/

































































































































































































Автор: Штакельберг А.А. Гуцевич А.В. Мончадский А.С.
Теги: insecta hexapoda насекомые энтомология фауна ссср комары
Год: 1970




Текст
ФАуНА
СССР
ДВУКРЫЛЫЕ
III
выпуск
4»
АК АДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
НОВАЯ СЕРИЯ № 100
ФАУНА СССР
НАСЕКОМЫЕ ДВУКРЫЛЫЕ
Том III, вып. 4
А. В. ГУЦЕВ ИЧ, А. С. МОНЧАДСКИЙ, А. А. ШТАКЕЛЬБЕРГ
КОМАРЫ
СЕМЕЙСТВО CULICIDAE
е
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
Ленинград 1970
УДК Ш.171
Компвы (Семейство Culicidae). Гуцевич А. В., М о нч ад-
ский Л. С., Штакельберг А. А. В серии: Фауна СССР,
Насекомые двукрылые, том III, вып. 4. 1970. Изд-во «Наука»,
Лопипгр. отд., Л. 1—384.
Монографическое описание важнейших кровососов фауны СССР,
приносящих вред, кроме непосредственного всасывания крови,
передачей тяжелых заболеваний человека и животных, из кото-
рых на первом месте стоит малярия. Книга содержит введение
с необходимыми для целей определения сведениями но морфоло-
гии комаров, очерком их биологии, значения и географического
распространения. Специальная часть содержит определительные
таблицы и описания как взрослых комаров, так и их личинок.
Илл.—262, табл. — 1, библ. — 210 назв.
Главный редактор
директор Зоологического института АН СССР
академик В. Е. Выховский
Р е д-а кционная коллегия:
И. М. Громов, А. С. Мончадский, О. А. Скарлато, А. А. Стрелков
(редактор тома) и А. А. Штакельберг
Александр Васильевич Гуцевич,
Александр Самойлович Мончадский,
Александр Александрович Штакельберг
КОМАРЫ. СЕМЕЙСТВО CULICIDAE
Фауна СССР. Насекомые двукрылые, т. III, вып. 4.
Новая серия № 100i
г Утверждено к печати^
Зоологическим институтом Академии наук СССР
Редактор издательства Л. В. Шоренкова
Технический редактор Е. Н. В о лк о в а
Корректоры Л. М. Бова, Н. В. Лиха р е в а и Т. Г. Эвелъмна
Сдано в набор'29/VIII 1969 г. Подписано к печати 15/IX 1970 г.
РИСо АН СССР № 28-96В .Формат бумаги 70xl08i/i6. Бум. л. 121/,.
Печ. л. 24 + 1 вкл. печ. л.). = 33.95 усл. печ. Уч.-изд. л. 31.74. Изд.
№ 3870» Тип. зак. № 455. М-10360. Тираж 1400. Бумага № 2. Цена 3 р. 37 к.
Ленинградское отделение издательства «Наука»
Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. издательства «Наука». Ленинград, В-34, 9 линия, д. 12
2-10-6; 4-9-3
430-69 (II)
ПРЕДИСЛОВИЕ
Книги, посвященные комарам, были изданы в серии «Фауна СССР» —
А. А. Штакельберг, 1937, Семейство Culicidae (Кровососущие комары
Палеарктики), и в серии «Определители по фауне СССР» — А. С. Мон-
чадский, 1936 и 1951, Личинки кровососущих комаров СССР и сопредель-
ных стран. Для настоящего издания был использован материал этих
книг, а также обширные оригинальные и литературные материалы, по-
явившиеся за годы, прошедшие после выхода упомянутых изданий.
В основу работы положено изучение обширных коллекционных ма-
териалов Зоологического института Академии наук СССР (ЗИН). Эти
материалы накапливались в течение десятилетий при широком участии
очень многих лиц. Не имея возможности назвать всех, кто предоставил
свои материалы для пополнения коллекций института, мы считаем долгом
выразить им чувства глубокой признательности.
Чтобы избежать значительного увеличения объема книги и не за-
труднять пользование определительными таблицами, в работу включено
85 видов, найденных в Советском Союзе; к ним добавлено девять пале-
арктических видов Aedes, представляющих интерес как переносчики воз-
будителей болезней, или таких, обнаружение которых на территории
СССР более или менее вероятно.
Авторы старались не перегружать текст материалами, представляю-
щими узко специальный интерес, и не стремились дать подробные описа-
ния видов; в их характеристики включены признаки, имеющие главным
образом диагностическое значение. В отношении синонимики авторы
ограничились указанием лишь немногих наиболее распространенных
синонимов, в отношении литературы — необходимым минимумом ссы-
лок преимущественно на капитальные и относительно более новые ра-
боты. В частности, мы не имели возможности назвать авторов тех весьма
многочисленных работ, на основании которых дана характеристика
географического распространения каждого из видов комаров.
Что касается рисунков, то значительная, их часть взята из упомяну-
тых выше книг (Штакельберг, 1937; Мончадский, 1951), часть изготовлена
заново, часть заимствована из различных литературных источников, что,
конечно, во всех случаях оговорено.
Пользуемся случаем выразить глубокую благодарность Е. А. Афана-
сьевой, Т. А. Ляхтинен и К. Г. Федоровой за большую многолетнюю
работу по изготовлению препаратов, постановке и хранению коллекции
комаров Зоологического института.
Январь 1968 года
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
Сем. Culicidae — Кровососущие комары
I. Подсем. Anophelinae
1. Род A nopheles Mg. - Малярийные комары
1. Подрод Anopheles Mg.
Стр.
1. An. (An.) algeriensis Theob........................................ 80
2. An, (An.) plumbeus Steph........................................... 82
3. An. (An.) claviger Mg.............................................. 86
4. An. (An.) marten Sen.-Prun......................................... 88
An. m. marteri Sen.-Prun....................................... 90
An. m. sogdianus Kesh. . ...................................... 90
5. An. (An.) maculipennis Mg.......................................... 90
An. m. sacharovi Favre........................................ 99
6. An. (An.) lindesayi Giles ...................................... 100
An. I. lindesayi Giles ...................................... 103
An. I. japonicus Yam.......................................... 103
7. An. (An.) hyrcanus Pall.......................................... 103
2. Подрод Myzomyia Blanch.
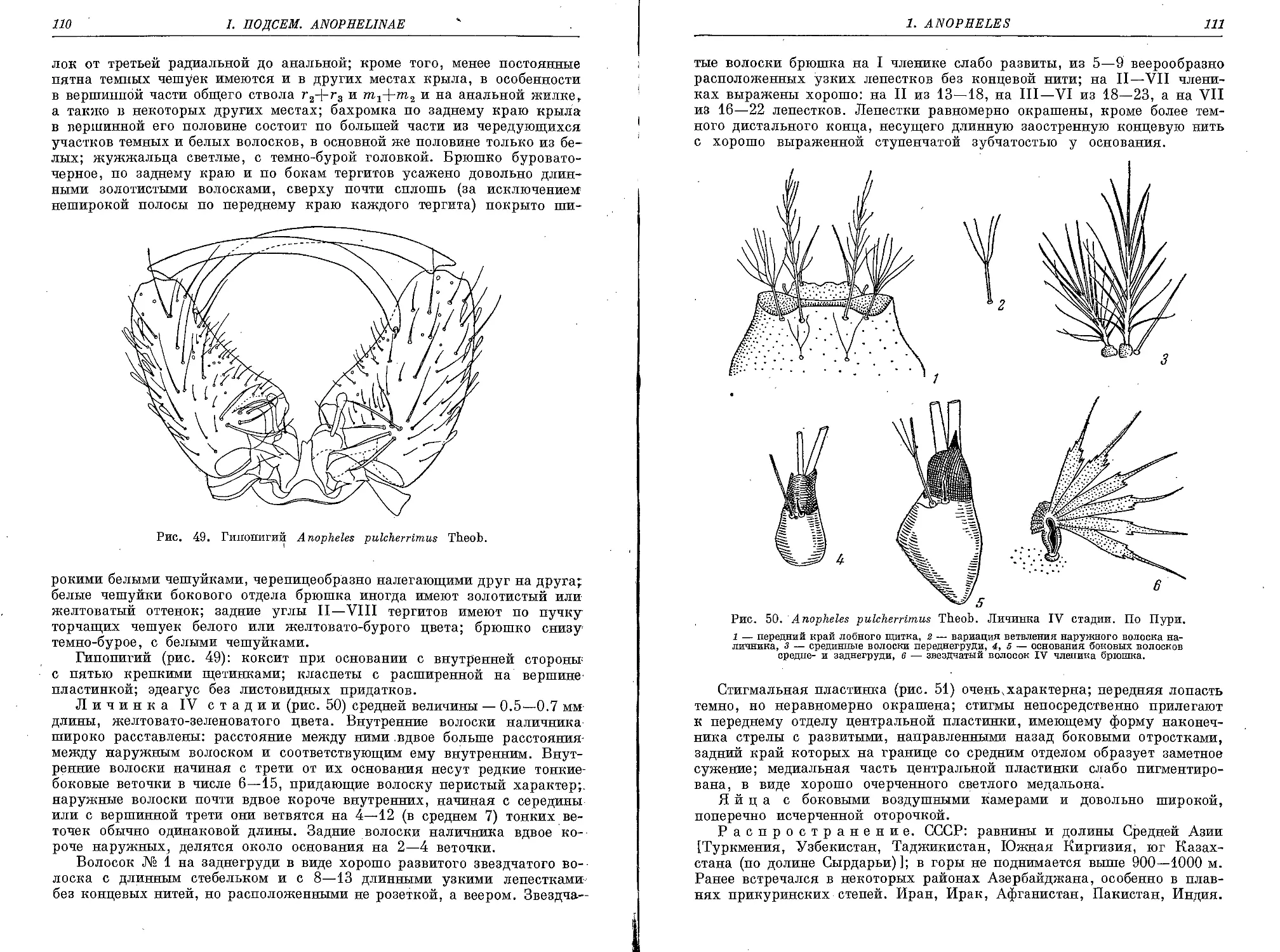
8. An. (M.) pulcherrimus Theob....................................... 108
9. An. (M.) superpictus Grassi ..................................... 113
II. Подсем. Toxorhynchit.inae
2. Род Toxorhynchites Theob.
1. T. christophi Portsch............................................. 118
111. Подсем. Culicinae
3. Род V ranotaenta Arrib.
1. U. unguiculata Edw................................................ 126
4. Род Orthopod от у ia Theob.
1. O. pulchripalpis Rond. ........................................ . 130
5. Род С и I i s e t a Felt
1. Подрод Allotheobaldia Brol.
1. C. (All.) longiareolata Macq...................................... 138
2. Подрод Culiseta Felt
2. С. (C.) glaphyroptera Sehin...................................... 142
3. С. (C.) alaskaensis Ludl.......................................... 144
C. a. alaskaensis Ludl......................................... 147
C. a. indica Edw............................................... 147
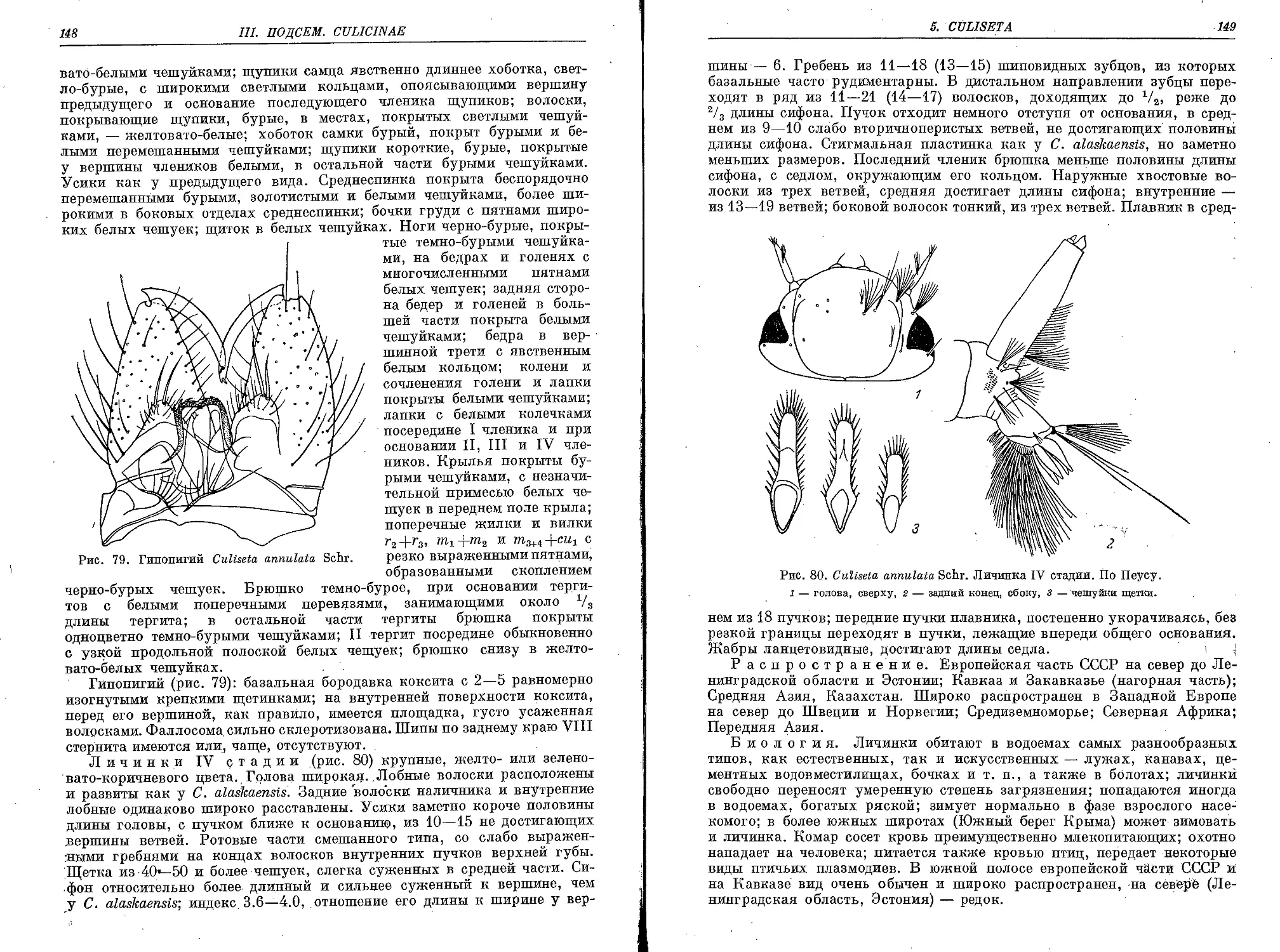
4. С. (C.) annulata Schr............................................ 147
C. a. annulata Schr........................................... 150
C. a. subochrea Edw........................................... 150
5. С. (C.) bergrothi Edw............................................. 151
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
5
3. Подрод Culicella Felt
Стр.
6. С. (С.) morsitans Theob.................................................... . 153
7. С. (С.) ochroptera Peus ....................................................... 156
8. С. (С.) fumipennis Steph...................................................... 159
9. С. (С.) setivalva Masi.......................................................... 160
6. Род Mansonia Blanch.
1. М. richiardii Fic............................................................... 164
2. M. buxtoni Edw.................................................................. 167
7. Род A e d e s Mg.
1. Подрод Ochlerotatus Arrib.
1. A. (0.) caspius Pall.......................................................... 191
A. c. caspius Pall.......................................................... 192
A. c. dorsalis Mg........................................................... 195
2. A. (0.) mariae Serg.......................................................... 196
3. A. (0.) pulchritarsis Rond.................................................... 199
A. p. pulchritarsis Rond................................................... 200
A. p. asiaticus Edw......................................................... 201
4. A. (0.) cantans Mg.............................................................. 202
5. A. (O.) riparius D. К.............................. . . ........................ 205
A. r. riparius D. К........................................................ 208
A. r. ater Guts............................................................. 208
6. A. (O.) behningi Mart........................................................ 208
7. A. (O.) excrucians Walk..................................................... 211
8. A. (O.) beklemishevi Den..................................................... 214
9. A. (O.) annulipes Mg........................................................ 216
10. A. (O.) flavescens Mii.ll................................................... 218
11. A. (O.) cyprius Ludl. .................................................... 221
12. A. (O.) rusticus Rossi .................................................... 223
13. A. (O.) reiiki Med........................................................ 225
14. A. (O.) lepidonotus Edw..................................................... 227
15. A. (O.) subdiversus Mart.................................................... 230
16. A. (O.) communis Deg....................................................... 232
17. A. (O.) pionips Dyar....................................................... 235
18. A. (O.) punctor Kirby..................................................... 237
19. A. (O.) hexodontus Dyar ........................'............................. 240
20. A. (O.) sticticus Mg........................................................... 243
21. A. (O.) nigrinus Eck.................:......................................... 245
22. A. (O.) hungaricus Mih...................................................... 246
23. A. (O.) diantaeus H. D. К................................................... 247
24. A. (O.) intrudens Dyar...................................................... 250
25. A. (O.) pullatus Coq........................................................ 252
26. A. (O.) nigripes Zett....................................................... 255
27. A. (O.) impiger Walk. ..........................................".............. 258
28. A. (O.) cataphylla Dyar..................................................... 261
29. A. (O.) leucomelas Mg....................................................... 263
30. A. (O.) detritus Hal........................................................ 264
31. A. (O.) simanini Guts....................................................... 267
32. A. (O.) kasachstanicus Guts........................................... . 269
2. Подрод Aedimorphus Theob.
33. A. (A.) vexans Mg. . . ........................................................ 270
A. v. vexans Mg. . .................................................... . 273
A. v. nipponii Theob........................................................ 273
3. Подрод Finlaya Theob.
34. A. (F.) geniculatus 01...................................................... 274
35. A. (F.) echinus Edw.......................................................... 278
36. A. (F.) nipponicus LaCasse et Yam............................................ 280
37. A. (F.) seoulensis Yam....................................................... 283
38. A. (F.) alektorovi Stack..................................................... 284
6
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
. Стр.
39. A. (F.) togoi Theob.......................................................... 288
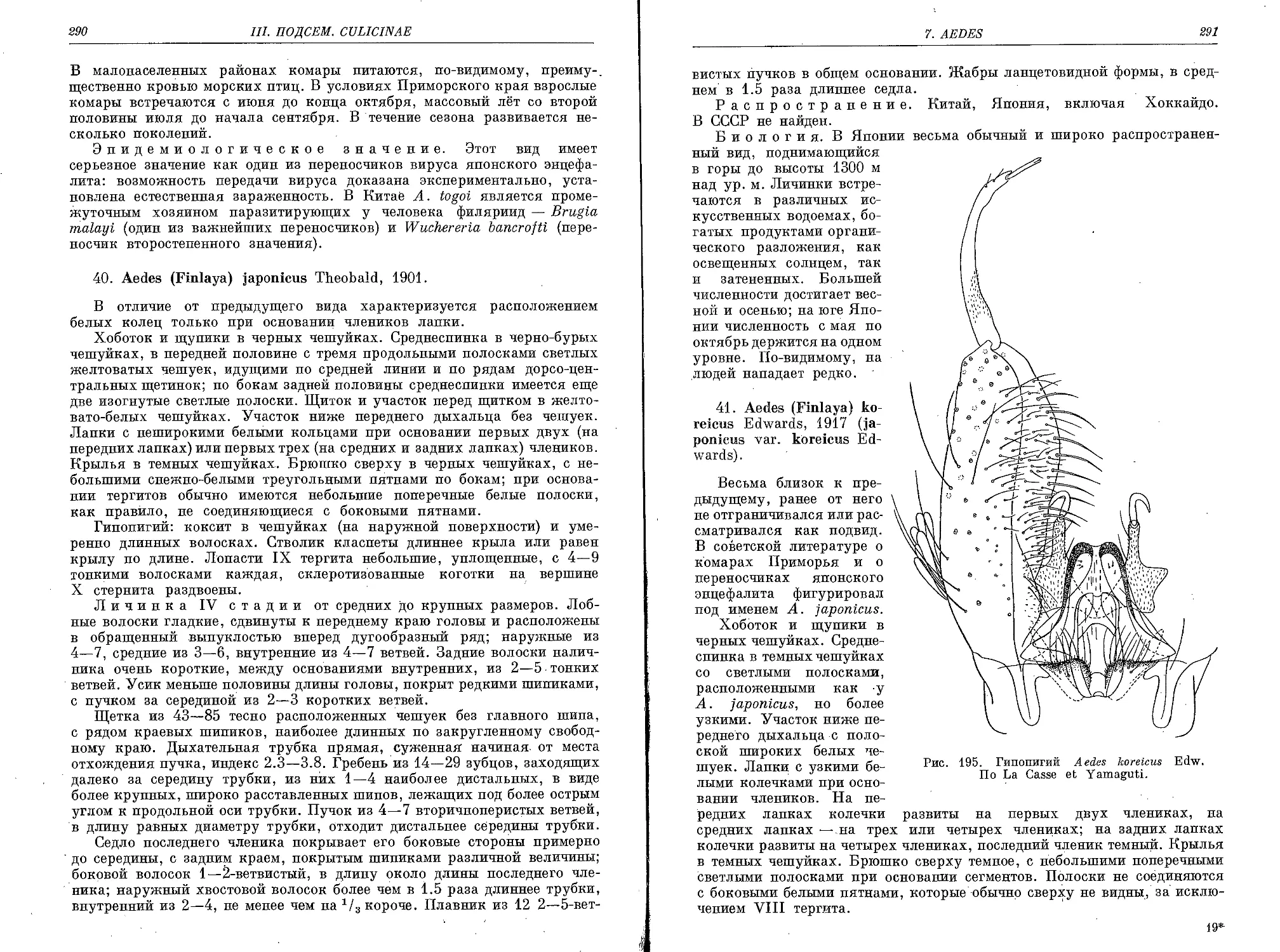
40. A. (F.) japonicus Theob...................................................... 290
41. A. (F.) koreicus Edw......................................................... 291
4. Подрод Stegomyia Theob.
42. A. (S.) aegypti L........................................................... 294
43. A. (S.) cretinus Edw....................................................... 298
44. A. (S.) vittatus Big......................................................... 299
45. A. (S.) galloisi Yam........................................................ 300
46. A. (S.) albopictus Sk....................................................... 303
47. A. (S.) flavopictus Yam..................................................... 305
48. A. (S.) chemulpoensis Yam.................................................... 306
5. Подрод Aedes Mg.
49. A. (A.) cinereus Mg......................................................... 308
A. c. cinereus Mg...................................................... 310
A. c. rossicus D. G. M.................................................. ЗГ1
A. c. esoensis Yam...................................................... 312
50. A. (A.) nobukonis Yam......................................................... 313
51. A. (?) aureus Guts............................................................ 313
8. Род C ulex L.
1. Подрод Lutzia Theob.
1. C. (L.) fuscanus Wied.......................................................... 322
2. C. (L.) vorax Edw............................................................ 324
2. Подрод Barraudius Edw.
3. С. (B.) modestus Fic. . ...................................................... 325
4. С. (B.) pusillus Macq......................................................... 327
3. Подрод Neoculex Dyar
5. C. (N.) territans Walk........................................................ 331
6. C. (N.) hortensis Fic......................................................... 333
7. C. (N.) martinii Med.......................................................... 335
8. C. (N.) hayashii Yam......................................................... 336
4. Подрод Culex L.
9. С. (C.) bitaeniorhynchus Giles................................................ 341
10. С. (C.) sinensis Theob....................................................... 345
11. С. (C.) whitmorei Giles...................................................... 345
12. С. (C.) mimeticus Noe ........................................................348
13. С. (C.) jacksoni Edw......................................................... 350
14. С. (C.) orientalis Edw....................................................... 352
15. С. (C.) tritaeniorhynchus Giles.............................................. 354
16. С. (C.) univittatus Theob..................................................., 358
17. С. (C.) theileri Theob....................................................... 360
18. С. (C.) pagans Wied.......................................................... 362
19. С. (C.) torrentium Mart...................................................... 364
20. С. (C.) pipiens L............................................................ 365
Добавление (Aedes)
A. (O.) montchadskyi Dubitzky.................................................. 375
A. (O.) stramineus Dubitzky...................................................... 376
A. (O.) fitchii Felt et Young.................................................... 377
A. (O.) rempeli Vock............................................................ 378
ВВЕДЕНИЕ
КРАТКИЙ МОРФОЛОГИЧЕСКИЙ ОЧЕРК
ВЗРОСЛОЕ НАСЕКОМОЕ
Тело комара, как у любого насекомого, разделяется на 3 основных
отдела: голову, грудь и брюшко (рис. 1).
Голова
Голова комара имеет в общем
шарообразную форму. Боковые
поверхности головы заняты почти
нацело большими фасеточными
глазами, сближенными или даже
соприкасающимися в верхней и
нижней частях головы и имею-
щими в общем почковидную форму.
Количество омматидий (фасе-
ток) одного глаза — от 300 до 1000.
Размеры каждой омматидии у са-
мок в среднем немного больше,
чем у самцов; соответственно ко-
личество омматидий у самцов не-
сколько больше, чем у самок.
Переднюю часть головы за-
нимает наличник — перед осно-
ваниями усиков, и лоб — над
усиками, между глазами; задняя
часть головы (за глазами) носит
название затылка.
Усики, сближенные своими ос-
нованиями, причленяются к пе-
редней части головы (рис. 2). Ко-
личество члеников усика у обоих
полов равно 15. Первый членик
(скапус) имеет вид кольца или
пластинки. Второй членик (то-
рус)— относительно крупный, осо-
бенно у самцов, более или менее
шаровидный. В нем помещается
Рис. 1. Строение тела комара.
1 — голова и ее придатки, II — грудь, III —
брюшко; 1—8 — сегменты брюшка.
сложно устроенный джонстонов
орган, представляющий собой орган слуха; детали его строения различны
у комаров разных родов (Risler, 1955). Остальные 13 члеников образуют
Рис. 2. Голова и ее придатки комара.
А — голова с расщепленным хоботком, Б — хоботок в момент
начала кровососания (по Павловскому), В — усики (по Кристо-
ферсу); г — голова, н — наличник, щ — щупик, у — усик, вг —
верхняя губа, вч — верхняя челюсть (мандибула), нч — нижняя
челюсть (максилла), п — подглоточник, нг — нижняя губа, х —
комплекс колющих частей хоботка.
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ВЗРОСЛЫЕ 9
жгутик усика. Членики жгутика построены в общем по одному типу,
приближаясь по форме к цилиндру и обладая более или менее одинаковой
длиною. Исключением являются лишь последние членики усиков самца,
отличающиеся значительной длиной. Волосяной покров усиков состоит
большей частью из розеток волосков, значительно более густых и длин-
ных у самца, нежели у самки (рис. 2, В). У самцов некоторых групп (подрод
Lophoceraomyia рода Culex) волоски на некоторых члениках усиков пре-
образованы в роговидные придатки.
Различные по строению и по функции волоски на усиках представляют
собой сенсиллы разных типов. Так, на усиках Aedes aegypti можно раз-
личить 5 типов волосков (Slifer a. Sekhon, 1962). Толстостенные во-
лоски — механорецепторы; тонкостенные со множеством мельчайших
отверстий — хеморецепторы. Отверстия в стенках волосков видны при
электронномикроскопическом исследовании ультратонких срезов.
Ротовые органы комара имеют форму хоботка (рис. 2). Хоботок самки
представляет собой сложное образование, состоящее из верхней губы,
пары верхних челюстей (мандибул), пары нижних челюстей (максилл),
подглоточника (гипофаринкс) и нижней губы. Верхняя губа образует
канал, по которому жидкая пища в момент сосания поступает в пище-
варительный тракт. Подглоточник прободен тончайшим каналом, по
которому в момент кровососания течет слюна. В спокойном состоянии
все упомянутые части хоботка вложены в желобок нижней губы, вслед-
ствие чего хоботок представляется как бы цельным.
Самки всех палеарктических комаров, за исключением Т oxorhynchites,
питаются как растительными соками, так и кровью позвоночных. При
кровососании пучок колющих частей погружается в покровы позвоноч-
ного. Образующая футляр нижняя губа как бы складывается вдвое,
оставаясь снаружи (рис. 2). В толще покровов хозяина пучок колющих
частей изгибается в разных направлениях и проникает в один из капил-
ляров, после чего начинается кровососание. Возможно и поглощение
крови, излившейся из капилляра при его повреждении в толщу ткани.
В этом случае акт кровососания длится несколько дольше.
Самцы комаров не питаются кровью. Хоботок самца отличается от
хоботка самки отсутствием или редукцией челюстей. Степень редукции
верхних и нижних челюстей различна у самцов комаров разных родов
(Marshall, 1938).
По бокам хоботка при его основании к нему причленяются нижнече-
люстные щупики, или щупальцы, —членистые придатки, достигающие
довольно значительной длины у самцов и самок Anopheles и, как правило,
у самцов прочих родов, но весьма короткие у всех самок Culicinae и у сам-
цов Uranotaenia и подрода Aedes (из палеарктических родов). Щупики
состоят из пяти члеников, но у самок V (а иногда и IV) членик может
отсутствовать или быть представленным в виде небольшого рудимента.
Грудь
Грудной отдел (рис. 3) распадается на три части: переднегрудь,
среднегрудь и заднегрудь. Каждая из поименованных частей представляет
собою в общем подобие кольца, состоящего из нескольких пластинок.
Спинные пластинки образуют спинку, боковые пластинки — бочки груди.
В семействе Culicidae, как и во всем отряде двукрылых, наиболее круп-
ным отделом груди является среднегрудь, к которой причленяются
крылья. Дорсальная поверхность груди — среднеспинка — представ-
ляется равномерно выпуклой. По средней линии среднеспинки тянется
10
ВВЕДЕНИЕ
ряд щетинок, носящих название акростихальных; кнаружи от послед-
них, примерно на половине расстояния от средней линии тела до боковых
краев среднеспинки, последняя несет также продольный ряд щетинок,
называемых дорсоцентральными. Сзади со спинкой граничит щиток
(scutellum), трехлопастный у Culicinae, цельнокрайний у Anopheles.
Под щитком и кзади от него расположен так называемый postscutellum.
Части груди, видимые сбоку, также состоят из нескольких склеритов
(склеротизованных пластинок), отделенных друг от друга швами. Боковые
пластинки каждого сегмента разделяются на два отдела: эпистерны
(часть, лежащая впереди) и эпимеры (задняя часть). Сбоку груди видны
Рис. 3. Грудь комара, сбоку.
шс — шейный склерит. Переднегрудь: псп — переднеспинка,
пам — проэпимеры, пас — проэпистерны. Среднегрудь: срсп —
среднеспинка, щ — щиток, иск — постскутеллум, 19 — перед-
нее дыхальце, лэс — мэзэпистерны (верхний отдел), сп — мез-
эпистерны (нижний отдел — «стерноплевры»), к — место при-
крепления крыла, мэп — меззпимеры, сг — среднегрудка. Зад-
негрудь: 29 — заднее дыхальце, мтас — метаэпистерны, мтап —
метаэпимеры. Ноги: 1к, 11к, 111к — коксы. Брюшко: 1т — тер-
гит, 1с — стерпит.
следующие части (рис. 3): переднеспинка, эпистерны и эпимеры.передне-
груди и эпистерны среднегруди. Последние у двукрылых, в частности
у комаров, разбиты на два отдела: меньший — передне-верхний и боль-
ший — нижний. Нижний отдел вклинивается в виде треугольника между
основаниями передних и средних ног и носит условное название «стерно-
плевра». Далее кзади идут эпимеры среднегруди (мезэнимеры) и эпи-
стерны и эпимеры заднегруди. Перепончатый участок между основанием
передних ног и передне-нижним краем стерноплевр носит название пост-
коксального участка. Расположение чешуек на боковой поверхности
груди, в частности на проэпимерах, стерноплеврах и мезэпимерах, играет
большую роль в систематике комаров, особенно рода Aedes.
Между, эпимерами переднегруди, эпистернами среднегруди и боковым
краем среднеспинки лежит переднее дыхальце (стигма). На границе
эпимер среднегруди и эпистерн заднегруди расположено заднее дыхальце.
Дыхальца играют важную роль в жизнедеятельности насекомого, так
как через них не только поступает воздух в трахейную систему, но и
испаряется вода. Экспериментально доказано, что, например у Anopheles
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ВЗРОСЛЫЕ
11
Рис. 4. Расположение щетинок на бочках груди.
А — Culiseta, Б — Aedes', пн — пронотальные, пэс —
проэпистернальные, пэм — проэпимерные, спр — ды-
хальцевые (спиракулярные), пспр — задыхальцевые
(постспиракулярные), пк — предкрыловые, сп — стер-
ноплевральные, вмэп — верхние мезэпимерные, нмэп —
нижние мезэпимерные.
maculipennis, испарение осуществляется в основном с поверхности трахей
(Виноградская, 1953). Размер дыхалец связан с интенсивностью испаре-
ния. У комаров, обитающих в зоне1 сухого климата, размер дыхалец
меньше, чем у комаров, живущих в условиях высокой влажности. Пока-
зателем размера дыхальца служит дыхальцевый индекс — отношение
длины передней грудной
стигмы к длине груди, выра-
женное в процентах (Вино-
градская, 1950).
Вблизи дыхалец и в дру-
гих определенных местах бо-
ковой поверхности груди
располагаются группы ще-
тинок. Их расположение яв-
ляется постоянным и хоро-
шим отличием для многих
групп семейства. В соответ-
ствии с положением щетинки
носят следующие наименова-
ния (рис. 4): пронотальные
(на pronotum, т. е. на перед-
неспинке); проэпистерналь-
ные; проэпимерные; дыхаль-
цевые, или спиракулярные
(непосредственно перед пе-
редним дыхальцем, между
ним и краем проэпимер);
задыхальцевые, или постспи-
ракулярные (позади перед-
него дыхальца, в верхнем
отделе мезэпистерн); стерно-
плевральные; верхние и ниж-
ние мез эпимерные.
Ноги
Вентральные придатки
груди, как у всех насекомых,
имеются в числе 3 пар —
передней, средней и задней
и причленяются соответст-
венно к передне-, средне-
и заднегруди. Ноги имеют
следующие отделы: корот-
кий тазик, сочлененный с грудью; вертлуг в виде небольшого колечка;
бедро, представляющее собой длинный цилиндр, иногда в основной части
слегка утолщенный; голень, имеющую также форму длинного и тонкого
цилиндра; пятичленистую лапку,, на конце которой сидят парные ко-
готки (рис. 5).
Коготки лапки с нижней, вогнутой, стороны могут быть вооружены
одним или двумя зубцами. Вооружение коготков представляется сокра-
щенно в виде формулы, например: 2,1; 2,1; 1,1. Формула эта расшифро-
вывается так: один из коготков передней лапки имеет два зубца, другой —
один зубец; строение коготков средней лапки повторяет строение перед-
1»
ВВЕДЕНИЕ
ПОЙ; па задних лапках оба коготка несут по одному зубцу. Формула
0,0; 0,0; 0,0 обозначает, что все коготки на лапках простые. Строение когот-
ков у разных видов (в частности рода Aedes) неодинаково, поэтому этот
признак играет роль при определении некоторых комаров (Vockeroth,
1950; Сазонова, 1958). Форма коготков лапки разных пар ног почти оди-
накова, но размеры коготков убывают от передних ног к задним. Поэтому
при определении обращают внимание в первую очередь на коготки перед-
ней лапки.
У некоторых родов семейства под коготками имеются присоски (pul-
villi) — небольшие овальные пластинки, усаженные нежными волосками
Рис. 5. Концевые придатки лапки комара. Из Е. Н. Павловского.
А — последний членик лапки Culex, Б — коготки самки Aedes, В — коготки
самки Culex', п — присоски (пульвиллы).
(рис. 5). Между коготками располагается непарное образование, так
называемый эмподий, чаще в виде перистой или разветвленной щетинки.
В отношении топографии свободного отдела конечности необходимо
помнить следующее. Сочленение тазика, вертлуга и бедра является много-
осным, в силу чего при поворотах в этом сочленении свободный отдел
конечности может принимать разнообразные положения по отношению
к основным осям тела насекомого. По этой причине удобнее ориентиро-
вочным центром для рассмотрения топографии конечности выбрать одно-
осное сочленение бедра и голени, так называемое коленное сочленение.
При согнутой в коленном сочленении конечйости бедро и голень будут
обращены друг ,к другу всегда одной и той же поверхностью, которая
носит название вентральной (при разогнутой конечности бедро и голень
обращены ею вниз), или нижней. Противоположная и параллельная ей
поверхность будет носить название дорсальной, или верхней. Перпенди-
кулярные же к поименованным сторонам поверхности будут называться
передней и задней.
Крылья
Дорсальные придатки грудного отдела комара, крылья, имеют в общем
удлиненно-овальную форму и снабжены продольными и поперечными
жилками, поддерживающими пластинку крыла. Место прикрепления
крыла к грудному отделу носит название основания. Противоположный,
дистальный, конец крыла называется вершиной. Край крыла, при рас-
простертых крыльях (т. е. тогда, когда длинная ось крыла перпенди-
кулярна продольной оси тела) обращенный кпереди, называется перед-
ним, направленный назад — задним.
Продольные жилки носят следующие наименования (рис. 6): косталь-
ная, или краевая, жилка (с), идущая по наружному краю крыла (у кома-
ров она развита как по переднему, так и по заднему краю); следующей
по направлению спереди назад продольной жилкой является субкосталь-
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ВЗРОСЛЫЕ
13
пая’жилка (sc), начинающаяся в основании крыла, идущая почти парал-
лельно переднему его краю и соединяющаяся в вершинной трети крыла
с костальной жилкой. Далее идет сектор радиальных жилок, число кото-
рых у комаров равно четырем (тх, г2, г3, г4+Б; формула ri+& обозначает,
что эта жилка в процессе эволюции данной систематической группы обра-
зовалась из двух жилок — г4 и г5, которые имеются в виде самостоятель-
ных образований у некоторых генетически связанных с комарами и,
как правило, у более примитивных форм). Начинаясь в основании крыла
общим стволом, радиальные жилки постепенно разветвляются, причем
от г4 отходит сначала ствол r2+г3+ г4+Б; далее от него ответвляется г4+Б;
наконец, ветви г2 и г3 образуют переднюю, или радиальную, вилку крыла;
отрезок г2+3 от поперечной жилки до основания вилки называется рукоят-
Рис. 6. Крыло Culex.
Жилки: с — костальная (краевая), sc — субкостальная, r2, r3, ri+s — радиальные,
mi, т2, m3+t — медиальные, си, — кубитальная, ап — анальная, тт — передняя попе-
речная (радиомедиальная), тт — задняя поперечная, h — плечевая.
кой радиальной вилки. Следующая жилка носит название медиальной (т);
у представителей сем. Culicidae она имеет 3 ветви — m1, т2 и т3+4; пер-
вые две образуют среднюю, или медиальную, вилку крыла; последняя
вместе с кубитальной жилкой (си) в вершинной части крыла образует
заднюю вилку. Наконец, последняя продольная жилка носит название
анальной (ап).
Поперечные жилки на крыле комара имеются в числе трех, а именно:
плечевая (й), соединяющая костальную и субкостальную жилки и распо-
ложенная близ основания крыла; радиомедиальная, или средняя (гт),
соединяющая системы радиальных и медиальных жилок и расположен-
ная близ середины крыла; медиокубитальная, или задняя (тси), соединяю-
щая медиальную и кубитальные жилки.
Продольные и поперечные жилки, ограничивая отдельные участки
крыла, образуют так называемые ячейки, которые обозначаются по назва-
нию жилки, лежащей перед соответствующей ячейкой. В отличие от жи-
лок, обозначаемых строчными буквами, ячейки обозначаются прописными
буквами, например 2?х, М, Си, Ап.
Крыловые жилки комаров покрыты чешуйками, более густо располо-
женными у самок, более редкими — у самцов. В большинстве случаев
чешуйки, покрывающие жилки, узкие — линейные или ланцетовидные;
реже, например у рода Mansonia, чешуйки более широкие. Пластинка
крыла покрыта мельчайшими волосками, различимыми только под микро-
скопом, — микротрихиями.
Дорсальными придатками заднегруди являются весьма характерные
для отряда двукрылых колбовидные придатки — жужжальца, пред-
14
ВВЕДЕНИЕ
ставляющие собой рудименты задней пары крыльев. Жужжальца функ-
ционируют как гироскопический орган равновесия. На ножке жужжалец
располагаются две группы сенсилл.
Брюшко
Брюшко комара имеет в общем удлиненно-цилиндрическую форму
и состоит из 10 сегментов, причем два последних, будучи приспособлен-
ными к выполнению половых функций, сильно модифицированы. Каждый
из восьми первых сегментов состоит из двух склеротизованных пластинок:
дорсальной, носящей название тергита, и вентральной — стернита; обе
.пластинки соединены тонкой плевральной перепонкой, на которой, по
бокам II—VII
сегментов, помещаются
1Хт
Рис. 7. Гипопигий Aedes.
к — гоаококсит (коксит), с — гоностиль (стиль),
пс — придаток стиля, еб— вершинная бородавка,
бб — базальная бородавка, си — стволик клас-
петы, ии — крыло класпеты, 1Хт — лопасти
IX тергита.
брюшные дыхальца — стигмы.
У голодной самки с неразви-
тыми яичниками боковые края
тергитов загнуты на брюшную'
сторону и при рассматривании
комара сверху не видны; пере-
понка, соединяющая тергиты са
стернитами, образует многочис-
ленные складки; такой же вид
имеет и брюшко высохшего ко-
мара. При насыщении комара и
при развитии яичников тергиты
распрямляются, а плевральная
перепонка растягивается.
Два концевых сегмента
брюшка (IX и X), как сказана
выше, приспособлены к выполне-
нию функции размножения и но-
сят название гениталий. Генита-
лии самца (рис. 7—9), так назы-
ваемый гипопигий, устроены
довольно сложно, причем, как
правило, их строение сильно
изменяется от вида к виду.
Поэтому строение гипопигия —
признак, играющий весьма важ-
ную роль в систематике кома-
ров, как и других насекомых.
Во многих случаях вид комара с наибольшей достоверностью может быть
определен по строению гипопигия, а в некоторых случаях определение
возможно только по этому признаку. В номенклатуре частей гипопигия
нет единообразия. Одни и те же части различными авторами обозначаются
по-разному.
У самцов гипопигий после выхода из куколки поворачивается на 180&
вокруг продольной оси тела так, что тергит становится на место стер-
нита, и наоборот. Таким образом, например, IX тергит брюшка будет
виден снизу, а IX стернит — сверху. Названия «тергит» и «стернит»
сохраняются, конечно, за дорсальной (морфологически) и вентральной
пластинками IX сегмента, несмотря на то что топографически они зани-
мают обратное положение.
IX сегмент, как и предыдущие, представляет собой подобие кольца,,
состоящего из дорсальной и вентральной пластинок. Истинный IX тергит
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ВЗРОСЛЫЕ
15
имеет вид неширокой пластинки, в боковых отделах заднего края кото-
рой более или менее развиты выросты, носящие название лопастей
IX тергита; вершина последних в большинстве случаев усажена щетинками
или волосками. Иногда, например у некоторых видов подрода Stegomyia,
помимо боковых лопастей, IX тергит имеет еще среднюю, чаще конусо-
видную или языковидную
Рис. 8. Гипопигий Culex.
с — стиль, к — коксит, пб — предвер-
шинная бородавка, Ig5, 2g5 — 1-й и
2-й отделы фаллосомы, Хс — X стернит,
Хбп — базальный придаток X стернита.
Истинный IX стернит обыкновенно
имеет вид узкой полоски, иногда
с боковыми выростами.
Наиболее крупным отделом гипо-
пигия являются латеральные при-
Рис. 9. Гипопигий Anopheles. По Сое, Free-
man a. Mattingly.
с — стиль, к — коксит, кл — класпеты, а —
эдеагус.
датки IX сегмента — гоноподиты (вальвы), представляющие собой пар-
ные образования. Каждый гоноподит состоит из двух члеников — гоно-
коксита и гоностиля. Эти части для краткости ниже обозначаются просто
«коксит» и «стиль». Коксит представляет собой массивное образование.
Он имеет с внутренней стороны в большинстве случаев выросты в виде
бородавок — вершинной и основной, или базальной (у большинства
Aedes; рис. 7). Иногда имеется одна бородавка (Culex, Culiseta), которая
располагается у середины коксита или, чаще, несколько ближе к его
вершине (предвершинная бородавка, характерная для рода Culex;
рис. 8). Внутренняя сторона коксита имеет обыкновенно шипы или креп-
кие щетинки (чаще в числе 1—3), расположение, строение и число кото-
рых характерно для отдельных видов, в частности рода Anopheles (рис. 9).
У Aedes шипы, если имеются, обычно располагаются на базальной боро-
давке. Снаружи, сверху и снизу коксит усажен щетинистыми волосками,
а иногда и чешуйками.
На вершине коксита или несколько перед нею (у подрода Aedes и
некоторых других) прИчленяется стиль, имеющий чаще узколанцетовид-
ную форму; иногда стиль бывает расширен. В вершинной части стиля
16
ВВЕДЕНИЕ
к нему причленяется пальцевидный или шиповидный придаток, иногда
весьма короткий.
Ковнутри от кокситов расположены так называемые класпеты (claspet-
tes), представляющие собой выросты основного отдела коксита. Класпеты
выражены вполне только у некоторых групп, особенного развития дости-
гая в роде Aedes у подродов Ochlerotatus и Finlaya, у которых класпеты
представляются двучленистыми образованиями, состоящими из основ-
ного отдела — стволика — и вершинного отдела — крыла, форма кото-
рого различна у разных видов. На выпуклой стороне крыла класпеты
может быть прозрачное пластинчатое расширение. Отсутствуют класпеты
у Culex и некоторых других родов.
Рис. 11. Брюшко самки комара.
По Сое, Freeman a. Mattingly.
А — Anophelesj Б — Culex, В '— Aedes.
Рис. 10. Внутренние склериты
гипопигия Aedes, сбоку.
По Matheson.
IXm — IX тергит, IXc — IX стернит,
Хт — X тергит, Хс — X стернит,
д5 — фаллосома, п — парамеры, бп —
базальная пластинка.
Ближе к средней линии тела расположены более или менее изменен-
ные склериты X (анального) сегмента, которые окружают анальное от-
верстие. X тергит представляется обыкновенно в виде двух незначитель-
ной величины пластинок, связанных на вершине с X стернитом. X стер-
нит (парапрокт) состоит из двух, у большинства форм семейства ясно
выраженных удлиненных пластинок, в некоторых случаях усаженных
короткими щетинистыми волосками; последние особенно развиты у рода
Culex, для которого характерно наличие шипиков или зубчиков на вер-
шине X стернита и та или иная степень развития его базального отростка.
В некоторых случаях, например у Aedes, склеротизованная вершина
каждой пластинки X стернита образует простой или зазубренный коготок.
Собственно генитальным органом комаров является эдеагус (aedeagus;
рис. 10) — сложное образование, состоящее из нескольких склеритов,
а именно: латеральных пластинок, парамер и фаллосомы (мезосомы).
В роде Culex фаллосома достигает значительной степени сложности и,
как правило, распадается на два отдела, которые в свою очередь бывают
разделены на несколько зубцов. Строение фаллосомы — признак, имею-
щий большое систематическое значение.
Концевые сегменты брюшка самки, по сравнению с предыдущими
сегментами, представляются относительно слабо измененными. VIII стер-
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЯЙЦО 17
пит у большинства форм представляется в виде простой пластинки; у не-
которых групп (особенно подроды Stegomyia и Finlaya рода Aedes) он
достигает значительного развития. На конце брюшка бывают заметны
парные образования, так называемые церки (рис. 11, В). В некоторых
случаях они бывают вполне явственно развиты, выдаваясь в виде двух
небольших эллипсоидальных пластинок, в других они более или менее
скрыты. Степени развития VIII стернита и церок для групп рода Aedes
являются признаками, связанными между собою по типу обратной кор-
реляции.
На просветленных препаратах заднего конца брюшка самки в области
VII—VIII сегментов видны сперматеки в виде склеротизованных капсул.
Количество сперматек различно — одна {Anopheles, Uranotaenia и как
исключение некоторые Aedes), две (некоторые Mansonia) или три (по-
давляющее большинство Culicinae).
Строение гениталий самки не играет большой роли в систематике
комаров, но довольно характерно для отдельных родов и подродов
(La Casse a. Yamaguti, 1955).
Следует добавить, что при определении комаров учитывается в ка-
честве важных систематических признаков строение разнообразных
придатков кожных покровов. Эти придатки обозначаются как чешуйки
(удлиненные, реже широкие, продольно исчерченные пластинки), шипы
(относительно толстые образования, обычно развитые в том или ином
месте, например на базальной бородавке коксита, в определенном для
данного вида количестве), щетинки (более тонкие, чем шипы, чаще их
количество неопределенно), волоски (еще более тонкие образования).
ЯЙЦО
Откладка яиц самками комаров производится различно. Самки Ano-
pheles откладывают яйца на поверхность воды поодиночке, хотя впослед-
ствии они, как не смачивающиеся водой, могут собираться вместе, обра-
зуя различные геометрические фигуры. Самки родов Culex, Mansonia
и подрода Culiseta откладывают яйца в виде компактных кладок — лодо-
чек, форма которых обусловлена пространством между скрещенными
задними ногами комара, куда производится откладка. Самки рода Aedes
и подрода Culicella рода Culiseta откладывают яйца в большинстве слу-
чаев в понижениях на влажную почву, где они могут сохраняться дли-
тельное время в сухом состоянии. После затопления мест откладки (при
выпадении осадков или при таянии снега) происходит частичный или пол-
ный выход из них личинок. Диапауза на фазе яйца широко распростра-
нена, особенно у видов рода Aedes, но еще недостаточно исследована.
Покровы яйца состоят из трех слоев. Внутренний слой — тонкая
желточная оболочка — непосредственно окружает богатую желтком яй-
цевую клетку. Следующая, наружная оболочка — хорион — образована
двумя слоями: средний — эндохорион — твердый и непрозрачный и
наружный — экзохорион — в большинстве случаев более мягкий и про-
зрачный. Экзохорион на поверхности несет различного рода скульптур-
ные образования, характерные для отдельных родов и видов.
Терминально {Culex, Aedes) или слегка субтерминадьно {Anopheles)
на переднем конце яйца находится сложно устроенный микропилярный
аппарат, обеспечивающий попадание спермиев внутрь яйца и его оплодо-
творение. Он состоит из розетковидной мембраны, окруженной кольцевым
валиком разросшейся наружной оболочки — экзохориона. В центре,
розетки находится отверстие с ведущим внутрь каналом, устье которого
2 Фауна СССР, А. В. Гуцевич и др.
18
ВВЕДЕНИЕ
закрыто изнутри особой пробочкой и открывается только при прохожде-
нии спермиев.
Форма яиц у Culicinae продолговато-овальная или почти цилиндри-
ческая, с более расширенным передним концом и более суженным, при-
Рис. 12. Личинка Anopheles. Общий
вид со спинной стороны.
Римские цифры — членики брюшка
(в скобках — первичные членики).
тупленным — задним. Поверхность ча-
сто (Aedes и др.) имеет сетчатую по-
лигональную структуру. У яиц Culiseta
ячеи такой сети более мелкие, а у Мап-
sonia — значительно крупнее.
Яйца Anophelinae имеют форму ло-
дочки с слабо вогнутой верхней (брюш-
ной) и выпуклой нижней (спинной)
поверхностью. Боковая поверхность
обычно окружена узким плавательным
пояском, который прерывается у сере-
дины различно развитыми более круп-
ными плавательными камерами с из-
менчивым у разных видов числом со-
ставляющих их секций. У Anopheles
maculipennis sacharovi яйца летних по-
колений имеют только плавательный
поясок, а зимние — кроме того, и слабо
развитые плавательные камеры. Рису-
нок экзохориона верхней поверхности
яиц, особенно для подвидов An. macu-
lipennis, имеет важное систематическое
значение (см. стр. 96 и вклейку между
стр. 96 и 97).
ЛИЧИНКА
Тело личинок состоит из трех резко
отграниченных друг от друга отде-
лов (рис. 12 и 13): заключенной в
сплошную склеротизованную капсулу
головы, груди, состоящей из трех сли-
тых вместе члеников, и 9-членикового
брюшка. Последний членик отогнут на
брюшную сторону; на его конце откры-
вается заднепроходное отверстие,
окруженное 2 парами жабр. На брюш-
ной стороне последнего членика нахо-
дится орган движения личинки — плав-
ник. На спинной стороне VIII членика
брюшка у личинок Anophelinae откры-
вается единственная функционирующая
задняя пара стигм, окруженная клапанным аппаратом, образующим
стигмальную пластинку; у всех личинок Culicinae задняя пара стигм со
стигмальной пластинкой находится на конце дыхательной трубки —
сифона, отходящей от спинной стороны VIII членика брюшка. Грудь —
наиболее широкий из отделов тела. Членики брюшка более узкие.
Размеры личинок IV стадии колеблются от 0.5—0.7 см у мелких видов
(Uгanotaenia, некоторые Culex) до 1.5 см и больше у крупных (Culiseta,
Aedes, Toxorhynchites и др.).
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
19
Окраска личинок разнообразна. Их кутикула, за исключением твер-
дых склеротизованных отделов (голова, дыхательная трубка и др.),
прозрачна и бесцветна. Поэтому об-
щая окраска тела личинок зависит
от окраски просвечивающих наружу
их органов и тканей, главным обра-
зом жирового тела. Пигмент сосре-
доточен преимущественно на верхней
поверхности спинной стороны лопа-
стей наружного слоя жирового тела.
Лопасти внутреннего жирового тела
почти лишены его. Соответственно
окраска спинной стороны личинок
всегда темнее, чем брюшной. Это
очень важно, особенно для видов,
развивающихся весной при низкой
температуре, так как пигмент способ-
ствует поглощению тепловых лучей
и развитие личинок происходит при
более благоприятном температурном
режиме. Поэтому на севере личинки
одного и того же вида пигментиро-
ваны сильнее, чем на юге.
Личинки Toxorhynchites окрашены
в винно-красный цвет, так же как
и личинки Orthopodomyia. Личинки
Ochlerotatus и Culiseta окрашены
от светло- до темно-коричневого, чер-
новатого цвета; Mansonia — желто-
коричневого цвета. Личинки Culex
окрашены в светло-коричневые или Рис. 13. Личинка Culicinae (Aedes cine-
серо-зеленые тона, многие виды, про- reus). Общий вид.
зрачные на младших стадиях, к сере-
дине IV стадии приобретают сине-зеленую окраску (С. territans). Зеленые
тона (от желто- до коричнево-зеленого) характерны для IV стадии личи-
нок Anopheles, тогда как младшие стадии темного, почти черного цвета.
Волоскитела
Тело личинок покрыто многочисленными разнообразными волосками.
Более длинные расположены так, что в своей совокупности образуют
вокруг личинки достаточно обширную чувствительную зону. Волоски
улавливают малейшие колебания воды, возникающие при приближений
хищников и заставляющие личинок менять свое местоположение. Осо-
бенно сильное удлинение части волосков наблюдается у личинок (Aedes
alektorovi, Orthopodomyia pulchripalpis и др.), живущих в небольших
скоплениях воды в дуплах деревьев, где в условиях скученности и не-
достатка пищи часто наблюдается явление каннибализма. У личинок
других дупловых видов, наоборот, происходит укорачивание волосков
и превращение их в шипы (личинки Toxorhynchites), часто придающие
личинкам вид ежа (Aedes geniculatus, A. echinus, A. galloisi и др.). Для ли-
чинок, обитающих у поверхностной пленки воды (Anopheles), характерны
укороченные или специально видоизмененные волоски на спинной сто-
роне тела и сильно развитые боковые волоски, тогда как у личинок,
2*
20
ВВЕДЕНИЕ
ведущих погруженный образ жизни, волоски развиты во все стороны
относительно равномерно.
Из различных кутикулярных образований, встречающихся у личинок,
наибольшее значение имеют следующие 2 типа.
К первому типу относятся различного рода скульптурные придатки
поверхности кутикулы: мелкие шипики (хетоиды), часто густо покры-
вающие тело личинки (Aedes cyprius, Culex hortensis); короткие заострен-
ные шипы на поверхности усиков или вдоль заднего края седла послед-
него членика брюшка, образующие гребенчатые ряды; микроскопические
шипики на поверхности дыхательной трубки и др. К этого же типа обра-
зованиям следует, вероятно, отнести и чешуйки щетки по бокам VIII чле-
ника брюшка и зубцы гребня на дыхательной трубке, так как в ряде
случаев можно проследить все переходы между ними и типичными мел-
кими шипиками хетоидного типа. Как правило, все эти образования
отличаются значительной количественной изменчивостью, меньшей у бо-
лее специализированных зубцов гребня и чешуек щетки.
Ко второму типу относятся настоящие волоски, строго обособленные
от окружающих частей наружных покровов. Каждый волосок имеет
внутри полость, слегка расширенное в виде головки основание, сидящее
в особой чашевидной сочленовной ямке, стенки которой образуют вокруг
головки базальное кольцо. Кутикула вокруг последнего может утол-
щаться в виде более темной пластинки, охватывающей иногда группу
сидящих рядом волосков (Toxorhynchites, Anopheles и др.), или в виде
бугорка с различного рода отростками (боковые волоски на груди и чле-
никах брюшка). Волоски второго типа часто имеют важное систематиче-
ское значение. Их положение и число постоянны не только у личинок
одного вида, но в большинстве случаев и одного рода, а во многих слу-
чаях и всего семейства. Однако строение каждого отдельного волоска и
степень его ветвления очень разнообразны и часто изменчивы. При этом
у одного и того же вида более сильное ветвление какого-либо волоска
связано с большей тонкостью его ветвей, и наоборот.
В простейшем случае волоски второго типа имеют вид длинного утон-
чающегося к свободному концу стержня, поверхность которого может
быть гладкой или нести вторичную перистость; при этом вторичные во-
лоски могут быть или короткими, или длинными и расположенными
в одной плоскости (у Anopheles). Стержневые волоски могут быть в раз-
личной степени расщеплены на конце. Другую группу составляют вет-
вящиеся волоски. Они могут отходить от базального кольца одним ко-
ротким стержнем, а затем начинают ветвиться дихотомически, древовидно,
беспорядочно (наружные волоски наличника у Anopheles) или в виде
веера на коротком основании. В ряде случаев отдельные ветви отходят
не в одной плоскости, а по кругу в виде ребер многогранной пирамиды
(Aedes geniculatus И др.). Наконец, волоски могут уплощаться, и тогда
каждая ветвь получает вытянуто листовидную форму (звездчатые во-
лоски Anopheles; рис. 22). Во всех случаях за самостоятельный волосок
надо считать все, что отходит от одного базального кольца.
Помимо упомянутых волосков и других образований, на теле личинок
могут иметься более темноокрашенные пластинки разнообразной формы.
Часто они приурочены к местам прикрепления мышц (спинные пластинки
•брюшка у Anopheles) или имеют опорное значение (седло или кольцо
на последнем членике брюшка, пластинки по бокам VIII брюшного чле-
ника у Uranotaenia и др.). Часто такие пластинки сами по себе или в комби-
нации с видоизмененными в шипы волосками имеют защитное значение, пре-
дохраняя личинок от нападения хищников (Orthopodomyia, Toxorhynchites).
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
21
Голова
Хитиновая капсула головы имеет более плоскую брюшную и вы-
пуклую спинную стороны. Общая форма колеблется от удлиненной в про-
дольном направлении — яйцевидной (Anopheles', рис. 14) — до широкой,
вытянутой поперечно овальной головы (Mansonia, Culex и др.; рис. 15).
У Toxorhynchites форма ее приближается к прямоугольной. Эти различия
связаны с типами питания личинок и особенностями функционирования
и строения их ротовых частей.
На спинной стороне головы находится крупный лобный (фронталь-
ный) щиток, обычно слегка вытянутый продольно и достигающий наи-
Рис. 14. Голова личинки Anopheles, со спинной стороны. По Пури.
А — I стадия, Б — IV стадия, лщ — лобный щиток, лги — лобный шов, ащ —
затылочный щиток («воротничок»), зги — затылочный шов, тги. — теменной
шов, яз — яйцевой зуб;. 2,3 — внутренний и наружный волоски наличника,
4 — задний волосок наличника, 5—7 — внутренний, средний и наружный
лобные волоски, s — теменной волосок, 9 — транссутуральный волосок,
12 — волосок у основания усика, 14 — глазной волосок.
большей ширины впереди глаз. У личинок Toxorhynchites он четырех-
угольной формы (рис. 57). Лобный щиток с боков и сзади отделен лобным
швом. Спереди к лобному щитку примыкает наличник (клипеус), отде-
ленный от него эпистомальным швом. Форма наличника связана со строе-
нием и функцией верхней губы: он сужен посредине и сильнее развит по
бокам. Морфологически передний отдел лобного щитка следует считать
наличником, так как там прикрепляются мышцы верхней губы, а эпи-
фарингеальная мускулатура прикрепляется в заднем отделе. Поэтому
в действительности лобный щиток является фронтоклипеальным, а на-
личник — преклипеусом.
С боков к лобному щитку примыкают щечные щитки. В заднем отделе
головы они подходят к средней линии и внутренними краями образуют
короткий теменной (корональный) шов. Щечные склериты на брюшной
стороне головы доходят до внутренних углов оснований нижних челюс-
тей, откуда начинаются направленные назад швы, отделяющие щеки от
брюшного срединного щитка — горла. Передний край последнего грани-
чит с основанием нижней губы.
В срединной части горла у личинок IV стадии Anopheles и некоторых
других, особенно в заднем отделе, имеется продольный шов, не доходящий
82
ВВЕДЕНИЕ
, пии лш
до основания нижней губы. Во время линьки головная капсула расхо-
дится по лобному шву на три части: лобный щиток приподымается, а оба
боковых щечных щитка расходятся в стороны, сгибаясь по упомянутому
срединному шву горлового щитка (рис. 14). Сквозь образовавшееся
отверстие вытаскивается голова линяющей личинки.
Задний край головы образует затылочное отверстие, более широкое
у Anopheles и Т oxorhynchites и более узкое у остальных личинок. Оно
окружено темноокрашенным узким кольцевым затылочным щитком (ворот-
ничком), отделенным от головной капсулы затылочным швом. Затылочный
шов прерывается сверху у теменного
шва, а с брюшной стороны — у сре-
динного линочного шва.
У Anopheles затылочный склерит
претерпевает в течение личиночного
развития ряд изменений. У вышедших
из яйца личинок он очень узкий, но в
течение I стадии его длина постепенно
увеличивается и перед линькой дости-
гает 70% длины головы. То же повторя-
ется и на II стадии. В течение III ста-
дии прирост длины воротничка мень-
ший — 30—32% длины головы, а на
IV стадии —11%. У других личинок
такие изменения не наблюдаются.
У всех личинок I стадии посредине
у заднего края лобного щитка нахо-
дится яйцевой зуб (рис. 14, 4), име-
ющий значение при выходе сформи-
ровавшейся личинки из яйца.
Окраска головной капсулы из-
менчива: от светлой полупрозрачной
(Culex) до желтовато-коричневой, темно-коричневой (многие Aedes) и
черной (Culiseta longiareolata, Uranotaenia). В случаях более темной
пигментации голова окрашена сравнительно равномерно, при светлой —
на лобном и щечных щитках, особенно в заднем отделе головы, имеются
закономерно расположенные более темные пятна, образующие своеобраз-
ный рисунок. Это — точки прикрепления мышц, приводящих в движение
ротовые придатки и усики, а также расширителей глотки (рис. 14). Эти
места, подвергаясь наибольшему механическому воздействию, склероти-
зуются более интенсивно. Дальнейшее укрепление головной капсулы
достигается слиянием отдельных пятен между собой с образованием
темных дугообразных поперечных полос в виде двух арок, укрепляющих
задний отдел головы. Учитывая, что такой рисунок головы в течение
стадии становится все более интенсивным, он не имеет значения как
диагностический признак.
Рис. 15. Голова личинки Aedes,
со спинной стороны.
лщ — лобный щиток, тш и лш — темен-
ной и лобный швы; 1 — щетинки налич-
ника, 2 — задние волоски наличника,
з, 4, 5 — внутренние, средние и наруж-
ные лобные волоски, в, 7 — теменной и
транссутуральный волоски.
Волоски головы
Голова личинок несет около 20 пар волосков, число, расположение
и особенности развития которых относительно постоянны и характерны
для большинства видов. Приводим описание только главнейших из них,
имеющих значение для определения вида личинок.
У переднего края наличника на его боковых выступах имеется пара
различной длины и толщины крепких, изогнутых внутрь заостренных
преклипеальных щетинок (рис. 15).
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
23
Следующий за ними ряд из двух пар клипеальных волосков располо-
жен на переднем крае лобного щитка — пара внутренних и пара наруж-
ных. Они имеют большое значение для определения вида у личинок
Anopheles и отсутствуют у других родов, за исключением Тoxorhynchites.
У личинок Anopheles имеет значение их взаимное расположение, развитие
и характер ветвления (рис. 14).
На некотором расстоянии позади передних волосков наличника по-
мещается их задняя пара. У личинок Anopheles она развита слабее перед-
них, у большинства других видов они очень короткие и отодвинуты зна-
чительно более назад, обычно между основаниями следующего ряда
более сильно развитых лобных (фронтальных, или постантеннальных)
волосков. Последних три пары: наружные, средние и внутренние. Они
могут быть расположены или в один изогнутый дугообразно назад ряд
(Anopheles, Culiseta, Mansonia и некоторые виды Aedes и Culex), или сред-
ние могут лежать впереди задних, образуя два треугольника с направлен-
ными наружу вершинами. При этом средние обычно более широко рас-
ставлены, чем задние (Ochlerotatits, некоторые виды Culex и др.). У личи-
нок Anopheles лобные волоски — перистые; только у An. plumbeus они
очень короткие и простые. У личинок Culiseta, Mansonia, многих Aedes и
Culex они сильно развиты, ветвятся от основания в виде плоского веера
и часто несут вторичную перистость. У других видов — они простые или
слабо ветвистые. Наружные, как правило, ветвятся сильнее остальных.
Следующий за лобными ряд волосков состоит из трех пар значительно
более коротких волосков: наружные — глазные — лежат позади простых
глазков, средние — транссутуральные — между внутренним краем слож-
ных глаз и лобным швом на щечных щитках и внутренние — теменные —
по бокам заднего отдела лобного щитка. Волоски брюшной стороны
головы не имеют систематического значения.
Глаза
У личинок IV стадии имеются две пары глаз. Одна из них — простые
глазки — в виде маленьких округлых пигментных пятен, расположенных
по бокам головы на уровне наибольшей ее ширины. Глазки вполне раз-
виты, функционируют уже у только вышедших из яйца личинок I стадии
и остаются у взрослых комаров, хотя и прикрыты у них чешуйками.
Вторая пара — сложные глаза, лежащие по бокам головы впереди
глазков. Они отсутствуют у личинок I стадии и постепенно развиваются
в процессе метаморфоза. У личинок, живущих в слабо освещенных водое-
мах (дупла деревьев), развитие и пигментация сложных глаз начинаются
позднее, часто на IV стадии. Сложные глаза начинают функционировать
еще у личинок, так как на сброшенных шкурках с темно пигментирован-
ной головой участок щечного щитка, прикрывающий глаза, всегда остается
не пигментированным.
Усики, или антенны (рис. 16), одночленистые и относительно мало
подвижные. Их длина очень изменчива: от */4—V3 длины головы (неко-
торые виды Finlaya и Stegomyia) до величины, в 1.5—2 раза ее превышаю-
щей (Mansonia, рис. 94; Aedes diantaeus, рис. 155). Наиболее просто
устроенные короткие усики имеют палочковидную форму, слегка су-
жаются к концу, на котором сидят чувствительные органы: волоски и
щетинки. Тело усика гладкое, без мелких шипиков, а около его середины
всегда имеется короткий и простой волосок. Такие усики в длину
не заходят вперед за границу волосков боковых лопастей верхней
губы.
84
ВВЕДЕНИЕ
Более длинные усики имеют слегка изогнутую форму и более ясное
разделение на базальный и более тонкий концевой отделы. На границе
между ними отходит волосок, состоящий из более или менее длинных и
многочисленных веерообразно расположенных ветвей, часто несущих
вторичную перистость. Тело усика, особенно в его базальной части,
Рис. 16. Различные типы усиков личинок Culicinae.
1 — Culiseta morsitans, 2 — Aedes, 3 — Culex pipiens.
покрыто редкими или более густо сидящими шипиками. Концевые во-
лоски и щетинки разделяются на две группы: вершинные — на самом
конце усика и предвершинные — на различном от него расстоянии
(рис. 16). Обе группы могут достигать значительной длины. Такое строе-
ние наиболее характерно для личинок Culex, Mansonia и некоторых дру-
гих. Усики пигментированы или однородно, или у их основания, а часто
и у вершины (многие Culex и др.).
У личинок Toxorhynchites вместо одного волоска на усике— три;
кроме того, на его теле имеются округлые чувствительные органы, сход-
ные с кольцевыми органами некоторых личинок Tendipedidae.
Ротовые органы и питание личинок
Строение ротовых частей у отдельных групп личинок тесно связано
с особенностями их типа питания и биологии. Всех личинок Culicidae по
этому признаку можно разделить на две группы: растительноядных
(всеядных) и хищных. Первичным является растительное питание.
У растительноядных личинок известно три различных типа питания.
1. Питание с поверхностной пленки воды, характерное для личинок
Anopheles и отчасти Ur сто taenia. 2. Питание с субстрата, путем слизыва-
ния или соскребывания налетов и обрастаний на поверхности погружен-
ных в воду предметов — перифитона, состоящего из различных микро-
организмов растительного или животного происхождения. Этот тип
свойствен ряду видов Aedes, Culiseta и др. 3. Фильтрационно-планктон-
ный тип питания, при котором личинки питаются находящимися во
взвешенном состоянии частицами (бактериями, водорослями, инфузо-
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
25
риями и др.). Этот тип питания встречается главным образом у личинок
Culex, Mansonia, некоторых Aedes и Culiseta. Наконец, есть немногие
виды, специально приспособившиеся к питанию нитчатыми водорослями
{Culex bitaeniorhynchus, отчасти С. sinensis).
Между этими типами существуют всевозможные переходы, и личинкам
каждого вида не исключительно присущ тот или иной из них, но обычно
один из типов питания превалирует, а другие лишь дополняют его. Так,
питающиеся с поверхностной пленки воды личинки Anopheles могут
питаться и с субстрата, а у некоторых из них наблюдалось и явление
каннибализма (An. plumbeus). Личинки с фильтрационным типом пита-
ния могут питаться и с субстрата, и с поверхностной пленки. То же наблю-
дается и у типичных обскребывателей.
Рис. 17. Верхняя губа. По Везенберг-Лунду.
1 — Culiseta morsitans, 2 — Culiseta annulata.
Активное хищничество и каннибализм у личинок Culicidae — явление
вторичное, более позднее, возникшее на базе уже развившегося приспособ-
ления к растительноядному питанию в условиях существования в водной
среде. У представителей разных подсемейств и родов семейства переход
личинок отдельных видов или групп к хищному типу питания происходил
самостоятельно и разновременно. В нашей фауне хищные личинки имеются
у Toxorhynchites и Lutzia.
Тип питания обусловливает не только ряд особенностей строения
и функции ротовых частей, но отражается и на строении и развитии уси-
ков, лобных и некоторых других волосков головы, общей формы послед-
ней и даже на строении дыхательной трубки. Все эти морфологические
особенности в своей совокупности образуют единый трофический функ-
циональный комплекс признаков, коррелятивно связанных между собой
и характеризующих приспособление личинок к тому или иному веду-
щему типу питания. Не приводя подробного описания ротовых органов
(см.: Мончадский, 1936, 1951), рассмотрим кратко их основные модифи-
кации и их связь с особенностями строения других элементов трофиче-
ского функционального комплекса.
Ротовые органы личинок, как и у всех насекомых, состоят из следую-
щих частей. 1. Верхняя губа (рис. 17) образована срединной
лопастью и двумя более сильно развитыми боковыми лопастями — ве-
ерами, состоящими из большого числа гибких длинных волосков. Это
основной орган, подгоняющий пищу к ротовому отверстию, а у хищни-
26
ВВЕДЕНИЕ
ков — захватывающий добычу. 2. Надглоточник (эпифаринкс),
ограничивающий сверху предротовую полость, является продолжением
Рис. 18. Верхняя челюсть.
1 — Anopheles (по Пури), 2 — Culiseta annulata, 3 — Culiseta morsitans (по Везенберг-Лунду).
основания внутренней поверхности верхней губы. Он несет симметричные
группы направленных назад волосков, щетинок и шипов, в совокупности
несущих функцию счесывания с лопастей верхней губы пищевых частиц
Рис. 19. Нижняя челюсть.
1 — Anopheles (по Пури), 2 — Culiseta morsitans, 3 — Culiseta annulata
(по Везенберг-Лунду).
и препятствующих их обратному току при отведении лопастей.
3. Верхние челюсти (мандибулы) (рис. 18) сильно склеротизо-
ваны, с различно развитыми темными зубами на дистальном конце внут-
реннего края и с системой шипов и волосков. При движении верхних
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА 27
челюстей их концевые зубы не соприкасаются и жевательной функции
не имеют. При накоплении пищевого комка в предротовой полости перед
подглоточником концевые зубы ударяют в него, размельчая небольшое
количество пищевых частиц. Подглоточник при этом служит как бы на-
ковальней. Системы волосков и шипов функционируют так же, как сход-
ные образования у эпифаринкса. Основания верхних челюстей ограни-
чивают предротовую полость с боков. 4. Нижние челюсти (мак-
силлы, рис. 19) устроены проще верхних; по своей форме, развитию
щупика, волосков и по степени подвижности варьируют сильнее у раз-
ных видов. Они ограничивают задне-боковые отделы предротовой по-
лости. 5. Нижняя губа помещается посредине между основаниями
нижних челюстей и вместе с подглоточником ограничивает с брюшной
стороны предротовую полость. Она состоит из трех последовательно нале-
гающих друг на друга треугольных пластинок, вершиной направленных
вперед, из которых две внутренние — ментум и субментум — имеют
зубчатые края и более крупный вершинный зуб. 6. Подглоточник
помещается непосредственно под ротовым отверстием и отделяется от
основания нижней губы отверстием протока слюнных желез. Это сильно
склеротизованная сложного строения пластинка, покрытая разного рода
бугорками и выступами. Над гипофарниксом накапливается пищевой
комок, по мере образования подвергающийся ударам зубцов верхних
челюстей, а затем попадающий через ротовое отверстие в глотку.
Видоизменения трофического
функционального комплекса
Для личинок, питающихся с поверхностной, пленки (Anopheles),
характерна удлиненная форма головы как следствие продольного располо-
жения мышц верхней губы, боковые лопасти которой при подгоне пищи
двигаются спереди назад. Во время питания личинки Anopheles повора-
чивают голову брюшной стороной кверху на 180°. При этом они упираются
снизу в поверхностную пленку воды двумя парами специальных образо-
ваний, отсутствующих у личинок с иным типом питания: 1) двумя спе-
циальными плоскими ножевидными щетинками на конце каждого усика
и 2) крупными щупиками нижних челюстей, несущими для увеличения
площади упора плоско ветвящийся боковой волосок, а на вершине —
листовидно уплощенные чувствительные органы. Пучки боковых ло-
пастей верхней губы не дифференцированы и состоят из волосков одного
типа и относительно одной длины. Верхние и нижние челюсти служат
для пассивного приема пищевых частиц, активно подгоняемых верхней
губой, снятия их с волосков последней и препровождения их в задний
отдел предротовой полости. В связи с этим их способность к движению
ограничена. Ряд перистоветвящихся лобных волосков, дополняемый
древовидными (у видов, их имеющих) наружными волосками наличника,
имеет значение для направления притока пищевых частиц. У личинок
An. plumbeus с развитой способностью к питанию с субстрата лобные и
клипеальные волоски простые и короткие. Отсутствие дыхательной
трубки, при наличии основных ее элементов, обеспечивает возможность
тесного контакта тела личинки с поверхностной пленкой. У личинок
Uranotaenia, живущих и питающихся непосредственно под поверхностью
воды, дыхательная трубка расположена под очень тупым углом к про-
дольной оси тела.
При питании с субстрата, не плоского, как поверхностная пленка
воды, а имеющего самую различную форму, движения лопастей верхней
88':
ВВЕДЕНИЕ
губы становятся более сложными, а верхние и нижние челюсти, хотя
еще и в небольшой степени, начинают принимать более активное участие
в непосредственном захвате пищи. Поэтому мышцы верхней губы при-
обретают диагональное направление, а мускулатура челюстей усили-
вается.
В голове сильнее развиваются боковые части щечных склеритов,
особенно в заднем отделе, вследствие чего голова сильнее развивается
в ширину, чем в длину. В строении волосков боковых лопастей верхней
губы наблюдается дифференциация. Концы внутренних волосков более
уплощены и несут 10—25 зубчиков, образующих гребень, с помощью
которого . личинки обскребывают налеты с поверхности субстрата. Чем
сильнее выражен этот тип питания, тем большее число волосков имеет
гребенчатые окончания и тем сильнее они развиты, в крайних случаях
образуя более Длинный внутренний пучок. Средняя лопасть более узкая,
с густо расположенными в виде щетки шипиками. Верхние и нижние
челюсти, несущие ряды коротких, более толстых щетинок, не только
участвуют в снятии пищевых частиц с лопастей верхней губы, но и в об-
скребывании субстрата. Зубцы мандибул сильно развиты, а максил-
лярный щупик очень мал. Антенны всегда палочковидные и короткие,
никогда не заходят за концы волосков верхней губы; более длинные усики
мешали бы питанию. Средние и внутренние лобные волоски относительно
короткие, простые или слабо разветвленные. Дыхательная трубка обычно
короткая, длина в среднем в 4—5 раз больше ширины основания. Этот
тип питания свойствен ряду видов подрода Ochlerotatus и рода Culiseta,
но степень приспособления к нему очень различна и не исключает воз-
можности планктонного типа питания.
Фильтрационно-планктонный тип питания связан с еще более сильным
развитием головы в ширину, становящейся поперечно овальной. Сложные
сгибательные и ротационные движения, совершаемые верхней губой
и нижними челюстями при облавливании толщи воды и подгоне взвешен-
ных в ней пищевых частиц в предротовую полость, вызывают общую
диагональную направленность их мускулатуры под большим углом к про-
дольной оси тела. Боковые лопасти верхней губы удалены друг от друга
на значительное расстояние. Они состоят из двух резко различных пуч-
ков! внутреннего, более широкого и короткого, направленного вперед
и внутрь, и наружного, более длинного, направленного по диагонали
вбок и вперед. Волоски внутреннего пучка не имеют гребенчатых кон-
цов. Срединная лопасть широкая, полукруглой формы, а не узкая, как
у личинок, питающихся с субстрата или с поверхностной пленки воды.
Зубы мандибул более тонкие, а ряды волосков более густые и длинные.
Сильно изменены максиллы: они конической, слегка суженной к основа-
нию формы с сильно редуцированным щупиком. На вершине максиллы
лежит пучок длинных, прямых или легко S-образно изогнутых волосков,
а ее внутренняя сторона густо покрыта более короткими волосками.
Максиллы очень подвижны и активно участвуют наряду с верхней губой
в подгоне пищи (вершинный пучок) и в ее отфильтровывании (пучок
внутренне-боковых волосков). Усики обычно длинные или очень длинные,
изогнутые; группа апикальных щетинок часто лежит далеко от субапи-
кальных, все они длинные. Пучок в виде сильно развитого веера из вто-
ричноперистых ветвей. Лобные волоски длинные, обычно веерообразно
ветвящиеся. Они вместе с пучком на усике ограничивают сверху и с боков
поток пищевых частиц, подгоняемый лопастями верхней губы. Дыхатель-
ная трубка обычно тонкая и длинная, что увеличивает радиус сферы
облавливаемого пространства.
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
29
Как вторичное приспособление хищный тип питания возник у личинок,
уже достаточно глубоко приспособленных к питанию с субстрата или
планктоном. Соответственно у подавляющего большинства видов и групп
хищных личинок органом схватывания добычи стала верхняя губа,
у всех растительноядных форм активно подгоняющая пищу к ротовому
отверстию. У встречающихся в СССР хищных личинок Т oxorhynchites
и Lutzia, таким образом, для схватывания добычи видоизменены боковые
лопасти верхней губы. При этом они сильно раздвинуты в стороны, сред-
няя лопасть редуцирована, а боковые отделы наличника увеличены и
выдвинуты вперед. Волоски лопастей резко уменьшаются в числе (у Тохог-
hynchites до 10, у Lutzia до 30—40) и превращаются в крючкообразно
загнутые щетинки. У Тoxorhynchites на конце они имеют грубые зубцы,
а у Lutzia — гребни хитиновых зубчиков, значительно более мощные,
чем у личинок, питающихся с субстрата. Верхние челюсти служат для
схватывания уже пойманной щетинками верхней губы добычи и для
проталкивания ее в глотку. Волоски на их теле частично редуцированы,
а зубы развиты сильнее. У нижних челюстей волоски превратились
в более короткие щетинки, их тело уменьшилось и приобрело прямо-
угольную форму. Усики сравнительно короткие, а голова (у Toxorhynchi-
tes) прямоугольная, с широким затылочным отверстием.
Грудь
Все три членика груди (передне-, средне- и заднегрудной) слиты
вместе настолько полно, что отделы ее, соответствующие отдельным из
них, можно внешне различить только по группировке волосков тремя
последовательными поясами.
Грудь уплощена в спинно-
брюшном направлении, с
брюшной стороны сильнее,
чем со спинной. Форма ее
колеблется, в зависимости
от стадии и степени зрелости
личинки, от округло-6-уголь-
ной до округло-овальной.
Сквозь прозрачную кутикулу
у старших стадий просвечи-
вают, особенно на брюшной
стороне, зачатки придатков
груди взрослого комара —
имагинальные диски в числе
5 пар. 3 пары на брюшной
стороне — зачатки ног — ле-
жат по бокам и разрастаются
к средней линии по напра-
Рис. 20. Волоски груди личинки А порholes.
По Пури:
А — передний край переднегруди с плечевым клапаном:
2, в — средний и наружный срединные волоски, 4—в —
плечевые волоски. Б — основание боковых волосков
переднегруди: 9, ю — брюшной и спинной волоски перед-
ней пары, и, 12 — спинной и брюшной волоски задней
пары.
влению к грудному отделу
брюшной нервной цепочки. На боках, ближе к спинной стороне, помещаются
зачатки крыльев, а позади них — небольшие зачатки жужжалец. Кроме
того, на спинной стороне по бокам переднего края груди расположены
зачатки рожков куколки, окончательно развивающиеся к концу IV ста-
дии. Перед линькой в куколку они входят в контакт с дыхательной си-
стемой личинки, наполняются воздухом, темнеют и сильно преломляют
свет. Последнее — верный признак готовой к окуклению личинки.
30
ВВЕДЕНИЕ
У всех личинок Anopheles по бокам переднего края спинной стороны
груди помещаются плечевые клапаны (рис. 20). Это полые прозрачные
выросты, направленные диагонально вперед и вбок, состоящие из двух
лопастей, сидящих на широком основании. От него отходит мускул-
ретрактор, прикрепляющийся на брюшной стороне, способный втягивать
клапан внутрь. Расправление происходит за счет упругости кутикулы
под влиянием внутреннего давления гемолимфы. Функционально пле-
чевые клапаны, отсутствующие у всех других личинок, тесно связаны
Рис. 21. Волоски груди личинки Anopheles. По Пури.
д __брюшная сторона: S—12 — боковые волоски, 13, 14 — прочие волоски. Б —
спинная сторона — переднегрудь: 1—3 — срединные волоски (внутренний, средний
и наружный) 4—7 — плечевые волоски; среднегрудь: 1 — внутренний срединный
волосок- заднегрудь: срединный волосок, часто звездчатого типа.
с жизнью личинок Anopheles на поверхности воды. Они поддерживают
передней конец их на поверхностной пленке. При поворотах головы
на 180° они обеспечивают телу личинок необходимую устойчивость.
Плечевые клапаны не имеют отношения ни к рудиментам переднегрудных
стигм, ни к дыхательной системе, вообще.
Многие из волосков груди имеют значение для видового определения
личинок Anopheles. Значение имеют следующие волоски (рис. 21): сре-
динные (7, 2, 3) на переднегруди, боковые волоски всех трех поясов (9—
12) и волосок 1 на заднегруди, часто видоизменяющийся в звездчатый.
У личинок Culicinae для определения вида также может иметь значение
группа срединных (медиальных) волосков переднегруди, у личинок
Тoxorhynchites — боковые волоски всех трех поясов. Однако эти при-
знаки обычно не используются, так как существуют более удобные на
последних члениках брюшка.
Брюшко
Брюшко состоит из девяти хорошо отграниченных члеников. Пер-
вично их было 10, но VIII и IX членики слились вместе, образовав вто-
ричный сложный VIII членик брюшка (рис. 12). Соответственно послед-
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
31
ний X членик стал вторично IX. Это слияние обусловлено образованием
стигмальной пластинки, как единого функционирующего органа, в со-
став которого вошли элементы VIII (стигмы, передняя и боковые лопасти,
или клапаны) и IX (задние лопасти, или клапаны, и центральная пла-
стинка, или рычаг с его отростками) члеников. При этом задний отдел
IX членика у большинства видов может сохраняться в виде промежуточного
кольца между VIII и последним члениками, а у некоторых Uranotaenia
(рис. 65) и других исчезает и этот рудимент. Последний X членик стано-
вится, таким образом, IX.
Так как общее строение и развитие волосков у первых семи члеников
брюшка относительно сходно, а у двух последних — значительно откло-
няется, их описание в дальнейшем будет проводиться отдельно.
Рис. 22. VI членик брюшка личинки Anopheles. По Пури.
А — спинная сторона, Б —• брюшная сторона, пт и зт — передняя и задняя
тергальные пластинки; 1—13 — волоски членика (1 — звездчатый и 6 — бо-
ковой). В — звездчатый волосок при большом увеличении.
Каждый из члеников первой группы несет до 13 пар волосков, рас-
положенных в один неправильный зигзагообразный ряд (рис. 22). Наи-
большего развития достигают боковые (плевральные) волоски.
У личинок Anopheles наибольшее значение для видового определения
имеют звездчатые волоски, находящиеся на спинной стороне, ближе
к заднему краю. Они варьируют как по форме, так и по степени развития
на передних члениках брюшка. На I членике у личинок подрода Anophe-
les они рудиментарны — в виде коротких, обычно простых волосков.
Только у An. lindesayi они из 3—4 ветвей. У большинства видов подрода
Myzomyia они хорошо выражены. Начиная со II членика они более или
менее развиты, а с III по VII членик достигают полного развития, за
исключением An. turkhudi, у которого они развиты на IV—VI члениках.
Звездчатые волоски (рис. 22) состоят из короткого округлого стер-
женька,- от которого веером отходят листовидно уплощенные ветви,
расположенные в параллельной поверхности тела плоскости. Форма ле-
пестков, интенсивность их пигментации, зазубренность краев и т. д.
варьируют у разных видов. Значение этих волосков заключается в под-
держании, совместно с плечевыми клапанами и стигмальной пластинкой,
личинки в подвешенном состоянии на поверхностной пленке воды. Во-
лоски в раскрытом состоянии лежат на поверхности воды. При погруже-
нии личинки эластичные лепестки смыкаются своими концами и захва-
тывают внутрь пузырек воздуха, с помощью которого при последующем
32
ВВЕДЕНИЕ
подъеме они преодолевают упругость поверхностной пленки и ложатся
на нее в раскрытом состоянии.
Грудь с плечевыми клапанами и стигмальная пластинка на VIII чле-
нике брюшка несколько возвышаются над поверхностью брюшка. По-
этому при подъеме, когда плечевые клапаны и стигмальная пластинка
касаются поверхности воды, у звездчатых волосков ближайших к груди
(I и II) и к стигмальной пластинке (VII и VIII) члеников возможность
контакта с поверхностной пленкой меньше, чем у расположенных на
средних члениках брюшка (III—VI). Тот, факт, что на IV—VI члениках
Рис. 23. Задний конец личинки Culicinae (А), сбоку.
VII, VIII — членики-брюшка; в—13 — волоски позади щетки (в, 13—
крайние верхний и нижний, 9 — средний, 7, и — промежуточные). В —
чешуйка щетки, Б — зубец гребня, Г — дыхательная трубка (сифон)
личинки Culiseta, сбоку. Стрелками показаны линии промеров длины сифона
и ширины его основания.
звездчатые волоски развиты у всех личинок Anopheles, на I членике —
лишь как исключение, а на VIII они отсутствуют, объясняет большой
размах изменчивости в их развитии на I—III члениках брюшка. Поэтому
полностью особенности их строения выражены только на средних (IV—
VI) члениках.
Боковые (плевральные) волоски 6 (рис. 22) — другая пара волосков
брюшка, имеющих значение для определения вида у личинок Anopheles.
У них важны особенности их ветвления и относительного развития.
Волоски на первых семи члениках брюшка у личинок Culicinae не
используются в систематических целях. Очень большое значение имеют
волоски и другие образования на двух последних члениках. VIII членик
брюшка у личинок Culicini и Тoxorhynchitini (рис. 23) несет в заднем
отделе дыхательную трубку (сифон). Под основанием трубки, по бокам,
ближе к заднему краю, помещается щетка, состоящая из различного
числа чешуек, свободными концами обращенных назад. Число их ко-
леблется от 5—7 до многих десятков. Каждая чешуйка имеет основание,
от которого назад отходит неплотно прилегающее к кутикуле тело, часто
заканчивающееся заостренным шипом. По бокам его основания почти
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
33
всегда находятся шипики или шипы, уменьшающиеся к основанию.
Иногда эти боковые шипы мало уступают по величине главному. У мно-
гих личинок (Culiseta, Culex, немногие Aedes) главный шип отсутствует,
а все тело чешуйки или только ее свободный конец усажены рядом шипи-
ков. Строение, число и взаимное расположение чешуек у разных видов
различно, но для определенных стадии и вида относительно постоянны.
Чешуйки в щетке, если их мало, могут располагаться в один прямой,
дугообразный или зигзагообразный ряд, если много — в виде пятна
различных очертаний. Часто строение крайних чешуек (верхних, ниж-
них или задних) иное, чем срединных. У личинок Uranotaenia чешуйки
своими основаниями отходят от заднего края хитинового склерита. У To-
xorhynchites вместо чешуек имеются 1—3 сильно развитых шипа, отходя-
щих от боковой пластинки, окружающей их основание. У личинок Ortho-
podomyia удлиненные чешуйки расположены в 2 правильных ряда и
имеют сложное строение.
Щетка сзади и сверху дугообразно окружена пятью волосками. Из
них крайние верхний и нижний и особенно средний — наиболее сильно
развиты и веерообразно ветвятся, а два промежуточных между ними —
обычно простые и более короткие.
Большое значение для систематики личинок имеет дыхательная трубка,
или сифон. У большинства видов начиная с III стадии она склеротизована
до основания, а у многих — сильно пигментирована. У личинок I стадии
склеротизован только ее дистальный конец. На II стадии склеротизация
приближается к основанию. Сифон редко имеет правильную цилиндри-
ческую форму. Обычно он сужен к вершине, редко (Culex territans) —
слегка расширен. У ряда видов наибольшая ширина приходится на сере-
дину трубки. У вершины сифон имеет в поперечном сечении форму круга
или овала, вытянутого продольно. Основание обычно более темно пигмен-
тировано и слегка утолщено спереди й с боков, а сзади имеет более или
менее глубокий вырез. По бокам, ближе к заднему краю, от кольца осно-
вания отходят два небольших отростка — ушки — точки прикрепления
мышцы, проходящей поперек основания, при сокращении которой сифон
отгибается назад. Вырез у его основания облегчает это движение, имею-
щее значение при отрыве личинки от поверхности воды. У личинок Stego-
myia и Toxorhynchites и некоторых других ушки отсутствуют.
Одним из существенных признаков при видовом определении (Culex,
некоторые Aedes) следует считать сифональный индекс — отношение
длины сифона к ширине его основания. В зависимости от способа изме-
рения этих величин можно получить различные значения индекса, часто
несравнимые друг с другом у разных видов и по данным различных авто-
ров. Поэтому необходимо придерживаться единого способа измерения:
1) измерять сифон сбоку, 2) измерять длину без клапанов стигмальной
пластинки, от основания ушковидных отростков базального кольца до
заднего края вершины сифона у основания задних клапанов стигмальной
пластинки, 3) ширину основания измерять по линии, идущей от перед-
него его края, через основание ушков до места ее пересечения с задним
краем основания, или с его продолжением, если вырез заднего края
заметно выражен (рис. 23, Г). В некоторых случаях (подрод Culiseta)
имеет значение сравнение отношений длины сифона не только к ширине
у основания, но и у вершины.
Большое систематическое значение имеет строение гребня сифона,
а также положение, относительное развитие и число пучков на его по-
верхности.
3 Фауна СССР, А. В. Гуцевич и др.
34
ВВЕДЕНИЕ
Из личинок нашей фауны гребень на сифоне отсутствует толька
у Orthopodomyia и Тoxorhynchites. У всех остальных он имеется в виде
двух симметричных продольных рядов зубцов, шипов или даже волосков
по бокам задней стороны сифона, начинающихся от его основания. Зубцы
гребня неодинаковы по строению; несколько ближайших к основанию —
рудиментарны; чем дальше зубцы от основания, тем они крупнее.^ Каж-
дый зубец имеет вид заостренного шипа, как правило, с небольшими
добавочными зубчиками у основания, из которых наиболее удаленный
от него крупнее других. Зубцы в гребне расположены более или менее
тесно на равном расстоянии друг от друга, или наиболее дистальные
более широко расставлены и крупнее остальных. В таких случаях гре-
бень может заходить за середину сифона. Иногда (Culiseta longiareolata)
Рис. 24. Задний онец брюшка личинки Anopheles, сбоку.
к По Мартини.
0~13 — волоски.
гребень состоит из ряда сильно расставленных по всей длине сифона
зубцов. У некоторых личинок подрода Culicella короткий гребень про-
должается в виде ряда крупных шипов, а у подрода Culiseta только не-
сколько ближайших к основанию имеют форму зубцов, а затем они удли-
няются и превращаются в длинные тонкие волоски.
У большинства личинок сифон, кроме пары коротких апикальных
щетинок на передней стороне вершины, имеет только одну пару пучков
волосков (Aedes, Culiseta), которые лежат или около середины (Aedes
и др.), или у основания трубки (Culiseta, Тoxorhynchites). Иногда, кроме
этой пары, на передней стороне сифона или у его вершины имеются 2—4
пары различно развитых волосков (личинки Aedes из группы rusticus,
A. cinereus). У личинок Culex сифон несет 4—8 или более пар пучков
волосков, расположенных симметрично или смещенных на заднюю по-
верхность. Кроме числа, расположения и характера ветвления пучков на
сифоне, систематическое значение имеет их длина, сравнительно с шири-
ной дыхательной трубки у места отхождения волосков (некоторые виды
Culex и Aedes).
У личинок Anopheles волоски на VIII членике брюшка по числу и
расположению в общих чертах сходны с волосками у Culicinae, но систе-
матического значения не имеют. У личинок Anopheles I стадии на боках
VIII членика, как у Culicinae, имеется щетка, состоящая из 5—6 про-
долговатых чешуек. Но уже на II стадии как самостоятельное образова-
ние она отсутствует.
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
35
Одна из характерных особенностей личинок Anopheles — отсутствие
дыхательной трубки. Вместо нее у личинок имеется специальное образо-
вание, поддерживающее стигмальную пластинку и гомологичное си-
фону.
Оно состоит из темной хитиновой дуги, сзади и с боков окружающей
основание задних лопастей, которые опираются на нее с помощью двух
боковых отростков (рис. 24). По бокам концы дуги переходят в треуголь-
ную пластинку. Задний край пластинки
несет ряд сильно развитых шипов. Верх-
ние шипы этого ряда соответствуют греб-
ню сифона Culicinae, а нижнего ряда —
их щетке по бокам VIII членика брюшка.
Относительное развитие, строение и число
шипов в гребне варьируют у разных ви-
дов, а также по стадиям. У личинок I
стадии боковые хитиновые пластинки еще
не развиты. Имеется только тонкая хи-
тиновая дуга и самостоятельно лежащие
группы чешуек, соответствующие гребню
и щетке.
Строение стигмальной пластинки име-
ет в ряде случаев важное систематичес-
кое значение, а для определения личи-
нок Anopheles дает ряд хорошо заметных
отличительных признаков.
У личинок Anopheles (рис. 25) стигмы
окружены пятью лопастями: непарной
передней, парой боковых и парой задних
лопастей. У середины основания передней
лопасти имеется эллипсоидное прозрач-
ное образование, обеспечивающее ее по-
ворот на 180° назад. По бокам ее основа-
ния сидят три пары коротких волосков.
Верхняя поверхность ее может быть цели-
ком темно пигментирована (все личинки
II стадии; An. claviger, An. plumbeus,
An. superpictus, An. pulcherrimus трех
последних стадий) или темная пигмента-
ция ограничивается основанием,, а перед-
ний отдел окрашен более светло (Ап.
maculipennis, An. hyrcanus).
У боковых лопастей на верхней поверхности лежит удлиненная тем-
ная хитиновая опорная пластинка, несущая на суженной вершине чув-
ствительный волосок, более тонкое основание которой Направлено к
стигмам.
Задние лопасти своими верхними поверхностями слиты с центральной
пластинкой в одно целое. Последняя состоит из трех отделов: переднего,
сужающегося в лежащий между стигмами передний отросток, основание
которого может продолжаться назад в передний и даже в средний отделы;
среднего и более широкого заднего. По характеру скульптуры в централь-
ной пластинке можно различить боковые и срединную части. Последняя’
служит местом прикрепления дорсовентральных мышц, чем и определя-
ются ее ячеистая структура, расположение ячей и их пигментация. От пе-
реднего отростка отходят направленные вперед продольные спинные;
3*
Рис. 25. Стигмальная пластинка
личйнок Anopheles (схематизиро-
вано).
ал, бл и пл — задняя,-«боковая и перед-
няя лопасти, цп — центральная пла-
стинка, зо, со и по — ее задний, сред-
ний и передний отделы, ст — стигмы,,
сто — стигмальные отростки, вп и
нп — верхняя и нижняя (основание)-
поверхности задней попасти; а, в,
с, d, f, 1—5 — волоски стигмальной
пластинки.
-36
ВВЕДЕНИЕ
мышцы VIII членика. Так выглядит раскрытая стигмальная пластинка
личинок Anopheles-, когда они находятся на поверхности.
При погружении передняя лопасть поворачивается назад, прикрывая
собой стигмы и центральную пластинку до среднего отдела. Стигмы
при этом смещаются вперед к основанию передней лопасти. Боковые
лопасти, связанные своими основаниями со стигмами, но не имеющие
собственной мускулатуры, поворачиваются при этом назад и становятся
на ребро передним краем вверх. Одновременно центральная пластинка
при комбинированном сокращении мышц — продольных, отходящих от пе-
А
Рис. 26. Строение стигмальной пластинки (схематизировано).
£ fyC— стигмальная пластинка личинок Culicinae: зк, бк и пк — задний, боковой и перед-
1 ний клапаны, р — рычаг, пл и зл — передняя и задняя луки рычага, зо — задний отро-
сток, ст — стигма, п — плечо, сто — стигмальный отросток; а, с, /, 1—5 — волоски.
15 — сагиттальный разрез через конец сифона личинки Culiseta с раскрытой стигмальной
пластинкой.
реднего отростка, и дорсовентральных, отходящих от ее срединной
части, — движется вперед и вниз, углубляя образовавшуюся полость,
открытую сверху в заднем отделе. Эта полость заполнена воздухом,
захваченным при погружении. При подъеме этот воздух служит для
пробивания пленки поверхностного натяжения и установления контакта
.стигм с атмосферой.
У всех личинок Anopheles I стадии (рис. 14) строение стигмальной
пластинки одинаково и сильно упрощено. У личинок II стадии все ее эле-
менты уже выражены, но видовые особенности проявляются еще очень
слабо. Только к III стадии они развиваются достаточно резко, а у ли-
чинок An. hyrcanus и An. maculipennis это происходит только в IV стадии.
Наличие дыхательной трубки у личинок Culicinae ограничивает дей-
ствие мышц только в одном направлении» параллельном продольной оси
сифона. Вследствие этого хитиновые элементы стигмальной пластинки
приобретают более дифференцированное, чем у Anopheles, строение,
обеспечивающее в этих условиях точную координацию их движения.
В стигмальной пластинке Culicinae (рис. 26) стигмы окружены пятью
клапанами, гомологичными лопастям пластинки Anopheles: непарным пе-
редним, парой боковых и парой задних. Между задними клапанами и
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА 37
стигмами находится гомологичный центральной пластинке Anopheles ры-
чаг, соединенный при помощи специальных отростков с большинством;
элементов пластинки и координирующий их движение.
Рычаг состоит из рукоятки, лежащей в полости сифона между тра-
хейными стволами. При рассматривании сбоку она хорошо заметна.
Рукоять к вершине сифона расширяется в передне-заднем направлении
и переходит в сжатое с боков тело рычага, имеющее внутри открытую
сверху полость. Эта полость образуется вследствие закладки рычага по-
следующей личиночной стадии, окружая старый подобно футляру. У ли-
чинок I стадии, таким образом, полость в рычаге отсутствует. Рычаг
заканчивается продольным седлообразным расширением. Соответственно
его концы образуют переднюю и заднюю луку. На передней луке часта
имеется передний отросток для соединения с передним клапаном. Зад-
няя лука продолжается в задний отросток, форма которого характерна
для многих родов и видов. От боков седла отходят отростки к стигмам,
часто охватывающие их задне-боковые края. Стигмы овальной, редко
округлой формы лежат по бокам тела рычага. Их наружные края лежат
в плоскости пластинки, а внутренние скошены, следуя за изгибом седла
рычага.
При погружении личинок рычаг сокращением прикрепляющихся к ру-
кояти мышц ретракторов втягивается вниз. Одновременно приводятся
в движение все части стигмальной пластинки. Стигмы своими отверстиями
обращаются навстречу друг к другу и несколько погружаются вглубь;
все клапаны поворачиваются внутрь и смыкаются над стигмами, образуя
полую, внутри неправильную пирамиду. В образовавшуюся полость
захватывается воздух. Замыкание клапанов не бывает полным, так как
поверхность стигмальной пластинки не смачивается водой и последняя
по этой причине не может попасть в полость и вытеснить оттуда воздух.
При подъеме, когда направленные вверх волоски задних и боковых
клапанов касаются поверхностной пленки, ретракторы расслабляются и
сокращаются их антагонисты — пара леваторов рычага, отходящих от его
тела к верхнему краю сифона. Пластинка начинает раскрываться, а на-
ходящийся в полости воздух своим выпуклым мениском выступает над
полураскрывшимися клапанами и пробивает пленку поверхностного на-
тяжения. Устанавливается контакт трахейной системы с атмосферным
воздухом, и клапаны ложатся на поверхность воды.
Некоторые особенности строения трахейной системы имеют система-
тическое значение. От задней пары стигм отходят два главных трахей-
ных ствола, проходящих вперед до переднегруди. Главные трахейные
стволы, особенно в дыхательной трубке и в задних члениках брюшка,
у большинства личинок на поперечном сечении имеют форму удлинен-
ного овала, а сбоку кажутся лентовидными. У некоторых личинок Culex
(особенно подрода Neoculex), Culicella и других они тонкие и округлого
сечения. У личинок Toxorhynchites и Orthopodomyia главные трахейные
стволы образуют пару пузыревидных расширений, находящихся в задне-
груди и частично в I членике брюшка. У личинок Mansonia подобные
пузыри находятся на особых ответвлениях главных трахейных стволов.
Все эти расширения имеют гидростатическое значение.
Последний членик брюшка несколько уже остальных. Он отогнут
под углом на брюшную сторону, на его спинной стороне и на боках поме-
щается хитиновая пластинка — седло. Седло занимает, особенно у мо-
лодых личинок, только спинную часть членика или, чаще, заходит на бо-
ковые стороны. В ряде случаев оно, разрастаясь на брюшную сторону,
охватывает членик в виде кольца, имеющего назади вырез, огибающий
38
ВВЕДЕНИЕ
основание плавника. Если такой вырез отсутствует, то часть передних
пучков плавника своими основаниями оказывается заключенной внутри
кольца, которое, так же как и седло, имеет опорную функцию и при-
дает последнему членику необходимую при движении личинки жесткость.
Задний край седла несет различного рода зубцы и шипы, достигающие
особо сильного развития у Toxorhynchites.
Сбоку на седле всегда имеется умеренно или слабо развитый боковой
волосок. На спинной стороне заднего конца последнего членика посре-
дине, непосредственно позади седла, находится комплекс из двух пар
волосков — хвостовых волосков (рис. 23). Их наружная пара отходит
от небольших самостоятельных склеритов и состоит из более длинных
и менее разветвленных волосков. Внутренние хвостовые волоски отхо-
дят от общего склерита, они более короткие, ветвятся сильнее и часто
имеют вид асимметричных вееров. У личинок Anopheles (рис. 12) волоски
наружной пары имеют изогнутые крючкообразно концы, служащие для
неподвижного закрепления личинки во время питания к плавающим
на поверхности воды предметам. Без этого ритмические движения лопастей
верхней губы вызывали бы поступательное движение личинки.
Вокруг анального отверстия расположены две пары жабр; их отно-
сительное развитие, форма и окраска часто служат для видового опре-
деления личинок. Вопреки своему названию, участие жабр в процессах
газообмена не большее, чем любого из участков кожных покровов ли-
чинок. Экспериментально доказано, что их основная функция — осмо-
регуляторная. Длина жабр у разных видов сильно колеблется: от трети
длины тела (Aedes pulchritarsis, A. galloisi) до едва заметной шаровидной
формы (Aedes detritus). При этом у личинок одного вида длина жабр уве-
личивается с понижением концентрации солей в воде их биотопов и умень-
шается с ее повышением. Форма жабр очень разнообразна: от колбасо-
видной с округлыми или приостренными концами до ланцетовидной,
листовидной или шарообразной.
По своему строению жабры представляют собой тонкостенные мешко-
видные выросты, покрытые тонкой бесцветной кутикулой, легко прони-
цаемой для воды и растворенных в ней веществ. Подстилающие ее гипо-
дермальные клетки часто сильно вакуолизированы и у ряда видов содер-
жат пигментные включения. Полость жабр сообщается с полостью тела
и заполнена гемолимфой, движение которой направляется специальной
мембраной, продольно разделяющей полость жабры, но не доходящей '
до ее конца. В мембране проходит трахея, дающая редкие ответвления
к слою гиподермальных клеток. Жабры личинок Culicidae принадлежат,
таким образом, к типу кровяных, а не трахейных жабр.
На брюшной стороне последнего членика находится главный орган
движения личинок — плавник. Он состоит из ряда сильно развитых
изогнутых волосков, основания которых расположены зигзагообразно
по средней линии. В своей совокупности они образуют плоский веер,
асимметрично загнутый назад. Соответственно каждый волосок обычно
ветвится несимметрично, назад от главного стержня. Только у личинок
Anopheles и Toxorhynchites плавник имеет вид симметричного веера, в со-
ответствии с чем наблюдается и двустороннее ветвление его волосков.
При движении личинок максимум давления на плавник приходится
сбоку. Поэтому основание каждого волоска укреплено поперечными бо-
ковыми отростками. Вследствие зигзагообразного расположения основа-
ний волосков плавника боковые отростки развиты несимметрично: если
основание волоска помещается вправо от средней Линии, то левый бо-
ковой отросток длиннее, а правый короче, и наоборот. Концы отростков
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. ЛИЧИНКА
39
загибаются, сливаются друг с другом и образуют общее для всего плав-
ника основание, придающее ему особую устойчивость. В переднем на-
правлении волоски плавника укорачиваются, а боковые отростки у осно-
вания развиваются слабее, и концы их остаются свободными. У ряда
видов (многие Aedes) образуется ряд из 2—40 волосков, более коротких,
слабее ветвящихся, отходящих впереди от общего основания.
Плавник отсутствует у личинок I стадии большинства видов. У мно-
гих видов наблюдается уменьшение числа и ветвления волосков плав-
ника, сопровождаемое редукцией их общего основания и отростков. Та-
кое упрощение строения плавника связано с сильным развитием жабр
(например, у Aedes pulchritarsis, A. galloisi, A. aegypti). При этом хво-
стовые волоски во время движения личинок отгибаются книзу, в про-
межуток "между верхней парой жабр, а волоски плавника, загибаясь
кверху, становятся между нижней парой жабр, приобретая опорную
-функцию. Жабры при этом становятся органами движения.
Рост личинок, возрастные изменения
отдельных признаков, определение стадии
ивозраставнутристадии
В течение личиночной фазы — от выхода из яйца до окукления —
личинки линяют 3 раза, т. е. проходят 4 стадии. За этот период их ли-
нейные размеры увеличиваются более чем в 8 раз. При этом увеличение
размеров тела происходит так, что все твердые хитиновые образования
(голова, дыхательная трубка, седло и др.) имеют максимум прироста
во время линек, а минимум — в течение стадий, тогда как все остальное
тело личинки, покрытое мягкой кутикулой, наоборот, имеет максимум
прироста в течение стадий, а минимум — во время линек.
Голова, дыхательная трубка и другие твердые участки покровов ли-
чинки, сразу после линьки прозрачные и мягкие, в течение первых ча-
сов отвердевают и утрачивают прозрачность вследствие отложения .в них
органических и неорганических включений. Только в это время происхо-
дит небольшое увеличение их размеров за счет расправления хитина.
Исключение составляют личинки Anopheles (в отдельных случаях и ды-
хательная трубка немногих Culicinae), у которых в течение стадии про-
исходит постепенное увеличение «воротничка» (стр. 22 и рис. 27). Все
остальное, покрытое мягкой кутикулой тело, 90% своего прироста полу-
чает во время стадий и лишь 10% при линьках. В течение 6—16 часов
после линьки всегда происходит заметное увеличение размеров личинки
за счет расправления кутикулы, затем увеличение прекращается и лишь
к началу вторых суток начинается рост, проходящий равномерно до сле-
дующей линьки.
В процессе роста личинок не только увеличиваются их размеры,
но происходит и ряд приуроченных к линькам морфологических изме-
нений многих признаков, позволяющих с большей или меньшей точностью
установить стадию личинки или ее возраст внутри стадии.
Недавно слинявшие личинки, независимо от стадии, кроме светлой
пигментации головы, дыхательной трубки и других твердых хитиновых
образований, отличаются от более старших по возрасту личинок той
же стадии относительно крупной головой по сравнению с грудью. Во-
лоски тела, особенно боковые, имеют не прямые, а закругленно изогнутые
концы. Близкие к линьке в следующую стадию или к окуклению личинки
всегда более темно пигментированы, голова их по сравнению с грудью
всегда относительно мала, волоски, особенно боковые, с прямыми кон-
40
ВВЕДЕНИЕ
цами; кроме того, сквозь прозрачную кутикулу у них просвечивают
уже начинающие темнеть волоски следующей стадии, особенно длинные
боковые волоски каждого членика, волоски плавника и хвостовые во-
лоски, которые под старой кутикулой плотно обвивают несущий их чле-
ник, создавая впечатление, что вся личинка запеленута и обвязана.
Рис. 27. Контуры головы и груди всех стадий личинок
Anopheles maculipennis Mg. после линьки (7) и перед линь-
кой в следующую стадию (2).
I, II, III и IV — личиночные стадии (III и IV стадии изображены
в 4 раза меньшем масштабе, чем I и II).
У готовых окуклиться личинок на боковых углах переднегруди просве-
чивают темные, от заполняющего их воздуха, куколочные рожки.
Как правило, с каждой последующей стадией ветвление одноименных
волосков увеличивается. То же происходит и с числом чешуек щетки
и с зубцами гребня, при этом у последних увеличивается и число допол-
нительных зубчиков у основания. Если чешуек в щетке мало (Aedes ri-
parius, A. pulchritarsls), то начиная со II стадии их число не увеличи-
вается. Для каждого отдельного вида по совокупности количественных
изменений всех признаков внешней морфологии возможно определение
вида на любой из стадий развития личинок. Однако все эти признаки
настолько скрупулезны, что для Определения массового материала они
непригодны.
МОРФОЛОГИЧЕСКИЙ ОЧЕРК. КУКОЛКА 41
Имеется небольшое число признаков, объективно характеризующих
некоторые из личиночных стадий. Так, для всех личинок I стадии ха-
рактерно наличие в заднем отделе лобного щитка яйцевого зуба (рис. 14),
отсутствие плавника; кроме того, за редчайшими исключениями, все
волоски их тела — простые. У личинок IV стадии, как уже отмечалось,
развиваются зачатки куколочных рожек, которые перед окуклением всту-
пают в прямой контакт с трахейной системой и их полость заполняется
воздухом. Для всех личинок II и III стадий подсемейства Culicinae ха-
рактерно наличие пузыревидного утолщения заднего или задне-бокового
края стигм, хорошо заметного на раскрытой пластинке и хуже — на
закрытой.
Наиболее удобным является следующий метод определения стадии
личинок. Проба с личинками выливается в белую эмалированную та-
релку, около которой ставят в ряд четыре небольших сосуда (солонки,
часовые стекла и т. п.). При помощи пипетки прежде всего отбирают
самых крупных личинок с наибольшей шириной головы и отсаживают
в крайний из сосудов. Далее, самых мелких личинок с наименьшей ши-
риной головы отсаживают в другой крайний сосуд. Таким образом, вы-
является наибольшая и наименьшая ширина головы, что, в случае присут-
ствия в пробе всех четырех личиночных стадий (что обычно и бывает
на практике с личинками Anopheles вследствие растягивания генерации
и захождения одной генерации за другую), соответствует IV и I стадиям.
Оставшихся личинок — их большинство и притом всех стадий — вни-
мательно просматривают на предмет вылова личинок для IV стадии
более мелких, а для I стадии соответственно более крупных, имеющих
ширину головы, равную таковой у прежде отсаженных личинок. Та-
ким путем будут выловлены из пробы все личинки I и IV стадий. Осталь-
ные личинки, независимо от размеров их тела, на глаз разбивают
на две группы: одну с шириной головы, большей, чем у личинок I стадии,
другую с меньшей, чем у личинок IV стадии, и соответственно размещают
в двух средних сосудах. Ошибки, весьма обычные вначале, будут сразу
же бросаться в глаза, так как каждая отсаженная не в свой сосуд ли-
чинка будет резко выделяться по ширине головы от остальных. Если какая-
либо стадия в пробе отсутствует, то в двух соседних сосудах могут ока-
заться личинки с одинаковой шириной головы. Их сливают вместе. Когда
все личинки рассажены, проверяют правильность разделения и подсчи-
тывают соотношение между стадиями. При очень небольшом навыке этот
способ дает возможность абсолютно безошибочно определять стадии ли-
чинок и отнимает при этом минимальное количество времени.
КУКОЛКА
Зрелая личинка IV стадии после линьки переходит в фазу куколки.
На этой фазе происходят сложные процессы гистолиза и гистогенеза,
заканчивающиеся образованием внутри куколочной шкурки готового
к вылету вполне сформированного взрослого комара. Куколки не пи-
таются и с внешней средой связаны непосредственно только процессами
газообмена. Нормально они свешиваются вниз с поверхностной пленки
воды и их дыхательная система находится в прямом контакте с атмо-
сферным воздухом. Куколки способны к очень быстрым активным дви-
жениям и опускаются на дно, водоема при внешних раздражениях. При
отсутствии последних они могут находиться на поверхности неопределенно
долгое время. Длительность куколочной фазы в среднем не превышает
2—2.5 суток. Первыми всегда вылетают самцы.
42
ВВЕДЕНИЕ
Куколка сбоку имеет вид запятой, а сверху — неправильного овала.
Ее тело состоит из двух отделов: массивной головогруди и более тонкого,
уплощенного дорсовентрально брюшка. Верхняя и передняя ‘ части го-
ловогруди заняты грудным отделом, нижняя часть переднего отдела — го-
ловой. Развивающиеся придатки головы и груди занимают нижнюю и
заднюю части головогруди. Сильно пигментированные глаза просвечи-
вают сквозь прозрачную кутикулу. Над ними по бокам спинки отходят
куколочные рожки, суженные у основания и 'расширяющиеся к свобод-
ному концу, у Anophelinae сильнее, чем у Culicinae. Их отверстия скошены
внутрь, но при нахождении куколок на поверхности они лежат в плоско-
сти поверхностной пленки. От основания рожек отходит трахея, соеди-
няющая их полость с переднегрудной стигмой развивающегося комара.
Прямой контакт куколочных рожек с его трахейной системой устанавли-
вается еще у личинок IV стадии за несколько часов до окукления. При
этом полость рожек заполняется воздухом. У куколок Mansonia, в связи
•с особенностями их дыхания, рожки дистально заострены и сходятся
на концах.
Брюшко куколки, как и у личинки, состоит из девяти члеников,
из которых последний — морфологически X, а предшествующий ему
VIII — сложный, состоящий из слившихся вместе VIII и IX члеников
брюшка. На заднем крае спинной стороны VIII членика находятся два,
заходящих внутренними краями друг за друга плавника — органы дви-
жения куколки. Каждый из них имеет вид неправильно овальной пла-
стинки, сидящей на суженном основании. Пластинка укреплена продоль-
ным срединным ребром, не доходящим до ее свободного края, часто не-
сущего ряд зубчиков или мелких шипов. У конца ребра отходит воло-
сок, немного ковнутри от основания которого находится значительно
более короткий добавочный волосок. Интересно отметить, что парные
зачатки плавников куколки закладываются у личинок в заднем отделе
дыхательной трубки (Culicinae) или между поддерживающими стигмаль-
ную пластинку образованиями (Anophelinae), гомологичными сифону.
Последний членик брюшка имеет вид узкого кольца, от брюшной
дуги которого отходит пара терминальных лопастей, лежащих между
основаниями плавников. Это половые придатки, очень короткие у самок
и значительно более длинные у самцов. Таким образом, по степени их раз-
вития можно определять пол куколки.
На спинной стороне I членика брюшка находится пара звездчатых
волосков, напоминающих аналогичные волоски на спинной стороне I—
VII члеников брюшка личинок Anopheles и выполняющих ту же функцию.
У личинок Mansonia они отсутствуют. Относительное развитие и характер
ветвления волосков III—VIII члеников брюшка имеют значение для
видового определения куколок. III—VII членики имеют на спинной сто-
роне восемь пар волосков, VIII членик — 3 пары.
