненты могут иметь относительную самостоятельность: вы-
раженные субъективные переживания могут сопровождаться небольшими
периферическим проявлениями, напротив, при псевдоэмоциях яркие пери-
ферические проявления (например, мимические и вегетативные реакции при плаче
или смехе) могут протекать без существенных субъективных переживаний.
❖ Периферические (соматические и вегетативные) проявления эмойций.
• Соматические проявления эмоций запускаются от лобной коры через
экстрапирамидную и пирамидную системы в форме тонических (гипер-
тонус, скованность, тремор) и фазических компонентов двигательного
сопровождения ЭМОЦИЙ (мимика, жест, поза, интонации голоса, мигание).
• Вегетативные проявления эмоций запускаются от теменно-височных
отделов правого полушария через активацию гипоталамуса и проявля-
ются повышением ЧСС, АД, частоты дыхания, потоотделением, расши-
рением зрачков, выбросом в кровь гормонов и другими изменениями
функций внутренних органов.
1.7. Эмоциональное напряжение (стресс) и психосоматическиезабояевания, их
профилактика (см. также тему 34: 8.1).
❖ Величину эмоционального напряжения можно оценить по формуле: СН = Ц
(Ин, Ви, Эн - Ис, Вс, Эс), где СН - степень напряжения; Ц - цель; Ин, Вн, Эн -
необходимые информация, время, энергия; Ис, Вс, Эс -имеющиеся в распоряже-
нии организма информация, время, энергия. СН возникает, если Ин, Вн, Эн >
Ис, Вс, Эс. Степень напряжения пропорциональна при этом величине иера-
451
венства: CH-1 - состояние мобилизации внимания, работоспособности;
СН-2 - стеническая отрицательная эмоция с повышением работоспособно-
сти, но снижением эффективности деятельности; СН-3 - астеническая от-
рицательная эмоция с максимальной мобилизацией ресурсов организма,
которая не может компенсировать недостаток информации, энергии, вре-
мени; СН-4 — невроз (лсихологичесое состояние - тревожность, страх, депрессия).
❖ Периферические проявления эмоций (соматические и висцеральные) явля-
ются мишенью для управления эмоциональным напряжением.
• Соматический канал разрядки у современного человека ограничен в
связи с гиподинамией, этическими н правовыми отношениями.
• Преимущественная разрядкаэмоционального напряжения через вегета-
тивный канал ведет к «вегетативной буре», может приводить к дис-
функции внутренних органов, психосоматическим заболеваниям.
❖ Профилактика эмоционального стресса и психосоматических заболева-
ний.
• Мышечная система как регулятор эмоционального напряжения (роль
мышечной активности н релаксации).
• Антистрессорная роль сна.
• Адаптогены и повышение неспецифнческой резистентности.
• Самовнушение и аутотренинг («Бедняками нас делают неосуществимые жела-
ния». Восточная мудрость).
2. Физиология сна («Когда проснешься, не вскакивай с постели словно ужаленный. Сначала поду-
май о том, что тебе приснилось». Аварская мудрость).
2.1. Сон - периодический физиологический процесс, характеризующийся выключе-
нием сознания и значительным ослаблением связи организма с внешней средой.
2.2. Цикл «сон - бодрствование» является циркадианным биоритмом. Супрахиаз-
матические ядра гипоталамуса и эпифиз служат ритмоводителем (пейсмекером)
этого цикла.
2.3. Физиологическая роль сна.
❖ Восстановительная функция сна (преобладание процессов анаболизма).
❖ Антистрессорная функция сна (сон служит одним из механизмов психиче-
ской защиты личности).
❖ Адаптивная функция сна (синхронизация с циклом смены дня и ночи обес-
печивает оптимальное взаимодействие организма со средой, подготовку
организма к деятельности во время бодрствования).
❖ Роль сна в переработке информации: осуществление процесса консолида-
ции - перевода информации из кратковременной в договременную память.
2.4. Виды сна.
❖ Физиологический сон:
• периодический ежесуточный (моно- и полициклический);
• периодический сезонный;
Нефизиологический сон (наркотический, патологический).
2.5. Структура сна.
❖ Формула сна: Сон = (ФМС + ФБС) 4 - б ши
452
• Продолжительность цикла около 90 мин, количество циклов у коротко-
спящих 3 - 4, у долгоспящих 6-7.
• Фаза медленного сна (ФМС) занимает - 80% времени сна.
• Фаза быстрого сна (ФБС) занимает - 20% времени сна.
• Доля каждой фазы в разных циклах ночного сна неодинакова: в первых
циклах максимальна длительность ФМС, перед пробуждением увеличи-
вается длительность ФБС.
❖ Профиль сна: сон начинается с ФМС, переход в ФБС происходит из 2-й стадии ФМС (по-
верхностный сон), из ФБС переход осуществляется во 2-ю стадию ФМС, пробуждение
происходит из 1-й стадии (дремота) ФМС.
❖ Особенности структуры сна обусловлены:
• возрастом человека (в течение жизни укорачивается общая длитель-
ность сна, уменьшается глубина сна, происходит редукция дельта-сна,
сокращается доля ФБС);
• половой принадлежностью (у женщин средняя продолжительность сна
больше, чем у мужчин: среди мужчин больше «сов», среди женщин -
«жаворонков»).
2.6. Характеристика фаз сна.
❖ Фаза медленного сна.
• Стадии ФМС сна по ЭЭГ- критериям:
• ф- 1-я стадия (засыпание, дремота), на ЭЭГ преобладают а- и 0-ритмы,
в конце стадии возникают К-комплексы (серии высокоамплигудных мед-
ленных потенциалов длительностью 3-5 с);
- ф- 2-я стадия (поверхностный сон), на ЭЭГ имеются К-комплексы и
появляются веретена сна (частота - 15 Гц, вариант a-ритма), что совпа-
дает с отключением сознания; фаза занимает около 50% сна и по
длительности нарастает от первого к последнему пиклу;
• ф- 3-я стадия (глубокий сон) характеризуется наличием 8-ритма часто-
той 3 - 3,5 Гц, который занимает до 30% ЭЭГ;
4-я стадия (очень глубокий сон) - 8-ритм занимает свыше 30% ЭЭГ,
его частота - 1 Гц; 3-я и 4-я стадии имеются в первых циклах сна и
отсутствуют в последних.
• Вегетативные и соматические изменения во время ФМС: снижение
частоты дыхания, сердечного ритма, мышечного тонуса, усилена секре-
ция СТГ, мелатонина, пролактина, паратгормона и АДГ, преобладают
образование АТФ и анаболические процессы.
• Психические процессы во время ФМС:
•ф- сновидения преимущественно логического характера, связаны с ак-
тивностью левого полушария, происходит переработка информа-
ции, находящейся в промежуточной памяти (более характерны для ста-
дий б-сиа);
-ф- во время ФМС отмечаются сноговорение, снохождения, кошмары и
ночные страхи, скрежетание зубами.
❖ Фаза быстрого или парадоксального сна (Ю. Азеринский, Н. Клейтман, 1953).
453
• Характеризуется парадоксальным состоянием организма: по сравне-
нию с физиологическим покоем во время бодрствования одни функции
организма активированы, другие - заторможены.
• Активированные функции организма во время ФБС.
ф- «Вегетативная буря»: увеличены ЧСС, АД, частота дыхания, крово-
ток мозга, кровенаполнение органов малого таза и половых орга-
нов.
Ф’ Усилена секреция АКТГ и кортизола, пролактина.
ф- Регистрируются быстрые движения глазных яблок, обусловленные
активацией холинергических нейронов покрышки моста.
ф- На ЭЭГ отмечается появление высокочастотных ритмов, характер-
ных для бодрствования (а- и (3-ритмы, пилообразный 9-ритм) в свя-
зи с усилением активирующих влияний ретикулярной формации
среднего мозга (десиихронизация ЭЭГ).
Ф’ Психические процессы:
у 80 % испытуемых в эту фазу имеются яркие, образные, эмо-
циональные сновидения, связанные с активацией правого по-
лушария;
осуществляется перевод обработанной в предыдущей фазе сна
информации в долговременную память.
• Заторможенные функции организма во время ФБС.
"Ф* Отмечается максимальное падение тонуса скелетных мышц конеч-
ностей и туловища. Оно связано с активацией небольшой части мо-
ноаминергических нейронов голубого пятна среднего мозга, оказы-
вающих тормозное влияние иа спинальные мотонейроны.
ф- Снижение температуры тела (уменьшен терморегуляторный тонус).
Ф’ Снижена секреция СТГ и мелатонина.
ф- Повышены пороги возбуждения сенсорных систем (разбудить че-
ловека в ФБС труднее, чем в ФМС).
ф’ Резко снижена секреторная и перистальтическая активность пище-
варительного тракта.
2.7. Гуморальные механизмы сна (эндогенные факторы сна).
❖ Способствуют развитию ФМС пептид 8-сна, гликопептид S, МСГ и мела-
тонин, который стимулирует ГАМК-рецепторы (а также интерлейкин-1, интер-
ферон-а2, ангиотензин, соматолиберин, инсулин, простагландин Dj).
♦> Способствуют развитию ФБС холецитокннин-8, ВИП, соматостатин.
2.8. Нервные механизмы обеспечения цикла сон-бодрствование.
♦ > Сон как пассивный процесс связан со снижением активности механизмов
поддержания бодрствования (теория деафферентацин, по Ф. Бремеру): для под-
держания бодрствующего состояния коры необходим приток сеисориых
стимулов, при уменьшении этого притока развивается сои.
❖ Сои как активный процесс связан с торможением центров обеспечения
бодрствования и активацией центров сиа.
454
• Подкорковая ретикулярная теория сна и бодрствования (В. Гесс, Г. Мо-
руцци, X. Мегун).
ф- Центр бодрствования: его раздражение приводит к пробуждению,
при его разрушении - постоянный сон. Пробуждающие влияния
обеспечивает восходящая активирующая ретикулярная система
ствола, которая получает сигналы по неспецифическим экстралем-
нисковым путям, поэтому их перерезка приводит к развитию дли-
тельного сна и потере сознания. (Помимо нее существуют другие активи-
рующие системы: таламическая, заднего гипоталамуса.)
-ф- Центр сна - это совокупность взаимодействующих структур го-
ловного мозга, обеспечивающих циклическое развитие двух разных
процессов - медленного и парадоксального сна.
Сомногенная или синхронизирующая система медленного сна
включает в себя передний гипоталамус, прозрачную перего-
родку, ядра шва, моста, одиночного пути. Она вызывает тор-
можение восходящей активирующей ретикулярной формации,
синхронизацию таламокортикальной активности и появление
на ЭЭГ «веретен» сна, 5-волн, развитие коркового торможения.
В ФМС увеличена концентрация нейромедиаторов серотонина
. н ГАМК. Существуют реципрокные отношения между систе-
мами бодрствования (активирующей ретикулярной формацией) н син-
хронизирующей системой.
Десинхронизирующая система парадоксального сиа включает в
себя нейроны голубого пятна, тормозящие ядра шва синхрони-
зирующей системы, нейроны каудального и среднего оральных
ядер моста. В ФБС увеличен уровень нейромедиаторов норад-
реналина и ацетилхолина.
• Корковая теория сна.
-ф- Сон является разновидностью внутреннего торможения (И.П. Пав-
лов).
ф- Координация корковых и подкорковых структур в единую функ-
циональную систему (П.К. Анохин): ослабление тормозных влияний
коры на гипоталамические центры сна приводит к их активации,
что угнетает в таламусе передачу активирующего влияния ретику-
лярной формации на кору. В результате этого усиливается процесс
коркового торможения.
2.9. Физиологические основы и роль сновидений, гипнотические состояния.
(«Жизнь и сновидения - страницы одной и той же книги» А. Шопенгауэр.)
❖ Сновидения - следствие жизненного опыта человека, «небывалая комби-
нация бывалых впечатлений» (И.М. Сеченов); возврат к филогенетически
более древнему способу мышления (Ф. Ницше); осуществления подавлен-
ных желаний, королевская дорога к бессознательному (3. Фрейд).
• > Роль сновидений: защищают от пробуждения, являются одним из механиз-
мов психологической защиты личности («В сновидениях хорошие люди делают
455
то, что плохие наяву». Платон), осуществляют творческую, поисковую актив-
ность.
❖ Гипноз - это состояние ограниченного сознания, вызванное внушением из-
вне.
• Гипнабельность (подверженность гипнозу) зависит:
•ф- от возраста: дети и старики хуже поддаются гипнозу из-за ослаб-
ленного внутреннего торможения;
от пола: женщины более гипнабельны по сравнению с мужчинами;
от интеллекта: прн очень низком или высоком интеллекте гипна-
бельность низкая;
от типа ВНД: у представителей художественного типа она выше,
чем у представителей других типов.
• Возможные механизмы гипноза:
по И.П. Павлову: снижение контроля второй н увеличение активно-
сти первой сигнальной системы; для гипноза характерно наличие
«сторожевых» пунктов в коре, через которые осуществляется связь
гипнотизируемого с внешним миром (рапбрт); по мере гипнотизации
нарастает электрическая активность правого полушария прн сохра-
нении активности в левом речевом полушарии;
по 3. Фрейду: введение инструкции, минуя сознание, перевод ее из
сферы бессознательного через цензуру в систему сознания и преоб-
разование ее в соответствующее поведение;
по П.В. Симонову: специфическая разновидность бодрствования,
моральная капитуляция и некритическое отношение человека к
приказам гипнотизера.
3. Физиология памяти.
3.1. Нейрологическая иамять - это способность нейронных сетей фиксировать,
хранить и воспроизводить информацию. Совокупность этих процессов обеспе-
чивает формирование адаптивного поведения с учетом прошлого опыта. След
сформированной памяти называется энграммой. Емкость памяти составляет
около полумиллиарда единиц памяти (понятий, команд). (На процессы обеспечения
памяти затрачивается около 15% образующейся в нервной ткани АТФ.)
3.2. Некоторые виды нейрологической памяти.
❖ По психологической организации различают смысловую или словесно-
логическую память, которая оперирует с представлениями, и образную па-
мять: зрительную, слуховую, вкусовую, обонятельную, двигательную, ко-
торая оперирует с ощущениями н восприятиями.
❖ По временной организации различают сенсорную (доли секунды), кратко-
временную (секунды - часы), промежуточную (часы - сутки) и долговре-
менную память (дни - годы).
Есть и другое мнение: вся память рассматривается как долговременная, часть которой ста-
новится активной в двнной момент времени и в нее записывается энграмма вновь приоб-
ретенного опыта.
456
3.3. Современные представления о механизмах памяти (основные события развива-
ются в синапсах н связаны с внутриклеточными реакциями, формирующими в ннх долговре-
менную потенциацию. См. также тему 4; задание 1: 4.4 н тему 30; задание 1: 4.3).
❖ Следовые потенциалы («хвосты») нервного импульса являются основой
формирования сенсорной памяти. С ее помощью сенсорная кора головного
мозга на короткое время (доли секунды) удерживает в виде ощущений до-
вольно точную н полную картину внешнего мира, воспринимаемую орга-
нами чувств. (Сенсорная память имеет большую, почти неограниченную емкость. В бо-
лее длительный вид памяти переходит только незначительная часть информации, которая
имеет новизну и большую значимость для организма. Важную роль в фильтрации инфор-
мации играют внимание и эмоции.)
❖ Синаптической основой кратковременной памяти (КП) является ранняя фа-
за долговременной потенцнации (до трех часов). Повышение синаптической
проводимости в нейронных цепях при этом связано с многократной и вы-
сокочастотной передачей возбуждения в их синапсах, что через вторые по-
средники и Са2+ увеличивает экзоцитоз медиаторов и формирует длитель-
ные высокоамплитудные ВПСП. Причину многократной высокочастотной
стимуляции синапсов многие исследователи связывают с развитием ревер-
берации возбуждения в нейронных цепях. КП практически независима от
белкового синтеза, так как она осуществляется с участием уже сущест-
вующих в клетке белков. (Некоторые исследователи считают, что механизмы памя-
ти, Приписываемые реверберации, создаются специальными нейронами, имеющими пейс-
мекерную активность. Пейсмекерный цикл создают последовательные волны: деполяриза-
ции, обусловленной потенциалзависимыми Са2*-каналами, и гиперполяризации, обуслов-
ленной Са2*-завнсимыми К‘-каналами. Частота пейсмекерной активности может превы-
шать 100 Гц.)
Промежуточная память (или процесс консолидации, упрочения памяти) соответ-
ствует поздней фазе долговременной потенцнации. Ее механизмы связаны
с ббльшей степенью участия вторых посредников (цАМФ, ИФ3, ДАГ, NO и др.),
которые приводят к экзоцитозу медиаторов, активации рецепторов и ион-
ных каналов. Кроме этого, вторые посредники через ранние гены и их бел-
ки активируют транскрипцию генов, запускающих синтез белков, улуч-
шающих синаптическую передачу («белки памяти», модулирующие пептиды, фер-
менты синтеза медиаторов, белки рецепторов, ионных каналов и др ). Поэтому проме-
жуточная память чувствительна к нарушению белкового синтеза. В консо-
лидации памяти большую роль играют структуры лимбической системы,
особенно гиппокамп. Инерционность нервных процессов (например, у флегма-
тиков), сохраняя след памяти, способствует ее консолидации. Подавляют
долговременную потенциацию и консолидацию памяти ИЛ-lp н ИЛ-6.
❖ Долговременная память связана с механизмами стойкой активации локусов
генома, кодирующих белки, повышающие эффективность синаптической
передачи (напрнмер, в геноме при этом отмечено увеличение метилирования нуклеино-
вых кислот). Важную роль в долговременной памяти играют ультраструктур-
ные изменения, улучшающие синаптическую передачу: миелинизация пре-
синаптических окончаний, увеличение числа аксошипиковых синапсов и
площади синаптической поверхности, увеличение дендритного едерева»
нейронов.
457
❖ Роль сна в механизмах памяти: в фазе медленного сна происходит логиче-
ская переработка информации с «очисткой» регистра промежуточной па-
мяти; в фазе быстрого сна - перевод обработанной информации в долго-
временную память.
❖ Изучение механизмов памяти на уровне совокупности нейронов сформировало понятие
нейронных кодов памяти (А.Н. Лебедев, 1985). Их формируют группы содружественно
работающих иейроиов, генерирующих циклически повторяющиеся волны импульсов. Эти
пакеты волн составляют единицу памяти (единицу хранения информации). Минимальное
число нейронов в ансамбле составляет около 100 клеток. Каждый ансамбль за 1 цикл ак-
тивности образует от 1 до 10 залпов нервных импульсов. Максимально возможное число
нейронных кодов памяти около 0,5 миллиарда.
4» Физиология восприятия и ощущении («Мир - изменение, жизнь - восприятие». Марк Авре-
лий).
4.1. Восприятие — психофизиологический процесс, формирующий целостный чув-
ственный образ предметов, явлений объективного мира н осознание его значи-
мости. (Наряду с ощущениями и представлениями формирует конкретно-чувственное отра-
жение действительности.)
4.2. Физиологические осповы восприятия.
❖ На рецепторном уровне специфика ощущений и восприятий формируется
специализацией (модальностью) рецепторов, их высокой чувствительно-
стью к адекватному раздражителя в результате эволюционного приспособ-
ления (Д.А. Бирюков).
❖ Нейрофизиологические механизмы ощущений и восприятии связаны с
формированием вызванных потенциалов (ВП) и включают три последова-
тельных этапа ПО 100 МС каждый (А.М. Иваницкий, 1999).
• Сенсорный этап (первые 100 мс, ранние волны ВП) связан с обработкой фи-
зических характеристик сигнала в корковых проекционных зонах анали-
затора. Этот этап не сопровождается субъективным переживанием
(ощущение как психологический феномен отсутствует).
• Этап информационного синтеза (вторые 100 мс, промежуточные волны ВП)
отражает синтез на нейронах сенсорной коры двух видов информации:
наличной информации о физический свойствах раздражителя и инфор-
мации о его значении, хранящейся в памяти (диалог памяти с настоя-
щим). На этом этапе появляются ощущения. В основе синтеза лежит
кольцевой путь возбуждения от сенсорной коры —» в ассоциативные зо-
ны коры —»в лимбико-гиппокампальный комплекс —»в мотивационные
структуры гипоталамуса —» возврат возбуждения в кору, включая сен-
сорную.
• Этап принятия решения (последние 100 мс, поздние волны ВП) - происходит
опознание, принятие решения и категоризация раздражителя, т.е. фор-
мируется восприятие. В этом процессе ведущую роль играет лобная ко-
ра.
❖ Участие нейронов-детекторов в восприятии.
• Нейроны-детекторы простых признаков (например, детекторы чистых тонов,
шума, начала звука, конца звука и др.) позволяют произвести анализ сложного
раздражителя, разложив его на более простые компоненты.
458
• Нейроны-детекторы сложных признаков (например, детекторы лица) позво-
ляют воспринимать объект целиком.
• Имеется критический период в формировании нейронов-детекторов (на-
пример, детекторы различения цаета формируются до 14-16 лет, детекторы положения
прямых линий - в более раннем периоде).
❖ Роль движений и обратной связи в формирование восприятия (например, бо-
лее адекватное формирование зрительного восприятия у «возницы» по сравнению с «на-
ездником» в опытах Хелда).
❖ Полушария головного мозга и организация восприятия у человека.
• Правое полушарие осуществляет неосознаваемое восприятие, зритель-
но-пространственный анализ стимулов, обработку информации с более
высокой скоростью, целостное восприятие (алкоголь, травма, заболевания за-
медляют скорость переработки сигналов прежде всего в правом полушарии).
• Левое полушарие осуществляет осознание вербальных и невербальных
раздражителей; смысловой (семантический) анализ раздражителей, анали-
тическое восприятие, последовательный анализ сигналов.
4.3. Особенности восприятия человека.
Осознание восприятия (является результатом речевой функции и процессов абстракт-
ного мышления; при осознании устанавливается электрическая связь между восприни-
мающими центрами и речевыми моторными центрами).
•5* Большая степень обобщенности восприятия (объект воспринимается как принад-
лежащий к определенному классу).
4.4. Характеристики восприятия как психологической функции.
• ** Константность - относительная независимость от физических условий восприятия (из-
менений расстояния, освещенности, поворота объекта и т.д.).
* ** Предметность - выделение из фона отдельного объекта.
❖ Целостность - объединение разных элементов в целое.
❖ Обобщенность - каждый объект относится к определенному классу.
5. Физиология внимания.
5.1. Внимание - это избирательная направленность познавательной деятельности
человека на определенный объект (предмет, событие, образ, суждение). По В. Джейм-
су, внимание - «это овладение умом в четкой и яркой форме одним из несколь-
ких наличествующих объектов. Суть этого процесса - фокусирование, концен-
трирование сознания».
5.2. Физиологическая роль внимания-, ограничивает поле восприятия, направляет
психическую деятельность, обеспечивает отбор и дробление информации, оп-
ределяет последовательность анализа информации.
5.3. Направленность внимания зависит от внешних факторов (физических пара-
метров сигнала: интенсивности, частоты и др.) и внутренних факторов (смы-
словой значимости сигнала для человека).
5.4. Виды внимания.
•> Непроизвольное внимание (биологическое происхождение).
• Есть у человека и животных.
• Базируется на активности правого полушария.
• Зависит от силы и новизны раздражения.
459
• Оперирует событиями только в настоящем времени.
• Осуществляется автоматически на основе ориентировочного рефлекса
(у человека обеспечивает включение механизмов произвольного внимания).
Произвольное внимание (социальное происхождение).
• Появляется у ребенка только после начала освоения речи, базируется иа
активности левого речевого полушария.
• Оно может быть обращено как в прошлое, так и в будущее.
• Обработка информации осуществляется последовательно, требует уси-
лий на выделение и обработку информации, определяет приоритеты в
последовательной обработке информации.
53. Физиологические механизмы и индикаторы внимания.
❖ Роль ретикулярной формации в реакциях активации и организации внима-
ния.
• Генерализованная активация коры служит основой непроизвольного
внимания (связана с восходящим активирующим влиянием РФ ствола мозга).
• Локальная активация лобной коры является основой произвольного се-
лективного внимания (связана с активацией иеспецифического таламуса).
❖ Роль ориентировочного рефлекса (ОР) в организации внимания (рефлекс
«Что такое?», открытый И.П. Плановым).
• ОР определяет значимость сигнала для организма. Специфическим сти-
мулом для возникновения ОР является абсолютная или относительная
новизна раздражения.
• Компоненты ОР.
Ф" На корковом уровне ОР представлен десинхронизацией ЭЭГ (де-
прессией a-ритма, активацией 0- и у-ритмов), которая связана с актива-
цией иейронов-детекторов новизны в иеспецифических ядрах тала-
муса и гиппокампе.
ф- В вегетативной сфере ОР представлен урежением ритма сердца,
расширением зрачков, увеличением кожной проводимости, расши-
рением сосудов головы и др.
ф В моторной сфере ОР сопровождается поворотом головы, движени-
ем глаз в сторону нового раздражителя, повышением мышечного
тонуса («рефлекс прицеливания» по Конорскому).
ф В сенсорной сфере ОР представлен увеличением чувствительности
анализаторов.
• ОР тормозит сопутствующие реакции, особенно двигательную актив-
ность и условные рефлексы (феномен отрицательной индукции).
• ОР сопровождается выделением эндорфинов, что ведет к самоподкреп-
леиию исследовательской деятельности.
• Угасание ОР происходит при повторных действиях (от 10 до 30 раз) того
же раздражителя. Гиппокамп осуществляет сличение новых и старых
раздражителей и угасание реакции иа уже привычные длительно повто-
ряющиеся раздражения.
460
❖ Доминанта (А. А. Ухтомский) обеспечивает:
• устойчивость внимания;
• «фильтрацию» сигналов, избирательно подавляя одни и усиливая дру-
гие сигналы на пути к высшим отделам мозга.
• Лобная кора тормозит реакции на побочные раздражения, сохраняет
направленность и запрограммированность поведения.
❖ ЭЭГ-корреляты произвольного внимания:
• условно-негативная волна (волна ожидания продолжительностью около 1 с), ко-
торая выявляется на ЭЭГ (методом усреднения) после действия предупреж-
дающего сигнала;
• усиление у-ритма (частота - 40 Гц), который возникает на дендритах
нейронов иеспецифических ядер таламуса, имеет холинергическую
природу и реализуется через М-холннорецепторы нейронов новой коры.