.Так как точное определение вида лучше проводить по личинкам и
взрослым комарам, мы не даем описаний родовых и видовых отличий
по куколочной фазе, как не имеющих значения в практике определения,
особенно при краткости существования куколок.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Палеонтологические данные по семейству Culicidae настолько бедны
и фрагментарны, что базироваться на них при выяснении закономерностей
современного географического распространения группы совершенно не-
возможно. Геологический возраст группы с точностью неизвестен, од-
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
43
нако целый ряд примитивных признаков, свойственных Culicidae, как
в имагинальной, так и в личиночной фазе, говорит о значительной древ-
ности группы. Кровососущие комары появились едва ли позднее горского
периода (Edwards, 1923). Почти полное отсутствие в то время эвритерм-
ных позвоночных не исключало возможности эволюции группы. И в совре-
менных нам условиях некоторые виды комаров, например Culex territans,
явно предпочитают кровь рептилий и амфибий.
Достоверные палеонтологические данные по Culicidae имеются лишь
начиная с эоцена (именно из эоцена Северной Америки), откуда описаны
2 вида, относимые к роду Culex. Олигоценовая фауна комаров пред-
ставляется более богатой, однако интересно отметить, что в нижнеолиго-
ценовом янтаре Прибалтики кровососущие комары не найдены. Янтарь
дает лишь представителей других близких семейств, среди которых
кровососов нет, а именно Dixidae и Chaoboridae. Из олигоценовых отло-
жений известно около 10 видов комаров, относимых к современным родам:
Culex, Mansonia, Aedes, Anopheles.
Современная нам фауна кровососущих комаров (семейство Culicidae)
не может быть признана особенно богатой* всего к настоящему времени
описано около 2500 видов, относящихся к 30 родам (Stone, Knight
.a. Starcke, 1959). Эта цифра отнюдь не исчерпывает, конечно, всего бо-
гатства форм группы, которые имеются в природе. Но, принимая во вни-
мание чрезвычайно интенсивные исследования фауны комаров, произ-
водившиеся в течение последних десятилетий во многих странах земного
шара, нет оснований ожидать значительного и быстрого увеличения этой
цифры.
По своему географическому распространению комары в основном
группа тропическая. В тропиках наблюдается не только богатство видо-
вого состава, но и глубокая древняя дифференциация отдельных фаун,
которая выражается в наличии в каждой зоогеографической области
тропической зоны резко обособленных эндемических групп.
Особенно резко своеобразие фауны комаров сказывается в Неотро-
пической области, которая заключает в себе характерные эндемичные
роды и подроды, причем некоторые , из них достаточно богаты видами.
К таким принадлежат прежде всего роды трибы Sabethini, характерной
преимущественно для Неотропической области: Trichoprosopon (с 32 ви-
дами), Phoniomyia (22 вида), Sabethes (21 вид), Limatus (8 видов). Принад-
лежащий к той же трибе довольно обширный род Wyeomyia (около 90
видов) тоже ограничен в своем распространении преимущественно пре-
делами Неотропической области. Лишь немногие виды данного рода
проникают в южные районы Неарктики. Кроме того, эндемичными в Не-
отропической области являются некоторые группы подсемейства Ano-
phelinae: род Chagasia и подроды (рода Anopheles) Stethomyia, Kerteszia,
Lophopodomyia. He строго эндемичны, но весьма характерны для Неотро-
пической области такие группы, как роды Psorophora и Deinocerites,
подроды Melanoconion (Culex) и Nyssorhynchus (Anopheles).
Большим богатством в видовом отношении отличаются также фауны
комаров Эфиопской и Ориентальной областей, однако дифференциация
этих фаун не так резко выражена, как в Неотропической области. Фауна
Эфиопской области характеризуется наличием только одного эндеми-
ческого рода, именно Eretmapodites с 23 видами, а также значительным
развитием следующих групп: подродов Myzomyia (Anopheles), Stegomyia
:и Aedimorphus (Aedes), родов Ficalbia, Mansonia и Uranotaenia.
В Ориентальной области в качестве эндемических родов выступают,
^помимо некоторых мелких групп, роды Торотуга (21 вид), Heizmannia
44
ВВЕДЕНИЕ
(18 видов) и в особенности Armigeres (41 вид); по одному виду из двух
последних родов проникает в Палеарктику. В Ориентальной области
большого развития достигают подрод Myzomyia, род Toxorhynchites
el подроды Aedes, Finlaya л Stegomyia рода Aedes.
Фауна Австралии в отношении Culicidae является в сильной степени
обедненной, эндемизм ее выражен слабо. В качестве примеров послед-
него можно назвать род Optfex с одним видом, а также некоторые мелки©
и достаточно слабо дифференцированные подроды рода Aedes.
Что касается фауны комаров Голарктического отдела в целом (Неарк-
тики и Палеарктики), то по сравнению с фауной тропиков она, как и
фауна Австралии, является (в абсолютных цифрах видов) сильно обед-
ненной. Ее значительно легче охарактеризовать отрицательными при-
знаками — отсутствием тех или иных групп, нежели положительными —
их наличием. К наиболее характерным чертам фауны комаров этого>
отдела относятся: значительное видовое богатство подрода Ochlerotatus
(род Aedes), заметная роль в фауне родов Culiseta и Anopheles. К наиболее
существенным отрицательным чертам ее фауны могут быть отнесены:
слабое развитие (в Неарктике — полное отсутствие) подрода Myzomyia,
чрезвычайная незначительность роли в фауне таких групп, как роды
Toxorhynchites, Tripteroides, Uranotaenia, Armigeres, подроды Aedes el
Aedimorphus, относительная бедность видами рода Culex, наконец, пол-
ное отсутствие многих тропических групп комаров, упомянутых выше
при характеристике фаун тропических зон.
В качестве общей характерной черты фауны комаров Голарктического'
отдела приходится прежде всего отметить факт широкого распростране-
ния очень многих форм группы. Распространение имеет часто зональный
характер. Рассматривая с этой точки зрения распространение комаров,
мы можем наметить следующие большие разделы Голарктического отдела.
Бореальная область Северной Суши включает в себе Канадскую об-
ласть Неарктики, тундру й лесную зону Палеарктики (за исключением
подзоны широколиственных лесов Дальнего Востока). Основной фон
фауны комаров области составляют виды подрода Ochlerotatus (рода
Aedes), которые преобладают и по количеству видов, и по «живой массе».
Значительную роль в фауне этой зоны играют также некоторые Anopheles,
Culiseta, отдельные виды прочих подродов Aedes, немногие виды Culex.
Отрицательные черты — полное отсутствие Toxorhynchites, Tripteroides,
Uranotaenia, Orthopodomyia, Armigeres, подродов Myzomyia и Stegomyia-,
весьма слабое в видовом отношении развитие рода Culex.
В Бореальную область входит Арктическая подобласть Северной
Суши (ПалеарктикиД-Неарктики), характеризующаяся бедностью видо-
вого состава исключительно из родов Aedes (подрод Ochlerotatus) el Culiseta.
и полным отсутствием всех других групп.
Средиземноморская подобласть Северной Суши, обнимающая Среди-
земноморье, Малую Азию и прилегающие страны, Южный берег Крыма,
Закавказье и большую часть Средней Азии, характеризуется наличием
в составе фауны группы Myzomyia, которая в южных частях области
играет значительную роль, затем родов Uranotaenia, Orthopodomyia,
большим значением рода Culex, который по числу видов явно преобладает
над Ochlerotatus-, последний представлен очень небольшим числом видов.
Палеархеарктическая (Маньчжурская) подобласть Северной Суши
занимает восточную и северо-восточную часть Китая, п-ов Корею, Японию,
Приморский край и Южное Приамурье. Характерными чертами фауны
комаров этой области являются наличие Toxorhynchites, богатство Finlaya,
Stegomyia el ориентальных Culex. р.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
45
Недостаточно ясным представляется характер фауны комаров Монго-
лии, горных хребтов и нагорных пустынь Центральной Азии, так как
материалы по фауне комаров этих районов слишком фрагментарны.
Возможно, что количество видов Culicidae этой области окажется значи-
тельно большим, чем это известно в настоящее время.
Что касается фауны комаров южных частей Неарктики (Сонорской
области), то отметим лишь ее весьма своеобразный характер и влияние
на ее фауну неотропических элементов.
В отношении генезиса фауны комаров Голарктического отдела и,
в частности, Палеарктики, принимая во внимание полное отсутствие
в ней эндемизма даже подродового, не только родового порядка, прихо-
дится признать ее в общем значительную молодость. Несколько спорным
такое предположение делает то обстоятельство, что при относительном
единообразии условий обитания комаров темпы эволюционного про-
цесса могут быть несколько замедленными по сравнению с организмами,
подвергающимися боле? разнообразным влияниям внешней среды.
Формирование современной фауны Culicidae Палеарктики произошло,
конечно, неодновременно в разных частях области и не в одном центре.
На основании анализа мировой фауны Culicidae можно построить сле-
дующую схему постепенного ее создания. В доледниковое время имело
место наступление южных элементов на север. В отношении фауны ко-
маров это сказалось в иммиграции, с одной стороны, эфиопских элементов
в западной части Палеарктики, с другой — ориентальных элементов —
в ее восточной части.
Фауна комаров современного нам Средиземноморья складывалась под
определенным и, по-видимому, продолжительным влиянием эфиопской
фауны. Эти отношения находят свое отражение и в характерных чертах
видового состава комаров территории СССР; здесь представлена большая
группа видов (23), характерных для Средиземноморской подобласти;
некоторые из них распространены и в Эфиопской области. Средиземно-
морские виды занимают значительное место среди комаров юга Украины,
Кавказа и Средней Азии. Ареалы некоторых видов данной группы про-
•стираются далеко на север. Так, Anopheles algeriensis найден в Эстонии
.(Ремм, 1957), Aedes geniculatus — в Чувашии и Татарии (Волкова, 1956).
Замечательно, что некоторые из эфиопских видов проникли довольно
.далеко в северном и северо-восточном направлениях, например Culex
dheileri — до Харьковской области, a Culiseta longiareolata — даже до
южных районов Западной Сибири.
Аналогичное явление — проникновение южных элементов на север,
.но, пожалуй, еще в более резко выраженной степени, наблюдалось на
’востоке Палеарктики — в пределах Палеархеарктической (Маньчжур-
ской) подобласти: ее фауна является в основе производной фауны ориен-
'тальной. На крайнем юго-востоке СССР обнаружено 18 видов комаров,
характерных для Маньчжурской подобласти (включая ориентальные
виды). Распространение комаров данной группы в пределах СССР, как
правило, ограничивается Приморьем и югом Хабаровского края; некото-
рые виды найдены только на крайнем юге Приморья, как например То-
. xorhynchites' christophi, Aedes nipponicus, A. flavopictus, A. nobukonis,
A. aureus, Culex hayashii. Заслуживает внимания обнаружение здесь
довольно большой группы ориентальных видов рода Culex: С. йогах,
С. fuscanus, С. tritaeniorhynchus, С. bitaeniorhynchus, С. sinensis, С. whit-
тог ei, С.jacksoni.
Фауна бореальной и арктической зон с ее богатством и разнообразием
.видов подрода Ochlerotatus не имеет корней в фауне тропических зон
46
ВВЕДЕНИЕ
восточного полушария. Бореальная (и арктическая) фауна комаров
Палеарктики имеет тесные генетические связи с Неарктикой, где группы,,
характерные для бореальной зоны, достигают особенно пышного разви-
тия; так, 23 вида Culicidae являются общими для Палеарктики и Неарк-
тики; все они найдены в Советском Союзе. Мы вправе связывать генезис
нашей бореальной и арктической фауны комаров с Неарктикой.
Пути расселения бореальных комаров из неарктического центра
в Палеарктику не вполне ясны. Иммиграция, по-видимому, осуществля-
лась и западным, и восточным путем. Более ранний (до отделения Америки
от Европы) западный путь послужил для обмена некоторых форм, свой-
ственных более южным широтам западной Палеарктики. Такова, на-
пример, группа видов рода Orthopodomyia, близких к О. pulchripalpis.,
В Палеарктике эта группа представлена одним видом — О. pulchripalpis,
который распространен в Средиземноморье и доходит до южной Англии,,
а на восток до Азербайджана. Два близких вида этой группы свойственны
восточным штатам Северной Америки. Хорошо морфологически обо-
собленная группа rusticus подрода Ochlerotatus, распространенная в боль-
шей части Средиземноморья и далее на север и северо-восток до Поволжья,.
Западной Сибири и Забайкалья, имеет своего представителя также в Се-
верной Америке — A. trichurus Dyar.
Помимо западного, несомненно существовал еще и восточный путы
обмена фауны Неарктики и Палеарктики, за что говорит факт преиму-
щественного распространения некоторых общих для обоих областей видов
в западной части Неарктики, а также большое сходство видового- состава
комаров Северо-Восточной Азии и Северо-Западной Америки. Так, из
27 видов комаров, найденных на Аляске (Gjullin, Sailer, Stone a. Travis,.
1961), 17 видов обнаружены также в Сибири.
Относительно геологического времени, к которому можно приуро-
чить расселение в Палеарктике тех или иных элементов фауны комаров,
сказать что-либо трудно. С несомненностью можно констатировать лишь,
одно, а именно, что заселение бореальной зоны Палеарктики основным,
контингентом форм комаров произошло в верхнетретичное время и во
всяком случае до плейстоцена. Об этом говорит наличие в горных цепях
юга Палеарктики (Пиренеи, Кавказ, система Тянь-Шань) ряда форм
Culicidae, свойственных лесной зоне; таковы Aedes pullatus и Culiseta
alaskaensis. Распространение южных — эфиопских и ориентальных —
элементов до наступления бореальной фауны, осуществлялось, вероятно,,
на значительно большем пространстве, нежели теперь. Показателем
такого более широкого распространения южных форм являются приве-
денные выше факты наличия в фауне СССР довольно большого количества
эфиопских и особенно ориентальных видов.
При рассмотрении вопросов географического распространения кома-
ров в Палеарктике необходимо учесть, что ареалы большинства видов-.
Culicidae огромны и эндемизм отдельных географических районов отно-
сительно слабо выражен. Как пример обширности ареала часто приводят
всесветное распространение Culex plpiens. Но это типично синантропный
вид и его широкое расселение произошло, вероятно, в недавнее время
вслед за человеком. Более показательным примером может служить,
«дикий» Aedes vexans, ареал которого охватывает Европу, Азию, Австра-
лию, Океанию, Африку и Северную Америку.
С другой стороны, мы знаем лишь очень мало палеарктических видов ко-
маров с незначительными по площади ареалами. Возможными примерами
являются Culiseta setivalva (Крым, Западное Закавказье, Болгария,.
Анатолия), Aedes alektorovt (Приморье, Южное Приамурье). Но и в этих
ЗНАЧЕНИЕ КОМАРОВ ДЛЯ ЧЕЛОВЕКА 47
немногих случаях впечатление о малой протяженности ареала может
быть обусловлено лишь недостаточностью наших знаний.
Характерно, что в такой хорошо изученной энтомологами области,
как Кавказ, где известно много эндемичных видов насекомых разных
групп, нет ни одного эндемика среди комаров. Вообще для комаров не
характерен эндемизм, ограниченный горными районами. По крайней
мере среди комаров фауны СССР не известно ни одного вида, характер-
ного только для горных ландшафтов, тогда как среди других кровососу-
щих двукрылых — мошек, мокрецов и слепней — таких видов немало.
Единственный вид комара, который обычно характеризуется как гор-
ный — Aedes pullatus, в более северных частях своего ареала встре-
чается, и на равнине.
Всего по сведениям, имеющимся в настоящее время (1966 г.), список
комаров фауны СССР включает 85 видов восьми родов; половину общего
количества составляют виды рода Aedes. За 30 лет, прошедших со вре-
мени выхода предыдущего издания настоящей книги, список видов уве-
личился на 17 единиц. Увеличение произошло как за счет описания
новых видов (всего пять), так в особенности в результате обнаружения
17 видов, ранее неизвестных в СССР. С другой стороны, получены данные,
говорящие против признания видовой самостоятельности 4 видов, кото-
рые либо исключены из списка как сомнительные (Aedes duplex Mart.,
Culiseta silvestris Shing.), либо сведены в подвиды: Aedes esoensis, A. ros-
sicus. Один вид (Aedes lepidonotus'), впредь до получения достоверных
данных о его наличии, исключен из списка комаров, встречающихся
в СССР. Конечно, указываемое в настоящей работе количество видов
комаров фауны Советского Союза нельзя считать окончательным. Оно-
несомненно увеличится, но это увеличение вряд ли будет значительным..
ЗНАЧЕНИЕ КОМАРОВ ДЛЯ ЧЕЛОВЕКА
КОМАРЫ КАК КРОВОСОСЫ 1
Комары являются существенным элементом гнуса, каковым термином
обозначают всю совокупность крылатых насекомых, являющихся бичом
многих мест в тайге, тундре и в других ландшафтных зонах. Объектом
вредоносного действия этих кровососов является как сам человек, так
и домашние животные. Экономическое значение комаров определяется
обоими этими моментами. Непосредственное вредоносное действие ко-
маров связано с тем, что самки их наряду с различными растительными
и иными соками пьют кровь человека, млекопитающих, птиц и других
животных, которую добывают, прокалывая кожные покровы своих хо-
зяев колющими ротовыми органами.
Рассмотрим значение комаров как кровососущих паразитов. Комары —
временные паразиты, так как их контакт с хозяином бывает весьма крат-
ковременным; срок контакта определяется временем, потребным для
прокалывания кожи и насыщения кровью. Самая процедура прокалыва-
ния кожи ротовыми частями малоощутима или вовсе неощутима, что-
объясняется очень малой толщиной их: диаметр хоботка малярийного
комара — около 0.055 мм. Тотчас же при начале сосания крови через
канал подглоточника в покровы хозяина впрыскивается слюна комара,
являющаяся секретом пары лежащих в его груди слюнных желез. В общем
вводится ничтожное количество слюны, составляющее, по данным опы-
1 Составил Е. И. Павловский.
48
ВВЕДЕНИЕ
тов с Aedes aegypti, от 1 до 13 мкг, в среднем 4.7 мкг (Devine, Venard
a. Myser, 1965). Однако эффект укола (считая весь период насасывания
крови) во многих случаях весьма ощутим. Степень и характер эффекта
зависят от вида комара, от чувствительности субъекта и от дозировки
уколов (одиночное или массовое нападение комаров).
Как и по отношению к другим кровососущим паразитам, люди обла-
дают различной чувствительностью к уколу комаров определенного
вида — от полной нечувствительности, через реакцию средней силы до
большой степени чувствительности. Такие вариации могут зависеть от
конституциональных особенностей субъекта и от степени его аллергиче-
ского состояния (анафилаксия). Не касаясь более этих причин, отметим,
что многие малочувствительны к уколам малярийных комаров — на
месте укола образуется лишь небольшое покраснение кожи и маленький
узелок. В то же время после укола различных Aedes быстро образуется
волдырь, достигающий через 10—20 Минут размера до 12 мм в диаметре.
Субъективные ощущения проявляются в виде зуда и жжения. На уколы
Aedes резко реагируют различные птицы, тогда как уколы комаров рода
Culex для них, видимо, малоощутимы. Иногда уколы комаров ведут
к местным нагноениям. Такое осложнение является вторичным. Объяс-
няется оно тем, что ранее комар сосал какой-либо зараженный материал
и механически инокулировал микробы в покровы человека. Грубые
расчесы кожи также могут вести к вторичным осложнениям.
При массовых уколах общая реакция кожных покровов, равно как
и субъективные ощущения, соответственно возрастают. Волдыри могут
сливаться вместе, кожные покровы отекают, причем эти явления дер-
жатся до суток. Если одиночный укол комара вполне терпим, то массовые
их нападения повергают человека и домашних животных в весьма тягост-
ное и даже непереносимое состояние. В сезон массового лёта комаров
(в особенности Aedes) невозможны полевые работы, по крайней мере
в часы активного состояния, комаров; могут быть затруднены заготовки
леса, строительство плотин, мостов, дорог и т. д.
Страдают от комаров и домашние животные. Скот не может работать
и спокойно пастись на пастбище. У крупного рогатого скота наблю-
дается уменьшение нагула и удойности. Все эти явления зависят не от
потери крови, выпиваемой комарами, а от токсического действия слюны,
вводимой в организм, и от побочных воздействий уколов. Именно токси-
ческие свойства слюны комаров как таковой являются первопричиной
кожных и общих реакций (Павловский, Штейн и Перфильев, 1928; Hecht,
1929).
КОМАРЫ КАК ПЕРЕНОСЧИКИ ВОЗБУДИТЕЛЕЙ БОЛЕЗНЕЙ
Роль- комаров как переносчиков возбудителей болезней человека
исключительно велика. Передаваемые комарами болезни наиболее харак-
терны для тропических стран (филяриатозы, желтая лихорадка, денге),
Но некоторые из болезней данной группы распространены и в странах
умеренного пояса (малярия, комариные энцефалиты и др.). В профилак-
тике болезней, возбудители которых передаются комарами, видную роль
играет борьба с переносчиками. Ее успех в значительной степени зависит
от знания видового состава переносчиков и их видовых биологических
особенностей.
Весьма велика, хотя слабо изучена, роль комаров как переносчиков
возбудителей болезней сельскохозяйственных и домашних животных.
Кроме того, комары несомненно передают множество возбудителей болез-
ЗНАЧЕНИЕ КОМАРОВ ДЛЯ ЧЕЛОВЕКА
49
ней диких животных. Но эта проблема представляет собой почти нетро-
нутое поле исследований.
Возбудители, в распространении которых комары играют ту или
иную роль, относятся к паразитам животной природы (плазмодии, филя-
рии), к бактериям и вирусам.
Комары и малярийные плазмодии
Хорошо известна роль комаров рода Anopheles как переносчиков
всех четырех видов плазмодиев — возбудителей малярийных заболева-
ний человека: Plasmodium vivax, Р. ovale, Р. malariae и Р. falciparum.
Источником заражения комаров служат люди с гамонтами плазмодия
в крови. В организме переносчика происходит оплодотворение плазмо-
диев и совершается часть цикла их развития и размножения — спорого-
ния, заканчивающаяся появлением спорозоитов в слюнных железах ко-
мара. Заражение человека происходит в момент укола через слюну,
содержащую спорозоиты (специфическая инокуляция).
По-видимому, большинство видов Anopheles в условиях опыта в той
или иной степени способны передавать малярийных плазмодиев от чело-
века человеку. Но в естественной обстановке эпидемиологическое значе-
ние различных видов весьма неодинаково, что зависит от многих причин,
как-то: восприимчивости к заражению плазмодиями того или иного вида,
продолжительности жизни комара, частоты нападения на человека,
длительности сезона активности и т. д. (Беклемишев, 1941).
Лишь относительно небольшое количество видов в различных геогра-
фических областях земного шара может быть отнесено к важным (факти-
чески или потенциально) переносчикам малярии1:
Области Виды Anopheles
Европа (кроме Средиземно- морья), Сибирь An. maculipennis Mg.
Южная Европа, Закавказье An. maculipennis Mg. An. superpictus Gr.
Средняя Азия An. maculipennis sacharovi Favre An. superpictus Gr. An. pulcherrimus Theob.
Восточная Азия An. hyrcanus Pall. (s. 1.)'
An. pattoni Christ.
Юго-Восточная Азия An. minimus Theob. An. maculatus Theob.
An. sundaicus Rod. An. jeyporiensis James An. leucosphyrus Don. An. letifer Sand. An. umbrosus Theob. An. barbirostris V. d. Wulp An. aconiius Don. An. hyrcanus Pall. (s. 1.)
Австралия, Новая Гвинея An. punctulatus Don. An. farauti Lav.
Индостан An. stephensi List. An. culicifacies Giles An. minimus Theob. An. sundaicus Rod. An. fluviatilis James An. philippinensis Ludl.
1 Комбинировано по разным источникам: Russell et al., 1946; Horsfall, 1955;
Foote a. Cook, 1959, и др. Данные относятся ко времени широкого распространения
малярийных заболеваний. Теперь во многих странах малярия ликвидирована или
количество заболеваний резко сократилось.
4 Фауна СССР, А. В. Гуцевич и др. ;
50
ВВЕДЕНИЕ
Передняя Азия
Северная Африка
Африка к югу от Сахары
Южная Америка
Центральная Америка
Северная Америка
An. maculipennis Mg. (s. 1.)
An. superpictus Gr.
An. stephensi List.
An. maculipennis Mg. (s.fl.)
An. sergenti Theob.
An. multicolor Camb.
An. pharoensis Theob.
An. gambiae Giles
An. funestus Giles
An. nili Theob.
An. pharoensis Theob.
An. rufipes Gough
An. darlingt Root
An. albimanus Wied. .
An. aquas alts Curry
An. pseudopunctipennis Theob
An. albitarsis Arr.
An. albimanus Wied.
An. pseudopunctipennis Theob
An. aquasalis Curry
An. quadrimaculatus Say
An. freeborni Aitk.
Так, в прошлом, когда малярия была широко распространена, и»
девяти встречающихся в СССР видов Anopheles два рассматривались как
наиболее важные переносчики: An. maculipennis — почти на всей тер-
ритории, где встречались малярийные заболевания, и An. superpictus —
в Средней Азии и Восточном Закавказье. Еще три или четыре вида могли
быть отнесены к переносчикам, имевшим местное или второстепенное
значение, — An. pulcherrimus, An. hyrcanus, An. claviger, возможно,,
местами и An. plumbeus. Остальные три вида (An. algeriensis, An. marteri^
An. lindesayi) не играли никакой роли в распространении малярии.
В Индии из 42 встречающихся там видов Anopheles только семь относят
к важным переносчикам малярии (Roy, 1946). В Малайе в качестве важ-
ных переносчиков малярии рассматриваются шесть из 47 местных видов
Anopheles (Sandosham, 1959). В Бразилии из 50 видов Anopheles четыре
считаются основными переносчиками и пять видов — второстепенными;
(Ferreira, 1964).
Продолжительность гонотрофического цикла у малярийных комаров,
как правило, значительно короче, чем длительность спорогонии плазмо-
дия при той же температуре. Поэтому спорозоиты могут появиться только-
у самок, проделавших 3—4 гонотрофических цикла (Беклемишев, 1944).
Если продолжительность жизни комаров в данных условиях невелика,
то преобладающая часть комаров отмирает, прежде чем у них может
завершиться спорогония плазмодия (Детинова, 1962).
Когда в данной местности ведется борьба с комарами путем обработки
помещений стойкими инсектицидами, сокращается численность комаров
и уменьшается продолжительность их жизни. Самки не доживают до-
эпидемиологически опасного возраста, т. е. до того времени, когда у них
могут появляться спорозоиты в слюнных железах. Это ведет к прекраще-
нию циркуляции плазмодия. Обработка помещений стойкими инсектици-
дами служила (и служит) методом, при помощи которого (в чистом виде
или в сочетании с другими методами) малярия ликвидирована во многих
странах, включая и Советский Союз.
В некоторых странах малярия ликвидирована на большей части тер-
ритории. Но в отдельных районах, несмотря на тщательное проведение
мероприятий, Цепь передачи плазмодия полностью разорвать не удается,
заболевания продолжают регистрироваться. Это так называемые проблем-
ные районы (Комитет экспертов по малярии, 1965). Причины возникаю-
ЗНАЧЕНИЕ КОМАРОВ ДЛЯ ЧЕЛОВЕКА
51
щих затруднений могут быть связаны с «энтомологическим фактором»:
с появлением у комаров устойчивости к инсектицидам, с изменением
поведения переносчиков, с появлением новых для данной местности ви-
дов или подвидов комаров, с заменой эндофильных Anopheles экзофиль-
ными и т. д. Преодоление возникающих трудностей возможно только
на основе глубокого изучения видовой экологии малярийных комаров.
Борьба с малярийными комарами, которая велась на протяжении
длительного времени, привела не только к сокращению численности
комаров, но и к полному их истреблению во многих пунктах и даже в от-
дельных районах. Иллюстрацией могут служить данные (Калмыков,
1965) относительно постепенного сокращения области распространения
An. maculipennis sacharovi и An. pulcherrimus в Таджикистане.
Комары, как известно, являются также переносчиками плазмодиев,
паразитирующих у животных. Кажущееся исключение составляет Нера-
tocystis kochi — паразит обезьян Африки, который передается мокрецами,
как, вероятно, и другие виды того же рода. Но род Hepatocystis, по-види-
мому, должен быть исключен из семейства плазмодиев и отнесен к се-
мейству Haemoproteidae.
Что касается плазмодиев обезьян, то их переносчиками служат раз-
личные виды Anopheles. Так, паразит макак Plasmodium cynomolgi bastia-
nellii (который может паразитировать и у человека) в условиях опыта
развивался до стадии спорозоитов у Anopheles hyrcanus, An. kochi Don.,
An. maculatus, An. philippinensis Ludl. и An. sundaicus (Warren et al.,
1963). Естественным переносчиком данного вида плазмодия в Малайе
считают An. balabacensis Bais., An. leucosphyrus и An. hackeri Edw. Через
комара An. freeborni была успешно осуществлена передача Р. brasilianum
от обезьяны A teles geoffroyi другим обезьянам того же вида, а также
людям-добровольцам (Contacos et al., 1963).
Переносчиками плазмодиев грызунов также оказались комары рода
Anopheles. Наиболее изучен Plasmodium berghei, который широко исполь-
зуется для экспериментального изучения малярийных паразитов. В усло-
виях опыта удалось осуществить передачу этого паразита от грызуна
грызуну через комаров рода Anopheles (в частности An. stephensi,
An. quadrimaculatus; см.: Yoeli, 1965), а также через Aedes aegypti (см.: Raf-
faele, 1965). В естественных условиях, в Конго (Киншаса), переносчиком
данного вида плазмодия является Anopheles dureni Edw. (Vincke, 1964).
Беспозвоночными хозяевами плазмодиев птиц, как известно, служат
преимущественно комары рода Culex, а для некоторых видов (Plasmodium
gallinaceum, Р. lophurae, Р. fallax и др.) — комары рода Aedes. Получены
данные о возможности развития некоторых птичьих плазмодиев в орга-
низме малярийных комаров, например Р. gallinaceum у An. quadrimacu-
latus. Подробности можно найти в обзорной работе Хаф (Huff, 1965).
Таким образом, комары рода Anopheles оказались переносчиками не
только плазмодиев человека, но и плазмодиев животных — обезьян,
грызунов, птиц и др. В некоторых случаях плазмодии животных пере-
даются комарами, почти не нападающими на людей, как например
An. hackeri — переносчик нескольких видов плазмодиев обезьян в Ма-
лайе (Wharton et al., 1964). Но там же An. letifer оказался переносчиком
как человеческих плазмодиев, так и Р. traguli, паразитирующего у мел-
кого копытного — оленька (Wharton et al., 1963). При такой ситуации,
конечно, затрудняется оценка результатов исследования комаров на
естественную зараженность плазмодиями.
Получены также предварительные данные о возможной роли комаров
как переносчиков плазмодиев пресмыкающихся. У небольшого числа
4*
52
ВВЕДЕНИЕ
экземпляров некоторых видов (Culex territans и др.), пивших кровь яще-
риц, зараженных Р. floridense, были обнаружены ооцисты (Jordan, 1964).
Для полного выяснения вопроса о переносчиках плазмодиев рептилий
нужны дальнейшие исследования.
Комары и филярии
Хорошо известна роль комаров как промежуточных хозяев некоторых
нематод подотряда Filariata (по традиции обозначаемых ниже как филя-
рии). Комары инвазируются, поглощая с кровью позвоночного хозяина
циркулирующих в его крови микрофилярий. В теле комара личинки
филярий развиваются в грудных мышцах, в полости тела и жировом
теле или в мальпигиевых сосудах. Развитие в насекомом завершается
появлением зрелых инвазионных личинок, которые мигрируют в хоботок
комара и при кровососании внедряются в кожу позвоночного. В орга-
низме переносчика филярии не размножаются. Инвазионного состояния
достигает лишь незначительная часть личинок, поглощенных переносчи-
ком при кровососании. .
В результате длительной сопряженной эволюции паразита и его
промежуточного хозяина выработались замечательные приспособления.
Наиболее известное из них — периодичность появления микрофилярий
в периферической крови позвоночного соответственно суточному ритму
активности насекомого-переносчика. Так, микрофилярии вухерерии
Банкрофта (Wuchereria bancrofti) появляются в периферической крови
йнвазированных людей ночью, что в общем соответствует суточному
ритму активности большинства видов комаров-переносчиков. Исключение
составляет распространенная в Полинезии так называемая апериодиче-
ская разновидность паразита, промежуточным хозяином которой слу-
жит Aedes polynesiensls Маг., активный и днем (Mattingly, 1962).
Известно свыше 30 видов переносимых комарами филярий — парази-
тов человека и животных; подробности можно найти в обзорных работах
(Lavoipierre, 1958; Hawking a. Worms, 1961; Nelson, 1964). При этом
надо учесть, что жизненные циклы расшифрованы не более чем у десятой
части известных видов подотряда Filariata.
Из филярий, паразитирующих у человека, комарами передаются
Wuchereria bancrofti и Brugia malayi (об их переносчиках см.: Инвазии,
вызываемые Wuchereria и Brugia, 1963; Edeson a. Wilson, 1964). В орга-
низме комаров личинки обоих видов филярий развиваются в грудных
мышцах.
Особое внимание привлекает проблема вухерериоза; Wuchereria ban-
crofti инвазирует десятки миллионов людей в тропических и субтропиче-
ских странах обоих полушарий. Список комаров, в организме которых
личинки этой нематоды могут развиваться до инвазионной стадии, вклю-
чает значительное количество видов нескольких родов, преимущественно
Culex и Anopheles. Но у многих видов комаров тех же родов личинки
W. bancrofti либо вовсе не развиваются, либо их развитие замедляется
или приостанавливается на какой-либо промежуточной стадии. «Хоро-
шими» переносчиками могут служить виды различных родов, в то время
как у близких к ним видов тех же родов личинки паразита не развиваются.
Значительные различия могут быть и между подвидами одного вида.
Так, Culex pipiens fatigans известен как «классический» переносчик
W. bancrofti, тогда как в организме Culex pipiens pipiens личинки этой
нематоды не находят благоприятных условий для развития. Различия
в степени восприимчивости к заражению паразитом могут быть между
ЗНАЧЕНИЕ КОМАРОВ ДЛЯ ЧЕЛОВЕКА
53
разными популяциями одного вида комаров. Наблюдаются и индивидуаль-
ные отличия, что относится не. только к заражению комаров филяриями,
но и другими паразитами — плазмодиями, вирусами.
Аналогично тому что было отмечено выше в отношении малярийных
плазмодиев, фактическая роль комара того или иного вида как пере-
носчика филярий зависит от многих факторов. В естественных условиях
лишь немногие виды комаров играют важную роль в распространении
филяриозов человека. В большинстве стран, где эндемичен вухерериоз
Банкрофта, главнейшим переносчиком является циркумтропический си-
нантропный комар С. pipiens fatigans. Значительную роль в распростра-
нении паразита в тех или иных странах, по-видимому, играют также
Anopheles gambiae (Африка), An. hyrcanus и Aedes togoi (Восточная Азия),
An. minimus та An. nigerrimus Giles (Юго-Восточная Азия), An. farauti
и An. bancrofti Giles (Новая Гвинея и Соломоновы острова) и некоторые
другие виды. До сего времени Wuchereria bancrofti известна только как
специфический паразит человека; естественно, что в распространении
этого паразита главную роль играют синантропные виды комаров или
во всяком случае такие, для которых кровь человека служит обычным
источником питания. л
Для другого переносимого комарами вида филярии человека —
Brugia malayi, паразита, широко распространенного в странах Юго-
Восточной Азии, важнейшими переносчиками являются Aedes togoi,
Anopheles hyrcanus, An. nigerrimus и виды рода Mansonia. Возможно
заражение человека комарами, получившими микрофилярий от живот-
ных, так как данная филярия паразитирует не только у людей, но и у до-
машних и диких животных (кошки, обезьяны и др.). Переносчиками
возбудителя от животного животному служат в основном виды рода
Mansonia.
В Советском Союзе известны только завозные случаи вухерериоза.
Но нельзя считать исключенной возможность появления и местных слу-
чаев, поскольку в фауне комаров СССР представлены виды, являющиеся
доказанными переносчиками филярий, а температурные условия, на-
пример в Закавказье и Средней Азии, допускают возможность развития
личинок филярий в комарах. Заслуживает внимания факт обнаружения
местного очага вухерериоза в одном из горных районов Восточной Анато-
лии (Yucel et Deschiens, 1960).
Для других видов рода Brugia, паразитирующих у животных, проме-
жуточными хозяевами также, по-видимому, являются комары. Так,
детально изучено развитие паразита собак, Brugia ceylonensis, в орга-
низме Aedes aegypti', личинки локализуются в грудных мышцах комара
(Jayewardene, 1963).
Выявлена также роль комаров как промежуточных хозяев некоторых
филярий других родов, паразитирующих у животных. Лучше изучены
комары как переносчики дирофилярий, паразитирующих у собак, —
Dirofilaria immitis и D. repens. Личинки названных паразитов разви-
ваются в мальпигиевых сосудах комаров разных видов — Anopheles
maculipennis, An. claviger, An. hyrcanus, Aedes aegypti, A. albopictus,
A. vexans и др. Дирофиляриоз собак широко распространен на юге СССР
(Украина, Кавказ, Средняя Азия). Дирофилярии обезьян также пере-
даются комарами. Но личинки Dirofilaria aethiops, паразитирующей
у обезьян, развиваются не в мальпигиевых сосудах, а в полости тела и
в жировом теле комаров-переносчиков (Webber, 1955).
Комары оказались промежуточными хозяевами и некоторых филярий,
паразитирующих у сельскохозяйственных животных, в частности нематод
54
ВВЕДЕНИЕ
рода Setaria — S. equina, S. marshalli, S. digitata, S. labiatopapillosa;
относительно последнего вида есть сведения о возможности развития
личинок и в кровососущих мухах.
Наши знания о переносчиках филярий, паразитирующих у диких
животных, весьма отрывочны. Все же установлена роль комаров многих
видов (родов Aedes, Armigeres, Culex, Mansonia, Anopheles) как промежу-
точных хозяев филярий — паразитов диких млекопитающих, птиц,
а также рептилий и амфибий. Особенно интересен тот факт, что комары
оказались промежуточными хозяевами пяти видов рода Foleyella, кото-
рые паразитируют у лягушек. Прослежено развитие личинок этих нема-
тод в полости тела и жировом теле Culex pipiens fatigans, С. р. molestus,
Aedes aegypti. Следовательно, питание комаров некоторых видов на
земноводных — не исключительный факт, а обычное явление.
Комары и арбовирусы
К группе арбовирусов (сокращение от слов arthropod-borne, т. е.
переносимые членистоногими) относятся вирусы человека и животных,
которые размножаются в организме кровососущих членистоногих и
передаются позвоночным насекомыми или клещами при кровососании
черезЖпюну. Известно очень много таких вирусов: по данным, относя-
щимся к 1963 г., их 155, из них 117 были выделены из комаров (Gordon
Smith, 1964). Эти цифры ежегодно возрастают. В распространении транс-
миссивных вирусных инфекций комары играют главную роль (см. обзор-
ные работы: Mattingly, 1960; Chamberlain a. Sudia, 1961; Reeves, 1962,
1965; Сазонова, 1962; Gordon Smith, 1964; Гуцевич, 1964; Pratt, 1964;
см. также: Вирусы, переносимые членистоногими, 1962).
Ниже перечислены наиболее важные виды комаров-переносчиков тех
вирусных инфекций, которые относительно хорошо изучены:
Болезни
Желтая лихорадка
Денге (возбудители — вирусы
денге и чикунгунья)
Японский энцефалит
Энцефалит долины Муррея
Западнонильская лихорадка
Энцефалит Сан-Луи
Восточный энцефалит (Аме-
рика)
Западный энцефалит (Америка)
Венесуэльский лошадиный
энцефалит
Калифорнийский энцефалит
Лихорадка о’ньон-ньон (Аф-
рика)
Важнейшие переносчики
В городах — Aedes aegypti', в при-
родных очагах — А. africanus
Theob., A. simpsoni Theob. и дру-
гие виды подрода Stegomyia (Аф-
рика), Haemagogus spp., Aedes
(Finlaya) leucocelaenus D. et Sh.,
Sabethes chloropterus Humb. и дру-
гие виды (Южная и Центральная
Америка)
Aedes aegypti, A. albopictus и неко-
торые другие виды подрода Ste-
gomyia
Culex tritaeniorhynchus, С. gelidus
Theob., C. vishniii Theob.,
C. pipiens, Aedes togoi и др.
Culex annulirostris Sk. и др.
Culex pipiens molestus, C. univlttatus,
C. antennatus Beck., Aedes aegypti
и др.
Culex pipiens fatigans, C. tarsalis Coq.
Culiseta melanura Coq., Aedes sollici-
tans Walk., A. vexans, Culex res-
tuans Theob. и др.
Culex tarsalis
Mansonia perturbans Walk., M. titil-
lans Walk, и др.
Aedes melanimon D., Culex tarsalis
(предположительно)
Anopheles funestus, An. gambiae
ЗНАЧЕНИЕ КОМАРОВ ДЛЯ ЧЕЛОВЕКА
55
Роль комаров как переносчиков определяется возможностью интен-
сивного размножения вируса в организме насекомого и накопления
вируса в слюнных железах в количестве, достаточном для инфицирования
позвоночного хозяина.
Для большинства арбовирусов характерен широкий круг беспозво-
ночных хозяев. Потенциальными переносчиками того или иного вируса
чаще являются комары разных родов, реже — виды одного рода или
даже подрода. Так, переносчиками вируса денге, насколько известно,
служат лишь немногие виды подрода Stegomyia. Обычно тот или иной
вирус при одинаковых условиях интенсивно размножается в организме
комаров определенных видов, но слабо размножается или вовсе не раз-
множается у комаров близких видов. Таких фактов известно очень много.
Примеры можно найти в упомянутых выше обзорных работах.
Все или почти все вирусные инфекции, передаваемые комарами,
характеризуются природной очаговостью. Виды комаров, которые редко
нападают (или вовсе не нападают) на людей, могут играть заметную
роль в циркуляции вируса в природных очагах. Примером служит Culi-
seta melanura Coq. — важный переносчик вируса восточного энцефалита
от птицы птице. Синантропные же виды комаров могут обеспечивать
распространение инфекции среди людей, передавая вирус от человека
человеку. Такова роль Aedes aegypti и A. albopictus как переносчиков
вируса денге. Наконец, виды комаров с широким кругом хозяев, питаю-
щиеся как на людях, так и на различных животных, могут распростра-
нять вирус среди животных, а также передавать его человеку. Таковы,
например, Culex tarsalis — переносчик западного энцефалита в Северной
Америке, С. tritaeniorhynchus и С. gelidus — переносчики вируса япон-
ского энцефалита в странах Восточной и Юго-Восточной Азии.
При наличии комаров-переносчиков, питающихся как на диких жи-
вотных, восприимчивых к инфекции, так и на людях, вирус может ирра-
диировать из природных очагов в населенную зону. Примером может
служить вспышка желтой лихорадки в Эфиопии в 1960—1962 гг., когда
заболело свыше 15 000 человек при летальности около 85% (Serie et al.,
1964). Связующим звеном между инфицированными животными (обезья-
нами) и человеком служил, по-видимому, комар Aedes simpsoni, который
обитает как в ненаселенной местности (в лесах), так и в селениях, и вблизи
селений, нападая и на обезьян, и на людей.
Роль комаров в распространении вирусов среди диких животных
изучена еще очень слабо. Поскольку одни вирусы циркулируют преиму-
щественно среди птиц, а Другие среди млекопитающих (например, гры-
зунов), возможная роль комаров как переносчиков в значительной сте-
пени определяется их пищевыми предпочтениями.
Человек, как правило, является побочным необязательным звеном
в циркуляции передаваемых комарами вирусов. Но в природе такая
циркуляция может осуществляться в очень широких масштабах, о чем
свидетельствуют результаты серологических исследований животных.
При обсуждении вопроса о переносчиках вирусов должны быть приняты'
во внимание такие биологические особенности комаров, как их подвиж-
ность, частота кровососания и повторяемость гонотрофических циклов,
возможность массового размножения в относительно короткое время.
С учетом этих особенностей могут быть объяснены факты огромных вспы-
шек некоторых передаваемых комарами вирусных инфекций. Таковы
имевшие место в прошлом эпидемии желтой лихорадки в городах Европы
и Америки, а в недавнее время эпидемия лихорадки о’ньон-ньон в 1959—
I960 гг. в Восточной Африке (свыше миллиона заболеваний), эпидемии
56
ВВЕДЕНИЕ
денге и особой формы геморрагической лихорадки (также вызываемой
в основном вирусом денге) в Юго-Восточной Азии. В этих эпидемиях
переносчиками возбудителей были тесно связанные с человеком виды
комаров — Aedes aegypti (денге), Anopheles funestus и An. gambiae (ли-
хорадка о’ньон-ньон).
Передаваемые комарами вирусные инфекции наиболее характерны
для тропиков, но распространены и. в странах умеренного пояса, как
например несколько форм комариных энцефалитов в Северной Америке,
японский энцефалит в Восточной Азии.
Особого внимания заслуживает проблема переносчиков западно-
нильской лихорадки; ее возбудитель распространяется в основном, по-
видимому, комарами, но, вероятно, и клещами — аргасовыми и иксодо-
выми. Очаги западнонильской лихорадки известны в Африке, Передней
Азии, Индии, а также в Южной Европе, в частности в Южной Франции
(где вирус выделен из комаров Culex modestus) и в дельте Волги.
В Чехословакии изучена роль комаров в передаче вируса тягиня
(из группы калифорнийского энцефалита), выделенного от Aedes caspius
и A. vexans (Бардош и Даниелова, 1959). Экспериментально показана
возможность передачи данного вируса комарами A. vexans и A. sticticus,
но получен отрицательный результат в опытах с A. cinereus (Шимкова
и др., 1960; Danielova, 1962).
В Чехословакии же из Anopheles maculipennis выделен вирус, полу-
чивший название чалово (Bardos a Cupkova, 1962); этот вирус относится
к группе буньямвера. Весьма близкие, если не идентичные, вирусы выде-
лены в Малайе из Culex, gelidus (вирус батаи) и в Индии из Anopheles
barbirostris, An. tesselatus Theob., An. subpictus Gr. и Culex bitaeniorhyn-
chus (вирус читтур; Singh a. Pavri, 1966). И на территории Советского
Союза, по-видимому, распространены передаваемые комарами вирусы
групп калифорнийского энцефалита и буньямвера.
На западе Украины при исследовании комаров на зараженность
было выделено несколько штаммов вируса лимфоцитарного хориоме-
нингита из Aedes communis, A. diantaeus, A. cantans и других комаров
(Глущенко и др., 1957; Виговский и Гуцевич, 1961). Названный вирус
также может быть отнесен к группе арбовирусов, так как доказана воз-
можность его размножения в тканях членистоногих (Ржегачек, 1965).
Впоследствии в различных республиках и областях СССР (Азербайджан,
Казахстан и др.) из комаров разных видов выделено много штаммов
вирусов, изучение которых несомненно даст интересные результаты
(Ильенко и др., 1962; Мирзоева и др., 1964; Ананян, 1965).
Комары как механические переносчики
возбудителей болезни
При этой форме передачи возбудитель болезни в организме пере-
носчика не развивается и не размножается; распространение возбудителя
возможно и без участия переносчиков.
Наилучше изученный пример — передача комарами и другими крово-
сосущими насекомыми туляремийного микроба. В организме комаров,
питавшихся на инфицированных туляремией грызунах, микробы сохра-
няются длительное время (прослежено до месяца; Олсуфьев и Голов,
1938). Передача происходит при кормлении зараженного комара — по
типу механической инокуляции. Чем меньше промежуток, времени между
питанием на зараженном и на здоровом животном (человеке), тем болыпе-
шансов, что передача микроба осуществится. Передача возможна и при
МЕТОДЫ РАБОТЫ
57
раздавливании зараженного комара на скарифицированной коже живот-
ного (человека). Комары несомненно играют видную роль в циркуляции
микроба в природных очагах туляремии, а также в заражении людей
при трансмиссивных вспышках инфекции (Олсуфьев и Руднев, 1960).
Переносчиками могут быть различные виды комаров, особенно рода
Aedes: А. caspius, А. vexans, А. cinereus, A. excrucians, А. flavescens, а также
Mansonia richiardii и др. Зараженных комаров неоднократно удавалось
находить в природе. В распространении туляремийного микроба участ-
вуют главным образом те виды комаров, которые охотно нападают
на грызунов, что особенно характерно для Aedes.
Комары являются также механическими переносчиками ряда виру-
сов — возбудителей болезней животных: оспы птиц, миксоматоза кроли-
ков и др.
КРАТКИЕ СВЕДЕНИЯ О МЕТОДИКЕ СБОРА,
МОНТИРОВКИ И СОХРАНЕНИЯ КОЛЛЕКЦИОННЫХ
МАТЕРИАЛОВ ПО КОМАРАМ
ВЗРОСЛЫЕ КОМАРЫ
Сбор комаров можно производить почти повсюду и во всякое время
года. Зимой самки некоторых видов (особенно из родов Culex, Culiseta,
Anopheles) могут быть добыты в подвалах домов, погребах, пещерах,,
помещениях для животных. Начиная со времени таяния снега комары
начинают покидать места зимовок. Конец весны ознаменовывается мас-
совым вылетом комаров, зимовавших в фазах яйца (Aedes) или личинки
(некоторые Anopheles, Culiseta и др.). В начале лета появляется новое
поколение комаров, зимовавших во взрослом состоянии.
Методика ловли взрослых комаров проста: нападающих самок отлав-
ливают эксгаустером или накрывают пробирками во время сосания;
можно ловить комаров сачком (лучше небольшим, 20—25 см диаметром),
из которого насекомых выбирают пробирками. Помимо ловли кровосо-
сущих самок, весьма важна также ловля самцов, которых следует искать
на цветах (особенно белых цветах зонтичных, рябины, ревеня и т. и.),
на листьях кустарников и деревьев, или же ловить их во время роения.
К середине лета численность комаров обыкновенно несколько падает,
однако к этому времени появляются некоторые новые формы. Конец
лета следует признать благоприятным для ловли большинства Culex,
Anopheles, некоторых Aedes и др.
Широкое распространение получили методы лова комаров, обеспечи-
вающие полноту сборов и возможность количественного учета. При сбо-
рах нападающих кровососущих двукрылых, в частности комаров, наибо-
лее точные результаты дает метод учетного колокола (Мончадский и
Радзивиловская, 1947). Матерчатый колокол подвешивают на высоте
около 2 м; под колоколом располагается наблюдатель, служащий «при-
манкой»; после 5-минутной экспозиции колокол опускают и вылавливают
эксгаустером насекомых, оказавшихся внутри колокола. Менее точные
и полные результаты дает простой способ, заключающийся в отлове
эксгаустером насекомых, нападающих на наблюдателя в течение опреде-
ленного промежутка времени; при малом количестве нападающих кома-
ров для учета вместо эксгаустера можно воспользоваться пробиркой.
Для количественного учета применяют также лов сачком. Единицей
учета может служить, например, количество комаров, пойманных 10Q
взмахами сачка вблизи человека (Олсуфьев, 1939). Во избежание повре-
58
ВВЕДЕНИЕ
ждения материала комаров надо извлекать из сачка почаще, например
после каждых 10 взмахов. Менее удовлетворительные результаты дает
отлов сачком путем кошения по траве; комары при этом сильно повре-
ждаются. Применяют и различные варианты ловушек в виде небольшого
домика, палатки или полога (Чинаев, 1959); приманкой служит человек
или животное, например теленок.
Широкое распространение получили разные модели световых лову-
шек, которые могут быть использованы и для количественного учета.
Источником света могут служить обычные электрические лампы, а также
лампы ультрафиолетового или поляризованного света (Бреев, 1958, 1963;
Жоголев, 1959; Ковров и Мончадский, 1963; Жоголев и Щербина, 1966).
Для лова комаров пригодны световые ловушки, снабженные засасываю-
щим устройством (вентилятором). Хотя не все виды комаров привлекаются
искусственным светом в одинаковой степени, сбор на свет дает ценный
материал, так как, помимо самок, отлавливаются и самцы.
Наилучшие результаты, в смысле получения экземпляров хорошей
сохранности, дает не ловля комаров, а вывод их из личинок и куколок;
при таком способе обеспечивается наличие достаточного количества
самцов, которые при обычных энтомологических методах ловли взрослых
комаров попадаются в общем не столь часто.
Для умерщвления комаров употребляют обычные в энтомологической
практике вещества — цианкали, хлороформ, серый и уксусный эфир.
Комаров помещают в банку с парами названных веществ или вводят
слегка смоченный в этих жидкостях тампон ваты в пробирку или эксгау-
стер с комарами. Долго оставлять комаров в парах анестезирующих
реактивов не следует; 5—10 минут при нормальной концентрации паров
следует признать совершенно достаточным сроком. Комаров, вышедших
из куколок, рекомендуется умерщвлять не в день окрыления, а на сле-
дующий день, когда покровы насекомого достаточно окрепнут.
Умерщвленного комара лучше всего сразу наколоть на булавку. Для
этого берут энтомологические булавки средних номеров (например,
№№ 0, 00) или же короткие тонкие булавки — минуциш Наколка на
булавку производится следующим образом: комара кладут на белую
бумагу спинной стороной кверху и, слегка придерживая его пальцами
левой руки или мягким пинцетом, правой рукой вводят булавку в сред-
нюю часть груди, между основаниями крыльев, лучше несколько отступя
от средней линии; когда это удалось, втыкают б!улавку в пластинку торфа
(или пропускают через бумагу) и, постепенно вводя булавку все глубже,
доводят объект примерно до 2/3 длины булавки, так, чтобы над насеко-
мым осталось около 1/3 ее длины. Обычный в энтомологической практике
способ наколки — помещение накалываемого насекомого между концами
собранных «щепоткой» пальцев левой руки — для комаров совершенно
неприменим ввиду крайней ломкости их ног. При наколке на минуции
поступают, как описано выше, с той только разницей, что после наколки
комара минуцию вкалывают в небольшой (удобные размеры 11x6 мм) прямо-
угольник бристольского картона (хорошей, без линеек, библиотечной
карточки), который уже накалывают на обыкновенную энтомологическую
булавку, лучше всего №№ 1—2. Весьма быстрым и удобным способом
является наколка комаров в бок; в этом случае булавка или минуция
пропускаются примерно посредине между основанием крыла и основа-
нием средних ног. Для придания материалам научной ценности необхо-
дима точная их этикетировка: географическое название местности, точ-
ная дата, условия нахождения, фамилия собирателя. Для предохранения
наколотых комаров от насекомых, повреждающих коллекции, в коробку
МЕТОДЫ РАБОТЫ
59
помещают пакетик с нафталином. Наколотых комаров рассматривают
под бинокулярным микроскопом (бинокуляром) при падающем свете;
рекомендуется увеличение в 20—25 раз.
В случае невозможности произвести немедленную наколку материалов
можно применять следующий способ их сохранения: раскладывают кома-
ров на слои ваты, сверху прикрывая их тонкой бумагой, на которой от-
мечаются необходимые для этикетировки данные. Слои ваты, перело-
женные листами бумаги, накладываются по мере заполнения материалом
.друг на друга в соответствующую размерам слоев коробку (неметалличе-
скую). В коробку кладут нафталин. Перед наколкой сухих комаров по-
мещают во влажную камеру (где не должно быть избытка водяных паров)
на несколько (3—6) часов. Затем с комарами поступают, как со свеже-
умерщв ленными.
Для определения вида комара часто бывает необходимо детальное
исследование генитального аппарата самца (гипопигия), которое про-
изводится следующим образом. У наколотых экземпляров отрезают тон-
кими ножницами последние сегменты брюшка, погружают их на минуту
в 96° спирт, а затем на несколько часов в 10%-й раствор едкого кали.
После промывания объекта в воде проводят его через спирты возрастаю-
щей крепости, обезвоживают 96° спиртом, просветляют в гвоздичном
масле или карбол-ксилоле и заключают в канадский бальзам. Препарат
исследуют под микроскопом. Чтобы можно было сопоставить результат
определения вида по гипопигию и по другим признакам насекомого,
один и тот же номер ставят на этикетке наколотого комара, от которого
приготовлен препарат, и на препарате гипопигия.
Более подробные данные о методике сбора и сохранения комаров
можно найти в работах Е. Н. Павловского (1935, 1959), А. С. Мончад-
ского (1952) и П. А. Петрищевой (1964).
ЛИЧИНКИ
Сбор личинок — обязательное звено всякого общефаунистического
обследования местности на комаров, дающее наиболее полные данные
об их видовом составе, сроках развития и типичных местах выплода.
Обследовать необходимо все типы встречающихся на исследуемой терри-
тории естественных и искусственных водоемов, постоянных и временных.
Обследование надо начинать ранней весной, до окончания таяния
снега. Тогда можно уже найти личинок младших стадий ранневесенних
видов рода Aedes, вышедших из перезимовавших яиц. Зимующих личи-
нок подрода Cullcella рода Culiseta, Anopheles claviger, Mansonia richiardii
можно обнаружить не раньше середины или конца весны, когда обитае-
мые ими водоемы освободятся от льда и температура воды в них подни-
мется до 5—12°. До этого они находятся на дне в неподвижном состоя-
нии. К середине весны, в начале развития листвы у березы, в водоемах
появляются личинки средневесенних видов рода Aedes. К началу цвете-
ния черемухи окукляются ранневесенние виды и появляются личинки
поздневесенних видов. В конце весны, когда зацветает сирень, начинают
попадаться личинки, вышедшие из яиц, отложенных перезимовавшими
самками родов Anopheles, Culiseta и Culex.
Весна — наиболее напряженный период обследования, особенно на
севере и в таежной, лесной и лесостепной зонах СССР, связанный с разно-
образием фауны комаров рода Aedes. На юге СССР лето и осень дают
наибольшее разнообразие видов родов Culex и Anopheles. Из всего разно-
образия водоемов необходимо выбрать наиболее типичные, которые надо
во
ВВЕДЕНИЕ
обследовать каждые 10—14 дней, а на юге (Кавказ и Средняя Азия) —
еженедельно. Помимо проведения таких регулярных обследований,
каждые 3—4 недели следует, для проверки полученных данных, обследо-
вать другие, однотипные с выбранными водоемы. Обследование личинок
можно прекратить в середине—конце сентября, а на юге — в начале
ноября.
К обследуемому водоему надо приближаться без шума, а также так,
чтобы тень не падала на его поверхность. Поэтому в ясные дни следует
подходить к нему против солнца. Личинки ряда видов {Anopheles claviger,
An. superpictus, Aedes excrucians, Culiseta morsitans и др.) очень чувстви-
тельны к внешним раздражениям, быстро опускаются на дно, где и .
остаются в течение 10—15 минут, что может существенно изменить ре-
зультаты лова. В небольших водоемах взмучиванием воды в них можно
заставить опустившихся на дно личинок быстро подняться на поверх-
ность.
При обследовании сачок — наилучшее из орудий лова. Диаметр
обруча 10—15 см. Проволока для него должна быть толщиной 3—5 мм,
негнущаяся и нержавеющая, лучше оцинкованная. Палка должна быть
легкая, 2 м длиной. Материя для сачка должна быстро фильтровать
воду, не пропускать личинок I стадии и быть настолько плотной, чтобы
не рваться, если сачок заденет в воде за сучья, палки и т. п. Таким усло-
виям удовлетворяют мельничный газ крупных и средних номеров (№№ 8—
15) и мелкоячеистая конгресс-канва, служащая для вышивания «кре-
стом». Марля для сачка не годится. Форма сачка должна быть закруг-
ленно-коническая. При конической форме сачка личинки, скопляясь
на дне острого конуса, при быстром движении испытывают значительное
давление воды и сильно повреждаются. Глубина сачка не должна пре-
вышать его диаметр более чем в-1.5 раза.
Наиболее полные сборы можно получить при следующем способе лова.
Сачок погружают на две трети в воду под острым углом по направлению
движения и быстро проводят им вдоль поверхности на протяжении 2—
3 м. Затем сачок быстро поворачивают на 180°, погружают на глубину
10—15 см и проводят им в обратном направлении, навстречу образо-
вавшемуся току воды. Таким путем в сачке окажутся личинки и куколки,,
находившиеся на поверхности воды и покинувшие поверхность перед,
началом лова.
Для лова личинок в очень мелких или небольших по площади водое-
мах удобнее пользоваться насаженным на короткую рукоятку проволоч-
ным обручем диаметром около 10 см, обтянутым марлей или мельничным
газом. Пойманных личинок смывают или в кювету, или в пробирку с во-
дой, прикладывая ее отверстие к личинке и поворачивая пробирку вверх
дном, заткнув отверстие пальцем с противоположной стороны ткани.
Можно пользоваться также ковшиком, половина дна которого имеет ряд
мелких отверстий.
Водоемы в дуплах деревьев можно облавливать или маленьким сачком
диаметром около 5 см, или пользоваться специальным выкачивателем.
Необходимо несколько повторных выкачиваний; при этом часть выка-
чанной воды надо выливать обратно в дупло.
Содержимое сачка выворачивается в белую эмалированную тарелку
или кювету, наполовину заполненную водой из водоема. Сачок тщательно
в ней ополаскивают и удаляют все попавшие туда посторонние предметы
(листья, ветви и пр.), а затем всех хищников. Пробу сливают в банку и.
этикетируют. Банку надо защищать от прямых солнечных лучей,
а в жаркие дни ее обертывают влажной тряпкой или надевают смоченный
МЕТОДЫ работы
61
ъодой специальный чехол, сшитый из двух слоев марли с простеганной
между ними ватной прослойкой.
В лаборатории пробу с личинками выливают в белую эмалированную
тарелку или кювету и разбирают по видам и стадиям, если позволяет
иремя. При недостатке его отсаживают всех куколок и часть личинок
IV стадии на выплод взрослых комаров, остальных личинок консерви-
руют, а разбор откладывают на зиму.
Для выведения взрослых комаров личинок, и куколок рассаживают
поодиночке или по несколько особей одного вида в небольшие цилиндры
или эрленмейеровские колбы, заполненные не более чем наполовину
водой, лучше из того же водоема, а на поверхность кладут свежий зеле-
ный лист. Сосуды защищают от солнца, закрывают ватным тампоном и
-ставят наклонно отверстием к источнику света. Средний срок длитель-
ности куколочной фазы — 2 суток. Первыми окукляются и вылетают самцы.
Личинок Anopheles для определения необходимо исследовать со спин-
ной стороны. Большинство признаков при этом лучше всего заметны.
Поэтому определение живых личинок в их естественном положении на
поверхности воды не представляет затруднений, если пользоваться при
этом предметным стеклом с углублением. Лишь немногие признаки (гре-
бень поддерживающих пластинок на VIII членике, плавник, хвостовые
волоски и жабры) видны при рассматривании заднего конца личинок
сбоку. Эти признаки, однако, имеют при видовом определении второсте-
пенное значение.
Личинок Culicinae для определения следует рассматривать таким
образом: передний конец (голова, грудь и первые членики брюшка)
сверху, а задний конец — сбоку. Поэтому личинок необходимо разрезать
острыми препаровальными иглами на уровне IV—V члеников ' брюшка
и соответственно ориентировать передний и задний концы тела. Препа-
раты; изготовленные из целой личинки, для определения непригодны.
При консервировании личинок для систематических целей надо упо-
треблять 96° спирт, нагретый до 50—55° (появление на стенках пробирки
со спиртом первых пузырьков). Это обеспечивает лучшее сохранение
волосков тела и раскрывание стигмальной пластинки. Предварительно
надо отсосать пипеткой воду, а нагретый спирт лить в сосуд с личинками
не сверху, а с края, чтобы он попадал на личинок сбоку и снизу. Через
30 минут наливают свежую порцию 96° спирта, в котором и сохраняют
личинок.
Для длительного хранения и особенно транспортировки личинок
помещают в небольшие пробирки, предварительно снабженные внутрен-
ней этикеткой, написанной мягким карандашом, с указанием географи-
ческого пункта, типа водоема, даты и фамилии сборщика. Писать на
этикетке только номера сборов не рекомендуется. Пробирку с материалом
заливают 96° спиртом и плотно закупоривают ватным тампоном, чтобы
внутри не было пузырька воздуха, который при тряске может механи-
чески повредить личинок — обломать у них жабры и волоски. Пробирки
ставят вертикально в материальную банку, лучше ватным тампоном
вниз, предварительно подстелив слой ваты и залив ее 96° спиртом. Свобод-
ное пространство между пробирками плотно забивают ватой. Следующий
слой пробирок отделяется от нижнего прокладкой из ваты. Прост-
ранство между верхним слоем пробирок и пробкой тоже плотно запол-
няется ватой. Часто несоблюдение правил упаковки и хранения мате-
риала делает невозможным его точное определение и исследование.
При изготовлении постоянных препаратов для длительного хранения
и исследования нельзя пользоваться жидкостью Фора—Берлезе. Этот
62
ВВЕДЕНИЕ
способ пригоден только для быстрого определения личинок в полевых
условиях в целях общей ориентировки, для чего он и был предложен..
Введение его в практику исследовательской работы ничем не оправдано.
Равным образом ничего, кроме резкого ухудшения качества препаратовг
не приносит обработка личинок в щелочи.
Необходимо изготовление бальзамных препаратов. Для этого личинок,
хранящихся в 96° спирте, переносят в свежую его порцию на 1—2 часа,
после чего, проколов их, помещают их на такой же срок в абсолютный
спирт для полного обезвоживания. Затем личинок, предварительно над-
резав их (Anopheles) или разрезав в области IV—V члеников брюшка
(Culicinae), переносят в просветляющую среду (гвоздичное масло, эвге-
нол, терпинеол) на 2—3 часа. После просветления из личинок делают
постоянный препарат в бальзаме (пихтовом, канадском и других), раство-
ренном в ортоксилоле. Личинок переносят на предметное стекло, пра-
вильно ориентируют передний конец спинной стороной кверху, задний,
естественно, ложится правильно (если личинки Anopheles не ложатся
точно, необходимо препаровальными иглами удалить у них плавник),
затем наносят на них каплю бальзама и покрывают покровным стеклом
с восковыми ножками (к 10 частям натурального пчелиного воска при-
бавляют 1—2 части скипидара, расплавляют, перемешивают и дают
остыть) высотой, соответствующей толщине личинок, слабо прижимают
последнее и сбоку подливают бальзам. Стекло без восковых ножек, давя
на объект, обычно обламывает волоски.
Часто бывает необходимо изготовить препарат из сброшенной шкурки
личинки. Для этого шкурку из воды помещают на несколько минут в 96е"
спирт, откуда ее переносят в молочную кислоту, где шкурка размягчается.
Ее помещают на предметное стекло и под бинокуляром тщательно рас-
правляют иглами. Затем отсасывают молочную кислоту и заменяют ее-
96° спиртом. При этом фиксируется приданная шкурке форма. Далее-
поступают, как при изготовлении бальзамных препаратов.
При всех манипуляциях с личинками необходимо соблюдать большую
осторожность, чтобы не обломать их волоски, жабры и т. д. Поэтому их
нельзя брать пинцетом. Живых личинок ловить и отсаживать можно-
только с помощью пипетки с достаточно широким отверстием. Фиксиро-
ванных личинок наиболее удобно переносить при помощи небольшой,
петли, сделанной из энтомологической булавки и насаженной, как пре-
паровальная игла, на деревянную рукоятку.
ЛИТЕРАТУРА
Литература по комарам огромна. Чтобы представить ее сколько-нибудь полно,,
хотя бы в. масштабе Палеарктики, понадобилась бы отдельная книга. Мы ограничи-
ваемся относительно небольшим списком. Для подробных справок о более старой ли-
тературе, касающейся СССР, можно рекомендовать: Н. С. Штакельберг.
Библиография малярии (эпидемиология, переносчики, борьба с малярией, организа-
ция противомалярийного дела в СССР). 1771—1935. 1940. Изд. АН СССР, М_.—Л. : 1—
963. Вспомогательные указатели. 1941 : 1—136. Работа содержит полный перечень .
отечественной литературы по комарам, с аннотациями. За более новой и текущей
литературой можно следить по реферативному журналу «Биология» (выпуск «Зоопа-
разитология»), а также по спискам, ежегодно публикуемым в журнале «Медицинская
паразитология и паразитарные болезни» (составитель Н. Н. Духанина).
Наиболее полные списки иностранной литературы можно найти в книгах:
A. Stone, К. L. Knight, Н. S t а г с k е. A synoptic catalog of the mosquitoes
of the World (Diptera, Culicidae). 1959; библиография — 38 страниц, преимущественно
систематика и фаунистика. W. R. Horsfall. Mosquitoes. Their bionomics and.
relation to disease. 1955; библиография — 79 страниц, преимущественно биология г»
ЛИТЕРАТУРА
63
медицинское значение комаров. М. Bates. The natural history of mosquitoes. 1949;
библиография — 46 страниц, также преимущественно по биологии и медицинскому
значению. A. N. Clements. The physiology of mosquitoes. 1963; библиография —
56 страниц, преимущественно физиология и морфология.
Обширные списки литературы приведены в монографиях, перечисленных ниже.
За текущей иностранной литературой можно следить по реферативным журналам:
The Review of the Applied Entomology. Ser. В (Medical and Veterinary), London; Tro-
pical Diseases Bulletin, London.
А. МОНОГРАФИИ И РЕГИОНАЛЬНЫЕ СВОДКИ
Беклемишев В. Н. 1944. Экология малярийного комара {Anopheles maculi-
pennis Mgn.). Медгиз, M. : 1—299.
Беклемишев В. Н. и Шипицина Н. К. (ред.). 1957. Сезонные явления
в жизни малярийных комаров в Советском Союзе. М. : 1—527.
Маслов А. В. 1967. Комары рода Culiseta. Изд. «Наука», М.—Л. : 1—182.
Мончадский А. С. 1936. Личинки комаров (сем. Culicidae) СССР и сопредель-
ных стран. Определители по фауне СССР, 24. Изд. АН СССР, М.—Л. : 1—379.
Мончадский А. С. 1951. Личинки кровососущих комаров СССР и сопредельных
стран (подсем. Culicinae). Определители по фауне СССР, 37. Изд. АН СССР,
М.—Л. : 1—290.
Мончадский А. С. и Штакельберг А. А. 1943. Малярийные комары
Таджикистана и меры борьбы с ними. Сталинабад : 1—96.
Штакельберг А. А. 1927. Кровососущие комары (сем. Culicidae) Союза ССР
и сопредельных стран. Определители по фауне СССР, 1. Изд. АН СССР, М.—Л. :
1—170.
Штакельберг А. А. 1937. Семейство Culicidae. Фауна СССР, Насекомые дву-
крылые, 3, 4. Изд. АН СССР, М.—Л. : 1—257.
В ar г and Р. I. 1934. Family Culicidae. Tribes Megarhinini and Culicini.
The fauna of British India, including Ceylon and Burma. Diptera, 5. London :
1—463.
Bates M. 1949. The natural history of mosquitoes. New York : 1—379.
Carpenter S. J. and La C a s s e W. J. 1955. Mosquitoes of North America
(north of Mexico). Berkeley a. Los Angeles : 1—360.
Christophers S. R. 1933. Family Culicidae. Tribe Anophelini. The fauna
of British India. Diptera, 4. London : 1—371.
Christophers S. R. 1960. Aedes aegypti (L.), the yellow fever mosquito. Its
life, history, bionomics and structure. Cambridge : 1—739.
Clements A. N. 1963. The phisiology of mosquitoes. International series of mono-
graphs on pure and applied biology, 17. Oxford : 1—393.
Edwards F. W. 1921. A revision of the mosquitoes of the Palaearctic region. Bull.
Entomol. Res., 12, 3 : 263—351.
Edwards F. W. 1926. Una revisione delle zanzare delle regioni Paleartiche. Riv..
malariol., 5, 3—6 : 1—152.
Edwards F. W. 1932. Fam. Culicidae. Genera insectorum, diriges par Wytsman.
Diptera, fasc. 194. Bruxelles : 1—258.
Edwards F. W. 1941. Mosquitoes of the Ethiopian region. III. Culicinae adults
and pupae. London : 1—499.
Evans A. M. 1938. Mosquitoes of the Ethiopian region. II. Anophelini adults and
early stages. London : 1—404.
Feng L. C. et al. 1958. (Комары Китая. Шанхай: 1—250; на китайском языке).
Foote R. Н. and Cook D. R. 1959. Mosquitoes of medical importance. Agric.
Handb. (U. S. Dep. Agric.), 152. Washington : 1—158.
Gjullin С. M., Sailer R. J., Stone A. and Travis В. V. 1961.
The mosquitoes of Alaska. Agric. Handb. (U. S. Dep. Agric.), 182. Washing-
ton : 1—98.
Hopkins G. H. E. 1952. Mosquitoes of the Ethiopian region. I. Larval bionomics,
of mosquitoes and taxonomy of Culicinae larvae. 2nd Ed. London : 1—355.
Horsfall W. R. 1955. Mosquitoes. Their bionomics and relation to disease. New
York : 1—723.
Kirkpatrick T. W. 1925. The mosquitoes of Egypt. „Cairo : 1—224.
Kramar J. 1958. Komaii bodavi — Culicinae. Fauna CSR, 13, Praha : 1—286.
La Casse W. J. and Yamaguti S. 1955. Mosquito fauna of J apan and Korea.
Kyoto : 1—268, 1—213.
Lang W. D. 1920. A handbook of British mosquitoes. London : 1—125.
Marshall J. F. 1938. The British mosquitoes. London : 1—341.
64
ВВЕДЕНИЕ
Martini Е. 1920. Uber Stechmiicken besonders deren europaische Arten und ihre
Bekampfung. Leipzig : 1—267.
Martini E. 1929—1931. Culicidae. In: E. Lindner, Die Fliegen der palaearktischen
Region, 11 u. 12. Stuttgart : 1—398.
Matheson R. 1944. Handbook of the mosquitoes of North America. 2nd Ed. It-
haca : 1—314. t
Mihalyi F. es Gulyas M. 1963. Magyaroszag csipo szunyogjai. Budapest : 1—229
(на венгерском языке).
Natvig L. R. 1948. Contributions to the knowledge of the Danish and Fennoscan-
dian mosquitoes — Culicini. Oslo : 1—567.
P e u s F. 1942. Die Fiebermiicken des Mittelmeergebiets. Leipzig : 1—150.
Puri J. M. 1931. Larvae of anopheline mosquitoes, with full description of those
of the Indian species. Ind. Med. Res. Mem., 21 : 1—225.
Rioux J. A. 1958. Les Culicides du «Midi» Mediterraneen. Encycl. Ent. (A),
35 : 1—303.
Seguy E. 1923. Les moustiques de France. Encycl. prat. Natural., 19 : 1—225.
Senevet G. 1958. Les Anopheles du Globe. Encycl. Ent. (A), 36 : 1—215.
Senevet G. et And ar elli L. 1955. Les AnophHes de 1’Afrique du Nord et du
Bassin Mediterraneen. Encycl. Ent. (A), 33 : 1—280.
Senevet G. et Andarelli L. 1959. Les moustiques de 1’Afrique du Nord
et du Bassin Mediterraneen les genres Culex, Uranotaenia, Theobaldia, Ortho-
podomyia et Mansonia. Encycl. Ent. (A), 37 : 1—384.
Snodgrass R. E. 1959. The anatomical life of the mosquito. Smithsonian Miscel-
laneous collections, 139, 8. Washington : 1—87.
Stone A., Knight K. L. and S t a r c k e H. 1959. A synoptic catalog of the
mosquitoes of the World (Diptera, Culicidae). The Thomas Say Foundation, 6.
Washington : 1—358.
Wesenberg-Lund C. 1921. Contributions to the biology of the Danish Culi-
cidae. Kgl. danske vid., selskab., 7 : 1—210.
Б. СИСТЕМАТИКА, ФАУНИСТИКА, МОРФОЛОГИЯ, БИОЛОГИЯ
Алекторов А. А. 1931. К фауне комаров Culicidae Дальневосточного края СССР.
Паразитол. сб. Зоол. ин-та СССР, 2 : 229(—248.
Башкарева А. Л. 1931. Некоторые данные о малярийных и других комарах
Сочинского района. Паразитол. сб. Зоол. ин-та АН СССР, 2 : 55—58.
Благовещенский Д. И. 1937. Материалы по фауне наружных паразитов
(A rthropoda) животных Казалинского и некоторых других районов южного
Казахстана. Сб. «О вредителях животноводства в Казахстане», Изд. АН СССР,
М.—Л. : 11—84.
БД) ж к о в Д. К. 1966. Кровососущие комары (Diptera, Culicidae) Болгарии.
М Энтомол. обозрение, 45, 3 : 570—574.
Брегетова Н. Г. 1946. Активность нападения на человека и ее суточный ритм
у комаров Aedes caspius caspius (Pall.) Edw. и Aedes vexans Meig. в природных
условиях южного Таджикистана. Изв. АН СССР, сер. биол., 2—3 : 251—279.
Б р е е в К. А. 1958. О применении ловушек ультрафиолетового света для определе-
ния видового состава и численности популяции комаров. Паразитол. сб. Зоол.
ин-та АН СССР, 18 : 219—238.
Бреев К. А. 1963. Влияние источников света на численность и видовой состав
комаров, собираемых в световые ловушки. Энтомол. обозрение, 42, 2 : 280—303.
Вальх С. Б. 1959. К познанию фауны Culicidae востока Украины. Мед. паразитол.
и паразитарн. болезни, 28, 6 : 687—695.
В е л и ч к е в и ч А. И. 1936. К фауне и экологии комаров Южного берега Крыма.
,! Паразитол. сб. Зоол. ин-та АН СССР, 6 : 137—145.
Виноградова Е. Б. 1965. Морфологическая и биологическая характеристика
некоторых природных популяций кровососущих комаров Culex pipiens L.
j S ; (Diptera, Culicidae) на территории СССР. Сб. «Экология вредных насекомых и
энтомофагов», Тр. Зоол. ин-та АН СССР, 36 : 31—57.
Вдг ноградова Е. Б. 1966. Кровососущие комары комплекса Culex pipiens L.
(Diptera, Culicidae), их практическое значение, систематика и биология. Энтомол.
обозрение, 45, 2 : 241—257.
Виноградская О. Н. 1950. Стигмальный аппарат Anopheles и его видоизме-
нения в зависимости от сухоустойчивости и влаголюбивости видов. Энтомол.
обозрение, 31, 1—2 : 151—154.
Виноградская О. Н. 1953. Участие трахейной системы в испарении воды
у Anopheles maculipennis messeae Fall, и сезонная изменчивость дыхальцевого
индекса у видов подсемейства Culicinae (Diptera, Culicidae). Энтомол. обозрение,
33 : 157—160.
ЛИТЕРАТУРА
65
Волкова М. И. 1956. Фауна кровососущих комаров (сем. Culicidae) долины
средней Волги в пределах Татарской АССР. Уч. зап. Казанск. гос. ун-та, 116,
5 : 153—156.
Вольфтруб А. М. 1963. К вопросу о видовом составе и динамике активности
кровососущих двукрылых насекомых в Таймырском национальном округе.
Тр. Н.-и. ин-та сельск. хоз. Крайнего Севера, 11 : 113—116.
Глаголева Е. М. 1944. Некоторые данные по экологии личинок Anopheles
в Таджикистане. I. Биотопы Anopheles algeriensis Theob. II. Биотопы Anophe-
les lindesayi Giles и Anopheles marteri Sen. Prun. III. Биотопы Anopheles bifur-
catus. IV. Биотопы Anopheles pulcherrimus Theob. Мед. паразитол. и пара-
зитарн. болезни, 13, 4 : 58—64 и 65—67; 13, 5 : 47—52; 13, 6 : 41—44.
Глаголева Е. М. 1946. Об экологии личинок Anopheles в Таджикистане. Мед.
паразитол. и паразитарн. болезни, 15, 5 : 60—67.
Гуцевич А. В. 1955. Новые и мало известные формы комаров. Тр. Зоол. ин-та
АН СССР, 18 : 320—324.
Гуцевич А. В. 1962. Новый вид кровососущего комара рода Aedes (Diptera,
Culicidae) из Казахстана. Энтомол. обозрение, 41, 4 : 886—888.
Гуцевич А. В. 1966. Новый вид.комара из Узбекистана — Aedes (Ochlerotatus)
simanini (Culicidae). Зоол. журн., 45, 3 : 457—459.
Гуцевич А. В., Донец 3. С., Ежова Г. Г. и Попов А. М. 1962.
Кровососущие комары (Diptera, Culicidae) Черновицкой области. Энтомол.
обозрение, 41, 2 : 355—358.
Денисова 3. М. 1940. Изменчивость гипопигиев Anopheles maculipennis. mes-
seae Fall, в популяции Марбумстроя (Марийская АССР). Сб. «Вопросы физио-
логии и экологии малярийного комара», 1 : ИЗ—119.
Денисова 3. М. 1948. Материалы по изучению вариаций гипопигиев самцов
Anopheles maculipennis atroparvus Thiel и Anopheles maculipennis sacharovi
Favre. Мед. паразитол. и паразитарн. болезни, 3, 17 : 221—227.
Денисова 3. М. 1955. Новый вид Aedes (Ochlerotatus). Мед. паразитол. и пара-
зитарн. болезни, 24, 1 : 58—61.
Денисова 3. М. 1964. Вариации гипопигиев самцов некоторых подвидов маля-
рийного комара Anopheles maculipennis Mg. (Diptera, Culicidae). Энтомол.
обозрение, 43, 4 : 815—822.
Ениколопов G. К. 1937. Об экологии Anopheles algeriensis Theob. Мед. пара-
зитол. и паразитарн. болезни, 6, 3 : 354—359.
Жоголев Д. Т. 1959. Световые ловушки как метод собирания и изучения насеко-
мых — переносчиков возбудителей болезней. Энтомол. обозрение, 38, 4 : 766—773.
Жоголев Д. Т. и Щербина В. П. 1966. Новая модель портативной свето-
вой ловушки для сбора кровососущих двукрылых насекомых. Мед. паразитол.
и паразитарн. болезни, 35, 5 : 619—621.
Здродовский П. Ф. 1926. Малярия на Мугани. Эпидемиология и план
борьбы. М.—Баку : 1—174.
Казанцев Б. Н. 1930. К вопросу об изменчивости рисунка и окраски крыльев
комаров рода Anopheles Средней Азии. Паразитол. сб. Зоол. ин-та АН СССР,
1 : 229—235.
Казанцев Б. Н. 1931. Цветовые вариации бухарских Aedes caspius. Паразитол.
сб. Зоол. ин-та АН СССР, 2 : 85—90.
Калмыков Е. С. 1965. Изменения в распространении малярийных комаров
в связи с хозяйственной деятельностью человека. Научи, тр. по паразитологии,
изд. «Ирфон», Душанбе : 55—61.
Кешишьян М. Н. 1941. Culicidae Таджикистана. Мед. паразитол. и парази-
тарн. болезни, 10, 1 : 77—80.
Ковров Б. Г. и Мончадский А. С. 1963. О возможности применения
поляризованного света для привлечения насекомых. Энтомол. обозрение,
42, 1 : 49—55.
Л и с о в а А. И. 1935. Материалы по фауне комаров сем. Culicidae города Ташкента
и его окрестностей. Паразитол. со. Зоол. ин-та АН СССР, 5 : 27—54.
Лобкова М. П. 1964. Некоторые данные по распространению комаров (подсе-
мейство Culicinae) в Карелии. Сб. «К природной очаговости паразитарных и
трансмиссивных заболеваний в Карелии», изд. «Наука», М,—Л. : 108—119.
Львов Д. К. 1956. О видовой самостоятельности комара-переносчика японского
энцефалита Aedes esoensis Yam. (Diptera, Culicidae). Энтомол. обозрение, 35,
4 : 929—934.
Маркович Н. Я. 1941. Новые данные по биологии Anopheles bifurcatus (наблю-
дения на Северном Кавказе). Мед. паразитол. и паразитарн. болезни, 10, 3—
4 : 410—413.
Мартини Э. (Martini Е.). 1926. Комары Среднего Поволжья. Изв. АН СССР,
VI серия, 20, 9 : 607—618.
5 Фауна СССР, А. В. Гуцевич и др.
66
ВВЕДЕНИЕ
М а с лов А. В. 1946. Изменчивость и систематика Палласова малярийного комара.
Мед. иаразитол. и паразитарн. болезни, 15, 6 : 39—46.
Маслов А. В. 1964. О систематике кровососущих комаров группы Culiseta {Di-
ptera, Culicidae'). Энтомол. обозрение, 48, 1 : 193—216.
Мончадский А. С. 1947. К познанию личинок комаров (Diptera, Culicidae).
II. Новые и малоизвестные личинки комаров Дальнего Востока. Паразитол.
сб. Зоол. ин-та АН СССР, 9 : 167—180.
Мончадский А. С. 1949. Личинки Finlaya {Diptera, Culicidae) из дупел деревьев.
Паразитол. сб. Зоол. ин-та АН СССР, 11 : 253—261.
Мончадский А. С. 1952. Летающие кровососущие двукрылые — гнус.
Изд. АН СССР, М,—Л. : 1—67.
Мончадский А. С. и Радзивиловская 3. А. 1947. Новый метод
количественного учета активности нападения кровососов. Паразитол. сб. Зоол.
ин-та АН СССР, 9 : 147—166.
Олсуфьев Н. Г. 1939. Видовой состав и сезонная динамика численности крово-
сосущих двукрылых в дельте Волги и их возможная роль в эпидемиологии
туляремии. Зоолог, журн., 18, 5 : 786—798.
Павловский Е. Н. 1935. Методы изучения кровососущих комаров {Culici-
dae). Изд. 2-е. Изд. АН СССР, М,—Л. : 1—176.
Павловский Е. Н. (ред.). 1959. Лабораторный практикум медицинской пара-
зитологии. Медгиз, Л. (Комары : 200—249).
Петрищева П. А. 1936. Фауна, экология и биология Culicidae Туркмении.
Паразитол. сб. Зоол. ин-та АН СССР, 6 : 49—115.
Петрищева П. А. 1947. О кровососущих комарах Приморья. В кн.: Парази-
тология Дальнего Востока, Медгиз, Л. : 144—159.
Петрищева П. А. 1964. Методы изучения природных очагов болезней человека.
Изд. «Медицина», М. (Комары : 12—35).
Прендель А. Р. 1966. Происхождение и характеристика фауны кровососущих
комаров УССР. Сб. «Краевая паразитология и природная очаговость трансмис-
сивных болезней», Киев : 91—96.
Ре мм X. Я. 1957. Материалы по фауне и экологии комаров {Diptera, Culicidae)
Эстонской ССР. Энтомол. обозрение, 36, 1 : 148—160.
Сазонова О. Н. 1956. Новый вид кровососущего комара рода Aedes Mg. {Dip-
tera, Culicidae) из европейской части СССР. Паразитол. сб. Зоол. ин-та АН СССР,
16 : 148—151.
Сазонова О. Н. 1958. Таблица для определения самок комаров рода Aedes Mg.
{Diptera, Culicidae) лесной зоны СССР. Энтомол. обозрение, 37, 3 : 741—752.
Сафьянова В. М. 1960. Материалы по экологии кровососущих комаров Влади-
мирской области. Зоолог, журн., 39, 2 : 236—243.
Соловей В. Я. и Лиходед В. Г. 1966. К фауне и экологии кровососущих
двукрылых {Diptera) северо-западной части Мурманской области. Энтомол.
обозрение, 45, 3 : 565—569.
Ф е д о р о в В. Г. 1946. К обнаружению Culex molestus Forskal в Ленинграде. Мед.
паразитол. и паразитарн. болезни, 15, 2 : 58—67.
Фридолин В. Ю. 1936. Животно-растительное сообщество горной страны Хибин.
Тр. Кольской базы АН СССР, 3 : 1—295.
Хелевин Н. В. 1958. Влияние внешних условий на возникновение эмбриональ-
ной диапаузы и на количество поколений у Aedes caspius dorsalis Mg. ’{Diptera.,
Culicidae) в течение сезона. Энтомол. обозрение, 37, 1 : 24—47.
Чагин К. П. 1943. Наблюдения над циклом развития Aedes (F.) togoi в лабора-
торных и природных условиях. Мед. паразитол. и паразитарн. болезни, 12,
2 : 44—52.
Ч и н а е в П. П; 1959. Методы количественного учета кровососущих комарок
{Diptera, Culicidae). Энтомол. обозрение, 38, 4 : 757—765.
Шингарев Н. И. 1928. Заметки по Culicidae. III. Русск. журн. тропич. мед.,
6 : 47—53.
Шленова М. Ф. 1959. Биология важнейших видов Aedes в средней полосе евро-
пейской части СССР. Мед. паразитол. и паразитарн. болезни, 28, 2 : 193—198.
Bates М. 1940. The nomenclature and taxonomic status of the mosquitoes of the
Anopheles maculipennis complex. Ann. Entomol. Soc. America, 33 : 343—
356.
В non о m ini G. a. Mariani M. 1953. World anophelines belonging to the
subgenus Maculipennia Buonomini e Mariani, 1946. Riv. malarial., 32 : 173—188.
C all о t J. et Rioux J.-A. 1965.' Liste sommaire des Culicides de France. Ann.
parasitol. humaine et comparee, 40, 2 : 242—245.
C oil ess D. H. 1958. Notes on the culicine mosquitoes of Singapore. IV. The Aedes
niveus subgroup {Diptera, Culicidae)-. introduction and description of five new-
species and of one new subspecies. Ann. Trop. Med. and Parasitol., 52, 4 : 468—483.
ЛИТЕРАТУРА 67
Coluzzi М. a. Contini С. 1962. The larva and pupa of Mansonia (Coquillet-
tidia) buxtoni (Edwards), 1923 (Diptera, Culicidae). Bull. Entomol. Res., 53,
2 : 215—218.
Corbet P. S. 1965. Reproduction in mosquitoes of the high Arctic. Proc. XII In-
tern. Congress of Entomology : 817—818.
Dqbrowska Prot E. 1964. Communities of mosquitoes in three types of forest
land. Ekol. polska A, 12, 36 : 737—783.
Dodge H. R. 1962. Supergeneric groups of mosquitoes. Mosquito News, 22,.
4 : 365—368.
Edwards F. W. 1923. Oligocene mosquitoes in the British Museum; with a summary
of our present knowledge concerning fossil Culicidae. Quart. J. Geol. Soc. London,
79 : 139-155.
Feng L. C. 1938. The tree hole species of mosquitoes of Peiping, China. Chin. Med.
Jour., Suppl., 2 : 503—525.
Gordon R. M. a. Lumsden W. H. R. 1939. A study of the behavoir of the .
mouthparts of mosquitoes when taking up blood from living tissue; together with
some observations on the ingestion of microfilariae. Ann. Trop. Med. and Parasi-
tol., 33 : 259—278.
Jobling B. 1938. On two subspecies of Culex pipiens. Trans. Roy. Entomol. Soc.
London, 87, 8 : 246—248.
Kramar J. 1957. Culex territans v CSR. Ceskoslovenska Parasitologie, 4 : 201—204.
Mariani M., Bruno-Smiraglia C. eCaravaglios N. 1964. Ri-
cerche sulla interspecificita labranchiae-atroparvus. Riv. malariol., 43, 1—3 :
37—50.
Martini E. 1928. Beitrage zur medizinischen Entomologie und zur Malaria-Epide-
miologie des unteren Wolgagebiets. Abhandlungen aus dem Gebiet der Ausland-
skunde, 29, Hamburg (Die Stechmiickenfauna : 21—69).
Mattingly P. F. 1953. A change of name among the British mosquitoes (Diptera,
Culicidae). Proc. Roy. Entomol. Soc. London B, 22 : 106—108.
Mattingly P. F. 1955. Le sous-genre Neoculex (Diptera, Culicidae) dans la sous-
region mediterraneenne. I. Espece, sous-espece et synonymie nouvelles. Ann.
parasitol. humaine et comparee, 30 : 375—388.
M a t t i n g 1 у P. F., Rozeboom L. E., Knight K. L., Shute P. G.,
Drummond F. H. a. Christophers S. R. 1951. The Culex pipiens
complex. Trans. Roy. Entomol. Soc. London, 102, 7 : 331—342.
P eus F. 1933. Zur Kenntnis der Aedes-Arten des deutschen Faunengebietes (Diptera,
Culicidae). Die Weibchen der Aedes communis-Grnppe. Konowia, 12 : 145—
159.
Reid J. A. 1953. The Anopheles hyrcanus group in south-east Asia (Diptera, Culi-
cidae). Bull. Entomol. Res., 44, 1 : 5—76.
Reid J. A. .a. К n i g h t K. L. 1961. Classification within the subgenus Anopheles
(Diptera, Culicidae). Ann. Trop. Med. and Parasitol., 55, 4 : 474—488.
Rioux J.-A. et A r n о 1 d M. 1955. Les culicides de Camargue (Etude systematique
et ecologique). Terre et Vie, 4 : 244— 286.
R i s 1 e r H. 1955. Das Gehororgan der Mannchen von Culexpipiens L., Aedes aegypti L.
und Anopheles stephensi Liston (Culicidae). Zool. Jahrb., Abt. 2, 74 : 478—
490.
Roubaud E. etGhelovitch S. 1956. Observations sur le moustiques anthro-
pophile mediterraneen du groupe pipiens, Culex berbericus Roub. C. r. Acad,
sci., 242, 25 : 2900—2903.
Senevet G. et An darell i L. 1963—1964. Les moustiques de 1’Afrique du Nord
et du bassin mediterraneen. Arch. Inst. Pasteur Algerie, 1963, 41 : 115—172;
1964, 42 : 94—192.
Senior-White R. 1927. Notes on Ceylon mosquitoes. II. The larvae of the com-
moner non-anopheline mosquitoes. Spolia Zeylanica Ceylon Jour. Sci. (B),
14 : 61—76.
Skierska B. 1963. Przeglad pismiennictwa dotyczqcego komarow (Culicidae)
z obszarow polski oraz reiestracja i rejonizacja tych owadow na terenie naszego
kraju. Wiadom. parazytol., 9,6 : 579—597.
S 1 i f e r E. H. a. S e к h о n S. S. 1962. The fine structure of the sense organs
of the antennal flagellum of the yellow fever mosquito Aedes aegypti (Linnaeus).
Jour. Morphol., Ill, 1 : 49—66.
StackelbergA. А. (Штакельберг A. A.). 1943. A new species of Finlaya,
Theob. (Diptera, Culicidae) from Ussuri land. Bull. Entomol. Res., 34, 4 : 311.
Trpis M. 1965a. Typologie der Biotope in der Ostslowakei gemass der Gesellschaften
von Larven und Imagines der Stechmiicken. Entomologicke Problemy. Brati-
slava, 5 : 9—69.
5*
68
ВВЕДЕНИЕ
Т г р i s М. 1965b. Verbreitung der Stechmucken (Diptera, Culicidae) in der Ostslowakei.
Entomologicke Problemy. Bratislava, 5 : 71—100.
Vockeroth J. R. 1950. Specific characters in tarsal claws of some species of Aedes
(Diptera, Culicidae). Canad. Entomologist, 82 : 160—162.
Vockeroth J. R. 1952. The specific status of Aedes pionips Dyar (Diptera, Culi-
cidae). Canad. Entomologist, 84 : 243—247.
Vockeroth J. R. 1954. Notes on northern species of Aedes, with descriptions of two
new species (Diptera, Culicidae). Canad. Entomologist, 86 : 109—116.
Seminar on the ecology, biology and control of the Culex pipiens complex. 1965.
World Health Organisation (Vector Control), 125 : 1—217.
В. КОМАРЫ КАК ПЕРЕНОСЧИКИ
Ананян С. А. 1965. Новые для СССР, передающиеся комарами, возбудители
вирусных инфекций. Сб. «Актуальные проблемы вирусных инфекций», Матер.
XII науч, сессии Инет, полиомиелита и вирусных энцефалитов, М. : 194—195.
Бардош В. и Даниелова В. (Bardos V., Danielova V.). 1959.
Вирус Tahyiia — вирус, выделенный из комаров в Чехословакии. Ж. гигиены,
эпидемиол., микробиол., иммунол. (Прага), 3, 2 : 182—193.
Бардош В. и Чуйкова Е. (Bardos V., С u р к о v а Е.). 1962. Вирус
Calovo — второй арбовирус, выделенный из комаров в Чехословакии. Ж. ги-
гиены, эпидемиол., микробиол., иммунол. (Прага), 6 : 255—260.
Беклемишев В. Н. 1941. О факторах, определяющих значение различных
видов Anopheles как переносчиков малярии. Мед. паразитол. и паразитарн.
болезни, 10, 1 : 5—8.
Беклемишев В. Н. 1944. Экология малярийного комара (Anopheles таси-
lipennis Mgn.). Медгиз, М. : 1—299.
Битовский А. И. и Гуцевич А. В. 1961. Предварительные результаты
изучения природных очагов лимфоцитарного хориоменингита на западе
S Украины. Докл. АН СССР, 1'40, 5 : 1223—1225.
Глущенко П. А., Гуцевич А. В. и Дудкина М. С. 1957. Исследова-
ние комаров как переносчиков вируса лимфоцитарного хориоменингита на за-
паде Украины. Докл. АН СССР, 113, 5 : 1181—1183.
Гуцевич А. В. 1964. Насекомые как переносчики вирусов — паразитов человека
и животных. Зоол. ж., 43, 3 : 429—442.
Детинова Т. С. , 1962. Методы установления возрастного состава двукрылых
насекомых, имеющих медицинское значение. Всемирн. организ. здравоохр.,
Женева : 1—220.
Ильенко В. И., Стерхова А. Н., Мирзоева Н. М., Д ю н и н а К. А.
иАхундов М. Г. 1962. Материалы по изучению трансмиссивных вирусных
инфекций в некоторых районах Азербайджанской ССР. Сб. «Вопросы эпидемио-
логии, бактериологии, гигиены, паразитологии и вирусологии», Тез. докл.,
Душанбе : 182—184.
Мирзоева Н. М., Ильенко В. И., Дюнина К. А. иАхундов М. Г.
1964. Данные о серологической и вирусологической разведке арбовирусов
в Кызылагачском заповеднике им. С. М. Кирова Азербайджанской ССР. В кн.:
Клещевой энцефалит, Кемеровская клещевая лихорадка, геморрагические лихо-
радки и другие арбовирусные инфекции. М. : 205—206.
Олсуфьев Н. Г. (и Голов Д. А.). 1938. Роль комаров в передачей хранении
туляремии. Тр. Отдела паразитологии ВИЭМ, 3 : 213—246.
О л с у ф ь е в Н. Г. 1941а. Новые экспериментальные данные по вопросу о роли
комаров в передаче и хранении туляремийной инфекции. Сб. работ, посвящ.
30-летию научн. деят. акад. Е. Н. Павловского, Л.—М. : 176—189.
Олсуфьев Н. Г. 19416. Роль комаров в передаче туляремийной инфекции ди-
‘ ким грызунам, птицам и домашним животным. Сб. работ, посвящ. 30-летию
научн. деят. акад. Е. Н. Павловского, Л.—М. : 190—197.
Олсуфьев Н. Г. и Руднев Г. П. (ред.). 1960. Туляремия. Медгиз,
М. : 1—459.
Орехов М. Д. 1952. Дипеталонематоз верблюдов в Туркменской ССР и меры
борьбы с ним. Ветеринария, 4 : 28—30.
Осипов А. Н. 1965. К выявлению промежуточных хозяев возбудителя сетариоза
крупного рогатого скота в СССР. Матер, к научной конференции Всесоюзного
общ. гельминтологов, 4 : 181—184.
Павловский Е. Н., Штейн А. К. и Перфильев П. П. 1928. Экспе-
риментальные исследования над влиянием действующих начал комара Culex
pipiens на кожу человека. Докл. АН СССР, А, 13 : 251—254.
ЛИТЕРАТУРА
69
Петрищева П. А. 1947. Комары — переносчики японского энцефалита в Южном
Приморье. В кн.: Паразитология Дальнего Востока. Медгиз, Л. : 106—143.
Сазонова О. Н. 1962. Кровососущие комары (Diptera, Culicinae). В кн.: Пере-
носчики возбудителей природноочаговых болезней. Изд. «Медицина», М. : 9—63.
Симанин П. И. 1930. К вопросу об экспериментальном заражении комаров тро-
пической малярией. Паразитол. сб. Зоол. музея АН СССР : 37—44.
Ржегачек Ю. (ЙеЬасек J.). 1965. Культивирование различных вирусов
в клещевых тканевых культурах. Acta virologica, 9,. 4 : 332—337.
Шимкова А., Даниелова В. и Бардош В. (Simkova A., Danie-
1 о v а V., Bardos V.). 1960. Экспериментальный перенос вируса Tahyiia
комарами Aedes vexans. Acta virologica, 4, 6 : 341—347.
Вирусы, переносимые членистоногими. 1962. Всемирн. организ. здравоохр.,
сер. техн, докл., 219, Женева : 1—87.
Инвазии, вызываемые Wuchereria и Brugia. 1963. Всемирн. организ. здравоохр.,
сер. техн, докл., 233. Женева : 1—59.
Комитет экспертов по малярии. Десятый доклад. 1965. Всемирн. организ. здра-
воохр., сер. vTexH. докл., 272. Женева :Д—62.
Bardos V. a. Cupkova Е. 1962. The Calovo virus — the second virus isola-
ted from mosquitoes in Czechoslovakia. J. Hyg. (Prague), 6, 2 : 186—192.
Bates M. 1949. The natural history of mosquitoes. New York : 1—379.
Chamberlain R. W. and Su di a, W. D. 1961. Mechanism of transmission
of viruses by mosquitoes. Ann. Rev. Entomol., 6 : 371—390.
Contacos P. G., Lunn J. S., CoatneyG. R., Kilpatrick J. W.
a. J о n e s F. E. 1963. Quartan-type malaria parasite of New World.monkeys
transmissible to man. Science, 142 : 676.
Danielova V. 1962. Experimentalni studie vztahu viru Tahyna k nekterym dru-
hfim котагй. Cs. epidemiol., mikrobiol., imunol., 11, 3 : 171—174.
Devine T. L., Ven ar d С. E. and M у s e r W. C. 1965. Measurement of sali-
vation by Aedes aegypti (L.) feeding on a living host. J. Insect Physiol., 11,
3 : 347—353.
Edeson J. F. B. and Wilson T. 1964. The epidemiology of filariasis due to
Wuchereria bancrofti and Brugia malayi. Ann. Rev. Entomol., 9 : 245—268.
Ferreira E. 1964. Distribuipao geografica dos anofelinos no Brazil e sua relapao
com о estado atual de erradicajao de malaria. Rev. brasil. malariologia, 16,
3 : 329—348.
Foote R. H. and Cook D. R. 1959. Mosquitoes of medical importance. Agric.
Handb. (U. S. Dep. Agric.), 152. Washington : 1—158.
Gordon Smith С. E. 1964. Factors influencing the behaviour of viruses in their
arthropodan hosts. «Host-parasite relationships in invertebrate hosts», Oxford: 1—31.
Hannoun C., Panthier R., Mouchet J. et EouzanJ. P. 1964;
Isolement en France du virus West-Nile a partir de malades et du vecteur Culex
modestus Ficalbi. C. r. Acad, sci., 259 : 4170—4172.
Hawking F. and Worms M. J. 1961. Transmission of filaroid nematodes.
Ann. Rev. Entomol., 6 : 413—432.
Hecht O. 1929. Die Hautreaktionen auf Insektenstiche als allergische Erscheinungen.
Arch. Schiffs-Tropenhygiene, 33 : 364—366.
Horsfall W. R. 1955. Mosquitoes. Their bionomics and relation to disease. New
York : 1-723.
Huff C. G. 1965. Susceptibility of mosquitoes to avian malaria. Exper. Parasitol.,
16, 1 : 107—132.
Jayewardene L. G. 1963. Larval development of Brugia ceylonensis Jayewar-
dene, 1962, in Aedes aegypti, with a brief comparison of the infective larva with
those of Brugia spp., Dirofilaria repens and Artionema digitata. Ann. Trop. Med.
and Parasitol., 57, 3 : 359—370.
Jordan H. B. 1964. Lizard malaria in Georgia. J. Protozool., 11, 4 : 562—566.
Lavoi pierre M. M. J. 1958. Studies on the host-parasite relationships of filarial
nematodes and their arthropod hosts. II. The arthropod as a host to the nematode:
A brief appraisal of our present knowledge based on a study of the more impor-
tant literature from 1878 to 1957. Ann. Trop. Med. and Parasitol., 52, 3 : 326—345.
Mattingly P. -F. 1960. Ecological aspects of the evolution of mosquito-borne
virus diseases. Trans. Roy. Soc. Trop. Med. and Hyg., 54, 2 : 97—112; discuss. :
130-134.
Mattingly P. F. 1962. Some considerations relating to the role of Culex pipiens
fatigans Wiedemann in the transmission of human filariasis. Bull. World Health
Org., 27 : 569—578.
Mattingly P. F 1965. The evolution of parasite-arthropod vector systems. «Evo-
lution of Parasites», Oxford : 29—46.
70
ВВЕДЕНИЕ
Mims С. A., D а у М. F. a. Marshall J. D. 1966. Cytopathic effect of Sem-
liki forest virus in the mosquito Aedes aegypti, Amer. J. Trop. Med. and Hyg.,
15, 5 : 775—784.
Nelson G. S. 1964. Factors influencing the development and behavoior of filarial
nematodes in their arthropodan hosts. «Host-parasite relationships in invertebrate
hosts», Oxford : 75—119.
Pratt H. D. 1964. Virus diseases transmitted by mosquitoes and other arthropods.
Mosquito News, 24, 2 : 91—103.
Raffaele G. 1965. Adattamento allo sviluppo nel polio di Plasmodium berghei.
Riv. malariol., 44, 1/3 : 1—8.
Reeves H. C. 1962. Mosquitoes and virus diseases. «Biological transmission of di-
sease agents», New York : 75—82.
Reeves W. C. 1965. Ecology of mosquitoes in relation to arborviruses. Ann. Rev.
Entomol., 10 : 25—46.
Roy D. N. 1946. Entomology (medical and veterinary). Calcutta : 1—358.
Russell P. F., West L. S. and M a n w e 11 R. D. 1946. Practical malario-
logy. Philadelphia : 1—684.
Sandosham A. A. 1959. Malariology with special reference to Malaya. Singa-
pore : 1—327.
Serie C., A n d r al L., Lindrec A. and Neri P. 1964. Epidemie de fievre
jaune en Ethiopie (1960—1962). Bull. World Health Org., 30, 3 : 299—319.
Singh K. R. P. a. P a v r i Kh. M? 1966. Isolation of chittoor virus from mosqui-
toes and demonstration of serological conversions in sera of domestic animals
at Manjri, Poona, India. Indian J. Med. Res., 54, 3 : 220—224.
V i n ck e I. H. 1964. Le paludisme des rongeurs en Afrique. Ann. Soc. beige med.
trop., 44, 3 : 579—586.
Warren M., Eyles D. E., Wharton R. H., Ow Yang С. K. 1963.
The susceptibility of Malayan Anophelines to Plasmodium cynomolgibastianellii.
Indian J. Malariol., 17, 1 : 85—105.
Webber W. A. F. 1955. Diro-filaria aethiops Webber, 1955, a filarial parasite
of monkeys. III. The larval development in mosquitoes. Parasitology, 45 : 388.
Wharton R. H., Eyles D. E., Warren M., Moorhouse D. E.
and Sandosham A. A. 1963. Investigations-leading to the identification
of members of the Anopheles umbrosus group as the probable vectors of mouse deer
malaria. Bull. World Health Org., 29, 3 : 357—374.
Wharton R. H., Eyles D. E., Warren McW. and C h e о n g W. H.
1964. Studies to determine the vectors of monkey malaria in Malaya. Ann. Trop.
Med. and ParasitoL, 58, 1 : 56—77.
Y о e 1 i M. 1965. Studies on Plasmodium berghei in nature and under experimental
conditions. Trans. Roy. Soc. Trop. Med. and Hyg., 59, 3 : 255—271; discuss. :
272—276.
Yucel A. et Deschiens R. I960. Depistage d’un foyer de filariose a Wuche-
reria bancrofti en Turquie Orientale. Bull. Soc. parthol. exot., 53, 5 : 885—891.
СПЕЦИАЛЬНАЯ ЧАСТЬ
Семейство Culicidae ранее рассматривалось в более широком объеме,
с включением трех подсемейств: Culicinae, Dixinae и Chaoborinae. В на-
стоящее время преобладает тенденция рассматривать названные группы
в качестве самостоятельных семейств длинноусых двукрылых. Каждое
из них достаточно четко характеризуется комплексом существенных
морфологических признаков, что относится как ко взрослым насекомым,
так и к личинкам.
Сем. CULICIDAE—КРОВОСОСУЩИЕ КОМАРЫ
Ротовые органы в виде хоботка, длина которого в несколько раз пре-
вышает диаметр головы (рис. 1, 2). Усики 15-члениковые, покрытые
густыми длинными (б) или более редкими короткими (5) волосками.
Крылья узкие (рис. 6), жилки в чешуйках. Костальная жилка развита
как по переднему, так и по заднему краю крыла; субкостальная жилка
длинная, доходит до костальной жилки; радиус четырехветвистый, при-
чем г2+3 с развилком, г4+5 простая; поперечные жилки между гг и г2
отсутствуют; медиальная жилка трехветвистая; поперечные жилки гт
и шеи развиты; си одноветвистая; анальная жилка длинная, доходит до
края крыла.
Личинки очень различны по внешнему виду. Все три грудных членика
слиты вместе в более широкий, чем брюшко, грудной комплекс. Брюшко
состоит из 9 члеников, из которых VIII — сложный, образован путем
слияния VHI и IX члеников, а последний, IX членик является морфоло-
гически X. У всех личинок имеется функционирующая задняя пара
стигм или непосредственно на спинной стороне VIII членика брюшка,
или на конце особой дыхательной трубки. Ряд личинок — хищники, но
ни у кого из них усики не видоизменяются для схватывания добычи.
Пузыревидные расширения главных трахейных стволов если и имеются,
то только в груди.
Семейство Culicidae подразделяется на три подсемейства: Anophelinae,
Т oxorhynchitinae и Culicinae. Первые два представлены в Палеарктике
одним родом каждое; поэтому их характеристика, применительно
к фауне СССР, совпадает с характеристикой рода, соответственно Ano-
pheles Mg. и Toxorhynchites Theob. (Megarhinus Rob,-Des.). Их отличи-
тельные признаки, а также признаки подсемейства Culicinae, представлен-
ного в фауне СССР шестью родами, приведены ниже в определительной
таблице родов.
72
СЕМ. CULICIDAE
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Щупики самки длинные, равные по длине хоботку (рис. 28).
Последние два членика щупиков самца образуют булавовидное
утолщение (рис. 29). Брюшко без чешуек или по меньшей мере стер-
ниты на более или менее значительном протяжении голые, без че-
шуек ................................. 1. Anopheles Mg. (стр. 76).
2 (1). Щупики самки короткие, в несколько раз короче хоботка (рис. 1).
Последние два членика щупиков самца не образуют булавовидного
утолщения. Как тергиты, так и стерниты брюшка на всем протяже-
нии покрыты чешуйками.
Рис. 28. Строение ротового аппарата самки Anopheles.
вг — верхняя губа, вч — верхние челюсти (мандибулы),
г — гипофаринкс (подглоточник), -ня — нижние челюсти
(максиллы), иг — нижняя губа, щ — нижнечелюстные
щупики.
3 (4). Хоботок утолщенный, в вершинной половине явственно утончаю-
щийся и изогнутый книзу. Щиток цельнокрайний. Крупные комары
с пучками ярких волосков на конце брюшка...........................
..........................2. Toxorhynchites Theob. (стр. 118).
4 (3). Хоботок тонкий, на всем протяжении более или менее одинаковой
толщины или слегка утолщающийся к концу, в вершинной части
нерезко изогнутый. Щиток трехлопастный (рис. 30). (Подсемейства
Culicinae).
5 (6). Анальная жилка упирается в. задний край крыла на уровне ответ-
вления общего ствола г2+3-|-г4+5 (рис. 31)..........................
...................................3. Uranotaenia Атт. (стр. 126).
6 (5). Анальная жилка упирается в задний край крыла на более или
менее значительном расстоянии за местом ответвления общего ствола
г2+з+г4+5 (ближе к вершине крыла).
7 (8). Дыхальцевые щетинки имеются (рис. 4)........................
.............. 5. Culiseta Felt (Theobaldia N.-Lem.) (стр. 133).
8 (7). Дыхальцевые щетинки отсутствуют (рис. 4).
9 (10). Задыхальцевые щетинки имеются. Коготки самки, как правило,
с зубчиком. Церки самки более или менее выступают (рис. 11) . . .
..........................................7. Aedes Mg. (стр. 169).
10 (9). Задыхальцевые щетинки отсутствуют. Коготки самки простые,
без зубчика. Церки самки не выступают.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
73
11 (12). I членик передних лапок длиннее четырех следующих члеников
тех же лапок, вместе взятых; IV членик передних лапок у обоих
полов сильно укороченный.. . 4. Orthopodomyia Theob. (стр. 130).
12 (11). I членик передних лапок не длиннее четырех следующих члени-
ков тех же лапок, вместе взятых; IV членик передних лапок у самок
не укороченный.
Рис. 30. Щиток комара.
А — Culicinae, Б — Anophelinae.
Рис. 31. Крыло Uranotaenia.
13 (14). Пульвиллы под коготками на лапках имеются (рис. 5). Чешуйки
крыла узкие...................................8. Culex L. (стр. 314).
14 (13). Пульвиллы под коготками на лапках не развиты. Чешуйки крыла,
как правило, широкие.................6. Mansonia Blanch, (стр. 162).
Примечание. В некоторых случаях определение облегчается тем обстоятель-
ством, что три рода представлены в фауне СССР одним видом каждый; эти виды легко
определяются по характерным особенностям окраски: Uranotaenia ungiculata —
голубоватые полоски по бокам груди, Orthopodomyia pulchripalpis — узкие белые
продольные полоски на фоне темных бурых чешуек среднеспинки, Toxorhynchites
christophi — пучки длинных ярко окрашенных волосков в задней части брюшка.
74
СЕМ. CULICIDAE
33
Особенности строения гипопигия самцов имеют характер видовых
признаков; родовые же отличия выражены менее отчетливо; поэтому
в руководствах по определению комаров таблица для определения рода
по строению гипопигия обычно не приводится. Настоящая таблица имеет
предварительный характер. Род Toxorhynchites в таблицу не включен,
так как гипопигий Т. christophi не описан.
1 (2). Коксит без выступов (бородавок). Стиль длиннее коксита, реже
примерно равен ему по длине.......................1. Anopheles Mg.
2 (1). Коксит у большинства видов (но не у всех) с бородавкой или
с двумя бородавками. Стиль короче коксита, реже почти равен ему
по длине.
,3 (4). Правый и левый кокситы тесно сближены. Коксит с одной, уса-
женной шипами и щетинками бородавкой, расположенной перед
вершиной, реже сразу за серединой. X стернит на вершине с много-
численными шипиками или поперечным рядом зубцов. . . 8. Culex L.
4 (3). Правый и левый кокситы, как правило, несколько раздвинуты.
Коксит с двумя бородавками (базальной и вершинной), а если с од-
ной, то она расположена у основания или у середины коксита; реже
бородавка отсутствует. X стернит на вершине без многочисленных
шипиков и без ряда зубцов (чаще с 1—3 крупными склеротизован-
ными зубцами).
5 (6). Придаток стиля относительно длинный, его длина превышает
ширину стиля (рис. 7), если длина придатка не больше ширины стиля,
то придаток причленяется перед вершиной стиля; если же придатка
вовсе нет, то стиль при основании раздвоен .... 7. Aedes Mg.
6 (5). Придаток стиля короткий; как правило, его длина меньше ширины
стиля.
7 (8). Бородавка коксита с одним толстым, сильно склеротизованным
тупым шипом (рис. 93).........................6. Mansonia Blanch.
8 (7). Бородавка коксита, если она имеется, по крайней мере с двумя
заостренными шипами или крепкими щетинками.
9 (10). Коксит сильно утолщенный, короткий, с небольшой плоской
бородавкой. Стиль широкий, пластинчатый (рис. 64)..............• .
......................................... 3. Uranotaenia Arr.
10 (9). Коксит не сильно утолщенный, более или менее удлиненный,
с конической бородавкой, реже без бородавки. Стиль относительно
узкий.
11 (12). Придаток стиля по длине примерно равен ширине стиля в наи-
более утолщенной его части (рис. 68) ... 4. Orthopodomyia Theob.
12 (11). Длина придатка меньше ширины стиля в наиболее утолщенной
его части (рис. 71 и 74); на вершине стиля может быть два при-
датка — у С. longiareolata .........................5. Culiseta Felt.
ЛИЧИНКИ
1 (2). Стигмы помещаются непосредственно на спинной стороне VIII чле-
ника брюшка; окружены стигмальной пластинкой (подсемейство
Anophelinae).............................Anopheles Mg. (стр. 76).
,2 (1). Стигмы, окруженные стигмальной пластинкой, помещаются
на конце дыхательной трубки — сифона, отходящей от спинной
стороны VIII членика брюшка; длина сифона по меньшей мере равна
его ширине у основания.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
75
3 (4). Боковые лопасти верхней губы приспособлены для схватывания
добычи и состоят каждая приблизительно из 10 расположенных
в один ряд мощных подвижных шипов с крючкообразно загнутыми
и зазубренными концами (подсемейство Toxorynchitinae)...............
...............................Toxorhynchites Theob. (стр. 118).
4 (3). Боковые лопасти верхней губы лишь в редких случаях приспособ-
лены для схватывания добычи и состоят каждая не менее
чем из 30 различно развитых волосков (подсемейство Culicinae).
5 (6). Сифон и клапаны стигмальной пластинки видоизменены в бура-
вящий аппарат............................Mansonia Blanch, (стр. 162).
6 (5). Сифон и клапаны стигмальной пластинки нормального строения.
7 (14). Сифон с одной парой пучков на задней поверхности.
8 (9). Пучки на сифоне помещаются у его основания.................
......................................Culiseta Felt (стр. 133).
9 (8). Пучки на сифоне помещаются у его середины или ближе к вершине.
10 (11). По бокам VIII членика брюшка имеется по хитиновой пластинке,
от заднего края которой отходят чешуйки щетки. Зубцы гребня
расщеплены на концах.......................Uranotaenia Arr. (стр. 126).
11 (10). По бокам VIII членика брюшка — только щетка из чешуек,
расположенных в один ряд или в виде пятна различной формы.
Если пластинки имеются, то чешуйки щетки лежат самостоятельно.
Зубцы гребня, при его наличии, шиповидно заострены на
конце.
12 (13). Гребень на сифоне отсутствует. На спинной стороне VI—VIII чле-
ников брюшка имеются хитиновые пластинки, развитие которых уси-
ливается к заднему концу .... Orthopodomyia Theob. (стр. 130).
13 (12). Гребень на сифоне имеется. Пластинки на VI—VHI члениках (
брюшка отсутствуют.............................. Aedes Mg. (стр. 169).
14 (7). Сифон с несколькими парами пучков на задней и боковых его
поверхностях . ..................................Culex L. (стр. 314).
I. Подсемейство ANOPHELINAE
Длина наличника, как правило, превосходит его ширину. Мандибулы
и максиллы самки хорошо развиты. Щупики у обоих полов в большинстве
случаев равны по длине хоботку, реже у самок несколько короче (у не-
которых экзотических видов). Среднеспинка слабо выпуклая; щиток про-
стой, дуговидный (за исключением тропического рода Chagasia). Ноги
очень длинные и тонкие, в мелких, плотно прилегающих чешуйках.
Пульвиллы не развиты. Крылья относительно узкие, часто с пятнами
темных или светлых чешуек; базальная часть г4+5 пересекает поперечную
радиомедиальную жилку и от нее на небольшом протяжении проходит
по направлению к основанию крыла, закрыловая пластинка (у основа-
ния заднего края крыла) по краю усажена узкими чешуйками. Брюшко
в волосках или (реже) в волосках и чешуйках. Гипопигий простой
(рис. 9); коксит короткий, как правило, без базальной бородавки; на
месте последней развиты шипы или крепкие щетинки в различном числе
(1—6); анальный сегмент нацело перепончатый или со слабо развитой
склеротизацией; эдеагус простой, трубчатый, с конечными листочками
или без них; самка, с одной сперматекой.
Личинки отличаются сильно развитыми боковыми волосками на чле-
никах тела, лежащими в плоскости, параллельной поверхности воды.
Характерен их перистый тип ветвления, веерообразный не встречается.
Голова сильнее вытянута в длину. На переднем крае две пары волосков
76
I. ПОДСЕМ. ANOP HELINAE
наличника - наружная и внутренняя, позади которых находится пара
задних волосков наличника. Позади усиков на лобном щитке лежит
ряд из трех пар, как правило, длинных перистых волосков. У личинок
тропи ноского рода Chagasia их две пары. Усики короткие, палочковид-
ной формы, с одним перистым, реже простым волоском; на конце несут
два крупных уплощенных шипа и чувствительные волоски. Голова,
в связи с особенностями питания с поверхностной пленки воды, может
поворачиваться в обе стороны более чем на 180°, брюшной стороной
кверху. Грудь по бокам переднего края имеет двухлопастные, способные
втягиваться плечевые клапаны, служащие для фиксации личинки на
поверхностной пленке. Волоски на груди расположены тремя поясами,
соответственно числу грудных члеников, по 13—14 пар различно разви-
тых волосков в каждом. Брюшко отличается наличием особых звездча-
тых волосков, по одной паре на каждом из семи передних члеников
брюшка; эти волоски на I—III члениках могут быть рудиментарными.
Звездчатые волоски, как и плечевые клапаны, служат для прикрепления
личинки к поверхностной пленке. Стигмы расположены непосредственно
на спинной стороне VIII членика брюшка и окружены стигмальной пла-
стинкой. Последняя поддерживается двумя боковыми пластинками
с гребнем из шипов, соединенными сзади хитиновой дугой.
Куколки характеризуются короткими, широко открытыми дыхаль-
цевыми воронками; боковые волоски брюшных сегментов расположены
непосредственно у заднего бокового угла сегмента; хвостовые плавники
с двумя волосками, один из которых расположен у вершины срединной
жилки, другой, более слабо развитый, — перед нею.
Яйца, как правило, с оторочкой и воздушными камерами.
Подсемейство заключает в себе три рода. Палеарктике свойственны
лишь виды центрального рода Anopheles.
1. Род ANOPHELES Meigen
С признаками подсемейства; щиток цельнокрайний, дуговидный;
ствол медиальной вилки прямой. Личинки палеарктических
видов рода Anopheles распадаются на три группы: 1) все виды подрода
Anopheles, 2) An. (An.) plumbeus и 3) виды подрода Myzomyla. Признаки
групп указаны в определительной таблице. Род Anopheles распадаетси
на шесть подродов, из которых четыре являются в основном неотропиче-
скими; Палеарктике свойственны виды подродов Anopheles Mg. и Myzo-
myia Blanch. (Cellia Theob.).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
22
1 (14). Поперечные жилки и основания вилок r2+r3, тг-\-т2 и ш3+4-|-сг/.1
покрыты темными чешуйками; костальная жилка по переднему
краю в одноцветных темных чешуйках или имеет не более двух
светлых пятен. (Подрод Anopheles Mg.).
2 (11). Крылья с темными чешуйками (темно-желтовато-бурыми или
бурыми), часто с пятнами, образованными скоплениями чешуек
еще более темного цвета, но без пятен из белых чешуек; передние
бедра тонкие, цилиндрические.
3 (10). Крылья с более или менее одноцветными чешуйками, без скопле-
ний (пятен) более темных чешуек; основание передней вилки крыла
1. ANOPHELES
77
(r2+3), как правило, расположено несколько ближе к основанию
крыла, нежели основание средней вилки (/и1+т2).
4 (5). Пучок узких белых чешуек на лбу отсутствует. Среднеспинка
коричнево-желтая, одноцветная, с довольно длинными темно-бу-
рыми волосками........................1. An. (An.) algeriensis Theob.
5 (4). Белый пучок на лбу явственно развит. Боковые отделы средне-
спинки темные; волоски, покрывающие среднеспинку, в некоторых
ее отделах светлые.
6 (7). Вершина хоботка светлая (беловатая) ........................
.................................4. An. (An.) marteri Sen.-Prun.
а (б). Бахромка крыла у его вершины белая...........................
......................... An. (An.) marteri marteri Sen.-Prun.
б (а). Бахромка крыла у его вершины темная..........................
............................An. (An.) marteri sogdianus Kesh.
7 (6). Вершина хоботка темная.
8 (9). Мельче и темнее; преобладающая окраска тела черно-серая
со свинцовым отливом. Пучок чешуек на переднем крае средне-
спинки сильно развит и чисто белого цвета .........................
..................................2. An. (An.) plumbeus Steph.
9 (8). Крупнее и светлее; преобладающая окраска тела желто-бурая
или бурая. Пучок чешуек на переднем крае среднеспинки слабее
развит и имеет желтоватый оттенок..................................
....................................3. An. (An.) claviger Mg.
40 (3). Крылья с пятнами, образованными скоплениями чешуек более
темного цвета; основания радиальной r2+r3 и медиальной иДг/ц
вилок на крыльях расположены на одинаковом расстоянии от осно-
вания крыла.............................5. An. (An.) maculipennis Mg.
а (б). Общая окраска тела темно-бурая; среднеспинка с широкой серой
продольной полосой и темно-бурыми боковыми отделами; темные
пятна на крыльях явственно выражены ............................ .
An. (An.) maculipennis maculipennis Mg.
An. (An.) maculipennis messeae Fall.
An. (An.) maculipennis melanoon Hack.
An. (An.) maculipennis labranchiae Fall.
An. (An.) maculipennis atroparvus van Tiel.
б (а). Общая окраска тела светло-желтовато-бурая; среднеспинка
на всем протяжении более или менее одноцветно светло-бурая; тем-
ные пятна на крыльях слабо выражены, особенно у самцов . . . .
...........................An. (An.) maculipennis sacharovi Favre.
11 (2). Крылья по переднему краю с белыми пятнами, Образованными
скоплениями белых чешуек (или с одним белым пятном); передние
бедра при основании утолщенные.
12 (13). Костальная жилка с одним белым пятном вблизи вершины. Зад-
ние бедра с явственным предвершинным белым кольцом. Передние
бедра при основании слабо утолщены..................................
..........................................6. An. (An.)'lindesayi Giles,
а (б). Белые чешуйки нижней поверхности задних бедер развиты при-
мерно на х/3 длины бедра от его основания...........................
......................... An. (An.) lindesayi lindesayi Giles.
б (а). Белые чешуйки нижней поверхности задних бедер развиты только
на */6 длины бедра от его основания................................
............................. An. (An.) lindesayi japonicus Yam.
43 (12). Костальная жилка с двумя светлыми пятнами. Задние бедра без
78
14
15
16
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
I. ПОДСЕМ. ANOPHELINAE
белого предвершинного кольца. Передние бедра при основании
явственно утолщены................7. An. (An.) hyrcanus Pall.
(1). Поперечные жилки и основания вилок г2+г3 и вщ+Шз покрыты
белыми чешуйками; костальная жилка по переднему краю с четырьмя
или более светлыми пятнами. (Подрод Myzomyia Blanch..).
(16) . Брюшко покрыто чешуйками, образующими с боков торчащие
в стороны пучки. V членик задних лапок белый. Щупики самки
в основной части покрыты торчащими чешуйками...................
..............................8. An. (М.) pulcherrimus Theob.
(15) . Брюшко покрыто волосками, без чешуек. V членик задних лапок
темный. Щупики самки в прилегающих чешуйках....................
.............................9. An. (М.) superpictus Grassi.
ss
(14). Коксит при основании с внутренней стороны по большей части;
с двумя (реже с одной или тремя) крепкими щетинками, из которых
по меньшей мере одна расположена на сосочковидном бугорке.
(Подрод Anopheles Mg.).
(3). Коксит при основании с внутренней стороны с одним толстым
шипом ............................ 1. An. (An.) algeriensis Theob.
(2). Коксит при основании с внутренней стороны с 2—3 крепкими
щетинками.
(5) . Коксит при основании с внутренней стороны с тремя крепкими
щетинками, из которых две разветвлены ...........................
.......................................3. An. (An.) claviger Mg.
(4). Коксит при основании с внутренней стороны с двумя простыми
(не разветвленными) крепкими щетинками.
(9). Щетинки (шипы) класпет сближены, но не слиты.
(8). Эдеагус короткий и широкий, на вершине без листовидных при-
датков ............................. 2. An. (An.) plumbeus Steph.
(7). Эдеагус длинный и узкий, на вершине с листовидными придат-
ками .................................5. An. (An.) maculipennis Mg.
(6). По крайней мере некоторые щетинки (шипы) класпет слиты, обра-
зуя прозрачную пластинку.
(13) . Лопасти IX тергита конические или трапециевидные, короткие
(не высокие); их высота примерно равна их ширине.
(12) . Коксит близ Середины своего внутреннего края с толстой щетин-
кой. (Крылья в одноцветно темных чешуйках)........................
.................................4. An. (An.) marteri Sen.-Prun.
(11) . Коксит близ середины своего внутреннего края с относительно-
тонкой щетинкой. (Крылья с пятнами белых чешуек)..................
............................... 6. An. (An.) lindesayi Giles.
(10). Лопасти IX тергита длинные; их длина (высота) в 3—4 раза пре-
восходит их ширину....................7. An. (An.) hyrcanus Pall.
(1). Коксит с внутренней стороны при основании с 4—6 крепкими ще-
тинками, сидящими непосредственно на теле коксита, а не на сосоч-
ковидных выростах его. (Подрод Myzomyia Blanch.).
(16) . Эдеагус на вершине с короткими листовидными придатками;
............................... 9. An. (М.) superpictus Grassi.
(15) . Эдеагус на вершине без листовидных придатков ..............
.................................8. An. (М.) pulcherrimus Theob.
1. ANOPHELES
79
1
2
3
4
5
6
7
8.
9
10
11
12
13
14
15
16
17
ЛИЧИНКИ IV СТАДИИ
(2) . Лобные волоски короткие и простые. Боковые волоски на IV—
VI члениках брюшка длинные, явственно перистые...................
................................ 2. An. (An.) plumbeus Steph.
(1). Лобные волоски длинные и перистые. Боковые волоски на IV—
VI члениках брюшка не перистые, а делятся недалеко от основания
на несколько ветвей.
(16). Основания внутренних волосков наличника сильно сближены.
(И). Наружные волоски наличника простые или слабо разветвленные
на концах или начиная с середины.
(6). Волоски наличника начиная от середины или концевой трети не-
сут тонкую вторичную перистость. Лепестки звездчатых волосков
длинные и узкие. Гребень на боках основания стигмальной пластинки
в среднем из 20—25 зубцов .... 1. An. (An.) algeriensis Theob.
(5). Волоски наличника гладкие, без тонкой вторичной неристости.
(8). Задние волоски наличника из 2—5 ветвей. Лепестки звездчатых
волосков брюшка без вытянутой концевой нити, широкие, с глубокой
зубчатостью по боковым краям. Стигмы не соединены с центральной
пластинкой.. Гребень на боках основания стигмальной пластинки
из 12—16 зубцов........................3. An. (An.) claviger Meig.
(7). Задние волоски наличника простые. Стигмы соединены с централь-
ной пластинкой, часто имеют для этого специальные отростки.
(10). Звездчатый волосок на заднегруди слабо развит, из 10—11 ле-
пестков. Концы лепестков звездчатых волосков брюшка с вытянутой
концевой нитью....................4. An. (An.) marteri Sen.-Prun.
(9). Звездчатый волосок на заднегруди хорошо развит, из 15—20 ле-
пестков. Концы лепестков звездчатых волосков брюшка постепенно
заостряются и не вытянуты в концевую нить........................
..................................6. An. (An.) lindesayi Giles.
(4). Наружные волоски наличника древовидно разветвлены.
(13). Волосок на усике длинный, около половины длины последнего;
отходит от его середины или лишь немного смещен к основанию
. . ................................. 7. An. (An.) hyrcanus Pall.
(12). Волосок на усике очень короткий, не превышает ширину послед-
него у места его отхождения, около основания.
(15). Наружные волоски наличника доходят до переднего края рас-
правленных боковых лопастей верхней губы или даже заходят
за них. Стигмальная пластинка меньших размеров (длина 0.38—
0.50, ширина между концами боковых лопастей 0.39—0.52 мм),
Слабо пигментирована; срединная часть заднего отдела центральной
пластинки светлая, ячеи и складки ее переднего отдела развиты
слабо .................. An. (An.) maculipennis sacharovi Favre.
(14). Наружные волоски наличника не доходят до переднего края
расправленных боковых лопастей верхней губы. Стигмальная пла-
стинка больших размеров (длина 0.50—0.57, ширина между концами
боковых лопастей 0.56—0.59 мм), темно пигментирована. Срединная
часть заднего отдела центральной пластинки, ячеи и складки ее пе-
реднего отдела темные, хорошо развиты............................
................................5. An. (An.) maculipennis Meig.
(3). Основания внутренних волосков наличника сильно удалены
друг от друга.
(18). Наружные волоски наличника тонковетвистые, с числом ветвей
от 6 до 12—15. Задние волоски наличника 2—4-ветвистые, в виде
80
I. ПОДСЕМ. ANOPHELINAE
редкого исключения простые. У стигмальной пластинки централь-
ная пластинка в передней части среднего отдела заметно сужена;
ее срединная часть слабо пигментирована, в виде резко ограни-
ченного светлого медальона. Передний отдел центральной пластинки
в форме наконечника стрелы; его боковые отростки вытянуты, от-
ростки у стигм отсутствуют ... 8. An. (М.) pulcherrimus Theob.
18 (1.7). Наружные и задние волоски наличника простые. У стигмальной
пластинки центральная пластинка широкая, не суженная в перед-
ней части среднего отдела; резко очерченный медальон в ее средин-
ной части отсутствует. Передний отдел центральной пластинки
не имеет боковых отростков; отростки у стигм имеются, реже от
сутствуют . ..........................9. An. (М.) superpictus Grassi.
1. Подрод ANOPHELES Meigen
Поперечные жилки и основание вилок г2 -|~г3 и в темных
чешуйках; костальная жилка по переднему краю крыла имеет, как пра-
вило, не более двух светлых пятен или вовсе без пятен. Коксит при осно-
вании с внутренней стороны по большей части с двумя (реже с одной
или тремя) крепкими щетинками, из которых по меньшей мере одна рас-
положена на сосочковидном бугорке.
Личинки подрода Anopheles, за исключением An. plumbeus, отли-
чаются тесно сближенными основаниями внутренних волосков налич-
ника, хорошо развитыми перистыми лобными волосками, перисто вет-
вящимся волоском на усиках и длинными боковыми волосками (№ 6)
на IV и V члениках брюшка, ветвящимися около основания на 2—5 вет-
вей, и короткими на VI членике. Личинки An. plumbeus отличаются
удаленными друг от друга внутренними волосками наличника, корот-
кими простыми лобными волосками и волоском на усиках и длинными
перистыми волосками на IV—VI члениках брюшка.
Подрод Anopheles (известно свыше 100 видов, из них в Палеарктике 10)
широко распространен; виды этого подрода представлены тем или иным
количеством видов во всех зоогеографических областях; что касается
палеарктических видов подрода Anopheles, то они в отношении своего
распространения распадаются на две группы: 1) группа видов, близких
к An. maculipennis, свойственна по преимуществу умеренной зоне, при-
чем отдельные ее представители идут достаточно далеко на север; 2) виды,
близкие к An. hyrcanus, имеют тяготение к Ориентальной области, где
эта группа представлена рядом видов; распространена на север эта группа
значительно меньше.
1. Anopheles (Anopheles) algeriensis Theobald, 1903.
Легко отличается от прочих видов рода отсутствием пятен на крыльях,
а также отсутствием пучка белых чешуек на лбу. Грудь и брюшко по
большей части одноцветно светло-бурые; среднеспинка с длинными
темными волосками. Задние лапки иногда с неясными светлыми колеч-
ками.
Гипопигий (рис. 32) имеет весьма характерное строение: при осно-
вании коксита расположен один крупный шип, сидящий на хорошо раз-
витом бугорке; эквивалентом наружных шипов базального отдела ко-
ксита, имеющихся у An. claviger, являются 1—2 непостоянных слабых
щетинки; у вершины внутренней стороны коксита расположена креп-
кая щетинка; класпеты трехлопастные; наружная лопасть имеет 2—3
1. ANOPHELES
81
близко сидящих крепких острых шипа; средняя лопасть усажена во-
лосками, внутренняя несет три более или менее одинаковых по величине
щетинки; эдеагус у вершины с 2—3 парами длинных и тонких листовид-
ных придатков.
Личинка IV стадии (рис. 33) сходна с таковой An. claviger.
Голова: сближенные внутренние волоски наличника длинные и простые
или, реже, слабо разветвленные на концах; начиная от середины или
ближе к вершине несут тонкую вторичную перистость; наружные —
вдвое короче внутренних, то же с вторичной перистостью, простые или
2—3-ветвистые на концах. Задние волоски наличника гладкие, простые,
Рис. 32. Гипопигий Anopheles algeriensis Theob.
реже 2-ветвистые на концах, обычно достигают основания передних.
Усики темные, покрыты более многочисленными, сильнее пигментиро-
ванными, чем у An. claviger, шипиками.
Звездчатые волоски брюшка на III—VII члениках из 16—18 равно-
мерно, но слабо пигментированных ланцетовидных лепестков со слабо
выраженной концевой нитью и явственно зазубренными краями.
Стигмальная пластинка не имеет темной пигментации и резко выра-
женной ячеистости в срединной части центральной пластинки и в сред-
нем ее отделе не выделяется от боковых отделов. Наоборот, срединная
полоса обычно светлее окружающих частей. Основание переднего отростка,
как правило, заходит назад не далее границы переднего и среднего от-
делов центральной пластинки. Гребень по бокам VIII членика брюшка
из 7—11 крупных зубцов (наиболее длинные из них крайние), между
которыми сидят по 1—3 мелких шипа. Шипы по верхнему краю у осно-
вания несут ряд мелких шипиков.
Яйца продолговато-овальные, в средней части сильно расширен-
ные, с хорошо развитыми боковыми воздушными камерами, но без ото-
рочки.
6 Фауна СССР, А. В. Гуцевич и др.
S2
I. ПОДСЕМ. ANOPHELINAE
Распространение. В основном — Средиземноморская под-
область. В Западной Европе на север проникает до Англии. Замечателен
факт нахождения An. algeriensis на острове Саарема у западного побе-
режья Эстонии (Ремм, 1957). В СССР вид распространен в Закавказье,
на Северном Кавказе, в Средней Азии. Как правило, встречается редко.
Массовое появление наблюдалось в плавнях рек, например в Дагестане
в плавнях р. Сулак (Ениколопов, 1937).
Рис. 33. Anopheles algeriensis Theob. Личинка IV стадии.
1 — передний край лобного щитка, 2 — левый усик, з — срединные волоски переднегруди
(правая сторона), 4 — стигмальная пластинка.
Биология. Личинки встречаются как в крупных заболочен-
ностях, густо поросших растительностью (из погруженных растений
для таких водоемов особенно характерна водяная сосенка), так и в не-
больших, но сильно затененных водоемах, преимущественно в стоячей,
иногда довольно сильно засоленной воде; в Таджикистане (Глаголева,
1944) личинки приурочены к родниково-сбросовым заболоченностям
с жесткой щелочной водой; личинки нередко попадаются совместно с ли-
чинками An. hyrcanus и Culex territans. Зимуют личинки. Комары на-
падают на человека и животных, под открытым небом; в условиях Сред-
ней Азии (Таджикистан) комары попадаются по преимуществу в холод-
ные сезоны — весной и осенью; комары держатся, как правило, в ка-
мышовых зарослях, изредка залетая в помещения для скота и еще реже
в жилые помещения.
2. Anopheles (Anopheles) plumbeus Stephens, 1828 (nigripes Staeger,
1839). — Дупловой, или черноногий, малярийный комар.
Отличается темной окраской и несколько меньшими по сравнению
с An. claviger размерами.
1. ANOPHELES
83
Голова с боков усажена черными чешуйками, на лбу между глазами
расположен направленный вперед пучок узких, чисто белых чешуек.
Хоботок и щупики черные. Грудной отдел черно-бурый; среднеспинка
с широкой, занимающей более 1/3
ширины среднеспинки, светлой пе-
пельно-серой продольной полосой;
посредине переднего края средне-
спинки развит пучок узких белых
чешуек; боковые отделы средне-
спинки и бочки груди черно-бу-
рые; щиток черноватый. Ноги
черные или черно-бурые, тазики
и основания бедер светлее. Крылья
без пятен, густо покрыты темно-
бурыми чешуйками; жужжальца
с черной головкой. Брюшко
сверху черное, блестящее, снизу
несколько светлее. Волоски груди
и брюшка светло-бурые, с золо-
Рис. 34. Гипопигий Anopheles plumbeus
Steph.
тистым оттенком.
Гипопигий (рис. 34): коксит с двумя крупными простыми щетинками
при основании; кроме того, крепкая щетинка расположена близ середины
внутренней стороны коксита; шипы на вершине класпеты сближенные,
но не слитые; эдеагус короткий и широкий, без придатков.
Рис. 35. Anopheles plumbeus Steph. Личинка IV стадии.
1 — голова, сверху, 2 — левый усик, 3 — стигмальная пластинка.
Личинка IV стадии (рис. 35) —• от темно-серого до темно-ко-
ричневого цвета. У недавно слинявших особей брюшко может иметь
кольчатый вид вследствие ослабления пигментации между члениками.
6*
84
I. ПОДСЕМ. ANOPHELINAE
Голова темно-коричневая, одноцветная, со слабо развитыми зачат-
ками сложных глаз. Волоски наличника тонкие, на концах могут слегка
ветвиться. У личинок An. plumbeus var. bartanensts они простые, глад-
кие и не ветвящиеся. Основания волосков расставлены приблизительно
на равном расстоянии друг от друга. Задние волоски наличника, лоб-
ные и другие волоски тонкие, короткие и простые. Усики темные, пря-
мые, лишенные шипов, с коротким простым волоском ближе к осно-
ванию.
Брюшко: звездчатые волоски развиты на II—VII члениках. На I
членике — короткие и простые. Число лепестков не меньше 15; лепестки
однородно пигментированы, ланцетовидной формы, с заостренным кон-
цом, но без концевой нити; их края начиная с х/3 от основания могут быть
слабо зубчатыми. Боковые волоски на IV—VI члениках брюшка длин-
ные и перистые. Стигмальная пластинка с темно пигментированной
передней лопастью. Стигмы соединены специальными отростками или,
реже, непосредственно с передним отделом центральной пластинки.
Последний имеет характерную форму лука или фигурной скобки с тон-
ким темным передним отростком. Грануляция срединного отдела цен-
тральной пластинки темная, резко выделяется от более светлых боковых
частей.
Яйца удлиненно овальной формы, с довольно широкой оторочкой
но без воздушных камер.
Распространение. Основная часть ареала находится в пре-
делах Средиземноморской подобласти. В Западной Европе на север
проникает до Англии и Южной Швеции, в СССР — до Эстонии (Ремм,
1957). Распространен на Украине (где встречается редко), на Северном
Кавказе, в Закавказье, в Туркмении (в лесах Копет-Дага) и в Таджи-
кистане. ;
Найденный в Индии и Пакистане дупловой малярийный комар боль-
шинством авторов рассматривается как особый вид — An. bartanensts
James, 1911. Никаких существенных отличий между An. plumbeus
и An. bartanensts не указано. Исследованные нами имаго и личинки из
Таджикистана являются вполне типичными An. plumbeus.
В наиболее южных частях ареала вид приурочен к горным и долин-
ным широколиственным лесам, где не поднимается в горы выше 1600—
2000 м.
Биология Личинки обитают в водоемах в дуплах различных
видов деревьев (на Украине — береза, берест, дуб, ясень; в Крыму —
граб, дуб, каштан, клен, орех, ясень, фруктовые деревья; на Кавказе —
бук, дуб, каштан, клен, орех, черноклен, чинар, ясень, яблоня; в Тад-
жикистане — грецкий орех, тут, чинар), преимущественно глубоких
и сильно затененных. В Абхазии наблюдались случаи обитания личинок
в различных, сильно затененных искусственных водоемах (заброшенные
колодцы, бассейны канализационной системы и т. и.). Встречаются ли-
чинки совместно с личинками Aedes geniculatus, A. pulchritarsis и Ortho-
podomyia pulchrtpalpis, а также с личинками-«крысками» Myiatropa flo-
rea L. (из сем. Syrphtdae). Вода в биотопах An. plumbeus, по исследо-
ваниям в Таджикистане (Глаголева, 1946), имеет ясно выраженный ще-
лочной характер за счет значительного количества солей щелочных
металлов (преимущественно калия и натрия), помимо которых она со-
держит гуминовые и ульминовые кислоты, дубильные вещества,' пиг-
менты и т. д., экстрагируемые из древесины дупла; по цвету вода в таких
водоемах приближается к цвету крепкого чая. Зимуют яйца и личинки
.всех стадий; при промерзании водоема могут выживать, причем
1. ANOPHELES
85
яйца выдерживают достаточно длительные сроки Промерзания. Является
типично лесным насекомым; в помещения почти не залетает; на человека
нападает относительно редко.
Рис. 36. Anopheles claviger Mg.
В дупловом малярийном комаре малярийные плазмодии могут про-
делывать полный цикл спорогонии, но значение этого вида в качестве
переносчика очень невелико вследствие свойственных ему экологиче-
ских особенностей; в прежние годы имел некоторое эпидемиологическое
значение в лесных районах курортной зоны Кавказа.
86
I. ПОДСЕМ. ANOPHELINAE
3. Anopheles (Anopheles) claviger Meigen, 1804 (bifurcatus auct.).1—
Родниковый (лесной) малярийный комар (рис. 36).
Общая окраска по большей части желтовато-бурая. Голова с боков
с узкими темно-бурыми чешуйками, сверху в затылочном отделе
с такими же чешуйками чисто белого цвета; лоб с торчащим вперед пуч-
ком кремовых чешуек и волосков; хоботок и щупики одноцветно темно-
бурые; усики бурые, основные членики светлее; пушок и волоски, по-
крывающие усики, как у предыдущего вида. Среднеспинка бурая, посре-
дине с широкой, кпереди суживающейся беловато-серой продольной
Рис. 37. Гипопигий A nopheles claviger Mg.
полосой, занимающей около 3/6 ширины среднеспинки, спереди (у го-
ловы), близ средней линии, с пучком желтовато-белых чешуек; волося-
ной покров среднеспинки в средней (серой) части беловато-желтого,
с боков буроватого цвета; щиток бурый, часто светло-бурый, по заднему
краю с буроватыми щетинками; бочки груди бурые, покрытые едва за-
метным налетом. Ноги бурые или темно-бурые, с нижней стороны бедер
и голеней, а также на сочленениях несколько более светлые; лапки од-
ноцветные темно-бурые. Крылья прозрачные, жилки покрыты одноцвет-
ными бурыми чешуйками; жужжальца с темно-бурой головкой. Брюшко
бурое, по заднему краю тергитов более темное, покрытое довольно длин-
ными светло-бурыми волосками.
Гипопигий (рис. 37) по внутреннему краю коксита с одной креп-
кой щетинкой, расположенной близ вершины членика; при основа-
нии коксита имеются три крупные и крепкие щетинки, из которых
две наружные сближены своими основаниями и в вершинной своей по-
1 Как выяснилось, под названием «Culex bifurcatus/) Линней описал самца Culex
pipiens', в связи с этим название Anopheles bifurcatus L. должно быть заменено
на An. claviger Mg.
1. ANOPHELES
87
ловине ветвятся; внутренняя же, сидящая на сосочковидном бугорке,
простая.
Личинка IV стадии (рис. 38) крупной величины и темной
окраски с сильно пигментированными головой и стигмальной пластинкой.
Внутренние волоски наличника длинные, почти одной длины с телом
усика, простые или 2—4-ветвистые у вершины, без вторичной перистости,
со сближенными основаниями. Наружные значительно короче внутрен-
них, гладкие, редко простые, чаще 2—4-ветвистые начиная от середины
или вершинной трети. Задние волоски наличника короткие, тонкие, из
Рис. 38. Anopheles claviger Mg. Личинка IV стадии.
1 — передний край лобного щитка (слева ветвящиеся на концах наружные волоски наличника),
2 — левый усик, з — стигмальная пластинка.
2—5 ветвей, отходящих почти от основания. Усик вдвое короче головы,
его тело с внутренней стороны покрыто редкими шипиками, волосок
короткий, из 4—7 ветвей, лежи!? близко к основанию.
Звездчатые волоски хорошо развиты на III—VII члениках брюшка;
лепестки ланцетовидной формы с слегка вытянутой вершиной, но без
концевой нити, с гладкими или слабо зазубренными краями; слабо,
но равномерно пигментированы. Боковые волоски на I—III члениках
перистые, на IV—VI — ветвистые.
Стигмальная пластинка с целиком темно пигментированной передней
лопастью, отсутствием соединения со стигмами переднего отдела централь-
ной пластинки, имеющего форму треугольника с направленной вперед
вершиной, и с отчетливо выраженной срединной пигментацией, целиком
темной в переднем и заднем отделах и ячеистой с темными контурами
в среднем отделе. Необходимо отметить, что личинки II и III стадий
An. algeriensis и An. hyrcanus и II стадии An. maculipennis имеют сход-
ное строение стигмальной пластинки.
Гребень по бокам VIII членика брюшка из 12—14 крупных и средней
величины шипов, чередующихся с более мелкими.
88
I. ПОДСЕМ. AEOPHELJNAE
Яйца продолговато-овальные, с боков с большим количеством
воздушных камер и с простой, без поперечной исчерченности, отороч-
кой, имеющейся только у концов яйца.
Замечания по систематике. Описано несколько подвидов A n. cla-
viger. Один из них, характерный для Средиземноморья, предложено рассматривать
в качестве самостоятельного вида — An. petragnani Del Vecchio, 1939; форма отли-
чается деталями хетотаксии личинки, строения яйца и куколки. В условиях лабора-
тории А п. claviger и An. petragnani не скрещиваются или не дают жизнеспособного
потомства. Для вполне убедительного обоснования видовой самостоятельности Ап.
petragnani нужны дополнительные данные.
Распространение. Европейская часть СССР на север до Ле-
нинградской области (60° сев. шир.); широко распространен в Крыму
и на Кавказе (Северный Кавказ, Дагестан, Грузия, Армения, Азер-
байджан); Средняя Азия (Туркмения, Узбекистан, Таджикистан,
Киргизия); в горах Средней Азии местами доходит до высоты 2000 м;
Казахстан, Западная Сибирь на восток до Томской области. Большая
часть Западной Европы от Средней Швеции и Норвегии до Италии;
Передняя Азия (Палестина, Малая Азия, Ирак, Иран); Северная Африка.
Биология. Личинки встречаются преимущественно в слабо про-
греваемых солнцем, умеренно или слабо минерализованных водоемах,
питаемых родниками, как стоячих (типа заболоченностей), так и про-
точных, затененных деревьями, кустами, скалами и т. п. В горных мест-
ностях личинки обитают в заводях горных ручьев или в родниковых
выключениях, а также в каналах орошения, питаемых водой роднико-
вого происхождения. В некоторых горных и предгорных районах места
обитания личинок иногда бывают приурочены к населенным пунктам
или их ближайшим окрестностям; в таких случаях комары нередко за-
летают в дома и помещения для животных. Зимуют личинки III и IV
стадии в непромерзающих до дна водоемах; в южных частях ареала раз-
витие не приостанавливается даже зимою, хотя и сильно замедляется.
Личинки очень подвижны и при малейшем беспокойстве опускаются
на дно. Дневки комаров располагаются среди растительности; весною
при слабом развитии растительности комары попадаются и в постройках.
Нападает по преимуществу на открытом воздухе. Самки некоторых по-
пуляций An. claviger способны к автогенному развитию яиц первой кладки
без сосания крови (Маркович, 1941).
An. claviger является потенциальным переносчиком малярийных плаз-
модиев человека; эпидемиологическое значение вида на большей части
ареала невелико, однако в местах, где личиночные биотопы приурочены
к населенным пунктам, оно может быть значительным. Установлена
естественная зараженность An. claviger возбудителем туляремии.
4. Anopheles (Anopheles) marteri Senevet et Prunelle, 1927.
По внешнему виду похож на An. claviger, от которого отличается
беловатой вершиной хоботка (самки), а также строением гипопигия;
последний имеет в основной части коксита две крепкие простые, не пе-
ристые щетинки; по строению гипопигия близок An. maculipennis, от ко-
торого его отличает характер придатков класпет, имеющих вид уз-
ких пластинок (у А п. maculipennis на класпетах имеются лишь щетинки
или же только на дорсальном отделе класпеты щетинки слиты в одну
узкую пластинку).
Затылок в передней части в беловатых чешуйках; между глазами
имеются длинные волосовидные чешуйки. Хоботок с беловатой вершиной,
1. ANOPHELES
89
без светлых колечек; при основании щупиков имеется незначительное
число (1—2) белых чешуек; длина щупиков самца примерно равна длине
хоботка. Среднеспинка с беловатой срединной полосою, которая развита
лишь в передней ее части (х/2—1/3); по бокам от нее имеется по две темные
полоски. Среднеспинка покрыта многочисленными узкими бурыми или
золотистыми чешуйками, которые в области срединной продольной по-
лосы окрашены светлее и являются более широкими; темные полоски
в передней части среднеспинки лишены чешуек или волосков. Щиток
с более развитыми в боковых отделах щетинками. Ноги: тазики желто-
вато-белые в светло-желтых волосках; бедра, голени и лапки черно-
бурые, с синеватым отливом; лапки без колечек. Крылья без пятен, в гус-
тых темных чешуйках; бахромка у вершины крыла белая.
Рис. 39. Гипопигий Anopheles marteri Sen.-Prun.
Гипопигий (рис. 39): коксит с двумя крепкими шиповидными щетин-
ками в основной части, из которых медиальная загнута на конце, одной
крепкой щетинкой в вершинной трети медиального края и многочислен-
ными мелкими волосками на дорсальной поверхности. Класпеты дву-
лопастные, причем иногда разделение на лопасти очень слабо выражено;
дорсальная лопасть класпеты с тремя узкими пластинчатыми выростами;
вентральная лопасть с расширяющейся к вершине пластинкой; эдеагус
с длинными листовидными придатками.
Личинка IV стадии близка к личинке An. lindesayi. Волоски
наличника простые, основания внутренних сближены; последние более
чем вдвое длиннее наружных; задние волоски одной длины с наружными
или длиннее. Лобные волоски перистые^ с 6—7 парами боковых ветвей,
немного короче внутренних волосков наличника; концы внутренних
лобных волосков едва достигают оснований задних волосков наличника.
Усики с темно пигментированной вершиной не менее чем на V5 их длины;
волосок из 3—4 умеренно длинных ветвей лежит у середины, ближе
к основанию.
Волосок № 1 на заднегруди видоизменен в звездчатый, из 10—11 ле-
пестков, более широких у основания и сужающихся к вершине резкими
уступами в концевую нить. Звездчатые волоски хорошо развиты на II—
VII члениках брюшка, из 15—19 удлиненных узких лепестков; начиная
с середины они 2—3 неправильными уступами переходят в концевую
нить, достигающую 1/3 общей длины лепестка.
90
I. ПОДСЕМ. ANOPHELINAE
Стигмальная пластинка с передней лопастью, темно пигментирован-
ной у основания и более светлоокрашенной у переднего края; стигмы
специальными отростками соединены с передним отделом центральной
пластинки; последняя очень широкая в среднем отделе, грануляция
которого в срединной части более светлая.
Распространение. Средиземноморье: Корсика, Балканский
полуостров, Алжир, Палестина. В Средней Азии и Северном Иране осо-
бый подвид — An. marteri sogdianus Kesh. В СССР данный вид обнаружен
только в горах Таджикистана (Кешишьян, 1938). >
Anopheles marteri marteri Senevet et Prunelle.
G признаками вида.
Anopheles marteri sogdianus Keshishian, 1938.
Отличается от основной формы отсутствием белого пятна на бахромке
крыла у его вершины. По личинкам отличия от основной формы отсут-
ствуют. По материалу из Таджикистана первоначально был описан в ка-
честве самостоятельного вида (Кешишьян, 1938).
Распространение. СССР: горные районы Таджикистана на
высотах более 1000 м. Северо-Западный Иран.
Биология. Личинки в большом количестве встречаются весною
(апрель—июнь) в затененных каменистых водоемах по течению горных
рек, ручьев и родников, часто совместно с личинками An. claviger и Ап.
lindesayi; экологически сходны с личинками последнего вида.
5. Anopheles maculipennis Meigen, 1818. — Обыкновенный малярий-
ный комар (рис. 40).
Вопрос о классификации «комплекса maculipennis» — один из наибо-
лее трудных и спорных в систематике комаров. Как известно, был опи-
сан ряд «рас» данного вида, которые затем обычно трактовались как под-
виды; из них шесть представлены в фауне Палеарктики: maculipennis
Mg., messeae Fall., melanoon Hack, (subalpinus Hack, et Lewis), labran-
chiae Fall., atrorparvus van Tiel, sacharovi Favre; последний был описан
в качестве самостоятельного вида. В монографии, специально посвя-
щенной виду An. maculipennis (Беклемишев, 1944), все перечисленные
формы рассматриваются как подвиды. .
Далее, было предложено каждый из названных подвидов возвести
в ранг вида (Bates, 1940). Это предложение принято многими авторами.
Однако такая «крайняя» точка зрения не получила общего признания.
Было замечено, что некоторые из форм комплекса maculipennis ближе
друг к другу, чем к другим формам; намечаются группы внутри ком-
плекса. Эти группы и было предложено рассматривать в качестве видов.
Такая «компромиссная» трактовка вопроса принята многими, в частности
авторами всесветного каталога комаров (Stone et al., 1959). В составе
фауны Палеарктики вместо одного вида — An. maculipennis — признаются
три: maculipennis (с подвидами maculipennis, messeae и melanoon), labran-
chiae (с подвидами labranchiae и atroparvus) и sacharovi. Североамери-
канский An. occidentalis D. К. также рассматривается в качестве само-
стоятельного вида. Эти четыре вида, вместе с несколькими другими близ-
кими видами, образуют группу maculipennis, входящую в состав одной
из шести групп видов, на которую предложено разделить подрод Anopheles
[Reid a. Knight, 1961; разделение подрода Anopheles на шесть групп
1. ANOPHELES
91
видов было предложено еще Эдвардсом (Edwards, 1932)]. Предлагалось
даже указанную группу выделить в особый подрод Maculipennia (Buo-
nomini е Mariani, 1953), но это предложение не получило поддержки.
Рис. 40. Anopheles maculipennis Mg.
Сторонники дробления комплекса maculipennis на большее или мень-
шее количество видов указывают на возможность их достоверного распоз-
навания по окраске дорсальной поверхности яйца (что зависит от строения
яйцевой оболочки) и на наличие существенных биологических отличий
между формами (см. ниже). Особо подчеркивается невозможность нор-
92
I. ПОДСЕМ. ANOPHELINAE
мального скрещивания отдельных форм и, следовательно, существова-
ние репродуктивной изоляции при их совместном обитании (в одной
местности нередко встречаются 2—3 формы комплекса).
По нашему мнению, при решении вопроса о классификации комплекса
maculipennis должны быть приняты во внимание следующие соображения.
Классификация комаров основывается главным образом на анализе
совокупности признаков имаго (9 и (?) и личинок. На этом же материале
базируется и практика определения видов, что находит свое выражение
в построении трех таблиц для определения видов каждого рода. Любой
из палеарктических видов комаров, самостоятельность которого обще-
признанна, надежно определяется хотя бы по одной из этих трех «фаз».
Соблюдение этого принципа дает твердую опору при классификации,
и мы не видим оснований, чтобы от него отказываться.
До сих пор не обнаружено никаких достоверных различий между
членами комплекса ни по личинкам, ни по имаго, ни, в частности, по
строению гипопигия; определение возможно только по яйцекладкам.
Известные отличия sacharovi от прочих форм комплекса несущественны
и имеют характер отличий подвидового ранга. Указывалось также на
некоторые отличия в строении гипопигия, в частности в форме шипов
на класпетах. Тщательное сравнительное исследование вооружения
класпет различных форм An. maculipennis показало, что по этому при-
знаку нельзя определять отдельные особи, но общая картина изменчи-
вости данного признака характерна для каждого подвида (Денисова,
1940, 1948, 1964).
Что касается биологических особенностей, то довольно существенные
внутривидовые различия выявлены у ряда видов комаров. Так, у многих
видов обнаружены неавтогенные и автогенные популяции. В этом слу-
чае мы имеем дело, видимо, с внутривидовыми группировками — био-
типами, иногда может быть, с подвидами. Весьма вероятно, что если бы
другие полиморфные, широко распространенные виды комаров изучались
бы столь же основательно, как изучается An. maculipennis, то и в других
видах была бы обнаружена такая же разнородность, и возник бы вопрос
о дроблении многих видов, что, по нашему мнению, нельзя признать
оправданным.
Мнение о существовании строгой репродуктивной изоляции между
членами комплекса maculipennis основано главным образом на опытах
с atroparvus, так как этот подвид легко культивируется в лаборатории.
Не вполне ясно, насколько полной и повсеместной является репродуктив-
ная изоляция всех членов комплекса. К тому же опыты скрещивания
определенных подвидов не всегда дают один и тот же результат; воз-
можность получения плодовитого потомства зависит от многих обсто-
ятельств. Так, в опытах гибридизации labranchiae/. atroparvus резуль-
тат зависел от того, были ли взяты комары (labranchiae) из природы или
из лабораторной культуры (Mariani et al., 1964). У других видов комаров,
например у Culex pipiens, также известны факты по крайней мере час-
стичной нескрещиваемости между различными подвидами и даже по-
пуляциями.
Так или иначе, в данном случае частичная или даже полная репро-
дуктивная изоляция подвидов свидетельствует лишь о начавшейся ди-
вергенции, которая, однако, еще не продвинулась достаточно далеко
и не привела к формированию новых видов. И лишь форма sacharovi
стоит на грани превращения в самостоятельный вид.
В силу изложенного мы считаем более правильным по-прежнему
рассматривать An. maculipennis как единый политипический вид с 6 пале-
1. ANOPHELES
93
арктическими подвидами: maculipennis, messeae, melanoon, labranchiae,
atroparvus и sacharovi. Последний, как более обособленный, в известном
смысле может быть противопоставлен остальным; только он выделен
в вышеприведенных таблицах для определения видов по самкам и по
личинкам. Мы полагаем, что признание за каждой из названных форм
видового статуса нарушает цельность и стройность классификации под-
рода Anopheles, так как различия между этими формами заведомо иного
характера и меньшего масштаба, чем различия между прочими видами
подрода Anopheles палеарктической фауны. Впрочем, вопрос о таксоно-
мическом ранге разновидностей An. maculipennis не имеет принципиаль-
ного характера, поскольку, как указывал еще Дарвин, разновидность —
это возникающий вид.
Ниже мы даем сначала общее описание An. maculipennis, относящееся
ко всем палеарктическим подвидам, кроме An. т. sacharovi, а затем
указываем отличительные особенности названного подвида. Важнейшие
морфологические и биологические особенности подвидов сопоставлены
в виде таблицы (см. вклейку между стр. 96—97).
Anopheles maculipennis — вид, подверженный очень большим коле-
баниям в окраске и величине; экземпляры более южного происхождения,
как правило, отличаются посветлением окраски и меньшими размерами.
Голова с боков усажена торчащими темно-бурыми чешуйками, сверху
с направленным вперед пучком довольно длинных беловатых чешуек
и волосков. Усики бурые, как у предыдущих видов. Щупики и хоботок
бурые, преимущественно в прилегающих чешуйках. Среднеспинка в сред-
ней части с широкой, кпереди суживающейся сероватой продольной
полосой, в передней половине которой намечаются обыкновенно 2—3
неясные буроватые полоски; по переднему краю среднеспинки у головы
имеется пучок беловатых чешуек; боковые отделы среднеспинки в пе-
редней своей части бурые, в задней — черно-бурые; волосяной покров
среднеспинки в средней ее части и на щитке — светло-бурый, в осталь-
ной части, в особенности над основаниями крыльев, — темно-бурый.
Ноги бурые, с нижней стороны бедер и внутренней стороны голеней
более светлые; лапки темные. Крылья покрыты буроватыми чешуйками,
с четырьмя пятнами черно-бурых чешуек, расположенными при основании
радиальной и медиальной вилок, на поперечных жилках и в основной
части общего ствола г2-(-г3; чешуйки, окаймляющие край крыла, на его
вершине, как правило, светлые, желтоватые. Жужжальца желтые, с бу-
роватой головкой. Брюшко бурое, иногда черно-бурое, часто с более
светлыми пятнами в боковых отделах тергитов; брюшко покрыто до-
вольно длинными золотисто-жёлтыми или буроватыми волосками.
Гипопигий (рис. 41) близ середины внутреннего края коксита несет
одну крепкую щетинку; в -основной части коксита сидят на бугорках
две крупные щетинки. Шипы на класпетах сближены, но никогда не
сливаются; форма шипов разнообразна (тупые, заостренные), их количество
также варьирует. Эдеагус длинный и узкий, с листовидными придатками
у вершины. Различия между подвидами An. maculipennis по личинкам
настолько малы, что пока не удалось найти признаков, позволяющих
с достоверностью их определять. Исследование массового материала
с последующей вариационно-статистической обработкой показало раз-
личия в среднем числе и относительном развитии ветвей некоторых во-
лосков (№ 1—6, по Мартини) на спинной стороне члеников брюшка,
при значительном захождении крайних вариантов друг за друга у всех
подвидов. Это делает индивидуальное определение подвидов по ли-
94
I. ПОДСЕМ. ANOPHELINAE
чинкам невозможным. Поэтому мы приводим только описание личи-
нок основного подвида An. maculipennis и подвида An. maculipennis
sacharovi, различия между которыми достаточно достоверны.
Личинки IV стадии (рис. 42), в связи с разнообразием
мест обитания и условий развития, очень сильно изменчивы по внешнему
виду, окраске и величине. Как правило, особи из более северных широт
крупнее, чем из более южных, с более темно пигментированными хити-
новыми образованиями (голова, стигмальная пластинка и т. п.). Общая
Рис. 41. Гипопигий Anopheles maculipennis Mg.
окраска тела, в зависимости от окраски окружающей личинку среды
и от освещенности водоема, колеблется от серо-желто-зеленой до черно-
зеленой.
Голова с очень изменчивой пигментацией: от светло-коричневых
пятен обычного числа и расположения до черно-коричневых полос на
общем темном фоне головной капсулы. Волоски наличника: внутренние
тесно сближены основаниями, ветвятся на концах (3—8ветвей); наруж-
ные древовидно ветвятся. Характер ветвления изменчив: чаще от основ-
ного стержня отходят две ветви, далее дихотомически или древовидно
ветвящиеся в горизонтальной плоскости и образующие двойную метелку;
реже от основного стержня отходят три или более вторичные ветви, вет-
вящиеся дальше и образующие одинарную метелку. Волоски направлены
под расходящимся кпереди углом. Наружные волоски наличника не
доходят до переднего края расправленных боковых лопастей верхней
губы, целиком лежат на фоне последних. Задние волоски наличника
короткие, слабо ветвистые (2—8 ветвей), лишь немного заходят за линию
оснований передних волосков. Усики почти прямые, до 2/3 длины головы,
Рис. 42. Anopheles maculipennis Mg. Личинка IV стадии.
наличника (дихотомическое0 ветвление) ~/СН°сильио наружного волоска
внутренних волосков Ha^J L?т
Яйца подвидов Anopheles maculipennis Mg. По Missiroli, Hackett, Martini.
1 — An. m. subalpinus (темная форма, melanoon); 2 — 6. — An. m. messeae; 7—8 — An.
m. maculipennis; 9—12 — An. m. atroparvus; 13—14 — An. m. labranchtae; 15 — An. m.
sacharovi.
Подвиды Anopheles maculipennis Mg.
Яйцо (см. рис. на стр. 96) Биологические особенности Географическое рас- пространение
окраска верхней поверхности воздушные камеры
Группа maculipennis An. m. maculipennis Mg. An. m. messeae Fall. An. m. melanoon Hack, (subalpinus Hack, et Lewis) Две темные поперечные по- лосы на уровне наружных концов воздушных камер; между полосами темных пя- тен нет (иногда 1—2 пятна). Помимо темных попереч- ных полос (иногда неотчет- ливых), в промежутке между ними имеются темные пятна. Окраска одноцветно темная или более светлая, с попереч- ными полосами и темными пятнами между ними (кон- трастный рисунок на светлом фоне). Хорошо развиты; по- верхность камер между ребрышками («межре- берная перепонка») мор- щинистая, реже гладкая. Хорошо развиты, длин- нее, чем у предыдуще- го; межреберная пере- понка морщиниста я. Хорошо развиты, но короче, чем у предыду- щего; межреберная пере- понка гладкая или слег- ка морщинистая. Копуляция на лету. Зимов- ка в холодных, реже в теп- лых помещениях; иногда пьет кровь зимой. Копуляция на лету. Зимов- ка в холодных помещениях; зимой крови не пьют. Копуляция на лету. Зимует чаще в теплых помещениях, может пить кровь зимой. Европа на север до 60—62°, на юг до Ис- пании, Италии, Греции; Западная Сибирь; не- которые горные районы Средней Азии (Копет- даг), Закавказье; Север- ный Иран. Особенно обилен на Кавказе. Северная и Средняя Европа, Северная Азия; на запад, север и восток до границы ареала вида; на юг до Испании, Гре- ции, а в СССР до Кав- казского хребта, Цент- рального и Юго-Восточ- ного Казахстана, до Се- верной Киргизии. Встре- чается в Монголии, захо- дит в Северо-Восточный Китай. Южная Европа, За- кавказье (где он явля- ется преобладающим или единственным подвидом); изредка встречается на Северном Кавказе.
Группа labranchiae An. m. labranchiae Fall. An. m. atroparvus Van Thiel Поперечных полос у концов камер нет; рисунок образован темными пятнами, отходящи- ми от краев и суживающимися к середине. Общая окраска более темная у An. т. atropa- rvus и более светлая у Ап. т. labranchiae. Хорошо развиты, но относительно короткие; межреберная перепонка гладкая или слегка мор- щинистая. Копуляция на лету. Обычно зимует в теплых помещениях, пьет кровь и зимой. Копуляция в сидячем поло- жении или на лету. Легко размножается в лаборатории. Зимует в теплых помещениях, зимой пьет кровь. Западное Средизем- номорье, на восток до Югославии и Ливии, на север не доходит до Пи- ренеев и Альп. В СССР не обнаружен. Европа на север до 55—56°, на юг до Испа- нии, Италии и Балкан. В СССР — Кавказ, Ук- раина, Белоруссия.
An. m. sacharovi Favre lel..ius Edw). Окраска одноцветная, се- рая. Отсутствуют или ру- диментарны. Копуляция на лету. Зимует чаще в теплых помещениях, зимой может пить кровь. Восточное Средизем- номорье — Италия, Бал- канский полуостров, Северная Африка, Пе- редняя Азия. В СССР— Средняя Азия (где, как правило, единственный подвид), Кавказ (вос- точные равнинные рай- оны). Иран, Афганистан. Западный Китай.
1. ANOPHELES
97
с более темным концом; волосок короткий (рис. 48), в среднем равен
ширине усика у места своего отхождения, лежит около основания и со-
стоит из 4—6 тонких ветвей.
Срединные волоски переднегруди: наружный — короткий и простой,
средний — перистый, с 3—7 боковыми ветвями с каждой стороны, внут-
ренний почти вдвое короче, слегка ветвится на конце. Звездчатый во-
лосок на заднегруди рудиментарный.
Звездчатые волоски брюшка развиты на III—VII члениках. Каждый
из них состоит из 16—24 лепестков, слегка расширенных в среднем от-
деле, который более сильно пигментирован. Концевая нить достигает
трети длины лепестка, нередко заострена на конце; ее основание несколь-
кими неправильными уступами переходит в тело лепестка в наиболее
широком его отделе.
Стигмальная пластинка крупная: длина 0.5—0.57 мм, ширина между
концами боковых клапанов 0.56—0.59 мм. Твердые хитиновые образования
обычно сильно пигментированы. Передняя лопасть с темным основанием,
остальная часть заметно светлее. Стигмы на некотором расстоянии от
центральной пластинки, не имеют отростков. Передний отросток централь-
ной пластинки своим основанием не доходит до границы между передним
и средним ее отделами, а переходит в складки и контуры ячей средин-
ной части. Последняя темно пигментирована от заднего края до середины
заднего отдела, затем кпереди переходит в гранулированную часть,
занимающую весь средний отдел и заднюю половину переднего отдела.
Она может быть сплошной или, чаще ячеи в среднем отделе расходятся
в стороны, оставляя более светлой срединную часть. При сплошной гра-
нуляции ячеи срединной части могут образовывать более темную полосу,
как у стигмальной пластинки An. claviger.
Гребень на боковых пластинках VIII членика состоит из 6—10 (чаще
7—9) крупных шиповидных зубцов, между которыми находятся в раз-
личном числе (1—4) значительно более мелкие.
Боковой волосок на седле длинный и обычно простой. Наружные
хвостовые волоски ветвятся несимметрично, их концы крючкообразно
загнуты, внутренние — ветвятся сильнее на верхней стороне. Плавник
из 18—20 пучков. Жабры в среднем равны длине седла, обе пары одной
длины.
Яйца An. maculipennis, за исключением An. т. sacharovi, имеют
занимающие среднюю треть яйца плавательные камеры и оторочку.
Структура и окраска наружной оболочки — экзохориона — различна,
но характерна для отдельных подвидов. Она зависит от местных изме-
нений строения экзохориона и состоит из темных и светлых пятен
и полос. Различия подвидов по этому признаку даны в Таблице и на
рисунке (см. стр. 96).
Распространение. Европа, Северная Африка, Северная
Азия (мы не касаемся вопроса о статусе североамериканской формы,
которая теперь большинством авторов рассматривается как самостоя-
тельный вид — An. occidentalis D. К.). Самое северное местонахождение
An. maculipennis — район юго-западнее Мурманска, 69° с. ш. (Соловей
и Лиходед, 1966). Далее граница ареала идет на юго-восток (в Карелии
у 66°) и на восток до р. Лены примерно на той же широте по долинам рек,
опускаясь значительно южнее по водоразделам и особенно в горах. От
Якутска (где вид обнаружен) граница ареала, плохо прослеженная на
этом участке, идет на юг и пересекает долину Амура примерно в 150 км
восточнее Благовещенска. Ареал вида включает также прилегающие
к Забайкалью районы Северо-Восточного Китая, Монголии, Синьцзян,
7 Фауна СССР, А. В. Гуцевйч и др.
98
I. ПОДСЕМ. ANOPHELINAE
Северный Афганистан и Иран, страны Передней Азии и отдельные районы
Северной Африки.
Биология. Вопросам экологии An. maculipennis посвящена ог-
ромная литература, в том числе специальная монография (Беклемишев,
1944). Ниже указаны лишь некоторые важнейшие черты экологии вида,
преимущественно общие для всех подвидов.
Личинки обитают в чрезвычайно разнообразных по типу, глубине
и величине постоянных и временных стоячих водоемах, хорошо осве-
щенных солнцем (на юге — в полу затененных), богатых скоплениями
нитчатых водорослей и высшей водной растительностью. В глубоких
постоянных водоемах они встречаются в местах скопления водной рас-
тительности, образующих у поверхности «второе дно». При летнем пере-
сыхании водоемов встречаются на мелководье и в заводях слабо проточ-
ных водоемов. На юге встречаются и в слабо солоноватой воде.
Зиму вид проводит во взрослой фазе, зимуют оплодотворенные самки,
обычно в различных нежилых помещениях, часто в помещениях для
сельскохозяйственных животных, реже в природе — в пещерах, норах.
В северной и восточной частях ареала комары, зимующие в необитае-
мых постройках, выносят многомесячное пребывание при очень низких
температурах (An. т. messeae).
Фенология вида хорошо изучена применительно к условиям различ-
ных ландшафтных зон (Беклемишев и Шипицина, 1957). На протяжении
теплого времени, когда температура благоприятствует размножению
(температурный минимум развития лежит около 10°), вид дает ряд
поколений: от 2 — на севере до 4—6 — на юге. Максимум численности
имаго обычно приходится на середину лета, но в районах с очень жар-
ким летом в июле—августе может наблюдаться снижение числен-
ности.
Главная масса комаров концентрируется в населенных пунктах
и вблизи них. Основной источник питания — кровь сельскохозяй-
ственных животных (коров, лошадей, свиней и др.), в меньшей степени —
людей. При малом количестве или отсутствии сельскохозяйственных
животных комары питаются преимущественно на людях; могут пить
также кровь домашних и диких птиц.
Обыкновенный малярийный комар — облигатный кровосос; питание
кровью необходимо для размножения; автогенного развития яиц у Ап. ma-
culipennis не наблюдалось. Для данного вида (как, вероятно, и для рода
в целом) летом характерна гонотрофическая гармония: одного полного
насыщения кровью достаточно для развития и откладки яиц.
Эпидемиологическое значение. В прошлом An. ma-
culipennis был важнейшим переносчиком всех трех основных видов маля-
рийных плазмодиев; на большей части Европы, в Казахстане, Сибири,
Забайкалье обыкновенный малярийный комар был практически един-
ственным переносчиком малярии. В южных частях ареала An. macu-
lipennis являлся (местами является и сейчас) важным переносчиком
малярии наряду с другими видами.
Обыкновенный малярийный комар известен как промежуточный хо-
зяин филяриид, паразитирующих у собак — Dirofilaria immitis и D. re-
pens.
В Чехословакии из An. maculipennis выделен вирус, получивший
название чалово.
По-видимому, An. maculipennis может играть роль в циркуляций
туляремийного микроба: установлена естественная зараженность.
1. ANOPHELES
99
Anopheles maculipennis sacharovi Favre, 1903 (elutus Edwards, 1921).
От других подвидов отличается более светлой окраской тела; боко-
вые отделы среднеспинки окрашены в светло-желтовато-бурый цвет,
более или менее одинаково со средним ее отделом. Темные пятна на кры-
льях, в особенности у самцов, более слабо выражены, а у потертых эк-
земпляров вовсе неразличимы. Чешуйки, окаймляющие край крыла,
одноцветные, без светлого пятна на вершине.
Указанные признаки не носят абсолютного характера. Надежное рас-
познавание подвида по имаго не всегда возможно.
Личинка IV стадии (рис. 43) очень сходна с An. maculi-
pennis, но в среднем меньшей величины. Имеющиеся отличия — только
количественного порядка — выражены слабой лежат в пределах изменчи-
вости. По общей окраске преобладают светлые желтовато-зеленые личинки
со слабо пигментированными головой и стигмальной пластинкой. Ри-
сунок на голове менее резкий, напоминает рисунок у недавно слиняв-
ших личинок An. maculipennis. Волосок на усике и наружные волоски
наличника в среднем относительно длиннее, чем у основной формы; со-
ответственно концы последних доходят до переднего края расправлен-
ных боковых лопастей верхней губы, их концы могут даже слегка вы-
даваться вперед; однако в ряде популяций личинок наблюдается боль-
шой процент исключений. В еще большей степени это относится к из-
менчивости числа лепестков в звездчатых волосках, которое у Ап. т.
sacharovi в среднем меньше, чем у основной формы, при одинаковом
их строении.
Стигмальная пластинка общими размерами и интенсивностью пиг-
ментации центральной пластинки отличается от An. maculipennis. Она
хотя и сильно варьирует в размерах, но заметно меньше и слабее пиг-
ментирована. Длина колеблется от 0.38 до 0.5 мм, ширина между кон-
цами боковых лопастей — от 0.39 до 0.52 мм. Центральная пластинка
с очень слабо выраженной светлой срединной грануляцией. Темный
участок в заднем отделе ограничен только ее задним краем; впереди она
прерывается и становится заметной только в среднем отделе. Основание
переднего отростка лишь слегка заходит в передний отдел, где рас-
ходится в виде складок, которые только у сравнительно редких, более
сильно пигментированных личинок достигают среднего отдела.
Яйца, отложенные самками весеннего, летних и осеннего поколе-
ний, лишены воздушных камер; имеется только оторочка. Однако на
юге ареала, где развитие в течение зимы не прекращается, яйца зимних
кладок имеют рудиментарные плавательные камеры, число и степень
развития которых» очень сильно изменчивы.
Распространение. Средиземноморская подобласть, преимуще-
ственно восточная ее часть: Италия, юг Балканского полуострова, Пе-
редняя Азия. В СССР — Средняя Азия на север до Аральского моря
и Киргизского хребта, Восточное Закавказье, Дагестан. Данный под-
вид встречается также в Иране, Афганистане и прилегающей к Средней
Азии части Синьцзяна (КНР).
Биология. Более характерен для аридных зон, где местами
достигает высокой численности. Личинки встречаются в открытых, хо-
рошо освещенных солнцем неглубоких водоемах, богатых водорослями
и негусто заросших высшей водной растительностью; выдерживают воду
с значительным содержанием солей. Часто, совместно с личинками Cu-
lex modestus Fic.; личинки малоподвижны, редко и на короткий срок
покидают поверхность воды.
7*
100
I. ПОДСЕМ. ANOPHELINAE
Во всех частях своего ареала An. т. sacharovi был (кое-где и теперь
остается) важным переносчиком малярии.
6. Anopheles (Anopheles) lindesayi Giles, 1900.
Окраска темно-бурая. Голова черная, с боков густо усажена черными
торчащими чешуйками, средняя часть затылка несет торчащие чешуйки
чисто белого цвета; лоб между глазами с направленным вперед пучком
Рис. 43. Anopheles maculipennis sacharovi
Favre. Стигмальная пластинка личинки
IV стадии.
желтовато-белых волосков; усики как у An. maculipennis, щупики и хобо-
ток черные. Среднеспинка в средней части светло-пепельно-серая, с боков
с темно-бурой, довольно широкой продольной полосой, которая снаружи
ограничивается узкой сероватой полоской; преобладающая окраска
бочков груди темная; щиток серый; волосяной покров грудного отдела
светло-желтоватого цвета. Ноги черно-бурые, тазики и вертлуги светлые;
задние бедра в третьей четверти своей длины с широким, чисто белым
кольцом. Крылья в большей своей части покрыты темными чешуйками;
на костальной жилке у вершины крыла имеется одно крупное белое пятно,
которое распространяется на и г2; участки белых чешуек имеются,
кроме того, в средней части г3, в основной части г4+5, в средней
части тг, в основной и вершинной третях пг3+4, кроме того, в белый
цвет обычно бывают окрашены чешуйки на концах жилок г4+5, т2,
си1 и ап- жужжальца светлые, с черной головкой. Брюшко чернобурое,
снизу сероватое, покрыто довольно длинными буроватыми волосками.
Гипопигий (рис. 44) при основании коксита с двумя щетинками;
шипы на конце класпеты слиты и образуют в совокупности длинную
и узкую пластинку; лопасти IX тергита короткие, конические.
1. ANOPHELES
101
Личинка IV стадии (рис.. 45) до 6 мм длиной, серого или
серо-зеленого цвета. Голова желто- или темно-коричневая. Посредине
лобного щитка от его заднего края лежит темное овальное пятно, за-
ходящее вперед за основание лобных волосков, по бокам от него около
глаз — два небольших овальных пятна. Волоски наличника простые,
как исключение задние могут быть 2-ветвистыми на концах; внутренние
сближены основаниями, очень длинные, наружные на х/3 — 1/2 короче;
задние волоски наличника одной длины с наружными. Лобные волоски
Рис. 44. Гипопигий Anopheles lindesayi Giles. По La Casse a.
Y ainaguti.
короткие, наиболее длинный внутренний едва заходит за основания
задних волосков наличника. Усик сильнее пигментирован у вершины,
волосок у основания из 6—9 веточек, короткий.
Волосок № 1 на заднегруди — звездчатый, хорошо развит, из 15—
22 лепестков без концевой нити; иногда рудиментарен. Звездчатые волоски
брюшка хорошо развиты на II—VII члениках: на II из 13—17, на III и
VI из 19—24 и на VII из 15—19 лепестков; лепестки темные, с хорошо
развитой длинной концевой нитью; у основания последней зубчатость
сильно растянута, часто переход в концевую нить очень постепен-
ный.
Стигмальная пластинка с'темной у основания передней лопастью,
светлее окрашенной к переднему краю. Стигмальные отростки обычно
Рис. 45. Anopheles lindesayi Giles. Личинка IV стадии. По Пури.
1 — передний край лобного щитка, 2 — усик (правый) с внутренней боковой стороны,
з — срединные волоски переднегруди, 4—6 — основания боковых волосков передне-,
средне- и заднегруди, 7 —- звездчатый волосок IV членика брюшка, 8 стигмальная
пластинка.
1. ANOPHELES
103
хорошо развиты, при их отсутствии передний отдел центральной пла-
стинки непосредственно прилегает к стигмам; его края резко очерчены,
грануляция среднего отдела ясно выражена, но лишь немного темнее
окружающих боковых его частей.
Распространение. СССР: горные районы Таджикистана,
на высоте 800—1600 м над уровнем моря (Кешишьян, 1938). Горные
районы Индии, Малайского полуострова, Китая, Филиппинских остро-
вов. В Японии особый подвид, An. lindesayi japonicus Yam.
Биология. Личинки встречаются в затененных, бедных расти-
тельностью горных ключах и ручьях поверхностного или снегового пи-
тания, с холодной (11—15°) чистой водой. Личинки приурочены к местам
с более слабым течением — излучинам и прибрежной зоне. Зимуют ли-
чинки. Комары нападают на человека преимущественно по вечерам и
вне помещений.
Anopheles lindesayi lindesayi Giles.
Характеризуется сильным развитием белых чешуек на нижней по-
верхности задних бедер, которые занимают около 1/3 бедра от его осно-
вания. Личинка — см. описание вида.
Распространение и биология — см. описание вида.
Anopheles lindesayi japonicus Yamada, 1918.
Отличается от типичной формы слабым развитием белых чешуек
на нижней поверхности задних бедер, которые занимают примерно
1/6 бедра от основания. Личинки не имеют существенных отличий от ос-
новной формы.
Распространение. Япония (Хоккайдо, Хонсю, Кюсю),
п-ов Корея, Китай.
Биология. Обитает в горных районах; спутниками личинок
вида являются личинки Anopheles - koreicus Yam., An. hyrcanus, Culex
hayashii и C. infantulus Edw.; предпочитают водоемы с чистой
водой.
7. Anopheles (Anopheles) hyrcanus Pallas, 1771 (? sinensis Wiedemann,
1828). — Палласов (камышовый) малярийный комар (рис. 46).
Весьма сильно варьирующий как по величине, так и по окраске
вид.
Голова темно-бурая, с боков с черно-бурыми, сверху с белыми тор-
чащими чешуйками, на лбу между глазами с направленным вперед пуч-
ком белых волосков; хоботок и щупики темно-бурые, последние у самки
с тремя белыми колечками и белой вершиной; кроме того, к бурым че-
шуйкам на щупиках примешиваются иногда в более или менее значитель-
ном числе белые чешуйки; щупики самки, особенно в основной половине,
покрыты торчащими чешуйками. Усики самки темно-бурые, покрытые
белым пушком, у основания члеников с розетками довольно длинных
бурых волосков; первые 5—7 члеников усиков несут некоторое количе-
ство белых чешуек. Среднеспинка бурая, в средней части с более или
менее ясно выраженной широкой серой полосой, часто разделенной
темными продольными полосками на две или четыре более узкие серые
полосы; иногда вся среднеспинка, за исключением упомянутых узких
Рис. 46. Anopheles hyrcanus Pall.
1. ANOPHELES
105
темных полос, покрыта серым налетом; в передней части среднеспинки
(у головы) имеется пучок белых чешуек; волосяной покров среднеспинки
светло-желтовато-бурый с золотистым оттенком; бочки груди бурые,
покрытые сероватым налетом. Ноги бурые, с нижней стороны бедер и
внутренней стороны голеней более светлые; основания передних бедер
явственно утолщены; лапки темно-бурые, с белыми колечками у вер-
шины первых трех или четырех (последнее особенно часто наблюдается
на задних лапках) члеников или же весь
IV членик задних лапок белый; иногда
белая окраска распространяется и на
часть V членика. Крылья на большей
части своего протяжения покрыты тем-
ными чешуйками; по переднему краю
крыла имеются два больших белых пят-
на — одно в начале дистальной трети,
другое
первое
первую
Рис. 47. Гипопигий Anopheles hyrcanus Pall. По La Casse a.
Yamaguti.
перед вершиной крыла, из них
занимает жилки костальную—
радиальную, второе —
косталь-
nyid—вторую радиальную; более постоянные участки белых чешуек
имеются в основной трети 1-й радиальной, на вилках r2+r3 и т^т^,
m3+i и в особенности на кубитальной и анальной жилках, которые иногда
в большей своей части покрыты белыми чешуйками; кроме того, в не-
которых случаях белые чешуйки в большем или меньшем количестве
примешиваются к бурым, не образуя сплошных участков; бахромка
крыла у вершины белая, в остальной части крыла буроватая; жужжальца
светлые, с бурой головкой. Брюшко бурое с довольно длинными и гу-
стыми бурыми или золотистыми волосками.
Гипопигий (рис. 47): коксит с внутренней стороны при основании
с двумя крепкими простыми щетинками; шипы класпеты слиты, образуя
узкую пластинку; IX тергит с длинными и узкими выростами.
Личинки IV стадии (рис. 48) очень изменчивы по окраске
и пигментации твердых хитиновых образований, в частности головы и
стигмальной пластинки.
106
I. ПОДСЕМ. ANOPHELINAE
Внутренние волоски наличника сближены, простые или чаще слабо
разветвленные на концах; наружные не менее 2/3 длины внутренних, '
могут доходить до переднего края боковых лопастей верхней губы, дре-
вовидно ветвистые: от главного ствола отходят 3—6 ветвей, далее много-
кратно ветвящихся, причем внутренние ветви развиты сильнее наруж-
ных; задние развиты слабо, имеют 2—3 или больше ветвей, заходящих
за основания передних волосков.
Рис. 48. Усики личинок.
1 — Anopheles maculipennis Mg., 2 — Anopheles hyrcanus Pall., 3 — стигмальная пластинка
личинки IV стадии Anopheles hyrcanus.
Усик с сильно развитым волоском, отходящим около середины, ближе
к основанию, и состоящим из 7—8 ветвей, достигающим не менее (часто
более) половины длины усика.
Волосок № 1 на заднегруди слабо развит, со сложенными длинными
лепестками без концевых нитей. Звездчатые волоски брюшка рудимен-
тарны на I—II и хорошо развиты на III—VII члениках, из 17—24 ле-
пестков, с нерезко выраженной концевой нитью, переходящей несколь-
кими ступенчатыми уступами в тело лепестка с темноокрашенной ба-
зальной половиной и светлой вершиной.
Стигмальная пластинка с темной только у основания передней ло-
пастью, со стигмами, не соединенными с передним отделом центральной
пластинки, передний отросток которой своим темным основанием вдается
в глубь пластинки и заходит за границу между его передним и средним
отделами; срединная темная пигментация заднего отдела центральной
пластинки слабо развита, только у заднего края, ячеистая грануляция
в среднем отделе светлая, без темных контуров.
1. ANOPHELES
107
Яйца характеризуются наличием умеренной величины боковых
воздушных камер и довольно широкой, в средней части яйца, гладкой
(без поперечной исчерченности) оторочкой.
Замечания по систематике. Объем данного вида различными авто-
рами трактуется по-разному. Описанные как подвиды А п. hyrcanus ориентальные формы
(nigerrimus Giles, 1900; pseudo sinensis Baisas, 1935; lesteri Baisas et Hu, 1936) теперь
обычно рассматриваются как самостоятельные виды. Возможно, что ориентальный
sinensis Wied., первоначально описанный по материалу из Кантона, представляет
собой особый вид, отличный от палеарктического An. hyrcanus (см.: Reid, 1953).
В пределах Палеарктики описан ряд внутривидовых форм (аберраций) An. hyr-
canus, в частности ab. pseudopictus Grassi (отличается белой окраской IV членика зад-
ней лапки) и ab. messopotamiae (со светлым расплывчатым рисунком крыла). Учиты-
вая весьма значительную изменчивость окраски An. hyrcanus, нет оснований призна-
вать за указанными формами ранг подвидов (Маслов, 1946). В частности «ab. mes-
sopotamiae», вероятно, представляет собой сезонную форму; светлая расплывчатая
окраска крыльев наблюдается по преимуществу у зимующих самок (Казанцев, 1930).
Что касается описанного по материалу из Ташкента А п. chodukini Mart., 1929, то по всей
вероятности он является аберрантным уклонением альбинистического ряда.
Распространение. Ареал простирается от Атлантического
до Тихого океана, охватывая юг и юго-восток Европы, запад, юг и восток
Азии, а также некоторые области Центральной Азии (Монголия, Синь-
цзян). Областью процветания вида (а также группы близких видов) яв-
ляется Юго-Восточная Азия (Индия, Зондские и Филиппинские острова),
откуда вид далеко проникает на север и запад в пределы Палеарктики.
На востоке ареал вида занимает Китай, Японию, п-ов Корею и Юж-
ное Приморье (вид особенно обилен в районе озера Ханка и на побережье
крайнего юга Приморья); северная граница вида здесь проходит по до-
лине Амура и Амгуни, затем по территории Монголии и Китая; от вер-
ховьев Иртыша (озеро Норзайсан) северная граница его распространения
идет в СССР по Центральному Казахстану к низовьям Урала, Волги
и Дона, южнее Харькова и Киева, немного севернее Кишинева. Вид
распространен также в Южной и Юго-Восточной Европе и в Передней
Азии. На юге СССР (Южный Казахстан, Средняя Азия, Кавказ) распро-
странен очень широко, в речных долинах и дельтах рек достигая местами
огромной численности; в горы поднимается до высот 1500—1600 м.
Биология. Личинки встречаются в хорошо прогреваемых солн-
цем стоячих, поросших зеленой растительностью водоемах, как неболь-
ших, так и крупных (типа заболоченностей и плавней), поросших ка-
мышом; встречаются на рисовых полях и в каналах оросительной си-
стемы, особенно в сбросных. В северных и северо-восточных (Дальний
Восток) частях ареала, особенно в более холодные периоды сезона, ли-
чинки встречаются по преимуществу в открытых (незатененных) водое-
мах. На юге ареала, а в остальных частях ареала — в наиболее жаркий
период, личинки обитают по преимуществу в затененных или сильно
заросших водоемах типа грунтовых заболоченностей и т. п., с мягкой,
средней степени минерализации водой повышенной окисляемости; со-
вместно с личинками вида попадаются, как правило, личинки Anopheles
pulcherrimus, An. maculipennis sacharovi, Culex modestus и др. Зимуют
самки; места их зимовок приурочены к тростниковым зарослям тугаев,
густому кустарнику и т. п. Период активности комаров продолжается
с весны до поздней осени (ноябрь), причем численность вида к осени
возрастает. В течение года — два (на севере)—четыре поколения.
Вид относительно слабо связан с человеком. Самки лишь изредка
залетают в жилые помещения, но в заметных количествах попадаются
в помещениях для скота. После кровососания комары вылетают из по-
108
I. ПОДСЕМ. ANOPHELINAE
мещений. Дневки — преимущественно в зарослях гигантолинеид. На-
падают по преимуществу на открытом воздухе, причем не только в су-
мерки и ночью, но и при ярком солнечном освещении; в тугаях в неко-
торые периоды попадаются в массе, весьма назойливы. В Южном При-
морье места выплода встречаются в ближайшем окружении человека,
а самки могут массами залетать в жилые помещения.
Эпидемиологическое значение вида весьма различно в зависимости
от условий местности; на Кавказе и в Средней Азии имел второстепен-
ное значение в качестве переносчика малярии, причем по преимуществу
в низменных частях долин крупных рек; в условиях эксперимента
An. hyrcanus не заражался плазмодием тропической малярии, но про-
являл восприимчивость к плазмодиям трехдневной малярии (Симанин,
1930). На Дальнем Востоке (Южное Приморье) являлся единственным
переносчиком малярии, хотя там последняя и не имела широкого рас-
пространения. В Восточной Азии (п-ов Корея, некоторые районы Китая и
Японии) являлся (а местами и теперь является) главнейшим перенос-
чиком малярии.
Вероятно, An. hyrcanus- играет роль в циркуляции возбудителя в не-
которых пойменных очагах туляремии; установлена естественная за-
раженность.
• 2. Подрод MYZOMYIA Blanchard (CELLIA Theobald)
Поперечные жилки и основания вилок крыла в белых чешуйках;
костальная жилка с четырьмя или более белыми пятнами. Гипопигий
в основной части коксита имеет 4—6 крепких щетинок.
Подрод Myzomyia имеет весьма резко выраженный ориентально-
эфиопский центр процветания; из Эфиопской области известно не менее
80 видов, из Ориентальной — не менее 45, из Австралийской — 14 видов;
для Нового Света подрод не констатирован. Палеарктические виды тес-
нейшим образом связаны с этим центром, причем западно-палеарктиче-
ские (средиземноморские в узком смысле) виды подрода тяготеют к аф-
риканскому сектору, восточные, в частности An. pulcherrimus, — к ин-
дийскому сектору этого центра.
Личинки подрода Myzomyia отличаются сильно расставленными ос-
нованиями внутренних волосков наличника, расстояние между которыми
больше расстояния между наружным волоском и соответствующим ему
внутренним; перистыми лобными волосками; коротким и простым во-
лоском на теле усика; длинными перистыми боковыми волосками на IV—
VI члениках брюшка.
8. Anopheles (Myzomyia) pulcherrimus Theobald, 1902. — Белый ма-
лярийный комар.
Очень характерный вид, отличающийся от всех прочих, за исключе-
нием африканского An. pharoensis, ясно выраженными пучками чешуек
по бокам тергитов брюшка, а также одноцветно белыми IV и V члени-
ками задних лапок.
Голова с боков усажена светло-бурыми, сверху чисто белыми торча-
щими чешуйками, на лбу между глазами с направленным вперед пучком
белых волосков. Усики самки темно-бурые, покрыты коротким и нежным
белым пушком; каждый членик несет розетку длинных белых волосков;
первые 7—8 члеников, кроме того, покрыты чисто белыми чешуйками,
которые закрывают весь членик, иногда же только некоторые его части
1. ANOPHELES
109
(вершину, внутреннюю или наружную поверхность); усики самца бурые,
с длинными золотистыми волосками. Хоботок покрыт светло-бурыми
прилегающими чешуйками, щупики бурые, покрыты светло-бурыми
чешуйками, в основной половине щупика торчащими, в вершинной по-
ловине преимущественно прилегающими; все членики щупиков на вер-
шине с белыми колечками, образованными скоплением белых чешуек;
белые колечки занимают от 1/6 до 2/4 длины членика, за исключением
последнего, на котором белое колечко занимает около 1/2 его длины.
Среднеспинка бурая или серовато-бурая, густо покрыта довольно ши-
рокими белыми чешуйками; лишены последних лишь неширокие полосы
в задней половине среднеспинки, а иногда также узкие полоски в перед-
ней ее половине; щиток одного цвета со среднеспинкой, покрыт белыми
чешуйками; бочки груди буровато-серые с сероватым налетом, густо
покрыты, в особенности в средних своих отделах, белыми чешуйками.
Ноги бурые, в большей части покрыты белыми чешуйками; передние
ноги с явственно утолщенными в основной трети бедрами, покрытыми бе-
лыми чешуйками, к которым часто примешиваются в более или менее
значительном количестве бурые чешуйки; передние голени покрыты
белыми чешуйками, с наружной стороны по большей части с двумя, иногда
сливающимися, узкими продольными полосками бурых чешуек; перед-
ние лапки черно-бурые, с белыми колечками у вершины I, II и III, а
иногда и IV члеников; средние бедра спереди в большей своей части
покрыты бурыми чешуйками, среди которых попадаются белые чешуйки,
иногда сгруппированные в виде продольной полосы, в вершинной трети
и у самой вершины с белыми пятнами, сзади с белыми чешуйками; сред-
ние голени спереди с бурыми, сзади с белыми чешуйками, у вершины
с белым кольцом; средние лапки бурые, с белыми колечками у вершины
первых 2—3 члеников; задние бедра окрашены, как и бедра средней
пары; преобладающая окраска чешуек, покрывающих задние голени,
белая; задние лапки со светло-бурым первым члеником, имеющим у вер-
шины белое колечко; II членик при основании с темно- или черно-бурым
колечком, занимающим от 1/4—1/2 длины членика; вершинная часть
II членика, а также членики III—V белые.
Крылья в большей части покрыты белыми чешуйками; по переднему
краю крыла имеется обыкновенно шесть черных пятен, из которых первые
два, расположенные в основной четверти крыла, незначительной вели-
чины, точкообразные, и развиты только на костальной жилке; третье
черное пятно, значительно более длинное (по длине равно примерно
х/2 своего расстояния от основания крыла) и широкое, распростра-
няется на костальную, субкостальную и первую радиальную жилки;
четвертое пятно, превосходящее в длину третье в 1.5 раза, распростра-
няется на жилки костальную, субкостальную, первую радиальную и
по большей части также на общий ствол г2+г3+г4+6; последний, однако,
в некоторых случаях в участке, прилегающем к четвертому пятну, по-
крыт белыми чешуйками; что же касается первой радиальной жилки,
то на ней четвертое пятно часто бывает разбито на три небольших участка,
покрытых белыми чешуйками; пятое пятно по большей части аналогично
по форме и расстоянию от вершины крыла третьему пятну и его расстоя-
нию от основания; шестое пятно в большинстве случаев короткое, рас-
положенное на изгибе к вершине крыла, занимает жилки костальную,
первую и вторую радиальные; кроме пятен по переднему краю крыла,
имеются более или менее постоянные пятна темных чешуек в основной
трети обоих или одной ветви вилки г2+гз> в средней трети г3, в основной
трети вилки /п1+/п2, а также в вершинных частях всех продольных жи-
по
I. ПОДСЕМ. ANOPHELINAE
лок от третьей радиальной до анальной; кроме того, менее постоянные
пятна темных чешуек имеются и в других местах крыла, в особенности
в вершинной части общего ствола г2+гз и ^i+^2 и на анальной жилке,,
а также в некоторых других местах; бахромка по заднему краю крыла
в вершинной его половине состоит по большей части из чередующихся
участков темных и белых волосков, в основной же половине только из бе-
лых; жужжальца светлые, с темно-бурой головкой. Брюшко буровато-
черное, по заднему краю и по бокам тергитов усажено довольно длин-
ными золотистыми волосками, сверху почти сплошь (за исключением'
неширокой полосы по переднему краю каждого тергита) покрыто ши-
Рис. 49. Гипопигий Anopheles pulcherrimus Theob.
рокими белыми чешуйками, черепицеобразно налегающими друг на друга;
белые чешуйки бокового отдела брюшка иногда имеют золотистый или
желтоватый оттенок; задние углы II—VIII тергитов имеют по пучку
торчащих чешуек белого или желтовато-бурого цвета; брюшко снизу
темно-бурое, с белыми чешуйками.
Гипопигий (рис. 49): коксит при основании с внутренней стороны1
с пятью крепкими щетинками; класпеты с расширенной на вершине
пластинкой; эдеагус без листовидных придатков.
Личинка IV стадии (рис. 50) средней величины — 0.5—0.7 мм
длины, желтовато-зеленоватого цвета. Внутренние волоски наличника
широко расставлены: расстояние между ними вдвое больше расстояния
между наружным волоском и соответствующим ему внутренним. Внут-
ренние волоски начиная с трети от их основания несут редкие тонкие-
боковые веточки в числе 6—15, придающие волоску перистый характер;,
наружные волоски почти вдвое короче внутренних, начиная с середины
или с вершинной трети они ветвятся на 4—12 (в среднем 7) тонких ве-
точек обычно одинаковой длины. Задние волоски наличника вдвое ко-
роче наружных, делятся около основания на 2—4 веточки.
Волосок № 1 на заднегруди в виде хорошо развитого звездчатого во-
лоска с длинным стебельком и с 8—13 длинными узкими лепестками
без концевых нитей, но расположенными не розеткой, а веером. Звездча—
1. ANOPHELES
111
тые волоски брюшка на I членике слабо развиты, из 5—9 веерообразно
расположенных узких лепестков без концевой нити; на II—VII члени-
ках выражены хорошо: на II из 13—18, на III—VI из 18—23, а на VII
из 16—22 лепестков. Лепестки равномерно окрашены, кроме более тем-
ного дистального конца, несущего длинную заостренную концевую нить
с хорошо выраженной ступенчатой зубчатостью у основания.
Рис. 50. Anopheles pulcherrimus Theob. Личинка IV стадии. По Пури.
1 — передний край лобного щитка, 2 — вариация ветвления наружного волоска на-
личника, з — срединные волоски переднегруди, 4, 5 — основания боковых волосков
средне- и заднегруди, в — звездчатый волосок IV членика брюшка.
Стигмальная пластинка (рис. 51) очень.характерна; передняя лопасть
темно, но неравномерно окрашена; стигмы непосредственно прилегают
к переднему отделу центральной пластинки, имеющему форму наконеч-
ника стрелы с развитыми, направленными назад боковыми отростками,
задний край которых на границе со средним отделом образует заметное
сужение; медиальная часть центральной пластинки слабо пигментиро-
вана, в виде хорошо очерченного светлого медальона.
Яйца с боковыми воздушными камерами и довольно широкой,
поперечно исчерченной оторочкой.
Распространение. СССР: равнины и долины Средней Азии
[Туркмения, Узбекистан, Таджикистан, Южная Киргизия, юг Казах-
стана (по долине Сырдарьи)]; в горы не поднимается выше 900—1000 м.
Ранее встречался в некоторых районах Азербайджана, особенно в плав-
нях прикуринских степей. Иран, Ирак, Афганистан, Пакистан, Индия.
Ш
I. ПОДСЕМ. ANOPHELINAE
Биология. Личинки встречаются по преимуществу в неглубоких
озерах, тугаях и больших заболоченностях, богатых плавающей на по-
верхности или близ поверхности водной растительностью — рдестами,
роголистником и т. п.; в отличие от личинок других комаров, попадаю-
щихся преимущественно в прибрежной зоне, личинки белого малярий-
ного комара приурочены главным образом к рдестовым полям открытой
зоны, где густые заросли рдестов заменяют личинкам дно водоема; ли-
чинки встречаются также в непересыхающих лужах, питаемых подпочвен-
ной водой или водой арыков, хаузах и т. п., реже — в клетках рисовых
полей. Водоемов с небольшой
Рис. 51. Anopheles pulcherrimus Theob. Стигмаль-
ная пластинка личинки IV стадии.
площадью водяного зеркала
(меньше 1—2 м2) или с во-
дой значительной степени
солености личинки опреде-
ленно избегают. В условиях
СССР зимует в стадии сред-
невозрастной личинки (в Ин-
дии, по-видимому, зимуют
взрослые комары), причем
нормально наблюдается зна-
чительная гибель личинок
на зимовках, чем объясняет-
ся обычно небольшая числен-
ность первого поколения ко-
маров; количество комаров
возрастает к осени, когда и
достигает максимума. Для
белого малярийного комара
характерны случаи резкого
колебания численности в од-
ном и том же месте в раз-
ные годы; особенно губи-
тельное действие на этот вид
оказывают более низкие, чем
средняя многолетняя, темпе-
ратуры зимних месяцев и холодная затяжная весна; наоборот, в теплые,
зимы и весны гибель личинок сводится к минимуму, что вызывает в бли-
жайшее лето возрастание численности вида; во многих случаях такие
колебания численности имеют узко .локальный характер.
Самки белого малярийного комара охотно залетают в дома и в по-
мещения для скота; часто встречаются под навесами и т. п. Нападают
и в открытой природе, и в помещениях, причем преимущественно в су-
мерки и ночью, хотя наблюдались случаи интенсивных нападений ко-
маров этого вида на человека и днем; в отличие от других Anopheles,
охотно летят на свет; по сравнению с другими видами рода белый маля-
рийный комар является более световыносливым и суховыносливым.
Роль белого малярийного комара как переносчика малярии в ми-
ровой литературе расценивается как второстепенная. Искусственное
заражение плазмодиями трехдневной и тропической малярии дало
положительные результаты. В природе были найдены естественно зара-
женные комары. В некоторых районах Средней Азии, где An. pulcher-
rimus появлялся в. большом количестве, этот комар рассматривался как
один из важных переносчиков малярии; в настоящее время область
его распространения местами сократилась.
1. ANOPHELES
113
9. Anopheles (Myzomyia) superpictus Grassi,
малярийный комар (рис. 52).
1899. — Украшенный
Довольно сильно варьирующий вид, в особенности в окраске ног и
крыльев; задние лапки или одноцветно бурые, или со слабо намечающи-
мися светлыми колечками.
8 Фауна СССР, А. В. Гуцевич и др.
114
1. ПОДСЕМ. ANOPHELINAE
Голова с боков с бурыми, сверху с белыми торчащими чешуйками;
усики с розетками бурых волосков; хоботок длинный и тонкий, со свет-
лой вершиной, щупики черные, с резкими белыми колечками и белой
вершиной; чешуйки на щупиках прилегающие. Среднеспинка светло-
бурая, с боков несколько темнее, усажена редкими светло-бурыми во-
лосками и густо покрыта, в особенности в передней своей части, узкими
беловатыми чешуйками; бочки светло-бурые, с сероватым оттенком. Ноги
темно-бурые, тазики светлые, на месте сочленения бедра и голени и го-
Рис. 53. Гипопигий Anopheles superpictus Grassi.
лени и лапки имеется узкое белое колечко; лапки по большей части од-
ноцветно бурые, реже с едва заметной кольчатостью. Крылья очень
сильно варьируют в окраске; по переднему краю крыла имеется обыкно-
венно четыре пятна белых чешуек (четвертое — у самой вершины крыла);
кроме того, в некоторых случаях наблюдается более или менее сильное
развитие белых чешуек в базальной части костальной жилки; промежу-
точные участки переднего края крыла покрыты черно-бурыми чешуй-
ками; как светлые, так и темные пятна переднего края крыла распро-
страняются на жилки костальную, субкостальную и первую радиальную;
жилки остальной части крыла (кзади от первой радиальной жилки)
в большей своей части покрыты белыми чешуйками; более или менее
постоянные пятна темных чешуек имеются на общем стволе Гг+гд, в ба-
зальной части г4-|-г5, на медиальной (два пятна на общем стволе тг-\-т2
и по одному пятну на каждой из ветвей вилки) ш3+4, кубитальной и аналь-
ной жилках; жужжальца бурые, с темной головкой. Брюшко бурое,
покрыто довольно длинными нежными светло-желтоватыми волосками.
1. ANOPHELES
115
Гипопигий (рис. 53): коксит относительно короткий и толстый, при
основании с пятью крепкими щетинками; класпеты с расширенной на вер-
шине пластинкой; эдеагус с короткими листовидными придатками.
Личинки IV стадии (рис. 54) мелких размеров, около 5 мм,
серого или серо-зеленого цвета. Внутренние волоски наличника широко
расставлены: расстояние между ними более чем превышает расстояние
Рис. 54. Anopheles superpictus Grassi. Личинка IV стадии. По Пури.
1 — передний край лобного щитка, 2 — срединные волоски переднегруди, з—5 — основания боковых
волосков передне-, средне- и заднегруди, 6— звездчатый волосок (№ 1) заднегруди, 7 — звездча-
тый волосок IV членика брюшка.
между наружным и соответствующим ему внутренним волоском. Послед-
ние тонкие, простые, с редкой вторичной перистостью; наружные вдвое
короче, простые и гладкие; задние волоски наличника одной длины с на-
ружными и такого же строения.
Волосок № 1 на заднегруди слабо развит, состоит из 6—8 тонких
узких и длинных ланцетовидных лепестков, сложенных веерообразно.
Звездчатые волоски на брюшке развиты на II—VII члениках брюшка.
На I членике они в виде обычных волосков, с 5—7 тонкими веточками,
отходящими часто почти совместно; на II членике состоят из 12—15;.
на III — из 14—18, на IV—VI — из 15—19 и на,VII —из 14—16 лепест-
ков; лепестки с хорошо развитыми концевыми нитями, достигающими
2/3 длины лепестка, резко очерченными и заостренными, с узким осно-
ванием и изменчивой зубчатостью.
8*
116
I. ПОДСЕМ. ANOPHELINAE
Стигмальная пластинка (рис. 55) с передней лопастью, темно, но не-
равномерно пигментированной; стигмы или непосредственно прилегают
к переднему отделу центральной пластинки, или соединены с ним при по-
мощи отростков, центральная пластинка широкая, не сужающаяся в пе-
редней части среднего отдела, боковые края которого параллельны друг
ДРУГУ, передний отдел не имеет боковых отростков, а сужается под тупым
углом; грануляция в срединной части центральной пластинки развита
слабо, имеет расплывчатые очертания и не образует хорошо очерченного
светлого медальона.
Яйца сильно варьиру-
ют; в норме яйца имеют до-
вольно широкую, поперечно
исчерченную оторочку, но
лишены воздушных камер;
яйца, откладываемые в хо-
лодное время года, часто
имеют небольшое количество
мелких воздушных камер,
сгруппированных в виде бо-
лее или менее правильного
эллипса у одного конца
яйца на его плоской стороне.
Р а с п р о с транение.
Вид характерен для Среди-
земноморской подобласти. До-
стигает высокой численности
в странах с сухим климатом,
встречается редко или отсут-
ствует в районах высокой
влажности. В СССР область
распространения включает
Рис. 55. Anopheles superpictus Grassi. Стигмальная Предкавказье (Дагестан), За-
пластинка личинки IV стадии. кавказье (Грузия, Армения,
Азербайджан), Среднюю Азию
(Туркмения, Узбекистан,
Таджикистан, Южная Киргизия, немногие южные районы Ка-
захстана). Южная Европа — Испания, южная половина Апеннинского
и Балканского полуостровов; Северная Африка (от Алжира до ОАР),
Малая Азия, Ирак, Иран, Афганистан, Пакистан, Северо-Западная
Индия.
Биология. Личинки являются обитателями проточных, слабо
проточных или стоячих водоемов родникового или грунтового (поверх-
ностно-фильтрационного) происхождения с прозрачной, хорошо про-
греваемой солнцем водой. Встречаются в водоемах высыхающих русел
рек, часто в небольших углублениях (следы от копыт и т. п.). В более
быстро текущих водах попадаются в скоплениях водорослей между кам-
нями или у берегов. Были находимы на рисовых полях, если вода их про-
точная и грунтового происхождения. Личинки отличаются широкой тер-
пимостью к степени минерализации воды и к качественному составу раство-
ренных в ней солей, но избегают мутной, загрязненной продуктами раз-
ложения органических веществ воды с высокой окйсляемостью. Личинки
исключительно подвижны и при малейшем беспокойстве (сотрясении,
затенении и т. п.) мгновенно опускаются в глубь водоема, где часто заби-
ваются под камни или другие предметы. При искусственном взмучива-
1. ANOPHELES
117
нии воды личинки поднимаются на ее поверхность. Зимуют взрослые
оплодотворенные самки и лишь в небольшом количестве личинки.
Зимовка самок в большинстве случаев не прерывает полностью их
жизнедеятельности: известный процент самок в течение зимы несколько
раз пьет кровь. Весеннее поколение личинок (в Средней Азии оно, как
правило, приурочено к апрелю) немногочисленно, причем развитие ли-
чинок идет сравнительно медленно; при средней температуре 21.5° оно
длится 24 дня, при 27° — 10 дней; средняя продолжительность разви-
тия комаров — 12—13 дней. Быстрое увеличение численности вида на-
чинается с конца мая—первой половины июня (в Средней Азии), после
спада паводковых вод; в середине лета (в Средней Азии — июль) числен-
ность вида достигает максимума; с августа начинается очень постепенное
уменьшение численности, заметно усиливающееся во вторую половину
сентября, что можно во многих случаях поставить в связь со спуском
воды с рисовых полей; этот период характеризуется также усилением
залета самок в жилые помещения, связанным с началом ночных похоло-
даний; в течение октября комары постепенно уходят на зимовку.
Самки украшенного малярийного комара в более или менее равной
мере сосут кровь как человека, так и животных. Самки часто в массовых
количествах залетают в жилые помещения на дневки; влет комаров
на дневки осуществляется, как правило, с восходом солнца, вылет —
после его заката; самки нападают на человека преимущественно в по-
мещениях; помимо жилых помещений, значительное количество комаров
концентрируется также на дневках в помещениях для скота. Местами
дневок комаров этого вида в жилых помещениях являются затененные
углы, пустая посуда, складки развешенного платья и т. п., вне жилья —
дупла деревьев, трещины лёссовых обрывов и глинобитных построек,
норы грызунов (сусликов, песчанок и др.), сюмы, пещеры и т. п. Наиболь-
шая активность самок наблюдается при закате солнца; она несколько ос-
лабевает с наступлением полной темноты, но не прекращается и днем.
Эпидемиологическое значение вида как переносчика всех форм маля-
рии чрезвычайно велико. Во многих местах Средней Азии этот вид имел
несомненно наибольшее значение по сравнению с другими видами ма-
лярийных комаров; последнему способствовали следующие особенности
биологии вида: его наибольшая численность совпадает с самым жарким
временем года (июль—сентябрь), вследствие чего цикл развития маля-
рийного плазмодия в комаре проходит в максимально сжатые сроки;
это усугубляется достаточно резко выраженной антропофильностью самок
и значительной скоростью процессов пищеварения, что вызывает более
частую, чем у других видов малярийных комаров, потребность в крово-
сосании; наконец, особенности экологии личинок из года в год поддержи-
вают численность вида на высоком уровне. В странах Ближнего и Сред-
него Востока An. superpictus в прошлом являлся (а местами и теперь
является) важнейшим переносчиком малярии.
II. Подсемейство TOXORHYNCHITINAE
Крупные, ярко окрашенные комары. Ширина наличника превосходит
его длину. Хоботок относительно толстый, в вершинной поло вине утончаю-
щийся, как правило, загнутый вниз. Максиллы и мандибулы доходят
примерно лишь до половины длины хоботка и лишены зубчиков. Дыхаль-
цевые щетинки имеются, задыхальцевые отсутствуют. Щиток дуго-
видный, цельнокрайний. Крылья с очень короткой радиальной вилкой
(г2+3); длина ствола вилки в несколько раз превосходит длину самой
118
И. ПОДСЕМ. TOXORHYNCIHTINAE
вилки; между m3+4 и си± имеется утолщение мембраны крыла в виде узко-
треугольной пластинки; у вершины си 4 задний край крыла образует не-
большую выемку. Брюшко покрыто широкими плоскими чешуйками,
у конца часто с пучками ярких волосков.
Личинки крупные, около 20 мм в длину, темного винно-красного
с фиолетовым оттенком цвета с прямоугольной головой, волоски которой
слабо развиты и смещены в переднюю ее треть, где расположены в ряд
двумя боковыми группами. Усики короткие, палочковидные. Ротовые
органы приспособлены к хищному типу питания. Для схватывания до-
бычи служат боковые лопасти верхней губы, находящиеся на боковых
выступах наличника и состоящие из небольшого числа (около 10) лежа-
щих в один ряд крепких, крючкообразно загнутых и зазубренных на
конце подвижных шипов. Верхние челюсти с сильно развитыми зубцами.
Грудь относительно узкая. В заднем отделе просвечивает пара тра-
хейных пузырей — расширений главных стволов трахей, имеющих гидро-
статическое значение. Волоски тела частично укорочены в виде грубых
шипов, несущих вторичные шипики. Боковые волоски развиты более
сильно, на спинной стороне т— наиболее слабо, что связано с жизнью
личинок непосредственно под поверхностной пленкой воды. Вокруг
оснований волосков и шипов характерно образование более сильно скле-
ротизованных участков кутикулы в виде хитиновых пластинок, чаще
всего по три пары на каждом членике, расположенных в три продоль-
ных ряда, с каждой стороны.
На VIII членике брюшка по бокам лежат более крупные пластинки.
На его спинной стороне расположена короткая дыхательная трубка
с парой пучков у основания; ушки и гребень отсутствуют. Короткий
последний членик окружен хитиновым кольцом, задний край которого
усажен шипами.
Биология. Личинки развиваются в небольших скоплениях воды
в пазухах листьев различных растений (бромелин), стволах бамбука и
дуплах деревьев, питаясь развивающимися там личинками других видов
комаров. Исключительно активные хищники, . уничтожающие и друг
друга. На ряде островов тропической зоны специально применяются
для уничтожения личинок дупловых видов кровососущих комаров.
Самки откладывают яйца поодиночке на поверхность воды. Встре-
чаются преимущественно в верхнем ярусе леса, питаются нектаром цветов
и не сосут кровь.
Подсемейство заключает в себе один род с тремя подродами.
2. Род TOXORHYNCHITES Theobald (Megarhinus Robineau-Desvoidy *)
С признаками подсемейства.
По своему распространению род является тропическим, лишь единич-
ные виды свойственны наиболее южным частям умеренной зоны. Всего
известно более 50 видов, из которых меньшая часть (подроды Ankylo-
rhynchusTzLynchiella) свойственна тропикам западного полушария, осталь-
ные (подрод Toxorhynchites) — преимущественно Ориентальной области;
палеарктические представители рода более близки к ориентальной
группе видов.
1. Toxorhynchites christophi Portschinsky, 1884 (? towadensis Matsumura,
1916) (рис. 56).
1 Имя Megarhinus теперь рассматривается как синоним наименования подрода
Lynchiella.
2. TOXORHYNCHITES
119
Относительно крупное насекомое; длина без хоботка 10—13 мм;
Почти все чешуйки прилегающие, с металлическим отливом, зеленого,
синего, фиолетового или серебристо-белого цвета.
Самка. Затылок в широких темно-бурых чешуйках, имеющих
металлический синий отлив. Задние края глаз в голубовато-белых (бирю-
Рис. 56. Toxorhynchites christophi Portsch.
зового цвета) чешуйках. Хоботок очень длинный (2/3—3/4 длины тела)»
загнутый вниз, в темно-бурых чешуйках, имеющих стальной отлив;
в средней части хоботка намечается неясное кольцо более светлых че-
шуек. Основание хоботка снизу в длинных черных волосках. Щупики
короткие в черно-бурых с металлическим отливом чешуйках. Усики бурые,
торус в белых чешуйках.
Среднеспинка в коричнево-бурых узких чешуйках; передний край и
боковые отделы среднеспинки в более широких светло-голубых чешуйках;
над основаниями крыльев — пучки желтых волосков. Боковая поверх-
ность груди в большей части покрыта широкими белыми, серебристо-
ISO
II. ПОДСЕМ. T0X0RHYNCHIT1NAE
блестящими чешуйками. Щиток в блестящих широких чешуйках — зеле-
новатых посредине, золотистых по бокам; последние более многочис-
ленны. Щетинки по краю щитка образуют непрерывный ряд.
Ноги: бедра и голени в черных со стальным отливом чешуйках; основ-,
ная часть бедер снизу покрыта белыми чешуйками. На лапках окра-
шены в белый цвет: вершинная половина I, весь II, иногда и основание
III членика передней лапки, основание (иногда и вершина) I и все после-
дующие членики средней лапки, а также II членик задней лапки. Осталь-
Рис. [57. Toxorhynchites christophi Portsch.
Личинка IV стадии. Голова, сверху.
ные членики лапок черные, с
синим отливом. Коготки лапки
простые.
Крылья прозрачные, слегка
затемненные; жилки в бурых
чешуйках. Поперечная жилка
между г2+з и г4+5 расположена
дистальнее радиомедиальной
жилки; последняя лежит на
одной прямой с задней попе-
речной жилкой (т=т).
Брюшко в черных чешуй-
ках, в передней половине брюш-
ка имеющих зеленоватый, в
задней половине — синеватый
металлический отлив. В перед-
ней половине V (а иногда и III)
тергита более или менее выра-
жена поперечная перевязь бе-
лых чешуек, такого же цвета
пятна имеются по бокам тер-
гитов. V тергит по бокам с бе-
лыми полосками, VI — с пуч-
ками более длинных густых
черных волосков, VII и VIII
тергиты с такими же пучками ярко-оранжево-желтых волосков.
Самец (ввиду отсутствия самцов в нашей коллекции приводим
описание самца Т. towadensts по: La Casse a. Yamaguti, 1955). Щупики
немного короче хоботка, в темных чешуйках, с двумя светлыми кольцами;
вершина щупиков загнута кверху й заострена. Окраска тела как у самки;
VI и VII тергиты по бокам с густыми пучками темных волосков, VIII тер-
гит с такими же пучками волосков золотисто-коричневого цвета.
Описание строения гипопигия в известной нам литературе отсутствует.
Л и ч.и нка IV стадии (рис. 57) около 16—20 мм длины, винно-
красно-фиолетового цвета, близка к личинкам Т. splendens Wied.
Голова прямоугольной, с округленными углами формы, с лобным
щитком, заметно суженным в среднем отделе, позади глаз. Передне-боко-
вые края щитка выдаются вперед и темно пигментированы; соответственно
передний край лобного щитка вогнут назад. Наличник сильно развит,
особенно его боковые отделы, несущие органы схватывания добычи, —
боковые лопасти верхней губы. Каждая из них состоит из' 10 утолщен-
ных когтевидно изогнутых темных крючьев с 2—3 короткими зубчиками
на концах. Остальные ротовые части сдвинуты к переднему краю головы,
вследствие чего на ее брюшной стороне область горла развита очень
сильно. Верхние челюсти с 2 крупными и 3—4 более мелкими зубцами на
концах. Усики короткие, меньше общей длины головы; их концы
2. TOXORHYNCHITES
121
находятся на одном уровне с концами крючков верхней губы при схва-
тывании добычи. Поверхность усиков лишена шипиков. В концевой
четверти отходят два войоска; из них наиболее удаленный от вершины —
простой и направлен внутрь, оба остальных более короткие и разветвлен-
ные.
На наличнике три пары сильно расставленных простых волосков.
Из них две наружные пары соответствуют клипеальным волоскам личи-
Рис. 58. Тoxorhynchites christophi Portsch. Личинка
IV стадии.
Хетотаксия груди (развертка на плоскость). Римские цифры —
членики груди, арабские — порядковые номера волосков каждого
членика.
нок Anopheles, а наиболее длинные внутренние — средним лобным во-
лоскам. На лобном щитке четыре пары волосков, смещенных в переднюю
его треть и лежащих двумя боковыми рядами. Из них две средние пары —
простые и наиболее длинные — соответствуют наружному и внутреннему
лобным волоскам, наиболее короткая, разветвленная внутренняя пара
соответствует задним клипеальным волоскам, а наружная пара — те-
менным; вблизи последних, кнаружи от лобного шва, находятся 2-ветви-
стые транссутуральные волоски.
Рис. 59. Toxorhynchites christophi Portsch. Личинка IV стадии.
Хетотаксия I, II и IV—VI члеников брюшка. Ряды спинно-боковых (сб), боковых (б) и брюшно-бо-
ковых (бб) волосков левой стороны.
Рис. 60. Toxorhynchites christophi Portsch. Личинка IV стадии. Задний конец,
f сбоку.
3. TOXORHYNCHITES
123
Волоски тела разнообразны по своему строению. Они бывают трех
родов: 1) крупные, в виде шипов, с грубой вторичной перистостью, про-
стые или 2-ветвистые, 2) длинные тонкие гладкие и простые волоски,
иногда слабо разветвленные на концах и 3) очень короткие простые или
слабо ветвящиеся на некотором расстоянии от основания. Отдельные
группы волосков сидят на склеротизованных пластинках.
Каждый из члеников груди имеет 14 пар волосков, образующих не-
сколько групп, расположенных тремя поперечными поясами на передне-,
средне- и заднегруди, а в продольном направлении — несколькими ря-
дами: спинным, спинно-боковым, боковым, брюшно-боковым и брюшным
(рис. 58). Расположение волосков и хитиновых пластинок на члениках
брюшка изображено на рис. 59 и 60.
По бокам VIII членика брюшка лежат сильно развитые пластинки,
от заднего края которых отходят два длинных вторичноперистых шипа,
выше сидят два тонких коротких волоска и вне пределов пластинки,
под основанием сифона, — один более длинный тонкий волосок. Эти
пять пар шипов и волосков соответствуют волоскам позади щетки личинок
Culicinae. Щетка, таким образом, отсутствует. Сифон короткий, имеет
форму усеченного конуса. Длина его немного больше ширины основа-
ния. Ушки и гребень отсутствуют. Два коротких волоска из 5—6 вто-
рично перистых ветвей находятся на задне-боковой стороне сифона,
немного отступя от его основания.
Последний членик короткий и широкий. Седло окружает его сплош-
ным кольцом, которое вдоль заднего края по бокам несет ряд тесно си-
дящих различной длины шипов. Боковой волосок в виде шипа, не дости-
гающего длины членика. Наружный и внутренний хвостовые волоски
из 6—7 ветвей каждый. Наружные несколько длиннее внутренних. Плав-
ник смещен назад, из 18—20 длинных вторичноперистых ветвей, в виде
асимметричного веера. Жабры короткие, шаровидные.
Биология. Личинки были найдены в скоплениях воды в дуплах
вяза и липы. Встречаются с конца июня до конца августа—начала сен-
тября. В кишечнике личинок найдены остатки личинок A. (St.) galloisi
Yam. и A. (F.) alektorovi. Очень редкий вид.
Замечания по систематике. Как уже указывалось (Штакельберг,
1937), японский вид towadensis Mats., по-видимому, следует считать синонимом вида,
описанного И. А. Порчинским. Никаких существенных различий между ними не отме-
чено. Некоторое сомнение вызывает лишь то обстоятельство, что в довольно подроб-
ном описании Т. towadensis (см.: La Casse a. Yamaguti, 1955) отмечены (и изображены)
золотисто-коричневые волоски на VIII сегменте брюшка самки, но не сказано о нали-
чии ярко окрашенных волосков на V—VII тергитах. Между тем именно пучки волос-
ков по бокам этих трех сегментов, соответственно белых, черных и желтых, при-
дают насекомому весьма своеобразный характерный облик. Возможно, эти волоски
не сохранились на экземпляре, послужившем для описания М. towadensis.
Распространение. Южное Приморье (АлектороВ, 1931; Мон-
чадский, 1951). Япония — о. Хонсю.
2. Toxorhynchites sp. (рис. 61 и 62).
Известны только личинки.
Личинка IV стадии похожа на личинку Т. christophi, от
которой отличается следующими признаками: 1) более развитой в ши-
рину, чем в длину головой, 2) более сильным развитием шипов на теле;
не 2-ветвистым, а простым шипом № 8 на боковой пластинке среднегруди;
почти цилиндрической, а не усеченно конической формой сифона, пучок
Рис. 61. Toxorhynchites sp. Личинка IV стадии. Передний конец, сверху..
Рис. 62. Toxorhynchites sp. Личинка IV стадии. Задний конец, сбоку.
2. TOXORHYNCHITES
125
на котором больше ширины сифона у места своего отхождения; более
длинным, чем последний членик, боковым шипом на седле; слегка заги-
бающимися вверх концами хвостовых волосков, слабее ветвящимся
наружным из них; плавником, состоящим не более чем из 15 волосков,
на которых вторичная перистость начинается от самого основания, и
более сильно развитыми жабрами эллипсоидальной формы.
Биология. Личинки были найдены в скоплениях воды в дупле
пня корейского кедра и в прикорневых дуплах кленов в июле и августе
совместно с личинками Л. (St.) galloisi, A. (F.) alektorovi и A. (F.) nip-
ponicus.
Распространение. Юг Приморской области, Барабашский
район, заповедник «Кедровая падь» (Мончадский, 1940; Амосова, 1951).
III. Подсемейство CULICINAE
Длина наличника превосходит его ширину; передний край наличника
закругленный. Хоботок относительно тонкий, редко к вершине слегка
утолщенный, не крючковидный; мандибулы и максиллы самки длинные,
за крайне редкими исключениями (некоторые экзотические роды) доходят
до конца нижней губы, в вершинной части с зубчиками. Среднеспинка
заметно выпуклая. Щиток явственно трехлопастный, с тремя пучками
щетинистых волосков; промежутки между лопастями голые. Крылья
с длинной радиальной вилкой; длина вилки редко менее длины ее ствола;
роговидная пластинка между пг3+4 и сиг отсутствует. Брюшко покрыто
чешуйками, как правило, прилегающими. Гипопигий весьма различного
строения; анальный сегмент всегда с хорошо развитым стернитом; эдеагус
без листовидных придатков. Самки, как правило, с 3 сперматеками.
Личинки имеют, в зависимости от типа питания, чаще более широкую,
реже округлую или удлиненную голову, не способную к повороту на
180°. Лобный щиток несет в переднем отделе три пары различно развитых
лобных волосков, расположенных или в дугообразный ряд, или двумя
симметричными треугольниками, пару задних волосков наличника;
ветвление их веерообразное, но не перистое. Усики разнообразной формы
и длины, у личинок с фильтрационным типом питания более длинные,
у питающихся перифитоном и хищников — более короткие, палочко-
видные, с различно развитым волоском на теле и с комплексом чувстви-
тельных щетинок на конце. Ротовые органы устроены различно в зави-
симости от типа питания.
Волоски тела ветвятся или веерообразнй, в одной плоскости, или
звездчато: ветви расположены, как ребра многогранной пирамиды; ветви
могут нести вторичную перистость.
Грудь широкая, без плечевых клапанов. Брюшко без звездчатых
волосков. VIII членик брюшка по бокам несет различное число чешуек,
образующих щетку, позади которой расположены пять волосков. На
•спинной стороне VIII членика у его заднего края находится дыхательная
трубка, имеющая гребень и несущая от одной до 5—6 пар различно распо-
ложенных пучков волосков. Последний членик, кроме седла, иногда
образующего сплошное кольцо, несет две пары хвостовых и одну пару
боковых волосков. Плавник, за редким исключением, имеется.
Обширное подсемейство Culicinae различными авторами подразде-
ляется на разное (от 2 до 12) количество триб. Автор одной из последних
работ о надродовых группах комаров (Dodge, 1962) предлагает разделить
подсемейство на четыре трибы: Sabethini, Aedini, Uranotaeniini, Culicini.
Мы придерживаемся более простой схемы классификации, принятой
126
III. ПОДСЕМ. CULICINAE
в каталоге комаров (Stone et al., 1959), согласно которой подсемейство-
Culicinae делится на две трибы: Sabethini с восемью родами (распростра-
ненными в тропиках обоих полушарий, преимущественно в Южной Аме-
,рике) и Culicini с 19 родами. Из них в Палеарктике известно восемь родов:
Uranotaenia, Culiseta, Orthopodomyia, Mansonia, Aedes, Culex, Heizmannia
и Armigeres-, в СССР распространены первые шесть. Триба Sabethini
в Палеарктике, насколько известно, представлена только одним видом —
Tripteroides bambusa Yam. (Rachionotomyia bambusa), обитающим в Япо-
нии и в Китае.
3. Род URANOTAENIA Arribalzaga
Мелкие виды темных цветов. Глаза соприкасаются как в верхней,
части головы над усиками, так и в нижней (за хоботком). Щупики у обоих
полов очень короткие, как правило, в длину не превышают х/в длины
хоботка. Среднеспинка с хорошо развитыми акростихальными и дорсо-
центральными щетинками. Лопасти переднеспинки широко расставлены,
как правило, с'тремя щетинками; дыхальцёвая щетинка одна, реже (у не-
которых экзотических видов) они отсутствуют вовсе; задыхальцевые
щетинки отсутствуют; кроме того, имеются примерно две предкрыловые,
несколько верхних стерноплевральных, как правило, две верхние мезэпи-
мерные и одна нижняя мезэпимерная щетинки. Ноги с несколько расши-
ренными к основанию средними (реже и передними) бедрами. Голени и
лапки самцов многих экзотических форм имеют характерные особенности:
вторичнополового порядка. Коготки у обоих полов простые, без зубцов;,
пульвиллы отсутствуют. Мембрана крыла с весьма мелкими волосками
(микротрихиями), почему при недостаточном увеличении кажется голой;
вилки крыла короткие; вилка ш1+2 явственно короче своего стебелька.
Анальная жилка вливается в край крыла на уровне или перед основанием
общего ствола г2+3 и г4+5; закрыловые пластинки без чешуек. Брюшко-
короткое, у самки на вершине притупленное. Гипопигий с коротким
кокситом, несущим умеренной величины базальную бородавку, усажен-
ную щетинками; стиль простой, с вершинным придатком; анальный
сегмент перепончатый, без склеротизованных зубцов; фаллосома разде-
лена на пластинки, иногда несущие зубцы.
Личинка небольших размеров, с темной, слабо удлиненной головой и
короткими усиками. По бокам VIII членика брюшка имеются хитиновые
пластинки, по заднему краю несущие немногочисленные зубцы щетки.
Зубцы гребня сифона с мелко расщепленными краями, не заостренные.
Волосков на дыхательной трубке одна пара около ее середины. Личинки
держатся горизонтально под поверхностью воды.
Род включает около 140 видов, из которых преобладающее большин-
ство свойственно тропической зоне обоих полушарий; в Палеарктике род.
представлен одним видом в Средиземноморье (в широком смысле, включая
Кавказ и Среднюю Азию) и одним (чисто ориентальным) видом в Японии-
1. Uranotaenia unguiculata Edwards, 1913 (рис 63).
Очень мелкий комар, характеризующийся наличием узких серебри-
стых полосок по бокам среднеспинки и по боковой поверхности груди;
общая окраска темно-бурая.