Увеличение локального мозгового кровотока является показателем актив-
ности мозговых структур (коры и подкорковых отделов), участвующих в
процессе внимания.
5.6. Психологические характеристики внимания.
Избирательность или селективность внимания - это направленность на физические или
лингвистические характеристики стимула («фокусировка»).
Устойчивость внимания определяется по длительности выполнения задания, требующего
непрерывного внимания; внешние факторы обеспечивают ее на 2 - 3 с, внутренние - мо-
тивация, интерес - иа более длительный срок.
*1' Концентрация внимания (определяется доминантой и латеральным торможением).
Переключаемостъ внимания (скорость перехода от одной деятельности к другой).
Объем внимания определяется количеством одновременно отчетливо осознаваемых объ-
ектов; он близок к объему кратковременной памяти 7 ± 2 стимула.
♦♦♦ Распределение внимания - способность использовать ограниченные ресурсы внимания для
одновременного выполнения двух заданий.
6. Физиология речи («Нет ничего сильнее и бессильнее слова». И.С. Тургенев).
6.1. Речь - форма общения с помощью звуковых и зрительных знаков. По И.П.
Павлову, слово является «сигналом сигналов», выполняет функцию «удвоения
мира», обеспечивает передачу опыта предыдущих поколений. (Критический пери-
од для освоения речи - до 10 лет. После утрачивается способность для построения нейронных
сетей, необходимых для формированию центра речи.)
6.2. Функции речи.
❖ Коммуникативная функция - речь как средство общения передает слуша-
телю:
• вербальную информацию (смысл словесного сообщения);
• невербальную информацию (тембр, высота голоса, пол и возраст, эмо-
циональное состояние говорящего).
❖ Регулирующая функция - внешняя и внутренняя речь как средство регуля-
ции функций организма и поведения:
• управление поведением других людей (например, во врачебной практи-
ке - внушение, гипноз);
461
• управление собственными действиями (внутренняя речь, развитие про-
извольного внимание, волевого поведения).
❖ Понятийная функция-, речь как орудие абстрактного мышления, средство
выражения мыслей, один из механизмов интеллектуальной деятельности.
❖ Программирующая функция речи состоит в построении смысловых схем
речевых конструкций, основанных на внутреннем программировании,
осуществляемом с помощью внутренней речи.
6.3. Периферические механизмы речи.
Механизм фонации локализован в гортани, его физическая основа - коле-
бания голосовых связок. Участвуют в фонации: генератор тонов - мышцы
гортани и голосовые связки; энергодатчики - межреберные мышцы, диа-
фрагма, мышцы брюшного пресса и спины, гладкие мышцы бронхов.
❖ Механизмы артикуляции обеспечивают образование звуков, физическая
основа артикуляции - резонанс полостей с постоянной или переменной
конфигурацией (полости глотки, рта, носа, придаточные пазухи).
❖ Развитие тонких манипуляторных функций правой руки (в том числе языка жестов) в ходе
эволюции привело к формированию особых функций левого полушария и передаче «спо-
собностей» руки артикуляционным органам (развитию речи).
6.4. Центральные механизмы речи.
❖ Механизмы восприятия речи.
• Слуховой центр речи Вёргике (поля 42 и 22 верхней височной извилины ле-
вого полушария):
ф’ Обрабатывает акустическую информацию из первичной слуховой
коры, обеспечивает понимание смысла слов, организацию их в
предложения, формирует внутреннюю речь (при его поражении воз-
никают затруднения в понимании устной речи н в письме под диктовку -
сенсорная афазия; речь при этом достаточно беглая, но обычно бессмыс-
ленная, т.к. больной не замечаег речевых ошибок);
ф- передает информацию через дугообразный пучок в двигательный
центр устной речи, хранящий программы артикуляции;
Ф Функциональной асимметрии полушарий:
левое вербальное полушарие обеспечивает анализ смысла слов,
организацию их в предложения, их осознание, формирование
внутренней речи;
правое полушарие обслуживает невербальный канал епчзи:
различение интонации, мужских и женских голосов, мелодий,
оно способно анализировать отдельные слова, но не в состоя-
нии организовывать их в предложения.
• Оптический центр речи (угловая извилина на границе затылочной, еигоч-
ной и теменной долей, поле 39 левого полушария) обеспечивает:
ф- обрабатывает зрительную информацию, поступившую из первич-
ной и вторичной зрительной коры, обеспечивает понимание пись-
менной речи; затем информация переносится в центр Вернике
(связь зрительной и акустической формы слова);
Ф* передает информации в двигательный центр устной речи;
462
ф- Функциональная асимметрия полушарий: аналогичичное поле 39
правого полушария значительно уступает левому в способности
понимать письменную речь и не может «озвучить» полученную
информацию. (Вместе с тем правое полушарие может не только читать
вопросы, но отвечать на низ с помощью левой руки, составляя слова из
буквенных карточек.)
❖ Механизмы воспроизведения речи.
• Двигательный центр устной речи (центр Брокй, поле 44 коры нижней лоб-
ной извилины) у большинства людей расположен в левом полушарии (у
правшей - 95%, у левшей - 70%):
Ф- Содержит командные нейроны, обеспечивающие программирова-
ние речевых конструкций;
Ф- Управляет через переднюю центральную извилину сокращениями
мышц языка, губ, гортани, дыхательными мышцами.
• Двигательный центр письменной речи (средняя лобная извилина в поле
б левого полушария, расположена рядом с зоной, управляющей движе-
нием правой руки).
• Функциональная асимметрия полушарий:
ф- При поражении центра Брокй нарушается устная речь больного в
результате затруднения построения слов, т.к. невозможно быстро
переключиться с одного слога на следующий (эфферентная моторная
афазия); нарушена способность формулировать высказывания, пере-
водить свои мысли в развернутую речь (динамическая афазия). При
этом больные свои ошибки осознают, понимание чужой речи со-
храняется. При письме больной хорошо выводит одну букву, но не
может написать слово.
-ф- Речедвигательная функция правого полушария значительно реду-
цирована: за счет него могут повторяться отдельные слоги, слова,
особенно жаргонные, пение, заученные тексты, но отсутствует спо-
собность организовывать слова в незаученные предложения.
ф- Устная речь до 10-летиего возраста контролируются обеими полу-
шариями. Имеется возможность «переноса» речевых функций из
доминантного полушария в субдомииантное. (Об этом говорят частые
случаи моторной афазии у детей при поражении правого полушария и
восстановление устной речи в течение года после поражения левого по-
лушария. У взрослых латервлизация центра устной речи резко выражена,
однако известны случаи, когда при потере правой руки и возникновении
леворукости моторная афазия возникала при поражении правого полуша-
рия.)
6.5. Речь как вторая сигнальная система.
*♦* Взаимоотношения второй и первой сигнальных систем.
• Связь двух сигнальных систем по типу вС —* Я» (словесный раздражитель -» непо-
средственная реакция) лежит в основе управления поведением как через внешнюю
речь, так и через внутреннюю речь.
463
* Связь двух сигнальных систем по типу *Н —* С» (непосредственный раздражитель —»
словесная реакция) обеспечивает функцию называния, в основе которой лежит связь
нейронов-детекторов непосредственных раздражителей с командными нейронами, ко-
торые управляют программой построения соответствующего слова.
* Взаимное торможение двух сигнальных систем способствует отвлечению слова от
конкретного явления, которое оно обозначает, создает относительную независимость
их действия.
♦♦♦ Способом изучения смысловой роли слова могут быть условные рефлексы на словесные
раздражители (Б.Н. Соколов, 1981). Вызывали условный оборонительный рефлекс на со-
четание слова с болевым раздражителем. Использование различных слов показало, что
центр оборонительного рефлекса образует множество связей с целым набором близких по
смыслу слов.
• Слова, близкие по смыслу, образуют ядро смысловых связей н вызывают оборони-
тельную реакцию.
• Слова, лежащие на границе смысловой близости, вызывают не оборонительную реак-
цию, а ориентировочный рефлекс.
* Применяя выработку дифференцировки (подкрепляя одно близкое по смыслу слово и
не подкрепляя другое) можно получить сужение смыслового поля: часть условных
оборонительных реакций будет замещаться на ориентировочные.
* Индифферентные слова не вызывают нн ориентировочной, нн оборонительной реак-
ций.
6.6. Контроль речеобразования (механизмы обратной связи).
♦♦♦ Контроль качества речи.
♦ Слухоречевой - контролирует соответствие речевого сигнала акустическому эталону,
сформированному в процессе обучения (восприятие собственной речи происходит
преимущественно через костную проводимость).
* Проприоцептивный - обеспечивает сравнение двигательного эталона, хранящегося в
памяти, с реальным исполнением (соматоречевой контроль).
Контроль конечного результата (смысла высказывания).
• Зрительный канал (оценка по реакции слушателя: принял ли'он сообщение, понял лн
смысл высказывания и т.д.).
• Слуховой канал (характер ответа слушателя, характеризующий разборчивость и гром-
кость речи).
7. Физиология сознания («Сознание - самое очевидное н самое непонятное свойство человека».
Оксфордский словарь).
7.1. Сознание - это форма отражения действительности, которая с помощью слов,
математических символов, художественных образов может быть передана дру-
гим людям (co-знание, совместное знание). Оно возникло в эволюции на основе
потребности членов сообщества к общению, объединению усилий, передачи
знаний.
7.2. Физиологические концепции сознания.
❖ Концепция «светлого пятна» (И.П. Павлов, 1913): сознание связано с динами-
чески образующимися в коре больших полушарий участками оптимальной
возбудимости, где легко образуются условные рефлексы. («Если бы можно
было видеть через черепную коробку и если бы место оптимальной возбудимости
светилось, то мы увидели бы иа думающем сознательном человеке, как по его
464
большим полушариям передвигается постоянно изменяющееся в форме и величи-
не причудливо меняющихся очертаний светлое пятно». И.П. Павлов)
❖ Концепция информационного синтеза (А.М. Иваницкий, 1997): для того чтобы
раздражитель был осознан, он должен действовать не менее 300 - 500 мс.
Вызванные потенциалы в сенсорной коре при этом формируют этапы ин-
формационного синтеза и принятия решений (см. выше 4.2).
❖ Вербальная (речевая) теория сознания говорит о решающей роли речи в
феномене сознания («Сознание вооружено речью». П.В. Снмоиов): для сознания
обязательна активации связей между сенсорными, гностическими (ассо-
циативными) структурами и двигательной речевой зоной, которые устанав-
ливаются как через корковый уровень, так и через таламус. (Однако связь соз-
нания с речью не следует понимать упрощенно. Больные с временным выключением речи
- например при инсульте, после восстановления речевой функции помнят события, проис-
ходившие во время утраты речи, т.е. у них не нарушались важнейшие функции сознания -
способность к абстракции и запоминание последовательности событий.)
❖ Сознание наиболее сильно зависит от функций речевого полушария (обыч-
но левого). При поражении левого полушария сознание утрачивается
раньше и восстанавливается хуже, чем при поражении правого («асиммет-
рия сознания»).
7.3. Подсознание, сверхсознание и интуиция.
••• Подсознание (синонимы: бессознательное, неосознанное) — это в широком смысле
все психические явления вне сферы сознания, связанные с восприятием
слабых, но мотивационно важных раздражителей, которые не осознаются
субъектом, но вызывают у него биоэлектрические, вегетативные и эмоцио-
нальные реакции, а также влияют на поведение и психические функции че-
ловека. (В узком смысле, по 3. Фрейду, бессознательное понимается как вытес-
ненные из сферы сознания автоматизированные навыки, усвоенные нормы пове-
дения, мотивационные конфликты.)
• Стимулы, вызывающие неосознанное восприятие, должны быть ниже
минимального порога для осознанного восприятия по силе (например, вы-
зывать только раннне волны вызванных потенциалов) или по длительности дей-
ствия (действовать меньше 300-500 мс). Однако ОНИ должны быть выше ми-
нимального порога для неосознаваемых раздражителей, т.е. вызывать
вегетативные, биоэлектрические и др. реакции организма.
• Повышение порога осознания эмоционально и мотивационно значимых
раздражителей является механизмом психологической защиты лично-
сти. Оно связано с тем, что эмоционально значимые неосознаваемые
слова вызывают возбуждение лимбической системы, которое по меха-
низму обратной связи оказывает восходящее тормозное влияние на но-
вую кору. Повышение порога осознания эмоционально значимых слов
свидетельствует о возможности их смыслового анализа на неосознавае-
мом уровне.
• Условнорефлекторная деятельность на неосознаваемом уровне.
•ф- Вырабатываются прочные условные рефлексы, если условный сиг-
нал сочетается с «подкрепляющим» неосознаваемым словом,
имеющим отношение к мотивационной или эмоциональной доми-
465
нанте. (Прочность этих рефлексов объясняет давние наблюдения относительно
консервативности подсознания, стойкости влечений, эмоциональных пережива-
ний в случаях, когда их повод остается неосознанным.)
Если же условные рефлексы вырабатываются с помощью подсозна-
тельных сигналов, не связанных с эмоциональной или мотивацион-
ной доминантой, то временная связь сохраняется только в кратко-
временной памяти и не переходит в долговременную память.
"Ф* Транквилизаторы на период своего действия подавляет условнорефлекторные
реакции, вырабатываемые иа осознаваемые раздражители, но не действуют на
условные рефлексы, выработанные на неосознаваемые сигналы, что говорит о
качественных различиях временных связей, образуемых на осознаваемом н не-
осознаваемом уровнях.
• Функциональная асимметрия больших полушарий в бессознательном.
Исследование людей с «расщепленным мозгом» (операция перерезки мозо-
листого тела у больных с эпилепсией) показало, что в осуществлении функций
бессознательного ведущую роль играет правое полушарие. Оно спо-
собно осуществлять сложную когнитивную деятельность, например,
выбрать правильный предмет, если его изображение илн слово посту-
пило в правое полушарие, анализировать отдельные предметы, но не
может их назвать. Правое полушарие может анализировать отдельные
слова н даже «произносить» их, но не в состоянии организовать их в
предложения
❖ Сверхсознание или интуиция - научное и художественное творчество, не
контролируемое сознанием. Это создание нового из старых элементов во
внутреннем мире человека, трансформация и рекомбинация следов (эн-
грамм) памяти и создание из них новых комбинаций. Критический анализ и
отбор наиболее правдоподобных гипотез протекает на уровне сознания с
включением речевого полушария.
8. Физиологические основы мышления («Мысль - ключ ко всем сокровищам». О. Баль-
зак).
8.1. Мышление - процесс познавательной деятельности, обобщенное н опосредо-
ванное отражение действительности (внешнего мира и внутренних пережива-
ний).
8.2. Основные формы мышления.
Ф Конкретно-образное мышление (ощущения, восприятия, представления) -
первая ступень развития мышления у ребенка.
❖ Абстрактное (словесио-логическое) мышление проявляется в виде понятий,
суждений, умозаключений н является более поздним этапом развития.
Формы использования понятий в суждениях н умозаключениях: индукция
(от частного к общему - левое полушарие сначала анализирует информа-
цию, а затем правое синтезирует); дедукция (от общего к частному - осу-
ществляется в правом полушарии).
83. Роль разных структур мозга в обеспечении мышления.
❖ Генерация стимулнезависимых (спонтанных) мыслей связана с активацией
передних зон лобной коры; этот отдел участвует также в волевом контроле
прн выполнении задания.
466
❖ В распознавании и принятии решения участвуют лобная и височная кора.
❖ Поиск стратегии решения задачи выполняют теменно-затылочные отделы
коры.
❖ Установление соответствия принятого решения выбранной стратегии вы-
полняют лобные, височные и лимбические отделы мозга при ведущей роли
лобной коры.
8.4. Физиологические механизмы мышления.
Вызванные потенциалы: принятие решения о выборе удлиняет латентный
период ВП и увеличивает амплитуду его поздних компонентов. .
❖ Сверхмедлеиная активность: при различных мыслительных операциях (на-
пример, принятии решения) на ЭЭГ можно зафиксировать отрицательную «вол-
ну ожидания» (амплитуда 3-20 мкВ, продолжительность 1-5 с).
❖ Концепция фокусов взаимодействия и информационного синтеза (А.М.
Иваницкий, 1977).
• Методом картирования внутрикоркового взаимодействия было показа-
но, что согласование ритмов двух и более структур характеризует обра-
зование связей между ними. При активации мышления связи начинают
сходиться к определенным областям ассоциативной коры, образуя узлы,
названные фокусами взаимодействия. При образном мышлении оии
расположены преимущественно в теменно-височных отделах, при абст-
рактно-вербальном мышлении - в лобных отделах. Нахождение реше-
ния сопровождалось образованием фокуса в речевой левой височной
области, даже если словесного ответа не требовалось.
• Фокус взаимодействия является местом конвергенции нервных связей.
Связи, подходящие к фокусу, устанавливаются на разных частотах. Со-
ответственно фокус состоит из групп нейронов с разной частотой ак-
тивности, настроенных на одинаковые с ними по частоте группы нейро-
нов на периферии. Внутри фокуса нейроны, работающие иа разных час-
тотах, объединены жесткими связями, основанными, вероятно, на
структурных изменениях в синапсах. Считается, что такая конструкция
имеет большую информационную емкость.
• Периферические по отношению к фокусу группы нейронов образуют с
ним кольцевые гибкие связи. Каждая связь приносят к фокусу из опре-
деленных участков коры или подкорковых образований свою информа-
цию.
• Таким образом, на нейронах фокуса происходит информационный син-
тез и сопоставление информации из оперативной памяти с информаци-
ей, извлекаемой из долговременной памяти и мотивационных центров.
Предполагается, что это лежит в основе конечной цели мыслительной
операции - нахождения решения. Субъективно это переживается как
процесс думания и нахождения ответа.
❖ Регистрация электрической активности малых популяций нейронов коры и
подкорковых структур с помощью погруженных в мозг многожильных
электродов позволяет обнаружить достоверные изменения частоты разря-
дов при выполнении мыслительных операций и различия реакций тех же
467
нейронов при выполнении разных проб (мозговые коды психической деятельно-
сти, по Н.П. Бехтеревой).
8.5. Функциональная асимметрия полушарий в процессах мышления.
♦ > Правое полушарие (особенно теменно-височная кора) обеспечивает конкретно-
образное мышление (первая сигнальная система), лучшее решение зрительно-
пространственных задач, одномоментную целостную обработку информа-
ции, интуитивное мышление.
❖ Левое полушарие (особенно лобная кора) обеспечивает абстрактно-логическое
мышление (вторая сигнальная система), лучшие возможности по оценке вре-
мени, аналитическую, поэтапную обработку информации, логическое
мышление, осознание информации («когнитивные» медиаторы - дофамин, ацетил-
холин, ГАМК - преобладают а левом полушарии).
❖ Взаимодействие полушарий при решении разных задач может осуществ-
ляться в форме антагонизма, синергизма, последовательной переработки
информации.
9. Профильный материал (см. тему 32; задание 1: 7-9).
Задание 2. Подготовить реферативное доклады иа тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющие-
ся у вас учебники и пособия): [1-16].
3.2. Дополнительная литература (см. с. 514 - 522): [1], [3], [10 ], [14], [15], [22], [23],
[28], [29], [30], [43], [48], [56], [59-61], [67], [94], [105], [111], [112], [114], [120],
[121], [132], [134], [136], [145], [152], [156], [161-164], [166], [168], [171], [173],
[175], [176], [177], [179], [188], [196], [200], [201], [213], [214], [222], [236], [242],
[243], [245], [251], [253].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ni/-nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» /В.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором н слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться иа практи-
ческом занятии по данной теме, итоговом занятии и экзамене.
468
Тема 32. Физиологические основы целенаправленного поведения чело-
века
Цель: а) знать основные механизмы поведения, ведущую роль потребностей и мотиваций в органи-
зации поведения, биологически детермнннроввнные виды целенаправленного поведения (пи-
щевое, половое, оборонительное и т.д.) и социально детерминированные виды поведения
(трудовая деятельность, обучение и т.д.);
б) уметь использовать эти знания для анализа физической и умственной работоспособности
человека, для рациональной организации труда, формирования здорового образа жизни.
Мотивация темы: теория функциональных систем позволила рассматривать поведенческий акт как
«единицу интегративной деятельности организма» (по П.К. Анохину), где полезный приспо-
собительный результат является главным системообразующим фактором, целью поведения.
Впервые цель поведения становится объективно изучаемой характеристикой, которая рас-
сматривается с материалистических позиций. Этот подход оказался успешным при изучении
трудовой деятельности, в спортивной, педагогической и клинической практике.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала. Общая характеристика поведения как
целенаправленной деятельности.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, н обозначены в «Графике и
содержанки самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции н подготовки рефератов.
1. Поведение - это активное взаимодействие организма с окружающей средой, на-
правленное на удовлетворение ведущей потребности. (В отличие от рефлекса оно заканчи-
вается не действием, а полезным приспособительным результатом, удовлетворяющим доминирую-
щую потребность.) Поведение обеспечивает адаптацию к изменяющейся среде и (у че-
ловека) ее целенаправленное изменение. Поведение представляет собой взаимосвя-
занный комплекс врожденных (генетически обусловленных) и приобретенных иа ос-
нове обучения компонентов.
2. Организатором поведенческого акта явлиются потребности.
2.1. Потребность - особое психологическое состояние неудовлетворенности тре-
бований организма, необходимых для нормальной биологической и социальной
жизнедеятельности. Возникновение потребности подчиняется механизму доми-
нанты. Потребность является движущей силой поведения и устраняется через
поведение.
2.2. Классификация потребностей.
❖ Витальные (биологические) потребности в пище, воде, сне и бодрствова-
нии, экономии сил и т.д. Это не любая «физиологическая нужда» организ-
ма, а только та, которая ие устраняется гомеостатическими механизмами.
Неудовлетворение этих потребностей ведет к гибели организма, поэтому
их называют витальными.
❖ Социальные потребности выражаются в стремлении человека принадле-
жать к определенной социальной группе и занимать в ней определенное
469
место. Они реализуются при взаимодействии с другими людьми, т.е. при
общественном поведении.
❖ Идеальные потребности (саморазвития) обращены к будущему, это ис-
следовательские, игровые, имитационные, «рефлекс свободы». Определяют
место человека в ноосфере (интеллектуальном освоении мира). (Две главные
идеальные потребности - это потребность в познании, создающая духовность, и
альтруистическое желание добра, создающее душевность. Человек духовен на-
столько, насколько он задумывается над великими и вечными вопросами о добре
и зле, истине, правде и лжи, смысле своего существования.)
3. Мотивация как актуализация потребности.
3.1. Мотивация - эмоционально окрашенное стремление человека совершать пове-
дение, направленное на удовлетворение своих потребностей.
3.2. Классификация мотиваций аналогична классификации потребностей (см. выше
2.2).
3.3. Нервный субстрат мотивации.
❖ Гипоталамус (центры голода, жажды, насыщения, полового поведения и
ДР-)-
•> Лобная кора играет важную роль в:
• формировании социальных и идеальных мотиваций, социализации био-
логических мотиваций;
• направленном поиске и выбор путей реализации мотивации;
• вероятностном прогнозировании результата;
• формирование эмоций по ходу возникновения и реализации мотиваций.
❖ Лимбическая система:
• организует иерархию мотиваций;
• формирует эмоцию как аппарат оценки возможности удовлетворения
мотиваций;
• активирует мозговые структуры, участвующие в реализации мотиваций:
информационные, соматические, управляющие вегетативными процес-
сами.
3.4. Физиологические механизмы трансформации потребностей в мотивационное
возбуждение.
Сигнализация от механорецепторов (например, «пустой желудок» и чувство голо-
да, эрогенные зоны и половая мотивация).
❖ Сигнализация ог хеморецепторов об изменениях гуморального состава
внутренней среды («голодная кровь» и активация центра голода, «метаболическое»
илн истинное насыщение при активации центра насыщения субстратами крови, повыше-
ние осмотического давления и возникновение жажды, уровень половых гормонов и поло-
вая активность, агрессивность и т.д).
❖ Центр мотивации обладает свойствами пейсмекера: его иейроны имеют
высокую возбудимость, спонтанную деполяризацию.
❖ При формировании мотивации у многих нейронов различных структур го-
ловного мозга регистрируется пачечный тип активности, регулярно сле-
дующий с интервалами - 150 мс, свидетельствующий об установлении
межнейронных связей.
470
❖ Центр мотивации обладает всеми свойствами доминантного очага (см. тему
6; задание 1: 1.7).
❖ Нейрохимические механизмы обеспечения мотивационного возбуждения.
• Эмоционально-мотивационные нейромедиаторы (серотоиии, норадреналин)
в большей концентрации находятся в правом полушарии.
• Нейропептиды обеспечивают модуляцию секреции нейромедиаторов:
торможение активности нейронов (осуществляют fl-эндорфии, соматостатин,
гонадолиберин и др.); активация нейронов (происходит под действием субстанции
Р, ангиотензина II); участие в формировании памяти (вазопрессин, фрагменты
АКТГ ускоряют запоминание н обучение, окситоцин вызывает амнезию).
• Гормоны и регуляция мотиваций (например, пентагастрии, холецистокинин,
лептин являются регуляторами пищевого поведения, тестостерон действует на гипота-
ламические центры регуляции полового поведения).
4. Врожденным компонентом поведения являются инстинкты.
4.1. Инстинкты - целенаправленное приспособительное поведение, обусловленное
врожденными механизмами и характеризующееся постоянством ответных ре-
акций на действие внешних н внутренних раздражителей (И.П. Павлов относил ин-
стинкты к сложнейшим рефлекторным комплексам). В инстинктах сохраняется весь
опыт эволюционного развития вида. Оии необходимы для сохранения индиви-
дуального организма, целиком вида и продолжения рода. Инстинкты включают
в себя все элементы поведения: потребность, мотивацию, целенаправленную
деятельность, подкрепление. Особенностью является то, что все компоненты
поведения генетически жестко детерминированы.
❖ Инстинкты в «чистом виде» у высших животных и человека могут быть
только сразу после рождения. Затем происходит индивидуализация видово-
го опыта, т.е. возникает сплав наследственных и приобретенных форм по-
ведения. (Инстинкты характерны для насекомых и животных, у которых потом-
ки, как правило, не встречаются с родителями.)