Голова в большей своей части покрыта темно-бурыми чешуйками,
сверху обыкновенно с продольной полоской серебристых чешуек. Хобо-
ток и щупики самца и самки черно-бурые, в такого же цвета! чешуйках-
Щупики самца короткие, как у самки. Усики и покрывающие их во-
S. URANOTAENIA
127-
лоски у обоих полов бурые; основные членики усиков в серебристых
чешуйках. Среднеспинка покрыта темно-бурыми, имеющими иногда
красноватый оттенок, чешуйками, с боков с неширокой продольной по-
Рис. 63. Uranotaenia unguiculata Edw.
лоской серебристых чешуек, тянущейся от переднего края среднеспинки
до основания крыла; щиток в бурых чешуйках; бочки груди в средней
части с продольной полосою серебристых чешуек. Ноги: тазики передней
и средней пары в серебристых чешуйках; бедра спереди в бурых чешуйках,
обыкновенно с резко выраженной продольной полоской белых чешуек,
128
III. ПОДСЕМ. CULICINAE
которая иа передней и средней паре ног вполне обособлена, на задней
же — сливается с покрытой белыми чешуйками задней поверхностью;
' задняя поверхность передних и средних бедер также в белых чешуйках;
голени спереди в черно-бурых чешуйках, посредине часто с расплывчатым
белым пятном; голени сзади в белых чешуйках; вершины бедер и голеней
покрыты белыми чешуйками; лапки черные; коготок передней лапки
самца с одним зубцом. Крылья прозрачные, слегка дымчатые; жилки
крыльев покрыты довольно
альных жилок в основной
Рис. 64. Гипопигий Uranotaenia
unguiculata Edw.
широкими бурыми чешуйками; ствол ради-
части в белых чешуйках. Брюшко сверху
покрыто темно-бурыми чешуйками, с
боков с треугольными пятнами сере-
бристых чешуек, иногда выраженными
только на последних тергитах брюш-
ка; снизу брюшко в белых чешуйках.
Гипопигий (рис. 64): коксит отно-
сительно толстый и короткий, непра-
вильно конический, в базальной трети
с внутренней стороны с небольшой
плоской бородавкой, несущей три круп-
ные щетинки; стиль широкий, в ос-
новной трети слегка суженный, перед
вершиной с шиповидным придатком.
Личинка IV стадии (рис. 65)
мелких размеров, желто-зеленого или
желто-коричневого цвета с темно пиг-
ментированной головой.
Голова немного сильнее вытянута в
длину. Ротовые органы сдвинуты впе-
ред, вследствие чего на брюшной сторо-
не область горла развита в 1.5—2 раза
сильнее, чем у личинок других родов,
а основание верхней губы смещено бо-
лее вперед, что связано с питанием с
нижней стороны поверхностной пленки
воды и сгибанием при этом головы назад при горизонтальном положе-
нии тела личинки.
Наружные лобные волоски из 4—7 коротких ветвей. Кнутри от них
находятся более длинные простые утолщенные средние волоски; внутрен-
ние лежат позади средних, но сильнее сближены, простые, а в редких
случаях из 2—3 ветвей. Задние волоски наличника короткие, простые
или слабо ветвистые, лежат впереди внутренних лобных волосков. Усики
очень короткие, прямые, слегка сужаются к вершине, темно пигменти-
рованы, почти или вовсе лишены шипов; волосок тонкий, очень короткий,
отходит у середины верхней поверхности.
Брюшко: VIII членик по бокам имеет по хитиновой пластинке, верх-
ний край которой может заходить на спинную сторону. Пластинки слабо
заметны у недавно слинявших личинок, темнеют к концу стадии. От
нижней части их заднего края отходят зубцы щетки, расположенные
в один неправильный ряд из 5—8, чаще шести темных, конически заост-
ренных шипов, бока которых покрыты тонкими, очень короткими во-
лосками, сидящими в один ряд.
Сифон с хорошо развитыми ушками у основания, умеренной длины
(индекс 3.2—3.8, чаще 3.5—3.6), с прямой передней стороной, несколько
сужается к вершине. Гребень из 13—18 (чаще 15—16) слабо пигментиро-
3. UR A NOT AENIA
129
Рис. 65. Uranotaenia unguiculata Edw. Личинка IV стадии.
1 — голова, сверху, г — задний конец, сбоку.
зубцов, расположенных, за исключением 2—4 базальных, на
расстоянии друг от друга. Каждый
ванных
равном
краям которой сидят в один
ряд короткие тонкие шипики.
Одна пара волосков из 8—
12 слегка вторичноперистых
ветвей отходит от середины
сифона у наиболее дисталь-
ных зубцов гребня.
Стигмальная пластинка
(рис. 66) отличается задним
отростком рычага, который
оканчивается двумя заострен-
ными и когтевидно загну-
тыми внутрь выростами, а
также наличием на. утолщен-
ных внешних боковых краях
стигм направленного кверху
слегка извитого волоска хе-
тоидного типа с сильно рас-
ширенным основанием.
Последний членик цели-
ком заключен в темное хитиновое
боковым волоском. Наружные хвостовые волоски 2-ветвистые, одной
зубец в виде чешуйки, по
unguiculata Edw. Личинка
Рис. 66.
IV
Uranotaenia
стадии. Стигмальная пластинка,-
кольцо с тонким 3-ветвистым
9 Фауна СССР, А. В. Гуцевич и др.
130
III. ПОДСЕМ. CULICINAE
длины с сифоном, внутренние из четырех более коротких ветвей. Плав-
ник в среднем из 10 пучков. Жабры ланцетовидные, с округленными
концами, значительно короче последнего членика.
Распространение. Средиземноморская подобласть. СССР:
юг Украины, дельта Волги, Кавказ, Средняя Азия, на северо-восток
до бассейна р. Или в Алма-Атинской обл. Южная Европа, Северная
Африка, Передняя Азия, Иран, Пакистан.
Биология. Личинки встречаются в небольших стоячих, заросших
высшей водной растительностью (иногда ряской) водоемах и держатся
в более затененных их участках; предпочитают пресную или почти прес-
ную воду, избегая воды с содержанием солей выше 0.1—О.2°/оо; личинки
встречаются совместно с личинками Anopheles maculipennis sacharovi,
An. hyrcanus, Culex pipiens, Aedes caspius caspius. Комары данного вида
на человека нападают очень редко.
4. Род ORTHOPODOMYIA Theobald
Род характеризуется отсутствием пульвилл на лапках, а также ды-
хальцевых щетинок на бочках груди; проэпимерных щетинок две; длина
I членика передних лапок превосходит длину остальных, вместе взятых.
Относительно крупные формы, по большей части черного цвета, с белым
рисунком.
Личинка отличается красноватой или розоватой окраской тела, перед
окуклением принимающей синевато-фиолетовый оттенок, очень длин-
ными, особенно боковыми, волосками груди и брюшка, наличием у боль-
шинства видов пластинок на VI—VIII члениках брюшка, отсутствием
гребня на сифоне и щеткой из двух прямых рядов сильно вытянутых
своеобразной формы чешуек. Главные трахейные стволы в заднегруди
расширены в виде пузырей, имеющих гидростатическое значение. Ли-
чинки развиваются в скоплениях воды в дуплах деревьев.
Небольшой род, включающий немногим более 20 видов, распростра-
ненных в Ориентальной области, в южной части Северной, в Центральной
и Южной Америке; в пределах Палеарктики — один средиземноморский
вид.
1. Orthopodomyia pulchripalpis Rondani, 1872 (albionensis MacGregor,
1919).
Вид отличается от прочих палеарктических комаров своеобразным
рисунком среднеспинки (рис. 67).
Голова черная, в черных чешуйках, с некоторой примесью белых
чешуек. Задние края глаз в верхней части в белых чешуйках. Хоботок
в черных чешуйках, близ середины с более или менее широким белым
кольцом; щупики самца явственно длиннее хоботка, черные, с белыми
колечками в местах сочленений и белым конечным члеником; щупики
самки по длине почти равны половине длины хоботка, черные, с белым
колечком посредине и белой вершиной. Усики черные, основной членик
в белых чешуйках. Среднеспинка черная, в узких черных чешуйках,
с очень узкими белыми продольными полосками, из которых две сред-
ние тянутся через всю среднеспинку от переднего ее края до щитка,
в передней половине среднеспинки будучи параллельными друг другу,
а в задней половине слегка дугообразно изогнутыми; две полоски, лежа-
щие латерально от первых, развиты лишь в задней половине средне-
спинки, над основанием крыльев; боковой край среднеспинки окаймлен
узкой белой полоской. Ноги в черных чешуйках, имеющих стальной
Рис. 67. Orthopodomyia pulchripalpis Rond.
9*
132
III. ПОДСЕМ. CULICINAE
отлив; основания бедер покрыты желтовато-белыми чешуйками; дорсаль-
ная поверхность бедер с более или менее значительным количеством
вкрапленных белых чешуек; колени с белым пятном; лапки черные, с бе-
лыми колечками, опоясывающими вершину предыдущего и основание
последующего членика; белые колечки на передних и средних лапках
слабо развиты и имеются обыкновенно лишь на основных члениках;
на задних лапках белые колечки явственно развиты на всех члениках;
V членик задних лапок белый.
Крылья прозрачные; жилки по-
крыты бурыми чешуйками, осно-
вание ствола радиальных жилок
в серебристо-белых чешуйках.
Брюшко черно-бурое, в такого же
цвета чешуйках, по переднему
краю тергитов с умеренно широ-
кими поперечными перевязями бе-
лых чешуек.
Гипопигий (рис. 68): коксит с
конической бородавкой, располо-
женной ближе к основанию кок-
сита; бородавка несет 4—5 креп-
ких щетинок; стиль относительно
узкий, в дистальной половине
слабо утолщенный, с придатком,
примерно равным ширине стиля
в наиболее широкой его части.
Личинки IV стадии
(рис. 69) красноватого цвета, при-
нимающего перед окуклением фи-
олетовый оттенок. Голова темная,
округлая. Лобные волоски вто-
ричноперистые, длинные. Наруж-
ные короче остальных; средние
(9—10 ветвей) и внутренние (5—
7 ветвей) своими концами да-
леко выдаются за передний край
головы. Немного впереди и между
отходят задние волоски налич-
Рис. 68. Гипопигий Orthopodomyia
pulchripalpis Rond.
внутренними лобными волосками
ника, по длине и ветвлению сходные с наружными лобными волосками.
Усики прямые, не покрыты шипиками; на расстоянии х/3 длины от осно-
вания отходит волосок, состоящий из семи коротких, веерообразно расхо-
дящихся ветвей. Ротовые органы — переходного к фильтрационному
типу питания.
Грудь широкая, особенно в заднем отделе, отличается очень длинными
боковыми волосками. Членики брюшка заметно сужаются к заднему
концу. Боковые волоски тоже очень длинные. На спинной стороне VI—
VIII члеников у зрелых личинок имеются темные пластинки; на VI чле-
нике — небольшая и узкая, на VII — занимает в длину большую часть
членика и заходит на боковые стороны, на VIII — более узкая, но дальше
заходит на боковые стороны. В промежутке между VIII и последним
члеником тоже узкие боковые пластинки.
Дыхательная трубка (индекс 3.5—4) лишена гребня и ушков у осно-
вания. У ее середины, ближе к основанию, на задней поверхности нахо-
дится волосок из 8—10 веерообразно расположенных ветвей. Щетка из
5. CULISETA
133
двух рядов длинных с острым шипом зубцов; часть зубцов заднего ряда
(16—20) помещается в промежутках между основаниями зубцов перед-
него (7—10). Из волосков позади щетки сильно развит только средний,
похожий на сифональ'ный волосок.
Последний членик брюшка с кольцевидным седлом, которое по ниж-
нему краю значительно короче верхнего. Наружные хвостовые волоски
Рис. 69. Orthopodomyia pulchripalpis Rond. Личинка IV стадии.
1 — задний конец, сбоку, г—4 — чешуйки щетки (2 — крупные, з — мелкие, 4 —• вид сбоку).
почти одной длины с брюшком, простые, внутренние в виде более корот-
кого асимметричного веера из 9—14 ветвей. Плавник из 14 пучков. Жабры
ланцетовидные; верхняя пара одной длины с седлом, нижняя вдвое
короче.
Распространение. В основном — Средиземноморская под-
область. СССР: Южный берег Крыма, Закавказье, от Сочи до Ленкорани.
Южная Европа, Северная Африка (Алжир, Тунис), Турция. В Западной
Европе на север доходит до Бельгии и юга Англии.
Биология. Личинки встречаются в скоплениях воды в дуплах
деревьев (бук, конский каштан и др.) или под корнями, часто совместно
с личинками Anopheles plumbeus, Aedes pulchritarsis и Aedes geniculatus",
зимуют личинки IV стадии. Комары днем в тенистых местах изредка
нападают на человека.
5. Род CULISETA Felt1 (THEOBALDIA Neveu-Lemaire)
Родовое имя Theobaldia Neveu-Lemaire, 1902 не может быть сохранено,
так как ранее (1885) оно было использовано для обозначения рода мол-
люсков. Поэтому оно должно быть заменено именем Culiseta Felt, 1904.
1 При составлении настоящей главы широко использованы данные работ А. В. Мас-
лова (1964, 1967).
134
III. ПОДСЕМ. CULICINAE
Комары данного рода обычно имеют средние или относительно круп-
ные размеры. Дыхальцевые щетинки имеются, задыхальцевые отсут-
ствуют. Коготки ног самки простые, пульвиллы не развиты. Ствол ради-
альной жилки в основной части с волосками, более многочисленными на
нижней поверхности крыла. Чешуйки на жилках узкие. Церки § не
выступают. Гипопигий: коксит, как правило, с бородавкой, стиль с очень
коротким придатком; класпеты отсутствуют.
Личинки от крупных до очень крупных размеров, с широкой, кроме
личинок Allotheobaldla, головой, с ротовыми частями, приспособленными
или к питанию с субстрата {Allotheobaldia), или к смешанному {Culiseta
s. str.), или к фильтрационному типу питания {Culicella)-, соответственно,
у них или короткие усики со слабо развитым пучком {Allotheobaldia), или
более длинные {Culiseta), или очень длинные с пучком в форме широкого
веера {Culicella)' аналогичны и изменения лобных волосков. Щетка на
VIII членике брюшка из большого числа чешуек, не имеющих главного
шипа. Сифон различной длины с относительно коротким гребнем, часто
продолжающимся к дистальному концу или в виде шипов (Л llotheobaldia,
частью Culicella), или в виде ряда волосков {Culiseta s. str.). Пучок всегда
у основания сифона. Стигмальная пластинка с задним отростком рычага
в виде более или менее массивной нерасщепленной пластинки. Главные
трахейные стволы или широкие, лентовидные, с овальным поперечным
сечением {Allotheobaldla, Culiseta,s. str.), или тонкие с округлым попереч-
ным сечением. Седло или слабо развито {Allotheobaldia), или кольцом
охватывает последний членик брюшка. Плавник сильно развит, часть
пучков лежит впереди общего основания и прободает кольцо седлав
Жабры различной длины.
Род с небольшим числом видов (около 30), из которых Палеарктике
свойственно 12—13. В своем большинстве виды приурочены к умеренной
зоне Палеарктики и Неарктики.
J ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
9?
1 (2). Среднеспинка с резко выраженными продольными белыми поло-
сами, образующими подобие лировидного рисунка (рис. 70). Бедра
и голени спереди с резко выраженными белыми пятнами. Костальная
жилка на большей части своего протяжения покрыта белыми чешуй-
ками. (Щупики самца немного короче хоботка). (Подрод Allotheo-
baldia Brol.)..............1. С. (Allotheobaldia) longiareolata Macq.
2 (1). Среднеспинка без белых полос. Бедра и голени спереди в одно-
цветно темных чешуйках или же с более или менее значительным
числом вкрапленных светлых чешуек. Костальная жилка покрыта
преимущественно или исключительно темными чешуйками. (Щу-
пики самца длиннее хоботка или равны хоботку).
3 (10). гт и тт расположены на одной прямой; если же они слегка
сдвинуты, то расстояние между ними, как правило, не превышает
длины тт. Крылья часто с темными пятнами. (Подрод Culiseta
Felt).
4 (7). Лапки одноцветно темные.
5 (6). Крылья с более или менее выраженными нерезкими темными
пятнами .......................... 5. С. (Culiseta) bergrothi Edw.1
1 С. bergrothi и С. glaphyroptera с трудом различаются по самкам, но хорошо отли-
чаются по строению гипопигия.
5. CULISETA
135
6
7
8
9
10
11
12
13
14
15
16
(5) . Пятна на крыльях отсутствуют или едва заметны...............
........................2. С. (Culiseta) glaphyroptera Sehin.1
(4). Лапки с белыми кольцами.
(9) . I членик задних лапок посредине без светлого кольца; бедра без
предвершинного светлого кольца...................................
...............................3. С. (Culiseta) alaskaensis Ludl.
а (б). Темные чешуйки на теле и ног ах темно-бурые или черные, а
светлые чешуйки — чисто белые, почему рисунок на теле и ногах кон-
трастный. Светлые чешуйки на крыльях отсутствуют или их немного.
Темные пятна на крыльях резко выражены. При основании терги-
тов брюшка имеются суживающиеся к средней линии перевязи
из белых чешуек; светлые чешуйки в задней половине тергитов
отсутствуют . . . . , С. (Culiseta) alaskaensis alaskaensis Ludl.
б (а). Темные чешуйки на теле и ногах светло-бурые или бурые, а свет-
лые чешуйки — охристые, почему рисунок на теле и ногах расплыв-
чатый. На крыльях имеется значительная примесь светлых чешуек.
Темные пятна на крыльях не резко выражены. При основании тер-
гитов светлые перевязи отграничены не резко; светлые чешуйки
в небольшом числе вкраплены на всей поверхности тергитов (и в зад-
ней их половине)...........С. (Culiseta) alaskaensis indica Edw.
(8). I членик задних лапок посредине с белым кольцом; бедра с пред-
вершинным светлым кольцом . . . 4. С. (Culiseta) annulata Schr.
а (б). Костальная жилка в темных (черных) чешуйках. Темные пятна
на крыльях резко выражены. При основании тергитов брюшка
имеются резко отграниченные перевязи светлых (белых) чешуек;
светлые чешуйки в задней половине тергитов отсутствуют ....
....................................С..(Culiseta) annulata annulata Schr.
б (а). Костальная жилка в темных и светлых перемешанных чешуй-
ках. Темные пятна на крыльях слабо выражены, расплывчатые. Свет-
лые перевязи при основании тергитов брюшка не резко отграничены
и образованы желтоватыми (а не белыми) чешуйками; такие же че-
шуйки вкраплены среди темных чешуек и в задней половине терги-
тов ......................... С. (Culiseta) annulata subochrea Edw.
(3). mm поперечная жилка расположена явственно ближе к основанию
крыла, нежели гт; расстояние между ними, как правило, не менее
длины тт. Крылья без темных пятен. (Подрод Culicella
Felt).
(14). Стерниты брюшка, как правило, с рисунком из темных чешуек
в виде буквы V, направленной острием вперед.
(13). Светлые колечки развиты на всех члениках передних, средних и
задних лапок......................8. С. (Culicella) fumipennis Steph.
(12). Светлые колечки развиты не на всех члениках лапок . . . .
...................................9. С. (Culicella) setivalva Masi.
(11). Стерниты брюшка без рисунка из темных чешуек в виде буквы V.
(16). Узкие перевязи светлых чешуек расположены только при осно-
вании тергитов. Чешуйки на крыле не образуют скопления (пятна)
................................6. С. (Culicella) morsitans Theob.
(15). Узкие, не резко отграниченные перевязи светлых чешуек распо-
ложены при основании, иногда, кроме того, и у вершины тергитов,
или же - они отсутствуют вовсе; иногда тергиты сплошь покрыты
светлыми чешуйками. В средней части крыла, при основании г4+5
может быть выражено неясное темное пятно...................... . .
................................7. С. (Culicella) ochroptera Peus.
136
III. ПОЦСЕМ. CULICINAE
dd
1 (2). IX тергит по бокам с длинными выростами; эдеагус массивный
...........................1. С. (Allotheobaldia) longiareolata Macq.
2 (1). IX тергит простой, без боковых выростов; эдеагус нормальный.
3 (4). Коксит с вершинной бородавкой, несущей пучок длинных чешуй-
чатых пластинок...................2. С. (Culiseta) glaphyroptera Sehin.
4 (3). Вершинная бородавка коксита усажена волосками или она вовсе
отсутствует.
5 (6). Коксит со слабо выпуклой вершинной бородавкой, густо усажен-
ной короткими волосками .... 3. С. (Culiseta) alaskaensis Ludl.
6 (5). Коксит без вершинной бородавки.
7 (8). На внутренней поверхности коксита перед вершиной обычно
имеется участок со скоплением густых волосков. Фаллосома сильно
склеротизованная . . .............. 4. С, (Culiseta) annulata Schr.
8 (7). Скопления густых волосков перед вершиной коксита нет. Фал-
лосома, как правило, слабо склеротизованная.
9 (10). Коксит в очень длинных и крепких щетинках, которые
(в количестве 2—3) развиты и на внутренней поверхности коксита
.................................... 9. С. (Culicella) setivalva Masi.
10 (9). Коксит в умеренно длинных и тонких щетинках.
11 (12). Базальная бородавка коксита несет две щетинки. VIII стернит
посредине заднего края с гребнем шипов, обычно их не менее десяти
.................................... 5. С. (Culiseta) bergrothi Edw.
12 (11). Базальная бородавка коксита несет не менее трех крепких щети-
нок. Задний край VIII стернита с меньшим количеством шипов
(не более десяти) или вовсе без шипов.
13 (14). VIII стернит по заднему краю без шипов.......................
...................... 7. С. (Culicella) ochroptera Pens (частью).
14 (13). Задний край VIII стернита с гребнем плотно сидящих шипов или
с несколькими разбросанными шипами.
15 (16). Дистальные членики щупиков (или один последний) отчетливо
утолщенные ........................ 6. С. (Culicella) morsitans Theob.
16 (15). Дистальные членики щупиков самца не утолщенные, такой же
толщины, как предыдущие.
17 (18). Базальная бородавка коксита с 3—4 крепкими щетинками
....................................8. С. (Culicella) fumipennis Steph.
18 (17). Базальная бородавка коксита с 5—8 крепкими щетинками (как
исключение, их 3—4)...................................................
................... 7. С. (Culicella) ochroptera Peus (частью).
ЛИЧИНКИ
1 (12). Сифон относительно короткий и толстый (индекс не более 4).
Главные трахейные стволы широкие, лентовидные. Усики короче
головы, со слабо развитым пучком.
2 (3). Гребень сифона из небольшого числа хорошо развитых зубцов,
широко расставленных по всей длине сифона. На поверхности
усиков шипики отсутствуют; пучок на усике у его середины, ближе
к дистальному концу, короткий, не более чем из трех ветвей (подрод
Allotheobaldia Brol.) . . 1. С. (Allotheobaldia) longiareolata Macq.
3 (2). Гребень сифона из более тонких зубцов, сидящих у основания,
продолжается в дистальном направлении в виде ряда длинных
волосков. Поверхность усика покрыта редкими шипиками; пучок
5. CULISETA.
137
4
5
6
7
8
9
10
11
12
13
14
15
16
на усике-у его середины, ближе к основанию, короткий,, более чем
из пяти ветвей (подрод Culiseta Felt).
(7). Усики в длину достигают не менее половины длины головы.
Средние лобные волоски из 4—8 ветвей. Щетка в среднем более
чем из 60 чешуек. .
(6). Усики достигают 2/3 длины головы. Гребень сифона занимает
не менее 3/4 его длины; волоски в нем численно преобладают над
зубцами. Впереди плавника пять пучков, три.из них своими основа-
ниями прободают кольцо седла ..............................
........................ 2. С. (Culiseta) glaphyroptera Sehin.
(5). Усики достигают половины длины головы. Гребень сифона за-
нимает 2/3 его длины; зубцы в нем или равны числу волосков, или
немного преобладают над ними. Впереди плавника 3—4 пучка,
из которых два своими основаниями прободают кольцо седла . . .
...............................5. С. (Culiseta) bergrothi Edw.
(4). Усики менее половины длины головы. Средние лобные волоски
не более чем из трех ветвей. Щетка в среднем менее чем из 50 чешуек.
(9). Сифон более короткий и толстый, слабо суженный к вершине:
отношение его длины к ширине у основания — 2.4—3.0 (2.7),
а к ширине у вершины — 3.4—3.9 (3.7). Чешуйки щетки узкие, удли-
ненные, с параллельными боковыми краями, без заметного расшире-
ния у основания ............... 3. С. (Culiseta) alaskaensis Ludl.
(8). Сифон более длинный и тонкий, сильнее суженный к вершине:
отношение его длины к ширине основания 3—4, а к ширине
у вершины — 5—6. Чешуйки щетки слегка сужены посредине,
с заметно расширенным основанием.
(11) . Расстояние между задними волосками наличника равно таковому
между внутренними лобными волосками.............................
.................................4. С. (Culiseta) annulata Sohr.
(10). Расстояние между задними волосками наличника меньше расстоя-
ния между внутренними лобными волосками ........................
............... С. (Culiseta) annulata subochrea Edw.
(1). Сифон относительно длинный и тонкий (индекс выше 5). Главные
трахейные стволы тонкие, округлой формы. Усики длиннее головы,
с хорошо развитым пучком (подрод Culicella Felt).
(16). Сифон, помимо гребня у основания, на задне-боковой поверх-
ности имеет более длинные, чем зубцы гребня, шипы, расположен-
ные без определенного порядка. Волосок у основания боковых
клапанов стигмальной пластинки в виде полусложенного веера,
сидящего на хитиновом основании.
(15). В щетке около 50 чешуек. Средний из волосков позади щетки
достигает длины последнего членика брюшка. Наружные хвостовые
волоски в виде асимметричного веера, ветвящегося на спинную
сторону, в среднем из шести ветвей. Передние пучки плавника,
не объединенные общим основанием, почти вдвое короче задних
..............................8. С. (Culicella) fumipennis Steph.
(14). В щетке около 100 или больше чешуек. Средний из волосков
позади щетки заметно короче последнего членика брюшка. Наруж-
ные хвостовые волоски из трех ветвей. Передние пучки плавника,
не объединенные общим основанием, одной длины или только не-
много короче задних...............9. С. (Culicella) setivalva Masi.
(13). Добавочные шипы, помимо гребня на сифоне, отсутствуют.
Волосок у основания боковых клапанов стигмальной пластинки
обычного строения.
138
III. ПОДСЕМ. CULICINAE
17 (18). Внутренние лобные волоски из 5—9 ветвей. Чешуйки у заднего
края щетки с более темной продольной заостренной к концу полосой.
Волосок на задних клапанах стигмальной пластинки крючкообразно
изогнут только на самом конце. Жабры в 1.5—2 раза длиннее седла
....................................7. С. (Culicella) ochroptera Pens.
18 (17). Внутренние лобные волоски из 2—3 ветвей. Чешуйки у заднего
края щетки без темной продольной полосы. Волосок на задних
клапанах стигмальной пластинки крючкообразно изогнут по всей
длине. Жабры короче седла . . . 6. С. (Culicella) morsitans Theob.
1. Подрод ALLOTHEOBALDIA Brolemann
Комары этого подрода характеризуются короткими щупиками самца,
длина которых равна примерно 3/4 длины хоботка. Коксит без вершинной
бородавки, IX тергит с двумя длинными отростками, эдеагус массивный.
Личинки с небольшой головой, ширина которой заметно менее чем
в 1.5 раза превышает ее длину, с ротовыми органами смешанного типа
питания, но с преобладанием типа питания с субстрата над фильтрацион-
ным. Усики короткие, лишенные шипиков, с коротким, слабо ветвя-
щимся волоском. Относительно короткий сифон обычно не склеротизован
до основания и не имеет ушков; имеется пара пучков у основания и гре-
бень, состоящий из небольшого числа шиповидных, широко расставлен-
ных зубцов, беспорядочно расположенных по всей длине трубки. Главные
трахейные стволы широкие, лентовидные. Последний членик брюшка
со слабо развитым седлом, сильно разветвленными пучками плавника и
короткими жабрами.
Известен только один вид подрода — С. (All.) longiareolata.
Недавно было предложено (Маслов, 1964) выделить из рода Culiseta
монотипический подрод Allotheobaldia Brol. и рассматривать его в ка-
честве самостоятельного рода. Действительно, С. (All.) longiareolata Macq.
отличается от прочих представителей рода довольно существенными
признаками — строением гипопигия, длиной щупиков самца и др.
Но при решении данного вопроса необходимо соблюдать «единый масштаб»
в трактовке различных групп семейства. В некоторых родах (например,
Aedes, Culex) различия между видами во всяком случае не менее значи-
тельны, чем в роде Culiseta. Дробление же многих родов комаров, по на-
шему мнению, не представляется целесообразным. Обособленность
С. (АП.) longiareolata достаточно подчеркивается выделением вида в осо-
бый подрод.
1. Culiseta (Allotheobaldia) longiareolata Macquart, 1838.
Резко отличается от всех прочих палеарктических видов рода белым,
имеющим вид лиры, рисунком среднеспинки (рис. 70), а также белым
цветом чешуек, покрывающих костальную жилку по переднему ее краю.
Голова сверху близ средней линии с довольно широкими белыми
прилегающими чешуйками, далее в латеральном направлении с отсто-
ящими бурыми щетинками; на боках головы и на затылке к последним
вновь примешиваются белые чешуйки; края глаз окаймлены густо ле-
жащими белыми чешуйками. Щупики самца короче хоботка, бурые;
длинный членик щупиков при основании и у середины белый; предпослед-
ний членик щупиков у своего основания, последний также и у вершины,
в белых чешуйках; последний членик сильно утолщенный, короткий.
Щупики самки короткие, темно-бурые, с примесью белых чешуек; вер-
Рис. 70. Culiseta longiareolata Macq.
140
111. ПОДСЕМ. CULICINAE
Рис. 71. Гипопигий Cu-
liseta longiareolata Macq.
шина щупиков самки в белых чешуйках. Хоботок черно-бурый. Усики
самца черно-бурые, с белыми колечками при основании члеников и длин-
ными бурыми волосками; усики самки одноцветно черно-бурые с покры-
тыми белыми чешуйками основными члениками. Среднеспинка покрыта
бурыми или желтовато-бурыми узкими чешуйками; вдоль средней линии
от переднего края среднеспинки до щитка тянется полоса белых чешуек;
аналогичные белые полосы тянутся в передней половине среднеспинки
по ее боковому краю; загибаясь далее кнутри (по поперечному шву
среднеспинки), они идут до щитка, располагаясь кнаружи от линии дорсо-
центральных щетинок; среднеспинка у основания крыльев, щиток и
бочки груди покрыты белыми чешуйками. Ноги черные, с многочислен-
ными белыми пятнами, часто сгруппирован-
ными в продольные полосы; бедра и голени
на задней поверхности в большей своей
части покрыты белыми чешуйками, лапки
черные с разбросанными пятнами белых
чешуек и с неширокими белыми кольцами
при основании первых двух, трех или че-
тырех (на задних лапках) члеников; коготки
самки простые, у самца на передних и сред-
них лапках с зубцом. Крылья по жилкам
покрыты бурыми чешуйками; костальная
жилка на всем своем протяжении по перед-
нему краю в белых чешуйках. Брюшко очень
сильно варьирует в окраске покрывающих
его чешуек; сверху в большинстве случаев
оно покрыто бурыми чешуйками; при осно-
вании тергитов имеются довольно широкие
поперечные перевязи чисто белых чешуек; зад-
няя треть тергитов покрыта желтоватыми че-
шуйками, которые, в виде выдающегося впе-
ред языка, часто доходят до белых чешуек переднего края тергитов или же
покрывают (что особенно часто наблюдается на последних тергитах)
всю оставшуюся свободной от белых чешуек часть тергита, замещая,
таким образом, бурые чешуйки; брюшко снизу покрыто белыми чешуй-
ками. VIII стернит самки с двумя широкими лопастями и выемкой между
ними.
Гипопигий (рис. 71) очень своеобразной формы с сильно развитыми
пальцевидными выростами IX тергита и весьма массивным склеротизо-
ванным эдеагусом.
Личинки IV стадии (рис. 72) крупных размеров, серого
цвета, с небольшой темной головой и коротким сифоном. Голова менее
чем в 1.5 раза больше в ширину, чем в длину. Лобные волоски слабо
развиты: наружные из 3—4 ветвей, средние (впереди внутренних) —
простые, внутренние — простые, реже 2-ветвистые. Пара простых задних
волосков наличника находится между средними лобными волосками.
Усики короткие, лишены шипиков, с коротким, не более чем в 2 раза
превышающим ширину усика, 1—3-ветвистым волоском.
Щетка из 40—75 чешуек, очень изменчивых по величине и форме —
от имеющих хорошо выраженный шип до окаймленных только рядом
шипиков. Из волосков позади щетки очень сильно развит средний.
Сифон (индекс 1.5—2.1) темно пигментирован, но его склеротизация
обычно не доходит до основания; ушки отсутствуют. Гребень состоит из
4—7 крупных гладких шипов, широко расставленных по всей длине
5. CULISETA
Ш
трубки; в ее базальном, не склеротизованном отделе имеется 3—7 более
мелких рудиментарных зубчиков. Можно наблюдать все переходы между
последними и чешуйками щетки. Пучок лежит у основания сифона, на
границе его склеротизованной части, состоит из 10—15 вторичноперистых
ветвей, более тонких и коротких, чем у среднего волоска позади щетки.
Стигмальная пластинка как на рис. 73.
Рис. 72. Culiseta, longiareolata Macq. Личинка IV стадии.'
1 — голова, сверху, г — задний конец; сбоку, з —. чешуйки щеткив
Последний членик брюшка со слабо развитым седлом, покрывающим
не более 3/4 его спинной стороны и едва заходящим на боковые стороны:
задний край густо покрыт короткими шипами. Наружные хвостовые
волоски одной длины с сифоном, 1—2-ветвистые; внутренние — в виде
веера из 10—16 ветвей, одной длины с плавником, состоящим из 16—18
сильно ветвящихся пучков. Длина жабр сильно изменчива, в зависимости
от степени солености водоема, от 1/2 Д° 1х/г длины последнего членика.
Распространение. В Палеарктике — в основном Средиземно-
морская подобласть. СССР: степная и пустынная зоны — Средняя и
Южная Украина на север до Харькова, Нижнее Поволжье, Северный
Кавказ, Закавказье (Грузия, Армения, Азербайджан), Казахстан и не-
которые южные районы Западной Сибири (Омск), Средняя Азия (Туркме-
ния, Узбекистан, Таджикистан, Киргизия). Западная Европа на север
до Южной Англии, Африка, Передняя Азия и далее на восток до Ирана,
Индии и Пакистана. *
Биология. Личинки обитают в различных мелких водоемах, как
искусственных (неглубокие колодцы, шурфы, бочки), так и естественных,
142
HI. ПОДСЕМ. CULICINAE
с пресной или слабо солоноватой водой; переносят значительную степень
загрязнения воды; большую часть жизни личинки проводят у поверхности
воды, редко опускаясь на дно; часто попадаются совместно с личинками
Culex pipiens, реже с Aedes caspius caspius', зимует в фазе личинки;
в Средней Азии развитие может продолжаться и в относительно холод-
ной воде, в связи с чем возможны вылеты комаров и в зимнее время.
Комары на человека нападают очень редко; они являются переносчиками
Рис. 73. Culiseta longiareolata Macq. Личинка
IV стадии. Стигмальная пластинка.
кровепаразитов птиц; в этом
комаре некоторые виды плаз-
модиев птиц могут проделы-
вать полный цикл спорого-
нии. У некоторых популя-
ций установлена возможность
откладки первой порции яиц
без кровососания.
2. Подрод CULISETA Felt
Щупики самца длиннее
хоботка или равны хоботку,
с утолщенными двумя вер-
шинными члениками. Коксит
часто с предвершинной бо-
родавкой или резко отгра-
ниченным участком густо
сидящих волосков в вершин-
ной трети медиальной поверх-
ности; IX тергит без длинных
придатков; эдеагус нормаль-
ный; фаллосома у большин-
ства видов сильно склеро-
тизованная, с крючковидно изогнутыми вершинными отделами.
Личинки с умеренно развитой головой, ширина которой в среднем
в 1.5 раза превышает ее длину, с ротовыми органами преимущественно
смешанного типа питания, с короткими усиками, лишь в единичных
случаях (С. glaphyroptera'} превышающими половину длины головы,
несущими относительно короткий пучок, с короткими предвершинными
и вершинными щетинками, расположенными двумя группами. Щетка из
большого числа тесно расположенных чешуек. Сифон умеренной длины
(индекс менее 4), с парой пучков у основания, с гребнем, продолжаю-
щимся в дистальном направлении в виде ряда длинных тонких, тесно
сидящих волосков. Главные трахейные стволы широкие, лентовидные.
Седло последнего членика окружает его кольцом. Впереди плавника не-
сколько (2—4) пучков прободают своими основаниями седло. Наружные
хвостовые волоски из нескольких ветвей.
Подрод представлен в Палеарктике шестью видами, из них один
в основном ориентальный.
2. Culiseta (Culiseta) glaphyroptera Schiner, 1864.
Вид характеризуется, помимо особенностей в строении гипопигия,
одноцветно темно-бурыми, без светлых колечек, лапками, а также отсут-
ствием или слабым развитием темных пятен на крыльях.
Голова с боков и сверху по краю глаз покрыта беловатыми чешуй-
ками, верхняя часть затылка в торчащих бурых чешуйках. Хоботок и
5, CULISETA
143
щупики у обоих полов темно-бурые, у самца щупики явственно длиннее
хоботка с расширенным, но плоским последним члеником. Усики самца
бурые, со светлыми колечками на основных и
средних члениках и одноцветно бурыми конеч-
ными члениками; усики самки одноцветно бу-
рые. Среднеспинка покрыта бурыми или золо-
тисто-бурыми чешуйками, последние образуют
неясные полосы и пятна; бочки груди местами
покрыты прилегающими белыми чешуйками.
Ноги покрыты бурыми чешуйками, задняя
поверхность бедер и голеней в желтовато-белых
чешуйках; лапки темно-бурые, без светлых
колец. Крылья с одноцветно темно-бурыми
чешуйками, иногда с едва намечающимися пят-
нами более темных чешуек при основании ви-
лок г и т и на поперечных жилках. Брюшко
сверху темно-бурое, по переднему краю тер-
гитов с поперечными перевязями белых че-
шуек, занимающими около 1/3 длины тергита;
брюшко снизу в значительной своей части по-
Рис. 74. Гипопигий Cu-
liseta glaphyroptera Sehin.
крыто белыми чешуйками, по заднему краю стернитов с перевя-
зями бурых чешуек.
Рис. 75. Culiseta glaphyroptera Sehin. Личинка IV стадии. По Пеусу.
1 — голова, сверху, г — задний конец, сбоку, з — чешуйки щетки.
Гипопигий (рис. 74): IX тергит без боковых выростов; коксит перед
вершиной с выступом, несущим пучок длинных чешуйчатых пластинок.
ш
III. ПОДСЕМ. CULICINAE
Личинки IV стадии (рис. 75) крупных размеров, светлого
желто-коричневого цвета, полупрозрачные, с широкой головой. Лобные
волоски сильно развиты, вторичноперистые; наружный — из 8—12,
средний — из 5—7, внутренний — из 7—9 ветвей, в длину достигающих
верхней губы. Задние клипеальные волоски впереди внутренних и сильнее
сближены. Усики длинные, заметно превышают половину длины головы;
пучок у середины, немного ближе к основанию, в длину не меньше поло-
вины длины усика, в среднем из 10 ветвей. Ротовые части фильтрацион-
ного типа питания, без гребенчатых окончаний даже на внутренних воло-
сках верхней губы, но с относительно короткими ее наружными лопа-
стями.
Щетка из 60—70 или более удлиненных чешуек с расширенным осно-
ванием, суженной средней частью и широким, закругленным дистальным
концом; края чешуек окаймлены рядом различно развитых шипиков.
Сифон (индекс 3.4—3.5) в вершинной трети слабо сужен. Гребень состоит
в среднем из 12—24 (15—21) шиповидных зубцов, далее, по направлению
к вершине, продолжающихся в виде ряда из 17—29 (19—22) более длин-
ных волосков до начала вершинной четверти сифона. Сифональный пу-
чок помещается недалеко от основания, между 9—12-м зубцами гребня,
из восьми ветвей, заметно более длинных, чем ширина сифона у основа-
ния. Стигмальная пластинка сходна с таковой у С. alaskaensis, но зна-
чительно меньших размеров.
Последний членик брюшка окружен сплошным кольцом седла, имею-
щим глубокий вырез на брюшной стороне. Наружные хвостовые волоски
из 3—4 ветвей, внутренние в виде веера в среднем из 14 ветвей. Боковой
волосок короткий, из двух ветвей. Плавник из 13—16 пучков, объеди-
ненных общим основанием, и из пяти более коротких впереди него; из
них передние три прободают своими основаниями кольцо седла. Жабры
в 1.5—2.5 раза длиннее седла, прозрачные, заострены на концах.
Распространение. Горные районы центральной и юго-вос-
точной Части Западной Европы. В СССР: Закарпатская область; данные
для других районов СССР (Ленинградская область и др.) относятся,
по-видимому, к С. bergrothi.
Биология. Вид приурочен к гористым местностям. Личинки
встречаются в полузатененных, слабо освещаемых солнцем водоемах
в углублениях каменистого ложа горных речек и ручьев. В водоемах
с чистой прозрачной водой и чистым каменистым дном и соответственно
с бедной микрофлорой и фауной личинки отсутствуют, тогда как более
загрязненные водоемы с опавшей листвой и т. п., богатые детритом,
густо населены личинками этого вида.
3. Culiseta (Culiseta) alaskaensis Ludlow, 1906.
Вид отличается от прочих палеарктических видов, обладающих тем-
ными пятнами на крыльях, отсутствием белых колец перед вершиной
бедер и посредине I членика задних лапок.
Голова сверху и с боков покрыта желтовато-белыми прилегающими
и темно-бурыми торчащими чешуйками. Усики самца как у предыдущего
вида; усики самки темно-бурые, основные 2—3 членика, в особенности
с внутренней стороны, покрыты белыми чешуйками. Хоботок и щупики
у обоих полов покрыты бурыми чешуйками, с довольно многочисленной
примесью вкрапленных белых чешуек; щупики самца явственно длиннее
хоботка. Среднеспинка покрыта темно-бурыми и золотистыми чешуйками,
местами сгруппированными в одноцветные пятна, местами же беспоря-
дочно перемешанными; боковые отделы среднеспинки, как правило, свет-
5. CULISETA
145
Рис. 76. Гипопигий Culiseta alaskaensis Ludl.
лее; бочки груди с пятнами белых чешуек. Ноги: бедра и голени спереди
покрыты черными чешуйками с примесью одиночных, но довольно много-
численных белых чешуек; задняя поверхность бедер и голеней покрыта
белыми чешуйками; вершины бедер и голеней в белых чешуйках; лапки
черные с белыми кольцами при основании II—III или II—IV (на задних
лапках) члеников. Крылья покрыты темно-бурыми чешуйками с незна-
чительной примесью одиночных белых чешуек; скопления черных чешуек
в виде явственно выраженных темных пятен имеются вдоль поперечных
жилок и на развилках г2-|-г3,
TTll —j—777-2 ^"3+4 —j-^-^1*
Брюшко сверху покрыто
черными чешуйками, при
основании тергитов с пере-
вязями белых чешуек, зани
мающими около 1/3 длины
тергита и к средней линии
тела слегка суживающимися;
брюшко снизу почти на всем
своем протяжении покрыто
белыми чешуйками.
Гипопигий (рис. 76): VIII
стернит по заднему краю
близ середины с гребнем ко-
ротких шипиков; коксит пе:
ред вершиной со слабо вы-
пуклой бородавкой, густо
усаженной волосками; ба-
зальная бородавка коксита
с двумя, реже с тремя креп-
кими щетинками, иногда
изогнутыми под углом. Фал-
лосома склеротизована.
Личинки IV ста-
fl и и (рис. 77) очень круп-
ные, от желто-коричневого
до почти черного цвета;
брюшко кажется поперечнополосатым вследствие отложения пигмента
в лопастях наружного слоя жирового тела, отсутствующего между чле-
никами.
Голова широкая. Лобные волоски вторичноперистые, наружный —
наиболее короткий — из 9—11 ветвей, средний — из 2—3, внутренний —
из 5—6 ветвей. Задние волоски наличника короткие и тонкие, из 3 вет-
вей; расстояние между ними равно расстоянию между внутренними лоб-
ными волосками. Усики заметно короче половины длины головы; пучок
на расстоянии 2/5 длины усика от основания, из 8—11 ветвей, достигающих
его вершины. Ротовые органы смешанного типа питания.
Щетка из 35—55 (46) чешуек, изменчивых по форме, но без сужения
посредине, которые несут ряд шипиков, более длинных и тесно сидящих
на округленном дистальном конце. Сифон короткий и широкий, слабо
суженный к вершине, индекс 2.5—3.0, а отношение его длины к ширине
у вершины — 3.4—3.9. Гребень состоит из 6—8 длинных шиповидных
зубцов и из 3—6 рудиментарных зубчиков у основания; к вершине, про-
должается в виде ряда из 16—18 волосков, в среднем одинаковой длины,
кроме более коротких 2—3 наиболее дистальных. Волоски не доходят
10 Фауна СССР, А. В. Гуцевич и др.
часто, особенно у молодых личинок,
146
III. ПОДСЕМ. CULICINAE
до вершинной трети или четверти сифона. Пучок у основания из 7—10 слабо
вторичноперистых ветвей, немного превосходящих половину длины сифона.
Стигмальная пластинка очень крупная (как на рис. 78). Последний членик
около половины длины сифона, седло охватывает его кольцом. Наружный
хвостовой волосок из 3—5 ветвей, из них средняя достигает длины си-
фона; внутренний в виде мощного веера из 22—27 ветвей. Боковой во-
лосок короткий, из 2—3 ветвей. Плавник из 16—18 сильно развитых
Рис. 77. Culiseta alaskaensis Ludl. Личинка IV стадии. Задний конец,
сбоку.
пучков и из 3—4 более коротких впереди общего основания. Жабры
до 1.5 длины седла или короче, ланцетовидные, заострены на конце.
Распространение. Бореальная зона Палеарктики и Не-
арктики. В пределах СССР ареал вида охватывает тундровую, таежную
и лесную зоны; вид обнаружен и в наиболее северных районах европей-
ской и азиатской частей СССР (Кольский полуостров, мыс Челюскин).
Западная Европа; Северная Америка. В горных районах юга СССР,
в Иране, Пакистане и в Северной Индии — особый подвид, С. (С.) ala-
skaensts indica.
Биология. Личинки встречаются, как правило, в непересыхающих
в течение лета, полузатененных водоемах, бедных водной растительностью,
с дном, покрытым гниющей опавшей листвой, расположенных в широких
прогалинах преимущественно лиственного леса или кустарника или
на открытых местах по соседству с ними; в зоне тундры личинки оби-
тают в плоских заболоченностях. Совместно с личинками этого вида
5. CULISETA
147
Рис. 78. Culiseta alaskaensis Ludl. Личинка
IV стадии. Стигмальная пластинка.
часто попадаются личинки Aedes cinereus, A. caspius dorsalis, A. cantans,
A. excrucians тл. другие, реже — личинки Anopheles maculipennis и Cu-
lex territans. Вид зимует во взрослой фазе в дуплах деревьев, пещерах,
подвалах и т. д., часто совместно с Culex pipiens, Anopheles maculipennis,
представителями сем. Helomyzidae (Diptera, Acalyptrata'), зимними кома-
риками Petauristidae (Diptera, Nematocera') st грибными комариками My-
cetophilidae', весною вылетает
с зимовок, как правило, рань-
ше других видов кровососущих
комаров (в окрестностях Ле-
нинграда — в середине или во
второй половине апреля). Ак-
тивный кровосос, хотя напа-
дает на человека обыкновенно
в небольших количествах как
в сумерки, так и при ярком
солнечном освещении; в тундре
и лесотундре нападает на се-
верных оленей.
Culiseta alaskaensis alaska-
ensis Ludlow.
С признаками вида. Дохо-
дит. до южных границ лесной
зоны. Экземпляры из южных
районов окрашены светлее.
Culiseta alaskaensis indica
Edwards, 1920.
Окрашен значительно свет-
лее основной формы. Щупики
самца часто в светлых чешуй-
ках. Среднеспинка в золотисто-
желтых чешуйках. Брюшко ча-
сто с более широкими перевя-
зями белых чешуек при основа-
нии тергитов. Гипопигий в об-
щем как у основной формы,
но шипики по заднему краю
VIII стернита часто развиты
слабее (число их у некоторых
экземпляров сокращено до 3).
Личинка с достоверностью неотличима от личинки типичной формы.
Р аспро стр анение. Горные районы юга СССР: Кавказ, Сред1
няя Азия — Узбекистан, Таджикистан, Киргизия, Казахстан.
4. Culiseta (Culiseta) annulata Schrank, 1776.
Близок к предыдущему, от которого отличается наличием белого
кольца в вершинной трети бедер и в средней трети I членика задних
лапок.
Голова с боков в желтовато-белых прилегающих чешуйках, особенно
густо расположенных по заднему краю глаз; голова свефху усажена
желтовато-бурыми щетинками, сзади — торчащими такого же цвета че-
шуйками. Хоботок самца светло-бурый, по большей части покрыт желто-
10*
118
III. ПОДСЕМ. CULICINAE
вато-белыми чешуйками; щупики самца явственно длиннее хоботка, свет-
ло-бурые, с широкими светлыми кольцами, опоясывающими вершину
предыдущего и основание последующего членика щупиков; волоски,
покрывающие щупики, бурые, в местах, покрытых светлыми чешуй-
ками, — желтовато-белые; хоботок самки бурый, покрыт бурыми и бе-
лыми перемешанными чешуйками; щупики короткие, бурые, покрытые
у вершины члеников белыми, в остальной части бурыми чешуйками.
Усики как у предыдущего вида. Среднеспинка покрыта беспорядочно
перемешанными бурыми, золотистыми и белыми чешуйками, более ши-
рокими в боковых отделах среднеспинки; бочки груди с пятнами широ-
ких белых чешуек; щиток в белых чешуйках. Ноги черно-бурые, покры-
тые темно-бурыми чешуйка-
ми, на бедрах и голенях с
многочисленными пятнами
белых чешуек; задняя сторо-
на бедер и голеней в боль-
шей части покрыта белыми
чешуйками; бедра в вер-
шинной трети с явственным
белым кольцом; колени и
сочленения голени и лапки
покрыты белыми чешуйками;
лапки с белыми колечками
посередине I членика и при
основании II, III и IV чле-
ников. Крылья покрыты бу-
рыми чешуйками, с незначи-
тельной примесью белых че-
шуек в переднем поле крыла;
поперечные жилки и вилки
r2-j-r3, mj.-j-TTZa и с
Рис. 79. Гипопигий Culiseta annulata Schr. резко выраженными пятнами,
образованными скоплением
черно-бурых чешуек. Брюшко темно-бурое, при основании терги-
тов с белыми поперечными перевязями, занимающими около V3
длины тергита; в остальной части тергиты брюшка покрыты
одноцветно темно-бурыми чешуйками; II тергит посредине обыкновенно
с узкой продольной полоской белых чешуек; брюшко снизу в желто-
вато-белых чешуйках.
Гипопигий (рис. 79): базальная бородавка коксита с 2—5 равномерно
изогнутыми крепкими щетинками; на внутренней поверхности коксита,
перед его вершиной, как правило, имеется площадка, густо усаженная
волосками. Фаллосома. сильно склеротизована. Шипы по заднему краю VIII
стернита имеются или, чаще, отсутствуют.
Личинки IV стадии (рис. 80) крупные, желто- или зелено-
вато-коричневого цвета., Голова широкая. .Лобные волоски расположены
и развиты как у С. alaskaensis. Задние волоски наличника и внутренние
лобные одинаково широко расставлены. Усики заметно короче половины
длины головы, с пучком ближе к основанию, из 10—15 не достигающих
вершины ветвей. Ротовые части смешанного типа, со слабо выражен-
яыми гребнями на концах волосков внутренних пучков верхней губы.
Щетка из-40«—50 и более чешуек, слегка суженных в средней части. Си-
фон относительно более длинный и сильнее суженный к вершине, чем
у С. alaskaensis; индекс 3.6—4.0, отношение его длины к ширине у вер-
5. CULISETA
149
шины — 6. Гребень из 11—18 (13—15) шиповидных зубцов, из которых
базальные часто рудиментарны. В дистальном направлении зубцы пере-
ходят в ряд из 11—21 (14—17) волосков, доходящих до 1/а, реже до
2/3 длины сифона. Пучок отходит немного отступя от основания, в сред-
нем из 9—10 слабо вторичноперистых ветвей, не достигающих половины
длины сифона. Стигмальная пластинка как у С. alaskaensis, но заметно
меньших размеров. Последний членик брюшка меньше половины длины
сифона, с седлом, окружающим его кольцом. Наружные хвостовые во-
лоски из трех ветвей, средняя достигает длины сифона; внутренние —
из 13—19 ветвей; боковой волосок тонкий, из трех ветвей. Плавник в сред-
Рис. 80. Culiseta annulata Schr. Личинка IV стадии. По Пеусу.
1 — голова, сверху, г — задний конец, сбоку, 3 —чешуйки щетки.
нем из 18 пучков; передние пучки плавника, постепенно укорачиваясь, без
резкой границы переходят в пучки, лежащие впереди общего основания.
Жабры ланцетовидные, достигают длины седла. i
Распространение. Европейская часть СССР на север до Ле-
нинградской области и Эстонии; Кавказ и Закавказье (нагорная часть);
Средняя Азия, Казахстан. Широко распространен в Западной Европе
на север до Швеции и Норвегии; Средиземноморье; Северная Африка;
Передняя Азия.
Биология. Личинки обитают в водоемах самых разнообразных
типов, как естественных, так и искусственных — лужах, Канавах, це-
ментных водовместилищах, бочках и т. п., а также в болотах; личинки
свободно переносят умеренную степень загрязнения; попадаются иногда
в водоемах, богатых ряской; зимует нормально в фазе взрослого насе-
комого; в более южных широтах (Южный берег Крыма) может зимовать
и личинка. Комар сосет кровь преимущественно млекопитающих; охотно
нападает на человека; питается также кровью птиц, передает некоторые
виды птичьих плазмодиев. В южной полосе европейской части СССР и
на Кавказе вид очень обычен и широко распространен, на севере (Ле-
нинградская область, Эстония) — редок.
150 HI. ПОДСЕМ. CULICINAE
Culiseta annulata annulata Schrank.
С признаками вида. Гипопигий: базальная бородавка коксита обычно
с двумя крепкими щетинками; шипы по заднему краю VIII стернита
чаще отсутствуют, а если имеются, то их не более четырех.
Culiseta annulata subochrea Edwards, 1921.
Отличается от основной формы более слабым развитием темных пя-
тен на крыльях, имеющих вместе с тем большее количество белых
чешуек, в особенности в переднем поле. Брюшко почти на всем своем про-
тяжении, за исключением узких белых перевязей при основании терги-
тов, покрыто светло-охряно-желтыми чешуйками, среди которых вкрап-
лены обыкновенно единичные бурые чешуйки; белые колечки на лапках
значительно шире, нежели у основной формы. Гипопигий: базальная
бородавка коксита обычно с 3—4 крепкими щетинками; шипы по заднему
краю VIII стернита чаще имеются (в числе 4—8).
Личинки IV стадии крупные, от бледно- до темно-коричне-
вого цвета, с более светлыми головой и сифоном. Голова широкая.
Лобные волоски вторичноперистые, наружный из 7—12, средний из 3
(реже 2), внутренний из 5—8 ветвей; задние волоски наличника из 3—4
тонких ветвей. Расстояние между последними меньше расстояния между
внутренними лобными волосками. Усик около половины длины головы;
пучок у середины, ближе к основанию, из 9—14 тонких ветвей, почти
достигающих конца усика. Ротовые органы смешанного типа питания.
Щетка из 30—50 чешуек, сходных с чешуйками С. annulata, но с более
тесно сидящими щипиками и с менее расширенным основанием. Сифон
(индекс 3.3—3.6; отношение его длины к ширине у вершины — 4.5—5.3)
как у С. annulata. Гребень из 10—15 тонких шиповидных зубцов и 2—5
мелких рудиментарных шипиков у основания. Гребень продолжается
в ряд волосков (18—26), основания которых не доходят до вершины
сифона на 0.3—0.4 его длины. Пучок расположен немного отступя от осно-
вания, из 7—11 вторичноперистых ветвей, обычно не достигающих по-'
ловины длины сифона. Стигмальная пластинка как у С. annulata. Послед-
ний членик меньше половины длины сифона, с седлом, охватывающим
его кольцом. Наружные хвостовые волоски из 3—5 ветвей, из них брюш-
ная наиболее длинная, превосходит длину сифона; внутренние — в виде
вееров из 16—19 ветвей. Боковой волосок тонкий, из трех ветвей. Плав-
ник из 16—18 пучков и из 2—3 впереди общего основания; относительное
развитие пучков как у С. annulata. Жабры ланцетовидной формы с за-
остренными концами, несколько короче седла.
Распространение. В пределах СССР широко распространен
в различных местах Средней Азии. В Западной Европе известен из Южной
Норвегии и Финляндии, Дании, Англии, Франции и других стран. Се-
верная Африка, Передняя Азия (Палестина, Сирия, Иран, Ирак). В За-
падной и Центральной Европе ареалы обоих подвидов налегают друг
на друга.
Биология. Вид является типичным галофилом; личинки оби-
тают в лужах, как правило, с богатой водной растительностью; в более
южных широтах (Средняя Азия) личинки тяготеют к водоемам, хорошо
затененным древесной или тростниковой растительностью, или же к доста-
точно глубоким колодцам, куда солнечный свет не проникает. Зимуют
самки; иногда зимовка очень кратковременна; по-видимому, могут пере-
зимовывать и личинки, давая ранний вылет взрослых. В течение года
вид дает до 4—5 поколений. На человека нападает редко.
5. CULISETA
151
5. Culiseta (Culiseta) bergrothi Edwards, 1921 (borealis Shingarev, 1927).
Вид характеризуется одноцветно черными, без светлых колечек, лап-
ками и едва намечающимися темными пятнами на крыльях; гт распо-
ложена несколько дистальнее тт.
Голова в светло-золотистых прилегающих чешуйках и черно-бурых
торчащих чешуйках и щетинках. Хоботок в глубоко черных чешуйках,
с металлическим отливом. Щупики самки в черных чешуйках, с незна-
чительным числом вкрапленных светлых чешуек. Щупики самца длин-
ные, в черно-бурых чешуйках, в вершинной половине (начиная с вершин-
ной части длинного членика) в длинных желтовато-бурых волосках.
Среднеспинка сильно варь-
ирует по цвету покрываю-
щих ее чешуек; или средне-
спинка более или менее рав-
номерно, но не густо покрыта
узкими золотистыми чешуй-
ками, или. же среднеспинка
в темно-бурых, почти черных
чешуйках, с продольными
полосами золотистых чешу-
ек, идущими в задней поло-
вине среднеспинки по ря-
дам дорсоцентральных ще-
тинок, и такого же цвета,
но более широкими полоса-
ми, идущими по боковым
краям среднеспинки; боко-
вые края голого простран-
ства перед щитком и сам
щиток в узких белых чешуй-
ках; задний край щитка в
длинных черно-бурых щетин-
ках; бочки груди С ПЯТ- Рис. 81. Гипопигий Culiseta bergrothi Edw.
нами белых ланцетовидных
чешуек. Ноги: бедра и голени спереди в черно-бурых чешуйках, с за-
метной примесью белых чешуек, в особенности в основной части бедра;
бедра и голени сзади, а задние бедра и спереди, за исключением верхней
поверхности и неясного кольца перед вершиной, а также вершины всех
бедер и голеней, в белых чешуйках. Лапки одноцветно черные, без свет-
лых колечек; I членик передних лапок самца очень длинный, обычно
длиннее остальных члеников той же лапки, вместе взятых. Крылья по
жилкам в узких темно-бурых чешуйках, с едва намеченными темными
пятнами при основании радиальной и медиальной вилок, а также на по-
перечных жилках.
Брюшко в черных чешуйках, с умеренно широкими перевязями бе-
лых чешуек при основании тергитов; как светлые, так и темные чешуйки
верхней поверхности брюшка имеют иногда голубоватый или стальной
отлив.
Гипопигий (рис. 81): коксит длинный;’ базальная бородавка хорошо
развита, с двумя крепкими и длинными щетинками; вершинная бородавка
отсутствует; VIII стернит по заднему краю с гребнем шипов.
Личинки IV стадии (рис. 82) крупных размеров, серо- или
желто-коричневого цвета. Голова широкая. Лобные волоски вторичнопе-
152
III. ПОДСЕМ. CULICINAE
ристые, наружный из 8—16 (10—11), средний из 5—9 (6—7), внутрен-
ний из 9—13 (10—11) ветвей; задние волоски наличника из 2—6 (3—5) вет-
вей, отходят впереди внутренних лобных волосков, более сильно сбли-
жены. Усики достигают 1/, длины головы; пучок у середины, немного
Рис. 82. Culiseta bergrothi Edw. Личинка IV стадии.
1 — голова, сверху, г — задний конец, сбоку.
ближе к основанию, из 5—14 (9—11) вторичноперистых ветвей, концами
не достигающих вершины усика. Ротовые органы фильтрационного типа.
Щетка из 50—80 (60—70) узких длинных чешуек, с нерезко выра-
женным сужением ближе к основанию; края окаймлены рядом шипиков,
более тесным на дистальном конце. Сифон заметно сужается к вершине;
длина его варьирует: у дальневосточных личинок индекс 3.1—4.1 (3.6—
3.9), а у личинок из Карельской АССР — 2.8—3.6 (3.2). Гребень состоит
в среднем из 14 (9—22) умеренной длины зубцов с 2—3 дополнительными
зубчиками у их основания и из 2—8 (3—4) рудиментарных зубчиков
5. CULISETA
15S
у кольца основания сифона; он продолжается в ряд из 7—15 (10—12)
волосков, из которых 3—4 ближайших к основанию более короткие;
волоски доходят до 2/3 длины сифона от основания. Пучок расположен
у основания, из 4—9 (6—7) вторичноперистых ветвей, достигающих длины
последнего членика. Стигмальная пластинка как у С. annulata, более
сильно пигментирована, по размерам значительно меньше, чем у С. ala-
skaensis. Последний членик брюшка с охватывающим его в виде кольца
седлом, имеющим вырез на брюшной стороне. Наружные хвостовые во-
лоски из 2—4 ветвей, из них средняя длиннее сифона, внутренние — в виде
веера из 13—16 ветвей; боковой волосок из 2—6 коротких ветвей. Плав-
ник из 13—19 сильно развитых пучков и из 3—4 более коротких и менее
разветвленных впереди общего основания; из них 1—2 расположены
в брюшном вырезе, а два прободают седло своими основаниями. Жабры
узкие ланцетовидные, с заостренными концами, в среднем в 2 раза длин-
нее седла.
Распространение. В СССР вид широко распространен в
южной полосе зоны тундры (Кольский полуостров, Ненецкий националь-
ный округ), в таежной зоне и в северной полосе лесной зоны; на Даль-
нем Востоке заходит в зону широколиственных лесов. Север Западной
Европы (Швеция, Норвегия, Финляндия).
Биология. Личинки встречаются в различных стоячих водоемах
от тундровых заболоченностей до лесных луж и копанок близ жилья
человека; на юге ареала обитают в затененных водоемах; переносят зна-
чительную степень загрязнения воды. На севере .ареала — одно поко-
ление, на юге — 2—3. Зимуют самки. Сосут кровь млекопитающих,
в частности крупного рогатого скота; иногда залетают в жилые поме-
щения и помещения для скота.
3. Подрод .CULICELLA Felt
Щупики равны по длине хоботку или длиннее его; последние два чле-
ника щупиков не утолщенные или более или менее утолщенные, в явственно
развитых волосках; бедра и голени без резко выраженных светлых пятен;
крылья без темных пятен; тт расположена явственно перед rm; коксит
без предвершинной бородавки или резко ограниченного участка густо
сидящих волосков, IX тергит без отростков; фаллосома слабо склеро-
тизованная; ее вершинные отделы слабо изогнутые, не крючковидные.
Личинки крупные, с широкой головой, с ротовыми органами фильтра-
ционного типа питания, с длинными изогнутыми усиками, сильно раз-
витым пучком на них, расположенным далеко за серединой; две длин-
ные предвершинные щетинки сидят отступя от вершины усика, на кото-
рой только одна щетинка длинная. Сифон (индекс 5—7) с парой пучков
у основания и с гребнем из небольшого числа шиповидных зубцов. Послед-
ний членик брюшка длинный, с кольцеобразно охватывающим его сед-
лом. Впереди плавника несколько (около шести) пучков, прободающих
основаниями кольцо седла. Наружные хвостовые волоски разветвленные.
Жабры умеренной длины с заостренными концами. Главные трахейные
стволы тонкие, округлой формы.
В подроде 8 видов, из них 5—6 палеарктических.
6. Culiseta (Culicella) morsitans Theobald, 1901.
Голова покрыта желтовато-белыми чешуйками, сверху, кроме того,
усажена торчащими бурыми чешуйками, кпереди постепенно переходя-
щими в длинные бурые щетинки. Хоботок самца светло-бурый, к осно-
154
III. ПОДСЕМ. CULICINAE
ванию и вершине несколько темнее, в светло-желтоватых чешуйках; щу-
пики самца явственно длиннее хоботка, бурые, на основных члениках
с широкими, на двух конечных члениках — с узкими светлыми коль-
цами; конечный членик щупиков явственно утолщенный; хоботок и щу-
пики самки одноцветно темно-бурые, в черно-бурых чешуйках. Средне-
спинка с узкими бурыми и золотистыми чешуйками (последние сгруппи-
рованы главным образом по средней линии, по линии дорсоцентральных
щетинок и по боковому краю среднеспинки); бочки груди местами по-
крыты пятнами белых чешуек; щиток в золотистых чешуйках. Ноги бу-
рые; бедра и голени спереди покрыты одноцветно бурыми, сзади желто-
вато-белыми чешуйками; колени и сочленения голени и лапки в белых
чешуйках; лапки с узкими белыми колечками при основании II, III,
а иногда и IV члеников. Крылья с одноцветно бурыми чешуйками, без
пятен. Брюшко черно-бурое,
в большей своей части по-
крыто такого же цвета че-
шуйками, при основании
тергитов с узкими попереч-
ными перевязями белых че-
шуек; брюшко снизу в од-
ноцветных желтовато-белых
чешуйках.
Гипопигий (рис. 83): дли-
на коксита превосходит ши-
рину при основании не более
чем в 3 раза; базальная бо-
родавка коксита с 3—6 креп-
кими щетинками; VIII стер-
нит по заднему краю с не-
правильным гребнем шипов
или довольно длинных креп-
ких щетинок, числом 3—10,
Рис. 83. Гипопигий Culiseta morsitans Theob. Личинки IV с т а-
По Natvig. д и и (рис. 84) крупных
размеров, изменчивой ок-
раски — от полупрозрачных до желтовато-зеленого и коричневого
цвета, со светлой головой, ширина которой более чем в 1.5 раза превы-
шает длину. Лобные волоски длиннее головы, слабо вторичноперистые;
наружные из 6—7, средние и внутренние из 2—3 ветвей. Задние волоски
наличника отходят впереди от внутренних, сильнее сближены, короткие
и простые. Усики длиннее головы, S-образно изогнутые, с темными су-
женным концом и основанием; волосок в виде густого веера из 20—25 вто-
ричноперистых ветвей помещается на расстоянии 2/3 длины усика от осно-
вания; предвершинные щетинки отходят отступя от вершинных, из ко-
торых одна — длинная. Щетка более чем из 100 тесно расположенных
длинных и узких чешуек со слегка расширенными основанием и дисталь-
ным концом, окаймленным рядом тонких чешуек. Сифон (индекс 5—7)
прямой, слабо суженный к вершине, с гребнем из 5—8 длинных тонких
зубцов, из которых наиболее дистальный немного расставлен и расположен
на расстоянии около 1/6 длины сифона от основания. Пучок у основания,
из 4—6 ветвей, в длину достигающих 1/i длины сифона. Стигмальная
пластинка очень своеобразна (рис. 85). Главные трахейные стволы отно-
сительно тонкие, с округлым поперечным сечением. Последний членик
брюшка длинный, с седлом, охватывающим его кольцом; задний край
5. CULISETA
155
на спинной стороне покрыт шипиками, исчезающими в переднем и боко-
вом направлениях. Наружные хвостовые волоски из трех ветвей, из ко-
Рис. 84. Culiseta ochroptera Peus. По ПеуСу.
1 — задний конец, сбоку, 2 — Culiseta morsitans Theob., задний конец, сбоку.
торых средняя достигает длины сифона, внутренние — в виде веера из 14—
17 ветвей. Боковой волосок простой, вдвое короче седла. Плавник из 10-
Рис. 85. Culiseta morsitans Theob. Личинка IV стадии.
Стигмальная пластинка.
14 пучков в общем основании и около шести пучков впереди него; из них
один лежит в выемке седла, а остальные прободают своими основаниями
кольцо. Жабры ланцетовидные, почти вдвое короче седла; верхняя пара
может быть немного короче нижней.
156 III. ПОДСЕМ. CULICINAE
Распространение. Европейская часть СССР от Прибалтики
и Ленинградской области до Крыма и Северного Кавказа; Средний Урал;
Западная Сибирь. Западная Европа от Норвегии, Швеции и Финляндии
до Средиземноморья; Северная Африка; Передняя Азия.
Биология. Личинки развиваются в различного типа водоемах,
как стоячих, так и слабо проточных, характеризующихся более или менее
значительной затененностью, с дном, покрытым опавшими листьями;
часто попадаются во временных водоемах в лесу или по опушкам; зимуют
личинки в сравнительно глубоких водоемах постоянного типа; при про-
мерзании водоема личинки гибнут; личинки этого вида являются, как
правило, спутниками личинок Anopheles claviger, а весною — личинок
Aedes. Комары местами нередки в лесных стациях, в частности в широко-
лиственных лесах; дневки встречаются в густой траве, под навесами раз-
личных построек и т. и.; в северных частях ареала (Прибалтика, Ленин-
градская область) более обычен во вторую половину лета и в начале
осени (вторая половина июля—сентябрь). Нападает как на сельскохо-
зяйственных животных, так и на человека, в основном, по-видимому,
питается на диких животных, вероятно на птицах.
7. Culiseta (Culicella) ochroptera Pens, 1935.
Голова в прилегающих беловатых чешуйках, с торчащими черными
чешуйками; по краю глаз длинные темные щетинки. Щупики самки темно-
бурые, иногда со светлыми чешуйками у вершины III и IV члеников.
Щупики самца выступают за вершину хоботка на длину полутора вер-
шинных члеников; конечный членик щупика не утолщенный; щупики
покрыты преимущественно желтоватыми чешуйками, лишь I членик, осно-
вание II и кольцо при основании III членика темные.
Среднеспинка в ржаво-бурых или золотисто-бурых чешуйках; по бо-
кам и у заднего края среднеспинки развиты узкие серповидные бело-
вато-латунные чешуйки, которые преобладают в пространстве перед щит-
ком, а также покрывают щиток. Такие же чешуйки иногда образуют
на среднеспинке неясные продольные полоски. Ноги: бедра спереди тем-
ные, сзади бледно-желтоватые. Лапки с узкими светлыми колечками при
основании первых трех члеников лапок; несколько светлых чешуек мо-
жет быть и на вершине члеников; нередко светлые колечки на лапках
слабо выражены, едва различимы. Крылья по жилкам в бурых чешуйках;
иногда при основании г4+5 чешуйки расположены более густо, образуя
подобие темного пятна.
Окраска чешуек брюшка весьма изменчива. Обычно тергиты бурые,
со скоплениями светло-желтых чешуек, образующих поперечные пере-
вязи при основании, а иногда (более узкие) и у вершины тергитов; послед-
ний тергит на всем протяжении светлый. Стерниты в светлых и темных
чешуйках, которые в основном сгруппированы, как на тергитах. Дальне-
восточные популяции чаще характеризуются одноцветными желтоватыми
(иногда более темными, бурыми) чешуйками на тергитах; встречаются
экземпляры с узкими базальными светлыми перевязями на тергитах или
с единичными светлыми чешуйками, вкрапленными на темном фоне
брюшка.
Гипопигий (рис. 86): бородавка коксита с 3—9 слабо изогнутыми ши-
пами (крепкими щетинками). Задний край VIII стернита с немногочислен-
ными (не более 8) шипами, иногда без шипов. Фаллосома обычно яйце-
видная, слабо склеротизованная.
Личинки IV стадии (рис. 84 и 87) крупные, светло- или
темно-коричневого цвета, с желто-коричневой головой и сифоном (Мон-
5. CULISETA
157
чадский, 1947). Лобные волоски вторичноперистые; наружные — из 7—
13 ветвей достигают 2/3 длины усика; средние очень длинные, длиннее
усика, всегда из двух ветвей; внутренние одной длины с наружными,
из 5—9 ветвей. Задние волоски наличника из 2—4 тонких ветвей. Усики
немного короче головы, более темно пигментированы у основания и в су-
женном вершинном отделе, изогнутом у места отхождения длинных пред-
Рис.’ 86. Гипопигий
Culiseta ochroptera
Peus.
Рис. 87. Culiseta
ochroptera Peus. Ли-
чинка IV стадии.
1 — голова, сверху,
2 — стигмальная пла-
стинка.
вершинных щетинок, расположенных заметно отступя от вершины; пучок из
20—37 ветвей. Щетка из 60—95 чешуек, из которых часть имеет продоль-
ный темный гребень; сужение посредине их тела, если имеется, то выра-
жено слабее, чем у С. morsitans. Сифон длинный (индекс 5.7—7.0), слабо
сужающийся к вершине. Гребень из 2—7 зубцов, из которых 2—3 дисталь-
ных шиповидные, более широко расставлены, и из 2—4 рудиментарных
зубчиков у основания. Пучок у основания из 5—10 ветвей, достигающих
х/4—1/3 длины сифона; ветви расположены не веером, а расходящимся
пучком. Стигмальная пластинка своеобразного строения (рис. 87). Послед-
ний членик брюшка длинный, седло охватывает его кольцом. Наружные
хвостовые волоски из 2—3 ветвей, не достигающих 3/4 длины сифона;
158
III. ПОДСЕМ. CULICINAE
внутренние — в виде веера из 12—23 ветвей; боковой волосок длинный
и простой. Плавник из 10—12 пучков, объединенных общим основанием,
и из 6—8 более коротких впереди него; часть их прободает кольцо седла
своими основаниями. Жабры тонкие, ланцетовидные, длинные (от 1 до 2.6
длины седла).
Замечания по систематике. А. В. Маслов (1964) считает С. ochrop-
tera синонимом описанного ранее С. silvestris Shing. (Шингарев, 1928). Но если судить
по весьма неполному описанию, представленному Н. И. Шингаревым, то «не идентич-
ность этих двух видов несомненна» (Штакельберг, 1937). Голотип С. silvestris не сохра-
Рис. 88. Culiseta ochroptera Pens. Личинка IV стадии.
1.— задний конец, сбоку, 2— зубцы гребня, 3 — чешуйки щетки.
нился; А. В. Масловым были исследованы лишь переданные ему паратипы. Ввиду
несовершенства первоначального описания С. silvestris представляется более правиль-
ным сохранить наименование С. ochroptera.
Данный вид А. В. Маслов (1964) предлагает разделить на четыре подвида: С. sil-
vestris silvestris Shing. (Восточная Европа, Приуралье, Западная Сибирь), С. s. ochrop-
tera Pens (Центральная Европа, Прибалтика), С. s. amurensis Masi. (Приамурье и
Приморье) и С. s. minnesotae Barr. (Северная Америка); автор приводит таблицы для
определения подвидов по самкам, самцам и по личинкам. Однако ввиду слабой изу-
ченности вида в целом предлагаемое разделение на подвиды и их сравнительная харак-
теристика имеют предварительный характер.
Распространение. Лесная зона Палеарктики от Западной
Европы до Приморья и Северо-Восточного Китая [не исключено, что
японский вид — Culiseta (Culicella) nipponica La Casse et Yamaguti —
тоже относится к C. ochroptera', вид описан только по личинкам с острова
5. CULISETA
159
Хоккайдо]. Северная и южная границы ареала не прослежены; в СССР,
помимо Дальнего Востока, С. ochroptera найдена во Владимирской области
(Сафьянова, 1960), на Урале, в Западной Сибири и в немногих других
районах.
Биология. Личинки приурочены к крупным неглубоким водое-
мам болот, лесных вырубок и т. и., а также к заболачивающимся бере-
гам озер; зимуют, по-видимому, личинки. Но в условиях востока Украины
зимуют самки (Вальх, 1959). Комары обитают преимущественно в при-
родных условиях; питаются кровью птиц, реже диких млекопитающих,
иногда нападают на человека.
8. Culiseta (Culicella) fumipennis Stephens, 1825.
Очень близок С. morsitans, от которого отличается наличием белых
колечек на всех члениках лапок и присутствием образованного темными
Рис. 89. Гипопигий Culiseta fumi-
pennis Steph. По Natvig.
Рис. 90. Culiseta fumipennis Steph.
Личинка IV стадии. Задний конец,
сбоку. По Сегюи.
чешуйками рисунка в виде буквы V на большинстве стернитов брюшка,
а также наличием скопления светлых чешуек в средней трети хоботка.
160
111. ПОДСЕМ. CULICINAE
Гипопигий (рис. 89): длина коксита в 3—3.5 раза превышает его ши-
рину при основании; базальная бородавка коксита с 3—4 крепкими ще-
тинками; задний край VIII стернита с несколькими шипами.
Личинки IV стадии (рис. 90) крупные, полупрозрачные,
зеленоватого или желтовато-коричневого цвета, с широкой головой. Лоб-
ные волоски вторичноперистые, наружные из 5—6, средние из 2, внутрен-
ние из 2—4 ветвей. Усики длиннее головы, с пучком из 10—13 вторично-
перистых ветвей, отходящих около границы суженной вершинной трети
усика. Предвершинные щетинки на некотором расстоянии от вершины.
Щетка из 120—160 длинных узких чешуек, слегка расширенных на кон-
цах и окаймленных тонкими шипиками. Сифон расширен у основания
и сначала заметно, а затем постепенно сужается к вершине; индекс
(около 5) при расширенном основании не дает точного представления
о длине сифона. Гребень из 7—8 хорошо развитых и 4—6 рудиментарных,
тесно сидящих зубцов, расположенных в косой, огибающий основание
пучка ряд; его продолжением являются 2—5 (3—4) крупных гладких,
сильно пигментированных шипов, неравномерно расставленных и отсут-
ствующих в вершинной трети сифона. Пучок отходит у основания, из 4—
5 гладких ветвей, достигающих половины длины сифона. Стигмальная
пластинка с сильно развитыми крючковидно изогнутыми вцдосками на
задних клапанах и с крупными, в виде полусложенного веера из 8—12 вет-
вей, волосками на боковых клапанах. Главные трахейные стволы тонкие,
округлого поперечного сечения.
Последний членик длинный; седло охватывает его кольцом. Наружные
хвостовые волоски из трех ветвей, из них средняя утолщена и более длин-
ная, чем сифон; внутренние хвостовые волоски в виде веера из 14—15 вет-
вей. Плавник из 14 пучков и 6 более коротких, слабее ветвящихся пучков
впереди него. Жабры ланцетовидное, более чем вдвое короче последнего
членика.
Распространение. Европейская часть СССР (Эстония, Ленин-
градская область, Среднее Приуралье, Украинское’ Полесье, Северный
Кавказ). Все указания на нахождение вида в СССР, как основанные
на взрослых комарах, нуждаются в подтверждении. Личинки С. fumi-
pennis в СССР найдены не были. Большая часть Западной Европы на се-
вер до Южной Швеции и Норвегии; Средиземноморские страны, вклю-
чая Алжир.
Биология. Личинки встречаются начиная с весны, в неглубоких
водоемах, с зеленой травянистой растительностью, а также в водоемах,
покрытых ряской (Lemna minor и L. trisulca), часто совместно с личин-
ками Culiseta morsitans, Culex territans, C. hortensis (в южных частях ареала),
реже с личинками Anopheles claviger] личинки питаются планктоном,
большую часть времени проводя в глубине водоема.
9. Culiseta (Culicella) setivalva Maslov, 1937.
Вид по Цветовым признакам очень близок к С. fumipennis, от которого
его резко отличает строение гипопигия, а именно отсутствие шипов в сред-
ней части заднего края VIII стернита и наличие очень длинных и крепких
щетинок на коксите.
Голова в прилегающих беловатых чешуйках и торчащих черно-бурых
чешуйках и щетинках. Хоботок самца светло-бурый, к основанию и на вер-
шине несколько темнее, в светло-желтоватых чешуйках; щупики длинные,
выступают за хоботок примерно на длину вершинного их членика, в бу-
рых чешуйках, с более йли менее заметной примесью светлых чешуек;
последние образуют более или менее широкие кольца близ середины
5. CULISETA
161
длинного членика и у основания предвершинного и вершинного члеников;
волоски, покрывающие щупики самца, буроватые, причем они особенно
длинные в вершинной половине длинного членика; волоски при основании
предвершинного и вершинного члеников светло-желтые. Хоботок и щупики
самки в темно-бурых чешуйках, лишь с незначительной примесью оди-
ночно вкрапленных светлых чешуек. Среднеспинка и щиток в довольно
редких и узких светло-золотистых чешуйках; бочки груди с небольшими
пятнами широких белых чешуек. Ноги (бедра и голени) спереди, темно-
Рис. 91. Гипопигий Culiseta setivalva Masi.
бурые, без примеси или лишь с незначительной примесью светлых че-
шуек, сзади белые; задние бедра спереди иногда с продольной полоской
светлых чешуек; вершина бедер и голеней с желтоватыми чешуйками;
при основании I—III или I—V члеников лапок имеются узкие белые
колечки. Лапки передних ног самца короткие, явственно короче соответ-
ствующей голени; длина I членика передних лапок примерно равна или
едва менее длины остальных члеников той же лапки, вместе взятых.
Крылья по жилкам в густых темно-бурых чешуйках. Брюшко сверху
в темных чешуйках с перевязями желтовато-белых чешуек по переднему
краю тергитов, занимающими около х/3—1/1 длины тергита; брюшко снизу
в желтовато-белых чешуйках с более или менее значительной примесью
темных чешуек, которые иногда образуют рисунок в виде перевернутой
буквы V, наподобие такого же рисунка у С. fumipennis.
Гипопигий (рис. 91): VIII стернит самца по заднему краю близ се-
редины без шипов, с 2—4 очень длинными крепкими щетинками по бокам
11 Фауна СССР, А. В. Гуцевич и др.
162 III. ПОДСЕМ. CULICINAE
от срединного выроста (по одной с каждой стороны) и рядом таких же длин-
ных и крепких щетинок (числом около восьми), расположенных за се-
рединой стернита; коксит с очень длинными и крепкими щетинками,
из которых 2—3 расположены на внутренней стороне коксита; базаль-
ная бородавка незначительных размеров, но явственно развита, у вер-
шины с 2 крепкими щетинками; стиль длинный и тонкий.
Личинка IV стадии похожа на личинку С. fumipennis, от ко-
торой отличается рядом признаков, сводящихся главным образом к более
слабому развитию волосков заднего (а, возможно, и переднего) конца
тела. Число чешуек в щетке значительно больше 100. Средний из волосков
позади щетки заметно короче последнего членика брюшка. Дополнитель-
ные широко расставленные дистальные зубцы гребня на сифоне развиты
слабее; пучок у основания сифона развит более сильно. На спинной
стороне заднего края VIII членика брюшка (рудимент IX членика) име-
ется узкая поперечная хитиновая дуга. Число пучков впереди плав-
ника 7—8, причем они лишь немного короче передних пучков, окружен-
ных общим основанием. Наружные хвостовые волоски только из трех
ветвей, из которых одна длиннее остальных. Жабры более чем вдвое
короче последнего членика.
Распространение. Юг Европейской части СССР (Южный
берег Крыма), Черноморское побережье Кавказа. Малая Азия (Анато-
лия).
Биология. По данным Величкевич (1936), личинки встречаются
в неглубоких, поросших водной растительностью, илистых, богатых
детритом, стоячих или слабо проточных водоемах, вода которых богата
содержанием кальция (101.2 мг на 1 л). Летом эти водоемы сильно затенены
растущими по их берегам деревьями и кустарниками, а осенью и зимой,
после опадения листвы, хорошо прогреваются солнцем. В году одно
поколение. Зимуют личинки III—IV стадии; окукление в конце апреля.
Лёт с начала мая. В течение летних месяцев водоемы почти нацело пере-
сыхают. Яйца, откладываемые или в воду, или на влажную землю, про-
водят этот период в состоянии диапаузы. Осенью, при новом заполнении
водоемов водой от выпавших дождей, из яиц выходят личинки, которые
к декабрю достигают II—IV стадии, в которой и зимуют. Личинки встре-
чаются совместно с личинками Culiseta morsitans, С. annulata, Culex pipiens,
C. territans, C. hortensis, Anopheles maculipennis. Самки нападают на чело-
века и животных в течение всего дня и вечером, особенно в затененных
местах и в кустарниках. Комары не отлетают далеко от мест выплода,
прячась в зарослях камыша, ежеголовки, в прибрежных кустах, а также
в дуплах деревьев; в помещения не залетают.
6. Род MANSONIA Blanchard
Глаза соприкасаются или почти соприкасаются. Хоботок умеренной
длины и более или менее равномерной толщины. Щупики самца равны
по длине хоботку или длиннее его; щупики самки короткие, не
длиннее Ч4 длины хоботка. Среднеспинка с хорошо развитыми акрости-
хальными и дорсоцентральными щетинками. Дыхальцевые щетинки от-
сутствуют, задыхальцевые имеются или отсутствуют; верхние стерно-
плевральные щетинки хорошо развиты; верхние мезэпимерные щетинки,
как правило, имеются. Ноги: I членик задних лапок короче задней го-
лени; передние лапки самца с одним более развитым коготком, воору-
женным нормально двумя зубчиками, и одним слабее развитым коготком,
как правило, лишенным зубчиков; коготки самки простые; пульвиллы
6. MANSONIA
163
отсутствуют. Крылья по жилкам с относительно широкими чешуйками.
Вилки крыла относительно длинные. Закрыловые пластинки с чешуйками.
Коксит с хорошо развитой базальной бородавкой. VIII сегмент самки
короткий и широкий; церки короткие, не выступают.
Личинки отличаются очень широкой головой, длинными усиками,
со жгутообразно вытянутой вершинной частью, с далеко отстоящими от
конца предвершинными щетинками и с сильно развитым пучком.
Ротовые части резко выраженного фильтрационного типа питания.
Грудь с сильно развитыми боковыми пучками, на средне- и заднегруди
имеющими хитиновые основания. В заднегруди боковые ответвления
главных трахейных стволов пузыревидно расширены. Короткий сифон
с клапанами, видоизмененными в буравящий аппарат, служащий для
добывания воздуха из воздухоносных полостей водных растений. Послед-
ний членик удлинен, с йольцевидно охватывающим его седлом и сильно
развитыми плавником и хвостовыми волосками.
Всего в роде Mansonia известно свыше 60 видов, из которых боль-
шинство свойственно тропикам как восточного, так и западного полу-
шарий; в Палеарктике род представлен лишь тремя видами — одним
(М. richiardii Fic.), широко распространенным в СССР, одним редким
средиземноморским, найденным и на юго-западе СССР (М. buxtoni Edw.),
и одним тропическим (М. uni/ormis Theob.), заходящим в Палеарктику
на южные острова Японии (Кюсю, Хонсю). Род подразделяется на 4 под-
рода. Оба вида, представленные в фауне СССР, принадлежат к подроду
Coquillettidia.
Подрод COQUILLETTIDIA Dyar
Задыхальцевые щетинки отсутствуют. VII сегмент брюшка самца
большой, VIII сегмент незначительной величины, но все же явственно
развитый, без хитиновых крючков. Гипопигий: базальная бородавка
коксита с палочковидным или иным придатком, в длину равным боро-
давке или превосходящим ее; стиль с коротким вершинным придатком.
Подрод включает приблизительно 35 видов, из которых свыше по-
ловины свойственно тропической Африке, 5 — Ориентальной области,
5 — Австралии с Новой Зеландией, 1 — Америке и 2 — Палеарктике.
?$
1 (2). Лапки со светлыми колечками. Хоботок, как правило, со светлым
кольцом посредине. Чешуйки крыла относительно широкие . . .
• • . .............................. 1. М. (С.) richiardii Fic.
2 (1). Лапки без светлых колечек. Хоботок без светлого кольца. Чешуйки
крыла относительно узкие...................2. М. (С.) buxtoni Edw.
83
1 (2). Стиль при основании широкий, посредине с перетяжкой . . . .
• • • .............................. 1. М. (С.) richiardii Fic.
2 (1). Стиль при основании более узкий, без перетяжки............
.......................................2. М. (С.) buxtoni Edw.
ЛИЧИНКИ
1 (2). Из волосков позади щетки на VIII членике брюшка крайний верх-
ний — из 2—4 ветвей. Поверхность сеДла, особенно на спинной
стороне, покрыта одиночными хетоидными шипиками . .... ^
....................................... 1. М. (С.) richiardii Fic.
И*
IM
III. ПОДСЕМ. CULICINAE
2 (1). Из волосков позади щетки на VIII членике брюшка крайний верх-
ний — из 5—7 ветвей. Поверхность седла покрыта более короткими
хетоидными щетинками, сидящими группами по 2—8 в одном ряду
......................................... 2. М. (С.) buxtoni Edw.
Рис. 92. Mansonia richiardii Fic.
1. Mansonia richiardii Ficalbi, 1889.
Вид характеризуется относительно широкими чешуйками крыльев
и наличием у преобладающего большинства экземпляров белого кольца
на хоботке и посредине I членика задних лапок (рис. 92). ...
6. MANSONIA
165
Голова в желтовато-белых прилегающих и бурых торчащих чешуйках.
Хоботок в бурых чешуйках, в средней трети с кольцом беловатых чешуек,
более широким у самки, более узким у самца. Щупики самки в переме-
шанных бурых и желтовато-белых чешуйках; у самца последние группи-
руются обыкновенно в основной части члеников, образуя подобие свет-
лых колец. Среднеспинка в бурых и золотистых перемешанных чешуй-
ках; щиток покрыт золотистыми чешуйками; бочки груди с пятнами
Рис. 93. Гипопигий Mansonia richiardii Fic. По Сое, Freeman a. Mattingly.
Отдельно изображен стиль М. richiardii (А) и М. buxtoni Edw. (В). По Aitken.
широких желтовато-белых чешуек. Ноги: бедра и голени спереди пестрые,
в бурых и желтых перемешанных чешуйках; лапки бурые, как правило,
с широкими (особенно на задних лапках) белыми кольцами при основании
члеников; I членик задних лапок с белым кольцом посредине; реже кольца
слабо выражены или отсутствуют вовсе, и ноги в большей своей части
одеты светлыми чешуйками. Такие светлые варианты (var. nikolskyi
Shing.) распространены в Средней Азии и Казахстане. Коготки самца
с зубчиками (2.0; 2.0; 0.0), у самки простые. Крылья слегка дымчатые;
жилки крыла покрыты относительно широкими белыми и бурыми пере-
мешанными чешуйками. Брюшко в бурых чешуйках, с треугольными
пятнами желтовато-белых чешуек по бокам тергитов.
Гипопигий (рис. 93): коксит толстый, короткий, с бородавкой, несу-
щей крепкий склеротизованный шип, оканчивающийся тупо. Стиль
с неправильными расширениями в основной и вершинной трети. X стер-
166
III. ПОДСЕМ. CULICINAE
нит на вершине с несколькими зубцами. Фаллосома сильно склеротизован-
ная, по краям зазубренная. Выросты IX тергита с 8—10 тонкими щетин-
Рис.-94. Mansonia richiardii Fic.
Личинка IV стадии. Голова,
сверху.
ками. VIII стернит с группой из 10—15
тесно сближенных крепких прямых ще-
тинок.
Личинка IV стадии (рис. 94
и 95) желтоватого или желто-коричневого
цвета. Голова широкая, не сужена к ос-
нованиям усиков. Из лобных волосков
наружные (из 9 ветвей) и средние (из 4—
5 ветвей) длинные и вторичноперистые,
внутренние — короткие, сходны с лежа-
щими впереди от них задними волосками
наличника; обе пары в среднем из 5—6
ветвей. Усики в 1.2—1.8 раза длиннее
головы за счет сильного развития конце-
вой части. Соответственно предвершин-
ные щетинки удалены от конца; пучок
из 15—20 вторичноперистых ветвей;
Грудь широкая, с хорошо развитыми
трахейными пузырями в заднем отделе.
Волоски переднего края груди длинные,
так же как и боковые. Концы срединных
волосков выдаются за передний край
головы.
Брюшко сужается к заднему концу. Щетка из неправильного ряда
(более 25) чешуек, с хорошо развитым главным шипом. Волоски позади
Рис. 95. Mansonia richiardii Fic. Личинка IV стадии. Сифон,
сбоку. По Везенберг-Лунду.
щетки разделены на две группы: верхнюю из двух волосков, отходящих
около верхних чешуек щетки, и нижнюю из трех волосков, расположен-
ных около ее середины; крайний верхний волосок из 2—4, средний из
2—3 длинных упругих ветвей.
6. MANSONIA
167
Сифон очень короткий, в форме усеченного конуса. Ушки у основания,
гребень и сифональные пучки отсутствуют. Клапаны и рычаг стигмаль-
ной пластинки видоизменены в буравящий аппарат; ее волоски — в длин-
ные шипы.
Последний членик сильно удлинен. Седло охватывает его кольцом,
расширяющимся кзади. Его поверхность, особенно со спинной стороны
у заднего конца, покрыта короткими одиночными хетоиДными щетин-
ками. Хвостовые волоски в виде полусложенных вееров; наружные длин-
нее внутренних. Боковые волоски 2—3-ветвистые. Плавник смещен
назад, из 10—14 длинных пучков. Жабры ланцетовидные, с заострен-
ными концами, не длиннее наибольшей ширины последнего членика.
Распространение. Вид широко распространен в западной
части Палеарктики. В пределах СССР известен из Ленинградской об-
ласти (на север до Приозерска), Эстонии, Латвии, Московской области,
Урала, Украины, Северного Кавказа, Закавказья, Средней Азии (Турк-
мения, Узбекистан, Таджикистан, Киргизия), Казахстана и южной зоны
Западной Сибири. Широко распространен в Западной Европе от Южной
Швеции, Финляндии, Дании и Англии до Италии, Венгрии, Болгарии
и Румынии; Передняя Азия (Палестина).
Биология. Личинка обитает в различных по величине водоемах
постоянного типа, богатых высшей водной растительностью (Acorns,
Carex, Glyceria, Ranunculus, Sparganium, Typha и др.), из воздухонос-
ных полостей которых личинки получают воздух для дыхания. Личинки
ведут придонный образ жизни, малоподвижны; удельный вес их больше
воды; при плавании, в связи с наличием трахейных пузырей, держатся
горизонтально. На севере ареала одно поколение в году, на юге — 2—3
поколения. Вышедшие осенью из яиц личинки зимуют обычно в III—IV
стадии; окукление в мае—июне. Куколки при помощи видоизмененных
дыхательных рожек способны, подобно личинкам, вбуравливаться в стебли
и корни растений в придонном слое водоемов. Перед выходом комара
концы рожек обламываются и куколка всплывает на поверхность. На юге
(Средняя Азия) в массовых количествах встречается в постоянных во-
доемах поймы среднего и нижнего течения крупных рек, как пресных,
так и слабо засолоненных, часто совместно с личинками Anopheles pulcher-
rimus. В южных частях ареала, в частности в Средней Азии, комар яв-
ляется весьма активным кровососом, причем в некоторых районах (поймы
крупных рек) явно преобладает над другими видами кровососущих ко-
маров; нападения наблюдаются по преимуществу в условиях дикой при-
роды; в помещения комар залетает редко.
2. Mansonia buxtoni Edwards, 1923.
Отличается от М. richiardii отсутствием колец на лапках и одно-
цветно темной окраской хоботка и щупиков.
Голова в желтовато-белых прилегающих и бурых торчащих чешуйках.
Хоботок и щупики покрыты бурыми чешуйками, местами частично тор-
чащими. Хоботок по длине примерно равен передним бедрам. Средне-
спинка в золотисто-коричневых чешуйках, более светлых перед щитком
и у основания крыльев. Ноги: бедра и голени спереди в темных чешуйках,
с небольшой примесью тусклых светлых чешуек, образующих неясные
продольные полоски; вершины бедер и голеней в беловатых чешуйках;
лапки бурые, одноцветные; задняя голень явственно длиннее I членика
лапки; коготки самки простые. Крылья в одноцветных темных чешуйках,
более узких, чем у М. richiardii (но более широких, чем у Culex). Брюшко:
тергиты в бурых чешуйках с отчетливыми беловато-кремовыми пятнами
168
III. ПОДСЕМ. CULICINAE
по бокам; на некоторых сегментах пятна соединены узкими перевязями
у переднего края тергитов; стерниты со светлыми перевязями при осно-
вании. Гипопигий очень похож на гипопигий М. richiardii и отличается
только формой стиля (рис. 93): при основании он относительно широкий,
посредине без перетяжки, вершинная половина сильно вздута.
Рис. 96. Mansonia buxtoni Edw. Личинка IV стадии.
1 — голова, сверху, 2 — задний конец, сбоку, з — группы хетоидных
щетинок на последнем членике.
Личинка IV стадии (рис. 96) очень похожа на личинку
М. richiardii. Она бледно-зеленого, цвета, величиной до 5—7 мм, с го-
ловой в 1.4 раза большей в ширину, чем в длину. Лобные волоски: наруж-
ные умеренной длины, из девяти, средние почти вдвое длиннее, из 5—7
вторичноперистых ветвей; задние — короткие, из восьми простых вет-
вей, сходны с лежащими впереди задними волосками наличника. Усики
как у М. richiardii.
7. AEDES
169
Щетка на VIII членике брюшка из 16—22 расположенных в непра-
вильный ряд (частично двойной у спинной стороны) чешуек с остроко-
нечным главным шипом. Из волосков позади щетки крайний верхний
из 5—7 (в среднем 6) ветвей, средний и промежуточные — из двух ветвей,
у последних они вдвое короче.
Последний членик как у М. richiardii, но поверхность его покрыта
рядами коротких хетоидных щетинок, собранных рядами в группы по
2—8 штук. Боковой волосок из четырех ветвей.
Распространение. Израиль, Сирия, Сардиния, Корсика,
Марокко. В СССР найден только в Черновицкой области (Гуцевич,
Донец, Ежова и Попов, 1962).
Биология почти не изучена. Известно, что комары этого вида
активно нападают на людей под открытым небом. Личинки встречены
на корнях ближе не определенных видов родов Acorus и Typha.
7. Род AEDES Meigen, 1818
Дыхальцевые щетинки отсутствуют, задыхальцевые имеются. Коготки
лапок самки, как правило, с зубчиками на всех ногах; у некоторых ви-
дов зубчики имеются лишь на двух или на одной паре ног. Пульвиллы
(присоски) под коготками на лапках отсутствуют. На задних ногах I
членик лапки короче голени. Задний конец брюшка самки более или
менее заострен, церки в той или иной степени выступают.
Личинки с ротовыми частями, приспособленными для питания планк-
тоном или перифитоном. Исключение — хищные личинки тропического
подрода Mucidus. Усики палочковидные, обычно короткие, с пучком
у середины или перед нею. Срединные и боковые волоски переднегруди
умеренно развиты. Трахейные пузыри и главные трахейные стволы с ок-
руглым сечением отсутствуют. Спинные хитиновые пластинки, за исклю-
чением седла последнего членика, отсутствуют. Сифон относительно
короткий (индекс не более 4), всегда имеет гребень и пару пучков на задне-
боковой поверхности, редко с дополнительными парными волосками на
передней стороне и боках трубки. Стигмальная пластинка, как правило,
с двуветвистым задним отростком рычага.
Богатый видами род; описано около 700 видов. Из них в фауне Со-
ветского Союза представлено 42, что составляет половину общего числа
найденных в СССР видов комаров. В тундре и в зоне леса по числу видов
и по количеству особей Aedes превосходят остальные роды комаров,
вместе взятые.
Род подразделяется на много (более 20). подродов, из которых Пале-
арктике свойственно пять: Ochlerotatus, Aedimorphus, Finlay a, Stegomyia,
Aedes. Подроды хорошо отличаются по строению гипопигия самцов,
менее четко — по самкам.
Личинки группируются иначе, чем соответствующие им взрослые.
Личинки подродов Aedes, Ochlerotatus и Aedimorphus очень близки друг
другу. С другой стороны, личинки некоторых видов рода Ochlerotatus
сходны с личинками Stegomyia, а многие из последних — с личинками
Finlaya. Поэтому по личинкам дается общая определительная таблица
видов, а в характеристиках отдельных подродов личинки, как правило,
не упоминаются.
Комары рода Aedes причиняют огромный вред как массовые крово-
сосы и как переносчики многих возбудителей болезней человека и жи-
вотных.
170
III. ПОДСЕМ. CULICINAE
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
$?
(50). Лапки со светлыми кольцами, иногда очень узкими, более раз-
витыми на задних лапках (светлые кольца лучше видны на темном
фоне).
(13). Каждое светлое кольцо лапок опоясывает 2 членика: вершину
предыдущего и основание последующего (рис. 97, А).
(6). Жилки крыла покрыты светлыми и темными чешуйками.
(5). Брюшко сверху с продольной светлой полосой, иногда почти
сплошь в светлых чешуйках.......................................
. ........................... 1. A. (Ochlerotatus) caspius Pall.
а (б). Среднеспинка в золотистых или ржаво-желтых чешуйках с двумя
продольными светлыми полосками. Жилка крыла в перемешанных
светлых и темных чешуйках..............A. caspius caspius Pall,
б (а). Среднеспинка с коричневой продольной полосой; боковые отделы
среднеспинки в серебристо-серых или кремовых чешуйках. Светлые
чешуйки крыла сгруппированы на определенных жилках ....
.......................................A. caspius dorsalis Mg.
(4). Брюшко без продольной светлой полосы. (С большей достовер-
ностью вид определяется по личинкам и по самцам)................
..............................2. A. (Ochlerotatus) mariae Serg.
(3). Жилки крыла в одноцветных темных чешуйках.
(8). Светлые кольца лапок очень узкие, развиты только на задних,
иногда и на средних ногах. Щупики самки и самца в одноцветных
темных чешуйках.................38. A. (Finlaya) alektorovi Stack.
(7). Светлые кольца более широкие, развиты на всех лапках. Щупики
темные с белой вершиной у самки, с белыми кольцами у самца.
(10). Последний членик лапки белый. (Средиземноморская подобласть)
......................... 3. A. (Ochlerotatus) pulchritarsis Rond,
а (б). Среднеспинка без продольной светлой полосы...............
..............................A. pulchritarsis pulchritarsis Rond.
б (а). Среднеспинка с узкой продольной полоской белых чешуек
.................................A. pulchritarsis asiaticus Edw.
(9). Последний членик лапки целиком темный или тёмный со светлым
кольцом при основании (Дальний Восток).
(12). Среднеспинка с большим белым пятном........................
..............................37. A. (Finlaya) seoulensis Yam.
(11). Среднеспинка без белого пятна . . 39. A. (Finlaya) togoi Theob.
(2). Светлые кольца опоясывают лишь основание члеников лапки
(рис. 97, Б).
(15). Белые кольца на лапках очень широкие, затемнены только вер-:
шины члеников. На задних голенях, помимо прилегающих чешуек,
имеются темные торчащие чешуйки.................................
..................... 32. A. (Ochlerotatus) kasachstanicus Guts.
(14). Светлые кольца на лапках не столь широкие, не шире 2/3 длины
соответствующего членика лапки, обычно уже. Задние голени без
торчащих чешуек.
(37). Хоботок явственно длиннее передних бедер. Чешуйки щитка
желтоватые или беловатые, узкие, изогнутые.
(20). Церки короткие, едва выступающие. Светлые пятна по бокам
брюшка образованы серебристо-белыми чешуйками.
(19). На задних лапках белые кольца имеются лишь на первых трех
члениках........................40. A. (Finlaya) japonicus Theob.
7. AEDES
171
19 (18). Белые кольца развиты на четырех члениках задней лапки. . .
..................................... 41. A. (Finlaya) koreicus Edw.
20 (17). Церки более длинные, явственно выступающие. Светлые чешуйки
брюшка не имеют блестящего серебристого оттенка.
21 (22). Белые кольца на лапках очень узкие, как правило, не превы-
шающие V4 длины соответствующего членика (рис. 97, В) ... .
......................................33. A. (Aedimorphus) vexans Mg.
Рис. 98. Пятна чешуек на бочках груди
Aedes.
пап —’проэпимерное, г — гипостигмальное,
пар — парастигмальное, мет — метастигмальное,
пк — посткоксальное, мае — мезэпистернальное
(стерноплевральное), мэп — мезэпимерное.
Рис. 97. Лапки Aedes.
А — A. caspius Pall., Б — A. cantans Mg.,
В — A. vexans Mg.
а (б). Вершинная половина каждого тергита брюшка в одноцветных
темных чешуйках............................A. vexans vexans Mg.
б (а). Тергиты в вершинной части сегментов со светлыми пятнами,
иногда образующими прерывистую продольную полоску . . . .
. . . . .............................. A. vexans nipponii Theob.
22 (21). Светлые кольца на лапках более широкие, занимающие на сред-
них члениках задней лапки не менее х/3 длины членика (рис. 97, Б).
Последующие виды группы cantans далеко не всегда могут быть
с достоверностью определены по самкам.1
23 (26). Брюшко сверху в одноцветных светлых чешуйках; иногда среди
них вкраплены единичные темные.
1 При определении видов группы cantans и особенно группы communis большую
роль играют такие признаки, как расположение пятен чешуек на боковых склеритах
груди (рис. 98—101), а также форма коготков лапок (рис. 102).
172
III. ПОДСЕМ. CULICINAE
24 (25). Общая окраска тела самки охряно-желтая. Среднеспинка в золо-
тисто-желтых чешуйках. Бочки груди в кремовых чешуйках, не резко
отличающихся по окраске от чешуек среднеспинки. Нижние мез-
эпимерные щетинки имеются . . . 11. A. (Ochlerotatus) cyprius Ludl.
Рис. 99. Расположение пятен чешуек на боковых склеритах
груди Aedes. По О. Н. Сазоновой.
А — A. cataphylla Dyar, В — A. pullatus Coq., В — A. intrudens Dyar,
Г — A. punctor Kirby, Д — A. nigrinus Eek., Е — A. dian-
taeus Н. D. К.
25 (24). Общая окраска тела самки желтовато-серая. Среднеспинка
в мелких ржаво-коричневых чешуйках.. Бочки груди покрыты серо-
вато-белыми чешуйками, резко отличающимися по окраске от тем-
ных чешуек среднеспинки. Нижние мезэпимерные щетинки от-
сутствуют .................... . 10. A. (Ochlerotatus) flavescens Mtill.
7. AEDES
173
26 (23). Темные чешуйки на тергитах брюшка имеются в более или менее
значительном количестве (не единичные); иногда темные чешуйки
преобладают.
Рис. 100. Расположение пятен чешуек на боковых склеритах
груди Aedes. По О. Н. Сазоновой.
А — A. behningi. Mart., Б — A. beklemishevi Den., В — A. excrucians
Walk., Г — A. riparius D. К., Д — A. cyprius Ludl., Е — A. flavescens
мои.
27 (28). Брюшко сверху преимущественно в темных чешуйках, без светлых
поперечных перевязей; светлые чешуйки (у самки) в большинстве
случаев сгруппированы в расплывчатые пятна вдоль средней линии
тела. Среднеспинка, как правило, в одноцветных мелких бронзовых
или цвета ржавчины чешуйках................................... . . .
............................. 6. A. (Ochlerotatus) behningi Mart.
174
III. ПОДСЕМ. CULICINAE
28 (27). Брюшко сверху с более или менее выраженными светлыми по-
перечными перевязями; если их нет, то светлые чешуйки разбросаны
на тергитах, не образуя пятен. Среднеспинка иной окраски: в золо-
Рис. 101. Расположение пятен чешуек на боковых склеритах
груди Aedes. По О. Н. Сазоновой.
А — A. impiger Walk., Б — A. caspius dorsalis Mg., В — A. vexans Mg.,
Г—-A. dnereus Mg.t Д — A. geniculatus 01.
тистых чешуйках, в светлых чешуйках с темной продольной полосой
или в темных коричневых чешуйках с неясными светлыми пятнами.
29 (32). Общая окраска относительно темная. Среднеспинка преимуще-
ственно в темных коричневых чешуйках или в более светлых с тем-
ной продольной полосой. Бочки груди в белых чешуйках. Брюшко
7. AEDES
175
сверху с поперечными светлыми перевязями, обычно яснее выражен-
ными на передних сегментах.
30 (31). Среднеспинка преимущественно в шоколадно-коричневых чешуй-
ках с небольшими неясными светлыми пятнами; иногда намечается
продольная темная полоса. На средних члениках задней лапки
белые кольца, как правило, занимают менее половины длины чле-
ника. Коготок лапки круто изогнут..................................
..............................4. A. (Ochlerotatus) cantans Mg.
Рис. 102. Коготки лапок комаров рода Aedes. По О. Н. Сазоновой.
1 — A. cinereus Mg., 2 — A. impiger Walk., 3 — A. nigripes Zett., 4 — A. ca-
taphylla Dyar, 5 — A. leucomelas Mg., 6 — A. pullatus Goq., 7 — A. intrudens
Dyar, 8 — A. punctor Kirby, 9 — A. hexodontus Dyar, 10 — A. pionips Dyar,
11 — A. communis Deg., 12 — A. nigrinus Eek., 13 — A. diantaeus H. D. K.,
14 — A. sticticus Mg., 15 — A. caspius dorsalis Mg., 16 — A. vexans Mg., 17 —
A. behningi Mart., 18 — A. cantans Mg., 19 — A. riparius D. K., 20 — A. be-
klemishevi Den., 21 — A. excrucians Walk., 22—A. cyprius Ludl., 23 —
A. flavescens Mtill., 24— A. geniculatus 01.
31 (30). Среднеспинка с более или менее выраженной темной продольной
полосой; боковые отделы среднеспинки в золотистых чешуйках.
На средних члениках задней лапки белые кольца занимают не ме-
нее половины длины членика. Коготок лапки полого изогнут . . .
...................................5. A. (Ochlerotatus) riparius D. К.
а (б). На крыльях, хоботке и щупиках среди темных вкраплены светлые
чешуйки.................................A. riparius riparius D. К.
б (а). Крылья, хоботок и щупики в одноцветных темных чешуйках
......................................... A. riparius ater Guts.
32 (29). Общая окраска относительно более светлая. Среднеспинка, как
правило, преимущественно в золотистых или кремовых чешуйках,
иногда с темной продольной полосой. Бочки груди обычно в желто-
ватых чешуйках. Брюшко сверху чаще без выраженных светлых
перевязей; если они имеются, то образованы чешуйками желтоватого
оттенка.
176
Ш. ПОДСЕМ. CULICINAE
33 (36). Среднеспинка преимущественно в золотистых чешуйках, без ясно
выраженной продольной темной полосы.
34 (35). Коготок лапки круто изогнутый, крупный; зубчик направлен
почти параллельно вершинной части коготка............................
.......................... 7. A. (Ochlerotatus) excrucians Walk.
35 (34). Коготок лапки менее изогнутый, с большим, резко отставленным
зубчиком. (Вид с достоверностью определяется только по личинкам)
................................8. A. (Ochlerotatus) beklemishevi Den.
36 (33). Среднеспинка с четко отграниченной продольной темной полосой,
боковые части среднеспинки в серовато-желтых чешуйках ....
...........................9. A. (Ochlerotatus) annulipes Mg.
37 (16). Хоботок не длиннее передних бедер. Чешуйки щитка широкие,
прямые, серебристо-белые.
38 (41). Среднеспинка со снежно-белыми пятнами.
39 (40). Среднеспинка в средней части с четырьмя круглыми белыми
пятнами...............................44. A. (Stegomyia) vittatus Big.
40 (39). Среднеспинка в передней половине с двумя треугольными или
полулунными белыми пятнами по бокам..................................
..........................48. A. (Stegomyia) chemulpoensis Yam.
41 (38). Среднеспинка с одной или несколькими белыми продольными
полосками.
42 (43). Среднеспинка с четырьмя узкими продольными полосками;
из них более выражены латеральные, изогнутые кнаружи в передней
половине среднеспинки....................42. A. (Stegomyia) aegypti L.
43 (42). Среднеспинка с серебристо-белой срединной продольной полоской.
44 (45). Последний членик задней лапки на вершине затемненный или
целиком темный...................... . 45. A. (Stegomyia) galloisi Yam.
45 (44). Последний членик задней лапки белый.
46 (47). Коготки передних и средних лапок самки с зубчиком (Средизем-
номорская подобласть) .... 43. A. (Stegomyia) cretinus Edw.
47 (46). Коготки передних и средних лапок самки без зубчика (Даль-
ний Восток).
48 (49). Небольшое светлое пятно у края среднеспинки над основанием .
крыла образовано широкими прямыми серебристо-белыми чешуйками
.....................................46. A. (Stegomyia) albopictus Sk.
49^ (48). Неотчетливое светлое пятно над основанием крыла образовано
Р узкими изогнутыми беловатыми или желтоватыми чешуйками . . .
...............•...........47. A. (Stegomyia) flavopictus Yam.
50 (1). Лапки без светлых колец.
51 (96). Хоботок явственно длиннее передних бедер.
52 (57). Церки самки короткие, едва выступающие. Светлые пятна
на брюшке блестящие, серебристые.
53 (54). Среднеспинка с двумя большими серебристо-белыми пятнами,
иногда сливающимися в одно пятно . ............................. .
..................36. A. (Finlaya) nipponicus La Casse et Yamag.
54 (53). Среднеспинка с широкой продольной темной полосой, без сереб-
ристо-белых пятен.
55 (56). Щиток в узких желтоватых чешуйках ........................
...............................34. A. (Finlaya) geniculatus 01.
56 (55). Щиток в широких белых чешуйках............................
...................................35. A. (Finlaya) echinus Edw.
57 (52). Церки относительно длинные, явственно выступающие. Светлые
пятна на брюшке не имеют блестяще-серебристого оттенка.
7. AEDES 177
58 (69). Б вершинной половине тергитов имеются или преобладают свет-
лые чешуйки.
59 (62). Чешуйки верхней части проэпимер широкие, прямые, черные.
Среднеспинка с широкой темной продольной полосой или с двумя
сближенными темными полосками.
60 (61). Светлые перевязи на тергитах брюшка не резко отграничены и
не образуют расширения посредине; иногда брюшко сверху в пере-
мешанных светлых и темных чешуйках, первые обычно преобладают
.....................................13. A. (Ochlerotatus) refiki Med.
61 (60). Светлые перевязи на тергитах брюшка чаще довольно четко отгра-
ничены и, как правило, образуют выступы вдоль средней линии
тела.............................12. A. (Ochlerotatus) rusticus Rossi.
62 (59). Чешуйки верхней части проэпимер чаще узкие, изогнутые, а если
прямые, то не черные (желтоватые или светло-бурые). Среднеспинка,
как правило, без темной продольной полосы.
63 (66). Светлые чешуйки на тергитах брюшка не образуют поперечных
перевязей; брюшко сверху либо сплошь покрыто светлыми чешуй-
ками, либо среди светлых вкраплено большее или меньшее коли-
чество темных, иногда образующих неясные пятна.
64 (65). Постскутеллум (языковидный выступ позади щитка) с пучком
чешуек ......... 14. A. (Ochlerotatus) lepidonotus Edw.
65 (64). Постскутеллум без пучка чешуек................................
....................... 15. A. (Ochlerotatus) subdiversus Mart.
66 (63). Тергиты брюшка, как правило, с базальными перевязями свет-
лых чешуек (в задней половине тергитов также имеется большее
или меньшее количество светлых чешуек).
67 (68). Посткоксальное пятно чешуек отсутствует. Покровы груди ко-
ричневые. В вершинной половине каждого тергита светлые чешуйки
перемешаны с темными.................30. A. (Ochlerotatus) detritus Hal.
68 (67). Посткоксальное пятно чешуек имеется. Покровы груди черно-
бурые. Вершинная половина каждого тергита в темных чешуйках
с большей или меньшей (иногда незначительной) примесью светлых.
............................... 31. A. (Ochlerotatus) simanini Guts.
69 (58). Вершинные отделы тергитов в одноцветных темных чешуйках;
светлые чешуйки образуют перевязи или пятна при основании или
по бокам тергитов.
70 (73). Среднеспинка в длинных густых черных щетинках. Щетинки раз-
бросаны по всей задней половине проэпимер.
71 (72). Пятно светлых чешуек на стерноплеврах доходит до переднего
угла склерита (у основания передних ног). Коготок лапки слабо
изогнутый, удлиненный . . . 26. A. (Ochlerotatus) nigripes Zett.
72 (71). Пятно светлых чешуек на стерноплеврах не доходит до их перед-
него угла. Коготок лапки резко изогнутый..............................
........................... 27. A. (Ochlerotatus) impiger Walk.
73 (70). Щетинки среднеспинки менее длинные и густые, чаще коричневые
или золотистые. Щетинки на проэпимерах имеются лишь у их заднего
края.
74 (77). Передние бедра спереди пестрые: многочисленные светлые че-
шуйки перемешаны с темными. Светлые чешуйки вкраплены среди
темных в различных частях крыла, особенно на костальной и первой
радиальной жилках.
75 (76). Хоботок с более или менее значительной примесью светлых че-
шуек, особенно в его средней части. Светлые чешуйки в довольно
12 Фауна СССР. А. в. Гуцевич и др.
178
III. ПОДСЕМ. CULICINAE
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
значительном количестве разбросаны на различных жилках крыла
......................... 29. A. (Ochlerotatus) leucomelas Mg.
(75). Хоботок в одноцветных темных чешуйках. На крыле светлый
чешуйки вкраплены в небольшом количестве главным образом лишь
в передней части крыла . . 28. A. (Ochlerotatus) cataphylla Dyar.
(74). Передние бедра спереди, как правило, не пестрые, преимущест-
венно в темных чешуйках или с небольшой примесью светлых.
Светлые чешуйки на крыле имеются только при его основании или
их вовсе нет.
(81). Небольшое пятно светлых чешуек непосредственно книзу от пе-
реднего дыхальца (гипостигмальное пятно) имеется.
(80). Пятно белых чешуек на мезэпимерах доходит до их нижнего края.
Нижние мезэпимерные щетинки имеются................................
.............................25. A. (Ochlerotatus) pullatus Coq.
(79). Пятно белых чешуек на мезэпимерах явственно не доходит
до их нижнего края. Нижние мезэпимерные щетинки, как правило,,
отсутствуют....................24. A. (Ochlerotatus) intrudens Dyar.
(78). Гипостигмальное пятно отсутствует.
(83). Пятно белых чешуек на стерноплеврах не доходит до их переднего
угла.......................23. A. (Ochlerotatus) diantaeus Н. D. К.
(82). Пятно белых чешуек на стерноплеврах доходит до их переднего
угла (у основания передних ног).
(91). Пятно белых чешуек на мезэпимерах доходит до их нижнего-
края. Нижние мезэпимерные щетинки имеются.
(86). Посткоксальные чешуйки отсутствуют (т. е. нет чешуек на пере-
пончатом участке, расположенном между стерноплеврами и местом
сочленения переднегруди с коксами первой пары ног).................
.............................16. A. (Ochlerotatus) communis Deg.
(85). Посткоксальные чешуйки имеются.
(88). Основание костальной жилки в темных чешуйках. Светлые пере-
вязи II—V тергитов брюшка посредине резко сужены .....
..........................18. A. (Ochlerotatus) punctor Kirby.
(87). При основании костальной жилки, как правило, имеется большее
или меньшее количество светлых чешуек. Светлые перевязи II—
V тергитов брюшка равномерной ширины или незначительно сужены
посредине. (Последующие два вида по имаго различаются с .трудом,
с уверенностью они могут быть определены по личинкам).
(90). Среднеспинка в относительно светлых серовато-желтых чешуй-
ках, с двумя сближенными неясно отграниченными коричневыми про-
дольными полосками .... 17. A. (Ochlerotatus) pionips Dyar.
(89). Чешуйки среднеспинки темные, чаще более или менее одно-
цветные, ржаво-коричневые ... 19. A. (Ochlerotatus) hexodontus Dyar.
(84). Пятно белых чешуек на мезэпимерах явственно не доходит
до их нижнего края. Нижние мезэпимерные щетинки отсутствуют.
(95). Задние голени, как правило, снаружи покрыты преимущественно-
светлыми чешуйками.
(94). Жилки крыла в темных чешуйках. Светлые перевязи брюшка
посредине сильно сужены. Первый членик жгутика усиков при осно-
вании желтый...................20. A. (Ochlerotatus) sticticus Mg.
(93). При основании крыла имеются светлые чешуйки. Светлые пере-
вязи брюшка более или менее равномерной ширины. Первый членик
жгутика усиков целиком черный.......................................
. . . ................. .... .21. A. (Ochlerotatus) nigrinus Eck.
7. AEDES
179
95 (92). Задние голени снаружи в темных чешуйках ...................
............................22. A. (Ochlerotatus) hungaricus Mih.
96 (51). Хоботок не длиннее передних бедер или (у A. aureus) длина хо-
ботка лишь немного превышает длину передних бедер.
97 (98). Среднеспинка в золотистых чешуйках с продольной темной поло-
сой. Крылья со значительной примесью светлых чешуек. Яркие зо-
лотистые чешуйки образуют на затылке крупное пятно..................
.....................................51. A. (sgen. ?) aureus Guts.
98 (97). Среднеспинка без темной полосы. Крылья в темных чешуйках.
Затылок без пятна золотистых чешуек.
99 (100). Тергиты брюшка с белыми пятнами по бокам, без светлых пере-
вязей .................................50. A. (Aedes?) nobukonis Yam.
100 (99). Тергиты брюшка одноцветно темные или темные, со светлыми
перевязями................................49. A. (Aedes) cinereus Mg.
а (г). Брюшко сверху без светлых перевязей.
б (в). Чешуйки среднеспинки относительно темные, красновато-бурые,
иногда с золотистым или бронзовым оттенком. Голова преимуще-
ственно в темных чешуйках.................A. cinereus cinereus Mg.
в (б). Чешуйки среднеспинки более светлые, золотисто-бурые или
желтоватые. Голова в светлых беловато-серых чешуйках...............
............................... A. cinereus rossicus D. G. M.
г (а). Брюшко со светлыми перевязями при основании тергитов ....
................................... A. cinereus esoensis Yam.
<3<3 1
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ
1 (2). Стиль состоит из двух ветвей: медиальной и латеральной. Щупики
в несколько раз короче хоботка, как у самки........................
..................................Подрод Aedes Mg. (стр. 183).
2 (1). Стиль простой, т. е. не разделен на две ветви. Щупики по длине
приблизительно равны хоботку, иногда немного длиннее или немного
короче хоботка.
3 (6). Класпеты имеются.
4 (5). Коксит с более или менее развитыми базальной и вершинной
бородавками, или по меньшей мере с одной из них....................
.......................... Подрод Ochlerotatus Arr. (стр. 180).
5 (4). Коксит без бородавок (иногда имеется зачаточная базальная
бородавка в виде незначительного бугорка) ......................
.......................... Подрод Finlaya Theob. (стр. 182).
6 (3). Класпеты отсутствуют.
7 (8). Придаток стиля причленяется на некотором расстоянии от его
вершины. Стиль широкий цилиндрический, к вершине слегка расши-
ренный. Базальная бородавка обособлена в виде колбовидного обра-
зования, усаженного на конце волосками — единственный встре-
чающийся в СССР вид подрода — A. (Aedimorphus) vexans Mg.
8 (7). Придаток стиля причленяется у вершины или немного отступя
от вершины. Стиль иной формы. Бородавка расположена в средней
части коксита...................Подрод Stegomyia Theob. (стр. 183).
1 Самцы A. nobukonis и A. aureus неизвестны.
12*
180
HI. ПОДСЕМ. CULICINAE
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Подрод OCHLEROTATUS Arribalzaga
1 (8). Базальная бородавка коксита несет ряд длинных ланцетовидных
чешуек.
2 (3). Придаток стиля S-образно изогнут . . 12. А. (О.) rusticus Rossi.
3 (2). Придаток стиля прямой или почти прямой.
4 (5). Крыло Класпеты удлиненное, с поперечной исчерченностью . . .
.......................................... 13. А. (О.) refiki Med.
5 (4). Крыло класпеты более короткое, без исчерченности.
6 (7). В основной части коксита имеется лишь одна бородавка, несущая
группу ланцетовидных чешуек .... 14. А. (О.) lepidonotus Edw.
7 (6). Помимо бородавки, усаженной длинными ланцетовидными чешуй-
ками, имеется меньших размеров бородавка, несущая 1—3 длинных
и несколько более коротких щетинок ..............................
............................. 15. А. (О.) subdiversus Mart.
8 (1). На базальной бородавке коксита ланцетовидных чашуек нет.
9 (14). Коксит на внутренней стороне с 3 крупными шипами, из них два
обычно помещаются на базальной бородавке.
40 (И). Коксит с направленным внутрь пучком густых волосков . . .
....................................23. А. (О.) diantaeus Н. D. К.
11 (10). Коксит без направленного внутрь пучка волосков.
12 (13). Стволик класпеты близ середины с пальцевидным выростом.
Коксит у вершины с направленным дистально пучком волосков
.......................................24. А. (О.) intrudens Dyar.
13 (12). Стволик класпеты без выроста. Коксит без пучка волосков
.......................................... 25. А. (О.) pullatus Coq.
14 (9). Коксит с внутренней стороны с 1—2 шипами (на базальной бо-
родавке) или вовсе без шипов.
15 (16). Базальная бородавка коксита с двумя шипами.............
............................................ 1. А. (О.) caspius Pall.1
(Строение гипопигия у обоих подвидов весьма сходно, часто встре-
чаются промежуточные формы).
а (б.) Базальная бородавка слабо выпуклая, основания шипов сбли-
жены; передний шип на вершине крючковидно изогнут................
A. caspius caspius Pall.
б (а). Базальная бородавка сильно выпуклая, основания шипов удалены
друг от друга; передний шип слегка изогнут ......................
A. caspius dorsalis Mg.
16 (15). Базальная бородавка с одним шипом или без шипа.
17 (24). Вершинная бородавка коксита слабо развита, иногда вовсе не раз-
личима.
18 (19). Очень слабо развитая базальная бородавка усажена относительно
короткими и тонкими щетинками и волосками, без шипов или креп-
ких щетинок.................................2. А. (О.) mariae Serg.
19 (18). Более или менее развитая базальная бородавка несет довольно
длинные щетинки; обычно одна из них отличается большей толщиной.
20 (21). Вершинная бородавка отсутствует. Базальная бородавка слабо
выпуклая . . .........................3. А. (О.) pulchritarsis Rond.
21 (20). Небольшая вершинная бородавка имеется. Базальная бородавка
выпуклая, коническая.
1 Сходно строение гипопигия A. hungaricus Mill. (стр. 246); этот вид отличается
ют A. caspius отсутствием белых колечек на лапках, одноцветными темными чешуй-
ками крыльев и другими признаками.
7. AEDES
181
22 (23). Коксит в длинных волосках. Базальная бородавка без отчетливо
выраженного шипа. Фаллосома сильно склеротизована, раздвоена.
Выросты IX тергита с 14—20 щетинками.............................
......................................26. А. (О.) nigripes Zett.
23 (22). Коксит в относительно более коротких волосках. Базальная
бородавка с шипом, более или менее выделяющимся среди соседних
щетинок. Фаллосома слабо склеротизованная, двузубчатая. Выросты:
IX тергита с 4—12 щетинками................27. А. (О.) impigerWalk.
24 (17). Вершинная бородавка коксита хорошо развита.
25 (48). Базальная бородавка коксита с одним шипом или с крепкой ще-
тинкой, которая отчетливо выделяется среди более тонких щетинок
и волосков.
26 (29). Крыло класпеты узкое, без прозрачного пластинчатого расши-
рения.
27 (28). Стволик класпеты короткий; крыло класпеты обычно склероти-
зовано, посредине слегка расширено . . . 18. А. (О.) punctor Kirby.
19. А. (О.) hexodontus Dyar.
28 (27). Стволик класпеты длинный; крыло класпеты очень узкое, слабо
склеротизованное ......................... 16. А. (О.) communis Deg.
17. А. (О.) pionips Dyar.
29 (26). Крыло класпеты с прозрачным пластинчатым расширением,
иногда заметным лишь при определенном положении класпеты.
30 (33). Коксит в очень длинных, направленных медиально волосках;
вершины волосков левого и правого кокситов заходят друг за друга.
Стволик класпеты резко изогнутый.
31 (32). Лопасти IX тергита обычно с 6—10 короткими прямыми щетин-
ками ...................................28. А. (О.) cataphylla Dyar.
32 (31). Лопасти IX тергита обычно с 10—15 относительно длинными ще-
тинками, направленными несколько наружу.............................
..................................... 29. А. (О.) Ipucomelas Mg.
33 (30). Волоски на внутренней стороне кокситов, как правило, отно-
сительно более короткие, обычно не заходящие вершинами друг
за друга (исключение — A. detritus). Стволик класпеты прямой
или не резко изогнутый.
34 (37). Вершинная бородавка с широким основанием (в проксимальном
направлении оно доходит до середины коксита), покрыта короткими
волосками. Крыло класпеты короткое.
35 (36). Дистальная часть базальной бородавки суженная, тонкая . . .
............................. 20. А. (О.) stictions Mg.
36 (35). Дистальная часть базальной бородавки более широкая, закруг-
ленная ...................................21. А. (О.) nigrinus Eck.
37 (34). Вершинная бородавка, отграниченная дистальной третью кок-
сита, усажена относительно длинными волосками. Крыло класпеты,
как правило, относительно длинное.
38 (39). Базальная бородавка узкая, ее высота значительно превышает
ее ширину при основании. Крыло класпеты весьма широкое, ширина
не меньше длины..............................4. А. (О.) cantans Mg.
39 (38). Высота базальной бородавки не превосходит ее ширину при ос-
новании. Крыло класпеты более узкое, ширина меньше длины.
40 (41). Базальная бородавка уплощенная, в коротких густых волосках,
с толстым, сильно склеротизованным шипом........................ .
...................................10. А. (О.) flavescens Mull.
41 (40). Базальная бородавка коническая, с шипом или крепкой щетин-
кой умеренной толщины и длинными волосками.
182
42
43
44
a
б
45
46
47
48
49
50
51
52
53
54
1
2
3
4
5
III. ПОДСЕМ. CULICINAE
(45). Шип базальной бородавки хорошо развит, отчетливо выделяется
среди соседних щетинок и волосков. (Лапки со светлыми кольцами).
(44). Стволиккласпеты длинный, изогнутый ... 11. А. (О.) cyprius Ludl.
(43). Стволик класпеты относительно короткий, прямой, а если изо-
гнутый, то прозрачное расширение крыла класпеты начинается
отступя от основания крыла (имеется «рукоятка»)..................
....................................5. А. (О.) riparius D. К.
(б). Рукоятка крыла класпеты короткая или отсутствует, прозрачное
расширение выражено в основной и вершинной частях крыла . . .
.................................... A. riparius riparius D. К.
(а). Рукоятка крыла класпеты длинная, занимает около половины его
длины; прозрачное расширение выражено только в вершинной
половине крыла класпеты......................A. riparius ater Guts.
(42). Шип (крепкая щетинка) базальной бородавки не резко выделяется
среди соседних более мелких щетинок. (Лапки без светлых колец).
(47). Внутренняя поверхность коксита в густых, довольно длинных
волосках. Прозрачное расширение крыла класпеты развито в средней
и вершинной частях крыла. Фаллосома яйцевидная, без перетяжки
........................................30. А. (О.) detritus Hal.
(46). Внутренняя поверхность коксита в относительно редких и более
коротких волосках. Прозрачное расширение крыла класпеты раз-
вито на небольшом протяжении его средней трети. Фаллосома удли-
ненная, с легкой перетяжкой . . . . 31. А. (О.) simanini Guts.
(25). Базальная бородавка без шипа (крепкой щетинки).
(50). Базальная бородавка коническая, ее высота примерно равна
ширине при основании ....................6. А. (О.) behningi Mart.
(49). Базальная бородавка уплощенная, ширина значительно больше
высоты.
(52). Базальная бородавка слегка выпуклая, в длинных, очень густых
волосках. Крыло класпеты при основании с небольшим крючковид-
ным выступом на вогнутой стороне . . 32. А. (О.) kasachstanicus Guts.
(51). Базальная бородавка едва различимая, плоская, в коротких
волосках. Крыло класпеты при основании без крючковидного вы-
ступа.
(54). Стволик класпеты относительно тонкий, длинный, к вершине
явственно сужается ...................7. А. (О.) excrucians Walk.
8. А. (О.) beklemishevi Den.
(53). Стволик класпеты более толстый, к вершине не сужается . . .
........................................ 9. А. (О.) annulipes Mg.
Подрод FINLAYA Theobald
(2). Коксит с пучком весьма крупных чешуек в дистальной части
внутренней поверхности . . 36. A. (F.) nipponicus La Casse et Yamag.
(1). Коксит несет мелкие чешуйки главным образом с наружной
стороны.
(6). Крыло класпеты длиннее стволика.
(5). Крыло класпеты значительно длиннее стволика. Придаток стиля
относительно короткий, его длина составляет х/7—1/5 длины стиля
..........................................39. A. (F.) togoi Theob.
(4). Крыло класпеты немного длиннее стволика. Придаток стиля отно-
сительно более длинный, его длина составляет приблизительно
1/4 длины стиля.........................37. A. (F.) seoulensis Yam.
7. AEDES
183
6
7
8
9
40
11
42
13
44
1
2
3
4
-5
6
7
8
9
40
1
2
3
4
(3). Крыло класпеты не длиннее стволика.
(8). При основании внутренней поверхности коксита имеется неболь-
шой бугорок с двумя крепкими и одной более тонкой щетинкой
..................................... 38. A. (F.) alektorovi Stack.
(7). Бугорка с крепкими щетинками при основании внутренней поверх-
ности коксита нет.
(12). Лопасти IX тергита с 2—6 крепкими щетинками.
(11) . Коксит в очень длинных густых волосках ...................
....................................35. A. (F.) echinus Edw.
(10) . Волоски коксита менее длинные и густые ...................
.................................... 34. A. (F.) geniculatus 01.
(9). Лопасти IX тергита с 5—10 тонкими волосками.
(14). Лопасти IX тергита выпуклые, полушаровидные. Коготок X стер-
нита на вершине простой ....... 41. A. (F.) koreicus Edw.
(13). Лопасти IX тергита уплощенные. Коготок X стернита на вершине
раздвоен...............................40. A. (F.) japomcus Theob.
Подрод STEGOMYIA Theobald
(2). Стиль колбовидный, на вершине сильно расширенный . . . .‘
.........................................44. A. (S.) vittatus Big.
(1). Стиль цилиндрический или веретеновидный.
(6). Придаток расположен на вершине стиля. Задний край IX тергита
с вырезкой.
(5) . Бородавка коксита округлая, резко очерченная. Задний край
IX тергита с неглубокой дугообразной вырезкой ...................
................................48. A. (S.) chemulpoensis Yam.
(4). Бородавка коксита не резко очерчена. Задний край IX тергита
с глубокой вырезкой........................42. A. (S.) aegypti L.
(3). Придаток стиля расположен немного отступя от его вершины. Зад-
ний край IX тергита выпуклый или с выступом.
(8) . Бородавка коксита вытянута в поперечном направлении . . . .
................................. 45. A. (S.) galloisi Yam.
(7) . Бородавка коксита вытянута в продольном направлении.
(10) . Задний край IX тергита зазубренный, дугообразный . . . .
................(Дальний Восток) 47. A. (S.) flavopictus Yam.
(Средиземноморье) 43. A. (S.) cretinus Edw.
(9) . Задний край IX тергита гладкий, с выступом посредине . . . .
.....................................46. A. (S.) albopictus Sk.
Подрод AEDES Мeigen
Подвиды A. (Aedes) cinereus Meigen.
(2). Латеральная (более длинная) ветвь стиля на вершине двураздель-
ная .................................... A. cinereus cinereus Mg.
(1). Латеральная ветвь стиля простая или зазубренная, но не двураз-
дельная.
(4). Бородавка коксита двувершинная...............................
.................................A. cinereus rossicus D. G. M.
(3). Бородавка коксита простая, ступенчатая.......................
...................................... A. cinereus esoensis Yam,
184
III. ПОДСЕМ. CULICINAE
ЛИЧИНКИ IV СТАДИИ1
1 (2). Усик заметно длиннее головы...............................
...........................A. (Ochlerotatus) diantaeus Н. D. К.
2 (1). Усик меньше длины головы.
3 (10). Лобные волоски расположены в один дугообразный ряд.
4 (5). Лобные волоски не смещены к переднему краю головы, находятся
в средней части лобного щитка. Щетка на VIII членике брюшка
из небольшого числа (до 20) чешуек с остроконечным главным шипом.
Дистальные зубцы гребня более широко расставлены, заходят
за середину дыхательной трубки; пучок ближе к вершине из 3—5 ко-
ротких ветвей.............._.................Подрод Aedes Meig.
а. Типичные личинки: с признаками подрода....................
.............................A. (Aedes) cinereus cinereus Meig.
б. Типичные личинки: на передней поверхности дыхательной трубки,
кроме предвершинных щетинок, имеются две пары тонких коротких,
едва заметных волосков из 2—5 ветвей ........................
A. (Aedes) cinereus rossicus D. G. M.
в. Типичные личинки: на боковых поверхностях дыхательной трубки
у ее вершины имеются четыре пары дополнительных, очень коротких
и тонких волосков.........A. (Aedes) cinereus esoensis Yam.
5 (4). Лобные волоски смещены к переднему краю лобного щитка. Щетка
на VIII членике брюшка не менее чем из 40—50 чешуек, не имеющих
главного шипа.
6 (7). Дыхательная трубка короткая (индекс около 2), одной длины
с последним члеником; пучок около вершины трубки. Жабры ко-
роткие, тупоконические.................A. (Finlaya) togoi Theob.
7 (6). Дыхательная трубка более длинная (индекс около 3), заметно
длиннее последнего членика; пучок ближе к ее середине. Жабры
одной длины с последним члеником брюшка или длиннее, ланцето-
видной формы.
8 (9). Зубцы гребня расположены на приблизительно одинаковом рас-
стоянии друг от друга; из них не более двух дистальных заходят
за основание пучка.....................A. (Finlaya) koreicus Edw.
9 (8). Наиболее дистальные зубцы гребня в числе 1—4 более широко
расставлены в виде крупных шипов, лежащих под более острым
углом к продольной оси трубки ... A. (Finlaya) japonicus Theob.
10 (3). Лобные волоски с каждой стороны расположены треугольником:
средние впереди внутренних.
11 (12). Средний и внутренний лобные волоски смещены к переднему
краю головы; задние волоски наличника одной длины с ними, лежат
между основаниями средних; все они сильно ветвятся. Щетка из рас-
положенных в. один ряд 8—15 чешуек с острым главным шипом
........................ A. (Finlaya) nipponicus La Casse et Yamag.
12 (11). Средний и внутренний лобные волоски не смещены к переднему
краю головы;' задние волоски наличника значительно короче их;
если не короче, то лобные волоски слабо ветвятся.
13 (26). Ушки по бокам основания дыхательной трубки, ближе к ее зад-
нему краю, отсутствуют. Усики с небольшим простым волоском,
короткие, не покрыты шипиками или, как исключение (Л. vittatus)^
шипики очень слабо развиты..............Подрод Stegomyia Theob.
1 Личинки A. simanini и A. kasachstanicus описаны на стр. 378 и 379, личинки
A. aureus и A. nobukonis неизвестны.
7. AEDES
185
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
(19). Наружные лобные волоски простые.
(16). Волоски тела, включая боковой волосок последнего членика,
звездчатого типа ветвления. Жабры колбасовидной формы, в 5 раз
длиннее седла . .............A. (Stegomyia) chemulpoensis Yam.
(15). Волоски тела ветвятся в одной плоскости. Жабры не более чем
в 3 раза длиннее седла.
(18). Чешуйки щетки, кроме главного шипа, имеют 2—6 и больше
добавочных шипов по бокам основания, из которых 1—2 крупнее,
и иногда могут достигать размеров главного. Добавочные волоски
на боковых сторонах дыхательной трубки отсутствуют..............
....................................A. (Stegomyia) aegypti L.
(17). Чешуйки щетки не имеют крупных добавочных шипов по бокам
основания главного шипа, несут только один ряд тонких коротких
щетинок. На боковых сторонах дыхательной трубки, кроме пучка,
имеется по простому волоску одной длины с пучком................
................................A. (Stegomyia) cretinus Edw.
(14). Наружные лобные волоски не менее чем из 2—3 ветвей.
(21). Гребень занимает около 2/3 длины дыхательной трубки, кроме
него ближе к вершине лежит один крупный гладкий шип. Волосок
на трубке находится между ним и наиболее дистальным зубцом
гребня ......................... A. (Stegomyia) vittatus Big.
(20). Гребень занимает не более половины дыхательной трубки, без
более крупного шипа ближе к вершине.
(23). Жабры в 1.2—1.6 раза длиннее дыхательной трубки. Задние
волоски наличника только немного короче лобных, сильно ветвятся,
из 9—10 ветвей. Боковой волосок на последнем членике из 2—4 ко-
ротких одинаковых ветвей............A. (Stegomyia) galloisi Yam.
(22). Жабры короче дыхательной трубки. Задние волоски наличника
заметно короче лобных, слабо ветвятся — из 2—5 ветвей. Боковой
волосок из 2 неравных, более длинных ветвей.
(25). Волоски тела звездчатого типа ветвления .................
. ............... A. (Stegomyia) flavopictus Yam.
(24). Волоски тела ветвятся в одной плоскости .................
..............................A. (Stegomyia) albopictus Skuse.
(13). Ушки по бокам основания дыхательной трубки, ближе к ее зад-
нему краю, хорошо развиты. Усики различного типа, обычно с хо-
рошо заметными шипиками, реже гладкие, с простым или, чаще,
ветвящимся волоском.
(28). Средние лобные волоски очень длинные, длиннее головы и вдвое
длиннее разветвленных внутренних. Жабры длинные, заостренные
на концах, более короткая нижняя пара одной длины с дыхательной
трубкой ..........................A. (Finlaya) alektorovi Stack.
(27). Средние лобные волоски примерно одной длины с внутренними.
Жабры самой различной длины.
(38). Усики гладкие, лишены шипиков. Жабры длинные, длиннее
седла.
(35). Пучок на усике в виде простого волоска. Зубцы гребня в виде
длинных, сидящих на равном расстоянии острых шипов.
(32). Щетка из 55—60 чешуек, расположенных неправильно треуголь-
ным пятном. Все лобные волоски сильно ветвятся, не менее чем
из пяти ветвей. Жабры колбасовидные, верхняя пара длиннее дыха-
тельной трубки, нижняя короче ... A. (Finlaya) seoulensis Yam.
(31). Щетка из И—18 чешуек, расположенных в один дугообразный
186
III. ПОДСЕМ. CULICINAE
ряд. Лобные волоски слабо ветвятся, из 1—4 ветвей. Верхняя пара
жабр не длиннее дыхательной трубки, нижняя заметно короче.
33 (34). Гребень занимает от V4 до 2/5 длины дыхательной трубки. Сильно
развитые волоски тела звездчатого типа ветвления из меньшего
числа более тонких ветвей................A. (Finlaya) geniculatus 01.
34 (33). Гребень занимает не менее половины дыхательной трубки. Сильно
развитые волоски тела звездчатого типа ветвления состоят из боль-
шого числа более толстых ветвей ... A. (Finlaya) echinus Edw.
35 (30). Пучок на усике из 2—4 коротких ветвей. Зубцы гребня короткие,
не шиповидные, с широким основанием.
36 (37). Длина дыхательной трубки в 4—5 раз превышает ее ширину
у основания; последняя в 1.4—1.5 раза больше ее ширины у вер-
шины: трубка слабо, но равномерно сужена к концу. Колбасовид-
ные жабры с округленными концами в среднем равны длине дыха-
тельной трубки .......................................................
.............A. (Ochlerotatus) pulchritarsis pulchritarsis Rond.
37 (36). Длина дыхательной трубки в 3.0—3.5 раза превышает ее ширину
у основания; последняя в 1.5 раза больше ее ширины у вершины:
трубка более сильно сужена к концу. Колбасовидные жабры в сред-
нем вдвое длиннее трубки .............................................
............ . A. (Ochlerotatus) pulchritarsis asiaticus Edw.
38 (29). Усики докрыты хотя бы редкими шипиками. Если шипики раз-
виты очень слабо (4. mariae), то жабры очень короткие, шаровидные.
39 (46). На передней поверхности дыхательной трубки, кроме предвер-
шинных щетинок, имеются 2—3 пары грубых, хорошо развитых
волосков, а на боковых поверхностях, кроме пучка, около оснований
дистальных зубцов гребня находится одна пара тонких боковых
волосков.
40 (43). Зубцы гребня, ближайшие к вершине, заходят за основание
пучка дыхательной трубки.
41 (42). Пучок на дыхательной трубке из 6—8 ветвей. Наиболее удален-
ный из расставленных зубцов гребня не доходит до вершинной трети
трубки.................................A. (Ochlerotatus) rusticus Rossi.
42 (41). Пучок на дыхательной трубке в виде длинного простого волоска.
Наиболее удаленный из расставленных зубцов гребня расположен
в вершинной трети трубки, почти у ее конца . .........................
.............................A. (Ochlerotatus) subdiversus Mart.
43 (40). Наиболее дистальные зубцы гребня не заходят за основание
пучка дыхательной трубки.
44 (45). На передней поверхности дыхательной трубки три пары волосков.
Из волосков, расположенных позади щетки, крайний нижний наи-
более короткий..........................-A. (Ochlerotatus) refiki Med.
45 (44). На передней поверхности дыхательной трубки две пары волосков.
Из волосков, расположенных позади щетки, крайний нижний наи-
более длинный .....................A. (Ochlerotatus) lepidonotus Edw.
46 (39). На передней поверхности дыхательной трубки другие волоски,
кроме предвершинных щетинок, отсутствуют, на боковых сторонах,
кроме пучка, нет других волосков.
47 (50). Дистальные зубцы гребня широко расставлены и заходят за ос-
нование пучка дыхательной трубки.
48 (49). Седло кольцом опоясывает последний членик брюшка . . . .
...................................... A. (Ochlerotatus) nigripes Zett.
49 (48). Седло последнего членика брюшка заходит только за середину
его боковых сторон ..... A. (Ochlerotatus) cataphylla Dyar.
7. AEDES
187
50 (47). Дистальные зубцы гребня не заходят за основание пучка дыха-
тельной трубки.
51 (72). Впереди плавника отходят 4—6 более коротких пучков, не
объединенных общим основанием. Пучок у середины дыхательной
трубки (исключение — A. mariae с очень короткой дыхательной
трубкой) в длину не меньше диаметра трубки у места его отхождения.
52 (53). Вся поверхность тела покрыта мелкими темными шипиками,
лежащими густыми рядами. Голова и длинная дыхательная трубка
(индекс 4—5) охряно-желтого цвета; гребень с 2—3 более крупными
и шире расставленными зубцами ... A. (Ochlerotatus) cyprius Ludl.
53 (52). Поверхность тела голая, не покрыта мелкими шипиками.
54 (57). Щетка на VIII членике брюшка в среднем из 6—12 чешуек.
55 (56). Чешуйки щетки расположены в один неправильный, часто изо-
гнутый ряд. Средние, и внутренние лобные волоски из 2—3 ветвей.
Индекс дыхательной трубки 3.5—4.0......................................
.....................A. (Ochlerotatus) riparius Dyar et Knab.
56 (55). Чешуйки щетки расположены в два (реже три) неправильных
ряда. Средние и внутренние волоски простые. Индекс дыхательной
трубки 2.0..............................A. (Ochlerotatus) nigrinus Eck.
57 (54). Щетка на VIII членике брюшка из 15—45 чешуек.
58 (59). Седло последнего членика брюшка слабо развито, не полностью
покрывает его сверху и едва заходит за боковые стороны. Жабры
короткие, шаровидные. Индекс дыхательной трубки не более 2
........................... A. (Ochlerotatus) mariae Ed. et Et. Serg.
59 (58). Седло хорошо выражено. Жабры не менее половины длины седла.
Индекс дыхательной трубки не менее 3.
60 (63). Волоски на вершине задних клапанов стигмальной пластинки
крючкообразно изогнуты и утолщены.
61 (62). Щетка из 30—40 (чаще 32—36) чешуек. Задние волоски налич-
ника из 2—3 тонких коротких ветвей. Волоски на вершине задних
клапанов стигмальной пластинки сильнее утолщены........................
............................. A. (Ochlerotatus) excrucians Walk.
62 (61). Щетка из 20—28 (среднее 24) чешуек. Задние волоски наличника
из 6—8 тонких коротких ветвей.’ Волоски на вершине задних
клапанов стигмальной пластинки менее утолщены..........................
................ ...............A. (Ochlerotatus) hehningi Mart.
63 (60). Волоски на вершине задних клапанов стигмальной пластинки
обычной формы: только слегка изогнуты и не утолщены.
64 (67). Дистальные зубцы гребня расставлены.
65 (66). Наиболее дистальные 1—3-й зубцы гребня лежат за серединой
дыхательной трубки, ближе к вершине. Пучок из 2—4 ветвей . . .
.................................. A. (Ochlerotatus) heklemishevi Den.
66 (65). Наиболее дистальные зубцы гребня заметно не доходят до сере-
дины дыхательной трубки. Пучок на ней из 4—7 (чаще 5—-6) ветвей
....................... A. (Ochlerotatus) flavescens Miill. (частично).
67 (64). Дистальные зубцы гребня не расставлены; промежуток между
двумя последними зубцами в среднем равен таковому между двумя
предыдущими. ' /
68 (69). Впереди плавника 6—7 более коротких пучков, не окруженных
общим основанием ... A. (Ochlerotatus) flavescens Miill. (частично).
69 (68). Впереди плавника 4—5 более коротких пучков, не окруженных
общим основанием.
70 (71). Плавник из 18—19 пучков, окруженных общим основанием.
Задний отросток стигмальной пластинки, кроме обычных двух
188
III. ПОДСЕМ. CULICINAE
ветвей, имеет между ними различно развитую среднюю ветвь . . .
................................ A. (Ochlerotatus) cantans Meig.
71 (70). Плавник не более чем из 16 пучков, окруженных общим основа-
нием. Задний отросток стигмальной пластинки обычной Для подрода
формы...................................A. (Ochlerotatus) annulipes Meig.
72 (51). Впереди плавника не более трех пучков, не объединенных
общим основанием. Если их четыре (A. vexans), то пучок на дыха-
тельной трубке короткий, меньше ее ширины у места его отхож-
дения, и лежит в начале вершинной трети.
73 (78). Жабры, как правило, короче седла, реже не более чем на1/3 длин-
нее его, не пигментированы.
74 (75). Все чешуйки щетки без длинного главного шипа, имеют по краю
ряд шипиков, из которых апикальные — наиболее длинные. Рычаг
стигмальной пластинки не имеет плеч и отростков к стигмам, сильно-
смещен назад, в промежуток между задними клапанами. Средние
лобные волоски из 1—2, внутренние из 2—3 ветвей. Жабры шаровид-
ные ................................. A. (Ochlerotatus) detritus Hal.
75 (74). Хотя бы некоторые чешуйки щетки с длинным главным шипом.
Рычаг стигмальной пластинки имеет плечи и стигмальные отростки
и занимает приблизительно центральное положение. Средний и
внутренний лобные волоски обычно простые. Жабры удлиненной
формы, не менее трети длины седла.
76 (77). Ветвление пучков плавника начинается далеко от их основания:
длина неразветвленного стебелька средних пучков плавника в 1.5—
2 раза больше длины боковых отростков их основания. Некоторые
из ближайших к брюшной стороне чешуек щетки с длинным главным
шипом, остальные с 2—3 более длинными шипиками у вершины и
с 2—3 более короткими по бокам основания...............................
................................A. (Ochlerotatus) leucomelas Meig.
77 (76). Ветвление пучков плавника начинается близко от их основания::
длина неразветвленного стебелька не больше длины боковых отрост-
ков их основания. Чешуйки щетки с более или менее выражен-
ным главным шипом могут единично встречаться в любом месте
щетки..................................A. (Ochlerotatus) caspius Pall,
а. Пучок на дыхательной трубке лежит за ее серединой, ближе к вер-
шине, из 5—40 ветвей. Внутренние хвостовые волоски из 12—15 вет-
вей. Средний волосок позади щетки из 7—14 ветвей...................
............................A. (Ochlerotatus) caspius caspius Pall,
б,. Пучок на дыхательной трубке лежит у ее середины, из 3—5
(редко больше) ветвей. Внутренние хвостовые волоски из 4—12 вет-
вей. Средний волосок позади щетки из 5—8 ветвей....................
. . . ....................A. (Ochlerotatus) caspius dorsalis Meig.
78 (73). Жабры не менее чем в 1.5 раза длиннее седла, чаще пигменти-
рованные. 7
79 (84). Число чешуек в щетке более 40; все или большинство чешуек
лишены главного шипа, несут по краю ряд мелких шипиков.
80 (81). Средние и внутренние лобные волоски простые, как редкое
исключение, один или два из них 2—3-ветвистые...........................
............................A. (Ochlerotatus) communis De Geer.
81 (80). Средние и внутренние волоски из 3—7 ветвей.
82 (83). Усики тонкие, длинные, до 2/3 длины головы или немного длин-
нее. Все чешуйки щетки без главного шипа...............................
................................ A. (Ochlerotatus) pionips Dyar.
7. AEDES
189
•83 (82). Усики более толстые и короткие, около половины длины головы.
У некоторых из чешуек щетки имеется слабо выраженный главный
шип ..............................A. (Ochlerotatus) pullatus Coq.
84 (79). Число чешуек в щетке меньше 30.
85 (88). Седло опоясывает кольцом последний членик брюшка или на его
. брюшной стороне между боковыми краями седла имеется узкая
щель.
86 (87). Число чешуек в щетке 10—20; чешуйки мелкие — 0.07—0.08 мм
................................A. (Ochlerotatus) punctor Kirby.
87 (86). Число чешуек в щетке 5—9; чешуйки более крупные — 0.ll-
о.13 мм ........................A. (Ochlerotatus) hexodontus Dyar.
88 (85). Седло не опоясывает кольцом последний членик брюшка, может
доходить только до нижнего края его боковых сторон.
89 (92). Внутренние лобные волоски простые. Дистальные зубцы гребня
не расставлены.
90 (91). Средние лобные волоски простые. В щетке 7—16 (в среднем
13—14) чешуек. Жабры пигментированы....................................
........................... A. (Ochlerotatus) impiger Walk.
91 (90). Средние лобные волоски из 2 ветвей. В щетке 16—24 (в среднем 20)
чешуйки. Жабры не пигментированы ......................................
................................A. (Ochlerotatus) hungaricus Mih.
92 (89). Внутренние лобные волоски из 2—5 ветвей.
93 (96). Дистальные зубцы гребня (1—3) всегда широко расставлены,
заметно крупнее остальных.
94 (95). Пучок на дыхательной трубке за серединой, ближе к вершине,
всегда впереди наиболее дистального из зубцов гребня; из 3—8
(чаще 4—6) тонких ветвей, более коротких, чем ширина трубки
у места отхождения пучка..................A. (Aedimorphus) vexans Meig.
95 (94). Пучок на дыхательной трубке у середины или немного за ней,
из 3—8 (чаще 4—6) ветвей, равных ширине трубки у места его отхо-
ждения или длиннее. Если число широко расставленных зубцов
гребня больше двух, пучок лежит между их основаниями...................
................................A. (Ochlerotatus) intrudens Dyar.
96 (93). Зубцы гребня расположены на равном расстоянии друг от друга;
как исключение, один наиболее дистальный может быть немного
удален от остальных. В щетке 16—27 (чаще 22—24) чешуек ....
.....................................A. (Ochlerotatus) sticticus Meig.
1. Подрод OCHLEROTATUS L. Arribalzaga1
Гипопигий: коксит с двумя бородавками — базальной и вершинной —
или по крайней мере с одной из них; стили простые, не раздвоенные;
придаток стиля причленяется на его вершине; класпеты имеются; фалло-
сома простая (не разделенная на 2 пластинки). Щупики самца немного
длиннее хоботка или равны ему, реже немного короче хоботка, обычно
с небольшими утолщениями на III и IV или на III и V члениках. Самка:
брюшко кзади суживается, церки отчетливо выступают; хоботок явст-
венно длиннее передних бедер.
Определение видов по самкам встречает значительные трудности.
Желательно иметь хорошо сохранившиеся, не потертые экземпляры.
В ряде случаев достоверное определение возможно только по самцам
или по личинкам. Важный систематический признак — расположение
пятен чешуек на боковой поверхности груди (Peus, 1933). При опреде-
1 См. также стр. 375.
190
III. ПОДСЁМ. CULICINAE
лении некоторых видов учитывается форма коготков лапки (Vockeroth,.
1950). Ряд полезных систематических признаков указан О. Н. Сазоновой
(1958); ею составлена таблица для определения видов Aedes лесной зоны
по самкам.
Подрод включает около 140 видов, распространенных преимущественно
в Палеарктике и Неарктике; в СССР известно 29 видов. Подрод Ochlero-
tatus особенно характерен для лесной зоны и тундры. Ареалы некоторых
видов простираются до 75—78° с. ш. Комары данного подрода во многих
местах составляют основной компонент гнуса, а также играют большую
роль в циркуляции возбудителей туляремии и некоторых вирусных ин-
фекций.
Палеарктические виды Ochlerotatus для удобства ориентировки по
взрослым особям можно разделить на четыре группы (не придавая им
определенного таксономического ранга), которые условно обозначаются
по наименованию одного из видов: caspius, cantans, rusticus, communis.
7. AEDES
191
Первые две группы характеризуются наличием светлых колец на лап-
ках (рис. 97), последние две — отсутствием таких колец.
Виды группы caspius.
Лапки с белыми кольцами, которые опоясывают основание и вершину
члеников; границы члеников видны посреди белых колец (рис. 103).
Гипопигий характеризуется слабым развитием или отсутствием вершин-
ной бородавки коксита, что, однако, свойственно и некоторым видам дру-
гих групп.
К группе caspius относятся три палеарктических вида.
1. Aedes (Ochlerotatus) caspius Pallas, 1771.
Весьма сильно варьирующий в окраске вид, представленный в пре-
делах Палеарктики двумя подвидами, из которых один — А. с. caspius —
свойствен более южным и
сухим районам, другой — А. с.
dorsalis — более северным и
влажным. На Дальнем Востоке,
в Сибири, Казахстане и в не-
которых областях европейской
части СССР распространены
промежуточные формы.
Голова сверху в белых и жел-
товато-бурых чешуйках. Хобо-
ток и щупики в бурых и белых
чешуйках, соотношение кото-
рых весьма изменчиво. Окраска
груди и крыльев обоих подви-
дов различна. Лапки с белыми
или кремовыми колечками, опо-
ясывающими основания и вер-
шины члеников; иногда светлые
колечки выражены неотчетли-
во; они лучше видны на темном
фоне. Брюшко с не резко отгра- Рис. 104. Гипопигий Aedes caspius dorsalis Mg.
ниченными светлыми попереч-
ными перевязями и со светлой продольной полосой, которая может быть
развита не на всех сегментах. В общем на брюшке преобладают светлые
чешуйки; темные иногда образуют только пятна по бокам тергитов, реже
брюшко сплошь в светлых чешуйках.
Гипопигий (рис. 104): базальная бородавка коксита с двумя шипами;
один из них более длинный, изогнутый, другой — более короткий, пря-
мой. Крыло класпеты узкое, без прозрачного расширения.
По личинкам оба подвида отличаются только признаками количествен-
ного порядка; общие размеры тела, дыхательной трубки, стигмальной
пластинки, жабр, степень ветвления волосков и др. Количественные
значения этих отличий, даже у резко выраженных популяций обоих под-
видов, в своих крайних вариантах всегда соприкасаются или заходят
друг за друга, что явствует из приведенных ниже описаний.
Распространение. Ареал вида простирается от Атланти-
ческого океана до Тихого и от 64° с. ш. (в Финляндии) и района Якутска
до южных границ Палеарктики (Северная Африка, Иран); широко рас-
пространен в Северной Америке (только А. с. dorsalis).
192
III. ПОДСЕМ. CULICINAE
Биология. Наиболее характерен для речных долин, где часто
по численности занимает первое место среди комаров, нападающих на
людей и животных. При массовом появлении комары данного вида про-
никают в населенные пункты, залетают в дома и помещения для животных.
Характерен массовый выплод в пойменных водоемах после паводка.
После летних дождей может появиться новое поколение, обычно численно
уступающее весеннему.
Замечания по систематике. Подвиды — А. с. caspius Pall, и
А. с. dorsalis Mg. — многими авторами рассматриваются как самостоятельные виды.
Ареалы подвидов на огромном пространстве налегают друг на друга. В Западной и
Центральной Европе обе формы легко отличаются по окраске имаго, менее четко —
по строению гипопигия; особи с промежуточными признаками почти не встречаются.
Но чем дальше на восток, тем чаще встречаются особи, о которых трудно или невоз-
можно сказать, к какому подвиду они относятся. Переходные формы распространены
в некоторых районах европейской части СССР (Поволжье, Северный Кавказ), в Казах-
стане и Сибири.
Что касается дальневосточных A. caspius, то их признаки обычно имеют смешан-
ный, промежуточный характер. Среди A. caspius из Забайкалья, Приморья и Северо-
Восточного Китая преобладают особи, у которых среднеспинка имеет две продольные
широкие полосы беловатых или кремовых чешуек; между ними проходит узкая темная
каштановая или коричневая полоса; боковые края среднеспинки также покрыты кашта-
новыми чешуйками. От такого переходного между А. с. caspius а А. с. dorsalis варианта
наблюдаются различные отклонения: вместо продольных светлых полосок могут быть
светлые пятна неправильных очертаний, кремовые чешуйки могут преобладать, а тем-
ные образуют лишь продольную полосу в передней части среднеспинки. По строению
гипопигия комары из Приморья также занимают промежуточное положение,
а по окраске крыла они более сходны е А. с. dorsalis. Южные популяции (Квантун-
ский полуостров) стоят ближе к А. с. caspius.
В Средней Азии распространен А. с. caspius, хотя встречаются и переходные
формы. В высокогорных районах (Памир) обитает А. с. dorsalis.
Поскольку (по крайней мере в восточной части ареала вида) А. с. caspius и А. с. dor-
salis связаны между собой постепенными переходами, мы считаем более правильным
рассматривать их в качестве подвидов.
Детальное исследование географической изменчивости A. caspius, возможно,
даст основания разделить вид не на два, а на большее количество подвидов. С другой
стороны, некоторые вариации окраски, а также других признаков явно зависят от эко-
логических факторов, в частности от условий развития преимагинальных фаз. Поэтому
описание новых подвидов без учета их экологии нецелесообразно.
Aedes duplex Mart, описан по двум самцам из Саратовской области (Martini, 1926,
1928). Согласно описанию, A. duplex похож на A. caspius, но отличается одноцветной
светлой окраской чешуек среднеспинки и слабым развитием светлых колечек на лап-
ках, а также удвоенным числом шипов базальной бородавки коксита: на каждой боро-
давке имеются два сближенных прямых шипа и два сближенных изогнутых. Однако
подобные особенности окраски отмечаются нередко у А. caspius в различных частях
его ареала. Удвоение же количества шипов коксита изредка наблюдается как аберра-
ция у комаров различных видов. В коллекции ЗИН имеются самцы 4. caspius из г. Са-
репты Волгоградской области, у которых на базальной бородавке, помимо двух шипов,
развиты один, два или несколько дополнительных шипов, почти таких же по размерам
и строению, как и «основные» шипы. Между обычными и аберрантными экземплярами
имеются постепенные переходы.
По нашему мнению, для признания видовой самостоятельности A. duplex нет до-
статочных оснований.
Aedes caspius caspius Pallas.
Помимо указанных в определительных таблицах, отметим следующие
особенности. Хоботок и щупики самки преимущественно в светлых че-
шуйках. В основной четверти костальной жилки темные чешуйки вкрап-
лены среди светлых или темные чешуйки преобладают. Темные пятна
на тергитах брюшка коричневые, светлые пятна имеют желтоватый от-
тенок, местами — белый. Проксимальный шип базальной бородавки,
коксита резко изогнут, с тонкой крючковатой вершиной.
7. AEDES
193
Окраска среднеспинки исключительно изменчива. Светлые продоль-
ные полоски могут быть узкими, отчетливыми (наиболее обычный вари-
ант) или более широкими, расплывчатыми. Иногда они желтоватые и едва
выделяются на основном фоне чешуек среднеспинки. Наконец, нередко
среднеспинка покрыта одноцветными желтовато-белыми или песочного
оттенка чешуйками. Размеры темных пятен по бокам тергитов брюшка
Рис. 105. Aedes caspius caspius Pall. Личинка IV стадии. Задний конец,
сбоку.
варьируют вплоть до полного вытеснения темных пятен светлыми че-
шуйками. Б. Н. Казанцев (1931) показал, что в районе Бухары светлые
«песочные» A. caspius развиваются в водоемах с высокой концентрацией
солей, тогда как комары из пресных водоемов характеризуются кон-
трастной окраской. Описанные варианты окраски характерны не для
определенной географической области, а встречаются в различных час-
тях ареала подвида.
Личинки IV стадии (рис. 105) средней величины, желто-
коричневого цвета, с более светлой головой и дыхательной трубкой.
Лобные волоски слабо вторичноперистые, наружные из 7—10 ветвей,
средние впереди внутренних, обе пары простые, реже 2-ветвистые, как
редкое исключение, один из волосков 3-ветвистый; задние волоски на-
13 Фауна СССР, А. В. Гуцевич и др.
194
III. ПОДСЕМ. CULICINAE
личника из 3—5 коротких и очень тонких ветвей. Усик около 1/'2 длины
головы, покрыт редкими, слабо развитыми шипиками; пучок у середины,
ближе к основанию, в среднем из девяти ветвей, достигающих половины
длины усика.
Щетка из 18—28 (чаще 20—25) чешуек, расположенных в 2—3 не-
правильных ряда; чешуйки очень изменчивы по форме, начиная от ко-
ротких, лишенных главного шипа, несущих на вершине и по краю ряд
более мелких шипиков, укорачивающихся к основанию чешуйки, до более
удлиненных, имеющих один (или больше) ясно выраженный шип с более
короткими по бокам основания. Волоски позади щетки: крайние верхний
из 5—10, нижний из 6—7, средний из 7—14 тонких ветвей; промежуточные
простые. Дыхательная трубка прямая, начиная от середины слегка су-
женная к вершине (индекс 1.8—2.6), сильно изменчива в длину. Гребень
из 17—26 (чаще 20—22) зубцов, из которых 1—4 ближайшие к основанию
рудиментарны; зубцы сидят на равных расстояниях друг от друга;
гребень слегка заходит за середину трубки. Пучок из 5—10 слабо вто-
ричноперистых коротких ветвей, за серединой трубки, ближе к вершине.
Стигмальная пластинка типична для личинок подрода Ochlerotatus, ее раз-
меры: длина 0.30—0.36 мм, ширина у задних клапанов 0.27—0.32 мм,
между концами боковых 0.28—0.34 мм.
Седло доходит до середины боковых сторон; боковой волосок простой
и короткий, меньше половины длины седла; наружный хвостовой во-
лосок простой, превышает длину трубки, внутренний из 12—15 более
чем вдвое коротких ветвей. Плавник из 14—17 пучков, окруженных об-
щим основанием, и из 2—3 более коротких впереди него; количество
ветвей в каждом из средних пучков достигает 9—12, отдельные ветви
тонкие. Жабры короткие — 0.3—0.9 длины седла, листовидной или
ланцетовидной формы, нижняя пара короче верхней.
Распространение. Подвид характерен для степной зоны
и для сухих степей с солоноватыми водоемами. В Европе распространен
на север до Англии, юго-западной Финляндии, центральных областей
европейской части СССР. Ареал подвида включает также южные районы
Сибири, Казахстан, Среднюю Азию, а вне Советского Союза — Монго-
лию, Северный и Западный Китай, Западную Азию, Северную Африку.
В степной Украине, на Нижней Волге, на востоке Предкавказья и Закав-
казья, в Казахстане и Средней Азии А. с. caspius — преобладающий
по численности вид комаров, местами в огромном количестве нападаю-
щий на людей и животных, например, в Муганской степи (Здродовский,
1926), в ряде районов Туркмении (Петрищева, 1936), в низовьях Сыр-
дарьи (Благовещенский, 1937).
Биология. Личинки встречаются в открытых или затененных (на
юге) водоемах, постоянных или временных, возникших в результате
таянья снега, речных паводков или сбросов воды с полей орошения, бедных
растительностью, с глинистым дном, часто с большим содержанием солей
(до 5% и выше). Яйца откладываются на влажную почву недалеко от
уреза воды, устойчивы к высыханию. Могут иметь несколько поколений
в течение года и местами достигать высокой численности (Средняя Азия).
Самки нападают на людей и животных, особенно в сумеречные
часы.
Отличается тепловыносливостью и сухоустойчивостью. В Таджикистане
наибольшую активность проявляет при температуре 19—31° (Бреге-
това, 1946). Нередко нападает днем при ярком освещении. Из мест мас-
сового размножения может мигрировать на значительное расстояние
иногда на десятки километров.
7. AEDES
195
В некоторых очагах туляремии играет роль в циркуляции возбудителя,
который в организме А. с. caspius может сохраняться в течение месяца
(Олсуфьев, 1938). Является переносчиком вируса тягиня в Чехословакии
(Бардош и Даниелова, 1959).
Aedes caspius dorsalis Meigen, 1830.
Основные отличительные признаки указаны в определительных таб-
лицах. Хоботок и щупики самки преимущественно в темных чешуйках.
Костальная жилка в основной четверти покрыта белыми чешуйками,
без примеси темных. Темные пятна тергитов брюшка черно-коричневые,
Рис. 106. Aedescaspius dorsalisMg. Личинка IV стадии. Задний конец,
сбоку.
а светлые имеют беловато-серый оттенок. Проксимальный шип базаль-
ной бородавки коксита слегка изогнут, его вершина обычно не загнута
крючком.
Личинка IV стадии (рис. 106) несколько крупнее, чем у ос-
новной формы, и более темной, до коричнево-черной окраски. Лобные во-
лоски вторичноперистые, наружные из 4—8 (чаще 5—6) ветвей, средние
впереди внутренних, обе пары простые, редко 2-ветвистые; задние во-
лоски наличника короткие, из 2—5 тонких ветвей. Усик около х/2 длины
головы, с редкими шипиками и пучком у середины из 4—7 ветвей, не пре-
вышающих х/2 его длины.
Щетка из 13—34 (чаще 20—25) чешуек, расположенных в 2—3 непра-
вильных ряда; чешуйки очень изменчивы; характер изменчивости как
13*
'196
111. ПОДСЕМ. CULICINAE
уосновной формы. Волоски позади щетки: крайние — верхний из 3 —8
(•чаще 4—5), нижний из 4—6, средний из 5—8 ветвей, промежуточные
простые. Дыхательная трубка более стройная, чем у основной формы,
индекс 2.5—3.0, реже меньше. Гребень из 14—23 зубцов, из которых
1—4 у основания рудиментарные, не доходит до середины трубки; зубцы
сидят на равных расстояниях друг от друга, в виде исключения наиболее
удаленный может быть слегка отставлен; гребень не доходит до середины
трубки. Пучок из 3—8 (обычно 4—5) вторичноперистых ветвей у середины
трубки; их длина не превышает
ширину трубки у основания. Стиг-
мальная пластинка (рис. 107) от-
личается от таковой у основной
формы несколько более крупными
размерами и более темной пигмен-
тацией.
Седло как у основной формы,
но интенсивнее пигментировано и
резко очерчено по нижнему краю.
Внутренний хвостовой волосок из
4—12 (чаще 8—9) ветвей, более
чем вдвое короче наружного.
Плавник по числу пучков не от-
личается от основной формы, но
каждый пучок ветвится слабее,
в среднем из 7—9 ветвей. Жабры
несколько длиннее, чем у основ-
Рис. 107. Aedes caspius dorsalis Mg. ной формы; из пресных водое-
Личинка IV стадии. Стигмальная пла- мов — д0 f 3 длины седла, а в со-
стинка. лоноводных — не более 0.3—0.4
длины седла.
Р аспространение. Характерен для открытых стаций под-
зоны смешанных лесов и для лесостепи. На север доходит до северной
границы вида; на юг до Венгрии, Болгарии, степной Украины, Северного
Кавказа, Нижней Волги и гор Средней Азии, включая Памир. Распро-
странен в Сибири, на Дальнем Востоке (см. выше), а также в Северной
Америкё.
Биология. Личинки встречаются преимущественно в открытых
небольших водоемах и заболоченностях, постоянных или временных,
после таяния снега, паводка, выпадения дождей или питаемых почвен-
ными водами, как в пресной, так и в соленой воде. Зимовка на фазе яйца.
Как и у основной формы, может давать несколько поколений в течение
летнего сезона, особенно при периодическом затоплении водоемов.
^Местами достигает большой численности. В долинах рек, например Волги,
Днепра, является массовым кровососом. В Московской области наиболее
активно нападает при температуре 16—22°; сезонная кривая численности
двувершинная (Шленова, 1959). В средней полосе европейской части
СССР за сезон дает 2—3 поколения (Хелевин, 1958).
. Принадлежит к числу переносчиков вируса западного энцефалита
.(лошадиного энцефаломиелита) в США.
2. Aedes (Ochlerotatus) mariae Ed. et Et. Sergent, 1902.
Отличается от предыдущего вида отсутствием продольной светлой
полосы на брюшке и отсутствием шипов на базальной бородавке коксита.
Нижеследующее описание составлено преимущественно по работе Сеневэ
7. AEDES
19Г
и Андарелли (Senevet et Andarelli, 1964), отмечающих большую» измен-
чивость признаков данного вида.
Хоботок в коричневато-бурых чешуйках, иногда с примесью беловатых
чешуек в средней части хоботка. Среднеспинка покрыта ржаво-коричне-
выми чешуйками, среди которых обычно разбросаны немногочисленные бе-
ловатые чешуики, иногда образующие
неясно очерченные узкие продольные
полоски («var. zamittii» Theob.). Лапки
с белыми кольцами, каждое кольцо
опоясывает два членика; последний
членик задней лапки целиком белый.
Крылья в перемешанных темных и
светлых чешуйках, последние более
многочисленны на костальной, субко-
стальной и первой радиальной жил-
ках. Окраска брюшка сильно варь-
ирует (рис. 108). При основании
тергитов выражены неширокие попе-
речные перевязи светлых чешуек,
обычно расширяющиеся по бокам в
виде латеральных пятен. Большая
часть каждого тергита покрыта тем-
ными чешуйками; продольной полосы
Рис. 108. Брюшко Aedes marine
Serg., сверху (различные варианты
окраски). По Senevet et Andarelli.
светлых чешуек нет.
Гипопигий (рис. 109): вершинная бородавка коксита не развита, почти?
неразличима. Умеренно выпуклая базальная бородавка несет волоски,,
среди которых неотчетливо выделяются 4—5 относительно более крупных
щетинок. Класпеты с коротким прямым стволиком и относительно узким
Рис. 109. Гипопигий Aedes marine Serg. По Senevet
et Andarelli.
A — базальная бородавка, Б — класпета, В — лопасти
IX тергита.
изогнутым крылом, лишенным прозрачного расширения. Фаллосома
более или менее прямоугольная. Лопасти IX тергита с относительно
длинными (по сравнению с A. caspius) щетинками, по 4—6 щетинок с каж-
дой стороны.
Личинки IV стадии (рис. 110) очень изменчивы по окраске..
Лобные волоски: наружные в среднем из семи ветвей, обе внутренние пары
простые, на 1/3 длиннее наружной, средние впереди внутренних; задние
волоски наличника очень тонкие и короткие, ветвистые. Усики тонкие,
слегка изогнутые, со слабо развитыми шипиками; пучок у середины,
из 6—9 ветвей, достигающих половины длины усика. . »
.198
III. ПОДСЕМ. CULICINAE
Щетка из 16—25 чешуек, расположенных в 2—3 ряда; чешуйки
имеют или хорошо выраженный главный шип с сильно изменчивым чис-
лом различно развитых зубчиков у основания, или последние в числе
1—2 развиты до размеров главного шипа. Дыхательная трубка короткая
(индекс 1.4—2.0), слабо суженная, с очень изменчивой степенью хити-
низации основания; гребень доходит до середины трубки, из 15 или более
тонких удлиняющихся в дистальном направлении зубцов с рядом допол-
нительных зубчиков вдоль базальной х/3—х/2 зубца; пучок из 6 ветвей,
в длину равных ширине трубки, отходит у ее середины, ближе к вершине.
Рис. 110. Aedes mariae Serg. Личинка IV стадии. По Эдвардсу.
1 — голова, сверху, г — задний конец, сбоку.
Седло на последнем членике слабо развито, не полностью покрывает
его спинную сторону и лишь немного заходит на боковые; боковой во-
лосок простой. Наружные хвостовые волоски простые и длинные, внут-
ренние в 3—4 раза короче, из 11—14 волосков. Плавник сильно развит
из 12—13 пучков в общем основании и 4—5 впереди него. Жабры короткие,
шаровидной формы.
Распространение. Страны Европы, Африки и Азии, преиму-
щественно на побережье Средиземного моря, от Марокко до Сирии.
В СССР с достоверностью не найден.
Биология. Личинки встречаются в скоплениях воды в углубле-
ниях скал вдоль морского берега, часто у самой полосы прибоя. Обычная
концентрация солей в таких водоемах 2—4%, но личинки переносят
и более высокую концентрацию солей, до 18%.
Переносчик возбудителя малярии птиц, Plasmodium relictum.
Замечания по систематике. Вопрос о наличии в СССР А. mariae
яеясен. Имеется указание о нахождении его на Украине (Прендель, 1966), с оговор-
7. AEDES
199
кой о том, что определение требует подтверждения. В коллекции ЗИН хранится пре-
парат гипопигия комара (возможно, A. mariae), найденного в окрестностях Днепро-
петровска в мае 1928 г. (материал В. В. Горицкой). Гипопигий в общем сходен с гипо-
пигием A. caspius со следующими существенными отличиями: базальная бородавка
коксита, помимо многочисленных волосков, несет неправильный ряд из 7—8 прибли-
зительно одинаковых щетинок; шипов или крепких щетинок на базальной бородавке
нет; слабо развитая вершинная бородавка несет несколько волосков, более длинных,
чем у A. caspius.
Возможно, А. mariae встречается на юге СССР, но не отграничивается от А. cas-
pius.
3. Aedes (Ochlerotatus) pulchritarsis Rondani, 1872.
Отличается отсутствием светлой продольной полосы на брюшке и
одноцветно бурыми чешуйками крыльев. Распадается на два подвида.
Рис. 111. Гипопигий Aedes pulchritarsis Rond.
Голова в желтоватых и бурых чешуйках. Щупики самки бурые, с бе-
лой вершиной; щупики самца бурые, с белыми колечками на последних
члениках. Окраска среднеспинки различна у обоих подвидов. Щиток
в желтоватых чешуйках. Бочки груди с пятнами белых чешуек. Лапки
с очень контрастными белыми кольцами, опоясывающими вершину пре-
200
111. ПОДСЕМ. CULICINAE
дыдущего и основание последующего членика. Последний членик лапки
белый. Крылья в одноцветных темных чешуйках. Брюшко бурое, с пере-
вязями белых чешуек при основании тергитов.
Гипопигий (рис. 111) отличается отсутствием вершинной бородавки
коксита; слабо выпуклая базальная бородавка несет длинный, изогну-
тый на конце шип и несколько более тонких щетинок; крыло класпеты
узкое, длинное; лопасти IX тергита слабо выпуклые с 4—6 короткими
щетинками.
Ярко-белая окраска чешуек на лапках и брюшке A. pulchritarsis
придает ему некоторое сходство с комарами подрода Stegomyia, что может
служить источником ошибок. Но у A. pulchritarsis на среднеспинке отсут-
ствует рисунок в виде лиры, характерный для A. (Stegomyia) aegypti.
Кроме того, у комаров подрода Stegomyia белые кольца на лапках распо-
ложены при основании члеников, тогда как у A. pulchritarsis каждое
кольцо опоясывает вершину одного членика и основание следующего.
Личинки полупрозрачные, от молочно-белого до сероватого цвета,
с желто-коричневой головой и более темной дыхательной трубкой.
Подвиды в основном различаются окраской среднеспинки, а по ли-
чинкам — длиной жабр.
Распространение. Средиземноморская подобласть Палеарк-
тики, южная Азия. В СССР — Южный берег Крыма и другие области
Украины на север до Ужгорода и Харькова, Кавказ, Средняя Азия.
Биология. Выплаживается в дуплах деревьев. Обычен во влаж-
ных лесах и в зоне поливного земледелия. Не принадлежит к числу мас-
совых кровососов.
Aedes pulchritarsis pulchritarsis Rondani.
Среднеспинка в золотистых чешуйках, на фоне которых выделяются
бурые пятна; иногда- темная окраска преобладает. Бедра и голени спе-
реди темные, с небольшими пятнами белых чешуек или с разбросанными
светлыми чешуйками; нередко бедра и голени спереди сплошь темные
с белой вершиной. Лапки с кольцами чисто белого цвета.
Личинка IV стадии с головой почти округлой формы, с широ-
ким передним краем у лобного щитка. Размеры изменчивы, в зависи-
мости от величины дупла.
Лобные волоски смещены к переднему краю щитка, наружные из
6—13 (чаще 10—12), средние из 4—7 (чаще 6—7) ветвей лежат впереди
от более длинных внутренних из 8—10 (редко меньше), лежащих на одной
линии с наружными; задние волоски наличника из 11—18 ветвей, отходят
между средними и внутренними лобными волосками. Усики тонкие,
слабо изогнутые, лишенные шипиков; пучок у середины 1—4 (чаще 2—3)
тонких коротких волосков.
Щетка по бокам VIII членика из 6—10 (чаще 8) чешуек, расположен-
ных в один, иногда неправильный, ряд; каждая чешуйка с заостренным
концевым шипом, с очень тонкими короткими шипиками у основания.
Дыхательная трубка тонкая, прямая, индекс 4—5; гребень из 17—24
близко сидящих зубцов, относительно коротких, с широким основанием,
несущим несколько шипиков; пучок у середины из 3—4 ветвей. Стиг-
мальная пластинка с подкововидным задним отростком рычага; органы
на верхней поверхности боковых клапанов через свои основания функ-
ционально сильнее связаны со стигмальными отростками рычага, чем
со стигмами.
Последний членик со слабо развитым седлом, обычно только сверху
прикрывающим членик; боковой волосок простой и длинный; наружные
7. AEDES
201
хвостовые волоски простые, длинные, внутренние из 3—5 ветвей, более
длинных, чем жабры. Плавник развит очень слабо, из 6—8 (редко 9)
слабо ветвящихся (2—4, чаще 3 ветви) пучков, одной длины с внутрен-
ними хвостовыми волосками. Жабры достигают длины дыхательной
трубки, колбасовидной формы с округленными концами.
Р а с п р о с т ранение. Западная часть ареала вида, в СССР —
Украина, Кавказ.
Биология. Личинки встречаются в скоплениях воды в дуплах
деревьев (ясень, дуб, берест и др.), как в стволовых, так и в прикорневых
и пеньковых, часто совместно с A. geniculatus, An. plumbeus и Or. pulch-
ripalpis. Зимовка на фазе яйца. Вылупление личинок сразу после запол-
нения дупла водой. Начиная с марта—апреля несколько поколений.
Самки — второстепенные кровососы.
Замечания по систематике. В качестве особого вариетета был опи-
сан А. р. var. berlandi Seguy, 1921, который затем было предложено рассматривать
в качестве самостоятельного вида (Rioux et Arnold, 1955; Callot et Rioux, 1965).
У A. berlandi светлые пятна на бедрах и голенях отсутствуют или слабо выражены,
среднеспинка большей частью покрыта золотистыми чешуйками различных оттенков,
иногда с зачатками продольной белой полосы; в строении гипопигия различий нет.
Форма, обозначаемая как A. berlandi, распространена в странах Средиземноморья,
от Алжира до Ирана, часто встречается вместе с «типичным» А . pulchritarsis. По нашему
мнению, для признания видовой самостоятельности А. berlandi нет достаточных основа-
ний. Признаки, указываемые как отличительные, весьма изменчивы. Экземпляры
из Крыма и Закавказья обнаруживают различные варианты окраски среднеспинки
и разную степень развития светлых пятен на бедрах и голенях.
Aedes pulchritarsis asiaticus Edwards, 1926
(var. stegomyina Stackelberg et Montchadsky, 1926).
Отличается главным образом окраской среднеспинки. Голова в белых
и черных чешуйках, сгруппированных пятнами. Среднеспинка в черно-
бурых или темно-коричневых чешуйках, с продольной срединной поло-
сой белых чешуек и белыми попе-
речными полосками; иногда полоски
среднеспинки образованы желтова-
тыми чешуйками, неясно выделяю-
щимися на основном фоне; боковые
края среднеспинки в белых чешуй-
ках. Кольца на лапках кремовые.
Личинка (рис. 112 и 113)
этого подвида отличается от основ-
ной формы следующими признаками:
менее длинная дыхательная трубка
(индекс 3.0—3.3); меньшее в сред-
нем число зубцов в гребне (16—18);
жабры вдвое длиннее дыхательной
трубки и почти одной толщины с нею;
Рис. 112. Aedes pulchritarsis asiaticus
Edw. Личинка IV стадии. Голова,
сверху.
плавник с тем же числом пучков, но
ветвление каждого из них слабее —
не более 2 ветвей, что связано с
более сильным развитием жабр.
Распространение. Восточная часть ареала вида. В СССР —
Средняя Азия: Узбекистан, Туркмения, Таджикистан, где подымается
в горы до высоты 2100 м над ур. м.
Биология. Личинки встречаются только в скоплениях воды
в дуплах деревьев (тополь, грецкий орех, карагач и др.), как в прикор-
202
III. ПОДСЕМ. CULICINAE
невых, заливаемых при сбросе воды с полей орошения, так и в стволовых
на высоте 0.5—2.0 м, с марта до октября. Зиму и засушливый период
проводят на фазе яйца, которая очень устойчива к высыханию. В Таджи-
кистане встречается совместно с An. plumbeus, но численно преобладает
в дуплах с менее щелочной водой, содержащей меньшее количество солей.
Самки — кровососы второстепенного значения. Несколько поколений
в течение года.
Рис. 113. Aedespulchritarsis asiaticus Edw. Личинка IV стадии.
1 — стигмальная пластинка, г — задний конец, сбоку.
Виды группы cantans.
Лапки с широкими светлыми кольцами при основании члеников
(рис. 97, Б). Комары крупных размеров, реже средней величины. В строе-
нии гипопигия нет особенностей, характерных для всех видов группы.
Личинки отличаются наличием 4—7 более коротких, расположенных
впереди от объединенных общим основанием пучков плавника.
Комары этой группы нападают почти исключительно под открытым
небом, в дома проникают лишь в виде исключения.
К группе cantans, характерной для лесной зоны Палеарктики и Неарк-
тики, принадлежит восемь палеарктических видов; все они встречаются
в СССР. Распознавание некоторых из этих видов по самкам представляет
значительные трудности и не всегда может быть достоверным. Данные о
географическом распространении видов группы? нуждаются в уточнении.
4. Aedes (Ochlerotatus) cantans Meigen, 18181.
Крупный комар, отличается темной окраской; на хоботке, крыльях,
бедрах и брюшке среди темных чешуек вкраплено много контрастно
выделяющихся белых или желтоватых.
1 Во многих работах различных авторов вид фигурировал как А. maculatus MeiJ
gen, 1804; но это имя следует считать синонимом A. rusticus Rossi, 1790.
7. AEDES
203
Голова в желтовато-белых и бурых чешуйках; последние обычно
сгруппированы в виде двух крупных пятен. Щупики самки бурые, с при-
месью белых чешуек и белой вершиной; щупики самца бурые, с неотчет-
ливыми белыми кольцами при основании члеников. Среднеспинка, сильно
Рис. 114. Гипопигий Лейех cantans Mg.
варьирующая в окраске, покрыта шоколадно-коричневыми чешуйками;
беловато-серые или кремовые чешуйки образуют пятна неопределенных
очертаний в боковых частях среднеспинки или покрывают боковые от-
делы среднеспинки почти целиком. Щиток в белых или желтоватых че-
шуйках. Бочки с пятнами белых чешуек. Лапки с белыми кольцами при
основании члеников; на средних члениках задней лапки каждое кольцо
занимает обычно немного менее половины длины соответствующего чле-
ника. Коготок на лапке круто изогнутый (рис. 102). Крылья в темных
чешуйках, с более или менее значительной примесью светлых. Брюшко
сверху с нерезко отграниченными белыми или кремовыми перевязями
при основании тергитов; иногда перевязи очень узкие или почти отсут-
ствуют. Среди темных чешуек, покрывающих тергиты, вкраплено боль-
шее или меньшее' количество светлых.
Гипопигий (рис. 114) весьма своеобразен: базальная бородавка кок-
сита с одним толстым шипом, удлиненная, ее высота значительно превы-
шает ширину; крыло класпеты очень широкое.
Личинка IV стадии (рис. 115) крупных размеров, коричне-
вой окраски, с более светлой головой и дыхательной трубкой. Лобные
204
III. ПОДСЕМ. CULICINAE
волоски: наружный из 6—11 (чаще 7—8), средний из 2—3 (чаще 2), впе-
реди внутреннего (из 3—4), ветвей; задние волоски наличника из 2
(реже 3) тонких коротких ветвей, между основаниями средних. Усик
вдвое или более короче длины головы, слабо изогнутый, с хорошо замет-
ными шипиками и с коротким пучком из 6—9 ветвей.
Рис. 115. Aedes cantans Mg. Личинка IV стадии. Задний
конец, сбоку.
Щетка на VIII членике брюшка из 28—40 (чаще 35—36) чешуек, рас-
положенных в 2—4 неправильных ряда; каждая чешуйка имеет острый
главный шип с мелкими шипиками по бокам основания, ближайшие
к нему 1—2 могут быть заметно крупнее. Дыхательная трубка прямая,
начиная с 2/в от основания постепенно сужается к вершине (индекс 3.0
или немного меньше); гребень из 21—33 (в среднем 26—27), из которых
2—6 базальных рудиментарны, тесно сидящих шиповидных зубцов, не
доходит до середины трубки; пучок у середины, из 5—7 ветвей, не пре-
вышающих ширины основания трубки. Стигмальная пластинка (рис. 116)
характерна по строению заднего отростка рычага: между основаниями
направленных назад боковых ветвей имеется различно развитая (чаще
7. AEDES
205
Рис. 116. A edes cantans Mg. Личинка IV стадии.
Стигмальная пластинка.
слабо) срединная ветвь в форме продолговатого листка; кроме того, перед-
ний клапан сильнее, чем у других видов, развит в длину.
Седло последнего членика доходит до середины его боковых сторон;
боковой волосок длинный, простой; наружные хвостовые волоски длин-
ные и простые, внутренние из 7—9 ветвей, вдвое короче наружных. Плав-
ник из 18—19 пучков в общем
основании и из четырех (реже
пяти), более коротких, впереди
него. Жабры варьируют в дли-
ну: редко одной длины с сед-
лом, обычно в 1.5 раза
длиннее.
Распространение.
От Западной Европы до Даль-
него Востока. В западных ча-
стях ареала весьма обычен, на
востоке редок. Характерен для
подзоны смешанных лесов; про-
никает в тайгу. Сведения о на-
хождении в лесотундре и тундре,
вероятно, относятся к другим
видам той же группы. На юг
доходит до Северного Кавказа,
Крыма и стран Южной Евро-
пы — Италии, Испании.
Биология. Личинки
встречаются в разного рода
весенних водоемах — лужах,
ямах, канавах с большим ко-
личеством опавшей листвы,
лишенных травянистой растительности или с небольшим ее развитием,
расположенных по лесным опушкам или в разреженных участках леса,
часто совместно с A. communis и A. punctor. Средневесенний вид, вылетаю-
щий позднее перечисленных видов и имеющий одно поколение в году;
иногда после летних дождей бывает повторный небольшой выплод. Зи-
мовка на фазе яйца; последние откладываются на влажную землю по
берегам водоемов. Один из наиболее широко распространенных массовых
кровососов, нападающих на людей и животных; во многих районах лес-
ной полосы европейской части СССР занимает по численности одно из
первых мест. Нападает почти исключительно в природных условиях.
5. Aedes (Ochlerotatus) riparius Dyar et Knab, 1907 (semicantans Mar-
tini, 1920).
Близок к предыдущему. Достоверно отличается по самцам и личин-
кам. Комар средней величины, реже крупный.
Среднеспинка, как правило, с отчетливой продольной полосой темно-
бурых или бронзовых чешуек. Боковые отделы среднеспинки в более
светлых золотистых или кремовых чешуйках, иногда с узкими бронзо-
выми полосками одного цвета с широкой срединной полосой. Передняя
поверхность задних бедер чаще светлая. Коготок на лацке полого изогну-
тый. Белые перевязи при основании тергитов брюшка относительно ши-
рокие и четко отграниченные; иногда узкие светлые перевязи выражены
и у вершины тергитов.
206
III. ПОДСЕМ. CULICINAE
Окраска у данного вида весьма изменчива. Так, североамериканские
популяции характеризуются отсутствием темной полосы на среднеспинке.
Ввиду изменчивости окраски самки не всегда могут быть определены
с уверенностью. Более надежно определение по строению гипопигия
(рис. 117). У A. riparius по сравнению с A. cantans базальная бородавка
коксита менее удлинена, ее высота приблизительно равна ширине при
основании; крыло класпеты
более узкое.
Личинка IV ста-
дии (рис. 118) средних раз-
меров и темной окраски.
Лобные волоски вторичнопе-
ристые: наружный из 4—9
(среднее 5—6), средний впе-
реди внутреннего, обе пары
из двух (редко трех) ветвей;
задние волоски наличника
между основаниями средних
тонкие, короткие, 3—4-вет-
вистые. Усик немного превы-
шает длину головы, почти
прямой, с развитыми шипи-
ками и с пучком у середины
из 3—5 ветвей.
Щетка VIII членика
брюшка из 6—9 крупных че-
шуек, расположенных в один
неправильный ряд; каждая
чешуйка с сильно развитым
заостренным шипом, по бо-
кам основания которого си-
дят более мелкие шипики.
Дыхательная трубка прямая,
начиная с базальной трети
слегка сужающаяся к вер-
шине, индекс — 3.5—4.0; гре-
бень из 14—21 (среднее 16)
стройных, тонких зубцов, за-
нимающих немного более
Рис. 117. Гипопигий Aedes riparius riparius D. К. трети ДЛИНЫ трубки, 2 3 бли-
По Carpenter a. La Casse. жайших к вершине зубца
более широко расставлены;
пучок у середины, немного ближе к основанию, из 3—5 ветвей, со сла-
бой вторичной перистостью, достигает 7з длины трубки.
Седло последнего членика на 4/в или больше покрывает его боковые
стороны; боковой волосок простой, почти равен длине седла; наружный
хвостовой волосок простой, превосходит длину дыхательной трубки,
внутренний в виде веера из 5—8 вдвое более коротких ветвей. Плавник
из 15—18 пучков в общем основании и из 4—6 более коротких впереди
него. Жабры не менее чем в 1.5 раза длиннее седла, тонкие, пигментиро-
ванные, с заостренными концами.
Отличия по личинкам между обоими подвидами A. riparius не уста-
новлены.
Рис. 118. Aedes riparius D. К. Личинка IV стадии. По Car-
penter a. La Casse.
1 — передний конец, сверху, 2 — задний конец9 сбоку.
208
III. ПОДСЕМ. CULICINAE
Распространение. Лесная зона Европы и Северной Азии,
Северная Америка. На севере доходит до границ лесной зоны, местами,
по-видимому, встречается и в тундре; на юге проникает в лесостепь,
Границы ареала точно не установлены. Принадлежит к числу относи-
тельно редких видов.
Биология. Личинки встречаются в весенних водоемах открытых
ландшафтов, преимущественно в торфяных болотах со сфагновым дном,
вместе с A. punctor. Зимовка на фазе яйца. Как правило, одно поколение
в году. Самки — активные кровососы, но не встречаются в большой
численности.
Распадается на 2 подвида.
Aedes riparius riparius Dyar et Knab.
Хоботок темный, с примесью светлых чешуек, образующих в средней
части хоботка расплывчатое кольцо. Щупики самки в темных чешуйках,
с небольшим количеством светлых. Последний членик задней лапки при
основании с белым кольцом. Крыло в темных чешуйках, среди которых
разбросаны светлые. Второй тергит брюшка без продольной светлой
полосы или такая полоса слабо выражена и развита не по всей длине
тергита.
Гипопигий (рис. 117) в общем похож на гипопигий A. cantans: стволик
класпеты прямой, крыло с очень короткой рукояткой, пластинчатое рас-
ширение наиболее выражено в средней части или ближе к основанию
крыла.
Европа, Сибирь. Североамериканская форма, по-видимому, более
близка к европейской.
Aedes riparius ater Gutsevich, 1955.
Хоботок, щупики самки и крылья в одноцветных бурых чешуйках;
несколько светлых чешуек может быть при основании костальной жилки.
Последний членик задней лапки без белого кольца. Второй тергит брюшка
с отчетливой продольной белой полосой, доходящей до заднего края
сегмента.
Гипопигий (рис. 119) не похож на гипопигий A. cantans: стволик
класпеты изогнутый, крыло с очень длинной рукояткой, пластинчатое
расширение выражено только в дистальной половине, а иногда лишь
в вершинной трети крыла.
Личинка А. г. ater неизвестна.
Приморье, Хабаровский край, Сибирь, северо-восток европейской
части СССР.
Замечания по систематике. Между обоими формами (подвидами?)
имеются довольно существенные отличия как в окраске, так и, особенно, в строении
гипопигия. Весьма возможно, что А. г. ater правильнее рассматривать как самостоя-
тельный вид, близкий к североамериканскому A. squamiger Coquillett, 1902.
6. Aedes (Ochlerotatus) behningi Martini, 1926.
По размерам меньше прочих представителей группы cantans', пре-
обладающая окраска темная.
Хоботок и щупики бурые, с небольшой примесью светлых чешуек.
Среднеспинка й одноцветных узких бронзовых или цвета ржавчины чешуй-
ках, иногда (чаще у самцов) с едва намечающейся темно-коричневой про-
дольной полосой. Покровы среднеспинки черно-бурые. Лапки со свет-
лыми кольцами, занимающими на средних члениках задней лапки около
половины длины членика, иногда менее половины. Крыло в бурых чешуй-
7. AEDES
209
ках, среди которых вкраплены немногочисленные светлые. Брюшко
в бурых чешуйках с примесью светлых, обычно образующих расплывчатые
пятна вдоль средней линии тела; иногда одно пятно охватывает два смеж-
ных тергита. Отчетливых светлых перевязей на брюшке нет. У самцов
светлые чешуйки, как правило, разбросаны на тергитах беспорядочно,
не образуя пятен.
Рис. 119. Гипопигий Aedes riparius ater Guts.
Отдельно изображена класпета,
Точное определение по самкам не всегда возможно. По строению
гипопигия (рис. 120) легко отличается от других видов. Хорошо развитая
коническая базальная бородавка коксита лишена шипов и крепких щети-
нок. У других видов группы cantans базальная бородавка с шипом,
а если шипа нет, то бородавка слабо выражена, плоская.
Личинка IV стадии (рис. 121) очень сходна с личинкой
A. excrucians и во многих случаях практически неотличима. Лобные
волоски грубые, вторичноперистые, наружный из 6—8, средний, впереди
внутреннего, из 2—3, последний из 2—4 (чаще 2) ветвей; задние волоски
14 Фауна СССР, А. В. Гуцевич и др.
210
III. ПОДСЕМ. CULICINAE
наличника из 6—8 очень тонких коротких ветвей. Усик около половины
длины головы, с резко выраженной шиповатостью и с пучком из 8—9 вет-
вей у середины.
Щетка из 20—28 (чаще 24) чешуек, расположена в 2—3 неправильных
ряда; чешуйки с хорошо развитым острым главным шипом с несколькими
дополнительными шипиками по бокам основания. Дыхательная трубка
«ужена к вершине, но менее сильно, чем у A. excrucians (индекс 3.0—4.0,
среднее 3.5); гребень из 18—28 (чаще 22—24) зубцов, 1—3 более круп-
Рис. 120. Гипопигий Aedes behningi Mart.
ные и сильнее расставлены; однако, как не редкое исключение, расстояние
между ними может быть не большим, чем у соседних зубцов; пучок из
пяти ветвей не достигает длины трубки и находится у середины, ближе
к вершине. Волоски на задних клапанах стигмальной пластинки крючко^-
образно изогнуты, но относительно немного тоньше, чем у A. excrucians.
О строении самой пластинки данные отсутствуют.
Седло почти доходит до нижнего края последнего членика; боковой
волосок простой, равен длине седла; наружные хвостовые волоски прос-
тые, одной длины с дыхательной трубкой, внутренние в виде веера из
14—17 ветвей. Плавник из 14—17 пучков в общем основании и из 5—6
более коротких впереди него. Жабры одной длины с седлом или лишь
немного длиннее, непигментированные.
Распространение. Найден в различных пунктах европей-
ской части СССР и Сибири, на восток до Красноярска и Барнаула, на
север до южной границы тайги, на юг до степной части Украины вклю-
чительно; вне пределов Советского Союза A. behningi найден в Чехосло-
вакии (Tipis, 1965&); на западе граница ареала вида не установлена.
Биология. Местами, в поймах рек, появляется и нападает в зна-
чительном количестве. Дает одно поколение за сезон, весной.
7. AEDES
21г
7. Aedes (Ochlerotatus) excrucians Walker, 1856.
От трех предыдущих видов отличается более светлой окраской, а также
строением гипопигия. Очень сильно варьирующий в окраске вид.
Хоботок и щупики в бурых и желтовато-белых чешуйках. Средне-
спинка в ржаво-желтых или красновато-бурых чешуйках, по бокам обычно
чешуйки более светлые. Иногда намечается неясная широкая продольная
полоса более темных чешуек. Покровы среднеспинки коричневые, реже
черно-бурые в средней части, коричневатые по бокам. Чешуйки на боках
груди желтоватого оттенка. Передняя поверхность передних бедер пест-
Рис. 121. Aedes behningi Mart. Личинка IV стадии.
Задний конец, сбоку. По Martini.
рая. Лапки со светлыми кольцами, ширина которых на средних члениках,
задней лапки составляет около половины длины соответствующего чле-
ника. Коготок лапки большой, круто изогнутый; крупный зубчик на-
правлен почти параллельно коготку (рис. 122). Встречаются, впрочем,
экземпляры, относящиеся, по-видимому, к A. excrucians, с менее разви-
тым и направленным в сторону зубчиком коготка. Крылья в перемешан-
ных темно-бурых и желтовато-белых чешуйках, обычно темные чешуйки
преобладают. Брюшко в бурых и желтовато-белых чешуйках; последние
обычно образуют нечеткие перевязи при основании, а иногда и у вершины
сегментов. Нередко тергиты на всем протяжении пестрые. Вообще окраска
брюшка исключительно изменчива. Например, у части экземпляров из
Забайкалья и Дальнего Востока темных чешуек на брюшке мало или
вовсе нет, на крыльях светлые чешуйки преобладают. Такие экземпляры,
похожи на A. cyprius и A. flavescens.
Гипопигий (рис. 122): базальная бородавка плоская, едва возвы-
шающаяся над поверхностью коксита, густо покрытая короткими во-
лосками, без крепких щетинок или шипов; стволик класпеты длинный,
слегка изогнутый, к вершине немного суженный; длина крыла превы-
шает его наибольшую ширину приблизительно в 3 раза.
1'4.'*'
212
III. ПОДСЕМ. CULICINAE
Личинка IV стадии (рис. 123) крупных размеров, желтовато-
коричневого цвета. Лобные волоски с различно развитой вторичной
пористостью: наружные из 4—9 (чаще 7—8), средние из 1—3 (чаще 2),
внутренние — позади последних — из 2—6 (чаще 2—3) ветвей; задние
Рис. 122. Гипопигий Aedes excrucians Walk. По Carpenter a. La Casse.
1 — коготок лапки, 2 •— гипопигий.
волоски наличника из 2—3 тонких коротких ветвей. Усик около
х/2 длины голЬвы с покрытым шипиками телом, у середины которого,
ближе к основанию, отходит пучок из 3—8 (чаще 7—8) вторичнопери-
стых волосков, достигающих половины длины тела усика.
Щетка на VIII членике брюшка в среднем из 30—40 (чаще 32—36)
чешуек. Наблюдается сильная изменчивость числа чешуек, в редких
случаях их может быть около 20 и даже меньше. Такие личинки по этому
признаку определить невозможно. Каждая чешуйка с резко выраженным
7. AEDES
213
главным шипом, по бокам основания которого сидят более мелкие и
тонкие шипики. Дыхательная трубка прямая и стройная, начиная от
2/5 от основания заметно сужается (индекс от 3.2 до 4;4, иногда доходит
до 5); гребень из 16—27 шиповидных зубцов, удлиняющихся в дисталь-
ном направлении, 1—3 наиболее удалены, крупнее и более широко
расставлены; пучок за серединой, из 5—6 широко расходящихся вто-
ричноперистых ветвей.
Рис. 123. Aedes excrucians Walk. Личинка IV стадии.
1 — голова, 2 — задний конец, сбоку.
Стигмальная пластинка (рис. 124) очень характерна и отличается от
других близких видов: расстоянием между вершинами задних клапанов,
значительно превышающим продольный диаметр пластинки; крючко-
видно изогнутыми и утолщенными волосками на вершинах задних кла-
панов и узкой темной хитинизацией верхней поверхности переднего кла-
пана, слабо расширенной в вершинной части.
Седло заходит за середину боковых сторон последнего членика; боко-
вой волосок простой, достигает длины седла; наружные хвостовые во-
лоски очень длинные, длиннее дыхательной трубки, внутренние в виде
веера из 7—9 ветвей, вдвое более короткие, чем наружные. Плавник из
214
III. ПОДСЕМ. CULICINAE
16—20 пучков, объединенных общим основанием, и из 4—6 более корот-
ких впереди него. Жабры длинные, не менее длины седла, но могут дости-
гать длины дыхательной трубки, непигментированные.
Распространение. Европа, Казахстан, Сибирь, Дальний
Восток СССР, Монголия, Северо-Восточный Китай, Япония, Северная
Америка. На севере доходит до границ тундры и местами, особенно по
поймам рек, проникает в тундру (на севере Норвегии до 70°); на юге —
до Крыма, Кавказа и гор Малой Азии. В зонах леса и лесостепи встре-
чается повсеместно, иногда в большой численности.
Биология. Личинки встречаются во временных весенних водое-
мах, реже в постоянных, с травянистой растительностью, в открытых
луговых стациях и поймах, вдоль лесных опушек и в разреженных участ-
ках леса; иногда в слабо соленой воде на морском побережье. Нор-
Рис. 124. Aedes excrucians Walk. Личинка IV стадии. Стиг-
мальная пластинка.
мально — одно весеннее поколение из перезимовавших яиц; вылет на-
чинается на неделю или декаду позже первого появления Aedes — средне-
весенний вид. Личинки при внешних раздражениях опускаются на дно,
часто на длительное время; искусственное взмучивание воды заставляет
их подняться на поверхность. Самки — массовые кровососы; лёт про-
должается довольно долго, иногда и летом наблюдается значительное
нападение.
Играет роль в циркуляции туляремийного микроба: установлена
естественная зараженность.
8. Aedes (Ochlerotatus) beklemishevi Denisova, 1955 (grandilarva Sazo-
nova, 1956).
Весьма близок к предыдущему. С полной уверенностью может быть
определен только по личинкам.
Среднеспинка в золотистых или бронзовых чешуйках. Передние бедра
спереди светлые с небольшим количеством темных чешуек. Коготок
лапки (рис. 102, 20) крупный, с резко отставленным зубчиком (Сазо-
нова, 1958). Крылья преимущественно в темных чешуйках с примесью
желтоватых. Передние тергиты с четкими светлыми перевязями при
основании сегментов, обычно и с очень узкими — у вершины тергитов.
7. AEDES
215
В строении гипопигия каких-либо отличий по сравнению с A. excrucians
не отмечено.
Личинка IV стадии (рис. 125) очень крупных размеров,
в среднем более 1 см длины, темно пигментирована. Лобные волоски
вторичноперистые, грубые, наружные из 5—9 (чаще 7—8), средние из
1—3 (в 90% — 2), впереди внутренних из 2—4 (чаще 3) ветвей; задние
2
Рис. 125. Aedes beklemishevi Den. Личинка IV стадии.
По О. Н. Сазоновой.
1 — голова, сверху, 2 — задний конец, сбоку.
волоски наличника между оснований средних, тонкие и короткие, из
2—7 (чаще 3—6) ветвей. Усик короткий, меньше половины длины головы,
слегка изогнутый и покрытый мелкими шипиками с пучком у середины
из 2—7 (чаще 3—5) ветвей.
Щетка на боках VIII членика брюшка из 14—20 (чаще 16—18) че-
шуек, расположенных в 2—3 неправильных ряда, с длинным с острой
вершиной главным шипом, по бокам основания которого сидят неболь-
шие тонкие шипики. Дыхательная трубка длинная, прямая, заметно
суженная в вершинной трети (индекс 3.1—3.5); гребень из 16—28 (чаще
216
III. ПОДСЕМ. CULICINAE
22—26) зубцов, из которых 2—5 у основания рудиментарные, а 2—6
(чаще 3—4) ближайших к вершине — более крупные и широко расстав-
ленные, крайний из них находится в концевой трети трубки; около него
сидит пучок из 2—4 (чаще 3) тонких, слабо вторичноперистых ветвей,
не достигающих в длину ширины основания трубки.
Волоски на концах задних клапанов стигмальной пластинки не крюч-
ковидно изогнутые и утолщенные (как у A. excrucians), а обычного для
личинок Ochlerotatus вида.
Седло последнего членика почти на 3/4 заходит на его боковые сто-
роны; боковой волосок простой, около 2/в длины седла. Наружные хво-
стовые волоски простые, немного длиннее дыхательной трубки, внутрен-
Рис. 126. Гипопигий Aedes an-
nulipes Mg. По Martini.
ние в виде веера из 11—16 (чаще 14—16)
вдвое более коротких волосков. Плавник
из 17—22 (чаще 19—20) пучков, объеди-
ненных общим основанием и в среднем из
4—7 более коротких пучков впереди него.
Жабры длинные, почти одной длины с
дыхательной трубкой, слабо пигментиро-
ванные.
Распространение. Описан по
материалу из Курской, Ярославской и
Московской областей (Денисова, 1955).
Найден в ряде пунктов европейской части
СССР и Сибири, на восток до побережья
Охотского моря. Границы ареала неизве-
стны. Часто встречается вместе с А. excru-
cians, но, как правило, значительно
уступает ему по численности.
Биология. Личинки во времен-
ных открытых водоемах на лугах и вы-
рубках, реже в лесах. Поздневесенний вид,
имеющий одно поколение, но при выпаде-
нии дождей единично встречающийся и
осенью. Зимовка на фазе яйца. При ши-
роком распространении в массе не встре-
чается. Самки — активные кровососы, на-
падающие в дневное и в вечернее время.
9. Aedes (Ochlerotatus) annulipes Meigen, 1830 (quartus Martini, 1920).
Весьма близок к двум предыдущим видам. Отличается окраской груди
и брюшка; есть небольшое отличие и в строении гипопигия. Ввиду боль-
шой изменчивости окраски определение по этому признаку не вполне
надежно.
Среднеспинка с широкой темной продольной полосой шоколадно-
коричневых чешуек; боковые части среднеспинки в кремовых или серова-
тых чешуйках. Брюшко с четкими желтоватыми перевязями при основа-
нии тергитов; светлые чешуйки могут быть и у вершины сегментов.
Гипопигий (рис. 126) в основном как у A. excrucians, но стволик
класпеты относительно толстый, к вершине не суженный, иногда стволик
в вершинной части даже немного толще, чем при основании. у
Личинка IV стадии (рис. 127) очень близка к личинке А. can-
tans. Личинки последнего вида, несколько отклоняющиеся от типичных,
определялись как A. annulipes.
7. AEDES
217
Наружные лобные волоски в среднем из 7, средние впереди внутрен-
них, первые из 1—4 (чаще 2), вторые из 3—6 (чаще 4) относительно грубых
вторичноперистых ветвей; задние волоски наличника между основа-
ниями средних короткие, тонкие, из 2—3 ветвей. Усик слегка изогнут,
более темный к вершине, с хорошо заметными шипиками и пучком у сере-
дины из 4—7 ветвей.
Щетка на VIII членике брюшка из 29—45 (среднее 33—34) чешуек,
расположенных в 2—4 неправильных ряда; каждая чешуйка имеет остро-
Рис. 127. Aedes annulipes Mg. Личинка IV стадии. Задний конец,
сбоку. По Marshall.
конечный главный шип и ряд шипиков по бокам основания, из которых
1—2 ближайших к последнему могут быть развиты сильнее, но не до-
стигают размеров главного. Дыхательная трубка начиная от середины
умеренно сужается (индекс в среднем 3.0, до 3.5). Гребень из 17—26
(чаще 21—23) тесно сидящих зубцов и 1—4 рудиментарных зубчиков
у основания; занимает не более 2/в длины трубки от основания. Пучок
у середины, из 3—7 (чаще 5) ветвей со слабой вторичной перистостью,
более длинных, чем ширина трубки у его отхождения. Стигмальная
пластинка с двураздельным задним отростком рычага обычного для под-
рода строения.
Седло последнего членика заходит за середину его боковых сторон;
боковой волосок простой; наружные хвостовые волоски простые, немного
218
III. ПОДСЕМ. CULIC1/NAE
длиннее дыхательной трубки, внутренние почти вдвое короче, в среднем
из 8 ветвей. Плавник в среднем из 16 пучков в общем основании и 4—5
более коротких впереди него. Жабры обычно немного короче седла,
удлиненно ланцетовидной формы.
Распространение. Западная и Центральная Европа — до
юга Швеции на севере и Югославии на юге. Местами, например в неко-
торых районах Польши и Чехословакии, A. annulipes— массовый вид
(Skierska, 1963; Dabrowska-Prot, 1964; Trpis, 1965а). В литературе
имеется много указаний на нахождение данного вида в различных пунк-
Рис. 128. Гипопигий Aedes jlavescens Mull.
тах европейской и азиатской частей СССР, преимущественно в пределах
лесной зоны. Большинство этих указаний, вероятно, относится к другим
видам группы cantans. Определение по самкам нельзя считать надежным
при отсутствии самцов. То же отмечает и О. Н. Сазонова (1958). Ве-
роятно, A. annulipes в СССР распространен только в европейской части:
Эстония (Ремм, 1957), Украина (Вальх, 1959).
Биология. Личинки встречаются в открытых или частично за-
тененных кустарником весенних водоемах, в низинах на лугах и на лес-
ных опушках, часто совместно с A. cantans, но в меньшей численности.
Средневесенний вйд, имеющий одно поколение; зимовка на фазе яйца. Ли-
чинки по поведению похожи на Л. excrucians. Самки — активные кровососы.
10. Aedes (Ochlerotatus) flavescens Muller, 1764 (lutescens Fabri-
•cius, 1775).
Крупный рыжеватый комар относительно темной окраски. Отли-
чается от предыдущих одноцветным светлым брюшком, а также строе-
нием гипопигия.
7. AEDES
819
Хоботок и щупики самки в перемешанных бурых и желтовато-белых
чешуйках. На щупиках самца светлые чешуйки образуют широкие кольца.
Среднеспинка в мелких темных чешуйках ржавого или красновато-бурого
Рис. 129. Aedesflavescens Miill. Личинка IV стадии.
По Carpenter a. La Casse.
1 — передний конец, сверху, 2 — задний конец, сбоку.
оттенка. Бочки груди в серовато-белых чешуйках, сплошь покрывающих
дорсальные мезэпистерны. Нижние мезэпимерные щетинки отсутствуют.
Ноги со светлыми кольцами, занимающими на средних члениках задней
лапки около половины (или немного более) длины соответствующего
220
III. ПОДСЕМ. CULICINAE
Рис. 130. Aedes flavescens
Miill. Личинка IV ста-
дии. Дыхательная
трубка.
Седло последнего
членика. Крылья в бурых чешуйках, среди которых вкраплены желто-
вато-белые. Брюшко сверху в одноцветных желтовато-серых чешуйках;
иногда среди них вкраплены единичные темные чешуйки.
Гипопигий (рис. 128) характеризуется наличием на базальной боро-
давке коксита очень толстого темного шипа; бородавка слегка уплощен-
ная, густо покрытая короткими волосками, сидящими на маленьких
бугорках. Класпеты с коротким прямым стволиком, крыло относительно
короткое, с рукояткой и хорошо выраженным пластинчатым расшире-
нием в вершинной половине.
Личинка IV стадии (рис. 129) круп-
ных размеров, полупрозрачная, желто-коричне-
вого цвета. Лобные волоски вторичноперистые:
наружные из 6—9, средние из 2—4 (чаще 3) лежат
впереди внутренних из 2—4 ветвей; задние волоски
наличника очень малы, из 2—3 ветвей, лежат
между основаниями средних. Усик короткий,
менее х/2 длины головы, с хорошо развитыми
шипиками и с пучком из 5—8 ветвей со слабой
вторичной перистостью, отходящим у середины
ближе к основанию усика и достигающим не-
много менее половины его длины.
Щетка на VIII членике брюшка из 17—36
(чаще 20—27) чешуек, расположенных в три
ряда; каждая чешуйка несет длинный заострен-
ный шип с узким базальным отделом и с хорошо
развитыми шипиками по краю основания чешуйки.
Дыхательная трубка (рис. 130) прямая, на-
чиная с середины сужается к вершине; индекс
3.2—4.0. Гребень заметно не доходит до середины
трубки, состоит из 17—28 (чаще 19—22) зуб-
цов, из которых 1—3 — наиболее дистальных —
более широко расставлены; встречаются особи с
нерасставленными дистальными зубцами; точное
определение их затруднительно. Пучок у середины,
из 4—7 (чаще 5—6) вторичноперистых ветвей,
достигающих х/4—х/3 длины трубки,
леника заходит за середину его боковых сторон;
боковой волосок простой, равен длине седла; наружные хвостовые во-
лоски простые, длиннее дыхательной трубки, внутренние в виде вееров
из 12—13 ветвей, вдвое более коротких. Плавник из 18—19 пучков
в общем основании и 6—7, более коротких, впереди него. Длина жабр
изменчива: от более коротких, чем седло, до вдвое превышающих его
длину; жабры не пигментированы, верхняя пара длиннее нижней.
Распространение. Ареал простирается от Атлантического
до Тихого океана, на севере до Карелии и Якутска включительно, на юге
до Закавказья, Средней Азии, Северного Китая (Цайдам), Монголии.
Встречается в Северной Америке, на Аляске доходит почти до Полярного
круга.
Биология. Личинки встречаются в открытых небольших или,
крупных луговых или пойменных водоемах и заболоченностях, возник-
ших от таянья снега или весеннего разлива рек и озер, поросших травя-
нистой растительностью. Развитие личинок медленное, и вылет самок
происходит в конце весны; лёт продолжается до конца лета. Одно поколе-
ние в году. Зимовка на фазе яйца.
7. AEDES
221
Во многих районах подзоны смешанных лесов и лесостепи появляется
в значительном количестве, хотя обычно и не относится к преобладающим
по численности видам. Обычен в населенных пунктах, но в дома и другие
постройки залетает редко. Активно нападает на людей и сельскохозяй-
ственных животных.
В условиях опыта прослежено сохранение туляремийного микроба
в организме A. flavescens в течение 20 суток, возможность передачи ми-
кроба уколом — до 16 дней, выделение микробов с испражнениями —
до 14 дней после заражения комаров возбудителем туляремии (Олсу-
фьев, 1941а). *
И. Aedes (Ochlerotatus) cyprius Ludlow, 1920 (freyi Edwards, 1921).
Крупный рыжий комар, отличается от предыдущего более светлой
окраской самки; общая окраска самцов темнее.
Хоботок и щупики в перемешанных бурых и желтоватых чешуйках.
Среднеспинка в светлых золотистых или охряно-желтых чешуйках,
Рис. 131. Гипопигий A edes cyprius Ludl.
иногда со слабо выраженной более темной продольной полосой. Покровы
среднеспинки светло-коричневые у самок, темно-коричневые или бурые
у самцов. Бочки груди в желтовато-белых чешуйках. Чешуйки не покры-
вают целиком дорсальные мезэпистерны, а сгруппированы на этом скле-
рите в виде двух более или менее разобщенных пятен. Нижние мезэпи-
мерные щетинки имеются. Лапки, с широкими светлыми кольцами, желтая
окраска которых ярко контрастирует с темной окраской дистальной
части каждого членика; на средних члениках задней лапки ширина кольца
составляет около 2/3 длины членика. Крылья в перемешанных бурых
и желтых чешуйках, последние обычно преобладают. У самцов окраска
222
III. ПОДСЕМ. CULICINAE
крыльев и ног не столь пестрая, как у самой;- на крыльях преобладают
темные чешуйки. Тергиты брюшка покрыты одноцветными охристо-жел-
тыми чешуйками, среди которых могут быть вкраплены единичные темные.
Гипопигий (рис. 131): базальная бородавка коксита коническая с од-
ним шипом и тонкими щетинками; стволик класпеты довольно длинный,
слегка изогнутый; крыло класпеты без рукоятки, пластинчатое расши-
рение наиболее выращено в средней части крыла.
Личинка IV стадии (рис. 132) крупных размеров, темной
окраски, непрозрачная, с ярко-желтой головой и дыхательной трубкой.
Поверхность кутикулы на всем теле личинок начиная со II стадии густо>
inilH 11 '‘""ц., ’
flt I // иЧ Ч I 1 I I I I 1 Ч 1| v [*,'
г
з
Рис. 132. Aedes cyprius Ludl. Личинка IV стадии. ’
1 — сифон, сбоку; шипики на поверхности кутикулы личинок III (2)
и IV (3) стадий.
покрыта темными мелкими шипиками хетоидного типа, расположенными
неправильными поперечными рядами, тесно прилегающими друг к другу.
По этому признаку личинки этого вида резко отличаются от других пред-
ставителей рода Aedes.
Наружные лобные волоски из 2—3 ветвей, средние впереди внутрен-
них, обе пары из двух ветвей, реже простые; задние волоски наличника
тонкие, чаще простые. Усик короткий, слегка изогнут и утолщен у осно-
вания, с мелкими шипиками и пучком из 1—3 ветвей у середины.
Щетка VIII членика брюшка из 9—15 (чаще 10—12) чешуек, располо-
женных двумя неправильными рядами; каждая чешуйка с одним крупным
острым шипом и с несколькими более короткими шипиками по бокам
основания. Дыхательная трубка длинная, стройная, постепенно сужается
к вершине (индекс 4.0—4.6); гребень из 17—27 (чаще 20—21) зубцов,
из которых 2—3, наиболее удаленные от основания, крупнее остальных
и более широко расставлены; пучок лежит за серединой, ближе к вер-
шине, состоит из 3—4 тонких ветвей, в длину превышающих ширину
основания трубки.
Седло последнего членика доходит до середины его боковых сторон;
боковой волосок простой; наружные хвостовые волоски простые,, за-
7. AEDES
газ
метно длиннее дыхательной трубки, внутренние в виде веера из 9—10
ветвей, вдвое короче наружных. Плавник из 16—18 пучков, окружен-
ных общим основанием, и из 3—6 (чаще 4—5) коротких пучков впереди
него. Жабры одной длины с седлом, если длиннее, то не более чем в 1.5 раза,
удлиненно ланцетовидной формы, пигментированы.
Распространение. Лесная и лесостепная зоны Палеарктики
от Западной Европы до Хабаровского края и Приморья. На севере про-
проникает в южную тайгу (в Карелии до 63°), на юге — до степной
Украины и Центрального Казахстана.
Биология. Личинки развиваются в крупных по площади, глу-
биной 40—80 см водоемах, возникших или в результате таянья снега,
или разлива рек, на берегах, поросших кустарником и богатых травяни-
стой растительностью. Встречаются одновременно с личинками A.jla-
vescens, A. excrucians, A. cantans, A. riparius и др., но держатся не у бе-
регов, а в более глубоких частях водоема с более низкой температурой.
Средневесенний вид, имеющий одно поколение; зимовка на фазе яйца.
Самки интенсивно нападают на человека и животных и местами, осо-
бенно в Сибири, являются злостными кровососами.
Виды группы rusticus.
Группа включает четыре относительно редких вида — А. rusticus,
A. refiki, A. lepidonotus и A. subdiversus, для которых характерно стро-
ение гипопигия: базальная бородавка коксита несет группу удлиненных
прозрачных чешуек ланцетовидной формы. Самки не обладают столь
характерным отличительным признаком. От двух предыдущих групп
комары группы rusticus отличаются отсутствием светлых колечек на
лапках. Чешуйки на проэпимерах, как правило, относительно ши-
рокие, прямые (отличие от группы communis).
Личинки этой группы отличаются дополнительными волосками на
дыхательной трубке, помимо обычной для Aedes пары пучков у ее сере-
дины: 2—3 пар грубых волосков на ее передней поверхности и пары
более тонких и коротких волосков на ее боковых сторонах около наиболее
дистальных зубцов гребня.
12. Aedes (Ochlerotatus) rusticus Bossi, 1790 (diversus Theobald, 1901).
Крупный комар, характеризующийся темным цветом широких че-
шуек, покрывающих верхнюю часть проэпимер.
Хоботок и щупики в черных чешуйках. Среднеспинка в золотисто-
бронзовых чешуйках, с двумя продольными темно-коричневыми или
черными полосками, более выраженными в передней половине груди;
кроме того, могут быть развиты две, расположенные латеральнее, узкие
темные полоски в задней половине среднеспинки. Чешуйки проэпимер
очень широкие, в верхней части черные. Лапки преимущественно в тем-
ных чешуйках. На крыльях преобладают темные чешуйки, среди которых
вкраплены единичные светлые. Тергиты брюшка при основании со свет-
лыми перевязями, расширенными в средней части; иногда эти расширения
образуют почти сплошную продольную светлую полосу.
Гипопигий (рис. 133): базальная бородавка коксита с пучком ланце-
товидных чешуек, без шипов или крепких щетинок; придаток стиля
волнисто-изогнутый; стволик класпеты довольно длинный, крыло ко-
роткое, неправильной формы, без пластинчатого расширения.
Личинка IV стадии (рис. 134) крупных размеров, от корич-
невого до почти черного цвета. Наружные лобные волоски в среднем
224
III. ПОДСЕМ. CULICINAE
из восьми, средние из двух (реже трех), лежат впереди внутренних,
которые из трех (реже двух) ветвей. Усики короткие, около х/2 длины
головы, тонкие, почти прямые, покрыты шипиками; пучок около середины,
ближе к основанию, из 5—6 вторичноперистых ветвей, достигающих
х/2 длины усика.
Щетка из 12—18 (чаще 14—15) чешуек, расположенных в два непра-
вильных ряда; каждая чешуйка с длинным острым шипом, 1—2 более
короткими боковыми шипами и тонкими базальными шипиками. Дыхатель-
ная трубка прямая, длинная (индекс 3—3.5), начиная от базальной х/3
Рис. 133. Гипопигий Aedes rusticus Rossi. По Natvig.
Отдельно изображены, при большем увеличении, стиль (А), класпета (В), и базаль-
ная бородавка коксита (В).
постепенно сужается. Гребень занимает базальные 2/Б трубки и состоит
из 15—25 тесно сидящих зубцов, удлиняющихся в дистальном направле-
нии; его продолжают 1—3 более длинных, широко расставленных шипа,
не заходящих на вершинную треть трубки. Пучок у середины трубки
около дистальных зубцов гребня в среднем из 6—8 вторичноперистых
ветвей. На передней поверхности трубки сидят три (реже четыре) пары
длинных грубых вторичноперистых, иногда ветвящихся волосков; пара
более тонких 1—2-ветвистых волосков на боковых сторонах трубки
за серединой гребня.
Седло заходит за середину боковых сторон последнего членика; бо-
ковой волосок простой, почти достигает длины седла; наружные хвосто-
вые волоски длинные и толстые, внутренние в виде более чем вдвое ко-
ротких вееров из 7—9 ветвей. Плавник сильно развит, в среднем из
16 пучков в общем основании и 3—4 более коротких впереди него. Жабры
почти вдвое короче седла, удлиненно листовидной формы, верхняя пара
длиннее нижней.
Распространение. Западная, Центральная и Южная Европа,
на север до Шотландии и Дании, Северная Африка, Малая Азия (var.
7. AEDES
225
subtrichurus Martini, 1927; отличается тем, что чешуйки среднеспинки
обычно одноцветные золотисто-коричневые, по бокам беловатые). В С.ССР
найден на Карпатах. Данные о находке в Ленинградской области тре-
буют подтверждения.
Биология. Личинки встречаются в лужах и других пересыхаю-
щих лесных водоемах с травянистой растительностью, полузатенен-
ных кустарником и деревьями; более редки в открытых постоянных
водоемах типа прудов.
Встречаются совместно с
личинками А. leucomelas
и A. cantans. В связи с
распространением вида да-
леко на юг годичный цикл
жизни меняется: в зави-
симости от условий зимо-
вать могут яйца, личинки
и оплодотворенные самки;
при этом, вследствие вы-
сыхания водоемов, наблю-
дается одно поколение в
году. Самки интенсивно
нападают на человека, но
не относятся к числу мас-
совых кровососов.
13. Aedes (Ochlerotatus)
refiki Medschid, 1928.
Отличается от преды-
дущего окраской терги-
тов брюшка, а также
строением гипопигия.
Хоботок и щупики в
черных чешуйках, иногда
с вкраплениями светлых
чешуек. Среднеспинка в
желтоватых чешуйках, с
широкой темной продоль-
ной полосой или с двумя
сближенными полосками.
Рис. 134. A edes rustic us Rossi. Личинка IV стадии.
Задний конец, сбоку. По Marshall.
проэпимер прямые, двухцветные, в ниж-
в верхней темно-коричневые или черные.
Бочки груди с пятнами
белых чешуек; чешуйки
ней части — беловатые,
Посткоксальное и гипостигмальное пятна чешуек имеются. Передние
и средние бедра и голени спереди в бурых чешуйках с примесью светлых.
Лапки темные. Крылья по преимуществу в темных чешуйках, основание
радиальной жилки покрыто светлыми чешуйками. Окраска брюшка
сильно варьирует. Чаще тергиты в черных чешуйках с нерезко ограни-
ченными белыми перевязями при основании; нередко белые перевязи
редуцированы, иногда до степени небольших пятен по бокам; могут быть
выражены узкие светлые перевязи и у вершины тергитов; иногда на тер-
гитах преобладают светлые чешуйки, среди которых вкраплены немно-
гочисленные темные.
Гипопигий (рис. 135): базальная бородавка уплощенная, она рас-
положена почти у середины внутренней поверхности коксита; бородавка
15 Фауна СССР, А. В. Гуцевич и др.
226
III. ПОДСЕМ. CULICINAE
усажена ланцетовидными поперечно исчерченными чешуйками; рядом
с бородавкой помещается дополнительный бугорок, несущий 2—3 длин-
ные щетинки. Стволик класпеты длинный, крыло вытянутое, поперечно
исчерченное. Удлиненные выросты IX тергита несут по 7—10 крепких
коротких щетинок. У заднего края VIII стернита находится скопление
весьма длинных щетинок.
Рис. 135. Гипопигий Aedes refikl Med.
Личинка IV стадии (рис. 136) меньше и темнее пигментиро-
вана, чем A. lepidonotus. Наружные лобные волоски из 6—9, средние
простые, редко из 2—3, внутренние позади средних из 2—5 (средние 3)
грубых темных вторичноперистых ветвей; задние волоски наличника
короткие, тонкие, из 3 ветвей. Усик короткий, слабо изогнутый, с ши-
пиками; пучок из 12—15 коротких и тонких ветвей.
Щетка по бокам VIII членика брюшка из 6—11 чешуек, расположенных
в один неправильный ряд, имеющих хорошо развитый заостренный глав-
ный шип с несколькими уменьшающимися боковыми шипиками у осно-
вания. Дыхательная трубка прямая, слегка суженная к вершине (ин-
декс 3.0—4.0, редко больше); гребень из 12—21 (чаще 13—16) тесно
сидящих зубцов и 1—2 широко расставленных крупных гладких шипов,
лишь немного заходящих за базальную треть трубки. Пучок из 6—9
сильно развитых вторичноперистых ветвей у середины трубки, но ближе
7. AEDES
227
к основанию; на передней поверхности трубки три пары грубых вто-
ричноперистых волосков, а на боковых поверхностях пара тонких
2—5-ветвистых волосков около дистальных шипов гребня.
Последний членик с седлом, почти доходящим до нижнего края боковых
сторон, боковой волосок из 1—3 длинных вторичноперистых ветвей;
наружные хвостовые волоски длинные и простые, внутренние на 1/3 ко-
роче, в среднем из девяти ветвей. Плавник хорошо развит, из 15 пучков
в общем основании и из 2—3 впереди него. Жабры листовидные, с окру-
гленными концами, в длину достигают длины седла, слегка пигменти-
рованы. /
Распространение. Южная и Центральная Европа (Испа-
ния, Франция, ГДР и ФРГ, Чехословакия, Венгрия, Югославия);
///// 1111 2
Рис. 136. Aedes refiki Med. Личинка IV стадии. По Medschid.
1—голова, сверху, г — задний конец, сбоку.
Малая Азия. В СССР с достоверностью обнаружен только в Крыму:
Южный берег, Симферополь. Относится к числу редких видов.
Биология. Личинки (Крым) встречались ранней весной в водо-
емах и лужах, в местах выемки дерна, с травяным или глинистым дном,
покрытым опавшей листвой. Зимуют на фазе яйца. Развитие личинок
начинается с конца февраля, вылет в конце апреля. Летнее поколение
возможно только при наличии воды в водоемах, где были отложены яйца.
В Крыму отмечалось нападение на человека (Величкевич, 1936).
14. Aedes (Ochlerotatus) lepidonotus Edwards, 1920.
Вид отличается наличием чешуек на постскутеллуме.
Голова сверху покрыта желтоватыми чешуйками, кнаружи от ко-
торых имеются черные торчащие чешуйки; боковые отделы головы в бе-
лых чешуйках. Хоботок черный. Щупики самки по длине равны х/3 длины
хоботка, черные, с незначительной примесью белых чешуек. Щупики
самца в большей своей части покрыты белыми чешуйками; вершина,
а в некоторых случаях и основания члеников в черных чешуйках; послед-
ние членики явственно утолщены, с пучками длинных волосков, желтых,
с черными вершинами на предпоследнем членике, одноцветно черных
15*
228
III. ПОДСЕМ. CULICINAE
на последнем. Среднеспинка в светло-желтых чешуйках; бочки груди
густо покрыты белыми чешуйками; постскутеллум с пучком светло-желтых
чешуек. Ноги: бедра и голени, за исключением покрытой черными че-
шуйками вершины, в желтовато-белых чешуйках; лапки в черных че-
шуйках, с примесью белых чешуек при основании I членика. Крылья
Рис. 137. Гипопигий Aedes lepidono-
tus Edw.
Рис. 138. Aedes. lepidonotus Edw.
Личинка IV стадии. По Medschid.
1 ~ голова, сверху, з задйий койец,
сбоку.
в большей своей части покрыты желтовато-белыми чешуйками, за исключе-
нием костальной и первой радиальной жилок, которые покрыты темными
чешуйками. Брюшко в большинстве случаев покрыто светлыми серовато-
желтыми чешуйками, к которым в незначительном числе примешиваются
беспорядочно разбросанные чешуйки; в некоторых случаях, особенно
у самцов, средняя часть тергитов почти сплошь покрыта черными че-
шуйками.
Гипопигий (рис. 137): коксит со слабо развитой базальной бородав-
кой, несущей пучок узких чешуек; добавочного выступа с длинными
щетинками близ базальной бородавки нет; вершинная едва различима;
класпеты имеют прямой утолщенный стволик и короткое, неправильной
формы крыло.
7. AEDES
229
Личинка IV стадии (рис. 138) средней величины, темно-
окрашенная. Наружные лобные волоски из 8—10, средние из 1—3, внут-
ренние из 3—4 ветвей, позади средних; все они вторичноперистые; задние
волоски наличника короткие и тонкие, из 2—3 ветвей. Усики более
чем вдвое короче головы, покрыты шипиками; пучок отходит от середины,
немного ближе к основанию, из 5—6 вторичноперистых ветвей, дости-
гающих х/2 длины усика.
Щетка из 6—11 крупных чешуек, расположенных в один неправиль-
ный ряд; каждая чешуйка имеет или один заостренный главный шип
с боковыми шипиками у основания, или последние достигают размеров
главного, а последний может быть рудиментарным или отсутствовать.