❖ В инстинктах различают два главных этапа его осуществления: поисковый
и завершающий. На поисковом этапе поведение является пластичным, за-
висимым от внешней среды. Оно формируется на основе ориентировочного
рефлекса - врожденной формы ориентировочно-исследовательской дея-
тельности. Напротив, завершающий этап является жестко стереотипным.
❖ Запускают инстинкты как внутренние факторы (нейрогуморальные откло-
нения в организме, формирующие потребности и мотивации), так н внеш-
ние факторы (ключевые или пусковые факторы внешней среды).
❖ Классификация инстинктов базируется на классификации потребностей и
мотиваций: 1) витальные инстинкты (пищевой, питьевой, оборонительный,
регуляции сна - бодрствования, экономия силы); 2) ролевые или зоосоци-
альные (половой, родительский, территориальный, иерархический и др.); 3)
саморазвития (исследовательский, имитационный, преодоления сопротив-
ления н др.).
4.2. Роль этологии в изучении врожденных форм поведения. Этология (от греч. ethos -
обычай, нрав, характер) - наука, изучающая преимущественно генетически обу-
словленные компоненты поведения животных в естественных условиях среды.
Этологн собрали большой материал по наблюдениям за формами поведения
471
животных (К. Лоренц, Нобелевская премия, 1973), который используется для понима-
ния эволюционно-генетических основ поведения человека.
5. Приобретенные компоненты поведения как результат обучения.
5.1. Неассоциативное обучение.
❖ Подражание (имитация) - приобретение индивидуального опыта путем не-
посредственного наблюдения за действиями другого человека. Оно осо-
бенно характерно для раннего онтогенеза. (Л.А. Орбели считал подражание глав-
ным охранителем вида.)
❖ Ориентировочно-исследовательская деятельность основана на ориенти-
ровочном рефлексе (ОР). (Компоненты ОР изложены в теме 31, задание 1: 5,5). Вы-
делено три вида взаимоотношений ОР и текущей деятельности организма
(по П.К. Анохину): 1) конфликтные отношения - ОР тормозит деятельность
(открытое Павловым торможение условных рефлексов); 2) ОР усиливает текущую
доминантную деятельность (например, при оборонительной доминанте новый
раздражетель вызывает не ОР, а оборонительный рефлекс); 3) трансформация
ОР в поведенческий акт, связанная с реализацией скрытой доминанты. ОР
связан с деятельностью «нейронов новизны», расположенных преимущест-
венно в гиппокампе, на которые конвергирует возбуждение с нейронов-
детекторов сенсорной коры при действие новых раздражителей.
❖ Привыкание связано с неоднократном повторении раздражителя, вызы-
вающего ОР. Хотя ОР относится к безусловным рефлексам, он обнаружи-
вает способность к угашению, что сближает его с условным рефлексом, т.к.
в обоих случаях имеет место обучение. Предположено, что при неодно-
кратном действие стимула в гиппокампе создается «нейронная модель»
раздражителя. При повторном действии, если модель и раздражитель сов-
падают, происходит возбуждение «нейронов тождества», и ОР угасает, т.е.
возникает привыкание как особая форма научения.
5.2. Импринтинг (запечатление) - быстрый тип обучения, аналог натурального ус-
ловного рефлекса на экологически адекватный раздражитель. В качестве внеш-
него сигнала выступает движущийся объект, в качестве подкрепления - состоя-
ние беспокойства, страха в раннем постнатальном онтогенезе. Устанавливает
прочную связь ребенка после рождения с матерью и родственниками. У ребенка
проявляется либо в формах, обеспечивающих контакт, «привязанность к объек-
ту», либо в реакциях избегания, страха перед незнакомцем. (У человека импринтинг
слабо выражен, некоторые исследователи наличие его у человека отрицают.)
53. Ассоциативное обучение основано на образовании условных рефлексов на базе
безусловных рефлексов, на основе других условных рефлексов, в том числе об-
разованные на раздражители второй сигнальной системы, а также образован-
ных на основе целенаправленной деятельности организма (инструментальные
рефлексы).
5.4. Когнитивное (познавательное) обучение - это высшая форма обучения. Оно осно-
вано на познании закономерных связей между явлениями и предметами среды,
использует рассудочную деятельность (конкретно-чувственное и абстрактное
мышление) и способность мозга к вероятностному прогнозированию (предвос-
хищению будущего). На его основе осуществляются социально обусловленные
виды обучения (труд, образование и др.).
472
6. Функциональная система поведения (П.К. Анохин, К. В. Судаков).
6.1. Потребности, лежащие в основе мотиваций, являются системоорганизующим
фактором построения функциональных систем поведения (ее полезным резуль-
татом). Поведение в каждый данный момент не ответ на прошлое событие, а
подготовка и обеспечение будущего.
6.2. Афферентный синтез формируется при участии следующих физиологических
механизмов (рис. 22).
❖ Доминирующая мотивация обеспечивает избирательное активирование
мозговых структур, направляющих поведение иа удовлетворение господ-
ствующей потребности, т.е. определяет «что делать». Она мобилизует ге-
нетически обусловленные или индивидуально приобретенные программы
поведения на достижение полезного результата.
❖ Память создает информационное обеспечение поведения (.«как делать»); в
этом обеспечении участвуют как генетическая память (безусловные рефлексы,
инстинкты), так и приобретенные при обучении компоненты поведения (ус-
ловные рефлексы, динамические стереотипы и др.). Характер извлекаемой
из памяти информации определяется доминирующей мотивацией.
❖ Обстановочная афферентация дает информацию о состоянии окружаю-
щей организм среды и (вместе с таламо-темеииой ассоциативной системой) решает
вопрос «можно ли делать»; обстановочная афферентация и память могут
сменить доминирующую мотивацию.
❖ Предпусковая интеграция, готовность создается взаимодействием моти-
вации, обстановочной афферентацни и памяти (при ведущей роли таламо-лобиой
ассоциативной системы в планировании целенаправленного поведения);
❖ Пусковая афферентация реализует сложившуюся в ЦНС предпусковую
интеграцию и определяет «когда делать»; пусковыми факторами являются
время и различные условные раздражители, ориентировочный рефлекс.
63. Принятие решения.
❖ Выделение целей поведения и способов достижения цели (участвуют ней-
роны поискового поведения).
❖ Критерии выбора решения.
• Вероятность достижения успеха, которую оценивают информационные
структуры мозга (выбирается тот путь, который с максимальной вероятностью
приводит к цели). При этом гиппокамп оценивает сигналы с низкой веро-
ятностью подкрепления, а лобная кора - высоковероятностные события.
• Минимизация энергетических затрат (выбирается обычно тот путь к цели,
который экономнее для организма по энергетическим затратам).
❖ Выделению целей способствуют эмоции: включаясь в мыслительную дея-
тельность, они нарушают равнозначность целей в ситуации выбора, выде-
ляя лишь некоторые из них.
❖ Латеральное торможение ограничивает число степеней свободы (механизм
выбора единственного пути для достижения цели).
❖ Принятие решения сопровождается включением в функции речевых центров
коры.
6.4. Акцептор результатов действия (АРД) - нейронная модель будущего резуль-
тата.
473
❖ В АРД программируются:
• параметры полезного результата;
• параметры среды, способствующие и препятствующие достижению по-
лезного результата;
• способы достижения полезного результата.
❖ Физиологические механизмы формирования АРД.
• Реверберация возбуждения при участии пирамидных и вставочных ней-
ронов лобной коры, гиппокампа обеспечивает формирование и дли-
тельное удержание в нервной системе АРД.
• Кратковременная память и ее консолидация обеспечивает сохранение
АРД.
• Эмоции обеспечивают прогностическую и оценочную функцию (подкре-
пляющие системы мозга обеспечивают минимизацию или максимизацию поведенче-
ского акта по ходу удаления от цели или приближения к ней).
6.5. Эфферентный синтез формирует эфферентную программу действия.
❖ Эфферентная программа действия - это совокупность возбужденных
нервных центров, объединенных с использованием механизмов доминанты,
синхронизации электрической активности удаленных структур, усвоения
ритма.
❖ Спектр центров, составляющих эфферентную программу деятельности.
• Моторные центры пирамидной и экстрапирамидной систем, создают
двигательный компонент целенаправленного поведения.
• Вегетативные центры при участии коры, лимбической системы, гипота-
ламуса обеспечивают энергетическое и пластическое обеспечение пове-
дения.
• Активация психофизиологических процессов (восприятия, памяти,
внимания, мышления, воли и др.) обеспечивает переработку информа-
ции, которая необходима мозгу для контроля за адекватностью пове-
денческого акта поставленным целям.
6.6. Целенаправленные действия - деятельность исполнительных органов, форми-
рующая полезный результат, удовлетворяющий доминирующую потребность
организма.
6.7. Оценка результатов поведенческой деятельности.
❖ В оценке результатов поведенческой деятельности участвуют проприоре-
цепторы, интерорецепторы и экстерорецепторы, информация с которых
сравнивается в АРД.
❖ Обратная афферентация (обратная связь) участвует:
• в поэтапной коррекции поведения;
• в санкции на завершение данного поведенческого акта при достижении
полезного результата (формируется положительная эмоция удовлетво-
рения);
• в запуске ориентировочно-исследовательской реакции и отрицательной
эмоции при рассогласовании реального результата и его модели в АРД:
на их основе корректируются АРД и эфферентная программа действия.
474
❖ При невозможности достижения полезного результата усиливается отрица-
тельная эмоция и формируется эмоциональный стресс, что может вызвать у
предрасположенных людей психосоматическую патологию.
❖ Достижение полезного результата не выключает поведение, а дает санкцию
субъекту на реализацию следующего поведенческого акта, который также
будет компонентом непрерывного поведения (поведенческого континуума),
совершаемого человеком на протяжении всей жизни.
7. Профильный материал для студентов лечебного факультета.
7.1. Факторы, лежащие в основе возрастных изменений ВНД.
*♦* Постепенная гибель нейронов в коре (до 50 % от числа нейронов новорожденно-
го); преимущественно поражаются лобные и височные отделы коры, участвующие
в реализации высших психических функций.
Снижение кровотока в мозге, уменьшение доставки Оз, энергообеспечения клеток
мозга, особенно в лобных и височных долях.
Ф Снижение числа клеточных рецепторов и их активности при действии медиаторов
и гормонов.
7.2. Возрастные проявления изменений ВНД.
❖ Снижена скорость приема и переработки информации, ослаблена обратная связь и
подвижность нервных процессов (к 80 - 90 годам подвижность составляет 60 % от
уровня 20- летних).
❖ Ослаблена реактивность клеток коры и их работоспособность вследствие:
• ослабления процессов восстановления;
• снижения активирующих влияний РФ, в том числе из-за снижения двигатель-
ной активности и импульсации от проприорецепторов;
• ослабления всех видов торможения, снижения его координирующей н охрани-
тельной роли.
❖ Ослаблена подвижность нервных процессов:
• увеличивается время выполнения задания, количество пауз;
• динамические стереотипы переделываются с большим трудом (увеличивается
консерватизм);
• затруднена выработка новых условных рефлексов;
• легко возникают фазовые состояния с уравнительной, парадоксальной и ульт-
рапарадоксальной фазами.
Ф Возникают нарушения в мотивационной сфере, снижение инициативности, ослаб-
ление социальных связей.
❖ Невербальный интеллект снижается уже с 30 - 35 лет, тогда как вербальный со-
храняется гораздо дольше.
7.3. ЭЭГ и старение.
••• Стабильность характеристик ЭЭГ сохраняется до 50 лет.
После 50 лет наблюдается перестройка спектра ЭЭГ:
• уменьшение амплитуды и частоты а- и P-ритма, наиболее выраженное в лоб-
ной коре;
• нарастание доли 6-ритма;
• падение лабильности приводит к сужению диапазона усвояемых ритмов, уве-
личению латентного периода прн регистрации вызванных потенциалов.
475
7 А. ЭЭГ при наркозе.
❖ Постепенное замещение на ЭЭГ ритмов бодрствования (а- и (3-) на 0- и 5-ритмы.
♦ ♦♦ I уровень наркоза характеризуется полиморфной электрической активностью с
низкой амплитудой всех четырех типов ритма.
❖ И уровень характеризуется регулярной 0- н 3-активностью с амплитудой до 100
мкВ.
❖ Ш уровень сопровождается 3-активностью с периодами «электрического молча-
ния».
❖ IV уровень возникает при передозировке наркотического вещества и характеризу-
ется увеличением периодов «электрического молчания», что является признаком
глубокого нарушения функции стволовых структур мозга.
73. Возрастные изменения сна.
❖ Снижена общая продолжительность сна и длительность как фазы медленного сна,
так и фазы быстрого сна.
❖ Имеется возврат к полицикличности сна, частые пробуждения среди ночи.
7.6. Возрастные изменения памяти.
❖ После 70 лет в большей степени поражается чувственно-образная память по срав-
нению со смысловой памятью.
❖ Ослабляются процессы консолидации, в результате чего снижается память на те-
кущие события при относительно сохранной памяти на события далекого прошло-
го.
4 * Снижается помехоустойчивость при запоминании и воспроизведении информа-
ции.
8. Профильный материал для студентов педиатрического факультета.
8.1 • Особенности ВИД новорожденного.
❖ Формирование условных рефлексов осуществляется прежде всего на жизненно
важные стимулы - на натуральные раздражители.
• Скорость образования условных рефлексов вначале невысока - на первом ме-
сяце жизни требуется до 200 сочетаний сигнала и подкрепления.
• Условные ^рефлексы образуются на время кормления (четко проявляются с
8-го дня), на положение для кормления (с конца 2-й недели), на зрительные
сигналы - с 3-го мес.
• Внешнее торможение существует сразу после рождения.
• Внутреннее дифференцировочное торможение появляется к концу 1-го мес.
(например, к 20-му дню ребенок различает положения для кормления и пеле-
нания);
• Эмоциональные реакции имеют преимущественно отрицательный характер
8.2 . ВИД у детей разного возраста.
❖ Грудной возраст (2 -12 мес.).
• Образование условных рефлексов осуществляется быстрее, они более проч-
ные, вырабатываются не только на простые сигналы, но и на комплексные.
• С 5-го мес. возможно формирование условного рефлекса при раздражении
любой сенсорной системы.
• К концу первого года слово становится сигналом для выработки условного
рефлекса.
• Начинает формироваться внутреннее торможение:
476
"Ф запаздывающее торможение начинает формироваться только с 5 мес., за-
паздывание вырабатывается с большим трудом, под влиянием воспитания
и тренировки только к старшему школьному возрасту;
ф угасание происходит гораздо медленнее, поэтому ребенка трудно отучить
от вредных привычек, неправильных навыков.
• Образуются динамические стереотипы, облегчающие приспособление ребенка
к окружающей среде, так как сила и подвижность нервных процессов у ребен-
ка еще невелики.
• Появляются положительные эмоции: улыбка - со 2-го мес., смех - с 3-го мес.
• Начинается развитие речевой функции,
ф гуление и лепет в 2 - 4 мес.;
ф освоение сенсорной речи (понимания значения слов и запретов, названия
имен близких людей, игрушек, предметов) начинается с 7 - 8 мес.;
ф развитие моторной речи начинается с 10 - 12 мес., важную роль играет
имитационный рефлекс;
ф словарный запас к концу первого года составляет 10-12 слов.
Возраст 2-3 года.
• Морфофункциональное созревание мозга:
ф устанавливаются связи между полушариями, между проекционными и
непроекциоиными отделами полушарий (отражением этого процесса
служит становление a-ритма на ЭЭГ);
ф происходит миелинизация проводящих путей;
ф увеличивается площадь коры, специализация ее участков под влиянием
сенсорной информации (влияние обогащенной информационной среды).
• Активная исследовательская деятельность:
ф происходит становление взаимодействия анализаторов;
ф двигательный анализатор используется в освоении количественных и
пространственных соотношений: дальше-ближе, больше-меньше.
• Образующиеся динамические стереотипы очень прочны, поэтому нарушение
порядка, режима воспринимается болезненно; образование стереотипов важно
для правильного воспитания, так как переделать их будет чрезвычайно трудно.
• Развитие речи:
ф развитие речи связано со способностью совершать тонкие манипуляции
пальцами;
ф накапливается словарный запас - к 2 годам он составляет ~ 200 слов, ос-
ваивается фразовая речь и грамматика, развивается обобщающая функция
слова. К началу 2-го года развивается I ступень интеграции, где слово вы-
ступает как символ конкретного предмета; к концу 2-го года - II ступень,
когда слово замещает несколько однородных предметов (слово «кукла»
становится «сигналом сигналов»; для этого необходимо, чтобы на него
выработался пучок условных связей - не менее 15 ассоциаций, иначе сло-
во останется символом, замещающим лишь один конкретный объект).
Лишь в 3 года ребенок достигает III ступени, когда слово замещает ряд
разнородных предметов («игрушка»).
• Мышление остается конкретным, предметным.
477
Ф Возраст 4-6 лет.
• Происходит усиление межцентральных связей ассоциативных зон и появление
способности оценивать вероятность подкрепления (у детей старше 5 лет).
• Соотношение процессов возбуждения и торможения: отмечается преоблада-
ние генерализации условных рефлексов; постепенно происходит становление
и усиление внутреннего торможения.
• Эмоциональные реакции неустойчивы, бурно проявляются (аффекты).
• Развитие речи:
ф словарный запас в начвле 4-го года составляет 500 - 700 слов, появляется
новый тип связей: словесный раздражитель - словеснвя реакция;
ф формируется IV ступень интеграции: к 5 годам слово становится обоб-
щающим понятием для ряда ранее выработанных обобщений предыду-
щих степеней («вещи» как обобщение слов-понятий «игрушка», «ме-
бель», «одежда»);
ф к 6 годам развивается внутренняя речь и словесное мышление.
❖ Возраст 7-11 лет.
• Сформирована достаточная сила и уравновешенность нервных процессов.
• Выражены все виды внутреннего торможения, но механизмы активного вни-
мания недостаточно развиты, отсюда быстрая утомляемость и отвлечение.
• Более четко выявляются типологические особенности ВНД, все характеристи-
ки еще подвижны (сила, уравновешенность, подвижность), что создает воз-
можность направленной педагогической коррекции характера.
• Обобщающая функция слова приближается к уровню взрослого.
Ф Возраст полового созревания (пубертатный, подростковый).
• Характерна повышенная возбудимость ЦНС.
• Ослаблены процессы торможения, ухудшаются дифференцировки, возникает
широкая генерализация возбуждения.
• Затруднено формирование новых условных рефлексов.
• Характерна эмоциональная неустойчивость.
8.3. Особенности ЭЭГ и вызванных потенциалов в разные периоды онтогенеза.
Ф Особенности ЭЭГ в онтогенезе (см. тему 6. задание 1: 6.2).
Ф Вызванные потенциалы как отражение в онтогенезе функционального созрева-
ния разных отделов мозга.
• Отмечается гетерохронность формирования вызванных потенциалов:
' Ф раньше всего, к 4 годам, формируется соматосенсорный ВП (ответ на
стимуляцию кожи);
ф позже всего формируется зрительный ВП: окончательный вид он приоб-
ретает к 18 годам.
• Происходит укорочение латентных периодов ВП при азрослении, появление
всех компонентов ВП, в том числе и поздних, отражающих переработку ин-
формации с участием механизмов опознания сигнала в ассоциативных систе-
мах мозга.
8.4. Особенности змоциопалъпых реакций маленьких детей.
478
❖ Доминируют процессы эмоциональной активности, компенсируя недостаток ин-
формации для достижения цели.
❖ Отмечается неустойчивость эмоций из-за слабости контроля со стороны высших
отделов ЦНС, слабости процессов внутреннего торможения.
❖ Положительные эмоции подкрепляют и закрепляют социализацию потребностей.
8.5. Формирование фаз и периодов сна в онтогенезе.
* ♦* Сокращение времени сна: у новорожденного до 21 ч в сутки, в 3 месяца - 16 - 19
ч, к 1 году - 14 ч, в 3 - 5 лет - 13 ч, в 10 лет - 10 ч, к 16 годам продолжительность
сна, как и у взрослых.
❖ Переход от полицикличного сна к моноцикличному: смена сна и бодрствования
происходит у новорожденного до 7 раз за сутки, в 1 год - 3 раза, с 1 до 6 лет - 2
раза, с 7 лет сон становится моноцикличным, как и у взрослого.
❖ Изменяется структура сна: ФБС занимает у новорожденного до 60% общего вре-
мени сна, к 5 годам достигает, как и у взрослого, 20% общего времени.
При созревании механизмов регуляции сна и бодрствования раньше происходит
становление ЭЭГ, типичной для стадии глубокого сна, а с 3 лет на ЭЭГ выделяет-
ся стадия поверхностного сна.
8.6. Возрастные изменения памяти: у детей в раннем возрасте преобладает чувственно-
образная память; обобщающая функция слова и преобладание смысловой памяти по-
является к 5-6 годам, усиливается и развивается в процессе обучения.
8.7. Возрастные особенности восприятия.
* 1* Созрбвание разных областей коры происходит неодновременно (например, пер-
вичные сенсорные зоны уже к рождению находятся в определенной стадии зрело-
сти и осуществляют простой анализ).
❖ До 3 - 4 лет ассоциативные зоны дублируют функции сенсорных.
• •• К 5 - 6 годам развивается таламо-париетальная ассоциативная система, которая
участвует в опознании сложных образов, а сенсорные зоны осуществляют более
простой анализ.
❖ С 7 лет совершенствование восприятия идет за счет развития таламо-фронтальной
ассоциативной системы.
• 2* К 10 - 11 годам формируется произвольное избирательное восприятие.
8.8. Роль полушарий мозга в организации внимания у ребенка.
• > Произвольное внимание носит социальный характер и формируется с 10-го мес.
по мере освоения речи (по инструкции взрослого). Сначала взрослый принуди-
тельно организует внимание ребенка, выделяет объект из среды. По мере освоения
речи именно внутренняя речь становится средством программирования внимания.
До 5 лет произвольное внимание легко подавляется непроизвольным. Непроиз-
вольное внимание связано с активностью правого полушария.
♦ > К 6 - 7 годам отмечается появление реакции десинхронизации на ЭЭГ, возрастает
роль речевой инструкции в организации внимания.
❖ К 9 - 10 годам происходит созревание лобных долей и обеспечение локальной ре-
гулируемой активации - основы произвольного внимания.
❖ В 12 - 16 лет (пубертатный период) отмечено ослабление регулирующих корко-
вых влияний на активационные процессы и ослабление внимания, его произволь-
ной регуляции.
8.9. Развитие речевой функции (см. выше 8.2).
9. Профильный материал для студентов стоматологического факультета.
479
9.1. Влияние функционального состояния организма на деятельность слюнных желез
и сенсорную функцию полости рта.
♦ ♦♦ Функциональные состояния, приводящие к снижению слюноотделения. Психио-
эмоциональное возбуждение (например, стресс на экзамене) может сопровождать-
ся в случае избыточной активации симпатического отдела и усиленного выброса
катехоламиноа снижением секреции слюнных желез и ощущением сухости во рту,
что приводит к нарушениям вкусовой чувствительности и речевой функции.
Функциональные состояния, приводящие к повышению саливации.
• В начальной фазе адаптации к зубным протезам.
• Рефлекторное усиление секреции происходит при гингивитах, стоматитах, яз-
венной болезни двенадцатиперстной кишки, панкреатите.
♦ ♦♦ Изменение активности рецепторов и их плотности называется функциональной
мобильностью. Она зависит от нисходящих влияний ретикулярной формации на
рецепторный аппарат ротовой полости. Может проявляться в форме гастролин-
гаального рефлекса - снижения чувствительности вкусовых рецепторов ротовой
полости при введении в желудок глюкозы через зонд, изменениях чувствительно-
сти при голоде и насыщении, при стрессе и других состояниях.
9.2. Значение учения И.П. Павлова о ВНД в работе врача стоматолога.
♦ ♦♦ Условные рефлексы на интерьер стоматологического кабинета, звук бормашины,
стоматологическое кресло, подкрепляемые болевым раздражением, вырабатыва-
ются быстро и становятся очень прочными. Это приводит к формированию чувст-
ва страха перед посещением стоматологического кабинета, вызывает психоэмо-
циональное напряжение со сдвигами в вегетативной и соматической сферах и тре-
бует от врача психопрофилактической работы с пациентом.
♦ ♦♦ Наличие второй сигнальной системы, основанной на речи, позволяет врачу уста-
новить коммуникативный контакт с пациентом и воздействовать на него, про-
граммировать его поведение (выполнять по указанию врача действия, способст-
вующие проведению манипуляций в ротовой полости), контролировать его психо-
эмоциональное состояние (внушение, беседа, произвольный контроль дыхания,
напряжение скелетных и жевательных мышц), по болевой реакции и ее словесно-
му выражению судить о степени поражения зуба.
♦ ♦♦ У больных нарушения речевой функции часто связаны с периферическими при-
чинами: изменением секреции слюнных желез, нарушением функций мышц язы-
ка, целостностью зубных рядов, наличием зубных протезов, дефектами твердого
неба и др. Устранение косметических дефектов у пациента и восстановление пол-
ноценной речевой функции возвращает человека к профессиональной деятельно-
сти, восствнавливает его социальные связи.
9.3. Особенности психоэмоционального состояния больных с повреждениями и дефек-
тами челюстно-лицевой области.
Стоматологические заболевания, сопровождающиеся потерей зубов, травмами челю-
стно-лицевой области приводят к значительному изменению облика больного. Для ря-
да профессий, связанных с межличностным общением (артисты, педагоги и др.), фак-
тор внешнего облика играет важную роль. Наличие подобного рода дефектов может
вызывать снижение самооценки, привести к хроническим депрессивным состояниям и
развитию психосоматических нарушений.
9.4. Условно- и безусловно-рефлекторные изменения в деятельности внутренних орга-
нов при стоматологических вмешательствах.
Психоэмоциональный стресс (страх при стоматологических вмешательствах формиру-
ет оборонительные условные рефлексы на боль и звук бормашины) сопровождается
480
активацией симпатоадреналовой системы и выбросом катехоламинов, избыточной мо-
билизацией функциональных резервов организма («вегетативная буря»). Это может
проявляться в росте ЧСС, АД, частоты дыхания, активации потовых желез и другими
симпатоадреналовыми реакциями. На фоне сердечно-сосудистой патологии это может
привести к гипертоническому кризу, сосудистому коллапсу. Боль как безусловнореф-
лекторный фактор затрагивает все без исключения физиологические системы организ-
ма, вызывает нарушения их функционирования, повышает мышечный тонус (защитно-
оборонительные реакции), является мощным стрессором.
9.5. Психофизиологические методы подготовки стоматологических больных к лече-
нию.
❖ Уменьшение степени подверженности стрессорным воздействиям.
• Разные виды фармакологического воздействия для устранения страха и боле-
вой реакции.
• Снижение субъективной интенсивности восприятия (снятие страха перед
вмешательством в ротовой полости) путем использования психотерапии, от-
влекающих внимание больного бесед, функционаяьной музыки и т.д.
• Снятие чрезмерной мышечной скованности, мешающей манипуляциям в по-
лости рта.
♦ ♦♦ Использование навыков снижения чрезмерного уровня функционального напря-
жения (нервно-мышечная релаксация, дыхательный контроль - произвольная ре-
гуляция дыхания и снижение болевых ощущений).
♦ ♦♦ Повышение физической активности как средство профилактики психосоматиче-
ских повреждений внутренних органов при избыточном выбросе катехоламинов.
9.6. Адаптация к зубным нротезам как проявление пластичности нервных центров.
♦♦♦ Зубной протез вызывает в начальной стадии:
• усиление слюноотделения (безусловный рефлекс на отвергаемый раздражи-
тель);
• дискоординацию жевательного акта н глотвння;
• нарушение речи и процессов артикуляции;
• мощное раздражение структур головного мозга потоком импульсов из ротовой
полости, вызванных наличием в ней инородного тела.
❖ Прн привыкании к протезу происходят:
• механическая адаптация тканей десны;
• адаптация рецепторного, проводникового и центрального отдела ротового
анализатора;
• восстановление эффективности жевания, нормализация саливации, ослабление
рвотного рефлекса, нормализация речевой функции.
❖ Скорость адаптации зависит от следующих факторов:
• пластических способностей высших отделов ЦНС перестраивать свои функ-
циональные взаимосвязи под влиянием импульсов (обратная связь), информи-
рующих о результатах текущей деятельности;
• типа ВНД и функционального состояния: переутомления, волнения, психиче-
ской травмы и т.д.;
• характера распределения давления на рецепторы периодонта и слизистой ро-
товой полости, что зависит от величины, конструкции и способа фиксации
протеза.
481
Задаяяе 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). Прн под-
готовке реферата использовать список основной я дополнительной литера-
туры (задание 3; 3.1 н 3.2). Объем реферата 2 - 3 с., регламент 5 - 7 мин.
Задаяяе 3. Для выполнения заданий 1 н 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющие-
ся у вас учебники и пособия): [1-16].
ЗЛ. Дополнительная литерапура (см. с. 514 - 522): [1], [3], [10], [12], [14], [22], [23],
[28-30], [40], [43], [48], [59-61], [67], [90], [105], [111], [121], [132], [134], [136],
[145], [152], [161], [162], [163], [166], [168], [171], [173], [175-177], [179], [188],
[196], [200], [201], [213], [214], [226], [236], [241], [242], [243].
3.3. Дистанционные средапи обучения. Использование страницы кафедры в Интер-
нете - www.vstna.ac.ni/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» IB.Б. Брин. - Владикавказ: СОГМА. - 1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки. оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. /Сонщрояь самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по данной теме и экзамене.
482
Тема 33. Система цоспроизведеиия
Цель: а) знать физиологические основы воспроизведения, его нейрогуморальную регуляцию (безус-
ловно- и условнорефлекторные механизмы, половые гипофизотропные и гонадотропные гормо-
ны, гормоны гонад; формирование и механизмы половой мотивации, роль половых гормонов в
формировании полового поведения; фазы полового цикла у мужчин (половое влечение, эроген-
ные зоны, эрекция, копулятивная и эякуляционная фазы, оргазм, рефрактерная фаза), особенно-
сти полового цикла у женщин; роль социальных факторов в реализации половых функций чело-
века, физиологические закономерности беременности и родового акта;
б) использовать эти знания для понимания функциональной деятельности целостного организ-
ма
Мотивация темы: в физиологическом смысле о системе воспроизведения можно, перефразируя слова
Сократа, сказать, что воспроизведение - не все, но все без воспроизведения ничто. Функции
системы воспроизведения являются физиологическим базисом для ряда медицинских дисцип-
лин: акушерство, гинекология, андрология, сексопатология.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Воспроизведение - сложная функциональная система, обеспечивающая деторожде-
ние и продолжение рода. Состоит из ряда функциональный подсистем - половой мо-
тивации и полового поведения, полового акта, развития эмбриона и плода, грудного
вскармливания, воспитания потомства. Начало ее функционирования связано с про-
цессом полового созревания (у мальчиков с 13-16 лет, у девочек - с 12-15 лет), одна-
ко закладка и формирование системы начинается с первого дня развития организма.
Критериями полового созревания являются появление вторичных половых признаков
(половые особенности скелета, мышц, распределения жира, оволосения, развитие грудных желез, мен-
струации у женщин, эрекции и семяизвержение у мужчин и др.). Основными критическими пе-
риодами этой функциональной системы являются внутриутробный период, периоды
полового созревания и угасания.
2. Внутриутробный период. В это время происходит формирование генетического,
гаметного и гормонального пола.
2.1. Генетический пол определяется набором половых хромосом в оплодотворен-
ной яйцеклетке - 46 (XX) в женском и 46 (XY) в мужском организме.
2.2. Гонадный пол определяется формированием половых желез - янчек или яич-
ников. На 3-й неделе в стенке желточного мешка появляются первичные поло-
вые клетки - предшественники сперматогониев н овогониев. На 4-й неделе на
медиальной поверхности первичных почек появляются зачатки половых желез
- первичные гонады (половые тяжи), состоящие из мезенхимальных клеток
первичной почки и покрытые целомическим эпителием. На 5-6 неделе первич-
ные половые клетки перемещаются в половые тяжи - начинается становление
гонадного пола.
483
❖ Развитие яичек. Первичные гоиады дифференцируются в яички, если они
заселяются первичными половыми клетками с кариотипом 46 (XY). В муж-
ском плоде происходит активация SRY гена (sex-determining region) в Y-
хромосоме, кодирующего фактор развития яичка - ДНК-связывающего
белка, который индуцирует транскрипцию других генов, определяющих
дифференцировку яичек (формируются семенники). (В этом процессе участвует
также ген на Х-хромосоме, регулирующий экспрессию фактора развития яичка, и ген на
аутосоме, определяющий структуру рецептора клеточной мембраны к фактору развития
яичка.)
❖ Развитие яичников. Первичные гонады дифференцируются в яичники, если
они заселяются первичными половыми клетками с кариотипом 46 (XX).
Они размножаются и превращаются в овогоиии (около 7 млн). Часть из них
(около 15%) превращается без деления в овоциты I порядка, остальные деге-
нерируют. Овоциты вступают в мейоз, который блокируется иа стадии
профазы (деление будет возобновляться только в постнатальном периоде в зре-
лых преовуляторных фолликулах). Овоциты I порядка покрываются одним
слоем эпителиальных клеток - образуются примордиальные фолликулы. В
процессе созревания вокруг овоцита формируется прозрачная оболочка
(zona pellucida), многослойный эпителий (zona granulosa). Затем вокруг
фолликула образуется внешняя оболочка (teca folliculi). Структура яичника
в основном формируется к 20-й неделе. К моменту рождения имеется около
1 мли фолликулов. Необходимым условием развития женских половых ор-
ганов во внутриутробном периоде является отсутствие секреции тестосте-
рона и фактора редукции мюллеровых канальцев.
2.3. Гормональный пол.
❖ В мужском плоде формируется секрецией тестостерона (начинается с 7-8-й не-
дели).
• Клетки Лейдига семенников начинают образовывать тестостерон, кото-
рый действует на рецепторы клеток вольфовых каналов первичной поч-
ки, превращая их в семявыносящие протоки и простату. Геи рецептора
андрогенов локализован на Х-хромосоме. Уровень тестостерона мак-
симален на 20-й неделе и постепенно снижается к концу внутриутроб-
ного периода.
• Клетки Сертоли в это время образуют фактор регрессии мюллеровых
канальцев, которые служат закладкой для маточных труб, матки и верх-
ней трети влагалища. Поэтому в мужском плоде они не образуются (не
считая простатической маточки).
• Физиологическая роль тестостерона во внутриутробном периоде: 1) ин-
дуцирует дифференцировку первичных половых клеток в сперматого-
нии (после 22-х недель); 2) необходим для дифференцировки гонадо-
тропных клеток гипофиза; 3) превращается в клетках полового бугорка,
складок и валиков в дигидротестостерон, под влиянием которого фор-
мируется половой член и мошонка (к 12-16 неделям); 4) совместно с
фактором регрессии мюллерова протока обеспечивает опускание яичек
в мошонку.
484
• Регулятором секреции тестостерона с 13-й недели являются хориониче-
ский гонадотропин, секретируемый плацентой, и лютеинизирующий
гормон (пик секреции между 24 - ЗО-й неделями).
❖ В женском плоде при отсутствии образования тестостерона и фактора рег-
рессии мюллеровых протоков последние превращаются в маточные трубы,
матку и верхнюю часть влагалища. (Из полового бугорка образуется клитор, из по-
ловых складок - малые половые губы, из половых валиков - большие половые губы, из
урогенитального синуса - нижняя часть влагалища, бартолиниевые и скеииевы железы.)
❖ Плоды обоего пола защищены от высокого уровня эстрогенов крови мате-
ри, имеющимся в их крови белком а-фетопротеином, который связывает
эстрогены.
2.4. Половая дифференцировка гипоталамуса происходит непосредственно перед
и сразу после родов и состоит в формировании двух центров - тонического и
циклического, состоящих из мелкоклеточных ядер, нейроны которых образуют
гонадолиберин.
❖ Тонический половой центр локализован в аркуатном (дугообразном) ядре
медиобазального гипоталамуса. Этот центр функционирует и в женском, и
в мужском организме. Секреция гонадолиберина происходит в импульсном
режиме у мужчин с периодом 3-4 часа, у женщин в фолликулярной фазе
цикла около 1,5 часов, в лютеиновой фазе - 3 - 4 часа.
❖ Циклический половой центр расположен в преоптической зоне гипотала-
муса и функционирует только в женском организме, начиная с периода
полового созревания. Он действует в течение 24 - 48 часов в менструаль-
ном цикле, обеспечивая предовуляториый выброс ЛГ и ФСГ. У плода муж-
ского пола перед родами и в первые месяцы после рождения этот центр
выключается высоким уровнем тестостероном (точнее образующимися из
него эстрадиолом в специфических нейронах). (Если в плод женского пола перед
родами ввести тестостерон, то циклический центр в женском организме необратимо вы-
ключается.)
3. Периоды детства (до полового созревания).
3.1. Эти периоды характеризуются низкой концентрацией в организме половых
гормонов, что обеспечивается высокой чувствительностью гипоталамуса к по-
ловым гормонам (торможение по механизму отрицательной обратной связи).
При этом уровень женских и мужских половых гормонов примерно одинаков у
девочек и мальчиков. Они образуются преимущественно в надпочечниках.
3.2. В яичниках в это время имеется около 400 тысяч примордиальных фолликулов
с очень медленным независимым от гонадотропина ростом, в яичках - сперма-
тогонии.
33. Торможение полового созревания осуществляется также гормонами эпифиза и
тимуса, активность которых в этом периоде у детей высока.
4. Период полового созревания или пубертатный период (продолжается в течение 5 -
10 лет, половая зрелость у девочек наступает в 16 - 18 лет, у мальчиков - в 18 - 20 лет).
4.1. Снижается чувствительность гипоталамуса к тормозным влияниям половых и
эпифизарных гормонов и повышается секреция ЛГ, ФСГ и пролактина. Увели-
чивается концентрация половых гормонов в крови: у мальчиков за счет андро-
485
генов, у девушек - эстрогенов. Главным стимулятором их секреции становится
ЛГ.
4.2. Происходит интенсивное развитие внутренних и наружных половых органов
(например, объем яичек увеличивается от 5 до 20 см3). Возникает скачок соматического
роста.
43. Начинается интенсивный сперматогенез по следующей схеме: размножение
сперматогониев (митоз) —» образование сперматоцитов I порядка, вступающих
в мейоз, —» после 1-го деления образуются сперматоциты П порядка —»после 2-
го деления - сперматиды, которые без деления превращаются в сперматозоиды
(за сутки их образуется около 100 млн). У мальчиков появляются поллюции.
4.4. У девочек формируется овариальный цикл и менструации.
4.5. Формируются вторичные половые признаки: лобковое оволосение, набухание
грудных желез и др. Формируется соматический пол (фенотип субъекта: муж-
чина или женщина). !'
4.6. Формируется способность совершать половой акт, беременность и р<йы.
4.7. Формируется половое самосознание, определяющее половое поведение (прежде
всего выбор полового партнера), формируется гражданский пол (мужчина,
женщина), пол воспитания (форма одежды, прическа, игры и др.).
5. Период половой зрелости (у мужчин с 18-20 лет, у женщин - с 16-18 лет).
5.1. Мужской организм. Основные анатомо-физиологические особенности муж-
ского организма (первичные вторичные половые признаки) связаны с гормональ-
ной функцией его системы воспроизведения.
❖ Семенники (яички) имеют три вида клеток: интерстициальные клетки
Лейдига, клетки Сертоли и половые клетки: сперматогонни —» спермато-
зоиды.
• Клетки Лейдига расположены между извитыми семенными канальцами,
образуют основной гормон - тестостерон (около 5 мг/сутки, 95% всех андро-
генов организма), небольшое количество дигидротестостерона (100
мкг/сутки), эстрадиола (80% всех эстрогенов мужского организма) н прогестеро-
на. Роль двух последних гормонов у мужчин до настоящего времени не
выяснена. (Соотношение образования тестостерона и эстрогенов зависит от локаль-
ной температуры: при 5 - 10° С ниже температуры тела, например, в мошоике) обра-
зуется преимущественно тестостерон, при 37 °C резко возрастает продукция эстроге-
нов, нвпример, при неопущеиии яичка - крипторхизме). Тестостерон в крови
транспортируется в связанной форме с Р-глобулином (60%), альбуми-
ном (37%) и в свободной активной форме (1-3%). В клетки-мишени
проникает только свободный тестостерон и связанный с альбуминами.
Основной катаболизм осуществляется в печени (за один пассаж крови инак-
тивируется 80% гормона), экскреция с мочой осуществляется в виде 17-
кетостероидов (вндростерона и этиохолаиалоиа).
'у’ Физиологические эффекты тестостерона.
Участвует в сперматогенезе, чему способствует его высокая
концентрация в извитых семенных канальцах.
486
Участвует (после превращения его в дигидротестостерон) в разви-
тии наружных половых органов (мошонки, полового члена) как
во внутриутробном, так и в пубертантном периодах.
Обеспечивает развитие и сохранение вторичных половых при-
знаков
Имеет анаболический эффект: активирует синтез белка, рост
скелета и мышечной массы.
Превращаясь в эстрогены в нейронах гипоталамуса, выключает
циклический половой центр в конце внутриутробного разви-
тия.
Участвует в формировании половой мотивации, полового по-
ведения и степени агрессивности.
ф" Регуляция продукции тестостерона.
Стимулируют продукцию гонадолиберин гипоталамуса и лю-
теинизирующих гормон гипофиза, которые секретируются в
пульсирующем режиме у мужчин с периодом 3-4 часа. (Толь-
ко импульсное воздействие гонадолибернна стимулирует секрецию
ЛГ и ФСГ в гипофизе; напротив, непрерывное воздействие гонадо-
либерина или его пролонгированных аналогов приводит к десеисити-
, зацин его рецепторов и к подавлению секреции ЛГ и ФСГ - обрати-
мая бескровная кастрация.)
Тормозят секрецию высокие уровни тестостерона крови (отри-
цательная обратная связь) и пролактина, а также эстрогены.
• Клетки Сертоли эпителия извитых семенных канальцев имеют рецеп-
торы ФСГ и тестостерона.
ф" Образуют андргогенсвязывающий белок (АСБ), который создает
высокую концентрацию тестостерона в семенных канальцах.
ф" Образуют эстрогены путем ароматизации тестостерона.
ф" Выделяют ингибин, тормозящий синтез ФСГ.
ф" Образуют гематотестикулярный барьер.
ф- Имеют трофическую функцию.
* Ф* Участвуют в образовании семейной жидкости.
• Сперматогенез является результатом взаимосвязанного влияния на
сперматогонии клеток Лейдига, образующих тестостерон и клеток Сер-
толи, выделяющих АСБ, что создает высокую местную концентрацию
ТС, необходимую для сперматогенеза.
❖ Функция простаты.
• Вырабатывает секрет, содержащий фруктозу, цитрат цинка и протеазы,
разжижающий эякулят и инициирующий двигательную активность
сперматозоидов.
• Вырабатывает пептид FPP (fertilization promoting peptide), предупреждающий
преждевременную акросомную реакцию спермиев (акросома - апикальное
тельце, облегчающее проникновение сперматозоида через оболочку яйцеклетки при
оплодотворении). Недостаток этого пептида ведет к бесплодию.
487
• Образует из тестостерона более активный гормон - дигидротестостерон
(фермент 5а-редуктаза) и эстрогены (фермент - ароматаза), которые регулиру-
ют её функции. Образует простатический специфический антигеи (мар-
кер её опухолевого поражения).
• Вырабатывает (наряду с клетками Сертоли) ингибин, тормозящий секре-
цию ФСГ аденогипофизом.
• Вырабатывает простагландины, поддерживайте тонус гладкомышеч-
ных клеток в самой простате и участвующие в механизмах эрекции.
• Тонус гладкомышечных клеток простаты помогает удержанию мочи.
5.2. Женский организм. Основные физиологические особенности женского орга-
низма в репродуктивном периоде связаны с гормональной функцией его систе-
мы воспроизведения.
❖ Овариально-менструальный цикл (28 ± 7 дней) включает в себя коорди-
нированные изменения нейросекреторных структур гипоталамуса, яични-
ков, труб и матки. Состоит из двух фаз (фолликулярной и лютеиновой) и
двух последовательных процессов: овуляции, которая приходится пример-
но иа середину цикла, и менструации, завершающей овариальный цикл, ес-
ли не произошло оплодотворение и имплантация яйцеклетки. (Примечание: в
акушерстве н гинекологии для более удобного расчета продолжительности цикла н его фаз
началом цикла считают первый день менструации.)
• Фолликулярная фаза характеризуется развитием фолликулов по схеме:
примордиальный фолликул —> первичный —> вторичный —> третичный
фолликул.
Примордиальный фолликул.
К началу половой зрелости общее количество примордиальных
фолликулов 200 - 400 тыс. В течение репродуктивного периода
98% из них погибает, 2% (4 - 8 тыс.) достигает стадии первич-
ного и вторичного фолликулов, и только около 400 образуют
третичные фолликулы (граафовы пузырьки).
В каждом цикле от 3 до 30 примордиальных фолликулов всту-
пают в фазу роста и формируют первичный фолликул. (Все на-
чавшие развитие, но не достигшие стадии овуляции фолликулы подверга-
ются атрезии и превращаются в атретическне тела.)
Продолжительности жизни овоцита фолликула до его оплодо-
творения от 14 до 50 лет, что увеличивает риск генных дефек-
тов особенно на фоне интоксикации никотином и алкоголем.
• ф’ Первичные фолликулы. На фолликулярных клетках гранулезы
имеются рецепторы ФСГ, ЛГ, эстрогена и тестостерона. ФСГ инду-
цирует синтез ароматазы (зстрогенсинтетазы), под действием которой
из тестостерона образуется эстрадиол. Эстрадиол стимулирует про-
лиферацию фолликулярных клеток (рост фолликула) и экспрессию
в них новых рецепторов к ФСГ и стероидам (тестостерон тормозит про-
лиферацию фолликулярных клеток).
Вторичные фолликулы морфологически отличаются от первичных
более выраженными оболочками (текой): наружной - соединитель-
488
нотканной и внутренней - гранулезной, между фолликулярными
клетками которой появляются полости, заполненные жидкостью.
Среди вторичных фолликулов один (редко два) в росте опережает
остальные (он называется доминантным). В функциональном плане
вторичные фолликулы характеризуются тем, что на клетках их теки
появляются рецепторы к ЛГ. ЛГ начинает стимулировать синтез
тестостерона в клетках теки. Тестостерон диффундирует вглубь
фолликула в клетки гранулезы, где превращается под действием
ароматазы в эстрадиол (ФСГ усиливает этот процесс синтеза эстро-
генов). Часть тестостерона н эстрадиола секретируется в фоллику-
лярную жидкость. На этой стадии в крови резко увеличивается
концентрация эстрадиола. (Для осуществления этих процессов стимули-
рующее влияние ЛГ осуществляется только при импульсной секреции гоиадолн-
бернна с периодами в фолликулярной фазе около 1,5 часов, в лютеиновй фазе - 3
- 4 часов. При отсутствии импульсной секреции, например прн гипоталамиче-
ской аменорее, созревание фолликулов, овуляция не происходят.)
ф- Третичный фолликул (граафов пузырек) возникает из доминантного
вторичного фолликула, в котором образуется полость с быстрым
накоплением жидкости и увеличением размеров с 200 мкм до 2,5
см в диаметре. Высокое содержание эстрадиола в крови по меха-
ннзму положительной обратной связи (самоусиления) вызывает
предовуляторный выброс гонадолиберина нз циклического центра,
что обеспечивает резко выраженный предовуляторный пнк ЛГ н
менее выраженный пнк ФСГ.
ф- Эстрогены стимулируют образование рецепторов для прогестерона
в клетках яичника, матки, молочной железы. Они стимулируют
рост железистого эпителия эндометрия, гладких мышц матки, из-
меняют физические свойства шеечной слизи - она непроницаема
для сперматозоидов в начале цикла, но проницаема в периоде ову-
ляции.
ф- В матке в это время развивается пролиферативная фаза: пролифе-
рируют клетки базального слоя и восстанавливается функциональ-
ный слой.
Овуляция (происходит через -30 часов после достижения пика эстра-
диола и через -11 часов после пика ЛГ; обычно через 11-13 дней 28-
дневного цикла).
ф- Первое (редукционное) деление мейоза завершается в течение 36
часов от достижения пика ЛГ (второе деление завершается не ранее
оплодотворения).
Ф- ЛГ стимулирует синтез простагландинов в фолликулярных клетках,
простагландины стимулируют протеолитические ферменты грану-
лезы, что приводит к истончению и разрыву стенки фолликула. В
деполимеризации стенки фолликула и овуляции участвует также
гормон релаксин.
Лютеиновая фаза характеризуется образованием желтого тела
489
•ф- Менструальное желтое тело функционирует до завершения цикла
(если нет имплантации), под влиянием ЛГ его клетки синтезируют
н секретируют прогестерон и эстрогены (в стероидогенезе прогестерон
является предшественником вндрогенов и эстрогенов). Уровень прогестеро-
на достигает пика через 8-9 дней после овуляции, что соответствует
времени имплантации. Физиологические эффекты прогестерона.
Обеспечивает условия для имплантации оплодотво-
ренной яйцеклетки и ее последующего развития.
Тормозит секрецию гонадолибернна, ФСГ и созревание новых
яйцеклеток.
Усиливает секрецию желез эндометрия, увеличивает содержа-
ние гликогена в секрете.
Изменяет состояние шеечной слизи, создает условия для ми-
грации сперматозоидов (снижается вязкость слизи, pH становится ще-
лочным, через 1-2 дня после овуляции формируется непроницаемый барь-
ер из плотной слизи).
Снижает возбудимость мускулатуры матки в результате изме-
нений проницаемости миоцитов для ионов К+ и Na+, прн этом
уменьшается число низкоомных контактов между клетками.
Повышает температуру тела на 0,4 - 0,5 °C (этот пирогенный
эффект используется для диагностики овуляции и биологиче-
ского способа контрацепции).
Во время беременности вызывает структурно-функциональные
изменения матки, молочных желез, способствующие вынаши-
ванию плода и лактации.
• ф В матке в лютеиновую фазу цикла имеется секреторная фаза: ма-
точные железы разрастаются и секретируют гликоген, гликопро-
теины, липиды, муцин.
• Менструация - циклические кровянистые выделения из матки, обу-
словленные отторжением поверхностного слоя эндометрия; возникает,
если овариальный цикл не сопровождается беременностью. Ее начало
совпадает с началом гибели желтого тела. В механизмах менструации
важную роль играют сосудистые реакции. Перед началом менструации
возникает спазм спиральных артериол в основании эндометрия, кото-
рый приводит к некрозу и отслоению поверхностного слоя слизистой от
базального слоя. Локальное освобождение сосудорасширяющих мета-
болитов вызывает расширение артериол, наступает кровотечение.
❖ Эффекты эстрогенов вне системы размножения.
• На метаболизм костной ткани: избыток эстрогенов ускоряет закрытие
ростовых зон, недостаток эстрогенов замедляет окостенение в пубер-
татном периоде и вызывает остеопороз в постменопаузе;
• На анаболические процессы (действие слабее, чем андрогенов): усили-
вают синтез ряда белков в печени; липопротеинов высокой плотности,
тормозят синтез липопротеинов низкой плотности, снижают уровень
холестерола в крови, факторов свертывания крови (VII, IX, X) (до мено-
490
паузы риск возникновения инфаркта миокарда у женщин в несколько раз меньше, чем
у мужчин);
• На ионный баланс: эстрогены способствуют депонированию Са2+ в ске-
лете, вызывают задержку Na+, прогестерон - потерю Na+.
❖ Андрогены в женском организме.
• Источники образования: надпочечники и яичники (тестостерон у женщин
0,5 - 2,5 нмоль/л; у мужчин 13-30 нмоль/л).
• Физиологическая роль: формирование полового влечения, стимуляция
роста волос на лобке и подмышечных впадинах.