Дыхательная трубка прямая, изменчива по форме (индекс 2.5—4.0),
сужается начиная с базальной трети. Гребень из 9—21 (среднее 14—16)
зубца, число которых варьирует в прямом соответствии с изменениями
длины трубки; из них 1—2, наиболее дистальных, широко расставлены,
но не заходят за базальную половину; пучок из 8—13 вторичноперистых
ветвей, у середины трубки, обычно впереди расставленных зубцов гребня;
на передней поверхности трубки две пары грубых длинных вторично-
перистых волосков; на боковых сторонах трубки около наиболее удален-
ного зубца гребня отходит обычно простой гладкий волосок, длина ко-
торого превышает ширину трубки.
Седло последнего членика доходит до его нижнего края; боковой
волосок хорошо развит", из 3—5 длинных вторичноперистых ветвей;
наружные хвостовые волоски простые, превышают длину дыхательной
трубки; внутренние на х/3 короче, в среднем из 9 ветвей. Плавник из 16
пучков в общем основании и из двух значительно более коротких впе-
реди него. Жабры меньше длины седла, листовидной формы, верхняя
пара немного длиннее нижней. ,
Распространение. Греция, Турция (Анатолия).
Биология. Личинки встречаются в различных открытых ве-
сенних водоемах, в местах, затопленных весенним разливом рек, слабо
засоленных, часто совместно с A. detritus. В течение года одно поколение
ранней весной; зимовка на фазе яйца.
Замечания по систематике. По материалу из Сибири был описан
новый вид — A. albescens Edwards, 1921, который Мартини (Martini, 1931) идентифи-
цировал с A. lepidonotus. Приводим описание A. albescens по Эдвардсу. «Вид явно род-
ствен A. lepidonotus Edwards, но postnotum (постскутеллум) без чешуек. Почти все
чешуйки тела беловатые, но на хоботке и щупиках они большей частью коричневые.
Чешуйки на проэпимерах беловатые, довольно узкие, некоторые из них изогнутые.
Щупики довольно длинные, составляющие 1/3 длины хоботка. Мембрана крыла бело-
ватая; все чешуйки по переднему краю крыла беловато-желтые, на радиальной жилке
имеется несколько темных чешуек. Чешуйки на ногах беловато-желтые, на последних
члениках лапки коричневые; лапки без белых колец. Коготки (формула); 1 :1, 1 : 1,
1 : 1. Сибирь: Омск (Grano); одна самка в музее Хельсинки». Этот экземпляр был ис-
следован Мартини (Martini, 1931). По его заключению, у комара на постскутеллуме
явственно сохранилась одна чешуйка и заметно несколько точек — по-видимому,
следы прикрепления отпавших чешуек.
Основываясь на приведенном описании А . albescens и предложенной идентификации
albescens=lepidonotus, мы определяли относящихся к группе rusticus комаров из Се-
верного Казахстана как A. lepidonotus. Но ни личинок, ни самцов, соответствующих
описанию последнего, мы не находили, равно как не находили и комаров с чешуйками
на постскутеллуме. Поэтому теперь мы считаем более правильным найденных в Сибири
и Казахстане комаров группы rusticus отнести к A. subdiversis (см. ниже). Вопрос же
об A. albescens Edw. остается открытым: либо это самостоятельный вид, никем после
1920 г. не обнаруженный (что маловероятно), либо он идентичен одному из видов
группы rusticus—lepidonotus или subdiversus. Предварительно мы считаем, что
A. lepidonotus в СССР с достоверностью не обнаружен; A. albescens мы рассматриваем
как сомнительный вид.
330
Ш. ПОДСЕМ. CULICINAE
15. Aedes (Ochlerotatus) subdiversus Martini, 1926.
От прочих трех видов группы rusticus отличается строением гипопигия.
Голова сверху в светлых и темных чешуйках. Хоботок и щупики
самки в черных чешуйках, чаще с примесью светлых. Среднеспинка
с широкой, иногда неясно отграниченной продольной полосой бронзовых
или желтовато-коричневых чешуек, состоящей как бы из трех полосок —
более светлой срединной и более темных боковых. Боковая поверхность
груди с пятнами серебристых или беловато-кремовых чешуек. Верхняя
половина проэпимер в грязно-желтых, иногда более темных, бронзовых
Рис. 139. Гипопигий Aedes subdiversus Mart.
прямых или слегка изогнутых чешуйках. Посткоксальное и гипостигмаль-
ное пятна чешуек имеются; последнее составляет часть продольной по-
лосы, расположенной ниже переднего дыхальца. Ноги: бедра и голени
передних ног спереди пестрые; лапки бурые, без белых колечек, но со
значительной примесью светлых чешуек. Крылья преимущественно
в темных чешуйках, светлые сконцентрированы при основании и вдоль
некоторых жилок, главным образом в передней части крыла. Брюшко:
тергиты в беловато-серых или кремовых чешуйках, чаще с примесью
темных, иногда образующих неясные пятна; соотношение светлых и тем-
ных чешуек на брюшке сильно варьирует.
Гипопигий (рис. 139): стиль прямой или слегка серповидно (не волно-
образно)- изогнут, базальная бородавка коксита усажена узкими лан-
цетовидными чешуйками; близ нее имеется небольшой добавочный выступ,
несущий 2—3 длинные и несколько более коротких щетинок; стволик
класпеты длинный, с относительно коротким крылом, лишенным попе-
7. AEDES
231
речной исчерченности; фаллосома склеротизована, на вершине зазубрена,
с перетяжкой посредине; выросты IX тергита несут по 8—10 коротких
толстых щетинок; задний край VIII стернита близ середины густо уса-
жен длинными тонкими волосками.
Личинки IV стадии крупные, желто-коричневого цвета,
с немного более светлыми головой и сифоном.
Голова в ширину в 1.5 раза больше, чем в длину. Лобные волоски
слабо вторичноперистые, наружные из пяти, средние из 2—3, внутрен-
ние из двух ветвей; средние лежат впереди внутренних, немного более
широко расставлены; между их основаниями находятся 2-ветвистые
задние волоски наличника. Усики достигают половины длины головы,
покрыты шипиками; у их середины находятся пучок из 3 ветвей около
1/3 длины усика. Ротовые части смешанного типа питания — с субстрата
и планктоном, как у A. rusticus.
Щетка из 12—17 (чаще 14—15) чешуек с длинным и острым шипом
и только с мелкими зубчиками у основания, расположенных обычно
в два неправильных ряда. Волоски позади щетки как у A. rusticus. Сифон
(индекс 3—3.3) с гребнем, занимающим около 2/5 его длины и состоящим
из 11—16 зубцов, из которых 2—5, прилежащих к основанию, руди-
ментарные; 1—2 наиболее дистальных зубца более крупные и шире рас-
ставлены. Каждый зубец в виде шипа с дополнительными шипиками
у основания. Между дистальным зубцом гребня и вершиной сифона
расположены 3—4 гладких, более длинных шипа. Пучок отходит около
дистального зубца гребня или немного ближе к вершине, в виде длинного
простого волоска. Сбоку от средних зубцов гребня отходит тонкий 2-вет-
вистый добавочный волосок. На передней поверхности сифона имеются
3—4 пары длинных слабо вторичноперистых волосков; иногда часть их
расположена зигзагом.
Последний членик с седлом, охватывающим около 3/4 боковых сторон.
Наружные хвостовые волоски простые, одной длины с сифоном; внутрен-
ние — в форме веера из 11 ветвей. Плавник из 14—15 пучков в общем
основании и из 6—7 более коротких впереди него. Жабры и боковые
волоски в имеющемся материале обломаны.
Замечания по систематике. Вид характеризуется большой измен-
чивостью окраски. Светлые варианты, которые нами ранее определялись как А. 1е-
pidonotus (см. выше), отличаются одноцветно светлой окраской чешуек на тергитах,
значительной примесью светлых чешуек на щупиках самки и на хоботке, а также более
светлой окраской чешуек верхней половины проэпимер. Между «типичными» и свет-
лыми вариантами имеются постепенные переходы.
Распространение. От Волги (вид был описан по материалу
из Саратовской области; Martini, 1926, 1928) до Забайкалья. В Казахстане
и на юге Сибири довольно обычен. Границы ареала не установлены.
Биология. Дает одно поколение за сезон, встречается только
весной. Местами, например в Кустанайской области, отмечено массовое
нападение на людей и сельскохозяйственных животных.
Виды группы communis.
Как правило, комары средней величины. От видов, относящихся
к группам caspius и cantans, отличаются темными лапками без светлых
колец, а от комаров группы rusticus — строением гипопигия. Богатая
видами разнородная группа, включающая 16 палеарктических видов; не-
которые из них принадлежат к числу массовых кровососов. В зонах леса
и тундры среди комаров, нападающих на людей и сельскохозяйственных
232
III. ПОДСЕМ. CULICINAE
животных, виды группы communis обычно явно преобладают. Нападают
преимущественно в природных условиях под открытым небом. Но при
большой численности комаров они проникают и в населенные пункты,
нападают в домах и помещениях для животных.
Большинство видов группы дает за сезон только одно весеннее поко-
ление. После дождей может наблюдаться небольшой летний выплод
некоторых видов, количественно многократно уступающий весеннему.
Суммарная сезонная кривая активности комаров данной группы обычно
одновершинна, максимум приходится на конец весны и начало лета.
На севере тайги и в тундре комары, которые там представлены в основ-
ном видами группы communis, в наибольшем количестве появляются
в середине или во второй половине лета.
Важный систематический признак — расположение пятен чешуек на
боковой поверхности груди (Pens, 1933; рис. 99). Чешуйки легко отпа-
дают; поэтому самки группы communis с уверенностью могут быть опре-
делены только по экземплярам хорошей сохранности. Потертые экземп-
ляры, со сбитыми чешуйками, до. вида, как правило, неопределимы;
часто приходится ограничиваться определением до группы видов. Более
надежно определение по гипопигиям самцов и особенно по личинкам.
Рис. 140. Гипопигий Aedes communis Deg. По Natvig.
Отдельно изображены, при большем увеличении, базальная бородавка кок-
сита (А), лопасти IX тергита (Б) и крыло класпеты (В).
16. Aedes (Ochlerotatus) communis De Geer, 1776 (nemorosus Meigen,
1818).
Комар средней величины. От близких видов отличается отсутствием
посткоксального пятна чешуек, а также строением гипопигия.
Щупики самки и хоботок покрыты бурыми чешуйками. Последний
членик щупика самца утолщен. Среднеспинка в золотисто-желтых или
бронзовых чешуйках, обычно с двумя сближенными продольными тем-
ными полосками, которые могут быть слиты в одну; иногда они слабо
выражены или отсутствуют. Бочки груди в белых чешуйках, не имеющих
серебристого оттенка. Посткоксального пятна нет, т. е. нет чешуек на
небольшом перепончатом участке между основанием передних тазиков
и передним краем мезэпистерн. Пятно чешуек на мезэпистернах дохо-
7. AEDES
333
дит до их переднего угла. Пятно светлых чешуек непосредственно под
передним дыхальцем (гипостигмальное пятно) отсутствует. Чешуйки на
мезэпимерах доходят до их нижнего
края. Лапки в черно-бурых чешуйках,
иногда с небольшой примесью серых
или беловатых. Крылья в темных че-
шуйках, светлые лишь на небольшом
протяжении покрывают основание ко-
стальной жилки. Брюшко сверху с по-
перечными белыми перевязями у пе-
реднего края тергитов, остальная
часть каждого тергита в темных чешуй-
ках; перевязи на II—V сегментах брюш-
ка более или менее равномерной шири-
ны, без явственного сужения посре-
дине.
Гипопигий (рис. 140): коксит в длин-
ных волосках, с хорошо развитыми ба-
Рис. 141. Aedes communis Deg. Ли-
чинка IV стадии. Голова, сверху.
зальной и вершинной бородавками. Базальная бородавка полукруглая,
с одним длинным шипом; по внутреннему краю бородавки расположен
ряд коротких, изогнутых в одну сторону волосков. Стволик класпеты
длинный, прямой или слабо изогнутый, крыло очень узкое, длинное,
без расширения, слабо склеротизованное.
Личинка IV стадии (рис. 141—143) крупных размеров,
изменчивой, преимущественно темной окраски. Лобные волоски вторично-
перистые: наружные из 4—8, средние впереди внутренних, обе пары
234
III. ПОДСЕМ. CULICINAE
простые в редких случаях один из волосков из двух или, в виде исклю-
чения, из трех ветвей; задние волоски наличника тонкие, короткие,
из 3—4 ветвей. Усик около 1/2 длины головы, покрыт шипиками и слегка
изогнут, с пучком у середины, из 6—7 ветвей, не достигающих V2 длины
Рис. 143. Aedes communis Deg. Личинка
IV стадии.
1 — чешуйки щетки, 2 — стигмальная пластинка.
усика.
Щетка в среднем из 60—80 чешуек, расположенных неправильным
треугольным пятном; каждая чешуйка не имеет главного шипа, ее дис-
тальный конец усажен рядом
заостренных шипиков, иногда
заходящих и на боковые края.
Дыхательная трубка от середи-
ны сужается к вершине, индекс
2—3 (в среднем 2.7). Гребень
из 17—26 (чаще 21—22) тесно
сидящих зубцов, из которых
1—4 базальных рудиментар-
ны, остальные удлиняются по
направлению к вершине; гре-
бень не доходит до середины
трубки. Пучок у середины, из
5—9 (чаще 6—7) вторичнопе-
ристых ветвей, достигающих V3
длины трубки.
Седло доходит до середины
или нижнего края последнего
членика; боковой волосок ко-
роткий и простой; наружные
хвостовые волоски простые,
длиннее, чем трубка, внутрен-
ние в виде веера из 6—8 более
коротких ветвей. Плавник из
16—19 пучков в общем осно-
вании и двух более коротких,
впереди него. Жабры длинные,
в 1.5—2 раза длиннее седла,
пигментированные, с заострен-
ными концами.
Распространение.
Зоны леса и тундры обоих по-
лушарий: Европа, Сибирь, Даль-
ний Восток СССР, Северная
Америка. На севере доходит
до побережья Ледовитого океана (район Мурманска, устье Колы-
мы), на юге — до Восточного Казахстана, Северного Кавказа, степ-
ной Украины и Болгарии.
Биология. Личинки встречаются в различного рода постоянных
и временных водоемах, возникших от таянья снега, с дном, покрытым
опавшей листвой или другими растительными остатками. На юге пред-
почтительно встречается в затененных лесных стациях, на севере —
в открытых местах. Личинки могут появляться из перезимовавших яиц
еще подо льдом. Одно поколение в течение года, но во влажную осень
может быть небольшой дополнительный выплод личинок; в сухую весну
основная масса личинок может появиться при выпадении летних осад-
ков. В лесной зоне — один из наиболее обычных массовых видов. Во мно-
7. AEDES
235
гих местах среди комаров, нападающих на людей и животных, занимает
первое место. Вылет начинается раньше, чем у большинства других
видов Aedes. К середине лета численность комаров резко снижается.
Может, по-видимому, играть некоторую роль в циркуляции туляре-
мийного микроба (установлена естественная зараженность).
17. Aedes (Ochlerotatus) pionips Dyar, 1919.
Весьма сходен с A. communis, от которого отличается наличием пост-
коксального пятна чешуек, а также строением личинки.
Рис. 144. Гипопигий Aedes pionips Dyar. По Carpenter
a. La Casse.
Голова сверху преимущественно в относительно светлых желтовато-
коричневых чешуйках. Хоботок и щупики в одноцветных бурых чешуй-
ках. Последний членик щупика самца тонкий. Среднеспинка в желто-
вато-серых или золотисто-бронзовых чешуйках с не резко ограниченной
широкой продольной коричневой полосой или с । двумя сближенными
полосами, разделенными узкой полоской золотистых чешуек. В задней
половине среднеспинки обычно выражены и боковые продольные темные
полоски. Пятна чешуек на бочках груди расположены так же, как у А. com-
munis, с тем отличием, что чешуйки имеются и на посткоксальной пере-
понке. Крылья в темных чешуйках, единичные светлые обычно имеются
Pre. 145. Aedes pionips Dyar. Личинка IV стадии. По Carpenter
a. La Casse.
J — передний конец, сверху, 2 — задний конец, сбоку.
7. AEDES
237
при основании крыла. Брюшко сверху в бурых чешуйках, со светлыми
поперечными перевязями более или менее равномерной ширины.
Гипопигий (рис. 144) как у A. communis; отличия в строении гипо-
пигия обоих видов (Vockeroth, 1952) незначительны, и по-видимому, ле-
жат в пределах внутривидовой изменчивости. Практически оба вида
по гипопигиям неразличимы.
Личинка IV стадии (рис. 145) крупная, темно-коричневой
окраски, с головой, ширина которой в 1.5 раза превышает длину. Усики
тонкие, стройные, слегка изогнутые внутрь, в длину более 2/3 длины
головы; тело их покрыто шипиками; волосок отходит у середины, ближе
к основанию, из 7—13 (среднее 8—9) слабо вторичноперистых ветвей,
в длину достигающих около половины длины усика. Лобные волоски:
наружные из 5—9 (среднее 7—8), средние из 3—5 (в 50% случаев из 3,
как единичное исключение из 2), внутренние из 3—5 (среднее 4, как исклю-
чение 2) вторичноперистых ветвей; задние волоски наличника из 3—5
коротких тонких волосков, лежащих между основаниями средних.
Щетка на VIII членике брюшка из 61—78 (среднее 68, как редкое
исключение меньше 60) немного расширенных на конце, слегка вытянутых
чешуек, края которых усажены в один ряд иногда несимметрично рас-
положенными короткими, уменьшающимися к основанию шипиками.
Дыхательная трубка прямая, слегка сужается к вершине; индекс около 3.0.
Гребень из 18—24 тесно расположенных, заметно удлиняющихся в дис-
тальном направлении зубцов и из 1—6 рудиментарных зубчиков у осно-
вания, занимает от 1/3 до 2/5 длины трубки. Пучок у ее середины из 4—9
(среднее 6) вторичноперистых ветвей. Последний членик с седлом, на 2/3
заходящим на боковые стороны; боковой волосок тонкий и простой;
наружные хвостовые волоски заметно длиннее дыхательной трубки,
простые, внутренние из 9—14 (среднее 12) гладких ветвей одной длины
с последним члеником. Плавник из 17—21 (среднее 19) пучка, объеди-
ненных общим основанием, и из 2—3 коротких пучков впереди него. Жабры
удлиненно ланцетовидной формы, заостренные на концах и пигментиро-
ванные; их длина изменчива: равняется длине седла или вдвое превы-
шает ее.
Распространение. Аляска, Канада, Северо-Запад США.
В Палеарктике до недавнего времени (Сазонова, 1958) не был найден,
не отграничивался от A. communis. Распространение в СССР: северная
половина европейской части, Западная и Восточная Сибирь, Хабаров-
ский край, Камчатка. Границы ареала не установлены. Возможно, что
некоторые указания о широком распространении и высокой численности
A. communis в северных районах должны быть отнесены к A. pionips
и к Л. hexodontus (см. ниже).
Биология. Личинки из перезимовавших яиц ранней весной
появляются в лужах и других разнообразных небольших водоемах,
образовавшихся после таянья снега в открытых и лесных местностях,
не только в равнинных, но и в горных ландшафтах до высоты около
1000 м над ур. м.
18. Aedes (Ochlerotatus) punctor Kirby, 1837 (meigenanus Dyar, 1921).
По окраске весьма сходен с двумя предыдущими; отличается от них
строением гипопигия, отсутствием светлых чешуек на крыльях и формой
светлых перевязей брюшка (с перетяжкой посредине).
Хоботок и щупики в черно-бурых чешуйках. Среднеспинка в золо-
тисто-или красновато-бронзовых чешуйках, обычно с темно-коричневой
продольной полосой или с двумя сближенными темными полосками.
238
III. ПОДСЕМ. CULICINAE
Встречаются и экземпляры с одноцветной среднеспинкой. Посткок-
сальное пятно имеется (отличие от A. communis). Пятно белых чешуек
на мезэпистернах доходит до их переднего угла, а на мезэпимерах —
до их нижнего края. Гипостигмальных чешуек нет. Голени и лапки в тем-
ных чешуйках. Крыло в одноцветных темных, почти черных чешуйках;
как исключение, при основании костальной жилки может быть несколько
Рис. 146. Гипопигий Aedespunctor Kirby. По Carpenter a. La Casse.
желтоватых чешуек. Поперечные светлые перевязи на тергитах брюшка
явственно сужены посредине; иногда выражены лишь белые пятна по бокам
сегментов, соединяющиеся узкой светлой полоской при основании тергитов.
Гипопигий (рис. 146): вершинная бородавка коксита уплощенная,
покрытая короткими изогнутыми волосками; ее основание в прокси-
мальном направлении доходит почти до середины коксита; базальная
бородавка с одним шипом, сильно выпуклая, густо покрыта короткими
7. AEDES
239
волосками, сидящими на маленьких бугорках. Класпеты с коротким
прямым стволиком и слегка расширенным в средней части длинным лан-
цетовидным крылом; его длина превышает ширину не менее чем в 2—3
раза; на препаратах крыло класпеты обычно имеет одноцветнокоричне-
вую окраску, без прозрачного расширения, выраженного у большинства
видов Ochlerotatus.
Личинка IV стадии (рис. 147) от средних до крупных
размеров, темно-коричневой у зрелых личинок окраски. Лобные волоски:
наружные из 2—8 (среднее 5), средние и внутренние из 1—3 (чаще 2)
Рис. 147. Aedes punctor Kirby. Личинка IV стадии. Задний
конец, сбоку.
ветвей со слабо развитой вторичной перистостью; задние волоски на-
личника между основаниями средних, тонкие, короткие, из 2—4 ветвей.
Усики менее половины длины головы, слегка изогнуты, с пучком у се-
редины из 4—7 не достигающих длины V2 усика ветвей.
Щетка из 11—24 (чаще 14—15) сравнительно мелких (0.07—0.08 мм)
чешуек, имеющих главный шип с маленькими шипиками у основания,
расположенных в 2—3 неправильных ряда. Число чешуек очень измен-
чиво, даже у одной особи с левой и правой стороны. Дыхательная трубка
прямая, слабо суженная к вершине (индекс около 3.0); гребень заметно
не достигает ее середины, состоит из 14—26 (чаще 19—22) зубцов, из ко-
торых 1—6 — рудиментарные; зубцы тесно сближены и постепенно удли-
нены по направлению к вершине трубки; пучок из 3—9 (среднее 5) вет-
вей у середины трубки, часто ближе к основанию.
Седло кольцом охватывает последний членик; с брюшной стороны име-
ется глубокий вырез, в котором помещаются передние пучки плавника;
240
III. ПОДСЕМ. CULICINAE
редко нижние края кольца не сливаются вместе, а только сближены
друг с другом, образуя на брюшной стороне узкую срединную щель.
Боковые волоски простые, одной длины с седлом; наружные хвостовые
сильно развиты, простые, внутренние из 5—8 ветвей. Плавник из 16—
19 пучков в общем основании и из 1—2 пучков впереди него. Жабры
длинные, суженные к концу, пигментированные, в среднем в 1.5—2 раза
длиннее седла.
Распространение. Зоны леса и тундры обоих полушарий:
Европа, Сибирь, Дальний Восток, Северная Америка. На севере ареал
простирается до Ледовитого океана, на юге до Крыма, Северного Кавказа,
Восточного Казахстана; найден в горах Алжира. Вопрос о распростра-
нении на севере нуждается в уточнении ввиду обнаружения там близ-
кого вида — A. hexodontus.
Биология. Личинки встречаются в небольших открытых или
полузатененных водоемах, образованных после таянья снега в торфяниках
или лесных заболоченностях, с дном, покрытым разлагающейся опавшей
листвой, хвоей, мхами или травянистой растительностью. В тайге и
тундре, а также в лесах Приморья A. punctor — один из важнейших
компонентов гнуса; среди комаров, нападающих на людей и сельско-
хозяйственных животных, по численности нередко занимает первое место.
Относится к числу ранних весенних видов, появляется одновременно
с A. communis или несколькими днями позже. Летом после дождей иногда
дает небольшой второй выплод. Установлена естественная зараженность
A. punctor возбудителем туляремии.
19. Aedes (Ochlerotatus) hexodontus Dyar, 1916.
Весьма сходен с A. punctor, отличается светлыми чешуйками при
основании крыла. От A. communis отличается существованием посткок-
сального пятна чешуек, а от внешне весьма сходного A. pionips — более
темной общей окраской, особенно окраской среднеспинки. С уверенностью
определяется по личинкам.
Голова сверху преимущественно в темных коричнево-бронзовых че-
шуйках. Хоботок и щупики покрыты черными чешуйками. Среднеспинка,
как правило, в одноцветных ржаво-коричневых чешуйках; иногда неясно
намечается темная продольная полоса. Крыло в темных чешуйках; свет-
лые имеются лишь при основании крыла. Иногда их немного, чаще свет-
лые чешуйки покрывают основание костальной и радиальной жилок
до уровня плечевой поперечной жилки. Брюшко в бурых чешуйках,
со светлыми перевязями, которые обычно немного сужены посредине,
реже — равномерной ширины. Иногда бурые чешуйки тергитов имеют
светлый бронзовый оттенок и нерезко отличаются от серовато-белых че-
шуек, образующих перевязи при основании сегментов.
Гипопигий (рис. 148). Самцы A. hexodontus и A. punctor практически
неотличимы.
Личинки IV стадии (рис. 149) темно- или желто-коричне-
вого цвета, с головой, в ширину в 1.3—1.4 раза превышающей длину.
Лобные волоски грубые, с редкой вторичной перистостью; наружные
из 4 (3—6) ветвей, средние лежат впереди внутренних, обе пары, как
правило, простые, реже из двух ветвей; случаи 2-ветвистости чаще встре-
чаются в средней и в южной части ареала. Задние волоски наличника
тонкие, из 2—3 ветвей. Усики около половины длины головы покрыты
шипиками, с пучком из 3—4 ветвей, не достигающих вершины. Ротовые
органы смешанного типа, с преобладанием питания с субстрата.
7. AEDES
341
Щетка в среднем из 7 (6—9, исключительно редко 10—11) чешуек
с резко выраженным главным шипом и тонкими мелкими шипиками у осно-
вания. Сифон (индекс около 3) прямой, постепенно сужающийся к вер-
Рис. 148. Гипопигий Aedes hexodontus Dyar. По Carpenter a. La Casse.
шине. Гребень из 12—17 сравнительно тесно сидящих зубцов, занимаю-
щих от х/3 до 2/s длины сифона от основания; наиболее дистальный иногда
крупнее остальных и, как исключение, может быть шире отставлен.
Пучок дистальнее гребня, из 2—7 (4—5) ветвей.
Последний членик брюшка с кольцевидно охватывающим его седлом.
Наружные хвостовые волоски длинные, простые; внутренние — из 4—
16 Фауна СССР, А. В. Гуцевич и др( j
Рис. 149. Aedes hesodontus Dyar. ЛичинкаТ1У стадии.
По Carpenter a. La Casse.
I —, передний конец, сверху, 2 — задний конец, сбоку.
7. AEDES
243
10 (6—8) расположенных веером ветвей; боковой волосок простой. Плав-
ник из 16—18 пучков, окруженных общим основанием, с двумя (реже
с одним) более короткими пучками впереди него. Жабры ланцетовидные,
слабо пигментированные, в среднем вдвое длиннее седла.
Распространение. Вид до последнего времени смешивался
с другими видами группы
communis, вследствие чего
его распространение выясне-
но недостаточно. В СССР
обнаружен на севере Коль-
ского полуострова, в Карель-
ской АССР (Пряжинский
район), в окрестностях Ле-
нинграда (Петродворец) и
в Ненецком националь-
ном округе (Нижнепечерский
район), в Западной и Восточ-
ной Сибири. По-видимому,
широко распространен в суб-
арктической, тундровой и
таежной зонах, а южнее —
в горных местностях. Север-
ная Швеция (Vockeroth, 1954),
Канада, Аляска и горные
районы США.
Биология. Ранневе-
сенний вид. Личинки появ-
ляются с началом таянья
снега в различного рода мел-
ких водоемах, в горах —
с каменистым дном. Взро.слые
вылетают, по-видимому, рань-
ше, чем A. punctor, и явля-
ются злостными кровососами.
20. Aedes (Ochlerotatus)
sticticus Meigen, 1838.
От предыдущих видов от-
личается меньшими размера-
ми, расположением чешуек
на мезэпимерах, окраской
среднеспинки и голеней, а
также строением гипопигия. Рис. 150. Гипопигий Aedes sticticusMg. По Carpen-
Хоботок, щупики и кры- ter a. La Casse.
лья в одноцветных тем-
ных чешуйках. Среднеспин-
ка с широкой, четко отграниченной продольной полосой (или с двумя
сближенными полосками) темно-коричневых чешуек; кроме того, могут
быть выражены еще две продольные темные полоски, расположенные
латеральнее. Боковые отделы среднеспинки в серебристо-серых или кре-
мовых чешуйках. Пятно белых чешуек на мезэпимерах явственно не до-
ходит до их нижнего края. Передние и средние бедра спереди темные.
На наружной поверхности задних голеней светлые чешуйки образуют
продольную полоску; иногда эта полоска слабо выражена, едва различима.
16*
244
III. ПОДСЕМ. CULICINAE
Брюшко сверху в черных, с бронзовым отливом чешуйках, с белыми
треугольными пятнами по бокам, соединенными поперечными перевязями
при основании сегментов; перевязи посредине сужены, иногда прерваны.
Гипопигий (рис. 150) сходен с гипопигием А. punctor, но отличается
формой базальной бородавки коксита и крыла класпеты: базальная бо-
родавка в дистальной части отделена от поверхности коксита; крыло
класпеты с небольшим прозрачным расширением, короткое, его длина
не более чем в 2 раза превышает ширину.
Личинка IV стадии (рис. 151) крупных размеров, коричне-
вой окраски, с более светлыми головой и дыхательной трубкой. Лобные
волоски вторичноперистые: наружные в среднем из пяти, средние впереди
внутренних, обе пары из 2—3 ветвей, редко больше; как очень редкое
исключение, один из волосков может быть простым. Задние волоски
наличника между основаниями средних, короткие, 1—4-ветвистые. Усик
Рис. 151. Aedes sticticus Mg. Личинка IV стадии. По Пеусу.
1 — VIII членик брюшка, сбоку, 2 — голова, сверху.
короткий, 0.4—0.5 длины головы, покрыт редкими шипиками; пучок
у середины, ближе к основанию, из 4—5 коротких ветвей.
Щетка на VIII членике брюшка из 16—27 (чаще 20—24) чешуек,
расположенных в 2—3 неправильных ряда; каждая чешуйка имеет уме-
ренно развитый главный шип, с несколькими более короткими шипи-
ками по бокам основания; иногда ближайшие к главному шипы почти
достигают размеров последнего. Дыхательная трубка относительно ко-
роткая, прямая (индекс 2—2.5, редко 3.0), от базальной трети постепенно
сужается к вершине. Гребень заходит за середину, в среднем из 20—25 си-
дящих на равном расстоянии зубцов; иногда наиболее дистальный может
быть несколько шире остальных. Пучок за серединой, из 5—6 вторично-
перистых ветвей, в длину достигающих ширины сифона у основания.
Седло последнего членика доходит до его середины или нижнего края;
боковой волосок короткий и простой; наружные хвостовые волоски про-
стые, значительно длиннее дыхательной трубки; внутренние из 6—7 вет-
вей. Плавник имеет около 20 пучков в общем основании и 1—2 впереди
него. Жабры более длины седла, заостренные, не пигментированные.
Распространение. Европа — на севере до Финляндии и Ка-
релии, на юге до Югославии, юга Украины и Северного Кавказа; Сибирь,
Забайкалье, Хабаровский край, Приморье; Япония, Северная Америка.
Биология. Личинки встречаются в водоемах, возникших в ре-
зультате таянья снега или разлива рек, полузатененных деревьями,
с дном, покрытым прошлогодней листвой, часто совместно с A. vexans.
7. AEDES
345
Рис. 152. Aedes nigrinus Eck. Личинка IV стадии.
По Пеусу.
1 — голова, сверху, г — VIII членик брюшка, сбоку.
Зимует на фазе яйца; последние, при отсутствии воды, могут переживать
длительное время — до 2—3 лет. Характерен для подзоны смешанных
лесов и для лесостепи. Сообщения о нахождении A. sticticus на севере
тайги и в тундре, вероятно, основаны на неточном определении. Местами
нападает в значительном количестве (Западная Украина, Забайкалье).
Дает одно поколение в год. Появляется значительно позже, чем A. com-
munis и А. punctor. По сезону активности скорее летний, чем весенний вид.
21. Aedes (Ochlerotatus) nigrinus Eckstein, 1918.
Очень близок предыдущему, от которого отличается деталями окраски
усиков, ног и крыльев (Pens, 1933).
I членик жгутика усиков утолщенный, одноцветно черный (у A. sti-
cticus — не утолщенный, при основании желтый). Основание костальной
жилки, субкостальная и основание медиальной жилки в белых чешуйках.
Передние и средние бедра спереди пестрые. Светлые перевязи брюшка
к средней линии не сужива-
ются или суживаются незна-
чительно.
Гипопигий: базальная бо-
родавка коксита короче и
шире, чем у A. sticticus-, ее
наружный край равномерно
выпуклый; крыло класпеты
с небольшим пластинчатым
расширением; длина крыла
превосходит его ширину при-
мерно вдвое.
Личинки IV ста-
fl и и (рис. 152) сходны
с личинками А. sticticus. Лоб-
ные волоски слабо вторичноперистые: наружные из пяти вет-
вей, средние впереди внутренних, обе пары простые, очень редко один
из волосков может быть 2-ветвистым; задние волоски наличника корот-
кие, из нескольких ветвей. Усик немного меньше х/2 длины головы, покрыт
редкими шипиками, с пучком из 3—5 коротких ветвей, на расстоянии
2/5 длины усика от основания.
Щетка в среднем из 10—12 (редко до 15—17) чешуек, более удлинен-
ных, с сильнее вытянутым, чем у A. sticticus, главным шипом. Дыха-
тельная трубка короткая, прямая, несколько суженная к вершине
(индекс 2.0 или немного больше). Гребень почти доходит до середины
трубки, в среднем из 17 зубцов и нескольких рудиментарных у основания.
Пучок у середины или немного за нею, из 4—6 ветвей.
Седло последнего членика доходит почти до нижнего края его боков;
боковой и хвостовые волоски как у A. sticticus. Плавник в среднем из 13—
14 пучков в общем основании и из четырех или больше более коротких
пучков впереди него. Жабры очень изменчивы в длину, в крайних вариан-
тах могут достигать длины дыхательной трубки, у форм с более корот-
кими жабрами верхняя пара длиннее нижней, в случаях с длинными
жабрами они одной длины.
Распространение. Европа — ГДР и ФРГ, Дания, Южная
Скандинавия, Финляндия, Чехословакия. Найден в нескольких пунктах
европейской части СССР: Эстония, Ленинградская и Вологодская области,
Северный Урал, Западная Сибирь. Редкий вид, свойственный преиму-
щественно открытым луговым пространствам.
246
III. ПОДСЕМ. CULICINAE
Биология. Личинки встречаются в открытых, богатых травяной
растительностью луговых водоемах, образованных или в результате таянья
снега, или затопления паводковыми водами. Зимуют на фазе яйца. Дан-
ные о нескольких поколениях в течение лета нуждаются в подтверждении.
22. Aedes (Ochlerotatus) hungaricus Mihalyi, 1955.
Относительно недавно описанный вид, систематическое положение ко-
торого не вполне ясно.
Мелкий комар. Хоботок и щупики самки в темных чешуйках. Средне-
спинка покрыта беловато-серыми чешуйками с отчетливой темно-корич-
невой продольной полосой, разделяющейся в задней половине средне-
спинки на две полоски. Пят-
Рис. 153. Гипопигий Aedes hungaricus Mih.
Mihalyi.
но светлых чешуек на мез-
эпистернах почти доходит до
их переднего угла, а на мез-
эпимерах — не достигает их
нижнего края, как у A. sttcti-
cus и A. nigrinus. Гипостиг-
мального и посткоксального
пятен нет. Лапки темные,
светлая полоска на наруж-
ной поверхности задних го-
леней отсутствует. Крылья
в темных чешуйках, особенно
густых вдоль костальной «и
первой радиальной жилок.
Брюшко сверху в темных че-
шуйках, со светлыми пере-
вязями при основании тер-
По гитов; перевязи слегка су-
жены посредине; по бокам
тергитов имеются пятна бе-
лых чешуек.
Гипопигий (рис. 153): коксит относительно узкйй, в густых волосках;
вершины волосков левого и правого кокситов слегка заходят друг за друга.
Базальная бородавка коксита умеренно выпуклая, с двумя крепкими
щетинками: одна из них длинная, изогнутая, другая — тонкая, короткая,
прямая. Вершинная бородавка слабо развита. Класпеты с почти прямым
коротким стволиком и довольно длинным крылом, явственно расширен-
ным в средней части; нерасширенная часть основания крыла («рукоятка»)
хорошо выражена. Фаллосома короткая, яйцевидная. Выросты IX тер-
гита с 3—4 довольно длинными изогнутыми щетинками.
Личинка IV стадии желто-коричневого цвета с более темной
головой, со светлыми пятнами в местах прикрепления мускулатуры.
Лобные волоски: наружные из 6—8, средние из двух ветвей, внутренние —
простые, лежат позади средних; все ветви слабо вторичноперистые.
Задние волоски наличника между основаниями средних лобных волосков,
едва заметные. Усики немного больше 1/2 длины головы, покрыты редкими
шипиками, с пучком из 8—10 ветвей, лежащим на расстоянии х/3 длины
от основания.
Щетка на VIII членике брюшка из 16—24 (среднее 20) чешуек с длин-
ным острым шипом и с более короткими шипиками у основания; у край-
них снизу и сверху чешуек ближайшие к главному шипу могут быть
одной величины с ним. Дыхательная трубка цилиндрическая, с наиболь-
7. AEDES
. 247
шей шириной на расстоянии х/3 от основания; индекс 2.0—2.4 (сред-
нее 2.2); гребень из 19—24 (среднее 20) тесно сидящих зубцов, увеличи-
вающихся в дистальном нацравлении, заходит за середину трубки; каждый
зубец в виде тонкого, слегка изогнутого шипа, с несколькими зубчи-
ками у основания; пучок за серединой трубки, заметно удален от наибо-
лее дистального зубца гребня, из 3—5 ветвей со слабой вторичной по-
ристостью.
Последний членик с седлом, покрывающим бока до середины, боко-
вой волосок простой, достигает 1/2 длины седла; наружные хвостовые
волоски простые, внутренние в виде веера в среднем из 10 ветвей. Плав-
ник из 15 пучков, окруженных общим основанием, и из трех, реже че-
тырех (у молодых личинок) более коротких впереди него. Жабры лан-
цетовидные, не пигментированные; верхняя пара длиннее нижней; за-
метно превышают длину седла.
Распространение. Венгрия, долина Дуная (Mihalyi es Gu-
lyas, 1963). По-видимому, характерен для пойменных лесов. В СССР
не обнаружен.
Замечания по систематике. По окраске А. hungaricus сходен с дру-
гими видами группы communis, особенно с А. sticticus и А. nigrinus, от которых отли-
чается отсутствием светлой полоски на наружной поверхности задних голеней. Но строе-
ние гипопигия — наличие двух крепких щетинок на базальной бородавке коксита и
очень слабое развитие вершинной бородавки — более характерно для видов группы
caspius.
23. Aedes (Ochlerotatus) diantaeus Howard, Dyar et Knab, 1917.
В среднем несколько крупнее других комаров группы communis. Све-
жие экземпляры отличаются стальным отливом черных чешуек хоботка,
ног и брюшка.
Рис. 154. Гипопигий Aedes diantaeus Н. D. К. По Natvig.
Отдельно изображены, при большем увеличении, базальная бородавка кок-
сита (А), крыло класпеты (В) и попасти IX тергита (В).
Хоботок и щупики в черных чешуйках. Среднеспинка в золотистых
или серебристо-серых чешуйках, с широкой продольной темной полосой
или двумя сближенными полосками. Пятно белых чешуек на мезэпистер-
нах не доходит до их переднего угла (отличие от всех предыдущих видов
группы communis). Посткоксальные и гипостигмальные чешуйки отсут-
ствуют. Крылья, голени и лапки в одноцветных темных чешуйках. Брюшко
Рис. 155. Aedes diantaeus H. D. К. Рис. 156. Aedes diantaeus'H.. D. К. Личинка IV стадии. Задний конец, сбоку.
Личинка IV стадии. Голова, сверху.
7. AEDES
249
сверху в черных чешуйках, с треугольными белыми пятнами по бокам,
иногда соединенными узкими перевязями по переднему краю тергитов.
Гипопигий весьма своеобразного строения (рис. 154). Внутренняя по-
верхность коксита несет густой широкий пучок направленных медиально
волосков. Базальная бородавка с тремя шипами; два из них сближены
и помещаются на особом выступе. Класпеты с коленчато изогнутым ство-
ликом и широким крылом.
Личинка IV стадии (рис. 155—157) крупных размеров,
от светло- до темно-коричневого цвета; исключительно длинные усики
позволяют, особенно во время движения, отличать личинок этого вида
в водоемах невооруженным гла-
зом.
Лобные волоски вторичнопе-
ристые, длинные: наружные из
3—6 (чаще 4), средние из 2—5 (ча-
ще 3—4) впереди внутренних из
2—6 (чаще 3—4) ветвей; задние
волоски наличника между осно-
ваниями средних, короткие и
очень тонкие. Усики в 1.1—1.3
раза длиннее головы, почти пря-
мые и тонкие, усажены хорошо
заметными шипиками и около се-
редины несут пучок из 3—7 (чаще
4—5) вторичноперистых ветвей.
Щетка на VIII членике брюш-
ка из 6—13 (11—12) крупных че-
шуек, расположенных в два не-
правильных ряда; каждая чешуй-
ка с длинным заостренным глав-
ным шипом, по бокам основания Рис 157. Aedes diantaeus Н. D. К. Личинка
которого лежат короткие тонкие IV стадии. Стигмальная пластинка,
шипики. Дыхательная трубка пря-
мая, начиная с х/3 от осно-
вания постепенно сужается к вершине; гребень из 15—22 (чаще 16—18)
длинных шиповидных зубцов, укорачивающихся к основанию трубки,
а 1—2 наиболее дистальных более широко расставлены; пучок у середины,
ближе к вершине, из 6—10 (чаще 7—8) слабо вторичноперистых ветвей,
в длину достигающих ширины основания трубки.
Седло доходит до нижнего края боков последнего членика; боковой
волосок простой, короче седла; наружный хвостовой волосок длиннее
дыхательной трубки, простой, внутренний почти вдвое короче, из 10—
13 ветвей. Плавник из 20—21 пучка в общем основании и из 2—4 пучков
впереди него. Жабры узкие, ланцетовидные, немного длиннее седла.
Распространение. Ареал простирается от Западной Европы
до Приморья и Хабаровского края, и от севера Норвегии и Карелии
до юга Украины и Северного Кавказа, а также охватывает часть Север-
ной Америки.
Биология. Личинки развиваются в разного рода временных лес-
ных водоемах, возникших от таянья снега, — ямах, канавах, лужах
ит. п., затененных или расположенных в более открытых местах, но с хо-
лодной водой. Зимуют на фазе яйца и имеют, как правило, одно поколение
в году. Поздневесенний вид с медленным личиночным развитием. Вид
характерен для влажных смешанных лесов, но распространен и в тайге.
250
III. ПОДСЕМ. CULICINAE
Рис. 158. Гипопигий Aedes intrudens Dyar.
Чаще встречается в местах с разреженным лесным покровом, на лесных
полянах и опушках. Вылетает в конце весны, позже, чем ряд других
видов Aedes. Активный кровосос. Обычно встречается в небольшом или
умеренном количестве. Местами, например в некоторых районах При-
карпатья, относится к числу преобладающих видов.
24. Aedes (Ochlerotatus) intrudens Dyar, 1919.
Отличается от предыдущих видов наличием гипостигмального пятна,
т. е. небольшого количества чешуек, расположенных непосредственно
книзу от переднего дыхальца. Темные чешуйки хоботка, крыльев, ног
и брюшка в отличие от A. diantaeus не имеют стального оттенка.
Хоботок и щупики в бурых чешуйках, на щупиках разбросаны серо-
вато-белые чешуйки. Среднеспинка в золотисто-бронзовых чешуйках, бо-
лее светлых (беловатых) у
боковых краев; иногда на
среднеспинке выражена не-
ясно отграниченная продоль-
ная темная полоса или две
узкие полоски. Посткоксаль-
ного пятна нет, гипостиг-
мальное имеется. На мезэпи-
мерах чешуйки не доходят до
нижнего края склерита. Пе-
редние бедра спереди пре-
имущественно в темных че-
шуйках с большей или мень-
шей примесью светлых. Лап-
ки темные. Крылья в одно-
цветных темных чешуйках,
иногда при основании ко-
стальной жилки имеется не-
сколько светлых. Брюшко
сверху темное, с белыми пе-
ревязями при основании сег-
ментов; перевязи равномер-
ной ширины, реже сужены
посредине.
Гипопигий (рис. 158) как
и у предыдущего вида,
характеризуется наличием
и густого пучка волосков
трех шипов на базальной бородавке
на внутренней поверхности коксита. Этот пучок в отличие от A. diantaeus
меньше по размерам, расположен дистальнее — у вершинной бородавки,
причем волоски направлены медиально и назад. Стволик класпеты посре-
дине с пальцевидным выступом, на вершине которого находится волосок.
Личинка IV стадии (рис. 159) от средних до крупных раз-
меров от желто- до темно-коричневой окраски. Лобные волоски вторично-
перистые: наружные из 5—9 (чаще 6—7), средние из 3—4 (чаще 3) лежат
впереди внутренних из 3—5 (чаще 4) ветвей; задние волоски наличника
между основаниями средних, короткие, из нескольких тонких ветвей.
Усик немного более половины длины головы, покрыт редкими шипиками,
с пучком из 6—8 ветвей, отходящим от середины, ближе к основанию.
Щетка из 10—18 (чаще 12—16) чешуек, расположенных в два непол-
ных ряда; каждая чешуйка крупная, темная, с длинным остроконечным
7. AEDES
251
шипом с очень короткими тонкими шипиками по бокам основания. Ды-
хательная трубка прямая, начиная от середины заметно сужена к вер-
шине, индекс 2.8—3.5 (в среднем около 3.0). Гребень из 14—22 (чаще 18—
19) удлиняющихся в дистальном направлении зубцов, из которых 1—4
(чаще 2—3) более сильно расставлены в виде более крупных гладких
шипов, тогда как остальные имеют 1—2 дополнительных зубчика, нем-
ного отстуря от основания; ближайший к вершине шип лежит заметно
за серединой1 трубки. Пучок из 4—10 (среднее 7—8) тонких ветвей со сла-
бой вторичной перистостью, в длину равных ширине трубки у места
его отхождения; пучок лежит у середины или ближе к вершине, при
этом, если широко расставленных дистальных зубцов гребня 1—2, он ле-
жит от них ближе к вершине, если их больше, то между их основаниями.
Седло последнего членика доходит до середины или нижнего края
его боковых сторон и имеет очень неровные очертания; боковой волосок
простой и короткий; наружный хвостовой волосок простой, заметно длин-
252
III. ПОДСЁМ. CULICINAE
нее дыхательной трубки; внутренний в среднем из 6—8 ветвей, в длину
около 2/3 трубки. Плавник из 16—18 пучков в общем основании и 1—3 бо-
лее коротких впереди него; ветвление особенно наиболее крупных задних
пучков плавника начинается далеко от их основания и это расстояние
заметно превышает длину боковых отростков последнего. Жабры длин-
нее седла, сильно варьируют по этому признаку, заострены на концах
и не пигментированы.
Распространение. От Западной Европы до Хабаровского
края и Камчатки; на севере доходит до лесотундры, но, по-видимому,
не проникает в тундру. На юге известен до степной Украины. Северная
и Средняя Европа, север США и южная часть Канады.
Биология. Личинки встречаются в неглубоких/ лесных лужах
и заболоченностях, возникших в результате таянья снега или фильтрации
при повышении уровня грунтовых вод; характерны водоемы без травя-
нистой растительности, с дном, покрытым опавшей листвой. Встреча-
ется совместно с A. excrucians, A. cataphylla, A. communis, A. diantaeus
и A. cinereus. Самки активно, часто в значительном числе, нападают
на человека, преимущественно в лесах, предпочитая олыпанники. В лес-
ной зоне — обычный вид. Дает одно весеннее поколение, появляющееся
раньше, чем у большинства видов. Летом численность резко снижается.
25. Aedes (Ochlerotatus) pullatus Coquillett, 1904.
Близок к предыдущему. Отличается деталями окраски и строением
гипопигия.
Хоботок и щупики в темных чешуйках, на щупиках может быть при-
месь светлых. Среднеспинка в золотистых или беловатых чешуйках,
с двумя продольными узкими темными полосками; эти полоски покрыты
темно-коричневыми чешуйками или свободны от них. Хитин среднеспинки,
как правило, угольно-черный. Гипостигмальное пятно имеется, посткок-
i с х чешуек нет. На мезэпимерах чешуйки доходят до нижнего края
склерита. Нижние мезэпимерные щетинки имеются (1—5). Ноги: перед-
ние бедра спереди темные, лапки в бурых чешуйках. Крыло в темных
чешуйках; светлые в небольшом количестве имеются при основании крыла.
Брюшко сверху в темных чешуйках, со светлыми перевязями при осно-
вании тергитов; перевязи посредине иногда сужены.
Гипопигий (рис. 160): коксит в длинных густых волосках, при осно-
вании с тремя шипами, как у двух предыдущих видов, но в отличие от них
пучок густых волосков на коксите отсутствует; один из двух сближенных
шипов плоский, в виде узкой пластинки, на конце иногда с двумя неж-
ными веточками; стволик класпеты коленчато изогнутый, без пальце-
видного отростка.
Личинка IV стадии (рис. 161 и 162) средней величины,
светло-коричневой или более темной окраски. Лобные волоски сильно
ветвистые, вторичноперистые; наружные из 8—13, средние из 3—8 (сред-
нее.6), внутренние из 3—7 (среднее 5) ветвей; задние волоски наличника
короткие, тонкие, из 4—5 ветвей. Усики относительно короткие, лишь
немного больше половины длины головы, покрыты шипиками с пучком
в среднем из пяти вторичноперистых ветвей.
Щетка из 50—60 чешуек, варьирующих по строению: от лишенных
главного шипа и несущих по краю уменьшающиеся к основанию шипики
до имеющих более или менее ясно выраженный главный шип с более
короткими боковыми шипиками. Дыхательная трубка начиная от ба-
зальной трети равномерно сужается к вершине; индекс колеблется от 3.0
до 3.5. Гребень из 15—25 тесно сидящих зубцов, увеличивающихся в ди-
Рис. 160. Гипопигий Aedes pullatus Coq.
Рис. 161. Aedes pullatus Coq. Личинка IV стадии. Голова,
сверху. По Carpenter a. La Casse.
254
III. ПОДСЕМ. CULICINAE
стальном направлении, и 2—3 рудиментарных, далеко не доходит до се-
редины трубки; пучок у середины, заметно впереди наиболее дисталь-
ного зубца гребня, из 5—8 вторичноперистых ветвей, в длину превы-
шающих ширину трубки у места отхождения пучка.
Седло последнего членика более чем наполовину покрывает его бо-
ковые стороны; боковой волосок простой, в длину немного превышает
половину длины седла; наружный хвостовой волосок простой и длинный,
Рис. 162. Aedes pullatus Coq. Личинка IV стадии. Задний конец, сбоку. По Carpenter
a. La Casse.
внутренний в виде симметричного веера из 6—10 гладких ветвей. Плав-
ник в среднем из 15 пучков в общем основании и 1—3 более коротких
впереди него. Жабры пигментированные, удлиненно ланцетовидные, слегка
изогнутые; верхняя пара немного длиннее нижней, в 1.5—2 или более
раза длиннее седла, могут достигать длины дыхательной трубки. «Н
Распространение. Ареал вида прерывистый. В Западной
Европе и на юге СССР A. pullatus встречается в горах, севернее — и
вне горных районов. Найден в Пиренеях, Альпах, Шварцвальде, Гарце,
Татрах, Карпатах, на Кавказе и в горах Тянь-Шаня на юго-востоке
Казахстана, а также в Ленинградской, Московской и Саратовской обла-
стях, на Кольском полуострове, в Тувинской АССР, в Забайкалье
и в Приморском крае. В коллекции ЗИН имеются экземпляры A. pul-
latus с этикеткой: западное подножье Отхон-Тенгри, 2300 м, Козлов,
VII 1924. Распространен также в Северной Америке: Аляска, Канада,
США в области Скалистых гор, местами до высоты 3000—4000 м над ур. м.
7. AEDES
255
Биология. Вид на севере ареала встречается в низменных и
равнинных местах, на юге приурочен к горным районам до 2000 м над ур. м.
и выше. Личинки встречаются в разнообразных небольших весенних
водоемах, затененных (на юге) или открытых (на севере и в горах), ли-
шенных травянистой растительности; в горных местностях — в неболь-
ших озерах с песчаным или каменистым дном. В Хибинах — в торфяных
водоемах и заболоченностях вдоль рек с илистым, каменистым или покры-
тым опавшей листвой карликовой березы дном. В течение года одно по-
коление из перезимовавших яиц, которое появляется позже, чем у дру-
гих видов Aedes; преимущественно летний вид. В большинстве районов
обнаружения не достигает высокой численности. Местами отмечен как
массовый вид, например на Кольском полуострове, в Хибинах (Фри-
долин, 1936).
26. Aedes (Ochlerotatus) nigripes Zetterstedt, 1838 (alpinus Edwards, 1921).
Темноокрашенный комар средней величины. От всех видов группы
communis, кроме A. impiger, отличается волосками груди и расположе-
нием проэпимерных щетинок.
Хоботок и щупики в одноцветных темно-бурых чешуйках. Щупики
самца заметно короче хоботка, концевой членик щупиков без расширения.
Среднеспинка густо покрыта длинными буровато-черными щетинками.
Покровы среднеспинки черные, чешуйки — бурые или-коричневые, по бо-
кам среднеспинки — с золотистым оттенком. Бочки груди в беловато-
кремовых чешуйках. Посткоксальное пятно имеется, гипостигмальное
отсутствует. Щетинки на проэпимерах беспорядочно разбросаны в задней
половине и у верхнего края. На мезэпистернах чешуйки доходят до пе-
реднего угла склерита. Бедра спереди пестрые, в белых и бурых пере-
мешанных чешуйках, иногда преимущественно темные, с небольшой при-
месью светлых чешуек. Коготок лапки удлиненный, слабо изогнутый,
с коротким зубчиком; длина зубчика меньше половины длины коготка.
Крылья в темных чешуйках, светлые имеются лишь при основании крыла.
Брюшко сверху в темных чешуйках (нередко с легким красноватым отли-
вом), с белыми перевязями при основании тергитов; перевязи, особенно
на задних сегментах, иногда нерезко очерченные, с размытым краем.
Гипопигий (рис. 163): коксит в густых длинных волосках; концы
волосков левого и правого кокситов обычно слегка заходят друг за друга.
Вершинная бородавка коксита слабо развита; базальная бородавка со ще-
тинками, но без явственно выраженного шипа или выделяющейся крепкой
щетинки; крайняя щетинка обычно немного крупнее остальных. Стволик
класпеты в вершинной половине изогнут, реже прямой. Крыло класпеты
со слабо выраженным пластинчатым расширением в средней трети крыла.
Выросты IX тергита крупные, с 14—20 крепкими короткими щетинками.
Фаллосома сильно склеротизована, темная, на конце раздвоена.
Личинки IV стадии (рис. 164) от средних до крупных раз-
меров, темно- или желто-коричневого цвета, с умеренно широкой голо-
вой. Лобные волоски со слабой вторичной перистостью; наружные в сред-
нем из пяти ветвей, средние и лежащие позади них внутренние — простые;
задние волоски наличника из 3—5 тонких коротких ветвей. Усики ко-
роткие, меньше половины длины головы; пучок из 1—3 коротких ветвей,
ближе к вершине. Ротовые органы смешанного типа, с преобладанием
питания с субстрата.
Щетка из 10—20 остроконечных зубцов, с едва заметными мелкими
шипиками у основания. Сифон (индекс 2.5—3) прямой, слабо сужаю-
щийся к вершине. Гребень из 12—17 зубцов, из которых 1—4 наиболее
256
III. ПОДСЕМ. CULICINAE
дистальных, в виде более длинных широко расставленных гладких шипов.
Пучок из 2—6 тонких коротких, около диаметра сифона у основания,
ветвей.
Последний членик брюшка с седлом или кольцевидно окружающим
его, или не полностью смыкающимся, с узкой щелью на середине брюшной
стороны. Наружные хвостовые волоски длинные, простые; внутренние —
в среднем из 5—6 ветвей; боковой волосок 1—2-ветвистый. Плавник
из 16—18 пучков, окруженных общим основанием. Жабры длинные,
Рис. 163. Гипопигий Aedes nigripes Zett.
Распространение. Циркумполярный вид, характерный для
тундры. Распространен на Аляске, в Канаде (включая острова аркти-
ческой зоны), в Гренландии, на севере Скандинавии и Финляндии. В СССР
найден на Кольском полуострове, в Архангельской области, в Ненецком
национальном округе, в бассейне р. Кары, в Норильске, Обдорске, Средне-
колымске, в бассейне Хатанги, в устье Лены, на Камчатке и Коман-
дорских островах.
Биология. Ранневесенний вид, развивающийся в лужах и дру-
гих временных водоемах, возникших при таяньи снега, в тундре — в за-
росших травой и кустами карликовой ивы и березы заболоченностях.
В тундре местами — массовый кровосос. Экология изучалась на земле
Элсмир (остров к западу от Гренландии), в пункте, расположенном
на 81°49' с. ш. (Corbet, 1965). Личинки появляются в начале июня в во-
доемах, возникших при таяньи снега. Через 3—4 недели начинается вылет.
Рис. 164. Aedes nigripes Zett. Личинка IV стадии. По Car-
penter a. La Casse.
1 st- передний конеца сверху, 2 задний конец, сбоку.
17 Фауна СССР, А. В. Гуцевич и др.
258
III. ПОДСЕМ. CULICINAE
В первые сутки комары питаются нектаром цветов, в частности на аркти-
ческом растении Dryas integrifolia. Спаривание происходит во время
роения. Самки могут пить кровь начиная со второго дня после вылета.
Наблюдалась и автогенная откладка яиц. Яйца откладываются на осве-
щенных местах по краям водоемов, на 5—10 см выше уровня воды.
27. Aedes (Ochlerotatus) impiger Walker, 1848 (nearcticus Dyar, 1919;
parvulus Edwards, 1921).
Ранее A. nearcticus Dyar и A. parvulus Edw. рассматривались как
самостоятельные виды, различающиеся главным образом по строению
Рис. 165. Гипопигий Aedes impiger Walk,
гипопигия — наличием (A. nearcticus) или отсутствием (A. parvulus) шипа
на базальной бородавке коксита. Но этот признак в данном случае отли-
чается изменчивостью. Базальная бородавка у одних экземпляров несет
несколько почти одинаковых щетинок, у других — одна из щетинок не-
много крупнее остальных, у третьих — имеется шип, явственно отличаю-
щийся от более тонких щетинок. Поэтому A. nearcticus и A. parvulus
правильнее рассматривать как один вид (Natvig, 1948), причем оба имени
следует считать синонимами A. impiger Walk. Исследование типов Cu-
lex impiger Walk, и Aedes nearcticus Dyar обнаружило их идентичность
(Vockeroth, 1952).
7. AEDES
259
Aedes impiger, как и A. nigripes, отличаются от других видов группы
communis щетинками груди и расположением проэпимерных щетинок.
Оба названных вида различаются строением гипопигия, формой коготков
лапки, расположением чешуек на мезэпистернах, а также размерами
тела — A. impiger мельче.
Хоботок и щупики в темных чешуйках. Щупики самца по длине равны
хоботку или немного короче хоботка. Среднеспинка густо покрыта отно-
сительно длинными черными щетинками. Покровы среднеспинки черные,
чешуйки коричневато-бурые, иногда с примесью более светлых, желто-
вато-белых, образующих местами небольшие пятна. На проэпимерах ще-
тинки разбросаны в задней половине склерита. Чешуйки на мезэпистер-
нах не доходят до их переднего угла. Посткоксальное пятно имеется,
гипостигмальное отсутствует.
Бедра спереди пестрые или
преимущественно темные, с при-
месью светлых чешуек. Коготок
лапки по сравнению с A. nigripes
более изогнут, с относительно
длинным зубчиком (его длина бо-
лее половины длины коготка).
Крыло в темных чешуйках, не-
большое количество светлых че-
шуек имеется при его основании.
Брюшко сверху темное, с белыми
перевязями при основании терги-
тов.
Гипопигий (рис. 165): коксит
в умеренно длинных волосках;
концы волосков левого и правого
кокситов обычно не перекрещива-
ются. Вершинная бородавка сла-
бо развита. Базальная бородавка
со щетинками; "крайняя из них
довольно отчетливо выделяется
конце она изогнута. Класпеты:
Рис. 166. Aedes impiger JWalk. Личинка
IV стадии. Голова, сверху. По Carpenter
a. La Casse.
большей толщиной и длиной; на
стволик от середины изогнут,
крыло класпеты с хорошо выраженным пластинчатым расширением,
достигающим конца второй трети крыла. Выросты IX тергита относи-
тельно небольшие, с 4—10 щетинками каждый. Фаллосома слабо склеро-
тизована, на конце двузубчатая.
Л и ч и й к а IV стадии (рис. 166 и 167) средней величины, от
светло- до темно-коричневого цвета, с коричневой головой, ширина ко-
торой на четверть превышает длину. Лобные волоски: наружные из 2—
4 вторичноперистых ветвей, средние гладкие и простые, внутренние про-
стые, часто вторичноперистые; задние волоски наличника тонкие, корот-
кие, из 2—4 ветвей, лежат между основаниями средних. Усики корот-
кие — меньше половины длины головы, с редкими шипами; пучок из 2—
4 вторичноперистых ветвей, у середины, но ближе к основанию.
Щетка на VIII членике брюшка из 7—16 (среднее 13—14) чешуек,
расположенных в два неправильных ряда; каждая чешуйка с хорошо
развитым острым главным шипом и с мелкими дополнительными шипи-
ками у основания. Дыхательная трубка начиная от базальной трети
постепенно сужается к концу; индекс — около 3.0. Гребень из 10—20
(среднее 12—14) тесно сидящих зубцов, увеличивающихся в дистальном
направлении, занимает базальную треть трубки. Пучок у середины, ближе
17*
•260
HI. ПОДСЕМ. CULICINAE
к основанию, из 2—6 (среднее 4) вторичноперистых ветвей, длиннее, чем
ширина трубки у места его отхождения.
Седло последнего членика немного заходит за верхнюю половину
боковых сторон, с коротким простым боковым волоском. Наружные
хвостовые волоски простые и длинные, внутренние из 3—4 ветвей. Плав-
ник из 14—16 пучков, окруженных общим основанием, и из 1—3 ко-
ротких пучков перед ним. Жабры в 1.5 раза длиннее седла, удлиненно
ланцетовидные, заостренные и пигментированные.
[Распространение. Зона тундры обоих полушарий. Прони-
кает, обычно вместе с A. nigripes, на север далее других видов комаров.
Рис. 167. Aedes impiger Walk. Личинка IV стадии. Задний конец, сбоку.
По Carpenter a. La Casse.
Найден на острове Элсмир под 82°30' с. ш.; это наиболее северное место-
нахождение комаров на земном шаре. В южном направлении ареал вида
проникает в зону леса, особенно в горных районах. Известен на Аляске,
в Канаде, Гренландии, на севере Скандинавии и Финляндии. В СССР
имеются указания о нахождении вида на Кольском полуострове, в Архан-
гельской области и Ненецком национальном округе, на Таймыре, в устье
Колымы и на Новосибирских островах. Обычно встречается вместе
с A. nigripes. Из двух арктических видов в районе Норильска (70° с. ш.)
преобладает по численности A. impiger (Вольфтруб, 1963).
Биология. Личинки развиваются в открытых местностях в раз-
личной величины водоемах, образованных при таяньи снега в тундровой
зоне и в лесотундре. Местами (низовья Енисея) достигает высокой числен-
ности и активно нападает на человека и животных, особенно на северного
оленя. Весной появляются ранее других встречающихся в той же местности
видов.
7. AEDES
261
28. Aedes (Ochlerotatus) cataphylla Dyar, 1916 (rostochiensis Martini,
1920).
От всех предыдущих видов группы communis отличается наличием
светлых чешуек не только при основании, но и в других частях крыла.
Характерна также пестрая окраска покрывающих бедра чешуек.
Хоботок в одноцветных черных чешуйках. Щупики в темных чешуй-
ках, с примесью светлых. Среднеспинка в боковых отделах покрыта се-
ребристыми или светло-серыми чешуйками, посредине с широкой, обычно
Рис. 168. Гипопигий Aedes cataphylla Dyar. По Carpenter
a. La Casse.
Отдельно изображены лопасти IX тергита A. cataphylla Dyar (А)
и A. leucomelas Mg. (Б).
нерезко отграниченной полосой красновато-бурых или темно-бурых че-
шуек. Иногда почти вся среднеспинка в каштаново-коричневых чешуй-
ках, окаймленных по краям более светлыми. Посткоксальное пятно
имеется, равно как и гипостигмальное. Последнее полоской чешуек сое-
диняется с округлым белым пятном на парастигмах — небольшом скле-
рите между передним дыхальцем и передне-верхним краем мезэпистерн.
Бедра спереди пестрые, в белых и бурых перемешанных чешуйках. Голени
и лапки в темных чешуйках, среди которых вкраплены светлые. Крылья
в бурых чешуйках, с небольшой примесью беловатых, сконцентрирован-
ных при основании крыла, а также в небольшом количестве разбросан-
ных вдоль костальной, субкостальной и первой радиальной жилок.
Брюшко сверху в черно-бурых чешуйках, при основании тергитов с до-
вольно широкими белыми перевязями более или менее равномерной ши-
262
III. ПОДСЕМ. CULICINAE
рины; последние тергиты иногда на большей части поверхности покрыты
светлыми чешуйками.
Гипопигий (рис. 168) отличается длиной и густотой волосков на внут-
ренней поверхности кокситов; вершины волосков левого и правого кокси-
тов заходят друг за друга. Базальная бородавка коксита относительно
небольшая, с одним шипом (крепкой щетинкой); вершинная бородавка
сильно выпуклая. Класпета с резко изогнутым стволиком, крыло с пла-
стинчатым расширением. Выросты IX тергита узкие, с 4—13 (обычно 6—
8) относительно короткими крепкими шипами, на-
правленными назад.
Личинка IV стадии (рис. 169) сред-
них размеров, темно пигментирована. Лобные
волоски: наружные из 3—6 вторичноперистых
ветвей, средние впереди внутренних, обе пары
гладкие, простые; задние волоски наличника
между основаниями средних, из 2—3 коротких
и тонких ветвей. Усики более чем вдвое короче
головы, прямые, с умеренно развитыми шипиками
и с пучком у середины или немного ближе к вер-
шине, из 3—5 ветвей, не достигающих половины
длины усика.
Щетка из 10 (редко)—28 (среднее 25) чешуек,
расположенных в 2—3 неправильных ряда;
каждая чешуйка с длинным, заостренным шипом
и с рядом уменьшающихся шипиков у основания.
Дыхательная трубка прямая, суженная в вершин-
ной половине (индекс 2.5—3.0, редко больше);
гребень из 11—21 тесно сидящего зубца и из 2—4
более крупных гладких острых шипов, широко
расставленных и доходящих до границы третьей
четверти длины трубки от основания; пучок
, У середины трубки, ближе к основанию, около
Рис. 169. Aedes cataphyl- or -
la Dyar. Личинка IV ста- места окончания гребня, из 3—5 ветвей.
дии. Сифон. Седло последнего членика заходит за сере-
дину боковых сторон; боковой волосок короткий
и простой; наружные хвостовые волоски простые,
в среднем одной длины с дыхательной трубкой, внутренние вдвое короче,
из 5—8 ветвей. Плавник из 18 пучков в общем основании и 1—2 более
коротких впереди него. Жабры пигментированные, ланцетовидно заострен-
ные, варьируют в длину, равны длине седла или вдвое превышают ее.
Распространение. Ареал охватывает Европу, Казахстан, Си-
бирь, Дальний Восток, частично Северную Америку. На юге доходит
до степной Украины, Северного Кавказа, Киргизии (в горах). На севере,
по литературным данным, проникает в тундру. Вопрос о северной гра-
нице распространения нуждается в уточнении. В Скандинавии известен
до 64° с. ш., т. е. не доходит до северной границы леса (Natvig, 1948).
На. Аляске A. cataphylla распространен до 65° с. ш. (Gjullin et al., 1961).
Биология. Зимовка на фазе яйца. Личинки встречаются в. лу-
жах, образовавшихся от таянья снега или в результате весеннего разлива
рек, преимущественно в открытых местах или на опушках. В лесной
зоне весьма обычен, местами нападает в значительном количестве, но нигде
не отмечен в качестве преобладающего вида. Более характерен для откры-
тых стаций, мест с разреженным лесным покровом. Дает одно поколение
за сезон. Появляется весной на несколько дней ранее других видов или
7. AEDES
263
одновременно c A. communis и A. punctor. К середине лета лёт заканчи-
вается.
29. Aedes (Ochlerotatus) leucomelas Meigen, 1804 (salinellus Edwards,
1921).
По признакам имаго (окраска, строение гипопигия) очень близок
к предыдущему. Отличается примесью светлых чешуек на хоботке и
большим количеством светлых чешуек на крыльях.
Хоботок в темных чешуйках, с более или менее значительной при-
месью желтовато-белых, обычно преобладающих в средней части хоботка.
-Среднеспинка в бронзово-коричневых чешуйках, без выраженной темной
продольной полосы; по бокам среднеспинки чешуйки светлее — золоти-
стые или серовато-белые. Крылья в темных чешуйках, со значительной
примесью светлых, более многочисленных на костальной, субкостальной
и радиальных жилках. Бочки груди и ноги как у A. cataphylla. Брюшко
-с широкими белыми поперечными перевязями на тергитах, обычно четко
Рис. 170. Основание пучков плавника личинок IV стадии.
По Федорову.
1 —. у Aedes leucomelas Mg., 2 — у Aedes caspius Pall,
очерченными. Встречаются экземпляры с размытыми краями перевязей
и единичными (на задних тергитах иногда многочисленными) светлыми
чешуйками в задней половине тергитов. Такие особи имеют некоторое
-сходство с A. detritus.
Гипопигий как у A. cataphylla, с тем отличием, что выросты IX тер-
гита широкие и несут 6—17 (чаще 10—12) более тонких длинных щетинок,
направленных назад и в стороны.
Личинка IV стадии (рис. 170) средней величины разных
-оттенков коричневого цвета. Лобные волоски вторичноперистые: наруж-
ные из 3—6, средние впереди внутренних; обе пары простые, лишь редко
внутренние 2-ветвистые; задние волоски наличника между основаниями
-средних, тонкие и короткие, из 4 ветвей. Усик почти вдвое короче головы,
почти прямой, со слабо развитыми шипиками и с пучком у середины
из 3—6 ветвей, достигающих половины длины усика.
Щетка на VIII членике брюшка из 18—29 (в среднем 24) чешуек,
расположенных в 2—3 неправильных ряда, и сильно изменчивой формы:
большинство ближайших к спинной стороне чешуек более короткие, ли-
шены главного шипа, а несут 2—3 более длинных шипика с несколькими
более короткими по бокам; часть чешуек, ближайших к брюшной сто-
роне, с ясно выраженным главным шипом, с мелкими шипиками у основа-
ния. Дыхательная трубка прямая, начиная со второй трети сужается
к вершине,, индекс 2.5—3 (среднее 2.6). Гребень занимает базальную
треть трубки, из 15—24 (чаще около 20) тесно сидящих зубцов и несколь-
ких рудиментарных у основания. Пучок у середины трубки, ближе к осно-
ванию, из 3—8 ветвей со вторичной перистостью.
264
III. ПОДСЕМ. CULICINAE
Седло доходит до середины боковых сторон последнего членика; бо-
ковой волосок простой, одной длины с седлом; наружные хвостовые во-
лоски простые и длинные, внутренние из 5—9 вдвое более коротких
ветвей. Плавник из 15—18 пучков в общем основании и с 1—3 более
короткими впереди него; очень характерно строение каждого пучка:
ветвление его начинается далеко от основания, и длина базальной, не-
разветвленной, части пучка в 1.5—2 раза больше длины бокового отростка
у его основания. Жабры одной длины или короче седла, листовидной
формы, нижняя пара короче верхней.
Распространение. В Европе встречается от Южной Сканди-
навии и Карелии, где он распространен до 65° (Лобкова, 1964), до Венгрии,
степной Украины и Северного Кавказа. Известен также из Казахстана,
Сибири, Забайкалья (Улан-Удэ), Монголии (Улан-Батор). Границы аре-
ала, особенно в его азиатской части, нуждаются в уточнении.
Биология. Личинки встречаются в разного рода открытых или
полузатененных весенних водоемах, преимущественно в низинах, на опуш-
ках леса или в разреженных его участках, с дном, покрытым опавшей
листвой, часто около одиночных деревьев. Встречаются в водоемах
с слегка солоноватой водой. Совместно с A. detritus, A. cataphyIla, А. рипс-
tor. В зонах леса и лесостепи — широко распространенный, но не мас-
совый вид. Дает одно поколение за сезон. Весной появляется одновре-
менно с наиболее ранними Aedes, к середине лета лёт прекращается.
30. Aedes (Ochlerotatus) detritus Haliday, 1833 (salinus Ficalbi, 1896).
От всех прочих видов группы communis, кроме A. simanini, хорошо
отличается окраской брюшка (рис. 171).
Хоботок и щупики в бурых чешуйках, с более или менее значительной
примесью светлых. Среднеспинка в перемешанных желтовато-коричне-
вых и белых чешуйках. Покровы груди коричневые. Бочки груди в широ-
ких белых чешуйках. Посткоксальное пятно отсутствует, гипостигмальное
имеется. Бедра спереди пестрые; голени и лапки в темных чешуйках, с более
или менее значительной примесью светлых. Крылья пестрые, покрыты:
широкими темными и светлыми чешуйками. Брюшко сверху с нерезко
очерченными светлыми поперечными перевязями при основании сегмен-
тов. Задний отдел каждого тергита в перемешанных бурых и желтовато-
белых чешуйках, причем темные обычно преобладают на передних, а свет-
лые — на задних сегментах.
Гипопигий (рис. 172): волоски коксита густые и довольно длинные:
концы волосков левого и правого кокситов немного заходят друг за друга.
Базальная бородавка с одним шипом (крепкой щетинкой) и более корот-
кими щетинками. Стволик класпеты короткий, изогнутый; ее крыло с ру-
кояткой; пластинчатое расширение развито в средней и вершинной ча-
стях крыла. Фаллосома короткая, яйцевидная, без перетяжки.
Личинка IV стадии (рис. 173 и 174) средней величины, от
желтоватой до коричневой окраски; лобные волоски: наружные из 7—
12, средние из 1—3 (чаще простых) впереди внутренних из 2—3 ветвей;
задние волоски наличника 2—3-ветвистые, тонкие и короткие. Усик около
0.6 длины головы, тонкий и слегка изогнутый, покрыт редкими шипиками,
с пучком у середины из 5—8 (среднее 6) ветвей, достигающих 1/2 длины
усика.
Щетка очень изменчива по числу чешуек — от 25 до 60 или больше; каж-
дая чешуйка лишена главного шипа, ее короткое тело, отходящее от су-
женного основания, несет по краю ряд шипиков, из которых вершинные
наиболее длинные. Дыхательная трубка короткая, индекс 2.2—2.5, на-
7. AEDES
265
чиная от середины слабо сужена к вершине. Гребень из 18—27 зубцов, за-
нимает до 1 /2 длины трубки; зубцы равномерно расставлены, только наи-
более дистальный иногда удален от остальных. Пучок у середины или
ближе к основанию, из 6—10 (чаще 8) ветвей. Стигмальная пластинка
отличается рядом характерных признаков в строении рычага: его тело не
Рис. 171. Aedes detritus Hal.
находится, как обычно, в центре пластинки между стигмами, а сдвинуто
назад, к задним клапанам; соответственно задняя лука неразвита, а задний
отросток с сильно изрезанным задним краем и с тонкими рудиментарными
ветвями; плечи и отростки к стигмам отсутствуют; очень сильно развита
передняя лука рычага, далеко вдающаяся в промежуток между стигмами.
266
III. ПОДСЕМ. CULICINAE
Седло последнего членика брюшка доходит до середины, его боковой
волосок простой, достигает длины седла; наружный хвостовой волосок
Рис. 172, Гипопигий Aedes
detritus Hal.
длинный, превышает длину трубки, внутрен-
ний в виде веера из 8—11 ветвей. Плавник
из 16—18 пучков в общем основании и из
2—3 более коротких впереди него. Жабры
очень короткие, шаровидные.
Распространение. Западная и
Южная Европа на север до Англии и
Южной Скандинавии; Северная Африка,
Передняя Азия. В СССР распространен в
республиках Средней Азии и на юге Казах-
стана. Имеются также сообщения о нахо-
ждении A. detritus в Ленинградской об-
ласти, на Украине, на Нижней Волге и
даже в Западной Сибири. Некоторые из
этих находок, возможно, основаны на неточ-
ном определении. Сообщалось также об обна-
ружении A. detritus в Кашгаре (Синьцзян)
и на северо-западе Монголии (цит. по: Nat-
vig, 1948).
Биология. Личинки встречаются в не-
больших весенних временных стоячих водое-
мах и лужах преимущественно с сильно со-
лоноватой, реже пресной водой, на юге аре-
ала — исключительно в солоноватой воде,
с содержанием солей от 0.83 до 5.2%, часто в заболоченностях вдоль
морских берегов. В Таджикистане и Киргизии встречен в горных районах
Рис. 173. Aedes detritus Hal. Личинка IV стадии.
1 — голова, сверху, 2 — стигмальная пластинка.
на высоте 1500—2000 м над ур. м., в водоемах, возникших после таяния
снега. Иногда попадаются совместно с A. caspius caspius, С. longiareolata и др.
Для умеренных широт есть указания на два или больше поколений в году
7. AEDES
867
после выпадения летних дождей; в более южных широтах наблюдается
одно поколение, возможно, в связи с ранним пересыханием водоемов.
Ранневесенний вид, зимующий на фазе яйца. Самки весьма активно напа-
дают на человека, местами в заметной численности. Характерен для при-
морских районов и внутриконтинентальных областей с обилием солоно-
ватых водоемов. В СССР является массовым видом в некоторых пустын-
Рис. 174. Aedes detritus Hal. Личинка IV стадии. Задний
конец, сбоку. По Marshall.
ных и степных районах Туркмении, Таджикистана и Узбекистана. На боль-
шей части ареала (далеко не достаточно выясненного) относится к числу
относительно редких видов.
Предполагается, что A. detritus является промежуточным хозяином
филярииды Dipetalonema evansi, вызывающей тяжелое заболевание верб-
людов (Орехов, 1952).
31. Aedes (Ochlerotatus) simanini Gutsevich, 1966.
Близок к предыдущему. Отличается более темной окраской, наличием
посткоксального пятна чешуек, строением гипопигия.
Хоботок и щупики в перемешанных темных и светлых чешуйках, обычно
светлые более многочисленны. Покровы груди черно-бурые. Средне-
спинка в желтовато-бронзовых и беловатых чешуйках; последние преоб-
ладают по бокам среднеспинки, перед щитком и на щитке. Бочки груди
в белых чешуйках, за исключением верхней половины проэпимер, покры-
той прямыми и относительно широкими чешуйками сероватого цвета.
Посткоксальная перепонка густо покрыта чешуйками. Гипостигмальное
868
III. ПОДСЕМ. CULICINAE
пятно продолжается кзади в виде полоски. Ноги: передние и средние
бедра и голени спереди пестрые; лапки без светлых колечек, но белых че-
шуек много на всех члениках лапок, иногда они преобладают; сочлене-
ния члеников лапки темные. Крылья пестрые, в относительно крупных
перемешанных бурых и белых чешуйках; последние разбросаны на всех
жилках. Тергиты брюшка в черно-бурых чешуйках, с довольно широкими
Рис. 175. Гипопигий Aedes simanini Guts.
Отдельно, при большем увеличении, изображены базальная бородавка
коксита (А), лопасть IX тергита (Б) и крыло класпеты (В).
белыми поперечными перевязями при основании сегментов. Немногочи-
сленные белые чешуйки разбросаны среди темных, покрывающих боль-
шую часть каждого тергита; иногда скопления светлых чешуек образуют
неясные перевязи и у заднего края некоторых сегментов.
Гипопигий (рис. 175): коксит покрыт волосками умеренной длины,
вершины волосков левого и правого коксита не перекрещиваются. Ба-
зальная бородавка коксита с одной крепкой щетинкой и более мелкими
щетинками и волосками. Вершинная бородавка развита. Стволик кла-
спеты прямой; ее крыло с пластинчатым расширением, развитым только
на небольшом протяжении, ближе к основанию. Фаллосома удлиненно-
овальная, с легкой перетяжкой?
1 Описание личинки см. на стр. 379.
7. AEDES
269
Вид описан (Гуцевич, 1966) по материалу, собранному П. И. Симани-
ным в марте—апреле 1929 г. в Коканде (Узбекистан). Найден также в Ка-
захстане: пос. Илицск Алма-Атинской области, окрестности Алма-Аты
на высоте 1800 м над ур. м. Вероятно, распространен в Средней Азии,
но не отграничивался от А. detritus.
Добавление к подроду OCHLEROTATUS
32. Aedes (Ochlerotatus) kasachstanicus Gutsevich, 1962.
Отличается белой окраской лапок и торчащими чешуйками задних
голеней. Не может быть отнесен ни к одной из групп видов палеаркти-
Рис. 176. Aedes kasachstanicus Guts.
А — гипопигий, Б — крыло класпеты, В — область поперечных жилок крыла, Г —. часть »
задней голени.
ческих Ochlerotatus (Гуцевич, 1962). От других видов подрода A. kasach-
stanicus отличается более коротким хоботком; его длина приблизительно
равна длине передних бедер.
270
III. ПОДСЕМ. CULICINAE
Щупики самки и хоботок в темных и светлых чешуйках; последние
преобладают в средней части хоботка. Щупики самца со светлыми коль-
цами. Среднеспинка в темно-каштановых чешуйках, среди которых раз-
бросаны беловатые, образующие неясную каемку впереди и по бокам
среднеспинки, а также покрывающие щиток и площадку перед щитком.
Бочки груди в широких белых чешуйках. Посткоксальное пятно чешуек
имеется, гипостигмального пятна нет.
Бедра и голени спереди в перемешанных светлых и темных чешуйках.
На задних голенях, помимо прилегающих, имеются и торчащие чешуйки,
главным образом темные; у самца торчащих чешуек на голени больше,,
чем у самки. Лапки белые, затемнены только вершины члеников. Крылья
в относительно широких бурых и белых чешуйках; белые разбросаны
на всех жилках, но бурые преобладают. На брюшке сверху преобладают
светлые чешуйки; темные имеются преимущественно в средней части,
тергитов. Церки выступают отчетливо.
Такие признаки, как широкие чешуйки на бочках груди и торчащие-
чешуйки на задних голенях, указывают на известное сходство A. kasach-
stanicus с видами подрода Muctdus, распространенного в Эфиопской,
Ориентальной и Австралийской областях.
Гипопигий (рис. 176): коксит с базальной- и вершинной бородавками;
вершинная — умеренно развита, базальная — крупная, выступающая,
густо усажена довольно длинными щетинистыми волосками, располо-
женными в несколько рядов. Шипов или крепких щетинок, выделяющихся
своей толщиной, на базальной бородавке нет. Класпеты: стволик пря-
мой, крыло со слабо развитым пластинчатым расширением; вблизи осно-
вания крыла класпеты имеется небольшой крючковидный выступ.
Распространение. Найден в ряде пунктов по течению
р. Или (Алма-Атинская область, Казахстан).
Биология. Характерные биотопы — леса и заросли кустарни-
ков в пойме. Появляется в мае, в наибольшем количестве нападает в июне;,
к августу лёт заканчивается. Очевидно, дает только одно поколение-
за сезон. Активный кровосос.
Личинка A. kasachstanicus найдена А. М. Дубицким (стр. 378).
2. Подрод AEDIMORPHUS Theobald, 1903 (ECCULEX Felt)
Близок к подроду Ochlerotatus. Отличается строением гипопигия?
класпеты отсутствуют, базальная бородавка коксита обычно сильно раз-
вита, иногда обособлена в виде образования колбовидной формы; палочко-
видный придаток (если есть) причленяется перед вершиной стиля; эдеа-
гус разделен на две зазубренные пластинки. По личинкам не отличается
от подрода Ochlerotatus.
Подрод распространен в Эфиопской и Ориентальной областях, всего
известно свыше 90 видов. В Палеарктике подрод представлен двумя
видами; один из них широко распространен, другой — A. alboscutella-
tus Theob. — в ее пределах найден только на юге Японии.
33. Aedes (Aedimorphus) vexans Meigen, 1830.
Характерный отличительный признак — очень узкие светлые ко-
лечки при основании члеников лапок (рис. 97).
Хоботок и щупики в бурых чешуйках, с небольшой примесью белых.
Среднеспинка покрыта одноцветными красновато-бурыми или бронзо-
7. AEDES
271
выми чешуйками; небольшое пространство перед щитком и щиток в золо--
тистых чешуйках. Бедра и голени спереди покрыты темными чешуйками,
среди которых разбросаны светлые. Лапки темные, с беловатыми колеч-
ками; каждое из них обычно занимает не более 1/5—1/4 длины соответ-.
ствующего членика; в этом
отличие от видов группы can-
tans, у которых колечки
шире. Ширина светлых ко-
лечек у А. vexans сильно
варьирует. Экземпляры с от-
носительно более широкими
колечками похожи на А. can-
tans; но последние отлича-
ются окраской среднеспинки,
брюшка и наличием светлых
чешуек на различных жил-
ках крыла. У A. vexans
крылья в бурых чешуйках,
единичные светлые могут
быть только при основании
крыла. Окраска брюшка раз-
лична у двух подвидов, на
которые подразделяется А.
vexans.
Гипопигий очень своеоб-
разен (рис. 177): базальная
бородавка почти отделена от
тела коксита в виде колбо-
видного выступа, расширен-
ный конец которого густо
усажен волосками; стили ши-
рокие, пластинчатые, с па-
лочковидным придатком, при-
членяющимся далеко отступя
от вершины стиля; по этому
последнему признаку вид
можно определить по целым
экземплярам, без приготов-
ления препаратов.
Личинки А. vexans, в свя-
зи с очень широким распро-
странением вида и разнооб-
разием условий развития
преимагинальных фаз и оби-
тания взрослых, чрезвычай-
но изменчивы, особенно по
габитуальным признакам, со-
Рис. 177. Гипопигий Aedes vexans Mg. По Carpen-
ter a. La Casse.
здавая впечатление существования эколого-географических внутри-,
видовых различий. Однако сравнение описаний личинок у разных авторов,
из Европы, Азии, Африки и Северной Америки и просмотр материалов,
по СССР не позволяют различать их по личиночной фазе даже с минималь-.
ной степенью достоверности.
Личинка IV стадии (рис. 178) от средних (на юге) до круп^.
ных (на севере) размеров, от желтовато- (на юге) до темно-коричневого,.
272
III. ПОДСЕМ. CULICINAE
почти черного (на севере) цвета. Лобные волоски вторичноперистые:
наружные из 6—12 (чаще 7—9), средние впереди внутренних и немного
шире расставлены, из 1—2 (редко 3), внутренние из 2—5 (чаще 3—4)
ветвей. Задние волоски наличника из 2—5 коротких и тонких ветвей,
между основаниями средних. Усик около 1 /2 длины головы, с умеренной
шиповатостью, более развитой у северных форм, с пучком из 5—12 вет-
вей (достигающих 1 /2 дли-
ны усика), лежащим на
расстоянии 1 /3 его длины
от основания.
Щетка из 7—13 (чаще
8—10) чешуек с остроко-
нечным шипом и с рядом
тонких шипиков по бокам
основания. Дыхательная
трубка начиная со второй
трети постепенно сужается
к вершине, длина очень
изменчива: индекс 2.5—
4.3 (в среднем 3.0—3.5),
у южных форм меньше,
чем у северных. Гребень
доходит до середины труб-
ки или (у крупных личи-
нок) заходит за нее, со-
стоит из 12—25 (чаще
18—20) зубцов, из которых
1—3 у основания рудимен-
тарны, а 1—3 наиболее
дистальных широко рас-
ставлены и крупнее ос-
тальных. Пучок ближе
к вершине из 3—8 (чаще
4—6) коротких ветвей.
Седло на последнем
членике достигает середи-
ны или нижнего края его
Рис. 178. Aedes vexans Mg. Личинка IV стадии. боков; боковой ВОЛОСОК
Задний конец, сбоку. По Marshall. простой, реже из 2—3
ветвей; наружный хво-
стовой [волосок простой, длиннее дыхательной трубки, внутренний
из 4—9 вдвое более коротких ветвей. Плавник из 11—13 пучков, окру-
женных общим основанием, и из 3—4 более коротких впереди него. Жабры
в 1.5 раза больше длины седла или длиннее, ланцетовидной формы, не
пигментированы; обе пары одной длины или нижняя немного короче.
Распространение. Европа, Азия, Африка, Северная Аме-
рика. В Европе проникает на север примерно до 60—62° с. ш., местами,
по долинам рек, и дальше. Особенно широко распространен в Азии:
Средняя Азия, Казахстан, Сибирь, Дальний Восток СССР, Япония,
Китай, Индокитайский полуостров, Индия, Иран, Малая Азия. В Африке
встречается от средиземноморского побережья до самого юга материка;
в Америке — от юга Канады до Мексиканского залива.
Биология. Личинки на юге приурочены к пресноводным или
солоноватым пойменным водоемам, к полоям дельт, оставшимся после
7. AEDES
273
паводков, весной чаще бедных растительностью; севернее — в различных
весенних водоемах (лужи, канавы, ямы) в открытых местах или на опуш-
ках, но в пресной воде. Вид наиболее характерен для областей с теплым
или жарким влажным летом. Особенно высокой численности достигает
в поймах и дельтах рек. Весной вылет начинается несколько позже,
чем у большинства видов Aedes. Весеннее поколение обычно относительно
немногочисленно. После летних дождей или летнего паводка у рек, теку-
щих с юга, происходят повторные выплоды, вылет может продолжаться
до осени. Период массовой активности длится от двух до четырех меся-
цев; сезонная кривая численности чаще дву- или многовершинна, при-
чем максимум может приходиться на различные месяцы, в зависимости
от реяшма рек и от помесячного распределения летних осадков.
В различных частях ареала A. vexans наблюдалось его массовое раз-
множение (Приморье, Хабаровский край, низовья Дуная, Днепра, Дона,
Волги и Терека, некоторые районы Закавказья, долина Рейна, многие
местности в США и т. д.). Нередко A. vexans по численности занимает
первое место среди насекомых, нападающих на людей и животных.
Из мест массового выплода может мигрировать на 10—20 км и более.
Нападает преимущественно под открытым небом, но нередко комары этого
вида в большом количестве проникают в дома и помещения для животных.
Эпидемиологическое значение. В очагах туляремии
A. vexans может играть роль в циркуляции возбудителя; установлена
естественная зараженность, а также возможность передачи туляремий-
ного микроба комарами данного вида в условиях эксперимента. В За-
карпатье из A. vexans был выделен вирус группы лимфоцитарного хорио-
менингита, а в Словакии — вирус тягиня. В США A. vexans является
одним из переносчиков вируса восточного энцефалита (энцефаломиелита
лошадей). Личинки филярииды собак — Dirofilaria immitis — могут раз-
виваться в организме комаров данного вида до инвазионной стадии.
Aedes vexans vexans Meigen.
Брюшко сверху в бурых чешуйках, по переднему краю тергитов
с белыми, посредине слегка суженными поперечными перевязями; форма
перевязей напоминает лежащую букву В. Задний край последних терги-
тов нередко в белых чешуйках. Экземпляры из Закавказья и Средней
Азии обычно отличаются светлой окраской чешуек и покровов средне-
спинки.
Описание личинок см. в описании вида (стр. 271).
Распространение. Европа, Америка, Африка, Западная
Азия, Казахстан, Сибирь.
Aedes vexans nipponii Theobald, 1907.
Отличается несколько более крупными, в среднем, размерами и боль-
шим количеством светлых чешуек на брюшке, а также на хоботке, щу-
пиках, бедрах и при основании крыльев. На брюшке, помимо довольно
широких поперечных перевязей, светлые чешуйки образуют пятна либо
в виде продольной полоски (обычно на передних сегментах), либо в форме
треугольника, обращенного вершиной вперед, обычно на задних сег-
ментах брюшка. На задних тергитах иногда светлые чешуйки преобла-
дают. Светлые чешуйки имеются при основании крыла, как исключение —
и в других его частях.
По личинкам не отличается от основной формы.
Распространение. Подвид характерен для Дальнего Вос-
тока: Приморье, Хабаровский край, Забайкалье, Монголия,. Китай,
18 Фауна СССР, А. В. Гуцевич и др.
274
III. ПОДСЕМ. CULICINAE
Япония. Встречается в Сибири и в Казахстане. Оба подвида связаны по-
степенными переходами. В одной и той же местности могут встречаться
как «типичные формы», так и переходные варианты.
3. Подрод FINLAYA Theobald
Отличается от других подродов Aedes главным образом строением
гипопигия. Класпеты имеются. Базальная бородавка коксита отсут-
ствует или слабо развита, вершинной бородавки нет. Придаток стиля
причленяется на его вершине. Эдеагус простой, неразделенный. Щупики
самца, как правило, немного короче хоботка.
Характерно строение задних сегментов брюшка самки: VIII стернит
широкий, не втягивающийся в предыдущий; церки самки короткие (отли-
чие от двух предыдущих подродов). Хоботок явственно длиннее перед-
них бедер. Особенность окраски — снежно-белые пятна по бокам брюшка;
у видов Ochlerotatus светлые чешуйки брюшка обычно не имеют снежно-
белого цвета. Хоботок и крылья у палеарктических видов Finlaya по-
крыты одноцветными темными чешуйками.
Личинки подрода Finlaya не имеют специфических отличий от других
палеарктических подродов рода Aedes. Их усики обычно лишены шипи-
ков, реже последние слабо развиты; волосок на их теле простой или
2-ветвистый. Волоски на теле могут иметь звездчатое строение или в виде
сильно развитых шипов. Дыхательная трубка короткая, с хорошо раз-
витыми ушками у основания. Зубцы гребня часто расположены в дуго-
образный или слабо S-образный ряд. Жабры различной формы; в обрат-
ном отношении с их величиной развит плавник.
Личинки большинства видов развиваются в небольших скоплениях
воды (дупла деревьев, пазухи листьев, пустоты в стволах бамбука,
углубления в скалах и т. п.). Биотопы отдельных видов специфичны,
вследствие чего распространение большинства видов пятнистое и часто
узко локализованное.
Подрод широко распространен, всего известно около 190 видов. Наи-
большего развития достигает в Ориентальной области. Из видов, встречаю-
щихся в Палеарктической области, большинство связано с ориенталь-
ными группами. В СССР известно пять видов, лишь один из них распро-
странен в Европе; остальные встречаются только на Дальнем Востоке.
Комары подрода Finlaya, как правило, не относятся к числу массо-
вых кровососов, но некоторые виды имеют серьезное значение как пере-
носчики возбудителей болезней человека — японского энцефалита, фи-
ляриозов, а отчасти, по-видимому, и желтой лихорадки (Aedes leucocelae-
nus Dyar et Shannon в Южной Америке).
34. Aedes (Finlaya) geniculatus Olivier, 1791 (ornatus Meigen, 1818).
Относительно крупный комар с широкой темной продольной поло-
сой на среднеспинке и белыми пятнами на вершине бедер (рис. 179).
Темные чешуйки хоботка, ног и брюшка имеют фиолетовый оттенок.
Хоботок и щупики в черных чешуйках. Среднеспинка с широкой,
резко отграниченной продольной полосой черно-бурых или шоколадно-
коричневых чешуек, или с двумя сближенными темными полосками;
по бокам задней половины среднеспинки развиты еще две темные полоски.
Боковые отделы среднеспинки покрыты серебристо-серыми или кремо-
выми чешуйками. Щиток в узких желтоватых чешуйках. Бочки груди
с пятнами широких снежно-белых чешуек. Голени и лапки в черных че-
7. AEDES
275
шуйках. На вершине бедра, в месте сочленения с голенью, имеется не-
большое пятно белых чешуек, резко выделяющееся на фоне черной окраски
голеней и дистальных отделов бедер. На передних и средних лапках
Рис. 179. Aedes geniculatus 01.
самки коготки с зубчиком, на задних — простые. Чешуйки крыльев одно-
цветные, бурые. Брюшко сверху в черных чешуйках, с треугольными^,
чисто белыми пятнами по бокам каждого сегмента.
18*
376
III. ПОДСЕМ. CULICINAE
Рис. 180. Гипопигий Aedes geni-
culatus 01. По Natvig.
Гипопигий (рис. 180): коксит густо покрыт чешуйками (с наружной
стороны) и волосками; базальная бородавка в виде покрытого волосками
небольшого бугорка без шипов и ще-
тинок. Стволик класпеты прямой, длин-
нее крыла; волосок на стволике рас-
положен ближе к основанию стволи-
ка; крыло класпеты узкое, изогнутое.
Лопасти IX тергита с 3—6 щетинками.
Личинки IV стадии (рис.
181) крупных размеров, слабо пигмен-
тированные, грязно-серого или желто-
ватого цвета, с более темными голо-
вой и дыхательной трубкой; сильно
развитые звездчато ветвящиеся волоски
придают личинке шиповатый вид. Лоб-
ные волоски слабо развиты: наруж-
ные короткие и тонкие, из 2—4 вет-
вей, средние сдвинуты вперед, из 1—2
(чаще) более толстых ветвей, лежат
впереди внутренних, состоящих из
простого, но вдвое более длинного во-
лоска; между основаниями средних нахо
дятся задние волоски наличника, из
7—8 веерообразно расходящихся тонких коротких ветвей. Усик тонкий,
около х/2 длины головы, без шипиков, с пучком в виде простого, редко
2-вётвистого, короткого волоска.
Рис. 181. Aedes geniculatus 01. Личинка IV стадии.
1 — задний конец, сбоку (по Marshall), г — стигмальная пластинка,
7. AEDES
277
Щетка на VIII членике из 11—16 чешуек, расположенных в один
выпуклый назад дугообразный ряд; чешуйки с длинным шиповидным
зубцом, по бокам основания которого сидят тонкие короткие щетинки.
Дыхательная трубка прямая и короткая, индекс около 3.0. Гребень
из 14—20 увеличивающихся в дистальном направлении зубцов, тесно
расположенных в один слегка изогнутый ряд, заходящий за базальную
1/4 трубки; пучок из 4—5 ветвей, впереди наиболее дистального зубца
гребня.
, Седло последнего членика заходит за его боковые стороны, с покры-
тым мелкими шипиками задним краем; боковой волосок значительно
длиннее седла, из 2—3 ветвей; наружный хвостовой волосок простой,
более толстый, превышает длину дыхательной трубки, внутренний в сред-
нем из трех почти вдвое более коротких ветвей. Плавник из 7—10 2—4-
ветвистых пучков в общем основании и 1—2 более коротких впереди
него. Жабры колбасовидные, слегка суженные на концах, длиннее седла,
нижняя пара более короткая.
378
III. ПОДСЕМ. CULICINAE
Личинки из северного Ирана (рис. 182) отличаются меньшими раз-
мерами, более длинными и сильнее ветвящимися, но более тонкими во-
лосками на теле, с более заметно развитой вторичной перистостыо в их
базальной части, более длинными пучками плавника и более короткими
жабрами.
Распространение. Европа на север до Англии и юга Скан-
динавии (до 58°); Северная Африка; Передняя Азия на восток до Ирана.
В СССР распространен на западе и на юге: Литва, Белоруссия, Украина,
Кавказ. Известны единичные находки в Туркмении, а также в Чувашии,
Татарии, Куйбышевской области, Воронеже.
Биология. Личинки встречаются в скоплениях воды в дуплах
деревьев (бук, дуб, орех, тополь, ясень), как в сильно затененных (ство-
ловые дупла), так и "в лучше освещенных (скопления воды в пнях).
Изредка развиваются в открытых водоемах (лужи, ямы и т. п.). Совместно
с личинками Ай. plumbeus, A. pulchritarsis, Orthopodomyia. В северной
части ареала зимовка на фазе яйца, в южной — могут перезимовывать
и личинки, давая в теплые зимы вылет взрослых комаров. Относительно
высокой численности достигает в лиственных лесах областей с большим
количеством летних осадков и мягкой зимой; реже встречается в смешан-
ных и еще реже — в хвойных лесах. В лесах Карпат, Кавказа и Талыша
местами является массовым кровососом, численно преобладающим по
сравнению с другими видами комаров. В остальных районах его обнару-
жения в СССР редок. Нападает преимущественно в природных условиях,
изредка проникает в населенные пункты. Весной появляется позже боль-
шинства видов Aedes. Сезонный ход численности в основном зависит от лет-
них осадков, максимум обычно в середине лета.
В условиях эксперимента установлена возможность передачи кома-
рами A. geniculatus вируса желтой лихорадки; в очагах желтой лихо-
радки этот вид не встречается.
35. Aedes (Finlaya) echinus Edwards, 1920.
Очень близок к предыдущему. Отличается окраской груди, в частности
цветом и формой чешуек щитка.
Хоботок и щупики в черных чешуйках. Среднеспинка с двумя сбли-
женными каштаново-коричневыми продольными полосами, резко отгра-
ниченными по бокам белыми чешуйками. Щиток в относительно широких
белых чешуйках. Бочки груди с пятнами снежно-белых чешуек, резко
выделяющимися на черном фоне покровов. Бедра с небольшим белым
пятном на вершине; голени и лапки черные. Крылья покрыты темными
чешуйками. Брюшко сверху темное, с узкими беловатыми перевязями
при основании сегментов; по бокам тергитов перевязи расширяются
в виде треугольных белых пятен.
В строении гипопигия по сравнению с A. geniculatus существенных
отличий, по-видимому, нет. Можно лишь отметить, что у A. echinus коксит
покрыт очень длинными густыми волосками.
Личинка IV стадии (рис. 183 и 184) отличается исключи-
тельно сильным развитием волосков груди и брюшка, которые в виде
звездчато ветвящихся от основания толстых шипов, несущих вторичную
перистость, придают ей сходство с ежом, за что вид и получил свое на-
звание. Описание составлено по материалу из Болгарии, любезно пре-
доставленному Д. К. Божковым.
Лобные волоски длинные, без вторичной перистости: наружные
из 2—4 (чаще 3), средние из 1—2 ветвей, впереди внутренних — всегда
простых; задние волоски наличника из 7—10 (чаще 7) более тонких и
7. AEDES
279
коротких ветвей, делящихся у основания. Усик длинный (0.60—
0.75 длины головы), заметно изогнутый внутрь, лишен шипов, с пучком
в виде простого, редко 2-ветвистого волоска, отходящего на 3/6 длины
усика от основания; иногда у вершины волосок расщепляется на 2—
3 ветки.
Щетка из 12—18 (чаще 14) сильно вытянутых чешуек, расположенных
в обращенный выпуклостью назад слегка дугообразный ряд; каждая
чешуйка имеет сильно развитый главный шип, по бокам которого начи-
ная от его вершины или отступя от нее сидят мелкие шипики, уменьшаю-
щиеся к основанию чешуйки. Дыхательная трубка почти цилиндриче-
ская, индекс 3.0—3.5 или немного больше; с хорошо развитыми ушками
Рис. 183. Первый членик брюшка, сверху. По Edwards.
1 — у Aedes geniculatus 01., 2 — у Aedes echinus Edw.
у основания. Гребень из 15—27 (чаще 18—22) тесно сидящих длинных
шиповидных зубцов, длина которых слегка уменьшается к основанию
трубки, и ближайших к нему 2—4 рудиментарных; у основания каждого
зубца имеется один более крупный шипик и несколько, мелких; гребень
заходит за середину трубки. Пучок перед наиболее дистальным зубцом
гребня, из 2—4 (чаще 3) ветвей, в длину заметно превышающих диаметр
трубки.
Седло последнего членика почти доходит до нижнего края его боковых
сторон; боковой волосок из двух грубых ветвей, превышающих длину
седла; наружный хвостовой волосок простой, более чем в 2 раза длиннее
трубки, внутренний из трех (редко двух) более коротких ветвей. Плав-
ник из 9—13 2—3-ветвистых пучков в общем основании и 1—2 впереди
него. Жабры сильно развиты, верхняя пара в 3—4, нижняя в 1.5 раза
длиннее седла.
Распространение. Средиземноморская подобласть: Север-
ная Африка, Малая Азия, Южная Европа, включая юг Балканского
полуострова; найден в Болгарии (Божков, 1966). В СССР не обнаружен.
Биология личинок малоизучена, по-видимому, сходна с тако-
вой у A. geniculatus. В Анатолии личинки встречаются в прикорневых
280
1П. ПОДСЕМ. CULICINAE
дуплах олив, заполняющихся водой при весенних разливах. На черно-
морском побережье Болгарии достигает заметной численности.
36. Aedes (Finlaya) nipponicus La Casse et Yamaguti, 1948.
Подобно двум предыдущим видам, характеризуется отсутствием белых
коле^к на лапках. От других палеарктических видов с темными лапками
отличается наличием белых пятен на среднеспинке. В литературе ранее
фигурировал как A. niveus Ludlow, 1903. В 1948 г. по материалу из Япо-
Рис. 184. Aedes echinus Edw. Личинка IV стадии. По Edwards.
1 —• голова, сверху, г — задний конец, сбоку.
нии был описан новый подвид — A. niveus nipponicus La Casse et Yama-
guti, который в 1950 г. те же авторы предложили рассматривать в ка-
честве самостоятельного вида. Относящаяся к группе geniculatus под-
группа niveus представлена довольно большим количеством видов, рас-
пространенных в Ориентальной области (Colless, 1958). Различия между
видами в ряде случаев, по-видимому, незначительны. В частности, при-
знаки, по которым различаются A. niveus и A. nipponicus (литературные
данные; см. ниже), не кажутся вполне достаточными для обоснования
видовой самостоятельности A. nipponicus.
Хоботок и щупики в одноцветных бурых чешуйках. Среднеспинка
с двумя большими пятнами серебристо-белых чешуек, разделенными
посредине. продольной темной полосой, расширяющейся кзади. Щиток
7. AEDES
281
в светлых чешуйках. Крылья и лапки в одноцветных бурых чешуйках.
Брюшко покрыто черными чешуйками с белыми пятнами по бокам сег-
ментов, иногда соединенными узкими белыми перевязями при основании
тергитов; чаще перевязи развиты на задних сегментах. Сперматека одна
(La Casse a. Yamaguti, 1955), что является исключением; у видов Aedes,
как правило, три сперматеки.
Aedes nipponicus отличается
от A. niveus «тем, что белые пят-
на среднеспинки почти всегда
(хотя имеются исключения) от-
граничены с медиальной сторо-
ны вогнутой линией» (La Casse
a. Yamaguti, 1955). У комаров
этого вида из Приморья окраска
среднеспинки весьма изменчива.
В большинстве случаев темная
продольная полоса, разделяю-
щая белые пятна, доходит до
переднего края среднеспинки,
причем линия, разграничиваю-
щая светлый и темный участки,
Рис. 185. Гипопигий
La Casse et Yamag. По
guti.
Aedes nipponicus
La Casse a. Yama-
Рис. 186. Aedes nipponicus La Casse
et Yamag. Личинка IV стадии. Голова,
сверху.
темная полоса
пятна оказы-
передняя поло-
может быть прямой или (чаще) слегка вогнутой. Часто
не доходит до переднего конца среднеспинки, и белые
ваются соединенными. Иногда, особенно у самцов, вся
вина среднеспинки покрыта серебристыми чешуйками.
Гипопигий весьма своеобразен (рис. 185). Чешуйки покрывают не
только наружную, как у других видов Finlaya, но отчасти и внутреннюю
поверхность коксита: в дистальной части его поверхности расположен
пучок крупных чешуек. Базальная бородавка весьма слабо развита,
с двумя короткими щетинками. У основания коксита расположен пучок
густо сидящих коротких волосков. Стили с длинным придатком, со-
282
III. ПОДСЕМ. CULICINAE
ставляющим около 3/4 длины стиля. Сближенные лопасти IX тергита
несут по несколько тонких длинных щетинок.
Личинки IV стадии (рис. 186 и 187), как у всех дупловых
видов, изменчивы по величине, зеленовато-серого или серо-коричневого
цвета, с темными головой и дыхательной трубкой.
Лобные волоски сдвинуты к переднему краю головы, вторичноперис-
тые, с более длинными центральными ветвями по сравнению с боковыми;
наружные — наиболее длинные, из 6—16 (чаще 9—12), средние из 7—12
Рис. 187. Aedes nipponicus La Casse et Yamag. Личинка IV стадии. Задний
конец, сбоку.
(чаще 9—10), внутренние из 8—18 (чаще 9—12) ветвей. Задние волоски
наличника сильно развиты, сходны со средними, лежат между ними,
из 6—12 (чаще 9—11) ветвей. Усик длинный (0.6—0.65 длины головы),
тонкий, покрыт мелкими шипиками, с пучком из 5—11 (чаще 6—9) вет-
вей, достигающих х/2 длины усика.
Щетка из 8—15 (чаще 10—12) чешуек, расположенных в один ряд;
каждая чешуйка с прямым острым шипом, по бокам основания которого
сидят мелкие щетинки. Дыхательная трубка слегка S-образно изогнута,
индекс 2.5—3.8, чаще около 3; у основания имеет ушки, хотя и слабо
развитые. Гребень из 10—24 (чаще 16—18) зубцов и из 1—5 (чаще 2)
мелких зубчиков у основания; все зубцы лежат на равном расстоянии,
наиболее дистальные не крупнее остальных; каждый зубец у широкого
основания имеет один крупный добавочный зубчик и несколько едва за-
метных щетинок. Пучок из 3—7 (чаще 4—6) вторичноперйстых ветвей,
имеющих звездчатое расположение.
7. AEDES
283
Седло последнего членика заходит за середину его боковых сторон,
с темно пигментированным задним и покрытым шипиками передним
краем; боковой волосок из 1—5 (чаще из 2—4) ветвей; наружные хвосто-
вые волоски грубые и простые, внутренние из 2—5 (чаще 3—4) ветвей,
из которых нижняя самая длинная. Плавник из 5—8 (чаще 6) 2- или
3-ветвистых пучков и из 1—2 более коротких передних; пучки не объеди-
нены общим основанием. Жабры ланцетовидной формы, верхняя пара
в среднем достигает длины седла, нижняя почти вдвое короче.
Распространение. Япония, п-ов Корея, Северный Китай.
В СССР найден только на крайнем юге Приморской области.
Биология. Личинки встречаются исключительно в небольших
скоплениях воды в дуплах деревьев и пней, а на юге — в стволах бам-
бука. В пределах СССР личинки были находимы очень часто в дуплах
липы (Tilia amurensis) и кленов (Acer mono и A. manshuricum), реже
в дуплах березы (Betula ermani и В. dahurica), редко в дуплах бархата,
манчжурского ореха и кедра. Личинки встречались в прикорневых и
в стволовых дуплах до высоты 8 м, совместно с личинками A. galloisi,
A. alektorovi и Toxorhynchites sp. Зимовка на фазе яйца. Появление ли-
чинок в первой декаде июня. Длительность личиночной фазы 20—25,
куколочной — 4 суток. Вылет комаров первого летнего поколения в пер-
вой декаде июля. В течение сезона возможно 3—4 поколения.
Самки весьма активные кровососы, интенсивно нападают на человека.
Способны отлетать от мест выплода на расстояние до 1 км. Нападение
в лесу в течение всего дня. Численность к осени заметно увеличивается.
Залетают в помещения, где и нападают.
37. Aedes (Finlaya) seoulensis Yamada, 1921.
Характеризуется наличием белого пятна на среднеспинке и белых
колечек на лапках.
Хоботок и щупики в черных чешуйках. Среднеспинка в передней
своей половине покрыта узкими белыми чешуйками; задняя половина
среднеспинки в темно-бурых чешуйках. Задняя граница участка, Покры-
того белыми чешуйками, имеет вид дуги. Лишенное чешуек пространство
перед щитком окаймлено беловатыми чешуйками. Щиток в белых че-
шуйках. Лапки с белыми кольцами на первых трех члениках; каждое
кольцо охватывает вершину одного членика и основание следующего;
на передних лапках кольца развиты только на первых двух члениках.
Крылья в бурых чешуйках, с пятном светлых чешуек при основании
костальной жилки. Брюшко сверху в черных чешуйках со стальным отли-
вом; по переднему краю тергитов развиты узкие беловатые перевязи, по
бокам :— белые пятна; светлые чешуйки имеются и у заднего края терги-
тов на последних сегментах.
Гипопигий: коксит с чешуйками на наружной поверхности. Придаток
стиля равен примерно х/4 его длины. Класпеты с коротким стволиком
и узким крылом, крыло немного длиннее стволика. Лопасти IX тергита
несут по 6—8 волосков.
Личинки IV стадии описаны недостаточно полно (Feng,
1938). Лобные волоски не сдвинуты к переднему краю, средние впереди
внутренних; все три пары, особенно наружные и средние, длинные, не
менее чем из пяти ветвей. Задние волоски наличника не короче средних,
хорошо ветвятся, лежат между средними и внутренними, но более сильно
сближены. Теменной волосок простой и длинный, не короче лобных.
Усик немного превышает половину длины головы, с редкими шипиками
284
III. ПОДСЕМ. CULICINAE
и с простым или 2-ветвистым волоском, отходящим у середины, но ближе
к вершине, своим концом достигает последнюю.
Щетка из 55—60 тесно расположенных чешуек, имеющих главный
шип, бока которого от основания до 2/3 его длины покрыты рядом тонких
щетинок. Дыхательная трубка прямая, индекс 3.0. Гребень из 20—22
длинных зубцов, расположенных на равном расстоянии, наиболее ди-
стальный из них заходит за середину трубки. Пучок из 5—6 ветвей лежит
ближе к вершине.
Последний членик сильно развит в длину, с седлом, доходящим до се-
редины его боков; боковой волосок простой; наружный хвостовой волосок
простой, более чем в 1.5 раза длинее трубки, внутренний из шести более
коротких ветвей. Плавник из 10 пучков в общем основании. Жабры колба-
совидные, верхняя пара в 1.3 раза длиннее трубки, нижняя — 0.75 ее
длины.
Рас пр остр а пение. П-ов Корея, Северный и Северо-Восточный
Китай. В СССР не найден.
Биология. Личинки — весьма обычные обитатели водоемов
в дуплах деревьев (Ulmus pumila, Quercus acutissima и Sophora japo-
nica) в течение летнего сезона.
38. Aedes (Finlaya) alektorovi Stackelberg, 1943.
От двух предыдущих видов отличается отсутствием белого пятна на
среднеспинке; лапки с узкими белыми кольцами, развитыми не на всех
члениках (Штакельберг, 1943).
Рис. 188. Гипопигий Aedes alektorovi Stack.
Хоботок и щупики в одноцветных темных чешуйках. Щупики самца
одной длины с хоботком или немного короче. Среднеспинка в двухцвет-
ных чешуйках: каштаново-коричневые составляют основной фон, золо-
тистые образуют полосы и пятна; иногда, особенно у самцов, преобладают
золотистые чешуйки. Лапки передних и средних (или только передних)
ног темные, если не считать небольшого белого пятна на сочленении
7. AEDES
285
голени и лапки. Задние (иногда и средние) лапки с двумя, тремя или
четырьмя узкими белыми кольцами. Каждое из них опоясывает два чле-
ника: вершину одного и основание следующего. Крылья в темных чешуй-
ках. Брюшко темное, с белыми пятнами по бокам, иногда соединенными
узкими белыми перевязями при основании тергитов. Церки едва за-
метны.
Гипопигий (рис. 188): базальная бородавка коксита имеет вид не-
большого выступа с двумя крепкими изогнутыми, щетинками и одной
(или двумя) более тонкой щетинкой. Придаток стиля составляет около
четверти его длины. Стволик класпеты длиннее крыла, посредине слегка
изогнут, крыло класпеты в вершинной половине несколько расширено.
Лопасти IX тергита с 3—6 щетинками.
Личинка IV стадии (рис. 189—191), в зависимости от вели-
чины дупла и его населенности, варьирует в размерах, светло-коричне-
вого цвета, с более темными головой и особенно дыхательной трубкой
(Мончадский, 1949). Лобные волоски: наружные из 4—8 (чаще 6) вто-
ричноперистых ветвей, средние простые и гладкие, отличаются значи-
тельной длиной — около 1.25 длины головы; внутренние позади них,
из 3—5 вдвое более коротких вторичноперистых ветвей; задние волоски
наличника впереди средних, сильнее сближены, из 10—12 коротких
тонких ветвей. Усики около половины длины головы, покрыты единич-
ными шипиками. с пучком у середины из 1—2 ветвей.
Хетотаксия тела сильно развита в длину, особенно боковые волоски
брюшка. Щетка из 20—41 (чаще 25—30) чешуек, расположенных в 2—
3 ряда, не имеющих главного шипа, а несущих по свободному краю ряд
шипиков, укорачивающихся к основанию чешуйки. Дыхательная трубка
тонкая, слегка изогнутая, индекс 3.9—4.5. Гребень заметно не доходит
до ее середины, состоит из 19—27 (чаще 21—24) зубцов и из 1—3 руди-
ментарных у основания, тесно сидящих и увеличивающихся в дисталь-
ном направлении. Пучок у середины, из 3—4 ветвей одной длины с диа-
метром трубки.
Седло последнего членика не доходит до середины его боковых сторон;
боковой волосок короткий, из 2—4 (чаще 3) тонких ветвей; наружный
хвостовой волосок почти в 1.5 раза длиннее дыхательной трубки, про-
стой, внутренний из 2—3 более коротких ветвей. Плавник развит очень
слабо и боковые отростки у основания пучков не соединяются общим
основанием; состоит из 4—7 (чаще 5—6) пучков с развитыми боковыми
отростками оснований и из 3—4 более коротких впереди них. Жабры
отличаются значительной длиной, в 1.5 раза длиннее дыхательной трубки,
колбасовидной формы, слегка заострены на концах; нижняя пара на
15—25% короче верхней.
Распространение. Найден в ряде пунктов на Дальнем
Востоке СССР к югу от Хабаровска.
Биология. Относительно редкий вид, характерный для широко-
лиственных лесов Советского Приморья. Личинки встречаются только
в скоплениях воды в дуплах деревьев (липа — Tilia amurensis, клен —
Acer mono, Ac. manshuricum и Ac. pseudosieboldianum, березы — Betula
-ermani и В. dahurica, бархата — Phellodendron amurense и др.), сов-
местно с личинками A. galloisi, A. nipponicus и Toxorhynchites christophi.
Личинки обитают преимущественно в сильно затемненных дуплах с узким
отверстием и отсутствуют в открытых прикорневых дуплах, а внутри
них держатся в наиболее темных местах. Зимовка на фазе яйца. Развитие
в июле длится около 15 суток.
Рис. 189. Aedes alektorovi Stack. Личинка Рис. 190. Aedes alektorovi Stack. Личинка IV стадии. Задний
IV стадии. Голова, сверху. конец, сбоку.
Рис. 191. Aedes alektorovi
Stack. Личинка IV ста-
дии.
1 — чешуйки щетки,
2 — гребень сифона.
288
III. ПОДСЕМ. CULICINAE
39. Aedes (Finlaya) togoi Theobald, 1907.
Характеризуется наличием белых колец на лапках всех ног, причем
каждое кольцо опоясывает два членика (рис. 192).
Хоботок в темных чешуйках. Щупики самки в черных чешуйках,
с белой вершиной. Щупики самца, равные примерно-4/5 длины хоботка,
Рис. 193. Гипопигий Aedes togoi
Theob. По La Casse a. Yamaguti.
черные, с белыми колечками на
сочленениях. Среднеспинка в черно-
бурых чешуйках с нерезко отгра-
ниченными узкими продольными по-
лосками желтовато-белых чешуек,
идущими по средней линии, по рядам
дорсо-центральных щетинок и по бо-
ковым краям среднеспинки. Щиток и
небольшое пространство перед щит-
ком в желтовато-белых чешуйках.
Лапки черные, с узкими белыми
кольцами, опоясывающими вершину
предыдущего и основание последу-
ющего членика. Крылья в одноцвет-
ных бурых чешуйках. Брюшко в чер-
ных чешуйках, по бокам с треуголь-
ными белыми пятнами, соединенными
узкими перевязями при основании
тергитов.
Гипопигий (рис. 193) отличается
строением класпет: узкое крыло
явно превышает по длине укорочен-
ный стволик. Коксит сверху и из-
нутри в длинных, несколько упло-
щенных, продольно исчерченных ще-
тинках. На наружной поверхности
коксита много чешуек, иногда еди-
ничные чешуйки находятся и на
внутренней его поверхности. При-
даток стиля короткий. Лопасти IX
тергита несут по 6—9 тонких во-
лосков.
Личинки IV стадии
(рис. 194) от умеренных до круп-
ных размеров, со слабо окрашенным
телом, темной головой и немного
более светлой дыхательной трубкой.
Лобные волоски сдвинуты к пе-
реднему* краю головы и расположены
в выпуклый вперед дугообразный ряд; наружные из 6—7, наиболее длин-
ные средние из 7—8 и внутренние из 11—13 веерообразно расходящихся
ветвей. Задние волоски наличника между основаниями внутренних,
в среднем из 7 более коротких ветвей. Усик почти прямой, короткий,
с телом, неравномерно покрытым редкими шипиками, и с коротким воло-
ском из 2—3 ветвей.
Щетка из 80—125 тесно сидящих чешуек, изменчивых по величине,
с закругленным свободным краем, покрытым рядом шипиков, укорачи-
вающихся к основанию. Дыхательная трубка короткая (индекс 1.8—2.3),
7. AEDES
289
слегка бочонкообразной формы. Гребень далеко заходит за середину
трубки, из 18—25 тесно сидящих шиповидных зубцов, уменьшающихся
к основанию трубки; по нижнему краю зубца сидят 2—5 широко расстав-
ленных зубчиков. Пучок из 7—10 веерообразно расходящихся вторично-
перистых ветвей, лежит близко к вершине и в длину превышает диаметр
трубки.
Последний членик одной длины с трубкой, со слабо развитым седлом,
почти не заходящим на боковые его стороны; боковой волосок короткий
и простой; наружный хвостовой волосок простой, в среднем в 2.5 раза
Рис. 194. Aedes togoi Theob. Личинка IV стадии.
1 — голова, сверху, 2 — задний конец, сбоку.
длиннее трубки, внутренний на 1/2 короче, в виде веера из 9—11 ветвей.
Плавник из 1—14 пучков по 7—12 ветвей, впереди от общего основания
1—2 более коротких пучка. Жабры изменчивой неправильно листовидной
формы, в 2.5—3 раза короче последнего членика.
Распространение. Приморская полоса Китая, п-ова Кореи и
Японии, включая Хоккайдо. В СССР найден на побережье южной части
Приморского края, на Сахалине и на южных островах Курильской
гряды.
Биология. Личинки в большой численности встречаются в не-
больших водоемах (углубления или расщелины) скалистого морского
побережья у полосы прибоя, с водой часто очень значительной солености,
резкие колебания которой, вследствие выпадения осадков, прибоя или вы-
сыхания, личинки легко переносят. Иногда попадается совместно с ли-
чинками С. vagans и An. hyrcanus. Несколько поколений в течение се-
зона — с мая по октябрь. Как вторично заселяемые биотопы — камен-
ные бассейны, цистерны и другие искусственные водоемы в населенных
пунктах, иногда далеко удаленных от побережья, например в Пекине.
Местами принадлежит к числу массовых кровососов. Нападает как
в природных условиях, так и в населенных пунктах, включая города.
19 Фауна СССР, А. В. Гуцевич и др.
290
III. ПОДСЕМ. CULICINAE
В малонаселенных районах комары питаются, по-видимому, преиму-
щественно кровью морских птиц. В условиях Приморского края взрослые
комары встречаются с июня до конца октября, массовый лёт со второй
половины июля до начала сентября. В течение сезона развивается не-
сколько поколений.
Эпидемиологическое значение. Этот вид имеет
серьезное значение как один из переносчиков вируса японского энцефа-
лита: возможность передачи вируса доказана экспериментально, уста-
новлена естественная зараженность. В Китае A. togoi является проме-
жуточным хозяином паразитирующих у человека филяриид — Brugia
malayi (один из важнейших переносчиков) и Wuchereria bancrofti (пере-
носчик второстепенного значения).
40. Aedes (Finlaya) japonicus Theobald, 1901.
В отличие от предыдущего вида характеризуется расположением
белых колец только при основании члеников лапки.
Хоботок и щупики в черных чешуйках. Среднеспинка в черно-бурых
чешуйках, в передней половине с тремя продольными полосками светлых
желтоватых чешуек, идущими по средней линии и по рядам дорсо-цен-
тральных щетинок; по бокам задней половицы среднеспинки имеется еще
две изогнутые светлые полоски. Щиток и участок перед щитком в желто-
вато-белых чешуйках. Участок ниже переднего дыхальца без чешуек.
Лапки с неширокими белыми кольцами при основании первых двух (на
передних лапках) или первых трех (на средних и задних лапках) члеников.
Крылья в темных чешуйках. Брюшко сверху в черных чешуйках, с не-
большими снежно-белыми треугольными пятнами по бокам; при основа-
нии тергитов обычно имеются небольшие поперечные белые полоски,
как правило, не соединяющиеся с боковыми пятнами.
Гипопигий: коксит в чешуйках (на наружной поверхности) и уме-
ренно длинных волосках. Стволик класпеты длиннее крыла или равен
крылу по длине. Лопасти IX тергита небольшие, уплощенные, с 4—9
тонкими волосками каждая, склеротизованные коготки на вершине
X стернита раздвоены.
Личинка IV стадии от средних До крупных размеров. Лоб-
ные волоски гладкие, сдвинуты к переднему краю головы и расположены
в обращенный выпуклостью вперед дугообразный ряд; наружные из
4—7, средние из 3—6, внутренние из 4—7 ветвей. Задние волоски налич-
ника очень короткие, между основаниями внутренних, из 2—5 тонких
ветвей. Усик меньше половины длины головы, покрыт редкими шипиками,
с пучком за серединой из 2—3 коротких ветвей.
Щетка из 43—85 тесно расположенных чешуек без главного шипа,
с рядом краевых шипиков, наиболее длинных по закругленному свобод-
ному краю. Дыхательная трубка прямая, суженная начиная от места
отхождения пучка, индекс 2.3—3.8. Гребень из 14—29 зубцов, заходящих
далеко за середину трубки, из них 1—4 наиболее дистальных, в виде
более крупных, широко расставленных шипов, лежащих под более острым
углом к продольной оси трубки. Пучок из 4—7 вторичноперистых ветвей,
в длину равных диаметру трубки, отходит дистальнее середины трубки.
Седло последнего членика покрывает его боковые стороны примерно
до середины, с задним краем, покрытым шипиками различной величины;
боковой волосок 1—2-ветвистый, в длину около длины последнего чле-
ника; наружный хвостовой волосок более чем в 1.5 раза длиннее трубки,
внутренний из 2—4, не менее чем на х/3 короче. Плавник из 12 2—5-вет-
7. AEDES
291
вистых пучков в общем основании. Жабры ланцетовидной формы, в сред-
нем в 1.5 раза длиннее седла.
Распространение. Китай, Япония, включая Хоккайдо.
В СССР не найден.
Биология. В Японии весьма обычный и широко распространен-
ный вид, поднимающийся
в горы до высоты 1300 м
над ур. м. Личинки встре-
чаются в различных ис-
кусственных водоемах, бо-
гатых продуктами органи-
ческого разложения, как
освещенных солнцем, так
и затененных. Большей
численности достигает вес-
ной и осенью; на юге Япо-
нии численность с мая по
октябрь держится на одном
уровне. По-видимому, на
людей нападает редко.
41. Aedes (Finlaya) ko-
reicus Edwards, 1917 (ja-
ponieus var. koreicus Ed-
wards).
Весьма близок к пре-
дыдущему, ранее от него
не отграничивался или рас-
сматривался как подвид.
В советской литературе о
комарах Приморья и о
переносчиках японского
энцефалита фигурировал
под именем A. japonicus.
Хоботок и щупики в
черных чешуйках. Средне-
спинка в темных чешуйках
со светлыми полосками,
расположенными как у
A. japonicus, но более
узкими. Участок ниже пе-
реднего дыхальца с поло-
ской широких белых че-
шуек. Лапки с узкими бе-
лыми колечками при осно-
вании члеников. На пе-
Рис. 195. Гипопигий Aedes koreicus Edw.
По La Casse et Yamaguti.
редних лапках колечки развиты на первых двух члениках, на
средних лапках — на трех или четырех члениках; на задних лапках
колечки развиты на четырех члениках, последний членик темный. Крылья
в темных чешуйках. Брюшко сверху темное, с небольшими поперечными
светлыми полосками при основании сегментов. Полоски не соёдиняются
с боковыми белыми пятнами, которые обычно сверху не видны, за исклю-
чением VIII тергита.
19*
292
III. ПОДСЕМ. CULICINAE
Гипопигий (рис. 195) сходен с гипопигием A. japonicus со следую-
щими отличиями: лопасти IX тергита полушаровидные, выпуклые; ко-
ротки на вершине X стернита простые.
Рис. 196. Aedes koreicus Edw. Личинка IV стадии.
1— голова, сверху, 2 — концы волосков верхней губы.
Рис. 197. Aedes koreicus Edw. Личинка IV стадии. Задний конец, сбоку.
Личинка IV стадии (рис. 196 и 197) крупных размеров,
часто интенсивно пигментирована. Лобные волоски без вторичной пери-
стости, смещены к переднему краю головы и расположены в один вы-
пуклый вперед дугообразный ряд; более короткие наружные из 3—6
чаще 4—5), средние из 2—6 (чаще 3—5), внутренние из 4—6 (чаще 5—6)
7. AEDES
293
ветвей. Задние волоски наличника короткие, из 2—4 отходящих от сере-
дины общего стебелька тонких веточек. Усик менее половины длины
головы, с телом, неравномерно покрытым редкими шипиками, и с корот-
ким пучком из 1—4 (чаще 2—3) коротких ветвей.
Щетка из 49—70 (среднее 58) тесно сидящих чешуек, изменчивых
по величине и форме, не имеющих главного шипа, края которых усажены
рядом шипиков, более длинных у вершины чешуйки. Дыхательная трубка
от места отхождения пучка заметно сужается к вершине, индекс 2.8—
3.4 (среднее 3.1). Гребень около половины длины трубки или немного
более, из 18—27 (чаще 20—25) тесно расположенных зубцов, наиболее
дистальные из них крупнее и лежат под более острым углом к продольной
оси трубки; на уровне этих зубцов отходит пучок в виде веера из 4—7
вторичноперистых ветвей, достигающих 1/3 длины трубки.
Седло последнего членика слабо развито, с неправильно изрезанными
боковыми краями, доходящими до верхней трети боковых сторон, и с уса-
женным мелкими и более крупными шипами задним краем; боковой во-
лосок простой, реже 2-ветвистый; наружный хвостовой волосок простой,
в 1.5 раза длиннее дыхательной трубки, внутренний из 3—5 (чаще 4)
немного более коротких ветвей. Плавник из 12, реже 11, пучков в общем
основании и 1, реже 2, пучком впереди него. Жабры ланцетовидные,
узкие, в среднем достигают 1.75 длины седла.
Распространение. Северо-Восточный Китай, п-ов Корея,
Япония (о. Хоккайдо). В СССР найден в Приморском крае.
Биология. Личинки встречаются в большой численности в по-
жарных бочках, кадках с дождевой водой в течение всего лета, всегда
совместно с С. vagans. Вид довольно обычен в лесных районах Южного
Приморья. Встречается преимущественно в обжитых местах, в населен-
ных пунктах или вблизи них. Нападает под открытым небом, иногда
в заметном количестве, в дома залетает редко. Питается на людях,
сельскохозяйственных животных, нередко встречается в курятниках.
В условиях Приморского края взрослые насекомые попадаются с июня
по сентябрь, массовый лёт в июле—августе. За сезон развивается, ве-
роятно, не менее двух поколений.
По-видимому, принадлежит к числу потенциальных переносчиков
японского энцефалита; сообщалось о возможности передачи вируса
в условиях эксперимента.
4. Подрод STEGOMYIA Theobald, 1901
Комары мелких размеров, реже средней величины. Хоботок относи-
тельно толстый и короткий, по длине приблизительно равен передним
бедрам. Щупики самца немного длиннее хоботка, с тонкими, обычно
загнутыми вверх вершинными члениками, без пучков волосков. У самки
VIII стернит сильно развит, церки короткие.
Гипопигий: класпеты отсутствуют, базальная (или предвершинная)
бородавка' коксита хорошо развита, вершинж)й бородавки нет; эдеагус
разделен на две пластинки, у большинства видов несущих на вершине
мелкие зубчики.
Весьма характерна окраска: среднеспинка с рисунком, образованным
светлыми чешуйками, лапки с белыми кольцами при основании члеников.
Светлые пятна на голове, на щитке и бочках груди, а также на брюшке
образованы преимущественно широкими чешуйками снежно-белого цвета
с блестящим серебристым оттенком.
294
III. ПОДСЕМ. CULICINAE
Во избежание повторений укажем подробнее особенности окраски,
общие для описываемых ниже видов подрода Stegomyia. Хоботок в одно-
цветных темных чешуйках. Щупики самки темные в основной половине,
белые в вершинной. Щупики самца с белыми кольцами. Голова сверху
в темных и светлых чешуйках; последние обычно образуют продольную
полоску посредине и округлые пятна по бокам, а также окаймляют зад-
ний край глаз. Среднеспинка с характерным для каждого вида светлым
рисунком. Щиток с тремя сближенными белыми пятнами. Лапки с белыми
кольцами при основании члеников, более развитыми на задних ногах.
Крылья в темных чешуйках, иногда с небольшой группой белых чешуек
при основании костальной жилки. Тергиты брюшка при основании с бе-
лыми перевязями, по бокам с пятнами такого же цвета.
У личинок подрода Stegomyia лобные волоски не сдвинуты к перед-
нему краю головы, средний впереди внутреннего. Усик без шипиков
на теле, с маленьким, обычно простым волоском. Пластинки у основания
боковых волосков средне- и заднегруди обычно с шипом. Зубцы щетки
немногочисленны, расположены обычно в один ряд. Дыхательная трубка
не имеет ушков по бокам основания.
Подрод включает свыше 90 видов, распространенных преимущественно
в Эфиопской и Ориентальной областях; немногие виды заходят в Пале-
арктику на юго-западе (три вида) и юго-востоке (четыре вида). Из них
четыре вида встречаются (или встречались) в СССР.
Некоторые из видов подрода играют весьма важную роль как перенос-
чики возбудителей болезней, особенно вирусных инфекций — желтой
лихорадки, денге и других.
42. Aedes (Stegomyia) aegypti Linneus, 1762 (argenteus Poiret, 1787;
fasciatus Fabricius, 1805; calopus Meigen, 1818). —Желтолихорадочный
комар (рис. 198).
От других видов подрода отличается окраской среднеспинки (рисунок
в виде лиры) и строением гипопигия.
Общие для описываемых видов подрода особенности окраски см. выше
(стр. 294). Среднеспинка в темных коричневых чешуйках с характерным
рисунком, состоящим из двух отчетливо выраженных боковых белых
полосок, изогнутых в передней половине, и из двух менее четких узких
прямых полосок, проходящих продольно вблизи средней линии. Вер-
шины бедер белые, голени темные. Передние и средние лапки черные,
с белыми кольцами при основании первых двух члеников. Задние лапки
с белыми кольцами, занимающими 1/i—1/3 длины членика на первых
трех члениках и около 4/5 длины четвертого членика. Последний членик
задней лапки целиком белый. Ширина белых колец изменчива. Коготки
передних и средних лапок самки с зубцом, задних лапок — простые.
Крылья по жилкам густо покрыты бурыми чешуйками. Брюшко сверху
покрыто черно-бурыми чешуйками, обыкновенно имеющими краснова-
тый или стальной отлив. В передней части каждого тергита имеются
узкие белые перевязи, по бокам — по одному серебристо-белому пятну.
Гипопигий (рис. 199): коксит короткий, утолщенный; посреди его
внутренней поверхности расположен нерезко очерченный уплощенный
выступ (бородавка), густо усаженный короткими щетинистыми воло-
сками. Стили к вершине сильно сужены, с коротким придатком на вер-
шине. Задний край IX тергита с глубокой вырезкой.
Л и ч и нка IV стадии (рис. 200 и 201) очень изменчива по раз-
мерам и окраске, в зависимости от условий развития. Лобные волоски
длинные и простые; средние, более короткие, смещены к переднему краю
7. AEDES
295
головы, внутренние позади них на уровне наружных. Задние волоски
наличника между основаниями средних, из 4—6 коротких тонких ветвей.
Усик короткий, около х/2 длины головы, без шипиков на поверхности,
с простым коротким волоском за серединой.
Рис. 198. Aedes aegypti L.
Щетка из 8—12 (чаще 10, иногда больше — до 18) чешуек, распо-
ложенных в один дугообразный ряд; чешуйки изменчивы, чаще с более
или менее выраженным главным шипом, по бокам которых лежат обычно
более короткие шипы, уменьшающиеся к основанию. Дыхательная трубка
относительно короткая, за серединой сужающаяся (индекс 1.5—2.5), без
ушков^по бокам основания. Гребень из 12—22 (чаще 15—17) тесно сидя-
296
III. ПОДСЕМ. CULICINAE
гцих зубцов, наиболее дистальный из которых может быть слегка от-
ставлен и лежит за серединой трубки; каждый зубец с широким: основа-
нием и с одним более крупным и 1—3 более мелкими дополнительными
зубчиками. Пучок около наиболее дистального зубца гребня, из 2—5 ко-
ротких ветвей.
Седло последнего членика доходит до нижнего края его боковых
сторон, боковой волосок из 2—3 коротких ветвей; наружные хвостовые
Рис. 199. Гипопигий Aedes aegypti L. По Carpenter a. La Casse.
волоски простые, вдвое длиннее трубки, внутренние из 2—4 более корот-
ких ветвей. Плавник из 8—10 2—3-ветвистых пучков. Жабры колбасо-
видной формы, с округленными концами, немного меньше длины дыхатель-
ной трубки.
Распространение. Тропическая и субтропическая зоны
обоих полушарий. В Европе: Франция, Испания, Португалия, Италия,
Югославия (только юг Адриатического побережья), Албания, Греция.
Африка — большая часть континента. В Азии: Передняя Азия, Иран
(у Шатт-эль-Араба), Пакистан, Индия, страны Юго-Восточной Азии,
приморские районы Китая (в Северном и Северо-Восточном Китае не встре-
чается), Япония — только на крайнем юге. В Западном полушарии:
Южная, Центральная и Северная Америка, в США до штатов Иллинойс
и Индиана, т. е. почти до 40° с. ш. Примерно таково было распространение
7. AEDES
297
A. aegypti в недавнем прошлом. В ряде стран, где этот вид комара ранее
был широко распространен, теперь он истреблен или встречается редко.
Рис. 200. Aedes aegypti L. Личинка IV стадии. По Macfie.
1 — задний конец, сбоку, 2 — голова, сверху, 3 — второй членик брюшка, сверху,
4 — основание боковых волосков заднегруди.
В СССР этот вид ранее был распространен на Черноморском побережье
Кавказа от Сочи до Батуми. Нередко он встречался и в удаленных от Чер-
номорского побережья пунктах Закавказья:
Кутаиси, Тбилиси, Баку. В результате ра-
бот, проведенных по инициативе Н. П. Ру-
хадзе, численность A. aegypti резко снизи-
лась до единичных находок; в последние
годы вид вовсе не обнаруживается.
Биология. Личинки развиваются
в самых разнообразных искусственных
скоплениях воды вокруг жилищ человека и
внутри них. Переносят значительную сте-
пень загрязнения воды. Длительность раз-
вития личинок и куколок при 27—30° —
около 10 суток; ниже 20° развитие останав-
Рис. 201. Aedes aegypti ' L.
Личинка IV стадии. Стиг-
мальная пластинка.
ливается.
Первоначальная область обитания A. ae-
gypti — тропическая Африка, где он и теперь
встречается в природных условиях. Вне
этой зоны этот вид обитает почти исключительно в населенных пунктах
или вблизи них; он более тесно связан с человеком, чем любой другой
вид. Нападает главным образом в домах и около домов. Копуляция про-
исходит внутри помещений; в лаборатории комары могут спариваться
298
III. ПОДСЕМ. CULICINAE
в маленьких садках, благодаря чему данный вид легко культивировать
(при температуре не ниже 20—22°, оптимум 25—30°). В естественной
обстановке за сезон развивается несколько поколений.
Эпидемиологическое значение A. aegypti исключительно велико. Он
является важнейшим или единственным переносчиком вируса желтой
лихорадки вне природных очагов болезни, отсюда наименование — «желто-
лихорадочный комар». В настоящее время «городские» очаги желтой ли-
хорадки в основном ликвидированы, но при наличии желтолихорадочного
комара сохраняется угроза возникновения эпидемий в результате заноса
вируса из природных очагов в населенную зону. Желтолихорадочный
комар является также главнейшим переносчиком вируса денге; в условиях
опыта, возможно, и в естественных условиях может передавать ряд пато-
генных для человека вирусов — возбудителей комариных энцефалитов и
лихорадок, а также лимфоцитарного хориомениНгита. Передает вирус
миксоматоза кроликов (механическая передача). В организме Л? aegypti
личинки возбудителя малайского филяриоза, Brugia malayi, завершают
свое развитие, а личинки Wuchereria bancrofti, как правило, не достигают
инвазионной стадии.
По вопросам биологии A. aegypti и его медицинского значения имеется
огромная литература (см.: Horsfall, 1955); этому виду посвящена обшир-
ная монография Кристоферса (Christophers, 1960).
Рис. 202. Гипопигий Aedes cretinus Edw.
43. Aedes (Stegomyia) cretinus Edwards, 1921 (lindtropi Shingarev, 1927).
Отличается от предыдущего окраской среднеспинки и строением ги-
попигия.
7. AEDES
299
Среднеспинка в черно-бурых чешуйках, с продольной белой полосой
посередине и двумя небольшими округлыми белыми пятнами по бокам.
В остальном окраска и строение коготков лапки как у Л. aegypti.
Гипопигий (рис. 202): коксит удлиненный, уплощенный выступ на его
внутренней поверхности, густо усаженный волосками, вытянут в продоль-
ном направлении. Стили, суженные в средней части, с тонким, относи-
тельно длинным придатком, причленяющимся немного отступя от вер-
шины стиля. Задний край IX тергита с дугообразным выступом.
Личинка описана недостаточно. Сходна с таковой A. aegypti
и отличается от последней более тонкими усиками с грубым простым
волоском у середины; щеткой из расположенных в один ряд 10 чешуек
с длинным заостренным шипом, несущим короткие тонкие шипики по бо-
кам основания; дыхательной трубкой (индекс около 2.0) с гребнем из 11 зуб-
цов, более узких, чем у Л. aegypti, с более мелкими зубчиками у основания;
с пучком из трех ветвей, отходящим у середины трубки, и с простым во-
лоском, отходящим от ее боковой стороны в начале вершинной трети;
жабрами, не превышающими а/3 длины дыхательной трубки, и более сильно
развитым плавником.
Распространение. Крит, Македония. В пределах СССР
найден в единичных экземплярах на черноморском побережье Кавказа:
Сухуми, Гудауты, Сочи (Башкарева, 1931). Последняя по времени извест-
ная нам находка: Гульрипш (вблизи
Сухуми), бамбуковая роща, 20 VIII1939,
две самки, отловленные в момент нападе-
ния (сбор Т. Я. Авдеевой). Вероятно, к
этому же виду относятся два недостаточ-
но хорошо сохранившихся комара с Юж-
ного берега Крыма, бесспорно принад-
лежащих к подроду Stegomyia.
Биология не изучена. Личинки
были обнаружены в скоплениях воды
в дуплах деревьев совместно с личин-
ками An. plumbeus, Or. pulchripalpis и
Л. geniculatus.
44. Aedes (Stegomyia) vittatus Bigot,
1861.
Отличается окраской среднеспинки
и наличием белого кольца на голенях.
Голова и ее придатки как у Л. ae-
gypti. Среднеспинка покрыта черно-бу-
рыми чешуйками, с четырьмя сереб-
ристо-белыми круглыми пятнами в
средней части. Голени всех пар близ се-
редины с белым кольцом. Лапки с бе-
Рис. 203. Гипопигий Aedes vitta-
tus Big.
лыми кольцами при основании первых трех члеников; на передних и средних
лапках кольца узкие, на задних — широкие. Два последних членика
задней лапки белые, иногда у вершины затемненные.
Гипопигий (рис. 203) отличается весьма своеобразным строением стиля:
на вершине он сильно вздут; длинный искривленный придаток помещается
на внутренней стороне стиля за его серединой.
Личинка IV стадии (рис. 204). Лобные волоски длинные:
наружные из 3—6 ветвей, направлены в стороны, средние впереди внут-
ренних, обе пары простые.. Задние волоски наличника между основаниями
300
III. ПОДСЕМ. CULICINAE
средних, маленькие, разветвленные. Усик немного больше половины
длины головы с очень слабо развитыми шипиками, с маленьким волоском
из трех ветвей у середины.
Щетка из 6—9 (чаще 8) крупных чешуек с длинным острым шипом,
по бокам основания которого в небольшом числе сидят тонкие короткие
шипики. Дыхательная трубка короткая (индекс 2.0 или немного больше),
слегка суженная к вершине, без ушков по бокам основания. Гребень
из 20—34 шиповидных зубцов, занимает около а/3 длины трубки; кроме
них, один более крупный гладкий шип лежит значительно ближе к вершине.
Между ним и наиболее дистальным зубцом гребня помещается пучок из 3;—
6 ветвей, в длину равных ширине трубки у места его отхождения.
Рис. 204. Aedes vittatus Big. Личинка IV стадии.
По Barraud.
1 — усик, 2 — дыхательная трубка, сбоку, 3 — зубцы гребня,
4 — чешуйки щетки.
Последний членик длиннее, чем у других личинок Stegomyia. Седло
слабо развито, лишь немного заходит на его боковые стороны; боковой
волосок короткий и простой; наружные хвостовые волоски, простые, длин-
ные, внутренние из 4—6 ветвей. Плавник из 5—7 2-ветвистых пучков
в общем основании и из 3—4 впереди него. Жабры лишь немного короче
дыхательной трубки, более широкие у основания и заостренные на концах.
Распространение. Южная Европа, Африка, Южная и Юго-
Восточная Азия. В СССР не найден.
Биология, Личинки встречаются в скоплениях воды в углубле-
ниях камней и скал, а также в разного рода искусственных водоемах
вокруг жилья человека. Местами нападает на людей в заметном количестве.
Принадлежит, по-видимому, к числу переносчиков желтой лихорадки:
возможность передачи вируса доказана экспериментально.
45. Aedes (Stegomyia) galloisi Yamada, 1921.
Характеризуется окраской груди (среднеспинка с белой продольной
полосой) и задних лапок (последний членик целиком или частично за-
темненный), а также строением гипопигия.
Среднеспинка в каштаново-бурых чешуйках, с продольной светлой
полосой, слегка суживающейся кзади; по бокам развиты менее отчетли-
вые светлые полоски, изогнутые в передней половине среднеспинки и
7. AEDES
301
прямые — в задней; чешуйки срединной и боковых светлых полос в зад-
ней части среднеспинки имеют желтоватый оттенок. Лапки всех ног с уз-
кими белыми колечками на первых двух члениках. Последующие членики
передней и средней лапки темные; на задних лапках третий членик белый
в основной половине; четвертый и пятый членики белые, с затемненной
вершиной. Коготок передней и средней лапок с зубцом, задней лапки —
простой. Брюшко в черных чешуйках, с узкими белыми перевязями при
основании сегментов и с белыми пятнами по бокам тергитов.
У экземпляров из Хабаровского края и Приморья наблюдаются по срав-
нению с приведенным оригинальным описанием значительные отклонения
в окраске средних и задних лапок: последний членик задней лапки цели-
Рис. 205. Гипопигий Aedes galloisi Yam.
ком темный, а окраска второго членика средней лапки самки весьма из-
менчива: он может быть сплошь белым, белым с затемненной вершиной или
темным с белым кольцом при основании (как у комаров этого вида из
Японии); иногда этот членик покрыт белыми чешуйками с наружной сто-
роны и темными с внутренней. Очевидно, окраска лапок — признак,
играющий большую роль в систематике рода — у этого вида весьма
изменчива. Возможно, здесь мы имеем дело с наличием двух или не-
скольких форм (подвидов?).
Гипопигий (рис. 205): бородавчатый выступ на внутренней поверхности
коксита выпуклый, вытянут в поперечном направлении, по наружному
краю густо усажен волосками. Стиль с перетяжкой посредине, палочко-
видный придаток причленяется немного отступя от.вершины стиля. Задний
край IX . тергита в виде пологой дуги, обращенной выпуклостью назад;
иногда посредине заднего края IX тергита развит треугольный выступ.
302
III. ПОДСЕМ. CULICINAE
Личинка IV стадии (рис. 206) коричневой окраски, с сильно
развитыми звездчато ветвящимися волосками на теле. Величина личинок
варьирует в зависимости от условий обитания (величина водоема, степень
заселенности, наличие пищи).
Лобные волоски слабо развиты, средние впереди внутренних, сдви-
нуты к переднему краю; наружные и средние из 2—3 ветвей, внутренние
простые. Задние волоски наличника между основаниями средних, лишь
немного короче их, из 9—10 ветвей. Усик короткий, около трети длины
головы, с телом, лишенным шипиков, и с коротким простым волоском
у его середины.
Рис. 206. Aedes galloisi Yam. Личинка IV стадии.
1 — голова, сверху, 2 — задний конец, сбоку.
Щетка из 8—12 (чаще 9—10) чешуек, расположенных в один ряд и
имеющих хорошо развитый острый главный шип с короткими тонкими
щетинками по бокам его основания. Дыхательная трубка без ушков, темно
пигментирована, заметно сужается от середины к вершине, индекс 2.9—
3.1. Гребень из 10—16 маленьких тонких шиповидных зубцов с расши-
ренным основанием, несущим 1—3 базальных зубчика; наиболее дисталь-
ный зубец не доходит до середины трубки. Пучок у середины, из 4—8 вет-
вей, длина которых не превышает ширины основания трубки.
Седло доходит до нижнего края боков короткого последнего членика,
в редких случаях охватывает его кольцом; боковой волосок короткий,
из 2—4 ветвей; наружный хвостовой волосок простой, одной длины с труб-
кой, внутренний из двух ветвей, из которых спинная короче. Плавник
развит слабо, из шести простых (реже 2-ветвистых) волосков, одной длины
с жабрами, в общем основании и из двух более коротких и простых впереди
него. Жабры колбасовидной формы, с округленными концами, 1.2—
1.6 длины трубки.
Распространение. Северо-Восточной Китай, п-ов Корея, Япония.
В СССР — Хабаровский край, Приморье, Сахалин. Кроме того, известны
два местонахождения в Западной Сибири: окрестности Томска и Колыван-
скй район Новосибирской области. Возможно, что появление этого вида
в Западной Сибири — результат случайного завоза по железной дороге.
7. AEDES
303
Рис. 207. Гипопигий Aedes albopictus Sk.
По La Casse a. Yamaguti.
Биология. Личинки в большой численности встречаются в скоп-
лениях воды в прикорневых и стволовых дуплах липы, ряда видов клена.
и березы, бархата и кедра совместно с личинками Л. alektorovi, A. nippo-
nicus и Toxorhynchites. Нередки находки их в плохо закрытых пожарных
бочках и в других искусственных водохранилищах, при условии их пол-
ного или частичного затем-
нения; попадаются совместно
с личинками A. koreicus и
С. vagans. Личинки из таких
более крупных биотопов до-
стигают вдвое большей вели-
чины, чем из дупел. Несколь-
ко поколений в течение се-
зона — с мая но октябрь.
Зимовка на фазе яйца; лес-
ной вид, относительно редко
нападающий на человека.
46. Aedes (Stegomyia) al-
bopictus Skuse, 1895.
Среднеспинка с белой про-
дольной полосой; от других
видов со сходным рисунком
среднеспинки отличается
строением гипопигия и ок-
раской задних лапок.
Основной фон чешуек
среднеспинки каштаново-ко-
ричневый; белая продольная
полоса сужается кзади.
В задней половине средне-
спинки намечаются боковые
продольные белые полоски,
в передней половйне средне-
спинки их нет (отличие от
A. galloisi). У края средне-
спинки, непосредственно впе-
реди основания крыла, рас-
положено нерезко отграни-
ченное небольшое пятно се-
ребристых широких прямых
чешуек. Лапки передних и
средних ног с узкими белыми
двух члеников. На задних лапках первые четыре членика с более широ-
кими белыми кольцами; последний членик задней лапки целиком белый.
Коготки лапок без зубчиков. Брюшко с белыми пятнами по бокам терги-
тов; кроме того, на тергитах при основании сегментов иногда развиты
перевязи, суживающиеся или прерывающиеся посредине.
Гипопигий (рис. 207): бородавка коксита, ограниченная его средней
третью, вытянута в продольном направлении. Она усажена весьма длин-
ными простыми волосками. Задний край IX тергита посредине с выступом.
Личинка IV стадии (рис. 208) средних размеров. Лобные
волоски слабо развиты, наружные из Двух, реже одной и трех коротких
ветвей, средние сдвинуты вперед, из 1—2 ветвей, внутренние позади
колечками при основании первых
304
III. ПОДСЕМ. CULICINAE
последних, наиболее длинные, простые, со слабой вторичной перистостью.
Задние волоски наличника сильнее, чем средние, сдвинуты к переднему
краю и сближены, из многих (6-<-15) тонких ветвей, отходящих от корот-
кого стебелька. Усик короткий, тонкий, без шипиков на поверхности,
волосок у середины, короткий и простой.
Щетка из 6—13 (чаще 8—10) крупных чешуек, расположенных в один
ряд, имеющих хорошо развитый заостренный главный шип с рядом ко-
ротких тонких щетинок по бокам основания. Дыхательная трубка без уш-
Рис. 208. Aedes albopictus Sk. Личинка IV стадии.
1 — усик, г — дыхательная трубка, 3 — чешуйки Щетки, 4 — зубцы
гребня, 5 — голова, сверху, в — основание одного из боковых
волосков груди. 1—4 — по Barraud, 5—в — по Senior-White,
ков, короткая (индекс 1.7—2.5), начиная от середины заметно сущающаяся
к вершине. Гребень из 7—23 (чаще 9—16) зубцов, с широким основанием,
от которого отходят несколько дополнительных зубчиков, наиболее
дистальный из них у середины трубки. Впереди него отходит пучок из 2—
4 (редко 5—6) ветвей, в длину равных диаметру трубки.
Последний членик короткий, с седлом, доходящим до брюшного края
его боковых сторон; боковой волосок из двух (реже больше) ветвей, из ко-
торых одна длиннее седла, вторая — вдвое короче; наружный хвостовой
волосок простой, вдвое и более длиннее дыхательной трубки, внутренний
из 1—3 (чаще 2—-3) немного более коротких ветвей. Плавник в среднем
из 8 обычно 2-ветвистых пучков, задние одной длины с наружным хвосто-
вым волоском. Жабры в 1.5—2 раза длиннее седла, с несколько сужен-
ными, округленными концами.
Распространение. Характерен для Ориентальной области
и Океании. Проникает также в Африку и в Австралию. В Палеарктике
7. AEDES
305
Рис. 209. Гипопигий Aedes jlavopictus Yam.
По La Casse a. Yamaguti.
в лабораторных
миологиче-
ски е. В Восточ-
Океании A. albo-
перенос-
известен в Северном Китае и в Японии (кроме о. Хоккайдо). В СССР
не найден.
Биология. Личинки развиваются в большой численности в раз-
ного рода искусственных водоемах и заполненных водой сосудах в бли-
жайшем окружении человека и в его жилищах и только, как исклю-
чение, — в небольших естественных почвенных водоемах. В течение се-
зона (апрель—октябрь) несколько поколений. В странах Восточной Азии
A. albopictus принадлежит к
числу массовых видов. Местами
в городах и селениях встреча-
ется в большом количестве. По
наблюдениям в Юго-Восточ-
ной Азии, человек служит ос-
новным источником питания ко-
маров этого вида. Легко куль-
тивируется .
условиях.
Э п и д е
ское знач
ной Азии и
pictus — важнейший
чик вируса лихорадки денге.
Некоторые виды птичьих плаз-
модиев {Plasmodium lophurae,
Р. gallinaceum) в организме
A. albopictus развиваются до
стадии спорозоитов. К зараже-
нию филяриями человека не-
восприимчив, но филярии собак
{Dirofilaria immitis) в комарах
этого вида развиваются до ин-
вазионной стадии.
47. Aedes (Stegomyia) fla-
vopictus Yamada, 1921.
Очень близок к предыду-
щему виду, отличается дета-
лями окраски среднеспинки и
строения гипопигия.
Среднеспинка в каштаново-
коричневых чешуйках, с про-
дольной белой полоской, сужи-
вающейся кзади и разделяю-
щейся перед щитком на две узкие
полоски. Кроме того, имеются
две слабо выраженные боковые,
продольные полоски в задней половине среднеспинки. Непосредственно
кпереди и чуть выше основания крыльев располагается нерезко очер-
ченное пятно беловатых или желтоватых узких, слегка изогнутых чешуек.
Лапки с узкими белыми колечками при основании первых двух члеников
передней и средней лапки; на задней лапке более широкие белые кольца
развиты при основании четырех члеников; последний членик задней лапки
почти целиком белый. Коготки лапок простые. Крылья в темных чешуйках,
с небольшим белым пятном при основании костальной жилки. Брюшко
20 Фауна СССР, А. В. Гуцевич и др. ;
зов
III. ПОДСЕМ. CULICINAE
с белыми пятнами по бокам тергитов; при основании сегментов иногда раз-
виты узкие белые перевязи.
Гипопигий (рис. 209): бородавка коксита, вытянутая в продольном
направлении, удлиненная; она лишь немного не доходит до вершины
коксита; на бородавке располагаются волоски двух родов: одни простые,
несколько более короткие, чем у A. albopictus, другие уплощенные, лан-
цетовидные. Задний край IX тергита зазубренный, выпуклый, дуго-
образный.
Личинка IV стадии с сильно развитыми волосками на теле
звездчатого типа ветвления. Лобные волоски: наружные из 2—3 коротких
ветвей, средние впереди внутренних, обе пары из 1—2 ветвей, последние
длиннее остальных; задние волоски наличника смещены к переднему
краю головы и сближены, из 9—16 ветвей, с тонкой вторичной пери-
стостью, отходят от короткого стебелька. Усик около половины длины
головы, с телом без шипиков и с простым коротким волоском у середины.
Щетка из 6—12 чешуек, расположенных в один ряд с главным шипом,
по бокам основания которого сидят короткие тонкие щетинки. Дыхатель-
ная трубка короткая, индекс 2.2—2.5, за серединой сужается к вершине;
без ушков по бокам основания. Гребень с изменчивым числом зубцов
(4—16), имеющих ряд зубчиков у широкого основания; почти доходит до се-
редины трубки. Пучок у середины из 3—6 ветвей, в длину около ширины
основания,
Седло последнего членика доходит до нижнего края его боковых сторон,
короткое, с покрытым шипиками задним краем; боковой волосок из 2—
7 ветвей, превышающих длину седла; наружный хвостовой волосок про-
стой, вдвое длиннее трубки, внутренний из 2—3 немного более коротких
ветвей. Плавник из 6—8 1—2-ветвистых пучков одной длины с внутрен-
ними хвостовыми волосками. Жабры колбасовидные, изменчивой длины —
от длины седла до длины дыхательной трубки.
Р а с п р о странение. Япония (включая о. Хоккайдо), п-ов Корея,
Индия, Пакистан. В СССР найден только в заповеднике Кедровая падь
(Южное Приморье,, сборы А. С. Мончадского).
Биология. Личинки встречаются в скоплениях воды в срезанных
стволах бамбука, дуплах деревьев, различных искусственных водоемах
в ближайшем окружении человека. Самки — активные кровососы.
48. Aedes (Stegomyia) chemulpoensis Yamada, 1921.
Отличается окраской ног и среднеспинки (отсутствием белой продоль-
ной полосы), а также строением гипопигия.
Среднеспинка в темно-коричневых чешуйках со снежно-белым ри-
сунком, состоящим из следующих частей: а) трех небольших пятен у пе-
реднего края, б) двух больших треугольных или полулунных пятен по бо-
кам передней половины среднеспинки, в) небольших пятен перед основа-
нием крыльев, г) двух продольных полосок и небольшого срединного
пятна в задней половине среднеспинки; последнее может отсутствовать.
Передние и средние бедра спереди с белой полоской в основной части и
несколькими белыми пятнами в вершинной половине. Голени с пятном
или кольцом белых чешуек в конце первой трети. Лапки с белыми колеч-
ками при основании члеников; последний членик задней лапки целиком
белый или белый с темной вершиной. Коготки на лапках без зубчиков.
Тергиты брюшка с белыми пятнами по бокам и узкими базальными белыми
перевязями равномерной ширины.
Гипопигий (рис. 210): в средней части внутренней поверхности коксита
расположен резко очерченный округлый бородавчатый выступ, густо уса-
7. AEDES
307
женный волосками. Стили саблевидные, без перетяжки и без расширения
на конце; придаток причленяется на вершине стиля. Задний край IX тер-
гита с неглубокой вырезкой.
Личинка IV стадии описана недостаточно, очень сходна
с A. galloisi. Лобные волоски все простые, средние впереди внутренних,
сильно сдвинуты вперед, внутренние почти вдвое длиннее наружных.
Задние волоски наличника между основаниями средних, из 4—5 ветвей..
Усик короткий, гладкий, с коротким простым волоском за серединой.
Рис. 210. Гипопигий Aedes chemulpoensis Yarn.
Волоски тела звездчатого типа ветвления. Щетка на VIII членике
брюшка из 6—10 чешуек, расположенных в один ряд, имеющих по 2—4 до-
полнительных зубчика по бокам основания главного шипа. Дыхательная
трубка без ушков у основания, индекс около 3.0. Гребень доходит почти
до середины трубки, из 15 зубцов. Пучок из четырех ветвей, у середины,
немного ближе к вершине.
Седло доходит до нижнего края боковых сторон последнего членика;
боковой волосок звездчатый, многоветвистый; наружные хвостовые во-
лоски длинные, простые, внутренние из 2—3 ветвей, более короткие.
Плавник из шести слабо ветвящихся пучков. Жабры колбасовидные,
в 5 раз длиннее седла, обе пары одной длины. -
Распространение. П-ов Корея, Северный Китай. EHCGCP не
найден.
Биология не изучена. Личинки встречались в стволовых'и при-
корневых дуплах Salix matsudana, Quercus ^cutissima и Sophora japoriica.
20*
308
III. ПОДСЕМ. CULICINAE
5. Подрод AEDES Meigen, 1818
Наиболее характерная особенность — короткие щупики самцов; их
длина, как и у самок, составляет около J/e длины хоботка. Своеобразно
также строение гипопигия: стили при основании раздвоены, без придатка;
класпеты отсутствуют. Отличия самок менее выражены: хоботок по длине
обычно равен передним бедрам, пятна чешуек на бочках груди слабо раз-
виты, церки явственно выступают (отличие от двух предыдущих подро-
дов). Подрод включает свыше 70 видов, распространенных преимущест-
венно в Ориентальной области.
Личинки очень сходны с личинками подродов Aedimorphus и Ochlero-
tatus. Строение стигмальной пластинки — единственный специфичный
личиночный признак, отличающий, как и по ряду имагинальных призна-
ков, подрод Aedes от других подродов этого рода.
49. Aedes (Aedes) cinereus Meigen, 1818.
Комар небольших размеров. Хоботок и щупики темно-бурые. По длине
хоботок приблизительно равен передним бедрам. Бочки груди с неболь-
шими пятнами желтоватых или белых чешуек, не имеющих блестящего
серебристого оттенка. Лапки одноцветные, темные. Крылья в темных
Рис. 211. Гипопигий Aedes cinereus rossicus D., G., M.
чешуйках, без примеси светлых. Окраска головы, среднеспинки и брюшка
различна у подвидов, на которые распадается данный вид.
Гипопигий (рис. 211): коксит короткий, при основании толстый, к вер-
шине сильно сужающийся. Базальная бородавка коксита хорошо раз-
вита, густо усажена волосками; дорсальный отдел бородавки обособлен
в виде так называемой дорсобазальной бородавки. Стили, приклоняющиеся
к кокситу далеко отступя от его вершины, раздвоены, причем латеральная
ветвь длиннее и уже медиальной.
7. AEDES
309
Личинка IV стадии (рис. 212) средних размеров, от полупро-
зрачного желтоватого до коричневого или темного цвета. Лобные волоски
вторичноперистые, расположены в один вогнутый назад дугообразный
ряд; наружные из 6—14 (чаще 7—8), средние из 3—7 (чаще 5-*-7), внутрен-
ние из 3—7 (чаще 4—6) ветвей. Задние волоски наличника короткие,
тонкие, из 3—7 ветвей. Усик длинный — около 0.7 длины' головы; тело
покрыто редкими шипиками, пучок из 4—7 ветвей отходит на 2/5 длины
усика от основания и достигает х/2 его длины.
Рис. 212. Aedes cinereus cinereus Mg. Личинка
IV стадии.
1 — голова, сверху, 2 — задний конец, сбоку.
Щетка на VIII членике из 10—19 (чаще 14—16) чешуек, расположен-
ных в 1—2 неправильных ряда; каждая чешуйка с остроконечным шипом,
с рядом мелких тонких щетинок по бокам основания. Дыхательная трубка
тонкая, стройная, индекс 3.0—4.0. Гребень занимает х/2—3/5 длины трубки,
состоит из 13—21 зубца, из которых 1—3 ближайших к основанию руди-
ментарны, а 1—3 наиболее дистальных широко расставлены и более круп-
ные. Пучок короткий, из 3—7 (чаще 4—5) ветвей, сдвинут к вершине,
заметно впереди наиболее дистального зубца гребня. На передней по-
верхности трубки у части личинок находятся две пары тонких коротких,
часто едва заметных пучков из 2—5 ветвей. Кроме того, в вершинной части
трубки, помимо имеющейся у всех личинок пары апикальных щетинок
310
III. ПОДСЕМ. CULICINAE
у ее переднего края, может быть четыре пары коротких тонких, слабо
ветвистых волосков, сидящих по краям боковых сторон; иногда они встре-
чаются в меньшем числе или отсутствуют. г
Стигмальная пластинка (рис. 213) отличается строением заднего от-
ростка рычага;, задняя лука последнего в виде длинного хитинового
стержня, расширяющегося
в дистальном направле-
нии, по бокам его зад-
него края сидят два когте-
видно загнутых навстречу
друг другу выступа, обра-
зующих задний отросток.
Седло последнего чле-
ника доходит до сере-
дины его боковых сторон
или заходит за нее; боко-
вой волосок из двух, реже
одной или трех ветвей; на-
ружные хвостовые воло-
ски длиннее дыхательной
трубки, простые; внут-
Рис. 214. Стиль (слева) и базальная бородавка
коксита Aedes cinereus Mg. По Д. К. Львову.
А — A. cinereus cinereus, Б, В, Г — переходные формы,
Д — A. cinereus esoensis Yam.
Рис. 213. Aedes cinereus Mg.
Личинка IV стадии. Стиг-
мальная пластинка.
ренние более чем вдвое короче, из 4—8 (чаще 7) ветвей. Плавник
из 9—10 пучков в общем основании и из 2—4 более коротких впереди него.
Жабры длинные, не менее чем вдвое длиннее седла, узкие, заостряющиеся,
не пигментированные.
Распространение. Европа, Сибирь, Дальний Восток СССР
(включая Сахалин и Камчатку), Северо-Восточный Китай, п-ов Корея,
Япония, Канада, США. На север в качестве редкого вида проникает до
69° с. ш. (в Норвегии), до Кольского полуострова и низовьев Печоры.
На юге распространен до Закавказья и Средней Азии включительно.
Aedes cinereus cinereus Meigen.
Голова преимущественно в темных чешуйках. Чешуйки среднеспинки
красновато-бурые, иногда с золотистым или бронзовым оттенком. Брюшко
сверху без светлых перевязей, по бокам тергитов с продольной желтовато-
белой полосой.
7. AEDES
311
Гипопигий (рис. 214): латеральная ветвь стиля на вершине раздвоена,
медиальная ветвь по длине составляет 2/2—J/3 латеральной. Дорсобазаль-
ная бородавка состоит из двух ветвей — латеральная длиннее медиальной.
Личинки IV стадии характеризуются признаками, приве-
денными при описании вида. Типичные формы не имеют дополнительных
волосков на передней поверхности дыхательной трубки и в ее вершинной
части. Переходные имеют на передней поверхности трубки одну или две
пары дополнительных волосков. Длина жабр увеличи-
вается по направлению к востоку.
Распространение. Европа, Кавказ, Сред-
няя Азия, Казахстан, Сибирь, Дальний Восток СССР,
Северная Америка.
Биология. Личинки развиваются как в по-
стоянных, сравнительно глубоких, затененных или
полузатененных водоемах в разреженном лесу или
на опушках, так и в покрытых кустарником луговых
заболоченностях или лужах, бедных растительностью.
Во многих географических районах А. с. cinereus при-
надлежит к числу массовых видов; нередко занимает
одно из первых мест среди комаров, нападающих на
людей. Большая часть ареала вида лежит в пределах
лесной зоны, но он более характерен для открытых био-
топов: лугов, разреженного леса, участков с преоблада-
нием кустарников. Комары укрываются в траве, у земли,
и при появлении людей подымаются снизу, колют глав-
ным образом ноги. Активны не только утром и ве-
чером, но нередко и днем.
Aedes cinereus — полисезонный вид. Весной комары
появляются позже ранневесенних Ochlerotatus, летают
до наступления осенних холодов. Кроме первой,
поздневесенней генерации обычно (но не повсеместно)
бывает вторая, после летних дождей. Если второй
выплод обилен, максимальная численность вида наблю-
дается в августе—сентябре (Ремм, 1957).
При некоторых трансмиссивных вспышках туля-
ремии отмечался как один из важнейших переносчи-
ков (механическая передача).
Рис. 215. Aedes ci-
nereus rossicus D.,
G., M. Личинка
IV стадии. Дыха-
тельная трубка,
сбоку.
Aedes cinereus rossicus Dolbeshkin. Gorickaja et Mitrofanova, 1930
(=4. tarnogradskii Martini, 1931).
Отличается строением гипопигия, а также окраской. Голова преиму-
щественно в беловато-серых чешуйках. Среднеспинка покрыта золотисто-
бурыми чешуйками. Вдоль боковых краев среднеспинки тянется расплыв-
чатая полоска желтовато-белых чешуек, более выраженная в задней по-
ловине груди. Брюшко сверху в черно-бурых чешуйках, без светлых
перевязей, по бокам со светлой продольной полосой.
Гипопигий (рис. 211): стили на вершине не раздвоены; медиальная
ветвь стиля по длине составляет 1/3—х/2 латеральной. Дорсобазальная
бородавка состоит из двух ветвей.
Типичные личинки IV стадии сходны по всем признакам
с основной формой, но всегда имеют две пары добавочных коротких во-
лосков на передней поверхности дыхательной трубки, которые у пере-
ходных форм могут частично или полностью отсутствовать (рис. 215).
312
III. ПОДСЕМ. CULICINAE
Распространение. Найден на Украине (в Днепропетровской,
Киевской, Закарпатской и Волынской областях), на Кавказе (Орджо-
никидзе, Теберда), в Чувашии, на Урале (Пермь, Оренбург) ив Якутии.
За пределами СССР — в Венгрии, Чехословакии и в Японии.
Биология. Личинки встречаются в разного типа небольших
временных открытых водоемах с травянистой растительностью. Имеют
несколько поколений в году. По-видимому, А. с. rossicus в основном оби-
тает в поймах рек. Местами он встречается в значительном количестве.
Комары весьма назойливы, нападают и днем.
Aedes cinereus esoensis Yamada, 1921.
Отличается окраской брюшка и строением гипопигия. Голова и грудь
как у А. с. cinereus. Брюшко сверху в черно-бурых чешуйках, иногда
с бронзовым оттенком. При основании тергитов развиты перевязи желто-
вато-бурых чешуек, по бокам — пятна такого же цвета.
Гипопигий (рис. 214): латеральная ветвь стиля на вершине не раздво-
ена; медиальная ветвь по длине составляет приблизительно 1/3 латераль-
ной. Дорсальная бородавка простая, т. е. состоит из одной медиальной
ветви.
Личинки IV стадии этого подвида сходны по всем признакам
с основной формой, но отличаются от нее наличием четырех пар дополни-
тельных, очень маленьких коротких волосков у вершины дыхательной
трубки, сидящих по краям боковых сторон. У переходных форм они могут
быть развиты не в полном числе, на одной из боковых сторон, или даже
отсутствовать. Жабры длиннее дыхательной трубки.
Распространение. Дальний Восток СССР, Казахстан, Се-
веро-Восточный Китай, п-ов Корея, Япония.
Биология. Каких-либо существенных отличий от А. с. cinereus
не отмечено. Во многих районах Приморья и Хабаровского края А. с. eso-
ensis принадлежит к числу массовых кровососов. Рассматривается как
один из возможных переносчиков вируса японского энцефалита (Петри-
щева, 1947).
Замечания по систематике. Формы, трактуемые нами как подвиды
A. cinereus, чаще рассматриваются как самостоятельные виды. Действительно, они
различаются не только окраской, но и довольно существенными особенностями строе-
ния гениталий. Но существуют постепенные переходы, причем серию промежуточных
вариантов можно подобрать в отношении каждого из отличительных признаков.
Так, у форм, переходных между cinereus и esoensis, светлые перевязи тергитов развиты
не на всех сегментах; иногда перевязи очень узкие или неполные, иногда заменены
небольшим пятном, состоящим всего из нескольких чешуек. Характерная для А. с. ci-
nereus вилка на конце латеральной ветви стиля нередко образована двумя неравными
выступами, один из них может быть очень коротким, едва заметным. Латеральная
ветвь дорсобазальной бородавки иногда укорочена или имеет вид маленького бугорка
и т. д.
Специальное исследование (Львов, 1956) показало, что из названных двух подвидов
в европейской части СССР распространен только А. с. cinereus', в Забайкалье наряду
с ним часто встречаются и переходные формы, весьма обычные в Хабаровском крае и
в Приморье, где распространены также «типичные» А. с. cinereus и А. с. esoensis,
причем последние преобладают. Точно так же связаны постепенными переходами
А. с. cinereus и А. с. rossicus. По окраске они различаются менее отчетливо, чем А. с. eso-
ensis от А. с. cinereus. Гипопигий А. с. rossicus по строению занимает промежуточное
положение между А. с. cinereus и А. с. esoensis.
То же наблюдается и у личинок. Все основные личиночные признаки, включая и
строение стигмальной пластинки, оказываются общими для всех трех описанных его
подвидов и ни у одного не выходят за пределы изменчивости основной формы. В типич-
ных случаях личинки А. с. cinereus не имеют дополнительных двух пар волосков на пе-
редней поверхности дыхательной трубки, но в той же пробе встречаются личинки
с одной или двумя парами этих волосков, хотя и в меньшей численности. Обратные
/
7. AEDES
313
отношения наблюдаются у А. с. rossicus. Типичные А. с. esoensis с четырьмя парами
дополнительных волосков в вершинной части дыхательной трубки встречаются у нас
на Дальнем Востоке сравнительно редко. Большинство личинок имеет их 1—2, реже
3 пары, часты случаи непарного развития, а у одной личинки наряду с полным разви-
тием волосков у вершины были и обе пары волосков на передней поверхности дыхатель-
ной трубки. Для Японии же (La Casse a. Yamaguti, 1955) описываются только типич-
ные А. с. esoensis' о наличии там упомянутых выше переходных форм или типичных
А. с. cinereus не упоминается.
Приведенные данные говорят против мнения о видовой самостоятельности А. esoen-
sis и A. rossicus.
50. Aedes (Aedes?) nobukonis Yamada, 1932.
К подроду Aedes этот вид относят предположительно, так как самцы
неизвестны. От других видов подрода отличается наличием белых пятен
по бокам сегментов брюшка при отсутствии светлых перевязей на тер-
гитах.
Голова сверху преимущественно в бурых чешуйках. Широкие серо-
вато-белые чешуйки образуют пятна по бокам головы. Узкие светлые
чешуйки вкраплены посредине затылка и образуют каемку вдоль заднего
края глаз. Хоботок и щупики бурые. Длина хоботка примерно равна
длине передних бедер. Покровы среднеспинки коричневые, чешуйки
узкие, золотисто-бронзовые. Золотистые чешуйки более светлого от-
тенка образуют неясную, едва заметную продольную полоску, которая
перед щитком раздваивается, окаймляя небольшой свободный от чешуек
участок. Щиток в узких золотистых чешуйках. Бочки груди с пятнами
белых чешуек. Ноги:/бедра спереди темные, сзади светлые; голени и
лапки сплошь в темных чешуйках. Коготок передних и средних ног
с зубчиком, задних ног — простой. Крылья в узких бурых чешуйках.
Брюшко сверху покрыто бурыми чешуйками. На первом тергите
нерезко выделяется пятно более темных чешуек с металлическим от-
ливом. По бокам тергитов базально располагаются белые пятна; свет-
лых перевязей на тергитах нет, поэтому брюшко сверху кажется сплошь
темным.
Личинка неизвестна.
Распространение. Япония (о. Кюсю). В коллекции ЗИН име-
ется экземпляр с этикеткой: Кедровая падь Приморской обл., 26 VII1940,
берег Кедровки, яма под корнем дерева, сбор А. С. Мончадского.
Экземпляр обнаруживает лишь небольшие отличия по сравнению с ориги-
нальным описанием. По описанию светлые чешуйки на голове образуют
две субмедиальные и две латеральные полоски; среднеспинка со срединной
и двумя субмедиальными полосками.
Добавление к роду AEDES
51. Aedes aureus Gutsevich, 1955.
Так как самцы этого вида неизвестны, вопрос об его принадлежности
к тому или иному подроду остается открытым.
Комар средней величины, отличается яркой золотистой окраской
чешуек боковых частей среднеспинки, наличием золотистого пятна на
голове и светлых чешуек на крыльях.
Голова сверху с резко очерченным срединным пятном из торчащих
золотистых чешуек и волосков. Хоботок и щупики в одноцветных бурых
чешуйках; в средней трети хоботка могут быть вкраплены единичные
светлые чешуйки. Хоботок немного длиннее передних бедер. Средне-
спинка с продольной, расширяющейся кзади, полосой шоколадно-ко-
ричневых чешуек. Боковые отделы среднеспинки в золотистых чешуйках.
314 III. ПОДСЕМ. CULICINAE
Лапки темные. Крыло: светлые кремовые чешуйки преобладают на
субкостальной, а также при основании радиальной, медиальной и куби-
тальной жилок; остальные жилки целиком или преимущественно в тем-
ных чешуйках. Брюшко сверху преимущественно в бурых чешуйках,
более светлые желтовато-серые чешуйки образуют расплывчатые пятна
в средней части и у переднего края тергитов. Церки очень короткие.
Личинка неизвестна.
Такой признак, как короткий хоботок (длина его едва превышает
длину передних бедер), позволил при описании вида высказать предпо-
ложение о его принадлежности к подроду Aedes.
Найден К. П. Чагиным в небольшом числе экземпляров на юге При-
морского края в Хасанском районе (Гуцевич, 1955).
8. Род CULEX Linnaeus
Средней величины или мелкие формы. Щупики самца, как правило,
длиннее хоботка или равны ему, реже (С. hayashii, а также многие экзо-
тические виды) короче хоботка; щупики самки короткие. Дыхальце-
вые щетинки отсутствуют; стерноплевральные и верхние мезэпимерные
щетинки хорошо развиты; нижних мезэпимерных щетинок, как правило,
одна, реже они отсутствуют вовсе; еще реже их 2—3; лишь у подрода
Lutzia не менее четырех нижних мезэпимерных щетинок. Большая часть
боковой поверхности груди лишена чешуек; имеется лишь несколько
пятен немногочисленных чешуек. Ноги: I членик задних лапок, за редким
исключением (подрод Barraudius), равен по длине задней голени или
длиннее ее. У самок коготки лапки простые. Крылья с длинной радиаль-
ной вилкой. Волоски на радиальной жилке при ее основании отсутствуют.
Закрыловые пластинки с узкими чешуйками. Церки самки не выступают.
Гипопигий: базальная бородавка коксита и класпеты отсутствуют;
предвершинная бородавка обыкновенно хорошо развита и несет более
или менее метаморфозованные щетинки (пластинки, пальцевидные вы-
росты и т. п.); X стернит на конце с пучком шипиков или с поперечным
гребнем зубцов; фаллосома, как правило, состоит из двух парных пла-
стинок, часто по краю изрезанных и несущих вторичные зубцы.
Личинки отличаются большой широкой головой с длинными усиками,
несущими хорошо развитый пучок в виде веера вторичноперистых вет-
вей и длинные предвершинные щетинки, часто сидящие на некотором
расстоянии от вершины. Ротовые части фильтрационного типа питания,
с длинными пучками простых волосков на боковых лопастях верхней
губы и на верхних челюстях. Исключение составляют хищные личинки
подрода Lutzia с верхней губой, приспособленной для схватывания до-
бычи. Волоски груди обычно длинные и разветвленные. Хитиновые
пластинки и отростки у их основания умеренно развиты. Пластинки на
члениках брюшка отсутствуют, за исключением последнего членика,
седло которого опоясывает его кольцом. Щетка по бокам VIII членика
брюшка обычно из большого числа чешуек. Сифон, за редким исклю-
чением (подроды Lutzia, Barraudius), длинный или очень длинный и тон-
кий, с хорошо развитым гребнем и с четырьмя или более парами сифо-
нальных волосков или пучков, расположенных или на боковых и задней
сторонах сифона, или смещенных на заднюю сторону и сидящих в один
зигзагообразный или прямой ряд. Главные трахейные стволы двух типов:
тонкие, с округлым сечением, и широкие, лентовидные, с продолговато-
овальным сечением. Трахеям первого типа соответствует строение стиг-
мальной пластинки как у С. territans, трахеям второго типа — как у С. pi-
piens.
8. CULEX 31S
Род, весьма богатый видами (известно более 400 видов), большинство
которых свойственно тропической зоне западного и восточного полушарий.
Род подразделяется на 16 подродов, из числа которых свойственные
фауне СССР охарактеризованы ниже.
Подразделение личинок на подроды не полностью отражает существую-
щие отношения по взрослой фазе. Личинки подрода Lutzia образуют
хорошо обособленную группу по ряду признаков, сочетающихся с их
хищническим типом питания, но имеют некоторые признаки (располо-
жение и число сифональных пучков, детали строения стигмальной пла-
стинки), связывающие их с личинками подродов Culex и Barraudius.
Личинки обоих последних подродов, а также подрода Lophoceraomyia
не имеют признаков, позволяющих их отделить друг от друга. Личинки
подрода Neoculex имеют тонкие с округлым поперечным сечением глав-
ные трахейные стволы и особый тип строения стигмальной пластинки,
но личинки С. mimeticus и С. jacksoni, принадлежащие к подроду Culex,
имеют такие же признаки. Таким образом, личинки рода Culex, представ-
ляя хорошо очерченную группу, внутри рода распределяются иначе,
чем соответствующие им взрослые комары.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
9?
1 (4). I членик задних лапок явственно короче задней голени (длина
его равна 4/5—®/7 голени). Мелкие комары (Подрод Barraudius
Edw.).
2 (3). Брюшко с боков с продольной полосой светлых чешуек, иногда
образующих более или менее развитые треугольные пятна по перед-
нему краю тергитов ...... 3. С. (Barraudius) modestus Fic.
3 (2). Брюшко с боков при основании тергитов с более или менее раз-
витыми обособленными пятнами светлых чешуек ........
....................х.......... 4. С. (Barraudius) pusillus Macq.
4 (1). I членик задних лапок немного длиннее задней голени или равен
ей по длине.
5 (8). Нижние мезэпимерные щетинки многочисленны, их не менее че-
тырех, обычно 6—10. Комары крупных размеров. (Подрод Lutzia
Theob.).
6 (7). Все тергиты брюшка по заднему краю со светлыми перевязями
более или менее одинаковой ширины. Основная половина передней
поверхности задних бедер с многочисленными темными чешуй-
ками ...................................2, С. ( Lutzia) vorax Edw.
7 (6). II—IV тергиты брюшка одноцветно темные или с очень узкими
светлыми перевязями по заднему краю; V—VIII тергиты на всем
протяжении покрыты светлыми чешуйками или же имеют очень
широкие перевязи светлых чешуек по заднему краю. Основная по-
ловина передней поверхности задних бедер в белых чешуйках, без
вкраплений темных чешуек . . . . 1. С. (Lutzia) fuscanus Wied.
8 (5). Нижние мезэпимерные щетинки чаще в количестве одной, реже
отсутствуют вовсе, еще реже в числе 2—3. Обычно комары средней
величины или мелкие. (Подроды Culex L. и Neoculex Dyar).
9 (20). Хоботок и лапки с белыми колечками, иногда очень узкими.
10 (13). Крылья, особенно по костальной жилке, с большими светлыми
пятнами, образованными скоплениями белых чешуек.
11 (12). Вершинная часть жилки сщ в светлых чешуйках..........'.
............................... 14. С. (Culex) orientalis Edw.
316
111. ПОДСЕМ. CULICINAE
12 (11). Вершинная часть жилки сих в темных чешуйках...............
............................... 12. С. (Culex) mimeticus Noe.
13. С. (Culex) jacksoni Edw.
13 (10). Крылья без пятен, иногда с беспорядочно вкрапленными в более
или менее значительном числе белыми чешуйками.
14 (15). Чешуйки на крыльях бурые, с более или менее многочисленными
вкраплениями белых чешуек .........................................
.......................... 9. С. (Culex) bitaeniorhynchus Giles.
15 (14). Крылья в одноцветных бурых чешуйках.
16 (19). Чешуйки среднеспинки резко двуцветные: передние 2/3 средне-
спинки покрыты светлыми (светло-желтыми или беловато-серыми)
чешуйками, задняя треть среднеспинки в темно-бурых чешуйках.
17 (18). Перевязи светлых чешуек расположены по заднему краю терги-
тов брюшка. Передняя часть среднеспинки в светло-желтых чешуй-
ках. Крылья в узких чешуйках . . .10. С. (Culex) sinensis Theob.
18 (17). Перевязи светлых чешуек расположены по переднему краю тер-
гитов брюшка. Передняя часть среднеспинки в белых чешуйках.
Чешуйки крыльев более широкие . . 11. С. (Culex) whitmorei Giles.
19 (16). Среднеспинка на всем своем протяжении покрыта более или
менее одноцветными темными чешуйками ..............................
........................ 15. С. (Culex) tritaeniorhynchus Giles.
20 (9). Хоботок и лапки без светлых колец.
21 (24). Брюшко сверху одноцветно бурое, без поперечных перевязей
или боковых пятен, образованных светлыми чешуйками.
22 (23). Брюшко сверху покрыто одноцветно черно-бурыми чешуйками,
иногда с едва намечающимися перевязями белых чешуек по заднему
краю тергитов. (Щупики. самца значительно длиннее хоботка).
Средиземноморская подобласть . 7. С. (Neoculex) martinii Med.
23 (22). Брюшко сверху покрыто одноцветно красновато-бурыми чешуй-
ками. (Щупики самца значительно короче хоботка). Дальний Вос-
ток ................................8. С. (Neoculex) hayashii Yam.
24 (21). Тергиты брюшка с более или менее развитыми поперечными пере-
вязями, образованными скоплениями белых или желтоватых чешуек
(как исключение, у отдельных экземпляров С. pipiens тергиты без
светлых перевязей).
25 (28). Передние и средние бедра и голени спереди темные, со светлой
продольной полосой; реже лишь передние бедра имеют такую полосу.
26 (27). Светлые поперечные перевязи брюшка посредине заднего края
обычно с треугольным выступом. Задние бедра снизу в вершинной
трети с темной полосой.................17. С. (Culex) theileri Theob.
27 (26). Светлые поперечные перевязи брюшка более или менее равно-
мерной ширины, без выступов. Задние бедра снизу на всем протя-
жении светлые............................18. С. (Culex) vagans Wied.
28 (25). Передние и средние бедра и голени спереди без светлой полосы.
29 (32). Светлые поперечные перевязи расположены по заднему краю
тергитов брюшка. Проэпистерны покрыты широкими светлыми че-
шуйками.
30 (31). Щупики в темных чешуйках с примесью светлых. Светлые пере-
вязи брюшка относительно широкие, иногда с выступом посредине.
Задние голени у вершины с белым пятном. (Последние членики щупи-
ков самца голые или почти голые) . . 6. С. (Neoculex) hortensis Fic.
31 (30). Щупики в одноцветно темных чешуйках. Светлые перевязи
брюшка узкие, без выступа посредине. Задние голени у вершины
8. CULEX
317
32
33
34
1
2
3
4
5
6
7
8
9
10
И
12
13
14
15
16
без белого пятна. (Последние членики щупиков самца в длинных
волосках)..................... 5. С. (Neoculex) territans Walk.
(29). Светлые поперечные перевязи или светлые боковые пятна распо-
ложены по переднему краю тергитов. Проэпистерны покрыты узкими
светлыми чешуйками.
(34). Светлые поперечные перевязи брюшка образованы чешуйками
чисто белого цвета..............16. С. (Culex) univittatus Theob.
(33). Светлые поперечные перевязи брюшка образованы чешуйками
желтоватого цвета; в некоторых случаях светлые перевязи брюшка
редуцированы или имеют вид пятен по бокам брюшка...............
.....................................20. С. (Culex) pipiens L.
19. С. (Culex) torrentium Mart.
(4). Коксит на наружной поверхности в мелких чешуйках. Предвер-
шинная бородавка расположена едва за серединой коксита.
(3). Стиль относительно длинный и тонкий; фаллосома короткая, ее зад-
ний край не выступает за вершину X стернита....................
................................3. С. (Barraudius) modestus Fic.
(2). Стиль относительно короткий и толстый; фаллосома длинная,
ее задний край заходит за вершину X стернита...................
................................4. С. (Barraudius) pusillus Macq.
(1). Коксит на наружной поверхности без чешуек. Предвершинная
бородавка расположена явственно за серединой коксита.
(14). Предвершинная бородавка коксита без пластинчатых придатков;
X стернит на вершине с одним рядом относительно крупных зубцов
или с несколькими рядами коротких шипиков.
(9). X стернит на вершине с многочисленными шипиками.
(8). Зубчики фаллосомы (иногда они едва заметны) расположены не-
посредственно на теле фаллосомы ... 1. С. (Lutzia) fuscanus Wied.
(7). Зубцы, обычно хорошо развитые, расположены на треугольном
выступе наружного края фаллосомы . . . 2. С. (Lutzia) vorax Edw.
(6). X стернит на вершине с поперечным рядом зубцов.
(11). Коксит на вершине с широким пальцевидным склеротизованным
выростом, выступающим на значительное расстояние за основание
стиля .......................... 6. С. (Neoculex) hortensis Fic.
(10). Коксит на вершине без пальцевидного выроста.
(13). Стиль относительно короткий, за серединой сильно расширен-
ный ............................7. С. (Neoculex) martiniiMed.
(12) . Стиль к вершине явственно и постепенно суживающийся . . .
. . . ..........................5. С. (Neoculex) territans Walk.
а (б). Фаллосома на вершине с многочисленными тупыми зубчи-
ками ........................... С. territans territans Walk.
б (а). Фаллосома на вершине почти ровная, без зубчиков . . . .
.......................... С. territans judaicus Edw.
(5). Предвершинная бородавка коксита несет одну или несколько
прозрачных пластинок овальной или ланцетовидной формы; реже
пластинки узкие, ножевидные; X стернит на вершине с шипиками,
расположенными в несколько рядов. (
(16). Предвершинная бородавка коксита несет несколько ножевидных
пластинок........................8. С. (Neoculex) hayashii Yam.
(15). Предвершинная бородавка коксита несет одну пластинку.
318
III. ПОДСЕМ. culicinae
17 (18). Фаллосома с каждой стороны образует одну простую (не зубча-
тую и не расщепленную) пластинку. Пластинка на предвершинной
бородавке коксита ланцетовидная ... 10. С. (Culex) sinensis Theob.
18 (17). Фаллосома с каждой стороны состоит не менее чем из двух скле-
ритов, часто вторично расщепленных или зубчатых.
19 (20). Фалллосома с каждой стороны состоит из двух простых (не рас-
щепленных) заостренных на концах серповидно изогнутых склери-
тов. Пластинка предвершинной бородавки коксита узкая, ноже-
видная ........................ 9. С. (Culex) bitaeniorhynchus Giles.
20 (19). Фаллосома с каждой стороны состоит из трех склеритов, если же
из двух, то один из них расщеплен или по краю зубчатый. Пластинка
предвершинной бородавки коксита более широкая, листовидная.
21 (24). X стернит без базального придатка или он имеется в рудимен-
тарном состоянии.
22 (23). Второй отдел фаллосомы с 4—5 длинными пальцевидными вы-
ростами ...............................И. С. (Culex) whitmorei Giles.
23 (22). Второй отдел фаллосомы простой . . . 20. С. (Culex) pipiens L.
а (б). Второй отдел фаллосомы узкий, крючковидный.................
.....................................С. pipiens pipiens L.
С. pipiens molestus Forsk.
б (а). Второй отдел фаллосомы широкий, лопастевидный ....
................................... С. pipiens fatigans Wied.
24 .(21). X стернит с явственно развитым, обычно достигающим довольно
значительной величины базальным придатком.
25 (26). Коксит у вершины близ основания стиля с пучком густых на-
правленных дистально волосков . . . 14. С. (Culex) orientalis Edw.
26 (25). Пучок волосков у вершины коксита отсутствует.
27 (32). Фаллосома относительно простая, с небольшим числом зубцов.
28 (31). Стиль постепенно суживается от основания к вершине. Пластинка
предвершинной бородавки яйцевидная.
29 (30). Фаллосома в вершинной части разделена на две пластинки, из ко-
торых одна образует 2—3 направленных латерально зубца ....
...................................17. С. (Culex) theileri Theob.
30 (29). Фаллосома в вершинной части разделена на 3 пластинки, из ко-
торых одна направлена дистально . . . 18. С. (Culex) vagans Wied.
19. С. (Culex) torrentium Mart.
31 (28). Стиль в вершинной трети расширен. Пластинка предвершинной
бородавки широкая, асимметричная . . 16. С. (Culex) univittatus Theob.
32 (27). Фаллосома сложная, многораздельная.
33 . (34). Второй отдел фаллосомы с 2—3 направленными латерально зуб-
цами .................................. 12. С. (Culex) mimeticus Noe.
13. С. (Culex) jacksoni Edw.
34 (33). Второй отдел фаллосомы с 4—5 зубцами, из которых медиальные
направлены дистально ... 15. С. (Culex) tritaeniorhynchus Giles.
ЛИЧИНКИ IV СТАДИИ
1 (2). Боковые лопасти верхней губы из 30—40 крепких, S-образно
изогнутых щетинок, несущих на дистальной половине гребень из
ряда коротких шипиков. Сифон короткий, не длиннее последнего чле-
ника брюшка, резко сужен на конце, с гребнем, занимающим всю
его длину до сужения (подрод Lutzia) ... С. (Lutzia) vorax Edw.
С. (Lutzia) fuscanus Wied.
2 (1). Боковые лопасти верхней губы обычного строения. Сифон длин-
8. CULEX 319
3
4
5
6
1
8
9
10
И
12
13
14
15
16
17
18
нее последнего членика брюшка, не резко сужен на конце, с гребнем,
занимающим не более половины его длины.
(4). Сифон короткий, длина в среднем в 3 раза больше ширины у ос-
нования. Пучки на сифоне расположены зигзагом на его задней
поверхности ...................... С. (Barraudius) pusillus Macq.
(3). Длина сифона не менее чем в 4 раза превышает ширину его осно-
вания.
(12). Щетка по бокам VIII членика брюшка из 4—8 крупных чешуек
с острым главным шипом.
(7). Нижнегубная пластинка по краю с многочисленными мелкими
зубчиками одинаковой величины. Волоски боковых лопастей верх-
ней губы из меньшего, чем обычно, числа тонких упругих коленчато
или крючковидно изогнутых волосков. Длинный тонкий сифон
с 4—5 парами коротких, слабо ветвящихся пучков.................
...........................С. (Culex) bitaeniorhynchus Giles.
(6). Нижнегубная пластинка из небольшого числа увеличивающихся
к середине зубцов.
(9). Нижнегубная пластинка с каждой стороны состоит из 10—^мел-
ких зубчиков у основания и 6—8 увеличивающихся к середине зуб-
цов. Предвершинные щетинки на усике отходят от середины рас-
стояния между концом и основанием пучка. Гребень из 2—4 малень-
ких зубцов у основания сифона .... С. (Culex) sinensis Theob.
(8). Нижнегубная пластинка только с более крупными зубцами. Гре-
бень из 8—14 зубцов, занимает не менее V4 длины сифона.
(И). Пучки на сифоне короткие, в длину около его диаметра у мест
их отхождения, каждый из 3—5 ветвей. Жабры не больше половины
длины сифона .........................*С. (Culex) vishnui Theob.1
(10). Пучки на сифоне очень длинные, более чем вдвое превышают
его диаметр у мест их отхождения, каждый из двух ветвей. Жабры
больше половины длины сифона ... С. (Culex) whitmorei Giles.
(5). Щетка по бокам VIII членика брюшка из большого числа (не ме-
нее 20) чешуек, у большинства видов не имеющих главного шипа,
а несущих по краю ряд в различной степени развитых зубчиков
или шипиков.
(14). Сифон, кроме зубцов гребня и предвершинных щетинок, имеет
на задней стороне в дистальной четверти дополнительные шипы
в числе от двух до девяти..............С. (Culex) jacksoni Edw.
(13). Дополнительные шипы в дистальной четверти сифона отсутствуют.
(28). Все (или кроме одного или двух у вершины) сифональные пучки
расположены на задней поверхности сифона зигзагообразно или
сильно сближенными к средней линии парами.
(21). Главные трахейные стволы узкие, меньше половины диаметра
сифона, округлого сечения. Стигмальная пластинка тип а «С. territans».
(18). Внутренний и средний лобные волоски короткие, не менее чем
вдвое короче наружных. Зубцы гребня в виде длинных тонких
шипов, несущих вдоль всего заднего края ряд острых тонких зуб-
чиков. Лопасти жирового тела темно пигментированы и придают
личинке поперечнополосатый вид ... С. (Neoculex) hayashii Yam.
(17). Внутренний и средний лобные волоски длинные, одной длины
с наружными волосками или длиннее их. Лопасти жирового, тела
не содержат темного пигмента и личинки не имеют поперечнополо-
сатого вида.
1 Знаком * отмечены виды, включенные только в определительную таблицу
личинок; в СССР они не встречаются.
320
III. ПОДСЕМ. CULICINAE
19 (20). Ближайшие к основанию 2—3 пучка волосков на сифоне располо-
жены между более широко расставленными дистальными зубцами
гребня, не менее чем в 3 раза длиннее диаметра сифона, последую-
щие пучки укорачиваются по направлению к вершине.................
.................................С. (Neoculex) hortensis Fic.
20 (19). Ближайший к основанию пучок волосков на сифоне расположен
дистальнее последнего из зубцов гребня; пучки на сифоне в среднем
в 2 раза длиннее диаметра сифона . . С. (Culex) mimeticus Noe.
21 (16). Главные трахейные стволы лентовидные, широкие, больше по-
ловины диаметра сифона, овального сечения. Стигмальная плас-
тинка типа «С. pipiens».
22 (25). Ближайшие к основанию 1—3 пучка волосков на сифоне лежат
между более широко расставленными дистальными зубцами гребня.
23 (24). Кроме пучков на задней поверхности сифона, расположенных
зигзагом, в концевой четверти сифона есть две пары более коротких
пучков, из которых одна пара, более удаленная от вершины, распо-
ложена на боковой стороне, а другая — на границе боковой и зад-
ней ...................................*С. (Culex) laticinctus Edw.
24 (23). Все пучки на сифоне расположены на задней его стороне . . .
............................... С. (Barraudius) modestus Fic.
25 (22). Ближайший к основанию пучок волосков на сифоне расположен
дистальнее последнего из зубцов гребня.
26 (27). Предвершинные щетинки на усике лежат у вершины суженного
концевого отдела усика. Пучков на сифоне пять пар, из них пред-
последняя от вершины расположена на боковой поверхности сифона
.......................................С. (Culex) theileri Theob.
27 (26). Предвершинные щетинки на усике расположены на расстоянии
х/4—х/3 длины от вершины суженного концевого отдела усика.
Пучков на сифоне 6—8 пар, расположенных или попарно, или зиг-
загом, из них две пары, из числа ближайших к вершине, располо-
жены на боковой поверхности сифона ... С. (Culex) orientalis Edw.
28 (15). Сифональные пучки волосков расположены парами по краям
задней поверхности сифона; часть их может быть на боковой его
поверхности.
29 (32). Сифон наиболее широк в средней трети, к вершине сужен силь-
нее, чем к основанию.
30 (31). Четыре пары пучков расположены по бокам задней поверхности
сифона в одну линию; длина их меньше ширины сифона у мест
их отхождения. Ширина сифона у вершины вдвое меньше, чем
у основания ...........................*С. (Culex) gelidus Theob.
31 (30). Из четырех пар пучков на сифоне предпоследняя пара от вершины
лежит на боковой его стороне; длина пучков не меньше ширины
сифона у мест их отхождения. Ширина сифона у вершины больше
половины ширины основания ... С. (Culex) pipiens fatigans Wied.
32 (29). Сифон наиболее широк у основания.
33 (36). Главные трахейные стволы узкие, меньше половины диаметра
сифона, округлого сечения. Стигмальная пластинка типа «С. terri-
tans».
34 (35). Сифон (индекс около 7) явственно расширен у вершины. Зубцы
гребня с 1—2 дополнительными зубчиками на заднем крае . . . .
.................................. С. (Neoculex) territans Walk.
35 (34). Сифон (индекс 7.5—11) не расширен у вершины, или это расши-
рение едва заметно. Зубцы гребня с 3—5 длинными дополнительными
зубчиками на заднем крае...............С. (Neoculex) martinii Med.
8. CULEX.
321
36 (33). Главные трахейные стволы лентовидные, широкие, больше по-
ловины диаметра сифона, овального сечения. Стигмальная пластинка
типа «С. pipiens».
37 (38). Пучки на сифоне очень короткие, менее половины его диаметра
у мест их отхождения.....................*С. (Culex) laurenti Newst.
38 (37). Пучки на сифоне более половины его диаметра у мест их отхо-
ждения или длиннее.
39 (42). Все пучки на сифоне одной длины.
40 (41). Пучков на сифоне 6—7 пар. Внутренний хвостовой волосок
из трех ветвей. Предвершинные щетинки на усике расположены
на расстоянии х/4—х/3 длины от вершины суженного концевого отдела
усика ............................ С. (Culex) tritaeniorhynchus Giles.
41 (40). Пучков на сифоне 4—5 пар. Внутренний хвостовой волосок
из двух ветвей. Предвершинные щетинки на усике расположены
непосредственно у вершины суженного концевого отдела усика . .
................................... С. (Culex) univittatus Theob.
42 (39). По меньшей мере две первые от основания пары пучков на си-
фоне длиннее остальных и больше его диаметра у мест их отхождения.
43 (44). Наиболее боковое положение на сифоне занимает ближайшая
к вершине пара сифональных пучков ... С. (Culex) torrentium Mart.
44 (43). Наиболее боковое положение на сифоне занимает вторая от вер-
шины пара сифональных пучков.
45 (48). Сифональных пучков четыре пары. Ближайшая к основанию
сифона пара лежит на заметном расстоянии ближе к вершине от наи-
более дистального зубца гребня. Боковой волосок на последнем
членике, как правило, простой.
46 (47). Сифон относительно более длинный (индекс больше 4.5). Жабры
заметно длиннее седла, обе пары одинаковой длины. Концы задних
пучков плавника на одном уровне с концами жабр......................
..................................С. (Culex) pipiens pipiens L.
47 (46). Сифон относительно более короткий (индекс не больше 4.5).
Жабры не длиннее седла, нижняя пара немного короче верхней.
Концы задних пучков плавника значительно выступают за концы
жабр...............................С. (Culex) pipiens molestus Forsk.
48 (45). Сифональных пучков пять пар. Ближайшая к основанию сифона
пара лежит на одном уровне или между первым и вторым из наи-
более дистальных зубцов гребня. Боковой волосок на последнем
членике, как правило, двуветвистый ... С. (Culex) vagans Wied.
1. Подрод LUTZIA Theobald
Виды подрода Lutzia отличаются от видов других подродов рода
Culex наличием четырех или более нижних мезэпимерных щетинок.
Личинки характеризуются небольшой округлой головой с видоизме-
ненными для хищничества ротовыми органами. Для схватывания добычи
служат боковые лопасти верхней губы, волоски которой видоизменены
в небольшое число (около 40) крепких изогнутых щетинок, сидящих
на сильно развитом широком основании. Зубы нижних челюстей крупные,
удлиненные. Лобные волоски длинные и обычно простые. Сифон короткий,
на задней поверхности почти по всей длине несет срединный ряд пучков;
гребень сильно развит, далеко заходит за середину сифона. Последний
членик брюшка очень длинный, часто длиннее сифона; кольцо седла
на спинной стороне этого членика значительно шире, чем на брюшной.
Небольшой подрод (семь видов), свойственный тропической зоне
как Нового (два вида), так и Старого (четыре вида в Ориентальной об-
21 Фауна СССР, А. В. Гуцевич и др.
322
III. ПОДСЕМ. CULICINAE
ласти, один — в Эфиопской области) Света; два вида из Ориентальной
области заходят в пределы Палеарктики.
Личинки встречаются в различных небольших естественных и искус-
ственных водоемах, чаще вблизи жилья человека, и питаются развиваю-
щимися совместно личинками других видов комаров (A. koreicus, С. va-
gans и др.). Личинки встречаются с июня до осени. Взрослые самки в нор-
ме не являются кровососами.
1. Culex (Lutzia) fuscanus Wiedemann, 1820.
Крупный комар, легко отличающийся окраской брюшка.
Голова покрыта узкими беловатыми чешуйками, с большим числом
торчащих* темных чешуек. Хоботок в темных чешуйках у основания
и у вершины; в средней части хоботка имеются, иногда преобладают,
светлые чешуйки. Щупики самки в перемешанных темных и светлых
чешуйках, щупики самца с неясными светлыми кольцами и светлой вер-
Рис. 216. Гипопигий Culex fuscanus Wied.
шиной. Среднеспинка покрыта золотисто-коричневыми узкими чешуйками,
с неясно выраженными пятнами или полосами, образованными скопле-
ниями светлых чешуек. Ноги: передняя поверхность передних и средних
бедер в темных чешуйках, с примесью многочисленных светлых чешуек;
передняя поверхность задних бедер в основной половине, за исключе-
нием верхнего края, покрыта белыми чешуйками; светлые чешуйки
в вершинной половине той же поверхности задних бедер образуют линию,
идущую до вершийы бедра. Лапки без светлых колец. Крылья по жил-
кам в одноцветных бурых чешуйках. Брюшко: вершинные тергиты (V—VIII)
покрыты желтыми чешуйками или последние образуют широкие пере-
вязи по заднему краю сегментов; тергиты II—IV на всем протяжении
темные или имеют лишь очень узкие перевязи светлых чешуек по заднему
8. CULEX
383
краю тергитов. Таким образом, брюшко самки сверху кажется отчетливо
двухцветным: передняя половина брюшка темная, задняя — светлая;:
у самца контрастность окраски брюшка менее выражена.
Гипопигий (рис. 216): коксит умеренно утолщенный; предвершинная
бородавка коксита несет три толстых шипа и несколько волосков; стиль
к вершине равномерно суживается. Фаллосома сильно склеротизована,
на ее узкой изогнутой части нет зубцов или расположено несколько не-
больших зубчиков. Пластинки X стернита с многочисленными шипиками
Рис. 217. Culex fuscanus Wied. Личинка IV стадии. Голова, сверху,
и щетинка верхней губы.
на вершине; базальный отросток X стернита хорошо развит, но слабо
склеротизован. Лопасти IX тергита слабо выпуклые, с рядом. длинных
щетинок.
Личинки IV стадии1 (рис. 217 и 218) крупных размеров,
с желто-коричневым телом и желто- или коричнево-красной головой
и более светлым сифоном.
Голова слегка вытянута в длину. Верхняя губа видоизменена для
схватывания добычи, с боковыми лопастями, состоящими из небольшого
числа (около 40) крепких, слегка S-образно изогнутых щетинок, на конце
имеющих гребень из тонких шипиков. Зубы верхних челюстей крупные
и удлиненные. Усики короткие, с коротким волоском у основания и с креп-
кими острыми щетинками на конце. Лобные волоски длинные, простые
или двуветвистые. Сифон короткий, с резко суженной вершиной за счет
изгиба его задней поверхности, на которой по всей ее длине располо-
жено 14—15 длинных пучков волосков, состоящих из 2—3 вторично-
перистых ветвей. Кроме того, на боках сифона за его серединой имеется
пара тонких брлее коротких волосков. Гребень из 8—11 зубцов, сидя-
1 Виды подрода Lutzia с достоверностью по личинкам неразличимы; возможно,
что настоящее описание относится и к личинке Culex (Lutzia) vorax.
21*
384
III. ПОДСЕМ. CULICINAE
щих по всей длине сифона, вплоть до вершинного сужения. Щетка из 35
или более чешуек, варьирующих по форме от удлиненных, с хорошо
выраженным центральным шипом, до более коротких, окаймленных
рядом шипиков. Последний членик длинный, заметно длиннее сифона.
Седло кольцом опоясывает членик и по спинному краю более чем в 2 раза
длиннее, чем по брюшному; задний край, таким образом, резко скошен
на брюшную сторону. Поверхность седла покрыта мелкими шипиками.
Хвостовые волоски
длинные и простые. Жабры короткие.
Распространение. СССР: юг
Приморского края. П-ов Корея, Япония,
Китай, страны Юго-Восточной и Южной Азии.
Рис. 219.'Гипопигий Culex vorax Edw. По Ваг-
raud.
Рис. 218. Culex fuscanus
Wied. Личинка IV стадии.
Дыхательная трубка, сбоку.
Биология. Личинки в небольших стоячих водоемах, как естест-
венных, так и искусственных; попадается в скоплениях воды близ жилья
человека; личинки питаются личинками различных Culicidae.
2. Culex (Lutzia) vorax Edwards, 1921.
Близок предыдущему. Отличается от него следующими признаками.
Бедра и голени спереди преимущественно в темных чешуйках; среди
которых светлые чешуйки образуют пятна, довольно правильно чере-
дующиеся с темными участками. Передняя поверхность задних бедер
в основной половине с многочисленными вкрапленными темными чешуй-
ками; вершинная половина той же поверхности задних бедер без про-
дольной линии, образованной белыми чешуйками. Все тергиты брюшка
(II—VIII) у вершины с неширокими перевязями светлых чешуек. Вер-
шина щупиков самца темная. Зубцы фаллосомы расположены на явст-
венно развитом треугольном выступе тела фаллосомы с боковой ее сто-
роны (рис. 219).
Личинка см.: С. (L.) fuscanus.
Распространение. СССР: юг Приморского края. Япония
(Хоккайдо, Хонсю, Сикоку, Кюсю), п-ов Корея, Китай, Юго-Восточная
и Южная Азия.
8. CULEX
325
Биология. Личинки обитают (в Японии) во всевозможных ско-
плениях воды, в частности в деревянных или металлических бочках
и т. п.; в естественных водоемах попадаются реже; переносят значитель-
ную степень органического загрязнения воды; чаще сопутствуют ли-
чинкам Culex pipiens и С. tritaeniorhynchus, которых уничтожают в за-
метных количествах. Нападает на человека крайне редко.
2. Подрод BARRAUDIUS Edwards
I членик задних лапок явственно короче голени (длина его примерно
4/5 длины голени); щупики самца длиннее хоботка, без длинных волосков;
голова по краю глаз в узких чешуйках. Коксит снаружи покрыт чешуй-
ками; предвершинная бородавка, расположенная едва за его серединой,
без пластинчатых придатков.
Личинки см. стр. 315.
Подрод включает в себя лишь два вида.
3. Culex (Barraudius) modestus Ficalbi, 1889.
Комар мелких размеров. Голова в желтовато-бурых чешуйках.
Хоботок и щупики бурые. Щупики самца кажутся очень тонкими, ните-
видными, так как они лишены длинных волосков. Среднеспинка в бурых
чешуйках, щиток покрыт беловатыми чешуйками. Бедра и голени спереди
в бурых, сзади в белых чешуйках. I членик задних лапок явственно
короче задней голени. Крылья по жилкам в бурых чешуйках; брюшко
в одноцветно бурых чешуйках, с боков с продольной полосой желтовато-
белых чешуек.
Гипопигий (рис. 220): коксит снаружи в более или менее густых че-
шуйках; предвершинная бородавка, расположенная едва за серединой
коксита, без пластинчатого придатка; бородавка несет 2—3 толстых
тупых шипа и несколько волосков; стиль в вершинной половине изо-
гнутый, относительно тонкий и длинный, его длина превышает поло-
вину длины коксита, реже равна ей; фаллосома с каждой стороны в виде
простой, почти прямой пластинки, не выступающей за усаженную ши-
пиками вершину X стернита.
Личинки IV стадии (рис. 221) слабо пигментированы, почти
прозрачные, со светлыми головой и сифоном. Голова в ширину почти
в 1.5 раза превосходит длину. Ротовые органы фильтрационного типа.
Усики умеренно шиповатые, светлые, с более темным основанием; к вер-
шине сужены начиная от точки прикрепления пучка (около 3/5 длины
усика от основания), состоящего из 15—25 вторичноперистых ветвей,
достигающих х/2 длины усика. Лобные волоски вторичноперистые, на-
ружные из 7—8, средние из 3—4 (реже 2), внутренние из 3—5 (реже 2)
ветвей.
Щетка VIII членика брюшка из 50 и более тесно сидящих мелких
продолговатых чешуек с рядом шипиков на вершине и отчасти на боках.
Сифон прямой, постепенно сужающийся к вершине; длина в 3.8—5 раз
превосходит ширину основания. Гребень занимает около 2/5 длины сифона,
состоит из 12—13 сравнительно сильно расставленных тонких зубцов
с 3—5 дополнительными шипиками по заднему краю. В среднем 10 пуч-
ков расположены посредине задней поверхности сифона зигзагообраз-
ным рядом начиная от уровня 2—3 дистальных зубцов гребня и почти
до вершины. 2—4 ближайших к ней пучка могут быть короче остальных
и слабее ветвятся. Остальные в длину равны или больше ширины сифона
Рис. 221. Culex modestus Fic. Личинка IV стадии. Задний конец,
сбоку.
8. CULEX
327
у мест их отхождения и состоят в среднем из 5—7 ветвей. Главные тра-
хейные стволы широкие, лентовидные. Стигмальная пластинка как
у С. pipiens. Последний членик с кольцевидно опоясывающим его сед-
лом, в длину немного превосходит ширину. Боковой волосок из 2—3
коротких ветвей. Наружные хвостовые волоски длинные и простые, внут-
ренние короче наружных, из 3—4 ветвей, из которых нижняя — наиболее
длинная. Плавник из 11—13 хорошо развитых пучков из 5—7 ветвей
в каждом. Жабры заметно короче седла, обе пары одной длины.
Распространение. СССР: Европейская часть на север до
Московской и Свердловской областей, Кавказ и Закавказье, Средняя
Азия, Казахстан, Южная Сибирь на восток до Приморского края. Юж-
ная Европа, Передняя Азия, Северная Индия.
Биология. Личинки встречаются с конца весны до поздней осени
в различных размеров пресноводных или слегка солоноватых водоемах
и заболоченностях, богатых зеленой растительностью и хорошо осве-
щенных солнцем, совместно с личинками An. maculipennis, An. т. sacha-
rovi, An. hyrcanus и реже с An. pulcherrimus. Комары встречаются осо-
бенно часто на лугах и тому подобных открытых местах. .
Установлена естественная зараженность С. modestus возбудителем
туляремии.
4. Culex (Barraudius) pusillus Macquart, 1850.
Очень мелкий комар. Вместе с С. modestus отличается от прочих пале-
арктических видов рода более коротким I члеником задних лапок, длина
которого равна 4/5 длины голени. От предыдущего вида отличается более
темной окраской тела и наличием белых латеральных пятен при осно-
вании каждого тергита брюшка.
Гипопигий (рис. 222): стиль прямой или слегка равномерно изогну-
тый, относительно короткий (отличие от С. modestus), его длина меньше
или равна половине длины коксита; сильно выпуклая бородавка коксита
несет два неодинаковых по величине толстых шипа с тонкой изогнутой
вершиной, а также много щетинок и волосков - фаллосома с каждой стороны
в виде нерасчлененной узкой длинной пластинки, изогнутой на конце; вер-
шина фаллосомы, как правило, выступает за густо усаженную ши-
пиками вершину X стернита и почти достигает уровня предвершинной бо-
родавки.
Личинки IV стадии (рис. 223) имеют почти прозрачное
тело желтоватого или светло-жеПто-зеленого цвета, с бледно пигменти-
рованными головой и сифоном. Голова поперечно овальной формы, ши-
рина в 1.4 раза превосходит длину. Усики слабо шиповатые, длинные:
до.0.8—0.9 длины головы, светлые, за исключением основания и суженной
вершинной части; пучок из 15—25 вторичноперистых ветвей, в длину пре-
восходящих половину усика; предвершинные щетинки сближены с вершин-
ными. Лобные волоски длинные, наружные из 7, средние и внутренние
из 2—3 ветвей. Щетка VIII членика брюшка из 50 и более мелких тесно
сидящих чешуек, окаймленных рядом шипов. Сифон короткий, длина
в 2.6—3.2 раза превосходит ширину основания. Гребень занимает около
половины длины сифона, состоит из 11—13 зубцов. В среднем 10 пучков
расположены зигзагом или по прямой линии посредине задней поверх-
ности сифона; около трех ближайших к основанию пучков лежат между
дистальными зубцами гребня. Пучки из 8—10 ветвей, длиной не меньше
ширины сифона. Главные стволы трахей широкие, лентовидные. Стиг-
мальная пластинка как у С. pipiens. Последний членик лишь немного
328
III. ПОДСЕМ. CULICINAE
короче сифона. Боковой волосок короткий, из 1—2 ветвей. Наружный
хвостовой волосок более чем в 1.5 раза длиннее сифона, внутренний —
короче, имеет одну, реже две более короткие ветви у своего основания.
Плавник из 12 хорошо развитых пучков. Жабры около х/4 длины седла,
верхняя пара лишь немного длиннее нижней.
Рис. 222. Гипопигий Culex pusillus Macq.
Распространение. Средиземноморская подобласть, преи-
мущественно ее восточная часть. СССР: Средняя Азия (Туркмения, Узбе-
кистан, Таджикистан). Северная Африка (Алжир, ОАР), Западная
Азия (Анатолия, Иран, Ирак).
Биология. Личинки встречаются в стоячих водоемах и заболо-
ченностях с повышенным содержанием солей (до 3%), часто совместно
с личинками A. caspius, An. pulcherrimus и др.; места выплода лежат
преимущественно близ морских побережий, соленых озер или оазисов.
Не относится к числу массовых видов.
3. Подрод NEOCULEX Dyar
Коксит без чешуек, с предвершинной бородавкой, несущей более
или менее развитые шиповидные придатки, как правило, без листовидной
пластинки; X стернит у большинства видов без базального отростка,
на вершине с пучком шипиков или гребнем тупых пластинчатых отрост-
ков; фаллосома простая, образованная одной парой пластинок.
Рис. 223. Culex pusillus Macq. Личинка IV стадии.
По Edwards.
1 — голова, сверху, г — задний конец, сбоку.
330
Ш. ПОЦСЕМ. CULICINAE
Личинка отличается длинным и тонким сифоном, иногда слегка рас-
ширяющимся к концу, тонкими, с округлым сечением, главными тра-
хейными стволами и стигмальной пластинкой типа С. (N.) territans.
Рис. 224. Гипопигий Culex territans Walk. По Carpenter
a. La Casse.
Личинки C. (Culex) mimeticus и C. (Culex) jacksoni очень сходны с ли-
чинками Neoculex.
Подрод включает около 60 видов, из которых 30 свойственны тропи-
ческой Африке, 9 — Ориентальной области, 10 — Австралийской,
5 — Центральной Америке, 8 — Палеарктике (из них один свойствен
также Неарктике); из числа палеарктических видов один (С. territans)
имеет голарктическое распространение, пять свойственны средиземно-
8. CULEX
331
морской подобласти в широком смысле (из них два доходят до Средней
Азии) и два характерны для Дальнего Востока.
5. Culex (Neoculex) territans Walker, 1856 (apicalis auct., nec Ad.).
Описан по материалу из США. Там же позднее был описан близкий
вид С. apicalis Adams, 1903. Европейский вид подрода Neoculex перво-
начально был определен как С. apicalis и под этим именем фигуриро-
вал во многих работах (Edwards, 1921, 1926; Martini, 1931; Мончадский,
1936, 1951; Штакельберг, 1937, и
широко распространенный в Па-
леарктике комар идентичен
С. territans, а не С. apicalis.
Последний распространен на
юго-западе США и в Мексике,
а в Палеарктике вовсе не встре-
чается (Mattingly, 1953, 1955;
Kramar, 1957). Следовательно,
во всех отечественных работах,
где упоминается С. apicalis,
соответствующие указания дол-
жны быть отнесены к С. terfl-
tans.
Описанный в Японии С. ги-
bensis Sasa et Takahashi, 1948,
вероятно, представляет собой
дальневосточный подвид С. ter-
ritans. По имаго и личинкам
обе формы неразличимы (La
Casse a. Yamaguti, 1955); воз-
можно, С. rubensis правильнее
рассматривать как синоним
С. territans.
Culex territans отличается от
близких форм, в частности от
С. hortensis (см. ниже), общей
более темной окраской, опуше-
нием щупиков самца, а также
. В дальнейшем выяснилось, что
Рис. 225. Culex territans Walk. Личинка
IV стадии.
1 — дыхательная трубка, сбоку, 2 — стигмальная
пластинка.
строением гипопигия.
Голова в белых и желтоватых чешуйках. Хоботок и щупики темно-
бурые в такого же цвета чешуйках, без примеси светлых. Щупики самца
с длинными волосками на последних двух члениках. Среднеспинка в бу-
рых чешуйках; передний и боковой края среднеспинки, а также щиток
в беловато-серых чешуйках. Бочки груди серые, с пятнами белых чешуек.
Ноги темно-бурые; бедра и голени спереди, лапки на всем своем протя-
жении покрыты темно-бурыми чешуйками; сзади бедра и голени в бе-
ловатых чешуйках; вершина задней голени без белого пятна. Жилки
крыла покрыты темно-бурыми чешуйками. Брюшко сверху в черно-бурых
чешуйках, по заднему краю тергитов с узкими белыми поперечными
перевязями более или менее равномерной ширины. Снизу брюшко, как
правило, зеленоватое, в бурых и белых чешуйках, сгруппированных
обыкновенно в широкие поперечные полосы;
Гипопигий (рис. 224): предвершинная бородавка коксита с двумя
толстыми прозрачными шипами, загнутыми на конце, и несколькими
более тонкими щетинками — прямыми или слегка загнутыми или за-
332
III. ПОДСЕМ. CULICINAE
зубренными. Фаллосома состоит из двух простых пластинок, соединен-
ных поперечными перемычками при основании и перед вершиной
(у С. apicalis поперечной пластинки в вершинной части фаллосомы нет,
в этом его главное отличие от С. territans). Степень развития вершинной
поперечной пластинки фаллосомы сильно варьирует, судя по материалу
из различных областей СССР; эта пластинка может быть широкой или
узкой, иногда весьма узкой.
Личинки IV стадии (рис. 225), вначале прозрачные, перед
окуклением утрачивают прозрачность и становятся желто- или сине-зе-
леными. Голова и сифон тоже приобретают коричневую окраску.
Голова очень широкая: ширина в 1.6 раза превосходит длину. Ротовые
органы фильтрационного типа. Усики почти одной длины с головой
с пучком в виде веера из 25—32 вторичноперистых ветвей, достигающих
3/4 длины усика; предвершинные щетинки у самого конца усика. Лоб-
ные волоски длинные, вторичноперистые наружные из 6—8, средние-
наиболее длинные из одной, внутренние из двух ветвей.
Щетка по бокам VIII членика брюшка из 50 или более продолговатых
мелких чешуек, край которых усажен рядом из мелких шипиков. Сифон
тонкий, длина в 7 раз превосходит ширину основания; слабо, но заметно
расширенный у самой вершины. Гребень из 13—16 зубцов занимает
около х/4—х/3 длины сифона от основания. 4—6 пар более длинных, чем
диаметр сифона, пучков из 2—4 ветвей помещаются вдоль границ бо-
ковых и задней сторон сифона. Базальные пары более сближены, чем
дистальные. Главные трахейные стволы тонкие, округлого сечения.
Стигмальная пластинка характерного строения (рис. 225) с сильно раз-
витыми задними клапанами, тонким длинным задним отростком рычага
и маленькими, глубоко погруженными округлыми стигмами. Последний,
членик брюшка длинный, с опоясывающим кольцевидно седлом. Хвосто-
вые волоски: наружный длиннее сифона простой, внутренний из четырех
более коротких ветвей. Плавник из 13—14 пучков. Жабры почти одной
длины с седлом, нижняя пара короче верхней.
Замечания по систематике. Вид распадается на два подвида —
С. t. territans и С. t. ju/iaicus', последний иногда рассматривается как самостоятельный
вид (Mattingly, 1953); но для признания его самостоятельности нужны дополнительные
обоснования.
Распространение. СССР: европейская часть от Ленинград-
ской и Свердловской области на севере до Украины, Крыма, Предкав-
казья и Нижнего Поволжья на юге, Кавказ и Закавказье, Средняя Азия,
Казахстан, Сибирь, Дальний Восток. Большая часть Западной Европы,
Япония, Северная Африка, Северная Америка.
Биология. В умеренных широтах личинки встречаются с июня
по сентябрь преимущественно в небольших, не пересыхающих в течение-
лета водоемах, хорошо освещенных солнцем и богатых зеленой расти-
тельностью, совместно с личинками An. maculipennis, A. cinereus,
С. alaskaensis и др. На юге личинки приурочены к хорошо затененным,
иногда родниковым водоемам и заболоченностям. Комары питаются
кровью рептилий и амфибий; на людей и сельскохозяйственных живот-
ных, как правило, не нападают.
Culex territans territans Walker.
С признаками вида.
Распространение и биология: см. оцисание вида.
8. CULEX
333
Culex territans judaicus Edwards, 1926.
Отличается от основной формы более короткими щупиками самца,
длина двух вершинных члеников которых примерно вдвое менее длины
третьего длинного их членика (у основной формы та же величина равна
примерно 2/3); длинный членик щупиков самца без длинных волосков;
конечные членики щупиков в менее густых волосках; фаллосома на вер-
шине с едва заметными бугорками. Мелкий комар, длина крыла около
3 мм (у С. t. territans около 4 мм).
Распространение. Передняя Азия (Палестина).
6. Culex (Neoculex) hortensis Ficalbi, 1889.
Близок к C. (N) territans-, отличается от него светлым пятном на вер-
шине задней голени, голыми или почти голыми вершинными члениками
щупиков самца, а также строением гипопигия. Щупики самки сверху
с вкраплением светлых чешуек. Поперечные светлые перевязи на тер-
Рис. 226. Гипопигий Culex hortensis Fic.
Отдельно показана дистальная часть коксита, сбоку.
гитах брюшка, как правило, более широкие, нежели у предыдущего
вида, и имеют в некоторых случаях по середине переднего края более
или менее значительный выступ, вдающийся в расположение темных
чешуек. Общая окраска тела и покрывающих его чешуек более светлая.
Гипопигий (рис. 226): коксит на вершине с широким склеротизован-
ным пальцевидным выростом; шиповидные придатки предвершинной
бородавки коксита относительно короткие; пластинчатый придаток бо-
родавки отсутствует.
Личинки IV стадии (рис. 227) чаще непрозрачные, желто-
вато-коричневого цвета. Поверхность их кутикулы покрыта мелкими
хетоидными волосками, более густо сидящими на средне- и заднегруди.
Голова в ширину более чем в 1.5 раза превосходит длину. Усики од-
ной длины с головой, густо покрыты шипиками, часто темно пигменти-
рованы. Пучок из 25—35 вторичноперистых ветвей, достигающих 3/4
длины усика. Предвершинные щетинки на небольшом расстоянии от
вершины. Лобные волоски: наружные из 7—12 ветвей; средние впе-
реди внутренних, расставлены немного шире, обе пары из 2—4 более
длинных и толстых ветвей.
Щетка по бокам VIII членика брюшка из 35—45 удлиненных чешуек
с одним рядом шипиков, окаймляющих слегка расширенный концевой
отдел. Сифон длинный, длина в среднем в 8 раз превышает диаметр у за-
334
III. ПОДСЕМ. CULICINAE
метно расширенного основания. Гребень занимает около l/s длины си-
фона, из 11—14 зубцов. 9—12 пучков расположены на задней поверхности
сифона в один зигзагообразный ряд; из них 2—4 ближайших к основанию
Рис. 227. Culex hortensis Fic. Личинка IV стадии.
Дыхательная трубка, сбоку.
лежат между дистальными зубцами гребня. Первые 6—8 пучков очень
длинные, последующие постепенно укорачиваются. Кроме них, на
боковых сторонах сифона ближе к вершине лежат 1—2 пары более ко-
ротких боковых пучков. Главные трахейные стволы тонкие, округлой
8. CULEX
335
формы; стигмальная пластинка как у С. territans. Последний членик
удлинен, с кольцевидно опоясывающим его седлом. Боковой волосок
из двух ветвей. Наружные хвостовые волоски длиннее сифона, внутрен-
ние — из 3—4 более коротких ветвей. Плавник из 12—14 пучков. Жабры
равны или превосходят длину седла.
Распространение. В основном — Средиземноморская под-
область. В СССР: Крым, Кавказ и Закавказье, Средняя Азия (Туркме-
ния, Узбекистан, Таджикистан), Южный Казахстан. Западная Европа
на север до Берлина, западная Азия (Малая Азия, Палестина, Сирия,
Иран), Северная Африка.
Биология. Личинки обитают в водоемах с достаточно чистой
водой, богатых водорослями и вообще зеленой растительностью, часто
совместно с личинками Anopheles maculipennis, Culiseta longiareolata,
C. annulata, Culex laticinctus Edw. и др. Комары проводят большую
часть дня в пещерах и тому подобных укрытых местах. На человека
нападают неохотно.
7. Culex (Neoculex) martinii Medschid, 1930.
Очень мелкий комар. Общая окраска тела желтовато-бурая. Брюшко
сверху в одноцветных чешуйках. Хоботок и лапки без светлых колечек.
Голова светлая; скулы в беловатых чешуйках. Хоботок бурый, в вершин-
ной трети слегка утолщенный. Щуг
щупики самца длиннее хоботка при-
мерно на длину последнего их чле-
ника, на вершине не утолщенные.
Среднеспинка желтоватая, с очень
мелкими золотисто-бурыми чешуй-
ками и довольно длинными бурыми
волосками. Бедра и голени спереди
в темных чешуйках, сзади светлые.
Крылья по жилкам в бурых чешуй-
ках. Брюшко сверху в одноцветно
бурых чешуйках, по бокам тергитов
с треугольными беловатыми пят-
нами, расширяющимися к заднему
краю сегментов.
Гипопигий (рис. 228): коксит ко-
роткий и толстый; бородавка распо-
ложена за серединой коксита, близко
к вершине; бородавка несет два
умеренно длинных, но очень толстых шипа. Стиль короткий и широкий,
у середины явственно расширенный. X стернит с широкими лопастевид-
ными боковыми выростами, усаженными мелкими волосками; на вершине X
стернит несет ряд узких тупых зубцов, образующих как бы гребень или
бахрому.
Личинки IV стадии (рис. 229): голова в ширину в 1.5 раза
превосходит длину. Усики достигают не менее 3/4 длины головы, с пучком
из 22—26 вторичноперистых ветвей, в длину достигающих 0.7—0.8 длины
усика. Предвершинные щетинки одной длины с пучком, расположены
на небольшом расстоянии от вершины. Наружные лобные волоски в сред-
нем из пяти ветвей, средние простые, реже 2-ветвистые, очень длинные:
достигают вершины усика или даже заходят за нее; лежат впереди значи-
тельно более коротких 2-ветвистых внутренних и больше расставлены,
чем последние. Щетка из 35—40 мелких сильно удлиненных чешуек,
336
III. ПОДСЕМ. CULICINAE.
несущих по краям и на конце ряд шипиков. Сифон очень тонкий и длин-
ный, длина в 7.5—11 раз превосходит ширину заметно расширенного
основания; иногда немного расширен у вершины. Гребень из 12—16 зуб-
цов, занимающих около базальной х/6 сифона и несущих 3—5 дополни-
тельных зубчиков. Сифональные пучки в числе 10—12 расположены сна-
чала зигзагом по бокам задней стороны сифона, а далее к вершине непра-
вильными парами, часть которых находится на боковых сторонах. Пучки
Рис. 229. Culex таг-
tinii Med. Личинка
IV стадии. Задний
конец, сбоку.
короткие, из 2—5 ветвей, одной длины с диаметром сифона или только
немного длиннее; ближайшие к вершине более короткие. Трахеи тонкие,
с округлым сечением; стигмальная пластинка типа С. territans.TLocnej^nm
членик удлинен, с кольцеобразно опоясывающим его седлом. Боковой
волосок 2-ветвистый. Наружные хвостовые волоскй простые, в среднем
одной длины с сифоном; внутренние — 4-ветвистые. Плавник из 11—12
пучков. Жабры около половины длины седла.
Р аспространение. Средиземноморская подобласть. СССР:
Средняя Азия: Узбекистан (Лисова, 1935 — «С. sp.»), Таджикистан,
Киргизия. Балканский полуостров (Югославия), Малая Азия (Анатолия).
Биология неизвестна.
8. Culex (Neoculex) hayashii Yamada, 1917.
Мелкий комар. Отличается от прочих видов Culex относительно ко-
роткими щупиками самца, а также строением гипопигия. Хоботок и
щупики темные. Щупики самца составляют 2/3—i/i длины хоботка.
Среднеспинка коричневая, в узких красно-бурых чешуйках бронзового
оттенка. Голени и лапки темные'без светлых колец. Крылья в одноцветных
8. CULEX
337
темных чешуйках. Брюшко сверху в темно-бурых чешуйках без светлых
перевязей.
Гипопигий (рис. 230): предвершинная бородавка коксита с 4—5 тесно
сближенными узкими пластинками и 2—3 более длинными палочковид-
ными придатками с крючковидной вершиной. Правая и левая пластинки
фаллосомы ближе к основанию соединены поперечной перемычкой; каж-
РиС. 230. Гипопигий Culex hayashii Yam. По La Casse
a. Yamaguti.
дая пластинка расширена, с разбросанными на ее поверхности зубцами
различной величины и короткими склер.отизованными выступами, направ-
ленными вперед и внутрь. X стернит на вершине с многочисленными ши-
пиками, расположенными в несколько рядов. Лопасти IX тергита слабо
развиты, каждая несет 4—6 волосков.
Личинки IV стадии (рис. 231—233) прозрачные, но с просве-
чивающим коричневым жировым телом, придающим им характерную
поперечную полосатость, видимую простым глазом.
Голова в ширину в 1.6—1.7 раза превосходит длину. Усики, покрыты
шипиками, почти одной длины с головой с темным основанием и сужен-
22 Фауна СССР, А, В. Гуцевич и др.
338
III. ПОДСЕМ. CULICINAE
ной вершинной частью. Пучок состоит из 25—30 ветвей, предвершинные
щетинки у конца усика. Лобные волоски: наружные длинные, из 5—
Рис. 231. Culex hayashli Yam. Личинка IV стадии. Распределение пигмента в жировом
теле.
2 — грудь и первые два членика брюшка (й — со спинной и v — с брюшной стороны),— I—
VIII членики брюшка, сбоку.
Рис. 232. Culex hayashli Yam. Личинка IV стадии. Голова,
сверху.
8 ветвей, средние и внутренние короткие, из 2—4 ветвей, почти одной
длины с задними волосками наличника. Щетка из 34—46 узких удли-
ненных чешуек с рядом шипиков. Сифон сравнительно короткий: длина
8. CULEX
339
в 5.5—6 раз превосходит ширину основания; слабо расширен у вершины.
Гребень из 10—13 зубцов, расположенных на одинаковом расстоянии
друг от друга, в виде тонких шипов, несущих по всей длине ряд тонких
Рис. 233. Culex hayashli Yam. Личинка IV стадии.
1 — задний конец, сбоку, 2 — стигмальная пластинка.