6. Половая мотивация и поведение. Половая мотивация (половое влечение, либидо) -
эмоционально окрашенное стремление человека совершать поведение, направленное
на удовлетворение половой потребности. Она относится к биологической, но сильно
социализированной мотивации; эмоциональный компонент в ней также резко выра-
жен, достигая во время полового акта степени аффекта и наслаждения. Эта мотива-
ция формируется в процессе полового созревания под влиянием двух групп факто-
ров.
6.1. Гормональные факторы (андрогены и эстрогены) создают «темное» половое
влечение без избирательной направленности. Решающую роль, как у мужчин,
так и у женщин играет уровень андрогенов, эстрогены придают половому вле-
чению жейский тип. У мужчин суточный ритм продукции андрогенов имеет
максимум в 2 - б ч (пик либидо), у женщин эстрогены увеличивают либидо в
середине менструального цикла (во время овуляции). Прогестерон в низких
концентрациях способствует формированию полового влечения, в высокой
концентрации (лютеиновая фаза цикла, беременность) тормозит половое влече-
ние. Ведущей мишенью для этих гормонов при формировании половой мотива-
ции является гипоталамус, исключительное значение при этом имеют преопти-
ческая область и передний гипоталамус (введение тестостерона только в эти области
гипоталамуса восстанавливает половое поведение у кастрированных животных, их разрушение
у здоровых животных прекращает половое поведение.) Другие области гипоталамуса,
влияющие на половое поведение, действую преимущественно через преоптиче-
скую область и передний гипоталамус.
6.2. Психические и социальные факторы формируют избирательное половое вле-
чение, т.е. определяют «идеал» полового партнера. Психические, социальные и
соматические характеристики партнера фиксируются в виде нейронной модели
в акцепторе результата действия половой функциональной системы. Совпаде-
ние качеств полового партнера с нейронной моделью вызывает усиление поло-
вого влечения вплоть до возникновения «чувства любви». Таким образом, по-
ловая мотивация и поведения формируются сложным комплексом физиологи-
ческих и психосоциальных факторов, определяемых общественной средой:
нормами морали, общением с родителями и сверстниками, разными видами ис-
кусств, средствами массовой информации и др.
6.3. Этапы (формы) полового влечения: платоническое влечение - интерес к противоположному по-
лу, не носящий сексуального характера (например, детская влюбленность); эротическое влече-
ние - желание не только духовного, но и телесного контакта (нежность, ласки); сексуальное
влечение - влечение к половому акту.
491
6.4. Формы полового поведения: ритуальное поведение (танцы, ласки и др.), направленный поиск
полового партнера, взаимодействие в поповом акте, сохранение и воспитание потомства.
7. Половой акт, фазы и механизмы (условно-физиологический ритм 1 - 3 раза в неделю).
7.1. Психическая фаза - осознание полового влечения, направленное на конкретно-
го человека. Ее физиологическая основа - возникновение половой доминанты, в
формировании которой участвуют условнорефлекторные стимулы, связанные с
личным опытом человека: зрительные раздражители (особенно у мужчин), слухо-
вые раздражители (особенно у женщин), и безусловно-рефлекторные стимулы: раз-
дражение эрогенных зон - генитальных (наружные половые органы) и экстра-
генитальных (губы, грудные железы и др.).
7.2. Фаза эрекции (увеличение кровенаполнения и плотности наружных половых
органов: полового члена, малых половых губ, клитора).
❖ Центр эрекции расположен в нижнепоясничных и крестцовых сегментах
спинного мозга (особенно в S2.4), имеет высокую возбудимость (низкий по-
рог) по сравнению с центром эякуляции. Раздражается рефлекторным и
психогенным (с корковых центров) путем, имеет хорошую способность к
суммации раздражения. (При разрушении крестцового центра психогенная эрекция у
25% людей обеспечивается пояснично-грудными сегментами; прн разрыве спинного мозга
в шейном и грудном отделах рефлекторная эрекция сохраняется.)
❖ Эфферентная часть представлена импульсацией по парасимпатическим пп.
erigentes (медиаторы ацетилхолин, ВИП), вызывающей увеличение крове-
наполнения наружных половых органов с увеличением их объема и плот-
ности: венозных синусов пещеристых тел и губчатого тела полового члена,
малых половых губ и клитора у женщин. При этом артерии расширяются,
кровоток увеличивается, а венозный отток резко уменьшается.
❖ Увеличение секреторных процессов: желез простаты, уретры, влагалища,
транссудация жидкости во влагалище.
7.3. Фрикционная стадия (введение и поступательно-возвратные движения поло-
вого члена во влагалище).
❖ Ее физиологической основой является суммацией возбуждения с механо-
рецепторов головки полового члена, влагалища и клитора в центре эякуля-
ции н лнмбнко-коргикальных структурах.
❖ В наружной трети влагалища за счет застойной венозной гиперемии обра-
зуется оргастическая манжетка (обеспечивает более выраженную стимуля-
цию эрогенных зон, затрудняет вытекание спермы).
❖ Происходит увеличение молочных желез и «эрекция» соска.
7.4. Эякуляторная фаза (семяизвержение) у мужчин.
❖ Центр эякуляции расположен в нижнегрудных и поясничных сегментах,
имеет низкую возбудимость нейронов. В регуляции эякуляции участвует
парацентральная долька, соединяющая пре- и постцентральную извилины
на медиальной стороне больших полушарий. (При разрушении крестцовых сег-
ментов эякуляция может быть, если есть эрекция; прн разрыве спинного мозга в шейных н
грудных отделах эякуляция отсутстаует - предполагается тормозное влияние центра эрек-
ции на центр эякуляции.) Оказывает эфферентное влияние:
• симпатическое влияние вызывает сокращение гладких мышц придатка,
семявыносящего протока, семенных пузырьков и простаты (семенная
492
жидкость поступает в задний отдел уретры, при этом внутренний
сфинктер мочевого пузыря сокращается);
• соматические влияния от спинальных моторных центров вызывают
мощные сокращение мышц тазового дна, что обусловливает толчкооб-
разный выброс семенной жидкости.
7.5. Фаза оргазма - соответствует по времени фазе эякуляции, выражается высшей
степенью эмоций и формированием сладострастного ощущения. Оргазм, как
эмоциональное ощущение, включается в АРД половой функциональной систе-
мы и становится мощным стимулятором половой мотивации.
❖ В формировании оргазма принимает участие сенсорная кора больших по-
лушарий, их парацентральных долек, лимбической системы, а также гипо-
таламус, вегетативные и двигательные центры ствола и спинного мозга.
❖ Вегетативные реакции во время оргазма - увеличение сердечного ритма до
180 уд. в мин, систолического и диастолического давления на 50-100 мм рт.
ст., учащение дыхания, расширение зрачков, секреция пота и др.
7.6. Рефрактерная фаза завершает половой акт и имеет два периода: абсолютной и
относительной половой невозбудимости. В периоде абсолютной рефрактерио-
сти никакие раздражители не могут вызвать повторную эрекцию, в периоде от-
носительной рефрактерности при действии сверхсильных раздражителей воз-
можен повторный половой акт.
7.7. Особенности фаз полового цикла у женщин.
• > Более длительная психогенная фаза, ведущую роль играет не сексуальное
влечение (стремление к половому акту), а эротическая (стремление к лас-
кам), тесно связанная с оценкой личностных качеств партнера.
❖ Сексуальное удовлетворение менее связано с оргазмом, чем у мужчин.
❖ Половой акт не имеет абсолютного рефрактерного периода.
8. Физиология беременности. Во время беременности формируется функциональная
система мать-плод (комплекс мать-плацента-плод).
8.1. Яйцеклетка в лютеиновой фазе цикла поступает в фаллопиеву трубу, где она
пребывает примерно четверо суток, в течение которых чаще всего и происходит
ее оплодотворение. Перистальтические сокращения яйцеводов (главный регу-
лятор - эстрогены) доставляют яйцеклетку в матку, где оиа и имплантируется
в эндометрий, подготовленный к этому процессу прогестероном и эстрогенами.
8.2. Циклическое желтое тело трансформируется в желтое тело беременности,
которое функционирует в течение первых 7-8 недель беременности, образуя
прогестерон и эстрогены. Стимуляция секреции его гормонов вначале осущест-
вляется хорионическим гонадотропином, в дальнейшем - хорионическим сома-
томаммотропином.
8.3. Формирующаяся плацента с 4-го месяца берет на себя гормональное обеспе-
чение развития плода (в опытах на животных гнпофизэктомия беременных на этом этапе
уже не приводит к прерыванию беременности).
❖ Стероидные гормоны плаценты (прогестерон, эстрогены, андрогены) ока-
зывают локальное действие на уровне матки; их синтез происходит только
в системе мать-плацеита-плод, так как в плаценте отсутствует ряд фермен-
тов, необходимых для стероидогенеза. Высокие уровни прогестерона и эст-
493
рогенов подавляют у беременной и у плода трансплантационный (клеточ-
ный) иммунный ответ (но увеличивается риск вирусных инфекций) и способству-
ют развитию трофобласта и плаценты.
❖ Белково-пептидные гормоны плаценты.
• Хорионический гонадотропин имеет эффекты, сходные с ЛГ гипофиза.
(Его синтез начинается в примитивном трофобласте уже через день после импланта-
ции и достигает пика в плаценте на 8-10-й неделе беременности; появление ХГ в моче
служит ранним индикатором беременности - биологическая проба Ашгейм - Цоиде-
ка.)
Способствует вынашиванию беременности, поддерживает функцию
желтого тела н стимулирует продукцию прогестерона, тормозит
продукцию ФСГ.
Активирует продукцию тестостерона в яичках плода и этим обес-
печивает дифференцировку гипоталамуса и полового тракта по
мужскому типу (тонический тип секреции гонадолнберина).
• ф Стимулируя образование прогестерона и эстрогенов, участвует в
образовании иммунотолерантности.
Ф По современным данным ХГ образуется не только в плаценте, но некоторыми
органами плода - почками и яичками.
• Хорионический соматомаммотропин имеет эффекты, подобные СТГ
(его секреция прямо пропорциональна сроку беременности).
В организме матери тормозит использование глюкозы, активирует
глюконеогенез, липолиз с образованием кетоновых тел, которые
(глюкоза н кетоновые тела) используется для питания плода.
• ф Прн действии на плод стимулирует синтез белков и рост плода.
• ф Поддерживает продукцию прогестерона в плаценте.
• ф Снижает чувствительность к инсулину.
• ф Возможно, способствует росту н дифференцировке молочных же-
лез.
• Релаксин плаценты.
• ф Обеспечивает деполимеризацию тазовых связок и расхождение
лонного сочленения (подготовка родовых путей).
• ф Размягчает шейку матки и расслабляет гладкие мышцы матки.
• Другие гормоны, выделенные из плаценты (хорионические тиреотропин н кортико-
тропин, пептиды, подобные гипоталамическим лнберннам и статинам, 0-зндорфин н
др).
9. Физиология родов. Роды - физиологический процесс изгнания из матки через вла-
галище плода н последа (плаценты, плодных оболочек н пуповины)
9.1. Факторы, инициирующие роды.
❖ Преодоление прогестеронового блока, снижающего возбудимость и сокра-
тимость матки: в конце беременности (с ЗО-Зб-й недели) количество обра-
зующегося прогестерона уменьшается, к тому же часть его превращается в
эстрогены. Соотношение эстрогены/прогестерон увеличивается.
494
❖ В конце беременности в организме женщины увеличиваются тормозные
процессы в высших отделах головного мозга, что приводит к повышению
возбудимости спинного мозга и снижению порога возбудимости матки.
❖ Увеличение размеров плода и его двигательной активности, а также отно-
сительное уменьшение объема околоплодных вод приводит к увеличению
афферентной импульсации с интерорецепторов матки, что способствует
началу родовой деятельности.
❖ Непосредственным инициатором родов являются прогестероны Fja н Ег,
которые стимулируют сокращение матки. Продукцию простагландинов в
матке увеличивают эстрогены, кортикостероиды плода, окситоцин.
9.2. Биомеханика родов. Имеется три периода родов: раскрытие шейки матки, из-
гнание плода и послеродовой период.
❖ Раскрытие шейки матки. Показателем начала родов являются схватки -
непроизвольные волнообразные, ритмические сокращения матки. Они сна-
чала кратковременные (около 10 с) с периодом 10-15 мин, далее они удли-
няются (до 1 мин), а период сокращается до 2 мин. Повышение внутрима-
точного давления во время схваток постепенно приводит к укорочению и
расширению шейки матки. Раскрытию шейки способствует плодный пу-
зырь, который перемещается во время схваток в эту зону. Во время очеред-
ной схватки происходит разрыв плодного пузыря и отхождение вод.
❖ Период изгнания плода. К схваткам присоединяются потуги - сокращения
мышц брюшного пресса, диафрагмы и промежности. Они также возникают
непроизвольно, но могут усиливаться и ослабляться роженицей. Давление в
матке при этом резко увеличивается, что приводит к продвижению плода
через родовой канал. После рождения перевязывают пуповину, отделяя но-
ворожденного от последа.
• > Послеродовой период продолжают схватки, которые уменьшают размер
плацентарной площадки. Плацента, не способная к сокращению, начинает
отслаиваться. Послеродовой период завершается выделением последа.
Объем кровопотери при нормальных родах равен около 200 мл.
10. Физиология лактогенеза и лактации.
10.1. Процесс лактогенеза требует участия многих гормонов.
❖ Во время беременности повышается уровень пролактина, кортизола, эстро-
генов, плацентарного лактогена, стимулирующих развитее секреторного
аппарата молочной железы. Однако, их эффект минимальных, так как вы-
сокие концентрации эстрогенов и прогестерона блокируют действие про-
лактина на железу.
❖ После родов резкое снижение уровня эстрогенов и прогестерона снимают
этот блок, и начинается лактогенез. Синтез казеина, a-лактоглобулина, жи-
ра и секрецию молока реализует пролактин при содействии кортизола и
инсулина. Один из механизмов секреции связан с синтезом простагланди-
нов, которые активируют образование РНК и белков.
10.2. Нейроэндокринный механизм лактации (сосательный рефлекс). Механиче-
ское раздражение сосков при сосании вызывает афферентную пульсацию, кото-
495
• В первые годы менопаузы уменьшение массы костей составляет 2,5%
ежегодно. В дальнейшем оно составляет около 0,75% в год.
• Причиной остеопороза является снижение уровня эстрогенов, которые
способны тормозить резорбтивные процессы в костной системе, и за
счет небольшого анаболического действия (образование белкового мат-
рикса кости) способствует задержке Са2+ в костной ткани.'
11.2. Возрастные изменения у мужчин.
❖ Продукция тестостерона прогрессивно падает с 18 до 80 лет и составляет в
старческом возрасте 50-65 % от уровня в молодом возрасте.
❖ Растет продукция тестикулярных эстрогенов и соотношение свободные эс-
трогены/свободный тестостерон.
❖ Уменьшается масса яичек, снижается интенсивность сперматогенеза и чис-
ло сперматозоидов.
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). При под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3; 3.1 и 3.2). Объем реферата 2 - 3 с., регламент 5-7 мин.
Задание 3. Для выполнения заданий 1 и 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющейся у
вас учебники и пособия): [1-9], [12], [14], [16].
3.2. Дополнительная литература (см. с. 514 - 522): [27], [30], [35], [43], [47], [63],
[107], [147], [150], [152], [196], [222], [225], [229], [248], [251], [253].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интер-
нете - www.vsma.ac.ru/~nphys: Слайды для компьютерной демонстрации из «Атла-
са по физиологии человека» / В.Б. Брин. - Владикавказ: СОГМА. -1999; тесты ос-
таточного уровня знаний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диа-
проектором и слайдами (по графику кафедры).
3.5. Копсультацип преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практи-
ческом занятии по д аниой теме и экзамене.
497
Тема 34. Физиология адаптации
Цель'. а) знать понятие об адаптации, индивидуальную адаптацию организма, лежащие в ее основе
биологические и социальные факторы, виды, фазы и критерии адаптации, механизмы развития
адаптивных реакций, резистентность, понятие о перекрестной резистентности и адаптации, тре-
нирующие режимы, роль биоритмов в адаптации, социальный аспект в адаптации.
б) уметь использовать эти знания для понимания деятельности человека в различных условиях
окружающей среды.
Мотивация темы: «Адаптация - итоговая проблема физиологии и медицины» К. Бернар.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логиче-
скую структуру учебного материала.
Внимание: фрагменты задания 1, предназначенные для самостоятельного изучения, оп-
ределяются лектором в конце читаемой по данной теме лекции, и обозначены в «Графике и
содержании самостоятельной работы студентов». Остальные фрагменты задания 1 могут
быть использованы для коррекции материала записанной лекции и подготовки рефератов.
1. Общая физиологическая характеристика адаптации.
1.1. Адаптация - это приспособление организма к изменению природных, произ-
водственных и бытовых условий. Стратегической целью адаптации является
восстановление гомеостаза организма в изменившихся условиях среды. Адап-
тационные возможности организма являются мерой его здоровья. С этой точки
зрения физиологическая норма - это максимальные адаптационные возможно-
сти при минимальном расходовании функциональных резервов.
Адаптация является неотъемлемым свойством всего живого, она осуществ-
ляется на различных уровнях: клеточном, тканевом, органном, организменном,
популяционном. В данной теме рассматривается адаптация на организменном
уровне, генетические и фенотипические механизмы которой обеспечивают ин-
дивидуальное приспособление путем изменения функций организма.
1.2. В основе адаптации лежит методологический принцип единства организ-
ма и среды обитания. Организм как открытая термодинамическая система
осуществляет обмен веществ, энергии и информации со средой, обеспечивая
поддержание гомеостаза. В действительности происходит непрерывное нару-
шение н восстановление гомеостаза под влиянием факторов среды, которое
имеет ритмический колебательный характер и обозначается термином гомеоки-
нез. Длительное отклонение констант от границ нормы ведет к нарушению
функций и создает угрозу жизни. Эти отклонения возникают под влиянием
факторов среды.
1.3. Классификация факторов среды (Н.А. Агаджанян, 2003).
❖ Экзогенные: природные, техногенные, экологические.
❖ Эндогенные: психоэмоциональное напряжение, болезни, вредные привыч-
ки.
• > Социальные: плохое качество жизни (работа, семья, жилье), чувство стра-
ха (преступность, терроризм), утрата жизненных ценностей.
❖ Действующие факторы помимо качественных различий отличаются и ко-
личественно. Существуют пределы адаптивных возможностей при дейст-
498
вии разных по интенсивности факторов. Если воздействие по интенсивно-
сти отклоняется (увеличивается или уменьшается) от оптимума, вызываю-
щего норму реакций, то такие факторы получили название экстремальных
или чрезвычайных раздражителей (по Павлову), нли стрессоров (по Селье).
❖ Необходимо учитывать, что, как правило, в естественных условиях на ор-
ганизм действует не отдельный фактор, а комплекс факторов среды; эти
воздействия носят прерывистый (ритмический) характер.
1.4. Выделяют два главных вида адаптации - фенотипическую и генотипиче-
скую.
Генотипическая адаптация осуществляется на основе мутации, наследственности и естест-
венного отбора. Она является основой эволюции, в наиболее «чистом» виде проявляется у
новорожденного ребенка. Генотипическая адаптация в дайной теме не рассматривается.
❖ Фенотипическая (или индивидуальная) адаптации формируется в процессе
взаимодействия организма со средой. Уровень фенотипической адаптации,
характерный для человека, достигается только в человеческом обществе.
Фенотипическая адаптация не передается по наследству. Различают сле-
дующие типы фенотипической адаптации.
• По основным стратегиям осуществления различают:
• ф'. Пассивную адаптацию или толерантность (гипобиоз, гибернация,
гипометаболизм). Она сопровождается минимизацией функций с
некоторыми нарушениями гомеостаза - зимняя спячка, оцепенение
при недостатке пнщи и воды, охлаждении, голодании, гипоксии.
Характерными ее признаками являются понижение уровня метабо-
лизма и снижение потребления кислорода.
• ф- Активную адаптацию или резистентность, она сопровождается ак-
тивацией катаболизма и увеличением потребления кислорода, не-
экономична, может давать патологические последствия (острые и
хронические болезни адаптации, в том числе психосоматические).
• По времени развития различают два вида адаптации.
"Ф' Срочная или кратковременная адаптация, при которой с помощью
нервных и эндокринных механизмов мобилизуются готовые налич-
ные резервы организма. Она осуществляется физиологическими и
функциональными системами организма при кратковременном дей-
ствии факторов среды или на начальном этапе адаптации при дли-
тельном воздействии стрессоров. Приспособление достигается пу-
тем мобилизации функциональных резервов организма, осуществ-
ляется на пределе физиологических возможностей, сопровождается
активацией не только специфических, но и неспецифических реак-
ций. Срочная адаптация дает организму возможность «продержать-
ся» до развития долговременной адаптации, она не бывает полной и
экономичной. Во время развития этой фазы возможны поврежде-
ниями систем организма.
"Ф' Долговременная адаптация, при которой реализуется связь между
функцией и генетическим аппаратом клеток и формируется струк-
турный след адаптации, позволяет расширить диапазон функцио-
499
нальной мощности систем организма, участвующих в данной адап-
тации. Она возникает при многократном или длительном действии
фактора и обеспечивает выживание организма в экстремальных ус-
ловиях.
2. Общий адаптационный синдром (стресс-синдром, Г. Селье, 1936) является вариантом
преимущественно срочной адаптации, ио и долговременной адаптации, если при этом
сохраняется напряженность регуляторных систем.
2.1. Общая характеристика.
❖ Стресс-синдром - совокупность защитных физиологических реакций в от-
вет на действие экстремальных факторов: экзогенных, эндогенных, соци-
альных. Стресс - экстренная адаптационная программа, включающая в се-
бя как неспецнфнческне, так н специфические реакции, осуществляющаяся
в результате взаимодействия стресс-реализующей и стресс-лимитирующей
систем. Соотношение стресс-синдрома и адаптации следующее: стресс-
синдром прекращается при достижении устойчивой долговременной адап-
тации, при которой снижается напряженность регуляторных систем.
❖ Стадии стресса (по Г. Селье) (рис. 64).
• Стадия тревоги (или аварийная) разделяется на две фазы: шока и проти-
вотока. Непродолжительная фаза шока следует сразу после действия
раздражителя. Для нее характерны торможение ЦНС, снижение АД,
мышечного тонуса, температуры тела, уровня глюкозы н лейкоцитов
крови н др. Резистентность организма прн этом снижена. Если сила раз-
дражителя велнка, то в этой фазе может наступить смерть (этому способ-
ствует недостаточность коры надпочечников). Если сила стрессора невелика, то
фаза шока отсутствует н сразу наступает фаза противотока, во время
которой повышается резистентность организма. Стадия тревоги соот-
ветствует срочной адаптации н развивается в результате нейро-
гуморальной мобилизации существующих резервов физиологических
систем организма.
• Стадия резистентности характеризуется максимальной устойчивостью
организма к стрессору, которая может сохраняться длительное время. В
этой фазе срочная адаптация переходит в долговременную адаптацию,
формируется структурный след в адаптации (см. ниже 3), уменьшается
напряженность регуляторных систем.
• Стадия истощения возникает прн действии сильных н длительных
стрессоров. При этом снижается резистентность организма (и уровень
глюкокортикоидов в крови), возникают болезни адаптации. Эта фаза изуча-
ется в патофизиологии.
2.2. Стресс-реализующая система - регуляторный комплекс, активирующий н ко-
ординирующий все изменения в организме, составляющие адаптивную реакцию
на стрессоры. Состоит нз центрального звена и периферических (эфферентных)
звеньев, соединяющих центральное звено со всем организмом.
500
❖ Центральнее звено находится в гипоталамусе и других отделах ствола моз-
га. Гипоталамус получает информацию о действии стрессоров от провод-
никового (через таламуса) и коркового отделов сенсорных систем н запус-
кает работу стресс-снстем. В этом участвуют три главные группы нейро-
нов.
• Нейроны паравентрикулярных ядер вырабатывают кортиколибернн (КЛ-
нейроны), активирующий ось АКТГ-глюкокортнконды коры надпочеч-
ников. Кортиколибернн реализует поведенческие и гормональные реак-
ции прн стрессе, влияет
на иммунную систему.
• Нейроны паравентрику-
лярных ядер, вырабаты-
вающих гормон арги-
нин-вазопрессин (АВ-
нейроны), который акти-
вирует образование
АКТГ, симпатический
Рис. 64. Стадии стресс-сиидрома. I - стадия
тревоги (в - фаза шока, б - фаза противотока),
П - стадия резистентности, Ш - стадия исто-
щения, сплошная линия - уровень резистент-
ности, прерывистая линия - уровень глюко-
кортикоидов в крови.
ртдел нервной системы
н тормозит иммунные
реакции.
Нейроны гипоталамуса
н голубого пятна сред-
него мозга, вырабаты-
вающие норадреналин (НА-нейроны).
• В последние десятилетия выявилась роль и других гормонов: иейропептида Y, выде-
ляющегося из НА-иейроиов гипоталамуса, ои активирует выработку кортиколиберина,
угнетает нейроны голубого пятна, усиливает вазоконстрикторное действие катехола-
минов; цитокинов, опосредующих действие стресс-системы иа иммунную ситему; ан-
гиотензина П; вещества Р и др.
Эфферентные звенья соединяют центральное звено с исполнительными
органами.
• Гипоталамо-гипофизарно-надпочечникое звено, действующим продук-
том которого является глюкортикоиды коры надпочечников.
• Менее изучена роль гипоталамо-гипофизарно-тиреоидного и гипота-
ламо-гипофизарно-гонадного звеньев.
• Симпатико-адреналовое звено включает в себя возбужденные спиналь-
ные центры симпатической нервной системы и мозговой слой надпо-
чечников, конечными продуктами которых являются норадреналин и
адреналин, действующие на различные типы а- н Р-адренорецепторов.
* Парасимпатическое зеено изучено плохо (известно, что оио участвует в развитии ги-
периисулинемии).
23. Стресс-лимитирующие системы осуществляют по механизму саморегуляции
и отрицательной обратной связи ограничение интенсивности ответа на стрес-
сорные воздействия н предупреждают развитие повреждений, в том числе пси-
хосоматических заболеваний. К стресс-лимитирующим системам относятся
501
следующие факторы.
❖ Мелатонин - гормон, вырабатываемый эпифизом, координирует взаимо-
действие нервной, эндокринной и иммунной систем в ответ иа поврежде-
ние, оказывает модулирующее, прежде всего тормозящее действие на вы-
работку кортиколиберина в гипоталамусе, АКТГ в гипофизе. Он тормозит
стероидогенез в надпочечниках при очень высокой секреции глюкокорти-
коидов. Мелатонин участвует в активации других стресс-лимитирующих
систем (см. ниже), является самым мощным эндогенным антиоксидантом,
ингибирует перекисное окисление липидов и защищает от повреждения
свободными радикалами белки и генетический аппарат клеток, снижает ги-
бель клеток как по механизму апоптоза, так и по механизму некроза.
❖ ГАМК-ергическая система оказывает свое влияние и в ЦНС, и на перифе-
рических нервных окончаниях, ограничивает секрецию кортиколиберина в
гипоталамусе, АКТГ в гипофизе, освобождение норадреналина и адрена-
лина
❖ Опиоидергическая система. Помимо освобождения из центральных нейро-
нов ЦНС, опиоидные пептиды (эндорфины, энкефалины, динорфины) вы-
деляются из аденогипофиза и мозгового слоя надпочечников, они ограни-
чивают повреждающие эффекты избытка катехоламинов. Опиоиды тормо-
зят выработку кортиколиберина, вазопрессина, окситоцина, глюкокорти-
коидов и катехоламинов. Одновременно подавляется секреция гонадотро-
пинов и ТТГ.
❖ Оксид азота (NO) выделяется одновременно с норадреналином из оконча-
ний симпатических нейронов и предупреждает спазм сосудов. Ои модули-
рует выброс кортиколиберина, повышает активность антиоксидантных
ферментов и усиливает синтез белков теплового шока (стресс-белков), пре-
дупреждающих денатурацию белков клетки.
❖ Локальные тканевые стресс-лимитирующие системы представлены веще-
ствами, образующимися в самих тканях: простагландины, аденозин, анти-
оксиданты. К антиоксидантам организма относятся белки церулоплазмин,
миоглобин, трансферрин, ферменты - супероксиддисмутаза, каталаза, глю-
татионпероксидаза, малые молекулы - глютатион, витамины С, Е, бета-
каротины.
❖ Важную роль в формировании резистентности клеточных структур играет
накопление стресс-белков (белков теплового шока), предотвращающих де-
натурацию белков. Белки теплового шока обеспечивают стабилизацию
белковых структур при действии широкого спектра повреждающих факто-
ров от гипоксии до ионизирующей радиации. Эти белки могут ингибиро-
вать рецепторы кальмодулина и тем самым препятствовать активации кле-
точных протеаз.
2.4. Системные механизмы развития стресс-синдрома осуществляют быстрый
ответ и резко преобладают в стадии тревоги и в начале стадии резистентности.
❖ Вначале реализуются поведенческие механизмы адаптации с помощью
врожденных и приобретенных реакций выбора предпочитаемых условий
для оптимального существования (температура, газовый состав среды,
пищевых компонентов и др.: «превращение нужд тела в поведение»), По-
502
веденческне механизмы осуществляются с участием ориентировочных ре-
акций и поиска решения проблемной ситуации. Во время бодрствования
происходит поиск путей изменения ситуации с помощью различных,
вплоть до социальных форм поведения. В фазе быстрого сна происходит
поиск путей внутренней адаптации к самой ситуации, в фазе медленного
сна происходит селекция информации по критерию значимости.
❖ Реализация адаптивного поведения осуществляется с участием активации
симпатоадреналовой и гипоталамо-гипофизарио-иадпочечниковой систем,
которые мобилизуют физиологические и метаболические резервы организ-
ма. Характерными признаками этой стадии являются гетерохронность, сла-
бая координированность вовлеченных физиологических систем, работа на
пределе возможностей, неадекватно большое число вовлеченных в ответ
систем, возникновение эмоций и вегетативных компонентов реакции, на-
рушение гомеостаза.
❖ Катехоламины и кортизол, активируя катаболические реакции, обеспечи-
вают «стирание» структурных следов предшествующих адаптаций в клет-
ках систем, не участвующих в данной адаптивной реакции. Освободившие-
ся структурные и энергетические резервы (аминокислоты, глюкоза, жирные
кислоты и другие вещества) перебрасываются к тем системам, которые ак-
тивно участвуют в процессах адаптации для формирования в них нового
структурного следа. Эта направленная передача структурных и энергетиче-
ских ресурсов к физиологическим и функциональным системам, осуществ-
ляющим адаптацию, становится возможной благодаря рабочей гиперемии
органов, обусловленной усиленным образованием NO, влияние которого
противостоит сосудосуживающему эффекту, обусловленному при стрессе
массивным выбросом катехоламинов, вазопрессина и ангиотензина П. Дли-
тельный спазм сосудов может стать фактором, вызывающим повреждения
в ряде органов, например, образование язв в желудочно-кишечном тракте.
2.5. Клеточные механизмы реализации стресс-синдрома обусловлены действием
медиаторов и гормонов на клетки органов и тканей.
❖ Наиболее ранним эффектом является действие норадреналина и адрена-
лин через а- и /3-адренорецепторы клеточной мембраны, включающих сис-
темы вторых посредников - аденилатциклазную (цАМФ) и фосфоинози-
тидную (ИФз и ДАГ) и Са2+. Через соответствующие протеиикиназы осу-
ществляется фосфорилирование (активация или инактивация) уже сущест-
вующих в клетке функционально активных белков (ферментов, насосов,
каналов, переносчиков, рецепторов) (подробно в теме 8; задание 1:2.3).
❖ Вторые посредники и их протеиикиназы являются активаторами ранних ге-
нов (протоонкогенов), обеспечивающих первичную реакцию генома клетки
на действие раздражителей. Активация ранних генов начинается через 5-7
мин после действия раздражителей и достигает максимума через 60-90
мин. Ранние гены являются универсальными регуляторами транскрипции
генома. Кодируемые ими белки обеспечивают выборочную активацию от-
дельных генов в геноме. В связи с этим они являются ключевым звеном в
осуществлении приспособительных реакций организма и перехода кратко-
временной адаптации в долговременную.
503
❖ Более длительное действие на геном имеют гормоны, проникающие в ядро
клетки - глюкокортикоиды, тиреоидные и половые гормоны, играющие
главную роль в переходе срочной в долговременную адаптацию.
3. Долговременная адаптация.
3.1. Долговременная адаптация возникает при длительном действии стрессоров. Ее
формирование начинает осуществляться в фазе резистентности стресс-
синдрома. Характерным признаком ее является образование структурного следа
в адаптации, приводящего к уменьшению напряжения регуляторных систем в
организме.
3.2. Механизмы образования структурного следа в адаптации связаны с действием
гормонов, проникающих в клетку (стероидных и тиреоидных), являющихся ре-
гуляторами экспрессии генов, а также с влиянием анаболических гормонов -
СТГ, соматомединов, факторов роста. В результате их действия в клетках тех
органов и тканей, которые входят в состав формирующейся функциональной
системы адаптации, активируется синтез ключевых функциональных белков,
необходимых для адаптации: ферментов энергетического обеспечения, белков
насосов, каналов, ферментов, сократительных мышечных белков.
33. Регуляция по типу «всем, всем», характерная для срочной адаптации, сменяется
иа регуляцию по типу «только для главных участников адаптивного процесса»,
т.е. для имеющих отношение к формированию структурного следа, что приво-
дит к снижению напряжения регуляторных систем. Этот феномен виден на рис.
64: в стадии резистентности адаптационный эффект сохраняется, несмотря на
снижение до нормы уровня глюкокортикоидов крови. Адаптированная система
организма отличается высокой экономичностью функционирования. (Например,
адаптированное к физической нагрузке сердце на равное количество работы потребляет на 1/3
меньше кислорода и субстратов окисления, чем не адаптированное сердце.)
3.4. Структурный след может быть локализован в ограниченном числе органов. На-
пример, при адаптации к действию ядовитых веществ увеличивается мощность
систем микросомального окисления в печени. Но может формироваться и раз-
ветвленный след, например, при адаптации к физической нагрузке он формиру-
ется в нервных центрах, обеспечивающих обучение и память (долговременная
потенциация), в скелетных и дыхательных мышцах, эндокринной и вегетатив-
ной системах и т.д.
4. Особенности психоэмоционального стресса (см. также тему 31; звдвние 1:1.7).
4.1. Психоэмоциональный стресс - состояние ярко выраженного психоэмоциональ-
ного переживания человеком конфликтных жизненных ситуаций, которые остро
или длительно ограничивают удовлетворение его социальных или биологиче-
ских протребностей (БМЭ, 1986, т.28, с. 152). Психоэмоциональный стресс возни-
кает при действие сильных психогенных факторов. Особенно патогенна ситуа-
ция, когда цель не достигнута, и отрицательная завершающая эмоция поддержи-
вает состояние мобилизации, что не дает результата и, в свою очередь, порожда-
ет отрицательную эмоцию. Индивидуальная значимость стрессора определяется
индивидуальным опытом, личностными особенностями («важно не то, что с Вами
происходит, а то, как Вы к этому относитесь» Г.Селье), силой действующего стрес-
сора (индивидуальная зона оптимума или пессимума).
504
4.2. В соответствие с двумя стратегиями адаптации (активной и пассивной) выделя-
ют два типа эмоционального стресса-. 1) активный эмоциональный стресс, при
котором преобладают процессы возбуждения, способствует развитию сердечно-
сосудистых заболеваний и язвенной болезни; 2) пассивный эмоциональный
стресс, при котором преобладают процессы торможения, способствует подавле-
нию иммунитета и развитию злокачественных опухолей и инфекционные забо-
леваний.
4.3. Особенности личности, влияющие иа действие психических стрессоров.
❖ Уровень притязания личности: завышенный уровень способствует психо-
эмоциональному стрессу.
❖ Иерархия потребностей и мотиваций: затруднение реализации идеальных и
социальных потребностей способствует возникновению психоэмоциональ-
ного стресса.
❖ Эмоциональная стабильность - чем выше она, тем более стрессоустойчив
человек, противоположное состояние - иейротизм способствует развитию
психического стресса.
❖ Состояние душевного здоровья как способности справиться с требования-
ми, предъявляемыми из внешней и внутренней среды (разные уровни адап-
тационных возможностей).
❖ Выносливость - зависит от типологических особенностей ВИД, прежде
всего силы нервных процессов.
❖ Преобладающие механизмы психологической защиты личности: активные
и гибкие стратегии поведения более эффективно защищают от развитя
стресса, пассивные и ригидные - менее эффективно.
❖ Характера взаимодействия между информационными структурами мозга
(лобная кора - гиппокамп) и мотивационными (гипоталамус - миндалина)
(П.В.Симонов).
4.4. Роль коры больших нолушарий и их функциональной асимметрии.
❖ Структура психоэмоционального стресса включает в себя, кроме эмоцио-
нального компонента, перцептивный (искажение восприятия), мотиваци-
онный (изменение эмоций), снижение интеллектуального потенциала и
способности принятия адекватных решений.
❖ Внешний информационный стрессор активирует преимущественно правое
полушарие и несколько снижает активность левого. Такое перераспределе-
ние функциональной активности полушарий в начале процесса адаптации
происходит и при действии физических стрессоров. Правое полушарие
лучше оценивает обстановку в условиях неопределенности среды и прогно-
зирует маловероятные события.
5. Физиологическая роль адаптации.
3.1. Общие свойства всех адаптаций.
❖ Повышают вероятность выживания индивидуума, организм приобретает
отсутствовавшую ранее устойчивость к действующим факторам среды.
❖ Повышают эффективность и экономичность использования ресурсов.
❖ Предупреждают повреждение организма не только действующими экстре-
мальными факторами, ио и другими воздействиями (перекрестная рези-
стентность).
505
❖ Функциональные улучшения в одной системе могут сопровождаться ухуд-
шением в других системах организма (сенсибилизация или цена адапта-
ции).
3.2. Критерии адаптации (по В.П. Казначееву).
❖ Термодинамический - восстановление стационарного состояния, т.е. ра-
венства прихода и расхода энергии. При развитии активной адаптации рас-
ход энергии, как правило, возрастает даже в условиях покоя, например, в
горах иа 25%.
❖ Кибернетический - качество или эффективность систем управления, полез-
ный приспособительный результат при сформированной адаптации дости-
гается не любой, а минимальной ценой.
❖ Физиологический - восстановление иа новом уровне нарушенного гомео-
стаза, перенастройка показателей гомеостаза. Адаптированный организм -
это функционально другой организм.
❖ Биологический - прогрессивная эволюция и оптимальная численность ви-
да, при этом восстанавливается репродуктивная способность индивидуума.
3.3. Перепрограммирование адаптации. В процессе эволюции стресс возник как
средство перепрограммирования адаптаций. С одной стороны, с помощью ката-
болического звена он «стирает» структурные следы ставших ненужных адапта-
ций, разоружает эти системы (отсюда феномен сенсибилизации, ослабления од-
них систем при развитии адаптивных процессов в других). С другой стороны,
он обеспечивает передачу освободившихся структурных и энергетических ре-
зервов в активно работающие системы и «закрепление» результатов процесса
адаптации в виде нового структурного следа в данных системах с помощью ана-
болического звена.
3.4. Антимутагенный эффект адаптации. В опытах было показано, что введение
химического мутагена дноксиднна адаптированным к болевым стрессорным
воздействием животным вызывало значительно меньшее количество хромосом-
ных повреждений, чем не адаптированным животных (или адаптированным к гипок-
сии). (При этом адаптация к болевому раздражителю или к гипоксии ие увеличивала число хро-
мосомных повреждений.)
3.5. Концепция адаптационной медицины (Ф.З. Меерсои, 1993). Содержанием этого
нового научного направления является изучение фундаментальных механизмов
адаптации, адаптационная терапия и адаптационная профилактика. При этом
прямые и перекрестные защитные эффекты адаптации специально воспроизво-
дятся не только у здоровых людей (адаптационная профилактика), но и на фоне
протекающей болезни (адаптационная терапия). Например, адаптация к физиче-
ской нагрузке создает возможность осуществления перекрестных защитных эф-
фектов: антигипоксических, антигипертензиоииых, антидиабетических, анти-
стрессорных. Это позволяет использовать физические нагрузки для профилак-
тики возникновения атеросклероза, диабета, гипертонии.
6. Повреждающие эффекты стресс-реакции (цена адаптации). Болезни адаптации.
6.1. Стрессогенная патология касается в основном инсулинзависимых органов и тканей, т.к глюко-
кортикоиды и адреналин подавляют секрецию инсулина, ослабляют его действие на инсулин-
зависимые ткани и уменьшают потребление ими глюкозы, сажают на «голодный паек» иммун-
ную систему, желудочно-кишечный тракт, опорно-двигательный аппарат. Диабетоподобная пе-
506
рестройка обмена обеспечивает снабжение глюкозой ЦНС, надпочечников, сердечной мышцы,
печени - ведущих систем, обеспечивающих защиту от опасности.
6.2. Хронический стресс является фактором риска развития язвенной болезни желудка и двенадца-
типерстной кишки, иммунодефицитов, что в свою очередь повышает риск развития онкологи-
ческих заболеваний, ожирения, сахарного диабета, атеросклероза и гипертензий. Стресс нару-
шает репродуктивные функции, вызывает задержку роста и физического развития. Истощая
опиоидергическую стресс-лимитирующую систему мозга, он способствует формированию за-
висимости от биоактивных веществ (алкогольной, никотиновой и наркотической зависимости).
Избыток внутриклеточного Са2* может вызывать активацию липаз и ПОЛ (см. выше) с после-
дующим повреждением тканей
6.3. Структурное обеспечение доминирующих в адаптации систем сопровождается торможением
развития других систем - феномен сенсибилизации или структурная цена адаптации. Например,
при тренировке к физической нагрузке увеличение структурных резервов в сердце, скелетных
мышцах может сопровождаться одновременным снижением числа нефронов, клеток печени и
надпочечников на 20-30%, что снижает резервные возможности этих органов (детренирован-
ность этих систем - цена адаптации). Особенно опасно это в детском организме, т.к. может не-
обратимо снизить мощность заторможенных систем и соответственно этому снизить резистент-
ность к действию некоторых факторов среды
Задание 2. Подготовить реферативные доклады на тему данного занятия. Тема рефера-
та выбирается с использованием учебных элементов (задание 1). Прн под-
готовке реферата использовать список основной и дополнительной литера-
туры (задание 3,3.1 и 3.2). Объем реферата2- 3 с., регламент5-7 мнн.
Задание 3. Для выполнения заданий 1 н 2 использовать следующие средства для са-
моподготовки студентов.
3.1. Основной учебно-методический комплекс (см. с. 514, использовать имеющийся у вас
учебники и пособия): [1-9], [14], [15].
3.2. Дополнительная литература (см. с. 514 - 522): [1-3], [15], [21], [30], [58], [75], [76],
[109], [186], [197], [222], [230], [243], [245], [247], [253].
3.3. Дистанционные средства обучения. Использование страницы кафедры в Интернете -
www.vsma.ac.ru/-nphvs: Слайды для компьютерной демонстрации из «Атласа по физиоло-
гии человека» / В.Б. Брнн. - Владикавказ: СОГМА. - 1999; тесты остаточного уровня зна-
ний по данной теме.
3.4. Учебная комната для самоподготовки, оснащенная учебными таблицами, диапроекто-
ром и слайдами (по графику кафедры).
3.5. Консультации преподавателей еженедельно по индивидуальному графику.
3.6. Контроль самостоятельной работы студентов будет осуществляться на практическом
занятии по данной теме, итоговом занятии н экзамене.
507
Ответы к ситуационным задачам
Задача 2-1. Снижения уровня АТФ в нейронах в этих условиях будет увеличивать внутриклеточ-
ную концентрация Na* в результате энергетической необеспеченности функции K*,Na*-Hacoca.
Поэтому будут активироваться К*-каналы, зависимые от АТФ и от ионов натрия, что будет уве-
личивать диффузию К* из клетки и препятствовать деполяризации.
Задача 2-2. 1) Снижение активности K*,Na*-Hacoca плазмолеммы приведет к повышению уровня
Na* в кардиомиоцитах, что вызовет деполяризацию мембраны, поэтому пороговый потенциал
уменьшится, а возбудимость увеличится.
2) Снижение градиента концентрации Na* между гиалоплазмой и внеклеточной жидкостью
уменьшит активность №*/Са2*-ионообмениика в результате чего выход Са2* во внеклеточную
среду уменьшится, а уровень его в гиалоплазме будет увеличиваться. 3) Повышение уровня Са2*
в гиалоплазме будет активировать зависимые от него К*-каиалы и диффузию К* из клетки, по-
этому скорость фазы реполяризации будет увеличиваться, а время потенциала действия
уменьшаться.
Задача 2-3. 1) Накопление высокой концентрации К* в межклеточной жидкости (следствие его
выхода из клетки в фазе реполяризации) свидетельствует о недостаточности K*,Na*-Hacoca, сле-
довательно, концентрация Na* в гиалоплазме будет также повышаться. Оба процесса будут вызы-
вать деполяризацию мембраны, что уменьшает пороговый потенциал и повышает возбудимость
клетки. 2) После интенсивной активности нейрона высокая концентрация Na* внутри клетки и К*
вне клетки будут продолжать активировать K*,Na*-Hacoc, электрогенная функция которого может
вызвать гиперполяризацию клеточной мембраны, (за один цикл удаляется из клетки 3 Na* и пере-
мещается в клетку 2 К*).
Задача 2-4. Концентрация Na* в межклеточной жидкости 90 ммоль/л представляет собой сниже-
ние его уровня по сравнению с нормой на 35-40%. В этих условиях даже прн полном открывании
активационных m-ворот в №*-каиалах входящий в клетку №*-ток становится настолько слабым,
что не может запустить регенеративную деполяризацию, поэтому генерация потенциалов дейст-
вия прекратится.
Задача 2-5. Увеличение градиента ионов Na приведет к повышению их диффузии в клетку (на-
пример, по каналам утечки). Это вызовет деполяризацию мембраны, снижение порогового потен-
циала, повышение возбудимости, и генерация ПД станет возможной при действии подпороговых
раздражителей. При развитии ПД увеличится входящий в клетку Na*-TOK и, следовательно, уве-
личится скорость фазы деполяризации ПД.
Задача 3-1. Поскольку закрывание ииактивациоииых h-ворот в натриевых каналах вызывает про-
лонгированная деполяризация, для их открывания необходимо применить гиперполяризующее
действие анода.
Задача 3-2. Для нервного волокна лабильность равна: 1с/1мс = 1000 Гц, для мышечного волокна
- 200 Гц. При действии сильных раздражителей она может быть несколько выше, так как в конце
спайка ПД имеется фаза относительной рефрактерности.
Задача 3-3. Продолжительность импульса тока в этих условиях равна 1 с/100000 Гц = 0,01 мс, что
в 10 раз меньше, чем хронаксия этой мышцы. Такой кратковременный раздражитель не может
вызвать возбуждение и сокращение мышцы.
Задача 4-1. Резкое снижение синтеза АТФ нарушит функцию калий-натриевого насоса плазмо-
леммы нейронов. Ионы калня, вышедшие из нейрона в фазе реполяризации ПД, не будут полно-
стью транспортироваться обратно из-за нарушения функции насоса. Накопление внеклеточного
калия затруднит диффузию ионов калия из клетки, что является главным механизмом формиро-
508
вания мембранного потенциала покоя. Наряду с этим блокада насоса уменьшает его электроген-
ную функцию, которая также участвует в создании потенциала покоя. Следовательно, произой-
дет деполяризация плазмолеммы.
Постепенно нарастающая деполяризация вызовет инактивацию натриевых каналов в результате
закрывания h-ворот. Натриевая инактивация приведет к полной невозбудимости нейрона, абсо-
лютной рефрактерности, а его импульсная активность прекратится.
Задача 4-2. Замена ионов натрия иа ионы холина с учетом полной непроницаемости мембраны
для холина уменьшит диффузию положительных ионов внутрь клетки. Возникнет сдвиг мем-
бранного потенциала в сторону равновесного калиевого потенциала, т.е. гиперполяризация мем-
браны. Потенциал действии не может возникнуть, так как будет нарушена фаза деполяризации из-
за отсутствия входящего в клетку натриевого тока. Следовательно, в данных условиях мембран-
ный потенциал покоя увеличится (гиперполяризация мембраны), и исчезнет способность к гене-
рации потенциала действия.
Задача 5-1. Это процесс торможения, который ограничивает возбуждение в ЦНС, придавая ему
нужную интенсивность и направление.
Задача 5-2. Клетки Реншоу обеспечивают возвратное торможение мотонейрона, при его сильном
возбуждении. Если бы в пресинаптических окончаниях этих клеток стал секретироваться глута-
мат, который является возбуждающим медиатором, по механизму положительной обратной связи
произошло бы перевозбуждение мотонейронов и неконтролируемое по силе сокращение иннер-
вируемых мышц.
Ответ к задаче 5-3. Известно, что самое медленное проведение возбуждения происходит в цен-
тральном звене рефлекса и оио связано с наличием многочисленных синапсов, в каждом из кото-
рых имеется синаптическая задержка в проведении возбуждения. Поэтому более длительное ла-
тентное время второго рефлекса, несмотря на более короткий рефлекторный путь и более скоро-
стные его волокна, можно объяснить более многочисленными синаптическими переключениями в
центре этого рефлекса.
Ответ к задаче 6-1. В этих экспериментах А.А. Ухтомский открыл новый принцип координаци-
онной и интегративной деятельности мозга - принцип доминанты. Доминантный очаг был обу-
словлен рефлекторным влиянием с рецепторов переполненного дистального отдела толстой киш-
ки. Афферентная импульсация из задней конечности поступила в доминатный очаг и увеличила
его возбуждение, что привело к акту дефекации.
Задача 9-1. Длительный прием глюкокортикоидов в высокой дозе по механизму обратной связи
ингибировал образование как кортиколиберина в гипоталамусе, так и АГТГ в аденогипофизе, что
ликвидировало их стимулирующее влияние на пучковую зону коры надпочечников. Развиваю-
щаяся при этом ее атрофия приводит к резкому снижению образования ферментов синтеза глюко-
кортикоидов с почти полным прекращением их образования. Через двое суток после отмены
приема глюкокортикоидов концентрация принятых глюкортикоидов снизилась до нулевого уров-
ня терапии. Ингибированная инертная система гипоталамус-гипофиз-кора надпочечников не мо-
жет в первые сутки обеспечить нормальную продукцию эндогенных гормонов. Сниженное со-
держание глюкокортикоидов в организме резко снижает его устойчивость к стрессовым факто-
рам.
Задача 9-2. У данного больного имеется феохромоцитома «адреналинового» типа, о чем свиде-
тельствуют гипергликемия и глюкозурия.
Задача 9-3. Рост у мужчин ниже 135 см называется карликовостью. В данном случае это гипофи-
зарный нанизм. Возможные механизмы при этом следующие: а) снижена продукция соматотроп-
ного гормона аденогипофизом; б) нарушена рецепция СТГ органами и тканями (особенно опорно-
509
двигательным аппаратом и печенью —> снижен синтез в ней соматомединов); в) нарушена рецеп-
ция соматомединов тканями и органами.
Задача 10-1. Для сокращения мышцы в режиме зубчатого тетануса очередной импульс должен
поступать в фазу расслабления мышцы, т.е. в промежутке 50-75 мс. Для сокращения в режиме
гладкого тетануса очередной импульс должен поступать в промежутке между фазой абсолютной
рефрактерности и фазой расслабления мышцы, т.е. в промежутке 5-50 мс.
Задача 10-2. Для улучшения синаптической передачи в этих условиях можно использовать сле-
дующие лекарства:
1) ингибиторы фермента холинэстеразы, разрушающей ацетилхолин;
2) активаторы фермента ацетилхолинтрансферазы, синтезирующего ацетилхолин;
3) активаторы кальциевых каналов пресинаптических окончаний для усиления экзоцитоза медиа-
тора;
4) активаторы синтеза рецепторов постсинаптической мембраны;
5) ингибиторы образования антител к рецепторам пресинаптической мембраны.