острых зубчиков. 11—13 сифональных пучков расположены зигзаго-
образным рядом на задней поверхности сифона; парное расположение
встречается в виде исключения. Пучки из 3—6 гладких ветвей, более
длинных, чем диаметр сифона у мест их отхождения. Главные трахейные
22*
340
Ill. ПОДСЕМ. CULICINAE
стволы тонкие, округлого сечения. Стигмальная пластинка (рис. 233)
по типу строения сходна с таковой С. (TV.) territans, но отличается от нее
рядом деталей (наличие на конце заднего отростка направленного вверх
хетоидного волоска, строение органов на верхней поверхности боковых
клапанов и др.). Седло кольцом опоясывает последний членик. Короткий
боковой волосок из 3—4 ветвей. Наружный хвостовой волосок одной длины
с сифоном, внутренний более короткий, из 2—5 последовательно укорачи-
вающихся к спинной стороне волосков. Плавник из 12—13 пучков. Жабры
ланцетовидной формы, в среднем одной длины с седлом; нижняя пара
длиннее верхней. '
Распространение. СССР: юг Приморского края. П-ов Корея,
Китай, Япония (Хоккайдо, Хонсю, Кюсю).
Биология. Личинки встречаются обычно в небольшой числен-
ности с конца весны до осени в различных освещенных солнцем мелких
водоемах, богатых гниющей опавшей листвой и поросших травянистой
растительностью, а также в различных искусственных водовместилищах
и цистернах, совместно с An. hyrcanus, A. communis (весной), С. modes-
tus, С. bitaeniorhynchus, С. tritaeniorhynchus, С. infantulus Edw. и С. vish-
nui Theob. Приурочены к разреженным лесам предгорных районов.
Самки не являются серьезными кровососами. Редкий вид.
4. Подрод CULEX Linnaeus
Коксит без чешуек; стиль, как правило, простой, без предвершинного
гребня шипиков; X стернит у вершины с пучком густых волосков или
шипиков, при основании у большинства видов с более или менее раз-
витым отростком; фаллосома сложного строения, часто с зубцами или
выростами. IX тергит в виде узкой поперечной полоски. Лапки, как
правило, темные, реже со светлыми колечками; последние в некоторых
случаях опоясывают вершину предыдущего и основание последующего
членика.
Личинки видов подрода разнообразны по своему строению и могут
быть охарактеризованы только немногими, притом отрицательными чер-
тами: отсутствием у ротовых частей каких-либо приспособлений к хищ-
ному образу жизни и расположением чешуек щетки, которые никогда
не образуют одного правильного ряда. Разделению видов по признакам
взрослой фазы на группы не соответствует разделение по личинкам,
которые изучены еще недостаточно. Личинкй некоторых видов этого
подрода имеют сходство с личинками видов подрода Barraudius, а ли-
чинки С. mimeticus — с личинками видов подрода Neoculex.
В подроде насчитывается более 180 видов; по своим морфологическим
особенностям (хетотаксия бочков груди, строение гипопигия) виды под-
рода разделяются на две большие группы; из них первая группа, вклю-
чающая из палеарктических видов все виды со светлыми колечками на
лапках, имеет определенно ориентальный центр процветания; лишь неко-
торые виды этой группы свойственны Африке; в ^Палеарктике эта группа
представлена преимущественно на крайнем ее юго-востоке (в СССР,
в Приморском крае), в меньшей степени — на юге (С. mimeticus). Вторая
группа видов этого подрода, палеарктические представители которой
внешне характеризуются одноцветно темными, без светлых колечек,
лапками, имеет явное тяготение к Эфиопской области: 1 в пределах по-
1 В эту группу входит также значительное количество неотропических видов,
однако по своим морфологическим особенностям они являются достаточно резко обо-
собленными, чтобы их можно было рассматривать как одно целое с эфиопскими пред-
ставителями группы.
8. CULEX
341
следней эта группа достигает наибольшего видового разнообразия, при
значительной близости многих видов между собой; интересно отметить,
что к этой группе принадлежит также обычный в большей части СССР
С. pipiens.
9. Culex (Culex) bitaeniorhynchus Giles, 1901.
Близок C. sinensis, от которого отличается наличием на крыльях
светлых чешуек, вкрапленных среди темных чешуек, а также строением
гипопигия.
Рис. 234. Гипопигий Culex bitaeniorhynchus Giles.
А — внутренние склериты гипопигия, Б — коксит и стиль,
В — базальная бородавка коксита, Г — второй отдел фалпо-
сомы.
Голова в средней части темени с узкими светло-золотистыми приле-
гающими и торчащими вильчатыми чешуйками; боковые края глаз в ши-
роких светлых прилегающих чешуйках. Щупики самки короткие, рав-
ные х/6 длины хоботка, в темных чешуйках; вершина щупиков самки
в светлых чешуйках. Щупики самца явственно длиннее хоботка (почти
на длину двух вершинных члеников); вершинные членики с кольцом
светлых чешуек при основании; длинный членик щупиков с двумя свет-
342
III. ПОДСЕМ. CULICINAE
лыми кольцами; вершинная часть (х/2— а/5) концевого членика щупиков
светлая. Хоботок за серединой с белым кольцом. Среднеспинка темно-
бурая, в передних 2/3 покрыта светло-желтоватыми или золотистыми
узкими чешуйками; близ середины покрытой светлыми чешуйками части
среднеспинки расположены два хорошо выраженных округлых пятна
темных чешуек; задняя треть среднеспинки покрыта темными чешуй-
ками. Бочки груди с белыми чешуйками на стерноплеврах и мезэпимерах.
Рис. 235. Culex bitaeniorhynchus Giles. Личинка IV стадии.
1 — голова, сверху, 2 — нижнегубная пластинка.
Ноги бурые; бедра и голени спереди в бурых чешуйках с более или менее
значительной примесью беспорядочно разбросанных белых чешуек,
сзади в большей своей части в белых чешуйках; лапки темно-бурые,
с белыми колечками при основании 1—4-го члеников. Крылья: жилки
покрыты бурыми и желтоватыми перемешанными чешуйками. Брюшко
бурое, с более или менее широкими перевязями белых чешуек у вершины
тергитов; иногда при основании тергитов. имеются также следы перевя
зей, но значительно более узких.
Гипопигий (рис. 234) своеобразной формы; пластинка на предвершин-
ной бородавке коксита узкая, ножевидная; фаллосома с каждой стороны
состоит из двух заостренных на концах, серповидно изогнутых пласти-
нок; по бокам фаллосома образует выступ с четырьмя или пятью пальце-
видными зубцами; базальный отросток X стернита слабо развит.
Личинки IV стадии (рис. 235, 236), вначале прозрачные,
к концу становятся зеленого или коричнево-зеленого цвета. Голова не-
большая и сравнительно неширокая. Основания усиков выдвинуты вперед.
8. CULEX
343
Усики палочковидной формы, у середины, за местом прикрепления пучка
из 15—25 ветвей, сужаются; предвершинные щетинки у вершины, на-
правлены внутрь. Ротовые органы видоизменены для питания нитчатыми
водорослями. Волоски боковых лопастей верхней губы состоят из мень-
шего, чем обычно, числа упругих и крючковидно изогнутых волосков;
Рис. 236. Culex bitaentorhynchus Giles. Личинка IV стадии. Задний
конец, сбоку.
обе пары челюстей тоже видоизменены, а нижнегубная пластинка в виде
равнобедренного треугольника, с очень мелкой, ровной зубчатостью по
краям. Лобные волоски вторичноперистые, наружные из 4—7, наиболее
длинные средние из двух и внутренние из трех ветвей. Щетка из 4—7 круп-
ных шиповидных чешуек. Сифон длинный и тонкий, длина в 6—8 раз
превосходит ширину основания. Гребень из 7—9 тонких зубцов у самого
основания трубки. 4—5 пар пучков на задней поверхности сифона распо-
ложены в один зигзагообразный ряд, каждый из 2—5 коротких, меньше
344
III. ПОДСЕМ. CULICINAE
диаметра сифона, ветвей. Седло кольцом опоясывает последний членик
брюшка. Боковой волосок короткий, 1—3-ветвистый. Наружные хвосто-
вые волоски простые, короче сифона, внутренние — из 3—4 ветвей.
Плавник из 12 пучков. Жабры немного длиннее седла, ланцетовидной
формы.
Распространение. СССР: юг Приморского края на север
примерно, до реки Имана, где широко распространен в низменных и
Рис. 237. Гипопигий Culex sinensis Theob. По La Casse
a. Yamaguti..
прибрежных районах. Япония, на север до Хонсю, п-ов Корея, Китай,
Юго-Восточная и Южная Азия, Африка, Северная Австралия.
Биология. Личинки обитают в открытых, хорошо прогреваемых
солнцем стоячих или слабо проточных водоемах с чистой водой, богатой
зеленой растительностью и скоплениями нитчатых водорослей, где они
преимущественно и концентрируются. Совместно с личинками этого вида
попадаются личинки An. hyrcanus, реже — С. tritaeniorhynckus, С. vishnui,
С. hayashii и др. Активный кровосос; нападает по преимуществу ночью
в районах рисосеяния.
8. CULEX
345
Рис. 238. Culex sinensis Theob.
Нижнегубная пластинка личинки.
По Barraud.
. Эпидемиологическое значение. По японским дан-
ным (Yamaguti a. La Casse, 1955), предположительно является перенос-
чиком японского энцефалита; доказана восприимчивость особей этого
вида к заражению вирусом японского энцефалита (Петрищева и Шуб-
ладзе, 1940).
10. Culex (Culex) sinensis Theobald, 1903.
Весьма близок по цветовым признакам предыдущему виду, от которого
отличается одноцветно бурыми чешуйками крыльев (без вкраплений
светлых чешуек), а также строением гипопигия.
Белые чешуйки на ногах обыкновенно сгруппированы в небольшие,
несколько беспорядочно расположенные пятна (правильного ряда пятен
никогда не образуется). Лапки с узкими белыми колечками при основа-
нии члеников; брюшко с перевязями
светлых чешуек при основании тер-
гитов .
Гипопигий (рис. 237) близок к тако-
вому предыдущего вида; пластинка
предвершинной бородавки коксита уз-
кая, ланцетовидная; фаллосома состоит
из пары простых (не расчлененных и
не зубчатых) слабо изогнутых пласти-
нок; базальные отростки X стернита
крупные, сильно склеротизованные.
Личинка близка к личинкам
С. bitaeniorhynchus. Отличается от ли-
чинок этого вида следующими при-
знаками: предвершинными щетинками,
отходящими на значительном рассто-
янии от вершины усика; нижнегуб-
ной пластинкой (рис. 238) с значительно более крупными зуб-
цами, увеличивающимися к середине; щеткой из 4—6 крупных чешуек
с острыми шипами; очень слабо развитым гребнем из 2—4 очень мелких
зубцов у самого основания сифона; крупным, слегка изогнутым шипо-
видным волоском на передней поверхности конца сифона; жабрами,
в 2.5 раза превышающими длину седла, и внутренними хвостовыми во-
лосками из двух ветвей.
Распространение. Большая часть ареала лежит в пределах
Ориентальной области. СССР: юг Приморского края. Япония (Хонсю,
Кюсю), п-ов Корея, Китай, Юго-Восточная и Южная Азия.
Биология. Личинки встречаются в чистой воде на рисовых полях
и в водоемах на заболоченных лугах.
11. Culex (Culex) whitmorei Giles, 1904.
Вид характеризуется резко двуцветной среднеспинкой, большая
часть которой покрыта белыми чешуйками, а задняя треть — темными
чешуйками; хоботок и лапки с белыми колечками; крылья с широкими
темными чешуйками на передних продольных и кубитальной жилках,
а также с пятном таких же чешуек при основании г4+5.
Большая часть головы покрыта белыми чешуйками. Хоботок в основ-
ной и вершинной части темно-бурый, в средней части с резко выражен-
ным желтовато-белым кольцом. Щупики самки в длину равны длины
хоботка, в бурых, с примесью белых, чешуйках; основание самих щупи-
ков и основание 3-го их членика в белых чешуйках; щупики самца в длину
346
111. ПОДСЕМ. CULICINAE
превосходят длину хоботка почти в I1/2 раза, черно-бурые; вершина и
основание 5-го членика щупиков и основание 4-го их членика в белых
чешуйках; более или менее широкое, но нерезко ограниченное кольцо
посредине 3-го членика и вершина 2-го членика светлые; волоски щупи-
ков многочисленные и длинные, темно-бурые. Среднеспинка бурая;
передняя часть среднеспинки (2/3) покрыта белыми чешуйками, близ
середины с золотистым почковидным пятном или тремя небольшими пят-
нами; задняя часть (Vg) среднеспинки в темно-золотистых чешуйках
и таких же щетинках с вкраплением белых чешуек. Ноги: бедра и голени
спереди в бурых чешуйках с незначительными вкраплениями белых че-
шуек; лапки при основании 1—4-го члеников с узкими белыми колечками.
Крылья в бурых чешуйках; костальная, субкостальная, первая радиаль-
ная, задняя радиальная (г4+3) и кубитальная жилки покрыты широкими
темными чешуйками. Брюшко со светлыми перевязями или с несколько
вытянутыми назад треугольными пятнами белых чешуек при основании
тергитов; тергиты V—VIII с боков с более или менее развитыми пятнами
белых чешуек; нижняя поверхность брюшка бурая, с перевязями светлых
чешуек.
8. CULEX
347
Гипопигий (рис. 239): предвершинная бородавка коксита с одной
умеренно широкой пластинкой, двумя палочковидными выростами,
из которых один тонкий и острый, другой толстый и тупой, и щетин-
ками; X стернит с относительно коротким и узким базальным отростком;
верхний отдел фаллосомы с хорошо развитым медиальным отделом в виде
крючка и относительно широкой треугольной латеральной пластинкой;
нижний отдел фаллосомы — нормально с четырьмя более или менее оди-
наковой величины зубцами.
Личинка IV стадии (рис. 240): голова почти в 1.5 раза силь-
нее развита в ширину, чем в длину. Лобные волоски вторичноперистые;
наружные из 6—10 ветвей, сред-
ние лежат немного кзади и ко-
внутри от наружных, обе пары
длинные, 2-ветвистые, реже
один из них из трех ветвей.
Задние волоски наличника ко-
роткие, тонкие, из двух ветвей,
реже простые. Теменной и транс-
сутуральный немного длиннее,
из трех или больше ветвей.
Усики длинные, достигают дли-
ны головы, с телом, покрытым
шипиками. Пучок из 10 или
более ветвей отходит за сере-
диной усика, тело которого бо-
лее сужено от места отхождения
пучка. Две предконцевые ще-
тинки лежат субтерминально.
Щетка из 5—8 чешуек с хо-
рошо развитым главным шипом,
расположенных неправильным
зигзагообразным рядом; боко-
вые шипики у их основания
очень слабо развиты, практи-
чески незаметны. Волоски по-
Рис. 240. Culex whitmorei Giles. Личинка
IV стадии. По Barraud.
1 — дыхательная трубка, сбоку, г — усик
зади щетки: крайние верхний и нижний соответственно из 3—5 и 2—3 вет-
вей, средний из 6—8 ветвей, промежуточные — простые. Сифон у осно-
вания расширен, затем постепенно сужается, индекс 5.0—6.0. Гребень
из 6—10 тонких зубцов, из которых 1—2 наиболее дистальных крупнее
и шире расставлены, занимает базальную часть сифона и далеко не дохо-
дит до трети от основания. Сифональные пучки расположены попарно
в один ряд, занимают среднюю треть задней поверхности трубки, в числе
4—6 пар. Каждый пучок из двух длинных слабо вторичноперистых вет-
вей. Кроме них, на боковых сторонах сифона лежат две пары значительно
более коротких 2-ветвистых пучков, один у середины, другой— дисталь-
нее последнего, из брюшных пучков. Главные трахейные стволы тонкие,
округлого сечения.
Последний членик заметно больше в длину, чем в ширину, седло
кольцом окружает его. Боковой волосок короткий, из двух ветвей. Хвосто-
вые волоски длиннее сифона, простые или 2-ветвистые. Плавник из 12 пуч-
ков. Жабры ланцетовидной формы, изменчивы: от длины седла до длины
сифона.
Распространение. Крайний юг Приморского края. Япония
(Кюсю, Хонсю — редок), п-ов Корея, Восточная и Юго-Восточная Азия.
348
III. ПОДСЕМ. CULICINAE
Биология слабо изучена; личинки попадаются в стоячейТили
слабо проточной воде,.на рисовых полях и т. п. Массовых нападений|на
человека и домашних животных не наблюдалось.
12. Culex (Culex) mimeticus Noe, 1899.
Очень характерный вид с пятнистыми крыльями, несколько напоми-
нающий по общему облику An. superpictus (рис. 241).
Голова в желтовато-белых и бурых чешуйках. Хоботок черно-бурый,
у самки с широким, у самца — несколько более узким белым кольцом
близ середины; щупики самки в черно-бурых чешуйках, вершина щупи-
ков обыкновенно покрыта белыми чешуйками; щупики самца черно-
бурые, в такого же цвета чешуйках, как правило, с 3 белыми колечками;
проксимальное кольцо, расположенное на длинном членике щупиков,
явственно шире остальных; вершина щупиков самца светлая. Усики
у обоих полов бурые, у самца со светлыми колечками. Среднеспинка в сред-
ней части в светло-бурых или золотисто-бурых, по краям в сероватых
или беловатых чешуйках; бочки груди с пятнами белых чешуек. Ноги:
бедра и голени спереди в черно-бурых чеЩуйках, часто с примесью белых
чешуек, иногда сгруппированных в виде узкой продольной полоски;
бедра и голени сзади покрыты белыми чешуйками; лапки с узкими бе-
лыми колечками при основании 1—4 члеников лапок; колечки частично
охватывают и вершины члеников. Крылья по жилкам в черно-бурых
чешуйках, по переднему краю с 3 большими белыми пятнами, располо-
женными в начале и конце средней трети переднего края крыла, а также
перед его вершиной, и занимающими жилки: с, sc и гх; более значитель-
ные участки, покрытые белыми чешуйками, имеются, кроме того, в сле-
дующих местах: на разветвлениях r2-]-rs и т^А-т^, на г4+5 (в большей
части своего протяжения эта жилка покрыта белыми чешуйками), т3+4
и ап\ жилка шх целиком в черно-бурых чешуйках. Брюшко темно-бурое,
сверху в черных чешуйках, при основании тергитов с поперечными пере-
вязями белых чешуек, занимающими чаще всего около х/3 длины тергита;
брюшко снизу в беловатых чешуйках.
Гипопигий (рис. 242): коксит в относительно коротких волосках;
предвершинная бородавка коксита несет широкий листовидной формы
пластинчатый придаток и обыкновенно пять крупных щетинок; стиль
к вершине постепенно сужающийся; X стернит с базальным придатком;
нижний отдел фаллосомы имеет 3—5 направленных наружу зубцов.
Личинки (рис. 243) по ряду признаков сходны с личинками
подрода Neoculex.
Голова очень широкая, часто темная; средние лобные волоски впе-
реди внутренних, более широко расставлены; обе пары из двух вторично-
перистых ветвей. Усики светлые, с более темной вершиной, покрыты ши-
пиками на 2/з от основания; пучок из многочисленных вторичноперистых
ветвей, у середины усика или ближе к вершине достигает половины его
длины. Предвершинные щетинки одной длины с пучком у середины су-
женной концевой части усика. Брюшко: щетка из 20—35 чешуек, с хо-
рошо развитым острым концевым шипом и рядом мелких шипиков по
бокам его основания. Сифон прямой, расширен у основания и постепенно
сужается к вершине; сифональный индекс — 4.5—7, варьирует в зави-
симости от ширины основания. Гребень занимает около х/3 длины сифона,
из 12—13 зубцов, из которых 1—2 наиболее дистальные могут быть круп-
нее и более широко расставлены. Пучков 5—6 пар. Первые 3—4 пары
более длинные, из 4—5 ветвей, часто могут располагаться зигзагом на
неравном расстоянии друг от друга; следующая пара из двух тонких
Рис. 241. Culex mimeticus Noe.
350
III. ПОДСЕМ. CULICIDAE
более коротких ветвей занимает более латеральное положение. Главные
стволы трахей тонкие, с округлым поперечным сечением. Стигмальная
пластинка типа С. territans. Последний членик с простыми наружными
хвостовыми волосками, примерно одной длины с сифоном, с внутренними
из двух ветвей, из которых верхняя вдвое короче нижней, и с коротким
2-ветвистым боковым волоском. Плавник из 12 пучков. Жабры в 1.5—
2 раза длиннее седла, заострены на концах.
Рис. 243. Culex mi-
meticus Noe. Ли-
чинка IV стадии.
Дыхательная
трубка, сбоку.
Рис. 242. Гипопигий Culex mimeticus Noe.
Распространение. Юг Палеарктики и Ориентальная область.
СССР: Северный Кавказ, Дагестан, Закавказье, Средняя Азия (Туркме-
ния, Таджикистан), Приморский край. Южная Европа, Малая Азия,
Иран, Индия, Китай, Япония и другие страны Азии.
Биология. Личинки встречаются в небольших скоплениях
дождевой воды в углублениях скал, а также в поросших растительностью
заводях вдоль русел быстро текущих горных ручьев, иногда совместно
с An. marteri, An. claviger. An. superpictus, Culiseta longiareolata.
13. Culex (Culex) jacksoni Edwards, 1934.
Комар внешне (по имаго) весьма сходен с С. mimeticus, но хорошо
отличается строением личинки.
8. CULEX
351
По описанию, среднеспинка в красновато-коричневых чешуйках.
По бокам среднеспинки и на пространстве перед щитком имеются более
светлые чешуйки. В остальном (включая строение гипопигия), как С. mi-
meticus.
Личинка IV стадии от зеленоватого (у молодых) до желто-
коричневого (у зрелых) цвета, средних размеров (до 1 см).
Голова в среднем в 1.5 раза больше в длину, чем в ширину. Лобные
волоски с хорошо развитой вторичной перистостью, наружные из 6—12
(в среднем 8), более длинные средний и внутренний соответственно из 2—6
(в среднем 3) и 3—7 (в среднем 5) ветвей. Задние волоски наличника ко-
роткие и тонкие, в среднем из 2—3 (1—5) ветвей; теменной и транссутураль-
ный немного длиннее — из 3—4 (1—5) нежных ветвей.
Усики немного короче головы, лишь слегка изогнуты; базальная
их часть слабо пигментирована, густо покрыта тонкими волосковидными
щетинками; дистальный отдел темно пигментирован, более тонкий, покрыт
редкими более короткими и толстыми шипиками. Пучок в виде веера из
11—26 (в среднем 18) вторичноперистых ветвей, достигающих конца
усика. Две длинные предвершинные щетинки; вершинная одной длины
с ними, но немного тоньше.
Щетка VIII членика брюшка из 19—40 (среднее 32) остроконечных
чешуек, расположенных неправильным пятном. Каждая чешуйка по
бокам основания несет ряд краевых шипиков, из которых ближайшие
к главному шипу всегда короче последнего. Волоски позади щетки в сред-
нем: крайний верхний из пяти, нижний из четырех, средний из семи вто-
ричноперистых ветвей; промежуточные гладкие и простые.
Сифон длинный, несколько расширен у основания (индекс около 6).
Гребень из 8—20 (в среднем 14) зубцов и 2—4 рудиментарных у основа-
ния. Зубцы в виде тонких стройных шипов с несколькими шипиками
у слабо расширенного основания; все зубцы слабо пигментированы. Гре-
бень занимает от трети до половины длины сифона, редко больше. 1—5,
чаще 3, наиболее дистальных зубцов длиннее остальных и значительно
более широко расставлены. В концевой четверти сифона, на задне-боко-
вой его поверхности, как продолжение гребня находятся значительно
более крупные дополнительные гладкие шипы, направленные острием
к вершине трубки. Это — характерный признак личинок этого вида.
Общее их число колеблется от девяти до двух (в среднем четыре, т. е. две
пары). При небольшом числе шипов некоторые из них могут быть руди-
ментарными: от широкого основания отходит маленький короткий шипик.
Число пучков на сифоне относительно постоянно: три пары на его
задней поверхности расположены неправильным зигзагом; ближайшие
к основанию 1—2 пучка лежат около или между наиболее дистальных
1—2 зубцов гребня, а ближайшие к вершине — между дополнительными
шипами. Пучки короткие, обычно равны ширине сифона у места их от-
хождения, состоят из 2—6 гладких ветвей. На боковых сторонах конце-
вой четверти сифона лежат две пары коротких 2-ветвистых (редко простых)
волосков.
Главные трахейные стволы широкие, лентовидные. Стигмальная
пластинка типа «pipiens».
Последний членик короткий: длина лишь немного превышает его
ширину. Седло кольцом охватывает его. Боковой волосок тонкий и ко-
роткий, из 2—3 ветвей. Хвостовые волоски: наружный простой, в сред-
нем одной длины с сифоном, внутренний на х/3 короче, из двух, редко трех
ветвей; как исключение — простой. Плавник сдвинут к заднему краю
членика, из 12 пучков. Жабры ланцетовидные, заострены на концах;
352
III. ПОДСЕМ. CULICINAE
обе пары почти одной длины, составляют 1.25—1.5 длины седла, редко
длиннее или короче.
Биология. Личинки были встречены в богатых нитчатыми водо-
рослями мелководных заболоченностях (Сучанский район; А. И. Лисова),
в мелких лужах и в заводи ручья (о. Фуругельм; В. К. Трасис), преиму-
щественно в предгорных с всхолмленным рельефом местностях. Най-
дены совместно с личинками An. hyrcanus, A. vexans, С. bitaeniorhynchus,
С. mimeticus, С. modestus, С. vagans и др.
14. Culex (Culex) orientalis Edwards, 1921.
Очень близок к С. mimeticus, от которого отличается следующими
признаками: крыловые чешуйки более длинные, вследствие чего крылья
кажутся более густо покрытыми чешуйками; сих у вершины в светлых
чешуйках; покрытый светлыми чешуйками участок находится и на общем
стволе т^зА-сис, вершина щупиков самца с многочисленными светлыми
чешуйками, которые в некоторых случаях преобладают над темными.
Гипопигий (рис. 244): коксит широкий и несколько утолщенный,
густо покрытый волосками, в особенности в вершинной части и близ
предвершинной бородавки; последняя несет прозрачную пластинку и
ряд шипов, слегка зазубренных или изогнутых на конце; вокруг них
имеются длинные волоски; стиль у середины сильно расширен, к вер-
шине заострен; второй отдел фаллосомы имеет три крупных и несколько
мелких зубцов; от С. mimeticus гипопигий отличается наличием густых
волосков в вершинной части коксита.
8. CULEX
353
Личинки IV стадии (рис. 245—247) полупрозрачны и лишь
перед окуклением приобретают желтоватую или зеленую окраску.
Голова широкая, светло-желтая. Лобные волоски длинные, вторично-
перистые, наружные из 5—10 ветвей, средние и внутренние сдвинуты
назад; средние более широко расставлены, из двух, внутренние из 3—
4 ветвей. Усики длинные, более 3/4 длины головы, пигментированы у осно-
Рис., 245. Culex orientalis Edw. Личинка IV стадии.
Голова, сверху.
вания и в концевом отделе; пучок из 20—35 достигающих 1/2 длины усика
вторичноперистых ветвей, явственно за серединой; предвершинные ще-
тинки длиннее пучка, помещаются на расстоянии 1/3 длины концевого
отдела от вершины, вершинная щетинка более короткая.
Щетка из 20—40 (среднее 30) чешуек, с рядом шипов по краю, у вер-
шины заметно более крупных. Сифон длинный, тонкий (индекс 6.2—7.9).
Число и расположение пучков изменчиво; на задней стороне 10—13 пуч-
ков, сидящих или попарно, или зигзагом; ближайшие к основанию,
более длинные, немного превышают ширину сифона, из 2—5 ветвей;
на боковых поверхностях ближе к вершине имеются 1—2 пары пучков;
все_ближайшие к вершине пучки укорочены. Гребень из 9—14 хорошо
23 Фауна СССР, А. В. Гуцевич и др.
354
Ш. ПОДСЕМ. CULICINAE
развитых зубцов и из 1—4 рудиментарных зубчиков у основания. Глав-
ные трахейные стволы овального поперечного сечения. Стигмальная
пластинка типа С. pipiens. Последний членик с длинными (1.3—1.5 длины
сифона) простыми наружными хвостовыми волосками и на 1/4—1/3 более
короткими внутренними, из 2—4 ветвей. Плавник из 11—13 пучков.
Жабры удлиненно-ланцетовидной формы, равны или длиннее седла
(до 1.7 его длины); нижняя пара более короткая.
Рис. 246. Culex orientalis Edw. Личинка IV стадии. Задний конец, сверху.
Распространение. Страны Восточной Азии. СССР: Хаба-
ровский и Приморский края, за исключением горно-таежных районов.
Япония, п-ов Корея, Китай, Филиппины.
Биология. Личинки встречаются преимущественно в открытых,
освещенных солнцем, богатых высшей растительностью и водорослями
водоемах, в канавах, на рисовых полях, часто совместно с личинками
An. hyrcanus, С. bitaeniorhynchus, С. tritaeniorhynchus, A. vexans и др.
Комар дает два, реже — три поколения. Как кровосос на человеке и
домашних животных не зарегистрирован.
15. Culex (Culex) tritaeniorhynchus Giles, 1901.
Вид характеризуется наличием светлых колечек на хоботке и лап-
ках, а также отсутствием светлых чешуек на крыльях (рис. 248).
8. CULEX
355
Голова в красновато-бурых чешуйках. Хоботок с белым кольцом,
которое у самки, в особенности на нижней поверхности хоботка, дохог
дит иногда почти до его основания. Щупики самки темные. Щупики
самца на длинном членике, с двумя белыми кольцами, из которых одно
явственно шире.другого; вершина щупиков самца темная. Среднеспинка
в одноцветных темных красновато-бурых чешуйках. Бедра и голени
в бурых чешуйках; нижняя поверхность бедер покрыта беловатыми
чешуйками. Лапки бурые, с узкими бе-
лыми колечками при основании члеников;
колечки иногда слабо заметны, они более
выражены на передних лапках.
Гипопигий (рис. 249) похож на гипо-
пигий С. mimeticus. Предвершинная боро-
давка коксита с листовидной пластинкой
и 3—4 шипами. Нижний отдел фаллосо-
мы несет с каждой стороны 4—5 зубцов,
из них некоторые (обычно медиальные)
направлены назад (отличие от С. mimeti-
cus') . Базальные придатки X стернита
хорошо развиты.
Личинки IV стадии (рис. 250)
почти прозрачные, слабо пигментирован-
ные.
Голова умеренно широкая: ширина
в 1.3 раза превышает длину. Лобные
волоски длинные, вторично перистые; на-
ружные из 6—8, средние из двух, внут-
ренние из трех ветвей; последние сбли-
жены больше средних. Усики около 0.7
длины головы, более темно пигментирова-
ны у основания и в суженной концевой,
части; пучок на расстоянии около а/3 дли-
ны усика от основания, в среднем из 30
вторичноперистых ветвей. Предвершинные
щетинки длиннее пучка, отстоят на 1/3
длины суженной концевой части от вер-
шины; вершинные — более короткие.
Щетка в среднем из 50 относитель-
Рис. 247. Culex orientalis Edw.
Личинка IV стадии. Измен-
чивость в расположении во-
лосков на дыхательной трубке.
но коротких чешуек, окаймленных не-
большим числом щетинок, увеличивающихся к вершине. Сифон длин-
ный (индекс 6.5—7.8), тонкий, с заметно расширенным основанием; гре-
бень |щз 9—15 зубцов с более широко расставленными 2—3 наиболее-
дистальными из них. Каждый зубец вдоль всей нижней поверхности
имеет 5—9 тонких острых зубчиков, увеличивающихся к вершине. Пять-
пар пучков по бокам задней поверхности сифона начиная со второй трети
его длины от основания. Пучки из 3—4 (реже из 2 — ближайший к вер-
шине) ветвей, не длиннее ширины сифона, за исключением ближайшего
к основанию; одна пара более коротких 2—3-ветвистых боковых пучков
в вершинной трети сифона. Трахейные стволы овального поперечного
сечения. Стигмальная пластинка типа С. pipiens. Последний членик:
брюшка с более длинными, чем сифон, простыми наружными хвостовыми'
волосками, с внутренними — из 3 ветвей, из которых нижняя одной,
длины с сифоном, а каждая последующая вдвое короче предыдущей ш
с 2—3-ветвистым коротким боковым волоском. Плавник из 12 пучков^
23*
356
111. ПОДСЕМ. CULICINAE
Жабры короткие, до половины длины седла, заострены на концах, верх-
няя пара длиннее нижней.
Распространение. Огромный ареал вида охватывает Ориен-
тальную область, южные и юго-восточные районы Палеарктики и зна-
чительную часть Эфиопской области. В СССР найден на юге Азербайд-
жана, в Туркмении и Узбекистане, на юге Приморского края (на север
Рис. 249. Гипопигий Culex tritaeniorhynchus Giles.
A — внутренние склериты гипопигия, Б — коксит и стиль, В —>
базальная бородавка коксита, Г — лопасти IX тергита.
Рис. 250. Culex tritaeniorhynchus Giles. Личинка IV стадии
По Kirkpatrick.
1 — головд, сверху, 2 — задний конец, сбоку.
358
111. ПОДСЕМ. CULICINAE
распространен примерно до реки Имана). Из сопредельных с Советским
Союзом стран известен в Турции, Иране, Китае, КНДР, Японии.
Биология. Личинки обитают по преимуществу в мелких, хорошо
прогреваемых водоемах, в некоторых случаях лишь периодически за-
топляемых, а также в озерах-старицах, в которых заселяется личинками,
как правило, лишь прибрежная, мелководная, лишенная растительности
полоса; заселение личинками луговых водоемов происходит обычно
только после сенокоса, когда водоемы прогреваются более сильно; на-
иболее обильны личинки с середины июля до конца сентября. Активный
кровосос; самки нападают преимущественно под открытым небом, как
правило, начиная с захода солнца до 24 ч.—1 ч. ночи и в утренние часы
(6—8 ч.); при пасмурной погоде комары могут нападать в течение круглых
суток; проникает в ближайшие к местам выплода населенные пункты,
нередко залетает в дома и помещения для сельскохозяйственных животных.
Эпидемиологическое значение. Вид является одним
из основных переносчиков японского энцефалита, причем в отношении
этого вида доказана как возможность передачи вируса через укол, так и
наличие естественно зараженных комаров в природных условиях.
16. Culex (Culex) univittatus Theobald, 1901 (perexiguus Theobald, 1903).
Отличается от близких видов незначительной величиной, узкими,
чисто белыми поперечными перевязями брюшка, а также наличием более
или менее развитой белой полосы на передней (наружной) поверхности
задних голеней.
21 — коксит и стиль, Б .— внутренние склериты гипопигия.
Голова сверху покрыта бурыми и золотистыми чешуйками; края глаз
в белых чешуйках. Хоботок бурый, иногда в основной половине с белыми
чешуйками. Шипики в бурых чешуйках, иногда с примесью белых чешуек
на вершине щупиков (самки). Среднеспинка в золотисто-бурых чешуйках;
боковые отделы среднеспинки, пространство перед щитком и щиток в се-
рых чешуйках; бочки груди с пятнами белых чешуек. Ноги в бурых
чешуйках; бедра снизу, задние бедра в основной половине, также спе-
реди, покрыты белыми чешуйками; задние, а иногда и средние голени
8. CULEX
359
снаружи с более или менее выраженной белой продольной полосой;
лапки черно-бурые. Жилки крыла покрыты бурыми чешуйками. Брюшко
темно-бурое в такого же цвета чешуйках; при основании тергитов имеются
узкие поперечные перевязи чисто белых чешуек.
2
Рис. 252. Culex univittatus Theob. Личинка IV стадии. По Edwards.
1 — голова, сверху, 2 — задний конец, сбоку.
Гипопигий (рис. 251): пластинка предвершинной бородавки широкая,
асимметричная; стиль за серединой явственно расширен; базальный при-
даток X стернита хорошо развит; второй отдел фаллосомы широкий,
простой.
Личинки IV Стадии (рис. 252) слабо пигментированы, почти
прозрачные. Ширина головы в 1.4 раза превышает длину. Лобные во-
лоски вторичноперистые, наружные из 7—9, средние из 2—3, внутрен-
ние из 3—4 ветвей; средние впереди внутренних, более широко расстав-
лены. Усики менее 3/4 длины головы, с пучком из 24—28 вторичноперистых
360
111. ПОДСЕМ. CULICINAE
ветвей, отходящим на расстоянии 2/3—3/4 длины усика от основания.
Предвершинные щетинки сближены с вершинными.
Щетка из 30—40 узких вытянутых чешуек, дистальный конец которых
окаймлен рядом тесно сидящих шипиков. Сифон (индекс 5.9—7.6) варьи-
рует в длину, более крупным особям соответствует большая величина
индекса. Гребень из 10—12 крупных зубцов, занимающих базальную
х/4 сифона; наиболее дистальные могут быть шире расставлены. Пять
пар пучков на задне-боковой поверхности сифона из 2—4 ветвей, в длину
не превышающих его ширину; длина пучков и степень их ветвления
убывают к вершине. Каждый последующий из ближайших к основанию
трех пучков занимает более боковое положение. Из ближайших к вер-
шине двух пучков один смещен на боковую, а другой — на заднюю по-
верхности сифона. Главные трахейные стволы широкие, овального се-
чения. Стигмальная пластинка типа С. pipiens. Последний членик брюшка
с простым, одной длины с сифоном наружным хвостовым волоском,
с 2-ветвистым внутренним и с хорошо развитым 2—3-ветвистым боковым
волоском. Плавник из 12 пучков. Жабры от 3/4 до 11/2 длины седла,
удлиненно-яйцевидной формы.
Распространение. Эфиопская область и Средиземноморская
подобласть Палеарктики, Иран, Пакистан, Индия. В пределах СССР
найден в Туркмении.
Биология. Личинки обитают в различного типа стоячих водое-
мах с пресной или слабо солоноватой водой с содержанием солей до 0.2°/^
и pH от 7.2 до 9.7. Совместно с личинками этого вида чаще встречаются
личинки С. laurenti Newst., несколько реже — С. pipiens pipiens, A. cas-
pius caspius и An. pharoensis Theob. Самки питаются преимущественно
на птицах.
Эпидемиологическое значение. Является одним из
переносчиков вируса западнонильской лихорадки.
17. Culex (Culex) theileri Theobald, 1903.
Относительно крупный вид, характеризующийся светлыми продоль-
ными полосками на передней поверхности бедер и голеней.
Голова в белых или желтоватых прилегающих и бурых торчащих
чешуйках. Хоботок в бурых чешуйках с более или менее значительной
примесью белых чешуек. Щупики самки в бурых чешуйках, сверху с не-
большой примесью белых чешуек. Щупики самца в бурых чешуйках с бо-
лее или менее значительными пятнами белых чешуек, имеющими иногда
кольцевидную форму. Среднеспинка в бурых чешуйках, довольно сильно
варьирующих в окраске; пространство перед щитком и щиток покрыты
беловатыми чешуйками. Ноги: бедра и голени спереди в бурых чешуйках
с резко выраженной продольной полосой белых чешуек; бедра и голени
сзади в белых чешуйках. Крылья по жилкам покрыты бурыми чешуйками.
Брюшко в черно-бурых чешуйках, при основании тергитов с перевязями
белых чешуек, обыкновенно образующими по середине своего заднего
края более или менее значительные выступы, которые в своей совокуп-
ности представляются в виде светлой срединной продольной полосы.
Брюшко снизу в светлых чешуйках.
Гипопигий (рис. 253): предвершинная бородавка коксита е относи-
тельно широкой эллипсоидальной формы пластинкой; стиль серповидный;
X стернит с хорошо развитым базальным отростком; фаллосома с большим
числом мелких зубцов.
Личинки IV стадии (рис. 254) прозрачные или желтоватого
цвета с часто более пигментированной головой. Лобные волоски вторично-
8. CULEX
361
перистые, длинные, наружные из 7—9, средние (впереди внутренних и
немного сильнее расставлены) из 2—3, внутренние из 3—5 ветвей. Усики
до 3/4 длины головы с более темными осно-
ванием и концевой частью. Пучок из 25—30
ветвей; предвершинные щетинки близко у кон-
ца усика, одной длины с пучком и с вершин-
ной щетинкой. Щетка из 30 и более чешуек,
с слабо развитым вершинным шипом. Сифон
почти прямой, постепенно сужающийся к вер-
шине, индекс 5.5—6. Гребень немного заходит
за базальную х/4 сифона, из 6—11 широко
расставленных шиповидных зубцов с 3—5 ко-
роткими зубчиками у основания. Пять пар пуч-
ков на сифоне начиная со второй трети его
длины от основания; три базальные пары бо-
лее длинные, из 4—8 ветвей на задней поверх-
Рис. 253. Гипопигий Culex
theileri Theob. Внутренние
ности сифона, часто расположены зигзагом; склериты.
четвертая пара на боковой поверхности трубки,
из 2—4 более коротких ветвей; пятая, ближайшая к вершине пара из 2—
4 коротких ветвей, на задне-боковой поверхности трубки. Главные тра-
Рис. 254. Culex theileri Theob. Личинка IV стадии. По Edwards.
1 — задний конец, сбоку, 2 — голова, сверху.
хейные стволы овального сечения. Стигмальная пластинка типа С. pipi-
ens. Последний членик брюшка с простыми, более длинными, чем сифон,
наружными хвостовыми волосками, с 4-ветвистыми внутренними и с ко-
ротким и простым хвостовым волоском. Плавник в среднем из 14 хорошо
362
III. ПОДО JIM. CULICINAE
развитых пучков. Жабры одной длины с седлом или немного длиннее,
с заостренными концами; обе пары примерно одной длины.
Распространение. Ареал вида включает Эфиопскую область,
Средиземноморскую подобласть Палеарктики и западные районы Ориен-
тальной области (Индия). В СССР: степная Украина, Крым, Кавказ и
Закавказье, Средняя Азия.
Биология. Личинки обитают в различных заросших раститель-
ностью стоячих водоемах и каналах, часто в сильно загрязненной воде,
в большинстве случаев в пресной или слабо солоноватой (до 0.2% соле-
ности) воде, но нередко и в соленой (0.5—1.0% солености, с pH 9.5).
Весной личинки можно найти в чистой воде на затопленных лугах. Комары
данного вида нападают преимущественно под открытым небом, местами
в довольно значительном количестве (например, в некоторых районах
Армении). Иногда залетают в дома и другие постройки.
18. Culex (Culex) vagans Wiedemann, 1828 (exilis Dyar, 1924).
По общему облику несколько напоминает С. theileri, а экземпляры
со слабо развитой светлой полосою на передних и средних бедрах и голе-
нях — С. pipiens-, от обоих названных видов хорошо отличается по строе-
нию гипопигия. Описанного по материалу из Владивостока С. exilis Dyar,
1924 следует рассматривать как ..синоним С. vagans Wied.
Рис. 255. Гипопигий Culex vagans Wied.
А — коксит и стиль, Б — внутренние склериты.
Голова в желтоватых или светло-буроватых чешуйках. Хоботок бурый,
в средней части, как правило, с примесью светлых чешуек; щупики самки
темные, с небольшой примесью’светлых чешуек с верхней стороны; щупики
самца длиннее хоботка примерно на длину последнего их членика; длин-
ный членик щупиков самца темный, с примесью светло-желтых чешуек;
конечные членики щупиков самца не утолщенные. Среднеспинка бурая,
покрытая буроватыми с бронзовым оттенком или желтоватыми чешуйками,
более светлыми по краям среднеспинки. Ноги с дорсальной поверхности
темные, с вентральной — светлые; передние и средние бедра и голени
Всех пар ног спереди с белой продольной полосою; реже белая продольная
8. CULEX
363
полоса развита лишь на передних бедрах. Брюшко в бурых чешуйках,
С резко выраженными перевязями желтовато-белых чешуек при основании
тергитов; у некоторых экземпляров эти перевязи имеют посредине заднего
края более или менее значительные треугольные выросты (такие экземп-
ляры внешне напоминают С. theileri).
Гипопигий (рис. 255) с относительно широкой пластинкой на пред-
вершинной бородавке коксита; фаллосома в вершинной части разделена
на 2—3 пластинки, из которых
одна направлена дистально; X
-стернит с базальным придатком.
Личинка IV стадии
(рис. 256) желто-коричневого
цвета, с пигментированными
головой и сифоном. Голова в
среднем в 1.5 раза превышает
ширину. Лобные волоски вто-
ричноперистые; наружные из
'6—11, средние из 4—6, внут-
ренние из 5—7 ветвей; обе по-
следние пары сближены, внут-
ренние сильнее. Усики около
а/з длины головы, коричневые,
более темные у основания. Пу-
пок на расстоянии 2/3—3/4 дли-
ны усика от основания, из 25—
30 ветвей, с более сильно разви-
той вторичной перистостью у
ветвей, обращенных внутрь.
Предвершинные щетинки у са-
мого конца, немного короче
вершинной.
Щетка из 37—40 (24—47)
чешуек, окаймленных тонкими
шипиками, более длинными у
вершины чешуйки. Сифон (ин-
декс 5—6) со второй трети
длины постепенно сужается к
вершине, почти вдвое более
узкой, чем основание. Гре-
бень из 7—13 зубцов с 2—4
рудиментарными зубчиками, за-
нимает около х/3 длины сифона; наиболее удаленный сильнее отставлен.
Пучков пять пар, из которых ближайшие к основанию три пары наиболее
длинные, не менее чем в 1.5 раза превышают ширину сифона из 3—4 (2—
б) ветвей, расположены на задне-боковой поверхности; первая пара часто
на уровне оснований наиболее дистального из зубцов гребня; четвертая
пара на боковой поверхности сифона, из 3 (2—4) более коротких ветвей;
пятая пара на задне-боковой поверхности, короче остальных, из 3—4 вет-
вей. Главные трахейные стволы широкие, овального сечения. Стигмаль-
ная пластинка типа С. pipiens. Последний членик брюшка с простыми,
более длинными, чем сифон, наружными хвостовыми волосками, с 2-вет-
вистыми внутренними и с боковым волоском из двух ветвей. Плавник
из 12 пучков, немного длиннее жабр. Последние вдвое длиннее седла,
заострены на концах.
Рис. 256. Culex vagans Wied. Личинка IV ста-
дии. По Barraud.
1 — дыхательная трубка, сбоку, г — усик, з — по-
следний членик, сбоку.
364
III. ПОДСЕМ. CULICINAE
Распространение. СССР: Забайкалье, Хабаровский и При-
морский края. Япония (Хоккайдо, Хонсю, Сикоку, Кюсю), п-ов Корея,
Китай, Индия (преимущественно горные и предгорные районы).
Биология. Личинки встречаются в различных естественных и
искусственных стоячих водоемах (ямы, канавы, пожарные бочки и т. п.),
преимущественно вблизи жилищ человека, вслед за которым распро-
страняются в неосвоенные районы тайги. На человека нападает редко.
19. Culex torrentium Martini, 1925.
Автором данного вида следует считать Мартини, хотя в большинстве
литературных источников комар фигурирует под именем С. exilis (Dyar,
1924); но этим именем был обозначен вид, описанный по материалу из Вла-
дивостока, где С. torrentium, насколько известно, не встречается; имя
exilis Dyar, как указывалось выше, следует считать синонимом С. vagans
Wied. Вопрос об отношениях обеих названных форм друг к другу и к комп-
лексу С. pipiens (см. ниже) неясен. По строению гипопигия С. torrentium
и С. vagans практически неотличимы. Но по окраске оба вида хорошо
отличаются: у С. vagans спереди на бедрах и голенях развиты продольные
полоски белых чешуек, отсутствующие у С. torrentium. Степень развития
этого признака варьирует. Встречаются экземпляры с едва выраженными
полосками, почти неотличимые от С. torrentium. Возможно, что torrentium
следовало бы рассматривать как подвид С. vagans, тем более что обе формы
сходны экологически.
Но тогда возникает вопрос об отношении этого вида к С. pipiens-, из-
вестно, что С. torrentium и С. pipiens надежно отличаются только по строе-
нию гипопигия: у С. torrentium хорошо развиты базальные отростки X стер-
нита. Встречаются, правда весьма редко, экземпляры с относительно слабо
развитыми отростками, как бы промежуточные между С. torrentium и
С. pipiens. Что касается других указываемых в литературе отличительных
признаков обоих видов (окраска среднеспинки и брюшка, длина рукоятки
радиальной вилки), то они, судя по нашему материалу, не позволяют
с уверенностью отличать обе формы.
Возможно,следовательно,троякое решение вопроса: а) считатьpipiens—
torrentium—vagans за один вид, б) рассматривать torrentium в качестве
западного подвида С. vagans-, в) признавать, как обычно принимают,
существование трех самостоятельных видов: С. pipiens (всесветное распро-
странение), С. torrentium (Европа, Сибирь), С. vagans (Сибирь, Дальний
Восток СССР, Китай, Япония). Впредь до специального исследования во-
проса (что наиболее удобно провести в Восточной Сибири, где, по-види-
мому, встречаются все три формы) нам представляется более целесообраз-
ным принять последний вариант.
По внешнему виду С. torrentium чрезвычайно близок С. pipiens (см.
ниже), от которого с достоверностью может быть отличен лишь по строе-
нию гипопигия. Общая окраска тела и покрывающих его чешуек у С. tor-
rentium более темная; среднеспинка в темных желтовато-бурых чешуйках,
как правило, без золотисто-красного оттенка, характерного для С. pi-
piens. Бедра и голени спереди темно-бурые, без примеси светлых чешуек.
Темные чешуйки брюшка более темные, почти черно-бурые; чешуйки
светлых перевязей брюшка белее, нежели у С. pipiens.
Гипопигий по строению чрезвычайно близок таковому С. vagans
(ср. рис. 255); X стернит с довольно длинным базальным отростком; фал-
лосома на вершине с тремя пластинками, из которых одна направлена
дистально, а две — латерально.
8. CULEX
365
В литературе указывались и другие отличительные признаки, позво-
ляющие, якобы, с уверенностью различать и самок: окраска стернитов,
расположение поперечных жилок и особенно длина радиальной вилки
крыла: у С. torrentium рукоятка более длинная, вилка всего в 3—4 раза
длиннее рукоятки, тогда как у С. pipiens рукоятка более короткая, вилка
в 5—6 раз длиннее рукоятки. Просмотр обширного материала из различ-
ных географических районов показал, что широкая изменчивость, свой-
ственная С. pipiens, «перекрывает» указанные отличия и они не являются
достоверными. Можно отметить, что хранящийся в коллекции ЗИН эк-
земпляр, определенный автором вида («С. torrentium, Saratov, IV 1925,
Е. Martini»), имеет очень короткую рукоятку радиальной вилки, и по этому
признаку экземпляр должен быть определен как С. pipiens.
Личинка IV стадии желто-коричневого цвета, с умеренно
широкой головой. Средние лобные волоски впереди внутренних, значи-
тельно сильнее расставлены, в среднем из 5 ветвей. Усики с пучком из 20 и
более ветвей, достигающих а/3 его длины. Предвершинные щетинки
сближены с вершинными.
Щетка из 35—40 чешуек, окаймленных рядом шипиков, более длинных
на конце чешуйки. Сифон (индекс 6—7) начиная со второй трети постепенно
сужается к вершине. Гребень из 12 зубцов, занимает около 1/5 длины
сифона от основания. Четыре пары пучков из 2—3 ветвей лежат на задне-
боковой поверхности сифона; ближайшая к вершине пара занимает наи-
более боковое положение. Первые три пары в длину в 1.5 раза превосходят
ширину сифона у основания; четвертая заметно короче. Трахейные стволы
широкие, овального сечения. Стигмальная пластинка типа С. pipiens.
Последний членик брюшка с длинными и простыми наружными хвосто-
выми волосками и 2-ветвистыми внутренними. Плавник из 11—12 пучков.
Жабры ланцетовидной формы, вдвое длиннее седла.
Распространение. Вид распространен в СССР очень широко;
известен из Ленинградской, Пермской, Свердловской, Оренбургской,
Уральской областей, Нижнего Поволжья, Крыма, Северного Кавказа,
Западной Сибири и других областей. Средняя Европа, Малая Азия.
Биология. Вид до последнего времени смешивался с С. pipiens,
в связи с чем экология его изучена недостаточно. Личинки обитают в во-
доемах различного типа; в Нижнем Поволжье были находимы в небольших
водоемах, богатых растительностью, в пойменной полосе рек, в Средней
Европе (Шварцвальд) в водоемах в углублениях скал вдоль русла горных
речек совместно с личинками Culiseta glaphyroptera. В отличие от С. pi-
piens не является синантропом. На человека, по-видимому, нападает очень
редко.
20. Culex (Culex) pipiens Linnaeus, 1758 (рис. 257).
Голова в бурых чешуйках; боковые отделы головы и задние края
глаз, как правило, покрыты белыми чешуйками. Хоботок бурый. Щупики
самки в бурых чешуйках, иногда с примесью белых чешуек близ середины
и у вершины. Щупики самца заметно длиннее хоботка, в бурых чешуйках;
с нижней стороны двух конечных члеников в белых чешуйках; темно-
бурые щетинистые волоски на вершине 3-го, на 4-м и 5-м члениках щупика
хорошо развиты; более или менее выраженные колечки белых чешуек
имеются на длинном членике щупиков. Среднеспинка покрыта красновато-
бурыми или желтовато-бурыми чешуйками; боковые края среднеспинки и
пространство перед щитком покрыто более светлыми чешуйками; бочки
груди с незначительными пятнами белых чешуек на проэпистернах,
на стерноплеврах и на мезэпимерах. Ноги в бурых чешуйках, бедра снизу
366
III. ПОДСЕМ. CULICINAE
покрыты желтовато-белыми чешуйками. Крылья по жилкам в бурых
чешуйках. Брюшко в темно-бурых чешуйках, по переднему краю тергитов
Рис. 257. Culex pipiens L.
с более или менее широкими перевязями светло-желтых или беловатых
чешуек; в редких случаях перевязи могут отсутствовать,
Гипопигий (рис. 258): предвершинная бородавка коксита несет до-
вольно широкую, на вершине закругленную пластинку; второй отдел
8, CULEX
367
фаллосомы простой — не расщепленный и без зубцов; базальный отросток
X стернита отсутствует или представлен в виде незначительного палочко-
видного или треугольного выступа; в редких случаях он более развит,
но в отличие от С. torrentium не изогнут и не склеротизован.
Личинка IV стадии (рис. 259) габитуально очень изменчива,
от светлого серо-желтого до желто-коричневого цвета, с желтоватой или
коричневой головой и сифоном. Средних, редко крупных размеров.
Рис. 258. Гипопигий Culex pipiens L. По Сое, Freeman a. Mattingly.
А — внутренние склериты С. р. pipiens L., Б — то же С. р. fatigans Wied.
Голова в 1.3 раза больше в ширину, чем в длину. Лобные волоски
хорошо развиты, вторичноперистые; наружные из 6—10, средние впереди
внутренних, более широко расставлены, обе пары из 3—7 (в среднем 4)
ветвей. Задние волоски наличника короткие и простые, между основаниями
средних лобных волосков. Усики около 0.6—0.7 длины головы, более густо
покрыты шипиками в базальной части и единичными в вершинной трети,
у начала которой отходит пучок из 20—30 вторичноперистых ветвей,
достигающих трети длины усика. Предконцевые щетинки близко от вер-
шины, короче пучка.
Щетка из 30—55 тесно лежащих полукруглым пятном продолговатых
чешуек, изменчивых по величине, не имеющих главного шипа, а несущих
по jKpaHM и на свободном конце ряд шипиков. Волоски позади щетки:
крайние верхний из 3—7, нижний из 3—6, наиболее длинный средний из 6—
10 вторичноперистых ветвей; промежуточные более короткие и простые.
Сифон изменчивой длины (индекс 4—6.5) и формы: от почти прямой трубки,
наиболее широкой у основания, до слегка изогнутой, более широкой в сред-
ней трети. Гребень занимает около 1/5—1/4 длины сифона из 9—18 близко
сидящих зубцов, из которых 1—3 ближайших к основанию рудиментарные.
368
111. ТЮДСЕМ. CULICINAE
Пучков на сифоне четыре пары (как очень редкое исключение — три
или пять пар), лежащих по бокам его задней поверхности. Ближайшая
к основанию первая пара лежит немного отступя от дистального зубца
гребня, из 3—8 ветвей; основания пучков первой пары наиболее сближены.
Вторая пара — у середины сифона, более смещена на боковые стороны,
из 2—7 ветвей; третья пара, из 2—7 более коротких ветвей, занимает
наиболее боковое положение; четвертая — наиболее короткая, из 2—
Рис. 259. Culex pipiens pipiens L. Личинка IV стадии.
1 — задний конец, сбоку, 2 — стигмальная пластинка.
5 ветвей, занимает сходное со второй парой положение. Длина пучков
изменчива — от более коротких, чем ширина сифона, у мест их отхожде-
ния до заметно более длинных.
Главные трахейные стволы широкие, лентовидные, занимают не меньше
половины ширины сифона. Стигмальная пластинка (рис. 259) немного
больше в ширину, чем в длину. Тело рычага широкое, с большой полостью,
между стигмами сужается, образуя переднюю луку, заходящую далеко
за середину стигм. Задняя лука имеет закругленный край, от середины
которого отходит вверх длинный волосок хетоидного типа, имеющий
более широкое основание. Позади основания волоска лежит задний от-
росток в виде неправильно овальной, слабо пигментированной пластинки,
более темной на дистальном конце, где прикрепляются оторочки задних
клапанов. Плечи и стигмальные отростки рычага слабо развиты.
Стигмы с очень сильно скошенным внутрь отверстием и соответственно
с глубоко погруженными внутренними краями, следующими за изгибом
рычага, и с утолщенным и сильно вытянутым вверх передне-боковым краем.
8. CULEX
869
Обе поверхности переднего клапана более сильно хитинизированы
только в срединной части в виде двух полосок на верхней и нижней его
поверхностях. Органы на боковых клапанах с расширенным основанием,
заканчивающимся отростком, направленным к стигмам.
Псследний членик короткий, лишь немного больше в длину, чем в ши-
рину. Седло охватывает его кольцом, с простым коротким боковым во-
лоском. Наружные хвостовые волоски 2-ветвистые (редко простые), в 1.5—
2 раза длиннее жабр, внутренние — из 2—4 более коротких ветвей,
из которых нижняя самая длинная. Плавник в среднем из 12 пучков, из них
более длинные задние больше или одной длины с жабрами. Последние
одной длины с седлом или длиннее, могут более чем вдвое превышать его
длину.
Замечания по систематике.1 Комары комплекса Culex pipiens
представляют политипический вид, включающий несколько внутривидовых форм,
в разной степени морфологически и биологически обособленных друг от друга (Jo-
bling, 1938; Mattingly et al., 1951; Seminar on the ecology, biology and control of Culex
pipiens complex, 1965). Первоначально в состав комплекса включались С. р. pipi-
ens L., С. р. fatigans Wied, и С. р. molestus Forsk.; по мере более детального изучения
в разных частях ареала были обнаружены популяции, характеризующиеся проме-
жуточными морфологическими признаками и разным сочетанием биологических осо-
бенностей, в отдельных случаях получившие свое самостоятельное название, напри-
мер С. р. pallens Coq. Сравнительно недавно из Австралии был описан — С. р. aust-
ralicus Dobr. и близкий к комплексу С. globocoxitus Dobr.; таксономический уровень
и систематическое положение последнего- еще недостаточно ясны. С. р. australicus
встречается только в Австралии, главным образом в сельской местности; биологи-
чески он сходен с С. р. pipiens, а морфологически напоминает промежуточные формы,
возникающие при скрещивании С. р. fatigans и С. р. pipiens, хотя последний и не
отмечен в Австралии.
С. р. fatigans (в американской литературе чаще встречается его синоним —
С. р. quinquefasciatus Say) наиболее четко отличается от других форм шириной второго
отдела фаллосомы самцов и строением сифона личинок; для спаривания имаго доста-
точно небольшого пространства (стеногамность), в сезонном цикле развития отсутствует
диапауза (гомодинамность). Морфологические различия между С. р. pipiens и С. р. mo-
lestus носят количественный характер, а биологические значительны: С. р. molestus
является гомодинамным, стеногамным и способным к формированию первой яйцекладки
без кровососания (автогенность); С. р. pipiens имеет диапаузу (гетеродинамность),
для копуляции требует большого пространства (эвригамность), созревание яйцу самок
происходит только после приема крови. Морфологические различия между С. р. pi-
piens и С. р. molestus незначительны; это справедливо как для имагинальной фазы
(окраска, сравнительная длина хоботка и первых четырех члеников щупиков самца),
так и для личиночной (величина сифонального индекса, число ветвей в сифональных
пучках, степень ветвления волосков и др.). Наиболее удобный диагностический при-
знак личиночной фазы — величина сифонального индекса (у С. р. molestus он не бо-
лее,4.5 и ниже, а у С. р. pipiens более 4.5 и выше) — подвержен значительной измен-
чивости, которая является непрерывной, но не клинальной; часто в пределах одного
места в разных водоемах встречаются микропопуляции, сильно различающиеся сред-
ней величиной сифонального индекса. Незначительное модифицирующее влияние
на величину сифонального индекса могут оказывать температурные условия развития;
возможна и комбинативная форма изменчивости (при естественной гибридизации рас-
сматриваемых форм), что, вероятно, наблюдается в случае некоторых промежуточных
популяций. Существование значительной изменчивости морфологических признаков
сильно затрудняет и часто делает невозможным правильное определение подвидовой
принадлежности одной особи; степень надежности определения увеличивается при ис-
следовании серии экземпляров (получение средних величин в случае количественных
признаков и дополнение морфологической характеристики биологической, имеющей
главное значение при распознавании С. р. pipiens и. С. р. molestus).
На протяжении истории изучения комплекса С. pipiens таксономический уровень
составляющих его форм трактовался различно. С. р. fatigans долгое время считался
самостоятельным видом; в настоящее время его принято рассматривать в качестве
подвидовой формы комплекса. Сложнее вопрос с С. р. molestus (некоторые ученые,
в частности французские, предпочитают называть его С. р. autogenicus Boubaud).
1 Составила Е. Б. Виноградова; подробности см. в работах автора (Виноградова,
1965, 1966, 1969).
24 Фауна СССР, А. В. Гуцевич и др.
370
111. ПОДСЕМ. CULICINAE
Существование значительных биологических различий между С. р. pipiens и С. р. mo-
lestus явилось причиной того, что некоторые авторы были склонны возводить послед-
ний в ранг самостоятельного вида; сейчас эта крайняя точка зрения большинством ис-
следователей отклоняется. Зато в литературе в последнее время все больше утвер-
ждается противоположная тенденция — рассматривать С. р. pipiens и С. р. molestus
в рамках одного подвида С. pipiens, включающего автогенные и неавтогенные формы;
это явилось прежде всего следствием объективной трудности дифференцировать
их по одним морфологическим признакам и существованием в природе многочисленных
промежуточных форм (более подробно они будут рассмотрены далее). Разделяя эту
точку зрения, мы все же считаем целесообразным привести ниже биологическую
характеристику крайних типичных форм С. р. pipiens и С. р. molestus.
Кроме перечисленных подвидов, в пределах комплекса обнаружено несколько
промежуточных форм, встречающихся в разных частях ареала и характеризующихся
разной степенью стабильности. С. р. pattens, сочетающий признаки С. р. pipiens и
С. р. fatigans, найден в широкой зоне соприкосновения ареалов упомянутых подвидов
в Северной Америке (между 39° и 36° с. ш.) и на Дальнем Востоке. Он отличается про-
межуточными морфологическими признаками, в частности умеренно широким вторым
отделом фаллосомы. Его дальневосточные формы представляют собой длительно
существующие устойчивые популяции, а североамериканские, возможно, носят се-
зонный характер.
Гибридные формы, промежуточные между С. р. molestus и С. р. fatigans, известны
из Австралии; появление их в некоторых местах приурочено к летне-осеннему периоду,
а элиминация — к зиме.
Широко распространены в природе формы, промежуточные между С. р. pipiens
и С. р. molestus. Они известны преимущественно из южных районов Палеарктики —
Северной Африки, Франции, Италии, СССР (Крым, Северный Кавказ, Закавказье).
Как правило, они характеризуются промежуточными морфологическими признаками
(средняя величина сифонального индекса личинок варьирует между 4 и 5) и разным
сочетанием биологических особенностей. Некоторые авторы, в частности французские
(Roubaud et Ghelovitch, 1956; Rioux, 1958, и др.), обозначают такие биотипы специаль-
ными названиями: С. р. berbericus Roubaud (стеногамный, неавтогенный, антропо-
фильный и гомодинамный), С. р. calloti Rioux et Peek, (стеногамный, неавтогенный,
орнитофильный и гомодинамный). Нам это кажется нецелесообразным, так как возмож-
ное число сочетаний биологических признаков достаточно велико и может привести
к нежелательному усложнению терминологии. Особый практический и научный ин-
терес представляют антропофильные популяции С. pipiens на территории СССР.
Массовое нападение С. pipiens на людей отмечено в Москве, Ростовё-на-Дону, Одессе,
Краснодаре, Ставрополе, Новороссийске и других городах.
Заслуживает внимания вопрос о происхождении популяций, характеризующихся
промежуточными признаками. Большинство авторов склонно приписывать если не всем,
то некоторым из них гибридное происхождение. Б пользу этого свидетельствуют много-
численные успешные опыты по скрещиванию внутривидовых форм, приводящие к по-
явлению плодовитого потомства, в том числе экспериментальное получение С. р. pal-
lens. Закономерности наследования основных морфологических и биологических при-
знаков (сифональный индекс личинок, строение гипопигия самцов, автогенность,
стеногамность) также находятся в соответствии с этой концепцией.
Очевидно, в процессе становления С. р. pipiens и С. р. molestus большое значение
имело приспособление последнего к существованию в ближайшем окружении чело-
века, в городах. Действительно, в условиях умеренного климата с достаточно суровой
зимой возможно существование либо формы с диапаузой (С. р. pipiens), либо гомо-
динамной формы (С. р. molestus), приуроченной к такой специфической экологической
нише, как теплые подвалы и тому подобные места, и размножающейся там круглого-
дично. Существование в замкнутой нише также способствует и отбору на автогенность
и стеногамность, что подтверждено экспериментальным путем и природными наблю-
дениями. Степень морфологической и биологической обособленности обоих подвидов
по мере движения к северу увеличивается, что вероятнее всего связано с действием
климатического фактора.
Culex pipiens pipiens L.
Среднеспинка, как правило, в красновато-бурых чешуйках. Второй от-
дел фаллосомы узкий, крючковидный (рис. 258, Д).
Личинка IV стадии (рис. 259) с очень изменчивыми призна-
ками как по внешнему виду, так и по степени ветвления и развития отдель-
8. CULEX 371
ных волосков. Как правило, у основной формы ветвление в среднем слабее,
что возможно установить не на отдельных особях, а при исследовании
массового материала. Средние и внутренние лобные волоски из 3—5 (сред-
нее 4) ветвей. Средний из волосков позади щетки из 6—8 ветвей. Пучки
на сифоне (считая от основания): 1-й — из 3—4, 2-й и 3-й — из 2—3 и 4-й —
из 2—4 ветвей.
Сифон прямой, в среднем отделе очень слабо изогнут, но не расширен,
постепенно сужается к вершине, более длинный, чем у других подвидов
(индекс 4.5—6.4, среднее 5.3—5.4).
Последний членик относительно короткий, только немного больше
в длину, чем в ширину. Наружные хвостовые волоски в среднем в 1.5 раза
длиннее, чем жабры, из двух ветвей. Наиболее длинные задние пучки
плавника своими концами достигают концов жабр, но не выступают за них.
Жабры ланцетовидные, с заостренными концами, заметно длиннее седла;
обе пары одинаковой длины.
Распространение. Встречается преимущественно в зоне
умеренного климата. Палеарктика, северная половина Северной Америки,
северная часть Африки; в Западной Африке встречается редко; нарушение
нормального процесса спаривания, предположительно, мешает его распро-
странению в тропиках. На территории СССР распространен повсеместно,
по долинам рек и в населенных районах, примерно до Полярного круга.
К востоку от Байкала встречается редко.
Биология. Личинки встречаются в самых разнообразных естествен-
ных и искусственных водоемах со стоячей водой — заболоченностях раз-
ного происхождения, мелководной части крупных водоемов, канавах,
ямах, мелких лужах, оросителях, арыках, бочках, цементированных
водоемах разного назначения. Личинки предпочитают сравнительно
чистую воду, хотя в некоторых случаях развиваются и в загрязненной орга-
ническими веществами воде. Места выплода могут быть не связаны с жильем
человека. Отличается широким вертикальным распространением, подни-
маясь в Талыше и Западном Памире до высоты 2000—2500 м над ур. м.
Личинки встречаются с мая, а в более южных областях — с марта до сен-
тября—ноября. Максимум численности приурочен ко второй половине
лета. В течение сезона возможно два или большее число (на юге) поколе-
ний. Зимуют в основном оплодотворенные самки в состоянии имагиналь-
ной диапаузы, используя в качестве мест зимовки всевозможные естест-
венные убежища, а в населенных пунктах — неотапливаемые подвалы и
погреба; в последних плотность зимующих самок может быть очень высокой.'
В южных частях ареала, вероятно, возможно непрерывное развитие в те-
чение всего года. Имаго отличаются эвригамностыо — для копуляции
требуется большое пространство. Созревание яиц у самок происходит
только после кровососания. Этот подвид принято считать строго орнито-
фильным; действительно, многие авторы, наблюдавшие большую числен-
ность их в природе, отмечали почти полное отсутствие их в сборах на лю-
дях. Случаи нападения на человека чаще встречаются в южных районах,
однако достоверное определение подвидовой принадлежности нападающих
особей затруднено. Наблюдения за выведенными в лаборатории комарами
также подтверждают большую агрессивность по отношению к человеку
южных форм, чем северных.
Роль как переносчика не совсем ясна. Есть данные о том, что С. р. pi-
piens является переносчиком западного лошадиного энцефалита и энце-
фалита Сан-Луи в Северной Америке, однако точность определения его
подвидовой принадлежности в данных случаях вызывает некоторые сом-
нения.
24*
372
111. ПОДСЕМ. CULICINAE
Culex pipiens molestus Forskal, 1775.
По морфологическим признакам и по окраске с достоверностью не от-
личается от С. р. pipiens.
Личинка IV стадии (рис. 260), как и основная форма, очень
изменчива, но степень ветвления волосков, как правило, в среднем силь-
нее. Средние и внутренние лобные волоски из 4—6 (средние 5) ветвей.
Средний волосок позади щетки из 6—10 ветвей. Пучки на сифоне (считая
от основания): 1-й — из 3—8, 2-й и 3-й — из 2—7 и 4-й — из 2—5 ветвей.
Сифон относительно более короткий, чем у основного подвида (индекс
не более 4.5), и немного сильнее сужен к вершине, не расширен в среднем
отделе.
Рис. 260. Culex pipiens molestus Forsk. Личинка
IV стадии. Задний конец, сбоку.
Последний членик более длинный, чем у основного подвида. Наружные
хвостовые волоски вдвое, или более, длиннее, чем жабры, хотя абсолютно
они короче, чем у основного подвида. Жабры одной длины с седлом или
короче, верхняя пара длиннее нижней.
Расп р о с т р а н е н и е. С. р. molestus является формой, симпатри-
ческой с С. р. pipiens- единственным исключением является Австралия,
где встречается только С. р. molestus (он был ввезен сюда морским путем).
К настоящему времени С. р. molestus обнаружен во многих городах Ев-
ропы,, а также Азии, Африки и Америки; в Англии, Швеции, Норвегии,
Польше, во Франции; в ГДР и ФРГ, в Чехословакии, Италии, в ОАР,
в. Израиле, в Судане, в Японии. В СССР он отмечен в Ленинграде (Федо-
ров,, 194.1), Вологде, Москве, Киеве, Харькове, Днепропетровске, До-
нецке, Красном Лимане, Николаеве, Баку. Нападает на людей в жарких
странах и в годы массового размножения в городах умеренного пояса.
Биология. В зоне умеренного климата личинки встречаются
в разнообразных искусственных скоплениях воды в подвалах и на чердаках
жилых домов и предприятий (преимущественно в городах, где размноже-
ние продолжается в течение круглого года), а также в бочках и других
искусственных водоемах вблизи домов (летом и ранней осенью). В тропи-
8. CULEX
373
ческих и субтропических странах возможно круглогодичное развитие в от-
крытых природных водоемах. Личинки предпочитают сильно загрязнен-
ную, богатую продуктами распада органических веществ воду, вплоть
до сточных вод канализационной системы, хотя могут развиваться и в ме-
нее загрязненной воде. В течение года наблюдается развитие нескольких
поколений; самки неспособны диапаузировать. Имаго отличаются стено-
гамностыо; минимальный объем, необходимый для копуляции, составляет
27 см3. Для изучения проявления стеногамности чаще используются садки
Фриззи (5 дм), садки Рубо (26 дм) и садки Бейтса (337 дм). Самки способны
к автогенному развитию яичников (формирование по крайней мере первой
яйцекладки без кровососания) за счет личиночных запасов питательных
веществ. В случае бедного личиночного питания эта способность реали-
зуется лишь у некоторых самок. Характерным признаком является строе-
ние яичников у молодых, недавно отродившихся особей: первые фолли-
кулы находятся, как правило, на разных фазах развития, представляя
мозаичную картину. Автогенность является наследуемым признаком.
Эпидемиологическое значение. С. р. molestus яв-
ляется одним из переносчиков вируса западнонильской лихорадки.
Culex pipiens fatigans Wiedemann, 1828 (quinquef asciatus Say, 1823).
Очень близок по окраске к С. р. pipiens, от которого отличается желто-
вато-бурой, а не красновато- или темно-бурой окраской чешуек средне-
спинки; щупики самца менее густо покрыты волосками; поперечные пе-
Рис. 261. Culex pipiens fatigans Wied. Личинка IV стадии.
По Barraud.
1 — дыхательная трубка, сбоку, г — усик, 3 — последний членик брюшка,
сбоку.
ревязи на тергитах брюшка беловатые, к боковым краям они обыкновенно
суживаются и имеют в общем полулунную форму. Хорошо отличается
от С. р. pipiens строением фаллосомы (рис. 258, Б), второй отдел которой
имеет лопастевидную форму и отличается значительною шириною (уС.р. pi-
piens — узкий, крючковидный). Самки обоих подвидов с достоверностью
неотличимы.
374
Ш. ПОДСЕМ. culicinae
Личинка IV стадии (рис. 261) отличается от остальных под-
видов прежде всего формой сифона, имеющего заметно выпуклую переднюю
сторону, а заднюю.сторону выпуклую в среднем отделе и слегка вогнутую
(реже прямую) при сужении к основанию и вершине. В связи с этим наи-
большая ширина сифона не у основания, а в средней части трубки; при этом
ширина у вершины сифона несколько больше половины ширины у основа-
ния. Пучки на сифоне в среднем сильнее ветвятся, чем у двух других под-
видов: обе ближайшие к основанию пары из 6—7 ветвей, в длину или до-
стигают ширины сифона у места их отхождения, или немного длиннее.
Как правило, и другие волоски тела, в частности лобные, ветвятся силь-
нее,чем у личинок других подвидов (средние и внутренние из 5—7 вторично-
перистых ветвей). Жабры или равны длине седла, или длиннее.
Распространение. Встречается повсеместно в тропических
и субтропических областях земного шара. В пределах СССР достоверно
не отмечен.
Биология. Личинки встречаются в разнообразных искусственных
водоемах в городах и небольших населенных пунктах; в естественных во-
доемах встречаются реже. Способны развиваться в сильно загрязненной
продуктами распада органических веществ воде, что дает им явные пре-
имущества перед другими видами комаров, входящими в данный биоценоз.
Очень быстрое увеличение численности в последнее время в тропических
и субтропических странах связано с процессом урбанизации (быстрый рост
небольших городов и укрупнение поселений сельского типа), создающей
благоприятные условия для размножения комара. Имаго являются стено-
гамными; на протяжении года дают несколько поколений, развиваясь
без диапаузы. Для яйцекладки необходимо кровососание. Круг пищевых
объектов достаточно широк, разные популяции в этом отношении разли-
чаются; в некоторых частях ареала комары этого подвида питаются пре-
имущественно на человеке, являясь назойливыми кровососами.
Эпидемиологическое значение. Важнейший пере-
носчик филяриоза, вызываемого Wuchereria bancrofti; является одним
из переносчиков Brugia malayi и Dirofilaria immitis, а также ряда видов
птичьих плазмодиев. В Америке — важнейший переносчик вируса энце-
фалита Сан-Луи и один из переносчиков вируса западного (конского)
энцефалита. В Восточной Азии — один из переносчиков вируса японского
энцефалита.
ДОБАВЛЕНИЕ
За время нахождения рукописи в производстве получены новые данные по кома-
рам, встречающимся в СССР: описаны 2 новых вида, Aedes (Ochl.) montchadskyi Dub.,
1968 и Aedes (Ochl.) stramineus Dub., 1970, найдено 2 новых для фауны СССР вида —
Aedes (Ochl.) jitchii Felt et Young и A. (Ochl.) rempeli Vocker., а также 2 вида — Aedes
(Ochl) kasachstanicus Guts, и A. (Ochl.) simanlnt Guts., личинки которых были ранее
неизвестны. Авторы особо благодарны А. М. Дубицкому (Алма-Ата) и П. Е. Поляковой
(Новосибирск) за предоставление материалов по личинкам, и взрослым комарам и дан-
ных по их биологии, послуживших основой для добавления.
Aedes (Ochlerotatus) montchadskyi Dubitzky, 1968.
Самка. Хоботок и щупики в перемешанных темных и светлых чешуйках; вер-
шина хоботка темная. Чешуйки среднеспинки преимущественно золотисто-желтые;
беловатые чешуйки расположены по краям среднеспинки, а также образуют 2 неболь-
ших пятна в ее средней части. Бочки груди в белых чешуйках, которые на стерноплев-
рах доходят до их переднего угла, а на мезэпимерах — до их нижнего края. Гипостиг-
мальное пятно имеется, посткоксапьное отсутствует. Бедра и голени спереди в переме-
шанных темных и светлых чешуйках. Лапки без выраженных светлых колечек; перед-
няя поверхность первых двух или трех члеников лапки в тёмных чешуйках, задняя —
в светлых; на боковых поверхностях лапки светлые чешуйки расположены преиму-
щественно в основной части члеников, создавая впечатление наличия светлых колечек;
вершинные членики лапки темные. Крылья в темных чешуйках со значительной при-
месью светлых вдоль костальной и первой радиальной жилок. Брюшко сверху темное
со светлыми перевязями при основании тергитов, расширяющимися по бокам; перевязи
занимают около четверти каждого тергита; на темном фоне светлых чешуек нет.
Самец. Окраска в основном как у самки, но светлые перевязи брюшка шире,
они занимают около трети каждого тергита. Гипопигий: коксит покрыт волосками
умеренной длины; базальная бородавка крупная, несет изогнутый шип, ряд из не-
скольких щетинок и несколько рядов волосков: вершинная бородавка умеренной ве-
личины. Стволик класпеты слегка изогнутый, ее крыло узкое, с небольшим пластинча-
тым расширением. Выросты IX тергита с 5—8 щетинками каждый.
Личинка IV стадии серо-бурого цвета, с хорошо развитой вторичной
пористостью на волосках тела.
Голова немного больше в ширину, чем в длину. Лобные волоски: наружные из
8—10, средние из 2—3, внутренние из 3—4 ветвей; внутренние длиннее средних. Зад-
ние волоски наличника короткие, 2—3-ветвистые. Теменные волоски простые, транс-
сутуральные более короткие, из 2 ветвей. Усики не более половины длины головы,
с редкими шипиками; пучок относительно короткий, из 6—8 ветвей, лежит за середи-
ной, ближе к вершине.
Срединные волоски переднегруди: передние, более тонкие и короткие, доходят
до уровня середины сложных глаз, задние толще, заметно длиннее; обе пары от основа-
ния 3-ветвистые; промежуточные — простые, одной длины с передними; все несут
редкую вторичную перистость.
Щетка в среднем из 27 (22—33) чешуек, расположенных в 2—3 ряда выпуклым
назад полулунным пятном; чешуйки изменчивы по размерам и форме: более короткие
с рядом шипов на дистальном конце и удлиненные с главным шипом, по бокам основа-
ния которого лежат редкие более короткие шипики. Волоски позади щетки сильно
развиты: крайний верхний из 4—7, средний из 5—7, крайний нижний из 3 ветвей;
промежуточные гладкие и простые.
Сифон постепенно сужается к вершине; индекс 3.3—3.5. Пучок у середины, не-
много ближе к основанию, из 5—8 ветвей, в 1.5—2 раза превышающих ширину трубки
у места отхождения. Гребень из 19—20 тонких тесно расположенных зубцов, несущих
2—5 зубчиков у основания; наиболее дистальный зубец не доходит до основания
пучка. ’ ’
Длина седла заметно превышает его ширину; его боковые стороны почти доходят
до брюшного края членика. Боковой волосок очень характерен, из 3 (редко из 2 или
4) вторичноперистых ветвей, в длину заметно превышающих длину седла. Наружные
376
ДОБАВЛЕНИЕ
хвостовые волоски простые, в 1.5 раза превышают длину сифона; внутренние —
из 8—10 вторичноперистых более чем двое коротких ветвей. Плавник из 17—18 пучков
в общем основании и одного более короткого впереди него. Жабры маленькие, овальные,
в 4—6 раз короче седла.
Распространенне й биология Юго-Восточный Казахстан. Пойма
верхнего и среднего течения р. Или. В небольших временных водоемах в тугайных
зарослях или около них, с различной степенью минерализации. Поздневесенний вид,
имеющий одно поколение в году.
Замечания по систематике. Принадлежит к группе communis',
по расположению светлых чешуек на лапках имеет некоторое сходство с видами группы
cantans. Пестрая окраска бедер и голеней, а также наличие светлых чешуек на крыле
сближают данный вид с A. leucomelas, от которого A. montchadskyi отличается окраской
груди, а также строением гипопигия.
Aedes (Ochlerotatus) stramineus Dubitzky, 1970.
Самка. Хоботок преимущественно в светлых чешуйках, вершина хоботка тем-
ная. Щупики в перемешанных светлых и темных чешуйках. Среднеспинка без полос,
в желтоватых чешуйках, более темных по краям, более светлых в задней трети средне-
спинки. Бочки груди в белых чешуйках, доходящих до переднего угла стерноплевр и
до нижнего края мезэпимер. Гипостигмальное и посткоксальное пятна чешуек имеются.
Бедра и голени спереди в перемешанных светлых и темных чешуйках, лапки с неясными
светлыми кольцами, опоясывающими вершину одного и основание следующего членика.
Крылья преимущественно в светлых чешуйках, темные в большем или меньшем коли-
честве вкраплены на костальной и других жилках. Мембрана крыла с желтоватым
оттенком. Брюшко сверху в одноцветных желто-белых чешуйках.
Самец. Окраска в основном как у самки. Гипопигий: коксит в относительно
коротких волосках и только у вершины коксита имеется несколько толстых длинных
волосков; базальная бородавка коксита выпуклая, в ее дистальной части располо-
жены 2 шипа — больший, в вершинной половине изогнутый крючком, и меньший,
прямой; кроме того, бородавка несет ряд щетинок, по толщине почти не уступающих
шипам, и несколько рядов волосков; вершинная бородавка слабо развита, едва за-
метна. Стволик класпеты прямой, короткий; крыло класпеты по длине равно ство-
лику, умеренно склеротизованное, несколько расширенное в дистальной половине.
Выросты IX тергита с 6—8 короткими щетинками. Фаллосома широкая, с легкой пере-
тяжкой.
Личинка IV стадии. Средней величины (до 1 см) от грязно- до темно-бурого
цвета с темной головой и более светлой дыхательной трубкой.
Голова немного больше в ширину, чем в длину. Со спинной стороны в срединной
части лобного щитка имеется темная пигментация, выделяющаяся на фоне более
светлых боковых склеритов. При умеренной степени пигментации в затылочном от-
деле она наиболее широка, далее кпереди, после небольшого сужения, она расширяется,
достигая наибольшей ширины между внутренними лобными волосками; после короткого
сужения она на уровне средних лобных волосков вновь расширяется. С возрастом
пигментация усиливается, ее контуры становятся менее четкими вследствие общего
потемнения головной капсулы. Лобные волоски: наружные из 9—11 вторично-
перистых ветвей, средние — простые, а внутренние из 5, реже из 3—6 ветвей. Задние
волоски наличника — между основаниями средних лобных, короткие, из 3—4 ветвей.
Теменной волосок короткий и простой, транссутуральный — значительно длиннее,
2-ветвистый. Усики достигают 2/3 длины головы, покрыты мелкими шипиками; пучок
у середины, из 5—8 ветвей.
Внутренние медианные волоски на переднегруди: передние, более короткие,
достигающие уровня заднего края глаз, из 3—4 ветвей, задние немного длиннее,
из 3—6 ветвей; обе пары ветвятся от основания; промежуточные — простые, одной
длины с задними. Все волоски лишены вторичной перистости.
Щетка по бокам VIII членика в среднем из 30 (24—36) чешуек, расположенных
треугольным Пятном. Чешуйки изменчивы по размерам и форме, от коротких и более
широких с гребенчатым рядом шипиков на дистальном конце до более узких и длин-
ных, заостренных на свободном конце, но без выраженного главного шипа, с рядом
уменьшающихся к основанию шипиков, вдоль боковых краев Волоски позади щетки:
крайний верхний из 7—9, средний из 9—10, крайний нижний из 6—7 вторичноперистых
ветвей; промежуточные гладкие и простые.
Сифон начиная от середины постепенно сужается, индекс 3.1—З.З/ Пучок из 7—9
вторичноперистых ветвей, в длину превышающих не более чем в 1.5 раза ширину си-
фона у места его прикрепления, находящегося за серединой, немного ближе к вершине.
Гребень из 29—33 зубцов, лежащих на равном расстоянии друг от друга, занимает
базальную половину трубки и на длину одного зубца не доходит до основания пучка:
зубцы тонкие и длинные со слабо выраженным рядом зубчиков вдоль основания.
Длина седла почти в 2 раза превышает его ширину; его боковые стороны доходят
до середины боков последнего членика. Боковой волосок простой, тонкий, немного
ДОБАВЛЕНИЕ
377
более 1/2 длины седла. Хвостовые волоски: наружные простые, почти в 1.5 раза длин-
нее сифона, внутренние из 12—14 ветвей, достигающих х/3 длины наружных. Плавник
из 14—16 пучков в общем основании и из 1—2 (редко 3) более коротких пучков
впереди него. Жабры короткие, в 5—6 раз меньше длины седла, яйцевидные, нижняя
пара немного короче верхней.
Ра.спространение и биология. Описан по материалу из Казах-
стана (Дубицкий, 1970). Комары, относящиеся вероятно к данному виду, встречаются
в ряде районов Средней Азии, Казахстана и Западной Сибири. Выплаживается весной
во временных больших открытых солоноватых водоемах совместно с A. detritus и
A. montchadskyi. За сезон дает только одно поколение.
Замечания по систематике. Весьма близок к A. caspius, от которого
ранее не отграничивался.
Aedes (Ochlerotatus) litchii Felt et Young, 1904.
Довольно крупный комар относительно темной окраски.
Самка. Хоботок темный, щупики в темных чешуйках с небольшой примесью
светлых. Среднеспинка в желтовато-бронзовых чешуйках с продольной темно-коричне-
вой полосой и небольшими неясно очерченными беловатыми пятнами. Бочки груди
в белых чешуйках, которые на стерноплеврах доходят до их переднего угла, а на мез-
эпимерах не доходят до их нижнего края. Гипостигмального пятна чешуек нет, пост-
коксальное имеется. Бедра и голени спереди пестрые. Лапки с белыми кольцами при
основании члеников; последние два членика передней лапки и один членик средней
лапки темные; на средних члениках задней лапки кольца занимают около половины
длины членика. Крылья преимущественно в темных чешуйках с большим или мень-
шим количеством светлых, вкрапленных на разных жилках. Брюшко сверху со свет-
лыми перевязями — широкими при основании тергитов и узкими у их вершины;
на задних тергитах преобладают светлые чешуйки.
Самец. Окраска в основном как у самки. Гипопигий: коксит в относительно
коротких волосках; базальная бородавка крупная, закругленная или тупо коническая,
слегка асимметричная; бородавка несет один шип и множество густых довольно
длинных волосков; вершинная бородавка сильно выпуклая. Стволик класпеты слегка
S-образно изогнутый или почти прямой; крыло класпеты короче стволика, слабо скле-
ротизованное, относительно узкое, при основании с небольшой выемкой. Выросты
IX тергита с 7—9 длинными щетинками.- Фаллосома цилиндрическая, удлиненная,
с глубокой выемкой на вершине.
Личинка IV стадии средней величины. Голова не менее чем в 1.5 раза
более развита в ширину, чем в длину. Лобные волоски вторичноперистые; внутренние
из 3—4, как исключение из 2 ветвей, средние впереди внутренних, из 2—3 более корот-
ких ветвей, наружные в виде многочисленного пучка. Задние волоски наличника между
основаниями средних лобных волосков короткие, из 3—4 ветвей. Теменной и транс-
сутуральный простые. Усики около половины длины головы, покрыты шипиками,
с пучком, отходящим около середины и не достигающим конца усика.
Срединные волоски переднегруди вторичноперистые, внутренний простой или
2—3-ветвистый, средний длинный и простой, наружный длинный, 2-ветвистый или
простой.
Щетка из 12—28 чешуек с хорошо развитым главным шипом и с рядом в 3—4 раза
более коротких шипиков у основания. Волоски позади щетки длинные, средний и край-
ние верхний и нижний из 3—4 вторичноперистых ветвей, промежуточные — гладкие
и простые.
Дыхательная трубка (индекс 4—5) начинает постепенно сужаться несколько от-
ступя от основания, а в концевой четверти становится цилиндрической. Гребень из
15—24 равномерно расположенных зубцов, немного не доходит до середины трубки;
зубцы увеличиваются по направлению к вершине. Пучок из 3—6 ветвей со слабой
вторичной пористостью лежит около середины трубки, за наиболее дистальным из
зубцов гребня. Волоски на задних клапанах хорошо развиты и изогнуты.
Последний членик удлинен, с седлом, немного заходящим за середину его боковых
сторон. Боковой волосок простой и гладкий, в длину достигает длины седла. Хвосто-
вые волоски наружные длинные и простые, внутренние в виде веера из 6 ветвей.
Плавник с одним или двумя пучками впереди общего основания. Жабры заостренные,
ланцетовидные, в 1.5—2 раза длиннее седла.
Распространение и биология. Найден в Магаданской области
на р. Анадырь и в зоне среднего течения Колымы на широте около 64° с. ш. (Поля-
кова, 1970). Появляется в июне, летает до конца августа; нападает на людей в довольно
большом количестве. Широко распространен в западном полушарии: Аляска, Канада,
США.
Замечания по систематике. Принадлежит к группе cantans; по сам-
кам вид не всегда может быть определен с уверенностью ввиду сходства с другими
представителями группы, в частности с A. riparius.
378
ДОБАВЛЕНИЕ
Aedes (Ochlerotatus) rempeli Vockeroth, 1954.
Описание имаго даем по литературным источникам (Vockeroth, 1954; Carpenter
a. LaCasse, 1955); до настоящего времени в Советском Союзе найдены только личинки;
взрослых комаров, которых с уверенностью можно отнести к данному виду, в нашем
распоряжении нет.
Самка. Хоботок темный, щупики в темных чешуйках с небольшой примесью
светлых. Среднеспинка в серовато-желтых чешуйках с продольной темно-коричневой
полосой. Бочки груди в беловатых чешуйках, которые на мезэпимерах достигают
их нижнего края. Гипостигмального и посткоксального пятен нет. Брюшко сверху
темное со светлыми перевязями при основании тергитов. Бедра и голени спереди
темные с небольшой примесью светлых чешуек. Лапки темные. Крылья в темных че-
шуйках, немногочисленные светлые имеются лишь при основании костальной жилки.
Самец. Окраска как у самки. Гипопигий: коксит в довольно длинных волосках,
базальная бородавка крупная, коническая, без шипа, с несколькими щетинками,
образующими ряд, в котором щетинки постепенно укорачиваются; вершинная боро-
давка небольшая, несет лишь несколько коротких волосков. Стволик класпеты изо-
гнут, крыло класпеты немного короче стволика, с расширением в базальной трети,
к вершине изогнуто и сужено. Ширина фаллосомы примерно равна ее длине.
Личинка IV стадии от темно-бурого до черного цвета, средних размеров
с очень темными головой и дыхательной трубкой.
Голова лишь немного больше в ширину, чем в длину. Лобные волоски с редкой
вторичной перистостью: средние впереди внутренних, обе пары — простые; наружные
из 4—6 более тонких ветвей с легкой вторичной перистостью. Задние волоски налич-
ника — между основаниями средних лобных волосков, короткие, из 4—5 ветвей.
Теменные тонкие и простые, транссутуральные, более короткие, двуветвистые. Усики
короткие, 0.5—0.6 длины головы, с телом, покрытым шипиками; пучок из 5—7 тонких
ветвей, не достигающих конца усика, отходит у середины, немного ближе к основанию.
Внутренние медианные волоски переднегруди гладкие, два передних простые и
более тонкие, задний 2-ветвистый от основания.
Щетка из 32—46 (среднее 38) чешуек, расположенных слегка выгнутым назад
пятном. Чешуйки изменчивы по величине и форме: от более длинных и узких до широ-
ких и коротких, чаще встречающихся у ее брюшного края. Чешуйки без главного
шипа или, как исключение, он очень слабо выражен; их передний край в виде гребня
из шипов, иногда заходящих на боковые. Волоски позади щетки: крайние верхний
из 2—4, нижний из 2—6, средний из 4—7 вторичноперистых ветвей, промежуточные —
гладкие и простые.
Дыхательная трубка в концевой трети сужается к вершине; индекс 3.5—4. Пу-
чок из 8—12 вторичноперистых ветвей отходит около середины трубки, заметно ближе
к основанию. Гребень из 20—23 тесно сидящих зубцов занимает лишь немного больше
базальной четверти трубки. Предконцевые шипы на трубке хорошо развиты. Волоски
на задних клапанах стигмальной пластинки крючковидно изогнуты.
Последний членик относительно короткий, его длина лишь немного превышает
высоту. Седло кольцом охватывает его; при этом со спинной стороны оно значительно
длиннее, чем с брюшной. Боковой волосок простой, заметно длиннее седла. Наружные
хвостовые волоски простые, гладкие, в 1.5 раза длиннее дыхательной трубки; внутрен-
ние — из 4—5 более коротких ветвей. Плавник из 14—16 пучков, объединенных общим
основанием, и из 1—2 более коротких впереди последнего; длина неразветвпенного
основания каждого пучка в среднем в 2—3 раза превышает длину отростков у их ос-
нования, при этом длина неразветвленного основания увеличивается от передних
пучков к задним. Жабры узкопанцетовидной формы, в 1.7—2 раза превышают длину
седла, слабо пигментированные.
Распространение и биология. Взрослые описаны по материалу
из Канады (Vockeroth, 1954). В СССР найден в Якутии (Аксенова и Ануфриева, 1969),
в различных других пунктах Сибири (Барабинская степь, бассейн Колымы — мате-
риал П. Е. Поляковой и Л. П. Кухарчук), а также на Алтае в Восточно-Казахстанской
области (Дубицкий и Тупицин, 1970). Вероятно указание о нахождении А. (О.) nigri-
pes под Саратовом относится к А. (О.) rempeli.
Aedes (Ochlerotatus) kasachstanicus Guts.
Личинка IV стадии средних размеров, темно-коричневой окраски, с тем-
ной дыхательной трубкой и более светлой головой (по А. М. Дубицкому).
Голова в 1.4 раза больше в ширину, чем в длину. Лобные волоски: наружные
в среднем из 8 (6—11), средние из 3 (2—4), внутренние из 5 (4—7) ветвей с редкой вто-
ричной перистостью, задние волоски наличника между основаниями средних лобных,
из 3 (2—7) очень коротких ветвей. Теменной волосок из 2 (1—3), транссутуральный —
из 3 (2—4) тонких ветвей. Усик короткий, около половины длины головы, с телом,
покрытым редкими мелкими шипиками и с пучком из 7—8 (6—11) ветвей, отходящим
у середины усика, немного ближе к основанию.
ДОБАВЛЕНИЕ
379
Внутренние срединные волоски переднегруди относительно короткие: передний
и средний из 1—2 ветвей, задний из 2—3, как редкое исключение из 4, более длинных,
но не достигающих уровня заднего края глаз ветвей с редкой вторичной пери-
стостью.
Щетка в среднем из 31 (27—34) чешуек, расположенных в. 2—3 ряда выпуклым
назад полулунным пятном. Каждая чешуйка имеет хорошо выраженный главный
шип с рядом более тонких и коротких шипиков по бокам основания. Волоски позади
щетки: крайний верхний из 4 (3—6), средний из 6 (5—10), крайний нижний из 5 (4—7)
ветвей с хорошо развитой вторичной перистостью; промежуточные гладкие и простые.
Сифон длинный, цилиндрический в базальной трети и постепенно сужающийся к вер-
шине; индекс 3.9—4.0. Гребень состоит в среднем из 20 (16—21) зубцов, расположен-
ных на равном расстоянии друг от друга, и из 2—6 мелких рудиментарных зубчиков
у основания. Гребень занимает базальную треть сифона. Каждый зубец имеет 2—4 зуб-
чика у слегка расширенного основания. Пучок в среднем из 4 (3—5) вторичноперистых
ветвей, в длину немного превышающих ширину сифона у места его отхождения, лежит
дистальнее конца гребня на расстоянии длины зубца.
Седло последнего членика в длину вдвое больше, чем в ширину; его боковые края
заходят за середину членика. Боковой волосок простой, немного короче длины седла.
Хвостовые волоски: наружный простой, немного длиннее сифона, внутренний более
чем вдвое короче, из 9 (8—10) ветвей. Плавник из 16 (12—18) пучков, объединенных
общим основанием, и из 2—3 более коротких впереди последнего. Жабры короткие,
около трети длины седла, яйцевидной или листовидной формы.
Биология. Места выплода приурочены к весенним водоемам, возникшим
в результате разлива рек или таянья снега, преимущественно в тугайной зоне с большим
количеством опавшей листвы, со слабо минерализованной водой. Поздневесенний вид,
имеющий одно поколение.
Aedes (Ochlerotatus) simanini Guts.
Личинки IV стадии средней величины, желтовато-коричневого цвета
с более темно пигментированной головой и более светлым сифоном|(по А. М. Дубицкому).
Голова почти в 1.5 раза больше в ширину, чем в длину. Лобные волоски: наруж-
ный в среднем из 5 (3—8), средние простые, внутренние простые (у 60%) или 2-ветви-
стые (у 40%), все с едва заметной вторичной перистостью. Задние волоски наличника
между основаниями средних лобных из 2, как исключение из 3—4, коротких тонких
ветвей. Теменные и транссутуральные длиннее последних, 1- или 2-ветвистые. Усики
короткие, около 0.4 длины головы, с телом, покрытым редкими шипиками; пучок
у середины, ближе к основанию, из 4—5 (3—6) ветвей, заметно не достигающих конца
усика.
Срединные волоски переднегруди со слабой вторичной перистостью: передний и
средний простые или 2-ветвистые, задний из 2—3 более длинных, доходящих до уровня
заднего края глаз ветвей.
Щетка в среднем из 22 (16—36) чешуек, расположенных в 2, реже в 3 ряда выпук-
лым назад полулунным пятном.. Каждая чешуйка имеет хорошо выраженный главный
шип и небольшое число шипиков по бокам его основания. Основания крайних спинных
и брюшных чешуек в пятне заметно сильнее расширено, чем у остальных. Волоски
позади щетки: крайний верхний из 5 (4—7), средний из 7 (6—9), крайний нижний
из 5 (4—8) вторичноперистых ветвей; промежуточные гладкие и простые; вокруг осно-
ваний крайнего верхнего и среднего волосков имеются неправильной формы более
темные склеротизованные пластинки. Сифон (индекс около 3.4) слабо сужается к вер-
шине. Гребель в среднем из 13 (12—17) равномерно расставленных зубцов и из 2—4 мел-
ких рудиментарных зубчиков у основания: занимает около 1/6, не более 1/i базальной
части трубки. Каждый зубец умеренной длины, с более широким основанием, край
которого обычно несет один более крупный и 2—3 более мелких зубчика. Пучок из
5 (4—7) вторичноперистых ветвей, в длину немного превышающих ширину трубки
у места его отхождения, находящегося ближе к ее основанию, но заметно отстоящего
от наиболее дистального зубца гребня.
Последний членик в 1.5 раза сильнее развит в длину, чем в ширину. Боковые края
седла у середины членика и только в передней его половине заходят за нее. Боковой
волосок простой, редко 2-ветвистый, не более половины длины седла. Хвостовые во-
лоски: наружные простые, в среднем одной длины с дыхательной трубкой, внутренние
вдвое короче, из 5 (4—8) ветвей. Плавник из 17 (14—18) пучков, окруженных общим
основанием, и из 1, редко 2 более коротких пучков впереди него. Жабры короткие,
листовидные, не более 14 длины седла.
Биология. Личинки развиваются в различных временных неглубоких во-
доемах, сильно загрязнейных органическими остатками, различной степени минера-
лизации, открытых и затененных, в зоне тугаев и вблизи населенных пунктов. Личинки
и куколки экологически очень пластичны. Выдерживают промораживание до —5, —7е
в течение 3—4 дней; при повышении температуры до 4—5° продолжают развитие.
Ранневесенний вид, имеющий 1 поколение в течение года. Личинки встречались с конца
380
ДОБАВЛЕНИЕ
2-й декады марта, комары — с середины—конца 1-й декады апреля; лет и нападение
заканчивались к концу апреля.
ЛИТЕРАТУРА
Аксенова А. С. иАнуфриева В. Н. 1969. Фауна и некоторые вопросы биоло-
гии гнуса в районе алмазных разработок в Якутской АССР. Мед. паразитол. и
паразитарн. бол., 47, 1 : 8—16.
Дубицкий А. М. 1968 Новый вид комара Aedes montchadskyi sp. п. из бассейна
Или. Паразитология, 2, 3 : 218—224.
Дубицкий А. М. 1970. Новый вид комара Aedes stramineus sp. n. (Diptera, Culi-
cidae). Паразитология, 4, 5 : 408—413.
Дубицкий А. М.иТ у пици нЮ. H. 1970. Описание личинок нового для фауны
СССР комара Aedes (О.) rempeli Vockeroth. Паразитология, 4, 2 : 171—174.
Полякова П. Е. 1970. Новый для фауны СССР вид комара (Diptera, Culicidae)
Aedes fitchii Felt and Young. Паразитология, 4, 4 : 389—391.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ КОМАРОВ 1
aconitus, Anopheles 49
Aedes 9, 10, 11*, 12*, 14*, 15, 16*, 17,
18, 23, 24*, 25, 33, 34, 39, 43, 44, 47,
48, 51, 54, 57, 59, 72, 74, 75, 169—314
Aedes (subgenus) 179, 183, 308
Aedimorphus (subgenus) 43, 44, 171, 270
aegypti, Aedes 9, 39, 48, 51, 53—56, 176,
183, 185, 294, 295*, 296*, 297*
africanus, Aedes 54
alaskaensis, Culiseta 46, 135—137, 144,
145*, 146*, 147*
alaskaensis alaskaensis 135, 147
alaskaensis indica 135, 147
albescens, Aedes 229
albimanus, Anopheles 50
albionensis, Orthopodomyia 130
albitarsis, Anopheles 50
albopictus, Aedes 53—55, 176, 183, 185,
303*, 304*
alektorovi, Aedes 19, 46, 170, 183, 185,
284*, 286*
algeriensis, Anopheles 45, 50, 77, 78, 79,
80, 81*, 82*
Allotheobaldia (subgenus) 134, 136, 138
alpinus, Aedes 255
annulata, Culiseta 25*, 26*, 135—137,
147, 148*, 149*
annulata annulata 135, 150
annulata subochrea 135, 137, 150
annulipes, Aedes 176, 182, 188, 216, 217*
annulirostris, Culex 54
Anopheles 9, 10, 15*, 16*, 17, 18*, 19,
20, 21*, 22*, 23—25, 26*, 27, 29*,
30*, 31*, 32, 34*, 35*, 36—40, 42—
44, 49—52, 54, 57, 59, 61, 62, 72,
74, 76—117
Anopheles (subgenus) 76, 78, 80
Anophelinae 18, 42, 43, 75
antennatus, Culex 54
apicalis, Culex 331
aquasalis, Anopheles 50
Armigeres 44, 54
aureus, Aedes 45, 179, 313
argenteus, . Aedes 294
balabacensis, Anopheles 51
bancrofti, Anopheles 53
barbirostris, Anopheles 49, 56
Barraudius (subgenus) 325
behningi, Aedes 173, 182, 187, 208, 210*,
211*
beklemishevi, Aedes 176, 182, 187, 214,
215*
bergrothi, Culiseta 134, 136, 137, 151*,
152*
bifurcatus, Anopheles 86
bitaeniorhynchus, Culex 25, 45, 56, 316,
318, 319, 341*, 342*, 343*
borealis, Culiseta 151
buxtoni, Mansonia 163, 164, 167, 168*
calopus, Aedes 294
cantans, Aedes 56, 175, 181, 188, 202,
203*, 204*, 205*
caspius, Aedes 56, 57, 170, 180, 188, 191
caspius caspius 170, 180, 188, 190*, 192,
193*
caspius dorsalis 170, 180, 188, 191, 195*,
196*
cataphylla, Aedes 178, 181, 186, 261*,
262*
Chagasia (subgenus) 43
Chaoboridae 43
chemulpoensis, Aedes 176, 183, 185, 306*
307*
chloropterus, Sabethes 54
christophi, Toxorhynchites 45, 118, 119*,
120, 121*, 122*
cinereus, Aedes 19*, 34, 56, 57, 179, 183,
308, 310*
cinereus cinereus 179, 183, 184, 309*,
310*
cinereus esoensis 179, 183, 184, 312*
cinereus rossicus 179, 183, 184, 308*, 311*
claviger, Anopheles 35, 50, 53, 59, 60,
77—79, 85*, 86*, 87*
communis, Aedes 56, 178, 181, 188,
232, 233, 234*
Coquillettidia (subgenus) 163
cretinus, Aedes 176, 183, 185, 298*
Culex 9, 12*, 13*, 15*, 16*, 17—19,
21—25, 33, 34, 37, 43—45, 48, 51, 52,
54,-57, 59, 73-75, 314—374
Culicella (subgenus) 17, 34, 37, 59, 153
Culicidae 9, 13, 24, 25, 38, 42—46, 71
culicifacies, Anopheles 49
Culicinae 9, 10, 17, 18, 24*, 30, 32*,
34—36, 39, 40, 42, 61, 62, 125
Culicini 32, 125, 126
Culiseta 11*, 15, 17—19, 23—25, 28,
33, 34, 44, 57, 59, 72, 74, 75, 133—162
cyprius, Aedes 20, 172, 182, 187, 221,
222*
darlingi, Anopheles 50
Deinocerites 43
1 Полужирным шрифтом набраны страницы с описанием вида, курсивом выде-
лены синонимы, а звездочка при цифре обозначает, что на этой странице помещен
рисунок данной формы.
382
УКАЗАТЕЛЬ
detritus, Aedes 38, 177, 182, 188, 264,
265*, 266*, 267*
diantaeus, Aedes 23, 56, 178, 180, 184,
247*, 248*, 249*
diversus, Aedes 223
Dixidae 43
dup lex, Aedes 192
dureni, Anopheles 51
echinus, Aedes 19, 176, 183, 186, 278,
279*, 280*
Eretmapodites 43
excrucians, Aedes 57, 60, 176, 182, 187,
211, 212*, 213*, 214*
exilis, Culex 362, 364
farauti, Anopheles 49, 53
fasciatus, Aedes 294
Ficalbia 43
Finlaya (subgenus) 16, 17, 23, 44, 170,
179, 274
fitchii, Aedes 377
flavescens, Aedes 57, 172, 181, 187, 218,
219*, 220*
flavopictus, Aedes 45, 176, 183, 185,
305*
fluviatilis, Anopheles 49
freeborni, Anopheles 50, 51
freyi, Aedes 221
fumipennis, Culiseta 135—137, 159*
funestus, Anopheles 50, 54, 56
fuscanus, Culex 45, 315, 317, 318, 322*,
323*, 324*
galloisi, Aedes 19, 38, 39, 176, 183, 185,
300, 301*, 302*
gambiae, Anopheles 50, 53, 54, 56
gelidus, Culex 54—56, 320
geniculatus, Aedes 19, 20, 45, 176, 183,
186, 274, 275*, 276*, 277*
glaphyroptera, . Culiseta 135—137, 142,
143*
grandilarva, Aedes 214
hackeri, Anopheles 51
hayashii, Culex 45, 316, 317, 319, 336,
337*, 338*, 339*
Heizmannia 43
hexodontus, Aedes 178, 181, 189, 240,
241*, 242*
hortensis, Culex 20, 316, 317, 320, 333*,
334*
hungaricus, Aedes 179, 189, 246*
hyrcanus, Anopheles 35, 36, 49—51, 53,
78, 79, 103, 104*, 105*, 106*
impiger, Aedes 177, 181, 189, 258*, 259*,
- 260*
intrudens, Aedes 178, 180, 189, 250*,
251*
jacksoni, Culex 45, 316, 318, 319, 350
japonicus, Aedes 170, 183, 184, 290
jeyporiensis, Anopheles 49
kasachstanicus, Aedes 170, 182, 269*, 378
Kerteszia (subgenus) ,43
kochi, Anopheles 51
koreicus, Aedes 171, 183, 184, 291*, 292*
laticinctus, Culex 320
laurenti, Culex 321
lepidonotus, Aedes 47, 177, 180, 186,
227, 228*
letifer, Anopheles 49, 51
leucocelaenus, Aedes 54
leucomelas, Aedes 178, 181, 188, 263*
leucosphyrus, Anopheles 49, 51
Limatus 43
lindesayi, Anopheles 31, 50, 77, 78,
100, 101*, 102*
lindesayi lindesayi 77, 103
lindesayi japonicus 77, 79, 103
lindtropi, Aedes 298
longiareolata, Culiseta 22, 34, 45, 134,
136, 138, 139*, 140*, 141*, 142*
Lophoceraomyia 43
Lophopodomyia 43
lutescens, Aedes 218
Lutzia (subgenus) 25, 29, 321
maculatus, Aedes 202
maculatus, Anopheles 49, 51
maculipennis, Anopheles 11, 18, 35, 36,
40*, 49, 50, 53, 56, 77—79, 90, 91*,
94*, 95*, 96*, 97, 106*
maculipennis maculipennis 77, 90, 96*
maculipennis atroparvus 77, 90, 96*
maculipennis elutus 99
maculipennis labranchiae 77, 90, 96*
maculipennis melanoon 77, 90, 96*
maculipennis messeae 77, 90, 96*
maculipennis sacharovi 77, 79, 90, 98,
99, 100*
maculipennis subalpinus 90, 96*
Mansonia 13, 17—19, 21, 23—25, 37,
43, 53, 54, 73—75, 162—169
mariae, Aedes 170, 180, 187, 196, 197*,
198*
marteri, Anopheles 50, 77, 79, 88, 89*
marteri marteri 77, 90
marteri sogdianus 77, 90
martinii, Culex 316, 317, 320, 335*, 336*
Megarhinus (subgenus) 76, 118
meigenanus, Aedes 237
melanimon, Aedes 54
Melanoconion (subgenus) 43
melanura, Culiseta 54, 55
mimeticus, Culex 316, 318, 320, 348,
349*, 350*
minimus, Anopheles 49, 53
modestus, Culex 56, 315, 317, 320, 325,
326*
montchadskyi, Aedes 375
morsitans, Culiseta 24*, 25*, 26*, 60,
135, 136, 138, 153, 154*, 155*
multicolor, Anopheles 50
Myzomyia (subgenus) 31, 43, 44, 108
nearcticus, Aedes 258
nemorosus, Aedes 232
Neoculex (subgenus) 37, 328
nigerrimus, Anopheles 53
nigrinus, Aedes 178, 181, 187, 245*
УКАЗАТЕЛЬ
383
nigripes, Aedes 177, 181, 186, 255, 256*,
257*
nigripes, Anopheles 82
nili, Anopheles 50
nipponicus, Aedes 45, 176, 182, 184,
280, 281*, 282*
niveus, Aedes 280
nobukonis, Aedes 45, 179, 313
Nyssorhynchus (subgenus) 43
Ochlerotatus (subgenus) 16, 19, 23, 28,
44—46, 170, 179
ochroptera, Culiseta 135, 136, 138, 155*,
156, 157*, 158*
Opifex 44
orientalis, Culex 315, 318, 320, 352,
\ 353*, 354*, 355*
<ornatus, Aedes 274
Orthopodomyia 19, 20, 33, 34, 37, 44,
46, 73—75, 130—133
parvulus, Aedes 258
pattoni, Anopheles 49
perexiguus, Culex 358
perturbans, Mansonia 54
petragnani, Anopheles 88
pharoensis, Anopheles 50
philippinensis, Anopheles 49, 51
Phoniomyia 43
pionips, Aedes 178, 181, 188, 235*, 236
pipiens, Culex 24, 46, 54, 317, 318, 365,
366*, 367*
pipiens pipiens 52, 318, 321, 368*, 370
pipiens fatigans 52—54, 318, 320, 373*
pipiens molestus 318, 321, 372*
pipiens quinquefasciatus 369, 373
plumbeus, Anopheles 23, 25, 27, 35,
50, 77—79, 82, 83*
polynesiensis, Aedes 52
pseudopunctipennis, Anopheles 50
Psorophora 43
pulcherrimus, Anopheles 35, 49—51, 78,
80, 108, 110*, 111*, 112*
pulchripalpis, Orthopodomyia 19, 46, 130,
131* 132* 133*
pulchritarsis, Aedes 38, 40, 170, 180, 199*
pulchritarsis pulchritarsis 170, 186, 200
pulchritarsis asiaticus 170, 186, 201*, 202*
pulchritarsis berlandi 201
pullatus, Aedes 46, 47, 178, 180, 189,
252 253* 254*
punctor, Aedes 178, 181, 189, 237, 238*,
' 239*
punctulatus, Anopheles 49
pusillus, Culex 315, 317, 319, 327, 328*,
329*
quadrimaculatus, Anopheles 50, 51
quartus, Aedes 216
refiki, Aedes 177, 180, 186, 225,226*
rempeli, Aedes 378
restuans, Culex 54
richiardii, Mansonia 57, 59, 163, 164*,
165*, 166*
riparius, Aedes 40, 175, 182, 187, 205,
207*
riparius riparius 175, 182, 206*, 208
riparius ater 175, 182, 208, 209*
rostochiensis, Aedes 261
rufipes, Anopheles 50
rusticus, Aedes 177, 180, 186, 223, 224*,
225*
Sabethes 43
Sabethini 43, 125, 126
salinellus, Aedes 263
salinus, Aedes 264
semicantans, Aedes 205
seoulensis, Aedes 170, 182, 185, 283
sergenti, Anopheles 50
setivalva, Culiseta, 46, 135, 136, 137,
160, 161*
Silvestris, Culiseta 47, 158
simanini, Aedes 177, 182, 267, 268*, 379
simpsoni, Aedes 54, 55
sinensis, Anopheles 103
sinensis, Culex 25, 45, 316, 318, 319,
344*, 345*
sollicitans, Aedes 54
Stegomyia (subgenus) 15, 17, 23, 33, 43,
44, 54, 55, 176, 179, 293
stephensi, Anopheles 49—51
Stethomyia 43
sticticus, Aedes 56, 178, 181, , 189, 243*,
244*
stramineus, Aedes 376
subdiversus, Aedes .177, 180, 186, 230*
subpictus, Anopheles 56
sundaicus, Anopheles 49, 51
superpictus, Anopheles 35, 49, 50, 60,
78, 80, 113*, 114*, 115*, 116*
tarnogradskyi, Aedes 311
tarsalis, Culex 54, 55
territans, Culex 19, 33, 43, 52, 317, 320,
330*, 331*
territans territans 317, 332
territans judaicus 317, 333
tesselatus, Anopheles 56
theileri, Culex 45, 316, 318, 320, 360,
361*
Theobaldia 72, 133
titillans, Mansonia 54
togoi, Aedes 53, 54, 170, 182, 184, 287*,
288*, 289*
Topomyia 43
torrentium, Culex 317, 318, 321, 364
Toxorhynchites 9, 18, 19—25, 29, 30,
33, 34, 37, 38, 44, 72, 75, 118-125
Toxorhynchitinae 117
Toxorhynchitini 32
Trichoprosopon 43
trichurus, Aedes 46
Tripteroides 44
tritaeniorhynchus, Culex 45, 54, 55,
316, 318, 321, 354, 356*, 357*
turkhudi, Anopheles 31
umbrosus, Anopheles 49
unguiculata, Uranotaenia 126, 127*, 128*,
129*
univittatus, Culex 54, 317, 318, 321,
358*, 359*
384
УКАЗАТЕЛЬ
Uranotaenia 9, 17, 18, 20, 22, 24, 27,
31, 33, 43, 44, 72, 74, 75, 126—130
vagans, Culex 316, 318, 321, 362*, 363*
vexans, Aedes 46, 53, 54, 56, 57, 171,
179, 189, 270, 271*, 272*
vexans vexans 171, 273
vexans nipponii 171, 273
vishnui, Culex 54, 319
vittatus, Aedes 176, 183, 185, 299*, 300*
vorax, Culex 45, 315, 317, 318, 324*
whitmorei, Culex 45, 316, 318, 319,
345, 346*, 347*
Wyeomyia 43
СОДЕРЖАНИЕ
Стр.
Предисловие ....................................................... 3
Систематический указатель видов ................................... 4
Введение .......................................................... 7
Краткий морфологический очерк...................................... 7
Взрослое насекомое .......................................... 7
Яйцо......................................................... 17
Личинка ..................................................... 18
Куколка...................................................... 41
Географическое распространение ................................... 42
Значение комаров для человека..................................... 47
Комары как кровососы ........................................ 47
Комары как переносчики возбудителей болезней................. 48
Краткие сведения о методике сбора, монтировки и сохранения коллекционных
материалов по комарам....................................... 57
Литература ....................................................... 62
Специальнаячасть.................................................. 71
Семейство Culicidae.............................................. 71
Определительная таблица родов.................................... 72
I. Подсемейство Anophelinae....................................... 75
1. Род Anopheles ............................................ 76
II. Подсемейство Т oxorhynchitinae .............................. 117
2. Род Toxorhynchites....................................... 118
III. Подсемейство Culicinae.................................... 125
3. Род Uranotaenia......................................... 126
4. » Orthopodomyia........................ 130
5. » Culiseta............................................. 133
6. » Mansonia............................................. 162
7. » Aedes................................................ 169
8. » Culex.............................................. 314
Добавление....................................................... 375
Указатель латинских названий комаров............................. 381