Задача 12-1. При длительном голодании уменьшение поступления белков с пищей приводит к
снижению белков в плазме крови (гипопротеинемии). Снижение онкотического давления крови
при этом приводит к перемещению воды из крови в межклеточное пространство наиболее рыхлой
ткани - подкожной жировой клетчатки. Если человек при этом не находится в условиях постель-
ного режима, то отеки локализуются в области ног, так как здесь наибольшее давление крови в
сосудах за счет гидростатического давления столба крови при вертикальном положении.
Задача 12-2. Более низкую величину pH имеет венозная кровь. Это связано с выходом из клеток
тканей и органов в кровоток кислых продуктов обмена, преимущественно СОг (превращающегося
в угольную кислоту) и молочной кислоты.
Задач 13-1. В 1-м анализе имеется нормальное содержание эритроцитов и гемоглобина, но уве-
личенное количество лейкоцитов, что соответствует пищеварительному лейкоцитозу. Во 2-м ана-
лизе увеличены все три показатели, что соответствует физической нагрузке. В 3-м анализе все по-
казатели в диапазоне нормы и соответствуют физиологическому покою.
Задача 13-2. Главный механизм увеличения количества эритроцитов и гемоглобина у постоян-
ных жителей высокогорья - стимуляция через повышенную продукцию эритропоэтина пролифе-
рации эритроидных клеток костного мозга, о чем свидетельствует и увеличение ретикулоцитов
крови. У только что прибывших в высокогорные зоны людей главная причина увеличения коли-
чества эритроцитов и гемоглобина - мобилизация крови из депо (особенно из селезенки).
Задача 13-3. Истощение депо железа в организме первично приведет к нарушению синтеза гемо-
глобина. Поскольку пролиферация эритроидных клеток при этом прямо не нарушена, то количе-
ство эритроцитов будет снижаться в меньшей степени, чем гемоглобина, однако насыщение эрит-
роцитов гемоглобином (цветовой показатель) будет снижено. Концентрация эритропоэтина (ак-
тиватор его синтеза - гипоксия) будет компенсаторно увеличиваться.
Задача 13-4. Лейкоцитарная формула изменится за счет увеличения содержания эозинофилов
(эозинофилия). Физиологический смысл этой реакции заключается в фагоцитозе гранул тучных
клеток эозинофилами с последующем разрушением гистамина ферментом гистаминазой.
Задача 14-1. Время свертывания крови будет резко увеличено, т.к. недостаток фактора УШ при-
ведет к замедлению фазы I коагуляционного гемостаза. Время кровотечения будет нормальным,
т.к. эта методика отражает состояние сосудисто-тромбоцитарного гемостаза, а количество тром-
боцитов и их функция у пациента не изменены.
510
Задача 14-2. Избыток тромбина является раздражителем хеморецепторов каротидного и аорталь-
ного телец. Рефлекторный ответ при этом завершается выбросом из тучных клеток гепарина, уве-
личение в крови антитромбина Ш, активаторов фибринолиза, что характеризует стимуляцию про-
тивосвертывающей системы крови. Этот опыт позволили предположить существование в орга-
низме, кроме свертывающей, противосвертывающей системы крови.
Задача 14-3. Агглютинация не произойдет ни в одной из капель, т.к. в сыворотке крови IV груп-
пы нет агглютининов аир.
Задачи 14-4 и 14-5. Врач нарушил требование переливать только одногрупную кровь и восполь-
зовался старым правилом разведения. Однако переливание большого количества крови на фоне
уменьшенного объема циркулирующей крови привело к реакции а-агглютинина переливаемой
крови с агглютиногеном А эритроцитов пациента. Эритроциты перелитой крови не будут агглю-
тинированы, так как не содержат агглютиногены А и В. Действия врача в этой ситуации являются
иллюстрацией мысли Авиценны: «Невежество ближе к спасению, чем полузнание».
Задача 16-1. Длительный прием приведет к компенсаторному увеличению числа р-
адренорецепторов или (и) чувствительности их к медиатору. Поэтому при отмене препарата эндо-
генные адреналин и норадреналин вызовут увеличение частоты и силы сердечных сокращений.
Задача 19-1. При отравлении СО происходит образование карбоксигемоглобина, который не
может присоединить кислород, в связи с этим содержание кислорода в крови снижается. По-
скольку показатели внешнего дыхания существенно не изменены, то Ро? и количество физически
растворенного Оз в артериальной крови, отражающие уровень легочной вентиляции, тоже не из-
меняться. (В этом трагедия отравления СО: нарастающее снижение поступления кислорода в головной
мозг некоторое время субъективно не воспринимается, т.к. основной раздражитель для сосудистых хеморе-
цепторов - Ро2 крови - остается в норме. Внезапно возникающая потеря сознания не позволяет человеку
покинуть место поражения.)
Задача 19-2. Выдыхаемый воздух представляет собой смесь альвеолярного воздуха и воздуха
мертвого пространства, последний из них по содержанию Ог и СО2 равен атмосферному воздуху,
т.е. в нем значительно больше Оз и меньше СОз, чем в альвеолярном воздухе.
Задача 20-1. При данной травме сохранение моторных центров диафрагмы (сегменты С2.5) и
мышц рук (С5.7) и выключение моторных центров межреберных мышц свидетельствует о пораже-
нии серого вещества и проводящих путей на уровне сегмента С$.
Задача 23-1. Поскольку аноспинальный центр находится в S|.2,T.e. ниже места повреждения, акт
дефекации будет осуществляться при поступлении кала в прямую кишку и повышении в ней дав-
ления. Однако разрыв пирамидных путей приведет к отсутствии произвольного контроля его на-
чала (попечечно-полосатый мышечный сфинктер ануса). Поскольку моторные центры диафрагмы
(С2.5) и мышц брюшного пресса (Th? - L3) находятся выше места повреждения спинного мозга
произвольное сокращение мышц брюшного пресса (натуживание) возможно
Задача 23-2. Повышение трипсина в крови свидетельствует о воспалении поджелудочной желе-
зы. В основе увеличение его активности в крови лежит процесс инкреции пищеварительных фер-
ментов, усиленный повышением мембранной проницаемости клеток при воспалении.
Задача 25-1. Токсическое вещество поражает белки-переносчики для глюкозы в клетках прокси-
мальных извитых канальцев, в которых происходит реабсорбция глюкозы.
Задача 25-2. Фильтрация в нефронах не прекратится, так как при этом будет осуществляться ре-
абсорбция.
Задача 25-3. Уменьшение концентрации Na* в крови вызывает увеличение секреции альдостеро-
на в коре надпочечников. Альдостерон в почках не только увеличивает реабсорбцию Na*, но и
511
увеличивает секрецию К+ в мочу, активируя образование молекул K+,Na+-Hacoca базальной мем-
браны и К*-каналов апикальной мембраны клеток собирательных трубок.
Задача 26-1. Прн погружении человека с перегреванием организма в холодную воду у него про-
изойдет стимуляция холодовых рецепторов кожи, что приведет к рефлекторному возбуждению
центра теплопродукции, и еще большему повышению температуры тела.
Задача 26-2. Охлаждение наступит раньше у первого человека, так как теплоотдача через кожу в
условиях конвекции воды при плавании у него будет выше.
Задача 26-3. Теплая, неплотная, свободного покроя одежда препятствует воздействию воздуха,
температура которого выше температуры кожи и в то же время не мешает испарению пота с ко-
жи, секреция которого стимулирована приемом горячего чая.
Задача 26-4. Блокада симпатических ганглиев резко снижает иесократительный термогенез в от-
вет иа стимуляцию холодовых рецепторов при развивающейся гипотермии, а блокада синапсов
скелетных мышец - резко снижает сократительный термогенез. Смысл этих мероприятий состоит
в сохранении энергетических резервов организма, необходимых для выхода из гипотермии с уче-
том послеоперационного периода.
Задача 27-1. Неполный анальгезнрующий эффект перерезки задних корешков по сравнению с
разрушением спинальных ганглиев свидетельствует о том, что часть аксонов нейронов спиналь-
ных ганглиев, проводящих болевые сигналы, поступает в спинной мозг не через задние корешки,
а через передние, что подтверждено морфологическим исследованиями.
Задача 27-2. Причины две: при ощупывании включаются новые тактильные рецепторы, а также
проприорецепторы; следовательно, в головной мозг поступает бблыиая информация о предмете.
Задача 28-1. Блокада М-холинорецепторов круговой мышцы радужной оболочки выключит па-
расимпатические влияние на нее. Зрачок расширится, и исследование глазного дна будет облегче-
но. Однако блокада при этом М-холииорецепторов миоцитов цилиарной мышцы выключит и
рефлекс аккомодации хрусталика, что нарушит восприятие близко расположенных предметов.
Задача 28-2. Возбуждение симпатического отдела нервной системы при болевом воздействии
приведет к выбросу адренвлина из мозгового слоя надпочечников. Адреналин будет стимулиро-
вать а-адреиорецепторы миоцитов радиальной мышцы радужной оболочки, что приведет е ее со-
кращению. Диаметр зрачка увеличится.
Задача 28-3. Лучи света от близко расположенного к глазу предмета имеют расходящееся на-
правление, что увеличивает размер сфокусированного изображения предмета на сетчатке. Сохра-
нение фокуса изображения на сетчатке при этом сохраняется в результате рефлекса аккомодации
хрусталика (его утолщением с увеличением преломляющей способности).
Задача 28-4. Целостное восприятие зрительных образов осуществляется правым полушарием.
При этом передача возбуждения в корковые зоны, регулирующие произвольные движения доми-
нантной руки, у левши (в правом полушарии) осуществляется быстрее, чем у правши (в левом по-
лушарии)
Задача 29-1. Причиной втяжения барабанной перепонки было снижение давления в барабанной
полости в результате всасывания кислорода из ее воздуха в кровь сосудов слизистой оболочки
полости.
Задача 29-2. В правом ухе воздушная проводимость превышает костную, что соответствует нор-
ме. В левом ухе костная проводимость превышает воздушную проводимость (отрицательный
опыт Риние), что свидетельствует о нарушение воздушной проводимости.
512
Задача 29-3. У первого пациента в бальном ухе нарушена воздушная проводимость, и звук в
нем воспринимается фоторецепторами по кратчайшему пути через костную проводимость. У
второго пациента нарушено звуковосприятие в кортиевом органе, поэтому смещение звуковос-
приятия происходит в сторону здорового уха.
513
Литература для самоподготовки студентов
Основной учебно-методический комплекс
1. Атлас по нормальной физиологии/Подред. Н.А. Агаджаняна. ~ М.: Высшая школа, 1987.-351 с.
2. Начала физиологии: Учебник для ВУЗов /Под ред. акад. А.Д. Ноздрачева. -СПб.: Изд-во «Лань»,
2002. -1088 с.
3. Основы физиологии человека: Учебник/Н.А. Агаджнанян, -М.: РУДН.-468 с.
4. Основы физиологии человека: Учебник: В 3 т. / Под ред. акад. РАМН Б.И. Ткаченко. - СПб., 1994.
- Т. 1. - 567 с.; - СПб., 1994. - Т. 2. - 413 с.; - М.: Литера, 1998. - Т. 3. - 474 с.
5. Физиология человека. Compendium / Под ред. Акад. РАМН Б.И. Ткаченко, проф. В.Ф. Пятима -
Самара: Дом печати, 2003. -496 с.
6. Физиологические основы здоровья человека: Учебник / Под ред. Б.И. Ткаченко, - СПб; Арха-
гельск: Издательский центр СГМУ, 2001. - 728 с.
7. Физиология человека: Учебиик / Под ред. В.М. Покровского, Г.Ф. Коротько. - М.. Медицина,
2003. - 656 с.
8. Физиология человека: Учебник/ Под ред. акад. РАМН Н.А. Агаджаняна, проф. В.И. Циркина. -
СПб.: Сотис, 2003. - 528 с.
9. Физиология человека: Учебник /Под ред. В.М. Смирнова. - М.: Медицина, 2001. -608 с.
10. Физиология челюстно-лицевой области: Учебник / Под ред. С.М. Будылиной, В.П. Дегтярева. - М.:
Медицина, 2000. - 352 с.
11. Дегтярев В.П., Коротич В.А. Фенькина Р.П. Нормальной физиологии: Учеб, пособие. - М.: Ака-
демиздатцентр «Наука» РАН, 2002. - 302 с.
12. Нормальная физиология: Курс физиологии функциональных систем / Под ред. К.В. Судакова. - М.:
МИА, 1999.-718 с.
13. Нормальная физиология: Учеб, пособие / Под ред. В.А. Полянцева. - М.: Медицина, 1989. - 239 с.
14. Нормаяьная физиология: (Учебные модули для самостоятельной работы студентов): Учеб, пособие:
- 3-е изд. / Под ред. В.Н. Яковлева. - Воронеж: Изд-во им. Е.А. Болховитинова, 2005. - 522 с
15. Особенности физиологии детей: Учеб, пособие / Под ред. В.М. Смирнова. - М.: РГМУ, 1993. -
168 с.
16. Физиология. Основы и функциональные системы: Курс лекций / Под ред. К.В. Судакова. - М.: Ме-
дицина, 1999. - 784 с.
17. Яковлев В.Н, Есауленко И.Э. Общая физиология возбудимых тканей: Учебное пособие. 2-е изд. -
Воронеж: ВГМА, 2003. - 73 с.
18. Яковлев В.Н, Есауленко И.Э. Физиология головного мозга: Учеб, пособие: - 2-е изд. - Воронеж:
ВГМА, 2001.-75 с.
Дополнительная литература
1. Агаджанян НА. Познай себя, человек! - М. - Астрахань, 1995. - 339 с.
2. Агаджанян НА., Губин Д.Г. Десинхроноз: механизмы развития от молекулярно-генетического до ор-
ганизменного уровня // Успехи физиол. наук. - 2004. - Т. 35, № 2. - С. 57 - 72.
3. Агаджанян НА., Марачев А.Г., Бобков ГА. Экологическая физиология человека. - М.: «КРУК»,
1998.-416 с.
4. Адрианов О.С. Организованный мозг//Успехи фнзиол. наук. - 1995. -Т. 26, № 1 (Сообщение I). - С.
25 - 45; № 2 (Сообщение II). - С. 18 - 29.
5. Акмаев ИГ Современные представления о взаимодействиях регулирующих систем: иераной, эндок-
ринной и иммунной И Успехи физиол. наук. - 1996. - Т. 27, № 1. - С. 3 -17.
6. Акоев ГН, Алексеев Н.П. Функциональная организация механорецепторов. - Л.: Наука, 1985. - 223 с.
7. Алипов НН Пейсмекерные клетки сердца: Электрическая активность и влияние вегетативных ней-
ромедиаторов // Успехи физнол. наук. - 1993. - Т. 24, № 2. - С. 37 - 64.
8. Алюхин Ю.С. Энергетический «бюджет» сердца: (Литературный обзор) // Успехи физиол. наук. -
2000.-Т.31,№ 1.-С. 47-53.
9. Анисимов В.Н. Эволюция концепций в геронтологии: Достижения и перспективы // Успехи геронто-
логии. - 1999. - Вып. 3. - С. 32 - 53.
514
10. Анохин К.В. Молекулярные сценарии консолидации долговременной памяти И Журнал высшей нерв-
ной деятельности им. И.П. Павлова. - 1997. - Т.47, №2. - С. 261 - 279.
11. Анохин П.К. Биология и нейрофизиология условного рефлекса. - М.: Медицина, 1968. -160 с.
12. Анохин П.К. Очерки по физиологии функциональных систем. - М.: Медицина, 1975. - 327 с.
13. Апанасенко ГЛ., Попова Л.А. Медицинская валеологня. - Ростов н/Д.: Феникс, 2000. -248 с.
14. Арушанян Э.Б. Вклад неостриатума в ритмическую организацию мозговой деятельности и приспосо-
бительное поведение животных // Успехи физиол. наук. - 1992. - Т. 23, № 1. - С. 58 - 73.
15. Арушанян Э.Б. Участие эпифиза в антистрессовой защите мозга И Успехи физиол. наук. - 1996. - Т.
27,№3.-С. 31-45.
16. Ата-Муратова Ф.А. Развивающийся мозг: Системный анализ. - М.: Медицина, 1990. - 296 с.
17. Бабичев В.Н. Нейроэндокринная регуляция секреции гонадотропинов и пролвктинв и роль нейроме-
диаторов в ней И Успехи физиол. наук. - 1995. - Т. 26, № 2. - С. 44 - 57.
18. Бажанова Е.Д. Роль апоптоза в разаитии нервной системы млекопитающих // Успехи совр. биоло-
гии. - 1999. - Т. 119, № 4. - С. 368 - 374.
19. Баевский Р.М. Концепция физиологической нормы и критерии здоровья // Рос. физиолог, жури. им.
И.М. Сеченова. - 2003. - Т. 89, № 4. - С. 473-487.
20. Балашов А.М. Эндогенные аллостерические регуляторы рецепторов И Успехи физиол. наук. - 2004. -
Т. 35,№2.-С. 73-91.
21. Барбараш Н.А. Периодическое действие холода и устойчивость организма // Успехи физиол. наук. -
1996.-Т. 27, №4.-С. 116-131.
22. Бассин Ф.В. Проблема бессознательного. - М.: Наука, 1980. - 486 с.
23. Батуев А.С. Высшая нервная деятельность. - М.: Высшая школа, 1991.-256 с.
24. Батуев А.С., Таиров О.П. Мозг и организация движений. - Л.: Наука, 1978. - 140 с.
25. Беркинблит М.Б., Глаголева Е.Г. Электричество в живых организмах. - М.: Наука, 1988. - 288 с.
26. Бернштейн Н.А. О ловкости и ее развитии. - М.: Физкультура и спорт, 1991. - 288 с.
27. Биология старения: Руководство по физиологии. - Л.: Наука, 1982. - 618 с.
28. Биохимия мозга: Учеб, пособие / Под ред. И.П. Ашмарина, П.В. Стукалова, Н.Д Ещенко. - СПб.: С-
Петерб. ун-т, 1999. - 328 с.
29. Блум Ф., Лейзерсон А., ХофстедтерЛ. Мозг, разум и поаедение. - М.: Мир, 1988.-248 с.
30. Большая медицинская энциклопедия. - М.: Советская энциклопедия. - 3-е изд. - Т. 1 (1974) - Т. 28
(1986). Использовать тематические статьи.
31. Боровский Е.В., Леонтьев В.К. Биология полости рта. - М.: Медицина, 1991. - 304 с.
32. Брагин ЕО. Нейрохимические механизмы регуляции болевой чувствительности. - М.: УДН, 1991. -
248 с.
33. Бредбери М. Концепция гематоэнцефалического барьера. - М.: Медицина, 1983. -480 с.
34. Бреслав И.С. Как управляется дыхание человека. - Л.: Наука, 1985. - 265 с.
35. Быков В.Л. Цитология и общая гистология: (Функциональная морфология клеток и тканей человека).
- СПб.:СОТИС, 1998. - 520 с.
36. Вандер А. Физиология почек. - СПб.: Питер, 2000. - 256 с.
37. Вартанян Н.А. Физиология сенсорных систем: Руководство. Серия «Мир медицины». - СПб.: Лань,
1999.-224 с.
38. Василенко В.Ю. Механизмы температурной чувствительности нейронов гипоталамуса // Успехи фи-
зиол. наук. - 1995. - Т. 26, № 2. - С. 62 - 73.
39. Вашкинелъ В.К., Петров В.М. Ультраструктура и функции тромбоцитов человека. - Л.: Наука, 1982.
-86 с.
40. Введенский Н.Е. Избранные произведения - М.: Изд. АН СССР, 1951. - Ч. П. - С. 409 - 679.
41. ВейнА.М., Авруцкий М.Я. Боль и обезболивание. -М.: Медицина. - 280 с.
42. Воейков В.В. Физико-химические и физиологические аспекты реакции оседания эритроцитов // Успе-
хи физнол. наук. - 1998. - Т. 29, № 4. - С. 55 - 73.
43. Возрастная физиология: Руководство по физиологии. - Л.: Наука, 1975. - 691 с.
515
44. Газенко О.Г., Григорьев А.И., Егоров АД. Физиологические эффекты действия невесомости иа чело-
века в условиях космического полета//Физиология человека. - 1997. -Т. 3,№ 3. - С. 138 -146.
45. Гареев Р.А. Транскапиллярный обмен и лимфообразование. - Алма-Ата: Наука, 1989. -144 с.
46. Геннис Р. Биомембраны: Молекулярная структура н функция: Пер. с англ. - М.: Мир, 1997. - 624 с.
47. Гистология: (Введение в патологию) / Под ред. Э.Г. Улумбекова, Ю.А. Челышева. - М.: ГЭОТАР,
1997.-960 с.
48. Годухин О.В., Щипакина Т.Г. Механизмы синаптической пластичности: Роль фосфорилирования си-
наптических белков и экспрессии генов // Успехи физ иол. наук. -1995. - Т. 26, № 3. - С. 41 - 53.
49. Гальберг ЕД, Дыгай А.М., Провалова Н.В., Скурихин ЕГ, Суслов Н.И. Роль нервной системы в регу-
ляции кроветворения. - Томск: Изд-во Том. ун-та, 2004. -146 с.
50. Гомозков О.А. Молекулярные и физиологические аспекты молекулярной дисфункции. Роль эндоген-
ных химических регуляторов //Успехи физиологических наук. -2000. Т. 31, №4. -С. 48-62.
51. Гормональная регуляция размножения у млекопитающих / Пер. с англ.; Под ред. К. Остина и Р.
Шорта. - М.: Мир, 1987. - 305 с.
52. Григорьев А.И., Ларина И.М. Принципы обмена кальция // Успехи физиол. наук. - 1992. - Т. 23, № 3.
-С. 24 - 52.
53. Гриппи М.А. Патофизиология легких / Пер. с англ. - М.: Вост, книжная компания, 1997. - 344 с.
54. Громыхина НЮ., Крымская Л.Г., Козлов В.А. Роль макрофагов в процессе формирования регулятор-
ных связей между иммунной, нервной и эндокринной системами в ходе иммунного ответа // Успехи
физиол. наук. - 1993. -Т. 24, № 1. - С. 59-73.
55. Групповые системы крови человека и гемотрансфузионные осложнения / Под ред. М.А. Умновой. -
М.: Медицина, 1989. - 160 с.
56. [урин А.В. Функциональная роль оксида азота в центральной нервной системе И Успехи физнол. на-
ук. - 1997. - Т. 28, № 1. - С. 53 - 59.
57. Гурфинкель В.С., Левик Ю.С. Скелетная мышца: Структура и функция. - М.: Наука, 1985. -143 с.
58. Давтян Т.К, Аванесян Л.А. О взаимоотношении иммунного н адаптивного ответов //Успехи совр.
биологии. -2001. - Т. 121, №3. - С. 275 - 286.
59. Данилова НН. Психофизиология. - М.: Аспект Пресс, 1999. - 373 с.
60. Данилова НН, Крылова АЛ. Физиология высшей нервной деятельности: Учебник. - М.: МГУ, 1997.
-432 с.
61. Данилова Р.А., Ашмарин И.П. Инверсная иммунорегуляция поведения и проблема существования ре-
гуляторных аутоантител // Успехи физиол. наук. -1994. - Т. 25, № 1. - С. 3 - 21.
62. ДворецкийД.П., Ткаченко Б.И. Гемодинамика в легких. - М.: Медицина, 1987. - 288 с.
63. ДедовИ.И., Мельниченко ГА., Фадеев В.В. Эндокринология: учебник. -М.: Медицина, 2000.-632 с.
64. Джонсон П. Периферическое кровообращение / Пер. с англ. - М.: Медицина, 1982. - 440 с.
65. Држевецкая НА. Эндокринная система растущего организма. - М.: Высшая школа, 1987. - 207 с.
66. Дыхательная недостаточность: Руководство для врачей / Под ред. А.Н. Зильбера. - М.: Медицина,
1989.-446 с.
67. Емельянов НА., Самойлов НО. Молекулярно-клеточные механизмы долговременной потенциации //
Успехи физнол. наук. - 1996. - Т. 27, № 3. - С. 12 - 27.
68. Заморский И.И., Мыслицкий В.Ф., Пишак В.П. Латеральное ядро перегородки мозга: Морфологиче-
ская н функциональная организация, роль в формировании хроноритмов // Успехи физнол. наук. -
1998. - Т. 29, № 2. - С. 68 - 87.
69. Зефиров АЛ., Черанов С.Ю. Молекулярные механизмы квантовой секреции медиатора в синапсе
//Успехи физиол. наук. -2000. -Т.31, №3. -С. 3-22.
70. Зилов В.Г., Судаков К.В., Этитейн О.И. Элементы информационной биологии и медицины. - М.:
МГУ, 2000.-248 с.
71. Зинчук В.В., Борисюк М.В. Роль кислородсвязывающих свойств крови в поддержании прооксидант-
ио-антноксидантного равновесия организма // Успехи физиол. наук. -1999. - Т. 30, № 3. - С. 38 - 45.
516
72. Зинчук В.В. Деформируемость эритроцитов: физиологические аспекты И Успехи физиол. наук. -2001.
-Т. 121, №3. -С. 275-286.
73. Иванов ЕП. Руководство по гемостазиологии. - Минск: Беларусь, 1991. - 302 с.
74. Иванов КП. Основы энергетики организма: Теоретические и практические аспекты. Том I. Общая
энергетика, теплообмен и терморегуляция. - Л.: Наука, 1990. - 307 с.
75. Иванов КП. Изменения физиологических функций, механизмы их восстановления н температурные
границы жизни при гипотермии // Успехи физиол. наук. -1996. - Т. 27, № 3. - С. 84 -103.
76. Иванов КП. Пределы физиологической адаптации человека к последствиям современных катастроф
// Физиология человека. - 1997. - Т. 23, № 3. - С. 109 - 121.
77. Иванов КП. Создание искусственной крови и уроки физиологии для практической медицины // Ус-
пехи физиол. наук. - 1996. - Т. 27, № 4. - С. 133 -139.
78. Иванов КП. Физиология системы крови и иммунной системы // Успехи физиол. наук. -1994. - Т. 25,
№2.-С. 75-81.
79. Иванов Л.Н. Физиология почки и водно-солевой обмен: Основные направления исследований в Рос-
сии // Успехи физиол. наук. -1995. - Т. 26, № 3. - С. 1 -10.
80. Ивашкин В.Т., Минасян ГА.. Угалев А.М. Теория функциональных блоков и проблемы клинической
медицины. - Л.: Наука, 1990. - 303 с.
81. ИржйкЛ.И. Гемоглобины и их свойства. - М.: Наука, 1975.-240 с.
82. Калюжный Л.В. Физиологические механизмы регуляции болевой чувствительности. - М.: Медици-
на, 1984.-216 с.
83. Кассиль В.Г. Регуляция потребления пищи (онтогенетические аспекты) //Успехи физиол. наук. -1995.
-Т.26, №1.-С. 46-75.
84. Кисляков Ю.А., Бреслав И.С. Дыхание, динамика газов и работоспособность при гипербарии. -Л.:
Наука, 1981.-237 с.
85. Клеточная биология легких в норме и при патологии / Под ред. В.В. Ерохина и Л.К. Романовой. - М.:
Медицина, 2000. - 496 с.
86. Клиническая нейрофизиология: Руководство по физиологии. - Л.: Наука, 1972. - 720 с.
87. Кобрин В.И. Гетерогенность миокарда и аритмии сердца //Успехи физиол. наук. -1993. -Т. 24, №4. -
С. 47-59.
88. Коркушко О.В., Коваленко А.Н. Система свертывания крови при старении. - Киев: Здоров’я, 1988. -
215 с.
89. Корнева ЕА., Шхинок Э.К Гормоны и иммунная система. - Л.: Наука, 1988. - 250 с.
90. Королева С.В., Ашмарин Н.П. Нейропептид Y: Многообразие и кажущаяся противоречивость функ-
ций. Анализ возможных опосредованных эффектов // Успехи физиол. наук. - 2000. - Т. 31, № 1. - С.
31-46.
91. Коротько Г. Ф. Введение в физиологию желудочно-кишечного тракта. - Ташкент: Медицина УзССР,
1987.-221 с.
92. Коротько ГФ. Регуляторная роль ферментов экзо- и эндосекретируемых пищеварительными желе-
зами // Успехи физиол. наук. -1996. - Т. 27, № 4. - С. 96 -111.
93. Костюк П.Г Кальций и клеточная возбудимость. - М.: Наука, 1986. - 254 с.
94. Кратин Ю.Г., Сотничвнко Т.С. Неспецифические системы мозга. - Л.: Наука, 1987. -159 с.
95. Кузник БИ., Васильев Н.В... Цыбиков Н.Н. Иммуногенез, гемостаз и неспецифическая резистентность
организма. - М.: Медицина, 1989. - 320 с.
96. Куприянов В.В, Бородин Ю.И, Караганов Я.Л. Выренков Ю.Е Микролимфология, - М.: Медицина,
1983.-288 с.
97. Куффлер С., Николс Дж. От иейроиа к мозгу. - М.: Мир, 1979. - 439 с.
98. Лебедев Н.Н. Биоритмы пищеварительной системы. - М.: Медицина, 1987. - 256 с.
99. Левтов В.А., Регирер С.А., Щадрин HJC. Реология крови. - М.: Медицина, 1982. - 272 с.
100. ЛейкокДж.Ф., Вайс П.Г. Основы эндокринологии / Пер. с англ. - М.: Медицина, 2000. - 504 с.
517
101. Леонов А.Н. Элементы научной теорнн гипербарической медицины. // Журнал теоретической и прак-
тической медицины. Т.1.-К» 1.-2003. С. 7 - 15.
102. Лиманский Ю.П. Рефлексы головного мозга. - Киев: Наукова думка, 1987. - 240 с.
103. Лиманский ЮЛ. Физиология боли. - Киев: Здоров’я, 1986. - 96 с.
104. Макаров А.Ю. Концепция интегративной функции ликвора в деятельности центральной нервной сис-
темы // Успехи физиол. наук. - 1992. - Т. 23, № 4. - С. 40 - 47.
105. Мак-Фарленд Д. Поведение животных: Психобиология, этология и эволюция / Пер. с англ. - М.:
Мир, 1988.-520 с.
106. Марков ХМ. Окснд азота и сердечно-сосудистая система //Успехи физиол. наук. -2000. -Т.32, №3. -
С. 49 - 65.
107. Марри Р., ГренерД, Менее П., Родуэлл В. Биохимия человека. - М.: Мир, 1993. - Т. 2. - 415 с.
108. Мартинчик А.Н., Маев И.В., Петухов А.Б. Питание человека (основы нутрициологии). /Под ред. А.Н.
Мартинчнка. - М.: ГОУ ВУНМЦ МР РФ, 2002. - 576 с.
109. Медведев В.И. Адаптация человека. - СПб.: Ин-т мозга человека РАН, 2003. - 584 с.
110. Меерсон Ф. 3. Адаптационная медицина: концепция долговременной адаптации. - М.: «Дело», 1993.
-138 с.
111. Механизмы деятельности мозга человека. Ч. 1: Нейрофизиология человека: Основы современной фи-
зиологии / Ред. Н.П. Бехтерева. - Л.: Нвука, 1988. - 677 с.
112. Механизмы памяти: Руководство по физиологии / И.П. Ашмарни, Ю.С. Бородкин, П.В. Бундзен и др.
-Л.: Наука, 1987.-432 с.
113. Морман Д, Хеллер Л. Физиология сердечно-сосудистой системы. - СПб.: Питер, 2000. - 256 с.
114. Наатанен Р. Внимание и функции мозга: Учеб, пособие / Пер. с англ.; Под ред. Е.Н. Соколова. - М.:
МГУ, 1998.-560 с.
115. Наследов ГА. Тоническая мышечная система позвоночных. - Л.: Наука, 1981. - 187 с.
116. Натан Д.Г., Зифф К.А. Регуляция кроветворения // Гематология и трансфузиология. - 1994. - № 2. -
С. 3 -10.
117. Наточин Ю.В. Проблемы эволюционной физиологии водно-солевого обмена. - Л..* Наука, 1984. - 38
с.
118. Нейрофизиология дыхания / Под ред. В.А. Сафонова, А.А. Чумаченко. - М.: Медицина, 1984. - 224 с.
119. Нефрология: Руководство / Под ред. И.Е. Тареевой. - М.: Медицина, 1995. - Т. 1. - 495 с.
120. Никитин В.П., Судаков К.В. Механизмы интегративной деятельности нейронов // Успехи фнзнол.
наук. - 1997. - Т. 28, № 1. - С. 27 - 43.
121. Николаева Е.Н. Психофизиология. Психологическая физиология с основами физиологической пси-
хологии. Учебник, - М.: ПЕР СЭ; Логос, 2003. - 544 с.
122. Ноздрачев АД. Химическая структура периферического автономного (висцерального) рефлекса //
Успехи физиол. наук. - 1996. - Т. 27, № 2. - С. 28 - 59.
123. Ноздрачев АД, Вуколова Р.П. Симпатический ганглий - периферический нейроэндокринный центр
//Успехи физиол. наук. - 1993. -Т.24,№ 1.-С. 80-95.
124. Ноздрачев АД, Чернышова М.П. Висцеральные рефлексы. - Л.: Наука, 1989. - 326 с.
125. Общая анатомия лимфатической системы / Бородин Ю.И., Сапин И.Р., Этннген Л.Е. и др. - Новоси-
бирск: Наука, 1990. - 243 с.
126. Общая физиология нервной системы: Руководство по физиологии. - Л.: Наука, 1979. - 717 с.
127. Общая и частная физиология нервной системы: Руководство по физиологии. - Л.: Наука, 1969. - 555
с.
128. Общая физиология возбудимых мембран: Руководство по физиологии / Б.И. Ходоров. - М.: Наука,
1975.-406 с.
129. Общая физиология пищеварения: Руководство по физиологии. - Л.: Наука, 1974. - 762 с.
130. Орбели Л.А. Избранные труды. Т.1. Вопросы эволюционной физиологии. - М-Л.: Изд-во АН СССР,
1961.-457 с.
131. Орлов И.В. Вестибулярная функция. - СПб.: Наука, 1998.-248 с.
518
132. Основы пснхофнзиологнн: Учебник/Отв. ред. Ю.И. Александров. - М.: ИНФРА-М, 1997.-432 с.
133. Отеллин В.А., Арушанян Э.В. Нигростриоиигральная система. - М.: Медицина, 1989. - 272 с.
134. Охотин В.Е., Калиниченко С.Г. Морфофункцнональная характеристика нейронов и их связей в новой
коре человека и животных // Успехи физиол. наук. - 1997. - Т. 28, № 2. - С. 40 - 52.
135. Павлов АД., Моршакова Е.Ф. Регуляция эритропоэза: Физиологические и клинические аспекты. -
М.: Медицина, 1987.-272 с.
136. Павлов И.П Двадцатилетний опыт объективного изучения ВИД (поведения) животных // Поли. собр.
соч,- М.; Л.: АН СССР, 1949. - Т. 3. - 605 с.
137. Павлов И.П. Лекции о работе главных пищеварительных желез. - М.; Медицина, 1951.-592 с.
138. Пальцев М.А., Иванов А.А. Межклеточные взаимодействия. - М.: Медицина, 1995. - 224 с.
139. ПетровР.В. Иммунология: Учебник для мед. вузов.-М.: Медицина, 1987.-414с.
140. Полунин ИА. Ритмогеиез сердца. - Астрахань, 1997. - 285 с.
141. Почки и гомеостаз в норме и патологин / Под ред. С. Клара. - М.: Медицина, 1987.-448 с.
142. Прокоп О., Геллер В. Группы крови человека / Пер. с нем. - М.: Медицина, 1991. -512 с.
143. Прошева В.И. Функциональная специфичность пейсмейкерной системы сердца //Успехи физиол. на-
ук. -1998. -Т.29, №3. -С.79-91.
144. Пытель Ю.А., Борисов В.В., Симонов В.А. Физиология человека: Мочевые пути. - М.: Высшая шко-
ла, 1992.-286 с.
145. Раевский КС.. Сотникова Т.Д, Гайнетдинов Р.Р. Дофаминергические системы мозга: Рецепторная
гетерогенность, функциональная роль, фармакологическая регуляция И Успехи физиол. наук. - 1996.
-Т. 27, № 4. - С. 3-26.
146. Регуляция висцеральных функций: Закономерности и механизмы. - Л.: Наука, 1987. - 255 с.
147. Репродуктивная эндокринология: В 2 т. / Пер. с англ.; Под ред. С. Йена, Р. Джаффе. - М.: Медицина,
1998.
148. Рецепторы клеточных мембран для лекарств и гормоиоа: Междисциплинарный подход/ Пер. с англ.;
Под ред. Р. Штрауба. - М.: Медицина, 1986. - 365 с.
149. РивинД.Л. Физиология и патология органа слуха. -М.: Радио и связь, 2000. -112 с.
150. Розен В.Б Основы эндокринологии: Учебник. - М.: МГУ, 1994. - 384 с.
151. Ройт А. Основы иммунологии / Пер. с англ. - М.: Мир, 1991. - 328 с.
152. Руководство по геронтологии / Под ред. Д.Ф. Чеботарева, Н.Б. Мнньковского, В.В. Фролькиса. - М.:
Медицина, 1978. - 503 с.
153. Руководство по кардиологии: Структура и функция сердечно-сосудистой системы в норме и патоло-
гии / Под ред Е.И Чазова. - М.: Медицина, 1982. - Т. 1. - 667 с.
154. Руководство по физиологии дыхания: Руководство по физиологии. - Л.: Наука, 1973.-251 с.
155. Рут Г. Кислотно-основное состояние и электролитный баланс. - М.: Медицина. 1978. - 118 с.
156. Самойлов В.О. «Рефлекс свободы» Ивана Петровича Паалова И Успехи физнол. наук. - 1999. - Т. 30,
№3.-С. 93-102.
157. СапинМ.Р., Этинген Л. Е. Иммунная система человека.-М.: Медицина, 1996.-300 с.
158. Селье Г. На уровне целого организма. - М.: Наука, 1972. - 122 с.
159. Семененя И.Н. Функциональное значение щитовидной железы И Успехи физиол. наук. - 2004. - Т.
35, №2.-С. 41-56.
160. Сенсорные системы: Обоняние н вкус / Под ред. Г.В. Гершуни. - Л.: Наука, 1980. - 183 с.
161. Сеченов И.М. Рефлексы головного мозга. - М.: АН СССР, 1961. - 99 с.
162. Сеченов И.М. К 150-летию со дня рождения / АН СССР, История естествознания и техники. - М.:
Наука, 1980.-607 с.
163. Симонов П.В. Мотивированный мозг. - М.: Наука, 1987. - 348 с.
164. Симонов П.В. Эмоциональный мозг. - М.: Наука, 1981. - 324 с.
165. Скок В.М., Шуба М. Ф. Нервно-мышечная физиология. - Киев: Вища школа, 1986. - 224 с.
519
166. Скребицкий В.Г, Чепкова А.Н. Синаптическая пластичность в аспекте обучения и памяти // Успехи
физиол. наук. -1999. - Т. 30, № 4. - С. 3 -13.
167. Слуховая система: (Основы современной физиологии) / Под ред. Я.А. Альтмана. - Л.: Наука, 1990. -
620 с.
168. Смирнов В.М. Нейрофизиология и высшая нервная деятельность детей и подростков: Учеб, пособие.
- М.: Академия, 2000. - 400 с.
169. Смирнов В.М., Яковлев В.Н. Физиология центральной нервной системы. - М.: Академия, 2002. - 352
с.
170. СмолярВ.М. Рациональное питание.-Киев: Наукова думка, 1991.-368с.
171. Спрингер С., Дейч Г. Левый мозг, правый мозг/Пер. с англ.-М.: Мир, 1983.-256 с.
172. Современная нефрология / Под ред. С. Клара, С.Г. Массари. - М.: Медицина, 1986. - 432 с.
173. Старение мозга / Под ред. В.В. Фролъкиса. - Л.: Наука, 1991. - 227 с.
174. Стародуб Н.Ф., Назаренко В.И. Гетерогенная система гемоглобина: Структура свойства, синтез,
биологическая роль. - Киев: Наукова думка, 1987. - 200 с.
175. Судаков КВ. Голографический принцип системной организации процессов жизнедеятельности // Ус-
пехи физиол. наук. - 1997. - Т. 28, № 4. - С. 3-31.
176. Судаков КВ. Информационный принцип в физиологии: Анализ с позиций общей теории функцио-
нальных систем // Успехи физиол. наук. -1995. - Т. 26, № 4. - С. 3 - 26.
177. Судаков КВ. Системная организация функций человека: Теоретические аспекты // Успехи физиол.
наук. - 2000. - Т. 31, № 1. - С. 81 - 96.
178. Судаков КВ. Физиологические активные вещества в организме // Успехи физиол. наук. - 1988. - Т.
19, №2.-С.45 - 60.
179. Тель Л.З. Валеология: Учение о здоровье, болезни и выздоровлении. В 3 т. - М.: «Изд-во АСТ»; «Ас-
трель», 2001. ТЛ - 432 с., Т.2 - 480 с., Т.З - 416 с.
180. ТепперменД, Теппермен X. Физиология обмена веществ и эндокринная система. - М.: Мир, 1989. -
656 с.
181. Титов В.Н. Транспорт в крови жирных кислот липопротеинами как макромолекулами: Факты и ги-
потезы // Успехи физиол. наук. - 1999. - Т. 30, № 3. - С. 23 - 37.
182. Ткаченко Б.И., Куприянов В.В., Орлов Р.С, Гуревич М. И. и др. Физиология кровеносных сосудов:
(Некоторые итоги и перспективы исследований) // Успехи физиол. наук. - 1989. - Т. 20, № 4. - С. 3 -
26.
183. Ткачук В.А., Авакян А.Э. Молекулярные механизмы сопряжения G-белков с мембранными рецепто-
рами и системами вторичных посредников. - Рос. физиолог, жури. нм. И.М. Сеченова. - 2003. - Т.
89, №12.-С. 1478-1490.
184. Тотолян А.А., Фрейдлин И.С. Клетки иммунной системы. - СПб.: Наука, 2000. - 231 с.
185. УголевА.М. Эволюция пищеварения и принципы эволюции функций: Элементы современного функ-
ционализма.-Л.: Наука, 1985.-543 с.
186. Умрюхин /7.Е. Ранние гены в церебральных механизмах эмоционального стресса // Успехи физиол.
наук. - 2000. - Т. 31, № 1. - С. 54 - 70.
187. Уразаев AJC., Зефиров АЛ. Физиологическая роль оксида азота // Успехи физиол. наук. - 1999. - Т.
30,№1.-С. 54 - 72.
188. Ухтомский А.А. Избранные труды /Под ред. Е.М. Крепса.- Л.: Наука, 1978.-358 с.
189. Фабер ДА., Безруких М.М. Морфологические аспекты изучения физиологии развития ребенка
//Физиология человека. -2001. -Т. 27, №5. -С. 8-16.
190. Фаллер ДИ., Шилдс Д Молекулярная биология клетки. Руководство для врачей. - М.: БИНОМ -
Пресс. 2003.-273 с.
191. Фанарджян В.В. Тормозные механизмы мозжечка: Функциональные особенности И Успехи физиол.
наук. - 1992. - Т. 23, № 4. - С. 3 - 22.
192. Федин А.Н., Ноздрачев АД., Бреслав И.С. Физиология респираторной системы: Учеб, пособие. -
СПб.: С-Петерб. уи-т, 1997. -188 с.
520
193. Федоров В.И. Холинергическое влияние на гемодинамику и экскреторные функции почки // Успехи
физнол. наук. - 1998. - Т. 29, № 4. - С. 42 - 50.
194. Федосеев Г.Б. Механизмы обструкции бронхоа. - СПб.: Мед. информ, агентство, 1995. - 336 с.
195. Фермилен Ж., Ферстате М. Гемостаз. - М.: Медицина, 1984. - 192 с.
196. Физиологические основы здоровья человека/ Под ред. Б.И. Ткаченко. - СПб; Архангельск: Издат.
центр СГМУ. 2001. - 728 с.
197. Физиология адаптационных процессов: Руководство по физиологии. - М.: Наука, 1986. - 635 с.
198. Физиология вегетативной нервной системы: Руководство по физиологии.-Л.: Наука, 1981. -752 с.
199. Физиология всасывания: Руководство по физиологии. - Л.: Наука, 1977. - 668 с.
200. Физиология высшей нервной деятельности. Ч. 1: Руководство по физиологии. - М.: Наука, 1970. -
612 с.
201. Физиология высшей нервной деятельности. Ч. 2: Руководство по физиологии. - М.: Наука, 1971. -
392 с.
202. Физиология гистогематических барьеров: Руководство по физиологии. - М.: Наука, 1977. - 575 с.
203. Физиология движений: Руководство по физиологии. - Л.: Наука, 1976. - 376 с.
204. Физиология дыхания: Руководство по физиологии. - Л.: Наука, 1973. - 352 с.
205. Физиология и патофизиология желудочно-кишечного тракта / Пер. с англ.; Под ред. Дж.М. Полак и
др. - М.: Медицина, 1989. - 496 с.
206. Физиология и патофизиология легочных сосудоа / Пер. с англ.; Под ред. К.Е. Уэйра, Дж.Т. Ривса. -
М.: Медицина, 1995. - 672 с.
207. Физиология и патофизиология сердца / Под ред. Н. Сперелакиса. - М.: Медицина, 1990. - Т. 1. - 532
с.; Т. 2. - 624 с?
208. Физиология кровообращения. Физиология сердца: Руководство по физиологии. - Л.: Наука, 1980. -
598 с.
209. Физиология кровообращения. Физиология сосудистой системы: Руководство по физиологии. - Л.:
Наука, 1984. - 652 с.
210. Физиология кровообращения. Регуляция кровообращения: Руководство по физиологии. - Л.: Наука,
1986.-640 с.
211. Физиология питания / Л.Ф. Павлоцкая и др. - М.: Высшая школа, 1989. - 368 с.
212. Физиология пищеварения: Руководство по физиологии. - Л.: Наука, 1974. - 762 с.
213. Физиология поведения. Нейрофизиологические закономерности: Руководство по физиологии / Под
ред. А.С. Батуева. - Л.: Наука, 1986. - 767 с.
214. Физиология поведения: Нейробиологические закономерности: Руководство по физиологии / Под ред.
А.С. Батуева. - Л.: Наука, 1987. - 736 с.
215. Физиология почки: Руководство по физиологии. - Л.: Наука, 1972. - 398 с.
216. Физиология сенсорных систем. 4.1: Физиология зрения: Руководство по физиологии. - Л.: Наука,
1971.-416с.
217. Физиология сенсорных систем. Ч. 2: Руководство по физиологии. - Л.: Наука, 1972.-703 с.
218. Физиология сенсорных систем. Уч. пособие двя вузов / Под ред. Я.А. Альтмана. - СПб.: Паритет,
2003. - 352 с.
219. Физиология сердца: Учеб, пособие / Под ред. акад. РАМН Б.И. Ткаченко. - СПб.: Специальная лите-
ратура, 1998. -128 с.
220. Физиология системы крови. Физиология эритропоэза: Руководство по физиологии. - Л.: Наука, 1979.
-360 с.
221. Физиология терморегуляции: Руководство по физиологии. - Л.: Наука, 1984. - 470 с.
222. Физиология человека: В 3 т. / Пер. с аигл.; Под ред. Р. Шмидта и Г. Тевса. - М.: Мир, 1996. - Т. 1. -
323 с.; Т. 2. - 313 с.; Т. 3 - 198 с.
223. Физиология эндокринной системы: Руководство по физиологии. - Л.: Наука, 1979. - 680 с.
224. Филаретов А.А. Функциональное значение многозвенного построения гипоталамогипофизарных
нейроэндокринных систем // Успехи физиол. наук. -1996. - Т. 27, № 3. - С. 3 -11.
521
225. Фундаментальная н клиническая физиология: Учебник для студентов вузов / Под ред. А.Г. Камкина
и А.А. Каменского.- М.: Академия, 2004. -1047 с.
226. Функциональные системы организма: Руководство / Под ред. К.В. Судакова. - М.: Медицина, 1987. -
432 с.
227. Хазипов Р.Н., Зефиров А.Л., Бен-Ари Е. ГАМК - основной медиатор возбуждения на ранних этапах
развития гиппокампа // Успехи физиол. наук. -1998. - Т. 29, № 2. - С. 55 - 67.
228. Хендерсон Дж.М. Патофизиология органов пищеварения / Пер. с англ. - М.; СПб.: Бином - Невский
Диалект, 1997. - 287 с.
229. Хеффнер Л. Половая система в норме н патологии: Учеб, пособие. - М.: ГЭОТАР-МЕД, 2003. - 128
с.
230. Хочачка Д, СомероДж. Биохимическая адаптация. - М.: Мир, 1988. - 568 с.
231. ХьюбелД Глаз. Мозг. Зрение/Пер. с англ. -М.: Мир, 1990.-239 с.
232. Частная физиология нервной системы: Руководство по физиологии. - Л.: Наука, 1979. - 241 с.
233. Частная физиология нервной системы: Руководство по физиологин. - Л.: Наука, 1983. - 734 с.
234. Чернух А.М., Александров П.Н., Алексеев О.В. Микроциркуляция. - М.: Медицина, 1975. - 456 с.
235. Шейман Дж.А. Патофизиология почкн / Пер. с англ. - М.: Восточная книжная компания, 1997. - 224
с.
236. Шеперд Г. Нейробиология: В 2 т. - М.: Мнр, 1987. - Т. 1. - 454 с.; Т. 2 - 368 с.
237. Шик ЛЛ. Основные черты управления дыханием // Успехи физиол. наук. - 1998. - Т. 29, № 2. - С. 3
-11.
238. Шиффман Ф. Дж. Патофизиология крови / Пер. с англ. - М.; СПб.: Бнном - Невский Диалект, 2000.
-448 с.
239. Шиффман Х.Р. Ощущение и восприятие. - СПб.: Питер, 2003. - 928 с.
240. Шишкина ГТ, Дыгало Н.Н. Молекулярная физиология адренергических рецепторов // Успехи физи-
ол. наук. - 1997. - Т. 28, № 1. - С. 59 - 73.
241. Шуваев В.Т. Амигдало-каудатная система н поведение // Успехи физиол. наук. - 1993. - Т. 24, № 2. -
С. 84-108.
242. Шулъговский В.В. Физиология центральной нервной системы: Учебник. - М.: МГУ, 1997. - 397 с.
243. Эверли Дж., Розенфельд Р. Стресс, природа и лечение. - М.: Медицина, 1985. - 224 с.
244. Эволюционная физиология: Руководство по физиологии. Ч. 1. - Л.: Наука, 1979. - 603 с.; Ч. 2. - Л.:
Наука, 1983. - 507 с.
245. Эккерт Р., РэнделлД., Огастин Дж. Физиология животных: Механизмы и адаптация: В 2 т. - М.:
Мир, 1991. - Т. 1. - 424 с.; Т. 2. - 344 с.
246. Экологическая физиология человека. Адаптация человека к климато-географическим условиям: Ру-
ководство по физиологии. - Л.: Наука, 1980. - 549 с.
247. Экологическая физиология человека. Адаптация человека к экстремальным условиям среды: Руково-
дство по физиологии. - Л.: Наука, 1979. - 704 с.
248. Эндокринология / Пер с англ.; Под ред. Н. Лавина - М.: Практика, 1999. -1128 с.
249. Эрман М.В. Нефрология детского возраста в схемах н таблицах: Справ, руководство. - СПб.: Спец,
лит., 1997.-413 с.
250. Ярилин А.А. Основы иммунологии: Учебник. - М.: Медицина, 1999. - 606 с.
251. Bullock J., Boyle J., Wang M. Physiology: 3rd edition / By Williams & Wilkins. Baltimore, 1994. - 641 p.
252. Costanzo L S. Physiology: 2nd edition / By Williams & Wilkins. Baltimore, 1998. - 326 p.
253. Guyton A.C., Hall J.T. Human Physiology and Mechanisms of Disease: 6th td. Saunders Company. Phy-
ladelphia, 1997.-737 p.
254. Silbemagl S., Despopoulos A. Taschenatlas der Physiologic. G. Thieme Verlag, Stuttgart; New York, 1991.
- 371 s.
522